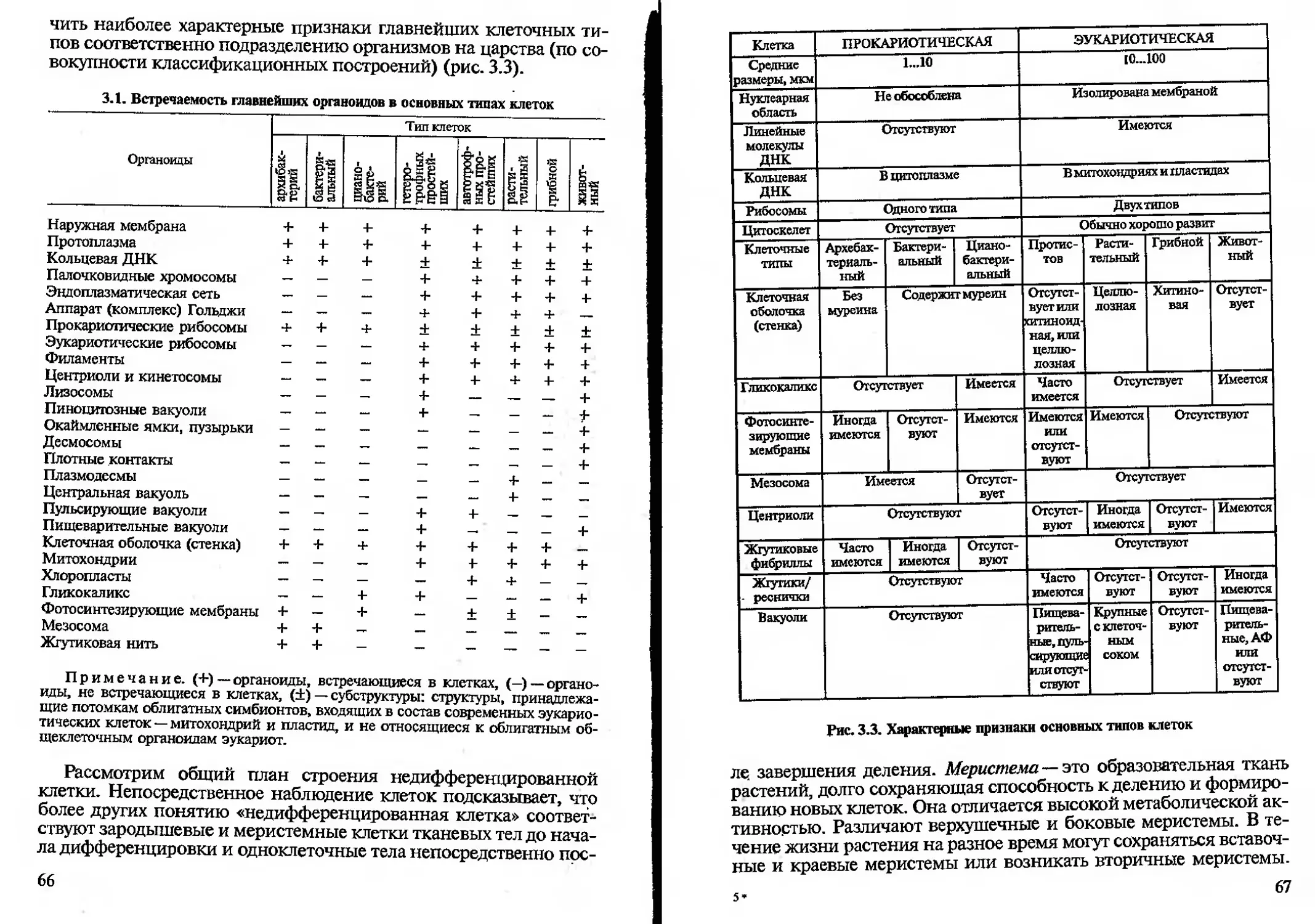
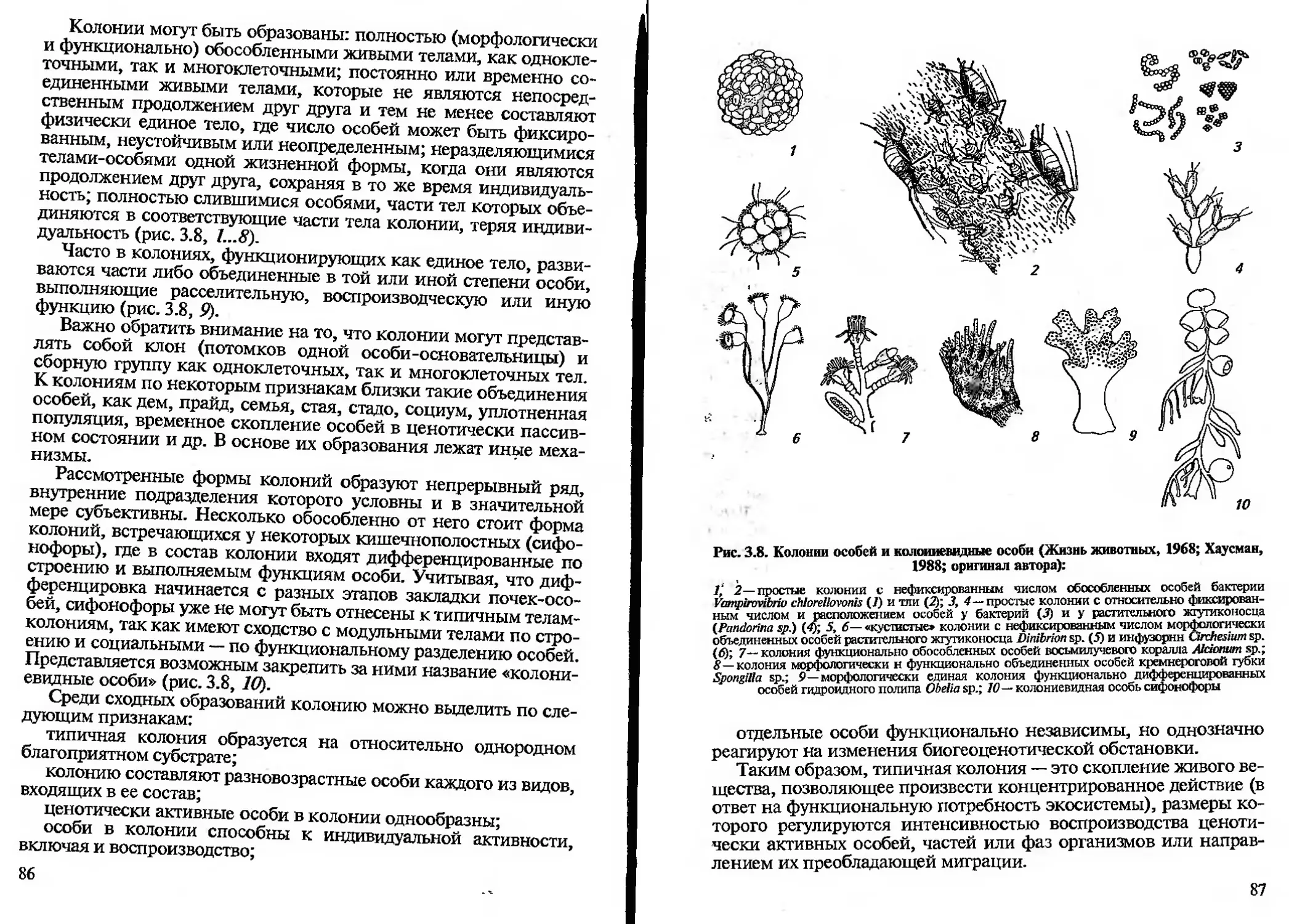
/








































































































































































Автор: Присный А.В.
Теги: общая экология биоценология гидробиология биогеография общая биология
ISBN: 978-5-9532-0669-3
Год: 2009




Текст
КЛАССИЧЕСКОЕ УНИВЕРСИТЕТСКОЕ ОБРАЗОВАНИЕ
А. В. ПРИСНЫМ
ОБЩАЯ БИОЛОГИЯ
ДУАЛИСТИЧЕСКАЯ
И МАТЕРИАЛИСТИЧЕСКАЯ
КОНЦЕПЦИИ ЖИЗНИ
НА ЗЕМЛЕ
«КолосС»
УДК 574/577(075.8)
ББК 28.0я73
П77
Редактор М. И. Толмачева
Рецензенты: доктор педагогических наук, профессор
И. Н. Пономарева (РПГУ им. А. И. Герцена), доктор биологичес-
ких наук, профессор Л. Н. Хицова (ВГУ)
На обложке: фрагмент картины М. К.Чюрлениса из цикла
«Сотворение мира»
Присный А. В. Общая биология. Дуалистическая и мате-
П77 риалистическая концепции жизни на Земле. — М.: КолосС,
2009. - 351 с.: ил.
ISBN 978-5-9532-0669-3
Изложены современные представления о сущности жизни и ее прояв-
лении в условиях Земли в виде живой системы — биосферы. Рассмотрены
особенности вещественных, энергетических и информационных процессов
на уровнях функционирования биосферы. Особое внимание уделено опи-
санию живых тел. Приведены основные гипотезы и теории, касающиеся
происхождения и эволюции земной формы жизни. С системных позиций
даны представления о человеке. Значительная часть материала излагается в
авторской трактовке.
Для студентов университетов, обучающихся по специальности 020201
«Биология» и направлению 020200 «Биология», а также для студентов ву-
зов, обучающихся по естественно-научным специальностям.
УДК 574/577(075.8)
ББК 28.0я73
Оригинал-макет книги является собственностью издательства «КолосС»,
и его воспроизведение в любом виде, включая электронный,
без согласия издателя запрещено.
ISBN 978-5-9532-0669-3
© Издательство «КолосС», 2009
Глава 1
ВВЕДЕНИЕ В БИОЛОГИЮ
Современную биологию можно определить как отрасль зна-
ния, как направление науки, как сферу деятельности людей. И
хотя эти три аспекта биологии взаимно проникают друг в друга,
между ними существуют и некоторые различия.
Биология как наука предполагает продуктивную деятель-
ность, то есть получение новых знаний (добычу новых фактов,
построение логических конструкций), запись их в форме, при-
годной для хранения и воспроизводства. Здесь взаимодействуют
изучающий субъект — ученый-биолог и объект изучения — жи-
вая природа.
Биология как отрасль знания предполагает прежде всего реп-
родуктивную деятельность, то есть передачу знаний (установ-
ленных фактов и гипотез) «по горизонтали» —- распростране-
ние, передачу их среди одновременно живущих людей и «по
вертикали» — от поколения к поколению. Эта отрасль требует
обязательного наличия Учителя и Ученика, где учитель являет-
ся связующим звеном между биологией-наукой и людьми, ис-
пользующими получаемые знания в своей практической дея-
тельности.
Биология как сфера деятельности одновременно и входит в
Предыдущие аспекты, и обособлена от них. В этом случае на пер-
вый план выступает человек, занимающий определенное функци-
ональное положение в структуре общества: в науке, в образова-
нии, в производстве и пр. Функциональность же определяется
формой (биосоциально зрелым человеком, связанным с набором
специализированных усилителей телесных органов — орудий тру-
де), способной реализовать заданную функцию, и дефицитом
функции, то есть существующей в обществе потребностью реали-
8ВЦИИ данной функции.
Нам предстоит усвоить некоторую сумму выработанных чело-
Мчеством биологических знаний, и, следовательно, мы имеем
ДМО со вторым аспектом биологии.
3
1.1. ПРЕДМЕТ И ЗАДАЧИ БИОЛОГИИ
Биология, как и любая наука, имеет свой предмет, использу-
ет характерные для нее методы и решает свои задачи. Традици-
онно считается, что биология — это комплексная наука, вклю-
чающая множество частных наук, сгруппированных в разделы.
Буквальный перевод ее названия — наука о живом (bios + logos).
В соответствии с названием в качестве предмета биологии долж-
на рассматриваться жизнь во всем ее многообразии, то есть яв-
ление жизни, живые системы как его материальное воплоще-
ние, живое вещество — активная часть живых систем, живые
тела — элементарные подразделения видовых форм живого ве-
щества, процессы, идущие в живом веществе на разных уровнях
его организации. Более широко известна иная формулировка:
биология изучает живые организмы, их многообразие, идущие в
них жизненные процессы, индивидуальное развитие организ-
мов и историческое развитие органического мира. Как можно
видеть, в ней предмет биологии не только сокращен в объеме,
но и упрощен. Человек, на уровне его биологической составля-
ющей, также относится к объектам, изучаемым биологией. Со-
циальная составляющая человека изучается комплексом гума-
нитарных наук.
Применительно к живым объектам биология изучает их внут-
ренние характеристики и отношения, в то время как выделившая-
ся в самостоятельную науку экология изучает их внешние отноше-
ния.
Задачи, решаемые биологией, многочисленны и разноплано-
вы, но в целом они состоят в познании жизни во всех ее прояв-
лениях для выработки таких форм отношений человека с ос-
тальной биосферой, которые позволяли бы ему неопределенно
долго и эффективно использовать ресурсы биосферы и разви-
ваться вместе с ней, то есть эффективно реализовать видоспе-
цифичную биосферную функцию. В приведенную формулиров-
ку заложена бесконечность (все проявления), делающая бес-
конечным и процесс их познания. Разумеется, можно опреде-
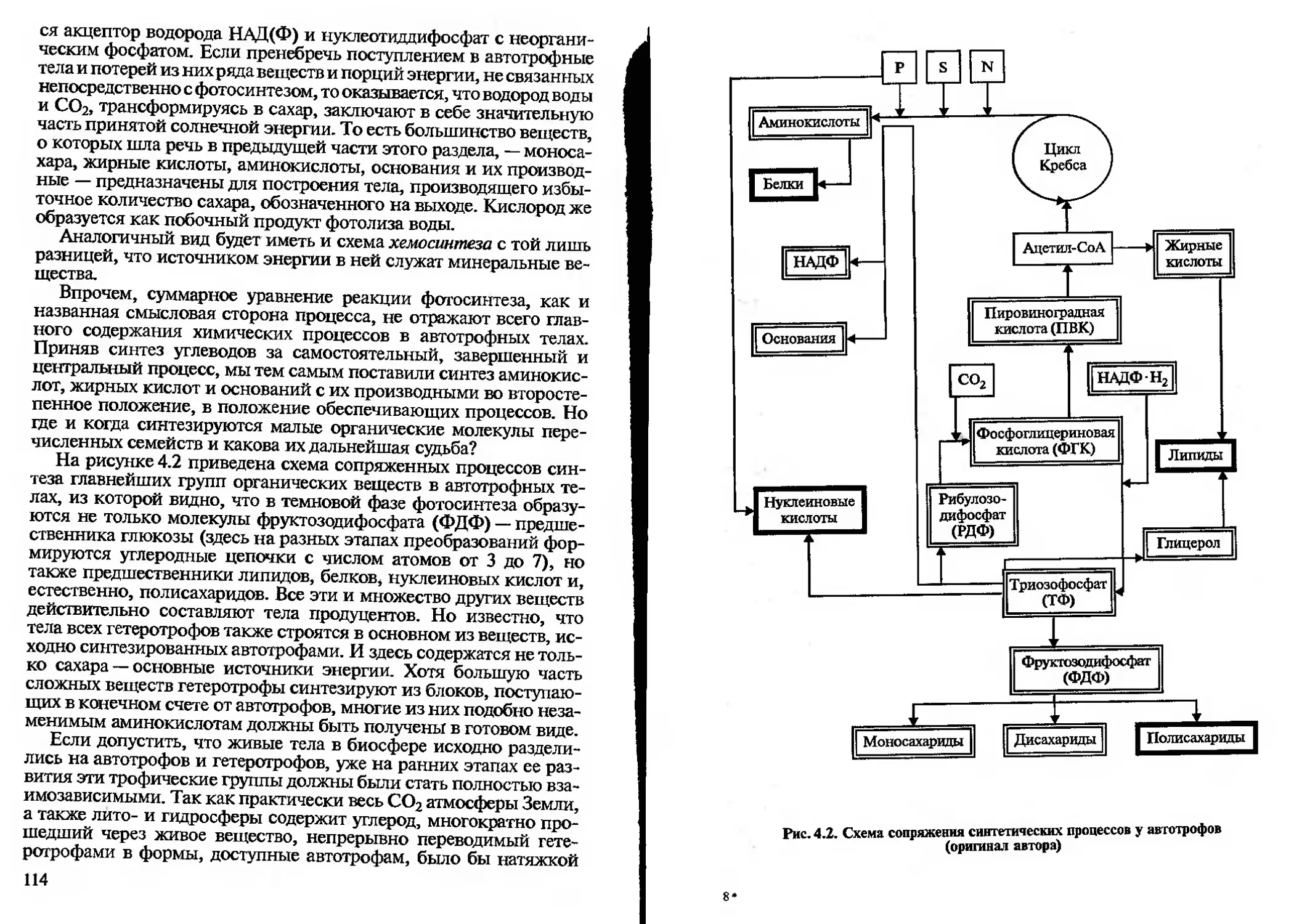
лить, обозначить и конкретные задачи биологии, но число
таких задач будет произвольно определяться их емкостью и ха-
рактером.
1.2. МЕТОДЫ БИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Биология сочетает в себе признаки и эмпирической (идущей от
опыта), и теоретической науки. Как эмпирическая наука «есте-
ственной» группы биология пользуется ее классическими метода-
4
ми — наблюдением, экспериментом, сравнением (в том числе из-
мерением), а как теоретическая — анализом, синтезом, индукцией
(восхождением от частного к общему) и дедукцией (восхождением
от общего к частному).
В современной биологии наблюдение и эксперимент, если
следовать утвердившимся определениям этих методов, факти-
чески неразделимы. Так, наблюдение исходно предполагает по-
лучение информации об объекте без воздействия на него со сто-
роны исследователя, а эксперимент предполагает получение
информации об объекте хотя бы частично в измененных и кон-
тролируемых экспериментатором условиях. Но наблюдение,
как правило, сопутствует эксперименту, а объект наблюдения
часто требует специальной обработки для облегчения наблюде-
ния.
Наблюдение в силу своей уникальности и непременной субъек-
тивности наблюдателя поставляет менее точную информацию об
объекте. Связано это с тем, что органы чувств отфильтровывают с
индивидуальной чувствительностью из всего диапазона внешних
раздражителей лишь значимую для существования вида часть ин-
формации. Она дешевле энергетически, а потому с помощью на-
блюдения можно получить большее количество, хотя и менее цен-
ной, информации.
Основное условие эксперимента — его повторимость. Изуче-
ние объекта в контролируемых условиях позволяет разным ис-
следователям в разных местах и в разное время, переводя с помо-
щью стандартного оборудования снимаемые параметры в унифи-
цированную форму, в конечном счете получить сведения, не за-
висящие или мало зависящие от индивидуальных особенностей
органов чувств самих исследователей. Следовательно, получае-
мая информация здесь точнее, объективнее, но дороже энергети-
чески.
Сравнение — способ оценки отдельных параметров изучаемого
объекта путем сопоставления их с известными характеристиками
другого объекта. Такими известными характеристиками обладают
и специальные измерительные инструменты, позволяющие про-
изводить более или менее точную количественную оценку изучае-
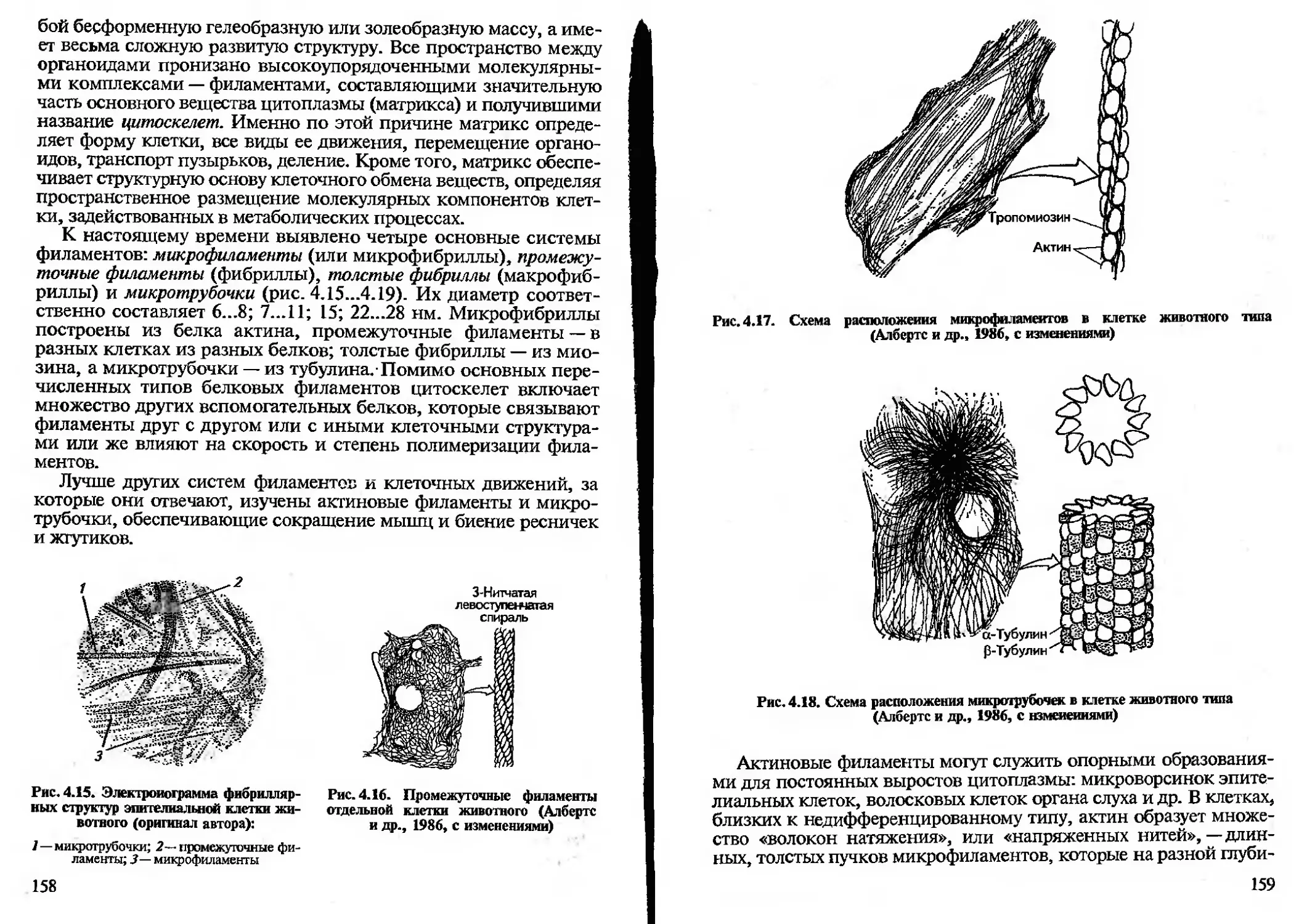
мого объекта.
В учебной деятельности используют все названные методы как
в плане получения навыков их применения, так и в качестве спо-
собов получения знания.
И наблюдение, и эксперимент, и сравнение могут проводиться
как в естественных или приближенных к ним, так и в искусствен-
но созданных условиях. Соответственно их разделяют на полевые
и лабораторные. Эксперимент, кроме того, может быть мыслен-
ным, и эту его форму называют «моделирование».
5
Общие методы, как уже отмечалось, едины для всего комп-
лекса естественных наук, но частные методы, или методики, ис-
пользующиеся в биологическом исследовании, весьма специ-
фичны.
В большинстве случаев научное исследование проходит не-
сколько стандартных этапов: формулирование рабочей гипотезы,
наблюдение (предварительный сбор данных), постановку экспе-
риментов, проверку и корректировку гипотезы, постановку кон-
трольных экспериментов, обобщение результатов, описание
факта.
Накапливаемые факты сами по себе обычно не имеют высокой
информационной ценности, но, будучи организованными в ка-
кую-либо теоретическую конструкцию (гипотезу, теорию, кон-
цепцию), они могут иметь то ли мировоззренческое значение, то
ли хозяйственно-практическое, а то и послужить руководством к
непосредственному действию. Даже один и тот же факт может
быть истолкован по-разному или служить аргументом в противо-
поставляемых теориях.
Говоря о фактах или теоретических конструкциях вроде гипо-
тезы, теории, следует всегда ясно представлять относительность
их объективности, истинности. До сих пор человечество не приду-
мало другого критерия для оценки истинности факта, кроме его
повторимости (или воспроизводимости), а для оценки истинности
гипотезы — кроме практики. А это означает, что не существует аб-
солютной истины, есть лишь некий объем динамичной, все более
увеличивающейся и упорядочивающейся информации. На каж-
дый момент времени эта информация уже хоть незначительно, но
обновлена, и потому то, что «вчера» было чуть ли ни истиной, «се-
годня» может рассматриваться как заблуждение. Из сказанного
следует, что нет особого смысла в делении логических конструк-
ций на гипотезы и теории. (Гипотеза — логическая конструкция,
имеющая характер предположения, а теория — гипотеза, подтверж-
денная практикой.) Правильнее говорить лишь о гипотезах, а уж
гипотезы могут быть более или менее аргументированными. Ви-
димо, ни одно теоретическое положение в биологии не может
быть ни доказано, ни опровергнуто потому, что мы имеем дело (в
логических конструкциях) не с самими явлениями, а с нашими
представлениями об этих явлениях. Представления же формиру-
ются на основе показаний органов чувств. Ну а органы чувств,
как известно, лишь фильтруют раздражения, производимые яв-
лениями. Комплекс органов чувств поставляет нам только часть
информации о явлениях. Тем не менее любое живое тело (вклю-
чая человека) получает преимущественно ту часть информации,
которая позволяет ему действовать в определенном смысле целе-
сообразно.
6
Именно поэтому мы можем говорить о значении биологичес-
ких знаний, биологического поиска для практической деятельнос-
ти человека. Наиболее тесно биология связана с теми сферами де-
ятельности человека, которые непосредственно имеют дело с жи-
вой природой, а именно: с производством продуктов питания и
биологического сырья; с взаимоотношениями человека и других
живых тел. К таким сферам относятся сельское, лесное и охотни-
чье хозяйство, звероводство, рыбоводство, промысел биоресурсов,
биотехнологическая промышленность, медицина, прикладная
экология (в том числе охрана природы), повседневный быт и не-
которые другие.
1.3. СИСТЕМА БИОЛОГИЧЕСКИХ НАУК
Биология — наука о живом — чрезвычайно многогранная на-
ука. Ее комплексный характер определяется, с одной стороны,
многообразием проявлений жизни, а с другой стороны, разнооб-
разием и многоплановостью практического приложения биологи-
ческих знаний.
Возникнув как частное направление естествознания, биоло-
гия сначала существует в виде двух классических подразделений:
зоологии и ботаники, каждое из которых включает весьма емкие
аспекты — систематику, морфологию, физиологию и позже эко-
логию, историческое и индивидуальное развитие. Колоссальный
объем описаний живых организмов уже в прошлом столетии
привел к обособлению, выделению из ботаники и зоологии част-
ных наук, предметом которых являются отдельные систематичес-
кие группы растений и животных. Такие частные науки опять-
таки носят комплексный характер, поскольку включают и систе-
матический, и морфологический, и физиологический, и другие
аспекты.
Исторически очень рано обособляется от общего ствола биоло-
гии наука о человеке — антропология — с частным подразделени-
ем — учением о строении и функциях человеческого тела — анато-
мией и физиологией человека, развитие которой стимулируется
прежде всего потребностями медицины.
Деление биологии на частные науки возможно и в ином ра-
курсе: на систематику, морфологию, физиологию с этологией,
биологию развития с генетикой, историческое развитие с пале-
онтологией и экологию. В рамках каждого из направлений мож-
но выделить частные разделы по систематическим группам: сис-
тематика водорослей, физиология насекомых, поведение живот-
ных и т. д.
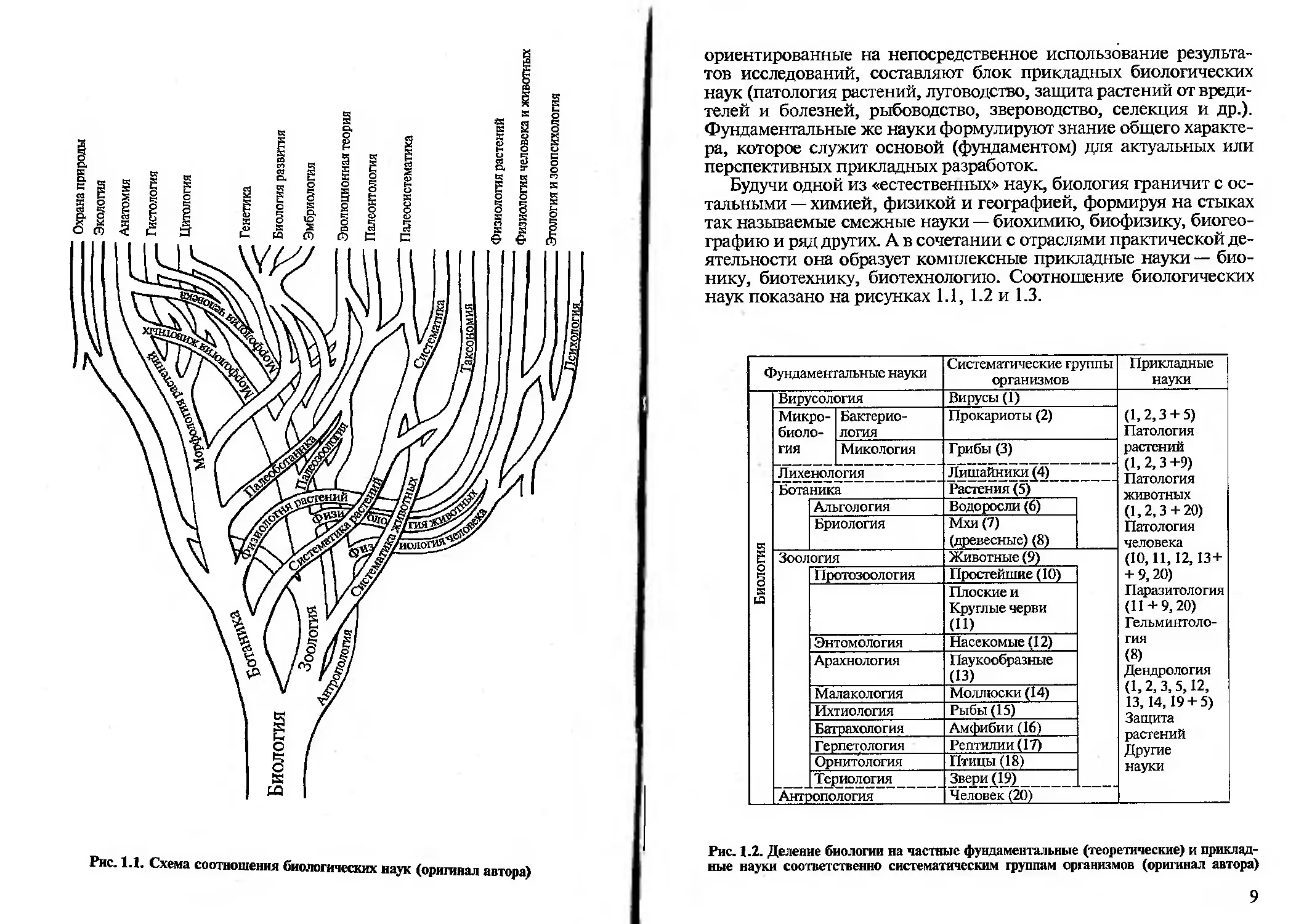
Отдельные отрасли биологических знаний или их комплексы,
7
Охрана природы
Экология
Анатомия
Рис. 1.1. Схема соотношения биологических наук (оригинал автора)
ориентированные на непосредственное использование результа-
тов исследований, составляют блок прикладных биологических
наук (патология растений, луговодство, защита растений от вреди-
телей и болезней, рыбоводство, звероводство, селекция и др.).
Фундаментальные же науки формулируют знание общего характе-
ра, которое служит основой (фундаментом) для актуальных или
перспективных прикладных разработок.
Будучи одной из «естественных» наук, биология граничит с ос-
тальными — химией, физикой и географией, формируя на стыках
так называемые смежные науки — биохимию, биофизику, биогео-
графию и ряд других. А в сочетании с отраслями практической де-
ятельности она образует комплексные прикладные науки — био-
нику, биотехнику, биотехнологию. Соотношение биологических
наук показано на рисунках 1.1, 1.2 и 1.3.
Фундаментальные науки Систематические группы организмов Прикладные науки
I Биология I Вирусология Вирусы (1) (1,2,3 + 5) Патология растении (1,2,3+9) Патология животных (1,2,3+20) Патология человека (10,11,12,13+ + 9,20) Паразитология (11 + 9,20) Гельминтоло- гия (8) Дендрология (1,2, 3,5,12, 13,14,19 + 5) Защита растений Другие науки
Микро- биоло- Бактерио- логия Прокариоты (2)
ГИЯ Лихе Микология Грибы (3)
нология Лишайники (4)
Бота ника Растения (5)
Альгология Водоросли (6)
Бриология Мхи (7) (древесные) (8)
Зоол огня Животные (9)
Протозоология Простейшие (10)
Плоские и Круглые черви (И)
Энтомология Насекомые (12)
Арахнология Паукообразные (13)
Малакология Моллюски (14)
Ихтиология Рыбы (15)
Батрахология Амфибии (16)
Герпетология Рептилии (17)
Орнитология Птицы (18)
Териология Звери (19)
Антропология Человек (20)
Рис. 1.2. Деление биологии на частные фундаментальные (теоретические) и приклад-
ные науки соответственно систематическим группам организмов (оригинал автора)
9
1\tv \
а\ '''W
Ч\ 7'""
* \Ч, / йго. , ..._
^/Лй1я ! куля?^
ТехноЛОг
"" , \ Биоин- ,
Р^' \ жене- / Ь*с
₽™ 2
\ I
БИОЛОГИЯ ,----
"^шя
Mol®' '
6ИО1К**’
si
tQ А?
Рис. 1.3. Схема образования комплексных научных дисциплин на стыках биологии с
другими науками и сферами знаний (оригинал автора)
Разделяя биологию на науки общие и частные, мы действуем
субъективно, определяя «емкость» составных частей относительно
произвольно выбранных подразделений отсчета. Например, сис-
тематика растений выступает как общая наука относительно сис-
тематики Мхов, но как частная — относительно ботаники.
1.4. ОСНОВНЫЕ ЭТАПЫ РАЗВИТИЯ БИОЛОГИИ
Первоначально знания о живой природе формируются у чело-
века на основе противопоставления себя окружающему органи-
ческому миру. Уже само противопоставление предполагает опоз-
нание себя и подобных себе и всего отличающегося от «я» и «мы».
На таком уровне биологического знания, возможно, находится
большинство животных. По некоторым косвенным данным, мож-
но предположить, что у людей, собственно уже у первых предста-
вителей рода Человек, на определенном этапе познания биогичес-
10
кого окружения формируются представления о единстве всего жи-
вого, включая и его — человека, а также о непрерывности в воз-
никновении живого из неживого. До сих пор «наивные» представ-
ления такого рода имеют место у народов, которые принято счи-
тать отставшими в своем экономическом и социальном развитии,
где биологическое в человеке преобладает (?)* над социальным, а
единство с природой более гармонично.
Характерной чертой этих представлений является отождествле-
ние себя с животными. Так, коренные жители Таймыра считали,
что животные ничем не отличаются от людей, приписывали им
способность к речи и мышлению. У якутов еще в XIX в. бытовало
мнение, что медведь умен, как человек, даже умнее (!)**: он все
может, все понимает и знает, а не разговаривает только потому,
что не хочет. Племена Западной Африки, Южной Америки и дру-
гих мест, где обитали обезьяны, как правило, считали их такими
же людьми, как и они сами. У многих народов Африки и Азии су-
ществует стойкое представление о единстве всего живого через ду-
ховную сферу (как в возникновении, так и в индивидуальной жиз-
ни).
Для них, как и для человека, необходимо правильно соотно-
сить себя с окружающими объектами (что представляет пищевую
ценность; что, наоборот, воспринимает тебя как пищу; что может
представлять реальную опасность; как вести себя по отношению к
телам, подобным себе и остальным, в конкретных ситуациях). Но,
как принято считать, животные в отличие от человека не способ-
ны осознавать себя и, следовательно, не применяют абстрактные
логические конструкции для всестороннего соотнесения себя с
другими живыми телами, других живых тел между собой.
Не ставя себя над окружающей природой, эти народы и воз-
никновение человека объясняли своеобразно. Жители ряда эква-
ториальных стран сохраняют поверье, что человек, так же как рас-
тения или черви, «произошел» прямо из влажной земли. Преда-
ния жителей острова Самоа гласят, что люди произошли от земля-
ных червей. Жители одного из индейских племен на территории
Калифорнии считают своими предками койотов. Подобные при-
меры многочисленны и хорошо освещены в специальной литера-
туре.
С развитием культуры, то есть социальной формы записи на-
капливаемой информации, расстояние между животными и чело-
веком в его представлениях все более увеличивается. Растут и от-
*3десь и далее вопросительным знаком автор подвергает сомнению традици-
онное представление.
••Восклицательным знаком автор показывает эмоциональное восприятие су-
шествующей легенды.
U
личительные черты животного-предка или покровителя, все более
отдаляющие его от животного мира и приближающие к человеку.
Его родословная обрастает все большим числом легенд и мифов.
Он становится Культурным героем (то есть мифическим лицом,
которому приписывают введение обычаев, обрядов, обучение тру-
довым навыкам), создателем людей, творцом мира. Так, претерпе-
вая сложный путь развития, тотем постепенно превращается в бо-
жество.
Следующим шагом можно считать «появление» высших су-
ществ-героев или Великих духов, прямо превращающих тотемных
предков в людей племени. Позже они становятся богами-покро-
вителями и богами стихийных сил.
Противопоставление себя растениям и другим животным без
вычленения из живой природы определяет в этот период накопле-
ние в традициях и запретах (табу) жизненно важных сведений об
окружающем мире, воспринимаемых как бы изнутри.
С появлением представлений о едином Боге этот Бог начинает
играть роль творца не только людей, но и всего мира или хотя бы
всех животных и растений. Но сотворение человека Богом отлича-
ется уже тем, что именно в него «Бог вдохнул душу» или же сотво-
рил его по «образу и подобию своему». Представляя себя венцом
творения, человек одновременно ставит себя над природой и фак-
тически вне ее. Познание природы переходит на другую основу —
созерцание, а позже — исследование «высшим» «низшего». Живые
тела становятся преимущественно неодушевленными объектами
изучения, а само познание уподобляется описанию шестеренок
сложного механизма и поиску скрытых пружин, приводящих его в
движение.
Становление и распространение религиозных догм о соотно-
шении Бога, Человека и Природы резко тормозили процесс само-
познания человека и изучения живых организмов, начавшийся с
работ Аристотеля. «Единый акт творения» жизни легко разрешал
проблему единства человека и остальной живой природы, утверж-
дая одновременно непознаваемость всего сущего.
Тем не менее научный подход к миропониманию развивался
во многих центрах древней цивилизации и прежде всего в Сре-
диземноморье, где собственно и сформировалась биология как
наука.
В определенной степени рождение научных методов познания
обязано представлениям об отдельности Природы и Бога и бого-
подобности человека, так как в Природе оставлено только матери-
альное, осязаемое, разделяемое на части, оно оказывается подда-
ющимся познанию через опыт.
История развития биологической науки приведена в табли-
це 1.1.
12
1.1. Хронология главнейших биологических открытий и разработок (основные
положения учений, главнейшие труды и открытия)
Периоды и I даты 1 Ученые, их доли и опфьпяя |
530—470 гг. до н. э. Гераклит Эфесский: единство всех природ- Умосозерцатель- ных явлений, порождаемых движением и ная натурфилосо- взаимодействием элементов космоса (огня, фия воды, воздуха и земли) Фалес, Анаксимандр, Анаксимен: природа возникает из первичного материала — воды в ходе естественного развития; все возника- ет из воздуха, способного разрежаться и уплотняться, а человек и животные проис- ходят из земной слизи
490-430 гг. 460-370 гг. Эмпедокл: развитие идей Гераклита Демокрит: мир состоит из неделимых ато- мов в бесконечном пространстве, которые постоянно соединяются и разъединяются; они однородны по качеству, но различны по величине, массе, форме; вся природа возникает из различных сочетаний атомов
384—322 гг. Аристотель заложил принципы классифика- Начало системати- ции животных, основы сравнительной ческого изучения анатомии и сравнительной эмбриологии: природы разрабатывал идею «биогенетического закона» и эпигенеза; сформулировал закон о ступенчатом расположении природных объектов в системе Природы; обосновал единство происхождения животных
IV в. дон.э.— 0-XIV в. н.э. Развиваются только анатомия и физиоло- Ведутся только гия животных и человека в связи с по- прикладные ис- требностями медицины. Некоторое рас- следования пространение получили «травники»
1514—1564 гг. А. Везалий; научное описание строения Расширение изу- органов и систем человека чения природы,
1564-1642 гг. 1583 Г. Г. Галилей: изобретение микроскопа углубление изуче- А. Чезальпино: попытка создания класси- ния человека фикации растений на основе строения цвет- ков, плодов и семян
1628 г. У. Гарвей: учение о кровообращении; от- крытие яйцеклеток
1660-1694 гг. Р. Гук, М. Мальпиги, Н. Грю, Р. Камерра- Господствующие риус: изучение тонкого строения растений гипотезы в биоло- и их половых различий гии: «овизм»,
1669 г. М. Мальпиги, Я. Сваммердам: изучение «анималькулизм»,
1673 г. строения и развития насекомых «эпигенез», «пре- А. Левенгук: описание микроорганизмов, формизм» эритроцитов, сперматозоидов
1686-1704 гг. Дж. Рей описал свыше 18 тыс. видов расте- Описательная бо- ний, сгруппировал их в 19 классов; ввел таника и зоология понятие «вид»; создал классификацию по- звоночных животных
13
Продолжение
Периоды и
даты
Ученые, их труды и открытия
1707-1778 гг.
1707—1788 гг.
1745, 1764 гг.
К. Линней обосновал реальность вада;
установил соподчинение систематических
подразделений; разработал классификацию
животных и растений
Ж. Бюффон предложил гипотезу истории
Земли, в которой появление животных и
растений отнес к последним периодам
Ш. Бонне разрабатывал идею «лестницы
существ»
1759, 1768 гг. К. Вольф вел эмбриологические исследо-
вания
1782, 1804 гг. Ж. Сенбъе, Н. Соссюр: изучение роли сол-
нечного света в способности листьев погло-
щать углекислый газ и выделять кислород
Конец XVIII в. Л. Спалланцани: обоснование невозмож-
Преобладающие
направления
Изучение строе-
ния, развития и
функций организ-
мов; первые фун-
даментальные
обобщения
1797-1808 гг.
1743—1805 гг.
1744-1829 гг.
1769-1832 гг.
1817 г.
1827 г.
1830 г.
1838—1839 гг.
1831—1882
1859
ности самозарождения жизни
Т. Роозе, Ж.Б. Ламарк, Г. В. Тревиранус
вводят в научную литературу термин
«биология»
У. Паллей: аргументация представлений о
«сотворении» живых организмов, становле-
ние «креационизма»
Ж. Б. Ламарк: эволюционная трактовка
«лестницы существ»
Ж. Кювье: исследования по сравнительной
анатомии и систематике; разработка теории
корреляции органов н частей тела, «теории
катастроф»
X. Пандер: исследования по сравнительной
эмбриологии позвоночных животных
К. Бэр: разработка принципов сравнитель-
ной эмбриологии.
Э. Жоффруа Сент-Илер: установление
единства плана строения животных; обос-
нование возможности эволюционных пра-
образований
М. Шлейден, Т. Шванн: разработка «кле-
точной теории»
Ч. Дарвин: исследования по географии рас-
тений и животных, их родственным свя-
зям и приспособительным изменениям;
создание «эволюционной гипотезы» и «ги-
потезы происхождения человека»
Середина Ю. Либих, Ж. Б. Буссенго: изучение особен-
XIX в. ностей питания растений и животных
1845 г. К. Бернар: изучение роли секреторных орга-
нов в пищеварении
1847 г. Г. Гельмгольц, К. Людвиг: разработка мето-
дов изучения нервно-мышечной системы и
органов чувств
14
Продолжение
Периода и 1 даты | Ученые, их труды и открытия | Преобладающие направления
1829-1905 гг. 1863 г. 1880-1920 гг. 1880-1936 гг. 1887—1891 гг. 1892 г. 1875, И. М. Сеченов: изучение высшей нервной деятельности. Л. Пастер: опыты по невозможности самозарождения живых организмов К. А. Тимирязев: изучение фотосинтеза И. П. Павлов: изучение безусловных и условных рефлексов и высшей нервной деятельности С.Н. Виноградский: открытие и изучение хемосинтеза у бактерий Д. И. Ивановский: открытие вирусов Э. Страсбургер, В. Флеминг: изучение Изучение меха-
1882 гг. сложных процессов клеточного деления низмов наслсд-
1875—1888 гг. О. Гертвиг, Г. Фоль, Э. Бенеден, Т. Бо- ственности; раз-
1865, вери: изучение процессов созревания по- ловых клеток и оплодотворения Г. Мендель: описание закономерностей на- витие экологии; оформление син- тетической теории
1900 гг. следования признаков у растений эволюции (СТЭ)
1875-1899 гг. 1902-1907 гг. 1903 г. 1910 г. и позже 1924 г. 1926 г. 1927 г. 1927, 1935, 1945 гг. 1920-1940 гг. 1935 г. 1942 г. 1944 г. Ф. Гальтон, К. Негели, Э. Страсбургер, А. Вейсман, X. Де Фриз: поиски носите- лей наследственности и формулирование теорий наследственности и изменчивости Т. Бовери, У. Сеттон: оформление хромо- сомной теории наследственности В. Иогансен: учение о чистых линиях; фор- мулирование понятий «ген», «генотип», «фенотип» Т. Морган (и его школа): развитие хромо- сомной теории наследственности А. И. Опарин: гипотеза происхождения жи- вых тел В. И. Вернадский: становление науки био- геохимии; разработка учения о биосфере Н. К. Кольцов: формулирование представ- лений о наследственных молекулах матрич- ного типа Э. С. Бауэр и А. Г. Гурвич: представление об устойчивой неравновесности как глав- ной характеристике живых систем С. С. Четвериков, Дж. Холдейн, Р. Фишер, С. Райт, Дж. Хаксли, Ф. Г.Добжанский, Э. Майр, И. И. Шмальгаузен, А. Н. Север- ное, Н. В. Тимофеев-Рессовский, И.А. Ра- попорт: разработка синтетической теории эволюции А. Тенсли: разработка учения об экосис- темах В. И. Сукачев: становление науки биогео- ценологии О. Эйвери: доказательство связи ДНК с на- следственностью
15
Продолжение
Периоды и
даты
Ученые, их труды и открытия
Преобладающие
направления
1945—1985 гг. Э. Шредингер, И. Пригожин, Ж. Николис, Начало разработки
М. Эйген, Б. П. Астауров, М. В. Волькен- общей теории
штейн, Б. М. Медников и др.: изучение,. жизни
процессов самоорганизации в физико-хи-
мических системах, моделирование пред-
биологических этапов зарождения жизни
на Земле, поиск подходов к разработке тео-
ретической биологии
С 1950-х годов развернулись широкие исследования в таких
областях биологии, как молекулярная биология, иммунология, ге-
нетика и селекция, экология популяций и биогеоценология.
Мощное развитие получили прикладные аспекты биологии: био-
технология, защита растений от вредных организмов, космичес-
кая и радиационная биология, экологическое моделирование.
Новые факты нанесли более или менее сильные удары по клас-
сической генетике, вызвали необходимость корректировки основ-
ных положений синтетической теории эволюции.
В восьмидесятые годы прошлого столетия появляется множе-
ство гипотез, противопоставляемых утвердившимся догмам об-
щей биологии (И. Пригожин, М. Эйген, С. Гоулд, М. Кимура,
В. А. Кордюм, Э. М. Галимов, В. Я. Савенков и др.). В теоретичес-
кой биологии наметился кризис, попытки выхода из которого в
настоящее время выражаются в разработке общей теории жизни.
Приведенная хронология биологических открытий показывает,
что развитие биологии идет неравномерно, как бы рывками, раз-
деляемыми все более короткими промежутками, а периоды на-
копления фактов сменяются периодами их анализа и обобщения.
Побудительные же стимулы и условия развития биологии здесь
невидны. Поэтому кратко остановимся на их характеристике.
360—330 гг. до н. э. — эпоха расцвета античной культуры. На-
копление знаний о природе, не имеющих прямого хозяйственного
значения.
Перерыв в активном познании природы длится более 15 столе-
тий. Этот период характеризуется (в европейских центрах цивили-
зации) утверждением религиозных догматов, подавлением экс-
пансии человека в природу. В практической деятельности преоб-
ладает ручной оседлый труд. Немногочисленные исследования рас-
тений и тела животных и человека ведутся в связи с
потребностями медицины и в поисках вместилища души (у чело-
века).
1540—1590 гг. — начало эпохи Возрождения. Пробуждается по-
знавательная активность. Развиваются техника, ремесла, искусст-
16
ва. Углубляется изучение человека. Обобщаются знания по бота-
нике, накопленные лекарями.
Почти столетний спад в накоплении фактов сопровождается
первыми попытками использования инструментария в биологи-
ческих исследованиях.
1665—-1705 гг. — усовершенствование микроскопов. Начало
эпохи путешествий и географических открытий. Расширение сфе-
ры доступного наблюдению позволяет вести изучение микроорга-
низмов и обусловливает всплеск описаний растений и животных.
1730—1840 гг.— зарождение капиталистических форм произ-
водства. Формирование материалистических идей. Наблюдается
общее раскрепощение сознания, что позволяет вести эмбриологи-
ческие исследования и создает предпосылки для формирования
эволюционных представлений.
1840—1860 гг. — период систематизации и обобщения фактов и
совершенствования методов изучения процессов, идущих в телах
растений и животных.
1860—1895 гг. — бурное развитие капиталистического способа
производства. Развитие химии, техники. Расцвет материалисти-
ческой философии. Утверждается эволюционная теория. Новые
методы исследований позволяют на новом качественном уровне
изучать процессы, идущие в живых организмах и даже в клетке.
Накопленные факты вызывают кризис в некоторых общепри-
нятых концепциях индивидуального и исторического развития
организмов.
1900—1920 гг. — социально-политические кризисы и военные
конфликты. Резкое усиление экспансии человека в природу. В не-
которых научных школах прорыв в области изучения механизмов
наследования признаков. Осмысление новых подходов к изуче-
нию природы.
1920—1940 гг. — бурный научно-технический прогресс, связан-
ный прежде всего с экономической конкуренцией государств раз-
ной социально-политической ориентации. Переход к интенсив-
ным технологиям в сельском хозяйстве и промышленности. От-
крытия в химии и физике и разработка на их основе новых мето-
дов изучения живых организмов.
С 1950-х годов вооруженная борьба за мировое господство пе-
реходит в форму «устрашения». Лидерство в науке, технике, тех-
нологии, образовании отчасти становится самоцелью. Ряд откры-
тий в химии и физике позволяет резко расширить границы позна-
ваемого, в том числе и в биологии. Некоторые разработки в облас-
ти генетики, физиологии, биохимии, микробиологии получили
успешное прикладное развитие, в связи с чем практические по-
требности стали эффективнее направлять развитие биологических
наук.
2 Присный А.В
Глава 2
ОБЩАЯ ХАРАКТЕРИСТИКА ЖИЗНИ
2.1. МАТЕРИАЛЬНЫЕ СИСТЕМЫ
Выдвинутые в разное время гипотезы, касающиеся проблемы
сущности и происхождения жизни, многочисленны и противоре-
чивы. Связать их воедино, не изменив при этом существенных
сторон, практически невозможно. Более того, анализ даже наибо-
лее аргументированных из этих предположений с позиций сис-
темного подхода показывает если не ошибочность, то по меньшей
мере их ограниченность, односторонность. В данном курсе «Об-
щей биологии» произведено согласование некоторых представле-
ний на основе дуалистической концепции жизни. Предлагаемый
учебник позволяет устранить лишь некоторые явные противоре-
чия между «признанными» и «непризнанными» гипотезами и од-
новременно показать через разнообразие мнений сложность само-
го предмета биологии.
Поскольку несовпадение «взглядов» начинается уже с различ-
ного смысла, вкладываемого в основные понятия и термины, це-
лесообразно начать именно с них. Нам потребуется разобраться в
том, что такое система, материя, движение, энергия, информа-
ция, пространство и время, и лишь после этого мы сможем при-
ступить к изучению некоторых существенных сторон земной
формы жизни.
2.1.1. СИСТЕМА
Существует около десятка значений термина «система». Его ис-
ходный смысл — целое, составленное из частей. В контексте изла-
гаемых представлений о сущности жизни под системой понимают
некоторую целостность, составные части (элементы) которой пря-
мо или косвенно определяют состояние друг друга, или же систе-
ма — это некоторое множество элементов, связанных взаимодей-
ствием в единое целое (рис. 2.1). В таких определениях, в общем
адекватных исходному значению слова, подчеркивается проис-
хождение целостности. В качестве систем при некоторых допуще-
18
Тела
А
Системы
Тела
Однородные мате-
риальные тела
(не взаимодействуют)
Тела взаимодействуют,
определяя состояние друг
друга и всего множества
Г
Разнородные мате-
риальные тела (не
взаимодействуют)
Группировка элементов в
системе делает взаимодейст-
вие в группах и между группа-
ми неравноценными, но более
':4- упорядоченным
Исходная неравноценность и взаимодопол-
нительность элементов во взаимодействии
связывает систему единым энергетическим
потоком и удаление даже одного из них
разрушает всю систему
Определенность отношений, связей, взаимодействий, то есть объем информации
, в системах на схеме: И>Б, К>В, К>И
Рис. 2.1. Тела и системы (оригинал автора)
2*
ниях можно, например, рассматривать: условно ограниченный
объем газа, молекулы которого находятся в тепловом движении;
живое тело; дубраву; горно-металлургический комбинат и т. п.
Системы могут составляться из однородных или разнородных эле-
ментов при связывании их единым энергетическим процессом
или образовываться разделением целого на взаимосвязанные час-
ти. Целостность системы будет определяться прежде всего тем,
насколько высока степень взаимной дополнительности (или ком-
плементарности) элементов. Так, одноименно заряженные эле-
ментарные частицы могут взаимодействовать, образуя систему
лить на короткое время, а разноименно заряженные, наоборот,
попадая в поле взаимного действия, образуют устойчивое един-
ство (до определенного значения их полной энергии). Одинако-
вые живые клетки способны сформировать лишь колонию, с не-
которыми допущениями — таллом, но многоклеточное тканевое
живое тело составляется только из морфологически и функцио-
нально дополняющих друг друга клеток
Системы могут быть открытыми и замкнутыми. Для первых ха-
рактерен обмен материей, энергией и информацией с окружаю-
щей средой, в том числе и с другими системами, а для вторых та-
кой обмен исключается. Реально в природе не существует замкну-
тых систем. Выделяют их условно для различных теоретических
построений. Особенно важны они для термодинамических расче-
тов.
Всякая система обладает протяженностью в пространстве и
длительностью во времени. При этом, какую бы часть Вселенной
мы ни выделили мысленно как систему, ее всегда можно предста-
вить как элемент системы более емкого уровня. Поведение систе-
мы имеет внутренний и внешний аспекты и может быть определе-
но соответственно через поведение элементов этой системы или
же как поведение некоторой целостности в системе более емкого
уровня. Главнейшими характеристиками поведения элементов
считаются соотносительное изменение их положения и состояния
и проявление связей, отношений, сопровождающиеся взаимодей-
ствием и перераспределением энергии внутри системы. Поведе-
ние системы тем более предсказуемо, а прогнозируемые ее состоя-
ния тем более вероятны, чем большее число элементов входит в
систему и чем с большими (до определенных значений) скоростя-
ми они движутся и определеннее соотносятся друг с другом: в
этом случае проявляется динамическая инерционность. В системе
с небольшим числом медленно движущихся и неопределенно со-
относящихся элементов увеличивается частота возникновения ма-
ловероятных состояний — флуктуаций.
Поведение элементов системы и самой системы может быть
выражено посредством пространственных и временных харак-
20
теристик. Через пространство описывается взаимное положе-
ние или изменение взаимного положения элементов, и, как не-
трудно заметить, это будет внешнее отношение, то есть положе-
ние или изменение положения относительно чего-либо внешне-
го к элементу или системе. Пространственная система координат
находится вне рассматриваемого объекта, и характеристики ее
осей не зависят от него. Поэтому, например, мы используем
одинаковые измерительные инструменты для оценки размеров
различных объектов, тем самым приводя их условно к одной
«точке отсчета», предполагая одновременно, что мерная шкала
в промежутках между измерениями не растягивается и не сжи-
мается. Измерительные инструменты здесь должны быть уни-
фицированной частью внешней среды по отношению к измеря-
емым объектам.
В трехмерной (неискривленной) пространственной системе ко-
ординат число направлений, по которым можно соотнести по-
ложение элементов в системе, три (оси здесь взаимно перпенди-
кулярны). Состояние или изменение состояния системы, то есть
взаимоотношения элементов в системе или динамику распределе-
ния энергии в системе, в отличие от предыдущего нельзя выразить
через что-то внешнее. Выразить состояние — это прежде всего со-
отнести настоящее системы с ее прошлым или будущим, иными
словами, показать ее в длительности, дать временную характерис-
тику. Позже будет показано, что время можно трактовать как
трехмерное относительно «стрелы времени», а конкретное его на-
правление выводится аналогично результирующему направле-
нию перемещения в трехмерном пространстве. Временная система
координат, следовательно, находится в самом рассматриваемом
объекте, который определяет характеристики ее осей. По этой
причине в каждой системе время «течет» по-своему и «уплотняет-
ся» или «растягивается» относительно внешнего времени при уве-
личении изменчивости системы.
2.1.2. МАТЕРИЯ
Предполагается, что системы материальны, то есть элементы
системы образованы каким-либо видом материи. Но что такое ма-
терия? На самом ли деле во Вселенной нет ничего, кроме движу-
щейся материи?
Считается вполне корректной общеизвестная формулировка
Ф. Энгельса, согласно которой «вся окружающая нас неживая
природа, начиная от звезд и кончая элементарными частицами, и
живая природа с ее качественно многообразными формами — от
неклеточных форм до многоклеточных организмов — все это ма-
21
терия». Философское понятие «материя» не выражает ничего ино-
го, кроме объективной реальности (существующей вне сознания и
независимо от него), и его нельзя отождествлять с естественно-на-
учным. И все же между философским понятием материи и есте-
ственно-научными знаниями о ней имеется не только различие,
но и неразрывная связь: естественные науки изучают различные
формы движения одной и той же объективной реальности — мате-
рии.
К числу основных видов материи относят космический вакуум
(базовый вид), поле и вещество (производные от вакуума), а также
плазму (вид, производный от вещества). В физике вакуум рассмат-
ривается как невозбужденная сверхплотная (1093г/см3) среда с
мелкозернистой структурой, представляющая смесь элементарных
частиц, поле трактуется как возбужденный неразреженный вакуум,
вещество — как возбужденное разреженное состояние вакуума, а
плазма—как предельно возбужденное однородное вещество,
представляющее собой «сгусток энергии».
Каждый вид материи представлен несколькими формами.' Ив-
вестны электромагнитное (условно разделяемое на электрическое
и магнитное) и гравитационное поля. Вещество мы разделяем на
неживое неорганическое, неживое органическое, живое неоргани-
ческое и живое органическое, но можно определять формы веще-
ства и по составу элементарных частиц. Формы и виды материи в
определенных условиях утрачивают различия и способны во мно-
гих случаях к прямому или опосредованному взаимному превра-
щению. ,
Вещество во Вселенной образует скопления нескольких раз-
мерных уровней: элементарные частицы, атомы, молекулы, веще-
ственные частицы и тела различного масштаба. Каждый из них
включает предыдущие, но все они конечны в пространстве, то
есть вещество прерывисто (дискретно). Промежутки между веще-
ственными частицами и телами заполнены вакуумом. И частицы,
и вакуум как бы пронизаны различными полями. Таким образом,
материя в целом действительно непрерывна.
2.1.3. ДВИЖЕНИЕ
Когда мы говорим о движении материи, в понятие « движение»
мы вкладываем его философское значение. Одно из полных и дос-
таточно точных определений движения принадлежит Ф. Энгельсу.
Оно звучит так: «Движение, рассматриваемое в самом общем
смысле слова, то есть понимаемое как способ существования ма-
терии, как внутренне присущий ей атрибут, обнимает собой все.
происходящие во Вселенной изменения и процессы, начиная от
22
простого перемещения и кончая мышлением». В настоящее время
принято различать пять форм движения материи: механическую
(перемещение частиц и тел), физическую (движение элементар-
ных частиц и атомов, включая взаимодействие элементарных час-
тиц), химическую (взаимодействие атомов и молекул), биологи-
ческую (рост, онтогенез, эволюция и др.) и социальную (мышле-
ние, общественные движения, смена общественных систем и др.).
Важно помнить, что движение включает не только перемещение в
пространстве, но также изменение состояния, то есть движение в
«объеме времени». Здесь уместно отметить, что покой, неизмен-
ное состояние — это также движение, но с постоянной скоростью
относительно движения в системе отсчета или сохранение взаимо-
связи с движущейся точкой отсчета.
Частной формой движения является развитие — такая форма
Движения, при которой на основе старого возникает новое, низ-
шее заменяется высшим, простое — сложным. Развитие объединя-
ет, суммирует в себе прогресс, регресс и круговорот. Круговорот в
Жйвых системах занимает особое место: он лежит в основе боль-
шинства жизненных процессов на всех уровнях от клетки до био-
сферы.
В процессе движения различные материальные образования
взаимодействуют друг с другом либо приобретают способность
к такому взаимодействию. В физике принято выделять четыре
формы взаимодействия: сильное, электромагнитное, слабое и
гравитационное. Но это взаимодействия элементарных частиц
к других вещественных тел через переносчиков взаимодей-
ствий — частицы полей. Базовые формы взаимодействий непос-
редственно обусловливают важнейшие внутриатомные и галак-
тические процессы, а опосредованно — процессы на промежу-
точных уровнях, где обнаруживаются иные формы взаимодей-
ствий. Совокупность взаимодействующих объектов ранее мы
условились называть системой, а сами объекты — элементами
системы. Можно сказать, что любая система обладает некото-
рым количеством движения, обусловливающим взаимодействие
ее элементов или самой системы с внешней средой (либо эле-
ментами надсистемы).
2.1.4. ЭНЕРГИЯ
Общая мера различных форм движения материи в системе
обозначается понятием энергия. Энергию материальной систе-
мы нельзя измерить непосредственно, и поэтому ее оценивают
ПО действию, оказываемому на внешнюю среду. Для практичес-
кихрасчетов принимают, что энергия системы равна сумме всех
23
действий, которые производятся вне системы, когда она каким-
либо способом спонтанно (самопроизвольно) переходит из оп-
ределенного состояния в произвольно выбранное («нулевое»)
состояние. Измеряется энергия в единицах механической рабо-
ты.
Запасенная системой внешняя (полученная из внешней сре-
ды) энергия может быть превращена в механическую энергию.
Часть теряемой внутренней энергии способна превращаться в
немеханические формы и составляет свободную энергию систе-
мы. За счет свободной энергии система может совершать работу.
Другая часть внутренней энергии выделяется в виде тепла, рассе-
иваемого в окружающем пространстве, она не может быть пре-
вращена в работу. Это связанная энергия, характеризуемая энт-
ропией — мерой рассеяния, хаоса. Соотношение между приведен-
ными формами энергии выражается формулой Н (полная энер-
гия системы, или свободная энтальпия) =pV (запасенная
внешняя энергия) + F (свободная энергия) S (энтропия). Энер-
гия, затрачиваемая системой на работу, направленную на внешнюю
среду, по ряду причин не может достигать pV+ F. Работа систе-
мы связана с направленным движением составляющих ее эле-
ментов, а придание направления уже само по себе требует расхо-
да энергии.
Считается, что в живых системах в работу превращается энер-
гия структуры живой материи. В качестве упрощенной аналогии
можно рассматривать превращение в работу потенциальной энер-
гии взведенного курка. Энергия же питательных веществ, потреб-
ляемых живой системой, затрачивается на возобновление структу-
ры; в приведенной аналогии — на взведение курка. При этом при-
ведение и освобождение энергии сопряжено с изменением кон-
формации рабочей структуры.
Потеря системой энергии в результате спонтанных процессов
приводит к термодинамически более выгодному состоянию. Но
такая потеря возможна лишь в том случае, если высвобождаемая
энергия больше удерживающей ее. Завершается переход энергии
нагреванием или уменьшением упорядоченности внешней среды.
Известно, что движущиеся час тицы и тела обладают кинети-
ческой энергией, а частицы и тела, преодолевающие какие-либо
поля, накапливают потенциальную энергию. В некоторых случаях
энергия имеет гибридный характер, а в системах, построенных из
полярных (комплементарных) элементов, заключена особая энер-
гия взаимного притяжения. Классификация известных видов
энергии приведена на рисунках 2.2...2.5.
Поток частиц поля обусловливает динамические формы энер-
гии, а частицы или тела, перемещающиеся против потока частиц
поля, накапливают статические формы энергии (см. рис. 2.5).
24
Рис. 2.2. Размерные уровни проявления основных видов взаимодействий (оригинал
автора)
Невозбужденные
и возбужденные
кварки и анти-
по
3
кварки разных Г
«ароматов» сши- \ Г
ваются сильным <
взаимодействием
по
2
адроны
барионы
протоны нейтроны
| мезоны |
Взаимодействующие
частицы
лептоны
мюоны нейтрино электроны
""'"""'"‘'в
Участвуют во взаимодействии [ гравитационном | [ сильном | слабом [
Рис. 2.3. Образование элементарных частиц н их участие во взаимодействиях
Рис. 2.4. Проявление взаимодействий в природных процессах
Частицы Энергия
нейтрино нейтрвнодинамическая, нейтриностатическая
— Поле
заряженные частицы и тела ►1 электрическое 1- электродинамическая, электростатическая
фотоны ►электромагнитное!- электроиодинамическая (световая), магнитостатическая
гравитоны ►j гравитационное гравидинамическая, гравистагическая
пионы мезонодинамическая {
Рис. 2.5. Виды энергии, связанные с движением их носителей
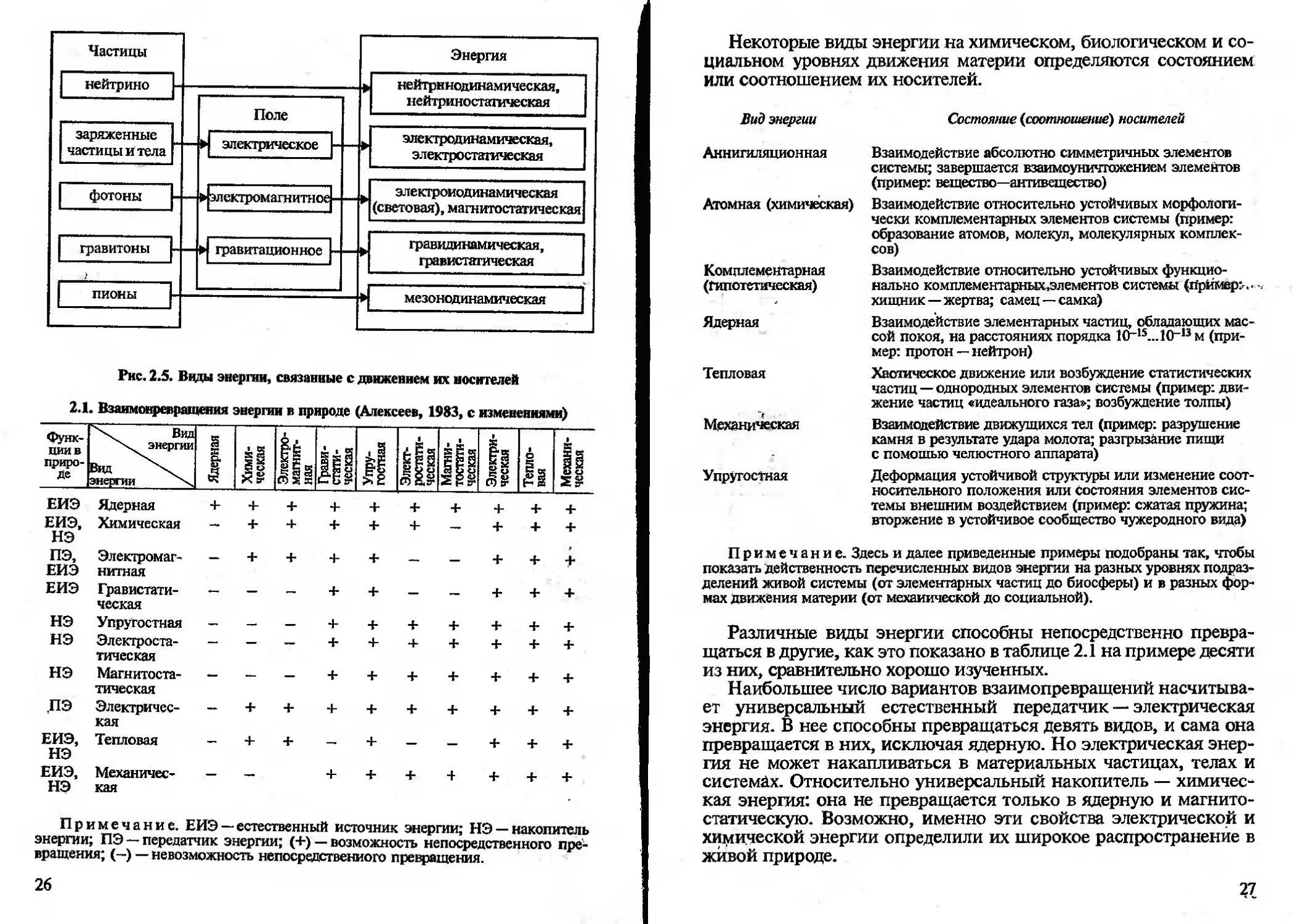
2.1. Взаимонреяращения энергии в природе (Алексеев, 1983, с изменениями)
НЭ
ПЭ,
ЕИЭ
ЕИЭ
НЭ
НЭ
НЭ
ПЭ
ЕИЭ,
НЭ
ЕИЭ,
НЭ
Электромаг-
нитная
Гравистати-
ческая
Упругостная
Электроста-
тическая
Магнитоста-
тическая
Электричес-
кая
Тепловая
Механичес-
кая
Примечание. ЕИЭ — естественный источник энергии; НЭ — накопитель
энергии; ПЭ — передатчик энергии; (+) — возможность непосредственного пре-
вращения; (—) — невозможность непосредственного превращения.
26
Некоторые виды энергии на химическом, биологическом и со-
циальном уровнях движения материи определяются состоянием
или соотношением их носителей.
Вид энергии Состояние (соотношение) носителей
Аннигиляционная Взаимодействие абсолютно симметричных элементов системы; завершается взаимоуничтожением элементов (пример: вещество—антивещество)
Атомная (химическая) Взаимодействие относительно устойчивых морфологи- чески комплементарных элементов системы (пример: образование атомов, молекул, молекулярных комплек- сов)
Комплементарная (Гипотетическая) Взаимодействие относительно устойчивых функцио- нально комплементарных,элементов системы (примеры- хищник —жертва; самец —самка)
Ядерная Взаимодействие элементарных частиц, обладающих мас- сой покоя, на расстояниях порядка 1СГ15...10-,3м (при- мер: протон — нейтрон)
Тепловая Хаотическое движение или возбуждение статистических частиц — однородных элементов системы (пример: дви- жение частиц «идеального газа»; возбуждение толпы)
Механическая Взаимодействие движущихся тел (пример: разрушение камня в результате удара молота; разгрызание пищи с помощью челюстного аппарата)
Упругостная Деформация устойчивой структуры или изменение соот- носительного положения или состояния элементов сис- темы внешним воздействием (пример: сжатая пружина; вторжение в устойчивое сообщество чужеродного вида)
Примечание. Здесь и далее приведенные примеры подобраны так, чтобы
показать действенность перечисленных видов энергии на разных уровнях подраз-
делений живой системы (от элементарных частиц до биосферы) и в разных фор-
мах движения материи (от механической до социальной).
Различные виды энергии способны непосредственно превра-
щаться в другие, как это показано в таблице 2.1 на примере десяти
из них, сравнительно хорошо изученных.
Наибольшее число вариантов взаимопревращений насчитыва-
ет универсальный естественный передатчик — электрическая
энергия. В нее способны превращаться девять видов, и сама она
превращается в них, исключая ядерную. Но электрическая энер-
гия не может накапливаться в материальных частицах, телах и
системах. Относительно универсальный накопитель — химичес-
кая энергия: она не превращается только в ядерную и магнито-
статическую. Возможно, именно эти свойства электрической и
химической энергии определили их широкое распространение в
живой природе.
27
2.1.5. ИНФОРМАЦИЯ
Степень определенности, предсказуемости состояний и отно-
шений, в широком значении принято обозначать термином ин-
формация. В кибернетике (в узком значении) информация — это
мера определенности связи, сообщения. Информацию можно так-
же определить как количественный эквивалент меры организо-
ванности, упорядоченности системы, то есть отрицательной энт-
ропии (негэнтропии, или неэнтропии). А это означает, что инфор-
мация противоэквивалентна энтропии.
Различают три вида неорганизованности: движения, строения
и отношений. В соответствии с этим выделяют и три вида энтро-
пии: тепловую, структурную и информационную. Понятиями,
противоположными им по смыслу, будут тепловая, структурная и
информационная негэнтропии (меры упорядоченности, организо-
ванности). Но негэнтропии и информация — понятия эквивалент-
ные, а в некоторых случаях и тождественные. Структурная инфор-
мация — понятие вполне конкретное. Оно характеризует опреде-
ленность пространственных и временных отношений элементов в
системе. Увеличению объема этого вида информации соответству-
ет усложнение строения системы. Таким образом, наименьшим
объемом информации обладают максимально упорядочивающие-
ся системы с предельно симметричными элементами (сферичес-
кими, тетрагональными, кубическими, икосаэдрическими и т. п.)
одного вида и одним уровнем структурной иерархии. Структурная
негэнтропия эквивалентна разнообразию элементов в системе и
организации их в иерархические объединения.
Тепловая негэнтропия как мера организованности движения в
определенном смысле эквивалентна работе, и мы условимся ис-
пользовать это понятие («работа») вместо понятия «тепловая ин-
формация». Единицей измерения работы является джоуль.
Наконец, противоэквивалент информационной энтропии —
информационную негэнтропию можно обозначить как собствен-
но информацию. Она выражает количественно меру определен-
ности отношений, связей, взаимодействий, порождаемых движе-
нием (перемещением и изменением) элементов системы, и изме-
ряется в байтах (bite). Объем информации эквивалентен отрица-
тельной энтропии, несвободе движения элементов.
Ценность информации не может быть определена через самое
себя. Она не абсолютна, а относительна. Более того, ценность вы-
ступает как некая характеристика используемого низшего для ис-
пользующего высшего. Некоторые типы систем (стационарные
неравновесные) способны продуцировать информацию (отрица-
тельную энтропию) в процессе притока энергии или при переходе
из неустойчивого состояния в устойчивое, в том числе строить ка-
28
чоственно новую или более сложную информацию на основе ис-
пользования информации внешней среды. При этом объем ин-
формации в среде уменьшается, а сама «поглощаемая» информа-
ция обесценивается.
Таким образом, объем информации в системе возрастает с уве-
личением сложности ее строения, количества и разнообразия на-
правленного движения, а также определенности связей и отноше-
ний между элементами.
2.1.6. ПРОСТРАНСТВО И ВРЕМЯ
Из традиционной физики известно, что мы живем в четырех-
мерном мире: три измерения характеризуют пространство и
одно — время. Существуют гипотезы, представляющие наш мир
семи- и даже одиннадцатимерным. В них увеличивается число
пространственных координат за счет учета «направлений» искрив-
ления пространства.
В реальном мире из всех форм движения наиболее понятным
для нас представляется перемещение в пространстве. Траекторию
движущегося тела можно описать линией, и в зависимости от
сложности этого движения линия будет: прямой, лежащей по
меньшей мере в двух плоскостях; кривой, лежащей в одной плос-
кости; кривой, лежащей в объеме. В каждый отдельный момент
времени движение описывается вектором, который может быть
разложен по направлениям (на три взаимно перпендикулярные
оси). Проще всего произвести разложение вектора, совместив его
с одной из координатных осей, когда значения по другим осям
превратятся в 0. Но тогда сама система координат становится не-
постоянной, подвижной. Более полно движение будет описано,
если одну из координатных осей совместить с вектором движения
системы, к которой относится описываемый движущийся эле-
мент. Тогда, естественно, «идеальной» следует считать систему ко-
ординат, ориентированную на Вселенную.
Пространство мы определяем как меру протяженности матери-
альных объектов, их соотносительного положения и изменения
протяженности и положения (безотносительно времени). Движе-
ние во времени —это самовоспроизведение состояния. В такой
трактовке сохранение состояния, покой можно рассматривать как
полное воспроизводство прежнего состояния, что аналогично пе-
ремещению в пространстве по прямой линии. Прямолинейность
состояния характерна для «стационарных» (а это неустойчивое,
промежуточное состояние) систем, то есть систем, воспроизводя-
щихся полно (воспроизводящих собственное состояние на каж-
дый следующий момент времени в неизменном виде). Примени-
29
тельно к ним мы говорим об условно равномерном течении време-
ни, а на практике некоторые из них используем в качестве хроно-
метров. Изменение состояния есть отклонение его от прямоли-
нейности (полного самовоспроизводства). Чем сильнее отклоне-
ние состояния от предшествующего, тем соответственно меньше
доля полно воспроизводимого состояния. Направления отклоне-
ний определяются в первую очередь выбранной точкой отсчета
(характером изменений), образуя в сумме плоскость, перпендику-
лярную линии неизменного состояния (рис. 2.6).
В замечании о понятности перемещения тела в пространстве
более отражена иллюзия, чем действительное положение, и преж-
де всего потому, что приходится одновременно касаться и про-
странственных, и временных характеристик. Вне пространства нет
изменения положения, но нет его также и вне времени. Траекто-
рия — производное пространства и времени — плод нашего вооб-
ражения. Она представляет собой графическое выражение или
мысленное отображение всего множества последовательных поло-
жений объекта в пространстве как оставшихся следов, рассматри-
ваемых в один момент времени. Восприятие движения возможно
лишь при сохранении в памяти предшествующих положений или
состояний объекта.
Попробуем расчленить перемещение на пространственную и
временную составляющие. В пространственной системе'коорди-
нат без учета координат времени (вне четвертого измерения) «Дви-
жущееся тело» займет некоторый объем, как бы присутствуя во
всех его частях одновременно. То есть тело становится более про-
тяженным в пространстве, чем если бы оно было покоящимся.
Так происходит потому, что, снимая время, мы позволяем'Делу
Рис. 2.6. Схема проявления лрехмериости времени в системе (оригинал автора)
30
быть в каждом месте, не исчезая в других. Но что произойдет, если
мы снимем пространство? Во-вторых, будет невозможно фикси-
ровать последовательные положения тела. А во-первых, исчезнут
пространственные характеристики самого тела. Длительность же
существования тела останется в виде его непрерывного исчезнове-
ния — появления. Этот процесс можно представить как спонтан-
ное превращение в себя, самовоспроизводство без существования
в пространстве. Самопревращение материальных образований —
это проявление течения времени. Сделаем несколько ходов назад:
если повторение себя происходит каждый раз в новой точке или в
новом объеме пространства, реконструируется перемещение или
изменение объема или формы тела.
Независимо от характера движения в пространстве в каждый
последующий момент времени тело в большей или меньшей сте-
пени сохраняет или изменяет свое состояние. В случае с равно-
ценным, полным самовоспроизводством тела в одной и той же
точке пространства мы говорим об абсолютном покое. Но если у
тела, перемещающегося в пространстве, непрерывно меняются
параметры состояния, то для описания его движения уже недоста-
точно трех измерений пространства и одного измерения време-
ни — к ним следует прибавить еще две координатные оси време-
ни. Суммируя направления движения, мы получим вектор пере-
мещения и вектор изменения состояния. Связь времени с состоя-
нием указывает на то, что время характеризует перераспределение
энергии в системе и отношений между ее элементами. На различ-
ных размерных (структурных) уровнях организации материи вре-
мя проявляется и соотносится с пространством по-разному. Так,
на уровне элементарных частиц, по крайней мере не обладающих
массой покоя, пространство и время «сливаются».
Итак, еще раз подчеркнем, что время существует только в сис-
темах. На «срезе» время системы — это некоторая плоскость, огра-
ниченная замкнутым контуром, каждая точка которой характери-
зуется вектором. При таком изображении площадь плоскости эк-
вивалентна запасу информации в системе. В системах, спонтанно
деградирующих (неживых), векторы от контура направлены
внутрь, образуя конус. Их суммарное направление — это направ-
ление «стрелы» времени, совпадающее с направлением роста энт-
ропии. Крутизна конуса зависит от интенсивности деградации си-
стемы: чем быстрее развиваются энтропийные процессы, тем
ниже конус. Стационарное, или «0»-состояние у системы факти-
чески отсутствует, так как векторы, образующие поверхность ко-
нуса, в этом случае должны приобрести параллельное направле-
ние и «расположиться» на поверхности цилиндра. Поверхность же
цилиндра соответствует противодействию энтропийным процес-
сам и активному сохранению состояния — это стационарно актив-
31
ная (развивающаяся) система в отличие от стационарно пассив-
ной (деградирующей). Типичная развивающаяся система имеет
вид конуса, направленного основанием вперед (относительно на-
правления «стрелы» времени), что характеризует общую антиэнт-
ропийную направленность процессов, происходящих в ней
(рис. 2.7). Суммарное направление векторов развивающейся сис-
темы также совпадает с направлением «стрелы» времени.
Изолированный вектор направлений изменений в развиваю-
щейся или деградирующей системе своей вершиной потенциаль-
но может описывать полусферу, основание которой перпендику-
лярно оси «прошлое — будущее». Изменение состояния системы
прямо связано с усилением взаимодействия элементов всех уров-
ней ее структурной организации. При этом происходят сжатие
или деформация пространства и уплотнение времени. Механиз-
мом уплотнения времени служит отклонение вектора времени от
центральной оси, сопровождающееся увеличением его проекции
Рис. 2.7. Схема поведения времени в системе:
Л —механизм искривления плоскости настоящего (времени); Б, В— направление векторов
времени на внешнем контуре развивающейся (Б) и деградирующей (В) систем (оригинал
автора)
32
на основание полусферы и укорочением проекции на ось. Это од-
новременно означает, что в устойчиво неравновесных системах,
активно сохраняющих отклоненный вектор, время непрерывно
«ускоряется» (уплотняется по оси) относительно внешнего време-
ни и процесс этот самоусиливающийся. Из сказанного следует от-
носительная независимость характера течения времени в разных
системах. Усиление взаимодействия элементов в системе вызыва-
ет деформацию пространства — его сжатие с одновременным рас-
тяжением в ближайшей внешней среде. Уплотнение времени в
зоне сжатия пространства также сопровождается искривлением
плоскости времени, а именно в сторону, противоположную на-
правлению вершины конуса, в прилежащей зоне (см. рис. 2.7, А)
для развивающихся систем и в сторону вершины конуса для уско-
ренно деградирующих систем.
Выше приведенная трактовка времени как меры относительно-
го состояния системы не противоречит пониманию времени как
меры длительности в направлении «прошлое — настоящее — буду-
щее», но позволяет понять механизм перехода системы из устой-
чиво равновесного состояния в неустойчиво равновесное или ус-
тойчиво неравновесное, то есть из устойчивого «неживого» состо-
яния, характеризующегося медленным ростом энтропии, в дегра-
дирующее с быстрым ростом энтропии или в «живое» состояние,
характеризующееся ростом негэнтропии.
2.1.7. МАТЕРИАЛЬНОЕ И НЕМАТЕРИАЛЬНОЕ (ИДЕАЛЬНОЕ)
В СИСТЕМЕ
Итак, уточнив понятийный аппарат, мы можем вновь обра-
титься к материальным системам. Первое, что нам предстоит вы-
яснить: является ли догмой представление о материальности сис-
тем и действительно ли идея вторична по отношению к материи.
Помня о том, что материя весьма многообразна, и о том, что в
предшествующих рассуждениях мы опирались на вещественные
системы, примем изначально как догму положение о веществен-
ности элементов, образующих любую систему.
Элементы находятся в непрерывном движении и взаимодей-
ствуют друг с другом, определяя состояние друг друга и системы в
целом. Взаимодействие элементов делает систему единым целост-
ным образованием, где каждый из видов элементов уникален и
производит только ему присущее действие. Такую систему можно
наблюдать, измерять: она вполне осязаема. Например, участок
суши с приземным слоем воздуха, группой растений, растительно-
ядных животных, хищников и почвенными микроорганизмами
может быть условно представлен как вещественная система. Ее
3 Присный А.В
33
элементы — взаимодействующие живые и неживые тела. Вычле-
нив отдельную пару живых тел и проследив за их движением, мы
обнаруживаем, что они отличаются некоторыми особенностями
строения и характера движения, а также то, что их взаимодей-
ствие привело к разрушению одного из них и поглощению дру-
гим. В нашем восприятии: одно животное съело другое. Повторив
наблюдение над аналогичными элементами той же системы, мы
получим тот же результат. Кроме того, при достаточной длитель-
ности наблюдения мы увидим, что в системе на месте разрушен-
ных элементов появляются новые, такие же, и все повторяется
сначала. Система при всем этом остается устойчиво целостной.
Мы предполагаем, что наблюдаемые процессы, а они вполне ма-
териальны, являются нормой для данного типа системы, частью ее
содержания. Материальные процессы здесь складываются в круго-
ворот веществ.
Из приведенного примера следует, что элементы рассматривае-
мой системы дополнительны (комплементарны) к другим и не
столько вещественно, сколько по содержанию, в функциональном
аспекте. Отмечая одинаковость поведения однородных элементов,
мы фиксируем их «предназначение», то есть функцию в системе.
Если мы обозначаем вычлененные элементы до их взаимодей-
ствия как «животные», то по результатам взаимодействия уже мо-
жем разделить их на «хищника» и «жертву». Анализ вещественной
стороны элементов или выяснение их пространственных отноше-
ний лишь в малой степени позволяют судить об их функциональ-
ной стороне: полноценно это возможно лишь на основании выяс-
нения взаимоотношений элементов. Степень определенности от-
ношений — информация — это уже характеристика не материаль-
ных элементов или процессов, а идеальной составляющей этой
системы. Но можно ли считать отношения и функции лишь не су-
ществующим в природе отражением процессов и тел в нашем со-
знании? Если системы существуют реально и наши представления
о них не ошибочны, то мы должны признать как факт, что строе-
ние и функция элемента взаимоопределяют друг друга, а вместе
они зависят от положения элемента в системе. И так же, как мате-
риальные элементы определяют материальные процессы в систе-
ме, отношения устанавливаются именно между функциональны-
ми сторонами элементов, то есть между идеальными элементами.
Функции и отношения могут быть как частными, так и общими,
дополняющими и исключающими остальные, что позволяет допу-
стить существование сложно структурированной нематериальной
сферы систем. А из этого следует двуединость материальной и
идеальной составляющих в любой реальной системе.
Материальные системы с дискретными вещественными эле-
ментами— это только логическая конструкция, которую мы-ис-
34
пользовали для удобства в рассуждениях. Она помогла нам позна-
комиться с основной терминологией и некоторыми схемами пост-
роения и функционирования самих систем. Поэтому к сказанно-
му ранее добавим, что: а) любая система включает не только веще-
ственные элементы, но также элементы и внутреннюю среду, по-
строенные из других видов материи; б) каждый элемент в той или
иной форме и степени взаимодействует не только с «родственны-
ми», но и с другими материальными элементами и средой (мате-
рией, не оформленной в элементы) данной системы, если такое
взаимодействие возможно; в) идеальная сфера системы также долж-
на быть неоднородной (аналогично материи), а отношения между
идеальными элементами — многообразными.
Сформулируем кратко основные положения раздела «Матери-
альное и идеальное в системе» и следствия из них.
1. Любая система включает упорядоченные в пространстве и во
времени элементы.
2. Элементы имеют двойственную природу: их материальная
составляющая во взаимодействии подчинена идеальной составля-
ющей, но в то же самое время определяет ее реализацию в отно-
шениях.
3. Система удерживается как целостность благодаря заключен-
ной в ней энергии и в ряде случаев способна высвобождать часть
энергии, совершая при этом работу.
4. Мы принимаем Вселенную за наиболее емкую по уровню из
всех известных нам систем.
5. Системы образуют иерархическую соподчиненность, и эле-
менты «низших» систем являются одновременно элементами сис-
тем всех и каждого из более емких уровней. Так, из элементарных
частиц состоят не только атомы, но также молекулы, веществен-
ные частицы, тела, планеты, солнечные системы, галактики и
Вселенная, а человек является членом семьи, жителем населенно-
го пункта, гражданином государства, представителем человече-
ства.
6. Многоплановость и иерархичность систем обусловливают
истинность всеобщих законов сохранения массы, энергии и ин-
формации.
7. Многообразие видов материи предопределяет равновероят-
ную возможность существования систем, элементы которых «по-
строены» не из вещества (в основе), а из других видов материи,
например плазмы. Исходя из этого, допускается возможность су-
ществования «параллельных миров» со своими независимыми
формами жизни.
8. Поскольку возможны превращения одних видов материи в
другие, возможны также переходы между параллельными мирами
и разными формами жизни.
35
9. Как разные виды и формы материи образуют единую много-
плановую систему — Вселенную, так разные виды и формы идеи
образуют единую многоплановую Вселенскую (Космическую)
идею.
2.1.8. ФУНКЦИОНИРОВАНИЕ СИСТЕМ
Ранее было отмечено, что системы подразделяются на замкну-
тые и открытые. Строго говоря, замкнутых систем в природе не
существует. Любая реальная система обменивается с окружающей
средой материей, энергией и идеей (информацией). В зависимос-
ти от направления потоков этих трех составных частей неразрыв-
ной триады (в систему, в системе или из системы) и соотношений
их количества и качества системы могут быть разделены на дегра-
дирующие, стационарные (как неустойчивое промежуточное со-
стояние) и саморазвивающиеся. В деградирующей системе разру-
шается структура, рассеивается энергия, уменьшается запас ин-
формации. Причиной деградации служат исходная неспособность
неживых систем и нарушение способности живых систем контро-
лировать внутренние потоки энергии. В деградирующей системе
нарушается соответствие между строением и функцией (структу-
рой и информацией) при невозможности восполнять (развивать)
недостающее. Замкнутые (в пространстве) системы, если бы они
существовали, должны неизбежно деградировать.
Для саморазвивающихся систем характерны подчинение их
подсистем и элементов единой системной идее, формирование
все более сложной внутренней расчлененности и накопление ин-
формации. Здесь значительная часть энергии расходуется на орга-
низацию энергетических потоков, приводящих элементы и всю
систему в неравновесное состояние. При этом система становится
работоспособной, подчиняет себе прилежащую часть среды, из-
влекает из нее новые порции материи, энергии и информации.
Одновременно система с большей или меньшей полнотой воспро-
изводит собственное состояние и состояние приграничной внеш-
ней среды. Различают неполное, полное и расширенное воспроиз-
водство. Последнее предполагает увеличение длительности удер-
жания энергии в системе, повторное (многократное) использова-
ние вовлекаемой в систему материи и наращивание информации
через усложнение структуры материальной и идеальной сфер.
Саморазвивающиеся системы всегда имеют сложную структур-
ную организацию. В качестве их элементов выступают так называ-
емые функциональные системы (правильнее — функциональные
подсистемы, элементы), которые могут формироваться или неза-
висимо от системы, в которую включаются, или же в результате
разделения системы на комплементарные части. Именно к «функ-
36
циональным системам» приложимо в полной мере понятие сте-
пени воспроизводства. Как правило, расширенное воспроизвод-
ство одних идет за счет неполного воспроизводства других «функ-
циональных систем», и в итоге общее количество материи, дви-
жения и идеи в объединяющей их системе остается почти неиз-
менным.
Введение понятия функция для характеристики сложных само-
развивающихся (живых) систем и их элементов неизбежно. Но что
такое функция? В логической схеме соотношения понятий «функ-
ция» устанавливается рядом с понятием «строение» (элемента) и
между понятиями «идеальные элементы» и «отношения». Это сле-
дует понимать так, что каждый элемент системы в силу своей дво-
якой сущности не только определенным образом устроен, но так-
же и функционален, то есть выполняет специфическую функцию,
взаимодействуя с другими элементами не только определенным,
но и предопределенным образом. В силу этого строение элемента
адекватно той функции, которую он выполняет. Строение, форма
элемента определяются его системной функцией, то есть функци-
ей, задаваемой ему системой, реализация этой функции зависит от
наличия соответствующего элемента.
Если понимать развивающуюся систему как самоорганизующу-
юся целостность, то и поведение ее элементов должно быть под-
чинено «идее» системы, хотя наличные элементы накладывают ог-
раничения на функционирование и развитие самой системы. Вза-
имоотношения элементов и системы можно выразить в следую-
щих положениях.
1. Материальные элементы (не сама материя) порождаются си-
стемой в соответствии с развитием ее идеальной сферы.
2. Идея не способна к обособленному саморазвитию. Это воз-
можно лишь через ее реализацию материальными элементами.
3. Направление развитию системы задает внешняя информа-
ция, а пути развития определяются структурой самой системы.
4. Максимальной ценностью для элемента обладает информа-
ция того уровня системы, в который он входит непосредственно,
хотя поведение элемента в конечном счете подчиняется идее Все-
ленной.
5. Элементы не существуют вечно, но порождаются системой,
когда в ней развивается дефицит функции, и более не воспроизво-
дятся, когда потребность в них исчезает.
6. Система существует, пока ее элементы взаимодействуют. Си-
стема зависит от элементов, но и определяет их существование.
7. Идея системы реализуется через индивидуальное поведение
элементов.
8. В функционирующей системе под воздействием потока
внешней информации и реорганизации внутренней информации
37
развивается дисбаланс материальной и идеальной сфер, стимули-
рующий новообразования. Невозможность таких новообразова-
ний и чрезмерные флуктуационные новообразования могут разру-
шить систему.
9. Расширенное воспроизводство включает и сохранение суще-
ствующего, и новообразования на его основе. В устойчивых систе-
мах существует предельная мера новообразований.
10. Каждый элемент в сложных системах или род элементов в
простых системах уникален и обеспечивает их полноту и целост-
ность.
И. Каждый элемент в сложных системах строго индивидуален,
его строение и функция согласованы между собой и с положением
среди других структурных элементов.
12. Устойчивость сложных систем пропорциональна заключен-
ной в них информации — определенности отношений между эле-
ментами.
13. Структурные и энергетические преобразования в системах
определяются законами их функционирования и развития. Любые
флуктуации нарушают устойчивость системы.
14. Информационный шум (ложная информация) нарушает
функционирование системы.
Приведенные следствия в определенной мере могут служить и
правилами поведения человека (как элемента) в обществе (в сис-
теме).
2.2. СУЩНОСТЬ ЖИЗНИ В ОПРЕДЕЛЕНИЯХ
К пониманию сущности земной формы жизни наука подходит
только сейчас, хотя попытки найти границу между живым и нежи-
вым предпринимались давно. Именно к концу XXв. оказалось,
возможным объединить факты, накопленные в экспериментах, с
теоретическими разработками в астрономии, физике, кибернети-
ке, биологии, социологии. Жизнь как явление можно охарактери-
зовать, дав ей более или менее удовлетворительное определение.
Такое определение должно содержать характеристику не только
субстрата жизни, но также и субстанции, то есть не только мате-
риальной основы, но и сущности процессов, происходящих в жи-
вой системе. Эволюция формулировок определений жизни отра-
жает процесс накопления знаний о живом веществе и представле-
ний о главном содержании происходящих в нем процессов. Поче-
му речь вдет именно о веществе, если существуют и другие виды
материи? Земная форма жизни (как мы ее понимаем) — это пре-
имущественно вещественная форма. Активные элементы живой
системы Земли образованы в основном органическим веществом,
38
мигирое в сочетании с некоторыми минеральными веществами со-
игииляет каркас биосферы.
Первое приемлемое с позиций науки толкование специфики
жизни принадлежит Ф. Энгельсу: «Жизнь — это способ существо-
НАИия белковых тел, существенным моментом которого является
Постоянный обмен веществ с окружающей их внешней природой.
Причем с прекращением этого обмена веществ прекращается и
жизнь, что приводит к разложению белка».
Позже с открытием материальной основы наследственности
(информационных матричных молекул) в определение жизни на-
ряду с белками были включены нуклеиновые кислоты, а развитие
кибернетики (науки об оптимальном управлении и связи) создало
основу для рассмотрения живых тел как кибернетических систем,
что нашло отражение, например, в определении М. В. Волькен-
штейна: «Живые тела, существующие на Земле, представляют со-
бой открытые саморегулирующиеся и самовоспроизводящиеся си-
стемы, построенные из биополимеров белков и нуклеиновых кис-
лот».
На следующем этапе оказалось возможным вообще опустить
указание материального носителя жизни. Так, Б. М. Медников
считает, что «жизнь — это активное, идущее с затратой энергии,
поддержание и воспроизведение специфической структуры».
Наконец, отметим взгляд Онзагера и Моровица на данную про-
блему. Рассматривая жизнь как экологическое свойство, они счи-
тают, что именно экологические процессы отражают единство
разнородных сложных биологических систем, непрерывно вступа-
ющих друг с другом в самые различные, чрезвычайно сложные от-
ношения. Эти процессы объединяет общая тенденция: как можно
полнее использовать то, что окружающая среда поставляет им в
виде сырья и энергии. Полагая, что экологический подход к оп-
ределению жизни позволит выяснить главное, они характеризуют
жизнь как «свойство материи, приводящее к сопряженной цирку-
ляции биоэлементов в водной среде, движимой в конечном счете
энергией солнечного излучения по пути увеличения сложности».
К биоэлементам относят углерод, водород, азот, кислород,
фосфор и серу. Все они широко распространены в космосе. Все,
кроме фосфора, первоначально были газообразными, то есть на
них в первую очередь действовало солнечное излучение. Суще-
ствующая гипотеза о возможности построения органических ве-
ществ на основе кремния отвергается прежде всего потому, что
кремний не имеет газообразной формы и потому не может заме-
нить углерод.
Суммируя сведения о системной организации Вселенной с
приведенными определениями, можно сделать вывод о том, что
жизнь и нежизнь представляют собой нечетко разграниченные со-
39
стояния материи, где жизнь есть расширенное воспроизводство
информации в материальных системах. При этом соотношение
между этими состояниями определяется характером перераспре-
деления материи, энергии и информации и общими энтропийно-
негэнтропийными процессами, протекающими во Вселенной. Со-
ответственно состояниям отдельные подразделения Вселенной
обозначаются как живые и неживые системы, или живое и нежи-
вое состояния систем. Одной из живых систем, единственной из-
вестной нам, является биосфера Земли. Специфика земной фор-
мы жизни состоит в том, что она развивается преимущественно на
основе одного вида материи — вещества. Здесь вещественная ос-
нова образует совокупную пассивную неживую среду — неживое
вещество и активные живые тела — результат морфофункциональ-
ной дифференцировки совокупного живого вещества.
2.3. КРИТЕРИИ ЖИВЫХ СИСТЕМ
Хотя между неживым и живым состоянием систем нет четкой
границы ни в истории, ни в любой момент их одновременного су-
ществования, а функционирование неживых и живых систем на
молекулярном уровне подчиняется единым, наиболее общим за-
конам природы, все же можно выделить некоторые характерные
признаки (критерии) живых систем. Понятно, что при этом мы
ориентируемся на единственную известную нам живую систему —
биосферу Земли.
Перечень критериев, предложенных разными авторами, вклю-
чает: специфический химический состав, обмен веществ, самовос-
производство, наследственность и изменчивость, рост и развитие,
раздражимость, дискретность, саморегуляцию, ритмичность,
энергозависимость. Этот перечень вполне может быть принят, но
при условии, что в качестве живых систем могут рассматриваться
все девять (от молекулярного до биосферного) уровней (см. раз-
дел 2.5).
Среди приведенных критериев отсутствуют системообразую-
щие. Их введение обнаруживает, что критерии необходимо разде-
лить на относящиеся к системам и относящиеся к элементам сис-
тем.
Одним из основополагающих разграничительных критериев
между «живым» и «неживым» состояниями систем служит их спо-
собность или неспособность к поддержанию асимметрии, нерав-
новесное™. В частности, на эту особенность указывает Э. С. Бау-
эр в следующей формулировке: «Как в неживых системах устойчи-
во их равновесное состояние, характеризующееся минимальным
значением свободной энергии и максимальным энтропии, так в
40
живых системах устойчиво их стационарно неравновесное состоя-
ние, характеризующееся запасами свободной энергии и негэнтро-
пии, носителем которых является их структура». В земной, веще-
ственной, форме живые системы «привязаны» к узкому диапазону
состояний вещества. По определению Э. М. Галимова, живые сис-
темы—это «итеративные (повторяющиеся, воспроизводящиеся)
стационарные системы необратимых процессов, протекающих в
линейной области соотношений сил и процессов» — «между рав-
новесием и нелинейностью».
Рассмотрим характерные признаки биосферы как живой систе-
мы.
1. Специфическую химическую основу биосферы образуют
асимметричные линейные и глобулярные молекулы органических
веществ. Соотношение в них химических элементов резко отлича-
ется от соотношения этих же элементов в космических телах и
земной коре. Так, содержание углерода в живом веществе в
7800 раз выше, чем в земной коре, азота —- в 160 раз, водорода — в
70 раз, а кислорода — в 1,5 раза (табл. 2.2).
2.2. Содержание, %, наиболее распространенных элементов в космических телах
на Земле и в земной коре
Элементы | Хондриты | Земля 1 Земная кора | Живое вещество
Кислород 33,2 29,5 46.-47 65-75
Железо 27,2 34,6 4,6...5,7 0,01-0,10
Кремний 17,1 15,2 25,9-29,5
Магний 14,3 12,7 1,9.„2,4 0,01-0,10
Сера 1,93 1,93 Нет данных 0,1—1,0
Никель 1,64 2,39
Кальций 1,27 1,13 2,9-4,3 0,1...1.0
Алюминий 1,22 1,09 8,0-8,1 Нет данных
Натрий 0,64 0,57 2,3-2,8 0,01-0,10
Хром 0,29 0,26 #
Марганец 0,25 0,22 0,1
Фосфор 0,11 0,10 о,1 0,1-1,0
Кобальт 0,09 0,13 Нет данных
Калий 0,08 0,07 1,9-2,6 0,1-1,0
Титан 0,06 0,05 0,4
Углерод * * 15...18
Водород * * 0,01 8...10
Азот * * * 1,5-3,0
Хлор * • 0,01-0,10
•Менее 0,03 %.
2. Биосфера образована условно дискретными в пространстве-
времени, активно взаимодействующими с внешним окружением
органическими живыми телами и неживыми телами (неживой
41
средой), связанными в динамическое единство потоками веще-
ства, энергии и информации.
3. Для биосферы характерна способность к саморегуляции —
особой форме двигательной (в широком понимании) ответной ре-
акции на раздражения.
4. Циклические процессы и способность к саморегуляции по-
рождают в биосфере специфическую форму динамически устой-
чивого существования — ритмичность.
5. Постепенная реорганизация — эволюция — не является ха-
рактеристикой только лишь биосферы, но в ней реорганизация
целого достигается направленными эволюционными преобразова-
ниями частей.
6. Как и другие открытые системы, биосфера энергозависима,
но ее внутренняя организация образует сеть накопителей, транс-
форматоров и передатчиков энергии. При этом в живых телах на-
капливается преимущественно химическая энергия, а передает-
ся — электрическая. Часть свободной энергии живые системы ис-
пользуют для активизации энергетического «входа».
7. Будучи открытой системой, биосфера имеет не только внут-
реннюю, но и внешнюю среду. Ее атрибутным свойством следует
считать общий положительный баланс в обмене веществом, энер-
гией и информацией с внешней средой.
8. Важнейшая особенность, отличающая биосферу от извест-
ных нам неживых систем, заключается в том, что каждый вид
ее элементов функционально комплементарен сумме остальных.
В неживых системах элементы обычно равноценны и взаимозаме-
няемы, а если комплементарны, то морфологически.
9. Биосфера целостна: для нее характерна относительная завер-
шенность строения.
10. В биосфере время имеет растущую негэнтропийную состав-
ляющую, что обеспечивает устойчивый рост объема информации.
Приведенные особенности биосферы можно принять в каче-
стве базовых характеристик особой группы систем, обозначаемых
нами как «живые». Возможно, существуют и другие признаки, до-
полняющие отличия живых и неживых систем. В то же время, и
это необходимо отметить особо, живые системы—лишь разно-
видность материальных систем.
Следующие четыре признака характеризуют живые тела как ак-
тивные элементы живой системы:
1) воспроизводство в поколениях. Непрерывность живых сис-
тем во времени при дискретности (на каждый отдельный момент)
входящих в них живых тел достигается путем формирования но-
вых поколений этих тел. В основе такого воспроизводства лежат
реакции матричного синтеза;
2) рост и развитие. Индивидуальная жизнь живого тела вклю-
42
чает рост и развитие, посредством которых достигается и в про-
цессе которых реализуется функционально активное состояние
тола;
3) наследственность и изменчивость. Новые поколения живых
ТОЛ сохраняют большую часть признаков родительских поколе-
ний, но несут и новообразования. Наследственность и изменчи-
вость обеспечивают динамическую устойчивость всей системы;
4) раздражимость. Живые тела, образующие активную часть
живой системы, обладают разнообразными формами раздражи-
мости, которые проявляются в ответных двигательных реакци-
ях.
Сложно организованные тела-элементы могут обладать отдель-
ными характеристиками систем, но они не являются настоящими
системами, поскольку образованы лишь живым веществом и обла-
дают функциональностью.
2.4. ХАРАКТЕРНЫЕ ПРИЗНАКИ ЖИВОГО ВЕЩЕСТВА
При всей кажущейся очевидности отличий живой материи от
неживой, следующих из приведенных определений жизни и кри-
териев живых систем, между ними невозможно провести резкую
границу. Хотя наиболее просто устроенные «живые тела» — виру-
сы — вне клеток живых организмов, то есть в неактивной форме,
проявляют свойства кристаллов и в определенном смысле сближа-
ются с неживыми телами, их нельзя рассматривать в качестве пе-
реходной ступени между неживым и живым веществом. Это
крайне специализированные живые тела, которые могли появить-
ся в биосфере не ранее, чем полноценные клеточные организмы,
и уж в любом случае — после появления нуклеиновых кислот (ге-
нетических матриц).
Мы не можем точно воспроизвести детали переходов вещества
от химической к предбиологической и далее к биологической эво-
люции, тем более что этот процесс был чрезвычайно длительным
и, вероятнее всего, неодноактным. Поэтому попробуем понять су-
щество отличий живого вещества от неживого, ориентируясь на
современное состояние природы.
1. Живое вещество биосферы характеризуется огромной сво-
бодной энергией. Это, по выражению В. И. Вернадского, форма
активированной материи. В отличие от неживой материи, осуще-
ствляющей пассивное взаимодействие, живая материя активно
взаимодействует с окружающей средой, чем, с одной стороны,
обеспечивает поддержание собственного существования, а с дру-
гой — одновременно изменяет среду, делая невозможным прежнее
неизменное существование в ней.
43
2. В живом веществе резко (в 103... 105 6 раз) ускоряется протека-
ние химических реакций, что объясняется действием ферментов.
При этом реакции носят в большей или меньшей степени контро-
лируемый характер.
3. Индивидуальные химические соединения, слагающие живое
вещество, устойчивы только в живых телах, в том числе и минера-
лы, входящие в живое вещество. Эту особенность подметил еще
Ф. Энгельс, указывая, что «смерть есть разрушение органического
тела, ничего не оставляющего после себя, кроме химических со-
ставных частей, образовавших его субстанцию».
4. Общим признаком всякого живого естественного тела в био-
сфере является в значительной степени регулируемое произволь-
ное движение. Выделяют две специфические формы движения
живого вещества: пассивную, которая создается размножением и
присуща всем живым организмам, и активную, которая осуществ-
ляется за счет направленного перемещения организмов и особен-
но наглядно проявляется у животных.
Живое вещество стремится заполнить собой все доступное пространство. Свой-
ство к максимальной экспансии (распространению, расширению) присуще живому
веществу так же, как свойственно теплоте переходить от более нагретых тел к менее
нагретым, растворяемому веществу находиться в растворе, а газу — рассеиваться в
пространстве. Экспансия живого вещества в неживой среде проявляется в подчине-
нии этой среды самому существу живого. Подчинить — означает сделать своим, ча-
стью себя или условием собственного существования. Как живое вещество вообще,
так и каждое живое тело в частности, осуществляя активное взаимодействие с внеш-
ней средой, тем самым реализует свое атрибутное свойство — способность и стрем-
ление к экспансии. В простейшем случае экспансия проявляется в способности жи-
вых тел воспринимать раздражения, воздействия внешней среды. «Опознание» сре-
ды для живого тела — начальный этап ее освоения, использования для собственного
воспроизводства. Проявлением экспансии можно считать и питание, то есть извле-
чение из среды порций вещества, энергии и информации для самовоспроизводства
и реализации системной функции. При этом часть внешней среды становится час-
тью тела. К проявлению экспансии относится и средообразующая деятельность жи-
вых тел и всего живого вещества, заключающаяся в создании условий для существо-
вания. Наиболее интересные формы экспансия приобретает у человека: общение,
познание, творчество, власть и др.
Движение живого вещества в его специфической форме, обозначаемой как
экспансия, предполагает захват нового при сохранении старого, что совпадает с
содержанием «расширенного самовоспроизводства». Об образовании нового при
неполном сохранении прежнего в предшествующих разделах речь шла уже неод-
нократно: криволинейное перемещение, трехмерность времени и др. Из достаточ-
но хорошо известных примеров этого рода приведем наследственную изменчи-
вость, сукцессию (последовательную смену) биоценозов, эволюцию биосферы.
Видимо, все это различные формы экспансии, а сама экспансия есть специфичес-
кое проявление трехмерности времени в особой группе неравновесных веще-
ственных систем — живых системах.
5. Живое вещество обнаруживает значительно большее морфо-
логическое и химическое разнообразие, чем неживое. Известно
более 2 млн различных органических соединений, входящих в со-
44
став живого вещества. В то же время количество природных со-
единений неживого вещества — минералов — составляет всего
около 2 тыс. Кроме того, в отличие от неживого, не связанного
своим происхождением с живыми организмами (абиогенного) ве-
щества живое вещество не бывает представлено исключительно
жидкой или газовой фазой. Тела организмов построены из ве-
ществ, находящихся во всех трех фазовых состояниях.
6. Органические вещества биогенного и абиогенного проис-
хождения имеют существенные отличия. В биоорганическом мире
полностью нарушена зеркальная симметрия (закон хиральной,
или киральной, чистоты). Что под этим подразумевается? Моле-
кулы, в которых имеются атомы углерода, связанные своими че-
тырьмя валентностями с разными «соседями», существуют в двух
зеркально противоположных формах. Их называют левыми и пра-
выми оптически активными зеркальными стереомерами. Так, все
природные аминокислоты — левые, а сахара — правые зеркальные
изомеры. При искусственном же синтезе этих веществ образуются
в равных отношениях и левые, и правые.
Открытие молекулярной асимметрии живой природы принадлежит Луи Пас-
теру. По его словам «асимметрия — единственная четкая демаркационная линия,
которую ... можно провести между химией живой и неживой природы». Сегодня
считается установленным, что все важнейшие вещества, играющие в организме
первостепенную роль, имеют определенный знак асимметрии, всегда один и тот
же д ля всех живых тел на нашей планете. Оптически активное вещество вне орга-
низма в течение определенного времени Становится нейтральным. Чтобы предот-
вратить это, требуется непрерывная затрата энергии. Между тем живой организм
поражает чистотой асимметрии, которая является непременным условием жизни
(Акопян).
7. Живое вещество представлено в биосфере в виде дисперсных
(отдельных, рассеянных) тел — индивидуальных живых организ-
мов, размеры которых лежат в пределах от 2,5 10-9 до 1,5 • 102 м, а
неживое вещество, как непрерывное неживое тело, может иметь
протяженность порядка 10'м. Самые крупные в геологической
истории организмы встречаются в настоящее время: из живот-
ных — это киты, а из растений — секвойи и эвкалипты.
Непрерывная протяженность ряда неживых веществ на многие тысячи кило-
метров имеет стационарный характер и не зависит от времени. Живое вещество,
несмотря на дисперсность организации, при рассмотрении его в пространстве-
времени, то есть в динамике, также оказывается непрерывным. Это имеет место и
в случае с каждым отдельным видовым живым веществом, и в случае с разнород-
ными смесями от биоценоза до биосферы.
8. Будучи дисперсным, живое вещество никогда не находится
на Земле в морфологически чистой форме, то есть в виде популя-
ций организмов одного вида. Оно всегда представлено смесями —
45
комплексами популяций разных видов. Такие комплексы популя-
ций принято называть сообществами живых организмов или биоце-
нозами.
9. Живое вещество существует на Земле в форме непрерывного
чередования поколений. Благодаря этому современное живое ве-
щество, характеризуясь непрерывным обновлением, оказывается
генетически связанным с живым веществом всех прошлых геоло-
гических эпох. Краткая формулировка этой особенности: «Все
живое из живого», обозначается как «принцип Реди». В противо-
положность этому неживое вещество абиогенного происхождения
поступает в биосферу из космоса или вносится порциями из ни-
жележащих оболочек земного шара. Отдельные такие порции мо-
гут образовываться в результате протекания одинаковых процес-
сов и, таким образом, быть аналогичными по составу, но генети-
ческой связи между собой в общем случае они не имеют.
10. Характерным для живого вещества является наличие эволю-
ционного процесса. Воспроизводство живого вещества идет не по
типу точного копирования предшествующих поколений, а путем
новообразований на их основе. Общее направление эволюции жи-
вого вещества состоит в увеличении упорядоченности живых сис-
тем при накоплении свободной энергии. При этом упорядочива-
нию подвергаются прежде всего динамические отношения, про-
цессы. Способность неживых систем и элементов живых систем
переходить в более упорядоченное состояние, рассеивая свобод-
ную энергию, не является аналогом эволюционного процесса, так
как она реализуется в отдельных живых телах, а эволюция —в
ряду поколений живых тел.
11. А. В. Лапо (1987) включает в число отличительных призна-
ков живого и неживого вещества их массу за геологическое время.
Общая масса вещества живого происхождения — это сумма нарас-
тающим итогом массы живых тел за все время их существования,
что во много раз превышает массу ныне живущих на Земле орга-
низмов. В то же время масса неживого абиогенного вещества зем-
ного происхождения является постоянной величиной в геологи-
ческой истории. С учетом сказанного в пунктах 7 и 9 различие
здесь сводится к тому, что масса живого вещества на Земле увели-
чивается настолько, насколько уменьшается масса неживого ве-
щества: часть планетного вещества постепенно охватывается жиз-
нью.
2.5. УРОВНИ ОРГАНИЗАЦИИ ЖИВОЙ МАТЕРИИ
Выделение уровней организации живой материи до настоящего
времени представляет предмет дискуссий. При этом существую-
щие разногласия зачастую обусловлены неопределенностью Под-
46
хода: выделяются ли уровни организации живого вещества или
живых систем, или речь идет об уровнях, на которых протекают
специфические жизненные процессы, или же о субъективно выде-
ляемых уровнях рассмотрения, изучения живого. Наиболее пол-
ный перечень, включающий молекулярный, органоидный, ком-
партментный, клеточный, тканевый, органный, организменный,
популяционный, биоценотический/биогеоценотический и био-
сферный уровни, отражает ориентацию на структурную иерархию
живого вещества, хотя и не полностью соответствует ей. Эти уров-
ни не представляют однородной группы, не всегда соподчинены.
Далеко не все они соответствуют и критериям систем. Рассмотрим
каждый из них в отдельности.
1. Молекулярный уровень. Отдельные молекулы вещества, в
том числе и сложные полимерные, не обладают многими систем-
ными характеристиками. В то же время молекулярные комплексы,
объединенные сопряженными циклическими процессами (реак-
циями), в некоторых случаях можно рассматривать как простей-
шие живые системы. Это относится прежде всего к первичной мо-
лекулярной биосфере. Оказавшись «плодородным» субстратом
предбиологической эволюции, молекулярные комплексы сыграли
важнейшую роль в становлении и развитии жизни на Земле. Име-
ются сведения о существовании самовоспроизводящихся комп-
лексов и в настоящее время. В современной биосфере сложные
комплексы молекул-организаторов биохимических процессов об-
разуют так называемые «неклеточные формы жизни» и лишь ус-
ловно могут быть отнесены к живым телам, будучи тем не менее
облигатными элементами биосферы. В то же время, начиная
именно с молекулярного уровня, проявляется вещественная не-
равновесность, столь характерная для материальной составляю-
щей живых систем.
2. Органоидный уровень, или уровень молекулярных комплек-
сов. Органоиды, в том числе включающие биологические мембра-
ны, — это функциональные структуры клеток. Они представляют
собой высокоупорядоченные комплексы молекул, условно диск-
ретные, редко обладающие способностью к самовоспроизводству
и, следовательно, не реализующие наследственность и изменчи-
вость в матричном варианте. Только митохондрии и пластиды мо-
гут рассматриваться как дискретные самовоспроизводящиеся
тела, но именно их происхождение связывают с переходом клеток
к облигатному симбиозу. Органоиды не дифференцированы на
комплементарные части и в целом не могут быть отнесены к сис-
темам. Они скорее представляют уровень интеграции молекул жи-
вого вещества.
3. Компартментный уровень. Компартменты — это функцио-
нальные подразделения эукариотических и отчасти прокариоти-
47
ческих клеток, в которых поддерживаются специфические физи-
ко-химические параметры и протекают специфические процессы.
Они, как и органоиды, не существуют как обособленные «самодо-
статочные» живые тела и потому могут рассматриваться только
как промежуточный уровень организации живого вещества.
4. Клеточный уровень организации удовлетворяет некоторым
критериям систем. Клетка достаточно четко дифференцирована
на дополняющие друг друга функциональные части, имеет специ-
фичную внутреннюю среду и достаточно определенно соотносит-
ся с внешней средой. Уже поэтому становится возможным свобод-
ное существование одноклеточных живых тел. В активном состоя-
нии клетка является базовым элементом всех более емких уровней
организации живого вещества в биосфере. Она обладает хорошо
выраженным жизненным циклом, способна к самовоспроизведе-
нию, повторяя себя в следующем поколении, подвержена онтоге-
нетическим, а в длинном ряду поколений и эволюционным изме-
нениям. Строго говоря, на собственно клеточном уровне органи-
зации сейчас находятся только тела дробянок и протестов. Клетки
же, составляющие многоклеточные живые тела с тканевой орга-
низацией, более интенсивно выполняют часть функций, характер-
ных для одноклеточных тел, и не способны существовать обособ-
ленно (в естественных условиях). Следовательно, клетки в одних
случаях выступают в качестве самостоятельных живых тел, облада-
ющих свойствами систем, а в других случаях — в качестве струк-
турно-функциональных элементов таких тел.
5. Группы клеток общего происхождения, имеющие сходное
строение и выполняющие сходные функции, вместе с выделяе-
мым ими межклеточным веществом обозначают понятием ткани.
Как следует уже из определения, ткани — это специализирован-
ные многоклеточные образования, которые могут быть только ча-
стью системы или ее элемента. Они с необходимостью предпола-
гают наличие других, дополняющих частей. Из этого следует, что
тканевый уровень — это уровень интеграции элементарных под-
разделений многоклеточного живого тела, что не может счи-
таться полноценной системой и не должно считаться самодоста-
точным уровнем организации живого вещества. В то же время тал-
ломные или слоевищные тела по характеру организации подобны
тканям.
6. Органы — это функциональные части многоклеточного жи-
вого тела. Как и ткани, они не могут образовывать обособленного
завершенного целого, отвечающего критериям системы. Даже го-
воря о системах органов (системах жизнеобеспечения), вводя в
формулировку понятие «система», мы подразумеваем разделение
функциональной части на специализированные более дробные
части, но не придаем им значения систем определенного иерархи-
48
ческого уровня. Представляется, что органы — результат диффе-
ренцировки многоклеточного тканевого живого тела.
7. Центральным и одновременно элементарным уровнем орга-
низации живого вещества биосферы признается организменный.
Организмы — это отдельные простые или комплексные живые тела
(особи) или группы живых тел, объединенные общим жизненным
циклом (онтогенезом), реально участвующие в биогеоценотичес-
ких процессах и воспроизводящиеся в поколениях. Организмен-
ный уровень реализуется через живые тела, которые могут быть
образованы одиночной клеткой, талломом, многоклеточным те-
лом с обратимой или необратимой дифференцировкой клеток.
Организация более сложных тел — модульных, социальных и мо-
дульно-социальных — также соответствует организменному уров-
ню.
8. Правомочность выделения популяционного и видового уров-
ней до настоящего времени не определена. Есть, однако, несколь-
ко аргументов, не позволяющих рассматривать популяции и виды
аналогично разновидностям и типам тканей в качестве настоящих
систем. Во-первых, они включают (объединяют) однородные эле-
менты, которые не могут образовывать завершенное единство. Во-
вторых, размеры видовых популяций чаще всего устанавливаются
произвольно. В-третьих, жизнедеятельность популяций и видов,
их состояние в основном определяются тем биогеоценозом, в ко-
торый они входят. В-четвертых, между популяцией и видом нет
четкой границы, как в большинстве случаев нет границы между
организмом и популяцией, организмом и видом (см. главу 3). 77о-
пуляция и вид — объединение организмов, живое однородное ве-
щество, то есть уровень организации живого вещества.
9. В структурной иерархии вещества следует выделить биоце-
нотический уровень. Биоценоз составлен генетически разнород-
ным живым веществом — комплексом видовых популяций, орга-
низованных в пищевую цепь. В пределах пищевой цепи происхо-
дит передача вещества и энергии от звена к звену, сопровождаю-
щаяся устойчивыми биотическими отношениями (ценотической
информацией) между последовательными и параллельными зве-
ньями. Однако в биоценозе не образуется замкнутый вихревой по-
ток вещества, как и в организме, изолированном от внешней сре-
ды. Для завершения цикла необходима неживая составляющая —
часть внешней среды, которую населяет сообщество. То есть в от-
личие от организма и биоценоза как уровней организации веще-
ства в этом случае формируется биогеоценоз, а соответствую-
щий — биогеоценотический уровень уже является уровнем орга-
низации системы.
10. Биосфера — оболочка Земли, включающая все живое веще-
ство—биоту (биотический уровень организации живого веще-
4 Присный А В
49
Рис. 2.8. Схема соотношений уровней организации жизни на Земле
ства), часть газовой оболочки, всю водную оболочку и незначи-
тельную часть твердой оболочки. Для земной формы жизни это
наиболее емкий и полноценный уровень организации систем, ко-
торый может быть представлен как глобальный биогеоценоз и в
пределах которого замыкаются все биогеохимические циклы.
Таким образом, системная организация в полной мере присуща
лишь биосферному или глобальному молекулярному уровню, с
небольшими ограничениями — биогеоценотическому и условно —
организменному уровню (рис. 2.8). Отдельный живой организм
функционирует как система, создает определенную направлен-
ность вещественно-энергетического потока, но неспособен к по-
вторному использованию переработанного вещества и, следова-
тельно, длительному обособленному существованию даже при
притоке энергии. Биогеоценоз, будучи составленным из компле-
ментарных популяций организмов и некоторого объема среды,
обеспечивает не только поэтапную передачу вещества в пищевых
цепях, но вовлекает часть отработанных веществ в повторные
циклы —части общего круговорота, происходящего в биосфере,
то есть глобального круговорота химических элементов. Уже на
этом основании можно утверждать, что биосфера возникла рань-
ше оформленных организмов.
Обращает на себя внимание тот факт, что биосфера, обладаю-
щая всеми признаками систем, воспроизводится, самообновляясь
не в идентичных поколениях, а через новые поколения составля-
ющих ее активных элементов. Чем «выше» уровень подразделения
биосферы, тем менее явно проступает матричное воспроизводство
поколений, сглаживается прерывистость. Это заставляет думать,
что так называемые уровни организации живых систем представ-
ляют'уровни функционирования (реализации процессов) единой
и единственной планетной живой системы — биосферы, которая в
своем развитии не составляется из частей, а дифференцируется на
функциональные части.
Глава 3
ЖИВЫЕ ТЕЛА
В предыдущем разделе мы отметили, что содержание земной
формы жизни можно свести к накоплению упорядоченной нерав-
новесное™ в вещественных системах, сопряженному с каскадом
переносов и превращений энергии в них. В наиболее общем виде
последовательность энергетических процессов в живом веществе
включает: поглощение и связывание энергии; перенос и удержа-
ние ее до высвобождения; придание направления и формы высво-
бождаемой энергии; внесение энергии в окружающую среду; воз-
буждение среды (вовлечение ее в жизненные процессы). При этом
поглощается рассеянная, а выделяется концентрированная на-
правленно высвобождаемая энергия. Одновременно значительная
часть энергии расходуется на самоподдержание живого вещества
как особой формы неравновесное™.
Все энергетические процессы происходят в материальных сис-
темах при прямом участии их элементов, в данном случае — .веще-
ственных элементов. В рамках нашего рассмотрения живое веще-
ство образует живые тела, условно ограниченные в пространстве и
во времени. На биосферном уровне живое вещество неоднородно,
но едино. Единство разнородности просматривается также на всех
уровнях функционирования биосферы: на молекулярном — это
единство химических элементов, малых молекул, фундаменталь-
ных процессов, протекающих в элементарных структурно-функ-
циональных единицах живых тел — клетках; на клеточном — по-
вторяемость строения клеток и участие клеток одинаковых типов
в построении генетически разнородных живых тел. Клетки — эле-
ментарные единицы видового живого вещества — образуют реаль-
ные живые тела — особи или объединения особей.
Собственно дискретные реально существующие вещественные
живые тела и являются предметом рассмотрения настоящего раз-
дела. К живым телам мы будем относить отдельные, обособленно
существующие клетки, клеточные ассоциации и их производные,
многоклеточные тела как в активном, так и в покоящемся состоя-
нии. Вирусы и близкие к ним образования — плазмиды, эписомы
52
и прионы не отнесены к живым телам, поскольку вне живых кле-
ток они не проявляют признаков жизни, а, включаясь в них, упо-
добляются активным функциональным частям клеток.
Одновременно нам предстоит рассмотреть один из узловых
уровней организации живого вещества, а именно — организмен-
ный. Именно этот уровень представлен не просто реальными жи-
выми телами, но и элементарными структурно-функциональными
подразделениями активной подсистемы биосферы. Организмен-
ный уровень организации живого вещества предполагает непос-
редственную реализацию элементарных системных функций в
конкретных условиях и способность к полноценному воспроиз-
водству путем образования дочерних поколений живых тел.
В качестве ключевых понятий, рассматриваемых в данном раз-
деле, выделим: организм, особь и индивидуум как обобщенные
типы живых тел. Каждый из этих типов может быть представлен
простыми (одиночными) и сложными (сборными) телами, а слож-
ные, в свою очередь, непрерывными и дискретными. Дискрет-
ность сложных тел проявляется в пространстве, во времени или в
пространстве — времени. В конкретном выражении живые тела
имеют разнообразные планы строения. Основные из них: прока-
риотическая клетка (Архебактерии, Бактерии и Цианобактерии);
эукариотическая клетка (Протисты, то есть одноклеточные, Гри-
бы, Растения и Животные); плазмодий (часть Протистов); псевдо-
плазмодий (часть Протистов); таллом (часть Растений, Лишайни-
ки и Грибы); бестканевое тело (Пластинчатые и Губки); тканевое
тело (Сосудистые растения и Эуметазои); клеточная группа (Про-
кариоты); колония (Прокариоты, Протисты, Паразои, Эумета-
зои); колониевидное тело (Инфузории, Сифонофоры); модульное
тело (часть Семенных растений, часть теплокровных Позвоноч-
ных); социальное тело (Термиты, часть Перепончатокрылых); мо-
дульно-социальное тело (Человек).
3.1. ОРГАНИЗМ, ОСОБЬ, ИНДИВИДУУМ (В ОПРЕДЕЛЕНИЯХ)
Один из величайших парадоксов современной науки о живом
состоит в отсутствии единого понимания таких базовых биологи-
ческих терминов, как организм, особь, индивидуум. Какие бы ас-
пекты биологического знания мы ни взяли в качестве примера,
всегда обнаружим смешение понятий «особь» и «организм»,
«особь» и «индивидуум», «организм» и «индивидуум». Даже про-
исхождение этих терминов трактуется неодинаково. Сравним со-
держание соответствующих статей в Словаре биологических поня-
тий и терминов (Реймерс, 1988) и Биологическом энциклопеди-
ческом словаре (1989):
53
«Организм (от гр. органон, лат. организмус — орудие, инстру-
мент):
1) любое живое существо, целостная система, реальный носи-
тель жизни, характеризующийся всеми ее свойствами; происходит
от одного зачатка: семени, споры, зиготы и т. д.; индивидуально
подвержен факторам эволюции и экологическим воздействиям;
2) отдельность живого как индивид с характерными для него
симбионтами;
3) биохимическая отдельность как составляющее живого веще-
ства биосферы (В. И. Вернадский);
4) любая биологическая и биокосная система, состоящая из
взаимосвязанных элементов, функционирующих как единое це-
лое (от особей-индивидов и колоний до популяций и биогеоцено-
зов)».
«Организм (от среднелат. organizo — устраиваю, придаю строй-
ный вид). В широком, самом общем смысле живой организм —
любая биологическая или биокосная целостная система, состоя-
щая из взаимодополняющих и соподчиненных элементов, взаимо-
отношения которых и особенности строения детерминированы их
функционированием как целого. В этом смысле в понятие «орга-
низм» входят не только особи (индивиды), но и колонии (колони-
альные организмы), семьи (у общественных животных), популя-
ции, биогеоценозы и т. д. В узком смысле организм — особь, ин-
дивидуум, живое существо».
«Особь — элементарная (неделимая) единица живого. Характе-
ризуется строгой взаимозависимостью непосредственно соприка-
сающихся функциональных частей. Отделение любой такой части
ведет к потере целостности и гибели особи. Как правило, особь
составлена индивидом со значительным числом неотрывных от
него видов других организмов. Понятие «особь» наиболее прило-
жимо к высшим неколониальным видам живого. Для колониаль-
ных вегетативно размножающихся и симбиотических форм оно
относительно: их морфологические части могут существовать не-
зависимо от целого и восстанавливать недостающие функцио-
нальные части (например, восстановление гидры из нескольких
клеток)».
«Особь — индивид, индивидуум, неделимая единица жизни.
Самый существенный признак особи — строгая взаимозависи-
мость отдельных частей: разделить особь на части без потери ин-
дивидуальности невозможно. В полной мере применимо к выс-
шим неколониальным организмам. Для колониальных вегетатив-
но размножающихся и симбиотических организмов понятие
«особь» относительно. С эволюционной точки зрения особь —
морфофизиологическая единица, происходящая из одной зиготы
или гаметы (в случае гаметофита, партеногенеза), от одной споры
54
или почки (при вегетативном размножении), индивидуально под-
лежащая действию элементарных эволюционных факторов».
«Индивид(отпат. индивидуум — неделимое). Элементарная еди-
ница жизни, экземпляр живого, имеющий все признаки, свой-
ственные виду, к которому он принадлежит, и вместе с тем отлича-
ющийся специфическими (генетическими и фенотипическими)
особенностями. В этом смысле — существо, происходящее от од-
ной зиготы, гаметы, споры, почки или другого зачатка и индивиду-
ально подверженное действию эволюционных факторов. В эколо-
гическом смысле ивдивид — живое существо в стерильной среде.
Пока вопрос об общебиологическом определении индивида у коло-
ниальных, симбиотических, а также размножающихся бесполым
путем организмов однозначно не решен. В качестве рабочей гипо-
тезы у этих форм можно принять за индивид (и одновременно
особь) отдельность, существующую самостоятельно или способную
жить в течение какого-то отрезка времени (от деления до деления,
от почкования до индивидуальной гибели), или минимальную
часть колонии и симбионта, способную существовать без явно вы-
раженных процессов дегенерации. Синоним индивидуум чаще
употребляется для обозначения человека-личности, субъекта».
Поскольку в понятиях «особь» и «организм» в той или иной
форме отражаются уровни организации живого вещества и живых
систем, попытаемся соотнести их именно в таком ракурсе. В каче-
стве исходной посылки примем одну из ранее отмеченных особен-
ностей живого вещества, а именно существование в виде дискрет-
ных тел-организмов. Данная посылка обязывает нас принять орга-
низменный уровень в качестве базового в последующих рассужде-
ниях-. Именно на организменном уровне обеспечивается и
реализуется текущее самовоспроизводство биосферы как полно-
ценной системы. Воспроизводство же организмов как элементов с
признаками системы включает и внутреннюю составляющую —
текущее самовоспроизводство через обновление собственных
структурных элементов, и внешнюю — образование нового, заме-
щающего поколения. Первое характерно для организма как систе-
мы, а второе — как для элемента системы. Полное замещение по-
колений живых тел имеет непосредственный выход на жизненные
циклы организмов или их полные онтогенезы. Будучи характер-
ным для организмов, замещающее воспроизводство не находит
выражения на надорганизменных уровнях подразделений биосфе-
ры, где воспроизводство реализуется только через воспроизвод-
ство базовых элементов-организмов в поколениях.
Для дальнейшего анализа содержания понятий «особь» и «орга-
низм» мы должны разобраться в типах воспроизводства («беспо-
лом» и «половом»), а также охарактеризовать пространственно-
временные отношения особи и организма.
55
Когда мы говорим об особи как морфологической отдельности,
фактически мы учитываем лишь пространственные отношения
вне временных, вне длительности. То есть это размерные, струк-
турные и тому подобные характеристики дискретных живых тел
на некоторый произвольно выбранный момент времени. Здесь
важно подчеркнуть, что снятие временных отношений устанавли-
вает запрет на функциональность тела, ибо процессы реализуются
в единстве пространственно-временных отношений. Тем не менее
мы можем «прослеживать» дискретное живое тело в последова-
тельности «срезов времени» в течение всего периода его существо-
вания. Но и в этом случае мы будем лишь фиксировать простран-
ственные, хотя и неодинаковые состояния. Видимо, можно при-
нять, учитывая сказанное, что особь — это дискретная единица
живого вещества на каждый конкретный момент времени от ее
образования или обособления от другой дискретной единицы до
индивидуальной смерти или разделения ее соматической части на
дочерние.
Отдельное тело может образовываться при слиянии других тел
как в случае с типичным оплодотворением или образованием
псевдоплазмодия и плазмодия, так и в результате разделения ро-
дительского тела на два или более дочерних — при митозе, фраг-
ментации, почковании и др. Судьба каждого тела также может
быть разной: деление, образование гамет, гибель до или после до-
стижения естественной «старости». В случае полного прохожде-
ния организмом отдельного жизненного цикла умирает только так
называемая соматическая часть тела, в то время как генеративная
часть оказывается «бессмертной», продлеваясь в генеративных те-
лах, на основе которых строится новое поколение тел с соматичес-
кой частью.
Следовательно, производя «срезы» на разных этапах индивиду-
ального цикла, мы потенциально можем обнаружить дискретные
тела: гаметы, споры, зиготы, личинки, семена, развитые живые
тела и т. д. Каждое из них с полным основанием может быть на-
звано особью, хотя и очевидна разнокачественность этих тел, при-
надлежащих одному организму.
Включение всех тел, характерных для полного жизненного
цикла, в единую непрерывную последовательность обязывает нас
принять во внимание (а иначе мы не получим непрерывности)
временной аспект. В пространстве—времени все особи, составля-
ющие видовое живое вещество, связываются в сеть, моменталь-
ный «срез» с которой при асинхронности развития поколений
включает более или менее полный набор характерных для него
тел-особей (рис. 3.1).
Единое пространственно-временное отношение предполагает
наличие причинно-следственного ряда, то есть взаимоопределяю-
56
Рис. 3.L Пространственно-
временная сеть видового жи-
вого вещества (оригинал
автора):
• — момент среза; <? — мужские
гаметы; 9 — женские гаметы;
3 — зигота; Э—эмбрион; Л —
личинка; БР — бесполое раз-
множение; НО—неполовозре-
лая особь; ПО — половозрелая
особь; ПР — половое размноже-
ние; СО —старая особь; стрел-
кой показано направление тече-
ния времени
щих материальных и идеальных процессов, или функциональнос-
ти дискретных единиц видового живого вещества. В приведенных
ранее определениях функциональный аспект связывался с поня-
тием «организм», поэтому в определении организма сделаем ак-
цент на то, что это функционирующее живое тело. Но, говоря об
уровнях организации живых систем и живого вещества, мы выде-
ляли в качестве базового подразделения организменный уровень,
заметим, уровень особей. В этом случае организмы рассматрива-
ются как элементы системы. Элементы же наделяются не только
строением (формой), но также и функцией (содержанием). Кроме
того, они функционально комплементарны сумме остальных эле-
ментов в системе. Следовательно, организм как элемент биосферы
(или ее некоторого подразделения) должен выполнять определен-
ную элементарную системную функцию.
Организмы, составляя видовое живое вещество, непосред-
ственно входят в биогеоценозы и выполняют в них определенные
ценотические функции, то есть они функционально активны. Раз-
ные этапы их полного жизненного цикла (поколения или фазы
индивидуального онтогенеза) неравноценны по характеру актив-
ности. Активность тел на отдельных этапах жизненного цикла мо-
жет быть направлена на другие элементы биогеоценоза, и тогда
они реализуют специфические биогеоценотические функции, или
же активность тел замкнута на данное видовое живое вещество для
выполнения определенных внутривидовых функций. Последнее
делает целесообразным разделение тел-особей на биогеоценотичес-
ки (далее — ценотически) активные и ценотически пассивные. Но
тогда мы были бы вынуждены исключить ценотически пассивные
тела-особи из живой системы, так как исходно предполагается
обязательная функциональная активность всех элементов в си-
стеме.
Возникшая проблема легко разрешается, если в состав организ-
ма мы включим все множество функционирующих тел-особей как
57
дискретных единиц видового живого вещества, составляющих за-
вершенный онтогенез (или полный жизненный цикл). Именно
такой организм способен реализовать ценотическую функцию в
относительно полном объеме и полноценно воспроизводиться в
поколениях по «замещающему» типу. При данном подходе снима-
ется проблема полярности тел, участвующих в половом воспроиз-
водстве, так как они рассматриваются в качестве свободных, но
необходимых комплементарных частей единого организма. Орга-
низм же строится на диффузно-комплементарном принципе, уни-
версальном для разных уровней организации вещества и систем в
целом. Примеры схем построения организмов приведены на ри-
сунке 7.4,
В свете излагаемых представлений организм может быть оди-
ночным телом (у части простейших, для которых неизвестен по-
ловой процесс в вице направленного переноса генетического ма-
териала от одного тела к другому), двумя телами (у прокариот и
части простейших с полярными типами тел-клеток без учета пере-
мещающихся из клетки в клетку хромосом или ядер), большим
числом тел, включающим последовательно или одновременно су-
ществующие однородные или разнородные функционирующие
особи. Далеко не всегда отдельное тело в длительности и протя-
женности представляет собой «чистый» ценотически активный
или пассивный тип. Чаще тело может включать и активную (сома-
тическую «рабочую»), и пассивную (генеративную и соматичес-
кую «обеспечивающую») части одновременно или же, трансфор-
мируясь, приобретать то или иное состояние (фазу). Следователь-
но, организм как диффузно-комплементарное образование может
последовательно или одновременно включать ценотически актив-
ные и пассивные тела, части и состояния.
Разделение функционирующих особей на ценотически актив-
ные и ценотически пассивные и принятие их за части организма
позволяют несколько иначе трактовать и формы воспроизводства.
Мы уже обозначили текущее самовоспроизводство и замещающее
воспроизводство в поколениях как понятия разного порядка. По-
скольку первое фактически обеспечивает существование тел в
длительности (во времени) и присуще всем без исключения жи-
вым телам, оно принимается как нечто само собой разумеющееся
и традиционно не связывается с понятием «воспроизводство». Го-
воря же о воспроизводстве, подразумевают прежде всего замеща-
ющее воспроизводство в поколениях ценотически активных жи-
вых тел, то есть формирование дочерних тел, подобных воспроиз-
водящимся. И именно к этому явлению относят так называемые
бесполую и половую формы размножения. В общем случае под
бесполым размножением понимают увеличение числа особей в ре-
зультате разделения материнского тела на два или более дочерних,
58
нс сопровождающееся комбинированием генетических матриц.
При половом размножении, как считается, должны сначала сфор-
мироваться, обычно многочисленные, половые клетки полярных
типов, которые затем, попарно сливаясь и составляя при этом но-
вые сочетания наследственных матриц, образуют увеличенное
Число дочерних особей.
Учитывая, что термин «размножение» по смыслу означает уве-
личение числа (особей), было бы более правильным в качестве об-
щего использовать термин «воспроизводство» в сочетании с уточ-
няющими терминами «половое» и «бесполое». Тогда такие явле-
ния, как половой процесс (конъюгация), вегетативное размноже-
ние (в частности — возобновление из зимующих органов) и
другие, не сопровождающиеся увеличением числа потомков, бу-
дут лишь частными случаями воспроизводства и не вступят с ним
В смысловое противоречие. Одновременно заметим, что половое
воспроизводство всегда предполагает комбинирование генетичес-
ких матриц, формирующихся в генеративной составляющей орга-
низма (его части или состоянии), а бесполое воспроизводство ос-
новывается на копировании прежде всего соматической составля-
ющей организма. Следовательно, с разными типами воспроизвод-
ства должны быть связаны и разные формы изменчивости:
комбинативная (комбинационная) — с половым, вариационная —
с бесполым, мутационная — это лишь информационный шум, со-
провождающий и то и другое.
Организм — элементарная полноценная функциональная еди-
ница биосферы, и в этом смысле он является носителем элемен-
тарной биосферной информации, то есть элементарной информа-
ционной единицей. Через организмы внешняя информация пре-
образуется во внутреннюю информацию живого вещества и вос-
производится в преобразованном виде путем тиражирования
реорганизованных матриц (молекул нуклеиновых кислот). Но
преобразование информации осуществляется в ходе участия орга-
низмов в биогеоценотических процессах. Значит, воспроизводству
предшествуют реализация, воплощение функции через матери-
альную структуру организмов. Биогеоценотическая среда, в кото-
рой действуют организмы, крайне неоднородна, что в основном и
определяет внутривидовое разнообразие организмов. Поэтому от-
дельные организмы, представляющие видовое живое вещество,
для обеспечения видовых приспособлений к изменяющейся сре-
де, то есть для выполнения функций в развивающейся системе,
«обязаны» суммировать, объединять частные вариации. Следова-
тельно, в интеграции видовой информации заключается смысл
воспроизводства, предполагающего комбинирование преобразо-
ванных генетических матриц.
Комбинирование матриц возможно лишь при условии их пере-
59
мещения (активного или пассивного) в ядре, в клетке, между
клетками, между частями тела, между комплементарными телами
организма в пределах видовой популяции. Матрицы в транспорт-
ном положении принято называть гаметами. Гаметы могут быть
активно подвижными (с помощью фибрилл цитоскелета, клеток-
носительниц, роста или движения сопутствующих или вспомога-
тельных клеток) или же неподвижными. Это хромосомы, переме-
щающиеся из одной хромосомной группы в другую или из одной
клетки в другую, — хромогаметы, подвижные и неподвижные,
сливающиеся в синкарион ядра — нуклеогаметы, гаметы клеточ-
ного строения — цитогаметы и многоклеточные гаметные тела —
сомогаметы. Кроме того, фрагменты матриц могут пассивно
транспортироваться специфическими переносчиками типа виру-
сов. В большинстве случаев хромосомный набор гамет, являю-
щихся ценотически пассивными особями организма, уменьшен
вдвое по сравнению с клетками ценотически активных особей,
хотя в принципе нет разницы: будет ли гамета диплоидной или
гаплоидной, — в любом случае она должна обеспечить комбини-
рование матриц. Для тиражирования матриц формируются специ-
альные внутренние или внешние обособленные или необособлен-
ные тела различного строения и относительных размеров: начиная
с превращения клетки-особи в предшественник гаметы (напри-
мер, гамонт у споровиков) и заканчивая длительно существующи-
ми сложными гонадами или кратковременно существующими це-
нотически пассивными полоносными особями. Все эти ценоти-
чески пассивные тела (состояния, органы и особи организма) долж-
ны иметь надежную информационную связь, морфологическую
или хотя бы дистанционную коммуникацию с ценотически актив-
ными состояниями, органами или особями организма, которая
обеспечивала бы прохождение потока информации к матрицам.
Эта связь может осуществляться через внутренние среды тела, по-
средством диффузно распространяющихся веществ, физических
взаимодействий. И если ценотически активные особи организма
стерильны (бесплодны), как у общественных насекомых, то они
одновременно служат и «выносными рецепторами» ценотически
пассивных (плодущих) особей, и имеют развитый язык химичес-
кой, электромагнитной, зрительной, звуковой, механической или
иной формы коммуникации.
Не участвуя непосредственно в биогеоценотических процессах,
генеративные составляющие организмов тем не менее оказывают-
ся столь же необходимыми, как и соматические. С одной стороны,
они обеспечивают отзывчивую на внешнюю информацию непре-
рывность (во времени) видового живого вещества, а с другой —
входя в состав конкретного тела, обусловливают морфологичес-
кие, физиологические и поведенческие признаки половой комп-
60
лементарности или полярности. Число комплементарных типов в
пределах вида может составлять от одной до двадцати (у некото-
рых инфузорий) пар. Но благодаря чему у организмов имеются
особи комплементарных типов? Прежде всего благодаря возмож-
ности комбинирования матриц с накопленной и реорганизован-
ной внешней информацией и соответственно возможности созда-
ния совокупной динамичной видовой информации. Во-вторых,
при образовании гамет обеспечивается более полная выбраковка
мутаций путем перевода их в гаплоидное, то есть «гомозиготное»,
состояние. Наконец, диффузно-комплементарный принцип пост-
роения организма является единственным способом существова-
ния популяции, а в пределе — и вида как единого целого. В силу
отсутствия четких границ между организмом и популяцией/видом
можно говорить об организменно-видовом уровне подразделения
живого вещества.
Традиционный подход к живому телу как системе в рамках раз-
виваемой гипотезы не может быть принят, поскольку даже функ-
ционирующее живое тело не удовлетворяет некоторым критериям
живых систем (эволюционному развитию, устойчивому росту ин-
формации и др.). Отдельное тело часто не является даже полно-
ценным элементом живой системы. Так, оно может не обладать
способностью к реализации полной элементарной биоценотичес-
кой функции, как правило, не может воспроизводиться в дочер-
нем поколении с реорганизацией накопленной информации соот-
ветственно внешней (системной) информации, в определенной
части индивидуального развития не имеет морфологической за-
вершенности. В большей мере системообразующие критерии про-
являются у организма, составленного из ценотически активных и
пассивных тел-особей. Однако и здесь мы видим комплексное
тело, образованное только живым веществом — частью одной из
подсистем полноценной живой системы — биосферы. Именно по-
этому и функционирующую особь, и организм, и популяцию, и
вид, и биоценоз, и биоту в целом следует рассматривать лишь как
уровни подразделения живого вещества, в большей или меньшей
степени обладающие отдельными свойствами систем.
Приведенные рассуждения позволяют трактовать организм как
простое (образованное одной особью) или сложное (состоящее из
двух или более особей), компактное (непрерывное) или диффуз-
но-комплементарное (дискретное) тело во временных границах
полного онтогенеза (жизненного цикла), способное выполнять
определенные биогеоценотические функции, усваивать внешнюю
информацию и обеспечивать воспроизводство следующего поко-
ления аналогичных тел всеми характерными для вида способами.
Тогда особи — это ценотически активные и ценотически пассив-
ные простые или сложные обособленные или независимые отдру-
61
гих в индивидуальной жизни тела одного организма. Каждая
особь на определенном этапе жизнедеятельности выступает по от-
ношению к части остальных особей своего вида как морфофунк-
циональный или функциональный комплементарный тип. Дис-
персностью живых тел определяется диффузно-комплементарный
принцип построения организмов, то есть принцип, соответствен-
но которому каждая из функциональных особей вида потенциаль-
но может занять строго определенное место дополнительно к осо-
бям других типов в составе организма. Обычно отдельная особь
способна воспроизводиться лишь одним из характерных для вида
способов.
Рассмотрим некоторые из основных типов особей. Первую
группу образуют особи, связанные, хотя бы часть времени инди-
видуального существования, с организацией потоков вещества,
энергии и информации соответственно биогеоценотической функ-
ции вида. Они могут быть одноклеточными, псевдоплазмодиаль-
ными, плазмодиальными, многоклеточными с одинаковыми или
разными клетками, модульными и т. д. Во вторую группу входят
так называемые ценотически пассивные особи. Это клеточные га-
меты, споры, цисты, плоды и семена, отделяющиеся почки, опор-
ные, запасающие и непитающиеся полоносные особи. Нередко
пассивные особи не обособлены от активных особей, а являются
их частью или временным состоянием. В качестве примеров мож-
но привести: расселительно-проникающую стадию (спорозоит,
мерозоит) у споровиков, плавающие «бродяжки» у сосущих инфу-
зорий или личинки губок, кишечнополостных, некоторых плос-
ких и кольчатых червей, моллюсков, (условно) взрослые полонос-
ные особи муравьев и термитов, половые железы ценотически ак-
тивных особей многоклеточных животных, гаметофиты высших
растений, плодовые тела грибов и др.
Пассивные особи (части, состояния) выполняют вполне опре-
деленные функции в организме: расселение, переживание небла-
гоприятных условий, интенсивную реорганизацию строения, за-
мыкание жизненного цикла, производство и комбинирование ге-
нетических матриц, временное накопление питательных веществ
И др.
Наиболее простым вариантом построения организма является
пара комплементарных особей с последовательной сменой трех
фаз активности (ценотически пассивной расселительной или мор-
фогенетической, ценотически активной, ценотически пассивной
репродуктивной) и ядерными или хромосомными гаметами. В бо-
лее сложных вариантах обособляются репродуктивное поколение
и поколение клеточных гамет или гаметных многоклеточных тел.
Особой сложности достигают организмы, выполняющие сложные
биогеоценотические функции. Здесь «активное» поколение может
62
включать две различные фазы или же вообще распадаться на два
(или более) поколения. Есть и другой путь предельного усложне-
ния строения организма: развитие опосредованно (через взрослую
фазу) ценотически активной «детской» фазы (при проявлении за-
боты о потомстве) с разделением взрослых функционирующих
особей на различные типы с телесными рабочими органами и вне-
телесными усилителями рабочих органов.
Систематические группы, традиционно обозначаемые как
Простейшие и Многоклеточные, объединяют весьма близкие ва-
рианты строения организмов, но при таком рассмотрении раз-
деляются без труда на основании лишь одного признака —ко-
личества типов клеток, слагающих ценотически активную часть
или фазу, или особь организма. Если это один тип, одна жизнен-
ная форма клеток, организм — одноклеточный, простейший; если
два или более — многоклеточный. Первые могут дать только не-
сколько десятков жизненных форм, зато вторые—по меньшей
мере несколько тысяч.
Мы также должны принять во внимание, что природные жи-
вые тела клеточного строения всегда (а другого пока не отмечено)
содержат в себе генетически инородные матрицы или тела клеточ-
ного строения. Бактерии содержат различного рода вирусные час-
тицы и свободные фрагменты нуклеиновых кислот. В клетках во-
дорослей, грибов, простейших, как правило, обитают прокариоти-
ческие клетки и вирусы, реже — эукариотические клетки. В мно-
гоклеточных организмах поселяются, кроме того, грибы, другие
многоклеточные и более обильно — представители бактерий и
простейших. Далеко не всегда такое сожительство носит антаго-
нистический характер. Чаще это облигатный и факультативный
симбиох Следовательно, можно утверждать, что организм всегда
неоднороден генетически, но целостен функционально и подчи-
нен программе, записанной на всех генетических матрицах, вхо-
дящих в его состав.
3.2. КЛЕТКА
Клетки современных живых тел представляют собой результат
длительной эволюции живого вещества и, участвуя в построении
подразделений живых систем, обязательно выполняют строго оп-
ределенные функции. Поэтому недифференцированные или не-
специализированные клетки в реальных живых телах отсутствуют.
Иными словами, недифференцированная клетка —это абстрак-
ция, включающая наиболее общие признаки клеток.
Отвлекаясь от специфики конкретных доступных наблюдению
клеток, мы можем вести речь о неких генерализованных типах
63
прокариотической и эукариотической клеток, названия которых
уже подчеркивают специфику их строения и функциональных
особенностей. Возникает парадоксальная ситуация: накопив дос-
таточно большой объем сведений о самых разнообразных видах
клеток, мы испытываем затруднения в формулировке емкой ха-
рактеристики единого плана строения для всех клеток. Причина
затруднения заключается, видимо, в том, что мы не имеем факти-
ческих данных о строении протоклетки. Хотя и здесь уместен воп-
рос: а была ли единая протоклетка? — то есть имели ли первые
живые тела единый план строения? От ответа на этот вопрос зави-
сит правильность многих наших представлений о живом веществе.
Будет менее ошибочным допущение исходного разнообразия про-
тобионтов, сформировавшихся в биосфере на ее предбиологичес-
ком этапе становления. Но, относя клетки разных планов строе-
ния к единому уровню организации живого вещества, попробуем
все же дать им единую характеристику, которая должна быть при-
годной и для современных клеток как элементарных живых тел.
Произвести такую операцию можно путем выделения общих при-
знаков клеток организмов, относящихся к разным царствам (с
учетом разнообразия классификационных построений).
Известные функциональные части различных клеток составля-
ют внушительный список. Мы ограничимся перечислением наи-
более регулярных из них, исключив, кроме того, органические и
минеральные включения. Но прежде условимся, что мы будем по-
нимать под названием «клетка». Клетка может быть определена
как минимальный сгусток живого вещества, способный организо-
вать пропуск через себя потока вещества и энергии, усваивать,
хранить, новообразовывать и воспроизводить информацию о соб-
ственных строении и функции. Перечисленные свойства клетки
обеспечиваются ее внутренней дифференцировкой на функцио-
нальные части — компартменты и органоиды, в силу чего клетка
приобретает многие характерные черты живой системы.
Традиционно клетка определяется как основная структурно-
функциональная единица всех живых организмов, элементарная
живая система. Клетка может существовать как отдельное живое
тело (архебактерии, бактерии, цианобактерии, простейшие, водо-
росли и грибы), в составе тел бестканевых животных и тканей
многоклеточных растений и животных или же участвовать в пост-
роении талломов грибов и растений.
Содержимое клетки — протоплазма. Каждая клетка, кроме не-
которых специализированных в зрелом состоянии, содержит ге-
нетический аппарат в нуклеоиде (прокариотическая) или в ядре,
отделенном от цитоплазмы мембранами (эукариотическая). Раз-
меры клеток варьируют в пределах от 0,1 мкм до 155 мм (редко —
более). Многообразные функции клеток выполняют специализи-
64
рованные структуры — органоиды. Универсальные органоиды: на-
ружная мембрана, рибосомы и хромосомы. Для эукариотических
клеток, кроме того, характерны митохондрии (хондриосомы), эн-
доплазматическая сеть, аппарат (комплекс) Гольджи и лизосомы.
Во многих клетках присутствуют также структуры, способствую-
щие поддержанию формы и ее изменению: микротрубочки и фиб-
риллы. Все клетки эукариот имеют сходный набор (общеклеточ-
ных) органоидов, сходно регулируют метаболизм, запасают и рас-
ходуют энергию, сходно с прокариотами используют генетичес-
кий код для синтеза белков. У них принципиально сходно
функционируют мембраны.
Сравнение наборов функциональных частей в разных умозри-
тельно выделенных типах клеток показывает, что наиболее сильно
отличаются про- и эукариотические клетки. Среди прокариот
большей морфологической обособленностью обладают цианобак-
терии. У эукариот можно выделить три четко определяемых кле-
точных типа: растительный, грибной и животный (рис. 3.2). Гра-
ницы между ними частично сглаживаются клеточными типами
простейших. Пользуясь данными таблицы 3.1, мы можем обозна-
Рис. 3.2. Главнейшие типы клеток (оригинал автора):
/ — бактериальный; 2—цианобактерий: 5—грибной; 4—животный; 5—растительный
5 Присный А В
чить наиболее характерные признаки главнейших клеточных ти-
пов соответственно подразделению организмов на царства (по со-
вокупности классификационных построений) (рис. 3.3).
3.1. Встречаемость главнейших органоидов в основных типах клеток
Органоиды Тип клеток
архибак- I торий бактери- альный циано- бакте- рий гетеро- трофных простей- ших автотроф- ных про- стейших расти- тельный грибной |
Наружная мембрана + + + + + + + + Протоплазма +4-4- + + + + + Кольцевая ДНК ++ + ± + ± ± ± Палочковидные хромосомы — — — + + ++ + Эндоплазматическая сеть _ _ _ + + + + +- Аппарат (комплекс) Гольджи _ _ _ + + + + — Прокариотические рибосомы + ++ ± ± ± ± ± Эукариотические рибосомы _ _ _ 4- + + + + Филаменты — — — + + ++ + Центриоли и кинетосомы — — — + + + + + Лизосомы — — - + — — — + Пиноцигозные вакуоли — — - + — - — 4- Окаймленные ямки, пузырьки — — — — — — — + Десмосомы — — — _ — _ _ + Плотные контакты - — — — - - — + Плазмодесмы — — — _ — + _ — Центральная вакуоль — — — _ _ +- _ _ Пульсирующие вакуоли — — - + + — — — Пищеварительные вакуоли _ _ _ + _ ’ _ _ + Клеточная оболочка (стенка) + ++ + + + + — Митохондрии — — — + + + + + Хлоропласты — — — — + + — — Гликокаликс — — + + — — — + Фотосинтезирующие мембраны + — + — ± ± — — Мезосома ++— — _ __ _ _ Жгутиковая нить ++— — — — — —
Примечание. (+)—органоиды, встречающиеся в клетках, (^ — органо-
иды, не встречающиеся в клетках, (±) — субструктуры: структуры, принадлежа-
щие потомкам облигатных симбионтов, входящих в состав современных эукарио-
тических клеток — митохондрий и пластид, и не относящиеся к облигатным об-
щеклеточным органоидам эукариот.
Рассмотрим общий план строения недифференцированной
клетки. Непосредственное наблюдение клеток подсказывает, что
более других понятию «недифференцированная клетка» соответ-
ствуют зародышевые и меристемные клетки тканевых тел до нача-
ла дифференцировки и одноклеточные тела непосредственно пос-
66
Клетка ПРОКАРИОТИЧЕСКАЯ ЭУКАРИОТИЧЕСКАЯ
Средние размеры, мкм 1...10 I0...100
Нуклеарная область Не обособлена Изолирована мембраной
Линейные молекулы ДНК Отсутствуют Имеются
Кольцевая ДНК В цитоплазме В митохондриях и пластидах
Рибосомы Одного типа Двух типов
Цитоскелет Отсутствует Обычно хорошо развит
Клеточные типы Архебак- териаль- ный Бактери- альный Циано- бактери- альный Протес- тов Расти- тельный Грибной Живот- ный
Клеточная оболочка (стенка) Без муреина Содержит муреин Отсутст- вует или китиноид- нам, или целлю- лозная Целлю- лозная Хитино- вая Отсутст- вует
Гликокаликс Отсутствует Имеется Часто имеется Отсутствует Имеется
Фотосинте- зирующие мембраны Иногда имеются Отсутст- вуют Имеются Имеются или отсутст- вуют Имеются Отсутствуют
Мезосома Имеется Отсутст- вует Отсутствует
Центриоли Отсутствуют Отсутст- вуют Иногда имеются Отсутст- вуют Имеются
Жгутиковые фибриллы Часто имеются Иногда имеются Отсутст- вуют Отсутствуют
Жгутики/ - реснички Отсутствуют Часто имеются Отсутст- вуют Отсутст- вуют Иногда имеются
Вакуоли Отсутствуют Пищева- ритель- ные, пуль- сирующие иля отсут- ствуют Крупные с клеточ- ным соком Отсутст- вуют Пищева- ритель- ные, АФ или отсутст- вуют
Рис. 3.3. Характерные признаки основных типов клеток
jiq завершения деления. Меристема — это образовательная ткань
растений, долго сохраняющая способность к делению и формиро-
ванию новых клеток. Она отличается высокой метаболической ак-
тивностью. Различают верхушечные и боковые меристемы. В те-
чение жизни растения на разное время могут сохраняться вставоч-
ные и краевые меристемы или возникать вторичные меристемы.
67
Иногда термин «меристема» применяют и по отношению к неспе-
циализированным клеткам животных, способным к дифференци-
ровке. Но на основе зародышевых и меристемных клеток можно
охарактеризовать обобщенный тип только эукариотической клет-
ки. Генерализованный же тип живой клетки будет включать лишь
несколько наиболее общих признаков, характерных как для кле-
ток, содержащих оформленные ядра, так и для прокариотических
клеток.
Любая живая клетка независимо от того, в образовании какого
живого тела она участвует, обязательно включает (см. рис. 3.3)
клеточную мембрану и протоплазму, образующие протопласт. По
определению протопласт — это клетка, лишенная клеточной
стенки или оболочки и имеющая только клеточную мембрану, ко-
торая ограничивает цитоплазму с различными органоидами и
включениями. Протопласту соответствуют клетки многоклеточ-
ных животных и некоторых бактерий (микоплазмы), а также ли-
шенные клеточных оболочек или стенок любые другие клетки.
Клеточная мембрана (цитоплазматическая мембрана, плазмати-
ческая мембрана, плазмалемма, наружная плазматическая мемб-
рана) — мембрана, отделяющая цитоплазму клетки от внешней
среды или от надмембранных образований. Толщина мембраны
составляет 7... 10 нм. Формируется она в гранулярной эндоплазма-
тической сети (гЭПС), затем модифицируется в аппарате (комп-
лексе) Гольджи. Клеточная мембрана играет определяющую роль
в обмене веществ между цитоплазмой и внешней средой, в движе-
нии клеток и в образовании межклеточных контактов. «Таможен-
ные» свойства мембраны обусловлены ее ограниченной избира-
тельной проницаемостью (полупроницаемостью) для различных
веществ. Так, вода проходит через нее свободно против градиента
концентрации веществ. Скорость диффузии других веществ об-
ратно пропорциональна их молекулярной массе. Полярные моле-
кулы и ионы переносятся через мембрану с помощью специаль-
ных белковых молекул-переносчиков. Аминокислоты, глюкоза, а
также ионы Na+ и К+ переносятся через мембрану с затратами
энергии. Клеточная мембрана обеспечивает способность клетки к
фагоцитозу и пиноцитозу, образованию выростов и впячиваний.
Обычно мембрана заряжена: ее электрический потенциал состав-
ляет 20... 100 мВ.
Строение клеточной мембраны рассмотрено в разделе 4.3.2.
Протоплазмой называют содержимое живой клетки, то есть ее
цитоплазму и нуклеоид/ядро.
Цитоплазма — эго обязательная составляющая любой клетки,
заключенная между клеточной мембраной и нуклеоидом/ядром.
Структурно цитоплазма представляет собой высокоупорядочен-
ный многофазный коллоид — гиалоплазму с находящимися в ней
68
немембранными и мембранными органоидами. Для цитоплазмы
характерны непрерывное перемещение и изменение состояния ее
компонентов. Цитоплазма пронизана микротрубочками и белко-
выми фибриллами, полимеризация и распад которых обеспечива-
ют обратимые переходы отдельных участков цитоплазмы из золе-
образного состояния в гелеобразное. Совокупность микротрубо-
чек и фибрилл составляет цитоскелет — опорно-двигательную си-
стему клетки. В цитоплазме осуществляются все процессы
клеточного обмена веществ, кроме синтеза нуклеиновых кислот в
эукариотических клетках. В цитоплазме клеток Простейших при-
нято выделять два слоя: эктоплазму (наружный слой), лишенную
большинства органоидов, более однородную и вязкую, и эндо-
плазму (внутренний слой), содержащую различные органоиды,
сильно структурированную и менее вязкую.
Нуклеоид — это ДНК-содержащая область прокариотической
клетки, не отграниченная мембранами. Он представляет собой
кольцевую молекулу ДНК, не соединенную с гистоновыми белка-
ми. ДНК закреплена в одной точке на внутренней поверхности
клеточной мембраны. Некоторые прокариоты имеют по несколь-
ку нуклеоидов в одной клетке. Иногда молекулы ДНК, содержа-
щиеся в нуклеоидах, называют бактериальными хромосомами.
По наличию или отсутствию в клетках ядра все организмы при-
нято делить соответственно на эукариоты и прокариоты. Ядро —
это область клетки, отграниченная двухмембранной оболочкой и
включающая сложные ДНК-содержащие структуры — хромосомы.
Ядро’обычно имеет сферическую или эллипсоидную форму. Его
размеры чаще варьируют в пределах от 1 мкм до 1 мм. Клетки мо-
гут иметь несколько, иногда много ядер, одинаковых или отлича-
ющихся по строению и функциям. Ядерная оболочка пронизана
порами, на краях которых наружная мембрана переходит во внут-
реннюю. Ядро содержит кариоплазму и оформленные элементы:
хроматин, перихроматиновые фибриллы и гранулы. Путем реали-
зации заключенной в генетическом материале информации ядро
управляет пластическими и энергетическими процессами в клет-
ке. Ядра могут делиться митотическим или мейотическим путем,
перешнуровкой на две или более дочерние клетки или почковани-
ем. При этом могут разделяться целые хромосомные группы
(скрытый митоз).
Общее разделение клеток на прокариотические и эукариоти-
ческие далеко не исчерпывает всего разнообразия планов их стро-
ения. В пределах первого надцарства — Прокариот — различают
по меньшей мере две группы клеток. Гетеротрофные с хемоавто-
трофными и частью фотоавтотрофных — архебактерии и эубакте-
рии составляют одну из них, а фотоавтотрофные — цианобакте-
рии—другую. В числе общих для различных прокариотических
69
клеток структурно-функциональных элементов отметим помимо
клеточной мембраны, цитоплазмы и кольцевой ДНК клеточную
стенку и рибосомы. Для клеток архебактерий и эубактерий, кроме
того, характерно наличие мезосом (впячиваний мембраны) и час-
то жгутиков, а для клеток цианобактерий — надмембранного же-
латинозного слоя (гликокаликса), фотосинтезирующих мембран с
бактериальным хлорофиллом, белковых и фосфатных гранул и ли-
пидных глобул.
Эукариотические клетки еще более разнообразны. Условно
можно выделить три-четыре главные группы: животный тип, рас-
тительный тип, грибной тип и клеточный тип протистов. Грибные
клетки и клетки простейших имеют ряд признаков, повторяю-
щихся у двух первых групп, но в иных сочетаниях, а кроме того,
собственные характерные признаки. Тем не менее можно выде-
лить несколько структур, объединяющих их и одновременно отли-
чающих от прокариотических клеток. К таким структурам отно-
сятся: ядро с хроматином (нуклеопротеиновым комплексом) и яд-
рышком (областью ядра, где происходит синтез рибосомных РНК
и сборка прерибосомных гранул); фибриллярные структуры, обес-
печивающие движение цитоплазмы и органоидов; мембранные
комплексы, разделяющие цитоплазму на функциональные облас-
ти (компартменты); рибосомы, отличающиеся от прокариотных;
митохондрии. Одновременно, и это следует помнить, эукариоти-
ческие клетки содержат в митохондриях, а растительные клетки
также и в хлоропластах кольцевые ДНК и рибосомы прокариотно-
го типа.
В перечень функциональных частей клетки входит клеточная
стенка. В данном случае это сборное понятие, обозначающее
плотные надмембранные образования: клеточные стенки бакте-
рий, клеточные оболочки растительных и грибных клеток, панци-
ри, раковинки и домики простейших. Кроме них клетка может
иметь в качестве надмембранного образования гликокаликс или
его усиленную форму цисту. Наконец, покров может быть образо-
ван комплексом плазматических мембран и белковыми пластин-
ками, расположенными во внешнем слое эктоплазмы, и тогда он
называется пелликулой.
Толщина клеточной стенки бактерий варьирует от 10 до 50 нм
и может составлять 10...50 % сухой массы клеток. У большинства
бактерий, то есть у грамположительных, зеленых фотосинтезиру-
ющих, цианобактерий, пурпурных фотосинтезирующих и нефото-
синтезирующих грамотрицательных бактерий, которых иногда
объединяют в царство эубактерий, в состав клеточной стенки вхо-
дит полимерное соединение пептидогликан (или муреин), состоя-
щее из дисахаридов, связанных короткими цепочками аминокис-
лот. У микобактерий в состав клеточной стенки входят липиды.
70
Клеточная стенка грамотрицательных бактерий кроме муреиново-
го слоя содержит липопротеиновую мембрану. Стенки архебакте-
рий не содержат муреина: они состоят из кислого гетеросахарцца
или белка.
Большинство бактерий способно образовывать на поверхности
клеточной стенки особый слой переплетенных углеводных моле-
кул — гликокаликс, а многие — непрочно связанную с «телом» по-
лисахаридную оболочку — капсулу (рис. 3.4).
Клеточная оболочка — это структурное образование на перифе-
рии клетки над клеточной мембраной, придающее ей прочность,
сохраняющее ее форму и защищающее протопласт. У растений
клеточная оболочка способна к одревеснению, образуя скелет рас-
тения, выполняющий опорную функцию. Основу клеточной обо-
лочки растений составляют целлюлозные фибриллы, погружен-
ные в матрикс, образованный гемицеллюлозой и пектином. В со-
став клеточной оболочки могут входить и другие органические и
неорганические вещества. Различают первичную и вторичную
клеточную оболочку. Меристемные и другие растущие клетки
имеют первичную оболочку, образованную в основном матриксом.
Фибриллы целлюлозы здесь расположены неупорядоченно. На
более тонких участках образуются канальцы, через которые про-
ходят плазмодесмы. Вторичная оболочка образуется после дости-
l uni ihi tn и iniHiiiiiHf iiiiiin:
— Капсула---------
Гликопротеид------
Внешняя мембрана \
•Пептидогликан (муреин)-
IIIII111 IIII Illi III UllllllillUII
U Hill 11 III 111111IIIII Hill HIIII
Плазматическая мембрана-
_______Цитоплазма ——____
Рис. 3.4. Строение клеточной стенки грамположительных (2) и грамотрицательных
• (2) бактерий н клеточной оболочки растений (3) я грибов (4) (оригинал автора)
— Оболочка
71
жения клеткой окончательного размера и накладывается на пер-
вичную изнутри. Здесь преобладает целлюлоза с более мощными
и упорядоченно расположенными фибриллами. В каждом слое на-
правление фибрилл меняется. Вторичная клеточная оболочка
пронизана порами.
Оболочка грибов образована в основном полисахаридом хити-
ном. Кроме грибов хитин в составе клеточных оболочек имеют
хитридиомицеты и некоторые группы саркодовых, причисляемые
к протестам (в некоторых классификациях — царство). У ряда
протестов в число основных компонентов оболочки входят цел-
люлоза, другие полисахариды, иногда с включением кремнезема
или карбоната кальция. Диатомовые водоросли имеют кремнезем-
ную оболочку. Интересно, что раковинные амебы и фораминифе-
ры также включают кремнезем и карбонат кальция, но ими инк-
рустируется хитиновая основа оболочки. Эти образования называ-
ются раковинками и домиками.
Пелликула характерна для трех групп протистов: эвгленовых,
инфузорий и споровиков. У них под наружной плазматической
мембраной находятся плоские мембранные мешочки с белко-
выми пластинками, или свободные белковые пластинки, или же
комплекс из двух плазматических мембран. Как правило, на-
ружная поверхность пелликулы бывает скульптурирована. При
этом изгибы наружной плазматической мембраны строго упо-
рядочены.
Надмембранные образования — клеточная стенка, оболочка,
гликокаликс — выполняют ряд функций: формообразовательную,
механической и химической защиты, опорную и коммуникацион-
ную. В случае накопления в них минеральных веществ организмы
выполняют и важнейшую биогеоценотическую функцию — средо-
образующую, переводя растворенные минералы в осаждаемое
кристаллическое состояние.
3.3. ОСОБЕННОСТИ СТРОЕНИЯ СВОБОДНОЖИВУЩИХ
КЛЕТОК
К свободноживущим мы будем относить те клетки, которые
представляют отдельные ценотически активные тела хотя бы в ча-
сти жизненного цикла. Такие клетки-особи обладают некоторыми
особенностями в структурной и динамической организации, по-
скольку в них, как и в свободноживущих истинно многоклеточ-
ных функционирующих особях, протекают все процессы индиви-
дуального и видового жизнеобеспечения, что, собственно, и явля-
ется воплощением биосферной функции живого вещества того
вида, к которому они принадлежат.
72
С «внешней» стороны жизнедеятельность активных особей
выглядит как результирующая деятельность «черного ящика». На
его «входе» имеются вещество в определенной форме и опреде-
ленном фазовом состоянии и энергия, заключенная в этом веще-
стве или приходящая от внешнего источника в усваиваемой фор-
ме, а на «выходе» — преобразованное вещество, обогащенное или
обедненное энергией. Внутренняя же сторона жизнедеятельности
полностью подчинена внешней стороне. По этой причине строе-
ние особи, включая его изменение в процессе развития, должно
обеспечивать выполнение элементарной биосферной функции.
Собственно, совокупность процессов, протекающих в живом теле
между «входом» и «выходом», с некоторой условностью мы и бу-
дем называть индивидуальным жизнеобеспечением, а внутренние
подразделения тела, предназначенные для реализации процес-
сов, — его функциональными частями или функциональными
структурами.
Комплексы специфичных функциональных структур свобод-
ноживущих клеток связаны преимущественно со средой обита-
ния, из которой они извлекают вещество и энергию, и их местом,
занимаемым в конкретных экосистемах. И для автотрофных, и для
гетеротрофных одиночных клеток важна способность к движению
или, наоборот, к сохранению стационарного положения в про-
странстве. В подвижной или перемешивающейся среде, обеспе-
чивающей непрерывный приток ресурсных веществ, если ско-
рость «обновления» среды не превышает значительно скорости
движения самой клетки, развивается два основных способа ло-
комоции: псевдоподиальный и ресничный. Они определяются
видом потребляемого вещества (плотные частицы или раствор),
и соответственно типом покровов (эластичные или жесткие).
Если же эффективность поглощения вещества и улавливания
рассеянной энергии не зависит от собственной скорости пере-
движения или скорость обновления среды значительно превы-
шает миграционные способности клеток, то такие клетки будут
сидячими (прикрепленными), парящими или пассивно под-
вижными.
Прокариотические клетки не имеют опорно-двигательных фи-
ламентов типа микротрубочек и микрофибрилл. Кроме того,
большинство их покрыто жесткой клеточной стенкой. Бактерии,
преобразующие световую энергию в энергию связей органических
веществ, могут обойтись без двигательных органоидов, если они
обитают на открытой дневной поверхности и в прозрачной среде.
Но те из них, которые поглощают органические (фоторедуценты)
или неорганические вещества (серобактерии), должны иметь воз-
можность менять положение в пространстве, если среда их обита-
ния не перемешивается.
73
Хемогетеротрофные бактерии аналогично могут быть или ак-
тивно подвижными, или пассивно подвижными. Одна из наибо-
лее общих особенностей прокариот состоит в том, что все они
представляют собой свободноживущие тела-клетки и никогда не
образуют истинно многоклеточных тел. Поэтому и механизмы
движения тел этой группы организмов достаточно однообразны:
пассивное движение в среде; скользящее движение с помощью ве-
щества, выдавливаемого через клеточную стенку; движение с по-
мощью жгутика. Жгутик прокариот имеет мало общего в строении
со жгутиком эукариот. Принципиально иной и способ его враще-
ния. Однако достигаемые эффекты сходны. Жгутиков у бактерий
может быть один или несколько, но они никогда не покрывают
всю поверхность тела, как у опалин и инфузорий.
Из других специализированных функциональных структур,
связанных с одиночным состоянием прокариотических клеток,
отметим характерные для палочковидных, кокковидных и мице-
лиальных бактерий эндоспоры — внутриклеточные покоящиеся
репродуктивные образования, предназначенные для пассивного
расселения и для переживания неблагоприятных условий среды.
Они очень устойчивы к высушиванию, воздействию высоких тем-
ператур, растворителей, действию радиации и пищеварительных
ферментов.
Эукариотические клетки-тела устроены сложнее других типов
клеток. Строение их цитоскелета и покровов обусловлено особен-
ностями образа жизни. Пассивно подвижные и парящие протисты
часто имеют выступающие за общий контур тела выросты панци-
ря (при жесткой оболочке) или внутреннего минерального скелета
(при эластичных покровах) (рис. 3.5, В некоторых случаях
парение в воде облегчается благодаря наличию аксоподий — спе-
циального типа псевдоподий с опорными микротрубочками
(рис. 3.5, 4) или простых длинных тонких псевдоподий (рис. 3.5, 5, б).
Клетки, которые эффективно перемещаются в пространстве с
Микротоками воды, вызываемыми более крупными животными,
вполне могут обходиться без двигательных органоидов (рис. 3.5, 7).
Но большинство свободноживущих протистов способны к актив-
ному движению.
Механизм движения клеток амебоидного типа основан на
сложном взаимном скольжении цитоплазматических филаментов,
сокращении подмембранной фибриллярной сеточки и переходе
гиалоплазмы из одного физико-химического состояния в другое.
К образованию псевдоподий способны преимущественно «пол-
зающие» по плотному или рыхлому субстрату одноклеточные
(рис. 3.5, 8, 9).
Протисты-гетеротрофы, собирающие пищу в толще воды, ис-
пользуют в качестве движителей реснички и жгутики — образова-
74
пия одинакового строения, но разных длины и формы движения
(рис. 3.5, 10... 19). Вероятно, развитие одного или другого типа
двигательных органоидов определяется требуемым сочетанием
скорости и точности перемещения. Реснички, жгутики и псевдо-
подии могут присутствовать и у половых особей протистов
(рис. 3.5, 20...22). Эти органоиды обеспечивают лишь транспорти-
ровку генетических матриц, расходуя энергию, ранее запасенную
ценотически активными особями или фазами.
Передвижение в среде может осуществляться или с помощью
перемещения тела, или благодаря перемещению среды относи-
тельно тела. Вторая форма реализуется, как и первая, посредством
ресничных образований и используется как прикрепленными (си-
дячими) одноклеточными, так и подвижными. Примерами могут
служить воротничковые жгутиконосцы, кругоресничные и равно-
ресничные инфузории (рис. 3.5, 23..30).
У инфузорий фибриллы образуют и еще один специализиро-
ванный органоид, отсутствующий в клетках истинно многокле-
точных тел, — палочковый аппарат, который служит для втягива-
ния пищевых частиц в цитоплазму и называется клеточным ртом
или клеточной глоткой (рис. 3.5, 31).
Необходимость выбора оптимальных условий для течения фо-
тосинтеза определяет развитие у подвижных автотрофов и мик-
сотрофов специализированного светочувствительного органо-
ида — глазка {стигмы). Глазок представляет собой гранулу свето-
чувствительного пигмента (рис. 3.5, 32, 33). У парящих и непод-
вижных протистов, как и в клетках многоклеточных живых тел,
соответствующих органоидов нет.
Подавляющее большинство свободноживущих клеток обитает в
воде или в средах, насыщенных водой. Тем из них, которые связа-
ны с минеральными или органическими растворами относительно
низкой концентрации, то есть обитателям гипотонических сред
(по отношению к концентрации цитоплазмы), приходится решать
проблему удаления избытка воды во внешнюю среду. Прокарио-
тические клетки имеют мощную клеточную стенку, которая, с од-
ной стороны, уменьшает проникновение воды в протопласт, а с
другой стороны, выдерживает повышенное осмотическое давле-
ние, что освобождает их от проблемы осморегуляции при неразви-
той цитоплазматической мембранной сети. Для простейших же,
постоянно обитающих в пресной или слабосоленой воде (амеб,
жгутиконосцев, инфузорий), очень характерен специальный ос-
морегуляторный органоид — сократительная (пульсирующая) ва-
куоль (рис. 3.5, 34). Сократительные вакуоли могут быть подвиж-
ными и с переменным положением, как у саркодовых, или стаци-
онарными, как у клеток с относительно постоянной формой, оди-
ночными или множественными (у некоторых инфузорий —до
75
24
42
43
44
Рис. 3.5. Специализированные функциональные структуры одноклеточных живых
тел (Жизнь животных, 1968; Хаусман, 1988):
/—выросты панциря у пирвдинеи (Ceratium sp.); 2, J—выросты внутреннего скелета у золо-
тистой водоросли (Distephanus sp.) (2) w радиолярии (Acantharea sp.) (J); 4—аксоподии у сол-
нечника (Acanthosphaerium sp.); 5, б—тпевканые псевдоподии у фораминиферы (Globigenna
sp.) (5) к моноталамиды (Monothalamida) (б); 7—утолщенные покровы пассивно подвижой
цисты простейшего (Echinosphaerium sp.); 8, 9— простые псевдоподии у раковинной (Arcella
sp.) (8) и голой (Amoeba sp.) (9) амеб; 10, //—псевдоподии и жгутики у голой амебы (Naegleria
sp.) (10) и растительного жгутиконосца (Silicoflageilidd) (II); 12— разные жгутики у животного
жгутиконосца Bodo sp. (Kinetopbstida); /5—видоизмененный жгутик — «ундулирующая мемб-
рана» у животного жгутиконосца (Trypanosoma sp.);/4,15— бахромчатые жгутики у эвгленовых
(Euglenida) (14) и золотистых водорослей (Cluysomonadida) (15); 16... 19— многочисленные жгу-
тики у гипермастигиды (Teratonympha sp.) (16) w опалины (Opalina ranarum) (17) и реснички у
«бегающей» (Stylonychia sp.) (18) и «плавающей» (Paramecium sp.) (19) инфузорий; 20...22—
двигательные жгутики ценотически пассивных особей — зооспор миксомицета (20) и водорос-
ли (Chlamodomonada sp.) (21) и мужской гаметы споровика (22); 23...27— прикрепительные до-
мики, стебельки и цилиарный двигательный аппарат простейших с обращенной формой дви-
жения у прикрепленных «подгонятелей» инфузорий Eufolliculina spp. (23, 24) и Stentorsp. (27) и
жгутиконосцев Salpingoeca sp. (25) и Poterioochromonas sp. (26); 28— стебелек и улавливающие
псевдоподии прикрепленного «собирателя» Desmothoracida (саркодовые); 29— стебелек и лов-
чие щупальца прикрепленной хищной инфузории (Discophrya sp.); 30— «скребные» пластинки
и реснички подвижной паразитической инфузории (Trichodina sp.); 31— палочковый аппарат
хищной инфузории; 32,33—светочувствительный глазок эвглены (32) и хламидомонады (33);
34— сложная пульсирующая вакуоль инфузории; 35— кинетопласт (видоизмененная мито-
хондрия) трипанозомы; 36— стрекательная нить споры микроспоридии; 37— апикальный
комплекс (коноид, роптрии и микронемы) споровика; 38...44— защитные структуры разового
действия: трихоциста (38); мукоциста (39); токсициста (40); гаптоциста (41); эжекгосома (42);
дискоболоциста (43); нематоциста (44)
100) . Сократительная вакуоль окружена более или менее заметны-
ми приводящими канальцами.
Иногда к специализированным органоидам простейших отно-
сят и пищеварительные вакуоли. Но эти мембранные структуры
формируются у любых клеток, способных к фагоцитозу, включая
и ряд клеток «высших» многоклеточных животных. Мембрана пи-
щеварительных вакуолей во время их образования отшнуровыва-
ется от наружной клеточной мембраны, а при опорожнении ваку-
оли вновь встраиваются в нее.
Таким образом, свободноживущие клетки кроме полного набо-
ра общеклеточных органоидов и структур, характерных для того
или иного их типа, могут иметь временно или постоянно несколь-
ко специализированных функциональных частей, необходимых
для обеспечения индивидуальной или видовой жизнедеятельнос-
ти. В число таких частей входят: у прокариот — клеточная стенка,
жгутиковая фибрилла, оболочка эндоспоры; у эукариот — жгутики
и реснички, выросты, с помощью которых осуществляется паре-
ние в воде, светочувствительные гранулы, пульсирующие вакуоли,
палочковые аппараты. Функциональные части клетки обозначают
термином «органоиды».
Органоиды — это постоянные клеточные структуры, обеспе-
чивающие выполнение специфических функций в процессе
жизнедеятельности клетки. Специальные органоиды однокле-
точных живых тел (жгутики, реснички, кинетопласт — комп-
78
лсксная митохондриальная структура (рис. 3.5, 35), сократи-
тельные вакуоли, глазки, палочковый аппарат) принято назы-
вать органеллами.
Кроме органоидов непрерывного индивидуального жизнеобес-
печения в живых телах-клетках встречаются и структуры разового
действия. Как правило, такие клетки не являются ценотически ак-
тивными, а обеспечивают возможность реализации активности.
Например, книдоспоридии имеют на определенной стадии жиз-
ненного цикла многоклеточные споры со стрекательными капсу-
лами, заякоривающимися в кишечнике рыб и обеспечивающими
проникновение в тело рыб ценотически активных паразитических
амебоидных клеток. У микроспоридий — внутриклеточных пара-
зитов насекомых — стрекательный аппарат является частью клет-
ки, отбрасываемой после проникновения активной спороплазмы
в клетку хозяина (рис. 3.5, 36). У споровиков ценотическая пас-
сивность связана не с отдельными клетками или частями клеток, а
с временным состоянием клеток. Это инвазионные стадии, на-
значение которых состоит в заселении здоровых клеток хозяина.
У них развивается комплекс специальных органоидов проникно-
вения через клеточную мембрану хозяина, не восстанавливаю-
щийся после срабатывания (рис. 3.5, 37).
Иногда простейшие (например, инфузории) имеют структуры
разового действия и в ценотически активном состоянии. Это пре-
имущественно однородные по составу и простые по строению за-
щитные образования: трихоцисты, экструсомы и др. (рис. 3.5,
38...44).
Свободноживущие эукариотические клетки в силу необходи-
мости непосредственно взаимодействовать с внешней средой ин-
дивидуально, как правило, имеют более сложное строение, чем
клетки многоклеточных тел-особей, что обусловливает их относи-
тельную функциональную завершенность.
3.4. ЯДЕРНО-ЦИТОПЛАЗМАТИЧЕСКИЕ КОМПЛЕКСЫ
ПРОТИСТОВ И МНОГОКЛЕТОЧНЫХ
Существует несколько форм живых тел, которые только на ос-
новании внешних признаков трудно однозначно отнести к «одно-
клеточным» или «многоклеточным». Это плазмодии, псевдоплаз-
модии и многоклеточные тела, клетки которых образуют синци-
тии и симпласты (рис. 3.6). Трудность состоит в том, что, имея
внешнее сходство, они связаны постепенными переходами как с
одиночными свободноживущими клетками, так и с истинно мно-
гоклеточными (тканевыми) телами, а в некоторых случаях — и с
типичными колониями.
79
7
Рис. 3.6. Ядерно-цитоплазматические комплексы (оригинал автора):
/—агрегирующие амебоидные особи миксомицета образуют псевдоплазмодий; 2—слияние
амебоидных особей миксомицета приводит к образованию плазмодия; 3 — совокупность веге-
тативных клеток вольвокса, соединенных ллазмодесмами, представляет собой симпласт; 4—
гифы грибов с неполным разделением цитоплазмы впячиваниями оболочки организованы по
типу симпласта; 5—клетки многоклеточных растений, соединенные многочисленными плаз-
модесмами, образуют симпласт; 6 — сливающиеся клетки тела губок образуют синцитий; 7—
частичное разрушение наружных клеточных мембран в эпителии плоских червей приводит к
образованию синцития
В зоологической литературе плазмодиями принято называть ве-
гетативные тела слизевиков (миксомицетов), представляющие
многоядерную массу протоплазмы. Однако известно, что, напри-
мер, у малярийного паразита вегетативное тело также называется
плазмодием. Иногда говорят, как о плазмодиях, о многоядерных
телах-клетках инфузорий, некоторых жгутиконосцев, опалин.
Принятие во внимание механизма образования данной жизнен-
ной формы живых тел и их дальнейшей судьбы в жизненном цик-
ле позволяет считать плазмодий сборным понятием. Так, у слизе-
виков плазмодиальная масса в типичном случае образуется в ре-
зультате полного слияния отдельных особей миксамеб, предвари-
тельно собирающихся в огромный, площадью до нескольких
сотен квадратных сантиметров, клеточный агрегат. Такой плазмо-
80
дий способен быстро передвигаться в поисках подходящей пищи.
В последующем он может снова распасться на отдельные клетки
или сформировать аналог плодового тела грибов, состоящий из
инцистированных амеб. У некоторых акразиевых (класс в типе
Саркомастигофоры) и слизевиков «объединяющиеся» амебы со-
храняют клеточные мембраны, образуя псевдоплазмодий. По строе-
нию и механизму образования псевдоплазмодий более соответ-
ствует колонии одноклеточных живых тел, а по особенностям ос-
новных моментов жизнедеятельности и последующих преобразо-
ваний — это обособленное живое тело, которое онтогенетически
предшествует (но не обязательно) плазмодиальной фазе или сле-
дует за ней.
У споровиков термином «плазмодий» обозначают обычно
многоядерный шизонт, который сам является лишь приступив-
шей к размножению трофической фазой ценотически активно-
го поколения особей. В дальнейшем такой шизонт распадается
на отдельные клетки соответственно числу образовавшихся
ядер. Здесь при ином механизме образования судьба плаз-
модия в определенной мере аналогична судьбе плазмодия сли-
зевиков.
Более устойчивое состояние плазмодия наблюдается у ряда
саркомастигофор и других простейших. Так, весь класс опалины,
большая часть видов класса акарпомиксы, класс плазмодиофоры,
взрослые фазы миксоспоридий представлены плазмодиальными
формами. Кроме того, многие грибы в вегетативной части образо-
ваны, по сути, одной сильно ветвящейся клеткой, в цитоплазме
которой расположено множество ядер. Грибы дают и переходную
форму к телам, составленным из многих клеток: мицелий может
быть разделен перегородками разной степени полноты так, что
цитоплазма оказывается все же непрерывной.
Во всех этих случаях ядерно-цитоплазматические комплексы
образуются в результате незавершенного деления, то есть за
расхождением дочерних ядер не следует разделение цитоплаз-
мы. К этой группе тел не следует относить простейших с двумя
(или более) разнокачественными ядрами (фораминиферы, инфу-
зории), для которых ядерный дуализм является устойчивой нор-
мой, а в обеспечении вегетативных процессов участвует только
одно ядро или один из типов ядер.
Итак, простейшие и грибы способны образовывать многоядер-
ные амебоидные тела — устойчивую или кратковременную жиз-
ненную форму — плазмодий или незавершенный ее вариант —
псевдоплазмодий. Такая морфофункциональная реорганизация
позволяет синхронизировать и, следовательно, активизировать де-
ятельность значительной по объему цитоплазматической массы, а
иногда и войти в новую ценотическую группировку.
6 Присный A JB.
81
Многоклеточные живые тела также могут образовывать посто-
янные или временные ядерно-цитоплазматические комплексы^,
обозначаемые терминами «синцитий» и «симпласт». Четкого
смыслового разделения этих терминов нет, но чаще «симпласт»
употребляется применительно к тканям растений, а «синцитий» —
к тканям животных. В этом случае симпласпгом называют много-
ядерный протопласт многоклеточного растения, у которого утра-
чены или не развиваются оболочки клеток, а также совокупность
протопластов, соединенных протоплазматическими нитями —
плазмодесмами. Из последнего следует, что большинство живых
тел, построенных из множества клеток, окруженных клеточной
оболочкой, имеют симпластный тип организации. Но диапазон
вариаций этого типа довольно широк: от совершенно нерасчле-
ненного симпласта у сифоновых водорослей (каулерпа) до почти
полностью обособленных клеток с вторичными оболочками, со-
единенных лишь тонкими цитоплазматическими тяжами у древес-
ных сосудистых растений.
Симпласту растений у животных соответствует синцитий — та-
кой вариант строения ткани, при котором клеточные мембраны
частично или полностью утрачиваются или не формируются изна-
чально и соответственно границы между клетками не выражены.
Встречается синцитий в телах животных разных систематических
групп: у стеклянных губок большая или меньшая часть клеток тела
может сливаться в общую цитоплазматическую массу, облегаю-
щую скелетные элементы; у плоских червей клетки кожного эпи-
телия часто сливаются в синцитий — цитоплазматическую плас-
тинку; по некоторым свидетельствам, полость тела нематод пред-
ставляет собой продукт полного слияния паренхимных клеток, то
есть синцитий; к этому же типу образований относятся много-
ядерные мышечные клетки, зародышевая соединительная ткань —
мезенхима, костная ткань, ретикулярная соединительная ткань.
Обратимые превращения в синцитий может претерпевать кишеч-
ный эпителий моллюсков и насекомых.
Ядерно-цитоплазматические комплексы, как следует из при-
веденных примеров, формируются и как объединения однокле-
точных тел, и как специализированные части многоклеточных
живых тел. По-видимому, это явление — один из путей сумми-
рования однородных функций с исключением индивидуальнос-
ти. Одновременно следует заметить, что такой путь не может
рассматриваться в рамках гипотезы возникновения многокле-
точности.
Рассуждая последовательно, мы должны признать, что и тал-
ломные тела водорослей и грибов, и усложненные талломы мохо-
видных, и истинно многоклеточные тела сосудистых растений
имеют единый принцип построения. Различия же между синцй-
82
тием и симпластом, несмотря на внешнее сходство между ними и
также между ними и плазмодиальными телами «одноклеточных»,
позволяют говорить о трех формах ядерно-цитоплазматических
комплексов: плазмодиях простейших, симпластах грибов и расте-
ний и синцитиях животных (рис. 3.7).
Б
В
Рис. 3.7. Механизмы образования ядерно-цитоплазматических комплексов (оригинал
автора):
А — слияние ценотически активных одноклеточных особей у протистов; Б — образование мно-
гоклеточного тела с последующим полным или частичным разрушением смежных наружных
клеточных мембран у животных; В—формирование неполных перегородок из врастающей
оболочки у грибов; Г—диффузное образование первичной клеточной оболочки растений,
пронизанной плазмодесмами
83
3.5. МНОГОКЛЕТОЧНЫЕ ЖИВЫЕ ТЕЛА
Существовавшее долгое время разделение организмов на одно-
клеточные и многоклеточные нельзя признать сколь-нибудь удов-
летворительным, и потому мы говорим об одноклеточных и мно-
гоклеточных живых телах. При этом, как было показано в преды-
дущем разделе, ядерно-цитоплазматические комплексы плазмо-
диального типа (плазмодии, псевдоплазмодии, синцитии и
симпласты) являются фактически многоклеточными образования-
ми. Сразу же отметим, что в рассматриваемую группу попадают
такие неодинаковые тела, как колонии одноклеточных особей,
колониальные особи одноклеточных, многоклеточные бесткане-
вые и многоклеточные тканевые особи, организмы. Поэтому по-
пробуем придать более конкретное содержание широко употреб-
ляемым, но «расплывчатым» терминам: одноклеточный, колони-
альный, колониеподобный, талломный, многоклеточный и мо-
дульный, помня, что все они характеризуют строение не
организма, а особи — отдельного, не всегда полностью обособлен-
ного в пространстве живого тела.
Отдельная обособленная свободноживущая клетка — однокле-
точное живое тело. Целесообразно сюда относить ценотически ак-
тивные живые тела, хотя одноклеточными могут быть и ценоти-
чески пассивные тела (гаметы), и стадии жизни особей (споры,
цисты). Далеко не все простейшие одноклеточны и не все одно-
клеточные тела относятся к простейшим. (Простейшие — систе-
матическая группа, но не уровень организации.)
3.5.1. КОЛОНИИ И КОЛОНИЕВИДНЫЕ ТЕЛА
Основное значение слова колония ~ поселение. Соответствен-
но колониальный — относящийся к колонии, поселению- В био-
логической литературе широко употребляются такие понятия, как
«колония организмов» и «колониальный организм». При всей ви-
димой простоте они оказываются весьма неопределенными. Рас-
смотрим их трактовку в терминологических словарях.
По Реймерсу, «колония организмов — поселение, группа со-
вместно живущих особей одного или нескольких видов, способ-
ных жить самостоятельно, но иногда выполняющих различные
функции или даже служащих как бы органами целого. Особи в
колонии извлекают выгоду от совместного питания, близости к
местам добычи корма, коллективной защиты от хищников, созда-
ния особого химизма среды и т. д. Различают 4 оттенка понимания
термина «колония»: 1 — объединение организмов одного вида,
возникшее вследствие того, что дочерние особи при бесполом раз-
84
множении не отделяются от материнского организма и иногда,
специализируясь, становятся как бы органами целого; 2 — объеди-
нение специализированных особей типа общественных насеко-
мых; 3 — совокупность аналогичных особей, выигрывающая от
объединения в борьбе за существование; 4 — скопление микроор-
ганизмов одного вида на богатой питательной среде».
Согласно БЭС «колониальные организмы — организмы, у ко-
торых особи дочерних поколений при бесполом размножении ос-
таются соединенными с материнским организмом, образуя более
или менее сложное объединение — колонию. Колониальные орга-
низмы встречаются главным образом среди одноклеточных, водо-
рослей, губок, книдарий (кишечнополостных), оболочников,
мшанок. Одни колониальные организмы ведут прикрепленный
образ жизни на неподвижном субстрате и имеют более или менее
развитый скелет, другие обитают в толще воды, обычно полупроз-
рачны, скелета не имеют. В колониях отдельные особи занимают
определенное место и выполняют специальные функции, важные
для всей колонии. Для многих колониальных организмов характе-
рен метагенез: чередование колониального бесполого и одиночно-
го полового поколений. Колониальными организмами иногда на-
зывают организмы, ведущие постоянно или временно скучен-
ный образ жизни: ряд бактерий, тлей, птиц».
Легко заметить, что понятия «колония» и «колониальный орга-
низм» понимаются как целое и часть, но неоднозначно определя-
ются. «Колония», кроме того, фактически является синонимом
«колонии организмов», а приведенные оттенки термина не совпа-
дают с основным его значением. Придадим этим понятиям более
конкретный смысл. Колония — относительно устойчивое компакт-
ное поселение особей, условно ограниченное резким спадом
плотности населения (числа особей на единицу объема). Колония
может быть образована идентичными особями, но не включать
полные организмы, поэтому правильнее говорить именно о коло-
нии особей или колонии живых тел (в минералогии используется
аналогичное понятие — «колония кристаллов» как колония нежи-
вых тел).
Колония может состоять из особей одного вида или особей раз-
ных видов и соответственно называться одновидовой (простой)
или многовидовой (сложной) колонией.
В основе образования колонии лежат прежде всего субстратные
отношения (пищевой субстрат или часть объема среды, более дру-
гих отвечающая потребностям вида). Второй важной побудитель-
ной причиной колониальности можно считать усиление функцио-
нального эффекта при пространственном объединении ценотичес-
ки активных особей, частей или фаз организмов, что уже упомина-
лось в отношении ядерно-цитоплазматических комплексов.
85
Колонии могут быть образованы: полностью (морфологически
и функционально) обособленными живыми телами, как однокле-
точными, так и многоклеточными; постоянно или временно со-
единенными живыми телами, которые не являются непосред-
ственным продолжением друг друга и тем не менее составляют
физически единое тело, где число особей может быть фиксиро-
ванным, неустойчивым или неопределенным; неразделяющимися
телами-особями одной жизненной формы, когда они являются
продолжением друг друга, сохраняя в то же время индивидуаль-
ность; полностью слившимися особями, части тел которых объе-
диняются в соответствующие части тела колонии, теряя индиви-
дуальность (рис. 3.8, 1...8).
Часто в колониях, функционирующих как единое тело, разви-
ваются части либо объединенные в той или иной степени особи,
выполняющие расселительную, воспроизводческую или иную
функцию (рис. 3.8, 9).
Важно обратить внимание на то, что колонии могут представ-
лять собой клон (потомков одной особи-основательницы) и
сборную группу как одноклеточных, так и многоклеточных тел.
К колониям по некоторым признакам близки такие объединения
особей, как дем, прайд, семья, стая, стадо, социум, уплотненная
популяция, временное скопление особей в ценотически пассив-
ном состоянии и др. В основе их образования лежат иные меха-
низмы.
Рассмотренные формы колоний образуют непрерывный ряд,
внутренние подразделения которого условны и в значительной
мере субъективны. Несколько обособленно от него стоит форма
колоний, встречающихся у некоторых кишечнополостных (сифо-
нофоры), где в состав колонии входят дифференцированные по
строению и выполняемым функциям особи. Учитывая, что диф-
ференцировка начинается с разных этапов закладки почек-осо-
бей, сифонофоры уже не могут быть отнесены к типичным телам-
колониям, так как имеют сходство с модульными телами по стро-
ению и социальными — по функциональному разделению особей.
Представляется возможным закрепит!, за ними название «колони-
евидные особи» (рис. 3.8, 10).
Среди сходных образований колонию можно выделить по сле-
дующим признакам:
типичная колония образуется на относительно однородном
благоприятном субстрате;
колонию составляют разновозрастные особи каждого из видов,
входящих в ее состав;
ценотически активные особи в колонии однообразны;
особи в колонии способны к индивидуальной активности,
включая и воспроизводство;
86
Рис. 3.8. Колонии особей и колоииевидаые особи (Жизнь животных, 1968; Хаусман,
1988; оригинал автора):
1‘ 2— простые колонии с нефиксированным числом обособленных особей бактерии
Vampirovibrio chlorellovonis (7) и тли (J); 3, 4— простые колонии с относительно фиксирован-
ным числом и расположением особей у бактерий (3) и у растительного жгутиконосца
(Pandorina sp.) (4); 5, 6— «кустистые» колонии с нефиксированным числом мор^юлогически
объединенных особей растительного жгутиконосца Dinibrion sp. (5) и инфузория Circhesium sp.
(б); 7— колония функционально обособленных особей восьмилучевого коралла Aicionutn sp.;
колония морфологически н функционально объединенных особей кремнероговой губки
Spongilla sp.; 9—морфологически единая колония функционально дифференцированных
особей гидроидного полипа Obelia sp.; 10— колониеввдная особь сифонофоры
отдельные особи функционально независимы, но однозначно
реагируют на изменения биогеоценотической обстановки.
Таким образом, типичная колония — это скопление живого ве-
щества, позволяющее произвести концентрированное действие (в
ответ на функциональную потребность экосистемы), размеры ко-
торого регулируются интенсивностью воспроизводства ценоти-
чески активных особей, частей или фаз организмов или направ-
лением их преобладающей миграции.
87
Перечислим преимущества и ограничения колониальное™:
колония способна произвести интенсивное локальное дей-
ствие;
в колонии в силу более выраженного средообразующего дей-
ствия уменьшаются индивидуальные энергетические затраты на
реализацию ценотической функции;
образование колонии — это способ перехода в иную пищевую
цепь, ведущий к усложнению и повышению устойчивости экосис-
темы, в которую входит колония;
в колонии увеличивается интенсивность информационных
процессов;
в колонии предопределяется индивидуальная неравноценность
особей, связанная с их пространственным распределением и кон-
центрацией в пространстве индивидуальных изменений призна-
ков;
в колонии ограничивается индивидуальная свобода отношений
с абиотической и биотической средой;
в пищевых цепях колония выступает как целостное образова-
ние, что приводит в экстремальных ситуациях к гибели большей
части особей одновременно.
Эволюционные изменения коснулись и колониального уровня
организации, вызвав появление форм, сходных с талломным и
даже типичным многоклеточным уровнем. Так, в прикрепленных
колониях некоторых инфузорий (Zoothamniori) по достижении ими
предельных размеров способность особей к размножению утрачи-
вается и вся колония начинает работать на небольшое число осо-
бей, способных к активной миграции, — будущих основателей но-
вых колоний. У низших талломных форм водорослей, традицион-
но относимых к растительным жгутиконосцам, имеющим клеточ-
ную оболочку, в ряде случаев вегетативные клетки изначально
утрачивают способность к размножению, делегируя ее отдельным
более крупным клеткам, питаемым через цитоплазматические
тяжи. Сходство колоний, например, воротничковых жгутиконос-
цев с такими талломными водорослями можно считать конвергент-
ным. Поскольку индивидуальная жизнь каждой из особей в такой
колонии ограничивается жизнью колонии как единого тела, то
половое воспроизводство также обеспечивается колонией как це-
лым. Аналогично половое воспроизводство талломного тела Volvox
и некоторых других растительных жгутиконосцев, относимых к
протистам, обеспечивается специализированными клетками, в то
время как их ценотически активные клетки не дифференцирова-
ны.
Все сказанное выше о колониях относится только к прокарио-
там, одноклеточным эукариотам (протистам) и многоклеточным
животным.
88
3.5.2. ТАЛЛОМНЫЕ ЖИВЫЕ ТЕЛА
По сложившейся традиции талломными или слоевищными назы-
вают тела водорослей, грибов, лишайников и некоторых моховид-
ных, то есть тела части организмов, ранее объединявшихся в цар-
ство Растения и не имеющих дифференцировки на настоящие
ткани и органы. Сюда же обычно относили и вегетативные тела
слизевиков, заростки высших споровых растений. При этом тал-
ломами называли самые разные живые тела, начиная с одиночных
клеток и заканчивая телами, дифференцированными на функцио-
нальные части (листовидные, стеблевидные и корневидные)
(рис. 3.9).
Типичный таллом — это многоклеточное тело, все клетки кото-
рого относятся к одной и той же жизненной форме, то есть диф-
ференцировка на ткани отсутствует. Это тело включает ядерно-
цитоплазматический комплекс — симпласт и непрерывную обо-
лочку, покрывающую его, — апопласт. Наличие апопласта и от-
сутствие цитоскелета препятствуют активному движению, но
обеспечивают ростовое движение. Способностью к размножению
в талломе обладают преимущественно краевые клетки, что харак-
терно для тел-прорастателей. Будучи поверхностными или глу-
бинными прорастателями без механических тканей, талломные
тела не способны к существенному воздействию на неживую среду
и, в частности, вертикальному транспорту веществ.
Талломный тип построения живых тел обеспечивает освоение
и заполнение среды, оказывая минимальное противодействие гра-
витационным силам. При этом автотрофы с талломным типом
строения полностью (водоросли) или в сильной степени (мохо-
видные) связаны с водной средой, а гетеротрофы (грибы)—с
плотным органическим субстратом. Симбиоз авто- и гетеротро-
фов дает новую форму талломных тел, характерную для лишайни-
ков.
Как и колонии, талломы претерпевают изменения и в сторону
упрощения, и в сторону усложнения строения. Например, вегета-
тивные тела некоторых водорослей и грибов могут быть представ-
лены одиночными клетками, а печеночники и мхи часто имеют
сходство с сосудистыми растениями.
3.5.3. МНОГОКЛЕТОЧНЫЕ ЖИВЫЕ ТЕЛА (С ТКАНЕВОЙ
ОРГАНИЗАЦИЕЙ)
Рассмотренные до сих пор живые тела образованы разным чис-
лом клеток, но их ценотически активные части сложены однотип-
ными, однообразно дифференцированными клетками. По этому
89
Рис. 3.9. Талломные тела цеиотически активных особей грибов, лишайников,
водорослей, печеночников, мхов и цеиотически пассивных особей хвощей и папорот-
ников (Рейвн и др., 1990; оригинал автора):
7—дрожжевые трибы; 2, 3—хитрвдиомицеты Myiophagus sp. {2) и Coelomycidium slmulii (5);
4— оомицеты; 5— базидиомицеты; 6— схема строения лишайника (а, в, г—слои, образован-
ные гифами гриба, б— слой с клетками водоросли); 7— накипной лишайник диплосхистес;
8— кустистый лишайник летария; 9...11—листовые формы цетрария {9) и кладония {10, 77);
12... 15— зеленые водоросли Hydrodiction sp. {12), Utothrix sp. {13), Cladophora sp. {14),
Cokochaete sp. (75); 16— красные водоросли Polysiphonia sp.; 17, 18~Ъууы,е водоросли
Laminaria sp. {17) и Sargassum sp. {18); 19, 20— печеночники Marchantia sp. {19) и Clasmatocolea
sp. {20); 21— мох Pofytrichium sp.; 22—заросток хвоща; 23— заросток папоротника
признаку разделение тел на одно- и многоклеточные произвес-
ти невозможно: здесь имеются множественные постепенные
переходы. В то же время большинство видов природных живых
тел построено из четко дифференцированных по строению и
функциям клеток. Исходная специализация предполагает раз-
деление клеток на два основных типа: клетки, связанные с пи-
танием (фотосинтезирующие или пищеварительные), и клетки,
связанные с характером движения (опорно-механические или
двигательные). Сочетание фотосинтезирующих и опорно-меха-
нических тканей дало начало сосудистым растениям, а пищева-
рительных и двигательных — эуметазоям, или настоящим мно-
гоклеточным животным (рис. 3.10). Кроме основной дифферен-
цировки ценотически активные части «истинных многоклеточ-
ных» часто дают дополнительную — на покровные, проводящие,
запасающие, соединительные, нервные и другие ткани. Указан-
ные особенности позволяют растениям и животным переме-
щать вещества против направления действия силы тяжести и в
горизонтальном направлении. Отметим также, что истинно
многоклеточные автотрофы — это вертикальные прорастатели,
способные формировать каркас вертикальной структуры биоце-
нозов, прежде всего наземных. Истинно многоклеточные гете-
ротрофы способны активно проникать в различные ярусы сооб-
ществ.
Многоклеточные животные и растения способны активно
воздействовать на минеральную составляющую биосферы, со-
здавая условия для вовлечения или вовлекая непосредственно
новые порции неживого вещества в биологический круговорот.
Необходимое уточнение сделаем для некоторых групп водных
многоклеточных животных и растений. Поскольку водная среда
уменьшает прямое действие гравитационных сил, механические
ткани могут замещаться различными производными покровных
тканей, минеральными и органическими опорными образова-
ниями или вместо них используется «гидростатический скелет».
У наземных животных также наблюдается сочетание использо-
вания полостных жидкостей, производных эпителия и соб-
ственно соединительных тканей для формирования опорно-
двигательных структур. В некоторых случаях функциональная
дифференцировка охватывает не группы клеток, а их части.
Так, эпителиальные клетки кишечнополостных разделяются на
пищеварительное тело и мускульные отростки или покровное
тело и мускульные отростки. Возможно, такие уклонения от ти-
пичной дифференцировки следует считать специализирован-
ными новообразованиями именно полипов и медуз, а исходное
состояние характерно для планулы — личинки кишечно-полост-
ных, равно как для пластинчатых и амфибластулы губок, у ко-
92
G
Рис. 3.10. Многоклеточные тела с тканевой организацией (Рейвн и др., 1990;
оригинал автора):
А, 5—опорно-проводящие клетки ксилемы (7) и фотосинтезирующие клетки флоэмы (2) в
стебле Psilotum (А) и листе Pinus (Б); В— пищеварительно-двигательные (3) и покровно-двига-
тельные (4) клетки тела гидры; Г— пищеварительные (5) и двигательные (6) клетки тела бески-
шечной турбеллярии
торых клетки разделяются на пищеварительные и двигательные
(ресничные или сократимые).
Нередко тканевые живые тела могут не быть целыми особями,
а представлять временно существующие живые части особей. Это,
например, регенерирующие части плоских червей, кишечнополо-
стных или иглокожих, части растений, половые щупальца некото-
рых головоногих моллюсков и др. (рис. 3.11). В одних случаях та-
кие живые фрагменты могут дать начало полноценной особи, то
есть быть ее фазой, а в других, выполнив собственную специаль-
ную функцию, погибнуть.
Рассмотрим преимущества и ограничения многоклеточнос-
ти. Многоклеточность позволяет резко увеличить функцио-
нальную эффективность живых тел посредством умножения од-
нородных специализированных элементов рабочих органов, а
также стабилизации внутренней среды и сравнительно высокой
степени независимости каждой клетки от изменений во внеш-
ней среде. Наверное, будет правильным полагать, что много-
клеточность — это закономерный этап эволюционного разви-
тия живых тел, позволяющий биосфере и обязывающий ее вый-
ти на новый структурный уровень и усиливающий интенсив-
ность активного взаимодействия живого вещества с неживой
средой. Этот мезоуровень обладает большей информационной
емкостью.
В то же время истинные многоклеточные тела обладают ка-
чественно иной реакцией на изменение внешних и внутренних
условий. Их численная реакция обычно сильно замедлена, а
функциональная — ускорена, что связано с развитием двух пос-
ледовательных уровней реакций (клеточного и организменно-
го). Они обладают меньшей устойчивостью к действию экстре-
мальных неблагоприятных факторов. Клетки, составляющие их,
обычно утрачивают полифункциональность и, обеспечивая вы-
полнение преимущественно одной специальной функции, ста-
новятся полностью зависимыми от комплементарных типов
клеток.
Рис. 3.11. Живые тела-части многоклеточных особей (оригинал автора):
/—обособленная плодущая часть тела многощетинкового червя палоло отмирает после раз-
множения; 2— обособленное половое щупальце самца аргонавта отмирает после переноса
сперматофора в мантийную полость самки; 5—части тела морской звезды восстанавливаются
до более или менее полноценных особей; 4— из частей тела поврежденной гидры восстанавли-
ваются полноценные особи (справа — неповрежденная особь); 5—фрагмент тела лишайни-
ка—самостоятельное живое тело; 6—фрагмент корневища пырея дает начало новой особи;
7— сломанная ветка может, сохраняя жизнеспособность, дать начало новой полноценной осо-
би; 8,9— мясистый стержневой корень и стеблевой клубень после отмирания надземных час-
тей растения временно существуют как самостоятельные живые тела; W— изолированный
столон земляники — полноценное живое тело
94
95
3.5.4. МОДУЛЬНЫЕ И СОЦИАЛЬНЫЕ ЖИВЫЕ ТЕЛА
В последние годы применительно к многоклеточным сосудис-
тым растениям вошел в употребление термин модульный организм.
Этот термин (правильнее — «модульное живое тело») применяют
для обозначения тел, имеющих постоянную, устойчивую часть, на
которой в различных сочетаниях и соотношениях надстраиваются
необходимые функциональные части-органы (сменяемые моду-
ли). Целесообразно к модульным относить не только раститель-
ные тела, но также тела животных, активно дополняемые времен-
ными внешними усилителями телесных структур. Например, пти-
цы используют длинные тонкие палочки для добычи личинок на-
секомых из укрытий, а каланы — камешки для разбивания
раковин моллюсков.
В перечень основных типов тел входит «социальный организм».
Каки большинство несистематизированных терминов, этоттермин
понимается весьма неопределенно. Им пользуются и в биологии, и
в социологии, но всегда подчеркивается какое-либо коренное отли-
чие общественных животных от человека (отсутствие сознания, аб-
страктного мышления, способности трудиться и т. п.).
Учитывая, что «социальный» переводится на русский язык как
«общественный», социальный организм следует понимать прежде
всего как организм, для которого характерна надындивидуальная
морфофункциональная комплементарность. Каждый такой орга-
низм включает ряд типов (жизненных форм) особей и обладает
рядом свойств систем. В частности, у многих перепончатокрылых,
у термитов ценотически активные функционирующие особи, спе-
циализирующиеся на выполнении вполне конкретных функций,
имеют отличия от других типов особей из этой же семьи, от кото-
рых полностью зависят (рис. 3.12). Семья же функционирует как
целое и даже может воспроизводиться половым способом через
разлетающихся полоносных особей и бесполым способом —от-
почковыванием части семьи. Комплементарные полоносные осо-
би у этих насекомых ценотически пассивны.
Человек как социальное животное имеет три существенных от-
личия от других социальных животных: генеративная составляю-
щая здесь образует ценотически пассивную часть (систему орга-
нов) в каждой ценотически активной особи; морфофункциональ-
ная дифференцировка у человека происходит преимущественно
не на телесном уровне, а на уровне объединения тел со сменяемы-
ми внешними усилителями (орудиями труда); запись информации
у человека производится не только на естественных генетических
матрицах, но и на искусственных носителях, хотя и основана на
принципе биологического кодирования. Такой тип устройства тел
следует обозначать как модульно-социальный.
96
Рис. 3.12. Морфофункциональная дифференциация особей социального организма
(Энтомология, 1985; Брайен, 1986, с изменениями):
1.-6—социальные касты у термитов: плодущая самка (7), самец (3, солдаты (J, 4), рабочие
особи (5, 6); 7—12— социальные касты у муравья Pheidole sp.: самец (7), плодущая самка (8),
рабочие особи {9, 10), солдаты {11, 12)
Ранее указывалось, что живые тела не бывают генетически од-
нородными. Поэтому в реальных условиях отдельные особи орга-
низмов могут представлять комплексные тела: истинное много-
клеточное тело, например, содержит в себе другие многоклеточ-
ные, талломные и одноклеточные тела. Распространенность тако-
го явления позволяет считать, что это не аномалия, а
норма — специфика построения многовидовых сообществ, форма
существования биосферы.
3.6. ТИПЫ ЦЕНОТИЧЕСКОЙ АКТИВНОСТИ
Все живые организмы биосферы Земли с некоторой условнос-
тью могут быть разделены на автотрофов и гетеротрофов. Однако
важно помнить, что это деление применимо в основном к ценоти-
7 Пр»|сный А.В.
97
чески активным особям, частям и фазам организмов, так как це-
нотически пассивные особи почти всегда гетеротрофны.
К автотрофным мы будем относить тела, способные усваивать
электромагнитную или минеральную химическую энергию, пере-
водя ее в энергию химических связей органических соединений.
Но органические соединения производятся автотрофами в избыт-
ке, то есть в количестве, превышающем их собственные нужды.
Более того, после естественной гибели автотрофов слагающая их
органика становится мертвой и недоступной для повторного ис-
пользования новыми поколениями автотрофов.
Для объяснения сущностной стороны избыточной продуктив-
ности существует несколько различающихся подходов. Согласно
одному из них потребители живой биомассы находят здесь обиль-
ную пищу и автотрофы как бы компенсируют потери, синтезируя
органику с запасом. Упрощенный системный подход предполага-
ет, что автотрофы «платят дань» потребителям живых органичес-
ких веществ за то, что они осуществляют регуляцию масс и чис-
ленностей в популяциях фото- и хемосинтетиков.
Целесообразнее все же рассматривать избыточную продукцию
как часть общей продукции органических веществ, выражение восхо-
дящей части биогенного цикла. В одно и то же время фото- и хемо-
синтез предполагают обеспечение как самовоспроизводства живых
тел-элементов системы, так и воспроизводства всей живой системы.
Термины «автотрофы» и «продуценты» не являются синонима-
ми. Первый отражает источник и механизм образования живого
вещества, способ питания, а второй — направление процессов в
живых телах и место в экосистемах.
Гетеротрофы — это не только целые организмы, использующие
готовые органические вещества для энергетических и пластичес-
ких нужд, но также большинство ценотически пассивных частей,
фаз или особей автотрофных организмов, обеспечивающих инди-
видуальную или видовую жизнь. Гетеротрофы составляют нисхо-
дящую часть биогенного круговорота веществ, переводя в конеч-
ном итоге органику в формы, доступные для повторного исполь-
зования автотрофами.
Судьба органических веществ, синтезированных продуцента-
ми, неодинакова: часть их становится мертвым веществом с инди-
видуальной смертью живых тел, а другая часть изымается гетеро-
трофами в «живом» состоянии. Как и в предыдущем случае,
объяснить существование потребителей органических веществ
можно с помощью, по крайней мере, трех подходов. Распростра-
нена гипотеза, согласно которой гетеротрофы появились раньше
автотрофов и используют их как возобновимый источник веще-
ства и энергии. Второе объяснение предполагает обратное отно-
шение: гетеротрофные организмы (не гетеротрофность вообще!)
98
формируются позже автотрофных и буквально обеспечивают воз-
можность их существования, получая взамен некоторые выгоды
(кислородное дыхание, озоновый экран, среду обитания и др.).
Остановимся подробнее на третьем подходе. В истории био-
сферы обе группы формируются практически одновременно как
составные части целого, как исходная дифференцировка протобио-
сферы на комплементарные элементы. Тела современных орга-
низмов очень часто построены именно так. К ним относятся и
упомянутые фотосинтезирующие растения с гетеротрофными
клетками, и огромное число животных с симбиотическими водо-
рослями, и животные погонофоры с хемосинтезирующими бакте-
риями, и типичные миксотрофные простейшие, и комплексные
тела лишайников и др. При таком подходе отпадает необходи-
мее™ в экосистемах
Продуценты ] L । [ Редуценты i [ Деструкторы и 1 редуценты*
Источники питания
Хемоавто- Фотоавто- Фотогетеро- Хемогетеро-
трофы трофы трофы трофы
Источники энергии
Неорганические соединения Свет Органические соединения
Источники углерода
Диоксид углерода Органические соединения
Источники водорода
Вода Серово- дород Органические соединения
Вода Органические соединения
Рис. 3.13. Общая классификация трофических групп организмов
* Миксотрофы (эвгленовые) способны переходить к фотоавтотрофному пита-
нию.
Т
99
мость объяснять существование одного элемента через наличие
другого.
Гетеротрофы подразделяют по месту, которое они занимают в
цепях питания, на консументов первого, второго и т. д. порядка, а
по характеру действия на потребляемую органику — на редуцентов
и деструкторов. Редуценты — гетеротрофы, понижающие запас
энергии в используемых веществах и упрощающие структуру этих
веществ. К ним относятся биотрофы — потребители живой орга-
ники и сапротрофы — потребители мертвой органики. Деструк-
торы— последнее звено. Они окончательно разрушают органи-
ческие молекулы, минерализуют их, переводя в состояние, дос-
тупное для повторного использования автотрофами и частично ге-
теротрофами в биогенном цикле.
Среди гетеротрофов есть небольшая труппа микроорганиз-
мов — несерные пурпурные бактерии, которые используют одно-
временно и солнечный свет как источник энергии, и органичес-
кие вещества как источник углерода. В связи с этим фактом в об-
щей классификации трофических групп организмов на первое ме-
сто выступает новый аспект — отношение к источникам энергии,
углерода и водорода (рис. 3.13).
Глава 4
БИОЛОГИЯ КЛЕТКИ
4.1. КЛЕТОЧНАЯ ТЕОРИЯ
История изучения клетки во многом отражает историю разви-
тия микроскопической техники и методов исследования. В это
трудно поверить, но большая часть сведений о строении и жиз-
недеятельности клетки получена лишь в последние три десятиле-
тия. Тем не менее регистрация ряда фактов, касающихся клетки
и воспринимаемых сейчас как очевидные, стала в свое время не
просто серией крупных открытий, а фундаментом для изучения
жизни на молекулярном и клеточном уровнях организации, без
чего познание организменного, биоценотического и биосферно-
го уровней было бы если не невозможным, то бесконечно ущерб-
ным.
В XVII в. стеклянные линзы и простейшие микроскопы, изоб-
ретенные Захарием Янсеном еще в 1590 г., использовались чаще
всего для забавы. В числе первых создателей микроскопов был Ро-
берт Гук. Круг профессиональных интересов английского ученого
отличался разнообразием: метрология, физика, инженерное дело,
архитектура и биология. Любопытно, что замечательное открытие
сделано им как бы мимоходом. В 1665 г. Р. Гук опубликовал аль-
бом рисунков, изображающих детали различных предметов и ма-
териалов, наблюдаемых под микроскопом. Среди них был и рису-
нок среза пробковой ткани, фактически мертвых клеточных обо-
лочек, заключающих крошечные камеры. Клетками Гук назвал
собственно перегородки между полостями, полагая, что это и есть
живое вещество. Сейчас же под термином «клетка» понимают
именно содержимое полостей (в живом состоянии).
Далее приведены последующие этапы изучения клетки.
1650—1700 гг.— голландец Антони ван Левенгук при помощи
хорошо отшлифованных линз и усовершенствованного микроско-
па с увеличением до 270 раз наблюдал клетки крови, сперматозои-
ды, простейшие и даже бактерии.
1700—1800 гг.— расширение наблюдений с помощью микро-
скопов, увеличивающих объекты в 100...200 раз. Накопление опи-
саний одноклеточных организмов и растительных тканей.
101
1827 г. — итальянский физик Джованни Баттист Амичи усовер-
шенствовал технологию изготовления линз для микроскопов, уст-
раняющую оптические аберрации, что дало толчок распростране-
нию микроскопов и проявлению интереса к изучению микрообъ-
ектов.
1828 г. — немецкий химик Фридрих Велер синтезировал моче-
вину.
1831 г. — англичанин Роберт Браун описал ядро в клетках рас-
тений.
1837 г. — немецкий ботаник Маттиас Шлейдеп сформулировал
теорию, согласно которой все растения состоят из клеток.
1838—1839 гт. — М. Шлейлен и его друг — физиолог живот-
ных — Теодор Шванн обобщили свои идеи с результатами иссле-
дований других ученых и сформулировали клеточную теорию. В
ней они утверждали, что все растения и животные состоят из од-
ной или множества клеток; клетка является основной единицей
структуры и функции во всех живых организмах; животные и рас-
тения имеют единый принцип роста, основанный на клеточном
делении.
1840 г. — Я. Пуркинье ввел термин «цитоплазма» для обозначе-
ния клеточного содержимого.
1855 г. — немецкий патологоанатом Рудольф Вирхов доказал,
что все клетки образуются из других путем клеточного деления.
1865 г. — чешский настоятель монастыря Грегор Мендель уста-
новил основные закономерности наследования признаков и их
изменчивости.
1886 г.— Эрнст Геккель установил, что хранение и передачу
наследственных признаков осуществляет ядро.
1886—1888 гг. — в разных странах изучали клеточное деление,
подробно описали его различные формы, было сделано описание
морфологии метафазных хромосом.
1880, 1888 гг. — открыты пластиды растений.
1890 г. — открыты митохондрии.
1898 г. — открыт мембранный комплекс, получивший название
«аппарат Гольджи».
1887—1900гг.—усовершенствована микроскопическая техни-
ка: разрешающая способность микроскопов доведена до физичес-
кого предела — до половины длины волны света. Развивались ме-
тоды изготовления микропрепаратов.
1880—1900 гг. — развитие химии малых органических молекул:
синтезированы сахара, аминокислоты, липиды, пурины, пирими-
дины; составлено представление об обмене веществ в клетке.
1900 г. — переоткрытие «законов Менделя».
1900—1940 гт. — замедление исследований в области морфоло-
гии и химии клетки. Область между разрешающей способностью
102
светового микроскопа и молекулами, доступными для химическо-
го изучения (0,001...0,1 мкм), остается абсолютно неисследован-
ной.
1930-е годы — изобретение электронного микроскопа.
1945 г. — Кейт Р. Портер, Альбер Клод и Эрнст Ф. Фуллам при-
менили электронный микроскоп для изучения клетки.
1950—1970 гг. — велись интенсивные работы по изучению ульт-
раморфологии клетки.
1930—1970 гг. — введены новые методы изучения биохимии
клетки: хроматография, электрофорез, изотопное мечение, высо-
коскоростное улырацентрифугирование.
С 1950-х годов объединяются морфологические, биохимичес-
кие, биофизические и генетические методы изучения клетки.
Соизмерять важность открытий с этого времени становится все
труднее. Отметим лишь «построение» Дж. Уотсоном и Ф. Криком
модели структуры ДНК. Впрочем, открытия последних четырех
десятилетий мало изменили содержание клеточной теории, кото-
рую можно выразить следующими пунктами.
1. Клетка — это структурная единица живых организмов, пред-
ставляющая собой определенным образом дифференцированный
участок цитоплазмы, окруженный клеточной мембраной.
2. Клетка — элементарная структурная и функциональная еди-
ница живого.
3. Клетки различных организмов гомологичны по своему стро-
ению.
4. Основные метаболические процессы в клетках всех живых
систем принципиально близки.
5. Размножение клеток происходит путем деления исходной
клетки.
6. Многоклеточные организмы представляют собой сложные
ансамбли клеток, объединенные в целостные, интегрированные
системы тканей и органов, взаимоподчиненные и связанные меж-
ду собой межклеточными и нервными формами регуляции.
4.2. ВВЕДЕНИЕ В БИОХИМИЮ КЛЕТКИ
4.2.1. ХИМИЧЕСКИЙ СОСТАВ ЖИВОЙ КЛЕТКИ (НЕОРГАНИЧЕСКИЕ
И МАЛЫЕ ОРГАНИЧЕСКИЕ МОЛЕКУЛЫ)
Разделение всех организмов, составляющих живое вещество
биосферы, на продуцентов и консументов (редуцентов и деструк-
торов) одновременно означает и то, что живые организмы, а точ-
нее — ценотически активные живые тела, выстраиваются в цикли-
ческую последовательность. В рамках цикла завершается поэтап-
103
ная передача отдельных порций вещества — круговорот химичес-
ких элементов, а также рассеяние части энергии в виде тепла при
протекании экзотермических реакций и при совершении работы.
Круговорот элементов в масштабе биосферы представляет со-
бой незамкнутый цикл: в него непрерывно вовлекаются новые
порции неживого вещества, а часть биогенного вещества (в целом
несколько меньшая) выходит из цикла преимущественно в виде
неорганических соединений. Именно поэтому единство химичес-
ких элементов в живом и неживом веществе составляет в одно и то
же время и причину, и следствие. Но круговорот элементов не яв-
ляется единственной и тем более главной сущностью жизненных
процессов. Как ранее было показано, трансформация веществ
идет параллельно со связыванием, передачей и высвобождением
энергии при общем ее накоплении в подразделениях живой систе-
мы — биосферы. Накопление же энергии усиливает неравновес-
ность, асимметрию структур, состояний и процессов и в конечном
счете увеличивает объем информации.
Общую вещественно-энергетическую картину живой составля-
ющей биосферы можно представить в виде схемы глобальной со-
пряженности взаимно обусловливаемых процессов синтеза и рас-
пада (рис. 4.1). Обособленные живые тела, в которых протекают
Рис. 4.1. Схема сопряженности процессов синтеза п распада в живом веществе
(оригинал автора)
104
отдельные фрагменты биосферных энергетических процессов, в
этом случае несложно свести к трансформаторно-конденсатор-
ным схемам, на выходе которых энергия либо накапливается (у
автотрофов-продуцентов), либо высвобождается с совершением
работы (у гетеротрофов-консументов) или непосредственно в виде
тепла (у гетеротрофов-редуцентов). Но энергетические процессы,
лежащие в основе жизнедеятельности живых тел, — это в конеч-
ном счете процессы взаимодействия молекул, составляющих их.
Именно на молекулярно-биосферном уровне происходит сопря-
жение всех звеньев в условно замкнутые циклы.
Если мысленно вычленить, изолировать живое естественное
тело, устранить его биоценотическую сущность, иными словами,
представить его не элементом живой системы, а собственно от-
дельным телом, то не составит особого труда сделать его физико-
химический «портрет». Это будет несколько сотен повторяющихся
реакций, сопряженных в единый комплекс каскадов и циклов, в
которых участвует около 70 (а всего до 100) химических элемен-
тов.
Химические элементы, входящие в состав живого вещества,
широко распространены в земной коре и в космосе в целом. Од-
нако соотношение долей каждого из них в живом и неживом ве-
ществе резко различается. Часть элементов: углерод, кислород, во-
дород и азот — имеют особенно высокое относительное предста-
вительство в живом веществе, то есть обладают высокой биофилъ-
ностью (С — 7800, О — 1,5, Н — 70, N — 160). На долю этих так
называемых органообразующих элементов приходится около 97 %
всей массы живого вещества. Вместе с фосфором, серой, калием,
кальцием, натрием, хлором и железом перечисленные элементы
входят в число макроэлементов, содержание каждого из которых в
живом веществе превышает 0,01 %.
Группу микроэлементов образуют химические элементы, содер-
жание которых в живом веществе колеблется в пределах
0,001...0,01 %. Еще меньше содержание элементов, относимых к
группе ультрамикроэлементов.
Самым распространенным соединением в живом веществе яв-
ляется вода. В клетках живых тел она составляет в среднем около
70 % массы при вариациях от 60 до 95 %. Почему именно вода?
Это вещество имеет целый ряд уникальных свойств аномального
характера, связанных с полярностью ее молекул. Так, вода плавит-
ся и кипит при более высоких температурах, чем это должно быть
в отсутствие водородных связей между молекулами. В твердом со-
стоянии ее плотность меньше, чем в жидком, из-за чего лед не то-
нет в воде. Кроме того, у воды высокая теплоемкость и большое
поверхностное натяжение. Она практически не сжимаема и не ра-
стяжима.
105
Молекула воды, имея равное число протонов и электронов, в
целом электронейтральна. Но из-за того что электроны располо-
жены несимметрично, она оказывается полярной. Так происходит
потому, что ядро атома кислорода несколько оттягивает электро-
ны от ядер атомов водорода, оставляя на них суммарный положи-
тельный заряд. Области же с небольшим отрицательным зарядом
располагаются в двух других углах воображаемого тетраэдра.
Поляризованные соседние молекулы воды способны образовы-
вать водородные связи, прочность которых максимальна, когда
два атома кислорода и атом водорода лежат на одной прямой.
Каждая молекула, объединяясь с четырьмя другими, участвует в
построении пространственной решетки. Когезивная (основанная
на способности частиц к слипанию) природа воды определяет и ее
высокое поверхностное натяжение, и высокую удельную теплоем-
кость, и большую теплоту испарения.
В силу своей полярности молекулы воды группируются вокруг
других полярных молекул и стабилизируют их водородными свя-
зями. Поэтому вода выступает в качестве универсального раство-
рителя гидрофильных веществ. Неполярные молекулы, вклинива-
ясь между молекулами воды, разрушают ее структуру, усиливают
беспорядок. Система из молекул гидрофобного вещества и воды
стремится прийти к более энергетически выгодному состоянию,
то есть к более упорядоченному состоянию, что выражается в вы-
талкивании «неудобных» молекул, их сближении и образовании
меньшего числа более крупных частиц. При этом структурная
упорядоченность смеси увеличивается.
Молекулы воды обладают слабовыраженной тенденцией к
ионизации, приводящей к временному переходу протона из одной
молекулы в другую и, следовательно, к проявлению одновременно
и кислотных, и щелочных свойств. Концентрация таких полусво-
бодных протонов в воде составляет 10-7моль/л (pH 7), что приня-
то обозначать выражением «нейтральная среда».
Когда в воде растворяются соединения, молекулы воды окру-
жают их и ослабляют взаимное притяжение анионов и катионов.
Подвижность ионов при этом увеличивается, что вызывает возра-
стание их реакционной способности.
Среди перечисленных особенностей воды нет несущественных
для развития земной формы жизни, но наиболее значимые из них
связаны с тем, что молекулы воды малы и полярны.
В биосфере функции воды неисчислимы. Вода в жидком и па-
рообразном состояниях является и внутренней, и внешней средой
жизни. Она переносит и концентрирует различные вещества, рас-
средоточивает или упорядочивает молекулы веществ, участвует в
поддержании и изменении формы живых тел, обеспечивает спо-
собность к терморегуляции и в целом стабилизирует температуру
106
живых тел. Вода участвует в химических реакциях. Она является
основным источником протонов для фотосинтеза. Распределение
и сочетание жизненных форм организмов в пределах биосферы
прежде всего определяется обилием и состоянием воды. С водой
связано явление, получившее название «осмос», которое заключа-
ется в том, что через мембраны, разделяющие два водных раствора
и пропускающие только молекулы воды, вода движется из гипото-
нического раствора в гипертонический, повышая в последнем ос-
мотическое (гидростатическое) давление.
Только воде уступают по распространенности в живом веще-
стве соединения углерода — наиболее биофильного среди хими-
ческих элементов. Углерод отличается от всех элементов способ-
ностью образовывать большие молекулы. Имея на внешнем уров-
не четыре электрона, он может образовывать четыре прочные ко-
валентные связи с другими атомами, в том числе и с атомами
углерода. В последнем случае строятся цепочки или кольца —
структурная основа всех органических молекул. Потенциальное
число различных органических молекул бесконечно, но в клетках
живых тел используется лишь небольшая их часть —всего не-
сколько миллионов. Правда, это число будет уменьшено на три
порядка, если произвести оценку числа так называемых малых
органических (или биологических) молекул, которые находятся в
растворе и из которых могут строиться, как из отдельных блоков,
разнообразные сложные — большие — биологические молекулы.
Молекулярная масса малых органических молекул варьирует от
100 до 1000, а число углеродных атомов в них, как правило, не
превышает тридцати. В таких молекулах имеется четыре типа
комбинаций атомов, определяющих их физические и химичес-
кие свойства: метильная группа (—СН3); гидроксильная группа
(—ОН); карбоксильная группа (—СООН) и аминогруппа (—NH2).
Содержащиеся в клетках малые органические молекулы обра-
зуют четыре основных семейства: простые сахара, жирные кисло-
ты, аминокислоты и нуклеотиды. Естественно, имеются молеку-
лы, не относящиеся к перечисленным семействам, но их общая
масса в клетке крайне мала.
Если рассматривать основные органические вещества в плане
особенностей соединения и сочетания атомов углерода с атомами
других химических элементов, нетрудно заметить, что они образу-
ют серию однородных групп.
Углеводороды — соединения, в которых углеродный скелет как
бы одет в «чехол» из водородных атомов. Это устойчивые непо-
лярные молекулы, неспособные к образованию водородных свя-
зей и нерастворимые в воде. Углеводородные цепи могут быть ча-
стями других более сложных веществ.
Так называемые С—О~соединения содержат один или несколько
107
атомов, связанных с кислородом. Это спирты, содержащие гидро-
ксильную группу; альдегиды и кетоны, содержащие карбониль-
ную группу; карбоновые кислоты, имеющие карбоксильную груп-
пу; эфиры — соединения, образующиеся при реакции кислот со
спиртами.
Углерод может быть соединен с атомом азота. В этом случае
образуются C-N-соединения: амины, амиды и азотистые основа-
ния. Амины способны присоединять в водном растворе протоны и
потому являются основаниями. Амиды же образуются в процессе
реакций кислот с аминами и в отличие от последних неполярны.
Азотистые основания разделяются на две группы: пуриновые и
пиримидиновые. В их основе лежит шестичленное пиримидино-
вое кольцо, сконденсированное с пятичленным кольцом или сво-
бодное.
Отметим также соединения углерода с фосфатом. Фосфат не-
органический — это стабильный ион фосфорной кислоты. При
взаимодействии фосфата со свободной гидроксильной группой
образуются фосфатные эфиры, а при взаимодействии с карбо-
ксильной группой или с другой фосфатной группой — кислые ан-
гидриды, которые легко гидролизуются с освобождением энергии.
Перечисленные группировки атомов в различных сочетаниях
связываются друг с другом, образуя большую часть разнообразия
органических веществ, которые мы уже отнесли к четырем основ-
ным семействам.
Моносахариды — это часть альдегидов и кетонов, имеющих не
менее двух гидроксильных групп. Их общая формула (СН2О)„. Уг-
леродная цепь моносахаров содержит от 3 до 7 атомов, и потому
наиболее простыми сахарами являются триозы глицеральдегид и
дигидроксиацетон:
о ОН о
। и
'с-с~сн2он сн2он-с-сн2он
Н Н
Глицеральдегид
Дигидроксиацетон
В природе очень широко распространены моносахариды, со-
держащие 5 и 6 углеродных атомов. Это пентозы и гексозы. При
длине углеродной цепочки более 4 атомов альдегидная и кето-
группа сахара могут реагировать с гидроксильной в пределах од-
ной молекулы, что приводит к образованию 5- или 6-членного
кольца, — явление, очень характерное, например, для рибозы и
глюкозы.
Возможность изменения положения отдельных атомов, связан-
108
них с углеродным скелетом, например обмен направлениями ори-
ентации водорода и гидроксильной группы, обусловливает суще-
ствование у сахаров множества стереомеров. Так, циклическая
глюкоза дает путем изомеризации моносахара маннозу и галакто-
зу. Сама же она существует в двух зеркально противоположных
стереоформах (обозначаются как D-глюкоза и L-глюкоза).
Гидроксильные группы в молекулах моносахаридов могут быть
заменены другими группами (аминогруппой, карбоксильной
и др.) или их комплексами, что сопровождается и изменением
свойств сахаров — образуются производные моносахаридов. Но
гидроксил сахара может замещаться и аналогичной молекулой, то
есть может происходить образование веществ, молекулы которых
включают два моносахарида, а связь между ними называют глико-
зидной. Подобным способом образуются, в частности, сахароза,
лактоза и др.
Если одинаковые моносахариды связываются в линейные или
разветвленные молекулы, то образуются олигосахариды и полиса-
хариды (крахмал, парамил, гликоген, целлюлоза и т. д.). Впрочем,
сложные сахара могут включать и разные моносахариды, а «одно-
образные» полимерные молекулы могут образовываться из произ-
водных сахаров (например, хитин).
Молекулы моносахаридов могут быть ковалентно связаны с
молекулами белков (в гликопротеидах) и жиров (в гликолипидах)
с образованием важнейших сигнальных, регуляторных и комму-
никационных веществ.
Основная, но далеко не единственная функция сахаров в клет-
ках живых тел — энергетическая. В результате последовательных
сопряженных реакций сахар постепенно расщепляется до более
коротких производных и, в конце концов, до неорганических ве-
ществ, из которых, он был образован. В этом процессе освобожда-
ется энергия, в большей или меньшей степени накапливаемая в
связях нуклеофосфатов (большей частью АТФ), и формируется
восстановительная способность в виде протонов, ассоциирован-
ных с их акцептором — НАД или НАДФ. Особенно легко окисля-
ются сахара-альдегиды.
Моносахара — не только источники энергии. Их функции в
живых телах многообразны. Так, глицеральдегид и дигидроксиа-
цетон выступают в качестве промежуточных продуктов в углевод-
ном обмене (в фотосинтезе, в дыхании). Глицеральдегид, кроме
того, используется в качестве строительного блока в синтезе липи-
дов. Пентозы участвуют в образовании нуклеотидов — макроэрги-
ческих соединений и мономеров нуклеиновых кислот, кофермен-
тов (НАД, НАДФ, ФАД, ФМН, кофермента А). Рибулезодифос-
фат связывает СО2 в темновой фазе фотосинтеза. Гексозы — наи-
более распространенные моносахара — основа для синтеза
109
резервных строительных и энергетических веществ ди-, олиго- и
полисахаридов.
Главнейшие широко распространенные производные моноса-
харидов: глицерол — предшественник липидов; аскорбиновая
кислота; глюкозамин — аминосахар, используемый в синтезе хи-
тина и ряда других полимеров.
Жирные кислоты — это углеводородные цепочки, оканчиваю-
щиеся карбоксильной группой. Такое строение молекулы придает
ей интересные свойства: углеводородная цепь химически малоак-
тивна и гидрофобна, а карбоксильная группа очень легко ионизи-
руется в воде и, следовательно, очень гидрофильна и реакционно-
способна. Жирные кислоты, имеющие в цепочке двойные связи,
называются ненасыщенными, углеводродная цепь у них искрив-
лена. Ионизированная карбоксильная группа легко связывается
с другими группами, в результате чего образуются эфиры или
амиды.
В клетках жирные кислоты обычно связываются с глицеролом,
присоединяясь к двум или трем его гидроксильным группам. Ког-
да третья гидроксильная группа связывается с фосфатом, а тот, в
свою очередь, с другими небольшими гидрофильными группами
(этаноламином, серином или холином), образуются фосфолипи-
ды — главнейшие структурные компоненты клеточных мембран.
Способность к формированию мономолекулярных поверхностных
пленок или погруженных в воду сфер — это прямое следствие раз-
личного отношения к воде «хвоста» и «головы» молекулы липида
(жира) или отдельной жирной кислоты. Фосфолипиды способны
в воде самоорганизовываться в бислои (бимолекулярные слои),
связываясь гидрофобными «хвостами». Главная роль в этом про-
цессе принадлежит взаимодействию гидрофобной части молекулы
с нарушаемой ею структурой воды. Аналогичное поведение обна-
руживают и гликолипиды — молекулы, содержащие два длинных'
гидрофобных «хвоста» и полярную область, состоящую из остат-
ков сахаров.
Триглицеролы, как правило, образуют резерв энергонасыщен-
ного вещества (жира). При их расщеплении в макроэргические
связи АТФ переводится почти в два раза больше энергии, чем при
расщеплении такого же количества глюкозы. Так происходит из-
за высокого содержания в молекуле углерода и водорода и низко-
го — кислорода.
Кроме жиров к липидам относят и некоторые другие вещества,
растворимые в органических растворителях. Это, например, сте-
роиды и полиизопреноиды — компоненты мембранных комплек-
сов и биологически активные вещества.
Аминокислоты — органические вещества, к первому углеродно-
му атому в цепочке которых одновременно присоединены карбо-
ПО
ксильная (СООН) и амино- (NH2) группы. Всего известно около
170 аминокислот.
Боковые цепи природных аминокислот, входящих в полипеп-
тидные цепи, представлены всего 20 видами, которые подразделя-
ют по своим свойствам на четыре семейства: кислотные, основ-
ные, полярные незаряженные и неполярные. Аминокислоты в
водном растворе при pH 7 всегда ионизированы из-за потери про-
тона карбоксильной группой и приобретения его аминогруппой.
Боковые цепи также несут больший или меныпий заряд, что зави-
сит от pH раствора.
Кислые боковые цепи имеют аспарагиновая и глутаминовая
кислоты; основные — лизин*, аргинин* и гистидин*; полярные
незаряженные — глицин, аспарагин, глутамин, серин, треонин*,
тирозин, цистеин (образует дисульфидные мостики, поддержива-
ющие третичную структуру белков); неполярные боковые цепи
имеют аланин, валин*, лейцин*, изолейцин*, пролин, фенилала-
нин*, метионин* и триптофан* (их боковые цепи в белках стре-
мятся сгруппироваться внутри трехмерной структуры молекулы).
Аминокислоты способны соединяться в длинные цепочки-по-
лимеры — белки посредством пептидных связей между карбо-
ксильной группой одной аминокислоты и аминогруппой другой
аминокислоты. Кроме 20 «магических» (или «волшебных») амино-
кислот в белках встречается несколько редких — производных от
них, которые не кодируются в ДНК, но образуются путем моди-
фикации аминокислот, уже включенных в полипептидную цепь.
Около 150 аминокислот никогда не встречается в белках, хотя
присутствует в клетках в свободном или связанном виде.
Нуклеотиды — сложные вещества, которые включают основа-
ния, пятичленные сахара и один или несколько фосфатных остат-
ков (фосфорную группу). Основаниями называют азотсодержа-
щие, циклические соединения — пурины и пиримидины. В пури-
нах, представляющих как бы спаренные циклы, имеется четыре
постоянных атома азота в 1, 3,7 и 9-м положениях и дополнитель-
но к углероду во 2-м (у гуанина) или в 6-м (у аденина) положении
присоединены аминогруппы. В пиримидинах «общие» атомы азо-
та занимают только 1-е и 3-е положения в одиночном цикле, а
аминогруппа связана с углеродом в 6-м положении только у цито-
зина. Урацил и тимин в этом положении имеют атом кислорода.
Кроме того, у всех пиримидиновых оснований к углероду во 2-м
положении присоединен атом кислорода.
К пятичленным сахарам нуклеотидов относятся только рибоза
и дезоксирибоза. Они отличаются друг от друга лишь наличием
*Так называемые незаменимые аминокислоты, синтезируемые только расте-
ниями.
Ill
или отсутствием атома кислорода при углероде во 2-м положении:
гидроксил — у рибозы; водород — у дезоксирибозы. При соедине-
нии основания с сахаром N-гликозидная связь образуется между
первым атомом углерода в сахаре и атомом азота в 1-м положении
у пиримидинов и в 3-м положении у пуринов. Фосфатная группа
присоединяется всегда к 5-му углеродному атому сахара.
Нуклеотиды (основание + сахар + фосфат) называют соответ-
ственно образующим их основаниям: аденозин, гуанозин, уридин,
цитидин и тимидин. Каждый из них может иметь от одной до трех
фосфатных групп, благодаря чему может служить временным на-
копителем энергии в связях между фосфатами. Трифосфатный
эфир аденина (АТФ) выступает в качестве энергетического по-
средника чаще, чем другие нуклеотиды. Его концевые (второй и
третий) фосфаты легко гидролизуются и образуют ковалентные
связи с другими, самыми разнообразными, молекулами. При этом
энергия, высвобождаемая при гидролизе, может сразу же исполь-
зоваться в любой эндоэргической реакции. Гуанозинтрифосфат
(ГТФ) служит источником энергии для синтеза белка на рибосо-
мах высших хордовых. В растительных клетках в этом качестве ис-
пользуются инозиндифосфат (ИДФ) и инозинтрифосфат (ИТФ),
у низших хордовых и иглокожих — креатинфосфат, а у кольчатых
червей, членистоногих и моллюсков — аргининфосфат.
Некоторые нуклеотиды способны служить переносчиками от-
дельных химических групп (атомов водорода, остатков сахаров
и др.) с одной молекулы на другую. Циклический аденозинмоно-
фосфат является универсальным внутриклеточным сигналом, ре-
гулирующим скорость протекания многих реакций в клетке.
Нуклеотиды в форме трифосфатов легко соединяются в поли-
мерные цепи (образуя фосфорные эфиры) ковалентными связями
между 3-гидроксильной группой остатка сахара одного нуклеоти-
да и 5-фосфатом другого. Поскольку в образовании полимерных
нуклеиновых кислот принимают участие четыре рибонуклеотида
или четыре дезоксирибонуклеотида, последовательность их соеди-
нения может варьировать. Вследствие этого существует возмож-
ность образования многочисленных уникальных последователь-
ностей. Чем длиннее полимерная цепь, тем больше уникальных
вариантов. Такая особенность отчасти определяет участие нуклео-
тидов в информационных процессах. Второй особенностью нук-
леотидов, обеспечивающей их участие в передаче генетической
информации, следует считать способность к нековалентному вза-
имодействию между определенными парами оснований. Без двух
названных особенностей нуклеотидов самосборка цепей РНК и
ДНК была бы невозможна.
Помимо отмеченного нуклеотиды и их производные служат
важными коферментами. К их числу относятся кофермент А
112
(СоА), никотинамидадениндинуклеотид (НАД), НАД-фосфат
(НАД-Ф), флавинадениндинуклеотид (ФАД), упоминавшиеся ра-
нее АМФ, АДФ, АТФ, циклический АМФ и др.
4.2.2. МАКРОМОЛЕКУЛЫ И ОСНОВНЫЕ ХИМИЧЕСКИЕ ПРОЦЕССЫ,
ИДУЩИЕ В ЖИВОЙ КЛЕТКЕ
Накопление энергии фотоавтотрофами в процессе фотосинтеза и
хемосинтеза и использование ее гетеротрофами. Теперь, когда мы
рассмотрели основные группы органических веществ на уровне
малых молекул, мы можем, хотя и очень схематично, изобразить
общую картину физико-химических процессов, проходящих в
природных органических телах.
Вся организация тел автотрофов, от плана строения до соотно-
шения комплексов молекул, подчинена одной «задаче» — перевес-
ти поступающую извне рассеянную энергию в «конвертируемую»
(обращаемую) форму. Но решается эта задача у современных жи-
вых тел чрезвычайно сложным путем, включающим текущее само-
воспроизводство, поддержание постоянства внутренней среды в
меняющейся внешней среде, воспроизводство в поколениях и
производство избыточной продукции малых и больших молекул.
Как отмечалось в разделе 2.1.4, оптимальной формой накапливае-
мой и хранимой энергии следует считать химическую, которая по
обратимости уступает только электродинамической, но последняя
не может быть накоплена и сохранена.
Рассмотрим смысловую сторону трансформаций вещества и
энергии в фотоавтотрофных телах. Упростив до предела схему
процесса фотосинтеза, ее можно представить в следующем виде:
(ЭМЭ) “> Хл°Р°Филл
Световые НАД(Ф) Н
реакции АТФ
АДФ+ФН
НАД(Ф)
В схеме есть вход, цикл и выход. На входе обозначены вода, СО2
и энергия электромагнитного поля. В цикле неизменными остают-
8 Присный А.В.
113
ся акцептор водорода НАД(Ф) и нуклеотиддифосфат с неоргани-
ческим фосфатом. Если пренебречь поступлением в автотрофные
тела и потерей из них ряда веществ и порций энергии, не связанных
непосредственно с фотосинтезом, то оказывается, что водород воды
и СО2, трансформируясь в сахар, заключают в себе значительную
часть принятой солнечной энергии. То есть большинство веществ,
о которых шла речь в предыдущей части этого раздела, — моноса-
хара, жирные кислоты, аминокислоты, основания и их производ-
ные — предназначены для построения тела, производящего избы-
точное количество сахара, обозначенного на выходе. Кислород же
образуется как побочный продукт фотолиза воды.
Аналогичный вид будет иметь и схема хемосинтеза с той лишь
разницей, что источником энергии в ней служат минеральные ве-
щества.
Впрочем, суммарное уравнение реакции фотосинтеза, как и
названная смысловая сторона процесса, не отражают всего глав-
ного содержания химических процессов в автотрофных телах.
Приняв синтез углеводов за самостоятельный, завершенный и
центральный процесс, мы тем самым поставили синтез аминокис-
лот, жирных кислот и оснований с их производными во второсте-
пенное положение, в положение обеспечивающих процессов. Но
где и когда синтезируются малые органические молекулы пере-
численных семейств и какова их дальнейшая судьба?
На рисунке 4.2 приведена схема сопряженных процессов син-
теза главнейших групп органических веществ в автотрофных те-
лах, из которой видно, что в темновой фазе фотосинтеза образу-
ются не только молекулы фруктозодифосфата (ФДФ) — предше-
ственника глюкозы (здесь на разных этапах преобразований фор-
мируются углеродные цепочки с числом атомов от 3 до 7), но
также предшественники липидов, белков* нуклеиновых кислот и,
естественно, полисахаридов. Все эти и множество других веществ
действительно составляют тела продуцентов. Но известно, что
тела всех гетеротрофов также строятся в основном из веществ, ис-
ходно синтезированных автотрофами. И здесь содержатся не толь-
ко сахара — основные источники энергии. Хотя большую часть
сложных веществ гетеротрофы синтезируют из блоков, поступаю-
щих в конечном счете от автотрофов, многие из них подобно неза-
менимым аминокислотам должны быть получены в готовом виде.
Если допустить, что живые тела в биосфере исходно раздели-
лись на автотрофов и гетеротрофов, уже на ранних этапах ее раз-
вития эти трофические группы должны были стать полностью вза-
имозависимыми. Так как практически весь СО2 атмосферы Земли,
а также лито- и гидросферы содержит углерод, многократно про-
шедший через живое вещество, непрерывно переводимый гете-
ротрофами в формы, доступные автотрофам, было бы натяжкой
114
Рис. 4.2. Схема сопряжения синтетических процессов у автотрофов
(оригинал автора)
8'
считать такую взаимообусловленность жизни различных органи-
ческих тел «эволюционной притиркой». На первый взгляд гете-
ротрофы «изымают» у автотрофов лишь небольшую часть биомас-
сы. Но это чаще всего прижизненное разовое отторжение. Если же
учесть многократное или постоянное питание биотрофов, напри-
мер, на растениях, сезонную переработку органики после индиви-
дуальной смерти автотрофов и отмирания частей их тел сапротро-
фами, то окажется, что практически вся органика, образованная
фото- и хемосинтетиками, в конечном счете переходит в тела
животных, грибов, бактерий и, растекаясь по живому веществу,
рано или поздно вновь возвращается в тела автотрофов. В этом
круговороте следует отмерить малозаметную, но исключительно
важную деталь: общая масса живого вещества и сложность его
динамической организации увеличиваются с геологическим вре-
менем. А это означает, что в биогенный круговорот вовлекаются
все новые порции земного вещества, непрерывно накапливается
химическая энергия, в прогрессии увеличиваются энергетичес-
кие затраты на структурную и динамическую организацию био-
сферы.
Теперь рассмотрим схему процессов освобождения энергии из
веществ-накопителей, например из полисахаридов:
ДЕЙСТВУЮЩАЯ
ЭНЕРГИЯ
Как и следовало ожидать, последовательность событий в схеме
в целом противоположна изображенному на схеме связывания
энергии автотрофами. Значит ли это, что разнонаправленные про-
цессы полностью уравновешивают друг друга? Лишь отчасти, а
именно количественно, в строгом соответствии с законами сохра-
нения масс и энергии (с учетом медленного увеличения массы ве-
щества и запаса энергии в совокупном живом веществе биосферы
в геологической истории). В качественном отношении процессы
не абсолютно равноценны: на входе в живую систему (или в жи-
вые тела автотрофов) энергия не носит столь концентрированного
и дифференцированного характера, как на выходе из системы
116
(или из тел гетеротрофов), где она становится направленно дей-
ствующей (см. рис. 4.1). Значительное количество энергии живые
тела теряют в виде тепла, излучаемого в окружающую среду. Такое
рассеяние энергии сопровождается увеличением хаоса в среде и
одновременно увеличением порядка в живых телах, в живом ве-
ществе в целом.
Далее нас будут интересовать структуры, которые обеспечива-
ют работу живых тел, и особенно структуры, связанные с транс-
формацией энергии и пространственно-временной организацией
энергетических процессов. Здесь нам не обойтись без предвари-
тельного знакомства с большими органическими молекулами,
строительными блоками для которых служат уже рассмотренные
малые органические молекулы.
Полисахариды. Эти молекулы большей частью составляют пи-
щевой пластический и энергетический резерв, но выполняют и
строительную функцию. Они плохо растворимы в воде и потому
не оказывают на клетку ни химического, ни осмотического дей-
ствия. В зависимости от вида мономеров полисахариды могут на-
зываться пентозами или гексозами. К числу наиболее распростра-
ненных полимерных сахаров относят крахмал, гликоген и целлюло-
зу. Все три этих вещества образованы глюкозой. Крахмальные мо-
лекулы включают два компонента: амилозу и амилопектин.
Первый имеет простую цепь, сворачивающуюся в спираль, а вто-
рой образован ветвящейся цепью. Крахмал — основное резерв-
ное вещество растений. Гликоген подобен амилопектину, но его
молекулы еще более разветвлены. Синтезируется он в клетках
животных. Целлюлоза — самое распространенное в природе
органическое вещество. Цепь у нее насчитывает до 10 тыс. глю-
козных остатков. Выступающие за контуры молекулы ОН-груп-
пы образуют в параллельных молекулах водородные связи, креп-
ко сшивающие их в микрофибриллы. Пучки сплетающихся мик-
рофибрилл называют макрофибриллами. В клеточных оболочках
макрофибриллы расположены перекрещивающимися слоями.
Такие оболочки тем не менее оказываются проницаемыми и для
света, и для различных веществ, растворенных в воде. Лишь не-
многие виды живых тел способны расщеплять целлюлозу до мо-
номеров.
К полисахаридам близки такие соединения, как хитин, глико-
протеины и гликолипиды. Хитин входит в состав клеточной обо-
лочки у большинства грибов, а также в состав кутикулы кольчатых
червей, членистоногих и некоторых других групп животных.
Это — полимер глюкозамина. Молекулы хитина, как и целлюло-
зы, образуют фибриллы. Гликопротеины и гликолипиды образуют
гликокаликс — оболочку клеток животного типа. С их присутстви-
ем связаны процессы межклеточного узнавания, а также антиген-
117
ные свойства клеток. Пектины и гемицеллюлоза входят в состав
матрикса клеточной оболочки у растений и способны образовы-
вать гели. Мукополисахариды в качестве мономеров имеют диса-
харидные остатки, в которых один из сахаров — обязательно ами-
носахар. Они входят в состав соединительных тканей животных
(хряща, костной ткани, роговицы глаза и др.).
Липиды. В дополнение к сведениям об этой группе веществ,
приведенным ранее, отметим, что к ним кроме триаглицеролов
(жиров и масел) относят фосфолипиды, воска, стероиды и терпе-
ны, гликолипиды и липопротеины. Если жиры и масла — это ти-
пичные энергетические депо, которые, скапливаясь в специализи-
рованных запасающих тканях и компартментах, могут выполнять
и другие дополнительные функции, то остальные липиды чаще
монофункциональны.
Воска часто включаются в покровные образования у растений и
животных и участвуют в терморегуляции, а также в защите от вы-
сыхания. Фосфолипиды — главные компоненты клегочных мемб-
ран. Они образуют бислои, поверхности которых составлены гид-
рофильными концами молекул. Стероиды, хотя и относятся к ли-
пидам, не содержат жирных кислот. Большинство стероидов регу-
лирует вегетативные функции животных, являясь их гормонами. В
эту же группу входят гликозиды растений. К терпенам относят не-
которые ароматические вещества растительного происхожде-
ния, ростовые вещества — гиббереллины, ряд фотосинтезирую-
щих пигментов (фитол, каротиноиды). Липопротеины, как и фос-
фолипиды, участвуют в построении мембран и в то же самое время
представляют собой транспортную форму липидов. Гликолипи-
ды — компоненты мембран хлоропластов и миелиновых оболочек
нервных клеток.
Белки. Это полимерные соединения, молекулы которых постро-
ены из аминокислот, о которых речь шла в начале раздела. Каждому
белку свойственна своя аминокислотная последовательность, и,
поскольку число мономеров в них может достигать нескольких
миллионов, потенциальное разнообразие белков бесконечно. Даже
отдельные клетки живых тел мотут содержать до десяти тысяч раз-
личных белков. Чрезвычайно многообразны и функции белков.
В зависимости от количества аминокислотных остатков белки
подразделяют на пептиды (до четырех аминокислот) и полипепти-
ды. Каждая молекула может состоять из одной или нескольких
полипептидных цепей и, кроме того, образовывать комплексы с
другими молекулами.
Для каждого белка характерна своя собственная трехмерная
структура, которая в значительной степени определяет его функ-
циональные свойства. Общая конформация молекулы обусловле-
на четырьмя уровнями ее организации.
118
Первый уровень описывается как первичная структура. Факти-
чески это последовательность аминокислот, определяемая генети-
ческим кодом. Второй уровень, или вторичная структура, устанав-
ливается при образовании водородных связей между водородом
аминогруппы и кислородом карбоксильной группы, отстоящих
друг от друга на четыре аминокислотных остатка. Поскольку это
регулярно повторяющиеся связи, молекула прочно удерживается в
спирально скрученном состоянии, хотя на отдельных участках мо-
лекулы такая спираль может нарушаться. Изгибание цепи может
происходить при образовании дисульфидных мостиков (S—S-свя-
зей) между серосодержащими аминокислотами. Деспирализован-
ные участки содержат аминокислоту пролин, которая не способ!®
образовывать водородные связи. Ко вторичной структуре относят
и p-конформацию (или складчатую структуру), которая возникает
вследствие образования водородных связей между параллельными
или антипараллельными полипептидами (например, в фиброи-
не — белке шелка гусениц бабочек). К разновидности р-конфор-
мации можно отнести и спирально-параллельную упаковку белко-
вых цепей в коллагене.
Третичная структура белковых молекул поддерживается ион-
ными, водородными и дисульфидными связями и, кроме того, с
помощью гидрофобных взаимодействий. В воде белки, имеющие
третичную структуру, укладываются в комок (глобулу) таким спо-
собом, что все их гидрофобные части упрятаны внутрь, а гидро-
фильные — вывернуты наружу.
Многие белки образуют молекулярные комплексы, в которых
несколько полипептидных цепей удерживается ионными и водо-
родными связями и благодаря гидрофобным взаимодействиям.
Как правило, это глобулярные молекулы. Такие белковые агрега-
ты описываются четвертичной структурой.
По составу белки принято делить на простые, содержащие
только аминокислоты, и сложные, содержащие дополнительно
небелковую, так называемую простетическую, группу. К простым
белкам относят альбумины, глобулины, гистоны и склеропротеи-
ны; к сложным — фосфопротеины, гликопротеины, нуклеопроте-
ины, хромопротеины, липопротеины, флавопротеины и металло-
протеины.
В структурной классификации белков выделяют фибрилляр-
ные, глобулярные и промежуточные. В фибриллярных белках тре-
тичная структура почти не выражена. Они не растворяются в воде,
обладают большой механической прочностью. Как правило, моле-
кулы фибриллярных белков длинные и, скрепляясь с параллельно
расположенными молекулами, образуют волокна или слоистые
структуры.
В глобулярных белках цепи свернуты в глобулы, описываемые
119
третичной структурой. Эти белки в среде, отклоняющейся от изо-
электрической точки, приобретают положительный или отрица-
тельный заряд и потому легко растворяются в воде, образуя колло-
идные суспензии.
Промежуточные белки характеризуются развитием только вто-
ричной структуры, но, как и глобулярные, они растворимы в воде.
При воздействии на белки некоторых жестких факторов, таких,
например, как высокая температура, сильные кислоты и щелочи,
излучения, тяжелые металлы, органические растворители, третич-
ная структура может нарушаться, но вторичная структура часто
сохраняется.
Поскольку вторичная, третичная и четвертичная структуры
белковых молекул полностью или почти полностью определяются
последовательностью конкретных аминокислот, то замена хотя бы
некоторых из них в полипептидной цепочке обязательно приведет
к изменению конформации молекулы, а следовательно, и к изме-
нению ее свойств.
Отдельные белковые молекулы могут в результате нековалент-
ных взаимодействий образовывать агрегаты с новыми свойствами.
Такое явление очень широко распространено в клетках живых
тел, и его считают одним из простейших механизмов эволюции и
глобулярных белков, и белковых фибрилл. Фибриллы могут обра-
зовываться не только как вариант 0-конформации фибриллярных
белков. Глобулярные белки, соединяясь последовательно, форми-
руют цепочки или кольца. Если каждая глобула соединяется с че-
тырьмя другими, образуется двухцепочечная спираль, как, напри-
мер, в случае с фибриллами актина — основного белка цитоскеле-
та. Глобулы, связанные с шестью такими же, упаковываются в
плоские слои, которые, впрочем, легко сворачиваются в трубочки.
Примером последнего служат микротрубочки — фибриллярные
структурные элементы клеток, состоящие из молекул белка тубу-
лина и участвующие в построении жгутиков, ресничек, нитей ве-
ретена деления и др. Сложные функциональные белковые агрега-
ты строятся преимущесгенно путем самосборки и при действии на
них определенных химических сигналов легко распадаются на
глобулы — субъединицы.
В клетке белки выполняют различные функции. Большинство
клеточных белков относится к группе глобулярных и обладает
способностью ускорять течение химических реакций, то есть яв-
ляется ферментами, или биокатализаторами. В основе каталити-
ческой активности белков лежит поверхностная конформация
глобул. Общий изгиб полипептида создает участки поверхности,
так называемые центры связывания, на которых сосредоточива-
ются группы, способные образовывать водородные связи. Распо-
ложение и количество таких групп очень индивидуальны для
120
каждого конкретного белка. Молекулы, конформация которых и
расположение участков, способных образовывать водородные
связи, комплементарны центрам связывания белков — фермен-
тов, взаимодействуют с ними. При этом электронные облака мо-
лекул смещаются настолько, что протекание определенных реак-
ций становится как бы неизбежным. Но любая реакция, катали-
зируемая белками, может идти как в прямом, так и в обратном
направлении, и «уход» от точки химического равновесия дости-
гается несколькими способами: созданием разности концентра-
ций реагирующих веществ и продуктов реакции путем быстрого
использования последних в дальнейших превращениях; разделе-
нием веществ внутриклеточными мембранами; образованием
мультиферментных комплексов; включением в ферментные сис-
темы аллостерических белков. Во всех этих случаях в клетке под-
держивается химическая неравновесность.
Аллостерические белки заслуживают особого внимания. Так
называют группу полипептидов, глобула которых может суще-
ствовать в двух или более устойчивых формах при неустойчивых
промежуточных состояниях. Аллостерические белки способны:
направлять течение реакций; служить регуляторами метаболизма,
запуская и останавливая реакции; участвовать в клеточной сигна-
лизации, то есть связывать обменные процессы с какими-либо хи-
мическими стимулами; совершать полезную работу в виде соб-
ственного направленного перемещения, как это делают, напри-
мер, белок миозин и белок ДНК-геликаза, или в виде переноса
малых молекул через мембраны, например белок натрий-калий-
зависимая АТФаза (Na-K-насос), глюкозный насос и др., затрачи-
вая энергию гидролиза АТФ; синтезировать АТФ, мобилизуя
энергию градиента протонов, как это происходит на внутренних
мембранах митохондрий у гетеротрофов.
Перечисленные основные свойства и функции белковых моле-
кул обусловливают рад производных от них функций белков как
группы органических соединений: строительную, каталитичес-
кую, двигательную, транспортную, защитную (механическую и
иммунную), регуляторную, сигнальную, информационную и
энергетическую. Это почти полный комплекс функциональных
свойств живых тел, и именно поэтому главные функциональные
элементы живых тел построены преимущественно из белков, а
жизнь как явление часто связывается с белковыми комплексами.
Нуклеиновые кислоты. Ранее мы уже обсуждали строение нук-
леотидов, их функций и отметили способность к образованию
крупных полимерных молекул нуклеиновых кислот. Совокуп-
ность различных нуклеиновых кислот составляет сложную систе-
му преобразователей структурной информации в динамическую и
наоборот. По своей сути нуклеиновые кислоты —это простран-
121
ственно-временные организаторы процессов, реализуемых по-
средством белков. В такой трактовке главное свойство нуклеино-
вых кислот может быть представлено как многократно усиленная
способность мононуклеотидов выступать в качестве энергетичес-
ких посредников. Традиционное толкование функции этих поли-
меров как хранение и передача наследственной информации долж-
но пониматься без упрощения: здесь имеет место и непрерывное
постоянное самовозобновление нуклеиновых кислот, и непрерыв-
ное постоянное копирование каждого нового их состояния.
Нуклеиновые кислоты принято разделять на дезоксирибонук-
леиновые (ДНК) и рибонуклеиновые (РНК) по виду сахара в нук-
леотидах. Однако это общее их деление весьма условно. Напри-
мер, хлоропластная ДНК содержит не только дезоксирибонуклео-
тиды, но и рибонуклеотиды. Вместе с митохондриальной ДНК
они составляют группу так называемых прокариотных ДНК. Об-
щими для этой группы признаками будут: относительно неболь-
шая молекулярная масса (до 107); включение в цепь уридилового
нуклеотида; связь с белками-протаминами в нуклеопротеиновых
комплексах. В отличие от них эукариотная, или ядерная, ДНК об-
ладает на 2...3 порядка большей молекулярной массой, имеет
характерный нуклеотидный состав, содержит значительное число
участков с обращенными последовательностями и некодирующи-
ми генами, связывается с белками-гистонами.
Особенности структурной организации ДНК изучены сравни-
тельно хорошо. В настоящее время имеется описание трех основ-
ных конформационных типов молекулы (структурные формы
ДНК): В, А и Z (рис. 4.3). Первая из них — она же основная, рас-
шифрованная еще в 1953 г., — это правозавитая двойная спираль с
десятью парами спрятанных внутрь оснований в каждом витке.
Форма А близка к предыдущей, но ее основания наклонены отно-
сительно оси и вывернуты наружу, практически заполняя собой и
малую, и большую борозды спирали. Z-форма — это левозавитая
спираль с зигзагообразным сахарофосфатным остовом и двенад-
цатью парами оснований, приходящимися на каждый виток. Ре-
альные молекулы ДНК лишь короткое время, часто на отдельных
участках, соответствуют той или иной структурной форме: они на-
ходятся в состоянии непрерывного превращения одной конфор-
мации в другую. При этом с помощью специальных ферментов
достигается положительная или отрицательная суперспирализа-
ция и, следовательно, большая или меньшая напряженность дуп-
лекса (двойной спирали). В результате молекула изгибается, мес-
тами расплетается, скручивается в крестообразные фрагменты
и т. д., меняя конформацию поверхности, сближая поочередно то
одни, то другие участки, формируя тот или иной уровень упаков-
ки хроматина. (Хроматином называют нуклеопротеиновый комп-
122
Рис. 4.3. Структурная ортанизаштя ДНК (Албертс и др., 1986; оригинал автора).
-нековалентное взаимодействие между комплементарными н^е5"^$к2-вариаНТЬ’
изображения двойной цепочки ДНК; 3— структурные формы ДНК
леке.) В митохондриях, хлоропластах и в клетках прокариот ДНК
часто образует кольцо с большей или меньшей степенью супер-
спирализации. В ядрах эукариот в разные периоды жизни клетки
хроматин ведет себя по-разному.
В деспирализованном (интерфазном) состоянии хроматин
представляет собой структуру, напоминающую «бусы на нитке»,
это третий уровень упаковки — нуклеосомная нить.
Здесь двойная спираль (2-й уровень) «намотана» на блоки,
каждый из которых составлен из восьми молекул белков-гистонов.
Диаметр нуклеосомной нити 11 нм, но сама она скручивается в
спиральную хроматиновую фибриллу диаметром около 30 нм (4-й
уровень). Фибрилла на очередном (5-м) уровне упаковки образует
серию динамичных петель с общей шириной (толщиной) до
300 нм. При подготовке клетки к делению фибриллы скручивают-
ся в так называемые конденсированные хромосомы (6-й уровень)
и далее в метафазные хромосомы (рис. 4.4). Начиная с третьего
уровня, упаковка хроматина стабилизируется белками и разблоки-
ровка цепей ДНК происходит только на период считывания с них
информации, то есть при синтезе мРНК и редупликации.
Установлено, что количество ДНК в клетках непостоянно: оно
меняется не только в клетках разных особей одного вида, но зави-
сит также от возраста и функциональной активности клетки или
Рис. 4.4. Уровни упаковки хроматина (Албертс и др., 1986, с изменениями):
/—двойная спираль; 2—нуклеосомная нить; 3~ хроматиновая фибрилла; 4—петельные до-
мены; 5— конденсированный участок хроматина; 6 — метафазная хромосома
124
живого тела. Всю ДНК клетки предложено разделять на облигат-
ный (ОК) и факультативный (ФК) компоненты. К ОК относят
основную часть ядерной ДНК, упакованную в нуклеосомную
нить. ФК представляет собой фрагменты ДНК, находящиеся в
кариоплазме, цитоплазме и органоидах цитоплазмы. Сюда же в
рвде случаев включают свободные фрагменты вирусных и бакте-
риальных ДНК. Факультативная ДНК способна встраиваться в об-
лигатную, давать транскрипты, переходить из одного участка ДНК
в другой, изменять сочетания регуляторных генов. По назначению
факультативный компонент предположительно является посред-
ником между цитоплазмой и облигатным компонентом.
Рибонуклеиновые кислоты (РНК) в общем плане отличаются
от дезоксирибонуклеиновой кислоты, как указывалось, видом са-
хара-пентозы и набором оснований в нуклеотидах. Основные
типы РНК: информационная, или матричная (мРНК), несущая
код белка от ДНК; транспортная (тРНК), образующая комплексы
с аминокислотами и переправляющая их к месту синтеза белка;
рибосомная (рРНК), связывающая группу белков в полипептид-
ный комплекс и связывающаяся с мРНК; гетерогенная (встречает-
ся в ядрах эукариот, имеет смешанный нуклеотидный состав).
Функциональные особенности РНК достаточно конкретны и в
целом более понятны. К ним мы вернемся позже при рассмотре-
нии механизмов синтеза белка в клетке.
4.3. СТРУКТУРНЫЕ КОМПОНЕНТЫ КЛЕТКИ И ГЛАВНЕЙШИЕ
ЖИЗНЕННЫЕ ПРОЦЕССЫ, ПРОХОДЯЩИЕ В НЕЙ
4.3.1. ЦИТОПЛАЗМА
В главе 3 приведена краткая характеристика цитоплазмы как
обязательной части живой клетки, заключенной между наружной
клеточной мембраной и ядром, указаны ее основные свойства и
функции.
Поскольку в цитоплазме пространственно сосредоточены глав-
нейшие метаболические процессы, именно она представляет фун-
кционально активную часть живого вещества, которое существует
в форме дискретных живых тел. В то же время важно помнить, что
цитоплазма — эго сложно структурированная часть любой клетки,
и ее органоиды, представляющие собой элементарные преобразо-
ватели вещества, энергии и информации, составляют обычно ме-
нее 10 % массы живой клетки. Все остальное — это вода — внут-
ренняя среда, вне которой большинство химических реакций рез-
ко замедляется либо останавливается вовсе, а также растворенные
в ней вещества.
125
Если представить цитоплазму всех живых тел как единое обра-
зование, можно заметить, что меньшая часть растворенных ве-
ществ в виде последовательности продуктов реакций, сначала ус-
ложняющихся, а затем упрощающихся, связана с накоплением,
переносом и высвобождением энергии. Другая же, большая, часть —
это вещества — строительные блоки функциональных и вспомога-
тельных структур, обеспечивающих течение этих энергетических
процессов и направляющих часть энергии на внешнюю среду.
Рассмотрим главные структурные компоненты цитоплазмы под
этим углом зрения.
Основное водянистое вещество цитоплазмы — цитоплазмати-
ческий матрикс, или гиалоплазма, или цитозоль, заполняет все про-
странство между клеточными органоидами, что составляет около
55 % объема всей клетки. Гиалоплазма содержит в виде истинных
растворов неорганические ионы, сахара, аминокислоты, жирные
кислоты, нуклеотиды и витамины. В ней растворены и некоторые
газы. Полимерные белки и нуклеиновые кислоты образуют колло-
идные растворы невязкой (золь) или вязкой (гель) консистенции.
Гель обычно занимает периферические области цитоплазмы. Гиа-
лоплазма способна к взаимным переходам гелеобразного и золеоб-
разного состояний. Поэтому, а также из-за различий в составе свой-
ства гиалоплазмы в разных частях клетки отличаются.
Общее коллоидное состояние гиалоплазмы определяет не-
сколько ее важных свойств:
гиалоплазма удерживается в плазматической мембране, так как
крупные молекулы не способны проникать через мембрану само-
произвольно;
будучи гидрофильным коллоидом, гиалоплазма имеет замет-
ное, но небольшое осмотическое давление, что позволяет клетке
«мягко» регулировать содержание воды;
коллоидная гиалоплазма при изменении температуры, давле-
ния, pH, состава ионов металлов может переходить из состояния
золя в состояние геля и обратно. Фазовые переходы гиалоплазмы
входят в число механизмов клеточного движения. Это же явление
наблюдается и в межклеточном веществе, например, при сверты-
вании крови, загустении спермы и т. п.;
при поглощении воды коллоиды набухают (как, например, же-
латин в воде) и увеличение объема используется живыми телами
для локального повышения давления, в частности для выталкива-
ния гамет, семян и спор.
В основном веществе, кроме того, всегда присутствуют типич-
ные функциональные структуры: микрофиламенты, промежуточ-
ные и толстые фибриллы и микротрубочки. В нем протекают не-
которые обменные процессы. Это, в частности, гликолиз, синтез
жирных кислот, нуклеотидов и некоторых аминокислот.
126
Живая цитоплазма непрерывно движется, и это движение
проявляется прежде всего в упорядоченном перемещении боль-
шинства органоидов. Иногда в живых клетках обнаруживаются
ясно выраженные круговые токи, получившие название «цик-
лов».
Гиалоплазма обладает буферными свойствами, и в ней посто-
янно поддерживается слабощелочная реакция. Химический состав
гиалоплазмы определяет ее осмотические свойства, усиливаемые
внешней и внутренними мембранами.
4.3.2. ОРГАНОИДЫ КЛЕТКИ
Содержащиеся в цитоплазме эукариотической клетки органо-
иды принято разделять на мембранные и немембранные. К пер-
вым относят наружную плазматическую мембрану (НПМ), эндо-
плазматическую сеть (ЭНС), аппарат (комплекс) Гольджи (дикти-
осомы), митохондрии, пластиды, лизосомы, пероксисомы (или
микротельца), вакуоли; ко вторым — рибосомы, иногда центрио-
ли, кинегосомы (базальные тельца) и цитоскелет. Отдельную
группу составляют поверхностные клеточные структуры: микро-
ворсинки, ложноножки (псевдоподии), микрошипы, реснички,
жгутики, межклеточные контакты (плазмодесмы, десмосомы, ще-
левой, химический и плотный контакты).
Рибосомы. Это крупные нуклеопротеиновые частицы, пред-
ставляющие собой мультиферментные комплексы.
У прокариот и эукариот рибосомы сходны по строению и функ-
циям. Каждая из них состоит из двух субъединиц (субчастиц):
большой и малой. Наибольший линейный размер рибосомы эука-
риот составляет 32 нм. Молекулы рибосомной РНК образуют кар-
кас, к которому самопроизвольно в определенном порядке присо-
единяются белки. Большая субчастица включает три молекулы
рРНК и 45 различных рибосомных белков, а малая — одну моле-
кулу рРНК и 33 различных белка. Рибосомы прокариот, митохонд-
рий и пластид несколько мельче: длина 29 нм; в большой субчас-
тице две молекулы рРНК и 34 белка, в малой — одна молекула
рРНК и 21 белок. Общая молекулярная масса эукариотической
рибосомы 4,5 млн, прокариотической — 2,8 млн. В рибосомах
обоих типов имеются бороздки, удерживающие синтезируемую
молекулу белка и молекулу матричной РНК (мРНК). Кроме того,
«собранная» рибосома имеет два участка связывания транспорт-
ных РНК (тРНК) (рис. 4.5). Первый из них называется пептидил-
тРНК-связывающим участком (P-участок), а второй — аминоа-
цил-тРНК-связывающим участком (A-участок). Все три РНК свя-
зываются с рибосомной РНК лишь в том случае, если их соотает-
127
Рис. 4.5. Рибосома (Албертс и др., 1986, с изменениями):
Л..5— внешний вид рибосомы в проекциях; 4— схема расположения участков связывания на
рибосоме (стрелками показаны участки нековалентного связывания рРНК с иРНК и тРНК
с мРНК)
ствующие триплеты нуклеотидов (кодоны) окажутся комплемен-
тарными.
Рибосома — это белоксинтезирующий агрегат, работу которого
мы рассмотрим несколько позже. Скорость синтеза полипептид-
ных цепей на рибосомах весьма высока. Так, на синтез одной
средней молекулы белка (около 1000 аминокислот) требуется око-
ло 1 мин. Тем не менее такая скорость явно недостаточна для
обеспечения клетки необходимым количеством белков. Поэтому
новая рибосома связывается с молекулой мРНК как только пре-
дыдущая сместится от ее «переднего» конца на 80 нуклеотидов,
что приводит к образованию полирибосомных комплексов, или
полисом.
Среди рибосом различают свободные и мембранно-связанные.
Свободные рибосомы располагаются в гиалоплазме и синтезируют
некоторые ферменты, необходимые для метаболических процес-
сов, идущих здесь же, белки хроматина (гистоны и др.), направля-
емые впоследствии через ядерные поры в ядро, большинство бел-
ков хлоропластов и митохондрий и часть белков микротелец.
Мембранно-связанные рибосомы — это рибосомы, самопроизвольно
связывающиеся со специальными белками на уплощенных участ-
ках ЭПС (цистернах) и образующие ее «шероховатые» области.
Такие рибосомы синтезируют белки для разнообразных органо-
идов и их мембран, а также все известные секреторные белки.
Образование рибонуклеопротеинов — предшественников ри-
босом — происходит в ядре, в области, называемой ядрышком.
Здесь на определенных локусах хромосом (ядрышковых организа-
торах), где находятся серии генов, кодирующих рибосомную РНК,
или на их внехромосомных копиях синтезируются молекулы пре-
рибосомной РНК (прРНК). В периферической области ядрышка
прРНК связывается с поступающими сюда из цитоплазмы белка-
ми и образует прерибосомные гранулы. Перед выходом в цито-
плазму гранулы расщепляются на части и образуют рибосомные
128
субчастицы. Окончательная сборка рибосом происходит только в
цитоплазме и только при наличии мРНК, формилметионин-
тРНК, АТФ и белковых факторов инициации. В некоторых случа-
ях в рибонуклеопротеиновый комплекс дополнительно встраива-
ются (в большую субчастицу), сменяя друг друга, белок инициа-
ции, белок элонгации (продления синтеза) и белок терминации
(остановки) синтеза полипептида.
Цитомембраны, или клеточные мембраны. Цитомембраны, а точ-
нее, связанные с ними эффекты, были известны еще в прошлом
веке. Но оптические микроскопы не позволяли не только изучить
детали их строения, но и увидеть вообще, так как толщина мемб-
ран на два порядка меньше разрешающей способности световой
оптики. Только к 1972 г. удалось с применением электронных
микроскопов и сложных комбинированных методов исследований
(например, метод замораживания — скалывания) составить более
или менее ясное представление о тонком строении, точном хими-
ческом составе мембран и механизмах главнейших мембранных
процессов.
Мембраны — едва ли не самый многофункциональный тип
клеточных структур: они обеспечивают химическую целостность
клетки; регулируют обмен веществ между клеткой и внешней сре-
дой; разделяют клетку на хорошо обособленные участки (компарт-
менты), формируя сеть внутренних барьеров-ограничителей; слу-
жат прикрепительными поверхностями для сложных ферментных
комплексов; сами обеспечивают протекание некоторых физико-
химических процессов (таких как световые реакции фотосинтеза,
окислительное фосфорилирование при дыхании и др.); на мемб-
ранах образуется разность потенциалов, и они же проводят элект-
рические заряды; с мембранами связаны процессы клеточного уз-
навания, а следовательно, дифференцировка тканей, питание,
опознание клеток, оплодотворение и многое другое.
Многофункциональность клеточных мембран при единстве их
плана строения обусловила возможность развития в клетке их спе-
циализированных участков, образующих относительно дискрет-
ные, но динамически единые мембранные органоиды.
В настоящее время большинством биологов принята «жидко-
стно-мозаичная» модель строения клеточных мембран, предло-
женная С. Сингером и Г. Николсоном в 1972 г. Согласно этой мо-
дели основу мембран образуют два слоя липидных молекул, обра-
щенных гидрофильными частями наружу, а гидрофобными —
друг к другу (рис. 4.6). Липидный бислой строится в воде самопро-
извольно. Его толщина 4...5 нм. Отдельные молекулы не перехо-
дят из одного слоя в другой, хотя в принципе это возможно с ве-
роятностью один раз в неделю для каждой из них. Но в плоскости
«своего» слоя они весьма подвижны: с соседними молекулами ме-
9 Присный А В | ПО
Рис. 4.6. «Жкткоство-мозаичвая» модель наружной плазматической мембраны
(оригинал автора)
няются местами примерно 10 000 000 раз в секунду и потому быст-
ро перемещаются (на суммарное расстояние до 2 мкм в секунду), а
также быстро вращаются вокруг собственной оси. Иными слова-
ми, липидная основа — это крайне текучее образование.
Мембранные липиды относятся к трем классам: это фосфоли-
пиды, холестерол и гликолипиды. Соотношение их в мембранах
разных типов клеток, в разных мембранах одной клетки и даже в
слоях одной мембраны различно. Разные липиды имеют разную
текучесть и по-разному ведут себя при разных температурах. Как
общая закономерность, мембраны животных клеток содержат
больше холестерола и отличаются большей текучестью, легче сли-
ваются, отшнуровываются и восстанавливаются при повреждени-
ях. В клетках живых тел с непостоянной температурой (пойкило-
термных) состав жирных кислот, входящих в липиды мембран,
меняется с изменением температуры так, что текучесть мембран
остается постоянной.
Считается, что липидный бислой служит двумерным (плоско-
стным) растворителем для белковых молекул, находящихся в со-
ставе мембраны. Более того, многие мембранные белки проявля-
ют специфические свойства только в присутствии определенных
фосфолипидов.
Гликолипиды обнаруживаются исключительно в наружном
слое, их сахарные группы ориентированы всегда к поверхности
клетки. Олигосахариды здесь могут быть очень сложными, но их
функции не вполне ясны.
Непроницаемый для растворимых в воде веществ липидный
бислой определяет большую часть общих для всех клеточных мем-
бран свойств и функций. Специфические же свойства и функции
130
снизаны главным образом с белками. Содержание белков в мемб-
ранах варьирует от 25 % в миелиновой «изолирующей» мембране
нервных волокон до 75 % во внутренних мембранах хлоропластов
и митохондрий.
Белковые молекулы имеют гидрофильные и гидрофобные уча-
стки. Гидрофобными участками они могут быть погружены в ли-
пидную основу с внешней или с внутренней стороны и удержи-
ваться в ней гидрофобными взаимодействиями. Белки могут так-
же ковалентно связываться с двумя жирными кислотами и встраи-
ваться в липидный бислой «на общих основаниях» или же
пронизывают его насквозь, выставляя гидрофильные части нару-
жу, а гидрофобные пряча внутрь. При этом внутренняя часть мо-
лекулы, как правило, представлена или сс-спиралью, или р-слоем.
Третичная структура белка при растворении его в липидах не из-
меняется.
В клеточных мембранах отмечены тысячи различных белков.
Среди них есть структурные белки, белки, образующие транспорт-
ные комплексы с внутренними или межмолекулярными гидро-
фильными каналами-порами (через такие поры могут проходить
полярные молекулы), специфические рецепторные белки, пере-
носчики электронов и преобразователи энергии, часть мембран-
ных белков является ферментами.
Белки, как и липиды, хотя и в меньшей степени, способны пе-
ремещаться в плоскости мембраны, если они не связаны в жест-
кие структуры с подмембранными белками или надмембранными
углеводами. Таким образом, мембраны в целом оказываются теку-
чими, в значительной степени способными к самосборке, само-
восстановлению, слиянию и отшнуровыванию.
Так же как и к липидам, к белкам мембраны могут ковалентно
присоединяться олигосахаридные боковые цепи (с образованием
гликопротеинов). Считается, что подавляющее большинство бел-
ков, выступающих на поверхности наружной плазматической
мембраны, связано с остатками сахаров, а белки, обращенные к
цитоплазме, с олигосахаридами не связываются. Аналогично
внутренние клеточные мембраны несут гликопротеины лишь на
стороне, обращенной внутрь обособленных специализированных
компартментов.
Кроме мембранных гликопротеинов и гликолипидов на внеш-
ней стороне плазматической мембраны большинства клеток рас-
полагаются адсорбированные гликопротеиды и протеогликаны,
секретируемые клеткой в межклеточное пространство. Поэтому,
строго говоря, резкой границы между клеткой и внешней средой
не существует. Предполагают, что часть гликолипидов может уча-
ствовать в процессах клеточного узнавания. Некоторые сильно
разветвленные олигосахариды, возможно, стабилизируют мемб-
9*
131
ранные белки, особенно те из них, которые осуществляют транс-
порт веществ через мембрану.
Транспорт веществ через мембраны. Мы уже отмечали, что мем-
браны непроницаемы для ионов и полярных молекул, поскольку
липиды, входящие в состав мембран, отталкивают их. Тем не ме-
нее они непрерывно поступают в клетку и выводятся из нее. И это
не только питательные вещества, но также ионы, необходимые
для стабилизации других химических параметров гиалоплазмы,
конечные продукты обмена веществ, биологически активные ве-
щества, секреты и др.
Известно четыре механизма транспорта веществ: диффузия, ос-
мос, активный транспорт и экзо-эвдоцитоз (рис. 4.7). Диффузия и
осмос — процессы самопроизвольные, пассивные и не требуют
энергетических затрат в отличие от активных процессов.
Рис. 4.7. Механизмы транспорта веществ через мембрану (оригинал автора):
1 — диффузия; 2— котранспорт; 3— мембранный транспорт; 4— образование «окаймленной
ямки»
132
Диффузия. Под диффузией понимают распространение ве-
ществ из области большей концентрации в область меньшей кон-
центрации. Если градиент концентрации пересекает биологичес-
кая мембрана, то диффузия идет и через мембрану, но липофиль-
ные (жирорастворимые) вещества проникают через нее гораздо
быстрее, чем гидрофильные (водорастворимые). Разновидностью
диффузии считается «облегченная диффузия», при которой опре-
деленные вещества проходят через мембранные поры, образован-
ные гидрофильными частями амфипатических (проникающих
через всю толщину мембраны) белков. Такие белки называют ка-
налообразующими или туннельными. Диффузный транспорт
вдет в том случае, если концентрация вещества с одной стороны
мембраны выше концентрации того же вещества с другой сторо-
ны мембраны. Правда, здесь имеет значение и электрический
градиент, поскольку заряженные молекулы не могут пассивно
двигаться в направлении одноименного заряда. Поэтому приня-
то объединять два явления и говорить об общем электрохимичес-
ком градиенте.
В других случаях облегченная диффузия обеспечивается фер-
ментоподобным поведением мембранных белков: переносимое
вещество связывается с активным центром белка, вызывая его
конформационное изменение, приводящее к «протаскиванию»
транспортируемой молекулы через мембрану.
Осмос. Осмосом называют пассивную диффузию молекул рас-
творителя из области с более высокой их концентрацией в область
с более низкой концентрацией через полупроницаемую мембрану.
Особенность осмоса в живых телах заключается в том, что в каче-
стве растворителя почти всегда выступает вода, а мембраны клеток
являются избирательно проницаемыми. По отношению к клетке
внешняя водная среда может быть: изотонической, когда вода по
обе стороны мембраны находится в динамическом равновесии
(например, клетка в морской воде); гипертонической, когда вода
диффундирует из клетки (клетка в насыщенном солевом раство-
ре); гипотонической, когда вода диффундирует в клетку (клетка
в пресной воде). Но осмотическое поступление воды в клетку
можно предотвратить, приложив к клетке гидростатическое дав-
ление. Это и будет осмотическое давление. Оно тем выше, чем
больше концентрация растворимых веществ в гиалоплазме.
Клетка может регулировать осмотическое давление, переводя,
например, нерастворимые полисахариды в растворимые моно- и
дисахариды.
Проблемы, решаемые живыми телами, в связи с явлением ос-
моса различны для сухопутных, пресноводных, морских и гало-
фильных обитателей.
Осмотическое давление поддерживает тургор клетки, обеспе-
133
чивает передвижение веществ внутри клетки и между клетками,
движение самих клеток, явления обезвоживания и лизиса.
Активный транспорт. Диффузия~процесс обратимый,
поскольку направление диффузии определяется направлением
электрохимического градиента. В отличие от диффузии активный
транспорт представляет собой перемещение молекул против элек-
трохимического градиента, то есть в направлении, противополож-
ном естественному, приводящему к уменьшению их потенциаль-
ной энергии. Такое «насильственное» перемещение требует энер-
гетических затрат и приводит к увеличению неравновесности со-
стояния.
В обобщенном виде активный транспорт малых молекул через
мембраны принято разделять на унимпорт — независимый транс-
порт однородных молекул в одном направлении и котранспорт —
сопряженный транспорт двух веществ. Последний включает сим-
порт (вещества накачиваются в одном направлении) и антипорт
(вещества переносятся в противоположных направлениях) (см.
рис. 4.7).
Наиболее распространенным в клетках живых тел считается
транспортный мембранный комплекс, так называемый
(Na++ К+)-насос (натрий-калиевый насос), который поглощает
из внешней относительно мембраны среды ионы калия и протал-
кивает их в клетку, одновременно выкачивая из клетки ионы на-
трия. В результате концентрация ионов калия внутри клетки ста-
новится выше, чем снаружи, а концентрация ионов натрия, на-
оборот, в клетке становится ниже. (В едином комплексе с натрий-
калиевым насосом работает второй транспортный белок — «канал
утечки калия», позволяющий ионам калия выходить из клетки по
градиенту концентрации, то есть посредством облегченной диф-
фузии.)
Натрий-калиевый насос работает по принципу антипорта: на-
трий выходит наружу, а калий входит внутрь клетки. На каждые
три иона натрия, выкачанных из клетки, в клетку накачиваются
два иона калия. Дополнительно выходящие из клетки ионы калия
через «канал утечки» создают избыточный потенциальный заряд
на внешней стороне мембраны, то есть мембрана заряжается. Пе-
рекачка ионов против градиентов концентрации требует посто-
янного притока энергии. Источником энергии служит АТФ, а
ферментом, расщепляющим АТФ, служит сам белок, образующий
«насос». Гидролиз одной молекулы АТФ необходим для заверше-
ния цикла переноса трех ионов натрия и двух ионов калия. Ежесе-
кундно одна молекула этого вида АТФазы расщепляет 100 моле-
кул АТФ. В принципе возможно заставить «насос» работать на-
оборот — на синтез АТФ, но для этого надо создать высокие гра-
диенты концентрации натрия и калия по обе стороны мембраны.
134
Натрий-калиевый насос, обеспечивая перенос ионов через
мембрану, не только создает мембранный потенциал-основу элек-
трической активности клетки. От концентрации этих ионов зави-
сит осмотическое давление в клетке. Градиент концентрации не-
которых ионов способствует транспорту в клетку глюкозы, амино-
кислот и других малых органических молекул. Поэтому клетки
расходуют на работу натрий-калиевого насоса примерно третью
часть всей синтезируемой АТФ.
Мембранно-связанные белковые транспортные комплексы, об-
ладающие АТФазной активностью, способны осуществлять пере-
нос и других ионов, например Са2+, Н+. Для обеспечения энерге-
тических процессов в живых телах особенно важной оказалась об-
ратимость работы протонных насосов. В клетках аэробных бакте-
рий и в митохондриях градиент протонов возникает на отдельных
этапах окислительного фосфорилирования, в хлоропластах — на
световой фазе фотосинтеза, а у галобактерий градиент протонов
формируется при участии фотоактивируемого пигмента родопси-
на. Во всех этих случаях выход протонов через насосы по градиен-
ту концентрации сопровождается синтезом АТФ.
Мембранный транспорт макромолекул и час-
тиц. Для переноса макромолекул и частиц вещества в клетке об-
разуются окруженные мембраной пузырьки-везикулы. Если, на-
пример, клетка какой-либо железы продуцирует секрет, то снача-
ла выделяемое вещество заключается во внутриклеточный пузы-
рек, затем пузырек передвигается в цитоплазме, подходит к
плазматической мембране, сливается с ней и открывается во вне-
клеточное пространство, высвобождая свое содержимое. Транс-
портирующие пузырьки такого типа встречаются практически у
всех эукариотических клеток, а явление слияния внутриклеточно-
го пузырька с плазматической мембраной называется экзоцитоз.
Клетки способны не только выделять высокомолекулярные ве-
щества и частицы, но и поглощать их. Это процесс, противопо-
ложный экзоцитозу с соответствующим названием — эндоцитоз.
Последовательность событий в этом процессе противоположна
последовательности событий в экзоцитозе. Поглощаемое веще-
ство как бы следует за впячивающейся мембраной, которая посте-
пенно смыкается над ним, окружая его, а затем отшнуровывается,
образуя внутриклеточный пузырек. В зависимости от размеров пу-
зырьков, а они, в свою очередь, зависят от консистенции погло-
щаемого вещества, различают две формы эндоцитоза: пиноци-
тоз — поглощение жидкости (здесь пузырьки мельче) и фагоци-
тоз — поглощение частиц вроде микробов или обломков клеток,
или плотных органических частиц (с образованием более крупных
пузырьков-вакуолей).
Остается до сих пор непонятным: почему и как многочислен-
135
ные разнородные, с различным содержимым пузырьки переме-
щаются к строго определенным пунктам назначения и в конце
концов сливаются только с определенными мембранными струк-
турами?
Экзоцитоз веществ, производимых клеткой, может происхо-
дить непрерывно через поток внутриклеточных пузырьков, на-
правленный непосредственно к плазматической мембране, но из-
вестен и другой путь: производимые вещества накапливаются в
специализированных секреторных пузырьках и высвобождаются
за пределы клетки после соответствующего, чаще всего химичес-
кого, сигнала извне.
В большинстве эукариотических клеток почти непрерывно
идет поглощение внеклеточной жидкости (жидкостный эндоци-
тоз). По этой причине наружная клеточная мембрана постоянно
уходит в цитоплазму. Скорость такого втягивания мембраны раз-
лична в разных клетках и при разном состоянии клеток, но поря-
док составляет 100 % площади наружной мембраны за несколько
десятков минут. Площадь же поверхности и объем клетки при
этом остаются постоянными. Следовательно, значительная часть
мембраны из внутриклеточных пузырьков должна быстро возвра-
щаться в наружную плазматическую мембрану.
Обычно пузырьки в цитоплазме сливаются друг с другом и
(или) с лизосомами и образуют пищеварительные вторичные ли-
зосомы, от которых позже отшнуровываются мелкие пузырьки,
сливающиеся в конце концов с наружной плазматической мемб-
раной (НПМ). Но лизосомы образуются путем отшнуровывания
от комплекса Гольджи. Забегая несколько вперед, отметим, что
мембраны комплекса Гольджи связаны своим происхождением с
мембранами эндоплазматической сети (ЭПС), а та, в свою оче-
редь, непосредственно продолжается в ядерную мембрану.
Некоторые эндоцитозные пузырьки не сливаются с лизосома-
ми и движутся через всю цитоплазму на противоположную сторо-
ну клетки, где освобождаются от содержимого путем экзоцитоза.
Такой механизм транспорта веществ встречается в ряде специали-
зированных клеток, например в клетках эндотелия (эпителий, вы-
стилающий кровеносные капилляры). Иногда здесь пузырьки
сливаются в непрерывные трубочки и образуют временные
транспортные канальцы. У саркодовых (тип Плазможгутиковые), в
частности у амеб, эндоцитозные пузырьки с непищевым содержи-
мым также быстро проходят через все тело.
Поглощение клетками специфических питательных веществ
носит еще более упорядоченный характер. Сначала рецепторные
молекулы мембраны связываются с ними, затем с внутренней сто-
роны мембраны формируется сеточка из белковых фибрилл, к ко-
торой подходят филаменты цитоскелета. Впячивание мембраны
136
приводит к образованию окаймленной ямки и далее окаймленного
пузырька. Погрузившись в цитоплазму, а это происходит намного
быстрее, чем в ранее рассмотренном случае, пузырьки сбрасывают
окаймление и сливаются с лизосомами. В процессе образования
окаймленной ямки происходит концентрирование молекул погло-
щаемого вещества за счет смещения рецепторных молекул мемб-
раны к общему центру.
В некоторых случаях поглощение макромолекул может произ-
водиться и без образования пузырьков. Например, передача ДНК
у бактериальных клеток, поступление белков в шероховатую эн-
доплазматическую сеть, проникновение токсинов в клетки проис-
ходят через мембранные межклеточные каналы, или каналы
трансмембранных белков, или же временные межмолекулярные
мембранные поры. В этих видах транспорта часто бывают задей-
ствованы мембранные рецепторные белки, но точный механизм и
последовательность событий в них во многом остаются пока неиз-
вестными.
Механизм слияния и отшнуровывания мембран весьма прост и
сводится к слипанию и последующему объединению липидных
бислоев или образованию и разрыву тонкой мембранной перетяж-
ки. И в том и в другом случае молекулы липидов стремятся «зак-
рыть» свои гидрофобные «хвосты» гидрофильными «головами».
Важным следствием из рассмотренных механизмов поглоще-
ния больших молекул и частиц является динамическое родство
всех мембран клетки, совершающих непрерывный и очень интен-
сивный круговорот. Лишь небольшая часть клеточных мембран в
каждый отдельный момент располагается на поверхности клетки.
Площадь мембран, сосредоточенных во внутриклеточных пузырь-
ках, превышает их в 10...30 раз. Например, в ацинарных клетках
поджелудочной железы площадь НПМ составляет около 30 мкм2,
а площадь везикулярной мембраны — около 900 мкм2.
На подвижности клеточных мембран основана концепция эндо-
мембранной системы. Согласно этой концепции мембраны клетки,
за исключением мембран митохондрий и пластид, представляют
собой единое динамическое целое и берут начало от эндоплазма-
тической сети.
Наружная плазматическая мембрана. Характеризуя цитомембра-
ны, мы фактически довольно подробно описали строение и функ-
ции наружной плазматической мембраны. Перечень главнейших
функций НПМ включает:
поддержание физической целостности и определенности клет-
ки; обеспечение в значительной степени химической определен-
ности клетки;
избирательный пассивный и активный транспорт веществ в
клетку и из нее (в том числе эвдоцитоз и экзоцитоз);
137
регуляцию осмотического давления клетки и основанных на
нем движений;
создание мембранного электрического потенциала и проведе-
ние электрического импульса;
участие в ряде случаев в синтезе АТФ; опознание внеклеточно-
го вещества (сигнальных молекул, пищевых частиц, клеток своего
и чужого типов и т. д.);
обеспечение контакта с соседними клетками в многоклеточных
телах;
образование поверхностных структур специального назначе-
ния.
Остановимся подробнее на двух последних функциях, которые
мы пока рассматривали.
Большинство живых тел многоклеточны, и НПМ клеток в них
является пограничной или контактной поверхностью. А это озна-
чает, что мембрана каждой клетки в зоне постоянного контакта с
мембранами смежных клеток должна обеспечивать механическую
и химическую связь с окружением (рис. 4.8).
Удержание клеток в тесной связке происходит благодаря десмо-
сомам — структурам, образованным пучками поперечно располо-
женных волокон-тонофиламентов, подходящих из глубины ци-
топлазмы к электронно-плотному веществу на внутренней поверх-
пластинка пространство
Рис. 4.8. Клеточные контакты (оригинал автора)
138
ности мембраны, и пластинками из плотного вещества (или пучка
фибрилл, поперечных перегородок, или же сотовидной перего-
родчатой структуры) в межмембранном пространстве толщиной
около 30 нм. Площадь десмосомы в плоскости мембраны состав-
ляет около 0,1 мкм2.
Щелевые контакты позволяют малым молекулам беспрепят-
ственно проходить из одной клетки в другую. Щелевой контакт
образован двумя гексагональными структурами, погруженными в
контактирующие мембраны точно напротив друг друга и тесно со-
прикасающимися в щели между клетками. Через каналы щелевых
контактов могут проходить молекулы размером не более 2 нм, а
это ионы, аминокислоты, сахара и нуклеотиды.
У клеток эпителиальных тканей обнаружен еще один тип кон-
тактов: так называемый плотный контакт. Обычно это кольцевой
выступ, сопровождаемый пучком подмембранных филаментов,
связанных проникающими через плотно прижатые друг к другу
мембраны замыкающими элементами. Назначение плотных кон-
тактов состоит в изоляции веществ, расположенных над апикаль-
ными (вершинными) и под базальными (основными) частями
эпителиальных клеток.
Следуя логике изложения, сейчас можно было бы дать характе-
ристику поверхностным клеточным образованиям: микроворсин-
кам, псевдоподиям, ресничкам, жгутикам и сложным мембран-
ным покровам. Но поскольку плазматическая мембрана в них не
имеет новых функциональных проявлений (они связаны с фиб-
риллярным аппаратом), рассмотрим их далее при описании фила-
ментов и микротрубочек. Здесь же обратим внимание на несколь-
ко-специализированных типов плазматической мембраны: миели-
новую оболочку, мембраны фоторецепторных клеток, пурпурные
мембраны галобактерий (рис. 4.9) и синаптические мембраны.
Миелиновая оболочка — это плазматическая мембрана «шван-
новской клетки»-. Ее специфическая особенность — обеднен-
ность белками. Соотношение белков и липидов в ней составляет
25 : 75. Такая мембрана почти не пропускает ионы и может слу-
жить изолятором для специализированных, проводящих заряд
участков нервных клеток. Усиление эффекта изоляции достигает-
ся многократным накручиванием плоской части шванновской
клетки на аксон.
Собственно пурпурная мембрана — это четко очерченные участки
плазматической мембраны, на которых имеется бакгериородопсин
(белок, входящий в состав плазматических мембран фотоавтотроф-
ных и фотогетеротрофных пурпурных бактерий), связанный с
простетической группой или так называемым «хромофором»,
идентичным хромофору родопсина из палочек сетчатки глаза у
позвоночных животных. При попадании на хромофор кванта све-
139=
Рис. 4.9. Производные наружной плазматической мембраны (Албертс и др.. 1986,
с изменениями):
У —«пурпурная мембрана» бактерии Halobacterium halobium: о—внешний вид, б—фрагмент;
2—миелиновая оболочка шванновской клетки: а —внешний вид, б—фрагмент; 3— фоторе-
цепторная мембрана: а—в дисках палочки сетчатки млекопитающего, б— в микровиллах ре-
тинальной клетки омматвдия насекомого
та он активируется и вызывает конформационные изменения в
белке. Вследствие этого белок переносит на внешнюю сторону
мембраны два протона, создавая градиент протонов и разность
потенциалов. Накапливающаяся на мембране электростатическая
энергия используется для синтеза АТФ.
У некоторых бактерий плазматическая мембрана впячена
внутрь клетки и образует примитивные органоиды — мезосомы и
фотосинтезирующие мембраны. Последние содержат пигменты
бактериохлорофилл или бактериородопсин, участвующие в транс-
формации энергии фотонов в энергию химических связей АТФ.
Отметим также, что сходные мембранные образования участвуют
в фиксации азота.
Фоторецепторные клетки могут иметь различное строение, но
их световоспринимающие части имеют принципиальное сходство.
Принято считать, что наружный сегмент фоторецепторной клетки
представляет собой видоизмененный жгутик, в котором плотно
уложены столбиком плоские мешочки — «диски». Например, в
«палочке» глаза человека их около 1 тыс. В мембранах дисков на-
ходятся молекулы светочувствительного пигмента родопсина.
Плотность укладки этих молекул достигает 100 000 на 1 мкм2. На-
ружная плазматическая мембрана фоторецепторной клетки в тем-
ноте деполяризована, поскольку все ее натриевые каналы полнос-
тью открыты. При попадании на родопсин даже одного фотона
140
света он распадается: его простетическая ipynna отделяется, а оп-
син изменяет свою конформацию и каким-то образом стимулиру-
ет закрытие натриевых каналов в наружной мембране. Из-за этого
мембрана поляризуется и мембранный потенциал достигает 1 мВ.
На первый взгляд «диски» следовало бы отнести к внутренним
мембранным органоидам фоторецепторной клетки, однако сопос-
тавление строения палочки глаза человека и, например, зритель-
ной клетки сложных глаз насекомых показывает, что плоские ме-
шочки дисков и впячивания между «микровиллами» — образова-
ния идентичные (см. рис. 4.9, 3). Одинаковы и светочувствитель-
ные пигменты мембран, родственные бактериородопсину из
плазматической мембраны пурпурных бактерий, хотя они могут
иметь структурные вариации с максимумами поглощения света в
разных частях спектра.
Синапсами называют специализированные функциональные
контакты между возбудимыми клетками (нервными, мышечными,
секреторными), служащие для передачи и преобразования нервных
импульсов. Синапс состоит из окончания пресинаптического ак-
сона (пресинаптическая часть); синаптической щели (простран-
ство, разделяющее мембраны контактирующих клеток) и участка
клетки, к которому подходит пресинаптическое окончание (пост-
синаптическая часть). Собственно мембраны, заключающие си-
наптическую щель, имеют лишь незначительные структурные
особенности, которые выражаются в том, что на внутренней
стороне пресинаптической мембраны расположены парные ряды
внутримембранных белковых частиц. В их области при открытии
«кальциевых каналов» под воздействием изменения мембранного
потенциала происходит залповое слияние пузырьков, содержащих
ацетилхолин, с мембраной. При этом ацетилхолин выбрасывается
в синаптическую щель, а белки мембран синаптических пузырь-
ков перемешиваются с белками наружной мембраны. В последую-
щие 10 с белки пузырьков перемещаются, собираются в крупные
частицы, захватываются окаймленными ямками и отшнуровыва-
ются в составе мембран, идентичных мембранам синаптических
пузырьков. То есть в рассматриваемом случае особенным является
не столько строение, сколько поведение мембран — сочетание
электрического и химического воздействий на мембрану в процес-
се передачи нервного импульса.
Внутренние мембраны клетки. Многие биохимические процес-
сы, протекающие в клетке, приурочены к мембранам, поскольку
мембраны обеспечивают создание градиента веществ или усло-
вий, упорядочение ферментных систем, разделение исходных и
конечных продуктов реакций и т. д. В клетках прокариот такие
процессы сопряжены обычно с единственной наружной плазма-
тической мембраной. Исключение составляют сложно устроен-
141
ные синезеленые водоросли. Считается, что сильное развитие
внутриклеточных мембран в клетках эукариот обусловлено малым
значением отношения между площадью плазматической мембра-
ны и объемом клетки. Для расположения всех молекул мембран-
но-связанных белков-ферментов требуются дополнительные
площади. Но, по-видимому, следует принять во внимание и не-
обходимость разграничения в пространстве участков цитозоля с
разными параметрами (например, локальное увеличение кон-
центрации исходных продуктов для смещения направления реак-
ции и т. п.). Важна компартментализация (разделение на «отсе-
ки») цитозоля и для повышения эффективности процессов, свя-
занных с внутриклеточным транспортом веществ. Более того, мы
не должны останавливаться только на определениях «для чего»,
так как их невозможно отделить от определений «в результате
чего». Так, если мы говорим, что появление лизосом связано с не-
обходимостью изолировать мембраной транспортируемые агрес-
сивные вещества-ферменты, мы учитываем й то, что транспорти-
ровка отсортированных молекул от стационарного места их синте-
за и сортировки к относительно устойчивому или временному,
подвижному пункту назначения через подвижную среду наиболее
эффективна при использовании транспортных пузырьков, что ве-
дет к образованию особого рода компартмента. Вспомним также,
что при разделении функционирующего тела на взаимосвязанные
комплементарные части увеличивается объем всех видов его ин-
формации.
В большинстве эукариотических клеток принято выделять семь
основных компартментов, или ограниченных мембранами объем-
ных отделов, промежутки между которыми заполняет цитозоль:
эндоплазматическую сеть (или эндоплазматический ретикулум);
комплекс Гольджи, включающий диктиосомы (или аппарат Голь-
джи); митохондрии; пластиды; лизосомы; пероксисомы и ядро.
Эндоплазматическая сеть (ЭПС), или эндоплазматический рети-
кулум (ЭР). Это обязательный мембранный органоид эукариоти-
ческих клеток (рис. 4.10, /, 2). На его долю обычно приходится
более половины общего количества цитомембран. Мембрана ЭР
образует единую поверхность, отграничивающую полость этого
органоида от гиалоплазмы, причем полость занимает около 10 %
общего объема клетки. Мембрана ЭР составляет непрерывное об-
разование с наружной ядерной мембраной. В трехмерном изобра-
жении ЭР выглядит как система плоских цистерн, покрытых ри-
босомами и соединенных гладкими трубочками. Соответственно
его разделяют на шероховатый ЭР и гладкий ЭР. В клетках разных
функциональных типов соотношение шероховатой и гладкой
мембраны может сильно отличаться. Клетки, продуцирующие в
большом количестве белки, закономерно имеют особенно разви-
142
Каналы гладкой ЭПС
Рис. 4.10. Мембранные органоиды клетки (Албертс и др., 1986, с изменениями):
1 — электронограмма эндоплазматической сети; 2—реконструкция объемного изображения
ЭПС; 3— электронограмма аппарата (комплекса) Гольджи; 4— реконструкция объемного
изображения аппарата Гольджи; 5—элек тронограмма различных типов лизосом; о электро-
нограмма пероксисом
тую сеть мембран, покрытых рибосомами, а клетки, секретирую-
щие липиды, наоборот, — мембраны, свободные от рибосом.
Главные функции ЭР — синтез и транспорт белков и липидов.
Известно, что органоидами, непосредственно обеспечивающими
синтез белков, являются рибосомы. Ранее уже упоминалось, что
свободные рибосомы синтезируют белки для цитозоля, а мемб-
ранно-связанные рибосомы направляют синтезируемые белки в
цистерны ЭР, откуда они транспортируются в другие мембранные
органоиды или за пределы клетки. Но свободные и мембранно-
связанные рибосомы — это лишь два состояния одних и тех же
рибосом. Прикрепление рибосомы к мембране происходит после
синтеза сигнальной последовательности аминокислот, связываю-
щейся с рецепторным белком мембраны. Область ЭР, покрывае-
мая рибосомами, сплошь усеяна такими рецепторными белками,
которые, собственно, и определяют протяженность шероховатой
зоны. Рецепторный белок направляет сигнальную последователь-
ность в цистерну через собственный канал, а в это время большая
субчастица рибосомы связывается с мембранными белками и
фиксируется на мембране ЭР. С этого момента рибосома считает-
ся мембранно-связанной. В цистерне от синтезируемой белковой
молекулы мембранным белком-ферментом отрезается сигнальная
последовательность и готовый полипептид подвергается гликози-
лированию, то есть к нему присоединяется олигосахарид, впо-
следствии сильно трансформируемый. Только в таком виде белок
может быть направлен к месту назначения.
Гладкие участки ЭР синтезируют липиды (фосфолипиды и хо-
лестерол) для формирования новых клеточных мембран. Особен-
ность этого синтеза состоит в том, что липиды образуются только
на внешней половине липидного бислоя, встраиваясь в него.
Транспортировка липидов к другим органоидам происходит в
виде мелких мембранных пузырьков, и только некоторые липиды
митохондриальных мембран доставляются на место специальны-
ми транспортными белками, растворимыми в воде. В число липи-
дов, синтезируемых мембранами ЭР, входят и стероидные гормо-
ны. Поэтому, например, клетки коры надпочечников и половых
желез имеют хорошо развитый гладкий ЭР.
Из других функций ЭР отметим участие его в процессах деток-
сикации некоторых конечных продуктов обмена веществ, лекар-
ственных препаратов (у человека и домашних животных) и ряда
иных веществ.
Особую разновидность эндоплазматического ретикулума пред-
ставляет саркоплазматический ретикулум мышечных клеток (мы-
шечных волокон). Он образует систему уплощенных цистерн, ко-
торая охватывает каждую миофибриллу. При переходе потенциала
действия с Т-трубочек плазматической мембраны на мембрану
144
саркоплазматического ретикулума с ней мгновенно сливаются пу-
зырьки с накопленными ионами кальция и выход кальция в цито-
золь приводит к разблокировке миозина тропомиозином и индук-
ции мышечного сокращения.
Аппарат (комплекс) Гольджи. В большинстве клеток эукариот
выявляется сложный мембранный комплекс, получивший свое
название по фамилии исследователя, впервые наблюдавшего его в
1898 г., благодаря применению в качестве красителей оксида ос-
мия (VIII) и дихромата рубидия (рис. 4.10, 5, 4).
В животных и растительных клетках аппарат Гольади выглядит
не совсем одинаково, и в клетках растений его принято называть
диктиосомой. Наверное, можно согласиться с мнением цитологов,
предлагающих называть диктиосомой отдельную стопку плоских
цистерн. Таких цистерн в стопке может быть от двух до восьми,
редко — до тридцати, а в типичном случае — шесть. Тоща все дик-
тиосомы клетки (от одной до нескольких сотен) можно было
бы обозначать как аппарат Гольджи.
В непосредственной близости с диктиосомами находятся много-
численные мелкие (диаметром около 50 нм) и крупные (до 1000 нм)
мембранные пузырьки. Основная масса мелких пузырьков сосре-
доточена около цистерны, расположенной со стороны ЭР, и по
периферии стопки цистерн. Многие из этих пузырьке» имеют
окаймление, а образуются они отпочковыванием от ЭР. Круп-
ные — секреторные пузырьки расположены рядом с диктиосомой
со стороны плазматической мембраны и отпочковываются от на-
ружной цистерны.
Внутренняя и наружная стороны аппарата Гольджи отличаются
не только динамическими характеристиками. Мембраны цистерн,
приближенных к мембранам ЭР, равны им по толщине
(2,5—4,0 нм), но и те и другие тоньше мембраны, находящейся с
противоположной стороны, и плазматической мембраны (7,5 нм).
Различно и содержимое цистерн в пределах одной стопки. Пред-
полагается, что пузырьки, идущие от ЭР к аппарату Гольджи, ис-
ходно дифференцируются и сливаются с разными цистернами.
Тем не менее отдельные секреторные белки обнаруживаются
последовательно во всех цистернах, хотя перед поступлением в
секреторные пузырьки они накапливаются только в наружной
цистерне.
По-видимому, аппарат Гольджи управляет распределением
большинства синтезируемых макромолекул по клетке. Через его
цистерны и пузырьки проходят секретируемые белки, гликопро-
теины и протеогликаны, липопротеины и гликолипиды плазмати-
ческой мембраны, белки лизосом, компоненты матрикса клеточ-
ных оболочек. Однако во время переноса молекулы подвергаются
значительным модификациям, которые преимущественно сводят-
10 Присный А.В
145
ся к изменениям олигосахаридов. Кроме того, некоторые глико-
протеиды синтезируются на мембранах диктиосом, например му-
цин, образующий в водном растворе слизь и встречающийся в са-
мых разных живых телах. Преобразования олигосахаридов, свя-
занных с белками и липидами, идут по огромному числу путей,
«ветвящихся» и не приводящих к одинаковым конечным результа-
там. Продукты этих преобразований отличаются друг от друга
олигосахаридными частями, которые, как считается, служат сиг-
нальными участками, или маркерами, по которым определяется
направление дальнейшего движения или преобразования той или
иной молекулы.
Своеобразную роль играет аппарат Гольджи в растительных
клетках в телофазе их деления. Секреторные пузырьки, направля-
емые микротрубочками в область «клеточной пластинки», достав-
ляют сюда материал матрикса клеточной оболочки, а мембрана
этих пузырьков идет на построение плазматической мембраны до-
черних клеток.
Помимо перечисленного аппарат Гольджи выполняет еще одну
важную функцию: в нем образуются лизосомы.
Считается, что аппарат Гольджи образуется из гладких (аграну-
лярных) мембран ЭР, которые, в свою очередь, происходят от те-,
роховатых (гранулярных) мембран. Следуя этой точке зрения,
примем, что цистерны диктиосом представляют собой слившиеся
транспортные пузырьки, дающие начало секреторным пузырькам
и лизосомам.
Лизосомы. Эти органоиды присутствуют в большинстве типов
эукариотических клеток, однако наиболее регулярны и многочис-
ленны они в фагоцитирующих клетках: у простейших, низших
многоклеточных животных, в некоторых тканях высших живот-
ных. В таких клетках лизосомы имеют типичную округлую форму
и размеры до 0,5 мкм (рис. 4.10, 5).
У растений органоиды, сходные с типичными лизосомами, вы-
явлены только в погибающих клетках. В остальное же время роль
лизосом играют крупные центральные вакуоли. Не исключается,
что центральные вакуоли своим происхождением связаны именно
с пузырьками, отшнуровывающимися от аппарата Гольджи.
Лизосомы — простые мембранные мешочки, наполненные гид-
ролитическими ферментами (протеазами, нуклеазами, липазами,
гликозидазами, фосфолипазами, фосфатазами и сульфатазами),
которые служат для контролируемого внутриклеточного расщеп-
ления макромолекул. Содержимое лизосом имеет кислую реак-
цию, и их ферменты обнаруживают наибольшую эффективность
при низких значениях pH (около 5). Учитывая, что цитозоль име-
ет pH несколько выше 7, в случае попадания в него ферментов из
лизосом ферменты оказываются не в оптимальных условиях и их
146
действие в небольших количествах не имеет катастрофических по-
следствий для клетки. Если же лизосома по какой-то причине раз-
рушится и все ее содержимое выйдет в цитоплазму, произойдет
значительное переваривание структурных элементов клетки.
Функциональные особенности лизосом проявляются и в строе-
нии их мембраны: она проницаема для продуктов расщепления
макромолекул; в ее состав входит специальный транспортный бе-
лок, который, используя энергию гидролиза АТФ, накачивает в
полость лизосомы протоны, благодаря чему достигается необходи-
мое значение pH; мембрана содержит белки-акцепторы, обеспе-
чивающие «стыковки» со специфическими транспортными пу-
зырьками.
Образование лизосом можно наблюдать как на внешних цис-
тернах диктиосом аппарата Гольджи, так и на шероховатой части
эндоплазматического ретикулума, в которой синтезируются их
ферменты. В настоящее время принято считать, что пузырьки,
образующиеся на диктиосомах, — это и есть собственно лизосо-
мы, или первичные лизосомы, а от ЭР отпочковываются транспорт-
ные пузырьки, обычно имеющие клатриновое окаймление. Когда
первичные лизосомы сливаются с вакуолями, в которых содержат-
ся подлежащие перевариванию частицы или вещества, их называ-
ют вторичными лизосомами или пищеварительными вакуолями. По
окончании переваривания в мембранных пузырьках остаются не-
переваренные или непереваримые вещества и частицы — образу-
ются «остаточные тельца». Остаточные тельца освобождаются от
своего содержимого, сливаясь с плазматической мембраной (ти-
пичный механизм экзоцитоза).
Лизосомы могут переваривать внутриклеточный материал или
выводить ферменты за пределы клетки. Но основная функция ли-
зосом состоит в переваривании веществ, поступающих в клетку
извне. Именно поэтому они более характерны для гетеротрофов.
Амебоидные клетки крови и лимфы способны переваривать захва-
тываемые бактерии и вирусные частицы. Лишь немногие микро-
организмы обладают устойчивостью к действию лизосомных фер-
ментов. Это, например, туберкулезная палочка, способная даже
размножаться во вторичных лизосомах.
В щитовидной железе млекопитающих ферменты лизосом рас-
щепляют предшественник гормонов тироксина и трийодтирони-
на, освобождая их перед выходом в кровяное русло.
Способность ферментов лизосом к разрушению различных
биологических структур и макромолекул используется клетками
для уничтожения ненужных им органоидов. Подвергаемая разру-
шению структура сначала окружается одиночной мембраной глад-
кого эндоплазматического ретикулума. Такой замкнутый мемб-
ранный мешочек сливается с первичными лизосомами, в резуль-
147
10*
тате чего образуется вторичная лизосома, или аутофагирующая ва-
куоль. Переваривание собственных органоидов особенно хорошо
проявляется в период клеточных дифференцировок и при резком
снижении функциональной активности клеток. Пока неизвестно,
как первичные лизосомы распознают специфику веществ, подле-
жащих перевариванию, которые заключены в мембранных ме-
шочках. Такое распознавание необходимо для доставки соответ-
ствующих ферментов.
Поскольку вторичные лизосомы неоднородны в морфологи-
ческом отношении, им дают специальные названия: при фагоци-
тозе крупных частиц (бактерий, водорослей и т. п.) образуются
крупные пищеварительные вакуоли; при захвате лизосомами цели-
ком мелких (около 50 нм) пузырьков, отшнуровывающихся от
комплекса Гольджи, образуются мулыпивезикулярные тельцу при
слиянии лизосом с пузырьками, содержащими собственные орга-
ноиды клетки, подлежащие перевариванию, образуются аутофа-
гирующие вакуоли, или аутофагосомы.
Пероксисомы (или микрозельца). Это округлые пузырьки, обра-
зованные одиночной мембраной (рис. 4.10, 6). Их диаметр состав-
ляет от 0,3 до 1,5 мкм. Мембрана пероксисом обладает необычно
высокой проницаемостью для молекул, не превышающих размера
молекул глюкозы. Такие крупные пероксисомы встречаются лишь
в некоторых типах клеток. Гораздо чаще, по крайней мере во всех
типах клеток млекопитающих, встречаются более мелкие микро-
тельца диаметром 0,15...0,25 мкм, получившие название микропе-
роксисомы. В крупных пероксисомах обычно хорошо заметна кри-
сталловидная сердцевина — трубчатая структура, содержащая фер-
менты в чрезвычайно концентрированном виде.
Пероксисомы содержат несколько ферментов. Известно, что
три из них участвуют в образовании пероксида водорода, а один —
в его разложении. Содержание последнего — каталазы — составля-
ет около 40 % массы всех белков пероксисомы. Пероксид водоро-
да образуется в клетках как побочный продукт некоторых окисли-
тельно-восстановительных реакций. Он очень токсичен и должен
быстро разлагаться. Возможно, поэтому каталаза относится к чис-
лу наиболее быстродействующих ферментов.
Крупные пероксисомы печени и почек человека играют важ-
ную роль в обезвреживании различных веществ. Например, почти
половина выпиваемого спирта окисляется именно в пероксисомах
(до ацетальдегида). Обычно эти органоиды используют около 10 %
кислорода, поступающего в печень, где происходит также превра-
щение жиров в углеводы. По-видимому, в пероксисомах происхо-
дит, кроме того, распад жирных кислот до ацетил-СоА (далее по-
ступающего в митохондрии).
Существует гипотеза, согласно которой пероксисомы — это ру-
148
диментарные остатки органоидов, выполнявших в примитивных
проэукариотических клетках все функции, связанные с обменом
кислорода, когда он появился в атмосфере. Образовавшиеся позже
митохондрии приняли на себя осуществление части окислитель-
ных реакций, связав их с реакциями синтеза АТФ. Примитивным
считается и механизм формирования крупных пероксисом в клет-
ках современных живых тел. Так, некоторые мембранные белки,
характерные только для них, синтезируются в ЭР, затем группиру-
ются в мембране ЭР с образованием проорганоида, который от-
почковывается от гладкой области ЭР. Главные ферменты перок-
сисом синтезируются в цитозоле и транспортируются в них через
мембраны. Зрелые пероксисомы часто не отшнуровываются пол-
ностью от эндоплазматического ретикулума, а остаются связанны-
ми с ним тонким трубчатым отростком.
Количество пероксисом в клетках изменяется в зависимости от
характера вешеств, поступающих в них; оно различно и в клетках
разных типов. Более того, в разных клетках присутствуют и раз-
ные пероксисомы, отличающиеся, в частности, набором ферментов
и протекающими в них реакциями. В листьях растений, напри-
мер, пероксисомы катализируют окисление побочных продуктов
фотосинтеза с использованием кислорода. Этот процесс называет-
ся фотодыханием. В прорастающих семенах совершенно другие
пероксисомы превращают жирные кислоты в сахара. Эти перо-
ксисомы называют глиоксисомами, поскольку они обеспечивают
(только в клетках растений) протекание реакций глиоксилатного
цикла.
Вакуолярный аппарат растительных клеток (рис. 4.11). Длитель-
ное противопоставление растений и животных как единственных
царств живых тел отразилось и на терминологии, принятой для
обозначения их внутриклеточных структур. Но в последние годы
были проведены тщательные исследования, позволяющие во мно-
гих случаях унифицировать названия
как органоидов, так и процессов,
протекающих в клетках самых раз-
ных живых тел.
В ботанике вакуолями принято на-
зывать ограниченные мембраной ча-
Рис.4.11. Вакуолярный аппарат растительной
клетки (оригинал автора):
1— центральная вакуоль; 2—вакуоли (сливающиеся
пузырьки Гольджи, отшнуровываюшиеся от ЭПС)
149
ные в мембраны, обнаруживаются в клетках животных. Правиль-
нее считать, что это пиноцитозные капли, которые еще не сли-
лись с первичными лизосомами.
2. Углеводы. Основными формами углеводных включений яв-
ляются: крахмальные зерна в растительных клетках; парамиловые
зерна в клетках автотрофных простейших; гликогеновые гранулы
в клетках бактерий, грибов и животных. Крахмальные и парами-
ловые зерна образуются в некоторых фотосинтезирующих органо-
идах, а затем выходят в цитозоль. Например, крахмальные зерна
образуются по одному в амилопластах. Клетки запасающих орга-
нов растений могут накапливать это вещество в значительных ко-
личествах. То же можно сказать и о гликогене, откладывающемся
в клетках запасающих органов животных. В печени млекопитаю-
щих он может заполнять клетки практически целиком, причем его
хранулы сливаются в более крупные агрегаты.
Углеводы могут запасаться в клетках в виде растворов. Если это
раствор — компонент цитозоля, то его не относят к включениям.
Но если, например, сахароза накапливается в вакуолях, то не бу-
дет ошибкой считать их включениями.
3. Белки. К накоплению белков в виде гранул способны циано-
бактерии, растения и животные. Для растений характерно отложе-
ние белковых гранул в непигментированных пластидах, а осталь-
ные образуют их в цитозоле.
4. Фосфат. В клетках неорганический фосфат способен запа-
саться в виде минеральных солей. Он регулярно присутствует, по
крайней мере, у цианобактерий.
Липиды, белки, углеводы и фосфат — типичные включения,
образуемые резервными веществами. Включения — это времен-
ные структуры, которые в зависимости от функционального со-
стояния клетки могут растворяться, использоваться, а через неко-
торое время появляться вновь. Отложение запасных веществ в хи-
мически видоизмененной форме и в твердом виде для клетки
предпочтительнее, поскольку они становятся химически неактив-
ными, инертными и при длительном хранении не опасны для са-
мой клетки; они занимают малый объем, даже накапливаясь в от-
носительно больших количествах.
Некоторые запасаемые в вакуолях клеток растений раствори-
мые вещества по достижении высоких концентраций образуют ти-
пичные, иногда достаточно крупные кристаллы. Так ведет себя,
например, оксалат кальция. Наряду с этим кристаллы, присут-
ствующие в цитоплазме, могут быть и типичными экскретами, то
есть продуктами обмена, далее не используемыми данной клеткой
и подлежащими удалению. Мочевая кислота у животных — одна
из форм удаляемых конечных продуктов азотного обмена. Иногда
она быстро выводится из клетки в жидком состоянии. В других
152
случаях, в частности в клетках мальпигиевых сосудов насекомых,
она переводится в кристалловидное состояние, быстро выталкива-
ется в просвет сосудов и далее выводится через заднюю кишку за
пределы тела. Наконец, как временное включение мочевая кисло-
та ведет себя у насекомых с полным метаморфозом в фазе кукол-
ки. Когда разрушаются личиночные органы, а имагинальные (ха-
рактерные для взрослых) органы еще не образовались, это веще-
ство накапливается в кристаллическом состоянии в клетках жиро-
вого тела (запасающего органа).
Одним из конечных продуктов азотного обмена у насекомых (у
прямокрылых и чешуекрылых) является птерин — смесь солей мо-
чевой кислоты. Птерин не выводится за пределы тела, но направ-
ляется в клетки кожных покровов и обеспечивает белую или жел-
тую окраску покровов- То есть его зерна выступают одновременно
и в качестве экскрета, и в качестве пигмента. Тот же птерин у мно-
гих насекомых включается в пигментные клетки органов зрения,
где наряду с другим пигментом, оммохромом, обеспечивает повы-
шение направленной чувствительности к свету отдельных глазков
сложного глаза. В окрашивании покровов у членистоногих кроме
птерина и оммохрома принимают участие биливердины, мелани-
ны и другие пигменты, представленные гранулами или зернами в
цитоплазме клеток. Упомянутый меланин — по-видимому, самый
распространенный пигмент животных (хотя встречается и у расте-
ний). Образуется он в особых клетках — меланоцитах и выполняет
в живых телах множество функций: защищает клетки от жесткого
ультрафиолета, способствует накоплению тепла, обеспечивает
зрительную коммуникацию, участвует в электрических процессах
ит.д.
Продукты выделения включают вещества, подлежащие удале-
нию из клетки. Их подразделяют на экскреты, секреты и инкреты
опять-таки весьма условно. И если к экскретам одновременно
относят и продукты ассимиляции, и неиспользованные, и не-
нужные, и вредные вещества, то понятно, что часть из них попа-
дает в группу секретов, так же как часть секретов — это типичные
инкреты.
Нет достаточно четких различий и между продуктами выделе-
ния и включениями. Многие минеральные вещества, поступаю-
щие в клетки простейших и низших многоклеточных животных в
растворенном виде, концентрируются в них до кристаллического
состояния и выводятся из цитоплазмы на поверхность клетки или
в межклеточное пространство или же образуют внутриклеточные
опорные элементы.
Целесообразно к экскретам относить те выделяемые клеткой
вещества, которые образуются как побочный или конечный про-
дукт метаболизма и данным живым телом более не используются.
153
В этом случае к секретам можно было бы отнести продукты спе-
циального синтеза, действие которых направлено на внешнюю по
отношению к клетке, выделяющей их, среду и выводящиеся за
пределы тела (в том числе в среду, проникающую в глубокие впя-
чивания — полости).
Особой формой секретов следовало бы считать инкреты — био-
логически активные вещества, выделяемые клетками многокле-
точных тел для регуляции вегетативных процессов, протекающих
в клетках того же самого живого тела.
Этот вариант деления позволяет обособить те минеральные и
органические вещества, которые выделяются клеткой на свою по-
верхность или в качестве межклеточного вещества и являются
обязательной частью данного живого тела.
Клеточная оболочка. В разделе 3.2 была приведена краткая ха-
рактеристика клеточной стенки бактерий и клеточной оболочки
растений и грибов. Отметим еще раз, что клеточная оболочка осо-
бенно развита у живых тел — глубинных прорастателей и, как пра-
вило, отсутствует у активно подвижных живых тел.
Поверхностный взгляд на надмембранные образования дает
повод рассматривать их как разновидность экскретов или секре-
тов. Однако известно, что клеточные оболочки выполняют специ-
фические активные функции, важные не только для клеток или
тканей, но для всего тела растения или гриба: они участвуют в
транспорте, поглощении и выделении веществ, в них может быть
сосредоточена лизосомальная активность.
Остановимся подробнее на строении клеточной оболочки рас-
тений. Основным ее компонентом является целлюлоза. Длинные
молекулы этого вещества объединяются в микрофибриллы толщи-
ной 10...25 нм. Упорядоченное расположение молекул в каждом
отдельном участке микрофибриллы придает целлюлозе кристал-
лические свойства. Пучки скрученных микрофибрилл образуют
нити, которые, в свою очередь, взаимно свиваются в макрофиб-
риллы. Толщина макрофибрилл достигает 0,5 мкм, а прочность
соответствует прочности стальной проволоки такого же диаметра.
Целлюлозные фибриллы образуют каркас, все промежутки кото-
рого заполнены молекулами матрикса (гемицеллюлозами, пекти-
новыми веществами и гликопротеинами) (рис. 4.12).
Вторым по распространенности (после целлюлозы) компонен-
том клеточной оболочки является лигнин. Он увеличивает жест-
кость оболочки и в основном содержится в клетках механических
тканей. Заполнение промежутков между фибриллами лигнином
лежит в основе одревеснения клеточных оболочек. Из одревеснев-
ших клеток состоят древесина и склеренхима. В оболочках защит-
ных тканей откладываются кутин, суберин и воски. Их наличие
предотвращает чрезмерную потерю воды растением.
154
Рис. 4.12. Схема молекулярной организации клеточной оболочки растительной
клетки (Албертс и др., 1986, с изменениями)
В типичном случае оболочка образована двумя слоями: сере-
динной пластинкой (межклеточным веществом) и первичной кле-
точной оболочкой. Многие клетки дополнительно формируют
вторичную клеточную оболочку (рис. 4.13, 1). Серединная плас-
тинка состоит большей частью из пектиновых веществ. Пектин
содержится и в первичной оболочке, что делает ее эластичной и
растяжимой по мере роста клетки. Именно поэтому в активно де-
лящихся клетках вторичная оболочка не развита. Такие клетки со-
храняют способность к дифференцировке жизненной формы и
участвуют в регенерационных процессах. Первичные оболочки в
отдельных местах имеют тонкие участки — первичные поровые
поля; здесь обычно проходят плазмодесмы (о которых речь пойдет
далее).
Вторичную оболочку протопласт начинает откладывать только
после того, как рост клетки прекращается. В ней больше целлюло-
зы и гемицеллюлозы, а пектины и гликопротеины отсутствуют.
Рис. 4.13. Оболочка растительной клетки (оригинал автора):
1 — схема относительного расположения слоев оболочки растительной клетки;2— хаотическое
расположение целлюлозных фибрилл в первичной оболочке; 3— упорядоченное расположе-
ние целлюлозных фибрилл во вторичной оболочке
155
Поэтому наличие вторичной оболочки придает клеточной оболоч-
ке в целом требуемую прочность и жесткость. Дополнительное
увеличение прочности достигается перекрещивающимся располо-
жением целлюлозных фибрилл в слоях вторичной оболочки. Вто-
ричная оболочка не образуется на первичных поровых полях, бла-
годаря чему в ней остаются углубления — поры. Иногда поры об-
разуются вне связи с поровыми полями. В оболочках смежных
клеток они располагаются друг против друга. Известно два типа
пор: простые и окаймленные.
Рост клеточных оболочек — весьма сложный, но точно органи-
зованный процесс. Волокна (фибриллы) целлюлозы синтезируют-
ся плазматической мембраной, а их расположение определяется
ориентацией микротрубочек, находящихся под мембраной. Но-
вообразованные фибриллы могут откладываться поверх образо-
ванных ранее фибрилл или между ними. В оболочке клеток, рас-
тущих во всех направлениях, они располагаются хаотически, а в
клетках, вытягивающихся в одном направлении, — поперек оси
(рис. 4.13, 2, 5). По мере роста клетки фибриллы меняют ориента-
цию, становясь почти параллельными направлению роста. Веще-
ства, входящие в состав матрикса оболочки, транспортируются к
плазматической мембране от диктиосом в секреторных пузырьках
и выделяются в результате экзоцитоза.
При делении клетки механизм разделения цитоплазмы зависит
от наличия клеточной оболочки и ее жесткости. Клетки большин-
ства живых тел разделяются путем втягивания плазматической
мембраны или эластичной оболочки от периферии к центру с об-
разованием борозды деления, которая постепенно углубляется,
вплоть до полного разделения дочерних клеток. У сосудистых рас-
тений, мохообразных и некоторых водорослей клетки разделяются
благодаря образованию клеточной пластинки. Еще в телофазе из
микротрубочек образуется волокнистая структура — фрагмопласт.
В его экваториальную плоскость от диктиосом собираются мелкие
пузырьки, выстраивающиеся в ней, не разрушаясь. Затем пузырь-
ки сливаются, выделяя пектиновые вещества, из которых и фор-
мируется серединная пластинка. Мембраны пузырьков, сливаясь,
включаются в плазматические мембраны, разделенные середин-
ной пластинкой. На следующем этапе каждый из смежных прото-
пластов начинает выделять вещество первичной оболочки, покры-
вающее серединную пластинку, а также новый слой оболочки,
продолжающийся вокруг всего протопласта.
Во время образования клеточной серединной пластинки из
фрагментов трубчатого эндоплазматического ретикулума заклады-
ваются и проникающие через нее плазмодесмы (рис. 4.14).
Плазмодесмами называют тонкие, диаметром 20...40нм, ци-
топлазматические тяжи, соединяющие практически все живые
156
Рис. 4.14. Плазмодесмы (Албертс и др., 1986; оригинал автора):
/ — электронотрамма плазмодесм растительной клетки; 2—схема разделения тканей расте-
ний на компартменты, определяемого наличием плазмодесм
клетки растения со смежными клетками. В области каждой плаз-
модесмы плазматическая мембрана одной клетки без перерыва
продолжается в мембрану другой. Сама плазмодесма представляет
собой цилиндрический выстланный мембраной канал. По оси ка-
нала из одной клетки в другую проходит десмотубула — мембран-
ная трубочка, соединяющая эндоплазматические ретикулумы со-
седних клеток. Пространство между плазматической мембраной и
десмотубулой заполнено цитозолем. У концов канала отмечены
сужения с кольцевыми структурами, которым приписывают роль
клапанов, регулирующих переход молекул из клетки в клетку.
Благодаря плазмодесмам тело растения оказывается непрерыв-
ным единством всех составляющих его протопластов. Это дает по-
вод для разделения такого тела на два крупных компартмента:
симпласт (внутриклеточный компартмент) и апопласт (внекле-
точный компартмент, включающий все клеточные оболочки и
мертвые пустые проводящие клетки).
Помимо веществ, образующих оболочки, клетки растений вы-
деляют и типичные межклеточные вещества, близкие к пектинам
и гемицеллюлозе. Это слизи и камеди, которые накапливаются в
слизевых ходах и железках. Слизи, растворяясь в воде, приобрета-
ют вязкую консистенцию. Особенно обильно они накапливаются
у суккулентов, поскольку способствуют удержанию воды. Камеди,
выступая на поверхности поврежденных частей растений, засты-
вают в стекловидную массу, препятствующую проникновению па-
тогенной микрофлоры и стеблевых ксилофагов (потребителей
древесины).
Фибриллярные структуры цитоплазмы. Исследования последних
20...25 лет показывают, что цитоплазма клетки не представляет со-
157
бой бесформенную гелеобразную или золеобразную массу, а име-
ет весьма сложную развитую структуру. Все пространство между
органоидами пронизано высокоупорядоченными молекулярны-
ми комплексами — филаментами, составляющими значительную
часть основного вещества цитоплазмы (матрикса) и получившими
название цитоскелет. Именно по этой причине матрикс опреде-
ляет форму клетки, все виды ее движения, перемещение органо-
идов, транспорт пузырьков, деление. Кроме того, матрикс обеспе-
чивает структурную основу клеточного обмена веществ, определяя
пространственное размещение молекулярных компонентов клет-
ки, задействованных в метаболических процессах.
К настоящему времени выявлено четыре основные системы
филаментов: микрофиламенты (или микрофибриллы), промежу-
точные филаменты (фибриллы), толстые фибриллы (макрофиб-
риллы) и микротрубочки (рис. 4.15...4.19). Их диаметр соответ-
ственно составляет 6...8; 15; 22...28 нм. Микрофибриллы
построены из белка актина, промежуточные филаменты — в
разных клетках из разных белков; толстые фибриллы — из мио-
зина, а микротрубочки — из тубулина.-Помимо основных пере-
численных типов белковых филаментов цитоскелет включает
множество других вспомогательных белков, которые связывают
филаменты друг с другом или с иными клеточными структура-
ми или же влияют на скорость и степень полимеризации фила-
ментов.
Лучше других систем филаментов и клеточных движений, за
которые они отвечают, изучены актиновые филаменты и микро-
трубочки, обеспечивающие сокращение мышц и биение ресничек
и жгутиков.
Рис. 4.15. Электроиограмма фибрилляр-
ных структур эпителиальной клетки жи-
вотного (оригинал автора):
7 — микротрубочки; 2- промежуточные фи-
ламенты; 3— микрофиламенты
Рис. 4.16. Промежуточные филаменты
отдельной клетки животного (Албертс
и др., 1986, с изменениями)
158
Рис. 4.17. Схема расположения микрофиламеитов в клетке животного типа
(Албертс и др., 1986, с изменениями)
Рис. 4.18. Схема расположения микротрубочек в клетке животного типа
(Албертс и др., 1986, с изменениями)
Актиновые филаменты могут служить опорными образования-
ми для постоянных выростов цитоплазмы: микроворсинок эпите-
лиальных клеток, волосковых клеток органа слуха и др. В клетках,
близких к недифференцированному типу, актин образует множе-
ство «волокон натяжения», или «напряженных нитей»,—длин-
ных, толстых пучков микрофиламентов, которые на разной глуби-
159
не в цитоплазме идут параллельно клеточной мембране, а также
тонкую сеть микрофиламентов непосредственно под наружной
клеточной мембраной. Волокна натяжения особенно развиты в
ползущих и распластанных клетках и почти не обнаруживаются в
неподвижных округлых клетках, а подмембранная сеть наблюда-
ется преимущественно на переднем крае ползущей клетки. В спе-
циализированных мускульных клетках или сократимых отростках
других типов клеток актиновые филаменты уложены в параллель-
ные пучки, обычно перемежаясь с толстыми (миозиновыми) фиб-
риллами. Скольжение актиновых филаментов относительно мио-
зиновых лежит в основе механизма мышечного сокращения. Дру-
гие виды клеточного движения могут осуществляться благодаря
направленным процессам полимеризации — распада микрофиб-
рилл.
Молекулы немышечного миозина участвуют в построении ра-
нее упомянутых «напряженных нитей» и, кроме того, входят в со-
став «сократимого кольца» делящихся клеток и «опоясывающего
кольца» эмбриональных эпителиальных клеток. Во всех этих слу-
чаях взаимное относительное скольжение актиновых микрофила-
ментов связано с присутствием образующих агрегаты молекул
миозина.
Промежуточные филаменты в клетках разных тканей распреде-
ляются по-разному, а в некоторых типах клеток их вообще нет.
Обычно промежуточные филаменты пронизывают всю цитоплаз-
му, но в одних клетках они образуют пучки, а в других — суще-
ствуют в виде отдельных нитей. В клетках многих типов распреде-
ление этой системы фибрилл похоже на распределение микротру-
бочек. Промежуточные филаменты — самые устойчивые элементы
цитоскелета и самые труднорастворимые компоненты клетки.
Именно эти волокна первоначально обозначались термином «ци-
тоскелет». Наиболее богаты промежуточными филаментами те ча-
сти клеток, которые подвергаются механическим нагрузкам: отро-
стки нервных клеток; области контактов между эпителиальными
клетками; большая часть цитоплазмы мускульных клеток гладкого
типа.
К толстым фибриллам относят нитевидные агрегаты молекул
миозина (см. рис. 4.19). Наиболее крупные агрегаты образуются
при полимеризации миозина скелетных мышц.
Строение и функции фибрилл. У подавляющего
большинства эукариот присутствуют множественные гены, коди-
рующие белок актин. Известно шесть различных типов актина, но
отличия в строении их молекул настолько незначительны, что
даже для далеких в систематическом отношении организмов раз-
новидности этого белка оказываются функционально взаимозаме-
няемыми.
160
Отдельные молекулы актина — это глобулы, прочно связанные
с ионами Са и нековалентно — с молекулой АТФ. При полимери-
зации концевой фосфат АТФ отщепляется. Хотя высвобождающа-
яся энергия не требуется для образования фибриллы, гидролиз
аденозинтрифосфата значительно повышает скорость полимери-
зации.
Актиновые филаменты состоят из двух цепей глобул и закруче-
ны в двойную спираль. Кроме них в образовании микрофиламен-
тов принимают участие и некоторые другие белки, например ста-
билизирующий актиновые цепи тропомиозин, молекулы которого
уложены в бороздки спирали филамента (см. рис. 4.17).
Клетки многих типов способны образовывать на своей поверх-
ности микроскопические пальцевидные выросты — микроворсинки
и микрошипы. При этом длина выростов в зависимости от состоя-
ния клетки может изменяться. Опорными образованиями таких
выростов служат актиновые филаменты. Изменение длины или
направления смещения опорного пучка определяется соотноше-
нием скоростей образования и разрушения филамента на его кон-
цах. Известно несколько примеров так называемой «взрывной»
полимеризации, когда скорость удлинения микрофиламентов
Рис. 4.19. Толстые фибриллы (Албертс и др., 1986, с изменениями):
1— молекула миозина; 2—расположение молекул миозина в макрофибрилле; 3— волокно
поперечно-полосатой мышцы; 4— схема расположения миозиновых и актиновых фибрилл
в мышечном волокне
11 Присный А.В.
161
превышает норму в сотни раз и достигает 20 мкм в секунду, что
соответствует связыванию примерно 5 тыс. актиновых глобул в
каждой из двух цепочек филамента.
В неполимеризованном состоянии значительная доля актина
связана в нестойкие комплексы с каким-либо белком, что служит
механизмом поддержания необходимой концентрации его моле-
кул для установления нормальной скорости полимеризации. Эти
же белки могут служить своеобразными включателями образова-
ния микрофиламентов.
Актиновые филаменты — это полярные структуры. Полярность
заключается в неодинаковости характера взаимодействия фила-
мента с молекулами миозина в разных направлениях от центра.
Полярность проявляется и в различной скорости их роста с раз-
ных концов. Однако направление нарастания или укорачивания
может регулироваться и прикреплением к тому или иному концу
фибриллы блокирующих белков.
Белок миозин присутствует практически во всех клетках, по
крайней мере, позвоночных животных. Он встречается в несколь-
ких различных формах. Одна из них — миозин скелетных (попе-
речно-полосатых) мышц — способна самопроизвольно полимери-
зоваться в гигантские комплексы — толстые фибриллы.
Отдельные молекулы миозина на электронных микрофотогра-
фиях выглядят как длинные палочковидные структуры с двумя
глобулярными головками на одном конце. В состав каждой из та-
ких молекул входят шесть полипептидных цепей: две одинаковые
тяжелые и две пары легких. Альфа-спирали тяжелых цепей закру-
чены одна вокруг другой в суперспираль, образующую палочко-
видный хвост. Каждая головка образована глобулярной частью тя-
желой цепи и включает по одной молекуле обоих типов легких
цепей.
Молекулы миозина ассоциируют друг с другом за счет палочко-
видных хвостов: несколько сотен молекул, располагаясь парал-
лельно со сдвигом по д лине, формируют пучок, из которого в сто-
роны выступают миозиновые головки, отсутствующие только на
небольшом серединном участке, где соединяются два противопо-
ложно направленных пучка миозиновых хвостов (см. рис. 4.19).
Глобулярные головки миозина способны взаимодействовать с
актином, и это взаимодействие порождает силу, реализующуюся в
клеточном сокращении. КПД этого преобразователя достигает
50...80 %. Необходимая для смещения фибрилл энергия получает-
ся при гидролизе АТФ. Миозин сам по себе обладает АТФазной
активностью, то есть обладает гидролитической ферментной ак-
тивностью по отношению к аденозинтрифосфату. Однако в комп-
лексе с актином скорость расщепления АТФ увеличивается в
150...300 раз.
162
Клетки гладкой мускулатуры содержат актин и миозин особого
типа. Их АТФазная активность почти в 10 раз ниже, чем в попе-
речно-полосатых мышцах, а взаимодействие миозина с актином
возможно только тогда, когда его легкие цепи фосфорилированы.
В гладких клетках тонкие и толстые фибриллы расположены мно-
го менее упорядоченно, чем в скелетных мышцах.
Актиновые филаменты, будь то центральный пучок микровор-
синок или напряженные нити, или сократимое кольцо, должны
иметь опору и потому, как правило, прикреплены одним концом
к клеточной мембране. Правда, механизм прикрепления пока не
ясен.
С актиновыми филаментами связано изменение фазового со-
стояния цитоплазмы (например, у движущихся саркодовых): загу-
стение, или переход в гелеобразное состояние, сопровождается
образованием поперечных сшивок между нитями, а разжижение,
или переход в золь, — фрагментированием филаментов.
Промежуточные филаменты — это прочные и относительно
долговечные фибриллы, имеющие вид прямых или слегка изогну-
тых волокон. Такие волокна образованы нитевидными молекула-
ми; Скрученными наподобие каната (см. рис. 4.16). Считается, что
процесс сборки промежуточных филаментов в клетке необратим.
Поэтому их деполимеризация осуществляется специальными фер-
ментами, избирательно расщепляющими ту или иную группу фи-
ламентов. Клетки разных типов обычно содержат только одну из
групп промежуточных филаментов: нейроны — нейрофиламенты;
эпителиальные клетки — кератиновые филаменты — «тонофила-
менты»; в большинстве других клеток имеются виментиновые
(как в фибробластах) или виментинсодержащие филаменты (как в
глиальных и мышечных клетках).
Основная функция промежуточных филаментов — противодей-
ствие растягивающим силам, то есть механическая. При этом фила-
менты могут находиться в клетках в виде рыхлых или плотных тя-
жей или же после отмирания образующих их клеток накапливаться
в межклеточном веществе. Так же образуются кератиновые чешуи,
волосы, ногти, то есть поверхностные защитные образования. В
других случаях функции промежуточных филаментов оказываются
менее ясными. Не получило объяснения и многообразие их групп в
плане неоднородности химического состава.
Микротрубочки и центриоли. Микротрубочки — это
нитевидные полые структуры, расположение и формирование ко-
торых тесно связаны с центром клеточной организации (центрио-
лями) (рис. 4.20). Особенно развиты они в клетках животного
типа. Отдельные микротрубочки отходят от клеточного центра и
тянутся к клеточной мембране. В отличие от пучков микрофила-
ментов радиально расположенные микротрубочки не исчезают,
п, 163
когда клетка принимает округлую форму, хотя их длина при этом
изменяется. При делении клетки микротрубочки практически
полностью перестраиваются, что указывает на их важную роль в
организации этого процесса. Более устойчивое состояние микро-
трубочки сохраняют в специализированных двигательных струк-
турах клеток—ресничках и жгутиках.
Микротрубочки состоят из глобулярного белка тубулина, об-
разующего димеры из слегка отличающихся полипептидов (а- и
р-тубулинов). При формировании микротрубочек молекулы тубу-
лина соединяются друг с другом в нитевидные комплексы — про-
тофиламенты. Обычно 13 таких протофиламентов, тесно примы-
кающих один к другому по всей длине, образуют цилиндрическую
структуру — собственно микротрубочку. Протофиламенты слегка
смещены относительно друг друга, так что тубулиновые димеры в
стенке цилиндра образуют спиральные ряды.
Сборка микротрубочек в клетке сходна со сборкой актиновых фи-
ламентов: она происходит самопроизвольно и сопровождается гид-
ролизом нуклеотидфосфата, правда, не АТФ, а ГТФ. Полимеризация
и деполимеризация тубулина идут в клетке постоянно и с участием
преимущественно одних и тех же молекул. Например, микротрубоч-
ки цитоплазмы интерфазной клетки на раннем этапе ее деления
разрушаются, а тубулин используется для постройки ахроматино-
вого веретена. В конце митоза происходит обратный процесс.
В ресничках и жгутиках микротрубочки связаны в единый пу-
чок—аксонему посредством специализированного образования,
получившего название «базальное тельце» (или «кинетосома»).
Эта структура служит матрицей для упорядочения системы мик-
ротрубочек по типу «9 + 2» (9 периферических дуплетов микро-
трубочек и 2 центральные микротрубочки) и инициирует построе-
ние дуплетов. Вся аксонема дополнительно стабилизируется вспо-
могательными белками, а кроме того, в ней присутствуют другие
белковые структуры, взаимодействие которых обеспечивает рес-
ничку (или жгутик) энергией и преобразует энергию в механичес-
кую работу волнообразного или штопорообразного движения ор-
ганоида. Наиболее важными из них считают короткие боковые
выросты дуплетов, направленные к соседним дуплетам и располо-
женные по всей длине каждой полной микротрубочки через рас-
стояния, примерно равные ее толщине. Они состоят из белка ди-
неина. Другой белок — нексин через несколько большие расстоя-
ния связывает соседние дуплеты. От каждого периферического
дуплета внутрь аксонемы отходят «радиальные спицы», заканчи-
вающиеся глобулярными образованиями, находящимися в непос-
редственной близости от «центральной капсулы». Капсула образо-
вана тонкими дуговидно загнутыми отростками центральной пары
микротрубочек (см. рис. 4.20, /).
164
Рис. 4.20. Ультратонкое строение жгутика (/) н центриоли (5). Механизм цилиарно-
го движения (2) (Албертс и др., 1986; Хаусман, 1988; Догель, 1991, с изменениями)
Базальное тельце имеет форму короткого цилиндра — продол-
жения аксонемы, но здесь к ней прибавляются девять неполных
микротрубочек, из-за чего периферийное кольцо составляется из
девяти триплетов слившихся микротрубочек. В центре цилиндра
микротрубочки с центральной капсулой отсутствуют.
Доказано, что смежные периферийные дуплеты способны
скользить друг относительно друга. Динеин в этом процессе дей-
ствует, как АТФаза. Каждая динеиновая ручка развивает пример-
но такое же усилие, как одна молекула миозина в сокращающейся
мышце. Сила прикладывается к соседнему дуплету микротрубо-
чек, с которым контактируют ручки, и стремится сдвинуть его по
направлению к вершине аксонемы (ем. рис. 4.20, 2). Считается,
что радиальные спицы и центральная капсула регулируют работу
динеиновых ручек таким образом, что получается волна движе-
ния, распространяющаяся вдоль органоида. Если бы все динеино-
вые ручки проявили активность одновременно, то аксонема свер-
нулась бы в тугую спираль. Регуляция работы ресничек и жгути-
ков, по-видимому, обеспечивается потоками ионов, но детали
этого процесса пока не изучены.
Большинство микротрубочек в клетке закреплено одним из
своих концов: аксонема — в базальном тельце, а микротрубочки
цитоскелета интерфазной клетки —в клеточном центре (откуда
они отрастают после разрушения). Поэтому базальные тельца и
центриоли называют центрами организации микротрубочек.
В клетках животного типа в середине клеточного центра почти
всегда имеется пара цилиндров диаметром около 0,1 мкм и дли-
ной около 0,3 мкм. Их называют центриолями. Так же, как и ба-
зальные тельца, центриоли образованы девятью триплетами мик-
ротрубочек, расположенными по окружности (см. рис. 4.20, 3).
В делящихся клетках растений и грибов, а также в ооцитах жи-
вотных центриоли не обнаруживаются.
Центриоли и базальные тельца совершенно одинаковы по
строению и в ряде случаев функционально взаимозаменяемы. Так,
например, у хламидомонады в начале деления жгутики исчезают,
а их базальные тельца мигрируют к ядру, где организуют построе-
ние митотического веретена, превращаясь в центриоли, а после
завершения деления перемещаются на периферию клетки, где
вновь становятся базальными тельцами, от которых «отрастают»
жгутики. Более того, в одном из экспериментов базальные тельца
хламидомонад и инфузорий-тетрахимен служили центриолями
яйцеклетки лягушки.
Центриоли почти во всех эукариотических клетках расположе-
ны перпендикулярно друг другу, и до начала деления они удваива-
ются. Дочерние центриоли образуют полюсы митотического вере-
тена. Каждая из новообразованных центриолей состоит из девяти
166
одиночных микротрубочек, а остальные, неполные, трубочки
триплетов достраиваются несколько позже.
В клетках с множеством ресничек или жгутиков базальные
тельца представляют собой потомство центриолей, которые обра-
зуют необходимое множество предшественников, распределяю-
щихся под мембраной, где они дозревают и инициируют рост ак-
сонем. Существует гипотеза, согласно которой центриоли являют-
ся саморегулирующимися органоидами, имеющими собственный
геном.
Интерфазное ядро. Ядро — самый крупный из органоидов клет-
ки. Его диаметр обычно колеблется в пределах от 1 мкм до 1 мм.
Большинство эукариотических клеток имеет одно ядро. Изредка
встречаются двуядерные и многоядерные клетки. Многоядерность
не всегда имеет одинаковое происхождение: в одних случаях это
нормальное состояние интерфазных клеток, где число ядер может
достигать нескольких десятков; в других случаях число ядер уве-
личивается в течение индивидуальной жизни клетки. Внешнее
сходство с многоядерными клетками имеют ядерно-цитоплазма-
тические комплексы (плазмодии, симпласты и синцитии), образу-
ющиеся в результате незавершенного деления или слияния от-
дельных одноядерных клеток. Обычно ядра многоядерных клеток
однообразны по строению и функциям. Но у некоторых простей-
ших наблюдается так называемый ядерный дуализм — разделение
ядер по размерам, хромосомному набору, функциям (у некоторых
инфузорий, фораминифер и др.).
Форма ядер чаще всего округлая, овальная или линзовидная.
Реже встречаются ядра лопастные, четковидные, другой формы.
Ядра способны изменять свою форму и внутреннюю структуру в
зависимости от состояния клетки. В большинстве клеток ядра пе-
ремещаются с токами цитоплазмы, но иногда они совершают са-
мостоятельные амебоидные движения.
Типичное ядро окружено двухмембранной ядерной оболочкой,
пронизанной порами, на краях которых наружная мембрана пере-
ходит во внутреннюю. Содержимое неделящегося ядра включает
кариоплазму с оформленными элементами: хроматином, перихро-
матиновыми фибриллами (толщиной 3...5 нм), перихроматиновы-
ми гранулами (40...50 нм), интерхроматиновыми гранулами
(20...25 нм) и функциональной областью — ядрышком (рис. 4.21).
Ядерные мембраны разделены перинуклеарным простран-
ством-полостью шириной 20...40 нм. Наружная мембрана со-
ставляет единое целое с эндоплазматической сетью, и ее внешняя
поверхность часто усеяна рибосомами. Поскольку перинуклеар-
ное пространство сообщается с полостью ЭПС, его можно рас-
сматривать как специализированный отдел эндоплазматической
сети. Хотя наружная и внутренняя мембраны являются продолже-
167
Рис. 4.21. Интерфазное ядро (оригинал автора):
1— ядерная пора; 2— ядерная пластинка
нием друг друга, они сильно отличаются по биохимическим свой-
ствам и функциям. Связь ядерной мембраны с мембраной ЭПС
позволяет ей быстро увеличивать и сокращать свою площадь.
К внутренней мембране со стороны нуклеоплазмы примыкает
слой, который получил название «фибриллярная ламина» или
«ядерная пластинка». Этот слой принимает участие в формирова-
нии ядерной оболочки и в организации ниже расположенного
хроматина (см. рис. 4.21). Прикрепление хроматиновых фибрилл
к ядерной оболочке обеспечивает возможность их дальнейшей
упаковки и блокирует репликацию ДНК в области будущей цент-
ромеры.
Транспорт веществ через ядерную оболочку осуществляется че-
рез ядерные поры. Поры окружены большими кольцевыми струк-
турами (с внутренним диаметром около 80 нм) — ядерными поро-
выми комплексами. Оболочка ядра типичной животной клетки
содержит несколько тысяч поровых комплексов, расположенных
очень упорядоченно: рядами или гексагоналями. Расчеты показы-
вают, что ежеминутно через канал каждого порового комплекса
проходит около 100 белковых молекул, до трех рибосомных час-
тиц и множество мелких молекул. Однако проницаемость пор для
разных веществ сильно отличается, и транспорт молекул через по-
ровые комплексы носит организованный характер.
Благодаря избирательной проницаемости двойная мембрана
изолирует «центральные» химические процессы, связанные с
репликацией ДНК и синтезом РНК от цитоплазмы, где реализу-
ется наследственная информация. Известно, что у прокариот та-
кого разделения этапов реализации генетического кода нет. И,
следовательно, эукариоты должны обладать неким преимуще-
ством, разделяя в пространстве и во времени транскрипцию
(ДНК РНК) и трансляцию (РНК -> белок). У прокариот синтез
белка начинается еще до того, как новосинтезируемая мРНК от-
соединяется от ДНК. Поэтому она транслируется полностью и без
168
изменений. В эукариотических клетках появляется возможность
«откорректировать» мРНК до выхода ее в цитоплазму, пока она не
связалась с рибосомными частицами. Этот дополнительный этап
между транскрипцией и трансляцией назвали «процессинг» или
созревание мРНК. Различают несколько видов процессинга, по-
зволяющих не только производить необходимую корректировку
матриц, но и оптимизировать форму записи генетического кода.
ДНК — это очень длинный неразветвленный линейный поли-
мер, содержащий у эукариотических клеток от 107 до 10п нуклео-
тидов. Каждый миллион нуклеотидов умещается на отрезке пря-
мой длиной 0,034 см и занимает объем Ю-15 см3. Так, гаплоидный
геном клетки человека содержит 3 • 109 нуклеотидов, а значит,
имеет протяженность более 13 см и может быть упакован в куб с
ребром 1,5 мкм.
У большинства организмов только очень небольшая часть всей
ДНК кодирует белки и важнейшие нетранслируемые РНК. Эту
ДНК называют кодирующей. Среди некодирующих участков име-
ются такие, которые обеспечивают компактную укладку ДНК в
составе хромосом. Компактная укладка ДНК важна для эукарио-
тических клеток, поскольку позволяет расположить очень длин-
ные молекулы в небольшом объеме клеточного ядра, а также кон-
тролировать функциональное состояние (активность) генов.
Центральную роль в структурной организации ядерной ДНК
играют специфические белки: гистоновой и негистоновой групп.
Гистоны — структурные белки, и по сравнению с остальными
белками их количество в клетке очень велико — почти равно по
массе ДНК, содержащейся в ядре. Считается, что вся ядерная
ДНК ассоциирована с гистонами, и этот нуклеопротеиновый
комплекс получил название хроматин. Можно сказать также, что
хроматин — это дисперсное (иногда говорят «диффузное») со-
стояние хромосом в интерфазе клеточного цикла. Тогда в хро-
матин включаются и негистоновые белки, а процентное соотно-
шение — ДНК: гистоны : негистоновые белки лежит в пределах
30...45:30...50:4...33 (в среднем 40:40 : 20).
Белки обеспечивают высокоорганизованную многоуровневую
упаковку ДНК, позволяющую в любой момент освободить, раз-
блокировать необходимые участки-гены. Полный перечень уров-
ней упаковки включает:
двойную спираль ДНК;
нуклеосомную нить (нуклеосомы — структурные единицы хро-
матина: участки ДНК длиной около 200 пар оснований, уложен-
ные на дисковидные гистоновые частицы диаметром около 11 нм,
каждая из которых состоит из 8 молекул белка);
хроматиновую фибриллу (парноспиральная упаковка нуклео-
сом в нитевидную структуру диаметром около 30 нм). В хромати-
169
новую фибриллу регулярно встроены негистоновые белки, спо-
собные сближаться, изгибая хроматиновую фибриллу в петли,
средняя длина которых независимо от видовой принадлежности
составляет около 400 нм);
«петельную фибриллу». Негистоновые белки в сближенном со-
стоянии образуют «осевой скелет» — диффузную фибриллу, в сто-
роны от которой отходят петли хроматиновой фибриллы. Такая
структура особенно хорошо наблюдается на хромосомах типа
«ламповых щеток», характерных для ооцитов, и политенных хро-
мосом насекомых. На петельном уровне ДНК еще достаточно лег-
ко освобождается от «упаковывающих» ее белков и на соответ-
ствующих участках становится возможной транскрипция.
Перечисленные четыре уровня упаковки хроматина имеют мес-
то в интерфазном ядре и называются эухроматином. Однако не
весь хроматин ядра находится в диффузном состоянии. Отдельные
его участки плотно упакованы — конденсированы и не активиру-
ются на протяжении всего клеточного цикла. Они получили на-
звание гетерохроматин. Протяженность гетерохроматиновых уча-
стков может быть различной. Такие инактивированные фрагмен-
ты ДНК сосредоточены в центромерах, теломерах, ядрышковых
организаторах. Аналогичную плотную упаковку имеет и одна из
Х-хромосом у самок млекопитающих.
В начале митоза происходит еще более плотная упаковка хро-
матина, прежде всего эухроматина: формируется еще один уро-
вень суперспирализации, при котором «петельная фибрилла»
скручивается в спираль, а сами петли сжимаются в компактные
комочки. Так образуются метафазные хромосомы, видимые в све-
товой микроскоп. Но еще до начала клеточного деления, в конце
интерфазы, хроматин полностью деспирализуется для обеспече-
ния репликации ДЖ—образования дочерних двойных цепей —
основы хроматид, из которых впоследствии формируются хромо-
сомы дочерних клеток.
Полимеризация нуклеотидов дочерних цепочек идет на обеих
антипараллельных цепях спирали в направлении от 5'-конца к
З'-концу, благодаря чему весь синтез осуществляется при учас-
тии одного фермента — самокорректирующей ДНК-полимера-
зы. Из-за антипараллельности цепей одна из них (ведущая)
синтезируется непрерывно, вторая (отстающая) синтезируется в
виде коротких фрагментов в противоположном направлении
(рис. 4.22).
Репликация ДНК начинается во многих точках одновременно
и от этих точек идет в обе стороны, что позволяет довольно быст-
ро построить дочерние хроматиды. В краткой записи репликация
включает: освобождение ДНК от упаковочных белков, разрыв
комплементарных связей между нуклеотидами двух цепочек, раз-
170
3’
3’
5'
Новосинтезированный фрагмент на «ведущей» цепи
ДНК-полимераза
ДНК-топоизомераза
РНК-затравки
Новосинтезированный фрагмент
на «отстающей» цепи
ДНК-полимераза
3'
5'
ДНК-геликаза
Дестабилизирующие
белки
РНК-проймаза
Рис. 4.22. Схема репликации ДНК (оригинал автора)
Матрицы
ведение цепочек, удержание их в разведенном состоянии, устра-
нение суперспирализации, синтез дочерних цепочек ДНК, снаб-
жение «упаковочными» белками одной или обеих цепей, форми-
рование нуклеосомной нити. Процесс репликации обеспечивает
целая серия белков: ДНК-полимераза и РНК-праймаза катализи-
руют полимеризацию нуклеотидтрифосфатов; ДНК-геликаза и де-
стабилизирующие белки расплетают и удерживают спираль; ДНК-
лигаза и фермент, разрушающий РНК-затравки, сшивают фраг-
менты Оказаки; ДНК-топоизомераза устраняет суперспираль пе-
ред репликационной вилкой; инициаторные белки образуют
новые репликационные вилки.
Ядрышко. Хроматин интерфазного ядра имеет несколько круп-
ных петель. ДНК в них содержит многочисленные повторяющие-
ся гены рибосомных РНК, которые транскрибируются с необы-
чайно высокой скоростью. Эти петли носят название «ядрышко-
вые организаторы» и занимают одну или несколько более или ме-
нее компактные области — ядрышко (или ядрышки). Ядрышко —
это не отдельный органоид, а диффузная структура, в которой
происходит сборка рибосомных частиц (в виде прерибосомных
гранул). Рибосомные белки доставляются сюда из цитоплазмы,
где синтезируется большинство клеточных белков. Почти готовые
рибосомные частицы покидают ядро через поры. В цитоплазме
происходит их окончательное «дозревание», когда они приобрета-
ют способность связываться с мРНК.
Размер ядрышка варьирует в разных клетках и в зависимости от
степени его функциональной активности. Иногда оно может за-
нимать до 25 % всего объема ядра. Основными химическими ком-
понентами ядрышка считают ДНК ядрышкового организатора,
предшественники рибосомных частиц — прерибосомные гранулы
и РНК-транскрипты.
171
Митотическое ядро. Как известно, процесс деления клетки ус-
ловно подразделяется на шесть этапов: профазу, прометафазу, ме-
тафазу, анафазу, телофазу и цитокинез. Последняя из них обозна-
чает разделение цитоплазмы, а остальные описывают последова-
тельные события, происходящие в ядре. Строго говоря, ядро в ха-
рактерном виде присутствует только в профазе и цитокинезе.
Поэтому митотическим ядром следует называть профазное ядро.
На первом этапе митоза еще сохраняется ядерная оболочка, а
хроматин завершает конденсацию, приобретая вид компактных
хромосом, каждая из которых состоит из двух дочерних хроматид,
соединенных в области центромеры и прикрепленных здесь по-
средством ядерной пластинки к внутренней мембране ядерной
оболочки. На этом участке вплоть до начала анафазы блокируется
завершение репликации ДНК, что обеспечивает правильное рас-
хождение хроматид в анафазе. К концу профазы каждая хромосо-
ма достигает наибольшей степени конденсации (упаковки).
У большинства простейших митоз совершается в неразрушаю-
щемся ядре и в этом случае митотическим можно называть ядро
на любом этапе деления. Иногда микротрубочки митотического
веретена проникают через ядро, в котором формируются специ-
альные каналы, из-за чего форма ядра резко усложняется. В то же
время хромосомы имеют типичный вид (рис. 4.23). Такой «скры-
тый» митоз (эндомитоз) завершается распределением хроматид по
полусферам ядра и последующим его разделением на дочерние
ядра экваториальной перетяжкой.
Число хромосом в клетках эукариот сильно варьирует — от двух
до нескольких сотен. Каждый вид обладает характерным и, как пра-
вило, постоянным набором хромосом в клетке — кариотипом. Ви-
доспецифично не только число хромосом, но также их размер, фор-
ма, положение на них центромеры и ядрышкового организатора.
Хроматиды профазных хромосом на следующих этапах разде-
ляются и, собираясь у полюсов веретена деления, образуют дочер-
ние кариотипы, лишь незначительно отличающиеся или практи-
Рис. 4.23. Схема протекания эндомитоза (без разрушения ядерной оболочки)
(оригинал автора)
172
чески не отличающиеся друг от друга и от материнского кариоти-
па. Точно, до мельчайших деталей организованный процесс рас-
пределения хроматид в дочерние ядра служит механизмом реали-
зации наследственности — способности к передаче признаков от
родительского поколения к дочернему.
Пластиды. К органоидам, характерным только для растительных
клеток, относят пластиды: хлоропласты, хромопласты, лейкоплас-
ты, этиопласты, амилопласты и другие, способные к взаимопревра-
щениям. Все они окружены оболочкой, состоящей из двух мембран.
Внутри пластид различают более или менее однородное вещество-
строму и мембранную систему. Между собой пластиды отличаются
прежде всего по содержащимся в них пигментам.
Хлоропласты — пластиды, в которых протекает фотосинтез.
Обычно они дисковидные, реже — овальной, чашевидной или
иной формы. Диаметр этого органоида в среднем составляет
4...5 мкм. В одной растительной клетке может помещаться до не-
скольких десятков хлоропластов, в то время как у водорослей и
автотрофных простейших хлоропласт чаще один.
Строма хлоропласта пронизана густой сетью мембран. Часть
мембран образует плоские пузырьки, уложенные стопками. Это
тилакоиды, собранные в граны. Все тилакоиды отдельного хло-
ропласта объединены посредством тилакоидов стромы, или меж-
гранных тилакоидов (рис. 4.24, 4.25).
Пигменты, характерные для этих пластид, — хлорофиллы и ка-
ротиноиды — встроены в мембраны тилакоидов гран. Иногда хло-
ропласты содержат временные запасы продуктов синтеза: крах-
мальные зерна и мелкие липидные капли.
Хлоропласты подобно прокариотам имеют собственную коль-
цевую ДНК, размещенную в нуклеоиде, и способны размножаться
независимо от готовности к делению всей клетки. В то же время и
синтез пигментов, и образование дочерних хлоропластов каким-
то образом контролируются хромосомами ядра.
Рис. 4.24. Схема строения хлоропласта (оригинал автора)
173
-Граны
•Тилакоид
Рис. 4.25. Схема развития внутренней сети мембран хлоропласта (оригинал автора)
Основные функции хлоропластов: синтез углеводов и предше-
ственников других групп органических веществ, включая амино-
кислоты и жирные кислоты, а также временное хранение продук-
тов синтеза.
Хромопласты — разнообразные по форме пигментированные
пластиды. Способны синтезировать и накапливать каротинои-
ды — пигменты желтого, оранжевого и красного цвета. Хромопла-
сты могут развиваться из хлоропластов, которые при этом теряют
хлорофиллы и внутренние мембраны. Точная функция хромопла-
стов неизвестна, но в некоторых случаях, присутствуя в клетках
цветков и плодов, они оказывают привлекающее действие на жи-
вотных с развитым цветным зрением. В значительных количествах
каротиноиды накапливаются в клетках старых листьев и часто
корней растений.
Лейкопласты — это непигментированные органоиды расти-
тельных клеток. Одна из их разновидностей — амилопласты —
синтезируют крахмал. Некоторые лейкопласты способны к обра-
зованию и накоплению белков, липидов и др. На свету лейкоплас-
ты могут превращаться в хлоропласты, развивая сложную внут-
реннюю мембранную систему.
Пропластиды- В меристематических (делящихся) клетках со-
держатся предшественники наиболее дифференцированных плас-
тид хлоропластов, хромопластов и амилопластов. Обычно они
бесцветны или имеют бледно-зеленую окраску. В темноте пропла-
стиды формируют полукристаллические скопления трубчатых
мембран и превращаются в этиопласты, которые на свету, в свою
очередь, упорядочивают мембраны в тилакоидную структуру и
дают начало хлоропластам (явление, характерное для клеток заро-
дышей семян и листьев, оказавшихся в темноте) (рис. 4.26).
Митохондрии. Подавляющее большинство эукариотических
клеток содержат митохондрии (рис. 4.27) — органоиды, где проис-
ходит аэробное окисление органических веществ со связыванием
выделяющейся энергии. Число, размеры и форма митохондрий
зависят от вида живого тела, жизненной формы клетки и ее функ-
циональной активности. Высокоактивные клетки некоторых тка-
ней могут содержать до 100 000 митохондрий. Они могут быть ок-
руглыми, овальными, палочковидными, чашевидными, изогнуты-
ми, спиральными и разветвленными. Их длина варьирует от 1,5 до
10 мкм, а ширина — от 0,2 до 1 мкм. Митохондрии растут, увели-
174
Рис. 4.26. Схема взанмонревращения пластид в растительной клетке
(оригинал автора)
Рис. 4.27. Схема строения митохондрий (оригинал автора)
чиваясь в размерах, делятся поперечной перетяжкой, могут изме-
нять свою форму, а некоторые — перемещаться в клетке.
Митохондрия образована двумя несколько отличающимися
замкнутыми мембранами. Наружная, гладкая, состоит преимуще-
ственно из фосфолипидов (85%), неспецифически проницаема
для веществ с молекулярной массой менее 10...20 тыс. Внутренняя
мембрана отделена от наружной межмембранным пространством
в 6... 10 нм. Она образует многочисленные пластинчатые, призма-
тические или трубчатые впячивания — кристы и заключает полу-
175
жидкое содержимое — матрикс. В ее составе преобладают белки —
около 70 %, а фосфолипидов только 20 %. Проницаемость этой
мембраны строго избирательна и обеспечивается белковыми ком-
плексами. Особые комплексы-транслоказы переносят через мемб-
рану АДФ и АТФ, причем АТФ синтезируется при непосредствен-
ном участии шаровидного фрагмента комплекса, выступающего
на внутренней поверхности мембраны (в сторону матрикса). В ос-
новании комплекса расположены ферменты цепи переноса элект-
ронов.
В митохондриальном матриксе содержатся ферменты, участву-
ющие в окислении пировиноградной кислоты и жирных кислот,
кольцевая ДНК, специфические мРНК и тРНК, а также рибосо-
мы прокариотного типа. Часто в матриксе присутствуют гранулы
солей кальция и магния.
4.3.3. ГЛАВНЕЙШИЕ ЭНЕРГЕТИЧЕСКИЕ ПРОЦЕССЫ В КЛЕТКЕ
Количество солнечной энергии, достигающей поверхности
Земли, по разным оценкам, составляет (5...13)1023кал/год, что эк-
вивалентно энергии примерно 100 млн атомных бомб средней
мощности. Около 0,2 % этой энергии преобразуется клетками рас-
тений и других фотосинтезирующих организмов и обеспечивает
все процессы, проходящие в живом веществе биосферы.
Связывание электромагнитной энергии происходит преимуще-
ственно на мембранах хлоропластов растений. В них встроены и
фотосинтетические пигменты, и молекулы, образующие цепи пе-
реноса электронов.
Хлорофилл и другие пигменты собраны в функциональные
единицы-комплексы, получившие название «фотосистемы». Каж-
дая фотосистема содержит 250...400 молекул пигментов. Все моле-
кулы фотосистемы способны поглощать фотоны света, но только
одна молекула хлорофилла в комплексе может использовать по-
глощенную энергию в фотохимических реакциях. Эту молекулу
называют реакционным центром, а остальные молекулы — антен-
ными, так как они улавливают, поглощают энергию попадающих
на них фотонов и передают ее на центральную молекулу хлоро-
филла. Одновременно антенные молекулы расширяют и диапазон
световых волн, участвующих в фотосинтезе (хлорофилл поглощает
свет в красной и синей частях спектра, а каротиноиды, кроме
того, — в зеленой).
Пигмент хлорофилл обладает особым свойством: когда он по-
глощает энергию фотона, один из его электронов «отрывается» от
своей молекулы и затем передается от одного переносчика к дру-
гому в электронно-транспортной цепи. На конечном участке он
176
соединяется с параллельно идущим другим электроном и ионом
водорода (протоном) из стромы хлоропласта, куда они (протоны)
выводятся из полостей тилакоидов. Хлорофилл восполняет убыль
электронов электронами, освобождающимися при фотолизе воды,
где одновременно образуются ионы водорода. При выходе прото-
нов из полости тилакоида в строму, по градиенту концентрации,
сквозь ферментный комплекс АТФазу, синтезируются молекулы
АТФ (рис. 4.28).
Протоны, ассоциированные с акцептором (НАДФ), и энергия,
запасенная в АТФ, поступают в цикл фиксации диоксида углерода
(последовательность реакций темновой фазы). Здесь акцептор —
5-углеродный сахар рибулозодифосфат связывается с диоксидом
углерода и образует нестабильную 6-углеродную молекулу, быстро
распадающуюся на две 3-углеродные молекулы. В последующем
триозофосфаты могут использоваться для синтеза 6-упперодных
моносахаридов либо служат предшественниками других групп ма-
лых органических молекул, либо вступают в длинный ряд превра-
щений, главный смысл которых заключается в образовании трех
пентоз из пяти триоз, то есть происходит воссоздание дибулозоди-
фосфата. В итоге в шестиуглеродной молекуле сахара связывается
энергия света, выталкивающая электроны из хлорофилла и вызы-
вающая фотолиз воды. Суммарное уравнение фотосинтеза (у зеле-
ных растений) можно записать в следующем виде:
световая фаза
24Н+ + 240Н + 24НАДФ + 12АДФ + 12Ф„ + энергия ->
-» 24НАДФ • Н + 12АТФ + 6О2 + 12Н2О
Рис. 4.28. Схема синтеза АТФ в хлоропластах (Албертс н др., 1986, с нзмевенмями)
12 Присный А.В
177
темновая фаза
6СО2 + 24НАДФ • Н + 12АТФ ->
-э С6Н!2О6 + 12АДФ + 12ФН + 24НАДФ
С учетом энергетических затрат на восстановление РДФ (6 мо-
лекул АТФ) синтез одной молекулы глюкозы требует энергии
18 макроэргических связей, или 252 ккал.
Не все растения используют одинаковый способ фиксации ат-
мосферного углерода. Рассмотренный вариант включает образо-
вание триоз после присоединения диоксида углерода к РДФ и
называется «С3-путь» фотосинтеза. У многих растений тропичес-
ких широт СО2 связывается с триозой фосфоенолпируватом, об-
разуя четырехуглеродную молекулу оксалоацетата, которая здесь
же, в клетках мезофилла, превращается в молекулу малата или
аспарата. На следующем этапе малат (или аспарат) декарбокси-
лируется и образует диоксид углерода и пируват. Далее диоксид
углерода связывается с РДФ, а пируват фосфорилируется и
возвращается за очередной молекулой СО2. Этот вариант, или
«С4-путь», более энергозатратен, но он не требует высокой кон-
центрации диоксида углерода, а следовательно, обеспечивает те-
чение фотосинтеза при кратковременно или слабоогкрытых ус-
тьицах, что становится выгодным в условиях высоких температур
и низкой влажности, но при мощном световом потоке. В це-
лом скорость фотосинтеза у «С4-растений» в 2...3 раза выше,
чем у «С3-растений».
Известен еще один путь фиксации диоксида углерода: «кислот-
ный метаболизм толстянковых». Он характерен большей частью
для пустынных толстянковых, кактусов и молочаев. Устьица этих
растений открываются ночью, и СО2 фиксируется в форме орга-
нических кислот, от которых отщепляется и вовлекается в обыч-
ный путь фиксации днем, при наличии солнечного света, когда
устьица закрываются в целях экономии влаги.
Ежегодно в результате фотосинтеза в биосфере образуется око-
ло 150 млрд т органических веществ и выделяется около 200 млрдт
свободного кислорода. В результате фотосинтеза сформировалась
атмосфера, предотвращающая перегрев поверхности Земли и пре-
пятствующая проникновению жесткого ультрафиолета. Однако
самое существенное явление в фотосинтезе — это улавливание
рассеянной электромагнитной энергии и превращение ее в накап-
ливаемую концентрированную форму, доступную для дальнейше-
го использования в биосфере.
Помимо зеленых растений солнечную энергию для синтетичес-
ких процессов используют и некоторые прокариоты. У цианобак-
178
терий механизм и схема фотосинтеза такие же, что и у растений:
Свет--->Хлорофили--->1
С6Н12О6
х- МФ<К ф НАД >. f Н2О
ч>АТФ J I НАД НчК ^О;
со2
Фотосинтезирующие зеленые и пурпурные бактерии использу-
ют в качестве источников водорода не воду, а сероводород или
жирные кислоты:
Свет—>БаК^й°--
хлорофилл
со2
h2s,h2r
soJ?r
Альтернативный источник энергии — окисление неорганичес-
ких соединений — используют хемоавтотрофные бактерии. Тем не
менее схема сопряжения процессов в синтезе гексоз практически
не изменяется:
Следовательно, общее уравнение фотосинтеза и хемосинтеза
может быть записано в таком виде:
СО2 + 2Н2А + энергия -> (СН2О)„ + Н2О + 2А
Одновременно с синтезом органических веществ хемоавтотро-
фы преобразуют химические и физические параметры субстрата,
на котором они поселяются и который используют в качестве ис-
точника энергии. Например, нитрифицирующие бактерии нитро-
зомонас и нитробактер последовательно окисляют аммиак до
азотной кислоты:
2NH3 + ЗО2 -> 2HNO2 + 2Н2О + 158 ккал
2HNO2 + О2 -> 2HNO3 + 38 ккал
179
12-
Серобактерии бежиатоа и тиотрикс окисляют сероводород до
серной кислоты:
2H2S + О2-> 2Н2О + 2S
2S + ЗОг + 2Н2О -> 2H2SO4 + 115 ккал
Железобактерии кренотрикс и лептотрикс окисляют двухва-
лентное железо до трехвалентного, образуя «болотную железную
руду»:
4FeCO3 + О2 + 6Н2О -> 4Fe(OH)3 + 4СО2+ 81 ккал
Высвобождение энергии из химических связей органических
веществ, прежде всего сахаров, в общих чертах напоминает обрат-
ную последовательность процессов ее связывания, где построе-
нию углевода предшествует создание градиента концентрации
протонов. Иными словами, процесс дыхания противоположен
процессу фотосинтеза и в схематическом виде он распадается на
три этапа: гликолиз, цикл лимонной кислоты и цепь переноса
электронов.
На первом этапе, который является общим и для дыхания, и
для различных типов брожения, шестиуглеродная молекула глю-
козы распадается на две трехуглеродные молекулы пировиноград-
ной кислоты с образованием четырех атомов водорода, ассоции-
рованных с их переносчиками (НАД), и четырех молекул АТФ.
В этом процессе две молекулы АТФ расходуются, и поэтому «чис-
тый» выход АТФ из гликолиза составляет две молекулы. Многие
прокариоты, грибы и некоторые беспозвоночные полностью об-
ходятся энергией, получаемой таким способом. Но это приспо-
собление развивается в связи с их существованием в бескислород-
ной среде. Такие организмы называют анаэробными.
У большинства животных, грибов и гетеротрофных клеток ра-
стений полученные на первом этапе молекулы пирувата транс-
портируются из цитоплазмы в митохондрии для дальнейшего
окисления. Аэробные бактерии используют в окислительных
процессах ферменты, расположенные на внутренней поверхнос-
ти мембран.
В митохондриях межмембранное пространство служит «резер-
вуаром протонов» аналогично полости тилакоидов хлоропластов.
Протоны накачиваются в резервуар при помощи цепи переноса
электронов. На внутренних мембранах митохондрий расположе-
ны кроме электронно-транспортной цепи ферменты цикла ли-
монной кислоты.
180
Пировиноградная кислота попадает в цепь преобразований
после отщепления диоксида углерода и соединения с кофермен-
том А (СоА) в виде ацетилкофермента А (ацетил-СоА). Последо-
вательно ее углеродные атомы отрываются от цепочки в виде ди-
оксида углерода, а водородные атомы, отщепляясь, соединяются
с акцепторами НАД и ФАД и поступают на электронно-транс-
портную цепь. В цикле лимонной кислоты образуется лишь одна
молекула АТФ; вся остальная энергия пирувата остается практи-
чески заключенной в нагруженных электронами переносчиках:
НАД - Н Н+ и ФАД Н.
На третьем этапе атомы водорода присоединяются к транс-
портной молекуле на внутренней стороне внутренней мембраны
митохондрии и переносятся на наружную ее поверхность, где пе-
редаются следующему переносчику. Но этот второй переносчик
«отнимает» для дальнейшей транспортировки электроны, а про-
тоны поступают в Н+-резервуар. Электроны направляются об-
ратно через мембрану во внутреннюю полость и там соединяются
с протонами диссоциированной воды. Так повторяется несколь-
ко раз и, в конце концов, ионы водорода, электроны и кислород
соединяются с образованием воды. Протоны из Н+-резервуара
по каналу АТФазы движутся в направлении внутренней полости,
где их концентрация ниже и при этом синтезируется АТФ
(рис. 4.29). Общий энергетический выход кислородного окисле-
Н2О (метаболическая)
J‘ Н*+ОН «Н2О
Строма
*v Межмебранное
\ пространство
‘ НГ-резервуар
Мембрана’’-.
АТФ ФН+АТФ
Рис. 4.29. Схема синтеза АТФ в митохондриях
(Албертс н др., 1986, с изменениями)
181
Рис. 4.30. Общая последовательность химического измельчения и окисления пищи
в живом теле (Албертс и др., 1986, с изменениями)
ния веществ в митохондриях значительно выше, чем при глико-
лизе.
Общую последовательность химического измельчения и окис-
ления пищи в живом теле, включая клеточное анаэробное и
аэробное дыхание, можно представить в виде схемы, приведенной
на рисунке 4.30.
Окисление основного энергетического материала — глюкозы у
гетеротрофов вдет разными путями и суммарные уравнения, сле-
довательно, имеют разный вид:
(Молочнокислые
бактерии и
высшие
животные)
(Дрожжи)
(Уксуснокислые
бактерии)
(Аэробные
бактерии и
митохондрии)
Молочная
кислота
Этиловый
спирт
Уксусный
альдегид
Диоксид
углерода
и вода
Гликолиз:
С6Н12О6 + 2 АДФ + 2ФН -> 2С3Н60з + 2АТФ + 2Н2О
Спиртовое брожение:
С6Н12О6 + 2АДФ + 2ФН -> 2СО2 + 2С2Н5ОН + 2АТФ + Н2О
Кислородный процесс:
2С3Н60з + 6О2 + 36АДФ + 36ФН -> 6СО2 + 42Н2О + 36АТФ
Полное расщепление глюкозы:
С6Н12О6 + 6О2 + 38АДФ + 38Ф„ -> 6СО2 + 44Н2О + 38АТФ
Глава 5
НАСЛЕДОВАНИЕ И РАЗВИТИЕ ПРИЗНАКОВ
5.1. ПРИЗНАКИ И ГЕНЕТИЧЕСКАЯ СИСТЕМА КЛЕТКИ
Природные живые тела — элементы живой системы — биосфе-
ры образованы живым веществом, основную массу которого, за
вычетом воды, составляют органические вещества. Наибольшим
морфологическим и функциональным разнообразием среди орга-
нических веществ отличаются белки. У автотрофов — накопите-
лей энергии, продуцирующих большинство групп органических
веществ, белки в основном составляют ферментные комплексы, а
у гетеротрофов — преобразователей накопленной энергии в «дей-
ствующую» энергию, белки — это не только ферменты, но и глав-
нейшие двигательные структуры, непосредственно использующие
высвобождаемую энершю для совершения работы или реализа-
ции отношений. Поэтому, хотя и с некоторой условностью, мож-
но ‘сказать, что строение и жизненные процессы природных жи-
вых тел определяются преимущественно белками. Таким образом,
именно белки в конечном счете определяют развитие и выраже-
ние практически всех признаков и свойств живого вещества как
целого и отдельных обособленных тел его видовых форм, через
которые живая система реализуется.
Несколько ранее мы вели речь о том, что первичная структура
белков, то есть последовательность расположения аминокислот в
полипептиде, полностью подконтрольна последовательности нук-
леотидов в дезоксирибонуклеиновых кислотах. А ДНК, как извест-
но, содержится не только в ядрах клеток, но также в пластидах и
митохондриях. Поэтому принято различать ядерную и цитоплаз-
матическую наследственность. Учитывая, что с белками связаны
конкретные проявления живых тел, которые мы можем описать,
формализовать, то есть признаки и свойства живых тел, молекулы
ДНК, собственно, и представляют собой компактную запись всех
признаков и свойств. Такое отношение можно показать направ-
ленной последовательностью: ДНК -> белокпризнак.
Известно, что число мономеров в ДНК на несколько порядков
превышает число мономеров в полипептидах. Причиной тому
служат три явления: каждая аминокислота кодируется тремя нук-
184
леотидами (триплетом); одна молекула ДНК кодирует сотни и ты-
сячи белков; только часть молекулы ДНК несет информацию о
структуре белковых молекул. Принято представлять молекулу
ДНК в виде последовательности фрагментов определенного функ-
ционального назначения — генов. Ген — это нуклеотидная после-
довательность, либо кодирующая полипептид, либо определяю-
щая транспортную или рибосомную РНК, либо необходимая для
правильной транскрипции какого-то другого гена. (Ген —это
нуклеотидная последовательность, либо определяющая одну из
РНК, либо необходимая для правильной транскрипции какого-то
другого гена.) Гены, с которых происходит считывание информа-
ции (транскрипция), называют структурными, а нетранскрибиру-
емые — регуляторными.
У прокариот структурные гены непрерывны и организованы в
группы, на которых синтезируется единая мРНК. В сочетании с
геном-регулятором, промоторным участком (куда присоединяется
РНК-полимераза) и геном-оператором (запускающим транскрип-
цию) структурные гены образуют «оперон». Оперон считается наи-
меньшей, регулируемой одним репрессором, единицей хромосомы.
В ядрах эукариот подобных генных комплексов нет, но их
структурные гены содержат транслируемые и нетранслируемые
участки. Первые названы «экзонами», а вторые — «интронами».
Экзон-интронная структура генов характерна для грибов, расте-
ний, животных и их вирусов. Структурные гены прокариот и их
вирусов интронные участки не содержат или содержат небольшое
их число.
Во второй половине XX в. представление о генетической систе-
ме получило новое развитие: были открыты перекрывающиеся
последовательности на ДНК —участки, кодирующие фрагменты
различных белков; обнаружены так называемые «прыгающие» или
мобильные гены, способные перемещаться по хромосоме, встраи-
ваясь то в один ее участок, то в другой; некоторые из повторяю-
щихся генов могут быть заблокированными на все время их суще-
ствования.
В целом генетическую организацию клетки можно представить
в виде схемы, изображенной на рисунке 5.1. Отметим лишь две ее
особенности: 1) генетическая система клетки очень неоднородна и
мобильна; 2) реальное живое тело представляет собой открытый
генетический комплекс.
Все гены, содержащиеся в генетической системе живого тела,
обозначаются понятием генотип. По одному из определений гено-
тип — это ансамбль взаимодействующих генов.
Учитывая сложность организации генетической системы живо-
го тела, лишь с крайней степенью условности можно допустить
существование простых признаков, то есть признаков, контроли-
185
Рис. 5.1. Организация генетической системы эукариотической клетки
(оригинал автора)
руемых одним геном. Правильнее говорить о существовании более
или менее сложных признаков, так как обычно каждый ген уча-
ствует в контроле нескольких признаков, а каждый признак конт-
ролируется несколькими генами.
Аналогично генотипу совокупность всех признаков и свойств
живого тела составляет его фенотип. А отдельные, дискретные, да-
лее неделимые без потери качества признаки и свойства получили
название фены. Не будет грубой ошибкой считать, что фены конт-
186
ролируются небольшими, относительно обособленными группами
генов и относятся прежде всего не к количественным, а к каче-
ственным признакам. Общепризнано, что фенотип развивается на
основе генотипа в непрерывном его взаимодействии с внешней
(по отношению к генотипу) средой. На каждый момент времени
фенотип отдельного функционирующего живого тела будет но-
вым: фактически это реализованная на текущий момент времени
программа развития. Фенотип есть реализованный генотип. Впро-
чем, если строго следовать такому подходу, оказывается, что гено-
тип, как открытый комплекс, сам непрерывно возобновляется. Но
возобновляется с «опечатками» и последующей репарацией (ис-
правлением), с новыми положениями отдельных «прыгающих» ге-
нов, с изменением активности генов, с встраиванием и выходом
различных мигрирующих фрагментов ДНК (МГДЭ — мобильные
генетические диспергированные элементы), иными словами —
развивается в непрерывном взаимодействии с внешней средой.
Следовательно, каждое новое состояние генотипа представляет
фенотип по отношению к предшествующему состоянию. Исход-
ный же генотип — это состояние комплекса генов, характерное
для начала первой интерфазы в онтогенезе живого тела.
Генотип и фенотип строго индивидуальны в своей онтогенети-
ческой динамике для каждого отдельного организма. Организмам
одного вида свойственна относительная общность и генетической
системы, и развивающихся на ее основе признаков. Их полная
идентичность условно допускается только для вегетативных по-
томков одной особи, формирующихся в одинаковых условиях сре-
ды, и в реальности невозможна. Поэтому естественные популяции
организмов полиморфны и по генотипу, и по фенотипу. Если
принять, что генотип — это получаемый (или передаваемый) ком-
плекс матриц, а динамическое состояние матриц отнести к фено-
типу, то генотипический полиморфизм выражается в разнородно-
сти индивидуальных генотипов на «входе» в популяцию, а фено-
типический полиморфизм — в разнородности фенотипов особей
популяции на каждый конкретный момент времени.
Во временной протяженности, в длительности, фенотип харак-
теризуется изменчивостью, которую принято разделять на геноти-
пическую и фенотипическую. Генотипическая изменчивость
предполагает изменения в генотипе, которые, в конце концов,
должны проявиться в фенотипе, чаще в дискретном альтернатив-
ном выражении, реже в некоторых вариантах взаимодействия ге-
нов — в альтернативно-количественном выражении. Ее подразде-
ляют на комбинативную и мутационную. Первая из них связана с
комбинированием генетических матриц в процессе мейоза и при
оплодотворении, то есть имеет отношение к собственно геноти-
пам и представляет собой нормальное явление, а вторая связана с
187
возникновением аномальных изменений в развивающемся (фено-
типическом) генотипе. С комбинативной изменчивостью связано
формирование новых сочетаний аллельных генов, резких ново-
образований в фенотипе здесь не происходит. Мутации же, буду-
чи более или менее резкими, могут проявляться или в наруше-
нии сбалансированности генетической системы, или в ненор-
мальном развитии признаков и тогда выбраковываются отбором,
или же при их нейтральности и рецессивном характере накапли-
ваться в геноме. У многоклеточных живых тел мутации накапли-
ваются прежде всего в соматической части, и тогда они могут пе-
редаваться дочернему поколению при бесполом размножении и
реже — в генеративной составляющей, откуда «перейти» в дочер-
ний генотип, как правило, не могут, поскольку в большинстве
случаев элиминируются на стадии гамет, где, не имея соответству-
ющих (парных) аллелей, проявляются в фенотипе. В целом мута-
ции правомерно трактовать как генетический шум.
Фенотипическая изменчивость основана на существовании
«широких» признаков, то есть признаков, генотипическая основа
которых предполагает возможность развития того или иного ко-
личества признака в различных внешних условиях. Поскольку ин-
тервал развития признака наследуется, утвердившееся понимание
этой формы изменчивости как ненаследственной не совсем верно.
Можно говорить о ней как об изменчивости, не передаваемой по
наследству потомкам в конкретном фенотипическом выражении.
Требуют уточнения и такие понятия, как «неопределенная» и «оп-
ределенная» изменчивость (соответствующие генотипической и
фенотипической, или индивидуальной и групповой изменчивос-
ти). Если оценивается возможность развития признака на основе
взаимодействия конкретных условий внешней среды с генотипом,
то соотношение понятий будет верным, если же на основе конк-
ретного механизма изменения генотипа, в последующем взаимо-
действующего с внешними условиями, то их соотношение будет
противоположным. Современное понимание фенотипической из-
менчивости несколько отличается от утвердившегося классичес-
кого понимания. Новое, заменяющее ее понятие «онтогенетичес-
кая», или «вариационная», изменчивость, связанное с динамикой
формирования фенотипа и отражающее выработку индивидуаль-
ных адаптаций (приспособлений) в конкретных условиях среды,
предполагает и иной механизм приобретения изменений. Здесь
ведущая роль отводится факультативному компоненту ДНК (см.
раздел 4.2.2), который трансформируется под влиянием устойчи-
вых изменений в цитоплазматических процессах и, встраиваясь в
облигатную ДНК, фактически производит перезапись внешней
информации на генетические матрицы. Данная гипотеза известна
под названием «генетический неоламаркизм», она не является об-
188
щепризнанной, но хорошо объясняет и индивидуальную изменчи-
вость фенотипов, и механизм «внезапного», или «сальтационно-
го», видообразования, и информационные процессы, протекаю-
щие в экосистемах.
В целом же изменчивость следует связывать с образованием
новых аллельных состояний генов и новых комбинаций аллелей,
но не с образованием новых генов.
Для характеристики генетической системы популяции исполь-
зуют понятие «генофонд» — вся совокупность генов (с учетом их
аллельных состояний), которые имеются у особей той или иной
популяции. Целостность генофонда популяции поддерживается
через комбинирование генетических матриц при половом воспро-
изводстве и некоторых формах бесполого воспроизводства в ряду
поколений организмов. Здесь также отметим, что часть факульта-
тивного компонента ДНК можно рассматривать как мобильную
интегрирующую часть генетических матриц, в определенной сте-
пени противостоящую дифференцирующему действию облигат-
ного компонента. Эта часть представлена свободными фрагмента-
ми вирусных и бактериальных ДНК.
При всех существующих различиях в деталях организации ге-
нетической системы все живые организмы имеют единую систему
записи генетической информации в молекулах нуклеиновых кис-
лот — генетический код. В узком смысле генетический код—зто
«словарь» триплетов мРНК (кодонов), кодирующих те или иные
аминокислоты и остановки процесса белкового синтеза. Реализа-
ция генетического кода выражается в синтезе белка в клетке по-
средством двух последовательных матричных процессов: транс-
крипции и трансляции. Направление переноса информации в
них, как правило, описывается «центральной догмой молекуляр-
ной биологии»: ДНК-> РНК -> белок. Известное нарушение «дог-
мы» — механизм воспроизводства РНК-содержащих вирусов
(РНКДНК).
5.2. РЕАЛИЗАЦИЯ ГЕНЕТИЧЕСКОГО КОДА
Транскрипцией называют процесс переноса генетической ин-
формации от ДНК к РНК, что выражается в построении разных
видов РНК (матричной, или информационной, рибосомной,
транспортной, гетерогенной) на одной из цепей ДНК, которую
называют смысловой или +-цепыо. У прокариот и эукариот
транскрипция отличается многими деталями.
В бактериальных клетках синтез РНК катализируется комплек-
сным ферментом ДНК-зависимой РНК-полимеразой, состоящим
из пяти субъединиц. Для начала транскрипции необходимо нали-
189
чие фермента, АТФ или ГТФ и специального участка ДНК, на-
зываемого промотором. Промотор содержит около 40 пар осно-
ваний и расположен непосредственно перед участком начала
транскрипции. В его составе выделяют последовательности узна-
вания (около 30 пар нуклеотидов) и связывания (около 10 пар
нуклеотидов).
Считается, что мРНК для каждого отдельного гена строится
только на одной из цепей ДНК. Следовательно, цепь считывания,
как и направление считывания, для соседних генов могут не со-
впадать. (Направление определяется по вакантным связям со сто-
роны третьего или пятого углеродных атомов сахара нуклеотида на
концах цепи.) Движение фермента РНК-полимеразы происходит
по считываемой цепи в направлении от 3'- к 5'-концу, а цепь РНК
при этом наращивается в направлении от 5'- к З'-концу. Во время
движения фермент расплетает ДНК на протяжении одного витка,
удерживает в разведенном состоянии ее цепи и выстраивает пос-
ледовательность рибонуклеотидов, комплементарную последова-
тельности смысловой цепи. Связь новосинтезированной цепи с
матрицей менее стабильна, чем связь двух соответствующих цепей
ДНК; поэтому происходит вытеснение РНК из репликационной
вилки при восстановлении двойной спирали за проходящим фер-
ментом. Уже после связывания первых восьми рибонуклеотидов
от фермента-комплекса отсоединяется одна из субъединиц, и с
этого момента транскрипция переходит из стадии инициации (на-
чала синтеза) в стадию элонгации (удлинения цепи). Терминация
(прекращение роста цепи) происходит на особых участках ДНК,
где содержатся обращенные последовательности нуклеотидов:
преимущественно гуанозина и цитозина. Транскрипт с такого уча-
стка самопроизвольно изгибается с образованием «шпильки»,
препятствующей дальнейшему росту цепи (рис. 5.2).
У эукариот транскрипцию обеспечивают три ДНК-зависимые
РНК-полимеразы: 1-я катализирует синтез трех рРНК; 2-я —
Промотор (старт-сигнал для РНК-полимеразы)
Обращенные последо-
вательности (стоп-сигнал
для РНК-полимеразы)
Фактор узнавания
РНК-полимераза (а,а,Э, Р’, а — субъединицы) Освободившийся фактор узнавания
Рис. 5.2. Транскрипция (оригинал автора)
190
мРНК; 3-я — одну из рРНК и тРНК. В инициации, элонгации и
терминации принимают участие и другие ферменты.
Первичные мРНК-транскрипты часто представляют собой бло-
ки, состоящие из нескольких функциональных молекул, они мо-
гут содержать нетранслируемые вставки, лишние концевые пос-
ледовательности или нуждаться в присоединении дополнитель-
ных фрагментов. Поэтому новосинтезированные молекулы под-
вергаются посттранскрипционному процессингу («созреванию»),
при котором с помощью ферментов осуществляются необходимые
модификации: разрезания, удаления, присоединения и др. В про-
цессинге особое место занимает самостоятельное, без участия бел-
ков-ферментов вычленение не кодирующих белки последователь-
ностей (интронов) и сращивание {сплайсинг, или сплэйсинг) оста-
ющихся кодирующих фрагментов (экзонов). Сплайсинг позволяет
сформировать разные мРНК из одинаковых первичных транс-
криптов. В дополнение к этому созревающие мРНК эукариот до
выхода из ядра получают ковалентно связываемые: с 5'-кон-
цом — «шапочку», образованную тремя фосфатами и метили-
рованным нуклеотидом, а с З'-концом — «хвост», состоящий из
100...200 адениловых нуклеотидов. Считается, что по «шапочке»
опознается передний конец мРНК при прохождении ею ядерной
поры и при связывании с рибосомными субчастицами, а «хвост»
стабилизирует концевую последовательность кодирующих трип-
летов нуклеотидов.
Трансляция — это перевод последовательности оснований
мРНК в последовательность аминокислот полипептида. Для реа-
лизации кода, то есть построения белковой молекулы соответ-
ственно последовательности нуклеотидов, существует система де-
кодирования — так называемый «словарь кода». Четыре нуклеоти-
да ДНК и четыре комплементарных им нуклеотида мРНК, груп-
пируясь в триплеты, дают 64 варианта сочетаний. Известно, что
три сочетания не кодируют аминокислоты и являются терминиру-
ющими, то есть останавливающими синтез полипептида, а 61 —
кодируют 20 аминокислот, включаемых в полипептиды. При не-
подвижной «рамке считывания» код не перекрывается, а на от-
дельные аминокислоты приходится от одного до шести кодовых
сочетаний нуклеотидов (табл. 5.1).
Принято считать, что избыточное кодирование представляет
собой «вырожденное» состояние кода, то есть отставание формы
записи информации от ее смысла, что, впрочем, может иметь и
другое объяснение. Если исходить в объяснении избыточного ко-
дирования из принципа минимальной достаточности и допустить,
что 20 аминокислот обеспечивают всю необходимую полноту
свойств белков, то для их кодирования оптимальным вариантом
было бы построение триплетов из трех различных нуклеотидов
191
(27 сочетаний). Зная, что комплементарное спаривание происхо-
дит между определенными пуринами и пиримидинами, мы смог-
ли бы подобрать для построения ДНК либо два, либо четыре ос-
нования, либо же три, но с условием, что один из них способен
образовывать пары с двумя другими, а это равноценно двум ком-
плементарным основаниям. Мы, следовательно, вынуждены со-
гласиться с минимальной достаточностью именно четырех нукле-
отидов и неизбежностью избыточного кодирования. Одного
взгляда на приведенную таблицу достаточно, чтобы заметить: ко-
дирование идет преимущественно по первым двум «буквам-осно-
ваниям» и реже — по третьей. В механизме посредничества тРНК
в сборке белковой молекулы на этот случай заложено нестрогое
соответствие третьего основания кодона (триплета на мРНК) пер-
вому основанию антикодона тРНК. Тем более, это первое поло-
жение часто занимает инозин, способный образовывать водород-
ные связи и с аденозином, и с цитидином, и с уридином.
5.1. Кодирование аминокислот триолетами нуклеотидов мРНК
Первое | Второе положение Третье
(5‘- конец) ’ У Ц 1 1 А 1 Г положение (У- конец)
У УУУ-Фен УЦУ-Сер , УАУ-Тир УГУ-Гис У
УУЦ-Фен УЦЦ-Сер УАЦ-Тир УГЦ-Гис Ц
УУА-Лей УЦА-Сер УАА-Терм УГА-Терм А
УУГ-Лей УЦГ-Сер УАГ-Терм УГГ-Три Г
Ц ЦУУ-Лей ЦЦУ-Про ЦАУ-Гис ЦГУ-Арг У
ЦУЦ-Лей ЦЦЦ-Про 1(АЦ-Гис ЦГЦ-Арг Ц
ЦУА-Лей ЦЦА-Про ЦАА-Гли ЦГА-Арг А
ЦУГ-Лей ЦЦГ-Про ЦАГ-Гли ЦГГ-Арг Г
А АУУ-Илей АЦУ-Три ААУ-Асп АГУ-Сер У
АУЦ-Илей АЦЦ-Три ААЦ-Асп АГЦ-Сер Ц
АУА-Илей АЦА-Три ААА-Лиз АГА-Арг А
АУГ-Мет АЦГ-Три А АГ-Лиз АГГ-Арг Г
Г ГУУ-Вал ГЦУ-Ала ГАУ-Асп ГГУ-Гли У
ГУЦ-Вал ГЦЦ-Ала ГАЦ-Асп ГГЦ-Гли Ц
ГУА-Вал ПДА-Ала ГАА-Глу ГГА-Гли А
ГУГ-Вал ГЦГ-Ала ГАГ-Глу ПТ-Гли Г
Конформация транспортной РНК весьма сложна из-за нали-
чия нескольких обращенных последовательностей нуклеотидов и
их комплементарного спаривания. Она получила название «три-
листник», но напоминает его лишь отдаленно на плоском изобра-
жении (рис. 5.3). Функционально важными ее частями являются:
акцепторный З'-конец, который при помощи фермента амино-
ацил-тРНК-синтетазы и энергии двух связей АТФ присоединяет-
ся к аминокислоте и образует аминоацил-тРНК с активирован-
ным аминокислотным остатком; антикодоновый участок на соот-
ветствующей петле с триплетом, комплементарным триплету
192
Рис. 5.3. Строение тРНК (Албертс и др., 1986, с изменениями)
мРНК, который кодирует аминокислоту, связанную с данной
тРНК (каждая аминокислота имеет свою или свои тРНК).
Синтез белка начинается с того момента, когда образуется ком-
плекс из мРНК, субчастиц рибосомы, АТФ или ГТФ и определен-
ной аминоацил-тРНК (у прокариот — формилметионин-тРНК, у
эукариот — метионин-тРНК или валин-тРНК). Последний, соб-
ственно, и реагирует на установление в Р-связывающий участок
рибосомы сигнального кодона. В общем виде процесс синтеза по-
липептидной цепи сводится к тому, что к активируемому триплету
мРНК в A-участке связывания присоединяется антикодон тРНК,
связанной с аминокислотой. Аминокислота при этом устанавли-
вается рядом с другой, удерживаемой в P-участке связывания
тРНК. Один из белков рибосомы, расположенный между обозна-
ченными аминокислотами, — фермент пептидилтрансфераза, ка-
тализирует образование пептидной связи. Одновременно связь
аминокислоты с тРНК в P-участке разрывается и рибосома пере-
мещается на три нуклеотида по информационной РНК в направ-
лении 5' —> 3'. Такое смещение приводит к тому, что свободная
тРНК из P-участка «выталкивается» транспортной РНК, удержи-
вающей аминокислотную цепь, а A-участок освобождается для
приема следующей аминоацил-РНК. При активировании в А-уча-
стке терминирующего кодона синтез белка обрывается. В это вре-
мя «фактор освобождения белка» связывает последнюю амино-
кислоту с гидроксилом воды и освобождает полипептид от тРНК в
P-участке рибосомы (рис. 5.4).
Среди рибосом, как это уже отмечалось, имеются свободные и
мембранно-связанные. Первые взвешены в цитозоле, а вторые об-
разуют шероховатые области ЭПС. Специальные исследования
показали, что различий между рибосомами двух типов нет и каж-
дая рибосома в разные промежутки времени может работать то в
свободном, то в прикрепленном состоянии. Положение рибосомы
зависит от белка, синтез которого начат: если это белок секрета-
13 Присный А.В.
193
Рис. 5.4. Трансляция (оригинал ангора)
руемый, то первые его аминокислоты составляют «сигнальную»
последовательность, или «лидерный пептид». С сигнальной пос-
ледовательностью связывается «белокузнающий сигнал» и блоки-
рует продолжение синтеза до тех пор, пока мРНК с рибосомой и
новосинтезированным фрагментом белка не окажутся прикреп-
ленными этим узнающим белком к мембранному белку-рецепто-
ру. Осевший на мембрану комплекс вновь начинает работать, ког-
да сигнальный пептид, пройдя через канал, образованный рецеп-
торным белком, отщепляется ферментом (уже в полости ЭПС).
Длительность жизни белков в среднем составляет около 2 сут,
хотя отдельные белки не разрушаются в течение нескольких меся-
цев. Концентрация белковых молекул в клетке определяется отно-
шением скоростей их синтеза и разрушения.
5.3. МОРФОФУНКЦИОНАЛЬНЫЕ ТИПЫ КЛЕТОК
S.3.1. ПРЕДСТАВЛЕНИЕ О ЖИЗНЕННОЙ ФОРМЕ
В современной биологической литературе в качестве синони-
мов употребляются такие понятия, как «жизненная форма», «био-
морфа», «экобиоморфа» и «морфоэкологический тип».
В ботанике под жизненной формой понимают внешний облик
растения, отражающий его приспособленность к условиям среды,
или же группу растений со сходными приспособительными при»
знаками как единицу экологической классификации. Например,
194
деревья, кустарники, кустарнички, полукустарники, полукустар-
нички, наземные и водные травы и др. — это один из вариантов
подобной классификации.
Применительно к животным жизненной формой называют
группу особей, имеющих сходные морфоэкологические приспо-
собления. В основу классификации жизненных форм животных
могут быть положены способы передвижения и добывания пищи,
характер пищи, степень активности, приуроченность к определен-
ной среде обитания. И в этом случае различные стадии онтогене-
за, половые формы, касты и другие отличающиеся по строению и
функционально особи одного вида могут представлять разные
жизненные формы.
Экобиоморфа по определению — тип приспособительной струк-
туры растений и связанных с ней физиологических особенностей,
характеризующий группу растений, обычно обитающих в сходных
условиях среды. Принято считать, что экобиоморфа включает до-
полнительный по отношению к жизненной форме параметр —
физиологический аспект.
Как было показано в разделе, посвященном организации жи-
вых систем и элементов систем — живых тел, строение и функ-
ция<—это их стороны, взаимосвязанные, взаимоопределяющие,
не существующие отдельно друг от друга и одновременно связыва-
ющие элемент с системой. Поэтому говорить о внешнем облике
живого тела уже само по себе означает — связывать процессы,
проходящие в живом теле, с системными процессами, в которых
тело участвует, занимая ту или иную пространственную и функци-
ональную нишу (то есть экологическую нишу). В таком случае
можно было бы принять за основное значение понятия «жизнен-
ная форма» выражение единства строения, функции и условий,
или морфофункциональный тип. Тогда оно и характеризовало бы
различающиеся фазы онтогенеза, кастовые группы общественных
организмов и совпадало бы по значению с понятиями-синонима-
ми (экоморфы, экобиоморфы и морфоэкологические типы).
Морфофункциональные типы обнаруживаются на разных
уровнях организации живых тел и в разных пространственных и
временных подразделениях: на клеточном, на уровне отдельных
многоклеточных особей, стадий их онтогенеза и отдельных функ-
циональных частей, целых организмов.
5.3.2. ЖИЗНЕННЫЕ ФОРМЫ КЛЕТОК
Все свободноживущие клетки-особи, а также клетки, составля-
ющие тела колоний, талломов и многоклеточные тела, могут быть
отнесены к какой-либо жизненной форме. Именно на этом уров-
13*
195
не морфофункциональные типы выражены особенно четко. Один
из вариантов подобного разделения клеток мы уже обсуждали,
когда обозначили главнейшие трофические типы живых тел: фо-
тоавтотрофы, фотогетеротрофы, хемоавтотрофы и хемогетеротро-
фы. Возможность эффективного выполнения функций зависит от
сочетания характера источников вещества и энергии, их доступ-
ности и характера результирующих процессов, протекающих в
клетке со специальными приспособительными структурами, ори-
ентированными на взаимодействие с внешней средой. Поэтому
отражением функциональной ниши клетки будут ее внешнее и
внутреннее строение, особенность течения физиологических про-
цессов.
К числу узловых характеристик клетки, которые могут быть по-
ложены в основу разделения отдельных клеток или их фазовых
состояний на морфофункциональные типы, относятся: характер
движения (пассивное или активное, свободное или прикреплен-
ное, прорастание или перемещение); характер питания, или спо-
соб импорта веществ (диффузия и осмос, активный трансмемб-
ранный перенос, фаго- и пиноцитоз, включая образование окайм-
ленных пузырьков); преобладающие энергетические процессы
(накопление или освобождение энергии); преобладающие пласти-
ческие процессы (синтез или распад веществ); главнейшие экс-
портируемые вещества и способ экспорта (секреты, инкреты, экс-
креты, вещества надмембранных образований, белки, липиды, уг-
леводы, минеральные вещества, диффузия, активный транспорт,
мембранный транспорт); форма высвобождения энергии (выделе-
ние тепла, совершение механической работы, электрический раз-
ряд, свечение, выделение химически активных веществ; тип воз-
будимости (мембранная, органоидная, цитоплазматическая, ци-
тоскелетная). Сочетание узловых характеристик дает большое, но
ограниченное число морфофункциональных типов из-за «запре-
та» на некоторые комбинации.
Одиночные клетки вне водной среды встречаются в ценотичес-
ки пассивном, покоящемся состоянии. Для них характерны округ-
лая или овальная форма, отсутствие поверхностных цитоплазма-
тических выростов, почти полное прекращение пластических и
энергетических процессов и обмена с внешней средой, развитие
мощной защитной оболочки. Такая жизненная форма типична и
для прокариотических, и для свободноживущих эукариотических
клеток.
Для функционирующих пассивно подвижных клеток основным
условием существования должно быть равномерное распределе-
ние в среде источников вещества и энергии с регулярным возоб-
новлением ресурсов при перемешивании среды. Фотоавтотрофы
из этой группы часто способны регулировать плавучесть, накапли-
196
вая пузырьки газа. Обычно они имеют сферическую или овальную
форму. Их оболочки более или менее уплотнены, но проницаемы
для воды и газов. Увеличение площади поверхности в необходи-
мых случаях достигается развитием постоянных выростов покро-
вов или минерального скелета. Данная жизненная форма встреча-
ется у различных прокариот и эукариот. Близка к ней организация
фотоавтотрофных клеток, ведущих прикрепленный образ жизни.
Таковы, например, цианеи и некоторые водоросли с клейкими ос-
лизненными оболочками.
Клеткам тел автотрофов-прорастателей свойственно активное
(ростовое) движение, так как проникновение в глубь используе-
мого субстрата сопряжено с затратами энергии и вещества на пре-
одоление постоянных и регулярно действующих сил. Они также
могут быть свободными или прикрепленными, но для них особен-
но характерна способность к образованию фибрилл из полимер-
ных углеводов, откладываемых над клеточной мембраной в виде
оболочки. Общее направление обменных процессов, как и у двух
предыдущих жизненных форм, — связывание энергии в синтези-
руемых органических молекулах. Целлюлозные фибриллы и близ-
кие к ним полимеры выделяются на поверхность мембраны без
посредства вакуолей.
Сходную жизненную форму образуют и некоторые хемогете-
ротрофы, например грибы, также образующие прочную оболоч-
ку, но не из целлюлозы, а из хитина. Однако направленность
процессов у них иная: преобладает расщепление энергонасы-
щенных органических веществ с высвобождением энергии в виде
тепла.
Ряд жизненных форм клеток связан с различными формами ак-
тивного перемещения в пространстве. Для части одноклеточных —
это прямое перемещение, как у активно подвижных, а для другой
части — прикрепленных — это обращенное перемещение (переме-
щение среды относительно тела). У одноклеточных автотрофов из-
вестно два способа перемещения, связанного с выбором оптималь-
ных условий: созданием бегущих волн на участках поверхности тела
(как у диатомовых водорослей) и цилиарный механизм, то есть с
помощью жгутиков (каку растительных жгутиконосцев, многих во-
дорослей). У гетеротрофов перемещение, как прямое, так и обра-
щенное, должно обеспечивать сбор пищевых частиц или выбор сре-
ды с более концентрированным раствором органики. Сбор будет
тем эффективнее, чем болыпе (до определенных пределов) разница
в скоростях движения хемогетеротрофной клетки и поглощаемых
частиц и тел. У свободно перемещающихся гетеротрофов известно
четыре способа движения: скольжение, связанное с выделением
слизи задней или боковыми частями клетки; перемещение создани-
ем бегущих микроволн на поверхности тела при упорядоченных со-
197
крашениях фибрилл кортекса (эктоплазматической фибриллярной
сети); амебоидное движение, основанное на работе актин-миози-
новых комплексов цитоскелета и фазовых превращений гиалоплаз-
мы; цилиарное движение, обеспечиваемое работой сложных комп-
лексов микротрубочек. К последнему близок способ перемещения
прокариот, снабженных жгутиковыми фибриллами. Каждый из
способов движения предполагает особую организацию покровов,
подчиненную механизмам поглощения пищи. Движение, основан-
ное на работе актиновых и миозиновых фибрилл, дает дополни-
тельные жизненные формы клеток сократимого типа, а на основе
работы микротрубочек формируются движители прикрепленных
подгонятелей и клеток, способных создавать направленный ток
жидкости. Перечисленные приспособления пригодны для тех слу-
чаев, когда оказывается энергетически выгодным «догонять» пищу
прямым или обращенным способом. Если же пища в рассеянном
виде сама совершает медленное равномерное направленное движе-
ние, то оказывается выгодным развивать жизненную форму паря-
щего или сидячего «собирателя» (как у радиолярий и солнечников)
с резко увеличенной площадью поверхности за счет постоянных
или устойчивых выростов цитоплазмы или разветвленного мине-
рального скелета.
Варианты постоянной формы клеток, обусловленной строени-
ем покровов, аналогичны у прокариот и эукариот как свободно-
живущих, так и отдельных клеток многоклеточных организмов.
Постоянство формы тела исключает амебоидный способ движе-
ния, но является основой для эффективной работы жгутиковых
фибрилл, жгутиков и ресничек. В целом можно говорить о суще-
ствовании и аналогий, и гомологии в рядах жизненных форм у це-
нотически пассивных и ценотически активных свободных клеток-
тел (прокариот и эукариот), клеток-особей и клеток-структурных
единиц тел многоклеточных организмов (рис. 5.5).
Основные жизненные формы клеток, участвующих в построе-
нии многоклеточных (с тканевой организацией) тел, формируют-
ся путем усиления одной или немногих морфофункциональных
сторон «недифференцированной» клетки. При этом важнейшими
узловыми характеристиками, определяющими тип строения цено-
тически активных тел, видимо, являются способ импорта веществ
и способ движения в сочетании со строением покровов (рис. 5.6).
Оказалось что многоклеточные (с тканевой организацией) тела
могут быть составлены только из двух типов клеток: 1) с постоян-
ной формой, обусловленной наличием целлюлозной оболочки,
диффузным импортом веществ и ростовым движением; 2) с не-
постоянной формой, везикулярным импортом и движением, ос-
нованным на работе цитоскелета. Первые образуют тела растений,
а вторые — тела животных.
198
Рис. 5.5. Жизненные формы клеток (Хаусман, 1988; оригинал автора):
/...б—прикрепленные и пассивно подвижные авто- и гетеротрофные клетки (7 —циста
Entamoeba hlstofytika, 2—инкапсулированная бактерия, 3— мегаспора растений, 4— однокле-
точная зеленая водоросль Chlorella, 5— одноклеточная цианобактерия, б— яйцеклетка рыбы);
7, 8— клетки с активным амебоидным движением (7—голая амеба, 8—мужская гамета аска-
риды); 9—15— клетки с активным цилиарным движением (9—панцирный жгутиконосец,
10—двужгутиковая бактерия, 11—типичный сперматозоид животных, 12— зооспора хлами-
домонады, 13—опалина, 14— многожгутиковая бактерия, 15— многожгутиковая гамета опа-
лины); 16, /7—активно подвижные изгибающиеся клетки (16— спирохета, 17— мужская га-
мета малярийного плазмодия); 18— радиолярия Circospatis sexfurca, «парящая» в воде
Клетки с оболочками могут быть автотрофными и гетеротроф-
ными. Автотрофные клетки, как правило, имеют тонкую оболоч-
ку, проницаемую для света и неорганических молекул, и занима-
ют поверхностное или открытое положение. При этом у парен-
химных клеток оболочка однородна по толщине, а у покровных —
толще с внешней стороны. Гетеротрофные клетки могут иметь как
тонкую, так и толстую оболочку и занимают глубинное или скры-
тое (от света) положение. Основные функциональные группы та-
ких клеток с тонкой оболочкой — резервные, запасающие и желе-
зистые, а с толстой одревесневающей оболочкой — механические
и проводящие. Толстые оболочки мертвых клеток обеспечивают
199
I Одноклеточные тела , |
|Талломные тела [ j | i| Бестканевые тел а~|
Ткане- I
вметала'
Тканевые
тела
Форма клеток
| Постоянная или ограниченно изменчивая | | Непостоянная ]
Покровы
Оболочка
Клеточная
стенка
| Пелликула j
Мембрана (с
гликокал иксом)
Движение
Ростовое П I Пассивное и цилиарное
Цитоскелетное
и цилиарное
Способ импорта веществ
Диффузия
Активный транспорт
Везикулярный
транспорт
Рис. 5.6. Признаки, определяющие тип строения ценотически активных тел
различные формы защиты тела растения при поверхностном рас-
положении и транспорт веществ при внутреннем. Если учесть, что
многоклеточность у растений — это способ освоения и заполне-
ния среды по вертикали, особенно в условиях суши, то главней-
шими классами жизненных форм их клеток следует признать ас-
симилирующие и механические.
У многоклеточных животных первичная дифференцировка
клеток основана на одностороннем усилении способности или к
транспорту веществ, или к изменению формы. У них выделяют
200
две главные исходные формы: 1) клетки с относительно постоян-
ной формой, поддерживаемой преимущественно тонкими и про-
межуточными фибриллами, способные к эндо-экзоцитозу; 2)
клетки с изменчивой формой, определяемой взаимодействием
тонких и толстых фибрилл, способные преимущественно к актив-
ному транспорту молекул через наружную мембрану. Это соответ-
ственно пищеварительные и двигательные клетки. Одновремен-
ная специализация клетки в двух указанных направлениях приво-
дит к развитию амебоидной формы.
Дальнейшая дифференцировка на жизненные формы идет в
обеих группах: пищеварительные клетки или поглощают вещества
и переваривают их внутриклеточно, или выделяют пищеваритель-
ные ферменты и поглощают продукты пищеварения; двигатель-
ные клетки специализируются или на увеличении числа независи-
мых сократимых комплексов (возможность растяжения и сокра-
щения в разных направлениях), или на развитии единого линей-
ного сократимого комплекса (сокращение в одном направлении,
значительное растяжение без повреждений невозможно).
Многоклеточные тела инициируют развитие и такой жизнен-
ной формы, как покровные клетки. Их основная особенность —
способность противодействовать химической и физической дезор-
ганизации посредством усиления транспортных свойств мембра-
ны и развития промежуточных фибрилл цитоскелета. Специали-
зация на экспорте органических молекул основаниями клеток
позволяет им формировать базальную мембрану, а вершинами —
кутикулу, что в целом усиливает их функциональную эффектив-
ность. Специализация же на транспортировке неорганических
ионов создает основу для появления «мембрановозбудимых» нерв-
ных клеток.
Приведение примеров жизненных форм клеток можно было бы
продолжить, но их полный перечень составляет несколько сотен,
а общепринятая классификация отсутствует. Нам же важно усво-
ить следующее:
1) в построении многоклеточных тел растений и животных уча-
ствуют группы разных жизненных форм клеток;
2) строение и функции клеток взаимно определяют друг друга;
3) чем сильнее выражена односторонность специализации жиз-
ненной формы какой-либо клетки в живом многоклеточном теле,
тем в большую зависимость от комплементарных жизненных
форм в данном теле она попадает;
4) в телах разных организмов могут формироваться одинаковые
жизненные формы клеток, а в одном теле —- разные.
Жизненные формы могут быть временными, обратимыми и
постоянными, а это зависит от внешних и внутренних механизмов
регуляции дифференцировки клеток в многоклеточном теле.
201
Количество жизненных форм свободноживущих клеток исчис-
ляется десятками. Несколько сотен жизненных форм насчитыва-
ют клетки многоклеточных тел. Комбинирование групп жизнен-
ных форм клеток в пределах многоклеточных тел позволяет сфор-
мировать бесконечное число жизненных форм на этом более ем-
ком уровне организации, из которых в конкретных условиях
биосферы реализовано лишь несколько тысяч. Одна из них — че-
ловек — постоянно использует преобразованное неживое веще-
ство биосферы для создания сменяемых структур — внешних уси-
лителей телесных органов, еще более увеличивая разнообразие
жизненных форм.
5.3.3. ДИФФЕРЕНЦИРОВКА КЛЕТОК
Известно, что любая клетка, хотя бы на коротком этапе инди-
видуальной жизни, имеет характерное строение, определяемое
прежде всего той функцией, которую выполняет эта клетка в эко-
системе, в теле особи или же в составе организма (рис. 5.7).
16
202
Рис. 5.7. Жизненные формы клеток многоклеточных тел (оригинал автора):
1... 1O— клетки тканей растений: 1 — покровная эпидермиса, 2— покровная пробковой ткани,
3— покровно-всасываюшая корня, 4— фотосинтезирующая паренхимы, 5—запасающая па-
ренхимы, б—механическая колленхимы, 7—проводящая сосуда, 8— проводящая трахеиды,
9— склереида склеренхимы, 10— волокно склеренхимы; 11...33— клетки тканей животных:
11—недифференцированная клетка, 12— железистая эпителия, 13—покровная призматичес-
кая эпителия, 14— покровная цилиндрическая эпителия, 15— покровная плоская эпителия,
16— покровная всасывающая эпителия, 17— клетка мускульной ленты аскариды, 18— клетки
гладкой мышечной ткани, 19— волокно поперечно-полосатой мышечной ткани, 20— фибро-
бласт соединительной ткани, 21— фиброцит соединительной ткани, 22— хондроцит соедини-
тельной ткани, 23—жировая клетка соединительной ткани, 24— гранулоцит соединительной
ткани, 25— эритроцит соединительной ткани, 26— фагоцит соединительной ткани, 27— мер-
цательная выделительная клетка, 28— глиальные клетки нервной ткани, 29— униполярный
нейрон нервной ткани, 30— мультиполярный нейрон нервной ткани, 31 — шванновская клет-
ка нервной ткани, 32— чувствительная нервная клетка органа слуха насекомого, 33— чувстви-
тельная нервная клетка органа обоняния позвоночного; 34...36— половые клетки: 34— спер-
матозоид рака, 35— сперматозоид млекопитающего, 36— яйцеклетка млекопитающего
Жизненные циклы клеток, образующих одноклеточные тела
простейших, часто называют простыми, что должно означать от-
сутствие реорганизаций в промежутке между двумя делениями
(завершением одного и началом другого). В то же время хорошо
известно, что после завершения деления и при подготовке к деле-
нию в ядре и цитоплазме клетки происходят чрезвычайно слож-
ные процессы, хотя внешне клетка может претерпевать лишь не-
значительные изменения: у нее формируются или исчезают двига-
тельные органоиды, структуры, связанные с питанием, реоргани-
зуются покровы, приобретается или утрачивается специфичная
форма и др., то есть клетка или переходит в ценотически активное
состояние, или выходит из него. Начальные изменения в интер-
фазе направлены на достижение определенной жизненной формы
для возможности реализации ценотической функции, а конеч-
ные — на обеспечение репродукции.
Можно привести многочисленные примеры, подтверждающие
существование у одноклеточных тел явления дифференцировки, то
есть перехода от менее специализированного состояния к более
специализированному, или от ценотически пассивного к ценоти-
чески активному состоянию. Очень сложные морфогенетические
процессы проходят у кокцидиеобразных споровиков (рис. 5.8).
У этих организмов в ряду поколений.каждая особь претерпевает
переход от инвазионной стадии с постоянной формой, набором
органоидов проникновения и другими особенностями к амебоид-
ной ценотически активной стадии, а часть активных клеток-осо-
бей в одном из поколений, в свою очередь, преобразуется в женс-
кую половую клетку (оогамету). Имеет место у простейших и
дифференцировка клеток на разные жизненные формы, составля-
ющие многоклеточное тело, правда, с единственным морфофунк-
циональным типом ценотически активных клеток.
Рис. 5.8. Разные жизненные формы клеток на основе одного генотипа у кокцидиеоб-
разных (тип Apicomplexa) (оригинал автора):
1—инвазионно-расселитепьная форма — спорозоит; 2—трофическая форма —шизонт; 3—
инвазионная форма—мерозоит; 4— покоящаяся форма — гамонт; 5 — неподвижная макрога-
мета; 6— подвижная микрогамета
204
Многие виды простейших образуют идентичные жизненные
формы (лишь с незначительными различиями). Одновременно от-
дельные клетки-особи — потомки одной материнской клетки или
клетки, образующиеся в последовательных митозах, с идентичны-
ми генотипами развивают разные жизненные формы (см.
рис. 5.6...5.8). Отсюда можно заключить, что генотип как целое не
является единственным и определяющим фактором направления
дифференцировки.
У паразитических жгутиконосцев — трипаносом в жизненном
цикле могут последовательно сменять друг друга четыре морфоло-
гически отличающиеся формы, связанные с разными хозяевами и
средами обитания (рис. 5.9). Это позволяет думать, что внешние
сигналы служат одним из факторов-включателей дифференциров-
ки. Но должна быть и некая программа развития той или иной
жизненной формы, предопределенность (или детерминация) на-
правления формообразования.
Детерминация развития и дифференцировка клеток изучаются
преимущественно на многоклеточных (с тканевой организацией)
животных. Получены многочисленные свидетельства комбиниро-
ванного действия ядерных генов, цитоплазмы, клеточного окру-
жения, предшествующих поколений клеток и внешних стимулов
на развитие той или иной жизненной формы клеток.
В опытах с крупной одноклеточной водорослью ацетабулярией
по пересадке безъядерной части клетки, отличающейся у разных
видов, показано, что форма «шляпки» всегда соответствует тому
из видов, которому принадлежит ядро (рис. 5.10). При пересадке в
яйцеклетки лягушки Rana pipiens на место разрушенных ядер из
клеток гаструлы (того же вида), уже проявляющих признаки диф-
ференцировки, зачастую развиваются нормальные взрослые осо-
би. Эти опыты указывают на определяющую роль ядра в целом в
специализации клеток, но не его состояния на конкретный мо-
мент времени.
1 2 3 4 5
Рис. 5.9. Детерминация жизненных форм клеток условиями среды у трипаносомы
(оригинал автора):
1 — инцистироваиная покоящаяся форма; 2— лейшманиальная: у мухи цеце — вне клеток, у
человека —в клетках; 3— лептомонадная и 4—критидиальная —трофические формы
в мухе цеце; 5—трипаносомная—трофическая форма в крови человека
205
Рис. 5.10. Детерминация формы «шляпки» у одноклеточной водоросли ацетабулярии
(Албертс и др., 1987, с изменениями)
Бластомеры зародыша амфибий, образующиеся после первых
меридиональных делений, при их разделении, способны к нор-
мальному развитию. У них цитоплазма яйцеклетки однородна и
потому первые бластомеры пока еще одинаковы. Но у животных,
у которых цитоплазма яйцеклеток до начала деления дифферен-
цирована на слои и зоны, отдельные бластомеры могут развивать-
ся только в определенные части зародыша. Можно было бы ду-
мать, что ядра на плазмодиальной стадии развития яйца, напри.т
мер, у насекомых уже имеют разное состояние и тем определяют
дальнейшую судьбу формирующихся клеток. Но тщательное изу-
чение цитоплазмы яйца позволяет обнаружить структурные и хи-
мические отличия в ее отдельных частях. Правда, пока непонятны
механизмы исходной дифференцировки цитоплазмы в так назы-
ваемых мозаичных яйцах (рис. 5.11).
В 20-е годы прошлого столетия результаты опытов по пересад-
ке частей гаструлы амфибий послужили экспериментальной осно-
вой для гипотезы эмбриональной индукции. Гипотеза предполагает
наличие в зародыше определенным образом расположенных кле-
ток — первичных организаторов, которые определяют течение
дифференцировки клеток соответствующей зоны зародыша. Пер-
вичные организаторы детерминируют положение оси зародыша и
расположение групп клеток или отдельных клеток — вторичных и
Рис. 5.11. Последовательность развития «мозаичного яйца» насекомого
(дрозофиллы) (Албертс и др., 1987, с изменениями)
206
третичных организаторов. Существуют косвенные доказательства
того, что организаторы выделяют вещества белковой природы,
распространяющиеся диффузным путем. Отдельные группы кле-
ток имеют разные пороги чувствительности к этим веществам и
потому по-разному реагируют на одинаковые их концентрации.
Данное явление объясняет наличие резких границ между группа-
ми клеток разных жизненных форм.
Воздействие внешней среды на клеточную дифференцировку,
как правило, оказывается опосредованным. Свет, газовый состав
окружающей среды, химический состав пищи изменяют течение
процессов в цитоплазме, что, в свою очередь, определяет экспрес-
сию генов, задействованных в регуляции морфогенеза. Часто меж-
ду факторами внешней среды и клеткой-мишенью имеется по-
средник-клетка или группа клеток, или ткань, реагирующая на
внешние воздействия с помощью выработки и выделения биоло-
гически активных веществ типа гормонов, способных участвовать
в регуляции развития клеток.
По-видимому, все же ключевым регулятором дифференциров-
ки следует считать генотип в его конкретном состоянии. Хотя
клетки одного вида организмов, а тем более одной особи имеют
практически идентичный генотип, наборы активных (экспресси-
руемых) генов в каждом типе клеток отличаются. Последователь-
ность преобразований в клетке за период ее развития от неспециа-
лизированного типа до функционально активного морфологичес-
кого типа определяется последовательностью включения и вык-
лючения генов, но механизмы динамики активности генов пока
еще изучены недостаточно.
Специализация клеток многоклеточного тела ведет к усилению
немногих функциональных свойств за счет ослабления других.
Поэтому дифференцировка клеток развивающегося тела одновре-
менно ведет к усилению взаимозависимости клеточных типов друг
от друга (рис. 5.12). Каждая жизненная форма клеток комплемен-
тарна сумме остальных. Должна существовать и количественная
мера соотношений разных типов специализированных клеток в
пределах одного живого тела, определяемая прежде всего генети-
ческой программой развития. Тем не менее, как известно, оконча-
тельный фенотип формируется в процессе взаимодействия разви-
вающегося генотипа и внешней информации. Одним из проявле-
ний корректирующего действия среды на живое тело является ус-
тановление конкретных соотношений морфофункциональных
типов клеток. Суммарный информационный сдвиг, образующий-
ся при усиленном размножении некоторых типов клеток, — это
сдвиг, компенсирующий изменения в экосистеме.
Аналогично описанному выше механизму действуют механиз-
мы естественной динамики численности в сообществах однокле-
207
= Гиподерма
Мускулатура
Нервная система
Первичные половые клетки
Соматические элементы гонады
Кишечник
Рис. S.12. Дифференцировка клеток нематоды Caenorhabditis elegans (Албертс и др.,
1987, с изменениями):
А — последовательность и направления дифференцировки клеток; Б—участие клеток, проис-
ходящих из разных «зачатков», в формировании гиподермы, мускулатуры и нервной системы
точных живых тел. То есть в биоценотическом аспекте динамика
численности протистов и бактерий в определенном смысле подоб-
на суммарной динамике численности жизненных форм клеток у
особей одного вида в конкретной живой системе.
Дифференцировка зародышевых клеток, клеток образователь-
ных тканей и неспециализированных клеток при регенерации на
разных этапах развития живого тела указывает на то, что направ-
ление дифференцировки задается не одним стимулом, а комплек-
сом стимулов и неодноактно. Обычно это происходит задолго до
того, как программируемые изменения проявляются. Те клетки, у
которых программа дифференцировки уже определена, называют
детерминированными. По определению клетка считается детерми-
нированной, если в ней произошли стойкие изменения, делаю-
щие ее потомков отличными от других клеток. Изменения долж-
ны быть стойкими, то есть сохраняться после устранения внешне-
го стимула, вызвавшего эти изменения. Устойчивость, впрочем,
не предполагает необратимости. Множество подтверждений тому
208
можно найти у низших многоклеточных. У губок, например, вы-
ход амебоидной клетки во внутренний клеточный пласт неизмен-
но ведет к формированию жгугико-воротничкового типа, а воз-
вращение ее в мезоглею сопровождается переходом в амебоидное
состояние, которое по одному из путей дальнейшей дифференци-
ровки может привести к необратимому превращению в склеро-
бласт.
При сравнении многоклеточного живого тела с телом, образо-
ванным единственной клеткой, обнаруживаются некоторые общие
их черты. И в одном и в другом случае тело разделено на компарт-
менты для обеспечения условий, необходимых для эффективного
течения процессов. В обоих случаях тела включают функциональ-
ные части, каждая из которых комплементарна сумме остальных.
Количественные соотношения частей определяются состоянием
тела во внешней среде. Устойчивая интенсификация какой-либо
функции достигается увеличением соответствующей части через
умножение ее элементов. Например, увеличение числа первичных
лизосом у инфузории туфельки и увеличение числа секреторных
клеток в пищеварительной системе собаки при длительном устой-
чивом интенсивном их кормлении следует считать явлениями го-
мологичными, так же как и размножение хлоропластов в клетке ли-
ста, фотосинтезирующих клеток в листе, листьев на дереве при со-
здании устойчивых благоприятных условий для фотосинтеза.
Любые изменения в состоянии живого тела и его структурных
подразделений, выражающиеся в дифференцировке клеток или
внутриклеточных структур, можно рассматривать на разных функ-
циональных уровнях живых систем, но всегда надо учитывать вза-
имозависимость и взаимоподчиненность таких уровней: на сколь-
ко внутриклеточные процессы определяются в конечном счете
биосферными, на столько же последние в реализации зависят от
первых.
5.4. ФУНКЦИОНАЛЬНЫЕ ОБЪЕДИНЕНИЯ КЛЕТОК —ТКАНИ
В процессе размножения и дифференцировки клетки живого
тела, сходные по строению и функциям и с общей предысторией,
образуют группы. Такие клеточные группы вместе с выделяемым
ими межклеточным веществом называют тканями. Обычно клет-
ки, составляющие ткань, являются потомками одной или немно-
гих клеток, определивших направление их специализации. В пе-
риод эмбрионального развития родоначальные клетки проделыва-
ют строго определенный путь перемещений и преобразований.
У животных различают четыре типа тканей: эпителиальную,
соединительную, мышечную и нервную.
14 Присный А.В.
209
Эпителий —это объединения клеток, покрывающие поверх-
ность тела и выстилающие его полости. Эпителиальные клетки
обычно полярны и плотно прилегают друг к другу. Межклеточное
вещество выделяется в незначительном количестве (у «трехслой-
ных»): основания клеток выделяют тонковолокнистую базальную
мембрану, а вершины могут выделять волокнистое вещество кути-
кулы. Эпителий быстро изнашивается и потому содержит недиф-
ференцированные замещающие клетки. Основные функциональ-
ные виды эпителия: покровный, защитный, рецепторный, всасы-
вающий, железистый и экскреторный. Кроме них существуют
особые морфологические формы: погруженный эпителий, сим-
пласт и миоэпителий. По форме слагающих клеток различают
плоский, кубический, призматический и цилиндрический эпите-
лий; по числу клеточных слоев — однорядный и многорядный.
Клетки эпителия могут нести микроворсинки (простые или перис-
тые), реснички, жгутики или образовывать псевдоподии. В онто-
генезе эпителиальные ткани развиваются раньше других — из эк-
тодермального и эктодермального зародышевых листков.
Соединительная ткань образуется подвижными или неподвиж-
ными клетками, выделяющими, как правило, большое количество
внеклеточного материала (бесструктурного и волокнистого). Раз-
личают трофические (кровь и лимфа), опорно-трофические (кро-
ветворная, жировая, пигментная, рыхлая и плотная) и опорные
(хрящи и кости) соединительные ткани. Их основные функции:
опорно-механическая, защитная и трофическая. Развивается этот
тип ткани из мезодермы (среднего зародышевого листка).
Мышечная ткань обеспечивает различные виды движения и
образована сократимыми клетками или их производными. Основ-
ные виды мышечной ткани — гладкая, сердечная и поперечно-по-
лосатая— различаются по степени упорядоченности фибрилл в
клетках, физиологическим особенностям и локализации в теле.
Развивается, как и предыдущий тип ткани, из мезодермы.
Нервная ткань состоит из плотно соприкасающихся нервных
клеток — нейронов, специализирующихся на формировании и
проведении мембранного электрического потенциала (рецептор-
ные и проводящие), и нейроглии — клеток, поддерживающих
нейроны. Межклеточное вещество плохо развито. Формируется из
наружного зародышевого листка (эктодермы). Основная функция
нервной ткани — преобразование накопленной химической энер-
гии в электрическую, возбуждающую клетки-мишени, под воз-
действием различных видов внешней энергии.
Единая детальная классификация тканей животных отсутствует.
Разные ткани в теле животных образуют комплексы — рабочие
органы, которые, в свою очередь, в процессе развития преобразу-
ются или объединяются в системы органов различной степени
210
сложности. Среди систем органов следует различать системы ин-
дивидуального жизнеобеспечения (они формируют основу цено-
тически активной части тела) и видового жизнеобеспечения (об-
разуют ценотически пассивную часть тела). К первым относят си-
стемы: покровов, опоры и движения, питания и пищеварения, га-
зообмена, химической регуляции, транспорта веществ, запасания
веществ и регуляции функций, а ко вторым — системы воспроиз-
водства новых поколений тел.
Ткани растений имеют несколько вариантов классификации.
Выделяют простые (состоящие из клеток одного типа) и сложные
(состоящие из клеток нескольких типов) ткани.
Простые ткани — паренхима, колленхима, склеренхима. Па-
ренхима — выполняющая ткань, образованная живыми метаболи-
чески активными клетками. У травянистых растений она же вы-
полняет опорную функцию. Важнейшие разновидности паренхи-
мы—это мезофилл (фотосинтезирующая ткань листьев) и эпи-
дерма (однослойный клеточный пласт, покрывающий тело
растения). Колленхима — механическая ткань. Ее живые клетки
образуют средние жилки листьев и опорные пучки в местах ветв-
ления стеблей. Склеренхима — механическая ткань, образованная
оболочками мертвых клеток.
К сложным тканям относят ксилему и флоэму, включающие и
живые клетки, и оболочки мертвых клеток. Эти ткани формируют
проводящую систему растений.
По сравнению с животными растения имеют более просто уст-
роенные органы, хотя в их образовании принимают участие комп-
лексы тканей. Представляется целесообразным выделение у расте-
ний следующих систем жизнеобеспечения: индивидуального —
покровов, опоры, газообмена, химической регуляции (минераль-
ного, в том числе водного, обмена), питания, запасания веществ,
транспорта веществ, регуляции функций; видового — воспроиз-
водства поколений.
Тела многоклеточных животных и растений, как правило, име-
ют сложную форму из-за сосредоточения отдельных органов и си-
стем органов в тех местах, где они оказываются наиболее ценными
в функциональном отношении. Такие функциональные объеди-
нения называют частями тела, хотя применительно к сосудистым
растениям их принято называть вегетативными органами. Набор и
соотносительное положение частей тела определяют планы строе-
ния живых тел, которые могут сильно отличаться у представите-
лей разных систематических групп организмов. Например, у птиц
это голова, шея, туловище, хвост, конечности, а у цветковых рас-
тений — корень, стебель, лист, цветок (плод).
14*
Глава 6
РЕАЛИЗАЦИЯ БИОГЕОЦЕНОТИЧЕСКИХ ФУНКЦИЙ
В ИНДИВИДУАЛЬНЫХ ЖИВЫХ ТЕЛАХ
&
6.1. ИНДИВИДУАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ КОМПЛЕКСЫ
(СИСТЕМЫ ЖИЗНЕОБЕСПЕЧЕНИЯ)
Говоря о биогеоценотических функциях организмов, мы пред-
полагаем, что вид заключает в себе элементарную биосферную ин-
формацию, он комплементарен сумме всех остальных видов и реа-
лизуется в конкретных живых телах. Поэтому индивидуальная
жизнь любого простого или сложного ценотически активного жи-
вого тела может быть с определенной степенью условности при-
равнена к воплощению видовых биосферных (или биогеоценоти-
ческих) функций в материальные и информационные (идеальные)
процессы. Развитие организма в течение индивидуальной жизни
представляет собой смену состояний, фаз, специализированных
частей и поколений, но обязательно включает период ценотичес-
кой активности, то есть участия в тех процессах, которые можно
считать общесистемными и которые определяют воспроизводство
всей системы. И воспроизводство живого вещества (материальной
составляющей), и воспроизводство информации (идеальной со-
ставляющей) требуют непрерывного потока энергии через систе-
му, а следовательно, через каждое живое тело. Ценотически пас-
сивные состояния, фазы, части и поколения в силу своего неучас-
тия в жизни системы непосредственно, представляя подготовку к
специфической активности или обеспечивая поддержание непре-
рывности видового живого вещества, могут иметь строение, отли-
чающееся в той или иной степени от строения ценотически актив-
ных тел. И если в онтогенезе организма выделяются, например,
эмбриональная, личиночная и взрослая фазы, где личинка будет
ценотически активной, а взрослая особь — пассивной, с упрощен-
ной организацией, традиционное диагностическое описание сис-
тем индивидуального жизнеобеспечения по взрослой фазе не по-
зволяет определить место вида в экосистеме.
В ценотически активных телах можно выделить несколько
функциональных комплексов, которые направляют через себя
энергетический поток в виде множества русел.
Часть энергии используется на поддержание и возобновление
самих функциональных комплексов, а часть направляется во внеш-
212
нюю (биотическую и абиотическую) среду для реализации непи-
щевых отношений.
Пока не принято единой терминологии для обозначения функ-
циональных подразделений тел растений и животных, более того,
понятие «система органов» применительно к растениям вообще не
употребляется, а вегетативными органами растений называют час-
ти тела, содержащие наборы органов из разных систем и постро-
енные из комплекса тканей.
Ранее мы приняли, что и растения, и животные, а также моне-
ры и грибы — это лишь условно выделяемые группы организмов,
составляющих единое живое вещество биосферы. Следовательно,
будучи клеточными организмами, все они должны характеризо-
ваться и сходными, пусть даже разнонаправленными, процессами.
Как известно, протофиты, протозои и часть одноклеточных гри-
бов оказываются иногда настолько близкими, что бывает невоз-
можно однозначно отнести их к тому или иному царству. Единой
здесь стала и терминология, используемая для обозначения кле-
точных структур. Сложное строение истинных многоклеточных не
должно служить препятствием для выработки единого подхода к
рассмотрению родственных образований и процессов у организ-
мов, относимых к царствам растений и животных.
Любое живое тело должно иметь физическую и химическую
определенность, целостность, что обеспечивается наличием по-
кровных образований. У разных тел покровы представлены следу-
ющими основными морфологическими вариантами: наружной
плазматической мембраной, НПМ, усложненной надмембранны-
ми образованиями (включая раковинки, домики, панцири), пел-
ликулой, НПМ с клеточной оболочкой или клеточной стенкой,
покровами клеточного строения (эпидермисом, корой, кожным
эпителием, сложной кожей-дермой), эпидермисом или кожным
эпителием со слизистыми выделениями или кутикулой, кожей с
костными или роговыми надкожными образованиями — чешуей,
чешуйками, пластинами, перьями, волосами. Покровы целиком
или на отдельных специализированных участках избирательно
проницаемы для различных веществ. Представляя пограничную и
контактную с внешней средой поверхность, они обычно несут ре-
цепторные структуры и коммуникационные образования.
Поддержание формы тела, расположения его частей в про-
странстве и обеспечение движения связаны с опорно-двигательны-
ми системами. В некоторых случаях опорные образования одно-
временно являются или двигательными, или покровными. На кле-
точном уровне организации это: опорные промежуточные фиб-
риллы и тонкие фибриллы цитоскелета, опорные микротрубочки,
внутриклеточные минеральные скелеты, органические и мине-
ральные надмембранные образования, сократимые актин-миози-
213
новые комплексы, двигательные комплексы микротрубочек. В не-
которых типах клеток формируются сократимые отростки, сте-
бельки и т. п. В многоклеточных телах образуются опорные соеди-
нительные ткани (клеточные массы), полостные жидкости (в том
числе вода в пищеварительной полости некоторых кишечнопо-
лостных), кутикулы, внешние и внутренние скелеты, скопления
оболочек живых или мертвых клеток, системы опорных пучков,
жилки, двигательные отростки клеток, мышечные клетки, специ-
ализированные мышцы, мышечные массы. Опорные и двигатель-
ные системы в живых телах не следует противопоставлять друг
ДРУГУ, поскольку через них осуществляется движение тела или его
частей со скоростью, равной нулю или отличной от нуля, что и
обозначается как положение или перемещение.
Системы питания (в узком смысле) обеспечивают живые тела
веществами и энергией, то есть преобразуют поступающие из
внешней среды вещества и энергию в формы, доступные к ис-
пользованию данными телами. При этом автотрофы и гетеротро-
фы, будучи лишь функциональными составляющими единого
живого вещества биосферы, реализуют разнонаправленные со-
ставляющие единого вещественно-энергетического процесса. В
автотрофных телах преобладают процессы преобразования посту-
пающей извне энергии в энергию химических связей органичес-
ких веществ, синтезируемых из неорганических веществ. Это
процессы, сопровождающиеся последовательным укрупнением
молекул. Поэтому к системам питания у автотрофов на разных
функциональных уровнях следует относить фото- и хемосинтези-
рующие мембраны, фотосинтезирующие органоиды (пластиды),
фотосинтезирующие клетки, совокупность клеток мезофилла, где
каждая более емкая функциональная структура включает менее
емкие. Свет улавливается, а основной источник углерода — СО2
поглощается только открытой поверхностью, из-за чего у много-
клеточных тел системы питания имеют диффузное строение, а ча-
сти тела, несущие их фрагменты, имеют вид пластинок или плос-
костей. Все продуценты синтезируют органические вещества из-
быточно, то есть сверх того, что необходимо им для текущего са-
мовоспроизводства и воспроизводства следующего поколения.
Этот избыток и составляет сущностную сторону продуцентов.
В организмах гетеро трофов органические вещества, синтезиро-
ванные автотрофами (или гетеротрофами предшествующего уров-
ня консументов), постепенно измельчаются, высвобождая энер-
гию, используемую для собственных нужд и воздействия на внеш-
нюю среду, и образуя малые молекулы, включаемые в пластичес-
кие процессы или выделяемые за пределы тела. Такое
измельчение производят пищеварительные структуры, которые
могут быть представлены следующими морфологическими форма-
214
ми: пищеварительные органоиды (вакуоли и митохондрии), пи-
щеварительные клетки, пищеварительная «плоскость», пищевари-
тельная паренхима, пищеварительная полость, кишечная труб-
ка, кишечник с дополнительными пищеварительными железа-
ми. У многоклеточных тел пища обычно подвергается обработке
ферментами, выделяемыми железистыми пищеварительными
клетками. Поэтому для создания высокой концентрации фермен-
тов пищеварительные структуры должны быть представлены ком-
пактными полостями.
Поступление энергии и исходных веществ в системы питания у
автотрофов носит пассивный характер, и для них оказывается дос-
таточным эффективное расположение фрагментов систем пита-
ния в пространстве. Гетеротрофы, как правило, активно добывают
пищу, расположенную или перемещающуюся в разных точках
пространства, отдельными порциями. При этом пища может быть
малодоступной или непригодной к эффективному химическому
измельчению. Тогда гетеротрофы используют имеющиеся у них
специальные структуры для захвата, извлечения и предваритель-
ного механического измельчения пищи (реснички, жгутики, кле-
точный рот, «палочковый аппарат», или клеточную глотку, перед-
нюю кишку, щупальца, кутикулярные утолщения и челюсти, ро-
товые аппараты и др.). Такие структуры чаще всего морфологи-
чески сопряжены с пищеварительными системами, предваряя их,
но могут располагаться на конечностях и других частях тела (ког-
тях, копытах, клешнях и др.).
Живое тело, будь то автотрофное или гетеротрофное, обяза-
тельно расходует органические вещества на энергетической ста-
дии обмена. Потребности в энергии не всегда совпадают по вре-
мени с возможностью непосредственного ее получения извне. По
этой причине живые тела накапливают резервы питательных ве-
ществ, которые могут иметь вид внутриклеточных включений,
специализированных клеток, запасающих тканей или же запасаю-
щих органов и частей тела.
Если исключить из рассмотрения переход органического веще-
ства от одного живого тела к другому, выстроенный в простую пи-
щевую цепь, то обмен веществами с внешней средой сводится для
всей последовательности к поглощению и выделению воды, мине-
ральных веществ и СО2, а для отдельных живых тел — дополни-
тельно кислорода. Поскольку для автотрофов вода, диоксид угле-
рода и минералы — это исходные вещества, включаемые во внут-
ренние обменные процессы, а для гетеротрофов они являются ко-
нечными продуктами обмена, то и системы жизнеобеспечения,
отвечающие за их поступление в тело или выведение из него, от-
личаются и строением, и направленностью реализуемых процес-
сов. Соответственно составным частям обмена выделяются систе-
215
мы газообмена, водного и минерального обмена, причем две пос-
ледние обычно объединяются в систему химической регуляции,
включающей осмотическую регуляцию.
Газообмен — процесс перехода газообразных веществ из среды в
живое тело и обратно через всю пограничную поверхность или ее
специализированные участки. Автотрофы поглощают диоксид уг-
лерода, а большинство их выделяют кислород. Гетеротрофы, на-
оборот, выделяют СО2, а кислород не поглощают только анаэро-
бы. Газы движутся по градиенту концентрации и через покровы
проникают диффузным путем, даже будучи растворенными в воде.
Непосредственно с газообменом сопряжены два процесса: дыха-
ние и вентиляция. Под дыханием принято понимать окисление ве-
ществ со связыванием части освобождающейся энергии в клетке,
и поскольку окисление может идти по типу гликолиза в цитоплаз-
ме или по типу кислородного окисления в митохондриях, то и ды-
хание разделяют на анаэробное и аэробное. Вентиляция — это
процесс смены объема воздуха в газообменных полостях или у
скрытых газообменных поверхностей. Особенно отчетливо она
выражена у трахейных и легочных животных, где совершается
принудительно за счет сокращений специализированных мышц.
Водно-минеральный обмен, или химическая регуляция, — про-
цесс весьма многоплановый и непосредственно, как и дыхание,
сопряжен с питанием и пищеварением. На характер, механизм и
назначение водно-минерального обмена прежде всего влияют тип
и способ питания, условия существования живого тела и особен-
ности его строения. От содержания воды в теле зависит большин-
ство жизненных процессов и прежде всего сама возможность про-
текания (био)химических реакций.
Автотрофы нуждаются (в период активности) в постоянном
притоке воды в ассимилирующие органоиды, клетки и ткани, по-
скольку вода для них — один из исходных продуктов фото- и хе-
мосинтеза. Гетеротрофы же, по крайней мере аэробные, непре-
рывно образуют воду, окисляя водород органических веществ. Та-
кая вода, называемая метаболической, должна непрерывно или
порциями выводиться за пределы тела. Непрерывное выделение
воды в парообразном состоянии часто обеспечивает терморегуля-
цию, но у автотрофов транспирация в первую очередь обусловли-
вает «сосущую силу» — один из механизмов подъема воды по про-
водящим путям. Поглощение воды происходит через неспециали-
зированные (покровы в целом) или специализированные (участки
покровов, зона всасывания, системы питания и пищеварения)
структуры. То же можно сказать и о выделении воды: через покро-
вы, через их специализированные участки (в том числе газообмен-
ные), через выделительные (в том числе осморегуляторные) и пи-
щеварительные системы. Регуляция содержания воды в клетках
216
живого тела может основываться на изменении физико-химичес-
кого состояния гиалоплазмы, содержания ионов в цитоплазме, па-
раметров межклеточного вещества, интенсивности работы специ-
ализированных осморегуляторных органоидов, клеток и органов.
К числу осморегуляторных структур относятся пульсирующие ва-
куоли простого или сложного строения, соленоциты, «мерцатель-
ные» клетки протонефридиев, отчасти метанефридии, коксальные
железы и другие специализированные органы регуляции мине-
рального обмена.
Многие вещества, а особенно неорганические и низкомолеку-
лярные органические соединения, легко растворяются в воде.
Вода, поступая из внешней среды в тело или выводясь из него,
способна перемещать и растворяемые вещества. Так, поглощение
воды клетками талломов, ризоидами и корнями фотоавтотрофов
сопряжено с механизмом всасывания необходимых минеральных
и некоторых органических веществ. Обычно такие вещества по-
ступают в живые тела из водной среды или из почвы, причем от-
ток воды и переносимых ею веществ из пространства, непосред-
ственно прилежащего к зоне поглощения, вызывает их приток из
более удаленного окружающего пространства. Гетеротрофы ис-
пользуют выделение части метаболической и поглощаемой воды
для удаления из тела минеральных и малых органических моле-
кул — конечных продуктов обмена, особенно азотсодержащих.
Соотношение воды и растворенных в ней веществ в клетке и,
как следствие, осмотическое давление зависят не только от содер-
жания воды в теле, но также и от ее содержания во внешней среде.
Телам, обитающим в гипотонической, изотонической и гиперто-
нической (в том числе воздушной) среде, приходится по-разному
решать проблему водно-солевого обмена. В первом случае для
поддержания химического постоянства необходимо непрерывно
удалять избыток воды, а в последнем — образующиеся раствори-
мые отходы обмена при недостатке воды. Специализированные
органы химической регуляции (органы выделения) достигают
наибольшей сложности у многоклеточных животных — обитате-
лей воздушной среды. Часто их нефридии, почки и мальпигиевы
сосуды работают по принципу «противоточного умножителя», чем
достигаются высокая концентрация или кристаллическое состоя-
ние удаляемых из тела веществ и, следовательно, уменьшение по-
терь воды. В живых телах имеют место и явления, противополож-
ные отмеченным. Так, автотрофы могут выделять избыток мине-
ральных веществ (например, кремнезема или карбоната кальция),
а гетеротрофы, наоборот, — избирательно поглощать в концент-
рированном виде недостающие им минеральные вещества (хлорид
натрия, соли кальция и др.).
В любом живом теле вещества непрерывно перемещаются.
217
Скорости и механизмы движения веществ, естественно, зависят от
плана организации тела. У свободноживущих одиночных клеток
доставка веществ к местам назначения полностью обеспечивается
с помощью типичных клеточных процессов: трансмембранного
переноса, транспорта в мембранных органоидах, вакуолярного
транспорта, диффузии и токов цитоплазмы. У водорослей и гри-
бов благодаря талломной организации перемещение веществ меж-
ду клетками происходит с участием плазмодесм. На тканевом
уровне организации усиливается роль межклеточных контактов,
транспорт веществ осуществляется при посредничестве межкле-
точного вещества и жидкостей, заполняющих анатомические и
функциональные полости.
Специализированные транспортные системы, где они имеют
место, состоят из каналов, связывающих место входа или образо-
вания транспортируемого вещества с местом его выхода или ис-
пользования. Перемещаются вещества по каналам или с помощью
специальной транспортирующей жидкости (воды, полостной
жидкости, гемолимфы, крови, лимфы), или же одно из транспор-
тируемых веществ (вода) одновременно играет роль транспорти-
рующего. В первом случае транспортная система должна быть «за-
кольцованной», а во втором — «сквозной». У сосудистых растений
система структурных образований для транспорта воды, мине-
ральных и органических веществ формируется из специализиро-
ванных клеток ксилемы и флоэмы. Поглощаемые вещества пере-
носятся от корневых волосков до сосудов ксилемы и от них до ас-
симилирующих клеток по трем путям: симпластному, апопластно-
му и вакуолярному. Мертвые клетки ксилемы (трахеиды и
членики сосудов) образуют перегородчатые или сплошные кана-
лы, заполненные водой — основным транспортируемым веще-
ством и в то же время транспортирующим растворенные мине-
ральные вещества от корней к листьям. Движущая сила складыва-
ется из разности осмотического давления в ксилемном соке и поч-
венном растворе — «корневого давления» и сосущей силы верхних
клеток сосудов, из которых вода переходит в транспирирующие
клетки по направлению водного потенциала. Этой силы достаточ-
но, чтобы вода по проводящим пучкам могла подниматься непре-
рывным потоком на высоту более 100 м.
Транспорт продуктов ассимиляции осуществляется по живым
клеткам флоэмы, образующим ситовидные трубки. В отличие от
воды органические вещества (преимущественно сахароза) пере-
двигаются и вверх и вниз. Они также образуют водные растворы, и
их движение носит характер «массового потока». Механизмы дви-
жения связаны с работой клеток-спутников и в общих чертах мо-
гут быть сведены к сочетанию гидростатического давления, разви-
вающегося из-за разности осмотического давления в клетках на
218
концах сосудов, и хемиосмоса, в котором принимают участие и
оболочки, и ситовидные пластинки, и фибриллярные белки кле-
ток.
У многоклеточных животных сосудами и полостями транспорт-
ной системы связываются, как правило, активные клетки и ткани,
контактирующие с внешней средой и расположенные в глубине
тела. При этом внешняя среда может проникать глубоко в тело в
виде впячивания или сплошного канала. Жидкость, заполняющая
каналы, приводится в движение посредством сокращения стенок
оформленных сосудов или их специализированных частей (сер-
дец) или скелетной мускулатуры. Морфологически оформленные
транспортные системы многоклеточных животных разделяют на
кровеносные и лимфатические. В том случае, если кровеносная
система образована сочетанием сосудов и полостей, ее называют
«открытой» или «незамкнутой», а если только оформленными со-
судами — «закрытой» или «замкнутой». В общей схеме транспорт-
ная система включает каналы с пульсирующим участком сосуда и
несколько капиллярных сетей (общую тканевую, газообменную,
кишечную и выделительную).
Вегетативные (и генеративные) функции живого тела коорди-
нируются и синхронизируются благодаря наличию специальных
регуляторных систем. Жизнедеятельность тела проходит в конк-
ретных изменчивых условиях среды, и потому значительное место
в регуляции занимает реакция на внешние раздражители. Клеточ-
ный уровень организации живых тел позволяет каждой отдельной
клетке самостоятельно реагировать на изменения во внешней сре-
де. Но у одноклеточных и талломных тел все клетки непосред-
ственно граничат со средой, а у многоклеточных тел с тканевой
организацией внешнюю среду каждой отдельной клетки полнос-
тью или частично составляют другие клетки или межклеточное ве-
щество. Свободноживущие и функционально близкие к ним клет-
ки принимают раздражения с помощью мембранных рецепторных
комплексов и преобразуют накопленную энергию в энергию дви-
гательной реакции, позволяющей оптимизировать отношения со
средой. Форма двигательной реакции на раздражение может быть
местной (на уровне отдельных специализированных клеточных
структур) или общеклеточной (мембранной, цитоскелетной, ци-
топлазматической). Например, неадекватная скорости собствен-
ного движения инфузории сила контакта с внешним объектом вы-
зывает выстреливание соответственно расположенных трихоцист,
а попадание в область солевого раствора — движение всей клетки
от гипертонической среды. Двигательная реакция предполагает не
только перемещение, но также трансмембранный перенос ионов,
экспорт или импорт веществ, изменение формы, изменение осмо-
тических свойств и др. Разнообразию внешних раздражителей со-
219
ответствует и разнообразие специфических двигательных ответов
на них.
В многоклеточных телах, где ответ на внешние раздражения
реализуется на надклеточном уровне, но непосредственного дей-
ствия на большинство клеток среда не производит, развиваются
единые клеточные системы, опосредованно связывающие глубоко
расположенные клетки с внешней для тела средой. Такие системы
называются регуляторными и подразделяются на нервные и гумо-
ральные.
Нервная система представляет собой единство множества спе-
циализированных клеток-преобразователей запасенной химичес-
кой энергии в энергию трансмембранного переноса ионов калия и
натрия. Под действием внешних раздражителей типичные или ви-
доизмененные рецепторные нервные клетки возбуждаются и пе-
редают возбуждение в виде электрического или химического сти-
мула следующим, проводящим нервным клеткам, которые, в свою
очередь, непосредственно или через посредничество других таких
же клеток возбуждают эффекторы—двигательные клеточные
структуры, осуществляющие ответ на исходное раздражение. При
посредничестве нервной системы тело производит быстрый,
обычно локальный, двигательный ответ на внешний стимул.
Иначе действуют гуморальные регуляторные системы. Под воз-
действием диффузного (ненаправленного) фактора, внешнего или
внутреннего, часть клеток тела вырабатывает и выделяет во внут-
реннюю среду биологически активные вещества (гормоны, нейро-
гормоны, фитогормоны), действующие очень специфично на оп-
ределенные клетки-мишени, медленно изменяя в них течение об-
менных процессов. В растениях это прежде всего регуляторы рос-
та: ауксины, гиббереллины, цитокинины, абсцизовая кислота и
этилен. Они вырабатываются большей частью в зонах интенсив-
ного деления клеток. Регулируют клеточное деление, ростовые
процессы, покой почек и семян, старение и опадание листьев, со-
зревание плодов. У животных гормоны вырабатываются в специа-
лизированных и неспециализированных эндокринных железах и
нервных клетках особого рода (нейросекреторных). Это могут
быть вещества, регулирующие активацию и покой, направлен-
ность обменных процессов, морфогенетические процессы, поло-
вое созревание, половую активность, формы поведения и другие
процессы, охватывающие, как правило, все тело.
Двигательные реакции на внешние воздействия имеют место
как у животных, так и у растений. У одноклеточных движения
проявляются преимущественно в форме перемещений относи-
тельно действующего фактора — таксисов. Таксисы принято раз-
делять на положительные (движение к источнику раздражения) и
отрицательные (движение от источника раздражения). Классифи-
220
кация таксисов основана на видах раздражителей (фототаксис, хе-
мотаксис и др.). Многоклеточные растения проявляют: направ-
ленные ростовые или тургорные движения на одностороннее дей-
ствие фактора—тропизмы (фототропизм, геотропизм и др.), ко-
торые также могут быть положительными или отрицательными;
ненаправленные ростовые или тургорные движения, осуществляе-
мые в ответ на диффузно действующий фактор, — настии (напри-
мер, открывание и закрывание венчиков цветков при изменении
освещенности); круговые или маятникообразные движения росто-
вого или тургорного характера, не связанные с действием опреде-
ленного фактора, — нутации (движение стеблей лиан, усиков го-
роха и винограда и др.). Многоклеточные животные имеют в той
или иной степени развитую нервную систему и на ее основе спо-
собны к рефлекторным двигательным реакциям.
Ростовое и активное движение — две формы, каждая из кото-
рых связана с живыми телами характерного строения. Движение в
форме перемещения имеет общую морфологическую основу на
разных уровнях организации живых тел. Амебоидный и цилиар-
ный способы перемещения проявляются как у одноклеточных тел,
так и у специализированных клеток многоклеточных тел. Однако
наибольшее развитие в двигательных структурах многоклеточных
получает движение, основанное на работе акгин-миозиновых
комплексов цитоскелета. Оно позволяет сформировать особый
морфофункциональный тип клеток — мышечные клетки, из кото-
рых строятся весьма эффективные преобразователи химической
энергии в механическую. Одно из наиболее замечательных явле-
ний, связанных с развитием мышц у животных, — это работа эле-
ментов системы — живых тел, направленная на внешнюю по от-
ношению к ней среду, и, следовательно, способность к производ-
ству информации.
6.2. ПОДДЕРЖАНИЕ ПОСТОЯНСТВА ВНУТРЕННЕЙ СРЕДЫ
(ГОМЕОСТАЗ)
Химические и физико-химические процессы, протекающие в
живых телах, имеют разную степень эффективности в разных ус-
ловиях. К важнейшим параметрам условий для протекания внут-
риклеточных процессов относятся: концентрация веществ в гиа-
лоплазме, pH среды, температура и др. Живые системы и функци-
ональные комплексы, обладающие свойствами систем, характери-
зуются способностью в той или иной степени регулировать
состояние внутренней среды в соответствии с изменением пара-
метров внешней среды по принципу отрицательной обратной свя-
зи. Способность систем противостоять изменениям и сохранять
221
динамическое постоянство состава и свойств называется гомеос-
тазом. Принято различать две формы гомеостаза: физиологичес-
кий и генетический, относящиеся соответственно к уровням от-
дельных живых тел и популяций тел.
У свободноживущих клеток поддерживаются специфические
физико-химические параметры цитоплазматического матрикса,
отличающиеся от условий окружающей среды. Но из-за очень вы-
сокого значения отношения поверхности клетки к ее объему ком-
пенсаторные изменения оказываются слишком энергозатрат-
ными, и потому клетки-особи быстро и значительно меняют свое
функциональное состояние при отклонении внешних условий от
оптимальных значений. Однако было бы неправильно говорить о
полном отсутствии гомеостаза у одноклеточных тел.
Многоклеточные живые тела имеют внутреннюю среду, окру-
жающую клетки, что создает более устойчивые условия для каж-
дой из них. Усложнение внутренней дифференцировки таких тел
сопровождается повышением сложности и эффективности меха-
низмов гомеостаза. Формируются специализированные системы
индивидуального жизнеобеспечения, участвующие в процессах
поддержания постоянства внутренней среды: покровы, системы
газообмена, транспорта веществ, химической и функциональной
регуляции. Многие наземные и некоторые водные животные спо-
собны активно регулировать внутренний температурный режим,
используя экзогенное или эндогенное тепло.
Наибольшего совершенства гомеостаз достигает у млекопитаю-
щих животных. У них, в частности, обеспечиваются постоянство
объема внеклеточных жидкостей, концентрация в тканевой и по-
лостных жидкостях ионов, включая протоны (pH), осмотически
активных веществ, состава белков, липидов и углеводов, газов,
температуры тела. Такое постоянство поддерживается и в преде-
лах внутренней среды отдельных органов и тканей. В обеспечении
гомеостатических реакций у этих животных задействованы все
функциональные системы индивидуального жизнеобеспечения и
особенно эндокринная и нервная системы.
У растений для поддержания гомеостаза на клеточном уровне
особое значение имеют наружная плазматическая мембрана и то-
нопласт вакуолей. На тканевом уровне динамическое постоянство
параметров тела обеспечивают межклеточное вещество и клеточ-
ные оболочки (в том числе и мертвых клеток).
Гомеостаз как явление, характерное для отдельных живых тел,
не имеет четкой границы с адаптациями, или приспособлениями.
Если гомеостаз рассматривать в качестве оперативной (быстрой и
кратковременной) реакции тела на изменение условий существо-
вания, то адаптации могут быть представлены как совокупность
относительно устойчивых морфофизиологических, поведенческих
222
и других особенностей, характерных для множества живых тел,
составляющих видовую популяцию, которые обеспечивают воз-
можность специфического образа жизни в определенных условиях
внешней среды. То есть гомеостатические реакции можно рас-
сматривать как динамичные индивидуальные приспособительные
изменения, не выходящие за рамки популяционно-видовых адап-
таций. Комплекс же устойчивых индивидуальных адаптаций —
это фактически жизненная форма, или морфофункциональный
тип.
Терминологическая граница между индивидуальным и популя-
ционно-видовым гомеостазом еще более стирается при выделении
его разновидности — генетического гомеостаза, обозначающего
способность популяции поддерживать относительную стабиль-
ность и целостность генетической структуры в изменяющихся ус-
ловиях среды. Способность отдельных живых тел противостоять
изменениям и в том числе внедрению в них чужеродной генети-
ческой информации (других живых тел или выделяемых ими ве-
ществ) — иммунитет — можно рассматривать как частное прояв-
ление генетического гомеостаза.
В настоящее время принято различать три формы иммунитета:
конституциональный, фагоцитарный и лимфоидный. Эта класси-
фикация основана на группировке явлений устойчивости по при-
знаку их формирования и по принципам защитного действия.
Конституциональный иммунитет — это наследственная форма ус-
тойчивости. Он обусловлен особенностями устройства, конститу-
ции живого тела. Фагоцитарный иммунитет также можно отнести
к наследственному, так как он связан с определенным, наслед-
ственно закрепленным типом клеток тела (хотя и не всего тела),
способных к фагоцитозу. (Ранее мы говорили о том, что способ-
ность к развитию определенных жизненных форм клеток наслед-
ственно детерминирована.) В лимфоидной же системе наследует-
ся только потенциальная способность к развитию иммунитета, но
не сам иммунитет как таковой. То есть эта форма устойчивости
приобретается телом в течение его индивидуальной жизни.
Набор защитных механизмов у каждого конкретного биологи-
ческого вида зависит от его эволюционно сложившегося типа
организации, выражающейся в таксономическом (систематичес-
ком) положении. Система конституциональной устойчивости
действует у всех живых существ от прокариот до высших позво-
ночных. У подавляющего большинства многоклеточных живот-
ных, кроме конституциональной, присутствует устойчивость, свя-
занная с фагоцитами. И только у позвоночных животных имеется
дополнительно к названным система индивидуально приобретае-
мого реактивного лимфоидного иммунитета. Эта третья форма
обеспечивается специфической активностью особых веществ бел-
223
ковой природы — иммуноглобулинов, или антител, вырабатывае-
мых клетками лимфоидной системы в ответ на появление в теле
антигенов. Антигенами могут быть любые вещества, способные
вызывать образование соответствующих антител и связываться с
ними.
На основе естественного лимфоидного иммунитета человек
разработал способы искусственной иммунной защиты. Искусст-
венный иммунитет подразделяют на активный и пассивный. Ак-
тивный иммунитет вырабатывается в теле при введении в него ос-
лабленных или убитых возбудителей заболеваний или их ослаб-
ленных токсинов. Он проявляется не сразу, но сохраняется дли-
тельное время. Пассивный иммунитет возникает, когда в живое
тело вводят готовые антитела в виде сыворотки. Этот иммунитет
проявляется сразу же после прививки, но сохраняется кратковре-
менно.
Глава 7
ВИДОВОЕ ЖИЗНЕОБЕСПЕЧЕНИЕ
Ранее рассмотренные системы жизнеобеспечения составляют
ценотически активную часть живого тела, или комплекс структур,
обеспечивающих индивидуальную жизнь, а также реализацию
экосистемной информации. Участие в биогеоценотических про-
цессах развивающихся отдельных тел можно представить и как
«разворачивание» индивидуальной генетической программы в
конкретных условиях. Процессы, идущие в биосфере и ее подраз-
делениях, изменяют как внутреннюю среду этой системы, так и
связи, отношения между составляющими ее элементами. Поэтому
внутренняя информация видового живого вещества подлежит по-
стоянной корректировке в соответствии с новообразованиями,
возникающими во внешней для него экосистемной информации.
Частные вариации видовой генетической системы у отдельных
особей, кроме того, должны быть интегрированы в генофонд по-
пуляции и распространены в ней, чтобы стать видовым достояни-
ем. Одновременно вариации, заведомо неблагоприятные, не спо-
собные развиться в полноценный фенотип, должны быть элими-
нированы.
В онтогенезе каждое тело приобретает определенную жиз-
ненную форму на клеточном или надклеточном уровне. И в том
и в другом случае клетки детерминированы в своей дифферен-
цировке. Но клетки одного тела должны иметь одинаковую ге-
нетическую систему, чтобы реализовать единую генетическую
информацию. В случае недостаточности функционального дей-
ствия, производимого сформировавшимся телом, соответствую-
щие его части должны увеличиться, сохранив специализацию
элементов-клеток.
Почти всегда ценотически активное тело растет. И если сво-
бодноживущие клетки-особи завершают период собственного ро-
ста, связанный с синтезом веществ протоплазмы, разделением на
самостоятельные дочерние клетки, то многоклеточные особи уве-
личивают свою прежде всего соматическую часть посредством ро-
ста числа собранных воедино клеток.
15 Присный А.В.
225
В разных случаях воспроизводство генотипа требует разного
конечного результата: дочерние генотипы должны быть или иден-
тичными материнскому, или отличными от него, содержащими
новообразования, накопленные материнским поколением. В пер-
вом случае воспроизводство наследственных макромолекул идет
по принципу тиражирования копий, а во втором — включает ре-
комбинацию, выбраковку и составление новых комбинаций алле-
лей. Первый процесс носит название «митоз» и служит для увели-
чения числа одинаковых хромосомных групп, ядер или клеток с
постоянным числом хромосом; второй — «мсйоз», служит для пе-
ревода генов в «гомозиготное» гаплоидное состояние, позволяю-
щее элиминировать большинство вредных мутаций и «запрещен-
ных» комбинаций и одновременно избежать прогрессивного уве-
личения числа хромосом в кариотипе в ряду поколений.
7.1. ЖИЗНЕННЫЙ ЦИКЛ КЛЕТКИ
Митоз и мейоз — это лишь этапы в жизни клеток. Весь же
жизненный цикл, или последовательность состояний между
окончаниями двух делений, подразделяется на интерфазу (вклю-
чающую дифференцировку, период специфической для данной
жизненной формы клетки активности и период подготовки к де-
лению) и митоз или мейоз (приводящие к распределению хрома-
тид в дочерние кариотипы или хромосомные группы попарно
или поодиночке). Обычно деление завершается или сопровожда-
ется цитокинезом (разделением цитоплазмы между дочерними
клетками) (рис. 7.1).
Интерфаза. Переход от недифференцированного состояния
клетки, которым заканчивается предшествующее деление, к оп-
ределенному морфофункциональному типу сопровождается ин-
тенсивными процессами биосинтеза, формообразованием, уве-
личением размеров, упорядочиванием органоидов, построением
типичного цитоскелета. Ядрышко начинает продуцировать раз-
личные РНК, образуются рибосомы, клетка синтезирует и струк-
турные, и функциональные белки. С этого момента начинается
интенсивный обмен веществ, контролируемый ферментами.
Некоторое время клетка осуществляет организацию потока ве-
щества и энергии в специфичной для нее форме, что является
главным содержанием ее деятельности как элемента экосистемы
или структурной единицы элемента экосистемы. Этот период сле-
дует считать основным в жизненном цикле клетки.
На следующем этапе рост клетки замедляется, характер метабо-
лических процессов смещается в сторону синтеза мономеров нук-
леиновых кислот и гистоновых белков (у прокариот — протами-
226
Рис. 7.1. Жизненный цикл клетки (оригинал автора):
Д—дифференцировка; ФА — функциональная активность; РЯ — репликация ядерной ДНК;
РЦ — репликация ДНК цитоплазматических органовдов
нов). Происходят репликация ДНК и образование хроматид (до-
черних хромосом).
Заключительный этап интерфазы характеризуется подготовкой
к делению клетки: идут интенсивные синтетические процессы,
делятся митохондрии и хлоропласты, центриоли, если они есть,
удваиваются, форма клетки упрощается, замедляется движение,
затормаживается эндоэкзоцитоз.
Длительность интерфазы обычно составляет не менее 90 % всей
длительности клеточного цикла, которая продолжается от 15 мин
у некоторых прокариот до 100 дней и более у эукариот. Некоторые
клетки (нейроны, эритроциты и волокна поперечно-полосатых
мышц) в зрелом (дифференцированном) состоянии не делятся со-
всем.
Окончание интерфазы одновременно является и началом ми-
тоза или мейоза. Реорганизация ядра при делении идет с участием
сложных динамических структур, выстраиваемых из микротрубо-
чек в митотическое веретено. У многоклеточных растений и жи-
вотных во время митоза ядерная оболочка разрушается, у боль-
шинства протистов разделение хроматид на дочерние группы про-
исходит в четко оформленном ядре.
Митоз, или непрямое деление клетки. Весь процесс митотичес-
кого деления принято разделять на несколько этапов, или фаз:
профазу, прометафазу, метафазу, анафазу, телофазу и цитокинез.
Иногда первые две фазы объединяют в единую — профазу.
227
В профазе диффузный хроматин постепенно конденсируется
в видимые хромосомы, каждая из которых состоит из двух сест-
ринских хроматид. Ядрышко дезорганизуется и исчезает. Из ту-
булиновых молекул разрушенного цитоскелета строятся микро-
трубочки митотического веретена. Сначала они образуются в виде
звездообразных фигур вокруг каждой из рядом расположенных
дочерних центриолей. Часть микротрубочек — полюсные нити —
расталкивают митотические центры, выстраивая их в биполярное
митотическое веретено.
Прометафаза начинается с разделения ядерной оболочки на
фрагменты, мало отличимые от цистерн ЭПС. В область, ранее
занимаемую ядром, проникают нити веретена. В хромосомах с
каждой стороны центромеры образуются особые структуры, кото-
рые прикрепляются к специальной группе микротрубочек. Это
«тянущие» нити, взаимодействующие с «опорными» нитями и
приводящие хромосомы в движение.
В метафазе все хромосомы выстраиваются так, что их центро-
меры лежат в одной плоскости, перпендикулярной оси веретена
деления на равном расстоянии от его полюсов.
Переход от метафазы к анафазе связан с резким одновремен-
ным разделением хроматид в области центромер. (Здесь заверша-
ется репликация ДНК на участке, объединявшем хроматиды.) Ос-
вобожденные хроматиды медленно расходятся под тянущим дей-
ствием скользящих нитей к полюсам. При этом тянущие нити
укорачиваются, а полюса расходятся еще дальше.
В телофазе хроматиды подходят к полюсам, тянущие нити
исчезают и вокруг каждой группы хроматид выстраивается но-
вая ядерная оболочка. Хроматиды, собственно теперь хромосо-
мы, начинают деспирализоваться, превращаясь в типичный
хроматин, образуются ядрышки. На этом деление ядра заканчи-
вается, и клетка вступает в завершающую стадию митоза — ци-
токинез.
Разделение цитоплазмы начинается еще в поздней анафазе или
в телофазе. В клетках животного, грибного и бактериального ти-
пов на наружной мембране в средней части клетки образуется
кольцевая перетяжка — борозда деления. Она углубляется в плос-
кости экватора до тех пор, пока дочерние клетки полностью не
отделятся друг от друга. Механизм разделения цитоплазмы в рас-
тительных клетках был рассмотрен в разделе 4.3.2.
Митоз в силу особенностей прохождения не может внести но-
вой генетической информации в геном дочерней клетки. Его ре-
зультатом является образование двух дочерних клеток, генетичес-
ки идентичных родительской. Основное назначение митоза — ум-
ножение однородных элементов (клеток, ядер, хромосомных
групп) для усиления реализуемой функции.
228
Мейоз, или редукционное деление клетки. В отличие от митоза
эта форма деления приводит к уменьшению числа хромосом в до-
черних клетках по отношению к родительской в два раза. Факти-
чески мейоз приводит к обособлению каждой из хроматид и, сле-
довательно, к «чистому», гомозиготному состоянию каждой алле-
ли в хромосомах. Мейотическое деление ядра проходит в два эта-
па, в каждом из которых наблюдаются процессы, аналогичные
фазам митоза. В интерфазе, предшествующей мейозу, происходят
репликация ДНК и интенсивный синтез гистоновых белков. К
концу интерфазы каждая из гомологичных хромосом представле-
на парой хроматид.
В профазе первого этапа, или первого деления, хромосомы
конденсируются, гомологичные хромосомы тесно сближаются
друг с другом по всей своей длине и через некоторое время час-
тично расходятся, оставаясь соединенными в нескольких точ-
ках — хиазмах. В каждой хиазме происходит обмен участками хро-
матид путем разрывов и последующих перекрестных соединений.
Этот процесс получил название кроссинговер. При кроссинговере
осуществляется обмен аллелями или блоками аллелей между го-
мологичными хромосомами, происходящими от комплементар-
ных родительских особей. После кроссинговера хромосомы оста-
ются прочно связанными до начала анафазы. К концу профазы
первого деления центриоли в клетках, где они имеются, расходят-
ся к полюсам, ядрышки исчезают, ядерная оболочка разрушается
и формируется веретено деления.
В метафазе первого деления мейоза пары гомологичных хромо-
сом выстраиваются в экваториальной плоскости клетки. К цент-
ромерам каждой из них прикреплены нити веретена, тянущиеся к
полюсам. Следующая фаза — анафаза I. В ней хромосомы, состоя-
щие из двух хроматид каждая, нитями веретена растаскиваются в
противоположные стороны. Телофаза завершает первое деление
мейоза. У животных и части растений в конце телофазы хромосо-
мы деспирализуются, восстанавливаются ядрышки и вокруг каж-
дой из новых хромосомных групп образуется ядерная оболочка.
Затем у животных происходит разделение цитоплазмы перетяж-
кой, а у растений — клеточной оболочкой. У некоторых растений
сразу же за анафазой I следует профаза второго деления.
Клетки животных, сформировав ядерные оболочки, вступают в
интерфазу, обычно короткую и без синтетического периода. Удво-
ения ДНК здесь не происходит. В остальном же второе деление
сходно с митозом и включает фазы, аналогичные по содержанию.
В профазе снова разрушается ядро, хроматиды укорачиваются и
утолщаются, центриоли перемещаются к противоположным по-
люсам, формируется веретено деления. Во второй метафазе цент-
ромеры хромосом выстраиваются в экваториальной плоскости
229
веретена и прикрепляются к его нитям. В анафазе нити тяну!' хро-
матиды за центромеры к полюсам, и с этого момента разделивши-
еся хроматиды становятся хромосомами. Образование дочерних
хромосомных групп во второй телофазе сопровождается разруше-
нием веретен, деспирализацией хромосом, восстановлением ядер-
ной оболочки и ядрышек. Завершается мейоз цитокинезом.
В зависимости от положения в жизненном цикле организма
мейоз или восстанавливает нормальное гаплоидное состояние
ядра с дополнительным перекомбинированием аллелей (зиготи-
ческая редукция при образовании гамет через митоз или транс-
формацию клетки), или же служит механизмом образования гап-
лоидных гамет, слияние которых приводит к восстановлению нор-
мального диплоидного состояния ядра.
При митозе из каждой родительской клетки образуются две до-
черние с неизменными набором хромосом и сочетанием аллелей,
а при мейозе — четыре клетки с уменьшенным вдвое набором хро-
мосом и новым сочетанием аллелей в каждой из них.
7.2. ВОСПРОИЗВОДСТВО
Экологическая ниша вида — это его место в пространственной
и функциональной структуре живой системы. Специфическая
функция вида обеспечивается всем множеством ценотически ак-
тивных частей, состояний, фаз и особей его организмов. Объем же
множества в большей или меньшей степени соответствует потреб-
ностям системы, или объему функции, и при устойчивом измене-
нии объема функции наличный объем множества испытывает
адекватную численную реакцию. Результатом реакции может быть
уменьшение или увеличение суммарного числа функциональных
элементов путем элиминации (уничтожения) некоторой их доли
или, наоборот, их размножением. Механизм реакции включает не
только прямое уничтожение и размножение, но также и измене-
ние соотношения между скоростями рождаемости и смертности.
Ответ видана изменение потребности может выражаться в прямой
численной реакции и в опосредованной — через усиление функ-
циональной эффективности. На субклеточном, клеточном и над-
клеточном уровнях механизмы численной реакции принципиаль-
но сходны.
Увеличение числа функциональных частей без изменения их
качественной стороны реализуется через матричный принцип
воспроизводства по типу штамповки. Этому принципу соответ-
ствуют механизм транскрипции — трансляции и механизм репли-
кации-митоза. Именно по этой причине простое увеличение
числа клеток-особей, клеток многоклеточных тел и многоклеточ-
230
ных особей базируется на митозе. Тела клеточного строения долж-
ны быть способны к увеличению собственной численности без
подключения механизмов генетической рекомбинации, если из-
менения в экосистеме требуют только количественного усиления
функции, выполняемой ими. Процесс увеличения числа особей в
дочернем поколении без реорганизации генома на надхромосом-
ном уровне, то есть без формирования специальных гаметных тел
и последующего оплодотворения, получил название «бесполое
размножение». Но размножение — это частный случай воспроиз-
водства, которое может и не сопровождаться увеличением числен-
ности. Поэтому в общем случае целесообразно пользоваться поня-
тием «бесполое воспроизводство».
Бесполое воспроизводство реализуется на разных
этапах онтогенеза организма, но, как правило, приурочено к его
ценотически активной составляющей. Его формы весьма разнооб-
разны и реализуются как через тела особей, так и через их специа-
лизированные части.
Рассмотрим основные формы бесполого размножения
(рис. 7.2). Деление — способ размножения, характерный для одно-
клеточных живых тел. Бинарное деление — образование двух одина-
ковых дочерних клеток из одной родительской. Множественное
деление (шизогония) — образование нескольких дочерних клеток из
одной родительской одновременно путем серии последователь-
ных митотических делений ядра и разделением такого «много-
ядерного плазмодия» на дочерние особи (характерно для простей-
ших типа Apicomplexa).
Спорообразование — образование одноклеточным или много-
клеточным родительским телом (обычно в специализированных
органах — спорангиях) путем митоза или мейоза одноклеточных
дочерних покоящихся, расселительных или инвазионных тел, в
дальнейшем прорастающих или трансформирующихся в вегета-
тивные или генеративные тела или гаметы. Различают несколько
форм спор:
зооспоры — подвижная фаза в одном из поколений особей про-
стейших, грибов и водорослей, как правило, трансформирующая-
ся в гаметы;
спорозоиды (и мерозоиды) — расселительно-инвазионная фаза в
поколениях у Apicomplexa, снабженная комплексом органоидов
проникновения (апикальным комплексом), трансформирующаяся
в вегетативную фазу — шизонт или генеративную фазу — гамонт;
зигоспоры — снабженные питательными веществами и толстой
клеточной оболочкой споры некоторых водорослей и грибов,
обеспечивающие переживание неблагоприятных условий и прора-
стающие в вегетативные тела;
споры (типичные) — одноклеточные, снабженные защитной
231
Рис. 7.2. Формы бесполого размножения (Хаусман, 1988; Рейвн и др., 1990;
оригинал автора):
7— бинарное деление у инфузории Tetrahymena sp.; 2— шизогония у споровика Plasmodium
sp,; 3— образование зооспор в процессе двух последовательных митозов у водоросли
Chlamidomonada sp.; 4 — образование зигоспоры после слияния гамет у грибов; 5 — спорогония
у споровика Eimeria sp. (образование споробластов в результате редукционного деления и пос-
ледующий митоз с образованием спор); <5— спора плауна; 7— спора сосны (Ми—микроспо-
ра, Me —мегаспора); 8— внутреннее почкование (эндодиогения) у кокцидий (Я —делящееся
ядро); 9— внутреннее почкование у сосущей инфузории Tokophrya cyclopwn (Ма—делящийся
макронуклеус, ВП—внутренняя почка); 70—наружное почкование у сосущей инфузории
Ephelota sp. (Ма—делящийся макронуклеус, НП —наружная почка); 11— внутренняя почка
(геммула) губки бадяги; /2—внутреннее почкование у личинки (мирацидия) сосальщика
fasciola bepatica (ЗК — зародышевые клетки); 13— наружное почкование у гидры (НП — поч-
ка); 14— внутреннее почкование личинки (ценура) ленточного червя; 15— почкование у по-
крытосеменного растения Bryophyllam sp.; 16— полиэмбриония (личинки хальцидоидного на-
ездника, развившиеся в гусенице яблонной плодожорки из одного отложенного яйца); 77-
фрагментация (эпитокия) у многощетянкового кольчатого червя Autolytus comutus; 18— меха-
низм полиэмбрионии у растений (с— двухклеточная стадия развития предзародыша, б— нор-
мальный ив - аномальный трехклеточные предзародыши, г — нормальный ид—аномальный
пятиклеточные предзародыши); 19—механизм полиэмбрионии у животных (а— стадия двух
клеток, б— разделение бластомеров, в — образование двух морул, г— стадия морулы, д—об-
разование бластоцисты); 20...30— органы вегетативного размножения (20...27) и возобновле-
ния жизнедеятельности (28...30): 20—луковица, 27 — клубнелуковица, 22—стеблевой клу-
бень, 23— корневище, 24 — столон, 25—плеть, 26— черенок, 27—отводок, 28— корневые
клубни, 29— эндоспоры, 30— мясистый стержневой корень
оболочкой расселительные и расселительно-инвазионные тела,
устойчивые к неблагоприятному действию внешних факторов у
грибов, моховидных и папоротникообразных, прорастающие в ве-
гетативные или генеративные тела;
микроспоры (пыльцевые зерна) и мегаспоры (зародышевые меш-
ки) — споры семенных растений, прорастающие в сильно редуци-
рованные мужской и женский гаметофиты, которые обеспечивают
половое воспроизводство.
Почкование — неравноценное деление родительской особи пу-
тем образования на теле нароста, выпячивания или обособления в
теле одиночных или собранных в группы специализированных
клеток, из которых развиваются функционально полноценные до-
черние особи. Почкование может быть завершенным и приводить
к формированию морфологически обособленных тел дочернего
поколения или незавершенным, приводящим к разрастанию
морфологически единого колониального тела с функционально
обособленными или необособленными особями материнских и
дочерних поколений. Выделяют две формы почкования:
внешнее — дочерняя особь формируется на открытой поверхно-
сти материнского тела. Встречается у одноклеточных грибов, се-
менных растений, инфузорий, губок, кишечнополостных и плос-
ких червей;
внутреннее—дочерние особи формируются внутри материнс-
кого тела и представляют собой покоящиеся, расселительные или
активные тела-особи, выход которых из материнского тела пред-
шествует его гибели или следует за ней. Эта форма воспроизвод-
ства близка к вегетативному размножению сосудистых растений.
Встречается у споровиков (эндодиогения), губок (образование
геммул и соритов) и плоских червей (педогенез у сосальщиков).
Полиэмбриония — образование однояйцевых близнецов при раз-
делении первых бластомеров в эмбриогенезе многоклеточных жи-
вотных. Встречается у паразитических насекомых отряда Пере-
пончатокрылые и у млекопитающих.
Фрагментация — разделение талломного или многоклеточного
родительского тела на части, восстанавливающиеся до завершен-
ных идентичных дочерних особей. Этот способ бесполого размно-
жения особенно характерен для нитчатых водорослей, грибов, ли-
шайников, ресничных плоских червей, многощетинковых кольча-
тых червей, немертин и морских звезд. Размножение высших рас-
тений искусственной фрагментацией широко распространено в
практической деятельности человека.
Вегетативное размножение — размножение специализирован-
ными вегетативными органами у сосудистых растений. Во многих
случаях оно напоминает внешнее почкование, но отличается от
него тем, что от материнского тела не отделяется вполне сформи-
234
рованная дочерняя особь, а только развивается специализирован-
ное тело, из которого позже при определенных условиях может
сформироваться тело дочерней особи. Часто эта форма воспроиз-
водства служит и для переживания неблагоприятного периода.
Выделяют следующие главнейшие органы вегетативного размно-
жения:
луковица — укороченный стебель с мясистыми листьями и моч-
коватой корневой системой, состоящей из придаточных корней.
Обычно она содержит одну или несколько дочерних луковиц —
«деток», каждая из которых может развиться в новое растение;
клубнелуковица — укороченный утолщенный вертикальный
подземный стебель с пленчатыми листьями и придаточными кор-
нями. Из пазушных почек также могут развиваться детки;
корневище — подземный, редко надземный, горизонтальный
побег с листьями и придаточными корнями, отходящими от каж-
дого узла;
столон — надземный горизонтальный стелющийся побег, обра-
зующий придаточные корни только в отдельных узлах и несущий
развитые листья. Разновидностью столонов считают плети, или
усы. Развитие дочерних особей происходит на быстро растущих
побегах с пленчатыми листьями. Укореняющиеся узлы чередуют-
ся с узлами, не образующими придаточные корни;
стеблевые клубни — утолщенные вершинные части корневищ с
пазушными почками, каждая из которых может дать начало ново-
му растению;
черенки и отводки — искусственно отделяемые части побегов с
последующим или предварительным укоренением, используемые
человеком для размножения древесных и кустарниковых растений.
К бесполому воспроизводству правомерно относить и такие его
формы, которые не сопровождаются увеличением численности
особей в дочернем поколении. Фактически это формирование
специализированных частей или органов, которые предназначены
для переживания неблагоприятного периода и за счет которых
происходит возобновление жизнедеятельности того же самого жи-
вого тела. Отметим следующие из них:
образование корневых клубней — накопление запасных пита-
тельных веществ в придаточных корнях. Возобновление тела рас-
тения происходит из почек, расположенных в основании старого
стебля;
образование «мясистых» стержневых корней. Возобновление
тела растения происходит из почек, находящихся в основании ста-
рого стебля, на верхней стороне утолщенного корня;
образование эндоспор бактерий. Эндоспоры — это покоящиеся
тела, формирующиеся по одной внутри материнских клеток и
прорастающие в новые вегетативные клетки.
235
Учитывая, что бесполое воспроизводство основывается прежде
всего на митотическом делении хромосомной группы ядра мате-
ринской клетки, не будет ошибкой отнести к нему и формы неза-
вершенного митоза: образование полиплоидных ядер (митоз без
кариокинеза и цитокинеза) и образование многоядерных клеток
(митоз с кариокинезом, но без цитокинеза).
Организмы, как было отмечено ранее, обладают свойствами
систем, и в частности способностью к самовоспроизведению в по-
колениях. Отдельные особи организма, если они представлены
обособленным комплементарным (половым) типом, обычно ока-
зываются способными только к бесполому воспроизводству-раз-
множению, что выражается в увеличении численности особей в
дочернем поколении при их генетической идентичности роди-
тельским особям.
Половое воспроизводство имеет качественно иной
характер: в его процессе реализуется интеграция той части внеш-
ней информации, которая была накоплена, «усвоена» отдельными
ценотически активными особями диффузно-комплементарных
организмов и записана в новых состояниях их генетических мат-
риц. Именно половое воспроизводство обеспечивает генетичес-
кую целостность видового живого вещества.
Полный онтогенез подавляющего числа видов организмов
включает и бесполое, и половое воспроизводство. Половое вос-
производство — это этап генетических реорганизаций, сводящий-
ся к комбинированию аллелей в хромосомах и комбинированию
хромосом при образовании дочерних кариотипов. Этот этап пред-
полагает образование специальных гаметных тел (гаметогенез), в
которых каждая из гомологичных хромосом представлена обособ-
ленно, и оплодотворение, центральным моментом которого в ти-
пичном варианте является слияние гамет (сингамия). В полноцен-
ном оплодотворении можно выделить три функциональных мо-
мента: 1) активацию женской гаметы (механическую, химичес-
кую, тепловую или иную); 2) объединение гаплоидных наборов
хромосом при слиянии ядер гамет, происходящих от комплемен-
тарных родительских особей, или в процессе первого незавершен-
ного митоза; 3) инициацию деления образующейся зиготы.
Гаметные тела, или гаметы, могут быть различными: кроме
наиболее известных — цитогамет (гаметных тел-клеток) известны
многоклеточные гаметные тела (сомогаметы), гаплоидные под-
вижные и стационарные ядра (нуклеогаметы) и пассивно подвиж-
ные хромосомы и фрагменты ДНК (хромогаметы). Гаметы могут
быть состоянием, фазой жизни клеточного тела, ядра или хромо-
сомы и образовываться путем трансформации, могут образовы-
ваться митотическим делением, если предшествующее тело гапло-
идно, или мейотическим делением, если оно диплоидно.
236
Цитогаметы могут быть одинаковыми морфологически, и тогда
их называют изогаметами. Изогаметы могут отличаться по биохи-
мическим и физиологическим свойствам. Такие гаметы встреча-
ются у грибов и простейших. Неодинаковые морфологические
признаки или черты поведения, если они не контрастны, свой-
ственны так называемым анизогаметам. Если же гаметы резко от-
личаются размерами и подвижностью, их называют спермием
(мужской комплементарный тип) и яйцом (женский комплемен-
тарный тип). Слияние гамет в этих случаях называют соответ-
ственно изогамией, анизогамией и оогамией (или овогамией).
У протестов клетки-особи, несущие нуклеогаметы, в пределах
одного вида образуют полярные комплементарные типы, которых
может быть от одной до 14 пар (например, у инфузории
Paramecium aurelia) или даже до 20 пар (у Paramecium bursaria).
При этом система спаривания (или конъюгации) может быть би-
полярной, как у большинства, когда особи могут спариваться
только с одним комплементарным типом, или мультиполярной,
когда каждая особь спаривается с любым типом особей, кроме
одного — к которому принадлежит сама. Цитогаметы обычно со-
ставляют два комплементарных типа, но иногда могут не иметь
мужского.
Особи с талломной и тканевой организацией тела образуют
гаметы в специализированных органах размножения: архегони-
ях, антеридиях и оогониях — низшие растения и ряд грибов; ре-
дуцированных гаметофитах — высшие растения; гонадах и слож-
ных половых железах — тканевые животные. Органы размноже-
ния могут быть частью тела ценотически активной особи, частью
тела ценотически пассивной особи или отдельным поколением
особей.
В зависимости от того, какие гаметы продуцирует особь, ее на-
зывают женской, мужской или гермафродитной. Гермафроди-
тизм — явление весьма сложное и в определенной мере комплекс-
ное. Так, применительно к плоским червям оно означает наличие
у каждой ценотически активной особи полного комплекта орга-
нов и мужской, и женской половой системы, а применительно к
моллюскам — наличие единого комплекта органов, производяще-
го разные типы гамет в разное время жизни. Хотя у некоторых
рыб, например у морского окуня, устойчивая смена половой ори-
ентации уже не связывается с гермафродитизмом. Этим же поня-
тием иногда обозначают и половые аномалии у животных, выра-
жающиеся в смешанном проявлении и мужских, и женских при-
знаков, даже если такие особи не способны продуцировать гаметы
(гинандроморфизм). У высших растений к гермафродитным осо-
бям относят однодомные растения, то есть растения с раздельны-
ми однополыми (мужскими и женскими) цветками, но гермафро-
237
дитными же называют и обоеполые цветки. Двудомность расте-
ний соответствует раздельнополости животных, а явления, анало-
гичного многодомности растений, у животных не отмечено. Мно-
годомные растения наряду с обоеполыми цветками имеют и одно-
полые, причем сочетание их на одном растении может быть раз-
личным: обоеполые и мужские (у чемерицы); обоеполые и
женские (у смолевки); обоеполые, мужские и женские (у конского
каштана); на одних растениях обоеполые, на других мужские, а на
третьих женские (у ясеня, винограда).
Оплодотворение может протекать в форме моноспермии, когда
на втором его этапе с яйцом сливается один сперматозоид, и в
форме полиспермии, когда в яйцеклетку проникают несколько
сперматозоидов. Чтобы соединиться с яйцом, сперматозоид дол-
жен проникнуть через яйцевые оболочки. Для этого у некоторых
типов яиц имеется специальное отверстие — микропиле. В других
случаях сперматозоид, вступив в контакт с яйцом, осуществляет
акросомальную реакцию: ферменты, содержащиеся в сперматозо-
иде, выделяются наружу, размягчают участок оболочки яйца и об-
легчают проникновение его головки к плазматической мембране.
Мембраны гамет сливаются, и одновременно происходит актива-
ция яйца. Объединение хромосомных групп в типичном случае
происходит при слиянии ядер и ведет к инициации деления зиго-
ты.
В случаях, когда сперматозоид не сливается с яйцеклеткой, оп-
лодотворение сводится только к активации яйца и инициации его
деления, что и соответствует партеногенезу. Такое оплодотворе-
ние может вызываться и сперматозоидами других видов организ-
мов и даже у некоторых видов различными абиотическими воз-
действиями. Партеногенез встречается в облигатной и факульта-
тивной формах. В первом случае самцы в популяциях отсутствуют
или очень редки. При циклическом партеногенезе развитие до-
черних поколений из женских гамет чередуется с типичным опло-
дотворением. Партеногенезу может предшествовать мейотическая
редукция числа хромосом, или же гаметы остаются диплоидными.
Из яйцеклеток при партеногенезе могут развиваться и самцы, и
самки (амфитокия), или только самцы (арренотокия), или только
самки (телитокия). Одна из разновидностей партеногенеза — пе-
догенез — способ размножения, при котором у личинок развива-
ются неоплодотворяемые яйца, дающие начало новому поколе-
нию. Педогенез встречается у двукрылых, жуков и ветвистоусых
ракообразных.
У цветковых растений оплодотворение имеет свои особенности
и называется двойным. Оно заключается в том, что при формиро-
вании семени оплодотворяется не только яйцеклетка, но и цент-
ральное ядро зародышевого мешка. Из диплоидной зиготы разви-
238
вается зародыш семени, а из трипловдной центральной клетки —
питательная ткань эндосперм. Сперматозоиды в этом случае не
способны к активному движению и передвигаются по пыльцевой
трубке. Пыльцевая трубка проникает в семязачаток через микро-
пиле, и ее ядро разрушается. Если пестик содержит несколько се-
мязачатков, то каждый из них оплодотворяется отдельным пыль-
цевым зерном (микроспорой).
Считается, что половое воспроизводство возникает на опре-
деленном этапе эволюции, когда митотическое деление клетки
уже было достаточно «отшлифованным», то есть половое вос-
производство вторично по отношению к бесполому. Важней-
шим приобретением, связанным с половым процессом, соглас-
но традиционным представлениям, является возможность ком-
бинирования генетической информации (путем слияния клеток
особей и последующего восстановления нормального хромо-
сомного набора с помощью редукционного деления). Образова-
ние специализированных гаметных тел относят к более поздне-
му периоду.
Первичность воспроизводства поколений особей вида, осно-
ванного на митозе, спорна: обособление и длительное воспро-
изводство вида при сохранении его целостности в этом случае
невозможно, так как многократное митотическое деление ведет
к формированию «веера» генетических линий, не образующих
однородной обособленности, без чего невозможна функцио-
нальная определенность вида. Исходное половое воспроизвод-
ство в отсутствие бесполого также невозможно, в том числе из-
за необходимости обеспечения чисто количественной реакции
видов на изменения в экосистемах. В качестве логически воз-
можного варианта можно допустить одновременное развитие и
митоза, и мейоза как равноценных специализированных спосо-
бов деления клетки из более примитивного предшествующего
им способа при становлении жизненных циклов организмов
(рис. 7.3).
Дифференциация способов деления клетки имела бы смысл
только в том случае, если бы каждый из них исходно имел само-
стоятельное значение. Ранее было показано, что митоз обеспечи-
вает не только построение многоклеточного тела, но также увели-
чение числа ценотически активных клеток или вспомогательных
клеток, идентичных исходным клеткам в данном поколении орга-
низма. И то и другое направлено на усиление ценотической ак-
тивности видового живого вещества. Мейоз же в сочетании с оп-
лодотворением обеспечивает генетическую рекомбинацию, обоб-
щение информации индивидуальных тел и выбраковку «запре-
щенных» генетических комбинаций. Следовательно, только
сочетание обеих форм воспроизводства в полном жизненном цик-
239
Рис. 7.3. Гипотетическая схема одновременного становления митоза и мейоза
(оригинал автора):
1— последовательность расхождения хроматид у прокариот; II— разделение длинной кольце-
вой хромосомы на два фрагмента может вести к одинаковому гомозиготному результату в ми-
тотическом и мейотическом путях воспроизводства гаплоидного организма; III— разделение
длинной кольцевой хромосомы на четыре фрагмента увеличивает относительное число комби-
наций гомозиготного генотипа в мейотическом пути воспроизводства гаплоидного организма;
IV— разделение д линной кольцевой хромосомы на два фрагмента с последующим образовани-
ем дублирующих гомологичных хромосом позволяет резко усилить комбинативные возможно-
сти мейотического пути воспроизводсгва диплоидных организмов с образованием гетерозигот-
ных генотипов
ле организма обеспечивает морфофункциональную целостность и
определенность биологического вида. И тогда проблема становле-
ния форм воспроизводства сопрягается с проблемой становления
биологических видов или дифференцировки живого вещества
биосферы на видовые составляющие.
16 Присный А В.
241
7.3. ОНТОГЕНЕЗ
Под онтогенезом понимают индивидуальное развитие особи от
зарождения (деления материнской клетки, оплодотворения яй-
цеклетки или начала обособленной жизни органа вегетативного
размножения) до конца жизни (индивидуальной смерти или ново-
го деления).
Существует несколько гипотез индивидуального развития организмов.
«Овизм» — система взглядов, сформировавшаяся к концу XVII в. в основном бла-
годаря работам Ш. Бонне и А. Валлиснери. Считалось, что в женской половой
клетке в микроскопическом виде содержится взрослый организм, а его развитие
сводится лишь к увеличению в размерах. Овизм был распространен более широ-
ко, чем «анималькулизм» — противоположная система взглядов, сторонником ко-
торой был А. Левенгук. Анималькулисты считали, что яйцо служит лишь пита-
тельной средой для развивающихся из мужских половых клеток животных, кото-
рые в сперматозоидах уменьшены до невидимых размеров. Овизм и анимальку-
лизм — разновидности более емкого учения о преформации, или «преформизма».
Преформизм—это учение о наличии в половых клетках организмов материаль-
ных структур, предопределяющих развитие зародыша и признаки организма, раз-
вивающегося из зародыша.
Преформистские взгляды складывались в течение длительного времени. До
начала XVIII в. господствовало мнение о наличии в половых клетках полностью
сформированного зародыша. Теоретическая основа преформизма заключена в
креационистских представлениях, то есть в догме изначального творения живых
существ и всех будущих поколений, заложенных в них.
Во второй половине XVIII в. под влиянием данных о нарушениях развития, о
наследуемости как материнских, так и отцовских признаков и о способности
организмов к регенерации в биологии развития получило преобладание учение о
последовательных новообразованиях — «эпигенез». Особенно активно это учение
развивали К. Ф. Вольф и Ж. Л. Бюффон.
Когда была выяснена сущность процессов оплодотворения и клеточного деле-
ния (во второй половине XIX в.), возникли теории наследственности, что дало
новый толчок к развитию преформистских взглядов. Была доказана несостоятель-
ность представлений о развитии организмов под влиянием лишь внешних усло-
вий, что отстаивали сторонники «эктогенеза» (разновидность «эпигенеза»), или
каких-то нематериальных факторов, что пропагандировалось «виталистами».
Учение об индивидуальности хромосом и их роли в процессах оплодотворения
и наследования признаков породило в конце XIX в. ряд гипотез о закономернос-
тях наследственности и изменчивости, носивших в той или иной мере преформи-
стский характер. С возникновением генетики эти гипотезы получили серьезное
научное обоснование. В середине XXв., когда была выяснена химическая природа
генов, механизмов хранения и передачи наследственной информации, префор-
мизм практически слился с генетикой. Не было тем не менее забыто и эпигенети-
ческое учение, и современная теория онтогенетического развития обязательно
учитывает воздействие на развивающийся фенотип факторов внешней среды.
У многоклеточных животных в онтогенезе принято выделять
следующие основные этапы: предзародышевый (гаметогенез и оп-
лодотворение), зародышевый (до выхода особи из яйцевых или за-
родышевых оболочек), послезародышевый (до достижения поло-
вой зрелости), взрослое состояние, включая старение. Такая пери-
одизация непригодна для описания развития животных со слож-
242
ным жизненным циклом, например кишечнополостных, дигене-
тических сосальщиков и др. Но и в случаях с простыми жизнен-
ными циклами схема онтогенеза может выглядеть по-разному, в
связи с чем различают три типа онтогенеза животных: личиноч-
ный — когда после раннего выхода из яйцевых оболочек особь не-
которое время живет в форме личинки, существенно отличаю-
щейся от взрослой формы; яйцекладный — когда зародыш дли-
тельное время развивается внутри яйца и личиночная стадия от-
сутствует; внутриутробный — когда оплодотворенное яйцо
задерживается в яйцевых путях материнской особи и при этом
возникает связь тканей зародыша и материнской особи через пла-
центу.
У сосудистых растений, размножающихся половым путем, он-
тогенез также начинается с предзародышевого периода. Далее сле-
дуют периоды: эмбриональный, ювенильный, зрелости, размно-
жения и старости. Поскольку из оплодотворенной яйцеклетки
развивается спорофит, а в жизненном цикле растений присутству-
Рис. 7.4. Примеры дифференциации организмов на особи (организмы выделены пунк-
тирными линиями, особи —сплошными) (оригинал автора):
7—прокариоты; 2— диплоидные протесты с половым воспроизводством; 3—споровики; 4—
многоклеточные животные с бесполым и половым воспроизводством; 5—человек
ет еще и гаметофит, причем часто как отдельное поколение осо-
бей, то здесь следует дополнительно выделить периоды спороге-
неза, развития гаметофита и формирования органов полового раз-
множения.
Индивидуальные онтогенезы, или онтогенезы особей, пред-
ставляют собой лишь фрагменты полных онтогенезов организмов,
обычно обозначаемых как жизненные циклы. Полный онтогенез
можно определить как последовательность морфогенетических
процессов в телах, составляющих организм. Набор таких тел огра-
ничивается жизненным циклом организма, куда входят, как пол-
ностью или полуобособленные тела, фазы или части: гаметы, зи-
Интеграция накопленной информации
и передача ее новому поколению организмов
Реализация полученной генетической информации
и накопление новой ценотической информации
Рис. 7.5. Объединенная схема жизненных циклов организмов (оригинал автора)
244
готы, эмбрионы, ценотически активные (вегетативные) тела и
гаметообразующие тела всех комплементарных типов (рис. 7.4).
В жизненном цикле происходит чередование половой и бесполой
форм воспроизводства. Несмотря на чрезвычайное разнообразие
конкретных жизненных циклов организмов в разных системати-
ческих группах, представляется возможным изобразить их в ваде
единой схемы (рис. 7.5), в которой отчетливо прослеживаются
главнейшие закономерности онтогенезов, назначение их этапов,
параллельность развития внутренней информации вида и ее веще-
ственного воплощения и сопряженность вида с экосистемой, в
которую он входит.
7.4. ОСОБЕННОСТИ ОНТОГЕНЕЗА ЧЕЛОВЕКА
Этапы, периоды и фазы онтогенеза. В общем случае в онтогенезе
человека принято выделять два главных этапа: пренатальный (до-
родовой) и постнатальный (послеродовой), разделенные родами.
Роды представляют собой процесс смены среды обитания дочер-
него тела и переход его от полной зависимости от материнского
тела как посредника между ним и внешней средой к относитель-
ной самостоятельности в осуществлении питания, газообмена,
движения, регуляции параметров внутренней среды.
Иной вариант периодизации онтогенеза человека отражает его
способность в определенном биологическом возрасте к половому
воспроизводству. Здесь выделяют дорепродуктивный, репродук-
тивный и пострепродуктивный периоды, которые отличаются ха-
рактером морфогенетических процессов, степенью устойчивости
функционирования отдельных органов и систем органов, чувстви-
тельностью к внешним воздействиям, приспособительными воз-
можностями.
Дорепродуктивный период, в свою очередь, разделяют на эмб-
риональный (зародышевый), личиночный, период метаморфоза и
ювенильный период. Их понимание применительно к человеку
неоднозначно. Так, в одних случаях считается, что у большинства
млекопитающих, включая человека, эмбриональный период уко-
роченный и соответствует промежутку времени от оплодотворе-
ния до начала гаструляции, когда еще сохраняется яйцевая обо-
лочка. Следующий за ним личиночный период с наиболее интен-
сивными морфогенетическими процессами и характерными для
него чертами эмбрионизации и временными органами завершает-
ся с началом родов, которые принимаются за метаморфоз. Мета-
морфоз захватывает и первую послеродовую неделю. Далее идет
продолжительный ювенильный период, предшествующий репро-
дуктивному периоду, когда завершаются развитие скелета, уста-
245
новление гормонального баланса, развитие вторичных половых
признаков.
По другим представлениям развитие млекопитающих идет по
«прямому» типу, который характеризуется отсутствием периодов
личинки и метаморфоза. В этом случае продолжительность и со-
держание эмбрионального периода практически соответствуют
внутриутробному развитию.
Специфические особенности человека, связанные с его соци-
альностью, несколько усложняют общую периодизацию онтогене-
за из-за совмещения его двух аспектов: биологического и социаль-
ного. Так, постнатальная часть дорепродуктивного периода до-
полнительно разделяется на младенчество, детство, отрочество и
юность —фазы, отличающиеся психофизиологическими и мор-
фоанатомическими особенностями, характером усваиваемой со-
циальной информации и способом ее усвоения.
Старение и смерть как биологические явления. В индивидуаль-
ном развитии человека принято выделять специфическую фазу,
сопровождающуюся закономерными изменениями в физическом
состоянии и эмоциональной сфере и предшествующую смерти.
Эта фаза обозначается как старость и имеет ряд характерных внеш-
них проявлений. Хронологически старость соответствует возрасту,
превышающему 75 лет, хотя соответствующие ей отдельные био-
логические признаки могут появиться в зрелом и даже молодом
возрасте (поседение волос, появление морщин, специфическое
соотношение половых гормонов и др.). Состояние старости дости-
гается благодаря изменениям на всех структурных уровнях: моле-
кулярном (уменьшение интенсивности репарации повреждений
ДНК, снижение транскрипционной активности, утрата части ге-
нов рибосомной РНК и др.), субклеточном (снижение количества
и активности митохондрий, сокращение объема шероховатой
ЭПС), клеточном (наличие предельной кратности клеточных де-
лений), тканевом (замещение соединительной тканью мышечной
и других тканей, атрофия тканей) и органном (функциональные
расстройства сердечно-сосудистой, дыхательной, выделительной,
нервной, гуморальной систем и др.).
Суммарный результат частных проявлений старения выра-
жается в нарастающем снижении жизнеспособности особи,
уменьшении эффективности приспособительных и гомеоста-
тических механизмов. В целом старение приводит к прогрес-
сивному повышению вероятности смерти, делает ее неизбеж-
ной. Наступлению биологической смерти нередко предше-
ствует состояние клинической смерти, когда клетки и ткани
еще сохраняют уровень жизнеспособности, позволяющий при
определенных внешних воздействиях возобновить жизнедея-
тельность организма.
246
Существует несколько гипотез, объясняющих механизмы ста-
рения. Среди них достаточно аргументированы две: 1) гипотеза
возрастного накопления повреждений биологических структур и
механизмов под действием спонтанных внутренних, а также внеш-
них факторов; 2) гипотеза генетической программы, включающая
гипотезу биологических часов.
Индивидуальную смерть можно рассматривать как проявление
разрушительной фазы (другая фаза — восстановительная) в обще-
биосферном процессе обновления видового живого вещества.
Глава 8
ОСНОВЫ СИСТЕМАТИКИ
ЖИВЫХ ОРГАНИЗМОВ
8.1. ВВЕДЕНИЕ В ТАКСОНОМИЮ
В своей практической деятельности человек прямо или косвен-
но взаимодействует с остальным живым веществом биосферы. В
настоящее время известно 1,5...2,0 млн видов организмов («чис-
тых» подразделений живого вещества), а по некоторым оценкам,
на Земле существует от 30 до 80 млн видов. Ориентироваться в та-
ком многообразии несистематизированного материала без специ-
ального «рабочего инструмента» практически невозможно. В ка-
честве такого инструмента выступает классификация — условное
распределение живых организмов по иерархически соподчинен-
ным группам в соответствии с какими-либо признаками.
Любой вариант распределения организмов предполагает их
тщательное предварительное изучение. Существует специальный
раздел биологии — систематика, который занимается описанием,
обозначением (приданием названия) и классификацией по таксо-
нам, или систематическим группам, всех известных существую-
щих и вымерших организмов, установлением родственных связей
между отдельными видами и группами видов. Цель системати-
ки — создание всеобщей естественной классификационной систе-
мы органического мира с установлением соподчиненности таксо-
нов различного ранга. Под естественностью здесь следует пони-
мать правильное отражение существующей в природе дифферен-
цировки живого вещества как результата его эволюционных
преобразований. Все разработанные до настоящего времени клас-
сификационные системы в большей или меньшей степени искус-
ственны, так как они основаны на учете ограниченного числа ди-
агностических признаков, и сама классификация предполагает
обозначение условных, искусственных границ в едином, непре-
рывном в пространстве и во времени, живом веществе.
Составной частью систематики, устанавливающей ее методоло-
гию, является таксономия — теория научной систематики и клас-
сификации, учение о соподчинении таксонов.
В качестве узловых систематических единиц — таксонов —
приняты вид, род, семейство, отряд или порядок, класс, тип или
248
отдел, царство и империя или доминион. Кроме них устанавлива-
ются промежуточные таксоны: подвид, надвид, подрод, триба,
подсемейство, надсемейство, подотряд, надотряд, подкласс, над-
класс, подтип, раздел, подцарство и некоторые дополнительные
подразделения и объединения.
Основным таксоном считается вид. Остальные же таксоны,
вплоть до типа, устанавливаются на принципе предполагаемого
или доказанного родства. Более емкие таксоны лишь подчеркива-
ют элементы сходства морфологической организации представи-
телей типов.
До настоящего времени нерешенной остается проблема раз-
мерности одноименных таксонов в разных группах организмов. Ее
следствием стало появление «бестаксонных» систем, отличающих-
ся от традиционных прежде всего отсутствием строгого количе-
ства ступеней — узловых таксонов в «лестнице существ». В значи-
тельной мере проблемы систематики отражают проблемы, суще-
ствующие в теории вида.
8.2. ПРЕДСТАВЛЕНИЕ О ВИДЕ
Проблема вида — одна из сложнейших в биологии. Ее нельзя
решать в отрыве от проблемы жизни как явления вообще и про-
блемы живого вещества как составной части живой системы-био-
сферы в частности. Тем не менее в науке сложилось определенное
представление о виде, относительно удобное для использования в
теоретических построениях и на практике.
Впервые термин «вид» применен Аристотелем для обозначения
группы сходных между собой организмов. Но он рассматривал вид
как частную категорию по отношению к более общей — роду, то
есть понятие «вид» носило у Аристотеля чисто логический харак-
тер. Представление о виде как о биологической категории ввел
Дж. Рей. Он же ввел бинарную номенклатуру — двойное название
видов. Однако основателем и науки систематики, и теории вида
считается К. Линней. Линней первым сформулировал представле-
ние о виде как об универсальной, дискретной и объективно суще-
ствующей в природе, то есть реальной, категории. Основные кри-
терии вида, по Линнею, морфологический и репродуктивный. (Он
придерживался креационистских взглядов и лишь в конце жизни
признавал возможность возникновения новых вариаций при гиб-
ридизации.) В практической систематике теоретические взгляды
К. Линнея преломлялись в так называемую «типологическую мор-
фологическую концепцию вида». Этот подход сохраняется в сис-
тематике и до настоящего времени.
Ж. Б. Ламарк, исходя из представлений об изменяемости ви-
249
дов, разработал основы эволюционной гипотезы, но в то же вре-
мя отказался от концепции дискретности видов и отрицал их ре-
альность. Согласно такому «номиналистическому» подходу вид
существует как номинальная (определяемая), а не как реальная
категория.
Ч. Дарвин поставил вопрос о виде как исторически возникаю-
щей и изменяющейся совокупности организмов. Кыда было уста-
новлено, что вид обладает сложной структурой, были предприня-
ты попытки найти «элементарный» вид. В 1873 г. Д. Жордан со-
здал учение об элементарном виде. Возникло представление о
двух типах видов: «линеоне» — виде со сложной структурой и
«жорданоне» — виде с простой структурой.
В 1908 г. С. П. Семенов-Тян-Шанский сформулировал концеп-
цию «политипического вида», в соответствии с которой вид — си-
стема, не только возникающая и изменяющаяся, но и непрерывно
развивающаяся путем внутренней дифференцировки.
Н. И. Вавилов (1931) показал несостоятельность «жорданона»,
установив, что внутривидовое разнообразие определяется не толь-
ко степенью становления вида, но также разнородностью условий
среды: в этом проявляется адаптивность вида к среде обитания.
Доказательство реальности вида Н. И. Вавилов видел в реальном
участии его в (вещественно-энергетических) биосферных процес-
сах. К середине XX в. оформилась «политипическая морфологи-
ческая концепция вида».
В 1968 г. Э. Майр предложил «биологическую концепцию»
вида, суть которой заключается в следующем: виды определя-
ются не различием, а обособленностью; виды состоят не из не-
зависимых особей, а из популяций; виды репродуктивно изоли-
рованы друг от друга. Биологическая концепция не применима
для выделения видов в случаях с бесполым размножением и са-
моопылением, вымерших форм и при рассмотрении вида во
времени.
На сегодняшний день не сформулировано единого общеприня-
того определения вида, но, суммируя основные из них, можно
заключить, что вид — это множество организмов, сходных морфо-
логически, занимающих одинаковую экологическую нишу, имею-
щих общую эволюционную судьбу, образующих замкнутую гене-
тическую систему и занимающих определенный ареал. Такое оп-
ределение отражает характерные особенности вида, сформулиро-
ванные на основе его «критериев». Последние принято разделять
на первичные и вторичные (производные от первичных).
К первичным критериям вида относят:
эволюционный: вид как система популяций имеет единую эво-
люционную судьбу (Шмальгаузен, 1946);
генетический: вид представляет собой генетически замкнутую,
250
репродуктивно изолированную систему, состоящую из генетичес-
ки открытых (потенциально способных обмениваться между со-
бой генами) подсистем-популяций;
экологический (правило ниш): два вида не могут занимать одну
и ту же экологическую нишу на данной территории (Гаузе, 1934);
географический: вид обладает ограниченным ареалом террито-
рией с определенными абиотическими и биотическими условия-
ми, к которым вид оказывается адаптированным (Коржинский,
1892).
На практике для выделения видов пользоваться первичными
критериями трудно, поэтому обычно применяют производные
критерии. В их основе лежит соответствие формы и функции, вы-
раженное в определенных признаках.
Вторичные критерии вида включают:
морфологический: как правило, разные ввды имеют внешние и
внутренние морфологические отличия;
кариологический: генетическое своеобразие вида проявляется в
особенностях организации его кариотипа, количестве и строении
хромосом;
иммунологический: каждый вид специфичен по составу бел-
ков, что может быть выявлено серологическими тестами;
физиолого-биохимический: вид характеризуется особенностя-
ми протекания жизненных процессов на уровне организма (этот
критерий на практике используют редко).
В предыдущих разделах отмечалось, что каждый вид обладает
характерным для него жизненным циклом, в рамках которого
происходят определенные морфогенетические процессы, смена
фаз, состояний, поколений тел-особей, изменения в проявле-
нии взаимоотношений со средой, чередование способов вос-
производства. При этом подчеркивался диффузно-комплемен-
тарный принцип построения организма, исключающий нали-
чие резких границ между организмом и популяцией (видом).
Сказанное позволяет трактовать вид как множество живых тел,
объединяемых определенным устойчивым типом онтогенеза.
Каждый такой онтогенетический тип — уникальное выражение
элементарной биосферной информации, записанной на мате-
риальных носителях — генетических матрицах, а также в сово-
купности отношений и связей, составляющей идеальную сферу
живой системы.
Любому виду, получившему научное описание, присваивают
название, отвечающее «принципу бинарной номенклатуры». Вид
должен иметь название на латинском языке, состоящее из двух
слов: первое — название рода, к которому отнесен вид — обычно
существительное, второе — видовое название, чаще прилагатель-
ное. Естественно, не исключается придание видам названий на
251
«живых» языках, как научных, так и бытовых. Например, насеко-
мое с латинским научным названием Gryllotalpa gryllotalpa Linne,
1758 (буквально — сверчок + крот повторяется и в названии рода,
и в названии вида) имеет русское научное название «медведка
обыкновенная», а одно из местных бытовых названий — «волчок».
После названия вида записывают фамилию автора, сделавшего его
описание, и год описания.
Вид признан единственной систематической категорией, суще-
ствующей реально. В концепции вида выделено несколько крите-
риев его реальности.
1. Обособляющаяся популяция характеризуется общностью
происхождения от материнской популяции и имеющимся у мате-
ринской популяции типом генома. Общность организации генома
определяет единство потенциальных возможностей преобразова-
ния данной группировки, а конкретные условия среды — направ-
ление преобразования.
2. Все популяции вида занимают одну и ту же экологическую
нишу (пространственное и функциональное место в экосистеме),
поэтому все они взаимозамещаемы. Популяции разных видов не
могут заменить друг друга: они либо сосуществуют, занимая раз-
ные экологические ниши, либо вытесняют одни других. Экологи-
ческая ниша предполагает непосредственное участие всей сово-
купности организмов вида в определенных вещественных и энер-
гетических процессах.
3. Вид состоит из популяций, и унаследованная от предков общ-
ность видоспецифичных генов поддерживается через обмен между
популяциями.
В пределах ареала особи вида распределены неравномерно.
Разнородность абиотических условий и разнообразие подразделе-
ний биосферы, в каждом из которых создаются специфические
биотические условия, с течением времени приводят к появлению
в пределах вида устойчивых, относительно обособленных друг от
друга группировок. Это популяции. Степень изолированности по-
пуляций зависит от степени расчлененности объема среды в гра-
ницах ареала и экологической пластичности вида. Межпопуляци-
онные барьеры могут быть «прозрачными», то есть легко проница-
емыми и не препятствовать обмену аллелями, или же, наоборот,
трудно преодолимыми, изолирующими отдельные популяции и
приводящие к накоплению в них оригинальных комплексов гене-
тических новообразований. Изолированные популяции часто
имеют собственную эволюционную судьбу, отличную от судьбы
основных популяций вида, Микроэволюционные процессы в слу-
чае наличия потребности в биогеоценозе (свободной экониши) и
достаточной пластичности популяции обусловливают развитие
адаптивной формы, сохраняющей связь с материнским видом, но
252
уже имеющей морфологические или биоэкологические отличия.
Так образуются экоморфы, географические расы, подвиды. Когда
же устойчивые отличия охватывают и генетическую систему попу-
ляции, дочерние группировки вида могут претендовать на статус
полувидов.
В иерархии внутривидовых структурных подразделений — по-
лувид, подвид, географическая раса, экоморфа, популяция —
главным подразделением является популяция. Буквальное русское
значение термина «популяция» — население — отражает скорее
демографический аспект. Определение популяции как «совокуп-
ности особей одного вида с общим генофондом, обитающих в
пространстве с относительно однородными условиями», не позво-
ляет сколь-нибудь четко обозначать границы таких внутривидо-
вых подразделений в реальных условиях. Поэтому этот термин ча-
сто используется произвольно для обозначения населения условно
ограничиваемого объема среды, хотя фактически оказывается
пригодным лишь для обозначения группировок «островного
типа», то есть изолированных популяций. Для преодоления не-
определенности и в случаях, когда надо подчеркнуть структурно-
иерархический аспект, используют понятие «биологическая попу-
ляция». В этом случае под популяцией понимают способную к са-
мовоспроизведению, существующую в длительном ряду поколе-
ний совокупность организмов одного вида, входящую в состав
экосистемы некоторого уровня и реализующую здесь видоспеци-
фичную функцию, то есть занимающую собственную экологичес-
кую нишу. Генофонды разных биологических популяций одного
вида отличаются устойчиво поддерживающимися частотами от-
дельных аллелей или групп аллелей, что является отражением
своеобразия соотношения функциональной формы вида и локаль-
ных особенностей внешних условий.
Популяция имеет собственную сложную структуру: в ней при-
нято выделять группы особей по возрастному и половому призна-
кам, степени и характеру активности, размеру, генетическим осо-
бенностям, признаку родства. Иногда в популяциях выделяют
временные устойчивые объединения особей, такие, как стая, ста-
до, косяк, колония, скопление и др.
Не противоречит общим представлениям о популяции и пред-
ставление об организме как сложном подразделении видового жи-
вого вещества с диффузно-комплементарным типом строения.
Более того, только так можно объяснить целостность популяции.
Отсутствие четких понятийных границ между организмом, попу-
ляцией и видом отражает реальную взаимозамещаемость одно-
именных особей (гамет, половых комплементарных особей, деис-
тически активных особей) в популяции, а популяций в пределах
вида.
253
8.3. ПРОСТРАНСТВО И ВРЕМЯ ВИДА
Существует проблема, касающаяся объяснения исходного сти-
мула к образованию новой формы, превращающейся в новый вид.
Как различающиеся, противоположные скорее по форме, чем по
содержанию, можно принять во внимание следующие гипотезы.
1. Стимулом является конкуренция, которая через микроэво-
люционные процессы увеличивает приспособленность популяций
или внутрипопуляционных групп к меняющимся условиям, что
позволяет таким популяциям или группам «завоевывать собствен-
ное место под солнцем».
2. Стимулом являются изменения в устойчиво развивающейся
экосистеме, которая из материала, поставляемого микроэволюци-
онными процессами, «отбирает» удовлетворяющие ее формы.
Соответственно этим объяснениям по-разному представляется
и последовательность процессов и событий, приводящая к оформ-
лению нового вида.
I. Индивидуальная конкуренция вызывает расчленение вида на
группы -> группы расходятся по формируемым или собственным,
экологическим нишам -> межгрупповая конкуренция ведет к спе-
циализации групп и далее к образованию новых подвидов и ви-
дов -> конкуренция снижается при стабилизации видов.
2. Изменения в экосистеме порождают потребность в новой
функции (развивается «дефицит функции») -> оформляется фун-
даментальная экологическая ниша — комплекс экосистемных тре-
бований к новому функциональному элементу—> одна или не-
сколько группировок существующих видов на основе микроэво-
люционных процессов претендуют на занятие свободной ни-
ши -> группировка или группировки, способные дать требуемые
жизненные формы, заполняют нишу, превращая ее в реализован-
ную,-> новая жизненная форма закрепляется генотипом -> зак-
репление взаимообусловленности функции и формы в онтогенезе
свидетельствует о становлении нового вида.
Вымирание отдельных видов в рамках этих гипотез связывается
или с конкурентным подавлением, или с превращением в другие
виды, или же с исчезновением соответствующей системной функ-
ции в процессе развития биосферы. Причинами массовых выми-
раний могут быть глобальные или крупные катастрофические яв-
ления.
На каждый конкретный момент времени видовое живое веще-
ство занимает некоторый объем пространства. Но вид на таком
срезе времени не существует, поскольку вид — это морфофункци-
ональная единица. Функция же немыслима без процессов, как ма-
териальных, так и идеальных, которые выражаются в динамике
состояний живого вещества. Живое вещество в процессе реализа-
254
ции биосферной информации непрерывно самовозобновляется.
Каждое новое состояние вещества возникает на основе предыду-
щего и служит основой для последующего. Формируется причин-
но-следственный рад, который представляет собой проявление
времени. Таким образом, вид существует только в единстве про-
странства и времени.
Видовые популяции в реальных экосистемах устойчиво суще-
ствуют лишь при определенных сочетаниях большого числа так
называемых «благоприятных» условий. Важнейшие аспекты этих
условий определяют в каждом конкретном случае и возможность,
и необходимость существования данной популяции в данной эко-
системе, то есть наличие экологической ниши.
Термин ниша был введен в науку в 1917 г. Дж. Гриннеллом для
обозначения самой мелкой единицы распространения вида и оп-
ределял потенциальный характер распространения отдельного
вида в условиях отсутствия взаимодействия с другими видами. В
1927 г. Ч. Элтон трактовал нишу как место определенного вида в
биотической среде в смысле его пищевых связей и взаимодей-
ствий с врагами, подчеркивая реальный функциональный аспект.
В 20—40-е годы XX в. Лотка, Вольтерра, Гаузе и Парк в своих ра-
ботах сформулировали представление о конкурентном исключе-
нии, следствием из которого явилось «правило экологических
ниш» (два разных вида в одном сообществе не могут занимать
одну и ту же экологическую нишу). В следующее десятилетие по-
лучило распространение понимание ниши как «профессии» орга-
низма в конкретном сообществе.
Г. Е. Хатчинсон (1958) дал формальное, потенциально количе-
ственное трактование ниши. По его мнению, нишу следует опре-
делять с учетом всего диапазона физических, химических и биоти-
ческих переменных среды, к которым должен быть адаптирован
(приспособлен) вид и под действием которых видовая популяция
живет и возобновляется бесконечно долгое время. Градиент каж-
дого фактора среды в этом случае можно представить как некое
измерение пространства. Если ниша имеет несколько значимых
измерений, то ее можно описать в терминах многомерного про-
странства (гиперпространства).
Хатчинсон различает два состояния видовой ниши: фундамен-
тальную и реализованную. Первая предполагает все множество
оптимальных условий, в которых вид может обитать в отсутствие
врагов; вторая — фактический комплекс условий, в которых этот
вид обычно существует.
Если ниши дискретны, то есть если между фундаментальными
нишами нет перекрывания, видовое богатство сообщества зависит
от общего занимаемого ими пространства и от среднего размера
каждой ниши. Поскольку все разнообразие ресурсов, используе-
255
мых видовой популяцией, с их градиентами учесть на практике
невозможно, нишу стали отождествлять с распределением актив-
ности вида по одной или нескольким наиболее важным или наи-
более доступным для измерения из «осей» ниши. Ширина ниши
по каждой из осей слагается из двух составляющих: внутрифено-
типической компоненты, характеризующей уровень изменчивос-
ти в использовании ресурса отдельными особями, и межфеноти-
пической компоненты, определяющей изменчивость по этому па-
раметру среди особей всей видовой популяции.
В реальных сообществах происходит частичное перекрывание
фундаментальных ниш, то есть на каждый ресурс претендует не-
сколько видов. Предполагается, что существует обратная зависи-
мость между перекрыванием ниш и конкуренцией. Перекрывание
ниш по нескольким градиентам ресурсов приводит к диффузной
конкуренции и, как следствие, сильному сокращению реализо-
ванной ниши.
Ниша представляется как изменчивая в пространстве — време-
ни. При этом изменения, постепенные или резкие, могут проис-
ходить в пределах единичного онтогенеза (индивидуального раз-
вития) или в пределах последовательного ряда онтогенезов.
Существуют различные мнения о механизме формирования
ниш. Согласно одному из них ниша порождается тем, кто ее зани-
мает, другому — ниша представляет свойство данного сообщества
и вне его лишена смысла. В последнем случае ниша формируется
абиотическими и биотическими компонентами данного сообще-
ства и заполняется в результате адаптации видов в ходе эволюции.
Этим объясняются и возникновение похожих ниш в разных эко-
системах с близкими условиями, и сходства в адаптациях популя-
ций, занимающих аналогичные ниши. (Экологическая эквивален-
тность, конвергенции доказывают порождение ниши сообще-
ством.)
В целом в настоящее время экологическую нишу определяют как
пространственное и функциональное место, которое может быть
занято или занимается конкретным видом в конкретном сообще-
стве. Учитывая, что пространственное место —это фактическое
сочетание условий и прежде всего ресурсов, использование кото-
рых представляет функциональную сущность видовой популяции,
экониша отражает потребность в реализации определенной функ-
ции в экосистеме, а следовательно, возможность и необходимость
включения в сообщество видового живого вещества с соответству-
ющей жизненной формой ценотически активных особей.
Становление реализуемой функции может быть объяснено с
двух разных позиций: как активное, через уход от конкуренции
с новообразованием собственной функции (популяционный
подход), и как пассивное, через заполнение «готовой ячейки»
256
(экосистемный подход). Эти позиции отражают две крайние
точки зрения на механизм формирования живых систем:
1) объединение, интеграция независимых (самостоятельных)
элементов в целое; 2) дифференциация целого на взаимосвя-
занные элементы.
Если мы допускаем функциональную определенность вида, то
длительность его исторического существования соответствует
длительности существования функциональной ниши в системе.
Реализуемая функция предполагает определенную реализую-
щую форму. Поскольку функциональная ниша качественно од-
нозначна, а количественно изменчива и существует на фоне
неоднородных в пространстве и во времени условий, вид подвер-
жен пространственной и временной динамике. Это выражается
в волнах численности (количественная реакция на изменение
условий) и изменчивости (качественная реакция). Генотипичес-
кий полиморфизм и фенотипическая изменчивость представляют
собой компенсаторные реакции вида, обеспечивающие возмож-
ность устойчивого выполнения экосистемных функций в меняю-
щихся условиях. Описание пространственной динамики вида
вполне укладывается в традиционную трехмерную схему, но для
описания качественной динамики «традиционного» одномерного
времени недостаточно.
Изменение во времени объема пространства, занимаемого ви-
довым живым веществом, имеет характер накладывающихся друг
на друга разнопериодических пульсаций. Ритмы пульсаций опре-
деляются взаимодействием внешних (экзогенных) и внутренних
(эндогенных) ритмов. Наиболее выраженными и потому извест-
ными из них являются ритмы солнечной активности, годовые и
суточные ритмы движения Земли, лунные, геомагнитные, онтоге-
нетические ритмы и ритмы клеточного деления. Выраженность
онтогенетических пульсаций зависит от средней длительности
жизни особей каждого поколения, синхронности развития поко-
лений, скорости нарастания биомассы и соотношения между рож-
даемостью и смертностью.
Длительные направленные изменения условий в простран-
ственных подразделениях биосферы и связанная с ними динамика
биогеоценозов вызывают изменения пространства, занимаемого
видовым живым веществом. Это могут быть смещение, увеличе-
ние или уменьшение ареала и переход в другую пространственную
экологическую нишу. Особую форму пространственной динамики
представляют устойчивые сезонные миграции в тех случаях, когда
вид заполняет две пространственно-функциональные ниши.
17 Присный АЛ.
Глава 9
МНОГООБРАЗИЕ ЖИЗНИ НА ЗЕМЛЕ
9.1. ФОРМЫ ЖИЗНИ
Долгое время в биологической литературе упоминались две
формы жизни: доклеточная (или неклеточная) и клеточная. За-
частую эта традиция сохраняется и поныне. Но чем больше све-
дений накапливается о вирусах, фагах, плазмидах, эписомах и
прионах, не имеющих клеточного строения, тем менее понят-
ными оказываются их природа и место в биосфере. Теперь уже
мало кто считает вирусы и близкие к ним образования эволю-
ционными предшественниками клеточных организмов. Чаще
их представляют как особые молекулярные комплексы, реали-
зующие способность к самовоспроизводству в клетках с их соб-
ственной генетической системой, то есть, буквально, как пара-
зитические молекулярные комплексы. И в одном и в другом
случае принимается, что такие комплексы — это самостоятель-
ные тела или даже организмы, занимающие пограничное с не-
живыми телами положение. Такой подход базируется на пони-
мании живого вещества биосферы как суммы всех живых тел.
Противоположный подход, согласно которому живые тела —
это дискретные части, подразделения единого живого вещества
биосферы, позволяет рассматривать нуклеопротеиновые, нук-
леиновые и протеиновые комплексы в качестве функциональ-
ных элементов, не связанных непосредственно с преобразова-
нием вещества и энергии, но являющихся информационными
посредниками между клеточными телами или дезорганизатора-
ми клеточных процессов.
Существуют формы, занимающие как бы промежуточное поло-
жение между клетками и вирусами. Это микоплазмы, риккетсии и
хламидии. Все они по существующим представлениям отнесены к
клеточным телам, поскольку имеют собственную липопротеино-
вую мембрану, рибосомы и, кроме того, размножаются бинарным
делением.
Согласно клеточной теории клетка — это основная структурная
и функциональная единица живого вещества и поэтому любое
природное живое тело построено из клеток и их производных.
258
В главе 3 были рассмотрены основные варианты построения из
клеток тел особей и построения из особей сложных организмов и
приняты следующие положения:
особи могут быть образованы одной клеткой или множеством
клеток одной или нескольких жизненных форм;
особи могут быть ценотически активными или ценотически
пассивными в течение всей индивидуальной жизни или изме-
нять состояние на отдельных стадиях или фазах, или же иметь в
своем составе ценотически активные и ценотически пассивные
части;
и одноклеточные, и многоклеточные особи могут образовывать
колонии, сохраняя морфологическую и функциональную обособ-
ленность, или колониальные тела, объединяясь только морфоло-
гически или, кроме того, функционально;
особи одного вида, но разных функциональных типов, суще-
ствующие и одновременно, и последовательно, объединяются
полным онтогенезом и образуют диффузно-комплементарный
организм;
организм обладает элементарной биосферной информацией и
реализует ее в конкретной экосистеме, организуя вещественно-
энергетический поток в направлении среда -> ценотически актив-
ная собственная составляющая -> среда.
Все вещественно-энергетические процессы в конечном счете
протекают в живой цитоплазме клеток или организуются ею, а так
называемые «неклеточные формы» лишь корректируют эти про-
цессы, чаще подавляя специфическую ценотическую активность
организмов. Следовательно, биосфера представляет именно кле-
точную форму жизни, хотя живое вещество биосферы функцио-
нирует на нескольких уровнях: клеточном, многоклеточных тел,
Сложных организмов, популяционно-видовом, биоценотическом
и биоты.
9.2. НЕКЛЕТОЧНЫЕ ФОРМЫ ЖИВОГО ВЕЩЕСТВА
(ПОЛУАВТОНОМНЫЕ МОЛЕКУЛЯРНЫЕ КОМПЛЕКСЫ)
9X1. ВИРУСЫ И БАКТЕРИОФАГИ
(ВИРУСЫ, ПОРАЖАЮЩИЕ КЛЕТКИ ПРОКАРИОТ)
Вирусы — это сложные молекулярные комплексы, включающие
нуклеиновые кислоты и белки, размеры которых составляют от 20
до 300 нм. Они не способны к автономному самовоспроизведению
и самостоятельной реализации вещественно-энергетических про-
цессов, поскольку не имеют полного набора необходимых фер-
17*
259
ментных комплексов и в свободном состоянии представляют без-
водные органические кристаллы. Следовательно, они не могут
быть отнесены к живым организмам. Попадая в живые клетки,
вирусы становятся фактически их частью и работают как полу-
автономные паразитические генетические системы, направляя
клеточные процессы на воспроизводство собственных структур-
ных элементов. Транскрипция вирусных нуклеиновых кислот у
разных вирусов происходит в разных частях клетки-хозяина — в
цитоплазме, в кариоплазме без разрушения ДНК хозяина или с
ее разрушением, в ядре с включением в ДНК хозяина. В послед-
нем случае часто происходит обмен генами между вирусом и
клеткой.
По строению вирусы принято разделять на четыре группы:
спиральные, икосаэдрические, сложные без оболочки и сложные с
оболочкой (рис. 9.1). В каждой группе есть ДНК-содержащие и
РНК-содержащие вирусы. И те и другие могут иметь или одноце-
почечную, или двухцепочечную нуклеиновую кислоту, могут по-
ражать прокариот или эукариот.
Многие вирусы изучены сравнительно хорошо. Среди них
можно назвать бактериофаг Pfl (спиральный с одноцепочечной
ДНК); бактериофаг <рХ174 (икосаэдрический с одноцепочечной
ДНК); аденовирус (икосаэдрический с двухцепочечной ДНК);
бактериофаг Т2 (сложный без оболочки с двухцепочечной
ДНК); вирус осповакцины (сложный с оболочкой с двухцепо-
чечной ДНК); вирус табачной мозаики (спиральный с одноце-
почечной РНК); реовирус (икосаэдрический с двухцепочечной
РНК); вирус гриппа (сложный с оболочкой с одноцепочечной
РНК).
Общую схему строения вирусов можно представить в следую-
щем виде. Нуклеиновая кислота образует сердцевину вирусной
частицы (вириона). Она окружена оболочкой из фибриллярных
или глобулярных белков — капсомеров. Вся совокупность капсо-
меров образует капсид с высокой степенью симметрии. Некото-
рые вирусы имеют дополнительную липопротеиновую оболочку.
Эта оболочка образуется путем отпочковывания липидного бис-
лоя от клеточной мембраны хозяина, а белки принадлежат самому
вирусу.
Спиральные капсиды состоят из скрученных вокруг общей
оси фибрилл или из плотно спирально упакованных в цилиндр
глобул. Количество капсомеров здесь определяется прежде все-
го длиной нуклеиновой кислоты. Икосаэдрические капсиды
имеют по 20 треугольных граней, 12 вершин и 30 ребер. Их кап-
сомеры пяти- или шестиугольной формы у разных вирусов
представлены различным числом. На каждой из вершин обычно
расположены «выступы» — белковые молекулы, обеспечиваю-
260
9
Рис. 9.1. Вирусы и фаги (оригинал автора):
/ — РНК-сод ержаший спиральный вирус табачной мозаики (/)= 18 нм); 2— вирус #1, содер;
жащий одноцепочечную ДНК (£>=6нм); 3— реовирус, содержащий двухцепочечную РНК
(Д = 55...6О нм); 4— бактериофаг <рХ174, содержащий одноцепочечную ДНК (Л - 25 нм); э—
вирус гриппа, содержащий одноцепочечную 8-фрагаентаую РНК (23-80...120
вирус, содержащий кольцевую двухцепочечную ДНК (2) — 60...90 нм); /—вирус rllv (вин),
содержащий одноцепочечную 2-фрагментную РНК (D = 100 нм) (внешний вид и внутреннее
строение: ЛБ —липидный бислой, БС —белковые слои); 8— вирус осповакцины, содержа-
щий двухцепочечную палочковидную ДНК (D— 160x250 нм); 9— бактериофаг Т2, содержа-
щий двухдепочечную ДНК (L = 100 нм)
щие прикрепление к мембране клетки хозяина. В случае отсут-
ствия в капсиде шестиугольных кампсомеров, как, например, у
бактериофага (рХ174, образуется видоизмененная структура —
«додекаэдр».
Сложные’ капсиды включают икосаэдрическую и спиральную
части. Среди вирусов со сложной формой хорошо изучен бактери-
офаг Т2. Он имеет икосаэдрическую головку, в которой располо-
жена ДНК; хвостовой отросток, состоящий из чехла со спираль-
ной упаковкой капсомеров, в котором проходит полый стержень,
соединяющий головку с пластинкой основания, или базальной
пластинкой; отходящие от пластинки шесть коленообразно изог-
нутых хвостовых нитей с шипами у их оснований. Сложно устро-
енный капсид обеспечивает введение в клетку бактерии (в данном
случае — кишечной палочки) вирусной ДНК. Для этого хвостовые
нити связываются с рецепторными гликопротеинами клеточной
стенки, изгибаются и прижимают базальную пластинку хвостово-
го отростка к поверхности бактериальной клетки. Затем чехол от-
ростка сокращается, полый стержень прокалывает клеточную
стенку и мембрану, а ДНК буквально впрыскивается в цитоплазму
бактерии. Другие вирусы имеют подобные или иные механизмы
введения генетического материала в клетку хозяина. При этом в
одних случаях капсид проникает в цитоплазму, а в других — оста-
ется на поверхности клетки.
Как уже указывалось, воспроизводство вирусных частиц про-
исходит в клетках хозяев с участием клеточных ферментных ком-
плексов, исходных веществ и макроэргических соединений. На
рисунках 9.2, 9.3 приведены основные схемы размножения виру-
сов.
Специальные исследования показали, что в природе не су-
ществует естественных живых тел, клетки которых абсолютно
свободны от вирусных частиц. Поведение вирусов по отноше-
нию к клетке хозяина складывается из трех этапов: проникно-
вение в клетку, воспроизводство собственных частиц и выход из
клетки. На втором этапе генетическая программа вируса может
инактивировать геном клетки, не разрушая или разрушая его, и
направлять биосинтетические процессы по пути, нехарактерно-
му для данной клетки, и, следовательно, «выключать» клетку из
нормальной ценотической активности. Некоторые вирусы,
встраивающие свою ДНК в ДНК клетки хозяина, лишь частич-
но или на непродолжительное время подавляют ее функцио-
нальную активность. Изменение направлений главных процес-
сов, протекающих в клетках, выражается в развитии различных
заболеваний. Выход новых вирусных частиц из клеток хозяев во
многих случаях завершается гибелью клеток. И если поражают-
ся одноклеточные живые тела (прокариоты и протисты), то ви-
262
Фаг
Икосаэдричес-
кий вирус
Вирусе
оболочкой
Прикрепление к клеточной поверхности
I-----------------1 « 1
J Сокращение стержня| j I
|«Впрыскивание» Д Нк| j { £
Слияниес
мембраной клетки
Проникновение ДНК
Репликация вирусной ДНК Синтез мРНК на вирусной ДНК
(ДНК-полимеразой клетки хозяина) (РНК-полимеразой клетки хозяина)
Процессинг (в клетках эукариот),
присоединение мРНК к рибосомам,
синтез вирусных белков
I ДНК |
Самосборка вирусных частиц
Выход вирусов
| Разрушение клетки хозяина~
Рис. 9.2. Схема размножения ДНК-содержащих вирусов
русы выступают в роли регуляторов их численности. Заражение
же многоклеточных тел далеко не всегда заканчивается их гибе-
лью, даже если многие клетки будут разрушены. Но в любом
случае такие тела заметно снижают свою основную функцио-
нальную активность.
Вирусы, не разрушающие клетку хозяина во время выхода из
нее, меньше влияют на активность клетки, но способны менять
программу ее деятельности, вплоть до нарушения механизмов
генной регуляции вегетативных и генеративных процессов. Встра-
ивание в ДНК клетки и выщепление из нее ДНК-копий вируса
сопровождаются возникновением мутаций в структурных и регу-
ляторных генах, включая и перенос целых генов. Этот процесс,
263
Прикрепление к клеточной поверхности
Проникновение РНК вируса в клетку
Синтез ДНК-копии на вирусной РНК с помощью
обратной транскриптазы — фермента вируса
Синтез двухцепочечной ДНК-копии с помощью
клеточного фермента ДНК-полимеразы
Встраивание двухцепочечной ДНК-копии в геном
клетки хозяина
Синтез мРНК на генах клетки и вируса
Синтез белков на рибосомах
Самосборка вирусных частиц
Выход вирусов
(Иногда пораженные клетки начинают быстро
делиться, образуя опухоль)
Рис. 9.3. Схема размножения РНК-содержащих вирусов
названный «горизонтальным переносом генов», играет важную
роль в эволюции живых организмов.
9.2.2. ПЛАЗМИДЫ
В современной биологической литературе плазмидами называ-
ют короткие фрагменты ДНК, ранее разделяемые на плазмиды (не
способные встраиваться в ДНК клетки хозяина) и эписомы (обла-
дающие такой способностью). Плазмиды очень широко распрост-
ранены в природе, являясь, как принято считать, паразитами или
симбионтами.
Плазмиды представляют собой внсхромосомные факторы на-
264
следственности, генетические элементы, способные устойчиво
существовать в клетке в автономном (?) состоянии. К плазми-
дам относят генетический аппарат митохондрий и пластид и,
кроме того, группы генов, не являющихся жизненно важными
для существования клеток. К настоящему времени сравнитель-
но хорошо изучены плазмиды бактерий. Они часто определяют
устойчивость последних к антибиотикам или являются поло-
вым фактором, кодирующим белки своеобразных трубочек, не-
обходимых для обеспечения полового процесса (через них ДНК
передается от одной бактериальной клетки к другой). Плазми-
ды структурно и функционально близки к интронам, с которы-
ми (а также с вирусами) образуют «рассеянный генетический
материал» биосферы.
9.2.3. ПРИОНЫ
В середине XX в. в клетках нервной системы высших живот-
ных, а впоследствии и в клетках других организмов обнаружены
совершенно необычные воспроизводящиеся паразитические бел-
ковые тельца, названные прионами (протеиновыми инфекцион-
ными частицами). Это волокнистые структуры, состоящие из при-
оновых палочек, каждая из которых представляет комплекс при-
мерно из 1000 белковых молекул и имеет диаметр 10...20 нм и дли-
ну 100...200 нм.
Накопленные данные свидетельствуют о том, что в прионах во-
обще нет нуклеиновой кислоты и поэтому вероятными механиз-
мами их размножения могут быть или управляемый клеточным
белком синтез прионового белка, или сборка на полипептидной
цепи приона с помощью фермента обратной транскриптазы,
мРНК и последующая транскрипция с использованием клеточных
структур и механизмов, или же активизация белком-прионом
«скрытых генов», ответственных за их сборку.
Большинство известных прионов вызывают заболевания нерв-
ной системы млекопитающих, получившие общее название «мед-
ленные инфекции». Эти заболевания имеют длительный, от не-
скольких месяцев до нескольких десятков лет, инкубационный
период без проявления каких-либо симптомов. Когда же начина-
ется собственно болезнь, она неуклонно прогрессирует и обычно
приводит к гибели.
Самым распространенным заболеванием человека, вызывае-
мым прионами, является болезнь Альцгеймера — форма старчес-
кого слабоумия. Специальные исследования, проведенные в США
(в конце 80-х годов XX в.), показали, что среди причин смертнос-
ти она занимает четвертое место. Из других «прионовых» заболе-
265
ваний человека отметим «куру» (встречается у горных племен Но-
вой Гвинеи) и болезнь Крейтцфельда—Якоба (широко распрост-
ранена), проявляющиеся в неврологических расстройствах с не-
благоприятным исходом. Наиболее известным заболеванием жи-
вотных, вызываемым этими возбудителями, является губчатый эн-
цефалит, или бешенство коров.
Прионы могут также провоцировать активность генов, сфор-
мировавшихся при участии интронов, и этим вызывать появление
в фенотипе ранее отсутствовавшего признака.
9.3. КЛЕТОЧНАЯ ФОРМА ЖИЗНИ
Живое вещество биосферы, будучи организовано в виде от-
дельных живых тел, имеющих клеточное строение, весьма не-
однородно в морфофункциональном отношении. Ранее упоми-
налось, что систематики выделяют более 1,5 млн его видовых
форм. Но среди таких элементарных единиц можно выделить
Рис. 9.4. Основные систематические подразделения живого вещества
(отделы и типы) (оригинал автора)
266
растительным
растительным
СЛОЖНЫЕ
МНОГОКЛЕТОЧНЫЕ
бактериотил
растительным тип
грибной тип
животным тип
ТАЛЛОМНЫЕ;
грибном тип
ПРОСТЫЕ
животным тип
МНОГОКЛЕТОЧНЫЕ
Рис. 9.5. Главнейшие морфологические и клеточные типы живых тел
(оригинал автора)
ОКЛЕТОЧ
(МОНЕРЬ
группы, обладающие единством планов строения, обусловлен-
ным генетическим родством, характеризующиеся общностью
базовых и преобладающих внутриклеточных процессов, выпол-
няющих сходные биосферные функции (рис. 9.4, 9.5). Учиты-
вая, что относительно непротиворечивой в современной систе-
ме живых организмов на сегодняшний день является группи-
ровка видов в таксоны не выше типа/отдела и именно эти под-
разделения живого вещества имеют эволюционную основу,
примем их в качестве наиболее емких и хорошо отличающихся
выражений клеточной формы жизни и приведем их краткие ха-
рактеристики.
9.3.1. МОНЕРЫ, ИЛИ ПРОКАРИОТЫ
К монерам относят диффузно-комплементарные организмы с
одноклеточными особями, способные при размножении образо-
вывать скопления-колонии или устойчивые группы соединен-
ных особей. Известно более 3600 видов. Для них характерно не-
большое число жизненных форм, выделяемых на основе прежде
всего функциональной специализации. Протопласт монер наи-
менее дифференцирован, область расположения хромосомы не
267
обособлена от цитоплазмы. Классификация монер окончательно
не установлена. Наиболее емким подразделениям этой группы,
обозначаемым как архебактерии, бактерии и цианобактерии, в
последнее время придается таксономический ранг царств. Часто в
классификационной системе монер первые два подразделения
объединяют в группу бактерий, к которой относят следующие
подгруппы:
эубактерий (или настоящие бактерии) — фотоавтотрофныс,
фотогетеротрофные, хемоавтотрофные и хсмогетеротрофные бак-
терии разнообразного строения;
актиномицеты — хемогетеротрофные палочковидные или нит-
чатые без клеточных «перегородок» бактерии;
хламидобактерии (или нитчатые железобактерии) — палочко-
видные или нитчатые хемоавтотрофные бактерии;
нитчатые серобактерии — хемоавтотрофные бактерии с нитча-
тыми телами;
миксобакгерии (или слизистые бактерии) — палочковидные с
тонкими эластичными клеточными стенками хемогетеротрофные
бактерии;
спирохеты — длинные, тонкие, спиралевидные, с эластичными
клеточными стенками хемогетеротрофные бактерии;
микоплазмы — различной формы неподвижные мелкие без
клеточной стенки хемогетеротрофные бактерии;
риккетсии — очень мелкие упрощенного строения палочковид-
ные хемогетеротрофные бактерии, имеющие сходство с вирусами
из группы «сложные с оболочкой».
Бактерии реализуют свои биосферные функции во всех про-
странственных частях биосферы и при широком диапазоне усло-
вий: они живут и в кислородсодержащих, и в бескислородных сре-
дах, в воде, в почве, в телах других организмов, при высоком дав-
лении и температуре свыше 100 еС (некоторые — свыше 360 °C), в
щелочной среде (при pH до 11,5) и в среде, насыщенной пищева-
рительными ферментами. Некоторые из них при определенных
условиях сохраняют жизнеспособность более 10 000 лет.
Автотрофные бактерии вносят большой вклад в круговорот уг-
лерода. Особенно важна для биосферы способность многих видов
бактерий фиксировать азот. Большинство бактерий является гете-
ротрофами: как биотрофами, так и сапротрофами. Биотрофы —
важнейшие регуляторы активности жизнедеятельности эукариот,
а сапротрофы — основные переработчики мертвого органического
вещества, возвращающие химические элементы в биологический
круговорот, переводя их в состояние, доступное для повторного
использования. В морских экосистемах на долю бактерий прихо-
дится до 90 % всей биомассы. Известны сложные морские экосис-
темы, полностью построенные на первичной продукции хемоав-
268
тотрофных бактерий. Освобождая энергию преимущественно в
виде тепла, бактерии влияют на особенности климата. Некоторые
бактерии концентрируют в среде отдельные химические эле-
менты, выделяя их из растворенных в воде веществ (например,
железо).
Отдельные виды бактерий человек использует в своей практи-
ческой деятельности. Одни из них находят применение в биотех-
нологиях (в пищевой промышленности), другие — в сельском хо-
зяйстве (как агенты биологической борьбы с вредными организ-
мами или в качестве живых удобрений), третьи имеют медико-ве-
теринарное и фитосанитарное значение (как возбудители
заболеваний человека и домашних животных или болезней куль-
турных растений).
Цианобактерии, или синезеленые водоросли, — это фотосинте-
зирующие, способные связывать азот прокариоты с одноклеточ-
ными и нитчатыми телами. Распространены в водоемах, в почве,
на голых минеральных субстратах, на поверхности живых тел и
как симбионты в их клетках или межклеточном веществе. Циано-
бактерии — единственные прокариоты, образующие газообразный
кислород в процессе фотосинтеза. Считается, что формирование
кислородсодержащей атмосферы шло, по крайней мере на первых
этапах, за счет этой группы организмов. Со способностью некото-
рых цианобактерий к накоплению и кристаллизации карбоната
кальция связано образование отдельных разновидностей извест-
няковых отложений. Цианобактерии входят в пионерные группи-
ровки, заселяющие безжизненные субстраты. Многие из них мо-
гут вступать в симбиотические отношения с губками, амебами,
жгутиконосцами, диатомовыми и зелеными водорослями (лишен-
ными хлорофилла), мхами, сосудистыми растениями и грибами-
оомицетами. При этом они теряют оболочку и функционируют
как хлоропласты.
9.3.2. ЭУКАРИОТЫ
Эукариоты — организмы с телами клеточного строения. Их
клетки имеют сложную внутреннюю дифференцировку. Область
компактного расположения хромосом в них отделена от цитоплаз-
мы ядерной оболочкой. Для эукариот характерно большое число
жизненных форм клеток и многоклеточных особей. Эукариоти-
ческие организмы принято делить на несколько царств. Большин-
ство биологов признают выделение царств: Растения, Грибы и
Животные. Но известен и вариант разделения эукариот на один-
надцать царств: I) Багрянки, 2) Низшие грибы, 3) Высшие грибы,
4) Растения (включают Высшие растения, Зеленые и Харовые во-
269
доросли), 5) Животные (включают Воротничковые жгутиконосцы,
Губки, Многоклеточные), 6) Эвгленовые, 7) Криптомонады,
8) Золотистые + Желто-зеленые + Бурые + Диатомовые водорос-
ли, 9) Перидинеи, 10) Хлоромонадовые + паразитические жгути-
коносцы + амебы + Споровики, 11) Инфузории + Опалины (Ста-
робогатов, 1984). В 2005 г. принята (бестаксонная) система эукари-
от, насчитывающая более 30 групп, эквивалентных рангу «Царство»
(Карпов, 2005). Характеристики групп эукариот, приведенные
ниже, ориентированы прежде всего на ценотически активные
формы.
Царство Растения объединяет в общей сложности более
400 000 описанных видов.
Водоросли. Группа водных, реже наземных эукариотических
организмов, клетки которых способны к фотосинтезу и имеют
оболочки, построенные из углеводов. В настоящее время известно
около 33 000 видов. Тела водорослей не имеют внутренней диффе-
ренцировки на системы жизнеобеспечения, даже если построены
из множества клеток, нет у них и многоклеточных органов раз-
множения. В морских экосистемах водоросли — основные проду-
центы органического вещества и кислорода, и именно эта сторона
жизнедеятельности определяет их место в биосфере. Под общим
названием «водоросли» объединяют несколько естественных так-
сонов — отделов одноклеточных и многоклеточных организмов с
талломными телами.
К одноклеточным водорослям относят золотис-
тые, желто-зеленые, диатомовые водоросли, пиррофиты и др.
Золотистые водоросли распространены большей частью в пре-
сных водоемах. Многие из них лишены развитой полисахаридной
оболочки, но имеют наружные кремнеземовые чешуйки или внут-
ренние скелетные элементы. Большинство золотистых водорос-
лей — одноклеточные со жгутиками, а некоторые — безжгутико-
вые — могут иметь сходство с амебами.
Желто-зеленые водоросли преимущественно неподвижны,
распространены и в пресной, и в соленой воде. От золотистых
отличаются набором фотосинтезирующих пигментов (отсут-
ствует фукоксантин) и наличием в жизненном цикле жгутико-
вых гамет.
Диатомовые водоросли в пресноводных и морских экосистемах
часто являются доминирующим компонентом фитопланктона.
Большинство диатомовых лишены жгутиков, но снабжены дву-
створчатым панцирем, состоящим из полимерного кремнезема.
Эти водоросли передвигаются посредством тонко регулируемой
секреции веществ через поры и токов цитоплазмы в поровых ка-
налах. Кремнеземные панцири диатомей, накапливаясь в донных
отложениях, образуют рыхлый порошок — диатомовый ил, кото-
270
рый человек использует как абразивный материал для тонкой по-
лировки.
(В некоторых классификациях золотистые, желто-зеленые и
диатомовые водоросли рассматриваются как классы в отряде Хри-
зофиты.)
Пиррофитпы, или динофлагелляты, — отдел двужгутиковых од-
ноклеточных водорослей, имеющих обычно целлюлозную оболоч-
ку, образующую пластинчатый панцирь. Большинство из них —
автотрофы, но есть среди динофлагеллят и гетеротрофы. Очень
часто эти водоросли содержат симбиотические цианобактерии и, в
свою очередь, сами вступают в симбиоз с многоклеточными жи-
вотными: губками, кишечнополостными, оболочниками, свобод-
ноживущими плоскими червями, брюхоногими и головоногими
моллюсками. Одна из групп динофлагеллят — зооксантеллы — ти-
пичные симбионты кораллов, они почти полностью обеспечивают
потребности своих «хозяев» в питательных веществах. Как прави-
ло, это облигатный симбиоз. Некоторые динофлагелляты, вызы-
вающие в период массового размножения так называемые «крас-
ные приливы», обладают сильнодействующими нервными токси-
нами.
К одноклеточным водорослям часто относят отдел Эвгленовые.
Однако эти организмы лишены клеточной оболочки и имеют ряд
признаков, сближающих их с животными. В этой классификации
мы относим эвгленовых к одноклеточным животным, а именно —
к миксотрофным жгутиконосцам.
К многоклеточным водорослям относят красные,
бурые и зеленые водоросли.
Отдел красные водоросли, или багрянки, включает организмы
(около 4000 видов), тела которых построены из сложно организо-
ванных безжгутиковых клеток, лишенных центриолей. Это пре-
имущественно морские многоклеточные формы, прикрепленные
к стабильному субстрату. Красную окраску им придают вспомога-
тельные пигменты, обеспечивающие поглощение зеленых, фиоле-
товых и синих лучей, проникающих на большую глубину. Багрян-
ки, как правило, имеют нитчатые нечленистые или членистые
тела, а иногда образуют корковые наросты. Красные водоросли
распространены на разных глубинах, вплоть до 250 м, и считаются
наиболее глубоководными фотосинтезирующими продуцентами.
Бурые водоросли — крупные морские водоросли (около 1500 ви-
дов) умеренных и холодных областей земного шара. Чаще встреча-
ются на каменистых прибрежных субстратах или образуют плот-
ные плавающие массы (род Sargassum). Имеют обычно нитчатые
или листоподобные тела от микроскопических до гигантских раз-
меров (до 60 м и более). Некоторые крупные бурые водоросли
имеют простейшую дифференцировку клеток, напоминающую
271
дифференцировку клеток у сосудистых растений. Многие виды
использует человек.
Отдел зеленые водоросли — наиболее разнообразная (не менее
7000 видов) из всех водорослей группа. Ее представители распрост-
ранены во всех типах водоемов, в почве, некоторые способны рас-
ти на снегу, на стволах деревьев, есть симбионты простейших и
кишечнополостных. По ряду признаков зеленые водоросли напо-
минают растения: по набору пигментов, по строению и составу
клеточной оболочки. Некоторые виды человек культивирует для
получения пищевых веществ или использует как энергетическое
сырье.
Растения. Это многоклеточные с талломной или тканевой орга-
низацией сухопутные, полуводные и вторичноводные автотроф-
ные, редко гетеротрофные (паразитические) организмы с много-
клеточными генеративными органами. В их жизненном цикле
обычно чередуются в разной степени выраженные спорофигное и
гаметофитное поколения и соответственно способы воспроизвод-
ства — бесполое и половое. Клетки тела имеют развитые целлю-
лозные оболочки.
Отдел Печеночники и мхи включает наиболее просто устроен-
ные наземные и полуводные растения, обитающие во влажных за-
тененных местах. В современной флоре насчитывается более
26 000 видов. Опорная и проводящая ткани у них отсутствуют или
слабо развиты. Тела представлены талломами или дифференциро-
ваны на аналоги листьев, стеблей и корней. Корневидные вырос-
ты-ризоиды могут быть одноклеточными или многоклеточными.
Их назначение состоит в закреплении тел на субстрате, а вода и
минеральные вещества поглощаются всей поверхностью таллома.
Печеночники и мхи легко теряют воду, поскольку не имеют раз-
витой кутикулы, но так же легко переживают безводные периоды.
В современных экосистемах они занимают преимущественно под-
чиненное положение. На минеральную среду оказывают незначи-
тельное действие.
В отдел Папоротники, плауны и хвощи включают сосудистые
растения (более 15000 видов), размножающиеся спорами (хотя
имеет место и половое воспроизводство) и формирующие сравни-
тельно развитые опорную (состоящую из ксилемы и склеренхи-
мы) и транспортную (образуемую трахеидами и ситовидными
клетками) системы. Тела этих растений дифференцированы на
стебель, мелкие или крупные листья и обычно корень. Гаметофит
редуцированный, автотрофный. Современные представители от-
дела — преимущественно травянистые формы с невысокой цено-
тической активностью.
Наиболее процветающая в настоящее время группа расте-
ний— это (отдел) Семенные растения, насчитывающая более
272
236 000 видов. В их телах хорошо развиты механические и прово-
дящие ткани, а сами тела ясно дифференцированы на корень,
стебель и листья. Размножаются семенами. Гаметофит крайне ре-
дуцирован, для полового размножения вода из внешней среды не
нужна. Семенные растения в подавляющем большинстве входят в
наземные экосистемы, где являются основными продуцентами и
важнейшими средообразователями. Их тела образуют сложную
объемную мозаику, позволяющую разнообразному живому веще-
ству максимально полно использовать потенциальную емкость
среды.
Водоросли, моховидные, папоротникообразные и семенные
растения объединяются в царство Растения, наиболее общими
признаками которого являются: наличие оформленных ядер в
клетках тела, развитие углеводной оболочки, обязательный ком-
понент которой — целлюлоза, построение многоклеточных тел по
симпластному типу и способность к фотосинтезу.
Грибы. Важное место в биосфере занимают грибы, выделенные
в самостоятельное царство. Грибы — это гетеротрофные эукарио-
ты с хитиновой клеточной оболочкой, не способные к активному
перемещению в пространстве. Тело грибов построено из тонких
ветвящихся нитей — гифов, которые в совокупности образуют ми-
целий. В отличие от растений тело гриба не имеет типичного кле-
точного строения: гифы, как правило, не разделены перегородка-
ми (смежными участками клеточных оболочек) или частично раз-
делены перфорированными перегородками, так что цитоплазма
может перетекать вдоль гифов, а ядра не имеют строгого соответ-
ствия промежуткам между перегородками. В отличие от животных
грибы сначала переваривают пищу вне тела, выделяя ферменты,
а затем всасывают ее всей поверхностью. Большинство грибов —
сапрофиты, реже встречаются паразиты и еще реже — симбион-
ты. Сапрофитные грибы составляют группу деструкторов (по
иной терминологии — редуцентов), и одна из важнейших их
функций — разложение целлюлозы. Кроме них это самое рас-
пространенное в биосфере органическое вещество могут рас-
щеплять лишь некоторые прокариоты и жгутиконосцы. Пара-
зитические грибы могут вступать со своими хозяевами в обли-
гатные или факультативные отношения. Облигатные паразиты,
как правило, не приводят своих хозяев к гибели, а факультатив-
ные, убивая хозяина, продолжают развитие на мертвых органи-
ческих остатках.
В современной биосфере царство представлено примерно
90 000 видов. Разделение грибов на отделы основано на особенно-
стях их жизненного цикла, в частности полового воспроизводства,
и в меньшей степени на особенностях строения. В царстве Грибы
принято выделять пять отделов: хитридиомицеты — микроскопи-
18 Присный А.В
273
ческие, обычно одноклеточные, паразитические грибы; оомице-
ты — большая группа преимущественно паразитирующих на рас-
тениях грибов с тонкими ветвящимися нечленистыми гифами;
зигомицеты — большей частью сапрофитные грибы с хорошо раз-
витым нечленистым мицелием; аскомицеты — самая многочис-
ленная группа высокоорганизованных грибов с членистыми гифа-
ми и иногда с крупными плодовыми телами (сморчки и трюфели),
включающая и сапрофитов, и преимущественно облигатных пара-
зитов; базидиомицеты — высшие грибы с членистыми гифами и
характерными плодовыми телами, в подавляющем большинстве —
сапрофиты с долговечным мицелием, некоторые (ржавчинные и
головневые грибы) — паразиты растений.
Аскомицеты и базидиомицеты вступают в характерные отно-
шения с цианобактериями, водорослями и высшими растениями.
Известно две формы таких отношений: образование особой груп-
пы живых тел — лишайников и образование микоризы. Первая фор-
ма рассматривается или как проявление симбиотических отноше-
ний, или как облигатный контролируемый (самим грибом) пара-
зитизм гриба аскомицета на водоросли или на цианобактерии.
Морфообразование у лишайников регулируется с помощью взаи-
модействия слагающих их организмов. Лишайники как живые
тела — это автотрофы, способные существовать на чистом мине-
ральном субстрате, растущие чрезвычайно медленно и живущие
тысячелетия. Микориза — взаимополезное объединение ряда ба-
зидиомицетов с корнями сосудистых растений, в котором грибы
разлагают гуминовые кислоты и получают от дерева питательные
и биологически активные вещества.
Животные. Гетеротрофные живые тела, построенные из клеток,
лишенных жестких оболочек, и способные к активному переме-
щению в пространстве, относят к животным. Животные — это са-
мая многочисленная группа организмов с наибольшим числом
жизненных форм и планов строения. В настоящее время всех из-
вестных животных принято разделять на 27 типов: 7 типов объеди-
няют в подцарство Простейшие и 20 — в подцарство Многокле-
точные.
Простейшие — это клеточные, но далеко не всегда одно-
клеточные организмы. Многоклеточными могут быть и отдельные
особи простейших. У простейших все функции индивидуального
жизнеобеспечения выполняются каждой клеткой отдельно, а це-
нотически активные клетки у каждого из видов представлены од-
ной жизненной формой. Все простейшие в активном состоянии
обитают в воде или в среде, насыщенной водой. Они являются
неотъемлемой составной частью главнейших биокосных систем.
В водных экосистемах и в почве свободноживущие простейшие
выступают как основные регуляторы численности бактерий.
274
Практически все многоклеточные животные и многие организмы i
растительного типа связаны с простейшими как с комменсалами :
(«нахлебниками»), симбионтами и паразитами. В водных экосис-
темах простейшие выполняют средообразовательную функцию •
более эффективно, чем многоклеточные животные. Подцарство
включает около 17 000 современных видов. Недавно предложена
классификационная система простейших, предполагающая выде-
ление не менее 13 типов: Воротничковыс жгутиконосцы, Эвглено- I
зои, Многожгутиковые, Опалины, Лабиринтоморфы, Форамини-
феры, Солнечники, Амебозои, Динофиты, Инфузории, Апикомп-
лексы, Микроспоридии, Миксоспоридии. Мы будем рассматри-
вать типы животных соответственно классификации, принятой в
основных руководствах по зоологии.
Тип Саркомастигофоры включает разнообразных простейших с
простым или сложным жизненным циклом, включающим одно-
клеточные, реже многоклеточные (плазмодиальные и псевдоплаз-
модиальные) тела. Ценотически активные клетки амебоидные или
жгутиковые, гетеротрофные, редко миксотрофные, большинство
свободноживущие водные или почвенные, некоторые — парази-
ты. Половой процесс, если имеется, осуществляется в форме копу-
ляции.
Тип Лабиринтулы характеризуется колониальными телами, об-
разованными живой эктоплазматической сетью, внутри которой
перемещаются веретоновидные клетки. Размножаются путем де-
ления клеток или распада агрегатов. Известно менее 40 видов.
Тип Апикомплексы представлен организмами со сложно орга-
низованным жизненным циклом, включающим несколько поко-
лений клеток и чередование бесполого и полового воспроизвод-
ства. Клетки ценотически активных поколений обычно меняют
жизненную форму с расселительно-инвазионной на трофичес-
кую. Все представители — около 5000 видов — паразиты живот-
ных.
Тип Микроспоридии включает более 1000 видов простейших па-
разитов животных. Их тела-клетки не имеют митохондрий. Обра-
зуют сложно устроенные споры со спирально скрученной полой
стрекательной нитью и инвазионным споробластом.
Тип Асцетоспоровые объединяет паразитические организмы, об-
разующие многоклеточные споры без выстреливающего аппарата.
Тип Миксоспоридии представлен преимущественно паразитами
рыб. Ценотически активное тело — плазмодий. Споровые тела с 1,
2 или многими стрекательными капсулами. Тип насчитывает око-
ло 1200 видов.
К типу Инфузории относятся одноклеточные со сложным ядер-
ным аппаратом, включающим не менее двух разнородных ядер
(микронуклеус и макронуклеус), организмы. Хотя бы на одной из
275
стадий онтогенеза они имеют локомоторные реснички. Половой
процесс проходит в форме конъюгации. Большинство — свобод-
ноживущие, но встречаются симбиотические и паразитические
формы. Известно более 7500 видов.
Многоклеточные в отличие от простейших, а точнее —
их ценотически активные тела, построены не менее чем из двух
жизненных форм клеток: пищеварительных и двигательных. Кро-
ме того, хотя бы отдельные функции индивидуального жизнеобес-
печения у них реализуются клеточными кооперациями.
Наиболее просто организованными многоклеточными живот-
ными считаются Пластинчатые. Их ценотически активное тело
образовано по меньшей мере тремя разными жизненными форма-
ми клеток с обратимой дифференцировкой. Питаясь колониями
водорослей, они образуют лишь функционально замкнутую пище-
варительную полость, чего не наблюдается у других животных.
Пока известно только два вида Пластинчатых.
Губки организованы несколько сложнее, чем Пластинчатые.
Морфологически близки к взрослым Пластинчатым их ценоти-
чески пассивные фазы — расселительные личинки. Дифференци-
ровка клеток у Губок обратима, и настоящих тканей они не обра-
зуют. Часть индивидуальных функций у этих животных реализует-
ся на уровне отдельных клеток. Губки — обращение подвижные
животные-фильтраторы. Многие из них вступают в облигатный
симбиоз с водорослями. В водных экосистемах Губки входят в
число важнейших средообразователей. В современной фауне на-
считывается около 2600 видов.
Кишечнополостные — двуслойные радиально-симметричные с
замкнутой кишечной полостью хищники (и микрофаги), способ-
ные образовывать различные жизненные формы особей в разных
поколениях в пределах одного жизненного цикла (полип и меду-
зу) и сложные колониальные тела. В морских, особенно тропичес-
ких, экосистемах они выполняют и средообразующую, и ценозо-
образующую (специфические сообщества коралловых рифов)
функции. Тип представлен более чем 5000 видов.
Плоские череп—двусторонне-симметричные трехслойные па-
ренхиматозные животные, ведущие паразитический, реже хищни-
ческий образ жизни. Часть клеток тела длительное время сохраня-
ются недетерминированными, из-за чего черви обладают хорошо
развитой способностью к регенерации и вегетативному размноже-
нию, в том числе и на личиночных стадиях. Большинство парази-
тических видов — специфические регуляторы активности и чис-
ленности хордовых животных на высоких уровнях плотности их
популяций. Известно около 45 000 видов.
Первичнополостные, или Круглые черви, — двусторонне-симмет-
ричные трехслойные первичнополостные сапрофаги, паразиты
276
растений и животных и потребители микроорганизмов (микрофа-
ги). Клетки тела дифференцируются на основе ранней (преиму-
щественно в эмбриогенезе) детерминации. Для большинства
Круглых червей характерно отсутствие жгутиковых клеток. Обра-
зование жизненных форм ценотически активных особей в значи-
тельной степени замещено дифференцировкой жизненных цик-
лов, особенно у паразитических видов. В водных экосистемах и в
почве Круглые черви — обычно микроконсументы. Паразиты вхо-
дят в число важнейших регуляторов активности и численности
многоклеточных растений и животных в биоценозах. В настоящее
время известно до 20 000 видов, но предполагается, что тип вклю-
чает не менее 1 млн видов.
Кольчецы, или Кольчатые черви, в типичном виде это двусто-
ронне-симметричные трехслойные вторичнополостные водные
или наземные животные. Основные трофические группы — детри-
тофаги, фитофаги, хищники и наружные паразиты. Виды, исполь-
зующие в пищу мертвую, преимущественно растительную, орга-
нику — важнейшие функциональные элементы биокосных систем
(донных илов и почв). Жизненные формы ценотически активных
особей очень разнообразны. Системы индивидуального жизне-
обеспечения в значительной степени сопряжены с целомом как
морфологически, так и функционально. К данному типу относят
около 13 000 видов.
Наибольшего разнообразия жизненные формы особей достига-
ют у Членистоногих — двусторонне-симметричных трехслойных
со смешанной полостью тела организмов, освоивших всевозмож-
ные среды обитания в биосфере, входящих во все типы и уровни
экосистем и участвующих во всех трофических группировках.
Строение, биологические и экологические особенности Членис-
тоногих во многом определяются способностью их эктодермаль-
ного эпителия к выделению хитина и образованию на его основе
кутикулы — внешнего скелета. По существующим оценкам,
1 650 000... 1 700 000 видов современных членистоногих получили
научные описания. Вероятно, их разнообразие в биосфере превы-
шает 30 млн видов.
К Моллюскам относят около 133 000 видов двусторонне-сим-
метричных или с нарушенной симметрией трехслойных вторич-
нополостных с сильно замещенной полостью тела животных,
использующих в пищу мелкодисперсную или тонкоизмельчае-
мую органику. Среди Моллюсков есть растительноядные, хищ-
ники и фильтраторы. Большинство способны к концентрирова-
нию и кристаллизации карбоната кальция, являясь срсдообра-
зователями. Участвуют в регуляции биомассы водорослей, сосу-
дистых растений, а в морских сообществах — и некоторых
животных.
277
Иглокожие — это радиально-симметричные трехслойные вто-
ричнополостные вторичноротые хищные и детритоядаые дон-
ные морские животные. Жизненные формы немногочисленны.
Как и моллюски, способны к концентрированию и кристаллиза-
ции в покровах карбоната кальция. Тил объединяет около
6600 видов.
Хордовые — двусторонне-симметричные трехслойные целоми-
ческие вторичноротые животные с развитым осевым опорным
образованием. Имеют наибольшее число жизненных форм кле-
ток, но сравнительно немного жизненных форм особей. Прояв-
ляют высокую геохимическую активность на мезоуровне биосфе-
ры, а один из видов — человек — и на макроуровне. Известно
примерно 51700 видов, из которых 49 350 видов относят к позво-
ночным.
Глава 10
ОРГАНИЗМ—ПОПУЛЯЦИЯ—ВИД В ЕДИНСТВЕ
СО СРЕДОЙ
10.1. ЭКОЛОГИЧЕСКИЕ ФАКТОРЫ
Биосфера в том виде, в каком мы ее знаем, в основном обра-
зована взаимодействующими вещественными элементами. До
сих пор мы рассматривали преимущественно лишь одну часть ве-
щества, а именно — живое вещество, из которого построены жи-
вые тела. Подчеркивая его специфический химический состав,
мы одновременно отмечали, что и живое, и неживое вещество
состоят из одних и тех же химических элементов. Учитывая, что
органическое вещество — основа живых тел — образуется из не-
органического вещества и после разрушения снова становится
неорганическим, биосферу можно предствить как непрерывно
сменяющуюся часть потока планетного вещества, «охватываемо-
го» жизнью лишь на короткое время. То есть живое вещество —
это лишь временное состояние неживого вещества, что со всей
очевидностью проявляется в биогеохимических циклах отдель-
ных элементов.
Живое вещество биосферы в целом и каждое отдельное живое
тело в частности пропускает через себя поток вещества, непрерыв-
но самовозобновляясь, и это движение возможно лишь при не-
прерывном сопряженном потоке энергии. По своей сути круго-
ворот вещества и поток энергии и есть проявление той неравно-
весности, которая устойчиво поддерживается самой живой систе-
мой и отличает ее от неживых систем, где приток энергии выводит
взаимодействующие элементы из состояния равновесия, заставля-
ет их совершать работу, после чего система вновь приходит в рав-
новесное состояние.
Итак, каждое отдельное живое тело поддерживает некоторый
уровень неравновесности по отношению к окружающей его среде,
взаимодействуя с этой средой. Такое взаимодействие проявляется
на всех уровнях функционирования живого вещества: от молеку-
лярного до биотической составляющей биосферы. Все многообра-
зие отношений живого вещества со средой изучает одна из общих
биологических наук — экология. Но живое вещество в реальных
экосистемах выступает прежде всего в виде дискретных живых
279
тел, разнообразие отношений которых можно условно разделить
на отношения индивидуального тела (особи, организма) с нежи-
вым и живым окружением и взаимодействия на надындивидуаль-
ных уровнях — популяционно-видовом и биоценотическом, что
изучают соответствующие разделы: аутэкология, демэкология и
синэкология.
Различные параметры среды, действующие на живые тела или
живое вещество и значимые для их существования, называют эко-
логическими факторами. Существует несколько вариантов класси-
фикации экологических факторов. Например, принято выделять
абиотические, биотические и антропогенные (порожденные чело-
веком) факторы. Они могут быть положительными или отрица-
тельными в зависимости от того, способствуют или препятствуют
выживанию и размножению организмов.
К абиотическим факторам относят действие температуры, све-
та, влажности воздуха и почвы, давления, радиоактивного излуче-
ния, электрического и магнитного полей, силы притяжения Зем-
ли, солевого состава почвы и водной среды, газового состава ат-
мосферы, движения воздуха и воды и т. п.
Биотические факторы — это все формы воздействия живых тел
друг на друга. Такие воздействия могут быть прямыми (непосред-
ственными) или косвенными (опосредованными). Биотические
факторы разделяют на пищевые, половые и средовые отношения,
антагонистические и неантагонистические.
Выделение антропогенных (порожденных человеком) факто-
ров в самостоятельную группу связано с тем, что действие на по-
пуляции и среду их обитания производится посредством создавае-
мых и управляемых человеком внешних усилителей его телесных
органов или культивируемых человеком организмов, то есть в це-
лом связано с практической деятельностью человека как социаль-
ного организма. Антропогенные факторы направлены большей
частью на экосистемы в целом и реже на отдельные живые тела
или их популяции.
Человек, представляя собой живое тело — дискретную единицу
видового живого вещества, является одновременно (в совокупнос-
ти со специфическим набором внешних усилителей) единицей
функционального подразделения социума (особью социальной
популяции локальной социальной системы). Аналогично экосис-
темным факторам в социуме действуют социальные факторы. Они
направлены как на функциональные группы, так и на социальные
индивидуумы.
По своему виду факторы разделяются на вещественные,
энергетические и информационные, а по месту происхождения —
на экосистемные, планетные и космические. Любой фактор
можно охарактеризовать по разным аспектам одновременно.
280
Например, солнечный свет — это абиотический, энергетичес-
кий, космический, а свет, получаемый от искусственного ис-
точника, — антропогенный, информационный, эко(социо)сис-
темный.
Некоторые факторы оказываются относительно постоянными
в течение длительного времени: сила тяготения, свойства атмос-
феры, температура и состав океанов, солнечная постоянная. Дру-
гие факт оры очень изменчивы в пространстве и во времени. При-
чем одни и те же факторы в разных средах обитания организмов
изменяются по-разному. Изменения факторов во времени могут
быть регулярно-периодическими, ритмическими, нерегулярными
и длительно направленными. Существуют и разово действующие
(разовые) факторы.
Действие факторов на организмы и их отдельные тела также
различно. Они могут действовать: как внешние раздражители,
вызывающие временные приспособительные изменения; как
ограничители возможности существования в данном объеме
среды; как модификаторы, вызывающие стойкие приспособи-
тельные изменения; как сигналы изменения других факторов.
Иногда все используемые факторы объединяют в группу «ре-
сурсные», а непригодные к использованию, чуждые — «ограни-
чивающие».
Различен и результат действия конкретного фактора или сово-
купности факторов на отдельную особь и на популяцию организ-
мов одного и того же вида. Например, для особи-жертвы взаимо-
действие с хищником — это разовый катастрофический фактор, а
для популяции жертвы — один из механизмов регуляции соб-
ственной численности и биомассы и реализация видоспецифич-
ной биогеоценотической функции.
Значение факторов среды для живого тела обычно зависит от
силы их проявления. Благоприятная сила называется зоной оп-
тимума в пределах диапазона проявления. Отклонения силы за
пределы оптимума образуют зоны пессимума. Крайние макси-
мальное и минимальное значения силы фактора, выдерживае-
мые живым телом, обозначаются как критические точки. Диа-
пазон силы проявления фактора между критическими точками
называют экологической валентностью. Естественно, этот диа-
пазон для вида и популяции шире, чем для отдельной особи.
Сумма экологических валентностей по различным факторам
образует экологический спектр вида. Спектры разных видов не
совпадают: каждый вид специфичен по своим возможностям,
что является следствием из механизма образования системы,
где каждый вид-элемент комплементарен сумме остальных и
уже с момента возникновения представляет морфофункцио-
нальную индивидуальность.
281
Наибольшее значение для популяции имеет фактор, находя-
щийся либо д минимуме, либо в максимуме зоны оптимума, то
есть имеет тенденцию выхода в зону пессимума и прибли-
жения к критическим точкам. Пределы выносливости и зона
оптимума по отдельно взятому фактору могут изменяться в за-
висимости от того, в каком сочетании и с какой силой в том же
объеме пространства и в то же самое время действуют другие
факторы.
Сочетания факторов определяют необходимость и возмож-
ность реализации конкретными видовыми популяциями элемен-
тарных биосферных функций, то есть фактически они определяют
возможность и необходимость существования тех или иных попу-
ляций в определенном объеме пространства.
Популяции, организмы и особи, в свою очередь, используют те
или иные факторы, проявляя к ним неоднозначное отношение,
что предполагает:
сохранение устойчивости своих количественных и качествен-
ных характеристик к виду фактора или силе его проявления, сни-
жающей эффективность реализации видоспецифичной функции
(невосприятием, пассивным отражением фактора или активным
противодействием ему);
избирательное восприятие (фильтрацию и дозировку) и усвое-
ние необходимого качества и количества фактора в порядке реа-
лизации функции и для повышения эффективности ее реализа-
ции;
использование фактора (в процесс реализации функции) без
его накопления, в том числе дифференцированное изменение ко-
личественных и качественных параметров в ответ на различные
виды и количества факторов. •
Позитивное использование ресурсной среды популяциями
включает избирательное извлечение и накопление вещества для
построения и непрерывного воспроизводства популяционного
живого вещества, накопление преобразуемых форм энергии,
хранение и новообразование интегральной (для всей популя-
ции) информации. То есть популяция как пространственно-
функциональное подразделение видового живого вещества на-
правляет через себя потоки вещества и энергии, занимая опре-
деленное место в стационарной и динамической структуре эко-
системы.
Вещественно-энергетические и информационные процессы,
проходящие внутри популяции, реализуются преимущественно
через составляющие ее организмы. Поэтому, собственно, организ-
мы имеют видоспецифичные и популяционно-специфичные функ-
циональные (или полифункциональные) признаки разноуровне-
вого назначения:
282
индивидуальные — признаки, обеспечивающие существование
биогеоценотически активного или биогеоценотически пассивного
живого тела в конкретных условиях среды в длительности, то есть
при данном комплексе используемых и неиспользуемых составля-
ющих среды;
видовые — признаки, обеспечивающие воспроизводство поко-
лений видоспецифичных живых тел или непрерывность видового
живого вещества;
экосистемно-популяционные — признаки, непосредственно
связанные с реализацией простой или сложной видовой функции
в конкретной экосистеме.
10.1.1. АБИОТИЧЕСКИЕ ВЕЩЕСТВЕННЫЕ, ЭНЕРГЕТИЧЕСКИЕ
И ИНФОРМАЦИОННЫЕ ФАКТОРЫ
Рассмотрим основные факторы и требования к ним видовых
популяций. Вещественные факторы. Эти факторы должны опреде-
лять возможность расширенного или, по крайней мере, полного
воспроизводства некоторого множества популяций, способных
составить устойчиво функционирующий биоценоз, в конкретном
биотопе. Надо сказать, что экологические спектры видов, суще-
ствующих на Земле в настоящее время, настолько разнообразны,
что почти любые реальные сочетания условий в пределах эубио-
сферы позволяют сформироваться биоценозу, хотя бы даже про-
сто устроенному.
В перечень вещественных факторов входят вещества, распрост-
раненные в биосфере, независимо от того, являются ли они экс-
плуатируемым ресурсом или препятствуют жизненным процес-
сам: вода, минералы, органические вещества и газы природного
происхождения, а также вещества антропогенного происхожде-
ния. Жизненно важными для популяций организмов будут их от-
носительные и абсолютные количества, состояние, характер рас-
пределения в пространстве, доступность, усваиваемость, хими-
ческая и осмотическая активность по отношению к живому ве-
ществу и др. При этом рассматриваются два условно выделяемых
аспекта значимости вещественных факторов: метаболический и
средовый.
Определенные соотношения веществ формируют так называе-
мые среды обитания. Например, рыхлая минеральная основа, в
которой доминируют кремнезем, карбонат кальция, соли магния
и алюминия, насыщенная некоторыми органическими вещества-
ми (гуминовыми кислотами, целлюлозой, хитином и др.), в кото-
рую проникают жидкая и парообразная вода и газы, составляю-
283
щие атмосферный воздух, образует почвы. Те же вещества, но с
преобладанием органики и воды, при минимуме газов образуют
илы. Газовая основа, включающая прежде всего азот, кислород,
аргон, диоксид, углерода, метан, сероводород, насыщенная водя-
ными парами, содержащая минеральную и органическую пыль и
летучие органические соединения, образует атмосферный воз-
дух. Водная основа с растворами минералов, газов и органичес-
ких веществ представляет еще одну среду обитания. То же можно
сказать и о живых телах с органическими веществами, составля-
ющими их основы. Вещества, нехарактерные для природных
сред, попадающие в них в результате деятельности человека или
аномальных природных процессов, составляют группу загрязни-
телей.
По приуроченности к средам обитания организмы в их конк-
ретном состоянии относят к аэробионтам, эпигеобионтам, эда-
фобионтам, гидробионтам и обитателям живых тел. Каждая из
групп имеет свои подразделения. В пределах пространства с од-
нородной средой популяция входит в локальные пространствен-
ные группировки, связанные со специфическими сочетаниями
конкретных проявлений факторов, вещественных, а также энер-
гетических и информационных. При этом экологические спект-
ры, благоприятные для одних популяций, для других оказывают-
ся экстремальными или губительными. Так, например, среди ра-
стений существуют экологические группы, отличающиеся по
требованиям к свойствам почвы. Ацидофилы (сфагнум, хвощ)
растут на кислых почвах, нейтрофилы — на нейтральных почвах,
базифилы (мордовник, ветреница) — на почвах с щелочной реак-
цией, индифферентные растения (ландыш, купена) — на разных
почвах. По требованию к количеству зольных элементов выделя-
ют: эвтрофные виды (дуб, сныть), мезотрофные (большинство
луговых растений) и олиготрофные (растения бедных почв: со-
сна, молодило, бессмертник). На засоленных почвах находят
благоприятные условия так называемые галофильные растения
(солянки, солеросы, кермеки), на каменистых — петрофильные,
на песчаных — псаммофильные, на обнажениях известняков —
кальцефильные. Все эти группы характеризуются определенны-
ми морфологическими и физиологическими особенностями. По-
пуляция как целое оказывается более терпимой к вариациям со-
четаний вещественных факторов, чем отдельные составляющие
их организмы. Более того, разные популяции одного вида могут
функционировать в средах с отличающимися параметрами, имея
при этом и разные качественные и количественные характерис-
тики.
Энергетические факторы. Биосфера — типичная открытая энер-
гозависимая система. Ее избыточная энергия — результат отсро-
284
ченного высвобождения воспринимаемой, связываемой и накап-
ливаемой энергии. С увеличением массы и сложности живого ве-
щества в геологической истории количество удерживаемой энер-
гии возрастает. Именно эта энергия определяет наличный объем
информации в системе.
Источниками энергии являются поля, плазма, движение веще-
ства, взаимодействие полей, плазмы и вещества и перераспределе-
ние движения в системе. Наиболее мощными источниками энер-
гии для биосферы являются Солнце с его полями, движение Зем-
ли в координатах Солнечной системы, разогретое ядро Земли,
гравитационное и магнитное поля Земли.
Рассмотрим подробнее электромагнитное излучение (ЭМИ)
Солнца. Общий диапазон волн составляет от 0,001 нм (гамма-
лучи) до нескольких километров (радиоволны). Это излучение
легко проходит через межпланетную среду и геомагнитное поле,
но поглощается и трансформируется в ионосфере и нейтральной
атмосфере. В спектре ЭМИ выделяют: гамма-лучи (0,001.-0,01 нм),
рентгеновские лучи (0,01—5,0 нм), ультрафиолет (5—380 нм),
невидимый фиолетовый диапазон (380...397 нм), видимые лучи
(397...760 нм), инфракрасные лучи (760 НМ...1 мм), радиоволны
(1 мм ... п км). Участки спектра ЭМИ, достигающие поверхнос-
ти Земли, называют «окнами прозрачности». Выделяют следую-
щие окна прозрачности: «оптическое» (290—5200 нм), «инфра-
красные» (8—14 и 17—330 мкм), «радиоастрономическое»
(1 мм...30 м).
Интенсивность радиоволн, достигающих поверхности Земли,
примерно в 1 000 000 раз меньше интенсивности излучения в оп-
тическом диапазоне. Ультрафиолетовое излучение (УФИ) с вол-
нами короче 290 нм практически не достигает земной поверхнос-
ти. Оптическая часть спектра излучается Солнцем на постоянном
уровне. Она включает часть ультрафиолета, видимые лучи и часть
инфракрасного излучения. В оптическую часть включается физи-
ологическая радиация (ФР) в диапазоне 300...800 нм. Ее основную
часть составляет физиологически активная радиация (ФАР) —
380—710 нм. (Хотя пурпурные и серные бактерии используют для
фотосинтеза инфракрасное излучение в диапазоне 800... 1100 нм.)
Проницаемость атмосферы Земли для ЭМИ в «окнах прозрачнос-
ти» зависит от сложного взаимодействия земных и космических
факторов, но, в свою очередь, определяет световой, гидротерми-
ческий и тепловой режим в экосистемах, подчиненный прежде
всего суточному, лунному и годичному циклам. Взаимодействие
ЭМИ с (газовой) атмосферой приводит к возбуждению и иониза-
ции ее атомов.
Солнечный ветер — это непрерывный поток ионизированного
газа (плазмы) от Солнца. Он служит переносчиком «вморожен-
285
I,
ных» магнитных полей солнечного происхождения. Для потока
солнечной плазмы магнитное поле Земли служит некоторым пре-
пятствием, и перед магнитосферой образуется ударная волна, где
движение плазмы замедляется, и часть ее кинетической энергии
превращается в тепловую энергию. Уплотненная горячая плазма,
обтекая геомагнитное поле, деформирует его: силовые линии сно-
сятся на ночную сторону Земли и образуют сильно вытянутый
хвостам агнитосферы с нейтральным слоем. Через полярные каспы
и нейтральный слой возможны прорывы солнечной плазмы к
верхним слоям атмосферы, а в остальных областях магнитосферы
идет лишь медленное просачивание плазмы.
На высоте около 500 км (силовые линии магнитного поля
Земли на такой высоте замкнуты) происходит захват частиц си-
ловыми полями и образуются протонный и электронный радиа-
ционные пояса. Находясь в непрерывном конвекционном дви-
жении, частицы порождают электрические токи: экваториаль-
ную и полярную электроструи. Втекание электрической энергии
в верхние слои атмосферы в высоких геомагнитных широтах вы-
зывает свечение атомов и молекул кислорода и азота на высоте
95... 120 км (полярное сияние). Экваториальный кольцевой ток
(течет с востока на запад) имеет радиус, равный 3...5 радиусам
Земли, и образует небольшую постоянную составляющую гео-
магнитного поля.
При усилении солнечного ветра магнитосфера сжимается и на-
пряженность магнитного поля у поверхности Земли увеличивает-
ся. Одновременно увеличение плотности частиц плазменного слоя
усиливает кольцевой ток и соответственно его магнитное поле,
что заставляет магнитосферу принять прежние размеры и форму,
а также уменьшает напряженность магнитного поля. Формируется
единичный импульс геомагнитного возмущения. Длительное уси-
ление солнечного ветра порождает серию последовательных им-
пульсов.
Кроме макровозмущений в геомагнитном поле формируются
короткопериодические колебания (от долей секунд до нескольких
десятков минут), индуцирующие электрические токи в верхних
слоях земной коры, и другие типы и формы электромагнитных
колебании (ЭМК) в диапазоне низких и очень низких частот (в
том числе ЭМК звуковой частоты, возникающие в результате мол-
ниевых разрядов).
Вспышки на Солнце влияют на поведение коллоидных систем
(например, происходит увеличение скорости осаждения оксихло-
рида висмута), кристаллизацию воды, парафина, образование во-
дородных связей в молекулярных группах, проницаемость биоло-
гических мембран, содержание ионов в жидкостях живых тел,
свертываемость крови, возбудимость и проводимость нервных
286
клеток, деятельность сердечно-сосудистой и нервной систем в це-
лом, скорость осаждения белков, концентрацию лейкоцитов, ди-
намику заболеваний и несчастных случаев, возникновение эпиде-
мий, эпизоотий и эпифитотий, динамику численности популяций
и ритмику биоценозов.
Динамика активности Солнца определяет разнопериодические
ритмические процессы на всех уровнях организации биосферы и
ее живого вещества. Даже часть эндогенных ритмов, например,
человека синхронизирована с экзогенными, прежде всего солнеч-
ными, ритмами.
Солнечно-земные (экзогенные) Эндогенные ритмы
ритмы человека
_ 0,5 суг
1 сут 1 суг
— 3 суг
_ 7 суг
27 суг (солнечные сутки),
28 суг (лунный ритм) 26...29 суг
— 0,5 года
1 год 1 год
_ 3 года
_ 7 лет
11,13 года ~
22 года —
90 лет (вековой) —
180 лет (178 лет — парад планет Меркурия,
Венеры, Земли и Юпитера)
Более длительные периоды
Наиболее сильное воздействие Солнца на биосферу происхо-
дит в годы наложения друг на друга солнечных возмущений, опре-
деляемых ритмами с разными периодами.
Влияние ближайшего Космоса на биосферу отчетливо прояв-
ляется в динамике популяций. Популяционные волны и дли-
тельные направленные изменения популяционных ареалов в
значительной степени связаны с погодными и климатическими
ритмами и в конечном счете с ритмикой поступления энергии в
биосферу.
Участие популяций в построении локальных подразделений
биосферы, где каждая из них представляет собой специфическое
звено вещественно-энергетического потока, предопределяет и
разную значимость для них различных видов и форм энергии. Для
популяций фотоавтотрофов это электромагнитная и тепловая
энергия, для хемогетеротрофов — химическая, гравитационная и
тепловая.
287
Абиотические космические и планетные факторы составляют
основные фоновые материально-энергетические условия суще-
ствования экосистем. Но те же факторы (например, с выраженной
ритмикой силы проявления), будучи способными влиять на сте-
пень определенности связей и отношений между элементами эко-
систем, являются и информационными факторами (в частности,
сигнальными).
10.1.2. БИОТИЧЕСКИЕ ФАКТОРЫ
Основную группу информационных факторов образуют непи-
щевые и пищевые биотические факторы, поскольку именно они
формируют потребность включения каждой видовой популяции в
экосистему, доводя ее до завершенности и целостности. Устойчи-
вые пищевые сети складываются в экосистеме, где каждая популя-
ция выступает как целое и как целое вступает в конкретные отно-
шения с другими видовыми популяциями. Классификацию попу-
ляционно-значимых биотических факторов можно представить в
следующем виде.
I. Пищевые антагонистические факторы.
1. Прямые пищевые антагонистические взаимоотношения:
а) хищничество — способ воздействия популяции консумента-
биотрофа второго или третьего уровня на популяцию предыду-
щего уровня, заключающийся в систематическом использовании
его живой массы в качестве источника вещества и энергии и яв-
ляющийся механизмом регуляции численности популяции жерт-
вы;
б) паразитоидизм — способ воздействия популяции консумен-
та-биотрофа второго или третьего уровня на популяцию предыду-
щего уровня, заключающийся в систематическом использовании
его живой массы в качестве среды обитания и источника вещества
и энергии одновременно и являющийся механизмом регуляции
численности популяции хозяина;
в) паразитизм — способ воздействия популяции консумента-
биотрофа второго или третьего уровня на популяцию предыду-
щего уровня, заключающийся в систематическом использовании
его живой массы в качестве среды обитания и источника веще-
ства и энергии одновременно и являющийся механизмом регуля-
ции ценотической и репродуктивной активности популяции хо-
зяина;
г) полупаразитизм (факультативное хищничество) — способ
воздействия популяции консумента-биотрофа второго или третье-
го уровня на популяцию предыдущего, того же или следующего
уровня, заключающийся в систематическом ограниченном ис-
288
пользовании ее живой массы в качестве источника вещества и
энергии и в норме не являющийся механизмом регуляции ее чис- i
ценности и активности;
д) собирательство — способ воздействия популяции консумен-
та-биотрофа на популяцию автотрофа или другого биотрофа, |
заключающийся в систематическом использовании его живой j
массы в качестве источника вещества и энергии и являющийся
механизмом регуляции численности этой популяции (от хищни-
чества отличается тем, что относительные размеры и подвиж- I,
ность особей собирателей имеют значительное превосходство
над жертвами);
е) пастьба — способ воздействия популяции консумента-био-
трофа первого порядка на популяцию автотрофа или талломного 1
гетеротрофа, заключающийся в частичном использовании живой
массы осеней в качестве источника вещества и энергии и являю-
щийся механизмом регуляции биомассы эксплуатируемой попу-
ляции. |
2. Косвенные пищевые антагонистические взаимоотноше-
ния: 1
а) конкуренция — взаимное действие популяций одной трофи-
ческой группы друг на друга при использовании одного и того же .
пищевого ресурса, недостаточного для расширенного воспроиз-
водства всех конкурирующих популяций одновременно. (
3. Неантагонистические пищевые отношения:
а) мутуализм — прямое или опосредованное положительное
влияние двух или более популяций разных видов друг на друга на
основе пищевых отношений (когда каждый из симбионтов обес-
печивает другого или других элементами питания, включая взаим-
ное влияние на основе пищевых отрицательных связей части из
них с популяциями-посредниками);
б) комменсализм (нахлебничество) — зависимость популяции i
одного вида — консумента — от популяции другого вида, принад-
лежащей к той же трофической группе, возникающая в результате
совместного использования пищи, добываемой особями второй
популяции. I
II. Непищевые средовые факторы.
1. Субстратные стационарные:
а) квартирантство — субстратная зависимость одной видовой
популяции от другой, возникающая на основе совместного ис- I
пользования участков среды, преобразуемых второй популяцией;
б) конкуренция — соперничество популяций разных видов за I
ограниченное пространство для поселения.
2. Субстратные динамические: I
а) форезия — зависимость популяции одного вида от популя-
ции другого вида, используемой в качестве транспортного сред- i
19 Присный А В
289
ства для расселения особей в пределах ареала по дискретным участ-
кам специфической среды;
б) кумулятивное негативное воздействие одной популяции на
другую через трансформацию общей для них среды обитания в
процессе жизнедеятельности;
в) кумулятивное негативное воздействие одной популяции на
другую путем активной целенаправленной трансформации общей
среды обитания.
3. Фабрические: зависимость одной популяции от другой, мерт-
вые или живые тела (или части тел) которой используются для
трансформации своей среды обитания.
Существуют и другие формы межпопуляционных отношений
промежуточного или комплексного характера, не включенные в
приведенную классификацию.
Глава 11
БИОСФЕРА
11.1. ОБЩИЕ СТАЦИОНАРНЫЕ И ДИНАМИЧЕСКИЕ
ХАРАКТЕРИСТИКИ БИОСФЕРЫ
Впервые термин «биосфера» появился в научной литературе в
1875 г. Его предложил Эдуард Зюсс — лауреат золотой медали Рус-
ского географического общества, известный австрийский геолог.
Но, вводя в науку новое понятие, Э. Зюсс не дал ему никакого
определения. Предтечей современного учения о биосфере счита-
ют Василия Васильевича Докучаева, главная заслуга которого со-
стоит в том, что он первым среди естествоиспытателей понял
опасность дробления науки о природе на множество частных не-
взаимосвязанных дисциплин. Идеи Докучаева подхватил и блес-
тяще развил его ученик, в последующем академик, Владимир Ива-
нович Вернадский — основоположник новой науки — биогеохи-
мии. (Биогеохимия связывает химию Земли с химией жизни и ус-
танавливает роль живого вещества в преобразовании земной
поверхности.)
В современном понимании биосфера Земли представляет со-
бой глобальную открытую систему со своим «входом» и «выхо-
дом». Ее «вход» — это поток солнечной энергии, поступающей из
космоса, и поток химической энергии, поступающей из литосфе-
ры, вовлекаемое в биогенный круговорот вещество, наличная
внутренняя информация и поток внешней информации; «выход» —
рассеиваемая и излучаемая преимущественно тепловая энергия,
уходящее из круговорота вещество, реорганизованная внутренняя
информация и поток исходящей информации.
Биосфера как кибернетическая система обладает свойством са-
морегуляции. Одно из наиболее характерных проявлений органи-
зованности биосферы заключается в наличии озонового экрана
(максимальная концентрация озона находится на высоте около
40...45 км), поглощающего губительные для живого вещества жест-
кие ультрафиолетовые лучи. Более того, состав газовой оболочки
планеты в целом практически полностью регулируется биотой.
Другой пример саморегуляции представляет Мировой океан. Реки
ежегодно вносят в него около 1,5 млн т только карбоната кальция,
а состав океанической воды существенно не меняется. Связано
19*
291
это со способностью многих организмов концентрировать и крис-
таллизовать в покровных образованиях, внешних и внутренних
скелетах минеральные вещества, осаждающиеся после их гибели
на дно океана и формирующие осадочные горные породы. Таким
образом, саморегуляция биосферы Земли обеспечивается живыми
организмами, что позволяет считать биосферу централизованной
кибернетической системой, то есть системой, в которой один из
элементов или одна из подсистем играет главенствующую роль в
функционировании системы в целом. Такой элемент называют ве-
дущей частью системы или ее центром. Живое вещество в биосфе-
ре и является ведущей частью системы.
В биосфере выделяют поля устойчивости жизни и поля суще-
ствования жизни. Первые определяют условия, которые живое ве-
щество выдерживает, не прекращая своих функций, а вторые —
условия, при которых живое вещество может увеличивать свою
массу (посредством роста и размножения). Пределы (собственно)
биосферы обусловлены полем существования жизни и в первую
очередь достаточным количеством влаги, обеспечивающим нор-
мальный ход ферментативных процессов, достаточным количе-
ством диоксида углерода и кислорода, обеспечивающим пласти-
ческие и энергетические процессы, благоприятным температур-
ным режимом, исключающим высокие температуры, вызывающие
свертывание белков, и низкие температуры, прекращающие рабо-
ту ферментов.
Где проходят границы биосферы? Если к биосфере относить те
зоны Земли, в которых длительное время существуют постоянные
сообщества живых организмов, то верхнюю границу биосферы
(эубиосферы) следует проводить по высоте рекордных активных
залетов летающих животных — до 10 км. Выше этой границы до
озонового экрана располагается парабиосфера. По данным, полу-
ченным с помощью геофизических ракет, бактерии в состоянии
анабиоза встречаются на высоте до 70 км, в стратосфере. Эта зона
получила название «апобиосфера». Мощность эубиосферы по вер-
тикали составляет менее 20 км: нижний — до 10 км — слой атмос-
феры, вся толща воды в океане и незначительная донная пленка
жизни — до 11 км, а на континентах — тонкий наземный и мощ-
ный подземный слой (гипобиосфера) более 3 км. Ниже в пределах
нескольких десятков километров земная кора охватывает ряд гео-
логических оболочек, сформированных прежними состояниями
биосферы. Это метабиосфера. Все названные оболочки: метабиос-
фера, эубиосфера, парабиосфера и апобиосфера — образуют мно-
гослойную мегабиосферу, под которой находится базальтовый
слой земной коры (рис. 11.1).
Живые организмы являются структурными единицами живого
вещества и активными элементами биосферы как системы. Они
292
Рис. 11.1. Схема строения биосферы (Реймерс, 1994, с изменениями)
составляют неотъемлемую часть этой системы и выполняют в ней
определенные функции. Живые организмы — это и аккумуляторы
солнечной энергии, и трансформаторы для превращения ее в за-
пасаемую химическую и выделяемую действующую энергию, и ус-
тройства, выполняющие работу, и хранители, преобразователи и
передатчики информации. Живые организмы не изолированы от
неорганического «косного» вещества; вместе с ним они образуют
особые функциональные подсистемы, которые В. И. Вернадский
назвал биокосными. Это подсистемы биосферы, в которых живое и
неживое вещество проникают друг в друга. Основные биокосные
подсистемы — почвы, илы, водоемы.
293
Характерной особенностью биосферы является мозаичность ее
строения. Жизнь, будучи организована в планетарном масштабе,
функционирует в пределах структурных подразделений биосферы —
биогеоценозов. Биогеоценоз — сообщество организмов, связанных
с определенными почвенно-климатическими условиями. Сейчас
этот термин приобрел более широкое значение: им обозначают не
только наземные, но и «водные» сообщества, связанные с опреде-
ленными физико-химическими условиями.
В структуре биогеоценоза выделяют два главных элемента: био-
ценоз — сообщество живых организмов и биотоп — территорию с
однородными климатическими условиями, заселенную более или
менее однородными по составу организмами. Биоценоз можно ус-
ловно разделить на структурные единицы в соответствии с деле-
нием организмов на систематические группы: фитоценоз — сооб-
щество растений; зооценоз — сообщество животных; орнитоценоз —
сообщество птиц; энтомоценоз — сообщество насекомых и т. д. В
биотопе также могут быть выделены более мелкие элементы, об-
ладающие какими-либо специфическими условиями — стации.
Например, в дубраве это кроны деревьев, стволы, полог леса, под-
стилка, гниющие стволы и пни, почва.
Каждый биогеоценоз характеризуется следующими основными
показателями: видовым разнообразием; численностью или плот-
ностью видовых популяций (общее число особей одного вида или
число особей, приходящихся на единицу площади, объема); био-
массой (общее количество органического вещества, расположен-
ное на единице площади); взаимоотношениями между видами и
взаимодействием между организмами и средой.
Для характеристики вещественных, энергетических и инфор-
мационных процессов в биосфере используют особые, логические
ее подразделения. Английский ученый А. Тэнсли назвал их экоси-
стемами. Сейчас экосистему определяют как комплекс взаимосвя-
занных организмов разных видов и изменяемой ими абиотической
среды, обладающий способностью к саморегуляции и полному са-
мовозобновлению биоты (живой составляющей). В структуре эко-
системы выделяют несколько функциональных блоков: ресурсную
среду, продуцентов,, консументов нескольких последовательных
порядков и редуцентов. Значимыми характеристиками экосисте-
мы являются структурная сложность, биологическая продуктив-
ность (количество биомассы, образуемой за единицу времени), в
том числе первичная продуктивность (продуктивность автотро-
фов) и вторичная продуктивность (продуктивность гетеротрофов),
скорость и пути миграции химических элементов, веществ, интен-
сивность энергетического потока, накопление и расход энергии в
отдельных блоках и др.
Сопоставление содержания понятий «биогеоценоз» и «экосис-
294
тема» показывает, что первое из них имеет отношение к конкрет-
ным структурно-функциональным подразделениям биосферы, а
второе — к абстрактным моделям произвольно выделенных частей
биосферы или биосферы в целом.
Биогеоценозы находятся в постоянном взаимодействии друг с
другом, и все вместе организуют гигантский круговорот вещества
в пределах биосферы. Основными элементами, участвующими в
нем, являются водород, углерод, кислород, азот, калий, кальций,
кремний, фосфор, сера, стронций, барий, цинк, молибден, медь и
никель. Биогеохимический круговорот в биосфере не является
замкнутым. Степень воспроизводства циклов достигает 90...98 %.
В масштабе геологического времени неполная замкнутость биогео-
химических циклов приводит к разделению элементов и накопле-
нию их в атмосфере, гидросфере или осадочной оболочке Земли.
В свою очередь, для существования этого круговорота необходимо
постоянное поступление диоксида углерода и других веществ из
недр Земли, что, в свою очередь, лежит в основе геохимического
принципа сохранения жизни: «Жизнь на Земле и других планетах,
при прочих равных условиях, возможна лишь до тех пор, пока эти
планеты активны и происходит обмен энергией и веществом меж-
ду их недрами и поверхностью».
Непрерывному круговороту в биосфере подвергаются только
вещества, в то время как в отношении энергии можно говорить
лишь о направленном потоке. Поступающая в биосферу солнеч-
ная энергия частично расходуется на синтез органического веще-
ства. Запасенная таким образом энергия, передаваясь с одного пи-
щевого уровня на другой, постепенно рассеивается, совершая ра-
боту и выделяясь в виде тепла (рис. 11.2). После окончательного
разложения органических остатков энергия частично накаплива-
ется в земной коре в алюмосиликатах.
Обновление всего живого вещества биосферы Земли осуществ-
ляется в среднем за 8 лет. При этом вещество наземных растений
обновляется примерно за 14 лет. В океане циркуляция вещества
происходит быстрее: вся биомасса обновляется в среднем за
33 дня, в то время как фитомасса — каждый день. Процесс полной
смены вод в гидросфере осуществляется за 2800 лет. В атмосфере
смена кислорода происходит за несколько тысяч лет, а диоксида
углерода — за 6,3 года.
Миграция химических элементов в биосфере осуществляется
или при непосредственном участии живого вещества, или же она
протекает в среде, геохимические особенности которой преиму-
щественно обусловлены живым веществом в течение всей геоло-
гической истории. На миграцию элементов в биосфере влияют
также и абиогенные факторы, однако их роль второстепенна по
сравнению с суммарным влиянием живого вещества.
295
Рис. 11.2. Динамика вещества и энергии в биосфере. (Повторение цикла соответ-
ствует новому пространственно-временному состоянию биосферы.) (оригинал автора)
В общей динамике вещества, энергии и информации в биосфе-
ре живое вещество выполняет несколько функций, поддающихся
формализации. Отметим наиболее очевидные.
1. Энергетическая функция включает поглощение и связыва-
ние солнечной энергии при фотосинтезе, а химической — при
разложении энергонасыщенных веществ, передачу энергии по пи-
щевой цепи разнородного 'живого вещества и высвобождение
энергии во внешнюю среду.
2. Концентрационная функция связана с избирательным на-
коплением в ходе жизнедеятельности определенных видов веще-
ства, используемых для построения живых тел и удаляемых из них
при химической регуляции.
3. Деструктивная (разрушительная) функция предполагает: ми-
нерализацию отмирающего живого вещества; разложение неживо-
го неорганического вещества; вовлечение образовавшихся ве-
ществ в биогенный круговорот.
4. Транспортная функция заключается в переносе вещества
против силы тяжести и в горизонтальном направлении.
5. Средообразующая функция связана с преобразованием жи-
вым веществом физико-химических параметров среды.
6. Информационная функция включает накопление, хранение,
реорганизацию и передачу информации о структурных и динами-
296
ческих характеристиках активной подсистемы в пространственно-
временной протяженности.
Такая работа живого вещества оказывается единственным спо-
собом обеспечения собственного длительного существования и
развития в условиях ограниченности доступных минеральных ве-
ществ, необходимых для этого. Придать «конечному» свойства
«бесконечного» можно, лишь организовав его движение по замк-
нутому циклу. Каждый вид организмов представляет специализи-
рованное звено в этом круговороте. Причем один из главных па-
радоксов жизни заключается именно в том, что непрерывность и
развитие обеспечиваются прерывистостью организации самого
живого вещества и процессами распада и деградации.
В восходящей части биогенного цикла главную роль играют
фото- и хемосинтезирующие организмы: бактерии, цианобакте-
рии, водоросли, талломные и сосудистые растения. Нисходя-
щая часть цикла обеспечивается гетеротрофными частями тел
автотрофов и гетеротрофами: бактериями, грибами и животны-
ми.
По современным данным, например, наземная растительность
без ущерба для нормального воспроизводства переносит изъятие
до 75 % своей биомассы, в то время как растительноядные живот-
ные не могут прокормиться при снижении запасов корма больше
чем на 50...70 %. В экосистемах, не затронутых влиянием челове-
ка, в принципе невозможно чрезмерное выедание растительности
травоядными: до того как размер выедания станет катастрофичес-
ким для растительности, начнется уменьшение популяции траво-
ядных. Подобные механизмы саморегуляции в локальных экосис-
темах обусловливают динамическую устойчивость глобального
биогенного круговорота веществ в биосфере.
11.2. биосфера как эволюционирующая система
11.2.1. ГИПОТЕЗЫ ЗАРОЖДЕНИЯ БИОСФЕРЫ
Существующие гипотезы возникновения жизни на Земле так
или иначе связаны с попытками объяснить происхождение живо-
го вещества, а точнее — живых организмов и в конечном счете
происхождение человека. С учетом современного уровня знаний о
системной организации жизни эти гипотезы можно трактовать
как гипотезы зарождения земной формы жизни в виде живой сис-
темы — биосферы.
Все многообразие гипотез появления живого вещества на на-
шей планете объединяют в три основные группы: 1) живое веще-
ство на Земле сотворено Высшим началом; 2) живое вещество по-
297
падает на Землю из космоса в «готовом» виде; 3) живое вещество
возникает естественным путем на Земле.
Система представлений, объединяемая гипотезами первой
группы (гипотезы «сотворения»), носит общее название «креацио7'
низм». Большинство верующих в Бога людей фактически могу'г
быть причислены к креационистам. Факт Творения присутствует
в разных религиозных учениях, но его сроки, последовательность
и механизм трактуются по-разному.
По-иному представляют возникновение жизни на Земле сто-
ронники гипотез биогенеза. Общим для них является постулирова-
ние возможности появления живого только из живого. Сама фор-
мула «Все живое из живого» (Omne vivum е vivo) известна с XVII в.
И принадлежит Франческо Реди. Каждое опровержение абиогене-
за усиливало позиции сторонников биогенетических гипотез. Ло-
гика биогенеза устанавливает вечность и широкую распространен-
ность жизни в космосе. Наиболее известна из представлений этой
группы гипотеза «панспермии», или повсеместности «зародышей
жизни», сформулированная С. Арениусом (Швеция) в начале
XX в. Ее сторонником был и В. И. Вернадский. Другая гипотеза
этой группы — гипотеза «стационарного состояния» — утверждает
вечность жизни на вечно существовавшей Земле, но в настоящее
время ее серьезно не рассматривают. Близкая по форме гипотеза,
содержательно граничащая с гипотезами предыдущей группы,
высказана Ф. Криком и Л. Оргелом (США): на Землю жизнь по-
пала извне, но не случайно, в виде «семян жизни», а в результате
преднамеренного внесения высокоразвитой внеземной цивилиза-
цией. Подобные представления существуют и в ненаучном зна-
нии, по крайней мере, касательно происхождения человека. Так,
например, люди африканского Племени Дагонов считают своими
предками «пришельцев» с Сириуса.
Гипотезы третьей группы — гипотезы абиогенеза, или «самоза-
рождения жизни», — появились еще в XV—XVI вв. В XVIII в. идеи
самозарождения развивали и пропагандировали Г. Лейбниц (Гер-
мания), Ж. Бюффон (Франция), Дж. Нидхэм (Ирландия); в XIX в. —
Э. Дарвин (Англия) и Ж.Б. Ламарк (Франция). Главнейшими ар-
гументами в гипотезе признавались факты появления живых орга-
низмов на закрытых (как считалось) питательных субстратах. При
этом допускалось существование «жизненной силы», возбуждаю-
щей жизнь в мертвом субстрате в любое время. Данная гипотеза
основывалась на ошибках субъективного восприятия явлений.
Научное обоснование абиогенез получил в 20-е годы прошлого
столетия в работах А.И. Опарина. В опытах с концентрированны-
ми растворами органических веществ были получены устойчивые
сгустки — коацерватные капли, способные поглощать, преобразо-
вывать и выделять некоторые вещества. Коацерваты рассматрива-
298
ют как аналоги предбиологических систем, давших начало живым
телам. Возможность абиогенного синтеза органических веществ в
условиях Земли, обоснованная Опариным, была показана в лабо-
раторных экспериментах С. Миллера (США) в 1955 г. Позже та-
кие эксперименты с вариациями по исходным условиям были по-
вторены многократно. Гипотеза «коацерватов» сначала не каса-
лась передачи наследственной информации и потому была допол-
нена в 1929 г. Д. Холдейном гипотетическими механизмами
матричных процессов с участием нуклеиновых кислот.
В рамках обсуждения гипотезы абиогенеза состоялась дискус-
сия по вопросу о месте появления живого вещества. Обсуждались,
причем весьма аргументированно, следующие варианты: 1) жизнь
зародилась в океане, в «первичном бульоне»; 2) жизнь зародилась
в океане, на подводных вулканах; 3) жизнь зародилась на при-
брежных глинах; 4) жизнь зародилась в наземных условиях среди
частиц грунта. Во всех этих вариантах предполагалось, что жизнь
моложе планеты, на которой она развивается.
По современным представлениям, жизнь на Земле существует
практически столько же, сколько существует и сама Земля
(4,65 млрд лет). По-видимому, фотоавтотрофные прокариоты
были уже достаточно развитыми около 3,5 млрд лет назад, а клет-
ки появились не менее чем на 0,4 млрд лет раньше.
До сих пор наука не располагает сведениями, необходимыми
для достоверной реконструкции обстановки, сложившейся на по-
верхности Земли на начальных этапах ее существования. Первич-
ная атмосфера состояла, по одним расчетам, преимущественно из
диоксида углерода и выходящих с глубинными газами воды, сер-
ных соединений и азота, а по другим — из метана, аммиака, окси-
да углерода, воды, возможно, диоксида углерода и азота. В любом
случае атмосфера должна была иметь восстановительный или, по
крайней мере, нейтральный характер. Хотя отсутствие аммиака,
метана или свободного водорода противоречит условиям экспери-
ментов Миллера по абиогенному синтезу органических веществ,
это, как доказано, не является препятствием для синтеза органики
в естественных условиях.
Предполагается, что основная масса органических соединений
образовалась за пределами Земли в период, предшествующий ее
формированию. Доказательством возможности внеземного абио-
генного синтеза служит обнаружение в кометах и на метеоритах
углеводородов, содержащих до 16 атомов углерода в молекуле,
спиртов, карбоновых кислот, пуринов, пиримидинов, аминокис-
лот, органических полимеров. Все они не проявляют оптической
активности, а следовательно, имеют абиогенное происхождение.
В последние годы доказана принципиальная возможность неорга-
нического синтеза в открытом космосе и на Земле не только
299
«симметричной», но также и «асимметричной» органики. Счита-
ется, что образование органических соединений в Солнечной сис-
теме на ранних этапах ее формирования (более 4,5 млрд лет назад)
было обычным явлением. Что же касается нуклеотидов и поли-
мерных белков, то их синтез проходил скорее всего уже в специ-
фических условиях Земли.
Общую схему становления живой системы на Земле можно
представить в следующем виде.
На первом, «химическом», этапе зарождения органического ве-
щества существует множество хаотических реакций абиогенного
(фотохимического и др.) эндоэргического синтеза органических
молекул и их экзоэргического разложения, что в целом при дефи-
ците кислорода ведет к накоплению продуктов синтеза на мине-
ральных адсорбентах. В этих протоциклах участвуют прежде всего
биоэлементы, в том числе и фосфор, некоторые соединения кото-
рого отличаются более легкими связыванием и освобождением
энергии.
На втором этапе пространственно объединенные экзо- и эндо-
эргические реакции становятся сопряженными и во времени че-
рез «цикл фосфора» (некоторые фосфаты становятся энергетичес-
кими организаторами), и, кроме того, через общие конечные про-
дукты и исходные вещества различных реакций. Такие сопряжен-
ные процессы ведут к увеличению длительности удержания
энергии, упорядочению молекулярных комплексов и, следова-
тельно, накоплению информации. Этот этап, характеризующийся
становлением в условиях Земли процессов самоорганизации,
можно назвать «предбиологическим».
На третьем, «протобиологическом», этапе осуществляется аби-
огенный синтез пятиуглероднь£х моносахаридов, которые входят и
в состав нуклеотидов и в цикл связывания углерода. Предположи-
тельно, на этом этапе энергетическими организаторами становят-
ся нуклеофосфаты, что приводит к увеличению организованности
химических процессов, появлению реакций спонтанной полиме-
ризации, по сути — биохимических процессов. Идет оформление
органо-минеральных структурных элементов протобиосферы.
Первичными организаторами процессов матричного синтеза мог-
ли служить некоторые органические вещества, а также твердые и
жидкие кристаллы минералов (особенно графит, пирит и силика-
ты).
Перечисленные этапы могли иметь место и в открытом космо-
се, и на поверхности Земли, и на подводных вулканах. Поэтому
первичное накопление органических молекул на Земле скорее
всего было комплексным процессом.
Конец третьего этапа характеризуется, в частности, появлени-
ем двумерных мономолекулярных органических слоев в форме
300
пленок на границе раздела фаз вода—-минералы, способных к ав-
токаталитическому метаболизму и фиксации углерода, а также ме-
ланоидинов и протеиноидов, обладающих гидролитической ак-
тивностью и способностью к переносу электронов. Фактически на
этом этапе формируются диффузно организованные предавто-
трофный и предгетеротрофный биосферные блоки, пока еще не
разделяющиеся на неживое и живое вещество. Общее или локаль-
ное накопление органики при определенных условиях приводит к
появлению трехмерных органических пленок и капель, где разно-
родные органические молекулы и сформировавшиеся простейшие
химические системы получают дополнительные возможности ди-
намического сопряжения.
Дифференцировка вещества на «неживое» и «живое» происхо-
дит на четвертом, «биологическом», этапе, который почти одно-
значно связывают с нашей планетой, с земной формой жизни.
Близкую схему начальных этапов становления живой системы
на Земле дает В. Я Савенков (1991). Уже на ранней стадии геоло-
гической истории в атмосфере происходило образование простей-
ших органических молекул, а в литосфере — предшественников
нуклеопротеидов на нитевидных кристаллах графита с одной вин-
товой дислокацией. В первичных неглубоких водоемах, в припо-
верхностном слое воды, накапливалось значительное количество
неорганических и органических веществ и частиц, включая пред-
шественники нуклеопротеидов, что привело к возникновению
квазикристаллического состояния воды, близкого к состоянию ги-
алоплазмы. Здесь образовалась единая динамическая сложная ге-
терогенная (химическая) система, которая, взаимодействуя с
внешней газообразной и жидкой средой, развивалась в направле-
нии усложнения структуры. В ней под действием ультрафиолета и
видимого солнечного излучения происходило преобразование
электромагнитной энергии в химическую, развивались сопряжен-
ные окислительно-восстановительные реакции на минеральных
частицах, а позже — на мембранах. На следующем этапе в квази-
кристаллической среде сформировались «микросистемы типа ги-
перциклов», способные к самовоспроизведению. Все эти процес-
сы формировали интегративную (общую) информацию единой
развивающейся макросистемы в виде огромного поверхностного
слоя воды первичных водоемов. Когда возникающие^ ней и ус-
ложняющиеся микросистемы достигли вещественной, энергети-
ческой и информационной самообеспеченности и приобрели спо-
собность к воспроизводству, они образовали примитивные одно-
клеточные организмы, а квазикристаллический слой постепенно
разрушился.
Несколько иначе химический этап становления биосферы ри-
сует Э. М. Галимов (2001). Прежде всего в его гипотезе накопле-
301
ние на поверхности Земли и в первичной атмосфере азота, воды и
соединений углерода связывается с интенсивным осаждением
здесь метеоритного вещества (углистых хондритов). Расчеты пока-
зали, что масса органического углерода, поступавшего на Землю с
метеоритами и образовывавшегося при метеоритных бомбарди-
ровках, могла составлять около Ю12 г/год, а первичный океан в
этот период представлял «разрозненную цепь отдельных водо-
емов».
В первичном бульоне были широко распространены разнонап-
равленные реакции синтеза (понижение энтропии) и распада (по-
вышение энтропии) органических молекул, в том числе фотохи-
мические и с участием катализаторов, конечные продукты кото-
рых могли служить исходными реагентами для противоположных
реакций. На основе микроскопического химического сопряжения
образуются химические циклы и каскады — равновесные хими-
ческие системы с постоянной энтропией. При непрерывном дос-
тупе (электромагнитной) энергии в область прохождения реакций
происходит переход равновесной системы в стационарную систе-
му, способную к производству низкоэнтропийного продукта, то
есть к упорядочению. Вовлечение низкоэнтропийного продукта в
какой-либо последующий процесс вызывает компенсирующее его
воспроизводство системой и формирование еще более низкоэнт-
ропийного продукта и т. д. Переход от микроскопической упоря-
доченности к макроскопической осуществляется через «итерации»
(повторения, воспроизводство) до макроскопически заметного
количества. Итерации реализуются через катализ (в том числе ав-
токатализ) и репликацию.
В этот начальный период синтезируются самые разнообразные
группы органических молекул, включая аминокислоты, сахара,
жирные кислоты и основания, простейшие полимеры. В связи с
этим обсуждались различные сценарии «химического этапа», по-
лучившие названия «мир белков», «сахарная модель», «мир липи-
дов», «мир РНК». По мнению Э.М. Галимова, на роль молекулы,
образующейся в ходе микроскопического химического сопряже-
ния, обладающей способностью к организации стационарной рас-
ширяющейся микроскопической системы и одновременно обла-
дающей свойствами, необходимыми для обеспечения перехода к
макроскопической стационарной расширяющейся химической
системе, может претендовать только молекула, близкая к адено-
зинтрифосфату. Аденин легче других нуклеотидов образуется при
полимеризации молекул HCN (в эксперименте — из исходного
соединения NH4CN), а рибоза — при последовательной конденса-
ции формальдегида (НСОН). Аденин, обладая выраженными
электродонорными и элекгроакцепторными свойствами, легко
соединяется с другими молекулами, в том числе и с рибозой, а об-
302
разевавшийся комплекс, в свою очередь, способен связываться с
фосфатными группами. Молекула, близкая к АТФ, должна была
обеспечить реакции полимеризации, прежде всего образование
полинуклеотидов и полипептидов.
На следующем этапе центральное функциональное место в хи-
мических системах занимают молекулы РНК. С ними связано ста-
новление генетического кода — основного механизма итерации
при переходе к макроскопическим химическим системам, пред-
ставляющим многофазные истинные и коллоидные растворы
органических веществ. Саморазвивающаяся химическая система
полностью основывается на разнонаправленности вещественно-
энергетических процессов, лежащих вблизи химического равнове-
сия, но смещенных от него, непрерывном притоке «усваиваемой»
энергии и способности устойчиво неравновесной системы к про-
изводству упорядоченности.
Обсуждая вероятные «сценарии» происхождения жизни на
Земле, нельзя не коснуться и обобщений, проведенных в самом
конце XX в. И. А. Резановым. Поводом для этого послужило обна-
ружение в метеоритах (углистых хондритах) окаменевших остан-
ков бактерий. Подобные метеориты в Солнечной системе образу-
ют пояс астероидов — обломки разрушившейся около 4 млрд лет
назад планеты, получившей название Фаэтон.
Имеющиеся в распоряжении специалистов данные позволяют
предполагать значительное сходство начальных этапов формиро-
вания Земли и Фаэтона и условий на их поверхности 4,6...4,0 млрд
лет назад. Первичная высокоплотная водородная атмосфера экра-
нировала солнечное излучение и обусловливала давление на по-
верхности этих планет до 10 тыс. атм, а температуры — до 300...
400 °C. Одновременно обширные излияния базальтовой магмы
поставляли на поверхность в огромных количествах вулканичес-
кие газы (Н2О, СО, СО2, NH3, СН4, НС1). В таких условиях очень
интенсивно образуются и накапливаются углеродсодержащие со-
единения. В углистых хондритах их содержание достигает не-
скольких процентов по массе. Даже частичное воспроизведение
(170 °C, 1 атм) подобных условий позволило С. Фоксу и его колле-
гам получать из сухой смеси аминокислот полимерные цепочки,
содержащие до 23 мономеров. Промывка их горячей смеси водой
приводила к образованию микросфер, реагирующих на изменение
осмотического давления.
Простейшие химические системы, образованные органически-
ми молекулами, способны в ряде случаев противодействовать пе-
регреву путем включения эндотермических реакций. Предполага-
ется, что одной из таких реакций могла быть реакция, приводя-
щая в одно и то же время к образованию и формальдегида (исход-
ное вещество для синтеза рибозы), и кислорода (его продукция
303
сильнее других окислительно-восстановительных реакций пони-
жает температуру) (2Н2+2СО2->2СН2О+О2). Вероятно, некоторые
уже синтезированные протобелки служили катализаторами данно-
го процесса. На этом этапе могли сформироваться универсальный
энергетический посредник — аденозинтрифосфат (АТФ) и меха-
низм повторного синтеза протобелков-катализаторов после их
разрушения в виде комплементарной авторепродукции. Совер-
шенствование последнего завершилось возникновением генети-
ческого кода.
В соответствии с рассматриваемой гипотезой бактериальная
жизнь на основе термосинтеза появилась на Земле уже через
100 млн лет после ее образования. Все древнейшие осадочные
породы, графит и даже кварциты образовались при участии бак-
терий, и в них часто обнаруживают формы, не укладывающиеся
в современные классификации. Концентрация и мощность раз-
личных биогенных соединений, сопутствующих друг другу, сви-
детельствуют о сгущениях разнообразных форм жизни в первич-
ной биосфере. Из горных пород были выделены останки циано-
бактерий, перидиней, жгутиконосцев и даже многоклеточных
прокариотов.
После исчезновения водородной атмосферы биогенное осадко-
накопление почти прекратилось, в горных породах того времени
останков бактериальных клеток мало, а разнообразие их резко
снижено. В период 4,1...3,5 млрд лет назад происходила смена тер-
моавтотрофной жизни на фотоавтотрофную. Другие гипотезы свя-
зывают возникновение жизни на Земле именно с этим периодом.
11.2.2. БИОЛОГИЧЕСКИЙ ЭТАП РАЗВИТИЯ БИОСФЕРЫ
Появление оформленных в клетки дискретных единиц разно-
родного живого вещества характеризует становление биоты про-
кариотного типа. На этом этапе биосфера существует как отно-
сительно простая, с небольшой информационной емкостью,
трехкомлонентная система: продуценты (хемоавтотрофы) — дес-
трукторы (хемогетеротрофы) — неорганическое вещество. С воз-
никновением фотоавтотрофных клеток, высвобождающих свобод-
ный кислород, и гетеротрофных аэробных клеток сложность и
«работоспособность» живого вещества существенно возрастают.
В дальнейшем морфологическое и функциональное объединение
аэробных прокариот без цитоскелета и анаэробных прокариот с
цитоскелетом позволило: резко увеличить количество энергии,
используемой для воздействия на внешнюю среду; сформировать
группу редуцентов, связывающую разрозненные и неорганизован-
ные вещественно-энергетические циклы; создать условия для ста-
304
I
новления основных клеточных типов эукариот (животного, гриб-
ного, растительного).
Гипотезы происхождения эукариотических клеток. До настояще-
го времени не предложено непротиворечивой гипотезы происхож-
дения клеток эукариотического типа. Тем не менее среди биоло-
гов признанием пользуются две гипотезы: аутогенная и симбиоти-
ческая.
Аутогенный путь происхождения эукариот предполагает разви-
тие сети внутренних мембран в крупной прокариотической клетке
путем впячивания наружной плазматической мембраны. Эта ги-
потеза хорошо согласуется с данными о динамической непрерыв-
ности клеточных мембран, с гипотезой о монофилии эукариоти-
ческих организмов, опирается на хорошо аргументированный ме-
ханизм внутренней дифференцировки клетки на компартменты.
В остальном же аутогенная гипотеза оказывается спорной или не-
приемлемой.
Симбиотическую гипотезу сформулировала в 1970 г. Л. Марге-
лис, и с тех пор представление об эукариотах как организмах ком-
плексного происхождения широко распространилось в среде био-
логов. Согласно этой гипотезе крупная амебовидная гетеротроф-
ная прокариотическая клетка, получающая энергию путем анаэроб-
ного окисления органических веществ, на определенном этапе
включает мелкую аэробную бактерию — предшественник мито-
хондрии, что в условиях дефицита кислорода оказывается энерге-
тически выгодным; возникает клетка животного типа, захватыва-
ющая спирохетоподобные прокариотические клетки, дающие на-
чало жгутикам. Наконец, внедрение в жгутиковую аэробную клет-
ку прокариот, клеток, подобных современным цианобактериям,
приводит к появлению автотрофной эукариотической клетки.
Симбиотическая гипотеза лучше других объясняет происхож-
дение митохондрий и пластид, окруженных двойными мембрана-
ми и имеющих, как и бактерии, кольцевые ДНК и рибосомы про-
кариотного типа. Она сравнительно хорошо экологически обосно-
вана. Размножение энергетических органоидов вне зависимости
от фазы жизни клетки также согласуется с представлениями о
комплексности первичных эукариот. В то же время нельзя объяс-
нять происхождение жгутиков эукариот следствием их симбиоза
со спирохетоподобными бактериями. Не укладывается в гипотезу
и то, что митохондрии и хлоропласты не вполне автономны и на-
ходятся отчасти под контролем ядерных генов. Требует доказа-
тельств и повышенное содержание кислорода в цитоплазме аме-
боидного прокариота, без чего симбиоз с аэробным организмом
теряет смысл.
Аутогенная и симбиотическая гипотезы при всей их противо-
положности имеют весомые аргументы в свою пользу, и, учитывая
305
20 Присный А В.
это, можно было бы принять к рассмотрению гибридную гипоте-
зу, включающую положительные элементы каждой из них. В этом
случае мы получили бы следующую картину. На ранних этапах
биологического этапа эволюции биосферы существовали различ-
ные по биоэкологическим свойствам группы прокариот. Среди
них наиболее.«полезными» на перспективу свойствами обладали
автотрофные неподвижные и пассивно подвижные прокариоты,
близкие к цианобактериям, гетеротрофные пассивно подвижные
аэробные бактерии и крупные активно подвижные амебообразные
прокариоты. Последняя группа обладала хорошо развитым ци-
тоскелетом и эластичным покровом, но окислительные процес-
сы у ее представителей шли по гликолитическому пути. Способ-
ность подобных клеток к активному движению граничит со спо-
собностью активно воздействовать на внешнюю среду и в мень-
шей степени зависеть от наличных условий. Высокая
информационная емкость крупной подвижной клетки должна
была реализоваться как через усложнение генетического аппара-
та, так и через внутреннюю дифференцировку. В клетке с под-
вижным цитоскелетом и изменчивой формой несколько хромо-
сом, прикрепленных к мембране, требуют компактного размеще-
ния и относительной изоляции от других структурных элемен-
тов. Немаловажной для нее была бы и возможность обеспечения
процессинга мРНК. При названных условиях клетка — предше-
ственник эукариотической клетки должна была иметь несколько
фрагментов ДНК, удерживаемых в глубоком впячивании наруж-
ной мембраны, основные мембранные органоиды, обеспечиваю-
щие фагоцитарный способ питания и сравнительно хорошо раз-
витый цитоскелет.
Развитие симбиоза с аэробными гетеротрофами могло бы выз-
вать революцию в биосфере, так как непосредственное обеспече-
ние цитоскелета макроэргическими соединениями дает мощный
толчок к совершенствованию движения и механического воздей-
ствия на абиотическую и остальную биотическую часть локальных
биогеоценозов, а в сумме всей глобальной экосистемы Земли. Но
такой симбиоз требует общего повышения содержания кислорода
во внешней среде. В связи с этим дополнительные аргументы дает
гипотеза функционального замещения пероксисом митохондрия-
ми. Поэтому одновременно должны были развиться и иные вари-
анты симбиоза: прокариот без клеточной стенки с фотоавтотроф-
ными и аэробными гетеротрофными прокариотами, которые
впоследствии приобрели углеводные оболочки; гетеротрофных
анаэробных прокариот с аэробными гетеротрофами, которые при-
обрели хитиноидную клеточную стенку’. Жгутики эукариот разви-
ваются из цитоскелета и потому не требуют симбиогенного объяс-
нения. Принятие обозначенной последовательности постулирует
306
независимое происхождение клеток растительного, грибного и
животного типов.
Уже с начальных этапов своего становления биосфера пред-
ставляет единство активной (живое состояние вещества) и пассив-
ной (неживое состояние вещества) подсистем. Ее развитие изна-
чально определяется интенсивностью взаимодействия этих подсис-
тем. Но живое вещество, представленное одноклеточными телами,
не способно активно трансформировать даже непосредственно
прилежащую среду для извлечения из нее новых порций вещества
и энергии. Поэтому на определенном этапе эволюции биосферы
появляются многоклеточные бестканевые и талломные, а позже и
тканевые живые тела. Обладая способностью к ростовому и актив-
ному движению, они способствуют резкому расширению и уско-
рению биогенного круговорота веществ, потока энергии и нара-
щиванию объема информации.
Гипотезы происхождения многоклеточности. Мы рассмотрим че-
тыре гипотезы, устанавливающие разные переходные формы меж-
ду протестами и низшими многоклеточными организмами:
гипотезу «гастреи» Э. Геккеля;
гипотезу «фагоцителлы» И. И. Мечникова;
гипотезу «сидячей колонии» А. А. Захваткина;
гипотезу «целлюряризации» И. Хаджи.
Первые три гипотезы связывают возникновение истинно мно-
гоклеточных организмов с колониальными жгутиконосцами, а
последняя — с многоядерными инфузориями.
Геккель считал, что шаровидная колония жгутиконосцев с од-
ним слоем клеток могла подобно тому, как это происходит с блас-
тулой многоклеточных животных, путем впячивания образовы-
вать живые тела, эволюционное развитие которых привело к по-
явлению низших многоклеточных с двуслойной организацией
(подобных кишечнополостным).
Гипотеза Мечникова отличается от предыдущей лишь тем, что
образование внутреннего слоя клеток шло не путем впячивания
наружного пласта, а путем внедрения пищеварительных клеток
внутрь колонии. Такие клетки были названы фагоцитами, их
скопление — фагоцитобластом, а переходная форма — фагоцител-
лой. По строению и образу жизни гипотетическую форму должны
напоминать личинка современных губок и кишечнополостных и
животные, выделяемые в тип Пластинчатые.
По мнению А. А. Захваткина, переходная форма должна быть
сходной не с личинками современных многоклеточных, а с их
взрослыми формами. Поэтому, считает он, у гипотетического
предка имелись черты современных губок, а начало ему дали ко-
лониальные сидячие жгутиконосцы.
С так называемыми «колониальными» жгутиконосцами, сход-
307
ними с видами рода Вольвокс, связывают и происхождение мно-
гоклеточных растений.
Гипотеза «целлюряризации» основывается на таких явлениях,
как полимеризация органоидов и сложная дифференцировка ци-
топлазмы у инфузорий, а также образование поколения клеток
при делении многоядерного плазмодия. Обособление разнород-
ных участков цитоплазмы инфузории вокруг ядер с образованием
наружных разделяющих мембран, по мнению Хаджи, может сразу
привести к образованию трехслойных многоклеточных, близких к
Плоским червям.
Для того чтобы оценить каждую из гипотез, надо прежде всего
сформировать собственное представление о сути проблемы. Когда
мы пытаемся разобраться в эволюционной цепи «клетка — много-
клеточное тело», невозможно обойти такие вопросы: был ли еди-
ный предок для всех многоклеточных живых тел?; что происходит
сначала — дифференцировка клеток или умножение числа одно-
родных элементов (клеток) живого тела?
В главе 3 мы условились вкладывать разное содержание в поня-
тия «колония клеток» (у прокариот и протистов), «многоклеточ-
ное талломное тело» (у грибов и низших растений), «многоклеточ-
ное бестканевое тело» (у Пластинчатых и Губок) и «истинно мно-
гоклеточное (тканевое) тело» (у сосудистых растений и эуметазо-
ев). Следовательно, нам предстоит выяснить происхождение
талломных, бестканевых и тканевых многоклеточных тел.
Анализ жизненных циклов современных Простейших показы-
вает со всей очевидностью, что в большинстве систематических
групп в процессе эволюции развивается тенденция к формирова-
нию аналогов многоклеточности, то есть к умножению однород-
ных ценотически активных элементов в пределах одиночных жиз-
ненных циклов. При этом имеют место все мыслимые направле-
ния такой тенденции. А именно: умножение числа хромосомных
групп в ядре — высокоплоидные ядра; умножение числа одинако-
вых ядер в клетке, в том числе и полиплоидных ядер — много-
ядерные клетки; умножение числа одиночных ценотически актив-
ных клеток — бесполое размножение; умножение числа ценоти-
чески активных клеток с образованием псевдоплазмодия — по
форме — образование многоклеточного тела; умножение числа
ценотически активных клеток с образованием простых (недиффе-
ренцированных) колоний — образование многоклеточного тела
как компактного скопления множества одинаковых клеток; умно-
жение числа ценотически активных клеток с образованием слож-
ной (с частичной дифференцировкой клеток) колонии, объеди-
ненной в единое многоклеточное тело.
В ряде случаев встречается образование истинных плазмодиев
из агрегатов одноклеточных особей. В то же время имеет место и
308
противоположный процесс — образование клеток-особей из плаз-
модиев и многоядерных клеток.
Часто жизненные циклы Простейших организованы сложно и
включают несколько разнородных поколений, но ценотически ак-
тивные поколения, даже если их более одного, представлены оди-
наковыми жизненными формами клеток. Если в жизненном цик-
ле появляются последовательно или одновременно различные
жизненные формы клеток помимо основных, то они не являются
ценотически активными особями, а представляют собой вспомо-
гательные (покоящиеся, инвазионные, транспортирующие и др.)
или половые, ценотически пассивные особи, или их состояния,
фазы. В этом случае наиболее существенной характеристикой
Простейших будет способность формировать только однородные
ценотически активные видоспецифичные клетки, то есть клетки
одной жизненной формы. Соответственно для тканевых много-
клеточных будет определяюще важной способность к развитию
различных морфофункциональных типов ценотически активных
клеток на этапе гистогенеза, каждый из которых комплементарен
сумме остальных. Строго говоря, и Простейшие, и Многоклеточ-
ные способны формировать клетки одинаковых жизненных форм,
но у первых жизненная форма ценотически активных клеток бу-
дет одной из видовых характеристик, а у вторых видовой характе-
ристикой будет служить жизненная форма ценотически активной
части тела или фазы, или особи организма, образованной группа-
ми клеток разных жизненных форм.
Что касается гипотез происхожения многоклеточности, то их
слабость состоит в том, что каждая из них постулирует уникаль-
ность какого-либо одного из предполагаемых путей усиления
функциональной активности с одновременным установлением
монофилии многоклеточных. Логичнее было бы допустить, что
все тенденции к полимеризации активных структур равноценны
и, признавая возможность развития многоклеточности на основе
одного из них, мы автоматически обязаны признать такую воз-
можность и для каждого из остальных, что означает допустить по-
лифилетическое происхождение многоклеточности. Не исключе-
но также, что появление предков многоклеточных животных и ра-
стений не следует связывать с колониальными одноклеточными.
Принцип морфофункциональной дивергенции клеток должен
быть основополагающим в этом процессе: только он обеспечивает
неизбежность и целесообразность взаимозависимости клеток раз-
ных жизненных форм друг от друга.
Итак, ни «гастрея», ни «фагоцителла», ни «сидячая колония»,
ни «многоядерные инфузории» не могут в достаточной степени
универсально или хотя бы удовлетворительно объяснить причину,
механизм и путь возникновения истинной многоклеточности. Ус-
309
танавливаемые же гипотетические формы выводятся из ценоти-
чески пассивных форм, а это требует признания возможности
сальтационной смены ценотической активности целых этапов он-
тогенеза организма.
Интересная идея содержится в гипотезе Г. А. Югая. Возникно-
вение многоклеточное™ он связывает с необходимостью выноса
избыточной информации на надклеточный уровень организации
живого вещества. - Развивая данное предположение, можно уточ-
нить, что избыточность информации сопряжена с мощным пото-
ком внешней биоценотической информации, выражающейся в
дефиците функции в сообществе в период становления верти-
кальной структуры биосферы. Именно потребности системы
могли вызвать вынос жизненной формы на надклеточный уро-
вень, где морфофункциональная дифференцировка клеток спо-
собна дать широкий спектр новых жизненных форм, отсутствую-
щих у одноклеточных, для более мощного воздействия на вне-
шнюю среду.
Вполне можно согласиться с утверждением, что одноклеточные
организмы появились раньше многоклеточных. Но такая последо-
вательность связана не с постепенным развитием «более совер-
шенных форм»: одноклеточные и многоклеточные — не разные
уровни совершенства, а вершины разных ветвей специализации.
Одноклеточные совсем не проиграли в борьбе за существование —
их суммарная биомасса составляет более половины всего живого
вещества биосферы. Последовательность становления: однокле-
точные прокариоты, одноклеточные эукариоты, многоклеточные
эукариоты — определяется последовательностью развития новых
уровней биосферы соответственно требованию усиления функций
при ее расширенном воспроизводстве.
К истинно многоклеточным организмам следует относить
организмы, тела которых состоят из большого числа клеток, диф-
ференцированных на различные ценотически активные морфо-
функциональные типы. С такой оговоркой из числа (истинно)
многоклеточных мы исключаем водоросли, грибы, лишайники,
печеночники и даже мхи, оставляя сосудистые растения и эумета-
зои.
Гипотетические переходные формы при ближайшем рассмот-
рении оказываются не вполне одинаковыми. Так, сидячие жгути-
коносцы в гипотезе Захваткина — это типичные животные клетки,
а «колониальные» жгутиконосцы рода Вольвокс формируют рас-
тительное талломное тело (их клетки соединены плазмодесмами).
Представляется неправомерным выводить происхождение много-
клеточных животных из аналога современного талломного расти-
тельного живого тела.
Учитывая сказанное, становление многоклеточности у грибов,
310
растений и животных следует связывать с разными предковыми i
формами. ।
Многоклеточные талломные тела грибов и растений сформи-
ровались в результате сочетания линейного и плоскостного разра- !
стания клеток соответствующих типов и незавершенного их мито-
тического деления (без завершения цитокинеза). Здесь реализо- •
вался характерный для таких тел ростовой способ движения в од- >
нородном непрерывном субстрате без смены характера
ценотической активности. ।
Становление тканевой организации у растений шло от пассив- '
но подвижных одноклеточных водорослей-обрастателей через (
талломный тип, а колониальный тип организации фитомастигин
следует считать параллельной ветвью. Ткани растений — это лишь 1
значительно усложненный таллом, в котором дифференцировка
клеток достигается в основном с помощью видоизмененной обо- >
лочки и пластид. |
Развитие многоклеточное™ у животных предположительно ।
шло тремя независимыми путями, но всегда было связано с выде-
лением из полифункционального целого олиго- или монофункци-
ональных частей, сохранявших взаимозависимость и надклеточ- I
ную, корпоративную целостность. Установление в качестве пред- f
ковой формы для животных-паразоев примитивных зоомастигин с
колониальным типом организации не объясняет развития обрати-
мой морфофункциональной дифференцировки клеток-особей на
два основных типа: амебоидный пищеварительный и жгутиковый
двигательный.
Появление у животных многоклеточное™ с тканевым типом
организации могло определяться двумя механизмами: 1) исходной
дифференцировкой клеток на покровные и нервные через специа-
лизацию на уровне клеток и одновременно покровных на покров-
но-пищеварйтельно-двигательные и покровно-двигательные че-
рез специализацию на уровне частей клеток; 2) исходной необра-
тимой дифференцировкой на пищеварительные, двигательные,
покровные и нервные на уровне клеток. Первый путь становления
эуметазоев должен был привести к линии, заканчивающейся дву-
слойными, а второй — к линии трехслойных. В отличие от паразо-
ев у эуметазов совершенствуются не наружные двигательные
структуры цилиарного типа (свойственные подгонятелям и филь-
траторам), а внутренние сократимые комплексы (характерные дая
активно подвижных) и клеточные мембраны.
В заключение отметам, что в большинстве групп многоклеточ-
ных животных обнаруживается как одно из направлений специа-
лизации образование синцитиев. Это, впрочем, не дает повода вы-
страивать эволюционную линию Простейших из Многоклеточных
через синцитии и плазмодии, как и наоборот, но показывает сход-
311
ные способы усиления функции через однонаправленные про-
цессы.
Представления об истории живых организмов на Земле. В период
бурного развития капиталистических форм производства, то есть в
конце XVIII начале XIX в., в общественном (массовом) созна-
нии еще господствовали религиозные представления, утверждав-
шие постоянство, незыблемость всего существующего, созданного
Великим Творцом. Но становление новых форм общественных
отношений, развитие материального производства и успехи есте-
ственных наук послужили толчком к развитию материалистичес-
ких идей в философии, где в это время господствовала идеалисти-
ческая концепция мирового порядка. Противоположные фило-
софские течения оказали влияние и на биологию. С одной сторо-
ны, консервативная система взглядов нашла свое отражение в
«креационизме» — учении об устойчивости, неизменности приро-
ды как конечной цели ее сотворения, а с другой стороны, резуль-
таты морфологических, физиологических, палеонтологических,
других биологических, а также геологических исследований, пере-
ложенные на материалистическую основу, вызвали развитие «на-
учного трансформизма» — учения об историческом непостоянстве
природы, ее изменяемости и даже закономерной эволюции.
Наиболее известными сторонниками креационизма были
К. Линней, Ж. Кювье, Ч. Лайель. Как система взглядов на приро-
ду креационизм с течением времени значительно видоизменялся.
Первоначально считалось, что все виды живых организмов созда-
ны Богом в едином акте творения и с тех пор не подвергались ни-
каким изменениям. Более тщательные исследования строения ра-
стений и животных, а особенно находки форм, существовавших в
прошлые геологические эпохц, вынуждали ученых признавать,
что виды изменчивы не только на каждый момент времени, но
также и в длительности своего существования. Но не укладываю-
щиеся в общее миропонимание факты воспринимались весьма
своеобразно. Линней за четыре года до своей смерти в одном из
писем профессору Российской академии наук И. Г. Гмелину, от-
мечая факты отдаленной гибридизации растений, затрагивал воп-
рос видообразования. В частности, он писал: «Первоначально Бог
создал столько различных растений, сколько есть естественных
отрядов (по современной классификации — семейств). ... он сам
же затем эти растения отрядов перемешал скрещиванием ... по-
явилось столько растений, сколько существует разнообразных от-
четливых родов. Затем Природа эти родовые растения перемешала
между собой и умножила в существующие виды, все какие воз-
можно». Фактически, по Линнею, сотворены только семейства,
виды же образуются естественным путем.
Ж. Кювье, виднейший морфолог животных, не признавая эво-
312
люционных преобразований живого населения планеты, под дав-
лением фактов палеонтологии и геологии сформулировал компро-
миссную «теорию катастроф». Согласно этой теории перерывы в
палеонтологической летописи и резкие смены фаун и флор связа-
ны с периодическими глобальными катастрофами на поверхности
Земли и проникновением на освобождающиеся территории слу-
чайно сохраняющихся растений и животных. Позднее появление
новых видов в последовательных геологических напластованиях
объясняли дополнительными актами Творения.
Принятие катастроф в истории Земли как научного факта мог-
ло служить аргументом к доказательству направленных измене-
ний, то есть эволюции Земли. Но и катастрофы были логично
встроены в концепцию «Постоянства» стараниями авторитетней-
шего геолога Ч. Лайеля. Используя законы ньютоновской косми-
ческой механики для объяснения земных процессов, он сформу-
лировал теорию «динамического постоянства Земли» — изменяю-
щейся, но вечной планеты. Согласно представлениям Лайеля, ис-
тория Земли и геологические процессы должны подчиняться двум
главным принципам: постепенности и ненаправленности, а в це-
лом — принципу единообразия, или «униформизма». Унифор-
мизм как научная концепция противопоставлялся «дирекциона-
лизму» — концепции направленных изменений, куда в качестве
частного случая входил и чистый катастрофизм. Так называемые
катастрофы, или перерывы в постепенности, Лайель объяснял
вторичными локальными выпадениями пластов и изменениями
направленности процессов. И все же к концу жизни, когда свиде-
тельства направленного развития живой природы привели Лайеля
к принятию эволюционной теории, он отказался от своего убеж-
дения, что Земля находится в неизменном состоянии.
Современная форма креационизма весьма сложна для прямого
противопоставления трансформизму или эволюционной теории,
поскольку она вновь вернулась из биологии в философию, где
есть возможность признавать и эволюцию материи, и неизмен-
ность, сотворимость идеи одновременно. Так, например, креаци-
онисты допускают, что человек произошел от обезьяноподобных
предков, но душе и разуму человека приписывают божественное
начало.
Идеи изменяемости природы возникали еще у античных фило-
софов и проповедовались в средние века, но с конца XVII в. для
доказательства преобразований в природе привлекаются научные
факты. Среди известных натуралистов, в той или иной степени
придерживавшихся идей трансформизма, были Р. Гук, Ж. Ламет-
ри, Д. Дидро, Ж. Бюффон, Э. Дарвин, Э. Жоффруа Сент-Илер,
К.Ф. Рулье. Механизмы преобразований живых организмов свя-
зывались с так называемыми целесообразными изменениями в от-
313
вет на действие внешней среды. Предполагалось, что такие при-
способительные изменения могут передаваться дочерним поколе-
ниям.
Первое достаточно полноценное обобщение основных поло-
жений трансформизма как учения и их доказательств сделал
Ж.Б. Ламарк. Его система взглядов уже перерастает традицион-
ный трансформизм, но еще не становится настоящим эволюцион-
ным учением. В научной литературе она получила название «ла-
маркизм». Основные положения эволюционной гипотезы Ламар-
ка следующие:
живые организмы изменчивы;
эволюционирующие виды постепенно превращаются в другие;
общее направление эволюции — усложнение, совершенствова-
ние организации;
движущая сила эволюции — внутреннее, изначально существу-
ющее стремление к прогрессу.
Впоследствии два первых положения позволили Ламарку отри-
цать реальность вида. «Правильное» совершенствование, идущее
от стремления к прогрессу, Ламарк назвал «градацией». Внешние
условия могут вызывать отклонения от градации, что выражается
в приобретении организмами приспособительных изменений. Из-
менения, по Ламарку, наследуются.
Со временем не принятая большинством натуралистов из-за
недостаточности доказательств гипотеза Ламарка модифицирова-
лась в различные формы «неоламаркизма». «Автогенез» и «эктоге-
нез» — одни из этих форм — представляют скорее исторический
интерес, но «генетический неоламаркизм», развиваемый в послед-
ние десятилетия российскими учеными, может стать достаточно
аргументированной гипотезой.
Эволюционное учение. Объективная невозможность доказать
экспериментально эволюцию живого вещества была преодолена в
XIX в. благодаря «открытию» ее движущих сил. Почти одновре-
менно и независимо друг от друга Э. Блите, П. Метью, А. Уоллес и
Ч. Дарвин выдвинули предположение о действующем в живой
природе естественном отборе, то есть о переживании или преоб-
ладающем размножении форм, наиболее приспособленных к внеш-
ним условиям. Заслуга Дарвина состоит прежде всего в том, что он
нашел убедительные аргументы из биогеографии и области искус-
ственного отбора для доказательства существования неопределен-
ной наследственной изменчивости и борьбы за существование,
приводящей к естественному отбору, повышающему общую при-
способленность организмов и формирующему новые их «разно-
видности».
Собственно гипотеза эволюции, сформулированная Чарльзом
Дарвином, в настоящее время в первозданном виде почти не рас-
314
сматривается. За последние 50...60 лет сильно изменилось ее сло-
весное оформление, уточнено и дополнено содержание основных
положений. Как определенный этап в развитии «дарвинизма»
можно рассматривать «синтетическую теорию эволюции» (СТЭ),
по сути — синтез эволюционной теории и генетики. СТЭ оформи-
лась в 40-е годы прошлого столетия и, в свою очередь, также пре-
терпела изменения. Сейчас основные положения СТЭ, а точнее —
синтетической гипотезы эволюции (СГЭ), можно сформулировать
в следующем виде.
1. Материалом для эволюционных преобразований живых
организмов служат модификации и комбинации генов, мутации
же в основном представляют «генетический шум».
2. Модификации носят направленный характер, комбинации —
ненаправленный, а мутации могут быть как ненаправленными,
так и направленными.
3. Направляющим фактором эволюции служит дефицит функ-
ции в системе, а ее механизмом — естественный отбор.
4. Наименьшая эволюционная единица — популяция.
5. Эволюция обычно носит дивергентный характер, но может
быть и филетической. Новый вид образуется, как правило, из од-
ного материнского, при гибридизации — из двух, а при горизон-
тальном переносе генов — не менее чем из двух материнских ви-
дов.
6. Темпы эволюции меняются. Эволюционные преобразования
могут быть как постепенными, так и скачкообразными.
7. Границы между популяцией и видом, популяцией и организ-
мом условны. Неоднородность среды обитания вида обусловлива-
ет создание внутривидовых подразделений надпопуляционного
уровня: полувидов, подвидов, рас и морф.
8. Генетическая система вида открыта, но целостна.
9. Репродуктивная изоляция как критерий вида применима
только для видов с половым процессом.
10. Макроэволюция может идти как через микроэволюцию, так
и минуя традиционные микроэволюционные пути.
11. Эволюция не направлена к некоей конечной цели, но с уче-
том прошлой истории, генетического окружения и очевидного
влияния среды может быть прогнозируема.
12. Направленная изменчивость во взаимодействии с отбором
служат механизмом корректировки морфофизиологической орга-
низации существующих видов и выделения новых видов, соответ-
ствующих свободным эконишам. Отбор идет одновременно на
организменном, видовом и экосистемном уровнях.
13. Наиболее общими путями эволюционных преобразований
являются дифференциация, интеграция и новообразование.
14. Образование новых видов сопровождается прямым запол-
315
нением, дроблением или объединением экологических ниш, по-
рождаемых системой.
11.3. ПРЕДСТАВЛЕНИЯ О МЕХАНИЗМАХ ЭВОЛЮЦИИ
ЖИВОГО ВЕЩЕСТВА
Общее наращивание информации в биосфере характеризуется
опережающими значениями упорядочения функциональных от-
ношений между структурными элементами (собственно информа-
ции). Поскольку эволюционирующая биосфера и есть земная
форма жизни, как частный вариант расширенного воспроизвод-
ства информации общее направление эволюции биосферы — ее
прогрессивное развитие — совладает с направлением антиэнтро-
пийных процессов во Вселенной (противоположным энтропий-
ной направленности «стрелы времени»). В этом смысле прогрес-
сивная эволюция биосферы необратима. Регресс возможен только
в функциональных подразделениях биосферы. Со снижением
иерархического уровня он идет легче, а на глобальном уровне мо-
жет заменяться лишь деградацией, разрушением. Рассуждая об
эволюции, мы должны разграничивать несколько явлений на пер-
вый взгляд одного порядка: эволюцию биосферы как живой сис-
темы, эволюцию живой материи (в том числе живого вещества),
эволюцию элементарных подразделений живого вещества — орга-
низмов, эволюцию функций (экологических ниш). В традицион-
но употребительном толковании эволюцию рассматривают как
процесс формирования адаптации биологических систем к усло-
виям окружающей среды. Корректнее было бы представлять эво-
люцию биосферы как процесс повышения эффективности ис-
пользования доступной энергии для накопления информации ее
материальными элементами.
11.3.1. МИКРОЗВОЛЮЦИЯ
Изменчивость. По сложившейся традиции принято разграничи-
вать определенную и неопределенную изменчивость. Другие ис-
пользуемые в литературе понятия, такие как фенотипическая и ге-
нотипическая, ненаследственная и наследственная, групповая и
индивидуальная изменчивость, не имеют прямого соответствия
первым.
«Определенная изменчивость — это те внутривидовые разли-
чия, которые возникают под влиянием факторов внешней среды и
условий воспитания; они одинаково выражены у всех особей дан-
ного вида и могут быть предсказаны заранее. Неопределенная из-
316
менчивость проявляется в разнообразных слабых различиях, кото-
рыми отличаются особи одного и того же вида» (Тыщенко, 1992).
Иными словами, определенная изменчивость обусловлена внеш-
ними факторами, а неопределенная — зависит от этих факторов,
но носит случайный характер и обусловлена наследственными
Свойствами самих организмов.
Строго говоря, разграничение наследственной и ненаслед-
ственной изменчивости неправомерно, поскольку «определен-
ные» индивидуальные изменения также имеют наследственную
природу, хотя здесь наследуются не готовые признаки, а возмож-
ный диапазон развития того или иного признака, или норма реак-
ции (см. раздел 5.1).
Норма реакции — это обусловленная генотипом индивиду-
альная программа развития. Конечное выражение признака в
пределах нормы реакции определяется сочетанием экологичес-
ких факторов и потому может отличаться у разных особей даже
при идентичности их генотипов. Такую изменчивость называют
Модификационной, а соответствующие факторы — модифика-
торами.
Неопределенная изменчивость объясняется несовпадением
генотипов и установленных ими норм реакций у разных особей
одного вида при действии одинаковых модификаторов. Причи-
ной несовпадений являются генотипические отличия особей в
пределах видоспецифичной генетической системы, передавае-
мой в ряду поколений при воспроизводстве. Генетические от-
личия особей могут возникать в результате мутаций (спонтан-
ных или индуцированных изменений в структуре генетического
кода), комбинаций (образования новых сочетаний аллелей в
пределах видоспецифичного генотипа) и вариаций (изменений,
связанных с так называемыми «мобильными диспергированны-
ми генетическими элементами»). В формировании дочерних ге-
нотипов при половом и бесполом воспроизводстве эти способы
генетических новообразований принимают неодинаковое учас-
тие. Мутационная, комбинативная и вариационная изменчи-
вость служат источниками новых признаков и свойств, получае-
мых и передаваемых по наследству и потому служащих материа-
лом для эволюционного процесса. При этом особая роль в ре-
организации геномов отводится интронам. Доказано, что после
самовычленения из первичных РНК-транскриптов они могут
встраиваться непосредственно в безинтронные участки ДНК
того же самого гена или в РНК-транскрипт другого гена и на
следующем этапе через механизм обратной транскрипции со-
здавать новые участки ДНК. Одновременно может происходить
и перетасовка экзонов, с которых транслируются белки. Подоб-
ные генные преобразования могут происходить одновременно и
317
однонаправлено у большого числа особей в популяциях. Со-
гласно одной из гипотез вариационный механизм лежит в осно-
ве «скрытого» и быстрого формирования новых генов, способ-
ных определять появление новых фенотипических признаков в
готовом виде. Комбинативная же изменчивость оперирует су-
ществующим набором генов.
Эволюционные процессы в популяциях. Морфофункциональной
единицей видового живого вещества является особь. Именно осо-
би подвергаются действию внешних факторов, благоприятствую-
щих или препятствующих их индивидуальной жизнедеятельности
и воспроизводству, то есть действию естественного отбора. Вместе
с тем особь не является элементарной эволюционной единицей,
поскольку эволюция предполагает долговременное направленное
изменение генотипов (определяющих развитие жизненных форм,
которые способны реализовать экосистемные функции) в ряду
поколений. Поэтому в качестве элементарной эволюционной еди-
ницы принимаются популяции, имеющие относительную генети-
ческую обособленность. Генетическая изоляция не тождественна
репродуктивной изоляции, так как последняя предполагает обяза-
тельное наличие полового воспроизводства с участием компле-
ментарных половых типов особей.
В пределах популяционного генофонда осуществляется посто-
янный обмен аллелями при незначительном притоке аллелей из
других популяций того же вида. В процессе обмена непрерывно
формируются и испытываются новые комбинации аллелей, часть
из которых, прошедшие «сито» отбора, становятся достоянием по-
пуляции. В результате с течением времени популяция меняется и
приобретает генетическое своеобразие. Таким образом, популя-
ция как статистическая целостность обеспечивает взаимодействие
и преемственность генотипов и изменение генофонда, что опреде-
ляет ее эволюционное значение.
Механизмы изменения генофонда популяции можно понять,
анализируя процессы, нарушающие его равновесное состояние.
Внутренне обусловленная устойчивость генофонда формализова-
на в законе Харди-Вайнберга: «если на бесконечно большую пан-
миксную (со свободным скрещиванием) популяцию не действуют
возмущающие процессы, то частоты отдельных аллелей в ряду по-
колений в такой популяции не изменяются». В комплекс факто-
ров, нарушающих «идеальное равновесие», входят: изоляция по-
пуляций, колебания численности популяций, «дрейф генов» (слу-
чайное однонаправленное смещение частот аллелей) и дифферен-
цированное размножение (включая избирательную элиминацию
особей). При этом приспособительное направленное изменение
генофонда популяции обусловливает только дифференцирован-
ное размножение, или естественный отбор.
318
Борьба за существование. Содержание явлений, обозначаемых д
понятием «борьба за существование», в общем смысле не соответ- :
ствует ему. Здесь предполагается, что любому видовому живому
веществу присуще внутреннее стремление к неограниченному ро-
сту («давление жизни», «стремление к максимальной экспансии»),
которое сдерживается всей совокупностью экологических факто- i
ров. В результате такого встречного давления численность попу- b
ляции в течение длительного времени колеблется в определенных I
пределах с более или менее устойчивым средним значением. Это ;
означает, что начальные численности каждого условно синхрони-
зированного поколения или численности размножающихся осо- ।
бей должны быть равны. Следовательно, рост популяций, опреде- |
ляемый рождаемостью, компенсируется соответствующим уров- I
нем смертности. [•
Для статистического показателя — численности безразлично,
что происходит в популяции на уровне жизнедеятельности осо-
бей. Особи же, обладая рядом признаков автономных живых сис- I
тем, ведут себя подобно таким системам, то есть стремятся к со-
хранению постоянства своих внутренних параметров и распрост- I,
ранению себя на прилежащую часть внешней среды. Поскольку t
особи одной популяции представляют однородные морфофунк-
циональные биогеоценотические единицы, их взаимодействие в
условиях ограниченности вещественно-энергетических ресурсов |;
сводится к конкуренции, растущей вместе с ростом плотности I
(или численности) популяции. Межвидовая конкуренция прояв-
ляется только в «зонах перекрывания» экологических ниш. Про- ।
тиводействие неблагоприятным абиотическим факторам в рамках I,
борьбы за существование оказывается значимым только в том слу-
чае, когда сила проявления фактора лежит между порогами чув- I
ствительности и устойчивости к ним всех особей популяции. Ре-
зультат взаимодействия с консументами в каждом конкретном I;
случае определяется состоянием рассматриваемой особи и ее по- I.
едателя, а также ценотической ситуацией. Генотипическая и, еле- [
довательно, фенотипическая гетерогенность особей в популяции Г
предопределяет разные индивидуальные возможности дожить до |
репродуктивного периода и оставить потомство. 1
Естественный отбор. Вследствие мутаций популяционных волн, j
изоляций и дрейфа генов изменяется частота аллелей в популяци- р
ях независимо от того, вызывает это или нет изменение присособ- У
ленности организмов к комплексу экологических факторов. Борь- |
ба за существование сама по себе также не способствует развитию [
адаптаций. К повышению приспособленности может вести только i
естественный отбор. |
Естественный отбор можно рассматривать как процесс и как |
результат. Как процесс он выражается в дифференцированной [
319 f
элиминации генотипов (комбинаций аллелей), понижающих жиз-
неспособность особей в конкретных складывающихся условиях, и
в неравноценном участии особей в процессе воспроизводства но-
вого поколения в популяции. Как результат естественный отбор
представляет собой увеличение в популяции частот аллелей и
комбинаций аллелей, которые обеспечили родительским особям
относительно эффективное участие в воспроизводстве и, следова-
тельно, динамическое поддержание приспособленности к реаль-
ным условиям существования популяции. Сами условия при их
непрерывной изменчивости на значительных промежутках време-
ни характеризуются смещением средних параметров, что ведет и к
направленным изменениям биоэкологических параметров попу-
ляции.
В настоящее время принято различать четыре формы есте-
ственного отбора: движущий, стабилизирующий, дестабилизиру-
ющий и дизругггивный (рассекающий).
Движущий отбор вызывает однонаправленное изменение по-
пуляции за счет предпочтения редких или новых генотипов од-
ной группы и элиминации редких генотипов другой группы.
Проявляется при длительных направленных изменениях условий
среды.
Стабилизирующий отбор выражается в увеличении частот ге-
нотипов, определяющих среднее выражение признака за счет
крайних уклонений. Особенно отчетливо он проявляется в долго-
временно устойчивой среде.
Дестабилизирующий отбор, наоборот, подхватывает уклонения
от «стандарта» популяции и увеличивает их частоты за счет сред-
ней нормы. Разнообразие генотипов и размах изменчивости при-
знаков при этом увеличиваются, что является условием для начала
действия движущего и дизруптивного отбора. Дестабилизирующе-
му отбору подвергаются популяции, на которые уменьшается дав-
ление лимитирующих факторов.
Сочетание движущего, стабилизирующего и дестабилизирую-
щего отбора может привести к дизруптивной форме, когда сохра-
няются и увеличиваются в частотах генотипы, определяющие
крайние выражения фенотипов, и при этом элиминируются про-
межуточные.
Понятие «отбор» отражает содержание по меньшей мере трех
явлений:
1) многоуровневый отбор на способность к поддержанию гоме-
остаза в меняющихся условиях среды. Он обеспечивает функцио-
нальную устойчивость вида. Завершается выработкой организмен-
ных и популяционных адаптаций;
2) отбор по отдельным признакам фенотипа, не имеющим пря-
мого адаптивного значения для особей. Обеспечивает непрерыв-
320
ность видового живого вещества во времени и конкретизацию по-
токов вещества, энергии и информации в системе, дифференци-
рует систему;
3) отбор онтогенетических новообразований на соответствие
нише. Обеспечивает относительную полноту и целостность систе- 11
мы, интегрирует систему. Завершается становлением комплекса
системных адаптаций вида, «утверждает» тот или иной тип онто- 1
генеза, необходимый данной системе.
Все три названные формы отбора сходятся на организмах и со- •
ответствующих им уровнях функционирования экосистем и идут 1
одновременно. Действие комплексного отбора на отдельные при- L
знаки фенотипа ведет к развитию их полифункциональности.
В таком виде отбор не является фактором (движущей силой) эво- '
люции, а представляет корректирующий фильтр — часть механиз-
ма поддержания динамической устойчивости системы, ее морфо- i
функциональной полноты.
Избыточное размножение организмов традиционно считается I
необходимой предпосылкой отбора. Но в этом явлении есть и дру- [
гой аспект. Потоки вещества, энергии и информации не могут ,
быть реализованы при устойчивой численности особей на всем 1 j
протяжении жизни поколения. Часть видового живого вещества [
непрерывно направляется в пищевые цепи, часть особей, не успе- .!
вая реализовать биогеоценотические функции, гибнет от случай- ।
ных факторов, некоторые фенотипы элиминируются на началь-
ных стадиях развития из-за генетических аномалий. По этим и р
другим причинам в устойчивой системе замещение поколений ।
организмов обязательно сопровождается увеличением числа осо- .
бей. ;
Видообразование. Рассматривая проблемы вида, мы отмечали, 11
что вид как устойчивая целостность и обособленность существует (
в пространстве более или менее длительное время. Поэтому каж-
дый вид следует выделять и среди видов, существующих одновре- .
менно с ним, и в эволюционном ряду материнских и дочерних
форм. В соответствии с принципом «все живое — из живого» но- “
вый вид может сформироваться только на основе уже существую-
щего или существующих. При этом возможно несколько вариан- |
тов видообразования: 1) два новых вида или более образуются пу- (
тем разделения материнского вида с сохранением последнего; 2)
два или более новых видов образуются путем разделения материн-
ского вида без сохранения последнего; 3) один новый вид образу-
ется из материнского путем медленного или быстрого его преоб-
разования; 4) один новый вид образуется на основе двух или более I
материнских. j
Первые два варианта объединяются общим названием «дивер-
гентное видообразование». Многократно повторяющаяся дивер-
21 Присный А В ^21 |
I
генция представляет собой так называемую адаптивную радиа-
цию. Эволюцию, основанную на дивергенции и адаптивной ради-
ации, обозначают понятием «кладогенез».
Развитие таксона (в том числе вида) без образования «боковых
ветвей» называют анагенезом. Этот способ видообразования при-
водит к исторической смене видов без увеличения видового мно-
гообразия.
Последний из приведенных вариантов видообразования связан
с образованием гибридных форм, имеющих статус вида, а потому
не требует подготовительных микроэволюционных процессов.
Соответственно механизму образования этот способ получил на-
звание «гибридогенез».
Соотнесение пространственного распределения материнских и
дочерних видов позволяет считать вероятным видообразование
как на основе изоляции родительских группировок расчленением
общего исходного видового ареала (аллопатрическое, или геогра-
фическое, видообразование), так и на основе экологической изо-
ляции форм в пределах ареала общей родительской группировки
(симпатрическое, или экологическое, видообразование). Объеди-
няющим началом для аллопатрического и симпатрического видо-
образования служат экологические ниши (их фундаментальный и
реализованный аспекты).
Аллопатрическое и симпатрическое видообразование, вероят-
но, нс представляют собой чисто пространственные формы, но
связаны со сложностью или зрелостью экосистем. Первое должно
быть более характерно для молодых и обедненных видами сооб-
ществ и в механизмах образования основывается на пульсациях
ареалов, сопровождающих жизненные волны, а также на пассив-
ных естественных и в последнее время искусственных заносах ро-
доначальных форм. Если такое переселение приводит к попада-
нию уже существующего вида в адекватную ему, но новую для
данного сообщества нишу, то вид-вселенец испытывает лишь
всплеск гомеостатических реакций на индивидуальном и популя-
ционном уровнях. Если свободная экониша частично соответству-
ет функциональному содержанию вида, то такой вид может закре-
питься в сообществе и со временем дать новообразование, частич-
но или полностью устраняющее дефицит функции. Отсутствие
равнозначной ниши в сообществе вызывает быструю гибель чуже-
родной группировки. Наконец, если в сообществе соответствую-
щая функция имеется, но реализуется «собственным» видом, сле-
дует ожидать вступления вселенца в конкурентные отношения,
исход которых зависит от сравнительной эффективности конку-
рирующих видов. Таким образом, аллопатрическое видообразова-
ние — это, во-первых, заполнение ниши, а во-вторых, интенси-
фикация функции.
322
Симпатрическое видообразование, как предполагается, более
характерно для сложных насыщенных сообществ, в которых уже
имеется достаточное количество онтогенетических типов. Запол-
нение ниш в них отступает на второй план, в то же время устой- ((
чивость сложных сообществ создает предпосылки для интенси- 1
фикации функций путем их дробления с соответствующей диф-
ференцировкой наличных функциональных элементов. Конку- ।
рентные отношения здесь имеют некоторую специфику: в
первую очередь они возникают в группах новых функционала- I
ных единиц, претендующих на одну нишу, в период их становле-
ния. Но поскольку они не могут быть полностью тождественны-
ми, такая конкуренция должна приводить к дроблению самой
экониши и формированию множества узкоспециализированных I
и полифункциональных видов. Следует подчеркнуть, что, даже
несмотря на допущение малочисленности исходной (материне- ||
кой) группировки при аллопатрическом видообразовании, ста- I
новление нового вида — это выделение некоторого множества, 1
уже реализующего функцию, а следовательно, значительного
множества. В зависимости от степени однородности экосистемы
в пространстве новый вид может быть исходно мономорфным I
или полиморфным. !
Известно, что генотип на каждый конкретный момент времени
имеет различное состояние по соотношению активных и «молча-
щих» генов, как структурных, так и регуляторных. Активность ге-
нов выражается в построении фенотипа особи, который может из-
меняться в течение полного онтогенеза очень резко, давая в том
числе ценотически активные и ценотически пассивные жизнен-
ные формы. Исходя из этого, можно предположить возможность
нейтральных для вида изменений в генотипах организмов, если
они происходят в «молчащих» генах или затрагивают ценотически
пассивную фазу онтогенеза. Единообразность таких изменений у
многих особей материнской популяции-вида под воздействием
направленной внешней информации, при синхронном их «вклю-
чении» могла бы служить механизмом сальтационного видообра-
зования (рис. 11.3).
Поскольку движущей силой и направителем эволюции видо-
вых форм живого вещества является дефицит функции в развива-
ющейся системе, охватывающий, пронизывающий ее целиком,
пространственно-временная изоляция не должна быть необходи-
мым условием видообразования.
Видимо, можно различать три основных направления видооб-
разования: дифференцирующее — разделение материнской груп-
пировки на несколько специализированных дочерних — ведет к
интенсификации функции; интегрирующее — усложнение онто-
генеза у дочерней группировки, сопровождающееся развитием нс-
323
21*
Онтогенезы
Конвергенция
(развитие близких
жизненных форм
на основе разных
генотипов)
Полувиды
Генотип
Дефицит
функции
Дивергенция
с «переходными»
формами
Модифицирующее
давление внешней
среды
Интродукция
Частоты вариантов
морфофункци-
онального типа
(фенотипа)
Вымирание
Рис. 11.3. Модель «эволюционного ландшафта» (оригинал автора)
скольких различных ценотически активных фаз, — ведет к расши-
рению и объединению функций; заполняющее — образование но-
вой формы, полноценно соответствующей новой экологической
нише, — направлено на обеспечение полноты экосистемы.
11.3.2. МАКРОЭВОЛЮЦИЯ
Макроэволюция, или надвидовая эволюция, прослеживается в
нескольких аспектах. Это и упомянутая коэволюция, и эволюция
емких подразделений живого вещества, обозначаемых как надви-
довые таксоны, и эволюционные изменения биогеоценозов, и, на-
конец, эволюция биосферы. Предметом наибольших дискуссий и
разногласий среди них является эволюция надвидовых подразде-
лений живого вещества, что связано с существованием его в виде
«абстрактной» совокупности видовых форм. Последние доступны
наблюдению, сопоставлению, анализу и на современном материа-
ле, и в палеонтологической летописи, чего нельзя сказать, напри-
мер, о типе или классе, объединяющих разнородное вещество.
Утвердившийся метод анализа эволюции надвидовых таксонов
и стоящих за ними надвидовых подразделений живого вещества
позволяет проследить лишь процесс исторической дифференци-
ровки более емких подразделений на менее емкие и противопо-
ложный ему интеграционный процесс (рис. 11.4). Но стоит только
изменить плоское изображение эволюционных траекторий на
объемное, та же схема позволит получить дополнительную, ранее
324
Collembola
Protura
Diplura . ।
Ротовые части погружены в кармановидное вгшчивание
Наружные ротовые части
Класс Entognatha
—-----------Mikrocoryphia ;
------------Thysanura [_________________
Появление крыльев ____________
------------Ephemeroptera Г Инфракласс
-------------Odonata 1_^_р3!£а__
} Подкласс Apterigona
Появление складывающихся крыльев_
______________«----Dictyoptera
"I-----------------Isoptera
----------Orthoptera
।— Phasmoptera
1— Dermaptera
----------Grylloblattodea
—---------Embioptera L
----—--------Plecoptera r.
---------- Zoraptera J
~~ ।-----Psocoptera ;
I-----Phtiriaptera J
!------- Thysanoptera '
I------Hemiptera '
Появление личинок и куколок
Rhaphidioptera ;
Megaloptera J
Neuroptera ;
Появление надкрыльев !
Coleoptera j
Hymenoptera |
Mecoptera I
Siphonaptera
Diptera
Trichoptera
Lepidopteja
о
£
о о.
г
о
I
S Щ
S £
е
s
О
Z
3
h
° £
&g
Ё &
о о
£
а>
X
1
о.
е
X
S
Рис 11.4. Схема предполагаемого филогенеза отрядов энтогиат и насекомых
(Ч. Росс, Д. Росс, 1985, с изменениями)
скрытую информацию (рис. 11.5). На обеих схемах сравнительно
хорошо просматриваются группировка отрядов в естественные ге-
нетические комплексы, возраст отрядов и скорость эволюцион-
ных преобразований. Вторая же схема, хотя и основана на учете в
основном морфологических признаков взрослых особей, высвечи-
вает экологическую эволюцию: отряды обнаруживают тенденцию
Рис. 11.5. Схема предполагаемой морфофункциональной эволюции отрядов энтогнат
и насекомых (оригинал автора):
Расстояния между вершинами ветвей примерно соответствуют числу отличий по 18 учтенным
морфологическим и биологическим признакам. Выделяются группировки: ценотически актив-
ные во всех фазах подстилочно-почвенные сапрофаги и ризофаги (коллемболы — Coll, проту-
ры — Pro, липлуры — Dip, микрокорифии — Mic, щетинохвостки — Thy, сеноеды — Pso, зорап-
теры — Zor, эмбии — ЕтЬ), со смешанным питанием (уховертки — Der) и ксилофаги (терми-
ты — Iso); ценотически активные во всех фазах открыто живущие грызущие фитофаги (палоч-
ники — Pha и прямокрылые — Ort); ценотически активные во всех фазах многоядные и
хищники (таракацосверчки — Gry, тараканы — Bia и богомолы — Man); ценотически активные
во всех фазах сосущие фитофаги (трипсы — Thys, равнокрылые — Нот и клопы — Het); цено-
тически активные в личиночной фазе хищники (верблюдки — Rap, скорпионницы — Мес и
сетчатокрылые — Neu); ценотически активные в личиночной и частично во взрослой фазе па-
разиты (пухоеды — Mai, вши—Апо, весрокрылые — Str и блохи — Sip); ценотически активные
в личиночной фазе грызущие фитофаги (чешуекрылые — Еер); ценотически активные в личи-
ночной фазе водные детритофаги, фитофаги и хищники (стрекозы — Odo, поденки — Eph, вес-
нянки — Pie, вислокрылые— Meg, ручейники — 7и); группа отрядов, дающая дополнительно
экологическую дивергенцию с повторением предыдущих на уровне семейств (перепончато-
крылые — Нут, жуки — Col, двукрылые — Dip) (оригинал автора)
326
к группировке в емких экологических нишах по стволовым харак-
теристикам именно ценотически активных фаз онтогенеза. Воз-
раст отрядов и скорость эволюционных преобразований здесь свя-
зываются с периодами крупных реорганизаций в биосфере и реак-
цией наличных подразделений живого вещества на появление ем-
ких экологических ниш.
При рассмотрении эволюционных изменений биогеоценозов,
общие закономерности которых излагаются в курсах «Экология»
и «Эволюционная теория», важно учитывать, что биогеоценозы —
это структурно-функциональные подсистемы биосферы, и пото-
му их пространственно-временная динамика не может быть пол-
но описана через закономерности эволюции изолированных сис-
тем. Так, развитие одних биогеоценозов может компенсаторно
подавлять развитие других или идти за их счет. Одновременно
биогеоценоз сам выступает в качестве системы по отношению к
составляющим его живым и неживым элементам, через которые
реализуются экосистемные изменения и которые накладывают
собственные ограничения на возможность реализации измене-
ний.
Изучение палеонтологических и морфологических рядов в над-
видовых таксонах позволило вывести некоторые общие законо-
мерности и сформулировать их в виде правил макроэволюции.
1. Правило необратимости эволюции Л. Долло (1893): эволю-
ция является процессом необратимым, и организм не может вер-
нуться к прежнему состоянию, уже «осуществленному» в ряду его
предков. ,
2. Правило происхождения новых групп от неспециализиро-
ванных предков Э. Копа (1904): обычно крупные группы организ-
мов берут свое начало не от высших представителей предковой
группы, а от сравнительно примитивных, неспециализированных
предков. .
3. Правило Э. Копа (1904): во многих группах позвоночных
животных в течение филогенеза (исторического развития) наблю-
дается увеличение размеров тела.
4. Правило прогрессивной специализации Ш. Депре (1907):
группа организмов, вступившая на путь специализации, в даль-
нейшем будет все дальше идти по тому же пути, приспосабливаясь
к крайним условиям существования.
5. Правило филогенетического предварения признаков Л. С. Бер-
га (1922): среди низкоорганизованных растений и животных могут
появиться признаки высокоорганизованных групп, стоящих выше
в системе или соответствующих более поздним эпохам палеонто-
логической летописи.
Эволюция фенотипических признаков в надвидовых система-
тических ipynnax также характеризуется некоторыми закономер-
327
ностями. Дивергенция как механизм видообразования ведет одно-
временно к прогрессивному увеличению форм признака, развива-
ющихся на основе исходной формы. Во многих случаях в близко-
родственных группах один и тот же признак может не только
развиваться в одном направлении, но также давать сходные ряды
индивидуальной изменчивости (закон гомологических рядов
Н. И. Вавилова). Такое явление обозначается понятием «паралле-
лизм». Не редки примеры и появления сходных признаков и их
комплексов у групп неродственных организмов. При этом в эво-
люционных радах отмечается прогрессивное усиление внешнего
сходства. Подобные явления демонстрируют так называемые
«конвергенции». Можно отметить, что механизм развития дивер-
генций, параллелизмов и конвергенций хорошо согласуется с оп-
ределяющей ролью «дефицита функции» в процессе видообразо-
вания: реализация сходных функций в сходных условиях требует
сходства формы.
Выделение «микроэволюции» и «макроэволюции» соответству-
ет рассмотрению эволюции видовых форм живого вещества и его
надвидовых подразделений. Ранее указывалось, что живое веще-
ство не существует в морфологически чистой форме: оно всегда
представлено смесями. Поэтому и эволюцию видовых форм мы
обязаны рассматривать не изолированно, а только в преломлении
через эволюцию разнородного, но единого живого вещества. Тог-
да отпадает необходимость в введении такого понятия, как «ко-
эволюция», или сопряженная эволюция. Внутривидовая эволю-
ция, или микроэволюция, — это лишь условно выделяемый нами
уровень изменений в живом веществе биосферы.
11.3.3. ТЕМПЫ'эволюции
Палеонтологический материал, имеющийся в распоряжении
эволюционистов, со всей очевидностью доказывает, что темпы
эволюции значительно отличаются как в разных систематических
группах, так и в разные периоды развития биосферы. Так, напри-
мер, некоторые роды пластинчато-жаберных моллюсков возникли
более 350 млн лет назад, а среди современных хищных млекопита-
ющих нет ни одного рода старше 15 млн лет. Большинство таксо-
нов за период своего существования проходили один или несколь-
ко периодов взрывной адаптивной радиации, сменяющихся дли-
тельными периодами эволюционного застоя.
В морфологических радах неравномерность темпов макроэво-
люции проявляется в неодинаковой скорости исторического пре-
образования разных систем органов и тканей. В результате возни-
кает разный уровень развития некоторых частей организма.
328
Для объяснения неравномерности темпов надвидовой эволю-
ции было предложено несколько гипотез. По сальтационистской
гипотезе новая таксономическая группа возникает очень быстро, в
течение смены нескольких поколений (путем «скачка»). Экзоге-
нистская гипотеза объясняет ускоренную эволюцию резкими из-
менениями условий внешней среды, например климата, деятель-
ности вулканов, геологической обстановки, космических факто-
ров, преимущественно катастрофического характера Концепция
квантовой эволюции (Симпсон, 1948) предполагает закономерное
периодическое кратковременное ускорение эволюции при пере-
ходе таксона в «новую адаптивную зону», остававшуюся до того
незанятой. В настоящее время вопрос о причинах неравномернос-
ти эволюции остается открытым.
Для количественного измерения темпов эволюции предлага-
лись такие единицы: «Дарвин» — изменение параметров тела
или его части на одну метрическую единицу за единицу
времени (Холдейн, 1949); число новых родов за 1 млн лет (Симп-
сон, 1949); число нуклеотидных замен в структурном гене (или
аминокислотных замен в белке) — молекулярно-генетические
часы. Тем не менее пока ни одна из них не принята к использо-
ванию.
11.3.4. ПРОГРЕСС И РЕГРЕСС В ЭВОЛЮЦИИ
Делавшиеся неоднократно попытки определить содержатель-
ную сторону эволюционного прогресса или регресса каждый раз
оказывались несостоятельными, поскольку ни сложность строе-
ния, ни количество ДНК в клетках, ни какие-либо другие пока-
затели не характеризуют степень приспособленности системати-
ческой группы к условиям существования. Более того, следует
принять точку зрения, согласно которой все реальные существу-
ющие и существовавшие прежде виды (равно как и надвидовые
таксоны) обладают одинаковым уровнем адаптивности. В то же
время введенные в 1925 г. А. Н. Северцовым понятия морфофи-
зиологического и биологического прогресса и регресса характе-
ризуют определенные эволюционные явления и имеют четкие
критерии.
Биологический прогресс проявляется в стремлении живых
организмов к увеличению видового многообразия, нарастанию
численности видов и более широкому распространению в про-
странстве. В конечном счете биологический прогресс ведет к про-
цветанию систематической группы. Биологический регресс харак-
теризуется противоположными явлениями и ведет к вымиранию
групп.
329
Известные способы достижения биологического прогресса
включают: морфофизиологический прогресс, или ароморфоз;
идиоадаптации; морфофизиологический регресс.
Организмы, вступившие на путь ароморфоза, повышают уро-
вень жизнедеятельности и увеличивают объем потенциальных на-
правлений дальнейшей эволюции за счет постепенно формирую-
щегося комплекса новообразований, не связанных с частными
приспособлениями к конкретным условиям.
Идиоадаптации — это такие ограниченные приспособления к
среде обитания, которые связаны не с изменением уровня морфо-
физиологической организации, а с развитием признаков специа-
лизации.
Морфофизиологический регресс означает переход организмов
к более простым отношениям со средой обитания, что сопровож-
дается частичной или полной редукцией органов и частей тела,
имевшихся у родоначальной формы или группы.
11.3.5. СООТНОШЕНИЯ ОНТОГЕНЕЗА И ФИЛОГЕНЕЗА
Соотношения между последовательностью этапов индивиду-
ального развития и последовательностью этапов эволюционного
становления вида или иной систематической группы, законо-
мерности этих соотношений объясняются «теорией филэмбрио-
генезов Северцова». Под филэмбриогенезами понимают такие
изменения в онтогенезе, которые переходят во взрослое состоя-
ние и служат материалом для новых направлений филогенеза.
Северцов выделяет несколько способов филэмбриогенетических
изменений: анаболию, или надставки к конечным стадиям заро-
дышевого развития; девиацию — отклонения от типичного раз-
вития на одной из средних стадий эмбриогенеза; архаллаксис —
отклонения от типичного развития на начальных стадиях эмбрио-
генеза.
При анаболии и девиации в новом онтогенетическом типе пол-
ностью или частично повторяются последовательные этапы родо-
начальных форм онтогенеза. Процесс повторения филогенетичес-
ких признаков получил название «рекапитуляция».
Теория рекапитуляций, разработанная Э. Геккелем и рас-
сматриваемая как частный случай теории филэмбриогенезов,
предполагает два типа онтогенетических признаков: палингене-
зов — признаков филогенетически далеких предков, и ценоге-
незов — вторичных признаков, возникших в результате приспо-
собления к определенным условиям эмбриональной или личи-
ночной жизни. Центральным ядром теории рекапитуляций счи-
тается биогенетический закон Э. Геккеля и Ф. Мюллера:
330
«Онтогенез является кратким и быстрым повторением филоге-
неза». Идея биогенетического закона просматривается еще в
трудах К. Бэра в вице общего положения: «В каждой большой
группе общее образуется раньше, чем специальное», и его конк-
ретизации в виде закона зародышевого сходства: «Чем более
ранние стадии индивидуального развития исследуются, тем
больше сходства обнаруживается между различными организ-
мами».
Важное место в эволюционных преобразованиях занимают
филэмбриологические изменения, известные под названиями «ге-
теротопии» и «гетерохронии»: эволюционные изменения в распо-
ложении эмбриональных зачатков и смещения закладки органов
во времени (акцелерации и ретардации).
Кроме появления новых признаков в онтогенезе могут проис-
ходить и так называемые отрицательные филэмбриогенезы, кото-
рые приводят к частичной или полной редукции органа или его
частей. Различают два типа редукций: рудиментацию — упроще-
ние или недоразвитие органа и анафазию — полное исчезновение
органа у взрослой формы при сохранении его зачатка у эмбриона
(или у личинки).
11.3.6. СПОСОБЫ ФИЛОГЕНЕТИЧЕСКОГО
ИЗМЕНЕНИЯ ОРГАНОВ
Любые филогенетические изменения органов основываются на
известном принципе полифункциональности: любой орган вы-
полняет, как правило, несколько, хотя и неравнозначных функ-
ций. Особенности эволюционных преобразований функций обус-
ловлены различными принципами филогенетического изменения
органов. .Важнейшие из них следующие:
принцип интенсификации функции связан с усилением глав-
ной функции органа;
принцип расширения функций — добавление к первичным
функциям сложно устроенного органа одной или нескольких но-
вых функций;
принцип смены функций — интенсификация одной из второ-
степенных функций органа, приобретающей значение первосте-
пенной;
принцип активации функции — переход пассивной функции в
состояние активной функции;
принцип иммобилизации функции — переход от активной
функции к пассивной;
принцип филогенетической компенсации органов: при ос-
лаблении в эволюции функции одного органа происходит ком-
331
пенсаторная интенсификация другого органа, не гомологично-
го первому, но находящегося с ним в биологической координа-
ции;
принцип субституции органов — замещение в эволюции утра-
ченного органа другим без изменения его функции.
Важными и ’широко распространенными способами эволюци-
онных преобразований являются полимеризация и олигомериза-
ция органов или иных структур.
Не только знание, но прежде всего понимание закономернос-
тей и способов эволюционных преобразований живых тел в про-
цессе появления новых функциональных элементов в биосфере
необходимо и для соотнесения существующих гипотез о проис-
хождении человека.
Глава 12
ЧЕЛОВЕК В БИОСФЕРЕ
Предпосылками для появления человека как вида (первый
представитель рода — Человек умелый) были уменьшение количе-
ства доступного к использованию вещества (некоторых элементов
и соединений), снижение темпов накопления информации в био-
сфере и стабилизация потока энергии. Среди групп живых орга-
низмов, сформировавшихся к периоду 5...2 млн лет назад
(рис. 12.1), перспективными для последующих реорганизаций
свойствами и признаками телесной организации обладали австра-
лопитеки. Они имели сравнительно крупное тело, освобожденные
от локомоции передние конечности с хватательной кистью, боль-
шой мозг, были многоядными, вели стадно-родовой образ жизни,
проявляли заботу о потомстве, использовали различные предметы
в качестве внешних усилителей, обладали мимической и звуковой
коммуникацией, не боялись огня. Род австралопитеков включал
несколько существовавших одновременно и сменявших друг друга
видов, большинство из которых претендовали на занятие сформи-
ровавшейся экологической ниши, а некоторые существовали «па-
раллельно» с видами рода Человек (рис. 12.2). Следы поздних ав-
стралопитеков теряются в эволюционной истории в последние
тысячелетия. Непосредственным эволюционным предком Челове-
ка считают Австралопитека афарского, а оформление первого
вида Человека относят к периоду 3...2 млн лет назад.
Ключевыми новообразованиями, сопровождавшими выделе-
ние из ствола австралопитековых Человека умелого {Ното habitis),
считаются изменение строения кисти и зубной системы и резкое
увеличение, почти в 1,5 раза, объема мозга (до 540...700 см3) и его
нейроморфологической сложности.
Переход к следующему историческому виду — Человеку пря-
моходящему, или архантропу (сборный вид Homo erectus), связан с
развитием резко выраженной социальности на основе развития
второй сигнальной системы (объем мозга достигает 800... 1000 см3,
что выше минимального достаточной для развития речи). В этот
период (2,0...0,5 млн лет назад) бурно развивается материальная
333
Рис. 12.1. Схема экологической эволюции приматов (оригинал автора)
Рис. 12.2. Схемы, иллюстрирующие современные представления о происхождении
человека (Ламберт, 1991; Пилбим, 1984, с изменениями):
А — последовательность отхождения ветвей высших приматов от общего ствола гоминоидок
7—семейство людей (Hominidae): люди (Ното), 2...5— семейство высших узконосых обезьян
(Pongidae): 2—шимпанзе (Pan), J—гориллы (Gorilla), 4—орангутаны (Pongo), 5—гиббоны
(Hylobates); 6...10— низшие узконосые обезьяны (Cercopithecidae): 6— колобусы (Colobus), 7—
Шимпанзе Орангутан
*1 ‘Горилла/
iPongidae t
I ц/ОР4™™
! . */Hominidae
ГигаМпек Ак^ралопитек
20
О
Ё
§
Б
О
1
2
3
4
5
6
Homo sapiens Неоантроп
_____Homo neandertaliensis Палеантроп
( Homo erectus Питекантроп
Homo habitis Архантроп
В
тонкотелые обезьяны {Presbytis}, Я—мартышки (Cercopithecus), 9—макаки {Масаса), /О—па-
вианы {Рар'ю)', надсемейство широконосые обезьяны (Platyrrhini); Б—схема «путаницы» в
связях между труппами антропоидов; В — взгляды на родословную человека: 1—человек про-
изошел от австралопитека; 2—человек произошел не от австралопитека: а—Австралопитек
афарский, 6— Австралопитек африканский, в—Австралопитек могучий, г—Австралопитек
Бойса, д—Человек умелый, е—Человек прямоходящий, ж—Человек разумный, з— афри-
канские человекообразные обезьяны
культура и происходит быстрое широкое расселение человека в
Африке, Европе и Азии.
Выделенный ранее вид Человек неандертальский {Ното
neandertalensis),jfaM палеантроп, в настоящее время признается не
всеми антропологами: предлагается рассматривать его в качестве
исторического подвида Человека разумного {Homo sapiens). По
раду признаков (объем мозга около 1500 см3, сложные орудия тру-
да и др.) эти две формы трудно разграничить. Следы материаль-
ной и духовной культуры свидетельствуют о том, что выделение
неоантропа как вида произошло на основе окончательного ста-
новления социальности.
ПоД социальностью понимают дифференцировку ценотически
активной фазы на несколько специализированных функциональ-
ных групп тел-особей со своими жизненными формами, каждая
из которых комплементарна сумме остальных (в рамках одного
социума — относительно завершенной и обособленной совокуп-
ности комплементарных социальных элементов).
Для того чтобы понять масштабность возникновения человека
как эволюционного события, нам необходимо выяснить характер
и оценить значительность отличий человека от других видов орга-
низмов. Человека включают в группу социальных животных, а к
его существенным отличительным признакам относят способ-
ность к абстрактному мышлению и трудовой деятельности. Под-
робное обоснование постепенного превращения «обезьяны в че-
ловека» приводится в трудах Ч. Дарвина и Ф. Энгельса.
Кроме человека к социальным животным относят ряд видов
перепончатокрылых и термитов (класс Насекомые). Для них ха-
рактерно образование более или менее крупных поселений семей-
ного типа с морфофункциональной дифференцировкой особей.
Ценотически пассивные полоносные особи, как правило, сохра-
няют комплекс признаков рода, семейства и отряда, а ценотичес-
ки активные стерильные самки часть таких признаков утрачива-
ют, но приобретают специальные морфологические, физиологи-
ческие и поведенческие признаки, ориентированные на выполне-
ние специальных функций. То есть у этих животных происходит
«разделение труда», что и является первым и основным признаком
социальной организации.
По имеющимся данным, австралопитеки единообразно ис-
пользовали и изготавливали простейшие орудия труда и в этом
смысле не отличались от других приматов и даже некоторых птиц.
Но уже Человек умелый отличался от всех остальных животных
тем, что у него было разделение труда на основе не телесной диф-
ференцировки, а различных орудий труда — внешних (внеорга-
низменных) усилителей телесных структур. Разделение труда как
частный случай дифференциации сопровождается интеграцией
336
родового стада. Аналогично принципу построения .многоклеточ-
ного тела усиление специализации каждого структурного элемен-
та ведет к усилению его зависимости от суммы комплементарных
ему элементов.
Развитие разных функциональных социальных групп у челове-
ка происходит путем дифференцировки одной биологической по-
пуляции на основе внешних усилителей. Популяция человека с
этих позиций представляет сообщество, составленное из профес-
сиональных групп, что и в структурном, и в функциональном ас-
пектах аналогично биоценозу, образованному видовыми популя-
циями. Собственно социальными популяциями у человека явля-
ются группы ценотически активных биологических индивидов с
однородным набором используемых ими внешних усилителей,
представляющие ту или иную профессию. Только в единстве с
внешними усилителями тела людей образуют социальные жизнен-
ные формы, реализующие определенные социальные функции в
определенных социальных условиях. Каждая социальная популя-
ция в процессе жизнедеятельности производит избыточную про-
дукцию, отторгаемую от нее вышерасположенным уровнем соци-
альных консументов. В итоге формируется характерная социаль-
но-трофическая пирамида с типичными локальными трофически-
ми цепями и общей сетью.
Социальность на основе внешних усилителей требует дополни-
тельной энергии. Основного энергетического канала хватает толь-
ко на воспроизводство биотической составляющей человеческого
социума с небольшим запасом. (В пищевых цепях человек высту-
пает как хемогетеротроф, биосапротроф, редуцент.) Организуе-
мый человеком второй энергетический канал для воспроизводства
внешних усилителей представляет собой освобождение трансфор-
мированной и накопленной в органических веществах солнечной
энергии — целенаправленное использование огня. (Человек ста-
новится дополнительно сапротрофом-деструктором.)
Быстрое наращивание объема информации, и прежде всего со-
циальной, требует новых форм ее записи, но записи, пригодной
для хранения, тиражирования, передачи и корректировки. Перво-
начально запись производится в мимических, звуковых, поведен-
ческих символах, в виде предметов различного назначения. И если
хранение информации еще, до определенных пределов, возможно
в индивидуальной памяти, то эффективная ее передача дочерним
поколениям требует специального рода деятельности — обучения.
В социуме появляется уникальное новообразование — компле-
ментарная пара «Ученик—Учитель». Со временем появляются и
внеорганизменные органические и минеральные матрицы для за-
писи информации (символы, рисунки, письменность, магнитная,
голографическая запись и др.).
22 Присный А.В. 337
Воспроизводство внешних усилителей и их использование тре-
буют развитого опережающего отражения, способности опериро-
вать образами, понятиями, представлениями, оторванными от
конкретных материальных носителей. Трудно согласиться с тем,
что такой способностью обладает только Человек, но у Человека
она получает наибольшее развитие.
Итак, Человека в биосфере выделяют: социальная организация
на основе внешних усилителей (модульно-социальный тип тел),
организация дополнительного потока энергии, внеорганизменные
формы записи информации, специальный канал передачи инфор-
мации и абстрактное мышление. Биосоциальная сущность Чело-
века предполагает качественно новый вариант его эволюционных
преобразований: морфологическая эволюция реализуется через
эволюцию внешних усилителей, то есть через эволюцию той части
неживой среды, которая является внетелесным продолжением ви-
дового живого вещества Человека. На определенном этапе разви-
тия социума Человек включает в состав усилителей собственной
мускульной силы и усилителей орудий труда некоторые виды жи-
вотных, одомашнивая их, или же себе подобных. Позже вторич-
ные усилители заменяются тепловыми, электрическими и иными
двигателями: мощность усилителей непрерывно возрастает, а с
ней растет и воздействие Человека на минеральную и биотичес-
кую среду.
Ценотически активные жизненные формы у Человека пред-
ставляют собой единство живого тела и определенных наборов
внешних усилителей и обозначаются как профессиональные
типы. Эти «социальные виды» аналогичны биологическим видам,
и их эволюция идет по законам, общим для живых систем. Онто-
генетические особенности Человека состоят в том, что на онтоге-
нез живого тела накладывается социальный онтогенез, в процессе
которого индивид на основе внеорганизменной информации при-
обретает способность использовать определенные внешние усили-
тели для выполнения определенной функции в социуме, то есть
объединяет телесную и внетелесную части. Только такое сопряже-
ние онтогенезов позволяет индивиду занять функциональное мес-
то в человеческом обществе.
Растущие энергетические потребности человеческого общества
обусловили выделение из биоты некоторых видов организмов, для
которых создаются контролируемые условия развития и воспроиз-
водства. Частичное блокирование связей, этих искусственных аг-
роэкосистем с остальной биотой и внесение в них дополнитель-
ной энергии из «второго энергетического канала» позволяют ин-
тенсифицировать их биопродукгивность и производить изъятие
биомассы большее, чем из аналогичных естественных экосистем.
Таким способом увеличиваются интенсивность и объем веще-
338
ственно-энергетического потока на глобальном биосферном уров-
не (рис. 12.3).
С появлением Человека в биосфере выделяется крупное струк-
турно-функциональное подразделение — социальная сфера, вклю-
чающая живое вещество человека и связанную с ним непосред-
ственно как его продолжение часть биоты и неживого вещества.
Эта подсистема характеризуется не только высокой геохимичес-
кой активностью, но также стремительным наращиванием инфор-
мации с особыми формами и уровнями ее записи, хранения и пе-
редачи. Часто ее называют техносферой, иногда — ноосферой.
Термином «техносфера» правильнее обозначать всю совокупность
разнородных внешних усилителей, построенных из неорганичес-
кого и мертвого органического вещества, как часть социосферы.
Термин «ноосфера» не имеет однозначного толкования. По-
явившись в конце XIX в., он первоначально использовался для
обозначения совокупности земных оболочек, трансформирован-
ных и направляемых в развитии Человеком. В. И. Вернадский по-
нимал под «ноосферой» сферу «разума». В первом случае считает-
ся, что ноосфера формируется одновременно с появлением Чело-
века, а во втором — должна сформироваться в будущем, когда Че-
ловек станет гармоничным элементом биосферы.
{
Внешние
4. Выделение в культуру
растений и животных —
ноаесности используют-
ся вторичные внешние
усилители.
изготовление сложных
усилителей, расширение
искусственных экосистем.
7. Техносфера подавляет
биоту и человека (пред-
кризисное состояние).
3. Часть биоты регуляр-
но используется для
получения пищи и сырья, основы агроценозов.
Огонь добывается и при- Для усиления их нерав-
меняется (в том числе) ------------’---------
для изготовления внеш-
них усилителей.
2. Изготовление простейших орудий труда. Регу-
лярное использование огня. Фсрмирующтйся
«второй энергетический канал» связан с разви-
тием внешних усилителей большей частью опо-
средованно.
1. Регулярное использование «природных»
предметов в качестве внешних усилителей. Огонь
используется большей частью для обработки
пищи и защиты.
Рис. 12.3. Эволюция сопряжения человека с внешними усилителями телесных
структур (оригинал автора)
тельной энергии для
внешних усилителей
служат рабы, животные,
сжигаемое топливо.
339
Возникнув в биосфере в ответ на появление свободной экони-
ши, Человек к настоящему времени в основном выполнил свою
видовую функцию: извлек из недр и рассеял по поверхности ог-
ромное количество минерального вещества, ранее недоступного к
вовлечению в биогенный круговорот; выделил в атмосферу избы-
точное количество углерода; увеличил количество связываемой и
удерживаемой в биосфере энергии; создал новые емкие уровни и
формы записи информации. Значит ли это, что человек как вид,
как функциональный элемент биосферы должен исчезнуть? На
такой вопрос следует дать утвердительный ответ, но с дополни-
тельными пояснениями. Исчезновение вида далеко не всегда есть
прямое вымирание. Чаще это образование другого исторического
вида (филетическая эволюция) или нескольких новых дочерних
видов (дивергенция). Если мы принимаем социальные виды как
аналоги биологических видов, то вымирание грозит только боль-
шей или меньшей части существующих в настоящее время соци-
альных видов. Основу дифференцировки биосоциального вида
Человека составляют внешние усилители, они же составляют и ха-
рактерные видовые признаки. Следовательно, и исчезнуть долж-
ны те наборы усилителей, для которых исчез дефицит функции.
Структурная реорганизация социосферы должна сопровождаться
и новым информационным наполнением, то есть изменением
массового сознания, что, возможно, следует отождествлять с фор-
мированием ноосферы в понимании В. И. Вернадского.
ЛИТЕРАТУРА
Основная
Албертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки: В 5 т. —
М.: Мир, 1986, 1987.
Алексеев Г. Н. Энергоэнтропика. — М.: Знание, 1983.
Антипчук Ю. П. Гистология с основами эмбриологии. — М.: Просвещение,
1983.
Бернал Дж. Возникновение жизни. — М.: Мир, 1969.
Бигон М., Харнер Дж., Таунсенд К. Экология. Особи, популяции и сообщества:
В 2 т. - М.: Мир, 1989.
Биологический энциклопедический словарь/Гл. ред. М С. Гиляров. — М.: Сов.
энциклопедия, 1989.
Воронцов Н. И., Сухорукова Л. Н. Эволюция органического мира. — М.: Про-
свещение, 1991.
Второв П. П., Дроздов Н. Н. Рассказы о биосфере. — Мд Просвещение, 1981.
Галимов Э. М.Феномен жизни: между равновесием и нелинейностью. Проис-
хождение и принципы эволюции. — М.: Едиториал УРСС, 2001.
Ганты Т. Жизнь и ее происхождение. — М.: Просвещение, 1984-
Грии Н., Стаут У., Тейлор Д. Биология: В 3 т. — М,: Мир, 1990.
Девис П. Пространство и время в современной картине Вселенной. — М.:
Мир, 1979.
Де Дюв К. Путешествие в мир живой клетки. — М: Мир, 1987.
Жизнь животных. В 6 т./Гл. ред. Л. А. Зенкевич. — М.: Просвещение, 1968—
1971.
Иорданский Н. Н. Развитие жизни на Земле. — М.: Просвещение, 1981.
Камшилов М. М. Эволюция биосферы. — М.: Наука, 1974.
Карпенко ML Universum sapiens (Вселенная Разумная). — М.: Мир географии,
1992.
Карпов С. А. Система простейших: история и современность. — СПб.: ТЕССА,
2005.
Катастрофы и история Земли: Новый униформизм/Под ред. У. Берггрен,
Дж. Ван Кауверинг. — М.: Мир, 1986.
Кемп П., Армс К. Введение в биологию. — М.: Мир, 1988.
Кордюм В. А. Эволюция и биосфера. — Киев: Наукова думка, 1982.
Ламберт Д. Доисторический человек: Кембриджский путеводитель. —М.: Нед-
ра, 1991.
Лапо А. В. Следы былых биосфер. — М.: Знание, 1987.
Медавар П., Медавар Дж. Наука о живом. — М.: Мир, 1983.
Медников Б. М. Аксиомы биологии. — М.: Знание, 1982.
Микляев И. Ю. Чудеса мазерной медицины. — Харьков: Основа, 1992.
Мирошниченко Л. И. Солнечная активность и Земля. — М.: Наука, 1981.
Моисеев Н. Н. Экология человечества глазами математика (Человек, природа
и будущее цивилизации). — М.: Молодая гвардия, 1988.
341
Моисеев Н. Н. Человек и ноосфера. — М.: Молодая гвардия, 1990.
Моисеев Н. Н., Александров В. В., Терко А. М. Человек в биосфере. — М.: На-
ука, 1985.
Мухин Л. М. Планеты и жизнь. — М.: Молодая гвардия, 1980.
Назаретян А. П. Интеллект во Вселенной: истоки, становление, перспективы.
Очерки междисциплинарной теории прогресса. — М.: Недра, 1991.
Николис Г., Пригожин И. Самоорганизация в неравновесных системах. — М.:
Мир, 1979.
Опарин А. И. Происхождение жизни. — М.: Московский рабочий, 1924.
Опарин А. И. Жизнь, ее природа, происхождение и развитие. — М.: Наука,
1968.
Перельман А. И. Изучая геохимию... (о методологии науки). — M.i Наука, 1987.
Пригожин И., Стенгрес И. Порядок из хаоса. — М.: Прогресс, 1986.
П. Тейяр де Шарден. Феномен человека. — М.: Наука, 1987.
Резанов И. А. Происхождение и эволюция биосферы/Эл. ресурс: http://
www.ihst.ru/~biosphere/03-2/Rezan.htm.
Рейвн П., Эверт Р., Айкхорн С. Современная ботаника: В 2т. — М.: Мир, 1990.
Реймерс Н. Ф. Основные биологические понятия и термины. — М.: Просвеще-
ние, 1988.
Реймерс Н. Ф. Экология (теории, законы, правила, принципы и гипотезы). —
М.: Журнал «Россия Молодая», 1994.
Рис Э., Стенберг М. От клеток к атомам: иллюстрированное введение в моле-
кулярную биологию. — М.: Мир, 1988.
Роллер Э. Открытие основных законов жизни. — М.: Мир, 1978.
Савенков В.Я. Новые представления о возникновении жизни на Земле.—
Киев: Выща школа, 1991.
Свенсон К., Уэбстер П. Клетка. — М.: Мир, 1980.
Северцов А. Н. Главные направления эволюционного процесса. Морфобиоло-
гическая теория эволюции. — М.: Изд-во МГУ, 1967.
Спенглер О. Л. Слово о воде. — Л.: Гидрометеоиздат, 1980.
Ферхуген Дж., Тернер Ф.» Вейс Л. и Др. Земля. Введение в общую геологию:
В 2 т. — М.: Мир, 1974.
Фолсом К. Происхождение жизни. — М.: Мир, 1982.
Фоули Р. Еще один неповторимый вид. Экологические аспекты эволюции че-
ловека. — М.: Мир, 1990.
Франтов Г. С. Геология и живая природа (Уровни организации вещества, био-
ника и геоника, клетки и газово-жидкие включения). — Л.: Недра, 1982.
Хаусман К. Протозоология. — М.: Мир, 1988.
Холдейн Дж. Происхождение жизни//Планета Земля. — М.: Изд-во иностран-
ной литературы, 1961.
Шкловский И. С. Вселенная, жизнь, разум. — М.: Наука, 1984.
Шредингер Э. Что такое жизнь? — М_: Атомиздат, 1972.
Эйген М. Молекулярная самоорганизация и ранние стадии эволюции//Успехи
физических наук. — 1973. — Т. 100. — Вып. 3.
Энгельс Ф. Диалектика природы. — М.: Политическая литература, 1982.
Эткинс П. Порядок и беспорядок в природе. — М.: Мир, 1987.
Югай Г. А. Философские проблемы теоретической биологии. — М.: Мысль,
1976.
Специальная
Акифьев А. П. Этюды об эволюции//Препр. Черноголовка: Ин-т хим.-физ. —
1988.
Альштейн А. Д., Ефимов А. В. Физико-химическая основа происхождения ге-
нетического кода: стереохимический анализ взаимодействия аминокислот и нук-
леотидов иа основе гипотезы прогенов//Молекулярная биология. — 1988.—
Т. 22.— № 5.
342
Брайен М. Общественные насекомые: Экология и поведение. — М.: Мир,
1986.
Белый У., Шторх Ф. Введение в цитологию и гистологию животных. — М.:
Мир, 1976.
Венер Р. А. Состав и свойства меланоидинов//ХТТ. — 1967. — № 6.
Волькениггейн М. В. Молекулярные основы эволюции//Молекулярная биоло-
гия. - 1989. — Т. 23. — № 1.
Глотов Н. В., Тараканов В. В. Норма реакции генотипа и взаимодействие гено-
тип-среда в природной популяции//Журнал общей биологии. —1985. — Т. 46.—
№ 6.
Голубовский М. Д. Некоторые аспекты взаимодействия генетики и теории эво-
люции//Методологические и философские проблемы биологии. — Новосибирск:
Наука, 1981.
Голубовский М.Д. Организация генотипа и формы наследственной изменчи-
вости эукариотов//Методологические проблемы медицины и биологии. — Ново-
сибирск, 1985.
Голубовский М.Д. Неравномерность темпа мутирования — закономерность
микроэволюции//Проблемы микроэволюции. — М., 1988.
Дарвинизм: история и современность: Матер, конф.. Ленинград, 19—21 иояб.,
1984. - Л., 1988.
Иорданский Н. Н. Механизмы квантовой эволюции//Проблемы микроэволю-
ции.—М., 1988.
Каллиникова В. Д. Индивидуальное развитие одноклеточных и воспроизведе-
ние свойств вида в онтогенезе//Вид и его продуктивность в ареале: Матер. 5 Всес.
совещ., Тбилиси, 10—12 нояб., 1988. — Вильнюс, 1988.
Каллиникова В. Д. Возможные механизмы генотипической изменчивости
агамных простейших семейства трипаносоматид/Проблемы микроэволюции. —
М., 1988.
Картавцев Ю. Ф. Популяционно-генетические аспекты видообразования у
животных//Проблемы микроэволюции. — М., 1988.
Красилов В. А. Видообразование в палеонтологической летописи//Филогене-
тические аспекты палеонтологии. Тез. докл. 35-й сес. Всес. палеонтол. о-ва, 23—
27янв., 1989.—Л., 1989.
Левченко В.Ф., Старобогатов Я. И. Сукцессионные и эволюционирующие
экосистемы//Проблемы микроэволюции. — М., 1988.
Медников Б. М. Молекулярные механизмы видообразования//Проблемы мик-
роэволюции. — М., 1988.
Мирабдилаев И.М. Проблема происхождения эукариот//Успехи современной
биологии. — 1989. — Т. 107. — № 3.
Михайловский Г. Е. Организация времени в биологических системах//Журнал
общей биологии. — 1989. —Т. 50. — № 1.
Оиотфиеико Ю. И. Наследственная информация и эволюция//Эвопюц. исслед.
Вавилов, темы. — Владивосток, 1988.
Отрощенко В. Л., Васильева Н.В., Стригункова Т.Ф., Шведова Т.А. Матрич-
ный синтез полинуклеотидов на минеральной поверхности//Изв. АН СССР. Сер.
биол. —1989.
Ошмарин П. Г., Ошмарин А. П. Мультиорганность функций как одна из основ
филогенетических изменений органов//Эволюц. исслед. Вавилов, темы. — Влади-
восток, 1988.
Петрашов В. В. Глаза и мозг эволюции. — М.: Прометей, 1988-
Риклефс Р. Основы общей экологии. — М.: Мир, 1979.
Росс Ч., Росс Д. Энтомология. — М.: Мир, 1985.
Рувинский А. О- Эволюция пола и происхождение многоклеточности//Про-
блемы макроэволюции. — М., 1988.
Руденко А. П. Общая теория прогрессивной химической эволюции и биогене-
за//Проблемы макроэволюции. — М., 1988.
343
Савостьянов Г. А. Различия направлений эволюции многоклеточности//Про-
блемы макроэволюции. — М., 1988.
Северцов А. С. Современные концепции вида//Бюл. Моск, о-ва испыт. приро-
ды. Отд. биол. —1988. — Т. 93. — № 6.
Стадничук И. Н._ Эволюция хлоропластов как эндосимбионтных органелл
эукариотов//Биологические науки. —1989. — № 6.
Тийвель Т. Приспособление клеток к эндосимбиозу в эволюции//Журнал эво-
люционной биохимии и физиологии. — 1988. — Т. 24. — № 4.
Телегина Т. А., Бехоев .И. Д., Павловская Т. Е. Состав и функциональные свой-
ства абиогенно синтезированных мелановдиновых пигментов//Изв. АН СССР.
Сер. биол. — 1988- — № 5.
Телегина Т. А., Масиновский 3. Б., Мохнаткина Г. В., Павловская Т.Е. Протеи-
ноиды как комплексы белковоподобных соединений с меланоидиновыми пиг-
ментами//Изв. АН СССР. Сер. биол. —1989. — № 4.
Augros R., Stanciu G. Systematic differentiation. A new evolutionary synthesis//BioL
Forum. — 1987. — V. 80. - № 4.
Ayres R. U. Selforganisation in biology and economics//Res. Repts 1st. Appl. Syst.
Anal. —1988. - № 1.
Ballantyne J. S., Chamberfine M.E. Adaptstion and evolution of mitochondria:
osmotic and ionic consideration: Pap. Lake Louise Biochem. Adapt. Symp., Lake Louise.
Febr. 7—9, 1987//Can. J. Zool. - 1988. - V. 66. - № 5.
Bara I. J., Gavrila L., Tnita E., Garaghin M. A new theory of evolution going to be
bom?//EvoL and Adapt. — 1988. V. 3.
Beloussov L. V. Ontogenesis and self-organization theory//Biol. Forum. —1988. —
V. 81. —№2.
Ferris J. P. Prebiotic synthesis: problems and chal!enges//Cold Spring Harbor Symp.
Quant. Biol. Vol. 52. — Cold Spring Harbor (N. Y.), 1987.
Glaser H.-J. Neue Aspekte der Evolutionstheorie. T. 1. HZ. Klin. Med. —1988. —
V.43. — №24.
Glaser H.-J. Neue Aspekte der Evolutionstheorie. T 2 HZ. Klin. Med. —1988. —
43. - № 24.
Joyce G. F. RNA evolution and the origins of life//Nature. — 1989. — V. 338. —
№6212.
Lenk R., Crespi P., Greppin H. Evolution de fentropie et de la neguentropie ед
biologie//Arch. set — 1987. — V. 40. — № 3.
Lewin R. A heresy in evolutionary biology//Science. —1988. —V. 241. — № 4872.
Lovtrup S. The theoretical basis of evolutionary throught//Sci Natur. Zool. et BioL —
1987.-V. 8.-№4.
Me Clintock B. Mechanisms that rapidly reorganise the genome. — Stadler
Symposium, 1978.—V. 10.
Rakosy-Tican L., Soran V., Dragos N. Gene and genome transfer during evolution//
Evol. and Adapt. —1988. —V. 3.
ScholTeniels E. Adaptation and finality or a Boeing is not a submarine//ReguL
endocrinienn et adapt, environm.: Colloq. int. Cent. etud. biol. anim. sauvag., Chize, 1—
5 juill, 1985. —Paris, 1986.
Stebbins G. L., Hartl D. L. Comparative evolution: Latent potentials for anagenetic
advance//Proc. Nat. Acad. Sci. USA. —1988. — V. 85. — № 14.
Ulanowicz R. E., Hannon В. M. Life and the production of entropy//Proc. Roy. Soc.
London. - 1987. —V. 232. - № 1267.
Walker J. Compartmentalization and niche differentiation: Causal patterns of
competition and coexistence//Acta biotheor. — 1987. — V. 36. — № 4.
Wittenberger C. A functional view on biogenetic law//Evoh and Adapt. — 1988. —
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Австралопитеки 313—315
Автотрофы 98
Аксонема 164
Аксоподии 74
Аминокислоты ПО
Анималькулизм 242
Антипорт 134
Апикомплексы 275
Апопласт 89
Аппарат (комплекс) Голвджи 145
Асцетоспоровые 275
Базальное тельце 164
Бактерии 267—269
Белки 118
Биогеоценоз 294
Биосфера 49, 279, 291
Биофильность 105
Биоценоз 46, 49, 294
Брожение 180, 183
Вакуоли(ь) аутофагирующие 148
— пищеварительная 78, 148
— сократительная 75
Вакуум космический 22
Валентность экологическая 281
Вегетативные органы 211
Вещество 22
—живое 40, 43, 301
— неживое 40, 301
Взаимодействие 23
Вид 49, 249, 250
Видообразование 319—321
Вирусы 259
Витализм 242
Включения 151
Внешние усилители 96, 336—339
Вода 105
Водоросли 270
— бурые 271
— диатомовые 270
— желто-зеленые 270
— зеленые 272
— золотистые 270
— красные 271
Воспроизводство 42
— бесполое 231
— половое 236
Время 29
Гаметы 60, 236
Ген 185
Генотип 185
Генофонд 189
Гетеротрофы 98
Гетерохроматин 170
Гиалоплазма 126
Гипотеза абиогенеза 298
— аугогенеза 305
— биогенеза 298
— гастреи 307
— сидячей колонии 307
— симбиогенеза 305
— фагоцителлы 307
— целлюряризации 307
— эволюционная 314
— эктогенеза 242
— эмбриональной индукции 206
Глазок 75
Гликоген 117
Гликолиз 180, 182, 183
Гомеостаз 221, 222
Граны 173
Грибы 273
Губки 276
Движение материи 22
Двигательные реакции 220
Деление 231
— бинарное 231
— множественное 231
Десмосома 138
Деструкторы 100
345
Детерминация 208
Диктиосома 145
Динофлагелляты 271
Дифференцировка 204
Диффузия 133
ДНК 121—125, 170
Дыхание 180—183
Жгутик 74, 164
Животные 274
Жизненная(ый) форма 194
— цикл 244
— — клетки 226—230
----организма 244
Жизнь 38—40
Жирные кислоты ПО
Зигоспоры 231
Зооспоры 231
Иглокожие 278
Изменчивость 187, 316—318
Иммунитет 223
Индивид 55
Инкреты 153
Интроны 185, 191
Информация 28, 34
Инфузории 275
Квартирантство 289
Кинетосома 164
Кислородный процесс 183
Кишечнополостные 276
Классификация 248
Клетка 64
Клеточная(ый) глотка 75
— оболочка 71, 154—157
----вторичная 71, 155
----первичная 71, 155
— стенка 70
—теория 103
—сок 150
Клубнелуковица 235
Код генетический 189
Колония 84, 85
Комменсализм 289
Компартменты 47
Конкуренция 289
Контакты клеточные 138, 139
Консументы 100
Концепция «эндомембраниой
системы* 137
Корневище 235
Корневые клубни 235
Котранспорт 134
Крахмал 117
Креационизм 298, 312
Кристы 176
Критерии вида 250, 251
Лабиринтулы 275
Лейкопласты 174
Лизосомы 146—148
— вторичные 147
—первичные 147
Липиды 118
Лишайники 274
Луковица 235
Макроэволюция 324—328
Макроэлементы 105
Материя 22
Мегаспора 234
Мейоз 229, 230, 241
Мембрана клеточная 68, 129
— наружная плазматическая 137
—пурпурная 139
— синаптическая 141
— фотосинтезирующая 140, 176
Меристема 67
Методы 4, 5
Микориза 274
Микроворсинки 161
Микропероксисомы 148
Микроспора 234
Микроспоридии 275
Микротельца 148
Микротрубочки 163
Микрошипы 161
Микроэволюция 316
Микроэлементы 105
Миксоспоридии 275
Митоз 227, 228, 241
Митохондрии 174—176
Многоклеточные 63, 276, 309
Моллюски 277
Монеры 267—269
Моносахариды 108
Мультивезикулярные тельца 148
Мутуализм 289
Мхи 272
Мясистые стержневые кории 235
Негэнтропия 28
Неживое вещество 22, 43—46
Ниша 255
— функциональная 256, 257
— экологическая 255
Ноосфера 339
Нуклеиновые кислоты 121
Нуклеоид 69
Нуклеотиды 111
346
Оболочка клеточная 71
— миелиновая 139
Овизм 242
Окаймленная (ый) ямка 137
— пузырек 137
Онтогенез 242, 330
Оперон 185
Оплодотворение 238
— двойное 238
Органеллы 79
Организм 49, 54, 58, 59, 61
— модульный 96
— модульно-социальный 96
—социальный 96
Органоиды 47
Органообразующие элементы 105
Органы 48, 210
Осмос 133
Особь 54
— ценотически активная 57
---пассивная 57
Отбор 319—321
Отводки 235
Палочковый аппарат 75
Папоротники 272
Паразитизм 288
Паразитоидизм 288
Пастьба 289
ПеЛликула 70, 72
Пероксисомы 148
Печеночники 272
Плазма 22
Плазмиды 264
Плазмодесмы 156
Плазмодий 80
Пластиды 173, 174
Пластинчатые 276
Плауны 272
Плотный контакт 139
Покровы 213
Поле 22
Полисахариды 117
Полиэмбриония 234
Полупаразитизм 288
Популяция 49, 252
Почкование внешнее 234
— внутреннее 234
Преформизм 242
Прионы 265
Продуценты 98
Пропластиды 174
Простейшие 63, 274, 309
Пространство 29
Протоплазма 68
Протопласт 68
Процессинг 191
Псевдоплазмодий 81
Развитие 23
Размножение вегетативное 234
— бесполое 58
— половое 59
Растения 270, 272
— семенные 272
Регуляторные гены 185
Редуценты 100
Резервуар протонов 177, 181
Репликационная вилка 171, 190
Репликация ДНК 170
Реснички 164
Рибосомы 127—128
— мембранно-связанные 128,
— свободные 128
Ритмы 287
Сальтационизм 323
Саркомастигофоры 275
Секреты 153
Симпласт 82, 89
Симпорт 134
Синапс 141
Синтетическая теория эволюции 313
Синцитий 82
Система(ы) 18
— биокосная 293
— водно-минерального обмена 216
— газообмена 216
—живая 40
—регуляторная 219
— питания 214
— пищеварительная 214
— опорно-двигательная 213
— органов 211
—транспортная 218
Систематика 248
Слоевище 89
Собирательство 289
Социальность 96, 336
Спорозоиды 231
Спорообразование 231
Споры 231
Среды обитания (жизни) 283
Стеблевые клубии 235
Стигма 75
Столон 235
Структура белка 118—120
-ДНК 122, 185
Структурные гены 185, 186
— формы ДНК 122
Таксономия 248
Таллом 89
347
Теория катастроф 313
— эволюции 314
Техносфера 339
Тилакоиды 173
Ткани 48, 209
Тонопласт 150
Транскрипция 189
Трансляция 191—194
Транспорт активный 134
— мембранный 135
Трансформизм 312
Трихоцисты 79
Углеводороды 107
Углеводы 152
Унимпорт 134
Фен 186
Фенотип 186
Фибриллы 158—163
— промежуточные 158
—толстые 158
— тонкие 158
Фибриллярная ламина 168
Форезия 289
Фосфолипиды 118, 130
Фотосинтез 113
Фрагментация 234
Фрагмопласт 156
Функция 134, 137, 296
Хвощи 272
Хемоавтотрофы 98, 99, 179
Хемогетеротрофы 98, 99
Хемосинтез 114
Хищничество 288
Хлоропласты 173
Хлорофилл 176
Хордовые 278
Хроматиды 228, 229
Хроматин 122, 169
Хромопласты 174
Хромосомы 228, 229
Целлюлоза 117
Центриоли 166
Цианобактерии 269
Цитозоль 126
Цитомембраны 129
Цитоплазма 68
Цитоскелет 158
Человек 333—340
Черви кольчатые 277
— круглые 276
— плоские 276
Черенки 235
Членистоногие 277
Шизогония 231
Шелевой контакт 139
Экзоны 185, 191
Экзоцитоз 135
Экобиоморфа 195
Экологический(ие) спектр 281
— факторы 280
Экология 279
Экосистема 294
Экскреты 153
Экструсомы 79
Эктогенез 242
Элементы (системы) 18
Эндоплазматическая сеть (эндоплаз-
матический ретикулум) 142—145
Эндоспоры 74, 235
Эндоцитоз 135
Энергия 23
Энтропия 24
Эпигенез 242
Эписомы 264
Эргастические вещества 151
Эукариоты 65, 269, 305
Эухроматин 170
Ядерная(ые) мембрана 167
— оболочка 167
— пластинка 168
— поры 168
Ядро 167—173
Ядрышко 171
С—N-соединения 108
С—О-соединения 107
ОГЛАВЛЕНИЕ
Глава 1. ВВВДЕНИЕ В БИОЛОГИЮ...................................... 3
1.1. Предмет и задачи биологии................................. 4
1.2. Методы биологических исследований......................... 4
1.3. Система биологических наук........................... 7
1.4. Основные этапы развития биологии....................... 10
Глава 2. ОБЩАЯ ХАРАКТЕРИСТИКА ЖИЗНИ.............................. 18
2.1. Материальные системы.................................... 18
2.1.1. Система----------................................ 18
2.1.2. Материя............... ...._______......____________ 21
2.1.3. Движение................................. ........ 22
2.1.4. Энергия........................................ 23
2.1.5. Информация....................................... 28
2.1.6. Пространство и время............................... 29
2.1.7. Материальное и нематериальное (идеальное) в системе. 33
2.1.8. Функционирование систем............................ 36
2.2. Сущность жизни в определениях........................... 38
2.3. Критерии живых систем................................ 40
2.4. Характерные признаки живого вещества................... 43
2.5. Уровни организации живой материи.............-.......... 46
Глава 3. ЖИВЫЕ ТЕЛА.................................. ........ 52
3.1. Организм, особь, индивидуум (в определениях)............ 53
3.2. Клетка.................................................. 63
3.3. Особенности строения свобод ноживущих клеток............ 72
3.4. Ядерно-цитоплазматические комплексы протистов и многоклеточных.... 79
3.5, Многоклеточные живые тела............................... 84
3.5.1. Колонии и колониевидные тела________________________ 84
3.5.2. Талломные живые тела..................... ........ 89
3.5.3. Многоклеточные живые тела (с тканевой организацией). 89
3.5.4. Модульные и социальные живые тела.................. 96
3.6. Типы ценотической активности............................ 97
Глава 4. БИОЛОГИЯ КЛЕТКИ.......................................101
4.1. Клеточная теория.......................................101
4.2. Введение в биохимию клетки........................... 103
4.2.1. Химический состав живой клетки (неорганические и малые
органические молекулы)............................... 103
349
4.2.2. Макромолекулы и основные химические процессы, идущие
в живой клетке......-.................... -.........ИЗ
4.3. Структурные компоненты клетки и главнейшие жизненные
процессы, проходя шие в ней..............................125
4.3.1. Цитоплазма......................................125
4.3.2. Органоиды клетки................................127
4.3.3. Главнейшие энергетические процессы в клетке.....176
Глава 5. НАСЛЕДОВАНИЕ И РАЗВИТИЕ ПРИЗНАКОВ...................184
5.1. Признаки и генетическая система клетки...............184
5.2. Реализация генетического кода........................189
5.3. Морфофункциональные типы клеток......................194
5.3.1. Представление о жизненной форме.................194
5.32. Жизненные формы клеток...........................195
5.3.3. Дифференцировка клеток....................... 202
5.4. Функциональные объединения клеток—ткани.......... ...209
Г л ав а 6. РЕАЛИЗАЦИЯ БИОГЕОЦЕНОТИЧЕСКИХ ФУНКЦИЙ
В ИНДИВИДУАЛЬНЫХ ЖИВЫХ ТЕЛАХ.................................212
6.1. Индивидуальные функциональные комплексы (системы жизне-
обеспечения) ....................................... 212
6.2. Поддержание постоянства внутренней среды (гомеостаз).221
Глава 7. ВИДОВОЕ ЖИЗНЕОБЕСПЕЧЕНИЕ _.................-......225
7.1. Жизненный цикл клетки.....................—........226
7.2. Воспроизводство.................................. 230
7.3. Онтогенез......................................... 242
7.4. Особенности онтогенеза человека....................245
Глава 8. ОСНОВЫ СИСТЕМАТИКИ ЖИВЫХ ОРГАНИЗМОВ.....................248
8.1. Введение в таксономию —---..------„...—...-------...—........... 248
8.2. Представление о виде.——............................ 249
8.3. Пространство и время вида ....,..........................254
Глава 9. МНОГООБРАЗИЕ ЖИЗНИ НА ЗЕМЛЕ.............................258
9.1. Формы жизни..........»..................... .......... 258
9.2. Неклеточные формы живого вещества (полуавтономные
молекулярные комплексы).................„............~........259
9.2.1. Вирусы и бактериофаги (вирусы, поражающие клетки
прокариот).................................................259
9.2.2. Плазмиды............................................264
9.2.3. Прионы..............................................265
9.3. Клеточная форма жизни....................................266
9.3.1. Монеры, или прокариоты............................ 267
9.3.2. Эукариоты................. ..........................269
Глава 10. ОРГАНИЗМ - ПОПУЛЯЦИЯ - ВВД В ЕДИНСТВЕ
СО СРЕДОЙ...................................279
10.1. Экологические факторы.......................................279
10.1.1. Абиотические вещественные, энергетические и информа-
ционные факторы................„........................283
10.1.2. Биотические факторы.................~...............288
350
Глава 11. БИОСФЕРА ............„............................. 291
11.1. Общие стационарные и динамические характеристики биосферы.291
11.2. Биосфера как эволюционирующая система................297
11.2.1. Гипотезы зарождения биосферы.......................297
11.2.2. Биологический этап развития биосферы...............304
11.3. Представления о механизмах эволюции живого вещества...316
11.3.1. Микроэволюция......................-............316
11.3.2. Макроэволюция............-.................... 324
11.3.3. Темпы эволюции-------------------------------------- 328
11.3.4. Прогресс и регресс в эволюции----------------------329
11.3.5. Соотношения онтогенеза и филогенеза.............330
11.3.6. Способы филогенетического изменения органов.....331
Глава 12. ЧЕЛОВЕК В БИОСФЕРЕ...................................333
Литература................... -——-----------.....---------------—341
Предметный указатель---......--.........----------.........-.....-345