/
























































































































































































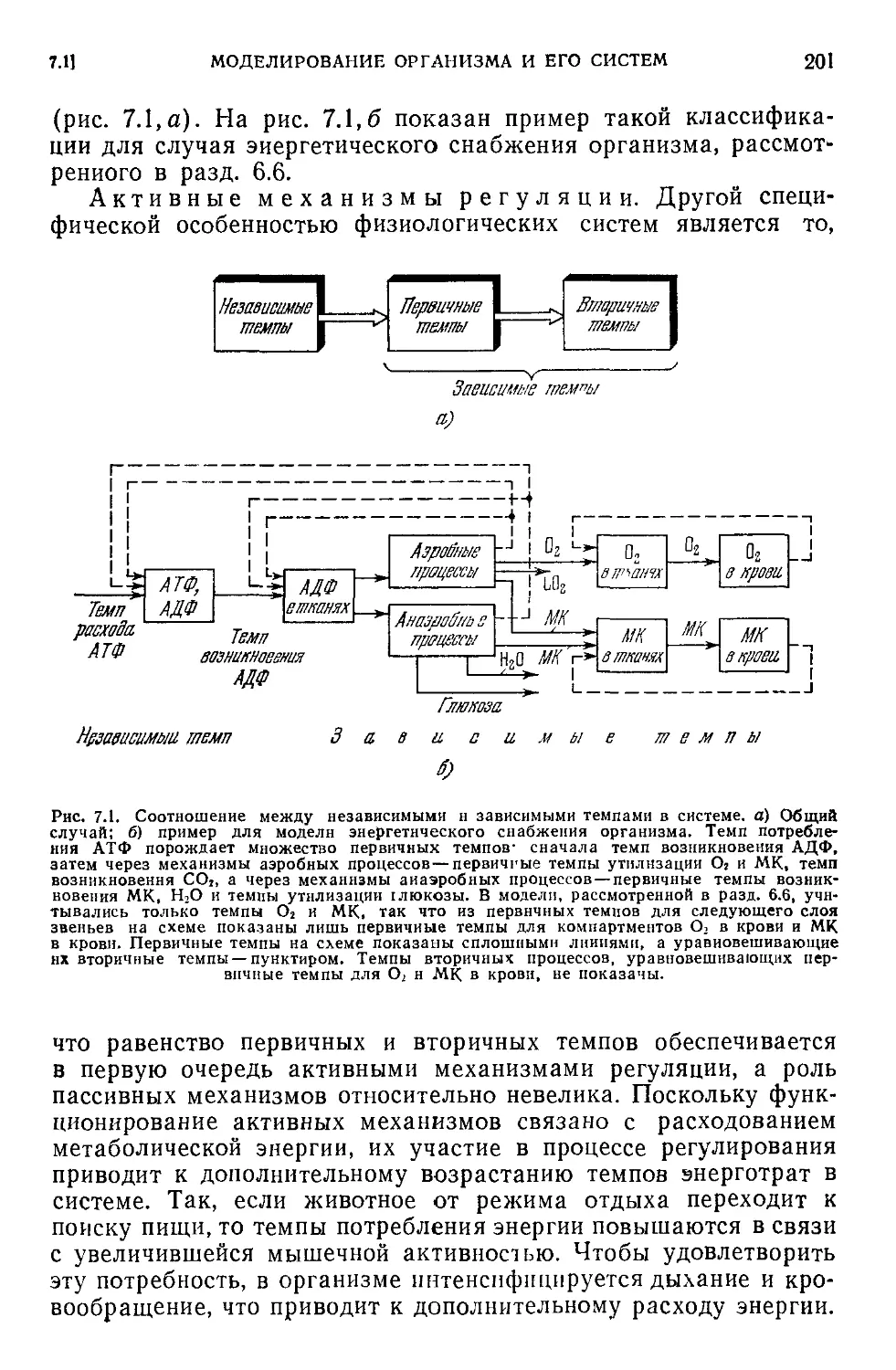
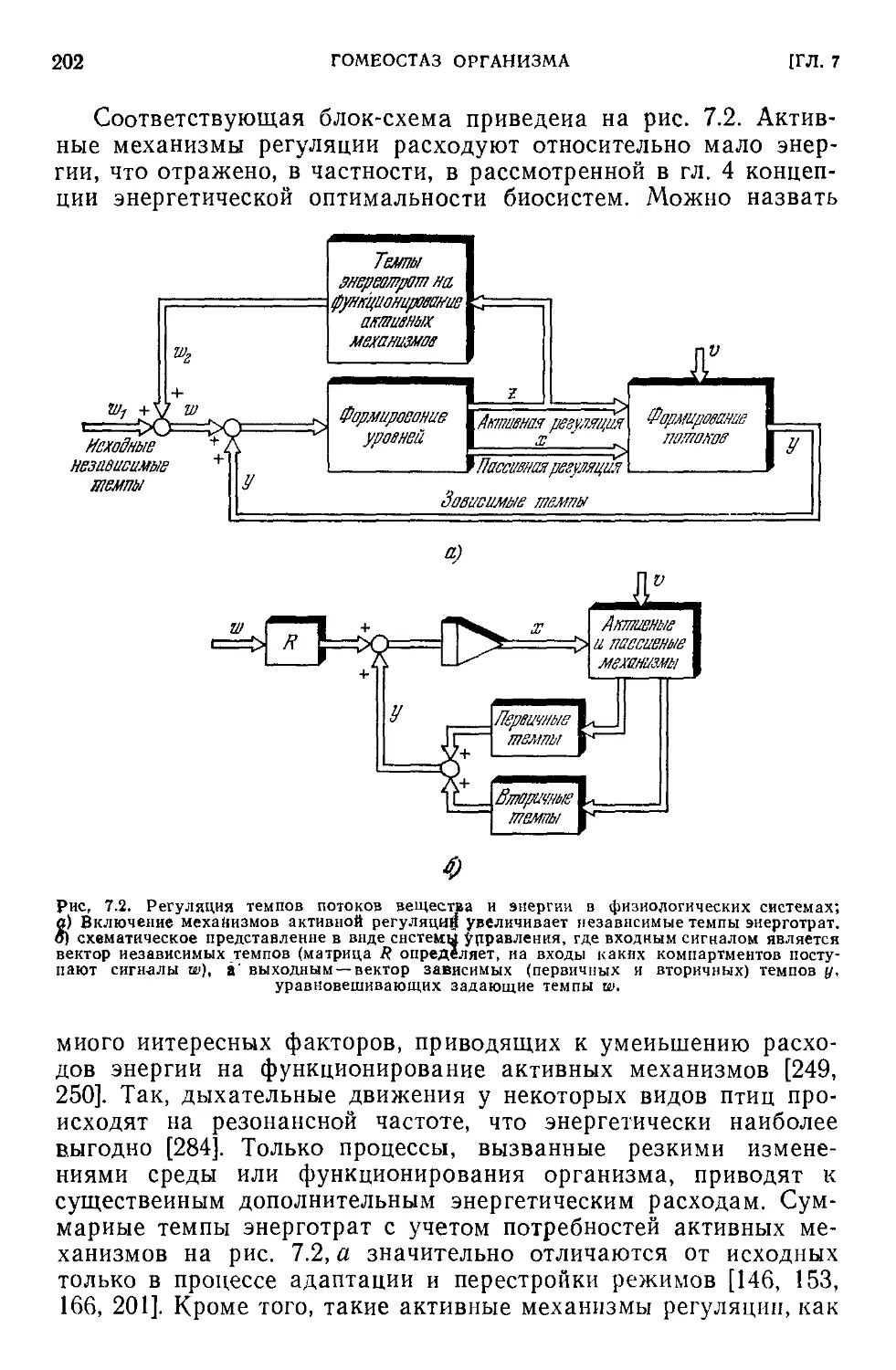






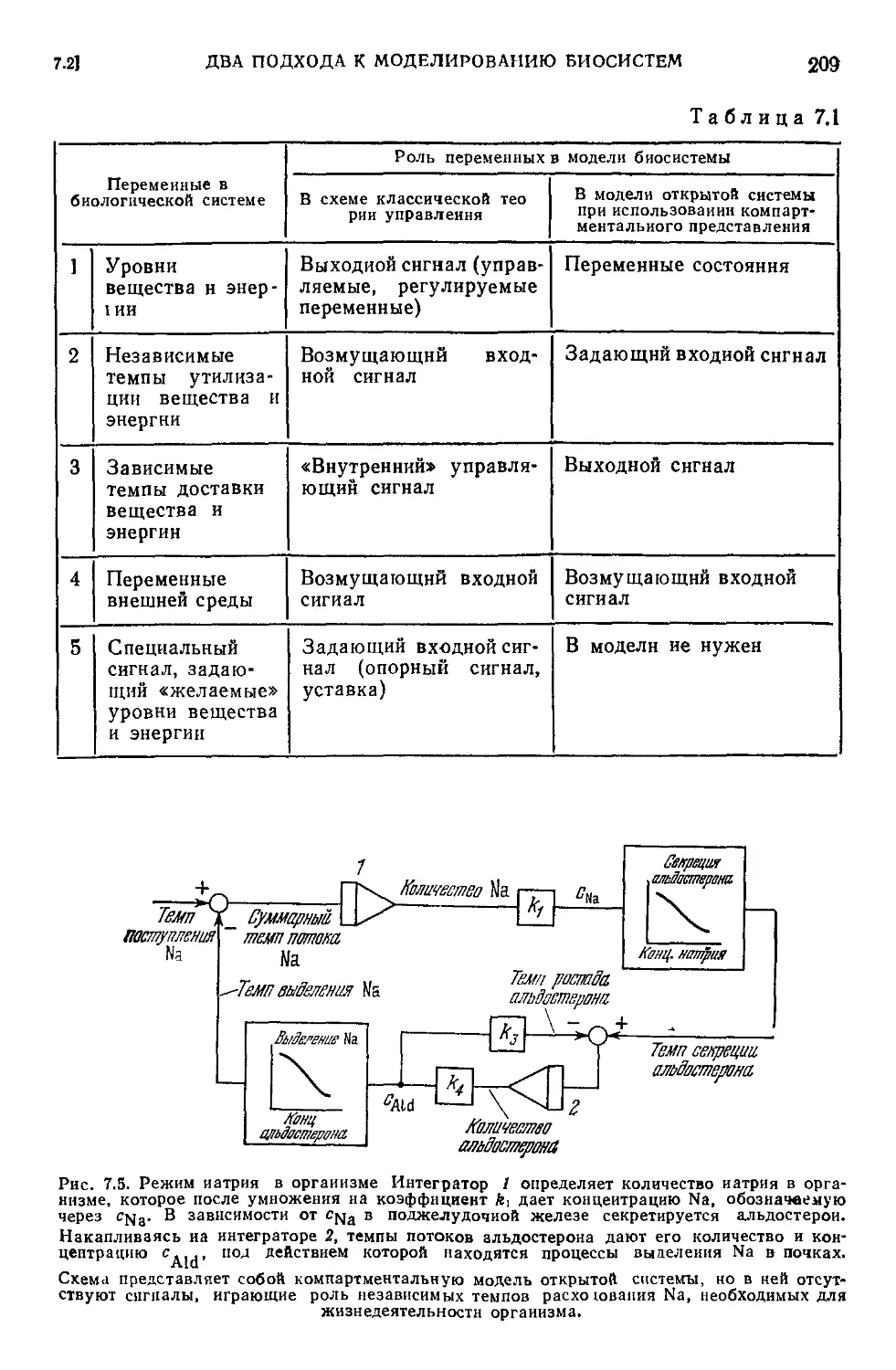



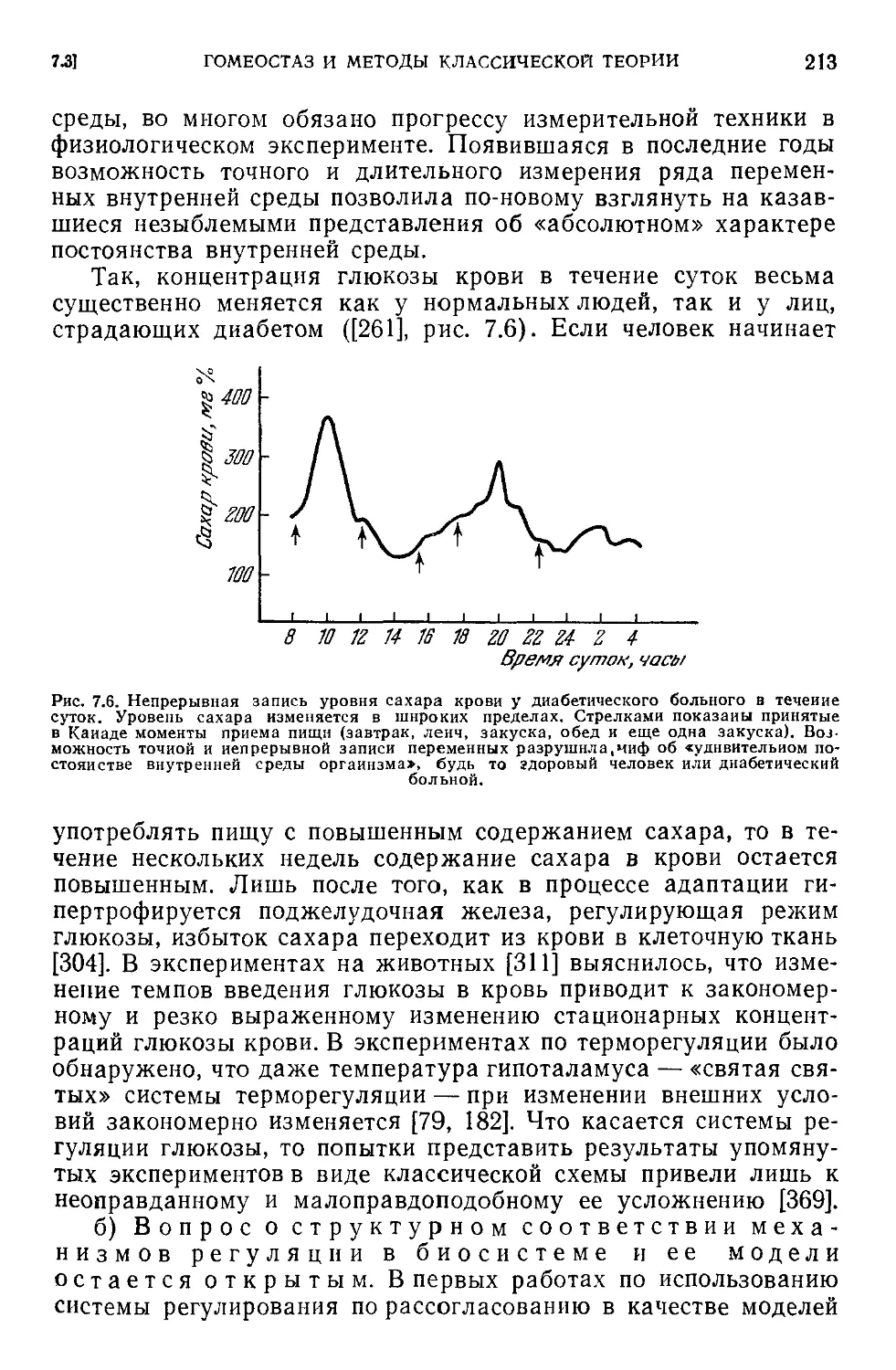





















































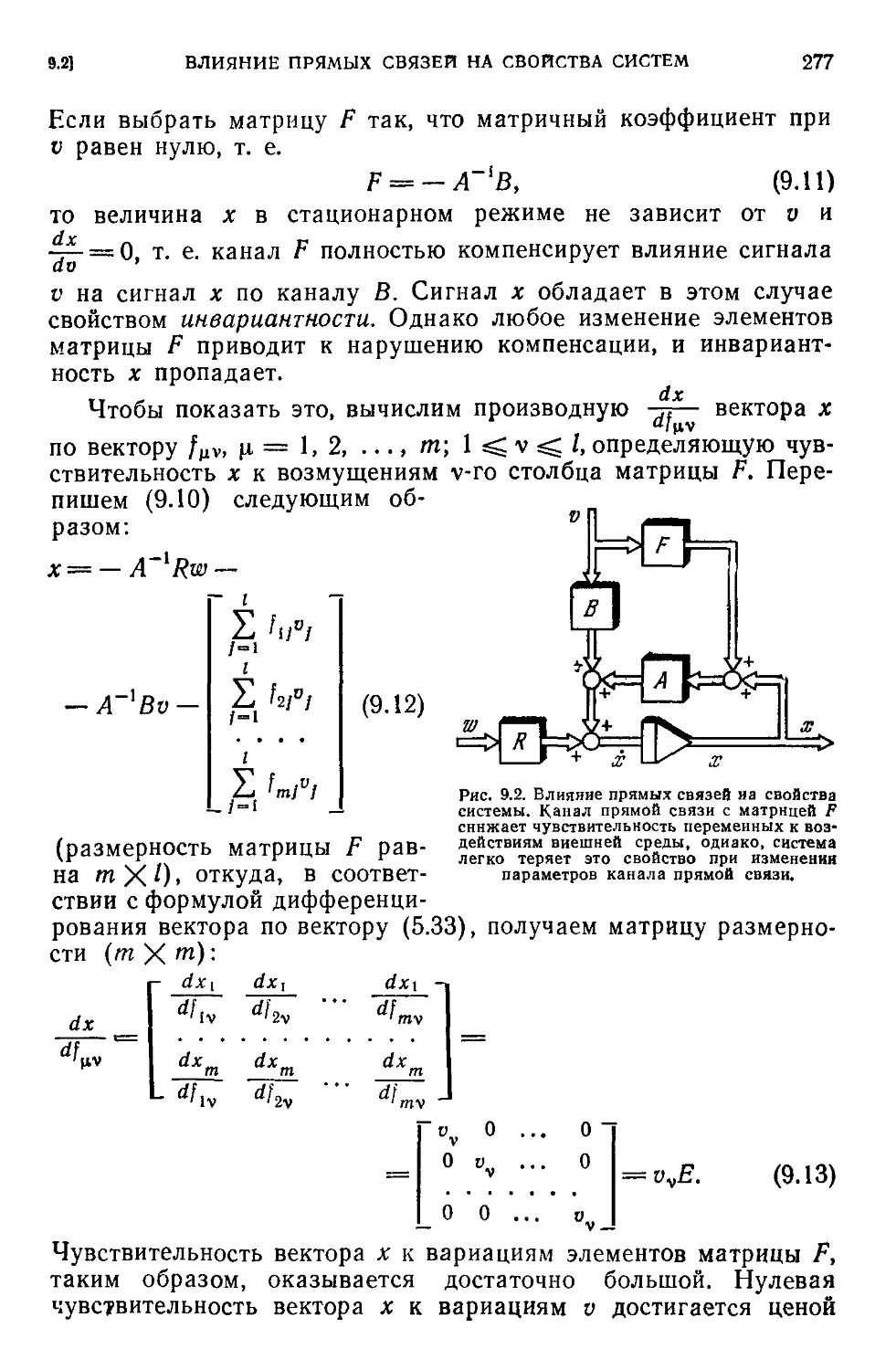







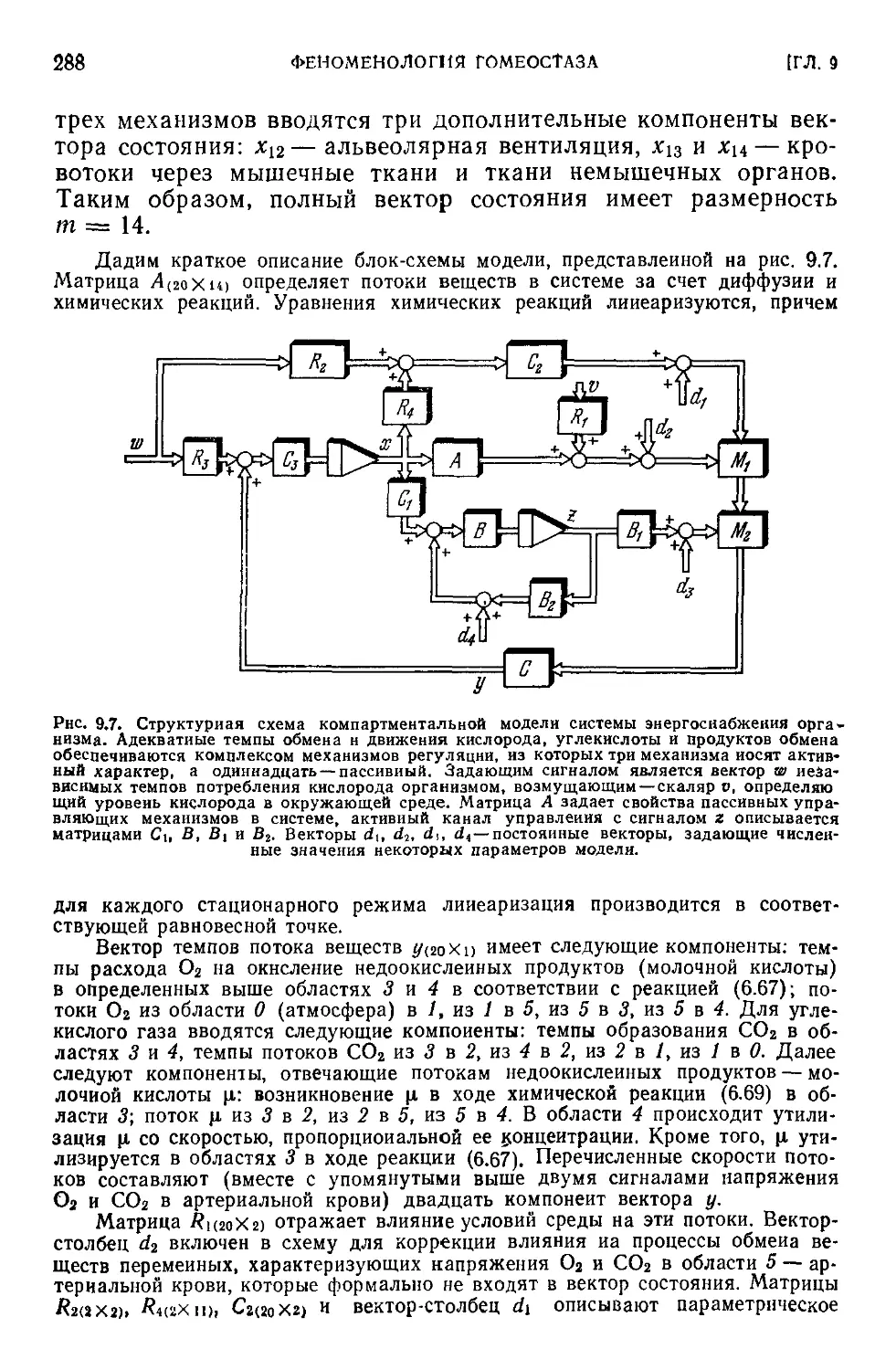




































Текст
В. Н. НОВОСЕЛЬЦЕВ
ТЕОРИЯ УПРАВЛЕНИЯ
И БИОСИСТЕМЫ
АНАЛИЗ
СОХРАНИТЕЛЬНЫХ СВОЙСТВ
МОСКВА «НАУКА»
//// \ ГЛАВНАЯ РЕДАКЦИЯ
ГП1 ФИЗИКО-МАТЕМАТИЧЕСКОЙ ЛИТЕРАТУРЫ
*===* 1978
2П2.154
Н 76
УДК 519.95
Теория управлении и биосистемы. Анализ сохранительных свойств. Ново-
сельцев В. Н. Главная редакция физнко-математнческой литературы изда-
тельства «Наука», М., 1978, 320 стр.
Теория управления в последние годы все шире применяется для изучения
процессов в биологических системах, и книги в этой области находят новых
читателей в лице биологов и медиков.
Книга рассказывает о современных методах теории управления и их при-
менении для исследования живых систем. В ней рассматриваются основные
идеи метода пространства состояний н компартментального моделирования
биосистем. Эти методы использованы для анализа сохранительных свойств
биологических систем. Изложение сопровождается примерами из области фи-
зиологии (системы терморегуляции, кислородного режима, энергетического
баланса) и других областей биологии.
Книга рассчитана на специалистов в области биокибернетики, бионики,
биофизики, а также биологов, медиков, физиологов и инженеров.
Илл. 91, библ. 369.
21004—1169
И 053(02)-78
166-78
© Главная редакция
физико-математической литературы
издательства «Наука», 1978
ОГЛАВЛЕНИЕ
Предисловие .........................................................6
ЧАСТЬ I
ТЕОРИЯ УПРАВЛЕНИЯ И ЖИВЫЕ СИСТЕМЫ
Глава 1. Специфика живых систем.....................................15
1.1. Живая система..............................................15
1.2. Структура биосистемы.......................................17
1.3. Принцип биологического эпиморфизма.........................20
1.4. Открытые системы...........................................21
1.5. Пассивное и активное управление в живых системах...........23
1.6. Темпы н уровни.............................................27
1.7. Иерархия целей в живых системах............................30
1.8. Термодинамические модели живых систем......................37
Глава 2. Сохраиительные способности живых систем....................42
2.1. Концепция самосохранения и живые системы..................42
2.2. Гомеостаз.................................................43
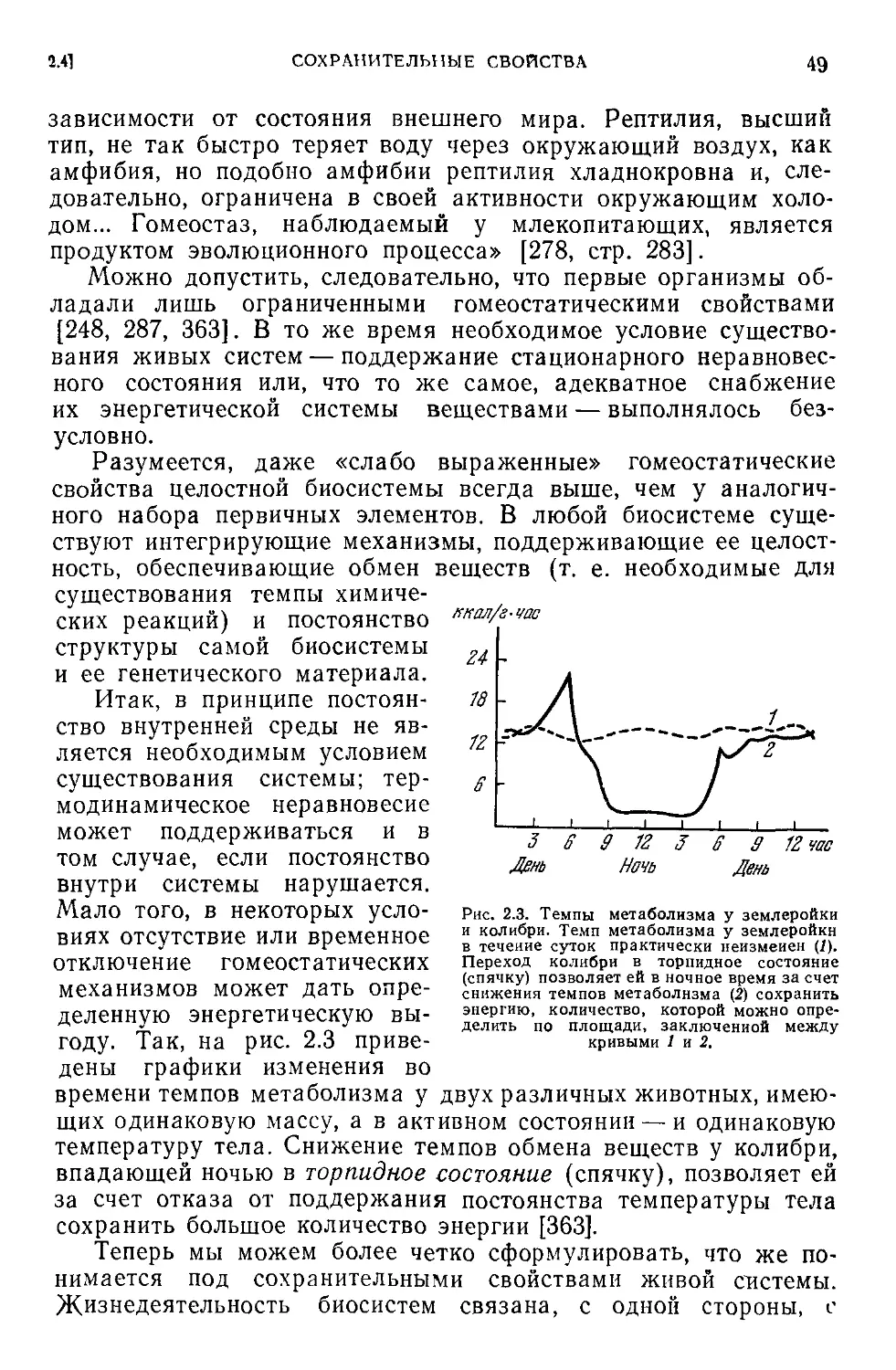
2.3. Гомеостатическая кривая...................................45
2.4. Сохраиительные свойства...................................48
2.5. Гомеостаз на разных уровнях организации жизни.............50
2.6. Некоторые общие свойства гомеостатических механизмов на раз-
ных уровнях организации биосистем..............................63
2.7. Гомеокииез, гомеорез и генетический гомеостаз..............66
Глава 3. Классическая теория управления и биосистемы................68
3.1. Методология теории автоматического управления..............68
3.2. Динамические системы. Схемы моделирования..................74
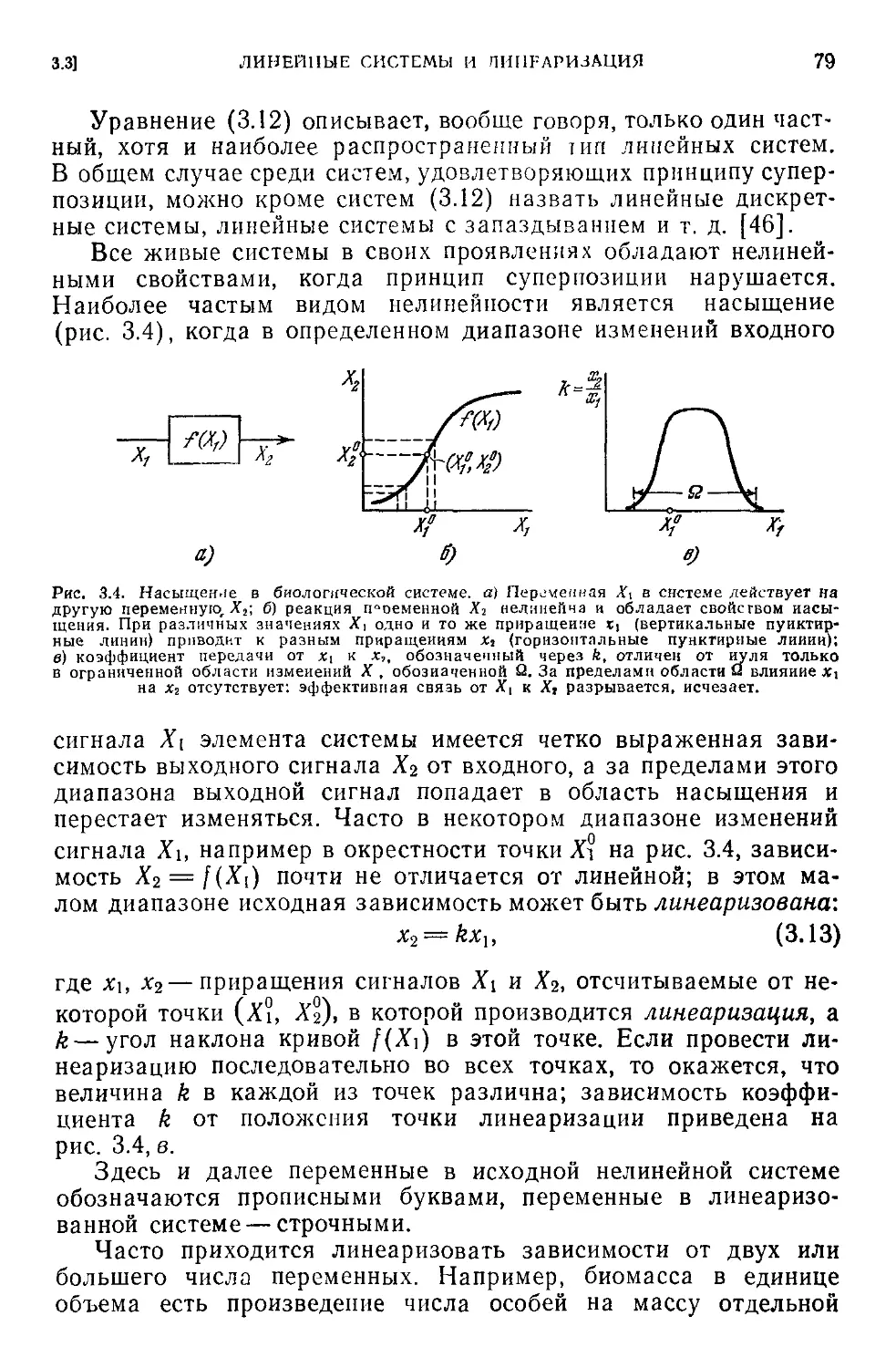
3.3. Линейные системы и линеаризация............................78
3.4. Обратная связь.............................................80
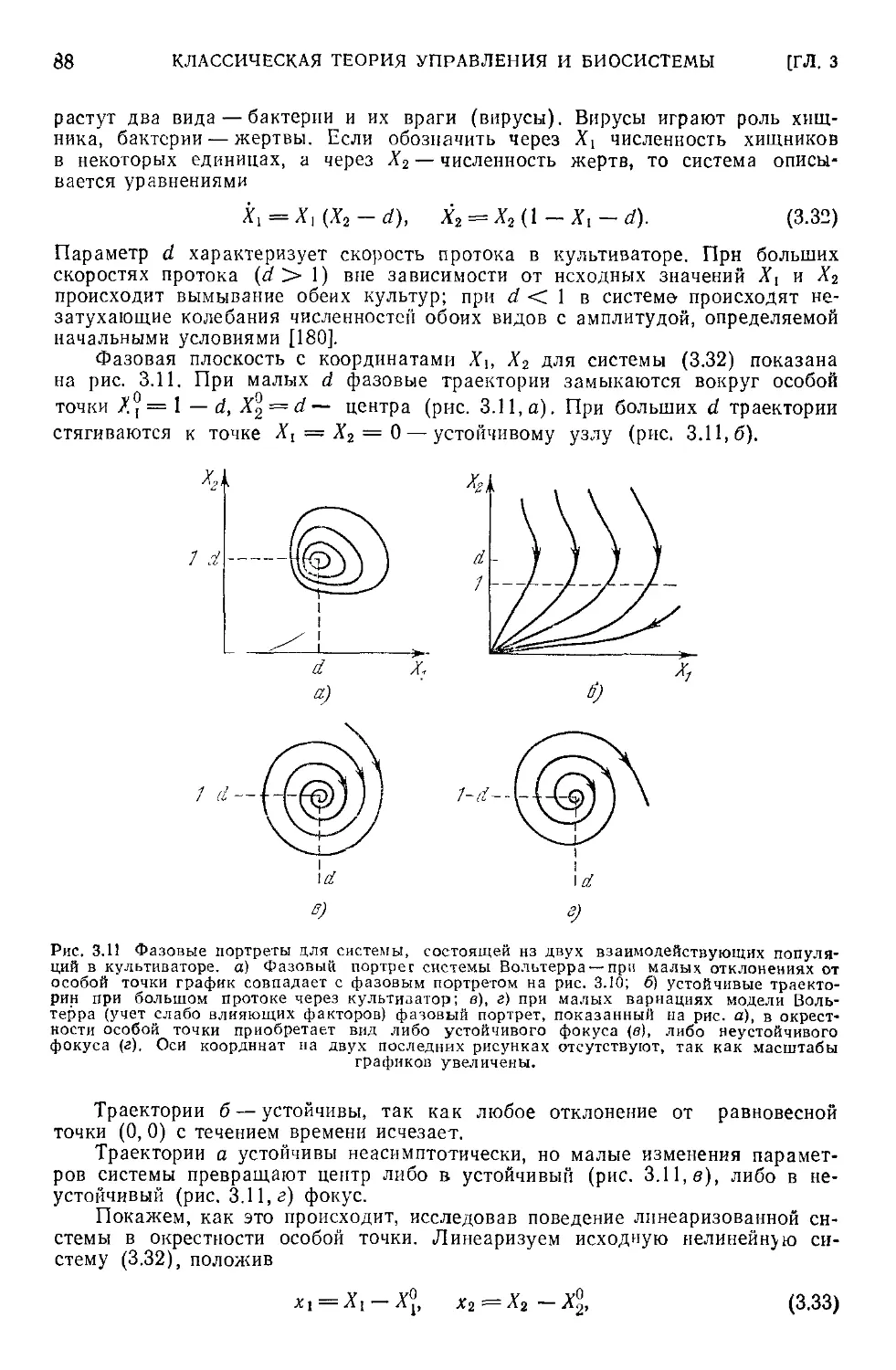
3.5. Устойчивость ..............................................85
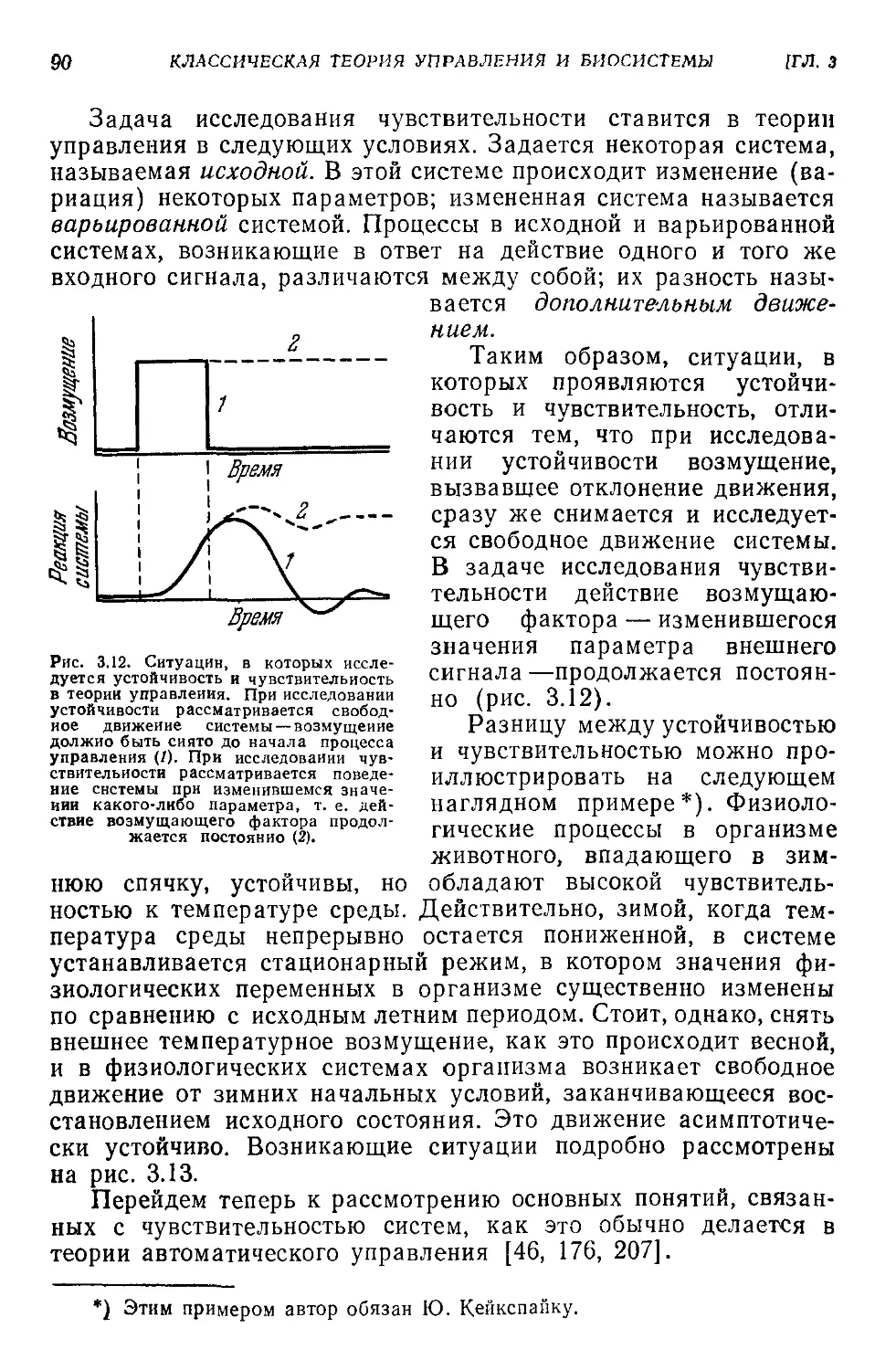
3.6. Чувствительность..........................................89
Глава 4. Оптимальность и адаптация в биосистемах....................97
4.1. Оптимальные системы........................................97
4.2. Адаптация .................................................99
V 4.3. Концепция оптимальности биосистем........................101
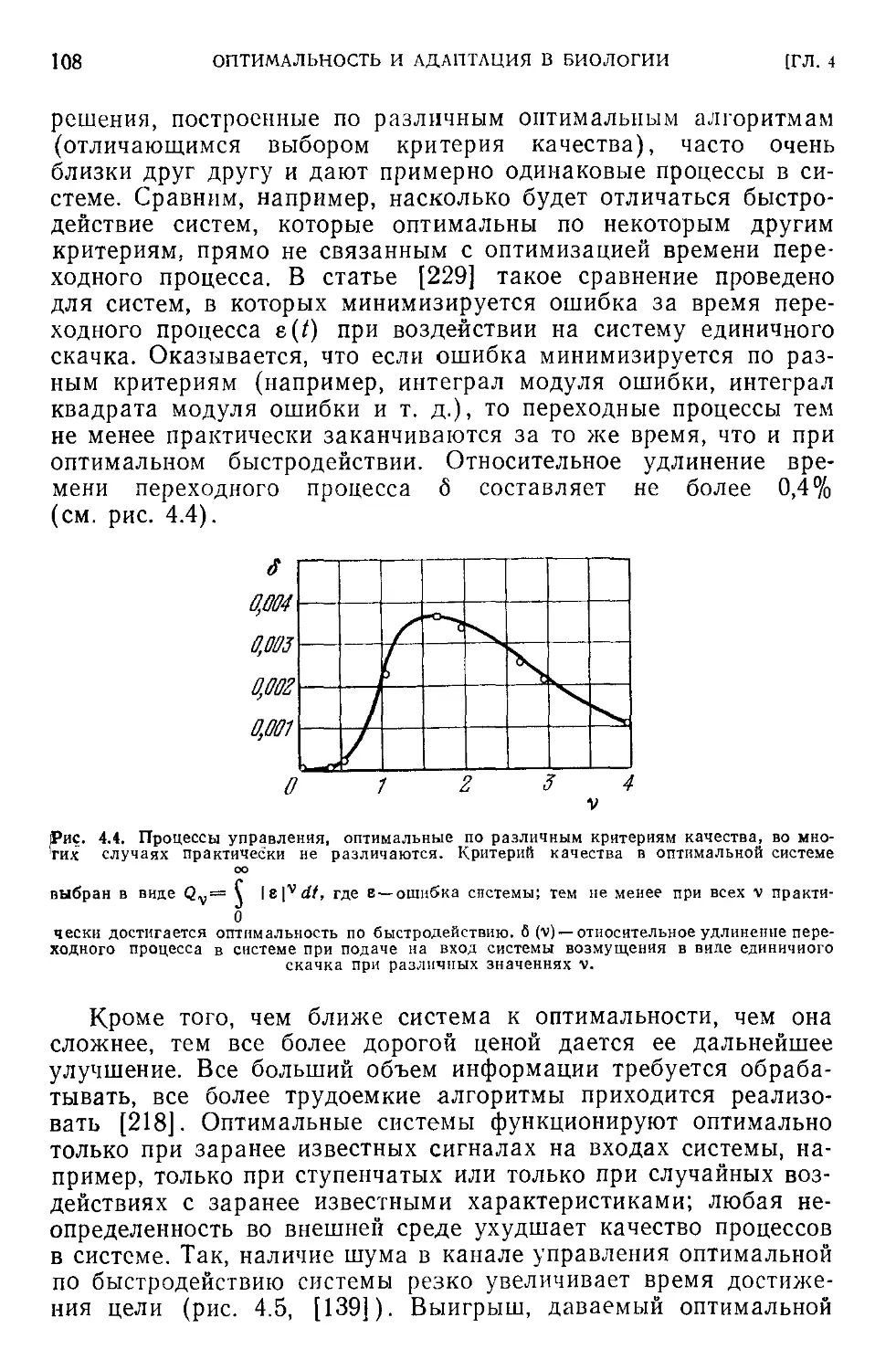
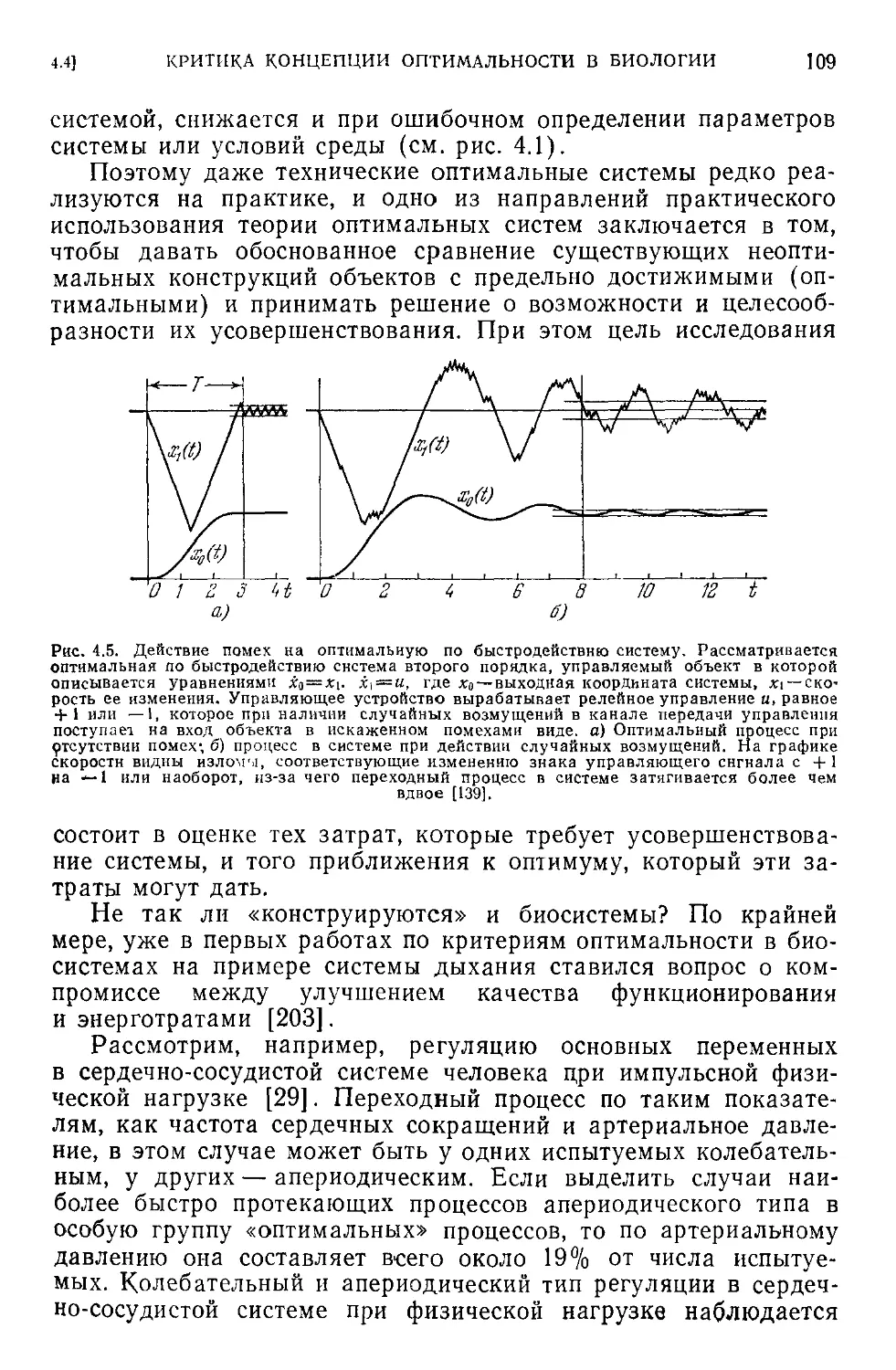
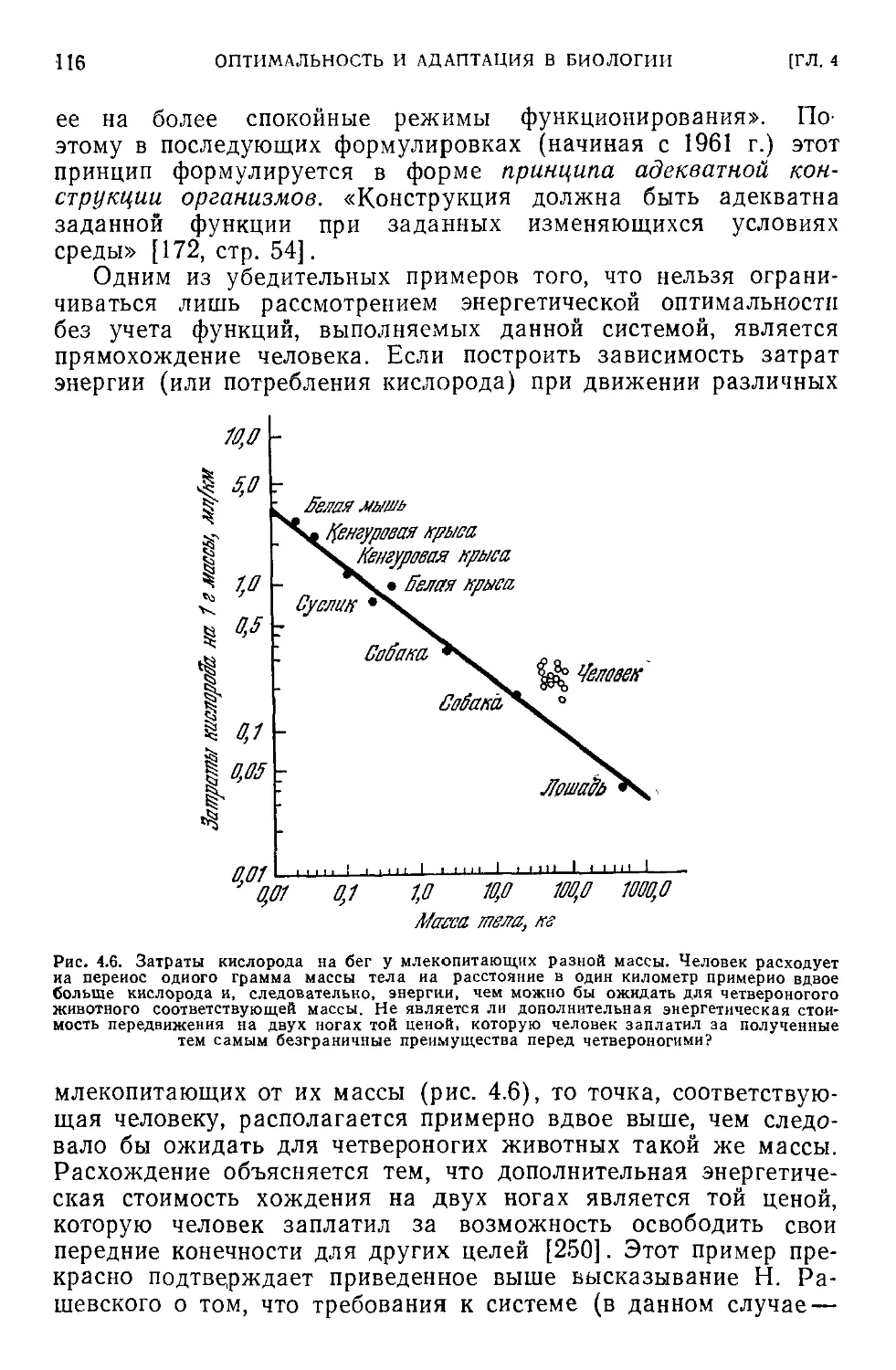
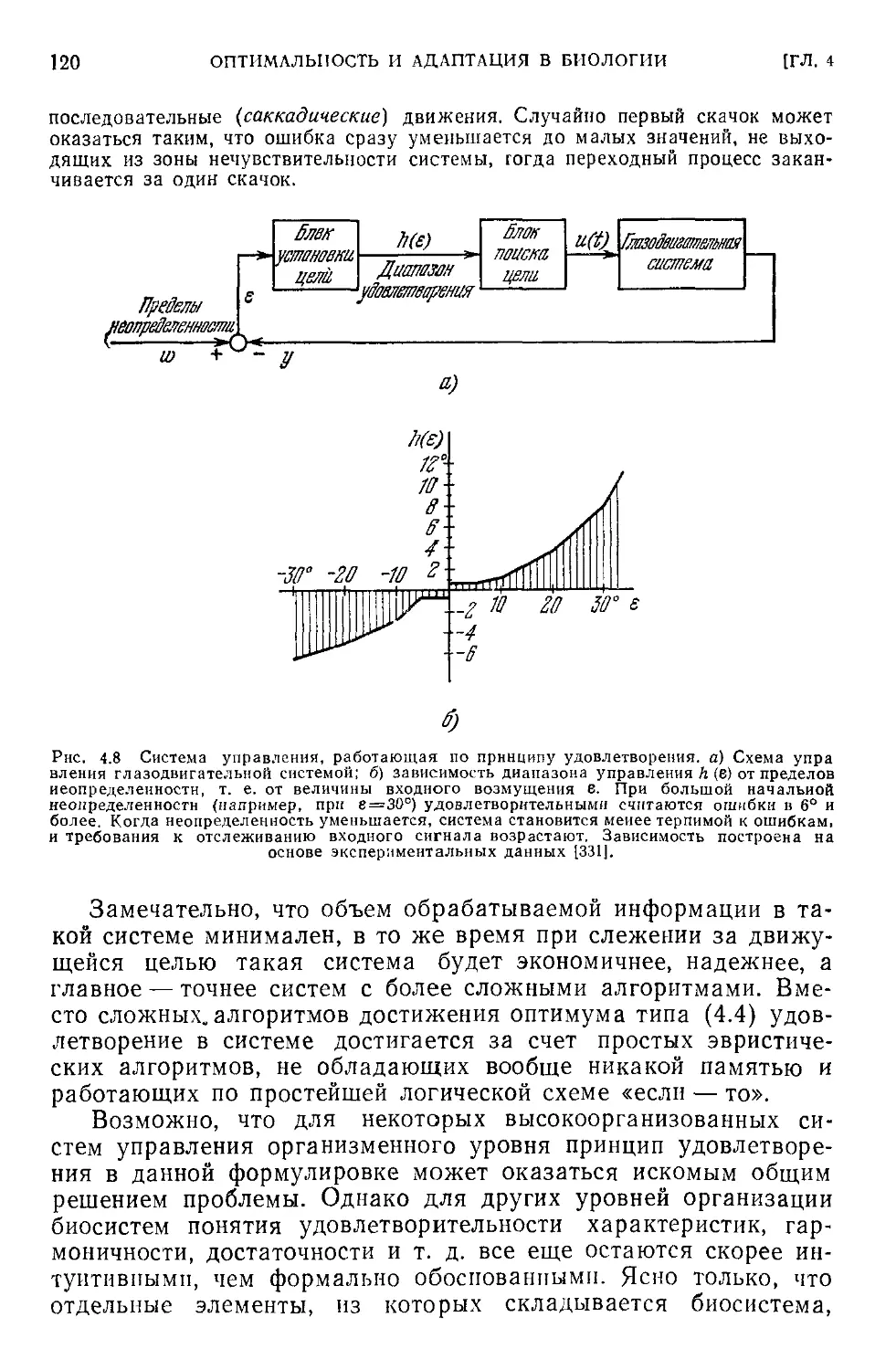
< 4.4. Критика концепции оптимальности в биологии...............104
4.5. Принцип удовлетворения....................................113
4.6. Хороший, плохой или оптимальный?..........................121
4
ОГЛАВЛЕНИЕ
Глава 5. Метод пространства состояний...............; . . . s > 124
5.1. Векторы и матрицы........................................124
5.2. Понятие состояния в биологических науках и в теории управ-
ления .......................................................133
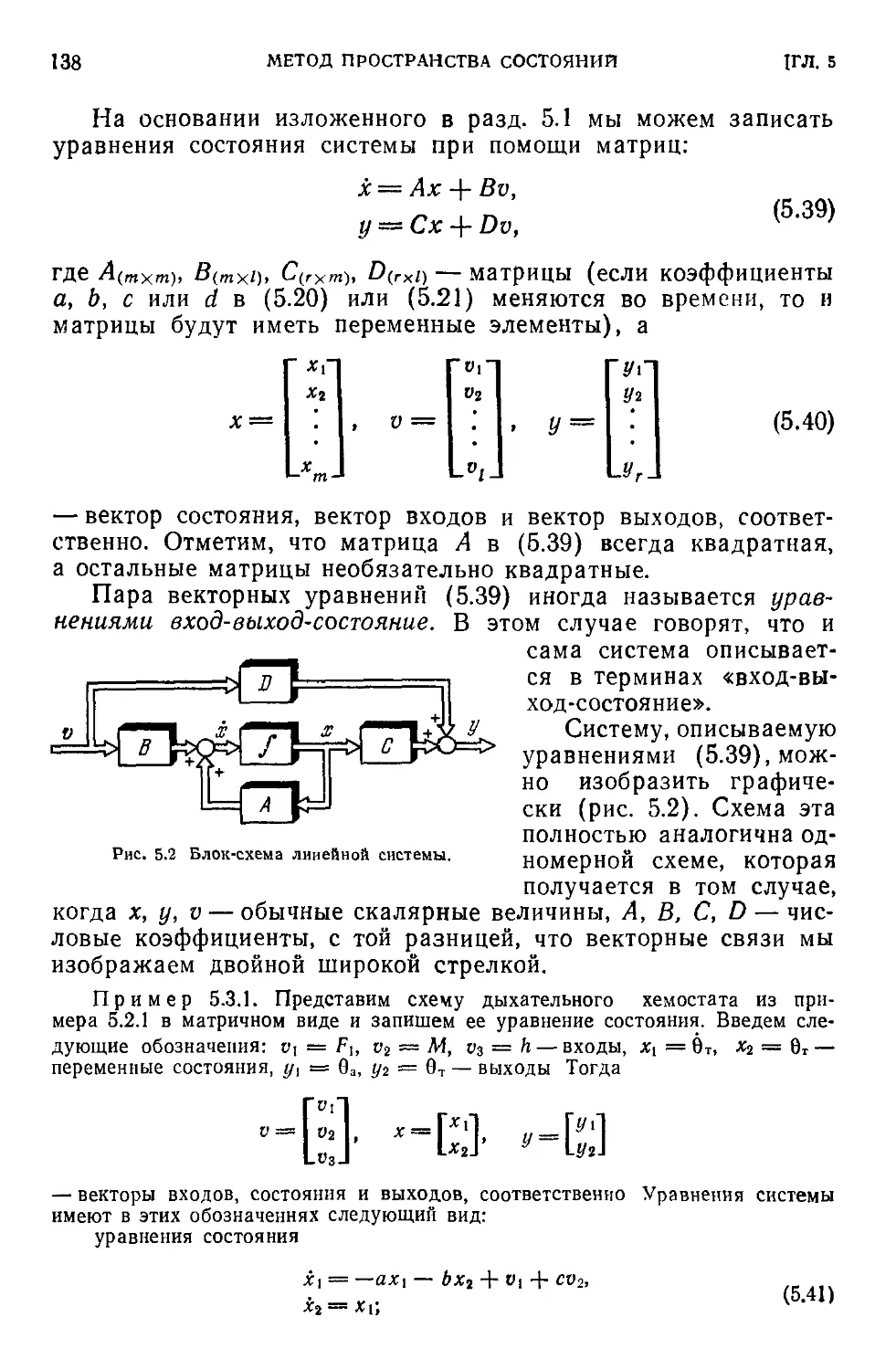
5.3. Уравнения состояния......................................136
5.4. Решение уравнений. Стационарный режим системы............140
5.5. Устойчивость системы и переходные режимы.................142
5.6. Чувствительность.........................................144
5.7. Наблюдаемость и управляемость............................146
5.8. Уравнения состояния для системы пассивного теплообмена . . 150
Глава 6. Компартментальные модели живых систем....................159
6.1. Определения..............................................159
6.2. Перемещение компонент....................................164
6.3. Взаимодействие и превращение компонент...................170
6.4. Утилизация компонент.....................................177
6.5. Общая схема компартментальной модели.....................182
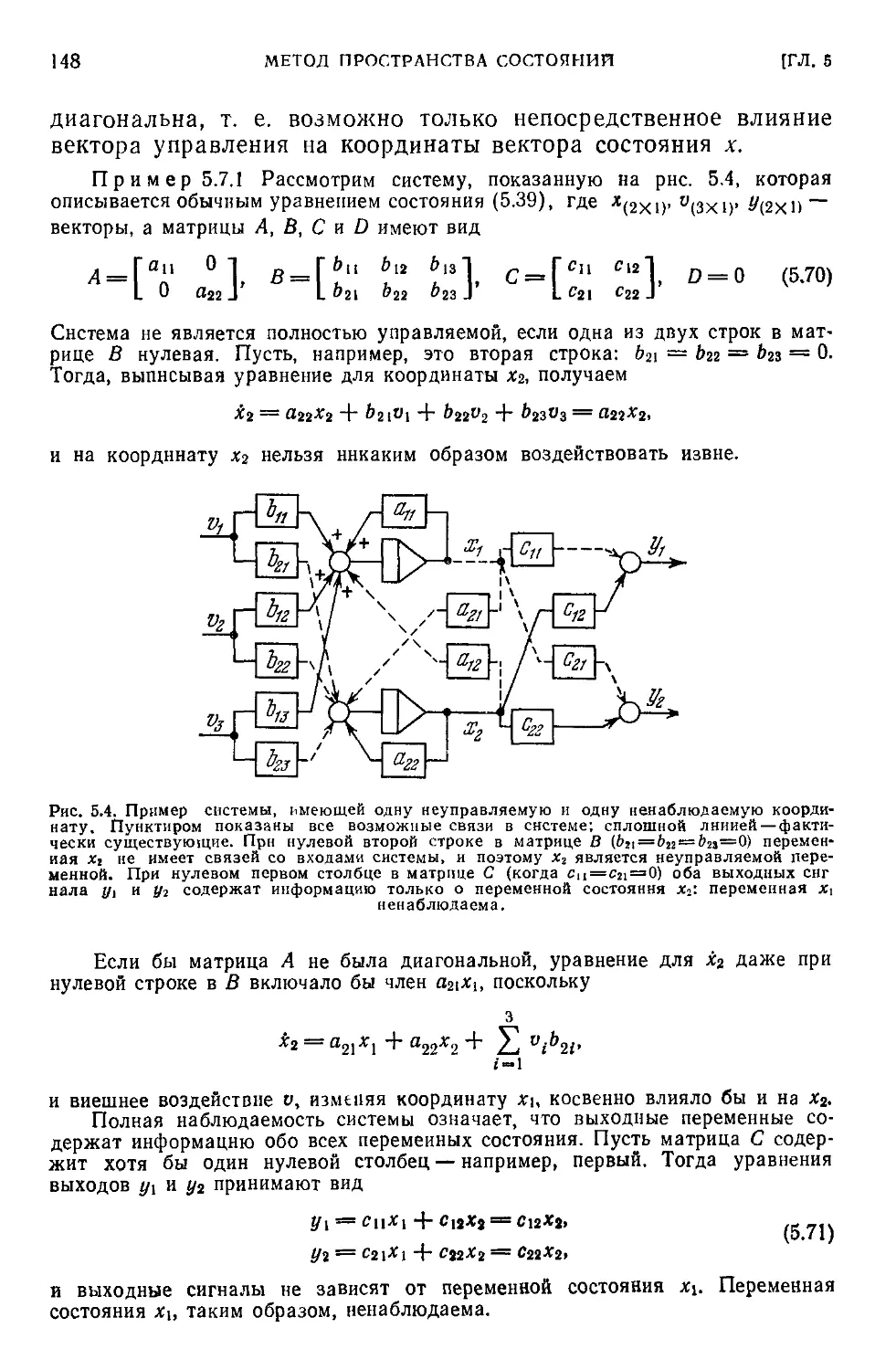
6.6. Компартментальная модель энергетической системы организма . 186
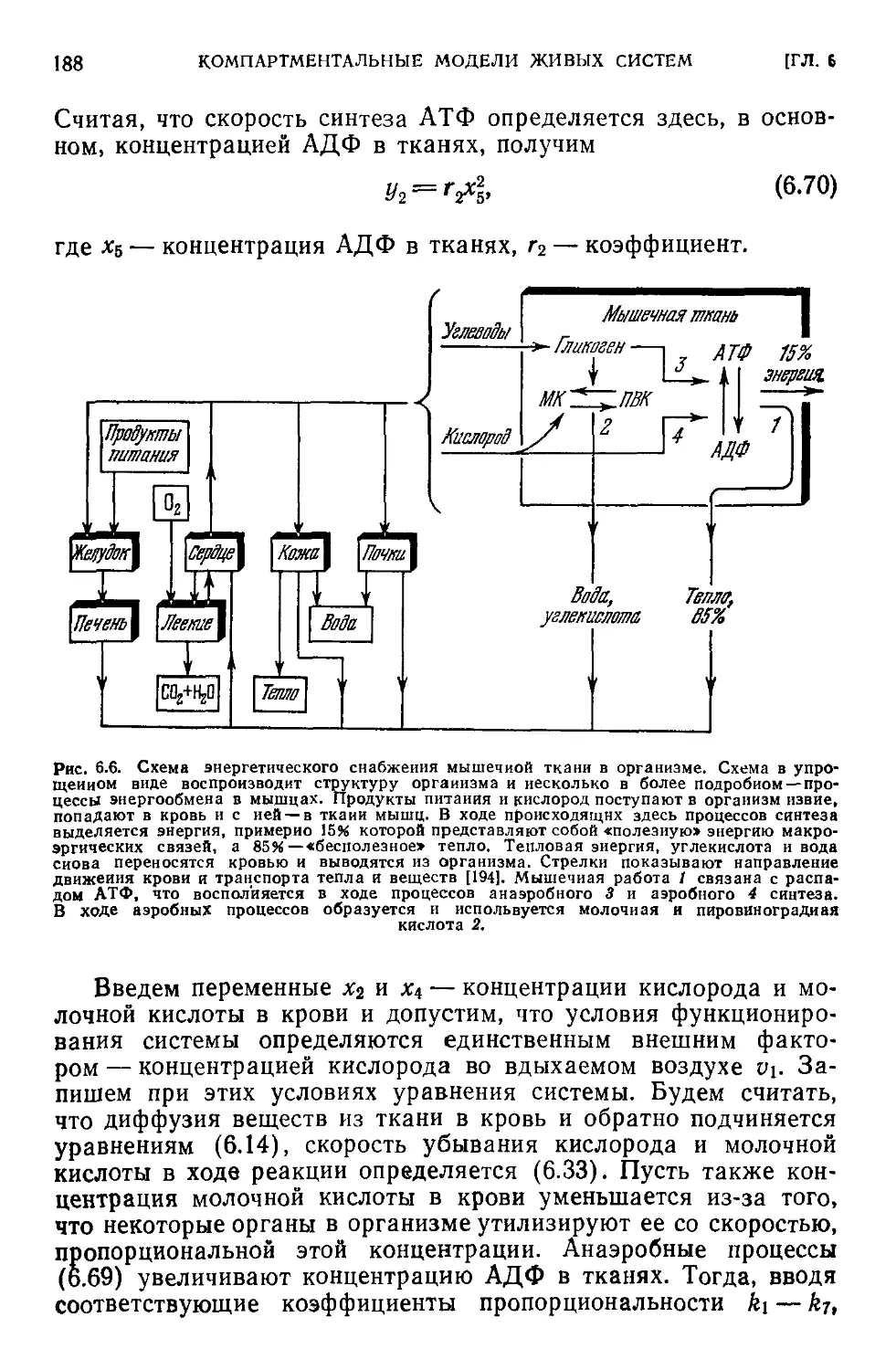
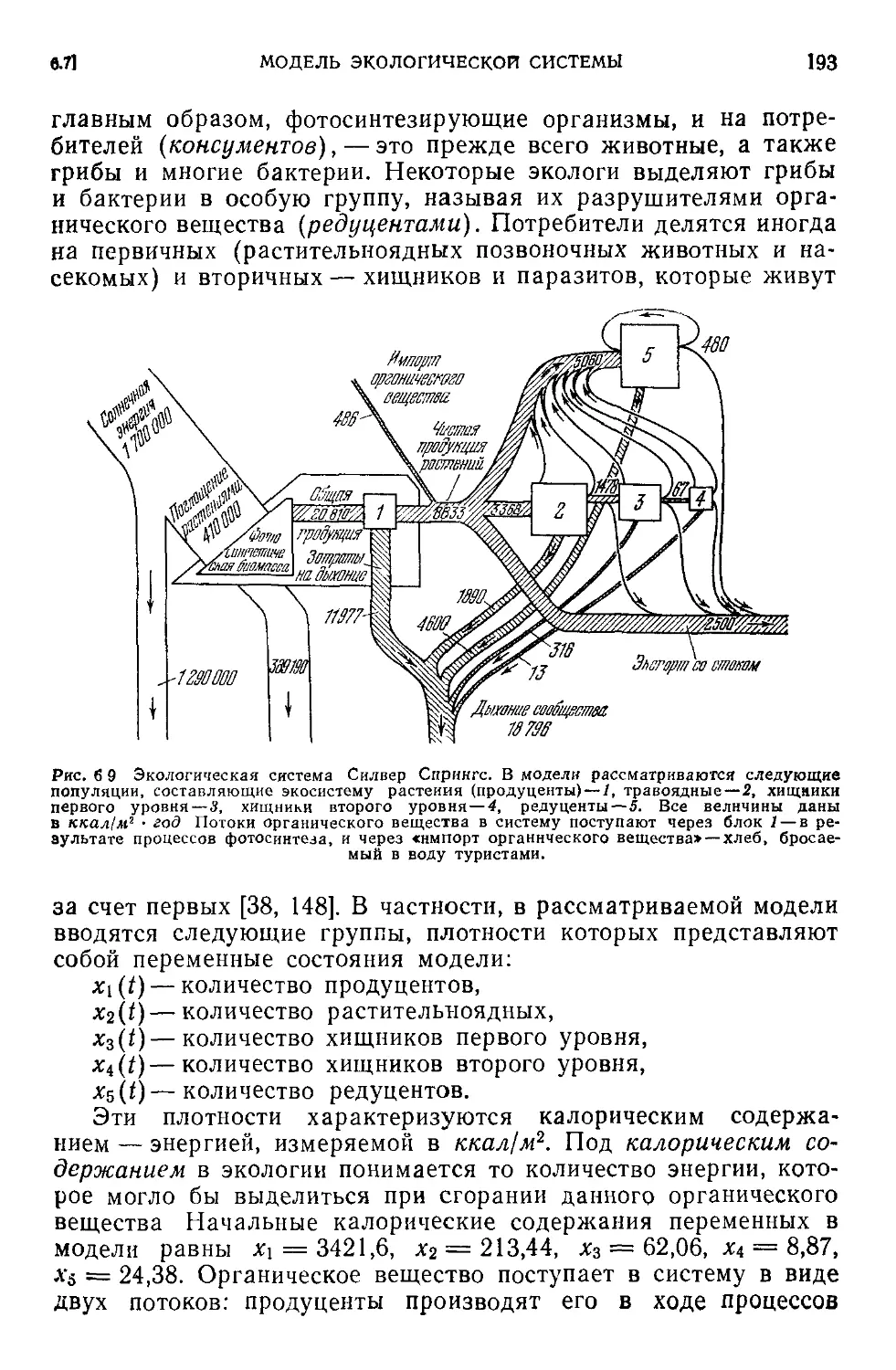
6.7. Компартментальная модель экологической системы...........192
ЧАСТЬ и
АНАЛИЗ СОХРАНИТЕЛЬНЫХ СВОЙСТВ БИОСИСТЕМ
Глава 7. Моделирование гомеостатических свойств организма .... 199
7.1. Замечания о моделировании сохранительных свойств организма
и его систем..................................................199
7.2. Два подхода к моделированию бносистем....................205
7.3. Моделирование гомеостаза методами классической теории ав-
томатического регулирования ................................. 210
7.4. Моделирование гомеостаза методами пространства состояний . 215
7.5. Влияние структуры компартментальной системы на ее гомеоста-
тические свойства.............................................220
7.6. Сочетание параллельных каналов регуляции.................225
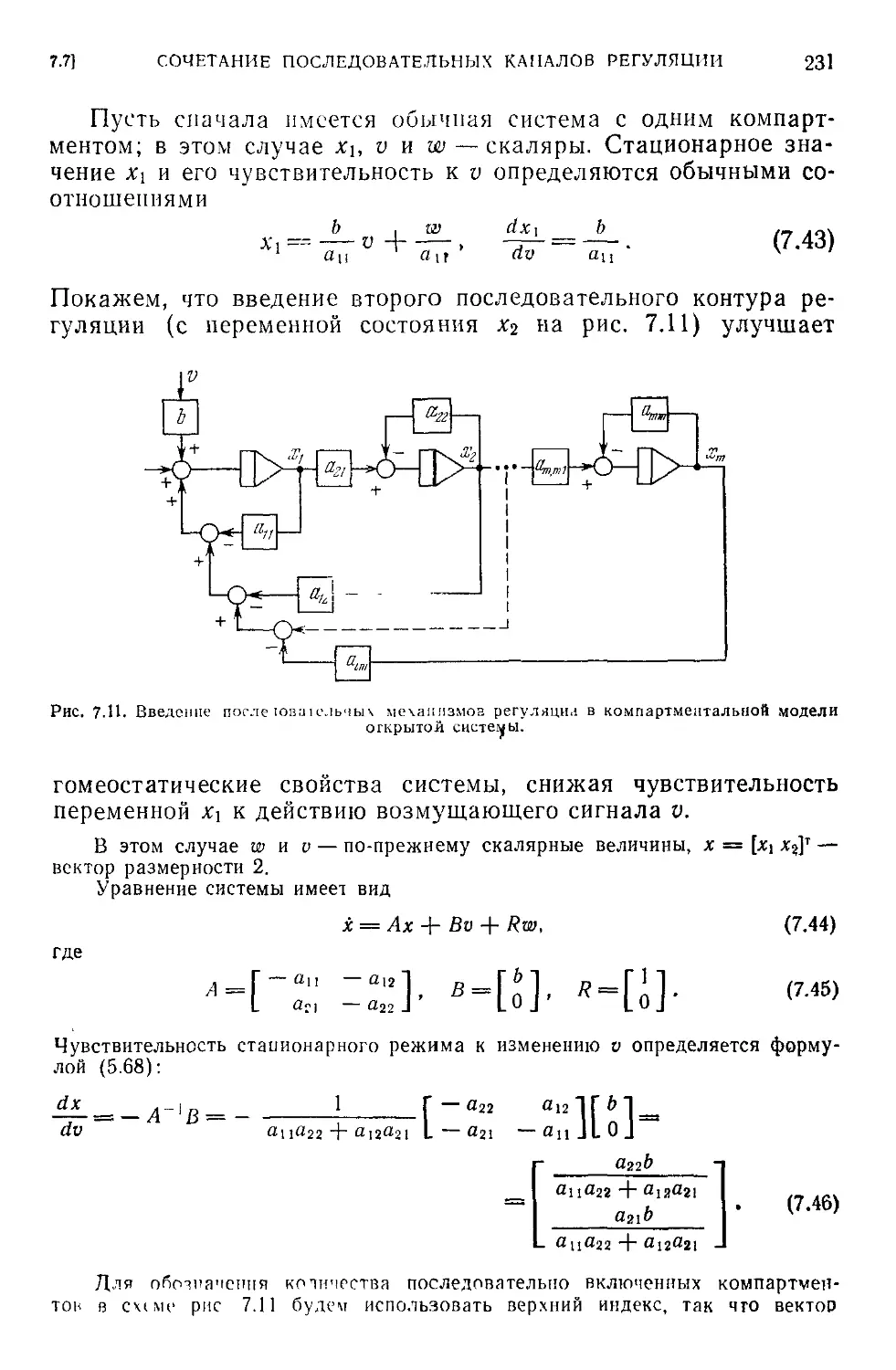
7.7. Сочетание последовательных каналов регуляции.............230
7.8. Гомеостаз в физиологических системах с активной регуляцией . 235
7.9. Стабилизация уровней вещества и энергии..................238
Глава 8. Количественная оценка гомеостатических свойств систем . . . 245
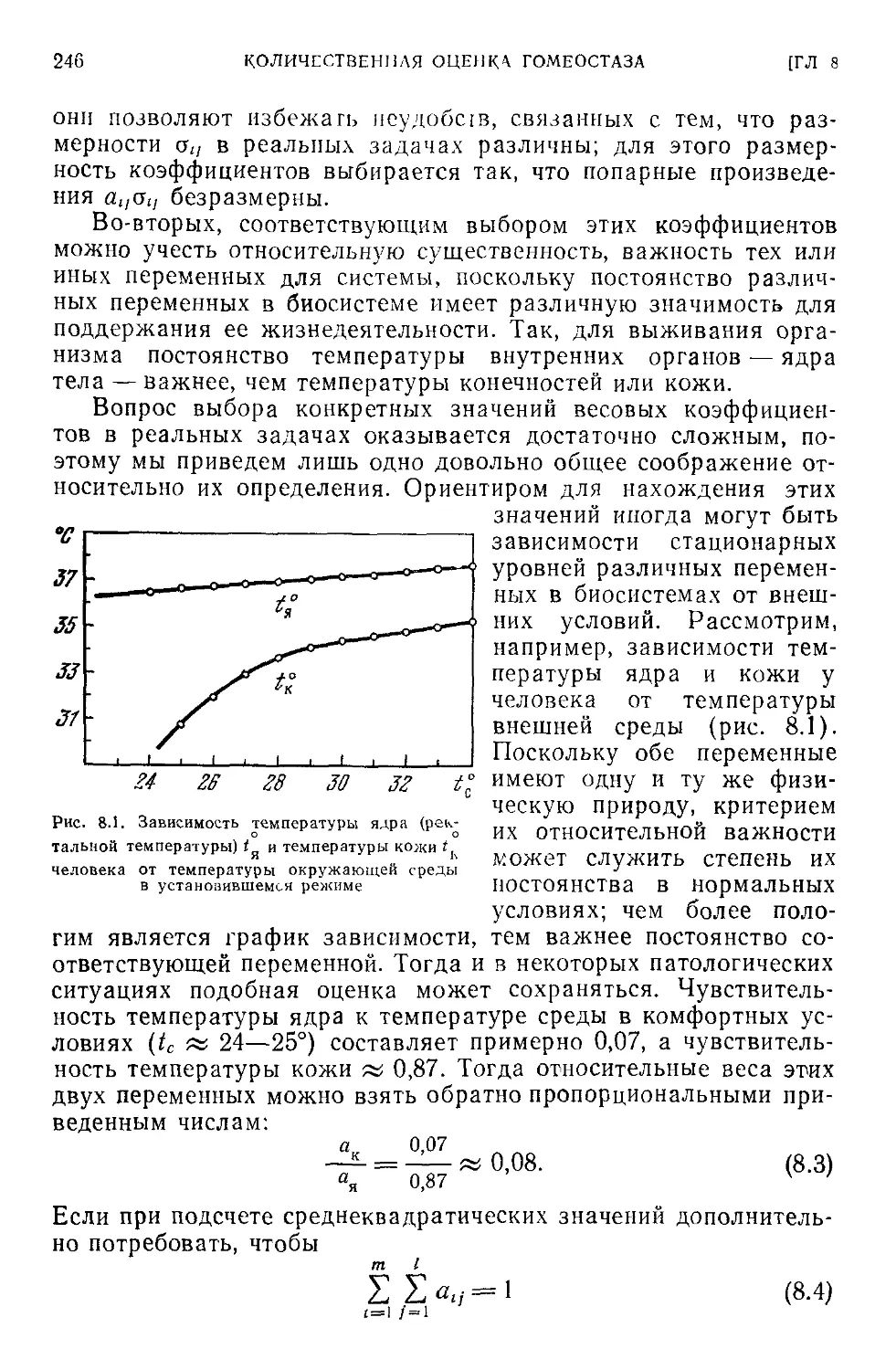
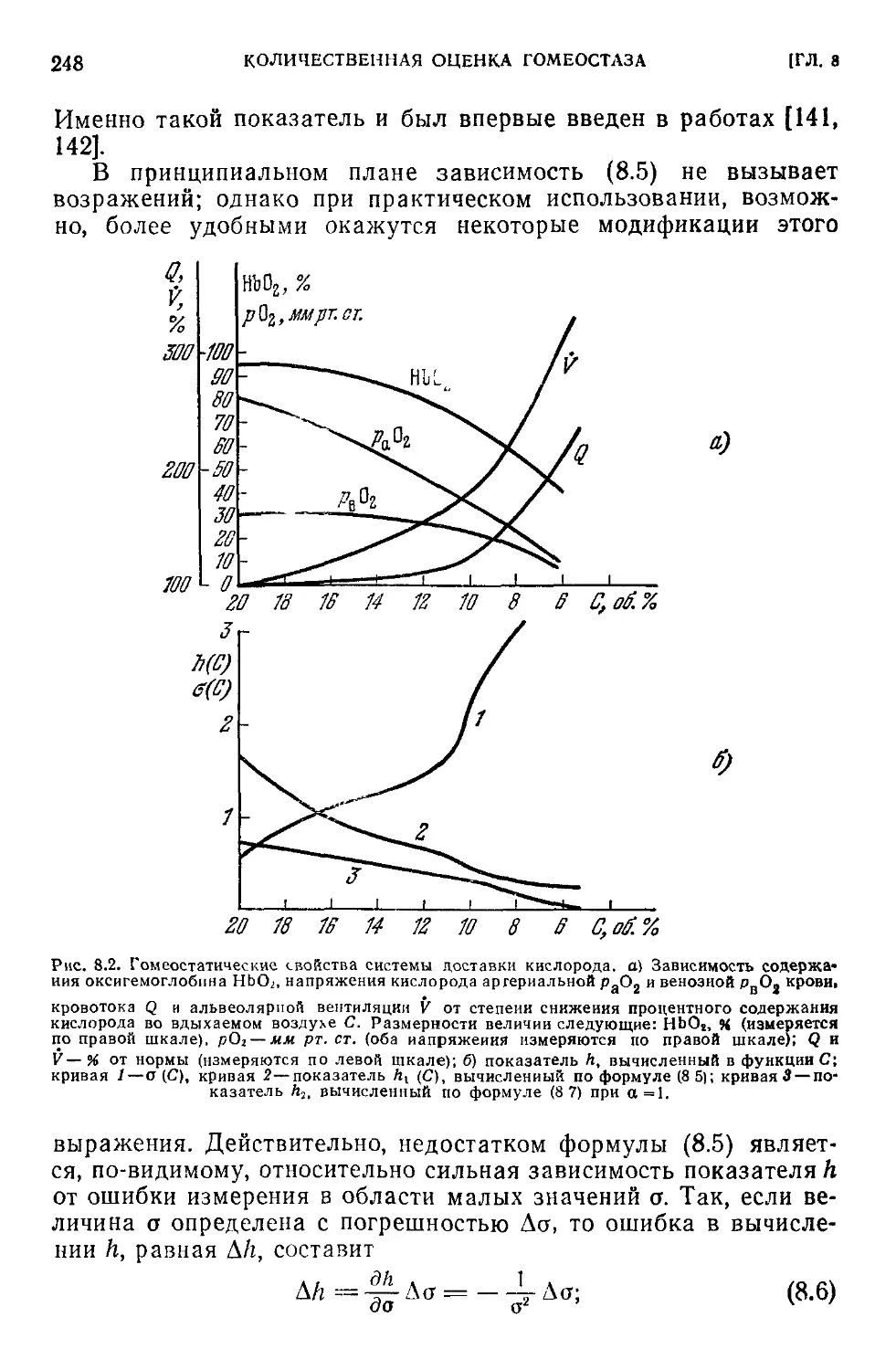
8.1. Показатель гомеостатической способности системы..........245
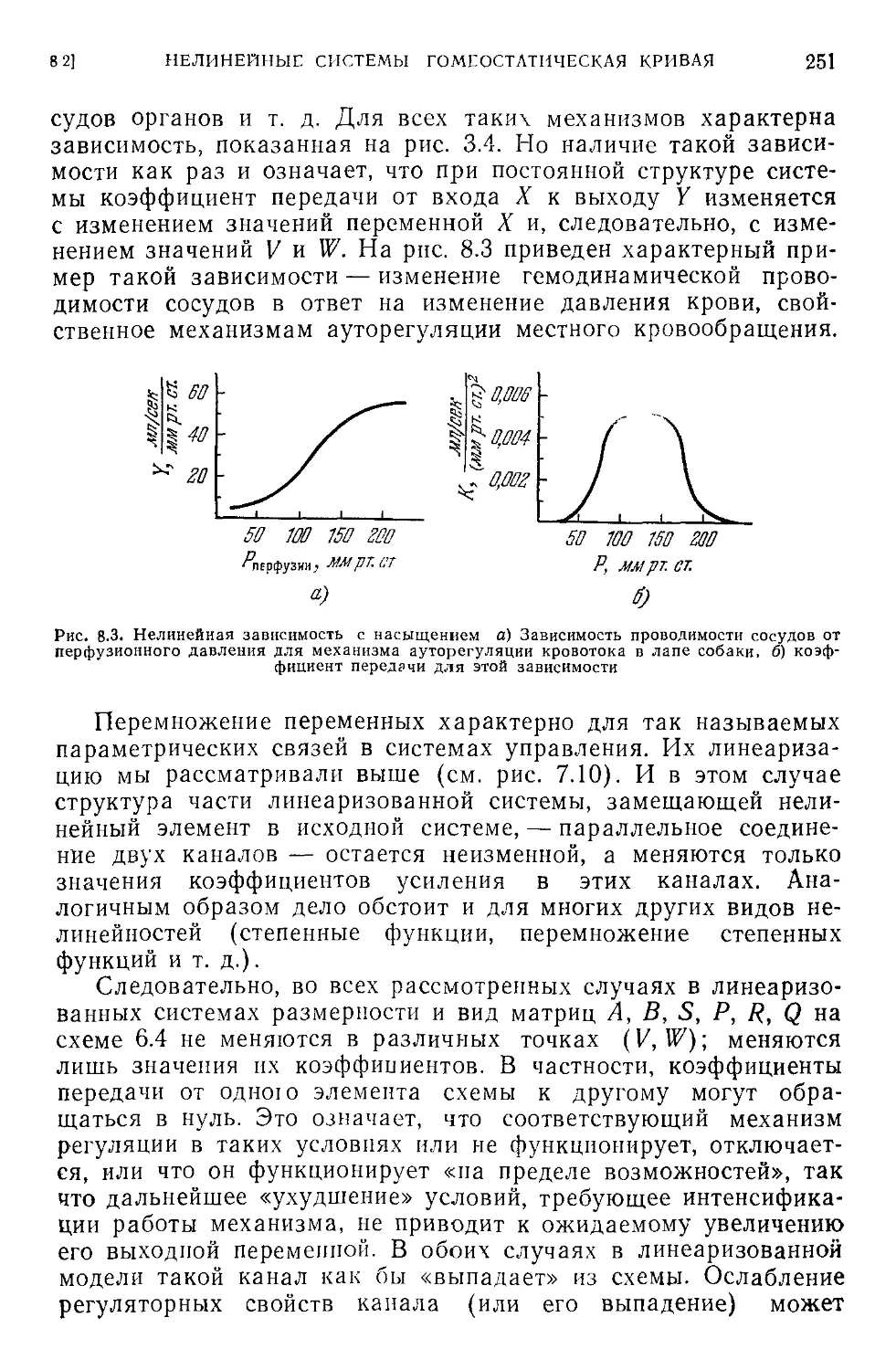
8.2. Гомеостаз нелинейных систем. Гомеостатическая кривая . . . 250
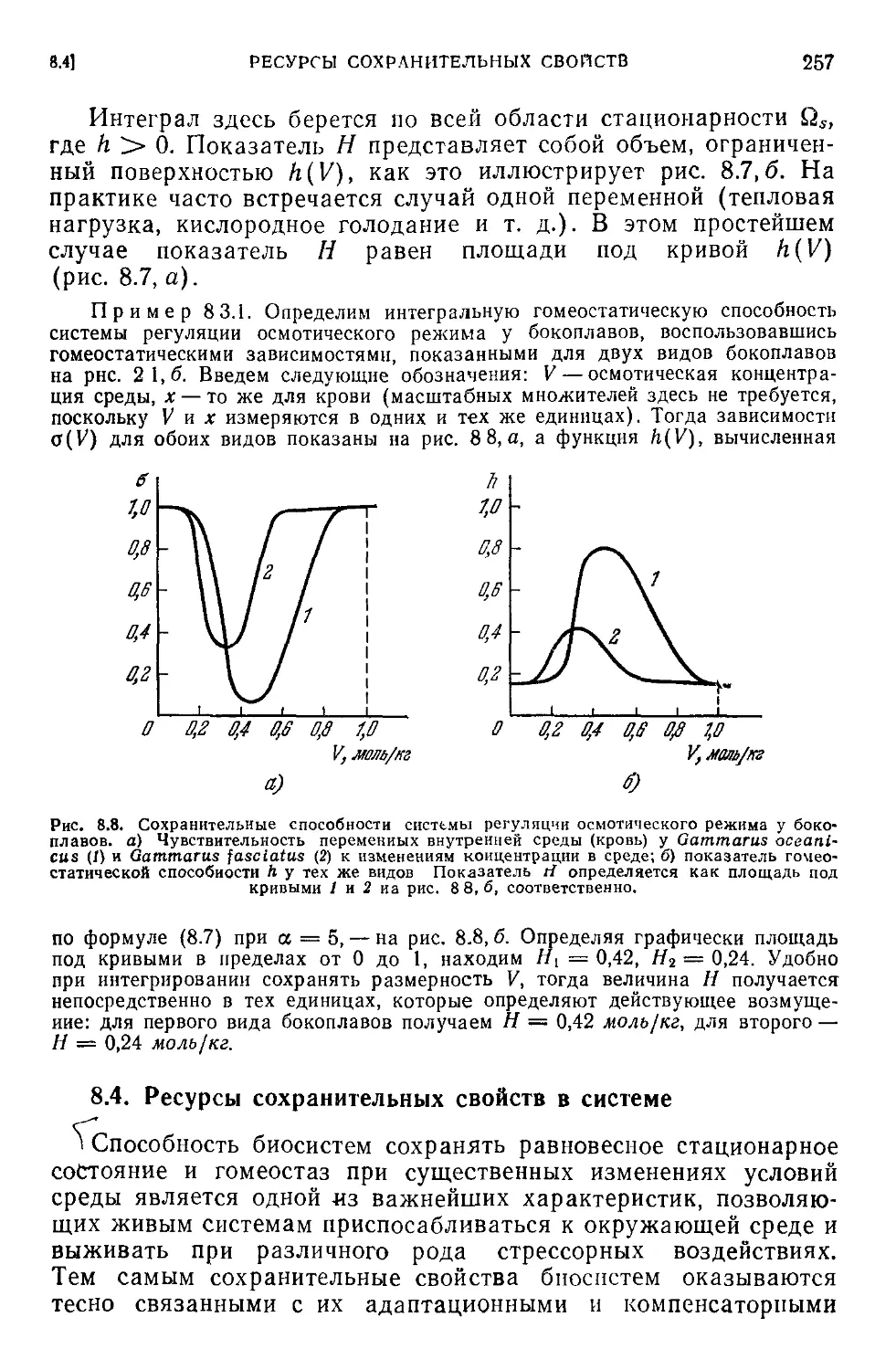
8.3. Интегральные оценки гомеостатических свойств системы . . . 256
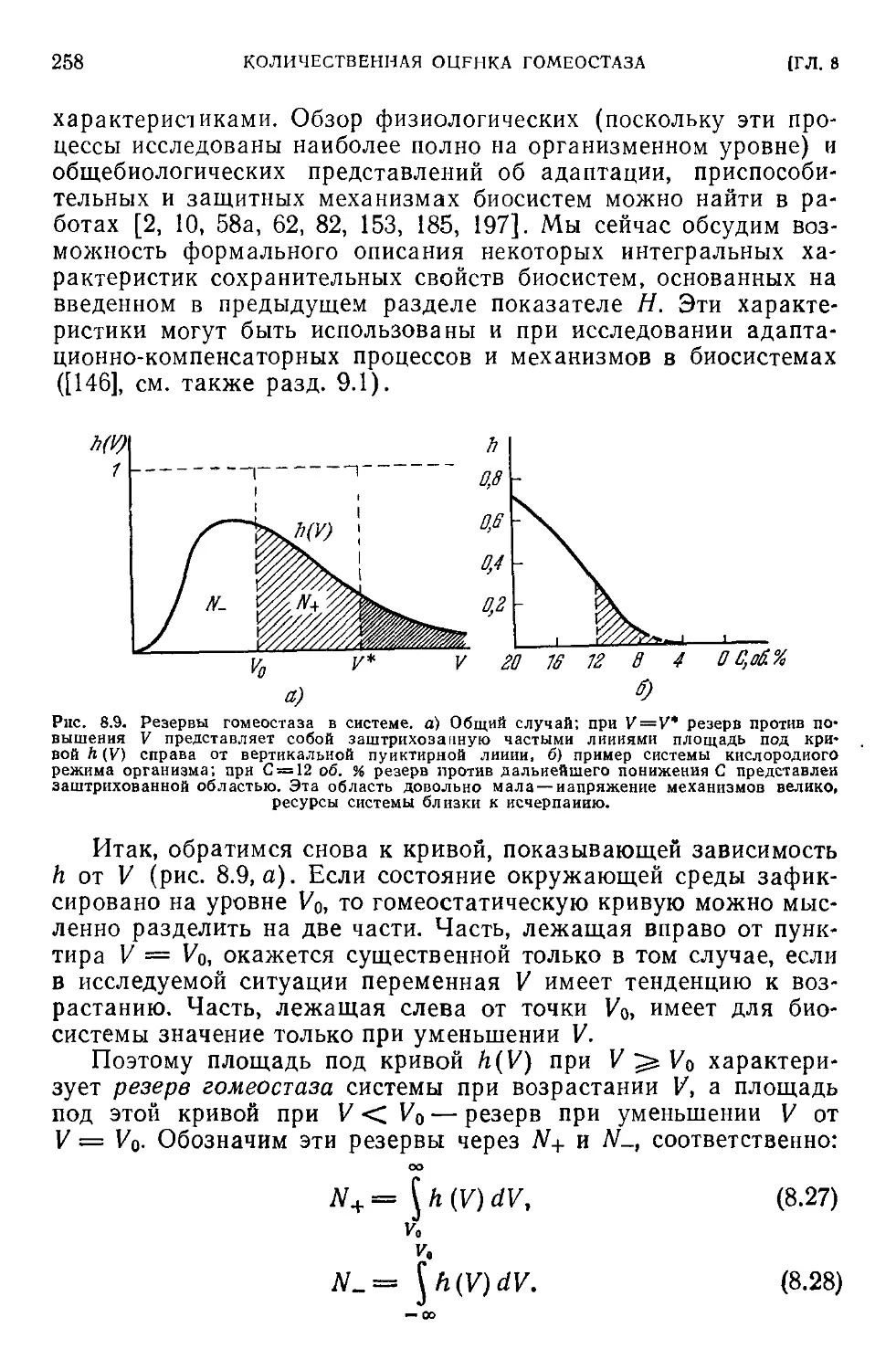
8.4. Ресурсы сохранительных свойств в системе.....................257
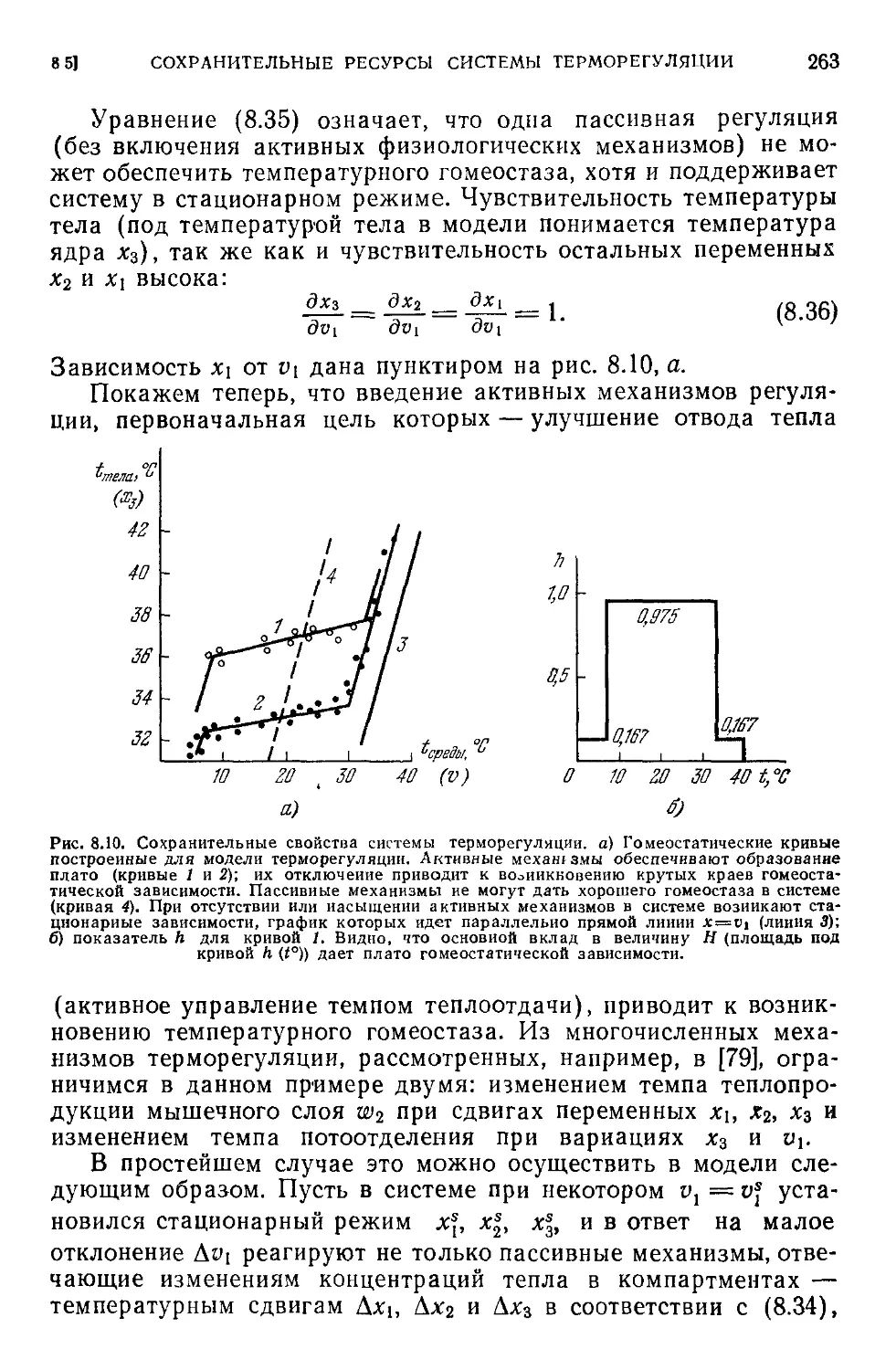
8.5. Сохранительные ресурсы системы терморегуляции................260
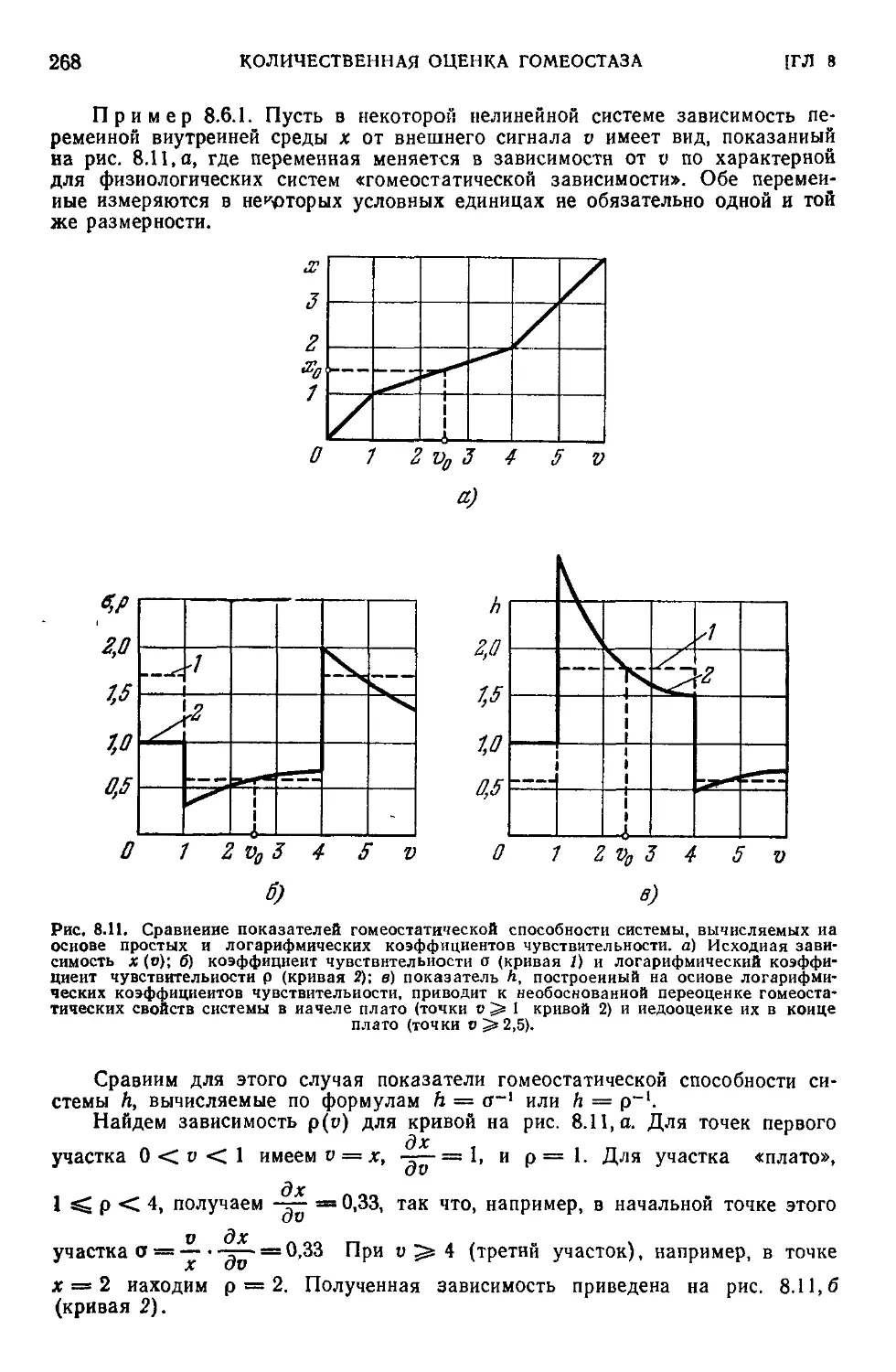
8.6. Логарифмические коэффициенты чувствительности................267
8.7. Формализованное представление показателя гомеостатической
способнасти...................................................269
Глава 9. Феноменология гомеостаза............................... . 272
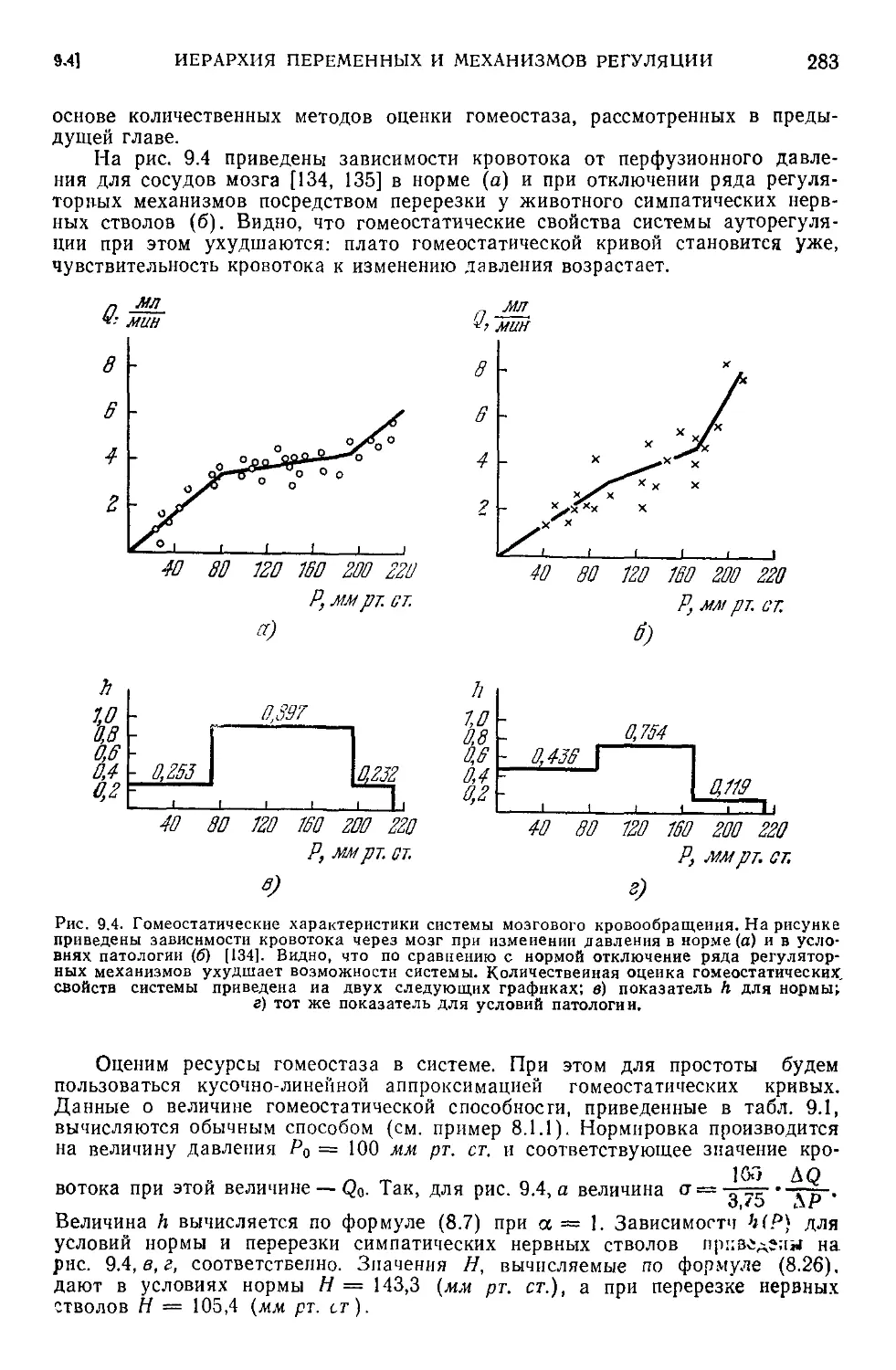
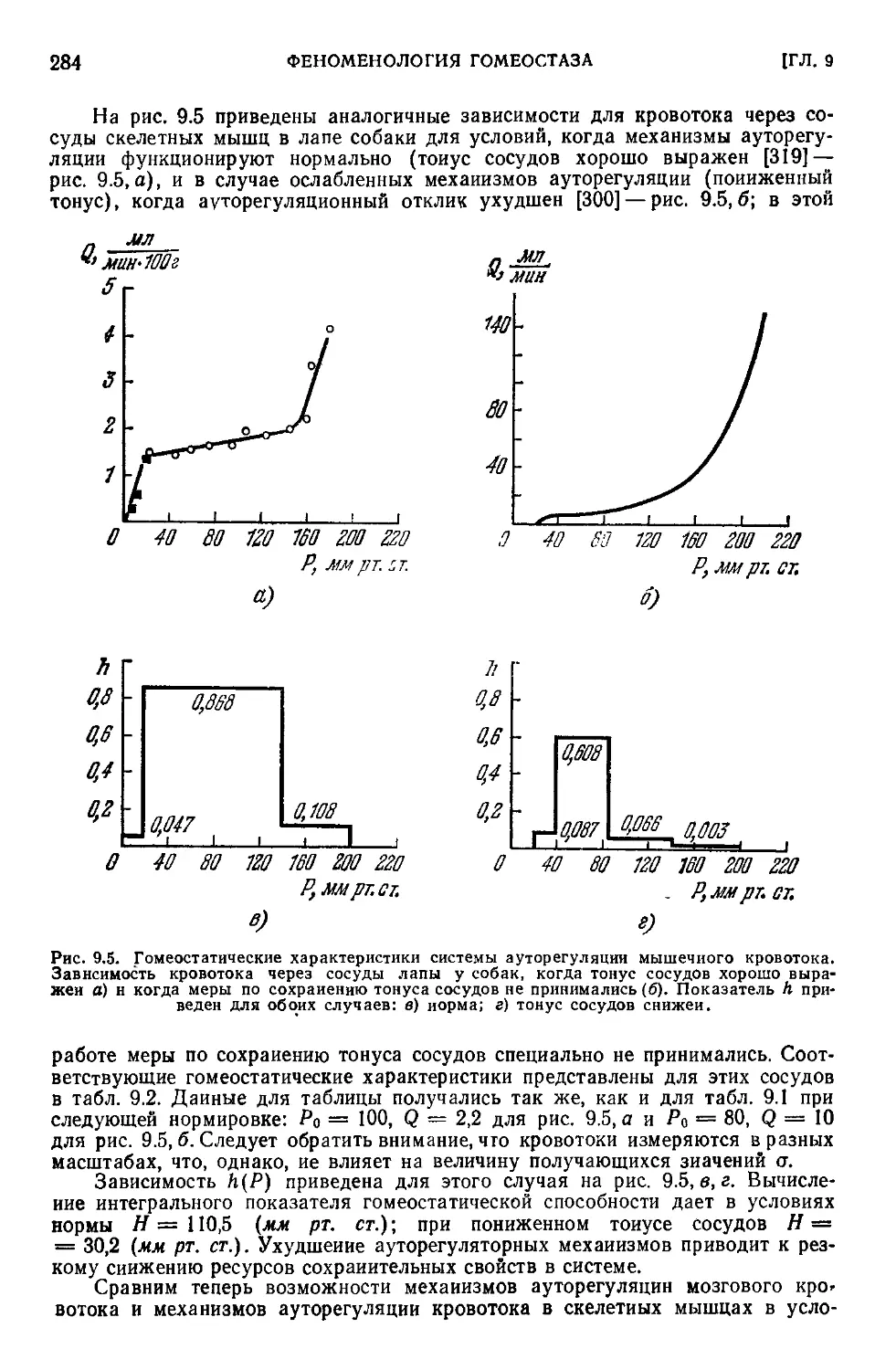
9.1. Типовые ситуации в гомеостатических системах.................272
9.2. Влияние прямых связей на свойства систем.....................276
ОГЛАВЛЕНИЕ Б
9.3. Двойственный характер переменных состояния в системе . . . 279
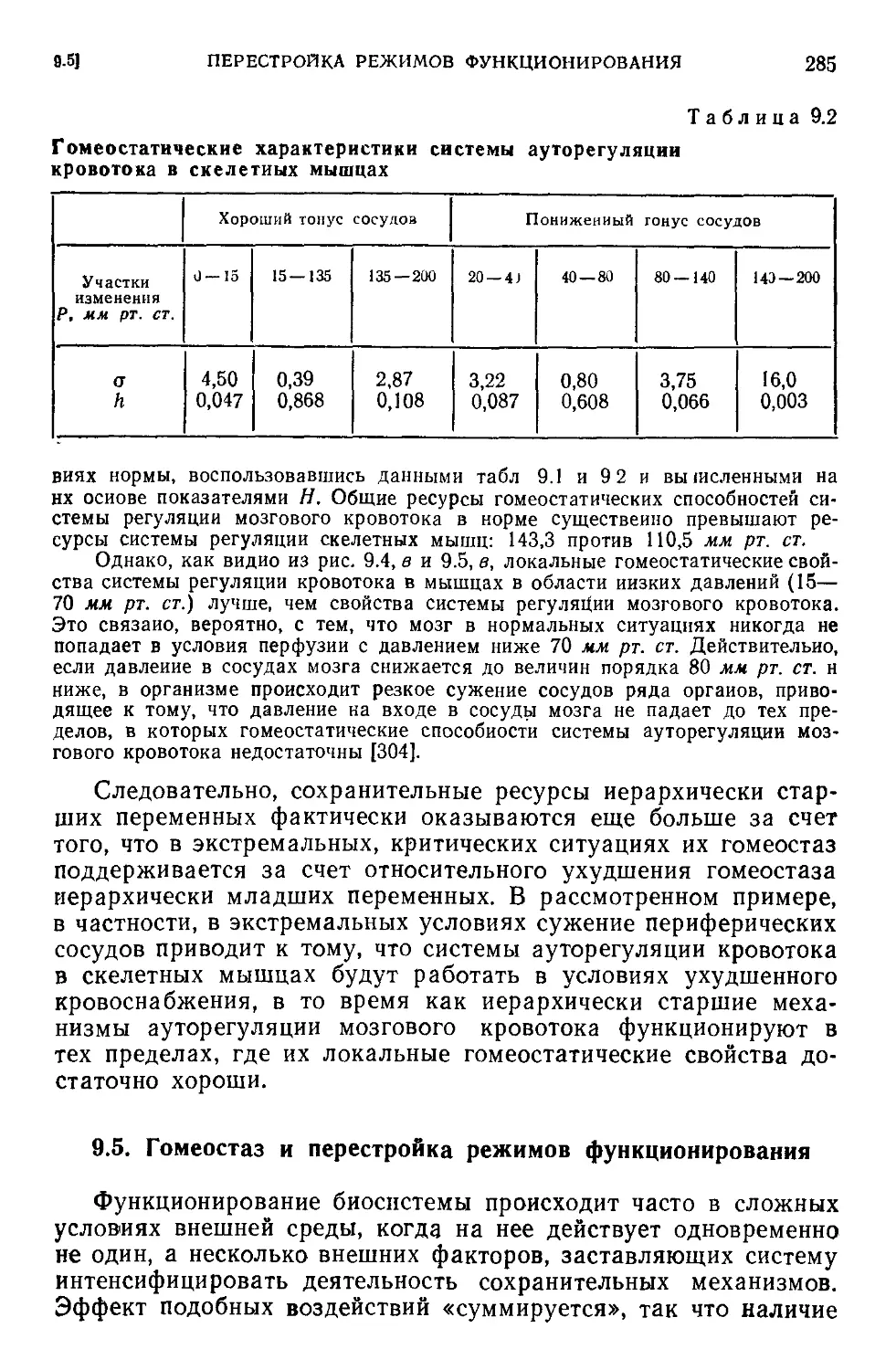
9.4. Иерархия переменных состояния н механизмов регуляции . . . 281
9.5. Гомеостаз и перестройка режимов функционирования .... 285
9.6. Нестационарные режимы в системе........................291
9.7. Моделирование биоснстем и принцип простоты.............293
Список обозначений и сокращений.................................296
Литература.................................................... 299
Предметный указатель.......................................... 315
ПРЕДИСЛОВИЕ
Проблема изучения тех свойств биосистем, которые позво-
ляют им в изменяющихся условиях окружения сохранять свою
жизнедеятельность, поддерживая стационарное неравновесное
состояние и обеспечивая постоянство внутренней среды, в силу
целого ряда причин приобретает в последние годы все большее
значение^ В данной книге для обозначения этой характерной
особенности биологических систем мы будем использовать тер-
мин сохраиительные свойства.
(Причины интереса к сохра нательным свойствам объектов
живой природы различны для разных уровней организации
жизниЗНа уровне организма и его систем интенсивное внимание
к анализу сохранительных механизмов объясняется прежде
всего освоением человеком новых сфер обитания — космоса, под-
водной среды, поскольку способность организма к самосохране-
нию непосредственным образом связана с процессами его при-
способления, адаптации к условиям обитания.ГНе мрнцшее зна-
чение имеет и прогресс медицинской науки —'разработка
и применение новых интенсивных методов лечения и-нротезиро-
вания все настойчивее требует точных количественных пред-
ставлений о ресурсах и возможностях организма и его систем.
Современные методы анестезиологии и реанимации, клиниче-
ские методы интенсивной терапии, все большее освоение спосо-
бов управляемого жизнеобеспечения призывают к анализу
целостных свойств организма во всей сложности составляющих
его систем. Целостность организма, его способность к самосо-
хранению, обеспечивается механизмами регуляции, поддержи-
вающими равновесие между отдельными системами организма
и обеспечивающими его стационарное функционирование.
Аналогичная картина наблюдается и на уровне сообществ
организмов. Сохраиительные механизмы экологических систем
в последние годы все чаще оказываются недостаточно мощными
перед давлением неблагоприятных факторов среды, все возра-
стающих по мере развития человеческой цивилизации. Сохране-
ние живой природы становится насущной задачей развивающе-
гося общества. Возникающие при этом проблемы оценки
допустимых пределов вмешательства человека требуют внима-
ПРЕДИСЛОВИЕ
7
тельного изучения сохранительных свойств различных биосистем
надорганизменного уровня — популяций, экосистем, ландшафтов.
В перспективе возникает уже и вопрос о сохранительных
свойствах и ресурсах биологических и социальных систем
в масштабах планеты. «Биосферный мониторинг... должен обе-
спечивать наблюдения, контроль и прогноз возможных измене-
ний уже не в региональном (экосистемном), а в глобальном
масштабе, т. е. в отношении биосферы в целом как среды жизни
всего человечества и ее изменений, вызванных деятельностью
общества», — пишет И. П. Герасимов [52].
Разными путями решаются задачи анализа сохранительных
способностей биологических систем на различных уровнях орга-
низации жизни, но по мере развития биохимии, физиологии,
экологии, разработки все более и более мощных и совершенных
методов исследования постепенно проявляется определенная
общность механизмов самосохранения объектов живой природы
от клеточного и субклеточного до популяционного и экологиче-
ского уровня. Все большую роль при анализе процессов само-
сохранения начинают играть количественные методы исследо-
вания, в том числе и методы теории управления, применению
которых для изучения сохранительных свойств биосистем по-
священа настоящая книга.
Самосохранение объектов живой природы является одним из
основных свойств, лежащих в основе жизненных явлений. «Со-
храняются только такие составляющие живой природы, в кото-
рых стремление к самосохранению выражено достаточно
ярко», — писал А. А. Ляпунов [ПО].
Понятие самосохранения имеет-в применении к биологиче-
ским системам два аспекта. Прежде всего самосохранение био-
системы означает ее способность поддерживать в изменяющихся
условиях функционирования и окружающей среды стационар-
ное неравновесное состояние, т. е. обеспечивать энтропийный
баланс в системе и баланс вещества и энергии. Другой, не ме-
нее важный аспект сохранительных свойств биосистем связан
с поддержанием гомеостаза — постоянства существенных для
жизнедеятельности системы переменных при наличии возмуще-
ний во внешней среде.
Живые системы — это необычайно сложные формы, возник-
шие в результате долгого эволюционного процесса, с сильно
выраженными способностями к адаптации, приспособлению
и эволюции, обладающие иерархической структурой и представ-
ляющие собой интеграцию многих гетерогенных элементов
и подсистем. В этой книге мы будем касаться далеко не всех
вопросов, связанных со сложностью, иерархией, гомеостазом
и адаптацией живых систем, ограничиваясь в основном лишь
теми их сторонами, где современные методы теории управления
8
ПРЕДИСЛОВИЕ
позволяют понять или уточнить механизмы функционирования
биосистем, выяснить, как образуются определенные свойства
в биологических структурах, дать количественные методы и кри-
терии их оценки.
Книга написана специалистом по теории управления и с по-
зиций этой теории. Рассчитанная на читателей-физиологов, био-
логов, медиков, инженеров и специалистов по теории управле-
ния, книга содержит разделы, по-разному интересные для
специалистов разных дисциплин.
Первая часть книги в основном носит обзорный характер
и посвящена рассмотрению общебиологических представлений
о гомеостазе и сохранительных способностях биосистем, с одной
стороны, и изложению идей и методов теории управления —
с другой.
В гл. 1—2 рассказывается о специфических характеристиках
биологических систем на разных уровнях организации жизни.
Материал этих глав излагается на биологическом языке, но
цель их написания состоит не только в том, чтобы суммировать
содержательные сведения о сохранительных свойствах био-
систем на различных уровнях организации жизни. Эта часть
написана под углом зрения специалиста по теории управления,
отражающим идею целостности биосистемы, мысль о единстве
управляющих механизмов биосистемы. Особое внимание здесь
уделяется представлениям о сохранительных свойствах объек-
тов живой природы, развиваемым классиками биологической
науки — взглядам У. Кэннона на гомеостаз живых систем
и И. М. Сеченова — на регуляцию потоков вещества и энергии
через биосистемы.
Гл. 3—5 посвящены идеям и методам теории управления.
Здесь следует заметить, что конкретные методы классической
теории управления, связанные, например, с анализом переда-
точных функций, не всегда дают адекватное описание сложных
биологических систем. Будучи приспособленными для решения
относительно простых задач регулирования со сравнительно
небольшим числом управляемых переменных, эти методы подчас
малоприменимы для исследования биологических проблем, где
даже при грубом упрощении приходится иметь дело с десятками
и сотнями переменных и параметров. Таковы, например, задачи
анализа биохимических или экологических систем, где рассмат-
ривается взаимодействие сотен компонент — различных веществ,
участвующих в биохимических реакциях, или видов растений
и животных, образующих экологические системы. Несколько
проще дело обстоит с анализом систем физиологического
уровня, где для описания функционирования таких систем, как
терморегуляция, регуляция кислородного режима, требуется
обычно не более двух-трех десятков переменных.
ПРЕДИСЛОВИЕ
9
Поэтому, хотя процессы управления в живых системах
в своей основе имеют много общего с управлением в техниче-
ских системах, сами конкретные методы классической теории
управления должны применяться для анализа биосистем
с большой осторожностью. Так, использование простой следя-
щей системы в качестве модели гомеостаза в физиологических
системах не позволяет дать сколько-нибудь полной картины
процессов адаптации и приспособления. Применение такой про-
стой модели для моделирования процессов в нервной системе
может дать описание только самой элементарной ее функции —
функции слежения у человека-оператора, и непригодно для
описания других ее свойств. Можно ожидать, что новые методы,
разработанные в теории управления, в ряде случаев будут
более адекватным аппаратом исследования биосистем. В част-
ности, применение метода пространства состояний, разработан-
ного в теории управления в последние годы, позволяет по-но-
вому осветить вопросы гомеостаза в физиологических системах.
Если классические методы теории управления достаточно
хорошо известны специалистам, работающим в области биоло-
гических применений кибернетики, в том числе и специалистам-
биологам и физиологам по вышедшим на русском языке в сере-
дине 1960-х гг. монографиям Ф. Гродинза и Дж. Милсума [60,
130], то разработанные к началу 1970-х гг. в теории управления
методы пространства состояний до сих пор малоизвестны ши-
роким кругам специалистов, интересующихся медико-биологи-
ческими применениями теории управления.
Метод пространства состояний требует использования мат-
ричного способа описания и исследования систем, с которым
знакомы сейчас далеко не все специалисты, работающие в этой
области. Поэтому описание метода пространства состояний
в книге сопровождается изложением элементарных основ тео-
рии матриц. Впрочем, изложение теории матриц включается
и в книги по методу пространства состояний, написанные для
инженеров. В этой же части обсуждается и проблема оптималь-
ности (гл. 4)—концепция, целиком пришедшая в биологию из
кибернетики и теории систем, разбираются ее основные черты
и особенно пределы применимости.
Гл. 6 посвящена теории компартментальных моделей.
Вторая часть книги, начиная с седьмой главы, посвящена
собственно анализу сохранительных свойств живых систем.
Анализ этих свойств проводится с применением метода про-
странства состояний, который, по-видимому, наиболее приспо-
соблен для исследования целостных характеристик биосистем.
Эффективность применения этого метода для анализа живых
систем определяется двумя обстоятельствами. Прежде всего
при использовании методов пространства состояния происходит
10
ПРЕДИСЛОВИЕ
отказ от привычных для кибернетических публикаций 1960-х го-
дов категорий «черного ящика», когда исследователь судит
о системе только на основе двух классов сигналов, входов и вы-
ходов системы, и не интересуется процессами внутри системы.
Новые методы вместо терминов «вход — выход» дают анализ
системы в терминах «вход — выход — состояние». Именно вве-
дение в рассмотрение понятия состояния — описания «внутрен-
него» поведения системы при данных входах и выходах — и поз-
воляет расширить биологические применения методов теории
управления.
Второе обстоятельство, позволяющее более широко приме-
нять метод пространства состояний для анализа многокомпо-
нентных биологических систем, состоит в том, что используемые
в нем матричные методы описания систем в равной степени
применимы к линейным системам как малой, так и большой
размерности.
Во второй части метод пространства состояний используется
для анализа сохранительных свойств биосистем на организмен-
ном уровне, в основном — для физиологических систем. Хотя
разработанные методы в принципе применимы и для анализа
других уровней организации живого, их конкретное использова-
ние для биосистем других типов требует учета особенностей
биосистем — своеобразия их структуры и функционирования,
наличия специфических целей систем и механизмов управления
на популяционном, экологическом и биохимическом уровнях.
Разнообразие биосистем на разных уровнях организации
жизни — при всем несомненном их сходстве — не позволило ав-
тору провести такого исследования.
Даже на организменном уровне анализ тех или иных систем
и структур открывает перед исследователем практически не-
ограниченное поле деятельности. Так, в отдельную ветвь давно
выделилось исследование принципов и механизмов управления
движениями. Другую не менее мощную ветвь составляет изуче-
ние моделей нервной системы — нейронов, нейронных систем
и структур. Эти важнейшие направления исследований на орга-
низменном уровне остаются за рамками изложения в этой
книге, в которой внимание читателя концентрируется на «веге-
тативном» уровне процессов регулирования и управления в ор-
ганизме. Это — низший этаж процессов самосохранения, свя-
занный со снабжением организма, его систем и органов веще-
ствами и энергией и с поддержанием постоянства условий
жизнедеятельности во внутренней среде организмов — го-
меостаза.
Конечно, и механизмы высшего уровня (в частности, пове-
денческие реакции животного организма) участвуют в поддер-
жании гомеостатического режима организма, накладываясь
ПРЕДИСЛОВИЕ
11
на механизмы вегетативного уровня. По словам К. Вилли [38],
мотивированное поведение служит ответом на то или иное
нарушение гомеостаза. Изучение взаимодействия вегетативных
механизмов управления и механизмов, связанных с процессами
в коре головного мозга животного организма, также осталось
за пределами изложения.
В гл. 7 обсуждаются вопросы моделирования сохранительных
свойств биологических систем в рамках классической теории
управления и с использованием метода пространства состояний.
Главная мысль этой главы состоит в том, чтобы продемонстри-
ровать единство открытого характера живых систем и их го-
меостатических свойств. Единая система управляющих меха-
низмов в целостной биосистеме может, как показано в этой
главе, не только поддерживать стационарный режим в открытой
системе, но при определенных условиях обеспечивать и ее го-
меостаз. Этот результат позволяет по-новому подойти к взгля-
дам Л. фон Берталанфи, противопоставлявшего теорию откры-
тых систем и кибернетические методы описания гомеостаза.
Гл. 8—9 посвящены развитию количественных представлений
и выработке численных характеристик для описания гомеоста-
тических свойств биологических объектов.
В книге рассматривается только одна характеристика живых
систем — их способность к самосохранению. Конкретное приме-
нение методы анализа сохранительных свойств в книге находят
для систем организменного уровня. Поэтому и список литера-
туры, в основном ограниченный системами этого уровня, ни
в коем случае не претендует на полноту.
В то же время современный аппарат теории управления,
применяемый при таком анализе, — и прежде всего метод про-
странства состояний — применим для описания значительно бо-
лее широких классов биологических систем на всех уровнях
организации жизни. Чтобы подчеркнуть эту особенность методов
теории управления, было выбрано двойное название книги:
«Теория управления и биосистемы. Анализ сохранительных
свойств».
Эта книга рассчитана на широкий круг читателей, интере-
сующихся проблемами управления в биологических системах,
хотя используемый в ней аппарат и нельзя всегда считать про-
стым. Изложение построено так, что все идеи даются не столько
на языке формул, сколько на языке, понятном специалистам-
биологам, не имеющим специальных знаний в области теории
управления.
Терминология книги во многом тяготеет к физиологическим
представлениям. Для обозначения механизмов, действующих
в биосистемах и обеспечивающих их существование и достиже-
ние адекватных условий функционирования, мы используем
12
ПРЕДИСЛОВИЕ
термин «управление», допуская его физиологический синоним
«регуляция». Конечно, процессы управления в биосистемах
почти всегда являются процессами самоуправления и саморе-
гуляции, за исключением случаев вмешательства человека.
Изложение материала сопровождается примерами. Чаще
всего примеры берутся из области физиологических систем —
как потому, что эти системы наиболее знакомы автору, так
и потому, что сохранительные свойства биологических систем,
возможно, наиболее ярко проявляются именно на этом уровне
организации жизни.
Книга, касающаяся широкого круга вопросов теории управ-
ления, биокибернетики, физиологии и других биологических
дисциплин, не была бы написана, если бы ие многочисленные
обсуждения й помощь многих специалистов в самых различных
областях, выражавшаяся к тому же в самой разной форме.
Автор глубоко признателен Я. 3. Цыпкину, который во мно-
гом способствовал формированию его научных взглядов и пред-
ставлений и тем самым появлению настоящей книги. Своим
интересом к физиологии организма автор обязан В. И. Шума-
кову и Е. Ш. Штенгольду, многолетнее сотрудничество с кото-
рыми оказалось для него таким плодотворным. С удовольствием
отмечает автор благожелательное внимание А. М. Петровского.
Чрезвычайно полезными в работе над рукописью оказались со-
веты и критические замечания Р. А. Полуэктова. Весьма цен-
ными для автора были контакты с А. Д. Армандом и К- П. Ива-
новым, прочитавшими ряд разделов рукописи, а также ее
обсуждение с М. П. Сахаровым и Л. А. Дартау. Особенно необ-
ходимым автор считает отметить содержательные и полезные
замечания, сделанные Ю. М. Свирежевым.
Большую работу по улучшению рукописи проделал редактор
книги А. А. Петров.
Всем им автор выражает глубокую благодарность. Однако
взгляды, выраженные в этой книге, — это взгляды ее автора,
и он принимает ответственность за ее недостатки, которых, воз-
можно, не удалось избежать.
В. Новосельцев
Ноябрь 1977 г.
ЧАСТЬ I
ТЕОРИЯ УПРАВЛЕНИЯ
И ЖИВЫЕ СИСТЕМЫ
В этой части книги излагаются основные представ-
ления о биологических системах и их сохранительных
свойствах на различных уровнях организации жизни
и описываются методы теории управления, применяе-
мые для исследования биосистем.
Под сохранительными свойствами живых систем
здесь понимается их способность поддерживать ста-
ционарное неравновесное состояние и постоянство
внутренней среды при изменении условий окружаю-
щей среды и режимов функционирования. Интерес
к самосохранению объектов как живой, так и неживой
природы восходит еще ко времени Возрождения. Ис-
следование сохранительных свойств биосистем пред-
ставляет собой общую проблему, включающую как
частную задачу изучение гомеостаза и гомеостатиче-
ских механизмов животного организма. Гомеостаз
и открытый характер биологических систем часто вы-
ступают как два противоречивых способа описания
свойств объектов живой природы. Анализ сохрани-
тельных свойств биосистем с позиций современной
теории управления позволяет во многом снять это
противоречие.
Для исследования сохранительных свойств биоло-
гических объектов в этой книге применяется аппарат
теории управления — традиционного метода исследо-
вания живых систем на всех уровнях их организации:
субклеточном, клеточном и органном, организменном
и надорганизменном.
Теория управления находит все более широ-
кую сферу применения в биологии и медицине, но
14
ТЕОРИЯ УПРАВЛЕНИЯ И ЖИВЫЕ СИСТЕМЫ
специалисты, работающие в области биокибернетики,
не всегда успевают за развитием идей и методов тео-
рии управления. Поэтому в главах данной части не
только дается обзор методов теории управления, уже
нашедших применение в биокибернетических и биоме-
дицинских исследованиях, но и излагаются современ-
ные разделы теории управления, которые еще недоста-
точно известны широким кругам исследователей
в этой области, и прежде всего — метод пространства
состояний.
Особое значение метод пространства состояний
приобретает в связи с появлением и распростране-
нием методов компартментального моделирования,
завоевавших уже прочные позиции при исследовании
экологических, физиологических, биохимических
(прежде всего — фармакокинетических) систем.
Таким образом, в этой части книги закладываются
основы, — как общебиологического, так и теоретико-
управленческого плана, — для количественного ана-
лиза сохранительных свойств объектов живой при-
роды.
ГЛАВА I
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
1.1. Живая система
Под биологической (или живой) системой обычно пони-
мается некоторая совокупность взаимодействующих элементов,
которая образует целостный биологический объект.
Примерами биологических объектов, которые можно рас-
сматривать в качестве живых систем, являются организмы; на
суборганизменном уровне — это клетки, ткани, органы, а на
надорганизменном — популяции организмов, экосистемы, био-
геоценозы, биосфера. Очевидно, что один и тот же биологиче-
ский объект может как выступать в виде целостной живой си-
стемы, так и входить в эту систему в качестве подсистемы или
элемента. Однако перечисление примеров живых систем, как
бы наглядно оно ни было, не снимает вопроса о том, чем же
собственно является «живая система». Четкого различия между
живым и неживым указать не удается.
«Жизнь есть сложная совокупность процессов, протекающих
в организованной материальной системе и характеризуемых со-
хранением специфичности системы в потоке обмена веществ
и наперекор разнообразным воздействиям наружной среды, если
эти воздействия не превышают регулятивную способность си-
стемы». Это определение, данное В. Н. Беклемишевым [28], не
только сохраняет дух уважения к живому, свойственный высо-
кой биологической науке, но и содержит все основные признаки,
которые в разных комбинациях входят в определение живой,
или биологической, системы, организма или жизни вообще,
предлагаемые для различных уровней организации живой
природы.
Обычно обсуждение требований, которым должна удовле-
творять живая система, сводится к трем основным вопросам
[44, НО, 248, 251, 257].
1. Живые системы являются открытыми системами. Откры-
той системой называют систему, обменивающуюся с окружаю-
щей средой веществом и энергией. Процессы обмена веществ
(метаболизм) должны всегда находиться в центре внимания
наук, связанных с изучением живых систем.
16
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
1ГЛ. I
2. Эти системы являются самоуправляемыми, саморегули-
рующимися, т. е. они могут обмениваться со средой информа-
цией и перерабатывать эту информацию для поддержания своей
структуры и управления процессами метаболизма.
3. Живые системы должны быть самовоспроизводящимися.
Впрочем, легко видеть, что в полном своем объеме это по-
следнее свойство может быть отнесено только к некоторым
уровням организации жизни (клетки, организмы). Почка, ожи-
дающая пересадки в аппарате искусственного жизнеобеспече-
ния, является примером живой системы, не обладающей этим
свойством [254].
Появляющиеся время от времени определения живых си-
стем, в которых делается попытка дать их исчерпывающие ха-
рактеристики, все же не решают вопроса о сущности жизни.
Приведем одну из наиболее развернутых формул живой системы
[333, 334].
Живая система представляет собой частичное подмножество
на множестве реальных систем, составленных из растений
и животных. Все живые системы обладают следующими харак-
теристиками:
'>а) они являются открытыми системами;
\б) они используют входы — пищу или горючее — чтобы вос-
станавливать собственную энергию и исправлять повреждения
в своей организационной структуре;
, в) они обладают уровнем сложности, превышающим некото-
рый минимум;
, г) они содержат генетический материал, состоящий из
дезоксирибонуклеиновой кислоты (ДНК), предположительно
полученной из некоторой изначальной ДНК, общей для всей
жизни;
д) они в значительной степени состоят из протоплазмы, со-
держащей белки и другие специфические органические ком-
поненты;
е) они содержат в себе решающую систему, которая контро-
лирует всю систему и организует взаимодействие ее подсистем;
ж) они содержат также другие специфические подсистемы
или находятся с таковыми в отношениях симбиоза или парази-
тирования, так что последние выполняют для них такие про-
цессы, в которых живые системы нуждаются;
з) эти подсистемы объединены в целостную систему, обла-
дающую свойствами саморегуляции, развития и самовоспроиз-
цедения и имеющую свои цели и задачи;
и) они могут существовать только в определенных условиях
окружения. Любое изменение в их окружении, такое как вариа-
ции температуры, давления, влажности, концентрации кисло-
рода атмосферы или интенсивности радиации, выходящее за
1.2J
СТРУКТУРА ВИОСИСТЕМЫ
17
пределы относительно узкого диапазона, существующего на по-
верхности Земли, вызывает стресс, к которому они не могут
приспособиться.
«Общеизвестно, как трудно определить понятие живого»,—
лучшего комментария к приведенным выше определениям жи-
вых систем, чем это высказывание, принадлежащее известному
американскому специалисту по молекулярной биологии Ф. Крику
[285], дать нельзя. Тем не менее ясно, что многие основные,
фундаментальные свойства живых систем связаны с процессами
управления (самоуправления, саморегуляции), протекающими
в открытых системах — системах, обменивающихся с окружаю-
щей средой веществами и энергией. С этими процессами мы и
будем иметь дело в данной книге.
1.2. Структура биосистемы
Для понимания процессов, протекающих в биосистеме, не-
обходимо учитывать две стороны ее функционирования. Одна
из них связана с тем, что система — открытая: это процессы
получения, накопления, передачи и использования энергии. Эги
процессы обеспечивают возможность сохранения структуры,
рост и выполнение всех специфических функций биологической
системы.
Другая сторона функционирования связана с управлением
энергетическими процессами; она включает восприятие, хране-
ние, переработку и использование информации. Информацион-
но-управляющие механизмы в системе определяют, какие энер-
гетические процессы и с какой скоростью происходят в ней.
Наиболее общей задачей управляющих систем организма
является сохранение его энергетической основы, создание бла-
гоприятных условий для постоянства ее функционирования при
меняющихся условиях внешней среды. «Кибернетические меха-
низмы для того и существуют, чтобы обеспечить стабилизацию
и сохранение энергетической части организма» [216, стр. 8].
Представление биологической системы в виде двух взаимо-
действующих компонент — энергетической и управляющей —
представляет собой основу системного подхода к анализу струк-
туры биосистем.
На рис. 1.1 показана схема потоков энергии и информации
в организме животного [216]. Энергетическая компонента орга-
низма обозначена как МС (метаболическая система), а управ-
ляющая часть представлена в виде блока регуляторных меха-
низмов (генетическое и физиологическое управление) и блока
эффекторов. Основным элементом схемы здесь является, безус-
ловно, метаболическая система — совокупность процессов об-
мена веществ, происходящих в клетках животного организма.
18
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
(ГЛ. 1
Одной из главных функций метаболической системы является
снабжение систем и органов организма энергией. Ниже
(разд. 1.4) мы приведем обычно даваемое определение метабо-
лизма, а еще ниже — в разд. 6.4 и 6.7 — коснемся ряда важных
свойств метаболических процессов.
На этом рисунке, как и во всем дальнейшем изложении,
мы используем широкие двойные стрелки для обозначения пере-
дачи одновременно не одного, а целой группы сигналов, ве-
ществ и т. д.
Субстраты, отбресв/
I Энергетическая
i—‘ L- потребность
мс F; .. zzzr
Реакции
Метаболиты
Энергоснабжение
Управление
(иены, молекулы, нервные импутсы)
Стимулы
Рис. 1.1. Схема потоков энергии и информации в организме животного. МС—метаболиче-
ская система, Р —регуляторные механизмы, включающие элементы генетического и физио-
логического управления. Все части организма потребляют метаболическую энергию, по-
ставляемую МС. Целью управления является поддержание стационарного неравновесия
в энергетической системе организма и сохранение ее структуры.
Поскольку эта схема содержит одновременно множество
управляющих цепей, выделим в ней три главных контура свя-
зей. Структура организма поддерживается механизмами генети-
ческого управления. Получая от остальных систем энергию
и информацию (в виде метаболитов — продуктов обмена ве-
ществ, а в период становления организма — в виде гормонов
роста), генетическая система управляет процессами синтеза не-
обходимых веществ и поддерживает жизнедеятельность осталь-
ных систем организма (рис. 1.2, а). Процессы в генетической
части протекают достаточно медленно: характерными дЛя нее
временами являются долгие интервалы времени, связанные
с процессами роста, становления организма и его старения,
процессы регенерации тканей и т. п.
Поведенческие реакции организма осуществляются системой
физиологического управления (рис. 1.2,6).
Функционирование эффекторов и других систем, потребляю-
щих энергию (мышцы, органы обоняния, осязания, работающие
1Л
СТРУКТУРА БИОСИСТЕМЫ
19
ткани внутренних органов), приводит к увеличению расхода
вещества и энергии. Этот расход должен компенсироваться
увеличением темпов синтеза веществ и энергии в метаболиче-
ской системе. На этом уровне физиологическое управление обе-
спечивает адекватное снабжение всех подсистем в соответствии
с возникающими потребностями (рис. 1.2,в).
Иты, молекулы,
(синтез, поЗдеужание жизнеЯеяшыюспнф
а.)
Стимулы РварферешсЩиЯ
б)
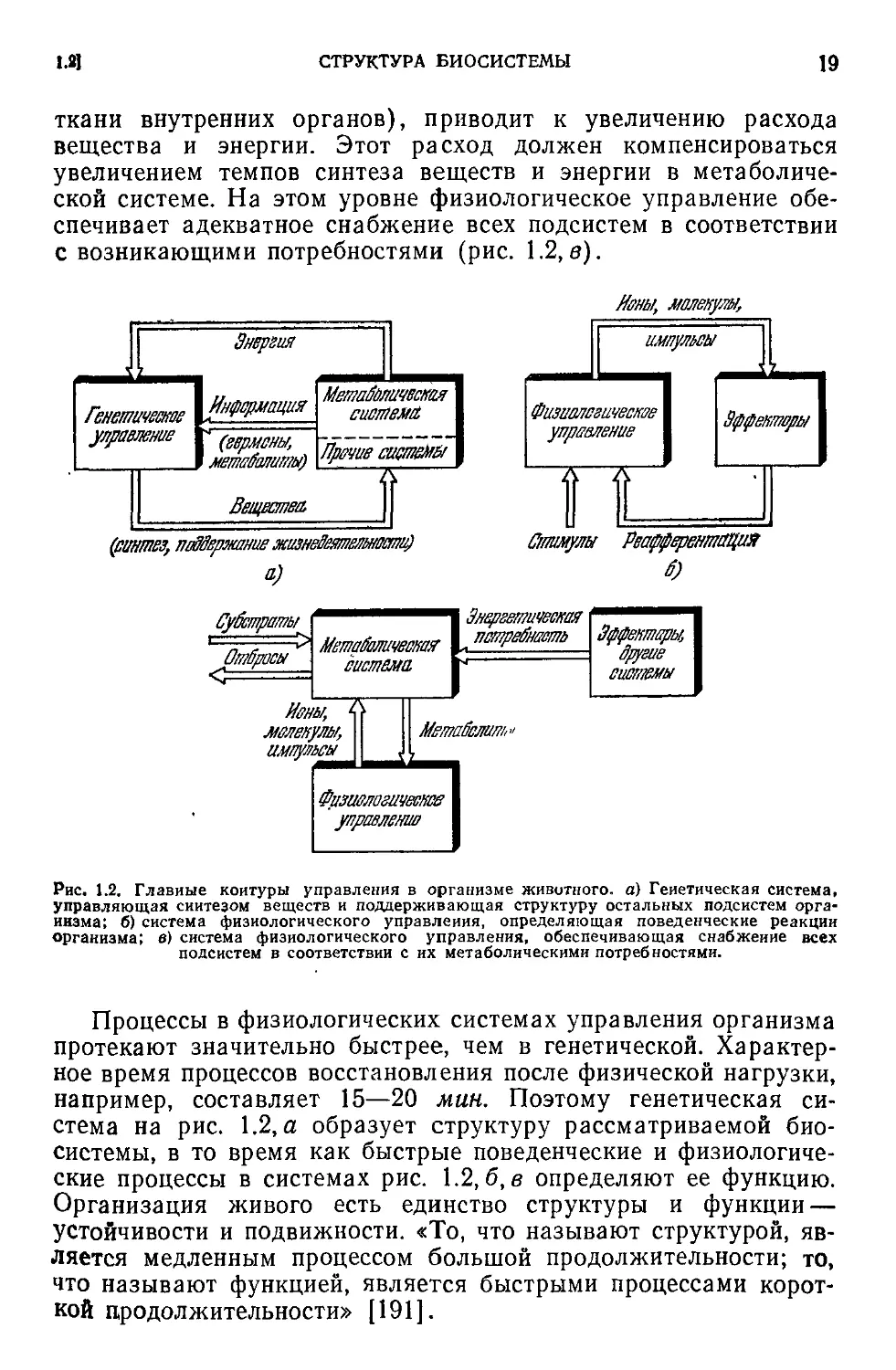
Рис. 1.2, Главные контуры управления в организме животного, а) Генетическая система,
управляющая синтезом веществ и поддерживающая структуру остальных подсистем орга-
низма; б) система физиологического управления, определяющая поведенческие реакции
организма; в) система физиологического управления, обеспечивающая снабжение всех
подсистем в соответствии с их метаболическими потребностями.
Процессы в физиологических системах управления организма
протекают значительно быстрее, чем в генетической. Характер-
ное время процессов восстановления после физической нагрузки,
например, составляет 15—20 мин. Поэтому генетическая си-
стема на рис. 1.2, а образует структуру рассматриваемой био-
системы, в то время как быстрые поведенческие и физиологиче-
ские процессы в системах рис. 1.2, б, в определяют ее функцию.
Организация живого есть единство структуры и функции —
устойчивости и подвижности. «То, что называют структурой, яв-
ляется медленным процессом большой продолжительности; то,
что называют функцией, является быстрыми процессами корот-
кой продолжительности» [191].
20
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
[ГЛ. 1
Таким образом, само строение биосистемы отражает здесь
две главнейшие ее характеристики — процессы обмена веществ
(открытый характер) и процессы управления.
1.3. Принцип биологического эпиморфизма
Как бы ни отличались организмы по своей физико-химиче-
ской конструкции, отношения между их биологическими свой-
ствами (такими как перемещение организма, процессы получе-
ния и использования пищи, выделительные процессы) остаются
всегда одними и теми же, т. е. инвариантны для всех организ-
мов. Однако перечисленные функции у высших организмов по
сравнению с такими же функциями у низших включают намного
больше элементарных компонент, элементарных процессов.
«Значит, природа соответствия между процессами или биологи-
ческими свойствами высших и низших организмов такова, что
одному элементарному процессу у низшего организма соответ-
ствует несколько элементарных процессов у высшего» [172].
Поскольку такое соответствие называется в математике эпи-
морфизмом, сам принцип эпиморфного отображения организмов
друг на друга был назван принципом биологического эпимор-
физма.
Принцип биологического эпиморфизма был предложен Н. Ра-
шевским [172, 347, 348] для сравнительного анализа аналогич-
ных процессов или свойств у разных типов организмов.
Нас интересуют сейчас процессы управления в биосистемах.
Поэтому мы можем сформулировать принцип биологического
эпиморфизма применительно к управляющим механизмам био-
логических систем. Механизмы управления, обеспечивающие
основные биологические свойства высших и низших организмов,
таковы, что они находятся в эпиморфном отношении: одному
элементарному механизму управления у низших организмов
соответствует множество элементарных механизмов у высшего.
В дальнейшем изложении этот принцип окажется для нас
очень и очень важным. Дело в том, что в процессе накопления
сложности сами биосистемы могут приобретать новые регуля-
тивные свойства. Рассмотрим, например, способность различных
биосистем к адаптации. В простой системе адаптация связана
с изменением относительно небольшого числа элементарных
процессов, и приспособительные возможности системы ограни-
чены тем, что эти процессы поочередно выходят на некоторые
граничные, предельные режимы. Когда «последний» из процес-
сов выйдет на такой режим, способность системы к дальнейшей
адаптации окажется исчерпанной. Очевидно, чго в сложной
эпиморфной системе возможности к адаптации могут быть зна-
чительно большими, чем в простой.
I.4J
ОТКРЫТЫЕ СИСТЕМЫ
21
Аналогичным образом можно связать с биологическим эпи-
морфизмом и возникновение гомеостатических свойств в си-
стеме, понимаемых как малая чувствительность ее управляющих
механизмов к вариациям внешней среды. В соответствующих
разделах (7.4—7.7) мы подробно рассмотрим этот вопрос. Сей-
час же мы лишь вкратце коснемся его. Когда некоторая функ-
ция в простой системе обеспечивается малым количеством ме-
ханизмов, на долю каждого из них приходится относительно
большая нагрузка. В ответ на внешнее возмущение реакция
каждого механизма оказывается ощутимой — механизмы чув-
ствительны к изменениям, происходящим в среде. В эпиморфной
системе, где таких механизмов много больше, их одновременное
включение может привести к уменьшению нагрузки, приходя-
щейся на каждый элементарный процесс, каждый регулятор.
В результате чувствительность каждого из регулирующих меха-
низмов к возмущениям становится меньше — возникают гомео-
статические свойства.
Ниже мы будем говорить о позднем (в эволюционном плане)
возникновении гомеостаза у организмов (разд. 2 4), а при рас-
смотрении вопросов моделирования гомеостаза специально оста-
новимся на соотношении «сложность — качество». Сложные про-
цессы возможны только в системе, богатой связями между эле-
ментами Эпиморфизм, возникающий в ходе эволюции, как раз
и способствовал возникновению сложной системы связей между
элементарными компонентами организмов; на определенном
этапе развития этих связей «хватает» не только на то, чтобы
поддерживать жизнедеятельность системы, обеспечивая нужные
ей темпы потребления вещества и энергии, но и на достижение
малой чувствительности переменных внутренней среды к внеш-
нему окружению.
Интересно, что аналогичные процессы наслоения качеств по
мере накопления структурной сложности происходят и в слож-
ных технических системах. Сначала возникает обычная устой-
чивость, затем способность системы хорошо функционировать
при помехах — помехоустойчивость, затем управляемость и, на-
конец, способность системы к самоорганизации [220].
1.4. Открытые системы
Понятие открытой системы впервые было введено в обиход
биологической науки Л. фон Берталанфи в 1932 г. [271]. Для
такой системы характерно, что в нее постоянно извне вводятся
вещества, которые внутри системы подвергаются различным ре-
акциям. В результате процессов анаболизма в системе возни-
кают компоненты более высокой сложности, утилизируемые ор-
ганизмом. Одновременно идут процессы катаболизма, конечные
22
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
(ГЛ. 1
продукты которого выводятся из системы. Совокупность процес-
сов анаболизма и катаболизма называется метаболизмом.
Одной из наиболее характерных черт открытых систем яв-
ляется то, что в них достигается состояние подвижного равно-
весия. При этом структура системы остается постоянной, но это
постоянство сохраняется в процессе непрерывного обмена и дви-
жения составляющих ее веществ.
На рис. 1.3 показана простая схема такой открытой системы.
На ее входе — поток
Рис. 1.3. Простая схема от-
крытой системы. Входной
поток веществ обозначен че-
рез yi, выходной — через у2.
поступающих веществ — субстратов у\, на
выходе — поток продуктов катаболизма
У2- Внутри системы вещества А в ходе
обратимых реакций преобразуются в ве-
щества В. Разумеется, в любой биологи-
ческой системе пути метаболических ре-
акций гораздо сложнее: они идут парал-
лельно друг другу, расходятся или схо-
дятся. Сама система метаболических ре-
акций обладает в значительной степени ауторегуляторными
свойствами, которые возникают из-за наличия в ней цепей об-
ратных и прямых связей [72, 98].
Пример такой более сложной биохимической системы при-
веден на рис. 1.4 [189]. Обратная связь состоит в угнетении
Рис. 1.4. Система метаболических реакций, обладающая ауторегуляторными свойствами.
В системе имеется обратная (/) и прямая (2) связи. Прямая связь представляет собой
активацию предшественником субстрата, которая приводит к увеличению скорости по-
требления продукта Sn в спобочных» реакциях, катализируемых ферментом Е^. Обратная
связь — угнетение фермента Е, конечным продуктом Sn. Изменение скорости у^ образова-
ния исходного субстрата So является возмущением в системе.
ключевого фермента Е\ конечным продуктом Sn, образующимся
из исходного субстрата So через ряд промежуточных форм
Si, ..., Sn-i- Ферменты, катализирующие промежуточные реак-
ции, обозначены через Еь Ег, •••> Е„; М\ — вещества, способные
активировать или угнетать фермент Ef, Mk — такие же вещества
для фермента Е*.
Прямая связь представлена механизмом активации фер-
мента веществом — предшественником субстрата. Соединение S„
1.5]
ПАССИВНОЕ И АКТИВНОЕ УПРАВЛЕНИЕ
23
служит субстратом для фермента Ek, чувствительного к активи-
рующему действию исходного субстрата So. Изменение скоро-
сти у0 образования исходного субстрата So действует как внеш-
нее возмущение на Sn — при увеличении скорости уо возрастает
и концентрация Sn. Однако активация фермента Ек субстратом
So компенсирует 'возмущающее действие уо за счет увеличения
скорости потребления вещества Sn в реакциях, катализи-
руемых Ek.
В биохимических системах эти механизмы дублируются, «за-
параллеливаются» более инерционными механизмами. Таковы,
например, репрессия синтеза ферментов Е\, ..., Еп конечными
продуктами (обратная связь) и индукция синтеза этих фермен-
тов исходным субстратом So.
1.5. Пассивное и активное управление
в живых системах
Совместное действие большого числа механизмов прямых
и обратных связей, регулирующих активность и скорость син-
теза ферментов, позволяет поддерживать концентрации конеч-
ных продуктов почти на неизменном уровне при изменении
внешних возмущений в довольно широких пределах. Механизмы
рассмотренного типа как бы «встроены» в регулируемую си-
стему, поэтому для них используется термин «внутреннее управ-
ление'».
Внутреннее управление является пассивным. Это означает,
что в реально существующей системе поддержание стационар-
ного равновесного состояния или возникновение характерных
ответов системы на внешнее возмущение не требует какой-либо
метаболической работы. Пассивные механизмы регуляции свой-
ственны не только биохимическому уровню организации биоси-
стем. Поддержание нормальной пространственной ориентации
у рыб обеспечивается пассивным механизмом регуляции — центр
плавучести и центр тяжести не совпадают, так что при отклоне-
нии осевой плоскости рыбы от вертикальной возникает крутя-
щий момент, возвращающий тело в нормальное положение.
Пассивное управление в системе возникает как результат
взаимодействия элементов', составляющих «саму систему».
Л. фон Берталанфи предложил для такого типа управления
термин «динамическое взаимодействие» или «первичная регу-
ляция». Типичным примером пассивного управления в системах
является взаимодействие между веществами, находящимися в
системе, и потоками этих веществ, циркулирующими внутри
системы и пересекающими границы системы со средой.
Пассивные механизмы управления лежат на грани, отделяю-
щей управляемые системы от неуправляемых.
24
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
1ГЛ. 1
В теории управления решение о том, относить или не отно-.
сить данную систему к классу управляемых систем, выносится
на основании наличия в описании этой системы обратных свя-
зей. Если обратная связь существует, система считается управ-
ляемой. Пассивные системы в теории управления обычно отно-
сятся к неуправляемым. Рассмотрим, например, такую техниче-
скую систему, как мост. При действии на него нагрузки мост
прогибается, и в результате деформации возникают силы, урав-
новешивающие действие нагрузки. Можно сказать, что мост
обладает механизмами сохранения своей формы. Поскольку,
однако, механизмы упругой деформации работают пассивно,
только за счет энергии, поступающей извне, подобные объекты
в теории управления не представляются в виде систем с обрат-
ной связью и считаются поэтому неуправляемыми. Простое
и подробное освещение этого вопроса имеется во вводных раз-
делах книги [235].
В теоретической биологии складывается противоположная
традиция: пассивные механизмы относятся к системам управле-
ния. Термин «пассивная система управления» введен У. Р. Эшби
[259] и вошел в учебники по теории управления для биологов
[130]. Пассивный характер управления в живых системах озна-
чает, что «если заданы всеобщие химические и физические за-
коны, а также компоненты и организация живой системы..., то
для поддержания стационарного состояния или возникновения
характерных ответов системы не требуется какой-либо метабо-
лической работы» [216, стр. 15].
Одна из причин того, что пассивные регулирующие меха-
низмы в живых системах относятся к системам управления, со-
стоит в следующем. Если в неживой природе можно указать
множество объектов, в которых присутствуют одни лишь пас-
сивные механизмы, как в упомянутом выше примере с мостом,
то в живых системах пассивные механизмы управления сплошь
и рядом составляют только «нижний этаж» иерархических
управляющих структур, на который накладываются следую-
щие этажи, составленные уже из активных механизмов управ-
ления.
Как мы увидим, многие интересные черты гомеостатического
поведения биологических систем получают простое объяснение,
если биосистема представляется в виде элемента, охваченного
«пассивной» связью, которая затем дублируется активными об-
ратными связями (см. разд. 7.4, 7.8). Для физиологических си-
стем, например, взаимодействие пассивной и активной регуля-
ции приводит к возникновению характерной зависимости пере-
менных внутренней среды от окружающих условий — так
называемой регуляционной или гомеостатической кривой
(см. разд. 2.3 и 8.2).
1.5J
ПАССИВНОЕ И АКТИВНОЕ УПРАВЛЕНИЕ
25
Если пассивное управление в равной мере присутствует
в живой и неживой природе, то активное управление в природе
специфично только для биосистем.
Активное управление требует от биосистемы метаболических
затрат, поскольку управление осуществляется специальными
механизмами, обособленными от элементов системы, которыми
они управляют. В этом смысле активное управление является
внешним, подобно тому, как пассивное управление является
внутренним.
Внешние управляющие механизмы «накладываются» на
внутренние, образуя вместе с ними единую систему замкнутых
контуров управления, комплекс прямых и обратных связей
в системе. В живых системах, в отличие от рассмотренных
выше технических систем, пассивное управление также может
быть представлено в виде обратной связи.
Внешние механизмы управления организованы так, что лю-
бое возмущение и на систему, изменяющее ее состояние, обна-
руживается (либо непосредственно, либо по тем эффектам в си-
стеме, которые обусловлены этим возмущением) при помощи
специальных чувствительных элементов. Сигнал с чувствитель-
ного элемента поступает на внешние элементы контура, где об-
рабатывается, в результате чего вырабатывается ответный
управляющий сигнал, противодействующий влиянию возмуще-
ния на систему.
Рис. 1.5. Наложение контура внешнего активного управления на внутренние механизмы
управления. Возмущение v изменяет реакцию А < * В; эффект D воспринимается чув-
ствительным элементом S, сигнал с которого поступает в устройство обработки информа-
ции М. Исполнительный элемент (эффектор) Л формирует управляющий сигнал, парирую-
щий действие возмущения v; у\—входной поток веществ, Уг— выходной поток.
На рис. 1.5 показано, как простой внешний контур актив-
ного управления накладывается на внутренний механизм пас-
сивного управления в метаболической структуре биосистемы
[216]. В реальных биосистемах имеется множество механизмов
пассивного управления, на которые накладываются многочис-
ленные механизмы активного управления. На рис. 1.6 представ-
лена схема наложения активных механизмов регуляции на пас-
сивные механизмы окислительных процессов в системе
26
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
(ГЛ. 1
физиологического уровня — системе кислородного снабжения
организма. Показанное на рисунке изображение является ин-
терпретацией в терминах активного и пассивного управления
схемы, приведенной в [9, 10].
Активные механизмы регуляции, по всей видимости, широко
распространены и на клеточном уровне. Хотя фактически о тех
специфических стадиях регуляторных процессов, где происхо-
дит потребление метаболической энергии, известно очень не-
много, нет сомнений в том, что в метаболизме клетки «должны
Активные
механизмы
регуляции,
>2
ЦНС
(кора
головного
мозга)
Втша/м
процессы
в тканях
(пассивная регуляция)
Хвморецепторы
Рис. 1.6. Наложение активных механизмов регуляции на пассивные механизмы окисли-
тельных тканевых процессов. Система окислительных процессов в тканях обладает само-
регуляторными свойствами, основу которых составляют пассивные механизмы регуляции
Механизмы активной регуляции составляют высший этаж управляющей системы. В схеме
рассматриваются следующие активные механизмы регуляции: целенаправленные изменения
дыхательного объема, жизненной емкости легких, объема резервного воздуха, глубины
вдоха, частоты дыхания, ударного объема сер ща, частоты пульса, объемной скорости
кровотока, величины сопротивления сосудов, количества гемоглобина в крови, величины
кислородной емкости крови и, наконец, эритропоэза.
существовать и будут, несомненно, обнаружены какие-то спе-
циальные пункты ' подачи энергии в регуляторный процесс»
[98]. Возможно, одним из технических аналогов активных ре-
гулирующих процессов клеточного и субклеточного уровня яв-
ляется механизм случайного поиска [169]. Своеобразная модель
механизма активной регуляции в субклеточных структурах была
исследована в работе [59]. Этот механизм представляет собой
реализацию случайного поиска на уровне митохондриальных
структур нейронной клетки, целью которого является «оптими-
зация энергетического режима нервной клетки».
Иногда считается, что активные процессы управления в био-
системах реализуются единственным видом отрицательной об-
ратной связи — отрицательной обратной связью по рассогласо-
ванию. Такой точки зрения придерживается, в частности,
Л. фон Берталанфи [31, 271]: «динамическое взаимодействие
в открытых системах применяется к первичным регуляциям, как
в клеточном метаболизме», а «модель обратной связи отражает
I в)
ТЕМПЫ И УРОВНИ
27
то, что может быть названо вторичной регуляцией в метабо-
лизме и других областях, т. е. регуляцию путем предварительно
установленных механизмов и фиксированных связей» [271,
стр. 163].
Из рассмотренных примеров, однако, уже сейчас ясно, что
такая простая модель механизмов регуляции, как отрицатель-
ная обратная связь по рассогласованию в одноконтурной си-
стеме, может быть лишь самым первым приближением при ис-
следовании активной регуляции в биосистемах. Ниже мы по-
дробнее рассмотрим этот вопрос — он будет в центре внимания
при моделировании гомеостаза и открытых систем (см. разд. 7.4).
Сейчас же мы еще раз отметим, что эта модель является не
единственным и не всегда лучшим способом описания слож-
ных систем и механизмов управления в биологических объ-
ектах.
1.6. Темпы и уровни
При описании живых систем все время приходится иметь
дело с двумя типами величин. Один из них связан с количе-
ствами веществ, имеющимися в различных частях биосистемы
или окружающей ее среды. Примерами здесь могут быть кон-
центрации различных субстратов или ферментов в клетках
(биохимические системы), содержание кислорода, концентрация
молочной кислоты в тканях, содержание гемоглобина в крови,
температура тела (физиологические системы), содержание
биомассы в некотором объеме или количестве животных дан-
ного вида на единице площади (экологические системы). Пере-
менные, определяющие эти величины, называются уровнями'.
уровень сахара крови, уровень численности популяции и т. д.
Другой тип переменных связан с изменением уровней; эти
переменные характеризуют динамику процессов накопления, по-
требления, усвоения, переработки и элиминации веществ.
Иногда они обозначаются как скорости процессов — скорость
синтеза ферментов, скорость выделения веществ и т. д. Мы бу-
дем употреблять для их описания общий термин темп-, темп
синтеза ферментов, темп поступления кислорода в ткани, темп
роста популяции.
Термины «темпы» и «уровни» становятся в последнее время
общепризнанными при моделировании не только биологических,
но и экономических и промышленных систем. Их английские
аналоги — flow rate и level — подробно рассматриваются в об-
зоре [344] применительно к моделированию биосистем, в част-
ности, экологических. Определение этих же терминов при мо-
делировании систем произвольного типа дано Дж. Форресте-
ром [224].
28
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
1ГЛ. I
Обе группы терминов — уровни и темпы — мы далее будем
твердо употреблять только для обозначения двух упомянутых
групп переменных величин. Уровни отражают достигнутое со-
стояние системы; темпы — ее активность, интенсивность проте-
кания в ней процессов. При таком использовании терминов не
метаболизма», «уровень синтеза ве-
е «темп обмена веществ», или, еще
более строго, «темп потребления
кислорода», «темп выделения угле-
кислоты». Близким аналогом темпа
является установившийся термин
«скорость», поэтому будем говорить
и так — «скорость химической ре-
акции», «скорость синтеза про-
дукта».
Размерности переменных, харак-
теризующих уровни вещества и
энергии, отражают либо их количе-
ства (г; мг; л; см3 и др.), либо ко-
личества, приходящиеся на единицу
площади или объема (г/см3; мэкв/л;
единиц на гектар; мг% и т. д.). Раз-
мерности темпов обязательно вклю-
чают размерность времени [/“']:
г/сек, л/мин, мэкв/л-час и т. д.
Темпы отражают изменение уров-
ней. Если какое-то вещество возни-
кает и элиминирует в данной систе-
ме в результате нескольких процес-
сов (безразлично какой природы •—
химической или физической), так
что у\, yz, ..., yr-s — темпы поступ-
ления этого вещества в систему или его синтеза внутри нее, а
yrs+i, ..., уг — темпы потребления или удаления его из си-
стемы, то уровень х связан с темпами у\, ..., уг простым урав-
нением
г—S г
Е <1Л>
fe=I s+1
Темпы как бы порождают уровни.
В свою очередь уровни в силу присущих системе закономер-
ностей оказывают регулирующее действие на темпы. Так, ско-
рость диффузии (темп у переноса вещества из одной среды
в другую) зависит от разности уровней Xi и х2 в этих средах:
у = а(х2 — х1),
следует говорить
щества». Точнее
«уровень
80
ВО
40
го
о
ю
го
SO N
-го
-40
-во -
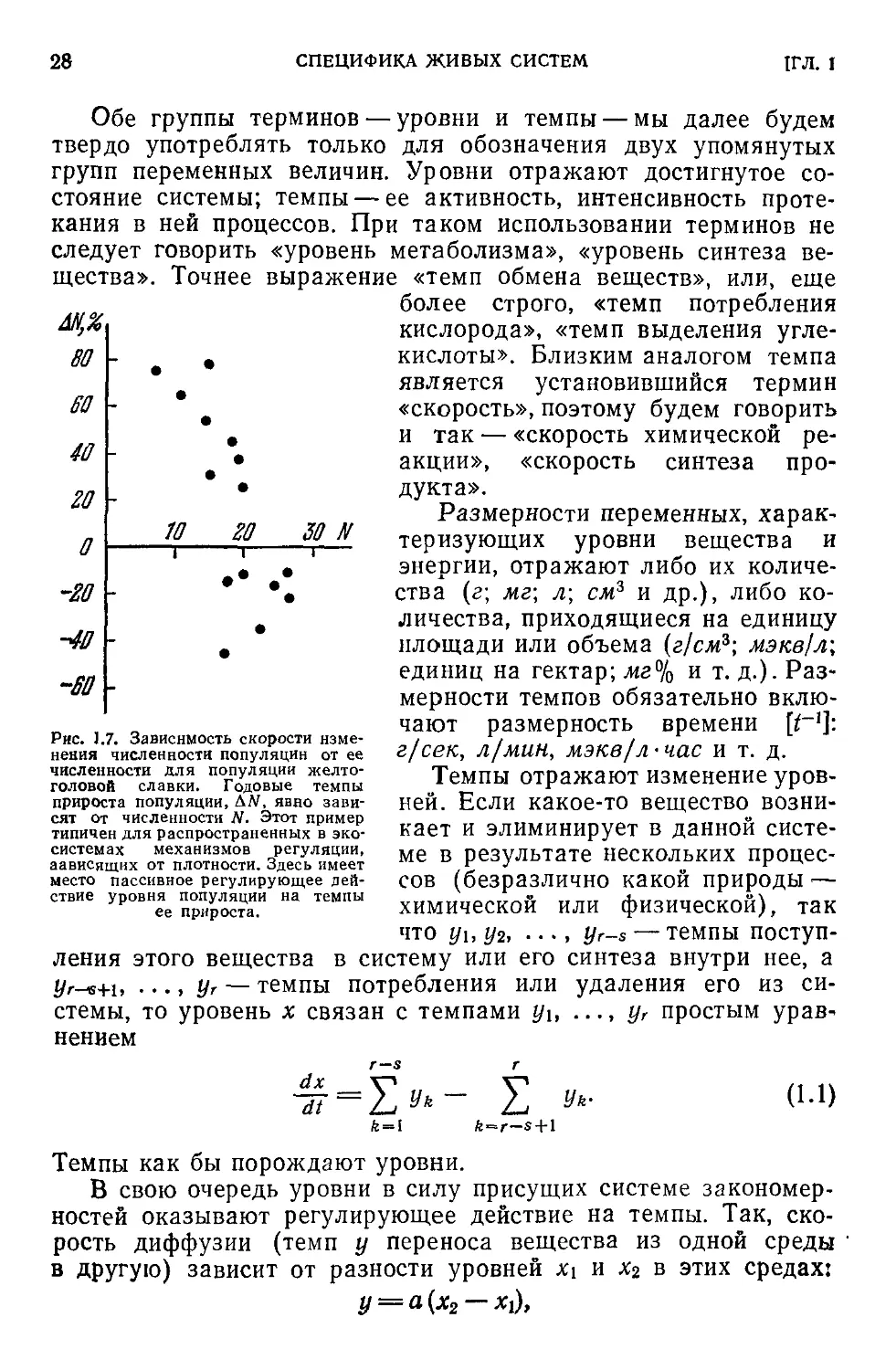
Рис. 1.7. Зависимость скорости изме-
нения численности популяции от ее
численности для популяции желто-
головой славки. Годовые темпы
прироста популяции, АЛТ, явно зави-
сят от численности ЛГ. Этот пример
типичен для распространенных в эко-
системах механизмов регуляции,
зависящих от плотности. Здесь имеет
место пассивное регулирующее дей-
ствие уровня популяции на темпы
ее прироста.
1.61
ТЕМПЫ И УРОВНИ
29
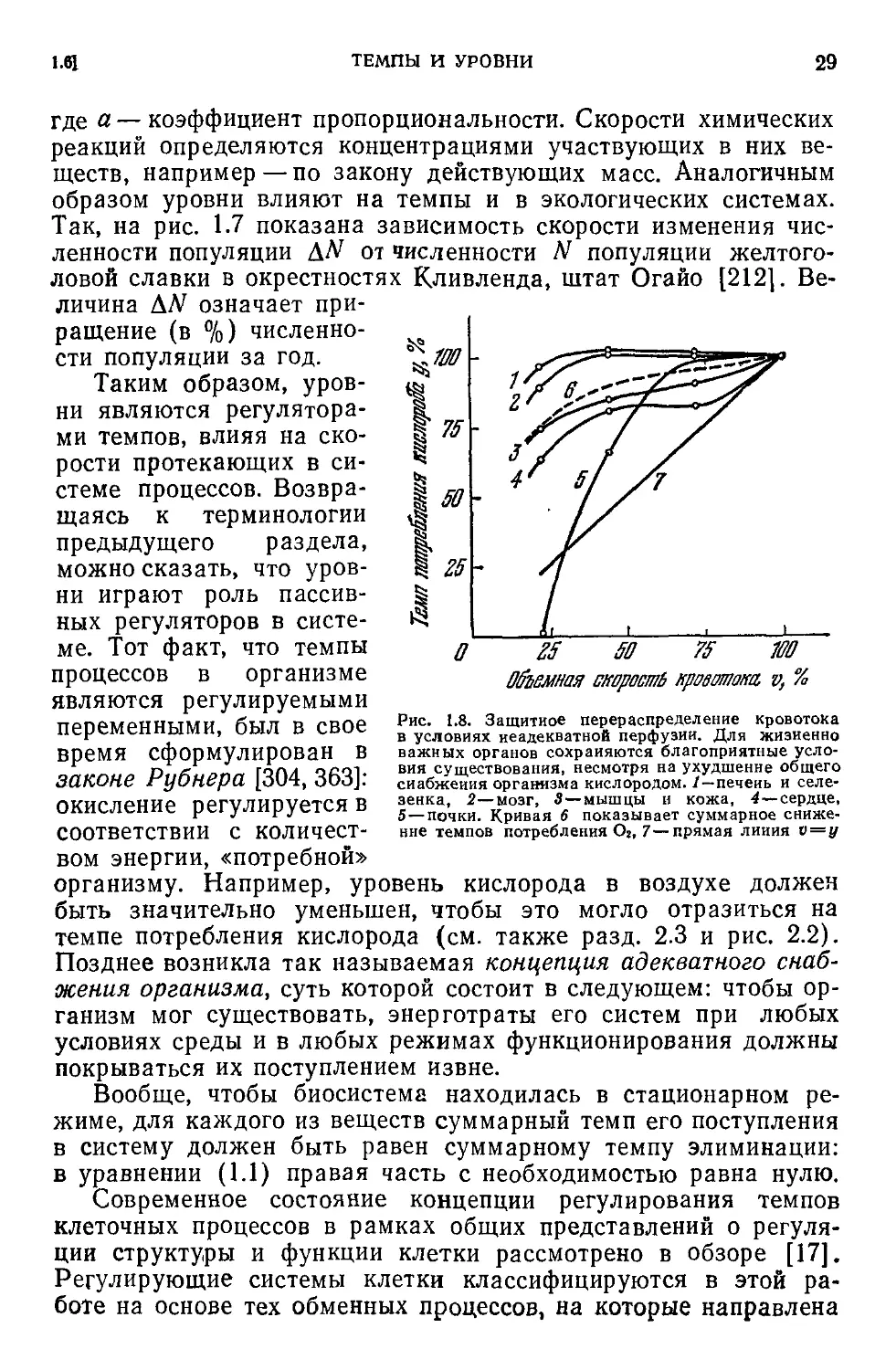
Объемная скорость кровотока, ц %
Рис. 1.8. Защитное перераспределение кровотока
в условиях неадекватной перфузии. Для жизненно
важных органов сохраняются благоприятные усло-
вия существования, несмотря на ухудшение общего
снабжения организма кислородом, /—печень и селе-
зенка, 2—мозг, 3—мышцы и кожа, 4—сердце,
5—почки. Кривая 6 показывает суммарное сниже-
ние темпов потребления Os, 7—прямая линия v=y
где а — коэффициент пропорциональности. Скорости химических
реакций определяются концентрациями участвующих в них ве-
ществ, например — по закону действующих масс. Аналогичным
образом уровни влияют на темпы и в экологических системах.
Так, на рис. 1.7 показана зависимость скорости изменения чис-
ленности популяции Д2У от численности N популяции желтого-
ловой славки в окрестностях Кливленда, штат Огайо [212]. Ве-
личина Д# означает при-
ращение (в %) численно-
сти популяции за год.
Таким образом, уров-
ни являются регулятора-
ми темпов, влияя на ско-
рости протекающих в си-
стеме процессов. Возвра-
щаясь к терминологии
предыдущего раздела,
можно сказать, что уров-
ни играют роль пассив-
ных регуляторов в систе-
ме. Тот факт, что темпы
процессов в организме
являются регулируемыми
переменными, был в свое
время сформулирован в
законе Рубнера [304, 363]:
окисление регулируется в
соответствии с количест-
вом энергии, «потребной»
организму. Например, уровень кислорода в воздухе должен
быть значительно уменьшен, чтобы это могло отразиться на
темпе потребления кислорода (см. также разд. 2.3 и рис. 2.2).
Позднее возникла так называемая концепция адекватного снаб-
жения организма, суть которой состоит в следующем: чтобы ор-
ганизм мог существовать, энерготраты его систем при любых
условиях среды и в любых режимах функционирования должны
покрываться их поступлением извне.
Вообще, чтобы биосистема находилась в стационарном ре-
жиме, для каждого из веществ суммарный темп его поступления
в систему должен быть равен суммарному темпу элиминации:
в уравнении (1.1) правая часть с необходимостью равна нулю.
Современное состояние концепции регулирования темпов
клеточных процессов в рамках общих представлений о регуля-
ции структуры и функции клетки рассмотрено в обзоре [17].
Регулирующие системы клетки классифицируются в этой ра-
боте на основе тех обменных процессов, на которые направлена
30
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
{ГЛ. I
регуляция: регуляция скорости протекания процессов синтеза,4
процессов распада веществ, энергообразования и, наконец, дви-
жения (транспортных процессов).
При ухудшении внешних условий в биосистеме активизи-
руются механизмы, направленные на поддержание темпов по-
требления необходимых веществ. Так, на рис. 1.8 приведены
кривые, характеризующие темп потребления кислорода у раз-
личными системами организма при искусственном кровообра-
щении при снижении темпов кровоснабжения (так называемого
объема перфузии) от значения 4,5 л/мин, принятого за 100%.
Согласно концепции защитного перераспределения кровотока
при снижении объема перфузии жизненно важные органы в ор-
ганизме находятся в избирательно благоприятных условиях по
сравнению с другими тканями [48], темп потребления кислорода
в них поддерживается на относительно высоком уровне.
Мы специально остановились на том, что темпы в биосисте-
мах можно рассматривать как управляемые переменные, что
цель системы управления можно формулировать в терминах
поддержания постоянства темпов. Существуют два математи-
чески формализованных подхода, при которых основное внима-
ние уделяется темпам процессов, происходящих в живых систе-
мах, потокам веществ, проходящих через биосистему. Один из
них связан с рассмотрением термодинамических моделей откры-
тых систем, другой — с так называемыми компартментальными
моделями открытых систем. Мы кратко ознакомимся с первым
из иих в разд. 1.8; компартментальным моделям посвящена гл. 6.
1.7. Иерархия целей в живых системах
Процессы, происходящие в реальных биологических объек-
тах, представляют собой управляемые (или самоуправляемые)
процессы в том смысле, что важнейшую роль в них играют
различного рода механизмы регуляции. Для содержательного
описания результата управления в биосистемах в биокибернети-
ческой литературе часто применяется понятие о цели управ-
ления.
Термин «цель механизмов управления в биосистеме», исполь-
зуемый в таком контексте, не предполагает каких-либо созна-
тельных или намеренных действий в анализируемых системах
(даже говоря о целостном организме, мы ограничиваемся иссле-
дованием физиологического уровня регуляции, не включая в
рассмотрение его поведенческие реакции).
Под целью в биологической системе мы будем подразумевать
то конечное состояние, в которое она приходит в силу своей
структурной организации [168] или ожидаемый результат ее
функционирования [И, 127]. Как обычно, говоря о достижении
1.7]
ИЕРАРХИЯ ЦЕЛЕЙ В ЖИВЫХ СИСТЕМАХ
31
цели в биосистеме, мы имеем в виду выполнение некоторых
соотношений между ее переменными. Если, например, меха-
низмы регуляции в организме обеспечивают равенство темпов
потоков веществ, с одной стороны, поступающих в его системы,
а с другой — расходуемых в них, то можно сказать, что цель
этих механизмов регуляции — обеспечение адекватного снабже-
ния или, другими словами, сохранение стационарного неравно-
весного состояния систем организма.
Таким образом, проблема анализа целей в биосистемах в на-
шем контексте оказывается весьма близкой задаче изучения
характеристик и свойств биосистем в их «конечных» состояниях.
Однако, характеристики конечного состояния, достигаемого в
живых системах, настолько разнообразны, что вопрос о целях
в теории биосистем все еще недостаточно ясен.
Живые системы — многоцелевые в том смысле, что при их
анализе можно указать множество полезных свойств, которые
достигаются в различных ситуациях и при разнообразных внеш-
них воздействиях. Анализ целей в живых системах усложняется
и тем, что эта проблема может обсуждаться на разных уров-
нях организации, и рассмотрение часто не связывается с конк-
ретными условиями окружения биосистем и условиями их
функционирования. Относительно мало внимания уделяется и
таким вопросам, как иерархия целей в живой природе, хотя
в сложных многоуровневых структурах, характерных для био-
систем, анализ глобальных целей системы и целей ее подсистем
может оказаться полезным и интересным. При построении «де-
рева целей» живой природы можно было бы с успехом исполь-
зовать идеи и результаты исследований сложных технических
систем [73, 93, 127].
Здесь мы рассмотрим только один аспект проблемы целей
в живых системах, связанный с зависимостью результата функ-
ционирования системы от условий окружающей среды. Биоси-
стема при этом будет рассматриваться как целостное структур-
ное образование на данном этапе эволюционного процесса. По-
этому, если считать, что целью живой природы является
получение «эволюционной пластичности и организменной устой-
чивости» [248], то мы будем иметь дело только со второй компо-
нентой этой цели — устойчивостью сложившейся биосистемы.
В частности, мы не будем касаться представлений о наилучшей
приспособленности биосистем к среде как максимальной ско-
рости размножения организмов, что восходит еще к идеям
Дарвина (см., например, [77, 161, 190, 233]).
Остановимся сначала на некоторых основных представлениях
о целесообразных свойствах биосистем в рассматриваемом
аспекте. В наиболее общем виде цель биосистемы можна пред-
ставить, вероятно, как «сохранение и продолжение ее жизни и
32
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
[ГЛ. 1
жизни коалиции аналогичных систем» [82]. «Нам представ'-
ляется, что в качестве такой цели живой природы в целом
можно принять стремление к самосохранению», — писал
А. А. Ляпунов [НО].
Но достижение цели самосохранение определяется, во-пер-
вых, способностью обеспечить свою жизнь необходимыми для
этого средствами и, во-вторых, защитой выполняемых жизнен-
ных функций от неблагоприятных внешних воздействий [248].
Эти две стороны сохранительных свойств тесно связаны между
собой и лежат в основе всех классических представлений о само-
сохранении биосистем.
Первая сторона сохранительных свойств биосистем обычно
связывается с представлениями Клода Бернара. Клод Бернар
писал, что организм имеет внешнее окружение и внутреннюю
среду, и подчеркивал, что «именно постоянство «внутренней
среды» является условием свободной и независимой жизни,...
все жизненные механизмы, как бы разнообразны они ни были,
имеют только одну цель, сохранение постоянства условий жизни
во внутренней среде...» ([270], см. также [3]). Эти представле-
ния легли в основу концепции постоянства внутренней среды
организмов и биосистем вообще — концепции гомеостаза.
Другая сторона сохранительных свойств биосистем связана
с необходимостью обеспечить адекватный приток веществ и
энергии в биосистему извне, чтобы уравновесить их расход
внутри системы, обеспечивая тем самым стационарное неравно-
весное состояние.
Оперируя понятиями темпов и уровней, рассмотренными в
предыдущем разделе, можно уточнить, что речь идет, с одной
стороны, о сохранении постоянства условий внутренней среды —
о поддержании неизменных уровней вещества и энергии внутри
системы, а с другой — о регулировании темпов потоков вещества
и энергии, которыми система и ее элементы обмениваются со
средой.
В биокибернетике обычно основное внимание уделяется по-
стоянству внутренней среды; регулированию потока веществ
отводится относительно малая роль, и зачастую проблема регу-
лирования скоростей и темпов потоков в биосистемах выпадает
из поля зрения исследователей.
«Сохранение высокой степени организации у живых систем
возможно лишь благодаря их способности извлекать энергию
извне и управлять ее расходованием внутри», — этими словами
начинается книга Дж. Милсума [130]. Следующая фраза, каза-
лось бы, должна содержать сведения о том, что живые системы
могут эффективно регулировать эти потоки энергии, должным
образом обеспечивая свои потребности. Однако вместо этого мы
читаем: «В связи с этим естественно ожидать, что все живые
17)
ИЕРАРХИЯ ЦЕЛЕЙ В ЖИВЫХ СИСТЕМАХ
33
системы «умеют» эффективно регулировать свою внутреннюю
среду». Недооценка процессов управления темпами вообще ха-
рактерна для многих работ в области биокибернетики.
В то же время в классической физиологии регуляции темпов
поступления веществ придавалось большое значение, начиная
с работ И. М. Сеченова. «Посмотрим теперь на регуляцию при-
хода с расходом... Механизмов, поддерживающих равновесие
между этими двумя величинами, в животном теле очень
много», — писал он в 1860 г. [193, стр. 529].
Большое внимание в биокибернетике уделяется еще одному
желаемому результату живых систем — высокому качеству их
функционирования — эффективности, экономичности, надежно-
сти. Часто для этих свойств биосистем используется терминоло-
гия оптимальных систем [82, 175, 230, 231]. Анализу проблемы
оптимальности биосистем ниже посвящена отдельная глава,
сейчас же мы рассмотрим только некоторые соотношения иерар-
хического порядка между тремя выделенными выше целями —
обеспечением стационарного неравновесного состояния или, что
то же самое, адекватного снабжения системы (регуляция тем-
пов потоков вещества и энергии), обеспечением постоянства
внутренних условий — гомеостаза (регуляция уровней вещества
и энергии) и, наконец, достижением высокого качества функ-
ционирования биосистем.
Для простоты мы ограничимся только стационарными режи-
мами работы биосистем, когда условия окружения фиксируются
на различных уровнях, а система достигает уравновешивания
со средой. Рассмотрим теперь, какие цели и в какой степени
могут быть достигнуты в тех или иных условиях окружающей
среды.
Впервые сопоставление двух целей биосистем, связанных с
управлением темпами потоков вещества и с регуляцией постоян-
ства внутренней среды, было сделано, вероятно, И. М. Сечено-
вым в 1860 г. Обсуждая «регуляцию прихода с расходом», он
писал: «Приход и расход минеральных веществ равны между
собой, и это есть, конечно, ручательство за то, что содержание
этих веществ в организме остается постоянным» [193, стр. 527—
529].
Конечно, постоянство внутренней среды достигается с боль-
шим трудом, чем просто равенство притока и расхода, и пред-
ставляет поэтому более высокий результат в системе. Совокуп-
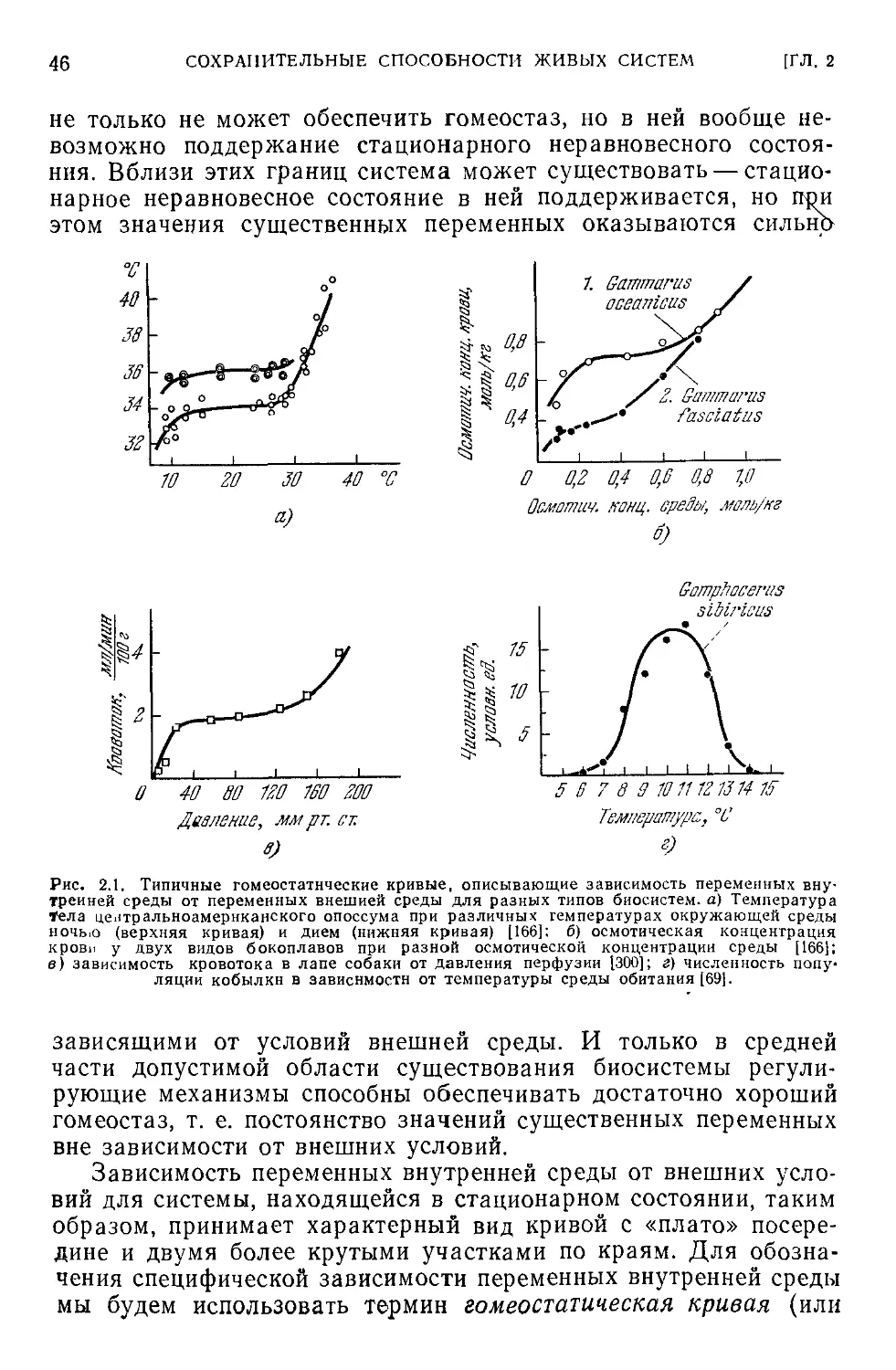
ность точек равновесия, построенная для некоторой переменной
внутренней среды ( какого-либо уровня вещества) в зависимости
от внешних условий, имеет характерный вид платообразной
кривой с пологим участком посередине и двумя более круты-
ми— по краям. Эта кривая целиком располагается в области,
где равенство притока и расхода (стационарность) обеспечено.
34
специфика живых систем
|ГЛ I
Такая кривая называется иногда регуляционной характеристи-
кой [166]. Ниже мы специально рассмотрим эту зависимость,
которую будем называть также гомеостатической кривой
(разд, 2.3, рис. 2.1). Платообразный характер этой кривой сви-
детельствует, что постоянство внутренней среды обеспечивается
в системе в относительно узком диапазоне внешних условий по
сравнению со всем диапазоном, в котором поддерживается ра-
венство «притока с расходом», т. е. при ухудшении условий
окружающей среды уравновешивание со средой наступает при
все больших и больших сдвигах внутреннего состояния био-
системы. И нарушается гомеостаз уже тогда, когда равенство
притока и оттока еще сохраняется. Можно сказать поэтому, что
сохранение равенства темпов притока и расхода веществ яв-
ляется целью первого порядка в биосистеме; эта цель дости-
гается в первую очередь и поддерживается в более широком
диапазоне внешних условий. Поэтому приведенное выше выска-
зывание И. М. Сеченова можно перефразировать следующим
образом: равенство темпов прихода и расхода вещества является
необходимым условием постоянства уровней вещества в системе.
В свою очередь постоянство уровней является достаточным усло-
вием равенства темпов притока и расхода.
Так, в биохимических системах регуляция направлена на
поддержание скоростей протекающих в них реакций [17, 72, 103,
289]. Однако эта регуляция приводит к тому, что и концентра-
ция веществ часто оказывается постоянной в некотором диапа-
зоне изменения условий внешней среды — система оказывается
«эластичной» в изменяющихся внешних условиях [72].
Нарушение энергетического снабжения биосистемы (невоз-
можность обеспечить равенство темпов прихода и расхода ве-
ществ), а не нарушение условий внутренней среды является
пределом жизни биосистем. «Необходимым критерием границы
физиологического приспособления служит нарушение законов
обмена», — писал Э. С. Бауэр [26].
Когда баланс темпов притока и расхода веществ обеспечен,
появляется возможность достижения цели второго порядка —
создания таких условий функционирования внутри системы, ко-
торые не зависели бы от изменения окружения — гомеостаза. По
словам С. Лема, «гомеостаз — так ученые называют стремление
к равновесию, т. е. к существованию вопреки изменениям,—
создал известковые и хитиновые скелеты, противодействующие
силе тяжести, обеспечил подвижность посредством ног, крыльев
и плавников, облегчил пожирание с помощью клыков, рогов, че-
люстей и пищеварительных систем и в то же время защитил от
пожирания панцирями и камуфляжами и дошел на этом пути
освобождения от внешней среды до регуляции, обеспечивающей
постоянство температуры тела...» [102, стр. 24].
1.71
ИЕРАРХИЯ ЦЕЛЕЙ В ЖИВЫХ СИСТЕМАХ
35
Наличие гомеостаза открывает (в эволюции), возможность
дальнейшего совершенствования свойств управляющих механиз-
мов в живых системах. В частности, гомеостаз позволяет внут-
ренним клеткам и органам живых систем работать в наилучших
возможных условиях и не расходовать энергию и управленче-
ские ресурсы на создание и функционирование специальных
сохранительных систем внутри организмов [117].
Нарушение гомеостаза автоматически влечет за собой изме-
нение режимов внутренних органов и систем и, возможно, ухуд-
шение их характеристик. Таким образом, цель «гомеостаз», ве-
роятно, оказывается проще достижимой, чем получение наилуч-
ших свойств биосистемы. Гомеостаз является необходимым усло-
вием высокого качества биосистем.
Этот вопрос, однако, не был до последнего времени предме-
том сколько-нибудь широкого обсуждения, хотя на отдельных
уровнях организации биосистем он и поднимался. Так, в эколо-
гии «зрелое сообщество... очень часто способно буферить
(тормозить) физическое окружение в гораздо большей степени,
чем молодое. Таким образом, с эволюционной точки зрения
достижение уровня стабильности или гомеостазиса более важно,
чем просто повышение продуктивности» [148].
Подводя итог сказанному, можно установить следующий по-
рядок достижения целей в биосистеме для случая установив-
шихся режимов, возникающих при различных фиксированных
условиях окружающей среды. Наиболее важной и достигаемой
в первую очередь целью является поддержание стационарного
неравновесного состояния. Эта цель может быть сформулиро-
вана как поддержание равенства темпов потоков вещества и
энергии, поступающих в систему и покидающих ее. После этого '
возникает возможность поддержания постоянства внутренней
среды — гомеостаза, что является целью второго порядка. На-
конец, если обе эти цели достигнуты, возможно прогрессирую-
щее улучшение качества процессов в системе. В этой ситуации
можно говорить об энергетической эффективности, оптимальной
конструкции, получении максимальной надежности функциони-
рования и т. д. Однако достижение максимально высоких пока-
зателей в биосистемах является уже целью третьего порядка по
сравнению с поддержанием стационарного неравновесия и
гомеостаза.
По крайней мере, очевидно, что такие качества, как макси-
мальная надежность системы, означающая способность к безот-
казному функционированию в широком интервале условий окру-
жения, относится именно к наилучшим внешним условиям (эти
условия часто' называют адекватными [82]). В неадекватных
условиях система вынуждена адаптироваться к среде, и это
ведет к определенному напряжению регуляторных механизмов
36 специфика ЖИВЫХ СИСТЕМ фл. 1
и перестройке их функционирования [2, 20, 219]. Аналогичное
рассуждение применимо не только к надежности, но и к другим
свойствам биосистем, в обеспечении которых принимает участие
множество механизмов. Исчерпание ресурсов этих механизмов
при ухудшении условий среды может повлечь за собой снижение
качества процессов в системе. Жизнедеятельность в неадекват-
ных условиях требует включения дополнительных механизмов
реакций, и система переходит сначала в состояние напряжения,
а затем — патологии [185, 197, 201]. В отличие от нормального
режима, который «обеспечивает оптимальные условия жизне-
деятельности (в некоторых пределах, выработанных в процессе
эволюции), аварийный режим преследует цель прежде всего
обеспечить самосохранение системы при действии патологиче-
ского фактора» [22].
Рис. 1.9 иллюстрирует иерархию трех типов целей в живых
системах. Пусть vt и Уг — некоторые переменные, описывающие
Рис. 1.9. Иерархия целей в биосистеме. Самосохранение системы связано с обеспечением
стационарного неравновесия — равенства темпов расхода и притока веществ, что является
иерархически наиболее важной целью, обеспечиваемой в системе в первую очередь.
Другой целью, иерархически менее важной, является поддержание постоянства внутренней
среды. Первая цель достигается в более широкой области изменения условий окружающей
среды, вторая — в более узкой, Заштрихована область Й в которой система мо-
жет иметь иаилучшие (оптимальные) характеристики. Пунктиром показана область, в кото-
рой система может существовать только ограниченное время; tii н Oj — переменные внешней
среды.
стационарные свойства окружающей среды. Эти сигналы будем
рассматривать как постоянные возмущения. Тогда на плоскости
(У1, v2) можно выделить область й5, где регуляторные меха-
низмы системы обеспечивают существование стационарных ре-
жимов, и меньшую область Йй, в которой поддерживается гомео-
стаз. Наконец, условно можно выделить и еще более узкую
область Й„, для которой характерно наиболее высокое качество
функционирования биосистемы в указанном выше смысле. Ра-
зумеется, границы между этими областями можно выделять
лишь приближенно.
1.8]
ТЕРМОДИНАМИЧЕСКИЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
37
По мере ухудшения условий в системе происходит «отказ»
от иерархически менее важных целей, связанных с получением
«оптимальных» характеристик (область Й^). Дальнейшее ухуд-
шение условий приводит и к потере гомеостатических свойств
(выход изображающей точки за пределы области Й^), а затем
и к потере способности системы обеспечить стационарный режим
при выходе за пределы области й8. В этом случае жизнедеятель-
ность системы может поддерживаться лишь некоторое ограни-
ченное время за счет запасов вещества и энергии, имеющихся
в системе, и расход которых временно позволяет сохранять
равенство темпов расходования веществ в местах их траты и
темпов поступления веществ к этим местам из «депо» внутри
системы. Эта область показана на рис. 1.9 пунктиром. Отметим,
что под свойствами среды могут пониматься не только ее физи-
ческие или химические характеристики, но и переменные, описы-
вающие биологическое окружение данной биосистемы.
В дальнейшем будут рассматриваться в основном два типа
целей биосистем — достижение равенства темпов потоков при-
хода и расхода и обеспечение постоянства внутренней среды.
В биокибернетической литературе чаще всего рассматри-
ваются цели второго и третьего порядка — гомеостаз и получе-
ние наилучших характеристик. Цель первого порядка — управ-
ление темпами потоков, достижение стационарного неравнове-
сия — исследуется в рамках теории термодинамических систем,
а в последнее время — в компартментальных моделях открытых
систем. Этим моделям посвящена ниже гл. 6, а сейчас мы крат-
ко рассмотрим некоторые идеи термодинамического подхода к
исследованию живых систем.
1.8. Термодинамические модели живых систем
Для описания биосистем с помощью термодинамических мо-
делей используются основные понятия термодинамики — такие,
как термодинамическое равновесие и неравновесие, а также
термодинамические функции, характеризующие состояния си-
стемы и их возможные изменения. Достаточные для наших це-
лей сведения по термодинамике живых систем можно почерп-
нуть, например, в [98, 202].
Из термодинамических функций для описания биосистем
наиболее часто используется энтропия S, определяемая с по-
мощью дифференциального уравнения
dS = J£_, (1.2)
где dS — приращение энтропии, dQ — приращение количества
тепла в системе при бесконечно малом квазистатическом
38
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
[ГЛ. 1
изменении ее состояния, а Т — абсолютная температура. Из вто-
рого принципа термодинамики следует, что в изолированной си-
стеме могут протекать только такие процессы, которые сопро-
вождаются возрастанием (или сохранением постоянного значе-
ния) энтропии.
В статистической физике используется другая трактовка
энтропии, определяемая как мера термодинамической вероят-
ности w состояния системы:
5=/г1пау, (1.3)
где k — постоянная Больцмана, £=1,38-10-16 эрг/град. При
этом под термодинамической вероятностью понимается число
микросостояний системы (например, число различных распреде-
лений молекул в газе), при которых реализуется данное макро-
состояние. Замкнутая термодинамическая система приходит в
состояние термодинамического равновесия с максимальной эн-
тропией S:
AS = О, S = max,
где AS — изменение энтропии системы.
Эффективность описания процессов в открытых системах с
помощью методов термодинамики открытых систем [162а] зави-
сит от того, насколько открытая система близка к термодинами-
ческому равновесию. Если рассматриваемая открытая система
близка к термодинамическому равновесию, а значения ее пере-
менных мало отличаются от равновесных, то поведение энтропии
определяется теоремой Пригожина:
В стационарном состоянии прирост энтропии, обусловленный
протеканием необратимых процессов внутри системы, имеет
минимальное из возможных и положительное значение.
Баланс энтропии в открытой системе представляется в виде
AS = ASf + ASe, (1.4)
где AS/, ASe — прирост энтропии за счет процессов внутри си-
стемы и приток энтропии извне, соответственно. В зависимости
от соотношения AS, и AS« величина AS может быть как поло-
жительной, так и отрицательной. В стационарном состоянии
AS = 0.
Согласно теореме Пригожина прирост энтропии внутри си-
стемы в единицу времени в стационарном состоянии принимает
максимальное значение.
Если потоки вещества и энергии через открытую систему
велики, а динамические значения переменных сильно отли-
чаются от равновесных, то в общем случае такие системы
нельзя описать термодинамическими методами; отдельные части
1.8]
ТЕРМОДИНАМИЧЕСКИЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
39
таких систем могут составлять подсистемы, близкие к равнове-
сию, но наряду с ними в системе действуют и «нетермодинами-
ческие» силы [180]. Ответ на вопрос, к какому классу открытых
систем относятся те или иные биологические системы, может
дать только эксперимент. Применение неравновесных термоди-
намических моделей в ряде интересующих нас задач рассмот-
рено, например, в монографии [96].
Для практически важных задач организменного и более вы-
соких уровней методы термодинамики открытых систем при-
меняются довольно редко. Более распространены методы моде-
лирования, при которых биологический объект представляется
в виде нескольких относительно изолированных и взаимодей-
ствующих между собой количеств вещества — компартментов.
Сейчас мы проанализируем простую открытую систему, состоя-
щую из двух пространственно разделенных отсеков и представ-
ляемую в виде двух компартментов, используя для ее описания
энтропийные характеристики. Этот пример позволит читателю
почувствовать близость таких внешне столь далеко отстоящих
друг от друга методов описания биосистем, как компартменталь-
ный анализ и методы термодинамики открытых систем.
Рассмотрим сначала замкнутую систему—ящик с двумя
отделениями. В перегородке имеется отверстие, через которое
находящиеся в ящике частицы
в отделение (рис. 1.10, а).
Такие модели используют-
ся иногда для объяснения
статистического характера
энтропии [202]. Эту систему
можно рассматривать и как
простой аналог компартмен-
тальной модели с двумя
компартментами, если сово-
купность частиц в каждом
отделении представить как
компартмент. Пусть в ящи-
ке находится всего шесть
частиц.
Определим энтропию, от-
могут переходить из отделения
а) б)
Рис. 1.10. Простая модель системы, содержа-
щая два компартмента. а) Закрытая система;
б) открытая система. Равновесное состояние
в системе а) не совпадает со стационарным
неравновесием в системе б). Эта модель может
рассматриваться как простейший случай ком-
партмеитальной системы, имеющей два ком-
партмеита; первый компартмент — совокупность
частиц в левом отделении системы, второй —
в правом.
вечающую каждому из воз-
можных состояний системы. Обозначим состояние символом
(т, п — т), где т— число частиц в левом отделении (первом
отсеке), а п — общее число частиц.
, Тогда удобно воспользоваться статистической трактовкой
энтропии (1.3), определяя число различных комбинаций частиц,
при которых реализуется данное макросостояние системы. Коли-
чество комбинаций, приводящее к состоянию (т, п — т), есть
40
СПЕЦИФИКА ЖИВЫХ СИСТЕМ
(ГЛ. I
число сочетании из п по т:
fl t
п — I / \ । •
и т ! (п — т) I
В табл. 1.1 приведены значения w и In w для всех возможных
состояний системы на рис. 1.10, а. Энтропия системы S полу-
чается для каждого состояния умножением величины, стоящей
в последнем столбце, на постоянную Больцмана.
Таблица 1.1
(1-5)
Состояние системы Число комбина- ций W In w Состояние системы Число комбика ЦИЙ W In w
(6,0) 1 0,00 (2,4) 15 2,71
(5,1) 6 1,79 (1,5) 6 1,79
(4,2) 15 2,71 (0,6) 1 0,00
(3,3) 20 2,99
Допустим, что частицы внутри системы движутся не полно-
стью случайно: перемещение их происходит от более «населен-
ного» отсека к менее «населенному». Пусть выравнивание числа
частиц подчиняется закону диффузии; поток частиц пропорцио-
нален разности концентраций в первом и втором отсеках, т и
(п — т), соответственно:
У\2 — а[т — (п — т)] = a (2m — п), (1.6)
где а — коэффициент пропорциональности. Для простоты можно
положить а = 0,5.
Тогда движение частиц будет приводить к тому, что в си-
стеме реализуются все более вероятные комбинации — энтропия
системы возрастает. В стационарном режиме система предпочти-
тельно находится в состояниях с максимальной энтропией, когда
п
т=-^.
Дополним теперь систему двумя внешними потоками частиц:
в первый отсек ящика извне поступает поток с заданным темпом
2 частицы в единицу времени; из второго отсека частицы с та-
ким же темпом уходят в окружающую среду (рис. 1.10,6). Обо-
значим потоки символом у с двойным индексом; среде присвоим
индекс 0. Тогда потоки из среды в отсек 1 и из отсека 2 в среду
равны, соответственно,
Ут ~ 2, у2о = 2.
Рассмотрим дискретную последовательность событий в си-
стеме при исходном состоянии (4,2). После притока двух частиц
1 81 ТЕРМОДИНАМИЧЕСКИЕ МОДЕЛИ ЖИВЫХ СИСТЕМ 41
извне в первый отсек состояние системы будет (6,2). Затем
согласно (1.6) из первого отсека во второй перейдут две ча-
стицы— система окажется в состоянии (4,4), а после оттока
двух частиц из второго отсека в среду — в состоянии (4,2). Пе-
реходы (4,2)->(6,2) и (4,4)—>(4,2) вызваны обменом со средой,
переход (6,2)->(4,4) — внутренними необратимыми процессами
в системе.
Вычислим приращение энтропии AS, и ASe в (1.4). Приток
энтропии извне составляется двумя компонентами: ASel и ASe2,
отвечающими указанным двум переходам системы (4,2)->(6,2)
и (4,4)->(4,2). Тогда
ASel = k In Ct - k In Сб = 0,626,
ASe2 = 61nCe- k In Cs = 6(2,71 - 4,25) =-1,546
и
ASe = ASel + ASe2 = —0,926.
Найдем теперь AS,. Для перехода, вызванного внутренними
процессами, имеем
AS,- = 6 In Се - 6 In Cl = 0,926.
Следовательно, общее изменение энтропии за цикл AS =
= ASe + AS, = 0, т. е. система находится в стационарном ре-
жиме. Начальное и конечное состояния системы совпадают:
четыре частицы слева, две справа. Это стационарное состояние
является неравновесным, оно поддерживается за счет непре-
рывного протекания через систему потока частиц. Энтропия ста-
ционарного неравновесного состояния (4,2), равная 2,716, мень-
ше максимального значения 2,996, отвечающего стационарному
равновесию (3,3). Система, предоставленная самой себе, спо-
собна совершить работу за счет выравнивания числа частиц в
обоих отсеках ящика.
В этом простом примере темп производства энтропии внутри
системы определяется только физическим процессом переноса
частиц; в более сложных случаях такой прирост энтропии яв-
ляется следствием также и необратимых химических реакций.
Приток же извне и в сложных системах обеспечивается прежде
всего за счет физических процессов перемещения вещества.
ГЛАВА 2
СОХРАНИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
2.1. Концепция самосохранения и живые системы
Обычно в литературе по теоретической и математической
биологии начало анализа сохранительных свойств биосистем
связывается с принципом Ле-Шателье-. «Если на систему, нахо-
дящуюся в устойчивом равновесии, подействовать извне, изме-
няя какое-нибудь из условий, определяющих положение равно-
весия, то равновесие смещается в том направлении, при котором
эффект произведенного воздействия уменьшается». Этот прин-
цип был сформулирован французским химиком А. Ле-Шателье
в 1887 г. для термодинамических систем. После того, как немец-
кий физик К. Браун показал, что это положение является след-
ствием второго начала термодинамики, иногда стали упо-
треблять название «принцип Ле-Шателье—Брауна».
Однако идея самосохранения объектов реального мира зна-
чительно старше, и ее возникновение относится к временам
Возрождения. Великий философ древности Аристотель учил, что
каждому элементу отвечает свое естественное место во Вселен-
ной, к которому тела стремятся по природе с тем, чтобы там
достигнуть покоя. Например, тела падают на землю потому, что
именно там их естественное место. По Аристотелю «движение
всего направлено к одному и тому же, если ничего не помешает»
114, стр. 45].
После открытия Ньютона законов движения стало ясно, что
такого естественного места не существует; все тела сохраняют
свое состояние, пока и поскольку внешние силы не вынуждают
к его изменению. Изменение состояния, каким бы оно ни было,
осуществляется внешними силами. Тела «сопротивляются» лю-
бому насильственному изменению своего состояния (см., напри-
мер, [174]).
В этих условиях Спиноза сформулировал свою концепцию
самосохранения-. «...Никакая вещь не имеет в себе ничего, через
что она могла бы уничтожиться..., наоборот, она... противодей-
ствует всему тому, что может уничтожить ее существование.
Следовательно, насколько возможно и насколько от нее зависит,
она стремится пребывать в своем существовании» ([196, стр. 88],
см. также [292]).
2.2]
ГОМЕОСТАЗ
43
Принцип Ле-Шателье — Брауна, таким образом, можно
трактовать как применение общего принципа самосохранения
к термодинамическим системам.
В сходных терминах был сформулирован американским фи-
зиологом У. Кэнноном и принцип самосохранения живых систем-.
«В открытой системе, такой как наши организмы, состоящие
из нестабильного материала и подверженные непрерывному
воздействию возмущений, само постоянство служит доказатель-
ством существования агентов, действующих или готовых к дей-
ствию, чтобы поддержать это постоянство. Если состояние
остается устойчивым, то это происходит потому, что любая
тенденция к его изменению автоматически вызывает увеличение
эффективности фактора или факторов, противодействующих
этому изменению» [278, стр. 281].
Важность концепции самосохранения для понимания процес-
сов в живых системах, таким образом, очевидна. Подобные пред-
ставления в различных сферах исследования биосистем исполь-
зуются в различных формах. Так, часто принципы сохранения
принимают гомеостатический характер, в той или иной мере
близкий принципу Ле-Шателье [28, 278]. В других случаях
связь концепций гомеостаза с принципом Ле-Шателье отри-
цается [271]. Интересная связь идей самосохранения с фило-
софскими концепциями отражения и отображения рассмотрена
в работе [192].
2.2. Гомеостаз
Одним из центральных элементов при анализе естественных
защитных сил биосистем, обеспечивающих их самосохранение,
является понятие гомеостаза?? Первоначально термин гомеостаз
был введен У. Кэнноном для описания способности организмов
поддерживать постоянство внутренней среды.
«Постоянные условия, которые поддерживаются в организме,
можно было бы назвать равновесием. Это слово, однако, имеет
довольно точное значение для относительно простых физико-
химических состояний в замкнутых системах, где уравновеши-
ваются известные силы. Координированные физиологические
процессы, которые поддерживают большинство установившихся
состояний в организме, настолько сложны и специфичны для
живых существ, включая, возможно, мозг и нервы, сердце, лег-
кие, почки и селезенку, работающие совместно, что я предложил
специальное обозначение для этих состояний — гомеостазис. Это
слово не предполагает чего-то установленного и неподвижного,
застывшего. Оно означает состояние — состояние, которое мо-
жет меняться, но которое относительно постоянно» [278,
стр. 24].
44
сохрапительные способности живых систем
[ГЛ. 2
Согласно концепции У. Кэннона при наличии угрозы со сто-
роны внешней среды в системах организма включаются агенты,
уменьшающие эту опасность. Корректирующие механизмы дей-
ствуют в основном через специальную часть нервной системы,
функционирующую как регуляторный механизм. Вследствие
этой регуляции в организме обеспечивается, «во-первых, запа-
сание веществ как средство обеспечения соответствия между
спросом и предложением и, во-вторых, изменяются скорости не-
прерывно протекающих в организме процессов» [278, стр. 285].
Эта концепция была последовательно применена У. Кэнноном
для анализа процессов сохранения постоянства содержания
воды и солей в крови, поддержания адекватного кислородного
снабжения, гомеостаза сахара, белков, жиров и кальция крови,
гомеостаза нейтральности крови и постоянства температуры
тела.
Позже понятие гомеостаза стало формулироваться в более
компактной форме: гомеостазом обычно называют сохранение
постоянства внутренней среды организма при наличии возмуще-
ний во внешней среде.
Отметим также, что термином «гомеостаз» иногда обозна-
чают и саму систему, обладающую гомеостатическими свой-
ствами. Такое использование этого термина восходит к работе
[258]. В [269], например, гомеостаз понимается как «определен-
ного рода буферирующие или защитные устройства внутри био-
логических систем, предохраняющие их от внешних дезоргани-
зующих сил».
Приведем также определение гомеостаза, данное в стандарт-
ном английском медицинском словаре [359]: «Гомеостаз (1)
состояние равновесия в живом организме, относящееся к раз-
личным функциям и химическому составу жидкостей и тканей,
например, к температуре, частоте пульса, давлению крови, содер-
жанию воды, сахара крови и т. д., (2) процесс, посредством
которого это равновесие поддерживается».
Согласно определению У. Кэннона гомеостаз — это свойство
целостного организма, целостной биосистемы. Другую позицию
занимает Л. фон Берталанфи: «Следует уяснить, что термин
«гомеостаз» может употребляться двояко. Он используется либо
в его первоначальном смысле, предложенном Кэнноном и иллю-
стрируемом примерами поддержания температуры тела и других
физиологических переменных с помощью механизмов обратной
связи, либо в другом смысле, который имеют в виду, а именно,
как синоним для органической регуляции вообще» [31, стр. 43].
Далее Берталанфи настаивает на использовании термина «го-
меостаз» как синонима термина «отрицательная обратная
связь».
2.3]
ГОМЕОСТАТИЧЕСКАЯ КРИВАЯ
45
Мы придерживаемся другой точки зрения, считая понятие
гомеостаза более широким и связанным с сохранительными спо-
собностями целостной биосистемы. «Клод Бернар, привлекший
внимание к стабилизации «внутренней среды», и Уолтер Кэннон,
исследовавший гомеостатические функции вегетативной нервной
системы, черпали силу своих концепций из того, что в наши
дни рассматривалось бы как анализ систем», — писал Т. Уотер-
мен [216].
Это целостное свойство биологических объектов обеспечи-
вается сложной системой управляющих механизмов — прямых
и обратных связей. Комплекс таких связей поддерживает отно-
сительное постоянство одновременно всех существенных пере-
менных в организме. Трудно представить себе ситуацию, когда
переменные одной системы поддерживались бы на постоянном
уровне, обеспечивая жизнедеятельность соответствующей систе-
мы, а гомеостаз других систем был бы полностью нарушен.
Ведь согласно определению У. Р. Эшби [259] под существен-
ными переменными понимаются такие переменные организма,
которые имеют важное значение для выживания организма и
тесно связаны между собой, так что значительное отклонение
одной из них рано или поздно приведет к значительным откло-
нениям остальных и к гибели организма. Ясно поэтому, что
равновесное состояние в биосистеме может быть достигнуто
только при одновременном поддержании относительного по-
стоянства всех ее существенных переменных. В этом смысле
и вся система регулирующих механизмов должна рассматри-
ваться как единый регулятор, обеспечивающий сохранение и
гомеостаз целостной системы. Когда же мы говорим о поддер-
жании постоянства какой-либо отдельной переменной — напри-
мер, о температурном гомеостазе или гомеостазе сахара кро-
ви, — мы должны отдавать себе отчет в том, что такой гомеостаз
возможен лишь в рамках общего гомеостаза целостной биологи-
ческой системы.
2.3. Гомеостатическая кривая
Как бы хорошо ни были организованы регуляторные меха-
низмы биосистемы, возможности их тем не менее ограничены.
Для каждой биосистемы существует область условий внешней
среды, в которой эти механизмы могут обеспечивать гомеостаз
системы. Температура, влажность, интенсивность радиации, со-
держание газов в атмосфере, наличие в окружающей среде
пищи и воды — вот некоторые из факторов, определяющих об-
ласть гомеостаза системы. Для каждого типа биосистем суще-
ствуют некоторые предельные — минимальные и максималь-
ные — значения этих факторов, при выходе за которые система
46
СОХРАПИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
не только не может обеспечить гомеостаз, но в ней вообще не-
возможно поддержание стационарного неравновесного состоя-
ния. Вблизи этих границ система может существовать — стацио-
нарное неравновесное состояние в ней поддерживается, но при
этом значения существенных переменных оказываются силыю
Осмотич. конц. среЯы, мт/кг
Девление, мм рт. ст.
в)
Рис. 2.1. Типичные гомеостатические кривые, описывающие зависимость переменных вну-
тренней среды от переменных внешней среды для разных типов биосистем, а) Температура
Тела центральноамериканского опоссума при различных температурах окружающей среды
ночью (верхняя кривая) и днем (нижняя кривая) [166]: б) осмотическая концентрация
кров» у двух видов бокоплавов при разной осмотической концентрации среды [166];
в) зависимость кровотока в лапе собаки от давления перфузии [300]; г) численность попу
ляции кобылкн в зависимости от температуры среды обитания [69].
Температуре, °C
Д
зависящими от условий внешней среды. И только в средней
части допустимой области существования биосистемы регули-
рующие механизмы способны обеспечивать достаточно хороший
гомеостаз, т. е. постоянство значений существенных переменных
вне зависимости от внешних условий.
Зависимость переменных внутренней среды от внешних усло-
вий для системы, находящейся в стационарном состоянии, таким
образом, принимает характерный вид кривой с «плато» посере-
дине и двумя более крутыми участками по краям. Для обозна-
чения специфической зависимости переменных внутренней среды
мы будем использовать термин гомеостатическая кривая (или
2.3]
ГОМЕОСТАТИЧЕСКАЯ КРИВАЯ
47
гомеостатическая зависимость). Разумеется, степень выражен-
ности «плато», его ширина сильно варьирует у разных биоси-
стем; чем выше организация биосистемы, чем лучше ее регуля-
тивные способности, тем шире плато и тем более полого может
идти гомеостатическая кривая.
На рис. 2.1 приведены характерные гомеостатические зави-
симости ряда переменных внутренней среды от переменных
внешней среды для разных типов биологических систем. Можно
видеть, что положение кривой, вообще говоря, зависит от ре-
жима функционирования системы. Так, на рис. 2.1, а приведена
зависимость температуры тела центральноамериканского опос-
сума от температуры окружающей среды ночью и днем. Это жи-
вотное ведет ночной образ жизни, и в режиме активности
(ночью) гомеостатическая кривая идет выше.
Заметим, что зависимость кровотока от давления перфузии
(рис. 2.1,в), строго говоря, не является гомеостатической кри-
вой; обычно на таких графиках изображается зависимость пере-
менной типа уровня для внутренней среды от переменной такого
же типа для внешней среды.
В этом смысле не являются го-
меостатическими кривыми и кри-
вые, приведенные на рис. 1.8.
В сравнительной физиологии
для описания платообразных за-
висимостей рассмотренного типа
(рис. 1.8,2.1) употребляются тер-
мины регуляция и конформация.
Эти термины относятся как к
уровням, так и к темпам; поло-
гие и слабо наклонные участки
кривых связываются с регуля-
цией; прямая пропорциональ-
ность трактуется как конформа-
ция [166]. На рис. 2.2 приведен
график, иллюстрирующий кон-
формацию и регуляцию темпа
Напряжение 02, мм рт. ст.
Рис. 2.2. Конформация и регуляция темпа
потребления кислорода у животных с не-
зависимым дыханием (карась) при раз-
ных значениях температуры и напря-
жения кислорода в среде [169].
потребления кислорода у животных с независимым дыханием.
При снижении напряжения кислорода рО2 в среде организм уве-
личивает интенсивность функционирования системы дыхания,
сохраняя в некотором диапазоне изменения внешних условий
темп потребления О2 неизменным (регуляция); далее темп по-
требления кислорода падает (конформация).
Характер гомеостатических кривых, так же как и вопросы
регуляции и конформации, оказываются чрезвычайно важными
для понимания процессов управления и построения моделей ме-
ханизмов управления в живых системах.
48
СОХРАПИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
2.4. Сохранительные свойства
Является ли гомеостаз необходимым свойством живых си-
стем? В свое время задавался вопрос, могут ли вообще суще-
ствовать «негомеостатические» системы [276]. Ответ на этот
вопрос, по-видимому, не столь прост, как полагает Дж. Миллер
[333], считающий, что негомеостатических систем существовать
не может, и «по крайней мере одна переменная должна поддер-
живаться на постоянном уровне в стационарном режиме». Рас-
смотрение сохранительных свойств биосистем на разных уровнях
организации Позволяет более обоснованно судить о гомеостазе
и его роли в сохранении различных типов биосистем.
Большинство живых организмов имеют хорошо выраженные
гомеостатические свойства по отношению к основным, опре-
деляющим факторам внешней среды. Однако имеется и множе-
ство форм, не обладающих хорошим гомеостазом вообще. Так,
в отношении теплового режима говорят о гомойотермных и
пойкилотермных животных в зависимости от того, насколько
хорошо выражен у них эффект постоянства температуры. Анало-
гично, регуляция водного обмена и осмотического баланса у жи-
вотных, обитающих в воде, проходит практически все града-
ции — от пойкилоосмотического типа животных до гомойоосмоти-
ческого (см. рис. 2.1,6).
Известно, что гомеостатические свойства организмов не
оказывают существенного влияния на численность видов или
ареал их расселения. Исключением, впрочем, являются случаи
освоения новых территорий, когда еще не сложилось зрелое
экологическое сообщество, и существование в экстремальных
условиях—в «тяжелые» годы или на краях ареалов (пустын-
ные территории, тундры). В этих случаях индивидуальные го-
меостатические свойства могут оказаться существенными для
данного вида животных. В нормальных же условиях выжива-
ние, численность и распространение вида зависит прежде всего
от других факторов — отношений хищничества, соревнования за
жизненные ресурсы и т. д. [266].
Выше при обсуждении целей управления в биосистемах мы
указали на необходимость обеспечения адекватного снабжения
веществами и энергией как на первую, наиболее важную их
цель. Во многих случаях в ходе эволюции биосистемы достигли
дополнительно и другой цели — приобрели свойство относитель-
ной независимости внутренней среды от внешних условий, свой-
ство гомеостаза [278, 356]. Эта точка зрения принадлежит
автору концепции гомеостаза У. Кэннону. «Не предпола-
гается,— пишет он, — что все гомеостатические механизмы бу-
дут найдены у всех форм животных... Амфибия неспособна
сохранять содержание воды и постоянство температуры вне
2.4]
СОХРАПИТЕЛЬНЫЕ СВОЙСТВА
49
зависимости от состояния внешнего мира. Рептилия, высший
тип, не так быстро теряет воду через окружающий воздух, как
амфибия, но подобно амфибии рептилия хладнокровна и, сле-
довательно, ограничена в своей активности окружающим холо-
дом... Гомеостаз, наблюдаемый у млекопитающих, является
продуктом эволюционного процесса» [278, стр. 283].
Можно допустить, следовательно, что первые организмы об-
ладали лишь ограниченными гомеостатическими свойствами
[248, 287, 363]. В то же время необходимое условие существо-
вания живых систем — поддержание стационарного неравновес-
ного состояния или, что то же самое, адекватное снабжение
их энергетической системы веществами — выполнялось без-
условно.
Разумеется, даже «слабо выраженные» гомеостатические
свойства целостной биосистемы всегда выше, чем у аналогич-
ного набора первичных элементов. В любой биосистеме суще-
ствуют интегрирующие механизмы, поддерживающие ее целост-
ность, обеспечивающие обмен веществ (т. е. необходимые для
существования темпы химиче-
ских реакций) и постоянство
структуры самой биосистемы
и ее генетического материала.
Итак, в принципе постоян-
ство внутренней среды не яв-
ляется необходимым условием
существования системы; тер-
модинамическое неравновесие
может поддерживаться и в
том случае, если постоянство
внутри системы нарушается.
Мало того, в некоторых усло-
виях отсутствие или временное
отключение гомеостатических
механизмов может дать опре-
деленную энергетическую вы-
году. Так, на рис. 2.3 приве-
дены графики изменения во
времени темпов метаболизма у
День Ночь День
Рис. 2.3. Темпы метаболизма у землеройки
и колибри. Темп метаболизма у землеройки
в течение суток практически неизменен (/).
Переход колибри в торпидное состояние
(спячку) позволяет ей в ночное время за счет
снижения темпов метаболизма (2) сохранить
энергию, количество, которой можно опре-
делить по площади, заключенной между
кривыми 1 и 2.
двух различных животных, имею-
щих одинаковую массу, а в активном состоянии — и одинаковую
температуру тела. Снижение темпов обмена веществ у колибри,
впадающей ночью в торпидное состояние (спячку), позволяет ей
за счет отказа от поддержания постоянства температуры тела
сохранить большое количество энергии [363].
Теперь мы можем более четко сформулировать, что же по-
нимается под сохранительными свойствами живой системы.
Жизнедеятельность биосистем связана, с одной стороны, с
50
СОХРАНИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
необходимостью обеспечить энтропийный баланс в системе, т. е.
поступление необходимых системе веществ и энергии в темпе,
равном темпу их расходования, а с другой — с необходимостью
обеспечить постоянство внутренних условий в системе. Выше
мы старались показать, что эти две стороны жизнедеятельности
тесно связаны между собой, но далеко не идентичны.
В организме животных, способных впадать в спячку, в тече-
ние всего периода активного поведения поддерживается как
баланс вещества и энергии, так и постоянство внутренней среды,
и прежде всего — температура тела. При впадении в спячку
гомеостаз системы нарушается — температура не только уста-
навливается на более низком уровне, но и становится более
вариабельной под влиянием условий среды. Равенство же тем-
пов потребления и поступления между собой при этом сохра-
няется, хотя обе величины — и темп потребления веществ, на-
пример, кислорода, и темп их поступления, — падают вместе до
существенно более низких значений. Аналогичные явления
можно указать и для экологических систем, когда при изменении
условий в системе происходит сохранение как видового состава
системы, так и сохранение темпов потока энергии через систему,
причем при прогрессирующем ухудшении окружающих условий
сначала будет нарушен видовой состав сообщества (в опреде-
ленном смысле такое нарушение соответствует нарушению го-
меостаза в системах организменного уровня), и лишь затем—
энергетические потоки через систему.
Способность биосистемы к самосохранению определяется
тем, насколько хорошо при наличии внешних возмущений в ней,
с одной стороны, поддерживаются темпы поступления веще-
ства и энергии и, с другой, обеспечивается постоянство внутрен-
ней среды. Таким образом, мы приходим к следующему опре-
делению.
Понятие «сохрапительные способности биосистемы» яв-
ляется объединением двух понятий — способности системы обес-
печивать гомеостаз и способности поддерживать стационарное
неравновесное состояние при изменяющихся условиях внешней
среды.
Поэтому ниже мы будем говорить о гомеостазе и гомеоста-
тических свойствах системы как о части ее сохранительных
свойств.
2.5. Гомеостаз на разных уровнях организации жизни
«Допущение о том, что жизненные явления основаны на фи-
зико-химической активности, хотя и справедливо, но не выяв-
ляет существа проблемы, поскольку невозможно обнаружить
физико-химических явлений, которые конструируют каждое су-
2.5]
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
51
щество по предсуществующему плану и производят удивитель-
ную субординацию и гармоническое единство органической ак-
тивности. В живом существе имеется приспособленность, своего
рода регулируемая деятельность, которая... является наиболее
впечатляющим качеством живых существ», — писал К. Бер-
нар [270].
Изучение процессов регуляции в организме и его системах
позволило классикам физиологической науки — К- Бернару,
И. М. Сеченову, У. Кэннону — сформулировать положение о том,
что «все жизненные механизмы, как бы разнообразны они ни
были, имеют тольку одну цель — сохранение постоянства уело*
вий жизни во внутренней среде» [270]. Эта концепция предстала
в окончательном виде после того, как И. М. Сеченов обратил
внимание на связь постоянства внутренней среды с адекватным
снабжением организма веществами и энергией, а У. Кэннон ввел
термин «гомеостаз».
В настоящее время общепризнано, что принцип гомеостаза
как таковой играет важную роль в жизненных процессах на
всех уровнях организации жизни [38, 41, 269]. Концепция гомео-
стаза оказалась настолько содержательной, что породила целый
ряд направлений в исследовании сохранительных свойств жи-
вых систем как на специфическом уровне, характерном для
различных типов живых систем, так и на уровне формальных
обобщений в биокибернетике. Так, на организменном уровне к
гомеостатическим реакциям относят обычно только реакции фи-
зиологических систем вегетативной регуляции, обеспечивающих
постоянство уровней вещества и энергии в организме, его ор-
ганах и тканях.
Многие целостные реакции, безусловно «предохраняющие ор-
ганизм от внешних дезорганизующих сил», по традиции не от-
носятся к гомеостатическим. Таковы, в частности, имунные ре-
акции, модели которых получили широкое распространение
[37, 131, 151]. Тем более термин «гомеостаз» не относится нами
к специфическим поведенческим реакциям, хотя такие реакции
играют важную роль, например, при поддержании теплового
режима организма [277], а гомеостатическая концепция была
распространена в свое время на соматическое поведение [65],
как, впрочем, и на экономические, политические и социальные
системы [32, 47, 65, 278].
Клеточный уровень. Устойчивость живых систем на
уровне клеточной организации (цитогенетический гомеостаз) яв-
ляется той основой, на которой базируются сохраиительные
свойства живых существ и, следовательно, живых систем всех
надорганизменных уровней организации. Для описания вну-
триклеточных механизмов управления биохимическими процес-
сами чаще всего используется термин ауторегуляция. Среди
52
СОХРАИИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
ауторегуляторных механизмов можно указать угнетение актив-
ности ферментов конечными продуктами, репрессию синтеза
ферментов конечными продуктами, активацию ферментов исход-
ными субстратами, наконец, индукцию синтеза ферментов соот-
ветствующими субстратами. Так, реакции гликолиза и окисли-
тельного фосфорилирования, являющиеся основой клеточного
метаболизма, угнетаются высокими концентрациями конечного
продукта — АТФ; повышение концентрации АДФ и АМФ, наобо-
рот, приводит к активации ферментативной активности, в резуль-
тате чего увеличиваются темпы синтеза конечного продукта.
Таким образом, клеточная активность хорошо координиро-
вана на ферментном уровне — лишь в весьма редких случаях
темпы синтеза и разрушения веществ отличаются от необходи-
мых для обеспечения нормального роста и поддержания метабо-
лизма клетки [68, 98]. Однако термин «гомеостаз» применяется
к подобным механизмам регуляции довольно ограниченно: гово-
рят, что клетка обладает гомеостатическими реакциями в том
смысле, что если ей наносится какое-либо «обратимое» повреж-
дение, то клетка сама его ликвидирует. Так, когда в ДНК
происходят определенные мутации, и вследствие этого синтез
некоторых ферментов становится невозможным, необходимые
метаболиты все-таки синтезируются, хотя и необычным для
нормальной клетки путем — «возможно именно в этом и состоит
значение альтернативных метаболических путей» [98, стр. 5541.
Чаще, однако, вместо гомеостаза говорят об устойчивости или
эластичности: устойчивость комплексов биохимических реакций
связывается со сложностью системы метаболических путей [72,
338]. «Тесно переплетаемая сеть реакций, поддерживаемых в
стационарном состоянии, обеспечивает химическую «эластич-
ность» клетки, от которой зависит ее выживание в изменяющих-
ся условиях среды» [72, стр. 12].
Интересная концепция анализа регуляторных свойств комп-
лексов ферментативных реакций изложена в [310]. Если огра-
ничиться рассмотрением стационарных состояний, то для ана-
лиза изменений в системе при воздействии возмущений в [310]
предлагается использовать понятие управляющей силы (control
strength — англ.) различных ферментов. Влияние фермента £)
с концентрацией х, на концентрацию с,- метаболита S,- в стацио-
нарном состоянии представляется элементом «матрицы управ-
ления»
Управляющая сила того же фермента Cj определяется как
мера его влияния на скорости реакций в системе. Если обозна-
чить через ys общий поток продукта в системе, а через yj —
2.5)
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
53
величину потока, связанного с ферментом Eh то
д In у
С. =_____~
1 d\nyj '
(2.2)
Очевидно, что показатели (2.1) и (2.2) могут быть исполь-
зованы для количественного описания не только регуляцион-
ных характеристик отдельных ферментов в комплексе реакций,
но и для определения сохранительных способностей всего
комплекса — возможности и степени поддержания основных по-
казателей его химических реакций, таких как концентрации суб-
стратов или скорость синтеза продуктов.
В многоклеточных организмах исключительно важную роль
играют механизмы надклеточного уровня регуляции, контроли-
рующие процессы клеточного деления — митотическую актив-
ность клеток. Проблема исследования гомеостаза на этом уров-
не связывается с влиянием внешних факторов на процессы кле-
точного роста и деления клеток [318, 324]. Регуляция клеточного
роста обеспечивает в организме постоянство количества диффе-
ренцированных клеток, выполняющих некоторую данную функ-
цию. Предметом обсуждения в этом случае служат механизмы,
обеспечивающие гомеостаз структуры органов и тканей в орга-
низме. При этом наиболее предпочтительными кажутся два ва-
рианта. Первый из них связан с возможностью регуляционных
принципов, основанных на сложном балансе влияний взаимодей-
ствующих клеток, другой — с регулирующим влиянием единст-
венного фактора — активности тканеспецифических митотиче-
ских ингибиторов, т. е. веществ, обладающих свойством подав-
лять митоз [269]. В обоих случаях речь идет о зависимости
темпов роста и деления от плотности клеточной популяции, в
которой находится данная клетка.
Модели гомеостатической регуляции роста эпидермиса были
разработаны в работах [315, 316]. Блок-схема такой системы
с обратной связью показана на рис. 2.4. Базальные клетки де-
лятся, новые клетки дифференцируются и переходят в роговое
вещество кератин. Дифференцирующаяся масса клеток произво-
дит вещества, регулирующие как скорость деления, так и ско-
рость дифференцирования клеток. На модели, реализованной
на ЦВМ [315], можно воспроизводить такие процессы, как одно-
кратное воздействие карциногена на кожу, влияние внешнего из-
носа кожи и т. д.
Гомеостатическая регуляция клеточного роста в тканях мле-
копитающих рассматривается в работе [297]. Регуляция клеточ-
ного роста обеспечивается за счет управления темпом образова-
ния новых клеток, причем рассмотрены четыре варианта дей-
ствия регулирующих веществ, производимых делящимися, неде-
лящимися, взрослыми неделящимися или, наконец, вновь обра-
54
СОХРАПИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
зуемыми клетками. Многие аспекты взаимоотношений процессов
в опухолевых тканях с проблемой ионного гомеостаза рассмо-
трены в книге [114].
Уровень организма и его систем. Гомеостаз орга-
низма является результатом одновременного действия много-
численных и сложно организованных регуляторных механизмов,
образовавшихся в процессе эволюционного развития.
Регуляторные механизмы клетки, о которых говорилось в
предыдущем разделе, составляют в этой системе, пожалуй, са-
мый нижний уровень. Этот уровень связан с наиболее древними
Рис. 2.4. Схема гомеостатической регуляции роста эпидермиса.
организменными механизмами — неспецифическими химически-
ми регуляциями, на которые наложились затем механизмы эндо-
кринной регуляции, действие которых также связано с влиянием
на активность ферментов, темпы их синтеза и, кроме того, на
проницаемость биологических мембран. Следующий уровень на-
слоения представляют собой механизмы гуморальной регуляции
с помощью активных веществ, вырабатываемых не в эндокрин-
ных железах, а в обычных рабочих органах [125, 304, 354]. На-
конец, самый верхний слой гомеостатических механизмов состав-
ляют регуляторные механизмы нервной системы, наиболее
быстродействующей управляющей системы организма. Сово-
купность этих механизмов позволяет не только обеспечивать не-
обходимые темпы поступления веществ и энергии ко всем рабо-
тающим органам и тканям, но и стабилизировать уровни внут-
риклеточных концентраций многих жизненно важных веществ.
Множественность регуляторных механизмов организма позво-
ляет ему достигать этих целей за счет относительно малого на-
пряжения каждого из участвующих в процессе регуляции меха-
низмов, расширяя тем самым область гомеостаза.
Новый этап в развитии идеи гомеостаза организма наступил
после того, как Н. Винер, ознакомившись с концепцией У. Кэн-
нона, предложил первую кибернетическую модель физиологиче-
2.5]
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
55
ской регуляции. Две страницы, которые он в 1949 г. в своей
«Кибернетике» [40, стр. 145—146] посвятил этому вопросу, ока-
зали глубокое влияние сначала на методы моделирования го-
меостаза, а затем и на физиологическую трактовку гомеостаза
вообще.' Изложив идею простого автоматического регулятора в
технике — схему отрицательной обратной связи по отклонению
от уставки (рис. 2.5), Н. Винер высказал предположение, что
Рис. 2.5. Схема простого автоматического регулятора по отклонению, предложенная Вине-
ром в качестве модели гомеостатической регуляции в организме.
таким же образом может обеспечиваться и постоянство внут-
ренней среды организма: «Итак, наше внутреннее хозяйство
должно включать набор термостатов, автоматических регулято-
ров концентраций водородных ионов, регуляторов давления и
тому подобных приборов — набор, которого было бы достаточно
для большого химического завода. Все это-в целом и составляет
наш гомеостатический механизм».
Высказанная Н. Винером идея о применении методов теории
управления в физиологии оказалась, несомненно, плодотворной
и породила целый ряд работ по моделированию гомеостаза в
физиологических системах. К этим работам мы еще вернемся в
главе о моделировании гомеостаза (см. разд. 3.4 и 7.3). С дру-
гой стороны, эта идея привела к тому, что возникла тенденция
вообще отождествлять гомеостаз организма с простой схемой на
рис. 2.5 [56, 322 и др.]. Дело в том, что сама методология клас-
сической теории управления, основанная на выделении конеч-
ного и небольшого числа регулируемых параметров, хорошо при-
способлена для изучения простых по конструкции автоматиче-
ских систем; для понимания же специфических процессов в
живом организме часто требуются методы, выходящие за рамки
классической теории регулирования [177]. Поэтому естественно,
что наибольшие успехи в понимании физиологических регуля-
торных механизмов животного организма с помощью простой
модели рис. 2.5 были получены при моделировании изолирован-
ных физиологических систем [60, 130, 253].
Отождествление концепции гомеостаза с простой однокон-
турной моделью следящей системы оказалось неприемлемым
при исследовании сложных физиологических систем. Л. фон Бер-
таланфи высказал мнение, что такая модель не может быть
56
СОХРАИИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
использована для анализа открытых систем: «В отличие от ки-
бернетики, занимающейся анализом механизмов обратной связи,
общую теорию систем интересует динамическое взаимодействие
внутри систем с многими переменными, причем для живых орга-
низмов наибольшее значение в этой связи имеет исследование
понятия открытой системы» [31, стр. 41]. Простая модель гомео-
стаза в форме отрицательной обратной связи осталась и вне ра-
мок теории функциональной системы П. К. Анохина в физиоло-
гии [9, 11]. Недостатки столь простой модели гомеостаза, необ-
ходимость разработки более адекватных конструкций, оЛисываю-
щих гомеостатические свойства организма, обсуждались и в ряде
других работ [153, 216, 245, 253].
Термин «гомеостаз» вначале использовался для характери-
стики устойчивости, постоянства внутренней среды организма в
целом. В дальнейшем, и прежде всего под влиянием винеров-
ской модели, это понятие было перенесено на отдельные функ-
ции, что позволило говорить о температурном гомеостазе [79,
149], гомеостазе сахара крови [13, 56, 70, 261], гомеостазе дыха-
тельной функции крови и т. д. Такой перенос центра тяжести с
целого организма на его отдельные функции основан на допу-
щении, что постоянство целого обеспечивается постоянством его
частей. Поскольку, однако, стабильность целого допускает за-
мену одного механизма или вещества другими, лишь бы не на-
рушалась цель регуляции, возможности организма по поддер-
жанию своего существования значительно богаче того, что пред-
лагает простая модель одноконтурной следящей системы [141,
200, 244],
Специфика гомеостаза различных биосистем организменного
уровня — растений, простейших одноклеточных организмов, бес-
позвоночных и, наконец, высших животных рассмотрена в моно-
графии [38]. Что касается организма человека, то проблема его
гомеостаза остается предметом внимательного изучения в са-
мых разных аспектах (см., например, [55, 186J).
Говоря о модели гомеостаза как о простой структурной схе-
ме, мы, разумеется, упрощаем рассмотрение вопроса. Ведь воз-
можно усложнение системы гомеостатической регуляции, на-
пример, образование цепей из таких контуров, параллельное
или последовательное их соединение, наличг.е логических це-
пей, переключающих управление с одного контура на другой
и т. д., как об этом будет говориться ниже (разд. 7.3). Однако
исходная идея поддержания абсолютного равенства некоторых
выходных сигналов (уровней вещества пли энергии) специально
заданным значениям^ усхавкам». остается неизменной и при та-
ком обобщенном '"понимании классической схемы гомеостаза.
Л именно идея «уставки» и становится подчас препятствием на
2.5]
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
57
пути понимания механизмов и функциональных свойств био-
систем.
Экологические системы. Для биологической систе-
мы, включающей животные и растительные популяции, а также
абиотические факторы и учитывающей взаимодействие между
ними, используется термин экологическая система или биогео-
ценоз [161, 187, 198, 204]. Сложные синтетические биосистемы
экологического уровня создаются в результате столкновения на
одном и том же пространстве разнообразных органических форм
и разделения сфер влияния, захвата экологических ниш. Состоя-
ние равновесия в экосистеме (биоценотический гомеостаз — см.,
например, [248]) поддерживается многочисленными механизма-
ми активной и пассивной регуляции.
Специфика экологических механизмов регуляции такова, что
исследование влияния условий внешней среды на свойства эко-
систем (и, в частности, на равновесные численности популяций
входящих в нее видов) довольно затруднительна. Не в послед-
нюю очередь эти трудности объясняются гем, что экосистемы
реагируют на малозаметные изменения, например, погодных ус-
ловий резкими колебаниями численности видов. Эти колебания
настолько велики, что обычно соответствующие графики строят-
ся в логарифмическом масштабе.
Наиболее изученными механизмами регуляции в экосистемах
являются, по-видимому, так называемые механизмы, зависящие
от плотности популяции [210, 212, 263, 329] (здесь очевидна ана-
логия с механизмами управления на клеточном уровне). Инте-
ресно отметить, что, в отличие от клеточного и организменного
уровня, аналогия механизмов экологической регуляции с про-
стыми системами автоматической стабилизации была подверг-
нута критическому разбору, и такие модели экосистем не полу-
чили распространения [336]. Однако механизмы регуляции, за-
висящие от плотности популяции, являются лишь одним из
довольно частных видов регуляции, существующих в экосистеме.
Регуляция в экосистемах связана и со структурой пищевых се-
тей, с механизмами конкретного взаимодействия отдельных ви-
дов (например, борьбой за места обитания) [148], а также гене-
тическими механизмами [170].
Способность экосистем сохраняться в условиях постоянной
или изменяющейся среды описывается в экологии с помощью
целой группы терминов. Часто для этой цели используются тер-
мины устойчивость или стабильность [210, 212, 328]. Однако эти
термины обладают рядом недостатков.
Во-первых, содержание самого термина «устойчивость» или
«стабильность» зачастую не является строго определенным:
устойчивость оценивается «по наблюдаемым во времени измене-
ниям численности» [212, стр. 248]. Очевидно, что наблюдаемые
58
СОХРАПИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
во времени изменения численности популяций определяются не
только свойствами экологической системы, но и свойствами
входного сигнала — величиной внешних воздействий на систему.
Ясно поэтому, что так определяемая устойчивость не является
характеристикой экосистемы, а характеризует сразу пару «эко-
система — внешняя среда».
Другой недостаток связан с тем, что по мере внедрения в
экологические исследования методов теории управления тот же
термин «устойчивость» все чаще используется и в другом — тео-
ретико-управленческом смысле (способность системы восстанав-
ливать численность при ее малых отклонениях от исходной, по-
ложение нулей характеристического уравнения системы в левой
полуплоскости [187, 256, 336]).
Поэтому в последнее время для описания слабой зависи-
мости состояния экосистемы от условий внешней среды предла-
гается ряд новых терминов. Было, в частности, предложено оп-
ределять поведение экосистем с точки зрения сразу двух
свойств, когда к понимаемому более или менее традиционно
свойству устойчивости добавляется новое свойство — упругость
или эластичность (resilience — англ., [280, 312, 313], см. также
[1866]). Под устойчивостью, в соответствии с обычными поло-
жениями теории динамических систем, тогда подразумевается
способность системы возвратиться в состояние равновесия после
возмущения. Упругость же определяет «сохранение взаимоотно-
шений внутри системы и является мерой способности системы
компенсировать изменение переменных состояния, переменных
внешней среды и в то же время сохраняться», — например, за
счет перехода в новое равновесное состояние. Аналогичный тер-
мин для описания сохранительных свойств биосистемы — поня-
тие инерции — было предложено У. Мардоком [336]: «тенденцию
популяции противостоять давлению сдвигов в среде я назвал
инерцией».
При анализе сохранительных свойств экосистем большое
внимание уделяется их открытому характеру [158, 187, 204, 307,
329]. В таких системах равновесные состояния могут смещаться
под влиянием внутренних изменений в системе и под воздейст-
вием внешних факторов. В этом случае устойчивость можно
трактовать на основе принципа Ле-Шателье: степень сохране-
ния состояния системы при действии возмущений зависит от
структуры потоков вещества и энергии в системе. (Блок-схема
одного из типов экосистем — луговой экосистемы — с учетом
структуры потоков приведена на рис. 2.6; потоки вещества, энер-
гии и информации рассматриваются в последнее время и при-
менительно к новому типу природных комплексов — так назы-
ваемым географическим системам [15, 16].)
2.5]
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
59
Проблема связи устойчивости экосистем с их структурой и
разнообразием состава давно является предметом обсуждения
в экологии. «Среди экологов считается почти аксиомой, что бо-
лее сложные по структуре сообщества и более устойчивы» [187,
стр. 132].- Однако конкретная зависимость, существующая между
разнообразием и сложностью экосистем и их устойчивостью, все
Рис. 2.6. Блок-схема экосистемы с учетом влияния человека.’ Модели такого рода, вклю
чающие наряду с природными и экологическими характеристиками и некоторые черты
влияния человека на среду, могут использоваться для анализа сохранительных свойств
экосистем. На схеме показана структура экосистемы (источник биомассы, популяции
продуцентов и консументов, сток биомассы) и структура населения региона (источник
населения, популяции сельского и городского населения, сток). Пятиугольниками пока-
заны внешние факторы, влияющие на систему, пунктиром — потоки информации, сплошными
линиями —материальные потоки. Темпы материальных потоков являются управляемыми
переменными; влияние управляющих переменных на темпы потоков показано пересече-
нием пунктирных и сплошных линий.
еще достаточно неясна и продолжает дискутироваться как в эко-
логических, так и в математических публикациях [101, 217, 329].
Современное состояние проблемы устойчивости в биологии
применительно к экологическим системам рассмотрено в содер-
жательном обзоре [101]. В этой работе предлагается разделить
термины «устойчивость» и «стабильность» экологических систем.
Устойчивость, в отличие от представлений теории динамических
систем, должна определяться как соотношение между измене-
нием характеристик системы и величиной вызвавшего их воз-
действия. Если при анализе поведения системы фиксируется
только изменение характеристик систем, безотносительно к ве-
личине вызвавшего его внешнего воздействия, то предлагается
использовать понятие стабильности.
Если, например, при воздействии на систему, обозначаемом
Ду, возникает изменение некоторой характеристики системы,
60
СОХРАИИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
обозначаемое Лу, то меру устойчивости S системы предлагается
определять отношением
5 = (2.3)
в то время как мера стабильности системы St должна характе-
ризоваться величиной
(2.4)
Что же является целью регуляции в экосистемах? В каком
соотношении с такой целью находится свойство устойчивости
или стабильности?
Впервые такая цель была, по-видимому, сформулирована в
работах А. Лотка [326а] в виде экстремального принципа — эво-
люция экосистем идет в сторону увеличения суммарного потока
энергии через систему, причем в стационарном состоянии дости-
гается его максимально возможное значение. Позднее К- Уатт
выразил это положение следующим образом: «Сообщество жи-
вотных и растений в любом месте земного шара представляет
собой ансамбль видов, который обеспечивает максимальное ис-
пользование падающей на Землю солнечной радиации при том
типе почв, который характерен для данного района» [210]. В со-
ответствии с изложенными в разд. 1.7 представлениями об
иерархии целей в биосистемах, можно рассмотреть и специфиче-
ские цели систем экологического уровня. Высшие цели в эко-
системах связаны с сохранением темпов производства органиче-
ского вещества: интенсивность использования падающей на
Землю солнечной энергии должна поддерживаться на макси-
мально возможном уровне. Сохранение же видового состава и
численности отдельных популяций целесообразно лишь постоль-
ку, поскольку оно способствует выполнению высших целей —
поддержанию максимального темпа- потока энергии через систе-
му. Сохранение видового состава поэтому представляет собой
цель второго порядка.
Действительно, если на экосистему действуют все более и
более жесткие условия среды, то сначала сохраняется и видо-
вой состав, и темп потока энергии через систему, затем — толь-
ко поток при измененном видовом составе, затем нарушается и
скорость потока энергии. Так, луговые системы на водоразде-
лах, подвергающиеся относительно слабым возмущениям
(дожди, холод, суховеи, но не поднятие грунтовых вод, приво-
дящее к кислородному голоданию корневой системы) обходятся
гомеостатическими механизмами отдельных растений, не вклю-
чая специфических экологических регуляторов, т. е. не изменяя
видового состава сообществ. Сообщества, обитающие на скло-
нах и в низменных участках, подвержены большим внешним
2 5]
ГОМЕОСТАЗ НА РАЗНЫХ УРОВНЯХ ОРГАНИЗАЦИИ
61
возмущениям (затопление корневой системы). Луговое сообще-
ство, включающее лишь незначительное количество болотных
растений, при повышении уровня грунтовых вод через несколько
лет отвечает существенной перестройкой видового состава, т. е.
включением специфических экологических регуляторов, обеспе-
чивающих неизменность темпов потребления энергии и постоян-
ство продукции сообщества. Лишь очень редкие в природе явле-
ния (например, извержения вулканов, приводящие к полному
изменению свойств среды обитания) или человек способны вы-
вести экосистему за пределы возможностей ее механизмов регу-
ляции — экосистема как целостное образование исчезает, на-
рушается энергетический баланс, сохраняется лишь жизнь от-
дельных видов, возможно, ранее даже отсутствовавших в данной
экосистеме *).
Механизмы, деятельность которых направлена на сохранение
экосистемы, в самом общем плане называются в экологии гомео-
статическими, хотя этот термин еще и не завоевал таких же
прав, как в физиологии организма, и иногда гомеостатической
регуляцией здесь называются механизмы регуляции, связанные
только с поддержанием численности видов.
В последнее время, однако, все большее признание получает
расширительное толкование гомеостатических регуляций в эко-
системах. Так, в работе [217] понятие степени гомеостаза при-
влекается для общего суждения о мере устойчивости экологиче-
ской системы. При этом рассматривается некоторая характери-
стика системы у, «близость которой к норме» существенна для
системы. При изменении внешнего воздействия v на величину
Ди характеристика у получает приращение Ду, а «степенью го-
меостаза в этом случае можно назвать отношение относитель-
ных изменений аргумента и функции»:
g=tl/v- (2-5)
Интересны и рекомендации по практической оценке величи-
ны G: когда зависимость у(и) неизвестна, но имеются дискрет-
ные измерения u(G) и y(tt), i — 1, 2, ..., п, величину G можно
найти как отношение средних отклонений ои и оу от их «нор-
мальных значений» и, и yt:
G = ^~,
°у
где
п . П ,
E^vi _ 1 V ^У{
] vi ' п L yt’
*) Этот пример принадлежит А А. Титляновой.
62
СОХРАНИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
Ранее идея оценки степени гомеостаза системы на основе
сравнения показателей о для воздействий среды и характеристик
сообщества высказывалась и в работе [343], хотя там использо-
вался более сложный способ определения величин ov и ov, свя-
занный с оценкой «благоприятных» и «неблагоприятных» тен-
денций в поведении системы (см. также [101]).
Другой аспект расширительного толкования гомеостатиче-
ских свойств экологических систем связан с работами Междуна-
родной биологической программы [291, 368]. В рамках этих ра-
бот, например, для луговых экосистем обсуждались следующие
положения:
1. В луговых экосистемах существуют гомеостатические ме-
ханизмы, которые стремятся удержать систему в состоянии
«луг», несмотря на действие возмущений. Однако существуют
пределы упругости системы: если возмущения достаточно силь-
ны, гомеостатические механизмы будут неадекватными, и систе-
ма будет изменяться от лугового состояния к некоторому дру-
гому.
2. Стресс в экосистеме встречается последовательными
«линиями обороны». Первая линия включает лишь немногие под-
системы целостной экосистемы. Увеличение стресса последова-
тельно приводит к включению большего и большего числа под-
систем.
3. Чем разнобразнее система, тем более она нечувствитель-
на к возмущениям.
Блок-схема луговой экосистемы и была представлена выше
на рис. 2.6.
Таким образом, понятие гомеостаза оказывается связанным
с кардинальными вопросами, на которые должна дать ответ
экологическая наука — что такое экосистема, чем определяется
ее тождественность самой себе, что такое «здоровье» экосистемы
и каковы механизмы, поддерживающие ее существование.
В этой книге мы ограничиваемся лишь теми аспектами со-
хранительных свойств биосистем, которые связаны с поддержа-
нием стационарных и не зависящих от времени состояний.
Нужно иметь в виду, однако, что в динамической теории
популяций и сложных экологических систем концепция сохране-
ния системы связывается не только (и даже не столько) со ста-
ционарными режимами, но и с проблемами роста популяций.
Широко распространенным понятием, характеризующим эти
свойства популяций, является приспособленность, отражающая
способность популяции к размножению.
Приспособленность популяции W определяется как удельная
скорость роста ее численности N
1 rfV
N di ’
(2.6)
2 6j
ОБЩИЕ СВОЙСТВА ГОМЕОСТАТИЧЕСКИХ МЕХАНИЗМОВ
63
Обсуждение этого понятия имеется, например, в [161, 233].
В [161] дается четкое определение приспособленности популя-
ции как целого и рассматриваются ее формализованные харак-
теристики.
Какие же выводы можно сделать из этого краткого обзора
по гомеостазу, устойчивости и стабильности экологических си-
стем?
Вероятно, задача исследования сохранительных свойств си-
стем надорганизменного уровня — одна из наиболее острых и
сложных в общей проблеме самосохранения живых систем.
Практические требования, связанные с необходимостью оценки
реальных возможностей человека по сохранению и целесооб-
разному изменению природных комплексов и систем [4, 52, 82,
115], вынуждают экологов и специалистов по математическому
исследованию экосистем выдвигать все новые критерии и поня-
тия для описания различных самосохранительных свойств эколо-
гических систем. Работа эта, вероятно, еще далека от совершен-
ства. Возможно, некоторые из предлагаемых в ходе такого
исследования терминов могут лучше и полнее отразить специфи-
ческие свойства природных экосистем, но работу по оценке со-
хранительных свойств систем надорганизменного уровня целесо-
образно начать с освоения уже разработанных в теории динами-
ческих систем понятий и представлений, таких как устойчивость
по Ляпунову и методы оценки чувствительности. Соответствую-
щие разделы теории управления будут изложены в следующей
главе.
2.6. Некоторые общие свойства
гомеостатических механизмов
на разных уровнях организации биосистем
Даже краткий обзор некоторых свойств гомеостатических
систем на различных уровнях живой природы позволяет выде-
лить ряд общих характеристик, присущих всем этим уровням.
С другой стороны, такой обзор дает нам возможность видеть ог-
раниченность некоторых приемов, используемых для описания
гомеостатических свойств биосистем на отдельных уровнях.
Попробуем вкратце сформулировать основные свойства го-
меостатической регуляции, в той или иной мере проявляющиеся
во всех рассмотренных выше сферах.
1. Гомеостатические качества являются, по-видимому, одним
из главных звеньев в системе сохранительных свойств биологи-
ческих объектов на всех уровнях организации жизни. Однако
при существенных изменениях условий внешней среды или
структуры системы гомеостатические свойства могут нарушать-
ся ради высшей цели — сохранения жизнедеятельности системы,
64 СОХРАИИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ [ГЛ. 2
ее стационарного неравновесного состояния. Следовательно, го-
меостаз биосистем не является обязательным условием жизни;
цель «гомеостаз» — иерархически младше цели «сохранение
жизни» (см. также разд. 1.7).
2. Гомеостатические свойства биосистем проявляются как
слабая зависимость некоторых существенных переменных био-
системы от условий внешней среды. В первую очередь понятие
гомеостаза связывается со стационарными режимами системы.
Хороший гомеостаз означает, что характеристики стационарно-
го режима мало зависят от окружающих условий, т. е. мало
чувствительны к факторам внешней среды.
3. При существенных сдвигах в условиях среды переменные
системы становятся все более чувствительными к этим изме-
нениям; возникает характерная зависимость внутренних пере-
менных от переменных внешней среды — гомеостатическая
кривая. z
4. Гомеостатические свойства бйосистем развиваются при
усложнении структуры системы, возникающем в эволюционном
процессе. Чем сложнее биосистема, тем лучше могут быть ее
гомеостатические свойства. К такому результату приводит нало-
жение активной регуляции на пассивные механизмы управления
в организме и его системах, разветвленная структура метаболи-
ческих путей в клетке, разнообразие и связанная с ним сложная
сеть регулирующих цепей в экосистеме.
5. При переходе к следующему, более высокому, уровню
организации биосистем гомеостатические механизмы каждого
данного уровня сохраняются в системе в качестве неспецифиче-
ского «.нижнего этажа», образующего «подсистемный слой» ме-
ханизмов регуляции. Клеточные механизмы регуляции сохра-
няются как подсистемные механизмы на органном и организ-
менном уровне. Физиологические регуляторы являются в свою
очередь неспецифическими подсистемными механизмами эколо-
гических систем.
6. Возможности каждого уровня гомеостатических механиз-
мов ограничены. Включаясь, регулирующий механизм начинает
изменять свои переменные вместе с изменением действующего
на систему возмущения. Достигнув предельных значений или
исчерпав наличные ресурсы, регулирующий механизм не ^спосо-
бен компенсировать дальнейшее возмущающее влияние среды.
Поэтому при действии стрессорных факторов внешней среды
происходит поочередное включение регуляторных механизмов
разных уровней, начиная, по-видимому, с неспецифических
«подсистемных». После исчерпания ресурсов подсистемного
уровня нагрузка ложится на специфические системные механиз-
мы; примером такого поведения могут быть реакции биосистем
экологического уровня.
2.61
ОБЩИЕ СВОЙСТВА ГОМЕОСТАТИЧЕСКИХ МЕХАНИЗМОВ
65
Поскольку, однако, каждая биосистема входит в качестве
элемента в биосистемы иерархически более старших уровней,
исчерпание ресурсов специфических гомеостатических механиз-
мов системы не обязательно ведет к ее гибели, прекращению ее
жизнедеятельности. Жизнь в этом случае поддерживают уже
надсистемные механизмы регуляции. Так, гомеостатические ме-
ханизмы целостного организма можно рассматривать как над-
системные в задачах, связанных с жизнедеятельностью отдель-
ных органов. Именно они поддерживают снабжение органов в
экстремальных условиях, например, при искусственном крово-
обращении и недостаточном объеме перфузии (см. рис. 1.8).
В какой-то мере о надсистемных механизмах можно говорить и
в случае действия сверхсильного стрессора на экологическую
систему.
7. Когда мы говорим об общих принципах самосохранения,
мы не должны упускать из виду преемственности многих основ-
ных черт сохранительных свойств у объектов живой и неживой
природы. Конечно, объекты живой природы — биосистемы — в
ходе эволюции приобрели и закрепили такие механизмы управ-
ления, которые позволяют им поддерживать стационарное не-
равновесное состояние во все более широком диапазоне условий
внешней среды. Не последнюю роль в этом расширении диапазо-
на сыграл процесс обогащения структуры регуляторов биосисте-
мы за счет возникновения в нем новых элементов и связей, спе-
цифических только для биосистем, процесс наслоения или соче-
тания управляющих механизмов. Но самый нижний слой сохра-
нительных механизмов в биосистемах — простейшие механизмы
пассивной регуляции — является общим как для живых, так и
для неживых объектов. Таковы, например, процессы пассивной
регуляции темпов диффузии, когда концентрации веществ иг-
рают роль пассивных регуляторов, пассивная устойчивость поло-
жения тел в пространстве или, наконец, сохранительные меха-
низмы молекулярного уровня, лежащие в основе устойчивости
генетического кода [251].
Известный пример пассивных механизмов поддержания нор-
мального положения у рыб (расположение центра плавучести
выше центра тяжести [216]) показывает, что вклад механизмов,
общих для живой и неживой природы, в процессы управления
в биосистемах может быть достаточно весомым. Молекулярная
биофизика уделяет большое внимание изучению физико-химиче-
ских закономерностей, играющих ведущую роль на уровне мо-
лекулярной организации биосистем и сохраняющих свое значе-
ние и на более высоких уровнях (тем, впрочем, меньшее, чем
этот уровень выше [44]). Такая позиция близка к представле-
ниям, высказанныхм в работе [156] о процессах «квазиуправле-
ния» в неживой природе.
66
СОХРАПИТЕЛЬНЫЕ СПОСОБНОСТИ ЖИВЫХ СИСТЕМ
[ГЛ. 2
2.7. Гомеокинез, гомеорез и генетический гомеостаз
К концепции гомеостаза близки и некоторые другие общебио-
логические представления. Хотя они и выходят за рамки нашего
рассмотрения, мы кратко упомянем их основные характеристики.
При рассмотрении гомеостаза биосистем основное внимание
уделяется стабильности, постоянству переменных внутренней
среды организма — свойству, изучение которого восходит еще
к Клоду Бернару и И. М. Сеченову. Более поздние физиологи-
ческие исследования показали, что все переменные в организме
и в биосистемах других уровней подвержены закономерным цик-
лическим изменениям. Этот факт лег в основу представлений
о живой системе как о компактной системе, образованной сово-
купностью сопряженных осцилляторов с ограниченными цикла-
ми и связанных с ними управляющих механизмов. Эти осцилля-
торы имеют множество степеней свободы — химических, элек-
трических, механических, гидравлических, так что величины
концентраций, потенциалов (уровней) и потоков вещества и
энергии (темпов) колеблются, совершая циклические движения.
Управление такими циклами осуществляется посредством инги-
бирования или освобождения от торможения, так что средние
значения частоты колебаний и их амплитуды поддерживаются
в некоторых пределах. «Систему регулирования, посредством ко-
торой эти колебательные системы модулируются в пределах их
нелинейного устойчивого рабочего диапазона, мы назовем го-
меокинезом», — пишут А. Ибералл и У. Мак-Каллок [78, стр. 56].
С нашей точки зрения понятие гомеокинеза и концепция го-
меостаза представляют собой очень близкие модели постоянства
внутренней среды биосистем; различие между ними состоит
единственно в том, что эти модели дают описание одного и того
же явления в отличающихся временных масштабах. Модель го-
меокинеза, обладая большей «степенью разрешения», улавли-
вает ритмические изменения переменных, в то время как модель
гомеостаза ограничивается рассмотрением лишь средних значе-
ний переменных внутренней среды. Концепция гомеокинеза но-
сит пока методологический характер; работающих моделей кон-
кретных биосистем на ее основе нет даже для организменного
уровня, на материале которого эта концепция была создана.'Что
касается колебательных процессов в биохимических и других
системах, то они не связаны с концепцией гомеокинеза и иссле-
дуются специфическими методами [21, 23, 57, 75, 131а, 132].
Чем выше организация биосистемы, тем сложнее процессы
ее становления и развития. Эти процессы должны быть скоорди-
нированы во времени и пространстве, т. е. должна существовать
интегрирующая управляющая система, формирующая и объеди-
няющая элементы, возникающие на каждой стадии развития
2.7]
ГОМЕОКИНЕЗ, ГОМЕОРЕЗ, ГЕНЕТИЧЕСКИЙ ГОМЕОСТАЗ
67
биосистемы на унаследованной основе генетической структуры.
Процессы формирования и объединения развивающихся систем
и структур в организме называются процессами эпигенетической
(или онтогенетической) интеграции [213].
Для сложных организмов система корреляции эпигенетиче-
ских процессов усложняется; создаются механизмы, обеспечи-
вающие устойчивость и целостность развивающегося организма
в его изменениях. Устойчивость путей развития особи или попу-
ляции в определенных условиях, т. е. поддержание внутреннего
равновесия на каждой стадии развития, смещение равновесия
при переходе от одной стадии к другой и восстановление равно-
весия на всех стадиях при его случайном нарушении, называет-
ся гомеорезом (или гомеорезисом) [213, 248]. Этот термин про-
исходит от греческих слов гомойос — сходный, одинаковый, и
рео — течь. Механизмы гомеореза обеспечивают сохранение нор-
мального течения процессов онтогенеза при наличии возмуще-
ний во внешней или внутренней среде.
Способность биосистемы сохранять свои индивидуальные
признаки в процессе развития, т. е. способность данного гено-
типа создавать в широком диапазоне условий среды один и тот
же фенотип, часто называется гомеостазом развития [129, 325].
Этот термин относится к одной особи, и его следует отличать
от генетического гомеостаза, который характеризует группу
особей. Генетический гомеостаз означает «устойчивость популя-
ции к изменению ее генетического состава под влиянием от-
бора» [129, стр. 267]. Гомеостатические свойства генетических
систем определяются их структурой; введение мутантных генов
может ухудшить гомеостаз. Так, в мутантных линиях введение
в генотип дрозофилы гена Ваг делает процесс образования фа-
сеток чувствительным к температуре среды.
Сохранение наследственной информации является одним из
кардинальных условий существования жизни. Однако надеж-
ность хранения генетического кода не исключает возможности
его изменения и использования таких изменений в эволюции.
Стабильность характеристик в особи сочетается таким образом
с пластичностью популяций и видов в эволюционном процессе.
Современные представления о молекулярно-генетических ме-
ханизмах и системах управления подытожены в монографии
[170], где рассматриваются специфические свойства генетических
систем управления во всем многообразии их свойств — в клет-
ках, организмах, сообществах. Сохранительные свойства систем
самовоспроизведения понимаются здесь как их устойчивость
в широком смысле, когда существенные состояния или режимы
системы'«устойчивы к достаточно широкому спектру возмуще-
ний, изменению параметров и конструктивных особенностей
системы». ([170]j£M. также [290]).
ГЛАВА 3
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ
И БИОСИСТЕМЫ
3.1. Методология теории автоматического управления
Методы теории управления в последнее время находят все
более широкое применение при анализе систем различного ро-
да — технических, биологических, медицинских, экономических.
В каждой из этих областей исследователь сталкивается с зада-
чами, обладающими специфическими особенностями. Так, мно-
гие технические системы имеют относительно небольшое число
переменных и параметров, а связи между ними почти всегда
можно определить в ходе экспериментального исследования си-
стемы или в ходе ее конструирования и разработки. В биологи-
ческих и медицинских задачах число существенных перемен-
ных, как правило, очень велико, а связи между ними, харак-
тер влияния одной переменной на другую, часто неясны. Если
учесть, что непрерывно совершенствуются и сами технические
системы, для которых собственно и был предложен и вместе с
которыми развивался и совершенствовался аппарат теории
управления, то ясно, что расширение области применения тео-
рии управления постоянно требовало как интенсивного разви-
тия идей и методов этой теории, так и разработки специфиче-
ских приемов анализа систем для различных областей при-
менения.
В развитии методов теории управления условно можно выде-
лить два основных периода. Различные специалисты по-разному
определяют характер и время перехода от одного периода к дру-
гому [237, 238, 293]. Однако общий смысл этого перехода состоит
в следующем: если на первом этапе развития теории управле-
ния — на этом этапе чаще использовался термин теория автома-
тического регулирования — интерес исследователей был со-
средоточен на простых системах, имеющих, например, один
вход и один выход и функционирующих в простых условиях, то
на втором этапе внимание специалистов привлекли сложные си-
стемы с большим числом переменных и параметров, имеющие
многообразные цели и решающие сложные задачи в меняющих-
ся условиях.
3 II
МЕТОДОЛОГИЯ ТЕОРИИ АВТОМАТИЧЕСКОГО УПРАВЛЕНИЯ
69
Первый период, условно называемый периодом передаточных
функций, продолжался примерно до 1960 г. В этот период были
разработаны в основном классические методы теории управле-
ния — понятие обратной связи, понятие передаточной функции
системы, методы преобразования и решения дифференциальных
и разностных уравнений управляемых систем. Главное внима-
ние в этот период уделялось исследованию систем с одним вхо-
дом и одним выходом, хотя определенные успехи имелись и в
изучении систем с несколькими входами и выходами [46, 260].
Второй период (1960—1970 гг.) охарактеризовался перено-
сом центра внимания исследователей на сложные системы уп-
равления со множеством входов и выходов. Главным достиже-
нием этого этапа принято считать формулирование понятия
состояния системы, а сам период поэтому часто называется пе-
риодом переменных состояния [293]. В этот период наибольшее
развитие получили матричные способы описания и исследования
систем, методы устойчивости по Ляпунову, методы исследования
систем в терминах «вход-выход-состояние».
Переход от первого этапа ко второму совпал с зарожде-
нием и развитием таких новых разделов теории управления, как
теория больших систем, теория иерархических структур, теория
адаптации и обучения, когда кругозор специалистов по теории
управления стал резко расширяться. По-новому стали формули-
роваться и представления о принципах организации и управле-
ния в биологических системах. Так, для системы управления
движениями были сформулированы такие «неклассические»!
принципы управления, как блочное управление — синергии,
принцип локального управления сложными системами, была по-
казана эффективность многоуровневого принципа организации
систем управления, преднастройки и обучаемости [223].
В последнее время возникает тенденция применять для ме-
тодов теории управления, разработанных на ранних этапах ее
развития, термин классическая теория управления, в отличие от
новых разделов и методов, появившихся и активно разрабаты-
ваемых на втором этапе. Хотя этот термин еще и не получил
всеобщего признания у специалистов по теории управления, он
широко используется в книгах по биологическим применениям
методов теории управления и математических методов [223]. Мы
также будем использовать его в данной книге.
Теория автоматического управления решает два типа задач.
Первый тип — задачи анализа. Задача анализа заключается в
том, чтобы уметь для заданной системы найти ее реакции при
том или ином входном воздействии и определить некоторые
обобщенные свойства системы.
Теория управления имеет дело не с реальными объектами, а
с их описаниями, называемыми моделями. Предполагается, что
70
классическая теория управления и биосистемы
[ГЛ. 3
описание объекта (иногда говорят — системы) дается в виде
некоторого количества стандартных элементов, связанных
между собой. Заданную таким образом систему будем называть
моделью реального объекта. При этом сама теория управления
вовсе не занимается проблемой построения моделей; считается,
что эта проблема лежит вне компетенции теории управления,
хотя некоторые оценки степени соответствия модели объекту
могут быть даны и в рамках теории управления. Действующие
на объект внешние факторы принято называть входными воз-
действиями или входными сигналами (входами), а ее реакции —
выходными сигналами (выходами).
В биологических системах будем различать два типа вход-
ных сигналов — возмущающие и задающие. Задающие сигналы
определяют некоторые требования, которые биологическая си-
стема должна выполнить. Остальные входные сигналы, в том
числе определяющие условия, в которых функционирует систе-
ма, относятся к возмущающим. На рис. 3.1 задающие входные
4)
6)
Рис. 3.1. Входные н выходные сигналы системы, a) w ш2, .... w^ —задающие входы,
Op t>2,..., —возмущающие входы, yv у2, уг—выходы; б) сигналы показаны обобщенно;
w, » и у—векторы размерности n, I иг, соответственно.
сигналы обозначены через Wi, w2, ..., wn, возмущающие — че-
рез ui, v2, , vi. Символами у\, у2, ..., уГ обозначены выход-
ные переменные.
Одна и та же реально существующая величина в зависи-
мости от желания исследователя и выбранного им способа опи-
сания системы может выступать в качестве различных сигналов
в модели. Так, при исследовании системы кровообращения в
организме часто в качестве выходной переменной выбирают.ар-
териальное давление. Величины, характеризующие другие си-
стемы организма и меняющиеся в процессе решения (например,
величина легочной вентиляции), играют в этом случае роль воз-
мущающих сигналов. Задающим входом в модели системы кро-
вообращения, построенной в соответствии со схемой классиче-
ского типа, является специальный опорный сигнал — уставка
(подробнее об этом сказано в разд. 7.3). При описании же ре-
акций системы дыхания (на вдыхание углекислоты) легочная
вентиляция играет уже роль выходного сигнала, входным сигна-
3.1]
МЕТОДОЛОГИЯ ТЕОРИИ АВТОМАТИЧЕСКОГО УПРАВЛЕНИЯ
71
лом является концентрация углекисло 1ы во вдыхаемом воздухе.
Если в ходе исследования меняются переменные в других си-
стемах (то же артериальное давление), то они будут выступать
в качестве внешнего возмущающего сигнала.
В простых случаях при использовании методов теории управ-
ления анализ систем ограничивается структурами с одной вход-
ной и одной выходной величиной. Системы же с несколькими
входами и выходами представляются в матричной форме, после
чего на них распространяются результаты классической теории.
Классическим методом описания линейных систем считается за-
пись связи между ее входом и выходом при помощи дифферен-
циального (или разностного) уравнения. Дифференциальные
уравнения стали привычным методом описания биологических
систем. Так, для организменного уровня классические методы
широко используются в известных монографиях Ф. Гродинза
[60] и Дж. Милсума [130].
Одним из главных методов анализа систем в классической
теории автоматического управления является решение уравне-
ний, описывающих систему. Для облегчения процесса получения
решения были разработаны эффективные методы преобразова-
ния — ряды и интегралы Фурье, преобразование Лапласа.
В классической теории управления большое внимание уде-
ляется тому, чтобы судить о поведении системы (устойчивости
и качестве), не решая описывающих ее уравнений. Отсюда воз-
никли различные критерии, позволяющие делать нужные выво-
ды о линейных системах на основании исследования коэффи-
циентов их уравнений (такие, как критерий Рауса — Гурвица
[46]).
Вторым типом задач, с которыми постоянно сталкивается
теория управления, является синтез систем с заданными свой-
ствами. Суть задачи синтеза состоит в следующем. Имеется ре-
альный объект, для которого, как и в задачах анализа, дается
готовая модель. Целью работы является выбор такого управ-
ляющего устройства, или регулятора, при котором исходный
объект вместе с присоединенным к нему регулятором дает си-
стему управления с нужными характеристиками.
Требования к синтезируемой системе обычно задаются сле-
дующим образом:
— система управления должна быть устойчивой, т. е. раз-
личные отклонения, возникающие под влиянием возму-
щающих сигналов, со временем должны затухать;
— ее реакции на входные сигналы должны обладать опре-
деленными свойствами, характеризующими качество си-
стемы — переходный процесс должен быть достаточно
быстрым; ошибки в установившемся состоянии должны
быть не очень велики и т. д.
72
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
(ГЛ. 3
Задача синтеза системы управления, обеспечивающей наи-
лучшие из возможных характеристик, известна как задача об
оптимальном управлении. Для создания оптимальной системы
управления необходима исчерпывающая информация о системе
и среде, которой фактически никогда нет, хотя бы потому, что
при расчетах используется неточное описание реальной системы
(ее модель), о чем говорилось в начале раздела.
Поэтому возникла задача синтеза систем управления, в ко-
торых процессы сохраняют высокое качество несмотря на то,
что параметры управляемого объекта и переменные внешней
среды могут быть неизвестными или меняться неизвестным об-
разом. Такие системы получили название адаптивных систем.
Их особенность состоит в том, что в ответ на изменение харак-
теристик объекта или среды происходит изменение параметров
в управляющей части системы. Конечно, адаптивность и опти-
мальность — два совершенно не связанных понятия. Адаптивная
система может быть не оптимальной, а оптимальная — не обяза-
тельно адаптивна.
Достоянием биокибернетики и других направлений исследо-
ваний биосистем в настоящее время являются, в основном,
лишь методы теории управления, полученные в первый период
ее развития. Одной из целей настоящей книги является пропа-
ганда использования в задачах исследования биосистем новей-
ших методов теории управления — методов пространства состоя-
ний. С новых позиций мы можем критически взглянуть и на те
методы, которые столь успешно использовались в задачах ис-
следования биосистем и используются при исследовании процес-
сов управления в биосистемах сегодня, хотя завтра, быть может,
новые задачи заставят нас применить те методы, о которых се-
годня мы только начинаем говорить.
Когда методы теории управления применяются для исследо-
вания систем живой природы, часто оказывается, что методоло-
гия этой теории не удовлетворяет специалистов-биологов. Это
происходит потому, что теория управления занимается в пер-
вую очередь задачами синтеза, а биологические задачи —
прежде всего задачи анализа. Поэтому возникает законный во-
прос: в какой мере инженерные решения, предложенные чело-
веком именно в ходе синтеза технических устройств, могут быть
использованы как образцы или модели при анализе живых си-
стем? По крайней мере, нужно отдавать себе отчет в том, что не
все методы и не все понятия теории автоматического управления
можно в чистом виде «пересаживать» на биологическую почву.
Понятия, методы и идеи теории управления — кирпичи, которые
могут органично укладываться в целостное здание наших пред-
ставлений о живом объекте; однако биологические представле-
3.1]
МЕТОДОЛОГИЯ ТЕОРИИ АВТОМАТИЧЕСКОГО УПРАВЛЕНИЯ
73
ния при этом не должны втискиваться в «прокрустово ложе» го-
товых теоретико-управленческих концепций [223, 253].
В последние годы все чаще появляются работы о теории уп-
равления, написанные для специалистов-биологов, начиная с ра-
боты Ф. Гродинза [60] и учебника Дж. Милсума [130], построен-
ных целиком на материале живых систем. Однако в таких рабо-
тах изложение, как правило, начинается с простейших методов
классической теории автоматики, они практически повторяют на
новом материале всю историю ее развития с самого начала
[136]. Поэтому со следующей главы мы начнем разговор о мето-
дах теории управления с того момента, на котором они обры-
ваются в указанных пособиях. А сейчас мы вкратце напомним
некоторые понятия этой теории, с которыми будем постоянно
сталкиваться в ходе дальнейшего изложения.
Поведение системы во времени описывается изменением не-
которых величин — например, входных или выходных. Величи-
ны, меняющиеся в процессе функционирования системы, назы-
ваются ее переменными. Реакция переменных, описывающих
систему, зависит от некоторого числа постоянных величин, опре-
деляющих свойства системы. Эти постоянные величины назы-
ваются параметрами системы.
Отличие переменной от параметра в системе состоит, вооб-
ще говоря, даже не в том, что параметры системы постоянны, а
переменные меняются. Один и тот же параметр может изменять
свою величину в серии экспериментов, принимая новое постоян-
ное значение в каждом из них. Мало того, в ряде случаев па-
раметры могут независимо изменяться во времени заранее за-
данным образом далее в течение одного эксперимента (в этом
случае говорят о системе с переменными параметрами). Разни-
ца между параметром и переменной состоит в том, что перемен-
ные в системе меняются под действием внутренних связей си-
стемы, а параметры, если и меняются, то только под влиянием
внешних обстоятельств, вне всякой связи с внутренними про-
цессами в системе.
Но даже такая попытка разделения описывающих систему
величин на параметры и переменные не может считаться стро-
гой. Ниже мы столкнемся с так называемыми параметрическими
обратными связями, когда в процессе адаптации в системе про-
исходит настройка некоторых параметров основного контура
управления (разд. 4.2). Если при этом удается разделить про-
цессы настройки, когда ищется нужное значение параметра, и
процессы функционирования, когда параметр остается на най-
денном фиксированном уровне, то приведенное толкование мож-
но сохранить как удовлетворительное. Если же, как это чэ.сто
бывает, например, в системах организма и в других биосисте-
мах, процессы настройки и функционирования системы нельзя
74
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
[ГЛ. 3
разделить, то разница между параметрами и переменными ста-
новится иллюзорной, и такое разделение основывается, пожалуй,
только на традициях и интуитивных представлениях исследова-
телей. Одна и та же величина, описывающая биосистему, может
выступать в одних случаях как переменная, в других — как па-
раметр: Например, масса животного является параметром в за-
дачах управления искусственным кровообращением и перемен-
ной— при исследовании онтогенетических процессов.
При описании биосистем с помощью математических моделей
переменные и параметры биосистемы являются переменными и
параметрами описывающих ее уравнений.
3.2. Динамические системы.
Схемы моделирования
Современные способы изображения биологических систем в
виде графических схем ведут свое происхождение, с одной сто-
роны, от традиционных изобразительных методов, используе-
мых в биохимии, физиологии-и других биологических дисцип-
линах [5, 9, 12, 49, 89, 234, 247], и, с другой, от структурных
схем, получивших в свое время широкое распространение для
программирования аналоговых вычислительных машин [42, 90,
228].
Применяемые в настоящее время схемы во многом сохра-
няют точность и строгость, присущие схемам аналогового моде-
лирования, но при этом не предназначены для узких целей ис-
следования систем с помощью тех или иных вычислительных
средств, а имеют общий смысл наглядного представления струк-
турных свойств и особенностей исследуемых моделей.
Для получения схемы моделирования какого-либо процесса
или системы требуется их математическое описание. Такое опи-
сание объектов часто дается в форме дифференциальных урав-
нений /г-го порядка, разрешенных относительно старших произ-
водных, т. е.
dk
(*ь ....^Г1’; • • •; > *(ГП; 4
dz
(3.1)
1=1, 2.....tn,
где х,— переменная системы (зависимая переменная), z — не-
зависимая переменная, х', х", . .., х^-1) — производные X/ по 2
от первого до (й — 1)-го порядка.
Систему уравнений (3.1) всегда можно представить в виде
системы уравнений первого порядка.
3.21
ДИНАМИЧЕСКИЕ СИСТЕМЫ. СХЕМЫ МОДЕЛИРОВАНИЯ
75
Для этого достаточно ввести новые переменные
Тогда уравнения (3.1) принимают вид
dyi, k— 1
dz 10, ^20, •••, ^i.fe-ь Ут0, У ml, •••> Уда,*-!; 2)-
(3.3)
Вместе с системой уравнений (3.2) уравнение (3.3) и представляет собой си-
стему, составленную из km уравнений первого порядка.
Поэтому всегда можно считать, что исследуемая система
описывается уравнениями вида
dx,
= х2, .хт; z), i=l, 2, .... т. (3.4)
Если независимой переменной z является время t, то система
дифференциальных уравнений (3.4), переписанная в виде
x — fi (хь х2, .... хт; I), i — 1, 2, ..т, (3.5)
называется динамической системой.
Динамические системы описывают протекание физических,
химических, биологических явлений и процессов во времени.
Если уравнения приведены к виду (3 4) или (3.5), то говорят,
что они представлены в так называемой нормальной форме
Коши, Решение системы (3.5) при некоторых ограничениях на
вид функций f, полностью определяется значениями функций
Xi, х2, ..., хт в некоторый момент времени /0, которые назы-
ваются начальными условиями системы (3.5). Начальные усло-
вия обычно записываются следующим образом:
X] (^о) = Х]0,
Х-2 (/0) = Х20, (3 6)
•*чп(А)) == ХтЬ'
76
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
1ГЛ. 3
Переменные %i, Х2...хт в (3.5) называются иногда фазо-
выми координатами системы. Задание совокупности фазовых
координат определяет состояние системы в текущий момент
времени.
Графические схемы, используемые для описания структуры
системы, должны с той или иной степенью полноты отражать
связи между ее фазовыми координатами. Различают несколько
Рис. 3.2. Символы, используемые в схемах моделирования. В левом столбце—обозначения,
наиболее часто используемые в схемах моделирования, в правой — обозначения, приме*
няемые для программирования аналоговых вычислительных машин, а) Суммирование (или
вычитание); б) умножение (или деление), во втором случае на схеме желательно указывать
делимое и делитель; в) интегрирующее звено; а) нелинейное преобразование; д) постоян-
ные коэффициенты.
типов графических представлений системы. Если в схеме отобра-
жаются лишь основные структурные связи между некоторыми
переменными, дающие представление о характере связей в си-
стеме, то такая схема называется блок-схемой системы. При
изображении блок-схемы можно не придерживаться каких-либо
строго установленных обозначений для отдельных элементов и
узлов. Наиболее приемлемыми символами блок-схемы можно
считать изображение отдельных частей схемы в виде прямо-
угольников произвольного размера с указанием операций, вы-
полняемых данным узлом [42]. Вообще говоря, в этом случае
3.2] ДИНАМИЧЕСКИЕ СИСТЕМЫ. СХЕМЫ МОДЕЛИРОВАНИЯ 77
можно ограничиваться и словесным описанием функций, вы-
полняемых блоком. Степень детализации элементов блок-схемы
может быть различной, что определяется только точкой зрения
исследователя.
Схема моделирования является строгим графическим отра-
жением уравнений системы. Символы, используемые для от-
дельных элементов, должны быть стандартизированы, а все эле-
менты структуры должны г меть один и тот же уровень детали-
зации. Обычно при построении схем моделирования исходят из
описания систем в нормальной форме Коши и используют набор
стандартных обозначений элементов, показанный на рис. 3.2.
Схема моделирования строится следующим образом: каждой
фазовой координате ставится в соответствие один интегрирую-
щий элемент (интегратор), а затем с помощью стандартных
символов графически изображаются связи между интеграторами
согласно правым частям уравнений (3.5). В различных областях,
где применяются схемы моделирования, формулируется ряд
специфических требований, более жестко регламентирующих
способы изображения систем (см., например, [42, 228]). Мы
этих требований придерживаться не будем. В наших схемах мы
будем использовать, в основном, элементы, приведенные в пер-
вом столбце рис. 3.2, хотя будем применять обозначение инте-
гратора в любой из двух возможных форм — в виде прямоуголь-
ника или в виде треугольника.
Пример 3.2.1. Изобразим блок-схему и схему моделирования для про-
стой динамической системы. Возьмем описание элемента кровеносного сосуда
в форме так называемого эластичного резервуара [253]. Пусть переменная х
описывает количество крови, находящееся в данном отрезке сосуда — резер-
вуаре. Тогда
х = w — у, (3.7)
где w и у — темп притока и темп оттока крови для данного резервуара, со-
ответственно. Будем считать приток крови независимым внешним сигналом.
Темп оттока прежде всего зависит от давления в резервуаре, которое опре-
деляется соотношением
Р = -Q- (х — а), (3.8)
где Р— давление в резервуаре, С — коэффициент, определяющий эластич-
ность кровеносного сосуда (емкость), а — константа, так называемый нена-
пряженный объем резервуара, т. е. такой объем крови, который может
содержаться в резервуаре без повышения давления Р. Тогда темп оттока
равен
У=-^-(Р~Ь), (3.9)
где R — сопротивление сосуда току крови, b — константа, равная давлению
в следующем резервуаре, куда оттекает кровь.
78
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
[ГЛ. 3
Система описывается тремя уравнениями (3.7) — (3.9). Чтобы привести
описание системы к форме Коши, в нашем случае достаточно подставить
в)
Рис. 3.3. Примеры блок-схемы системы и схем моделирования. Изображение системы в виде
блок-схемы позволяет использовать произвольные символы с разной степенью детализа-
ции (а). В схемах моделирования применяется изображение элементов только с помощью
стандартных символов; на рис. б) и в) показаны два возможных варианта схем моделиро-
вания эластичного резервуара для системы кровообращения.
выражения (3.8) и (3.9) в (3.7). Тогда получаем
1 . ь
х = ® — ~£>С (х — а) +-дГ (3.10)
Один из возможных вариантов блок-схемы и два варианта схем модели-
рования эластичного резервуара приведены на рис. .3.3.
3.3. Линейные системы и линеаризация
Система называется линейной, если для нее справедлив
принцип суперпозиции: если у (ufe) — реакция системы на воздей-
ствие Vk, то
саУЫ (3.11)
при любых I, Cl, ..., Ct И При любых функциях О1((), ...,
В задачах биокибернетики линейные системы часто описы-
ваются дифференциальными уравнениями вида
г I п
а.цУ1 + X bivVv + 2 CinWp, i = l, 2, ..., Г, (3.12)
/=1 V=1 Ц=1
где сигналы у, v и w имеют тот же смысл, что и на рис. 3.1, и
являются переменными описываемой системы. Коэффициенты
Цц, бы, — параметры системы.
3.3]
ЛИНЕЙНЫЕ СИСТЕМЫ И ЛИНЕАРИЗАЦИЯ
79
Уравнение (3.12) описывает, вообще говоря, только один част-
ный, хотя и наиболее распространенный тип линейных систем.
В общем случае среди систем, удовлетворяющих принципу супер-
позиции, можно кроме систем (3.12) назвать линейные дискрет-
ные системы, линейные системы с запаздыванием и т. д. [46].
Все живые системы в своих проявлениях обладают нелиней-
ными свойствами, когда принцип суперпозиции нарушается.
Наиболее частым видом нелинейности является насыщение
(рис. 3.4), когда в определенном диапазоне изменений входного
Рис. 3.4. Насыщение в биологической системе, а) Переменная в системе действует на
другую переменную, Х2; б) реакция племенной Х2 нелинейна и обладает свойством насы-
щения. При различных значениях Xi одно и то же приращение (вертикальные пунктир-
ные линии) приводит к разным приращениям х2 (горизонтальные пунктирные линии);
в) коэффициент передачи от х{ к х7, обозначенный через k, отличен от нуля только
в ограниченной области изменений X , обозначенной Q. За пределами области Q влияние Xi
на х2 отсутствует: эффективная связь от Х( к Х2 разрывается, исчезает.
сигнала Х{ элемента системы имеется четко выраженная зави-
симость выходного сигнала Х2 от входного, а за пределами этого
диапазона выходной сигнал попадает в область насыщения и
перестает изменяться. Часто в некотором диапазоне изменений
сигнала Xi, например в окрестности точки Х° на рис. 3.4, зависи-
мость Х2 = /'(Х|) почти не отличается от линейной; в этом ма-
лом диапазоне исходная зависимость может быть линеаризована:
х2 = kx\, (3.13)
где X], х2— приращения сигналов Х{ и Х2, отсчитываемые от не-
которой точки (Х°, X°), в которой производится линеаризация, а
k — угол наклона кривой /(Xi) в этой точке. Если провести ли-
неаризацию последовательно во всех точках, то окажется, что
величина k в каждой из точек различна; зависимость коэффи-
циента k от положения точки линеаризации приведена на
рис. 3.4, в.
Здесь и далее переменные в исходной нелинейной системе
обозначаются прописными буквами, переменные в линеаризо-
ванной системе — строчными.
Часто приходится линеаризовать зависимости от двух или
большего числа переменных. Например, биомасса в единице
объема есть произведение числа особей на массу отдельной
80
классическая теория управления и биосистемы
i [ГЛ. 3
особи; темп поступления кислорода в орган с кровью равен
произведению объемного кровотока в единицу времени на
количество кислорода, извлекаемого из единицы объема крови
а) б)
Рис. 3.5. Линеаризация произведения двух величии. После линеаризации нелинейный эле-
мент— множитель (а) —заменяется линейным элементом —сумматором входных сигналов
с соответствующими коэффициентами (б).
и т. д. В обоих случаях при рассмотрении небольших вариаций
приходится линеаризовать произведение
X3=feX1X2. (3.14)
На рис. 3.5 показано, как это делается: приращения всех сигна-
лов относительно исходных значений X°i, Х2, Х3 связаны соот-
ношением
х3 = к1х1 +/г2х2, (3.15)
причем ki = kX2, k2—kX°].
Подробнее о нелинейности и линеаризации рассказывается в
гл. 3 учебника [130], предназначенного для специалистов-био-
логов. Мы еще раз вернемся к этому вопросу в разд. 5.3.
3.4. Обратная связь
Передача сигнала в системе в направлении от входа к вы-
ходу называется прямой связью. Обратной связью называется
любая передача сигналов в обратном направлении, т. е. от вы-
хода системы к входу (рис. 3.6, а). В сложных системах может
возникнуть ситуация, когда та или иная связь между двумя
переменными может трактоваться как прямая (идущая в на-
правлении от входа к выходу) или обратная (рис. 3.6,6). Это
зависит от того, влияние какого входа на какой выход или на
какую внутреннюю переменную рассматривается. Поэтому при
исследовании сложных систем, имеющих множество входов и
выходов, такие термины, как «положительная» или «отрицатель-
ная» обратная связь, обычно не употребляются.
Однако не надо думать, что термин «обратная связь» по
мере перехода к исследованию все более и более сложных си-
стем полностью исчезнет. Нет, это понятие по-прежнему исполь-
зуется и будет использоваться при описании простых систем,
3.4]
ОБРАТНАЯ СВЯЗЬ
81
имеющих относительно небольшое число переменных, например,
одноконтурных систем.
Понятие отрицательной обратной связи удобно для понима-
ния и качественного осмысливания процессов в биологических
В)
Рис. 3.6. Обратная связь и сложная структура системы, а) В простой системе с одним
входам и одним выходом понятие обратной связи оказывается очень полезным: выделив
прямой канал acd, можно указать обратную связь fb, б) в сложной системе с несколькими
входами и выходами некоторая связь может рассматриваться как прямая или как обратная
в зависимости от того, какие вхоты и выходы берутся в качестве основных. Элемент d
играет роль прямой связи для капала aefl и обратной (cd)—для канала bgkh.
а;
системах; успешное применение этого понятия связано с необхо-
димостью выделения из сложной системы относительно простых
элементарных подсистем. Однако, вычленяя из системы ее от-
дельные компоненты и рассматривая их порознь, мы фактически
теряем возможность исследовать це-
лостный объект.
Пожалуй, наиболее распростра-
ненным видом обратной связи в био-
системах является так называемая
параметрическая обратная связь
(рис. 3.7). Здесь в прямом канале
от входа Vi к выходу Х\ имеется па-
раметр k, который может меняться
под действием некоторой перемен-
ной Х2, т. е. k = й(Х2). Тогда увели-
чение (или уменьшение) входного
сигнала Vi сначала по прямому ка-
налу вызывает увеличение (или
уменьшение) Хь но затем перемен-
ная Х2, изменившись под действием
Рис. 3.7. Параметрическая отрица-
тельная обратная связь. В прямом
канале ог входа Vi к выходу
имеется некоторый параметр —коэф-
фициент k. Увеличение Vi вызы-
вает увеличение Xit но возникаю-
щее вследствие этого изменение пере-
менной Л2 вновь приводит к умень-
шению — параметры прямого ка-
нала изменяются так, что k умень-
шается.
Xi, приведет к противопо-
ложно направленному изменению k (соответственно, к его
уменьшению или увеличению). Эффект действия такой отрица-
тельной связи — уменьшение изменения переменной Xi при дей-
ствии входа Уь
Пример 3.4.1. Типичным примером отрицательной обратной связи па-
раметрического типа в организме является изменение одного из наиболее су-
щественных параметров системы кровообращения, сопротивления сосудов, при
изменении кислородного режима тканей.
82
классическая теория управления и биосистемы
[ГЛ. 3
Рассмотрим процессы поступления и утилизации кислорода в ткани. Темп
потребления кислорода гО2 будем считать заданным и равным ш. Темп по-
ступления кислорода qO2 в клетки в первом приближении определяется сле-
дующими двумя факторами: градиентом напряжения кислорода гч = рО2 ме-
жду тканями и артериальной кровью и количеством крови Q, протекающим
через сосуды тканей. Количество крови Q зависит в свою очередь от пара-
метров системы — величины артериального давления Р и сосудистого сопро-
тивления R, величину которого можно связать с количеством кислорода в
тканях Х| [306] некоторой функцией /?(Х|). Взаимодействие перечисленных
выше переменных и параметров может быть представлено схемой на рис. 3.8, а.
Рис. 3.8. Система кислородного сложения как система с параметрической обратной связью,
а) Стрелкой в верхней части рисунка отмечен основной контур обратной связи; сопро-
тивление сосудов—параметр R—меняется в системе из-за наличия параметрического кон-
тура, показанного круглой стрелкой в нижией части рисунка; величина R изменяется под
действием сигнала х —внутренней переменной системы, б) увеличение интенсивности
параметрической обратной связи (коэффициент Ь) приводит к тому, что на одно и то же
изменение Ди система отвечает все меньшим и меньшим сдвигом Дх,.
Основной контур отрицательной обратной связи (обычный контур пассивной
регуляции) показан в верхней части рисунка. Нижний контур представляет
собой параметрическую отрицательную обратную связь.
Рассмотрение примера можно продолжить и дальше. Аппроксимируем
функциональную зависимость R(xt) линейной функцией R = a -j- bx, и про-
иллюстрируем влияние отрицательной параметрической связи на гомеостати-
ческие свойства системы.
Уравнение системы в стационарном режиме имеет вид
Xi = (vi — Xi) &1Q — ш = 0,
откуда после подстановки соотношения
Р _ Р
R а + bxi
получаем уравнение
ktP а
х' = kj> + b Vl ~ kiP + b w' (ЗЛ6>
Интенсивность параметрической обратной связи увеличивается при увеличе-
нии параметра Ъ. При отсутствии параметрической обратной связи b = 0, и
а
X; = Pi — f,~p~ W. (3.17;
Зависимость в этом случае представляет собой линейную функцию с
коэффициентом наклона, равным 1. При наличии параметрической обратной
3.41
ОБРАТНАЯ СВЯЗЬ
83
связи Ь > 0, и зависимость х^щ) по-прежнему остается линейной, однако
угол наклона ее к оси у1 с ростом b уменьшается. На рис. 3.6, б приведено
семейство Xi(Vi) для разных значений коэффициентов Ь. Увеличение х, приво-
дит к тому, что зависимость переменной от условий среды О| стаиовитси
меньше.
Для организма человека можно оценить среднее значение коэффициентов
а и Ь, подставив в формулу Q = Р(а -ф Ьх{) усредненные значения всех пере-
менных для двух стационарных режимов, например, покоя (и> = 12,5 мл/мин)
и физической нагрузки (и> = 1733 мл/мин) [253]. Тогда Q(l) = 1,4 л/мин,
рт — ЮО мм рт. ст., х*,1* = 40 мм рт. ст., Q<2> = 18,5 л/мин, Р<2> = 100 мм
рт. ст., х^ = 38 мм рт. ст. и я =—1330, b = 35. Угол наклона прямой
хДп,) в этом случае уменьшается по сравнению с 1:
ktP _ fef ЮО _ 74
ktP 4- b ~ kx • 100 + 35 109 — °’68- (3-18>
Однако наибольшее распространение при моделировании
биологических систем получила отрицательная обратная связь
по отклонению от уставки (рис. 3.9,а).
Задающий входной сигнал w сравнивается с выходом систе-
мы у; сигнал е = w — у называется сигналом ошибки или рас-
согласованием. Сигнал рассогласования поступает на вход ре-
гулятора, который вырабатывает управляющий сигнал и. Управ-
ление и поступает на вход объекта. На объект действует и
внешний возмущающий сигнал V.
Пример 3.4.2. Представим в виде схемы с отрицательной обратной
связью механизм пассивного регулирующего действия концентрации кисло-
рода в тканях некоторого органа на темп притока (рис. 3.9,6), с которым
мы столкнулись в примере 3.2.1.
Рис. 3.9. Обратная связь по отклонению от уставки в простой системе, а) Общая схема;
б) пассивная регуляция темпа поступления кислорода в ткани.
Пусть орган потребляет кислород с темпом и>, а целью управления бу-
дем считать обеспечение поступления кислорода с тем же темпом. Тогда темп
поступления у кислорода является выходом объекта. Разность между тем-
пами потребления и поступления, накапливаясь, дает напряжение кислорода
в тканях
t
* = J (У — ®) dt + х0, (3.19)
о
где х0 —исходное значение концентрации кислорода (при t = 0), У —объем
тканевого резервуара.
84
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
[ГЛ. 3
Уравнение регулятора (3.19) можно записать в виде
х = — w). (3.20)
Объект описывается уравнением
y = k(v — x), (3.21)
где k — коэффициент, v — напряжение кислорода в среде, окружающей рас-
сматриваемый тканевый резервуар.
Принцип обратной связи получил чрезвычайно широкое рас-
пространение при методологическом [1, 43, 112, 128, 137, 156]
и математическом описании биологических систем и явлений на
всех уровнях организации жизни и в свое время был признан
ведущим принципом саморегуляции и самоуправления в живой
природе [5, 33, 122, 154, 159, 226, 320]. Почти во всех случаях
применение идеи обратной связи проходило через этап использо-
вания простейшей одноконтурной модели отрицательной обрат-
ной связи по отклонению от уставки. Так, на организменном
уровне этот принцип был использован для моделирования прак-
тически всех сколько-нибудь важных систем — дыхания [60, 301
и др.], кровообращения [6, 83, 105, 107, 157, 214, 355, 367 и др.],
терморегуляции [74, 79, 84, 309, 361, 362 и др.], водно-солевого
обмена [305, 330 и др.], энергообразования [63, 243 и др.], си-
стемы сахара крови [13, 70, 225, 261]. Этим же принципом руко-
водствуются при рассмотрении процессов старения [118, 227],
лечения заболеваний [14, 155, 183, 199, 262] и при разработке
систем управления искусственными органами (см., например,
[35, 50, ИЗ, 252, 311]). Принцип обратной связи был использо-
ван и для объяснения протекания эволюционного процесса
[54, 242, 248], хотя, насколько известно автору, математических
моделей в форме простой отрицательной обратной связи разра-
ботано не было. Математический анализ эволюционного
процесса пошел по другому пути (см., например, [170, 186а,
255]).
Возвращаясь к вопросу о формах обратной связи и их ис-
пользовании в моделях биологических процессов и явлений,
нельзя не отметить, чтс многие критические замечания в адрес
математических методов исследования сложных явлений в жи-
вых системах по существу относились, да и относятся не столько
к принципиальным вопросам возможности и необходимости ис-
пользования этих методов, сколько к конкретным формам их
проявления, и прежде всего к использованию излишне упрощен-
ных способов описания сложных форм организации живой при-
роды — таких, как простейшая модель отрицательной обратной
связи по отклонению от уставки.
3.6]
УСТОЙЧИВОСТЬ
85
3.5. Устойчивость
В теории управления исследование устойчивости относится
к следующей ситуации. Сначала на систему действует некоторое
возмущение, нарушающее равновесие в ней. Затем это возмуще-
ние снимается, перестает действовать. Однако в системе уже
возникло некоторое неравновесное состояние. Это состояние
представляет собой начальные условия для свободного движе-
ния системы — процесса регулирования. Если после достаточно
малого возмущения в системе восстанавливается тот же режим,
который поддерживался в системе до начала действия возмуще-
ния, процесс называется сходящимся или устойчивым, в против-
ном случае — неустойчивым.
Строгое определение устойчивости дается обычно следующим
образом.
Пусть дана система обыкновенных дифференциальных уравнений
= h (xi, х2> • • •> хп), (3.22)
где функции fi таковы, что производные
вуют и непрерывны, и пусть ф;(/), i = 1,
удовлетворяющее при t = to условиям
dfi
dxk ’
i, k = 1, 2,
..., n, сущест-
2, ..., n, — решение этой системы,
<MZo)=<₽°’ l’ = 1-2.......n.
(3.23)
Решение ф,(/) называется устойчивым no Ляпунову, если для любого
е > 0 можно указать такое 6(e) > 0, что для всякого решения xi(t), началь-
ные условия х(’ которого удовлетворяют неравенствам
|х?-ф?| < б (е),
для всех t t0 справедливы неравенства
Iх/(0 — Ф;(0|<е, /=1, 2, ...,я.
(3.24)
(3.25)
Это означает, что решения, близкие по своим начальным значениям, остаются
близкими и в дальнейшем.
Если при сколь угодно малом б > 0 хотя бы для одного решения х<. это
неравенство не выполняется, то решение ф>(/), i = 1, 2..п, называется
неустойчивым.
Если решение q>i(t) не только устойчиво, но дополнительно удовлетворяет
условиям
lim | хД0-<р. {/) | = 0, » = 1, 2, ...,я, (3.26)
при | X; — <р° | < б, то решение Ф<(0 называется асимптотически устойчивым.
В популярном изложении иногда говорят, что устойчивые процессы, не
обладающие, однако, свойством асимптотической устойчивости, находятся на
грани устойчивости [179].
Наиболее просто понятие устойчивости для линейной систе-
мы с постоянными параметрами. Если корни ее характеристиче-
86
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
{ГЛ. 3
ского уравнения лежат в левой полуплоскости (см. разд. 5.5), то
все процессы, независимо от величины начальных отклонений,
вызванных внешними воздействиями, оказываются устойчивыми.
Тогда и сама система называется устойчивой.
Для нелинейных систем устойчивость поведения — понятие
более сложное. Движения в нелинейной системе могут быть
устойчивыми в одной области изменения переменных и неустой-
чивыми— в другой. Поэтому для нелинейного случая рассматри-
вают не устойчивость системы, а устойчивость движений или
траекторий.
Удобным способом наглядного изображения движений яв-
ляется метод фазового пространства. Описание этого способа на
примерах простых биосистем имеется в работе [179]. Суть ме-
тода заключается в следующем. Рассматривается пространство,
координатами в котором являются фазовые переменные системы
Xi, х2, ..., хт в соответствии с приведенным в разд. 3.2 опреде-
лением. Задание текущих величин фазовых координат опреде-
ляет в этом пространстве некоторую точку Р — изображающую
точку. При изменении фазовых координат во времени изобра-
жающая точка описывает в фазовом пространстве траекторию,
по виду которой мо^кно судить о свойствах системы. Наиболее
удобен этот метод для исследования систем второго порядка,
когда фазовых координат всего две, и фазовое пространство пре-
вращается в фазовую плоскость. Рассмотрим этот случай под-
робнее.
Линейная система второго порядка описывается уравнением
х + 2gcox + <о2х = 0. (3.27)
Пусть сначала | = 0. Введем переменные x\ = x(t), х2 = х (/)
и перепишем (3.27) в виде
х2 + co2Xi = 0. (3.28)
Для исследования системы методом фазовой плоскости надо
построить графическую зависимость, связывающую переменные
Xi и х2. Найдем связь между Xi и х2 следующим образом. Умно-
жим первое слагаемое (3.28) на х2, а второе — на равное ему xf.
х2х2 + <o2xix1 = 0 (3.29)
или
[2 Т
т-+4<|=о- (3-30)
Проинтегрировав (3.30), находим искомое соотношение между
X] и х2:
X2 + С^Х; = с2. (3.31)
3.5]
УСТОЙЧИВОСТЬ
87
Это — уравнение эллипса. Ряд решений при различных значе-
ниях с, соответствующих разным начальным значениям Xi и Хг,
показан на рис. 3.10. Совокупность траекторий в плоскости
(хь х2) образует фазовый портрет системы.
Время служит параметром при движении вдоль траектории;
рост времени соответствует движению по часовой стрелке;
t3 > h > Л- Это ясно из того, что при х2 > 0 (т. е. при it > 0)
величина Xi должна возра-
стать.
Аналогично можно по-
строить фазовые портреты для
системы (3.27) и при произ-
вольных | и со. В зависимости
от знака и величины £ и со в
линейных системах существует
несколько типов особых точек;
по характеру фазового портре-
та можно судить об устойчи-
вости системы — эти данные
можно представить в виде
табл. 3.1.
Рис. 3.10. Пример фазовой плоскости для
простой линейной системы второго порядка»
Фазовые портреты системы второго порядка приводятся во
многих книгах, например, в [66, 179].
Та блица 3.1
№ п/п Значения параметров Особые точки Свойства системы
Величина £ Знак при (д’
1 5 = о + Центр Устойчива, но не асимптотически
2 0<£ < I + Устойчивый фокус Устойчива асимптоти- чески
3 -1<5<0 + Неустойчивый фокус Неустойчива
4 1<5 + Устойчивый узел Устойчива асимптоти- чески
5 5< —1 + Неустойчивый узел Неустойчива
6 5 = 0 — Седловая точка Неустойчива
7 5 = 1 — Устойчивая седловая точка Устойчива асимптоти- чески
8 5 = -1 — Неустойчивая седловая точка Неустойчива Устойчива асимптоти-
9 5®>о о2 = 0 Нет чески
10 5®<о и2 = 0 Нет Неустойчива
Пример 3 5.1 Система двух взаимодействующих видов описывается
классической моделью Вольтерра. Пусть, например, в проточном культиваторе
88 КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ [ГЛ. 3
растут два вида — бактерии и их враги (вирусы). Вирусы играют роль хищ-
ника, бактерии — жертвы. Если обозначить через X] численность хищников
в некоторых единицах, а через Х2 — численность жертв, то система описы-
вается уравнениями
Xx=X}(X2-d), Х2 = Х2(\ — Х^ —d). (3.32)
Параметр d характеризует скорость протока в культиваторе. При больших
скоростях протока (d > 1) вне зависимости от исходных значений X,L и Хг
происходит вымывание обеих культур; при d < 1 в системе происходят не-
затухающие колебания численностей обоих видов с амплитудой, определяемой
начальными условиями [180].
Фазовая плоскость с координатами Хь Х2 для системы (3.32) показана
на рис. 3.11. При малых d фазовые траектории замыкаются вокруг особой
точки Х°= 1 — d, Х!~ = d— центра (рис. 3.11, а). При больших d траектории
стягиваются к точке X, = Х2 = 0— устойчивому узлу (рис. 3.11,6).
Рис. 3.11 Фазовые портреты для системы, состоящей нз двух взаимодействующих популя-
ций в культиваторе, а) Фазовый портрет системы Вольтерра —при малых отклонениях от
особой точки график совпадает с фазовым портретом на рис. 3.10; б) устойчивые траекто-
рии при большом протоке через культиватор; в), г) при малых вариациях модели Воль-
терра (учет слабо влияющих факторов) фазовый портрет, показанный на рис. а), в окрест-
ности особой точки приобретает вид либо устойчивого фокуса (в), либо неустойчивого
фокуса (г), Оси координат па двух последних рисунках отсутствуют, так как масштабы
графиков увеличены.
Траектории б — устойчивы, так как любое отклонение от равновесной
точки (0, 0) с течением времени исчезает.
Траектории а устойчивы неасимптотически, но малые изменения парамет-
ров системы превращают центр либо в устойчивый (рис. 3.11, в), либо в не-
устойчивый (рис. 3.11, г) фокус.
Покажем, как это происходит, исследовав поведение линеаризованной си-
стемы в окрестности особой точки. Линеаризуем исходную нелинейную си-
стему (3.32), положив
x^Xi-XO х2==Х2-Х°, (3.33)
3.6]
ЧУВСТВИТЕЛЬНОСТЬ
89
(3.36)
где X, = 1 — d, X?) — d. Тогда вместо (3.32) получаем
Xi = (l — d) х2, x2 = —dxi, (3.34)
или
Xj + d (1 — d)xi = 0. (3.35)
Это уравнение можно записать в виде (3.27), где 5 = 0, со2 = d(l —d). Си-
стема находится на границе устойчивости (строка 1 в табл. 3.1).
Стоит, однако, учесть какие-либо слабые зависимости в исходной нели-
нейной системе, как в линеаризованном уравнении (3.35) появится член с хь
и система будет соответствовать либо строке 2 (рис. 3.11, в), либо строке 3
(рис. 3.11, г) в табл. 3.1. Такая ситуация возникает, например, если предпо-
ложить, что взаимодействие хищник — жертва подчиняется закону Моно (ско-
рость размножения хищника пропорциональна концентрации жертвы при ма-
лых концентрациях, при больших концентрациях скорость размножения хищ-
ника не зависит от концентрации жертвы). Получающийся в этом случае фа-
зовый портрет показан на рис. 3.11, г.
Тогда система вместо уравнения (3.32) описывается уравнением
*1 = Т+л7~ dXb
X^XAl-d)--^-.
Вводя новые переменные
Х1=Х1—Х°1, х2 = Хг — Х°2
и линеаризуя затем правые части (3.36) в окрестности особой точки
л л kd
получаем уравнения, описывающие движение системы в окрестности
точки,
= — (1 — d) х2,
х2 = — dxt + 2 (1 — d) х2
или
х\ — d (1 — d) %! — d (1 — d)2 Xi = 0.
Поскольку положительные значения концентрации Х2 возможны только прн
0 < d < 1, видим, что в (3.37) значения коэффициентов 5 и <о2 всегда отве-
чают строке 3 табл. 3.1, т. е. особая точка представляет собой неустойчивый
фокус.
3.6. Чувствительность
Чувствительность системы в теории управления — свойство
системы изменять свой режим и характеристики поведения при
изменении какого-либо параметра. Чем больше меняется режим
системы при изменении какого-либо параметра, тем более чув-
ствительна система к его изменениям [36]. Чувствительностью
называют иногда также и показатель, количественно характери-
зующий это свойство.
особой
90
КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
[ГЛ. 3
Рис. 3.12. Ситуации, в которых иссле-
дуется устойчивость и чувствительность
в теории управления. При исследовании
устойчивости рассматривается свобод-
ное движение системы —возмущение
должно быть снято до начала процесса
управления (/). При исследовании чув-
ствительности рассматривается поведе-
ние системы при изменившемся значе-
нии какого-либо параметра, т. е. дей-
ствие возмущающего фактора продол-
жается постоянно (2).
Задача исследования чувствительности ставится в теории
управления в следующих условиях. Задается некоторая система,
называемая исходной. В этой системе происходит изменение (ва-
риация) некоторых параметров; измененная система называется
варьированной системой. Процессы в исходной и варьированной
системах, возникающие в ответ на действие одного и того же
входного сигнала, различаются между собой; их разность назы-
вается дополнительным движе-
нием.
Таким образом, ситуации, в
которых проявляются устойчи-
вость и чувствительность, отли-
чаются тем, что при исследова-
нии устойчивости возмущение,
вызвавшее отклонение движения,
сразу же снимается и исследует-
ся свободное движение системы.
В задаче исследования чувстви-
тельности действие возмущаю-
щего фактора — изменившегося
значения параметра внешнего
сигнала—продолжается постоян-
но (рис. 3.12).
Разницу между устойчивостью
и чувствительностью можно про-
иллюстрировать на следующем
наглядном примере*). Физиоло-
гические процессы в организме
животного, впадающего в зим-
обладают высокой чувствитель-
Действительно, зимой, когда тем-
остается пониженной, в системе
нюю спячку, устойчивы, но
ностью к температуре среды,
пература среды непрерывно
устанавливается стационарный режим, в котором значения фи-
зиологических переменных в организме существенно изменены
по сравнению с исходным летним периодом. Стоит, однако, снять
внешнее температурное возмущение, как это происходит весной,
и в физиологических системах организма возникает свободное
движение от зимних начальных условий, заканчивающееся вос-
становлением исходного состояния. Это движение асимптотиче-
ски устойчиво. Возникающие ситуации подробно рассмотрены
на рис. 3.13.
Перейдем теперь к рассмотрению основных понятий, связан-
ных с чувствительностью систем, как это обычно делается в
теории автоматического управления [46, 176, 207].
*) Этим примером автор обязан Ю. Кейкспайку.
3.61
ЧУВСТВИТЕЛЬНОСТЬ
9!
Пусть, например, исходная система описывается уравнением
(3.38)
при начальном значении Хо, а — параметр. Если в некоторый
г 20 10 а) 0 -10 -20 б) 0 40 20 е) о ч Зима Лето
i0 4 и ^3 U\
I Е п II t 11
40_ -20 11 11 .
: 1 : И ' — 1 1 1 1 II
tfl ^1^2 $$ ^4^5 $
Рис. 3.13. Сравнение понятий устойчивости н чувствительности на примере физиологических
систем организма животных. Показано условное изменение температуры среды в течение
года (а); температура тела торпидных (впадающих в спячку) животных (6); температура
тела животных, сохраняющих зимой активный образ жизни (в). Принято, что «анормальной»
является относительно высокая летняя температура среды, а зимнее снижение темпера-
туры рассматривается как действие постоянного возмущения — ДО • 1 (/). Кроме того, рас-
сматриваются кратковременные возмущения температуры в моменты времени ti и ti. Зимним
условиям соответствует интервал времени (to, t»), летним — интервал t tj. Анализ чув-
ствительности системы терморегуляции к изменению параметра ДО корректен и а времен-
ном интервале (to, t); анализ устойчивости свободного движения в системе —на интерва-
лах (t2, t3), (t", и прн t > ts. Все свободные движения, показанные на рисунке, устой-
чивы (это относится к переходным процессам, начинающимся в точках ti, t», t5). Система
терморегуляции на рис. 3.13, б чувствительна к изменению параметра ДО (процесс, начи-
нающийся в точке to), а система на рис. 3.13 в нечувствительна к этому параметру. К рас-
смотрению примера можно вернуться еще раз после прочтения всего текста данного
раздела.
момент времени величина а получила приращение Да, то варьи-
руемое движение описывается уравнением
= а + Да), (3.39)
а дополнительное движение
Дх(/) = х(/)-х(/). (3.40)
Если вариация Да настолько мала, что можно ограничиться ли-
нейным приближением при разложении x(t) и x(t) в ряд Тей-
лора по а, то из (3.40) можно получить уравнения первого при-
ближения
A,=O^«-A“- <ЗЛ1>
Функция времени
«(0 = ^- (3.42)
92 КЛАССИЧЕСКАЯ ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ [ГЛ. 3
называется функцией чувствительности координаты х к пара-
метру а.
С помощью функций чувствительности можно оценить сте-
пень влияния того или иного параметра на координаты системы
и найти отклонение процессов в варьированной системе от про-
цесса в исходной системе как функцию времени.
Ниже в этой книге мы, как правило, будем .интересоваться
зависимостью переменных х системы от изменяющихся условий
внешней среды, V. Покажем, как в этом случае можно сформу-
лировать задачу так, чтобы свести ее к анализу чувствительно-
сти системы. Допустим, что на систему действует внешний
сигнал в виде ступенчатого возмущения величиной и. Будем рас-
сматривать величину v в качестве параметра системы, на кото-
рую действует одно и то же внешнее воздействие — единичное
ступенчатое возмущение. Тогда
v (0 = v • 1 (/),
где символ 1 (/) означает единичный скачок:
fO, t < О,
/>0. (3,43)
Теперь мы можем сравнивать поведение исходной и возмущен-
ной систем, отличающихся величиной и, на одно и то же воз-
действие !(/). Если нас интересует чувствительность переменной
х к изменениям величины v, то теперь можно воспользоваться
формулой (3.42).
Пример 3.6.1. Вернемся к схеме пассивной регуляции темпа поступле-
ния кислорода в ткани, показанной на рис. 3.9, б. Переменная х согласно
(3.20) и (3.21) описывается уравнением
х = -рг [k (о — х) — а»), (3.44)
решение которого при нулевых начальных условиях имеет вид
х(/) = (о--у)[1-е"~']. (3.45)
Пусть нас интересует чувствительность переменной х к изменениям внешней
среды. Вычислим функцию чувствительности (3.42). Дифференцируя, напри-
мер, (3.45) по параметру о, получаем
График этой функции чувствительности приведен на рис. 3.14.
Чтобы аналитически построить функции чувствительности
в системах управления, надо решать так называемые уравне-
ния чувствительности, имеющие для случая (3.38) следующий
3.6]
ЧУВСТВИТЕЛЬНОСТЬ
93
вид:
du d / dx \ й/
di dt \ да) дх
да
(3.47)
Это линейные дифференциальные уравнения с переменными ко-
эффициентами, решение которых практически нецелесообразно
из-за большой сложности. Поэтому в теории управления разра-
ботан ряд методов получения функций чувствительности без
решения
тельности
уравнений чувстви-
([46], часть II,
[207]).
Для исследования биологи-
ческих систем сам вид функ-
ции чувствительности может
оказаться несущественным, до-
Рис. 3.14. Функция чувствительности для
системы регуляции темпа поступления кисло-
рода, показанной на рис. 3.9, б.
статочно лишь знать ее основ-
ные характеристики, такие как
время затухания или устано-
вившееся значение. Так, на
рис. 3.14 время затухания функции чувствительности составляет
от 3 до 5 постоянных времени, равных V/й; установившееся зна-
чение равно 1.
Особую роль при исследовании зависимости биосистем от
условий внешней среды играет установившееся значение функ-
ций чувствительности. Оно определяет сдвиги стационарного
режима системы при стационарном изменении условий внешней
среды.
Пусть х — стационарное значение внутренней переменной
системы, Ах— его изменение при изменившихся условиях среды,
и пусть возмущающее действие среды описывается ступенчатым
изменением возмущающего входа v на величину Ап. Тогда ста-
ционарное состояние системы смещается на величину Ах:
Ах =^~ • Ди,
dv ’
(3.48)
где -зг-= Нт «(/)— коэффициент передачи от внешнего входа —
/->00
возмущающего сигнала v — к переменной х. Ниже частную про-
дх
изводную , определяющую изменение стационарного значе-
ния переменной х при стационарном изменении входа v, мы бу-
дем называть статическим коэффициентом чувствительности или
просто коэффициентом чувствительности х к и и обозначать
символом <т.
Для получения коэффициентов чувствительности в устойчи-
вой системе достаточно рассмотреть только стационарные
режимы.
94
классическая теория управления и биосистемы
[ГЛ. 3
Продолжим рассмотрение предыдущего примера. Коэффициент чувстви-
тельности можно получить из (3.46):
дх дх (О
1.
Одиако проще рассмотреть непосредственно стационарный режим системы.
В таком режиме темп поступления вещества равен темпу его утилизации,
w = у. Поэтому из (3.20) и (3.21) имеем для стационарного режима
w дх ,
-т, откуда ¥»1.
В этом рассмотрении мы представили зависимость перемен-
ной х в стационарном режиме от входа v как предел функции
чувствительности (3.42) при /~>оо. При анализе биологических
систем, однако, можно воспользоваться и другой терминологией,
рассматривая в общем случае передачу сигнала от входа Vj к пе-
дх,
ременной хи Тогда можно величину считать коэффициен-
том передачи от входа системы Vj к переменной xt. Суть дела
при таком рассмотрении не меняется. Ниже мы будем пользо-
ваться терминолбгией чувствительности, поскольку при иссле-
довании гомеостатических свойств систем это позволяет сохра-
нить буквальную формулировку гомеостаза, принятую при опи-
сании физиологических систем и других биологических систем
организменного уровня (см. также разд. 8.1 и 7.4).
Важность исследования чувствительности для биологических
систем объясняется тем, что такая постановка задачи близка по
смыслу к задачам анализа сохранительных характеристик жи-
вых систем, зависимости переменных их внутренней среды от
внешних условий. В настоящее время можно указать целый ряд
работ, в которых исследуется чувствительность биосистем. Одно
из направлений таких поисков связано с определением факто-
ров, оказывающих наибольшее влияние на те или иные выход-
ные сигналы системы. Цель подобных работ обычно заклю-
чается в том, чтобы выбрать наиболее эффективные управляю-
щие переменные (например, для управления процессом
теплообмена в системе жизнеобеспечения в космосе [126]),
в оценке качества управления в системе (например, чувстви-
тельность процессов в сердечно-сосудистой системе человека
и животных [106, 246, 268, 342], чувствительность процессов
фармакокинетики [365], управление в ферментных системах
[310]), или, наконец, в том, чтобы решать вопросы прогнозиро-
вания поведения физиологических систем [323]. При таком
подходе о качестве системы можно судить только после упоря-
дочения функций чувствительности, организации способа оценки
степени влияния параметров системы на ее состояние. Одним
3.6]
ЧУВСТВИТЕЛЬНОСТЬ
95
из удобных способов такой оценки, такого упорядочения яв-
ляется суждение по статическому коэффициенту чувствитель-
ности. Общие вопросы упорядочения функций чувствительности
рассмотрены в [207]. Вопросы взаимосвязи чувствительности
системы и ее надежности обсуждаются в работе [176].
Другое направление (получившее определенное развитие,
в частности, в экологических и биохимических исследованиях)
связано с изучением способности систем сохранять свои харак-
теристики при перемене условий существования. Об этих рабо-
тах мы говорили выше, рассматривая гомеостатические харак-
теристики экосистем. Однако термин «чувствительность», как
правило, в подобных работах не используется (исключением
является, например, работа [332]). Вместо чувствительности
говорят о способности системы буферить среду [148], об устой-
чивости системы к изменению условий ее формирования [248],
условий обитания [28, 138, 170, 204], об инерции [336] или
упругости системы [72, 312]. Ряд авторов при этом особо
подчеркивает разницу в понятиях устойчивости в теории
управления и устойчивости в рассматриваемом смысле [312,
346].
Неточность формулировки задачи исследования зависимости
системы от внешней среды в экологии в терминах устойчивости
видна хотя бы из следующего примера. Мера устойчивости (или
мера изменчивости), традиционно используемая при анализе по-
пуляций — стандартное отклонение логарифма ее численности
[212]. Отклонение численности популяции происходит, как пра-
вило, под влиянием некоторых внешних факторов, и отклонение
зависит как от свойств системы (которые собственно и должны
интересовать исследователя), так и от свойств входного сигнала.
Например, отклонение численности в стационарном режиме
можно определить по формуле (3.48), но свойства системы оп-
ределяются в этой формуле не самим результатом — отклоне-
д зс
нием Дх, а коэффициентом чувствительности Результирую-
щее отклонение зависит как от этого коэффициента, так и от
величины возмущения Дщ связанного только со свойствами
внешней среды и не зависящего от свойств системы.
Из изложенного в настоящем разделе видно, что понятие
чувствительности системы по существу наиболее близко для ха-
рактеристики описываемых свойств экологических систем и мо-
жет явиться адекватным аппаратом исследования их сохрани-
тельных способностей. Фактически термин «устойчивость си-
стемы» в экологическом аспекте часто применяется как раз для
описания ее чувствительности — см., например, [101] или фор-
мулу (2.3) в разд. 2.5.
96 КЛАССИЧЕСКАЯ теория УПРАВЛЕНИЯ и БИОСИСТЕМЫ [ГЛ, 3
Рассматривая понятие чувствительности, принятое в теории
автоматического управления, следует иметь в виду, что в неко-
торых областях биологических исследований термин «чувстви-
тельность системы» используется иногда в смысле, не совпа-
дающем с указанным выше. Так, в физиологии говорят, что
система чувствительна (или нечувствительна) к действию не-
которого фактора, если пороговое значение этого фактора, на
которое система начинает реагировать, мало (соответственно,
велико). Ниже мы будем последовательно придерживаться трак-
товки чувствительности, принятой в теории управления,
ГЛАВА 4
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОСИСТЕМАХ
4.1. Оптимальные системы
Чтобы поставить и решить задачу об оптимальном управле-
нии в системе, нужно выполнить ряд условий. Прежде всего
необходимо иметь следующую информацию о задаче управ-
ления:
1) исчерпывающие сведения об управляемом объекте, т. е.
его математическое описание, например, в виде дифференциаль-
ных уравнений, связывающих между собой переменные, управ-
ляющие сигналы и возмущения;
2) информацию о среде, в которой объект функционирует,
в виде задания (как функций времени) всех действующих на
объект возмущений; если эти возмущения случайны, то тре-
буются их статистические характеристики;
"~ -3) сведения о цели управления, т. е. точно сформулирован-
ные требования к тому состоянию объекта, которое должно
быть достигнуто после окончания процесса управления;
4) информацию о качестве процесса управления, т. е. зада-
ние некоторого количественного показателя, который должен
быть максимизирован или минимизирован соответствующим вы-
бором управления;
5) данные об ограничениях, которые не могут быть нарушены
в ходе процессов управления.
Незнание или ошибки в количественном определении хотя бы
одного из перечисленных факторов приводят к ухудшению ка-
чества управления — система становится неоптимальной.
Любая задача оптимизации сводится к выбору лучшего в не-
котором смысле варианта из большого числа возможных ва-
риантов. Каждый из вариантов можно охарактеризовать ка-
ким-то числом (или набором чисел), определяющим близость
получаемого при данном варианте решения к поставленной
цели. Такое число называется показателем качества, и наи-
лучший вариант соответствует экстремуму показателя каче-
ства.
97
98
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
Задача оптимального управления в теории автоматического
управления формулируется следующим образом. Целью управ-
ления в общем случае является достижение экстремума (обычно
минимума) некоторого функционала Q — критерия оптимально-
сти системы:
Q (х, w, и, ()=min. (4.1)
Функционал — это величина, значение которой зависит от
вида некоторых функций. В формуле (4.1) величина Q зависит
от функций x(t), w(t), и (J) и времени t, где х — m-мерный век-
тор, описывающий состояние системы, w — l-мершлй вектор,
определяющий входные сигналы, которые система должна вос-
производить или преобразовывать заранее заданным образом,
и — г-мерный вектор, элементы которого описывают управляю-
щие сигналы в системе. Если на управляющие воздействия и
наложены ограничения, например, задана область Qu их допу-
стимых изменений, то минимум в (4.1) ищется при ограничении
и е Qu. (4.2)
Пример 4.1.1. В предыдущих примерах мы старались обращаться к те-
матике, связанной с биосистемами. Здесь мы, однако, выбираем пример тех-
нической систёмы. Дело в том, что для биологических объектов формулировка
перечисленных выше условий 1)—5) (предоставление информации о задаче
управления) сама по себе связана с большими трудностями, как об этом го-
ворится ниже в этой главе.
Возьмем в качестве объекта управления двигатель постоянного тока, ура-
внение которого имеет вид
Х1 + kiXi = k2U — V,
г — v I4-3)
*2 — *1,
где Xi — скорость вращения двигателя вала, х2 — угловое перемещение вала,
и — управляющий сигнал (напряжение, подводимое к якорю двигателя), v —
возмущение (нагрузка на вал), ki, k2 — постоянные коэффициенты. Таким об-
разом, требование 1) выполнено — уравнения объекта заданы.
Информация о среде состоит в том, что нагрузка v в процессе управле-
ния не меняется, v = const.
Цель управления заключается в том, что неподвижный двигатель надо
повернуть на некоторый угол и остановить; исходное положение двигателя
X! = 0, х2 — х21, конечное состояние должно быть Xi = 0, х2 = х22.
Показатель качества процесса управления — время Т, за которое дости-
гается заданное конечное состояние. Следовательно, в оптимальной системе
должно быть обеспечено минимальное время переходного процесса (оптималь-
ное быстродействие системы): Т — Тшп.
Ограничения могут быть заданы, например, в виде предельной величины
напряжения, которую можно подавать на якорь двигателя: |u| С umzx.
Ухудшение качества оптимальной по быстродействию си-
стемы при отсутствии полной информации об объекте и среде
можно проиллюстрировать на рассмотренном примере. Если при
4.2]
АДАПТАЦИЯ
99
расчетах величина ограничения ытах определена неверно, то
процесс будет идти дольше по сравнению с тем оптимальным
процессом, который можно было бы осуществить в системе при
точно известных значениях ограничений. Будем считать, что
процесс окончен, когда в достигну-
том состоянии значение координаты
х2 отличается от желаемого на 10%.
Тогда относительное удлинение вре-
мени переходного процесса в зави-
симости от ошибки при определении
ограничивающего значения umax
можно проиллюстрировать графи-
ком на рис. 4.1. График построен
для примера 4.1.1 при k\ = 0.
Оптимальные системы содержат
сложные решающие вычислитель-
ные устройства, реализующие опти-
мальный закон управления — зави-
1 2 3 4 5 G
^тпах
симость
иопт(/) = Ф(х(0,/), п(0,/)) (4.4)
оптимального управления в момент
времени t от измеряемых в этот же
момент времени значений всех пе-
ременных системы х2, ..., Хщ и
возмущений Vi, и2, ..., щ и их
«предыстории» на интервале време-
ни от начала управления до те-
кущего момента времени: х,(0, t),
Интерес к оптимальным систе-
Рис. 4.1. Ухудшение качества опти-
мальной системы при отсутствии пол-
ной информации о системе и среде.
При расчете оптимальной по быстро-
действию системы было принято,
что на ее вход можно подавать
максимально допустимый сигнал
ja|s=a(; фактически на вход посту-
пает сигнал, равный не Ир a uniax.
Оптимальное быстродействие дости-
гается, очевидно, только при
“тах=“1- При “тах>“1 в снстеме
возникает колебательный переход-
ный процесс, при «тах < и{ ~ апе-
риодический; в обоих случаях
быстродействие падает, а время
окончания процесса Т >
мам в теории управления возник в
результате работ А. А. Фельдбаума [218]. Позже был разра-
ботан и общий метод синтеза оптимальных систем управле-
ния— принцип максимума Л. С. Понтрягина [162, 240].
4.2. Адаптация
В последние годы терминология и основные идеи теории
оптимальных систем в технике начали широко использоваться
при анализе биологических систем. Оптимальные процессы
управления, оптимальные конструкции и характеристики орга-
низмов, наконец, оптимальные методы эксплуатации природных
объектов и систем — эти понятия пришли в биологические науки
из технических дисциплин. Так, идея оптимальной конструкции
биосистем организменного уровня целиком базируется на пред-
ставлениях об оптимальности, заимствованных из теории
100
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
управления. Термин же «адаптация» взят теорией управления
из биологии [169а, 237]. В биологии этим термином обозначается
любое приспособление организма к условиям существования.
Поэтому непосредственное применение столь широкого термина
к системам управления неудобно, так как любая, даже самая
простая, система с обратной связью обладает способностью ав-
томатически приспосабливаться к изменению внешних условий.
В результате в теории управления относят к. адаптивным систе-
мам только те, которые автоматически приспосабливаются
к непредвиденным изменениям параметров объекта и внешней
среды [46, часть III].
В наиболее простом случае адаптация системы управления
к изменениям внешних условий или параметров объекта состоит
в том, что в ответ на эти
Рнс. 4.2. Простая адаптивная система управления.
Кроме основного контура с отрицательной обрат-
ной связью, включающего регулятор Р и объект О,
в системе имеется дополнительная цепь адаптации.
Адаптивное устройство измеряет входные и выход-
ные переменные системы и на основании их ана-
лиза меняет параметры регулятора, поддерживая
работоспособность и качество системы.
изменения в системе про-
исходит изменение пара-
метров регулятора, на-
правленное на сохране-
ние работоспособности и
качества системы. Для
этого в систему управле-
ния, кроме основного кон-
тура, вводится дополни-
тельная цепь адаптации
(рис. 4.2). Совокупность
изменяемой части основ-
ного контура и цепи адап-
тации называется адап-
тивным регулятором.
Обычно цепь адаптации
характеризуется более медленным протеканием процессов, чем
основной контур управления. Так как рассмотренная выше в
разд. 3.5 параметрическая обратная связь имеет структуру, ана-
логичную приведенной схеме, то любая модель биосистемы,
включающая хотя бы одну параметрическую обратную связь,
является в терминах теории управления адаптивной системой.
В публикациях по использованию теории управления в био-
системах имеется тенденция считать адаптивные системы чем-то
вроде усовершенствованных оптимальных систем, когда допол-
нительный контур адаптации (самонастройки, приспосаблива-
ния) появляется только в оптимальных системах. Такая тенден-
ция является ошибочной. Адаптивность, вообще говоря, никак
не связана с оптимальностью; это — две самостоятельные ха-
рактеристики систем управления. Адаптация дает улучшение
качества системы, но не обязательно приводит к «идеалу» —
оптимальной системе. Однако в теории управления адаптация
4.31
КОНЦЕПЦИЯ ОПТИМАЛЬНОСТИ БИОСИСТЕМ
101
и в самом деле часто используется именно как средство дости-
жения оптимальности в заранее неизвестных условиях [237,238].
Достижение хорошего качества процессов в системе (или
даже оптимальности) за счет адаптации обычно связано с вклю-
чением в систему дополнительных более или менее сложных
устройств переработки информации. Поэтому сложные адаптив-
ные системы могут служить адекватной моделью адаптивного
поведения достаточно высокоорганизованных живых организ-
мов, которые могут с помощью хорошо развитого аппарата об-
работки информации производить оценку вероятностных харак-
теристик внешней среды и реагировать в соответствии с этими
оценками. В живых системах надорганизменного и суборганиз-
менного уровня процессы адаптации должны протекать на более
простом в смысле алгоритмического обеспечения уровне. Одним
из простейших возможных механизмов адаптации является ис-
пользование параметрических обратных связей.
Адаптивное поведение у живых организмов, осуществляемое
при помощи параметрической обратной связи, рассматривается,
в частности, в книге Р. Розена [175, гл. 11]. Адаптивный харак-
тер поведения системы при этом означает, что если ее реакция
на данное воздействие не является предпочтительной, то в си-
стеме постепенно происходят такие изменения, которые позво-
ляют достичь предпочтительного состояния. В нелинейных био-
логических системах адаптация осуществляется за счет того,
что характеристики отдельных звеньев системы начинают ме-
няться в зависимости от значений выходного сигнала системы
(или каких-либо внутренних сигналов).
Интересно, что Р. Розен, развивая идею оптимальности био-
логических систем, невольно наталкивает здесь читателя на
«еретическую» мысль — вместо того, чтобы создавать сложные
оптимальные конструкции и алгоритмы, биосистеме достаточно
иметь простые механизмы типа параметрической обратной
связи и, адаптируясь к условиям среды, достигать тех же внеш-
них характеристик поведения, что и в оптимальной системе, но
более простыми и надежными, хотя и более энергоемкими, спо-
собами. Оптимальность и адаптация при этом оказываются
двумя различными и даже конкурирующими вариантами до-
стижения одной цели — получения предпочтительного поведения
биосистемы.
4.3. Концепция оптимальности биосистем
До проникновения в биологию кибернетических представле-
ний и методов теории управления термины оптимум, оптималь-
ность применялись в различных биологических дисциплинах
для обозначения наиболее благоприятных условий обитания,
102
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
соответствующих желаемым условиям протекания процессов
жизнедеятельности, т. е. «для отражения факта зависимости всех
физиологических и биохимических процессов от условий среды»
[150]. Противоположным по значению термином было в этом
смысле понятие пессимума, характеризующее неадекватные ус-
ловия среды.
В последнее время, однако, термин «оптимальность» широко
используется при описании биосистем в другом смысле, когда
оптимальными являются свойства самих систем — их наилучшее
функционирование (оптимальные процессы, оптимальные алго-
ритмы) или их строение (оптимальная структура). Оптималь-
ность в биосистемах не только существенно по-разному пони-
мается у разных авторов (хотя прототипом оптимальной биоси-
стемы чаще всего является оптимальная система в технике), но
и встречает принципиальные возражения. Сейчас, ознакомив-
шись с основными представлениями теории управления, мы мо-
жем подробнее рассмотреть аргументы «за» и «против» этой
концепции.
Для биолога, вникающего в глубинные процессы, происходя-
щие в живой природе, характерно не только знание и понимание
многих основных и частных закономерностей жизненных явле-
ний. Содержательный язык, используемый им для описания
биосистем, не избегает эмоционально окрашенных терминов.
Изумительная приспособленность живых систем к условиям
обитания, удивительное постоянство их внутренней среды — тер-
мины, к которым естественные науки охотно прибегают для ха-
рактеристики процессов регуляции в биосистемах. Оптималь-
ность, понимаемая как высокое качество биосистемы, ее наи-
лучшая приспособленность, для естественных наук является
поэтому понятием едва ли не само собой разумеющимся. Инже-
нер, физик, а иногда и математик [366] со своей стороны часто
склонны более трезво смотреть на то, что безусловно восхи-
щает биолога.
Корни представлений об экстремальном характере процессов
и явлений в живых системах надо'искать прежде всего в экстре-
мальных принципах неживой природы. Леонарду Эйлеру при-
надлежит первая в истории формулировка этих принципов: «Так
как здание всего мира совершенно и возведено премудрым твор-
цом, то в мире не происходит ничего, в чем не был бы виден
смысл какого-либо максимума или минимума». «Если опустить
в этой цитате ссылку на бога, как это часто делается в попу-
лярных книгах об оптимальности, — пишет Я. 3. Цыпкин, — то
это утверждение, которое можно назвать принципом экстремаль-
ности или, если угодно, — принципом оптимальности, лежит
в основе многих фундаментальных законов естествознания»
4.31
КОНЦЕПЦИЯ ОПТИМАЛЬНОСТИ БИОСИСТЕМ
103
[239] . Такого рода принципы известны в оптике, механике,
электродинамике, теории относительности [167].
Аналогичным образом иногда делаются допущения и об
экстремальном характере процессов в живых системах. Таков,
например, «принцип наименьшего взаимодействия», предложен-
ный в работе [51]. Суть этого принципа заключается в следую-
щем. Для сложных управляющих систем типична структура, до-
пускающая выделение относительно автономных подсистем. Для
каждой из подсистем все остальные подсистемы относятся
к внешней среде, и целесообразность поведения подсистемы про-
является в том, что в устойчивых состояниях все они стремятся
функционировать как бы независимо, автономно. Система назы-
вается целесообразно работающей во внешней среде, если она
минимизирует свое взаимодействие со средой.
Возможно, однако, что в такой общей форме этот принцип
не будет достаточно конструктивным и потребуется его уточне-
ние для каких-то более конкретных типов живых систем; ведь
даже для неживой природы экстремальные принципы формули-
руются лишь для отдельных «классов» явлений.
Другим источником представлений об оптимальности живых
систем служит анализ эволюционных процессов живой природы.
Нет сомнений в том, что эволюционный процесс имеет глубокие
внутренние закономерности, приводящие к движению эволюции
по своего рода экстремальным траекториям (см., например,
[233]). В последних моделях эволюции биологических макро-
молекул отбор также интерпретируется как результат действия
некоторого экстремального принципа [255].
В наиболее развернутой форме различные аспекты принципа
оптимальности в биологии описаны, пожалуй, в книге [175],
имеющей, однако, тенденцию к популярности изложения. Основ-
ная идея этой книги может быть сформулирована следующим
образом: живые организмы, пребывающие совместно в течение
достаточно долгого времени в определенной совокупности
окружающих условий, постепенно приобретают признаки, опти-
мальные для этих условий. Такой процесс происходит при вы-
полнении двух следующих допущений: а) преимущества, имею-
щиеся у индивидуумов в борьбе за существование, передаются
потомству; б) данные условия среды остаются неизменными
достаточно долго.
Впервые гипотезу об оптимальной конструкции организмов
выдвинул Н. Рашевский [347, 348], впрочем, позже от нее от-
казавшийся (взгляды Н. Рашевского подробнее мы рассмотрим
ниже). В соответствии с этой гипотезой оптимальной является
конструкция, обеспечивающая наименьший расход метаболиче-
ской энергии на поддержание данного элемента структуры (ко-
торый необходим и достаточен для нужд организма). Примером
104
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
оптимальной конструкции служит модель ветвящегося сосуди-
стого русла (диаметры сосудов, места и углы ответвлений, рас-
считанные на основе гипотезы об оптимальной конструкции,
хорошо совпадают с экспериментальными данными Ц71.
173, 282]).
Биологические системы управления по Р. Розену также под-
вергаются действию естественного отбора, и поэтому из прин-
ципа оптимальной конструкции непосредственно следует, что
можно говорить и об оптимальных механизмах регулирования
их функций. В книге, однако, круг биосистем, где реализуется
принцип оптимальности, ограничен лишь организмами; по мне-
нию ее автора, в популяциях возникают хотя и хорошие, но не
оптимальные способы защиты от изменений условий среды
[175, стр. 119—120].
Хотя принципы оптимальности и распространяются на меха-
низмы управления в организме, в качестве примера в книге
приводятся, однако, не оптимальные системы управления, по-
добные рассмотренным выше в разд. 4.1, а механизмы мини-
мизации сигнала рассогласования в обычных моделях следящих
систем (терморегуляция, дыхательный хемостат, регуляция ве-
личины зрачка). В целом концепция оптимальности в форме,
предложенной Р. Розеном, вызывает возражения у целого ряда
авторов. Поэтому следующий раздел мы посвятим рассмотре-
нию недостатков этой теории.
4.4. Критика концепции оптимальности в биологии
Представления о том, что все живые системы могут рас-
сматриваться как оптимальные системы управления, подвер-
гаются критическому рассмотрению с различных исходных по-
зиций. Мы попытаемся проследить три таких подхода, суть
которых приблизительно можно выразить следующим образом:
1. Эволюция и естественный отбор не приводят к созданию
оптимальных биосистем.
2. Биосистемам не нужна оптимальность в том смысле, как
она понимается в теории управления.
3. Понятие оптимальности биосистемы не соответствует по-
становке задачи об оптимальных системах в теории управле-
ния.
Разумеется, порядок расположения этих подходов не связан
с их относительной важностью или значимостью, а определяется
лишь удобством изложения. Итак,
1. Эволюция не приводит к созданию опти-
мальных биосистем. Прежде чем обсуждать вопрос по
существу, отметим одно обстоятельство философского плана, ко-
торое при рассмотрении оптимальности биосистем обычно не
4.4]
КРИТИКА КОНЦЕПЦИИ ОПТИМАЛЬНОСТИ В БИОЛОГИИ
105
принимается в расчет. Дело в том, что экстремальные принципы
неживой природы и экстремальные принципы эволюции живых
систем — категории весьма разного порядка. Неизвестно, яв-
ляются ли экстремальные законы неживой природы результа-
том какого-либо процесса эволюции материи: по крайней мере,
в доступном человечеству масштабе времени эти законы высту-
пают как неизменные данности — стационарные свойства Все-
ленной. Если процессы эволюции этих данностей и представляют
собой реальность, то либо они давно «затухли», либо протекают
в том же темпе, в каком происходят другие процессы во Все-
ленной, например, ее расширение.
Эволюция живых систем, наоборот, протекает довольно бы-
стро. Самые медленные процессы эволюции происходят в гео-
логических масштабах времени, самые быстрые — на наших
глазах. Быстрая эволюция микроорганизмов и вирусов (в том
числе часто и благоприятная для человека), резкие изменения,
а иногда и дезинтеграция, эволюционно сложившихся биогео-
ценозов при интенсивном загрязнении среды — этими приме-
рами далеко не исчерпывается список быстрых эволюционных
сдвигов в биологических системах, которые происходят на гла-
зах человека.
Эволюционный процесс идет по своего рода экстремальным
траекториям. Но успевают ли эти траектории достичь стацио-
нарных точек? Пусть биосистемы проявляют тенденцию к улуч-
шению своей структуры, своего функционирования в процессе
эволюции, но на каком же этапе стремления к оптимальному
уравновешиванию со средой они находятся в каждый данный
момент эволюционного процесса? Так, принцип исключения
Гаузе гласит, что два вида не могут неограниченно долго суще-
ствовать в одной нише; один из них в конце концов должен
быть полностью вытеснен. Этот принцип, обсуждение которого
можно найти в [212], определяет лишь асимптотическое состоя-
ние модели системы, состоящей из двух видов; сам же процесс
вытеснения может длиться достаточно долго.
В палеонтологии не вызывает сомнений факт длительного
сосуществования в одном и том же ареале видов-аборигенов
и более высокоорганизованных видов-пришельцев, постепенно
вытесняющих первоначальное население: «...вытеснение орга-
низмов, уже занимающих известное местообитание, иммигран-
тами, в общем лучше вооруженными для борьбы за жизнь,...
требует более или менее значительного времени» [61]. Напом-
ним, что речь здесь идет о геологическом времени.
Ясно также, что в течение столь длительных периодов вре-
мени могут претерпевать существенные изменения сами условия
окружающей среды. Это обстоятельство учитывается в работах
по математической теории отбора и эволюции (см., например,
106
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
[231, 233]). Будь условия существования неизменными, стацио-
нарный режим в эволюирующей системе мог бы быть достигнут,
но ведь условия существования не определяются только отно-
сительно стабильными абиотическими факторами — в отличие
от предположения Р. Розена, популяция существует в условиях
биогеоценоза, организм — в популяции, биогеоценоз — в био-
сфере. Изменяясь в эволюции, каждая биосистема изменяет
и условия своего обитания — структуру биосистемы более высо-
кого уровня, элементом которой она является. Среда, в которой
эволюирует биосистема, меняется во времени с той же ско-
ростью, что и сама биосистема. Инженеры знают, что в таких
условиях оптимума достичь нелегко.
Палеонтологические данные, похоже, свидетельствуют о том,
что вновь возникшие виды наиболее приспособлены к своей
среде — они быстро распространяются в ней (рис. 4.3). Но, за-
крепляясь в сообществе живых организмов (биосе), новый вид
Рис. 4.3. Распространенность хвойных пород в различные геологические эпохи [152]. Воз-
никнув, новые виды, как правило, быстро увеличивают свою численность, что соответ-
ствует расширению ветвей на рисунке, но постепенно при изменении условий среды чис-
ленность видов начинает сокращаться, в вид может полностью исчезнуть. В правом
столбце —геологический масштаб времени, начиная от древнейшего девонского периода и
кончая современным четвертичным 1. Cordaltales. 2. Lebaihiaceae. 3. Voltziaceae. 4. Podo-
zamitaceae. 5. Araucariaceae. 6. Cheirolepidiaceae. 7. Palissyaceae. 8. Podocarpaceae
9. Cephalotaxaceae. 10. Taxaceae. 11. Pinaceae. 12. Sciadopityaceae. 13. Taxodiaceae.
14. Cupressaceae.
самим своим присутствием изменяет условия обитания — отно-
шения внутри биоса. Достигнув максимума, вид (или таксон)
постепенно теряет свойство «максимальной приспособленности»,
когда, возможно, он достигал наиболее благоприятного соотно-
шения с условиями окружения. Численность вида начинает со-
кращаться, и сам вид может полностью исчезнуть. Так вымерло
около 90% всех созданных природой к настоящему времени так-
сонов [61, 152]; «...основным фактором вымирания более или
менее широко распространенных групп форм... являются взаи-
моотношения внутри биоса» [61]. Были ли «оптимальными»
4.4}
КРИТИКА КОНЦЕПЦИИ ОПТИМАЛЬНОСТИ В БИОЛОГИИ
107
вымершие виды? Или, развиваясь в направлении оптимума, они
просто не успели достигнуть его? Оптимально ли трехкамерное
сердце рыб, которое «просто не доросло до четырехкамерного»
[116]? Человек как биологический вид, по крайней мере, явно
не успевает эволюировать вслед за изменяемой им же средой.
Уже сейчас звено «человек-оператор» является самым узким
местом в системе «человек-машина» [18]. Сохраняемая в эво-
люции как одна из основ самосохранения биохимическая инди-
видуальность оказывается сейчас едва ли преодолимым препят-
ствием для получения организмом помощи извне — пересадки
органов и тканей.
Эволюция близорука — она отдает предпочтение тем видам,
которые получают преимущество немедленно. Тем самым
в принципе достижимым является лишь ближайший локальный
экстремум за счет мелких изменений конструкции организма;
кардинальные мутации, как правило, детальны [102, 116, 248].
«Приспособленность, доведенная до максимума, представляет
собой относительный оптимум» [99].
Критический анализ эволюционного процесса с точки зрения
конструктора технических систем, — по выражению самого ав-
тора, «пасквиль на эволюцию» — принадлежит С. Лему [102,
стр. 456]. Подытоживая сказанное, можно согласиться с его
положением, что «эволюция оставляет своим творениям самые
примитивные органы и биохимическое оснащение, лишь бы с их
помощью они управлялись с сохранением вида». Согласиться,
помня, однако, что слова эти принадлежат скорее мыслителю,
чем специалисту в области систем.
2. Биологическим системам не нужна опти-
мальность. Оптимальное функционирование системы свя-
зано с необходимостью получения, хранения и переработки
большого количества информации о среде, состоянии и целях
системы и т. д. У многих типов биосистем — и прежде всего
у различных видов организмов — имеются достаточные для этого
возможности. Другие типы биосистем — многие органы и си-
стемы в организме, популяции, биогеоценозы — вряд ли обла-
дают соответствующими характеристиками. Но даже на орга-
низменном уровне вопрос о целесообразности использования
всех возможностей для получения строго оптимальных решений
весьма дискуссионен.
Начать с того, что оптимальность в технике не так уж при-
влекает конструкторов систем. Даже при учете всех факторов
(полная информация) оптимальность технических систем дает
обычно выигрыш, не превышающий 10—15% по сравнению
с грамотным, хотя и не оптимальным решением задачи. Таков,
например, выигрыш, который может дать оптимальность в де-
терминированной задаче быстродействия. Заметим также, что
108
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
решения, построенные по различным оптимальным алгоритмам
(отличающимся выбором критерия качества), часто очень
близки друг другу и дают примерно одинаковые процессы в си-
стеме. Сравним, например, насколько будет отличаться быстро-
действие систем, которые оптимальны по некоторым другим
критериям, прямо не связанным с оптимизацией времени пере-
ходного процесса. В статье [229] такое сравнение проведено
для систем, в которых минимизируется ошибка за время пере-
ходного процесса е(0 при воздействии на систему единичного
скачка. Оказывается, что если ошибка минимизируется по раз-
ным критериям (например, интеграл модуля ошибки, интеграл
квадрата модуля ошибки и т. д.), то переходные процессы тем
не менее практически заканчиваются за то же время, что и при
оптимальном быстродействии. Относительное удлинение вре-
мени переходного процесса 6 составляет не более 0,4%
(см. рис. 4.4).
Рис. 4.4. Процессы управления, оптимальные по различным критериям качества, во мно-
гих случаях практически не различаются. Критерий качества в оптимальной системе
00
выбран в виде Qv== I е |v dt, где 8 —ошибка системы; тем не менее при всех v практи-
0
чески достигается оптимальность по быстродействию, б (v) — относительное удлинение пере-
ходного процесса в системе при подаче на вход системы возмущения в виде единичного
скачка при различных значениях v.
Кроме того, чем ближе система к оптимальности, чем она
сложнее, тем все более дорогой ценой дается ее дальнейшее
улучшение. Все больший объем информации требуется обраба-
тывать, все более трудоемкие алгоритмы приходится реализо-
вать [218]. Оптимальные системы функционируют оптимально
только при заранее известных сигналах на входах системы, на-
пример, только при ступенчатых или только при случайных воз-
действиях с заранее известными характеристиками; любая не-
определенность во внешней среде ухудшает качество процессов
в системе. Так, наличие шума в канале управления оптимальной
по быстродействию системы резко увеличивает время достиже-
ния цели (рис. 4.5, [139]). Выигрыш, даваемый оптимальной
4.4]
КРИТИКА КОНЦЕПЦИИ ОПТИМАЛЬНОСТИ В БИОЛОГИИ
109
системой, снижается и при ошибочном определении параметров
системы или условий среды (см. рис. 4.1).
Поэтому даже технические оптимальные системы редко реа-
лизуются на практике, и одно из направлений практического
использования теории оптимальных систем заключается в том,
чтобы давать обоснованное сравнение существующих неопти-
мальных конструкций объектов с предельно достижимыми (оп-
тимальными) и принимать решение о возможности и целесооб-
разности их усовершенствования. При этом цель исследования
Рис. 4.5. Действие помех на оптимальную по быстродействию систему. Рассматривается
оптимальная по быстродействию система второго порядка, управляемый объект в которой
описывается уравнениями х0=Х1. Х|«в, где х0 — выходная координата системы, xi — cko’
рость ее изменения. Управляющее устройство вырабатывает релейное управление и, равное
+ 1 или —1, которое при наличии случайных возмущений в канале передачи управления
поступает на вход объекта в искаженном помехами виде, а) Оптимальный процесс при
отсутствии помех; б) процесс в системе при действии случайных возмущений. На графике
скорости видны излом'.!, соответствующие изменению знака управляющего сигнала с + 1
на или наоборот, из-за чего переходный процесс в системе затягивается более чем
вдвое [139].
состоит в оценке тех затрат, которые требует усовершенствова-
ние системы, и того приближения к оптимуму, который эти за-
траты могут дать.
Не так ли «конструируются» и биосистемы? По крайней
мере, уже в первых работах по критериям оптимальности в био-
системах на примере системы дыхания ставился вопрос о ком-
промиссе между улучшением качества функционирования
и энерготратами [203].
Рассмотрим, например, регуляцию основных переменных
в сердечно-сосудистой системе человека цри импульсной физи-
ческой нагрузке [29]. Переходный процесс по таким показате-
лям, как частота сердечных сокращений и артериальное давле-
ние, в этом случае может быть у одних испытуемых колебатель-
ным, у других — апериодическим. Если выделить случаи наи-
более быстро протекающих процессов апериодического типа в
особую группу «оптимальных» процессов, то по артериальному
давлению она составляет всего около 19% от числа испытуе-
мых. Колебательный и апериодический тип регуляции в сердеч-
но-сосудистой системе при физической нагрузке наблюдается
по
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
и у собак, хотя «оптимальный» тип при этом не выделяется
[364]. Само разнообразие результатов подобных экспериментов
свидетельствует о том, что биосистемы довольно «терпимы»
к свойствам отдельных своих элементов и подсистем.
Подробнее это свойство живых систем мы рассмотрим
в разд. 4.5.
3. Концепция оптимального управления не-
применима к биологическим системам по фор-
мальным причинам. Оптимальность в строгом смысле
этого слова требует выполнения довольно четко очерченного
круга условий. В биологических системах формулирование
и проверка этих условий связана с большими трудностями.
«Понятие оптимальности к живой природе во многих случаях
попросту неприложимо. Дело в том, что об оптимальности
можно говорить в условиях, когда высказаны жесткие ограни-
чения на то, что может меняться, когда ясно, что должно оста-
ваться неизменным, и когда имеется точно очерченная целевая
функция. В живой природе в общем случае эти обстоятельства
не имеют места», — писал А. А. Ляпунов [ПО].
Постановка задачи об оптимальности управления в технике,
рассмотренная в разд. 4.1, включает пять основных пунктов:
объект управления; среда, в которой объект функционирует; кри-
терий каиества системы; цель управления; наконец, ограничения
на систему управления. Обсуждение каждого из этих пунктов
позволяет не только оценить формальные трудности, стоящие на
пути анализа биосистем как систем оптимального управления,
но и более глубоко обсудить некоторые специфические черты
организации биосистем.
' Объект управления. Общеизвестны трудности, возникающие
при попытках разделить живую систему на «объект» и «регу-
лятор». Лишь на некоторых уровнях организации (в том числе
на организменном) существуют элементы системы (органы),
явно выполняющие регулирующие функции, такие как нервная
система. Но и на организменном уровне функции регулятора
реализуются не только нервной системой — регулирующие меха-
низмы включают в себя многие другие связи между его пере-
менными. Объектом управления можно считать, например, под-
множество переменных биосистемы, сохраняемых при вариа-
циях условий среды. Сохранение этих переменных «происходит
за счет изменения других; такие изменения мы будем называть
регулятивными, поскольку за их счет удерживаются специфиче-
ские признаки» биосистемы [28, стр. 24].
Другой подход к определению того, что считать объектом
управления, связан с разделением биосистемы на метаболиче-
скую компоненту и кибернетическую систему, обеспечивающую
сохранение энергетической основы системы ([216], см. также
4.4]
КРИТИКА КОНЦЕПЦИИ ОПТИМАЛЬНОСТИ В БИОЛОГИИ
111
разд. 1.2). Но и в этом случае нельзя забывать, что сама энер-
гетическая компонента в большой степени обладает свойствами
саморегуляции и самосохранения, т. е. имеет свойства как регу-
лятора, так и объекта одновременно.
Наконец, каждый целостный биообъект (внутри которого мы
сейчас пытались выявить «объект управления» и «регулятор»)
не является изолированным. Клетки являются частью орга-
низма, организм входит в популяцию, популяции формируют
биогеоценозы.
Поэтому, как бы ни была совершенна система управления
объектом данного уровня, она не самостоятельна в своем пове-
дении, в своих решениях. При воздействии факторов внешней
среды, возмущающих более высокие уровни организации био-
системы, механизмы регуляции подсистем подавляются, как бы
оптимальны они ни были с точки зренця этой подсистемы. Са-
мый совершенный хищник не может уничтожить до конца свою
«неоптимальную» по конструкции жертву — экологические ме-
ханизмы регуляции в биогеоценозе довлеют над регулирую-
щими механизмами организма и популяции. «Выживают не
виды, а ансамбли», — пишет по этому поводу М. Эйген [255].
Популяция, составленная из «более приспособленных» особей,
в целом может оказаться менее приспособленной [61].
Специфические трудности возникают на организменном
уровне при выделении в качестве объекта отдельных органов
и подсистем организма. Эти подсистемы в организме относи-
тельно автономны в нормальных условиях, так что выделение
их в качестве объекта исследования во многих случаях не вы-
зывает сложностей. Однако при ухудшении условий среды связи
между отдельными подсистемами становятся более тесными;
сильные стрессорные воздействия организм встречает не как
совокупность изолированных подсистем, а как целостная си-
стема [34, 164, 181, 188]. Таким образом, в сложных условиях,
когда в наибольшей степени должны проявляться оптимальные
свойства органов и систем организма, возникают и наибольшие
трудности с выделением отдельных «объектов регулирования»
в целостном организме. Аналогичные сложности могут возник-
нуть и при анализе экосистем.
Явно неоптимальные с точки зрения энергетики особенности
организма (петушиный гребень, например, [102, 192]) или про-
сто нецелесообразные на организменном уровне генетические
особенности вида (высокий процент генетических уродств [102,
248]) оказываются необходимыми для осуществления эволю-
ционного процесса. Само наличие по-разному приспособленных
особей и групп особей является необходимым условием отбора.
Существование «плохих» и, следовательно, неоптимальных кон-
струкций организмов в популяциях необходимо для процесса
112
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
улучшения, «оптимизации» самих популяций. Рассогласование
качеств отдельных особей является движущей силой эволюции.
«Материал для эволюции поставляют не победители в борьбе
за существование, а побежденные», — писал Э. С. Бауэр
[26, стр. 201].
Описание среды. С описанием среды в биосистемах, по-види-
мому, дело обстоит наиболее хорошо; по крайней мере, в первом
приближении среда должна описываться так же, как при иссле-
довании технических систем. .Однако определенные трудности
все же могут возникнуть из-за разделения целостной биоси-
стемы на подсистемы — «объект» и его окружение — «среду».
В частности, на организменном уровне для каждой подсистемы
в качестве среды выступают все остальные подсистемы [51],
и здесь мы сталкиваемся с теми же трудностями, что и при
определении объекта управления.
Критерий качества. Обычно улучшение качества биосистемы
связывается с уменьшением затрат энергии на ее функциониро-
вание [97, 171, 175, 230—232] или с улучшением протекания
процессов в системе (например, быстродействие [29, 108], ми-
нимизация флуктуаций переменных внутренней среды [34], до-
стижение экстремальных значений некоторых переменных со-
стояния [7]). Для экологических систем часто предполагается
максимальная приспособленность или максимальная скорость
размножения в среде [161, 231 и др-].
Однако выбор критерия оптимальности все же остается пол-
ностью делом вкуса исследователя. Интуитивная «ясность» кри-
терия служит недостаточным ориентиром хотя бы потому, что
в/стандартных условиях (например, при отработке ступенчатого
воздействия) процессы, минимизирующие энерготраты или
время переходного процесса, практически различить невозможно
([229], см. рис. 4.4). Следует иметь в виду, что в других слу-
чаях система может, наоборот, оказаться весьма критичной
к выбору критерия оптимальности. «Система, оптимальная
в некотором смысле, с другой точки зрения может оказаться
неоптимальной и даже бесполезной» [240].
Кроме того, при наличии упомянутых альтернатив у уссле-
дователя всегда остается выбор одной из двух постановок
задач: минимизация энергетического показателя при ограниче-
ниях на качество процесса или получение экстремума показа-
теля качества при ограничениях на энергетические траты. Ис-
черпывающей аргументации в пользу выбора того или другого
подхода в каких-либо конкретных ситуациях в литературе нет.
Зато, как уже отмечалось, имеется тенденция к рассмотрению
компромиссных критериев [175, 203].
Цели управления. Вопрос о целях, которые достигаются
в процессе развития биосистем — в эволюционном процессе,
4.5]
ПРИНЦИП УДОВЛЕТВОРЕНИЯ
113
в онтогенезе, — мы уже обсуждали (разд. 1.7). Здесь мы только
добавим, что биосистемы — многоцелевые системы, выполняю-
щие одновременно несколько разных функций; в различных ус-
ловиях среды непосредственные цели могут быть весьма раз-
личными. Для исследования многоцелевых систем разработаны
специальные методы [73, 93].
Трудность формирования цели является существенным воз-
ражением против концепции оптимальности биосистем: нельзя
одновременно обеспечить «оптимальные» процессы во всех слу-
чаях, если цели системы в каждой ситуации разные. На этом
основании многие авторы говорят о достаточности для системы
получения в каждом конкретном случае не оптимального, а лишь
удовлетворительного результата [136, 259, 278]. Для экологиче-
ских систем подобную мысль проводит и Р. Розен [175, стр. 120].
Подробнее об этом мы будем говорить в разд. 4.5.
Ограничения. В технических задачах ограничения на коор-
динаты системы и на управление возникают из чисто конструк-
тивных соображений — ручка управления может быть доведена
только до упора и не больше; элементы конструкции техниче-
ской системы рассчитаны на точно заданные напряжения и уси-
лия, запас топлива в баках строго ограничен.
В живых системах таких жестких ограничений нет — в про-
цессе приспособления и адаптации они могут менять уровень
ограничений. Нетренированный человек становится спортсменом-
разрядником, так что в задачах, аналогичных рассмотренным
в примере 4.1.1, предельные усилия «шах увеличиваются.
Вид, приспосабливаясь к условиям среды, приобретает необхо-
димые качества, в том числе путем морфологических измене-
ний. Вообще, структура биосистем (определяющая, как пра-
вило, ограничения) меняется при изменении функционирования
систем — выполняемая функция формирует структуру биоси-
стемы. Следовательно, постоянство ограничений, свойственное
обычной задаче оптимального управления, для биосистем, во-
обще говоря, не имеет места. Вместо того, чтобы усложнять ал-
горитмы управления, добиваясь оптимального процесса при дан-
ных ограничениях, биосистема может, адаптируясь, просто осла-
бить ограничения, и достичь лучшего качества за счет простых
и надежных средств, но это уже совсем другая задача оптими-
з'ации, не рассматриваемая в теории управления.
4.5. Принцип удовлетворения
По словам У. Р. Эшби, «требование найти оптимум может
быть излишним; в биологических системах достаточно того,
чтобы организм находил состояния или величины, лежащие
в заданных пределах» ([259, стр. 320]).
114
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
Подчеркивая высокую степень приспособленности живых
систем, многие авторы тем не менее считают термин «оптималь'
ность» не отвечающим существу положения. «Понятие опти-
мальности лучше заменить понятием гармоничности — на-
сколько все части организма участвуют в создании условий, не-
обходимых для существования каждой из них», — писал
В. Н. Беклемишев [28, стр. 30]. Другие же авторы, рассматривая
проблему приспособленности, подчеркивали, что в биосистеме
отсутствует стремление к достижению какого-то единственного
наилучшего состояния; уравновешивание со средой достигается
не в одной точке, а скорее в целой области возможных состоя-
ний системы. Идея пределов терпимости и близкие представле-
ния о пределах безопасности имеются еще у У. Кэннона [278]
и У. Р. Эшби [259]. Аналогичные идеи развиваются и в [102,
184]. Близким является и тезис о множестве вариантов дости-
жения цели [11].
Во всех этих случаях фактически происходит разрыв с пред-
ставлениями о единственном наилучшем из всех возможных,
оптимальном варианте достижения цели. Вместо единственного
решения в системе допускается целое множество возможных
вариантов — либо в виде непрерывного отрезка допустимых ве-
личин (пределы терпимости), либо в виде дискретного множе-
ства приемлемых решений.
Близкий аспект этой проблемы рассмотрен в [223] в связи
с так называемым принципом локального управления. Для
сложных биосистем, таких, например, как целостный организм,
обычно можно выделить множество относительно независимых
элементов. Они обладают относительной самостоятельностью
поведения в рамках «правил взаимодействия», и понятие опти-
мальности для такой системы взаимодействующих элементов
неудобно (можно говорить об оптимальности только в том слу-
чае, если есть следующий, более высокий уровень организации,
и задача, решаемая всей системой в целом). Более подходящим
термином в рассматриваемом случае оказывается область до-
пустимых режимов.
Определенным аргументом в пользу того, что биосистемам
не надо поддерживать строгую оптимальность, является наблю-
даемая в жизни переменность, вариабельность нх характерис-
тик. Один и тот же организм в онтогенезе проходит последова-
тельность состояний, и его регулятивные и приспособительные
характеристики сильно отличаются на разных этапах. Так, начи-
ная с шестнадцатилетнего возраста, время адаптации зритель-
ного аппарата к темноте у человека непрерывно увеличивается,
удваиваясь примерно каждые 10—15 лет. Понятие нормы как
наиболее часто встречающихся и поэтому наиболее адекватных
условиям существования числовых значений отдельных функций
4.5]
ПРИНЦИП УДОВЛЕТВОРЕНИЯ
115
организма также, оказывается, довольно сильно варьирует
с течением времени. Так, в XIX веке нормальным для человека
считалось содержание лейкоцитов 10—14 тыс. в куб. мм крови,
в начале XX века эта характеристика снизилась до 8—12 тыс.,
в 1920-х годах нормой считалось 6—10 тыс., в 1940-х —
6—8 тыс., лишь с конца 1950-х годов установилась современная
норма.
Характеристики организма существенно меняются и в ходе
процессов адаптации, тренировки. Организм тренированного
спортсмена обладает специфическим аппаратом регуляции
в сердечно-сосудистой системе и других системах организма
(см., например, [29, 86]). После окончания интенсивного тре-
нировочного процесса организм спортсмена не обязательно воз-
вращается к состоянию, характерному для нетренированных
людей. Аналогичные примеры легко можно привести и для
других типов биосистем.
Трудно сказать, к какому конкретно состоянию во всех пе-
речисленных случаях относилось бы понятие оптимальности.
Интересно в этой связи рассмотреть эволюцию взглядов
Н. Рашевского, впервые выдвинувшего в свое время мысль об
оптимальной конструкции организмов и позже отказавшегося
от нее [172].
Первоначальные положения были сформулированы следую-
щим образом. Любой организм осуществляет ряд биофизиче-
ских, биохимических и прочих функций, которые даже при оди-
наковой их интенсивности могут выполняться различными
структурами. Выдвинутый в 1943 г. общий принцип гласил, чю
«конкретная структура или конструкция, которую мы действи-
тельно находим в природе, является простейшей из возможных
структур или конструкций, способных выполнять данную функ-
цию. Иными словами, из возможных вариантов выбирается
тот, который удовлетворяет принципу максимальной простоты».
Заметим, что принцип максимальной простоты и в наше время
кажется привлекательным исследователям биосистем [180].
Однако, по словам Н. Рашевского, «простота — понятие не-
сколько неопределенное». Для математических применений этот
принцип оказался неудовлетворительным, и его пришлось
видоизменить, потребовав минимума затрачиваемого материа-
ла и энергии [171, 282]. В таком виде этот принцип и по-
лучил известность под названием принципа оптимальной кон-
струкции.
Оптимальность, однако, тоже понятие относительное. «Мо-
жет, например, оказаться, — говорит Н. Рашевский, — что опти-
мальность с точки зрения количества материала не соответ-
ствует оптимальности с точки зрения затрат энергии. Кроме того,
требования к конструкции можно снизить, если рассчитывать
116
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ. 4
ее на более спокойные режимы функционирования». По-
этому в последующих формулировках (начиная с 1961 г.) этот
принцип формулируется в форме принципа адекватной кон-
струкции организмов. «Конструкция должна быть адекватна
заданной функции при заданных изменяющихся условиях
среды» [172, стр. 54].
Одним из убедительных примеров того, что нельзя ограни-
чиваться лишь рассмотрением энергетической оптимальности
без учета функций, выполняемых данной системой, является
прямохождение человека. Если построить зависимость затрат
энергии (или потребления кислорода) при движении различных
Рис. 4.6. Затраты кислорода на бег у млекопитающих разной массы. Человек расходует
иа перенос одного грамма массы тела на расстояние в один километр примерно вдвое
больше кислорода и, следовательно, энергии, чем можно бы ожидать для четвероногого
животного соответствующей массы. Не является ли дополнительная энергетическая стои-
мость передвижения на двух ногах той ценой, которую человек заплатил за полученные
тем самым безграничные преимущества перед четвероногими?
млекопитающих от их массы (рис. 4.6), то точка, соответствую-
щая человеку, располагается примерно вдвое выше, чем следо-
вало бы ожидать для четвероногих животных такой же массы.
Расхождение объясняется тем, что дополнительная энергетиче-
ская стоимость хождения на двух ногах является той ценой,
которую человек заплатил за возможность освободить свои
передние конечности для других целей [250]. Этот пример пре-
красно подтверждает приведенное выше высказывание Н. Ра-
шевского о том, что требования к системе (в данном случае —
4.5]
ПРИНЦИП УДОВЛЕТВОРЕНИЯ
117
энергетические) можно существенно ослабить, если допустить
сужение возможностей системы, рассчитывать ее на более спо-
койные режимы функционирования.
Кстати, это не единственный случай резкого отличия энерге-
тических характеристик организма человека от животных.
В рамках теории Рубнера (эта теория касается связи темпов
обмена веществ с геометрическими размерами и массой орга-
низмов) было установлено, что на каждый килограмм массы
животного в течение жизни расходуется одно и то же количество
энергии — энергетический фонд, равный 191 000 ккал!кг. По-
этому мелкие животные с более высоким темпом метаболизма
имеют короткую продолжительность жизни, а крупные виды
с их сниженной интенсивностью обмена живут дольше. «Совер-
шенно непонятное исключение в свете этой теории — человек,
энергетический фонд которого в 4 раза выше, чем у остальных
млекопитающих» [19]. Если во всем придерживаться энергети-
ческого принципа оптимальности, то следовало бы признать,
что только человек с его уникальным энергетическим фондом
в 725 800 ккал!кг (по Рубнеру) обладает какими-то особенно-
стями структуры, делающими ее оптимальной. Все же остальные
представители животного мира не только не имеют наилучшей
возможной структуры, но поразительно далеки от возможного
оптимума.
Еще один, и тоже весьма показательный, пример энергети-
чески неоптимального решения был рассмотрен выше, в разд. 2.4.
Поддержание температурного гомеостаза в организме живот-
ного связано с расходованием дополнительного количества
тепла. Как видно из рис. 2.3, землеройка, например, чтобы до-
стичь лучшего качества процессов в системе терморегуляции
(можно ли говорить в этом случае об оптимальности?) поддер-
живает постоянные темпы метаболизма в течение всех суток.
Колибри же, отказавшись от постоянства температуры тела
и ослабив тем самым требования к системе терморегуляции,
получает очевидные энергетические выгоды. Спокойный режим
функционирования и в этом случае позволяет значительно сни-
зить энергетические требования к системе-—в тех условиях су-
ществования, которые характерны для жизни колибри, допол-
нительные энерготраты на температурный гомеостаз не оку-
паются.
Таким образом, применяя принцип адекватной конструкции,
следует учитывать не только функцию, подлежащую осуще-
ствлению, но и условия среды, с которыми биосистема может
встретиться в своей жизни; при этом выполняемые расчеты
подобны той работе, которую выполняет инженер при конструи-
ровании технических систем. Такой принцип допускает, оче-
видно, не единственное решение проблемы структуры и формы,
118
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
[ГЛ, 4
и в этом смысле он аналогичен допущению областей равно
приемлемых решений, пределов терпимости.
Эти представления на организменном уровне были подыто-
жены М. Месаровичем [331] в виде принципа удовлетворения.
Основным предположением при формировании этого принципа
является отсутствие жесткой определенности в понятии цели;
цель системы формулируется как достижение некоторого мно-
жества одинаково желаемых состояний. Система удовлетво-
ряется, достигнув любого из них. Такое ослабление требовании
к результату дает два рода преимуществ:
а)оно позволяет объяснить сильную вариабельность резуль-
татов некоторых экспериментов (это становится ясным после
рассмотрения, например, описанных ниже реакций глазодвига-
тельной системы);
б) дает возможность понять простые эвристические алго-
ритмы адаптации систем к среде.
Принцип удовлетворения формулируется следующим обра-
зом [331]. Организм характеризуется некоторыми переменными,
определяющими его функционирование (performance variab-
les— англ.) z/i, z/2, , Уг, и теми пределами, в которых воз-
можно их изменение без существенного ухудшения процессов
в организме ЙУ[, Эти пределы образуют диапазон
удовлетворения. На организм действуют внешние сигналы оц,
ау2, • • •, Wi, для которых задаются пределы неопределенности
Управляющие переменные обозначим через и.
Тогда при действии внешних воздействий организм выбирает
управление так, что переменные функционирования остаются
в диапазоне удовлетворения, пока и поскольку воздействия
остаются в пределах неопределенности. Любое управление, обе-
спечивающее такое состояние, одинаково приемлемо.
Математическая запись принципа удовлетворения такова.
Пусть рассматриваемая система описывается уравнениями
Dt (0 = ft («, w, t), i = l, 2, г, (4.5)
а управление выбирается в зависимости от текущих внешних
условий
«(/) = <p(w, 0, (4.6)
где t — время. Управление u(t) в каждый момёнт времени вы-
бирается так, чтобы обеспечить выполнение условий
f (и, w, /) е Qy, (4.7)
тогда и только тогда, когда
w(t)f=Qw. (4.8)
4.51
ПРИНЦИП УДОВЛЕТВОРЕНИЯ
119
Характерной особенностью этого принципа является то, что
диапазон удовлетворения прямо зависит от пределов неопре-
деленности; когда неопределенность возрастает, система де-
лается более «терпимой», и диапазон удовлетворения расши-
ряется. Когда неопределенность уменьшается, система стано-
вится более «честолюбивой», а диапазон удовлетворения
сужается.
Пример 4.5.1. В качестве примера в работе [331] рассмотрена глазо-
двигательная система. Испытуемый видит цель (светлое пятно на экране) и
следит за его смещениями. Качество системы оценивается по близости поло-
жения глаза и цели (рис. 4.7). Классические методы анализа глазодвигатель-
ной системы управления не могут объяснить следующих явлений: цель дости-
гается только после ряда последовательных движений — скачков; количество
Глаз
а)
б)
Рис. 4.7. Реакции глазодвигательной системы на смещение цели, а) Схема опыта;
/ — цель, 2—точка фиксации глаза; б) после ступенчатого смещения цели w (t) выходной
сигнал y{t) совершает саккадические движения — последовательность скачков. В данном
случае в ответ на оба ступенчатых изменения положения цели глазодвигательная система
ответила последовательностью из двух скачков, величина каждого из которых, однако,
никак не связана с величиной входного возмущения. В первом случае установившаяся
ошибка равна нулю, во втором—0,1°.
и величина последовательных скачков у данного испытуемого в повторных
экспериментах непредсказуемо варьируют и, в частности, цель может иногда
достигаться за один скачок.
Экспериментальные данные легко объясняются иа основе принципа удо-
влетворения. Схема системы управления представляется в виде, показанном
на рис. 4.8, а. Предел неопределенности есть начальное рассогласование —
ошибка е = w — у. Экспериментально удалось установить зависимость диа-
пазона удовлетворения от пределов неопределенности = й(е) (рис. 4.8,6).
Управляющий сигнал u(t) выбирается в каждый момент времени так, чтобы
перемещение глаза р(и) уменьшило ошибку до «допустимых» в этот момент
времени значений
w — р (и) s h (е). (4.9)
Тогда процесс идет последовательно, возможно, в несколько стадий. В на-
чальный момент времени неопределенность е велика, и выбранное управле-
ние и° может быть достаточно грубым — важно лишь перевести систему в
диапазон удовлетворения, который в начальный момент тоже достаточно ве-
лик. Тогда наступает второй этап: поскольку пределы неопределенности
уменьшились, требования к качеству достижения цели в системе возросли,
диапазон удовлетворения сузился. Теперь нужно новбе управление и1, пе-
реводящее систему в новый диапазон удовлетворения. Так возникают
120
ОПТИМАЛЬНОСТЬ И АДАПТАЦИЯ В БИОЛОГИИ
(ГЛ. 4
последовательные (саккадические) движения. Случайно первый скачок может
оказаться таким, что ошибка сразу уменьшается до малых значений, не выхо-
дящих из зоны нечувствительности системы, тогда переходный процесс закан-
чивается за один скачок.
я;
41(e)
12°~
-30° -20 -10
10 го го° s
—4
--о
$)
Рнс. 4.8 Система управления, работающая по принципу удовлетворения, а) Схема упра
вления глазодвигательной системой; б) зависимость диапазона управления h (в) от пределов
неопределенности, т. е. от величины входного возмущения в. При большой начальной
неопределенности (например, при е=30°) удовлетворительными считаются ошибки в 6° и
более. Когда неопределенность уменьшается, система становится меиее терпимой к ошибкам,
и требования к отслеживанию входного сигнала возрастают. Зависимость построена на
основе экспериментальных данных [331].
Замечательно, что объем обрабатываемой информации в та-
кой системе минимален, в то же время при слежении за движу-
щейся целью такая система будет экономичнее, надежнее, а
главное — точнее систем с более сложными алгоритмами. Вме-
сто сложных.алгоритмов достижения оптимума типа (4.4) удов-
летворение в системе достигается за счет простых эвристиче-
ских алгоритмов, не обладающих вообще никакой памятью и
работающих по простейшей логической схеме «если — то».
Возможно, что для некоторых высокоорганизованных си-
стем управления организменного уровня принцип удовлетворе-
ния в данной формулировке может оказаться искомым общим
решением проблемы. Однако для других уровней организации
биосистем понятия удовлетворительности характеристик, гар-
моничности, достаточности и т. д. все еще остаются скорее ин-
туитивными, чем формально обоснованными. Ясно только, что
отдельные элементы, из которых складывается биосистема,
4.6]
ХОРОШИЙ, ПЛОХОЙ ИЛИ ОПТИМАЛЬНЫЙ?
121
должны быть совместимы или «согласованы» друг с другом, об-
разуя вместе устойчивую открытую гомеостатическую систему.
Возможно также, что какие-то суждения о требованиях совме-
стности, предъявляемых к элементам биосистемы, можно полу-
чить, базируясь на принципе адекватной конструкции системы,
т. е. подходя к ее исследованию с позиций инженера, желаю-
щего достичь «целей живой природы» в соответствующей тех-
нической конструкции.
Концепция оптимальности, заимствованная из теории управ-
ления, оказывается слишком прямолинейной для описания ха-
рактеристик приспособленности биосистем, а адаптация в жи-
вых системах — не столько союзником оптимальности, сколько
ее соперником. В ряде случаев система достигает нужного для
своего существования уровня приспособленности не за счет со-
здания сложных оптимальных алгоритмов, а за счет целесооб-
разного (адаптивного) изменения параметров в простейших ал-
горитмах, что позволяет ей существовать в таких условиях ок-
ружения, для которых инженер не только не может рассчитать
оптимального решения, но пока даже не пытается дать достаточ-
но общую постановку задачи.
4.6. Хороший, плохой или оптимальный?
Вынесенный в заголовок настоящего раздела вопрос — на-
звание одной из рядовых статей по теории оптимальных систем
[351]. Авторы этой статьи с интригующим названием рассмот-
рели задачу синтеза оптимальной линейной системы управления
с квадратичным критерием качества (т. е. общепринятый в тео-
рии управления вариант постановки такой задачи) и показали,
что полученное в рамках строгой теории решение может ока-
заться неосуществимым при использовании на некоторых тех-
нологических объектах. Даже если решение, полученное для
рассматриваемых в теории идеализированных условий, и осу-
ществимо на практике, то оно обладает целым рядом серьезных
недостатков. Так, если в технологической системе, построенной
в соответствии с полученными рекомендациями, произойдет от-
каз одного из датчиков, то система не просто станет неоптималь-
ной, а потеряет устойчивость со всеми вытекающими отсюда
неприятными последствиями.
Выбором заголовка авторы рассмотренной статьи хотели под-
черкнуть, что термин «оптимальный» не всегда отвечает интуи-
тивным представлениям о хорошем или плохом функционирова-
нии систем. Требование хорошего, высококачественного функ-
ционирования практически означает не только (и не столько)
оптимальность в строго оговоренных условиях. В жизненных
ситуациях всегда существуют те или иные неучитываемые при
122 ОПТИМАЛЬНОСТЬ и адаптация в БИОЛОГИИ [гл. 4
теоретическом рассмотрении малые факторы, нарушающие ра-
боту системы, и реальная система должна при вариациях ус-
ловий сохранять работоспособность и качество.
Такие представления в теории управления связываются сей-
час с так называемыми робастными системами управления*),
суть которых состоит в следующем. Когда нет уверенности в
том, что идеальные условия, при которых получается теорети-
чески оптимальное решение задачи, могут быть реализованы на
практике, то надо конструировать не оптимальные системы или
структуры, а те, на которые меньше всего влияют небольшие
отклонения от идеальности. «Если какая-то вещь обладает при
идеальных условиях оптимальными характеристиками, то на
практике она окажется весьма далекой от оптимума», — в та-
кой форме высказал свое отношение к оптимальным системам
известный американский специалист в области прикладной ста-
тистики Дж. Тьюки [366]. «Многие ли... ясно понимают, в чем
единственное хорошее качество оптимума в реальном мире?
В чем ценность доказательства того, что меньше всего стекла
уходит на сферическую бутылку для молока?.. Единственное
хорошее качество его в том, что формальный оптимум будет
единственным, и о нем относительно легко писать доклады и
его сравнительно легко изучать»,— пишет он далее.
Идея абсолютной оптимальности, единственного из всего
многообразия решений, будь то в технических или живых си-
стемах— это абстракция. «Точная оптимизация не нужна для
практических целей ни в одной области. Везде требуется либо
получить просто достаточно хорошее решение, или решение,
которое лучше существующего. Герберт Саймон ввел для со-
циологических, в основном, задач глагол сатисфизировать — до-
биваться удовлетворительной работы. Слово мелиорировать
(улучшать) ничуть не хуже, чем оптимизировать» [366]. С прин-
ципом удовлетворения — satisfaction principle (англ.) — мы уже
сталкивались в предыдущем разделе.
Значит, система может быть хорошей, но не оптимальной,
или же оптимальной, но плохо работающей в неопределенных,
неадекватных условиях. Оптимальная система должна еще быть
и ...хорошей. Адаптация, обучение, использование векторных ме-
тодов оптимизации — по этим направлениям идет развитие про-
блемы оптимальности в теории управления [239].
Живые системы — безусловно «хорошие» системы. Их свой-
ства и характеристики, возникающие в процессе естественного
отбора и эволюции, таят в себе многие пока не понятые и тем
более не формализованные закономерности. Технические си-
стемы еще долго не достигнут совершенства в функционирова-
) От английского robust — крепкий, здоровый.
4.6, ХОРОШИЙ, ПЛОХОЙ ИЛИ ОПТИМАЛЬНЫЙ? 123
нии и работоспособности, свойственного живым системам. И дол-
го еще любые концепции, предлагаемые для усовершенствова-
ния качества технических систем, будут находить готовые про-
тотипы или аналоги в живой природе.
В таком положении находится сейчас концепция оптималь-
ности — одно из наиболее математизированных и формализо-
ванных направлений описания живых систем. В прошедшие
годы поиски строгой оптимальности были полезными и необхо-
димыми вехами исследования и конструирования технических
систем и, следовательно, одним из закономерных подходов к
изучению и описанию свойств живых систем.
ГЛАВА 5
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
5.1. Векторы и матрицы
^На том уровне сложности, который сейчас используется для
описания и анализа биосистем, как правило, возникает необхо-
димость привлекать большое количество информации о перемен-
ных в системе и связях между ними. По мере усложнения био-
логического объекта описывающая его система дифференциаль-
ных уравнений становится все более громоздкой.
Для линейных систем, однако, использование матричных ме-
тодов описания позволяет проводить исследование практически
одинаково компактно вне зависимости от количества перемен-
ных в системе. Считается, что аппарат матриц дает возмож-
ность более просто представлять различные математические
и физические операции с помощью операций над элементами
матриц.
Возьмем, например, систему линейных уравнений и покажем,
как удобно использовать понятие матрицы для ее компактного
представления. Пусть система уравнений имеет вид
X] = ЦцХ1 + #12*2 + • • • + а1тхт>
х2 — #21*1 Н- #22*2 Н- • • • Н- а2тхт> (Я. 1\
хт — ®т\х1 + ^т2х2 Н- • • • 4~ аттХт.
Эти уравнения описывают множество взаимосвязей, суще-
ствующих в некоторой реальной системе между ее перемен-
ными X], Х2, . . • , хт и скоростями ИХ ИЗМеНеНИЯ Х1, х2, ..., хт.
Все эти связи можно охарактеризовать упорядоченным набором
коэффициентов a,/, i, j = 1,2, ..., т, если обозначить этот набор
символом А и записать в виде прямоугольной таблицы
#11 #12
#21 #22
а1т
а2т
(6.2)
_#ml ат2
а
mm J
5.Ц
ВЕКТОРЫ И МАТРИЦЫ
125
Такая таблица, имеющая, как и исходное уравнение, т строк,
составленных из коэффициентов а1у-, называется матрицей.
Коэффициенты исходного уравнения составляют ее столбцы, ко-
торых в данном случае тоже т. Матрица (5.2) иногда обозна-
чается символом [а,/].
Линейные уравнения часто используются для описания био-
систем. Матрица коэффициентов в этом случае определяет сразу
все взаимосвязи, существующие между внутренними перемен-
ными системы х. Свойства системы, следовательно, определяют-
ся свойствами описывающей ее матрицы, V-
Поэтому мы рассмотрим терминологию, принятую в теории
матриц, и усвоим некоторые элементарные навыки обращения
с векторами и матрицами, чтобы читатель, ранее не сталкивав-
шийся с матрицами и векторами, мог понимать матричные обо-
значения, а при необходимости и самостоятельно продвигаться
дальше (в этом случае, например, можно рекомендовать чита-
телю книгу [66]).
Читатель, знакомый с матрицами, может вполне пропустить
этот раздел и продолжить чтение с разд. 5.2.
Матрицей называется прямоугольная таблица, элементами
которой могут быть числа или функции. Элементы матрицы
обозначаются при помощи двойного индекса: первый индекс
обозначает номер строки, второй — столбца. Так, элемент а32
стоит в матрице на пересечении третьей строки и второго столб-
ца. Если матрица имеет всего один столбец, она называется
вектором, или вектором-столбцом.
Пример 5.1.1. При исследовании биосистем удобно в качестве векто-
ров брать определенные группы сигналов. Так, все сигналы, характеризующие
внешнюю среду, можно представить в виде вектора следующим образом.
Пусть нас интересуют такие переменные среды, как температура О, концен-
трация кислорода сО2 и влажность ц. Обозначив эти величины через v2
и v3, соответственно, получим вектор
[f'1
I. (5.3)
1>3J
Поскольку вектор имеет всего один столбец, индекс у его
элементов одинарный, и обозначает он номер строки.
Матрица А, имеющая m строк и п столбцов, называется
(иг X н) -матрицей или матрицей размерности (или порядка) m
на п. В этом случае иногда пишут Л(тх«). Вектор (5.3) являет-
ся матрицей размерности три на один: v<3Xi)J о векторе гово-
рят — вектор размерности 3.
Если оба размера матрицы совпадают, то матрица Л(ЛХП) на-
зывается квадратной. Квадратная матрица имеет диагональ, со-
стоящую из элементов ап, аг2> .. ,апп. Если все элементы, кроме
диагональных, равны нулю, матрица называется диагональной.
126
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
Диагональная матрица иногда обозначается как [diaga;;]:
о
о
о
[diag ai(] =
Я11
О
О О
^22 О
О Лзз
(5.4)
ООО
апп
Если в диагональной матрице все диагональные элементы рав-
ны 1, матрица называется единичной и обозначается симво-
лом Е'.
1 о • • • 0-
0 1 • • • О
(5.5)
L0 0 • • • 1J
Если в матрице А все столбцы поменять местами со стро-
ками, то такая матрица называется транспонированной и обо-
значается 4Т.
Например, для матрицы А = ^транспонированная мат-
рица имеет вид = Транспонированный вектор-стол-
бец дает вектор-строку. Так, транспонируя вектор (5.3), полу-
чаем
oT = [oj v2 Ц3]. (5.6)
Транспонировать вектор, в частности, удобно для экономии ме-
ста при записи, если записывать его так:
v = [vi v2 w3]T. (5.7)
Для исследования сложных систем приходится обращаться
с векторами и матрицами примерно так же, как при исследова-
нии простых систем мы оперируем с переменными и коэффи-
циентами. Рассмотрим поэтому простейшие действия с матри-
цами.
Сложение и умножение матриц. Складывать
можно матрицы одной размерности, например (тХО- Суммой
матриц А и В одной размерности служит матрица С той же
размерности, каждый элемент которой есть сумма соответствую-
щих элементов матриц А и В:
= an -|- blf. (5.8)
Разность матриц D = А — В имеет элементы
dtj — aij — Ьц. (5.9)
5 Л
ВЕКТОРЫ И МАТРИЦЫ
127
Две матрицы одной размерности называются равными, если
их соответствующие элементы одинаковы, т. е.
А = В, (5.10)
только если ощ = biS для всех i, j.
Произведение матриц обозначается следующим образом:
С = АВ. Умножать можно матрицы, у которых внутренний раз-
мер совпадает. Так, если имеем Л(тхл), B^xk), то внутренний
размер этой пары есть п, и можно выполнить операцию умно-
жения. Элементы матрицы произведения С при этом получают-
ся посредством перемножения и суммирования элементов ис-
ходных матриц, а именно для получения элемента с номером
(i, /) надо по очереди перемножить пары элементов i-й строки
матрицы А и /-го столбца матрицы В (первый с первым, второй
со вторым и т. д.), а затем просуммировать их.
Найдем, например, элемент с номером (2, 3) матрицы С,
являющейся произведением двух матриц С = АВ, где
Г 1 3 h
Г1 2 0 3~1 -112
4(2X4) = [1 0 2 —1J’ 5(4X3)= 0 2 3
L 2 4 4.
(5.11)
Матрицы можно перемножить, так как их внутренние размеры
совпадают. Элемент Сгз, например, равен сумме попарных про-
изведений элементов второй строки матрицы А, на элементы
третьего столбца матрицы В:
с23=1 . 1+0-2 + 2-3 + (-1). 4 = 3.
Чтобы найти произведение матриц, надо поочередно найти
все элементы матрицы С. Размерность матрицы С будет опре-
деляться внешними размерами сомножителей:
4(тХл)5(пХА) = б?(тхА). (5.12)
Найдем, в частности, произведение матриц А и В из (5.11).
Имеем
4(2Х4)В(4ХЗ) = 0(2X3) =
_Г1-2 + 6 3+2+12 1+4+121 Г 5 17 174
~L 1-2 3 + 4-4 1+6-4J-L-1 3 з]'
Общее правило для определения элемента с номером (i, /)
матрицы С можно записать в виде формулы
п
Cii=T,aiyb4b (5.13)
v»l
где п — внутренний размер пары матриц А к В,
128
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
1ГЛ S
Легко видеть, что при умножении матриц их нельзя менять
местами (операция умножения матриц некоммутативна). Для
матриц А(тХП) и В(ПХт) существуют как произведение АВ, так
и произведение ВА, но размерность получающейся матрицы бу-
дет (т X т) в первом случае и (п X п)—во втором. В этом
случае говорят, что матрица А умножается на В справа и сле-
ва, соответственно. Даже для квадратных матриц, вообще го-
воря, АВ ВА.
Умножим, например, матрицу А(2х3) на матрицу В(зх2) снача-
ла справа, а потом слева. Пусть
Г1 ° 21 о q
Л(2ХЗ) = 10 1 11’ 5(3X2) = 2 3 •
Li oj
Имеем
[О -1
2 3
1 О
(5.14)
-11
7 . (5.15)
2J
Умножим теперь матрицу Л(2х3) из (5.14) справа на вектор
(5.3). Имеем
л Г1 0 21 Г^’1 Гр, + 2рз1
Ло:==10 1 1J [jj = bs + fs J •
Матрица Av имеет размерность (2 X 1)> т- е- является вектором.
При умножении матрицы или вектора на единичную матрицу
умножаемая матрица или вектор не изменяются. Умножим, на-
пример, Е(2х2) на матрицу Л(2хз):
Г1 01 Г1 3 21Г1 • 1+0-5 1-3+ 0-1 1 -2 + О- 11Г1 3 21
L0 iJlS 1 1J —Lo-1 + 1-5 0-3+bl 0.2+ 1 • 1J — [б 1 J’
При умножении матрицы произвольной размерности на ска-
лярный (не векторный) множитель k каждый элемент матрицы
умножается на величину k.
Определители. Определителем квадратной матрицы
Л(пХП) называется число, равное сумме п\ членов вида
(— 1)гащйщ ... ankfi, каждый из которых соответствует одному
из различных упорядоченных множеств ki, й2, ..., kn, получен-
ных г попарными перестановками элементов 1,2.....п Число
п есть порядок определителя.
Для квадратной матрицы А размерности (2 X 2)
А =
ап
<221
а 12
а22
(5 16)
определитель равен ацн22—<221012- Определитель любой мат-
рицы А(„х„) обозначается как det Л, или |Л |.
5 1]
ВЕКТОРЫ И МАТРИЦЫ
129
Определители квадратных матриц большей размерности вы-
числяются путем их сведения к определителям матриц (2 X 2)
с помощью метода Лапласа. Этот метод заключается в следую-
щем. Если в определителе | А | n-го порядка вычеркнуть i-ю
строку и /-й столбец, оставшиеся (п— 1) строк и столбцов об-
разуют определитель (п— 1)-го порядка. Обозначим его |М,/|.
Этот определитель называется минором элемента ач. Так, ми-
нор элемента Щг в определителе третьего порядка
я И Я12 Я13
| А | = а21 а22 а23
Л31 Й32 а33
равен 1^,2 1 = 12*,
Минор элемента а(/, взятый со знаком плюс, если (i 4~ /) четно,
и со знаком минус, если (i -ф /) нечетно, называется алгебраи-
ческим дополнением элемента аИ и обозначается через Сч:
Сг/ = (-1)1+/|М(/|. (5.17)
Алгебраическое дополнение элемента оцг в предыдущем при-
мере есть
£12= ( О3 ™ = (а21а33 а31а2з) = а31°23 — а21^33-
| изi изз|
Метод Лапласа состоит в том, что определитель произволь-
ного порядка раскладывается по элементам любой строки или
столбца. Например, для определителей третьего порядка разло-
жение по элементам первой строки дает
I А | = ЯцСп + О12С12 + Я13С13,
по элементам третьего столбца —
| А | = а13С13 + 023^23 + «ззСзз-
Определитель четвертого порядка сначала сводится к опре-
делителям третьего порядка, а затем — к определителям вто-
рого порядка. Аналогично вычисляются определители и высших
порядков при п 2
Таким образом, при разложении по /-му столбцу имеем фор-
мулу
п
I А I ' ^V/£v/>
а по й-й строке — формулу
Пусть дана матрица
п
I А | ^Ац£ац.
-1
А = 2
.1
О —2-|
3 О I
2 1J
(5.18)
(5.19)
(5.20)
130
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
Вычислим определитель |А| разложением, например, по эле-
ментам второго столбца:
det Л = 0 + 3- (+1) • |[ ~ J] + 2 • (-1) • (J “о| = 1.
Определитель удобно раскладывать по тем строкам или
столбцам, где имеется много нулевых элементов. Существуют и
другие способы вычисления определителей, на которых мы не
останавливаемся (см., например, [66]).
Обратная матрица. Если А — квадратная матрица,
Сц — алгебраическое дополнение а,/, то матрица, образованная
из алгебраических дополнений Сц, называется присоединенной
с А и обозначается Adj А:
AdjA = [CH]. (5.21)
Обратите внимание на порядок следования индексов в фор-
муле — матрица [С//] является транспонированной для матрицы,
составленной из Сц.
Найдем для примера присоединенную матрицу с матрицей
(5.20). Имеем Сц = 3, Сы = —2, С]3 = 1, С21 = —4, С22 ==13I
С23 — —2, С31 = 6, С32 = —4, С33 = 3. Следовательно,
Г 3
Adj А = [Сц] = -2
L 1
—4 6"
3 —4
—2 3. ’
(5.22)
Если квадратная матрица А имеет определитель det А, не рав-
ный нулю, то для нее можно построить обратную матрицу.
Матрица А~1 называется обратной матрицей матрице А в
том случае, если умножение матрицы А на А-1 справа или слева
дает единичную матрицу:
АА~' = А~,А = Е. (5.23)
Обратная матрица вычисляется по простой формуле
Л"1 = ^ТТА^Л- (5.24)
Умножение на обратную матрицу аналогично операции де-
ления обычных чисел, или скаляров; как нельзя делить на нуль,
так нельзя построить обратную матрицу для матрицы, у кото-
рой определитель равен нулю.
Матрица А, у которой det А = 0, называется особенной. Об-
ратная матрица существует для всех неособенных матриц.
Найдем матрицу, обратную матрице А из (5.20). Имеем
det А = 1. Следовательно, A-1 =-J^Adj А = Adj А, и А-1 оп-
ределяется соотношением (5.22).
5.11
ВЕКТОРЫ И МАТРИЦЫ
131
Проверим, чему равно произведение АЛ-1: из (5.20) и (5.22)
получаем
1 0 —21 Г 3 —4 6"|
2 3 0 —2 3 —4 1
.12 1JL 1 —2 з]
-3—2 —4+4 6—6 "I
6-6 -8+9 12-12
.3+4—1 -4+6-2 6— 8+з]
-1 0 0"|
0 10
.0 0 1J
— £(3X3)-
Собственные числа матрицы. Собственные (ха-
рактеристические) числа Ль .... Лп произвольной квадратной
матрицы А определяются как решения, или корни, характери-
стического уравнения для матрицы А:
det [ЛД — А] = 0. (5.25)
Матрица размерности (п X «) имеет п собственных чисел.
Найдем, например, собственные числа матрицы
Г2 —2 3-|
А = И 1 1
Li з -и
(5.26)
Характеристическое уравнение имеет ви.
2
det
-Л, —2
-1
Л — 1
—3
—3 "I
-1
Л + 1J
= Л3 — 2Л2 — 5Л + 6 = (Л — 1) (Л +- 2) (Л — 3) = 0
Определитель обращается в нуль при трех значениях Л : Xi = 1,
Хг — —2, Х3 = 3. Это — собственные числа матрицы А. Собствен-
ные числа могут быть действительными или комплексными.
Сумма собственных чисел называется следом матрицы и обо-
значается tr А;
tr А = Л] Х2 ... 4- Хп. (5.27)
След матрицы можно получить, не находя ее собственных чисел,
так как он равен сумме ее диагональных элементов:
п
tr А = У, а{{.
i-1
(5.28)
Ранг матрицы. Для матриц произвольной размерности
(п X т) минором 1-го порядка называется определитель, полу-
чающийся из исходной матрицы после вычеркивания любых
(п — I) строк и (т — I) столбцов. Максимальный размер от-
личных от нуля миноров матрицы называется ее рангом.
Например, ранг матрицы
Г1 0 0 01
[о 0 2 0]
132
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. в
равен двум, так как минор, составленный из первого и третьего
столбцов, отличен от нуля.
Блочные матрицы. Несколько матриц или векторов
можно объединить в одну матрицу или вектор, которые будут
в этом случае называться блочными. Наоборот, матрицу (век-
тор) можно свести к другой матрице (вектору), составленной
из элементов, которые являются подматрицами (подвекторами).
Например, проводя пунктирные линии на матрице А размер-
ности (3X3), можно записать матрицу (2X2), составленную
из подматриц Ль Аг, А3 и А^, следующим образом:
Л =
«II а 12 «13
«21 «22 «23
.аз. «32 «зз
(5.29)
где
Л1 = [йп],
А = К а13], Л4 = Гг “231.
1_аз2 «ззЗ
Действия над блочными матрицами выполняются так же, как
над обычными матрицами при условии, что подматрицы при вы-
полнении действий умножения и сложения согласуются (т. е.
имеют соответствующие размерности).
Некоторые операции над матрицами удобно производить,
представляя их в виде блочных. Так, операцию взятия обрат-
ной матрицы от блочной матрицы можно выполнить, восполь-
зовавшись следующей формулой. Пусть А(ПХП), В(пхт), С(тхП),
D(mxm') — подматрицы некоторой блочной матрицы. Тогда
гл i bi"1 Г(л - вр-'с)-1____j_- (а - во~|с)~1до~1~|
LcLdJ = L- (Р-СЛ-’вГ'Сл""1] (Р-’ся'-’вГ-1 ]’
Производная и интеграл от матрицы. Обычные
способы получения производных и интегралов по определению
переносятся на случай матриц при условии сохранения перво-
начального порядка сомножителей. Так, при дифференцирова-
нии и интегрировании по времени имеем
±Л(/) = Л(/) =
'ац(^) d12(0
a2l(/) «22(0
(5.31)
«1П(0“
a,2 (0 ••• \aln(t)dt
(5.32)
am2^dt *
5.2]
СОСТОЯНИЕ В БИОЛОГИИ И В ТЕОРИИ УПРАВЛЕНИЯ
133
Ниже нам понадобится брать производную вектора по век-
тору. Производная m-мерного вектора х по /-мерному век-
тору b есть матрица (тХО [104]:
dx db dxi dbl dx2 dbl dxi db2 ' dx2 db2 dxi ” db l dxj ' dbt
dx dx dx~
m tn
_ dbl db2 ’ dbi
(5.33)
Пример 5.1.2. Пусть вектор стационарных концентраций веществ в от-
секах некоторой открытой системы есть х = [х1 Ха]т, где х1 — концентрация
вещества в первом отсеке, х2 — во втором. Вектор внешних условий системы
тот же, что в примере 5.1.1. Чувствительность вектора стационарных концен-
траций х размерности 2 к внешним условиям v (вектор размерности 3) опре-
деляется производной dxjdv — матрицей размерности (2X3):
Г dxi dxi dxi -J
dx dvt dv2 dv3
dv dx2 dx2 dx2
. dv{ dv2 dv3 .
(5.34)
5.2. Понятие состояния в биологических науках
и в теории управления
В биологических науках интуитивно сформулировалось
представление о состоянии системы как об очень широком
круге показателей и характеристик, определяющих ее функцио-
нирование и реакции на различные внешние факторы. В физио-
логии, например, говорят о нормальном состоянии организма,
если все его переменные и их реакции на внешние стимулы
находятся в допустимых пределах. Понятие внутреннего состоя-
ния организма в сравнительной физиологии эквивалентно поня-
тию функционального уровня и учитывает скорости движения
веществ, скорости протекания различных процессов и такие ха-
рактеристики, как температура тела [166]. Еще более широко
термин «состояние» используется в биогеоценологии как аналог
некоторых интегральных характеристик биосистемы. Так, важ-
ным показателем является «состояние биоценотической системы
в целом и в особенности ее устойчивости во времени» [204].
Следует иметь в виду, что термин «состояние» в физиологии
и в ряде других биологических и медицинских дисциплин трак-
туется в двух различных плоскостях. В одних случаях этот тер-
мин означает конкретный набор числовых характеристик, опи-
сывающих структуру и функционирование организма или его
134
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. 5
систем. Например, состояние системы кровообращения в орга-
низме человека можно описать следующим образом: частота
сокращений сердца — 68 уд./мин, систолическое и диастоличе-
ское давление—150 и 110 мм рт. ст., минутный объем кро-
ви — 7 л1мин.
В других случаях понятие состояния используется в качестве
обобщающего, классифицирующего термина, который дает не-
которую оценку структуры и функционирования организма или
его систем. Термин «состояние» в этом случае относится к це-
лой группе мало отличающихся друг от друга наборов количе-
ственных характеристик. Именно в таком смысле следует пони-
мать клинические термины «удовлетворительное состояние» или
«состояние средней тяжести», относящиеся к целостному орга-
низму. Состояние системы кровообращения, характеристики ко-
торого приведены выше, может быть отнесено к классу гипер-
тензивных состояний, как и близкие к нему состояния, для ко-
торых, например, систолическое давление равно 160 или
170 мм рт. ст. Обзору и общей классификации состояний орга-
низмов — от нормальной жизнедеятельности через промежуточ-
ные состояния до анабиоза — посвящена монография [55а].
Таким образом, термин состояние биосистемы, хотя интуи-
тивно и ясен, не имеет строгого определения. Иногда он отно-
сится к переменным типа уровень, как в экосистемах, иногда —
к переменным системы типа темп, как в сравнительной физио-
логии (хотя в этом случае специальной разницы между тем-
пами и уровнями не делается), и, наконец, связывается с обоб-
щенными характеристиками биосистемы. В теории управления
имеется строгое определение понятия состояния системы [46,
66, 88].
Состояние динамической системы — это наименьший набор
чисел, который необходимо задать в данный момент времени t0,
чтобы была возможность в рамках математического описания
системы предсказать ее поведение в любой будущий момент
времени t > Со-
существует простой способ определения этого набора чисел
(называемого также вектором состояния системы), основанный
на использовании схем моделирования. Схемы моделирования,
рассмотренные выше в разд. 3.3, стали широко распространен-
ным методом исследования биосистем, поэтому для объяснения
смысла вектора состояния мы и воспользуемся этим способом.
Естественным вектором состояния в системе является век-
тор, составленный из выходных переменных всех интеграторов
в схеме моделирования [66, 344]. Легко видеть, что в этом слу-
чае координаты вектора состояния совпадают с рассмотренными
ранее фазовыми координатами системы (разд. 3.3). Поэтому мы
ниже будем говорить о состоянии системы только в этом смыс-
5.21
СОСТОЯНИЕ В БИОЛОГИИ И В ТЕОРИИ УПРАВЛЕНИЯ
135
ле. Состояние системы есть вектор, составленный из выходных
переменных всех ее интеграторов, или из ее фазовых координат.
Для биологических систем это определение является чрез-
вычайно удобным. Все живые системы есть открытые системы,
в которых протекают процессы обмена вещества и энергии. Су-
ществует широкий класс моделей биологических систем, в ко-
торых основное внимание направлено как раз на описание про-
цессов обмена, запасания и расходования веществ и энергии.
Такие модели называются компартментальными, и им будет
посвящена следующая глава. В компартментальных моделях
открытых систем выходные сигналы интеграторов всегда опи-
сывают запас веществ или энергии в системе, определяют их
уровни — количества или концентрации. Значит, вектор состоя-
ния биосистемы является вектором концентраций веществ в этой
системе. В терминах переменных типа темпов и уровней
(разд. 1.7) вектор состояния часто представляет собой вектор
уровней в системе.
Легко видеть, что задания уровней в системе достаточно для
определения всех темпов процессов, происходящих в ней, если
только известны законы функционирования системы и условия
среды, в которых она находится. Следовательно, состояние си-
стемы есть минимальный набор переменных, полностью опреде-
ляющий ее функционирование. Этот набор можно изменить, но
нельзя уменьшить.
Пример 5.2.1. В рассмотренном ранее в гл. 3 примере 3.4.1 приводится
модель системы кровообращения, в которой имеется шесть сигналов. Три из
них — внешние, это входы системы vh Wi, Р. Поэтому вектор входов имеет
размерность 3:
0=[01 wt Р]г.
Остальные три сигнала — внутренние: Q, Xt и R. Вектор состояния, однако,
имеет размерность единица, так как в схеме рнс. 3.8, а имеется только один
интегратор.
Действительно, достаточно задать одну из этих трех величин, как осталь-
ные находятся из уравнений системы и значений вектора входов. Естествен-
ной переменной состояния здесь является выход интегратора Х| в схеме
рис. 3.8. Задавшись значением Х|, легко находим сопротивление сосудов
R = а + 6xi
и величину кровотока
Р Р
Q R а + bxi
Метод пространства состояний в теории управления пришел
на смену классическому подходу, основанному на идее черного
ящика, когда система исследовалась только на основе анализа
соотношений между ее входами и выходами. Метод «черного
ящика» в сложных системах оказался неадекватным, поскольку
реакции выхода на один и тот же вход оказываются при таком
136
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
описании неоднозначными (факт, известный и в биологии). Опи-
сание системы с помощью передаточной функции не позволяет
учитывать влияние внутренних переменных и входов, имеющих-
ся в системе между рассматриваемыми входом и выходом (см.
[46], т. III, стр. 228). Когда эти переменные меняются, меняются
и свойства системы — реакции на один и тот же входной сигнал
зависят от их значений. Этот недостаток и устраняется введе-
нием в рассмотрение понятия состояния системы. Знание со-
стояния системы позволяет всегда однозначно определить лю-
бые ее характеристики и в этом смысле является исчерпываю-
щим описанием системы.
Пример 5.2.2. Дыхательный хемостат Ф. Гродинза ([60], уравне-
ния VII. 24 и VII. 26) имеет три входа и два выхода и описывается двумя
дифференциальными уравнениями второго и первого порядка:
От 4- лОт 4- ЬОт === 0j 4- сЛ4, 0а = б/От 4~ g&r 4- Л, (5.35)
где 0, — входной сигнал (концентрация углекислоты во вдыхаемом воздухе),
Л1— входной сигнал (темп выделения СО2 в процессе метаболизма; в работе
Ф. Гродинза считается постоянным), h — входной сигнал (вспомогательная
константа), 0т — выходной сигнал (концентрация СО2 в тканях), 0а — выход-
ной сигнал (концентрация СО2 в артериальной крови), a, b, с, d, g— пара-
Рис. 5.1. Схема моделирования дыхательного
хемостата Гродинза.
метры (коэффициенты, определяе-
мые структурой дыхательной си-
стемы).
Схема моделирования ([60],
стр. 164) приведена на рис. 5.1.
В схеме имеется два интегратора
с выходными переменными 0Т и
От. Они и образуют вектор состоя-
ния системы. Обозначив xt =0Т,
х2 = От, получаем вектор состоя-
ния в виде
X = [х, Х2]Т.
Модель Ф. Гродинза не отно-
сится к упомянутому выше классу
компартментальных моделей. Основной чертой таких моделей является выде-
ление каждого интересующего исследователя объема вещества, перемещающе-
гося и преобразующегося в системе, в ее независимый элемент — компартмент.
Тогда каждому компартменту отвечает своя переменная состояния. В рассма-
триваемом же случае своя переменная состояния отвечает только одному из
двух представляющих интерес объемов вещества, а именно количеству СО2 в
тканях. Поскольку модель Ф. Гродинза не является компартментальной, ее
вектор состояния не описывает уровней вещества в системе. Уровни вещества
представляют собой выходные сигналы модели.
5.3. Уравнения состояния
Поскольку достаточным описанием системы является зада-
ние в каждый момент времени ее вектора состояния и вектора
входов, то в современной теории управления динамические свой-
ства системы описываются с помощью уравнений, связываю-
6.3]
УРАВНЕНИЯ СОСТОЯНИЯ
137
щих между собой ее состояния и входы в различные моменты
времени. Что касается выходных сигналов системы, то они вы-
ражаются через вектор состояния с помощью отдельного урав-
нения.
В такой форме система описывается парой уравнений, из
которых первое — дифференциальное — определяет состояние
системы и тем самым задает ее динамические свойства. Второе
уравнение—алгебраическое — определяет выходы системы че-
рез ее текущее состояние.
Обозначив вектор состояния системы через х, вектор входов
через V, вектор выходов через у, можно записать уравнения не-
прерывной детерминированной системы в виде
х(/) = Ф[х(/), »(/)],
y(t) = W[x(t), и(0]. ( *
Первое уравнение в (5.36) называется уравнением состоя-
ния, а второе — уравнением выхода. Уравнение состояния обыч-
но бывает дифференциальным (в дискретных системах — раз-
ностным), а уравнение выхода—всегда алгебраическое.
Оба уравнения в (5.36) написаны для векторов. Когда по-
добные уравнения переписываются покоординатно, т. е. выпи-
сываются порознь для каждой координаты вектора состояния,
исходная динамическая система определяется уже не одним, а
несколькими уравнениями состояния и несколькими уравнениями
выхода. Число таких уравнений равно, очевидно, числу компо-
нент в векторе состояния и в векторе выхода.
Для системы, описываемой линейными дифференциальными
уравнениями, уравнения состояния записываются следующим
образом:
X] — ЛцХ1 -ф а12Х2 -ф • • • 4" U\mxm -ф &llff 4" ^12и2 4" • • • + b\iV[,
х2 = a2ixt -ф а22х2 + • • • 4- a2mxm 4* ^21^1 4* b22v2 + • • + b2tvi,
xm — amixi 4~ am2x2 4’ • • • 4" ammxm 4" bm}V\ 4" bm2V2 -ф . . . -ф bmlVlt
(5.37)
а уравнения выхода задаются системой алгебраических урав-
нений
У\ = ci 14 4- • • • 4- clmxm -ф йцУ1 4- ... 4- йцО/,
У2 — с21х1 4- • • • 4- C2mXm -ф d2\V! 4- . . . -ф d2[Vi, gg)
Уг — crixi 4* • • 4* crmxm 4- driVi 4* • •. 4- drivh
где xt, ..., xm — переменные состояния, щ, ..., Vi — входы,
г/1,..., уг — выходы, а все величины а, Ь, с, d — коэффициенты.
138
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
1ГЛ. 5
На основании изложенного в разд. 5.1 мы можем записать
уравнения состояния системы при помощи матриц:
х — Ах 4- Bv,
у = Сх-}- Dv,
(5.39)
где А(тХт), В(тХ1), C{rXm), D(rXi) — матрицы (если коэффициенты
а, Ь, с или d в (5.20) или (5.21) меняются во времени, то и
матрицы будут иметь переменные элементы), а
ственно. Отметим, что матрица А в (5.39) всегда квадратная,
а остальные матрицы необязательно квадратные.
Пара векторных уравнений (5.39) иногда называется урав-
нениями вход-выход-состояние. В этом случае говорят, что и
сама система описывает-
ся в терминах «вход-вы-
ход-состояние».
Систему, описываемую
уравнениями (5.39), мож-
но изобразить графиче-
ски (рис. 5.2). Схема эта
полностью аналогична од-
номерной схеме, которая
Рис. 5.2 Блок-схема линейной системы.
получается в том случае,
когда х, у, v — обычные скалярные величины, А, В, С, D — чис-
ловые коэффициенты, с той разницей, что векторные связи мы
изображаем двойной широкой стрелкой.
Пример 5.3.1. Представим схему дыхательного хемостата из при-
мера 5.2.1 в матричном виде и запишем ее уравнение состояния. Введем сле-
дующие обозначения: vt — Fh v2 — М, v3 = h — входы, xt = QT, x2 = QT —
переменные состояния, tp = 0a, y2 = 0T — выходы Тогда
— векторы входов, состояния и выходов, соответственно Уравнения системы
имеют в этих обозначениях следующий вид:
уравнения состояния
Xi = — axi — Ьх2 + сц + cv2,
х2 = хг,
(5.41)
УРАВНЕНИЯ СОСТОЯНИЯ
139
4 31
уравнения выходов
у 1 — dx 1 -|- gXi + о3,
Уг = хг.
При помощи матриц эти уравнения можно записать так
* = ^(2X2)* + 5(2хзЛ
У = С(2Х2)Х + D(2X3)°> ’
где матрицы имеют следующий вид:
-г ?]• «-к; а. с-к ?]•
ГО о 11
o = Lo О о} <5’43)
Эти матрицы получаются непосредственно из уравнения (5.41): их элементы
суть просто коэффициенты при соответствующих координатах вектора; если
некоторая координата вектора отсутствует в правой части уравнения, то в
матрице на этом месте ставится нуль.
Линеаризация нелинейных моделей биоси-
стем. Поскольку все биесистемы обладают нелинейными ха-
рактеристиками, построение для них линейных моделей осуще-
ствляется путем линеаризации исходных характеристик, о ко-
торой мы говорили в разд. 3.3.
Опишем процесс линеаризации для системы нелинейных
уравнений состояния (5.36). Обозначим в исходной нелинейной
системе векторы переменных входа, выхода и состояния пропис-
ными буквами X, Y, V, соответственно. Тогда
Х(О = Ф[Х(/), V(0L (544)
Y (!) = Т [X (/), И(/)].
Пусть нас интересует движение системы при малых отклоне-
ниях условий ее функционирования V от некоторых заданных
V* (например, поведение экосистем при небольшом изменении
условий обитания или поведение функций и органов организма
при изменении его режима функционирования — интенсивности
мышечной работы).
Если при V = V* в системе осуществлялось движение X —
= X*(t), К= Y*(t), то можно ввести обозначения
v (!) = V (!) - V* (!), х(1) = Х (!) - X* (!),
у (t) = Y (!) — Y* (!). (5.45)
Тогда, раскладывая функции в правых частях уравнений си-
стемы (5.44) в ряды Тейлора в точке (X*, V*) и ограничиваясь
140
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. 5
линейными членами, получаем
X- (/) + х и - Ф (Г, У) + • х + _я • V,
v-v* v^v* (5.46)
Г (0 + у и - Ч- (Г, Г) + | • у + 4^. | у.
'ЛаЛ А вА
у» у*
Производная вектора (функции) Ф по вектору X в (5.46), так
же как и остальные производные, согласно (5.33) являются мат-
рицами. Обозначим поэтому
, в=4®| ,
Ол у* Ov Ijf*, у*
с = i d=23LI (5‘47)
|^*. у*’ д И |^* у*
Поскольку Х*(/) = Ф(Х*, У*), Y*(t) = Т(Х*, И*), имеем с уче-
том (5.47)
х. (/) = А (I) х + В (/) v,
«/(/) = С (/)% + /)(/) и. (5,48)
Матрицы А, В, С и D имеют переменные во времени эле-
менты, поскольку линеаризация зависимостей в (5.46) произво-
дится каждый раз в различных точках X*(t),
Если нас интересует стационарный режим исходной системы
при V = const, когда переменные системы перестают изменять-
ся во времени, то с течением времени X*(t) сходится к стацио-
нарному значению X, а матрицы А, В, С, D в таком режиме ока-
зываются обычными матрицами с постоянными элементами
(5.39).
5.4. Решение уравнений. Стационарный режим системы
При исследовании биосистем мы часто интересуемся их ста-
ционарными состояниями. В терминах открытых систем, разу-
меется, следует говорить о стационарном неравновесном состоя-
нии, хотя в теории управления и в биокибернетических работах
можно встретить термин равновесный режим системы. Под рав-
новесием при этом, конечно, должны пониматься режимы, когда
уравновешены все силы, действующие в системе, уровни пере-
стают меняться, а темпы процессов сохраняются постоянными.
В стационарном режиме при v = const вектор состояния не
меняется, х=0, и дифференциальное уравнение состояния
(5.39) переходит в алгебраическое уравнение
Дх + Вп = 0. (5.49)
5.4]
РЕШЕНИЕ УРАВНЕНИЙ. СТАЦИОНАРНЫЙ РЕЖИМ СИСТЕМЫ
141
Это уравнение надо разрешить относительно х, т. е. записать
х в виде некоторой функции от и. Проделаем это следующим
образом.
Пусть <4 — неособенная матрица. Напомним, что Д в урав-
нениях системы — всегда квадратная матрица. Возьмем урав-
нение Ах Д- Bv — 0 и умножим все его члены на Л-1 слева.
Тогда А~хАх -f- A~xBv = 0, и так как А~хАх ~ Ех = х, полу-
чаем
х— — А~'Ви. (5.50)
Пример 5.4.1. Решим уравнения для дыхательного хемостата Гродинза
(5.42). В стационарном режиме имеем Ах + Bv — 0, т. е.
Г—а -61 , Г1 с °1
L 1 oj х + Lo 0 ojv — °’
где х — вектор размерности 2, v — вектор размерности 3. Найдем Л-1. Имеем
ГО 61
Лс1]Л = [_1 _aJ, det Л = 6.
Тогда
Стационарное состояние х определяется уравнением (5.50):
Можно записать решение для каждой из координат:
Г 0 0 0"1 Го)"
Р‘1 = 1 с Ь2 ,
l*2-1 Lt т 0JLd
откуда
лц = 0,
01 , CV2
х*=~т+—-
(5.51)
ОТ
0 V. (5.52)
(5.53)
(5.54)
Решив уравнение состояния, можно определить выходы си-
стемы, так как они связаны с состоянием алгебраическим урав-
нением (5.39). Подставляя х из (5.50) и помня о необходимости
соблюдения порядка сомножителей, получаем
y = -CA~'Bv + Dv = (p — CA~lB)v. (5.55)
Пример 5.4.2. Найдем стационарные значения выходных сигналов в
модели дыхательного хемостата. Из (5.42), (5.43) и (5.54) имеем
1
О
о. (5.56)
142
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. 5
предыду-
(5.57)
измене-
Расписав это векторное уравнение покоординатно, можно, как и в
щем примере, найти у\ и
S се
y\ = — ~vl----^-v2 + v,.
1 с
5.5. Устойчивость системы и переходные режимы
Переходный процесс возникает в системе либо при
нии входных сигналов, либо при постоянстве входных сигналов,
но при наличии начальных условий по х, отличающихся от рав-
новесных. При этом каждая координата системы претерпевает
некоторые изменения. Характер этих изменений в линейном слу-
чае полностью определяется свойствами матриц, описывающих
систему.
Если система является асимптотически устойчивой, как об
этом говорилось в разд. 3.6, то переходные процессы по всем
координатам, вызванные при неизменных условиях среды от-
клонениями некоторых координат х от стационарного состоя-
ния (5.50), с течением времени затухают, и переменные возвра-
щаются к этому состоянию. Устойчивость системы зависит ис-
ключительно от вида матрицы А. Динамическая система асимп-
тотически устойчива, если все собственные числа матрицы А
имеют отрицательные действительные части.
Пример 5.5.1. Система дыхательного хемостата Гродинза описывается
уравнениями (5.42). Имеем характеристическое уравнение
det — Д] — % (X + а) — Ь = 0,
откуда %i, 2 = -у (— а ± л/а? + 4b). Пусть, например, а = 1, Ь = —1. Тогда
матрица системы имеет два комплексных собственных числа с отрицательной
действительной частью, равной —%i, г = у (—1 ±1 л/Ю- В этом случае
система устойчива. В других случаях суждение об устойчивости или неустой-
чивости можно получить на основе табл. 3.1. В зависимости от значений а и
b в матрице А система дыхательного хемостата может оказаться как устой-
чивой, так и неустойчивой.
Обычно при исследовании реальных объектов, в том числе
и живых систем, приходится иметь дело с их нелинейными мо-
делями. Если в таком случае возникает необходимость исследо-
вания устойчивости, то часто совершается переход к линейному
случаю.
В теории динамических систем сформулированы условия, при
которых возможен переход от нелинейной системы к линеари-
зованному описанию — задача исследования устойчивости по
первому приближению,
6.5)
УСТОЙЧИВОСТЬ СИСТЕМЫ И ПЕРЕХОДНЫЕ РЕЖИМЫ
143
Пусть система описывается дифференциальными уравнениями
i=>F(x,t), (5.58)
где х = [xi, .... х„]т, a F(x) = [fi(x, t).f„(x, <)]т, причем fi — дифферен-
цируемые в окрестности начала координат функции, (ДО, 0,..., 0, t) 0.
В окрестности начала координат система описывается уравнениями
i{ = У, alfx + Rt .....хп, t), z = l, 2.....n, (5.59)
/-1
которые получаются при разложении правых частей (5.58) по х в окрестно-
сти начала координат по формуле Тейлора. Отметим, что это описание си-
стемы — не приближенное, а полное.
Вместо точки покоя системы (5.58) можно исследовать на устойчивость
точку покоя системы
i = Ах, (5 60)
где А = [а,;]. Система (5.60) называется системой уравнений первого при-
ближения для системы (5.58).
Если элементы матрицы А не зависят от времени, говорят, что система
(5.59) стационарна в первом приближении. Для стационарных в первом при-
ближении систем суждение об устойчивости исходной нелинейной системы
возможно в том случае, если все члены Ri в (5.59) ограничены (т. е. нигде
не обращаются в бесконечность при 0 =5 t <. оо) и разлагаются в ряды по
степеням хь хг, ..., хп в некоторой области У х2 е, причем разложения
i=t
начинаются с членов не ниже второго порядка.
Краткое изложение соответствующих теорем и простые примеры имеются,
например, в [94].
Аналитическое решение матричных дифференциальных урав-
нений в линейном случае получается следующим образом. Для
уравнения х = Ах -f- Bv имеем
t
x(f) = Ф (t) х (0) + Ф(/ — т) Ви(т) dx, (5.61)
о
где Ф(()— фундаментальная матрица системы, или переходная
матрица состояний. Для систем с постоянными коэффициентами,
у которых матрица А имеет п различных собственных значений,
фундаментальная матрица может быть вычислена по теореме
Сильвестра (см. [240], разд. V):
п / п \
ф«) = £р'Ц <5.62)
z=l\ /=1 ‘ ' I
X / =# i /
Вычислим, например, фундаментальную матрицу для системы с матрицей
144
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
Собственные значения матрицы А различны: /ч = —1, Х2 = —2. Тогда
в (5.62) под знаком произведения при i — 1 остается только член с j = 2,
а прн i = 2 — только член с / = 1. Поэтому
-U-HJ J]-
Выход системы получается, если выражение х(/) подставить
в уравнение выхода (5.38):
t
у (0 = СФх (0) + J СФВи (т) dx + Dv (t). (5.63)
о
Обычно в примерах и выкладках, которые включаются в
учебники и пособия по теории управления, рассматриваются
системы второго порядка. Для таких систем удобно пользовать-
ся выражением для фундаментальной матрицы, записанным в
явном виде. Если система второго порядка описывается уравне-
нием
*1 == а11*1 + Я12х2>
। (5.64)
Ха == ^22^2,
а собственные числа Ад и Хг, корни характеристического урав-
нения
Л2 — (ац -|- й12) Л -|~ (ацЙ22 — а12й21) — 0, (5.65)
различны, то
ф«-тД7ГХ
[(щ1 — Л2) — (flu — Xi) в22 (е^ * — в^) "|
(«Ц — Х2) (й|1 — X|) fKt „А,О Л \ АЛ г.. Л , АЛ I-
---- —--------(е — е ) (ап — Х2) е — (Оц — ЛЦ е |
Д12 J
(5.66)
О. Чувствительность
Стационарный режим в системе определяется ее внутренними
свойствами (структурой и параметрами) и условиями внешней
среды (входами). Состояние х и выход у в стационарном ре-
жиме определяются формулами (5.50) и (5.55):
х= — Д-1Ви,
. (5.67)
y = Dv-CA~'Bv.
Свойства системы задаются матрицами А, В, С и D. Их можно
трактовать следующим образом. Матрицы А и В определяют
внутренние свойства системы: А — основная матрица системы,
6.6)
ЧУВСТВИТЕЛЬНОСТЬ
145
от ее вида зависит, как было выяснено в предыдущем разделе,
устойчивость системы; В — матрица связи, эта матрица опре-
деляет структуру влияния входов на переменные состояния,
иногда она называется матрицей управления. Матрицы С и D
не определяют внутренних свойств системы; они лишь показы-
вают, как состояние системы отражается на ее внешних реак-
циях — выходах у. В технических задачах часто D = 0, по-
этому представительство переменных состояния в выходном
векторе определяется только матрицей С, которая в теории уп-
равления иногда называется матрицей наблюдения.
В биологических задачах, как мы увидим ниже, могут быть
ненулевыми как С, так и D (матрица D отражает, например,
свойства процессов обмена веществами и энергией между си-
стемой и средой; с таким случаем мы столкнемся ниже в при-
мере системы терморегуляции — см. (5.87)).
Чувствительность вектора стационарных значений перемен-
ных состояния к вектору стационарных значений входов v есть
d х
производная Согласно (5.34) и (5.67) имеем
^-=-Л“1В. (5.68)
Аналогично, чувствительность выходного вектора есть
^- = £)-СА_1В. (5.69)
Например, матрица —А~'В в дыхательном хемостате Ф. Гродинза (5.52)
имеет вид
ГО 0 0-1
Эта матрица в системе дыхательного хемостата определяет коэффициенты
чувствительности обеих переменных состояния ко всем трем входам. Устой-
чивость хемостата зависит от соотношения коэффициентов а и b (см. при-
мер 5.5.1); чувствительность состояния, кроме того, определяется и значе-
нием с.
Устойчивость и чувствительность системы, как мы уже го-
ворили раньше, представляют собой два ее различных свойства.
Устойчивость системы определяется свойствами одной лишь
матрицы А; чувствительность стационарных режимов — свой-
ствами матриц А и В. Устойчивость есть свойство системы воз-
вращаться в исходное состояние при наличии начальных усло-
вий, отличающихся от стационарных, независимо от причин,
приводящих к таким начальным условиям. Например, эти
условия могут возникнуть после кратковременного возмущения
состояния при постоянных условиях внешней среды. Чувстви-
тельность (5.68) характеризует способность системы сохранять
146
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
(ГЛ. 5
или изменять свои реакции на внешние возмущения (и сохра-
нять свои стационарные значения переменных) при изменении
параметров внешней среды. Предполагается, что однажды из-
менившись, внешний параметр должен оставаться на новом
уровне в течение всего времени рассмотрения системы.
Некоторые биологические аспекты понятия чувствительности
мы уже обсудили в разд. 3.7.
5.7. Наблюдаемость и управляемость
Как ясно из предыдущих разделов, основное методологиче-
ское различие между классическим подходом (вход-выход)
и методом пространства состояний (вход-выход-состояние) за-
ключается в следующем.
При классическом подходе исследователь выделяет из всего
объекта только те цепи, по которым проходит сигнал от различ-
ных входов до интересующего его выхода; все остальное в объ-
екте остается вне рамок рассмотрения. Поскольку такая цепь
может оказаться достаточно длинной, уравнения «вход-выход»,
описывающие ее свойства, имеют обычно высокий порядок.
В теории многосвязных систем этот метод распространяется на
случай нескольких входов и выходов, что усложняет рассмотре-
ние, но не меняет основной идеи.
При использовании метода пространства состояний исследо-
вателя прежде всего интересует целостная картина объекта.
Выписывая уравнения состояния в отличие от уравнения в тер-
минах «вход-выход», мы допускаем тем самым, что в объекте
могут существовать процессы и переменные, не проявляющиеся
в доступных исследователю выходных процессах. Объект, опи-
сываемый уравнениями состояния, «живет собственной жизнью»,
открывающейся для исследования только через наблюдение не-
которого количества выходных сигналов. Связь объекта с внеш-
ним миром достигается через его входы (в частности, управле-
ние таким объектом производится через вектор входов) и выходы
(только выходные переменные доступны наблюдению). Такой
подход должен глубоко импонировать специалистам-биологам.
Разница в постановке задачи исследования системы при
классическом подходе и методе пространства состояний условно
иллюстрируется рис. 5.3.
Понятия управляемости и наблюдаемости как раз и воз-
никли в связи с таким новым подходом к исследованию системы.
Интерес к этим понятиям объясняется желанием получить ответ
на следующие вопросы. При каких условиях можно восстано-
вить поведение вектора состояния системы, зная поведение вы-
ходного вектора на некотором интервале времени? При каких
условиях можно перевести систему из заданного начального со-
5.7]
НАБЛЮДАЕМОСТЬ И УПРАВЛЯЕМОСТЬ
147
стояния в требуемое конечное состояние за ограниченное
время?
Если структура системы (т. е. вид матриц А и С в (5.39))
такова, что по заданному выходному сигналу y(t) можно одно-
значно восстановить х(/), то говорят о наблюдаемости системы.
Точнее, состояние х (to) называется наблюдаемым в момент вре-
мени to, если можно указать интервал времени t t0 Т та-
кой, что по известным значениям v(t) и y(t), t to Т, можно
найти x(to). Если все состояния наблюдаемы в любой момент
времени, то говорят, что система полностью наблюдаема.
Рис 5.3. Разница между классическим подходом и методом пространства состояний.
а) При классическом подходе исследователя не интересует объект как целое: он выде-
ляет в нем только некоторые пути от входов v=[Vi п2]т к выходам у = [у\ у2]т. На «карте»
объекта —обширные «белые пятна»; б) при исследовании методом пространства состояний
объект рассматривается как целостная система во всей совокупности своих переменных,
описываемая матрицей А. Однако доступ воз-можен лишь к некоторым из внутренних пере-
менных х, н он осуществляется через внешние воздействия v; наблюдению доступны тоже
лишь некоторые нз переменных х через посредство вектора наблюдаемых выходных пере-
менных у. Обозначенные двойной лниией дуги В а С символизируют матрицы системы,
показанные на рнс. 5.2.
Управляемость системы означает, что при заданном виде
матриц Л и В в (5.39) существует такое управление, которое
за конечное время переводит систему из любого начального со-
стояния в некоторое заданное конечное состояние (например,
в состояние равновесия). Точнее, если заданное конечное со-
стояние х(Т) может быть достигнуто из начального состояния
x(to), то состояние x(to) называется управляемым в момент
времени to- Если все состояния системы управляемы в любой
момент времени, то система полностью управляема.
При рассмотрении вопросов управляемости следует иметь
в виду, что управление может действовать на некоторую рас-
сматриваемую координату как непосредственно (в этом случае
в правой части уравнения появляются координаты вектора
управления и), так и косвенно (в этом случае в правой части
уравнения для рассматриваемой координаты самих координат
вектора управления нет, но зато есть другие координаты век-
тора состояния, которые уже зависят от управления и). По-
этому сначала мы рассмотрим пример, в котором матрица А
148
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. 5
диагональна, т. е. возможно только непосредственное влияние
вектора управления на координаты вектора состояния х.
Пример 5.7.1 Рассмотрим систему, показанную на рнс. 5.4, которая
описывается обычным уравнением состояния (5.39), где °(3xi)’ ^(2x1) —
векторы, а матрицы А, В, С и D имеют вид
Д = Га'1 0 1 В = Р“ Р *131, С = РН С121, D = 0 (5.70)
L 0 ^22 J L 021 022 023 J L 021 с22 J
Система не является полностью управляемой, если одна из двух строк в мат-
рице В нулевая. Пусть, например, это вторая строка: 62i = ba = ba = 0.
Тогда, выписывая уравнение для координаты х2, получаем
^2 = £122-^2 + bz [СЦ + ^22^2 4" ^23^3 = ^22-^2»
и на координату х2 нельзя никаким образом воздействовать извне.
Рис. 5.4. Пример системы, имеющей одну неуправляемую и одну ненаблюдаемую коорди-
нату. Пунктиром показаны все возможные связи в системе; сплошной линией — факти-
чески существующие. Прн нулевой второй строке в матрице В (^2i=^22 = ^23==0) перемен-
ная хг не имеет связей со входами системы, и поэтому х2 является неуправляемой пере-
менной. При нулевом первом столбце в матрице С (когда сц=С21=0) оба выходных снг
нала {/j и у2 содержат информацию только о переменной состояния х2х переменная Х\
ненаблюдаема.
Если бы матрица А не была диагональной, уравнение для х2 даже при
нулевой строке в В включало бы член а21хь поскольку
з
*2 = “21Х1 +а22х2+ Е Vib2i’
< = 1
и внешнее воздействие и, изменяя координату Xi, косвенно влияло бы и на х2.
Полная наблюдаемость системы означает, что выходные переменные со-
держат информацию обо всех переменных состояния. Пусть матрица С содер-
жит хотя бы один нулевой столбец — например, первый. Тогда уравнения
выходов у\ и z/2 принимают вид
У\ = С11*1 + = С12Х2, (5 71)
Уг = C2iX| + с22х2 = СггХг,
и выходные сигналы не зависят от переменной состояния х,. Переменная
состояния хь таким образом, ненаблюдаема.
6 71
НАБЛЮДАЕМОСТЬ И УПРАВЛЯЕМОСТЬ
149
Вообще говоря, любую систему можно представить в виде
четырех подсистем, которые по-разному проявляют указанные
свойства. Такое представление иллюстрируется рис. 5.5.
Понятия управляемости и наблюдаемости имеют принципи-
альное значение при исследовании систем любой природы, в том
числе — биологических. Неучет неуп-
равляемых или ненаблюдаемых под-
систем в исследуемой системе часто
может привести к ошибочным выво-
дам. Например, наличие неуправляе-
мой части может при любых попытках
управления процессами в биосистеме
приводить к чрезмерному росту на-
блюдаемых переменных подсистемы
Si не за счет управления v, а только
за счет влияния неуправляемой части
S3 (см. рис. 5.5).
Мы рассмотрели понятия управляе-
мости и наблюдаемости только для
одного частного случая уравнений со-
стояния, имеющих вид (5.70). Для си-
стемы, записанной в общей форме
Рис. 5.5. Разбиение системы на
подсистемы, обладающие различ-
ными характеристиками с точки
зрения управляемости и наблю-
даемости. Система Si полностью
управляема и наблюдаема, S? —
управляема, но ненаблюдаема,
5з — неуправляема, но наблю-
даема, S4 — неуправляема и не-
наблюдаема.
(5.39), условия управляемости выглядят несколько более слож-
но. Это объясняется тем, что в рассмотренном случае диагональ-
ной матрицы А управление v может воздействовать на перемен-
ную х, только непосредственно (через i-ю строку матрицы В),
так как все переменные состояния «развязаны» и не влияют
друг на друга. В общем же случае матрица <4(ОТХт) недиаго-
нальна, и переменные состояния могут влиять друг на друга.
Поэтому, если даже нет влияния управления на данную коор-
динату состояния непосредственно через строку В, то такая
связь все-таки может возникнуть, хотя и более сложным обра-
зом: управление v влияет на какую-то другую координату, ко-
торая уже через матрицу А влияет на данную координату.
В таком случае роль матрицы В берет на себя произведение
матриц АВ. Мало того, если и в этом случае влияние v на рас-
сматриваемую координату состояния отсутствует, то может ока-
заться, что такое влияние осуществляется еще более опосред-
ствованным образом, через матрицу ААВ = (А)2В и т. д.
Строгое рассмотрение этого вопроса показывает, что условие
полной управляемости можно записать следующим образом.
Система является полностью управляемой, если ранг матрицы
[В АВ (А)2В ... (Л)т-1В] (5.72)
равен т.
150
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
Аналогичным образом выписывается и условие наблюдаемо-
сти. Система является полностью наблюдаемой если ранг
матрицы
[Ст АТСТ (Ат)2Ст ... (Ат)т"'ст] (5.73)
равен пг.
В следующем разделе мы покажем, как понятия наблюдае-
мости и управляемости применяются для анализа моделей био-
систем.
5.8. Уравнения состояния
для системы пассивного теплообмена
В этом разделе мы рассмотрим применение метода простран-
ства состояний для описания системы теплообмена в организме
животного, следуя работе [286]. Анализ этой работы в рамках
классической теории управления име-
ется в учебнике [130].
Представим ткани тела в виде трех-
слойной структуры (кожа, мышцы и
ядро) с температурами хь х2, х3, соот-
ветственно (рис. 5.6). Источники
тепла расположены в ядре (темп
притока тепла щ3 определяется ин-
тенсивностью обмена веществ) и в
мышцах (темп притока тепла w2 за-
висит от интенсивности мышечной ра-
боты). Потоки тепла от ядра к мыш-
Рнс. 5.6. Представление тела жи-
вотного в виде трехслойиой струк-
туры с внутренними источниками
тепла. Xi — температура слоя
кожи d\, Xj—температура слоя
мышц dit Xg — температура ядра
тела—слоя толщиной d$\ №t — по-
тери тепла с испарением. Источ-
ники потоков тепла ш2 н Wt рас-
положены в мышечном слое и
ядре. В формулах (5.74) коэффи-
циенты теплопроводности kit k\
определяются толщиной слоев
Аг=2А/(^1 + ^г).
Аз=2А/(</2
k —теплопроводность тканей
при нормальном кровоснабже-
нии.
цам, от мышц к коже и от кожи в ок-
ружающую среду определяются вели-
чиной перепада температур (х3— х2),
(х2— Xi) и (xi — ui), соответственно,
где Ui — температура окружающей
среды. Процессы теплообмена в орга-
низме определяются двумя видами ме-
ханизмов. Это прежде всего пассивные
механизмы, которые зависят от темпе-
ратурных перепадов между слоями
тела и от коэффициентов теплопровод-
ности k3, &2 и теплопередачи k\. На них
наложены активные терморегуляцион-
ные механизмы, которые реализуются, с одной стороны, через
изменение переменных ш3, ДО2 и теплопотерь с испарением гец,
а с другой стороны — через изменение коэффициентов &2, k\
(сосудистые реакции).
5.8]
СИСТЕМА ПАССИВНОГО ТЕПЛООБМЕНА
151
Если ограничиться рассмотрением пассивных механизмов
теплообмена, то их уравнения можно записать в виде
Т 1х1 = k2 (х2 — Х\) — (%! — Ui) — wb
Т2х2== k3(x3 —х2) — k2(x2 —Xi) + w2, (5.74)
7л = — k3 (х3 — х2) + о>з,
где Ti = clpldl, i — 1, 2, 3, — постоянные времени каждого из
слоев, ct — их удельная теплоемкость, р, — удельная плотность,
di—характеристика толщины слоев. Схема моделирования си-
стемы приведена на рис. 5.7.
Рис. 5.7 Схема моделирования пассивных механизмов теплообмена. Вектор состояния
x=[Xi х2 Хз]т имеет три компоненты: температура кожи» мышц и внутренних органов (ядра),
соответственно. Вектор входов w = [wj w2 представляет собой темпы потоков тепла,
возникающие из-за процессов метаболизма во внутренних органах, в мышцах, Wt, и
темпы теплопотерь от испарения влаги с поверхности кожи. Пассивные механизмы регу-
ляции представлены влиянием переменных состояния на темпы теплопередачи от ядра
к мышцам, от мышц к коже и от кожи в окружающую среду. Эти механизмы обеспечи-
вают равенство выходных переменных соответствующим входным величинам. В стацио-
нарном состоянии i==l, 2, 3. Однако пассивные механизмы регуляции теплового
режима не могут обеспечить постоянства внутренней среды (неизменность сигналов х* и
Xj) прн изменении температуры окружающей среды — возмущающего сигнала
Рассмотрим эту схему подробнее, положив для простоты
о>1 = 0. Прежде всего в системе три переменных состояния хь
х2, х3 — выходы трех интеграторов. Вектор состояния х, следо-
вательно. определяется как [х3 х2 х3]т. Внешних сигналов, дей-
ствующих на систему, тоже три: w3, w2 и vh Два из них, w$,
w2 — темпы; один, Vi,— уровень.
Здесь мы впервые сталкиваемся со спецификой биосистем:
вектор входов включает в себя две разнородные группы пере-
менных, темпы и уровни. В этом случае можно поступить одним
из двух следующих способов. Можно, не обращая внимания на
152
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
эту разницу, ввести общий вектор для всех внешних воздействий
w2
w3
(5.75)
Однако во многих случаях более оправдано представление внеш-
них воздействий в виде двух векторов, обозначенных, соответ-
ственно, через v и w. В данном случае можно было бы ввести
обозначения
у = [щ],
(5.76)
Переход от одного способа описания к другому производится
с помощью блочных матриц.
В нашем случае трехмерный вектор (5.75) можно предста-
вить как блочный вектор, составленный из векторов (5.76) раз-
мерности 1 и 2:
v = Г ^.1*’> 1. (5.77)
L w(2 X 1) J
Перепишем уравнения состояния (5.74) при помощи матриц и
векторов. Разделив обе части каждого из уравнений на соответ-
ствующую постоянную времени Tt, запишем получившуюся си-
стему в обычном виде, без использования блочных матриц:
х — Л(з х з>х + б(з х з)У,
(5.78)
где
Если мы хотим теперь выделить два типа входов v и w, то уравнения
(5.78) и (5.79) можно переписать в матричной форме, представив v в виде
(5.77), а матрицу В(ЗХз) в (5-79) —в виде блочной матрицы [S(3Xij-T?t3x2)J ,
Г ^1/Г. i 0 о -
В(3 X 3) — 0 ! 1/т2 0
. 0 i о 1/Гз -
(5.80)
Тогда уравнение состояния запишется в виде
* Азх 3)х + S(3 X 1)И + «(3 X 2)w
(5.81)
Вид матриц В и 7? в (5.81) ясен из (5 80). Ниже в этом примере мы будем
исходить из более простого описания системы (5 78) — (5.79).
5.8]
СИСТЕМА ПАССИВНОГО ТЕПЛООБМЕНА
153
Обсудим теперь вопрос о том, какие сигналы целесообразно
взять в данной системе в качестве выходных. В качестве выхо-
дов исследователь обычно рассматривает те сигналы, которые
могут его заинтересовать в процессе работы. В статье [286],
например, вводится переменная, представляющая средневзве-
шенную температуру тела. Обозначив ее через i/i,
= + (5.82)
yl di4-d2 + d3 v '
где di, di, d3 — характеристики толщины слоев, возьмем у\ в ка-
честве одного из выходов системы. Часто исследователя интере-
суют также значения температуры ядра и температуры кожи.
Поэтому в качестве двух других выходов, обозначенных через
z/2 и z/з, возьмем Xi и %з:
3/2 = *1, Уз = х3. (5.83)
С точки зрения анализа открытых систем важно, что в ста-
ционарном режиме темпы потоков тепла от ядра к мышцам и
от мышц к коже должны установиться на значениях, опреде-
ляемых интенсивностью метаболизма и интенсивностью мышеч-
ной работы w3 и W2, соответственно. Поэтому эти темпы можно
рассматривать как управляемые величины в системе регулиро-
вания теплового баланса системы, т. е. взять их в качестве вы-
ходов в системе управления (подробнее об этом говорится в
разд. 7.2). Значит, имеет смысл ввести еще две координаты
и 1/5 в вектор выхода системы, определив их как соответствую-
щие темпы потоков тепла
У4 = k3 (х3 — х2),
Z/5 = k2 (х2 — Xi).
Наконец, интерес может представить и общая теплоотдача
организма (темп потока тепла от кожи в среду). Введем по-
этому еще одну координату выхода
г/6= kx (xj — Oi). (5.85)
Таким образом, мы сформировали вектор выхода у =
— [f/i У2 Уз У4 У5 У<з\т, включив в него переменные, которые, по
нашему мнению, могут интересовать исследователя. Если бы мы
ошиблись и не рассмотрели еще каких-либо характеристик си-
стемы, вектор выхода можно было бы расширить и дальше.
Выпишем теперь уравнение выхода для системы терморегу-
ляции. В соответствии с (5.39) имеем
У — С(бхз)Х + D(gx3)V, (5.86)
154
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. S
где х = [х, х2 х3]т, v = [uj w2 о>3]т. Из (5.82) —(5.85) находим
вид матриц С и D:
С(6 х 3) —
______d2______
di + d2 + ds
О
О
— ks
k2
О
ds
di + d2 + ds
0
1
ks
0
0
0(6 X 3) =
0
0
0
0
kl
0 0-
0 0
о 0
о 0
о 0
о 0
(5.87)
0
Обратите внимание на то, как легко получаются элементы
матриц С и D из коэффициентов исходных уравнений — это
просто коэффициенты при соответствующих переменных в алге-
браических уравнениях (5.82) — (5.85).
Исследуем теперь уравнения состояния (5.78), рассмотрев
вопросы устойчивости и существования стационарного режима,
управляемости и наблюдаемости системы, а также чувствитель-
ность переменных состояния системы в стационарном режиме.
Устойчивость. Мы уже говорили, что исследование ус-
тойчивости для моделей естественных биосистем является вспо-
могательной задачей, помогающей оценить, насколько хорошо
модель воспроизводит объект (устойчивость реального биологи-
ческого объекта, естественно, сомнений не вызывает). В сложных
системах, исследование которых производится на вычислитель-
ных машинах, устойчивость проверяется, как правило, в про-
цессе отладки программы. Имеются и машинные процедуры для
непосредственного получения собственных значений матриц,
описывающих системы. В простых случаях возможно исследо-
вание устойчивости классическими методами (например, по упо-
минавшемуся выше критерию Рауса — Гурвица [46]). Для на-
ших целей сейчас достаточно убедиться, что система пассивного
теплообмена является устойчивой при относительно широком
изменении численных значений элементов матрицы А в (5.78).
Покажем, например, что система устойчива при k\ = k2 = k3 =
= Tl = T2 = T3 = 1.
В этом случае характеристическое уравнение (5.25) имеет
вид
[Л + 2 —1 от
-1 Л + 2 -1 |=0, (5.88)
о —1 А ч-2 J
5.8]
СИСТЕМА ПАССИВНОГО ТЕПЛООБМЕНА
155
откуда (%4-2)3 — 2(% + 2)=0, и корни характеристического
уравнения легко находятся: М — —2, %2,з = —1 ± ! Действи-
тельные части всех трех корней (—2 и —1) лежат в левой полу-
плоскости, и система асимптотически устойчива.
Стационарный режим системы. Поскольку система
устойчива, имеет смысл говорить об исследовании стационарного
режима х =—A~'‘Bv согласно (5.50).
Найдем обратную матрицу А~' (для сложных систем эта
операция также может быть поручена вычислительным маши-
нам). Для матрицы А, определяемой (5.79), имеем det Д —
= — - и, вычисляя Adj Л согласно (5.21), находим
I U 2* 3
11
kl
т\
kl
11
kl
Т2
kl
т2 (kt + kA
k | ^2
7~2 (fei 4 kA
kik2
ki
T3(kt+ kA
klk2
T3 (k>k2 -|- k\k3 k2k3}
kik2k3
(5.89)
Найдем теперь стационарное значение х, вычисляя сначала Bv
для В(зх1) из (5.80) и v из (5.77), а затем A~lBv. Получаем
Bv =
о
1
Т2
о
klVl ~
Г.
а>2
Т2
виз
Тз _
(5.90)
о
1
Т3 _
и трехмерный вектор-столбец х имеет вид
Vi
х = — A lBv =
Ol
fl
w2 виз
kl kl
kl + k2 _ , kl + k2
—n:— ®2 H-------rr— 01 з
ki«2 k}K2
k\ + #2 । &1&2 4” k\k$ + &2&3
--г—7--- +
k i«2
(5.91)
klk2k3
w3
Имея в явном виде вектор состояния х, можно по матрице
наблюдений состояния системы С(6х3) и матрице наблюдений
входов 0(6*1) (5.87) получить стационарное значение выхода
системы (5.86). Предоставим выполнение этой операции чи-
тателю.
Чувствительность. Чувствительность стационарного ре•
жима (5.91) к вектору стационарных значений входов показы-
вает, насколько смещается стационарный режим системы пас-
сивного теплообмена при переходе организма от одних условий
среды У1 к другим или от одной интенсивности физической
156
МЕТОД ПРОСТРАНСТВА СОСТОЯНИИ
[ГЛ. 5
нагрузки и>2 к другой. Считается, что после такого перехода
должно пройти время, достаточное для затухания переходных
процессов в системе.
Согласно (5.68) чувствительность вектора х размерности 3
к вектору v размерности 3 есть матрица — А~1В размерности
(3X3). Умножив А~1 из (5.89) на В из (5.79), получаем
ас
“ dXi дх\ dxi ~ 1 1
dvt dwj dw3 1 kl
дхг дх3 dxi ^1+^2
— dvi dwj dw3 — 1 k\kz
дх3 дх3 дх3 1 k\ + k3
dvt dwi dw3 1
1
fe 1 Ч~ fea
kikj -p k\k3 -p k3k3
£ 1&2&3
(5.92)
Легко видеть, что коэффициенты чувствительности, состав-
ляющие матрицу чувствительности (5.92), можно получить и не-
посредственно из (5.91), продифференцировав выражения, стоя-
щие в матрице в первой, второй и третьей строке, сначала по
Vi, потом по ш2 и, наконец, по ш3. Уравнение (5.92) свидетель-
ствует, в частности, о том, что при изменении температуры
окружающей среды температура всех слоев тела сильно ме-
няется:
Наблюдаемость и управляемость. Имея явное
решение уравнений системы в виде (5.91) для стационарного
режима, нет смысла проводить исследование управляемости по
методам, описанным в разд. 5.7. Действительно, из выражения
(5.91) очевидно, что каждая переменная состояния явно зависит
от всех входов, так что каждая переменная состояния управ-
ляема, и поэтому система полностью управляема.
Тем не менее для иллюстрации применения методов анализа
управляемости определим ранг матрицы (5.72). Для полной
управляемости требуется, чтобы ее ранг был равен трем. На-
чинаем составлять эту матрицу. Первые ее три столбца берутся
из матрицы связи В(3хз), определяемой (5.79), вторые три —из
матрицы (АВ)(зхз), имеющей вид
(fei ~b A _ki_ T\T, 0
k3
rf T2T3 (5.94)
k3 k3
и
T2T3 T~ 3 _
6.8]
СИСТЕМА ПАССИВНОГО ТЕПЛООБМЕНА
157
Тогда первые 6 столбцов матрицы (5.72) получаем в виде
fel 0 0 ki (k\ + ^2) А k2 т{т2 0
1 klk2 4” &3 k3
0 72 0 т,т2 1 2 т2т3 , (5.95)
0 0 1 0 ks
Т2 V3 т2 ' 3
и очевидно, что уже из них можно составить минор третьего
порядка, отличный от нуля (например, это три первых столбца
или второй, третий и четвертый столбцы) — система полностью
управляема.
Наблюдаемость системы в данном случае зависела от нас.
Выбором достаточного числа выходов, т. е. заданием матрицы
С(вхз) в (5.87) мы достигли того, что переменные выхода зависят
от всех переменных состояния. Тем самым все переменные со-
стояния представлены на выходе, и система полностью наблю-
даема. При желании, однако, можно убедиться и в выполнении
условий наблюдаемости, вычислив ранг матрицы (5.73) подобно
тому, как мы определили ранг матрицы (5.72).
В этом примере набор выходных переменных мы выбирали
сами. На практике, однако, дело часто обстоит иначе. Вектор
выходов не задается исследователем, как это было в рассмот-
ренном случае, а определяется возможностями его эксперимен-
тальной установки, например, наличием регистрирующей или
измерительной аппаратуры. Пусть, в частности, исследователя
интересует измерение всех переменных состояния, а имеется
возможность регистрации только одной температуры кожи
Выясним, наблюдаема ли система в этом случае.
Матрица С имеет вид
С(зх1) = [1 0 0]. (5.96)
Ее ранг равен единице, и поэтому придется выписывать и иссле-
довать всю матрицу (5.73). (Если бы ранг С был равен трем,
что возможно при трех и более выходных сигналах, мы бы сразу
сделали вывод, что система наблюдаема.) Имеем
4“ ^2 Tt k2 т2 0
Лт = k2 Tt 0 k2 + ks T2 ks T2 ks Ts ks Ts _ CT = 1 0 .0. . (5.97)
Умножая Ст слева на Ат, получаем второй столбец, а умножая
его еще раз слева на /Г, — и третий столбец искомой матрицы
158
МЕТОД ПРОСТРАНСТВА СОСТОЯНИЙ
[ГЛ. 5
(5.73). Она, следовательно, имеет вид
[Ст ДТСТ (дт)2ст]=
fe, + kz 1 —’ о А _ т-i 0 0 ^(*l+42+fe27’1
т\т^ Tik2 (kt + k2) — Т\k2 (k2 + &з) (5.98)
7-2Г2 &2&3 т,т2
Ее определитель
det [Ст ЛТСТ (дт)2ст] = 4^>0. х z -J Т^Т 1 Г 2 (5.99)
поэтому ее ранг равен трем, и система полностью наблюдаема.
Это означает, что, регистрируя процесс изменения во вре-
мени температуры кожи, исследователь может в пределах
выбранной модели определить также и текущие значения тем-
пературы мышц и ядра.
ГЛАВА 6
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
6.1. Определения
При описании биологических систем оказывается, что в них
можно выделить некоторые относительно независимые количе-
ства или объемы вещества, участвующие в кинетических про-
цессах как некое целое, легко выделяемое из остальной системы.
При математическом описании предполагается, что эти количе-
ства вещества равномерно распределены в некоторых объемах
пространства, так что их концентрация постоянна, т. е.
^- = 0, (6.1)
где с — концентрация рассматриваемого вещества, х,, i— 1, 2,
3, — пространственные координаты. Удобным способом изучения
таких систем стал компартментальный анализ [317], основным
элементом которого является понятие компартмента.
Обычно компартментом называется некоторое количество
вещества, выступающее в процессах транспорта и обмена как
самостоятельная единица. В общем случае компартментом мож-
но назвать «вещество, характеризующееся некоторой количе-
ственной мерой». Как правило, компартмент описывается массой
или концентрацией рассматриваемого вещества или объемом,
который оно занимает в пространстве.
Модели биологических систем, в которых используется цред-
ставление о компартментах, называются компартментальными
моделями. Такие модели, как правило, используются для иссле-
дования процессов переноса вещества и энергии внутри живой
системы и обмена их со средой. В этом случае компартментам
приписывается пространственная характеристика: компартменту
соответствует некоторая область или объем в биосистеме.
Так, в физиологических системах организма можно выделить
области, различающиеся по концентрации каких-либо веществ
или энергии (в модели системы дыхания Ф. Гродинза (см.
разд. 5.2) таких областей две — легочный и тканевый резер-
вуары, отличающиеся концентрацией кислорода; в рассмотрен-
ной в разд. 5.8 модели пассивного теплообмена — три области,
160
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
различающиеся температурой и т. д.). В водных экосистемах
можно производить разделение на области акватории или по
глубинным слоям. Например, в модели экосистемы Азовского
моря [58] таких областей семь. В экосистемах в каждой области
равномерно распределенными компонентами являются различ-
ные виды или возрастно-половые группы. В фармакокинетиче-
ских моделях (применяемых для исследования действия лекар-
ственных средств на различные органы) области определяются
тем, насколько равномерно концентрируются фармакологические
вещества в тканях [267], и их может быть довольно много.
В других случаях в качестве компартментов рассматри-
ваются взаимодействующие друг с другом вещества (или дру-
гие компоненты биосистемы), находящиеся в одном и том же
объеме. Тогда кинетика реакций также описывается с помощью
компартментальных моделей, а различным компартментам отве-
чают количества или концентрации различных веществ в одной
и той же пространственной области. Так, в тканях и жидкостях
тела происходят многочисленные реакции между химическими
веществами, и участвующие в них субстраты, ферменты и про-
дукты могут рассматриваться как различные компартменты.
В моделях наземных экологических систем часто делается допу-
щение, что все исследуемые процессы происходят на некоторой
ограниченной территории (иногда, как в классической модели
экологических систем — модели Вольтерра, такое допущение
делается неявно), и в качестве компартментов выступают чис-
ленности или плотности различных видов или возрастных групп
животных.
В общем случае компартментальная модель содержит не-
сколько связанных между собой компартментов, в которых
протекают три типа процессов — обмен компонентами между от-
дельными компартментами, а также между компартментами и
средой, превращение компонент друг в друга и, наконец, разно-
образные процессы, приводящие к исчезновению рассматривае-
мых компонент или веществ. Процессы последнего типа мы
будем обозначать общим термином утилизация.
Примерами процессов первого типа являются на организ-
менном и суборганизменных уровнях физические процессы —
диффузия, перенос веществ и энергии различного рода носи-
телями (кровь, воздух в легких), излучение энергии, испарение
воды с кожи и т. д. На уровне экосистем — миграции животных,
перенос биомассы потоком воды в проточном культиваторе, пе-
ремещение биомассы растительных и животных видов в вод-
ных экосистемах под действием течений и т. д.
К процессам превращения компонент относятся различного
рода химические реакции (в биосистемах от клеточного до орга-
низменного уровня), взаимодействия типа «хищник-жертва» на
6.11
ОПРЕДЕЛЕНИЯ
161
уровне популяций (когда одна компонента служит пищей для
другой), переход особей из одной возрастной группы в другую
в экологических моделях, где различаются половозрастные
группы.
Приведем и пример утилизации. Если в некоторой биоси-
стеме рассматриваются только процессы поступления и распре-
деления кислорода, то окислительные процессы, в результате
которых кислород переходит в молекулы СО2 и Н2О, можно
рассматривать как процесс утилизации кислорода. Утилизация
происходит в тех компартментах, где протекают соответствую-
щие процессы.
Различие между компартментом и пространственной обла-
стью легко видеть из следующего примера [317]. Пусть компо-
нента А переходит из одной пространственной области в другую
в некоторой физиологической системе посредством процесса
диффузии. Эта ситуация может быть описана двухкомпартмен-
тальной моделью, показанной на рис. 6.1, а, где Xi и х2 — коли-
чества вещества А в отсеках 1 и 2, соответственно. Пусть теперь
Рис. 6.1. Различие между компартментом и пространственной областью в компартменталь-
иоЙ модели. Если компонента А переходит из пространственной области / в область 2 и
при этом вступает в реакцию с некоторым другим веществом В, то в зависимости от темпов
протекания процессов переноса вещества А и комплекса АВ из области в область и тем-
пов реакции Л + В Л АВ модель может иметь два (а), три (в) или четыре (б) компарт-
мента.
вещество А вступает в реакцию с некоторым другим веществом
В, образуя комплекс АВ в каждой из двух пространственных
областей. Тогда система описывается моделью, содержащей
четыре компартмента: количества хг и х2 вещества А в отсеках
1 и 2 и количества Хз и х4 комплекса АВ в тех же отсеках
(рис. 6.1,6). Наконец, если обмен веществом А между простран-
ственными областями и реакция А + В АВ протекают относи-
тельно медленно, а выравнивание концентраций АВ между
этими областями происходит быстро, так что концентрация АВ
всюду в системе одинакова, то получаем трехкомпартменталь-
ную модель (рис. 6.1,в), где величины лц и х2 сохраняют преж-
ний смысл, а х3 — полное количество АВ в обеих пространствен-
ных областях вместе.
162
КОМПАРТМЕПТАЛЫ1ЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ 6
Подобные ситуации часто встречаются в физиологических
системах, когда в относительно небольшом числе пространствен-
ных областей происходят перемещение и химические превраще-
ния большого числа веществ. Легко видеть, что при таком под-
ходе число компартментов равно количеству дифференциальных
уравнений, описывающих модель, или порядку уравнений си-
стемы. Во многих работах, однако, разница между компартмен-
том и пространственной областью отсутствует. Так, в [81]
«компартмент рассматривается как определенное физиологиче-
ское пространство, в котором имеется то или иное количество
вещества, участвующего в процессах транспорта и обмена».
При описании экосистем компартментом считается обычно
любой элемент системы, с которым связано накопление био-
массы. Так, в обзоре по моделированию экосистем [344] рас-
сматривается компартментальная модель популяции коловра-
ток, в которой роль компартментов играют следующие величины:
количество молодых диплоидных самок, количество взрослых
диплоидных яиц, количество гаплоидных яиц и т. д.
Если некоторое вещество в компартментальной системе пе-
ремещается из одного компартмента в другой, темп изменения
количества этого вещества xt для i-ro компартмента опреде-
ляется уравнением
п п
Х(= 2L Uii— 2L Ум + Ую — Уо( + Si dh (6.2)
i=l у k=\
i ¥= i i
где Xi — количество вещества в i-м компартменте, ytj — темп по-
тока вещества из /-го компартмента в i-й, ую и yOi — темпы пото-
ков вещества из окружающей среды в i-й компартмент и из
компартмента в среду, соответственно, gt и dt — темпы возникно-
вения и исчезновения вещества в i-м компартменте.
Развитие компартментального анализа систем в течение
длительного времени было связано почти исключительно с изу-
чением так называемых индикаторных методов исследования
физиологических систем. Эти методы заключаются в том, что
для анализа потоков вещества в некоторой системе в нее вво-
дится извне специфическое вещество — индикатор, движение ко-
торого легко регистрируется. Обычно это радиоактивные или
окрашивающие вещества [352].
Пусть, например, в компартмент объема V при t — 0 введено
малое количество меченого вещества ро. Тогда изменение коли-
чества индикатора в компартменте с течением времени есть
функция p(i). Если обозначить плотность индикатора через
x(i), то
Р(0=Ух(/).
(6.3)
e.l]
ОПРЕДЕЛЕНИЯ
163
В случае радиоактивного вещества величина р есть общее
количество индикатора, измеряемое обычно в единицах мккюри,
делений в секунду или отсчетов в минуту (1 кюри = 3,7-1010 де-
лений в секунду). Величина х представляет собой количество
радиоактивного материала на единицу массы и называется
удельной активностью (мккюри/г).
В стационарном режиме темп притока вещества в данный
компартмент равен темпу его оттока. Обозначим этот темп
через г. Тогда темп уменьшения количества индикатора в ком-
партменте тем больше, чем больше темпы обмена вещества:
<=-«• <6-4>
Часто исследователя интересует отношение количества ве-
щества, покидающего компартмент в единицу времени, к общему
количеству V вещества в компартменте
L = (6.5)
Величина L называется клиренсом *) и определяет динамику
изменения количества индикатора
Процессы в компартментальных моделях описываются обык-
новенными дифференциальными уравнениями, что позволяет
широко использовать при их исследовании методы пространства
состояний, рассмотренные в гл. 5. Если система описывается
дифференциальными уравнениями в частных производных (на-
пример, если предполагается непрерывное изменение плотности
распределения каких-либо веществ в пространстве), то модель
не относится к компартментальным. Поскольку уравнения в
частных производных используются для описания потоков ве-
щества и энергии в пространстве, такие системы — в противовес
компартментальным моделям — иногда называют потоковыми
моделями (flow systems — англ.). Таковы, например, модель био-
аквоценоза [111], в которой рассматривается непрерывное изме-
нение характеристик системы — освещенности и концентраций
биомассы по глубине, модель терморегуляции в организме че-
ловека [298], где потоки тепла описываются уравнениями тепло-
проводности. Однако компартментальные модели и в этом слу-
чае могут служить хорошим приближенным описанием системы.
*) Английские термины clearance rate или turnover rate сохраняют слово
rate — «темп» и поэтому имеют буквальный смысл темпа изменения количе-
ства индикатора в компартменте.
164
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. в
6.2. Перемещение компонент
Законы, определяющие процессы перемещения компонент в
биосистемах, могут быть как простыми (в частности, таковы
физические законы, описывающие пассивный транспорт веще-
ства и энергии), так и чрезвычайно сложными (как, например,
законы, регулирующие активный транспорт в биохимических си-
стемах или расселение и перемещение видов и отдельных групп
животных). В компартментальных моделях живых систем в на-
стоящее время исследуются, однако, лишь простейшие зависи-
мости такого рода. Рассмотрим некоторые из них.
Диффузия. Наиболее распространенным механизмом
транспорта веществ в физиологических и биохимических систе-
мах является диффузия. Этот механизм представляет собой один
из способов, посредством которого могут перемещаться моле-
кулы вещества, растворенного в жидкости, и является след-
ствием теплового движения молекул — броуновского движения
[352, 357].
В 1855 г. Фиком был сформулирован первый закон диффузии,
связывающий плотность потока вещества через единичную пло-
щадь за единицу времени J с изменением концентрации этого
вещества с,
(6.7)
где D — коэффициент диффузии (см2/сек), а концентрация с по
предположению непрерывно меняется в пространстве и зависит
только от одной пространственной координаты х.
В более общем случае с — с(х, у, z, t), т. е. концентрация
может зависеть от пространственного положения точки
Р(х, у, z) и времени t. Тогда через бесконечно малый участок
плоскости в точке Р идет поток вещества с компонентами по
всем трем направлениям J = [/* /у /2]т, где
4 = J = — Jz = -D~. (6.8)
х дх ’ и ду г dz ' '
При этом обычно допускается, что раствор изотропен, т. е. обла-
дает одинаковыми свойствами по всем направлениям. Если во
всех уравнениях (6.8) величина D одна и та же, она называется
константой диффузии.
Поскольку в малом элементе раствора, окружающем точку
Р, справедливо уравнение непрерывности
дс dJx dJ dJ-
+ + —^- = 0, (6.9)
dt 1 дх ду 1 dz ' '
6,2]
ПЕРЕМЕЩЕНИЕ КОМПОНЕНТ
165
то после подстановки сюда (6.8) при D = const получаем соот-
ношение
4г=дДс> (бл°)
где оператор А определяется следующим образом:
Д = + <6Л1)
Цели концентрация имеет ненулевой градиент только вдоль
оси х, то получается второй закон диффузии Фика:
# = (6-12)
Очевидно, использованное выше уравнение непрерывности яв-
ляется лишь частным случаем закона сохранения массы. Если
обозначить через /щ и /от потоки вещества, входящие в некото-
рый объем пространства и выходящие из него, то получаем
обычное уравнение сохранения массы
dm , г
1Г~ ую — •'01-
(6.13)
Это принципиальное соотношение в различных источниках
называется по-разному. Часто для него используется название
принцип Фика, а в теории открытых систем оно известно как
уравнение Берталанфи.
При компартментальном анализе обычно используется сле-
дующее описание процессов диффузии. Пусть система содержит
п компартментов, и xt— уровень или концентрация некоторой
компоненты в i-м компартменте. Тогда процессы переноса ком-
поненты, вызванные различием концентрации (уровней вещества
или энергии) в разных компартментах и в окружающей среде,
описываются линейными дифференциальными уравнениями
1 fe.o
“ V7 Zj ki! (х’— + Т7 “ Xl^
j-i
i=l, 2.........n, (6.14)
где (x/ — xi) — разность уровней вещества или концентраций,
ky — постоянные неотрицательные коэффициенты, v — уровень
или концентрация рассматриваемой компоненты в окружающей
среде, Vi — объем i-ro компартмента.
Уравнения (6.14) можно переписать в векторном виде, если
ввести векторы х == [xi х2 ... х„]т и о =» [и]:
x^/ljx + BiV, (6.15)
166
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
где х О, v 0 (обычные условия неотрицательности концен-
траций), а матрицы Л1(„хп) и BI<rtXI) имеют следующие элементы:
(6.16)
Кроме процессов диффузии в модели можно учесть и компо-
ненты, появляющиеся в ее компартментах в силу других при-
чин: некоторое вещество (или энергия) может поступать или
Рис. 6.2. Схема компартментальной модели, а) Условное обозначение схемы, где
х2, •*•» хп — концентрации рассматриваемой компоненты в компартментах, у{, у2, ...,уп~-
приток компоненты извне, Ьц— коэффициенты, определяющие темпы диффузии; б) изо-
бражение при помощи матриц. Здесь х, у, v — векторы концентраций, притоков и условий
среды, соответственно.
удаляться из системы в ходе химических или физических про-
цессов. Так, в системе теплообмена (разд. 5.8) учитывается
тепловая энергия, выделяющаяся в ходе процессов метаболизма
в различных частях системы. Обозначая темп поступления рас-
сматриваемой компоненты в i-й компартмент через yt и вводя
вектор
У = [У\ У2 Уп]\ (6.17)
Иолучаем уравнение линейной компартментальной модели в виде
х — А[Х -|- B{v -|- R.{y, (6.18)
где /?1(пх») — диагональная матрица с элементами
ri( = V7. (6.19)
Матрица R позволяет по количеству компоненты, поступившей
извне, определить ее концентрацию в i-м компартменте.
Схема такой модели представлена на рис. 6.2.
6.2]
ПЕРЕМЕЩЕНИЕ КОМПОНЕНТ
167
Пример 6 2.1. В системе пассивного теплообмена (разд. 5.8) можно
разделить вектор входов (5.75) на два вектора, как это сделано в (5.77),
(5.81). Тогда, добавив компоненту Wi в вектор ш, получаем для у из (6.17)
и о из (6.14)
y = w(3x = w2 ®з]Т-
v= U(1 х 1)= [°1]’
и в уравнении (6 18) матрицы принимают следующий вид:
fei + fez fez «
Ti T\
ki (fez + fez) fez
T2 T2 T2
0 fe3 fea
T3 T3
(6.20)
(6.21)
MT,
О
. О
о
1/Г2
о
«Объем» пространственного элемента структуры в данном случае характери-
зуется коэффициентом пересчета накопленного тепла в изменение темпера-
туры, Ti = ctpd, (см. разд. 5.8).
Уравнение диффузии, записываемое в виде
z/12 = — £ (С] — с2), (6.22)
где z/12 — поток вещества в компартмент 1 из компартмента 2,
Ci и с2—концентрации вещества в этих компартментах, k—
коэффициент, определяемый свойствами «границы» между ком-
партментами, наиболее часто используется при моделировании.
Линейное уравнение (6.22) дает простейшую гипотезу о харак-
тере процессов транспортного обмена между компартментами
в тех случаях, когда природа и механизмы транспорта неиз-
вестны.
Нелинейные процессы переноса компонент.
Использование линейных уравнений диффузии в форме (6.18)
позволяет описать достаточно широкий класс явлений физиче-
ского переноса компонент биосистемы.
Однако во многих случаях физические законы переноса ком-
понент оказываются более сложными. Так, излучение тепловой
энергии от тела животного к окружающим предметам подчи-
няется закону Стефана — Больцмана
ух = aS (4 — vf), (6.23)
где у\—темп теплового потока (ккал/час), х, и Vi — темпера-
тура, соответственно, кожи и среды в градусах Кельвина, а —
постоянная Стефана — Больцмана (о ~ 0,49-10~7 ккал/ж2 • чао
град4), S — эффективная площадь излучающей поверхности.
168 КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ [ГЛ 6
Заметим, что уравнение (6.23) хорошо линеаризуется в обла-
сти тех температур, с которыми обычно имеет дело исследова-
тель (от —20 до +50°C). Можно привести примеры нелинейных
зависимостей переноса компонент, плохо поддающихся линеари-
зации.
Пример 6.2.2. Рассмотрим простейшую модель миграции саранчи под
действием феромона. Феромоны — особого рода «пахнущие» химические веще-
ства, выделяемые отдельными особями животных н используемые ими для хи-
мического общения с другими особями того же вида. Передача этих веществ
происходит по воздуху в соответствии с линейными законами диффузии, хотя
наличие ветра может усложнить условия их распространения. Транспорт фе-
ромонов подробно рассмотрен в [352]. Ниже мы будем интересоваться не
транспортом феромонов, а движением массы животных под влиянием изме-
нения концентрации феромона.
Пусть модель содержит две пространственные области — область, где
скапливается саранча, и остальное рассматриваемое пространство. Концен-
трацию особей саранчи в первой области (без разделения на личинок и
взрослых) обозначим через хь во второй — через х2. Поток саранчи (еди-
ниц/ед. времени} обозначим через у12. Перелет саранчи начинается при превы-
шении концентрацией феромона (которую обозначим символом хз) некото-
рого предельного значения Хз. Тогда уравнения системы имеют вид
Xi = ацХ — knt/12,
x2 = k2iyi2, (6.24)
Хз = #32* 1 — U33X3,
где а и k — положительные коэффициенты. Здесь принято для простоты, что
а) доля взрослых саранчуков в общей популяции постоянна (темп производ-
ства феромона пропорционален плотности популяции JCi); б) феромон исчезает
с темпом, пропорциональным своей концентрации Х3.
Чтобы отразить влияние условий среды и, надо ввести нелинейные зави-
симости аи = ац(у), а перелет определить условием
„ __Г 0» Хз < Хз,
9,2==Ьхь Хз>Хз. {6-25)
Катастрофический характер миграции саранчи объясняется существенно
нелинейным характером зависимости у{2 от концентрации х3, что воспроизво-
дится в модели уравнением (6.25). Модель имеет три компартмента при рас-
смотрении двух пространственных областей.
Конвективный транспорт (перенос компо-
нент носителями). Часто в биосистемах та или иная ком-
понента может переходить из компартмента в компартмент не
непосредственно, а с помощью некоторых переносчиков. Напри-
мер, в организме кислород из атмосферы в кровь попадает
с вздыхаемым воздухом, а из крови к органам переносится с
потоком крови. Тот же поток крови переносит и тепловую
энергию от компартмента к компартменту, а выдыхаемый воз-
дух уносит тепло в окружающую среду. На экологическом
уровне такие процессы имеют место, например, при переносе
семян растений ветром или животными. Уменьшение интенсив-
ности потока носителя в этом случае приводит к уменьшению
ПЕРЕМЕЩЕНИЕ КОМПОНЕНТ
169
e.SJ
темпов переноса рассматриваемых компонент. Иногда такое
сокращение может оказаться трагическим для переносимой ком-
поненты. Так, предполагают, что причиной вымирания водяного
ореха в эпоху голоцена было сокращение численности крупных
животных, которые служили переносчиками плодов ореха [236].
В эпидемических процессах перенос возбудителя происходит
также с носителем, роль которого иногда играют транспортные
потоки [24, 303].
Конвективный перенос в физических явлениях имеет место
в том случае, когда переносимое вещество находится в движу-
щейся жидкости или газе. Результирующее движение вещества
называется конвективным транспортом.
Если через некоторую поверхность идет нормальный поток
жидкости или газа с объемной скоростью и, то
Z/коив == (6.26)
где z/коив — темп перемещения вещества с конвекцией, ас — кон-
центрация вещества в носителе. Для трехмерного случая ана-
логично тому, как это имело место при рассмотрении диффузии,
можно записать уравнение конвективной диффузии
+ + + = (в.27>
где D — константа, и=\и v w]T — трехмерный вектор скоро-
стей вдоль осей х, у, г. Если вектор и — константа, то из
(6.27) получаем
Vc = D^c, (6.28)
где оператор u-V определяется следующим образом:
и • V = u-/- + v + w (6.29)
дх'ду'дг v '
т. е. конвективный транспорт описывается обычным уравнением
диффузии (6.10), в котором появляется добавочный член, отра-
жающий вклад конвекции.
В компартментальных моделях обычно достаточно использо-
вать уравнение переноса вещества с носителями, записанное в
виде
У = Qx, (6.30)
где у — темп (скорость) переноса компоненты с носителем, х —
уровень или концентрация рассматриваемой компоненты в носи-
теле (содержание компоненты в единице носителя), Q — темп
потока носителя.
170
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
Рассмотрим, например, процесс теплоотдачи с выдыхаемым воздухом.
Пусть удельная теплоемкость воздуха равна с0, температура тканей легких 0],
температура среды 0о, интенсивность дыхания V. Тогда потери тепла у вы-
ражаются соотношением
У = с0 (01 - 0о) V. (6.31)
6.3. Взаимодействие и превращение компонент
В процессе функционирования биосистем обращающиеся в
них компоненты не остаются неизменными. Сущность процессов
жизни в том и состоит, что компоненты, поступающие из окру-
жающей среды в биосистему и запасаемые в ней, взаимодей-
ствуют между собой, вступают в реакции, преобразуются и
включаются в кругооборот веществ и энергии, утилизируются,
а ненужные биосистеме вещества снова переходят в окружаю-
щую среду.
Так, вещества, поступившие в организм, подвергаются ряду
химических превращений, обеспечивая поступление энергии и
построение структур тканей и органов. В то же время происхо-
дит непрерывный распад тканей с образованием простых ве-
ществ. Эти два типа процессов (анаболизм и катаболизм) вме-
сте образуют то, что называется процессом метаболизма. Слож-
ные химические процессы метаболизма объединяются в единое
целое с помощью хорошо организованного управления, осуще-
ствляемого нервной и гуморальной системами.
При рассмотрении этих вопросов следует иметь в виду их
большую сложность и недостаточность наших знаний о функ-
ционировании систем и органов в целостном организме. По-
этому, как правило, нарисовать детальную и полную картину
химических процессов и процессов управления в организме не
удается. Однако имеется несколько типовых способов описания
превращений компонент в биохимических системах (закон дей-
ствующих масс, уравнения ферментативных реакций) и разра-
ботаны простейшие модели воздействия активных веществ на
органы и железы в организме. Кроме этих моделей мы кратко
рассмотрим вопросы превращения и взаимодействия компонент
в биосистемах экологического уровня.
Закон действующих масс. В качестве компонент
биосистем на всех уровнях организации можно прежде всего
рассматривать химические вещества, поступающие в организм
и утилизируемые в нем.
Если в компартменте системы происходит химическая реак-
ция с веществами А, В, ..., которые превращаются в вещества
М, N, ..., то закон действующих масс записывается для нее
в виде
аЛ +₽В+ ... + ..., (6.32)
в.З] ВЗАИМОДЕЙСТВИЕ И ПРЕВРАЩЕНИЕ КОМПОНЕНТ 171
где числа а, р, ..., ц, v, ... носят название стехиометрических
коэффициентов.
Важнейшей характеристикой реакции (6.32) является кон-
станта ее скорости k. Если концентрации веществ А, В, ..., М,
N, ... обозначить через ха, хв, . хм, xN, ..., то изменение
этих концентраций во времени определяется уравнением
</хЛ _ dxB dxM dx N
dt ~ dt — • ’ ’ dt ~ dt
... = k (x^x® ... x^x* ...). (6.33)
Часто для биохимических реакций оказывается, что некото-
рые из компонент, участвующих в реакции, имеются в избытке.
Тогда их изменением в ходе функционирования можно прене-
бречь, и в правой части (6.33) соответствующая концентрация
заменяется константой. Например, если все вещества, кроме
А и 2V, имеются в изобилии, то выражение в правой части (6.33)
заменяется на —k^x^. В таком случае закон действующих
масс принимает вид
dx. dx,.
-dT = --dT=-kM (6.34)
где kj — новая константа.
В биосистемах происходят сложные специфические комп-
лексы химических реакций. Их моделирование представляет со-
бой самостоятельную задачу [75,281]. Но при ориентировочных
грубых расчетах иногда вместо рассмотрения всей системы хи-
мических реакций пользуются уравнениями так называемой сум-
марной реакции, в которых учитываются лишь исходные и
конечные вещества этой реакции. Все промежуточные стадии и
возникающие при этом вещества не рассматриваются. Имеются
указания на то, что в биосистемах для оценочных расчетов
можно пользоваться законом действующих масс для суммарных
реакций [215, 281]. При моделировании процессов обмена .в
живых системах могут оказаться полезными методы математи-
ческого моделирования химических реакций, разработанные в
химической технологии [228].
Ферментативные реакции. Если уравнения, описы-
вающие закон действующих масс, одинаково характерны для
живой и неживой природы, то ферментативные реакции, в кото-
рых ферменты работают как катализаторы — ускорители реак-
ций, в основном специфичны для биосистем (хотя ферментатив-
ные реакции проходят не только в живом организме — in vivo,
но и в условиях химического эксперимента, в пробирке —
in vitro).
172
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
1ГЛ. 6
Ферменты — это различные белки, выполняющие каталитиче-
скую функцию в организме. Вещества, вступающие под дей-
ствием ферментов в реакцию, называются субстратами. Иногда,
кроме фермента и субстрата, для осуществления реакций необ-
ходимы еще и дополнительные вещества — коферменты. Дейст-
вие ферментов заключается во влиянии на скорость реакции, ко-
ферменты действуют либо как переносчики некоторых химиче-
ских групп, участвующих в катализируемой реакции, либо как
усилители действия (активаторы) ферментов [98, 279, 304].
При взаимодействии фермента Е с субстратом S образуется
комплекс ES, в котором реакционная способность субстрата
повышена — субстрат превращается в продукт реакции, а фер-
мент возвращается в исходное состояние:
E + S^=ES-^E + P. (6.35)
Обычно концентрация субстрата превышает концентрацию
фермента. В стационарных условиях концентрация свободного
фермента Х\ и концентрация связанного в комплексе фермента
Х2 постоянны.
Кинетические уравнения имеют вид
Xi = — х2 = — kiSxi + (/?_i + k2) х2 (6.36)
В стационарном режиме
х1 = х2 = 0. (6.37)
Общая концентрация фермента Хз есть Е:
Е = X] Ч- х2. (6.38)
Темп протекания процесса в стационарном режиме
у = — S — P = k2x2. (6.39)
Тогда, подставляя в (6.39) стационарное значение х2, легко
определяемое из (6.36) — (6.38), получаем уравнение для скоро-
сти (темпа) ферментативной реакции
где = Уравнение (6.40) называется уравнением
Михаэлиса — Ментен. При увеличении концентрации субстрата
S скорость ферментативной реакции растет, приближаясь к мак-
симальной
ym^ = k2E. (6.41)
Уравнение Михаэлиса — Ментен является одной из наиболее
часто используемых моделей при описании биохимических про-
6.3] ВЗАИМОДЕЙСТВИЕ И ПРЕВРАЩЕНИЕ КОМПОНЕНТ 173
цессов [352] и стало своего рода базовой моделью для различ-
ного рода исследований. В работе [294], например, статистиче-
ские проблемы построения кинетических моделей рассматри-
ваются применительно к модели Михаэлиса — Ментен.
Воздействие активных веществ на органы и
железы. Своеобразным механизмом превращения компонент
в организме является воздействие (стимулирующее или тормо-
зящее) некоторых веществ на железы и органы.
Так, если некоторое вещество (с индексом i) выделяется ка-
ким-либо органом под действием других веществ (с индексами
от 1 до k), то в общем случае можно записать
— .. хк), (6.42)
где yi — скорость выделения вещества с индексом I, X/, j = 1, ...
..., k, — концентрация вещества с индексом / в органе.
Простейшей формой уравнения (6.42) является линейная
зависимость, когда скорость синтеза некоторого вещества про-
порциональна концентрации некоторого другого вещества (гор-
мона, фермента). Такой способ описания применяется и в на-
стоящее время [164, 279], хотя чаще используются нелинейные
зависимости [30].
Пример 6.3.1. В щитовидной железе организма с участием йода синте-
зируется тироидный гормон ТГ, поступающий затем в кровь. Скорость синтеза
определяется концентрацией другого вещества — тиреотропного гормона ТТГ.
В работе [30] этот процесс описывается нелинейной зависимостью
У1 = Рх2х3 — <Xix2 — а2хь (6.43)
где Xi, г/i—концентрация ТГ в тканях щитовидной железы и скорость син-
теза ТГ, х2, х3— концентрация йода и ТТГ в тканях щитовидной железы,
аь «2, Р — коэффициенты.
Два последних слагаемых отражают ингибирование формирования и вы-
деления ТГ под влиянием йода и самого ТГ.
Превращение компонент в популяции. Если
популяция организмов рассматривается как целое без ее разде-
ления на какие-либо группы (возрастные или половозрастные),
то изменение ее численности часто описывается уравнением Фер-
хюльста— Пирла для однородной популяции
^ = (а-уУ)ЛГ, (6.44)
где У — численность особей в популяции в момент времени t,
а > 0 — коэффициент естественного прироста, у > 0 — коэффи-
циент, учитывающий конкуренцию внутри популяции (за пищу,
пространство и т. д.).
Во многих моделях, однако, рассматривается возрастной со-
став популяций. Так, в моделях водных экосистем обычно иссле-
дуется несколько компонент — в пределах каждой популяции —
174
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
группы сеголеток, рыб в возрасте от года до двух, от двух до
трех лет и т. д. [123, 124, 163, 222]. Обычно из-за сезонности
размножения популяций уравнения, описывающие переход осо-
бей из группы в группу, записывают в дискретной форме, счи-
тая, что в определенный момент низшая возрастная группа
переходит в высшую, а мальки поступают в нулевую группу
популяции.
Пусть популяция состоит из (п-)-1) компонент (п возраст-
ных групп плюс нулевая возрастная группа — мальки). Тогда
в год с номером k состояние популяции описывается вектором
х№) = [х(*) x\k> . . .
где Xi — численность i-й возрастной группы. Связь между со-
стоянием популяции в год с номером (&-]-1) и состоянием в
k-и год описывается уравнением
X(fe+1) = [/X«:)> (6.45)
где U — матрица размерности (k ф- 1)Х(& + 1),
г0 61 62 . . ега—1
Ро 0 0 . .. 0 0
(/ = 0 ₽1 0 . .. 0 0 (6.46)
_ 0 0 0 .. • ₽п—1 0 _
Здесь р,- — коэффициент выживаемости i-й возрастной группы,
е(-—коэффициент пополнения нулевой возрастной группы от осо-
бей i-й возрастной группы (средняя плодовитость особей i-ro
класса). Матрицы типа (6.46) называются матрицами Лесли
[212, 302]. Коэффициенты е,, р, могут сложным образом зави-
сеть от численности популяции и ее компонент, а также от
условий среды (абиотических факторов).
Если считать, что численности возрастных групп меняются
во времени непрерывно, то вместо модели (6.46) можно рас-
сматривать дифференциальные уравнения
Xi = Lxt. (6.47)
Здесь X, — численность i-й возрастной e2 0 — da — Pa ... группы, а матрица L
имеет вид ei — di — pi di
А = ~ — do — Ио d0 0 En—l 6n 0 0 0 0 , (6.48)
0 0 0 — an — Pn o _
где Ei имеют тот же смысл, что и в (6.46), а, — коэффициент,
определяющий вероятность перехода особей i-й группы в еле-
в.З] ВЗАИМОДЕЙСТВИЕ И ПРЕВРАЩЕНИЕ КОМПОНЕНТ 175
дующую возрастную группу, щ — коэффициент естественной
смертности в z-й группе.
Общая система уравнений динамической теории популяций,
объединяющая большинство известных математических моделей,
используемых в теории популяций для анализа динамики поло-
возрастной и генетической структур, предложена в [161]. На
основе этой системы уравнений удается определить и область
применения тех или иных частных моделей теории популяций.
Взаимодействие между популяциями в эко-
системе. Простейшей моделью взаимодействия популяций
является модель Вольтерра «хищник-жертва» [45], рассматри-
ваемая при следующих условиях: пища для жертвы имеется в
изобилии, хищник питается только жертвой. Прирост жертв за
малый промежуток времени пропорционален их численности,
прирост хищников пропорционален произведению количества
хищников на количество жертв, что соответствует частоте воз-
можных встреч хищника с жертвой. Естественная смертность
хищников пропорциональна их численности.
Модель имеет следующий вид:
= xl (е1 V12*2) >
. (6.49)
~dt~ — x2 (— e2 + У21Х1),
где Xi, X2 — количество особей жертв и хищников, соответствен-
но, ei — коэффициент естественного прироста жертв, е2 — коэф-
фициент естественной убыли хищников, у12, 721— коэффициенты,
определяющие «конкурентоспособность» жертв и хищников.
Уравнение Вольтерра получило широкое распространение
при моделировании биоценозов различного вида и неоднократно
обобщалось и дополнялось для исследования различных свойств
экосистем. Так, учет внутривидовой конкуренции жертв и хищ-
ников приводит к уравнению Костицына
= X] (б] у12Х2 У11Х1) ,
dx2 (6.50)
— хг (— е2 + YaiX, — У22хг)>
где ун, Y22 — коэффициенты, учитывающие внутривидовую кон-
куренцию (сокращение скорости прироста популяции при уве-
личении ее численности). В широком диапазоне значений коэф-
фициентов (а все коэффициенты этой системы уравнений поло-
жительны), система (6.50) имеет предельную устойчивую точку
(узел). Это означает, что колебания численности видов в эко-
системе отсутствуют, и имеется стационарный неравновесный
режим.
176
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
Другие типы обобщений даются, например, в работах: [160]—
для рассмотренных ниже систем Либиха, [187]—для проблемы
эксплуатации популяций. Поскольку решение уравнений Воль-
терра приводит к циклическим движениям, что является след-
ствием выбранной Вольтерра специальной формы уравнений, то
особое внимание при его обобщениях уделяется тому, чтобы
можно было получить все виды поведения системы — гибель
обеих или одной из популяций, устойчивое сосуществование и,
наконец, режим с периодическим увеличением и уменьшением
численности популяций [55, 91].
В настоящее время для математического описания экологи-
ческих систем чаще всего используется модель Ферхюльста —
Пирла [161, 187а], записанная для системы из п видов:
п
Xi = e.iXi + z=l, 2.......п, (6.51)
Pz /=i
где е(- — внутренние коэффициенты прироста, 0,- и ац — коэффи-
циенты, описывающие влияние популяций друг на друга. Совре-
менное состояние теории вольтерровских моделей рассмотрено
в [187а].
Часто уравнения взаимодействия популяций записываются в форме, от-
личной от (6.Й1), хотя существо процессов в таких уравнениях остается неиз-
менным. Так, в работе [221] исследуется водная экосистема, содержащая
10 популяцйй, которая описывается системой уравнений
ю
= <6-52)
где Bi — биомасса i-й компоненты, измеряемая в ккал/м3, Rt — суточный ра-
цион {ккал/м3), Hi и Ti — неусвоенная пища и траты на энергетический
обмен в тех же единицах, М, — отмирание (переход биомассы в мертвое
вещество в тех же единицах), с,у — доля i-ro вида в питании /-го вида. Легко
видеть, что если слагаемые Ri, Hi, Ti, Mi считать пропорциональными числен-
ности xi, а долю i-ro вида в питании /-го вида — пропорциональной численно-
сти i-ro вида, то четыре первых слагаемых в (6.52) аналогичны первому сла-
гаемому в (6.51). Остальные слагаемые обоих уравнений также одинаковы.
Системы с лимитирующими факторами. Про-
цессы в биосистемах часто протекают так, что на разных их
этапах наличие различных компонент системы оказывает ре-
шающее влияние на характер этих процессов. Так, для популя-
ции темп поглощения и обмена веществ на начальном этапе
развития (когда плотность популяции невелика, а продуктов пи-
тания достаточно) определяется только плотностью популяции.
Однако при тех же условиях внешней среды для разросшейся
популяции ограничивающим фактором дальнейшего ее роста
является наличие продуктов питания. Свойство систем, заклю-
чающееся в том, что на каждом этапе ее функционирования
в.4! УТИЛИЗАЦИЯ КОМПОНЕНТ 177
существует единственная переменная, ограничивающая темпы
протекающих в системе процессов, известно как принцип
Либиха.
В последние годы разработана формализованная теория си-
стем, функционирующих на основе принципа Либиха, — так на-
зываемая теория Л-систем [53, 160], в которой рассматри-
ваются системы с «обменом веществ» — их поглощение, пере-
работка и выделение. Примерами подобных систем являются
как биосистемы (организмы, биоценозы), так и многие системы
производства и экономики. Система Либиха описывается урав-
нениями
х = Ар, (6.53)
где х — n-мерный вектор концентраций веществ, х 0, А(п>(т) —
матрица, определяющая свойства системы. В правой части
(6.53) вместо обычного вектора х стоит вектор р размерности
т, р > 0, определяющий для любого текущего состояния в соот-
ветствии с принципом Либиха, какая переменная в системе
является лимитирующей; /-я компонента этого вектора задается
следующим образом:
где уц — поток компоненты с номером i на вход процесса с но-
мером /, и, — внешний лимитирующий фактор, а1;- — стехиомет-
рический коэффициент вещества i в процессе с номером /. Пред-
полагается, что протекающие в системе процессы имеют харак-
тер химических реакций.
6.4. Утилизация компонент
Чтобы жить, биосистема должна непрерывно расходовать
энергию, потребляя для этого нужные ей вещества. «Постоянный
приток свободной энергии — это только одно из условий поддер-
жания стационарного состояния. Другое условие — наличие ор-
ганизации, позволяющей поглощать и использовать эту энер-
гию» [98].
Рассмотрим, например, системы биохимического (клеточ-
ного) и физиологического (организменного) уровня. Каждая
клетка независимо от других клеток должна обеспечивать свои
энергетические потребности, получая необходимую энергию в
процессе обмена вещества. Потребности клетки можно выразить
через темп утилизации метаболической энергии — количества
распадающейся в единицу времени АТФ (например, грамм-мо-
лекул АТФ в минуту). Можно с уверенностью сказать, что
темпы энерготрат в клетке определяют и темпы протекающих
178
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ t
в ней процессов метаболизма, хотя интимные процессы клеточ-
ной регуляции темпов метаболизма изучены далеко не полно-
стью. «Каким именно образом химическая энергия подается к
местам ее утилизации (ретикулярным мембранам, актомиозину,
клеточным мембранам, рибосомам и т. д.) и какими средствами
обеспечивается регуляция направления движения и объема по-
токов, основанная на потребностях, в точности не известно. Но
что мощная внутриклеточная система (или системы) должна
существовать — это очевидно» [363, стр. 29].
Согласно так называемому энергетическому правилу поверх-
ности (или закону Рубнера) основной обмен организма опре-
деляется генетическими факторами и пропорционален поверх-
ности тела. Темпы обмена веществ тем интенсивнее, чем меньше
весовые и линейные параметры организмов. Хотя в современной
физиологии связь законов Рубнера с регулированием темпов
процессов в организме не выдвигается на первый план, для нас
важно следующее обстоятельство. Закон Рубнера численно
определяет заданную величину темпов метаболизма, которая за-
тем поддерживается физиологическими механизмами, несмотря
на колебания условий внешней среды (таких, например, как
снижение или повышение концентрации кислорода).
Темп энерготрат каждой клетки в конечном счете зависит
в организме только от генетически предопределенной реакции
высших координирующих центров. Каждая клетка должна вы-
полнять свои специфические функции в темпе, устанавливаемом
этими высшими координирующими центрами. Темпы всех кле-
ток, складываясь, дают то, что принято называть темпом обмена
веществ целостного организма. Интенсивность метаболизма
обычно выражается через темпы теплопродукции (например,
в ккал/час).
Минимально возможное значение темпов метаболизма в орга-
низме (так называемый основной обмен) достигается у человека
после сна и тридцатиминутного покоя при снятии всех психиче-
ских и эмоциональных факторов, при комфортной температуре
и при условии, что человек не ел в течение 12 часов. Факторами,
изменяющими темп метаболизма, в физиологии считаются физи-
ческая нагрузка, специфическое действие пищи (когда после
приема пйщи темп обмена веществ «автоматически» повы-
шается) , введение тироидных и некоторых других гормонов, воз-
раст, лихорадки любого происхождения, сон и нарушения пита-
ния. Наконец, темпы метаболизма у человека зависят от клима-
тических условий — темпы обмена веществ в тропиках на 10—
20% ниже, чем в Арктике [304].
Аналогичная ситуация имеется и для других типов биосистем.
Для каждого их типа существует некоторое минимальное
значение темпов обмена, отвечающее оптимальным условиям
6 4]
УТИЛИЗАЦИЯ КОМПОНЕНТ
179
внешней среды и условиям «покоя» биосистемы. В экологиче-
ских системах, например, аналогом основного обмена физиоло-
гических систем является обмен покоя [39, 123]. Ниже этого
уровня скорости обменных реакций в биосистеме уменьшиться
не могут. Любое ухудшение условие среды или увеличение
активности системы приводит к увеличению скорости обмена
веществ. Для большинства позвоночных животных суммарный
темп метаболизма (ккал/сутки) зависит от массы тела ш и ге-
нетически закрепленных характеристик организации, опреде-
ляющих коэффициент пропорциональности k,
W = kmb, (6.55)
где m — масса организма (кг). После «многолетних поисков
в десятках лабораторий мира» было принято k — 70, b = 3/4
[363]. Иногда, однако, можно встретить и отличающиеся от
(6.55) выражения. Так, у рыб траты на обмен определяются
формулой
W = ст0-8, (6.56)
где постоянная с учитывает как генетические характеристики,
так и поправку на отличие активного обмена от обмена покоя
[39]. В экосистемах темп потребления энергии измеряется
обычно в ккал/год.
Говоря о темпах утилизации вещества и энергии в биосисте-
мах, мы хотим отразить одну характерную особенность, которой
в литературе по биокибернетике уделяется недостаточно внима-
ния. Речь идет о том, что потоки вещества и энергии, входящие
в компартменты системы и покидающие их, не равноправны по
своей сути. Одна часть потоков обусловлена самим фактом
протекания процессов жизнедеятельности; эти потоки нарушают
равновесие в системе и ее компартментах.
Другая часть потоков, напротив, является реакцией системы
на исходное нарушение баланса вещества и энергии в компарт-
ментах, она носит восстановительный, регуляторный характер
и направлена на поддержание этого баланса. Темпы таких по-
токов целиком зависят от регуляторных механизмов системы, их
величина регулируется, подстраивается в соответствии с жиз-
ненными потребностями системы.
Чтобы подчеркнуть принципиальное различие между двумя
типами темпов, мы введем для них специальные обозначения.
Темпы процессов, непосредственно связанных с выполнением
биологических функций, мы будем называть первичными тем-
пами. Для процессов, направленных на восстановление непре-
рывно нарушаемого в ходе жизнедеятельности системы баланса
вещества и энергии, будет использоваться термин вторичные
темпы.
180
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
Понятие о первичных и вторичных темпах легко получается
при рассмотрении процессов расхода и восполнения энергии в
ходе функционирования живых клеток. Роль первичных тем-
пов здесь играют темпы энерготрат, например, темп обмена ве-
ществ или интенсивность энерготрат при основном обмене.
Расход энергии в каждой клетке должен в среднем покры-
ваться ее доставкой. Так, при резкой физической нагрузке в
первое время вторичные темпы отстают от увеличившихся пер-
вичных, и баланс веществ в компартментах системы нарушается.
Содержание гликогена в мышцах падает почти до нуля, резко
возрастает концентрация молочной кислоты в крови. Эти сдвиги
стимулируют регуляторные механизмы на всех уровнях орга-
низма и его систем, так что и по окончании работы вторичные
темпы остаются повышенными для восполнения энергетических
трат в клетках. В среднем темпы потоков энергии и веществ,
направленных на восстановление нарушенного баланса, должны
быть равны темпам первичных процессов во всех компартментах.
Темп утилизации кислорода в энергетической системе орга-
низма можно трактовать как первичный (именно он «задается»
высшими координирующими центрами, когда животное перехо-
дит от одного режима функционирования к другому, и энергети-
ческие потребности меняются — см. рис. 1.2, в в общей схеме
регуляции в организме животного). Темп поступления кислорода
в метаболическую систему должен в среднем следить за задан-
ным темпом утилизации, и темп поступления О2 можно считать
вторичным. Темп возникновения СО2 жестко связан с темпом
потребления О2 и не зависит от физиологических механизмов
регуляции. Поэтому и этот темп должен рассматриваться как
первичный. Темп удаления СО2 из организма, величина которого
определяется физиологическими механизмами, является вторич-
ным. Темпы теплопродукции при данном обмене веществ яв-
ляются первичными, темпы теплоотдачи, находящиеся во власти
физиологических регуляций, — вторичными. Величина вторичных
темпов с помощью механизмов физиологической регуляции в
организме должна поддерживаться на уровне первичных темпов.
В компартментальных моделях живых систем первичные
темпы расхода энергии связаны, с одной стороны, с поддержа-
нием структуры биосистемы (а в процессе развития — также с
образованием и ростом новых подсистем и структурных образо-
ваний), а с другой — с энергетическим обеспечением функцио-
нирования всех ее элементов. Темпы утилизации вещества и
энергии, идущих на поддержание структуры биосистемы, опре-
деляются ее массой и объемом, специфическими свойствами
составляющего ее биологического материала и, наконец, ее
пространственным строением. В частности, от пространственного
расположения компартментов системы зависит протяженность
ел]
УТИЛИЗАЦИЯ КОМПОНЕНТ
181
транспортных путей и, следовательно, расходы на транспорти-
ровку веществ между компартментами. Во многих случаях
можно считать, что темп потока энергии, расходуемой на под-
держание структуры, либо постоянен, либо пропорционален не-
которым переменным состояния биосистемы х,. Так, метаболиче-
ские траты на поддержание транспортных путей — перемещение
питательных веществ из почвы к надземным частям растений —
тем больше, чем крупнее растение [158].
Для /-го компартмента обозначим расходы на поддержание
структуры через w^.
Несколько сложнее дело обстоит с расходами энергии на
функционирование биосистемы. В предыдущих разделах мы
рассмотрели некоторые процессы переноса и превращения ком-
понент в биосистемах. Часть этих процессов происходит «пас-
сивно», т. е. механизмы, ответственные за их осуществление, не
расходуют метаболической энергии (см. разд. 1.6). Таковы про-
цессы диффузии (6.10) или (6.14), а также излучение тепловой
энергии (6.23). Если процессы переноса веществ (6.15) осу-
ществляются пассивно (например, в экосистемах — перенос се-
мян и биомассы потоками воздуха или воды), то такие процессы
также не связаны с активной затратой энергии в биосистеме.
Часто процессы преобразования компонент на основе закона
действующих масс (6.34) или в ходе некоторых ферментативных
реакций (6.40) также представляют собой пассивные про-
цессы, не приводящие к расходу метаболической энергии биоси-
стемы.
Другие процессы функционирования биосистем — перемеще-
ние компонент активным путем — миграция (6.24), активные
процессы конвекции — перенос веществ (6.30) в организме жи-
вотного с потоками крови или других веществ (лимфы, воздуха
в легких, воды в жабрах), активный синтез веществ (6.42) —
происходят с непосредственной затратой метаболической энер-
гии. Темпы расходования энергии в каждом компартменте про-
порциональны интенсивности активных процессов, происходя-
щих в нем. Обозначим через скорость потребления энергии
в /-м компартменте за счет некоторого активного процесса его
функционирования (переноса или преобразования компонент):
— v)> (6.57)
где х, v — векторы состояния и входов биосистемы.
Тогда, объединяя темпы расхода энергии в /-м компартменте
на поддержание структуры и на его функционирование, можно
определить первичные темпы расхода энергии как
wlв “4° + WT' (6.58)
182
К0МПАРТМЕ11ТАЛЫ1ЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
Фактически для всех уровней биосистем первичные темпы рас-
хода метаболической энергии, происходящего в живых клетках
организмов, оплачиваются «универсальной энергетической ва-
лютой», т. е. АТФ. Однако при исследовании процессов жизне-
деятельности с помощью компартментальных моделей часто
вместо процессов, определяющих баланс АТФ, рассматривают
процессы передачи, синтеза, накопления и утилизации тех ком-
понент, которые участвуют в энергетическом балансе (кисло-
рода, глюкозы, а в экосистемах — органического вещества).
Тогда вместо (6.57) и (6.58) в рассмотрение вводится вектор
темпов потоков веществ
w = [щ] w% ... w„]T, (6.59)
размерность которого определяется числом компартментов п.
Компоненты вектора w зависят в общем случае от вектора х и
от вектора внешних условий и. Для каждой комбинации усло-
вий среды v и состояний системы х вектор (6.59) определяет
необходимые для жизнедеятельности темпы утилизации веществ
и энергии в компартментах.
Таким образом, темпы утилизации вещества и энергии (или
темпы возникновения, появления веществ, аналогичные этим
темпам) играют особую роль в компартментальных моделях
биологических систем. Именно они выступают в качестве пер-
вичных темпов, определяющих потребности живых систем. Рас-
смотренные же в двух предыдущих разделах процессы транс-
порта и процессы превращения компонент чаще всего выступают
в роли генератора вторичных темпов. Изучение соотношения
между первичными и вторичными темпами лежит в основе
компартментального моделирования. Мы поэтому постараемся
представить схему компартментальной модели общего вида в
такой форме, чтобы первичные и вторичные темпы процессов
в ней были основными элементами структуры.
6.5. Общая схема компартментальной модели
Обычно считается, что в каждом из компартментов системы
процессы поступления, преобразования и утилизации компонент
протекают независимо друг от друга. Все три типа процессов,
рассмотренных выше, — перенос веществ, их превращения в
ходе «обмена веществ» и использование — происходят в ком-
партментах одновременно, так что изменение концентраций
веществ есть суммарный результат всех этих процессов, опре-
деляемый по закону сохранения веществ.
Общая схема мультикомпартментальной модели живой си-
стемы может поэтому быть изображена следующим образом
(рис. 6.3). Основной элемент схемы — вектор состояния системы
6.5]
ОБЩАЯ СХЕМА КОМПАРТМЕНТАЛЬНОЙ МОДЕЛИ
183
х, характеризующий уровни (концентрации или количества)
всех компонент, принимающих участие в обмене веществ в био-
системе.
Жизнедеятельность системы связана с процессами потребле-
ния веществ, темп или скорость которых определяется внешними
по отношению к системе факторами (например, мотивационными
и генетическими характеристиками системы, условиями внешней
Рис. 6.3. Общая схема компартментальной модели биосистемы. Скорость изменения век-
тора концентраций х равна сумме скоростей утилизации, переноса и синтеза компонент»
участвующих в обмене веществ. Поскольку величина темпов утилизации не может регу-
лироваться внутренними механизмами системы, эти темпы играют роль независимых пере
меииых—первичных темпов. Регулирующие механизмы должны уравновесить расход ком-
понент их синтезом и доставкой. Поэтому темпы процессов, направленных иа компенсацию
трат вещества и энергии, играют роль зависимых переменных —вторичных темпов. В ста-
ционарном состоянии вторичные темпы по величине должны быть равны первичным.
среды и) и некоторыми внутренними характеристиками (напри-
мер, биомассой). Выше эти темпы были обозначены как пер-
вичные.
Зависимость темпов потребления от вектора состояния л
показана стрелкой в нижней части рисунка. Напомним, что тер-
мин «темпы потребления» используется здесь условно, так как
соответствующий вектор скоростей описывает не только явления
утилизации, но также перемещения и синтеза компонент, необ-
ходимых для жизнедеятельности системы: скорость утилизации
кислорода и глюкозы в физиологических системах, скорость
утилизации солнечной энергии в экосистемах; скорость выделе-
ния углекислоты и тепла и скорость образования шлаков во
всех типах биосистем. Все эти скорости заданы для кажого из
компартментов биосистемы.
Процессы потребления уравновешиваются в биосистеме
преобразованием поступающих извне компонент в необходимые
Для жизнедеятельности вещества, что на схеме показано блоком
184
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
«процессы синтеза», и перемещением образовавшихся веществ
в те компартменты, где происходит их утилизация («процессы
переноса»). В соответствии с принятым выше условным разде-
лением рассматриваемые механизмы включают в себя и про-
цессы выведения из биосистемы продуктов ее жизнедеятельности
(отвод углекислоты и тепла из тех компартментов, где они обра-
зуются, выведение их в окружающую среду). Скорости про-
цессов синтеза и переноса зависят от вектора концентраций х и
от внешних условий v. Кроме того, возможно влияние на эти
процессы и вектора w.
Темпы доставки вещества в те компартменты, где происходит
их расходование, в биосистемах регулируются. Их величина за-
висит от темпов расходования и, разумеется, от возможностей
регуляторных механизмов, которые стремятся обеспечить баланс
вещества и энергии в системе. Выше эти темпы потоков веще-
ства и энергии мы обозначили как вторичные.
Тогда с точки зрения теории управления первичные темпы
могут играть в системе роль внешнего задающего сигнала, а
вторичные темпы — роль управляемого, выходного сигнала.
Ниже, в разд. 7.2, мы подробно остановимся на этом вопросе.
Полная скорость изменения концентраций х каждой компо-
ненты во всех компартментах определяется суммой скоростей
утилизации, переноса и синтеза этой компоненты в данном ком-
партменте. Если нас интересуют малые отклонения от некоторых
нормальных режимов в системе, то можно линеаризовать все
зависимости, указанные на рис. 6.3 в прямоугольниках. В этом
случае в соответствии с разд. 3.3 и 5.8 мы получим схему, пока-
занную на рис. 6.4, которая описывается линейными уравне-
ниями с переменными коэффициентами. Если при линеаризации
ввести следующие обозначения:
процессы переноса компонент
х(1) = Atx + B[V + Q\W, (6.60)
процессы синтеза компонент
х<2> = А2х + B2v + Q2w, (6.61)
скорость потребления компонент
х<з) = Rw + Рх + Sv, (6.62)
то вторичные темпы, как об этом говорилось выше, опреде-
ляются суммой (6.60) и (6.61), а первичные темпы — соотноше-
нием (6.62). Обозначив
Л = А + Л2, В = В1 + В2, Q = Qi + Q2. (6.63)
получим линейную схему, показанную на рис. 6.4.
6.6]
ОБЩАЯ СХЕМА КОМПАРТМЕНТАЛЬНОЙ МОДЕЛИ
185
Уравнения состояния системы в этом случае имеют вид
x=(X + P)x + (B + S) v + (/? + Q) да, (6.64)
где х—вектор состояния системы, и и да— входные сигналы,
А, Р, В, S, R, Q — матрицы соответствующих размерностей.
Уравнение (6.64) можно переписать в обычном виде уравнений
состояния для линейных систем (первое уравнение в (5.39)),
если должным образом переобозначить матрицы в (6.64).
Строго говоря, этот подход применим в том случае, если
выполнены условия, при которых о поведении исходной нели-
нейной системы можно судить по первому приближению
Рис. 6.4. Схема линеаризованной компартментальной модели. Первичные темпы в модели
зависят от режима работы системы, условий внешней среды и, наконец, от некоторых
переменных состояния системы, таких как общий объем или биомасса (матрицы R, S и Р,
соответственно). Вторичные темпы регулируются внутренними механизмами управления к
поддерживаются равными величинам первичных темпов (матрицы А, В и Q).
(см. разд. 5.5). Если в исходной нелинейной системе существует
стационарный режим, при котором переменные системы пере-
стают изменяться во времени, то и в линеаризованной системе
в конце концов устанавливается стационарное состояние, при
котором процессы утилизации и доставки (синтеза и переноса)
всех компонент уравновешены, так что
х = 0. (6.65)
Если матрица (А + Р) неособенная, то в соответствии с (5.50)
стационарное значение вектора состояния х определяется фор-
мулой
х = - {А + РГ1 [(В + S) v + (R + Q) да]. (6.66)
Уравнения (6.64) или (6.66) позволяют исследовать общие
свойства компартментальных моделей живых систем — их ди-
намические характеристики и поведение систем в стационарных
режимах. Такое исследование представляет собой основу ком-
партментального анализа живых систем.
186
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
В последние годы компартментальный анализ становится все
более распространенным методом исследования биологических
явлений и процессов. Хотя в настоящее время нельзя указать
достаточно полных источников, освещающих проблемы компарт-
ментального моделирования (книги [265, 317], к сожалению,
охватывают далеко не все интересующие специалистов в области
биокибернетики и исследования биосистем проблемы компарт-
ментального анализа), можно упомянуть ряд публикаций, ка-
сающихся как общих проблем компартментального моделиро-
вания (см., например, [296, 345, 350, 353]), так и ряда частных
применений. Среди последних есть как задачи традиционного
плана — биохимические, экологические или фармакологические
[314, 321, 346], так и задачи нового типа — такие, как модели-
рование клеточного цикла [318]. Наиболее близким к пробле-
мам компартментального моделирования справочным пособием
можно, по-видимому, считать обзор [344].
6.6. Компартментальная модель
энергетической системы организма
Все виды работы в организме — химическая, физическая,
электрическая — выполняются за счет расходования богатых
энергией (макроэргических) веществ и прежде всего аденозин-
трифосфата (АТФ). Отдав энергию, АТФ превращается в аде-
нозиндифосфат (АДФ). Расход АТФ в работающих органах
покрывается его синтезом за счет двух типов процессов: аэроб-
ных, протекающих с потреблением кислорода, и анаэробных,
проходящих в отсутствие кислорода. Основными веществами, из
которых в ходе протекания аэробных и анаэробных процессов
получаются макроэргические вещества, являются глюкоза и на-
капливаемый в печени гликоген. В упрощенном виде схему энер-
гетического снабжения живой клетки можно представить так,
как показано на рис. 6.5 [38, 98]. Блок-схема энергетических
процессов в организме животного и человека приведена на
рис. 6.6 [194].
Пусть расходование АТФ происходит со скоростью ш; синтез
АТФ в ходе аэробных и анаэробных процессов — со скоростью
Уу и z/2, соответственно. Составим простейшую модель энергети-
ческого снабжения организма [63, 253]. Рассмотрим модель, со-
стоящую из двух пространственных областей — функционирую-
щей ткани и омывающей ее крови.
Аэробные процессы восстановления АДФ до АТФ идут в со-
ответствии с суммарным уравнением
2С3Н6О3 + 6О2 + 180АДФ -> 6Н2О + 6СО2 + 180АТФ. (6.67)
молочная
кислота
6.6]
МОДЕЛЬ ЭНЕРГЕТИЧЕСКОЙ СИСТЕМЫ ОРГАНИЗМА
187
Считая, что скорость этой реакции определяется законом
действующих масс, причем определяют реакцию концентрации
кислорода и молочной кислоты, получим
(6.68)
где X] — концентрация кислорода в тканях, х3 — концентрация
молочной кислоты в тканях, rj — коэффициент.
Рис. 6.5. Схема энергетического снабжения мышечной клетки (тликолиз). При гликолити-
ческом расщеплении глюкозы чистый выход АТФ составляет 3 моль на 1 моль глюкозы.
Процесс протекает следующим образом: в результате нескольких ферментативных реак-
ций гликоген расщепляется до фруктозо-1,6-дифосфата (блок /), который затем расщеп-
ляется до двух трехуглеродных соединений (блок 2). Одно из них окисляется до 3-фосфо-
глицерииовой кислоты с образованием одной молекулы АТФ. Окисление происходит с уча-
стием иикотинамидадениндииуклеотида (НАД), который затем восстанавливается до НАД-Н
(пунктирный контур 3). В результате нескольких ферментативных реакций затем полу-
чается пировиноградная кислота (ПВК)» и образуется еще одна молекула АТФ (блок 4),
Далее процесс может развиваться двояко: в анаэробных условиях ПВК восстанавливается
до молочной кислоты (МК) за счет НАД-Н, в аэробных же условиях ПВК окисляется
в ходе так называемого цикла трикарбоновых кислот, так что в присутствии кислорода МК
не образуется. Возникшая в анаэробных условиях МК поступает с кровью в печень, где
включается в процессы синтеза гликогена печени (блок 5). Чистый выход в 3 моль АТФ
получается следующим образом: из одной молекулы шестиуглеродных соединений (блок/)
получается 2 молекулы трехуглеродных соединений (блок 2), каждая из которой вступает
в описанный выше процесс превращений, что дает 4 молекулы АТФ. Однако одна моле-
кула АТФ используется в ферментативных реакциях в блоке 1 [98].
Анаэробные процессы описываются суммарным уравнением
С6Н12О6 + 2АДФ 2С3Н60з + 2АТФ.
глюкоза молочная
кислота
(6.69)
188
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. Б
Считая, что скорость синтеза АТФ определяется здесь, в основ-
ном, концентрацией АДФ в тканях, получим
Z/2 = ^5> (6-70)
Рис. 6.6. Схема энергетического снабжения мышечной ткани в организме. Схема в упро-
щенном виде воспроизводит структуру организма и несколько в более подробном—про-
цессы энергообмена в мышцах. Продукты питания и кислород поступают в организм извне,
попадают в кровь и с ней —в ткани мышц. В ходе происходящих здесь процессов синтеза
выделяется энергия, примерно 15% которой представляют собой «полезную» энергию макро-
эргических связей, а 85%—«бесполезное» тепло. Тепловая энергия, углекислота и вода
снова переносятся кровью и выводятся из организма. Стрелки показывают направление
движения крови и транспорта тепла и веществ [194]. Мышечная работа / связана с распа-
дом АТФ, что восполняется в ходе процессов анаэробного 3 и аэробного 4 синтеза.
В ходе аэробных процессов образуется и испольвуется молочная и пировиноградная
кислота 2.
Введем переменные х2 и х4 — концентрации кислорода и мо-
лочной кислоты в крови и допустим, что условия функциониро-
вания системы определяются единственным внешним факто-
ром — концентрацией кислорода во вдыхаемом воздухе v\. За-
пишем при этих условиях уравнения системы. Будем считать,
что диффузия веществ из ткани в кровь и обратно подчиняется
уравнениям (6.14), скорость убывания кислорода и молочной
кислоты в ходе реакции определяется (6.33). Пусть также кон-
центрация молочной кислоты в крови уменьшается из-за того,
что некоторые органы в организме утилизируют ее со скоростью,
пропорциональной этой концентрации. Анаэробные процессы
(6.69) увеличивают концентрацию АДФ в тканях. Тогда, вводя
соответствующие коэффициенты пропорциональности ki — k-j,
в.6]
МОДЕЛЬ ЭНЕРГЕТИЧЕСКОЙ СИСТЕМЫ ОРГАНИЗМА
189
получим
Xi = ki (х2 — *1) — kfijx,
Х2 =k2(v — Х2) — kj (х2 — X;),
Хз = k5y2 — k3 (х3 — х4) — k6ylt
х4 = k3 (х3 — х4) — Й4Х4,
Х5 = W — Ух — у2,
(6.71)
где у\ и у2 определяются уравнениями (6.68) и (6.70). Послед-
нее уравнение описывает динамику АДФ в тканях; изменение
содержания АТФ в моде-
ли не рассматривается,
поскольку в рамках дан-
ного описания правые ча-
сти дифференциальных
уравнений для АТФ и
АДФ совпадают с точ-
ностью до знака. Если,
например, обозначить че-
рез х6 концентрацию АТФ
в тканях, то хе = —w +
Рис. 6.7. Блок-схема компартментальной модели
энергоснабжения организма. Задающий сигнал w —
темп потребления АТФ. Условия окружающей
среды v — концентрация кислорода во вдыхаемом
воздухе. Вектор состояния x=[xiXj ... Х|]т имеет
следующие компоненты: Xi и хз — концентрация
кислорода в тканях и кровн, Хз и х< — концентрация
молочной кислоты в тканях и крови, Х5—концен-
трация А.ТФ в тканях. Нелинейные уравнения,
описывающие аэробные и анаэробные процессы,
выделены в нелинейный блок F; все остальные
+ У1 + У2- Координату х6
мы поэтому рассматри-
вать в математическом
описании не будем.
Уравнение (6.71) мож-
но записать в компакт-
ной форме, используя
векторно-матричную за-
пись. Блок-схема энерге-
процессы в системе линейны.
тического снабжения организма в таком виде показана на
рис. 6.7.
Здесь х—вектор состояния (х = [xi х2 х3 х4 х5]т), v — вектор
возмущающих входов (и = [nJ), w — вектор задающих входов
(w = [®i]). Матрицы имеют следующий вид:
- kx
о
о
а =
о
о
— k3
k3
о
о
k3
— k3 — k4
kx
— kx — k3
0
0
0-
0
о
о
о _
(6.72)
190
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. f
Нелинейный функциональный преобразователь F изображен для
компактности записи, он образует из вектора состояния х век-
тор потоков у = [t/j у2]т, который поступает на входы компарт-
ментов в соответствии с матрицей R2. Выходной сигнал блока
F и матрица R2 задаются следующим образом:
Однако нас интересует не просто матричное представление,
а возможность такого описания системы, при котором в явном
Рис. 6.8. Уравновешивание первичных и вторичных
темпов в компартментальной модели энергоснаб-
жения организма. Вектор уровней вещества х по-
ступает на вход нелинейного блока F, описываю-
щего химические реакции в системе. Вектор темпов
этих реакций у (размерности 2) вместе с вектором х
(размерности 5) участвует в формировании первич-
ных и вторичных темпов потоков вещества; ма-
трицы А и Р поэтому имеют размерность (5X7).
Стационарное состояние в системе возникает в том
случае, когда вторичные темпы достигают величин,
равных первичным темпам в системе.
темп потребления кислорода в ходе
восстановления АТФ, — k7y\-,
виде выступает уравнове-
шивание первичных и
вторичных темпов пото-
ков.
Для каждого из пяти
компартментов (начиная
с ключевого — пятого)
первичные темпы можно
определить следующим
образом:
для компартмента 5
(уровня АДФ в тканях) —
темп образования АДФ,
непосредственно равный
темпу расхода АТФ в про-
цессах жизнедеятельности
организма, w\
для компартмента 1
(уровня О2 в тканях)—
аэробной реакции (6.68)
для компартмента 2 (О2 в крови)—темп исчезновения кис-
лорода в крови за счет физических процессов его перехода в
ткань, — k\ (х2 — Xt);
для компартмента 3 (МК в тканях) — темп образования мо-
лочной кислоты в ходе анаэробной реакции (6.69), k^y2;
для компартмента 4 (МК в крови)—темп поступления мо-
лочной кислоты в кровь из ткани, £3(х3— х<).
Вторичные темпы процессов изменения уровней вещества в
тех же компартментах определяются следующим образом:
для компартмента 5 — темп утилизации АДФ в ходе как
аэробных, так и анаэробных процессов, —гц — z/2;
для 1 — темп поступления О2 из крови, ki(x2 — Xj);
6 6]
МОДЕЛЬ ЭНЕРГЕТИЧЕСКОЙ СИСТЕМЫ ОРГАНИЗМА
191
для 2 — темп поступления О2 из легких, k2(v— х2);
для 3 — суммарный темп утилизации МК в аэробной реак-
ции,— k^Xi, и переноса МК из ткани в кровь, —й3(х3— хА\
для 4 — темп утилизации МК внутренними органами, —k^xi.
Таким образом, рассмотренная система может быть пред-
ставлена в виде блок-схемы, приведенной на рис. 6.8, где струк-
турные элементы определяются следующим образом. Нелиней-
ный блок химических реакций F по-прежнему задается уравне-
нием (6.73), матрица образования темпов первичных потоков Р
имеет размерность (5X7), так как она преобразует вектор
[xi x2x3x4X5 г/i г/2]т размерности 7 в вектор первичных темпов
размерности 5:
k-7 kt 0 — fel 0 0 0 0 0 0 0 0 0 - 0
р = 0 0 0 0 0 ks 0 — k-s 0 0 0 0 ks 0 (6.74)
0 0 0 0 0 0 0 _
а матрица R размерности (5X0 имеет вид
Я = [0 0 0 0 1]т. (6.75)
Вторичные потоки задаются матрицами А, В и Q размерностью
(5X7), (5X0 и (5 X 1), соответственно:
(6.76)
^2
О
о
В =
<2 = 0.
Из этого примера видно, что первичные и вторичные темпы
отражают химические и физические процессы, происходящие во
всех компартментах системы. Стационарное состояние может
быть достигнуто только в том случае, если все первичные тем-
пы будут уравновешены за счет вторичных.
А как происходит уравновешивание в энергетической си-
стеме организма? Строго говоря, уравновешивание в различных
компартментах наступает не одновременно, и в некоторых «глу-
бинных» компартментах такое уравновешивание может быть
отложено на довольно длительный срок, что связано с опреде-
ленной иерархичностью организации системы. В рассмотренном
192
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
(ГЛ 6
примере сначала равновесие темпов (т. е. стационарное состоя-
ние) достигается в компартменте 5 — темп синтеза АТФ должен
по возможности скорее достичь величины, равной темпу энерго-
трат организма, так как в противном случае будет невозможно
функционирование его систем и органов в заданном режиме.
После достижения равновесия темпов по АТФ процессы накоп-
ления МК и расхода Ог в тканях (компартмента 1 и 3) могут
не сразу достичь стадии уравновешивания. Что касается МК
крови, то здесь равновесие темпов может быть достигнуто еще
позже, так что уровень МК в крови возрастает в течение до-
вольно длительного отрезка времени.
Рассмотренная простая модель не позволяет, впрочем, дать
сколько-нибудь развернутого описания иерархии равновесий в
организме, когда стационарное состояние для более важных
компонент (веществ и энергии в тех или иных иерархически
важных компартментах) устанавливается при изменении усло-
вий среды или функционирования организма раньше, чем в
остальных элементах системы.
Но на одной особенности структуры потоков в энергетиче-
ской системе организма мы все же остановимся. Множество
первичных темпов в системе — вектор размерности 5 — опреде-
ляется по существу заданием лишь одного внешнего сигнала —
темпа w расхода АТФ. Поэтому, если не вдаваться в детали
происходящих внутри системы иерархических процессов «вклю-
чения» различных и все более «глубинных» компартментов (что
видно уже и из рассмотренного примера), то функционирование
энергосистемы организма направлено прежде всего на достиже-
ние заданной интенсивности энергообмена—уравновешивания
первичного темпа траты АТФ. В этом смысле можно сказать,
что энергетическая система является системой управления тем-
пом синтеза АТФ, в которой задающим сигналом является темп
расхода АТФ, определяемый только высшими регулирующими
структурами организма, а управляемым сигналом — темп син-
теза АТФ. Величина управляемого сигнала поддерживается
равной величине задающего сигнала с помощью физиологиче-
ских и биохимических механизмов регуляции.
6.7. Компартментальная модель экологической системы
Одной из ранних моделей реальных экосистем является мо-
дель водного экологического сообщества Силвер Спрингс во
Флориде, разработанная Ю. Одумом [148, 343]. Структура этой
системы иллюстрируется рис. 6.9.
Рассматривая энергетические и другие сложные аспекты су-
ществования экосистем, удобно все организмы делить на произ-
водителей (или продуцентов) органического вещества — это,
6.7]
МОДЕЛЬ экологической системы
193
главным образом, фотосинтезирующие организмы, и на потре-
бителей (консументов), — это прежде всего животные, а также
грибы и многие бактерии. Некоторые экологи выделяют грибы
и бактерии в особую группу, называя их разрушителями орга-
нического вещества (редуцентами). Потребители делятся иногда
на первичных (растительноядных позвоночных животных и на-
секомых) и вторичных — хищников и паразитов, которые живут
Рис. б 9 Экологическая система Силвер Спрингс. В модели рассматриваются следующие
популяции, составляющие экосистему растения (продуценты) — I, травоядные—2, хищники
первого уровня—З, хищники второго уровня—4, редуценты—5. Все величины даны
в ккал/м2 • год Потоки органического вещества в систему поступают через блок 1 — в ре-
зультате процессов фотосинтеза, и через «импорт органического вещества» —хлеб, бросае-
мый в воду туристами.
за счет первых [38, 148]. В частности, в рассматриваемой модели
вводятся следующие группы, плотности которых представляют
собой переменные состояния модели:
xi(Z) — количество продуцентов,
*20) — количество растительноядных,
x3(t)—количество хищников первого уровня,
xC(t)—количество хищников второго уровня,
x3(t)—количество редуцентов.
Эти плотности характеризуются калорическим содержа-
нием — энергией, измеряемой в ккал!м2. Под калорическим со-
держанием в экологии понимается то количество энергии, кото-
рое могло бы выделиться при сгорании данного органического
вещества Начальные калорические содержания переменных в
модели равны Х\ = 3421,6, х3 — 213,44, Хз = 62,06, х^ = 8,87,
х5 = 24,38. Органическое вещество поступает в систему в виде
двух потоков: продуценты производят его в ходе процессов
194
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
фотосинтеза, а растительноядные получают хлеб, бросаемый в
воду туристами. Эти потоки равны, соответственно, wi = 20810,
w2 = 486 (ккал/м2-год).
В [344] в иллюстративных целях предполагается, что обмен
энергией между элементами происходит по линейному закону.
Тогда уравнения системы можно записать в виде
х = Ах + Rw,
(6.77)
где
— Т12 — У 15—^-10—Рю 0 0 0 0 -
Т 12 —Тгз—Р25—Рго 0 0 0
— 0 Тгз — Тз4 — Рз5— Рзо 0 0
0 0 ^34 — Р-15— Рю 0
[115 Р25 И 35 Р-15 —Рзо -
(6.78)
7? = [1 1 0 0 0]т, w = [a>i w2]T, x = [xi х2 ... х5]т,
т — скорость потребления энергии для данного трофического
уровня, р — интенсивность
ность, Л — потери энергии
Рис. 6.10. Блок-схема экологической
системы Силвер Спрингс. В матрич-
Йдм виде система изображается
й компактной форме. Матрица Z?
определяет структуру потоков w
органического вещества, поступаю-
щего в систему, матрица А — все
внутренние свойства системы, вклю-
чая такие характеристики, как темпы
потребления энергии популяциями
экосистемы, темпы их дыхания,
коэффициенты естественной смерт-
ности и потери энергии со стоком
воды.
дыхания, у, — естественная смерт-
со стоком воды. Двойные индексы
использованы для удобства — они
указывают направление потоков
энергии; индекс 0 использован для
среды. Блок-схема системы в мат-
ричном виде приведена на
рис. 6.10.
В [344] эта модель рассматри-
вается как компартментальная мо-
дель экосистемы, включающая пять
компартментов. Здесь же приведены
программы на языке ФОРТРАН
для решения уравнений (6.77) —
(6.78).
Приведем теперь схему компарт-
ментальной модели экосистемы к
интересующему нас виду с явным
заданием первичных и вторичных темпов, как это было проде-
лано в разд. 6.5.
Выделим сначала первичные темпы утилизации энергии в
различных компартментах модели. Рассмотрим, например, ком-
партмент растительноядных организмов, х2. Процессы жизнедея-
тельности в нем в соответствии с уравнением (6.77) требуют
затрат на дыхание, равных —р20х2. Естественная смертность
увеличивает темп потока энергии через компартмент еще на
6.7]
МОДЕЛЬ ЭКОЛОГИЧЕСКОЙ СИСТЕМЫ
195
• -|Л25*2- Эти два слагаемых входят в состав первичного темпа
расхода энергии вне зависимости от того, в каких условиях су-
ществует популяция, хотя их величина и будет зависеть от ус-
ловий окружения. Поскольку популяция растительноядных вхо-
дит в конкретную экосистему, включающую хищников, то она
вынуждена еще увеличить темп потока энергии через компарт-
мент, чтобы выплатить «дань» в виде пищи для хищников пер-
вого уровня. В модели это третье слагаемое первичного темпа
энерготрат принято в виде —ггз^г- Таким образом, первичный
темп энерготрат для компартмента 2 равен (—т2з — р25— Рго)*2-
Аналогичные соображения для других компартментов дают
матрицу Р первичных темпов для общей схемы компартменталь-
ного моделирования, показанной на рис. 6.4, в виде
Р =
— Т12 — PlS—^lO — Р10 0 0 0 0 -
0 — Т23—Р-25 — Р20 0 0 0
0 0 — Тз4 — Р-35— Рзо 0 0
0 0 0 — Ц45 — Р40 0
0 0 0 0 —Рзо -
(6.79)
Для компартмента редуцентов первичный темп есть, очевид-
но, темп их энерготрат на дыхание.
Механизмы регуляции в экосистеме должны обеспечивать
темп поступления энергии в каждый из компартментов. Для
вторичных темпов в схеме рис. 6.4 матрица А тогда имеет вид
- k 0 0 0 0-
Т12 0 0 0 0
А = 0 т2з 0 0 0 (6.80)
0 0 T34 0 0
- Р-15 Ц25 Р35 Ц 45 0 _
Коэффициент k в первой строке определяет здесь продуктив-
ность в компартменте 1; численное значение его можно опреде-
лить, зная общую продукцию органического вещества и кало-
рическое содержание компартмента в стационарном состоянии.
В нашем случае k = 20810/3421,6 = 6,08. Вводя коэффициент k,
мы делаем более реалистическое предположение о величине
потока органического вещества, чем это было в исходном приме-
ре; величина Wi теперь зависит от количества фотосинтезирую-
щих организмов. Остальные элементы матрицы т(/ — темпы
потребления биомассы t-ro компартмента организмами /-го ком-
партмента, i — 1, 2, 3; / = 2, 3, 4. Компартмент, соответствую-
щий организмам-редуцентам 5, удовлетворяет свои энергетиче-
ские потребности за счет темпов отмирания организмов в ком-
партментах 1—4.
196
КОМПАРТМЕНТАЛЬНЫЕ МОДЕЛИ ЖИВЫХ СИСТЕМ
[ГЛ. 6
Кроме того, первичный темп в компартменте 2 частично
уравновешивается за счет темпа поступления органического ве-
щества W2- Конечно, растительноядные организмы экосистемы
могут употреблять в пищу и не весь хлеб, бросаемый в воду
туристами. Однако для простоты мы ограничимся случаем пол-
ного использования потока W2- Тогда матрица Q для вторичных
темпов в уравнении системы имеет вид
Q = [0 1 0 0 0]т- (6.81)
В этом случае блок-схема экосистемы принимает вид, показан-
ный на рис. 6.11, где матрицы А, Р и Q определяются (6.79) —
(6.81).
Система стремится к стационарному состоянию, с одной сто-
роны, изменяя биомассу в различных компартментах и тем са-
Рис. 6.11. Уравновешивание первичных
и вторичных темпов в системе, показан-
ной на рис. 6-9.
мым меняя величину потребно-
стей системы, т. е. ее первичные
темпы, а с другой стороны, стре-
мясь удовлетворить эти темпы
за счет регулирования вторич-
ных темпов.
Эта модель является, конеч-
но, простейшей моделью процес-
сов в экосистеме; ее можно рас-
сматривать как компартменталь-
ную модель распределения энер-
гии. В более сложных случаях
следует учитывать не только
«суммарную» характеристику процессов — калорическое содер-
жание компартментов, но и специфические компоненты си-
стемы. В экологических моделях часто используется специфиче-
ская терминология компартментального анализа. Так, в [58] го-
ворится о «камерных» моделях, где под камерами понимаются
пространственные области.
Модели реальных экосистем получили в последнее время ши-
рокое распространение. Разработаны компартментальные мо-
дели Перуанской системы, Атлантических вод Норвежского
моря, экологической системы Азовского моря [4, 58, 92, 221]. Наи-
более полной из них является, по-видимому, модель Азовского
региона [58]. Акватория региона в этой модели разбита на
7 пространственных областей, для каждой из которых вводится
120 переменных, так что размерность вектора состояния
равна 840,
ЧАСТЬ II
АНАЛИЗ СОХРАНИТЕЛЬНЫХ
СВОЙСТВ БИОСИСТЕМ
В этой части исследуются системы организменного
уровня, на котором сохранительные свойства биоси-
стем проявляются, по-видимому, наиболее полно.
Если условия среды совместимы с жизнью, то как
бы ни менялись характеристики процессов жизнедея-
тельности в физиологических системах, адекватное
снабжение организма, его систем и тканей в среднем
за некоторые достаточно большие промежутки вре-
мени не нарушается, и стационарное неравновесное
состояние сохраняется.
Нарушаться может лишь постоянство внутренней
среды — гомеостаз систем. Поэтому целью анализа в
этой части книги является исследование лишь одной
стороны сохранительных свойств биосистем — гомео-
стаза.
Мы будем иметь дело в основном с физиологией
животного организма — проблемами управления в це-
лостном организме, физиологических системах, в ор-
ганах и тканях. Для биосистем рассматриваемого
типа удобным аппаратом исследования является ме-
тод пространства состояний и компартментальное мо-
делирование. Специфические особенности функциони-
рования физиологических систем позволяют по-новому
взглянуть на вопросы построения их компартменталь-
ных моделей. Традиционные классы физиологических
сигналов получают при построении компартменталь-
ных моделей новые роли, так что формулируемая при
методологическом анализе основная цель биосистем —
поддержание стационарного неравновесного состоя-
ния — становится и формальной целью системы уп-
198
АНАЛИЗ СОХРАНИТЕЛЬНЫХ СВОЙСТВ БИОСИСТЕМ
равления в компартментальных моделях физиологиче-
ских систем.
Достижение и поддержание стационарного нерав-
новесия системы формулируется в терминах теории
управления как обычная задача обеспечения равен-
ства задающих и управляемых сигналов в ее компарт-
ментальной модели, но в роли задающего сигнала те-
перь выступают темпы потоков вещества и энергии,
необходимо расходуемых в процессах жизнедеятельно-
сти, а в роли управляемого сигнала — темпы потоков
вещества и энергии, направленных на восстановление
энергетических трат в системах и органах животного
организма.
Концепция гомеостаза в терминах метода про-
странства состояний получает теперь адекватную
трактовку как сохранение состояния системы при воз-
мущающих воздействиях окружающей среды. Из ка-
чественной категории гомеостаз превращается в ко-
личественную-. появляется возможность численно оце-
нить различные аспекты гомеостатических свойств
систем, их сохранительные ресурсы и приспособитель-
ные возможности.
Картина динамических процессов, протекающих в
компартментальных моделях физиологических систем
в изменяющихся условиях среды, по сравнению со
свойствами классических моделей теории автомати-
ческого регулирования оказывается значительно бо-
гаче и более полно воспроизводит многочисленные ас-
пекты процессов жизнедеятельности физиологических
систем.
ГЛАВА?
МОДЕЛИРОВАНИЕ ГОМЕОСТАТИЧЕСКИХ СВОЙСТВ
ОРГАНИЗМА
7.1. Замечания о моделировании сохранительных свойств
организма и его систем
Угроза животному организму связана со следующими тремя
видами опасности: химическими или физическими воздействиями
среды, нападением «микрохищников»— патогенных бактерий и
вирусов, грибов и простейших организмов, и, наконец, нападе-
нием различного рода «макрохищников». Для защиты от них
организм обладает соответствующими механизмами, начиная
от вегетативных реакций физиологических систем, фагоцитоза,
воспалительных реакций и процессов заживления, явлений им-
мунитета и кончая индивидуальными поведенческими реакциями
и коллективными механизмами защиты [87]. Мы будем иметь
дело только с первым видом опасностей — изменением условий
среды — ис низшим уровнем ответных реакций организма и его
систем — механизмами гомеостаза, осуществляемыми физиоло-
гическими регуляторными системами.
Целью физиологических механизмов управления является со-
хранение стационарного термодинамически неравновесного со-
стояния организма. Если процессы управления на рассматри-
ваемом уровне физиологических систем организованы доста-
точно хорошо, а условия среды не предъявляют чрезмерных
требований к ресурсам системы управления, то кроме обеспече-
ния стационарности возникает возможность поддерживать и го-
меостаз — постоянство условий во внутренней среде. Соотноше-
ние между этими двумя целями в биосистеме было подробно
рассмотрено в разд. 1.7.
Во всем дальнейшем изложении будет использоваться аппа-
рат компартментального описания биосистем. Блок-схема ком-
партментальной модели, в которой стационарное неравновесное
состояние поддерживается при равенстве первичных и вторич-
ных темпов потоков вещества и энергии, была приведена выше
на рис. 6.3 и 6.4. Для систем организма эту схему можно пред-
ставить в более подробном и удобном виде, воспользовавшись
специфическими особенностями систем организменного уровня.
Мы рассмотрим два таких аспекта физиологических систем —
200
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ 7
наличие у них заданного темпа функционирования и существо-
вание специфических активных механизмов регуляции, играю-
щих ведущую роль в процессах управления и самосохране-
ния.
Заданная интенсивность функционирова>
ния. При моделировании физиологических систем обычно ис-
следуется случай неизменности системы во времени, когда про-
цессы онтогенетического развития оказываются за пределами
рассмотрения. Тогда первичные темпы энерготрат определяются
только высшими регулирующими центрами организма и не за-
висят от характеристик самой системы (например, от ее раз-
мера). Физиологические механизмы регуляции не могут изме-
нить величин первичных темпов, установленных центральной ре-
гуляцией в организме.
Так, в условиях основного обмена темпы потребления кис-
лорода, выделения СОг, темп теплопродукции и т. д. опреде-
ляются только режимом функционирования организма. «Уро-
вень основного обмена не является физиологически регулируе-
мой величиной. Он установлен природой и необходим для
поддержания целостности и возбудимости тканей», — пишет
К- П. Иванов [79].
Изменение режима функционирования организма, его физи-
ческая активность при данных условиях среды однозначно опре-
деляет темп обмена веществ в организме и, следовательно, пер-
вичные темпы потоков веществ и энергии во всех компартмен-
тах модели.
Рассмотрим этот вопрос подробнее, вернувшись к принци-
пиальной схеме регуляции в организме животного, изложенной
6 разд. 1.2. Уровень физиологического управления, на котором
обеспечивается гомеостаз и энергетическое снабжение систем
и органов организма, отвечает рис. 1.2, в, где «энергетическая
потребность» определяет должные темпы протекания процессов
в системе.
Поскольку эти темпы являются для системы физиологиче-
ского управления внешними, а их роль состоит в том, чтобы
задать первичные темпы, рассмотренные в разд. 6.4 и 6.5, на-
зовем их задающими, или независимыми темпами. В соответ-
ствии с обозначениями, принятыми в разд. 6.4, для вектора не-
зависимых темпов (размерности пг) будем использовать обо-
значение w.
Тогда все темпы в компартментальных моделях систем ор-
ганизменного уровня можно классифицировать следующим об-
разом. Независимые темпы задают величину первичных темпов
потоков вещества и энергии в компартментах модели, а физио-
логические механизмы регуляции поддерживают величины вто-
ричных темпов равными соответствующим первичным темпам
7.U
МОДЕЛИРОВАНИЕ ОРГАНИЗМА И ЕГО СИСТЕМ
201
(рис. 7.1,а). На рис. 7.1,6 показан пример такой классифика-
ции для случая энергетического снабжения организма, рассмот-
ренного в разд. 6.6.
Активные механизмы регуляции. Другой специ-
фической особенностью физиологических систем является то,
Независимые
темпы
Первичные
темпы
Вторичные
темпы
Зависимые темпы
а)
Глюкоза
Независимый темп Зав и с и м ы в темпы
Рис. 7.1. Соотношение между независимыми н зависимыми темпами в системе, а) Общий
случай; б) пример для модели энергетического снабжения организма. Темп потребле-
ния АТФ порождает множество первичных темпов- сначала темп возникновения АДФ,
затем через механизмы аэробных процессов—первичные темпы утилизации О? и МК, темп
возникновения СО2, а через механизмы анаэробных процессов—первичные темпы возник-
новения МК, Н2О и темпы утилизации глюкозы. В модели, рассмотренной в разд. 6.6, учи-
тывались только темпы О2 и МК, так что из первичных темпов для следующего слоя
звеньев на схеме показаны лишь первичные темпы для компартментов О2 в крови и МК
в крови. Первичные темпы на схеме показаны сплошными линиями, а уравновешивающие
нх вторичные темпы —пунктиром. Темпы вторичных процессов, уравновешивающих пер-
вичные темпы для О2 и МК в крови, не показаны.
что равенство первичных и вторичных темпов обеспечивается
в первую очередь активными механизмами регуляции, а роль
пассивных механизмов относительно невелика. Поскольку функ-
ционирование активных механизмов связано с расходованием
метаболической энергии, их участие в процессе регулирования
приводит к дополнительному возрастанию темпов энерготрат в
системе. Так, если животное от режима отдыха переходит к
поиску пищи, то темпы потребления энергии повышаются в связи
с увеличившейся мышечной активностью. Чтобы удовлетворить
эту потребность, в организме интенсифицируется дыхание и кро-
вообращение, что приводит к дополнительному расходу энергии.
202
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
Соответствующая блок-схема приведена на рис. 7.2. Актив-
ные механизмы регуляции расходуют относительно мало энер-
гии, что отражено, в частности, в рассмотренной в гл. 4 концеп-
ции энергетической оптимальности биосистем. Можно назвать
wz
Темпы
знереотрот на
функционирование
активных
механизмов
w
г
v
Исходные
независимые
темпы
j. Формирование \дкцштая регуляща Формирование
уровней I я потоков
потоков
___^Пассивнаярегуляция
Зависимые темпы
У
У
0
Рис, 7.2. Регуляция темпов потоков вещества и энергии в физиологических системах;
а) Включение механизмов активной регуляций увеличивает независимые темпы энерготрат,
о) схематическое представление в виде системы Управления, где входным сигналом является
вектор независимых темпов (матрица /? определяет, на входы каких компартментов посту-
пают сигн-алы и»), а выходным —вектор зависимых (первичных и вторичных) темпов у,
уравновешивающих задающие темпы w.
много интересных факторов, приводящих к уменьшению расхо-
дов энергии на функционирование активных механизмов [249,
250]. Так, дыхательные движения у некоторых видов птиц про-
исходят на резонансной частоте, что энергетически наиболее
выгодно [284]. Только процессы, вызванные резкими измене-
ниями среды или функционирования организма, приводят к
существенным дополнительным энергетическим расходам. Сум-
марные темпы энерготрат с учетом потребностей активных ме-
ханизмов на рис. 7.2, а значительно отличаются от исходных
только в процессе адаптации и перестройки режимов [146, 153,
166, 201]. Кроме того, такие активные механизмы регуляции, как
7.П
МОДЕЛИРОВАНИЕ ОРГАНИЗМА И ЕГО СИСТЕМ
203
система дыхания и кровообращения, «выключить» нельзя, и
поэтому их вклад в суммарные темпы в какой-то мере суще-
ствует всегда. Поэтому во многих случаях анализа физиологи-
ческих систем достаточно рассматривать более простую схему
компартментальной модели, приведенную на рис. 7.2, б.
Эта схема представляет собой систему управления, в кото-
рой роль задающего сигнала играют независимые темпы w, а
роль регулируемого сигнала — зависимые темпы у. Вектор со-
стояния х есть совокупность уровней вещества и энергии в ком-
партментах модели, а сигнал и, описывающий условия внешней
среды, играет роль возмущающего сигнала.
Жизнедеятельность системы поддерживается в том случае,
если в схеме рис. 7.2 обеспечивается стационарный режим, т. е.
достигнуто равенство
z/ = w. (7.1)
В этой системе в принципе допустимы любые величины х, при
которых выполняется (7.1), так что множество значений х,
при которых возможен стационарный режим, можно трактовать
как диапазон удовлетворения в физиологической системе в со-
ответствии с концепциями принципа удовлетворения, рассмот-
ренными в разд. 4.5. Однако в физиологических системах в ста-
ционарном режиме реализуются далеко не все возможные зна-
чения х из этого диапазона. При изменении внешних условий
регуляторные механизмы стремятся поддерживать значение век-
тора состояния на относительно постоянном уровне. Лишь край-
не неблагоприятные условия, граничащие с возможностями
жизнедеятельности (в физиологии такие условия называются
экстремальными), приводят к выходу значений х на границы
диапазона удовлетворения.
Область изменения условий среды, в которой стационар-
ные значения существенных переменных системы малочувстви-
тельны к изменениям v, мы назвали в разд. 1.7 областью гомео-
стаза, а более широкую область, в которой существует стацио-
нарный режим — областью стационарности Qs. Теперь мы
проиллюстрируем связь, существующую между гомеостатической
кривой, рассмотренной в разд. 2.3, и расположением областей
Н5 И
Пусть на рис. 7.3 вектор v = [щ у2]т описывает условия внеш-
ней среды, а х — некоторая существенная координата вектора
состояния. Если зафиксировать значение v2 = v*2, то при изме-
нении значения щ в пределах от а до d в системе существует
стационарный режим; зависимость x = x(vi) в стационарном
режиме показана на рис. 7.3, а. Эта зависимость имеет типич-
ный вид кривой с плато; область плато как раз и соответствует
204
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
области гомеостаза — интервалу (Ь, с) на рис. 7.3, а и заштри-
хованной области на рис. 7.3, б.
Таким образом, для области гомеостаза характерна относи-
тельная независимость переменных системы от условий внешней
среды. Однако сам термин «относительная независимость» мож-
но трактовать по-разному. Поэтому при исследовании гомео-
Рис. 7.3. Область стационарности и
область гомеостаза системы. В обла-
сти £2^ поддерживается стационар-
ный режим системы, причем стацио-
нарное значение переменной х опре-
деляется величинами и и2: х—
—f (Oi, fs), и переменная х изменяется
в пределах удовлетворения (а). В об-
ласти гомеостаза переменная х
мало зависит от изменений вектора и*
Показана гомеостатическая кривая
для переменной х при v2==const (а)
и расположение областей £2 и (б).
кальные гомеостатические
стаза возникло несколько направле-
ний, основанных на различном по-
нимании того, что называется ела-
бой зависимостью переменных си-
стемы от условий среды. В следую-
щем разделе мы рассмотрим эти
трактовки более подробно, а пока
обратимся еще раз к гомеостатиче-
ской кривой на рис. 7.3, а. Можно
сказать, что эта зависимость харак-
теризует сохранительные способно-
сти системы с двух различных точек
зрения.
Локальные характеристики го-
меостаза должны отражать поведе-
ние системы при данных условиях
внешней среды и описывать реакции
системы в ответ на малые измене-
ния внешних условий. Локальные
гомеостатические свойства опреде-
ляются тем, насколько полого идет
график гомеостатической кривой при
данном значении v = v*. Чем более
полого идет гомеостатическая кри-
вая в данной точке, тем лучше ло-
свойства. Можно сказать поэтому,
что локальные гомеостатические свойства системы в точке
v = v* определяются чувствительностью стационарного значе-
ния переменной х к v в этой точке.
Интегральные характеристики гомеостаза системы связаны
с общим характером гомеостатической кривой. Они определяют
такие свойства системы, как общий вид и ширина участка плато
(размеры области, в которых система может поддерживать го-
меостаз и качество гомеостаза в этой области). Интегральные
характеристики отражают сохранительные свойства системы
при широком изменении условий среды, ее способность приспо-
сабливаться к таким изменениям. Ниже в этой главе мы не бу-
дем касаться интегрального аспекта гомеостатических свойств
системы, оставив его до гл. 8, а изложение в следующих разде-
лах главы относится к локальным гомеостатическим свойствам.
7 2]
ДВА ПОДХОДА К МОДЕЛИРОВАНИЮ БИОСИСТЕМ
205
По самому своему смыслу локальность гомеостатических
свойств означает, что исследователя интересуют лишь малые
сдвиги условий среды в окрестности заданной точки v = и*. Сле-
довательно, для устойчивых систем мы будем иметь дело и с
малыми сдвигами векторов состояния х и выходов у. Поэтому
при анализе локальных гомеостатических свойств системы мож-
но пользоваться линейным приближением, и векторные уравне-
ния «вход-выход-состояние» будут линейными уравнениями вида
(5.39).
7.2. Два подхода к моделированию биосистем
J Если рассматривать существующие в настоящее время мо-
дели биосистем на всех уровнях организации жизни под углом
зрения теории управления, то их условно можно разделить на
две большие группы.
' Первая группа моделей находится в непосредственной связи
с классической теорией управления. Наибольшее распростране-
ние такие модели получили при исследовании систем организ-
менного уровня. Общая схема такой модели представляет либо
непосредственно систему регулирования с отрицательной обрат-
ной связью по отклонению (см. разд. 2.2, рис. 2.5), либо ее обоб-
щение на случай многосвязной системы [5, 60, 130, 154, 259а].
Основная идея построения таких моделей состоит в задании
определенных требований, которым должны отвечать выходные
переменные системы. Обычно это требование равенства стацио-
нарных значений некоторых существенных переменных, выбран-
ных в качестве выходного сигнала, заранее заданным опорным
значениям или уставкам. Выбранные переменные при этом иг-
рают роль регулируемого или управляемого сигнала в системе
управления. *
В качестве выходного сигнала обычно выступают концентра-
ции веществ — глюкозы, кислорода, углекислоты, водородных
ионов, гормонов [13, 60, 70, 243, 261], температура (концентра-
ция тепловой энергии) [74, 84, 186] или некоторые другие био-
логические переменные в организме — например, артериальное
давление, давление в различных отделах сердца и т. д. [6—8,
105, 157, 214].
Таким образом, для первой группы моделей прежде всего
характерна идея о регуляции уровней вещества и энергии в си-
стеме или же непосредственно связанных с ними переменных
(таких, как давление). Если х— некоторый уровень вещества,
рассматриваемый как выход системы, х — его опорное значе-
ние — уставка, то для моделей первого типа в стационарном
режиме можно получить
(7.2)
Х = X.
206
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
Во вторую группу мы выделим модели, при построении ко-
торых интерес исследователя заключается в описании не столь-
ко уровней, сколько темпов изменения переменных, описываю-
щих биосистему.
Если вектор состояния биосистемы есть х, то внимание ис-
следователя сосредотачивается на поиске зависимостей
x = f(x). (7.3)
Такие модели широко распространены для всех видов ис-
следования биосистем — экологического, физиологического, био-
химического. Наиболее распространенным их типом являются
компартментальные модели (см., например, разд. 6.6 и 6.7).
Особо следует выделить модели второй группы для физиоло-
гических систем, хотя они составляют относительно малую долю
всех разработанных моделей систем организменного и суборга-
низменного уровня. Это, например, модели почечной функции
[330], кислородного снабжения [83], системы теплового режима
[2Н], модели поджелудочной железы [262]. Большинство же
моделей функционирования организма и его систем строится по
принципу классической схемы управления (см. разд. 3.4). »
v Для моделей второй группы основное внимание уделяется
темпам процессов в системе — скоростям обменных реакций на
клеточном уровне, потокам и их скоростям — на уровне орга-
низма и его систем, динамике изменения численности популя-
ций — на надорганизменном уровне [349]. В качестве цели, обес-
печиваемой механизмами регуляции в таких моделях, называет-
ся обеспечение стационарного режима, т. е. равенства темпов
поступления и расходования составляющего систему веще-
ства— первичных и вторичных темпов. Такое равенство может
достигаться или при постоянных во времени значениях перемен-
ных или «в среднем» — в ходе колебательных процессов. Что
касается переменных состояния, то при построении моделей по-
лучению каких-либо специфических свойств или характеристик
переменных состояния часто не уделяется достаточного вни-
мания.
• Впрочем, в последнее время в ряде работ по системам био-
химического уровня проводится мысль о том, что «хорошая»
регуляция темпов протекания процессов в сложных биохимиче-
ских комплексах обеспечивает не только стационарность состоя-
ния системы, но и постоянство концентраций при изменяющихся
условиях среды [72].
Разницу между двумя этими подходами при моделировании
систем подчеркивал еще Л. фон Берталанфи, считавший, что ме-
тоды кибернетики (и теории управления) применимы для по-
строения моделей первого типа, а модели второго типа должны
описываться в рамках теории открытых систем [31]. Из выше-
7.2]
ДВА ПОДХОДА К МОДЕЛИРОВАНИЮ БИОСИСТЕМ
207
изложенного, однако, ясно, что и открытые системы могут ус-
пешно исследоваться методами теории управления, хотя есте-
ственным аппаратом их описания являются не столько приемы
классической теории управления, сколько методы пространства
состояний.
Сравним теперь оба рассмотренных подхода с позиций тео-
рии управления, обращая внимание прежде всего на те роли,
которые играют в модели различные группы переменных, опи-
сывающих биосистему.
Напомним, что при описании физиологических систем мы
можем выделить следующие основные группы переменных:
1) уровни вещества и энергии в системе и ее компартментах
(концентрации или количества);
2) независимые темпы процессов — скорости утилизации ве-
щества и энергии в системе и ее компартментах;
3) зависимые темпы — первичные и вторичные темпы или
скорости транспорта веществ в системе и ее компартментах;
4) состояние окружающей среды.
В табл. 7.1 и на рис. 7.4 показано распределение ролей этих
групп переменных в модели классической теории автоматиче-
ского регулирования — схеме управления с отрицательной об-
ратной связью по рассогласованию—и в компартментальной
модели открытой системы. Напомним, что в классической схеме
используется представление «вход-выход», а компартментальная
модель строится в соответствии с методом пространства состоя-
ний и описывается уравнениями «вход-выход-состояние».
Принципиальное различие между двумя способами распре-
деления ролей переменных в моделях с точки зрения теории уп-
равления состоит в выборе задающих входных сигналов. Если
в классической модели задающий входной сигнал определяет
желаемые значения уровня вещества в системе и вводится до-
полнительно, образуя новую специфическую группу перемен-
ных, то в моделях открытых систем задающими сигналами счи-
таются необходимые для существования и жизнедеятельности
системы темпы потоков вещества и энергии. Остальные сиг-
налы распределяются в соответствии с этим главным положе-
нием.
Конечно, эта схема является лишь «скелетом», на котором
строятся конкретные модели биосистем, и поэтому она не мо-
жет отразить многих их характерных черт. Так, в схеме клас-
сической теории управления в качестве выходных переменных
выступают далеко не все уровни вещества и энергии; обычно
выбирается один из них (например, на физиологическом уров-
не— глюкоза, углекислота, кислород и т. д.), а остальные яв-
ляются «внутренними» переменными системы, т. е. в соответ-
ствии с классическим рецептом они не должны включаться в
208
ГОМЕОСТАЗ ОРГАНИЗМА
1ГЛ. 7
модель, а их суммарной характеристикой должна быть переда-
точная функция высокого порядка от рассматриваемого входа
к данному выходу. Однако в конкретных моделях физиологиче-
ских систем исследователи не пошли по этому пути, и их все
более сложные схемы постепенно отходили от простых класси-
ческих канонов. Поэтому, хотя методологически уставка все еще
продолжает играть принципиально определяющую роль в таких
моделях, фактически ее влияние на процессы в системе тем
меньше, чем больше «внутренних», незастабилизированных пере-
менных в модели.
ч) б)
Рис. 7.4. Роль различных переменных в моделях физиологических систем, а) В схеме клас-
сической теории управления: задающий входной сигнал—специально вводимые опорные
значениях (уставки), выходной регулируемый сигнал —значения соответствующих уровней,
концентраций х, возмущающие входные сигналы — состояние среды и и темпы потребления
вещества (иитеисивиость функционирования) w. Зависимые темпы у играют роль одного
из управляющих сигналов, 6) в схеме открытой системы задающий входной сигнал —не-
зависимые темпы w, выходной регулируемый сигнал — зависимые темпы у, возмущающий
входной сигнал —v. Уровни вещества и энергии —вектор состояния х — играют роль одного
из управляющих сигналов (пассивное управление) наряду с ие показанными на схеме
активными управляющими механизмами. На рисунках использованы обозначения: Р —ре-
гулятор, О —объект управления.
Другим фактором, не учитываемым в схематической табл. 7.1,
является наличие специальных активных регулирующих меха-
низмов, которыми должны быть дополнены обе схемы на
рис. 7.4. В табл. 7.1 в роли управляющих сигналов классиче-
ской схемы выступают зависимые темпы. При этом имеется
в виду, что их «управляющая функция» состоит в том, что они
участвуют в образовании регулируемых уровней. В компартмен-
тальной модели в качестве управляющего сигнала выступает
вектор состояния, выполняющий функцию пассивного регуля-
тора темпов.
Вопрос о том, какая из двух возможных схем дает более
адекватное описание процессов регуляции, составляет предмет
обсуждения специалистов по многим физиологическим систе-
мам. Во многих случаях современная физиология пока скло-
няется к выбору классической схемы, показанной на рис. 7.4, а.
Кроме традиции, здесь, вероятно, не последнее место занимает
и то, что в моделях классического типа проблема гомеостаза по-
лучает наиболее простое толкование.
Однако и схема компартментальной модели открытой си-
стемы, где в качестве регулируемого сигнала выступают темпы
7.21
ДВА ПОДХОДА К МОДЕЛИРОВАНИЮ БИОСИСТЕМ
209
Таблица 7.1
Переменные в биологической системе Роль переменных в модели биосистемы
В схеме классической тео рии управления В модели открытой системы при использовании компарт- ментальиого представления
1 Уровни вещества н энер- 1ИИ Выходной сигнал (управ- ляемые, регулируемые переменные) Переменные состояния
2 Независимые темпы утилиза- ции вещества и энергии Возмущающий вход- ной сигнал Задающий входной сигнал
3 Зависимые темпы доставки вещества и энергии «Внутренний» управля- ющий сигнал Выходной сигнал
4 Переменные внешней среды Возмущающий входной сигнал Возмущающий входной сигнал
5 Специальный сигнал, задаю- щий «желаемые» уровни вещества и энергии Задающий входной сиг- нал (опорный сигнал, уставка) В модели не нужен
Рис. 7.5. Режим натрия в организме Интегратор / определяет количество натрия в орга-
низме, которое после умножения на коэффициент ki дает концентрацию Na, обозначаемую
через <?Na. В зависимости от cj>ja в поджелудочной железе секретируется альдостерон.
Накапливаясь на интеграторе 2, темпы потоков альдостерона дают его количество и кон-
центрацию Сддд» под Действием которой находятся процессы выделения Na в почках.
Схема представляет собой компартментальную модель открытой системы, ко в кей отсут-
ствуют сигналы, играющие роль независимых темпов расхоювания Na, необходимых для
жизнедеятельности организма.
210
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
процессов, постепенно становится равноправной моделью физио»
логических систем. Характерной в этом отношении является по-
зиция, занятая известным американским физиологом А. Гайто-
ном, изложенная им в справочнике по медицинской физиологии
[304]. Описывая роль отрицательных обратных связей в орга-
низме человека, он подчеркивает: «Концентрация, темп процес-
са или количество вещества — вот те величины, которые долж-
ны регулироваться» [304, стр. 9].
В качестве примера систем управления в организме наряду
с классическими схемами регулирования по уставке в этом
справочнике приводится и схема режима натрия в организме
(рис. 7.5). Легко видеть, что по структуре она полностью соот-
ветствует модели открытой системы, а процессы управления в
ней направлены прежде всего на достижение равенства темпов
поступления и удаления натрия. Однако обозначение сигналов
в схеме все еще тяготеет к традиционной физиологической схе-
ме — классической системе регулирования по отклонению от
уставки. Темп поступления натрия рассматривается как возму-
щающий сигнал, а количество натрия в крови — как выходной,
управляемый. Таким образом, схема регуляции натриевого ре-
жима занимает «промежуточное» положение между классиче-
ской схемой и компартментальной моделью открытой системы,
хотя и ближе ко второй.
7.3. Моделирование гомеостаза
методами классической теории
автоматического регулирования
При моделировании гомеостаза в рамках классических пред-
ставлений делается следующее допущение: если некоторая пе-
ременная в биосистеме незначительно меняется при изменении
условий внешней среды, то она является выходной переменной
в некоторой системе регулирования, специально предназначен-
ной для поддержания постоянства этой переменной. Если в био-
системе, например в организме, поддерживается постоянство
множества переменных внутренней среды, то модель такой си-
стемы содержит множество систем регулирования, стабилизи-
рующих каждую из переменных внутренней среды [33, 40, 130].
Иногда все эти системы рассматриваются не порознь, а как
единая многосвязная система [259а].
Если система стабилизации (как называют систему автома-
тического управления с постоянным значением задающего сиг-
нала) устойчива, то в стационарном режиме значения всех вы-
ходных сигналов равны своим предписанным значениям; вне
зависимости от условий внешней среды и темпов расхода ве-
ществ в биосистеме выполняется условие (7.2).
7 3]
ГОМЕОСТАЗ И МЕТОДЫ КЛАССИЧЕСКОЙ ТЕОРИИ
211
В классической модели устойчивость системы автоматиче-
ского регулирования эквивалентна наличию гомеостаза выход-
ной переменной. Именно поэтому в литературе гомеостаз иногда
отождествляется с устойчивостью. В явной форме такое поло-
жение высказывается, например, в [175, 206]; в неявном же
виде оно встречается у многих авторов.
Условие (7.2) должно выполняться при вариациях v и изме-
нении режимов функционирования w. Впрочем, в некоторых мо-
делях в стационарном состоянии это условие не выполняется, и
система имеет установившуюся ошибку; такова, в частности,
рассмотренная выше модель дыхательного хемостата Ф. Гро-
динза.
Модель простой системы автоматического регулирования по
рассогласованию описывает лишь самый низкий уровень иерар-
хической системы управляющих механизмов [33, 34, 216]. По-
этому в работах С. Н. Брайнеса и В. Б. Свечинского [33, 34]
была предложена модель трехуровневой системы управления в
целостном организме, основанная на представлении гомеостаза
в рамках классической теории управления. Суть этого подхода
состоит в следующем.
Внутренняя среда организма характеризуется рядом физио-
логических показателей, обычно поддерживаемых в нем на при-
мерно постоянном уровне. В работе [34] эти показатели назы-
ваются параметрами; в терминах теории управления их обычно
называют переменными, что мы и будем делить ниже. Тогда в
пространстве значений этих переменных можно выделить об-
ласть, в которой еще возможно протекание жизненно важных
процессов в организме. Каждому состоянию организма в этой
области соответствует оптимальный набор значений переменных,
при котором достигает минимума или максимума некоторый
критерий оптимальности, учитывающий необходимый приток
энергии, расход ресурсов организма и т. д. Тогда для управле-
ния внутренней средой организма требуется следующая после-
довательность операций:
1. Определение критерия оптимальности с учетом внешних
условий и состояния организма.
2. Нахождение оптимальных значений переменных внутрен-
ней среды.
3. Поддержание требуемых значений физиологических пере-
менных в системах автоматической стабилизации.
В соответствии с решаемыми задачами структура системы
управления в целостном организме включает три этажа, выпол-
няющих указанные функции. Системы регулирования, находя-
щиеся на нижнем этаже, связаны между собой, так что факти-
чески на постоянном уровне должна поддерживаться не одна,
а сразу несколько переменных, что затрудняет работу системы.
212
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
Пусть, например, оптимальному режиму организма соответ-
ствует набор должных значений переменных (уставок в систе-
мах регулирования) х. Однако взаимное влияние переменных
не позволяет достичь одновременно всех должных значений.
Тогда цель нижнего уровня системы управления может быть
сформулирована как поиск минимума суммарной среднеквадра-
тической ошибки Q:
т
Q = min етЛе = min О/(х/— X;)2, (7.4)
х х i » 1
где е = х — х — вектор ошибок рассогласования, А — diag а, —
весовая матрица, учитывающая относительную важность по-
стоянства различных переменных для организма.
Схема моделирования гомеостаза в виде системы автомати-
ческого регулирования с отрицательной обратной связью по
рассогласованию, однако, обладает рядом недостатков, кото-
рые затрудняют ее использование во многих прикладных медико-
биологических задачах. Эти недостатки можно сформулировать
следующим образом [253].
а) Модель применима только в узкой области
изменения условий функционирования. В живых
системах постоянство переменных внутренней среды является
относительным; вариации внешних условий v и режима ра-
боты — вектора w изменяют стационарное состояние биоси-
стемы. Классическая модель в принципе обеспечивает «абсо-
лютное» постоянство выходных сигналов (7.2), так что модель
может быть адекватной исходной биосистеме только в обла-
сти гомеостаза, но не во всей области стационарности (см.
рис. 7.3).
Попытки приблизить эту модель к реальным условиям де-
лались в двух направлениях. С одной стороны, можно выбрать
такую структуру системы, что, несмотря на наличие опорного
значения, выходной сигнал не всегда ему равен. В этом слу-
чае не очень ясно, зачем вообще вводить опорное значение в
систему, и разработку модели можно оправдать лишь привер-
женностью к общепринятой схеме. С другой стороны, можно
попытаться ввести зависимость опорного значения от условий
среды и от режима функционирования биосистемы. Для орга-
низма такой прием в отдельных случаях оказался успешным
[50, 157, 214], но попытка найти общий метод определения такой
зависимости уводит нас от модели гомеостаза как системы ре-
гулирования по рассогласованию [253]. Не оправдала себя та-
кая попытка, например, в моделях системы терморегуляции
[31а].
Критическое отношение к уставке как величине, определяю-
щей неизменное во времени значение переменных внутренней
7.31
ГОМЕОСТАЗ И МЕТОДЫ КЛАССИЧЕСКОЙ ТЕОРИИ
213
среды, во многом обязано прогрессу измерительной техники в
физиологическом эксперименте. Появившаяся в последние годы
возможность точного и длительного измерения ряда перемен-
ных внутренней среды позволила по-новому взглянуть на казав-
шиеся незыблемыми представления об «абсолютном» характере
постоянства внутренней среды.
Так, концентрация глюкозы крови в течение суток весьма
существенно меняется как у нормальных людей, так и у лиц,
страдающих диабетом ([261], рис. 7.6). Если человек начинает
Время суток, часа
Рис. 7.6. Непрерывная запись уровня сахара крови у диабетического больного в течение
суток. Уровень сахара изменяется в широких пределах. Стрелками показаны принятые
в Канаде моменты приема пищи (завтрак, ленч, закуска, обед и еще одна закуска). Воз-
можность точной и непрерывной записи переменных разрушила,чиф об суднвительиом по-
стоянстве внутренней среды оргаинзма>, будь то здоровый человек или диабетический
больной.
употреблять пищу с повышенным содержанием сахара, то в те-
чение нескольких недель содержание сахара в крови остается
повышенным. Лишь после того, как в процессе адаптации ги-
пертрофируется поджелудочная железа, регулирующая режим
глюкозы, избыток сахара переходит из крови в клеточную ткань
[304]. В экспериментах на животных [311] выяснилось, что изме-
нение темпов введения глюкозы в кровь приводит к закономер-
ному и резко выраженному изменению стационарных концент-
раций глюкозы крови. В экспериментах по терморегуляции было
обнаружено, что даже температура гипоталамуса — «святая свя-
тых» системы терморегуляции — при изменении внешних усло-
вий закономерно изменяется [79, 182]. Что касается системы ре-
гуляции глюкозы, то попытки представить результаты упомяну-
тых экспериментов в виде классической схемы привели лишь к
неоправданному и малоправдоподобному ее усложнению [369].
б) Вопрос о структурном соответствии меха-
низмов регуляции в биосистеме и ее модели
остается открытым. В первых работах по использованию
системы регулирования по рассогласованию в качестве моделей
214
ГОМЕОСТАЗ ОРГАНИЗМА
1ГЛ. 7
биосистем четко отмечалось, что опорный сигнал вводится ав-
тором модели только для удобства описания. При этом не важ-
но, существует ли в реальной системе сигнал уставки, и если
существует, то чему он равен [60]. Поэтому, моделируя биоси-
стему, составленную из одних и тех же элементов, разные ав-
торы вводили уставки по разным переменным. Так, физиологи-
ческие модели, включающие элементы метаболизма тканей и
элементы, описывающие их снабжение кровью и кислородом,
были рассмотрены в работах [214, 301]. Но авторов работы
[301] интересует прежде всего система дыхания, и они вводят
уставку по концентрациям веществ в системе дыхания; в ра-
боте [214] главное внимание уделяется кровообращению, и ус-
тавка вводится по его переменной — артериальному давлению,
причем опорное значение меняется с изменением режима функ-
ционирования организма. Таким образом, в этих работах устав-
ка как бы заменяет немоделируемые части системы; «... уставка
является лишь концентрированным выражением всей совокуп-
ности воздействия организма на рассматриваемую систему»
[253].
С другой стороны, имеется группа работ, в которых делается
попытка осмыслить, как же образуется опорное значение для
тех или иных переменных в организме [216, 253, 295, 362]. В ча-
стности, считается, что при «более широком понимании уста-
новочная точка аналогична константе равновесия обратимой хи-
мической реакции» [216, стр. 28]. Тем самым подчеркивается, что
установочная точка — уставка — не столько является чем-то на-
вязанным системе извне, сколько формируется в самой системе
под влиянием взаимодействия ее компонент, а ее величина за-
висит от структуры системы. Например, для системы ориента-
ции у рыб установочная точка определяется функциональной
анатомией соответствующих органов чувств [216]. В системе тер-
морегуляции поддержание нормальной температуры обеспечи-
вается одновременным функционированием механизмов пото-
отделения, дрожи, сосудистых реакций и т. д., так что вся эта
система, благодаря своей структуре, автоматически смещается
к такому значению температуры, при котором суммарный темп
теплопродукции равен темпу суммарной теплобтдачи [295, 362].
Эта равновесная температура и играет роль установочной точки
в системе терморегуляции. Таким образом, в этих работах не
предполагается, что все элементы модели рис. 7.4, а реально су-
ществуют в биологическом прототипе. Наконец, в ряде работ
в свое время допускалось, что сравнивающие устройства (или
их аналоги) и опорные значения могут реально существовать
в организме [9, 34, 95].
Анализ недостатков схемы классической теории регулирова-
ния с уставкой можно закончить цитатой, принадлежащей из-
7 4]
ГОМЕОСТАЗ И МЕТОДЫ ПРОСТРАНСТВА СОСТОЯНИЙ
215
вестному американскому специалисту в области терморегуляции
К. Куперу. Обсуждая концепцию уставки в терморегуляции, он
задает вопрос, не являются ли подобные термины «превосход-
ным примером жаргонного словечка, которое закрывает путь
дальнейшим исследованиям и приводит к их свертыванию,
или же они на самом деле имеют какую-то реальную цен-
ность?» [283].
7.4. Моделирование гомеостаза методами
пространства состояний
В этом разделе мы рассмотрим, как можно описывать гомео-
статические свойства биосистем на основе компартментальных
моделей открытого типа (рис. 7.4, б).
Основное отличие этих моделей от систем, с которыми мы
имели дело в предыдущем разделе, состоит в том, что при выбо-
ре структуры модели никаких условий на «желаемое» поведение
переменных заранее не накладывается. Если в моделях с отри-
цательной обратной связью по рассогласованию заранее огова-
ривается, что интересующий нас сигнал будет постоянным во
всех стационарных режимах при любых изменениях внешней
среды (или, что непринципиально, будет меняться при таких ва-
риациях заранее оговоренным образом), то единственное свой-
ство, в котором мы уверены при исследовании компартменталь-
ной модели, — это то, что в стационарном режиме темпы
поступления веществ и энергии равны темпам их утилизации в
системе. Что касается стационарных значений переменных со-
стояния, то их поведение, вообще говоря, может быть самым
различным.
Поэтому главной задачей исследований на этом этапе яв-
ляется выявление таких структур компартментальных моделей,
в которых стационарные значения переменных в системе мало
зависят от изменений условий внешней среды, т. е. система об-
ладает гомеостатическими свойствами.
При моделировании гомеостатических свойств физиологиче-
ских систем мы интересуемся, в основном, стационарными ре-
жимами их работы. Это объясняется двумя обстоятельствами:
во-первых, именно такие режимы характерны для систем орга-
низма [271, 333, 334] и, во-вторых, с математической точки зре-
ния изучение стационарных режимов значительно удобнее.
Сформулируем условия, которым должна удовлетворять ис-
следуемая система, чтобы к ней можно было приложить разви-
ваемую концепцию анализа гомеостатических свойств в терми-
нах пространства состояний.
1. Исходная система уравнений должна быть линеаризуема
во всей области изменения входных сигналов и переменных
216
ГОМЕОСТАЗ ОРГАНИЗМА
1ГЛ. 7
состояния. (При строгом математическом рассмотрении считает-
ся, что система должна быть линеаризуема во всей этой области,
за исключением отдельных точек, в которых система может не
допускать линеаризации. В биологических системах обычно
нельзя проводить линеаризацию в тех точках, где происходит
«включение» механизмов активной регуляции, например, в си-
стеме терморегуляции организма — включение механизмов со-
судистых реакций или дрожи.)
2. Система должна допускать исследование на устойчивость
по первому приближению (как об этом говорилось в разд. 5.9).
3. Внешние сигналы, действующие на систему, стремятся к
постоянным значениям при t—> оо.
4. Во всех точках области устойчивости исследуемой систе-
мы существует стационарный режим, когда при t -> оо пере-
менные состояния стремятся к некоторым постоянным значе-
ниям.
Определим теперь, что будет пониматься под гомеостатиче-
скими свойствами системы. Рассмотрим два стационарных ре-
жима системы, возникающих при двух различных постоянных
значениях вектора входов v, обозначенных через а? и v2—
— vl Av.
В обоих случаях в системе при t -> оо устанавливаются ста-
ционарные значения вектора состояния х, так что потоки ком-
понент во всех компартментах уравновешены.
Однако поскольку условия среды v1 и v2 различны, будут
различны, вообще говоря, и векторы состояния х в обоих слу-
чаях. Обозначим стационарные значения вектора состояний,
отвечающие значениям v1 и v2, через хг и х2, соответственно,
причем положим х2 = х1 + Ах.
Рассмотрим какую-нибудь одну переменную вектора состоя-
ния, например х,-, и одну переменную вектора условий среды,
например V/. Тогда можно сказать, что при изменении условий
среды
Avy = v2 — v’ (7.5)
переменная состояния х, получила приращение
Ьх{ = х2 — х’, (7.6)
причем при достаточно малых Av/
Axy = o(/Av/, (7.7)
где
дх.
Т.4) ГОМЕОСТАЗ И МЕТОДЫ ПРОСТРАНСТВА СОСТОЯНИИ 217
представляет собой коэффициент чувствительности переменной
Xi к возмущению V/. Если величина а,/ мала,
< 1, (7.9)
то будем говорить, что переменная xt обладает локальными го-
меостатическими свойствами при изменении условий внешней
среды Vj.
Поскольку в нелинейной системе величина щ, различна при
разных значениях v, т. е.
= <yz/(v), (7.10)
то переменная xt обладает локальными гомеостатическими свой-
ствами в тех точках v, где выполняется условие (7.9).
Рассмотрим теперь, как можно охарактеризовать гомеоста-
тические свойства системы в целом. Коэффициенты чувствитель-
ности переменных системы составляют матрицу коэффициентов
чувствительности:
[о0] = - Л-’В, (7.11)
где А, В — матрицы линеаризованной модели системы. В идеа-
ле хороший гомеостаз системы мог бы означать, что все элемен-
ты матрицы (7.11) малы. Однако этого не приходится ожидать
для моделей реальных систем. Поэтому мы будем говорить, что
система обладает гомеостатическими свойствами в том случае,
когда условия (7.9) выполнены для существенных переменных
системы. Что понимать под существенными переменными, сле-
дует уточнить в каждом конкретном случае. Ниже мы сделаем
попытку охарактеризовать «существенность» переменных, т. е.
их вес в гомеостазе целостной системы, с помощью специально
выбираемых коэффициентов (см. ниже разд. 8.1).
Остановимся вкратце на вопросе о том, почему гомеостаз
системы связывается с малой чувствительностью вектора со-
стояния системы, а не вектора выходов. Дело в том, что внешне
поведение системы может быть стабильным, т. е. выходные пе-
ременные могут практически не изменяться, даже в том случае,
когда изменение окружающих условий вызывает резкие сдвиги
внутри системы. Часто эти сдвиги «компенсируются» внутри и
проявляются вовне не сразу. Так, в организме человека про-
исходит напряжение регуляторных механизмов (связанное, оче-
видно, с изменением переменных состояния), которое направ-
лено специально на поддержание выходных переменных (таких,
например, как артериальное давление [199]). И только уже весь-
ма значительные сдвиги переменных состояния проявляются как
изменение выходов. Вообще говоря, малая чувствительность
вектора состояния автоматически влечет за собой и малую
218
ГОМЕОСТАЗ ОРГАНИЗМА
(ГЛ. 7
чувствительность вектора выходов системы. В то же время малая
чувствительность вектора выходов может не означать малой чув-
ствительности переменных состояния.
Рассмотрим, например, случай, когда система описывается уравнением
состояния (5.39) при матрице D = 0. В этом случае х = Ах + Bv, у = Сх.
Равенство D = 0 имеет место во многих технических системах. Покажем
сначала, что если х не чувствителен к изменениям v, то и все компоненты
вектора у малочувствительны к этим изменениям. В соответствии с (5.69)
имеем
4^-=-СЛ_1В, (7.12)
dv '
откуда для fe-й компоненты выходного сигнала получаем
, т
<7|3>
Пусть с = max cki, а=тахо^, 7=1, 2,..., т. Тогда
С mcG. (7.14)
<Э0у
и при малых Gy вектор выхода также малочувствителен к v.
В живых системах матрица D в уравнении выхода, вообще говоря, не яв-
ляется нулевой. Однако, как видно из рассмотренных в разд. 5.8 примеров,
отличны от нуля элементы матрицы D, соответствующие в векторе выходов
компонентам типа темпов, причем только тем из них, которые определяют об-
мен веществами и энергией между системой и средой.
Для переменных типа уровней, к которым собственно и относится поня-
тие гомеостаза, все элементы матрицы D равны 0, и формула (7.14) остается
справедливой.
Наоборот, малая чувствительность вектора выходов к вектору v в общем
случае не означает малой чувствительности вектора состояния. Действительно,
пусть матрица С в уравнении состояния (5.49) имеет вид С = [—1 1], а ста-
ционарные значения вектора состояния [x'i х2]т чувствительны к скалярному
возмущению v. Чувствительность выходного сигнала в ста-
ционарном режиме тогда равна
A- (с.Г'Ъ-о.
dv do I L1 JJ
Специфический способ образования выходного сигнала не позволяет вну-
тренним сдвигам проявиться вовне; выходной сигнал системы будет оста-
ваться неизменным несмотря на то, что все переменные состояния будут изме-
няться под воздействием вариаций внешних условий.
Малая чувствительность выходного сигнала часто достигает-
ся за счет того, что изменения двух или нескольких сильно ме-
няющихся сигналов — переменных состояния, зависящих от
V, — компенсируют друг друга. Внешняя стабильность перемен-
ных системы скрывает в этом случае резкие сдвиги, которые
могут возникать во внутренней среде системы. Эти сдвиги не-
7.4) ГОМЕОСТАЗ И МЕТОДЫ ПРОСТРАНСТВА СОСТОЯНИЙ 219
наблюдаемы из-за специфического вида матрицы С. Стоит уве-
личить размерность выходного вектора или другим способом
изменить матрицу С, как внутренние сдвиги состояния прояв-
ляются вовне и становятся наблюдаемыми. Поэтому гомеостаз
системы мы будем связывать с малой чувствительностью именно
вектора состояния системы: в терминах пространства состояний
гомеостаз системы понимается как малая чувствительность ста-
ционарных значений существенных переменных состояния к пе-
ременным внешних воздействий.
Итак, система обладает хорошими гомеостатическими свой-
ствами, если для всех ее существенных переменных состояния
выполняется условие
дх
<£1, 1=1, ..., т; /=1, (7.15)
Если же чувствительность некоторых существенных переменных
состояния велика.
то система не обладает хорошим гомеостазом.
Например, система пассивного теплообмена животного орга-
низма, рассмотренная в разд. 5.8, не обладает хорошими го-
меостатическими свойствами. Действительно, для нее сигнал v
есть температура окружающей среды, и гомеостатические свой-
ства определяются первым столбцом матрицы чувствительности
(5.58), так что условие (7.15) не выполнено.
Если рассмотреть выходную переменную в простой схеме ре-
гулирования с отрицательной обратной связью с уставкой, то та-
кая переменная будет обладать хорошим гомеостазом. Разу-
меется, из этого не следует, что и все остальные существенные
переменные состояния в такой системе имеют хорошие гомеоста-
тические свойства. Таким образом, система автоматической ста-
билизации может не обладать выраженным гомеостазом, хотя
некоторые ее переменные (и прежде всего выходная) таким
свойством обладают. Ясно, что и компартментальная модель с
произвольной структурой также не обязательно обладает хоро-
шим гомеостазом.
Чтобы обеспечить гомеостаз, живая система должна обла-
дать какими-то специфическими структурными особенностями,
приобретенными в процессе эволюции. Можно связать возникно-
вение гомеостаза в эволюции, о котором говорил еще У. Кэннон,
с процессом постепенного усложнения живых систем от низших
форм к высшим, с принципом биологического эпиморфизма
(разд. 1.4). Наслоение сначала пассивных, а затем и актив-
ных связей делает структуру биосистемы сложнее и богаче и
220
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
позволяет за счет этого достигать все новых и новых свойств и
качеств. Гоместаз как раз и является одним из таких свойств.
Поэтому в следующих разделах ход наших рассуждений
будет таким. Сначала мы покажем, что в простой модели откры-
той системы с пассивным управлением, не обладающей доста-
точным богатством связей, можно обеспечить стационарный ре-
жим, но нельзя получить хороших гомеостатических свойств.
Затем мы увидим, что существуют такие способы усложнения
структуры открытых систем с пассивным управлением, которые
по мере увеличения сложности приводят к непрерывному улуч-
шению их гомеостатических свойств. И в заключение мы рас-
смотрим возникновение гомеостаза в системе с активным управ-
лением.
Заметим, что, хотя идея наслоения связей и обсуждается
в общей теории систем [3, 31, 334], речь там идет о другом.
Так, обычно предполагается, что гомеостаз возникает тогда,
когда на систему первично взаимодействующих компонент био-
системы накладываются «вторичные механизмы регуляции, уп-
равляемые фиксированными структурами преимущественно
типа обратной связи» [31, стр. 43]. Поскольку обратная связь
понимается здесь как классическая система автоматической
стабилизации с уставкой, такое наслоение как раз и представ-
ляет собой классический подход, рассмотренный в разд. 7.3.
Гомеостаз возникает скачком, мгновенно: пока есть только ме-
ханизмы первичного взаимодействия в открытой системе, гомео-
стаза нет. Но стоит на них наложить механизм отрицательной
обратной связи по рассогласованию, как немедленно появляется
гомеостаз.
При рассматриваемом ниже подходе механизм возникнове-
ния гомеостаза представляется другим. В отличие от подхода
Л. фон Берталанфи, наслаиваются друг на друга не два прин-
ципиально разных типа взаимодействий, а происходит постепен-
ное накопление однотипных связей; сами механизмы первичного
взаимодействия компонент биосистемы по мере увеличения их
количества в системе (в рамках определенных структур) посте-
пенно придают ей все лучшие гомеостатические свойства.
7.5. Влияние структуры компартментальной системы
на ее гомеостатические свойства
Сохранительные свойства открытых систем прежде всего на-
правлены на обеспечение стационарности — равенства темпов
поступления и расходования веществ в системе. В этом случае
именно темпы, скорости потоков являются объектом регулиро-
вания, а переменные типа уровней (в том числе концентрации
вещества и энергии) играют роль регулирующих сигналов. Цель
7.5] СТРУКТУРА МОДЕЛИ И ЕЕ ГОМЕОСТАЗ £21
настоящего раздела состоит в том, чтобы показать, как накоп-
ление пассивных механизмов регуляции, направленных первона-
чально на поддержание скоростей потоков через систему, при-
водит постепенно к возникновению в системе гомеостатических
свойств.
Для этого, как будет показано ниже, достаточно, чтобы
1) система была достаточно сложной, т. е. число ее элементов
было велико, и 2) эти элементы образовывали некоторые спе-
цифические структуры, при которых основная цель управле-
ния — обеспечение равенства темпов поступления и расхода
веществ — достигалась за счет сочетания многих механизмов
регуляции — совместного (параллельного или последовательно-
го) функционирования регулирующих элементов.
В дальнейшем нам придется неоднократно сталкиваться с
термином «механизм регуляции» в связи с введением новых
переменных состояния в компартментальную модель. Поэтому
следует уточнить, что мы будем понимать под этим термином.
В компартментальных моделях биосистем можно выделить
два типа влияний, направленных на регуляцию темпов потока
вещества и энергии. В первом случае мы имеем дело с непосред-
ственным влиянием концентраций веществ на скорость их пере-
носа; это обычная пассивная регуляция. В этом случае роль ре-
гулирующего механизма играет сама переменная состояния.
В других случаях регулирующее действие на темпы оказывают
не сами переменные состояния (которые мы отождествляем
с уровнями, концентрациями веществ и энергии в компартмен-
тах системы), а некоторые специфические сигналы. В физиоло-
гических системах, например, такими сигналами являются час-
тота сердечных сокращений или величина альвеолярной венти-
ляции. В терминологии пространства состояний это выходные
сигналы у системы, которые однозначо определяются через век-
тор состояния х. Формально их можно исключить из рассмот-
рения, заменив выходной сигнал ук соответствующей k-й строкой
уравнения выхода в (5.39):
т I
Ук = ^ CiXi + S djVf. (7.17)
7=1 /=1
Поэтому, говоря о сочетании механизмов регуляции при вве-
дении в систему новых элементов и новых переменных состоя-
ния, мы будем придерживаться следующего определения.
Механизмом регуляции в компартментальной модели мы бу-
дем называть любую структурно выделяемую комбинацию сиг-
налов, которая содержит, по крайней мере, одну переменную со-
стояния и оказывает влияние на темпы поступления вещества
или энергии в ее компартменты.
222
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
В частности, простейшим механизмом регуляции скорости
потока некоторого вещества является пассивное управление,
осуществляемое концентрацией этого вещества в компартменте.
На структурной схеме такой механизм представляет собой ин-
тегратор, от выходного сигнала которого зависит темп потока
вещества на его входе. Поэтому мы будем моделировать появ-
w^t) --------
а)
б)
Рис. 7.7. Простая система гормональной
регуляции. Введение регулирующего
сигнала — концентрации гормона г — при-
водит к появлению в модели системы
дополнительного компартмента. а) ис-
ходная схема движения вещества без
регулирующих механизмов; б) схема
с учетом регулирующего действия гор-
мона.
ление нового механизма регуля-
ции введением в схему дополни-
тельного компартмента с соответ-
ствующей переменной состояния
Xi и говорить о регулирующем
механизме (регулирующем ка-
нале) X;.
Пример 7.5.1. Рассмотрим простой
пример системы гормональной регуля-
ции [317]. Возьмем процесс движения
вещества, показанный на рис. 7.7, а.
Здесь q — количество некоторого веще-
ства, темп притока которого равен
а скорость элиминации пропор-
циональна количеству с коэффициен-
том k:
q = wi (/) — kq. (7.18)
Пусть теперь это вещество выступает в
роли компартмента в живой системе, и
увеличение его концентрации лц приво-
дит к ускорению темпа секреции неко-
торого гормона г, концентрация х2 кото-
рого в свою очередь увеличивает ско-
рость элиминации q. В линейном при-
ближении имеем
«1 = — аи«1 — ai2«2 + ®1,
Х2 = — #21-^1 — #22^-2 4“ ^2.
где w2—исходная скорость секреции гормона, ац, ai2, a2i. а22— положитель-
ные коэффициенты. Схема модели с регуляцией, имеющей теперь два ком-
партмента, показана на рис. 1.1,6.
Введение регулирующего сигнала — концентрации х2 гормона г — озна-
чает введение в модель дополнительной переменной состояния.
Остановимся еще на толковании понятия «нагрузка» или
«напряжение» регуляторных механизмов, часто используемого
в физиологических исследованиях. Этот термин описывает преж-
де всего внешнюю силу, внешнее воздействие на систему. Когда
тот или иной механизм «принимает нагрузку на себя», то это
значит, что меняется функциональное состояние органов и си-
стем, ответственных за выполнение его регуляторных функций.
Напряжение механизма регуляции означает, что координаты
вектора состояния в компартментальной модели, описывающие
его функционирование, Изменяются, приближаясь к своим пре-
7.5]
СТРУКТУРА МОДЕЛИ И ЕЕ ГОМЕОСТАЗ
223
дельным значениям. Для пассивных механизмов регуляции на-
пряжение означает изменение (часто сокращение) запасов ве-
щества в соответствующем компартменте, для активных — изме-
нение выходного сигнала активной регулирующей цепи (7.17),
вследствие чего его чувствительность к действию возмущающих
факторов уменьшается.
а) Модель с одним компартментом. Рассмот-
рим простую однокомпартментальную модель, показанную на
в)
Рис. 7.8. Простая модель открытой системы, а) схема с одним каналом регуляции. В си-
стеме обеспечивается стационарный режим, но отсутствуют гомеостатические свойства;
б) зависимость х (v) при различных значевшях коэффициента а в схеме на рис. 7.8, а;
в) схема открытой системы с двумя параллельными каналами регуляции.
рис. 7.8, а. Эта система при а >> 0 устойчива, и ее стационарный
режим определяется обычным уравнением (5.50):
Гомеостаз системы определяется чувствительностью х к и; за-
висимость (7.20) приведена на рис. 7.8, б. Хороший гомеостаз
в принципе может быть достигнут при а —► оо. Однако при боль-
ших коэффициентах усиления в системах управления обычно
возникает тенденция к неустойчивости, и требуются специаль-
ные усилия для преодоления этой тенденции [120, 121]. В реаль-
ных биосистемах, однако, величины а и & —' одного порядка; в
случае диффузии, например, а = Ь. Поэтому в простых линей-
ных системах гомеостаз практически обеспечен быть не может,
область гомеостаза отсутствует.
б) Модель с двумя компартментами. Рассмот-
рим, как изменятся свойства этой простой системы, если
усложнить ее структуру за счет введения параллельной цепи
224
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
регуляции, направленной, как и первая, на поддержание адек-
ватного темпа поступления веществ. Пусть схема имеет два ком-
партмента, соединенные так, как показано на рис. 7.8, в.
Уравнение системы в форме «вход-выход-состояние» имеет
вид
х = Ах + Bv + Rw, у = Сх, (7.21)
где
::]• в=[»;]- «-[:[].
Поскольку матрица А особенная (у нее две одинаковых
строки, и поэтому det А = 0), пользоваться обычным уравне-
нием (5.50) для определения стационарного режима системы
нельзя. Действительно, уравнение det [Л — ХЕ] =0 дает = 0,
^2 = —ai — а2. Это означает, что система с параллельными ре-
гулирующими каналами обладает стационарным режимом, но
стационарные значения переменных состояния зависят от на-
чальных условий.
Рассмотрим поведение системы (7.21) подробнее. Ограничимся для про-
стоты случаем а-, = а2 = а. Тогда фундаментальная матрица системы, в со-
ответствии с формулой (5.66), равна
,, 1 Г (1 4- e~2at) —fl — e~2at) 1
®<0“т[_;;д-J, (,+:-)]• (7-22)
Переходный процесс, вызванный начальными условиями х(0) и приложением
внешнего сигнала До, определяется формулой (5.61):
t
х (/) = Ф (/) х (0) + Ф (/ - т) В До dx. (7.23)
о
Вычислим первое слагаемое, определяемое начальными условиями х10 и х20:
Ф(Пх(0)=—Г (l+*-2o<)*>0 - (1 - e~2at) х201
Ф (0) 2 [ _ (1 _ е-2«9 Х[0 (1 + e-2at) Х2о J • <7-24)
Компоненты второго слагаемого в (7.23) одинаковы н также легко вы-
числяются:
i
у Ь} До J [(1 + е~2а ('"х>) - (1 - е~2а ('~'с))] dx = До [1 - е"2о(]. (7.25)
о
Окончательные уравнения переходного процесса для обеих координат имеют
вид
*i (0 = 4-Г*ю — *20 + e~2at (х10 + х20) + -Ь'^~ (1 — е~2а')1,
1 1л (7.26)
х» (/) = 2-[- *10 + *2о + e~2at (xw + x2Q) + la V (1 — е-2а')].
7.61
СОЧЕТАНИЕ ПАРАЛЛЕЛЬНЫХ КАНАЛОВ РЕГУЛЯЦИИ
225
В стационарном режиме при /-> оо
X10 — Х20 6 1 До
2 + 2а ’
х2
Ь\ Л.и
2а
(7.27)
Чтобы показать, что схема (7.21) может служить хорошей моделью соче-
тания механизмов регуляции в открытых системах, положим исходные значе-
ния Хю = х20 = 0 при о = 0. Тогда в ответ на любое приращение До пере-
менные и х2 получают приращения Дх ] = Дх2 =-^ Ди. Легко видеть, что
в системе с двумя каналами это приращение вдвое меньше того, которое по-
лучается в системе с одним каналом — однокомпартментальиой модели (7.20).
Это получилось потому, что нагрузку, приходившуюся ранее на одну регу-
лирующую переменную xt, теперь разделили между собой два параллельно
соединенных канала с двумя переменными xt и х2.
Такая система устойчива неасимптотически: возмущения, возникающие в
процессе ее функционирования, с течением времени не исчезают. Действи-
тельно, пусть начальные условия х10, х20 в (7.27) изменились под действием
какого-либо возмущения на Si и е2, соответственно. Тогда в системе устано-
вится новый стационарный режим:
81 — е2 , 6|До
Х‘ =-----2-----+~2^'
, А А (7.28)
— 61 Ч" 82 , Ь} ДР
Ха==-----2-----+ ~2^-
Хотя обе переменные состояния при этом изменяются, выходной сигнал у
остается неизменным:
у = — а (х] + х2) = b\hv (7.29)
Поэтому стационарный режим в системе поддерживается при произвольных
значениях X] и х2, лишь бы их сумма удовлетворяла (7.29).
7.6. Сочетание параллельных каналов регуляции
Живые системы — это сложные структуры, в которых можно
выделить множество управляющих цепей и механизмов, осуще-
ствляющих процессы регуляции. Из технических применений
теории управления известно, что усложнение системы, введение
дополнительных переменных в ее структуру, приводящее к по-
вышению качества системы, дает и нежелательные последствия,
поскольку возникающие побочные эффекты могут привести, в ча-
стности, к потере устойчивости системы. Поскольку высокая
устойчивость живых систем является общеизвестным фактом,
остается признать, что в их структурах природа использует
только такие способы усложнения, которые не приводят к по-
тере устойчивости и позволяют относительно просто реализовать
сам процесс управления. Некоторые способы такого упрощения
процессов управления в сложных живых системах рассмотрены в
[223] — принцип «блочного управления», иерархический принцип
226
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ 7
построения систем и некоторые друте. Дру1ие способы ус-
ложнения систем без ухудшения устойчивост, возможно, свя-
заны со введением механизмов управления, имеющих сильно
«разнесенные» постоянные времени.
Еще одно специфическое свойство процессов управления в
живых системах было впервые отмечено Дж. Тракселом [209]:
если в технических системах достижение нужного результата
обеспечивается, как правило, резкими изменениями одного уп-
равляющего сигнала, то в биологических объектах аналогичный
результат достигается за счет одновременного изменения мно-
жества регулирующих сигналов, каждый из которых при этом
получает лишь незначительные приращения. В этом и последую-
щем разделах мы рассмотрим именно такой способ управления
в физиологических системах, когда достижение должного ре-
зультата обеспечивается включением (одновременным или по-
следовательным во времени) множества управляющих механиз-
мов в системе.
Речь будет идти только об анализе систем, так что можно
не интересоваться вопросами их устойчивости. Наоборот, для
задач синтеза любые структурные преобразования должны вно-
ситься в схему лишь после проверки, не приведут ли они наряду
с ожидаемым улучшением качества системы к потере устой-
чивости.
Целью нашего рассмотрения является выяснение тех спосо-
бов усложнения структуры систем, которые можно наблюдать в
реальных биосистемах и которые за счет усложнения ее регули-
рующих каналов приводят к появлению малой чувствительности
переменных состояния этой системы к изменениям условий окру-
жающей среды. Мы рассмотрим ниже две простые схемы систем,
которые могут быть аналогами гомеостатических структур в жи-
вых системах.
Первая — это простое увеличение числа регулирующих пере-
менных, включаемых параллельно друг другу. Такое тривиальное
наслоение хорошо известно в теории управления. Простейшие
примеры биологических структур такого типа мы уже встре-
чали в книге: это система кислородного снабжения, рассматри-
ваемая как система с параметрической обратной связью
(рис. 3.8) или схема энергетической системы организма с двумя
параллельными каналами восстановления АТФ (анаэробным и
аэробным, разд. 6.6). Такой случай возникает и при линеариза-
ции многих типов нелинейных элементов в системах управле-
ния: выходной сигнал множительного звена представлялся в
виде суммы двух сигналов, включенных параллельно (рис. 3.5).
Поскольку нелинейные элементы широко распространены в мо-
делях биологических систем, ясно, что первый тип структур —
параллельное включение регулирующих механизмов — представ-
7 6]
СОЧЕТАНИЕ ПАРАЛЛЕЛЬНЫХ КАНАЛОВ РЕГУЛЯЦИИ
227
ляет собой весьма частое явление в управляющих структурах
биосистем.
Этот случай, впрочем, был бы достаточно тривиален и мог бы
не заинтересовать исследователя, если бы не одно обстоятель-
ство. Обычно в физиологических системах происходит парал-
лельное включение качественно неоднородных механизмов уп-
равления — пассивных и активных. При этом активные механиз-
мы настолько более эффективны, что в норме почти целиком
принимают на себя нагрузку от изменяющихся условий среды
и режимов функционирования, защищая переменные пассивных
регулирующих цепей. А поскольку регулирующие сигналы пас-
сивных цепей являются, в частности, концентрациями многих
важных для жизнедеятельности системы веществ, то тривиаль-
ное сочетание механизмов управления оборачивается нетриви-
альным фактом постоянства существенных переменных внутрен-
ней среды.
Вторая схема относится к случаю, когда суммируются не
сами управляющие сигналы, а результаты их действия. Вероят-
но, даже в простейших случаях регуляции в организме конечный
эффект обеспечивается дублирующим действием многих факто-
ров. Так, химическая регуляция местного кровообращения обус-
ловлена действием множества химических механизмов разного
назначения и разной эффективности [100]. Другими примерами
могут служить рассматриваемая ниже (разд. 8.5) система тер-
морегуляции с ее многочисленными механизмами теплоотдачи и
теплопродукции или схема множественных механизмов в систе-
ме дыхания (рис. 1.6).
Идея уменьшения чувствительности каждого из параллель-
ных каналов регуляции, обеспечивающих нужную скорость по-
ступления вещества или энергии, может быть сформулирована
следующим образом. Изменение переменных состояния х( ока-
зывает регулирующее действие на потоки г/*. При этом важен
лишь суммарный эффект всех приращений Ах,, а доля воздейст-
вия каждого регулирующего механизма тем меньше, чем боль-
ше регулирующих переменных в системе (чем больше компарт-
ментов включено параллельно). При многократном дублирова-
нии конечный результат (стационарное состояние системы или
равенство скоростей поступления и расходования веществ) обес-
печивается малыми одновременными сдвигами большого числа
переменных внутренней среды, что и означает хороший гомео-
стаз системы.
Рассмотрим влияние процесса сочетания параллельных каналов на го-
меостатические характеристики в системе, описываемой уравнением состояния
х — Ах + Bv + Rw, (7 30)
где х, и, w — векторы размерности m, I, п, соответственно, а матрицы А, В, R
имеют размерности (т\т), (тХ.1), (т\п). Будем вводить в систему
228
ГОМЕОСТАЗ ОРГАНИЗМА
(ГЛ. 7
последовательно новые переменные состояния, так что на каждом шаге раз-
мерность х увеличивается на единицу. Для удобства будем вводить измене-
ния в систему так, чтобы они касались только той части, которая обведена
пунктиром на рис. 7.9. Пусть сначала Qi = Q2 = ' Структура потоков,
которыми система обменивается со средой, остается при этом неизменной.
Рис. 7.9. Сочетание параллельных каналов регуляции в открытой системе, а) Общая схема.
Пунктиром показана та часть системы, в которой производятся изменения. Структура
внешних потоков при этом остается неизменной; б) введение параллельных каналов по
I-й переменной. Пунктиром по-прежнему показана часть системы, в которой происходят
изменения.
।--------------------------------------------------------------------1
Рассмотрим сначала сочетание параллельных каналов для одной из коор-
динат системы. Пусть происходит переход системы из одного стационарного
состояния в другое, вызванный малым изменением условий внешней среды
До. При таком переходе переменные состояния xi, i = 1, 2, ..., т, получают
приращения Дхь В соответствии с (7.30) приращения х и о связаны между
собой соотношениями
I т
- £ biv Kvv = S %дv <7-3i)
v=l U=I
Это условие можно трактовать как условие стационарности по i-й перемен-
ной состояния (суммарное приращение всех переменных на входе интегра-
тора равно нулю).
Для приращения i-й переменной состояния из (7.31) получаем
г- I т
Дх>=- т; £ biv д% + £ % ч • <7-32)
" Lv=l J
Заменим теперь единственный компартмент с переменной Xi параллель-
ным соединением новых переменных состояния хц, ..., xls (рис. 7.9,6). Вве-
дем также матрицу G, преобразующую координату х; в исходной системе в
вектор размерности s в новой системе:
G = [£i g2 ... gs]T. (7.33)
Уравнение части системы, обведенной пунктиром, теперь имеет вид
[т I -1
£ aiv.xi + Е • Л = 1, 2.................... (7.34)
ц-1 v-1 J
7.6]
СОЧЕТАНИЕ ПАРАЛЛЕЛЬНЫХ КАНАЛОВ РЕГУЛЯЦИИ
229
а переменная Xi представляет собой сумму переменных х{^.
S
xi=Fs^-Z (7.35)
К"° 1
где —скаляр, х^ — вектор [х(1 xi2 ... x(s]T, а ^ — матрица (1 X«)>
Fs=[l 1 ... 1]. (7.36)
Очевидно, что условие неизменности системы состоит в том, чтобы
= gv>0; v = l, 2......s. (7.37)
v=l
Из (7.35) и (7.32) для стационарного режима имеем
= (7.38)
V“»I
Пусть, например, gv = 1/s. Тогда приращения всех переменных состояния
уменьшились в s раз:
dxiv 1 dxi
dv s dv
(7.39)
При достаточно большом s все переменные xiv обладают хорошими гомеоста-
тическими свойствами.
Если вернуться к исходной системе на рис. 7.9, а, то проделанная операция
наслоения каналов по i-й переменной может быть представлена следующим
образом. Матрица Q1 теперь стала отлнчгой от единичной и «разверты-
вает» единственную переменную состояния х, в вектор x\s\ матрица Q2 «свер-
тывает» этот вектор снова в скаляр, оставляя прочие координаты вектора со-
где G и F определяются (7.33) и (7.36), а Е|, Е2— единичные матрицы раз-
мерности (1—1)XG— 1) и (m —i)X("i — 0, соответственно.
Уравнение состояния для системы, показанной на рис. 7.9, имеет вид
х = QtAQ2x + Bv + Rw. (7.41)
Легко видеть, что матрицы QI и Q2 — особенные из-за специфического вида
матриц F и G, поэтому анализ стационарных режимов такой системы должен
производиться аналогично тому, как в разд. 7.5 мы исследовали систему вто-
рого порядка. Это, безусловно, является недостатком рассмотренного способа
получения гомеостатических свойств в компартментальных моделях, так как
ие позволяет пользоваться обычной формулой (5.50) для вычисления стацио-
нарных режимов *).
*) Другим недостатком рассмотренного типа параллельного сочетания
механизмов регуляции является то, что, как легко показать, система оказы-
вается ненаблюдаемой. Однако, хотя вводимые в систему новые переменные
состояния и иеиаблюдаемы, это не мешйет системе улучшать свои гомеостати-
ческие свойства.
230
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
Рассмотренная структура в принципе позволяет обеспечить гомеостаз
всех переменных системы, если проделать рассмотренную выше операцию на-
слоения последовательно для всех координат системы. Матрицы Q, и Q2 в
этом случае будут иметь блочный вид:
(7.42)
Недостатком такой структуры с точки зрения моделирования
физиологических систем является то, что в принципе все парал-
лельные каналы при действии возмущающего фактора вклю-
чаются одновременно. Конечно, при малых коэффициентах пере-
Рис. 7.10. Появление параллельных ка-
йалов регуляции прн линеаризации ис-
ходной нелинейной системы. При лине-
аризации сложной зависимости у от
вектора переменных хт приращение Ху
определяется суммой приращений ком-
понент с соответствующими коэф-
фициентами передачи kit i = l, 2, т
дачи yv включение какой-то части
механизмов регуляции происхо-
дит достаточно медленно, и ста-
ционарное состояние достигается
при существенных сдвигах не по
всем переменным, но в рамках
описанной модели с наслоением
параллельных каналов не может
быть хорошо известного «эшело-
нированного» включения механиз-
мов регуляции [334].
Зато безусловным достоинст-
вом рассмотренной схемы явля-
ется ее близость биологическим
системам регуляции в следующем
смысле. Если темп поступления
вещества у в системе определяет-
ся множеством переменных х,, а уравнения, связывающие х и у,
сложны и нелинейны, то при исследовании реакций системы на
малые возмущения можно ограничиться линеаризованной моде-
лью. В этом случае вклад каждого из механизмов регуляции
представляется в виде слагаемого, а блок-схема системы имеет
вид параллельного соединения множества регулирующих кана-
лов, как показано на рис.7.10 (см. также [240], стр. 23).
7.7. Сочетание последовательных каналов регуляций
Рассмотрим теперь схему компартментальной модели откры-
той системы, в которой происходит сочетание последовательных
механизмов, каждый нз которых вносит свой вклад в регуляцию
темпов поступления вещества (рис. 7.11).
7.7]
СОЧЕТАНИЕ ПОСЛЕДОВАТЕЛЬНЫХ КАНАЛОВ РЕГУЛЯЦИИ 231
Пусть сначала имеется обычная система с одним компарт-
ментом; в этом случае хь v и w— скаляры. Стационарное зна-
чение Xi и его чувствительность к v определяются обычными со-
отношениями
___6 . w dx\ Ь
X1 a,i V ' ап ’ dv atl
(7.43)
Покажем, что введение второго последовательного контура ре-
гуляции (с переменной состояния х2 на рис. 7.11) улучшает
Рис. 7.П. Введение после юваюдьчых механизмов регуляции в компартментальной модели
открытой системы.
гомеостатические свойства системы, снижая чувствительность
переменной %i к действию возмущающего сигнала v.
В этом случае w и и — по-прежнему скалярные величины, х = [х, х3]т —
вектор размерности 2.
Уравнение системы имеет вид
х = Ах + Bv + Rw, (7.44)
где
л=Г-а1, -а>21 В = Р1, « = [']. (7.45)
L «;ч — <222 J L О J L О J
Чувствительность стапионарного режима к изменению v определяется форму-
лой (5.68):
— .У'В —_______________1 Г ~ «22 а‘2'1Р1==
dv <211«22 + «12«21 L— «21 — «lljLOj
«22^
____ «11«22 + «12«21
~ • (7Л6)
- « 11 «22 + «12«21 -
Для обозначения количества последовательно включенных компартмен-
тои в смме рис 7.11 будем использовать верхний индекс, так что вектоо
232
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
размерности т будет обозначен х(т>:
x("il _ х<т) x,m)]T.
Таким же индексом при необходимости будем обозначать размерность опре-
делителей и алгебраических дополнений матриц. Тогда нз (7.43) и (7.46) для
компоненты X] при п = 1 н п = 2 находим
b
о.. ’
b
, О12О21
он Н--------
022
Поскольку второе слагаемое в знаменателе (7.49) больше нуля,
dx<2> d^_
dv ’
(7.47)
dx1/’
dv
dx‘\2'
dv
(7.48)
(7.49)
dv
(7.50)
что мы и хотели показать.
Интересен предельный случай, когда коэффициент 022 = 0. В этом случае
система сохраняет устойчивость, поскольку характеристические числа матрицы
« __Г — 011 — 012 1
L 02. О J
имеют вид 1] 2 = -g-(— он ± ^О]! — 4о|2о21), ReX<0, и в стационарном
режиме согласно (7.46) получаем
dx(2’
dv
0 1
Ь .
а12 J
Чувствительность исходной переменной xj2) оказывается равной нулю, т. е.
в системе обеспечивается абсолютная инвариантность переменной х®.
Общий случай. Покажем теперь, что в общем случае —
при произвольном числе (m—1) ступеней в схеме рис. 7.11 —
добавление новой ступени снижает чувствительность стацио-
нарных значений всех переменных состояния х(^~Х}..............xm-il>-
Уравнение системы в общем случае имеет вид
где
S(m-1)=[6 0 0
---Он
021
О
д(т-1) __
вЛ(т-1'х(т-1) +В(т-1)р+ Kw
(7.51)
... 0]т,
Я|2
— 022
Оз2
— 013
0
— Озз
а1, т-2
О
О
171. т-1
0
о
am — 1, т—2 am—l, т — 1
(7.52)
0
о о
7.7)
СОЧЕТАНИЕ ПОСЛЕДОВАТЕЛЬНЫХ КАНАЛОВ РЕГУЛЯЦИИ
233
Чувствительность переменных состояния определяется обычным выражением
dx(m_1) Ь
—!------=---------Adj(m-1)a.., < = 1, 2, т — 1, (7.53)
dv detA^"1’ 1г
где Adj(m-1,an —алгебраическое дополнение элемента аи в А(т-,). Допустим
теперь, что в систему введен дополнительный канал регуляции с номером т,
и представим уравнение системы в блочном виде
^(т-1)
д(т)
лт
(7.54)
ГДе М (т X 1) — [ ш 0 . . . 0]т, A)(ix»n) — [0...0 dm, т~ 1], 7/(1 X1) ~ [ &тт]'
Тогда чувствительность вектора х<т> определяется формулой
dx(m>
dv
Г 41.
|_ и\l
][?т”
(7.55)
Для вычисления правой части (7.55) воспользуемся формулой (5.29), даю-
щей выражение обратной матрицы для блочных матриц. Найдем матрицу
(Д(т-1> — ML-'N} Поскольку в матрице ML~'N отличен от нуля только эле-
мент с номером (1, т), равный (— aimam: имеем
(А(л,-1) _ ML~XN) =
(7.56)
Тогда, обозначив элементы обратной матрицы через a;v> запишем
, Г Adi(m'« 1
(A*'"-1’ 1 = [a. 1 =------г—,—у---------j—r , (7.57)
1 ZvJ L det(A('”-1)-ML-'y)]
где Adj^'") avi — алгебраическое дополнение элемента с номером (v, i) в мат-
рице (А<'"-1) — ML~lN). Для элементов первой строки имеем
Adj<m>au = (- атт) Adj*"1-1»^, (7.58)
где Adj*m-1) da — то же, что и в (7.53). Кроме того, из (7.56) разложением
по элементам последнего столбца получаем
det (A(m-1) - ML~lN) = - атт det A'"1"1’ + аы Adj(m)aIv. (7.59)
Можно показать, что при нечетном т величины Adj(m)alm и det'm~[)A по-
ложительны, а при четном — отрицательны, т. е. их знаки всегда совпадают.
234
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ, 7
Из (7.55) — (7 59) тогда следует
дх1™ _ дх‘{т-''> 1
~v dv ' । । %,.-м,тЦт~
ammdet<m~')A
i=l, 2..............m-1. (7.60)
Последнее равенство означает, что введение m-го последовательного ре-
гулирующего элемента приводит к уменьшению чувствительности всех (т— 1)
переменных состояния Следовательно, по мере накопления связей в такой
системе можно достичь гомеостатических свойств всех переменных состояния.
Пусть, например, т = 3. Тогда из (7.60) получаем
dx? b
dv - = , (7.61) , <7|2<721 , <221 <713*232 fl|l -Г ~T ^22 #22^33
dxA dv b = . (7.62) aI1^22 । । ^13^32 _ 1 v. 12 + <221 <733
Выражение (7.61) меньше (7.48); в то же время уменьшилась и чувствитель-
ность хг в (7.62) по сравнению с (7.46).
Рассмотренная модель сочетания последовательных механиз-
мов регуляции лишена недостатков, свойственных модели соче-
тания параллельных механизмов. Стационарный режим в такой
модели определяется обычными соотношениями (5.50) при не-
вырожденной матрице А, т. е. стационарные значения не зави-
сят от начальных условий в системе, как это обычно и бывает
в компартментальных моделях открытых систем. При действии
возмущающих факторов механизмы регуляции включаются
последовательно во времени, так что система может достичь но-
вого стационарного состояния, включив только «первые эшело-.
ны» адаптивных процессов и имея неиспользованные резервы
адаптации. Динамические свойства такой модели достаточно
богаты и могут объяснить некоторые важные феномены гомео-
статической регуляции в биосистемах, связанные, например, с
замещением быстрых, но «слабых» механизмов регуляции более
медленными, но мощными регуляторными процессами.
Итак, мы рассмотрели два способа организации структуры
компартментальных моделей, при которых чувствительность пе-
ременных состояния системы может быть снижена. При этом
нам было важно показать принципиальную возможность обес-
печения малой чувствительности стационарных значений пере-
менных состояния за счет увеличения сложности системы путем
накопления пассивных каналов управления. В реальных моде-
лях, конечно, связи между переменными по всегда могут быть
представлены в виде относительно простых структур, подобных
изображенным на рис. 7.9 или 7.11. Хотя сам факт сочетания
7.8]
ГОМЕОСТАЗ В СИСТЕМАХ С АКТИВНОЙ РЕГУЛЯЦИЕЙ
235
регуляторных механизмов, безусловно, имеет место в сложных
биосистемах, конкретные формы этого сочетания могут отли-
чаться от рассмотренных выше. Кроме того, структуры, подоб-
ные исследованным в настоящем разделе, могут составлять
лишь часть взаимодействий, описываемых матрицей системы.
Важно, что гомеостатические характеристики физиологических
систем могут обеспечиваться за счет усложнения ее структуры,
что отражает принцип биологического эпиморфизма. Напом-
ним, что этот принцип, рассмотренный в разд. 1.3, подчеркивает
наличие у сложных биосистем множества механизмов для вы-
полнения тех функций, которые в более простых биосистемах
выполняются всего лишь одним механизмом.
В следующих главах мы на конкретных примерах рассмот-
рим, как при усложнении структуры систем в них будут возни-
кать гомеостатические свойства.
7.8. Гомеостаз в физиологических системах
с активной регуляцией
В двух предыдущих разделах было показано, что усложне-
нием структуры в компартментальных моделях открытых систем
можно достичь не только стационарного неравновесного состоя-
ния, но также и обеспечить гомеостаз всех переменных состоя-
ния. При таком рассмотрении в яРной или неявной форме дела-
лось допущение, что коэффициенты усиления всех пассивных ка-
налов в модели примерно одинаковы (так, в модели на рис. 7.8
считалось а = Ь, в модели рис. 7.9 все коэффициенты усиления
g'v выбирались равными 1/s).
Активные механизмы в физиологических системах обладают
значительно более высокими коэффициентами усиленйя, чем
механизмы пассивной регуляции. Хотя получить непосредствен-
ные оценки такого рода и затруднительно, некоторые представ-
ления о сравнительной эффективности пассивных и активных
механизмов в одних и тех же условиях могут дать следующие
цифры. Темп доставки кислорода в мышечные ткани под дей-
ствием пассивной регуляции (увеличение артерио-венозной раз-
ницы) может возрасти от 50 до 120—150 мл)л, т. е. в 2,4—3 раза.
Активная же регуляция кровотока через работающие мышцы
дает его увеличение примерно от 1 до 20 л!мин, что приводит к
увеличению темпов поступления кислорода в 20 раз. Пассивные
механизмы регуляции теплоотдачи с кистей рук (снижение тем-
пературы кожи) уменьшают темп отдачи тепла примерно на
25%, а активная регуляция (сосудистый спазм, например)
может снизить темпы теплоотдачи вдвое. Такая эффективность
активных механизмов регуляции при изменении условий среды
объясняется просто: переменные механизмов активной регуляции
236
ГОМЕОСТАЗ ОРГАНИЗМА
(ГЛ. 7
обладают весьма высокой чувствительностью к изменениям
внешних условий. Именно поэтому, когда в физиологии говорят
о гомеостазе, в число гомеостатируемых переменных никогда
не включаются переменные внутренней среды, описывающие ак-
тивные механизмы регуляции.
Рассмотрим теперь влияние активных механизмов регуляции
в схемах параллельного и последовательного соединения.
Параллельное соединение компартментов.
Возьмем схему двух каналов,
Рис. 7.12. Простая модель системы
с двумя каналами регуляции (одним
пассивным и одним активным).
показанную на рис. 7.8, и видоиз-
меним ее так, как изображено на
рис. 7.12. Пусть X! теперь опи-
сывает процессы накопления ка-
кого-либо вещества, необходи-
мого для жизнедеятельности си-
стемы (xi — существенная пере-
менная), а х2— процессы изме-
нения количества веществ, уча-
ствующих в активной регуляции.
Переменная х2 не входит в число
гомеостатируемых переменных
системы.
Повторяя выкладки, проделанные в разд. 7.5, находим урав-
нения переходного процесса, аналогичные (7.26):
1 Г а2 + а,е at
а [ -а,(1 — е~а!)
- а2 (1 — е at) I
а, a2e~at J
рю] , Г а' 1 6До
1х20 J + |_ а2 (1 — e~at) J а ’
(7.63)
где обозначено а = сц + а2; первое слагаемое определяет реак-
цию на начальные условия, а второе — на изменение окружаю-
щей среды Да. В стационарном режиме при t -> оо второе сла-
гаемое дает
= х* = 7Г%п-Ь^>- (7.64)
ai + а2 + а2 ' '
Если, как это характерно для активных механизмов регуляции,
О2 > «ь то
dxi „ . ^х2 1
dv dv ~ *’
(7.65)
т. е. введение параллельного канала активной регуляции обес-
печивает хороший гомеостаз существенной переменной. Ясно
также, что введение единственного канала активной регуляции
в более сложном случае может обеспечить гомеостаз сразу не-
скольких переменных, как, например, в схеме, показанной на
рис. 7.9, б.
7.8]
ГОМЕОСТАЗ В СИСТЕМАХ С АКТИВНОЙ РЕГУЛЯЦИЕЙ
237
Последовательное соединение компартмен-
тов. Покажем теперь, что если в схеме последовательного со-
единения (рис. 7.11) переменная хт отвечает активному меха-
низму, то все переменные хь х2, ..., хт_х обладают гомеостати-
ческими свойствами. Обратимся к формуле (7.60) и рассмот-
рим величину знаменателя. Поскольку Adj(m>alm получается из
определителя треугольной матрицы, его абсолютная величина
равна произведению всех коэффициентов а21, Дз2, • • •, am, m-i-
Для активных механизмов характерны большие отношения
Ctm, m—i/dmm (тогда переменная хт очень чувствительна к ц).
Кроме того, для таких механизмов можно считать большими и
величины aim. Тогда знаменатель в (7.60) достаточно велик, а
чувствительность всех переменных хь х2, ..., xm-i (кроме пере-
менной хт, отвечающей активному механизму) оказывается
малой, что и требовалось показать.
Таким образом, суть'активной регуляции в физиологических
системах можно кратко сформулировать следующим образом.
Целью первого уровня, которая достигается при функциониро-
вании механизмов активной регуляции, как и механизмов регу-
ляции вообще, является поддержание стационарного состояния
систем, т. е. обеспечение равенства темпов потоков (сначала не-
зависимых и зависимых, затем — среди зависимых — первичных
и вторичных, рис. 7.2). В этом смысле цели активных и пассив-
ных механизмов регуляции совпадают. Однако активные меха-
низмы регуляции обладают по сравнению с пассивными лучши-
ми регуляционными характеристиками: большими коэффициен-
тами усиления, большим быстродействием. Поэтому достигается
и цель второго уровня: при действии внешних возмущений
активные механизмы в основном принимают нагрузку на себя,
а на долю пассивных регуляторов остается лишь незначительная
часть нагрузки, и они оказываются нечувствительными к воз-
мущающим факторам. Что касается переменных состояния, вхо-
дящих в цепи активной регуляции, то их высокая чувствитель-
ность следует и из общих соображений — см. формулу (7.17).
Поскольку в компартментальных моделях все переменные со-
стояния описывают уровни вещества и энергии, то с точки зре-
ния теории управления возникает неожиданная ситуация: гомео-
стаз системы можно трактовать как постоянство части регули-
рующих (хотя и пассивных) переменных в системе. Это специ-
фическое свойство систем организма имеет, впрочем, аналогию в
технике. Пассивные механизмы физиологической регуляции на-
ходятся как бы в резерве, включаясь только в том случае, когда
активные механизмы уже теряют свою эффективность или выхо-
дят на предельные режимы функционирования. В этом смысле
их можно трактовать как средство повышения надежности
системы [118], и становится очевидной связь, существующая
238
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ 7
между гомеостазом и надежностью физиологических систем.
Взаимодействие активных и пассивных механизмов регу-
ляции оказывается, таким образом, одним из существенных
аспектов функционирования систем, что феноменологически про-
является во многих чертах их гомеостатического поведения. Не-
которые из таких свойств мы рассмотрим ниже при анализе воз-
никновения гомеостатических кривых в физиологических систе-
мах (разд. 8.5) и двойственного характера их переменных
(разд. 9.2).
7.9. Стабилизация уровней вещества и энергии
В этом разделе мы обсудим вопрос, каким образом опреде-
ляются в физиологических системах те уровни, на которых про-
исходит стабилизация переменных внутренней среды. Начнем с
того, что сам термин «стабилизация» не вполне точно опреде-
ляет существо дела:
Рис 7 13. Характерный вид
зависимости переменной вну-
тренней сречы от изменения
условий внешней среды,
и — диапазон изменения v,
в котором обеспечиваются
стационарность системы и ее
гомеостат ические свойства
соответственно
у многих животных, обладающих плохим
гомеостазом, при изменении условий ок-
оужения значения «стабилизируемых»
переменных могут изменяться в доволь-
но широких пределах.
Если рассмотреть изменение пере-
менной внешней среды v в достаточно
широком диапазоне, то для многих пере-
менных в различных организмах можно
обнаружить картину, аналогичную изо-
браженной на рис. 7.13. При больших
значениях v переменная состояния х при-
ближается к прямой х = v, затем при
уменьшении v система попадает в «нор-
мальный» диапазон внешних условий и
кривая x = x(v) образует изгиб (гомео-
статическая зависимость — см. разд. 2.3).
Дальнейшее изменение v снова приво-
дит к образованию пропорциональной
зависимости х от v, прямая х = х(у) идет параллельно прямой
х = v. Примеры такого поведения видны, в частности, на
рис. 2.1, а и б.
Плато, образуемое кривой х(ц), может быть довольно узким,
как на кривой 2 на рис. 2.1,6. Только по мере усложнения си-
стем и накопления в них регуляторных механизмов это плато
делается достаточно широким и пологим, так что выражение
«стабилизация переменных внутренней среды на некотором
уровне» становится оправданным. В организме человека, напри-
мер, можно говорить о стабилизации температуры внутренних
органов или напряжения кислорода в крови.
7 9]
СТАБИЛИЗАЦИЯ УРОВНЕЙ ВЕЩЕСТВА И ЭНЕРГИИ
239
В организме различаются два пути, приводящих к расшире-
нию гомеостатического плато. Первый из них связан просто с
накоплением пассивных регулирующих механизмов — увеличе-
нием числа механизмов, участвующих в регуляции (биологиче-
ский эпиморфизм, см. разд. 1.3), второй — с появлением специ-
фических физиологических механизмов активной регуляции.
Среди последних имеется большая группа регуляторных меха-
низмов, реагирующих на изменение самих переменных внутрен-
ней среды. Эти механизмы обладают специальными рецепто-
рами, которые позволяют им точно следить за текущим уровнем
своих переменных и предотвращать их большие изменения.
Здесь мы попадаем в область, где современная физиология
предоставляет в распоряжение исследователей обширный мате-
риал. Терморецепторы в системе теплового режима организма,
барорецепторы в системе кровообращения, хеморецепторы во
многих системах регуляции внутренней среды организма «сле-
дят» за протеканием процессов в этих системах и, если сущест-
венные переменные выходят на границы пределов безопасности,
включают мощные исполнительные механизмы, направленные на
предотвращение дальнейших изменений, а иногда, возможно, и
на возвращение отклонившихся переменных к их исходным
«нормальным» значениям. Впрочем, по мере совершенствования
измерительных методов все чаще,отмечается, что включение ре-
гуляторных механизмов не направлено на возвращение перемен-
ных к некоторому заданному уровню, а лишь эффективно пре-
пятствует их дальнейшему изменению (см., например, экспери-
менты по определению температуры гипоталамуса в [79, 182]).
Какую же роль играют рецепторы — датчики значений пере-
менных состояния — в концепции сохранительных с§ойств, как
укладываются схемы регуляции с рецепторами в модели ком-
партментальных систем? Эти вопросы достаточно сложны, и в
настоящем разделе мы можем изложить только некоторые прин-
ципиальные соображения. Начнем с того, что механизмы регуля-
ции с рецепторами внутренней среды в эволюционном плане
представляют собой довольно поздние образования, которые спо-
собны функционировать уже в условиях поддерживаемого ста-
ционарного неравновесного состояния. Хотя эти механизмы аб-
солютно необходимы для обеспечения нормального функциони-
рования организма, их отключение не приводит к прекращению
жизни. Хорошо известна, например, гипотермия — жизнь орга-
низма в условиях отключения нормальных механизмов терморе-
гуляции. Известны факты, когда при травмах черепа температу-
ра продолжает оставаться на нормальном уровне, но теряется
способность ос регулирования при изменении окружающей тем-
пературы [76] Жизнедеятельность организма продолжается и
при хирургических операциях под наркозом, когда многие
240
ГОМЕОСТАЗ ОРГАНИЗМА
(ГЛ 7
рецепторные механизмы регуляции отключаются. Денервация
отдельных систем организма не нарушает нормального снабже-
ния органов, хотя динамические свойства снабжающих систем
ухудшаются [252, 253]. Наконец, барорецептивные механизмы,
действие которых направлено на поддержание постоянства арте-
риального давления (быть может, даже в ущерб темпам крово-
тока в некоторых системах или органах), способны адаптиро-
ваться, и в экспериментальных условиях (перфузия изолирован-
ного каротидного синуса) их регулирующий эффект проявляется
только в течение 15—20 мин, после чего исчезает.
Жизнедеятельность организма связана с непрерывным изме-
нением независимых темпов расхода вещества и энергии, так что
в его системах постоянно протекают процессы, направленные на
поддержание равенства независимых и зависимых темпов. Тогда
можно сказать, что действие механизмов стабилизации внут-
ренней среды (термо-, баро-, хеморецепторов) обеспечивает та-
кое протекание этих процессов, чтобы существенные перемен-
ные не выходили за пределы безопасности. Поэтому в рамках
рассматриваемого подхода деятельность рецепторных регули-
рующих механизмов направлена на ограничение переменных
состояния в системе управления зависимыми темпами потоков
вещества и энергии.
Систему регуляции в организме тогда можно представить в
виде иерархической структуры, имеющей два уровня целей (см.
разд. 1.7) .^Система управления должна обеспечить равенство
зависимых и независимых темпов потоков, и при переходных
процессах в системе не должны нарушаться ограничения, нало-
женные на ряд ее переменных. Равенство темпов представляет
цель высшего уровня, выполнение ограничений, налагаемых на
существенные переменные, — цель низшего уровня.
Цель высшего уровня обеспечивается пассивными и многими
активными механизмами регуляции, цель второго уровня —
прежде всего механизмами рецепторного типа. Можно полагать,
что в случае конфликта между целями разных уровней ситуация
разрешается в пользу цели высшего уровня. Возможно, именно
так обстоит дело с адаптацией барорецепторов и исчезновением
рефлекса каротидного синуса, о котором говорилось выше.
Попробуем несколько более четко подчеркнуть аналогию с
обычной постановкой задачи синтеза системы автоматического
управления (см. разд. 3.1, 4.1). Если уравнение системы имеет
вид
х = f (х, и, t), (7.66)
то цель управления при синтезе системы может быть запи-
сана в виде задания желаемых значений переменных состоя-
ния в некоторый момент времени Т, отвечающий окончанию
7.9]
СТАБИЛИЗАЦИЯ УРОВНЕН ВЕЩЕСТВА И ЭНЕРГИИ
241
переходного процесса:
х(Т) — хт, (7.67)
причем качество самого переходного процесса должно удовлет-
ворять некоторым наложенным на систему условиям. Если, на-
пример, ищется оптимальное управление, то функционал каче-
ства (см. разд. 4.1) должен достигать минимального значения
Q (х, и, t) = min (7.68х
при наличии ограничений на управляющие воздействия, напри-
мер, вида
|м(/)|<М, (7.69)
и на область изменения фазовых координат — переменных со-
стояния х:
х‘<х(0<х**. (7.70)
Тогда в компартментальных моделях открытых систем соот-
ношению (7.67) отвечает условие стационарного неравновесия
(7.1). Введение ограничений (7.69), (7.70) ухудшает качество
переходного процесса, увеличивая показатель (7.68). Функциони-
рование рецепторных механизмов регуляции аналогично наложе-
нию на переменные состояния системы ограничения (7.70).
XСтремление системы к гомеостазу, приводящее к появлению
ограничений на фазовые переменные системы, ухудшает каче-
ство переходных процессов в ней. Две цели — высшего и низ-
шего уровня — вступают в противоречие: достижению равенства
первичных и вторичных темпов препятствуют механизмы, на-
правленные на недопущение значительных отклонений во внут-
ренней среде организма. Этот конфликт у разных видов живот-
ных решается по-разному. Так, темпы потоотделения у дикой
(гиеновой) собаки при изменении физической нагрузки (ско-
рости бега) меняются меньше, чем у домашней, а температур-
ный гомеостаз у нее соответственно хуже [250, стр. 85]. Вероят-
но, для живущей в более суровых условиях дикой собаки регу-
лирование темпов потоков веществ и энергии оказывается более
важным, чем для домашнего животного.
Стационарные значения переменных в компартментальных
моделях физиологических систем образуются как результат
уравновешивания многочисленных факторов. Это можно проил-
люстрировать схематически так, как показано на рис. 7.14. При
действии возмущающих факторов регулирующие механизмы в
соответствии с принципом Ле-Шателье способствуют восстанов-
лению стационарного состояния в системе. Изменение любой
переменной состояния х приводит к изменениям первичных и
вторичных темпов потоков в соответствующий компартмент, так
что новое стационарное состояние устанавливается при уравно-
вешивании первичных и вторичных потоков в компартменте
242
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ 7
(рис. 7.14, а). Действие возмущений приводит к тому, что за-
висимость либо первичных, либо вторичных темпов от х изме-
няется (пунктир на рис. 7.14, а), и стационарное значение х сме-
щается ОТ Xi ДО Х2-
Если же в системе имеются рецепторные механизмы актив-
ной регуляции, то зависимость от х вторичных темпов, регули-
руемых этими механизмами, может принять характерный поро-
говый вид (рис. 7.14,6). Тогда действие того же возмущающего
Рис. 7.14. Уравновешивание противоположно направленных факторов в компартментальных
моделях физиологических систем Влияние противодействующих сил приводит к разно-
направленному изменению первичных и вторичных темпов, стационарное значение к воз-
никает при уравновешивании этих темпов а) Схема с пассивными механизмами регуляции:
уравновешивание происходит при х — х}, при действии внешних возмущений (когда зави-
симость вторичных темпов смещается, как показано пунктиром) равновесие смещается
в точку Xi, б) схема с рецептивными механизмами ретутяцин, изменения стационарного
состояния при тех же внешних возмущениях практически нет.
фактора, что и в первом случае, не приведет к существенному
изменению стационарного значения переменной х. Изменение
характеристик системы под действием возмущений и соответ-
ствующее смещение х на обоих рисунках показано пунктиром.
В общем случае блок-схему образования стационарных со-
стояний в физиологических системах организма можно тогда
представить так, как показано на рис. 7.15, а. Изменение х при-
водит к изменениям первичных и вторичных темпов потоков,
уравновешивание которых и дает стационарные значения пере-
менных состояния. Схема формирования потоков включает в
себя все механизмы регуляции, в том числе и рецептивные ме-
ханизмы, обеспечивающие нормальное протекание переходных
процессов в системе.
7.9]
СТАБИЛИЗАЦИЯ УРОВНЕЙ ВЕЩЕСТВА И ЭНЕРГИИ
243
Рассмотрим, например, как 1Может происходить образование
стационарных значений переменных состояния в системе термо-
регуляции. Эта система принадлежит к числу наиболее изучен-
ных в физиологии, и поэтому она издавна служит как бы «базо-
вой системой» для проверки различных теоретико-управленче-
ских гипотез и биокибернетических концепций. Если в 1960-х гг.
исследователей вполне удовлетворяли модели системы терморе-
гуляции, построенные на принципах классической теории управ-
ления (см. разд. 7.3, а также [130]), то к началу 1970-х гг. на-
метился резкий перелом в отношении ведущих специалистов к
подобным моделям [295]. Авторы работ [274, 283, 335, 358] еди-
нодушны в том, что понятие уставки — концепция, целиком
заимствованная из теории управления, и в организме нет ана-
логичных ей управляющих структур. Более вероятно, что равно-
весное значение температуры получается в результате уравно-
вешивания противоположно направленных «холодовых» и «теп-
ловых» факторов, как было еще в 1927 г. предложено в работах
Базетта (см., например, [335]). Схема динамического баланса,
Рис. 7.15. Образование стационарных состояний в физиологических системах, а) Общий
случай, б) пример: система терморегуляции в трактовке Базетта. Схема а) представляет
собой общую схему, показанную на рис 7.2, причем для простоты в ней, как н в схеме б),
внешние воздействия v и w не указаны.
предложенная им, показана на рис. 7.15. Схема полностью совпа-
дает с приведенной здесь же общей схемой, если отождествить
обратную связь по возрастанию и уменьшению температуры с
первичными и вторичными темпами тепловых потоков.
В нескольких работах [274, 358] для системы терморегуляции
были рассмотрены и схемы нейронного уровня, в которых также
происходит уравновешивание противоположно направленных
факторов. В работе Дж. Блая [274] в качестве таких факторов
прямо указываются темпы теплопродукции и темпы теплопотерь
(рис. 7.16).
Таким образом, стационарные значения переменных состоя-
ния в физиологических системах возникают в результате
244
ГОМЕОСТАЗ ОРГАНИЗМА
[ГЛ. 7
уравновешивания двух противоположно направленных факто-
ров — первичных и вторичных темпов потоков вещества или
энергии, определяющих поступление веществ и их удаление из
компартментов. Управление темпами потоков в компартменталь-
ных моделях физиологических систем производится как пассив-
ными, так и активными механизмами регуляции, среди которых
Гипоталамус
6)
Рис, 7.16. Блок-схема структуры, регулирующей тепловой режим организма, а) Схема ней-
ронной модели. Изменение температуры кожи измеряется рецепторами, и информация пере-
дается в центр терморегуляции—гипоталамус, где имеется четыре типа нейронов. Резуль-
татом работы нейронной структуры является «баланс темпов теплопродукции и тепло-
потерь при некоторой температуре тела, близкой к оптимальной для некоторых функций
организма» 1274]; б) характеристика активности четырех основных типов нейронов, входя-
щих в схему а).
своеобразную роль ограничителей выполняют рецепторные ме-
ханизмы. Взаимодействие различных типов регуляции в физио-
логических системах и их компартментальных моделях может
быть достаточно сложным, чем и объясняется многообразие про-
цессов и явлений регуляции в физиологических системах.
ГЛАВА 8
КОЛИЧЕСТВЕННАЯ ОЦЕНКА
ГОМЕОСТАТИЧЕСКИХ СВОЙСТВ СИСТЕМ
8.1. Показатель гомеостатической способности системы
Гомеостаз физиологической системы в соответствии с кон-
цепцией, развитой в гл. 7, означает малую чувствительность су-
щественной части ее переменных состояния хг, i = 1, 2, ..., т,
к изменению входных возмущающих воздействий щ, / = 1 , 2, ...
...,/, т. е. малость соответствующих коэффициентов чувстви-
тельности о,-/ (7.8). Тогда гомеостатические свойства целостной
системы можно описать совокупностью всех значений 0,7, i =
= 1, 2, ..., т\ j = 1, 2....I, которую можно представить в
виде матрицы
°11 ° 12 • • °11
[о</] = °21 «22' • • а21 (8.1)
- °т' ° m2 • • ° ml -
Для линейных систем матрица (8.1) записывается в удобном
виде (7.11), а в случае нелинейных систем ее можно построить
для линеаризованных моделей. Эта матрица определяет количе-
ственные характеристики регуляторных реакций, возникающих
в системе. Чтобы описать некоторые обобщенные свойства систе-
мы, ее гомеостатическое поведение, целесообразно в качестве
меры этих свойств взять некоторые «усредненные» численные
характеристики элементов матрицы [<Т//], например, среднеквад-
ратическое значение коэффициентов чувствительности:
где ctij—весовые коэффициенты. Такая мера чувствительности
для оценки свойств переменных состояния была предложена
нами в [142]. В теории цепей аналогичные показатели, вводимые
для выходных переменных системы, рассматривались и ранее
(см., например, [272, 341]).
Весовые коэффициенты. Входящие в формулу (8.2)
весовые коэффициенты а,, выполняют две функции. Во-первых,
246
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ 8
они позволяют избежать неудобств, связанных с тем, что раз-
мерности о,/ в реальных задачах различны; для этого размер-
ность коэффициентов выбирается так, что попарные произведе-
ния ачоч безразмерны.
Во-вторых, соответствующим выбором этих коэффициентов
можно учесть относительную существенность, важность тех или
иных переменных для системы, поскольку постоянство различ-
ных переменных в биосистеме имеет различную значимость для
поддержания ее жизнедеятельности. Так, для выживания орга-
низма постоянство температуры внутренних органов — ядра
тела — важнее, чем температуры конечностей или кожи.
Вопрос выбора конкретных значений весовых коэффициен-
тов в реальных задачах оказывается достаточно сложным, по-
этому мы приведем лишь одно довольно общее соображение от-
носительно их определения. Ориентиром для нахождения этих
значений иногда могут быть
зависимости стационарных
уровней различных перемен-
ных в биосистемах от внеш-
них условий. Рассмотрим,
например, зависимости тем-
пературы ядра и кожи у
человека от температуры
внешней среды (рис. 8.1).
Поскольку обе переменные
имеют одну и ту же физи-
ческую природу, критерием
их относительной важности
может служить степень их
постоянства в нормальных
условиях; чем более поло-
гим является график зависимости, тем важнее постоянство со-
ответствующей переменной. Тогда и в некоторых патологических
ситуациях подобная оценка может сохраняться. Чувствитель-
ность температуры ядра к температуре среды в комфортных ус-
ловиях (tc 24—25°) составляет примерно 0,07, а чувствитель-
ность температуры кожи « 0,87. Тогда относительные веса этих
двух переменных можно взять обратно пропорциональными при-
веденным числам:
Рис. 8.1. Зависимость температуры ядра (рек-
о о
тальной температуры) / и температуры кожи
человека от температуры окружающей среды
в установившемся режиме
Если при подсчете среднеквадратических значений дополнитель-
но потребовать, чтобы
т I
(8.4)
<=1 /=»1
8.1)
ПОКАЗАТЕЛЬ ГОМЕОСТАТИЧЕСКОМ СПОСОБНОСТИ
247
(условие нормировки коэффициентов), то в нашем примере
вк + ая = 1, ииз (8 5) и (8.4) получаем ак — 0,075, ая = 0,925.
Такое рассмотрение может принести определенную пользу
при определении весовых коэффициентов и в других случаях,
хотя эта задача в общем виде оказывается значительно более
сложной и не имеет удовлетворительного решения несмотря на
то, что внимание ей уделяется уже достаточно давно: «Все, что
нам нужно, это возможность расположить переменные организ-
ма приблизительно в порядке их важности», — писал У. Р. Эшби
[259].
В некоторых практически интересных случаях используются
весьма специфические приемы классификации переменных со-
стояния по степени их важности для организма. Так, в системе
терморегуляции такая оценка может быть сделана на основании
того вклада, который вносится изменением температурного ре-
жима отдельных частей тела (и соответствующих переменных
состояния) в некоторый численный показатель, определяющий
включение центральных механизмов терморегуляции [298].
Напомним, что хотя функционирование активных механиз-
мов регуляции — таких, как дыхание и кровообращение — имеет
для организма кардинальное значение, описывающие эти меха-
низмы существенные переменные, за редким исключением, по
традиции не относятся к числу гомеостатируемых. Так, считает-
ся, что в системе кровообращения переменные, описывающие
давления в различных частях сердца и кровеносной системы,
должны поддерживаться на постоянном уровне, т. е. их харак-
теристики учитываются при анализе гомеостаза организма. В то
же время показатели энергетического баланса сердечной мыш-
цы, объемы крови в сосудистом русле и депо, частота сердеч-
ных сокращений, величина сопротивления сосудов при анализе
гомеостатических свойств остаются в тени. Поэтому для части
координат вектора состояния системы, описывающих пове-
дение активных механизмов регуляции, можно выбирать
ач ~ 0.
Средняя чувствительность системы о не всегда является
удобным мерилом гомеостатических свойств системы; хотелось
бы иметь «прямую» зависимость между качеством гомеостаза
и измеряющим его показателем. Поэтому целесообразно каким-
либо образом «обратить» показатель чувствительности о, введя
вместо него такую функцию h == f (ст), которая увеличивалась бы
с ростом гомеостатической способности системы. Простейший
вид такой функции — обратно пропорциональная зависимость
Л = (8.5)
248
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ. 8
Именно такой показатель и был впервые введен в работах [141,
142].
В принципиальном плане зависимость (8.5) не вызывает
возражений; однако при практическом использовании, возмож-
но, более удобными окажутся некоторые модификации этого
а)
20 18 18 14 12 10 8 в С,о&%
Рис. 8.2. Гомеостатические свойства системы доставки кислорода, а) Зависимость содержа-
ния оксигемоглобина НЬО2, напряжения кислорода артериальной раО2 и венозной РВО8 крови,
кровотока Q и альвеолярной вентиляции V от степени снижения процентного содержания
кислорода во вдыхаемом воздухе С. Размерности величии следующие: HbOt, Н (измеряется
по правой шкале), рО2 — мм рт. ст. (оба напряжения измеряются по правой шкале); Q и
V— % от нормы (измеряются по левой шкале); б) показатель й, вычисленный в функции С;
кривая /—о (С), кривая 2—показатель hv (С), вычисленный по формуле (8 5); кривая 3 — по-
казатель й2, вычисленный по формуле (8 7) при а=1.
выражения. Действительно, недостатком формулы (8.5) являет-
ся, по-видимому, относительно сильная зависимость показателя h
от ошибки измерения в области малых значений а. Так, если ве-
личина а определена с погрешностью До, то ошибка в вычисле-
нии h, равная Д/г, составит
Д/г =~Да =--^Да; (8.6)
да а2
8.1]
показатель гомеостатической способности
249
при малых а погрешность Д/г может быть очень большой. Если
учесть, что малые а соответствуют хорошему гомеостазу, то
ясно, что формула (8.6) будет давать большие ошибки именно
в наиболее важных для исследователя случаях.
Кроме того, обратно пропорциональная зависимость h от о
неудобна еще и потому, что при относительно больших значе-
ниях а в практически интересных случаях убывание h на гра-
ницах области гомеостаза оказывается недостаточно быстрым.
Поэтому для практических задач можно рекомендовать и
другие зависимости. Например, показатель
h — . J 2- (8.7)
1 4- аа2 ' '
лишен указанного недостатка и, кроме того, нормирован:
0</г<1 (8.8)
при изменении а от оо до 0.
Величина а в знаменателе позволяет по-разному оценивать
влияние а на показатель гомеостатических свойств. Значение
h = 1 достигается при о = 0.
Пример 81.1. Определим в качестве иллюстрации гомеостатическую
способность системы снабжения организма кислородом на основе данных,
приведенных в работе [95]. Зависимость пяти переменных этой системы от
процентного содержания кислорода во ’ вдыхаемом воздухе изображена на
рис. 8.2, а.
Предложенная выше методика определения h удобна для случая измере-
ния переменных состояния системы В данном примере модели системы нет, и
поэтому приходится сделать допущение, что величины, представленные
на рис. 8.2, а, служат переменными состояния некоторой модели системы кис-
лородного снабжения Тогда можно применить формулы (8.2) и (8.5) или
(8.7).
Введем безразмерные переменные, выбрав соответствующие масштабные
коэффициенты. В качестве масштабов выберем значения переменных при нор-
Q
мальных условиях (С = 20). Обозначим а = -^-,
, НЬО2 (С) _ НЬО2
11М НЬО2|с_20~ 95 ’
о О2 р 02
Хг (С)=-io- ’ Хз (С)=^о- ’ <8-9)
Xt = Too’’ Хз = Too •
д X
Тогда, например, для значения производной в точке v = 1 получаем
<?*! I 20 НЬО2 |с=20 — НЬО2 |с=16 _
dv Ip-] ~ <'5 IS-20 “ ’
Абсолютные значения производных в зависимости от v даны в табл. 8.1,
250
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ 8
I а б л и ц а 8.1
с дХ\ dv dv дх dv dv дх*. dv 1 1г h>
20 0,27 0,87 0,16 1,00 0,20 0,62 1,63 0,72
16 0,54 1,00 0,83 1,90 0,41 1,08 0,93 0,53
12 1,27 1,62 1,00 2,21 1,19 1,45 0,69 0,32
10 1,47 1,75 1,16 2,49 4,00 2,40 0,42 0,15
8 1,68 1,75 1,33 3,00 5,00 2,91 0,34 0,06
В таблице показатель вычисляется по (8 5), показатель /г2 — по (8 7)
при а = 1. Нормировочные коэффициенты в формуле (8 2) ач = 1. Зависи-
мости h\(C) и /г2(С) приведены на рис 8.2,6.
8.2. Гомеостаз нелинейных систем. Гомеостатическая кривая
При исследовании гомеостатических свойств нелинейных
компартментальных моделей мы можем провести линеаризацию
уравнений системы так, как это показано на рис. 6.4. Если в
исходной системе существует стационарный режим, то в рассма-
триваемом случае он существует и в линеаризованной си-
стеме. Наиболее важное для нас отличие нелинейного случая
от линейного состоит в том, что при линеаризации системы при
разных условиях внешней среды (различные V и У) матрицы
А, В, S, Р, R и Q на рис. 6.4 оказываются различными; в част-
ности,
A = A(V,W), B = B(V,W), (8.10)
где V » W означают, как и ранее, условия внешней среды и ре-
жим функционирования нелинейной модели биоснстемы, V и
W меняются в некоторой области рассматриваемых значений:
V s Qy, W с= Qu?.
Особенно часто в биосистемах приходится сталкиваться с
нелинейностями двух типов: параметрическими связями (напри-
мер, умножением переменных) и насыщением. Покажем, что при
линеаризации оба эти типа нелинейности приводят к тому, что
структура линеаризованной модели (вид описывающей ее мат-
рицы) остается неизменной во всех точках областей Q^,
но элементы матриц могут меняться в разных точках (V, W).
Нелинейности типа «насыщение» характерны для всех уров-
ней организации биосистем; часто встречаются они, например, в
механизмах активной регуляции физиологического уровня. Так,
ограничены пределы изменения переменных в активных меха-
низмах регуляции дых,ния и кровотоков, ограничены пределы
максимальной теплоотдачи через испарение, сопротивление со-
8 2]
НЕЛИНЕЙНЫЕ СИСТЕМЫ ГОМЕОСТАТИЧЕСКАЯ КРИВАЯ
251
судов органов и т. д. Для всех таких механизмов характерна
зависимость, показанная на рис. 3.4. Но наличие такой зависи-
мости как раз и означает, что при постоянной структуре систе-
мы коэффициент передачи от входа X к выходу У изменяется
с изменением значений переменной X и, следовательно, с изме-
нением значений V и W. На рис. 8.3 приведен характерный при-
мер такой зависимости — изменение гемодинамической прово-
димости сосудов в ответ на изменение давления крови, свой-
ственное механизмам ауторегуляции местного кровообращения.
а) 6)
Рис. 8.3. Нелинейная зависимость с насыщением а) Зависимость проводимости сосудов от
перфузионного давления для механизма ауторегуляции кровотока в лапе собаки, б) коэф-
фициент передачи для этой зависимости
Перемножение переменных характерно для так называемых
параметрических связей в системах управления. Их линеариза-
цию мы рассматривали выше (см. рис. 7.10). И в этом случае
структура части линеаризованной системы, замещающей нели-
нейный элемент в исходной системе, — параллельное соедине-
ние двух каналов — остается неизменной, а меняются только
значения коэффициентов усиления в этих каналах. Ана-
логичным образом дело обстоит и для многих других видов не-
линейностей (степенные функции, перемножение степенных
функций и т. д.).
Следовательно, во всех рассмотренных случаях в линеаризо-
ванных системах размерности и вид матриц А, В, S, Р, R, Q на
схеме 6.4 не меняются в различных точках (И, IF); меняются
лишь значения их коэффициентов. В частности, коэффициенты
передачи от одною элемента схемы к другому могут обра-
щаться в нуль. Это означает, что соответствующий механизм
регуляции в таких условиях или не функционирует, отключает-
ся, или что он функционирует «па пределе возможностей», так
что дальнейшее «ухудшение» условий, требующее интенсифика-
ции работы механизма, не приводит к ожидаемому увеличению
его выходной переменной. В обоих случаях в линеаризованной
модели такой канал как бы «выпадает» из схемы. Ослабление
регуляторных свойств канала (или его выпадение) может
252
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ. 8
привести к образованию гомеостатических кривых, характерных
для поведения внутренних переменных состояния организма.
Продемонстрируем, как возникает гомеостатическая зависимость
на примере простой компартментальной модели.
Рассмотрим схему с двумя компартментами, показанную на
рис. 8.4, которой можно дать следующее физиологическое тол-
кование. Независимый темп потока вещества W должен урав-
новешиваться зависимым темпом У, который регулируется
Рис. 8.4. Простая модель с параллельными каналами регуляции. Переменная модели*
рует пассивный канал регуляции темпа потока вещества У, переменная Xi—активный
канал.
двумя механизмами, активным и пассивным. Эти два механиз-
ма характеризуются переменными состояния и Х2 (например,
W — темп потребления О2 в тканях некоторого органа в ходе
процессов метаболизма, Xt — концентрация О2 в тканях органа,
Х2—кровоток через сосуды, питающие ткань). Возмущающий
сигнал V имеет смысл концентрации моделируемого вещества
в окружающей среде (в примере — напряжение О2 в крови).
Уравнения системы имеют вид
х,= ’ [_г + ^2(У-х,)],
71 (8.11)
Х2 = ^[-Г + ^2(У-Х,)1,
где обозначено k = kik2. Слагаемое
Y = kX2(V-Xi) (8.12)
имеет следующий смысл: по закону диффузии единица носителя
(единица массы крови) уносит из тканей ki(V — Xi) единиц ве-
щества, а весь носитель — в k2X2 раз больше.
Решение уравнения (8.12) в статическом режиме можно
получить следующим образом (напомним, что из-за вырожден-
ности матрицы системы обычное уравнение (5.50) непримени-
мо) . В статике, как обычно, У = W. Если на систему, находя-
щуюся в стационарном состоянии Xi, Х2, подействовало возму-
щение v, то все переменные перешли в новое стационарное со-
стояние, получив приращения Xi и х2:
(Х2+х2)'к‘(-Хх-хх+ V + v)~W, (8.13)
8.21
НЕЛИНЕЙНЫЕ СИСТЕМЫ. ГОМЕОСТАТИЧЕСКАЯ КРИВАЯ
253
откуда при малых хь х2 и v находим
X2x1-x2(V-Xi)-oX2 = 0. (8.14)
Сравним приращения Xi и х2. Как в исходном, так и в новом
стационарном режиме в = V — W = О, и поэтому
оо
xt = 8 (/) dt - ,
« (8-15)
х2 = -4- 8 (/) dt ,
1 2 J * 2
О
где символом е обозначен интеграл от в(0—«накопленная»
ошибка. Подставляя (8.15) в (8.14), получаем
Тогда
где
TiTA
Ti(Xi-V)+TA ’
Xi =? aT2v,
x2 = aT iv,
(8.16)
(8.17)
a — const Г1 (X[ _ yj + тл • (8.18)
Соотношения (8.17) имеют важный смысл: если активная
цепь регуляции обладает значительно большим быстродейст-
вием, чем пассивная, Т2 С Tt, то сдвиг х2 переменной Х2 актив-
ной цепи регуляции много больше, чем сдвиг Xi пассивной пере-
менной
х2>*1, (8.19)
и переменная Xi практически не меняется при изменении усло-
вий внешней среды:
*1 < <Эл, .. ,
— <1 или -^<1.
(8.20)
Обратим еще раз внимание читателя на то, что чувствитель-
ности переменных Xi и Х2, находящихся в параллельных каналах
регуляции, связаны обратным — илн, используя физиологиче-
ский термин, реципрокным — соотношением: если будет
уменьшаться в силу тех или иных причин, величина будет
возрастать.
254
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ 8
Для более общего случая аналогичный вывод получается с
помощью так называемых инвариантов чувствительности. Ин-
вариантом чувствительности называется некоторая комбинация,
составленная из функций чувствительности или коэффициентов
Рис. 8.5. Простая модель, демонстрирующая возникновение гомеостатической кривой
в системе. Постоянство Х{ нарушается в связи с тем, что у регулирующего механизма Х<
из-за насыщения падает коэффициент усиления и ухудшаются регуляторные свойства
чувствительности, остающаяся постоянной величиной при изме-
нении тех или иных условий в системе или внешней среде [362].
Для компартментальных моделей инварианты чувствительности
получаются, если продифференцировать по V условие стацио*
парности У = W. В нашем случае,
используя (8.12), получаем
-^\kX2 (/-Х1)] = О, (8.21)
или
дХг _ „
дУ Л2 дУ
(8.22)
Рис. 8.6. Гомеостатическая кривая
в простой модели, показанной иа
рис. 8.5. Пока переменная Х2 вклю-
чена параллельно переменной Xi,
она берет нагрузку на себя, так что
переменная Xi поддерживается на
относительно постоянном уровне.
После насыщения переменной Х2
(точки V* и V**) участок плато го-
меостатической кривой переходит в
более крутые участки.
где Xi, Х2, V — значения перемен-
ных для стационарных режимов
системы. Выражение (8.22) и пред-
ставляет собой инвариант чувстви-
тельности для нашей модели. От-
сюда
ЭХ, . дХ2
дУ дУ
(8.23)
где через b > 0 обозначен постоянный коэффициент, равныц
(К — Х1)/Х2. Из (8.23) ясно, что любое уменьшение чувстви-
тельности активного механизма Х2 приведет к ухудшению гомео-
стаза внутренней среды системы — возрастанию чувствитель-
ности стационарного значения концентрации Хр «Постоянство
Л4
8.2J
НЕЛИНЕЙНЫЕ СИСТЕМЫ. ГОМЕОСТАТИЧЕСКАЯ КРИВАЯ
255
одних переменных может быть связано с высокой активностью
других» [259, стр. 109].
Введем теперь ограничение в канал, моделирующий актив-
ный механизм регуляции, включив в схему нелинейный элемент
(рис. 8.5). При увеличении V на v в соответствии с (8.17) сигна-
лы У! и Х2 получают приращения X] и х2; точка равновесия Х2
смещается вправо и постепенно выходит из зоны линейности в
Рис. 8.7. Зависимость показателя гомеостатической способности системы. В наиболее
часто встречающемся случае на систему воздействует только один внешний фактор (а);
в этом случае показатель гомеостатической способности представляет собой плоскую кри-
вую. J3 более общем случае показатель h образует некоторую поверхность в пространстве
внешних возмущающих сигналов (б)
(8.24)
(8.25)
зону нелинейных эффектов, когда коэффициент передачи от
Х2 к Х2 падает. При этом, как следует из (8.23),
дХ, дХ,
дУ дУ ’
дХ,
и чувствительность начинает возрастать; при достижении
в точке Х2 = Х2 сигналом Х2 значения Х2 дальнейшее увели-
, дХ'2
чение Х2 не приводит к увеличению Х2, т. е. = 0. Поэтому,
начиная с этого момента,
дУ *'
Аналогичная последовательность событий происходит и при
уменьшении Х2 до Х2 . Таким образом, при изменении V сово-
купность стационарных точек системы на рис. 8.5 образует го-
меостатическую кривую, показанную на рис. 8.6. Относительное
постоянство (участок плато) переменной внутренней среды обес-
печивается за счет регулятивных изменений переменной Х2;
активный капал принимает нагрузку на себя. После выхода пе-
ременной Х2 на уровень ограничения (точки Х2 и х2 )
256 КОЛИЧЕСТВЕННАЯ ОЦГНКА ГОМЕОСТАЗА [ГЛ. 8
переменная Х2 перестает демпфировать переменную Xit и участок
плато переходит в более крутые участки кривой. Хотя рассмот-
ренный пример носит довольно абстрактный характер, он позво-
ляет понять основные принципы возникновения гомеостатиче-
ской кривой в биосистемах.
Если теперь построить зависимость показателя гомеостати-
ческой способности системы h от условий среды V, то получится
кривая, показанная на рис. 8.7, а. Области плато отвечают ма-
лые значения о и, следовательно, высокие значения й. Этот диа-
пазон обозначен на рисунке как £2л, а через Qs обозначена об-
ласть стационарности, включающая как участок плато, так и
участки с негомеостатическим поведением сигнала В более
общем случае зависимость h(V) показана на рис. 8.7,6, где
предполагается, что V — вектор размерности 2, т. е. V =
= [^ V2]\
8.3. Интегральные оценки
гомеостатических свойств системы
^Жизнеспособность организма сохраняется до тех пор, пока
регуляторные системы поддерживают адекватное снабжение его
энергетической системы. В живой системе «максимально воз-
можные механизмы защиты всегда сильнее максимально воз-
можных отклоняющих воздействий» [10].
Чем же можно охарактеризовать «силу» гомеостатических
механизмов?} В разд. 8.1—8.2 мы рассмотрели поведение систе-
мы при малых возмущениях условий среды, дав количественную
характеристику реакций системы, протекающих в соответствии
с принципом Ле-Шателье. Механизмы с лучшими локальными
гомеостатическими характеристиками дают в среднем меньшие
сдвиги состояния при тех же внешних возмущениях, причем для
наиболее существенных переменных сдвиги состояния минималь-
ны. Однако для общей оценки гомеостатической способности си-
стемы локальных характеристик гомеостаза недостаточно: важен
также и размер области гомеостаза, т. е. ширина плато гомео-
статических кривых для всех существенных переменных состоя-
ния системы.
’ Итак, «сила» гомеостатических механизмов определяется
двумя факторами: локальной способностью системы поддержи-
вать гомеостаз (величиной показателя h) в каждой точке, т. е.
наклоном плато, и шириной плато на гомеостатических кривых
переменных системы. Обе характеристики качества можно объе-
динить в едином интегральном показателе гомеостатической
способности системы
Н = J Л (V)dV. (8.26)
а
8.41
РЕСУРСЫ СОХРАНИТЕЛЬНЫХ СВОЙСТВ
257
Интеграл здесь берется по всей области стационарности
где h > 0. Показатель Н представляет собой объем, ограничен-
ный поверхностью h(V), как это иллюстрирует рис. 8.7,6. На
практике часто встречается случай одной переменной (тепловая
нагрузка, кислородное голодание и т. д.). В этом простейшем
случае показатель Н равен площади под кривой h(V)
(рис. 8.7, а).
Пример 8 3.1. Определим интегральную гомеостатическую способность
системы регуляции осмотического режима у бокоплавов, воспользовавшись
гомеостатическими зависимостями, показанными для двух видов бокоплавов
на рнс. 2 1,6. Введем следующие обозначения: V — осмотическая концентра-
ция среды, х — то же для крови (масштабных множителей здесь не требуется,
поскольку V и х измеряются в одних и тех же единицах). Тогда зависимости
a(V) для обоих видов показаны на рис. 8 8, а, а функция h(V), вычисленная
Рис. 8.8. Сохранительные способности системы регуляции осмотического режима у боко-
плавов. а) Чувствительность переменных внутренней среды (кровь) у Gamtnarus oceanh
cus (1) и Gamtnarus fasciatus (2) к изменениям концентрации в среде; б) показатель гомео-
статической способности h у тех же видов Показатель ti определяется как площадь под
кривыми 1 и 2 на рис. 8 8,6, соответственно.
по формуле (8.7) при а = 5, — на рис. 8.8, б. Определяя графически площадь
под кривыми в пределах от 0 до 1, находим = 0,42, Н2 — 0,24. Удобно
при интегрировании сохранять размерность V, тогда величина Н получается
непосредственно в тех единицах, которые определяют действующее возмуще-
ние: для первого вида бокоплавов получаем Н = 0,42 моль/кг, для второго —
Н == 0,24 моль/кг.
8.4. Ресурсы сохранительных свойств в системе
^Способность биосистем сохранять равновесное стационарное
состояние и гомеостаз при существенных изменениях условий
среды является одной из важнейших характеристик, позволяю-
щих живым системам приспосабливаться к окружающей среде и
выживать при различного рода стрессорных воздействиях.
Тем самым сохранительные свойства биосистем оказываются
тесно связанными с их адаптационными и компенсаторными
258
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
(ГЛ. 8
характеристиками. Обзор физиологических (поскольку эти про-
цессы исследованы наиболее полно на организменном уровне) и
общебиологических представлений об адаптации, приспособи-
тельных и защитных механизмах биосистем можно найти в ра-
ботах [2, 10, 58а, 62, 82, 153, 185, 197]. Мы сейчас обсудим воз-
можность формального описания некоторых интегральных ха-
рактеристик сохранительных свойств биосистем, основанных на
введенном в предыдущем разделе показателе Н. Эти характе-
ристики могут быть использованы и при исследовании адапта-
ционно-компенсаторных процессов и механизмов в биосистемах
([146], см. также разд. 9.1).
Рис. 8.9. Резервы гомеостаза в системе, а) Общий случай; при V=V* резерв против по-
вышения V представляет собой заштрихованную частыми линиями площадь под кри-
вой й (V) справа от вертикальной пунктирной линии, б) пример системы кислородного
режима организма; прн С = 12 об. % резерв против дальнейшего понижения С представлен
заштрихованной областью. Эта область довольно мала — напряжение механизмов велико,
ресурсы системы близки к исчерпанию.
Итак, обратимся снова к кривой, показывающей зависимость
h от V (рис. 8.9, а). Если состояние окружающей среды зафик-
сировано на уровне Ео, то гомеостатическую кривую можно мыс-
ленно разделить на две части. Часть, лежащая вправо от пунк-
тира V = Ео, окажется существенной только в том случае, если
в исследуемой ситуации переменная V имеет тенденцию к воз-
растанию. Часть, лежащая слева от точки Уо, имеет для био-
системы значение только при уменьшении V.
Поэтому площадь под кривой h(V) при V Ео характери-
зует резерв гомеостаза системы при возрастании V, а площадь
под этой кривой при V < Ео — резерв при уменьшении V от
V = Ео. Обозначим эти резервы через N+ и N-, соответственно:
оо
JV+ = \h(V)dV, (8.27)
Vo
v«
AL = ^h(V)dV. (8.28)
S.4J РЕСУРСЫ СОХРАНИТЕЛЬНЫХ СВОЙСТВ 259
Очевидно, N+ 4- N- = Н, т. е. интегральный показатель го-
меостатической способности дает количественную оценку сохра-
нительных ресурсов системы.
Биосистема находится в предпочтительном состоянии, если
она «настроена» так, что в нормальных, привычных для нее ус-
ловиях (точка Ио на рис. 8.9, а) резервы W+ и N- равны между
собой, т. е. площадь под кривой h(V) в этой точке делится
пополам.
^Сотя положение и форма кривой h(V) определяется прежде
всего генетическим аппаратом, унаследованным данной биоси-
стемой, известное влияние на нее оказывает и внешняя среда.
Для организмов, в частности, условия окружающей среды на
ранних стадиях развития «запечатлеваются», фиксируются в
структуре его регуляторных систем. Такое явление носит назва-
ние импринтинг (см., например, [125, 149]). Так, при помещении
животных в молодом возрасте в измененные условия (высокие
или низкие температуры) они приобретают приспособительные
изменения терморегуляции и шерстного покрова, которые спо-
собствуют смещению кривой h(V) в соответствующую сторону
[149]. У растений, акклиматизированных к условиям северного
климата, область гомеостаза также оказывается сдвинутой в
область низких температур [201].
Удобно считать, что еслц при некотором V = V* выполняет-
ся соотношение
М+ = М_, (8.29)
то регулирующие механизмы системы не напряжены; при сме-
щении точки V* регуляторные механизмы системы напрягаются.
Определим степень напряжения регуляторных механизмов систе-
мы как относительный резерв гомеостаза, истраченный системой
в точке V*. Следует, конечно, различать напряжение механиз-
мов при повышении или понижении V. Можно поэтому обозна-
чить степень напряжения механизмов системы при повышении
V через v+, а при понижении — через v“. Однако в реальных си-
туациях обычно не вызывает сомнений направление изменений
V, и знак «+» или «—» в этом случае можно опускать. Следо-
вательно, можно положить
—. 100%. (8.30)
Пример 8.4.1. Вычислим резерв гомеостаза системы регуляции кисло-
родного режима против понижения концентрации кислорода во вдыхаемом
воздухе (рис. 8 2,6). Построенная в более удобном масштабе зависимость
й(С) приведена на рис. 8.9,6;
20
N- = $ А (С) dC = 5,2 (об.%). (8.31)
о
260
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ. 8
У нас нет данных о гомеостатических кривых системы при С > 20, и мы
не можем определить ее другой половины и найти величину И в соответ-
ствии с (8.26). Однако, если допустить, что при С = 20 выполнено соотно-
шение (8.29), то можно определить, как увеличивается напряжение регуля-
торных механизмов системы при уменьшении С. Так, при С = 20 напряжение
по определению равно нулю, а при С = 12 оио составляет уже
V (12) = Ю0% = 69%, (8.32)
где N = 0,8 — площадь, заштрихованная на рис. 8.9, б. Напряжение механиз-
мов велико; гомеостатические способности системы близки к исчерпанию.
Разумеется, полученные оценки носят относительный харак-
тер, их величина зависит от выбора коэффициентов «ц в форму-
лах для о, от вида функции h(V) и, в частности, от значения
коэффициента а в формуле (8.7). Однако эти величины, вычис-
ляемые при одних и тех же предположениях для разных био-
систем (организмов, различных органов в одном и том же орга-
низме и т. д.), позволяют делать обоснованные выводы об их от-
носительных сохранительных возможностях. Пример такого рода
мы рассмотрим в следующей главе (разд. 9.2, пример 9.2.1).
8.5. Сохранительные ресурсы системы терморегуляции
В рамках общих предположений, развиваемых в настоящей
работе, главной целью системы регуляции теплового режима в
животном организме можно считать обеспечение адекватного
темпа рассеивания тепла, равного темпу теплопродукции. Темп
же теплопродукции — скорость образования тепла в ходе про-
цессов жизненной активности — определяется более высокими
уровнями управления в организме, включающими механизмы
мотивации и управления поведением животного. Поэтому задаю-
щим сигналом в блок-схеме системы терморегуляции, построен-
ной в виде компартментальной модели открытой системы, яв-
ляется темп теплопродукции, а управляемым — темп тепло-
отдачи.
Если обозначить вектор темпов теплопродукции в различ-
ных компартментах модели организма через w, а вектор вторич-
ных темпов, описывающих процессы теплоотдачи из этих же
компартментов, через у, то целью первого уровня в системе яв-
ляется выполнение равенства (7.1), т. е. у = w. В работах по
терморегуляции обычно предполагается, что такое равенство
должно выполняться для организма в целом, т. е. суммарный
темп теплопродукции должен быть уравновешен суммарным
темпом теплопотерь. Соотношение (7.1) является, таким обра-
зом, более строгим представлением или расшифровкой этого об-
щего положения.
8.5]
СОХРАИИТЕЛЬНЫЕ РЕСУРСЫ СИСТЕМЫ ТЕРМОРЕГУЛЯЦИИ
261
Соотношение (7.1) достигается при участии всех регулятор-
ных механизмов, как пассивных, так и активных. В частности,
у организмов, не обладающих выраженным температурным го-
меостазом, главную роль в обеспечении теплового режима иг-
рают пассивные механизмы теплопередачи, когда температура
каждого из органов в организме непосредственно влияет на темп
теплоотдачи. У гомойотермных организмов, однако, имеется
весьма развитый терморегуляторный аппарат, специфической
целью которого является, по-видимому, недопущение выхода не-
которых существенных температур за пределы своих диапазонов
безопасности.
Так, использование точных методов измерения температуры
показало, что понижение температуры окружающей среды
(до 5—2 °C) вызывает снижение температуры тканей мозга (ги-
поталамуса) примерно на 0,25°, а затем возникающая терморегу-
ляционная реакция дрожи препятствует дальнейшему падению
температуры мозга, но не возвращает ее к исходной величине
[79 стр. 139].
Такие опыты заставляют думать, что речь идет не столько
о стремлении системы терморегуляции вернуть температуру тела
к какой-то исходной установочной точке, сколько сделать внут-
реннюю температуру просто малочувствительной к изменению
температуры окружающей среды, поддерживать ее значения в
допустимых пределах.
В литературе по физиологии терморегуляции рассмотрение
часто ограничивается регуляторными механизмами именно этого
уровня. Вообще, основное внимание исследователей направлено
на трактовку регуляторной роли именно специфических термо-
регуляционных механизмов [79, 273, 286, 309, 361], хотя в при-
роде терморегуляция организмов обеспечивается не только и
даже не столько специфическими физиологическими механиз-
мами регуляции, сколько, с одной стороны, «подсистемными» не-
специфическими механизмами пассивной регуляции, а с дру-
гой — активными поведенческими механизмами «надсистемного»
уровня ([277, 287], см. также разд. 2.6).
При изменении условий окружения (вектор о) и условий
функционирования и режимов работы организма (вектор о>) в
нем возникают переходные процессы по всем переменным со-
стояния х = x(v,w,t), направленные на выполнение равенства
у = w. В ходе этих переходных процессов некоторые из пере-
менных х могут принять неприемлемые для нормальной жизне-
деятельности организма, значения. Чтобы предотвратить выход
переменных за пределы безопасности, во время переходных про-
цессов включаются рецепторные механизмы терморегуляции
(см. например, характеристики нейронов третьего и четвертого
типа на рис. 7.16).
262
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
1ГЛ. 8
Таким образом, функционирование терморецепторных меха-
низмов регуляции в организме можно считать направленным на
выполнение ограничений, налагаемых на переменные состояния
в системе. Во время действия сильных внешних или внутренних
возмущающих факторов, когда возникающие в системе переход-
ные процессы грозят выходом существенных переменных состоя-
ния за границы пределов безопасности, эти механизмы гаранти-
руют выполнение ограничений, налагаемых на температуру внут-
ренних органов тела, как об этом говорилось в разд. 7.9.
Описываемая ниже модель системы терморегуляции в орга-
низме построена именно на этой концептуальной базе и является
поэтому принципиальной: С целью лучшего понимания механиз-
мов, лежащих в ее основе, мы будем моделировать не организм
человека, а организм животного с относительно слабо выражен-
ными гомеостатическими свойствами — американского опоссума
[166]. Гомеостатические температурные кривые для этого живот-
ного приведены на рис. 2.1, а.
За основу модели возьмем рассмотренную в разд. 5.8 систему
пассивной регуляции темпов теплоотдачи механизмами типа
диффузии (см. уравнение (5.83) и рис. 5.7). Это уравнение с
учетом потерь тепла с испарением (—да;) можно записать в виде
Тi%! = k2 (х2 — Xi) — ki (Xi — t>i) — wi,
Т2х2 = k3 (х3 — х2) — k2 (х2 — Xj) + w2, (8.33)
Г3Х3 = — k3 (х3 — Х2) + w3.
Не имея точных данных о физиологии опоссума для иденти-
фикации модели на рис. 5.7, ограничимся иллюстративными зна-
чениями коэффициентов, положив k\ = k2 = k3 = 2. Кроме
того, будем считать, что темп теплопродукции от метаболизма
W3 = 8, от мышечной активности w2 = 4 и темп теплопотерь с
испарением Wi = 4 (все величины измеряются в условных без-
размерных единицах). При выбранных значениях задающих сиг-
налов и параметров системы решение уравнения (5.83) в стати-
ческом режиме имеет вид
w* -|- W2 — ш 1
ki '
W3+W2—W1 . Из + W2 /О
ki----------k2 ’ (8-34)
Шз + ! Шз + W2 W3
ki k2 k2
х2=ц1-ЬЮ, X3 = yi+14. (8.35)
При Vi — 23° уравнение (8.35) дает значение температур ко-
жи, мышечного слоя и ядра 27°, 33° и 37°, что довольно близко
соответствует режиму бодрствования животного.
Х1 = У1 +
х2 — Vi +
Хз = 1>1 +
или
Xi = Vi 4- 4,
8 5]
СОХРАНИТЕЛЬНЫЕ РЕСУРСЫ СИСТЕМЫ ТЕРМОРЕГУЛЯЦИИ
263
Уравнение (8.35) означает, что одна пассивная регуляция
(без включения активных физиологических механизмов) не мо-
жет обеспечить температурного гомеостаза, хотя и поддерживает
систему в стационарном режиме. Чувствительность температуры
тела (под температурой тела в модели понимается температура
ядра х3), так же как и чувствительность остальных переменных
х2 и Xi высока:
дхз дх2 dxi
dvi dvi dv!
(8.36)
Зависимость Xi от Vt дана пунктиром на рис. 8.10, а.
Покажем теперь, что введение активных механизмов регуля-
ции, первоначальная цель которых — улучшение отвода тепла
ъ
1,0 -
0,5 -
0,075
—\ 0,157
0,107
О 10 20 30 40t°C
Рис. 8.10. Сохранительные свойства системы терморегуляции, а) Гомеостатические кривые
построенные для модели терморегуляции. Активные механизмы обеспечивают образование
плато (кривые / и 2); их отключение приводит к вооиикновению крутых краев гомеоста-
тической зависимости. Пассивные механизмы ие могут дать хорошего гомеостаза в системе
(кривая 4). При отсутствии или насыщении активных механизмов в системе возникают ста-
ционарные зависимости, график которых идет параллельно прямой линии x=t’i (линия 3);
б) показатель Л для кривой 1. Видно, что основной вклад в величину И (площадь под
кривой h (t°)) дает плато гомеостатической зависимости.
(активное управление темпом теплоотдачи), приводит к возник-
новению температурного гомеостаза. Из многочисленных меха-
низмов терморегуляции, рассмотренных, например, в [79], огра-
ничимся в данном примере двумя: изменением темпа теплопро-
дукции мышечного слоя w2 при сдвигах переменных хц х2, х3 и
изменением темпа потоотделения при вариациях х3 и vt.
В простейшем случае это можно осуществить в модели сле-
дующим образом. Пусть в системе при некотором —vf уста-
новился стационарный режим xf, х®, х£, и в ответ на малое
отклонение Дщ реагируют не только пассивные механизмы, отве-
чающие изменениям концентраций тепла в компартментах —
температурным сдвигам Дхь Дх2 и Дх3 в соответствии с (8.34),
264
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ. 8
но и происходят малые сдвиги величин Wj и w2:
&Wl = — (х3 — хз) — k8 (и! — у!),
Аку2 = — k5 (х! — х9 — k6 (х2 — х2) — k7 (х3 — Хз),
т. е. увеличение потоотделения при повышении 1Ц, дрожь — при
понижении. Напомним, что при исследовании гомеостатических
свойств речь идет только о линейном приближении; исходная за-
висимость Wi и w2 от внутренних условий в организме и внешних
условий может быть достаточно сложной. Коэффициенты
— k% определяют чувствительность активных механизмов к из-
менению соответствующих переменных; мы будем здесь считать,
что вплоть до достижения переменными активных механизмов
Wi и w2 некоторых предельных значений (насыщения) величины
этих коэффициентов остаются постоянными.
Чтобы не увеличивать порядок системы, мы здесь не вводим
в модель новых переменных состояния при включении дополни-
тельных механизмов регуляции. Это допустимо потому, что в
системе терморегуляции гомеостаз традиционно рассматривается
как постоянство одних лишь температурных переменных; рас-
смотрение гомеостаза системы в более общем смысле потребо-
вало бы оценки и тех «усилий» системы, которыми обеспечивает-
ся постоянство температуры, и в показатель гомеостатической
способности системы в соответствии с (8.2) следовало бы
включать и переменные механизмов дополнительных каналов
регуляции. В этом случае пришлось бы ввести в систему соот-
ветствующее число дополнительных компартментов, как это ре-
комендовалось выше в общем случае (разд. 7.5). Поскольку, од-
нако, сейчас мы интересуемся только гомеостатическими свой-
ствами температурных переменных, достаточно рассмотрения
трех компартментов, с которыми мы имели дело в пассивной
системе в разд. 5.8.
Система с активными механизмами регуляции описывается
уравнением
Т1X1 = k2 (х2 — Xi) — ki (xi — У]) — wt +
Т2х2 = ^3(х3 — х2) — k2(x2 — Xi) + ay2 — Ада2, (8.38)
Т’зХз = — (х3 — Хг) + w3.
Стационарный режим этой системы находится аналогично
(8.34). Положив для простоты — 1, j = 4.........8, имеем
Х2 == 20,8 + 0,27щ, х2 = 31,2 + 0,08щ, х3 = 35,2 + 0,08иь (8.39)
Чувствительность х к v упала:
4^- = 0,27, = 0,08, 4е1 =0,08; (8.40)
ovi ’ ’ 001 1 ’ dvi v 7
гомеостатические свойства системы резко улучшились.
8.5]
СОХРАПИТЕЛЬНЫЕ РЕСУРСЫ СИСТЕМЫ ТЕРМОРЕГУЛЯЦИИ
265
Рассмотрим теперь, как образуется гомеостатическая кри-
вая. Для участка изменения Vi, на котором активные механиз-
мы регуляции не выходят на ограничение, стационарные значе-
ния переменных хь х2 и х3 изменяются в соответствии с (8.39),
но при этом значения w по мере увеличения Vi также увеличи-
ваются в соответствии с (8.37), где за Xs приняты значения 27°,
33°, 37°, a vf = 23°.
После достижения предельных значений переменных wi и
w2 механизмы (8.37) «насыщены», их дальнейшее изменение не-
возможно, и уравнения (8.37), определяющие значения прира-
щений и Дда2, заменяются на уравнения
Дш, — Дщ2 = 0, (8.41)
т. е. система, начиная с точки насыщения, снова описывается
уравнениями пассивной регуляции (8.33); чувствительность Xi,
х2 и х3 к изменению Vi снова возрастает до значения 1.
Из рис. 2.1, а видно, что при понижении температуры точка
излома (момент начала насыщения активных механизмов) рас-
полагается около значения v\ = 8 °C. Положим, что насыщение
достигается точно при ц* = 8°, а при vt v* значения kj = О,
j = 4, ..., 8. Аналогичное явление насыщения происходит и
при повышении температуры до 33 °C.
Тогда совокупность точек стационарного равновесия в ком-
партментальной модели терморегуляции состоит из трех участ-
ков: плато, описываемого уравнением (8.39), в диапазоне тем-
ператур примерно от 8° до 33 °C, и двух крутых участков при
щ < 8° и v2 > 33°, описываемых уравнениями типа (8.35).
Крайние участки соответствуют ситуациям, в которых из-за на-
сыщения активные механизмы не принимают участия в регуля-
ции тепловых потоков в организме.
Гомеостатическая кривая показана на рис. 8.10, а сплошной
линией /. Из изложенного ясно, что характер гомеостатической
кривой определяется структурой системы (количеством и видом
связей), значениями внутренних коэффициентов передачи и,
что является весьма существенным, величиной теплопродукции
w2, w3 при исходном значении щ.
Можно поэтому ожидать, что снижение уровня активности
животного приведет к изменению вида гомеостатической кривой.
Покажем, что это действительно так. Допустим, что днем, в пас-
сивный период жизнедеятельности, сниженная мышечная актив-
ность приводит к меньшей теплопродукции w2 = 1, т. е.
и> = [4 1 8]т (8.42)
(остальные тепловые потоки считаем неизменными). Решая си-
стемы уравнений (8.33) и (8.38) при новых значениях скоростей
тепловых потоков (8.42), находим зависимость 2, также
268
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ 8
Пример 8.6.1. Пусть в некоторой нелинейной системе зависимость пе-
ременной внутренней среды х от внешнего сигнала v имеет вид, показанный
на рис. 8.11, а, где переменная меняется в зависимости от и по характерной
для физиологических систем «гомеостатической зависимости». Обе перемен-
ные измеряются в некоторых условных единицах не обязательно одной и той
же размерности.
Рис. 8.11. Сравнение показателей гомеостатической способности системы, вычисляемых на
основе простых и логарифмических коэффициентов чувствительности, а) Исходная зави-
симость х(о); б) коэффициент чувствительности а (кривая /) и логарифмический коэффи-
циент чувствительности р (кривая 2); в) показатель h, построенный на основе логарифми-
ческих коэффициентов чувствительности, приводит к необоснованной переоценке гомеоста-
тических свойств системы в иачеле плато (точки о > 1 кривой 2) и иедооцеике их в конце
плато (точки о >2,5).
в)
Сравним для этого случая показатели гомеостатической способности си-
стемы й, вычисляемые по формулам h = или h — р“*.
Найдем зависимость р(п) для кривой на рис. 8.11, а. Для точек первого
участка 0 < v <Z 1 имеем v = х, 1» и р— 1- Для участка «плато»,
дх
1 р < 4, получаем — 0,33, так что, например, в начальной точке этого
участка а =— = 0,33 При и^4 (третий участок), например, в точке
х = 2 находим р = 2. Полученная зависимость приведена на рис. 8.11,6
(кривая 2).
8.71
ФОРМАЛИЗОВАННЫЙ ПОКАЗАТЕЛЬ ГОМЕОСТАЗА
269
Аналитическое выражение для каждого из трех участков этой зависимости
можно легко получить, воспользовавшись формулами статьи [299].
Построим теперь, как и прежде, зависимость коэффициента чувствитель-
ности а от V. Поскольку размерность х и v в принципе может быть раз-
лична, необходимо ввести некоторый масштабный множитель, имеющий раз-
[v 1 „ ,
— I. В качестве такого множителя удобно брать отношение значе-
ний и0 н хо в некоторой точке, соответствующей «нормальным» условиям
функционирования системы. В данном случае за такую точку можно взять
середину участка плато на кривой, положив = 2,5. Тогда х0 = 1,5 и
<т(и) = -^-^-(и); (8-46>
л о (JV
зависимость (8.46) также приведена на рис. 8.11,6 (кривая /). В точке о =
2 5
= ио, например, а = -jy • 1 = 1,67
На рис. 8.11, в приведены зависимости показателя гомеостатической спо-
собности системы h(y) для случая оценки по логарифмическому коэффи-
циенту чувствительности р (кривая 2) и по коэффициенту а (кривая /). Срав-
нивая эти кривые, видим, что при использовании логарифмического коэффи-
циента чувствительности получается резкое изменение показателя h в точках
v = 1 и v = 4, где имеются перегибы или изломы кривой х(и). Такое изме-
нение показателя, строго говоря, не является логическим следствием измене-
ния самих гомеостатических свойств системы—ее гомеостатической зависи-
мости x(v), а объясняется прежде всего выбором формулы логарифмической
чувствительности. Кривая 1, построенная на основе коэффициента чувстви-
тельности <т, дает более адекватную физиологическую картину.
Поэтому при анализе сохранительных свойств биологических
систем нам кажется более предпочтительным использование
коэффициентов о. Разумеется, выбор между о и р все-таки
остается делом вкуса исследователей; в принципе не исключает-
ся использование любого из двух способов описания.
8.7. Формализованное представление показателя
гомеостатической способности
В предыдущих разделах мы ввели понятие количественной
оценки гомеостатических свойств системы и показателя гомео-
статических свойств системы. Сейчас мы покажем, как эти по-
нятия можно изложить на несколько более формализованном
уровне [143].
Пусть при малых возмущениях вектора v, обозначаемых че-
рез е, стационарный режим в системе сохраняется, а все компо-
ненты вектора состояния получают приращение Дх:
Ax = Vox-e, (8.47)
где через Vox обозначена матрица (8.1) размерности (mXD:
Vox = [(Vox)I (Vox)2 ... (Vox)JT (8.48)
270
КОЛИЧЕСТВЕННАЯ ОЦЕНКА ГОМЕОСТАЗА
[ГЛ. 8
со строками
[дх. дх, дх. 1
••• L/=l, 2, .... tn. (8.49)
dv{ dv2 dvl J’ ’ ’ ’ v '
В качестве меры смещения состояния системы Дх возьмем
взвешенное среднеквадратическое значение Дх, обозначенное че-
рез р:
р2 = (Дх)т А (Дх), (8.50)
где А(тХт) = diaga( — матрица весовых коэффициентов, учи-
тывающая относительный вес переменных состояния, at 0. На-
пример, если значимость всех переменных состояния одинакова,
то все а, равны между собой, аг= 1/т/.
Определим матрицу чувствительности системы:
So= Vox)T A (Vox). (8.51)
Тогда
ц2 = eTSoe, (8.52)
а взвешенное среднеквадратическое значение оо коэффициентов
5х.
чувствительности определяется теперь по формуле
a» = tr (Vox)T A(V0x) = trS0. (8.53)
Тогда показатель гомеостатической способности системы h,
определяемый (8.5) или (8,7), запишется в виде компактной
формулы
h = (trS„)~'/! (8.54)
или, соответственно,
/г = (1 + atrS0)-1. (8.55)
Для линейных систем формулы, описывающие гомеостатиче-
ские свойства, можно выразить непосредственно через матрицы
А и В. Чувствительность переменных состояния тогда опреде-
ляется формулой (5.68), и можно записать
?ох=-А-1В. (8.56)
Таким образом, значения о не зависят от v; для линейной си-
стемы формулу (8.2) можно переписать в компактном виде, вос-
пользовавшись выражением для следа матрицы (5.28):
т I
tr[(- А“‘в)т(- А’*в)]= Ё Ёо?/, (8.57)
/-I 1
8.7] ФОРМАЛИЗОВАННЫЙ ПОКАЗАТЕЛЬ ГОМЕОСТАЗА 271
где 01/ — элемент матрицы (—А~1В) с номером (/, /). Следова-
тельно, формулу (8.2) можно (при atl — 1) записать как
tf2 = ~j-tr [(—А-1В)Т(—А-1В)], (8.58)
и зависимость h (о) выражается теперь через элементы матриц
А и В.
Поскольку для линейных систем величина h не зависит от v
и постоянна во всем диапазоне изменения v от — оо до + °°>
для линейных систем нет смысла вычислять интегральный по-
казатель Н, так как он будет бесконечно большим. Введение Н
целесообразно только для нелинейных систем, как это и было
отмечено в разд. 8.3.
ГЛАВА 9
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
9.1. Типовые ситуации в гомеостатических системах
В реальных ситуациях функционирование биосистем происхо-
дит в условиях непрерывных изменений как вне, так и внутри
системы. Если обратиться к схеме на рис. 7.2,6, то можно ска-
зать, что переходные процессы, приводящие к изменению пере-
менных состояния в физиологических системах, возникают в сле-
дующих случаях:
— при вариациях условий окружающей среды (воздействие
возмущающего входа v),
— при изменении режима функционирования системы (из-
менение задающего входа — независимых темпов потоков веще-
ства и энергии да),
— при изменениях параметров системы (вариации элементов
матриц А или В).
В ответ на эти изменения в биосистеме происходят сдвиги,
направленные прежде всего на сохранение стационарного нерав-
новесия, — система адаптируется, приспосабливается к новым
условиям существования. Приспособительные процессы при из-
менении v и да можно назвать адаптацией системы к внешним
возмущениям, а при изменении элементов матрицы А — процес-
сами компенсации. Эти термины достаточно хорошо соответ-
ствуют общебиологическим представлениям о процессах адап-
тации и компенсации [2, 58а, 82, 153, 165, 185, 197]. Здесь мы рас-
смотрим только процессы адаптации в системе — переходные
процессы, вызванные действием внешних возмущений. Процес-
сы, вызванные изменением v, мы будем называть адаптацией
системы, к внешним условиям. Для процессов, происходящих при
изменении да, будет использоваться термин перестройка режима
функционирования системы.
Адаптация физиологических систем. Адапта-
цию к изменениям условий среды можно представить себе как
переходный процесс в компартментальной модели, в результате
которого система переходит в новое стационарное состояние (со-
стояние адашированностиУ Обычно в физиологии считается,
что в состоянии адаптированности, во-первых, сохраняется
S.11 ТИПОВЫЕ СИТУАЦИИ В ГОМЕОСТАТИЧЕСКИХ СИСТЕМАХ 273
обычный или устанавливается повышенный уровень жизнедея-
тельности и, во-вторых, обеспечивается сохранение гомеостаза
на всех структурно-системных уровнях организма [82, 185, 197].
В наших терминах это означает, что независимые темпы w
после окончания процесса адаптации возвращаются почти к ис-
ходной величине, а переменные состояния сохраняются практи-
чески на постоянном уровне. Однако при адаптации может воз-
никнуть и новое состояние организма, существенно отличающее-
ся от исходного.
Обычно считается, что адаптация является полной, если все
системы организма сохраняют постоянство своих переменных, а
адаптационные возможности системы не уменьшаются. При не-
полной адаптации в некоторых системах организма происходят
уже существенные изменения значений переменных, но общие
показатели деятельности организма сохраняются, хотя и при
уменьшенных адаптационных возможностях. Крайняя степень
неполной адаптации иногда называется кажущейся адаптацией
[197].
Процесс адаптации. Наиболее характерной чертой
процесса адаптации является временное увеличение темпов ме-
таболизма— потребления кислорода и других веществ [166].
Так, у рыб и беспозвоночных при изменении концентрации со-
лей в воде потребление кислорода в период адаптации увели-
чивается в 9—10 раз [201]. Эффект увеличения темпов потребле-
ния веществ (независимых темпов) в ходе процесса адаптации
легко воспроизводится компартментальной моделью (рис. 7.2, а)
при соответствующем выборе ее элементов: в ходе переходного
процесса включаются активные механизмы регуляции, отчего
в период адаптации происходит увеличение независимых тем-
пов w.
Пример 9.1.1. Пусть на рис. 7.2, а все сигналы — скаляры, причем у =
= (х + г) — v, w2 = 2z, а переменные пассивного и активного каналов опи-
сываются, соответственно, уравнениями Тх + х = е, z + z = е, где рассогла-
сование между зависимыми и независимыми темпами потоков вещества и
энергии обозначено как е = у—w. Тогда исходный режим при v = 0, для
которого у = w = = 1, w2 = 0, достигается при значениях пассивного и
активного управляющих сигналов х = 1, z = 0. После ступенчатого возму-
щения и = 1 в системе возникает переходный процесс, и устанавливается но-
вый стационарный режим: z = 0, х = 2; w = 1, и = 1. Переходный процесс
иллюстрируется кривыми на рнс. 9.1, где видно характерное повышение «об-
щего уровня активности» — темпов потребления w.
Состояние адаптированност и. При описании
свойств адаптированной системы обычно оценивается степень
сохранения ее реакций на внешние возмущения и степень со-
хранения адаптационных возможностей. Поскольку под адап-
тационными возможностями обычно понимается способность си-
стемы приспосабливаться к широкому кругу внешних условий,
274
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
оба указанных свойства можно количественно охарактеризовать
с помощью введенных в гл. 7 показателей гомеостатической спо-
собности.
Действительно, если ограничиться рассмотрением действия
малых внешних возмущений, то сохранение реакций системы
означает, что и в новом состоянии система не выходит за пре-
Рис. 9.1. Процесс адаптации
как переходный процесс в ком-
партментальной модели открытой
системы. В ответ на действие
внешнего возмущения г (0 в фи-
зиологической системе регуляции
включаются активные, z(t), и
пассивные, х (0. механизмы.
Функционирование активных ме-
ханизмов приводит к увеличе-
нию энерготрат системы w2 (t),
вследствие чего во время про-
цесса адаптации темпы w (t) об-
мена веществ в системе повы-
шаются. I— исходное состояние,
II — период адаптации, /77—со-
стояние адаптированное™.
Перестройка
делы линейного диапазона по всем
своим переменным. В этом случае
коэффициенты чувствительности
сохраняются, и значение показателя h
в (8.7) остается неизменным. Что ка-
сается адаптационных возможностей,
то их следует связать с величиной ин-
тегрального показателя (8.26) — вели-
чиной Н. Но в этом случае ясно, что
любое приращение сигнала v, описы-
вающего рнешние условия (сдвиги
стационарной точки и0 на рис. 8.9, а),
приводит к изменению резервов гомео-
стаза У+ и N_. Поэтому, вероятно, по-
нятие полной (неполной) адаптации
должно связываться не столько с пол-
ным (неполным) сохранением адап-
тивных способностей (величин Н или
N), сколько с сохранением высоких
значений h и, следовательно, с незна-
чительным (существенным) измене-
нием резервов системы. Кажущаяся
адаптация соответствует случаю поло-
жения точки и0 на границе области
стационарности, когда резервы гомео-
стаза системы исчерпаны, N — 0.
режимов функционирования.
Переходные процессы в системе возникают и при изменении по-
ведения или активности животного. В этом случае причиной
сдвигов переменных состояния является изменение независимых
темпов w — задающего сигнала в системе управления.
С точки зрения поддержания постоянства внутренней среды
процессы адаптации при изменениях v и при изменениях w, хотя
и обладают существенными различиями, протекают во многом
аналогично. Поэтому рассмотренные выше методы исследова-
ния реакций системы на возмущающее действие сигнала v (го-
меостатические реакции системы) в принципе распространяются
и на случай изменения режима функционирования системы,
когда меняются требования к гомеостатическому регулятору —
величины независимых темпов потоков вещества и энергии ш.
9.1)
ТИПОВЫЕ СИТУАЦИИ В ГОМЕОСТАТИЧЕСКИХ СИСТЕМАХ
275
В работе [142] для определения реакций системы на изменение
w при сохранении постоянства условий среды v был применен
термин перестройка.
Определим для процессов перестройки количественные ха-
рактеристики, аналогичные введенным в предыдущих главах
показателям гомеостатической способности. Здесь мы ограни-
чимся формальным указанием на возможность проведения та-
кой аналогии в широком смысле и введем эти характеристики
так, как это было сделано в разд. 8.3, не используя для них ка-
кой-либо новой терминологии.
Если при малом изменении вектора w, которое будем обо-
значать через е, все компоненты вектора состояния получают
приращения Дх, то
Дх = Vwx • е, (9 1)
где V^x—матрица аналогичная (8.48):
Vasx = [(Vasx)1 (V№x)2 ... (?шх)„]т (9.2>
со строками
Г дх, дх, дх. Я
(^шх)г = ЬН- -Н- ... -Н- • (9-3)
' w dwl dw^ dwn J ' '
Введем no аналогии c (8.51) матрицу чувствительности си-
стемы к вариациям задающего входа
Sw = (Vfflx)T A (V^x), (9.4)
где Л(тхт) = diagfli — матрица коэффициентов, дающая относи-
тельный вес переменных состояния, 0. Если значимость по-
стоянства какой-либо переменной для биосистемы не зависит от
ситуации, в которой система находится, то матрица А совпа-
дает с матрицей весовых коэффициентов в показателях гомео-
статической способности системы.
Взвешенное среднеквадратическое значение коэффициентов
чувствительности dxjdwj теперь определится формулой
= tr (9.5)
а показатель, характеризующий способность системы сохранять
постоянство переменных состояния при перестройке, можно оп-
ределить по аналогии с (8.54)
g = (tr$j’2 (9.6)
или по аналогии с (8.55)
g = (1 + a tr SJ"1. (9.7)
276 ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА [ГЛ. 9
Интегральный показатель
G—^gdw (9.8)
а
определяет сохранительные ресурсы системы при изменении не-
зависимых темпов потоков w в области Й. Степень напряжения
механизмов и резервы гомеостаза можно теперь определить ана-
логично тому, как это сделано для гомеостатической способно-
сти системы в разд. 8.3—8.4.
Подобные оценки могут оказаться полезными, в частности,
при исследованиях способности организма приспосабливаться
к изменениям физической нагрузки и при определении его воз-
можностей по выполнению физической работы. Результаты таких
исследований излагаются, например, в [64, 133, 147]. Легко ви-
деть, что уменьшение приспособительных способностей системы
связано с выходом ее переменных состояния на нелинейный
участок; в этом смысле задача оценки адаптивных способностей
близка к технической задаче оценки степени нелинейности си-
стем [85].
9.2. Влияние прямых связей на свойства систем
Под прямой связью понимается непосредственное измерение
возмущающего фактора v и формирование на основе получен-
ной информации некоторого компенсирующего сигнала, предот-
вращающего изменение управляемых переменных у. С точки
зрения классической теории управления в этом случае имеет
место инвариантность при управлении по возмущению [212а].
В теории управления известны и сравнительные особенности
прямых и обратных связей при регулировании: в отличие от
обратных связей, которые эффективно работают при относи-
тельно больших коэффициентах усиления, прямые связи обес-
печивают хороший результат и при сравнительно малых коэф-
фициентах передачи. Но, с другой стороны, прямые связи не
могут точно компенсировать действие возмущений; качество
компенсации быстро ухудшается при внутренних возмущениях
в системе.
Действительно, рассмотрим случай приспособления к внеш-
ним возмущениям при наличии прямой связи F. Схема такой
биосистемы показана на рис. 9.2. Ее уравнение состояния имеет
обычный вид
x = Rw-Y Ах-\-(В-\- AF)v. (9.9)
Стационарный режим системы определяется из условия
х = - Л-1/?® - + F) V.
(9.10)
9.2}
ВЛИЯНИЕ ПРЯМЫХ СВЯЗЕЙ НА СВОЙСТВА СИСТЕМ
277
Если выбрать матрицу F так, что матричный коэффициент при
v равен нулю, т. е.
F = — A~'B, (9.11)
то величина х в стационарном режиме не зависит от и и
dx
— = 0, т. е. канал F полностью компенсирует влияние сигнала
v на сигнал х по каналу В. Сигнал х обладает в этом случае
свойством инвариантности. Однако любое изменение элементов
матрицы F приводит к нарушению компенсации, и инвариант-
ность х пропадает.
„ dx
Чтобы показать это, вычислим производную — вектора х
по вектору fnV, и = 1, 2, ...» m; 1 < v </, определяющую^чув-
ствительность х к возмущениям " "
пишем (9.10) следующим об-
разом:
х— —
v-ro
A~'Bv- (9.12)
1
Ё fmivi
столбца матрицы F. Пере-
R
Рис. 9.2. Влияние прямых связей на свойства
системы. Канал прямой связи с матрицей F
снижает чувствительность переменных к воз-
действиям внешней среды, одиако, система
легко теряет это свойство при изменении
параметров канала прямой связи.
(размерность матрицы F рав-
на m X I), откуда, в соответ-
ствии с формулой дифференци-
рования вектора по вектору (5.33), получаем матрицу размерно-
сти (т X т):
dx
dxi dxt
dfiv dl2v
dx\
ту
dxm dxm
m m
- dfiv df2V
dXm
di mv
°v
О
о
°v
= v„E. (9.13)
0 ”
0
о о
%_
Чувствительность вектора x к вариациям элементов матрицы F,
таким образом, оказывается достаточно большой. Нулевая
чувствительность вектора х к вариациям v достигается ценой
278
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
(ГЛ. 9
большой чувствительности системы к вариациям элементов но-
вого канала связи F.
Пример 9 2.1. Рассмотрим пример из области терморегуляции, демон-
стрирующий как достоинства, так и недостатки компенсации с помощью пря-
мой связи. На рис. 9.3 показана простейшая модель системы с каналом, изме-
Рис. 9.3. Простая модель компен-
сации в системе терморегуляции
по принципу прямой связи. Актив-
ные механизмы терморегуляции
могут обеспечить инвариантность
температуры тела лишь в малом
диапазоне изменений темпера*
туры среды. Большое изменение
окружающей температуры вы-
звало бы отклонение темпера-
туры ядра тела на одии-два гра-
дуса.
ряющим температуру внешней среды V. Пусть
приток тепла от процессов обмена веществ
W — 50 ккал!м2-час в сгационарном режиме
уравновешивается теплопогерями У, причем
Y-k(X-V), (9.14)
где X— температура тела, V—температура
среды, k — коэффициент. При k = k0 =
= 0,34 ккал!м2-час-град в стационарном режи-
ме при V = 20° температура тела X = 37°.
В схеме имеется активный механизм тер-
морегуляции, построенный по принципу прямой
связи. Терморецепторы, чувствительные к изме-
нению V, фиксируют изменения температуры
среды, что приводит к включению центральных
механизмов терморегуляции и последующему
изменению коэффициента теплоотдачи k.
Уравнение (9.14) при этом заменяется следую-
щим уравнением:
Г==*о[1 + f-(V-V0)](X-V), (9-15)
где значение коэффициента теплоотдачи k0
при температуре окружающей среды Vo = 20
принято за исходное, а коэффициент f определяет характеристики прямого
канала.
Величину f в соответствии с (9.11) можно выбрать так, чтобы обеспечить
инвариантность X в некоторых заданных условиях среды, т. е. чтобы измене-
ния V не сказывались на температуре тела X. Для этого надо линеаризовать
уравнение (9.15) и выбрать в точке линеаризации такое f, которое удовле-
творяет (9.11).
Линеаризация уравнения (9.15) в точке (Vi, X]) дает
х = ах + bv,
(9.16)
где х — X — Xb v — V — Уь а — £0(—1 — f Vi + f Vo), b — k$(—fXi + 1 +
+ 2fV, - fVa).
При l/1 = Vo = 20, X1 = 37 получаем a = —b0, b — b0(l—/XI+fV1).
Коэффициент b обращается в нуль, если
f = I, —0,059. (9.17)
При таком значении f переменная X в системе оказывается инвариантной
к изменению температуры внешней среды в точке V = 20°.
Стоит, однако, параметрам прямого канала несколько измениться, как
компенсация нарушается, и величина X начинает сильно зависеть от окру-
жающей температуры V. Эта зависимость определяется величиной производ-
ной в стационарном режиме. Действительно, имеем
<9Л”
9.3) ДВОЙСТВЕННЫЙ ХАРАКТЕР ПЕРЕМЕННЫХ СОСТОЯНИЯ В СИСТЕМЕ 279
т. е. изменение f, например, на 0,1 приведет к довольно заметному сдвигу
температуры Xj на 0,3 °C.
Посмотрим теперь, каким может быть диапазон изменений величины ко-
эффициента f в исходной нелинейной системе на рис. 9.3. Легко видеть, что
введенный в модель механизм теплоотдачи работает только при V < 37. При
изменении значения температуры среды VT, при котором происходит линеари-
зация, значение коэффициента f в линеаризованном уравнении (9.16), обеспе-
чивающее инвариантность переменной х, должно меняться. Например, при
Х1 = 37, Vi — 35 это значение оказывается равным f = 1/(У1 — Vi) = 0,500.
Таким образом, диапазон изменения f, обусловленный одним лишь нелиней-
ным характером исходной системы (9.15), существенно превышает взятую
выше величину 0,1, так что изменение температуры X при постоянном значе-
нии коэффициента f только из-за изменения температуры среды может соста-
вить один-два градуса.
Таким образом, простой канал прямой связи не может обес-
печить хороших гомеостатических свойств системы. В реальных
биологических системах прямой канал обычно является одним
из элементов единой системы управления. Роль его состоит в
том, чтобы уменьшить чувствительность переменных внутренней
среды, хотя полной компенсации действия возмущений может и
не быть. Окончательное устранение возмущений выполняется
механизмами обратной связи, обеспечивающими, в частности,
и адаптивные изменения в самом канале прямой связи.
9.3. Двойственный характер переменных состояния
в системе
Переменные состояния в компартментальных моделях откры-
тых систем выполняют двоякую роль. Прежде всего эти пере-
менные оказываются пассивными регуляторами, принимающими
участие в процессах поддержания равенства темпов поступления
и расходования вещества и энергии в биосистеме, т. е. в поддер-
жании ее стационарного неравновесного состояния. С другой
стороны, в сложных системах наличие множественности, дубли-
рования регуляторных механизмов приводит к тому, что они
оказывают стабилизирующее действие друг на друга. Включе-
ние каждого нового регулирующего механизма в систему (па-
раллельного, как в разд. 7.6, или последовательного, как в
разд. 7.7) приводит к тому, что нагрузка, приходящаяся на каж-
дую переменную состояния, уменьшается. Таким образом, на-
копление регуляторных механизмов, направленных первоначаль-
но на регуляцию темпов поступления вещества, приводит вторич-
но и к появлению новой регуляторной функции у системы — под-
держанию постоянства внутренней среды.
В этом смысле мы говорим о двойственном характере регу-
ляторных механизмов в системе: с одной стороны, это управляю-
щие переменные (цель управления — поддержание равенства
первйчных и вторичных темпов, т. е. притока и оттока вещества
280
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
в компартменты), с другой — управляемые (цель управления —
сохранение относительного постоянства переменных состояния,
т. е. гомеостаз системы).
Конечно, отсюда отнюдь не следует, что в физиологических
системах не может быть специфических механизмов, направлен-
ных исключительно на поддержание постоянства внутренней
среды или на постоянство некоторых наиболее важных для си-
стемы переменных. Наоборот, такие связи можно встретить на
разных уровнях организации систем. Так, на уровне целостного
организма примерами таких связей могут служить механизмы
аортального и каротидного рефлексов (регулирующее влияние
изменений давления в области дуги аорты или каротидного си-
нуса на сердечный выброс и тонус сосудистой системы [355]).
Эти связи в определенном смысле слова являются «вторичными»,
поскольку их функционирование не связано с непосредственным
обеспечением стационарности системы — равенства независи-
мых и зависимых темпов потоков вещества или энергии.
В разд. 7.9 мы пытались показать, что результат действия этих
механизмов можно трактовать как выполнение функции огра-
ничителей в обычной задаче синтеза теории автоматического
управления. Можно сказать, что и в этом случае мы сталки-
ваемся с двойственным характером поведения переменных в
системе. Действительно, недопущение больших отклонений пе-
ременных внутренней среды, выполняющих роль пассивных ре-
гуляторов, может привести к таким нежелательным, если ис-
ходить из общих предпосылок, последствиям, как увеличение
времени переходных процессов в системе, более позднее уста-
новление равенства первичных и вторичных темпов в компарт-
ментах системы и т. д. Ограничения, налагаемые деятельностью
таких механизмов в сложных по структуре физиологических си-
стемах, могут даже нанести ущерб адекватному снабжению от-
носительно маловажных органов и подсистем. Так, при стрес-
сорных воздействиях (см., например, рис. 1.8) происходит так
называемая централизация кровотока, которая может быть свя-
зана с ухудшением кровоснабжения — и, следовательно, с воз-
можными нарушениями равенства темпов снабжения и трат ве-
щества или энергии — ряда органов и периферических тканей
организма. Аналогичные явления имеют место и при охлажде-
нии организма, когда тепловые потоки к периферическим орга-
нам нарушаются ради сохранения постоянной температуры ядра
тела. Мы не будем подробно останавливаться на этих сложных
и многообразных явлениях, ограничившись только одним их
аспектом, связанным с пороговым поведением переменных си-
стемы.
Пороговые эффекты. Обычно наличие пороговых эф-
фектов связывается в системе с наличием нелинейности тина
9.4]
ИЕРАРХИЯ ПЕРЕМЕННЫХ И МЕХАНИЗМОВ РЕГУЛЯЦИИ
281
«порог». Присутствие таких нелинейностей в гомеостатических
системах приводит к тому, что переменные состояния (и вы-
хода) могут совершать так называемый нерегулируемый дрейф
в подпороговых пределах. Так, для сигналов активных цепей
регуляции это может означать, что соответствующая цепь
регуляции «включается» только после того, как величина
активирующего ее сигнала превысит некоторый пороговый
уровень.
Однако следует обратить внимание на одну особенность не-
линейных гомеостатических систем, связанную с образованием в
ней пороговых эффектов. Дело в том, что эффекты такого рода
могут возникать в системе и при формальном отсутствии нели-
нейностей типа «порог». Действительно, если какая-то перемен-
ная Xt в некотором диапазоне изменения условий внешней среды
v и задающих входов w достаточно хорошо защищена иерар-
хически низшими механизмами, то они в большой степени пре-
дотвращают изменение Xt вплоть до истощения своих запасов.
После этого дальнейшее изменение v или w приводит с точки
зрения внешнего наблюдателя к «внезапному» увеличению чув-
ствительности Xi к вариациям v или w. Зависимость х,(ц) или
х,(ву) в этом случае принимает вид, характерный для порого-
вых нелинейностей.
Пороговые эффекты связаны с известным при исследовании
биосистем явлением — необходимостью учета ненаблюдаемых
переменных состояния для объяснения наблюдаемого харак-
тера изменений выходных переменных системы [131а].
9.4. Иерархия переменных состояния
и механизмов регуляции
Для каждого уровня организации биосистем разработаны
свои достаточно разнообразные и многогранные представления
об иерархии, связанные со структурными или функциональными
их особенностями. Например, в экологических системах гово-
рят об иерархическом порядке, имея в виду место данного вида
или группы в трофической пирамиде. Свои представления об
иерархии развиты и для физиологических систем. Здесь мы бу-
дем ориентироваться только на одну сторону поведения физио-
логических переменных — их место и роль в регуляторных от-
ношениях в системе. Общие представления об иерархии в си-
стемах управления рассматриваются, например, в [127,360].
Иерархический порядок переменных, их важность для биоси-
стемы в рассматриваемом аспекте интуитивно связывается со
степенью их защищенности при неблагоприятных воздействиях
внешних факторов. На уровне физиологических систем, в ча-
стности, (Переменные можно расположить по степени их
282
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
важности, ориентируясь в каждой ситуации на то, как поддер-
живается постоянство этих переменных.
При этом высшую иерархическую группу переменных состав-
ляют выходные переменные у, определяющие потоки вещества
и энергии. Разумеется, в компартментальных моделях, как и
в реальных системах, переменные у меняются при изменении у,
т. е. коэффициенты чувствительности для стационарных режи-
мов системы отличны от нуля:
4? (9-19)
Это связано с тем, что при вариациях v могут изменяться сами
задающие сигналы — независимые темпы потребления вещества
и энергии в системе w:
w = w(v), (9.20)
а величины у «следят» за изменениями w (см. пример 9.1.1).
Переменные состояния, выполняющие в той или иной мере
роль регуляторов темпов потоков вещества и энергии, состав-
ляют вторую по важности группу переменных в биосистеме.
Однако двойственный характер переменных позволяет в свою
очередь произвести упорядочение переменных х по степени их
важности для системы в пределах этой группы. Иерархически
более старшими переменными можно считать те, которые в це-
лостной биосистеме сохраняются наилучшим образом. При этом
имеется довольно ясная связь между иерархической значи-
мостью переменной и ресурсами ее гомеостатических свойств,
хотя эта связь и не проста.
Пример 9.4.1. Механизмы ауторегуляции кровотока. Если давление Р,
при котором происходит кровоснабжение какого-либо органа в организме, ме-
няется, то относительное постоянство кровотока Q через сосуды органа под-
держивается так называемыми ауторегуляторными механизмами. Оценим го-
меостатические характеристики этих механизмов в различных органах и при
разных условиях с целью проиллюстрировать возможность их сравнения на
Таблица 9.1
Гомеостатические характеристики системы ауторегуляции
кровотока мозга
Норма Перерезка симпатических нервных стволов
Участки изменения Р, мм рт. ст. 0 — 70 70-190 190 — 230 0 — 85 85 — 170 170 — 205
а 1,715 0,334 1,825 1,132 0,566 2,720
й 0,253 0,897 0,232 0,436 0,754 0,119
9.4]
ИЕРАРХИЯ ПЕРЕМЕННЫХ И МЕХАНИЗМОВ РЕГУЛЯЦИИ
283
основе количественных методов оценки гомеостаза, рассмотренных в преды-
дущей главе.
На рис. 9.4 приведены зависимости кровотока от перфузионного давле-
ния для сосудов мозга [134, 135] в норме (а) и при отключении ряда регуля-
торных механизмов посредством перерезки у животного симпатических нерв-
ных стволов (б). Видно, что гомеостатические свойства системы ауторегуля-
ции при этом ухудшаются: плато гомеостатической кривой становится уже,
чувствительность кровотока к изменению давления возрастает.
h
1.0 - 0,397
_ ------------- ---------
0,0 -
0,4 - 0,253 \nZ5Z
0,2 ---------
-----1-----1----1----1-----1___L
40 80 120 160 200 220
P, мм рт. ст.
e)
0,436 f
—J____L_
40 80
0,754
0,119
_j_____i_- i ... Li
120 160 200 220
P, ммрт. CT.
Рис. 9.4. Гомеостатические характеристики системы мозгового кровообращения. На рисунке
приведены зависимости кровотока через мозг при изменении давления в норме (а) и в усло-
виях патологии (б) [134]. Видно, что по сравнению с нормой отключение ряда регулятор-
ных механизмов ухудшает возможности системы. Количественная оценка гомеостатических,
свойств системы приведена иа двух следующих графиках; в) показатель h для нормы;
а) тот же показатель для условий патологии.
Оценим ресурсы гомеостаза в системе. При этом для простоты будем
пользоваться кусочно-линейной аппроксимацией гомеостатических кривых.
Данные о величине гомеостатической способности, приведенные в табл. 9.1,
вычисляются обычным способом (см. пример 8.1.1). Нормировка производится
на величину давления Ро = 100 мм рт. ст. и соответствующее значение кро-
п _ _ . 100 AQ
вотока при этой величине — Q0- Так, для рис. 9.4, а величина ст — Q • г У .
Величина h вычисляется по формуле (8.7) при а = 1. Зависимости h(P) для
условий нормы и перерезки симпатических нервных стволов прквгдгиы на
рис. 9.4, в, г, соответственно. Значения Н, вычисляемые по формуле (8.26),
дают в условиях нормы Н = 143,3 (мм рт. ст.), а при перерезке нервных
стволов Н = 105,4 (л'..и рт. ст).
284
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
На рис. 9.5 приведены аналогичные зависимости для кровотока через со-
суды скелетных мышц в лапе собаки для условий, когда механизмы ауторегу-
ляции функционируют нормально (тоиус сосудов хорошо выражен [319] —
рис. 9.5,а), и в случае ослабленных механизмов ауторегуляции (пониженный
тонус), когда ауторегуляционный отклик ухудшен [300] — рис. 9.5, б; в этой
Р, ммрт. ст.
0,8
0,6
0,4
0,2
0,108
_i____
О 40 80 120 160 200 220
Р,ммрт.ст.
0,6 -
0,4 -
0,2 -
О
0,003
40 80 120 160 200 220
. Р,ммрт.от.
h
Рис. 9.5. Гомеостатические характеристики системы ауторегуляции мышечного кровотока.
Зависимость кровотока через сосуды лапы у собак, когда тонус сосудов хорошо выра-
жен а) н когда меры по сохранению тонуса сосудов не принимались (б). Показатель h при-
веден для обоих случаев: в) норма; г) тонус сосудов снижен.
работе меры по сохранению тонуса сосудов специально не принимались. Соот-
ветствующие гомеостатические характеристики представлены для этих сосудов
в табл. 9.2. Данные для таблицы получались так же, как и для табл. 9.1 при
следующей нормировке: Ро — 100, Q — 2,2 для рис. 9.5, а и Ро = 80, Q = 10
для рис. 9.5, б. Следует обратить внимание, что кровотоки измеряются в разных
масштабах, что, однако, ие влияет на величину получающихся значений а.
Зависимость h(P) приведена для этого случая на рис. 9.5, в, г. Вычисле-
ние интегрального показателя гомеостатической способности дает в условиях
нормы Н— 110,5 (лип рт. ст.)\ при пониженном тонусе сосудов Н —
= 30,2 (мм рт. ст.). Ухудшение ауторегуляторных механизмов приводит к рез-
кому снижению ресурсов сохранительных свойств в системе.
Сравним теперь возможности механизмов ауторегуляцин мозгового кро-
вотока и механизмов ауторегуляцин кровотока в скелетных мышцах в уело-
fl-5]
ПЕРЕСТРОЙКА РЕЖИМОВ ФУНКЦИОНИРОВАНИЯ
285
Таблица 9.2
Гомеостатические характеристики системы ауторегуляции
кровотока в скелетных мышцах
Хороший тонус сосудов Пониженный гонус сосудов
Участки изменения Р, мм рт. ст. 0—15 15—135 135 — 200 20 —4J 40 — 80 80 — 140 14Э —200
ст h 4,50 0,047 0,39 0,868 2,87 0,108 3,22 0,087 0,80 0,608 3,75 0,066 16,0 0,003
виях нормы, воспользовавшись данными табл 9.1 и 92 и вы шсленными на
их основе показателями Н. Общие ресурсы гомеостатических способностей си-
стемы регуляции мозгового кровотока в норме существенно превышают ре-
сурсы системы регуляции скелетных мышц: 143,3 против 110,5 мм рт. ст.
Однако, как видно из рис. 9.4, в и 9.5, в, локальные гомеостатические свой-
ства системы регуляции кровотока в мышцах в области низких давлений (15—
70 мм рт. ст.) лучше, чем свойства системы регуляции мозгового кровотока.
Это связано, вероятно, с тем, что мозг в нормальных ситуациях никогда не
попадает в условия перфузии с давлением ниже 70 мм рт. ст. Действительно,
если давление в сосудах мозга снижается до величин порядка 80 мм рт. ст. н
ниже, в организме происходит резкое сужение сосудов ряда органов, приво-
дящее к тому, что давление на входе в сосуды мозга не падает до тех пре-
делов, в которых гомеостатические способности системы ауторегуляции моз-
гового кровотока недостаточны [304].
Следовательно, сохранительные ресурсы иерархически стар-
ших переменных фактически оказываются еще больше за счет
того, что в экстремальных, критических ситуациях их гомеостаз
поддерживается за счет относительного ухудшения гомеостаза
иерархически младших переменных. В рассмотренном примере,
в частности, в экстремальных условиях сужение периферических
сосудов приводит к тому, что системы ауторегуляции кровотока
в скелетных мышцах будут работать в условиях ухудшенного
кровоснабжения, в то время как иерархически старшие меха-
низмы ауторегуляции мозгового кровотока функционируют в
тех пределах, где их локальные гомеостатические свойства до-
статочно хороши.
9.5. Гомеостаз и перестройка режимов функционирования
Функционирование биосистемы происходит часто в сложных
условиях внешней среды, когда на нее действует одновременно
не один, а несколько внешних факторов, заставляющих систему
интенсифицировать деятельность сохранительных механизмов.
Эффект подобных воздействий «суммируется», так что наличие
286
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
дополнительных неблагоприятных факторов снижает сохрани-
тельные ресурсы системы по отношению к некоторой рассмат-
риваемой переменной внешней среды. Так, тепловая нагрузка
хуже переносится организмом в условиях кислородного голода-
ния. Уменьшаются и резервы гомеостатических свойств.
Интуитивно ясно, что снижение гомеостатических способно-
стей системы при одновременном действии нескольких неблаго-
приятных факторов связано с перемещением изображающей
точки системы к границам области гомеостаза на рис. 1.9. Од-
нако до количественных оценок такое рассмотрение, как пра-
вило, не доводится [71]. Поэтому представляется важным тот
факт, что развитые в предыдущих разделах методы могут дать
количественное представление о характере одновременного дей-
ствия на систему несйольких факторов.
Особое место занимает здесь вопрос о связи гомеостатиче-
ских свойств системы с режимами ее функционирования. В био-
системе одни и те же механизмы управления обеспечивают как
зависимые темпы переработки вещества (тем большие, чем в
более интенсивном режиме находится система), так и ее гомео-
статические реакции. Поэтому естественно ожидать, что меха-
низмы интенсивно функционирующей системы в меньшей мере
способны поддерживать ее гомеостаз, а ресурсы сохранительных
свойств при увеличении интенсивности обмена веществ должны
сокращаться *).
Чтобы продемонстрировать это положение, мы рассмотрим
гомеостатические характеристики системы энергетического снаб-
жения организма на основе математической модели, разрабо-
танной в [253] и исследованной в [63]. В качестве двух факторов,
независимо вызывающих напряжение гомеостатических меха-
низмов, возьмем общий уровень энерготрат организма (оцени-
ваемый по независимым темпам потребления кислорода) и ус-
ловия внешней среды, определяемые напряжением кислорода
во вдыхаемом воздухе. Выбор таких факторов позволяет, в ча-
стности, моделировать ситуации, связанные с физической дея-
тельностью человека в неблагоприятных условиях среды.
В основе схемы лежит рассмотренная в разд. 6.6 компарт-
ментальная модель энергетической системы организма, допол-
ненная математическим описанием системы транспорта крови
*) Мы предполагаем здесь, что структура системы, работающей в отно-
сительно спокойном режиме и в режиме интенсивного функционирования,
остается одной и той же. Этот тезис, впрочем, не является очевидным для
физиологических систем организма: при интенсивном функционировании в
ием могут активизироваться механизмы, имеющие в состоянии покоя «нуле-
вые» коэффициенты усиления. В этом случае свойства системы следует опи-
сывать с помощью моделей с переменной структурой, но такие системы здесь
не рассматриваются.
9.5]
ПЕРЕСТРОЙКА РЕЖИМОВ ФУНКЦИОНИРОВАНИЯ
287
к тканям организма и системы обмена воздуха через легкие.
В модели пять пространственных областей: 1 — альвеолярный
воздух, 2 — венозная кровь, 3 — жидкости мышечной ткани,
4 — жидкости прочих (немышечных) тканей, 5 — артериальная
кровь. Между ними происходит обмен тремя компонентами —
кислородом, углекислым газом и недоокисленными продуктами
обмена, а между системой и средой — двумя, О2 и СО2 (рис. 9.6).
Среда описывается вектором v с двумя компонентами Wi и
и2—напряжением кислорода и углекислоты, при этом о2 = О
(у — возмущающий сигнал). Темп энерготрат задается вектором
w с компонентами Wi и w2 —
темпами потребления кисло-
рода тканями мышц и немышц
(ц> — задающий сигнал). Ос-
новная часть модели описы-
вает процессы диффузии ком-
понент и химические реакции
между ними согласно уравне-
ниям, приведенным в разд. 6.6.
Соответствующие компо-
ненты вектора состояния обо-
значим следующим образом:
%! — х4 — напряжение кис-
лорода в областях 1—4',
Хз — х8— напряжение угле-
кислого газа в этих же обла-
стях;
Хд — Хп — концентрация не-
доокисленных продуктов в об-
ластях 2, 3 и 4, соответ-
Рис. 9.6. Компартментальная модель системы
энергетического снабжения организма. Мо-
дель включает пять пространственных об-
ластей, показанных на рисунке цифрами от
1 до 5, и одиннадцать компартментов, со-
ответствующих уровням Ог, СОг и молочной
кислоты в этих областях. Режим работы
системы определяется двумя независимыми
темпами потребления Ог в областях 3 и 4;
внешняя среда описывается одной перемел-
иой — напряжением рО2 в атмосфере.
ственно.
Для упрощения процесса моделирования переменные арте-
риальной крови не представлены в векторе состояния; напряже-
ния О2 и СО2 в артериальной крови в модели являются компо-
нентами выходного сигнала. Таким образом, часть вектора со-
стояния, определяющая концентрации веществ в системе, имеет
размерность т = 11.
На систему пассивного обмена веществами между указан-
ными областями и средой в модели накладывается пять меха-
низмов управления более высокого уровня. Два из них пред-
ставляют собой пассивные механизмы увеличения отбора кисло-
рода из крови (для крови, протекающей через мышцы и ткани
прочих немышечных органов, соответственно), а остальные'
три — активные механизмы управления потоками веществ по-
средством изменения объемной скорости носителя. Для этих
288
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
трех механизмов вводятся три дополнительные компоненты век-
тора состояния: %i2 — альвеолярная вентиляция, Xi3 и — кро-
вотоки через мышечные ткани и ткани немышечных органов.
Таким образом, полный вектор состояния имеет размерность
т ~ 14.
Дадим краткое описание блок-схемы модели, представленной на рис. 9.7.
Матрица Д(2оХ14) определяет потоки веществ в системе за счет диффузии и
химических реакций. Уравнения химических реакций линеаризуются, причем
Рис. 9.7. Структурная схема компартментальной модели системы энергоснабжения орга-
низма. Адекватные темпы обмена н движения кислорода, углекислоты и продуктов обмена
обеспечиваются комплексом механизмов регуляции, из которых три механизма иосят актив-
ный характер, а одиннадцать —пассивный. Задающим сигналом является вектор w неза-
висимых темпов потребления кислорода организмом, возмущающим —скаляр о, определяю
щий уровень кислорода в окружающей среде. Матрица А задает свойства пассивных упра-
вляющих механизмов в системе, активный канал управления с сигналом z описывается
матрицами С,, В, Bt и Вг, Векторы d,, d2, d>, d4—постоянные векторы, задающие числен-
ные значения некоторых параметров модели.
для каждого стационарного режима линеаризация производится в соответ-
ствующей равновесной точке.
Вектор темпов потока веществ г/(2оХи имеет следующие компоненты: тем-
пы расхода О2 на окисление недоокислеиных продуктов (молочной кислоты)
в определенных выше областях 3 и 4 в соответствии с реакцией (6.67); по-
токи О2 из области 0 (атмосфера) в 1, из 1 в 5, из 5 в 3, из 5 в 4. Для угле-
кислого газа вводятся следующие компоненты: темпы образования СО2 в об-
ластях 3 и 4, темпы потоков СО2 из 3 в 2, из 4 в 2, из 2 в /, из 1 в 0. Далее
следуют компоненты, отвечающие потокам недоокислеиных продуктов — мо-
лочной кислоты р.: возникновение ц в ходе химической реакции (6.69) в об-
ласти 3; поток р. из 3 в 2, из 2 в 5, из 5 в 4. В области 4 происходит утили-
зация р со скоростью, пропорциональной ее концентрации. Кроме того, р ути-
лизируется в областях 3 в ходе реакции (6.67). Перечисленные скорости пото-
ков составляют (вместе с упомянутыми выше двумя сигналами напряжения
О2 и СО2 в артериальной крови) двадцать компонент вектора у.
Матрица Дц2оХ2) отражает влияние условий среды на эти потоки. Вектор-
столбец d2 включен в схему для коррекции влияния на процессы обмена ве-
ществ переменных, характеризующих напряжения О2 и СО2 в области 5 — ар-
териальной крови, которые формально не входят в вектор состояния. Матрицы
/?2<ах2), Яадхп), Сг(2оХ2) и вектор-столбец di описывают параметрическое
9.5) ПЕРЕСТРОЙКА РЕЖИМОВ ФУНКЦИОНИРОВАНИЯ 289
влияние сдвигов кривой диссоциации на темпы потоков вещества. Это влия-
ние определяется диагональной матрицей Л4Ь элементы которой зависят от
состояния системы.
Активные механизмы управления представлены вектором г(зХ1). Матрица
СцзХп) определяет связь z с состоянием х. Матрица ВгаХз; и вектор за-
дают характер влияния г на темпы потоков вещества между отдельными об-
ластями. Диагональная матрица В(3х з> задает временные характеристики ак-
тивных механизмов управления. Параметрическое влияние активных механиз-
мов управления определяется диагональной матрицей Мг, элементы которой
зависят от текущего состояния системы, а также матрицей ВцпХз) и век"
тором d3.
Матрица С<ц хзо; формирует потоки веществ внутри системы. Относитель-
ные объемы областей задаются диагональной матрицей Сз(цХп)-
Процессы моделируются так, что все переменные имеют размерности со-
ответствующих исходных физиологических данных [63, 253]. Мы не будем
здесь приводить конкретных выражений для матриц и векторов, указанных
на схеме рис. 9.7, тем более, что выше рассматривались вопросы линеаризации
и описания энергетической системы организма (разд. 6.6).
Рассмотрим теперь гомеостатические способности системы
в различных стационарных режимах, отличающихся темпом по-
требления кислорода, т. е. гомеостатические свойства организма
при различной физической нагрузке.
Будем исследовать зависимость показателя h от двух пере-
менных — от темпа потребления кислорода и от напряжения
кислорода в окружающей среде wu
h = h(vl,wi). (9.21)
Вычисление h проведем по формуле (8.5), причем в формуле
для определения чувствительности (8.2) выберем следующие
значения весовых коэффициентов: для переменных внутренней
среды yt, i = 1, 2, ..., 11, положим а, = 1, а для переменных
состояния, описывающих активные механизмы регуляции (i =
= 12, 13, 14), положим а, = 0.
Значения h вычисляются последовательно в стационарном
режиме при w\ = 72,5; 200; 400; 800; 1200; 1733 am/jwuh, v'j — 160;
145; 120; 100 мм рт. ст. Для нахождения коэффициентов чув-
ствительности в каждой точке величина v] получает прираще-
ние, равное AwJ = O, lw°, после установления нового стационар-
ного режима вычисляются значения
о? Дх,
^=4 — . г = 1, 2, ..., 11, (9.22)
X, Дсй
и затем находится величина
290
феноменология гомеостаза
[ГЛ. 9
В данном случае оказалось, что именно такой способ опре-
деления скорее всего приводит к цели; в общем случае, однако,
может потребоваться использование и существующих стандарт-
ных методов нахождения функций чувствительности с помощью
моделей чувствительности, как это сделано, например, в работе
[323].
Зависимость h от двух переменных (щ, wi) приведена на
рис. 9.8. В условиях покоя и при нормальном напряжении кис-
Рис. 9.8. Гомеостатические характеристики системы
энергоснабжения организма. Гомеостаз системы
ухудшается как при понижении содержания кисло-
рода во вдыхаемом воздухе, так и прн увеличении
темпов обмена веществ. При совместном действии
обоих факторов гомеостатические храктернстики
системы ухудшаются в значительно большей сте-
пени, Сигналы, действующие на систему, изме-
ряются в следующих единицах: задаваемый темп
потребления кислорода Wt — мл Ог/мин, напряже-
ние кислорода в воздухе th — мм рт. ст.
лорода в воздухе величи-
на показателя h макси-
мальна и равна 1,06.
Изменение каждой из пе-
ременных У], в небла-
гоприятную сторону при-
водит сначала к относи-
тельно медленному сни-
жению гомеостатических
свойств системы. Однако
при достижении активны-
ми управляющими пере-
менными — вентиляцией
Х12 и кровотоками Х1з и
х14 — их предельных зна-
чений 45 л/мин, 18 и
10 л1мин, соответственно,
начинается более суще-
ственное снижение гомео-
статических способностей
системы. Адекватное снаб-
жение тканей организма
кислородом при этом не
нарушается, но стацио-
нарный режим в системе
обеспечивается за счет
все больших и больших
сдвигов переменных внутренней среды x„ i — 1, 2..11. При
дальнейшем увеличении интенсивности функционирования (тем-
пов потребления кислорода wi), как и при снижении содержа-
ния кислорода во вдыхаемом воздухе, стационарный режим в
системе не достигается. Показатель гомеостатической способно-
сти в этой области равен нулю.
Физиологический смысл графика, приведенного на рис. 9.8,
очевидно, состоит в следующем. В нормальном режиме (покой
или физическая нагрузка невысокой интенсивности, достаточное
содержание кислорода во вдыхаемом воздухе), регулирующие
механизмы действуют достаточно эффективно в том смысле, что
9.6]
НЕСТАЦИОНАРНЫЕ РЕЖИМЫ В СИСТЕМЕ
291
стационарный режим достигается при незначительных сдвигах
переменных внутренней среды. Высокая чувствительность ак-
тивных механизмов управления Х12 — х14 позволяет при этом
поддерживать относительное постоянство переменных в цепях
пассивных управляющих механизмов — концентраций веще-
ства во всех компартментах биосистемы. Почти вся нагрузка
ложится на активные механизмы регуляции. Вследствие нели-
нейности характеристик активных управляющих механизмов
увеличение интенсивности функционирования или снижение со-
держания кислорода приводит к тому, что стационарное состоя-
ние достигается при все возрастающих сдвигах переменных
внутренней среды; нагрузка с активных регуляторов постепен-
но, хотя сначала и в незначительной степени, перекладывается
на пассивные регуляторы, так что гомеостатические характе-
ристики системы начинают ухудшаться. Наконец, при исчерпа-
нии ресурсов активных регуляторов вся нагрузка падает на пе-
ременные внутренней среды — гомеостаз резко ухудшается, а
затем система теряет и способность поддерживать стационар-
ное состояние.
Заметим, что даже при выходе системы за границы области
стационарности в течение некоторого времени она способна под-
держивать адекватное снабжение тканей кислородом за счет
веществ, находящихся в компартментах. Эти запасы веществ
определяют последний резерв системы — резерв пассивной ре-
гуляции.
9.6. Нестационарные режимы в системе
Все предыдущее рассмотрение в этой книге относилось к
гомеостазу, понимаемому как сохранение стационарных значе-
ний переменных состояния в системе. Однако полученные ре-
зультаты можно в определенном смысле обобщить и на тот
случай, когда биологическая система находится под постоян-
ным воздействием внешних возмущений, так что стационарный
режим в ней не достигается. Обратимся для этого вновь к
функции чувствительности [207].
При определении функций чувствительности различают два
случая. В одном случае возмущение параметров системы k не
зависит от времени, в другом — зависит. Тогда изменение пара-
метров в обоих случаях можно записать, соответственно, в виде
k = k0 + e или k — k0 + e • v (t).
Если возмущение не зависит от времени, то функция чувстви-
тельности u(k) системы, которая описывается уравнением
х — f (х, k, t),
(9.24)
292
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
определяется посредством обычной производной
u(kj) = hm--------------------------=
е->0 в
дх (О
dkj •
(9.25)
Поскольку переменные и, задающие характеристики внеш-
ней среды, являются параметрами в уравнениях системы, урав-
нение (9.25) определяет функции чувствительности и в этом
случае.
В предыдущем рассмотрении мы в качестве меры функции
чувствительности (9.25) брали смещение стационарных значе-
ний переменных состояния
, дх, (/) дх.
а и = lim — = -тг-
(~>оо
(9.26)
где kj, в частности, представляло собой j-ю компоненту век-
тора V.
Если возмущение, действующее на систему, зависит от вре-
мени, то функция чувствительности определяется следующим
образом:
х jk, + ev (О, Л — х (k„ t)
и (kh t) = lim ——---------------l-LZ., (9.27)
e->0 e
где v(<)—равномерно ограниченная интегрируемая функция,
a e — постоянная [207]. Функция (9.25) представляет собой ча-
стный случай (9.27). Функции чувствительности и в (9.27) те-
перь зависят от вида возмущающей функции v(t).
Упорядочим множество функций чувствительности, введя
норму
|| и (kj, v, /) || = X max | ui (kj, v, f) |. (9.28)
Здесь / = 1, 2, ..., l. Размерность вектора состояния по-преж-
нему равна tn.
Полагая
IIИ (Й/, V, t)\\ = Cj, (9.29)
получаем ряд чисел ci, ci, ..., Cd, который можно расположить
так, что
Ci < с2 < ... < cD, D I (9.30)
(одинаковым числам с/ в этом ряду соответствует один член).
Ряд (9.30) дает расположение параметров системы по степени
их влияния на состояние, когда все параметры возмущаются
одним и тем же сигналом v(i). Этот ряд носит название пара-
метрического ряда.
9.71
МОДЕЛИРОВАНИЕ БИОСИСТЕМ И ПРИНЦИП ПРОСТОТЫ
293
Нас интересуют сейчас соотношения, существующие между
параметрическим рядом для стационарной вариации v(t) — vo
и параметрическим рядом, соответствующим любой нестацио-
нарной вариации v(t), ограниченной условием
v(0<vo. (9.31)
Для таких рядов показано [207, гл. 2], что параметрический ряд
всех стационарных вариаций параметров представляет собой
верхнюю границу параметрических рядов, соответствующих не-
стационарным вариациям параметров, удовлетворяющих усло-
вию (9.31).
Отсюда получаем следующий вывод. Если система находится
в нестационарных условиях, когда внешняя среда непрерывно
меняется (однако амплитуда этих изменений не превосходит v0,
а форма изменений во времени всех параметров возмущений —
одна и та же — v(i)), то влияние этих возмущенй не больше
того, которое получается при действии на систему стационарных
возмущений vq.
Ясно поэтому, что все результаты, полученные для стацио-
нарных возмущений, могут быть распространены на случай не-
стационарных возмущений определенного вида.
9.7. Моделирование биосистем и принцип простоты
Целесообразность устройства живых систем сказывается- в
том, что биологические системы при всей их сложности надежны
и просты в управлении. Иногда это положение называют прин-
ципом простоты и формулируют его следующим образом: в ре-
зультате эволюции выжили и были отобраны наиболее простые
и надежные в управлении биологические объекты и процессы —
при условии, что они выполняют свои функции [128, 180].
Простота в управлении достигается в живых системах за
счет специфических форм организации элементов и подсистем,
таких, например, как блочная структура, синергии [223] или
временная иерархия [180]. При моделировании биологических
систем принцип простоты означает, что модель должна содер-
жать минимальное число переменных и уравнений. Вместе с
тем их число должно быть достаточно для описания главных
свойств и функций моделируемой системы или процесса. По-
этому большое значение при выборе математического описания
имеет задача упрощения моделей. Однако вопрос о том, что
считать главными свойствами и функциями системы, опреде-
ляется субъективно самим исследователем, интересы и пред-
почтения которого кладутся в основу модели [159]. Именно этим
последним обстоятельством и определяется тот факт, что с
294
ФЕНОМЕНОЛОГИЯ ГОМЕОСТАЗА
[ГЛ. 9
точки зрения моделирования сохранительных свойств биосистем
можно выделить два типа моделей.
В одном случае интересы исследователя направлены на вы-
яснение процессов и характера взаимодействия между отдель-
ными компонентами — процессов обмена веществ в открытой
системе. Принцип простоты в этом случае проявляется в стрем-
лении к упрощению моделей, например, на основе учета их вре-
менной иерархии [180]. Простота достигается путем сокращения
Числа элементов и, следовательно, числа управляющих механиз-
мов в системе. Легко понять поэтому, что в простых моделях
открытых систем невозможно возникновение гомеостатических
свойств за счет множественности или дублирования управляю-
щих механизмов. В этом случае исследователь обычно не ин-
тересуется зависимостью характеристик системы от внешних
условий — ее гомеостазом.
В другом случае во главу угла ставится задача воспроизве-
дения в первую очередь феномена постоянства некоторых пере-
менных в биологической системе — ее гомеостатических свойств.
Таковы, в частности, модели многих физиологических систем,
построенные по схеме классической теории управления. Про-
стота модели достигается в этом случае уже за счет того, что
для описания регуляции в системе берется готовая модель в
виде простейшей системы управления — отрицательная обрат-
ная связь по отклонению от уставки.
Можно, однако, сказать, что эта простота получена доста-
точно высокой ценой. Ведь для достижения в модели желатель-
ных свойств пришлось ввести в практику моделирования биоси-
стем новый класс сигналов — уставки, по всей видимости, от-
сутствующие во многих реальных живых системах. Уставка в
физиологических моделях — лишь более или менее удобный
способ описания всей совокупности воздействий организма на
рассматриваемую систему [253]. Однако модель с отрицательной
обратной связью получила такое распространение, что в био-
кибернетической литературе можно встретить мнение, что ус-
тавки представляют собой нечто реально существующее в жи-
вых системах.
Не противоречит ли такой способ моделирования гомеостаза
принципу Оккама — предшественнику современных принципов
простоты? «Бритва Оккама», сформулированный в первой по-
ловине XIV в. принцип лаконичности мышления (см., например,
[102]), требует от ученого, чтобы он старался объяснить каждое
явление возможно более просто, и гласит: «Entia non sunt mul-
tiplicanda praeter necessitatem»*).
) Сущностей не следует умножать сверх необходимости (лат.).
9.7)
МОДЕЛИРОВАНИЕ БИОСИСТЕМ И ПРИНЦИП ПРОСТОТЫ
295
Так или иначе, но переход к моделям гомеостаза открытых
систем, рассматриваемым в настоящей книге, означает не про-
сто усложнение модели за счет сочетания каналов управления.
Конечно, в классической модели гомеостаза как системы управ-
ления по рассогласованию постоянство некоторой переменной
может быть обеспечено всего лишь одним каналом обратной
связи, а в моделях открытых систем без введения специальных
уставок на переменные состояния гомеостаз обеспечивается
только в том случае, если имеется сочетание нескольких дубли-
рующих каналов управления. Даже в простейшем случае таких
каналов должно быть два — как в моделях физиологических
систем с пассивным и активным регуляторами темпов потоков
вещества.
В концептуальном же плане такие модели в определенном
смысле даже «проще» моделей с уставками — для описания тех
же классов явлений в них рассматривается не шесть типов пе-
ременных, а только пять, как это следует из разд. 7.2 и табл. 7.1.
Что касается математического аппарата исследования, то осо-
бого усложнения нет и здесь, так как в случае линейных систем
матричные методы позволяют в компактном виде описывать
системы произвольного порядка. Поэтому рассмотренный в
книге метод моделирования гомеостатических свойств открытых
систем лежит в общем русле развития работ по изучению био-
систем на основе принципа простоты и позволяет описывать и
исследовать сохраиительные свойства биосистем с помощью про-
стых и эффективных методов.
СПИСОК ОБОЗНАЧЕНИЙ И СОКРАЩЕНИЙ
А — величины сродства в феноменологических уравнениях,
Adj — присоединенная матрица,
А, В, С,... — матрицы,
а, Ь, с... — коэффициенты, константы,
С — управляющая сила фермента,
с — концентрация вещества,
с — удельная теплоемкость,
D — константа диффузии,
D — коэффициент диффузии,
d — скорость протока в культиваторе,
е — предел неопределенности (в принципе удовлетворения),
Е — единичная матрица,
Е — фермент и его количество, участвующее в ферментативных реакциях,
G — интегральный показатель сохранительных свойств системы при изме-
нении режимов функционирования,
g— показатель сохранительных свойств системы при перестройке режи-
мов функционирования,
Л — предел удовлетворения (в принципе удовлетворения),
h — показатель гомеостатической способности системы,
Н — интегральный показатель гомеостатической способности системы,
1 — плотность потока вещества в процессах диффузии,
k — константа Больцмана,
k — константа скорости химических реакций,
L — клиренс,
т — масса,
N — резерв гомеостаза системы,
N — численность популяции,
Р — давление крови в кровеносных сосудах,
Р — продукт в ферментативных реакциях и его количество,
Р — пространственная точка,
Q — критерий оптимальности системы,
Q — кровоток,
R — остаточный член в разложении Тейлора,
R — сопротивление кровеносных сосудов,
г — темп потока вещества,
S — матрица чувствительности,
S — площадь,
S — субстрат в ферментативных реакциях и его количество,
S — энтропия,
Т — момент окончания процесса,
Т — постоянная времени,
t — время,
U — матрица Лесли,
и—лимитирующий фактор в моделях Л-систем,
и — управляющее воздействие в системе автоматического управления,
СПИСОК ОБОЗНАЧЕНИЙ И СОКРАЩЕНИЙ
297
и — функция чувствительности,
V — внешнее воздействие (возмущающее) на нелинейную систему до ли-
неаризации,
V — объем резервуара, пространственной области, компартмента,
v — внешнее воздействие на произвольную систему (возмущающее),
и — то же в случае линеаризованной системы,
W — заданные темпы в нелинейной системе до линеаризации,
W — приспособленность популяций,
w — заданные темпы (первичные, независимые) в произвольной системе,
w — то же в линеаризованной системе,
w — фазовый объем,
X — переменная состояния, вектор состояния в нелинейной системе до ли-
неаризации,
х — переменная состояния, вектор состояния в произвольной системе,
х — то же в линеаризованной системе,
х—пространственная координата,
х — уровень вещества или энергии в системе и компартментах,
х — уставка по переменной х в системе управления,
У — выходная переменная, вектор выходов в нелинейной системе до ли-
неаризации,
у — выходная переменная, вектор выходов в произвольной системе,
у — то же в линеаризованной системе,
у — поток продукта,
у — темпы потоков вещества (вторичные, зависимые),
у — пространственная координата,
z — пространственная координата,
а, р, у,... — стехиометрические коэффициенты,
а — коэффициент в формуле показателя гомеостатической способности,
Р — коэффициент выживаемости в моделях экосистем,
у—коэффициент конкурентоспособности в моделях экосистем,
е — сигнал рассогласования в системах автоматического управления,
е — коэффициент пополнения в моделях экосистем,
Л — характеристическое число,
р — коэффициент естественной смертности в моделях экосистем,
v — степень напряжения геомеостатических механизмов,
р— удельная плотность,
р — логарифмический коэффициент чувствительности,
а — коэффициент чувствительности,
а — постоянная Стефана-Больцмана,
ц — влажность,
— температура,
О — концентрация СО2 в модели Ф. Гродинза,
£ — коэффициент затухания,
Ф — фундаментальная матрица линейной системы,
£2 — область изменения переменной,
® — собственная частота,
1 (/) — единичный скачок, ступенчатый сигнал единичной амплитуды в си-
стеме автоматического управления.
Сокращения
АДФ — аденознндифосфат,
АТФ — аденозинтрифосфат,
МК — молочная кислота,
ПВК — пировиноградная кислота,
ТГ — тироидный гормон,
ТТГ — тиреотропный гормон,
29S
СПИСОК ОБОЗНАЧЕНИЙ И СОКРАЩЕНИЙ
рОг — напряжение кислорода,
сОг — концентрация кислорода.
Индексы
а — артериальный,
в — венозный,
к — относящийся к коже,
опт — оптимальный,
с — относящийся к среде,
т — индекс, обозначающий транспонирование (верхний индекс)
т — тканевый,
я — относящийся к «ядру» тела,
е — внешний,
g — общий,
й — относящийся к гомеостазу,
i — внутренний,
q — относящийся к высокому качеству,
s — стационарный,
и — относящийся к управляющим воздействиям,
v — относящийся к внешним возмущающим воздействиям,
w — относящийся к внешним задающим воздействиям.
ЛИТЕРАТУРА
1 А б р а м о в а Н. Т., Целостность и управление, «Наука», М., 1974.
2 А в ц ы н А. П., Адаптация и дизадаптация с позиций патолога, Клиниче-
ская медицина, № 5, 1974.
3. Адольф 3., Развитие физиологических регуляций, «Мир», М., 1971.
4. А л е к с е е в В. В., Динамика водных биогеоценозов. Сб. «Человек и био-
сфера», изд. МГУ, М., 1975.
5. Амосов Н. М., Регуляция жизненных функций и кибернетика, «Нау-
кова думка», Киев, 1964.
6. Амосов Н. М., Палец Б Л., Агапов Б. Т., Ермакова И. И.,
Лябах Е. Г., Пацкина С. А., Соловьев В. П., Теоретические ис-
следования физиологических систем, «Наукова думка», Киев, 1977.
7. Амосов Н. М., Л и щу к В. А., Палец Б. Л., П а ц к и и а С. А., Ер-
макова И. И., Лябах Е. Г., Алгоритмическое обеспечение модели
внутренней сферы организма. Сб. «Управление и информационные про-
цессы в живой природе», «Наука», М., 1971.
8. А м о с о в Н. М., Остапов Ю. Г., Математическое моделирование ме-
' таболизма клетки. Сб. «Проблемы кибернетики», вып. 2£, «Наука», М.,
1972.
9. Айо хин П. К-, Теория функциональной системы как предпосылка к по-'
строению физиологической кибернетики. Сб. «Биологические аспекты ки-
бернетики», Изд. АН СССР, М., 1962.
10. А но хи и П. К-, Общие принципы формирования защитных приспособ-
лений организма, Вести. АМН СССР, № 4, 1962.
11. Анохин П. К-, Теория функциональной системы. Сб. «Общие вопросы
физиологических механизмов», «Наука», М., 1970.
12. Автомонов Ю. Г., Моделирование биологических систем. Справоч-
ник, «Наукова думка», Киев, 1977.
13. А ито мо но в Ю. Г., Ки фо ре нк о С. И., Микульская И. А.,
Пароконная Н. К., Математическая теория системы сахара крови,
«Наукова думка»,'Киев, 1971.
14. Аристотель, Физика, Гос. соц.-эк. Изд., М., 1937.
15. Ар м а и д А. Д., Теория поля и проблема выделения геосистем, Вопросы
географии. Сб. 98, «Мысль», М., 1975.
16 Армаид А. Д., Куприянова Т. П., Типы природных систем и фи-
зико-географическое районирование, Изв. АН СССР, сер. геогр., № 5,
1976.
17. Афанасьев Ю. И., Ноздрин В. И., Регуляция структуры и функ-
ции клетки, Успехи современной биологии, т. 83, вып. 3, 1977.
18. Ахутин В. М„ Неймарк Г. С., Иориш А. Е., Шиф М. И., Био-
нические принципы синтеза эргатнческих систем повышенной надежности,
изд. Научного совета по кибернетике, М.-Л., 1975.
19. Аршавский И. А., Физиологические механизмы продолжительности
жизни млекопитающих, Успехи современной биологии, т. 83. вып. 2.
1977.
300
ЛИТЕРАТУРА
20. Баевский Р. М., К проблеме прогнозирования состояния человека
в условиях космического полета, Физиологический журнал СССР, № 6,
1972.
21. Баевский Р. М., Геллер Е. С., О роли колебательных процессов в
механизмах адаптации биологических систем. Сб. «Методологические
проблемы биокибернетики», «Наука», М., 1974.
22. Б а е в с к и й Р. М., Геллер Е. С., Некоторые проблемы анализа энер-
го-информационных связей в живых системах. Сб. «Вопросы киберне-
тики», изд. Научного совета по кибернетике, вып. 22, М., 1976.
23. Баевский Р. М., Геллер Е. С., Нидеккер И. Г., Никули-
на Г. А., С е м е и о в а Т. Д , Биокибернетика: циклические процессы в
живом организме. Сб. «Общие вопросы физиологических механизмов»,
«Наука», М., 1970.
24. Б а р о я н О. В., Р в а ч е в Л. А., Ермаков В. В., Франк К. Д.,
Шашков В. А., Итоги машинного и математического моделирования
эпидемий гриппа в масштабе страны. Сб. «П Всесоюзный академия, сим-
позиум по применению математических методов и вычислительной тех-
ники в медико-биологических исследованиях (тезисы докладов)», изд.
Ин-та медицинской радиологии АМН СССР, Обнинск, 1971.
25. Б а р т о и А., Э д х о л м О., Человек в условиях холода, ИЛ, М., 1957.
26. Бауэр Э. С., Теоретическая биология, Всес. ин-т эксперим. медицины,
М.-Л., 1935.
27. Бахмутов В. А., Температура тела тетеревиных птиц Западной Си-
бири при разных состояниях активности, Экология, № 2, 1975.
28. Беклемишев В. Н., Об общих принципах организации жизни, Бюлл.
МОИП, т. LXIX, вып. 2, 1964.
29. Белецкий Ю. Б., Безуглов В. П., О возможности применения ме-
тодов теории автоматического управления для анализа изменений арте-
риального давления после физических нагрузок. Сб. «Проблемы управ-
ления функциями организма человека и животных», нзд. Научного со-
вета по кибернетике, М., 1973.
30. Беликова М. А., Ляпунов А. А., Старовойтова Э. П., Си-
стемный подход к математическому моделированию эндокринной си-
стемы и системы кровообращения. Сб. «Проблемы кибернетики», вып. 25,
М„ 1972.
31 БерталаифиЛ., Общая теория систем — критический обзор. Сб. «Ис-
следования по общей теории систем», «Прогресс», М., 1969.
31а . Б ё р н ш т е й н В. А., Критические замечания к теории «установочной
точки» в терморегуляции, Успехи физиологических наук, т. 6, № 4, 1975.
32. Бир С., Кибернетика и управление производством, Физматгиз, М., 1963.
33. Б р а й и е с С. Н., С в е ч и н с к и й В. Б., Проблемы иейрокибериетики и
нейробионики, «Медицина», М., 1968.
34. Брайн^с С. Н., Свеч и нс кий В. Б., К общей теории управления
в целостном организме в норме и патологии. Сб. «Биологическая и ме-
дицинская кибернетика», «Медицина», М., 1971.
35. Бурлаков Р. И., Юшкин А. В., Синтез управления ИВЛ на основе
стохастической модели, Медицинская техника, Xs 5, 1970.
36. Б ы х о в с к и й М. Л., Чувствительность и динамическая точность систем
управления, Изв. АН СССР, Техническая кибернетика, № 6, 1964.
37. Вериго В. В., Скотникова Г. А., Математическая модель коопе-
ративного иммунного ответа. С б. «Математическая теория биологиче-
ских процессов», изд. Научного совета по кибернетике, Калининград,
1976.
38. Ви лли К., Детье В., Биология, «Мир», М., 1974.
39. В и и б е р г Г. Г., Интенсивность обмена и пищевые потребности рыб,
изд. Белорусского гос. ун-та, Минск, 1956.
40. Винер Н., Кибернетика, «Сов. радио», М., 1968.
ЛИТЕРАТУРА
801
41. Винчестер А., Основы современной биологии, «Мир», М., 1967.
42. В и т е н б е р г И. М., Программирование аналоговых вычислительных
машин, «Машиностроение», М., 1972.
43. В о д о п ь я и о в П. А., Устойчивость в развитии живой природы, «Нау-
ка и техника», Минск, 1974.
44, ВолькенштейиМ. В., Молекулы и жизнь, «Наука», М., 1965.
45. Вольтерра В., Математическая теория борьбы за существование,
«Наука», М., 1976.
46. ВороновА-. А., Основы теории автоматического управления, ч. I—III,
«Энергия», Л., 1965-1970.
47 Г а в р и л е ц Ю. Н., Экономический гомеостаз. Сб. «Математика и ки-
бернетика в экономике», «Экономика», 1975.
48. Галетти П., Брич ер Г., Основы и техника экстракорпорального кро-
вообращения, «Медицина», М., 1966.
49 ГарфинкельД., Моделирование биохимических систем. Сб. «Вычис-
лительные устройства в биологии и медицине», «Мир», М., 1967.
50. Гельман В. Я., Тар а бу кин В. И., Анализ системы управления
периферическим артериальным давлением. Сб. «Автоматическое регули-
рование физиологических функций в условиях патологии», изд. Леиингр.
ии-та усоверш. врачей, Л., 1972.
51. Гельфанд И. М., Цетлин М. Л., О математическом моделировании
механизмов центральной нервной системы. Сб. «Модели структурно-
функциональной организации некоторых биологических систем», «Наука»,
М, 1966.
52. Г е р а с и м о в И. П., Научные основы современного мониторинга окру-
жающей среды, Изв. АН СССР, сер. геогр., № 3, 1975.
53. Гильдерман Ю. И., Кудрина К. Н., Полетаев И. А., Модели
Л-систем. Сб. «Исследования по кибернетике», «Сов. радио», М.,
1970.
54. Г и л я р о в М. С., Закономерности приспособления членистоногих к жиз-
ни иа суше, «Наука», М., 1970.
55. Гомеостаз. Под ред. П. Д. Горизонтова, «Медицина», М., 1977.
55а. Голдовский А. М. Основы учения о состояниях организмов, «Нау-
ка», Л., 1977.
56. Голдман С., К вопросу о кибернетических аспектах гомеостазиса.
Сб. «Самоорганизующиеся системы», «Мир», М., 1964.
57. Гольдэкр Р., Регуляция ритма и гомеостазиса в биологии и меди-
цине. Сб. «Кибернетика н живой организм», «Наукова думка», Киев,
1964.
58. Г о р с т к о А. Б , С у р к о в Ф. А., Математика и проблемы сохранения
природы, «Знание», М., 1975.
58а. Граве П. С., Об адаптации в живых системах. Сб. «Адаптивные си-
стемы», вып. I, «Зинатне», Рига, 1972.
59. Гринченко С. Н., Загускии С. Л., Загускииа Л. Д., Модель
оптимизации энергетики нейрона. Сб. «Проблемы бионики», вып. 15,
«Вища школа», Харьков, 1975.
60. Гродинз Ф., Теория регулирования и биологические системы, «Мир»,
М„ 1966.
61. Давиташвили Л. Ш., Причины вымирания организмов, «Наука»,
М„ 1969.
62. Давыдовский И. В., Приспособительные процессы в патологии,
Вести. АМН СССР, № 4, 1962.
63. Д а р т а у Л. А., Модель энергообмеиа организма как гомеостатическая
система управления. Сб. «Биологическая и медицинская кибернетика»,
изд. Научного совета по кибернетике, М.-Л., 1974.
64. Д а р т а у Л. А., Г у м е н е р П. И., Молчанова С. С., Новосель-
цев В. Н., Интегральная оценка эффективности гомеостатических
302
ЛИТЕРАТУРА
систем. Сб. «IFAC symposium on control mechanisms in bio and eco-
systems», v. 4, Leipzig, 1977.
65. Демпси Э. В., Гомеостаз. Сб. «Экспериментальная психология», т. 1,
ИЛ, М., 1960.
66. Деруссо П., Рой Р., Клоуз Ч., Пространство состояний в теории
управления, «Мир», М., 1970.
67. Д е ч е в Г., Матвеев М., Окислительно-восстановительный дебит в
биологических объектах как результат управляемых необратимых про-
цессов в открытых системах, Биофизика, т. XIV, № 6, 1969.
68. Диксон М., Уэбб Э„ Ферменты, ИЛ, М., 1961.
69. Дрё Ф., Экология, Атомиздат, М., 1976.
70. Дришель Г., Регулирование уровня сахара крови. Сб. «Процессы
регулирования в биологии», ИЛ, М„ 1960.
71. Дружинин В. В., Ко ит о ров Д. С., Проблемы системологии, «Сов.
радио», М., 1976.
72. Дэгли С., Ни колье о и Д., Метаболические пути, «Мир», М., 1973.
73. Емельянов С. В., Борисов В. И., Малевич Л. А., Черка-
шин А. М., Модели и методы векторной оптимизации. Сб. «Итоги науки
и техники», т. 5, Техническая кибернетика, изд. ВИНИТИ, М., 1973.
74. Ермакова И. И., Роль температуры артериальной крови в системе
терморегуляции человека (эксперименты на цифровой модели), Физиоло-
гический журнал СССР, № 11, 1973.
75. Жаботинский А. М., Концентрационные колебания, «Наука», М.,
1974.
76. Загер О., Межуточный мозг, Изд. АН РНР, Бухарест, 1962.
77. ' 3 о т и и А. И., Количественные теории роста. Сб. «Количественные
аспекты роста организмов», «Наука», М., 1975.
78. Ибера л л А. С., Мак-Калл ок У. С., Гомеокинез — организацион-
ный принцип сложных систем. Сб. «Общие вопросы физиологических
механизмов», «Наука», М., 1970.
79. Иванов К П., Биоэнергетика и температурный гомеостазис, «Наука»,
Л., 1972.
80. И в а х и е и к о А. Г., Долгосрочное прогнозирование и управление слож-
ными системами, «Техшка», Киев, 1975.
81. Измайлов С. Ф., Смирнов А. М., Функциональная компартмента-
ция бикарбоновых аминокислот в растительной клетке, Изв. АН СССР,
сер. биол., № 5, 1977.
82. Казначеев В. П., Биосистема и адаптация, изд. Сиб. филиала АМН
СССР, Новосибирск, 1973.
83. Каи Г. С., Кан Е. Л., А л ь б е р т и и с к и й Б. И., Эритрон как подси-
стема в сложной иерархической системе управления балансом кисло-
рода. Сб. «Общие вопросы физиологических механизмов», «Наука», М.,
ял 1976'
б4„ К а н д р о р И. С., Новосельцев В. Н., О возможном подходе к мо-
делированию системы терморегуляции человека. Сб. «Физиологические
адаптации к теплу и холоду», «Наука», Л., 1969.
85. К а пи то иен ко В.'В., Количественная оценка степени нелинейности
в задачах идентификации. Сб. «Третье Всесоюзное совещание по стати-
стическим методам в теории управления. Тезисы докладов», ч. I, изд.
Ин-та проблем управления, М., 1973.
86. К а р п м а н В. Л., Фазовый анализ сердечной деятельности, «Медйцина»,
М., 1965.
87. Карр Я., Механизмы биологической защиты, «Медицина», М., 1976.
88- К атков ник В. Я., Полуэктов Р. А., Многомерные дискретные
системы управления, «Наука», М., 1966.
89. Коган А. Б. (ред.), Биологическая кибернетика, «Высшая школа»,
М„ 1972.
ЛИТЕРАТУРА
303
90 К о г а н Б. Я., Электронные Моделирующие устройства .и их примене-
ние для исследования систем автоматического регулирования, Физмат-
гнз, М., 1963.
91. Колмогоров А Н., Качественное изучение математических моделей
динамики популяций. Сб. «Проблемы кибернетики», вып. 25, «Наука»,
М„ 1972.
92. Коняхина Л. В., Приц А. К., Фикс Б. В., К учету взаимных свя-
зей между коэффициентами удельного прироста биомассы различных
компонент ценоза. Сб. «Математическая теория биологических процессов»,
изд. Научного совета по кибернетике, Калининград, 1976.
93. К р а с н е н к е р А. С., Задачи и методы векторной 'оптимизации, Из-
мерения, контроль, автоматизация, № 1, 1975.
94 Краснов М. Л., Макаренко Г. И., Операционное исчисление.
Устойчивость движения, «Наука», М., 1964.
95. Лауэр Н. В., К о л ч и н с к а я А. 3., Ш к а б а р а Е. А., О регулиро-
вании кислородных режимов организма. Сб. «Кислородный режим орга-
низма и его регулирование», «Наукова думка», Киев, 1966.
96. Л а й ф у т Э., Явления переноса в живых системах, «Мир», М., 1977.
97. Л е в а д н ы й В. Г., Энергетически оптимальные структурные параметры
бронхиального дерева. Сб. «Вопросы кибернетики», изд. Научного Со-
вета по кибернетике, М., 1975.
98. Леви А., С и к е в и ц Ф., Структура и функции клетки, «Мир», М.,
1971.
99. Левине Р., Генетические последствия естественного отбора. Сб. «Тео-
ретическая и математическая биология», «Мир», М., 1968.
100. Левтов В. А., Химическая регулйцня местного кровообращения, «Нау-
ка», Л., 1969.
101. Левнч А. П., Понятие устойчивости в биологии. Математические ас-
пекты. Сб. «Человек н биосфера», вып. 1, изд. МГУ, М., 1976.
102. Лем С., Сумма технологии, «Мир», М., 1968.
103. Л ей ннгер А., Превращение энергии в клетке. Сб. «Живая клетка»,
ИЛ, М„ 1962.
104. Л и Р., Оптимальные оценки, определение характеристик и управление,
«Наука», М., 1966.
105. Л и щ у к В. А., Основные свойства саморегуляции нососной функции
сердца. Св. «Некоторые проблемы биокибернетики, применение электро-
ники в бнологнн н медицине», нзд. Ин-та кибернетики АН УССР, вып. 1,
Киев, 1969.
106. Л ищу к В. А., Палец Б. Л., Исследование математической моде-
ли сердечно-сосудистой системы (на укр. яз.), Автоматика, № 3,
1971.
107. Л ищу к В. А., Столяр В. Л., Мироненко В. И., Шидлов-
с к и й В. А., Теоретический анализ задачи идентнфикацнн математиче-
ской модели сердечно-сосудистой системы. Сб. «Математическая теория
биологических процессов», нзд. Научного совета по кибернетике, Кали-
нинград, 1976.
108. Л у кош ков а Е. В., Ха ют ин В. М., Шерман И. А., Об оптимиза-
ции быстродействия сердечно-сосудистой системы прн оборонительной
реакции. Сб. «Вопросы кибернетики», изд. Научного совета по киберне-
тике, М., 1975.
109. Л у не в ск а я Л. В., Сельков Е. Е., Чап лей и Р. А., Математи-
ческая модель кинетики АТФ-азной реакции. Сб. «Регуляция в биоло-
гических системах» (тезисы докладов), нзд. Ин-та биофизики АН СССР,
Пущнно-на-Оке, 1970.
ПО. Л я п у и о в А. А., О рассмотрении биологии с позиций изучения живой
природы как большой системы. Сб. «Проблемы методологии системного
исследования», «Мысль», М., 1970.
304
ЛИТЕРАТУРА
111. Ляпунов А. А., О кибернетических вопросах биологии. Сб. «Проб-
лемы кибернетики», вып. 25, «Наука», М., 1972.
112. Ляпунов А. А.. Багриновская Г. П., О методологических во-
просах математической биологии. Сб. «Математическое моделирование
в бнологнн». Материалы I школы по математическому моделированию
сложных биологических систем, «Наука», М., 1975.
113. Май стр ах Е. В., Автоматическое регулирование физиологических
функций в условиях патологии. Сб. «Автоматические системы для регу-
ляции жизнедеятельности в условиях патологии», Труды Ленингр. ин-та
усоверш. врачей, вып. 104, Л., 1972.
114. Маленков А. Г., Ионный гомеостаз и автономный рост опухоли,
«Медицина», М., 1976.
115. Малик Л. К., О сохранении качества поверхностных вод Западной
Сибири, Изв. АН СССР, сер. геогр., № 5, 1976.
116. М а л и н о в с к н й А. А., Типы управляющих биологических систем и их
приспособительное значение. Сб. «Проблемы кибернетики», вып. 4, М.,
1960.
117. Малиновский А. А., Общие вопросы строения систем и их значе-
ние для биологии. Сб. «Проблемы методологии системного исследова-
ния», «Мысль», М., 1970.
118. М а р к о с я н А. А., Надежность биологической системы — всеобщий за-
кон онтогенеза. Сб. «Ведущие проблемы советской геронтологии», «Нау-
кова думка», Киев, 1972.
119. Математическая теория биологических процессов (тезисы докладов
I конференции), нзд. Научного совета по кибернетике, Калининград,
1976.
120. Мееров М. В., Синтез структур автоматического регулирования вы-
сокой точности, «Наука», М., 1964.
121. Мееров М. В., Системы многосвязного регулирования, Фнзматгнз, М.,
’ 1967.
122. Меннцкнй Д. Н., Саморегуляция нервных механизмов. Сб. «Совре-
менные проблемы нейрокнбернетнкн», «Наука», Л., 1972.
123. М е и ш у т к и и В. В., Математическое моделирование популяций про-
мысловых рыб, нзд. Ин-та рыбного хозяйства н океанографии (ВНИРО),
М„ 1970.
124. Меншуткнн В. В., Моделирование процессов изучения и эксплуата-
ции озерной экологической системы. Журнал общей биологии, т. 32, № 2,
1971.
125. Меньшиков В. В., Гуморальные механизмы регуляции функций орга-
низма в норме и патологии, «Медицина», М., 1970.
126. Меньшов А. И., Космическая эргономика, «Наука», Л., 1971.
127. Месаровнч М., Мако Д., Такахара И., Теория иерархических
многоуровневых систем, «Мнр», М., 1973.
128. Методологические принципы физики, «Наука», М., 1975.
129. Меттлер Л., Грегг Т., Генетика популяций и эволюция, «Мнр», М.,
1972.
130. М н л с у м Дж., Анализ биологических систем управления, «Мнр», М.,
1968.
131. Молчанов А. М., Многобарьерный иммунитет, Биофизика, т. XVI,
вып. 3, 1971.
131а. М о л ч а и о в А. М., Релаксационная модель адаптации, Биофизика,
т. XVII, вып. 2, 1972.
132. Молчанов А. М., Математические модели в экологии. Роль критиче-
ских режимов. Препринт, изд. Ин-та биофизики АН СССР, Пущино-на-
Оке, 1973.
133. Молчанова С. С., Новосельцев В. Н., Математический способ
определения функциональных возможностей организма спортсмена при
ЛИТЕРАТУРА
305
дозированной мышечной работе. Сб. «Методы биокибернетического ана-
лиза функционального состояния спортсменов-подростков», изд. Ин-та
гигиены детей и подростков, М., 1977.
134. Москаленко Ю. Е., Закономерности регуляции кровоснабжения го-
ловного мозга, Вести. АМН СССР, № 11, 1974.
135. Москаленко Ю. Е„ Вайнштейн Г. В., Демченко И. Т. и др.,
Внутричерепная гемодинамика, «Наука», Л., 1975.
136. Наппельбаум Э. Л., Предисловие к книге Дж. Милсума «Анализ
биологических систем управления», «Мир», М., 1968.
137. Наумов Н. П., Бионика и надорганнзменные системы. Сб. «Проблемы
бионики», «Наука», М., 1973.
138. Нельзина Е. Н., Медведев С. И., Данилова Г. М., Географи-
ческая устойчивость бноценотнческого комплекса суслнчных нор, Билл.
МОИП, отд. биол., т. LXXII, вып. 1, М., 1967.
139. Новосельцев В. Н., Оптимальные по быстродействию системы упра-
вления при наличии случайных помех, Автоматика и телемеханика, т. 23,
№ 12, 1962.
140. Новосельцев В. Н., Об оценке гомеостатических свойств систем,
Автоматика и телемеханика, № 7, 1972.
141. Новосельцев В. Н., Гомеостаз систем управления, Автоматика и те-
лемеханика, № 12, 1973.
142. Новосельцев В. Н., Гомеостаз организма как система управления,
изд. Ин-та проблем управления, М., 1973.
143. Новосельцев В. Н., Гомеостаз в биологических системах управле-
ния. Сб. «IV Всесоюзное совещание по проблемам управления». Рефе-
раты докладов, ч. II, «Наука», М., 1974.
144. Новосельцев В. Н., Системный подход в биологии и моделирова-
ние гомеостаза. Сб. «Биологическая и медицинская кибернетика», ч. I,
Биокибернетика, изд. Научного совета по кибернетике, М.-Л., 1974.
145. Новосельцев В. Н., Бноснстема, Большая Медицинская Энциклопе-
дия, т. III, 1975.
146 Н о в о с е л ь це в В. Н., Адаптация и гомеостаз живых систем. Сб. Тру-
дой Ин-та проблем управления, вып. 8, М., 1976.
147. Новосельцев В. Н., Гуменер П. И., Понятие «функциональное
состояние организма» с биокибернетических позиций. Сб. «Методы био-
кибернетического анализа функционального состояния спортсменов-под-
ростков», нзд. Ин-та гигиены детей и подростков АМН СССР, М., 1977.
148. Одум Ю., Основы экологии, «Мир», М., 1975.
149. Ольнянская Р. П., О гомеостатических реакциях организма и его
физиологических состояниях, «Наука», Л., 1969.
150. Оптимум. Большая Советская Энциклопедия, т. 31, Изд. «Большая Со-
ветская Энциклопедия», М., 1955.
151. Орки и а Е. Л., Моделирование процессов в системе «организм-опу-
холь». Сб. трудов Ин-та проблем управления, вып. 8, М., 1976.
152. Основы палеонтологии. Голосеменные и покрытосеменные, Госгеолтехиз-
дат, М., 1963.
153. О х и я и с к а я Л. Г., Критерии оценки пределов адаптации человека при
длительном воздействии производственных веществ. Сб. «Адаптация и
компенсация при химических воздействиях», изд. ВИНИТИ, М., 1973.
154 Парии В. В., Баевский Р. М., Введение в медицинскую киберне-
тику, «Медицина», М., 1966.
15£. Петровский А. М., Сучков В. В., Шхвацабая И. К., Управле-
ние лечением заболеваний как проблема современной теории управле-
ния, Автоматика и телемеханика. № 5, 1973.
156. Пет рушен ко Л. А., Принцип обратной связи, «Мысль», М., 1967.
157. Пикеринг У. Д„ Н и к и ф о р у к П. Н., Мерриман Дж. Э.,
Исследование сердечно-сосудистой системы регулирования с помощью
806
ЛИТЕРАТУРА
аналоговой вычислительной машины. Сб. «Биоэлектрическое управление.
Человек в автоматические системы», «Наука», М„ 1970.
158. Полетаев И. А., О математических моделях элементарных процессов
в биогеоценозах. Сб. «Проблемы кибернетики», вып. 16, «Наука», М., 1966.
159. Полетаев И. А., О математическом моделировании. Сб. «Проблемы
кибернетики», вып. 27, «Наука», М., 1973.
160. Полетаев И. А., Модели Вольтерра «хищник-жертва» и их расшире-
ние с использованием принципа Либиха, Журнал общей биологии,
т. XXXIV, № 1, 1973.
161. Полуэктов Р. А. (ред.), Динамическая теория биологических попу-
ляций, «Наука», М., 1974.
162. Понтрягин Л. С., Болтянский В. Г., Гамкрелндзе Р. В.,
Мищенко Е. Ф., Математическая теория оптимальных процессов, Физ-
матгиз, М., 1961.
162а. П р н г о ж и и И., Введение в термодинамику необратимых процессов,
ИЛ, М, 1960.
163. П р и ц А. К., Принцип стационарных состояний открытых систем и ди-
намика популяций, нзд. Калинингр. гос. ун-та, Калининград, 1974.
164. Прогресс биологической и медипи некой кибернетики. Под ред. А. И. Бер-
га и С. Н. Брайнеса, «Медицина», М., 1974.
165. Прокопенко ГО. И., Уровни компенсации как показатель неблаго-
приятного действия факторов внешней среды, Вести. АМН СССР, № 8,
1974.
166. Проссер Л., Браун Ф., Сравнительная физиология животных,
«Мир», М., 1967.
167. Разумовский О. С., Современный детерминизм н экстремальные
принципы в физике, «Наука», М., 1975.
168. Рапопорт А., Математические аспекты абстрактного анализа систем.
Сб. «Исследования по общей теории систем», «Прогресс», М., 1969.
169. Растр игин Л. А., Статистические методы поиска, «Наука», М., 1968.
169а. Р а с т р и г и и Л. А., Проблема адаптации в технике, биологии и социо-
логии. Сб. «Адаптивные системы», вып. I, «Зинатне», Рига, 1972.
170. Ратнер В. А., Молекулярно-генетические системы управления, «Нау-
ка», Новосибирск, 1975.
171. Рашевски Н., Некоторые медицинские аспекты математической био-
логии, «Медицина», М., 1966.
172. Рашевский Н„ Модели и математические принпнпы в биологии. Сб.
«Теоретическая и математическая биология», «Мир», М., 1968.
173. Рашевский Н., Организмические множества: очерк общей теории
биологических и социальных организмов Сб. «Исследования по общей
теории систем», «Прогресс», М„ 1969.
174. Роджерс Э., Физика для любознательных, т. I, «Мир», М., 1969.
175. Розен Р., Принцип оптимальности в биологии, «Мир», М., 1969.
176. Розеивассер Е. Н., Юсупов Р. М., Методы теории чувствитель-
ности в автоматическом управлении, «Энергия», Л., 1971.
177. Розоноэр Л. И., Предисловие к книге Ф. Гродииза «Теория регули-
рования н биологические системы», «Мир», М., 1966.
178. Розоноэр Л И., Обмен н распределение ресурсов (обобщенный тер-
модинамический подход), ч. I, Автоматика н телемеханика, №'5, 1973.
179. Романовский Ю. М., Степанова Н. В., Ч е р и а в с к и й Д. С.,
Что такое математическая биофизика? «Просвещение», М., 1971.
180. Романовский IO. М., Степанова Н. В., Черпавский Д. С.,
Математическое моделирование в биофизике, «Наука», М., 1975.
181. Ростопшнн Ю. А., Математическое моделирование в физиологии
(одни метод выделения подсистем). Сб. «Математическая теория биоло-
гических процессов», изд. Научного совета по кибернетике, Калининград.
1976.
ЛИТЕРАТУРА
307
182. Румянцев Г В., Слепчук Н. А, Иванов К. П, Величина тепло-
рассеивания и ее регуляция при терморегуляторных сосудистых реак-
циях, Физиологический журнал СССР, № 8, 1973.
183. Саакян В. Г., Изучение некоторых процессов развития н лечения
хронических заболеваний с помощью математической модели. Сб. трудов
Ин-та проблем управления, вып. 8, М., 1976.
184. Саймон Г., Науки об искусственном, «Мир», М., 1972.
185. Саноцкнй И. В., Итоги и перспективы изучения физиологической
адаптации н временной компенсации хронического патологического про-
цесса при минимальных химических воздействиях. Сб. «Адаптация и
компенсация при химических воздействиях», нзд. ВИНИТИ. М., 1973.
186. Саркисов Д С., Очерки по структурным основам гомеостаза, «Ме-
дицина», М., 1977.
186а.Седых Е. И., Модель эволюции двух взаимодействующих популяций,
Автоматика и телемеханика, № 7, 1977.
1866. С в и р е ж ё в Ю. М., Логофет Д. О., Устойчивость биологических со-
обществ, «Наука», М., 1978.
187. Свнрежев Ю. М., Елизаров Е. Я., Математическое моделирование
биологических систем, «Наука», М., 1972.
187а. С в н р е ж е в ГО. М., Вито Вольтерра н современная математическая
экология. Послесловие в [45].
188. Се лье Г., На уровне целого организма, «Медицина», М., 1971.
189. Сельков Е. Е., Ауторегуляцня в биологической системе, Большая Ме-
дицинская Энциклопедия, т. III, 1975.
190. Семенов С. М., Семенова В. Э., Рост и развитие биологических
организмов в случайной среде. Сб. «Математическая теория биологиче-
ских процессов», нзд. Научного совета по кибернетике, Калининград,
1976.
191. Сетров М. И., Организация биоснстем, «Наука», Л., 1971.
192. Сетров М. И., Информационные процессы в биологических системах,
«Наука», Л., 1975.
193. Сеченов И. М., Рефлексы головного мозга. Избранные произведения,
т. I, Изд. АН СССР, М„ 1952.
194. Сн иг л тон В. Т„ Введение в эргономику, изд. Всемирной организации
здравоохранения, Женева, 1974.
195. Соколов Н. И., О построении всережнмных регуляторов. Сб, «Авто-
матическое управление и вычислительная техника», вып. 9, «Машино-
строение», М., 1968.
196. Спиноза Б., Этика, доказанная в геометрическом порядке, Гос. соц.-
эк. Изд., М.-Л., 1932.
197. Степанскнй Г. А., Адаптация человека к современному химическому
окружению. Сб. «Адаптация н компенсация при химических воздей-
ствиях», нзд. ВИНИТИ, М., 1973.
198. Сукачев В. Н., Биоценология и фитоценология, Докл. АН СССР,
т. 47, 1945.
199. Сучков В. В., Петровский А. М., Шхвацабая И. К., Эри-
на Е. В., Сидельникова Т. Я., Теоретические основы поиска инди-
видуальных границ колебаний н длительного поддержания артериаль-
ного давления в допустимых пределах при гипотензивной терапии, Кар-
диология, № 4, 1975.
200. Сушков Ю. Н„ Критериальный метод представления многомерных
уравнений регрессии в медико-бнологнческих исследованиях. Сб. «Мате-
матические методы в биологии и медицине», вып. I, изд. Ин-та медико-
биологических проблем, М., 1968.
201. Тарусов Б Н., Авторегуляторная роль антиоксидантов прн адапта-
ции организмов к условиям внешней среды, Биофизика, т. XV, вып. 2,
1970.
308
ЛИТЕРАТУРА
202. Taj) усов Б Н, Кольо О. Р. (ред.), Биофизика, «Высшая школа»,
М„ 1968.
203. Тенеибаум Л. А., О процессах управления параметрами внешнего
дыхания. Сб. «Общие вопросы физиологических механизмов», «Наука»,
М„ 1970.
204. Тимофеев-Ресовскнй Н. В., Яблоков А. В., Глотов Н. В.,
Очерк учения о популяции, «Наука», М., 1973.
205. Тихонов А. Н., О завнснмостн решений систем дифференциальных
уравнений, содержащих малый параметр, Математический сборник, т. 22,
№ 2, 1948.
206. Тома Р., Комментарий к статье К. Уоддингтона «Основные биологиче-
ские концепции». Сб. «На пути к теоретической биологии», «Мир», М.,
1970.
207. Томович Р., Вукобратович М., Общая теория чувствительности,
«Сов. радио», М., 1972.
208. Томович Р., Радановнч Л., Гомеостатическое управление дина-
мическими системами. Сб. «Самонастраивающиеся автоматические си-
стемы», «Наука», М., 1964.
209. Т ()' а к с е л Дж. Г., Некоторые специальные задачи проектирования си-
стем управления. Сб. «Приспосабливающиеся автоматические-снстемы»,
ИЛ, М„ 1963.
210. Уатт К., Экология и управление природными ресурсами, «Мир», М.,
1971.
211. - Уда,- Со в аж, Бонавентюр, Ледрю, Г ю й е, Управление тепло-
выми процессами в организме человека: регулирование температуры или
изменение теплообмена с помощью сервомеханизма? Сб. «Биологические
динамические системы», Труды американского общества инженеров-ме-
хаников (перевод с аигл.), т. 95, серия G, № 3, 1973.
212. Уильямсон М., Анализ биологических популяций, «Мир», М.,
1975.
212а. Уланов Г. М., Динамическая точность и компенсация возмуще-
ний в системах автоматического управления, «Машиностроение», М'.,
1971.
213. Уоддингтон К. X., Основные биологические концепции. Сб. «На пути
к теоретической бнологнн», «Мнр», М., 1970.
214. Уорнер X. Р., Некоторые методы исследования системы кровообраще-
ния с применением вычислительных машин. Сб. «Вычислительные устрой-
ства в биологии и медицине», «Мир», М., 1967.
215. Уотермэн Т., Проблема. Сб. «Теоретическая и математическая биоло-
гия», «Мир», М., 1968.
216. Уотермен Т., Теория систем и биология. Точка зрения биолога. Сб.
«Теория систем и биология», «Мнр», М., 1971. „
217. Федоров В. Д., Соколова С. А., Об устойчивости планктонного
сообщества, Океанология, т. XII, № 6, 1972.
218. Фельдбаум А. А., Основы теории оптимальных автоматических си-
стем, «Наука», М, 1966.
219. Философские проблемы теории адаптации, «Мысль», М., 1975.
220. Ф л е й ш м а н Б. С., Элементы теории потенциальной эффективности
сложных систем, «Сов. радио», М., 1971.
221. Флейшман Б. С., Крапив ин В. Ф., Математическая модель Пе-
руанской экосистемы. Сб. «Математические модели морских экологиче-
ских систем», «Наукова думка», Киев, 1974.
222. Флейшман Б. С., К р а п н в н н В. Ф., Руденко В. И., Исследова-
ние живучести экосистемы на основе ее математической модели. Сб. «Ин-
формационные методы в системах управления, измерений и контроля»,
т. II, изд. Ин-та рыбного хозяйства и океанографии (ВНИРО), Влади-
восток, 1972.
ЛИТЕРАТУРА
309
223. Фомин С. В., БеркннблитМ. Б., Математические проблемы в био-
логии, «Наука», М, 1973.
224. Форрестер Дж., Основы кибернетики предприятий, «Прогресс», М.,
1971.
225. Фостер, Соулднер, Тэн, Гайтон, Исследование гомеостаза глю-
козы у человека на коротких интервалах времени. Сб. «Динамические
системы и управление», Труды американского общества инженеров-ме-
хаников (перевод с англ.), т. 95, сер. G, № 3, 1973.
226. Франк Г. М., Саморегуляция клеточных процессов. Сб. «Биологические
аспекты кибернетики», «Наука», М, 1962.
227. Ф р о л ь к и с В. В., Регулирование, приспособление и старение, «Наука»,
Л., 1970.
228. Фрэнкс Р., Математическое моделирование в химической технологии,
«Химия», М., 1971.
229. Фуллер А. Т., Оптимизация релейных систем регулирования. Труды
I Международного конгресса ИФАК, т. 2 (теория дискретных, оптималь-
ных и самонастраивающихся систем), Изд. АН СССР, М., 1961.
230. Ханин М. А., Исследование некоторых вопросов механизма кровооб-
ращения человека. Сб. «Проблемы управления функциями организма
у животных», изд. Научного совета по кибернетике, М., 1973.
231. Ха нин М. А., О количественном критерии отбора. Сб. «Вопросы кибер-
нетики», вып. 12, нзд. Научного совета по кибернетике, М., 1975.
232. Ханин М. А., Бухаров И. Б., К феноменологической теории функ-
циональных параметров системы транспорта кислорода, Физиологический
журнал СССР, № 2, 1970.
233. Хании М. А., Дорфман Н. Л., Математические модели роста, осно-
ванные на эволюционном экстремальном принципе, Докл. АН СССР,
т. 212, № 3, 1973.
234. Ха юти и В. М., Сосудодвигательные рефлексы, «Медицина», М., 1964.
235. Хэммонд П., Теория обратной связи, «Наука», М., 1964.
236. Ц в е л е в И. Н., О способе распространения водяного ореха в прошлом
и вымирание его в историческое время, Ботанический журнал, № 9,
1964.
237. Цыпкин Я. 3., Адаптация и обучение в автоматических системах,
«Наука», М., 1968.
238. Цыпкин Я. 3., Основы теории обучающихся систем, «Наука», М„
1970.
239. Цыпкин Я- 3., Оптимальность в естествознании, технике и обществе.
Сб. «Проблемы управления», нзд. Ин-та проблем управления, М., 1975.
240. Чаки Ф., Современная теория управления, «Мир», М., 1975.
241. Чанс Б., Нестационарные метаболические процессы. Сб. «Теоретическая
и математическая биология», «Мир», М., 1968.
242. Шапошников Г. X., Направленность эволюции, Журнал общей био-
логии, т. 38, № 5, 1977.
243. Шарма Д. К., Управление биосинтезом в гормональных системах. Сб.
«Общие вопросы физиологических^ механизмов», «Наука», М., 1970.
244. Шидловский В. А., Динамические биологические системы. Сб.
«Динамические системы и управление». Труды американского общества
инженеров-механиков (перевод с англ.), т. 95, сер. G, № 3, М.,
1973.
245. Шидловский В. А., Особенности гомеостаза и гомеокннеза при мио-
госвязном мультифункцнональном и мультипараметрическом регулиро-
вании. Сб. «Биологическая и медицинская кибернетика», ч. I (Мате-
риалы Второй Всесоюзной конференции), нзд. Научного совета по ки-
бернетике, М.-Л., 1974.
246. [Пндловский В. А., Везеломский Д. В., Ганич кин ГО. Н.,
Л н щ у к В. А., Роль сердца в регуляции кровообращения. Сб. «Бноло-
310
ЛИТЕРАТУРА
гическая и медицинская кибернетика», ч. II, изд. Научного совета по ки-
бернетике М Л, 197!
247. Ш и к Л. Л., Физиолошя дыхания, «Наука», Л., 1973.
248. Шмальгаузен И. И., Кибернетические вопросы биологии, «Наука»,
Новосибирск, 1968.
249. Шмидт-Нильсен К., Животные пустынь. Физиологические проб-
лемы тепла и воды, «Наука», Л., 1972.
250. Шмидт-Нильсен К-, Как работает организм животного, «Мир», М.,
1976.
251. Шредингер Э., Что такое жизнь с точки зрения физики? Атомиздат,
М., 1972.
252. Штен гольд Е. Ш., Принципы и проблемы управления искусственным
сердцем. Сб. «Проблемы искусственного сердца и вспомогательного кро-
вообращения», «Медицина», М., 1970.
253. Шумаков В. И., Новосельцев В. Н., Сахаров М. П., Штен-
гольд Е. Ш., Моделирование физиологических систем организма, «Ме-
дицина», М., 1971.
254. Шумаков В. И., Штенгольд Е. Ш., Онищенко Н. А., Консер-
вация органов, «Медицина», М., 1975.
255. Э й г е и М., Самоорганизация материн и эволюция биологических макро-
молекул, «Мир», М., 1973.
256. Э м а н Т. И., О некоторых математических моделях биогеоценозов. Сб.
«Проблемы кибернетики», вып. 16, «Наука», М., 1966.
257. Энгельгардт В. А, Проблема жизни в современном естествознании.
Сб. «Ленин н современное естествознание», «Мысль», М., 1969.
258. Эшби У. Р., Введение в кибернетику, ИЛ, М., 1959.
259. Эшби У. Р., Конструкция мозга, ИЛ, М., 1962.
259а. Юматов Е. А., Проблема многосвязного регулирования дыхательных
показателей организма, Успехи физиологических наук, т. 6, № 4, 1975.
260. ЯнушевскнйР. Т., Теория линейных оптимальных многосвязных си-
стем управления, «Наука», М., 1973.
261. Ackerman Е., Gatewood L. С., Rosewear J. W„ Molnar G. D.,
Model studies of blood-glucose regulation, Bull. Math. Biophys., v. 27,
spec, issue, 1965.
262. Al bisser A. M„ Leibel B. S., Ewart T. G. et al., An artificial en-
docrine pancreas. Diabetes, v. 23, No. 5, 1974.
263. Amant J. L., The detection of regulation in animal population, Ecology,
v. 51, No. 5, 1970.
264. A m i n A. T., H a k i m i S. L., Graphs with given connectivity and indepen-
dence number of networks with given measures of vulnerability and sur-
vivability, IEEE Trans, on Circ. Theory, v. CT-20, No. 1, 1973.
265. Atkins G. L., Multicompartmental models for biological systems, Met-
huen, London, 1969.
266. Bartolomew G. A., The role of physiology and behavior in the main-
tenance of homeostasis in the desert environment, in: «Homeostasis and
feedback mechan sms», Cambridge Univ. Press, 1964. '
267. Bellman R., Topics in pharmacokinetics. 1. Concentration-dependent ra-
tes, Math. Biosci., v. 6, No. 1, 1970. (
268. Ben eken J„ Rideout V. C., The use of multiple models in cardiovas-
cular system studies: transport and perturbation rrtdthods, IEEE Trans.,
v. BME-15, No. 2, 1968.
269. В e г g e 1 F., General discussion, in: «Homeostatic Regulators», Churchill
Ltd., London, 1969.
270. Bernard C., An introduction to the study of experimental medicine, Do-
ver Publ., N. Y., 1957.
271. Bertalanffy L. von, General System Theory (Foundation, Develop-
ment, Application), G. Brazillier, N. Y., 1973.
ЛИТЕРАТУРА
311
272. Biswas R. N., Kuh E. S., A multiparameter sensitivity measure
for linear systems, IEEE Trans, on Circ. Theory, v. CT-18, No. 6,
1971.
273. В 1 i g h J., The thermosensitivity of the hypothalamus and thermoregula-
tion in mammals, Biol. Rev., v. 41, No. 3, 1966.
274. Bligh J., Neuronal models of mammalian temperature regulation, in:
«Essays on temperature regulation», North-Holland Publ., Amsterdam-
London, 1972.
275. Boling R. H., Toward state-space models for biological populations, J.
Theor. Biol., v. 40, No. 3, 1973.
276. В u с к R. C., On the logic of the general behavior system theory, in: «Min-
' nesota studies in the philosophy of science», v. 1, Univ. Minnesota Press,
1956.
277. Caban ас M., Thermoregulatory behavior, in: «Essays on temperature
regulation», North-Holland Publ., Amsterdam-London, 1972.
278. Cannon W. B., The wisdom of the body, Kegan Paul, Trench, Trubner
and Co., London, 1932.
279. Carson E. R., Finkelstein L., The dynamic and control of. chemi-
cal processes in man, Measurement and control, v. 3, No. 2, 1970.
280. Casti J., Connectivity, complexity and resilience in complex ecosystems,
IFAC Symposium on control mechanisms in bio- and ecosystems, Leipzig,
v. 1, 1977.
281. Chance B, Estabrook R. W., Williamson J. R. (eds.), Control
of energy metabolism, Academic Rress, N. Y., 1965.
282. Cohn D. L„ Optimal systems: 1. The vascular system, Bull. Math. Bio-
phys., v. 16, No. 1, 1954.
283. Cooper К. E., The body temperature «set-point» in fever, in: «Essays
on temperature regulation», North-Holland Publ., Amsterdam-London,
1972.
284. Crawford E. C., Kampe G., Resonant panting in pigeons, Comp. Bio-
chem. Physiol., v. 40A, No. 6, 1971.
285. Crick F. N. C., Of moleculas and men, Washington Univ. Press, Seattle-
London, 1966.
286. С г о s b i e R. J., Hardy J. T., Fessenden E., Electrical analog simu-
lation of temperature regulation in man, IRE Trans, on Bio-Med. Electron.,
v. BME-8, No. 4, 1961.
287. Davison T. J, Primitive mammals and patterns in the evolution of
thermoregulation, in: «Essays on temperature regulation», North-Holland
Publ., Amsterdam-London, 1972.
288. Deaking M. A. B., The steady states of ecosystems, Math. Bioscience,
v. 24, No. 3-4, 1975.
289. Dean A С., H i n s h e 1 w о о d C. N., Integration of cell reactions, Na-
ture, No. 201, 1964.
290. Demetrius L., On gene regulation, a mathematical model, I, Math. Bio-
sci. v. 14, No. 3-4, 1972.
291. De Wit С. T., Dynamic concept in biology, in: «Internat. Biol. Program:
prediction and measurement of photosyntetic productivity», Centre for
Agri. Publ. and Doc. Wageningen, Netherlands, 1970.
292. Dieter H., The basic structure of modern philosophy, Cultural Herme-
neutics, No. 2, 1974.
293. Dora to P., Book reviews, IEEE Trans, on Autom. Control, v. AC-18,
No. 6, 1973.
294. E n d r e n у i L., Statistical problems of kinetic model building, in: «Mathe-
matical models of metabolic regulation», Akademiai Kiado, Budapest,
1976.
295. Essays on temperature regulation. North-Holland Publ., Amsterdam-Lon-
don, 1972.
312
ЛИТЕРАТУРА
296. Estreicher J., Revillard C... Scherrer J. R., A new approach
to the problem of control in compartmental analysis, IFAC Symposium
on control mechanisms in bio- and ecosystems, v. 4, Leipzig, 1977.
297. Good P. I., Alternative mechanisms for homeostatic regulators of mam-
malian cell populations, J. Theor. B ol., v. 40, No. 3, 1973.
298. Gordon R. G., Roemer R. B., Horvath S. M., A mathematical mo-
del of the human temperature regulatory system — transient cold exposure
response, IEEE Trans, on BME-23, No. 6, 1976.
299. Gor$ki-Popel J., Classical sensitivity. A collection of formulas, IEEE
Trans, on Circ. Theory, v. CT-10, No. 2, 1963.
300. G r e e n H. D., R a p e 1 a G. E., Blood flow in passive vascular beds, Cir-
culation Res., v. XV, No. 2, suppl. 1, 1964.
301. Grodins F. S., James G., Mathematical model of respiratory regula-
tion, Ann. N. Y., Acad. Sci., v. 109, art. 2, 1963.
302. Gull P., Vogt A., Mathematical analysis of the Leslie population mat-
rix model, Bull. Math. BioL, v. 35, No. 5-6, 1973.
303. Gupta N. K., Rink R. E., Optimal control of epidemics, Math. Bioscien-
ce, v. 18, No. 3-4, 1973.
304. Guyton A. C., Textbook of medical physiology. 4th ed., W. B. Saunders
Publ., Philadelphia-London-Toronto, 1971.
305. Guyton A. C., Langston J. R.,'Navar G., Theory for renal auto-
regulation by feedback at the juxtraglomerular apparatus, Circulation
Res., v. 15, No. 2, suppl. 1, 1964.
306. Guyton A. C., Ross J. M., Carrier O., Walker J., Evidence for
tissue oxygen demand as the major factor causing autoregulation, Circu-
lation Res., v. 15, No. 2, suppl. 1, 1964.
307. Hannon B., The structure of ecosystems, J. Theor. Biol., v. 41, No. 3,
1973.
308. Hammel H. T., The set-point in temperature regulation, in: «Essays on
temperature regulations», Elsevier Publ. Co., Amsterdam, 1972.
309. Hardy J., Physiology of temperature regulation, Physiol. Rev., v. 41,
No. 3, 1961.
310. Heinrich R., Rapoport T. A„ The regulatory principles of the gly-
colysis of erythrocytes in vivo an.d in vitro, in: «Mathematical models of
metabolic regulation», Akademiai Ki ado, Budapest, 1976.
311. Hetenyi G. Jr., Norwich К. H., Zelin S., Analysis of the glucore-
gulatory system in dogs, Amer. J. Physiol., v. 224, No. 3, 1973.
312. Holling C. S., Resilience and stability of ecological systems, Annual
Review of Ecol. and Syst., v. 4, No 1, 1973.
313. Holling C. S., Ewing S., Blind man’s buff exploring the response
space generated by realistic ecological simulation models, Proc, of Inter-
nal. Symp. on Statistical Ecology, v. 2, Yale Univ. Press, New Haven, 1971.
314 Hubbel S. P., On sawbugs and systems: the ecological bioenergetics of
a terrestrial isopod. System analysis and simulation in ecology, v. 1, Aca-
demic Press, N. Y.-London, 1971.
315. Iversen О. H., A model of homeostas s, in: «Homeostatic Regulators»,
Churchill Ltd, London, 1969.
116. Iversen О. H., Evidence for chalone regulation of epidermis from ex-
periments with the fruit bat web, in: «Molecular mechanisms and control
of cell proliferation», Eighth Meeting of the European Study Group. Abs-
tracts, Moscow, 1973.
317. Jacquez J., A compartmental analysis in biology and medicine. Else-
vier Publ. Co., Amsterdam-London-N. Y., 1972.
318. Jansson B., Simulation of cell-cycle kinetics based on a multicompart-
mental model, Simulation, v. 25, No. 4, 1975.
319. Jones R. D., Berne R. M., Local regulation of blood flow in sceletal
muscle, Circulation Res., v. XV, No. 2, suppl. 1, 1964.
ЛИТЕРАТУРА
313
320. Jones R. W., Principles of biological regulation, Acad. Press, N. Y.-Lon-
don, 1973.
321. Julkinen R. J., Kekki M., Himberg J. J., Wai strom B., Non-
linear multcompartmental model for binding by extracellular proteins
with application to warfarin, Acta Pharmacol, et Toxicol., v. 38, 1976.
322. К e z d i P., Regulation of blood pressure, in: «Cardiovascular Disorders»,
Davis Comp., Philadelphia, 1968.
323. King R. E., Parametric sensitivity of physiological systems — prognostic
analysis, IEEE Trans., v. BME-14, No. 4, 1967.
324. Lamerton L. F., Cell population kinetics in relation to homeostasis,
in «Homeostatic Regulators», Churchill Ltd., London, 1969.
325. Lerner 1. M., Genetic homeostasis, John Wiley and Sons, N. Y., 1954.
326. L i с к о V., Some biochemical threshold mechanisms, Bull. Math. Biophys.,
v. 34, No. 1, 1972.
326a. Lot ka A. J., Elements of physical biology, Williams and Wilkins. Balti-
more, 1925 (new edition: Dover, N. Y., 1956).
327. Lubbers D„ Kessler M.,. Oxygen supply and rate of tissue respira-
tion in: «Oxygen transport in blood and tissue», Stuttgart, 1968.
328. McArthur R., Fluctuations of animal populations and a measure of
community stab lity, Ecology, v. 36, No. 3, 1955.
329. May R. M., Stability and complexity in model ecosystems, Princeton
Univ. Press, 1973.
330. M e r 1 e 11 i R., Weed H. R., Nonlinear model of body fluid volume and
osmolarity control systems, Abstracts of the 5th IFAC World Congress,
Instrumental Soc. of America, Pittsburgh, 1972.
331. Mesa ro vic M. D., Erlandson R., Mack о D., Fleming D., Sa-
tisfaction principle in modeling biological functions, Kybernetes, v. 2,
No. 1, 1973.
332. Miller D. R., W e i d h a u s D. E., Hall R. C., Parameter sensitivity
in insect population modelling, J. Theory. Biol., v. 42, No. 2, 1973.
333. Miller J. G., Living systems: basic concepts, Behavioral Sci., v. 10, No. 3,
1965.
334. Miller J. G., Living Systems, The Quart. Rev. Biol., v. 48, No. 2, 1973.
335. Mitchell D., Atkins A. R., Windham С. H., Mathematical and
physical models of thermoregulation, in: «Essays on temperature regula-
tion», North-Holland Publ., Amsterdam-London, 1972.
336. Murdoch W. M., Population regulation and population inertia, Ecology,
v. 51, No. 3, 1970.
337. Needham A. E., The organization of life, The Quart. Rev. Biol., v. 34,
No. 3, 1959.
338. Newman S. A., A source of stability in metabolic networks, J. Theor.
Biol., v. 35, No. 2, 1972.
339. Novoseltsev V. N., A new model of homeostatis in physiological sys-
tems, IFAC Symposium on control mechanisms in bio- and ecosystems,
v. 4, Leipzig, 1977.
340. Novoseltsev V. N., Weryhski A., Control theory and living systems,
Prace Inst, of Biocyb. i Inz. Biomed. PAN, Nr. 2, Warsaw, 1977.
341. P a г к e r S. R., Sensitivity: old questions, some new answers, IEEE Trans,
on Circuit Theory, v. CT-18, No. 1, 1971.
342. Patil M. K., G h i s t a D. N., Construction, simulation, clinical applica-
tion and sensitivity analysis of a human left ventricular control system
model, Bull. Math. Biol., v. 37, No. 5, 1975.
343. Patten B., Preliminary method for estimating stability in plancton,
Science, v. 134, No. 11, 1961.
344. Patten В. C., A primer for ecological modelling and simulation with
analog and digital computers, in: «Systems analysis and simulation in
ecology», v. 1; Academic Press, N. Y.-London, 1971.
314
ЛИТЕРАТУРА
345. Perl W., Lassen N. A., E f f e r s R. M., Matrix proof of flow, volume
and mean transit time theoiems for regional and compartrnental systems,
Bull. Math. Biol., v. 37, No. 6, 1975.
346. Powell T., A mathematical model for calcium homeostasis, Bull. Math.
Biophys., v. 34, No. 4, 1972.
347. Rashevsky N., Topology and life: in search of general mathematical
principles in biology and sociology, Bull. Math. Biophys., v. 16, No. 3,
1954.
348. Rashevsky N., Mathematical Biophysics. Third ed., v. 2, Dover Publ.,
N. Y„ 1960.
349. Rate control of biological processes. Cambridge Univ. Press, 1973.
350. Rosen G., Solutions to systems of nonlinear reaction-diffus.on equations,
Bull. Math. Biol-., v 37, No. 3, 1975.
351. Rosenbrock H. H., McMorran P. D., Good, bad or optimal? IEEE
'Trans, on automatic control, v. AC-16, No. 6, 1971.
352. R u b i n о w S., Mathematical problems in the biological sciences, SIAM
Publ., Philadelphia, 1973.
353. R u b i n о w S. 1., On closed or almost closed compartrnental systems,
Math. Bioscience, v. 18, No. 3-4, 1973.
354. Ruch T. C., Patton H. D. (eds.), Physiology and biophysics. 19th ed.,
Saunders Co., Philadelphia-London, 1965.
355. Sarnoff S. J., Mitchell J. H., The regulation of the performance of
the heart, Amer. J. Medicine, v. XXX, No. 5, 1961.
356. Segura E. T., A system approach to homeostatic evolution, 1FAC Sym-
posium on control mechanisms in bio- and ecosystems, v. 4, Leipzig, 1977.
357. Skilland A. H. P., Diffusional mass transport, John Wiley and Sons,
N. Y„ 1974.
358. Snellen J. W., Set point and exercise, in: «Essays on temperature regu-
lation», North-Holland Publ., Amsterdam-London, 1972.
359. Stedman’s Medical Dictionary. 20th ed., Bailliere, Tindall and Cassel, Lon-
don, 1967.
360. Simon H. A., The architecture of complexity, «General Systems», v. X,
No. 1, 1965
361. Stolwijk J. A. J., Mathematical model of thermoregulation, in: «Physio-
logical and Behavioral Temperature Regulation», Springfield, 111., 1970.
362. Swatny M. N. S., Bhushan C., Thulasiraman K-, Sensitivity in-
variants for linear time invariant networks, IEEE Trans., v. CT-20, No. 1,
1973.
363. Swan H., Thermoregulation and bioenergetics, Elsevier Publ. Co.,
N. Y.-London-Amsterdam, 1974
364. T о p h a m W. S., Warner H. R., The control of cardiac output during
exercise, in: «Physical bases of circulatory transport:, regulation and ex-
change», W. B. Saunders, Philadelphia-London, 1967.
365. Tuey D. B., Sensitiv.ty of pharmacokinetic control process models, Tech.
Rept. No. 72-35, Dept. Eiectr. Eng. Univ. South. Calif., Los Angeles, 1972.
366. T u к e у J. W., A practical statistician looks at the TRANSACTIONS, IEEE
Trans, on Information Theory, v. IT-12, No. 2, 1966.
367. Valentinuzzi M. E., Powell T., Hoff H. E., Geddes L.( A.,
Control parameters of the blood-pressure regulatory system, Med. ’and
Biol. Engineering, v. 10, No. 5, 1972.
368. Van Dyne G M., Organization and management of an integrated eco-
logical research program — with special emphasis on system analysis, uni-
versities and scientific cooperation, in: «Mathematical Models in Ecology»,
Blackwell Sci. Publ., Oxford, 1972.
369. ZelinS., Norwich К. H, Hetenyi G. Jr., The glucose control me-
chanism viewed as a regulator, Med. and Biol. Engineenng, v. 13, No. 6,
1975.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Адаптационные возможности 273
Адаптация 99
— и оптимальность 106
— кажущаяся 273
— неполная 273
— полная 273
—, процесс 273
— физиологических систем 258, 272
Адаптированность 272
Адекватное снабжение, концепция 29,
48
Активность митотическая 53
Анаболизм 21, 170
Аристотель, учение 42
Аррениуса закон 266
Ауторегуляция 51
— кровотока 251
---, гомеостатические свойства 282
---, иерархические соотношения 285
Базетта схема 243
Баланс энтропийный 50
Берталанфи уравнение 165
Биогеоценоз 57
Биос 106
Биосистема 17
—, качество функционирования 33
—.структура 17, 19
Блок-схема 76
Больцмана постоянная 38
Вектор 125
—.производная по вектору 133
— состояния 134
Вероятность термодинамическая 38 •
Взаимодействие динамическое 23
Воздействие возмущающее 70, 207
— задающее 70
— управляющее 70, 207
Вольтерра модель 87, 175
Гаузе принцип 105
Гомеокинез 66
Гомеорез 66
Гомеостаз 7, 34, 42—45, 51
— биоценотический 57
— на клеточном уровне 51
— на организменном уровне 54
— на разных уровнях организации 50
— на экологическом уровне 57
---- — —,степень 61
— , область 203
— , общие свойства 63
—,определение в терминах простран-
ства состояний 219
— , — Кэннона 43
— отдельных функций 56
— развития 67
—,резерв 258
—.трехуровневая схема 211
—, феноменология 272
—, характеристики интегральные 204
—,— локальные 204, 217
— цитогенетический 51
—.эволюционное происхождение 21,
49
Гомеостатическая кривая 24, 34, 45,
64
— —, в компартментальных моделях
250—255
Гомеостатические свойства 64, 216,
217
----интегральные 256
----локальные 245, 247
----, формализованное описание 269
Гомойоосмотия 48
Гомойотермия 48
Движение дополнительное 90
—,устойчивость 85
Движения саккадические 119
Действующих масс закон 170
Диабет 214
316
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
Диффузия 164
—,закон Фика второй 165
—,-----первый 164
— конвективная 169
—.константа 164
Дополнение алгебраическое 129
Допустимых режимов область 114
Дрейф 281
Иерархия в системах управления 281
— равновесий 192
— целей биосистем 30, 36, 63
Импринтинг 259
Инвариант чувствительности 254
Инвариантность 232, 277
Ингибитор 53
Индикаторные методы 162
Инерция биосистем 58
Интегратор 77
Интеграция эпигенетическая 67
Калорическое содержание 193
Канал регуляции, сочетание 225
Катаболизм 21, 170
Квазиуправление 65
Клиренс 163
Код генетический 65
Компартмент 159
Компартментальная модель, общая
схема 182
Компартментальный анализ живых
систем 185—186
Компенсация 272
Компоненты биосистемы 160
-----.перемещение 160, 164
-----.превращения 160, 170
— в популяциях 173
утилизация 160
Конвекция 168
Конструкция организмов адекватная
116
-----оптимальная 103, 115
-----простейшая 115
Консументы 193
Конформация 47
Координата фазовая 75
Костицына уравнение 175
Коэффициент весовой, выбор 246
Критерий оптимальности 211, 212
Ле-Шателье принцип 42, 58, 241
Либиха принцип 177
Лииеаризапи? 139, 184
Л-система 177
Матрица 125
— блочная 132, 152
— диагональная 125
— .дифференцирование 132
— единичная 126
— .интегрирование 132
— квадратная 125
— Лесли 174
— наблюдения 145
— обратная 130
—, определитель 128
— присоединенная 130
—, равенство 127
—, ранг 131
—, след 131
—, сложение 126
—,собственные числа 131
— состояний переходная 143
— транспонированная 126
—, умножение 127
— управления 145
— фундаментальная 143
Метаболизм 22, 170
Механизм кибернетический (регуля-
ции, управления) 17
— — активный 201, 235
---- в компартментальной модели
221
---- в экосистемах 57
----------, зависящий от плотности
57
----, двойственный характер 279
----молекулярно-генетический 67
----,нагрузка 222
----надсистемный 65
----.наложение (наслоение) 25,
65
—' —, напряжение 259
----нервной системы 54
----неспецифический 64
----пассивный 65
----, степень напряжения 259
---- физиологический 199
Михаэлиса — Ментен уравнение 172
Моделирование биосистем 205
----, два подхода 205
----классическими методами 210
----------.недостатки 56, 212
---- методом пространства состоя-
ний 150, 209, 215 ,
Модель компартментальная 159
----, схема 166
----физиологической системы 182—
185
— объекта 70
— потоковая 163
— термодинамическая 87—39
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
317
Модель физиологической системы
регуляции 55
---------- классическая 55
----------, недостатки 56
Наблюдаемость 146
— системы терморегуляции 156
Надежность 95
— физиологических систем 237—238
Насыщение 79
Нелинейность 250
Неопределеииости пределы 118
Непрерывности уравнение 164
Норма 114
Обмен основной 178
Объект управления 97, 110
Ограничения на систему 97—98
— в физиологических системах 240—
242
Оккама принцип 294
Опорное значение 205, 214
Определитель 128
— , порядок 128
Оптимальность биосистем, концепция
101
---.критика 104, 114, 122
— в технике 107—109
— и адаптация 100
— и эволюция 105—106
— .критерий 97, 112
— по быстродействию 108
Оптимизация 97
Оптимум 101
Осциллятор биологический 66
Ошибка в системе управления 83
— накопленная 253
Параметр 73
Переменная 73
— существенная 246
— физиологическая, распределение
ролей 207
Перестройка режима функционирова-
ния 272, 274, 285
Пессимум 102
Поведение адаптивное 101
Подсистема 111, 149
Поиск случайный 26
Помехоустойчивость 21
Пороговый эффект 280
Пределы безопасности 114, 239—240
— неопределенности 118
— терпимости 114
Пригожина теорема 38
Приспособленность 62, 106
Продукт 22
Продуценты 192
Простоты принцип 293
Прямохождение, энергетическая стои-
мость 116
Разнообразие экосистем 59
Рассогласование 83
Реакции поведенческие 18
— ферментативные 171
— химические 170
---.константа скорости 171
Регулятор адаптивный 100
Регуляция активная 237
— вторичная 27
— и конформация 47
— клеточного роста 53
— первичная 23, 26
— «прихода с расходом» 33
— физиологическая, классическая мо-
дель 55
Редуценты 193
Режим нестационарный 291
— переходный 142
— стационарный 64, 140, 215
Резервуар эластичный 77
Ресурс сохраиительный 259
Рецепторы 239
Реципрокное соотношение 253
Рубнера теория 29, 117, 178
Ряд параметрический 292
Самовоспроизведение 16
Самоорганизация 21
Саморегуляция 16
Самосохранение 32
— , концепция Кэннона 43
— , — Спинозы 42
Самоуправление 16
Связь обратная 23, 80, 84
--- отрицательная 81
—------по рассогласованию 26, 27,
44, 84
---параметрическая 81
---положительная 80
— прямая 22, 80, 276
Сила управляющая фермента 52
Система биологическая 15
— географическая 58
— динамическая 75
----, схема моделирования 74, 77
— живая 15
— метаболическая 17
318
предметный указатель
Система многосвязная 210
— наблюдаемая 150
— открытая 15, 17, 21, 58
— стабилизации 210
— управления 71
---- адаптивная 72
----, ограничения 113
---- робастная 122
---- с переменной структурой 286
----, характеристическое уравнение
131
— управляемая 24, 147, 149
— «человек-машина» 107
— энергетическая 110, 287
Скачок единичный 92
Состояние 134
— адаптированности 273
— биосистемы 133
— подвижного равновесия 22
— стационарное неравновесное 7, 33
— торпидное 49
Сохраиительные свойства 6, 42, 48,
50, 63
----.моделирование 199
----,преемственность 65
----, ресурсы 257
Среда внутренняя 32
— окружающая, описание 112
Стабилизация физиологических пере-
менных 238
Стабильность 57, 59
— , мера 60
Стационарности область 203
Стефана — Больцмана постоянная 167
Стресс 62, 64
Субстрат 22, 172
Суперпозиция 78
Темп 27
— вторичный 179
---- в экосистеме 19'
---- в энергетическо"
— зависимый 201
— заданный 200
— независимый 200
— первичный 179
---- в организме 200
-----в экосистеме 194
— ---в энергетической системе 190
— потребления 183
Теория автоматического регулирова-
ния 68
— управления, методология 68
----,задача анализа 69
----, — синтеза 71
----, период передаточных функций
69
Теория управления, период пере-
менных состояния 69
Терморегуляция, нейронные механиз-
мы 244
—,обзор моделей 243
—, пассивная система 150
—.роль прямых связей 278
— .сохраиительные ресурсы 260
—, уравновешивание факторов 243
—.установочная точка 214, 215
Точка изображающая 86
— особая 87
— установочная 214
---, в терморегуляции 214, 215
Транспорт конвективный 168
Удовлетворения диапазон 118, 203
— принцип 113, 118, 122
Управление активное 23, 25
— внешнее 25
— внутреннее 23
— генетическое 17, 19
— локальное, принцип 114
— оптимальное 72, 97
---, закон 99
— пассивное 23
— физиологическое 17, 19
Управляемость 21, 146, 147
Упругость 58
— , пределы 62
Уравнение выхода 137
— дифференциальное линейное 78
— —, линеаризация 79
---, начальные условия 75
---, нормальная форма Коши 75
— состояния 136
— характеристическое 131
Уравнения «вход-выход-состояиие»
138
Уравновешивание факторов 242
Уровень 27
—, регулирующее действие 28
Условия среды адекватные 35
---экстремальные 203
Уставка 56, 205, 214
—.концепция 210
—,—.недостатки 212
Устойчивость 85, 142
— асимптотическая 85 <
— биосистем 52, 57, 59, 95
--, мера 60
—, исследование по первому прибли-
жению 142
— по Ляпунову 85
Утилизация 177
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
319
Фазовая плоскость 86
Фазовое пространство 86
Фармакокинетика 94
Фермент 22, 170
Ферхюльста— Пирла уравнение 173,
176
Фика принцип 165
Фонд энергетический 117
Функционал 98
Функциональная система 56
Функция термодинамическая 37
— чувствительности 292
-----, упорядочение 292
Характеристика регуляционная 34
Хемостат дыхательный 136, 141, 142,
145
Целесообразность 103
Цель управления в живых системах
30, 112
----- второго порядка 35
----- в экосистемах 60
-----, иерархия 30, 36, 63
----- первого порядка 34
-----третьего порядка 35
Централизация кровотока 280
Цикл биологический 66
Человек-оператор 107
Чувствительности уравнение 92
— функция 92
Чувствительность 89, 144
— и устойчивость 91, 95
— , коэффициент 93
—, — логарифмический 267
Эволюция 84, 104—107, 112
Экосистема 57
—.компартментальная модель 192—
196
Экстремальности принцип 102
Эластичность 52
Энергетическое снабжение клетки 187
---- организма 188
--------, компартментальная модель
186—192
Энтропия 37
Эпиморфизм биологический 20, 219,
235, 239
---- и адаптация 20
---- и гомеостаз 21
Эффектор 17
Василий Николаевич
Новосельцев
ТЕОРИЯ УПРАВЛЕНИЯ И БИОСИСТЕМЫ
Анализ сохранительных свойств
М., 1978 г., 320 стр. с илл.
Художник Н. И. Максимов
Редактор А. А. Петров
Техн, редактор Е. В. Морозова
Корректор Е. В. Сндоркииа
ИБ № 11040
Сдано в набор 11.04.78. Подписано к печати 03.11.78.
Т-20131. Бумага 60 X 9O'/ir> тип. № 1. Литературная
гарнитура. Высокая печать. Услови. печ. л. 20.
Уч.-нзд. л. 21,36. Тираж 3000 экз. Заказ №2588 Цена
книги 2 р. 40 к.
Издательство «Наука»
Главная редакция физико-математической литературы
117071, Москва, В-71. Ленинский проспект, 15
Отпечатано с матриц Ордена Трудового Красного
Знамени Ленинградской типографии № 2 имени
Евгении Соколовой «Союзполиграфпрома» при Го-
сударственном комитете СССР по делам изда-
тельств, полиграфии и книжной торговли. 198052,
Ленинград, Л-52, Измайловский проспект, 29 в Ле-
нинградской типографии № 4 «Союзполиграф-
прома» прн Государственном комитете СССР по
делам издательств, полиграфии и книжной тор-
говли. Ленинград, ф-126 ул. Социалистическая,
«• 14.