/


































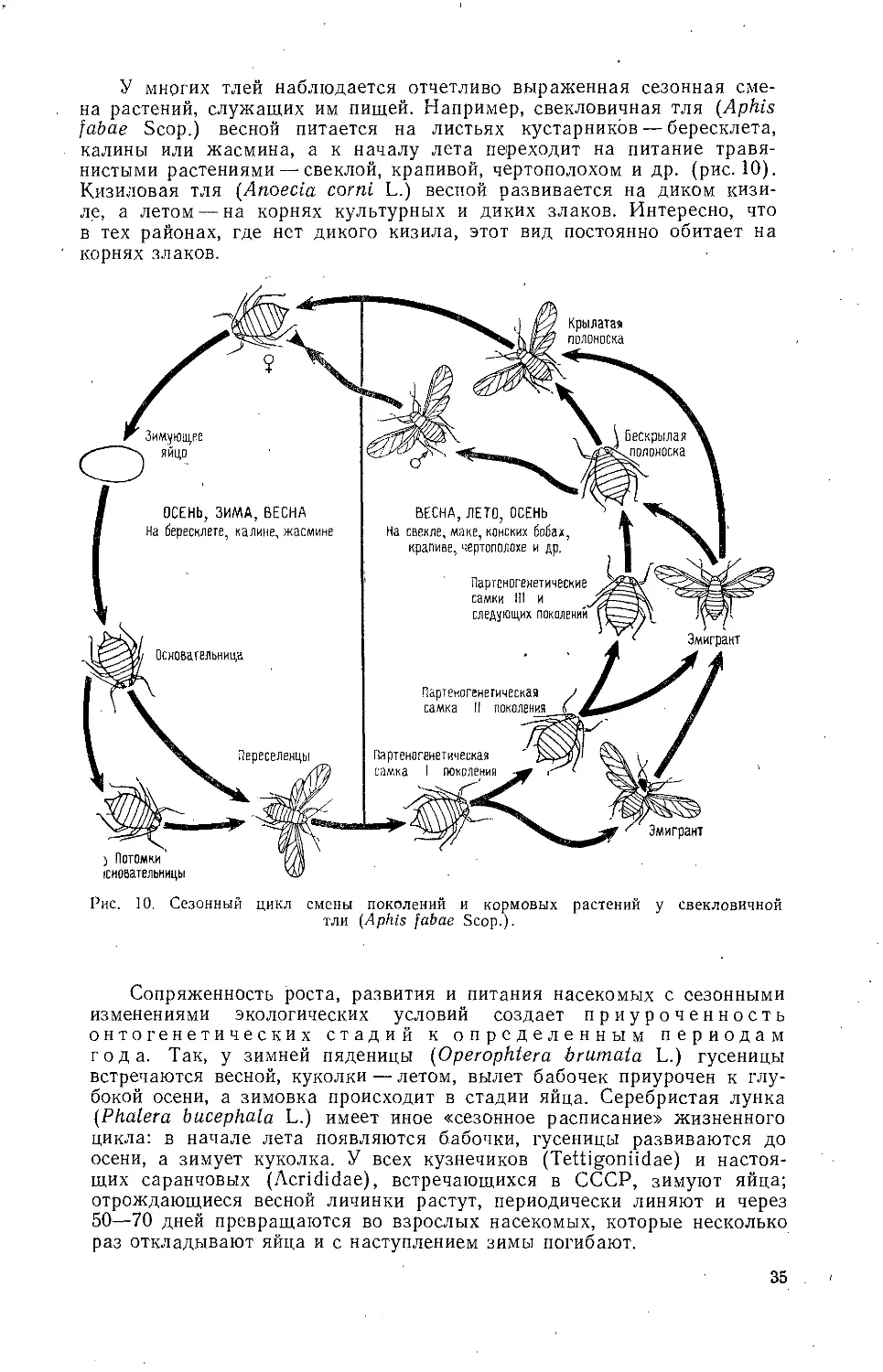














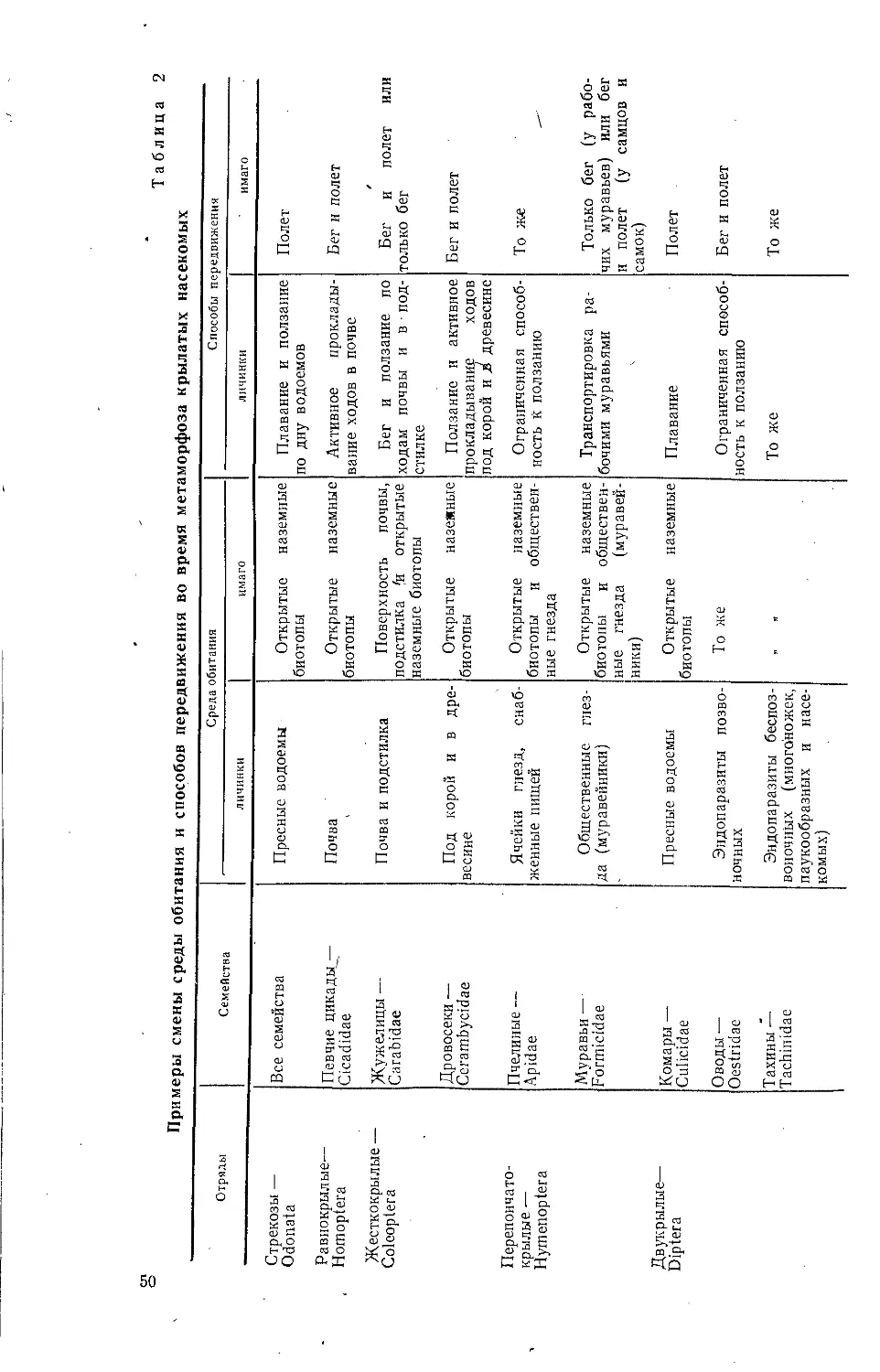





































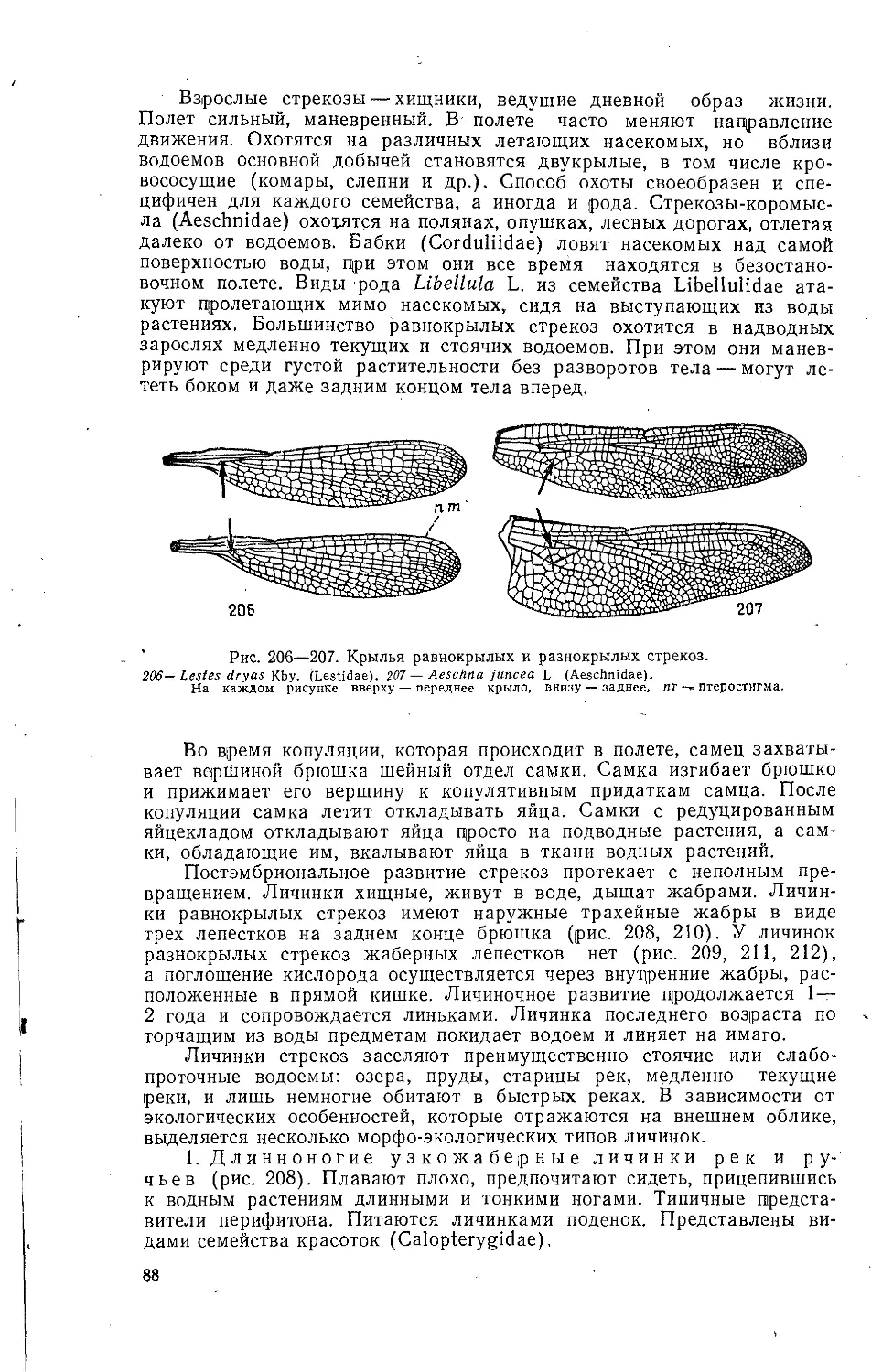










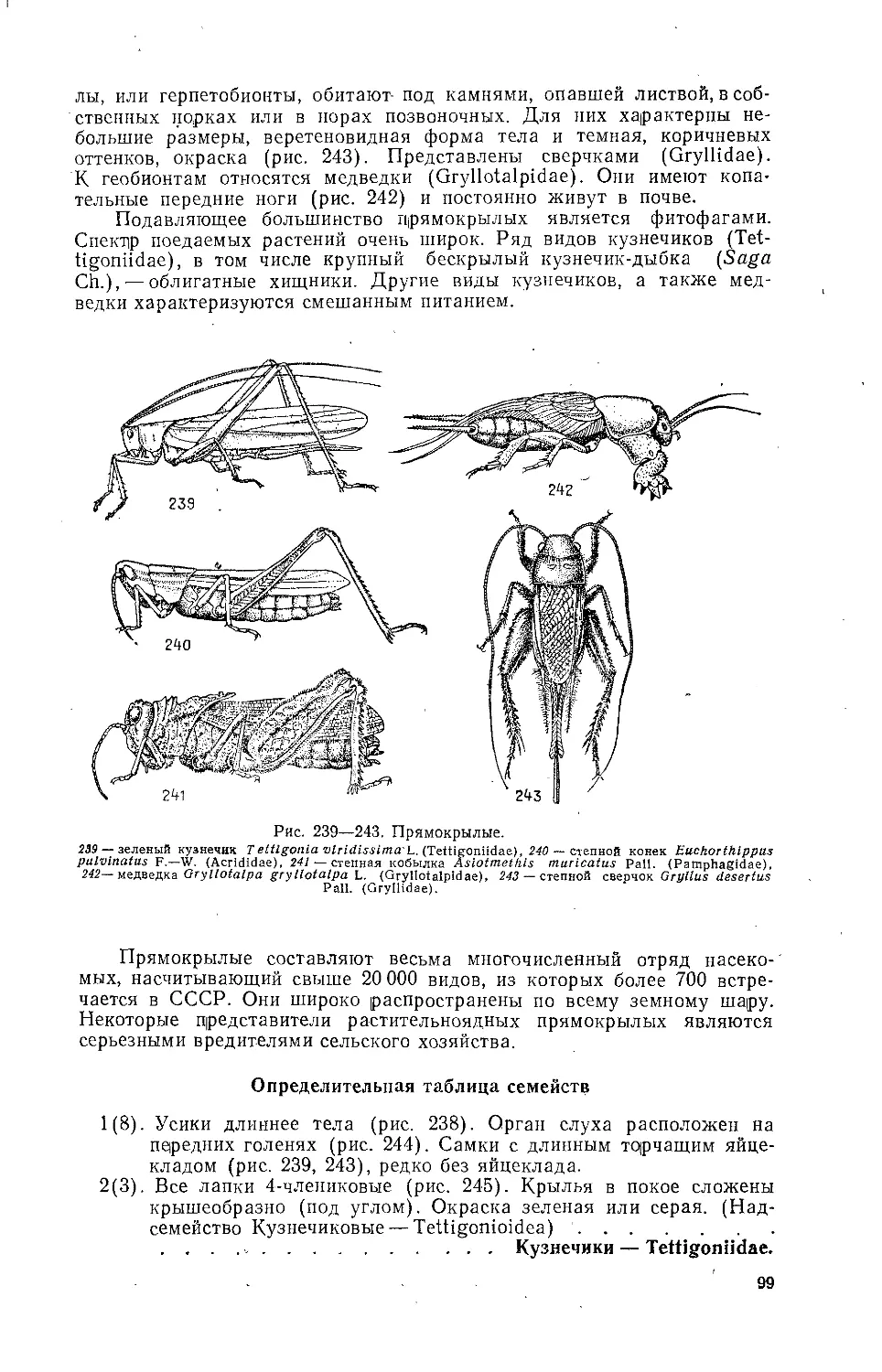






































































































































Текст
ЛЕНИНГРАДСКИЙ ОРДЕНА ЛЕНИНА
И ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ
ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ имени А. А. ЖДАНОВА
РУКОВОДСТВО
ПО ЭНТОМОЛОГИЧЕСКОЙ ПРАКТИКЕ
Под редакцией проф. В. П. Тыщенко
ЛЕНИНГРАД
ИЗДАТЕЛЬСТВО ЛЕНИНГРАДСКОГО УНИВЕРСИТЕТА
1983
Печатается по постановлению
Редакционно-издательского совета
Ленинградского университета
УДК 595.70 (076.5)
Руководство по энтомологической практике: Учеб. пособие/Под ред. В. П. Тыщен-
ко— Л.: Изд-во Лениигр. ун-та, 1983. — 230 с. Ил. — 587, табл. — 2, библиогр.—
48 иазв.
Руководство состоит из‘двух взаимосвязанных частей. В первой части освещаются
роль насекомых в наземных и пресноводных экосистемах, .пища и пищевые режимы,
поведение, сезонные явления, методы сбора и учета численности. Вторая часть содер-
жит эколого-морфологические очерки и определительные таблицы классов, отрядов
н семейств наземных членистоногих, встречающихся в европейской части СССР (24 от-
рядов насекомых, 4 отрядов паукообразных, наземных ракообразных—мокриц, мно-
гоножек — диплопод и хилопод).
Для студентов н преподавателей университетов, педагогических и сельскохозяй-
ственных институтов, а также для любителей природы и учителей при проведении
зоологических экскурсий со школьниками.
Авторы: А. К. Бродский, В. Е. Кипятков, И. А. Кузнецова, Е. Ф. Мартынова,
А. X. Саулич, А. А. Стекольииков, В. П. Тыщенко.
• Рецензенты: чл.-кор. АН СССР Ю. И. Полянский (ЛГУ), каид. биол. наук
А. А. Добровольский (ЛГУ), д-р биол. наук В. И. Кузнецов (Зоол. ии-т АН .СССР).
2005000000—189
Р 076(02)—83 ,,4—83
@ Издательство
Ленинградского
университета
1983 г.
ПРЕДИСЛОВИЕ
Полевая зоологическая практика составляет важную часть обще-
образовательной подготовки специалистов-биологов в университетах
и педагогических институтах. Можно (и нужно) тщательно штудиро-
вать учебники, читать специальную литературу, работать с коллекция-
ми и наглядными пособиями на практических занятиях, но достаточно
прочные знания в области зоологии должны быть обязательно закреп-
лены наблюдениями над животными в их, естественных местообита-
ниях.
Основная цель учебных пособий и руководств по полевой зоологиче-
ской практике заключается в том, чтобы помочь студентам и любите-
лям природы познакомиться с животным населением различных биото-
пов, научить понимать биологический смысл наблюдаемых-явлений, про-
демонстрировать многообразие экологических связей животных с ус-
ловиями их обитания. Обычно такие пособия и руководства написаны
по типу «зоологических путеводителей», в которых последовательно
рассматриваются тематические экскурсии в лес, поле, речную пойму, на
пруд, болото и т. д. Авторы настоящего «Руководства по энтомологи-
ческой практике» отказались от этой традиции, предоставляя возмож-
ность преподавателям и любителям природы самим выбирать конк-
ретные темы и составлять планы экскурсий. Эта книга является не
«зоологическим, путеводителем», а справочным пособием, предназна-
ченным для самостоятельной работы студентов с энтомологическими
объектами па летней практике. Авторы видели свою задачу в том, что-
бы научить студентов ориентироваться в сложном" мире наземных чле-
нистоногих, правильно оценивать и анализировать результаты своих
наблюдений над этими своеобразными созданиями живой природы.
В первой часди «Руководства» дается теоретическое освещение тех
аспектов биологйи и экологии насекомых, которые особенно наглядно
выявляются во время энтомологических экскурсий. Здесь приводятся
необходимые сведения о распределении насекомых по биотопам и об
их роли в природных экосистемах, освещаются вопросы питания, по-
ведения, сезонной цикличности насекомых, излагаются, основные ме-
тоды их сбора, количественного учета и хранения коллекционного ма-
териала.
Вторая часть содержит эколого-морфологические очерки отдельных
классов и отрядов, причем в число энтомологических объектов включены
не только насекомые, но и сопутствующие им другие наземные чле-
нистоногие— мокрицы, многоножки и паукообразные. Несмотря на так-
сономическую разобщенность насекомых с этими сопутствующими груп-
пами, все наземные членистоногие составляют четко очерченный при-
родный комплекс беспозвоночных животных, обладающих сходными
морфологическим^, экологическими и этологическими адаптациями. По-
3
этому мокриц, многоножек и паукообразных не следует исключать из
энтомологических объектов на том формальном основании, что они не
относятся к классу насекомых. В эколого-морфологических очерках
большое внимание уделяется не только взрослым (имагинальным), но
и личиночным стадиям, с которыми часто приходится встречаться во
время энтомологических экскурсий.
Определительные таблицы, которые следуют за эколого-морфологи-
ческими очерками, в большинстве случаев доведены до семейства. Ру-
ководствуясь этими таблицами, можно определить представителей
4 классов, 35 отрядов и 240 семейств наземных членистоногих. Опреде-
ление отрядов насекомых проводится и по имагинальным и по личи-
ночным стадиям.
«Вездесущность» наземных членистоногих в сочетании с постоянной
доступностью для наблюдений обусловливает их использование в ка-
честве показательных зоологических объектов, иллюстрирующих многие
явления живой природы. Нередко основная трудность заключается
в том, чтобы опознать встреченных насекомых, пауков, многоножек.
Авторы надеются, что издание настоящего «Руководства», снабженно-
го большим количеством рисунков с «опознавательными признаками»
наземных членистоногих, поможет преодолеть эту трудность, а эколого-
морфологические очерки позволят выяснить характерные особенности
«опознанных» объектов. На рисунках важнейшие морфологические при-
знаки, на которые следует обращать внимание при определении, либо
выделены черным цветом, либо указаны стрелками.
«Руководство» написано главным образом применительно к фауне
лесной и лесостепной зон европейской части СССР. В работе над кни-
гой принимали участие сотрудники кафедры энтомологии Ленинград-
ского государственного университета. Создавая «Руководство», ав-
rqpbi опирались на многолетний опыт проведения летней энтомоло-
гической практики со студентами биолого-почвенного факультета ЛГУ
на двух базах — в заповеднике «Лес на Ворскле» (Белгородская обл.)
и в пос. Кузнечное на берегу Ладожского озера (Ленинградская обл.).
Тотальные рисунки наземных членистоногих выполнены художниками
И. Г. Гай и Н. А. Флоренской.
Часть I
ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НАСЕКОМЫХ .
НАСЕКОМЫЕ В ЭКОСИСТЕМАХ*
В природе все организмы живут в тесном контакте друг с другом,
или, иначе говоря, живут в сообществах, которые именуются биоцено-
зами. Любой биоценоз взаимодействует с физической средой, образуя
с ней неразрывное единство, которое получило название экосистемы.
Экосистема обязательно включает два основных компонента: автотроф-
ный, для которого характерны фиксация световой энергии, использова-
ние простых неорганических веществ и построение сложных органиче-
ских молекул, и гетеротрофный, для которого характерны утилизация,
перестройка и разложение сложных органических соединений. Экосисте-
ма должна рассматриваться не как простая сумма слагаемых, т. е. от-
дельных, входящих в ее состав организмов, но как исторически сло-
жившаяся система, способная к устойчивости при относительной ста-
бильности внешней среды и к разнообразным изменениям в результате
перемен во внешней среде и в своем составе.
Размеры экосистем могут быть различными. В качестве таковых
иногда рассматривают сравнительно простые сообщества, как, напри-
мер, население кувшинки или разлагающегося пня. Однако подобные
сообщества мало автономны, сильно зависят от окружающей среды
и, кроме того, существуют непродолжительное время. Правильнее рас-
сматривать в качёстве экосистемы более крупные единства, обладающие
значительной степенью автономности и существующие достаточно дли-
тельное время, для того чтобы достичь равновесия с окружающей сре-
дой, например лес, луг, озеро и т. и. Независимо от абсолютных разме-
ров экосистемы организмы в ней всегда распределяются неравномерно,
образуя скопления или пятна. Наиболее хорошо бывают выражены вер-
тикальные подразделения, или ярусы. Так, в лесу растительность пред-
ставлена травой, кустарниками, древесными ярусами; почва также от-
четливо подразделяется на горизонты. Стратификация бывает хорошо
выражена не только в лесной или в водной экосистеме. Даже в такой,
казалось бы, «одномерной» экосистеме, какой представляется луг, мож-
но выделить несколько ярусов: почву, в которой проводят всю жизнь
дождевые черви, личинки жуков и другие животные; поверхность поч-
вы, к которой приурочены муравьи, жужелицы; слой мха, в котором оби-
тают ногохвостки и панцирные клещи; стебли и листья, с которыми свя-
заны прямокрылые, тли и другие насекомые; цветки, на которых про-
водят основную часть времени цветочные пауки, шмели (Bombus Latr.),
жуки и многие другие.
В основе способности экосистемы к самоподдержанию и саморегу-
лированию лежит поток энергии, пронизывающий ее. Лучистая энергия
солнца фиксируется зелеными растениями и далее передается от одйо-
* А. К- Бродский.
5.
го организма к другому с пищей. Это осуществляется путем поедания
одних организмов другими. Последовательность' поедания определяет
образование в биоценозе пищевых цепей. При каждом очередном пере-
носе большая часть (80—90 %) потенциальной энергии теряется, пере-
ходя в тепло. Это ограничивает возможное число этапов или звеньев
цепи обычно до 4—5. Пищевые цепи можно разделить на 2 основных'
типа: пастбищные цепи, которые начинаются с зеленого растения и ве-
дут далее к фитофагам и к их хищникам, и детритные цепи, которые ,
начинаются от мертвого органического вещества, ведут к сапрофагам,
а затем к их хищникам. Пастбищные пищевые цепи, в свою очередь,
делятся на пищевые цепи хищников и пищевые цепи паразитов. Первые -
ведут от растений к растительноядным, поедаемым мелкими плотоядны-
ми, которые, в свою очередь, поедаются более крупными хищниками
и т. д. По мере продвижения по цепи хищников животные все более
увеличиваются в размерах и уменьшаются численно. Вот пример обыч-
ной, часто встречающейся в сосновых лесах пищевой цепи хищников:
сосна^-тли—>-коровки—«-пауки-^насекомоядные птицы->хищные птицы.
Пищевые цепи паразитов, в отличие от пищевых цепей хищников, ведут
к организмам, все более уменьшающимся в размерах и увеличивающим-
ся численно. Примером может служить такая цепь: трава—«-травоядное
млекопитающее->блохи->-жгутиконосцы. Детритные цепи начинаются
с неживых органических веществ. В качестве потребителей выступают
сапрофаги, относящиеся к разным систематическим группам. Это мо-
гут быть мелкие животные, преимущественно беспозвоночные, которые
живут в почве, питаясь опавшей листвой, или же бактерии и грибы,
разлагающие органические вещества.
В экосистемах пищевые цепи образуют сложные переплетения, на-
зываемые пищевыми сетями. Организмы, получающие пищу от расте-
ний через одинаковое число этапов, считаются принадлежащими к од-
ному трофическому уровню. Зеленые растения занимают первый тро-
фический уровень (уровень продуцентов), растительноядные — второй
(уровень первичных консументов), хищники, поедающие растительнояд-
ных,— третий (уровень вторичных консументов), а вторичные хищни-
ки— четвертый (уровень третичных консументов). Популяция одного
вида может занимать один или более трофических уровней в зависи-
мости от используемых источников.
Членистоногие могут занимать второй, третий и четвертый трофи-
ческие уровни. Особенно обильно они представлены на втором уровне,
где по числу особей видное место принадлежит насекомым. Большая
группа насекомых питается вегетативными частями растений. В пищу
идет все — от корней до листьев. Особая группа — ксилофаги — питает-
ся древесиной. Генеративные части растений также имеют большое чис-
ло потребителей среди насекомых, таких, как цветочные мухи, пчелы,
жуки, чешуекрылые и мн. др. Связи насекомых с растениями весьма
разнообразны (рис. 1). Из-за ускорения прироста растений при изъятии
части этого прироста растительноядные насекомые способствуют увели-
чению продуктивности растений. Потребляя зеленую биомассу, фито-
фаги осуществляют поступление энергии к последующему звену — хищ-
никам, что представляет собой необходимое условие для направленного
движения веществ и энергии в экосистеме. При этом через большое
число видов насекомых-фитофагов от одного кормового растения берут
начало сразу несколько пищевых цепей. Возникшее множественное
дублирование пищевых цепей составляет важное условие стабильности
экосистемы. При дублировании пищевых цепей на видовом уровне зна-
чительно повышается устойчивость экосистем, так как выпадение (унич-
тожение) одного вида не прерывает движение веществ и перенос энер-
гии от продуцентов к редуцентам. ।
6
Рис. 1. Схема, показывающая многообразие связей насекомых с растениями (по Се-
ребровскому, с изменениями).
г. — галл; ж.с. — жук-семяед (Apion Hbst.) из семейства долгоносиков (Curculionidae) и его
личинка; к.б. — куколка бабочки; л. б. к. — личинка божьей коровки (Coccinellidae); л. ж. — ли-
чинка журчалки (Syrphidae); л. п. — личинка пилильщика (Tenthredinidae); л. п. ж. — личинка
пластинчатоусого жука (Scarabaeidae); м- — мины и растительноядный наездиик (Eurytomidae);
н. — наездник-энтомофаг (Ichneumonidae), заражающий гусеницу; п. — пчела (Apidae); т.— тли;
х. — хальцида; я.з. — яйца златоглазки (Chrysopidae).
7
Третий и четвертый трофические уровни представлены многочислен-
ными-'хищными насекомыми, паукообразными и губоногими многонож-
ками. Между фитофагами и хищниками существует тесная взаимосвязь:
чем больше первых, тем выше продуктивность вторых и, наоборот, па-
дение численности растительноядных форм вызывает уменьшение про-
дуктивности хищников. Как следствие этого, в экосистемах устанавли-
вается равновесие, когда выедание растений осуществляется в опти-
мальных размерах, численность фитофагов контролируется первичными
хищниками, которые, в- свою очередь, подвергаются -контролю со сторо-
ны вторичных хищников. Тем не менее в природе время от времени на-
блюдаются вспышки массового размножения растительноядных насеко-
। мых, что порой приводит к значительному уничтожению растительности.
Такие вспышки возникают потому, что энтомофаги (хищники и парази-
ты) не всегда в состоянии справиться с сильно размножившимися фи-
тофагами. Для предотвращения разрушительных колебаний числен-
ности и, следовательно, сохранения устойчивости экосистемы в целом
. необходимо, чтобы звено первичных хищников было представлено боль-
шим числом видов, что может обеспечить только самый богатый вида-
ми класс животных — насекомые. Существенное значение могут иметь
и другие хищные членистоногие, например паукообразные, однако для
нормального существования природных сообществ прежде всего необ-
ходимо присутствие в них большого числа видов насекомых. Не случай-
но насыщенность экосистемы видами насекомых возрастает по мере ее
развития: молодые стадии характеризуются обедненным видовым со-
ставов, тогда как зрелым, более устойчивым, свойствен богатый набор
видов. В этом заключается исключительно важная регулирующая роль
насекомых и других членистоногих в пресноводных и наземных1 экосис-
темах. Благодаря высокому видовому разнообразию насекомые обеспе-
чивают 'множественное дублирование пищевых цепей на видовом
уровне.
Важная роль, которую играют членистоногие в пресноводных и на-
земных сообществах, определяется высоким уровнем продуктивности
этих животных. По продукции (биомасса на единицу площади) членис-
тоногие значительно уступают позвоночным животным. Чем мельче ор-
ганизм, тем выше его удельный метаболизм (на единицу веса) и, сле-
довательно, тем ниже биомасса, которая может поддерживаться на дан-
ном трофическом уровне, и, наоборот, чем крупнее организм, тем выше
его продукция. Иначе обстоит дело с продуктивностью. Поскольку про-
дуктивность определяется как биомасса на единицу площади в едини-
цу времени, то преимуществами здесь обладают организмы мелких раз-
меров, которым благодаря более высокому уровню метаболизма присущи
и более высокие темпы размножения и обновления биомассы. Таким об-
разом, существенно уступая позвоночным животным по индивидуаль-
ным размерам и весу, членистоногие превосходят их по уровню продук-
тивности. В свою очередь, значительная продуктивность членистоногих
находит свое выражение в,высокой численности особей. Именно благо-
даря многочисленности членистоногие занимают главенствующее поло-
жение в энергетике экосистем — через них протекает основная часть
энергии. На втором, третьем и, в меньшей степени, четвертом трофиче-
ских уровнях они играют определяющую, доминирующую роль. Отсюда
становится понятным большое разнообразие жизненных форм, прису-
щее членистоногим, а также значительное влияние, которое они оказы-
вают иа среду обитания и на организмы, населяющие экосистему.
Важная особенность всех наземных сообществ — большое обилие
и разнообразие насекомых и других членистоногих. Они — обязательный
и важный компонент любого из наземных биоценозов, но тем не менее
для каждого типа экосистемы можно отметить специфику, свой харак-
8
терний набор видов, способных к совместному существованию именно
в данной экосистеме, и особые жизненные формы.
Для лесной зоны, где в основном проводится энтомологическая
практика, следует рассмотреть два основных типа экосистем — лес и луг.
Лесные экосистемы относятся к числу наиболее зрелых. Ис-
ключительно важной их особенностью можно считать то, что основная
масса органики в них утилизируется в пищевых цепях детритного типа.
За длительное время развития лесных сообществ происходит становле-
ние различных механизмов, препятствующих немедленному и непосред-
ственному выеданию зеленой биомассы. К таким механизмам можно
отнести возникновение грубых механических тканей растений, давление
хищников на фитофагов и ряд других. Использование органики в виде
листового опада и запасных питательных веществ в плодах, семенах,
корнях и т. д. происходит преимущественно в подстилке. Среди проду-
центов господствующее положение занимает древесная растительность.
В зависимости от доминирующего вида древесной растительности вы-
деляют сосняки, дубравы, различные смешанные леса и т. д. Взаимо-
отношения первого и последующих трофических уровней в лесу зависят
от типа леса, видового состава биоценозов и изменяются в зависимо-
сти от пространства и времени. Деревья создают ту органику, которая
в конечном итоге служит пищей остальным обитателям леса, пре-
доставляют убежища и укрытия для различных животных, а также ока-
зывают формирующее влияние на микроклимат. Травянистая расти-
тельность развита слабо, а ее видовой состав беден.
В лесу отчетливо выражена ярусйость, или вертикальные подраз-
деления. В кроне обитают различные листогрызущие и сосущие насе-
комые., Основную массу среди первых составляют жуки-листоеды
(Chrysomelidae), гусеницы и личинки пилильщиков (Tenthredinidae). <
Еще более многочисленны, хотя и представленные преимущественно
мелкими формами, сосущие насекомые: тли и цикадки. Среди вторич-
ных консументов преобладают в основном медленно летающие насеко-
мые, такие, как златоглазки (Chrysopidae) и верблюдки. Сюда же,
в кроны, проникают муравьи. Наибольшей активностью отличаются
лесные муравьи Formica L., которые играют значительную роль в ог-
раничении массового размножения пилильщиков, совок (Noctuidae)
и пядениц (Geometridae). В ветвях кроны часто строят ловчие сети
пауки-кругопряды (Araneidae) и пауки-линифииды (Linyphiidae).
Наиболее характерен для леса комплекс членистоногих, связанный '
со стволами и крупными ветвями деревьев. В зависимости от того, на-
сколько тесно членистоногие связаны с древесиной, можно выделить две
основные экологические группы. К первой группе относятся представи-
тели отрядов жесткокрылых (короеды — Ipidae, усачи — Cerambycidae,
златки — Buprestidae, долгоносики — Curculionidae и др.), перепончато-
крылых (рогохвосты — Siricidae) и чешуекрылых (древоточцы — Cossi-
dae и стеклянницы — Aegeriidae). Их личинки протачивают ходы раз-
личной конфигурации и глубины в коре, под корой и в древесине ослаб-
ленных, усыхающих и сухостойных деревьев, на неокоренной древеси-
не и свежих пнях. Питание древесиной является для них необходимым
условием развития и требует специальных приспособлений, одно из ко-
торых— мощно развитые челюсти — особенно характерно для всех пред-
ставителей этой группы. Более или менее цилиндрическая форма тела
личинок с опорными склеротизированными площадками хорошо служит
целям прокладывания ходов в древесине. Эти ходы позднее могут исполь-
зоваться различными обитателями щелей.и полостей. К числу специаль-
ных приспособлений к жизни в древесине относится особое строение
надкрылий жуков-короедов, у которых возникает так называемая «тач-
ка», помогающая жукам удалять из ходов буровую муку и опилки (см.
9
с. 128). За исключением короедов большинство насекомых-ксилофагов
характеризуется крупными размерами. В связи с этим хищники, питаю-
щиеся обитателями древесины, также достигают больших размеров.
Они-то и составляют основу второй экологической группы, связанной
с древесиной, и представлены личинками хищных жуков и двукрылых.
Местом охоты служат трещины древесины, полости под корой, а также
ходы, проделанные ксилофагами. Удлиненное гибкое, часто уплощенное
тело облегчает им передвижение в полостях и скважинах. К вторич-
ным консументам относятся также наездники (Ichneumonidae и Вгасо-
nidae) — преимущественно крупные формы, откладывающие яйца в ли-
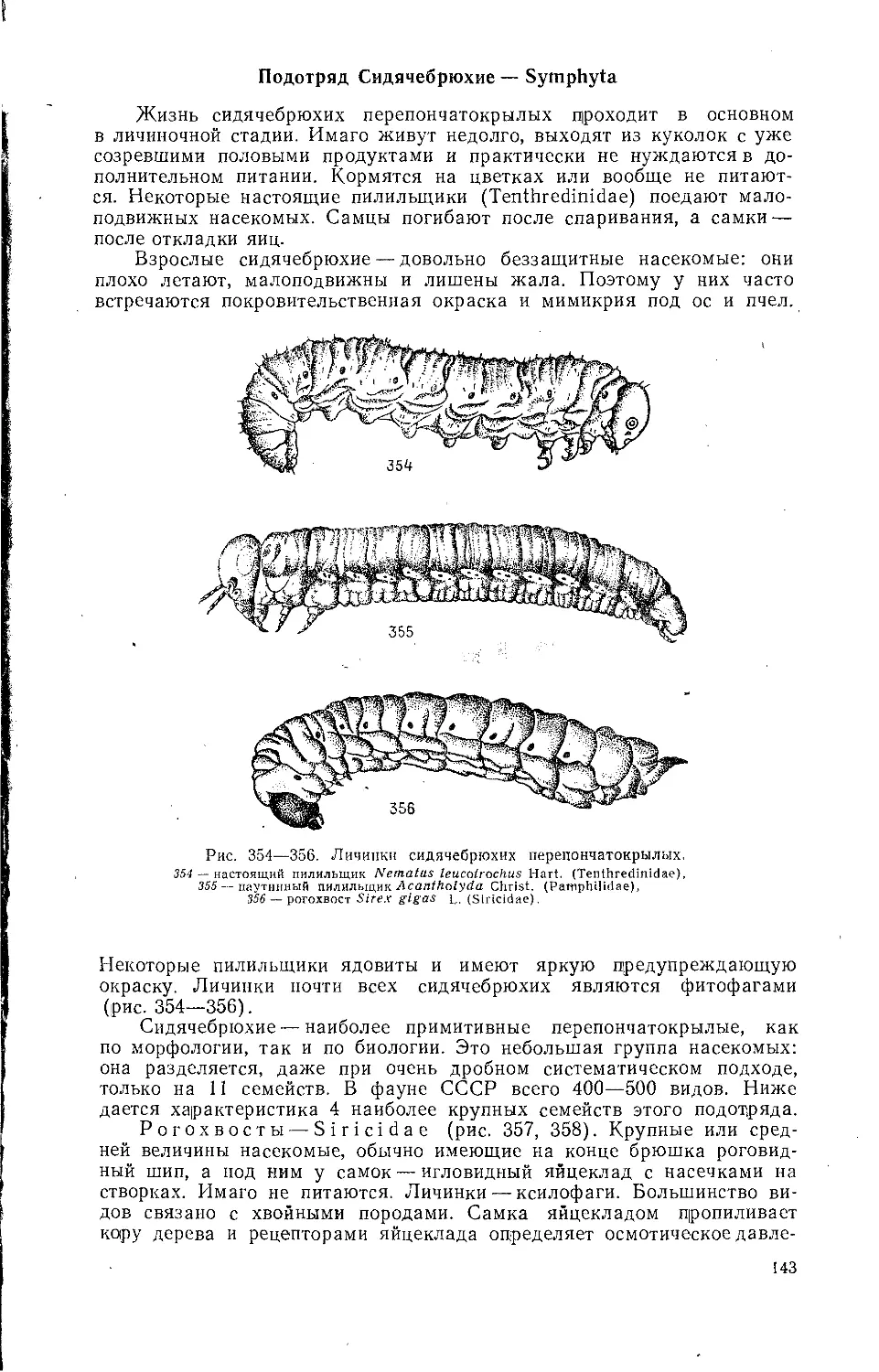
чинок насекомых-ксилофагов. В дуплах строит сложные картонные гнез-
да древесный черный муравей (Lasius fuliginosus Latr.), а муравьи-
древоточцы (Camponotu's herculeanus L. и C. vagus Scop.) поселяются
в ослабленных или мертвых елях. Под корой сооружают свои паутин-
ные гнезда и откладывают к.оконы пауки-скакунчики (Salticidae).
Сайи пауки часто охотятся на поверхности стволрв. Здесь же обычны
сенокосцы.
Насекомые-ксилофаги первыми заселяют ослабленные деревья. По
мере отмирания дерева увеличивается количество видов животных, жи-
вущих в конечном итоге за счет использования его древесины. Один
комплекс видов сменяет другой — так длится до тех пор, пока вся дре-
весина не будет использована. По наблюдениям Б. М. Мамаева, в евро-
пейской части СССР и на Северном Кавказе смена доминирующих групп
беспозвоночных, разрушающих древесину, может быть представлена
в таком виде:
Разрушение коры Разрушение древесины
Короеды (Ipidae) Усачи (Cerambycidae) (Усачи J (Cerambycidae) Жесткокрылые < ! (Рогачи (Lucanidae)
Жесткокрылые Плоскотелки X Перепончатокрылые Муравьи
(Cucujidae) Сверлилы (Lymexylonidae) (Formicidae) 1 ' Дождевые черви (Lumbricidae)
Ассоциации наиболее массовых и широко распространенных видов на-
секомых могут быть с успехом использованы в качестве индикаторов
той или иной стадии разрушения коры и древесины. Последними в раз-
лагающемся стволе или пне поселяются сапрофаги, представленные
в основном личинками двукрылых и жуков. Ими, в.свою очередь, пи-
таются губоногие многоножки, жужелицы (Carabidae), стафилиниды
(Staphylinidae) и другие хищные членистоногие. Сапрофаги и их хищ-
ники составляют специфический комплекс видов лесной подстилки,
в которой как раз и происходит утилизация большей части органики
леса. Большинство хищников подстилки, а также жуки-мертвоеды
(Silphidae) ведут ночной образ жизни. Светлое время суток они про-
водят, прячась в различных полостях, скважинах, под камнями и в
крупномерном детрите. Для их обнаружения необходимо применение
специальных методов (см. раздел «Сбор и хранение, насекомых»).
Большинство членистоногих, обитающих в лесу, характеризуется
темной однотонной окраской. Распознавание объектов с помощью зре-
ния имеет здесь подчиненное значение по сравнению с запаховой (хе-
морецепторной) и акустической ориентацией. Пахучие вещества, напро-
тив, играют существенную роль при различных биотических связях:
обнаружении жертвы, врага, партнера для спаривания и т. п. Этому бла-
гоприятствует небольшая по сравнению с открытыми местами подвиж-
ю
ность воздуха, что позволяет пахучим веществам подолгу сохраняться,
не рассеиваясь.
За редкими исключениями луга лесной зоны представляют собой
результат уничтожения лесов. Освобожденное от деревьев пространст-
во зарастает травянистой растительностью. Луга довольно разнообраз-
ны, они широко Используются как сенокосы и пастбища. Некоторые из
них сосредоточены вдоль рек — это заливные, или пойменные луга.
В луговых сообществах на первом трофическом уровне доми-
нирует травянистая растительность. Основная часть органики по-
требляется сразу, без задержки, поэтому детрита образуется мало. Пре-
обладают пищевые цепи пастбищного типа. Видовое разнообразие
растений весьма велико; что влечет за собой не меньшую насыщен-
ность видами второго трофического уровня. Одни растительноядные на-
секомые используют в пищу генеративные части растений, другие пи-
таются вегетативными частями. Большое число видов связано с цвета-
ми. Здесь находят пищу взрослые стадии многих жуков, чешуекрылых,
пчел (Apidae),' цветочных мух (Syrphidae), мух-тахин (Tachinidae)
и очень многие другие. Вторичные консументы представлены цветоч-
ными пауками (Thomisidae), бекасницами (Rhagionidae), ктырями
(Asilidae) и толкунчиками (Empididae). Членистоногие, так или иначе
связанные с цвеТами, образуют на лугу верхний ярус. Они наиболее
заметны, так как ярко окрашены и держатся открыто, перелетая с цвет-
ка на цветок. Ниже располагается комплекс видов, связанных с веге-
тативными' частыми травянистых растений. Примечательной чертой
сообщества луга в отличие от леса является обилие полужесткокры-
лых—сосущих растительноядных насекомых. Нет недостатка и в гры-
зущих фитофагах, среди которых -центральное место занимают прямо-
крылые, взрослые стадии жуков-долгоносиков (Ciirculionidae), личинки
жуков-листоедов (Chrysomilidae) и пилильщиков (Tenthredinidae).
Вторичные консументы представлены божьими коровками (Cocci-
nellidae), пауками-линифиидами (Linyphiidae) и пауками-тетрагнати-
1? дами (Tetragnathidae). Следует также отметить небольших размеров
наездников (Ichneumonidae, Braconidae, Chalcididae и др.), очень
многочисленных и потому играющих существенную роль в ограничении
численности фитофагов. К поверхности почвы приурочены ночные хищ-
ники— жужелицы (Carabidae) и стафилиниды (Staphylinidae). Их
добычей становятся растительноядные насекомые, мигрирующие в ноч-
ное время в приземный слой воздуха, где сохраняется наиболее высокая
f температура. Эти же насекомые, видимо, служат основной добычей и для
пауков-волков (Lycosidae). Виды жужелиц и стафилинид, встречаю-
щиеся на лугах, значительно мельче тех, которые занимают аналогич-
ную экологическую нишу в лесу.
Большинство Членистоногих луга характеризуется яркой контрастной
окраской. В биотических связях основная роль отводится зрению. Это-
му способствуют хорошая освещенность и отсутствие механических
» преград. Подчиненное значение имеет звуковая сигнализация, которая,
однако, преобладает у прямокрылых, занимающих видное место среди
членистоногих второго трофического уровня. Запаховая сигнализация
ослаблена или вообще невозможна из-за быстрого рассеивания паху-
чих веществ на открытых пространствах.
Лесные и луговые сообщества не исчерпывают всего разнообразия
природных комплексов лесной зоны, хотя и являются наиболее типич-
ными и характерными. Можно еще говорить о болотных экосисте-
мах, имея в виду верховые и низовые болота. Но наиболее обособлен-
ное положение занимают водные экосистемы. По территории
лесной зоны протекают крупные реки и имеется большое количество
O3eip. Несмотря на то, что пресноводные водоемы занимают небольшую
11
часть суши, они тем не менее характеризуются большим разнообразием
типов. Имеются стоячие .и проточные водоемы. Наиболее обычный и ча-
сто встречающийся тип водоема — пруды—небольшие стоячие водоемы
с относительно хорошо развитой литоральной зоной, т. е. мелководным
участком, в котором свет проникает до дна. В естественных прудах
и озерах для этой зоны характерны высшие растения, укореняющиеся
на дне водоема. Следующая зона — лимническая, т. е. толща воды, до
нижней границы которой доходит небольшое количество света, развита
в прудах гораздо слабее, и совсем отсутствует профундальная зона —
дно и толща воды, куда совсем не проникает солнечный свет. Две по-
следние зоны хорошо развиты в озерах. В реках и ручьях представлены
в общем две большие зоны: перекаты и плесы.
В животном населении любых пресноводных водоемов значитель-
ная доля видов приходится на членистоногих. Многие из них обитают
в воде постоянно, как в личиночной, так и во взрослой стадии. К этой
группе относятся ракообразные, паукообразные и часть насекомых. При
этом некоторые насекомые, например водные жуки и клопы, могут вре-
менно покидать водоем (например, при его пересыхании), отыскивая
другой, однако жизнь вне воды носит для них вынужденный и кратко-
временный характер. К нормальному существованию они способны
только в водной среде. К другой группе относятся насекомые, личи-
ночные стадии которых развиваются в воде, а взрослые стадии живут
вне воды, отлетают от водоемов иногда па большое расстояние и воз-
вращаются к ним для того, чтобы отложить яйца. Иными словами, они
являются своеобразными земноводными мира насекомых. Отсюда и на-
звание этой группы — амфибиотические насекомые. К ним относятся
представители разных отрядов: поденки, веснянки, стрекозы, вислокрыл-
ки, ручейники и многие двукрылые. После вылета из водоема взрослые
стадии этих насекомых становятся подчас важными компонентами лес-
ных и луговых сообществ. Так, на лугах, прилегающих к водоему, зна-
чительная доля всех вторичных консументов приходится на стрекоз. Как
существенный вклад в энергетику прибрежных экосистем можно рас-
сматривать массы погибших после спаривания веснянок, ручейников
и особенно поденок. Кровососущие комары (Culicidae), личиночные
стадии которых развиваются в воде, становятся важными компонентами
лесных сообществ, не уступая по числу особей и биомассе многим ис-
ходно лесным видам членистоногих.
В пресноводных водоемах, особенно стоячих, хорошо представлены
5 основных жизненных форм: бентос, перифитон, нектон, планктон и ней-
стон. Бентос формируют прикрепленные или покоящиеся на дне орга-
низмы, а также зарывающиеся в ил. Среди членистоногих к ним отно-
сятся личинки поденок, стрекоз, веснянок и ручейников. Они копошат-
ся на дне среди растительных остатков. Личинки поденок занимают
второй трофический уровень, а веснянки, стрекозы и ручейники — тре-
тий и четвертый. Личинки вислокрылок, а также некоторых стрекоз
и поденок зарываются в ил. Здесь же многочисленны личинки звонцов
(Chironomidae) и других двукрылых. Они получают свою пищу из дет-
рита. Дыхание бентосных членистоногих осуществляется при помощи
специальных органов, жабр различного строения, которые позволяют
использовать кислород, растворенный в воде. Весьма своеобразны кло-
пы, относящиеся к семейству водяных скорпионов (Nepidae). На заднем
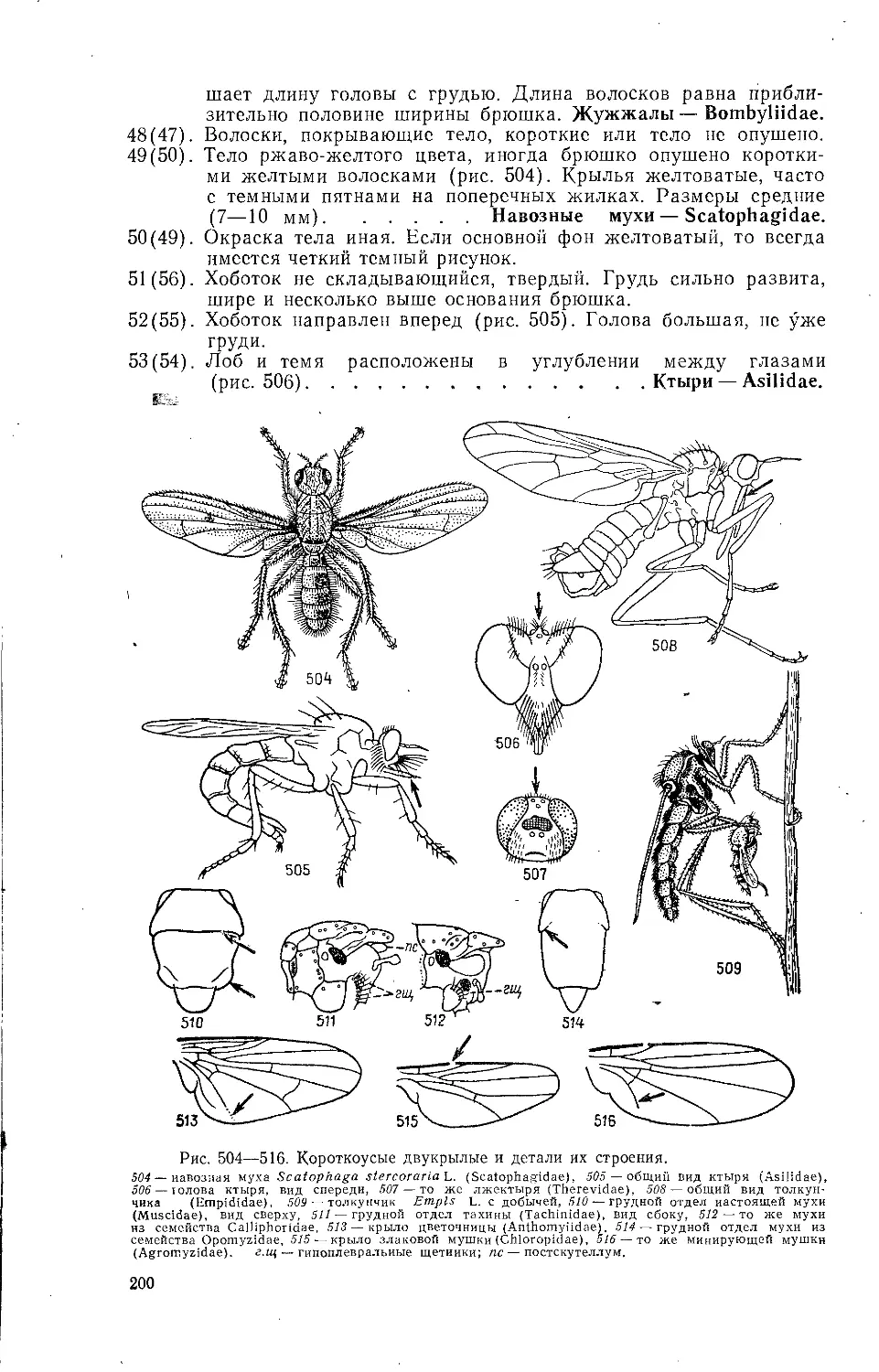
конце у них имеется длинный отросток, представляющий собой дыха-
тельную трубку. Конец этой трубки* клоп выставляет из воды и при
помощи ее набирает воздух.
К перифитону относятся животные, прикрепленные или уцепившие-
ся за стебли или листья высших растений. Распределение перифитон-
пых животных характеризуется, вертикальной зональностью. Среди них
12 ~
есть первичные и вторичные консументы. К первым относятся личинки
поденок — обитатели водной .растительности, некоторые водяные клопы
и жуки. Из вторичных консументов особой активностью выделяются ли-
чинки стрекоз. Выбрасывая вперед нижнюю губу (так называемую
маску), они ловят довольно крупных животных, проплывающих мимо.
Некоторые активно плавающие животные, цепляясь за высшие расте-
ния, могут на время переходить в состав перифитона. К примеру, клоп-
плавт (Ilyocoris cimicoides L.) не может быть отнесен исключительно
к нектону, так как часто и подолгу ползает по растениям.
В состав нектона входят плавающие, свободно перемещающиеся
в толще воды животные. Среди нектонных насекомых особенно замет-
ны личинки и взрослые стадии ныряющих жуков и клопов. Некоторые
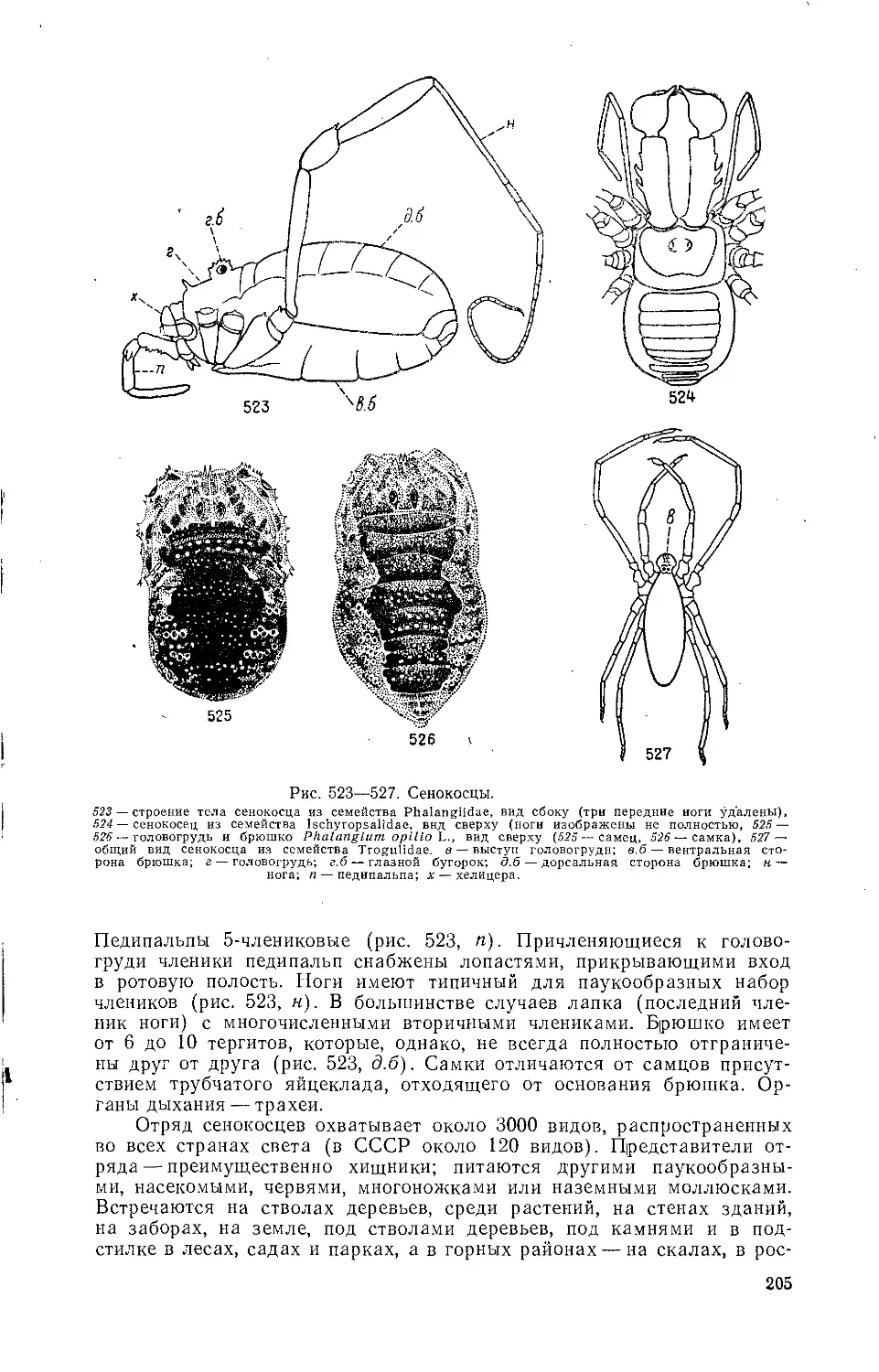
из них, в частности жуки-плавунцы (Dytiscidae) и клопы-гладыши
(Notonectidae), — активные хищники, другие — к примеру водолюбы
(Hydrophilidae) и гребляки (Corixidae) —занимают второй трофический
уровень. Большинство животных нектона обладает мощными гребными
конечностями. Различные личинки и куколки двукрылых остаются взве-
шенными в воде, часто у самой поверхности. Многие животные нектона
дышат атмосферным воздухом и несут с этой целью воздушный пузы-
рек на нижней поверхности тела или под надкрыльями.
К планктону относятся плавающие организмы, которые перемеща-
ются главным образом с помощью течений. Важные группы планктона
в прудах — крупные, плохо плавающие виды ветвистоусых и некоторые
виды веслоногих ракообразных.
Нейстон включает организмы, плавающие или бегающие по по-
верхности воды. Животные нейстона активно используют сйлы поверх-
ностного натяжения, что позволяет им быстро двигаться по водяной
пленке. Эта группа представлена клопами-водомерками (Gerridae)
и жуками-вертячками (Gyrinidae). Последние замечательны тем, что
глаза их разделены на 2 части, одна из которых используется для того,
чтобы видеть над водой, а вторая — для подводного зрения. Водомер-
ки передвигаются по. поверхности воды с не меньшей легкостью', чем
любые сухопутные насекомые по земле. У них .средние и задние ноги
смазаны снизу жировым веществом и поэтому йе смачиваются водой.
Клопы-водомерки — активные хищники. Стоит пролетающему мимо на-
секомому по той или иной причине упасть на воду, как оно тотчас
схватывается клопом.
Итак, пресноводные экосистемы характеризуются значительным
своеобразием по сравнению с лесными и луговыми. Не меньшее свое-
образие свойственно сообществу орг.анизмов, связанных
с почвой. Почва как среда обитания обладает целым рядом свойств,
присущих только ей. Физико-химические свойства почвы заставляют
говорить о ней, как об особой среде обитания. Хотя комплекс почвен-
ных членистоногих формально должен быть отнесен в качестве состав-
ной части к ранее рассмотренным типам биоценозов — лугу и лёсу,
его уместно рассмотреть отдельно. При этом важно подчеркнуть, что
именно в почве происходит основная утилизация мертвого органического
вещества. Следовательно, почвенные животные играют значительно бо-
лее важную роль в лесных экосистемах, нежели в луговых.
По числу особей, разнообразию жизненных форм и степени воздей-
.ствия на почву членистоногие занимают видное место среди почвенных
животных. Особенно. велика численность мелких почвенных и подсти-
лочных членистоногих — клещей и ногохвосток. Во многих типах почв
их плотность может достигать 300—400 тыс. особей на 1 м2. Многонож-
ки, лич-инки двукрылых, взрослые жуки и их личинки принадлежат
к более крупным почвенным беспозвоночным, которые встречаются
практически во всех типах почв, а их плотность составляет 100—300 осо-
13
бей на 1 м2. С почвой как средой обитания связаны и многие виды
муравьев.
По степени связей членистоногих с почвой можно выделить 3 эко-
логические группы. У геобионтов весь жизненный цикл протекает
в почве, и вне ее эти животные оказываются лишь случайно. Характер-
ными представителями геобионтов являются медведки (Gryllotalpa
Latr.), панцирные клещи, ногохвостки, некоторые многоножки — дипло-
поды и хилоподы. У геофилов только определенные стадии развития
проходят в почве, а на других стадиях эти членистоногие не относятся
к почвенным обитателям. Наиболее демонстративные примеры геофилии
дают многие насекомые, у которых метаморфоз нередко сопровожда-
ется сменой сред обитания. Так, у певчих цикад (Cicadidae) личинки
развиваются в почве, а взрослые цикады принадлежат к активно ле-
тающим, открыто живущим насекомым' У майских жуков (Melolon-
tha L.) все преимагинальные стадии обитают в почве и только имаго
живут и питаются на деревьях. Наконец, у геоксенов весь жизненный
цикл протекает вне цочвы, а почву они используют только в качестве
убежища. Живущие открыто наземные животные нередко зарываются
в землю при наступлении неблагоприятных условий в их естественной
среде обитания. Известно, что многие насекомые проводят в почве пе-
риод зимовки и летней засухи. К числу насекомых-геоксенов, живущих
открыто, но зимующих в почве, относятся такие важнейшие вредители
сельского хозяйства, как вредная черепашка {Eurygaster integriceps
Put.) и колорадский жук (Leptinotarsa decemlineata Say).
Геобионты и геофилы, как правило, имеют хорошо выраженные
приспособления к жизни в почве, а у геоксенов таких приспособлений
не бывает. Копательные передние ноги медведок {Gryllotalpa L.), ли-
чицок певчих цикад (Cicadidae) и жуков-навозников (Scarabaeidae)
могут служить яркими примерами морфологических приспособлений
к изготовлению нор и ходов путем разгребания почвы. Удлиненное гиб-
кое тело, свойственное многоножкам и почвенным личинкам насекомых,
облегчает передвижение этих животных по существующим в почве по-
лостям и скважинам. У многих членистоногих, заселяющих почву, так
же как и у пещерных животных, редуцируются органы зрения и по-
кровные пигменты.
Членистоногие оказывают многообразное'влияние на почву, что по-
зволяет говорить о их большой роли в процессах почвообразования.
Прежде всего следует упомянуть об участии членистоногих
в формировании первичных (пленочных) почв наскаль-
ных и осадочных породах. Когда на открытой породе появляются ли-
шайники, водоросли и другие первые поселенцы из мира растений,
одновременно порода заселяетс'я и животными, которые питаются этими
растениями. К таким животным принадлежат клещи, ногохвостки, мок-
рицы и многоножки-диплоподы. Экскременты этих членистоногих .со-
ставляют важнейшую часть формирующейся почвы, а их трупы, пере-
мешиваясь с растительными остатками, истлевают и превращаются
в перегной. Так зарождается первая почва, которая становится средой
обитания более крупных животных.
В почвенных биоценозах большое значение имеют членистоногие —
сапрофаги, которые питаются разлагающимися остатками растений
и животных и принимают активное участие в образовании гуму-
са. Отмирающие части растений, экскременты и трупы животных быст-
ро перерабатываются сапрофагами из мира членистоногих (многонож-
ками-диплоподами, почвенными клещами, ногохвостками, личинками
жуков и двукрылых, жуками-навозниками )н мертвоедами). Продукты
этой переработки поступают в почву и вовлекаются в дальнейшие про-
цессы гумификации, осуществляемые микроорганизмами. Органические
14
вещества заносятся почвенными сапрофагами из подстилки на всю глу-
бину перегнойного горизонта, создавая характерную' для него темную
окраску. В разных климатических зонах мощность этого горизонта сов-
падает с глубиной миграций почвообитающих личйнок насекомых, ко-
торые, следовательно, участвуют в формировании почвенного
профиля, в особенности в оформлении верхних горизонтов почвы.
Влияние членистоногих на химический состав почвы ска-
зывается не только в том, что они обогащают почву органическими ве-
ществами. Мокрицы и многоножки-диплоподы выступают в роли живот-
ных-концентраторов кальция, отлагающегося в их покровах. С ли-
ночными шкурками, экскрементами и трупами отмирающих животных
соли кальция поступают в почву и могут способствовать повышению ее
плодородия. Как показывают расчеты, при численности многоножек
и мокриц порядка 80 особей на 1 м2 образование ими линочных шкурок
равнозначно ежегодному внесению в почву 22—23 кг извести на 1 га.
Наряду с дождевыми червями некоторые почвенные членистоногие
создают благоприятную для роста растений зернистую структу-
ру почвы. Благодаря совокупной деятельности дождевых червей
и членистоногих-сапрофагов в почве возникают водопрочные струк-
турные агрегаты, содержащие органические и минеральные вещества,
Необходимые растениям. Прокладывая ходы и вырывая норки, почвен-
ные членистоногие увеличивают скважность почвы, а благодаря повы-
шенной скважности улучшается снабжение почвы воздухом и вбдой.
Участие членистоногих в почвообразовании основывается на их тес-
ных взаимоотношениях с другими компонентами экосистем. При пере-
работке растительных и животных остатков насекомые и другие чле-
нистоногие-сапрофаги выступают.в роли важнейшего связующего звена
между высшими растениями, позвоночными животными и микроорга-
низмами. Разлагая растительный опад, экскременты и трупы животных,
почвенные и подстилочные членистоногие подготавливают необходимые
субстраты для последующей деятельности микроорганизмов, обеспечи-
вают непрерывный круговорот веществ и направленный поток энергии
в наземных экосистемах.
•Без участия почвенных членистоногих экологическое ’ равновесие
в экосистемах нарушается и их дальнейшее существование ставится
под угрозу. Такая ситуация сложилась на Австралийском континенте
в период бурного развития скотоводства и освоения обширных терри-
торий под пастбища. Местная фауна австралийских жуков-навозников
способная питаться только экскрементами сумчатых, оказалась непри-
способленной к питанию навозом овец и крупного рогатого скота. На-
воз накапливался в огромных количествах на поверхности земли, за-
трудняя рост травы'и снижая призводительность пастбищ. Для ликви-
дации последствий нарушенного экологического равновесия пришлось
интродуцировать в Австралию несколько видов навозников из Южной
Африки. Интродуцированные навозники, усваивающие экскременты ко-
пытных, способствовали освобождению пастбищ от навоза и удобрению
почвы продуктами его переработки. Этот пример наглядно показывает
важное экологическое значение насекомых-сапрофагов в почвенных
биоценозах.
ПИЩЕВЫЕ РЕЖИМЫ И ПИЩЕВАЯ СПЕЦИАЛИЗАЦИЯ
НАСЕКОМЫХ*
Насекомые отличаются чрезвычайно широким разнообразием пи-
щевых режимов. В этом обширнейшем классе животных немало хищ-
ников, паразитов, кровососов и растительноядных видов (фитофагов).
* А. А. Стекольников.
15
Значительный удельный вес в питании насекомых занимает сапрофа-
гия — питание разлагающимися остатками отмерших организмов. Не-
которые насекомые используют в пищу продукты жизнедеятельности
животных (экскременты, шелк, воск, шерсть) или питаются низшими
растениями, микроорганизмами и грибами.
Одним из исходных режимов питания для насекомых является
хищничество. Сочетание морфологической примитивности и фило-
генетической древности с хищническим образом жизни свойственно
многим группам насекомых довольно высокого таксономического ран-
га. Так, все виды стрекоз — одного из наиболее древних отрядов крыла-
тых насекомых — являются активными хищниками как в личиночных,
так и в имагинальных стадиях. Среди прямокрылых хищничество ха-
рактерно для многих представителей древнего надсемейства кузнечи-
ковых (Tettigonioidea). Однако и среди типично растительноядных
групп иногда наблюдается вторичный переход к хищничеству (некото-
рые гусеницы, отдельные виды клопов-щитников — Pentatomidae и слеп-
няков — Miridae, жуки-мягкотелки — Cant'haridae и пестряки — С1е-
ridae).
Способы'захвата и поглощения пищи хищниками разнообразны
и связаны с рядом различных приспособлений как морфологического,
так и поведенческого характера. Многие хищники, активно преследую-
щие добычу, имеют хорошо развитые бегательные ноги (жужелицы —
Carabidae, скакуны Cicindela L.), либо обладают маневренным и стре-
мительным полетом (стрекозы, ктыри — Asilidae). Соответственно ме-
нее подвижные хищники нападают на личинок насекомых, дождевых
червей, моллюсков, тлей, пожирают яйца, членистоногих.
Особую группу составляют хищники-засадники, обладающие часто
великолепно развитой криптической внешностью и мощными хвататель-
ными конечностями. Таковы богомолы, обильно представленные в тро-
пических и субтропических областях. Сидящий среди листвы богомол
практически неподвижен, но стоит возле него появиться мухе или ба-
бочке, как совершается молниеносный бросок, и жертва оказывается
в цепких хватательных конечностях'хищника.
Необычный способ охоты наблюдается у личиной муравьиных львов
(Myrmeleontidae). Эти личинки выкапывают в песке воронки, работая
плоской и широкой головой как лопатой. На дне воронки хищные ли-
чинки и поджидают свою добычу — муравьев, 1 скатывающихся по на-
клонным стенкам песчаного кратера. Эффективность такой охоты уве-
личивается благодаря тому, что личинки муравьиных львов выкапывают
ловчие воронки вдоль муравьиных троп.
Своеобразными хищниками являются осы-парализаторы (сколии —
Scoliidae, дорожные и роющие осы — Pompilidae и Sphecidae), снаб-
жающие своих личинок' предварительно парализованными с помощью
жала насекомыми и пауками. Личинки ос в этом случае выступают как
пассивные хищники, пожирающие обездвиженную жертву. Некоторые
осы снабжают пищей своих личинок в течение всего их развития вплоть
до окукливания, как, например, обитающая на песках оса-бембекс
(Bembex F.). В этом случае личинкам доставляется убитая' добыча,
причем по мере роста личинки оса приносит ей все' более
крупную добычу. Одновременно самка выкармливает до 8 ли-
чинок.
Общественные складчатокрылые осы (Vespidae) не только убива-
ют добычу, но и переминают ее своими челюстями, снабжая личинок
своеобразно приготовленным свежим мясным фаршем из насекомых.
Создаются уникальные отношения, когда плотоядные личинки уже не
являются хищниками, поскольку пожирают обездвиженную, механиче-
ски переработанную пищу, а взрослые насекомые, питаясь нектаром
16
и пыльцой цветковых растений, обладают поведением специализирован-
ных хищников.
Значительное количество насекомых ведет паразитический образ
жизни. В отличие от хищничества паразитизм характеризует фи-
логенетически молодые группы.
Насекомые-эктопаразиты встречаются как среди отрядов с непол-
ным превращением (пухоеды, вши, некоторые клопы), так и в отрядах
с полным превращением (двукрылые, блохи). Эктопаразитическиена-
секомые с неполным превращением, обитая на птицах или млекопи-
тающих, питаются волосом, пером или эпидермисом. На основе такого
образа жизни легко возникает переход к питанию кровью — гемато-
фагии. Так возникла гематофагия у вшей — наиболее специализиро-
ванных эктопаразитов млекопитающих. Однако у полужесткокрылых —
паразитов птиц, летучих мышей, некоторых других млекопитающих
и человека — питание кровью возникает благодаря обитанию предков
кровососов в норах, гнездах и других убежищах птиц и млекопитаю-
щих.
Гематофагия широко распространена среди двукрылых и встреча-
ется в различных семействах этого отряда (мошки — Simuliidae, кома-
ры— Culicidae, слепни — Tabanidae, настоящие мухи — Muscidae, кро-
вососки— Hippoboscidae). Здесь наблюдаются чрезвычайно глубокие
морфологические приспособления к питанию кровью: возникают специа-
лизированные хоботки; Проявляются биохимические адаптации—-выде-
ление в ранку со слюной антикоагулянтов и анестезирующих веществ;
осуществляется поиск хозяина, требующий совершенствования органов
чувств; часто возникает гонотрофическая гармония—-зависимость со-
зревания яиц от степени насыщения кровью; иногда происходит потеря
крыльев и характерное для эктопаразитов уплощение тела (мухи-кро-
вососки). Однако наиболее специализированными эктопаразитами-кро-
вососами среди насекомых с полным превращением являются блохи.
В пределах этого отряда прослеживается четкий переход от временного
обитания на хозяине только в момент питания к стационарному пара-
зитизму. Некоторые гематофаги способны питаться не только кровью
теплокровных животных, но и нападают на других насекомых, питаясь
их гемолимфой, например мокрецы (Ceratopogonidae).
Необычный характер дополнительного имагинального питания,
необходимого для развития яиц, обнаружен у многих паразитических
перепончатокрылых. Самки этих насекомых слизывают капельку гемо-
лимфы, выступающую из ранки в покровах хозяина, нанесенной, яйце-
кладом при откладке яйца или при парализации жертвы. У ряда пара-
зитов наблюдается даже обязательное питание гемолимфой хозяина.
В этом случае самка • наносит специальный укол яйцекладом, без
откладки яйца.
Насекомые-эндопаразиты встречаются только среди от-
рядов, имеющих полное превращение, и преимущественно в таких про-
цветающих отрядах, как двукрылые и перепончатокрылые. В роли хо-
зяев для эндопаразитических насекомых выступают членистоногие
и позвоночные животные.
Все эндопаразитические насекомые обитают внутри хозяина лишь
в личиночной стадии. При этом характер паразитизма и пути его ста-
новления могут быть крайне разнообразными. Так, среди перепончато-
крылых-энциртид (Encyrtidae), развивающихся в ложнощитовках, встре-
чаются типичные эндопаразиты, у которых яйцо откладывается внутрь
хозяина, где и питается вышедшая личинка. В то же время личинки
самцов паразита Coccophagus lycimnia Wlk. развиваются внутри лож-
нощитовки, но за счет тканей ее паразита первого порядка. И, нако-
нец, у ряда паразитов типичная в морфологическом отношении эндопа-
2 208 ' 17
разитическая личинка питается как хищник, уничтожая одно за дру-
гим отложенные под тело ложнощитовки яйца.
У многих внутренних паразитов, особенно у перепончатокрылых,
в теле хозяина остается также и куколка и только вышедшее из нее
взрослое насекомое, прогрызая отверстие в покровах хозяина, перехо-
дит к свободному существованию.
Кроме паразитов-энтомофагов среди насекомых встречаются виды,
связанные с позвоночными животными (оводы — Oestridae и некоторые
мухи-саркофагиды — Sarcophagidae). У этих насекомых часто возника-
ют удивительно тонкие приспособления к паразитизму, особенно при
заражении хозяина. Муха Вольфарта (Wohlfartia magnified Sehin.) при
созревании яиц нападает на крупных позвоночных животных и выбрыз-
гивает из яйцевода сформировавшихся личинок на ссадины, ранки или
прямо на слизистую оболочку. Личинки активно внедряются в подкож-
ную клетчатку и мигрируют затем в лобные пазухи или гайморову по-
лость. Муха Dermatobia hominis L. использует для заражения своего
хозяина форезию. Ее самка откладывает яйца с готовыми к выходу
личинками на кровососущих двукрылых, в том числе и на комаров
(Culicidae), которые и переносят личинок дерматобии на их хозяина.
Типичными паразитами млекопитающих являются оводы (Oestri-
dae)— средней величины мохнатые и не питающиеся мухи. Подкожные
оводы (Hypoderma Latr.) откладывают яйца на шерсть крупного ро-
гатого скота, и личинки, внедряясь в подкожную клетчатку, двигаются
наверх, к спинной стороне тела, где начинают активно питаться и рас-
ти. Перед окукливанием зрелая личинка вылезает через свищ в коже
наружу, падает на землю и зарывается в почву. Желудочные оводы
(Gasterophilus Lea.), развивающиеся на лошадиных, откладывают яйца
либо на губы и щеки, либо на участки тела, которые животные выче-
сывают зубами. Во всех случаях личинки оказываются в пищеваритель-
ном тракте хозяина, где и проходит их развитие. Третья группа ово-
дов— носоглоточные оводы (Oestrus L.). Самки этих мух выбрызгива-
ют дичинок на слизистые оболочки позвоночных животных, обычно
копытных, и личинки развиваются в носовой полости или лобных
пазухах хозяина.
Трофические связи насекомых с позвоночными животными не огра-
ничиваются хозяино-паразитными отношениями. Ряд двукрылых и жест-
кокрылых развивается за счет экскрементов позвоночных, являясь коп-
рофагами. Личинки многих высших двукрылых, обитающие огром-
ными массами в экскрементах, выделяют наружу пищеварительные
ферменты, разжижающие пищевой субстрат, после чего и происходит
его заглатывание. Важной особенностью личинок таких мух является
их устойчивость к ядовитым веществам. Иначе используют для питания
экскременты животных жуки-навозники (Geotrupes Latr.-, Aphodius II.
и др.). Многие виды навозников затаскивают экскременты в специально
вырытые ими норы или камеры, где формируют из экскрементов массы
определенной формы, которыми и питаются их личинки. Однако у не-
которых навозников экскременты не заготавливаются впрок и не за-
таскиваются в норки. Взрослые жуки-навозники используют экскремен-
ты и для дополнительного имагинального питания.
Большой комплекс насекомых питается трупами позвоночных
животных (н е к р о ф а г и я). Это в основном личинки двукрылых и жу-
ки-мертвоеды (Silphidae). Для личинок двукрылых характерно внеки-
шечное пищеварение и быстрое развитие. В связи с этим у конкури-
рующих с ними за пищу жуков-могильщиков (Necrophorus F.) из се-
мейства мертвоедов (Silphidae) выработалось очень интересное поведе-
ние. Могильщики, вытаскивая из-под трупа землю, довольно быстро
закапывают его, избегая тем самым заражения своей добычи личинка-
18
Ми мух. По мере уничтожения мягких частей трупа происходит его за-
селение другими насекомыми, которые питаются шерстью и сухожилия-
ми. Это прежде всего жуки из семейства кожеедов (Dermestidae).
Не менее древним режимом Питания по сравнению с хищничеством
следует признать поллинофагию — питание пыльцой; Многие па-
леозойские насекомые, по-видимому, питались огромными скоплениями
на почве спор папоротников, плаунов, хвощей, хвойных. Затем произо-
шел переход ряда насекомых' к питанию пыльцой непосредственно на
генеративных органах растений и далее к питанию семенами. Доказано,
например, что палеозойские палеодиктиоптеры (Palaeodictyoptera) пи-
тались на кордаитах, прокалывая их семена мощным хоботком.
Сопряженная эволюция насекомых и растений привела к появлению
покрытосеменных и к возникновению у насекомых антофилии — пи-
танию на цветках. Насекомые, посещая цветки, получают для пита-
ния пыльцу и нектар, а растения — надежное перекрестное опыление.
То, что поллинофагия представляет собой один из первичных режимов
питания, подтверждается ее распространением среди современных групп
насекомых. Очень часто питание пыльцой приурочено к крайне архаич-
ным группам: первичные зубатые моли (Micropterygidae) среди чешуе-
крылых, некоторые кузнечики, уховертки, пилильщики и сетчатокрылые
являются поллинофагами. В то же время антофилия наблюдается у мо-
лодых и процветающих групп, таких как жалящие и паразитические
перепончатокрылые, двукрылые и чешуекрылые. У этих насекомых раз-
виты хоботки для поглощения нектара. Если же используется пыльца,
то возникают морфологические приспособления для ее сбора (корзи-
ночки пчелиных — Apidae) или наблюдаются своеобразные изменения
в ротовом аппарате для захвата и поглощения пыльцы (ложковидные
щетинки на ротовых частях жуков-малашек — Malachius F., хитиновая
пленка с бахромкой на мандибулах у горбаток — Mordellidae, необхо-
димая для загребания пыльцы и т. д.).
Довольно часто у антофильных насекомых параллельно с погло-
щением нектара наблюдается питание медвяной росой, или
падью — сахаристыми выделениями тлей и других равнокрылых. Пер-
востепенное значение пади, как источника углеводной пищи, проявля-
емся у муравьев (Formicidae), у которых на этой основе сложились сим-
биотические отношения с тлями и ложнощитовками. Важным резервом
углеводов служит медвяная роса и для многочисленной армии парази-
тических перепончатокрылых, а в период наибольшей активности тлей
падь собирают пчелы, перерабатывая ее в мед. Особенно возрастает
роль пади в питании насекомых в периоды засух, когда резко сокраща-
ется количество нектара в цветках.
Фитофагия — один из основных режимов питания насекомых.
У многих из них возникают самые разнообразные морфологические, по-
веденческие и физиологические приспособления к питанию на расте-
ниях. Фитофагов, как и хищников, можно подразделить на сосущих
и грызущих. Для сосущих растительноядных насекомых характерно
развитие хоботка, прокалывающего растительные ткани. У одних на-
секомых хоботок вводится в сосудистую систему, у других он погружа-
ется непосредственно в хлорофиллоносные клетки, разрушая их. Сосу-
щие фитофаги (полужесткокрылые, равнокрылые, трипсы) часто явля-
ются переносчиками вирусных заболеваний растений. Грызущие
фитофаги — более многочисленная группа насекомых, включающая
гусениц, личинок пилильщиков, многих жесткокрылых и прямокрылых.
Насекомые используют для питания все части растений — листья,
древесину, корни, генеративные органы. Так, например, в кроне дуба пи-
таются листьями гусеницы дубовой листовертки (Tortrix viridana L.),
непарного шелкопряда (Ocneria dispar L.), целого комплекса хохлаток
19
(Notodontidae) и пядениц (Geometridae), личинки и взрослые жуки
дубового листоеда (JPachybrachys tesselatus 01.), имаго майских жуков
(Melolontha L.) и долгоносиков (Curculionidae). В желудях развивают-
ся желудевая плодожорка (Laspeyresia splendana Hb.) и желудевый
долгоносик (Curculio glandium Маг.). Под корой дуба обитают личин-
ки дубового заболонника (Scolyttts intricatus Ratz.) и златок (Bupre-
stidae), а в древесине — целый ряд видов усачей (Cerambycidae). И, на-
конец, корнями дуба питаются личинки майских жуков и некоторых
щелкунов (Elateridae).
Рис. 2—5. Некоторые типы повреждения листьев в результате питания на-
секомых-фитофагов.
2 — скелетирование, 3 — полное объедание листвы, 4 — выгрызание отверстий в плас-
тинке листа, 5 —- минирование.
Среди фитофагов можно выделить насекомых, питающихся снару-
жи и обитающих внутри растительных тканей. Для филлофагов — на-
секомых, поедающих листья, характерны оба способа существования.
Открыто живущие фитофаги либо скелетируют листья, не затрагивая
сеть жилок (рис. 2), либо грызут лист сбоку, Оставляя часто один че-
решок (рис. 3), а иногда могут выгрызать одиночные окна в пластинке
листа (рис. 4). Скрытно живущие филлофаги обитают внутри листьев,
образуя в них ходы или обширные полости, — так называемые мины
(рис. 5). Минирование листьев вызывается личинками некоторых дву-
крылых и чешуекрылых, реже пилильщиками (Tenthredinidae) или жест-
кокрылыми. Так, личинки жуков-букарок (Coenorrhynus Thoms.) оби-
тают внутри черешков листьев яблони.
20
Среди карпофагов — насекомых, повреждающих плоды и семена,
также встречаются виды, обитающие внутри растений. Гусеницы яблон-
ной и грушевой плодожорок (Laspeyresia pomonella L. и L. pyrivora
Danil.) выедают семена, а затем протачивают ходы в мякоти плода.
Другие группы карпофагов поедают плоды и семена растений снаружи,
например, хлебные жуки (Anisoplia Serv.), выедающие семена злаков,
или же наносят хоботком уколы, высасывая содержимое плодов и се-
мян. Это прежде всего полужесткокрылые. Среди них наиболее извест-
ны своей вредоносностью черепашки (Eurygaster Lap.), высасывающие
зерна культурных злаков. К насекомым, питающимся на генеративных
частях растений, относятся также антофаги, поедающие бутоны
и цветки.
Особый тип фитофагии представляет собой ксилофагия— питание
древесиной. Для ксилофагов характерны мощные челюсти, прочная го-
лова и уплощенное тело. У многих личинок, живущих в древесине, осо-
бенно у жесткокрылых, редуцируются грудные ноги и появляются так
называемые опорные мозоли на грудных сегментах. В зависимости от
биологии вида или группы видов насекомые-ксилофаги развиваются
либо в древесине здоровых деревьев, либо на ослабленных и только что
поваленных стволах. Многие насекомые развиваются в мертвой древе-
сине, например жуки-точильщики (Anobiidae).
Корнями растений в основном питаются личинки жесткокрылых
и двукрылых, обитающие внутри корней или в почве вблизи корневой
системы.
Питание низшими растениями распространено в меньшей степени,
чем высшими. На хвощах и плаунах насекомые-фитофаги, видимо, во-
обще отсутствуют. На папоротниках питаются некоторые пилильщики.
На-мхах развиваются отдельные виды гусениц, ледничники (Boreidae)
из отряда скорпионниц. С лишайниками связаны некоторые сеноеды
и чешуекрылые. Довольно- значительный комплекс чешуекрылых пита-
ется водорослями. Сеноеды соскабливают водоросли со стволов де-
ревьев. Микроскопические пресноводные водоросли входят в рацион
личинок комаров (Culicidae), клопов-гребляков (Corixidae) и других
насекомых, обитающих в водоемах.
Довольно часто в отдельных группах фитофагов проявляется при-
уроченность к определенной жизненной форме растений. Имеются насе-
комые, обитающие только на травах (хортофилы) или же только на
древесных и кустарниковых породах (дендрофилы). При этом на тра-
вах развиваются более молодые и морфологически более специализи-
рованные группы.
В некоторых случаях отмечается прямая зависимость между уров-
нем морфологической специализации и филогенетического возраста на-
секомых-фитофагов и филогенетической древностью их кормовых рас-
тений. Так, обычно с хвойными связаны более архаичные
группы насекомых, чем фитофаги, обитающие на покрытосе-
менных. Например, среди тлей представители наиболее прими-
тивного семейства хермесов (Adelgidae) поселяются только на хвой-
ных деревьях.
Одним из наиболее древних режимов питания насекомых, возможно
даже более древним, чем хищничество, является с а пр о фаги я — пи-
тание разлагающимися остатками органического происхождения. Са-
профаги распространены среди самых различных групп насекомых, ве-
дущих обычно почвенный образ жизни. Так, их много среди личинок
двукрылых и жесткокрылых. В то же время детальное изучение пита-
ния насекомых показывает, что некоторые из них, считавшиеся ранее
типичными сапрофагами, питаются на самом деле грибками, дрожжами,
бактериями или другими микроорганизмами.
21
Довольно широко распространено среди насекомых питание
грибами, и мицетофаги (потребители грибов) встречаются в самых
различных отрядах этого класса. Личинки двукрылых обитают в пло-
довых телах шляпочных грибов, некоторые моли (Tineidae) выгрызают
ходы в трутовиках, сеноеды поедают мицелий паразитных грибов. От-
дельные виды короедов (Ipidae) заражают при яйцекладке древесину
спорами гриба Graphitlm, мицелием которого питаются их личинки,
проделывая ходы под корой. Разводят грибы в своих жилищах некото-
рые муравьи и термиты. Встречаются случаи, когда карпофаги в дей-
ствительности являются мицетофагами. Так, самка жука-казарки
(Rhynchites Schneid.) при яйцекладке заражает яблоко грибом Мо-
nilia, которым питаются ее личинки. Если заражения грибом не проис-
ходит, личинка погибает.
Многие насекомые, ведущие самый различный образ жизни, ис-
пользуют для питания микроорганизмы. Личинки кома-
ров-звонцов (Chironomidae) питаются микроорганизмами, которые за-
селяют придонный ил. Другие личинки двукрылых развиваются в бро-
дящем соке, вытекающем из пораженных деревьев, используя для пи-
тания дрожжи и бактерии. Личинки галлицы Miastor metraloas Mein.,
заселяющие гниющую древесину пней, также питаются бактериями.
Даже личинки дрозофил (Drosophila Fall.), живущие в гниющих фрук-
тах, развиваются за счет поглощения субстрата, нашпигованного мик-
роорганизмами.
Несомненно вторичен переход насекомых к некоторым уникальным
режимам питания — поглощению воска пчелиных сот вощинной огнев-
кой (Galleria mellonella L.) или шелка и шерсти некоторыми молями
(Tineidae) и кожеедами (Dermestidae). Известен также удивительный
пример питания личинок мухи Psylopa petrolei Coq. углеводородами
нефти. Личинки этой мухи обитают в нефтяных лужах. Во всех случа-
ях, которые указаны выше, развитие идет быстрее, если в основном пи-
щевом субстрате содержится примесь более питательной органики жи-
* вотного или растительного происхождения.
К перечисленным режимам питания следует добавить встречаю-
щуюся у насекомых с полным превращением афагию, при которой
имаго живет за счет резервных продуктов, накопленных личинкой и не
питается. Часто подвергается редукции и ротовой аппарат. Афагия рас-
пространена среди чешуекрылых, встречается у двукрылых, например
у оводов (Oestridae), очень характерна для самцов червецов и щитовок,
1 свойственна также- поденкам, веснянкам и другим насекомым. Афагам
присущи очень короткие сроки имагинальной жизни.
Для насекомых с полным превращением в целом характерно не-
полное совпадение или же значительное расхождение режимов питания
на имагинальной и личиночной стадиях развития. Так, большинство гу-
сениц— типичные фитофаги, а взрослые чешуекрылые выступают пре-
имущественно как потребители нектара. У двукрылых иногда к этим
различиям питания в онтогенезе добавляются и половые различия.
У слепней (Tabanidae), например, самки питаются кровью позвоночных
животных, самцы — нектаром, а личинки — прожорливые хищники, оби-
тающие в почве.
Нередко у насекомых отмечается смешанное питание. Уховертки спо-
собны питаться семенами растений и в то же время поедают ложно-
щитовок, тлей и других неподвижных или малоподвижных членистоногих,
выступая в роли хищников. Самки многих паразитических пере-
пончатокрылых по характеру питания принадлежат к типичным анто-
филам, потребляющим нектар с пыльцой, и одновременно к специали-
зированным гематофагам или даже к хищникам. Сочетание антофилии
и хищничества характерно для многих пилильщиков (Tenthredinidae)':
22
Пищевую специализацию, определяющую масштабы пище-
вого рациона, принято условно подразделять на 3 категории: полифа-
гию, олигофагию и монофагию. При полифагии насекомые используют
в пищу самые разнообразные, филогенетически удаленные виды
растений или животных, В качестве типичного примера поли-
фага можно привести непарного шелкопряда (Ocneria dispar
L.), гусеницы которого способны развиваться как на покрытосе-
менных, так и на хвойных. Олигофагия характеризуется более узким
набором пищевых объектов — в пищу используются растения или жи-
вотные одного семейства, близких семейств или нескольких родов. Клас-
сическим примером олигофагии служат белянки (Pieris L.), связанные
с крестоцветными растениями. Монофагия представляет собой крайнее
выражение олигофагии, когда питание происходит за счет одного вида
или нескольких близких видов растений .и животных, относящихся к од-
ному роду. Типичным монофагом является грушевая плодожорка (Las-
peyresia pyrivora Danil.), гусеницы которой живут внутри плодов гру-
ши, не повреждая яблоки.
Пищевая специализация у насекомых в различных экологических
группах проявляется неодинаково. Так, у хищников, где выбор жертвы
связан со способом и местом охоты, преобладает полифагия. Наоборот,
среди паразитических насекомых труднее найти полифага, чем олиго-
фага или монофага.. Глубокие приспособления паразита к обитанию
внутри хозяина или же на его теле часто затрудняют заражение других
видов хозяев. У паразитических насекомых возникают адаптации, на-
правленные на поиск хозяина и его заражение, на возможность нор-
мального дыхания и питания внутри животного организма, на подавле-
ние защитных реакций хозяина. Необходима также четкая синхрониза-
ция жизненных циклов хозяина и паразита.
Фитофагам свойственны самые разнообразные уровни пищевой спе-
циализации— от типичной полифагии (многие настоящие саранчовые —
Acrididae) до монофагии (некоторые червецы и щитовки). Для сосу-
щих фитофагов в большей степени, чем для грызущих, характерна оли-
гофагия. к резкому сужению пищевой специализации приводит также
обитание в плодах и древесине. Наоборот, почти все почвенные фито-
фаги, поедающие корни растений, относятся к категории полифагов.
Пищевую специализацию фитофагов определяют особые вещества,
содержащиеся в растениях. Вещества-аттрактанты обеспечивают при-
влекательность растения для насекомого, а реппеленты делают расте-
ния несъедобными. Разнообразные сочетания аттрактантов и реппелен-
тов поддерживают узкую пищевую специализацию многих насекомых-
фитофагов.
Пищевая специализация в пределах других режимов питания ис-
следована значительно менее детально. Интересные факты получены по
питанию некрофагов. У синей мясной мухи (Sarcophaga Mg.) была об-
наружена аномальная олигофагия (питание на нескольких далеко не
родственных организмах). Личинки этой мухи хорошо развиваются, пи-
таясь^ясом быка, гуся и лягушки. В то же время развитие личинок
невозможно на мясе свиньи, кролика и сазана. Истинная олигофагия
наблюдается у австралийских навозников, которые способны питаться
экскрементами только сумчатых животных. Известная степень пищевой
специализации характерна и для некоторых европейских навозников.
В пределах класса насекомых можно обнаружить отдельных пред-
ставителей, практически не проявляющих пищевую специализацию. Это
всеядные насекомые, или так называемые пантофаги. Типичными пан-
('офагами являются синантропные тараканы и фараонов муравей (Мо-
nomorium pharaonis L.), способные поедать любую пищу, как живот-
ную, так и растительную.
ПОВЕДЕНИЕ НАСЕКОМЫХ*
Поведение насекомых чрезвычайно разнообразно, что объясняется
исключительным многообразием их взаимоотношений со средой обита-
ния. В пределах этого класса мы встречаемся как с весьма примитив-
ным поведением, так и с такими удивительно сложными его проявле-
ниями, как забота о потомстве и социальное поведение.
В поведении насекомых, несомненно, преобладают врожденные,
инстинктивные реакции, не требующие для своего проявления какой-
либо тренировки или приобретения опыта. Прежде всего это простей-
шие безусловные рефлексы, такие, как отдергивание конечности при ее
раздражении, убегание или рефлекторная неподвижность (насекомое
«притворяется мертвым») при возникновении опасности и т. п. Все это
безусловные реакции на внешние стимулы.
В основе ориентации насекомых лежат довольно простые направ-
ленные движения, или таксисы. Они занимают важное место в поведе-
нии насекомых, поскольку те или иные формы ориентации составляют
основу любого поведения. В зависимости от фактора внешней среды, по
которому осуществляется ориентация, различают фото-, хемо-, гигро-,
термо- и геотаксисы. Таксис называют положительным, если движение,
насекомого происходит в направлении увеличения силы стимула, и от-
рицательным при противоположном направлении движения. Выделяют
также несколько типов таксисов, различающихся по механизму ориен-
тации. При тропотаксисе происходит сравнение стимулов, получаемых
парными рецепторами (глазами, усиками, органами слуха), и насеко-
мое движется так, чтобы уровень стимуляции обоих рецепторов был
одинаковым, и при этом, естественно, ориентируется прямо па источник
внешнего стимула. Именно так обычно насекомые находят источники
запаха или звука, ориентируются на источники света при прямолиней-
ном движении к ним. Если градиент концентрации стимула невелик
и его не могут уловить парные рецепторы, то насекомое совершает при
перемещении вперед движения из стороны в сторону (телотаксис), оце-
нивая концентрацию стимула и поворачивая, когда она снижается. Так
насекомые находят в полете источники запаха, подлетая к ним с под-
ветренной стороны. Менотаксис, или компасная ориентация, представ-
ляет собой движение под определенным углом к источнику света (солн-
це, луна) или силе тяжести. Такие таксисы особенно характерны для
насекомых, способных ориентироваться по солнцу или луне (медонос-
ная пчела — Apis mellifera L., многие муравьи — Formicidae). Прилет
насекомых ночью на искусственные источники света также в значитель-
ной степени объясняется менотаксисом: двигаясь под постоянным уг-
лом к источнику света, насекомое по спирали подлетает к нему все
ближе и ближе.
Многие двигательные реакции представляют собой рефлекторные
стереотипные движения, которые осуществляются в соответствии с врож-
денными программами поведения. Это, например, ходьба, полет, по-
следовательные движения при чистке тела, движения ротовых органов
при питании, яйцеклада при откладке яиц и т. и. Однако, как правило,
такие стереотипные движения находятся под контролем факторов вне-
шней среды, которые их ориентируют и определяют их характер. Так,
например, движения ног при ходьбе или ротовых органов при питании
согласуются с особенностями субстрата или пищевого объекта, полет
координируется целым комплексом внешних факторов и т. п.
Как правило, различные стереотипные движения объединяются
и вместе составляют отдельные поведенческие акты, или так называе-
* В. Е. Кипятков.
24
мые комплексы фиксированных действий (КФД), имеющие определен-
ное «смысловое», адаптивное значение (например, захват добычи, пи-
тание, передвижение, чистка тела, копуляция, откладка яиц). Каждый
КФД осуществляется в соответствии с врожденной программой и вклю-
чается обычно определенным специфичным для него стимулом, воспри-
нимаемым насекомым во внешней среде. Такие специфичные стимулы
получили в этологии название ключевых стимулов, или релизеров. Ре-
лизеры, действуя по принципу замка и ключа, вызывают специфичные
для них КФД, но, разумеется, только в том случае, если насекомое под-
готовлено к ним предшествующими КФД и находится в адекватном
физиологическом состоянии. Можно привести массу примеров такого
действия релизеров. Например, голодная бабочка или муха, наступив
ногой на каплю сахарного сиропа, сразу же вытягивает хоботок и на-
чинает питаться, так как хеморецепторы лапки восприняли вкус сахара,
являющийся релизером этого КФД. Хищная личинка жука-плавунца
рода Dytiscus L. совершает бросок и хватает челюстями любой появив-
шийся возле нее предмет небольшого размера. Релизером этого пове-
дения для очень плохо видящей личинки являются колебания воды
определенной интенсивности. Релизером копуляции является для сам-
ца запах или тактильные стимулы, воспринимаемые от самки, рели-
зером чистки тела — загрязнение усиков, тела и конечностей.
Нервная система насекомых тонко настроена на восприятие стиму-
лов-релизеров. Для этого во всех анализаторах (зрительном, слуховом,
обонятельном, тактильном) функционируют специальные механизмы,
воспринимающие релизеры, так называемые детекторы. Детекторы на-
строены на восприятие определенного стимула и реагируют только на
него. Например, во вкусовом анализаторе имеются специализирован-
ные хеморецепторы, реагирующие только на некоторые химические ве-
щества, имеющие для насекомого пищевую ценность (сахара, аминокис-
'лоты и другие соединения). Определенные хеморецепторы на усиках
самцов специфически реагируют на запах полового феромона самок.
В слуховом анализаторе насекомых, использующих акустические сиг-
налы, имеются детекторы, реагирующие на звуковые стимулы со стро-
го определенной спектральной характеристикой, т.-е. принадлежащие
особям того же вида и определенного пола. Таким образом, с помощью
детекторов насекомые выделяют из поступающих к ним внешних сти-
мулов только необходимую информацию, имеющую для них жизненно
важное значение.
Однако не все поведение насекомых объясняется реакциями на
внешние стимулы-релизеры. Ведь даже при полном их отсутствии на-
секомое не остается неподвижным, а обычно активно перемещается
в окружающей среде. Такая спонтанная активность возникает благодаря
существованию соответствующих эндогенных механизмов в нервной
системе и обычно проявляется в форме исследовательской активности,
т. е. целенаправленного обследования окружающей, среды. Исследо-
вательская активность имеет огромное значение, так как она направлена
прежде всего на отыскание в окружающем мире стимулов-релизеров, не-
обходимых для осуществления основных жизненных функций насеко-
мых (питания, размножения, расселения). Кроме того, спонтанно мо-
гут возникать и многие другие КФД, например, движения при дыха-
нии, выделении, чистки тела, линьке, строительстве кокона и выходе из.
куколки.
В большинстве случаев отдельные КФД в поведении насекомого
довольно жестко связаны в цепочки, или последовательности поведен-
ческих актов. Каждый член такой последовательности обычно не может
осуществиться, пока не будут реализованы все предшествующие КФД
и, в свою очередь, подготавливает условия для осуществления после-
25
дующего КФД. Рассмотрим, например, поведение одиночной осы фи-
ланта (Philanthus Fieb.) во время охоты на пчелу (рис. 6). Сидя на
каком-нибудь растении, филант поджидает пчелу (рис. 6, а). Он реаги-
рует на любое пролетающее насекомое сходного с пчелой размера (ре-
лизер), поворачивается и следит за ним (рис. 6,6). Если насекомое
садится на цветок (релизер), то оса' подлетает к нему с подветренной
стороны (рис. 6, s), воспринимая его запах. Если этот запах сходен
с запахом пчелы (релизер), то филант нападает на нее на цветке
(рис. 6) и, войдя в контакт с добычей, тактильно и по запаху (рели-
зеры) окончательно определяет, что это пчела, и жалит свою жертву
или же бросает ее в случае ошибки. Наездник, отыскивающий своего
хозяина, прежде всего ищет среду его обитания (например, растение
определенного вида) по запаху или зрительно, а затем уже обнаружи-
Рис. 6. Последовательность поведенческих актов при охоте роющей осы филанта
(Philanthus triangulum F.) на пчелу (по Тинбергену, с изменениями).
а — ожидание добычи; б — реакция иа пролетающее насекомое и слежение за ним; в ~ подлет
к опустившемуся на цветок насекомому с подветренной стороны, восприятие его запаха и
нападение иа добычу; г — направление ветра.
вает свою жертву и окончательно определяет ее пригодность для отклад-
ки яиц рецепторами яйцеклада. Такого рода цепная организация явля-
ется фундаментальной особенностью инстинктивного поведения насе-
комых.
Несмотря на явное преобладание врожденных форм поведения, на-
секомые способны и к приобретению опыта, т. е. к обучению. Про-
стейшей формой обучения является привыкание, т. е. угасание, безус-
ловной реакции при неоднократном предъявлении вызывающего ее
стимула. Так, например, постепенно исчезает реакция бегства или затаи-
вания при частом применении раздражающих воздействий. У всех ис-
следованных насекомых выражена способность к ассоциативному обу-
чению: у них легко образуются классические условные рефлексы, фор-
26
мирующиеся при совпадении условного стимула с безусловным, а так-
же инструментальные условные рефлексы, связанные обычно с обу-
чением методом проб и ошибок (обучение в лабиринте, обучение выбо-
ру различных предметов или стимулов, выработка реакции избегания).
Например, пчелиные (Apidae) и муравьи (Formicidae) легко обучают-
ся находить кормушки по определенному запаху, цвету или выбирать
геометрические фигуры определенной формы. У медоносных пчел
(Apis mellifera L.) вырабатываются условные рефлексы на время корм-
ления. Особой формой обуче-
ния является так называемое
скрытое обучение, для кото-
рого характерно запоминание
и обучение с одного раза при
отсутствии какого-либо под-
крепления. Примером такого
обучения является запомина-
ние места расположения гнез-
да многими насекомыми (пре-
жде всего жалящими пере-
пончатокрылыми). Экспери-
менты показывают, что
запоминание всех зрительных
ориентиров и расположения
гнезда дреди них полностью
осуществляется уже при пер-
вом вылете, а при возвраще-
нии насекомое легко и без-
ошибочно находит свое гнез-
Рис. 7. Зрительная ориентация роющей осы филанта (Philanthus triangulum F.) у ее
гнезда (по Тинбергену, с изменениями).
а —оса, вылетающая из гнезда, вход в которое окружен кольцом из шишек, совершает харак-
терный ориентировочный облет; б — возвращающаяся оса ищет вход внутри кольца из шишек
(вверху) и после его перемещения в сторону от гнезда (внизу); в — возвращающаяся оса ищет
вход внутри кольца из камешков, отличая его от треугольника.
27
до. Например, уже упоминавшаяся оса филант, вылетая из
своего гнезда, прежние ориентиры вокруг которого уничтожены и вы-
ложены новые в виде кольца из шишек, совершает характерные ориен-
тировочные облеты (рис. 7, а) и затем при возвращении безошибочно
выбирает круг, даже если он смещен в сторону от входа в гнездо
(рис. 7, б) или выложен не из шишек, а из камешков, и отличает его
от такого же треугольника (рис. 7, в).
Лучше всего выражена способность к обучению у общественных
насекомых (медоносные пчелы, муравьи), причем у них приобретение
опыта в течение онтогенеза играет немаловажную роль в организации
сообществ. Большое значение имеет способность к запоминанию ориен-
тиров у насекомых, сооружающих гнезда. Однако пока мы очень мало
знаем о том, как используют в естественных условиях свои способности
к обучению остальные насекомые, не обладающие столь сложными фор-
мами поведения, и имеют ли для них эти способности какое-либо адап-
тивное значение.
Медоносные пчелы и муравьи обладают даже некоторыми способ-
ностями к элементарному обобщению (абстрагированию). Например,
они могут опознавать простые геометрические фигуры (треугольник,
квадрат, четырехугольник, круг) вне зависимости от их цвета, размера,
конкретной формы и расположения. Возможно также опознавание
фигур по признаку двух- или трехцветности безотносительно к цветам
и их расположению, или карточек с 1, 2 или 3 черными пятнами вне
зависимости от конкретной формы и взаимного расположения этих пя-
тен. Таким образом, у этих насекомых могут формироваться простые
обобщенные понятия: треугольник, двухцветность, числа 1, 2’, 3 и т. д.
Фактически это элементарная рассудочная деятельность.
Все формы поведения насекомых можно разделить на 4 категории:
'1) индивидуальное поведение, 2) репродуктивное, 3) групповое и стад-
ное, 4) социальное. Индивидуальное поведение направлено на
удовлетворение основных жизненных потребностей данной особи и обыч-
но не предполагает взаимодействия с другими особями того же вида.
Это такие формы поведения, как покой, передвижение (локомоция, по-
лет, плавание), дыхание; выделение, поддержание чистоты тела, иссле-
довательская активность, избегание хищников, поиски убежищ, мигра-
ции и питание. Пищевое поведение нередко бывает достаточно слож-
ным и обычно состоит из отыскания пищи, определения ее пригодности
и собственно питания. Хищные насекомые имеют те или иные способы
отыскания добычи и ее поимки. У многих насекомых, развивающихся
с полным превращением, функцию отыскания пищи выполняет самка,
откладывающая яйца близ источника корма или непосредственно на
него. Миграционное поведение насекомых весьма разнообразно. Миг-
рации могут быть нерегулярными, суточными и сезонными. Многие на-
секомые в различные часы суток держатся в разных биотопах или
в различных участках одного биотопа. Например, комары (Culicidae)
по ночам мигрируют на поляны и просеки, где их мало днем; гусеницы
озимой совки \Agrotis segetum Den, et Schiff.) ночью питаются на рас-
тениях, а днем прячутся в почве. С миграциями нередко связаны подго-
товки насекомых к окукливанию и зимовке. Имаго насекомых с полным
превращением всегда обитают в иных местах, чем личинки, причем не-
редко это связано с дальними миграциями. Например, слепни (Tabani-
dae), комары (Culicidae), стрекозы развиваются в водоемах, а обитают
часто очень далеко от них. Некоторые виды насекомых совершают ре-
гулярные ’ дальние миграции, перелетая поодиночке (многие бабочки
семейства Nymphalidae) или стаями (перелетные саранчовые—Acridi-
dae, некоторые божьи коровки — Coccinellidae, стрекозы) на расстоя-
ния в десятки и даже сотни и тысячи километров,
28
Репродуктивное поведеййе складывается йз встречи насе-
комых противоположных полов и спаривания (половое поведение)
и той или иной формы заботы о потомстве (родительское поведение).
Половое поведение начинается с отыскания брачного партнера; при этом
активная роль принадлежит обычно самцу и значительно реже — самке.
Самцы поденок и комаров-звонцов (Chironomidae) образуют рои, кото-
рые самки отыскивают зрительно и по звуку. Самки короедов (Ipidae)
по запаху отыскивают самцов, начавших строительство гнезд в стволах
деревьев. При поисках самки самцы разных видов могут использовать
различные органы чувств. Так, многие насекомые (прямокрылые, цика-
ды, некоторые комары) используют для обеспечения встречи полов
акустические сигналы, обычно издаваемые самцами и привлекающие
самок. Самцы дневных насекомых обычно находят самок, руководст-
вуясь зрением (поденки, стрекозы, дневные бабочки, многие жуки,
двукрылые, перепончатокрылые). Такие насекомые обычно имеют ха-
рактерные рисунки или окраску, обеспечивающие взаимное узнавание
полов. Жуки-светляки (Lampyridae) используют для этого импульсы
света, испускаемые самками или особями обоих полов. Однако самцы
большинства насекомых пользуются при поисках самки в основном
обонянием, ориентируясь на запах веществ, выделяемых специальными
железами самок — половых феромонов, или аттрактантов.’ При этом
самка либо неподвижна и, выделяя феромон, ожидает самцов (у многих
ночных бабочек), либо самец находит самку по ее запаховому следу
(у многих пчелиных и термитов). Самцы некоторых ночных бабочек
могут прилетать на запах самки с расстояния в несколько километров,
ориентируясь с помощью телотаксиса и тропотаксиса в облаке феро-
мона, относимого от самки ветром. Во многих случаях в поисках самки
участвуют разные органы чувств. Так, у медоносной пчелы самец на-
ходит самку в полете по ее запаховому следу, а на более близком рас-
стоянии ориентируется зрительно.
Для некоторых'насекомых (стрекозы, кузнечики — Tettigoniidae,
сверчки — Gryllidae, некоторые тараканы, жалящие перепончатокрылые)
характерно интересное явление — территориальность: самцы занимают
определенные участки территории, которые охраняют от других самцов.
Занявший территорию самец спаривается с прилетающими к нему сам-
ками.
После встречи особей противоположных полов часто наблюдается
так называемое ухаживание: характерные движения самца перед сам-
кой, сопровождаемые демонстрацией рисунка на крыльях или теле, при-
косновениями к самке, вибрацией крыльев, звуковыми сигналами и т. д.
Во время ухаживания самец и самка окончательно устанавливают ви-
довую принадлежность партнера (в случае ошибки ухаживание пре-
рывается), а самец определяет, готова ли самка к копуляции. Обычно
это достигается благодаря восприятию феромонов самца и самки при
их контакте. Кроме того, стереотипные движения при ухаживании яв-
ляются ‘видовым признаком. Если самка готова к спариванию, то она
отвечает на ухаживание, принимая характерную позу...
Родительское поведение проявляется в заботе о потомстве со сто-
роны самки; крайне редко в ней участвует и самец (у короедов-—Ipi-
dae, жуков-могильщиков рода Necrophorus F. и у некоторых навозни-
ков из семейства пластинчатоусых жуков — Scarabaeidae). Только
некоторые насекомые совершенно не заботятся о потомстве и откладыва-
ют яйца в случайных местах. Большинство же видов откладывает яйца
поблизости от пищи или непосредственно на нее. Многие насекомые при
этом маскируют яйца, откладывая их в ткани растений, в почву, укры-
вая пушком или выделениями специальных желез (кубышки саранчо-
вых— Acrididae). Некоторые насекомые охраняют яйца и молодых
29
Лйчйнбк в специальной норке (уховертки, МбДведкй—
Gryllotalpidae, некоторые сверчки — Gryllidae) или укрывают своим
телом кладку (некоторые клопы-щитники — Pentatomidae), или вына-
шивают яйца в своем теле до появления личинок (некоторые тараканы,
многие короткоусые двукрылые). Наконец, ряд наиболее продвинутых
в этом отношении видов обеспечивает свое потомство и укрытием и пи-
щей, сооружая для него специальные гнезда, снабжаемые провизией
(многие жуки: пластинчатоусые — Scarabaeidae, могильщики рода
Necrophorus F., кравчики — Lethrus Scop., короеды — Ipidae; роющие
осы — Sphecidae и одиночные пчелы — Apidae). Наивысшим достиже-
нием на этом пути является постепенное выкармливание личинок еже-
дневно приносимым кормом (одиночные осы: Philanthus FieK, Вет-
bex F., Ammophila Kirby).
Групповое и стадное поведение связано с образованием
некоторыми насекомыми скоплений. Сюда не относятся временные
и непостоянные по составу скопления, вызванные локальным обилием
пищи или другими благоприятными факторами, или рои и подобные
группы, связанные с половым поведением, а только скопления, в кото-
рых наблюдается определенная групповая организация. Так, у ряда
насекомых (пилильщики-ткачи — Pamphilidae, -горностаевые моли —
Yponomeutidae) личинки из одной кладки живут вместе, сооружают
общее паутинное гнездо и совместно добывают корм. Некоторые насе-
комые (например, тараканы) постоянно живут тесными группами
вместе с личинками, иногда обнаруживая даже совместную заботу
о потомстве, сложную иерархическую организацию группы с наличием
доминирующих и подчиненных особей, или сигнализацию об опасности
с помощью феромонов тревоги (например, 'у многих тлей). Так'назы-
1ваемые перелетные саранчовые (азиатская перелетная саранча — Lo-
custa migratoria L., африканская перелетная саранча — Schistocerca
gregaria Forsk.) образуют огромные стаи, насчитывающие в своем со-
ставе миллионы и даже миллиарды особей и согласованно мигрирую-
щие на сотни километров.
Для насекомых с групповым и социальным поведением очень ха-
рактерно явление, получившее название эффекта группы. Обычно он
заключается в ускорении развития и созревания, интенсификации об-
менных процессов и других физиологических и поведенческих измене-
ниях у сгруппированных насекомых по сравнению . с находящимися
в одиночестве. В основе этого эффекта лежит восприятие других особей
своего вида с помощью зрения, обоняния и осязания. Среди необщест-
венных насекомых эффект группы достигает своего максимального вы-
ражения у перелетных саранчовых, у которых он выражается в форме
фазовой изменчивости: личинки, выросшие в группе,, превращаются
в имаго так называемой стадной формы, которая отличается от одиноч-
ной формы более темной окраской, более длинными крыльями и дру-
гими морфологическими признаками, а также ярко выраженной склон-
ностью к миграциям.
Социальное п о в е д е н и е — это замечательное явление, воз-
никшее независимо в нескольких группах насекомых. Социальными, или
общественными насекомыми являются термиты и некоторые перепонча-
токрылые: муравьи (Formicidae), бумажные осы (Vespidae) и многие
пчелиные (Apidae). Общественными называются насекомые, обитающие
большими сообществами, или колониями, в которых наблюдаются
кооперирование в выращивании потомства и других видах деятельно-
сти и репродуктивная дифференциация, т. е. разделение особей на
размножающихся и бесплодных, выкармливающих потомство, получен-
ное от плодовитых особей. Репродуктивная дифференциация приводит
к возникновению социального полиморфизма, выражающегося в разде-
30
ленйй особей на отдельные группы, йлй касты, которые различающей
по своей морфологии, физиологии и поведению. Разграничивают 2 ос-
новные касты — репродуктивную и бесплодную. У общественных пере-
пончатокрылых на касты разделяются только особи женского пола,
и репродуктивная каста у них представлена плодовитыми самками, или
царицами, а бесплодная каста — неразмножающимися самками — ра-
бочими. У термитов социальный полиморфизм характерен для обоих
полов: репродуктивную касту представляют и царицы и цари, а бес-
плодные рабочие особи могут быть как самками, так и самцами.
Рис. 8. Социальный полиморфизм у муравья Pheidole dentata Мауг (по Уилеру).
а — мелкий рабочий; б — крупный рабочий; в — солдат; г — царица, сбросившая крылья; д—~
крылатый самец.
Обычно царицы значительно крупнее рабочих и отличаются от них
по строению тела (рис. 8). В колонии общественных насекомых может
быть только одна царица {моногиния), и тогда колония в генетическом
отношении является семьей, т. ё. потомством одной матери, или же,
у других видов, цариц много {полигиния). Среди особей бесплодной
касты тоже может наблюдаться полиморфизм. В простейшем случае ра-
бочие просто сильно различаются по своим размерам. Такая изменчи-
вость часто сопровождается различиями в пропорциях тела: крупные
рабочие имеют увеличенную голову и-мощные челюсти. Нередко таких
крупноголовых рабочих выделяют в самостоятельную касту солдат.
У муравьев настоящие солдаты есть лишь у немногих видов (рис. 8),
а у термитов они имеются практически всегда.
31
Социальный' полиморфизм является основой разделения функций
между особями в колонии общественных насекомых — полиэтиэма, ко-
торый может проявляться в различных формах. Кастовый полиэтизм
выражается в различиях функций особей, относящихся к разным кас-
там. Так царицы выполняют в основном функцию размножения, а ра-
бочие— все остальные обязанности. Солдаты в отличие от рабочих
занимаются прежде всего охраной гнезда, а также могут выполнять
наиболее трудоемкие виды работ. В пределах одной касты различия
в функциях могут быть связаны с размерами особей (размерный поли-
этизм, см. с. 167). У большинства общественных насекомых хорошо
выражен возрастной полиэтизм, т. е. изменения функций и поведения
особи в зависимости от ее возраста. Обычно молодые рабочие выпол-
няют различные работы внутри гнезда, а с возрастом переходят к вне-
гнездовой деятельности (см. с. 163 и 167).
Социальное поведение у насекомых возникало в эволюции на осно-
ве усложнения родительского поведения. У целого ряда видов самка
выкармливает свое потомство в специальном гнезде и остается с ним
до выхода взрослых насекомых, которые в дальнейшем могут посе-
ляться в том же гнезде, приступая к размножению. Так формируется
ассоциация близкородственных особей живущих и размножающихся
в общем гнезде. В ходе дальнейшей эволюции возникают совместное
выращивание потомства, другие формы кооперирования и, наконец,
разделение на касты. Причины появления социального полиморфизма
очень сложны, до конца еще не поняты и их^рассмотрение выходит за
рамки данного пособия.
Забота о потомстве, явившаяся основой возникновения социально-
сти, достигает у высокоразвитых общественных насекомых наивысшей
•сложности и разнообразия. Яйца, личинки, куколки окружены постоян-
ным вниманием со стороны рабочих, которые их чистят, облизывают,
кормят личинок и удаляют их экскременты, помогают им при линьках.
Характерно кормление личинок изо рта в рот, т. е. отрыгивание ра-
бочими содержимого зобика или секретов специальных кормовых желез
непосредственно в рот личинки, нередко совершенно беспомощной и не-
способной питаться самостоятельно. Рабочие также постоянно обли-
зывают и кормят царицу (и царя у термитов). Кроме того, очень
характерно постоянно внимание рабочих особей друг к другу, они по-
долгу занимаются взаимным «туалетом», тщательно очищая и облизы-
вая тело, ноги и усики других особей, которые как бы «с удовольст-
вием» подставляют для этого то один, то другой участок своего тела.
Очень важной формой поведения является также трофаллаксис — пе-
редача жидкого корма путем отрыгивания изо рта в рот (у термитов
развит также проктодеальный трофаллаксис — поглощение жидкой
пищи, выделяемой из анального отверстия). Путем трофаллаксиса про-
исходит равномерное распределение пищи в колонии насекомых, а так-
же передаются специальные вещества (феромоны), имеющие сигналь-
ное значение.
Чрезвычайно сложное и й высокой степени координированное пове-
дение общественных насекомых базируется на многочисленных меха-
низмах коммуникации и передачи информации между особями. В этом
общении насекомыми используются различные типы сигналов. Акусти-
ческие сигналы имеют ограниченное распространение; они применяют-
ся для сообщения об опасности некоторыми термитами, муравьями,
осами, пчелами, а медоносные и безжальные пчелы используют их в пи-
щевой мобилизации. При трофаллаксисе и других формах взаимодей-
ствия и контакта между насекомыми широко используются тактильные
сигналы в форме прикосновений усиками, ногамй, ударов и вибрации
телом. Так, характерные прикосновения усиками являются релизерами
32
трофаллаксиса, и без них он невозможен. Не менее важными являются
химические сигналы — это выделяемые во внешнюю среду летучие ве-
щества — феромоны.
У общественных насекомых существуют 3 основных типа коммуни-
кации. Привлечение и узнавание обеспечиваются в основном выделением
феромонов-аттрактантов. Так, царицы выделяют специальные феромо-
ны, привлекающие к ним рабочих, стимулирующие облизывание и корм-
ление (кроме того, они подавляют плодовитость рабочих). Все особи
из одного гнезда у общественных насекомых имеют специфический об-
щий запах, который определяется феромонами и другими веществами,
находящимися на поверхности их тела и воспринимаемыми при контак-
те. Такие же поверхностные феромоны определяют узнавание личинок
и куколок и заботу о них, позволяют опознавать кастовую принадлеж-
ность, возраст, физиологическое состояние особи. Посторонние особи
в гнездо не допускаются или даже убиваются.
Для сигнализации об опасности обычно используются феромоны
тревоги, распространяющиеся из точки их выделения путем диффузии
и привлекающие к этому месту других особей, делая их, кроме того,
более агрессивными и готовыми к отражению опасности. Иногда для
сообщения об опасности применяются акустические сигналы (в сочета-
нии с феромонами и реже независимо).
Важнейшей формой коммуникации является мобилизация, т. е. воз-
буждение других особей и направление их в определенное место для
выполнения той или иной функции. Мобилизация может проводиться
для сбора корма, отражения опасности, ремонта гнезда и переселения.
Как правило, в ней участвует целый комплекс сигналов: тактильные,
химические, иногда акустические, или даже такие сложные формы сиг-
нализации, как танцы медоносных пчел (см. с. 163). У муравьев и тер-
митов основной формой мобилизации является мобилизация по следу:
рабочие особи возбуждаются разведчиками с помощью тактильных
сигналов и направляются в нужное место (например, к обнаруженному
источнику пищи) по следу, оставленному разведчиками. След прокла-
дывается с помощью следовых феромо'нов, выделяемых железами брюш-
ка и наносимых на субстрат при передвижении (см. рис. 395).
На основе коммуникации и передачи информации у общественных
насекомых сформировались разнообразные и сложные механизмы ре-
гуляции, обеспечивающие четкую организацию процессов жизнедея-
тельности колонии в целом. Эта регуляция затрагивает самые разно-
образные процессы: развитие и дифференциацию на касты в онтоге-
незе, разделение функций и распределение «рабочей силы» в за-
висимости от потребностей, поддержание определенной темпера-
туры и даже влажности в гнезде. Многие механизмы регуляции дей-
ствуют по принципу гомеостазиса, т. е. поддерживают величину того
или иного параметра на определенном необходимом уровне.
Для общественных насекомых очень характерен также эффект
группы, причем у наиболее высоко организованных из них присутствие
других особей не просто оказывает стимулирующее воздействие, но
является жизненной необходимостью, поскольку изолированные от
группы насекомые нередко не способны к какой-либо целенаправлен-
ной деятельности и быстро погибают.
СЕЗОННЫЕ ЯВЛЕНИЯ В ЖИЗНИ НАСЕКОМЫХ*
Сезонные изменения экологической обстановки оказывают силь-
нейшее влияние на численность и жизнедеятельность насекомых. Общим
правилом является увеличение численности какого-либо вида в наибо-
* В. П. Тыщенко.
3 208
33
лее благоприятные для роста, развития и размножения периоды года.
Так, по данным трехлетних наблюдений в Сухуми численность маля-
рийных комаров (Anopheles mactilipennis Mg.) достигает максимума
в июле и августе, снижаясь в 15—20 раз во время зимовки (рис. 9). По
всему ареалу малярийный комар имеет только один максимум чис-
ленности в течение года. Существует, однако, немало видов насекомых
с двумя или тремя максимумами. Регуляция сезонных колебаний чис-
ленности осуществляется через систему факторов, определяющих фено-
логию насекомых. Надежное соответствие фенологии сезонным изме-
нениям экологической обстановки служит гарантией высокой числен-
ности и оптимальной плотности популяции.
Рис. 9. Сезонные изменения численности малярийных комаров (Anopheles macull-
pennls Mg.) в Сухуми (по данным Авдеевой и Зимина).
Сезонные колебания численности являются итого-
вым результатом многих сезонных ритмов, отражающихся на темпах
размножения и уровне смертности. Назовем наиболее характерные для
насекомых проявления сезонной ритмики.
В районах с резко выраженной сезонностью климата отчетливо
проявляются сезонные изменения темпов роста и раз-
вития насекомых. Интенсивный рост личинок приурочен к теплому
периоду года, а зимой имеет место задержка роста или полное его
прекращение. Естественно, что в зимний период прекращаются линьки,
приостанавливается развитие яиц и всех постэмбриональных стадий.
Сезонные ритмы питания обычно сопряжены с изменения-
ми роста и развития насекомых. Как правило, в холодное время года
насекомые отказываются от питания или сокращают'его до минимума.
В летнее время интенсивность питания подвержена влиянию погодных
условий. В течение теплого сезона может изменяться и состав пищи
в зависимости от ее обилия и доступности. Так, у шведской му'хи
(Oscinosoma frit L.) в Нечерноземной зоне РСФСР первое поколение
развивается на всходах яровых хлебов, второе — на колосках овса
и ячменя, а третье —на всходах озимых злаков. Жуки обыкновенного
свекловичного долгоносика (Bothynoderes puctiventris Germ.) после
выхода из почвы, где они зимуют, вначале питаются марью и лебедой,
а затем переключаются на преимущественное потребление молодых
всходов сахарной свеклы.
34
У многих тлей наблюдается отчетливо выраженная сезонная сме-
на растений, служащих им пищей. Например, свекловичная тля (Aphis
fabae Scop.) весной питается на листьях кустарников — бересклета,
калины или жасмина, а к началу лета переходит на питание травя-
нистыми растениями — свеклой, крапивой, чертополохом и др. (рис. 10).
Кизиловая тля (Anoecia corni L.) весной развивается на диком кизи-
ле, а летом — на корнях культурных и диких злаков. Интересно, что
в тех районах, где нет дикого кизила, этот вид постоянно обитает на
корнях злаков.
Рис. 10. Сезонный цикл смены поколений и кормовых растений у свекловичной
тли (Aphis fabae Scop.).
Сопряженность роста, развития и питания насекомых с сезонными
изменениями экологических условий создает приуроченность
онтогенетических стадий к определенным периодам
года. Так, у зимней пяденицы (Operophtera brumata L.) гусеницы
встречаются весной, куколки — летом, вылет бабочек приурочен к глу-
бокой осени, а зимовка происходит в стадии яйца. Серебристая лунка
(Phalera bucephala L.) имеет иное «сезонное расписание» жизненного
цикла: в начале лета появляются бабочки, гусеницы развиваются до
осени, а зимует куколка. У всех кузнечиков (Tettigoniidae) и настоя-
щих саранчовых (Acrididae), встречающихся в СССР, зимуют яйца;
отрождающиеся весной личинки растут, периодически линяют и через
50—70 дней превращаются во взрослых насекомых, которые несколько
раз откладывают яйца и с наступлением зимы погибают.
35
Названные выше представители насекомых в течение года дают
одно полное поколение. Такой цикл развития называется моновольтин-
ным. Он характеризуется однократным пиком появления имаго, при-,
уроненным к определенному сезону (начало лета—-у серебристой лун-
ки, середина лета и осень — у саранчовых и кузнечиков, поздняя
осень — у зимней пяденицы).
При поливольтинном цикле, свойственном многим насекомым,
в течение года развиваются от 2 до 10—И поколений. В этом случае
обычно наблюдаются обособленные пики появления имаго, приходя-
щиеся на разные периоды года и соответствующие завершению разви-
тия каждого поколения. Например, в лесостепной зоне европейской
• З.К » «в « «в * «•
I I I I I । I । . 1 । I__| I_।___I-1—I___I-1__1---1--i-1---1-1--1—I
10 20 30 10 20 30 10 20 30 Ю 20 30 10 20
V VI VII VIII IX
Рис. 11. Фенология щавелевой совки (Acronycta rumicis L.) в условиях лесостепной
воны (по Данилевскому).
Кривые на графике — динамика лета бабочек в 1930 г. в Полтаве по учетам светоловушкой.
Под графиком — сроки развития гусениц и куколок. I и II — поколения; г — гусеницы; з.к — зи-
мующие куколки; к —куколки.
части СССР лет бабочек щавелевой совки (Acronycta rumicis L.)
происходит дважды: весной появляются бабочки, которые выходят из
перезимовавших куколок, затем происходит развитие следующего по-
коления, завершающееся вылетом бабочек в середине лета (рис. 11).
Бабочки летнего поколения откладывают яйца, из них вылупляются
гусеницы, которые к осени превращаются в зимующих куколок. В се-
верных районах европейской части СССР короткий вегетационный пе-
риод обеспечивает развитие только одного полного поколения, а на
юге щавелевая совка дает 3 поколения. Сходные сезонные ритмы об-
наруживаются у капустной совки (Barathra brassicae L.), капустной
белянки (Pieris brassicae L.) и у многих других чешуекрылых. Тли,
дрозофилы (Drosophilidae) и некоторые другие мелкие насекомые мо-
гут давать в течение года большое ^число (до 10—11) поколений, ко-
торые, однако, накладываются друг на друга, и периоды появления
имаго у них не так четко изолированы, как у щавелевой совки.
Некоторые насекомые завершают полный цикл развития в тече-
ние двух и более лет. Двухлетние и многолетние циклы особенно час-
то наблюдаются у обитателей почвы и древесины. Двухлетний цикл
с зимовкой личинок и имаго обнаружен у такого типичного геобионта
36
как обыкновенная медведка (Gryllotalpa gryllotalpa L.). У жука-
кузьки (Anisoplia austriaca Hrbst.), развитие которого тоже проте-
кает по двухлетнему циклу, зимуют личинки младших и старших воз-
растов (рис. 12). Многолетние циклы свойственны майским хрущам
Melolontha melolontha L. и М. hippocastani F. Развитие личинок у обо-
их видов хрущей происходит в почве и продолжается 3 года на юге
и 4 года на севере. Закончившие питание личинки на третий или чет-
вертый год окукливаются обычно в июне-июле. Жуки остаются в поч-
ве зимовать и вылетают весной следущего года. У личинок жука-оленя
(Lucanus cervus L.), обитающих в гниющей древесине лиственных пород,
развитие от яйца до куколки растягивается на 5—8 лет. Как правило,
при двухлетних и многолетних циклах каждый год наблюдается толь-
ко один период появления имаго, но численность взрослых насекомых
сильно изменяется по годам.
Апрель Май Июнь Июль Август Сентябрь зйг моб- ка
1 II in 1 II III 1 и in 1 II in 1 и ш 1 и in
40 40 40 0 40 9 tfi) • $ 40 • * 0 * е & • Ж 0 & 40 40 40 40 40
40 40 40 40 40 40 40 40 40 40 40 40 40 40 40 40
о» 0к
Рис. 12. Фенология жука-кузьки (Anisoplia austriaca Hrbst.) в условиях
степной зоны (по Добровольскому).
и — имаго; « — куколки; л —личинки; я —яйца.
Сезонные цикломорфозы наблюдаются только у поли-
вольтинных насекомых и проявляются в периодической смене поколений,
различающихся по морфологическим признакам. Примером относи-
тельно простого цикломорфоза могут служить весенние и летние ба-
бочки пестрокрыльницы Araschnia levana-prorsa L., различающиеся по
расцветке крыльев (рис. 13). Весенняя форма этого вида (levana)
имеет рыжевато-красные крылья с черным пятнистым рисунком и с не-
сколькими белыми пятнышками у вершины переднего крыла. Крылья
осенней формы (prorsa) буровато-черные, с белыми или желтоватыми
пятнышками и полосками.
У некоторых насекомых цикломорфозы сопровождаются перехо-
дами от обоеполого размножения к партеногенезу. Например, у тлей
в течение всего лета происходит только партеногенетическое размно-
жение самок, сочетающееся с живорождением и приводящее к быстрому
росту популяции (см. рис. 10). Время от времени в колонии пар-
теногенетических бескрылых тлей появляются крылатые самки-пере-
селенцы и эмигранты, перелетающие на новые растения. Осенью пар-
теногенетические самки-полоноски отрождают крылатых самцов и бес-
крылых самок обоеполого поколения. После спаривания осенние самки
откладывают крупные зимующие яйца, из которых весной выходят
крылатые самки-основательницы, дающие начало новому ряду парте-
ногенетических поколений.
37
Выбор насекомыми наиболее благоприятных экологических ниш
в период летнего развития и во время зимовки приводит к сезонным
миграциям и сезонной смене местообитаний. Окрылив-
шиеся клопы вредной черепашки (Eurygaster integriceps Put.) летом
питаются на посевах пшеницы и на дикорастущих злаках, осенью
мигрируют в лесные массивы и здесь зимуют под опавшей листвой,
а весной возвращаются на поля. Почвенные насекомые при неблаго-
приятных условиях (зимой и при засухе летом) совершают вертикаль-
ные миграции, уходя в глубокие горизонты почвы. Многие водные на-
секомые перед наступлением зимы или зарываются в ил и грунт на
дне водоемов или покидают водоемы и устраиваются на зимовку во
влажной земле.
gl_------1------1-------1------1-------1----__1-------1_------1------1------j--------1-----_J
I II III IV V VI VII VIII IX X XI XII
Месяцы
Рис. 13. Фенологическая схема смены сезонных форм у пестрокрыльницы (Araschnia
levana-prorsa L.) в Средней Европе (по Мюллеру).
/ — сезонные изменения длины дня; 2 — зимующие куколки; 3 — весеииие бабочки ’ (форма
levana); 4— яйца; 5 — гусеницы; £ —куколки; 7 — летние бабочки (форма prorsa).
Перечисленные выше сезонные ритмы иногда обусловливаются
прямыми влияниями экологических факторов па развитие, размноже-
ние и поведение насекомых. Недостаток полноценной пищи и пониже-
ние температуры перед наступлением зимы могут замедлить рост
и развитие, снижать двигательную активность, подавлять плодови-
тость насекомых, а в некоторых случаях даже создавать специфиче-
ские изменения окраски. Известно, что, если куколки чешуекрылых
содержатся при пониженных температурах, они могут дать темно-
окрашенных бабочек. Значительное понижение температуры (до +5
или 0°) вызывает полное обездвиживание и холодовое оцепенение на-
секомых. В засушливых районах сезонные изменения влажности тоже
могут индуцировать синхронные с ними изменения в поведении и раз-
множении насекомых.
Однако для регуляции сезонных циклов в природных условиях
более важное значение имеет не прямое (витаЛ|Ьное), а сигнальное
действие экологических факторов. Сигнальный фактор приводит к сме-
не сезонных биологических состояний, адаптированных к разным пе-
риодам года. У большинства насекомых сезонные ритмы основаны на
чередовании двух биологических состояний — активной жизнедеятель-
ности и диапаузы. Активная жизнедеятельность характеризуется вы-
38
соким уровнем энергетического обмена и синтеза органических соеди-
нений. Она сопровождается интенсивным ростом, развитием и размно-
жением, усиленным питанием и высокой подвижностью насекомых.
Для состояния диапаузы характерно торможение метаболизма, роста,
развития и ' размножения, полное или частичное подавление питания
и подвижности. Диапауза представляет собой специальную адаптацию
к перенесению .неблагоприятных условий, поэтому сезонный переход
от активной жизнедеятельности к диапаузе, как правило, сопровожда-
ется повышением устойчивости насекомых к отсутствию пищи, иссу-
шению, длительному действию низкой или высокой температуры. Сле-
довательно, активная жизнедеятельность и диапауза отражают при-
способление насекомых к двум основным периодам года: летнему
и зимнему в умеренных широтах, влажному и засушливому — в тро-
пических.
Таблица 1
Примеры приуроченности диапаузы к стадиям онтогенеза
у некоторых чешуекрылых
Виды Типы диапаузы Диапаузирующие стадии
Русские названия Латинские названия
Л истовертка дубовая Tortrix viridana L. Эмбриональная Яйцо
Шелкопряд сосновый Dendrolimus pint L. Личиночная Гусеница любого возраста
Плодожорка яблоневая Laspeyresia pomonella L. Пронимфальная Закончившая пита- ние гусеница перед окуклением
Белянка капустная Pieris brassicae L. Куколочная Куколка
Лимонница Gonepteryx rhamni L. Имагинальная Бабочка
У каждого вида насекомых диапауза приурочена к определенной
стадии (табл. 1). Некоторые насекомые имеют и зимнюю и летнюю
диапаузу. Например, у рыжего соснового пилильщика (Neodiprion
sertifer Geoffr.) диапаузируют зимой яйца, а летом—личинки в коко-
нах. Обычно диапауза заканчивается в течение неблагоприятного для
развития сезона, но иногда она переходит в суперпаузу, продолжаю-
щуюся более календарного года.
Важнейшим сигнальным фактором, определяющим наступление
зимней диапаузы, являются сезонные изменения длины дня {фотопе-
риод). Укорочение дня в конце лета и осенью запускает физиологиче-
ские процессы, которые позволяют насекомым своевременно сформи-
ровать диапаузу и уйти на зимовку в состоянии готовности к перене-
сению неблагоприятных условий зимнего сезона. Прекращение диапаузы
и обратный переход к активной жизнедеятельности обычно дости-
гаются благодаря длительному действию низких температур во время
зимовки. Этот процесс восстановления активной жизнедеятельности
у диапаузирующих насекомых получил название реактивации.
Длина дня регулирует не только переход от активной жизнедея-
тельности к диапаузе, но и некоторые другие проявления сезонной рит-
мики. Показано, что действие короткого дня вызывает сезонную сме-
ну окраски в разных поколениях бабочки Araschnia levana-prorsa L.
39
(см. рис. 13) и переход от партеногенеза к обоеполому размножению
у тлей (см. рис. 10).
Изучение сезонных ритмов и способов их регуляции представляет
не только теоретический, но и практический интерес. Только на осно-
ве конкретного познания сезонных ритмов можно составлять научно
обоснованные прогнозы массового появления вредных или полезных на-
секомых и разрабатывать систему мероприятий по защите растений
от вредителей.
СБОР, КОЛИЧЕСТВЕННЫЙ УЧЕТ И ХРАНЕНИЕ НАСЕКОМЫХ*
МЕТОДЫ СБОРА
Существует ряд универсальных методов сбора, при которых одно-
временно вылавливаются представители нескольких семейств и отря- ?
дов насекомых и другие членистоногие, такие, как пауки, клещи и мно- \
гоножки. Подобные методы дают возможность получить большой фау- ;
нистический материал, который позднее монтируется и определяется •
на лабораторных занятиях. В данном случае речь идет об укосах сач- J
ком, почвенных раскопках, отряхивании деревьев и кустарников и от- !
части лове на свет. Наряду с этим необходимо использование и спе- !
циальных методов сбора, рассчитанных на отлов определенных групп i
или даже отдельных видов насекомых (см. ниже). Полученные таким ;
способом данные о видовом составе насекомых и некоторых других j
членистоногих определенного биотопа в какой-то степени отражают i
и численность отдельных видов, так как редкие и малочисленные виды j
в сборах практически отсутствуют и, напротив, массовые встречаются j
лостоянно. Но в большинстве случаев их следует дополнить более объ- |
ективными данными о численности интересующих нас групп, которые ;
могут быть получены при специальных количественных учетах. Из-за ;
трудоемкости последних во время студенческой практики применяются ?
лишь некоторые из них, наиболее часто используемые при по.чвенно- j
биологических и биоценологических исследованиях.
Обычно уже при первом знакомстве с насекомыми в природе ста-
новится очевидным, что сбор их требует большого внимания и наблю- ;
дательности. Легче всего собрать насекомых в ясные солнечные дни, '
когда они активно питаются, перелетают и поэтому хорошо заметны. ,
Особенно много их на лесных полянах и опушках, в пойменных лугах ;
и по берегам водоемов. Насекомых легко поймать с помощью энтомо- ;
логического сачка, состоящего из металлического обруча и марлевого ;
мешка. Из сачка насекомые осторожно переносятся в морилку — в па- i
ры эфира или хлороформа. Сделав несколько (5—10) взмахов сачком !
по траве, кустарникам и ветвям деревьев, можно собрать большое ко- 1
личество насекомых и пауков, заселяющих эти ярусы растительности. )
Наряду с взрослыми формами в этом случае в сачок попадают и ли- j
чинки. Для большинства насекомых с полным превращением характер- '
на разобщенность мест обитания имагинальных и личиночных стадий. (
Поэтому места и методы сбора их совершенно различны. Приме- j
ром могут служить хорошо известные всем майские жуки (Melolon- >
tha L.), питающиеся листьями деревьев, в то время как их личинки)
живут на корнях в почве и могут встретиться лишь при почвенных '
раскопках. ;
Для укосов (в особенности количественных) применяются специ-
альные сачки с определенным диаметром обруча (30 или 31,5 см) '
* Е. Ф. Мартынова.
40
и мешком из бязи или мельничного газа. Мельничный газ является
очень хорошим материалом и для воздушных сачков.
Очень хорошие результаты могут быть получены и просто при
тщательном осмотре стволов деревьев, ветвей, листьев, в особенности
со следами погрызов, поврежденных плодов, ’семян и соцветий. Под
камнями, стволами и ветками, давно лежащими на земле, всегда мож-
но обнаружить многочисленных обитателей — различных жуков и их
личинок, муравьев рода Myrmica Latr., ногохвосток и многоножек.
Мелкие объекты собирают с помощью эксгаустера (рис. 14) или смо-
ченной водой кисточки и вместе с личинками помещают в пробирки
Рис. 14. Эксгаустер (по Тихомиро-
вой).
1 — стеклянный цилиндр; 2 — резиновая
пробка; 3 — стеклянные трубки; 4 — кол-
пачок из мельничного газа; 5 — резиновая
трубка; 6 — кольцо из изоляционной лен-
ты или лейкопластыря, укрепляющее края
цилиндра.
Рис. 15—16. Два типа эклектбров (по Ги-
лярову).
15 — Эклектор Тульгрена, 16 — Эклектор Тре-
горда.
1 — источник нагревания; 2 — пробы иа сите;
3—-воронка; 4 — сосуд с фиксирующей жид-
костью; 5 — цилиндр из жести; 6 — наружная
воронка с водой; 7 — термометр.
с 70%-ным этиловым спиртом (лучше с добавлением 2—3% глице-
рина). Очень разнообразна фауна подстилки и верхних горизонтов
почвы. При общих сборах подстилку, а затем и верхние слои почвы
переносят на светлую клеенку, ткань или в большую кювету и разби-
рают вручную или просеивают через специальные почвенные сита. При
таком способе выбираются только достаточно крупные объек-
ты— т$к называемая мезофауна. Для извлечения мелких членистоно-
гих используют специальные установки — эклекторы (рис. 15, 16),
основанные на свойственной всем мелким обитателям подстилки
и почвы высокой чувствительности к высыханию. Обитатели почвы по-
кидают верхние, подсыхающие слои и перемещаются вниз, где попа-
дают на сетку и проваливаются в воронку, соединенную с пробиркой,
наполненной фиксирующей жидкостью. Можно поместить подстилку
в полиэтиленовые мешки и проводить разборку в лаборатории.
Многие виды насекомых активны в сумеречные и ночные часы.
Поэтому необходимо вскоре после захода солнца осматривать цвету-
щие растения, используя в более поздние часы электрический фо-
нарик.
Широко применяются при сборе насекомых и различные ловушки.
Насекомые, активные в ночные часы, привлекаются любым источником
41
летают, а то и
Рис' 17. Свето-
ловушка.
света и могут отлавливаться или собираться специальными дветоло-
вушками. Светоловушка, изображенная на рис. 17, состоит из мощной
электрической лампы, абажура-отражателя и закрепленной под ним
металлической воронки, соединенной с широкогорлой банкой, напол-
ненной фиксирующей жидкостью (спиртом, бензином, керосином).
Прилетающие на свет насекомые ударяются о металлические части
прибора, падают в воронку и скатываются в ловчую банку. Ловушки
должны быть укреплены не ниже 2—3 м над землей. В теплые без-
лунные ночи на свет в массе прилетают чешуекрылые, жуки, двукры-
лые и перепончатокрылые. Зачастую из прилежащих участков при-
цриползают и дневные насекомые, привлеченные ярким
светом. В автоматических светоловушках часть ма-
териала повреждается (в особенности страдают-че-
шуекрылые), но при ежедневных утренних выборках
материал вполне пригоден для дальнейшей обработки.
Для некоторых насекомых привлекающими яв-
ляются различные сильно пахнущие приманки. Мерт-
воедов (Silphidae), жужелиц (Carabidae), ряд видов
жуков сем. Staphylinidae и мух привлекает запах
гниющего мяса. У трупов позвоночных животных их
всегда можно собрать в изобилии. Можно поместить
приманку на дно широкогорлого сосуда, погружен-
ного до краев в почву, и периодически проводить
выборку попавших туда насекомых. Пустые цилинд-
ры и стеклянные банки также могут служить для
сбора; в них чаще всего попадают виды, обитающие
на поверхности почвы и в подстилке. В цилиндры
иногда помещают воронку, прилегающую к краям
цилиндра, и банку с фиксатором (2—-4%-ным фор-,
малином).
некоторых групп чешуекрылых, главным образом со-
вок (Noctuidae), выставляют неглубокие корытца с бродящей смесью
из патоки, меда и пива, а также накладывают ловчие пояса и раз-
вешивают тряпки, пропитанные этой же смесью. Для фаунистических
сборов этот метод мало пригоден, так как бабочки часто тонут в рас-
творе или прилипают к нему, теряя при этом чешуйки. По таким эк-
земплярам трудно определить видовую принадлежность и приходится
использовать строение копулятивных придатков, что без специальной
подготовки достаточно сложно. Бродящие приманки иногда применяют
при изучении динамики лета отдельных, преимущественно вредяйхих
видов.
Для некоторых семейств чешуекрылых (волнянки — Lymantriidae,
павлиноглазки — Attacidae) и жуков-хрущей (Scarabaeidae) возможен
также вылов самцов на запах самки. Половые феромоны, выделяемые
самкой, воспринимаются самцами с большого расстояния, исчисляе-
мого иногда километрами. Самку при этом помещают в марлевый
мешочек, который, в свою очередь, прикрепляется внутри большого
садка, устроенного по типу рыболовной верши.
Для приманки вредителей древесины (усачей — Cerambycidae,
короедов — Ipidae) используют ловчие деревья, поваленные или час-
тично ошкуренные, но не спиленные. На таких ослабленных или мерт-
вых деревьях происходит интенсивная откладка яиц, а через неко-
торый промежуток времени здесь можно собрать личинок и взрослых
насекомых.
Для отлова насекомых, посещающих цветущие растения (пере-
пончатокрылые, двукрылые), используют привлекающую особенность
Для сбора
42
желтого цвета. Для того чтобы собрать прилетевших .насекомых, при-
меняют желтые ловчие чашки Мерике. Изготовить их несложно. Белые
эмалированные кюветы или неглубокие миски изнутри окрашивают
в желтый цвет так, чтобы краска покрывала дно и нижнюю половину
стенок. Миски заполняют водой до уровня окрашенного слоя и поме-
щают на участки с цветущей растительностью (разнотравные луга,
лесные опушки). Миски должны быть расположены на уровне голо-
вок цветков. Прилетевшие насекомые падают в воду, а наблюдатель,
переходя от одной чашки к другой, вынимает их из воды пинцетом или
кисточкой.
Во многих биотопах, в особенности лесных, постоянно можно
встретить различные виды муравьев (Formicidae) и их гнезда. Му-
равьи, которых мы находим в подстилке, на земле и лесных дорогах,
на стволах деревьев, кустарниках и травянистой растительности, явля-
ются рабочими особями. Других членов семьи —самцов и самок мож-
но собирать либо в момент роения, когда крылатые половозрелые
особи вылетают из куколок, либо при раскопках муравейников. Рас-
копки без крайней необходимости проводить не следует, учитывая
большую положительную роль муравьев в лесных биоценозах. Рабо-
чих муравьев лучше собирать вблизи муравейников, чтобы при опре-
делении видов муравьев представлять себе и типы их гнезд. Личинок
и куколок муравьев можно увидеть и собрать в насыпных муравейни-
ках рыжего лесного муравья (Formica rufa L.) и черного муравья
(Lasius niger L.), осторожно приподняв лопатой верхнюю часть ко-
нуса.
Личинки многих наземных насекомых развиваются в воде, а для
большого числа видов водная среда является постоянной средой оби-
таний. В проточной, быстро текущей воде, богатой кислородом, фауна
насекомых беднее, но некоторые группы, обитающие здесь, в стоячей
воде не встречаются. Это в первую очередь личинки многих видов по-
денок и веснянок, мелких кровососущих двукрылых — мошек (Simu-
liidae). В старицах медленно текущих рек, озерах и болотах число
обитающих там видов резко возрастает. Почти для всех таких водое-
мов характерны водные клопы — водомерки (Gerridae), гладыши (No-
tonectidae), гребляки (Corixidae), водяные скорпионы (Nepidae)
и плавты (Naucoridae); в них также часто встречаются жуки-плавун-
цы (Dytiscidae) и плавунчики (Haliplidae), водолюбы (Hydrophilidae),
вертячки (Gyrinidae), личинки стрекоз, ручейников, нескольких видов
мух. Вблизи воды в период яйцекладки и отрождения можно встре-
тить и взрослых насекомых, которых легко собрать с помощью воздуш-
ного сачка. Водных насекомых вылавливают специальным сачком из
плотного мельничного газа или канвы, погружая его в заросли водных
растений и на дно водоема. Пробы чаще всего содержат много ила
и растительных остатков и разбирать их нужно в специальных белых
кюветах или тазах. Насекомые выбираются пинцетом и раскладыва-
ются в банки с водой, так, чтобы хищные виды находились в отдель-
ных сосудах. Окончательная разборка проводится обычно в лаборато-
рии. Личинки (кроме полужесткокрылых) переводятся в спирт, а ос-
тальные— в морилку и на ватные слои. Многие водные насекомые,
в особенности полужесткокрылые, хорошо летят на свет и попадаются
в светоловушки даже на значительном удалении от водоемов.
МЕТОДЫ КОЛИЧЕСТВЕННОГО УЧЕТА f
Для оценки биологической роли отдельных видов или групп ви-
дов в определенных биоценозах необходимы конкретные данные о чис-
ленности, которые могут быть получены при специальных количест-
43
Рис. 18. Биоце-
нометр.
венных учетах. Мы познакомимся с двумя наиболее универсальными
методами количественной оценки энтомофауны.
Наиболее простой способ учета обитателей травянистого яруса —
кошение сачком стандартного размера (обруч диаметром 30 см,
глубина мешка 60—70 см и длина ручки 1,0—1,5 м). Движения сбор-
щика должны быть равномерными, чтобы при каждом взмахе сачком
захватывалось определенное расстояние. После каждого взмаха дела-
ется шаг вперед. Учеты проводятся на 50 или 100 взмахов. По окон-
чании кошения содержимое быстро стряхивается на дно сачка, мешок
перехватывается и завязывается шнурком, а нижняя часть мешка
с насекомыми помещается в широкогорлую морилку. Через несколько
минут неподвижные насекомые пересыпаются из сачка в банку или
в ту же морилку для последующей разборки и подсчета в лаборатории.
Каждая проба снабжается отдельной этикеткой. Этот метод не дает
так как из нижнего яруса захватывается лишь часть
населения, но он очень удобен для сравнительных •
целей, когда проводятся параллельные учеты в
различных биотопах или в одном и том же биото-
пе, но в разное время. Для того чтобы представить
себе численность насекомых на единицу площади,
можно воспользоваться формулой, предложенной
Л. Г. Динесманом (1966): x=NI2RLn, х —
количество насекомых на 1 м2, N — число насеко-
мых, пойманных при кошении, R — радиус сачка
в Метрах, L — средняя длина пути, проходимого
обручем сачка по травостою при каждом взмахе
сачка, п — число взмахов (ударов) сачка. Следу-
ет, однако, помнить, что полученные цифры дают
лишь приблизительное представление о численно-
сти насекомых и не являются исчерпывающими.
Наибольшая точность достигается при использовании б ио цен о-
метров. Принцип их работы сводится к изоляции участка почвы,
подстилки и травостоя с помощью металлической рамки определенно-
го размера, имеющей заостренные нижние края и закрепленный ме-
шок сверху (рис. 18). Проводится полная выборка субстрата вместе
с верхним слоем почвы, который затем тщательно просматривается.
На разбор одной пробы затрачивается иногда от нескольких часов до
1—2 дней. Из-за трудоемкости процесса биоценометры на учебной
практике не используются.
При работе с биоценозом учитывается население подстилки и са-
мого поверхностного слоя почвы. Далее можно продолжить эту ра-
боту, вынимая послойно почву и просматривая ее вплоть до гори-
зонта, в котором насекомые полностью отсутствуют. Данные учета
рассчитываются затем на единицу поверхности. В результате числен-
ность почвенных насекомых определяется количеством животных
в столбе почвы площадью 1 м2 на всю глубину их встречаемости. Та-
ким же образом можно рассчитать численность отдельных видов или
групп. Но чаще анализ почвенной энтомофауны проводится без пред-
варительного учета населения травостоя, тем более, что под кронами
деревьев и кустарников при сильном загущении и обйльной подстилке
травостой редок или практически отсутствует. При специальных поч-
венных раскопках выбирается ровная площадка размером 50X50 см
(площадью 0,25 м2). Вначале собирается и просматривается подстил-
ка, а затем на расстеленную сбоку клеенку или лист фанеры осторож-
но, небольшими порциями вынимается лопатой почва, перетирается
руками и внимательно просматривается. Учеты проводятся послойно
до глубины встречаемости насекомых. Как правило, выделяются сле-
44
дующие горизонты (в сантиметрах): 0—5, 5—10, 10—20, 20—30, 30—
50. Весной и в первую половину лета при достаточно высокой влажно-
сти в этих слоях и сосредоточены почти все почвенные обитатели.
Данные по каждому горизонту записываются отдельно.
ХРАНЕНИЕ СОБРАННОГО МАТЕРИАЛА
Взрослых насекомых и некоторых наземных личинок собирают
в морилки — различного размера банки с корковой пробкой или поли-
этиленовой крышкой. Внутрь морилки закладываются узкие полоски
фильтровальной бумаги, поглощающие влагу, а на дно — несколько
слоев из той же бумаги. Заряжается морилка серным эфиром или
хлороформом; последний не воспламеняется, менее летуч и поэтому
чаще применяется во время полевой практики. Желательно периоди-
чески освобождать морилку, перекладывая мертвых насекомых в дру-
гие банки, на вату или фильтровальную бумагу, так как вновь отлов-
Рис. 19—25. Схема наколки насекомых на энтомологические булавки (по Горно-
стаеву).
19 — прямокрылые, 20 — жесткокрылые, 21 — двукрылые, 22 — перепончатокрылые, 23 — клопы-щит-
ники, 24 — прочие полужесткокрылые, 25 — чешуекрылые. Точки обозначают места введения бу-
лавки у насекомых разных отрядов.
ленные насекомые, прежде чем погибнуть, могут сильно повредить
находящийся там материал. Чешуекрылых следует собирать в отдель-
ные морилки, а дневных бабочек, перед тем как положить в морилку,
нужно сильно сжать двумя пальцами в области груди, после чего
крылья остаются сложенными вместе, а чешуйки не стираются.
На лабораторных занятиях собранный материал следует до высы-
хания наколоть на энтомологические булавки или разложить на слои
ваты. При наколке булавку вводят в тело насекомого вертикально
на 2/3 ее длины в строго определенном месте (рис. 19—25). Наколотые
на булавки экземпляры размещаются в закрытых коробках с торфя-
ным или пробковым дном. Пауков и многоножек переносят в 70%-ный
спирт, но лучше это делать сразу же на экскурсии. Ногохвосток, тлей
и других мелких и ‘нежных насекомых помещают в спирт на месте
сбора или приносят в лабораторию с небольшой порцией влажного
субстрата. Волосистых гусениц и сильно склеротизованных личинок
жуков сразу помещают в спирт, а мясистых личинок с тонкими по-
кровами и гусениц с редкими и короткими волосками на 1—2 минуты
опускают в кипящую воду и только после этого переносят в спирт.
Мелких длинноусых двукрылых (комаров — Culicidae, мокрецов — Се-
ratopogonidae, мошек — Simuliidae), тлей, мелких муравьев также
лучше хранить в спирте, так как их тонкие структуры в сухом виде
чрезвычайно хрупки.
45
Часть 11
КРАТКИЙ ОПРЕДЕЛИТЕЛЬ
И ЭКОЛОГО-МОРФОЛОГИЧЕСКАЯ
ХАРАКТЕРИСТИКА КЛАССОВ,
ОТРЯДОВ И СЕМЕЙСТВ
НАЗЕМНЫХ ЧЛЕНИСТОНОГИХ
СТРОЕНИЕ И РАЗВИТИЕ НАЗЕМНЫХ ЧЛЕНИСТОНОГИХ*
Общая характеристика
Обширный тип членистоногих (Arthropoda) охватывает морских,
пресноводных, почвенных, свободноживущих наземных и паразитиче- v
ских животных с сегментированным (расчлененным) телом и членис-
тыми конечностями. В составе этого типа выделяют 8 ныне живущих
и несколько ископаемых классов, но к наземным членистоногим при-
надлежат только некоторые наземные и амфибиотические представи-
тели класса ракообразных (Crustacea), все многоножки (Myriapoda),
насекомые (Insecta) и паукообразные (Arachnida).
По хорошо обоснованным филогенетическим представлениям чле-
нистоногих следует выводить от аннелидных предков, имевших по
1 паре нерасчлененных придатков на всех сегментах (рис. 26). При
формировании тела членистоногого в процессе эволюции первые 5 или
6 сегментов слились вместе и образовали голову. Придатки этих сег-
ментов превратились в усики (антенны) и ротовые органы. Усики
служат основным местом размещения обонятельных и многих других
рецепторов. Ротовые органы используются для извлечения пищи из
кормового субстрата, ее захвата, удерживания, и размельчения. Ос-
тальные сегменты аннелидных предков у членистоногих вошли в со-
став груди и брюшка. У многоножек почти все туловищные сегменты
снабжены однотипно устроенными бегательными (ходильными.) нога-
ми. Поэтому тело многоножки подразделяют только на 2 отдела —
голову и туловище (рис. 27). У широко распространенных наземных
ракообразных, называемых мокрицами, бегательные ноги находятся
только на передних (грудных) сегментах, а придатки задних (брюш-
ных) сегментов сильно укорочены, видоизменены и выполняют иные
функции, не связанные с передвижением животного. Соответственно
в теле мокрицы выделяют 3 отдела — голову, грудь и брюшко (рис. 28).
У насекомых 3 передних сегмента туловища формируют грудной от-
дел, снабженный-тремя парами ног и, как правило, одной-двумя па-
рами крыльев, а сегменты брюшка лишёны конечностей (рис. 29).
Только на самых последних брюшных сегментах у насекомых могут
находиться парные или непарные придатки, резко отличающиеся от
грудных ног. У паукообразных отсутствуют усики, голова слита
с грудью в единый несегментированный отдел — головогрудь, а брюш-
ко бывает сегментированным или несегментированным и не имеет ко-
нечностей (|рис. 30). Отсутствие усиков у паукообразных в какой-то
степени возмещается педипальпами — второй парой конечностей голо-
вогруди.
Тело членистоногих покрыто твердой хитиновой кутикулой, обра-
зующей наружный скелет, к которому изнутри прикрепляются мышцы.
Окраска тела членистоногих в основном обусловлена пигментами, ло-
* В. П. Тыщенко.
46
кализованными в кутикуле. Особенности строения покровов нередко
используются в качестве «опознавательных признаков» при определе-
нии отрядов и семейств. Важное диагностическое значение могут
иметь своеобразные рисунки, создаваемые пигментами на поверхности
покровов, и различные модификации кутикулы — волоски, щетинки,
шипы, чешуйки и т. д.
Рис. 26—30. Схема сегментации отделов тела у наземных членистоногих и их анне-
лидного предка.
26 — аннелидный предок членистоногих, 27 — многоножка, 28 — мокрица — представитель на-
земных ракообразных, 29 — насекомое, 30 — паук,
б — брюшко; г —голова; гл — головогрудь; гр — грудь; к — крылья; п — педипальпы; с.г —
сегменты, вошедшие в состав головы; с.т — сегменты, вошедшие в состав туловища; г — туло-
вище; у — усики; щ —- щупики.
Наземные членистоногие не составляют единую таксономическую
группу, а их гомологичные признаки отвечают лишь общей исходной
организации всего типа. Однако при переходе к жизни на суше под
влиянием сходных условий обитания конвергентно выработались и не-
которые черты эколого-морфологического сходства между разными
группами наземных членистоногих. У открытоживугцих многоножек,
паукообразных и насекомых на поверхности тела формируется тонкий
восковой слой, который препятствует испарению воды и защищает жи-
вотное от иссушения. Этот слой может исчезать, когда личинки насе-
комых переходят к водному Или почвенному образу жизни. Типичные
органы дыхания наземных членистоногих представлены трахеями,
а у паукообразных — трахеями и легкими (легочными мешками). Для
47
многоножек, паукообразных и насекомых характерно развитие специ*
альиых органов выделения в виде открывающихся в кишечник труб-
чатых слепых отростков, названных мальпигиевыми сосудами. Пере-
ход к обитанию на суше привел к возиикновеиию внутреннего оплодо-
творения в большинстве групп наземных членистоногих и к ряду
сходных приспособлений для защиты яиц от высыхания.
Нервная система и органы чувств у наземных членистоногих до-
стигают высокого развития, что обусловливает весьма сложное ин-
стинктивное поведение этих животных. К числу наиболее совершенных
органов чувств, свойственных членистоногим, относятся фасеточные
(сложные) глаза насекомых, ракообразных и некоторых много-
ножек.
Определительная таблица классов
по имагинальным стадиям
1(6). Голова снабжена усиками (рис. 27—29, у).
2(3). Ног всегда больше 7 пар. Тело удлиненное, редко широкое
и короткое, состоит из многочисленных однородных члеников,
несущих по одной или две пары ног (рис. 27)....................
.......................Многоножки— Myriapoda (с. 55).
3(2). Ног 3 пары или 7 пар. Тело состоит из головы, груди, снабжен-
ной ногами, и членистого брюшка (рис. 28, 29)..................
4(5). Ног 7 пар. Тело уплощенное, крыльев нет (рис. 28)........
.......................Ракообразные— Crustacea (с. 52).
5(4). Ног 3 пары. Тело разнообразной формы, часто снабженное
крыльями (рис. 29) .'...................
............................Насекомые— Insecta (с. 61).
6(1). Голова без усиков, слита с грудью в нерасчленениую голово-
грудь. Ног 4 пары (рис. 30)................................ .
.....................Паукообразные— Arachnida (с. 201).
Постэмбриональное развитие и метаморфоз
Наземные членистоногие размножаются половым путем и откла-
дывают яйца, а некоторые из них способны к живорождению. Выхо-
дящие из яиц или отрождаемые самкой молодые животные растут
и периодически линяют. Перед каждой линькой старая кутикула от-
слаивается и под ней образуются новые кутикулярные слои. В момент
линьки отслаивающаяся кутикулярная шкурка растрескивается, а но-
вая поверхность покровов затвердевает и пигментцруется. Взрослая,
способная к размножению стадия членистоногих получила название
имагинальной стадии (имаго). Крылатые насекомые, достигшие взрос-
лого состояния, прекращают линять и более не растут, ио у многоно-
жек линьки и рост не прекращаются после наступления половозре-
лости.
Постэмбриальиое развитие наземных членистоногих может проте-
кать по типу эпиморфоза, анаморфоза или метаморфоза.
Эпиморфоз наблюдается у некоторых многоножек из подклас-
са хилопод, у большинства паукообразных и у мокриц — наземных
представителей класса ракообразных. При таком способе превращения
из яйца выходит нимфа, по своей организации вполне сходная с има-
го. Даже молодая нимфа имеет полный набор сегментов и конечно-
стей (рис. 31, Hi), а последующее ее развитие до имагинальной ста-
дии сводится только к процессам роста и формированию органов раз-
множения (рИС. 31, Hz, и).
48
При типичном анаморфозе вылупляющаяся из яйца личинка
отличается от имаго по расчленению тела и количеству конечностей,
а во время личиночного развития происходит новообразование туло-
вищных сегментов и ног. Например, у многоножек, относящихся к под-
классу диплопод, из яйца выходит личинка, имеющая только 7 туло-
вищных сегментов и 3 парц ног (рис. 32, Л]). После каждой линьки
число сегментов и ног постепенно увеличивается (рис. 32, л2), пока
не достигнет их полного набора, свойственного имаго (рис. 32, и).
У некоторых многоножек, относящихся к подклассу хилопод, из яйца
выходит личинка с 7 парами ног, а их полное число, равное 15 парам,
достигается на одном из старших личиночных возрастов еще до на-
ступления половозрелости. Разритие клещей можно рассматривать как
своеобразную модификацию такого превращения: из яйца выходит
Рис. 31—35. Основные типы постэмбрионального развития наземных членистоногих.
31 — эпиморфоз многоножек-хилопод, 32 — анаморфоз многоножек-диплопод, 33 ~~ стадии разви-
тия клещей, 34 — метаморфоз насекомых, развивающихся с неполным превращением, 35— мета'
морфоз насекомых, развивающихся с полным превращением, и— имаго; к, — куколка; л — личин-
ка; — личинка младшего возраста; л2 —личинка старшего возраста; я— нимфа; н, — нимфа
младшего возраста; я2 — иимфа старшего возраста.
шестиногая личинка (рис. 33, л), которая затем линяет на восьмино-
гую нимфу (рис. 33, н), а после следующей линьки переходит в ста-
дию имаго, тоже снабженную 8 ногами (рис. 33, и).
При метаморфозе насекомых наблюдаются более глубокие
морфофункциональные перестройки, чем при других способах постэм-
брионального развития наземных членистоногих. В большинстве случаев
метаморфоз приводит к формированию у имаго крылового аппарата,
отсутствующего у личинок. Способность имагинальной стадии к по-
лету значительно расширяет рассеЛительные возможности насекомых
и облегчает контакты между самцами и самками в период спаривания.
Метаморфоз многих насекомых сопровождается и сменой среды оби-
тания прн превращении бескрылой личинки в крылатую взрослую
форму (табл. 2).
У насекомых различают 2. главных типа метаморфоза — неполное
и полное превращение (рис. 34, 35). Это различие между основными
4 208
49
СП
Примеры смены среды обитания и способов передвижения во время метаморфоза крылатых насекомых
Таблица 2
Отряды Семейства Среда обитания Способы передвижения
личинки и.маго личинки имаго
Стрекозы — Odonata Все семейства Пресные водоемы Открытые биотопы наземные Плавание и ползание по дну водоемов Полет
Равнокрылые— Homoptera Певчие цикады — Cicadidae Почва Открытые биотопы наземные Активное проклады- вание ходов в почве Бег и полет
Жесткокрылые — Coleoptera Жужелицы — Carabidae Почва и подстилка Поверхность почвы, подстилка (и открытые наземные биотопы Бег и ползание по ходам почвы и в под- стилке Бег и полет только бег ИЛИ
Дровосеки — Cerambycidae Под корой и в дре- весине Открытые биотопы наземные Ползание и активное прокладывание ходов под корой и s древесине Бег и полет
Перепончато- крылые — Hymenoptera Пчелиные — Apidae Ячейки гнезд, снаб- женные пищей Открытые биотопы и ные гнезда наземные обществен- Ограниченная способ- ность к ползанию То же
Муравьи — Formicidae Общественные гнез- да (муравейники) Открытые биотопы и ные гнезда ники) наземные обществен- (муравей- Транспортировка ра- бочими муравьями Только бег чих муравьев) и полет (у самок) (у рабо- или бег самцов и
Двукрылые— Diptera Комары — Culicidae Пресные водоемы Открытые биотопы наземные Плавание Полет
Оводы-— Oestridae Эндопаразиты позво- ночных То же Ограниченная способ- ность к ползанию Бег и полет
Т ахины — Tachinidae Эндопаразиты беспоз- воночных (многоножек, паукообразных и насе- комых) То же То же
типами метаморфоза нашло свое отражение в выделении двух групп
крылатых насекомых: Hemimetabola — насекомые с неполным пре-
вращением и Holometabola — насекомые с полным превращением.
При неполном превращении в индивидуальном развитии
существуют только 2 постэмбриональные стадии — личинка и имаго,
а личинка характеризуется хорошо выраженными имагинальными при-
знаками (рис. 34). У таких личинок тело четко подразделяется на
голову, грудь и брюшко, имеются фасеточные глаза, грудные ноги и та-
кой же ротовой аппарат, как у имаго. Как правило, в конце личиноч-
ного развития появляются наружные зачатки крыльев, которые пре-
вращаются во вполне развитые и функционирующие крылья после
пдрньки на имаго. К группе Hemimetabola, охватывающей насекомых
с неполным превращением, относятся стрекозы, прямокрылые, равно-
крылые, полужесткокрылые и некоторые другие отряды.
При полном превращении существуют 3 постэмбриональ-
ные стадии — личинка, куколка и имаго, а личинка по своей морфо-
функциональной организации резко отличается от имаго (рис. 35).
Такие личинки лишены фасеточных глаз и дефинитивных грудных ко1
ценностей, часто они имеют червеобразную форму тела и иной ротовой
аппарат по сравнению с имагинальной стадией. Личинка в течение
своего существования претерпевает ряд линек, переходит из одного
возраста в другой, но ее организация не меняется, а наружные зачат-
ки крыльев всегда отсутствуют. Достигнув последнего возраста, ли-
чинка превращается в куколку, которая впоследствии линяет на имаго.
Куколка не питается и не растет, она обладает весьма ограниченной
подвижностью или совсем неподвижна, а ее биологическая функ-
ция сводится к процессам развития. Глубокие морфогенетические из-
менения, происходящие на куколочной стадии, обеспечивают пере-
стройку всей организации личинки и ее превращение во взрослое кры-
латое насекомое. В группу Holometabola, охватывающую насекомых
с полным превращением, входят жесткокрылые, перепончатокрылые,
ручейники, чешуекрылые, двукрылые и некоторые другие отряды.
Определительная таблица классов
по личиночным и нимфальным стадиям
1(2). Ног нет. Тело червеобразное, сегментированное .............
...................... Личинки насекомых, развивающихся
с полным превращением,— Insecta-Holometabola (с. 65).
2(1). Ноги имеются. Тело разнообразной формы, обычно сегменти-
рованное, но бывает и несегментированным.
3(10). Ног 3 пары На брюшных сегментах настоящих конечностей нет,
но могут присутствовать укороченные нерасчлененные ложнонож-
ки, а на конце брюшка иногда имеются членистые придатки —
церки (рис. 35, л\, л2).
4(9). Тело сегментированное, подразделенное на отделы — голову,
грудь и брюшко, или голову и туловище.
5(6), Первый сегмент, следующий за головой, лишен конечностей
(рис. 32, Л1). Длина тела не более 5 мм..........................
Анаморфные молодые личинки многоножек—Myriapoda (с. 55).
6(5). Конечности находятся на трех первых сегментах, следующих за
головой.
7(8). Фасеточные глаза имеются, усики длинные, обычно длиннее го-
ловы (рис. 34, ль л2). Брюшных ложноножек нет....................
Личинки насекомых, развивающихся с неполным превращением,—
Insecta-Hemimetabola (с. 64).
51
8(7). Фасеточные глаза отсутствуют, усики короткие, короче головы
или едва заметные (рис. 35, Ль лг). Часто имеются брюшные
ложноножки . . Личинки насекомых, развивающихся с полным
превращением,— Insecta-Holometabola (с. 65).
9(4). Тело несегментированное, не подразделенное на отделы (рис. 33,
л).................... Личинки клещей— Arachnida-Acarina (с. 221).
10(3). Ног больше 3 пар.
11(14). Усики имеются. Тело сегментированное. Ног больше 4 пар.
12(13). Тело уплощенное и широкое, подразделяется на голову, грудь
и брюшко. На груди 7 пар ног .......................................
Нимфы наземных ракообразных — Crustacea (с. 52).
13(12). Тело обычно удлиненное, редко укороченное и широкое, под-
разделяется на голову и туловище. Почти все туловищные сег-
менты (за исключением первого и последнего) снабжены ногами
(рис. 31, «1, «г; 32, лг) . . . Личинки и нимфы многоножек —
Myriapoda (с. 55).
14(11). Усики отсутствуют. Тело либо целиком несегментированное,
либо только брюшко имеет сегменты. Ног 4 пары.
15(16). Тело слитное, не подразделенное на отделы (рис. 33, «). . .
Нимфы клещей — Arachnida-Acarina (с. 221).
16(15). Тело подразделено на слитную головогрудь и слитное или сег-
ментированное брюшко . . . . Нимфы других паукообразных —
Arachnida (с. 201).
МОКРИЦЫ—НАЗЕМНЫЕ ПРЕДСТАВИТЕЛИ
КЛАССА РАКООБРАЗНЫХ — CRUSTACEA *
Общая характеристика
Ракообразные составляют обширный класс жабродышащих члени-
стоногих, распространенных главным образом в морях и континенталь-
ных водоемах. В трех отрядах ракообразных наблюдается выход на
сушу, но достаточно широко распространенными наземными животны-
ми среди них являются только мокрицы — сухопутные и амфибиотиче-
скне представители отряда равноногих (Isopoda).
Мокрицы имеют овальное, уплощенное тело, состоящее из головы,
груди и брюшка (рис. 36, 37). Слитная голова несет усики (антен-
ны), фасеточные глаза и ротовые органы, представленные верхними
и нижними челюстями. Главную роль в размельчении пищи играют
верхние челюсти (мандибулы). Многочлениковый усик подразделяется
на ножку и жгутик. Грудь у мокриц состоит из 7 обособленных сег-
ментов, снабженных бегательными ножками. У самок пластинчатые
выросты грудных ножек образуют крышечки выводковой камеры, рас-
положенной на нижней стороне четырех сегментов груди (рис. 37). Во
время размножения самки вынашивают в этой камере яйца и выходя-
щее из них молодое потомство. Брюшко состоит из 6 сегментов, а их
укороченные пластинчатые придатки выполняют дыхательную функ-
цию. Последний брюшной сегмент снабжен раздвоенными членистыми
придатками, которые называют уроподами.
Мокрицы имеют очень широкое распространение и встречаются
в различных ландшафтных зонах. Многие виды амфибиотических и наи-
более гигрофильных мокриц заселяют берега морей и внутренних во-
* В. П. Тыщенко.
52
доемов. В лесной зоне они живут в верхних горизонтах почвы, в под-
стилке, во мху, под камнями, под корой деревьев и пней, в гниющей
древесине. Известны пещерные мокрицы, а также синантропы, посе-
ляющиеся в сырых помещениях, сараях, подвалах и погребах.
Выбор местообитаний мокрицами определяется главным образом
их требованиями к условиям влажности, содержанию кальция, нали-
чию и доступности пищи. Все наземные ракообразные не могут в тече-
ние длительного времени жить в сухой атмосфере без доступа влаги,
так как они быстро теряют воду в результате испарения через прони-
цаемые покровы. Поэтому мокрицы в основном приурочены к влаж-
ным биотопам. В сухих биотопах они живут в таких местах, где во-
насыщенная влагой атмосфера (на-
Рис. 36—37. Строение тела мокриц (по
Катиеру и Шмёльцеру).
36 — самка мокрицы Porcellio scaber Latr.,
37 — то же, вид снизу, i—7 — грудные сегменты;
I—VI — брюшные сегменты. в.у — внутренняя
ветвь уроподы; г.н — грудные ноги; д.б.н,— ды-
хательные брюшные ножки; ж — жгутик усика;
к.в.к — крышечки выводковой камеры; н —
ножка усика; н.у— наружная ветвь уроподы;
у — усик; ф.г— фасеточный глаз.
круг тела животного создается
пример, в |норках, вырывае-
мых пустынными видами мо-
криц). У видов, населяющих
сухие местообитания, кутику-
ла утолщается и пропиты-
вается кальцием, что способ-
ствует повышению их устой-
чивости к дефициту влаги.
Такие вид(ы предпочитают
поселяться на субстратах,
обогащенных кальцием. На-
пример, Armadillidium offici-
nale Boer живет на сухих из-
вестняках южного берега
Крыма и имеет очень толстую,
сильно кальцинированную ку-
тикулу. Содержание кальция
у некоторых мокриц может
достигать 30—40 % от их су-
хого веса. Наряду с многонож-
ками-диплоподами (см. с. 57)
мокрицы принимают активное
участие в накоплении и миг-
рации этого элемента.
Основную пищу мокриц
составляют разлагающиеся
растительные остатки и силь-
но разрушенная древесина.
Многие мокрицы-сапрофаги охотно поедают кусочки моркови, картофе-
ля, яблок, а иногда могут переходить и к питанию живыми травянис-
тыми растениями. В парниках и теплицах они даже повреждают
выращиваемые растения. По-видимому, • у влаголюбивых мокриц пита-
ние сочными частями растений является одним из способов удовлетво-
рения потребностей в воде. Наряду с растительными тканями мокрицы
могут захватывать и частицы минерального грунта, восполняя тем са-
мым недостаток кальциевых соединений. ...
Мокрицы начинают размножаться весной и до наступления зим-
них холодов дают от 1—2 до 5—6 пометов. В каждом помете насчи-
тывается несколько десятков молодых рачков. Яйца вынашиваются
самками в выводковых камерах. Первые дни после завершения эмб-
рионального развития молодь тоже находится в выводковой камере,
а затем ее покидает. Начальный период самостоятельного существо-
вания молодых мокриц является наиболее критическим периодом в их
жизни. Молодые мокрицы крайне чувствительны к недостатку влаги,
и многие из них погибают в ранних возрастах.
53
В европейской части СССР известно около 50 видов мокриц, от-
носящихся к 5 семействам. Эти семейства образуют следующий эко-
логический |ряд, отражающий градуальное усиление адаптаций к на-
земному существованию: Ligiidae->Trichoniscidae->Oniscidae->Porcelli-
onidae->Armadillidiidae.
Один из самых обычных видов мокриц европейской фауны — Onis-
cus asellus L. из семейства Oniscidae имеет гладкие блестящие покро-
вы. У другого, тоже весьма обычного вида — Porcellio scaber Latr. из
семейства Porcellionidae грудные сегменты и голова со спинной сто-
роны покрыты мелкими бугорками, создающими матовую поверхность
покровов (рис. 36).
Определительная таблица семейств
1(2). Жгутик усиков с многочисленными члениками (рис. 38) . . .
.................................................. Ligiidae.
Рис. 38—41. Мокрицы.
38 — Llgiditim Brandt, 39 — Onisctis L , 40 — Trichoniscus Brandt, 41 — ArtnadiUidiutn Brandt.
2(1). Жгутик усиков с немногочисленными (не более 4) члениками
(рис. 36, 39—41).
3(8). На голове перед глазами имеются 2 боковых выступа. Уроподы
длинные, сверху хорошо заметные (рис. 36, 39, 40).
4(7). Жгутик усиков с 2—3 явственно обособленными члениками. Внут-
ренние ветри уропод заметно короче наружных (рис. 36, 39),.
Длина тела у взрослых особей от 6 до 18 мм (как правило, боль-
ше 10 мм).
5(6). Жгутик усиков с 3 обособленными члениками (рис. 39) . . .
Oniscidae.
6(5). Жгутик усиков с 2 обособленными члениками (рис. 36) . . .
.................................................Porcellionidae.
7(4). Жгутик усиков без явственно обособленных члеников. Внутрен-
ние ветви уропод не короче или едва короче наружных (рис. 40).
Длина тела у взрослых особей не превышает 7 мм . . . . . .
..................................................Trichonoscidae.
8(3). На голове перед глазами нет боковых выступов. Уроподы корот-
кие, сверху едва заметные или совсем незаметные (рис. 41).
Мокрицы способны свертываться в шар .... Armadillidiidae.
54
КЛАСС МНОГОНОЖКИ— MyRIAPODA*
Общая характеристика
К многоножкам относятся наземные членистоногие с трахейным
дыханием, имеющие слитную голову и вытянутое сегментированное ту-
ловище. Ротовые органы грызущего типа. Глаза устроены по типу
простых глазков и сгруппированы в 2 небольших скопления по бокам
головы. В редких случаях эти скопления образуют ложнофасеточные
глаза. Туловищные сегменты хорошо обособлены и снабжены конечно-
стями. Присутствие ног не только на передних (грудных), но и на зад-
них (брюшных) сегментах характеризует всех многоножек. Обычные
места обитания многоножек — поверхность почвы и подстилка, а их
распространение охватывает все климатические пояса европейской ча-
сти СССР.
В составе класса многоножек выделяют 4 подкласса, которые иног-
да возводят даже в ранг классов: симфилы (Symphyla), пауроподы
(Pauropoda), двупарноногие, или диплоподы (Diplopoda), и губоногие,
или хилоподы (Chilopoda). Симфилы и пауроподы составляют немно-
гочисленные группы крошечных и бледно окрашенных членистоногих.
Диплоподы и хилоподы отличаются более крупными размерами, заме-
чательным разнообразием жизненных форм и широким распростране-
нием в природе. Поэтому при проведении энтомологической практики
знакомство с классом многоножек целесообразно ограничить этими
двумя подклассами.
Определительная таблица подклассов
1(2). Очень мелкие (1—2 мм) многоножки с 9 парами ног (рис. 42).
Усики двуветвистые. Живут скрытно в почве и подстилке . . .
..........................................Пауроподы— Pauropoda.
2(1). Длина тела, как правило, более 2 мм. Число пар ног на имаги-
нальных стадиях более И. Усики обычные, одноветвистые.
3(4). Туловищные сегменты, начиная с
пятого, несут по 2 пары ног. Уси-
ки из 8—9 члеников...................
. . . . Диплоподы — Diplopoda,
4(3). Туловищные сегменты несут по
1 паре ног. Усики более чем из
10 члеников.
5(6). Первая пара ног превращена в
сильно утолщенные, изогнутые
крючкообразно, заостренные на
концах ногочелюсти. На имаги-
нальных стадиях число пар йог от
15 до 109. Хилоподы — Chilopoda.
6(5). Первая пара ног не отличается от
прочих. На имагинальных ста-
диях 12 пар ног (рис. 43). Мел-
кие (не более 8 мм), нежные, ли-
шенные глаз многоножки с длин-
ными многочлениковыми усиками.
Живут во влажной почве, под-
Рис. 42—43. Многоножки — пау-
роподы и симфилы.
42 — пауропода, 43 — симфила.
стилке, гнилых пнях, под камнями
и корой деревьев. ......
...............Симфилы — Symphyla.
* В. П. Тыщенко.
55
ПОДКЛАСС ДИПЛОПОДЫ — DIPLOPODA
Общая характеристика
Многоножки, относящиеся к этому подклассу, имеют темные
и плотные покровы, содержащие известковые включения. Голова дип-
лопод несет сравнительно короткие усики, глаза и ротовой аппарат,
приспособленный для поглощения твердой или полужидкой пищи. Наи-
более характерной особенностью диплопод является попарное слияние
туловищных сегментов. В результате такого слияния возникают двой-
Рис. 44. Строение тела диплопод (по Шубарту).
в.н — видоизмененная нога первого сегмента; г —голова; г.п — глазное поле; н — ноги; пр.с —
первый туловищный сегмент; пс.с — последний туловищный сегмент; т.с — туловищные сегменты;
у — усик.
ные сегменты, имеющие по 2 пары ног (рис. 44). Только первые 4 сег-
мента не сливаются друг с другом и, отличаясь от других сегментов,
образуют как бы грудной отдел тела. Туловищные сегменты бывают
округлыми или уплощенными, и основные семейства диплопод четко
различаются по очертаниям сегментов на поперечных срезах (рис. 45—
47). Для некоторых семейств диагностическими признаками служат
микроскульптурные особенности покровов, например расположение
продольных бороздок на туловищных сегментах (рис. 48, 49).
Многие диплоподы имеют пахучие железы, расположенные попар-
но в туловищных сегментах (рис. 48, 49). Выделения этих желез
содержат 2 основных компонента: 1) ароматические вещества, обладаю-
щие резким неприятным запахом и отпугивающие хищников; 2) веще-
ства контактного токсического действия, служащие для защиты от на-
павших врагов. Отпугивающими компонентами яда диплопод могут
быть камфара, хиноны и другие пахучие соединения, а тохсические
компоненты представлены серотонином, гиалуронидазой и цианидами.
Впрочем, яд европейских диплопод не опасен для человека.
Рис. 45—49. Туловищные сегменты диплопод.
Форма сегментов на поперечных срезах: 45— кивсяки (Julidae), 46— многосвязы (Polydesmidae),
41 — клубовидки (Glomeridae), 48—49 — расположение продольных бороздок на сегментах: 48 —
Julidae, 49 — Blaniulidae.
п.ж — пахучие железы; п.п—поры пахучих желез.
56
Постэмбриональное развитие диплопод протекает по типу анамор-
фоза. Из яйца выходит личинка, имеющая 7 туловищных сегментов
и 3 пары ног. После каждой линьки число сегментов и ног увеличи-
вается, пока не достигается их полное число, свойственное половозре-
лым особям. Во взрослом состоянии диплоподы тоже линяют.
Диплоподы обычно питаются перегнивающими растительными ос-
татками, реже живыми растениями. Подобно другим животным-сапро-
фагам они играют большую роль в разложении органического мате-
риала подстилки и вовлекают в почву вещества, необходимые для
образования гумуса. У некоторых видов важным компонентом пищи
является почвенный перегной. Обычно диплоподы заселяют поверхно-
стные слои почвы и лесную подстилку, но нередко встречаются также
Рис. 50—54. Многоножки-диплоподы.
50 — многосвяз (Polydesmidae), 51— Strongylosomidae, 52 — клубовидка (Glomeridae), 53 —- Glomeris
соппеха С. L. Koch, схема расположения светлых пятен на спинной поверхности туловищных
сегментов, 54— Glomeris sp. в свернувшемся состоянии.
под отставшей корой, в гнилой древесине и гнездах грызунов. Некото-
рые виды обнаруживают явную приуроченность к окультуренным ме-
стообитаниям— оранжереям, плодовым и ботаническим садам.
Подкласс диплопод охватывает 4 отряда и 10 семейств, встречаю-
щихся в европейской части СССР. Почти 90 % европейских видов это-
го подкласса относятся к 5 самым крупным семействам, перечислен-
ным ниже и включенным в определительную таблицу.
К и всяки (Julidae) и представители близкого к ним семей-
ства Blaniulidae имеют цилиндрическое тело, состоящее из боль-
шого числа сегментов (рис. 44, 45). У кивсяков отношение длины тела
к его толщине составляет 10 : 1, а у Blaniulidae — 20: 1. Глазки распо-
лагаются по бокам головы в виде скоплений разной формы, но могут
отсутствовать у некоторых Blaniulidae. В обоих семействах прикосно-
вение к телу многоножки вызывает ее рефлекторное свертывание
в спираль. При яйцекладке самки кивсяков сооружают сложные гнез-
да из смеси земли и экскрементов. Каждое гнездо содержит 60—70 яиц.
57
Оно имеет куполовидную форму и снабжено надстройкой с отверстием,
предназначенным для проветривания развивающихся яиц. Характер-
ным представителем этого семейства является весьма обычный в ле-
сах и на полях песчаный кивсяк (Schizophyllum sabulosum L.). Его
тело длиной от 15 до 47 мм имеет черную или черно-коричневую окрас-
ку с двумя светлыми продольными полосками на спине. В ряде райо-
нов европейской части СССР отмечено повреждение этими кивсяками
посевов кукурузы.
Многосвязы (Polydesmidae) имеют уплощенное вытяну-
тое в длину тело с боковыми крыловидными выступами (рис. 46, 50).
При яйцекладке самки многосвязов сооружают такие же гнезда, как
и самки кивсяков. Представители семейства Strongylosomi-
d а е похожи на многосвязов, но туловищные сегменты у них снаб-
жены боковыми вздутиями (рис. 51). В отличие от кивсяков все много-
связы и Strongylosomidae лишены глаз.
Клубовидки (Glomeridae) имеют необычное для много-
ножек короткое и широкое тело (рис. 47, 52, 53). По внешнему облику
они напоминают мокриц и, так же . как и некоторые мокрицы, могут
свертываться в шар (рис. 54). У европейских клубовидок тело обычно
коричневое или черное со светлыми пятнами. У наиболее часто встре-
чающегося вида Glomeris соппеха С. L. Koch каждый туловищный сег-
мент, за исключением первого и двух последних, украшен 4 светлыми
пятнами (рис. 53). Клубовидки питаются главным образом опавшими
листьями дуба, березы, осины и гниющей древесиной. Самки не делают
гнезд, а откладывают яйца поодиночке внутри округлых капсул, обра-
зованных частицами земли и сухими экскрементами. По-видимому, ма-
териал этих капсул служит первой пищей для вылупляющихся личи-
нок.
Определительная таблица отрядов и семейств
1(2). Тело короткое и широкое (рис. 52, 53), многоножки могут свер-
тываться в шар (рис. 54). Второй сегмент значительно больше
следующих за ним сегментов. (Отряд Oniscomorpha) . . . .
................................ Клубовидки — Glomeridae.
2(1). Тело узкое, удлиненное (рис. 44, 50, 51), многоножки в шар не
свертываются. Второй сегмент не больше следующих за ним сег-
ментов.
3(6). Туловищные сегменты округлые, без боковых выступов и взду-
тий (рис. 44), а общее число сегментов, снабженных ногами,
больше 20. (Отряд Juliformia).
4(5). Продольные бороздки имеются и на боковой и на спинной по-
верхности сегментов (рис. 48) ’...................Кивсяки — Julidae.
5(4). Продольные бороздки имеются только на боковой поверхности
сегментов (рис. 49)................................. Blaniulidae.
6(3). Туловищные сегменты снабжены вздутиями или крыловидными
выступами (рис. 50, 51), а общее число сегментов, снабженных
ногами, не больше 20. (Отряд Polydesmoidea).
7(8). Туловищные сегменты снабжены крыловидными выступами;
'спинная поверхность сегментов покрыта мелкими бугорками
(рис. 50) ......................Многосвязы—Polydesmidae.
8(7). Туловищные сегменты без крыловидных выступов, но с боковы-
ми вздутиями; спинная поверхность сегментов гладкая (рис. 51).
. . .......................... Strongylosomidae.
58
ПОДКЛАСС ХИЛОПОДЫ — CHILOPODA
Общая характеристика
однои парой конечностей (рис.
превращены в сильные хвататель-
рых коготков (рис. 55, н; 56). Каж-
Многоножки-хилоподы характеризуются равномерно расчленен-
ным туловищем и хорошо обособленной головой с длинными много-
члениковыми усиками (рис. 55). Ротовые органы состоят из верхней
губы, жвал и двух пар нижних челюстей. У всех хилопод туловищные
сегменты одиночные, снабженные
б. н). Конечности первого сегмент
ные ногочелюсти, имеющие вид ос
дая ногочелюсть имеет ядови-
тую железу, которая открывает-
ся на кончике коготка (рис. 57).
Хилоподы пользуются ногочелюс-
тями и при< нападении на чле-
нистоногих, которыми они пита-
ются, и при защите от врагов.
Крупные виды хилОпод способ-
ны прокусить кожу человека, а
их укусы могут вызвать местные
симптомы раздражения кожи и
небольшие воспаления. Послед-
ние туловищные сегменты, на-
зываемые генитальными сегмен-
тами, не имеют настоящих ног,
а сохраняют их рудиментарные
остатки, участвующие в ОплодО-
тв9ренйи. Последняя пара ног,
расположенная перед гениталь-
ными сегментами, длиннее ос-
тальных конечностей (рис. 55,
о.«). Эти длинные ноги выпол-
няют в основном не двигатель-
ную, а осязательную функцию.
Они густо усажены чувствитель-
ными щетинками и периодиче-
ски очищаются многоножкой при
помощи нижних челюстей. Ок-
раска тела у хилопод обычно
тусклая—желтая или коричневая.
В двух отрядах хилопод (Lith'obiomorpha и Scutigeromorpha) пост-
эмбриональное развитие протекает с анаморфозом, а личинки имеют
уменьшенное число ног и сегментов по сравнению с взрослыми ста-
диями. В остальных отрядах развитие протекает по типу эпиморфоза,
и личинки выходят из яиц с полным числом ног и сегментов.
Все хилоподы являются хищниками, нападающими на1 различных
членистоногих и червей, которых они умерщвляют ядом своих ного-
челюстей. Обитают под камнями, в щелях скал и почвы, под ствола-
ми деревьев, под корой пней, в гнилой древесине и подстилке, а так-
же в домах на стенах. В зависимости от типа местообитаний среди
хилопод можно выделить жизненные формы, охватывающие 4 основ-
ных семейства европейской фауны.
Геофилиды (Geophilidae) являются типичными обитателя-
ми почвы, Сильно вытянутое тело, составленное из большого числа
сегментов, служит специальной адаптацией геофилид к передвижению
в ходах и скважинах почвы (рис. 58). Подобно дождевым червям они
перемещаются в почве, раздвигая частицы земли головой и тулови-
Рис. 55—57. Строение тела и ногочелюсти
хилопод.
55— строение тела, 56—ногочелюсти сколопенд-
ры, 57 — коготок ногочелюсти с ядовитой желе-
зой.
б.н — бегательные ноги; г — голова; к — коготок
ногочелюсти; н — ногочелюсть; о.и. — осязатель-
ная йога; у — усики; я.ж.— ядовитая железа.
59
щем. Ноги в основном используются для отталкивания и фиксации
тела внутри почвенной скважины. Отсутствие глаз также характеризу-
ет этих многоножек, как представителей экологической группы геоби-
онтов. Почвенные геофилиды питаются главным образом клещами, пер-
вичнобеск|рылыми насекомыми и личинками насекомых.
Выбор местообитаний геофилидами предопределяется их высокой
чувствительностью к условиям- влажности. Снижение влажности воз-
духа вызывает уход геофилид в глубокие горизонты почвы. В степной
зоне они могут опускаться на глубину до 100—200 см. В сухих степях,
полупустынях и пустынях геофилиды часто поселяются в норах гры-
зунов. Наиболее разнообразная фауна и высокая численность геофи-
лид обнаруживаются в лесных почвах.
Рис. 58—62. Многоножки-хилоподы.
58 — геофилида (Geophilidae), 59— сколопендра (Scolopendridae), 60— костянка (Lithobiidae),
61 — неполовозрелая стадия костянки, имеющая неполное число ног и сегментов, 62 — мухоловка
(Scutigeridae).
Костянки (Lithobiidae) и сколопендры (Scolo-
pendridae) обитают в самых верхних горизонтах почвы, нередко
встречаются в подстилке, под камнями, в расщелинах скал, под корой
деревьев и в других укрытиях. Ночью они выходят на поверхность поч-
вы. В отличие от геофилид костянки и сколопендры неспособны пере-
двигаться в почве путем активного прокладывания ходов. В соответ-
ствии с новыми условиями обитания на поверхности земли у них раз-
виваются простые глазки, удлиняются конечности и уменьшается чис-
ло сегментов (рис. 59—61). Удлинение конечностей и уменьшение
числа сегментов отражают приспособление многоножек к передвиже-
нию по напочвенному субстрату. В период размножения самки сколо-
пендр проявляют заботу о потомстве: они свертываются спиралью во-
круг отложенных яиц и смачивают их секретом своих желез. Самки
60
костянок некоторое время носят яйца между придатками генитальных
сегментов, а затем откладывают их поодиночке, прикрывая сверху
мелкими частицами почвы.
В европейской части СССР особенно широко распространена обык-
новенная костянка (Lithobius forficatus L.). Ее тело окрашено в корич-
невый цвет и достигает в длину 30—35 мм. Эта многоножка живет- под
корой деревьев, среди камней и в лесной подстилке, охотно поселяет-
ся в развалинах и мусорных кучах. Сколопендры встречаются на юге
европейской части СССР. К ним относится кольчатая сколопендра
(Scolopendra cingulata) — самая крупная многоножка нашей фауны,
достигающая в длину 9—12 см. Тело этой многоножки бурого цвета
с зеленоватым оттенком. Укус кольчатой сколопендры вызывает у че-
ловека скоро проходящую боль, красноту и припухлость.
Мухоловкй (Scutigeridae) — наиболее приспособившиеся
к открытому обитанию хилоподы (рис. 62). Они бегают по поверхно-
сти почвы, по деревьям и стенам зданий не только ночью, но даже
и днем в сухих местностях. Такие морфологические признаки, как чрез-
вычайно длинные ноги и ложкофасеточные глаза мухоловок, несомнен-
но возникли под влиянием открытого и дневного образа жизни. Во
время охоты мухоловки захватывают летающих насекомых своими би-
чевидными лапками, обертывая их вокруг жертвы наподобие лассо.
Определительная таблица отрядов и семейств
1(2). Туловище длинное и узкое, снабженное более чем 30 парами ног
(рис. 58). Простые глазки и ложнофасеточные глаза отсутствуют
(рис. 63). (Отряд Geophilomorpha)....................................
Геофилиды— Geophilidae.
2(1). Ног не более 23 пар (рис.
59—62). Имеются простые
глазки (рис. 64) или ложно-
фасеточные глаза (рис. 65).
3(6). Последняя пара ног короче
тела (рис. 59—61). Имеются
простые глазки (рис. 64).
4(5). Ног от 21 до 23 пар (рис. 59).
(Отряд Scolopendromorpha).
Сколопендры — Scolopendridae.
5(4). Ног не более 15 пар (рис.60,
61). (Отряд Lithobiomorpha).
Костянки — Lithobiidae. иааш,
6(3). Последняя пара ног длиннее тела (рис. 62). Остальные ноги тоже
очень длинные, легко обламываются. Имеются ложнофасеточные
глаза (рис. 65). (Отряд Scutigeromorpha) ............................
............................... Мухоловки — Scutigeridae.
Рис. 63—65. Форма головы и располо-
жение глаз в разных семействах хи-
лойод.
63 — геофилида (Geophilidae), 64 — костянка
(Lithobiidae), 65 — мухоловка (Scutigeridae).
л. г — ложнофасеточные глаза; п.г — простые
КЛАСС НАСЕКОМЫЕ—INSECTA
Общая характеристика *
Насекомые составляют самый крупный класс животного мира, на-
считывающий более 800 000 видов. Они встречаются практически во
всех наземных и пресноводных биотопах, занимая в них все возмож-
ные экологические ниши. Освоение разнообразных местообитаний со-
* В. П. Тыщенко.
61
провождалось широкой адаптивной радиацией насекомых при сохра-
нении общего плана организации, характерного для данного класса.
Тело взрослого насекомого- (имаго) подразделяется на
голову, грудь и брюшко (рис. 66). На голове сосредоточены главней-
шие органы чувств и органы приема пищи. Основную часть головы
занимает слитная головная капсула; ее передняя поверхность назы-
вается лбом, верхняя — теменем, а задняя — затылком (рис. 67). По
бокам головной капсулы расположены крупные фасеточные глаза,
а между ними на темени могут находиться мелкие и одиночные дор-
зальные глазки. В большинстве случаев крылатые насекомые имеют
3 дорзальных глазка. К передней поверхности головы причленяются
усики (антенны), подразделенные на отдельные членики. Нижняя
часть головы занята на-
,. Рис. 66. Схема строения насекомого (по Мамаеву,
Медведеву и Правдину).
б—бедро; в — вертлуг; г — голень; д.г — дорсальный гла-
зок; з— затылок; з.н—задняя (прыгательная) нога; к —
крыло; л — лоб; лп — лапка; н — наличник; «к •—надкрылье;
п.н — передняя (бегательная) нога; р.а — ротовой аппарат;
т-—темя; тз — тазик; у — усик; ф.г — фасеточный глаз;ц —
церк; я — яйцеклад.
тивная радиация наблюдается в строении
мых. Исходным для этой группы считается
личником и ротовым ап-
паратом.
В разных отрядах и
семействах насекомых
модификации придатков
головы сильно отража-
ются на строении усиков
и ротовых аппаратов.
Усики могут варьиро-
вать по их относительной
длине, а количество чле-
ников изменяется от 3 до
нескольких десятков. В
зависимости от их фор-
мы усики могут быть
нитевидными, четковид-
ными, булавовидными,
гребенчатыми, перисты-
ми, коленчатыми или
пластинчатыми (рис. 68—
74). Значительная' адап-
ротовых аппаратов насеко-
грызущий ротовой аппарат,
в состав которого входят следующие ротовые придатки: 1) верхняя
губа (лабрум)—непарная цельная пластинка, соединяющаяся с на-
личником; 2) одна пара нерасчлененных и зазубренных верхних че-
люстей (мандибул), называемых также жвалами; 3) одна пара рас-
члененных нижних челюстей (максилл), несущих максиллярные щу-
пики; 4) слившаяся пара расчлененных придатков, составляющих
нижнюю губу (лабиум) с парой лабиальных щупиков (рис. 75). Гры-
зущий ротовой аппарат, снабженный сильными и толстыми жвалами,
приспособлен для питания твердыми продуктами. Некоторые насеко-
мые, обладающие грызущим ротовым аппаратом, могут перегрызать
корни деревьев. Приспособление насекомых к питанию жидкостями
привело к возникновению иных типов ротовых аппаратов, например ко-
лющего хоботка клопов и тлей, сосущего хоботка бабочек или слож-
ного грызуще-лижущего аппарата жалящих перепончатокрылых.
Грудь является ведущим локомоторным отделом насекомых, так
как па ней находятся органы передвижения — ноги и крылья. Грудь
состоит из трех сегментов, получивших название передне-, средне-
и заднегруди. Каждый грудной сегмент несет одну пару ног. В состав
ноги входят 2 коротких начальных членика—тазик и вертлуг, 2 более
длинных членика — бедро и голень, а за голенью следует состоящая из
нескольких члеников лапка. Обычно открыто живущие наземные на-
62
секомые имеют ноги бегательного типа (рис. 76), однако в некоторых
отрядах и семействах отдельные пары грудных конечностей могут пре-
образовываться в прыгательные, хватательные, плавательные или ко-
пательные ноги (рис. 77—80).
У большинства взрослых насекомых к спинной, поверхности сред-
не- и заднегруди причленяются 2 пары крыльев. Как правило, на по-
верхности крыла четко выделяются продольные и поперечные жилки,
образующие прочный каркас для прозрачной крыловой перепонки.
Однако у бабочек часто бывает трудно увидеть расположение жилок
Рис. 67—80. Придатки головы и ноги насекомых.
67—расположенно придатков на голове, 68—74 — усики (68 — нитевидный, 69— четковндный,
70 — булавовидный, 71 — гребенчатый, 72 — перистый, 73 — коленчатый, 74 — пластинчатый), 75 —?
придатки грызущего ротового аппарата, 76—80—ноги (76—бегательная, 77 — прыгательная, 78 —
хватательная, 79 — плавательная, 80 — копательная).
в.г —верхняя губа; в.ч —верхние челюсти; л.щ — лабиальные щупики; м.Щ — максиллярные щу-
пики; «.г —нижняя губа; н.ч — нижине челюсти; у — уснк.
из-за сплошного слоя окрашенных чешуек, покрывающих всю поверх-
ность передних и задних крыльев. Передние крылья могут быть зна-
чительно более твердыми, чем задние, и тогда они именуются над-
крыльями. Толстые qporoBeBiHHe надкрылья жуков не имеют явствен-
ных жилок; они прикрывают сверху брюшко, защищая его от механи-
ческих повреждений, и не участвуют в полете. В отряде двукрылых,
к которому относятся комары и мухи, хорошо развита одна передняя
пара крыльев, а задние крылья редуцированы до едва заметных жуж-
жалец. Есть насекомые и полностью лишенные крыльев. К первично-
бескрылым относятся некоторые примитивные представители насеко-
мых, составляющие особый подкласс (см. ниже), а вторичнобескры-
лыми могут быть названы такие насекомые, у которых крылья были
утрачены в связи с переходом к паразитическому образу жизни (вши
и блохи) или в силу каких-то иных причин.
Брюшко соответствует заднему отделу тела, который у большин-
63
ства взрослых насекомых состоит из 6—10 сегментов. Последний брюш-
ной сегмент несет анальное отверстие. Брюшко лишено истинных ко-
нечностей, а брюшные придатки, если они имеются, сосредоточены на
его конце. К типичным придаткам брюшка относятся церки, хвостовые
нити, яйцеклад самок, а также мужские и женские половые придатки.
Насекомые относятся к раздельнополым животным, и у многих из
них во взрослом состоянии проявляется явственный половой димор-
физм. Самцы и самки одного вида- могут различаться по окраске тела
или по относительным размерам и строению фасеточных глаз, усиков
и крыльев. У какого-либо пола могут быть развиты вторичные поло-
вые признаки, не свойственные представителям противоположного
пола. Например, у самцов жука-оленя (Lucanus cervus L.) верхние че-
люсти сильно увеличены, снабже-
ны зубцами и далеко выступают
впереди тела жука (рис. 81), а у
самок они имеют обычные размеры
(рис. 82). У некоторых чешуекры-
лых только самцы снабжены
крыльями, а самки бескрылые или
обладают сильно редуцированными
крыльями. В наибольшей степени
половой диморфизм проявляется у
равнокрылых из подотряда кокцид,
у которых маленькие самцы явля-
ются нормально развитыми кры-
латыми насекомыми, а самки во
много раз крупнее самцов, лише-
ны крыльев, имеют недоразвитые
Рис. 81—82. Половой диморфизм у жу- ноги и. почти утрачивают подраз-
ка-оленя (Lucanus cervus L.). деление тела на отделы. У терми-
81- самец, 82 -самка. тов общественных ПереПОНЧЗТО-
крылых половой диморфизм допол-
няется кастовым полиморфизмом. В таких случаях особи, относящиеся
к одному женскому полу, подразделяются на касты, существенно раз-
личающиеся по морфологическйм признакам и поведению.
Личинки насекомых, развивающихся с неполным
превращением (Hemimetabola), по общему плану организа-
ции сходны с имаго. Их тело всегда четко подразделено на голову, грудь
и брюшко, голова несет фасеточные глаза, грудь снабжена тремя па-
рами дефинитивных ног. Крылья могут быть представлены неболь-
шими парными зачатками на последних стадиях, а половые придатки
появляются только после имагинальной линьки. Многие характерные
признаки, свойственные имаго, присутствуют уже у таких личинок. На-
пример, личинки настоящих саранчовых (Acrididae) имеют прыгатель-
ные задние ноги (рис. 83), личинки богомолов характеризуются хва-
тательными, а личинки медведок (Gryllotalpa Latr.)—копательными
передними ногами (рис. 85, 86). Личинки уховерток на конце брюшка
снабжены характерными клещевидными церками (рис. 87), сохраняю-
щимися у имаго. Типичные личиночные (ценогенетические) признаки
при неполном превращении отчетливо проявляются только у личинок,
развивающихся в воде. К таким признакам относятся у личинок
стрекоз своеобразный выдвижной хватательный аппарат, называемый
маской и расположенный на нижней стороне головы (рис. 84), а у ли-
чинок равнокрылых стрекоз (Zygoptera), поденок и веснянок — на-
ружные трахейные жабры. Эти органы функционируют только на ли-
чиночной стадии и исчезают при превращении водной личинки в на-
земное взрослое насекомое.
64
Личинки насекомых, развивающихся с полным
превращением (Holometabola), весьма разнообразны по фор-
ме и сегментации тела. Их морфофункциональная организация цели-
ком определяется ценогенетическими признаками и всегда резко отли-
чается от организации имаго. Фасеточные глаза у таких личинок от-
сутствуют, а антенны бывают очень короткими (короче головы) или
вообще не выражены. Органы зрения представлены личиночными ла-
теральными глазками, но нередко глазки редуцированы вплоть до пол-
ного отсутствия. По наличию или отсутствию ног на грудных и брюш-
ных сегментах свободноживущих (непаразитических) личинок этого
типа можно разделить на 3 категории.
Рис. €3—87. Личинки насекомых, развивающихся с неполным превращением.
83 — саранча (Locusta LJ из отряда прямокрылых, 84—стрекоза, 85 — богомол, 86 — медведка
(Qryllotalpa Latr.) из отряда прямокрылых, 87 — уховертка.
з.л —зачатки крыльев; к.н — копательные ноги; м — маска; п.н — прыгательные ноги; х.н — хва-
тательные ноги; ц — церки.
Олигоподиальные личинки, свойственные ручейникам, сетчатокры-
лым и некоторым жесткокрылым, снабжены тремя парами вполне раз-
витых грудных конечностей при полном отсутствии брюшных ложно-
ножек (рис. 88). Тело личинок четко подразделяется на голову, грудь
и брюшко.
Полиподиальные личинки имеют 3 пары коротких грудных конеч-
ностей и несколько пар брюшных ложноножек (рис. 89). Тело черве-
образное, а сегменты груди и брюшка различаются слабо. К полипо-
диальным личинкам относятся гусеницы — личиночные стадии чешуе-
крылых и называемые ложногусеницами личинки растительноядных
перепончатокрылых.
Аподиальные личинки характерны для двукрылых, многих пере-
пончатокрылых и некоторых жесткокрылых. У них нет ни грудных ко-
нечностей, ни брюшных ложноножек (рис. 90—92). Обычно личинки
этой категории имеют червеобразное тело, в котором грудь не обособ-
лена от брюшка (рис. 90). Только у водных личинок комаров из се-
мейства Culicidae ширина грудного отдела заметно превышает шири-
ну брюшных сегментов (рис. 91). У многих личинок высших мух из
подотряда короткоусых двукрылых отсутствует головная капсула
(рис. 92), но остальные аподиальные личинки снабжены хорошо раз-
витой головой.
При полном превращении куколка соответствует промежуточ-
ной стадии между личинкой и имаго. Жесткокрылые, перепончатокры-
лые, сетчатокрылые и длинноусые двукрылые имеют свободную кукол-
ку, у которой ноги и другие придатки не прирастают к телу, а сочле-
5 208
65
няются с йиМ только своймй основаниями (рис. 93). У такой куколки
все части тела и его придатки имеют почти такое же строение, как
у взрослых насекомых. Свободные куколки сетчатокрылых и комаров
сохраняют даже ограниченную способность к перемещению. Чешуе-
крылым свойственна покрытая куколка, у которой придатки полностью
срастаются с телом и остаются заметными лишь их контуры (рис. 94).
У некоторых бабочек покрытая куколка находится в коконе, изготов-
ленном из выделений шелкоотделительных желез гусеницы. Высшие
представители короткоусых двукрылых окукливаются под покровом
затвердевшей личиночной шкурки, которая образует бочонковидный
Рис. 88—92. Типы личинок насекомых, развивающихся с полным превращением.
88 — олигоподиальная личинка жужелицы (Carabidae), 89 — полиподнал^ная личинка пилильщика
(Tenthredinidae), 90 — аподиальная личинка пчелы (Apidae), 91 — аподиальная водная личинка
комара (Culicidae), 92 — аподиальная личинка мухи (Muscidae).
ложный кокон, называемый пупарием (рис. 95). Куколки мух, имею-
щие вид пупариев, сохраняют следы сегментации, но придатки и от-
93 94 95
делы тела у них совсем не выражены.
Обычно насекомых подразделяют на 2 подкласса. К подклассу
первичнобескрылых (Apterygota) относят мелких нежных
насекомых, лишенных крыльев, но снабженных различными выроста-
ми или парными придатками на брюшных сегментах. Этот относитель-
но примитивный подкласс охватывает 4 небольших отряда, не имею-
щих, по-видимому, достаточно близкого
филогенетического родства. Подкласс
крылатых (Pt'er y.go.t а) -подраз-
деляется на 32 отряда и отличается
особенно широким видовым разнообра-
зием. К этому подклассу относят насе-
комых, испытывающих неполное или
полное превращение, снабженных крыль-
ями на имагинальной стадии или вто-
рично утративших крылья. В приведен-
ной ниже определительной таблице от-
рядов к первичнобескрылым насекомым
принадлежат ногохвостки (Collembola),
щетинохвостки (Thysanura), двухвост-
ки (Diplura), а к крылатым — все ос-
тальные отряды.
Рис. 93—95. Типы куколок (по
Веберу).
93 — открытая куколка долгоносика
(Curculionidac), 94 — покрытая кукол-
ка бабочки-ннмфалнды (t^Iymphalidae),
95 — пупарнй мухи (Calliphoridае).
66
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ОТРЯДОВ
ПО ИМАГИНАЛЬНОИСТАДИИ*
1(42). Крылья (иногда укороченные) имеются.
2(3). Брюшко заканчивается 2 или 3 членистыми хвостовыми нитя-
ми, длина которых превышает длину тела (рис. 96). Усики
обычно короче головы. Задние крылья меньше передних или
совсем отсутствуют. Ротовой аппарат редуцирован, не функцио-
нирует .......................Поденки — Ephemeroptera (с. 82).
3(2). . Брюшко без длинных членистых хвостовых нитей, иногда с ко-
роткими придатками, длина которых меньше длины тела.
4(5). Насекомые с одной передней парой крыльев, снабженных ясно
видимыми жилками (рис. 97—99). На задней груди пара була-
вовидных жужжалец (видоизмененные -задние крылья). Рото-
вые органы образуют хоботок, иногда недоразвиты ....
. .................................Двукрылые — Diptera (с. 189).
5(4). Насекомые с двумя парами крыльев, или же имеются лишь
крылья передней пары, превращенные в жесткие, лишенные
жилок надкрылья.
6(7). Крылья и тело покрыты легко стирающимися чешуйками,
крылья не ороговевшие. Форма тела и крыльев как на рис.
122—124. Ротовой аппарат часто в виде спирального закручен-
ного хоботка. Иногда хоботок редуцирован .......................
. Чешуекрылые — Lepidoptera (с. 176).
7(6). Крылья без чешуек (голые или покрыты волосками или муч-
нистым налетом) или передние крылья ороговевшие.
8(9). Передние и задние крылья с бахромкой длинных волосков на
' переднем и заднем крае, узкие и длинные, иногда недоразви-
тые (рис. 132). Очень мелкие (0,5—2 мм) насекомые с удлинен-
ным телом и короткими ногами ..............................- •
.........................Трипсы — Thysanoptera (с. 118).
9(8). По крайней мере передний край передних крыльев без бахром-
ки длинных волосков; если такая бахромка имеется, то тело
иной формы, не удлиненное.
10(13). Ротовой аппарат в виде членистого хоботка, без щупиков
(рис. 127).
11(12). Передние крылья однородные, перепончатые или кожистые
(рис. 130), в покое лежат кровлеобразно, т. е. прикрывают
брюшко не только сверху, но и с боков (рис. 131). Хоботок
причленяется отступя от переднего края головы и кажется
отходящим от ее заднего края.................................
.........................Равнокрылые — Homoptera (с. 103).
12(11). Передние крылья неоднородные: на вершине прозрачные, неж-
ные, в остальной части более плотные и часто прозрачные
(рис. 128, 129). Если передние крылья укороченные, они мо-
, гут состоять только из плотной части. В покое крылья лежат
плоско на брюшке. Хоботок причленен к передней части- го-
ловы (рис. 127) . . Полужесткокрылые — Hemiptera (с. НО).
13(10). Ротовой аппарат не в виде членистого хоботка: грызущий или
грызугце-лижущий с развитыми мандибулами (рис. 100, 101)
или рудиментарный грызущий с заметными нижнегубными
и нижнечелюстными щупиками. ,
14(17). Передние крылья представляют собой плотные ороговевшие
надкрылья, утратившие жилкование; в сложенном состоянии
они полностью или частично прикрывают брюшко, почти всег-
* В. П. Тыщенко, при участии Н. Ю. Клюге.
67
68
Рис. 96—145. Отряды насекомых.
96 — поденка (Ephemeroptera), 97, 98, 99 — различные представители двукрылых (Diptera), 100,
/9/— голова и расположение верхних челюстей (жвал) у насекомых, имеющих грызущий рото-
вой аппарат, 102 — уховертка (Dermaptera), 103, /94 — различные представители жесткокрылых
(Coleoptera), 105 — стрекоза (Odonata), 106, 107, 108, /99 — различные представители прямокрылых
(Orthoptera), //9-—богомол (Mantoptera), ///—таракан (Blattoptera), 112—скорпнонннЦа(Месор1ега).
113 — верблюдка (Raphidioptera), 114 — вислокрылка (Megaloptera), 115, 116 — сетчатокрылые (Neu-
roptera), 117 — термит (Isoptera), 118, 119 — сеноеды (Psocoptera), 120 — сидячебрюхое перепон-
чатокрылое (Hymenoptera), /2/— стебельчатобрюхое перепончатокрылое (Hymenoptera), 122, 123,
124 — различные представители чешуекрылых (Lepidoptera), 125, 126 — ручейники (Trichoptera),
127 — голова насекомого с членистым хоботком, 128, 129 — полужесткокрылые (Hemiptera), 130,
131 — равнокрылые (Homoptera), 132 — трипс (Thysanoptera), 133, 134 — веснянки (Plecoptera),
135, 136^- ногохвостки (Collembola). 137 — муравей (отряд Перепончатокрылые — Hymenoptera),
138 —• щДинохвостка (Tbysanura), 139, /49 — двухвостки (Diplura), /4/— термит (Isoptera), 142 —
бескрылая самка жука (Coleoptera), 143 — бескрылая самка бабочки (Lepidoptera), /44 — трипс
(Thysanoptera), 145 — сеноед (Psocoptera).
да не налегают друг на друга (за исключением жука-майки —
Meloe, см. рис. 323) и обычно сходятся по шву (рис. 102,
103, 104).
15(16). На конце тела имеются длинные клещевидные придатки
(рис. 102). Надкрылья укорочены и прикрывают только часть
брюшка...........................Уховертки — Dermaptera (с. 101).
69
16(15). На конце тела нет клещевидных придатков. Надкрылья при-
крывают все брюшко (рис. 103) или часть брюшка (рис. 104).
.............................. Жесткокрылые — Coleoptera (с. 120).
17(14). Передние крылья не ороговевшие, перепончатые или уплот-
ненные, кожистые с хорошо сохранившимся жилкованием.
18(19). Усики короче головы. Брюшко удлиненное (рис. 105). . . .
.............................Стрекозы — Odonata (с. 87).
19(18). Усики длиннее головы.
20(23). Тазики ног расставлены; между ними хорошо видны свобод-
ные участки грудных сегментов. Переднеспинка крупная
(рис. 106—109, 133).
21(22). Задние ноги прыгательные, с утолщенными бедрами (рис. 106—
108). Если расширение бедер на задних ногах недостаточно
выражено, передние ноги копательные с укороченными и рас-
ширенными члениками, снабженными острыми зубцами
(рис. 109).................Прямокрылые — Orthoptera (с. 97).
22(21). Задние ноги обычные, бедра всех ног не утолщенные. Слегка
уплощенные насекомые, в покое складывают крылья плоско
на спинке или облегающе (|рис. 133, 134).........................
................................... Веснянки — Plecoptera
23(20). Тазики ног каждой пары соприкасающиеся, крупные.
24(27). Передние крылья кожистые, более твердые, чем задние; по-
следние перепончатые, веерообразно складываются на брюшке
под передними крыльями. Крылья укладываются на брюшке
плоско или облегающе (рис. НО, 111).
25(26). Передние ноги хватательные, с острыми зубчиками на бедре
и голени (рис. ПО). Тело вытянутое, переднеспинка удлиней-
. ная...............................Богомолы — Mantoptera (с. 93).
26(25). Все ноги бегательные (рис. 111). Тело плоское, переднеспинка
расширенная, частично или полностью прикрывает голову
сверху..........................Тараканы — Blattoptera (с. 91).
27(24). Передние и задние крылья перепончатые.
28(29). Мембрана крыла покрыта мелкими волосками, придающими
ей матовый характер. Ротовой аппарат рудиментарный или
приспособлен для сосания. Насекомые похожи на бабочек
с относительно узкими крыльями (оис. 125). В покое складыва-
ют крылья кровлеобразно (рис. 126)...............................
...........................Ручейники — Trichoptera (с. 173).
29(28). Крылья без густых волосков на мембране.
30(33). Переднее крыло длиннее заднего и расширяется от основания
к вершине таким образом, что в расправленном состоянии его
наибольшая ширина находится около вершины заднего крыла
(рис. 118, 120, 121). Задние крылья способны сцепляться с пе-
редними и образовывать с ними.единую плоскость. Жилкова-
ние крыльев негустое. >
31(32). В покое крылья складываются кровлеобразно (рис. 119).
Мелкие (до 5 мм) насекомые с мягкими покровами и обычно
резко изогнутыми жилками на крыльях. Брюшко всегда сидя-
' чее (рис. 118, 119)...................................
..............................Сеноеды — Psocoptera (с. 102).
32(31). В покое крылья складываются плоско или облегающе. Насе-
комые различных размеров с твердыми покровами. Брюшко
сидячее (рис. 120) или стебельчатое (рис. 121)...................
................. Перепончатокрылые — Hymenoptera (с. 140).
33(30). Крылья иной формы; обычно передние и задние крылья сход-
ны по размерам и форме и не способны сцепляться.
70
34(35). Голова вытянута вниз наподобие клюва, на конце которого
находится грызущий ротовой аппарат (рис. 112)...................
.........................Скорпионницы — Mecoptera (с. 139).
35(34). Голова не вытянута наподобие клюва.
36(41). Жилкование сетчатое: крылья с многочисленными четкими
продольными и поперечными жилками. Крылья складываются
на спинке более или менее кровлеобразно (рис. ИЗ, 116). Оди-
ночные насекомые.
37(40). Усики расположены перед глазами (как на рис. 113, 114).
38(39). Переднегрудь удлиненная (рис. 113). Самка с длинным яйце-
кладом. Крылья прозрачные....................................
......................Верблюдки — Raphidloptera (с. 137).
39(38). Переднегрудь не удлиненная (рис. 114). Самка без яйцеклада.
Крылья буроватые . : . Вислокрылки — Megaloptera (с. 136).
40(37). Усики расположены между глазами (как на рис. 115, 116).
...................... Сетчатокрылые — Neuroptera (с. 137).
41(36). Передние и задние крылья без замкнутых ячеек, только с про-
дольными жилками, плоско складываются на спинке. Голова
крупная (рис. 117). Общественные насекомые......................
....................... Термиты—Isoptera (с. 94).
42(1). Крылья отсутствуют.
43(44). Усики очень короткие, короче головы. Тело плоское........
......................... , Двукрылые — Diptera (с. 189).
44(43). Усики длиннее головы.
45(48). В составе брюшка не более 6 сегментов.
46(47). Брюшко сидячее, причленяющееся к груди широким основа-
нием. Тело или вытянутое, цилиндрическое (рис. 135), или
укороченное с округлым брюшком (рис. 136).......................
...................... Ногохвостки—Collembola (с. 75).
47(46). Брюшко стебельчатое (имеется узкий перехват между грудью
и брюшком—рис. 137). Общественные насекомые (муравьи).
.................Перепончатокрылые—Hymenoptera (с. 140).
48(45). В составе брюшка более 6 сегментов.
49(52). Брюшко на конце снабжено 2 или 3 длинными хвостовыми
нитями или 2 клещевидными придатками.
50(51). Брюшко на конце снабжено 3 хвостовыми нитями (рис. 138).
........................Щетинохвостки — Thysanura (с. 80).
51(50). Брюшко на конце снабжено 2 хвостовыми нитями (рис. 139)
или клещевидными придатками (рис. 140)..........................
............................. Двухвостки — Diplura (с. 79).
52(49). Брюшко на конце без хвостовых нитей или клещевидных при-
9 датков.
53(54). Задние ноги прыгательные, с утолщенными бедрами (как на
рис. 106—108)....................Прямокрылые — Orthoptera (с. 97).
54(53). Задние ноги обычные, бедра на всех ногах не утолщенные.
55(64). Усики кцроче тела.
.56(59). Ротовой аппарат грызущего типа с хорошо развитыми жва-
лами (как на рис. 100, 101).
57(58). Покровы светлой окраски или неокрашенные. Голова крупная,
глаза небольшие, не выпуклые (рис. J41).........................
................................ Термиты — Isoptera (с. 94).
58(57). Покровы темные. Голова относительно небольшая с хорошо
заметными выпуклыми глазами (рис. 142)..........................
.........................Жесткокрылые — Coleoptera (с. 120).
59(56). Ротовой аппарат негрызущий, жвалы не развиты, а иногда
недоразвит весь ротовой аппарат.
60(61). Тело покрыто чешуйками и густыми волосками (рис. 143).
71
Ротовой аппарат в виде свернутого спиралью нечленистого хо-
ботка или отсутствует. ... ............................
..........................Чешуекрылые — Lepidoptera (с. 176).
61(60). Тело без чешуек. Ротовой аппарат иного типа.
62(63). Ротовой аппарат представлен длинным членистым хоботком
(как на рис. 127). . . .Равнокрылые—Homoptera (с. 103).
63(62). Ротовой аппарат иного типа, без членистого хоботка. Насеко-
мые очень мелкие (0,5—2 мм) с удлиненным телом (рис. 144).
.............................. Трипсы — Thysanoptera (с. 118).
64(55). Усики, равные по длине телу или длиннее его.
65(66). Средних размеров или крупные насекомые: длина тела боль-
ше 5 мм. Кончик брюшка с короткими церками......................
..........................Тараканы—Blattoptera (с. 91).
66(65). Мелкие насекомые: длина тела меньше 5 мм. Кончик брюшка
без церок (рис. 145). ..........................
............................. Сеноеды — Psocoptera (с. 102).
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ОТРЯДОВ
ПО ЛИЧИНОЧНЫМ СТАДИЯМ*
1(14). Личинки живут в воде.
2(3). Личинки живут в переносных чехликах из растительных остат-
ков и минеральных частиц илн в неподвижных домиках на
камнях, или в илу (рис. 146). Усики очень сильно редуцирова-
ваны, едва заметные..........................................
. ........................ Ручейники — Trichoptera (с. 173).
3(2). Личинки живут свободно, без чехликов и домиков. Усики обыч-
но хорошо заметные, но могут отсутствовать.
4(5). Ноги полностью отсутствуют; (рис. 147—149)...............
. . .....................4 Двукрылые — Diptera (с. 189).
5(4). Ноги имеются.
6(7). Нижняя губа очень большая, превращена в хватательный ап-
парат, способный далеко вытягиваться, а в покое складывать-
ся, прикрывая голову снизу в виде так называемой «маски»
(рис. 150, 151). Наружные жабры отсутствуют (рис. 152) или
имеют вид трех листовидных придатков на последнем сегмен-
те брюшка (рис. 153). . . Стрекозы — Odonata (с. 87).
7(6). Нижняя губа не удлиненная, не складывается в виде «маски».
8(11). Кончик брюшка с 2 или 3 длинными и членистыми хвостовыми
нитями (рис. 154, 155).
9(10). Кончик брюшка с 2 хвостовыми нитями, а брюшные сегменты
без трахейных жабр (рис. 154)...................................
............................ Веснянки — Plecoptera (с. 95).
10(9). Кончик брюшка с 3 хвостовыми нитями (рис. 155), если таких
нитей только 2, то на брюшных сегментах имеются трахейные
жабры.........................Поденки — Ephemeroptera (с. 82).
11(8). Кончик брюшка без хвостовых нитей, но иногда с одной длин-
ной дыхательной трубкой.
12(13). Ротовой аппарат в виде членистого хоботка (как на рис. 127),
фасеточные глаза имеются....................................
................. Полужесткокрылые—Hemjptera (с. ПО).
* В. П. Тыщенко.
72
Рис. 146—174. Личинки насекомых.
146 — ручейник (Trichoptera), 147, 148, 149 — различные представители двукрылых (Diptera), 150 —
передняя часть тела у личинки стрекозы с прижатой к голове нижней губой (маской), 151 — то
же с вытяиутой нижней губой, 152, 153—стрекозы' (Odonata), 154 — весняика (Plecoptera), 155 —
поденка (Ephemeroptera), 156— жук-водолюб^Й£?о1еор1ега), /57 — саранча (Orthoptera), 158 — та-
ракан (Blattoptera), 159 — голова равнокрылого (Homoptera), 160 — голова полужесткокрылого
(Hemiptera), 161 — пчела (Hymenoptera), 162— муха (Diptera), 163, 164 — жесткокрылые (Coleo-
ptera), 165 — галлообразующий комарик (Diptera), 166, /57— гусеницы (Lepidoptera), 168 — пи-
лильщик (Hymenoptera), 169, 170 — сетчатокрылые (Neuroptera), 171, 172, 173, 174 — различные
. представители жесткокрылых (Coleoptera).
73
13(12). Ротовой аппарат грызущий с хорошо развитыми жвалами, ор-
ганы зрения представлены одиночными глазками, сгруппиро-
ванными вместе, но не образующими фасеточных глаз. Форма
тела обычно, как на рис. 156....................................
.....................Жесткокрылые — Coleoptera (с. 120).
14(1). Личинки живут в наземных биотопах — в почве, подстилке,
древесине, под корой деревьев и пней, открыто на растениях,
в тканях растений (галлах и минах) или в качестве паразитов
на теле и внутри тела различных животных.
15(26). Фасеточные глаза имеются. Личинки старших возрастов могут
иметь зачатки крыльев (рис. 157).
16(23). Ротовой аппарат грызущего типа с хорошо развитыми жва-
лами (как на рис. 100, 101).
17(18). На конце тела имеются характерные длинные клещевидные
придатки (как на рис. 102)....................................
........................Уховертки — Dermaptera (с. 101).
18(17). На конце тела нет клещевидных придатков.
19(20). Передние ноги хватательные, удлиненные и расширенные, снаб-
женные острыми зубчиками и крепкими шипами (как на
рис. 110).......................Богомолы — Mantoptera (с. 93).
20(19). Передние ноги обычные, бегательные, иногда копательные.
21(22). Задние ноги прыгательные, с утолщенными бедрами, значи-
тельно длиннее передних и средних ног (рис. 157). Если рас-,
ширение бедер на задних ногах недостаточно выражено, то
передние ноги копательные, с укороченными и расширенными
члениками, снабженными острыми зубцами (как на рис. 109).
............................ Прямокрылые — Orthoptera (с. 97).
,22(21). Задние ноги бегательные, не длиннее или едва длиннее перед-
них и средних ног (рис. 158).....................................
...............................Тараканы Blattoptera (с. 91).
23(16). Ротовой аппарат в виде членистого хоботка (рис. 159, 160),
обычно подогнутого под грудные сегменты.
24(25). Лоб смещен на нижнюю поверхность головы (рис. 159).
............................Равнокрылые—Homoptera (с. 103).
25(24). Лоб не смещен на нижнюю поверхность головы (рис. 160).
................ Полужесткокрылые—Hemiptera (с. ПО).
26(15). Личинки без фасеточных глаз и даже в старших возрастах не
имеют зачатков крыльев. Органы зрения представлены одиноч-
ными глазками или отсутствуют.
27(36). Ноги полностью отсутствуют (рис. 161 —165).
28(31). Личинки паразитические или обитающие в сотах, гнездах, му-
равейниках, особых ячейках и камерах, устроенных взрослыми
насекомыми.
29(30). Головная капсула вполне четкая, обособленная от груди
(рис. 161). . . . Перепончатокрылые — Hymenoptera (с. 140).
30(29). Головная капсула полностью отсутствует (рис. 162).......
..............................Двукрылые — Diptera (с. 189).
31(28). Личинки непаразитические, обитают в почве, подстилке, гри-
бах, под кор^й деревьев и пней, в древесине и растительных
тканях.
32(33). Туловищные сегменты с продольными и поперечными склад-
ками (рис. 163, 164). Личинки живут в растительных тканях,
под корой, в древесине и почве ................... ..............
.....................Жесткокрылые—Coleoptera (с. 120).
33(32). Туловищные сегменты гладкие, без складок.
74
34(35). Личинки живут в галлах и имеют С-образное, заостренное
к обоим концам тело.........................................
.............. Перепончатокрылые—Hymenoptera (с. 140).
35(34). Личинки живут в почве, подстилке, грибах и растительных
тканях. У личинок, обитающих в галлах, тело не С-образное
(рис. 165)..........................Двукрылые — Diptera (с. 189).
36(27). Ноги имеются (рис. 166—174).
37(40). Личинки снабжены 3 парами грудных ног и брюшными лож-
ноножками (рис. 166—168). ,
38(39). Брюшных ложноножек от 2 до 5 пар (рис. 166, 167). . . .
................. Чешуекрылые—Lepidoptera (с. 176).
39(38). Брюшных ложноножек более 6 пар (рис. 168)............
.............. Перепончатокрылые — Hymenoptera (с. 140).
40(37). Личинки снабжены только грудными ногами, брюшные ложно-
ножки отсутствуют (рис. 169—174).
41(42). Ротовые части приспособлены для прокалывания и высасыва-
ния добычи. Жвалы тонкие, заостренные, обычно серповидной
формы, превышают длину головы. Форма тела — рис. 169, 170.
......................... Сетчатокрылые — Neuroptera (с. 137).
42(41). Ротовой аппарат грызущий, обычного типа. Жвалы имеют
разное строение, но более короткие. Форма тела — рис. 171 —
174........................Жесткокрылые — Coleoptera (с. 120}.
ОТРЯД НОГОХВОСТКИ, ИЛИ КОЛЛЕМБОЛЫ—COLLEMBOLA*
Размеры ногохвосток чаще всего составляют 1—2 мм, а наиболее
крупные представители европейской фауны не превышают 6 мм. Тело
ногохвостки состоит из головы, груди с тремя парами ног и брюшка,
1
снабженного своеобразными придат-
ками, присущими лишь этой группе
первичнобескрылых насекомых — вен-
тральной трубкой, прыгательной вил-
кой и зацепкой (рис. 175—177). Ха-
рактерной особенностью коллембол
является наличие у них лишь 6 брюш-
ных сегментов. Благодаря тенденции
к слиянию отдельных сегментов, об-
щее количество их во многих семейст-
вах еще более уменьшается, и у
большинства слитнобрюхих ногохвос-
ток тело почти однородное, нерасчле-
ненное (рис. 177, 178). Открытоживу-
щие формы ногохвосток имеют жел-
товато-коричневую, серо-зеленую,
сине-фиолетовую и коричнево-фиоле-
товую окраску. Часто пигмент распо-
лагается отдельными пятнами и пе-
ревязями, образуя яркий, контрастный
рисунок (рис. 178, 179). У обитателей
нижних ярусов подстилки окраска бо-
Рис, 175—177. Форма и строение
тела ногохвосток.
175 — Hypogastrura Born. (Hypogastruri-
dae), 176— Tomoceriis Nic. (Tomocerldae),
/77 — SminthuTtis Latr. (Sminthuridae).
в. т — вентральная трубка; з — зацепка;
п. в — прыгательная вилка.
лее светлая, а типичные почвенные
и пещерные формы вообще утрачива-
ют пигмент. С каждой стороны голо-
вы на темном поле расположены
8 простых глазков. У пещерных, поч-
* Е. Ф. Мартынова.
75
венных и почвенно-подстилочных форм наблюдается редукция числа
глазков, иногда до полного их исчезновения. Усики 4-члениковые, в се-
мействе Orchesellidae 6-члениковые, и снабжены многочисленными
сенсорными образованиями. У основания усиков находится по-
стантеннальный орган, служащий для регистрации уровня влаж-
ности и имеющий чаще всего кольцевидную или розетковидную
форму. Ротовой аппарат так называемого скрыточелюстного типа: все
Рис. 178—184. Ногохвостки.
178 —Heterosmintharus Stach (Sminthuridae), 179 — Еп omobrya Rond. (Entomobryidae), 180—
Tomocertis Nic. (Tomoceridae), 181 — Erlsea D.-Torre (Neanuridae), 182—Folsomia Willem (Isoto-
midac), 183— Onychiurus Gerv. (Onychiuridae), 184 — Isotomodes Axels. (Isotomldae).
76
его части находятся в ротовом конусе, погруженном в полость головы.
Все разнообразие типов ротового аппарата ногохвосток можно свести
к трем основным: 1) жующему, или перетирающему, 2) «отгрызающе-
му» и 3) колюще-сосущему. Ногохвостки с жующим ротовым аппара-
том используют в пищу гифы и споры грибов, водоросли, бактерии,
мертвые и разлагающиеся растительные остатки, а также пыльцу
хвойных и живые ткани растений. Для большинства «отгрызающих»
форм, не перетирающих пищевой субстрат, более характерно хищниче-
ство. В качестве объектов питания они используют мелких обитателей
почвы — коловраток, тихоходок, клещей, первичнобескрылых насеко-
мых, а также трупы беспозвоночных и позвоночных животных. Для
видов с колюще-сосущим ротовым аппаратом характерна сапрофа-
гия — высасывание продуктов разложения, содержащих микроорга-
низмы и простейших.
Тело ногохвосток покрыто многочисленными или более редкими
волосками и щетинками (хетами) различной длины. Часть простых
волосковидных хет опушена или приобретает ланцетовидную, головча-
тую, шиповатую и иную форму. Во многих семействах, наряду с раз-
личной формы хетами, имеются и многочисленные чешуйки. Строение
грудного отдела не однотипно. У части членистобрюхих ногохвосток
(подотряд Arthropleona) переднегрудной отдел редуцирован и заметен
• лишь сбоку в форме складчатого склеротизованного участка (рис. 176,
179, 180). Средне- и заднегрудь развиты нормально и часто близки по
размерам. Каждый грудной сегмент несет одну пару ног, у которых
лапка и голень сливаются, образуя единый тибиотарзальный членик.
Необычны в функциональном отношении упомянутые выше придатки
брюшка, являющиеся производными брюшных конечностей. От ниж-
ней поверхности первого брюшного сегмента отходит вентральная труб-
ка — мешковидное образование, из которого могут выдвигаться пу-
зырьки или длинные лентовидные тяжи, служащие, по-видимому, для
газообмена, а частично для прикрепления к гладким ' поверхностям
и пленке воды (рис. 175—177, в.т). Снизу четвертого брюшного сег-
мента, на непарном основании — манубрии, прикреплена прыгательная
вилка (рис. 175—177, п.в}. Концевые отделы вилки парные. В спокой-
ном состоянии вилка подогнута под брюшко и закреплена зацепкой,
расположенной на третьем сегменте брюшка (рис. 175, з). Вилка с по-
мощью сильной мускулатуры быстро отводится назад, и совершаемый
при этом прыжок может достигать нескольких сантиметров. На вод-
ной поверхности вилка используется в качестве нормального способа
передвижения, а в остальных случаях — при внезапно возникшей опас-
* ности. У обитателей нижних ярусов подстилки и почвы вилка укора-
чивается, а иногда и полностью исчезает, так как в узких почвенных
полостях она не может быть эффективно использована. Половое отвер-
стие находится на пятом брюшном сегменте. Копулятивный орган от-
сутствует и оплодотворение происходит путем захвата самкой пакетов
спермы — сперматофоров, отложенных на субстрат. Число линек не
фиксировано и варьирует от 3—4 до 12. Линьки могут продолжаться
I в течение всей жизни. Нимфы отличаются от взрослых особей мень-
шими размерами, числом хет, меньшим количеством зубчиков на ко-
готках и концевых частях прыгательной вилки, отсутствием или недо-
развитием генитальных структур.
Большинство видов ногохвосток принадлежит к мезофилам и гиг-
рофилам. Они боятся высыхания и могут длительно существовать лишь
в достаточно влажной среде. В лесостепной и степной зонах в наиболь-
шем количестве их можно встретить в байрачных лесах, дубравах,
поймах рек. При благоприятных условиях наблюдается массовое раз-
множение многих видов. Гигантские скопления ногохвосток можно
77
иногда наблюдать по берегам временных водоемов, озер, медленно
текущих рек (Podura aquatica L., Hypogastriwa Born., Isotomurus
Born.). Вызывает удивление, а иногда и беспокойство у наблюдавших
это явление людей, появление ранней весной на снегу.темных пятен,
состоящих из огромного скопления ногохвосток, которые поднимаются
во время оттепелей из подстилки на поверхность снега и нагретые, от-
таявшие участки почвы. Это чаще всего Isotoma viriais Bourl.,
I. fennica Rent, и виды рода Hypogastrura Born., сохраняющие актив-
ность при сравнительно низких температурах.
Благодаря высокой численности, даже при малых размерах тела,
ногохвостки играют существенную роль в процессах разложения орга-
нического вещества и в почве. В лесной подстилке и почве, где их чис-
ленность нередко достигает 30—60 тысяч особей на 1 м2, они активно
участвуют в переработке листового опада, животного и растительного
детрита. Отдельные виды имеют симбиотические связи с микроорганиз-
мами, что. расширяет возможности переработки растительной клетчат-
ки. Утилизацией экскрементов крупных сапрофагов ногохвостки за-
вершают окончательное разрушение растительных остатков. Для мно-
гих слитнобрюхих ногохвосток, обитающих в травостое, характерно
питание пыльцой хвойных и живыми тканями растений. Ранней весной,
в условиях повышенной влажности они иногда повреждают всходы
полевых и огородных культур. При этом сильнее поедаются те части
растений, которые находятся в контакте с почвой.
По внешнему облику ногохвосток легко разделить на 2 группы.
К первой принадлежат виды с удлиненным телом и ясными границами
между сегментами; лишь иногда у них сливаются 2—3 последних
сегмента. Это подотряд членистобрюхие — Arthropleona (рис. 175, 176).
Вторая группа объединяет виды с округлым, почти шаровидным те-
лом из плотно слитых сегментов, с почти неразличимыми межсегмент-
ными промежутками. Это подотряд слитнобрюхие — Symphypleona
(рис. 177). Каждый из подотрядов объединяет большое число семейств,
которые без изготовления специальных препаратов определить невоз-
можно. И только после просмотра материалов под микроскопом наи-
более крупных представителей можно «узнать» в природе.
В мировой фауне насчитывается брлее 4000 видов ногохвосток,
в СССР — около 700. Представители этого отряда распространены все-
светно—от арктических островов Северного полушария до Антаркти-
ки, встречаясь во всех ландшафтных зонах. На этой огромной
территории ногохвостки связаны главным образом с почвенными и поч-
венно-подстилочными ярусами, где численность и видовое многообра-
зие их максимальны. В травянистом ярусе, на кустарниках и стволах
деревьев обитает значительно меньшее число видов, относящихся пре-
имущественно к слитнобрюхим ногохвосткам и •представителям се-
мейств Sminthuridae (рис. 177, 178) и Entomobryidae (рис. 179). Это
относительно крупные, ярко окрашенные формьц часто с контрастным
рисунком, длинными усиками и ногами, хорошо развитой прыгатель-
ной вилкой, с 8 глазками и обычно лишенные постантеннального орга-
на. Эту группу жизненных форм чаще всего называют атмобиотиче-
скрй.
Очень обильна фауна ногохвосток верхнего слоя подстилки. Виды
этого комплекса (подстилочные ногохвостки) внешне напоминают ат-
мобиотические формы, отличаясь в целом меньшими размерами, часто
более короткими усиками, ногами и вилкой, присутствием у многих
видов постантеннального органа. Характерные представители подсти-
лочных форм, к которым относятся многие виды семейств Hypogastru-
ridae (рис. 175), Tomoceridae (рис. 176, 180), Neanuridae (рис. 181)
и Isotomidae (рис. 182), в значительной мере утрачивают пигмент;
78
усики, ноги и прыгательная вилка у них укорочены, а число глазков
уменьшено, иногда до одного с каждой стороны головы. Настоящие
почвенные формы, в особенности обитающие в глубоких горизонтах
почвы, характеризуются полной редукцией глаз и пигмента. Вилка от-
сутствует или сильно укорочена, постантеннальный орган сложного
строения. Последний признак' особенно характерен для семейства
Onychiuridae. Как правило, почвенным обитателям свойственны малые
размеры и суженное тело. Типичные представители этой группы — ви-
ды родов Onychiurus Gerv. из семейства Onychiuridae (рис. 183)
и Isotomodes Axels, из семейства Isotomidae (рис. 184). Следует отме-
тить также ряд узко специализированных жизненных форм — обитате-
лей пещер и гнезд общественных насекомых. У нейстонных ногохвос-
ток возникают своеобразные адаптации к обитанию на поверхности
воды, сопровождающиеся расширением концевых отделов вилки.
ОТРЯД ДВУХВОСТКИ, ИЛИ ВИЛОХВОСТКИ- DIPLURA*
Как и представители других отрядов первичнобескрылых насеко-
мых, двухвостки лишены крыльев,
тело имеет беловатую или жел-
товатую окраску. Глаза отсут-
ствуют, а на плоской, направ-
ленной ротовым отверстием впе-
ред голове сидят 1 длинные мно-
гочлениковые усики, превышаю-
щие иногда длину тела. Рото-
вые органы жующего типа, по-
груженные в полость головы;
снаружи они видны лишь час-
тично. Грудные сегменты несут
по одной паре ног, оканчиваю-
щихся одночлениковой лапкой
с . двумя коготками. На груди
расположены 3—4 пары дыха-
лец. Брюшко из 10 сегментов,
последний сегмент либо с длин-
ными многочлениковыми церка-
ми, напоминающими по форме
усики (рис. 185), либо с мощ-
ными клещевидными придатками
(рис. 186), которые используют -
ся двухвостками при захвате и
удержании добычи. На вентраль-
ной поверхности семи начальных
сегментов брюшка находятся
парные, относительно короткие
и покрытые волосками кониче-
ские придатки — грифельки, а
рядом с ними — втяжные кок-
сальные пузырьки (рис. 187).
Грифельки приподнимают брюш-
ко над субстратом. Наружные
половые придатки отсутствуют,
а половое отверстие расположе-
но между VIII и IX сегментами
брюшка. Перенос спермы у дву-
* Е. Ф. Мартынова.
а их узкое, длинное, червеобразное
Рис. 185—187. Двухвостки.
185— Campodea Westw. (Campodeidae), 186—
Japyx Halid. (Japygidae), 187 — грифельки и
коксальные пузырьки на вентральной поверх-
ности брюшного сегмента.
г — грифельки; к.п — клещевидные придатки;
п — коксальиые пузырьки; ц — церки.
79
хвосток происходит без контакта полов, и самка захватывает сперма-
тофоры, отложенные самцами на субстрат. Тело, ноги и придатки по-
крыты многочисленными волосками и щетинками. Их длина и распо-
ложение различны, и эти особенности шцроко используются при опре-
делении отдельных видов и родов. Большинство представителей этого
отряда не превышает 2—8 мм, но в семействе Japygidae встречаются
настоящие гиганты. Так, Japyx dux Skor. с Памира достигает 5 см.
Постэмбриональное развитие протекает по типу эпиморфоза. Нимфы
отличаются от взрослых особей, кроме размеров, меньшим числом
члеников в усиках и церках, недоразвитыми генитальными структура-
ми и более редкими щетинками. Линьки происходят не только на ним-
фальной стадии, но и в половозрелом состоянии.
Двухвостки питаются мелкими почвенными членистоногими и чер-
вями, а также мертвыми остатками животного и растительного проис-
хождения, гифами и спорами грибов, водорослями. Обитают в рыхлой,
достаточно влажной почве и подстилке, под камнями, стволами пова-
ленных деревьев, чаще под пологом леса. Распространены главным об-
разом в южных районах. Отечественная фауна двухвосток изучена не-
достаточно; они известны из горных и предгорных районов Средней
Азии и Кавказа, а также с Украины, из Поволжья и Прибалтики.
В центральных и южных районах европейской части СССР в лесной
подстилке можно встретить представителей семейства Campodeidae.
В мировой фауне насчитывается около 500 видов двухвосток;
в СССР немногим более 20 видов, принадлежащих двум семействам,
которых легко отличить по форме придатков на конце брюшка. У кам-
подей (Campodeidae) эти придатки длинные, многочлениковые
(рис.' 185), а у япиксов (Japygidae) они короткие, клещевидные
(рис. 186). В фауне СССР первое семейство представлено родом
'Campodea Westw., а второе —родом Japyx Halid.
ОТРЯД ЩЕТИНОХВОСТКИ —THYSANURA*
К этому отряду относятся довольно крупные (1,5—2 см) первич-
нобескрылые насекомые с удлиненным (рис. 188—190) или укорочен-
ным, иногда почти треугольным телом, у большинства видов плотно
покрытым чешуйками и в различной степени пигментированным. Голо-
ва направлена ротовым отверстием вперед (у Lepismatidae) или вниз
(у Machilidae). Глаза, как и у крылатых насекомых, фасеточные;
в семействе Machilidae они крупные и впереди почти всегда соприка-
саются (рис. 191), а в семействе Lepismatidae — небольшие и не со-
прикасаются (рис. 192). У почвенных щетинохвосток, а также у оби-
тателей муравейников и термитников глаза отсутствуют. Перед фасе-
точными глазами у видов семейства Machilidae расположены 3 простых
глазка. Усики длинные, многочлениковые и иногда в два раза и более
превышают длину тела. Ротовые части грызущие и расположены от-
крыто, поэтому щетинохвосток наряду с крылатыми насекомыми от-
носят к открыточелюстным насекомым. Грудной отдел состоит из трех
округленных по бокам сегментов, грудные ноги бегательные и бега-
тельно-копательные. Брюшко состоит из 11 сегментов, но последний
сегмент прикрыт предпоследним и не виден сверху. Его спинной учас-
ток вытянут в длинную хвостовую нить — каудальный филамент, имею-
щий тактильную или опорную функцию (рис. 188, к.ф). От нижней по-
верхности XI сегмента отходят длинные парные церки, состоящие из
многих члеников (рис. 188, ц). Они или заметно короче и тоньше кау-
дального филамента (рис. 189) или лишь незначительно отличаются
* Е. Ф. Мартынова, при участии В. Г. Каплина.
80
ES"
ЗйЙ
М"
,'K.tp
isa
Рис. 188—190. Щетннохвосткн (по Снодграссу, Стаху и орнг.).
188— махилис (Machilidae), 189 — чешуйница (Lepismatidae), 190 — синантропная
чешуйница Leplsma saccharina L. к Ф — каудальный филамент; ц — церки.
от него (рис. 188, 190). На нижней поверхности брюшка у щетино-
хвосток имеются парные грифельки и втягивающиеся мешочки, напо-
минающие таковые двухвосток (см. рис. 187). Мешочки у Lepismatidae
обычно отсутствуют, а грифельки чаще .всего имеются только на
VIII и IX сегментах. У Machilidae грифельки обычно сидят почти на
всех сегментах брюшка (со II по IX), но могут отсутствовать на не-
которых сегментах. При передвижении щетннохвосток слабо припод-
нятое брюшко скользит по субстрату, а грифельки при этом, выполняя
роль полозьев, смягчают трение его о субстрат. Длинные грифельки
IX сегмента используются видами семейства Machilidae для соверше-
ния прыжков, подобно тому, как ногохвостки используют прыгатель-
ную вилку. Но при этом во время прыжка они перемещаются не впе-
ред, а назад. Щетинохвостки семейства Lepismatidae быстро бегают,
опираясь на лапки, грифельки, каудальный филамент и опорные ще-
тинки. По наблюдениям в лаборатории домашняя лепизма (Lepismo-
des domestica Pack.) также
может совершать небольшие
прыжки. Самки щетннохвосток
имеют яйцеклад, отходящий
от нижней поверхности VIII
и IX брюшных сегментов. Кон-
чик яйцеклада часто снабжен
копательными шпорами.
Постэмбриональное раз-
витие щетннохвосток проте-
кает по типу эпиморфоза.
Рис. 191—192. Расположение глаз на голове
щетннохвосток.
191 — Machilidae, 192 — Lepismatidae.
6 208
81
Нимфы младших возрастов лишены чешуек, грифельков и гениталь-
ных структур. Половая зрелость наступает через 8—12 месяцев,
но и после этого взрослые щетинохвостки продолжают линять
и расти. В умеренном климате, по-видимому, один репродуктивный
цикл в году. Живут от 3 до 5 лет.
Щетинохвостки питаются отмершими растительными остатками,
лишайниками, микроскопическими грибами, напочвенными и наскаль-
ными водорослями. Синантропные виды повреждают пищевые продук-
ты, бумагу, книги, ткани, картины, написанные маслом. Среди синант-
ропных щетинохвосток наиболее известна чешуйница (Lepisma saccha-
ritia L., рис. 190), которая встречается в оранжереях, на складах и в
жилых домах, где ее легче всего отыскать в ванных комнатах. Это се-
ребристое, очень подвижное насекомое в немецких работах очень удач-
но называют «серебристой рыбкой» (Silberfischchen).
Щетинохвостки распространены главным образом в южных рай-
онах. Семейство Machilidae встречается преимущественно в горах,
среди каменистых россыпей, а семейство Lepismatidae — па равнинах.
Многие виды щетинохвосток обитают в подстилке и рыхлой почве, под
корой деревьев, в муравейниках. В СССР отмечено до 40 видов, от-
носящихся к трем семействам, которые встречаются в горах повсе-
местно, а в равнинных условиях преимущественно в южных районах
(в степях европейской части СССР, в пустынях Средней Азии, на
Дальнем Востоке и на юге Сибири). Из северных районов нашей
страны в настоящее время известен только 1 вид — Petrobius balti-
cus St. (семейство Machilidae), обнаруженный в Латвии.
В фауне европейской части СССР наиболее многочисленны пред-
ставители двух семейств — Machilidae и Lepismatidae, а в Крыму обыч-
ны представители еще одного семейства — Nicoletiidae. Указанные се-
мейства довольно легко различаются, так как у Nicoletiidae глаза
отсутствуют, у махилисов (Machilidae) они большие, занимают всю
переднюю часть головы и соприкасаются посередине (рис. 191), а че-
шуйницы (Lepismatidae) снабжены небольшими широко расставлен-
ными глазами (рис. 192).
ОТРЯД ПОДЕНКИ-EPHEMEROPTERA*
Общая характеристика
К отряду поденок относятся нежные крылатые насекомые с удли-
ненным телом и 2—3 длинными членистыми хвостовыми нитями на
конце брюшка (рис. 193). Ротовой аппарат редуцирован. Усики ко-
роткие. Крылья с богатым жилкованием (рис. 194—197), в покое не
складываются на брюшке. Передняя пара крыльев всегда значительно
больше задней; иногда задние крылья отсутствуют. Брюшко состоит из
10 сегментов, заполнено воздухом. Развит половой диморфизм. У сам-
цов глаза обычно крупные, чабто подразделены на 2 отдела. Передние
ноги самца очень длинные, на конце девятого стернита брюшка имеют-
ся членистые половые щипцы (форцепсы).
Превращение неполное. Личинки поденок живут в воде — от года
до 3 лет. Развитие сопровождается большим числом линек (до 25).
Из воды выходит субимаго — крылатая стадия с недозрелыми поло-
выми продуктами. На берегу субимаго линяет и превращается в има-
го. Крылатые поденки не питаются и живут недолго: субимаго от не-
скольких секунд до нескольких дней, имаго от нескольких часов до
10 дней. Субимаго отличается от имаго тусклой окраской, мутными
* А. К. Бродский.
82
' крыльями с бахромкой волосков по заднему краю и укороченными
передними ногами.
Самцы поденок для встречи с самками собираются в рой. В рое
они демонстрируют брачные танцы, состоящие из чередования актив-
ных взлетов и пассивных спусков. Для формирования роя на берегу
водоема выбирается место, по некоторым признакам отличающееся от
окружающей местности. Например, роение часто происходит над вы-
сокими тростниками, тропинкой, каким-нибудь камнем и т. д. Боль-
шинство поденок роится в сумерки и на рассвете. Ночью поденки
охотно летят на свет. Самки влетают в рой и после спаривания в воз-
духе летят к месту откладки яиц. Для некоторых поденок, например
Рис. 193—197. Строение тела и жилкование крыльев у поденок.
193— общий вид самца Sipfilonurus linnaeanus Etn. (Siphlonuridae), 194 — задний край
переднего крыла Heptagenia flava Rost. (Heptageniidae), 195 — переднее крыло Poiamanihtis lutetis
(Potamanthidae), 196 — то же Ephemerella igniia Poda (Ephemerellidae), 197 — задний край
переднего крыла LepiopHlebia marginata L. (Leptophlebiidae). Латинскими буквами обозначены
крыловые жилки.
для представителей семейства Palingeniidae, обитающих в больших
озерах, характерны лишь короткие взлеты над водой. У поденок, ко-
торым не свойственно брачное роение, часто наблюдается одновремен-
ный выход из воды многих тысяч особей. Во время массового лёта они
появляются в столь больших количествах, что над водой поднимается
«столб» снующих насекомых. Синхронный вылет и скученность поде-
нок облегчают встречу полов в течение короткого периода их имаги-
нальной жизни.
При яйцекладке самки во время полета откладывают кучки яиц
в воду. У некоторых видов, развивающихся в горных реках, самкам
для откладки яиц приходится погружаться в воду. Там они приклеи-
вают яйца к подводным предметам. Самки видов, обитающих в быст-
рых' и полноводных реках с илистым грунтом, перед яйцекладкой ле-
тят вверх по течению, что компенсирует снос течением яиц и личинок.
Личинки поденок (рис. 198—204) отличаются большой экологи-
ческой пластичностью; они заселяют водоемы самого различного типа
и разнообразные стации. Ротовой аппарат личинок сложного грызу-
щего типа, приспособленный для питания детритом и мелкими водо-
83
рослями. Очень редко личинки хищничают. В зависимости от условий
обитания и характера питания выделяются 6 морфо-экологических
типов личинок (по классификации О. А. Черновой).
1. Крупножаберные личинки зарослей (рис. 198).
Обитают в стоячих водоемах и тихих заводях рек. Общая поверхность
листовидных трахейных жабр, через которце поглощается растворен-
ный в воде кислород, велика. У личинок, обитающих в текучей неглу-
Рис. 198—204. Морфо-экологические типы личинок поденок.
198— Siphlonurus linnaeantis Etn. (Siphlonuridae), 199— Basils rhodani Pict. (Baetidae),
200— Ollgoneuriella rhenana Jmh. (Oligoneuriidae), 201— Caenis macrura Steph. (Caenidae),
202 — Ephemerella ignlta Poda (Ephemerellidae), 203— Ecdyontirus Etn. (Heptageniidae),
204— Ephemera L. (Ephemeridae). 4
бокой воде, жабры одиночные и с менее развитой трахеацией, а в не-
достаточно аэрированных водоемах живут личинки, снабженные двой-
ными жабрами. Хвостовые нити сильно опушены, что помогает ли-
чинкам держаться в спокойной воде среди зарослей растительности
и плавать. Плавают при помощи волнообразного, изгибания брюшка
в вертикальной плоскости; хвостовые нити при этом играют роль не-
парного плавника (рис. 205). Представлены видами семейства Siphlo-
nuridae и несколькими видами семейств Baetidae и Leptophlebiidae.
84
2. Мелкожаберные, быстр и иные, активно плаваю-
щие личинки (рис. 199). Обитают в быстро текущей воде, на по-
рогах и перекатах. Активно плавают среди камней. Плавание осуще-
ствляется так же, как у личинок предыдущего типа. Имеют небольшие
жаберные листки, обтекаемое тело, притупленную спереди голову,
сильные ноги с цепкими коготками. Представлены некоторыми видами
семейства Baetidae.
ПАССИВНОЕ ПОГРУЖЕНИЕ
Ecdyonurus
Рис. 205. Движения и позы личинок поденок.
Стрелки показывают возможные пути эволюции способов движения.
3. Крышечножаберные личинки илистых грунтов
(рис. 201). Обитают в низовьях рек, в прибойной зоне озер и местах,
где постоянно взмучивается ил. Личинки имеют видоизмененные жаб-
ры: первая жабра рудиментарна, вторая отсутствует, третья видоизме-
нена в плотную квадратную крышечку, которая прикрывает от оседаю-
85
щего ила нежные бахромчатые пластинки остальных жабр. Ноги хо-
дильного типа, покрыты волосками. Ползают по дну. Представлены
видами семейства Caenidae.
4. Плоскотелые или с цепкими ногами быстринные
личинки (рис. 200,202, 203). Обитают на каменистом грунте горных по-
токов и быстрых рек. У личинок, которые держатся на нижней поверхно-
сти камней, жабры образуют почти замкнутый диск — присоску, а сами
жаберные нити у основания отдельных пластинок присоски очень не-
велики (рис. 203). Такое строение жабр обеспечивает надежное при-
крепление к камням в условиях быстрого течения. Личинки имеют
уплощенную голову с острым передним краем. Уплощены также брю-
шко и бедра. Глаза расположены на дорзальной поверхности головы.
Плавают либо при помощи сильных волнообразных движений всего
тела (Heptagenia Walsch.), либо загребая ногами (Ecdyonurus Etn.)
(рис. 205). Эта группа представлена видами семейства Heptageniidae.
У личинок, которые держатся на боковой поверхности камней,
каждая жабра состоит из пластинок, прикрытых плотным листком.
Такие личинки имеют сильные, цепкие ноги (рис. 202). У некоторых
личинок изменена первая пара ног, которая образует подобие ного-
челюстей с сильно развитым фильтрующим аппаратом из длинных
щетинок на бедре и голени (рис. 200). Эта группа представлена вида-
ми двух семейств — Ephemerellidae и Oligoneuriidae.
5. Грунтовые, закапывающиеся личинки с «бивня-
ми» (рис. 204). Обитают в V-образных ходах глинистых и илистых
подмываемых берегах рек и озер. Имеют характерные приспособления
для закапывания: выступающий зубчатый лоб, клыки верхних челюс-
тей, копательные передние ноги. Средние и задние ноги приспособле-
•ны для опоры и проталкивания тела внутри хода. Жабры двураздель-
ные, перистые, загнуты на спинную сторону и предназначены для соз-
дания тока воды, промывающей ход личинки. Представлены видами
семейств Palingeniidae, Ephemeridae и Polymitarcidae.
6. Длиннокоготные и со «щетками» на голове личин-
ки песчаных грунтов. Закапываются вертикально в глубь пес-
ка. Представлены редко встречающимися видами семейств Ametropo-
didae и Behningiidae.
Всего известно более 1500 видов поденок, в фауне СССР — окол®
200 видов, относящихся к 15 семействам.
Определительная таблица семейств
1(4). Задних крыльев нет или они очень малы и не более чем с 3 про-
дольными жилками.
2(3). Поденки с 3 хвостовыми нитями. Задние крылья всегда отсут-
ствуют. Мелкие поденки с крупной грудью, относительно ко-
ротким брюшком. Широкие треугольные крылья в покое рас-
простерты в стороны.................................Caenidae.
3(2). Поденки с 2 хвостовыми нитями, стройные с овальными крыль-
ями..................................................Baetidae.
4(1). Задние крылья крупные; если они малы, то всегда со многими
жилками (рис. 193).
5(20). Жилкование богатое. Поперечные жилки расположены между
всеми продольными жилками (рис. 193—197).
6(19). Крылья не смяты в складки. Субкостральная жилка (Sc) на
переднем крыле до самого конца свободная (рис. 193, 195, 196).
7(18)-. Ширина переднего крыла составляет не более половины его
длины (рис. 193, 195, 196). Крылья имаго не замутнены белрш,
Глаза самцов крупные,
§6
8(11). Поденки с 2 хвостовыми нитями (рис. 193). ,
9(10). На передних крыльях в кубитальном поле (между жилками
СиА и СиР) от жилки СиА к заднему краю отходят многочис-
ленные волнисто-изогнутые жилки (рис. 193). . . .
........................................ , , Siphlonuridae.
10(9). В кубитальном поле 2 пары длинных интеркалярных (вставоч-
ных) жилок (рис. 194)...............................Heptageniidae.
11 (8). Поденки с 3 хвостовыми нитями.
12(17). Крылья без пятен. От жилки Ai к заднему краю крыла отхо-
дит не более 2 жилок или их нет совсем (рис. 195, 196).
13(14). Жилка МР% при своем отхождении от МР\ делает резкий из-
гиб, жилка СиА также с изгибом в основании (рис. 195) . .
................................................... Potamanthidae.
14(13). Все продольные жилки у основания прямые (рис. 196).
15(16). Между продольными жилками на наружном крае крыла ко-
роткие свободные краевые интеркалярные жилки (по крайней
мере в задней части крыла) (рис. 196). Форцепсы самца мас-
сивные, с 1 маленьким концевым члеником............................
....................... . . . ' . . . . , , Ephemerellidae.
16(15). На наружном крае крыла свободных интеркалярных жилок нет
(рис. 197). Форцепсы самца с 2 маленькими концевыми чле-
никами . . '................................... Leptophlebiidae.
17(12). Крылья обычно с темными пятнами. От жилки At к заднему'
краю крыла отходит несколько прямых параллельных жилок.
Жилка СиА с изгибом в основании. Крупные виды ....
...............................................Ephemeridae.
18(7). Ширина переднего крыла больше половины его длины. Крылья
‘имаго молочно-белые. Глаза самцов маленькие, как у самки.
Средние и задние ноги слабые, не функционирующие ....
................................ . . Polymitarcidae.
19(6). Передние крылья смяты в складки. Субкостальная жилка (Sc)
в дистальной части крыла слита с костальной. Длина тела 2—
3 см.......................................... Palingeniidae.
20(5). Продольные жилки слиты попарно, поперечные жилки только
в передней половине крыла..........................Oligoneuriidae.
ОТРЯД СТРЕКОЗЫ - ODONATA *
Общая характеристика
К этому отряду относятся крупные, ярко окрашенные насекомые,
имеющие большую подвижную голову с крупными фасеточными гла-
зами, короткими усиками и с грызущим ротовым аппаратом. Крылья
длинные, почти гомономные, с узелковыми структурами (изломом на пе-
реднем крае). Жилкование богатое. У представителей подотряда равно-
крылых (Zygoptera) передние и задние крылья одинаковые по форме
и размерам (рис. 206), а у видов, относящихся к подотряду разнокры-
лых (Anisoptera), задние крылья у основания расширены и по форме
значительно отличаются от передних (рис. 207). В покое крылья не
складываются на спинке, а держатся распростертыми или отводятся
назад. Тело стрекоз стройное, удлиненное. Брюшко заканчивается
короткими одночлениковыми придатками. Копулятивные придатки сам-
цов, в отличие от всех других насекомых, находятся на втором стер-
пите брюшка; половое отверстие расположено, как обычно, на девятом
сегменте.
* А. К. Бродский.
87
Взрослые стрекозы — хищники, ведущие дневной образ жизни.
Полет сильный, маневренный. В полете часто меняют направление
движения. Охотятся на различных летающих насекомых, но вблизи
водоемов основной добычей становятся двукрылые, в том числе кро-
вососущие (комары, слепни и др.). Способ охоты своеобразен и спе-
цифичен для каждого семейства, а иногда и рода. Стрекозы-коромыс-
ла (Aeschnidae) охотятся на полянах, опушках, лесных дорогах, отлетая
далеко от водоемов. Бабки (Corduliidae) ловят насекомых над самой
поверхностью воды, при этом они все время находятся в безостано-
вочном полете. Виды рода Libellula L. из семейства Libellulidae ата-
куют пролетающих мимо насекомых, сидя на выступающих из воды
растениях. Большинство равнокрылых стрекоз охотится в надводных
зарослях медленно текущих и стоячих водоемов. При этом они манев-
рируют среди густой растительности без разворотов тела — могут ле-
теть боком и даже задним концом тела вперед.
Рис. 206—207. Крылья равнокрылых и равнокрылых стрекоз.
206— Lestes drvas КЬу. (Lestidae), 207 — Aeschna juncea L. (Aeschnidae).
На каждом рисунке вверху — переднее крыло, внизу — заднее, пт -« птеростигма.
Во время копуляции, которая происходит в полете, самец захваты-
вает вершиной брюшка шейный отдел самки. Самка изгибает брюшко
и прижимает его вершину к копулятивным придаткам самца. После
копуляции самка летит откладывать яйца. Самки с редуцированным
яйцекладом откладывают яйца просто на подводные растения, а сам-
ки, обладающие им, вкалывают яйца в ткани водных растений.
Постэмбриональное развитие стрекоз протекает с неполным пре-
вращением. Личинки хищные, живут в воде, дышат жабрами. Личин-
ки равнокрылых стрекоз имеют наружные трахейные жабры в виде
трех лепестков на заднем конце брюшка (рис. 208, 210). У личинок
разнокрылых стрекоз жаберных лепестков нет (рис. 209, 211, 212),
а поглощение кислорода осуществляется через внутренние жабры, рас-
положенные в прямой кишке. Личиночное развитие продолжается 1 —
2 года и сопровождается линьками. Личинка последнего возраста по
торчащим из воды предметам покидает водоем и линяет на имаго.
Личинки стрекоз заселяют преимущественно стоячие или слабо-
проточные водоемы: озера, пруды, старицы рек, медленно текущие
реки, и лишь немногие обитают в быстрых реках. В зависимости от
экологических особенностей, которые отражаются на внешнем облике,
выделяется несколько морфо-экологических типов личинок.
1. Длинноногие узкожаберные личинки рек и ру-
чьев (рис. 208). Плавают плохо, предпочитают сидеть, прицепившись
к водным растениям длинными и тонкими ногами. Типичные предста-
вители перифитона. Питаются личинками поденок. Представлены ви-
дами семейства красоток (Calopterygidae).
88
2. Длиннобрюхие, активно плавающие личинки
стоячих вод (рис. 209). Движение осуществляется реактивным
способом: резко выталкивая воду из прямой кишки, личинки рывками
двигаются вперед. Активные хищники. Питаются различными водными
насекомыми, головастиками и мальками рыб. Представлены видами
семейства коромысл (Aeschnidae).
3. Широкожаберные личинки стоячих и медленно
текущих вод (рис. 210). Обитают в защищенных, неглубоких мес-
Рис. 208—212. Морфо-экологические типы личинок стрекоз.
208— Calopieryк vlrgo L. (Calopterygidae), 209— Aeschna juncea L. (Aeschnidae), 210— Lestes
sponsa Hans. (Lestidae), 211— Oph.iogotnph.us serpeniinus Charp. (Gomphidae), 212— Libellula
quad maculata L. (Libellulidae).
тах с богатой растительностью. Имеют удлиненное тело и листовидные
жабры на вершине брюшка. Плавают, загребая воду ногами и из-
гибая тело; хвостовые жабры при этом выполняют роль непарного
плавника. Питаются мелкими личинками насекомых и рачками. Пред-
ставлены видами всех семейств равнокрылых стрекоз, кроме красоток
(Calopterygidae).
4. 3 а р ы в а ю щ и е с я волосатые личинки (рис. 211).
Тело густо покрыто длинными волосками. Ноги сильные, с шипами
и часто с копательным зубцом на голени. Передвигаются очень медлен-
но. Большую часть времени проводят зарывшись в грунт, оставляя на
поверхности лишь усики и самый кончик брюшка. Типичные предста-
вители бентоса. Встречаются в реках и прибойных зонах озер. Связа-
89
ны с песчаными и галечниковыми участками дна. Питаются олигохе-
тами. Представлены видами семейства дедок (Gomphidae).
5. Короткобрюхие ползающие личинки стоячих
вод (рис. 212). Для движения не используют реактивный способ.
Медленно ползают по дну. Обитатели илистых грунтов. Питаются ли-
чинками комаров-хирономид и различными органическими частицами,
процеживая ил с помощью маски. Представлены видами семейств
бабок (Corduliidae) и настоящих стрекоз (Libellulidae).
Отряд стрекоз охватывает более 3000 видов, но большинство из
них приурочено к тропикам и субтропикам. Фауна СССР насчитывает
около 170 видов.
Определительная таблица семейств
1(8). Передние и задние крылья одинаковы по форме и жилкованию.
Имеется крыловой четырехугольник (рис. 206). (Подотряд Рав-
нокрылые стрекозы — Zygoptera).
2(3). Крылья темноокрашенные или дымчатые, их основание не сте-
бельчатое. Птеростигмы (рис. 206, пт) у самца нет, у самки на
ее месте находится светлое пятно ..............................
...............................Красотки — Calopterygidae.
Рис. 213—216. Детали строения стрекоз.
213 — основание переднего крыла у самца Coenagrlon. hastulattim Charp.. 214 — то же
у самца Platycnemls pennipes Pall., 215— голова стрекозы-бабки (Corduliidae),
вид сбоку, 216 — то же настоящей стрекозы (Libellulidae).
3(2). Крылья прозрачные, их основание стебельчатое (рис. 213, 214).
Птеростигма всегда имеется. ;
4(5). Птеростигма длинная, равная по длине по крайней мере двум
ячейкам (рис. 206). Тело обычно бронзово-зеленое..................
............................................ Лютки — Lestidae.
5(4). Птеростигма короткая, длиной в одну ячейку. Тело без метал-
лического блеска.
6(7). Крыловой четырехугольник правильный, длина его передней
стороны примерно равна длине задней стороны (рис. 213). Го-
лени двух задних пар ног у самца расширены........................
................... Плосконожки — Platycnemidae.
7(6).. Крыловой четырехугольник неправильный, передняя его сто-
рона значительно короче, чем задняя (рис. 214). Голени ног
у самца не расширены ...........................•...............
.................... . . . Стрелки — Coenagrionidae.
8(1). Передние и задние крылья не одинаковы по форме и жилко-
ванию. Имеется крыловой треугольник (рис. 207). (Подотряд
Разнокрылые стрекозы — Anisoptera).
9(14). Крыловые треугольники на всех крыльях расположены оди-
наково— вытянуты по длине крыла (рис. 207).
10(11). Глаза не соприкасаются. Самки без яйцеклада.................
. . ......................................Дедки — Gomphidae.
90
11(10). Глаза хотя бы частично соприкасаются. Самки с яйцекладом.
12(13). Глаза соприкасаются только в одной точке. Черные с желты-
ми пятнами стрекозы. В европейской части СССР один род
с одним видом — Cerdulegaster annulatus Latr.....................
....................................... Cordulegasteridae.
13(12). Глаза соприкасаются на некотором протяжении. Окраска дру-
гая..................................... Коромысла — Aeschnidae.
14(9). Крыловые треугольники расположены неодинаково: на перед-
них крыльях вытянуты поперек крыла, на задних — вдоль
крыла.
15(16). Задний край глаз посередине сильно изогнут (рис. 215). Тело
обычно с металлическим блеском ...................................
........................................Бабки — Corduliidae.
16(15). Задний край глаз прямой (рис. 216). Тело без металлического
блеска............................. Настоящие стрекозы — Libellulidae.
ОТРЯД ТАРАКАНЫ — BLATTOPTERA *
Общая^характеристика
К отряду тараканов относятся крупные или средних размеров на-
секомые с сильно уплощенным в дорсовентральном направлении телом
(рис 217). Голова характерной треугольной формы, сверху прикрыта
большой переднеспинкой. Ротовой аппарат грызущего типа. Усики
длинные, нитевидные. Крылья нередко укорочены или отсутствуют
(рис. 218). У крылатых форм передние крылья кожистые, жилкование
Рис. 217—223. Тараканы.
217 — общий вид таракана, 218 — самка с укороченными крыльями, 219 — самка Blatta orienialls
L. (Blattidae), 220— самец Blatella germanlca L. (Blatellldae), 221— самец Eciobtus lapponlcus
L. (Blatellldae), 222 — самка Eciobius duskei Adel., 223 ~ самка Phyllodromlca megerlei
Fieb. (Blatellldae).
♦ А. К. Бродский.
91
простое, почти исключительно продольное. Задние крылья перепонча-
тые, с богатым жилкованием и большой анальной лопастью (|рис. 217).
Анальная лопасть в покое складывается веером под плоско лежащими
на брюшке надкрыльями или подгибается лишь в одном месте и вееро-
образно не складывается. Ноги сильные, бегательные, вооружены ши-
пами. Лапки 5-члениковые. Конец брюшка с короткими членистыми
церками. Брюшко самцов часто с пахучими железами.
Превращение неполное. Яйца заключены в оотеку (плотный пакет,
покрытый общей оболочкой), которую самка вынашивает нй- конце
брюшка. Личинки претерпевают 5—9 линек до достижения имагиналь-
ной стадии. С каждой линькой помимо общего роста увеличивается
количество члеников в усиках и церках. Продолжительность жизни
тараканов — от 1 года до 7 лет. Крупные виды медленнее развиваются
и.дольше живут. Продолжительность онтогенеза у одного и того же
вида зависит от внешних факторов. Например, развитие черного та-
ракана (Blatta orientalis L.) заканчивается в среднем через год, но
при неблагоприятных условиях может растягиваться до 4—5 лет. Ос-
новные экологические особенности тараканов — скрытый образ жизни
и стенотермность.
Все тараканы активны в ночное время. Летают редко и неохотно,
предпочитая прятаться в различных полостях и скважинах. Бегают
быстро. Всеядны, но виды, живущие в природе, — либо детритофаги,
либо фитофаги.
Группа синантропных тараканов, связанных в своем распростра-
нении с человеком, охватывает несколько семейств. Самыми типичными
представителями этой цруппы являются черный таракан (Blatta orien-
talis L.) из семейства Blattidae и рыжий таракан, или прусак (Blatella
g-ermanica L.) из семейства Blatellidae. У черного таракана тело дли-
ной 18—30 мм, блестящее, как будто лакированное, черной или черно-
вато-бурой окраски (рис. 219); самки имеют укороченные крылья, не
достигающие первого брюшного сегмента. Прусак меньше по размерам
(10—30 мм), буровато-рыжего цвета с двумя темными полосками на
переднеспинке (рис. 220); надкрылья у самцов и самок прикрывают
все брюшко. Синантропные тараканы обычно живут в домах и лишь
в теплых странах встречаются и на воле.
Лесные тараканы обитают во влажных местах, часто по берегам
рек, в подстилке, в разлагающейся древесине. В европейской части
СССР эта экологическая группа представлена несколькими видами
рода Ectobius Steph, из семейства Blatellidae. Один из них —
Ectobius lapponicus L. распространен по всей лесной зоне, а на севере
заходит и в тундру. Для зоны степей типичен другой вид того же ро-
да— Е. duskei Adel. У самца Е. lapponicus в центре переднеспинки
находится темное пятно (рис. 221), а у самца и самки Е. duskei — чер-
ная дугообразная полоса (рис. 222). На юге Украины, в Крыму и на
Кавказе встречаются некоторые виды рода Phyllodrotnica Fieb.
(рис. 223), тоже относящиеся к семейству Blatellidae. Для зоны пус-
тынь характерны крупные тараканы-черепашки (Polyphagidae).
Определительная таблица семейств
1(2). Длина тела не более 13 мм. Самки крылатые, короткокрылые
или бескрылые................................... Blatellidae.
2(1). Длина тела от 18 до 40 мм. Самки с укороченными крыльями
или бескрылые.
3(4). Тело или хотя бы края переднеспинки и передний край надкрыль-
ев покрыты волосками.............................Polyphagidae.
4(3). Тело голое. Синантропы.........................Blattidae.
92
ОТРЯД БОГОМОЛЫ - MANTOPTERA
Богомолов легко опознать по их своеобразному внешнему облику.
Размеры средние или крупные, тело удлиненное. Голова треугольной
формы, ротовой аппарат грызущего типа. Усики многочлениковые,
обычно нитевидные, реже перистые. Переднегрудь шеевидно вытянута.
Обе пары крыльев хорошо развиты, иногда в различной степени уко-
рочены или отсутствуют. Передние крылья узкие, задние широкие
с веерообразной анальной частью. Все крылья с ясным продольным
жилкованием. В покое передние крылья ^рблегающе ч ложатся на
брюшко и прикрывают веерообразно сложенные задние. Передние но-
ги хватательные с очень длинными тазиками и сильными, увенчанными
крупным шипом голенями, которые могут наподобие перочинного ножа
~ задние ноги ходильного
и
Рис. 224—225. Богомолы.
224 — голова самки Empasa fasciata
Brulle (Empusidae)» 225— Mantis reli-
giosa L. (Mantidae), общий вид.
плоТно прикладываться к бедрам. Средние
типа. На вершине брюшка находятся мно-
гочлениковые церки.
Развитие с неполным превращением.
Яйца, как и у тараканов, заключены в
оотеку. Личинки первого возраста отлича-
ются от всех последующих возрастов; их
передние ноги короткие, тело покрыто ши-
пиками, брюшко заканчивается двумя
длинными хвостовыми нитями. Личиноч-
ное развитие сопровождается 7—8 линь-
ками.
Богомолы — теплолюбивые насекомые.
Они приурочены к сухим, хорошо прогре-
ваемом стациям и распространены глав-
ным образом в степях и пустынях. Зеле-
ный цвет степных богомолов по мере вы-
сыхания растительности постепенно пере-
ходит в бурый. Для пустынных богомолов
характерна серая окраска, прекрасно мас-
кирующая их на фоне песка и глины.
Взрослые богомолы и их личинки — это типичные хищники-засадники.
Подстерегая добычу, они могут подолгу сидеть неподвижно в харак-
терной позе с высоко поднятой на длинной переднегруди головой и сло-
женными как бы «в молитвен передними ногами. Нападение провоци-
руется движениями различных объектов в поле зрения богомола: стоит
какому-нибудь насекомому пролететь мимо, как ’богомол хватает его
одним молниеносным выпадом передних ног. Добычей могут стать до-
статочно крупные насекомые, такие как прямокрылые, бабочки, мухи
и перепончатокрылые. Личинки младших возрастов кормятся тлями
и другими мелкими насекомыми.
Отряд богомолов охватывает около 2000 видов. В СССР обнару-
жено 20 видов и 2 семейства. У представителей семейства
Empusidae голова украшена торчащим вперед коническим отро-
стком (рис. 224), а у видов семейства Mantidae голова без
такого отростка (рис. 225). В европейской части СССР к югу от 50—
54° с. ш. часто встречается обыкновенный богомол (Mantis religiosaL.)
из семейства Mantidae. Тело обыкновенного богомола достигает 50—
60 мм и окрашено в зеленый или буровато-желтый цвет; крылья и над-
крылья хорошо развиты.
* А. К. Бродский.
93
ОТРЯД ТЕРМИТЫ — ISOPTERA *
Термиты—это исключительно общественные насекомые мелких или
средних размеров. Ротовой аппарат грызущего типа. Усики нитевид-
ные, короче тела. Крылья имеются лишь у способных к размножению
самцов и самок. Крылья длинные, перепончатые, в сложенном состоя-
нии далеко выступают за конец брюшка. Задняя пара крыльев без
веерообразной анальной части, по форме и размерам напоминает пе-
реднюю. На конце брюшка имеются церки, состоящие из различного
числа члеников (от 1 до 8). х
Постэмбриональное развитие с неполным превращением. На базе
неполного превращения развился кастовый полиморфизм, который рас-
пространяется не только на имагинальные стадии, но и на личинкопо-
добных особей.
Личинка термитов'—это особая стадия развития без зачатков
крыльев и других признаков кастовой дифференцировки. Ее покровы
Рис. 226—229. Касты термита Anacanthoterm.es ahngerianus Jac.
226 — солдат, 227 — рабочий, 228 — крылатое имаго, 229 — самка, сбро-
сившая крылья (царица).
слабо склеротизованы, обычно белого цвета. Термиты всех каст про-
ходят, через стадию личинки, включающую несколько возрастов. '
Про со л дат — промежуточная стадия превращения личинки, ра-
бочего или нимфы в солдата. Морфологически эта стадия похожа на
солдата, но некоторые структуры головы еще не вполне сформирова-
ны, и все тело лишено пигментации. Функция солдат (рис. 226)
состоит в защите гнезда от врагов, особенно от других видов терми-
тов и муравьев. В качестве оружия используются мощные мандибулы,
затыкание головой отверстий гнезда и опрыскивание врага клейким
секретом лобной железьк Каждому способу^-- защиты соответствуют
специальные морфологические изменения головы.
Рабочие термиты (рис. 227)—бескрылые формы с недораз-
витыми мужскими или женскими половыми органами. Они занимаются
сбором и доставкой корма в гнездо, кормят половых особей, солдат
и личинок, заботятся о 'яйцах, ведут строительные работы.
* А. К. Бродский.
94
I
Нимфа — стадия развития с зачатками крыльев. После несколь-
ких линек она превращается в имаго. Крылатые имаго (рис. 228)
дружно вылетают из гнезда и вскоре после роения обламывают себе
крылья. Затем они отыскивают полового партнера и парами уходят
в укрытие, чтобы заложить новую семью. Брюшко яйцекладущей сам-
ки (царицы) разрастается до громадных размеров (рис. 229), а ее
плодовитость достигает 80 000 яиц в сутки.
Термиты ведут скрытый образ жизни. Живут колониями в почве
или древесине. Питаются древесиной, сухими растительными остатка-
ми и детритом, иногда разводят грибы. Многие тропические виды
строят надземные сооружения (термитники). Внутри термитника под-
держиваются оптимальные условия влажности и температуры. Однако
возможность регуляции физических условий внутри колонии ограни-
чена, поэтому большинство термитов обитает в тропиках и субтропиках.
Лишь немногие виды проникли в зону умеренного климата. В ев-
ропейской части СССР только 2 вида. Kalotermes flavicollis F. встре-
чается на Черноморском побережье Кавказа, a Ruticulitermes luci-
fugus Rossi. — в Молдавии и на юге Украины. Оба вида могут разру-
шать деревянные постройки и старые деревья.
На территории СССР обитают 7 видов термитов из 4 семейств.
Они встречаются помимо европейской части СССР в Средней Азии,
Закавказье и во Владивостоке (1 завезенный вид). Самый крупный
термит нашей фауны — Anacanthotermes ahngerianus Jac. (рис. 226—
229) распространен в Туркмении и на юге Казахстана.
ОТРЯД ВЕСНЯНКИ — PLECOPTERA*
Общая характеристика
К веснянкам относятся небольшие или средних
мые с удлиненным телом (рис. 230). Усики длинные,
Ротовой аппарат грызущего типа; нередко ротовые
дуцированы. Грудные сегменты оди-
наковых размеров. Обе пары .
крыльев перепончатые, с ветвящи-
мися продольными жилками.
Задние крылья отличаются от пе-
редних наличием анального веера.
В покое крылья складываются ли-
бо облегающе, либо трубкообраз-
но. У некоторых веснянок крылья
укорочены или вообще отсутству-
ют. Ноги ходильные, с 3-членико-
выми лапками (рис. 231—233). На
конце брюшка расположены цер-
ки — длинные, многочлениковые,
реже укороченные, нечленистые,
иногда сильно видоизмененные.
Превращение неполное. Ли-
Чинки водные, по внешнему облику
похожие на взрослых, отличаясь
от них отсутствием крыльев и на-
размеров насеко-
многочлениковые.
части сильно ре-
ружных половых придатков. Тра- Рис 230 Веснянка Perla Geoffr. (Рег.
хеиные жабры иногда имеют лис- lidae), общий вид.
товидную форму, но чаще пред- а.л — анальная лопасть.
* А. к. Бродский.
95
ставлены нитевидными выростами на разных сегментах груди и брюш-
ка. Развитие личинок продолжается от 1 года до 3 лет и сопровож-
дается большим числом линек (до 24).
Личинки веснянок живут в быстротекущих, холодных и богатых
кислородом водах. В целом они могут быть охарактеризованы как
холодолюбивые стенотермные оксибионты, приуроченные к горным ре-
кам и ручьям. Некоторые личинки хорошо плавают, но чаще ходят
по дну, цепляясь за подводные предметы. Ранние личиночные стадии
питаются детритом, поздние — преимущественно хищники. Они напа-
дают на личинок поденок и двукрылых, на ракообразных и себе по-
добных. В горных реках личинки веснянок наряду с личинками поде-
нок составляют основу кормовой базы многих рыб, в том числе фо-
релей.
Рис. 231—237. Детали строения веснянок.
231—233 — лапки (231 — Taeniopterygidae, 232— Nemouridae, 233 — Perli-
dae), 234—235 — передние (вверху) и задние (внизу) крылья (234—
Nemouridae, 235— Perlodidae). 236—237— передние крылья (236—
Arcynopteryx Klap., 237— Diura Bill.), а.л — анальная лопасть; С, R,
Ri, RS — крыловые жилки.
Личинки последнего возраста перед имагинальной линькой поки-
дают водоем. Только что перелинявшие взрослые веснянки длитель-
ное время остаются на месте, после чего перелетают на различные
предметы (стволы деревьев, травянистые стебли, камни и т. д.). Для
некоторых видов отмечены миграционные перелеты на расстояние в не-
сколько сот метров от водоема. Многие веснянки роятся: они быстро
летают у поверхности воды, часто меняя направление. Копуляция про-
исходит на субстрате, после чего самка летит откладывать яйца. Ле-
тящая самка легко контактирует с водой и освобождается от пакета
с яйцами, висящего на конце брюшка. Поэтому некоторые виды вес-
нянок имеют длиннокрылых самок и короткокрылых самцов. Взрослые
веснянки, за небольшим исключением, не питаются.
Отряд насчитывает примерно 2000 видов, из которых на террито-
рии СССР встречается около 200.
Определительная таблица семейств
1(8). Первый членик лапок длинный, приблизительно такой же дли-
ны, как третий (рис. 231). Церки одночлениковые или укоро-
ченные, иногда многочлениковые.
S6
2(3). Все 3 членика лапок приблизительно одинаковой длины
(рис. 231). Це|рки состоят не более, чем из 10 члеников. . . .
...............................................Taeniopterygidae.
3(2). Второй членик лапок самый короткий, менее половины длины
первого или третьего; первый и третий приблизительно равные
(рис. 232). ,
4(5). В вершинной прети крыльев поперечные жилки образуют
Х-образную фигуру (рис. 234). Крылья всегда развиты нор-
мально. Церки одночлениковые, часто видоизмененные. . .
.....................................................Nemouridae.
5(4). Крылья без Х-образной фигуры близ вершины (рис. 235).
Церки многочлениковые или одночлениковые, не видоизменен-
ные.
'•6(7). Церки одночлениковые. Крылья нормально развиты, склады-
ваются трубкообразно и выпукло лежат над брюшком. . . .
............................................. . Leuctridae.
7(6). Церки усиковидные; в церке не менее 7—8 члеников. Крылья
лежат на брюшке плоско, иногда укорочены.......................
...............................................Capniidae.
8(1). Первый и второй членики лапок короткие (рис. 233), третий
значительно длиннее, чем первый и второй вместе взятые. Цер-
ки длинные.
9(14). Крылья нормальной длины; достигают конца брюшка или за-
ходят за него.
10(13). Задние крылья с большой, хорошо развитой анальной ло-
пастью (рис. 230, 235, а.л) с многочисленными (4—10) аналь-
ными жилками.
11(12)'. Передние крылья с беспорядочной сетью жилок близ верши-
ны (рис. 236) или не более чем с 2 поперечными жилками
между Си/?! (рис. 235) или с темным пятном в области по-
перечной жилки между /? и /?5 (рис. 237)...........................
..............................................Perlodidae.
12(11). Передние крылья без сети жилок у вершины, более чем с 2 по-
перечными жилками между С и Ki, темное пятно отсутствует
(рис. 230)............................................. Perlidae.
13(10). Задние крылья со слабо развитой, небольшой анальной лопас-
тью, имеющей не более 3 анальных жилок......................... . .
............................................Chloroperlidae.
14(9). Крылья укороченные, не достигающие конца брюшка, часто
прикрывают лишь первые 4—5 брюшных сегментов.....................
. . „ . . . Perlodidae и Perlidae (короткокрылые самцы).
ОТРЯД ПРЯМОКРЫЛЫЕ-ORTHOPTERA*
Общая характеристика
К отряду прямокрылых относятся насекомые средних или крупных
размеров. Голова с отвесным или скошенным назад лбом. Ротовой ап-
парат грызущего типа. Усики длинные, многочлениковые; форма их
разнообразна, но преобладают нитевидные (рис. 238). Для всех пря-
мокрылых характерна крупная, нависающая на бока переднеспинка.
Передние и средние ноги бегательные, задние — прыгательные с утол-
щенными бедрами и длинными голенями. Передняя пара крыльев уз-
кая, кожистая, превращена в надкрылья. Задняя пара широкая, пере-
пончатая; в покое складывается веерообразно под узкими передними
* А. К. Бродский.
7 208
97
крыльями. Нередко встречаются короткокрылые формы, а иногда бес-
крылые. На конце брюшка расположены одночлениковые церки; они
обычно короткие, но у сверчков (Gryllidae) и медведок (Gryllotalpi-
dae) длинные и гибкие. У большинства прямокрылых самки имеют
яйцеклад — длинный мечевидный или серповидный (у кузнечиковых —
Tettigonioidea), копьевидный (у сверчковых — Grylloidea) или корот-
кий копательный (у саранчовых — Acridoidea). Многие прямокрылые
имеют специальные органы стрекотания, расположенные у кузнечико-
вых и сверчковых на надкрыльях, а у саранчовых на надкрыльях
и внутренней поверхности задних бедер. Органы слуха локализованы
на передних ногах (у сверчковых и кузнечиковых) или по бокам пер-
вого сегмента брюшка (у саранчовых).
Развитие с неполным превращением. Яйца откладываются в поч-
ву, реже — в ткани растений. Для саранчовых характерна откладка
яиц в почву с образованием
Рис. 238. Кузнечик Decticus verrucivo-
rus L.
кубышки (особой защитной
капсулы). За время личиноч-
ного развития саранчовых
происходит 4—5 линек. Кры-
ловые зачатки становятся за-
метными уже у личинок вто-
рого возраста и с каждой
линькой увеличиваются в раз-
мерах.
Прямокрылые —это в ос-
новном теплолюбивые насеко-
мые, населяющие открытые
местообитания (луга, поля-
ны, степи, пустыни). Сравни-
тельно немногие виды обита
ют под пологом леса. В за-
висимости от экологических
особенностей, которые отражаются на внешнем облике, выделяют 2 мор-
фо-экологических типа, называемых также «жизненными формами»
(по классификации Г. Я. Бей-Биенко).
1. Фитофилы — обитатели толщи растительного покрова
‘(рис. 239, 24,0). Наиболее многочисленны среди них хортобионты —
обитатели травянистой растительности (рис. 240). Для хортобионтов
характерно стройное удлиненное тело, скошенный лоб и защитная
окраска под цвет травы. Представлены главным образом многими ви-
дами семейства настоящих саранчовых (Acrididae). Другая группа
фитофилов — тамнобионты, или обитатели крон деревьев и кустарни-
ков (рис. 239). Представлены различными видами семейства ' кузне-
чиков (Tettigoniidae) и стеблевыми сверчками (Oecanthidae).
2. Геофилы — обитатели поверхности и толщи почвы (рис. 241 —
243). В свою очередь, подразделяются на открытых геофилов, скры-
тых геофилов и геобионтов. Представители первой группы на расте--
ния взбираются лишь на время питания. Основное время проводят на
поверхности почвы. Характеризуются более или менее уплощенным
телом; покровы испещрены различными морщинками и бугорками, что
делает их незаметными на фоне субстрата (рис. 241). Имеют крип-
тическую окраску под цвет гальки, песка, земли. Задние крылья не-
редко яркие — красные, голубые, с черной полосой. Они привлекают
внимание в момент взлета, который к тому же сопровождается тре-
ском крыльев. Преимущественно обитатели пустынь и полупустынь.
Представлены различными видами кобылок из семейств настоящих
и пустынных саранчовых (Acrididae и Pamphagidae). Скрытые геофи-
98
лы, или герпетобионты, обитают- под камнями, опавшей листвой, в соб-
ственных норках или в норах позвоночных. Для них характерны не-
большие размеры, веретеновидная форма тела и темная, коричневых
оттенков, окраска (рис. 243). Представлены сверчками (Gryllidae).
К геобионтам относятся медведки (Gryllotalpidae). Они имеют копа-
тельные передние ноги (рис. 242) и постоянно живут в почве.
Подавляющее большинство прямокрылых является фитофагами.
Спектр поедаемых растений очень широк. Ряд видов кузнечиков (Tet-
tigoniidae), в том числе крупный бескрылый кузнечик-дыбка (Saga
Ch.), — облигатные хищники. Другие виды кузнечиков, а также мед-
ведки характеризуются смешанным питанием.
Рис. 239—243. Прямокрылые.
239 — зеленый кузнечик Тeitigonla vlridissima L. (Tettigoniidae), 240 — степной конек Euchorthippas
pulvinatus F.—W. (Acrididae), 241— степная кобылка Asiotmethis muricaius Pall. (Pamphagidae),
242— медведка Gryllotalpa gryllotalpa L. (Gryllotalpidae), 243 — степной сверчок Gryllus desertus
Pall. (Gryllidae).
Прямокрылые составляют весьма многочисленный отряд насеко-'
мых, насчитывающий свыше 20 000 видов, из которых более 700 встре-
чается в СССР. Они широко распространены по всему земному шару.
Некоторые представители растительноядных прямокрылых являются
серьезными вредителями сельского хозяйства.
Определительная таблица семейств
1(8). Усики длиннее тела (рис. 238). Орган слуха расположен на
передних голенях (рис. 244). Самки с длинным торчащим яйце-
кладом (рис. 239, 243), редко без яйцеклада.
2(3). Все лапки 4-члениковые (рис. 245). Крылья в покое сложены
крышеобразно (под углом). Окраска зеленая или серая. (Над-
семейство Кузнечиковые — Tettigonioidea)..........................
....................................Кузнечики — Tettigoniidae.
99
3(2). Передние и средние лапки 3-члениковые (рис. 244). Крылья
в покое сложены плоско ('рис. 243). Окраска черная, коричне-
вая или желтовато-коричневая. (Надсемейство Сверчковые —
Grylloidea).
4(5). Передние ноги копательные (рис. 242). Самки без яйцеклада.
Крупные формы, более 35 мм . . Медведки — Gryllotalpidae.
5(4). Передние ноги обычного строения, бегательные (рис. 243).
Самки с копьевидным яйцекладом. Размеры тела не более
25 мм.
6(7). Задние голени сверху с мелкими шипиками между единичными
более крупными шипами (рис. 246). Голова плоская, ротовые
органы направлены вперед. Живут на растениях.......................
..........................Стеблевые сверчки — Oecanthidae.
Рис. 244—250. Детали строения прямокрылых.
244 — передняя левая нога сверчка (Gryllidae), 245 — задняя лапка кузнечика (Tettigoniidae).
246 — задняя голень стеблевого сверчка (Oecanthidae), 247 — правая задняя голень н лапка
сверчка (Gryllidae), 248—-голова, переднеспинка и надкрылья прыгунчика (Tetri'gidae), 249 —
заднее бедро представителя пустынных саранчовых (Pamphigidae), 250 — то же представителя
настоящих саранчовых (Acrididae).
7(6). Задние голени сверху без мелких шипиков (рис. 247). Голова
округлая, лоб отвесный, ротовые органы направлены вниз
(рис. 243). Живут на поверхности почвы и в норках . . . .
...........................................Сверчки — Gryllidae.
8(1). Усики короткие, не длиннее половины тела (рис. 240, 241).
Органы слуха размещаются по бокам, первого сегмента брюш-
ка. Самки с коротким яйцекладом. (Надсемейство Саранчо-
вые — Acridoidea).
9(10). Переднеспинка вытянута назад в длинный отросток, прикры-
вающий сверху все брюшко (рис. 248). Длина тела не превы-
шает 18 мм .............................Прыгунчики — Tetrigidae.
10(9). Переднеспинка короткая, не прикрывает брюшко или лишь
слегка заходит на него (рис. 240, 241). Длина тела от 10 до
80 мм.
11(12). Бедра задних ног снаружи между продольными килями в бу-
горках и ребрышках, не образующих правильной перистой
скульптуры (рис. 249). Поверхность тела сильно шероховатая,,
(рис. 241)................. Пустынные саранчовые—Pamphagidae.
12(11). Бедра задних ног снаружи между продольными килями с пра-
вильной перистой скульптурой (рис. 250). Поверхность тела
гладкая (рис. 240) или шероховатая.............................
...................... . Настоящие саранчовые — Acrididae.
100
ОТРЯД УХОВЕРТКИ, ИЛИ КОЖИСТОКРЫЛЫЕ —DERMAPTERA
Общая характеристика
Уховертки составляют сравнительно небольшой (около 1200 видов)
отряд насекомых, имеющих удлиненное тело и укороченные кожистые
надкрылья, под которыми спрятана сложенная вдоль и поперек вто-
рая, перепончатая пара крыльев (рис. 251, 252). Голова почти сердце-
видной формы. Ротовой аппарат грызущего типа. Усики нитевидные.
У ряда видов крылья могут отсутствовать. Ноги бегательные, -лапки
3-члениковые. В связи с укорочением надкрылий брюшные сегменты
плотные, сильно склеротизованные. На конце брюшка расположены
клещевидные нечленистые придатки, которые являются видоизменен-
ными церками. У самцов форма этих придатков изменчива, и внутри
одного вида встречаются особи как с укороченными, так и с длинны-
ми придатками (рис. 253, 254).
Рис. 251—258. Уховертки и детали их строения.
25/ — самец Forficala auricularia L. (Forficulidae), общий вид, 252 — заднее крыло, 253—254 —
клещевидные придатки самца (253— укороченные, 254— удлиненные), 255—256 — задние лапки
(255 — Forficulidae, 256 — Labiduridae), 257—258 — клещевидные придатки (257 — Labiidae, 258 —
Labiduridae).
Развитие с неполным превращением. Спаривание происходит в кон-
це лета. Яйца откладываются по 50—80 штук в специально вырытые
ямки или норки. В норках зимуют яйца, личинки, имаго. Самки про-
являют заботу о потомстве и охраняют кладки, прикрывая их головой
и передними ногами. \
Уховертки — это теплолюбивые и влаголюбивые насекомые. Ведут
ночной образ жизни, днем скрываются под камнями, опавшей листвой,
в гнилых пнях и т. д. Распространены в лесах. Пища уховерток чрез-
вычайно разнообразна. Они могут питаться гниющими органическими
остатками, водорослями, грибами или живыми частями высших расте-
ний. Некоторые виды хищничают, нападая на насекомых, пауков,
червей.
В СССР встречаются 26 видов и 3 семейства уховерток. В евро-
пейской части СССР самые массовые виды — Forficula auricularia L.
(рис. 251) и F. tomis Koi. из семейства Forficulidae. Они нередко по-
селяются па огородах, в садах и на бахчах.
* А. К. Бродский.
101
Определительная таблица семейств
1(2). Второй членик лапок расширен сердцевидно, шире третьего
(рис. 255)............................................. Forficulidae.
2(1). Второй членик лапок простой, цилиндрический, не шире третьего
(рис. 256).
3(4). При рассматривании насекомого сверху между основаниями кле-
щевидных придатков хорошо виден выступ — пигидий (рис. 257).
. , . , . ..................................................Labiidae.
4(3). Выступ между основаниями клещевидных придатков сверху не
виден (рис. 258).........................................Labiduridae.
ОТРЯД СЕНОЕДЫ-— PSOCOPTERA *
К этому отряду относятся в основном очень мелкие насекомые
длиной 1—5 мм с нитевидными усиками, бегательными ногами и двумя
парами перепончатых крыльев, складываемых крышеобразно (рис.
259); иногда крылья укорочены или отсутствуют (рис. 260). Ротовой
Рис. 259—260. Сеноеды.
259 — Amphigerontia Koi. (Psocidae), 260— Liposcelis divinatorius МОП.
(Liposcelidae).
аппарат своеобразный, грызущий. Верхние челюсти массивные, а внут-
ренняя лопасть нижних челюстей обособлена в виде подвижного и твер-
дого стилета, далеко вдающегося внутрь головы и снабженного на на-
ружном конце зубцами. Эти стилеты служат для раздробления пищи
перед едой. Уникальный дробящий ротовой аппарат ^позволил сено-
едам использовать сухую и твердую растительную пищу, недоступную
для большинства других насекомых. Это растительная труха, лишай-
ники, мицелий грибов, одноклеточные водоросли. Лишь немногие сено-
еды питаются живыми тканями растений.
Миниатюрность сеноедов позволила им заселить разнообразные
скрытые местообитания и мелкие полости различных субстратов. Они
обитают одиночно или группами в лесной подстилке, в трещинах коры
и под корой, в норах и гнездах, под камнями, в трещинах каменных
стен и т. п.
Размножение обоеполое, у части видов партеногенетическое. Спа-
риванию предшествуют ритуальные танцы самцов перед самкой. Сам-
ки откладывают яйца с помощью небольшого яйцеклада по одному или
кучками, прикрывая их железистыми выделениями задней кишки или
* В. Е. Кипятков.
102
паутиной. Паутинные железы у сеноедов открываются на нижней губе.
Развитие с неполным превращением. Личинки живут поодиночке или
группами, устраивая общее паутинное гнездо. Большинство видов
имеет несколько поколений в году. Зимуют яйца, реже личинки или
имаго.
Сеноеды широко распространены по всему земному шару, но
большинство видов обитает в тропиках и субтропиках. Известно более
1600 видов сеноедов, группируемых в 30 семейств и 3 подотряда;
в СССР встречается только 60 видов. В лесной зоне европейской части
СССР на стволах деревьев часто обитают снабженные крыльями виды
рода Amphigerontia Koi. (рис. 259) из семейства Psocidae. Среди си-
нантропных сеноедов заслуживают упоминания домовой сеноед, или
пыльная вошь (Trogium pulsatorium L.) из семейства Trogidae и книж-
ная вошь (Liposcelis divinatorius М.) из семейства Liposcelidae.
У Т. pulsatorium крылья рудиментарные, в виде чешуйчатых зачатков,
покрытых волосками, а у L. divinatorius отсутствуют даже зачатки
крыльев (рис. 260). Оба вида повреждают гербарии, коллекции насе-
комых, растительное лекарственное сырье, книги, портят и загрязняют
крупу, хлебные изделия, сахар, сухофрукты. Сеноеды, в массе встре-
чающиеся в стогах сена, могут являться промежуточными хозяевами
овечьих це(:тод.
ОТРЯД РАВНОКРЫЛЫЕ-HOMOPTERA*
Общая характеристика
Равнокрылые — внешне разнообразные насекомые (рис. 261—265)
с малоподвижной головой. Ротовой аппарат сосущего типа, представ-
лен членистым хоботком, отходящим от заднего края головы. Усики со-
стоят из 3—10 члеников, часто короче тела. Глаза обычно хорошо
развиты, иногда они редуцированы до трех фасеточных бугорков (у не-
которых тлей — Aphidinea) или вовсе отсутствуют (у некоторых кок-
цид — Coccinea). Крылья однородные перепончатые, в покое склады-
ваются кровлеобразно. У самцов кокцид развита лишь передняя пара
крыльев, а самки бескрылые. Жилкование крыльев явственное, обычно
обедненное, лишь у цикадовых (Cicadinea) имеются поперечные жил-
ки. Ноги бегательные, у листоблошек (Psyllinea) и большинства ци-
кадовых задние ноги прыгательные, некоторые тли обладают передни-
ми прыгательными ногами. Иногда ноги сильно укорочены (у некото-
рых тлей и кокцид) или недоразвиты (у самок кокцид).
Превращение неполное, у белокрылок (Aleyrodinea) и самцов кок-
цид усложненное неполное, у бескрылых форм упрощенное. Сезонное
развитие тлей часто сопровождается сменой кормовых растений и слож-
ными циклами с чередованием партеногенетических и обоеполых по-
колений, живорождения и яйцекладки, крылатых и бескрылых самок
(см. рис. 10).
Все равнокрылые являются фитофагами. Многие виды относятся
к вредителям сельского хозяйства, причем вред усугубляется способ-
ностью переносить возбудителей вирусных заболеваний растений. Мно-
гие виды выделяют сладкие экскременты, загрязняющие листья и дру-
гие органы растений, — так называемую медвяную росу, или падь, бла-
гоприятную для развития сажистых грибов.
Мировая фауна насчитывает более 30 000 видов равнокрылых, из
них для СССР отмечено 4000 видов. Отряд делится на 5 подотрядов:
* А. X. Саулич.
103
цикадовых (Cicadinea), листоблошек (Psyllinea), белокрылок (Aleyro-
dinea), тлей (Aphidinea) и кокцид (Coccinea). Во время энтомологи-
Рис. 261—265. Равнокрылые. ч
261 — цикада (Cicadinea), 262 — листоблошка (Psyllinea), 263 — белокрылка (Aleyrodinea)
264 — крылатая форма тли (Aphidinea), 265 — кокцида Coccus hesperldutn L. (Coccinea).
ческой практики более подробное знакомство с подотрядами равно
крылых целесообразно провести на примере двух подотрядов — цика
довых и кокцид.
Определительная таблица подотрядов
I 1(2). Лапки 3-члениковые. Усики очень короткие, 3-члениковые; тре-
тий членик в виде тонкой щетинки, иногда членистой. Крылья
с продольными и поперечными жилками (рис. 261).............
..................................Цикадовые—Cicadinea (с. 105).
2(1). Лапки 1—2-члениковые или отсутствуют. В составе усиков от 1
до 15 члеников; если усики 3-члениковые, то вершинный членик
не щетинковидный. Крылья, если есть, без поперечных жилок
(рис. 262—264).
3(8). Лапки обычно 2-члениковые, всегда с парой коготков; если лап-
ки отсутствуют, то тело покрыто белым восковым пушком, либо
брюшко с хвостиком или парой трубочек. Крыльев 2 пары или
их нет. Крылатые формы имеют хоботок.
104
4(5). Ноги короткие; задние ноги с утолщенными бедрами, прыгатель-
ные. Усики 10-члениковые (рис. 262)................................
...................................Листоблошки—Psyllinea.
>5(4). Ноги удлиненные, с тонкими бедрами (насекомые не прыгают).
В составе усиков не более ? члеников.
6(7). Усики 7-члениковые (рис. 263). Обе пары крыльев покрыты бе-
лым налетом, сходны по размерам и жилкованию.......................
...................................Белокрылки — Aleyrodinea.
7(6). В составе усиков от 3 до 6 члеников. Крылья без белого палета,
задние крылья меньше передних и с иным жилкованием (рис.
264).............................................. Тли — Aphidinea.
8(3). Лапки 1-члениковые, всегда с одним коготком. Крыльев 1 пара
или их нет. Крылатые формы без хоботка. Тело может быть по-
крыто сверху щитком (рис. 265). Кокциды — Coccinea. (с. 108).
Общая характеристика подотряда цикадовых — Cicadinea
Цикадовые — мелкие или средних размеров насекомые с чрезвы-
чайно разнообразной формой и окраской тела. Голова подвижно сочле-
нена с переднегрудью, несет по бокам большие фасеточные глаза и 2—
3 простых глазка, расположенных на темени. Большая часть лица
(нижняя часть головы) занята клипеусом, разделенным поперечным
швом на антеклипеус и постклипеус. У многих видов постклипеус сли-
вается со лбом, образуя фронтоклипеус. Лоб часто смещен на верх-
нюю половину головы, где образует резко очерченную лобную пла-
стинку. Хоботок 3-члениковый, прикреплен к нижней поверхности го-
ловы. Усики располагаготся под глазами или между ними. Они обычно
состоят из 3 члеников, последний из них несет длинную, иногда чле-
нистую щетинку.
Грудь состоит из трех сегментов: передне-, средне- и заднегруди.
Наиболее развита из них среднегрудь. Переднегрудь сверху прикрыта
переднеспинкой, форма и скульптурные образования которой часто
используются при определении видов. Крыльев 2 пары —’ передние бо-
лее плотные и задние перепончатые. Надкрылья разделены продоль-
ной складкой на кориум и клавус. Клавус в виде узкого треугольника
занимает внутреннюю часть крыла, а его остальная поверхность за-
нята кориумом. Наряду с полнокрылыми цикадами встречаются и ко-
роткокрылые формы. Передние и средние ноги обычно бегательные,
задние — у большинства прыгательные, хорошо развитые, с мощными
тазиками. У певчих цикад (Cicadidae) все 3 пары ног бегательные.
Размножение обычно половое, редко партеногенетическое. Яйца
цикадовых не отличаются большим разнообразием, овальные или вере-
теновидные. Они откладываются в ткани растений или свободно на по-
верхность стеблей и корней, группами или поодиночке. Количество от-
кладываемых яиц колеблется у разных видов. У певчих цикад самка
может отложить до 600 яиц.
Личинки имеют 5 возрастов и обычно похожи на имаго, только
у' певчих цикад они сильно отличаются от имагинальной стадии по
внешнему виду и образу жизни. Развитие личинок некоторых видов
происходит под землей, например, у личинок семейств Cicadidae
и Cercopidae. Для почвообитающих личинок характерен ряд морфоло-
гических и биологических особенностей. Их покровы слабо пигменти-
рованы, в ранних возрастах отсутствуют глаза, у личинок некоторых
видов тело сплющено в дорсовентралыюм направлении. Для облегче-
ния передвижения в почве передние Ноги у личинок певчих цикад пре-
вращаются в копательные конечности. На теле обычны торчащие ще-
105
тинки. Личинки остальных цикад живут открыто на земле или на рас-
тениях;. питание их связано с надземными частями растений. Личинки
пенниц (Aphrophoridae) живут на растениях в комке вырабатываемой
ими пены (кукушкины слезы).
Обычно цикады дают 1—2 поколения в год, на юге —3—4. У пев-
чих цикад многолетний цикл развития, а у ряда северо-американских
видов достигает 17 лет.
Все цикадовые являются фитофагами и при этом отличаются боль-
шой прожорливостью. Избыток поглощаемой воды и сахаров выводит-
ся из организма благодаря’ своеобразной" модификации кишечника
и образованию, фильтрационной камеры, которая возникает на основе
срастания начала и конца средней кишки. Вода, содержащаяся в пи-
ще, диффундирует сквозь стенки фильтрационной камеры, минуя сред-
ний отдел кишечника. Освобожденные от воды питательные продукты
в концентрированном виде проходят через среднюю кишку. Среди ци-
кадовых встречаются полифаги, олигофаги и монофаги, но наиболее
распространена олигофагия.
Рис. 266—272. Некоторые представители цикадовых и детали их строения.
266— певчая цикада Тiblcen plebe jus Scop. (Cicadidae), 267— пенница Aphrophora silicinaGz.
(Aphrophoridae), 268 — голова цикады-свинушки (Delphacidae), вид спереди, 269 — носатка
Dlctyophora europaea L. (Dictyopharidae), 270 — голова певчей цикады, вид спереди, 271— темя
пенницы, вид сверху, 272 — то же цикадки (Cicadellidae).
Цикадовые встречаются в самых разнообразных местообитаниях.
Особенно много их в травянистой растительности. Певчие цикады жи-
вут преимущественно на деревьях и кустарниках. В районах с жарким
климатом цикады наиболее активны в вечерние и утренние часы.
В умеренном климате они активны в течение всего дня.
Большинство видов цикад относится к вредителям сельского хо-
зяйства, лишь небольшая часть видов из-за малой численности и сла-
бой вредоносности индифферентна для человека. Большой вред расте-
ниям цикады причиняют в результате высасывания соков. В этом пла-
не особенно большие убытки от цикад несут хлопко- и рисосеющие
страны тропиков и субтропиков. Не менее сильный вред цикады нано-
сят растениям яйцекладом при откладке яиц. На месте повреждения
образуются язвочки и наплывы камеди, а сами ранки служат местом
вторжения бактерий и грибов. Но наибольший вред цикады наносят
как переносчики фитопатогенных вирусов.
В мировой фауне насчитывается, около 20 000 видов цикадовых из
40 семейств, распространенных в основном в тропиках. В СССР отме-
чено около 2000 видов. Ниже приводится характеристика наиболее
важных семейств, встречающихся на территории европейской части
СССР.
106
Певчие цикады — Cicadidae. Отличаются крупными раз-
мерами. Длина тела некоторых тропических видов достигает 6,5 см.
Самая крупная наша цикада — Tibicen plebejus Scop. (рис. 266), имеет
длину тела около 5 см, а размах крыльев достигает 10 см. Обе пары
крыльев хорошо развиты, прозрачные, с мощными жилками. На голо-
ве между фасеточными глазами — 3 простых глазка. Бедра передних
ног вздутые, с 2—3 зубцами. Свое название певчие цикады получили
за их удивительную способность к стрекотанию. В брюшке самцов
находится звуковой аппарат высокой сложности и эффективности. Ос-
новой его служат 2 мембраны, приводимые в колебание двумя корот-
кими, но очень толстыми мышцами. Резонаторами служат 2 огромные
полости, представляющие собой осрбые расширения трахей и запол-
няющие почти все брюшко. В непосредственной близости от звукового
аппарата находятся органы слуха. Самки тоже имеют органы слуха,
но лишены звуковых органов. Певчие цикады откладывают яйца
в ткани растений, надпиливая их яйцекладом. Личинки, выходящие из
яиц, падают на землю и зарываются в почву. Личинки внешне не по-
хожи па имаго, имеют копательные передние ноги. Питаются корнями
растений. Личиночное развитие продолжается несколько лет. Очень
большое семейство, распространенное главным образом в тропиках.
У нас около 15 видов из 5 родов.
Пенницы —Aphrophoridae. Довольно богатое видами се-
мейство. Обычно средней величины насекомые. Глазков 2. Передние
крылья кожистые, обычно пигментированные. Между передними
крыльями есть небольшой щиток (рис. 267). Задние ноги прыгатель-
ные. Личинки живут в комке пены, защищающем их от высыхания.
Большого хозяйственного значения не имеют. В СССР около 15 видов.
Горбатки — Membracidae. Очень 'богатое видами семей-
ство, распространенное преимущественно в тропиках. В мировой фауне
около 2500 видов, у нас только 3. Характерной чертой строения этих
цикадок является сильно развитая переднеспинка, несущая обычно
гребни, шипы и другие выросты нередко причудливой формы, значение
которых до сих пор не ясно. Обе пары крыльев перепончатые. Личин-
ки горбаток ведут такой же образ жизни, как и взрослые особи. Боль-'
шое хозяйственное значение у нас в стране имеет горбатка-буйвол
(Stictocephala. bubalus F.), завезенная в Европу из Северной Аме-
рики.
Цикадки — Cicadellidae. Самое большое семейство среди
цикадовых. В СССР насчитывается около 500 видов. Большей частью
мелкие очень подвижные насекомые. Надкрылья обычно плотные, встре-
чаются виды с укороченными надкрыльями. Обитают, главным образом,
на травянистых растениях. Личинки ведут такой же образ жизни, как
и взрослые особи. Цикадки имеют большое хозяйственное значение
как переносчики вирусных заболеваний растений. Один из наиболее
распространенных представителей этого семейства — зеленая цикадка
(Cicadella viridis L.). Тело этой цикадки длиной 5—9 мм, сверху
окрашено в зеленую или сизую окраску, а голова и нижняя сторона
тела оранжево-желтые.
Определительная таблица семейств цикадовых
1(4). На голове обычно развиты продольные кили. Усики прикреп-
лены под глазами (рис. 268).
2(3). На вершине задней голени имеется большая подвижная шпо-
ра, соизмеримая по величине с члениками лапки. Голова впе-
реди не вытянутая. Длина тела меньше 5 мм.......................
. , . ...........................Свинушки — Delphacidae.
107
3(2). Шпоры нет. Голова сильно вытянута вперед (рис. 269). Длина
тела больше 5 мм..................... Носатки — Dictyopharidae.
4(1). Продольных килей на голове обычно нет. Усики всегда при-
креплены ковнутри от глаза (рис. 270).
5(6). Задние ноги бегательные. На темени 3 простых глазка. Над-
крылья прозрачные, стекловидные (рис. 266)........................
. « . . ........................Певчие цикады — Cicadidae.
6(5). Задние ноги прыгательные. На темени 2 простых глазка. Над-
крылья часто плотные и окрашенные, иногда укороченные;
7(12). На темени есть ясно очерченная лобная пластинка (рис. 271).
8(9). Темя и передняя часть переднеспинки отвесные, переднеспин-
ка несет по крайней мере 1 отросток, направленный назад и ча-
стично прикрывающий брюшко . . Горбатки — Membracidae.
9(8). Темя и переднеспинка горизонтальные без отростков (рис. 267).
10(11). Переднеспинка шире головы с глазами, сильно расширенная
кзади. Надкрылья обычно ярко окрашенные.............................
................................................Cercopidae.
11(10). Переднеспинка примерно такой же ширины, как голова с гла-
зами (рис. 267), бока переднеспинки более или менее парал-
лельные. Надкрылья плотные, кожистые................................
...................................Пенницы — Aphrophoridae.
12(7). Темя без ясно очерченной лобной пластинки (рис. 272). . .
.............................................Цикадки — Cicadellidae.
Общая характеристика подотряда кокцид,
или червецов и щитовок — Coccinea
Самки большинства видов ведут
Рис. 273—274. Кокцида Кегтосос-
cus quercus L.
273 — самеп. 274 — самка.
Кокциды — самая обособленная и наиболее специализированная
группа среди сосущих насекомых, принадлежащих к отряду равнокры-
лых (Homoptera). Наиболее характерным морфологическим призна-
ком, отличающим кокцид от других равнокрылых, является одночлё-
никовая лапка,. заканчивающаяся одним коготком. Внешне самцы
и самки кокцид очень сильно отличаются друг от друга.
Самцы имеют тело с ясным расчленением на голову, грудь и брюш-
ко (рис. 273). Они очень мелкие, для многих видов вообще неизвестны.
Самцы питаются только в личиночной стадии, а во взрослом состоя-
нии лишены ротовых частей. Характерной чертой самцов является от-
сутствие задних крыльев. Ноги нормально развиты. Усики многочле-
никовые.
малоподвижный образ жизни.
В связи с этим тело их пре-
терпело сильную деградацию:
сегментация тела частично
утрачена, исчезло ясное де-
ление на голову, грудь и брю-
шко, утрачены крылья, ноги
претерпели различную степень
редукции, иногда они совсем
отсутствуют. Усики сведены
к одночлениковому бугорку,
глаза представлены пигмент-
ными пятнами или отсутству-
ют. Самки питаются как в
личиночной стадии, так и во
взрослой с помощью 1—3-
членикового хоботка. Личин-
ки и самки кокцид внешне
108
очень похожи па вздутия и наросты на коре, на лишаиники, комочки
мусора (рис. 265, 274). В связи с малой подвижностью у самок разви-
лись защитные приспособления в виде разнообразных твердых покро-
вов и приспособительная окраска. Наиболее распространены щитки из
воска, лака, сброшенных личиночных шкурок, утолщенной, сильно
склеротизованной кутикулы и т. д. Некоторые червецы живут в коре
деревьев и их можно обнаружить лишь по торчащим белым восковым
нитям.
Большинство кокцид имеет одно поколение в году, некоторые дают
3—4 поколения. Зимуют в стадиях яйца, личинки или взрослой самки.
Яйца откладываются в яйцевой мешок, под тело самки или под щи-
ток. Половые различия начинают проявляться у личинок второго-треть-
его возраста. Личинки самцов имеют 2 дополнительных возраста, в те-
чение которых они в отличие от самок претерпевают усложненное пре-
вращение и в этот период не питаются. Превращение осуществляется
в специальных ватообразных коконах или под щитком, который от-
личается от щитка самки меньшими размерами и удлиненной формой.
Личинки самок уже после второй линьки превращаются во взрослых
особей. Как у всех насекомых с неполным превращением, личинки са-
мок и взрослые самки очень похожи по внешнему облику.
С момента начала питания щитовки вообще не двигаются, а чер-
вецы перемещаются на небольшие расстояния в пределах одного рас-
тения. Перелеты самцов также ограничены. Несмотря на это, для кок-
. цид характерна высокая способность к пассивному расселению. Оно
осуществляется личинками первого возраста только в течение первых
2—3 дней жизни до начала питания. Эти личинки называются бро-
дяжками. Основным фактором, способствующим расселению бродя-
жек, -является ветер. Большую роль в распространении червецов и щи-
товок играет хозяйственная деятельность человека, связанная с пере-
возками растений.
Кокциды живут на большинстве древесных, кустарниковых и мно-
голетних травянистых растений, питаясь соками и повреждая все части
растений — корни, ветви, листья и плоды. Связи с кормовыми рас-
тениями различны. Многоядные червецы и щитовки (полифаги) встре-
чаются на всевозможных лиственных деревьях, кустарниках, травяни-
стых растениях и даже на хвойных породах. К таким многоядным кок-
цидам относятся акациевая ложнощитовка (Parthenolecanium corni
Bouche), калифорнийская щитовка (Diaspidiotes perniciosus Comst.)
и др. Обитатели близкородственных растений (олигофаги) охватывают
меньшее число видов. К ним, в частности, относятся червецы, живу-
щие на злаках, виды, живущие на грушах, яблонях и других расте-
, ниях семейства розоцветных. Большую группу образуют одноядные
червецы и щитовки (монофаги). Встречаются виды, роды и даже се-
мейства, приуроченные к определенному роду или виду растений. На-
пример, все представители семейства Acleridae живут на злаках, щи-
товки рода Leucaspis Targ. — на соснах и т. д. Многйе виды кокцид
поселяются не только на определенных растениях, но и на определен-
ных частях растений. Например, северный кермес (Kermococcus quer-
cus L.) живет только на стволах дубов с толстой корой (рис. 273,
274), а южный кермес (К. roboris Fourcr.)—исключительно на тонких
ветках дуба. Особенно сильно специализированы виды, живущие на
злаках.
Кокциды 'имеют громадное хозяйственное значение. Наибольший
вред они приносят в южных районах и в оранжереях. В северных об-
ластях СССР имеют меньшее значение, и массовые размножения на-
блюдаются у небольшого числа видов.. Среди вредителей сельского хо-
зяйства значительную часть составляют кокциды, завезенные из других
109
стран. На юге кокциды вредят цитрусовым, техническим культурам,
декоративным растениям, многим плодовым, винограду, чайному ку-
сту и другим сельскохозяйственным культурам. В северных районах
европейской части СССР постоянно вредят 2 вида щитовок—-запято-
видпая щитовка (Lepidosaphis ulmi L.) и ивовая щитовка (Chionaspis
salicis L.), поражающие древесные породы, ягодники и городские де-
коративные растения.
Повреждения растений кокцидами носят весьма разнообразный
xapaKTqp. При сильном заражении ствола щитовками происходит
омертвение тканей, нарушается деятельность проводящих сосудов.
При повреждении штамба молодые деревья погибают. Кокциды, жи-
вущие на листьях, вызывают опадение листвы. При сильном зараже-
нии деревьев наблюдается полная потеря листьев, которая нередко при-
водит к гибели деревьев. Кокциды, сосущие у плодоножки, вызывают
опадение плодов. При поражении плодов, особенно яблок, на них по-
являются красные пятна, сильно снижающие качество урожая. Пита-
ясь корнями злаков, они сильно задерживают рост хлебов. Вред чер-
вецов и щитовок усугубляется выделением медвяной росы — сладких
экскрементов, на которых развиваются сапрофитные грибы, сильно за-
грязняющие растения. Ряд вредных видов, отсутствующих в нашей
стране, внесен в список карантинных объектов.
Среди кокцид есть и полезные виды, например лаковый червец
(Laccifer lacca Kerr). Обильные выделения, покрывающие тело насе-
комых, перерабатываются в шеллак — ценный продукт, который имеет
хорошие электроизолирующие свойства и широко применяется в раз-
личных отраслях промышленности. Некоторые виды кокцид пользу-
ются известностью как насекомые, дающие красную краску кармин.
• Подотряд включает в себя 8 семейств. Из них щитовками
называют самое специализированное семейство Diaspididae. Тело са-
мок этого семейства покрыто щитком, состоящим из 1—2 личиночных
шкурок и секреторных продуктов самой самки. Подушечницами,
или ложнощитовками, называют семейство Coccidae (рис. 265),
а червецами (рис. 274) —всех остальных. Общее число видов кок-
цид, известных к настоящему времени, приближается к 4000, в СССР
обнаружено более 500 видов.
ОТРЯД ПОЛУЖЕСТКОКРЫЛЫЕ, ИЛИ КЛОПЫ— HEMIPTERA*
Общая характеристика
К этому отряду относятся наземные или водные насекомые с уп-
лощенным, реже цилиндрическим, телом (рис. 275, 276). Голова сво-
бодная, но обычно малоподвижная. У всех наших видов хорошо раз-
виты фасеточные глаза, расположенные по бокам головы (рис. 277,
278, ф.г). Форма глаз обычно округлая, овальная или почковидная.
Кроме сложных глаз для полужесткокрылых характерны простые глаз-
ки (рис. 278, п.г), которые располагаются обычно на темени. Они от-
сутствуют у личинок, хищников, у большинства водных клопов, а так-
же у представителей некоторых семейств, например у слепняков (Мд-
ridae).
На верхней поверхности головы различают лоб и темя (рис. 277,
л.т). Впереди лоб переходит в наличник и лежащие по бокам от него
щеки. Усики 4—5-члениковые, реже 3-члениковые, располагаются пе-
ред глазами (рис. 275, 276, у). Чаще всего они нитевидные, веретено-
видные или булавовидные. Неправильные усики встречаются у неко-
А. X. Саулич.
ПО
торых водных и паразитических клопов. Ротовой аппарат колюще-со-
сущего типа. Хоботок 3—4-члениковый, прикреплен к передней части
головы (рис. 277, 278, х). У растительноядных форм он обычно тон-
кий, в спокойном состоянии подогнут к нижней поверхности тела
(рис. 277). У хищников хоботок короткий, толстый, дугообразно изог-
нутый в виде клюва и далеко отстоит от нижней поверхности тела
(рис. 278). Переднегрудь сохраняет слабую подвижность, а средне-
грудь и заднегрудь тесно соединены друг с другом. Переднегрудь раз-
вита хорошо и прикрыта сверху крупной переднеспинкой (рис. 275,
276, пс). Края переднеспинки могут быть прямыми, выемчатыми или
вогнутыми. Они называются окаймленными, если отделены от основ-
ного диска вдавленной линией. Углы переднеспинки нередко заканчи-
ваются зубцами, шипами и другими скульптурными образованиями.
Рис. 275—278. Общий вид и детали строения полужесткокрылых.
275 — слепняк (Miridae), 276 — щитиик (Pentatomidae), 277 — голова и переднегрудь слепняка, вид
сбоку, 278 — голова хищнеца. (Reduviidae), вид сбоку.
г —голова; к —кориум; кл — клавус; кн — кунеус; л ~ лоб; н — ноги; п ~ перепоночка; п.г —
простой глазок; пс — переднеспинка; г — темя; у — усики; ф.г — фасеточные глаза; х— хоботок;
tu.K — шейное кольцо; щ— щиток; э — эмболиум.
Иногда переднеспинка разделена поперечным вдавлением на перед-.
нюю и заднюю доли. На передней доле обычны мозолистые возвыше-
ния. Передний край переднеспинки у ряда семейств (например, у слеп-
няков) отделен поперечной бороздкой в более или менее узкое шей-
ное кольцо (рис. 275, ш.к).
Среднегрудь представляет собой наиболее массивный грудной
сегмент, в котором находится сильная ножная и крыловая мускула-
тура. Сверху среднегрудь видна только в виде щитка (рис. 275,
276, щ), чаще небольшого треугольного, но иногда очень большого
и закрывающего всю спинную сторону брюшка, как, например, у щит-
ников-черепашек (Scutelleridae).
Заднеррудь сверху не видна. Снизу на боках ее между средними
и задними тазиками лежат отверстия пахучих желез. Наличие паху-
чих желез — характерная черта полужесткокрылых. Их секрет — не-
предельная цимициновая кислота с характерным неприятным запахом.
Отверстия пахучих желез отсутствуют у всех водных и хищных кло-
пов. Пахучие железы имеют, несомненно, защитное значение, а также
способствуют концентрации особей одного вида.
Среднегрудь и заднегрудь несут по одной паре крыльев. Передние
крылья (надкрылья) плотные, кожистые, чаще всего разделенные прр-
111
дольным швом на клавус — внутреннюю часть и кориум — наружную
(рис. 275, 276, кл, к). У некоторых видов от кориума отделена вер-
шина— кунеус и наружный край — эмболиум (рис. 275, кн, э). Вер-
шинная часть надкрыльев прозрачная, называется перепоночкой
(рис. 275, 276, п). На ней, как правило, жилкование ясное. Количе-
ство жилок на перепоночке различно у представителей разных се-
мейств и нередко используется при диагностике видов. Линия сопри-
косновения клавусов противоположных надкрылий позади щитка на-
зывается общим швом надкрылий. Задние крылья перепончатые.
Для полужесткокрылых характерно явление крылового полимор-
физма. Укорочение или полное исчезновение крыльев у имаго часто яв-
ляется видовым признаком, но широко распространено и явление фа-
культативного полиморфизма. Суть его заключается в том, что один
и тот же вид может существовать в природе в виде полнокрылых, ко-
роткокрылых и бескрылых форм. Чаще всего встречается половой ди-
морфизм, когда самки бескрылы, а самцы полнокрылые.
Ноги обычного строения. Лапка состоит из 2—3 члеников; на вер-
шине последнего членика находятся парные коготки. На бедрах, голе-
ни, члениках лапки бывают шипы, зубцы, щетинки, волоски. В зави-
симости от образа жизни, способа использования конечностей отдель-
ные части ног могут претерпевать различные модификации. Ноги чаще
всего бегательные, в этих случаях все 3 пары ног имеют одинаковое
строение (рис. 275, 276, «). У водных клопов ноги плавательные; они
снабжены разнообразными волосками, увеличивающими гребную по-
верхность. В некоторых случаях, например у гребляков (Corixidae),
каждая пара ног имеет своеобразное строение. В отдельных семей-
ствах ноги копательные, а у некоторых хищников передние ноги хва-
тательные.
Брюшко состоит из И сегментов, но первый и последний сегмен-
ты редуцированы. Генитальные сегменты сильно видоизменены. Верх-
няя сторона брюшка обычно скрыта под крыльями, чаще всего пло-
ская. Нижняя сторона сильно выпуклая. Боковые края брюшка обра-
зуют более или менее широкий брюшной ободок, отделенный от
остальной части брюшка вдавленной линией.
Полужесткокрылые — насекомые с неполным превращением. Яйца
клопов различаются по окраске и форме. Характерной их особенно-
стью является наличие крышечки на верхнем полюсе, которая откры-
вается при выходе личинки из яйца. Обычно яйца откладываются на
поверхность субстрата или иногда погружаются в ткань растения груп-
пами или поодиночке. Число откладываемых яиц варьирует от не-
скольких десятков до нескольких сот.
Личинки мало отличаются от имаго по морфологии и биологии.
Существует несколько признаков, по которым можно безошибочно от-
личить личиночную стадию от имагинальной. У личинки зачатки крыль-
ев появляются с третьего возраста, но они лежат по бокам тела, не
соприкасаясь по срединному краю, т. е. не имеют общего шва над-
крылий. Личинки лишены глазков. Усики у них 3- или 4-члениковые,
лапки 2-члениковые, отверстия пахучих желез открываются на спин-
ной стороне брюшка тремя непарными отверстиями. Личиночная ста-
дия имеет 5 возрастов. Продолжительность развития может коле-
баться у разных видов от нескольких недель до 2 лет.
Питание полужесткокрылых очень разнообразно. Большинство
видов относится к фитофагам, но .бывают среди них хищники и пара-
зиты. У растительноядных форм чаще всего проявляются олигофагия
и полифагия. Кормом служат .все части растений — вегетативные, гене-
ративные, содержимое семян. Корнями растений питаются земляные
щитники (Cydnidae). Говоря о фитофагии клопов, необходимо отметить
112
полную универсальность потребления растительной пищи. Низшими
водорослями питаются гребляки (Corixidae), с лишайниками и мхами
связана жизнь некоторых видов семейства Microphysidae, на папорот-
никах живут травяные клопы из семейства Miridae. Пищей для хищ-
ных клопов служат различные мелкие членистоногие.
Паразитизм достаточно широко распространен среди полужестко-
крылых. Постоянными паразитами летучих мышей являются виды тро-
пического семейства Polyctenidae — высоко специализированные пара-
зиты, приспособившиеся жить среди меха хозяина. Временными пара-
зитами теплокровных животных являются все виды семейства Cimicidae.
Они питаются на грызунах, птицах, человеке, сохраняя свои изначаль-
ные биотопы в дуплах деревьев, пещерах и норах животных.
Различные группы клопов сильно отличаются по своей биологии
и экологии. Большинство из них принадлежит к наземным формам, ио
многие относятся, к водным. Большая часть водных клопов живет
в толще воды, имея для этого разнообразные приспособления (напри-
мер, дыхательные трубки у водяных скорпионов — Nepidae). Некоторые
виды живут на поверхности воды; их тело покрыто густыми коротки-
ми несмачивающимися волосками. Водомерки (Gerridae) скользят по
поверхности воды, используя натяжение водяной пленки. Водомерки
рода Halobates живут на поверхности моря в сотнях километрах от
берега, питаясь плавающими остатками беспозвоночных и приклеивая
яйца к плавающим перьям птиц и другим предметам. Это единствен-
ный среди насекомых вид, постоянно обитающий в море.
Наземные полужесткокрылые заселяют разнообразные биотопы.
Очень богата фауна клопов древесных пород и кустарников. Одни кло-
пы питаются на лиственных, другие — на хвойных породах. Под ко-
рой встречаются виды, питающиеся ею, и хищники, охотящиеся за дру-
гими насекомыми, например за жуками-короедами. Здесь же живут
мицетофаги, потребляющие мицелий трутовиков. Зимой под корой
находят убежище многие виды, нередко образуя при этом большие
скопления. Очень много клопов на травянистых растениях. Наконец,
большая группа клопов живет в почве и на ее поверхности.
Полужесткокрылые широко распространены по всему свету. Но
наиболее богата ими фауна пустынь и полупустынь, что определяет-
ся большой термофильностью и ксерофильностью этой группы насе-
комых. Бедна фауна тундры, причем большая часть клопов этой зоны
связана с водой. Также небогата фауна лесной зоны.
Хозяйственное значение полужёсткодрылых очень велико. В пер-
вую очередь следует отметить их вредную деятельность. В этом отно-
шении на первом месте стоят фитофаги — вредители сельского хозяй-
ства. Одним из самых злостных вредителей зерновых культур во мно-
гих странах мира является вредная черепашка (Eurygaster integriceps
Put.) из семейства щитников-черепашек (Scutelleridae). Уколы от хо-
ботка вредной черепашки заметны невооруженным глазом; они бывают
окружены конусом беловатого вещества — засохшего сока растений.
От сосания стеблей молодых растений средний лист начинает усыхать,
и само растение постепенно погибает. Сосание ниже колоса вызывает
усыхание и побеление его. В период налива зерна черепашка полно-
стью его высасывает, в результате зерно становится легковесным, те-
ряет свои хлебопекарные свойства и всхожесть. Даже небольшие по-
вреждения‘зерна, не вызывающие сморщивания, а только оставляющие
следы укода, изменяют хлебопекарные свойства муки, обусловленные
разрушением зерен крахмала и значительным снижением количества
клейковины под действием ферментов слюнных желез клопов.
Вредителями зерновых культур являются многие виды семейства
слепняков, например хлебный клопик (Тrigonotylus ruficorni'i Geof.)
8 208
ИЗ
и странствующий слепняк (Notostira erratica L.). При массовом раз-
множении эти виды наносят существенный ущерб, вызывая задержку
роста и пожелтение растений. Кроме злаков названные виды повреж-
дают посевные травы — тимофеевку, житняк и др.
Основными вредителями кормовых трав являются также слепня-
ки— Adelphocoris lineolatus Gz., Lygus pratensis L. и др. Личинки лю-
цернового клопа (A. lineolatus) сосут преимущественно черешки моло-
дых растений. Взрослые особи повреждают все наземные части, но
предпочитают соцветия. Высосанные цветки быстро увядают и опа-
дают, что приводит к значительному снижению урожая семян.
Вредителями огородных культур являются клопы рода Eurydema
Lap. из семейства щитников (Pentatomidae). Сосание старых частей
растений не вызывает больших повреждений, но молодые растения за-
держиваются в росте и увядают. От этих клопов особенно сильно стра-
дает рассада капусты.
В южных странах, а также на юге СССР, большой вред приносит
щитник Nezara viridula L. — широкий полифаг, вредящий более чем
1000 видам растений из 35 семейств (хлопку, табаку, томатам, карто-
фелю, цитрусовым и др.). Многие растительноядные полужесткокры-
лые связаны с хлопком. На нем отмечено около 120 видов клопов, от-
носящихся к разным семействам.
В садоводстве некоторый вред причиняют кружевницы (Tingidae).
Питание этих клопов на плодовых, деревьях приводит к преждевремен-
ному опаданию листвы. При этом нарушается и нормальная закладка
почек будущего года. Большой вред кружевницы приносят в пи-
томниках грушам и яблоням, ослабляя молодые деревья. На цитру-
совых отмечено около 40 видов полужесткокрылых, но в СССР цитру-
совые практически не страдают от клопов. В лесном хозяйстве значи-
тельный вред приносят подкорники (Aradidae).
Помимо непосредственного вреда, вызываемого сосанием различных
частей растений, полужесткокрылые получают большое хозяйственное
значение как переносчики фитопатогенных вирусов.
Важным медицинским объектом является кровососущий постель-
ный клоп (Cimex lectularius L.) из семейства Cimicidae. В связи с по-
лупаразитическим образом жизни этот вид утерял способность к по-
лету и приобрел плоскую форму тела. Постельный клоп питается
кровью человека, а другие виды этого семейства являются гнездовы-
ми паразитами птиц и летучих мышей.
Положительное значение для человека имеют некоторые хищные
клопы из семейств Anthocoridae, Nabidae и Reduviidae, уничтожающие
многих вредителей — тлей, гусениц и т. д. В биологическом методе
борьбы с колорадским жуком в некоторых странах успешно использу-
ются хищные клопы рода Perillus из семейства щитников.
В мировой фауне о-писано более 40 000 видов полужесткокрылых,
из них для СССР указано свыше 2000 видов из 40 семейств. Подав-
ляющее большинство видов относится к нескольким семействам; крат-
кая характеристика некоторых из них приводится ниже.
Слепняки — Miridae (рис. 275). Одно из самых крупных се-
мейств полужесткокрылых. Средних размеров или мелкие клопы
с нежными покровами. Отличительной чертой семейства является от-
сутствие глазков у имаго. Фасеточные глаза очень большие. Усики
длинные, тонкие. Щиток небольшой треугольный. Перепоночка с 1 —
2 замкнутыми ячейками. Ноги тонкие, лапки 3-члениковые. Кроме ко-
готков они несут придатки двух типов — аролии, расположенные между
коготками, и псевдоаролии, прикрепленные к коготкам. Форма аролий
и их расположение используются при диагностике видов. Определение
видов этого семейства часто вызывает трудности, поэтому точная диаг-
114
ностика Т|ребует изучения строения гениталий. Слепняки — это в основ-
ном растительноядные формы, но среди них есть и специализирован-
ные хищники, играющие большую роль в снижении численности вред-
ных видов членистоногих. Хищные слепняки питаются яйцами насе-
комых, тлями, личинками мух, паутинными клещами. Зимуют яйца
или имаго слепняков. Число поколений в году варьирует у разных
видов и в разных зонах от 1 до 2—5. При массовом размножении
некоторые растительноядные виды могут причинять большой вред сель-
скому хозяйству.
Земляные клопы — Lygaeidae (рис. 279). Богатое в видо-
вом отношении семейство (в мире известно около 2000 видов,
в СССР — свыше 300). Мелкие и средней величины клопы. Хоботок
Рис. 279—287. Полужесткокрылые.
279 — представитель семейства Lygaeidae, 280 — ромбовик (Coreidae), 281 — щитник-черепашка
(Scutelleridae). 282 — водяной скорпион Nepa cinerea L. (Nepidae), 283 — гладыш Notonecta
L. (Notonectidae), 284— гребляк (Corixidae), 285— плавт Ilyocoris cimicoides L. (Naucoridae),
286 — кружевница (Tingldae), 287 — подкорник (Aradidae).
и усики 4-члениковые. Глазки обычно имеются. Перепоночка с 5 про-
дольными жилками. Часто встречаются короткокрылые формы. Боль-
шинство видов этого семейства живет в почве, под растениями и в под-
стилке. Обычно хорошо бегают. Растительноядны, но многие виды пи-
таются разлагающимися остатками растительного и животного
происхождения. В СССР важного значения как вредители не имеют.
Ромбовики, или краевики — Coreidae (рис. 280). Одно
из самых крупных семейств полужесткокрылых. Обычно крупные
и средних размеров. Окраска чаще всего бурая, желтоватая. Глазки
есть. Хоботок 4-члениковый. Усики 4-члениковые, место их прикреп-
ления видно сверху. Щиток маленький. Перепоночка с многочисленны-
ми жилками. Растительноядны. Являются второстепенными вредите-
лями, усиливая вредное значение других видов клопов.
Земляные гцитники — Cydnidae. Средних размеров или
мелкие клопы, обычно темноокрашенные, блестящие. Голени покрыты
115
сильными шипами, ноги копательные. Большей частью живут в почве
или у корней растений. Питаются корешками различных травянистых
растений. ,
Щитники — Pentatomidae (рис. 276). Крупные и средней
величины клопы с твердыми покровами. Окраска у многих видов яркая.
Усики 5-члениковые, очень длинные. Место их прикрепления видно
сверху. Глазки есть. Перепоночка с многочисленными жилками. Щи-
ток обычно большой, на 2/3 прикрывает брюшко. Большинство видов
растительноядны, среди них есть виды, имеющие большое экономиче-
ское значение как вредители сельского хозяйства. Среди щитников
много хищников, для которых обычную пищу составляют мелкие на-
секомые, их личинки и яйца.
Щ ит н и к и - ч е р е п а ш к и — S с u t е 11 е г i d а е (рис. 281). Сред-
ней-величины или крупные клопы с большим щитком, прикрывающим
почти все брюшко. Питаются на травянистых растениях, некоторые
виды сильно вредят. К числу опаснейших вредителей злаков относится
вредная черепашка (см. выше).
Хищнецы —Reduviidae. Крупные, реже средних размеров
клопы. Голова цилиндрическая, вытянутая в длину. Хоботок корот-
кий, толстый, дугообразно изогнутый (рис. 278). Глазки имеются. Боль-
шинство видов полнокрылые. Хищники, питаются различными насеко-
мыми; уколы крупных видов болезненны для человека. Большое се-
мейство. В мировой фауне насчитывается около 3000 видов, в СССР
около 90.
Водяные скорпионы — Nepidae (рис. 282). Крупные кло-
пы, живущие на дне водоемов. Передвигаются медленно. Добычу обыч-
но подкарауливают и ловят движением передних хватательных ног.
Дышат атмосферным воздухом с помощью дыхательной трубки.
Гладыши — Notonectidae (рис. 283). Средней величины
клопы. Тело продолговатое, выпуклое, гладкое. Усики 4-члениковые,
очень короткие, прикреплены под глазами. Глазков нет. Перепоночка
без жилок. Задние ноги пребные, прекрасно плавают. Дышат атмосфер-
ным воздухом, поднимаясь к поверхности воды и выставляя конец
брюшка с дыхательными отверстиями. Яйца откладывают в ткани вод-
ных растений. Хищники, могут нападать на мальков рыб. Укусы этих
клопов очень болезненны.
Водомерки — Gerridae. Тело удлиненное, густо покрыто не-
смачивающимися волосками. Усики 4-члениковые, очень длинные. Бы-
стро скользят по поверхности воды, используя натяжение поверхно-
стной пленки. Крылья обычно развиты, легко перелетают из водоема
в водоем. Хищники, питаются мелкими беспозвоночными.
Определительная таблица семейств
1(2). Ноги каждой пары своеобразного строения: передние короткие,
с 1-члениковой лопаткообразной лапкой, средние удлиненные
и тонкие с 1-члениковой лапкой, несущей 2 очень длинных за-
остренных коготка, задние веслообразные, с 2-члениковой
сплюснутой лапкой, по краям густо усаженной длинными во-
лосками (рис. 284). Хоботок очень короткий, нерасчлененный,
неподвижно соединен с передним концом головы. Живут в во-
доемах, плавают кверху спинкой, могут прилетать на свет.
........................................... Гребляки — Corixidae.
2(1). Ноги иного строения. Хоботок всегда отчетливо членистый, со-
единен с головой подвижно.
3(8). Усики короткие, гораздо короче головы, почти незаметны при
рассматривании насекомого сверху. Живут в водоемах.
116
4(5). На конце брюшка имеется длинная дыхательная трубка
(рис. 282). Хоботок короткий, направленный вперед . . . .
.............................Водяные скорпионы — Nepidae.
5(4). На конце брюшка нет дыхательной трубки. Хоботок подогнут
на брюшную сторону тела.
6(7). Тело широкое, овальное, заметно уплощенное. Надкрылья ле-
жат на спине почти плоско (рис. 285). Плавают кверху спин-
кой . . . ...............................Плавты — Naucoridae.
7(6). Тело сжато с боков, высокое, спинная сторона сильно выпук-
лая. Надкрылья лежат на спинет кровлеобразно. Задние ноги
длинные, веслообразные с густыми волосками (рис. 283). Пла-
вают на спине.........................Гладыши— Notonectidae.
8(3). Усики длинные, длиннее головы, хорошо видны при рассматри-
вании насекомого сверху. Живут на суше или на поверхно-
сти водоемов.
9(10). Нижняя сторона тела покрыта густыми и короткими серебри-
стыми волосками, не смачивающимися водой. Средние и зад-
ние ноги значительно длиннее передних. Тело узкое, удлинен-
ное. Живут на поверхности воды . . Водомерки — Gerridae.
10(9). Нижняя сторона тела без густых, коротких, прилегающих, се-
ребристо отливающих волосков, хотя часто бывает покрыта
более редкими и длинными волосками. Средние и задние ноги
не длиннее или едва длиннее передних. Живут на суше, иног-
да по берегам водоемов, но не на поверхности воды.
11(32). Усики 4-члениковые. Щиток обычно небольшой, прикрывает
меньше 1/3 длины брюшка.
12(17). Хоботок изогнут дугообразно и не прилегает к нижней сторо-
не тела (рис. 278), часто он короткий и толстый, клювовид-
ный.
13(16). Хоботок 3-члениковый (рис. 278).
14(15). Длина тела больше 6 мм. Перепоночка надкрылий с 2 ячей-
ками в основании и 2—3 отходящими от них жилками . . .
..........................................Хищнецы — Reduviidae.
15(14). Длина тела меньше 5 мм. Перепоночка с неявственным жилко-
ванием. Тело более или менее уплощенное. Живут по берегам
водоемов...............................................Saldidae.
16(13). Хоботок 4-члениковый. Первый членик его очень короткий.
Перепоночка у полнокрылых форм с 3—4 ячейками, от кото-
рых отходит 6—8 жилок..................................Nabidae.
17(12). Хоботок обычно не изогнут дугообразно, в покое прилегает
к нижней стороне тела, как правило, длинный и тонкий
(рис. 277).
18(19). Переднеспинка 5-угольная, задний край ее вытянут в большой
треугольный отросток, целиком или почти целиком прикры-
вающий щиток. Надкрылья и переднеспинка с ячеистой или
сетчатой структурой (рис. 286). Длина тела меньше 5 мм. .
...........................................Кружевницы — Tingidae.
19(18). Переднеспинка 4-угольная, редко 6-угольная, задний край ее
без треугольного отростка. Щиток всегда открыт сверху. Яче-
истой структуры нет.
20(23). Тело плоское, листовидное, крылья часто укорочены или от-
сутствуют. Глазков нет.
21(22). Голова сверху с шиповидным отростком и большим кониче-
ским выступом (наличником) между основаниями усиков
(рис. 287). Щиток большой треугольный или полукруглый.
Края брюшка фестончатые..........................................
.....................................Подкорники — Aradidae.
117
22(21). Голова без шиповидных отростков. Наличник обратно-треу-
угольный (расширяющийся к вершине). Щиток очень малень-
кий. Паразиты теплокровных......................................
, ........................Постельные клопы — Cimicidae.
23(20). Тело выпуклое. Крылья прикрывают все брюшко или большую
его часть. Глазки есть или отсутствуют.
24(27). Перепоночка с многочисленными продольными жилками
(рис. 280).
25(26). Отверстия пахучих желёз четкие, часто ушковидной формы.
.................................Ромбовики — Coreidae.
26(25). Отверстия пахучих желез нечеткие, а если четкие, то не ушко-
видные..............................................Rhopalidae.
27(24). Перепоночка с немногими (не более 8) жилками или совсем
не развита.
28(29). Перепоночка отсутствует или укорочена и не прикрывает
брюшко. Надкрылья красные; на каждом надкрылье находится
крупное черное пятно. . . . Красноклопы — Pyrrhocoridae.
29(28). Перепоночка развита, достигает конца брюшка.
30(31). Перепоночка с 5 продольными жилками (рис. 279). Глазки
имеются................... Земляные клопы — Lygaeidae.
31(30). Перепоночка с меньшим числом жилок, обычно образующих
замкнутые ячейки (рис. 275). Глазков нет........................
................................... Слепняки — Miridae.
32(11). Усики 5-члениковые, места их прикрепления прикрыты сверху
боковыми краями головы. Щиток большой, прикрывает не ме-
нее 1/3 длины брюшка, нередко он прикрывает почти всю
спинную сторону брюшка.
33(34). Голени с длинными толстыми шипами (ноги копательные).
............\.............Земляные щитники — Cydnidae.
34(33). Голени без длинных толстых шипов; если шипы имеются, то
тонкие и короткие.
35(36). Тело почти полушаровидное; целиком черные, мелкие формы
(короче 5 мм). . . . Полушаровидные щитники— Plataspidae.
36(35). Тело не полушаровидное, окраска тела иная.
37(38). Щиток очень большой, закрывает почти все брюшко и дохо-
дит до его вершины; основание щитка шире заднего края пе-
реднеспинки между задними (но не боковыми) углами
(рис. 281)....................Щитники-черепашки — Scutelleridae.
38(37). Щиток закрывает меньше 2/3 длины брюшка, если доходит до
вершины брюшка, то его основание уже переднеспинки между
задними углами (рис. 276). . . . Щитники — Pentatomidae.
I
ОТРЯД ТРИПСЫ, ИЛИ БАХРОМЧАТОКРЫЛЫЕ-THYSANOPTERA*
Трипсы — это очень мелкие насекомые с удлиненно-овальным те-
лом, обычно не превышающим в длину 1—2 мм, и с уникальным
асимметричным (правая мандибула редуцирована) колюще-сосущим
ротовым аппаратом. Бегательные ноги снабжены пузыревидной при-
соской на кончике каждой лапки. Крылья узкие, с редуцированным
жилкованием и бахромкой из1 длинных ресничек по краям (рис. 288).
У некоторых видов крылья отсутствуют. На конце брюшка самок
* В. Е. Кипятков.
118
имеется короткий яйцеклад (подотряд яйцекладные трипсы — Tereb-
rantia) или же у обоих полов брюшко заканчивается короткой вер-
шинной трубкой (подотряд трубкохвостые трипсы — Tubulifera).
Большинство трипсов живет в цветках или соцветиях растений,
реже на листьях, под корой и на коре деревьев, на лишайниках, во
мху, под опавшими листьями. Питаются обычно клеточным соком
растений. .Многие заглатывают зерна пыльцы или высасывают их.
Некоторые трипсы при развитии на растениях образуют галлы. Труб-
кохвостые трипсы могут также питаться гифами и спорами грибов,
лишайниками, мхами. Ряд трипсов, например виды семейства Aelo-
thripidae из подотряда Terebrantia,. являются хищниками и нападают
на других трипсов, тлей, сеноедов, паутинных клещей или поедают
яйца насекомых.
Рис. 288—290. Табачный трипс Thrips tabaci Lind.
288 — имаго, 289-— личинка, 290 — нимфа.
Яйца трипсов откладываются в ткани растений или в различные
микрополости; они окружены желеобразным веществом, выделяемым
самкой. Развитие сложное, приближающееся к полному превращению.
Личинки второго возраста (рис. 289) уходят в почву, подстилку, па-
зухи листьев и т. п., где обычно строят паутинный кокон, в котором
проходят непитающиеся, малоподвижные стадии пронимфы и нимфы
(рис. 290). У некоторых трипсов наблюдаются партеногенез и яйце-
живорождение. В год обычно развивается несколько (до 7—9) поко-
I лений.
Практическое значение трипсов связано прежде всего с тем, что
многие виды являются серьезными вредителями сельскохозяйствен-
ных культур и древесно-кустарниковой растительности. Они обычно
концентрируются на быстро растущих тканях (молодые листья, цветы,
верхушечные побеги), что вызывает усыхание и отмирание точек рос-
та у растений и- иногда гибель посевов. Трипсы являются также пере-
носчиками вирусных заболеваний растений. Хищные трипсы могут
иметь положительное значение, снижая численность паутинных кле-
щей, тлей, растительноядных трипсов и других вредителей. В ряде слу-
чаев трипсы могут выступать в роли опылителей растений, особенно
там, где крупные насекомые редки. В ряде районов Исландии, напри-
. мор, трипсы являются единственными опылителями некоторых ценных
кормовых трав.
119
В мировой фауне насчитывается свыше 5000 видов трипсов, отно-
сящихся к 165 родам и 5 семействам; в СССР 230 видов, 49 родов.
Распространены трипсы всесветно, но наиболее разнообразны в тро-
пиках и субтропиках.
ОТРЯД ЖЕСТКОКРЫЛЫЕ, ИЛИ ЖУКИ - COLEOPTERA *
Общая характеристика
Жуки легко узнаются по плотной и жесткой передней паре крыль-
ев, образующих так называемые надкрылья (рис. 291, «). Они обыч-
но прикрывают все брюшко сверху, но иногда укорочены, и часть
брюшка или все брюшные сегменты остаются неприкрытыми. У жуков,
Рис. 291—292. Жук Cicindela L., общий вид.
291 — вид сверху, 292 — вид снизу.
б — бедро; бр — брюшко; в — вертлуг; г — голень; гл — голова; ж— жвала; з. в — задний верт-
луг; зг— заднегрудь; з.т — задний тазик; к — крыло; к.ш— концевые шипы; л — лапка; «— над-
крылье; яг—нога; п—пигиднй; иг— переднегрудь; пс— переднеспинка; п-Т — передний тазик;
сг — среднегрудь; с.т—средний тазик; т.в — тазиковая впадина; // — усик; ф.г — фасеточные
глаза; Щ — щиток; э.н — эпиплевры надкрылий; э.п —- эпиплевры переднеспинки.
лишенных задних крыльев, надкрылья могут срастаться по шву, со-
ответствующему линии соприкосновения правого _ и левого надкрыль-
ев. На каждом надкрылье различают боковой (наружный) край, вер-
шину (кончик надкрылья), основание, плечевой угол, образованный
основанием с боковым краем. Обычно боковые края надкрылий более
или менее подогнуты на нижнюю сторону тела жука, образуя эпи-
плевры (рис. 292, э.н). Вторая пара крыльев (рис. 291, к) большей
частью прозрачная, с темными жилками, в покое прикрыта надкрылья-
ми. При полете активно действуют только задние крылья жуков.
Голова (рис. 291, гл) обычно округлая, у долгоносиков (Curculio-
nidae) впереди вытянутая в головотрубку, на конце которой помеща-
* И. А. Кузнецова при участии Н. Ю. Клюге.
120
ются рот и ротовые органы. Ротовой аппарат грызущего типа, все его
части хорошо развиты. Жвалы (рис. 292, ж) сильно склеротизованы,
у некоторых самцов достигают огромных размеров (см. рис. 81). Че-
люстные щупики 4-члениковые, нижнегубные — 3-члениковые. Фасе-
точные глаза (рис. 291, 292, ф.г) округлые, овальные или почковидные
(с выемкой на внутренней стороне). Иногда, например у жуков-вертя-
чек Gyrinidae, каждый глаз разделен на 2 части — верхнюю и ниж-
нюю. Усики с числом члеников от 2 до 40, прикреплены на лбу или
щеках. В зависимости от формы члеников они могут быть нитевидны-
ми, пильчатыми, четковидными, булавовидными неколенчатыми, була-
вовидными коленчатыми, пластинчатыми, гребневидно-булавовидными,
головчатыми, неправильными и т. д.
Из трех сегментов груди у жуков наиболее развита переднегрудь
(рис. 292, пг). На переднеспинке (рис. 291, пс) различают разнооб-
разные по форме передние и задние углы, передний край —- вершину
и задний край — основание. Переднеспинка .бывает поперечной, про-
дольной, округлой, квадратной, полукруглой, сердцевидной и т. д. Обыч-
но она снабжена более или менее резкими боковыми выступами, слу-
жащими границей между верхней стороной и подогнутыми книзу
боковыми краями ее; эти выступы называются эпиплеврами передне-
спинки (рис. 292, э.п). Среднегрудь (рис. 292, сг) в покое прикрыта
надкрыльями; сверху видна только часть ее — щиток (рис. 292, щ).
Заднегрудь (рис. 292, зг) разнообразной формы, в покое полностью
прикрывается надкрыльями.
На вентральной стороне каждого грудного сегмента имеется пара
тазиковых впадин (рис. 292, т.в), в которые входит тазик ноги
(рис. 292, п.т, с.т, з.т). Тазиковые впадины бывают замкнутые, когда
тазик окружен со всех сторон частями грудного сегмента, и открыты-
ми, когда между тазиком и прилегающими к нему частями груди оста-
ется щель. Передние тазики бывают шаровидными, конусовидными или
поперечными. Задние тазики обычно сильно поперечные; все тазики
могут быть соприкасающимися и разделенными. Ноги жуков могут
быть бегательными, копательными, плавательными или прыгательны-
ми. На каждой голени обычно имеется пара концевых шпор (рис. 292,
к.ш). Лапка состоит из 3—5 члеников (рис. 292, л).
Брюшко прочно соединено с заднегрудью (рис. 291, 292, б). В со-
ставе брюшка обычно различимы 5—7 сегментов с вентральной стороны
и 7—9 — с дорсальной. Вентральные участки сегментов соответствуют
стернитам, а дорсальные — терситам. Последний брюшной тергит на-
зывается пигидием (рис. 291, 292, п).
Размеры жуков колеблются от 0,2 мм до 20 см. Их окраска в зна-
чительной степени определяется условиями обитания и особенностями
поведения каждого вида. Многие жуки, живущие в почве или в пеще-
рах, лишены пигментов, имеют бледно-желтую окраску. Яркая окрас-
ка чащр всего свойственна дневным жукам. Она определяется не толь-
ко пигментами, но часто и особой структурой покровов, создающей так
называемую оптическую окраску блестящих металлических тонов.
«Наряд» многих жуков дополняется сложной и разнообразной скульп-
турой надкрыльев и переднеспинки.
По характеру питания имаго жуков можно разделить на 3 основ-
ные биологические группы: хищники, сапрофаги и фитофаги. Хищные
жуки питаются мелкими беспозвоночными, главным образом насеко-
мыми. К сапрофагам относят многочисленных жуков, потребляющих
разлагающиеся растительные остатки, навозников-копрофагов и не-
крофагов, питающихся на трупах и продуктах разложения позвоноч-
ных животных. Фитофаги охватывают собственно растительноядных
121
жуков и потребителей древесины, которые по типу питания прибли-
жаются к сапрофагам.
Развитие жуков протекает с полным превращением. В районах
северного и умеренного климата многие виды развиваются в течение
2 и более лет или же дают 1 поколение в год. Лишь некоторые листо-
еды (Chrysomelidae) успевают дать 2—3 поколения в течение года.
В период спаривания самцы жуков активно разыскивают самок.
В некоторых семействах встречаемость полов облегчается благодаря
скоплению большого числа особей на пищевом субстрате (на растени-
ях, трупах позвоночных, навозе и т. п.). У многих видов самцы находят
самок по запахам их половых феромонов. С помощью зрения отыскива-
ют друг друга жуки, активные в дневное время, а также светляки
(Lampyridae), имеющие органы свечения. Некоторые виды возможно
издают звуковые сигналы, служащие для привлечения полового парт-
нера.
Откладка яиц самками происходит, как правило, после копуляции,
однако у некоторых жуков встречается и партеногенез. Яйца разви-
ваются от нескольких часов до десятков суток. Многие жуки проявля-
ют заботу о потомстве. Прежде всего яйца откладываются в укрытия —
трещины, выемки, специально сделанные углубления (например, у дол-
Рис. 293—297. Личинки жуков.
293 — типичная камподеоидная личинка жужелицы (Carabidae), 294 — нетипичная камподеоидная
личинка мертвоеда (Silphidae), 295 — червеобразная, снабженная ногами личинка пластинчато-
усого жука (Scarabaeidae), 296 — червеобразная безногая личинка долгоносика (Curculionidae),
297 — червеобразная, снабженная короткими ногами личинка щелкуна (Elateridae).
122
гоносиков— Curculionidae) или в «заранее изготовленные более или
менее длинные ходы (например, у короедов — Ijpidae). Большинство
копрофагов, некрофагов и трубковертов заготавливают пищу для ли-
чинок, а некоторые из них охраняют потомство. В обеспечении потом-
ства кормом могут принимать участие и самки и самцы.
Личинки жуков олигоподиальные или аподиальные. Они могут
быть разделены на 2 основных морфо-экологических типа:
1. Камподеоидные личинки, получившие свое название
по внешнему сходству с двухвостками рода Campodea Westw. (см.
рис. 185), имеют вытянутое в длину тело с четким подразделением на
голову, грудь и брюшко; снабжены хорошо развитыми бегательными
ногами (рис. 293, 294). К этому типу относятся свободноживущие
обычно хищные личинки.
2. Червеобразные личинки, как правило, с мягкими белы-
ми покровами, слабо развитыми или отсутствующими ногами
(рис. 295—297). К этому типу относятся малоподвижные, скрытно жи-
вущие формы. По характеру питания они принадлежат к сапрофагам
или фитофагам. Среди личинок этого типа немало обитателей почвы,
потребителей корней, древесины, грибов или субстратов, переработан-
ных грибами.
Особую небольшую группу жуков составляют симбионты насеко-
мых (муравьев, пчел, ос, термитов). Личинки этих видов хищничают
или питаются остатками пищи и трупами своих хозяев.
Личинки, живущие открыто на растениях, в помете и трупах по-
звоночных, развиваются от 1 до 4 месяцев, а развитие в почве и дре-
весине протекает в течение года и более. Куколки жуков , открытого
типа, (см. рис. 93) развиваются от нескольких дней до месяца. Окукли-
вание происходит или в специальной камере, приготовленной личинкой
(например, у долгоносиков — Curculionidae и карапузиков — Histeri-
dae) или внутри последней личиночной шкурки (например, у божьих
коровок — Coccinellidae и у некоторых кожеедов — Dermestidae).
Жесткокрылые составляют самый крупный и широко расселив-
шийся отряд насекомых. Процветанию отряда способствовали следую-
щие особенности морфофункциональной организации жуков:
1) Развитие с полным превращением, позволившее личинкам
и имаго самостоятельно осваивать разнообразные экологические ниши.
2) Наличие прочных надкрылий, защищающих нежные задние
крылья при передвижениях насекомых в твердой, сыпучей и жидкой
среде.
3) Грызущий ротовой аппарат, как самый универсальный тип ро-
f товых аппаратов, позволяющий при незначительных модификациях ис-
' пользовать в пищу все вещества растительного и животного происхож-
дения независимо от их консистенции.
В настоящее время известно около 300 000 видов жуков. Их под-
' разделяют на 150 семейств. В СССР встречаются 25 000 видов и бо-
лее 100 семейств. Отряд делится на 4 подотряда. Основные подотряды,
представленные в фауне СССР, — это плотоядные жуки (Adephaga)
и разноядные жуки (Polyphaga).. В приведенной ниже (см. с. 133)
определительной таблице к подотряду плотоядных относятся вертячки
(Gyrinidae), плавунцы (Dytiscidae) и жужелицы (Carabidae), а все
остальные семейства принадлежат к подотряду разноядных.
В зависимости от особенностей обитания и питания жуков основ-
ные семейства этого отряда могут быть сгруппированы в 6 экологиче-
ских комплексов.
Комплекс водных жуков охватывает несколько специфиче-
ских водных семейств (например, плавунцы — Dytiscidae, водолюбы —
Ну drophilidae, вертячки — Gyrinidae) и отдельных представителей
123
таких семейств, которые в основном обитают на суше (например, род
Donacia F. из семейства листоедов — Chrysomelidae). Обычно водные
жуки заселяют только пресные водоемы, некоторые плавунцы и вер-
тячки могут жить в солоноватой воде, но обитателей морской воды
среди них нет.
Особенно заметны в природе водные жуки из подотряда плото-
ядных, составляющие наиболее многочисленную группу хищных на-
секомых, обитающих в водной среде. Жуки и личинки заселяют почти
все стоячие и текучие водоемы, уничтожая большое количество водных
беспозвоночных. Представители семейства плавунцов играют особенно
большую роль в ограничении численности личинок и куколок крово-
сосущих комаров. Некоторые крупные жуки, например виды рода
Dytiscus L., могут питаться даже мальками рыб.
Водные жуки, как правило, живут в зоне водорослей. Они исполь-
зуют растения для откладки яиц, устраивают здесь засады и спаса-
ются от врагов. Эти жесткокрылые тесно связаны с водной средой
в течение почти всего жизненного цикла. По степени требований к ус-
ловиям среды водные жуки проявляют высокую избирательность, и по-
тому многие формы являются индикаторами определенных условий,
что очень важно для разработки гидробиологической классификации
и комплексной оценки континентальных водоемов.
Комплекс наземных хищных жуков обитает главным об-
разом в почве, подстилке и открыто на поверхности земли или
на растениях. Эти жуки и их личинки играют важную роль в регу-
лировании состава и численности животного населения наземных
биоценозов. В комплекс входит несколько крупных семейств жестко-
крылых.
Жужелицы — Carabidae (рис. 291, 292). Большинство представи-
телей этого семейства принадлежат к активным, преимущественно
многоядным хищникам. Личинки жужелиц тоже хищные, имеют ти-
пичную камподеоидную форму тела (рис. 302). Обычно жужелицы ве-
дут ночной образ жизни, а днем прячутся под камнями, в подстилке
и почве. Наиболее известны крупные жужелицы двух родов — Calo-
soma Web. и Carabus Thoms. Это прекрасные бегуны, быстрые в пре-
следовании добычи. Они обладают удлиненным стройным телом, силь-
ными бегательными ногами. Хищные жужелицы известны как ведущий
биотический фактор, сдерживающий вредную деятельность многих
почвенных вредителей; они также входят в число естественных врагов
колорадского жука. Другой род — скакуны (Cicindela L.) объединяет
яркоокрашенных солнцелюбивых хищников (рис. 291, 292), при опас-
ности порывисто взлетающих с поверхности земли. Личинки скакунов
подкарауливают свою добычу, скрываясь в вырытых ими почвенных
норках.
Немногие представители семейства жужелиц питаются растения-
ми. К их числу относятся хлебные жужелицы рода Zabrus CI., кото-
рые в личиночных и имагинальных стадиях повреждают всходы и ко-
лоски злаков. В отличие от хищников жужелицы-фитофаги более ко-
ротконогие, с шаровидной головой, в движениях менее активны.
Карапузики — Histeridae (рис. 298). Небольшие жуки с компакт-
ным темноокрашенным телом, покрытым твердой гладкой кутикулой.
При опасности поджимают ноги и"усики, долго лежат неподвижно,
«притворяясь мертвыми». Практически все карапузики — хищники,
обитают в навозе, падали, отбросах, в гнездах и норах птиц и зверей,
особенно на песчаной почве. Некоторые виды живут под корой деревь-
ев или в муравейниках. Жуки и личинки питаются главным образом
личинками насекомых. Могут приносить пользу, уничтожая личинок
мух (в навозе), личинок блох (в норах) и короедов (под корой).
124
Рис. 298—303. Наземные жуки-хищннки.
298 — карапузик Hister L. (Histeridae), 299 — хищник Ocypus Leach. (Staphylinidae), 300 — Oxypo-
rus F. из того же семейства, 301— мягкотелка Cantharis obscura L. (Cantharidae), 302 — божья
коровка Hyppodamia Muis. (Coccinellidae), 303 — пчеложук Trlchodes aplarius L.
Хищники — Staphylinidae (рис. 299, 300). Легко опознаются по
вытянутому телу и сильно укороченным надкрыльям, прикрывающим
лишь первые 2 брюшных тергита. Имаго и личинки живут во всевоз-
можных гниющих органических веществах, под камнями, во мху, под
опавшей листвой, под корой деревьев, в грибах, в прибрежном песке,
в гнездах млекопитающих и птиц, в муравейниках, иногда на цветах.
Хищниками активно истребляются некоторые сельскохозяйственные
и стволовые вредители. Отдельные виды паразитируют на пупариях
мух. Жуки рода Paederus F., вызывающие у человека дерматит, инте-
ресны с медицинской точкц зрения.
Мягкотелки — Cantharidae (рис. 301). Умеренно подвижные жуки
с очень мягким удлиненным телом. Надкрылья мягкие, плоские. Жуки
подстерегают добычу на цветках. Хищные личинки мягкотелой живут
в почве. К типичным представителям этого семейства относится тем-
ная мягкотелка (Cantharis obscura L.), имеющая одноцветные, чер-
125
ные или буро-черные надкрылья (рис. 301). Ноги у этой мягкотелки
тоже че|рные, а переднеспинка черная с желтыми боками.
Божьи коровки — Coccinellidae (рис. 302). Хорошо заметны бла-
годаря яркой «предостерегающей» окраске взрослых жуков, личинок
и куколок. Надкрылья обычно красные или желтые с черными пятна-
ми. При прикосновении к их телу жуки выделяют из коленных сочле-
нений капельки едкой оранжевой гемолимфы. Несъедобны для боль-
шинства насекомоядных позвоночных. Личинки и жуки очень прожор-
ливы, питаются тлями, реже червецами, листоблошками, мелкими
гусеницами и клещами. В СССР используются для биологической борь-
Рис. 304—308. Растительноядные жуки.
304 — листоед Donacia F. (Chrysomelidae), 305 — Cryptocephalus Geoffr. из того же семейства.
306 — Agelastica alni L. из того же семейства, 307 — короткохоботный долгоносик (Curculionidae),
308 — длиннохоботный долгоносик.
бы с тлями и червецами. Сбор божьих коровок облегчается тем, что
они часто группируются в местах зимовки, образуя большие скопления.
Пестряки — Cleridae (рис. 303). Жуки средней величины, ярко-
окрашенные, надкрылья с цветными перевязями или пятнами. Личин-
ки хищные, живут в гнездах пчел и ос или под корой, где охотятся
за личинками и жуками короедов. Некоторые виды паразитируют на
саранчовых, а отдельные представители этого семейства развиваются
на сухой падали. Жуки' встречаются на цветах, реже на коре деревь-
ев. В европейской части СССР на цветах можно встретить обыкновен-
ного пчеложука (Trichodes apiarius L.) с надкрыльями, снабженными
тремя оранжевыми перевязями на черном фоне ,(рис. 303). ,
126
Жуки растительноядного комплекса и их личинки ис-
пользуют в пищу все части растений, начиная с почек и бутонов и кон-
чая корневищами и корнями. Для большинства представителей этого
комплекса характерна приуроченность к определенным растительным
органам; часто проявляется и строго избирательное отношение к раз-
ным видам растений. Подавляющее большинство жуков-фитофагов от-
носится к двум крупным семействам.
Листоеды—Chrysotnelidae (рис. 304—306). Мелкие или средней
величины жуки, похожие на усачей (Cerambycidae), но способные
подгибать усики на нижнюю сторону тела. Среди листоедов много вре-
дителей сельского и лесного хозяйства; вредят как имаго, так и личин-
ки. Жуки чаще всего живут открыто на листьях и стеблях; поврежда-
ют листья, бутоны, соцветия травянистых и древесных растений. Сре-
да обитания личинок более разнообразна. Они живут и питаются от-
крыто на листьях, в минах, образующихся во внутренних тканях листа,
внутри стеблей, в почве и на корнях растений или в своеобразных
чехликах, построенных из экскрементов и частиц почвы. Изящный, ме-
таллически блестящий жук — радужница (Donacia F.) обитает в воде
(рис. 304). Его личинки развиваются в корнях и стеблях водных
растений. Жуки обширного рода скрытоглавов (Cryptocephalus
Geoffr.), насчитывающего более 250 видов, грызут листья деревьев
и кустарников (рис. 305). Личинки и имаго козявки Agelastica alni L.
встречаются на ольхе, реже на тополях, березах и ивах (рис. 306).
К семейству листоедов относится также широко известный коло-
радский жук (Leptinotarsa decemlineata Say)—опасный карантинный
вредитель картофеля и других пасленовых. Тело этого жука достигает
в длину 9—11 мм, овальное, желтое, на каждом надкрылье по 5 чер-
ных .продольных полосок. Родина колорадского жука — Северная Аме-
рика. В Европе он был впервые обнаружен в 1922 г. В СССР сейчас
распространен почти во всей европейской части. В течение года разви-
ваются 1—2 поколения жука; зимует имагинальная стадия. Жуки спо-
собны совершать перелеты на 40-—300 км; обычно они проявляют
склонность к миграциям в середине лета и осенью перед зимовкой.
В СССР колорадский жук может легко акклиматизироваться практиче-
ски во всех зонах картофелеводства.
Долгоносики—Curculionidae (рис. 307, 308). Характеризуются вы-
тянутой в головотрубку передней частью головы, на конце которой на-
ходится типичный для жуков грызущий ротовой аппарат. Среди долго-
носиков обычно различают короткохоботных (рис. 307), у которых
головотрубка не длиннее основной части головы, и длиннохоботных
(рис. 308), у которых она намного превышает длину головы, а иногда
сравнима с длиной всего тела.
Большинство долгоносиков — фитофаги, хотя некоторые виды пи-
таются гниющей древесиной. Короткохоботные долгоносики, как пра-
вило, откладывают яйца в почву, а большинство длиннохоботных —
в ткани растений. Личинки безногие, С-образные (рис. 296), белые или
кремовые. Трудно найти вид растений, с которым не были бы связаны
долгоносики. Многие из них приурочены к питанию определенными ви-
дами и частями растений. Большинство долгоносиков развивается за
счет внутренних тканей растений. Так, развитие желудевого долгоно-
сика (Curculio glandium Marsch.) протекает внутри желудей. Реже
личинки питаются на поверхности листа или цветка. Значительное чис-
ло видов долгоносиков связано с травянистой растительностью. Поч-
венные личинки — часто полифаги, реже — олигофаги. Например ли-
чинки клубеньковых долгоносиков рода Sitona Germ, развиваются
только в клубеньках на корнях бобовых, а личинки свекловичного дол-
гоносика (Bothynoderes punctiventris Germ.) питаются корнями рас-
127
тений из семейства маревых. Среди долгоносиков очень много вреди-
телей растений; некоторые из них указаны выше.
К долгоносикам близки трубковерты — Atellabidae, которых в на-
стоящее время выделяют в отдельное семейство.* Личинки трубковертов
свертывают трубки из листьев или развиваются в побегах и пло-
дах. Жуки обычно имеют яркую металлическую окраску. К трубко-
вертам относятся казарка (Rhynchites bacchus L.) и букарка (Coenorr-
hinus pauxillus Germ.). Личинки казарки развиваются в плодах ябло-
ни, груши, абрикоса, персика, сливы и других фруктовых деревьев.
Они являются переносчиками возбудителя плодовой гнили. Личинки
букарки живут в черешках и листьях различных розоцветных.
Комплекс жуков-ксилофагов охватывает несколько се-
мейств, имеющих ряд общих признаков. Личинки ксилофагов питаются
древесиной и корой, поэтому они ведут скрытый образ жизни и в сво-
ем распространении тесно связаны с древесной растительностью. Од-
нако отдельные семейства этого обширного комплекса довольно сильно
различаются не только по таксономическому положению, но также по
их взаимоотношения^ с деревом, как объектом питания и средой оби-
тания.
Кброеды — Ipidae (рис. 309—311). Мелкие жуки (0,8—12 мм) с ко-
ленчатыми булавовидными усиками и головой, вытянутой в едва за-
метный хоботок. Тело цилиндрическое, бурого, черного или рыжего
цвета. Надкрылья у многих видов имеют на конце вдавление, так
называемую «тачку», которая служит для очистки ходов от опилок
и буровой муки (рис. 310, 311);. Личинки короедов похожи на личинок
долгоносиков. Все короеды обитают под корой, в коре, в древесине
или в корнях деревьев; очень редко поселяются в стеблях травянистых
растений. Биологически короеды связаны с определенными ’ породами
деревьев, а экологически — с определенными фитоценозами. Лишь
очень немногие виды многоядны. Короеды ведут' очень скрытный об-
раз жизни, обычно летают только в поисках мест для откладки яиц.
Ходы в коре и древесине характерны для каждого вида короедов.
В период размножения жуки прогрызают в коре входное отверстие,
которое переходит в «брачную камеру», где самец оплодотворяет одну
или несколько самок. Количество «маточных» ходов, отходящих от
«брачной» камеры, определяется числом яйцекладущих самок. Яйца
откладываются в стенки маточного хода, а личинки точат ходы в сто-
роны от его продольной оси. Каждый личиночный ход заканчивается
куколочной колыбелькой. Молодые жуки выгрызают круглые выход-
ные отверстия.
Короеды способны поселяться на здоровых деревьях, однако они
особенно опасны для ослабленного древостоя. Заселенные короедами
деревья быстро погибают, тем более, что многие короеды переносят
болезни деревьев. В лесах нашей страны хвойным деревьям особенно
сильно вредят виды рода Blastophagus Eichh. и шестизубый короед
(Ips sexdentatus Boerh.). Лиственные породы часто подвергаются на-
падениям непарного короеда (Xyleborus dispar Fabr.) и березового
заболонника (Scolytus ratzeburgi Jans.).
Усачи, или дровосеки — Cerambycidae (рис. 312—314). Жуки раз-
ной величины, большей частью крупные или средних размеров. Голо-
ва свободная. Тело продолговатой формы, обычно темноокрашенное
или с пестрым рисунком. Надкрылья без точечных бороздок. Усики
длинные, часто длиннее половины тела, складываются только поверх
надкрыльев. Большинство усачей способно издавать скрипучий звук
при трении среднегруди о переднегрудь («скрипуны»). Личинки мя-
* В определительной таблице иа с. 134 оии объединены с долгоносиками.
128
систые, с очень большой переднегрудью и особыми выпуклостями
(«двигательными мозолями») на брюшных сегментах. Ноги у личинок
в большинстве случаев не развиты. Личинки живут в древесине, ко-
торой они и питаются, реже поселяются в стеблях травянистых расте-
ний или в почве, где питаются корнями.
Распространение усачей тесно связано с кормовыми породами.
Хвойным породам особенно сильно вредят усачи рода Monochamus
Guer. (рис. 312). В средней полосе европейской части СССР самый
Рис. 309—315. Жуки-ксилофаги.
309 — короед Blastopnagus Eich. (Ipidae). 310 — Ips Deg. нз того же семейства, 311 — заболонник
Scolytus Geoifr. из того же семейства, 312 — усач Monochamus Guer. (Cerambycidae), 313 — Tetro-
pium castaneumL. из того же семейства, 314 — усач-кожевник Prionus coriartus L. нз того Же семей-
ства, 315 — златка Anthaxia quadripunctata L. (Buprestidae).
обычный вид этого рода — М. galloprovincialis 01. — развивается на
соснах. Личинки другого усача Tetropium castaneum L. (рис. 313)
живут преимущественно в засохших и здоровых старых стволах елей.
Усач-кожевник Prionus coriarius L. (рис. 314) развивается в древесине
старых хвойных и лиственных деревьев. Личинки домового усача
Hylotrupes 'bajulus L. повреждают деревянные постройки, телеграфные
столбы и другую техническую древесину. Многие взрослые усачи
встречаются на цветах.
Златки — Buprestidae (рис. 315). Жуки небольших или средних
размеров, реже крупные. Тело, суженное к концу, обычно с металли-
ческим блеском. Голова вертикальная, короткая и широкая, втянутая
в переднегрудь до уровня глаз. Усики короткие, пильчатые. Ноги тоже
короткие. Златки предпочитают тепло и яркое солнце. Встречаются на
9 208 1 29
стволах и бревнах (преимущественно на их южной стороне). Мелкие
виды более обычны на цветках и листьях. Полет отличается легкостью
и быстротой. Личинки слепые, безногие, имеют сильно развитую пе-
реднегрудь. Ходы личинок в древесине и лубе плоские, извилистые,
плотно забитые сыпучей буровой мукой. Молодые жуки при выходе
из древесины прогрызают летные отверстия в виде эллипса.
В качестве вредителей леса златки уступают короедам и усачам, /
но часто приобретают первостепенное значение среди жуков-ксилофа-
Рис. 316—320. Почвенные жуки и сапрофаги.
316 — узконадкрылка Oedemera 01. (Oedemeridae), 317 — навозник Geotrupes Latr. (Scarabaeidae),
318 — самец щелкуна Corymbltes Latr. (Elateridae), 319 — медляк Blaps F. (Tenebrionidae), 320 —
четырехточечный мертвоед Xylodrepa quadripunctata L. (Silphidae).
гов, повреждающих плодовые деревья. В европейской части СССР один
из наиболее распространенных видов — четырехточечная златка
(Anthaxia quadripunctata L.)—развивается в древесине хвойных по-
род (рис. 315). Мелкие виды рода Agrilus Curt, питаются на листвен-
ных породах, а взрослые жуки часто встречаются на цветках.
В комплекс почвенных обитателей и сапрофагов
входят семейства, связанные с почвой и подстилкой, но главным
образом во время личиночной жизни. Характерную особенность этого
комплекса составляет широкое разнообразие пищевых режимов и мно-
гоядность его представителей. Почвенные жуки и их личинки могут
питаться корнями растений, перегнивающими растительными и живот-
ными остатками, микроорганизмами почвы, навозом, трупами позво-
ночных животных. Ниже приводятся только основные семейства жуков-
сапрофагов, копрофагов и некрофагов.
130
Узконадкрылки— Oedetneridae (рис. 316). Средних размеров (от
8 до 20 мм) жуки с узким удлиненным телом и мягкими надкрыльями.
Длинные усики придают им внешнее сходство с усачами. Личинки
развиваются в гниющей или сухой древесине, иногда в лесной под-
жилке. По способу питания они занимают промежуточное положение
между ксилофагами и сапрофагами, но больше тяготеют к сапрофа-
гам. Взрослые жуки встречаются на цветах.
Пластинчатоусые— Scarabaeidae (рис. 317). Жуки различных
, размеров — от очень маленьких до крупных, обычно с выпуклым коре-
настым телом и копательными ногами. Усики с пластинчатой булавой
(см. рис. 74). Личинки мясистые, белые, С-образные, с хорошо разви-
тыми ногами и мощными челюстями (рис. 295); развиваются в почве,
навозе или гнилой древесине. В этом семействе выделяют 2 четко обо-
собленные группы — навозников и хрущей.
Навозники в личиночном и имагинальном состояниях питаются
пометом животных. Лишь немногие виды используют растительную
пищу. Взрослые жуки заготавливают пищу для личинок в особых нор-
ках, которые они выкапывают в земле. Навозники играют важную роль
в круговороте органических веществ в наземных экосистемах. В евро-
пейской части СССР к самым обычным крупным навозникам относят-
ся виды рода Geotrupes Latr. — овальные, сильно выпуклые жуки
с длиной тела от 16 до 27 мм, черного или синевато-черного цвета
(рис. 317).
Хрущи разиваются либо в почве, питаясь перегноем и корнями
растений, либо в гниющей древесине. Жуки ведут открытый наземный
образ жизни; питаются листьями, молодыми побегами и цветками,
а иногда вообще не принимают пищи. Многие виды хрущей принад-
лежав к опасным вредителям растений. Наиболее известным предста-
вителем этой группы бесспорно является майский жук (Melolontha F.).
В европейской части СССР встречаются 2 близких вида этого рода.
Западный майский жук (М. melolontha L.) распространен в Литве, Бе-
лоруссии и на Украине, а восточный майский жук (М. hippocastani F.)
заселяет всю лесную зону СССР. Оба вида летают весной по вечерам,
обгрызают листву деревьев. Яйца откладывают в почву, где и разви-
ваются личинки майских жуков.
Щелкуны. — Elateridae (рис. 318). Жуки средней величины или
мелкие. Тело длинное, на переднем и заднем концах суженное. Усики
пильчатые, у самцов иногда гребневидные (как на рис. 318). Только
жукам этого семейства присуща замечательная способность после пе-
реворачивания на спину подскакивать кверху с характерным щелчком.
Механизм «щелчка» обусловливается специальным отростком передне-
груди; этот отросток упирается в край ямки, расположенной на сред-
иегруди (см. рис. 341). Развитие щелкунов протекает в почве и в гни-
лой древесине. Личинки, получившие название проволочников, имеют
удлиненное цилиндрическое тело с гладкими плотными покровами
(рис. 297). Все 3 пары личиночных ног приблизительно одинаковой
длины. Большинство личинок щелкунов питается корнями и подзем-
ными частями растений; многие виды считаются опасными сельскохо-
зяйственными и лесными вредителями. Взрослые жуки живут открыто
на растениях, часто посещают цветки.
Чернотелки — Tenebrionidae (рис. 319). Представители этого се-
мейства очень обычны в степях, полупустынях и пустынях, а в лесной
зоне они немногочисленны. Покровы жуков плотные, обычно однотонно
черные или коричневые. Жуки не способны к полету; как правило, ве-
дут открытый напочвенный образ жизни, наиболее активны в вечерние,
ночные или утренние часы. Многие виды при опасности выделяют за-
щитные жидкости, обладающие резким неприятным запахом. Личинки
131
живут в почве, по внешнему виду похожи на проволочников и назы-
ваются ложноцроволочниками; передние ноги у них длиннее средних
и задних. Основную пищу ложнопроволочников составляют корни рас-
тений. Наряду с проволочниками они могут причинять серьезный вред
сельскохозяйственным культурам и лесопосадкам. В степях и лесо-
степях европейской части СССР на поверхности земли встречаются
крупные черные жуки —медляки Blaps F. (рис. 319). Некоторые чер-
нотелки вредят продовольственным запасам, развиваясь в зерне, муке
и мучных продуктах. Мелкие лесные виды этого семейства обитают
под корой деревьев и в древесных грибах.
Мертвоеды — Silphidae (рис. 320, 321). Разнообразные по внешне-
му облику жуки с длиной тела от 6 до 40 мм. Усики утолщаются
Рис. 321. Могильщик Necrophorus
vespillo L. (Silphidae).
к вершине, иногда с резко выраженной булавой. Личинки мертвоедов
плоские и широкие, похожие на мокриц (рис. 294). Жуки и личинки
питаются падалью, грибами, навозом; некоторые виды потребляют
растительную пищу или хищничают. Жуки-могильщики (Necropho-
rus F.) принадлежат к самым крупным европейским представителям
этого семейства. Они окрашены в черный цвет, а на надкрыльях име-
ют 2 оранжевые перевязи (рис. 321), питаются на трупах животных;
мелкие трупы птиц и грызунов зарывают в почву. К хищным мертво-
едам относится Xylodrepa quadripunctata L. (рис. 320); нападает глав-
ным образом на гусениц.
Своеобразный комплекс паразитоидов ограничивается одним
семейством нарывников — Meloidae, к которому относятся средней ве-
личины или крупные жуки с шеевидной перетяжкой позади фасеточ-
ных глаз (рис. 322). Наличие в теле жуков ядовитого вещества кан-
таридина защищает их от поедания многими насекомоядными живот-
ными. Личинки нарывников развиваются в гнездах пчел или в кубыш-
ках саранчовых. Превращение усложненное, включающее ряд форм
личинок, которые резко отличаются друг от друга. Из яиц выходят
подвижные камподеоидные личинки. Они отыскивают своих хозяев,
132
а затем превращаются в червеобразных личинок. Взрослые жуки ве-
дут активный, непаразитический образ жизни, питаются листьями
и цветками различных растений. Шпанская мушка (Lytta vesicato-
ria L.) имеет зеленую с металлическим блеском окраску тела и над-
крыльев (рис. 322). Жуки объедают листья растений и кустарников.
Личинки шпанской мушки живут в гнездах одиночных пчел, поедая
их яйца и запасы пищи. Крупные бескрылые жуки-майки (Meloe L.)
окрашены в черные или темно-синие тона (рис. 323). Их личинки так-
же паразитируют в пчелиных гнездах. На юге СССР часто встреча-
ются виды рода Mylabris F., имеющие красные или желтые надкрылья
с черными пятнами и перевязями. Все представители этого рода раз-
виваются в кубышках саранчовых.
Определительная таблица семейств
1(6). Тазики задних ног большие, неподвижно слиты с заднегрудью,
простираются назад до второго сегмента брюшка, так что пер-
вый брюшной сегмент снизу виден лишь по бокам от задних
Рис. 323—342. Некоторые жуки и детали их строения.
323 — майка Meloe L. (Meloidae), 324 — задние тазики и основание брюшка жужелицы (Carabidae),
вид снизу, 325— задние тазики и брюшко плавунца (Dytiscidae), вид снизу, 326— голова вер-
тячки (Gyrinidae), вид сбоку, 321 — задние тазики и брюшко мягкотелки (Cantharidae), вид снизу,
328 — то же щелкуна (Elateridae), 329 — лапка листоеда Cassida sabreticulata Suffг. (Chrysomelidae),
330 — лапка короеда (Ipidae), 331 — коготки нарывника (Meloidae), 332 — горбатка (Mordellidae),
вид сбоку, 333 — коготки пыльцееда (Alleculidae), 334— усик навозника Geotrupes L. (Scarabaeidae),
335— усик кравчика Lethrits L. (Scarabaeidae), 336—338 — разные типы передних тазиков жуков
(336 — конические торчащие, 337 — округлые, 338 — поперечные), 339 — лапка сверлильщика
(Lymexylonidae), 340 — лапка пестряка (Cleridae), 341 — щелкун (Elateridae), вид сбоку, 342 —
златка (Buprestidae), вид сбоку.
з.г—задний тазик, I—II—первый и второй сегменты брюшка, в.г — верхняя часть глаза, н.г—
нижняя часть глаза, б.п — бедренная покрышка, к.л —кожистые лопастинки.
133
тазиков (рис. 292, 324, 325). Если ноги бегательные, то верт-
луги задних ног большие, торчащие. (Подотряд Adephaga).
2(3). Ноги бегательные. Живут на суше...............................
........................................Жужелицы — Carabidae.
3(2). Хотя бы задние ноги плавательные, с длинными волосками.
Живут в воде.
4(5). Глаза разделены на верхнюю и нижнюю части (рис. 326). Пе-
редние ноги длиннее средних и задних..............................
.........................................Вертячки — Gyrinidae.
5(4). Глаза обычные. Передние ноги короче задних....................
.........................................Плавунцы — Dytiscidae.
6(1). Тазики задних ног обычно короткие, не достигают второго сег-
мента брюшка (рис. 328). Если же они длинные, конические
и способны доставать до второго брюшного сегмента, сочлене-
ние тазиков с грудью подвижное (рис. 327). Задние вертлуги
обычно небольшие, не торчащие. (Подотряд Polyphaga).
7(16). Все лапки 4-члениковые, их третий членик двулопастной
(рис. 329, 330).
8(11). Голова впереди вытянута в трубку (головотрубку); если она
неясная, то усики коленчато-булавовидные. Верхней губы нет.
9(10). Головотрубка ясная (рис. 307, 308)...........................
........................ . . . Долгоносики — Ctirculionidae.
10(9). Головотрубка очень короткая или ее нет. Усики коленчато-
булавовидные. Тело цилиндрическое, мелкое (рис. 309—311).
................................................Короеды — Ipidae.
11(8). Голова не вытянута. Усики не коленчатые и не булавовидные.
Верхняя губа явственная.
12(15). Тазики задних ног узкие, расставленные.
13(14). Голени с 2 шпорами. Усики длиннее ^половины тела, закиды-
ваются на спину. Тело обычно крупное, стройное и удлинен-
ное (рис. 312—314)...................................
....................Усачи, или дровосеки — Cerambycidae.
14(13). Голени без шпор или с 1 шпорой. Усики обычно короче поло-
вины тела, подогнутые вниз. Тело чаще коренастое, короткое
(рис. 305, 306), реже удлиненное (рис. 304).........................
......................................Листоеды — Chrysomelidae.
15(12). Тазики задних ног широкие, с приподнятым задним краем. Тело
округлое, маленькое (до 6 мм), в волосках. Надкрылья с то-
чечными бороздками..........................Зерновки—Bruchidae.
16(7). Третий членик лапок не двулопастиой; если двулопастной, то
лапки не 4-члениковые.
17(28). Лапки передних и средних пог 5-члениковые, задних — 4-чле-
никовые.
18(23). Переднеспинка с боковой каймой.
19(20). Голова за глазами резко сужена, с тонкой шеей, видной сни-
зу, не втянута в переднегрудь. Надкрылья к вершине сужи-
вающиеся, не прикрывают конец брюшка, который часто вы-
тянут и заострен (рис. 332). Тело длиной менее 10 мм, в при-
легающих волосках..............................Горбатки — Mordellidae.
20(19). Голова за глазами не сужена, втянута в переднегрудь.
21(22). Голова по бокам перед глазами с расширением, прикрываю-
щим основания усиков. Сегменты брюшка снизу неподвижно
сросшиеся. Жуки различных размеров — от мелких до круп-
ных.......................................Чернотелки — Tenebrionidae.
22(21). Голова перед глазами без расширения, основания усиков вид-
ны сверху. Коготки лапок гребенчатые (рис. 333). Жуки мел-
кие......................................... Пыльцееды — Alleculidae.
134
I:
23(18). Переднеспинка без боковой каймы.
24(25). Голова за глазами с резкой шеевидной перетяжкой (рис. 322,
323). Каждый коготок лапки расщеплен от основания на
2 части (рис. 331)............................Нарывники — Meloidae.
25(24). Голова за глазами без резкой шеевидной перетяжки. Коготки
не расщепленные.
26(27). Усики прикреплены на лбу. Тело узкое и длинное (рис. 316).
Надкрылья мягкие......................Узконадкрылки — Oedemeridae.
27(26). Усики прикреплены под боковым краем головы. Надкрылья
в беспорядочно разбросанных точках. Верхняя сторона тела
в длинных волосках.............................Мохнатки — Lagriidae.
28(17). Все лапки 5-, 4- или 3-члениковые или на передних лапках на
1 членик меньше, чем на средних.
29(30). Усики короче челюстных щупиков, булавовидные. Живут в во-
де или навозе...........................Водолюбы — Hydrophilidae.
30(29). Усики длиннее челюстных щупиков.
31(34). Усики коленчатые, с односторонней пластинчатой (рис. 74,
334), гребенчатой (рис. 73) или обволакивающей (рис. 335)
булавой. Передние ноги копательные (т. е. с расширенными
голенями), обычно с рыжим волосатым пятном у основания
бедра снизу. *
32(33). Булава усиков гребенчатая (рис. 73). Брюшко снизу с 5 сег-
ментами. Верхние челюсти (жвалы) ’обычно очень большие,
особенно у самца (рис. 81, 82)...............Рогачи— Lucanidae.
33(32). Булава усиков пластинчатая (рис. 74, 334) или обволакиваю-
щая (рис. 335). Брюшко снизу с 6 сегментами.......................
.............................Пластинчатоусые — Scarabaeidae.
34(31). Усики никогда не имеют односторонней пластинчатой или
гребенчатой булавы, лишь у могильщика — Necrophorus
(рис. 321) булава кажется пластинчатой, но членики ее на са-
мом деле имеют форму вогнутых чашек, а не пластинок. Осно-
вание переднего бедра всегда без рыжего волосатого
пятна.
35(36). Надцрылья сильно укорочены, обычно прикрывают лишь 2 пер-
вых сегмента брюшка (рис. 299, 300). Усики обычно ните-
видные..................................Хищники — Staphylinidae.
36(35). Надкрылья покрывают все брюшко; иногда остаются откры-
тыми лишь его последние сегменты или надкрылья и крылья
совсем отсутствуют.
37(46). Тазики передних ног торчащие, конусовидные, обычно сопри-
касаются (рис. 336).
38(39). Переднеспинка с распластанными краями. Усики булавовид-
ные или хотя бы слегка утолщающиеся на конце. Надкрылья
жесткие, с боковым кантом; либо надкрылья обрублены на
конце и не прикрывают 2—4 последних сегмента брюшка
(рис. 321), либо они покрывают все брюшко и имеют широ-
кий, обычно загнутый кверху боковой кант (рис. 320). Лапки
5-члениковые. Жуки средних размеров или крупные (не ме-
нее 1 см)....................................Мертвоеды — Silphidae.
г9(38). Форма переднеспинки, усиков или надкрыльев иная; либо
лапки нс 5-члениковые, либо размеры меньше 1 см.
40(45). Тазики задних ног конусовидно торчащие, сближенные
(рис. 327). Надкрылья мягкие. t
41(42). Лапки с длинными прямоугольными члениками (рис. 339).
Щиток с продольным килем. . . Сверлилы— Lymexylonidae.
42(41). Лапки с треугольными или сердцевидными члениками. Щиток
без киля.
135
43(44). Брюшко снизу с 6—7 сегментами (рис. 327). Вершины верх-
них челюстей (жвал) простые. Мягкотелки — Cantharidae.*
44(43). Брюшко снизу с 6 сегментами. Вершины верхних челюстей
раздвоенные............................Малашки — Melyridae.
45(40). Тазики задних ног не торчащие, отодвинуты друг от друга.
Подошва лапок с кожистыми лопастинками (рис. 340). Верх-
няя сторона тела обычно с ярким рисунком (рис. 303). . .
..................................... Пестряки — Cleridae.
46(37). Тазики передних ног не торчащие, округлые или поперечные
(рис. 337, 338).
47(52). Тазики задних ног с бедренными покрышками: часть тазика
нависает над бедром и может хотя бы частично прикрывать
его снизу (рис. 328).
48(49). Голени вкладываются в желобки на бедрах, а бедра — в уг-
лубления на нижней стороне тела.........................' . . . .
....................................Кожееды — Dermestidae.
49(48). Голени не вкладываются в желобки на бедрах.
50(51). Переднеспинка подвижна сверху вниз и не образует общую
выпуклость с надкрыльями (рис. 318, 341). Жуки могут под-
скакивать. Тело обычно матовое, без металлического блеска
...........................................Щелкуны—Elateridae.
51(50). Переднеспинка неподвижна, образует общую выпуклость с над-
крыльями (рис. 015, 342). Тело часто с металлическим блеском
...........................................Златки — Buprestidae.
52(47). Тазики задних ног без бедренных покрышек.
53(54). Усики коленчатые. Надкрылья не покрывают вершину брюшка
(рис. 298). Тело твердое, гладкое. Лапки 5-члениковые, редко
4-члениковые............................Карапузики — Histeridae.
54(53). Усики не коленчатые. Надкрылья покрывают все брюшко
(рис. 302). Тело сверху выпуклое, обычно с пятнами, снизу
плоское. Лапки 3-члениковые . . Коровки — Coccinellidae.
ОТРЯД ВИСЛОКРЫЛКИ — MEGALOPTERA **
Вислокрылки — темноокрашенные насекомые с широкой головой,
грызущим ротовым аппаратом и длинными нитевидными усиками. Фа-
сеточные глаза широко расставлены. Из грудных сегментов вы-
деляется широкая увеличенная переднегрудь. Крылья перепонча-
тые, темные, с сетчатым жилкованием, складываются кровлеоб-
разно, прикрывая 10-члениковое брюшко с короткими церками.
Яйцеклад отсутствует.
Превращение полное. Личинки водные, по внешнему облику очень
сходные со взрослыми насекомыми, однако имеют на первых семи сег-
ментах членистые подвижные придатки, похожие на ноги, и длинный
заостренный вырост на конце брюшка.
Взрослые вислокрылки встречаются в конце мая — начале июня
возле стоячих или медленно текучих водоемов с илистым или илисто-
песчаным грунтом, где обитают их личинки. Летают плохо и неохот-
но, не питаются; яйца прикрепляют крупными группами к листьям при-
брежной растительности. Личинки, выходя из яиц, падают в воду и за-
рываются в ил. Питаются донными беспозвоночными. Закончившие
развитие личинки выходят из воды и окукливаются без кокона во влаж-
ной почве. Куколки свободные, подвижные.
* К этому же семейству относятся светляки (Lampyrinae), иногда выделяемые
в отдельное семейство Lampyridae. Самки светляков совершенно бескрылы.
** А. А. Стекольников.
136
Вислокрылки — это небольшой отряд, включающий 2 семейства, из
которых только семейство Sialidae представлено в фауне СССР. В ев-
ропейской части СССР встречается 1 вид — Sialis lutaria Latr.
ОТРЯД ВЕРБЛЮДКИ — RAPHIDIOPTERA *
Верблюдки — насекомые с исключительно своеобразным обликом,
который создается узкой головой, вытянутой подвижной переднегрудью,
превышающей длину средне- и заднегруди вместе взятых, и прозрач-
ными, сложенными кровлеобразно крыльями с богатым сетчатым жил-
кованием и птеростигмой. Ротовой аппарат грызущий, глаза сравни-
тельно хорошо развиты, усики нитевидные. У самок присутствует длин-
ный и тонкий, изогнутый яйцеклад. Окраска обычно черная, ноги буро-
желтые.
Развитие с полным превращением. Личинки верблюдок внешне
сходны с имаго. Их сплющенное тело приспособлено к обитанию под
корой и в ходах короедов. Личинки нападают не только на короедов,
но и на мелких гусениц, поедают куколок и кладки яиц насекомых.
Многие виды живут под корой деревьев (преимущественно хвойных по-
род), некоторые заселяют поверхностные слои почвы, подстилку
и гниющую древесину. Развитие личинок продолжается 2 года. Окук-
ливаются личинки в колыбельках. Куколка подвижная. Взрослые верб-
людки— тоже активные хищники, встречаются на стволах сосен и дру-
гих деревьев, попадаются при кошении травостоя сачком в течение
всего лета.
Верблюдки — очень небольшой отряд, включающий несколько ви-
дов и 2 семейства. У представителей семейства Raphidiidae длина пе-
реднеспинки в 3—4 раза превышает ее ширину, голова треугольной
формы. Виды, относящиеся к другому семейству Inocellidae, имеют бо-
лее короткую переднеспинку (ее длина только в 2 раза превышает ши-
рину) и почти квадратную голову.
ОТРЯД СЕТЧАТОКРЫЛЫЕ - NEUROPTERA**
Общая характеристика
К сетчатокрылым относятся довольно разнообразные по внешнему
виду насекомые с двумя парами перепончатых крыльев (рис. 343—•
346). Иногда задние крылья уменьшены или значительно модифици-
рованы. Жилкование сетчатое ^рис. 347'—349), практически одинаковое
на передних и задних крыльях, причем продольные жилки в большин-
стве случаев с развилкой на конце. Усики разнообразного строения,
ротовой аппарат грызущий, фасеточные глаза хорошо развиты. Конеч-
ности сетчатокрылых бегательные, с 5-члениковыми лапками. Полет от-
носительно слабый. Сетчатокрылые активны преимущественно в су-
мерках и ночью, лишь некоторые виды ведут дневной образ жизни. Пи-
таются мелкими насекомыми и цветочной пыльцой.
Развитие с полным превращением. Личинки олигоподиальные,
хищные, с плоской головой, веретенообразным телом и развитыми
грудными ногами. Ротовой аппарат в виде парного колющего хоботка;
мандибулы несут на нижней стороне желоб, подстилаемый максилла-
ми. Через образованные между мандибулами и максиллами каналы
в жертву впрыскивается слюна и через эти же каналы происходит вы-
сасывание растворенного содержимого. Окукливаются личинки в шел-
* А. А. Стекольииков.
** А. А. Стекольииков.
137
ковистом коконе, материал для которого поставляется мальпигиевыми
сосудами через анальное отверстие.
Всего известно около 5000 видов сетчатокрылых. В европейской
части СССР встречается 45 видов из 9 семейств.
Наиболее широко представлены в европейской фауне зла-
тоглазки (Chrysopidae)—типичные обитатели лесов умеренно-
го пояса (рис. 343). Их яйца, снабженные стебельками, откладываются
на листья небольшими группами (рис. 350). Веретеновидные подвиж-
ные личинки златоглазок (рис. 351) живут открыто на деревьях и кус-
тарниках, реже на травах, пожирая тлей. Некоторые златоглазки об-
Рис. 343—349. Форма тела и жилкование крыльев сетчатокрылых.
343 — златоглазка (Chrysopidae), 344 — муравьиный лев (Myrmeleontidae), 345 ~ Mantispa styriaca
Poda (Mantispidae), 346 — Ascalaphus F. (Ascalaphidae), 347— переднее крыло златоглазки, 348 —
переднее крыло Hemerobius nitidulus L. (Hemerobiidae), 349 — переднее (вверху) и заднее (внизу)
крылья пыльцекрыла (Coniopterygidae).
ладают пахучими переднегрудными железами, которые выделяют сек-
рет с резким неприятным запахом. Близки по биологии к златоглаз-
кам представители семейств Hemerobiidae и Coniopterygi-
dae, личинки которых также обитают на растениях, высасывая тлей,
червецов и паутинных клещей.
Муравьиные львы (Myrmeleontidae) внешне напоми-
нают стрекоз-стрелок благодаря тонкому и длинному брюшку и узким
прозрачным крыльям (рис. -344). Личинки муравьиных львов имеют
плоское тело и серпообразные зазубренные мандибулы; встречаются
только на песках, где выкапывают ловчие воронки вдоль муравьиных
троп. Питаются муравьями, попадаю-
щими в эти воронки. Личинки пере-
двигаются, пятясь назад.
На юге европейской части СССР
можно встретить необычного вида
сетчатокрылое — Mantispa styriaca
Poda из семейства Mantispidae
(рис. 345). Эти хищные насекомые
обладают хватательными передними
ногами, а их личинки, обитая внутри
коконов пауков, питаются яйцами
и молодыми паучками. Другое южное
Рис. 350-351. Яйца и личинка семейство - A s с а 1 а р h i d а е пред-
златоглазки. ставлено на европейской части СССР
350 - яйца, 351 - личинка. ДВуМЯ ВИДЭМИ рОДЭ AsCalaphllS F,
138
(рис. 346). Эти насекомые обладают крупной головой с бу-
лавовидными усиками, коротким телом и пестро окрашенными
крыльями. Личинки аскалафусов живут на растениях или на
земле. Небольшая часть европейских сетчатокрылых, относящаяся к се-
мейству Sisyridae, в личиночной стадии ведет водный образ жизни,
развиваясь в губках или мшанках.
»
Определительная таблица семейств
1(12). Все ноги бегательные. Переднегрудь не длиннее среднегруди.
2(9). Усики на конце не утолщенные (нитевидные или четковидные).
3(8). Крылья прозрачные. Продольные жилки на конце с развилкой
(рис. 347, 348).
4(7). Усики нитевидные (рис. 343). Поперечные жилки в костальном
поле переднего крыла не ветвятся (рис. 347).
5(6). Тело зеленое или желтовато-зеленое. Крылья с легким радуж-
ным блеском. Насекомые средних размеров (в размахе крыль-
ев около 20 мм и более) . . Златоглазки — Chrysopidae.
6(5). Тело и крылья бурые. Мелкие насекомые (в размахе крыльев
около 10 мм) . . . . -..........................Sisyridae.
7(4). Усики четковидные. Поперечные жилки в костальном поле вет-
вятся (рис. 348)...........................Hemerobiidae.
8(3). Крылья матовые, белые. Продольные жилки на концах без
развилок (рис. 349).............................Coniopterygidae.
9(2). Усики утолщенные на конце—булавовидные или головчатые
(рис. 344, 346).
10(11). Усики во много раз короче тела (рис. 344). Глаза цельные.
Тело почти голое . . . Муравьиные львы — Myrmeleontidae.
11(10). Усики примерно равные длине тела (рис. 346). Каждый глаз
разделен глубокой продольной бороздой. Тело в длинных чер-
ных волосках.........................................Ascalaphidae.
12(1). Передние ноги хватательные, остальные — бегательные
(рис. 345). Переднегрудь длиннее среднегруди ....................
......................................... . Mantispidae.
ОТРЯД СКОРПИОННИЦЫ, ИЛИ СКОРПИОНОВЫЕ МУХИ — MECOPTERA*
Скорпионницы — небольшой отряд насекомых очень необычного
облика, который придает им голова, вытянутая в длинную головотруб-
ку. На ее конце расположены ножевидные или стилетообразные жвалы,
небольшая верхняя губа и членистые щупики. Головотрубка («клюв»)
образуется срастанием вытянутых основных члеников нижней губы
и максилл с пропорционально вытянутыми щеками и наличником. Фа-
сеточные глаза крупные, усики сравнительно длинные, нитевидные. Обе
пары перепончатых крыльев одинаковой формы и с одинаковым сет-
чатым жилкованием, удлиненные, нередко с бурыми пятнами. Иногда
крылья отсутствуют. Крылатые скорпионницы летают медленно и не-
уклюже.
Превращение полное. Личинки червеобразные, в большинстве слу-
чаев полиподиальные (с развитыми брюшными ложноножками, кото-
рые заканчиваются прочной щетинкой). Обитают в поверхностных
слоях почвы и в подстилке; питаются перегнивающими органическими
остатками, иногда поедают ризоиды мхов. Куколка свободная, в ко-
лыбельке.
* А. А. Стекольииков.
139
Отряд скорпионниц разделяется на 5 семейств. В СССР встреча-
ются 3 семейства — около 30 видов.
Представители семейства Panorpidae имеют вполне раз-
витые пестрые крылья, которые в покое распластываются в стороны.
У самцов утончающийся конец брюшка загнут кверху и направлен впе-
ред; последний, генитальный сегмент сильно вздут, что в целом соз-
дает поверхностное сходство с брюшком скорпиона. Семейство пред-
ставлено в нашей фауне несколькими широко распространенными ви-
дами рода Panorpa L. Эти насекомые ведут дневной и сумеречный об-
раз жизни, встречаясь в зарослях кустарников, на лесных полянах
и в других тенистых и сырых местах. Питаются мертвыми насекомыми
и другими органическими остатками, а также пыльцой и генеративны-
ми частями растений, нередко посещают экскременты. При питании вы-
деляют на пищевой субстрат секрет слюнных желез.
У видов семейства Bittacidae крылья без рисунка, в по-
кое складываются кровлеобразно. Длинные ноги придают им внешнее
сходство с комарами-долгоножками. Жвалы этих скорпионниц вытя-
нуты гораздо сильнее, чем у предыдущего семейства, и превращены
в мощные стилеты, которые, складываясь с длинной верхней губой
и частями максилл, формируют простейший хоботок. Все представи-
тели семейства — специализированные хищники. Свою добычу (в ос-
новном мелких двукрылых) они ловят длинными задними ногами, по-
сле чего вонзают в жертву хоботок и высасывают ее.
Для обоих семейств характерно довольно экзотическое поведение
при спаривании. У самцов сем. Panorpidae в период копуляции слюн-
ные железы выделяют коллоидное вещество, которое поступает через
рот наружу отдельными капельками и тут же поедается самкой. Сам-
цы сем. Bittacidae в период копуляции предлагают самкам вместо ка-
пелек слюны пойманную заранее добычу, которую самка поедает, пока
длится копуляция.
Семейство ледничников — Boreidae представлено в на-
шей фауне одним родом Boreus Latr. Это весьма своеобразные мелкие
насекомые, полностью лишенные крыльев. Они активны только в хо-
лодное время года — с октября до апреля; во время оттепелей быстро
бегают по снегу. Поздней осенью и ранней весной их можно встретить
на моховых подушках.
ОТРЯД ПЕРЕПОНЧАТОКРЫЛЫЕ — HYMENOPTERA*
Общая характеристика
По степени морфологической специализации и широте экологиче-
ской приспособленности перепончатокрылые уступают, например, отря-
ду двукрылых, но по сложности и разнообразию поведенческих реак-
ций они не имеют себе равных не только среди насекомых, но и среди
всех первичноротых животных. Именно у высших перепончатокрылых
мы встречаемся с самыми замечательными примерами так называемо-
го социального, или общественного, поведения.
Для перепончатокрылых характерно наличие 2 пар крыльев со
своеобразной системой жилок (рис. 352, к). Задние крылья значитель-
но меньше передних и в полете сцепляются с ними, образуя функцио-
нально единую крыловую поверхность. Число жилок на крыльях умень-
шается у мелких форм, и у многих мелких наездников они могут со-
всем отсутствовать. Во многих группах перепончатокрылых встреча-
ются совершенно бескрылые формы; как правило, это самки.
* В. Е. Кипятков.
140
Другой своеобразной морфологической чертой перепончатокрылых
является слияние нижней губы и нижних челюстей в способный вытя-
гиваться хоботок (лабио-максиллярный комплекс), приспособленный
для сосания и слизывания жидкой пищи, которой обычно является нек-
тар цветков (рис. 352, х). Это делает перепончатокрылых наиболее
важной группой среди насекомых-опылителей растений. Наилучшим об-
разом хоботок развит у высших, жалящих перепончатокрылых (Асп-
leata); он вторично отсутствует лишь у муравьев (Formicidae). В то
же время верхние челюсти (мандибулы) перепончатокрылых сохра-
Рис. 352—353. Общий план строения
перепончатокрылых.
352 — общий вид рабочей пчелы (Apis
mellifera L.)представителе стебельчато-
брюхих (Apocrita), 353—общий вид самки
пилильщика — представителя сидячебрюхих
(Symphyta); ноги и крылья удалены.
б — брюшко; г — глазки; ж. у — жгутик уси-
ка; зг — заднегрудь; к — крылья; кр — кор-
зиночка; м—мандибулы; н — ноги; пг —
переднегрудь; п.с — промежуточный сегмент
брюшка, вошедший в состав грудн; сг —
среднегрудь; с.у — стебелек усика; у — усик;
ф.г— фасеточные глаза; х— хоботок; ч.л —
первый членик задних лапок; щ —щеточка;
я—яйцеклад; I—IX — тергиты брюшиых
сегментов; 1~ 9 — стерниты брюшных сег-
ментов.
няют свою грызущую функцию и являются основным орудием в строи-
тельной деятельности (рис. 352, м), что и способствовало развитию
сложных форм поведения у высших перепончатокрылых. Таким обра-
зом, перепончатокрылые имеют полифункциональный ротовой аппарат,
который может быть назван грызуще-лижуще-сосущим.
Дорсальная часть переднегруди перепончатокрылых (передне-
спинка) неподвижно соединена со среднегрудью, тогда как вентраль-
ная ее часть (переднегрудка) соединена с ней и с переднеспинкой по-
движно. Переднегрудка вместе с причленяющимися к ней ногами и го-
ловой образует функционально единый аппарат (рис. 352). Этим
обеспечивается значительная подвижность ног и головы, что также
способствовало развитию сложных форм поведения, связанных со
строительством гнезд.
По строению брюшка (рис. 352, 353, б) перепончатокрылые раз-
деляются па 2 подотряда. У представителей сидячебрюхих пере-
пончатокрылых (Symphyta) брюшко сидячее: оно причле-
няется к груди без сужения или стебелька (рис. 353). У стебельча-
тобрюхих перепончатокрылых (Apocrita) либо имеет-
ся «талия» — сильное сужение брюшка к его основанию (висячее
брюшко — рис. 352), либо основание брюшка вытянуто в длинный
и тонкий стебелек (стебельчатое брюшко). Лишь у немногих мелких
наездников брюшко вторично становится сидячим. Такое строение
брюшка обеспечивает большую его подвижность, необходимую для
141
парализации добычи и откладки яиц. Кроме того, у стебельчатобрю-
хих морфологически первый сегмент брюшка вошел в состав груди-
(промежуточный сегмент), и брюшко фактически начинается со второго
сегмента (рис. 352, п.с). На конце брюшка у самок перепончатокры-
лых имеются яйцеклад (рис. 353, я) или жало.
Перепончатокрылые — насекомые с полным превращением. Личин-
ки почти всех видов имеют прядильные железы и окукливаются в па-
утинных коконах. Куколки открытого типа. Для всех перепончатокры-
лых характерен гапло-диплоидный механизм определения пола: самки
развиваются из оплодотворенных диплоидных яиц, а самцы — из не-
оплодотворенных гаплоидных путем партеногенеза (арренотокия).
Иногда (у некоторых рогохвостов — Siricidae, пилильщиков — Tenthre-
dinidae, орехотворок — Cynipidae и муравьев — Formicidae) из неопло-
дотворенных яиц могут развиваться самки (телитокия), что достигает-
ся за счет выпадения одного из делений мейоза и образования дипло-
идных яиц.
Многообразие перепончатокрылых и их обилие в различных био-
ценозах делают их весьма заметными в хозяйственной деятельности че-
ловека. Растительноядные перепончатокрылые вредят сельскому и лес-
ному хозяйству. Однако среди перепончатокрылых значительно больше
полезных видов, чем вредных. Прежде всего перепончатокрылые, в осо-
бенности пчелиные (Apidae), являются основными опылителями расте-
ний, в том числе возделываемых человеком. В настоящее время в свя-
зи с сокращением численности диких опылителей основную работу по
опылению культурных растений выполняют разводимые человеком ме-
доносные пчелы Apis mellifera L. Подсчитано, что экономический эф-
фект этой деятельности медоносных пчел во много раз больше, чем
.доходы от реализации основных продуктов пчеловодства — меда и во-
ска (к ним в последнее время прибавились ценные источники витами-
нов и лекарственных препаратов — прополис, маточное молочко и пыль-
ца). Разработаны и успешно используются методы искусственного раз-
ведения шмелей (Bombus Latr.) и пчел-листорезов (Megachile Latr.)
для опыления клевера и люцерны.
Огромное значение для человека имеет деятельность перепончато-
крылых-энтомофагов. Важнейшая роль в этом отношении принадлежит
паразитическим перепончатокрылым — наездникам и яйцеедам, которых
неоднократно использовали для организации биологической борьбы
с насекомыми-вредителями растений. В большинстве случаев успешного
применения биометода использовались перепончатокрылые и чаще все-
го наездники или яйцееды. Особенно перспективны яйцееды, уничто-
жающие вредителей в яйцах, т. е. до того, как они начали наносить
вред. Еще недостаточно изучена и оценена человеком деятельность оди-
ночных и общественных ос как энтомофагов, но она, несомненно, очень
существенна. А вот значение муравьев понято уже давно. В особенно-
сти полезны в лесах муравьи рода Formica L. Их искусственное раз-
множение и расселение позволяет успешно предотвращать массовые
размножения целого ряда вредителей и в ряде случаев даже отка-
заться от использования инсектицидов.
Перепончатокрылые — третий по величине отряд насекомых. По
числу известных видов (около 90 000) они уступают только жесткокры-
лым и чешуекрылым. Однако реальное число видов перепончатокрылых,
несомненно, гораздо больше и по оценкам разных авторов достигает
150—300 тысяч. В современной систематике отряд разделяется на
2 подотряда и 50—60 семейств. В СССР обнаружено около 10 000 ви-
дов перепончатокрылых.
142
Подотряд Сидячебрюхие — Syrnphyta
Жизнь сидячебрюхих перепончатокрылых проходит в основном
в личиночной стадии. Имаго живут недолго, выходят из куколок с уже
созревшими половыми продуктами и практически не нуждаются в до-
полнительном питании. Кормятся на цветках или вообще не питают-
ся. Некоторые настоящие пилильщики (Tenthredinidae) поедают мало-
подвижных насекомых. Самцы погибают после спаривания, а самки —
после откладки яиц.
Взрослые сидячебрюхие — довольно беззащитные насекомые: они
плохо летают, малоподвижны и лишены жала. Поэтому у них часто
встречаются покровительственная окраска и мимикрия под ос и пчел.
Рис. 354—356. Личинки сидячебрюхих перепончатокрылых.
354 — настоящий пилильщик Nematas leucoirochus Hart. (Tenthredinidae),
355 -- паутинный пилильщик Acantholyda Christ. (Pamphilidae),
356 — рогохвост Sirex gigas p. (Siricidae).
Некоторые пилильщики ядовиты и имеют яркую предупреждающую
окраску. Личинки почти всех сидячебрюхих являются фитофагами
(рис. 354—356).
Сидячебрюхие — наиболее примитивные перепончатокрылые, как
по морфологии, так и по биологии. Это небольшая группа насекомых:
она разделяется, даже при очень дробном систематическом подходе,
только на 11 семейств. В фауне СССР всего 400—500 видов. Ниже
дается характеристика 4 наиболее крупных семейств этого подотряда.
Рогохвосты — Siricidae (рис. 357, 358). Крупные или сред-
ней величины насекомые, обычно имеющие на конце брюшка роговид-
ный шип, а под ним у самок — игловидный яйцеклад с насечками на
створках. Имаго не питаются. Личинки — ксилофаги. Большинство ви-
дов связано с хвойными породами. Самка яйцекладом пропиливает
кору дерева и рецепторами яйцеклада определяет осмотическое давле-
143
ние сока флоэмы. Если оно понижено (дерево ослаблено), то рого-
хвост откладывает яйца. В основании яйцеклада имеется впячивание
кутикулы — «сумка», заполненная спорами симбиотического гриба, вы-
зывающего красную гниль древесины. При откладке яиц споры вво-
дятся в высверленное отверстие, и в дальнейшем личинки питаются дре-
весиной, пораженной грибом, без чего не могут нормально разви-
ваться.
Личинки рогохвостов имеют рудиментарные грудные ноги и ост-
рый шип на заднем конце тела, используемый при передвижениях
(рис. 356). Выгрызают в толще ствола цилиндрический ход, забитый
Рис. 357—358. Рогохвосты.
357 — самка большого соснового рогохвоста (Sirex gigas L.). 358 — самка ольхового рогохвоста
(Xiphydria camelus L.).
очень мелкой буровой мукой. Откладка яиц происходит в июне-июле.
Развитие личинок длится минимум 2 года, а при неблагоприятных
условиях может растягиваться на 5—6 лет. В конце развития личинка
окукливается без кокона в специальной колыбельке. Имаго выходят из
ствола, прогрызая отверстия своими мощными челюстями.
Стеблевые, или злаковые пилильщики — Cephidae
(рис. 359). Мелкие насекомые с узким, несколько сплющенным с бо-
ков телом. Самки с пиловидным, но обычно незаметным яйцекладом.
Личинки развиваются в стеблях злаков, реже — кустарников. В апре-
ле-мае самка откладывает 35—50 яиц по одному в каждое растение,
пропиливая яйцекладом верхние междоузлия. Личинки белые, с руди-
ментарными грудными ногами. Личинка питается и постепенно спуска-
ется вниз по стеблю. Затем она делает кольцевой надрез изнутри стеб-
ля на уровне почвы, отчего такой стебель скоро падает, а личинка зи-
мует в оставшёмся пеньке, затыкая его пробочкой из огрызков расте-
ния, и окукливается весной.
Настоящие пилильщики — Tenthredinidae (рис. 353,
360—362). Мелкие, средние, а иногда и довольно крупные насекомые
достаточно разнообразной внешности. У некоторых видов, связанных
с хвойными породами, самцы имеют гребневидные усики (рис. 360).
144
Рис. 359—363. Пилильщики. \
359 — самка малого хлебного пилильщика Cephas pygmaetis L. (Cephictae), 360 — голова самца сос-
нового пилильщика Diprion pint L. (Tcnthredinidae), 361 — самка тогохже вида, 362 — самка ело-
вого пилильщика Pachynematns scutellatas Лап из того же семейства,— самка паутинного
пилильщика Acantholyda hieroglyphica Christ. (Pamphilio^e).
У самок усики пиловидные (рис. 361) или нитевидные (риК 362), ред-
ко гребневидные. Самки с коротким пиловидным яйцекладам (рис.
353, я), который в покое втягивается в брюшко. Яйца всегда отклады-
ваются в сделанные яйцекладом надрезы в тканях растений, даже если
личинки развиваются открыто. При развитии происходит разбуха-
ние яиц.
Личинки (рис. 354) похожи на гусениц. Они называются ложно-
гусеницами и имеют 6—8 пар брюшных ложноножек (у гусениц не
более 5 пар); обычно ложногусеницы окрашены покровительственно,
иногда имеют яркие пятна для отпугивания врагов. У ряда видов
личинки снабжены ядовитыми железами, покрыты слизью, восковым
10 208
145
налетом или совершают отпугивающие врагов движения. По образу
жизни личинки настоящих пилильщиков также очень сходны с гусе-
ницами, причем на севере, где чешуекрылые немногочисленны, пи-
лильщики, напротив, обильны и экологически их замещают. Мы нахо-
дим среди личинок пилильщиков следующие экологические группы:
открыто живущие на листьях, обитающие в плодах, в свернутых листь-
ях, внутри стеблей, в тканях листьев (минирующие), образующие гал-
лы. Обычно .пилильщики—олигофаги и даже монофаги, редко поли-
фаги.
Размножение обоеполое, но у некоторых видов имеет место пар-
теногенез при почти полном отсутствии самцов (телитокия). Закон-
чившие питание личинки окукливаются в подстилке, в почве, в коко-
нах. Зимуют предкуколки. У многих видов развиваются 2 поколения
в год.
Паутинные пилильщики, или пилильщики-ткачи —
Pamphilidae (рис. 363). Небольшое семейство, развивающееся
в основном на хвойных породах. Самки приклеивают яйца к поверх-
ности растений. Личинки имеют очень оригинальную внешность: 3 па-
ры грудных ног, довольно длинные усики и членистые церки на зад-
нем конце тела (рис. 355). Паутинные железы личинок, в отличие от
большинства перепончатокрылых, функционируют в течение всей жиз-
ни и используются для строительства паутинных гнезд, которые мо-
гут иметь вид индивидуальных паутинных трубок (одиночные ткачи)
или же быть общими для всех личинок из одной кладки (обществен-
ные ткачи). Личинки передвигаются вне гнезд только по паутинкам.
Отгрызая хвоинки или кусочки листьев, они затаскивают их в гнездо
и там поедают. Окукливаются в почве без кокона. В год развивается
только 1 поколение.
Подотряд Стебельчатобрюхие — Apocrita
Среди стебельчатобрюхих перепончатокрылых доминируют виды,
личинки которых питаются животной пищей, главным образом насе-
комыми. Встречающиеся здесь растительноядные формы перешли к та-
кому образу жизни вторично.
Личинки стебельчатобрюхих ведут малоподвижный образ жизни,
питаясь пищей, запасенной для них взрослыми особями, или парази-
тируя в теле хозяина. Они лишены ног, глаз и усиков, имеют чаще
всего веретеновидную форму. У наездников личинки могут иметь весьма
своеобразную внешность, связанную с их приспособлением к парази-
тизму. В течение всей жизни личинки стебельчатобрюхих не выделяют
экскременты, что было бы губительно как для личинки-паразита,
так и для личинки, развивающейся в небольшой ячейке с заго-
товленной провизией. Только перед окукливанием происходит соеди-
нение среднего и заднего отделов кишечника и все накопившиеся за
период развития экскременты выводятся наружу. Обычно имаго вы-
ходят из куколок с незрелыми половыми продуктами, живут долго
и нуждаются в дополнительном питании, которое происходит прежде
всего на цветах; могут питаться также падью тлей. Многие наездни-
ки-эктопаразиты питаются гемолимфой хозяина, вытекающей из отвер-
стия, образованного яйцекладом при парализации. Характерна проте-
рандрия: самцы выходят из куколок раньше самок, что повышает их
репродуктивный успех и гарантирует своевременное оплодотворе-.
ние яиц.
Стебельчатобрюхих перепончатокрылых можно разделить на 2 боль-
шие группы: паразитические перепончатокрылые, или
наездники —Parasitica и жалящие перепончатокры-
146
л не — Aculeata. У Parasitica яйцеклад выполняет свою первичную
функцию откладки яиц, но используется также для парализации хозяи-
на, а иногда и для защиты от врагов; имеется только 1 ядовитая же-
леза, связанная с яйцекладом; вертлуги обычно состоят из 2 члени-
ков. У Aculeata яйцеклад преобразован в жало — орган защиты и на-
падения. С жалом связаны 2 ядовитые железы и оно ие используется
для яйцекладки. Яйца выводятся из полового отверстия на конце
брюшка. Вертлуги у жалящих всегда одинарные.
.Деление стебельчатобрюхих перепончатокрылых на паразитиче-
ских и жалящих условно, так как имеются группы, занимающие по
ряду признаков промежуточное положение. Такое деление не исполь-
зуется в современной систематике, но оно очень удобно в учебных це-
лях и поэтому сохранено в настоящем руководстве.
Подотряд Стебельчатобрюхие,
группа Паразитические перепончатокрылые,
или наездники — Parasitica
Обширная группа насекомых, разделяемая в настоящее время на
8 надсемейств и 25—30 семейств. Размеры тела весьма разнообразны:
от крупных до чрезвычайно мелких. Среди наездников-яйцеедов есть
формы, имеющие размеры около 0,2 мм, т. е. вообще самые мелкие
среди насекомых. У мелких форм жилкование на крыльях сильно ре-
дуцировано. Только для некоторых форм характерна вторичная фито-
фагия (см. ниже), большинство же паразитирует в личиночных стадиях
на насекомых и пауках. Некоторые типичные представители паразити-
ческих перепончатокрылых показаны на рис. 364—367.
Рис. 364—367. Паразитические перепончатокрылые.
364 — наездникApanteles glomeratus L. (Braconidae), 365 — яйцеед Trtchogramma evanescens Wstw.
(Chalcididae), 366 — розанная орехотворка Rhodites rosae L. (Cynipidae), 367 — яйцеед Telenomus
Hal. (Proctotrupidae).
147
Свое собирательное название наездники получили за присущую
многим из них характерную позу при откладке яиц: насекомое садит-
ся верхом на жертву и изгибает брюшко вниз, причем жертва часто
продолжает при этом двигаться (рис. 368—370). Самые крупные и наи-
более известные наездники относятся к обширному семейству Ichneu-
monidae. Тело этих наездников стройное, удлиненное; крылья обычно
с хорошо развитым жилкованием, а на передних крыльях имеется тем-
ный уплотненный участок — птеростигма (рис. 371, 372).
Взаимоотношения между личинкой наездника и ее хозяином значи-
тельно отличаются от паразитизма в классическом понимании, по-
скольку всегда приводят хозяина к гибели, и в этом отношении боль-
Рис. 368—370. Наездники и яйцееды.
368 — яйцеед Trichogramma evanescens Wstw. (Chalcididae), заражающий яйцо бабочки, 369 —
наездник Aphidias Nees (Braconidae), откладывающий яйцо в тело тли, 370 — наездник, отклады-
вающий яйцо под кору дерева на личинку жука-усача.
ше напоминают хищничество, чем паразитизм. Поэтому часто насеко-
мых, паразитирующих в беспозвоночных, называют не паразитами,
а паразитоидами.
По характеру паразитирования всех наездников делят на экто-
и эндопаразитов. Эволюционно первичен и более примитивен эктопара-
зитизм, когда личинка-паразит питается тканями хозяина через отвер-
стие в его покровах, оставаясь снаружи. При эндопаразитизме паразит
развивается внутри тела хозяина. Эктопаразитов отличают от эндопа-
разитов многие особенности морфологии, физиологии и поведения.
Эктопаразиты заражают, как правило, скрытно живущих хозяев,
обитающих в ходах, плодах, галлах, стеблях и других полостях, ко-
торые служат защитой личинке, питающейся на теле хозяина. Эндопа-
разиты же могут заражать как скрытно, так и открыто живущих хо-
зяев. Эктопаразиты перед откладкой яйца обычно парализуют хозяи-
на, что лишает его возможности сбросить с себя личинку паразита.
У эндопаразитов парализация встречается редко и всегда бывает вре-
148
меннои; ее цель иная: лишить хозяина возможности сопротивляться
в момент откладки яйца. Личинки эктопаразитов имеют высокую ско-
рость развития, что необходимо, так как парализованный хозяин мо-
жет погибнуть. Эндопаразиты развиваются дольше и могут диапаузи-
ровать в первом возрасте, что обычно сопряжено с диапаузой хозяина.
Эктопаразиты в физиологическом отношении менее специализиро-
ваны, чем эндопаразиты, часто они являются полифагами, заражаю-
щими широкий круг хозяев из разных систематических групп. У них
обычно наблюдается экологическая специализация, т. е. приурочен-
ность к определенным местообитаниям хозяев (например, под корой
деревьев, в подстилке, в плодах) или к общим типам повреждений рас-
тений (галлам, минам и т. п.). У эндопаразитов наряду с экологиче-
Рис. 371—372. Наездники семейства Ichneumonidae.
371 — Cryptus F., 372 Ophion F.
з — зеркальце; n—птеростигма; п.ж— остаток поперечной жилки; с.ж — склеротизоваиная жил-
ка 'на переднем крае крыла.
ской специализацией всегда выражена приуроченность к определенной
таксономической группе хозяев: к чешуекрылым, двукрылым, жукам,
щитовкам, или чаще к более узким группам — семействам, родам и да-
же отдельным видам хозяев.
Яйца эндопаразитов мелкие и почти не имеют желтка. При разви-
тии они активно поглощают питательные вещества из гемолимфы хо-
зяина и увеличивают свои размеры в десятки, а иногда в сотни раз.
Эндопаразитические личинки многообразны по внешнему и внутренне-
му строению, имеют различные приспособления к питанию, дыханию
и защите от иммунных сил хозяина. При питании они поедают вна-
чале гемолимфу и жировое тело хозяина, не затрагивая жизненно важ-
ных органов. Поэтому зараженные насекомые, хотя и становятся бо-
лее ослабленными, продолжают двигаться и питаться до конца жиз-
ни. Внутри тела хозяина у паразита проходит только 1 линька, вторая
же совпадает с выходом из хозяина, после чего нередко наблюдается
эктопаразитическое дополнительное питание и затем окукливание в ко-
коне. Развитие эндопаразитов обычно очень тесно связано с состоянием
хозяина и регулируется его эндокринной системой. Если у эктопарази-
тов регуляция сезонных циклов развития вполне самостоятельная, то
у эндопаразитов она зачастую обусловлена физиологическими влия-
ниями хозяина: например, диапауза у паразита наступает в ответ на
возникновение диапаузы у хозяина.
149
Эктопаразиты, как правило, являются групповыми: самка откла-
дывает на хозяина несколько яиц в зависимости от его размеров и плот-
ности его популяции. Эндопаразиты — чаще всего одиночные, т. е. в хо-
зяине может развиться только 1 паразит. Исключение составляют мно-
гие мелкие паразиты крупных хозяев, яйцо которых, испытывая полное
дробление, дает несколько, иногда сотни отдельных зародышей.
Это явление называется полиэмбрионией. Из хозяина, зараженного од-
ним яйцом, вылетают, таким образом, десятки взрослых наездников,
что сильно повышает репродуктивный потенциал таких видов. Это осо-
бенно важно, когда хозяева в природе редки и найти их нелегко.
Хозяевами эктопаразитов являются главным образом личинки на-
секомых, развивающихся с полным превращением (в основном жуков
и бабочек), причем обычно личинки старших возрастов. Значительно
реже хозяевами могут быть куколки. Хозяева эндопаразитов значи-
тельно более многообразны (хотя и здесь преобладают личинки жуков
и бабочек), причем заражаться могут любые стадии развития хозяев:
яйца, личинки, куколки и даже имаго. В яйцах насекомых развива-
ются так называемые яйцееды. Это наиболее многочисленные среди
самых мелких наездников представители семейств Chalcididae
(рис. 365, 368) и Proctotrupidae (рис. 367). Развитие паразита может
растягиваться на несколько онтогенетических стадий хозяина. Так,
у многих наездников яйцо откладывается в личинку, а выходит пара-
зит из куколки и даже иногда из имаго хозяина; может заражаться
яйцо, а выход паразита происходит из личинки и даже из куколки. Су-
ществуют также имагинальные эндопаразиты, заражающие имаго
и выходящие из имаго. Их хозяева — жуки, реже перепончатокрылые
и сетчатокрылые. Некоторые группы наездников развиваются в имаго
и личинках клопов, в тлях (рис. 369), щитовках, клещах и их нимфах.
У многих наездников наблюдается гиперметаморфоз: личинка пер-
вого возраста имеет специальные приспособления для нахождения сво-
его хозяина, а питаются личинки последующих возрастов. Так, самки
некоторых мелких наездников откладывают тысячи яиц на почву или
растения. Подвижные личинки первого возраста активно цепляются
к проходящим мимо насекомым. Если такими насекомыми оказывают-
ся рабочие муравьи, то с их помощью личинки добираются до своих
хозяев — личинок и куколок муравьев. Личинки многих наездников
семейства Ichneumonidae в первом возрасте активно ползают, отыски-
вая своих хозяев. Вообще же большинство эндопаразитов развивается
с гиперметаморфозом, так как личинки разных возрастов у них, как
правило, приспособлены к достижению разных целей.
Широко распространен среди наездников сверхпаразитизм, т. е.
паразитирование на паразитах (личинках наездников или паразитиче-
ских мух). Обычно наблюдается паразитизм второго порядка, реже—•
третьего и крайне редко — четвертого порядка. Самки наездников из
семейства Trigonalidae откладывают множество яиц на листья. Если эти
яйца будут проглочены питающимися листьями гусеницами, то из них
выходят личинки, для которых гусеница не является хозяином, а толь-
ко местом ожидания хозяина — паразитической личинки наездника из
семейства Ichneumonidae. Если гусеница окажется зараженной личин-
кой наездника, то личинка семейства Trigonalidae будет паразитиро-
вать на ней, являясь, таким образом, паразитом второго порядка.
Взрослые наездники обычно спариваются в местах дополнитель-
ного питания (на цветках); у некоторых видов наблюдается роение.
Оплодотворенная самка активно отыскивает хозяина, что обычно пред-
ставляет нелегкую задачу: сначала наездник по тем или иным призна-
кам отыскивает местообитание хозяина, а затем ищет его, ориентируясь
зрительно и по запаху. Установлено, что у многих наездников самка
150
запоминает запах того хозяина, в котором она развивалась, и затем
руководствуется им при отыскании хозяина для яйцекладки. Оконча-
тельно пригодность найденного объекта для откладки яйца определя-
ется рецепторами яйцеклада. Некоторые наездники обладают удиви-
тельными способностями к отысканию хозяев. Например, крупные пред-
ставители семейства Ichneumonidae каким-то образом отыскивают
развивающихся глубоко под корой личинок жуков и откладывают на
них яйца, просверливая кору своим длинным яйцекладом (рис. 370).
При откладке яйца в хозяина самка метит его специальным за-
пахом (феромоном), который сигнализирует другим самкам, что дан-
ный хозяин уже заражен, и те не откладывают в него свои яйца. Если
же это случится, то личинка, вышедшая из яйца первой, уничтожает
своих более мелких конкурентов. Для этого личинки многих эндопа-
разитов в первом возрасте имеют длинные серповидные мандибулы.
Во многих группах паразитических перепончатокрылых появились
растительноядные формы. Это характерно почти для всех видов семей-
ства орехотворок (Cynipidae), являющихся галлообразователями на
дубе (80% видов) и других растениях. Орехотворки — мелкие насе-
комые с коротким, обычно сжатым с боков телом (рис. 366). Самки
откладывают яйца в ткани растений, где при развитии личинок обра-
зуются галлы — разрастающиеся ткани растений, форма которых спе-
цифична для каждого вида орехотворок. Для орехотворок характерно
чередование обоеполого и партеногенетического поколений, причем
самцы и самки обоеполого поколения крылаты, а партеногенетические
самки бескрылы. Эти поколения связаны с разными сезонами года
и развиваются в галлах на разных частях растений. Некоторые орехо-
творки самостоятельно не образуют галлы, а развиваются как инкви-
лины в чужих галлах. Много видов галлообразователей также среди
наездников семейства Eurytomidae. Среди растительноядных наездни-
ков встречаются также семееды, развивающиеся в семенах растений.
Некоторые паразитические перепончатокрылые питаются пыльцой, раз-
виваясь как инквилины в гнездах одиночных пчел.
Подотряд Стебельчатобрюхие,
группа Жалящие перепончатокрылые — Aculeata
Самцы и самки жалящих перепончатокрылых значительно разли-
чаются по сложности своего поведения. Для самок, как правило, ха-
рактерны высокоразвитые проявления заботы о потомстве, связанные
со строительством гнезда, заготовкой корма для личинок или даже
непосредственно их выкармливанием. Самцы не участвуют в заботе
о потомстве и только у некоторых роющих ос (Sphecidae) они могут
охранять гнездо в отсутствие самки. Основная функция самцов сводит-
ся к оплодотворению самок. Механизмы, обеспечивающие встречу по-
лов, разнообразны. У ряда видов копуляция происходит на цветах;
у других самцы дежурят в местах гнездования, поджидая выходящих
позже них самок. Самцы многих пчел и ос занимают индивидуальные
территории, которые они охраняют от других самцов. На этих терри-
ториях самцы спариваются с прилетающими самками. У шмелей
(Bombus Latr.) самцы летают по замкнутым маршрутам, меченным
в определенных точках половым феромоном, и спариваются с привле-
каемыми феромонами самками. Самцы и самки многих мураевьев об-
разуют большие рои, где происходит спаривание. Самцы находят са-
мок (или самки самцов), ориентируясь зрительно и по запаху поло-
вых феромонов.
У одиночных видов жалящих перепончатокрылых каждая самка
самостоятельно выращивает свое потомство, сооружая для него гнез-
151
до, в котором может быть одна или чаще несколько ячеек. В каждой
ячейке развивается 1 личинка. Самка нередко строит в течение жизни
несколько гнезд. За год обычно развиваются 1—2 поколения. У многих
ос и пчел гнезда отдельных самок располагаются в непосредственной
близости друг от друга, образуя обширные поселения. У некоторых ви-
дов наблюдается коммунальное поведение, когда несколько самок
(обычно это сестры) обитают в одном совместно сооружаемом гнез-
де, но имеют в нем собственные ячейки и выкармливают своих личи-
нок самостоятельно. Коммунальное гнездование выгодно тем, что по-
зволяет самкам затрачивать меньше сил на строительство и более
эффективно охранять гнездо от хищников и паразитов. Многие одиноч-
ные осы и пчелы являются инквилинами (гнездовыми паразитами)
в гнездах других видов: они откладывают яйца в чужие гнезда, и их
личинки развиваются за счет провизии, заготовленной для своих ли-
чинок самкой вида-хозяина.
Во многих группах жалящих проявляется тенденция к возникно-
вению социального поведения. У целого ряда видов одиночных пчел
и ос самка остается в гнезде до выхода из куколок своего потомства.
Нередко ее дочери начинают размножаться в том же гнезде, выра-
щивая в нем своих личинок. В дальнейшем на основе такого поведения
может возникать кооперирование в выращивании расплода и других
видах деятельности, а затем и разделение на касты, т. е. переход к со-
374
Рис. 373—375. Некоторые представители эктопаразитических ос.
373 — самка блестянки Chrysis L. (Chrysididae), 374— Tlpfila femorata F. (Tiphiidae), 375 — самка
немки Mutllla L. (Mutillidae).
152
циальному образу жизни (см. раздел «Поведение насекомых»). Это
происходило неоднократно в эволюции жалящих перепончатокрылых:
существует более 10 групп, в которых общественное поведение возник-
ло самостоятельно и независимо. Это муравьи, общественные складча-
токрылые осы, медоносные пчелы и шмели и некоторые другие менее
известные группы.
Жалящие — обширная и разнообразная группа перепончатокры-
лых, разделяемая на 25—30 семейств. Ниже охарактеризованы только
важнейшие семейства, широко распространенные в европейской части
СССР.
Наиболее примитивные представители ос являются эктопарази-
тами, и их биология во многом сходна с биологией наездников. Так,
осы семейства Bethylidae паразитируют на гусеницах, личин-
ках жуков и других насекомых. Блестянки — Chrysididae
(рис. 373) являются специализированными паразитами скрытоживущих
Рис. 376. Сколия Scolia maculata Dr. (Scoliidae).
насекомых, в основном одиночных ос и пчел. Осы, относящиеся к не-
большому семейству Tiphiidae (рис. 374), охотятся на личи-
нок пластинчатоусых жуков и скакунов. Весьма своеобразные по вне-
шности немки — Mutillidae, у который самцы крылаты, а бескры-
лые самки напоминают муравьев, покрытых густыми волосками
(рис. 375), паразитируют в гнездах шмелей, одиночных ос и пчел, из-
редка встречаются в гнездах муравьев, а некоторые виды развиваются
на личинках жуков и мух.
Сколии—Scoliidae (рис. 376). Крупные осы с массивным
телом и крепкими ногами, обычно с рисунком из четырех светлых пятен
на брюшке. Парализуют личинок, пластинчатоусых жуков. Парализа-
ция производится уколом жала в единственный нервный ганглий ли-
чинки. Паралич постоянный. Парализованная добыча с яйцом осы на
брюшной стороне оставляется на месте поимки без сооружения ка-
кого-либо укрытия. Наиболее крупная наша сколия Scolia maculata
Dr. (рис. 376) парализует крупных личинок жука-носорога (Oryctes
nasicornis L.).
Дорожные осы — Pompilidae (рис. 377). О-бширное, но
слабо изученное семейство. Обычно темноокрашенные осы со строй-
ным, но коротким телом, очень длинными тонкими ногами и закручен-
ными в кольца усиками. Внешне несколько похожи на наездников.
153
случаиную полость, трещину в
Рис. 377. Дорожная оса Calicur-
gus hyalinatus F. (Pompilidae).
n — птеростигма; ю.о •— югальиая об-
ласть.
Чаще всего встречаются на открытых местах—полянах, просеках, доро-
гах, тропах, насыпях. При передвижениях для дорожных ос характер-
ны быстрые и резкие перебежки и броски из стороны в сторону. Все
дорожные осы охотятся на пауков, парализуя их уколом жала в един-
ственный нервный ганглий головогруди. Но перед этим оса обычно жа-
лит паука в рот, парализуя нервный центр, управляющий ядовитыми
хелицерами, что делает паука безопасным для осы. Провизия личин-
ки дорожной осы — всегда только один паук. Поэтому мелкие виды ос
охотятся на мелких пауков, а крупные — на крупных.
Самые примитивные дорожные осы оставляют парализованного
паука с приклеенным на грудь яйцом в месте поимки без всякого укры-
тия. Однако у большинства видов добыча затаскивается в какую-либо
почве, пустую норку, которые затем за-
брасываются почвой и камешками.
Некоторые дорожные осы не исполь-
зуют готовые полости, а самостоя-
) тельно выкапывают норку для каж-
дого добытого паука. Наконец, не-
многие виды сооружают гнездо еще
до охоты, а уже потом ловят паука,
доставляют в гнездо и откладывают
на него яйцо. Отдельные виды до-
рожных ос строят даже многоячей-
ковые гнезда или живут совместно
в одной норке (коммунальное гнез-
дование). Однако все это редкие
исключения, а у большинства до-
рожных ос поведение весьма прими-
тивное и ограничивается использо-
ванием готовых полостей для разме-
щения пойманной и парализованной
добычи.
Роющие осы — Sphecidae
(рис. 378—380). Обширное семейство,
разделяемое на целый ряд подсе-
мейств и триб. Очень разнообразны по внешнему облику и размерам,
которые могут быть от мелких до довольно крупных. Большинство ви-
дов гнездится в почве, выкапывая норки с одной или несколькими
ячейками, расположенными вокруг основного хода. Многие, в особен-
ности мелкие, виды гнездятся в древесине, используя для этого гото-
вые полости—ходы насекомых-ксилофагов. В таком ходе оса соору-
жает линейный ряд ячеек, разделяя его поперечными перегородками
из древесных опилок. Некоторые осы используют для этой цели пу-
стые соломинки или стебли и веточки, выгрызая у них сердцевину.
Только немногие роющие осы изготавливают гнезда из мокрой глини-
стой почвы (например, пелопей — Sceliphron К1.).
Все роющие осы являются парализаторами. Их добыча — самые
разнообразные насекомые и пауки, причем часто наблюдается четкая
специализация родов и даже видов ос на определенном типе добычи.
Обычно добыча представлена достаточно обширной группой, например,
мухами (для ос рода Bembex F. — рис. 380), пчелами (для рода Phi-
lanthus F.), долгоносиками и златками (для рода Cerceris Latr.—
рис. 379), гусеницами подгрызающих совок (для рода Ammophila
Kby. — рис. 381, 382), кузнечиками и саранчовыми (для рода Sphex L.—
рис. 378). Нередки также случаи более узкой специализации роющих
ос. Например, Sphex occidentalis охотится за кузнечиками рода Ephip-
piger Latr., a S. maxillosus L. — за полевыми сверчками (Gryllus сат-
154
pestris L.). Поведение осы во время охоты и способ парализации зави-
сят от характера добычи; в частности, количество уколов жалом опре-
деляется числом нервных ганглиев у жертвы. Так, для парализации
долгоносика или златки осе Cerceris Latr. нужно всего одно ужаление
в единственный туловищный ганглий, а оса Ammophila Kby. для пара-
лизации гусеницы должна ужалить ее последовательно во все сегмен-
тарные нервные узлы (рис. 381).
Среди роющих ос мы находим виды, соответствующие разным эта-
пам эволюции инстинктивного поведения. Наиболее примитивные из
них являются просто эктопаразитами и не прячут свою добычу, дру-
гие используют для этого любые случайные полости. Однако большин-
ство роющих ос строят специальные гнезда. Гнездо либо сооружается
уже после парализации добычи неподалеку от нее, как у многих видов
рода Sphex L., либо оно строится заранее и в него транспортируется до-
быча, как, например, у видов рода Ammophila Kby. (рис. 382). В ходе
дальнейшей эволюции осы переходят на парализацию более мелких
насекомых и снабжают каждую личинку не одной крупной, а несколь-
кими мелкими жертвами, что облегчает работу осы. Такое поведение
проявляется у большинства роющих ос. При дальнейшем усложнении
поведения осы переходят от массового провиантирования к постепен-
ному, т. е. приносят личинке парализованных насекомых не сразу,
155
а по-одному по мере их поедания. Так, например, многие виды рода
Ammophila Kby. имеют одновременно несколько гнезд и снабжают их
гусеницами поочередно. Гнездящиеся на песках осы-филанты (Philan-
thus F.) и бембексы (Bembex F.) выкармливают личинок, регулярно
принося‘им добычу (пчел у филантов и мух у бембексов). В данном
случае отпадает необходимость парализовать добычу, так как она не
хранится долго, и эти осы просто убивают свои жертвы. Необходи-
мость постоянного возвращения к гнезду выработала у ос блестящие
способности к ориентации (см. рис. 7); осы, имеющие несколько гнезд,
не только прекрасно помнят их расположение, но и точно знают, сколь-
ко и когда нужно принести корма каждой личинке в зависимости от
382
Рис. 381—382. Роющая оса Ammophila Kby.
во время парализации и транспортировки
добычи.
381 — парализация гусеницы озимой совки
Agrotis segetum Schiff.), 382 — транспортировка па-
рализованной добычи в гнездо.
ко па(пализовапных насекомых. Обычно
ее возраста.
Складчатойр ы л ы е
о с ы—V е s р i d а е (|рис. 383—
386). Способны складывать
в покое передние крылья
продольно вдоль их длины.
В отличие от остальных ос
всегда откладывают яйцо в
пустую ячейку, а затем уже
приносят в нее провизию.
Разделяются на 3 подсемей-
ства, из которых только
2 представлены в европейской
фауне.
Осы подсемейства
Eumeninae (рис. 383) яв-
ляются одиночными и строят
гнезда либо в почве (часто
с наружными пристройками
в виде башенок или трубочек,
как у стенных ос рода Ody-
nerus Latr.), либо в пустых
соломинках или внутри ходов
в древесине, либо сооружают
гнезда из смоченной слюной
глины в виде шариков и кув-
шинчиков, расположенных на
стеблях растений или на кам-
нях (рис. 387). В каждую
ячейку помещают по несколь-
это личинки жуков или гусе-
ницы. У некоторых видов наблюдается постепенное провиантирование.
Осы подсемейства Vespinae (рис. 384—386) называются
бумажными осами, потому что они строят свои гнезда из мелко пере-
жеванной и смоченной слюной древесины, которая после высыхания
напоминает грубую бумагу или картон. Почти все представители этого
подсемейства являются общественными насекомыми. Кастовый ди-
морфизм может быть выражен в разной степени. Так, у видов рода
Polistes Latr. (рис. 384) плодовитые самки-царицы почти не отличают-
ся по размерам от рабочих ос, а у шершней (род Vespa L.—рис. 385)
и представителей рода Vespula Th. (рис. 386) царицы значительно
крупнее рабочих.
Гнезда бумажных ос (рис. 388) состоят из одного или нескольких
сотов, расположенных обычно горизонтально друг над другом. Соты
формируются из шестигранных ячеек, обращенных отверстиями вниз.
Обычно соты окружены многослойной бумажной оболочкой с входным
156
Рис. 383—386. Складчатокрылые осы (Vespidae).
383 — стенная оса Odynerus poecilus Sauss.. 384 — оса-полист Pollstes gallicus L., 385 — шершень
Vespa crabro L., 386 — обыкновенная oca Vespula •vulgaris L.; д.я — дискоидальная ячейка.
Рис. 387. Одиночная складчатокрылая
оса Eumenes coarctatus L., доставляю-
щая парализованную гусеницу в гнездо.
Рис. 388. Гнездо бумажной осы Vespula
germanica F. в поперечном разрезе.
з. я —закрытые крышечками ячейки с куколка-
ми; л — леток; о.г — оболочка гнезда; с — соты.
157
отверстием внизу или сбоку, но у некоторых примитивных видов гнезда
без оболочки. Так, у видов рода Polistes Latr. гнездо состоит из одного
сота. Гнезда либо подвешиваются на бумажном стебельке к веткам
деревьев и под крышами зданий, либо строятся в дуплах и почвенных
норах. ' !
Бумажные осы ловят разнообразных насекомых, пережевывают
и скармливают их личинкам. Сами же питаются в основном углевода-
ми, получая их на цветках, а также в виде секрета лабиальных желез
личинок, которые отдают его осам в ответ на прикосновения усиками.
Некоторые виды накапливают в гнездах значительные запасы меда.
Личинки окукливаются в ячейках, заплетая их отверстия паутинными
крышечками.
Большинство бумажных ос имеет однолетний цикл развития. Гнез-
да основываются весной оплодотворенными осенью и перезимовавши-
ми молодыми царицами; иногда для этого могут объединяться не-
сколько цариц. Самостоятельно построив небольшое гнездо, царица
выращивает первое поколение рабочих ос, которые начинают выпол-
нять все основные работы в гнезде, а царица в дальнейшем лишь от-
кладывает яйца. У более примитивных видов, относящихся к роду
Polistes Latr., царица продолжает участвовать в строительстве и даже
в снабжении личинок кормом. Постепенно количество рабочих в гнез-
де возрастает, и во второй половине лета начинают появляться моло-
дые самцы и самки — будущие царицы. У ос родов Vespa L. и Vespu-
la Th. царицы выращиваются в более крупных ячейках, сооружаемых
в конце лета. К концу цикла развития численность ос в гнезде дости-
гает нескольких десятков или нескольких сот. Молодые самцы и самки
вскоре покидают гнездо, спариваются и самки уходят на зимовку,
а материнская колония постепенно распадается и погибает.
Несмотря на наличие резкого кастового диморфизма у наиболее
высокоорганизованных бумажных ос, они в целом имеют невысокий
уровень социальной организации. Разделение функций между рабо-
чими особями у них выражено довольно слабо, и каждая оса может
выполнять любые работы попеременно. Почти отсутствуют у них
и сложные формы передачи информации. Так, например, осы добыва-
ют корм всегда в одиночку и не могут сообщить о найденном источ-
нике корма другим особям. Обмен кормом между членами одной ко-
лонии (трофаллаксис) у ос наблюдается очень часто, но практически
отсутствует характерное для наиболее высокоорганизованных общест-
венных насекомых поведение взаимного облизывания и очистки тела.
В пределах этой группы мы находим все переходы от одиночного
к общественному образу жизни, что делает бумажных ос очень инте-
ресными объектами для исследования путей возникновения социаль-
ного поведения у насекомых.
Пчелиные — Apidae (рис. 352, 389). Одно из самых много-
численных семейств жалящих перепончатокрылых, насчитывающее бо-
лее 20 000 видов и разделяемое на 8—10 подсемейств, которые в на-
стоящее время обычно рассматриваются как отдельные семейства.
Пчелиные — это, по существу, осы, оставившие осиные повадки и пере-
шедшие к выкармливанию своих личинок пыльцой и нектаром. На-
стоящие пчелы возникли в меловом периоде вскоре после появления
цветковых растений; дальнейшее эволюционное развитие этих двух
групп тесно взаимосвязано и является прекрасным примером сопря-
женной эволюции. С тех пор и до настоящего времени пчелы являют-
ся основными опылителями цветковых растений. Приспособление к пи-
танию на цветках (антофилии) открыло перед пчелами совершенно
новую адаптивную зону и обусловило огромное разнообразие этих.на-
секомых. Только самые примитивные представители пчелиных соби-
158
рают пыльцу, проглатывая ее и перенося в гнездо в зобике; у боль-
шинства видов возникли для этой цели разнообразные собирательные
аппараты. Во-первых, тело пчел покрыто густыми перистыми волоска-
ми, в которых задерживается пыльца. Во-вторых, первый членик задних
лапок расширен и покрыт изнутри щеточкой волосков, предназначен-
ной для прочесывания тела и снятия с него пыльцы (рис. 352, щ).
В-третьих, пчелиные имеют так называемые «корзиночки», служащие
для накопления этой пыльцы и транспортирования ее в гнездо. Обычно
это скопление длинных волосков на задних ногах и на других частях
тела, щетка густых волосков на нижней стороне брюшка или окру-
женная длинными волосками блестящая площадка на задней голени,
где собирается комочек пыльцы — обножка. Именно по последнему
типу устроены корзиночки у шмелей (Bombus Latr.) и у медоносной
пчелы Apis mellifera L. (рис. 352, к).
Обычно одиночные пчелы собирают корм на растениях опреде-
ленной систематической группы, а иногда даже только с цветов одного
вида. При этом всегда имеет место точная синхронизация периодов
цветения кормовых растений и активности' посещающих их пчел. Об-
щественные пчелы, напротив, мало специализированы в отношении по-
сещаемых ими растений.
Гнезда пчелиных весьма разнообразны. Чаще всего они выкапы-
ваются в почве, иногда выгрызаются в древесине или в стеблях раз-
личных растений (рис. 390); у многих одиночных пчел гнезда устраи-
ваются в готовых полостях — пустых соломинках, ходах насекомых-
ксилофагов, чужих почвенных норках или даже в пустых раковинах
улиток. Такие гнезда могут быть двух типов: 1) линейные, когда нор-
ка или иная полость разделяется перегородками из почвы, древесины
и других материалов на отдельные ячейки; 2) гроздевидные, когда
ячейки независимо открываются в центральный ход или в его ответ-
вления. Пчелы-каменщицы (род Chalicodoma Lep). сооружают гнезда
на камнях или стенах из смоченной слюной почвы, застывающей
в очень твердую массу. Большинство пчел обмазывают стенки ячеек
изнутри секретом специальной железы, образующим водонепроницае-
159
мую пленку, или выделениями слюнных желез. У пчел подсемейства
Megachilinae такая обмазка отсутствует, и они изготовляют перегород-
ки или ячейки целиком из самых разнообразных приносимых извне
материалов: кусочков листьев, смолы, войлока, изготовленного из
пушка растений, смоченной слюны почвы и т. п. У видов подсемейства
Apinae, к которому относятся медоносная пчела, шмели и некоторые
тропические безжальные пчелы, ячейки строятся из воска, выделяе-
мого железами на тергитах или стернитах брюшка.
Многие одиночные пчелы являются инквилинами и откладывают
свои яйца в гнезда других видов пчелиных, где их личинки развива-
ются за счет чужих запасов провизии. Такие пчелы обычно не имеют
опушения и корзиночек, поскольку они сами не собирают пыльцу. Для
инквилинов всегда характерна специализация по хозяину, т. е. разви-
тие за счет пчелиных лишь определенной систематической группы.
Рис. 390. Гнездо одиночной пчелы Osmia Latr. в стебле ежевики.
' Среди пчелиных неоднократно возникал общественный образ
жизни. Так, в подсемействе Halictinae мы находим все переходы от
одиночного существования до примитивной социальности. Представи-
тели этого подсемейства — небольшие темноокрашенные пчелы, строя-
щие гроздевидные гнезда в почве. У общественных видов гнезда закла-
I дывают весной самки-основательницы. Среди их потомства, которое
остается в том же гнезде, только часть особей размножается, осталь-
1 ные бесплодны и являются рабочими. Однако у большинства видов это
разделение на касты половых и рабочих недостаточно четко выражено
и между ними почти нет морфологических различий. Только у немно-
гих видов первое поколение полностью представлено бесплодными ра-
бочими пчелами, имеющими несколько меньшие размеры, чем самка-
основательница (царица). Поведение этих пчел также примитивно:
отсутствуют сложные механизмы сигнализации и передачи информа-
ции, взаимное облизывание и обмен кормом.
Более высокий уровень социальной организации имеют шмели
(Bombus Latr.), относящиеся к подсемейству Apinae. Это крупные,
густо опушенные перепончатокрылые, распространенные в основном
в умеренном поясе и на севере. Чаще всего шмели гнездятся в почве,
нередко используя поры грызунов (рис. 391), на почве под прикрыти-
ем сена или мха или реже в дуплах. Личинки шмелей развиваются
группами по 5—10 особей в одной общей восковой ячейке, постепенно
растягивая ее по мере роста. Взрослые шмели дают им корм через
160
временные отверстия в стенке ячейки. Личинки окукливаются в бочон-
кообразных коконах, которые после выхода имаго используются для
хранения меда и пыльцы.
Гнезда шмелей существуют только одно лето, и их жизненный
цикл очень сходен с жизненным циклом бумажных ос. Весной выле-
тевшие после зимовки са-мки-основательницы (царицы), оплодотворен-
ные осенью, самостоятельно основывают гнезда и выращивают первое
поколение рабочих, которые начинают выполнять все необходимые ра-
боты, а царицы с этого времени только строят новые ячейки и откла-
дывают в них яйца. Число рабочих шмелей в гнезде постепенно увели-
чивается, а в конце лета начинают выращиваться самцы и плодовитые
Рис. 391. Гнездо шмеля Bombus lapidarius L., устроенное в покинутой норе мыши
(по Вильсону).
в.я — восковые ячейки с личинками (стенки двух из них вскрыты); к — коконы; м.г — медовые
горшки; п.к— пустые коконы с запасами меда и пыльцы; ц — царица.
самки — будущие царицы. К этому времени количество шмелей в гнез-
де может достигать нескольких сот. Самцы и самки вскоре покидают
гнездо для спаривания и зимовки, а материнская колония постепенно
погибает. Кастовые различия у шмелей всегда четко выражены в по-
ведении, физиологии и морфологии (у большинства видов царицы зна-
чительно крупнее рабочих). Однако у них нет сложных форм передачи
информации, например мобилизации на пищу. Отсутствуют также
взаимное облизывание и обмен кормом, слабо выражено разделение
функций среди рабочих. Шмели-кукушки (Psithyrus Lep.) являются
социальными паразитами (инквилинами) в гнездах шмелей; они утра-
тили собственных рабочих и собирательный аппарат. Самка кукушки
убивает царицу в гнезде вида-хозяина и замещает ее, а рабочие шмели
выращивают ее потомство.
11 208
161
Рис. 392. Участок сота в гнезде медоносной пчелы Apis mellifera L. (по Вильсону).
В ячейках видны яйца, личинки на разных стадиях развития, запасы пыльцы и меда (в пра-
вом верхнем углу); часть ячеек запечатана (в левом верхнем углу).
з.м — запечатанные маточники с куколками маток (один из них вскрыт); к.п — пчела, кормящая
личинку в ячейке; м — матка в окружении свиты рабочих пчел; о.к — обмен кормом между
пчелами; т — трутень.
Одной из вершин эволюции социального поведения у насекомых
является поражающее своей сложностью поведение медоносных пчел
рода Apis L. из того же подсемейства Apinae. Этот род представлен
всего 4 видами, из которых 3 обитают в тропиках, а 1 вид — медонос-
ная пчела Apis mellifera L. — разводится человеком с незапамятных
времен и распространился с ним по всей Земле (рис. 352). В естест-
венных условиях пчелы гнездятся в дуплах или трещинах скал, подве-
шивая к их потолку несколько висящих вертикально и строго парал-
162
лельно овальных восковых сотов (рис. 392). Соты состоят из обращен-
ных на обе стороны шестигранных ячеек двух размеров — мелких для
выращивания рабочих пчел и более крупных для выращивания трут-
ней (последние занимают нижнюю часть сота)' Центральная часть
средних сотов занята ячейками с расплодом, а их края и боковые со-
ты— запасами меда и пыльцы. Пчелы перерабатывают нектар в мед,
удаляя из него излишнюю влагу и расщепляя сахарозу на глюкозу
и фруктозу. Пыльца утрамбовывается в ячейках и под воздействием
молочнокислых бактерий превращается в длительно сохраняющийся
пчелиный хлеб, или пергу. Ячейки с медом, пергой и окуклившимися
личинками пчелы закрывают восковыми крышечками. В течение всего
активного сезона в гнезде поддерживается практически постоянная
температура около 35°С. Это достигается за счет выделения пчелами
физиологической теплоты, а при перегреве соты охлаждаются разбрыз-
гиванием мелких капелек воды при интенсивной вентиляции гнезда.
Зимуют пчелы в активном состоянии, поддерживая внутри зимнего
клуба (шарообразного скопления пчел) температуру 20—35° за счет
питания запасенным на зиму медом.
Колония пчел обычно состоит из 30 000—50 000 (иногда до 100 000)
рабочих пчел; она всегда моногинна, т. е. имеет только одну плодови-
тую самку (царицу, или матку) и, следовательно, в генетическом от-
ношении является семьей, т. е. потомством одной самки. Матка значи-
тельно крупнее рабочих, не имеет собирательного аппарата, восковых
и кормовых желез, живет до 5 лет (рабочие пчелы живут лишь 5 не-
дель летом и до 6 месяцев зимой), ее яичники гипертрофированы и она
может откладывать до 2000 яиц в сутки. Матка постоянно занята от-
кладкой яиц и движется по сотам, окруженная кольцом пчел свиты,
которые ее кормят (рис. 392, л). Яйцекладка начинается в январе
и прекращается в октябре. Личинок рабочих пчел и самцов (трутней)
пчелы кормят первые 3 дня секретом кормовых желез («пчелиным мо-
лочком»), а остальные 3 дня развития — смесью меда и пыльцы.
Новые матки выращиваются в специальных овальных ячейках
(маточниках), сооружаемых на краю сота (рис. 392, з.м), а их ли-
чинок кормят только молочком, что и приводит к развитию
матки. Рабочая пчела развивается 21 день, матка—16 дней, трутень —
24 дня.
Для рабочих пчел характерна возрастная смена функций. В тече-
ние первых 3 недель жизни пчела не покидает гнезда, выполняя внут-
ригнездовые функции. Самые молодые пчелы имеют развитые кормо-
вые железы и кормят личинок и матку; затем у них развиваются вос-
ковые железы, и они становятся строительницами. Параллельно они
выполняют и другие работы: перерабатывают нектар в мед, приготав-
ливают пергу, чистят ячейки и т. д. В течение 2 последующих недель
жизни пчела является фуражиром и вылетает из улья за кормом.
Фуражиры всегда специализируются (хотя бы временно) на посеще-
нии определенных видов растений.
Среди фуражиров имеется группа разведчиков, задача которых
найти новые источники корма и направить на них остальных пчел.
Эта мобилизация на пищу осуществляется пчелами с помощью слож-
ной формы сигнализации — виляющего танца'. Танцующая пчела, бы-
стро виляя брюшком, совершает прямолинейные пробеги под таким
углом к вертикали, который равен углу между направлением на солн-
це и на обнаруженный источник корма. После каждого пробега она
поворачивает то вправо, то влево, описывая фигуру, напоминающую
восьмерку (рис. 393). Скорость движения пчелы во время танца об-
ратно пропорциональна расстоянию до источника корма. Следующие
за танцовщицей пчелы, получив эту информацию, самостоятельно отыс-
163
кивают источник корма, ориентируясь, кроме того, еще и по запаху,
который они восприняли от танцовщицы.
У пчел существуют и другие сложные формы сигнализации. На-
пример, сигнализация о тревоге осуществляется с помощью выделения
феромона тревоги из железы, находящейся в сумке жала. Мандибу-
лярные железы матки синтезируют и выделяют так называемое ма-
точное вещество — оксидеценовую кислоту. Функции этого вещества
весьма разнообразны. Его воспринимают пчелы свиты и передают дру-
гим пчелам на своем теле. Маточное вещество сигнализирует пчелам
о присутствии матки, подавляет развитие их яичников, не допускает
строительства маточников и стимулирует все формы деятельности
Рис. 393. Информационное значение танцев у медоносной пчелы Apis mellifera L.
(по Вильсону).
Пчела (т.п) совершает виляющий танец на вертикальном соте; за ней следуют мобилизуемые
пчелы (м.п). Угол а между направлением виляющего пробега и вертикальной осью соответ-
ствует углу между направлениями на солнце и источник корма.
пчел. Все пчелы одного улья имеют общий запах, складывающийся из
собственного запаха пчел и запаха пыльцы, меда и других веществ
в улье. Этот запах сигнализирует пчелам о принадлежности данной
особи к их семье. Все посторонние пчелы распознаются пчелами-сторо-
жами у летка и не допускаются в улей. У пчел хорошо развиты обмен
кормом между особями (трофаллаксис), взаимное облизывание и очи-
стка тела.
Семьи медоносных пчел размножаются с помощью роения, кото-
рое происходит в мае-июне. Перед этим в улье выводятся трутни и за-
кладываются маточники. Матка прекращает откладывать яйца,
и в улье исчезает расплод. Летная деятельность пчел почти прекра-
164
щается, и они скапливаются на сотах в ожидании роения. Оно начи-
нается, когда в маточниках полностью созреют молодые матки. Раз-
ведчики возбуждают пчел зигзагообразными танцами, и примерно по-
ловина пчел вылетает из улья вместе со старой маткой. Рой привива-
ется где-нибудь на ветке и находится там до тех пор, пока развед-
чики не подыщут подходящее место для нового гнезда, о котором они
сообщают танцами на поверхности роя. Затем рой снимается, летит за
разведчиками и поселяется в новом помещении. В это время в старом
гнезде выходит из маточника первая матка, отыскивает и убивает
всех остальных, а затем несколько раз вылетает для спаривания
с трутнями и приступает к яйцекладке. При большом количестве пчел
иногда наблюдается выход второго и даже третьего роев (с молодыми
матками).
Муравьи — Formicidae. Все муравьи являются обществен-
ными насекомыми, за исключением ряда социально-паразитических ви-
дов, которые вторично утратили 'социальность. Это одно из самых круп-
ных семейств жалящих перепончатокрылых, охватывающее более
8000 видов и разделимое на 10—И подсемейств, из которых в СССР
встречаются в основном представители 2 подсемейств — Formicinae
и Myrmicinae. Наибольшее видовое разнообразие муравьев наблюда-
ется в тропиках; число видов резко сокращается с увеличением широ-
ты. Муравьи заселяют все наземные местообитания, за исключением
тундры, причем повсюду они встречаются в обилии, являясь в боль-
шинстве наземных биоценозов доминирующими хищниками среди бес-
позвоночных. Такое процветание муравьев обусловлено их социаль-
ностью и некоторыми специфическими чертами «муравьиного» образа
жизни. Рабочие муравьи бескрылы и имеют небольшие размеры, что
позволяет им свободно проникать в самые разнообразные микрополос-
ти и осваивать практически все участки биоценозов.
Обычно рабочие муравьи значительно меньше плодовитых самок-
цариц (см. рис. 8) и живут 2—4 года, тогда как царицы могут жить
до 15 и более лет. У большинства видов рабочие в одном гнезде не-
одинаковы по размерам; иногда эти различия очень велики. Крупные
рабочие, которых называют солдатами, могут иметь большую голову
с мощными мандибулами (см. рис. 8), а у мелких рабочих сохраняют-
ся обычные размеры головы и мандибул.
Численность рабочих в колонии у примитивных муравьев может
быть всего 10—20 особей, но обычно измеряется сотнями или тысячами.
В крупном муравейнике лесных муравьев рода Formica L. насчитыва-
ется до 1000 000 рабочих. В гнезде может быть одна царица (моноги-
ния) или несколько цариц (полигиния). У моногинных видов увеличе-
ние размеров колонии достигается за счет гипертрофии яичников ца-
рицы. Как правило, такие царицы имеют непомерно большое брюшко.
Самцы у муравьев (см. рис. 8) живут очень недолго и не выполняют
никаких функций, кроме участия в размножении.
Гнезда муравьев весьма разнообразны. В условиях умеренного
климата они обычно устраиваются в почве или связаны с ней. Гнезда
могут размещаться целиком в почве, где выкапываются ходы и камеры,
и открываться наружу отверстиями или насыпными кратерами, или же
имеют насыпные холмики из почвы и растительных остатков, также
поонизанные ходами и камерами. Насыпной холмик служит аккумуля-
тором солнечного тепла и сооружается тогда, когда почва недостаточно
прогревается из-за затенения травостоем. В горах многие муравьи гнез-
дятся под камнями, а в лесах — в живой или мертвой древесине. У ры-
жих лесных муравьев (Formica rufa L.) наземная часть муравейника
состоит из насыпного купола из растительных остатков. Купол погру-
жается в углубление почвы, окруженное земляным валиком; он имеет
165
плотную оболочку с выходами и рыхлую внутреннюю часть. Подзем-
ные помещения муравейника представлены ходами и камерами, ухо-
дящими на глубину 1,5 м. В центральной части гнездового купола
с весны до конца лета муравьи поддерживают почти постоянную тем-
пературу 27—30°.
Подавляющее большинство муравьев представлено хищниками; они
выкармливают личинок животной пищей (в основном насекомыми).
Некоторые виды являются зерноядными насекомыми. Обычно взрослые
муравьи питаются в основном углеводной пищей, которую они полу-
чают на цветах (некоторые даже принадлежат к специфическим опы-
лителям растений), или же потребляют экскременты сосущих насеко-
мых— тлей, щитовок, цикадок. В умеренной зоне для этой цели му-
равьи чаще всего посещают тлей (рис. 394). Этот симбиоз выгоден не
только муравьям, но и тлям, которых муравьи защищают от врагов,
Рис. 394. Рыжие лесные муравьи 'Formica polyctena Forst.,' посещающие
группу дубовых тлей Lachnus roboris L. и берущие от них падь (по Виль-
сону).
сооружают для них укрытия и даже сохраняют их зимующие яйца
в муравейниках. Установлено, что тли в присутствии муравьев сильно
увеличивают концентрацию сахара в пади (экскрементах) и выделя-
ют ее только в ответ на прикосновения муравьев. Как правило, му-
равьи не запасают пищу в гнезде, а накапливают на зиму питательные
вещества в жировом теле. Углеводную пищу рабочие могут некоторое
время хранить в зобиках.
Принесенная в гнездо жидкая пища распределяется между всеми
муравьями путем кормового обмена (трофаллаксиса). Пища, находя-
щаяся в зобике, не усваивается до попадания в среднюю кишку и не
принадлежит данному муравью, поскольку она может быть передана
другому члену колонии по первому его требованию. Совокупность зо-
биков всех муравьев является как бы «общественным желудком», пи-
ща в котором хорошо перемешивается благодаря трофаллаксису.
Муравьи проявляют постоянную заботу о потомстве: тщательно
облизывают яйца, личинок, куколок, переносят их в те камеры гнезда,
микроклимат которых более подходит для их развития. Эта постоянная
транспортировка расплода является характерной чертой поведения
муравьев. Нередко рабочие переносят и других (в особенности моло-
дых) муравьев; чаще всего это наблюдается при переселении в новое
гнездо. Молодых личинок муравьи кормят выделениями кормовых же-
лез и специальными бесплодными кормовыми яйцами, которые откла-
166
дываются рабочими особями. Более взрослые личинки получают со-
держимое зобика и кусочки насекомых. Рабочие помогают личинкам
при линьке освободиться от сбрасываемой шкурки, удаляют их экскре-
менты, образующиеся при окукливании. Молодые муравьи без помощи
взрослых не могут выйти из коконов или освободиться от куколочных
шкурок.
Царицу рабочие кормят только жидкой пищей, причем в основном
секретом кормовых желез. Наибольшим вниманием со стороны рабочих
окружены царицы моногинных видов: их постоянно ощупывают, обли-
зывают и кормят окружающие муравьи. У полигинных видов царицы
более самостоятельны и нередко сами выпрашивают пищу у рабочих.
Рабочие муравьи также очень внимательны и друг к другу: взаимное
облизывание и очистка тела являются важнейшими элементами в жиз-
ни муравейника.
У муравьев очень хорошо выражено разделение функций между
рабочими. Прежде всего это возрастная смена функций. Молодые му-
равьи занимаются в основном уходом за расплодом и кормлением
цариц; это муравьи-няньки. Более старшие рабочие обрабатывают
принесенную добычу, занимаются строительством, уборкой мусора
и прочими внутригнездовыми функциями. С возрастом они все чаще
выходят из гнезда и наконец переходят в последнюю возрастную груп-
пу, становясь фуражирами. Эти переходы из одной функциональной
группы в другую регулируются потребностями колонии: при недостат-
ке фуражиров муравьи переходят к деятельности вне гнезда в более
раннем возрасте, а если не хватает нянек, то у старых рабочих вновь
развиваются кормовые железы, и они начинают выкармливать личи-
нок. Среди фуражиров наблюдается специализация: одни становятся
охотниками, вторые — сборщиками пади, третьи доставляют строи-
тельный материал. Выделяется также группа активных фуражиров, или
разведчиков, занимающихся отысканием новых источников пищи. У ви-
дов с полиморфной кастой рабочих наблюдается распределение функ-
ций по размерам муравьев. Так, у рыжих лесных муравьев (Formica
rufa L.) в среднем более мелкие рабочие становятся сборщиками
пади, а более крупные — охотниками. У видов, имеющих солдат, они
специализируются обычно на охране гнезда и на выполнении наиболее
трудоемких функций (транспортировка и расчленение крупной добы-
чи, перемалывание зерен и т. п.).
Согласованность поведения муравьев в колонии обеспечивается
многочисленными механизмами сигнализации и передачи информации.
Огромную роль в общении муравьев играют разнообразные тактиль-
ные сигналы (прикосновения антеннами, ногами, удары головой, виб-
рации тела и т. п.), но они пока недостаточно исследованы. Без обме-
на тактильными сигналами не может начаться трофаллаксис, не про-
исходит кормление личинок, невозможны любые формы контактов меж-
ду муравьями. С помощью специфических тактильных сигналов раз-
ведчики активируют фуражиров при мобилизации их на найденный ис-
точник корма. Звуковые сигналы используются немногими муравьями
и обычно участвуют в сигнализации о тревоге. Потревоженные му-
равьи Camponotus Мауг. и Lasius F. ударяют брюшками о субстрат,
производя звуки, сигнализирующие о тревоге. Для той же цели обыч-
но используется стридуляция у муравьев, относящихся к подсемейст-
ву Myrmicinae.
Наилучшим образом изучены химические сигналы, имеющие у му-
равьев очень широкое распространение. Для сигнализации о возник-
шей опасности обычно используются феромоны тревоги, выделяемые
мандибулярными железами или железами жала. Они привлекают дру-
гих муравьев к источнику опасности, возбуждают их и приводят в аг-
167
рессивное состояние. Следовые феромоны выделяются в задней кишке
(у видов подсемейства Formicinae) или в железах жала (у видов под-
семейства Myrmicinae) и используются для мечения путей передви-
жения вне гнезда. Разведчик, обнаруживший пищу, возвращается
в гнездо и, прикасаясь кончиком жала или анальным отверстием к суб-
страту, оставляет на нем пахучий след из капелек следового феромо-
на (рис. 395). Затем по этому следу он приводит к корму группу мо-
билизованных в гнезде фуражиров, или же у многих муравьев они
приходят по следу' без разведчика-лидера.
Царица у муравьев, как и у медоносных пчел, выделяет специфи-
ческий феромон, привлекающий к ней рабочих, подавляющий их пло-
довитость и стимулирующий все формы их деятельности. Различные
феромоны, выделяемые на поверхности тела и воспринимаемые при
взаимных контактах муравьев, обеспечивают узнавание стадии разви-
тия, возраста, касты, физиологического состояния. Так, например, му-
равьи узнают личинок благодаря феромону на поверхности их тела,
стимулирующему поведение ухода за личинками. Этот феромон раз-
личается у личинок рабочих и цариц, у активно развивающихся и диа-
Рис. 395. Муравей Solenopsis Westw. прокладывает след, прикасаясь к субстрату
кончиком брюшка н оставляя на нем капельки следового феромона (по Вильсону).
паузирующих личинок. Все муравьи из одного гнезда имеют общий
запах, позволяющий отличать посторонних особей и не допускать их
в гнездо. Этот запах складывается из поверхностных феромонов,
а также запахов гнездового материала и пищи, абсорбированных ку-
тикулой.
Размножение у большинства муравьев происходит по следующей
схеме. В определенное время года (чаще всего во второй половине
лета) в муравейниках выращиваются крылатые самцы и самки. Вско-
ре (обычно после дождя) они дружно вылетают из муравейников,
причем у ряда видов образуют большие роевые скопления над высо-
кими деревьями. Самки спариваются с самцами, которые вскоре пос-
ле этого умирают. Оплодотворенные самки опускаются на землю
и сразу же обламывают себе крылья. Эти бескрылые молодые царицы
отыскивают подходящее место для нового гнезда и срроят небольшую
норку, в которой вскоре откладывают первые яйца. В дальнейшем
самка-основательница самостоятельно выращивает первое поколение
очень мелких рабочих, выкармливая личинок яйцами и выделениями
кормовых желез без поступления пищи извне только за счет ресурсов
собственного жирового тела и дегенерирующей Крыловой мускулатуры.
Появившиеся рабочие приносят пищу и кормят царицу, которая начи-
нает откладывать все больше яиц и сама уже не выкармливает личи-
нок. Численность рабочих постепенно увеличивается, но только через
8—10 лет колония достигает такого размера, когда начинается выра-
щивание крылатых самцов и самок.
Многие полигинные виды размножаются путем деления колонии
или почкования (отделения отводков). При этом часть рабочих уходит
168
из материнского гнезда в отводок, унося часть расплода и плодови-
тых цариц. Оплодотворенные молодые самки после брачного лёта ча-
ще не основывают новые гнезда, а возвращаются в гнезда своего ви-
да, увеличивая в них количество цариц. У рыжих лесных муравьев
(Formica rufa L.) отводки длительное время остаются тесно связанны-
ми с материнским гнездом: между ними происходят постоянные обме-
ны расплодом и молодыми рабочими, что обеспечивает единство
запаха и родственные отношения между этими муравейниками. Возни-
кает колония муравьев, обитающая в нескольких муравейниках. В со-
став такой сложной колонии может входить несколько десятков гнезд.
Некоторые виды муравьев утратили способность к самостоятель-
ному основанию гнезд молодыми самками и основывают гнезда путем
временного социального паразитизма в гнездах муравьев других видов..
Молодая царица такого вида либо проникает в гнездо, где погибла
царица, либо сама убивает ее. Рабочие-хозяева гнезда выкармлива-
Рис. 396. Муравьи-амазонки Polyergus rufescens Latr., совершающие набег, на гнездо
муравья Formica fusca L. (по Вильсону).
Слева — амазонки, сражающиеся с хозяевами у входа в их гнездо; справа — амазонки, унося-
щие похищенные коконы в свое гнездо.
ют потомство принятой ими чужой царицы, а сами постепенно выми-
рают, так как их царица отсутствует. Им на смену появляются рабочие
вида-паразита, и через несколько лет гнездо уже полностью принад-
лежит этому виду. Временный социальный паразитизм характерен для
рыжих лесных муравьев, которые паразитируют на мелких видах рода
Formica L., для ряда видов рода Lasius F. и многих других муравьев.
От временного социального паразитизма в эволюции муравьев,
неоднократно происходил переход к постоянному паразитизму. Самки
таких видов уже не убивают царицу вида-хозяина, а живут вместе
с ней в одном гнезде, причем обычно пользуются даже большим, чем
она, вниманием со стороны рабочих-хозяев, которые выращивают «чу-
жое» потомство, состоящее только из самцов и самок. Каста рабочих
у этих видов отсутствует, т. е. они утратили социальность, что является
следствием паразитического образа жизни.
Удивительнейшей формой социального паразитизма у муравьев
является рабовладение. Муравьи-рабовладельцы совершают набеги
на гнезда других видов муравьев и похищают их коконы, из которых
выводятся рабочие, становящиеся «рабами» в гнезде рабовладельцев.
Эти рабы ведут себя так же, как вели бы в собственном гнезде, т. е.
строят гнездо, добывают корм, выращивают личинок, но при этом их
усилия затрачиваются на благо чужого вида — муравьев-рабовладель-
цев. Некоторые виды являются факультативными рабовладельцами
(например, кроваво-красный рабовладелец Formica sanguinea Latr.)
169
и содержат рабов только на начальных этапах развития колонг
реходя впоследствии к независимому существованию. Другие ж
новятся облигатными рабовладельцами, и их строение и поведен
наруживают ряд регрессивных черт, являющихся следствием noi
ного паразитизма. Например, муравьи-амазонки Polyergus rufi
Latr. из подсемейства Formicinae утратили почти все формы пс
ния, свойственные муравьям, и даже не могут самостоятельно пит
(их кормят рабы). Острые серповидные челюсти Р. rufescens приг
только для сражений с хозяевами ограбляемых гнезд и для тран
тировки похищенных коконов (рис. 396). Единственным занятием
зонок являются набеги на гнезда хозяев, а все остальные функции
гнезде выполняют рабы.
Определительная таблица семейств*
1(8). Брюшко сидячее, т. е. не суженное или едва суженное у о
вания (рис. 353). (Подотряд Сидячебрюхие — Symphyta)
2(5). Голени передних ног с 1 вершинной шпорой. Самки обь;
с торчащим яйцекладом.
3(4). Задний край переднеспинки с глубокой выемкой. Брюшко
линдрическое с роговидным шипом на конце, или же с ост
ми боками, а к вершине постепенно заострено (рис. 357, ЗЕ
Размеры крупные или средние (8—40 мм). Самки с тор
щим яйцекладом............................Рогохвосты — Siricid
4(3). Задний край переднеспинки едва выемчатый. Тело тонк
стройное, немного сплющено с боков (рис. 359). Размеры 1
большие (7—10 мм). Яйцеклад заметен плохо....................
..........Стеблевые, или злаковые пилильщики — Cephid:
5(2). Голени передних ног с 2 вершинными шпорами (рис. 39"/
Самки с пиловидным яйцекладом, который обычно втян
в брюшко и не заметен.
6(7). Задний край переднеспинки прямой или едва выемчатый. Ус
ки к концу утоньшенные (рис. 363).........................
. , > ......................Пилильщики-ткачи— Pamphilida<
7(6). Задний край переднеспинки с глубокой выемкой. Усики ните
видные, булавовидные, пильчатые или перистые, но не утош
шейные к концу (рис. 361, 362)...............................
.................Настоящие пилильщики — Tenthredinidae.
8(1). Брюшко сильно сужено у основания, между ним и грудью име
ется резкий перехват (стебелек), обычно короткий (рис. 352)
иногда очень длинный (рис. 398). У очень мелких форм брюш-
ко бывает сидячим, или кажется таковым, но тогда крылья
с редуцированным жилкованием, без замкнутых ячеек (рис.
365, 367). (Подотряд Стебельчатобрюхие — Apocrita).
9(10). Стебелек брюшка состоит либо из 2 явственно обособленных
друг от друга члеников (подсемейство Myrmicinae — рис. 399),
либо из 1 членика с вертикально торчащей вверх пластин-
кой— «чешуйкой» (подсемейство Formicinae — рис. 400)
или с резко приподнятым бугорком. Общественные насекомые
с кастой бескрылых рабочих особей; крылатые самцы и самки
появляются лишь периодически. . . . Муравьи — Formicidae.
10(9). Стебелек брюшка простой, образован из передней части пер-
* Звездочками отмечены семейства, которые в современной систематике насе-
комых рассматриваются в ранге надсемейств и разделяются на ряд семейств, обычно
трудных для определения неспециалисту,
170
вого брюшного сегмента, без выступающих вверх пластинок
или резко приподнятых бугорков.
11(40). Крылья имеются.
12(15). Крылья с сильно редуцированным жилкованием, без замкну-
тых ячеек (рис. 365, 367). Мелкие и очень мелкие формы
(0,5—5 мм).
13(14). Тело обычно металлически блестящее. Яйцеклад отходит от
нижней строны брюшка далеко от его вершины. Усики всегда
коленчатые..........................................Chalcididae.*
14(13). Тело преимущественно черное, без металлического блеска.
Яйцеклад отходит от самой вершины брюшка. Усики коленча-
тые или простые..................................Proctotrupidae.*
15(12). Замкнутые ячейки на крыльях имеются. Средних размеров или
крупные, реже мелкие формы.
Рис. 397—404. Детали строения перепончатокрылых.
397 — часть передней ноги пилильщика (Tenthredinidae)c 2 вершинными шпорами на голени, 398 —
брюшко и длинный стебелек роющей осы Sceliphron KI. (Sphecidae), 399 — двухчлениковый
стебелек муравья из подсемейства Myrmiclnae, 400 — одночлениковый стебелек с вертикальной
чешуйкой у муравья нз подсемейства Formicinae, 401 — двухчлениковый вертлуг наездника
(Ichneumonidae), 402— одночлениковый вертлуг жалящего перепончатокрылого (Aculeata), 403 —
грудные сегменты дорожной осы (Pompilidae), - вид сверху, 404 — то же роющей осы (Sphecidae).
16(17). Передние крылья без глазка (птеростигмы), обычно лишь
с одной маленькой замкнутой ячейкой (рис. 366). Брюшко
сильно сдавлено с боков, тергиты его сильно развиты и скры-
вают стерниты. Грудь горбатая с сильно выпуклым щитком.
Окраска черная, желтоватая, красноватая. Размеры неболь-
шие.....................................Орехотворки—Cynipidae*.
17(16). Передние крылья, как правило, с птеростигмой (как на рис.
364, 371, 372). Если ее нет, то остальные признаки иные.
18(23). Вертлуги двучлениковые (рис. 401). Яйцеклад имеется, час-
то и в покое торчит наружу.
19(22). Передний край крыла перед птеростигмой и за нею с силь-
но склеротизова'нной жилкой (рис. 364, 371, 372).
20(21). В центре вершинной части крыла имеется маленькая ячейка
(«зеркальце»), которая гораздо меньше соседних ячеек
(рис. 371, з). Если зеркальца нет, то в крупной центральной
ячейке имеется остаток поперечной жилки (рис. 372, п.ж).
Тело тонкое, стройное. Самки обычно с торчащим яйцекла-
дом................................................ Ichneumonidae.
21(20). В центре вершинной части крыла ячейки близки по размерам
(рис. 364)............................................ Braconidae.
171
22(19). Сильно склеротизованной жилки на переднем крае крыла нет.
. . . Другие семейства паразитических перепончатокрылых.
23(18). Вертлуги одночлениковые (рис. 352, 402). Яйцеклад превра-
щен в жало, в покое всегда втянутое в брюшко.
24(25). Задние крылья без замкнутых ячеек.............Bethylidae.*
25(24). Задние крылья с замкнутыми ячейками; если изредка их нет,
то на переднеспинке имеются ' хорошо развитые плечевые
бугры (рис. 373).
26(27). Тело яркоокрашенное: синее, зеленое, фиолетовое, пурпурное
с сильным металлическим блеском. Брюшко состоит только из
3—4 (редко 5) видимых сегментов (рис. 373); остальные (вер-
шинные) сегменты сильно удлинены и телескопически втяги-
ваются в предыдущие. Обычно брюшко снизу вогнутое, и осы
могут сворачиваться в шарик. . . Блестянки — Chrysididae.
27(26). Окраска иная. Брюшко состоит более чем из 5 видимых сег-
ментов. Вершинные его сегменты иного строения.
28(37). Задний край переднеспинки выемчатый, его углы доходят до
крыловых крышечек и не образуют плечевых бугров (рис. 403).
29(30). Первая дискоидальная ячейка переднего крыла гораздо длин-
нее ячейки, лежащей между ней и основанием крыла (рис. 383—
386, д.я). Крылья в покое обычно складываются пополам вдоль
их длины. Задние углы переднеспинки заострены, налегают па
крыловые крышечки. Глаза с глубокой вырезкой на внутреннем
крае (рис. 383)....................Складчатокрылые осы — Vespidae*.
30(29). Первая дискоидальная ячейка короче ячейки, лежащей между
ней и основанием крыла (как на рис. 374, 376, 377). Крылья
в покое не складываются пополам. Задние углы переднеспин-
ки закругленные, не налегают на крыловые крышечки. Глаза
обычно без вырезки.
31(32). Бока среднегруди слабо выпуклые. Брюшко без поперечного
вдавления между первым и вторым сегментами. Передние та-
зики не короче переднегруди. Ноги длинные и тонкие. Усики
у живых особей обычно завиты в кольца (рис. 377). . . .
.........................................Дорожные осы — Pompilidae.
32(31). Бока среднегруди сильно выпуклые. Брюшко снизу с попереч-
ным вдавливанием между первым и вторым сегментами. Пе-
редние тазики короче переднегруди. Ноги относительно корот-
кие и утолщенные.
33(36). Тазики средних ног широко расставлены и не соприкасаются.
34(35). Передние крылья без глазка (птеростигмы). Глаза почковид-
ные. Размеры обычно крупные. Тело в довольно густых отстоя-
щих волосках. Второй и третий сегменты брюшка сверху
обычно с крупными пятнами (рис. 376)...............................
........................................Сколии — Scoliidae.
35(34). Передние крылья с птеростигмой. Глаза не почковидные. Раз-
меры средние. Тело слабо опушенное (рис. 374).....................
......................................... Тифии — Tiphiidae.
36(33). Тазики средних ног сближены, соприкасаются...............
........................... Немки—Mutillidae(caMnbi).
37(28). Задний край переднеспинки прямой, в виде валика, несущего
по бокам плечевые бугры, не доходящие до крыловых крыше-
чек (рис. 404).
38(39). Первый членик задней лапки цилиндрический (рис. 378—380),
не уплощенный с выемкой в основании. Эта выемка и внутрен-
няя поверхность большой шпоры задних голеней в густых во-
лосках..................................... Роющие осы — Sphecidae.
39(38). Первый членик задних лапок более или менее уплощенный, без
172
выемки в основании, обычно гораздо больше остальных чл<
ников (рис. 352, ч.л; 377) и на внутренней стороне целико;
покрыт густыми волосками в виде щеточки. Большая шпор
задних голеней без густых волосков. Тело обычно густо опу
шейное................................ Пчелиные — Apidae*
40(11). Крылья отсутствуют.
41(42). Брюшко сильно сдавлено с боков, его высота значительш
больше ширины. Тергиты брюшка очень сильно развиты, скры
вают стерниты. Окраска черная, желтоватая или Краснова
тая................................. Орехотворки — Cynipidae*
42(41). Признаки иные.
43(44). Брюшко снизу между первым и вторым сегментами с глубо-
ким поперечным вдавлением. Средней величины насекомые,
черные или черно-синие, обычно с красной грудью и перевя-
зями или пятнами из густых светлых волосков на брюшке
(рис. 375).......................... Немки — Mutiliidae (самки).
44(43). Брюшко обычно без поперечного вдавления между первым
и вторым сегментами. Окраска иная.
45(46). Стерниты брюшка склеротизованы значительно меньше, чем
тергиты; у коллекционных экземпляров брюшные стерниты
ссыхаются снизу в продольную складку или подтянуты под
тергиты и не видны сбоку. Вертлуги двучлениковые (рис. 401).
Самки часто с торчащим яйцекладом.................................
........................Наездники — Ichneumonidae, Braconidae.
46(45). Стерниты брюшка не образуют складку, обычно видны сбоку.
47(48). Длина головы больше ее ширины. Вертлуги одночлениковые
(рис.'402). Брюшко не сдавлено сверху вниз. Тело без метал-
лического блеска.................................... Bethylidae*.
48(47). Длина головы обычно не больше ее ширины. Вертлуги дву-
члепиковые, или же брюшко сдавлено сверху вниз и с острым
боковым краем, или же тело металлически блестящее. Мел-
кие и очень мелкие формы..........................................
.................Наездники—Chalcididae*, Proctotrupidae*.
ОТРЯД РУЧЕЙНИКИ - TRICHOPTERA **
Общая характеристика
К этому отряду относятся средних размеров, реже мелкие насе-
комые, по внешнему облику похожие -на бабочек. Тело и крылья по-
крыты густыми волосками. Ротовой аппарат мягкий, сосущий, но без
длинного хоботка, приспособлен для слизывания жидкостей. Многие
виды отличаются недоразвитием ротового аппарата. Усики длинные,
нитевидные. Крылья гетерономные: передние более или менее узкие,
задние либо широкие с хорошо развитой анальной лопастью, либо по-
хожи на передние, по короче их. В покое крылья складываются кров-
леобразно. Ноги бегательные, голени вооружены шпорами. Церки от-
сутствуют.
Развитие с полным превращением. Самки после спаривания от-
кладывают яйца в воду. Личинки развиваются в различного типа прес-
ных водоемах. Как у гусениц, у них имеется парная шелкоотделитель-
ная железа, выводное отверстие которой открывается на нижней губе.
Куколка свободного типа. Окукливание личинок происходит в постро-
енных ими домиках. Перед линькой на имаго куколка покидает домик
** А. К. Бродский.
173
и всплывает с помощью плавательных средних ног. Линька происходит
на береговых камнях или на растениях.
Взрослые ручейники сидят на растениях или летают вблизи водо-
емов, в которых проходит развитие личиночных стадий. Некоторые
виды появляются в массе, образуя рои. В течение короткой имагиналь-
ной жизни не питаются, лишь слизывают капли росы или дождя.
Развитие личинок протекает в разнообразных пресноводных водо-
емах. Выделяются 2 основных морфо-экологических типа личинок.
1. Личинки, живущие свободно и не строящие до-
миков. Форма тела камподеоидная (рис. 405, 406). Ротовые органы
Рис. 405—407. Морфо-экологические типы личинок
ручейников.
405 — Rhyacophila septentrionls F. (Rhyacophilidae), 406 —
Hydropsyche pellucidula Curt. (Hydropsychidae), 407 — Lim-
nophilus politus McLach (Limnophilidae).
направлены вперед, a
продольные оси головы
и тела совпадают. Тело
обычно с глубокими пе-
рехватами между сег-
ментами. Дышат при по-
мощи жабр кустистой
формы. Хищники. Для
ловли добычи часто ис-
пользуют выделения шел-
коотделительной железы,
сооружая ловчие сети
или воронки, направлен-
ные против течения.
Приурочены, как прави-
ло, к проточным водо-
емам, хотя отдельные ви-
ды могут встречаться
и в стоячей воде. Пред-
ставлены главным обра-
зом видами семейств
Hydropsychidae, Hydrop-
tilidae и Rhyacophilidae.
414
415
Рис. 408—415. Домики ручейников.
40S — Ooera pilosa F. (Sericostomatidae), 409—413 — разные виды семейства Limnophilidae (409 —
Limnophllus stigma Curt., 410 — L. rhombicus L., 411 — Anabolla soror Curt., 412 — Halesus
interpunctatus Zett., 413 — Limnophllus nlgriceps Zett.), 414 — Molanna an gustata Curtis
(Molannidae), 415 — Limnophllus borealis Zett.
174
2. Личинки, живущие в домиках (рис. 407). Домики, или
чехлики разнообразны как по избираемому материалу, так и по форме
и размерам (рис. 408—415). Песчинки или растительные частицы скреп-
ляются при помощи выделений шелкоотделительных желез. Личинки
имеют толстое цилиндрическое тело. Голова направлена ротовыми ор-
ганами вниз и образует с продольной осью тела прямой угол. Жабры
нитевидные. Личинки растительноядные. Населяют преимущественно
стоячие водоемы, хотя некоторые виды встречаются и в проточных во-
дах. Представлены видами семейств Limnophilidae, Phryganeidae
и многих других.
Определительная таблица семейств
1(2). Очень мелкие формы (длина тела не более 3,5 мм). Передние
крылья узкие, ланцетовидные, с бахромкой длинных волос-
ков.........................................: z Hydroptilidae.
2(1). Более крупные ручейники. Передние крылья не ланцетовидные,
без бахромки длинных волосков (рис. 416, 417).
Рис. 416—418. Детали
строения ручейников.
416 — крылья ручейника
из семейства Rhyacophi-
lidae (вверху--переднее
крыло, внизу — заднее),
417 — форма тела и
крыльев ручейника из
семейства Phryganeidae,
418 — ноги ручейника из
семейства Limnophilidae.
3(8). Простые глазки имеются, хотя могут быть иногда скрыты в во-
лосках.
4(5). Задние крылья сходны с передними по форме, но немного ко-
роче (рис. 416). Встречаются на кустах около горных ручьев
и водопадов........................................Rhyacophilidae.
5(4). Задние крылья широкие с хорошо развитой анальной областью
(рис. 417). Встречаются у озер и прудов.
6(7). Передняя голень с одной шпорой или вовсе без шпор, на сред-
ней голени не более 3 шпор (рис. 418). . . Limnophilidae.
7(6). Передняя голень с 2 шпорами, средняя — с 4. . Phryganeidae.
8(3). Простых глазков нет.
9(10). Усики тонкие и очень длинные, в 2—3 раза длиннее передних
крыльев. На средней и задней голенях по 2 шпоры. Вечерами
в массе роятся....................................... Leptoceridae.
10(9). Усики менее чем в 2 раза длиннее передних крыльев. На
задней голени 4 шпоры, на средней — 2 или 4.
11(12). На передней голени 3 шпоры. Встречаются около равнинных
рек. . ..............................................Polycentropidae.
12(11). На передней голени меньше 3 шпор или их нет вообще. Встре-
чаются около проточных водоемов.
13(14). Усики толстые и пильчатые.................... Arctopsychidae.
14(13). Усики тонкие и нитевидные.................... Hydropsychidae.
175
ОТРЯД ЧЕШУЕКРЫЛЫЕ ИЛИ БАБОЧКИ - LEPIDOPTERA*
Общая характеристика
В морфологическом отношении чешуекрылые составляют довольно
компактную группу крылатых насекомых. Все тело и 4 крыла густо
покрыты чешуйками и частично волосками. Голова (рис. 419, г) с круп-
ными фасеточными глазами, хорошо развитыми нижнегубными щупи-
ками и расположенным между ними длинным спирально скрученным
сосущим хоботком. Только у зубатых молей (Micropterigidae) рото-
вой аппарат грызущего типа. Усики (рис. 419, у) хорошо развиты, са-
мого разнообразного строения — от нитевидных до перистых или була-
вовидных.
Крылья обычно широкие, треугольные (рис. 420—422), реже узкие
(рис. 423) или даже ланцетовидные (рис. 424). Чаще всего передние
Рис. 419. Схема строения бабочки.
б— брюшко; г—голова; з. к — место
прикрепления задних крыльев; н — ноги;
н.щ — нижнегубные щупики; п.к. — место
прикрепления передних крыльев; сг—
среднегрудь; у — усик; ф.г — фасеточные
глаза; х— хоботок.
крылья несколько шире задних, но иногда (например, у видов семей-
ства Crambidae) наблюдаются обратные соотношения: задние крылья
гораздо шире узких передних крыльев. У низших чешуекрылых
(Micropterigidae, Eriocraniidae, Hepialidae) обе пары крыльев пример-
но одинаковы по форме и размерам.
Передние и задние крылья скреплены друг с другом специальным
оценочным аппаратом. Наиболее распространен френатный тип
сцепления крыльев (рис. 425). В этом случае сцепление осуществ-
ляется с помощью френулума (уздечки) и ретинанулума (зацепки).
Уздечка представлена одной или несколькими крепкими щётинками
в основании заднего крыла, а зацепка — либо рядом щетинок, либо
загнутым выростом в основании переднего крыла. У зубатых молей
и представителей небольшого семейства Eriocraniidae наблюдается
юго-френатный тип сцепления крыльев (рис. 426). В этом
случае уздечка зацепляется за специальную лопасть пластинки перед-
него крыла (югум). И наконец в семействе Hepialidae уздечка реду-
цирована, и сцепление крыльев осуществляется с помощью югума
(рис. 427). В некоторых группах френатный оценочный аппарат исче-
зает (например, у булавоусых чешуекрылых — Rhopalocera и коконо-
прядов— Lasiocampidae), и соединение крыльев обеспечивается нало-
жением переднего крыла на расширенное основание заднего крыла.
Такой тип сцепления крыльев называется аплексиформным.
* А. А. Стекольииков.
176
Жилкование крыльев чешуекрылых (рис. 428—432) характеризу-
ется значительной (редукцией поперечных жилок и незначительным вет-
влением основных продольных стволов. В пределах отряда выделяется
2 типа жилкования крыльев. Первый тип характерен для низших че-
шуекрылых, обладающих юго-френатным или югатным аппаратом
сцепления (рис. 428). В этом случае жилкование передних и задних
крыльев практически одинаковое (гомоневральный тип). Второй тип
Рис. 420—424. Чешуекрылые.
420 — многоцветница Vanessa polychloros L. (Nymphalidae), 421— бражник Smerinthus ocellatns
L. (Sphingidae), 422 — совка Pltisia gamma L. (Noctuidae), 423 — горностаевая моль Yponomeuta
mollnellus (Yponomeutidae), 424 — Lyonetia Hbn. (Lyonetidae).
жилкования (гетероневральный) характерен для чешуекрылых с фре-
натным и аплексиформным аппаратами сцепления крыльев. У этих ба-
бочек жилкование передней и задней пар крыльев различно за счет
упрощения жилкования задних крыльев (рис. 429—432).
Чешуйки на крыльях различно окрашены и образуют часто до-
вольно сложный рисунок. Нередко наблюдается структурная окраска
(пятна с металлическим блеском). По наружному и заднему краю
крыльев тянется бахрома, состоящая из нескольких рядов чешуек
и волосков.
В грудном отделе наиболее развита среднегрудь (рис. 419, сг).
Переднегрудь по бокам тергита несет лопастевидные придатки — па-
тагии. В среднегруди сходные образования расположены над основа-
нием передних крыльев и называются тегулами. Ноги бегательные
(рис. 419, н), часто со шпорами на голенях (рис. 433). У некоторых
121/2 208
177
чешуекрылых передние ноги сильно редуцированы, спрятаны в волося-
ном покрове, и бабочки передвигаются на четырех ногах.
Дневные чешуекрылые, образующие естественную группу Rhopa-
locera, в состоянии покоя поднимают и складывают крылья над спи-
ной (рис. 434). У большинства остальных бабочек обе пары крыльев
отводятся назад, складываются и вытягиваются вдоль брюшка
(рис. 435, 436); только некоторые пяденицы (Geometridae) и павлино-
глазки (Attacidae) не складывают крылья, а держат их распростерты-
ми в стороны (рис. 437).
Рис. 425—433. Детали строения чешуекрылых.
425 — френатный тип сцепления крыльев, 426 — юго-френатный тип сцепления крыльев, 427 —
сцепление крыльев с помощью югума у бабочек семейства Hepialidae, 428 — жилкование и фор-
ма крыльев у бабочек семейства Hepialidae (вверху — переднее крыло, внизу — заднее), 429 —
то же у белянок (Pieridae), 430 —то же у парусников (Papilionidae), 431 —то же у совок
(Noctuidae), 432 — то же у пядениц (Geometridae), 433 — нога бабочки со шпорами на голени.
Жилки крыльев: А — анальные; A4j, Мз, Л43—медиальные; R— радиальные; Sc — субкостальные.
Брюшко (рис. 419, б) состоит из 9 сегментов. Последний сегмент
резко модифицирован, особенно у самцов, у которых он формирует
копулятивный аппарат. Структурные особенности копулятивного аппа-
рата широко используются в систематике, позволяя четко разграничивать
даже близкие виды. У самок последние сегменты брюшка (обычно
с седьмого до девятого) преобразованы в телескопический мягкий яй-
цеклад. В большинстве случаев половая система самок у бабочек от-
крывается наружу двумя половыми отверстиями. Одно из них, терми-
нальное, служит только для откладки яиц, второе, расположенное
либо на конце седьмого сегмента, либо на восьмом сегменте, является
совокупительным отверстием. Этот тип половой системы назван дит-
ризным и характерен для большинства чешуекрылых. Однако
в архаичных семействах (Micropterigidae, Eriocraniidae и др.) половая
система построена по так называемому монотризному типу, при
котором имеется только одно половое отверстие. И наконец, в семействе
Hepialidae, хотя и развиты два половых отверстия, оба они занимают
терминальное положение (экз опорный тип половой системы).
Характерная черта бабочек — развитие у многих из них криптиче-
ских приспособлений, обеспечивающих их защиту от хищников. Слож-
ные рисунки на крыльях имитируют отдельные элементы окружающей
178
с изрезанным контуром крыльев
прошлогоднего листа.
Рис. 434—437. Положения крыльев
у бабочек в состоянии покоя.
434 — схема, показывающая типичное поло-
жение крыльев у дневных чешуекрылых
(Rhopalocera), 435 — то же у coBOK(Noctuidae),
436 — листовертка Ptycholoma lecheana L.
(Tortricidae), 437— схема, показывающая по-
ложение крыльев у пядениц (Geometridae).
среды. Так, у некоторых совок (Noctuidae), сидящих днем на стволах
деревьев, передние крылья по окраске и рисунку сходны с лишайни-
ками. Задние крылья, прикрытые сверху передними, не видны и не
имеют сложного рисунка. То же самое наблюдается и у дендрофиль-
ных пядениц (Geometridae), у которых на передних крыльях часто
воспроизводится изображение структуры коры. У некоторых нимфалид
(Nymphalidae) при складывании крыльев наружной оказывается их
нижняя сторона. Именно эта сторона окрашена у многих из них
в темно-бурые тона, что в сочетании
создает полную иллюзию засохшего
Часто параллельно с крип-
тической окраской у бабочек
наблюдаются рисунки с яркими,
броскими пятнами. Почти все
нимфалиды, обладающие крип-
тическим рисунком на исподе
крыльев, сверху окрашены ис-
ключительно эффектно. Много-
цветная яркая окраска исполь-
зуется бабочками для распозна-
вания особей своего вида. У
пестрянок (Zygaenidae), обла-
дающих ядовитой гемолимфой,
яркая контрастная окраска кры-
льев и брюшка выполняет иную
сигнальную функцию, указывая
на их несъедобность для хищ-
ников. У части чешуекрылых,
ведущих дневной образ жизни,
возникает замечательное внеш-
нее сходство с хорошо защищен-
ными насекомыми, такими как
жалящие перепончатокрылые. У
стеклянниц (Sesiidae) такое
сходство достигается окраской брюшка и прозрачностью узких крыль-
ев, на которых практически полностью редуцированы чешуйки.
Основным источником пищи для бабочек служит нектар. Пере-
летая при питании с цветка на цветок, бабочки, наряду с двукрылыми,
перепончатокрылыми и жуками активно участвуют в опылении расте-
ний. Примечательно, что бабочки, обладая достаточно длинным хо-
ботком, посещают цветки не только с открыто расположенными ис-
точниками нектара, но и с нектаром, глубоко скрытым в шпорцах
цветков или на дне трубчатого венчика и соответственно недоступным
для других насекомых. Цветки многих гвоздичных и орхидных благо-
даря их морфологии могут опыляться только чешуекрылыми. У неко-
торых тропических орхидей возникают специальные приспособления
к опылению цветков чешуекрылыми.
Помимо нектара многие бабочки охотно поглощают сок, вытекаю-
щий из пораненных деревьев или плодов. В жаркий летний день мож-
но наблюдать большие скопления белянок (Pieridae) возле луж. Сюда
же, привлеченные водой, прилетают и другие чешуекрылые. Многие
дневные бабочки часто кормятся на экскрементах позвоночных живот-
ных. Независимо в самых различных семействах чешуекрылых возника-
ет афагия: бабочки не питаются и их хоботок подвергается редукции.
Среди насекомых с полным превращением чешуекрылые — единствен-
ная крупная группа, в которой столь часто наблюдается переход
к афагии.
179
Большинство чешуекрылых ведет ночной образ жизни и только
некоторые группы активны днем. Среди последних ведущее место при-
надлежит булавоусым, или дневным чешуекрылым (Rhopalocera) —
группе, чрезвычайно обильно представленной в тропиках. Дневной
образ жизни характерен также для ярко окрашенных пестрянок (Zy-
gaenidae) и стеклянниц (Sesiidae). Среди других семейств чешуекрылых
палеарктической фауны виды с дневной активностью встречаются спо-
радически. Некоторые совки (Noctuidae), пяденицы (Geometridae),
огневки (Pyralidae), листовертки (Tortricidae) обладают круглосу-
точной активностью, однако днем эти бабочки чаще всего активны
в пасмурную погоду или в затененных местах.
Чешуекрылые — насекомые с четко выраженным половым димор-
физмом, который проявляется в строении усиков и сцепочного аппара-
та крыльев, в характере крылового рисунка и в степени опушения
брюшка. Наиболее демонстративный половой диморфизм в рисунке
крыльев наблюдается как у дневных, так и у ночных чешуекрылых.
Ярким примером половых различий может служить расцветка крыль-
ев у непарного шелкопряда (Ocneria dispar L.). Самки этого вида
крупные, со светлыми, почти белыми крыльями; они резко отличаются
от мелких и стройных самцов со сложным бурым рисунком на крыль-
ях. Усики у самок непарного шелкопряда слабо гребенчатые, у самцов
сильно гребенчатые. Половой диморфизм в окраске крыльев может
быть выражен в ультрафиолетовой части спектра и невидим для глаза
человека. Так, совершенно одинаковые белые бабочки боярышницы
(Aporia crataegi L.) на самом деле диморфны, и самцы отличаются
от самок ультрафиолетовым рисунком.
Крайним выражением полового диморфизма могут служить ме-
шочницы (Psychidae), некоторые пяденицы (Geometridae), отдельные
виды волнянок (Lymantriidae) и листоверток (Tortricidae), у которых
самки в отличие от самцов не имеют крыльев, либо обладают их ру-
диментами. Самки многих чешуекрылых выделяют пахучие вещества
(феромоны), запахи ' которых самцы улавливают обонятельными ре-
цепторами. Чувствительность рецепторов довольно высока, и самцы
улавливают запах самки с расстояния в несколько десятков, а иногда
и сотен метров.
У булавоусых и некоторых других бабочек половые феромоны вы-
деляются самцами; их источниками служат специальные андрокони-
альные железы, которые в разных группах расположены на крыльях,
на ногах или на брюшке. Видовая специфичность половых феромонов
обеспечивает репродуктивную изоляцию видов. В концентрированном
виде запах феромона легко улавливается человеком. Для этого необ-
ходимо продержать самца какого-либо вида в чистой закрытой про-
бирке всего несколько минут.
Яйца откладываются чешуекрылыми обычно непосредственно на
кормовое растение их гусениц. Самки приклеивают яйца к субстрату
с помощью секрета придаточных половых желез. Только у некоторых
бабочек отмечено рассеивание яиц прямо на почву возле кормовых
растений. Многие дендрофильные чешуекрылые откладывают яйца
в расщелины коры с помощью длинного телескопического яйцеклада;
лишь некоторые минирующие моли (например, виды семейства Eriocra-
niidae) способны помещать свои яйца в ткани листа. Яйца отклады-
ваются поодиночке или группой. Обычно кладка яиц остается незащи-
щенной, однако у волнянок (Limantriidae) яйца либо обволакиваются
пенистыми выделениями придаточных половых желез, застываю-
щими в виде зеленой стекловидной массы (как у ивовой волнян-
ки— Stilpnotia salicis L.), либо покрываются войлочком, образован-
ным из склеенных волосковидных чешуек брюшка самки (как у зла-
180
тогузки — Euproctis chrysorrhoea L. или непарного шелкопряда —
Ocneria dispar L.).
Развитие чешуекрылых сопровождается полным превращением.
Личинки (гусеницы) имеют цилиндрическое 10-члениковое туловище
и прочно склеротизованную голову, на которой расположены 6 пар
простых глазков и очень короткие 2- или 3-члениковые усики (рис. 438).
Ротовой аппарат грызущий с хорошо развитыми мощными мандибу-
лами. Максиллы. и нижняя губа образуют единый комплекс с прядиль-
ным сосочком на конце. С помощью прядильного аппарата гусеница
.выделяет шелк.
Грудные сегменты несут 3 пары расчлененных ножек, которые
используются только для опоры или захвата листьев во время пита-
ния. Для передвижения используются брюшные мясистые ложноножки
с рядом крючьев на их подошвах. В типичном случае (рис. 438) у гу-
сениц развиты 5 пар ложноножек (4 пары с III по VI сегменты брюш-
ка и 1 пара на конце туловища). В некоторых группах чешуекрылых
Рис. 438. Гусеница совки Laphygma exigua Hbn. (по Гера-
симову) .
происходит частичная редукция ложноножек. У пядениц (Geometridae)
обычно развиты брюшные ножки только на VI и последнем сегментах
брюшка, с чем связана своеобразная петлеобразная походка этих гу-
сениц. У некоторых совок (Noctuidae) в младших гусеничных возра-
стах тоже отсутствуют ложноножки на III и IV- сегментах брюшка.
У части хохлаток (Notodontidae) отсутствует последняя''пар а брюш-
ных ног, а у скрытно живущих гусениц могут иногда редуцироваться
все ложноножки.
Покровы гусениц усажены щетинками (хетами), имеющими строго
фиксированное положение у каждого вида. Расположение щетинок
(хетотаксия) широко используется в систематике для диагностики ви-
дов и таксонов более высокого ранга. С возрастом у некоторых гусе-
ниц появляется вторичный волосяной покров, густо покрывающий все
тело.
Куколки почти у всех чешуекрылых покрытые: придатки тела
плотно спаяны с покровами и неподвижны. Способность к движению
сохраняют только отдельные сегменты брюшка. Свободные куколки
с подвижными придатками обнаруживаются у архаичных чешуекры-
лых (Micropterigidae и Eriocraniidae). Куколки этих бабочек исполь-
зуют мандибулы либо для прогрызания кокона, либо для выхода из
почвы перед вылетом имаго. Кроме того, большая группа низших фре-
натных чешуекрылых обладает неполными куколками (промежуточ-
ный тип между свободными и покрытыми куколками). У этих куколок
все придатки тела свободны, но неподвижны, однако брюшные сег-
менты сохраняют подвижность в большей степени, чем у покрытых
куколок. Такие куколки способны перед линькой на имаго выбирать-
ся из укрытий. Этому способствует вооружение куколки (шипики на
поверхности кутикулы и шипы на конце брюшка).
.13 208 1 81
Гусеницы в подавляющем большинстве случаев являются фито-
фагами, причем можно обнаружить в Пределах отряда все степени
пищевой специализации. Примером. монофагии может служить дубо-
вая листовертка (Tortrix viridana L.)-, связанная с ранней расой дуба.
Капустная белянка (Pieris brassicae L.), развивающаяся на кресто-
цветных— классический пример олигофагии, а монашенка (Lymantria
monacha L.) может служить примером полифагии. Гусеницы монашен-
ки способны развиваться не только на самых разнообразных листвен-
ных древесных породах, но и на хвойных, таких, как ель, сосна и ли-
ственница.
По образу жизни гусеницы чрезвычайно разнообразны, хотя они
и уступают в этом отношении двукрылым или жесткокрылым. Ниже
приводятся основные жизненные формы гусениц.
1. Свободноживущие гусеницы. Для них характерно пи-
тание листьями непосредственно на растениях. Гусеницы, связанные
с древесной растительностью, часто покрыты вторичным волосяным
покровом и нцредко снабжены различными выростами (коконопряды—
Lasiocampidae, волнянки — Lymantriidae, хохлатки — Notodontidae).
Наоборот, гусеницы, связанные с травами, обычно лишены густого
волосяного покрова, за -исключением медведиц (Arctiidae). Возможно,
что густой волосяной покров открытоживущих гусениц выполняет за-
щитную функцию, предохраняя их от нападения позвоночных-энтомо-
фагов. Несомненно защитную функцию выполняют ядовитые волоски
некоторых гусениц. Например, у гусениц златогузки (Euproctis chry-
sorrhoea L.) кончики наполненных ядом волосков легко обламываются
и, попадая на слизистые оболочки или просто на кожу позвоночных
животных и человека, вызывают сильные воспалительные явления. .
Для некоторых свободноживущих гусениц известны специальные
защитные кожные железы. Так, у гусениц махаона (РарШо machaon
L.) при их раздражении из передней части груди выворачивается остро
пахнущая оранжевая железа в форме длинной вилки. У хохлаток Се-
гига Schr. и Dicranura Bsd. сходные железы ярко-красного цвета вы-
двигаются на концах заостренных хвостовых придатков.
Важной особенностью многих свободноживущих гусениц является
их криптическая внешность. В некоторых случаях в создании крипти-
ческого эффекта участвуют не только окраска и форма тела, но и по-
ведение. Так, гусеницы пядениц (Geometridae) способны вытягивать
тело под углом к ветке, на которой они сидят, за счет длительного
тонического сокращения туловищной мускулатуры. Подобная поза
в сочетании с покровительственной окраской и своеобразным рельефом
тела создает полную иллюзию торчащего сучка. . ।
2. Чехлоносцы. Эту жизненную форму представляет* гусени-
цы, обитающие внутри специальных чехликов, которые они строят из
тонких веточек, сухих листьев или песчинок, склеивая их шелком. Ти-
пичными примерами таких гусениц могут служить мешочницы (Psy-
chidae). Чехлики мешочниц можно часто увидеть на стволах деревьев,
на листьях или просто на стене дома и на заборе. В чехлика! проис-
ходит и окукливание гусениц, а у некоторых наиболее специализиро-
ванных мешочниц даже самка не покидает своего убежища, где со-
вершается копуляция и откладываются яйца.
3. Почвообитающие гусеницы. Среди гусениц обитателей
почвы сравнительно немного. Это преимущественно подгрызающие
совки из подсемейства Agrotinae. Для них характерны копательная
голова и полифагия.
4. Ли сто верты. К этой группе относят гусениц, обильно выде-
ляющих шелк и обитающих в свернутых или сплетенных шелковой
нитью листьях, которыми они питаются. Примерами листовертов мо-
182
гут служить некоторые листовертки (Tortricidae) и огневки (Pyrali-
dae).
5. Прядильщики. Биологически близкая группа к листовер-
там. Гусеницы-прядильщики оплетают шелковыми нитями ветви де-
ревьев и кустарников, образуя гнезда, в которых они обитают. Типич-
ными представителями прядильщиков служат горностаевые моли
(Yponomeutidae), которые не только питаются, но и окукливаются
в гнездах. При массовом размножении черемуховой горностаевой моли
(Y ponomeuta evonymelius L.) вес таких гнезд с куколками может до-
стигать нескольких килограммов.
6. Плодожорки. Эти гусеницы ведут типичный скрытый, образ
жизни внутри плодов и семян. Гусеницы-плодожорки лишены какого-
либо рисунка, обычно белого или желтого цвета, иногда розовые или
даже красные. Классическим примером подобного образа жизни мо-
жет служить яблоневая плодожорка (Laspeyresia pomonella L.), гу-
сеницы которой живут внутри плодов яблони, айвы, иногда груши.
В первых возрастах гусеницы выедают семена, а затем переходят к пи-
танию сочной мякотью плода.
7. Бурильщики. Четко очерченная жизненная форма гусениц,
обитающих внутри ветвей и стволов древесных пород, а также в кор-
нях. По режиму питания бурильщики являются ксилофагами. Для них
характерны многолетние циклы развития. По адаптивным морфологи-
ческим признакам бурильщики сходны с плодожорками. Внутри стеб-
лей и стволов обитают гусеницы древоточцев (Cossidae) и стеклянниц
(Sesiidae), а внутри корней травянистых растений — гусеницы тонко-
прядов (Hepialidae).
8. Минеры. К этой группе относятся гусеницы самых разнооб-
разных семейств. В отличие от бурильщиков минеры обитают внутри
зеленых частей растений, обычно внутри листьев. При выедании внут-
ренней части листа гусеницами образуется полость (мина), хорошо за-
метная снаружи в виде белых пятен (рис. 5). В зависимости от вида
чешуекрылых гусеницы могут выедать либо всю мякоть листа, образуя
двусторонние мины (Eriocraniidae, Gelechiidae), либо только поли-
садную паренхиму (Stigmellidae) и губчатую паренхиму (Lithocol-
letidae), образуя односторонние мины, либо повреждают клетки эпи-
дермиса. Эпидермальные минеры (сокоеды) способны высасывать сок
из растительных клеток. Минирующий образ жизни накладывает глу-
бокий отпечаток на общую организацию гусениц: тело уплощается, ча-
сто редуцируются ноги и появляются двигательные мозоли, видоизме-
няется ротовой аппарат, исчезает часть глазков и частично редуци-
руются усики.
9. Водные гусеницы не представляют собой единой жизнен-
ной формы, поскольку характер использования ими среды обитания
довольно разнообразен. В систематическом отношении все палеаркти-
ческие водные гусеницы относятся к огневкам (Pyralidae). Предста-
вители рода Schoenobius Dup. обитают внутри листьев водных расте-
ний и не соприкасаются непосредственно с водой. Гусеницы время от
времени строят чехлики, в которых они переносятся течением к но-
вому растению. Виды рода Hydrocampa и рясковая огневка (Cataclysta
lemnata L.) также минируют листья. Однако эти мины заливаются во-
дой и гусеницы дышат с помощью растворенного в воде кислорода не-
посредственно через покровы тела. У этих гусениц дыхальца редуци-
рованы. В старших возрастах гусеницы переходят от минирующего об-
раза жизни к чехликовому. Еще более приспособлена к подводному
образу жизни огневка Paraponyx stratiotata, гусеницы которой живут
в чехликах на водных растениях и обладают нитевидными трахейны-
183
ми жабрами. Иной образ жизни у Acentropus niveus 01. Гусеницы это-
го вида живут открыто под водой на водных растениях и полностью
лишены дыхалец; они используют для дыхания кислород, растворен-
ный в воде. Бескрылая самка A. niveus остается под водой и откла-
дывает яйца на растения.
Все перечисленные жизненные формы гусениц связаны с расте-
ниями. Однако среди чешуекрылых встречаются отдельные виды с ины-
ми режимами питания. Так, многие настоящие моли (Tineidae) пере-
шли к питанию трутовыми грибами, а некоторые виды этого семейства
специализировались на питании шерстью или волосом, развиваясь
в природной обстановке в норах и гнездах позвоночных животных. От-
дельные представители настоящих молей, например платяная моль
(Tineola biselliella Humm.), стали синантропными и повреждают шер-
стяные изделия в жилых домах и на складах. Специальный режим пи-
тания характерен для небольшой группы огневок (Pyralidae), кото-
рые живут в ульях медоносной пчелы или в гнездах шмелей, питаясь
вощиной. /
Особого внимания Заслуживают хищные гусеницы, дающие пред-
ставление о широких адаптивных возможностях отряда. Несомненно,
что хищный образ жизни возник у гусениц вторично. Среди многочис-
ленного семейства совОк (Noctuidae) нцредко наблюдается факульта-
тивное хищничество. Такие гусеницы могут поедать тлей, встретив-
шихся на растении, иногда нападают на более мелких или ослаблен-
ных гусениц. Питание животной пищей у перечисленных гусениц не
обязательно. Настоящих хищников среди чешуекрылых немного. Ши-
роко известен вид совки Coccidophaga scitula Rmb., гусеницы которой
специализировались на питании червецами. Спорадически встречаются
хищники и в других семействах. Крайне своеобразно поведение гусе-
ниц голубянки Lycaena alcon F. Ее гусеницы в первом возрасте пита-
ются на горечавке, но после первой линьки проникают’в гнезда му-
равьев, где уничтожают их личинок. Гусеницы этой голубянки обла-
। дают развитыми кожными железами, которые выделяют вещество,
привлекательное для муравьев.
. Гусеницы окукливаются либо в почве, либо открыто на расте-
ниях. В последнем случае гусеницы перед окукливанием прядут кокон
(например, коконопряды — Lasiocampidae, павлиноглазки—Attacidae,
волнянки — Lymantriidae и другие). Гусеницы булавоусых чешуекры-
\ лых (Rhopalocera) не прядут кокона и при окукливании многие из них
прикрепляются к растениям. Куколки этих бабочек обладают покро-
вительственной окраской, а их цвет часто зависит от цвета субстрата,
на котором происходит окукливание. У парусников (Papilionidae) и бе-
лянок (Pieridae) куколки прикрепляются к растениям шелковым по-
яском. У нимфалид (Nymphalidae) и бархатниц (Satyridae) куколки не
имеют пояска и повисают на растениях вниз головой. Нередко на-
блюдается окукливание гусениц в щелях коры или под отставшей
корой. В этом случае гусеницы изготавливают редкий кокон, либо фор-
мируют колыбельку.
В пределах отряда чешуекрылых наблюдается значительное раз-
нообразие жизненных циклов. В условиях умеренного климата встре-
чаются как моноциклические, так и полициклические виды. Зимуют
обычно гусеницы или куколки, реже наблюдается зимовка имаго
и яиц. Например, у совок (Noctuidae) 45,5 % европейских видов зи-
муют в стадии гусеницы, 32,5 %—в стадии _ куколки, 17 % видов об-
ладают эмбриональной диапаузой и только 5 % видового состава зи-
муют в стадии имаго. Как правило, у каждого вида стадия зимовки
приурочена к строго определенному этапу онтогенеза.
Циклы развития различаются и по времени размножения. У боль-
184
шинства видов имаго активны летом. Однако у некоторых бабочек
период размножения приурочен к осени (например, у зимней пядени-
цы— Operophtera brumata L., самки которой откладывают яйца в ок-
тябре или даже в ноябре). Встречаются виды и с ранневесенним раз-
множением, откладывающие яйца в конце марта или в апреле (неко-
торые листовертки рода Acleris и некоторые пяденицы — Geometridae).
У многих чешуекрылых — обитателей засушливых районов в цикле
развития помимо зимней диапаузы нЬблюдается и летняя остановка
развития. Летний покой проявляется и у северных видов, приступаю-
щих к размножению поздней осенью. Так, у зимней пяденицы, обла-
дающей эмбриональной диапаузой, имеет место летняя задержка в раз-
витии куколок, которые лежат в почве с июня до октября. Известны
у чешуекрылых двухлетние и даже трехлетние циклы развития, харак-
терные, в основном, для бурильщиков (например, для древоточца
Cossus cossus L.). У некоторых полициклических чешуекрылых наблю-
дается закономерная смена сезонных форм, определяемая длиной дня
(см. рис. 13).
Расселительную функцию у чешуекрылых выполняет имаго, за
исключением некоторых видов, у которых ветром на значительные рас-
стояния переносятся мелкие гусеницы первого возраста, обладающие
многочисленными длинными и тонкими волосками (например, у непар-
ного .шелкопряда — Ocneria dispar L.). Нередко ветром на десятки
и сотни километров переносятся и взрослые бабочки. Такие миграции
описаны для серьезного вредителя сельскохозяйственных культур — лу-
гового мотылька (Loxostege sticticalis L.). Перенос ветром в целом
характерен для мелких чешуекрылых со слабым полетом.
Удивительная способность к активным перелетам на значительные
расстояния известна для некоторых бражников. Так, олеандровый
бражник (Deilephila nerii L.), обитающий на Аппенинском полуостро-
ве и развивающийся на олеандре, каждый год появляется на Черно-
морском побережье Кавказа. Бабочки откладывают яйца на олеандр,
й гусеницы питаются до глубокой осени, однако они не способны бла-
гополучно перезимовать и вымерзают. Бабочек олеандрового бражни-
ка находили на севере Европы (под Ленинградом, в Финляндии и При-
балтике), куда они залетают, преодолевая тысячи километров. Точно
так же совершает дальние миграции другой бражник — мертвая го-
лова (Acherontia atropos L.), который залетает в центральную и север-
ную Европу из ее южных областей. Гусеницы этого вида развивают-
ся на пасленовых, в том числе и на картофеле, но, не достигая стадии
куколки, погибают в осенне-зимний период от холодов.
Среди насекомых с полным превращением отряд чешуекрылых за-
нимает одно из первых мест по богатству мировой фауны; в составе
отряда насчитывается более 140 000 видов. Большая их часть обитает
в тропических областях. К северу и югу от экватора' количество видов
постепенно уменьшается, однако даже в тундре еще сохраняется срав-
нительно обильная фауна чешуекрылых, а полярная перламутровка
{Boloria polaris В.) встречается севернее 82-й параллели.
Определительная таблица семейств
Г(6). Передние и задние крылья сходны по жилкованию и форме;
радиальная жилка разветвлена как на передних, так и на зад-
них крыльях (рис. 428, Р).
2(5). Передние и задние голени со шпорами (рис. 433). Мелкие ба-
бочки (длина крыла менее 10 мм).
3(4). Ротовой аппарат грызущий; средние голени без шпор. Бабоч-
185
ки встречаются днем в конце мая, часто сидят на цветках лю-
тика, где питаются пыльцой ....................................
..................................Зубатые моли — Micropterigidae.
4(3). Ротовой аппарат в виде хоботка (сосущий); средние голени
со шпорами. Бабочки летают в апреле, обычно среди берез
.....................................Первичные моли — Eriocraniidae.
5(2). Голени без шпор. Бабочки среднего и крупного размера, дли-
на крыла более 10 мм. Активны в сумерках и ночью . . .
............................................... . ... Тонкопряды — Hepialidae.
6(1). Передние и задние крылья различны по форме и жилкова-
нию; радиальная жилка разветвлена только на передних
крыльях (рис 429-—432, R).
Рис. 439—450. Детали строения чешуекрылых.
439 — переднее крыло бархатницы (Satyridae), 440 — голова настоящей моли (Tineidae), 441 —
заднее крыло огневки (Pyralidae), 442 — форма тела пестрянки (Zygaenidae), 443 — заднее крыло
хохлатки (Notodontidae), 444, 445 — варианты строения головы у волнянок (Lymantriidae), внд
сбоку, 446, 447, 448 — то же у коконопрядов (Lasiocampidae,), 449 — голова медведицы (Arctiidae),
450 — типичный рисунок переднего (вверху) и заднего (внизу) крыльев у совок (Noctuidae).
Жилки крыльев? А — анальные: R — радиальные: Sf — субкостальные.
7(18). Усики булавовидные (рис. 420). Крылья в покое складывают-
ся вертикально иад телом (рис. 434); зацепки нет. (Группа
Булавоусые, или дневные чешуекрылые—Rhopalocera).
8(9). Внутренний край заднего крыла с дугообразной выемкой
(рис. 430). Крупные яркие бабочки, часто с длинными хво-
стиками на задних крыльях или с полупрозрачными крылья-
ми .................................Парусники — Papilionidae.
9(8). Внутренний край заднего крыла без дугообразной выемки
(рис. 429).
10(11). Грудь широкая, только в 3—4 раза уже переднего крыла. Го-
186
лова лишь немного уже груди, с пучками чешуек на верхней
стороне. Задние голени с 2 парами шпор (рис. 433). Неболь-
шие бабочки буроватых или зеленовато-серых тонов . . . .
............................ Толстоголовки—Hesperiidae.
11(10). Грудь узкая, ее ширина меньше длины переднего крыла бо-
лее чем в 7 раз. Голова значительно уже груди, без пучков
чешуек. Задние голени с 1 парой шпор.
12(15). Передние ноги сильно укорочены и скрыты в волосяном по-
I крове груди.
13(14). Некоторые жилки передних крыльев в основании вздуты
(рис. 439). Бабочки обычно бурых тонов, часто с одним или
несколькими глазками на нижней стороне крыльев . . . .
.............................. Бархатницы, или сатиры — Satyridae.
14(13). Жилки передних крыльев в основании не вздуты. Бабочки
часто пестро и ярко окрашены, крылья нередко с изрезанны-
ми краями (рис. 420) . . . Нимфалиды — Nymphalidae.
15(12). Передние ноги не скрыты в волосяном покрове груди.
16(17). Глаза окаймлены белой полосой. Крылья с металлическим
блеском (голубые, огненно-красные, иногда фиолетовые) или
же бурые и без металлического блеска. На исподе крыльев
нередки перевязи из ртдельных глазков. Небольшие бабоч-
ки с четко выраженным половым диморфизмом в окраске
крыльев................................. Голубянки — Lycaenidae,
17(16). Крылья окрашены иначе (белые, желтые, оранжевые
и без металлического блеска). Глаза не окаймлены белым.
Средней величины бабочки, ^асто с половым диморфизмом
..............................................Белянки — Pieridae.
18(7). Усики не булавовидные, крылья в покое не складываются
вертикально над телом (рис 435—437); если же усики була-
вовидные или крылья складываются вертикально, то есть за-
.цепка.
19(22). Крылья прозрачные с бурым окаймлением, бабочки имитиру-
ют жалящих перепончатокрылых, активны днем.
20(21). Голени без шипов............................... . . . .
Бражники—Sphingidae. (Род Шмелевидки — HaemorrhagiaGr.).
21(20). Голени усажены короткими шипиками........................
.................................Стеклянницы — Sesiidae.
22(19). Крылья не прозрачные, густо покрыты чешуйками Активны
преимущественно ночью или в сумерках.
23(24). Голова в пучках торчащих волосков (рис. 440), широкая
(лишь немного уже груди) . . Настоящие моли — Tineidae.
24(23). Голова намного уже груди, без пучков торчащих волосков.
25(26). Задние крылья ланцетовидные, с бахромкой волосков, превы-
шающих ширину крыльев /рис. 424)................................
Более 20 семейств мелких чешуекрылых (Nepticulidae, Ti-
........................scheriidae, Gracillariidae и др.).
26(25). Задние крылья не ланцетовидные и с более короткой бах-
ромкой.
27(28). Задние крылья с вырезкой на внешнем крае перед вершиной
...............................Выемчатокрылые моли — Gelechiidae.
28(27). На задних крыльях перед вершиной нет вырезки.
29(38). Бахромка на задних крыльях длиннее 1/3 ширины крыльев.
30(31). Нижнегубные щупики короче челюстных щупиков или же уси-
ки длиннее крыльев.......................Минно-чехликовые моли —
.............................................Adelidae.
31(30). Нижнегубные щупики длиннее челюстных щупиков или по-
следние отсутствуют; усики всегда короче крыльев.
187
32(33). Нижнегубные щупики серпообразно загнуты вверх . . . .
................................Выемчатокрылые моли — Gelechiidae,
ширококрылые моли — Oecophoridae и другие семейства мел-
ких чешуекрылых
33(32). Нижнегубные щупики не загнуты вверх.
34(35). Голова мохнатая, хоботка нет. Самки бескрылые, самцы кры-
латые .....................................Мешочницы — Psychldae.
35(34). Голова покрыта чешуйками, хоботок есть.
36(37). Передние крылья обычно серебристо-белые с резкими черными
точками (рис. 423).........................Горностаевые моли —
Yponomeutidae.
37(36). Рисунок крыльев иной'(рис. 436)............................
..................................Листовертки — Tortricidae.
38(29). Бахромка на задних крыльях короче 1/3 ширины крыльев.
39(48). Задние крылья с 3 анальными жилками, доходящими до края
крыла (рис. 441, Д).
40(41). Субкостальная и радиальная жилки на задних крыльях слиты
за пределами срединной ячейки (рис. 441, Sc, 7?).................
......................................Огневки — Pyralidae.
41(40). Субкостальная и радиальная жилки на задних крыльях, если
и слиты, то не за пределами срединной ячейки (рис. 432,
Sc, R).
42(47). Срединных шпор на задних голенях нет, если есть, то очень
короткие.
43(44). В окраске тела и крыльев есть красные, синие или зеленые
цвета; если развиты пятна на крыльях, то только красные или
белые. Усики булавовидные (рис. 442). Бабочки часто встре-
, чаются днем на цветущих растениях...............................
..........................................Пестрянки — Zygaenidae.
44(43). Окраска крыльев иная, усики не булавовидные.
45(46). Крылья дымчатые, полупрозрачные, кроме чешуек покрыты
волосками; в размахе крыльев менее 30 мм. Самки бескры-
лые........................................Мешочницы — Psychldae.
46(45). Крылья плотные, покрыты чешуйками; тело массивное, в раз-
махе крыльев более 30 мм...........................
.....................................Древоточцы — Cossidae.
47(42). Срединные шпоры на задних голенях хорошо развиты. Бабоч-
ки в размахе крыльев не более 30 мм (рис. 436)...................
..................................Листовертки — Tortricidae.
48(39). На задних крыльях имеются только 1—2 анальные жилки, до-
ходящие до края крыла (рис. 431, Д).
49(56). На передних крыльях жилка М2 отходит от ячейки ближе
к ЛГ, чем к М3, или на равном расстоянии между М, и Л43
(рис. 431).
50(51). Усики веретеновидные (рис. 421). Крупные и среднего разме-
ра бабочки с мощным сигарообразным телом и быстрым по-
летом.......................................Бражники — Sphingidae.
51(50). Усики не веретеновидные.
52(53). На каждом крыле по одному срединному глазку. Наружные
края задних крыльев никогда не бывают волнистыми. Крупные
бабочки (размах крыльев от 50 до 150 мм).........................
.................................Павлиноглазки — Attacidae.
53(52). Крылья без срединных глазков; если с глазками, то только на
одной паре крыльев или же наружные края задних крыльев
волнистые.
54(55). Субкостальная жилка на задних крыльях резко изгибается
в основании, сближаясь или сливаясь на некотором расстоя-
188
нии с радиальной жилкой (рис. 432, Sc, R). Бабочки в покое
не складывают крылья вдоль тела, а держат их распростер-
тыми в стороны (рис. 437)...........Пяденицы — Geometridae.
55(54). Субкостальная жилка на задних крыльях в основании изогну-
та очень слабо и не сливается с радиальной жилкой (рис. 443,
Sc, R). Бабочки в покое складывают крылья кровлеобразно,
вытягивая их назад вдоль тела....................................
........................................ Хохлатки—Notodontidae.
56(49). На передних крыльях жилка М2 отходит от ячейки ближе
к Л43, чем к Л41 (рис. 431).
57(60). Грудь, голова и ноги, исключая лапки, скрыты в волосяном
покрове. Хоботок всегда редуцирован и скрыт в волосках.
58(59). Щупики тонкие и хорошо заметны (рис. 444, 445).' Зацепка
есть...................................Волнянки — Lymantriidae.
59(58). Щупики толстые или полностью скрыты в волосяном покрове
и не заметны (рис. 446—448). Зацепки нет.........................
..........................Коконопряды — Lasiocampidae.
60(57). Тело не опушено; если же опушение заметно, то голени не
скрыты в волосяном покрове груди и развит1хоботок (рис.449).
61(62). Передние крылья на вершине серповидно изогнуты. . . ..
.............................Серпокрылки — Drepanidae.
62(61). Передние крылья на вершине не изогнуты серповидно.
63(66). На задних крыльях имеется субкостальная жилка (рис. 431).
64(65). На задних крыльях субкостальная жилка слита с радиальной
на коротком расстоянии, менее 1/3 длины срединной ячейки
(рис. 431). Передние крылья обычно тусклой темной окраски
с характерным рисунком из поперечных полос и округлых или
• почковидных пятен (рис. 422, 450). . . Совки — Noctuidae.
65(64). На задних крыльях субкостальная жилка слита с радиальной
на большом протяжении, более чем на 1/3 длины срединной
ячейки. Крылья часто с ярким рисунком.............................
..........................................Медведицы — Arctiidae.
66(63). На задних крыльях субкостальной жилки нет. Черные или
коричневые бабочки с белыми пятнами на передних крыльях.
.........................Ложнопестрянки — Ctenuchidae.
ОТРЯД ДВУКРЫЛЫЕ, ИЛИ КОМАРЫ И МУХИ —DIPTERA*
Общая характеристика
К этому отряду относятся насекомые мелких и средних размеров
с одной парой крыльев (рис. 451). Вторая пара крыльев видоизменена
в короткие булавовидные придатки — жужжальца. Голова подвижная.
Глаза крупные. Ротовой аппарат представлен хоботком лижушего или
колющего типа. Усики либо многочлениковые, длинные (в несколько раз
длиннее головы), либо короткие, состоящие всего лишь из 3 члеников.
По этому признаку весь отряд подразделяется на 2 подотряда: длин-
ноусых (Nematocera) и короткоусых (В г а с hy се г а).
У короткоусых третий членик иногда имеет ряд насечек или перетя-
жек, что не следует путать с настоящей членистостью. Хорошо развит
грудной отдел, в котором преобладает среднегрудь. Крылья прозрач-
ные или окрашенные. Жилкование упрощенное, с изгибами продоль-
* А. К. Бродский.
189
них жилок и немногими замкнутыми ячейками. В покое крылья скла-
дываются плоско на брюшке. Ноги бегательные. Брюшко у длинно-
усых длинное цилиндрическое, а у короткоусых короткое овальное.
У самок часто имеется втяжной яйцеклад. Церки отсутствуют.
Развитие с полным превращением. Размножение обоеполое, реже
партеногенетическое. Известны случаи педогенеза. Яйца сигаровидные,
реже округлые (рис. 452). У некоторых видов яйца снабжены спе-
циальными фиксаторами; например, для прикрепления к телу хозяина
яйцо большеголовой (Gonbpidae) имеет особый четырехлапый якорек,
У некоторых высших представителей подотряда короткоусых наблюда-
ется живорождение. Иногда рождается личинка первого возраста, а у
кровососок (Hippoboscidae) наблюдается отрождение вполне зрелой
личинки, готовой к окукливанию.
Рис. 451—456. Комнатная муха Musca domestica L. (Muscidae)
и ее стадии развития.
451 — имаго, 452 — яйца, 453 — личинка первого возраста, 454 — взрос-
лая личинка, 455 — куколка, вынутая из пупария, 456 — пупарий.
Тело личинок чаще удлиненное, веретеновидное, червеобразное
(рис. 453, 454). Покровы слабо склеротизованы и непигментированные.
Редко тело окрашено: красное у звонцов (Chironomidae) или зелено-
ватое у открыто живущих видов журчалок (Syrphidae). Покровы могут
быть гладкими, но чаще они снабжены волосками, шипами, бугорками,
валиками и т. п. Основные особенности личинок двукрылы^ — это от-
сутствие ног и преимущественное развитие дыхалец на конце тела.
Отсутствие членистых грудных конечностей компенсируется двига-
тельными валиками, различными выростами тела; иногда имеются об-
разования, напоминающие ложные ножки личинок пилильщиков. Ли-
чинки мошек (Simuliidae), живущие в быстрых потоках, обладают
мощными присосками. Голова личинок очень изменчива. У многих длин-
ноусых она хорошо развита (Chironomidae, Bibionidae). Иногда голова
сильно уменьшается и частично втягивается в переднегрудь (Tipulidae,
Limoniidae, Empididae). У личинок высших короткоусых головы нет,
а имеются только черные ротовые крючки, просвечивающие сквозь по-
кровы головного сегмента тела.
Куколка свободная (рис. 455). Иногда перед окукливанием личин-
ка сплетает кокон (например, у мошек). Куколка многих мух' заклю-
чена в отставшую и затвердевшую линочную шкурку личинки послед-
него возраста — так называемый пупарий (рис. 456). При выходе
190
имаго.из куколки пупарий разрывается по кольцевому шву. На этом
основании часть короткоусых двукрылых получила название «кругло-
шовных». Другие короткоусые, у которых оболочка куколки разрыва-
ется при выходе имаго по прямому шву, получили название «прямо-
шовных».
Взрослые двукрылые, как правило, обитают открыто. Большин-
ство хорошо летает. Лучшими летунами среди всех насекомых явля-
ются журчалки (Syrphidae). В полете они часто используют зависание
на одном месте, резкие рывки вперед, назад или вбок. Ускорение,
развиваемое журчалками при взлете, в 18 раз превосходит ускорение
силы тяжести. Перед спариванием многие длинноусые двукрылые ро-
ятся. В рое самцы демонстрируют беспорядочный, хаотичный полет.
Самка обнаруживает рой и влетает в него. В свою очередь, самцы на-
Рис. 457—463. Личинки двукрылых, развивающиеся в почве.
457 — болотница Limonia macrostigma Schumm. (Limoniidae), 458 — древесница Coenomyia ferrugine
Scop. (Xylophagidae), 459 — ктырь Apoclea nelvipes Lw. (Asilidae), 460 — лжектырь Thereva
L. (Therevidae), 461 — траурный комарик Sciara Mg. (Sciaridae). 462 — бекаснн^а Rhagi oscolopaceus
L. (Rhagionidae), 463 — львинка Chloromyia Dune. (Stratiomyidae).
ходят самку либо с помощью зрения, либо по звуку, издаваемому ею
во время полета. Самцы толкунчиков (Empididae) во время брачного
полета таскают с собой «баллончик» с пенистыми стенками; внутри
«баллончика» находится убитая добыча, предлагаемая самке перед
спариванием.
После спаривания самка летит на яйцекладку. Способы откладки
яиц разнообразны. Многие виды мошек (Simuliidae) для откладки
яиц опускаются под воду. Пестрокрылки (Tephritidae) длинным яйце-
кладом прокалывают ткани растений, а некоторые тахины (Tachinidae)
могут прокалывать толстые покровы хозяина. Жужжалы (Bombyliidae)
вращательными движениями брюшка, вооруженного крючьями, дела-
ют воронку в почве, куда откладывают яйцо. Самка мухи Lucilia bu-
fotiivora Mon. из семейства Calliphoridae ползает перед лягушкой до
тех пор, пока та ее не проглотит — вылупление личинок из яиц проис-
ходит в кишечнике лягушки: У кровососущих длинноусых откладка яиц
сопряжена с питанием кровью. Так, у самок комаров рода Culex L.
после одноразового насасывания крови созревает до 250 яиц.
Большинство взрослых двукрылых питается росой, выступающим
соком растений, нектаром цветков, навозом. Довольно большая груп-
па мух (ктыри — Asilidae, толкунчики — Empididae, бекасницы — Rha-
191
gionidae) — хищники, нападающие на насекомых. Кровью позвоночных,
особенно млекопитающих, питаются некоторые длинноусые двукрылые
(настоящие комары — Culicidae, мокрецы — Ceratopogonidae, мошки —
Simuliidae), отдельные короткоусые прямошовные (слепни — Tabani-
dae) и круглошовные (настоящие мухи — Muscidae, кровососки —
Hippoboscidae). Небольшая часть двукрылых в имагинальном состоя-
нии не питается (бабочницы — Psychodidae, звонцы— Chironomidae,
галлицы — Cecidomyiidae, оводы — Oestridae и др.).
Экология личинок чрезвычайно разнообразна. Благодаря черве-
образной форме тела и редукции ног они заселили толщу самых раз-
личных субстратов: твердых и рыхлых, сухих и влажных, растительного
и животного происхождения, богатых и бедных органическим вещест-
вом. Двукрылые в личиночном состоянии, хотя и влаголюбивые, но
Рис. 464—470. Личинки двукрылых, развивающиеся в древесине.
464— мокрец Forcipotnyla nigra Winn. (Ceratopogonidae), 465 — KTbipbLaphrla flava L. (Asilidae),
466 — древесница Xyiophagus ater F. (Xylophagidae), 467— Synneuron Lund. (Hyperscelididae),
468 — львинка Pachygaster orbltalis Wahlb. (Stratiomyidae), 469 — болотница Epiphragma ocellarls
L. (Limoniidae), 470 — журчалка Brachyopa bicolor Fall. (Syrphidae). ,
главным образом сухопутные формы. В почве развивается большое
количество двукрылых (рис. 457—463). К наиболее характерным пред-
ставителям почвообитающих личинок относятся слепни (Tabanidae)
ктыри (Asilidae), лжектыри (Therevidae), бекасницы (Rhagionidae)
и многие долгоножки (Tipulidae). В лесной подстилке живут личинки
толстоножек (Bibionidae), долгоножек, галлиц и некоторых других
семейств. Гниющие остатки растительного и животного происхожде-
ния, в частности навоз, составляют основное место развития навозных
мух (Scatophagidae), бабочниц (Psychodidae), муравьевидок (Sepsi-
dae), горбаток (Phoridae), настоящих мух (Muscidae), а также многих
видов из семейств Calliphoridae и Sarcophagidae. Некоторые виды этой
группы переходят к обитанию в гнездах птиц и грызунов. Целый ряд
двукрылых связан с разлагающейся древесиной и развивается под
корой, либо в дуплах и пнях (рис. 464—470). Это преимущественно
представители длинноусых двукрылых (долгоножки, болотницы — Limo-
niidae, толстоножки и др.), личинки которых имеют крепкие жвалы,
приспособленные для разрушения древесины. Короткоусые двукрылые,
для которых более характерны безголовые личинки, чаще всего раз-
виваются под отставшей корой, где для передвижения они используют
192
естественные полости и ходы других насекомых. В тканях живых рас-
тений обитают личинки галлиц (Cecidomyiidae), злаковых мушек
(Chloropidae), цветочниц (Anthomyiidae) и др. Ряд двукрылых связан
с животными. Одни из них живут в трупах (Calliphoridae), другие яв-
ляются эндопаразитами млекопитающих (оводы — Oestridae, Sarcop-
hagidae) или насекомых и многоножек (тахины—Tachinidae, Sarcop-
hagidae).
Многие двукрылые перешли к развитию в водной среде (рис. 471—
479). При этом могут использоваться самые незначительные и времен-
ные скопления воды на дорогах, в дуплах и пазухах листьев. Подоб-
ные скопления становятся местом обитания форм с коротким периодом
развития. В стоячих водоемах развиваются представители многих се-
мейств (звонцов — Chironomidae, мокрецов — Ceratopogonidae, болот -
ниц — Limoniidae, настоящих комаров — Culicidae, долгоножек —
Tipulidae, зеленушек — Dolichopodidae, журчалок — Syrphidae и
др.). К быстротекущим водоемам — рекам, ручьям—’Приурочены
мошки (Simuliidae). В соленой воде прибрежной зоны морей и океа-
478
479
Рис. 471—479. Личинки двукрылых, развивающиеся в воде.
471 — комар Culex piplens L. (Culicidae), 472 — мошка Simulinm Latr. (Simuliidae), 473 — звонец
Chlronomus Mg. (Chironomidae), 474 — журчалка Eristalls tenax L. (Syrphidae), 475— мокрец
Cullcoldes Latr. (Ceratopogonidae), 476 — Dixa amphibia DeGeer из семейства Dixiidae, 477 — комар
Chaoborus Licht. (Culicidae). 47S — львиика Stlattomya chamaeleon Deg. (Stratiomyidae), 479 — сле-
пень Haematopota pluvialls L. (Tabanidae).
193
Рис. 480. Личинка бе-
лополосой журчалки
Syrphus albostriatus Fll.,
развивающаяся на рас-
тениях.
нов встречаются некоторые виды звонцов. Дыхание водных личинок
осуществляется либо за счет кислорода, растворенного в воде, либо за
счет атмосферного кислорода. В первом случае используются жабры, а во
втором на конце тела имеется дыхательная трубка. На вершине дыха-
тельной трубки открываются дыхальца, часто окруженные несмачивае-
мыми волосками (рис. 471, 474, 478). Первый способ дыхания свойст-
вен бентосным двукрылым, например звонцам; второй способ исполь-
зуется представителями нектона, к которым относятся настоящие
комары, львинки (Stratiomyidae) и некоторые другие семейства.
В дыхании водных двукрылых большое значение имеет также газооб-
мен через покровы.
Личинки некоторых двукрылых способны заселять, казалось бы,
совершенно непригодные для жизни насекомых местообитания. Так,
личинки некоторых видов семейства Ephydridae
живут в водоемах с высоким содержанием со-
лей, или в горячих, серных и даже нефтяных
источниках.
Личинок двукрылых, живущих открыто,
очень мало. На растениях обитают личинки
некоторых журчалок, например беловолосой
журчалки Syrphus albostriatus F11. (рис. 480).
Не менее велико разнообразие режимов пи-
тания личинок двукрылых. Разлагающимися
растительными остатками питаются сапрофаги,
к которым относится большинство толстоножек
(Bibionidae), долгоножек (Tipulidae) и многие
виды семейства Lauxaniidae. Живыми расти-
тельными тканями питаются фитофаги. Почво-
обитающие личинки долгоножек и толстоножек
подгрызают корни растений. Специализирован-
ные фитофаги,, проникая в ткани стеблей, листь-
ев и плодов, минируют их (злаковые мушки —
Chloropidae, минирующие мушки — Agro-
myzidae и др.) или вызывают образование
галлов (галлицы — Cecidomyiidae). Особую
группу среди личинок двукрылых образуют ми-
в плодовых телах съедобных грибов. Некоторые
галлицы, развивающиеся в гниющей древесине, под корой и в пнях,
•питаются мицелием грибов.
Огромное количество видов двукрылых питается животной пи-
щей. Личинки многих короткоусых прямошовных двукрылых — актив-
ные хищники. Личинки слепней (Tabanidae) и ктырей (Asilidae) в поч-
ве нападают на личинок насекомых. Личинки бекасниц (Rhagionidae)
и толкунчиков (Empididae) питаются дождевыми червями. Личинки
зеленушек (Dolichopodidae) уничтожают личинок короедов, а крупные
личинки ктырей рода Laphria Mg. (рис. 465) нападают на личинок
усачей. Личинки древесниц (Xylophagidae) прокалывают покровы
жертвы, выделяют внутрь тела пищеварительные соки и затем выса-
сывают содержимое. Они практически всеядны: поедают личинок ко-
роедов, усачей, проволочников, галлиц, долгоножек и дождевых чер-
вей. У личинок круглошовных короткоусых двукрылых хищничество на-
блюдается реже. Некоторые журчалки (Syrphidae) питаются тлями,
а личинки настоящих мух (Muscidae), обитающие в навозе, в старших
возрастах питаются личинками других копрофагов.
Многие водные личинки потребляют мельчайшие планктонные ор-
ганизмы. У малоподвижных личинок имеются особые приспособления
для приема пищи, например, веера мошек (рис. 472)', щетка верхней
цетофаги, обитающие
194
губы комаров, при помощи которых создается Мощный ток воды по на-
правлению ко рту для фильтрации частиц пищи.
Эндопаразитические личинки двукрылых развиваются в насекомых,
дождевых червях, моллюсках, пауках, мокрицах и многоножках. В ча-
стности, многие тахины (Tachinidae) паразитируют на гусеницах, не-
которые представители семейства Sarcophagidae развиваются на са-
ранчовых и их кубышках, большеголовки (Conopidae)—на жалящих
перепончатокрылых, жужжалы (Bombyliidae)—также на жалящих
перепончатокрылых и, кроме того, в кубышках прямокрылых. Двукры-
лые— паразиты позвоночных животных представлены немногими ви-
дами. В теле лягушек и жаб паразитирует Lucilia bufonivora Mon.
из семейства Calliphoridae, а оводы (Oestridae) являются специализи-
рованными паразитами млекопитающих.
Некрофагия (питание трупами позвоночных) широко распростра-
нена среди мясных мух, относящихся к семейству Calliphoridae. Зна-
чительно разнообразнее группа двукрылых-копрофагов. Под корой де-
ревьев экскрементами жуков питаются личинки некоторых мокрецов
(Ceratopogonidae). В помете позвоночных развиваются личинки му-
равьевидок (Sepsidae), навозных мух (Scatophagidae), некоторых ба-
бочниц (Psychodidae) и др. Существует характерный комплекс свя-
занных с навозом мух-синантропов, включающий представителей на-
стоящих мух (Muscidae), цветочниц (Anthomyiidae), а также семейств
Calliphoridae и Sarcophagidae. Мухи-синантропы обитают вблизи чело-
веческого жилья и скотных дворов.
Двукрылые составляют один из самых крупных, отрядов насеко-
мых; по количеству известных видов (более 80 000) они уступают толь-
ко жесткокрылым, чешуекрылым и перепончатокрылым. Широкое рас-
пространение двукрылых и их высокая численность обусловливают
важное практическое значение этого отряда. Как среди длинноусых,
так и среди короткоусых двукрылых немало кровососов, паразитов по-
звоночных животных и переносчиков возбудителей серьезных заболе-
ваний (например, малярийный комар Anopheles maculipennis Mg. из
семейства настоящих комаров — Culicidae). Немаловажное отрица-
тельное значение имеют двукрылые — вредители растений (например,
злаковые мушки — Chloropidae). Многие почвенные, подстилочные
и водные личинки двукрылых играют роль «природных санитаров»,
способствующих разложению органических остатков. Паразитические
двукрылые (например, многие виды семейства Tachinidae) снижают
численность вредных насекомых и могут быть использованы для био-
-* логической борьбы с вредителями сельского и лесного хозяйства.
Некоторые двукрылые легко разводятся в лабораторных условиях
и с большим успехом используются в качестве модельных объек-
тов генетических и цитологических исследований (например,
Drosophila melanogaster L. из семейства плодовых мушек — Drosop-
hilidae).
Определительная таблица семейств
1(18). Усики длинные, многочлениковые, обычно длиннее головы
/ (рис. 481). (Подотряд Длинноусые двукрылые — Nematocera).
2(5). Среднеспинка с V-образным швом между основаниями крыль-
ев (рис. 482).
3(4). Лицевая часть головы (рыльце) с «носом» или небольшим бу-
горком (рис. 483, 484). Крупные и среднего размера комары
с удлиненным телом и очень длинными тонкими Ногами, ко-
торые легко обламываются. . . Долгоножки — Tipulidae.
4(3). Рыльце без «носа» и бугорка. Мелкие, реже средних размеров
195
насекомые с длинными ногами. Встречаются в сырых лесах,
на болотах и заливных лугах. В зимние оттепели встречаются
на снегу..........................Болотницы — Limoniidae.
5(2). Среднеспинка без V-образного шва между основаниями
крыльев.
6(9). Ветвей жилок, упирающихся в край крыла, не менее 9
(рис. 487).
7(8). Мелкие (1,8—5 мм) двукрылые с густо опушенным телом
и крыльями, внешне напоминающие бабочек-молей. Летают
Рис, 481—491. Длинноусые двукрылые и детали их строения.
481 — общий вид самца из семейства настоящих комаров (Culicidae), 482— среднеспинка долго-
ножки (Tipulidae),4S3— голова самца долгоножки Prionocera turcica F. (Tipulidae), 484 — то же
Р. lapponica Tjed., 485— характерная поза малярийного комара (Culicidae), 486 — то же нема-
лярийного комара, 487 — крыло комара Culex pipiens L. (Culicidae), 488 — тоже толстоножки
Dilophus Mg. (Bibionidae), 489 — то же мокреца (Ceratopogonidae), 490 — то же мошки (Simuliidae),
491 — общий вид мошки.
плохо. Крылья в покое держат кровлеобразно или плоско на
брюшке (одно крыло налегает на другое). Хоботок короткий
и толстый.........................Б.абочницы — Psychodidae.
8(7). Насекомые средних размеров. Густого опушения нет. Крылья
по жилкам и ноги покрыты мелкими чешуйками. Хоботок длин-
ный и тонкий, длиннее головы и груди вместе взятых (рис. 481).
Самки, как правило, кровососы. Сидящие на субстрате маля-
рийные комары держат тело под острым углом к субстрату
(рис. 485), а у немалярийных комаров тело почти параллельно
субстрату (рис. 486). . . Настоящие комары — Culicidae.
9(6). Ветвей жилок, упирающихся в край крыла, менее 9 (рис. 488).
196
10(11). Костальная жилка обходит все крыло; на заднем крае крыла
она тоньше, чем на переднем. На крыле имеются только про-
дольные неветвящиеся жилки. Мелкие насекомые с длинными
усиками и стройными ногами. . . Галлицы—Cecidomyiidae.
11(10). Костальная жилка развита только по переднему краю крыла,
доходя лишь до его вершины.
12(13). Тело стройное, брюшко длинное, крылья далеко не достигают
его вершины. Ноги длинные, тонкие. Ротовые органы редуци-
рованы. Усики самцов длинноперистые. Звонцы — Chironomidae.
13(12). Тело коренастое, крылья заходят за вершину брюшка. Ноги
сравнительно короткие и толстые.
Рис. 492—498. Короткоусые двукрылые и детали их строения.
492 — кровососка Hippobosca equina L. (Hippoboscidae), 493 — крыло львинки (Stratiomyidae), 494 — то
же слепня (Tabanidae), 495 — златоглазик Chrysops relictus Mg. из того же семейства слепней,
496 — усик древесницы (Xylophagidae), 497 — бекасница Rhagio scolopaceus L. (Rhagionidae), 498—
ктырь'тонкобрюх Leptogaster cylindrlca DeGeer (Asilidae). Си, M — жилки крыла.
14(15). Насекомые средних размеров (5—10 мм). Бедра утолщенные.
На голове имеются простые глазки. Толстоножки—Bibionidae.
15(14). Насекомые мелкие (1—4 мм). Бедра не утолщены.
16(17). Кубитальная жилка образует развилку примерно в середине
крыла, т. е. имеется стебелек (рис. 489). . .....................
..............................Мокрецы — Ceratopogonidae.
17(16). Кубитальная жилка без стебелька (рис. 490). Брюшко широ-
кое и короткое (рис. 491)................. Мошки — Simuliidae.
18(1). Усики короткие, всегда состоят из 3 члеников (рис. 451). (Под-
отряд Короткоусые двукрылые — Brachycera).
19(20). Тело сильно уплощено, покровы кожистой или роговой консис-
тенции. Крылья плоско складываются на спинке (рис. 492)
197
или отсутствуют. Голова расположена в выемке переднего края
переднеспинки. Тазики, особенно средние, широко расставле-
ны. Кровососущие паразиты млекопитающих и птиц. . . .
...........................Кровососки — Hippoboscidae.
20(19). Тело более или менее выпуклое. Крылья всегда имеются.
21(22). Дискоидальная ячейка маленькая (рис. 493). Жилки собраны
у переднего края крыла. Брюшко, как правило, широкое, пя-
тиугольное. ............................Львинки — Stratiomyidae.
22(21). Дискоидальная ячейка крупная. Жилки не собраны у перед-
него края крыла.
23(30). Задних ячеек крыла (т. е. ячеек, ограниченных медиальной
и кубитальной жилками и открывающихся на заднем крае кры-
ла) 4—5 (рис. 494).
24(25). Тело плотное с широким сплюснутым брюшком; брюшко в сред-
ней части не уже груди (рис. 495). Глаза у живых и свежераз-
моченных экземпляров радужно переливаются, яркие. Хоботок
сильно склеротизованный.....................Слепни — Tabanidae.
25(24). Тело стройное, брюшко не сплюснуто, по крайней мере в сред-
ней части уже груди.
Рис. 499—503. Короткоусые двукрылые и детали их строения.
499 — общий вид большеголовки (Conopidae), 500 — зеленушка Dolichopus Latr. (Dolichopodidae),
501 — крыло журчалки (Syrphidae), 502 — журчалка Sphaerophoria St. Farg. et Serv. (Syrphidae).
503— пестрокрылка Urophora R.—D. (Tephritidae).
198
26(27), Третий членик усиков разделен на 8 колец (рис. 496). Мухи
с удлиненным цилиндрическим телом, черные, по облику напо-
минают мелких рогохвостов . . Древесницы — Xylophagidae.
27 (26). Усики иного строения.
28(29), Крылья длиннее брюшка, заходят за его вершину. Голова
обычно голая. Средних размеров мухи с длинным коническим
брюшком (рис. 497)......................Бекасницы — Rhagionidae.
29(28). Крылья не длиннее брюшка. На голове часто имеются длинные
волоски. Крупные или средних размеров мухи с длинным, но
не коническим брюшком (рис. 498). . . Ктыри — Asilidae.
30(23). Задних ячеек крыла не более 3.
31(32). Голова большая, пузыревидная (см. сбоку!). Глаза сильно сдви-
нуты вверх, широко расставлены и выступают по бокам го-
ловы. Конец брюшка обычно подогнут на вентральную сторо-
ну (рис. 499)......................, » Большеголовки — Conopidae.
32(31). Признаки иные.
33(34). Мелкие и средней величины (до 6 мм) мухи с удлиненно-ко-
ническим или сплюснутым с боков брюшком (рис. 500). Окрас-
ка металлически-зеленая. . . Зеленушки — Dolichopodidae.
34(33). Признаки иные. Если окраска металлически-зеленая, то
• брюшко не коническое, более или менее широкое и сплюснуто
в дорсовентральном направлении.
35(36). Крылья прозрачные с круглой темной точкой у вершины. Мел-
кие стройные мухи с круглой головой...............................
, . .............................Муравьевидки — Sepsidae.
36(35). Крылья с иным рисунком или бесцветные,
37(38). Поперечных жилок на крыле нет. Жилки на переднем крае
крыла утолщены, на остальной части едва намечены. Тело
бархатисто-черное.........................Горбатки — Phoridae.
38(37). На крыле есть поперечные жилки. На задней части крыла
жилки нормально развиты.
39(40). Жилкование крыла включает сверхкомплектную жилку
(рис. 501). Брюшко часто с желтыми пятнами (рис. 502).
.............................................Журчалки — Syrphidae.
40(39), Сверхкомплектная жилка отсутствует.
41(42). Ротовые части недоразвиты, хоботок отсутствует. Крупные му-
хи (9—16 мм) с телом, покрытым густыми волосками; реже
без густогб покрова из волосков. . , . Оводы — Oestridae.
42(41). Ротовые части вполне развитые, имеется мягкий втяжной или
твердый не складывающийся хоботок. Тело голое, покрытое
отдельными щетинками или густо опушенное.
43(46). Крылья пестрые, т. е. с хорошо различймыми интенсивными
пигментными пятнами.
44(45). Тело опушенное. Окраска однотонная, темная. Между глаза-
ми расположены густые очень короткие щетинки....................
................................. Жужжалы — Bombyliidae.
45(44). Тело не опушенное (рис. 503), длина тела не более 6 мм.
Встречаются на лугах, лесных полянах; приурочены главным
образом к различным сложноцветным . . '...........................
..................................Пестрокрылки — Tephritidae.
46(43), Крылья прозрачные или равномерно затемненные, без четких
пигментных пятен. (Могут лишь быть затемнены поперечные
жилки).
47(48). Тело покрыто очень густыми длинными волосками, которые
придают ему шаровидную форму. Между глазами располо-
жены густые очень короткие щетинки. Хоботок часто превы-
199
шает длину головы с грудью. Длина волосков равна прибли-
зительно половине ширины брюшка. Жужжалы — Bombyliidae.
48(47). Волоски, покрывающие тело, короткие или тело не опушено.
49(50). Тело ржаво-желтого цвета, иногда брюшко опушено коротки-
ми желтыми волосками (рис. 504). Крылья желтоватые, часто
с темными пятнами на поперечных жилках. Размеры средние
(7—10 мм)......................Навозные мухи — Scatophagidae.
50(49). Окраска тела иная. Если основной фон желтоватый, то всегда
имеется четкий темный рисунок.
51(56). Хоботок не складывающийся, твердый. Грудь сильно развита,
шире и несколько выше основания брюшка.
52(55). Хоботок направлен вперед (рис. 505). Голова большая, не уже
груди.
53(54). Лоб и темя расположены в углублении между глазами
(рис. 506).......................................Ктыри — Asilidae.
кы
Рис. 504—516. Короткоусые двукрылые и детали их строения.
504 — навозная муха Scatophaga stercoraria L. (Scatophagidae), 505 — общий вид ктыря (Asilidae),
506— 1олова ктыря, вид спереди, 507 — то же лжектыря (Therevidae), 508 — общий вид толкун-
чика (Empididae), 509 — толкунчик Erupts L. с добычей, 510— грудной отдел настоящей мухи
(Muscidae), вид сверху, 511— грудной отдел тахины (Tachinidae), вид сбоку, 512 — то же мухи
из семейства Calliphoridae, 513 — крыло цветочницы (Anthomyiidae), 514— грудной отдел мухи из
семейства Opomyzidae, 515 — крыло злаковой мушки (Chloropidae), 516 — то же минирующей мушки
(Agromyzidae). г.щ — гипоплевральиые щетинки; пс — постскутеллум.
200
54(53). Лоб и темя расположены на одном уровне с глазами
(рис. 507)..........................Лжектыри — Therevidae.
55(52). Хоботок направлен вниз (рис. 508). Голова маленькая, уже ,
груди (рис, 509)....................* . Толкунчики — Empididae. .
56(51). Хоботок складывающийся, более мягкий. Облик типично «му-
шиный» (рис. 451). На теле всегда различимы крепкие длин-
ные щетинки или группы жестких волосков.
57(66). Поперечный шов среднеспинки цельный, закрыловые бугорки
обособлены (рис. 510). Тело, как правило, больше 5 мм.
58(59). Постскутеллум хорошо развит, валикообразный (рис. 511; пс).
Тело темное в многочисленных крепких щетинках,’ брюшко
иногда с красноватым рисунком . . Тахины — Tachinidae.
59(58). Постскутеллум отсутствует или слабо развит, не валикооб-
разный (рис. 512).
60(61). Брюшко металлически-синее или зеленое . . Calliphoridae.
61(60). Окраска брюшка без металлического блеска.
62(63). Гипоплевральные щетинки над тазиками задних ног имеются
(рис. 511, 512, г.щ). Брюшко серое, со светлыми пятнами,
обычно образующими шахматный рисунок . . Sarcophagidae.
63(62). Гипоплевральные щетинки отсутствуют.
64(65). Анальная жилка, хотя бы в виде складки, доходит до заднего
края крыла (рис. 513) . . . Цветочницы — Anthomyiidae.
65(64). Анальная жилка не доходит до заднего края крыла ....
.............................Настоящие мухи — Muscidae.
66(57). Поперечный шов среднеспинки прерван посередине, закрыло-
вые бугорки не обособлены (рис. 514). Размеры мелкие
(меньше 6 мм).
67(68).. Костальная жилка на всем протяжении цельная. Тело часто
с металлическим блеском......................Lauxaniidae.^
68(67). Костальная жилка с перерывом или перетяжкой (рис. 515,
516).
69(70). Анальная жилка отсутствует (рис. 515). Спинка с контрастны-
ми темными продольными полосками................................
........................ . Злаковые мушки — Chloropidae.
70(69). Анальная жилка имеется (рис. 516). Тело серое, черное или
коричневое, иногда с желтым рисунком............................
...... .... Минирующие мушки — Agromyzidae.
КЛАСС ПАУКООБРАЗНЫЕ —ARACHNIDAE*
Общая характеристика
К этому классу относятся членистоногие с воздушным дыханием
и шестью парами конечностей. Усики отсутствуют. Глаза простые,
у мелких форм могут частично или полностью редуцироваться. Чаще
всего тело подразделено на 2 отдела: головогрудь, несущую конечно-
сти, и брюшко, лишенное конечностей. Сегменты головогруди слиты,
но у сольпуг (Solifugae) 2 задних грудных сегмента обособлены. Брюш-
ко у _ сольпуг, скорпионов (Scorpiones), ложноскорпионов (Pseudo-
scorpiones) и сенокосцев (Opiliones) сегментированное, а у большин-
ства пауков (Aranei) и клещей (Acarina) слитное, без каких-либо сле-
дов сегментации.
Первая пара конечностей головогруди называется хелицерами; они
состоят из 2—3 члеников и имеют вид клешней, крючков или стилетов.
* В. П. Тыщенко.
14 208
201
Вторая пара конечностей — педипальпы; они обычно состоят из 5—6
члеников. Педипальпы не участвуют в передвижениях животного; на
них располагаются разнообразные рецепторы. У скорпионов и ложно-
скорпионов они превращены в клешнеобразные органы, служащие для
захвата и удерживания добычи. Остальные 4 пары конечностей состоят
из 6—7 члеников; они играют роль ходильных (бегательных) ног. По-
следние членики всех ног заканчиваются маленькими коготками. Пост-
эмбриональное развитие паукообразных осуществляется по типу эпи-
морфоза, но у клещей наблюдается своеобразный анаморфоз (см.
с. 49).
В составе класса паукообразных насчитывается около 40 000 ви-
дов, заселяющих различные наземные биотопы. Только некоторые кле-
щи и паук-серебрянка (Argyroneta aquatica Cl.) вторично перешли
к жизни в водной среде. Многие паукообразные тесно связаны с поч-
вой или являются постоянными почвообитающими животными. Богатая
фауна пауков и клещей свойственна травянистым растениям, кустар-
никам и деревьям. Среди паукообразных немало синантропов, а некото-
рые клещи относятся к кровососам и паразитам позвоночных живот-
ных. Преобладающий тип питания паукообразных — хищничество.
Только в отряде клещей помимо хищников и паразитов есть типичные
сапрофаги и фитофаги.
Современных представителей паукообразных подразделяют на 10
отрядов. Фауна СССР насчитывает 7 отрядов. По всей европейской
части СССР широко распространены ложноскорпионы, сенокосцы, па-
уки и клещи. На юге нашей страны встречаются еще 2 отряда —
скорпионы и сольпуги. Отряд телифонов (Uropygi) на территории
СССР представлен одним редким видом, обитающим в подстилке хвой-
. но-широколиственных лесов Уссурийского края.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ОТРЯДОВ "
1(2). Педипальпы имеют вид клешней (рис. 517). Брюшко сегменти-
рованное. Длина тела не превышает 5 мм...........................
................ Ложноскорпионы — Pseudoscorpiones (с. 203).
2(1). Педипальпы не клешнеобразные.
3(4). Брюшко сидячее, сегментированное (рис. 518). Ноги длинные
и тонкие ....................Сенокосцы — Opiliones (с. 204).
4(3). Брюшко иесегментированное.
5(6). Тело подразделено на головогрудь и брюшко, соединенные уз-
Рис. 517—520. Отряды паукообразных.
517— ложпоскорпион (Pseudoscorpiones). 518 — сенокосец (Opiliones), 519— паук (Aranei), 520
клещ (Acarina).
202
ким стебельком (рис. 519). На конце брюшка имеются паутин-
ные бородавки...........................Пауки — Aranei (с. 206).
6(5). Тело цельное, стебелек отсутствует (рис. 520). Паутинных боро-
давок нет...............................Клещи — Acarina (с. 221).
ОТРЯД ЛОЖНОСКОРПИОНЫ - PSEUDOSCORPIONES
Общая характеристика
К ложноскорпионам относятся мелкие (обычно 2—5 мм) пауко-
образные с клешневидными педипальпами, которые придают им неко-.
торое внешнее сходство со скорпионами (рис. 521, 522). Хелицеры дву-
членистые; на их кончиках открываются протоки паутинных желез,
расположенных в головогруди. Клешня педипальпы имеет 2 пальца —
подвижный и неподвижный. На кончике неподвижного пальца откры-
Рис. 521—522. Ложноскорпионы.
521— Neobisiidae, 522 — Chernetidae.
вается проток ядовитой железы, а в некоторых семействах ядовитые
железы найдены в обоих пальцах клешни. Ноги состоят из 6 члени-
ков, иногда последний членик (лапка) подразделен на 2 вторичных чле-
ника. Головогрудь слитная; у большинства ложноскорпионов на пе-
реднем участке головогруди расположены 2 или 4 глаза. У предста-
вителей семейства Chernetidae и у пещерных видов других семейств
глаза отсутствуют. Брюшко широкое, сегментированное; с дорсальной
стороны оно несет 11 одиночных или парных склеротизованных пласти-
нок, соответствующих отдельным брюшным сегментам. Последний
(12-й) сегмент редуцированный, обычно имеет вид небольшого аналь-
ного конуса. Органы дыхания представлены трахеями. Окраска ложно-
скорпионов однотонная, от желтой до темно-коричневой.
Представители этого отряда широко распространены в природе,
I но мало известны из-за мелких размеров и скрытного образа жизни.
Они живут в подстилке, в сухой листве, в мусоре, под камнями, во мху,
под корой и в дуплах деревьев, в гнездах птиц и в муравейниках.
Встречаются в помещениях для скота и в норах грызунов. Некоторые
виды принадлежат к факультативным синантропам: они живут в книж-
ных шкафах, среди бумаг и книг, в коллекциях и гербариях. Иногда
во время экскурсий удается поймать муху, пчелу или другое крыла-
тое насекомое, к телу которого прицепился ложноскорпион, удержи-
ваясь на нем с помощью клешней педипальп. Это явление, получившее
203
название форезии, представляет собой частный случай комменсализма
и, по-видимому, служит средством расселения ложноскорпионов.
Все ложноскорпионы питаются мелкими членистоногими. Добыча
захватывается педипальпами и парализуется выделениями ядовитых
желез. Ложноскорпионы раздельнополы; их размножению предшест-
вует наружно-внутреннее осеменение. Самцы откладывают на суб-
страт сперматофор, представляющий собой каплю спермы на длинном
стебельке. Затем самец захватывает самку педипальпами и протаски-
вает ее над сперматофором, который попадает в половое отверстие
.самки. Вскоре после встречи с самцом самка сооружает гнездо из па-
утины, выделяемой хелицерами, и начинает откладывать яйца. Через
несколько дней из яиц выходят молодые нимфы, которые первое вре-
мя держатся в гнезде, а потом оставляют его и приступают к самосто-
ятельному существованию. В периоды линек и зимовки нимфы тоже
сооружают паутинные гнезда. Продолжительность развития у боль-
шинства ложноскорпионов — 1 год.
В настоящее время описано примерно 1300 видов этого отряда.
Миниатюрные размеры и большое внешнее сходство видов, относящих-
ся к разным семействам и родам, сильно затрудняют определение лож-
носкорпионов. В европейской части СССР особенно часто встречаются
4 семейства.
Определительная таблица семейств
1(4). Хелицеры крупные. Склеротизованные пластинки на дорсальной
поверхности брюшка цельные, не разделенные продольной лож-
бинкой (рис. 521). На головогруди 2 пары глаз или же (у пе-
щерных форм) глаза отсутствуют.
2(3). Головогрудь впереди более широкая, чем сзади. Хелицеры почти
достигают размеров головогруди. Живут под камнями и в лесной
подстилке..............................................Chthoniidae.
3(2). Головогрудь впереди имеет такую же ширину, как сзади. Хе-
лицеры заметно короче головогруди (рис. 521). Живут под кам-
нями, в подстилке и во мху.............................Neobisiidae.
4(1). Хелицеры сильно уменьшенные. Каждая склеротизованная пла-
стинка на дорсальной поверхности брюшка разделена продоль-
ной ложбинкой на 2 половинки — правую и левую (рис. 522).
5(6). На головогруди 1—2 пары глаз. Живут под камнями, в подстил-
ке, под корой деревьев, в домах и в гнездах птиц....................
............................................. . Cheliferidae.
6(5). Глаза отсутствуют (рис. 522). Живут во мху, под корой деревь-
ев и в муравейниках....................................Chernetidae.
ОТРЯД СЕНОКОСЦЫ —OPILIONES
Общая характеристика
Средних размеров паукообразные с длинными тонкими ногами
и сегментированным брюшком, которое всем своим широким основа-
нием причленяется к головогруди (рис. 523). Глаз 1 пара; иногда
(у пещерных видов) глаза могут отсутствовать. Обычно глаза распо-
ложены на приподнятых бугорках головогруди (рис. 523, г.б). Хелице-
ры состоят из двух члеников (рис. 523, х). Базальный членик простой;
апикальный членик снабжен клешней, приспособленной для захвата
и умерщвления добычи. У. представителей семейства Ischyropsalidae
очень крупные хелицеры являются специальным приспособлением для
питания наземными моллюсками, находящимися в раковинах (рис. 524).
204
Рис. 523—527. Сенокосцы.
523 — строение тела сенокосца из семейства Phalangiidae, вид сбоку (три передние иоги удалены),
524 — сенокосец из семейства Ischyropsalidae, внд сверху (ноги изображены не полностью, 525 —
526— головогрудь и брюшко Phalanglum opilio L., вид сверху (525—самец, 526—самка), 527 —
общий вид сенокосца из семейства Trogulidae. в — выступ головогруди; в.б — вентральная сто-
рона брюшка; г — головогрудь; г.б — глазной бугорок; д.б — дорсальная сторона брюшка; н —
нога; п — педипальпа; х — хелицера.
Педипальпы 5-члениковые (рис. 523, п). Причленяющиеся к голово-
груди членики педипальп снабжены лопастями, прикрывающими вход
в ротовую полость. Ноги имеют типичный для паукообразных набор
члеников (рис. 523, н). В большинстве случаев лапка (последний чле-
ник ноги) с многочисленными вторичными члениками. Брюшко имеет
от 6 до 10 тергитов, которые, однако, не всегда полностью отграниче-
ны друг от друга (рис. 523, д.б). Самки отличаются от самцов присут-
ствием трубчатого яйцеклада, отходящего от основания брюшка. Ор-
ганы дыхания — трахеи.
Отряд сенокосцев охватывает около 3000 видов, распространенных
во всех странах света (в СССР около 120 видов). Представители от-
ряда— преимущественно хищники; питаются другими паукообразны-
ми, насекомыми, червями, многоножками или наземными моллюсками.
Встречаются на стволах деревьев, среди растений, на стенах зданий,
на заборах, на земле, под стволами деревьев, под камнями и в под-
стилке в лесах, садах и парках, а в горных районах — на скалах, в рос-
205
сыпях камней и в пещерах. Нередки на паровых землях и среди кар-
тофельных кустов. Для большинства сенокосцев характерна способ-
ность к автотомии конечностей.
Сенокосцы — раздельнополые животные; их размножению предше-
ствует настоящее внутреннее осеменение путем копуляции. Оплодотво-
ренная самка откладывает яйца в почву. У большинства видов полный
цикл развития занимает 1—2 года, взрослые особи могут жить до
3 лет.
Фауна сенокосцев достигает наибольшего разнообразия в лесной
зоне. Особенно богато заселены ими лиственные и смешанные леса.
В европейской части СССР встречаются более 60 видов и 8 семейств,
в том числе наиболее многочисленное и широко распространенное се-
мейство Phalangiidae. Самый массовый вид, относящийся к этому се-
мейству,— Phalangium opilio L. На дорсальной стороне тела у Р. opi-
lio имеется темно-коричневая полоса с характерным расширением в ее
средней части (рис. 525, 526). Вентральная поверхность брюшка светло-
желтая или белая. Длина тела у взрослых особей составляет 5—7 мм.
Определительная таблица семейств
1(2). Последний членик педипальпы длиннее ее предпоследнего чле-
ника (рис. 523, п). Живут в траве, на поверхности земли, на
стенах домов и на стволах деревьев, встречаются также в почве
и в подстилке. Нападают на мелких членистоногих, но могут пи-
таться и на трупах животных или потреблять растительную
пищу............................................... Phalangiidae.
2(1). Последний членик педипальпы короче ее предпоследнего чле-
ника (рис. 524).
3(4). Вторая пара ног едва длиннее тела. Передний край головогруди
с двумя крючковидно изогнутыми выступами (рис. 527, в). Тело
уплощенное, часто покрытое приставшими к нему частицами
почвы. Живут в подстилке, во мху, в земле и под камнями.
Встречаются редко.................................... Trogulidae.
4(3). Вторая пара ног значительно (в 3 раза и больше) превышает
длину тела. Передний край головогруди без крючковидных вы-
ступов.
5(6). Хелицеры очень длинные и толстые, направлены вперед и закан-
чиваются крупными клешнями (рис. 524). Живут во влажных
местах, в подстилке и под камнями, питаются наземными улит-
ками и слизнями. Встречаются очень редко . . Ischyropsalidae.
6(5). Хелицеры относительно короткие, не выступающие вперед (как
на рис. 523). Живут в почве и подстилке, питаются мелкими на-
секомыми и клещами................................Nemastomatidae.
ОТРЯД ПАУКИ —ARANEI
Общая характеристика
Тело пауков состоит из двух отделов — головогруди и брюшка, ко-
торые соединяются узким стебельком (рис. 528). Головогрудь при-
крывается сверху одним сплошным щитом (рис. 528, г.в ее перед-
ней части расположено 6—8 глаз. Расположение и относительные раз-
меры глаз используются в качестве одного из самых важных диагно-
стических признаков, характеризующих отдельные семейства пауков.
У некоторых пещерных видов глаза могут отсутствовать. Стерниты
головогруди сливаются в единый стернальный щит, занимающий всю
нижнюю часть головогруди между базальными члениками ног. Стер-
206
нит сегмента педипальп остается свободным и образует нижнюю губу.
Хелицеры двучлениковые (рис. 528, х); второй членик превращен в ост-
рый коготок, связанный с ядовитой железой. Педипальпы короткие
(рис. 528, п); у половозрелых самцов они преобразованы в копулятив-
ный аппарат. Ноги сохраняют типичное для паукообразных строение
и состоят из 6 члеников (рис. 528, н). Лапки всех ног не имеют вто-
ричных члеников, за исключением пауков семейства Pholcidae, снаб-
женных особенно длинными ногами. Брюшко у европейских видов
несегментированное. Органами дыхания у большинства пауков служат
легочные мешки и трахеи.
Все пауки снабжены паутинными железами, которые открываются
на нижней стороне брюшка, ближе к его концу через паутинные бо-
родавки (рис. 528, п.б). У пауков европейской фауны, как правило,
имеются 3 пары паутинных бородавок. Паутина используется пауками
для постройки ловчих сетей, жилых трубок, гнезд, яйцевых коконов,
сеточек для сбора спермы самцами, иногда и для миграционных пере-
летов.
Рис. 528. Строение тела паука, вид
сбоку (три передние пары ног удале-
ны).
в.б — вентральная сторона брюшка: г —глаза; г.щ —
головогрудной щит; д.б—дорсальная сторона
брюшка; «—нога; н.г — нижняя губа; п— педи-
пальпа; /1.6--паутинные бородавки; с — стебелек;
х — хелицера.
Пауки приносят пользу, уничтожая вредных насекомых. Яд подав-
ляющего большинства видов безвреден для человека. Среди европей-
ских пауков только укус каракурта (Latrodectus tredecimguttatus Ros.)
может вызвать тяжелое отравление, а иногда бывает и смертельно
опасным. Каракурт заселяет сухие степные и йустынные районы, а в ев-
ропейской части СССР северная граница его распространения прибли-
зительно совпадает с северной границей степной зоны. Гораздо менее
токсичным для человека является укус тарантула (Lycosa singorien-
sis Laxm.), распространенного в пустынях, степях и в лесостепной
зоне.
Всего известно около 28 000 видов пауков. В СССР зарегистриро-
вано 1500 видов, относящихся к 36 семействам. В европейской части
СССР встречаются 30 семейств. Среди них можно выделить 5 биоло-
гических групп, различающихся по жизненным формам и способам лов-
ли добычи.
Бродячие пауки (рис. 529—536) охватывают семейства Sal-
ticidae, Oxyopidae, Pisauridae, значительную часть семейства Lycosi-
dae и отдельных представителей других семейств. Пауки этой группы
отличаются высокой подвижностью, не строят ловчих сетей и постоян-
ных убежищ, активно преследуют свою добычу в беге или прыжках.
Как правило, они ведут дневной образ жизни, имеют хорошо разви-
тые органы зрения и темную окраску тела (рис. 529).
Бродячие пауки-волки (Lycosidae) охотятся на поверхности поч-
вы. К ним относятся многочисленные виды рода Pardosa С. L. Koch
(рис. 537) и некоторые представители других родов, встречающихся
на всей территории европейской части СССР. В период размножения
самки этих пауков вынашивают яйцевые коконы, прикрепленные к па-
утинным бородавкам (рис. 530). После вылупления из яиц молодые
/
207
Рис. 529—536. Бродячие пауки.
529— паук из семейства Lycosidae в позе охоты, 530— самка паука из семейства Lycosidae с ко-
коном, 531— Dysdera Latr. (Dysderidae), 532 — Oxyopes Latr. (Oxyopidae), 533 — Alopecosa
Sim. (Lycosidae), 534 — Pisaura mirabilis Cl. (Pisauridae), 535—Heliophanus C. L. Koch (Salticidae),
536 — Myrmarachne Mac Leay (Salticidae).
паучки взбираются на брюшко самки и живут здесь некоторое время
под ее охраной.
Представители небольшого семейства Pisauridae, близкого к Lyco-
sidae, охотятся как на земле, так и на травянистых растениях и низ-
ких кустарниках. Широко распространенный в СССР крупный паук
Dolomedes fimbriatus Cl., снабженный двумя продольными светлыми
полосами на головогруди и брюшке, ведет амфибиотический образ жиз-
ни и может бегать по поверхности воды (рис. 538). Другой вид этого
семейства — Pisaura mirabilis CL, имеющий заостренное на конце брюш-
ко (рис. 534), встречается только на юге и заселяет остепненные су-
хие биотопы.
У видов семейства Oxyqpidae брюшко тоже заостренное, но тело
более компактное (рис. 532), ноги усажены длинными многочисленными
208
шипами. Пауки охотятся на растениях, могут совершать небольшие
прыжки. К этому семейству относится 1 европейский род Oxyopes
Latr., распространенный на юге в степях и других открытых сухих
биотопах.
К обширному семейству скакунчиков (Salticidae) относятся оби-
татели растений, стволов деревьев, стен и заборов, камней, скал и по-
верхности почвы. У большинства видов этого семейства тело компакт-
ное, а ноги короткие и относительно толстые (рис. 535). На переднем
крае головогруди находятся крупные глаза. Пауки передвигаются ко-
роткими перебежками и быстрыми прыжками. Заметив добычу на рас-
стоянии 10—15 см, паук осторожно подкрадывается и стремительно
прыгает на нее. В период размножения самки переходят к оседлому
Рис. 537. Головогрудь и брюш-
ко бродячего паука-волка Par-
dosa С. L. Koch (Lycosidae).
Рис. 538. Dolomedes fimbriatus Cl.
(Pisauridae) на поверхности воды.
образу жизни и изготавливают паутинные гнезда, в которых происхо-
дит откладка яиц. В гнезде помещаются один или несколько коконов
и самка, охраняющая яйца. К типичным и часто встречающимся пред-
ставителям скакунчиков относятся виды рода Evarcha Sim. (рис. 539).
Некоторые скакунчики обитают вблизи муравейников, а по форме
и расцветке тела великолепно имитируют муравьев (рис. 536).
Пауки-норники составляют обособленную биологическую
группу, которая в европейской части СССР представлена некоторыми
видами семейства Lycosidae. Пауки этой группы живут в изготовлен-
ных ими земляных норках и охотятся ночью, ориентируясь за счет не-
обычайно развитой тактильной чувствительности. Во время охоты паук
сидит у входа в норку и ловит пробегающую мимо добычу (рис. 540).
К наиболее известным норникам принадлежит самый крупный паук
нашей фауны — русский тарантул (Lycosa singoriensis Laxm.). В пе-
риод размножения самка держится в норе вместе с коконом, а после
вылупления паучков она покидает нору и отправляется странствовать
вместе с многочисленным потомством, размещенном на ее брюшке и го-
209
ловогруди. Во время продолжительных странствований самка посте-
пенно расселяет паучков.
Пауки-засадники (рис. 541—544) охватывают семейство
Thomisidae, а также часть семейств Clubionidae и Gnaphosidae. Они не
строят ловчих сетей, а подстерегают добычу на цветках, листьях, коре
деревьев, скалах и почве. Пауки этой группы могут иметь различную
расцветку и форму тела, обычно хорошо адаптированную к специфи-
ческим условиям их обитания.
Бокоходы (Thomisidae) являются типичными засадниками. Во вре-
мя охоты они выжидают добычу в состоянии почти полной неподвиж-
ности, широко раздвинув передние («хватательные») ноги' (рис. 545).
Рис. 539. Головогрудь и брюш-
ко скакунчика Evarcha Sim.
(Salticidae).
Рис. 540. Русский тарантул
Lycosa singoriensis Laxm. у
входа норы в ожидании до-
бычи (по Мариковскому).
Нередко бокоходы обладают великолепной покровительственной окрас-
кой, способствующей маскировке паука во время охоты. У цветочных
бокоходов покровительственная окраска особенно эффективна, потому
что в зависимости от цвета лепестков один и тот же паук может ме-
нять свою окраску — от белой и желтой до розовой и бледно-зеленой.
К самым обычным цветочным паукам европейской фауны принадле-
жат Mistimena vatia Cl. с округлым брюшком и Thomisus onustus
Walck., имеющий 2 угловидных выступа на заднем крае брюшка
(рис. 545). Основными объектами нападения для этих пауков служат
пчелы и другие насекомые, посещающие цветки. В травянистой расти-
тельности, в кронах деревьев и на почве обитают многочисленные
виды рода Xysticus С. L. Koch, имеющие темный рисунок на брюшке
и неспособные к изменениям окраски (рис. 543, 546). У охотящихся на
почве и в подстилке пауков рода Oxyptila Sim. тело покрыто булаво-
видными щетинками, которые удерживают частицы почвы и пыли, пре-
красно маскирующие паука. Виды рода Tibelltis Sim. имеют вытянутое
в длину тело (рис. 541), охотятся на стеблях травянистых растений
или на веточках деревьев и кустарников. Для большинства видов рода
Philodromtis Walck. характерно уплощенное и укороченное тело
210
(рис. 542) с расцветкой, имитирующей поверхность скал, крупных кам-
ней или коры деревьев.
Пауки, относящиеся к семейству Clubionidae, как правило, окра-
шены в светлые тона — зеленые, желтоватые, желто-зеленые или свет-
ло-коричневые. Широко распространенные в Европе виды рода Clubio-
па Latr. (рис. 544) живут на травянистых растениях, под корой и в ли-
стве деревьев или кустарников. Пауки охотятся в ночное время, а день
проводят в убежищах, которые изготавливаются из скрученных и за-
крепленных паутиной листьев.
Засадники из семейства Gnaphosidae имеют одноцветную темную
окраску (коричневую или черную), живут под камнями, в подстилке,
во мху, под корой деревьев и пней. Многие представители этого семей-
ства могут изготавливать убежища из паутины, но в отличие от видов
рода Clubiona никогда не скручивают листья растений.
Рис. 541—544. Пауки-засадники.
541 — TibellusS\m.. (Thomisidae), 542 — Phllodromus Walck. (Thomisidae), 543 — Xysticus C. L. Koch
(Thomisidae), 544 — Clubio Latr. (Clubionidae).
Пауки-тенетники (рис. 547—551) представлены следующими
семействами: Linyphiidae, Theridiidae, Agelenidae, Dictynidae и Phol-
cidae. Все они строят довольно плотные ловчие сети («тенета»), со-
ставленные из сухих и клейких паутинных нитей. У большинства пау-
ков этой биологической группы брюшко удлиненно-овальное или ок-
руглое и явственно приподнятое над головогрудью (рис. 549).
Мелкие науки, относящиеся к семейству Dictynidae, строят непра-
вильные ловчие сети из редких переплетающихся нитей'на стенах и за-
борах, на ветках и стволах деревьев или на отдельных широких листь-
ях. В европейской части СССР особенно часто встречаются виды рода
Dictyna Sund. (рис. 547).
Пауки семейства Linyphiidae располагают свои сети в углубле-
ниях почвы, в траве, на ветках деревьев и кустарников. Обычно сеть
состоит из куполообразного или горизонтального полога, который снизу
211
и сверху поддерживается вертикальными паутинными нитями (рис.
552). Во время охоты паук сидит под пологом брюшной стороной квер-
ху. Пролетающее над сетью насекомое задерживается верхними вер-
тикальными нитями и падает на поверхность полога, вызывая его со-
трясение. Паук подбегает к месту падения добычи и, прокусывая по-
лог, захватывает ее снизу коготками хелицер. Самые заметные и наи-
более распространенные представители этого семейства относятся к роду
Linyphia Latr. (рис. 553). Мелкие виды семейства Linyphiidae живут
в лесной подстилке, на почве и во мху.
Европейские представители обширного семейства Theridiidae имеют
2 типа ловчих сетей. Неправильная сеть, которая встречается в основ-
ном у широко распространенного рода Theridium Walck. (рис. 549),
Рис. 545. Цветочный паук Thomisus
onustus Walck. в позе охоты на
цветке.
Рис. 546. Головогрудь и брюш-
ко бокохода Xysticus
С. L. Koch (Thomisidae).
представлена перекрещивающимися паутинными нитями, идущими
в разных направлениях. В упорядоченной сети, характерной, напри-
мер, для рода Steatoda Sund., клейкие ловчие нити направлены к по-
верхности почвы, а ее полог образован сухими паутинными нитями
(рис. 554). Сеть этого типа приспособлена для ловли нелетающих чле-
нистоногих: пробегая по поверхности почвы, они приклеиваются к лов-
чим нитям и становятся жертвами паука. Главными объектами охоты
для этих пауков являются муравьи. У многих видов семейства Theri-
diidae ловчая сеть одновременно служит логовищем и в период, раз-
множения в сети можно обнаружить самку с яйцевыми коконами. Од-
нако иногда в сетях устраиваются специальные логовища, замаскиро-
ванные хвоинками, кусочками почвы и мелкими песчинками. К паукам,
строящим упорядоченную сеть, относится и каракурт (Latrodectus
tredecimguttatus Ros.). Взрослая самка каракурта достигает в длину
10—12 мм, а ее тело имеет интенсивно-черную окраску; брюшко округ-
лое (рис. 555). Самец в несколько раз меньше самки, брюшко у него
продолговатое с ярко-красными или желтыми пятнами, окруженными
белыми ободками. Опасен для человека только укус самки каракурта,
а самец никакой опасности не представляет.
212
Семейство Agelenidae в биологическом плане крайне разнородно.
К нему относятся как бродячие пауки, так и тенетники, обитающие
в траве, под камнями и корой деревьев. Водяной паук — серебрянка
(Argyroneta aquatica L.) приспособился к постоянной жизни под во-
дой, где он устраивает своеобразный паутинный колокол, заполненный
воздухом. Среди тенетников этого семейства один из самых примеча-
тельных объектов — лабиринтовый паук Agelena labyrinthica Cl.
(рис. 551), встречающийся на открытых солнечных местах по всей
территории европейской части СССР, за исключением ее- северных
районов. Воронковидная ловчая сеть A. labyrinthica располагается не-
посредственно над поверхностью почвы (рис. 556). При подкараулива-
нии добычи паук сидит у входа в трубку и скрывается в ней при ма-
лейшей опасности. Сходные по форме и небольшие по размерам лов-
Рис. 547—551. Пауки-тенетники.
547 Dlctyna Sund. (Dictynidae), 548 — Lepthyphantes Menge (Linyphiidae), 549 — Therldlum
Walck. (Theridiidae), 550 — Pholcus Walck. (Pholcidae), 551 — Agelena labyrinthica Cl. (Agelenidae).
[ чие сети строят виды рода Tegenaria Latr., обитающие в домах и под-
валах.
Синантропный образ жизни типичен и для видов рода Pholcus
$ Walck., относящихся к небольшому семейству Pholcidae. Все предста-
вители этого семейства имеют исключительно длинные ноги, что при-
дает им внешнее сходство с сенокосцами (рис. 550). Пауки нередко
! встречаются в домах и других постройках в средней полосе и на юге
европейской части СССР. Они строят неправильные ловчие сети, со-
стоящие из плотных переплетений паутинных нитей. В период размно-
жения самка изготавливает единственный кокон, который она посто-
янно охраняет и держит в хелицерах.
Пауки-кругопряды, к которым относятся 2 основных семей-
ства— Araneidae и Tetragnathidae, строят геометрически правильные
колесовидные сети среди травянистой и древесно-кустарниковой рас-
213
тительности. Колесовидная ловчая сеть состоит из многоугольной рамы
и расходящихся из центра радиальных нитей (рис. 557). Центр сети
или открытый или заплетенный паутинными нитями, образующими
центральную сеточку. Радиусы сети соединяются клейкими ловчими
нитями, расположенными по спирали. Между центральной сеточкой
и ловчими нитями находится свободная зона, которая может отсут-
ствовать. Ловчая сеть такого типа является наиболее совершенным
приспособлением для охоты пауков за летающими насекомыми, так
как в ней достигается максимальная эффективная поверхность ловчей
зоны при минимальных затратах паутины.
Рис. 553. Головогрудь и брюш-
ко паука-теиетника Linyphia
Latr. (Linyphiidae).
Рис. 552. Схема ловчей сети пауков се-
мейства Linyphiidae (по Виле).
I
Рис. 554. Упорядоченная ловчая сеть паука-тенетника Steatoda Sund.
(Theridiidae).
214
Рис. 555. Самка каракурта Latrodectus tredecimgutta-
tus Rossi (Theridiidae) в позе охоты на ловчей сети.
Рис. 556. Ловчая сеть лабиринтового паука Agelena labyrin-
thica Cl. (Agelenidae).
Кругопряды семейства Tetragnathidae имеют длинные тонкие ноги
и удлиненное, почти палочковидное тело (рис. 558). Они строят лов-
чую сеть, лишенную центральной сеточки, но снабженную хорошо вы-
раженной свободной зоной (рис. 559). У наиболее обычного в евро-
пейской части СССР рода Тetragnatha Latr. сети располагаются в тра-
ве вблизи водоемов. Плоскость сети имеет наклонное или почти
215
Рис. 558—559. Паук-кругопряд семейства Tetragna-
thidae и его ловчая сеть.
558 — Tetragnatha Latr., 559—ловчая сеть.
Рис. 557. Схема строения
ловчей сети паука-круго-
пряда (по Виле).
л.з — ловчая зона; н.р — нить
рамы; р — радиусы; с.з —
свободная зона; ц. с — цент-
ральная сеточка.
561
560
Рис.
560—561. Пауки-кругопряды семейства Araneidae.
560 — Araneus diadematus Cl., 561 — головогрудь и брюшко lygtella Plck-Cambr.
216
Рис. 562—’563. Ловчие сети пауков-кругопрядов семейства
Araneidae (по Виле).
562— Zyglella x-notata Cl., 563— Araneus alsine Walck. у — убежище.
Рис. 564. Ловчая сеть паука-кругопряда Cyclosa oculata Walck (Araneidae).
с — стабнлнмеит.
15 208
217
горизонтальное положение. Нередко эти пауки раскидывают свои сети
над водой на прибрежных растениях.
У пауков семейства Araneidae ноги более толстые, тело компакт-
ное, брюшко округлое или угловатое (рис. 560, 561). В подавляющем
большинстве случаев сети этих пауков снабжены хорошо выраженной
центральной сеточкой (рис. 562, 563). У видов рода Zygiella Pick.—
Cambr. сеть имеет свободный сектор, лишенный ловчих нитей (рис. 562).
В этом секторе проходит сигнальная нить, которая соединяет убежи-
ще паука с его сетью. У европейских видов рода Zygiella дорсальная
поверхность брюшка по краям более темная, чем в середине его
(рис. 561). К наиболее известным кругопрядам относятся многочислен-
ные виды крестовиков (род Araneus CL), а самым обычным видом это-
го рода является домашний крестовик Araneus diadematus Cl. Его лег-
ко опознать по характерному крестообразному рисунку на брюшке
(рис. 560). Другие виды рода Araneus различаются по рисунку брюш-
ка; у некоторых из них брюшко вообще без рисунка или только с от-
дельными темными точками и узкими полосками на светлом фоне. Лов-
чие сети крестовиков всегда изготавливаются по единому плану
(рис. 557), но отличаются у разных видов и поэтому могут служить
такими же надежными диагностическими признаками, как и рисунок
брюшка. Домашний крестовик не строит убежище, и во время охоты
паук сидит в центре сети, а у Л. alsine Walck. и у некоторых других
видов этого рода сеть снабжена убежищем (рис. 563, у). У отдельных
видов кругопрядов семейства Araneidae (например, у Cyclosa oculata
Walck.) сеть имеет специальное приспособление — стабилимент в виде
2 зигзагообразных лент или узких полосок, направленных вверх и вниз
от центра сети (рис. 564, с). Стабилимент служит для маскировки пау-
ка, сидящего в центре.
Определительная таблица семейств
1(2). Ноги очень длинные (вторая пара ног в 4—5 раз длиннее те-
ла). Пауки похожи на сенокосцев (рис. 550). Тенетники . .
..................................................... Pholcidae.
О 565 ОООо О о О О 566 со о о О 0 О О 567
о о О О О О О О 568 «о о® ° о о ° 569 оэ I Оо со 0° иэ: СО i I
О о о о о о о О 571 О О О о О О О О 572 8 00 8 О О 573
Рис. 565—573. Типичное расположе-
ние глаз в основных семействах па-
уков.
565 — Dysderidae, 566 — Salticidae, 567 —
Lycosidae, 568 — Oxyopidae, 569— Ageleni-
dae, 570 — Araneidae, 571— Pisauridae,
572 — Clubionidae, 573 — Linyphiidae.
Рис. 574—575. Расположение
передних паутинных борода-
вок на кончике брюшка пау-
ков (вид с вентральной сто-
роны).
574 — Gnaphosidae. 575 —Clubionidae.
218
2(1). Ноги не такие длинные (вторая пара ног менее чем в 4 раза
длиннее тела).
3(4). Глаз 6 (рис. 565). Форма тела — рис. 531. Бродячие пауки
и засадники. Живут под камнями, в подстилке и под корой
деревьев.............................................Dysderidae.
4(3). Глаз 8 (рис. 566—573).
5(10). Глаза расположены в 3 или 4 ряда (рис. 566—568). Бродячие
пауки, норники и засадники.
6(7). В первом ряду глаза очень крупные, намного крупнее осталь-
ных глаз (рис. 566). Форма тела—рис. 535, 536 ...................
.....................................Скакунчики — Salticidae.
7(6). Глаза первого ряда не крупнее остальных глаз (рис.
567, 568).
8(9). В первом ряду 4 глаза (рис. 567). Форма тела — рис. 533
. . .................................. Пауки-волки — Lycosidae.
9(8). В первом ряду 2 глаза (рис. 568). Форма тела —рис. 532
......................................................Oxyopidae.
10(5). Глаза расположены в 2 ряда (рис. 569—573).
11(12). Передние паутинные бородавки у основания широко раздви-
нуты (рис. 574). Бродячие пауки и засадники .......................
..............................................Gnaphosidae.
12(11). Передние паутинные бородавки у основания сдвинуты друг
к другу или соприкасаются (рис. 575).
13(14). Ноги последней пары длиннее всех остальных ног. У самок все
тело светло-зеленое, у самцов головогрудь зеленая, а брюшко
красное с 2 желтыми продольными полосами. Засадники . .
.............................Sparassidae (Род Micrommata Latr.).
14(13). "Ноги последней пары короче второй пары ног, если не короче,
то головогрудь и брюшко иной окраски.
15(16). Передние пары ног направлены в стороны. Форма тела —
рис. 541—543. Засадники .... Бокоходы — Thomisidae.
16(15). Передние ноги направлены вперед. Форма тела иная.
17(18). Передние паутинные бородавки длинные, выступающие поза-
ди брюшка (рис. 551). Тенетники, реже бродячие пауки и за-
садники. Расположение глаз — рис. 569 . . . Agelenidae.
18(17). Паутинные бородавки короткие, не выступающие позади
брюшка.
19(20). Тело сильно вытянутое, снабженное длинными ногами и круп-
ными, направленными вперед хелицерами (рис. 558). Круго-
пряды. ...........................Tetragnathidae (Род Tetragnatha
Latr.).
20(19). Тело округлое или слабо вытянутое, хелицеры направлены
вертикально вниз;
21(24). Ноги с многочисленными толстыми шипами.
22(23). Брюшко округлое или угловатое (рис. 560, 561). Передние
и задние боковые глаза соприкасаются друг с другом (рис.
570). Кругопряды.......................................Araneidae.
23(22). Брюшко удлиненно-овальное (рис. 534, 538). Передние и зад-
ние боковые глаза широко расставленные (рис. 571) . . .
......................................................Pisauridae.
24(21). Ноги с редкими шипами или без шипов.
25(26). Пауки мелкие (не более 5 мм), брюшко серое с темными пят-
нами. Форма тела — рис. 547. Тенетники . . . Dictynidae.
26(25). Пауки, как правило, крупнее 5 мм. Если длина тела меньше
5 мм, то окраска брюшка иная.
27(28). Ноги с отдельными толстыми шипами. Брюшко удлиненно-
го
овальное (рис. 544). Передние и задние боковые глаза явствет
но отставлены друг от друга (рис. 572). Засадники и бродячи
пауки.............................................Clubionidae
28(27). Ноги с отдельными щетинками и многочисленными волоскам!
но без толстых шипов, а иногда и без щетинок. Брюшко округ
лое или удлиненно-овальное. Передние и задние боковые гла
за соприкасаются друг с другом (рис. 573), реже отставлен
ные.
29(30). Глаза переднего ряда почти вплотную придвинуты к передне
му краю головогруди. Бродячие пауки ................................
.....................Tetragnathidae (Род Pachygnatha Sund.)
30(29). Глаза переднего ряда далеко отставлены от переднего кра;
головогруди. Тенетники. .
31(32). Основная окраска брюшка снизу темнее, чем сверху . .
.............................................. . Linyphiidae
32(31). Основная окраска брюшка снизу не темнее (часто, светлее)
чем сверху .............................................Theridiidae.
Определительная таблица ловчих сетей
семейства Araneidae -
•t-
1(6). Сеть со стабилиментом (рис. 564, с). Паук сидит в центре
сети; убежища нет.
2(3). Стабилимент в виде 2 зигзагообразных лент. Паук сидит
в центре сети с широко расставленными ногами. (Род Argiope
Sav. et And.)..............................A. bruennichi Scop.
3(2). Стабилимент не зигзагообразный, в виде 2 узких полосок, про-
ходящих по противоположным радиусам (рис. 564). Паук си-
дит в центре сети с плотно прижатыми к телу ногами. (Род
Cyclosa Menge). ;
4(5). Центр сети находится на высоте 1—2 м над поверхностью зем-
ли. В сети около 40 радиусов................., С. conica Pall)
5(4). Центр сети находится на высоте 20—50 см над поверхность^
земли. В сети всего 20—30 радиусов (рис. 564)...............
............... С. oculata Walck/
6(1). Сеть без стабилимента. •
7(12). Сеть имеет свободный секто1р, лишенный клейких ловчих нитей'
(рис. 562). (Род Zygiella Pick.-Cambr.). j
8(9). Сеть находится на стволе дерева, убежище паука под корой'
или в расщелине коры.........................Z. stroemi Thor.;
9(8). Сеть свободно провисает между ветками деревьев или строит-;
ся в траве.
10(11). Сеть без свободной зоны и ловчие нити начинаются сразу за;
центральной сеточкой....................... Z. atrica С. L. Koch.;
11(10). Сеть с явственной свободной зоной (рис. 562).........•
........................................: Z. x-notata Cl.i
12(7). Сеть без свободного сектора (рис. 557, 563). (Род Ara.neu.s~'
Cl.).
13(14). Сеть без свободной зоны и ловчие нити начинаются сразу за;
центральной сеточкой.................... A. ceropegius Walck.;
14(13). Сеть с явственной свободной зоной.
15(16). Сеть без убежища, с 30—42 радиусами (рис. 557). Брюшко па-:
ука с характерным крестообразным рисунком (рис. 560). . .'
...........................................A. diadematus Cl.;
220 i
/
I
16(15). Сеть с убежищем, соединенным с центром сети сигнальной
нитью (рис. 563).
17(18). Сети крупные: диаметр ловчей зоны 60—70 см. Убежище нахо-
дится непосредственно в сети вблизи от ловчей зоны . . .
...................................................A. cornutus Cl.
18(17). Сети средней величины и мелкие: диаметр ловчей зоны не бо-
лее 50 см.
19(20). Центр сети находится на высоте не более 30 см над землей.
Убежище расположено непосредственно в сети вблизи от лов-
чей зоны (рис. 563)..............................A. alsine Walck.
20(19). Центр сети находится на высоте 50—80 см над землей.
21(22). В сети не более 20 радиусов.............A. quadratus Cl.
22(21). В сети более 20 радиусов................A. marmoreus Cl.
ОТРЯД КЛЕЩИ - ACARINA
К отряду клещей относятся мелкие, иногда микроскопические на-
земные или вторичноводные членистоногие. Расчленение тела чрезвы-
чайно разнообразное: у примитивных форм сохраняется наружная сег-
ментация брюшка (рис. 576), у некоторых видов имеется нечетко обо-
собленная головогрудь (рис. 577, 578), но у большинства представи-
телей этого отряда тело вовсе не расчленено, а граница между грудью
и брюшком не выражена (рис. 579). Глаза имеются у одних клещей,
но отсутствуют у других. На брюшной и спинной стороне тела часто на-
Рис. 576—579. Разделение тела клещей на отделы.
576 — сохранение следов первичной сегментации у примитивных клещей, 577 — частичное обособ-
ление головогруди, 578 —- то же, вид сбоку (ноги удалены), 579 — полное слияние отделов тела,
г — гнатосома; с.б — сегментные бороздки; н первые членики ног.
ходится склеротизованные щитки, расположение и размеры которых
служат важными таксономическими признаками. Хелицеры и педипаль-
221
Рис. 580—587. Клещи.
580 — водяной клещ Hydrachna geographica Mull. (Hydrachnidae), 581 — почвообитающнй панцирный
клещ Orlbatella magntseta Ewing (Oribatellidae), 582 — паутинный клещ Tetranychus ulmi
C. L. Koch (Tetranychidae), 583— клеищ-краснотелка EuschGngastria indica Hirst (Trombiculidac),
584 — клещ-краснотелка Microtrombldium hystricinum Canest. (Trombidiidae), 585 — самец нксодового
клеща Ixodes persulcatus P. Sch. (Ixodidae) в позе ожидания хозяина, 586 — форма тела и рас-
цветка спинного щитка у нксодовых клещей рода Ixodes Latr. (Ixodidae), 587 — то же у нксодо-
вых клещей рода Dermacentor .С. L. Koch (Ixodidae).
222
пы сильно видоизменены; они входят в состав сложного ротового ап-
парата, называемого гнатосомой (рис. 579, г). Ноги состоят из 5—•
6 члеников. Большинство клещей обладает 3 парами ног на стадии ли-
чинки и 4 парами на стадиях нимфы и имаго. Иногда число ног у има-
го уменьшается до 2 пар. Дыхание клещей кожное или трахейное.
Постэмбриональное развитие протекает по типу анаморфоза и прохо-
дит через следующие стадии: яйцо, личинка, нимфа, имаго. Стадия ним-
фы может охватывать до 3 возрастов, разделенных линьками. Разви-
тие многих клещей представляет собой сложный и длительный процесс,
нередко сопровождающийся сменой свободноживущих и паразитиче-
ских или активных и покоящихся стадий.
Отряд клещей характеризуется крайне широким разнообразием
пищевых режимов. Он объединяет хищников, кровососов, эктопаразитов
и эндопаразитов, сапрофагов и фитофагов. Многие виды имеют важное
практическое значение в качестве вредителей растений, паразитов до-
машних животных и человека, переносчиков опасных инфекционных за-
болеваний. В зависимости от мест обитания клещей их можно разде-
лить на 4 экологические группы.
Водяные клеши встречаются в озерах, прудах, вдоль берегов
и в проточных водах. Взрослые формы—хищники, питаются пачками,
личинками насекомых и другими мелкими беспозвоночными. Как пра-
вило, свободпоживущие водяные клещи ярко окрашены в красный цвет,
а их ноги превращены в плавательные конечности, снабженные много-
численными длинными, густо сидящими волосками. Личинки обычно не
способны к самостоятельному существованию и являются специализи-
рованными паразитами насекомых и пресноводных моллюсков.
К числу самых крупных и наиболее заметных водяных клещей от-
носится Hydrachna geographica Mull. У взрослой самки этого вида дли-
на тела достигает 8 мм, а у самца — до 6 мм. Клеща легко узнать по
характерному черному рисунку на ярко-красном основном фоне брюш-
ка с его дорсальной стороны (рис. 580). Хищные взрослые клещи
и нимфы встречаются в мелких водоемах, прудах, канавах главным об-
разом весной. Личинки Н. geographica паразитируют на жуках-плавун-
цах. Иногда они в огромных количествах покрывают брюшную поверх-
ность плавунца.
Почвенные клещи охватывают большое число видов, относя-
щихся к разным семействам и надсемействам. Их численность может
достигать нескольких сот тысяч или даже миллионов особей на 1 м2.
Пищевые режимы клещей, заселяющих почву и подстилку, отличаются
широким разнообразием; среди них немало сапрофагов, хищников, фи-
тофагов и паразитов почвенных животных. Для некоторых видов ос-
новную пищу составляют микроорганизмы — бактерии и грибы.
Панцирные клещи-орибатиды (Oribatei) являются типичными пред-
ставителями почвенной фауны. Они встречаются во всех типах почв,
но предпочитают влажные и затемненные местообитания. Тело взрос-
лых орибатид обычно покрыто склеротизованным темноокрашенным
панцирем, снабженным различными выступами и шипами (рис. 581).
Неполовозрелые стадии орибатид лишены панциря. В большинстве
случаев орибатиды питаются разлагающимися растительными остатка-
ми и почвенными микроорганизмами. Наряду с другими почвенными
сапрофагами они принимают участие в процессах почвообразования. Пе-
рерабатывая растительные остатки, орибатиды способствуют их пре-
вращению в почвенный гумус и повышают плодородие почв. В таеж-
ных лесах большое значение имеет деятельность этих клещей как пер-
вичных разрушителей хвойного опада, который с трудом усваивается
другими сапрофагами.
223
Растениеобитающие клещи представлены главным об-
разом фитофагами и некоторыми хищниками, нападающими на рас-
тительноядных клещей и других мелких членистоногих. В ка-
честве вредителей растений особенно важное значение имеют
тетраниховые, или паутинные клещи (Tetranychidae). Длина тела
у этих клещей едва достигает 0,8 мм, а их окраска варьирует от
желтоватой и зеленоватой до оранжевой и красной. Некоторые клещи
приобретают красную окраску во время зимовки. В отличие от ориба-
тид тетраниховые клещи имеют мягкие покровы, лишенные панциря
и склеротизованных щитков. Овальное или яйцевидное тело клеща
снабжено длинными щетинками (рис. 582).
Тетраниховые клещи обычно живут колониями на нижней стороне
листьев, оплетая их тонкой паутинкой. Во время питания клещи про-
калывают хелицерами эпидермис листа и высасывают растительные
соки. При обильном заражении клещами ухудшается общее состояние
растений, а поврежденные ими листья отмирают и опадают. Особенно
сильно страдают от клещей плодовые деревья и хлопчатник.
Паразитические и кровососущие клещи весьма мно-
гочисленны и разнообразны. Во время экскурсий на растениях, во мху
или прямо на земле часто можно встретить клещей-краснотелок, отно-
сящихся к двум близким семействам — Trombidiidae и Trombiculidae.
Представители семейства Trombiculidae обычно имеют перетяжку по-
середине тела (рис. 583), а у Trombidiidae такой перетяжки никогда не
бывает (рис. 584). Клещи окрашены в ярко-красный цвет. Густой по-
кров щетинок на поверхности тела и ногах придает им бархатистый
вид. Хотя нимфы и взрослые краснотелки ведут непаразитический об-
раз жизни, их личинки принадлежат к специфическим паразитам насе-
комых и позвоночных. Личинки некоторых краснотелок из семейства
Trombiculidae являются тягостными паразитами человека: вгрызаясь
в кожу, они могут вызывать зуд и жжение.
Среди кровососущих клещей особого внимания заслуживают са-
мые крупные представители всего отряда—иксодовые клещи (Ixodidae).
Они относятся к временным кровососущим паразитам наземных поз-
воночных и пользуются широкой известностью в качестве переносчиков
возбудителей клещевого энцефалита, туляремии и других инфекцион-
ных болезней. В зависимости от стадии развития и упитанности раз-
меры иксодовых клещей колеблются от 0,5 до 20 мм. Цвет тела обыч-
но светло-желтый, коричневый или бурый. На юге СССР и в тропиках
встречаются клещи с беловато-эмалевой и разноцветной окраской. Те-
ло, как и у всех клещей, слитное и на больших его участках покрытое
мягкой кутикулой, растягивающейся при кровососании. Более твердые
и нерастяжимые участки покровов формируют щитки. Спинной щиток
самца прикрывает почти все тело (рис. 585), а у самок, нимф и личи-
нок он более короткий (рис. 586, 587). С нижней стороны тела самцы
снабжены брюшными щитками, отсутствующими у самок. С телом
клеща подвижно сочленены гнатосома (хоботок) и ноги.
Голодные иксодовые клещи взбираются на низкорослые растения
и принимают характерную позу в ожидании хозяев-прокормителей
(рис. 585). Обычно взрослые клещи питаются на крупных млекопитаю-
щих— копытных и хищниках, а многие виды нападают и на человека.
Типичными хозяевами личинок и нимф являются грызуны, насекомо-
ядные, мелкие хищники и птицы. Самцы характеризуются пониженной
пищевой активностью, а у некоторых видов они вообще не питаются.
Паразитирование клещей наблюдается в различные, но определенные
для каждого вида сезоны года.
Иксодовые клещи широко распространены по территории СССР.
В лесной зоне встречаются главным образом виды рода Ixodes Latr.
224
с однотонно окрашенным чернобурым спинным щитком (рис. 586), а для
степей и лесостепной зоны более характерны виды рода Dermacentor
С. L. Koch с беловатым эмалевым пигментом на спинном щитке, ногах
и хоботке (рис. 587).
Систематика клещей еще недостаточно разработана. Многие зоо-
логи даже полагают, что клещи составляют сборную группу, объеди-
няющую 2 или 3 самостоятельных отряда. Определение надсемейств
и семейств вызывает большие затруднения и доступно только специа-
листам. Поэтому в настоящем руководстве определительные таблицы
не приводятся, а знакомство с разными представителями клещей огра-
ничивается перечисленными выше экологическими группами.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Ажеганова Н. С. Краткий определитель пауков (Aranei) лесной и лесостеп-
ной зоны СССР. Л., 1968. 150 с.
Бей-Биенко Г. Я. Общая энтомология. М., 1980. 416 с.
Бей-Б иен ко Г. Я., Данилевский А. С., Иванов А. В. и др. Опреде-
литель классов и отрядов наземных членистоногих. М.; Л., 1957. 92 с.
Бродский А. К., Гейспиц К. А., Гринфельд Э. К. и др. Краткий опре-
делитель отрядов и семейств наземных членистоногих европейской части СССР. Л.,
1974. 102 с.
Бэкер Э., У а р т о и Г. Введение в акарологию. М., 1955. 475 с.
Гиляров М. С. Закономерности приспособлений членистоногих к жизни на
суше. М., 1970. 276 с.
Горностаев Г. Н. Насекомые СССР. М., 1970. 372 с.
Горностаев Г. Н., Левушкин С. И. Определитель пресноводных насеко-
мых европейской части СССР. М., 1973. 186 с.
, Добровольский Б. В. Фенология насекомых. М., 1969. 232 с.
Догель В. А. Зоология беспозвоночных. М., 1981. 606 с.
Жизнь животных. Т. 3. Беспозвоночные. М., 1969. 576 с.
Залесская Н. Т. Определитель многоножек-костянок СССР. М., 1978. 212 с.
Иванов А. В. Пауки, их строение, образ жизни и значение для человека. Л.,
1965. 304 с.
Козлов М., Нинбург Е. Ваша коллекция. М., 1971. 159 с.
Локшина И. Е. Определитель двупарноногих многоножек (Diplopoda) рав-
нинной части европейской территории СССР. М., 1969. 78 с.
Мамаев Б. М. Определитель насекомых по личинкам. М., 1972. 400 с.
Мамаев Б. М., Медведев Л. Н., Правдин Ф. Н. Определитель насе-
комых европейской части СССР. М., 1976. 304 с.
Мариковский П. И. Чем питаются насекомые. Алма-Ата, 1977. 256 с.
Методы почвенно-зоологических исследований. М., 1975. 277 с.
Определитель насекомых европейской части СССР. М.; Л., 1948. 675 с.
Определитель насекомых европейской части СССР. М.; Л„ Т. I. Низшие,
древнекрылые, с неполным превращением. 1964. 936 с.; Т. II. Жесткокрылые и вееро-
крылые. 1965. 668 с.; Т. III, первая часть. Перепончатокрылые. 1978. 584 с.; Т. IV,
первая часть. Чешуекрылые. 1978. 711 с.; Т. V, первая часть. Двукрылые, блохи.
1969. 809 с.; Т. V, вторая часть. Двукрылые, блохи. 1970. 943 с.
Определитель обитающих в почве личинок насекомых. М., 1964. 919 с.
Определитель сельскохозяйственных вредителей по повреждениям культур-
ных растений. Л., 1976. 696 с.
Павловский Е. Н., Лепнева С. Г. Очерки из жизни пресноводных жи-
вотных. М.; Л., 1948. 459 с.
Плавильщиков Н. Н. Определитель насекомых. М., 1957. 548 с.
Плавильщиков Н. Н. Краткая энтомология. М., 1961. 208 с.
Положенцев П. А., Козлов В. Ф. Малый атлас энтомофагов, М., 1971.
118 с.
Райков Б. Е., Р и м с к н й - К о р с а к о в М. Н. Зоологические экскурсии. Л.,
1956. 694 с.
Руководство к большому практикуму но энтомологии. I. Систематика на-
секомых. М., 1978. 160 с.
Р у п а й с А. А. Определитель вредителей декоративных и плодовых деревьев
и кустарников по повреждениям. Рига, 1976. 323 с.
Сердюкова Г. В. Определитель родов иксодовых клещей фауны СССР. М •
Л., 1963. 20 с.
Серебровский А. С. Биологические прогулки. М., 1973. 169 с.
226
Словарь-справочник энтомолога. М.; Л., 1958. 631 с.
Тыщенко В. П. Определитель пауков европейской части СССР. Л., 1971.282 с.
Тыщенко В. П., Горы шина Т. К.., Дольник В. Р. Сезонные ритмы.—
В кн.: Проблемы космической биологии, т. 41. Биологические ритмы. М., 1980,
с. 229—288.
Фабр Ж.-А. Жизнь насекомых. М., 1963. 459 с.
Фабр П. Насекомые. М., 1976. 192 с.
Фасулати К. К- Полевое изучение наземных беспозвоночных. М., 1971.
424 с
Фриш К. Из жизни пчел. М., 1980. 216 с.
X а л и ф м а н И. А. Пчелы. М., 1963. 398 с.
Халифман И. А. Муравьи. М., 1963. 303 с.
Халифман И. А. Шмели и термиты. М., 1972. 319 с.
Халифман И. А. Четырехкрылые корсары. М., 1978. 319 с.
Шевырев И. Я. Загадка короедов. М., 1969. 95 с.
Шовен Р. Жизнь и нравы насекомых. М., 1960. 246 с.
Шовен Р. Мир насекомых. М., 1970. 240 с.
Шумаков Е. М., Брянцева И. Б. Вредные и полезные насекомые. Л., 1968.
144 с.
Яхонтов В. В. Экология насекомых. М., 1969. 488 с.
ОГЛАВЛЕНИЕ
Предисловие (В. П. Тыщенко)........................................... . : 3
Часть I. ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НАСЕКОМЫХ
Насекомые в экосистемах (А. К. Бродский)................................... 5
Пищевые режимы и пищевая специализация насекомых (А. А. Стекольииков) 15
Поведение насекомых (В. Е. Кшгятков)....................................' 24
Сезонные явления в жизни насекомых (В. П. Тыщенко) ...................... 33
Сбор, количественный учет и хранение насекомых (Е. Ф. Мартынова) .... 4.0
Методы сбора.........................................................—
Методы количественного учета. :.....................................43
Хранение собранного материала. .....................................45
Часть II. КРАТКИЙ ОПРЕДЕЛИТЕЛЬ И ЭКОЛОГО-МОРФОЛОГИЧЕСКАЯ
ХАРАКТЕРИСТИКА КЛАССОВ, ОТРЯДОВ И СЕМЕЙСТВ
НАЗЕМНЫХ ЧЛЕНИСТОНОГИХ.
Строение и развитие наземных членистоногих (В. П. Тыщенко).................46
Общая характеристика.....................................; . —
Определительная таблица классов по имагинальным стадиям . . . 48
Постэмбриональное развитие и метаморфоз ........................48
Определительная таблица классов по личиночным и нимфальным
стадиям ........................................................51
Мокрицы — наземные представители класса ракообразных — Crustacea
(В. П. Тыщенко) 52
Общая характеристика.......................................... —
Определительная таблица семейств .............................. 54
Класс Многоножки — Myriapoda (В. П. Тыще:, ко) ...........................55
Общая характеристика.............................................—
Определительная таблица подклассов ............................. —
Подкласс Диплоподы — Diplopoda.......................................56
Общая характеристика........................................... —
Определительная таблица отрядов и семейств......................58
Подкласс Хилоподы — Chilopoda........................................59
Общая характеристика.............................................—
Определительная таблица отрядов и семейств................. . . 61
Класс Насекомые—Insecta 61
Общая характеристика (В. П. Тыщенко) ................................ —
Определительная таблица отрядов по имагинальной стадии (В. П. Ты-
щенко) 67
Определительная таблица отрядов по личиночным стадиям (В. П. Ты-
щенко) 72
Отряд Ногохвостки, или коллемболы — Collembola (Е. Ф. Мартынова) 75
228
Отряд Двухвостки, или вилохвостки — Diplura (Е. Ф. Мартынова) . . 79
Отряд Щетииохвостки — Thysanura (Е. Ф. Мартынова).......................80
Отряд Поденки — Ephemeroptera (А. К. Бродский)..........................82
Общая характеристика............................................. —
Определительная таблица семейств..................................86
••’Отряд Стрекозы — Odonata (А. К. Бродский).............................87
Общая характеристика......................................... . —
Определительная таблица семейств..................................90
•Отряд Тараканы — Blattoptera (А. К- Бродский)..........................91
Общая характеристика...............................................—
Определительная таблица семейств..................................92
Отряд Богомолы — Mantoptera (А. К- Бродский)............................93
Отряд Термиты — Isoptera (А. К. Бродский)...............................94
Отряд Веснянки — Plecoptera (А. К- Бродский)............................95
Общая характеристика.......................................; i —
Определительная таблица семейств..................................96
•Отряд Прямокрылые — Orthoptera (А. К. Бродский)........................97
Общая характеристика...........................................s —
Определительная таблица семейств..................................99
Отряд Уховертки, или кожистокрылые — Dermaptera (А. К. Бродский) . 101
Общая характеристика...............................................—
Определительная таблица семейств .......................... 102
Отряд Сеноеды — Psocoptera (В. Е. Кипятков).............................—
«Отряд Равнокрылые—Homoptera (А. X. Саулич).......................103
Общая характеристика...............................................—
Определительная таблица подотрядов.............................104
Общая характеристика подотряда цикадовых — Cicadinea , . . , 105
Определительная таблица семейств цикадовых.....................107
Общая характеристика подотряда кокцид, или червецов и щито-
вок— Coccinea....................................................108
«Отряд Полужесткокрылые, или клопы — Hemiptera (А. X. Саулич) . . ПО
Общая характеристика...............я..............................—
Определительная таблица семейств ...............................116
Отряд Трипсы, или бахромчатокрылые — Thysanoptera (В. Е. Кипятков) 118
•'Отряд Жесткокрылые, или жуки-—Coleoptera (И. А. Кузнецова) . . . 120
Общая характеристика............................................ —
Определительная таблица семейств ...............................133
Отряд Вислокрылки—Megaloptera (А. А. Стекольников)...................136
Отряд Верблюдки — Raphidioptera (А. А. Стекольников)..................137
'Отряд Сетчатокрылые—Neuroptera (А. А. Стекольников)....................—
Общая характеристика..............................................—
Определительная таблица семейств ............................... 139
• Отряд Скорпионницы, или скорпионовые мухи — Mecoptera (А. А. Сте-
«Отряд Перепончатокрылые — Hymenoptera (В. Е. Кипятков) .... 140
Общая характеристика.........................................„ _• —
Подотряд Сидячебрюхие — Symphyta.................................143
Подотряд Стебельчатобрюхие — Apocrita............................146
Подотряд Стебельчатобрюхие, группа Паразитические перепончато-
крылые, или наездники—Parasitica................................147
Подотряд Стебельчатобрюхие, группа Жалящие перепончатокры-
лые— Aculeata............................................. ... 151
Определительная таблица семейств .............................. 170
Отряд Ручейники — Trichoptera (А. К- Бродский).........................173
Общая характеристика........................................ г г —
Определительная таблица семейств ........................ ..... 175
«Отряд Чешуекрылые, или бабочки — Lepidoptera (А. А. Стекольников) . 176
Общая характеристика..................,......................... —
229
Определительная таблица семейств..................................185
• Отряд Двукрылые, или комары и мухи — Diptera (А. К- Бродский) . . 189.
Общая характеристика............................................; —
Определительная таблица семейств..................................195
Класс Паукообразные — Arachnida (В. П. Тыщенко) ........................... 201
Общая характеристика..................................... .- . —
Определительная таблица отрядов ............................... 202
Отряд Ложноскорпионы — Pseudoscorpiones.................................203
Общая характеристика................................................—
. Определительная таблица семейств ............................. 204
Отряд Сенокосцы — Opiliones............................................204
Общая характеристика...............................................—
Определительная таблица семейств...............................206
Отряд Пауки — Aranei...................................................206
Общая характеристика.............................................—
Определительная' таблица семейств...............................218
Определительная таблица ловчих сетей семейства Araneidae . . . 220
Отряд Клещи — Acarina . 221
Рекомендуемая литература.................................................... 226.
ИБ № 1593
РУКОВОДСТВО ПО ЭНТОМОЛОГИЧЕСКОЙ ПРАКТИКЕ
Редактор И. П. Дубровская
Художественный редактор А. Г. Голубев
Технический редактор Г. Л1. Матвеева
Корректоры С. К. Школьникова, Г. Н. Гуляева
Сдано в набор 18.01.83. Подписано в печать 20.10.83. М-19266. Формат 70X108'/ie. Бумага тип. № 2.
Гарнитура литературная. Печать высокая. Усл. печ. л. 20,3. Усл. кр.-отт. 20,56. Уч.-изд. л. 19,78.
Тираж 5000. Заказ № 208. Цена 85 коп.
Издательство ЛГУ имени А. А. Жданова. 199164, Ленинград, Университетская наб., 7/9.
Сортавальская книжная типография Госкомиздата Карельской АССР по делам издательств, полиграфии
и книжной торговли. Сортавала, Карельская, 42.
-
по
< 9/ -19
УКОВО,
энтомологической