/
























































































Теги: biology insects science war in ukraine science magazine insectology
Год: 2024




Текст
EDI TO R I A L
Genuine images in 2024
I
the pilot period, authors generally provided a satisfactory response and fixed the problems so that the paper
could proceed to further review or acceptance. However,
papers that should not be published were detected. Going forward, if the authors’ response is unsatisfactory
or raises additional concerns, we will probe further and
take steps that could include rejecting the paper. If image
integrity concerns are raised about a paper that a Science
journal already published, we will use Proofig to carefully
examine the suspicious images, which will inform subsequent actions (e.g., correction or retraction).
Among other changes at the journals, Ali Shilatifard,
the academic editor of Science Advances, has decided
to step down after 10 years of exemplary service. Because of the outstanding growth of
the journal, five new section editors
have been appointed to lead social
and interdisciplinary science; neuroscience; Earth, environmental, ecological, and space sciences; physical
and materials sciences; and biomedicine and life sciences.
Meanwhile, in this issue of Science,
the News staff highlights areas in
research and policy that are likely
to attract attention in 2024. From the
effects of El Niño on global temperatures to forging AI regulation across
the world to the outcome of a US presidential election, there will be no shortage of events and issues to keep an eye
on. Science will cover all of this and more, in its research,
news, and commentary sections. And Science’s newsletter, ScienceAdviser, will deliver exciting highlights from
the world of science and Science every weekday morning.
In addition, this month, the Science Press Package team—
the group responsible for presenting content from the
Science journals to thousands of journalists—will launch
a weekly press package called “Policy Pak” that will be
filled with policy-oriented content from across Science’s
six journals. It will be aimed at the producers of the many
newsletters relied on by policy-makers to keep abreast of
scientific issues.
As I wrote in my end-of-year editorial last month, 2024
will bring a lot of challenges. Through better monitoring for research errors, careful curation of content, and
sharing information with policy-makers, we hope to build
stronger trust and integrity in science in the coming year.
H. Holden Thorp
Editor-in-Chief,
Science journals.
hthorp@aaas.org
p
n recent years, the research community has become
increasingly concerned with issues involving the
manipulation of images in scientific papers. Some
of these alterations—involving images from experimental techniques such as microscopy, flow cytometry, and western blots—are inadvertent and may
not change the conclusions of papers. But in rare
cases, some are done deliberately to mislead readers.
Image sleuths who can detect these alterations, like the
scientific integrity consultant Elisabeth Bik, have risen
to prominence, as has the website PubPeer, where many
of the detected flaws are posted. High-profile incidents,
such as one involving the laboratory of former Stanford
University President Marc Tessier-Lavigne, have eroded public confidence in science and
harmed careers of investigators who
missed doctored images coming from
their own laboratories. To address
these problems, in 2024, the Science
family of journals is adopting the use
of Proofig, an artificial intelligence
(AI)–powered image-analysis tool, to
detect altered images across all six of
the journals.
Proofig screens images for duplication and other types of manipulation. It is similar to the iThenticate
plagiarism detection software (which
Science has been using for 7 years), but
it works on images rather than text. Science has been piloting Proofig for several months with clear evidence that problematic figures
can be detected prior to publication, so its use will expand
to all papers under consideration that present relevant
images. This should help identify both honest mistakes
and fraudulent activity before a decision is made on publication. Prior to the pilot phase, Science had been conducting “human-eye” image checks on a portion of papers,
so Proofig is a natural next step. This tool will enhance
Science’s review process and reduce the potential for human error because it captures many more alterations.
Proofig will be applied after a research paper is revised
by authors. After analyzing the images, the tool generates
a report flagging duplications and other abnormalities,
such as rotation, scale distortion, and splicing. The paper’s editor reviews the findings and determines whether
the AI-detected issues may be problematic. (In some
cases, figures may have intentional rotations or duplications that are explained in the paper.) If so, the editor
contacts the authors to request an explanation. During
y
y g
y
,
PHOTO: CAMERON DAVIDSON
g
“…we hope to
build stronger
trust and
integrity in
science in the
coming year.”
–H. Holden Thorp
10.1126/science.adn7530
SCIENCE science.org
5 JANUARY 2024 • VOL 383 ISSUE 6678
7
NEWS
p
g
y
Edited by Jeffrey Brainard
In October 2023, boats on
Puraquequara Lake in Brazil
were stranded after drought
lowered the water level.
y g
IN BRIEF
y
AREAS TO WATCH
P
olitical upheaval and policy disputes could create uncertainties for scientists this year as a momentous U.S. election
looms and European parties grapple over green policies, for
example. As the effects of the pandemic caused by SARSCoV-2 continue to recede, trials of Long Covid treatments
may serve up their first fruits. The Pacific Ocean warming
called El Niño will likely help boost global temperatures to new
records, providing a continual reminder that time is running out
for science-based interventions to keep the world from warming
beyond “safe” thresholds. Science’s news staff forecasts other areas
of research and policy likely to make news this year.
8
5 JANUARY 2024 • VOL 383 ISSUE 6678
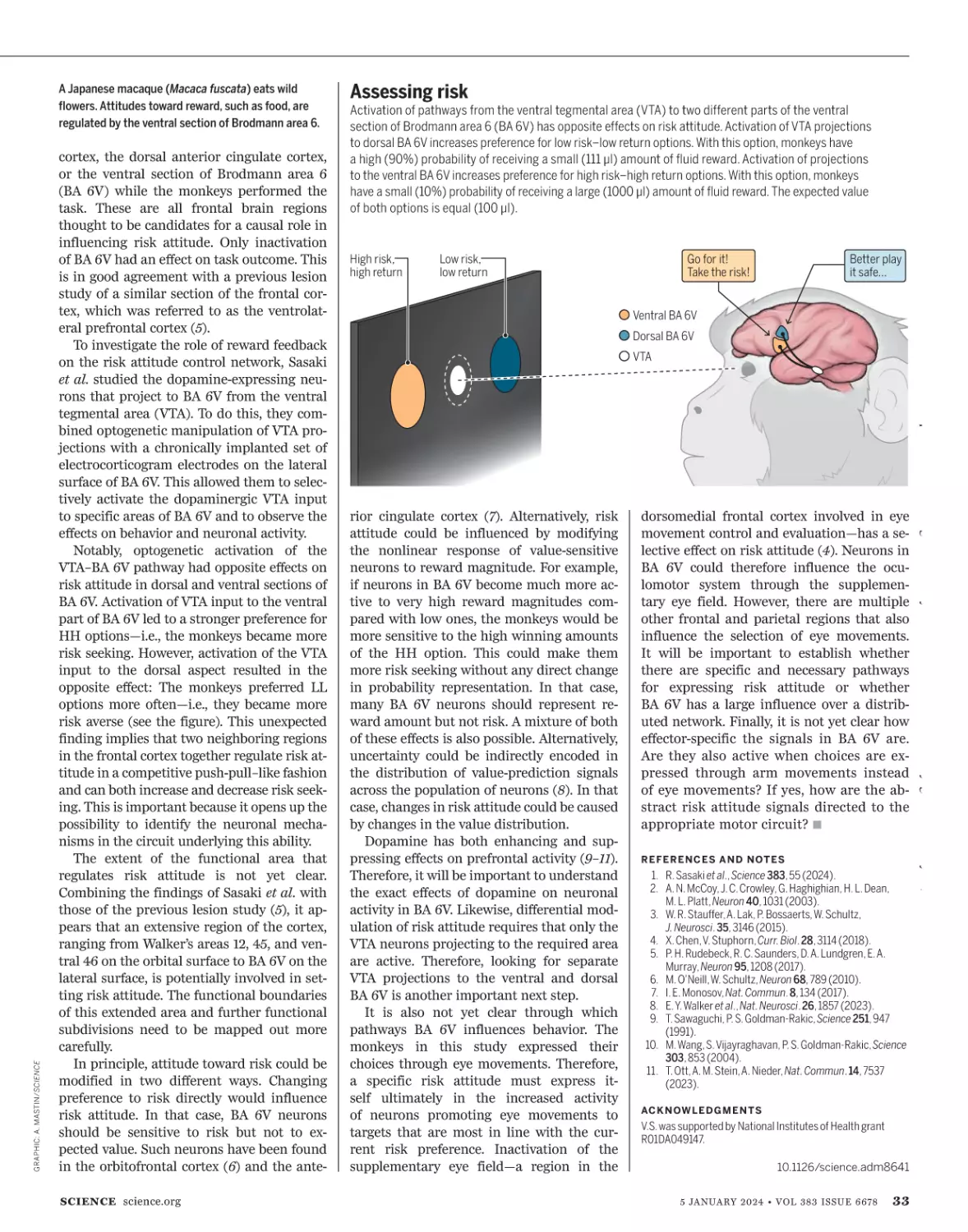
C L I M AT E S C I E N C E | An El Niño warming
in the eastern Pacific Ocean is likely to
grow stronger over the next few months
and might help push the average global
surface temperature 1.5°C above preindustrial levels for the first time. Part
of a regularly oscillating climate pattern,
the El Niño is expected to worsen drought
in the Amazon and Australia. The shift,
which began last year, is suspected of
helping make 2023 the hottest year in
modern history, as temperatures in the
first 11 months averaged more than 1.4°C
above preindustrial levels—higher than
rising emissions of greenhouse gases
alone can explain. El Niño has suppressed
sciencemag.org SCIENCE
,
What’s coming up in 2024
El Niño may extend record heat
calms a hyperactive immune system, and
vagus nerve stimulation recalibrates the
autonomic nervous system, potentially
ameliorating symptoms such as brain fog
and abnormal heart rate. Even if these
therapies aren’t successful, scientists hope
trial results will clarify the biology fueling
the condition, helping them identify what
to test next.
the ocean’s ability to absorb heat, and a
drop in Sun-blocking pollution also played
a role—effects that will continue this year.
U.S. politics shape science
PHOTO: JAIME SALDARRIAGA/AP
| Four years after
the pandemic caused by SARS-CoV-2
began, millions of people are debilitated
by Long Covid, a syndrome that includes
crushing fatigue, persistent headaches,
and shortness of breath. With no treatments clearly confirmed to help, patients
and their doctors are experimenting with
various drugs and dietary supplements.
This year, scientists hope some of the
first rigorous placebo-controlled clinical
trials of possible Long Covid treatments
will report outcomes, though they may
be preliminary. Some, such as Paxlovid,
other antiviral drugs, and monoclonal
antibodies, target the SARS-CoV-2 virus
itself, which may persist in the body.
Other candidates take aim at different
abnormalities identified in Long Covid
research: Intravenous immunoglobulin
C L I N I CA L R E S E A R C H
A N T H R O P O L O GY | Many Indigenous
people have been hesitant to engage with
researchers after centuries of colonial
war and decades of exploitation by some
scientists. But glimmers of reconciliation
are becoming more common, as more
Indigenous-led research projects and
genuine collaborations aim to incorporate
centuries of Indigenous knowledge of
the natural world into Western science.
This year, new partnerships may build on
high-profile examples from 2023, such as
a study of the ancestry of the horse in the
North America. The U.S. National Science
Foundation is supporting expanded
efforts with $30 million over 5 years for
a new Center for Braiding Indigenous
Knowledges and Science, and the National
Institutes of Health is providing $9 million to Stanford University and the Native
BioData Consortium, a biobank run by
Native geneticists, to create a COVID-19
database for Native people. Although
many points of tension remain, the new
5 JANUARY 2024 • VOL 383 ISSUE 6678
9
,
SCIENCE sciencemag.org
First fruits of Long Covid trials
Teaming with Indigenous experts
y
I N F E CT I O U S D I S E A S E S | A strategy
of releasing lab-modified mosquitoes to
prevent the spread of dengue fever is
poised to scale up this year after a series
of successes. The Aedes aegypti mosquitoes, developed and tested by the
nonprofit World Mosquito Program
(WMP), carry the bacterium Wolbachia
pipientis, which blocks them from
transmitting certain viruses and spreads
to their offspring when they mate with
wild insects. A trial in Indonesia showed
the strategy reduces dengue cases and
hospitalizations. And in the fall of 2023,
researchers reported that dengue cases
fell by at least 95% in treated areas in
the largest continuous release of the
modified mosquitoes yet, in Colombia’s
Aburrá Valley. WMP, which has projects
in 14 countries, is now expanding. It
plans to construct the world’s largest
Wolbachia mosquito production facility,
in Brazil, which will begin operations
this year. WMP says it also expects the
World Health Organization to issue formal guidance this year on deploying the
mosquitoes—a step that could lead more
countries to adopt the method.
y g
Antidengue mosquitoes take off
A mosquito modified to fight dengue lays eggs
at a World Mosquito Program factory in Colombia.
y
| Governments around the world
last year announced ambitious plans
to increase their oversight of artificial
intelligence—a race to regulate AI that will
likely speed up this year. U.S. agencies face
a daunting job to flesh out policy directives
announced in November 2023 by President
Joe Biden’s administration aimed at setting
standards for the responsible development
of AI. Members of Congress have also been
active, introducing more than 150 bills
designed to ensure that AI is a boon, not
a threat, to economic development, public
health, civil liberties, and national defense.
The European Union appears to be closer
to adopting ethical safeguards. But specific
rules may quickly become obsolete because
of the seemingly endless stream of powerful
new AI applications, such as chatbots based
on large language models.
POLICY
| Two experiments
working together could reveal how
the tiny masses of elusive particles called
neutrinos stack up. They come in three
types—electron, muon, and tau—that
morph into one another, a phenomenon
that might help explain how the universe
generated more matter than antimatter.
First, physicists have to flesh out their
theoretical model. They know that two
of the neutrinos have nearly the same
mass, but not whether there are two
light neutrinos and one heavier one, or
vice versa. Physicists working with the
T2K experiment in Japan and the NOvA
experiment in the United States study
neutrinos by shooting them hundreds
of kilometers through Earth to huge
detectors. This year, they plan to release
a joint analysis that could indicate which
of the two alternatives is correct. Finding
that the electron neutrino is light would
complicate planned experiments to search
for a type of nuclear decay that would
prove the neutrino is its own antiparticle.
PA R T I C L E P H YS I C S
g
The race to regulate AI
Bringing order to neutrino mass
p
D E M O C R AC Y | The U.S. national elections
in November will cap what promises to
be a bumpy political ride for scientists
this year. An ongoing partisan stalemate
in Congress over annual spending bills
could lead to a government shutdown
that would disrupt grantmaking at every
science agency. Most observers expect that
the final budget agreement will contain at
most small increases for individual agencies rather than the large boosts President
Joe Biden requested and Congress had
promised to deliver. The winner of the
race for president will shape policy on
myriad issues involving science, including
climate change, pandemic preparedness,
innovation, and immigration—topics on
which the likely major-party candidates,
Democrat Biden and his predecessor,
Republican Donald Trump, have starkly
different views.
NE WS | I N B R I E F
year will likely bring new models of collaboration, and more respect from some
Western scientists, in these and other
projects—for example in Native American
genetics and the effects of climate change
on Arctic ecosystems.
Pushback on EU green ambitions
POLICY
| With far-right, nationalist parties
Fusion megaproject faces delay
Costly Europa probe to launch
P L A N E TA RY S C I E N C E | NASA’s $5 billion
Europa Clipper probe is set to launch in
October aboard a SpaceX Falcon Heavy
rocket as the agency’s most expensive
planetary science mission since the
Viking Mars probes of the 1970s. Europa,
one of Jupiter’s large moons, has an icy
crust only kilometers thick, beneath
which swirls a vast salty ocean—a
potential nursery for life. When Clipper
arrives in 2030, it won’t land or sample
this ocean directly, but will swoop past
the moon 50 times, scanning its surface
and gathering clues about its interior.
Scientists had hoped the probe would
follow up on hints from telescopic surveys
of an active plume erupting from Europa
into space. But a recent campaign with
NASA’s JWST space telescope found no
evidence of it.
g
| Managers of ITER, the giant
experimental fusion reactor decades in
the making, are expected to announce this
year a new completion date, postponing
“first plasma” well beyond the longstanding target of 2025. The facility, being
built in France at a cost of tens of billions
of dollars to its international partners,
is meant to demonstrate the viability of
fusion as a carbon-free energy source,
but it has been beset with problems. The
COVID-19 pandemic slowed manufacturing of components, segments of the reactor
vessel were wrongly shaped and didn’t fit
together, cooling pipes were corroded, and
the French nuclear regulator was unconvinced of its safety. ITER Director-General
Pietro Barabaschi, who took control 1
year ago after the death of his predecessor, Bernard Bigot, is struggling to get
E N E R GY
construction back on track. He plans to
reveal the revised schedule this year but
told a fusion energy conference in October
2023, “It will not be good news.”
p
polling high for pan-European elections
in June, observers anticipate steppedup opposition to the bloc’s sprawling
green agenda. The European Green Deal,
approved in 2020, aims to make the bloc
“the first climate-neutral continent” and
reach zero greenhouse gas emissions by
2050, with new climate-friendly measures
planned across various policy areas, including energy, conservation, and transport.
Reflecting those goals, about one-third of
the funding under the massive Horizon
Europe program will go to research related
to climate change. Regardless of whether
far-right parties end up forming the third
biggest political force in the European
Parliament, the majority group—the
conservative European People’s Party—has
already cast itself as friendly to farmers
and industry by pushing back against
policies aimed at reducing pesticide use,
for example. Legislators could also seek to
limit the funding for the implementation
of green legislation that has to date largely
withstood conservative pushback—such
as the Nature Restoration Law, which sets
targets to restore degraded ecosystems.
SCIENCEMAG.ORG/NEWS
Read more news from Science online.
y
y g
y
,
Technicians at the Jet Propulsion Laboratory examine NASA’s Europa Clipper spacecraft, scheduled for launch in October to explore an icy moon of Jupiter.
10
5 JANUARY 2024 • VOL 383 ISSUE 6678
sciencemag.org SCIENCE
N E WS
IN DEP TH
p
A company called Running
Tide has released tons of woody
material off Iceland’s coast.
GEOENGINEERING
g
Plant waste buried at sea to fight climate change
Carbon in forestry or agriculture debris could remain locked on sea floor for centuries
y
By Saima Sidik
11
,
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
SCIENCE science.org
so the two layers don’t mix much at all—one
reason why very little oxygen makes it to the
sea floor. Without oxygen, microbes are limited in their ability to convert the carbon in
biomass back into greenhouse gases, such as
methane, and even if some methane is produced, chemical reactions in the sulfate-rich
waters will break it down. And because the
layers don’t mix, any trace greenhouse gases
that are produced will be locked in the depths
for hundreds or thousands of years. “There’s
all these additional processes that add more
layers of security,” Angel says.
The advantages are enough to lure investors hoping to sell credits for the carbon removed from the atmosphere. Carbon credit
marketplace Supercritical recently became
Rewind’s first customer, and this summer
the company plans to start sinking biomass
in burlap sacks—possibly including forestry
residue, river driftwood, and agricultural
waste. Bulgaria, Romania, Turkey, and Georgia have all shown interest in the project,
Angel says.
Frontier Climate—a group that makes
commitments to buy future credits from
carbon sequestration startups—recently
awarded $250,000 R&D grants to Rewind
and another firm, Houston-based Carboniferous, which hopes to sink sugarcane waste
in an oxygen-starved region of the Gulf of
Mexico known as Orca Basin. The waste is
abundant on Gulf Coast farms, says Morgan
y g
PHOTO: JENNIFER JOHNSON/RUNNING TIDE
D
ror Angel, a marine ecologist at the
University of Haifa, had for years
heard his archaeologist colleagues
talk about ancient shipwrecks on the
bottom of the Black Sea that were
perfectly preserved by the low-oxygen
environment. “You can see ropes,” Angel says.
“It’s something which is quite spectacular.”
Now, Angel wants to combat climate
change by purposefully adding to the wreckage, sinking waste wood to the sea floor,
where carbon that the trees stored up while
living can remain locked away for centuries.
Angel is a science lead for an Israeli company called Rewind, one of many companies
riding a wave of investment in technologies
that could help limit global warming by
drawing carbon out of the atmosphere and
locking it up. Whereas some carbon capture
schemes require expensive machines and
complex chemistry, burying terrestrial biomass at sea is exceedingly simple: All it takes
are tugboats, barges, and woody waste from
forestry and agriculture.
The approach has advantages over another
popular ocean-based carbon capture strategy: growing, and sinking, massive amounts
of seaweed or phytoplankton. Because the
plant material is grown on land rather than
in the ocean, it is less likely to rob nutrients
from the surrounding water and upset the
ecology. Industrial agriculture and forestry
have an extensive infrastructure for growing, processing, and transporting plants, in
contrast to marine farming, which has never
been attempted at scale. And because woody
plants are tough and unlikely to degrade, they
are good at hanging on to their carbon. “Decomposers don’t like to eat them—they don’t
get much out of it,” says Ning Zeng, a climate
scientist at the University of Maryland.
At the same time, the approach may
fall short of what’s needed to fight climate
change. To keep warming below 2°C, the
world needs to capture and store about
10 billion tons of carbon dioxide per year by
midcentury, according to the International
Energy Agency. But terrestrial biomass can
be sunk only where supplies of waste are located near suitable bodies of water. By one recent estimate, the approach could sequester a
few tens of billions of tons of carbon dioxide
in total—just a fraction of the need.
“The terrestrial biomass thing is not going to solve the full problem,” says ocean
engineer Kate Moran from Ocean Networks
Canada, a group that is assessing the efficacy
of carbon capture strategies. “It’s going to be
a small piece of the pie if it is deemed to be
more beneficial than risky.” But, she adds,
“We need all the tools in the toolbox.”
In the Black Sea, Rewind has one of the
world’s great carbon burial sites. The sea is
much saltier at the bottom than at the top,
NE WS | I N D E P T H
But researchers fear a plan to increase outside oversight of
six top schools could create new problems
By Dennis Normile
T
,
science.org SCIENCE
y
wenty years ago, five universities in
Japan were among the world’s top
100, according to an annual compilation by the ShanghaiRanking Consultancy. The University of Tokyo led
the contingent in 19th place, with
Kyoto University 30th.
But by 2023, Tokyo had tumbled to 27th
and Kyoto to 39th despite repeated efforts
to keep the universities globally competitive. The other three schools dropped out of
the top 100 entirely.
The decline has raised alarms, and Japan’s government has launched a new
effort to reverse it. Last month, lawmakers approved legislation that requires six
top-ranked universities to establish new
management policy councils designed to
give outside experts a greater voice in decisions. The councils, which will include
the university president and at least three
members independent of the school, will
weigh in on “significant operational policies and oversee the execution of the president’s duties,” the education ministry’s
higher education bureau said in a statement. And Masahito Moriyama, Japan’s education and science minister, has asserted
that the measure will “enhance education
and research.”
But critics fear the councils will only add
to already cumbersome university bureaucracies and push academic scientists to
emphasize applied research. And because
Moriyama’s ministry will vet council members, they worry the new law will erode university autonomy and open the door to political
meddling. The councils will lead to “increasingly complex and mysterious decisionmaking,” predicted sociologist Ryosuke
Nishida of the Tokyo Institute of Technology
in a critique published by The Asahi Shimbun, a prominent Japanese news outlet.
All sides agree that Japan’s research establishment has endured what some academics
call “the lost decades.” In addition to the slide
in university rankings, since the early 2000s
Japan has dropped from second to fifth in
the number of papers indexed in Clarivate’s
Web of Science, according to Japan’s National
Institute of Science and Technology Policy
(NISTEP). And it fell from fifth to 12th in the
number of papers ranking in the top 1% of
citations, trailing countries including Spain
and Iran.
Multiple factors have contributed to the
decline, says Masatsura Igami, a science policy researcher at NISTEP. But he singles out a
y g
5 JANUARY 2024 • VOL 383 ISSUE 6678
Japan tries, again, to boost
global ranking of its universities
y
12
SCIENCE POLICY
g
Saima Sidik is science journalist based in
Somerville, Massachusetts.
p
Raven, a biogeochemist at the University of
California, Santa Barbara and the company’s
chief science officer. “It’s already sitting in
piles,” she says. “The alternative for this material is essentially that it degrades, releases
methane, and requires tending so it doesn’t
light on fire.” Carboniferous is now applying
for permission to test its strategy from the
Environmental Protection Agency.
Portland, Maine–based Running Tide is
combining terrestrial and marine biomass
in one carbon capture strategy. The company
takes waste wood from a forestry operation in
Nova Scotia that would otherwise be burned
or left to decay and presses it together to create floating “buoys” no bigger than a basketball that are seeded with seaweed spores. The
buoys are released off the coast of Iceland,
where ocean currents carry them over a deep
region with little oxygen. Eventually they become waterlogged and sink, along with any
seaweed that has grown en route. Last summer, Running Tide sold its first carbon credits to Shopify, and the company says it has
sunk tens of thousands of tons of material
into the North Atlantic Ocean.
Marine scientist David Koweek of the
nonprofit Ocean Visions, which has previously supported Running Tide’s research,
lauds the simplicity of sinking terrestrial
biomass, because technology exists for almost every step in the process. That’s a
strong reason why “you might think about
doing this,” he says.
Beyond that, the benefits are murkier.
Even though boats are a climate-friendly
form of transportation (trucks emit at least
100 times more carbon per kilometer),
Angel says it wouldn’t make sense to ship
biomass around the world to get it to favorable sites. And although sunk terrestrial
biomass doesn’t steal nutrients from marine
life, removing it from land could deplete soil
of nutrients. “Over time we’re going to also
be losing some of the fertility that crops and
forests need,” says Charlotte Levy, a biogeochemist at Carbon180, which advocates for
scaling up carbon removal projects. Levy
also worries that as innovators find new
uses for scrap biomass—for example, as
sustainable building materials or biochar,
a charcoal-like soil additive—sinking the
biomass might not be the most environmentally friendly use.
Zeng agrees that sinking terrestrial biomass will be limited to a few areas of the
ocean for the foreseeable future. But the urgency of carbon removal demands that every possible scheme be explored thoroughly,
he says. “I think every idea deserves $1 billion of support to test it out.” j
Tohoku University, which hosts this advanced
synchrotron, is one of several Japanese schools to
fall off a list of the world’s top 100 universities.
Megaproject
will chart
human immune
diversity
Human Immunome
Project’s survey aims to
improve drugs and vaccines
By Mitch Leslie
T
y
,
13
y g
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
he hepatitis B vaccine is one of the
most potent immunizations, usually providing decades of protection
against the deadly liver virus. But
in about 10% of people it doesn’t
work, and in 2020, Amy Huei-Yi Lee,
a systems biologist at Simon Fraser University, and her colleagues set out to determine whether they could predict who
would benefit. The scientists found that
data on recipients’ immune systems such
as the abundance of certain proteins and
the activity patterns of a few genes foretold whether they would generate defenses
against the virus. “We got a sense of what
factors drive the vaccine response and what
[doesn’t],” Lee says.
She and her colleagues were only able
to take measurements from a handful of
patients, but an ambitious effort slated to
begin early this year will collect such data
from hundreds of thousands of volunteers
throughout the world. Called the Human
Immunome Project (HIP) and backed by
an international consortium of companies,
government agencies, and universities, the
effort will probe thousands of immune
variables in blood and tissue samples. The
result will likely be the world’s largest and
most comprehensive immunological database, a resource for scientists investigating immune system differences and how
they influence our responses to vaccines
and drugs and our vulnerability to illness.
“There’s a huge opportunity here in terms
of understanding human disease,” says
immunologist Mark Davis of Stanford University, who is not involved in the project.
And that’s just the start for the effort,
which currently operates on about $5 million a year in funding but could ultimately
cost billions. An offshoot of a previous effort
known as the Human Vaccines Project, HIP
g
SCIENCE science.org
IMMUNOLOGY
p
“stagnation” in research spending and a scientific workforce that has struggled to keep
pace with those found in other nations.
Many researchers trace Japan’s current
academic malaise to a 2003 law that turned
the nation’s 86 government-supported national universities into corporate entities
with greater independence. At the time,
policymakers argued that private entities
outperformed public institutions, and that
corporatization was a means of “improving efficiency and strengthening universities through competition,” says Kiyoshi
Yamamoto, an emeritus professor at the
University of Tokyo who specialized in university management.
While pursuing that strategy, however,
the government steadily cut operating funds
for the universities, which included some
support for research. Overall, those funds
dropped from $87 billion in 2004 to $76 billion in 2022, according to the Japan Association of National Universities. To compensate,
universities were allowed to raise tuition, increase revenues from affiliated hospitals, and
pursue contract research with industry.
The government also launched a number
of competitive grant programs intended to
promote promising areas of research and
strengthen top universities. In 2015, for example, it created the Japan Agency for Medical Research and Development (AMED) to
move basic biomedical research discoveries
into clinical trials. And just 2 years ago it
also launched an initiative that will award
up to $70 million annually to each university
deemed capable of becoming a world-class
institution (Science, 27 May 2022, p. 903). In
September 2023, Tohoku University won the
first such award.
Although the competitive programs have
helped some universities, they are “still far
from being able to make up for … the reduction in government funding,” says Futao
Huang, a higher education scholar at Hiroshima University.
The cost cutting, meanwhile, led to both
a dearth of entry-level research jobs and
“the creation of a career ladder of low-paid,
insecure jobs” that “drove talented young
researchers out of academia,” says Robert
Geller, a geophysicist and emeritus professor at the University of Tokyo. Enrollment in
Ph.D. programs has dropped precipitously.
Such developments are now undermining government efforts to boost innovation,
some scientists say. To cope with budget cuts,
for example, Japan’s national universities
increased the clinical duties of their hospital physicians. But that meant they had less
time for biomedical research, according to
an analysis reported by physician-scientists
Shotaro Kinoshita and Taishiro Kishimoto
of the Keio University School of Medicine
in an October 2023 letter in The Lancet. As
a result, “research output has declined,”
Kinoshita says—a trend that “may be hindering” AMED’s mission.
Critics of the latest tweak to university
governance don’t believe it addresses the
challenges universities face. Outsiders, they
note, already have a voice in managing most
universities. At the University of Tokyo, for
example, independent advisers serve on the
board and hold a majority of the seats on a
28-member Administrative Council.
Others worry about a provision in the new
law that allows the education minister to veto
council appointees, saying it could politicize
university governance. As an example, they
point to a recent controversy over appointees
to the Science Council of Japan (SCJ), which
plays a critical role in advising the government and is considered the nation’s foremost
association of researchers. Breaking with historic practice, in October 2020 then–Prime
Minister Yoshihide Suga blocked the SCJ appointments of six academics who had criticized the policies of his Liberal Democratic
Party (LDP), creating an uproar among researchers and the public.
Such concerns did not prevent the current LDP government from pushing the
new measure through the legislature in late
December 2023. Opponents take some solace, however, from resolutions added to the
bill that call for the protection of academic
freedom and respect for a university president’s council nominations.
The resolutions are not legally binding.
But Sayaka Oki, a historian of science at the
University of Tokyo, says they could give research advocacy groups that “had not seriously tried to cooperate before” a “means to
battle against possible harmful outcomes of
the new law.” j
NE WS | I N D E P T H
y
,
science.org SCIENCE
y g
5 JANUARY 2024 • VOL 383 ISSUE 6678
ect will then select a few variables that provide the clearest picture of how the immune
system is working. They will also provide the
basis for an immune monitoring kit, a standard set of assays that all the sites in the second part of the project will use.
In the end, HIP will generate nearly
2 trillion immune measurements, which will
be publicly available through a central database. With this data haul and other information, HIP will build a predictive AI model
that can forecast—based on immune profile,
ancestry, economic status, age, and other
information—how individuals will respond
to stresses or challenges, such as a particular drug or pathogen. The model could help
pharmaceutical companies identify opportunities for new treatments and drug reactions
to avoid. And by providing a much more
detailed view of a population’s health and
vulnerability to side effects, the model could
enable countries to better decide which
drugs are needed by and suitable for their
populations, thus allowing them to reduce
health care costs, Keirstead says.
What HIP is aiming for with its AI ambitions has “never been done before,”
Kierstead says, which is probably why this
part of the project draws more skepticism
from outside researchers. The project intends to generate not just predictive models, but also ones that replicate how the
immune system operates. Mathematical
biologist Reinhard Laubenbacher of the
University of Florida says the AI will detect patterns of responses but doubts it will
open a deeper understanding of the immune system. “Data collection efforts like
this are tremendously helpful, but we will
probably need more than that,” he says. A
priority is “building [a] theoretical framework” to understand the information the
project will accrue, he says.
Another challenge is money. To realize
its ambitions, HIP will require a supersize
budget, about $1 billion to $3 billion over
the next 10 years, Keirstead says. To raise the
needed funds, HIP now hopes to go beyond
its current partners to philanthropies, governments, and other pharmaceutical companies. “I am targeting everyone. There is not
going to be a stone left unturned,” he says.
Ensuring that HIP’s far-flung sites follow
the same procedures in collecting and analyzing the data will also be a challenge, Lee says,
adding that the immune monitoring kits will
be a big help in this regard. Thomas says that
attracting nonwhite participants could also
be difficult, given their mistrust of scientific
research like this. “They haven’t seen benefits
and have been exploited.”
Still, he and others are eager to see what
HIP produces. “If they pull this off, it will be
big,” Greenplate says. j
y
projects have collected limited
categories of information and,
in Project Baseline’s case, haven’t
made the data available publicly.
Another area where research
has fallen short is “the understanding of human immune variation and diversity,” says John
Tsang, a systems immunologist
at Yale University who helped
develop HIP’s scientific plan.
A litany of factors—including
age, sex, diet, living conditions,
previous disease exposure, and
genetics—shapes how the immune system functions. But
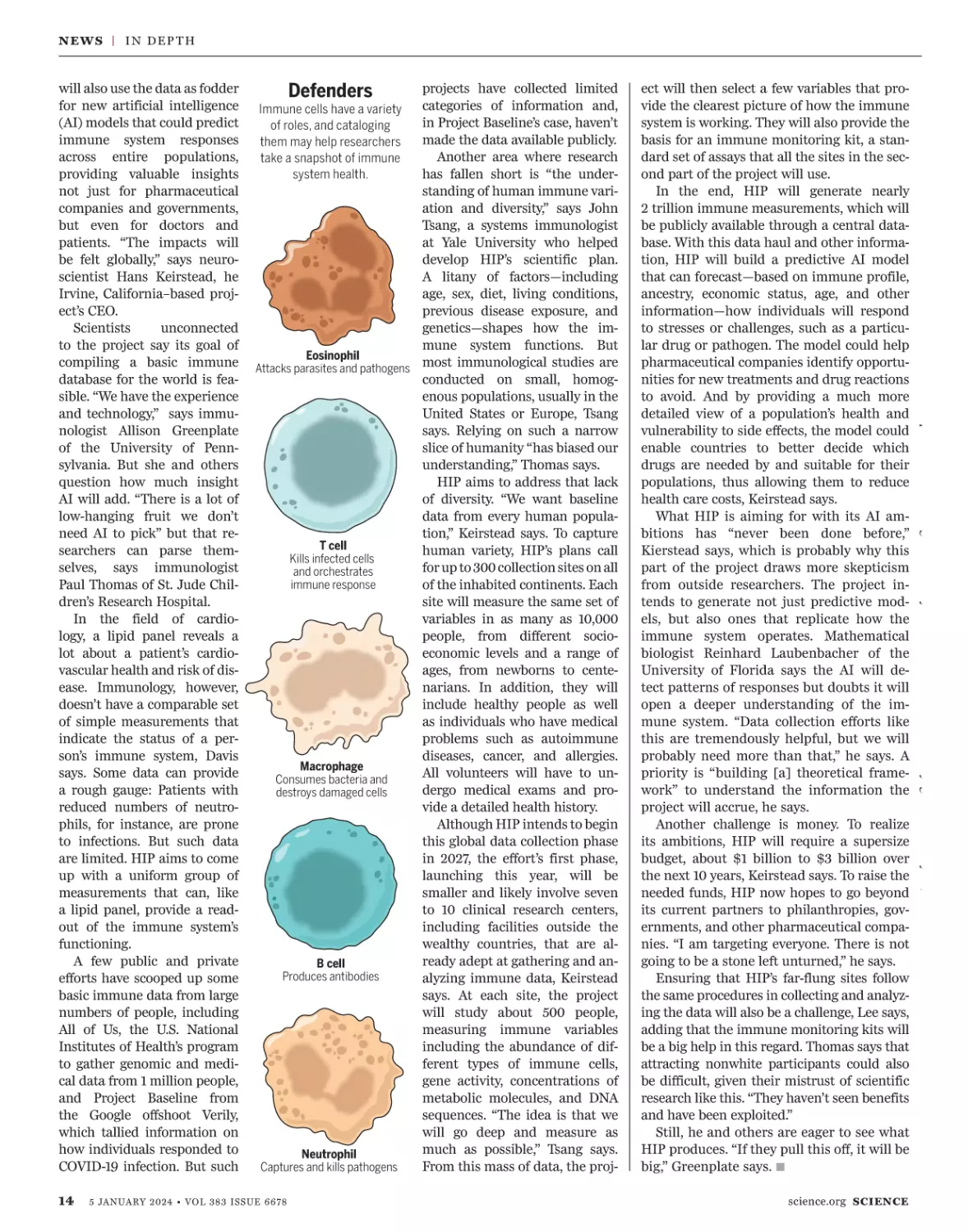
Eosinophil
most immunological studies are
Attackss p
parasites and p
pathogens
g
conducted on small, homogenous populations, usually in the
United States or Europe, Tsang
says. Relying on such a narrow
slice of humanity “has biased our
understanding,” Thomas says.
HIP aims to address that lack
of diversity. “We want baseline
data from every human population,” Keirstead says. To capture
T cell
human variety, HIP’s plans call
Kills infected cells
for up to 300 collection sites on all
and orchestrates
immune response
spo
of the inhabited continents. Each
site will measure the same set of
variables
in as many as 10,000
v
people, from different socioeconomic levels and a range of
ages, from newborns to centenarians. In addition, they will
include healthy people as well
as individuals who have medical
problems such as autoimmune
diseases, cancer, and allergies.
Macrophage
All
A volunteers will have to unConsumes bacteria and
dergo medical exams and prodestroys damaged cells
vide
a detailed health history.
v
Although HIP intends to begin
this global data collection phase
in 2027, the effort’s first phase,
launching this year, will be
smaller and likely involve seven
to 10 clinical research centers,
including facilities outside the
w
wealthy
countries, that are already adept at gathering and anB cell
Produces antibodies
alyzing immune data, Keirstead
says. At each site, the project
will
study about 500 people,
w
measuring immune variables
including the abundance of different types of immune cells,
gene activity, concentrations of
metabolic molecules, and DNA
sequences. “The idea is that we
will
w go deep and measure as
much as possible,” Tsang says.
Neutrophil
From this mass of data, the projCaptures and kills pathogens
g
14
Defenders
Immune cells have a variety
of roles, and cataloging
them may help researchers
take a snapshot of immune
system health.
p
will also use the data as fodder
for new artificial intelligence
(AI) models that could predict
immune system responses
across entire populations,
providing valuable insights
not just for pharmaceutical
companies and governments,
but even for doctors and
patients. “The impacts will
be felt globally,” says neuroscientist Hans Keirstead, he
Irvine, California–based project’s CEO.
Scientists
unconnected
to the project say its goal off
compiling a basic immune
database for the world is feasible. “We have the experience
and technology,” says immunologist Allison Greenplate
of the University of Pennsylvania. But she and others
question how much insight
AI will add. “There is a lot off
low-hanging fruit we don’t
need AI to pick” but that researchers can parse themselves, says immunologist
Paul Thomas of St. Jude Children’s Research Hospital.
In the field of cardiology, a lipid panel reveals a
lot about a patient’s cardiovascular health and risk of disease. Immunology, however,
doesn’t have a comparable set
of simple measurements that
indicate the status of a person’s immune system, Davis
says. Some data can provide
a rough gauge: Patients with
reduced numbers of neutrophils, for instance, are prone
to infections. But such dataa
are limited. HIP aims to come
up with a uniform group off
measurements that can, like
a lipid panel, provide a readout of the immune system’s
functioning.
A few public and private
efforts have scooped up some
basic immune data from large
numbers of people, including
All of Us, the U.S. National
Institutes of Health’s program
to gather genomic and medical data from 1 million people,
and Project Baseline from
the Google offshoot Verily,
which tallied information on
how individuals responded to
COVID-19 infection. But such
ASTRONOMY
Magellanic cloud may be two galaxies, not one
Rethink of familiar object may boost odds that its name, offensive to some, will be changed
By Daniel Clery
y
,
PHOTO: JUAN CARLOS MUNOZ-MATEOS/ESO
y g
15
y
5 JANUARY 2024 • VOL 383 ISSUE 6678
g
SCIENCE science.org
p
its parts, de los Reyes says, and relies on
“better data.” The question remains whether
the SMC’s two components are separate obhe Small Magellanic Cloud (SMC), a
jects that have drifted close together and are
hazy blob easily visible in the Southbound by their mutual gravity or whether
ern Hemisphere’s night sky, has long
one is debris drawn from the other by a close
been considered a lone dwarf galaxy
encounter with another galaxy such as the
close to the Milky Way. But a study
LMC. That the two parts seem to contain simposted online last month, and acilar masses of gas is “suggestive of two syscepted by The Astrophysical Journal, sugtems” without a connection, Murray says. But
gests the familiar site is not a single body,
de los Reyes counters that the level to which
but two, with one behind the other as
previous generations of stars have enriched
viewed from Earth.
the two gas clouds with heavier elements is
By tracking the movements of clouds
“relatively similar,” suggesting they are “not
of gas within the SMC and the young stars
entirely unrelated.”
recently formed within them, astronomer
If further studies confirm that the SMC
Claire Murray of the Space Telescope Sciis not one thing, but two, a new
ence Institute and her colleagues
name or names will be in order.
have found evidence of two stellar
De los Reyes argues that whatever
nurseries thousands of light-years
the clouds are called, astronomers
apart. If confirmed, the reassessneed to oust Magellan. She points
ment will likely amplify calls from
out that the objects had been
an increasing number of astronoknown and described by peoples
mers to change the SMC’s name
in the Southern Hemisphere for
and that of its neighbor, the Large
thousands of years, as well as reMagellanic Cloud (LMC).
corded by Italian and Arabic exSixteenth century Portuguese
plorers before Magellan’s voyage.
explorer Ferdinand Magellan, afThe astronomical community
ter whom the galaxies are named,
has long celebrated Magellan.
was not an astronomer, did not
There are Moon and Mars craters
discover them, and is recorded as
bearing his name, NASA’s Magelhaving murdered and enslaved
lan mission to Venus, and the twin
Indigenous people during his
Magellan Telescopes in Chile and
first-ever circumnavigation of the
the Giant Magellan Telescope curglobe. As a result, astronomer Mia
Magellanic clouds, seen here above Paranal Observatory in Chile,
rently under construction there.
de los Reyes of Amherst College
are due for a name change, some astronomers say.
De los Reyes says the atrocities
called for renaming the clouds in
an opinion piece for Physics magathat Magellan and his crews comzine last year. The idea has since “gotten a lot
Combining those two sets of data and asmitted in Argentina and elsewhere during
of informal support,” she says.
suming any stars that young would still be
their travels make him unsuitable for these
The two Magellanic clouds are in the gravimoving in concert with the clouds of gas
honors and for having his name in the more
tational sway of the much larger Milky Way
from which they formed, they identified two
than 17,000 peer-reviewed papers as a result.
and are on course to pass through it in the
distinct star-forming clouds. By measuring
De los Reyes is now working with the
future. The LMC is a disk galaxy, like the
how much light from the two clouds is abInternational Astronomical Union (IAU),
Milky Way in miniature. But the SMC is irsorbed by dust between them and Earth, they
which manages the naming of celestial obregular in shape, two-thirds of the LMC’s
calculated that one is more distant than the
jects, to present a resolution to its general
mass, and often gets overlooked in favor of
other. But determining the amount of sepaassembly in August. But what to call the
its heftier neighbor. “The SMC got the short
ration proved tricky. “Measuring distance to
familiar clouds? Murray suggests one of
end of the stick,” de los Reyes says. “It’s less
gas is very challenging,” Murray says. In the
their many Indigenous names. De los Reyes
well modeled.”
new study, her team describes constructing a
says the IAU resolution will include sevSome previous studies have suggested
simple computer model of the system. With
eral options for members to vote on. She
that the SMC may have multiple compothe available data plugged in, it predicts the
likes the idea of keeping the acronyms SMC
nents, but the truth has not been easy to pin
two clouds are 16,000 light-years apart—
and LMC—“to maintain continuity in the
down. The SMC “has suffered most” in its
more than half the distance between Earth
literature”—and just changing the “M” to
gravitational interactions with the LMC and
and the center of the Milky Way.
something “more physically meaningful”
the Milky Way, Murray says. “It’s simply full
This view of the SMC “is much more comsuch as “milky” or “meridional,” a reference
of disrupted gas. It’s a train wreck of sorts.”
pelling” than previous attempts to untangle
to the cloud’s southern location. j
T
So, she and her team set out to reinvestigate both the system’s stars and its gas, using
what Murray calls the “latest and greatest”
instruments available. To probe the SMC’s
gas they turned to the Australian Square
Kilometre Array Pathfinder, a radio telescope
made up of 36 dishes each 12 meters in diameter. With it, they zoomed in on radio waves
emitted by atomic hydrogen gas. They then
plotted the location and velocity of thousands
of stars in the SMC that are less than 10 million years old using Gaia, an orbiting observatory from the European Space Agency.
Over the past decade, Gaia has logged such
data for more than 1 billion stars across the
Milky Way with unprecedented accuracy.
NE WS | I N D E P T H
Monica Bertagnolli in her office at the National
Institutes of Health.
By Jocelyn Kaiser
science.org SCIENCE
,
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
16
y g
W
hen cancer surgeon Monica
Bertagnolli learned last year that
President Joe Biden wanted her to
lead the $47.5 billion National Institutes of Health (NIH), the world’s
largest funder of biomedical
research, “I didn’t embrace it,” she says with a
laugh. She was just a few months into heading NIH’s largest component, the National
Cancer Institute (NCI). “My reaction was:
‘But I’m the NCI director and I have plans.’
We were accomplishing some really great
things together. And it was just too soon.”
But the idea grew on her. She had gotten
to know the chiefs of NIH’s 26 other institutes and centers at weekly meetings and
found them “dynamic and talented and eager for innovation.” NIH’s acting director,
Lawrence Tabak, was lamenting the lack of a
permanent NIH chief since geneticist Francis
Collins stepped down in December 2021 after
12 years at the helm. And Bertagnolli saw the
post as a chance to advance “some things that
I am passionate about that I think will help
biomedical research overall.”
So, she took the job. A few weeks in, she sat
down recently for a 45-minute interview in
her office between a swirl of meetings, calls,
and the first town hall with NIH staff to fill
a nearly 500-seat room since the COVID-19
pandemic. She discussed her goal of making
NIH research more “equitable and accessible”
by expanding clinical trials to more rural and
minority patients. She also wants researchers
to bring more rigor to clinical care with the
help of artificial intelligence (AI). “We have
an opportunity to be able to deliver so much
more in terms of real high-quality, wellunderstood results that can help guide care
for people,” she says.
Colleagues say that although Bertagnolli,
age 64, has made a name as a lab scientist and
clinical trial leader, she is a physician first.
Raised on a ranch in Wyoming, she studied
engineering at Princeton University before
heading to the University of Utah for medical
school and a career as a physician-scientist at
Brigham and Women’s Hospital and the Dana
Farber Cancer Institute, where she was chief
of surgical oncology for 10 years. Dana Farber
colleague Suzanne George says Bertagnolli
was known to “take the hardest cases” of sarcomas, a rare cancer of bone and soft tissue,
offering hope to desperate patients.
At the same time, she was making her
name as a researcher. Until about 7 years
ago, Bertagnolli ran an NIH-funded tumor
immunology lab studying how inflammation
y
Improving patient care is top priority for head of world’s
largest biomedical research funder
g
Once reluctant, new NIH chief
Monica Bertagnolli embraces role
p
BIOMEDICAL RESEARCH
drives colon cancer in mice. The work led
to a landmark clinical trial testing whether
anti-inflammatory drugs called Cox-2 inhibitors can prevent precancerous colon polyps
in people prone to these growths.
Bertagnolli still remembers walking down
a hallway at Brigham when the trial’s results
came out in 2006. Cox-2 inhibitors reduced
polyps but also raised the risk of strokes and
heart attacks, putting the drugs off limits as a
preventive measure. “My oncology colleagues
said, ‘We’re so sorry,’” she says. “But cardiologists said, ‘That was great!’”—because
the large study firmed up the risks of Cox-2
drugs, which had already led regulators to remove some types from the market.
Bertagnolli’s trial experience led to a
job heading one of NCI’s large cooperative
groups, which run multiple cancer trials.
When a 2010 Institute of Medicine report
urged NCI to overhaul these groups, she
agreed to oversee a merger of three of them—
one run by researchers at powerhouse cancer
centers since the 1950s, another centered in
the Midwest with many community oncologists, and a third consisting of surgeons.
Bertagnolli gave these disparate groups of
nearly 10,000 cancer specialists what she
calls a “completely new identity” as the Alliance for Clinical Trials in Oncology. On its
committees, community oncologists from the
rural West now sit side by side with researchers from major academic centers.
Merging these groups was “extraordinarily
multifaceted and complex,” says Elad Sharon,
a former staffer in NCI’s clinical trials division who is now at Dana Farber. “She was
clearly head and shoulders the most innovative thinker running those shows.”
In October 2022, Bertagnolli became the
first woman to lead NCI. Two months later,
she announced that she had early breast cancer. (She has completed treatment and has a
good prognosis.) Within 6 months, she had
produced a document called the National
Cancer Plan that lays out eight strategies,
such as smoking cessation campaigns and
wider cancer screening, to achieve Biden’s
moonshot goal of cutting the U.S. cancer
death rate in half by 2047. She unveiled a new
unit at NCI aimed at working with the U.S.
Food and Drug Administration to develop innovative clinical trial designs. In May 2023,
she was nominated to become the second
woman to lead NIH.
It took until October for Senator Bernie
Sanders (I–VT), who heads the committee
that oversaw her nomination, to schedule
her confirmation hearing. She was confirmed
by a Senate vote of 62 to 36 after a hearing
where conservatives pressed her on NIH’s
Peru moves to crack down on
fraud in research publishing
Two new laws would make it easier to punish scientists
who buy authorship and commit other misdeeds
By María de los Ángeles Orfila
P
,
17
y
5 JANUARY 2024 • VOL 383 ISSUE 6678
y g
María de los Ángeles Orfila is a journalist in
Montevideo, Uruguay.
y
eru’s legislature is poised to approve
new laws that would make it easier
to investigate and punish researchers
who engage in fraudulent publishing
practices, including paying to have
their names added to a paper.
The move comes as Peru’s national science agency seeks to crack down on authorship buying and other unethical practices. It
recently removed two scientists accused of
violations from a national registry that is key
to receiving government grants, job promotions, and salary bonuses. And officials are
investigating more than 170 other researchers that a Peruvian media report alleged
were involved in publishing fraud, including
72 listed on the national registry who work at
14 universities in Peru.
The new legislation will empower universities and government officials to punish such
behavior, and would give Peru some of the
strongest measures against publishing fraud
in Latin America. Shady publishing practices
“transcend mere ethical violations” because
they enable researchers to obtain government
and private funding under false pretenses,
says Edward Málaga Trillo, a neurobiologist
and member of Congress who is the driving
force behind the bills, which lawmakers are
expected to finalize early this year. “These individuals are perpetrating fraud.”
As in many other nations, Peru’s academic
community has been struggling with a rising
tide of fake authorship and related problems.
One cause, some researchers say, is a 2014 law
that aimed to stimulate research by rewarding researchers who boost their publishing
output. For example, under a scoring system
used by universities, researchers can earn
five points for authorship in a high-impact
journal, and two points when the journal is
lower impact. Amassing points can bring bonus payments and career advancement.
The law creates perverse incentives, researchers say. In an October 2023 report,
journalists with the TV program Punto Final
reported that Peruvian researchers were paying up to $500 to add their names to papers
that they did not help write. The reward can
be substantial: Some for-profit universities in
Peru pay publishing bonuses of $2500, says
biologist Gisella Orjeda Fernandez of the
National University of San Marcos, a former
head of Peru’s science agency, the National
Council of Science, Technology and Technological Innovation (Concytec). “There is a
whole environment that incubates this [misconduct],” Orjeda Fernandez says.
The 72 researchers that Punto Final alleged were involved in publishing fraud
comprise an “alarming” 1% of all scientists
on Peru’s national registry, says Concytec
President Benjamín Marticorena Castillo.
One of the two researchers that Concytec has
already removed from the registry was found
to have republished articles by a Spanish researcher under their own name. The other
expelled researcher had assumed the identity
of a Venezuelan scientist.
Signs of publishing fraud can be obvious,
notes Nahuel Monteblanco, president of
Cientificos.pe, a Peruvian group othat investigates misconduct. Many of the papers cited
by Punto Final have numerous co-authors
who are based in many different nations and
have few prior publications on the same subject. “If your colleague consistently publishes
20 articles a year with collaborators from Nepal, Afghanistan, Kuwait, or Indonesia, that’s
highly suspect,” Monteblanco says.
Existing law, however, gives Concytec and
Peru’s 93 public and private universities
limited authority to investigate and punish
such misconduct. The two bills now before
Congress aim to change that. One would
amend a law covering universities, whereas
the other gives new powers to Concytec. The
legislation defines fraud to include “plagiarism, fabrication or falsification of information in publications, projects, reports and any
other academic product related to scientific
research.” Researchers who commit lesser
violations could be suspended from the national registry for 2 to 5 years. Those committing more serious infractions could be
permanently suspended and potentially face
criminal charges and imprisonment.
Peruvian science, Orjeda Fernandez says,
“should not hesitate to expel the guilty.” j
g
SCIENCE science.org
SCIENTIFIC MISCONDUCT
p
support of gender-affirming research, fetal
tissue studies, and NIH’s role in funding virus research in China that some claim led to
the pandemic.
Sanders had delayed her hearing because
he wants NIH to do more about drug prices.
He and others have called on NIH to use its
“march-in rights,” exercising a provision in
a 1980 law that some experts say allows the
agency to reassign a patent on a drug developed with taxpayer funding if a company is
charging an unreasonable price. The Biden
administration has since proposed that NIH
can use that mechanism to control prices—
a shift from previous administration policy.
The proposal is open for public comment.
“People should have fair and equitable
access” to NIH-funded drugs, but “the situation is complex,” Bertagnolli says. She is
now working to rebuild relationships with
members of Congress that frayed during the
pandemic, she says.
At NIH Bertagnolli plans to push for a goal
she’s championed for several years: building
a “learning health system.” The idea involves
folding patients’ electronic health records
into a research database and then using AI
tools to extract useful information. The approach could, for example, help diagnose a
condition or determine whether a cancer
drug tested in a clinical study is working in
the real world.
Bertagnolli wants NIH’s basic research
community to know that her interest in clinical research does not “diminish in any way
the importance of fundamental science.” But,
she adds, “The research arm needs to be able
to work with and embrace our entire health
system.” She expects to work across health
agencies to beef up standards for electronic
health records and promote their adoption
so patients can share their data for research.
Bertagnolli also faces immediate pressures.
A report delivered by her advisers last month,
for example, urges NIH-funded institutions
to raise the minimum salaries for postdocs
by 24% to $70,000, despite an agency budget that could stay level over the next 2 years.
“We can make a commitment to do all we can
with the extramural research community to
make [postdoc raises] possible,” she says. “But
sacrifices are going to need to be made. And
I don’t think we can entirely foresee how this
is going to work out. It’s just too complicated
and it’s so intertwined with the dynamics of
the academic community and environment.”
Colleagues think she is up to the challenges. Cell biologist and Huntsman Cancer
Institute CEO Mary Beckerle, who has known
Bertagnolli since her Utah medical school
days, compares her to the late Supreme Court
Justice Sandra Day O’Connor, who also grew
up on a ranch: A “bundle of wit, resilience,
and can-do spirit,” Beckerle says. j
NE WS
p
g
FEATURES
y
y g
LAID TO WASTE
Ukrainian scientists are tallying the grave environmental
consequences of the Kakhovka Dam disaster
y
,
By Richard Stone, in Kyiv and Odesa, Ukraine
I
n the predawn hours of 6 June 2023, a
pair of explosions rocked the Kakhovka
Dam, a 3-kilometer-long hydropower
facility on the Dnipro River in southern Ukraine. Waking up that morning to the unfolding catastrophe, “I
couldn’t believe it,” recalls Volodymyr
Osadchyi, director of the Ukrainian Hydrometeorological Institute
(UHMI). “I thought it had to be fake news.”
But footage captured by a Ukrainian military drone showed water from one of Europe’s largest reservoirs gushing through a
18
5 JANUARY 2024 • VOL 383 ISSUE 6678
gaping breach in the dam.
Over the next 4 days, 18 cubic kilometers of water surged downstream, inundating more than 620 square kilometers
and affecting 80 settlements. Scores of people died, and many more are unaccounted
for. Up to 1 million people lost access to
drinking water. In October, the Ukrainian
government pegged the cost of the disaster, which it blames on Russia, at roughly
$14 billion. Nearly half that figure—
$6.4 billion—is an estimate of lost ecosystem services due to chemical pollution and
habitat destruction along the Dnipro, one
of Europe’s largest rivers.
Assessing environmental harm in the
midst of a war in which the Dnipro itself
delineates more than 300 kilometers of
the front line is not easy. But Osadchyi and
other Ukrainian researchers have been sobered by what they’ve found so far. The toll
includes heavy damage to a unique sturgeon breeding facility, flooding of nature
reserves and agricultural land, and a death
blow to countless organisms adapted to
brackish estuaries near the confluence of
science.org SCIENCE
N E WS
p
g
PHOTO: ALEXEI KONOVALOV/TASS VIA ZUMA PRESS
long Dnipro (the Dnieper, for Russians)
5 JANUARY 2024 • VOL 383 ISSUE 6678
19
,
ORIGINATING IN RUSSIA, the 2200-kilometer-
flows across Ukraine’s northern frontier
with Belarus, near Chornobyl, wending
through Kyiv and the rest of the country
before spilling into the Dnipro-Bug Estuary and the Black Sea. After World War II,
Soviet engineers built or refurbished six
hydroelectric dams along the river. They
completed the Kakhovka Dam, the final
and largest one, in 1956.
Soon after Russia launched its fullscale invasion of Ukraine on 24 February
2022, it captured the Kakhovka Dam and
Nova Kakhovka, a city on the Dnipro’s left
bank built for the hydroelectric station’s
workers. In a TV address that October,
Ukrainian President Volodymyr Zelenskyy
claimed the Russian military had rigged
the dam with explosives and warned that
its destruction “would mean a large-scale
disaster.” At the time, Ukrainian troops
were on the verge of recapturing Kherson,
a major city downstream on the Dnipro,
triggering concerns Russia might blow up
the dam to unleash floodwaters that would
slow the Ukrainian advance.
It would not have been the first use of
water as a weapon during the conflict. In
March 2022, the Ukrainian army destroyed
a dam on the Irpin River, north of Kyiv, to
y
SCIENCE science.org
to thrive when humans are displaced. For
instance, because it has been too dangerous to fish in much of the Black Sea, fish
stocks are thought to be rebounding. With
hunting banned in a wintering ground for
migratory birds on the southern steppe,
near the Black Sea coast, “the situation for
many populations of birds is so much better
now,” says Vasiliy Kostiushyn, an ornithologist at the I.I. Schmalhausen Institute of
Zoology (IZAN), also part of NAS.
The ecological legacy of the Kakhovka
disaster is similarly nuanced, as new habitats emerge on terrain scoured by floodwater and in the former reservoir’s driedout lakebed. The Ukrainian government
has vowed to rebuild the dam after the
war. But some experts hope it will change
its mind and allow a natural recovery—and
perhaps even accelerate efforts to rewild
parts of the lower Dnipro Basin.
“Kakhovka is a tragedy,” says marine
ecologist Galyna Minicheva, director of
NAS’s Institute of Marine Biology (IMB).
“But it is also a huge and unprecedented
natural experiment.”
y g
the Dnipro and the Black Sea. Billions of
mussels are rotting on the former reservoir’s desiccated lakebed. And the Kakhovka’s destruction has added a new wrinkle
to a puzzle that arose earlier in the war:
the unexplained deaths of dolphins and
porpoises in the Black Sea off Ukraine’s
southern coast.
Such ecological miseries pale in comparison with the atrocities Russian forces
have committed against Ukrainians during
a nearly 2-year war that has claimed hundreds of thousands of lives. But when the
long and grinding war finally ends, Ukrainians will confront environmental damage
that extends well beyond the Dnipro to
widespread chemical contamination of agricultural fields and forests from shelling,
the wanton destruction of protected areas,
and the laying of innumerable mines that
experts say will take decades to clear.
“The environmental cost of the war has
been immense,” says Sergei Mosyakin, director of the M.G. Kholodny Institute of
Botany, part of the National Academy of
Sciences of Ukraine (NAS). “The ecocide
that Russia has inflicted on our country
will be studied for generations to come.”
Yet as in all wars, nature has found places
y
Billions of zebra and quagga mussels are rotting in the dessicated lakebed of the Kakhovka Reservoir (left),
which largely emptied after explosions tore apart the Kakhovka Dam in June 2023, unleashing a destructive flood.
NE WS | F E AT U R E S
The morning of the disaster, Sergiy
Afanasyev’s first thoughts were for colleagues at the S.T. Artyushchyk Production
Experimental Dnipro Sturgeon Breeding
Plant. Located southwest of Kherson in
Dniprovs’ke, the 40-year-old station annually stocked the Dnipro’s reservoirs with
some 1.5 million juvenile sturgeons adapted
to the region, including the critically endangered Danube sturgeon (Acipenser gueldenstaedtii). Ukrainian troops evacuated
its staff before a 4-meter wave of turbid,
polluted water flooded the station and its
breeding ponds. But the sturgeons, which
bog down Russian troops marching on the
capital. The next month it employed a similar tactic, punching a hole in a dam at the
Oskil Reservoir in the Kharkiv region. Then
in September 2022, Russia hit a dam on
the Inhulets River with cruise missiles, unleashing clayey floodwaters on Kryvyi Rih,
Zelenskyy’s hometown. Those and other
strikes on water infrastructure ruined cropland and wrecked local ecosystems.
But even though Ukraine clawed back
Kherson that November, the Kakhovka
Dam, still under Russian occupation, stayed
intact for another half a year.
Reservoir bogs
Dnipro River
Nikopol
Ukraine
Odesa
Shrubs and grasses
Marshy vegetation
Water bodies
Sand and silt
y
Zaporizhzhia
Nuclear Power
Plant
AROUND 9 A.M. on 4 October, Osadchyi and
Restoring the great meadow
Former lakebed below the embankment
would become steppe grassland.
Proposed
embankment
30
0
30
Steppe grassland site
When the Kakhovka blasts came, they
dwarfed the previous attacks. The breach
was wider than modeling of an explosion
had anticipated. “Something else made everything way worse,” says Amin Tavakkoli
Estahbanati, a remote sensing specialist at
the University of Houston. Analyzing synthetic aperture radar measurements from
2015 to 2023, he and his colleagues reported
at the American Geophysical Union meeting in San Francisco last month that the
dam had begun to deform months before
the blasts, possibly because of faulty operations and poor maintenance.
20
5 JANUARY 2024 • VOL 383 ISSUE 6678
are acutely sensitive to toxicants, could not
be rescued. “There is a very low possibility
that any survived,” says Afanasyev, director
of NAS’s Institute of Hydrobiology.
The breach would also have been devastating for the scarce wild sturgeons in the
Dnipro. In late spring, the fish swim upriver
from the Black Sea to spawning grounds
just below the Kakhovka Dam. Early June is
peak breeding season, Afanasyev says. “The
explosion wiped them out.”
Upstream of the dam, nearly 90% of
the Kakhovka Reservoir drained, exposing
1870 square kilometers of former lakebed,
science.org SCIENCE
,
km
y
0
Depth (meters)
Grygorii Derkach wriggled under a barbed
wire fence meant to keep civilians out of a
death zone along the Dnipro near Kherson,
less than 1 kilometer from Russian forces on
the opposite bank. Ukrainian army officers
urged the UHMI hydrologists to secure their
precious samples of river water quickly, before the Russians began their daily shelling. “We were in a sour mood,” Osadchyi
says—a lingering disquiet from the previous day, when the duo had driven through
the ruined village of Chornobaivka.
As the pair edged toward the Dnipro
that cool, sunny morning, they heard what
sounded like a motorbike engine. “Hit
the ground!” a soldier shouted. Osadchyi
dove under a tree as a Russian reconnaissance drone swooped in, just 20 meters
overhead. “It was terrifying,” he says. The
drone departed, and the rattled scientists
scrambled to fill their 40-liter plastic jug.
Lugging it back to their car in Kherson’s
River Port District, they heard shells explode near the area they’d occupied moments earlier.
Back at their laboratory, the UHMI researchers learned new details about water
quality in the Dnipro. Although the nasty
compounds in the reservoir sediments have
largely washed out of the water column, the
river is still freighted with nitrogen, phosphorus, and toxicants from sewage and
y g
Kakhovka
Dam
Nova Kakhovka
g
Kyiv
Dnipro River
p
The Kakhovka Dam breach on 6 June 2023 inundated settlements and unique ecosystems along the lower
Dnipro River. Nearly 90% of the reservoir drained, exposing 1870 square kilometers of former lakebed.
One proposal for postdisaster recovery calls for building a 50-kilometer-long embankment that would create
a narrower, deeper reservoir and allow for the return of steppe grassland.
including dense beds of zebra and quagga
mussels that once filtered and cleansed the
reservoir’s water. Now, as much as 500,000
tons of dead bivalves are rotting in the
desiccated lakebed. It could take a few
years for the soft body parts to decompose
fully, and their shells much longer, says
Volodymyr Yuryshynets, a parasitologist at
the Institute of Hydrobiology.
Normally, Afanasyev would have immediately dispatched scientists to assess
the Dnipro’s health. But because of the
“extreme danger of such investigations”
along the war’s front line, he says, his institute first enlisted volunteers—fishers
and others who remained along the river—
to report on fish stocks and send water
samples to Kyiv for analysis. Those samples enabled the scientists to verify that
sediments swept downstream were laden
with manganese and other heavy metals,
and long-lived organic compounds such as
polychlorinated biphenyls (PCBs) that had
accumulated over decades in sediments
behind the dam. “The colloidal particles
were quite toxic,” Afanasyev says.
To bring the picture into sharper focus,
however, the scientists knew there was
no substitute for seeing the disaster area
for themselves.
NE WS | F E AT U R E S
rine perch (Sander marinus). The species
had vanished from the region’s watersheds
until 2016, when fishers in the Dnipro-Bug
Estuary reported its surprising reappearance. “It’s very sensitive to toxicants and
lower salinity,” Afanasyev says. “Now it
only exists in a museum.”
y
,
science.org SCIENCE
y g
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
22
g
senic, and copper—and toxicants such as
petroleum byproducts and PCBs.
The double punch dealt a devastating
blow to marine life that couldn’t swim
away from the flood. Mollusks and other
near-shore sedentary creatures in the bay
were wiped out en masse, Minicheva says.
The surge of nutrients also ignited masULTIMATELY THE FLOODWATERS reached the
sive blooms of short-lived cyanobacteria,
sea. On a warm September day, 4 months
commonly known as blue-green algae.
after the breach, sunbathers lolled on OdeThen, bacteria that consume the algae rapsa’s beach and swimmers frolicked without
idly depleted dissolved oxygen, choking
fear in the Black Sea thanks to a metal net
coastal waters.
strung between piers that prevents RusIn August 2023, Ukraine’s military relaxed restrictions on Black
Sea research, hoping to glean
evidence for war crimes
charges.
IMB
scientists
now have permits to dive at
four locations in Odesa Bay.
They are chronicling what
Minicheva expects to be a
prolonged period of ecosystem recovery. And they are
warily eying an invasive sea
snail, the veined rapa whelk
DOWNSTREAM IN THE lower
(Rapana venosa). The predaDnipro Basin, the surge of
tor from the western Pacific
polluted floodwaters from
Ocean first appeared in the
the breach pummeled rare
Black Sea in the 1940s and
habitats, including Oleshky
in recent years has become
Sands National Nature Park,
a major pest in Ukrainian
an 80-square-kilometer prewaters, where it dines on enserve east of Kherson. The
demic oysters and other molsecond largest expanse of
lusks. “I worry that Rapana
Days after Russia launched its full-scale invasion in 2022, Galyna Minicheva risked
sand in Europe, it is home to
could be the big winner” as
conducting fieldwork in the highly saline Kuialnyk Estuary near Odesa, Ukraine.
the endangered sandy blind
creatures vie to recolonize
mole-rat (Spalax arenarius). “Many surely
sian mines from drifting into shore. On a
marine habitats damaged by the freshdrowned,” says IZAN zoologist Oleksiy
hillside patio overlooking the beach, howwater surge, says IMB marine biologist
Vasylyuk, who also leads the nonprofit
ever, Minicheva’s team was all business.
Mikhail Son.
Ukrainian Nature Conservation Group.
An IMB marine biologist in a wetsuit exResearchers are also hoping the marine
Other exceptional endemic species—
amined a basket of mollusks and other sea
measurements shed light on a mystery. In
including Nordmann’s birch mouse (Sicista
creatures. A colleague labeled a glass jar
March 2023, in a stunning claim that reloriger), a tree-dwelling ant (Liometopum
of seawater that will be analyzed for disceived widespread press coverage, a Ukraimicrocephalum), a sand gadfly, two spesolved oxygen and salinity. If it weren’t for
nian ecologist asserted that at least 50,000
cies of pearl knapweeds, and a rare birch
the war, a more salubrious place to do scidolphins died over 6 months in 2022, pri(Betula borysthenica)—inhabit nearby ecoence would be hard to imagine. “We have
marily because of underwater mine blasts
systems, including the 900-square-kilomea saying,” Minicheva said with a smile. “In
and acoustic damage from high-energy
ter Black Sea Biosphere Reserve that’s now
Kyiv, science is academic. In Kharkiv, it’s
submarine sonar. Other experts dismissed
in Russian hands. Vasylyuk fears some of
applied. In Odesa, science is shikarna.
that figure as a wild exaggeration—“simply
those populations are now extinct. But it’s
Luxurious.”
crazy,” says UkrSCES acting Director
too dangerous, he says—and in RussianFor more than a year, Ukraine’s miliViktor Komorin.
held areas, impossible—to get out into the
tary deemed research in Black Sea waters
The truth looks less alarming. In 2022,
field and check.
too perilous to undertake. But measureabout 900 porpoise and dolphin stranding
When the pulse of sludge-choked freshments close to shore in Odesa Bay, where
deaths were photo verified in the westwater reached the Dnipro-Bug Estuary, it
the Dnipro-Bug Estuary meets the Black
ern Black Sea—roughly twice the annual
hammered fish species adapted to brackSea, showed that the flood caused salinaverage recorded from 2018 to 2021, says
ish water. Taking heavy losses, according
ity to drop from 15 parts per million to
IZAN mammalogist Pavel Goldin. But in
to forecasts by Afanasyev’s team, were
4 parts per million—“practically fresh2023 there were only a handful. And necthe Black Sea roach (Rutilus frisii), the
water,” Minicheva says. Monitoring staropsies he and UkrSCES zoologist Karina
Dnieper barbel (Barbus borysthenicus),
tions in the bay run by the Ukrainian
Vishnyakova performed on several of those
and the Sarmatian bleak (Alburnus sarScientific Centre of Ecology of the Sea
dead animals have revealed no obvious
maticus). Particularly heartbreaking, he
(UkrSCES) then recorded high concentralinks to pathogens or toxicants released by
says, is the presumed demise of the estuations of metals—including cadmium, arthe floods or oil spills. They also did not
p
agricultural runoff. “The amount of wastewater pouring in is the same as before,”
says Osadchyi, who notes that determining
pollutant concentrations is a “complex and
meticulous task.” Compounding that bad
news is the loss of mussels that filtered the
water. “To a great degree, they purified the
reservoir” when alive, Afanasyev says. Until
the mussels rebound, wastewater entering
the Dnipro above Kakhovka Dam will wash
downstream largely unfiltered.
The emptied reservoir itself might also
be creating a health threat. There, the Dnipro now wends through a mucky patchwork of about 9000 newly
formed small lakes and
ephemeral ponds. Scientists
worry these wetlands are incubating disease-transmitting
mosquitoes and other bloodsucking insects, populations
of which could explode next
spring, Yuryshynets says.
“It’s a potential danger for
the region.”
p
find internal hemorrhaging—a hallmark
of exposure to loud sonar soundings or
underwater explosions. Further tests are
underway at the University of Padua and
the University of Veterinary Medicine Hannover. But so far, “The cause of the deaths
is not straightforward,” says Goldin, who
has been sharing the results with Ukrainian prosecutors investigating possible
war crimes.
A fuller picture of the harm inflicted on
the Black Sea will only come into focus after hostilities cease. On the eve of the fullscale invasion, UkrSCES had been gearing
up for the most ambitious Ukrainian expedition in years to monitor the Black Sea
environment. The war scuttled that voyage, and a missile strike on Odesa’s port in
July 2022 damaged its research vessel—the
Belgica, donated by Belgium in September
2021 and renamed the Borys Aleksandrov
after IMB’s previous director, who perished
in a fire at the institute in 2019. Restoration work must wait until after Ukraine’s
victory, says Komorin, who has already begun to plan an expedition with Ukraine’s
Black Sea allies: Bulgaria, Georgia, Romania, and Turkey.
g
After a centurieslong absence, wild asses called kulans are thriving in a Ukrainian reserve.
Reporting for this feature was supported in part by
the Richard Lounsbery Foundation.
5 JANUARY 2024 • VOL 383 ISSUE 6678
23
,
think the dam should be rebuilt,” Vasylyuk
says. Instead, he and others see great potential in letting nature take its course. “My
dream is that the lower Dnipro watershed
will be the next hot spot for rewilding,” says
Igor Studennikov, executive director of the
Centre for Regional Studies. As a model,
he points to a recent effort to restore the
Tarutino Steppe, southwest of Odesa near
the Danube delta region. A decade ago,
Oleg Diakov, an ecologist with Rewilding
Ukraine, and colleagues began to restore
native grasses and animals such as kulans
(Equus hemionus)—a native wild ass that
disappeared from the Ukrainian steppe
centuries ago—to a 5200-hectare sanctuary.
After the war, Diakov says, similar reserves
could be established around areas in the
lower Dnipro Basin where extensive minefields will deter agriculture and other land
uses for years to come. “We’d already identified this area before the war as having the
greatest potential for large-scale ecological
restoration,” he says. But those aspirations—
and a full accounting of the environmental
cost of the war—will remain unfulfilled,
Mosyakin says, until after Ukraine reclaims
territory under occupation.
“The sooner the war is over,” Goldin
says, “the sooner our ecosystems will get a
chance to recover.” j
y
SCIENCE science.org
basin soared, and in winter, when ice cover
suffocated fish.
This approach also offers “a chance to right
an historical wrong,” Osadchyi says. When
Soviet engineers impounded the reservoir,
he notes, they inundated a swath of steppe
and wetlands known as the Velykyy Luh, or
Great Meadow—an area prized by Cossacks
who have lived in southern Ukraine for centuries. A smaller reservoir would spare some
of that prized territory, preserving native
willows, poplars, and other vegetation that
are now colonizing hundreds of square kilometers of the drying basin.
Climatologist Svitlana Krakovska, head
of the Ukrainian delegation to the United
Nations Intergovernmental Panel on Climate Change, is skeptical of plans to reclaim the old lakebed. Before the disaster,
she notes, climate modeling forecasted
that the lower Dnipro Basin would become
much drier. Evaporation at the Kakhovka
Reservoir would have mitigated local
warming, she says. Without a reservoir,
“The region will only get hotter and drier.”
Krakovska also anticipates that the willows and poplars emerging in the former
reservoir’s lakebed will wither as the water
table recedes. “Sure, there will be a different ecosystem,” she says. “But I’m afraid it’s
impossible to go back to what it was like
there in the middle of the 20th century.”
Still, some experts would prefer to try
to rewind the clock even further. “I don’t
y g
sion on whether the Kakhovka Dam will be
rebuilt. For the Ukrainian government, a
paramount concern is economic recovery.
Reimpounding the reservoir could entice
residents back to abandoned homes, weekend dachas, and fishing boats along the former shoreline. And it would ease concerns
about a future restart of the Zaporizhzhia
Nuclear Power Plant, which drew cooling
water from the reservoir and, since the
dam’s destruction, has had to rely on hastily
dug wells for water to cool its reactor cores
and spent fuel.
Osadchyi, who is part of an expert group
from NAS that is assessing options for
future water infrastructure there, says a
failure to rebuild the reservoir “would be
another disaster.” Refilling it would allow
mussels to reestablish themselves and begin to filter the water again and provide
timely water releases for downstream
needs such as supplying municipalities, irrigating crops, and sustaining ecosystems.
Afanasyev favors an option he calls
“build back better.” It would involve creating a narrower and deeper reservoir by
repairing the Kakhovka Dam and confining the water within a 50-kilometer-long
barrier (see graphic, p. 20). The rationale
is that much of the Kakhovka Reservoir
was less than 2 meters deep. “It was essentially stagnant,” Afanasyev says. Fish kills,
he says, occurred frequently: in summer,
when water temperatures in the shallow
y
PHOTO: REWILDING UKRAINE
SCIENTISTS ARE ALSO awaiting a final deci-
Nuclear disasters–in–waiting
The Zaporizhzhia Nuclear
Power Plant, as seen
from a Ukrainian position
across the Dnipro River.
p
y
,
PHOTO: DMYTRO SMOLIENKO/SIPA VIA AP IMAGES
y g
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
SCIENCE science.org
The presence of IAEA observers at the Zaporizhzhia station since
September 2022 has deterred the theft of dirty-bomb ingredients.
But a major missile strike on one of its spent fuel repositories
could turn the plant itself into a dirty bomb, spreading radioactive
contamination in a radius of up to 30 kilometers, says Volodymyr
Borysenko, a nuclear engineer with the National Academy of Sciences of Ukraine’s Institute for Safety Problems of Nuclear Power
Plants (ISPNPP).
Even a smaller strike could contaminate the reactor complex.
And the spent fuel is also at risk from repeated electricity blackouts that have struck the plant, the latest in early December 2023.
Diesel-fueled generators can supply power for up to 10 days, but a
prolonged outage could be dangerous, as power is needed to pump
cooling water into the plant’s uranium reactor cores and pools holding spent fuel.
A lesser known radioactive risk is situated about 150 kilometers upstream from the Zaporizhzhia plant on the Dnipro River.
During the Cold War, the Prydniprovsky Chemical Plant was one
of Europe’s largest uranium ore processing facilities. The complex
accumulated some 40 million tons of tailings—leftovers of milling
uranium—and other foul residues before it closed in 1992. By early
2022, Ukraine, with help from the European Union, had fenced off
highly contaminated areas. But a missile or artillery strike on a
tainted building or dump could disperse radioactive dust over the
nearby city of Kamianske.
One relative bright spot is Chornobyl, where Ukrainian scientists
are restoring labs damaged early in the war. But large parts of the
exclusion zone remain off limits because of the threat of mines and
unexploded ordinance, says ISPNPP Director Anatolii Nosovskiy.
Complicating matters for radiation monitoring, he says, the Ukrainian army has built defensive fortifications in the zone, near the
border with Belarus.
Ukraine’s darkest fear is a Russian attack with a tactical nuclear
weapon, a saber that Putin and other officials rattle from time to
time. The State Emergency Service of Ukraine and other agencies need to develop plans for aid and evacuation, says Oleg
Voitsekhovych, head of radiation monitoring at the Ukrainian
Hydrometeorological Institute, which reports to the State Emergency Service. “To be honest, we are not yet ready.”
He’s fervently hoping for reason to prevail. “The most important
thing,” Voitsekhovych says, “is to stop this madness and prevent the
escalation of nuclear terrorism on the territory of Ukraine.” –R.S.
g
H
aving taken a heavy toll on Ukraine’s ecosystems and water
resources, the war with Russia threatens to create a another
environmental disaster: damage to the region’s extensive
nuclear infrastructure—including 15 power reactors and
three research reactors.
“There continues to be a highly precarious nuclear safety and
security situation across Ukraine,” International Atomic Energy
Agency (IAEA) Director General Rafael Mariano Grossi said in
a statement after explosions were heard near the Khmelnitsky
Nuclear Power Plant and its two Soviet-era reactors on 28 November 2023—the second near-miss in a single month at the site. “All of
Ukraine’s nuclear facilities remain vulnerable, either directly if hit by
a missile or indirectly if their off-site power supplies are disrupted.”
Russia’s assault on Ukrainian nuclear sites began on the very first
day of the full-scale invasion. On 24 February 2022, troops overran
the Chornobyl Nuclear Power Plant, infamous for the explosion
and fire there in 1986 that sent a plume of radioactive smoke into
Western Europe. During 5 weeks of occupation, Russian soldiers
ransacked labs and kicked up radioactive soil and dust as they dug
trenches and slogged through contaminated forests in the exclusion zone around the defunct plant. To the east that spring, Russian
troops frequently shelled the Kharkiv Institute of Physics and Technology, damaging a hall containing a subcritical nuclear reactor.
Shelling has also flared up repeatedly around the Zaporizhzhia
Nuclear Power Station, a complex of six reactors that constitutes
Europe’s largest nuclear power plant. Russia captured the plant
in March 2022 and the reactors were shut down 6 months later,
eliminating the risk of a core meltdown. Still, a prodigious amount
of nuclear material remains there: The reactor halls hold 1380
tons of fresh and spent uranium oxide fuel, and two repositories
store an additional 2100 tons of spent fuel laced with nasty longlived radionuclides—the ingredients, many Ukrainians fear, of a
“dirty bomb” that would use conventional explosives to spread
radioactive isotopes.
Russian propaganda often claimed that Ukraine plans to fashion a dirty bomb from such materials. After Russian President
Vladimir Putin amplified those accusations in October 2022,
“Russian hysteria over a dirty bomb was rampant,” says Maksym
Strikha, a physicist at the Academy of Sciences of the Higher
School of Ukraine who tracks Russian nuclear disinformation. For their part, Ukraine and its allies worry Russia
might detonate a dirty bomb and blame Ukraine.
21
p
PERSPECTIVES
g
EVOLUTION
y
Evolutionary paths
to new phenotypes
Ecological model systems
inform on innovative traits
in plants and animals
y g
By Kathryn R. Elmer
SCIENCE science.org
PHOTO: CHIEN C. LEE
plexity can coalesce has been a challenge
to explain since the time of Darwin (1). The
stepwise changes in phenotypes over long
times, involving many different forms and
parts with complex genetic bases, make
their existence seem vanishingly unlikely
(4). Indeed, during the evolution of these
composite traits, the intermediate states
might not be beneficial or even functional,
for example, as separate components or in
disparate combinations. However, advances
in quantitative and evolutionary genetics
have shown that the combinations of traits
and states that are beneficial for an organism can shift substantially and swiftly over
time and space. Although it is challenging
to predict, the history of selection and evo5 JANUARY 2024 • VOL 383 ISSUE 6678
27
,
School of Biodiversity, One Health & Veterinary
Medicine, University of Glasgow, Glasgow, UK. Email:
kathryn.elmer@glasgow.ac.uk
Springboard trapping is a composite trait that arose
in carnivorous pitcher plants through coincident
spontaneous evolution.
y
A
daptation by natural selection cannot
take any evolutionary path; it operates within the constraints of genetic
variation and environmental context,
with futures contingent on the past.
Therefore, how new suites of traits
arise is an enduring issue and is key to understanding the diversity of life (1). On pages 108
and 114 of this issue, Chomicki et al. (2) and
Stankowski et al. (3), respectively, investigate
two different cases of fascinating biological
complexity that arose through convergent
and convoluted evolutionary paths—one in
carnivorous pitcher plants (Nepenthes gracilis and Nepenthes pervillei) and another in
marine periwinkle snails (Littorina saxatilis). The studies use different approaches to
reconstruct evolution to reveal how complex
phenotypic traits arise in unexpected ways.
The results advance understanding not only
of the specific traits that are studied—feeding structures in plants and live-bearing (as
opposed to egg-laying) in snails—but also
how evolution in general might arrive at apparently unlikely combinations.
Highly complex organs or composite traits
are sets of biological features that, when
they come together, have a new synergy and
function. As highlighted by Chomicki et al.,
an example of a composite trait is that of
bird wings, which transformed from existing skin and forelimbs into new structures
that enable flight (1). Other classic examples
include animal eyes, for which many different parts evolved from different ancestral
forms to operate together. How such com-
I NS I GHTS | P E R S P E C T I V E S
10.
11.
12.
13.
14.
10.1126/science.adm9239
C
ontrolling the size and curvature of
materials with dimensions in the
nanometer scale is a challenge despite
the abundance of natural nanostructures, such as those that interfere with
light and produce color on butterfly
wings. Block copolymers are a class of materials that can spontaneously self-assemble into
ordered nanostructures with a range of potential applications, including photocatalysis,
nanomembrane separations, photonics, and
nanomedicine. However, such achievements
have been limited by the challenge of reproducibly synthesizing block copolymers that
form a desired network structure. On page 70
of this issue, Lee et al. (1) report a process for
reliably producing network nanostructures
by linking small functional groups to the end
of a diblock copolymer. By varying the chemistry of end groups and linking groups, the
authors demonstrate a simple set of design
rules for unlocking an unprecedented range
of network nanostructures using easily synthesized polymers.
A diblock copolymer comprises two
chemically distinct polymers that are covalently bound, forming a simple polymer
architecture (a single chain) that can selfassemble. A block is a segment of the polymer chain where all the monomers are
chemically identical. At equilibrium, diblock
copolymers exhibit a range of nanostructures that can be controlled by varying the
block fraction (2), which is the volumetric
fraction of the total chain that is occupied
by a single block. When one block is longer than the other in a diblock copolymer,
it is called the majority block and forms a
matrix surrounding the shorter, minority
block. Within a narrow range of composition
for such polymers, a bicontinuous network
Department of Chemical and Biological
Engineering, University of Sheffield, Sheffield, UK.
Email: a.nedoma@sheffield.ac.uk
science.org SCIENCE
,
8.
9.
By Alisyn J. Nedoma
y
5.
6.
7.
C. Darwin, The Origin of Species (Dent, ed. 6, 1872).
G. Chomicki et al., Science 383, 108 (2024).
S. Stankowski et al., Science 383, 114 (2024).
S. J. Gould, The Structure of Evolutionary Theory
(Harvard Univ. Press, 2002).
B. Riska, Evolution 43, 1172 (1989).
D. G. Blackburn, J. Morphol. 276, 961 (2015).
A. N. Ostrovsky et al., Biol. Rev. Camb. Philos. Soc. 91, 673
(2016).
A. J. Helmstetter et al., Nat. Commun. 7, 11271 (2016).
J. W. Sites Jr., T. W. Reeder, J. J. Wiens, Annu. Rev. Ecol.
Evol. Syst. 42, 227 (2011).
B. Crespi, C. Semeniuk, Am. Nat. 163, 635 (2004).
H. Recknagel et al., Nat. Ecol. Evol. 5, 1546 (2021).
B. Halliwell, T. Uller, B. R. Holland, G. M. While, Nat.
Commun. 8, 2030 (2017).
F. Racimo, S. Sankararaman, R. Nielsen, E. HuertaSánchez, Nat. Rev. Genet. 16, 359 (2015).
M. A. B. Haase et al., Genetics 217, iyaa012 (2021).
Chemically modifying
polymer ends enables
tailoring of nanostructured
materials
y g
1.
2.
3.
4.
Creating
the “plumber’s
nightmare”
y
5 JANUARY 2024 • VOL 383 ISSUE 6678
R E F E R E N C ES A N D N OT ES
CHEMISTRY
g
28
live-bearers. The diversity in these genetic
regions is highly consistent with natural selection driving the evolution of live-bearing,
rather than the pattern arising by chance. A
key message of the paper is that the genetics
of this complex trait, live-bearing, are not reflected in the history of most of the genome,
even though the trait is under selection and
fundamental to species’ biology.
There has been a long-standing debate
about whether the accumulation of small
stepwise changes or big leaps (saltation) is
more important in the evolution of diversity
(4). Chomicki et al. and Stankowski et al. did
not identify a single big evolutionary step or
large-impact mutation that moved the species to a new level of phenotypic innovation.
In pitcher plants, it is a case of spontaneous joining of different traits that happen to
have come together in the right way. In the
periwinkles, it was concluded that a gradual
accumulation of the right genetic combinations over long periods of time resulted in
live-bearing. This agrees with recent lessons
from genomic studies of organisms as varied
as humans and microbes: that changes to
relatively small genetic regions, not reflected
in the history of the overall genome, can have
outsized impacts on phenotypes (13, 14).
The genetic means by which innovative new traits evolve have been hard to
pin down in ecological model species. It is
notable that in the era of big genome data,
neither Chomicki et al. nor Stankowski et al.
pinpoint the genetic variants that cause the
traits they focus on. This leaves open the next
important question for both species: What
types of genetic changes are responsible
for these complex and composite traits? To
address this will require excellent genomelevel reconstructions and functional tests of
exactly which genes matter for phenotypic
outcomes. The amazing breadth of plant and
animal diversity across the globe has evolved
by circuitous paths, and resolving the complex history of genomes and traits unlocks
new depths for understanding evolution. j
p
lution can be highly flexible and variable
under realistic scenarios (5).
Chomicki et al. examined the evolution of
the so-called springboard-trapping feature
in carnivorous pitcher plants. This feature
consists of structural, chemical, and mechanical aspects: a flat, horizontal lid with
a waxy underside, which is triggered by rain
to fling insects into the pitcher. For springboard trapping to evolve once is an elegant
outcome, but the authors’ phylogenetic
reconstructions indicate that the feature
evolved separately in two different species
of pitcher plant. What type of evolutionary
path can result in the generation of such
a distinctive combination twice independently? The authors used advanced models
to predict how the traits evolved across the
phylogeny. One option was that selection
pushed the suite of traits together into their
current distinctive form. Although intuitive, this was not supported by the results.
Instead, the models strongly suggest that
the new composition arose by “spontaneous coincidence” of the independent components coming together, facilitated by high
levels of phenotypic variation.
Another complex composite trait that has
evolved through seemingly unlikely intermediate stages is that of females bearing
live offspring (viviparity) rather than laying
eggs (oviparity). This key innovation has
evolved in a discrete way independently
many times: in invertebrates (such as snails
and insects), anamniotes (such as fish and
amphibians), and amniotes (such as reptiles and mammals) (6, 7). At a macroevolutionary perspective, live-bearing tends to
be associated with higher rates of lineage
and species diversification, varying rates
of extinction, and expansion into new geographic areas (8, 9). At a microevolutionary
level, it has been linked to shifts in lifehistory traits that are important for daily
existence—for example, changes in reproductive output, mating and parental care
behavior, and using new habitats (10–12).
Otherwise indistinguishable egg-laying
and live-bearing forms of marine periwinkle snails are found on beaches across
many different shores of the northeastern
Atlantic Ocean. Stankowski et al. used the
DNA of live-bearing and egg-laying forms
to reconstruct the evolution of these populations of snails, with the aim of identifying the origin of live-bearing. By applying
an advanced analytical method, they found
that the genomic regions associated with
egg-laying or live-bearing have a different
evolutionary history than does the genome
overall. These reproductive mode–associated
regions involved the same genes that were
found to be expressed differently in the female reproductive tissues of egg-layers and
structure forms, known as a gyroid (3, 4). In
this structure, the minority block forms two
interlacing networks in which three tubes
intersect, whereas the majority block forms
a matrix surrounding this network (see the
figure). The junction between the two block
constituents is located at the interface where
the minority block extends into the tube formation and the majority block extends into
the matrix. Each diblock exhibits a pathway
of connected polymer, from one end of a
sample to the other, so that the structure is
continuous. Network structures can improve
the mechanical properties of the material
(5); simultaneously transport two quantities,
such as ions and electrons (6); and give rise
to specific optical and electronic properties
(7). They are notoriously difficult to synthesize because of the narrow range of block
nightmare” mesophase. The former has
four intersecting tubes that constitute the
minority block, whereas the latter has six
intersecting tubes (hence, the nightmare).
Thermodynamically equilibrated mesophases are attractive because they tend to
recover their equilibrium size and shape after a perturbation, which makes them more
robust to different processing conditions (12).
Lee et al. demonstrated thermal reversibility
by ramping up the temperature until the materials undergo a phase transition and then
cooling the sample down to room temperature. In all instances, the authors recovered
the original mesophase, which provides very
strong evidence that the room-temperature
network nanostructures are thermodynamically equilibrated. Lee et al. further showed
that the transformations between network
Two chemically distinct polymers that are covalently bound form a simple polymer architecture—the diblock
copolymer. The diblock can be modified with chemical groups appended to the ends of constituent polymer
chains. This enables the creation of structures never before realized, such as the diamond cubic and the plumber’s
nightmare cubic, which have four and six intersecting tubes, respectively, embedded in a polymer matrix.
Polymer 2 Linker
End-end chain
interactions
Chain 2
Tube
Matrix
Tube
GRAPHIC: N. BURGESS/SCIENCE
mesophases occur epitaxially, which could
lead to the application of these materials as
thermally actuated switches.
Lee et al. introduced two parameters to
qualitatively guide the design of network
structures: the strength of interactions between end groups and the degree of packing frustration, which is the compression of
polymer chains at the intersection between
tubes in the mesophase (and is modulated
by the linking group). At the intersection between tubes, the diblock copolymer chains
must compress to fit into the small volume
of the intersection node. Strong end-end interactions cause the polymer chains in the
majority block to stretch to accommodate the
close packing of the end groups. The resulting mesophase represents the equilibrium
balance between minimizing the interface
between blocks, minimizing chain stretching,
and constraining the system to fill space uni-
RE FE REN C ES AN D N OT ES
,
SCIENCE science.org
(Im3m)
Plumber’s
nightmare cubic
1. H. Lee et al., Science 383, 70 (2024).
2. F. S. Bates, G. H. Fredrickson, Annu. Rev. Phys. Chem. 41,
525 (1990).
3. D. A. Hajduk et al., Macromolecules 27, 4063 (1994).
4. M. W. Matsen, F. S. Bates, J. Chem. Phys. 106, 2436 (1997).
5. B. J. Dair et al., Macromolecules 32, 8145 (1999).
6. Y. Jung, S. Torquato, Phys. Rev. E Stat. Nonlin. Soft Matter
Phys. 72, 056319 (2005).
7. K. Hur et al., Angew. Chem. Int. Ed. 50, 11985 (2011).
8. J. Park, K. I. Winey, JACS Au 2, 1769 (2022).
9. A. C. Finnefrock, R. Ulrich, G. E. S. Toombes, S. M. Gruner,
U. Wiesner, J. Am. Chem. Soc. 125, 13084 (2003).
10. F. J. Martínez-Veracoechea, F. A. Escobedo,
Macromolecules 42, 1775 (2009).
11. M. Stefik et al., Chem. Mater. 21, 5466 (2009).
12. F. S. Bates, G. H. Fredrickson, Phys. Today 52, 32 (1999).
13. F. J. Martínez-Veracoechea, F. A. Escobedo,
Macromolecules 42, 9058 (2009).
14. P. F. W. Simon, R. Ulrich, H. W. Spiess, U. Wiesner, Chem.
Mater. 13, 3464 (2001).
15. H. Y. Hsueh et al., Adv. Mater. 23, 3041 (2011).
10.1126/science.adn0168
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
fractions in which they form (8). Gyroid
mesophases have been formed by blending
additional components into the diblock copolymer (9, 10) or by synthesizing a more
complex triblock polymer (11).
Lee et al. discovered a synthetically facile
route for linking end groups onto an existing diblock copolymer to induce the formation of equilibrium network nanostructures.
The method tolerates block fractions that
vary from 38 to 48 vol % for the minority
block, which extends the design space from
the conventional 37 to 39 vol % window in
which gyroids form (8). The ability to design
a single network-forming entity removes the
need for blending and simplifies the synthetic chemistry.
Beyond the gyroid mesophase, Lee et al.
observed two structures with diblock copolymers that had only been theorized: the
diamond cubic and the so-called “plumber’s
(Pn3m)
Diamond cubic
y g
(Ia3d)
Double gyroid
y
Chain 1
Chain packing
End
group
g
Polymer 1
p
A diversity of network structures
formly (13). Lee et al. found that the strength
of end-end interactions that is required to
induce network formation depends on the
equilibrium curvature of the unfunctionalized diblock copolymer. The authors’ design
rules provide a mechanistic basis for generating new block copolymer nanostructures.
Scaling up the use of network block copolymer nanomaterials remains a challenge
due to subtractive manufacturing techniques.
Polymer structures are used to template inorganic materials and then the polymer is
etched away or calcined (14, 15). The polymer
lost in these subtractive processes is generally unrecoverable, which leads to higher
manufacturing costs and greater embodied
carbon within the final product. The design
strategy of Lee et al. encourages a more efficient use of materials by enabling researchers
to use functional polymers that possess some
of the attributes of inorganic materials, such
as conductivity. Creating network structures
in which the polymer blocks perform the desired function reduces waste and is likely to
decrease the final cost of a technology.
Only a limited number of polymer pairs
have been used to form network structures
because self-assembly depends on the specific enthalpic interaction between the
polymer species (13). Lee et al. used linker
groups and end groups to introduce an additional degree of freedom that allows strong
interactions between the end groups to compensate for weaker interactions between
the polymer blocks. They demonstrate the
versatility of their technique by applying
end groups to different species of diblock
copolymers, thereby showing that network
structures form for both semicrystalline and
amorphous polymer blocks. The tunability
of end-group chemistry broadens the range
of potential polymer combinations that can
form network nanostructures, which will enable polymer chemists to select blocks based
on desired physicochemical properties rather
than their ability to self-assemble. j
29
I NS I GHTS | P E R S P E C T I V E S
IMMUNOLOGY
Restoring tolerance with antigen delivery
Strategies that modulate antigen delivery are being tested to reverse autoimmunity
By Bana Jabri1,2,3,4 and Valérie Abadie1,2,3
A
,
science.org SCIENCE
y
5 JANUARY 2024 • VOL 383 ISSUE 6678
y g
30
y
University of Chicago Department of Medicine, Chicago, IL,
USA. 2University of Chicago Celiac Disease Center, Chicago,
IL, USA. 3University of Chicago Committee on Immunology,
Chicago, IL, USA. 4University of Chicago Department of
Pathology, Chicago, IL, USA. Email: bjabri@bsd.uchicago.edu
g
1Section of Gastroenterology, Hepatology and Nutrition,
Although promising results with these
approaches have been reported in animal
models of autoimmunity, they have not yet
given rise to an efficacious antigen-specific
immunotherapy that provides a significant
clinical benefit. This may be for several reasons, including the degradation of antigens
before they reach their target, a failure to
delete pathogenic tissue-resident T cells, and
the emergence of de novo disease-driving antigens. To improve these approaches by promoting the differentiation of antigen-specific
Treg cells, delivery of the antigen by bacteria
that are genetically manipulated to secrete
regulatory cytokines, such as IL-10, has been
investigated. This improved the mucosal delivery of the antigen and induced more immunosuppressive Treg cells in recent-onset
nonobese diabetic (NOD) mice (9). The expansion of FOXP3+ Treg cells can also be augmented by coadministration of IL-2 muteins,
which are IL-2 variants specifically designed
to interact exclusively with the high-affinity
IL-2 receptor. This approach demonstrated
prolonged control of autoimmunity in the
NOD mouse model (10). However, if IL-2 muteins are provided alone, Treg cells expand in
a non–antigen-specific manner. Furthermore,
they might not be able to suppress effector
T cell responses because of the inflammatory
nature of the tissue environment targeted by
the autoimmune condition.
Another approach involves bioengineering that relies on either conjugating the
antigen to polymers (11) or nanotechnology
(6) to target different subsets of APCs. For
example, to better harness the tolerogenic
environment of the liver, antigens can be
conjugated to linear polymeric glycosylations, such as N-acetylgalactosamine and
N-acetylglucosamine, that are recognized by
C-type lectin receptors expressed by hepatic
dendritic cells, Kupffer cells, liver sinusoidal
endothelial cells, and hepatocytes. This results in increased uptake and presentation
of these synthetically glycosylated antigens
by liver APCs, which are poised to maintain
tolerance. This approach provided encouraging results in the BDC2.5 T cell adoptivetransfer mouse model of type 1 diabetes
(11). Intravenous administration of antigens
coupled to erythrocytes, which undergo sustained cell death called eryptosis, also leads
to continuous uptake of the coupled antigens
by splenic dendritic cells, hepatocytes, and
hepatic stellate cells. In animal models, these
p
utoimmune disorders encompass a
wide range of immunological diseases
that affect 4 to 10% of the population
worldwide. They occur when immunological tolerance toward self-antigens is broken, resulting in immune
responses against cells, tissues, or organs
that lead to tissue dysfunction and/or destruction. The goal of autoimmune disease
therapy is to restore tolerance to the self-antigen that causes the pathology by targeting
autoreactive T cells while preserving immune
competence to prevent infections and malignancies. The main challenge of this endeavor
resides in choosing the mode by which the
disease-driving antigen is delivered so that it
can initiate the removal or reprogramming
of the autoreactive T cells or the induction
and/or expansion of antigen-specific regulatory T (Treg) cells to suppress autoreactive
T cells. Although the potential of antigenbased immunotherapy approaches to restore tolerance have been demonstrated for
the treatment of immunoglobulin E (IgE)–
mediated peanut allergy (1), antigen therapies for autoimmune disorders are still being developed.
There are several key hurdles that a successful autoimmune disease antigen therapy
needs to overcome. For most autoimmune
disorders, there is absent or incomplete
knowledge of the disease-driving autoantigen. In addition, although one particular
disease-causing antigen triggers and drives
the autoimmune process, additional antigens
are exposed as tissue destruction takes place,
which causes a phenomenon called epitope
spreading, whereby the pathogenic T cell response widens to a diversity of antigens (2).
Furthermore, a successful therapy needs to
target functionally relevant subsets of both
tissue-resident and circulating pathogenic
memory T cells. This is not easily achieved
owing to the incomplete understanding of
the regulation of tissue-resident T cells and
given that there are important distinctions
in their mode of regulation (3) and transcriptional program (4). Moreover, even if Treg
cells are induced, they need to be functional
in an inflammatory environment. This is,
for example, problematic for forkhead box
P3–expressing (FOXP3+) Treg cells that fail to
control pathogenic effector T cells that have
been stimulated by the inflammatory cytokines interleukin-15 (IL-15), IL-21, or IL-7 that
are present in the autoimmune tissue environment (3).
To attempt to circumvent at least some of
these issues, diverse approaches have been
developed with the common goal of restoring immune tolerance by fine-tuning antigen
delivery to tolerogenic or resting antigenpresenting cells (APCs) that will induce Treg
cell differentiation, T cell deletion, or anergy
(a state of hyporesponsiveness). The rerouting of the T cell response involves two possible mechanisms. One results in pathogenic
T cells acquiring tolerogenic properties, such
as the ability to produce the immunosuppressive cytokine IL-10. The other depends
on the expansion and/or de novo generation
of inducible FOXP3+ Treg cells or type 1 regulatory T (Tr1) cells from naïve T cells. The
definition of Tr1 cells remains ambiguous because they can coproduce IL-10 and inflammatory cytokines or produce IL-10 only (5, 6).
Furthermore, it is important to note that the
beneficial effects of IL-10 are context dependent because it can have detrimental effects
in certain autoimmune diseases by promoting extrafollicular B cell responses and the
production of autoantibodies (7).
Several approaches are being tested in
clinical trials (see the table). One strategy
is based on delivering the disease-driving
antigens in their “naked” form (in the absence of any vehicle), leading to antigen
presentation in a tolerogenic environment.
This may be achieved by giving the antigen
orally, a strategy that has proven successful
in the context of peanut allergy, so that the
antigen is presented by intestinal tolerogenic
dendritic cells that promote the differentiation of FOXP3+ Treg cells (oral tolerance).
Alternatively, the disease-driving antigen
can be delivered outside of the site of the inflamed tissue to resting dendritic cells that
lack costimulatory molecules by injecting immunodominant peptides subcutaneously or
intramuscularly. Activating memory T cells
and naïve T cells through the T cell receptor
in the absence of costimulation by CD28 and
CD40 ligand leads to the deletion (and anergy) of the antigen-specific T cells (8).
Modes of antigen delivery
Systemic or mucosal administration of “naked” antigens targets resting or tolerogenic dendritic cells (DCs) to promote the deletion or induction of forkhead box
P3–expressing (FOXP3+) regulatory T (Treg) cells, respectively. To target additional types of antigen-presenting cells (APCs), several bioengineering approaches have
been developed. Antigens can be conjugated or delivered in nanoparticles. Nanoparticles can not only deliver antigens to many different types of APCs at different
locations but also deliver therapeutic cargo. Bioengineering approaches have the potential to promote more wide-ranging mechanisms of tolerance by inducing
regulatory type 1 and type 1–like cells and reprogramming pathogenic autoreactive T cells.
Naked antigen delivery
Approach
Low-dose oral antigen administration
Immune priming by
resting DCs (no signal 2)
Immune priming by intestinal
tolerogenic DCs
Deletion or anergy of autoreactive
T cells; de novo induction of Treg cells?
De novo induction of
Treg cells
Phase 1 for type 1 diabetes
(Cardiff University: NCT01536431)
Phase 2 for relapsing
remitting multiple sclerosis
(Medical University of Lodz)
Phase 2 for celiac disease
(ImmusanT: NCT03644069)
Phase 1/2 for type 1 diabetes
(Pre-POINT Study Group:
ISRCTN76104595)
Mechanism
Outcome
Deletion of
autoreactive T cells
De novo induction of
Treg cells
Reprogramming of
autoreactive T cells
Phase 1b/2a for multiple sclerosis
(Cellerys AG-2022-000801-28)
Phase 2 for celiac disease
(Anokion SA: NCT05574010)
5 JANUARY 2024 • VOL 383 ISSUE 6678
31
,
nature of its approach, mRNA therapeutics
may transform immune regulatory therapy
by enabling efficient delivery of antigen as
well as easier development and manufacturing. However, the inherent adjuvanticity
of mRNA-based vaccines that can activate
innate immune receptors may also lead to
the unwanted activation of APCs and the
enhancement of autoreactive immune responses. To tackle this issue, some modified
mRNA vaccines have been developed by introducing methylpseudouridine in place of
uridine. This nucleoside modification results
in an anti-inflammatory mRNA vaccine that
has reduced innate immune activation properties because it fails to activate Toll-like receptor 7 (TLR7) (15).
Despite the impressive development of
new technologies that offer the promise of
antigen-specific tolerogenic therapies, the
problems of epitope spreading and immune
regulation in an inflammatory tissue environment remain to be solved. Approaches
that can control both pathogenic circulating central memory and tissue-resident
effector memory T cells also need to be
identified. This will likely only be achieved
through combination therapies that are
adapted to each tissue and disease. As technologies for antigen delivery and therapeutic strategies with broader and stronger
tolerogenic potential are developed, the risk
of nonspecific immunosuppression also increases. Another critical issue to consider is
establishing the criteria for authorizing human trials and evaluating the feasibility of
acquiring pertinent preclinical data. To ensure the future success of these approaches,
it will be crucial to incorporate comprehensive and unbiased immunological and
multiomics investigations in clinical trials.
Although animal models are valuable for
y
increase the presentation of disease-driving
antigens by tolerogenic phagocytes and dendritic cells. This has been experimentally
achieved in mice by using negatively charged
microparticles engineered with polymeric
organic materials such as polylactide-co-glycolic acid (PLGA) that were passively taken
up by splenic marginal zone macrophages expressing the scavenger receptor MARCO (6).
Other approaches consist of using liposomes that contain peptide antigen and phosphatidylserine to facilitate their phagocytosis
by macrophages. By adding therapeutic cargo
that is delivered to different cell types and in
distinct anatomical locations, nanoparticles
have the capacity to induce all types of immune modulation and hence be adapted to
a given organ-specific autoimmune disorder.
This may be critical because each organ and
autoimmune disease has a particular immunological and tissue signature that requires
different modes of tolerance induction. For
example, KAN-101, a glycosylation signature
conjugated to deamidated gliadin peptides
that is specifically designed for liver targeting, is now advancing into phase 2 clinical
trials for the treatment of celiac disease (13).
Delivering antigens to anatomically distinct sites can also be achieved by altering
the nanoparticle chemistry. Selective organ
targeting has been developed, whereby manipulating multiple classes of lipid nanoparticles allows tissue-specific delivery of diverse
cargos, including mRNA, Cas9 mRNA–single
guide RNA (sgRNA), and Cas9 ribonucleoprotein complexes, which allows CRISPRCas–mediated gene editing in therapeutically
relevant cell types (14). Nevertheless, encapsulating many antigens might be necessary to
overcome epitope spreading.
Although nanoparticle technology is attractive because of the flexibility and holistic
y g
SCIENCE science.org
Targeting of and priming by resting DCs, tolerogenic DCs,
or nonprofessional APCs
y
approaches were shown to lead to deletion
of autoreactive T cells, de novo induction of
Treg cells, or reprogramming of autoreactive
T cells. Although phase 1 and 2 clinical trials
in patients with multiple sclerosis and celiac
disease are ongoing (2022-000801-28 and
NCT05574010, respectively), the efficacy of
these approaches in humans remains to be
demonstrated.
The use of nanoparticles offers distinct
ways to manipulate the immune system because they can be engineered to carry ligands
that target specific cell-surface receptors (e.g.,
DEC-205, which is highly expressed in dendritic cells) and therapeutic cargos (e.g., tolerance-promoting drugs, such as rapamycin,
or small interfering RNA) (6). Modulating the
amount of antigen delivered is also more easily achieved with nanoparticles than with naked antigen delivery. Delivering higher doses
of antigen may enhance tolerance induction
by promoting the gradual establishment of a
regulatory program in Tr1 cells that is characterized by the expression of specific negative
costimulatory molecules and transcription
factors—such as MAF, aryl hydrocarbon receptor (AHR), and nuclear factor interleukin-3–regulated protein (NFIL3)—in addition to IL-10 production and in the absence
of inflammatory cytokine production (12).
Activation of an immunosuppressive network
that involves IL-10–producing Tr1 cells might
be critical for establishing bystander suppression owing to the ability of IL-10 to downregulate the expression of costimulatory molecules and major histocompatibility complex
class II (MHCII) molecules on the surface of
APCs (5). Furthermore, the size, shape, and
surface chemistry of nanoparticles can be
manipulated to influence trafficking and the
functional program that is activated in the
targeted APCs. For instance, it is possible to
Nanoparticles
g
GRAPHIC: N. BURGESS/SCIENCE
Clinical trials
Bioengineering
Antigen coupling
p
Systemic antigen administration
I NS I GHTS | P E R S P E C T I V E S
32
5 JANUARY 2024 • VOL 383 ISSUE 6678
H
ow the nervous system deals with uncertainty in the outcome of actions is
a fundamental question in cognitive
neuroscience. Risky decisions represent a cost-benefit trade-off, whereby
the possibility of an outcome worse
than a safer alternative (i.e., cost) must
be weighed against the possibility of outcomes that are better than the safer alternative (i.e., benefit). This trade-off is entirely
subjective and depends on the individual’s
willingness to accept risk (i.e., uncertainty
about the outcome). Attitude toward risk
is strongly influenced by context, but very
little is known about the underlying neuronal circuitry. On page 55 of this issue,
Sasaki et al. (1) report that two neighboring regions in the frontal cortex of Japanese
macaques (Macaca fuscata) can influence
risk attitude in a competitive manner. This
1Department of Neuroscience, Johns Hopkins University
School of Medicine and Zanvyl Krieger Mind/Brain Institute,
Baltimore, MD, USA. 2Department of Psychological and Brain
Sciences, Johns Hopkins University, Baltimore, MD, USA.
Email: vstupho1@jh.edu
finding opens up the possibility to uncover
the neural circuit mechanism that underlies
attitude toward risk.
To study the influence of risk independently from the expected value (the probability-weighted sum of all possible outcome
values of the gamble), the authors designed a
choice task in which monkeys had to choose
between sets of gambles with a constant expected value but a changing probability of
winning the higher outcome. The gamble
options were presented as colored circles,
and the monkeys indicated their choice by
shifting their eyes to one or the other. Some
gambles had small probabilities of winning
but a high winning amount [high risk–high
return (HH)], whereas others had larger
winning probabilities but smaller winning
amounts [low risk–low return (LL)]. Because
the expected value was the same for HH and
LL options, any preference was the result of
risk attitude only. Six monkeys were tested,
and all showed a preference for HH options.
Thus, in line with previous findings, monkeys were overall risk seeking (2–4).
Sasaki et al. used muscimol injections to
bilaterally inactivate either the orbitofrontal
science.org SCIENCE
,
10.1126/science.adg7505
By Veit Stuphorn1,2
y
The authors thank C. Khosla and J. A. Hubbell for insightful
comments. The work of B.J. and V.A. is supported by grants
from the National Institutes of Health (NIH): R01 DK067180 to
B.J., R01 DK128352 to V.A., and Digestive Diseases Research
Core Center P30 DK42086 and R01 DK063158 to B.J. and V.A.
Specific brain pathways can lower or raise the willingness
of monkeys to take risks
y g
AC K N OW L E D G M E N TS
Dopamine regulates attitude
toward risk
y
1. G. Du Toit et al., N. Engl. J. Med. 372, 803 (2015).
2. C. J. Vanderlugt, S. D. Miller, Curr. Opin. Immunol. 8, 831
(1996).
3. B. Jabri, V. Abadie, Nat. Rev. Immunol. 15, 771 (2015).
4. L. K. Mackay et al., Science 352, 459 (2016).
5. M. G. Roncarolo, S. Gregori, R. Bacchetta, M. Battaglia,
N. Gagliani, Immunity 49, 1004 (2018).
6. P. Serra, P. Santamaria, Nat. Biotechnol. 37, 238 (2019).
7. S. Biswas, K. Bieber, R. A. Manz, Front. Immunol. 13,
970906 (2022).
8. M. K. Jenkins, R. H. Schwartz, J. Exp. Med. 165, 302
(1987).
9. S. Robert et al., Diabetes 63, 2876 (2014).
10. C. D. Moorman, S. J. Sohn, H. Phee, Front. Immunol. 12,
657768 (2021).
11. D. S. Wilson et al., Nat. Biomed. Eng. 3, 817 (2019).
12. B. R. Burton et al., Nat. Commun. 5, 4741 (2014).
13. J. A. Murray et al., Lancet Gastroenterol. Hepatol. 8, 735
(2023).
14. M. M. Żak, L. Zangi, Pharmaceutics 13, 1675 (2021).
15. K. Karikó, M. Buckstein, H. Ni, D. Weissman, Immunity
23, 165 (2005).
NEUROSCIENCE
g
R E F E R E N C ES A N D N OT ES
p
uncovering and defining general immune
concepts, pathways, and molecules, disparities exist between humans and animals in
terms of immune cell subsets, innate receptors, and exposure to environmental factors, such as diet, that might affect autoimmune pathogenesis. Consequently, reliably
predicting the response to antigen-delivery
therapy remains a challenge, even when using animal models that offer a higher degree of physiological relevance to humans
than laboratory mice.
Successfully navigating the complexities
of an individual’s immune system requires
a comprehensive approach, with the effectiveness of strategies being contingent upon
their alignment with the disease stage. This
challenge intensifies as tissue destruction advances, which introduces intricacies to tolerance restoration. Amid the myriad of technical and knowledge advances, it is crucial to
recognize the complexity of reinstating tolerance through antigen delivery. Consequently,
celiac disease, an autoimmune disorder that
targets the intestine, emerges as an exemplary human disease model. It provides a vital
platform for exploring the mechanisms that
underlie tolerance reinstatement through antigen delivery and understanding the factors
that influence therapeutic success or failure.
Notably, the strict human leukocyte antigen
(HLA) restriction of celiac disease coupled
with the knowledge of the disease-driving antigen (gluten) and accessibility of the tissue
targeted by the pathogenic immune response
make it a valuable focal point for advancing
therapeutic interventions aimed at tolerance
induction through antigen delivery. j
High risk,
high return
Go for it!
Take the risk!
Low risk,
low return
Better play
it safe...
Ventral BA 6V
Dorsal BA 6V
VTA
dorsomedial frontal cortex involved in eye
movement control and evaluation—has a selective effect on risk attitude (4). Neurons in
BA 6V could therefore influence the oculomotor system through the supplementary eye field. However, there are multiple
other frontal and parietal regions that also
influence the selection of eye movements.
It will be important to establish whether
there are specific and necessary pathways
for expressing risk attitude or whether
BA 6V has a large influence over a distributed network. Finally, it is not yet clear how
effector-specific the signals in BA 6V are.
Are they also active when choices are expressed through arm movements instead
of eye movements? If yes, how are the abstract risk attitude signals directed to the
appropriate motor circuit? j
y g
RE FE RE NC ES AN D N OT ES
AC KN OW LED G M E N TS
V.S. was supported by National Institutes of Health grant
R01DA049147.
10.1126/science.adm8641
5 JANUARY 2024 • VOL 383 ISSUE 6678
33
,
1. R. Sasaki et al., Science 383, 55 (2024).
2. A. N. McCoy, J. C. Crowley, G. Haghighian, H. L. Dean,
M. L. Platt, Neuron 40, 1031 (2003).
3. W. R. Stauffer, A. Lak, P. Bossaerts, W. Schultz,
J. Neurosci. 35, 3146 (2015).
4. X. Chen, V. Stuphorn, Curr. Biol. 28, 3114 (2018).
5. P. H. Rudebeck, R. C. Saunders, D. A. Lundgren, E. A.
Murray, Neuron 95, 1208 (2017).
6. M. O’Neill, W. Schultz, Neuron 68, 789 (2010).
7. I. E. Monosov, Nat. Commun. 8, 134 (2017).
8. E. Y. Walker et al., Nat. Neurosci. 26, 1857 (2023).
9. T. Sawaguchi, P. S. Goldman-Rakic, Science 251, 947
(1991).
10. M. Wang, S. Vijayraghavan, P. S. Goldman-Rakic, Science
303, 853 (2004).
11. T. Ott, A. M. Stein, A. Nieder, Nat. Commun. 14, 7537
(2023).
y
rior cingulate cortex (7). Alternatively, risk
attitude could be influenced by modifying
the nonlinear response of value-sensitive
neurons to reward magnitude. For example,
if neurons in BA 6V become much more active to very high reward magnitudes compared with low ones, the monkeys would be
more sensitive to the high winning amounts
of the HH option. This could make them
more risk seeking without any direct change
in probability representation. In that case,
many BA 6V neurons should represent reward amount but not risk. A mixture of both
of these effects is also possible. Alternatively,
uncertainty could be indirectly encoded in
the distribution of value-prediction signals
across the population of neurons (8). In that
case, changes in risk attitude could be caused
by changes in the value distribution.
Dopamine has both enhancing and suppressing effects on prefrontal activity (9–11).
Therefore, it will be important to understand
the exact effects of dopamine on neuronal
activity in BA 6V. Likewise, differential modulation of risk attitude requires that only the
VTA neurons projecting to the required area
are active. Therefore, looking for separate
VTA projections to the ventral and dorsal
BA 6V is another important next step.
It is also not yet clear through which
pathways BA 6V influences behavior. The
monkeys in this study expressed their
choices through eye movements. Therefore,
a specific risk attitude must express itself ultimately in the increased activity
of neurons promoting eye movements to
targets that are most in line with the current risk preference. Inactivation of the
supplementary eye field—a region in the
y
SCIENCE science.org
Activation of pathways from the ventral tegmental area (VTA) to two different parts of the ventral
section of Brodmann area 6 (BA 6V) has opposite effects on risk attitude. Activation of VTA projections
to dorsal BA 6V increases preference for low risk–low return options. With this option, monkeys have
a high (90%) probability of receiving a small (111 µl) amount of fluid reward. Activation of projections
to the ventral BA 6V increases preference for high risk–high return options. With this option, monkeys
have a small (10%) probability of receiving a large (1000 µl) amount of fluid reward. The expected value
of both options is equal (100 µl).
g
cortex, the dorsal anterior cingulate cortex,
or the ventral section of Brodmann area 6
(BA 6V) while the monkeys performed the
task. These are all frontal brain regions
thought to be candidates for a causal role in
influencing risk attitude. Only inactivation
of BA 6V had an effect on task outcome. This
is in good agreement with a previous lesion
study of a similar section of the frontal cortex, which was referred to as the ventrolateral prefrontal cortex (5).
To investigate the role of reward feedback
on the risk attitude control network, Sasaki
et al. studied the dopamine-expressing neurons that project to BA 6V from the ventral
tegmental area (VTA). To do this, they combined optogenetic manipulation of VTA projections with a chronically implanted set of
electrocorticogram electrodes on the lateral
surface of BA 6V. This allowed them to selectively activate the dopaminergic VTA input
to specific areas of BA 6V and to observe the
effects on behavior and neuronal activity.
Notably, optogenetic activation of the
VTA–BA 6V pathway had opposite effects on
risk attitude in dorsal and ventral sections of
BA 6V. Activation of VTA input to the ventral
part of BA 6V led to a stronger preference for
HH options—i.e., the monkeys became more
risk seeking. However, activation of the VTA
input to the dorsal aspect resulted in the
opposite effect: The monkeys preferred LL
options more often—i.e., they became more
risk averse (see the figure). This unexpected
finding implies that two neighboring regions
in the frontal cortex together regulate risk attitude in a competitive push-pull–like fashion
and can both increase and decrease risk seeking. This is important because it opens up the
possibility to identify the neuronal mechanisms in the circuit underlying this ability.
The extent of the functional area that
regulates risk attitude is not yet clear.
Combining the findings of Sasaki et al. with
those of the previous lesion study (5), it appears that an extensive region of the cortex,
ranging from Walker’s areas 12, 45, and ventral 46 on the orbital surface to BA 6V on the
lateral surface, is potentially involved in setting risk attitude. The functional boundaries
of this extended area and further functional
subdivisions need to be mapped out more
carefully.
In principle, attitude toward risk could be
modified in two different ways. Changing
preference to risk directly would influence
risk attitude. In that case, BA 6V neurons
should be sensitive to risk but not to expected value. Such neurons have been found
in the orbitofrontal cortex (6) and the ante-
Assessing risk
p
GRAPHIC: A. MASTIN/SCIENCE
A Japanese macaque (Macaca fuscata) eats wild
flowers. Attitudes toward reward, such as food, are
regulated by the ventral section of Brodmann area 6.
I NS I GHTS
P OLICY FORUM
BIODIVERSITY
Biodiversity monitoring for
a just planetary future
Data that influence policy and major investment decisions
risk entrenching social and political inequities
5 JANUARY 2024 • VOL 383 ISSUE 6678
science.org SCIENCE
,
34
y
1National Center for Ecological Analysis and Synthesis, University of California, Santa Barbara, Santa Barbara, CA, USA. 2Department of Environment Science, Policy, and Management, University
of California, Berkeley, Berkeley, CA, USA. 3School for Environment and Sustainability, University of Michigan, Ann Arbor, MI, USA. 4Department of Ecology and Evolutionary Biology, Yale
University, New Haven, CT, USA. 5Department of Society and Conservation, University of Montana, Missoula, MT, USA. 6Department of Global Development, Cornell University, Ithaca, NY, USA.
7Department of Computer Science, University of Cambridge, Cambridge, UK. 8Département de sciences biologiques, Universite de Montreal, Montreal, Canada. 9Department of Sociology, Tufts
University, Medford, MA, USA.10School of Engineering and Applied Sciences, Harvard University, Cambridge, MA, USA. Email: mchapman@nceas.ucsb.edu
y g
FROM DATA TO DECISIONS
Although the impact of data disparities
on decisions is central to discussions on
data governance throughout society—from
policing to finance to health care—the environmental domain has skirted many of
these critiques under the guise that its
data reflect and affect the natural world,
not people, politics, and histories. The ecological community agrees that data disparities exist but has yet to assess how those
disparities propagate through derived ecosystem indicators and policy and management decisions.
There are several ways in which data
disparities might be reflected in scienceinformed decisions in the context of global
biodiversity targets. For example, extensive
data collected within government-managed
parks compared to community-managed
and Indigenous lands (11) might lead to
systematic underestimates of biodiversity
y
E
A GLIMPSE INTO GLOBAL
BIODIVERSITY DATA
The systems that generate biodiversity
data are complex, uneven, and ultimately
human. Species observations reflect human processes across space and time: from
the decadal impacts of colonialism to the
weekly sway of work schedules in modern
society, from geopolitical strife to neighborhood-scale disparities.
Take, for example, the Global Biodiversity
Information Facility (GBIF). GBIF is a data
repository that synthesizes billions of species observations across the globe (see the
figure) and specifically aims to provide
global-scale biodiversity data to underpin
policy and inform decisions from invasive
species management to priorities for conservation investment.
Even at first glance, GBIF data do not
reflect latitudinal gradients of biodiversity,
but more closely trace macroeconomic patterns (see the figure). These data disparities are unsurprising to most ecologists
and, like the overrepresentation of population centers, roads, and protected areas
in global species observations (3), are increasingly adjusted for, even if imperfectly,
within existing modeling frameworks (4).
But digging deeper into these data, strikingly uneven patterns of data availability
reveal signatures of armed conflict (see
the figure) (5), the legacy effects of racist
policies (see the figure) (6), and changes in
political regimes (5).
Although descriptions of how biodiver-
cologists and conservation scientists
have long acknowledged that biodiversity data reflect legacies of social
inequity (see the figure). Although the
ramifications of these disparities were
easy to dismiss when the application
of large-scale biodiversity data was limited
to academic biogeography and theoretical conservation prioritizations, the stakes
have changed. Biodiversity data carry more
influence than ever before (1), guiding the
implementation of massive multilateral
commitments and global investments that
will affect nature and people for decades
to come—from informing priorities for
more than doubling the global area under conservation management to creating
international biodiversity offset markets.
We examine two contentious questions
that arise as we consider the disparities in
biodiversity data and their consequences
in the wake of contemporary biodiversity
policy: Are the best available data really a
suitable standard? Can more data and better statistical methods ensure that inequities aren’t entrenched when implementing
data-driven solutions?
With hundreds of billions of dollars being invested in conserving, restoring, and
sustainably managing ecosystems in the
wake of the post-2020 Kunming-Montreal
Global Biodiversity Framework (GBF) (2),
an understanding of the ways in which data
biases propagate through decision-making
is critical for the effective creation and
communication of data-driven solutions to
g
global biodiversity loss. The path forward
will require more than technocratic fixes.
Interdisciplinary collaboration and inclusive, bottom-up processes will be critical for
leveraging past, present, and future biodiversity data in a way that aligns with the
equity goals of global biodiversity policy.
p
By Melissa Chapman1,2, Benjamin R.
Goldstein2, Christopher J. Schell2, Justin S.
Brashares2, Neil H. Carter3, Diego Ellis-Soto4,
Hilary Oliva Faxon5, Jenny E. Goldstein6,
Benjamin S. Halpern1, Joycelyn Longdon7,
Kari E. A. Norman8, Dara O’Rourke2, Caleb
Scoville9, Lily Xu10, Carl Boettiger2
sity data disparities trace social and political inequity are notable (3, 5, 6), they rarely
provide the insights necessary to causally
attribute mechanisms of those disparities.
Human patterns captured by biodiversity
data undoubtedly include observational
biases but also reflect a reality of the
Anthropocene: People—and our politics,
economies, and histories—are major drivers of ecosystem composition. European
colonial history is still detectable in the
true distribution of alien floras worldwide
(7). Armed conflict affects underlying ecological processes in a variety of complex
ways (8). The legacy of residential segregation has influenced greenspace and
tree cover across neighborhoods, which in
turn affect habitat for and distribution of
urban wildlife (9). To add complexity, environments most degraded by extractive
infrastructure are often the most monitored—extractive infrastructures are often
also (biodiversity) knowledge infrastructures. For example, the Sacramento–San
Joaquin Delta in California is subject to a
tremendous amount of ecological monitoring, established as a political compromise
to assess the effects of building California’s
complex water infrastructure (10).
But will disparities in biodiversity data
really translate to ineffective and inequitable decisions for nature and people?
And if so, what can be done about it given
the urgency of the biodiversity crisis and
the immediacy of informing global policy
implementation?
Biodiversity data reflect legacies of social inequity
(1) The >2.6 billion species observations in the Global Biodiversity Information Facility (GBIF) database are disproportionately from high-income countries. (2) These
macroeconomic disparities in data density have become more pronounced through time. (3) There are fewer species observations in places and times of conflict (5).
For example, biodiversity observations notably declined during the Cambodian Civil War, which began in 1970, and especially the Cambodian genocide (1975–1979),
and remained low during the following decade of armed conflict. (4) In the United States, biodiversity observations unearth the legacy of the effects of racial and ethnic
discrimination in housing policy in the 1930s (“redlining”) (6). Neighborhoods that were redlined, or deemed “hazardous,” have approximately half the bird observation
density today of those neighborhoods that were deemed “safe” investments (6). (5) Human histories are reflected not only in where and when data are recorded but also
who collects, published, and owns data. In Nigeria, shifts can be seen in the country of data-publishing organizations following independence from Great Britain in 1960
(5). See https://github.com/milliechapman/humanDim-gbif for data, code, and further information about each panel.
1
100
101
Number of observations
102
103
104
2
105
106
0.30
0.25
0.20
p
Cumulative observations per hectare
0.35
High income
Upper middle income
Lower middle income
Low income
0.15
0.10
0.05
g
0.00
1960
1980
2000
2020
y
Battle-related deaths in Cambodia
<25
25–999
>1000
Cambodian
civil war
104
Khmer Rouge
regime
Cambodian-Vietnamese
war
103
10
2
101
1950
1960
1970
1980
1990
2000
2010
2020
Number of bird observations per hectare
105
United States
Urban areas
4
20
Not redlined
5
Country of data publisher
For observations recorded in Nigeria
Nigeria
Great Britain
Other
15
10
Redlined
5
0
Colonial
period
Post
independence
y g
Number of observations in Cambodia
3
Organizations based in Nigeria published 28% of
the country’s observations while Nigeria was
under colonial rule.
Since independence, they have published 70%.
y
GRAPHIC: M. HERSHER/SCIENCE
,
presence in the latter, misguiding ongoing
dialogues about the impact of different land
tenure, property rights, and management
regimes on biodiversity outcomes. Invasive
species might be detected earlier in more
intensely surveyed areas, driving investment toward removal and restoration in
areas most thoroughly monitored.
Without directly addressing and correcting for social and political disparities
in data, the conservation community will
likely fall into the same traps that other domains do—entrenching the inequities of the
past and present in future decision-making
through data. Quantitatively and qualitatively assessing data-derived decision biases
SCIENCE science.org
and the typologies of their impacts on people and communities is an important first
step to effectively mitigating the potential
negative impacts of these disparities.
MORE BIODIVERSITY DATA AND BETTER
MODELS MIGHT NOT SOLVE PROBLEMS
The past decade has marked a shift away
from labor- and resource-intensive specimen collection and field surveys and toward
a new generation of decentralized monitoring tools. Participatory science programs,
artificial intelligence–supported sensors,
and eDNA promise to substantially increase
the number of records per research dollar
and person-hour. More automated digitiza-
tion of natural history collections around
the world is increasing the capacity to understand long-term changes in ecosystems.
These technologies and their resultant
data streams will undoubtedly provide critical information to fill gaps in our knowledge about global biodiversity and inform
more robust global policy strategies. But
as biodiversity data become easier and
cheaper to collect, will sampling become
widespread enough that biases are an artifact of the past, buried under the massive
amounts of new information?
Although new monitoring technologies
continue collecting information about global
biodiversity and its degradation at finer
5 JANUARY 2024 • VOL 383 ISSUE 6678
35
I NS I GHTS | P O L I C Y F O RU M
,
R E F E R E N C ES A N D N OT ES
y
DATA AS SOCIAL INFRASTRUCTURE:
BIODIVERSITY MONITORING FOR
EQUITABLE DECISIONS
The realization that more data or better
models will never fully solve systemic bias
does not mean there are no solutions. It
means there are no shortcuts—no getting
around the need for local engagement, context-specific knowledge, and case-by-case
considerations when using this data. Investments in future monitoring should not
only prioritize new technologies that ease
the collection of massive amounts of biodiversity data, but also ensure that those data
include information about the local context
and social infrastructures.
Moving beyond quick technical fixes
will require connecting strategically to
y g
ing of those systems than global synthesis
data can provide.
Further, some human drivers of observational (and ecological) processes are not
digitizable or easily reduced into quantitative metrics. Although it might be possible
to investigate the impact of past residential
segregation policies in the United States
because there is geospatial information
on its history, dimensions that cannot
be reduced to polygons on a map, such
as human preferences, scientific funding
patterns, and industrial priorities, may
continue to be reflected in downstream
data products and decisions. “Datafication” can thus create another layer of bias:
between the social, political, and cultural
dimensions of data that are easy to digitize
and those that are not (14).
y
5 JANUARY 2024 • VOL 383 ISSUE 6678
“Moving beyond quick technical fixes
will require connecting strategically to
community-based partners…”
community-based partners and leveraging
expertise in social, ethical, cultural, and
political processes underlying data infrastructures and their histories. Communitybased monitoring and information systems
(CBMIS) provide a compelling framework
for locally engaged monitoring and are
highlighted in the GBF as one means of filling data gaps (2). Established networks of
CBMIS are already operational in several
countries and have proven effective at contributing to national and global-scale monitoring of ecosystems (15). Initiatives such as
the International Forestry Resources and
Institutions (IFRI) collect information on
institutional and social variables, alongside ecosystem data, through a network of
locally led Collaborating Research Centers
to understand the interrelationships among social and ecological
processes and outcomes in forest
systems around the world. There is
no technocratic solution for incorporating all relevant information
about ecosystems and their social
contexts into formal frameworks
for assessing biodiversity or devising policy
strategies at global scales. However, complementing global frameworks and synthesis databases with decentralized knowledge
collected as part of CBMIS (and programs
like IFRI) might help expose and ameliorate
data disparities that underpin biodiversity monitoring and mitigate the implications of these disparities on the distributional equity of downstream conservation
decision-making.
The success of the GBF, and the meaningfulness of its proposed indicators, requires
that policy-makers and scientists resist
technocratic shortcuts and instead assess
the equity implications of data disparities,
support local knowledge generation, and
work toward governance systems and monitoring frameworks that engage with biodiversity data as social infrastructure. j
g
36
izing unintended sociopolitical patterns in
data is an important step toward developing analytical methodologies that more accurately reflect true biodiversity patterns.
Although careful statistical models can
help identify and control for data disparities that can be quantified, they are not a
panacea. Quantitatively correcting socially
determined bias across spatial and temporal scales from the top down would require
a near-complete census of these multiscale
and interacting biases—an infeasible trap.
Even when such quantification reveals
statistically clear associations, conducting
inference on the multidirectional and interacting causal mechanisms that link social
infrastructure, monitoring, and biodiversity
is impossible without a deeper understand-
p
resolutions and with a broader scope, this
increasing amount of information has yet
to yield more representative data coverage
of biodiversity distributions. Instead, new
waves of biodiversity data have entrenched
the long-known overrepresentation of certain regions, taxa, and time periods in global
biodiversity data repositories (12).
Regardless of the volume or velocity of
data collection, where, when, how, and by
whom species are observed will always be
shaped by social, political, and economic
processes (13).
Collecting perfectly uniform global biodiversity data isn’t the only possible solution for addressing the gaps and disparities
in existing data. Ecologists and statisticians have worked extensively on methods
for bias correction of existing biodiversity data to infer how species
distributions and populations vary
in time and space despite imperfect data (4).
Nonrandom sampling effort can
be addressed in two ways: One is
by assuming that unobserved variation in sampling (e.g., geopolitical conflict–
associated differences in sampling effort) is
not confounded with the natural process of
interest (e.g., biodiversity distributions and
their change); another is by “correcting”
the bias in the process of interest with data
preprocessing or model-based inference.
In the case of social drivers of biodiversity
sampling at continental and global scales,
neither of these technical fixes is likely adequate to remove biases.
The first option—assuming that the sampling process in question is not related to
the natural process of interest—is baseless
in most cases. As outlined above, the drivers of data collection are often deeply intertwined with the natural processes that
scientists often seek to assess.
The second option—correcting for biases—is only as effective as the capacity
of quantifiable variables to explain the biases in the data. In ecology, bias correction
tends to focus on bioclimatic conditions,
latitudinal disparities, and simple accessibility metrics (e.g., population density,
proximity to roads) (4), meaning that the
other social infrastructures underlying
these data likely remain reflected in ecological insights (e.g., species distribution
models, metrics of community change) and
downstream decisions (e.g., conservation
priorities). Archiving and digitizing human societies’ darkest hours—from war to
colonialism to systemic racism—may allow
researchers to start to disentangle the past,
present, and future signatures of humans
on both biodiversity and the data capturing its distribution and change. Character-
1. A. Gonzalez, M. C. Londoño, Science 378, 1147 (2022).
2. Convention on Biological Diversity, Kunming-Montreal
Global Biodiversity Framework (2022).
3. A. C. Hughes et al., Ecography 44, 1259 (2021).
4. D. I. Warton, I. W. Renner, D. Ramp, PLOS ONE 8, e79168
(2013).
5. A. Zizka et al., J. Biogeogr. 48, 2715 (2021).
6. D. Ellis-Soto, M. Chapman, D. H. Locke, Nat. Hum. Behav.
7, 1869 (2023).
7. B. Lenzner et al., Nat. Ecol. Evol. 6, 1723 (2022).
8. K. M. Gaynor et al., Front. Ecol. Environ. 14, 533 (2016).
9. C. J. Schell et al., Science 369, eaay4497 (2020).
10. C. Scoville, Theory Soc. 48, 1 (2019).
11. S. T. Garnett et al., Nat. Sustain. 1, 369 (2018).
12. B. H. Daru, J. Rodriguez, Nat. Ecol. Evol. 7, 816 (2023).
13. J. E. Goldstein, H. O. Faxon, Environ. Plan. E Nat. Space 5,
39 (2022).
14. M. Chapman et al., One Earth 4, 790 (2021).
15. M. Ferrari et al., Biodiversity (Nepean) 2, 57 (2015).
10.1126/science. adh8874
science.org SCIENCE
B O OKS et al .
CLIMATE
By Erle C. Ellis
AC KN OW LED G M E N TS
The reviewer is a visiting fellow at the Oxford Martin School,
which provides funding for Our World in Data.
10.1126/science.adl5466
5 JANUARY 2023 • VOL 383 ISSUE 6678
37
,
1. Our World in Data; https://ourworldindata.org/.
y
SCIENCE science.org
RE FE REN C ES AN D N OT ES
y g
PHOTO: KATRIN SAUERWEIN/ISTOCK PHOTO
I
y
falsely, as a surprising number of educated
people do—that more people are starving,
n a time of daily environmental crisis
suffering, living in poverty, being exposed to
headlines and growing eco-anxiety, how
natural hazards, and dying young than ever
could anyone claim that a much better
before. The turning point for her came in
world is not only possible but that—in
the form of the late Swedish physician Hans
many ways—it already exists? The anRosling’s masterful, data-rich presentations
swer, according to data scientist Hannah
showing that the human world is almost uniRitchie in her first book, Not the End of the
versally changing for the better. Indeed, in
World, is that this is exactly what the data tell
many ways, Ritchie is building on Rosling’s
us. As the science outreach lead
legacy, although she goes much
of the widely cited project Our
further: Where Rosling focused
World in Data, she is certainly one
almost entirely on the successes
to know (1).
and challenges of human develNot the End of the World is built
opment, Ritchie takes on “the
on the same solid, data-driven
other side” of the sustainable fufoundations that characterize the
ture—the environmental damage
crisp, concise visuals produced by
that looms like a dark cloud over
Our World in Data, but it is a book
everything achieved in improving
Not the End
with a purpose beyond presenting
the human condition.
of the World:
facts about our planet: Ritchie’s
While the human world has
How We Can Be
goal is to convince young people
generally been getting better in
the First Generation
to shake off climate doom and get
critical ways for decades, the opto Build a
Sustainable Planet
motivated to address the unprecposite has largely been true for the
Hannah Ritchie
edented environmental challenges
rest of the planet. Yet, as Ritchie
Little, Brown Spark, 2024.
of our times. This mission is pershows in data-rich chapters
352 pp.
sonal, and she wraps the narrative
packed with illustrative charts,
of the book around her journey from econot all of the trends are going in the wrong
anxiety to “urgent optimism.”
direction, and we already have most, if not
Convinced that she “didn’t have a fuall, of the capabilities needed to address seture left to live for,” Ritchie long believed—
rious environmental challenges such as air
pollution, climate change, deforestation, inThe reviewer is at the Department of Geography and
sufficient food supply, biodiversity loss, ocean
Environmental Systems, University of Maryland, Baltimore
plastics, and overfishing.
County, Baltimore, MD 21250, USA, and is a visiting fellow
For each challenge, she describes how
at the Oxford Martin School, University of Oxford, Oxford
it was first identified and understood by
OX1 3BD, UK. Email: ece@umbc.edu
g
A data scientist offers an optimistic reality check
for the Anthropocene
scientists and how it has been addressed
to date. Most importantly, she highlights
and prioritizes a suite of specific solutions,
revealing how many seemingly intractable
threats to people and planet are, in fact, actionable and resolvable through strategies
available right now. However, she is careful
to show that even with solutions in hand,
many of the challenges we face are not being dealt with adequately and that, in many
cases, the damage is accelerating.
Ritchie also debunks two strategies that,
by her assessment, “won’t fix our problems”:
depopulation and degrowth. She argues that
the former is unnecessary—“peak child,” she
writes, has already happened—and more
rapid shrinkage is unachievable without
subjecting people to unconscionable harm
and that the latter would leave most people
on Earth unacceptably impoverished. True
believers in such propositions will likely remain unswayed, facts or no facts.
This brings us to the book’s greatest provocation—and subtitle—about how our generation can be the first to build a sustainable
planet. Recognizing the successes and future
potential of contemporary societies is certainly an important accomplishment of the
book. Nevertheless, making such a strong and
general claim about the superiority of today’s
societies over all that came before seems premature, problematic, and unnecessary and
exposes the absence of a deeper and more nuanced theoretical understanding of human–
nature relations in the book. Fortunately, this
and other oversimplifications do not detract
from the main aims of the work, which are
overwhelmingly pragmatic.
That this book represents “a synthesis of
nearly a decade of research and data” is clear
from its remarkable coverage of environmental evidence. Yet its main contribution may
be its ability to produce what Ritchie calls a
“perspective that helped me dig myself out
of a very dark place.” There is real peril in
our widespread failure to understand just
how much human lives have been improved
through societal efforts and in the level of
doom and helplessness that seems to be
spreading among young people. As Ritchie
demonstrates in Not the End of the World, a
better future for both people and planet is
possible and even achievable. j
p
Data-driven hope for the planet
Excessive plastic food packaging is not ideal, argues
Ritchie, but a little can help prevent food waste.
I N SI G H TS | B O O K S
SUSTAINABILITY
Water recycling goes mainstream
Purified:
How Recycled Sewage
Is Transforming Our Water
Peter Annin
Island Press, 2023. 248 pp.
Public health and public relations are key to successful
potable water reuse programs
By Sasha Harris-Lovett
y g
y
,
science.org SCIENCE
y
5 JANUARY 2023 • VOL 383 ISSUE 6678
g
38
p
contaminants (Orange County), do extra
monitoring in the sewer network (El Paso),
solicit additional surveillance from the
otable water reuse, also known as wapublic health department (Virginia), and
ter recycling or by the moniker “toilet
require specific training for potable water
to tap,” is the practice of deliberately
reuse treatment plant operators (El Paso).
incorporating treated wastewater
But regulations vary widely in ways that
into the drinking water supply. In Puhave the potential to compromise public
rified, journalist Peter Annin’s vivid
health and sour citizens on water reuse projand engaging new book, readers join Annin
ects. When Annin interviews potable water
on a tour of potable water reuse systems
reuse system engineers in Deep Spring,
across the United States, from California to
Texas, for example, he learns that the city
Texas to Florida. Annin’s meticulous reportdoes not have a monitoring system to deing reveals the technologies and individutect toxins, such as acetone, that can slip
als responsible for this new water supply,
untreated from sewage into drinking water
as well as the histories of the communities
through their reuse system. In much of the
that are—or soon will be—drinking it.
book, Annin seems to accept at
Annin sets the stage with a
face value the claims about water
portrait of water scarcity along
reuse technology made by the wathe Colorado River, which has
ter engineers he interviews, but
experienced record low water levI gave a silent cheer when he diels. Water levels in Lake Mead, we
rectly challenges one Deep Spring
learn, are nearing “dead pool”—
water engineer, asking “if it might
a state in which a reservoir conbe worthwhile for Big Spring watains so little water that it can no
ter officials to consider adding
longer flow downstream. Without
real-time monitoring for acetone
intervention, this will spell disasto their history-making plant?”
ter for the millions of people in
(The engineer cryptically replies,
the Southwest who depend on it.
“It may be.”)
“Enter purified sewage,” Annin
Annin contends that water
quips. “Thanks to climate change,
utilities, states, and the federal
never before has something so
government must ensure that we
foul looked so good.” This is cerhave the right regulations in place
tainly the case in parts of Texas.
to guarantee that potable water
Annin details how extreme
reuse projects maintain public
drought pushed cities such as Big
Spring, Wichita Falls, and El Paso
health. Such policies, he asserts,
to turn to potable water reuse
should include protecting sewPeople taste recycled wastewater in Segundo, California, in 2015.
when their reservoirs had almost
ers from chemical spills; creatrun dry. But drought is not the only driver
challenges. When the project’s engineers
ing real-time monitoring infrastructure for
for potable water reuse. In Florida, populadetected that their wastewater purification
early detection of problematic compounds;
tion growth is stressing freshwater supplies
technologies were creating a potent carand developing industry-standard regulato a breaking point, leading water engineers
cinogen, N-nitrosodimethylamine (NDMA),
tions governing treatment technologies,
to consider urban sewage as a source of
and adding it into the water supply, their
operator training, and emergency response
drinking water.
public relations team explained that the
plans. As potable water reuse comes into
Even when there are good reasons to
risk of ingesting NDMA in water was less
the mainstream, Annin warns that “there is
pursue potable water reuse, people are not
than the risk of eating hot dogs (which also
simply no margin for error.”
always excited about it. In the 1990s, San
contain NDMA) and described their techniAs fresh water supplies become increasDiego citizens roundly rejected a proposed
cal approach to ameliorating the problem.
ingly scarce and technologies for water
potable water reuse project, even though
This transparent communication paid off,
treatment improve, potable water reuse will
the city had already invested more than
and “no backlash occurred.”
likely become widespread. Annin’s timely
$15 million in it. Through interviews and
Annin’s case studies reveal how utilities
and important reporting empowers readanalysis of news articles, Annin forensithat have successfully implemented potable
ers to understand the critical issues at hand
water reuse go well beyond the regulatory
and offers an engaging introduction to poThe reviewer is at the San Francisco Estuary Partnership,
requirements to ensure that the water they
table water reuse. j
San Francisco, CA 94105, USA.
produce is safe. They do extra testing for
10.1126/science.adl2392
Email: sasha.harris-lovett@sfestuary.org
P
cally investigates why that project failed.
The engineers were clear: The technologies
used to treat sewage to drinking water standards were robust and would protect public
health. The problem, Annin concludes, is
that potable water reuse proponents in San
Diego did not have a strong enough communications and outreach strategy.
The importance of proactive outreach
and transparent communication to support potable water reuse projects is a
theme that recurs throughout the book.
In Orange County, for example, Annin reveals how the Groundwater Replenishment
System’s “sweeping public relations offensive” proved critical to its ability to weather
INSIGHTS
LET
LE T TTERS
ERS
p
g
y
NEXTGEN VOICES
Changing outdated expectations
y g
In the 3 November 2023 Working Life (https://scim.ag/WL_locationmatters),
Shanta Hejmadi questions the expectation that scientists should change institutions between their PhD and postdoc. We asked young scientists: What
traditional career expectation have you come across that seems outdated, counterproductive, or exclusionary? How should that expectation be adjusted? Read a selection of the responses here. Follow NextGen
Voices on social media with hashtag #NextGenSci. —Jennifer Sills
y
apply their skills in the innovation divisions
of large companies, at industry-led think
tanks, or as teaching faculty, and may later
choose another path or return to research.
These experiences deepen understanding
and generate new ideas. Academia can
improve inclusivity by encouraging innovation, reducing stigma around “alternative”
careers, and empowering individuals to
make career decisions that suit them.
Providing mentorship programs, career
workshops, and fellowships and grants for
diverse career stages and trajectories could
facilitate this flexibility.
Dequn Teng
Everyone who completes a PhD and postdoc
is expected to continue in academia. This
robs scientists of the flexibility to pursue less
conventional career paths such as funding,
science writing, science ethics, science journalism, or management. Information about
these options is not readily available, and
in some cases, there is a stigma attached to
changing careers. Choosing a career outside
of academia should be easy and accessible
at any point in the educational journey,
and people should be able to change course
without feeling like they have failed. Career
counseling should always be on offer, and
Garima Singh
24
5 JANUARY 2024 • VOL 383 ISSUE 6678
Fleming Fund South Asia, Mott MacDonald, Noida,
Uttar Pradesh, India. Email: singhg20@gmail.com
Scientists are expected to advance sequentially from PhD to postdoc to tenure-track
faculty. This expectation benefits prestigious
academic institutions, which can choose
from many applicants, but marginalizes
people who follow unconventional trajectories, such as transitions between academia,
industry, and teaching. Hiring teams in academia and beyond should embrace diverse,
nonlinear career paths. New PhDs may
Department of Engineering, Institute for
Manufacturing, University of Cambridge,
Cambridge, UK. Email: dt517@cam.ac.uk
A PhD is expected to have skills in grant
writing, teaching, industry, and entrepreneurship in addition to their subject area.
Universities benefit from this assumption
because it justifies the lack of help available
to specialists trying to accomplish tasks
outside of their area of expertise. Expecting
postdocs and faculty to immerse themselves
in tangential tasks slows down research
progress. Acknowledging that a PhD
indicates scientific but not administrative,
pedagogical, or business expertise would
free scientific experts to focus on research
science.org SCIENCE
,
Support realistic career paths
programs and scholarships at the university
level should help support career changes.
and justify hiring a specialist workforce to
conduct other tasks.
Houcemeddine Turki
Data Engineering and Semantics Research Unit,
University of Sfax, Sfax, Tunisia.
Email: turkiabdelwaheb@hotmail.fr
Assess experience equitably
Mayank Chugh
Department of Systems Biology, Harvard Medical
School, Boston, MA, USA.
Email: mayank_chugh@hms.harvard.edu
In China, students are expected to complete
their PhDs by age 35, after which they are
no longer given preference for positions at
universities. The age limit aims to encourage
younger candidates to apply for positions.
However, this requirement is unfair to those
who start their PhD late. When applying to
university positions, PhD graduates should
be judged by their achievement rather than
age. Awards, published papers, and patents
are more important and relevant indicators of ability than age at the time of PhD
completion.
exclusive commitment limits faculty’s ability
to provide job market insights to students
and lowers income potential compared with
private sector roles. Without exclusive commitment requirements, professors could help
students better navigate the job market, set
up science-related businesses, and engage in
other complementary activities that would
improve undergraduate teaching.
Paulo Bezerra
Department of Engineering and Technology,
Universidade Federal Rural do Semi-Árido, Pau dos
Ferros, RN, Brazil.
Email: paulo.bezerra@ufersa.edu.br
Doctors are expected to work long hours
to benefit patients and medical institutions, but the pressure leads to physical and
mental exhaustion. If institutions allowed
doctors to work reasonable schedules and
provide appropriate opportunities for vacation and self-care, doctors would be able
to better manage and balance their work
and personal lives, thereby enhancing their
work efficiency and quality.
Ju Wen
LiangBin Wu
Towards AI, Montreal, QC, Canada.
Email: bouchard.lf@gmail.com
School of Liberal Education, Chengdu Jincheng
College, Chengdu, Sichuan, China.
Email: jupiter@cdjcc.edu.cn
Peking University Shenzhen Hospital, Shenzhen
City, Guangdong, China.
Email: 864262711@qq.com
Standardize fair contracts
Incentivize quality research
Marine scientists who are just starting their
career are expected to accept unpaid work
opportunities. Although the positions often
require graduate-level education, numerous
certifications (such as SCUBA), and previous
experience, participants are rarely given
the option of advancing to a paid role. This
framework is exclusionary to those without
the financial means to support themselves
for weeks or months to gain what organizations tout as valuable experience. Instead,
marine conservation organizations should
adopt ethical programs that compensate
participants for their time and skills. Budget
allocations for stipends would be ideal, but
an alternative could be the creation of partnered internships with structured learning
objectives and benefits such as credits.
The “publish or perish” adage in academia
pressures postdoctoral researchers to prioritize quantity of publications over quality.
This paradigm, deeply ingrained in academic culture, benefits those who produce
a high volume of work, potentially at the
expense of thoroughness or innovation. The
expectation not only stifles creativity and
patience but also discourages investment in
high-risk and time-consuming but highreward and high-impact research. Academia
should pivot to valuing the quality and
impact of research rather than the number
of publications. This would cultivate an
environment that encourages innovative
and transformative science. Institutions can
promote this shift by redefining metrics of
success to include broader impacts such
as community engagement, teaching, and
policy influence, ensuring a holistic evaluation of medical researchers’ contributions.
SCIENCE science.org
Emily Yi-Shyuan Chen
Department of Marine Ecology, Institute of
Oceanology, Polish Academy of Sciences, Sopot,
Pomerania, Poland. Email: emily@iopan.pl
Jingtao Huang
In Brazil, many full-time faculty at public
universities are expected to agree to exclusive
commitment contracts, which prohibit
external employment. For example, as a
civil engineering professor, I’m barred from
authoring building designs. These contracts
are intended to reduce conflicts of interest,
ensure institutional loyalty, and provide
quality in teaching and research. However,
Shantou University Medical College, Shantou,
Guangdong Province, China.
Email: 19jthuang@stu.edu.cn
Scientific researchers are expected to
produce novel results. This focus, intended
to drive innovation, inadvertently fosters
an unhealthy culture that deters essential
replication studies. Excessively prioritizing originality, and neglecting or even
5 JANUARY 2024 • VOL 383 ISSUE 6678
25
,
Scientists are expected to train abroad
to build a competitive career. Working in
foreign countries undoubtedly provides
better international networks and collaborations; exposure to new methods, skills,
funding, and publishing landscapes; and a
diversity of ideas, people, and experiences.
However, expecting mobility is based on a
false assumption that global access is equal
to all. Some passports (mostly in the Global
North) provide more opportunities than others. The unwritten requirement of working
abroad, especially for postdocs, particularly
harms and excludes scholars from the
Global South. To provide equitable access
for everyone, institutions should reform
y
Shantou University Medical College, Shantou,
Guangdong, China.
Email: cherrychen66666@163.com
y g
Yan Chen
y
In the field of artificial intelligence, there
is an expectation that applicants with the
most prestigious institutional pedigrees will
have the most impressive skills. However,
this assumption overlooks individuals who
have gained valuable practical experience,
pursued alternative educational paths, or
come from underprivileged backgrounds
without access to top-tier education. To
remedy this inequity, institutions should
revise hiring criteria to focus on practical
experience, demonstrated expertise, and
portfolio quality rather than prioritizing
prestigious degrees.
g
Louis-François Bouchard
p
Artificial intelligence researchers are
expected to have PhDs. However, innovation in artificial intelligence often occurs in
industry settings, and industry experience
can rival academic credentials in driving
artificial intelligence advancements, thus
rendering the lengthy PhD process unnecessary. Therefore, industry experience should
be considered equivalent to a PhD. This
change would empower professionals to contribute without committing to the extended
time and narrow focus of a doctoral program. For those who do pursue a PhD, thesis
graduation criteria should include substantial industry achievements as an alternative
to academic certifications.
evaluation practices and metrics for early
career researchers and faculty during hiring,
tenure, promotions, funding, and potential
awards to prioritize a candidate’s research,
skills, background, and lived experiences
over their ability to secure international
opportunities.
I NS I GHTS | L E T T E R S
discouraging replication efforts, undermines the goal of establishing confirmed
and reliable knowledge. The scientific
community should acknowledge the value
of replication studies. To facilitate this shift,
funding bodies and academic journals need
to adjust their policies and reward systems
to recognize and support replication efforts.
Qianjun Wen
The Affiliated Hospital, Guizhou Medical
University, Guiyang City, Guizhou, China.
Email: wqjtmmu@126.com
Respect community ties
Many PIs spent their time as graduate
students and postdocs battling harsh (even
abusive) mentors in isolated, unsupportive
environments. They survived in academia by
accepting the circumstances and succeeding despite obstacles, and they rightly wear
their struggle as a badge of honor. However,
perpetuating the expectation that succeeding in academia requires struggling limits
the pool of future capable scientists. Instead,
institutions should strive to support each
scientist as the multifaceted individual they
are. Academic departments should normalize mentor-mentee discussions about work
styles, expectations, career plans, and project
management; provide training for careers
outside of academia; proactively address the
needs of scientists with disabilities, chronic
illnesses, and mental health challenges; and
allow for mid-career pauses. To see diversity
and equity in science, academia needs to be
a place where excellent scientists are encouraged, rather than challenged.
Madeline Klinger
Helen Wills Neuroscience Institute, University of
California, Berkeley, Berkeley, CA, USA.
Email: klinger@berkeley.edu
Scientists are expected to seek individual
achievement. Success is tied to earning
grants, authorships, and positions of power
such as PI. Overemphasizing these individualistic practices can stifle collaboration and
exclude or marginalize people whose cultures and communities prioritize collective
pursuits. I grew tired of the individualistic
environment I experienced while obtaining
a PhD, partly because it was a poor match
to my values as a Latina woman. Providing
more collective structures and practices,
such as mentoring the next generation
and encouraging individuals to help team
members, could prevent members of underrepresented groups from leaving science.
Emma Claudia Perez
Pasadena, TX, USA. Email: ecperez414@gmail.com
Encourage interdisciplinary work
Academic research conducted by business
students is expected to provide insight into
managerial challenges. Business schools
should expand their research objectives
to address global operational challenges
such as climate change and food insecurity.
Moving beyond the topic of management
will increase theoretical and interdisciplinary research and facilitate innovative
approaches that can tackle global problems.
Samuel Nathan Kirshner
University of New South Wales Business School,
University of New South Wales, Sydney, NSW,
Australia. Email: s.kirshner@unsw.edu.au
Applicants for lab positions are expected
to pursue projects closely aligned with the
group’s current research. Institutions benefit
from this strategy because new lab members
can integrate quickly and make immediate contributions. However, basing hiring
on narrow interests and goals hampers the
advancement of interdisciplinary studies.
Instead, institutions should recruit researchers with diverse disciplinary backgrounds
and interdisciplinary aspirations. Diverse
disciplinary teams and broad perspectives
are vital in emerging fields.
Chao Xu
School of Resource and Environmental Sciences,
Wuhan University, Wuhan, Hubei, China.
Email: xuchao@whu.edu.cn
Immunologists are expected to focus
exclusively on their own discipline, but an
interdisciplinary approach could be more
effective. For example, to decipher the
pathological changes responsible for autoimmune bone marrow failure, focusing only on
immunological aspects (lymphocytes) might
not lead to a comprehensive understanding.
Instead, immunologists need to collaborate
and look beyond immunology for the putative agent. The fields of genetics, molecular
genomics, and environmental and health
research could hold the answers.
Vandana Sharma
Scientists are expected to devote all of their
energy to their research to succeed, and
those raising children as they launch their
career are assumed to be less productive.
Department of Hematology, All India Institute
of Medical Sciences, New Delhi, India.
Email: sharmavandana.phd@gmail.com
10.1126/science.adn4211
science.org SCIENCE
,
5 JANUARY 2024 • VOL 383 ISSUE 6678
Create supportive environments
y
26
Climate-Energy Lab, Carnegie Institution for
Science, Stanford, CA, USA.
Email: evirguez@carnegiescience.edu
y g
As Shanta Hejmadi explains in her Working
Life, scientists are expected to change institutions between a PhD and postdoc, which
usually requires a long-distance move. The
goal is to expose scientists to different experiences, which can later inform their work as
faculty. I was so passionate about becoming faculty that I moved from Wisconsin to
Texas for a postdoc. My husband quit his job
to move with me. We missed our families
and the stronger support system we had in
our home state. Unlike Hejmadi’s “happy
ending,” the sacrifices we made ultimately
cost me my academic career. Instead of a
long-distance move, postdocs should be
given the option of working in a different
department, with a different faculty mentor,
at their PhD institution. Faculty who stay at
the same institution for a PhD and postdoc
could be an asset given their long-standing
knowledge of the institution’s history and
strong ties to the community. Not penalizing
Edgar Virguez
Boston Consulting Group, Minneapolis, MN, USA.
Email: bridget.stroup@gmail.com
y
Department of Aerospace Engineering, University
of Maryland, College Park, MD, USA.
Email: hartzell@umd.edu
Bridget Stroup
g
Christine Hartzell
Early-career scientists can be productive
researchers and responsible parents if they
can access supportive policies, such as
parental leave, flexible schedules, familysustaining wages, and advice on managing
time and setting boundaries.
p
Faculty are expected to take a sabbatical at
an institution in another state or country
about every 6 years. In theory, these experiences give faculty time to develop new ideas
and build collaborations with new people.
However, this expectation is unrealistic for
many families, including those with dual
incomes (if the spouse can’t work remotely
for months), health problems, children with
special needs, or local family obligations.
Institutions could address these challenges
by providing prestigious external funding
opportunities for multiple 1-week-long institutional visits during a sabbatical instead of
one months-long visit. I am fortunate to live
in a region with many nearby institutions, so
I can take a sabbatical without relocating my
family. However, any travel to institutions in
other states or countries currently would be
funded by my regular research grants, which
would not be as prestigious as a named sabbatical fellowship.
postdocs for staying at the same institution as their PhD would help postdocs with
spouses and families pursue their academic
dreams. This solution would especially benefit women and could even help increase the
rate of women in academic leadership.
RESEARCH
Photoluminescent inorganic
compounds embedded in a polymer
can be used to create dual-color
three-dimensional structures.
IN S CIENCE JOU R NA L S
Edited by Michael Funk
p
g
PEROVSKITES
y
A supramolecular boost
to emission
M
any inorganic perovskites can show strong
photoluminescence, including ones based
on hafnium and zirconium halides, but these
compounds are often air and water sensitive.
Zhu et al. extended a crown ether–assisted supramolecular assembly approach to guide and stabilize the
assembly of octahedral [HfBr6]2– octahedral centers. Solid
SCIENCE science.org
Science p. 62, 10.1126/science.adh8342
ORGANIC CHEMISTRY
Science p. 77, 10.1126/science.adk2312
Lighting the way
to amides
2D MATERIALS
Cobalt carbonyl complexes
catalyze the coupling of alkenes,
hydrogen, and carbon monoxide
to produce aldehydes through
Two-dimensional (2D) van der
Waals materials that exhibit
a moiré effect typically have
Mapping out moirés
complicated electronic band
structures. To study them,
researchers can apply magnetic
fields and look for oscillations in transport properties.
Thermodynamic properties
such as magnetization should
also exhibit quantum oscillations, but these are tricky to
detect in the very small 2D
material samples typically
available. Bocarsly et al. used a
very sensitive magnetometer—
a superconducting quantum
interference device—placed
on the apex of a sharp pipette
to scan a sample of moiré
graphene. The authors detected
patterns of magnetization
oscillations and used the data
to reconstruct an intricate band
structure. —JSt
Science p. 42, 10.1126/science.adh3499
5 JANUARY 2024 • VOL 383 ISSUE 6678
39
,
Cancer immunotherapy with
immune checkpoint inhibitors
blocks negative signals of T cell
activation to mount an immune
response against tumors but can
also lead to immunopathologies.
Colitis is a frequent and severe
adverse event in patients treated
with antibodies targeting the
checkpoint inhibitor cytotoxic
T-lymphocyte protein 4 (CTLA-4),
but the underlying mechanisms
leading to this reaction remain
unclear. Lo et al. demonstrate
that CTLA-4 blockade–induced
colitis in mice is dependent on
gut microbiota composition and
is driven by the unrestrained
activation of T cells and the
hydroformylation. Faculak et al.
report that by adding light to
the mix, they can swap out the
hydrogen for primary or secondary amines and produce amides
with the same catalyst. Adding
a silane afterward can reduce
these products to more highly
substituted amines if desired.
The authors propose that the
photoexcitation expels one
carbon monoxide ligand from
the catalyst, opening a critical
coordination site under mild
conditions. —JSY
y
PHOTO: ZHU ET AL.
Microbiota-reactive
T cells trigger colitis
concurrent depletion of a subset
of regulatory T cells in the gut
by receptors recognizing the Fc
domain of the anti-CTLA-4 antibodies. Anti-CTLA-4 nanobodies
lacking the Fc domain were found
to stimulate antitumor immunity without inducing colitis.
These findings may support the
development of next-generation
CTLA-4 inhibitors with reduced
inflammatory toxicities. —STS
Science p. 86, 10.1126/science.adi4196
y g
IMMUNOLOGY
powders of (18C6@K)2HfBr6 exhibited 96% photoluminescence quantum yield for blue emission,
and a zirconium analog had a photoluminescence
quantum yield of 83% for green emission. Addition
of a polymer created solution-processable inks for
printing luminescent thin films and structures. —PDS
R ES E ARCH
ALSO IN SCIENCE JOURNALS
NEUROSCIENCE
Brain circuits handle
risky decisions
The origins
of trait innovation
Traits are constantly changing in
populations, but the emergence
of a trait that provides a new
function is a rarer occurrence.
Two studies examined how such
traits arise. Stankowski et al.
identified the multiple genome
regions associated with the
transition from egg-laying to
live-bearing reproduction in a
clade of snails, demonstrating a
polygenic basis of live-bearing
reproduction with alleles that
accumulated in response to
selection over time. Chomicki et
al. found that the spontaneous
occurrence of three variable
traits created a novel mechanism for trapping insects in
pitcher plants, which arose separately in two different species.
Together, these studies provide
insight into the emergence of
complex, transformative traits
(see the Perspective by Elmer).
—BEL and CNS
Science p. 114, 10.1126/science.adi2982,
p. 108, 10.1126/science.ade0529;
see also p. 27, 10.1126/science.adm9239
Science p. 94, 10.1126/science.adj1962
IMMUNOLOGY
Double pi
The TAO(K3)
of T cell signaling
Carbenes contain a carbon atom
in which only two of the four
valence electrons are bonded
to other atoms. In general, at
least one of the two remaining
electrons resides in the same
plane as those bonds. Hu et al.
report an unusual carbene with
flanking phosphorus centers in
which both of the nonbonding
electrons occupy the π orbital
perpendicular to the bonding
plane. The carbene, isolated and
characterized in a complex with
each phosphorus coordinated to
T cell receptor signaling is
inhibited by the phosphatase
SHP-1, which dephosphorylates
and inactivates several effectors
downstream of the receptor.
Poirier et al. found that the
kinase TAOK3 promotes T cell
receptor signaling by phosphorylating SHP-1, leading to
the ubiquitylation and degradation of SHP-1. The loss of TAOK3
reduced both tonic and liganddependent T cell receptor
signaling in mouse CD4+ T cells
and in a human T cell line. Thus,
CARBENE CHEMISTRY
5 JANUARY 2024 • VOL 383 ISSUE 6678
,
41-B
Metal nanoparticles on oxide
surfaces used as catalysts for
industrial reactions tend to
become larger and less active
after prolonged exposure to
reaction conditions. Liu et al.
have shown that after exposure to methanol vapor at
200ºC, copper nanoparticles
in dealuminated beta zeolite
become smaller. The average
size decreased from ~5.6 to ~2.4
nanometers because copper
migrated and became trapped at
new surface sites. This catalyst maintained nearly 100%
conversion and selectivity for
ambient-pressure hydrogenation
of ethylene glycol to dimethyl
oxalate at 400ºC for 200 hours.
—PDS
EVOLUTION
y
Many block copolymers will
phase separate into distinct
regions, where the minority
component might form spheres,
cylinders, or layers inside the
majority component depending on the strength of the
interactions between the two
components and their relative
proportions. Some metastable
phases are also possible, including the bicontinuous “plumber’s
nightmare.” Lee et al. show
that it is possible to make
this phase a stable one in a
diblock copolymer by adjusting
the interactions of the chain
ends (see the Perspective by
Maintaining nanoparticles
with methanol
Sci. Signal. (2024)
10.1126/scisignal.adg4422
y g
The chemistry of
the chain ends matters
CATALYSIS
Science p. 81, 10.1126/science.adk6533
TAOK3 sets the threshold for
T cell activation by controlling
SHP-1 abundance. —AMV
y
POLYMERS
Science p. 70, 10.1126/science.adh0483;
see also p. 28, 10.1126/science.adn0168
rhodium, reacted as a Lewis acid
in the plane but as a Lewis base
above or below it. —JSY
g
Science p. 55, 10.1126/science.adj6645;
see also p. 32, 10.1126/science.adm8641
Nedoma). The strength of endgroup interactions was shown
to modulate the curvature of
the adjacent polymer, imposing
significant frustration when the
adjacent polymer is crystalline.
By developing a series of design
rules, the authors have found
a controlled way to reversibly make complex structures.
—MSL
p
Decision-making to select
between high risk–high return
and low risk–low return choices
is critical not only in economic
activities such as investing, but
also in many daily life situations. However, the underlying
neural circuits remain largely
elusive. Sasaki et al. identified
two subsystems of the brain’s
mesofrontal pathways that
control risk-dependent decision-making in macaques (see
the Perspective by Stuphorn).
Stimulation of the pathway
from the ventral tegmental
area to the ventral aspect of
the ventral Brodmann area 6
(area 6V) resulted in stronger
preference for high risk–high
return choices. By contrast,
activation of the circuit from the
ventral tegmental area to the
dorsal aspect of area 6V led to
low risk–low return inclination.
Repetitive stimulation of these
distinct pathways over time
resulted in stimulus-independent long-term enhancement or
reduction of preferences. —PRS
Edited by Michael Funk
science.org SCIENCE
R ES E A RC H | I N S C I E N C E J O U R NA L S
CANCER
Killing cohesin-mutant
MDS
Edited by Caroline Ash
and Jesse Smith
ELECTROCATALYSIS
Balancing Pd and Pt
for epoxidation
Propylene oxide is produced
at massive scale using dangerous and corrosive oxidants. A
potentially safer method uses
electrochemistry to oxidize
propylene with water, but
catalyst stability is a persistent
challenge. Chung et al. report
that by alloying palladium with
platinum, they could generate
an electrocatalyst that delivers >60% Faradaic efficiency
toward propylene epoxidation
by water oxidation. Initially
focusing on acetonitrile as a
cosolvent, the authors went
on to explore purely aqueous
media and discuss the prospects of improving propylene
throughput in the face of low
solubility. —JSY
y
Science p. 49, 10.1126/science.adh4355
Atypical models
of arrestin binding
Tissue-resident CD8+ T (TRM)
cells are constantly surveilling organs and tissues for the
presence of uninvited microbes.
Previous studies have shown
that TRM cell migration is triggered by chemoattractant and
adhesion molecule signaling,
which facilitates the rapid
detection of infected cells. More
recent evidence has indicated
that TRM cells within submandibular salivary glands display
different motility patterns
exclusive of chemosensing.
Ruef et al. now show that submandibular salivary gland TRM
cells from virally infected mice
display spontaneous retrograde
F-actin flow as a means of
force-generated translocation.
Similar patterns of locomotion
were detected in TRM cells from
other exocrine glands and were
dependent on the sensing of
changes in mechanical loads
Interactions between G protein–
coupled receptors and their
downstream signaling partners
are often weak, transient, or
mediated by short sequences
or structural motifs and are
thus difficult to study in detail.
Maharana et al. determined
cryo–electron microscopy
structures of the signaling and
regulatory protein b-arrestin in
complex with full receptors or
a phosphopeptide corresponding to the receptor C terminus.
Using refinement focused on
the arrestin component, the
authors visualized the peptidebound conformation. In one of
the structures, the protein’s C
terminus adopted an a-helical
conformation that the authors
propose is important for
dynamic recognition of different
receptors and phosphorylation
patterns. —MAF
5 JANUARY 2024 • VOL 383 ISSUE 6678
Science p. 101, 10.1126/science.adj3347
IMMUNODEFICIENCY
Immune imbalance
in rare disease
Combined immunodeficiencies
result from genetic mutations
that cause defective immune
responses. Lui et al. found that
human siblings homozygous for
a variant of lymphocyte-specific
protein tyrosine kinase (LCK), a
protein essential for T cell receptor (TCR) signaling, exhibited an
altered T cell compartment, suffered from recurrent infections,
and failed to thrive. The authors
created cell lines that expressed
the mutated LCK protein, which
had a single amino acid change,
and found that it disrupted TCR
signaling. In mice genetically
modified to carry the missense
mutation, some conventional
T cells could still mature, and
the development of intestinal
inflammation was attributed to
a deficiency in regulatory T cells.
—SHR
J. Exp. Med. (2024)
10.1084/jem.20230927
FLORAL EVOLUTION
Flower diversity
peaked early
The emergence of flowers
created new reproductive
modes, mutualistic interactions with pollinating animals,
and fodder for diversification
in angiosperms. Flower shape,
arrangement, color, and anatomy
vary widely between and within
some lineages. López-Martínez
et al. quantified disparities in
flower structure across lineages
and over time using morphological data from more than 1000
extant species and more than
100 fossil species and found that
floral disparity peaked in the
science.org SCIENCE
,
Mechanosensing
by T cells
y
SIGNALING COMPLEXES
y g
CELL MIGRATION
40
IN OTHER JOURNALS
g
Sci. Transl. Med. (2024)
10.1126/scitranslmed.ade2774
Sci. Immunol. (2023)
10.1126/sciimmunol.add5724
p
Splicing modulators have been
developed to treat cancers with
splicing factor mutations, such
as myelodysplastic syndromes
(MDS). Mutations in the cohesin
complex are also very common in MDS, but whether these
cohesin-mutant cancers would
respond to splicing modulation
is unknown. Wheeler et al. show
that cohesin-mutant cells are
particularly sensitive to splicing
modulators, which induce missplicing and down-regulation
of DNA damage repair genes,
leading to the accumulation of
DNA damage and sensitization
to talazoparib or chemotherapy
in vitro. In patient-derived
xenograft mouse models,
splicing modulators reduced
cohesin-mutant cancer burden,
and sequential treatment with
talazoparib or doxorubicin
and cytarabine significantly
improved survival. —MLN
through signals triggered by
nuclear deformation. —CNF
PARTICLE PHYSICS
Coming to terms with the
standard model
QUANTUM OPTICS
Straining diamond for
quantum control
T
y
Phys. Rev. X (2023)
10.1103/PhysRevX.13.041037
g
he optical and electronic properties
of color centers in diamonds make
them a promising platform for
quantum technologies, including
enhanced sensing and quantum
information processing applications.
However, coupling to vibrations in the
diamond crystal lattice can induce decoherence and typically requires operation
at millikelvin temperatures to mitigate
for the phonon-induced performance
loss. Guo et al. show that application of
uniform strain can enhance the properties of the color center for tin vacancy
centers in a diamond membrane. The
authors also show that the center can
be addressed with microwaves and the
operation temperature pushed up to 4
kelvin, conditions reachable with widely
available cryogenic systems. This work
demonstrates that strain-tuning provides a mechanism for the stable control
and coherence protection of color centers in diamond membranes. —ISO
p
The standard model (SM) of
particle physics is frequently
tested experimentally by
measuring some of its predictions. One such prediction is
the production rate of the W
boson along with a top and
antitop quark pair in protonproton collisions. Measurements
of this rate by the CMS and
ATLAS collaborations at the
Large Hadron Collider have
yielded values slightly higher
than, but still consistent with,
the SM prediction. However,
the SM prediction itself has an
uncertainty associated with the
complexities of the calculations
used to derive it. Buonocore et
al. improved the precision of the
SM value, using two different
methods to estimate a particularly difficult to calculate term.
The two approaches agree with
each other, giving confidence
in the result and enabling more
stringent tests of the SM. —JS
Phys. Rev. Lett. (2023)
10.1103/PhysRevLett.131.231901
Strain-tuning helps control the color centers
in diamond.
GLOBAL WARMING
Risky known unknowns
Inside a spider’s brain
Spiders are known to weave webs
to catch flying prey. However,
ancestors of spiders might have
been silk-lined burrow dwellers that adopted web building
during the Jurassic–Cretaceous
period, when flowering plants
and flying insects emerged. To
understand the evolution of their
distinctive hunting strategy,
Jin et al. looked into the brains
of spiders for clues. They took
SCIENCE science.org
Nat. Ecol. Evol. (2023)
10.1038/s41559-023-02238-y
VIROLOGY
Bat–virus arms race
Possibly because of the physiological demands of being a flying
mammal, bats have evolved to
trade off fever-inducing immune
responses for tolerance to a wide
As the severity of the global
warming crisis grows, a large
number of potential geoengineering responses have been
suggested. However, dangerous
unintended consequences of
approaches such as the injection of light-reflecting particles
(sulfate, for example) into the
stratosphere still are not understood. Vattioni et al. investigated
how stratospheric injections
of alumina particles might
perturb atmospheric composition, finding that the effects on
heterogeneous chemistry could
result in the destruction of more
than twice as much stratospheric
ozone as that eroded by chlorofluorocarbons in the late 1990s.
Once again, it is clear that there
is no silver bullet for curbing
global warming other than reducing the carbon dioxide content of
the atmosphere. —HJS
PLOS Biol. (2023),
10.1371/journal.pbio.3002398
Geophys. Res. Lett. (2023)
10.1029/2023GL105889
5 JANUARY 2024 • VOL 383 ISSUE 6678
41
,
EVOLUTION
range of viruses. Bat viruses,
however, rarely cause overt
disease in their hosts. Lytras et
al. have looked at the molecular
evolution of bat tolerance and
found that in horseshoe bats, a
prenylated residue is missing on
a potent mammalian antiviral
protein called 2′-5′-oligoadenylate synthetase 1 (OAS1). The
authors reconstructed in vitro
the most likely sequence of an
ancestral bat OAS1 and found that
it blocks severe acute respiratory
syndrome coronavirus 2 replication in A549-ACE2-TMPRSS2
cells. Protein structure predictions showed that the sites most
under selection in OAS1 lie near
the RNA-binding region at the
C terminus where the prenylation signal should be. What the
evolved OAS1 does for modern
horseshoe bats is a mystery. —CA
y
New Phytol. (2023)
10.1111/nph.19389
a high-throughput single-cell transcriptomic approach and built a
comprehensive atlas of cell types
for the adult spider brain. Aerial
web building and spatial niches
expose spiders to more environmental challenges and risks. The
authors found that genes that
have been positively selected in
the ancestral web-building spiders are preferentially expressed
in the integration center of the
brain and that these genes are
involved in learning and memory.
—DJ
y g
Early Cretaceous. This finding
may become even more apparent as more fossil flowers are
discovered and described. These
results provide quantitative support for several hypothesized and
qualitatively described patterns
in plant trait diversity. —BEL
RES EARCH
RESEARCH ARTICLE
◥
2D MATERIALS
De Haas–van Alphen spectroscopy and magnetic
breakdown in moiré graphene
Matan Bocarsly1†, Matan Uzan1†, Indranil Roy1†, Sameer Grover1, Jiewen Xiao1, Zhiyu Dong2,
Mikhail Labendik1, Aviram Uri2, Martin E. Huber3, Yuri Myasoedov1, Kenji Watanabe4,
Takashi Taniguchi5, Binghai Yan1, Leonid S. Levitov2*, Eli Zeldov1*
5 January 2024
1 of 7
,
Bocarsly et al., Science 383, 42–48 (2024)
y
*Corresponding author. Email: eli.zeldov@weizmann.ac.il (E.Z.);
levitov@mit.edu (L.S.L.)
†These authors contributed equally to this work.
y g
Department of Condensed Matter Physics, Weizmann
Institute of Science, Rehovot 7610001, Israel. 2Department of
Physics, Massachusetts Institute of Technology, Cambridge,
MA 02139, USA. 3Departments of Physics and Electrical
Engineering, University of Colorado Denver, Denver, CO
80217, USA. 4Research Center for Electronic and Optical
Materials, National Institute for Materials Science, 1-1 Namiki,
Tsukuba 305-0044, Japan. 5Research Center for Materials
Nanoarchitectonics, National Institute for Materials Science,
1-1 Namiki, Tsukuba 305-0044, Japan.
y
1
the studies of 2D electron systems mostly to
the nonthermodynamic Shubnikov–de Haas
(SdH) oscillations in transport coefficients.
Nevertheless, several studies have succeeded
in resolving magnetization oscillations in 2D
electron gas (2DEG) in gallium arsenide (GaAs)
heterostructures by using millimeter-sized samples (13–16), as well as in magnetically doped
zinc selenide (ZnSe) (17). By contrast, exfoliated clean van der Waals structures are typically limited to tens of micrometers in size, which
makes observation of the dHvA effect in atomically thin systems quite challenging. Furthermore, previous dHvA studies in 2DEG and in
bulk materials have been global, providing no
spatial information on the local BS and thermodynamic electronic properties.
We report the observation of dHvA effect in
a van der Waals structure and spatial mapping
of the thermodynamic QOs with resolution as
high as 170 nm. We observed very large magnetization oscillations in moiré flat bands in
Bernal-stacked bilayer graphene (BLG) aligned
to hexagonal boron nitride (hBN) (2, 18, 19).
The oscillations appear at low magnetic fields
and at carrier densities of a few electrons per
superlattice unit cell. In the integer quantum
Hall effect (QHE), the periodicity of the QOs is
tied in a universal manner to the carrier density
through the LL degeneracy. By contrast, the observed oscillations display characteristic frequencies that are an order of magnitude lower
than the fourfold-degeneracy frequency and
form complex spectra, revealing the coexistence
of multiple FSs and allowing accurate moiré
BS reconstruction.
When a number of FSs coexist, the QOs display several frequencies, reflecting the relative
Transport measurements of longitudinal resistivity (rxx) and Hall resistivity (ryx) (Fig. 1, A
and B) of the BLG sample (Fig. 2A and fig. S1)
were performed at a temperature T = 300 mK as
a function of applied out-of-plane magnetic
field Ba and carrier density n. Secondary peaks
in rxx (Fig. 1C) at n = 4n0 ≅ ±3.48 × 1012 cm–2
indicate that the BLG is aligned to the hBN
substrate with a twist of q ≅ 0.70°, forming a
moiré superlattice with unit cell size l ≅ 11.5 nm
(35), where n0 corresponds to one electron per
moiré unit cell. The weak rxx peaks at filling
factor n = n/n0 = ±4 reflect minima in the
density of states (DOS) and the absence of a
full gap between the low-energy flat bands
and the dispersive remote bands.
At low fields, ryx shows several sign reversals
in Fig. 1B. The Hall carrier density nH extracted
for Ba = 300 mT (Fig. 1D, green line) reveals a
van Hove singularity at n ≅ 3.5, accompanied by
a change in carrier type from electrons to holes.
Similar behavior is found at n ≅ –3.5. Several
additional nH sign reversals occur at higher jnj,
which is consistent with the presence of several
remote moiré bands. At higher fields, the sign
reversals in ryx in Fig. 1B disappear (Fig. 1D, blue
line), and nH follows nH = n (Fig. 1D, dashed
red line), a behavior characteristic of a single
band with no moiré potential.
At elevated Ba, a Landau fan originating from
the charge neutrality point (CNP) is visible at
all fillings (Fig. 1A), corresponding to fourfold
degenerate LLs stemming from spin and valley
degeneracies in graphene (fig. S4B). Additionally, Hofstadter patterns are visible as horizontal lines periodic with f0/Ba (Fig. 1A, dashed
black lines), arising from the interference of the
moiré unit cell with the area occupied by a flux
quantum f0 = h/e (where h is Planck’s constant
and e is the elementary charge), as reported
previously (1–3, 18, 36, 37). More complicated
Landau fans originating from close to n = ±4
are discerned at intermediate fields. In the
following, we describe local studies at fields
g
O
scillations in the thermodynamic and
transport properties of metals subject
to an external magnetic field are a fundamental quantum effect that originates
from the quantization of cyclotron orbit
areas. In two-dimensional (2D) systems, the
periodicity of quantum oscillations (QOs), explained by discrete Landau levels (LLs), has
a universal relation to the applied field and
Fermi surface (FS) geometry. The oscillations
are indispensable for resolving the band structure (BS) of moiré materials, in which the presence of a superlattice potential and enhanced
electron-electron interactions lead to the formation of narrow minibands with multiple FSs
and symmetry broken states (1–7). QOs can also
reveal the band topology (8, 9) and straininduced pseudomagnetic fields in graphene
(10–12).
In bulk materials, QOs are detected by measuring magnetization oscillations caused by
the de Haas–van Alphen (dHvA) effect. These
oscillations in thermodynamic properties, however, are usually experimentally inaccessible
in 2D electron systems because the signal scales
with the sample volume and is therefore extremely weak in two dimensions. This limits
Transport measurements
p
Quantum oscillations originating from the quantization of electron cyclotron orbits provide sensitive
diagnostics of electron bands and interactions. We report on nanoscale imaging of the thermodynamic
magnetization oscillations caused by the de Haas–van Alphen effect in moiré graphene. Scanning by
means of superconducting quantum interference device (SQUID)–on-tip in Bernal bilayer graphene
crystal axis-aligned to hexagonal boron nitride reveals large magnetization oscillations with amplitudes
reaching 500 Bohr magneton per electron in weak magnetic fields, unexpectedly low frequencies,
and high sensitivity to superlattice filling fraction. The oscillations allow us to reconstruct the complex
band structure, revealing narrow moiré bands with multiple overlapping Fermi surfaces separated by
unusually small momentum gaps. We identified sets of oscillations that violate the textbook Onsager
Fermi surface sum rule, signaling formation of broad-band particle-hole superposition states induced by
coherent magnetic breakdown.
size of the different Fermi pockets (FPs) encircled by the cyclotron orbits (17, 20–24). In
addition to these fundamental orbits, electron
orbits delocalized in k-space and supporting
coherently entangled states in different bands
can arise because of interpocket tunneling.
Such tunneling, which is hindered by momentum conservation at zero magnetic field, is
made possible at elevated fields through the
coherent magnetic breakdown (CMB) mechanism (25–32) that is expected to occur in the
vicinity of saddle points or Lifshitz transitions,
across which the topology of the FS changes
(23, 33, 34). Unlike conventional CMB, we found
a breakdown that occurs at low applied fields,
spans a wide range of energies, and displays
QO frequencies that are described by fractional Onsager quantization relations.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
below 350 mT, for which QOs in transport
measurements are barely resolved.
Imaging QOs
To study the magnetization oscillations, we
used a scanning superconducting quantum
interference device (SQUID) fabricated on the
apex of a sharp pipette [SQUID-on-tip (SOT)]
(38). An indium SOT (39) of about 170 nm in
Bocarsly et al., Science 383, 42–48 (2024)
above 500 Ω are saturated for clarity. (C) rxx versus n at Ba = 0 T showing
secondary peaks caused by hBN-BLG moiré. (D) Hall carrier density nH = Ba/(eryx)
versus n derived from ryx at Ba = 0.3 T (green), displaying several sharp sign
reversals that stem from the moiré BS. At Ba = 4.2 T (blue), these sign reversals are
absent. The dashed red line is drawn at a slope of 1 corresponding to nH = n in
absence of moiré flat bands that are washed out by the full CMB.
diameter (fig. S2) was scanned at a height of
h ≈ 200 nm above the sample surface (Fig. 2A)
dc
at T = 300 mK (35). The dc voltages Vtgdc and Vbg
applied to the top and bottom Pt gates, respectively, were used to control n. A small ac voltage
ac
of 5 to 20 mV root mean square was applied
Vbg
to the backgate, modulating the carrier density
by nac and the corresponding nac by 0.004 to
ac
0.016, and the resulting Bac
z ðx; y Þ ¼ n ðdBz =dnÞ
5 January 2024
was imaged across the sample. This signal reflects the induced ac modulation in the local
magnetization, mz(x, y) = dMz(x, y)/dn, which
can be reconstructed directly from the measured Bac
z ðx; yÞ (fig. S3) by a numerical inversion (40) [Mz is the magnetization per unit
area, and mz is the magnetization per excess
electron; both are dominated by orbital effects (35)]. A map is shown in Fig. 2B of the
2 of 7
,
Fig. 1. Transport measurements in BLG aligned to hBN. (A) rxx versus carrier
density n and magnetic field Ba at T = 300 mK shown on a logarithmic scale.
The oscillations at high Ba reflect the existence of fourfold degenerate LLs
(fig. S4) and display a Hofstadter’s butterfly (black dashed lines). (B) ryx versus
n and Ba featuring several sign reversals at low field that disappear at higher
Ba as the system enters the magnetic breakdown regime. Resistivity values
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
Fig. 2. Imaging the dHvA effect. (A) (Top) Optical image of the BLG-hBN
sample with indicated contacts for rxx and ryx measurements. The
dashed yellow rectangle indicates the area imaged in (B), and the dotted
black line indicates the line cut presented in Fig. 3. (Bottom) Schematic
sample structure indicating the applied top-gate and back-gate voltages,
dc
dc
ac
Vtg
and Vbg
þ Vbg
, and the corresponding ac magnetic field Bac
z imaged
BS of moiré bilayer graphene
To gain further insight, we performed continuummodel single-particle BS calculations of the BLGhBN moiré system (35, 41). The mismatch of
graphene and hBN’s lattice constants creates
a superlattice, causing band folding into the
moiré mini-Brillouin zone (mBz) (1–3, 41–45).
The calculated conduction C1 and valence V1
flat bands along with highly overlapping remote valence bands V2 and V3 are shown in
Fig. 4A. Generally, the inequivalent lattice sites
in the hBN substrate break inversion symmetry
and open a gap at CNP. However, remote bands
such as V2 and V3 are highly overlapping, creating complex FPs. Depending on BS parameters,
3 of 7
,
5 January 2024
than being restricted to integer fractions, f
varies continuously as a function of n. (iii) At
higher jnj, multiple characteristic frequencies
are present simultaneously. (iv) There are substantial spatial variations in the relative intensity of the QOs (Fig. 3A and movie S2). These
features reveal the presence of narrow moiré
bands with overlapping FSs.
y
Bocarsly et al., Science 383, 42–48 (2024)
was markedly different, characterized by strong
oscillations with substantially larger and variable periods Dn (Fig. 3, B and D). To quantify
the periodicity, we performed a fast Fourier
transform (FFT) over a narrow window of dn =
1.11 around a given n (35), for Bac
z ðnÞ as indicated with the white dashed line in Fig. 3A
(Fig. 3E; FFT at other positions are provided
in movie S2).
In the integer QHE, the frequency of the QOs
as a function of n is given by f ¼ N1 Bf0a , where N
is the spin-valley degeneracy. For a given N, f
is determined solely by Ba and should thus
be independent of the position and moiré
band filling n. For jnj ≲ 3:5, the FFT reveals
f0
, which is rather indepena peak at f ≅ 4B
a
dent of n and position (fig. S7 and movie S2).
Therefore, the QOs at these fillings originate
from the standard QHE, with fourfold degenerate LLs.
At jnj ≳ 3:5, the QOs show a very rich behavior (Fig. 3E), departing from the standard
QHE in a number of ways: (i) f is up to one
f0
. (ii) Rather
order of magnitude less than 4B
a
y g
mz(x, y) at n = –7.65 and Ba = 334 mT, displaying extremely large values reaching ±500 Bohr
magneton (mB)/electron and forming patches
of positive and negative mz with a characteristic size of about 1 mm. Upon varying n, the
patches move across the sample, and mz(x, y)
reveals quasi-periodic oscillations (Fig. 2C and
movie S1). The shape of the QOs and mz amplitude vary substantially with position (Fig. 2,
D and E). We observed these oscillations from
applied fields as low as 116 mT and up to Ba =
334 mT, at which our SOT has sufficient sensitivity (fig. S5). Similar behavior was found
at various values of displacement field D (fig.
S6) and in two additional samples (figs. S6
and S7).
To investigate the origin of the QOs, we
measured the evolution of Bac
z ðx Þ with n over
an extended range (Fig. 3A) by repeated scanning along the path indicated with the black
dotted line in Fig. 2A, while incrementing n
in 0.006 steps at Ba = 300 mT. For jnj ≲ 3:5,
weak periodic oscillations in Bac
z ðxÞ were discerned (Fig. 3C). For jnj ≳ 3:5, the behavior
with the scanning SOT. (B) Map of orbital magnetization at Ba = 334 mT,
T = 300 mK, and n = –7.65, showing domains of positive and negative local
magnetization mz(x, y) with amplitude of up to ±500 mB/electron. The color bar
applies to (B) to (E). (C) Tomographic rendering of mz(x, y, z) (movie S1).
(D) Slice of the tomographic data mz(y, n) at x = 1.36 mm. (E) mz(x, n) slice
along y = 1.22 mm.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
Bocarsly et al., Science 383, 42–48 (2024)
independent LLs, leading to QOs with multiple
fundamental frequencies. Traditionally, in bulk
materials QOs are measured versus 1/Ba, in
which case, each FP has fi proportional to the
FP area Si (20, 22). This conjecture, however,
does not hold for QOs measured versus n,
where the oscillation frequencies are given
f0
1 @Si
, where NX
by fi ¼ NNiððeeFFÞÞ 4B
i ðeF Þ ¼ 4p2 @e is the
a
DOS of pocket i, N ðeF Þ ¼ i N ðeF Þ is the total
DOS, and eF is the Fermi energy, and we consider fourfold degenerate bands (35). This stems
5 January 2024
from the fact that varying Ba affects the cyclotron motion of the entire Fermi sea, whereas
varying n affects behavior only near eF. Hence,
upon increasing eF by De, one LL is added to
a pocket when its Si has increased by DSi ¼
a
4p2 N i ðeF ÞDe ¼ 4B
X
f0 , leading to the Onsager
sum rule of fundamental frequencies, i fi ¼
f0
f0 ¼ 4Ba .
The experimental FFT data from Fig. 3E is
shown in Fig. 4D overlaid with color-coded lines
that indicate the fi (n) of each pocket calculated
4 of 7
,
either a full gap or a minimum in DOS occurs
between V1 and V2 at n = –4 (35).
For jnj ≲ 3:5, a single FP exists around the
K point (Fig. 4B). At jnj ≅ 3:5, a Lifshitz transition occurs [which is consistent with the
van Hove singularity at Ba = 0.3 T (Fig. 1D)],
forming two FPs centered around X and Y
points (movie S3). With increasing jnj, the FS
topology becomes more complicated, consisting of three or more FPs resulting from overlapping bands (Fig. 4C). Each FP accommodates
(D) Bac
z ðnÞ along the green segment in (A), revealing large-amplitude low-frequency
oscillations comprising multiple frequencies. (E) FFT of Bac
z ðnÞ at x = 4.97 mm
indicated with the white dotted line in (A) performed over a narrow window of dn = 1.11
around n (FFT at different locations is shown in movie S2). The frequency is in units
of f0/4Ba. At jnj ≲ 3:5, the QOs arise from conventional fourfold degenerate
f0
. For jnj ≲ 3:5, the low-frequency oscillations are governed
QHE LLs with f ¼ 4B
a
by multiple overlapping FSs. Positive and negative FFT frequencies are redundant
and are shown for clarity.
y
Fig. 3. Evolution of the dHvA QOs with n and position. (A) Bac
z ðx; nÞ measured
along the dotted black line in Fig. 1A at Ba = 300 mT (additional field values
are available in fig. S5). At jnj ≲ 3:5, the orbital magnetization and the corresponding
ac
Bac
z are weak, whereas for jnj ≳ 3:5, large Bz accompanied by pronounced lowfrequency QOs are present. (B) Zoomed-in cross-section of Bac
z ðnÞ along the
magenta segment in (A), showing low-frequency QOs with a period that gradually
varies with n. (C) Bac
z ðnÞ cross-section along the red segment in (A), showing
high-frequency periodic oscillations caused by conventional fourfold-degenerate LLs.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
Fig. 4. Calculation of BLG-hBN BS and of QOs. (A) Single-particle BS
calculation for a single valley (K) showing (top) the conduction flat band (C1),
(middle) the valence flat band (V1), and (bottom) two partially overlapping
remote valence bands (V2 and V3) in the moiré mBz of BLG aligned to hBN with
q = 0.75°. The bands are fourfold degenerate, with K′ bands rotated by 180°.
Tight-binding parameters were chosen to best fit (D) with rNB = 0.5 and w = 0.5
Magnetic breakdown
The BS calculations with enhanced lattice relaxation provide a good description of the observed
fundamental QO frequencies. Yet there are a
number of prominent lines in Fig. 4D that are not
accounted for by the calculated fi(n). Moreover,
these lines
Xdo not obey the Onsager band area
sum rule i fi ¼ f0 and cannot be explained by
simple harmonics of fi or by sample disorder (35).
These unaccounted-for lines indicate the presence
of electron orbits that encompass areas outside
the closed FS contours. Such trajectories are facilitated by interband electron tunneling caused
by the CMB mechanism, which has been widely
investigated in bulk metals (27) but has not
been identified in 2D vdW materials. When
two FPs are separated by a small momentumgap Dk, the magnetic field–induced interpocket
tunneling occurs with probability
!1=2
BMB
f
Dk3
f
≅ 0 Dk2
P ≅ e Ba ; BMB ¼ 0 1
2 R1 þ R12
2
5 of 7
,
5 January 2024
tion mechanisms and its effect on hBN-aligned
moiré heterostructures.
y
Bocarsly et al., Science 383, 42–48 (2024)
tunneling strengths between overlapping boron
and carbon atoms (tBC) and between overlapping nitrogen and carbon (tNC) to be equal, or
rNB ≡ tNC/tBC = 1. In (45), rNB ≅ 0.67, according
to ab initio density functional theory calculations. We found that both of these parameter
sets fail to fit our experimental data (fig. S8).
Experiments have revealed substantial lattice relaxation in magic-angle twisted bilayer
graphene (46), which has a substantial impact
on the BS (4, 47–49). Lattice relaxation is captured with the phenomenological parameter
w = tAA/tAB ≈ 0.8 (50). Recently, simulations on
aligned BLG-hBN heterostructures have also
shown substantial lattice relaxation, which was
proposed to affect band topology (19).
By correlating the high-resolution QOs with
simulations, we found rNB ≅ 0.5 and w ≅ 0.5
throughout the sample (movie S2). These values are substantially lower than estimated previously, leading to larger band overlaps with
smaller energy gaps (fig. S8) and to an enhancement of magnetic breakdown. This finding of
strong lattice relaxation in aligned BLG-hBN
calls for further exploration of lattice relaxa-
y g
from BS. For hole doping at 0 > n > –3.5, one FP
around K point (V1Κ) is present, resulting in a
single frequency fV1K = f0 (Fig. 4D, green). At
the Lifshitz transition at n ≅ –3.5, the FS breaks
into two pockets around the mBz corners, V1X
and V1Y (movie S3). As a result, two QO frequencies coexist for a small region of n, fV1X
and fV1Y, until V1 (Fig. 4D, green) overlaps V2
(Fig. 4D, light brown). At n < –5, V3 band (Fig.
4D, pink) starts to be occupied, forming two
FPs, V3X and V3Y, with increasing DOS with jnj
that coexist with the V2Κ FP with decreasing
DOS (Fig. 4C). As a result, the two V3 frequencies, fV3X and fV3Y, grow with jnj (Fig. 4D, pink),
whereas the fV2K frequency decreases (Fig. 4D,
light brown). The calculated behavior for –5 >
n > –10.5 closely follows the experimentally
derived frequencies and their evolution with
n. We observed similar behavior for electron
doping at n > 3.5.
With the above insight, the high sensitivity
to oscillation frequencies at fillings where multiple FPs coexist makes nanoscale magnetization imaging a particularly sensitive tool for
mapping the local BS. The work in (41) takes the
(fig. S8) (35). (B and C) Example of (B) simple and (C) complex FSs (solid
contours) at n = 2.054 and –10.14, respectively. The dashed contours indicate
the change in the areas of the FPs with small increase in n, reflecting the DOS
and the QO frequencies of each pocket. (D) The FFT of QOs from Fig. 3E, overlaid
with fundamental frequencies (lines coded by band colors) calculated by the
f0
relative DOS of each pocket, fi ðnÞ ¼ NNiððeeFFÞÞ 4B
.
a
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
5 January 2024
gives rise to extended equi-energy electron
orbits. The green dashed line in Fig. 5A indicates the shortest orbit that traces two-thirds
of the circumference of the two electron pockets
C3X and C3Y and one-third of the circumference of the hole pocket C2K. This results in a
QO frequency given by a fractional Onsager
relation
1
fMB ¼ ð2fC3X þ 2fC3Y þ fC2K Þ
3
where fi are the frequencies of the individual
FPs (35), in contrast to common CMB behavior
with integer Onsager relations (27, 32). Shown
6 of 7
,
Bocarsly et al., Science 383, 42–48 (2024)
(Fig. 5A, dark pink). The sharp touching points
between the C3Y and C2K pockets are characterized by a very small momentum gap Dk ≅
0.010 nm–1, and the gaps between the C3X
and C2K pockets have an even smaller Dk ≅
0.005 nm–1 (Fig. 5B, inset), leading to CMB at
low fields. Moreover, in contrast to the common situations in which close proximity of FPs
is limited to the vicinity of Lifshitz transitions,
the distinct highly overlapping BS of relaxed
BLG-hBN leads to small gaps extending over
almost the entire energy range of the remote
bands, with sharp ridges that closely follow each
other (Fig. 5B). The tunneling between the FPs
y
where BMB is the breakdown field and R1 and
R2 are the k-space radii of curvature of the two
FSs in the gap region (27). For BMB ≲ Ba ¼ 0:3 T,
this requires Dk ≲ 0:012 nm–1. Our BS calculations show very small gaps between FPs in remote bands (Fig. 5B).
To derive the MB orbits and their corresponding QO frequencies fMB, we analyzed two prominent unaccounted-for lines in the experiment
at 5 < n < 10.5 and –12.5 < n < –10 (Fig. 5, C and
E, bright green). The FS structure at n = 8.4 is
shown in Fig. 5A, displaying electron pockets
at X and Y originating from the C3 band (Fig. 5A,
red) and a hole pocket at K from the C2 band
of Bac
z ðnÞ at x = 2.04 mm, showing a pronounced frequency line for 4.5 < n < 10 that
does not obey the sum rule. This frequency is accounted for by the CMB orbit in (A),
resulting in fMB ¼ 31 ð2C3K þ 2fC3Y þ fC2K Þ (green line). (D) Calculated constant
energy BS cut at n = –12.28 showing three degenerate electron pockets from the
V4 band (brown) and one hole pocket from the V3 band (light purple). Scale bar
applies to both (A) and (D). The dashed green trajectory indicates the shortest CMB
orbit. (E) FFT of Bac
z ðnÞ at x = 4.44 mm with overlaid calculated fMB (green line)
(35). Positive and negative FFT frequencies are redundant and are shown for clarity.
y g
Fig. 5. Coherent magnetic breakdown. (A) Calculated constant energy BS cut
at n = 8.4 showing the occupied C3X and C3Y electron pockets (red, clockwise
black arrows) and the C2K hole pocket (dark pink, counterclockwise arrows). Scale
bar is shown in (D). The dashed green trajectory indicates the shortest magnetic
breakdown orbit. (B) Cut of the calculated BS along the dashed black line in
(A). Dashed line at e = 154.2 meV corresponds to the energy value in (A). The gap
between C2K and C3X pockets remains very small over a large range of energies.
(Inset) Zoom-in showing a gap of Dk ≅ 0.005 nm–1. (C) Expanded view of FFT
RES EARCH | R E S E A R C H A R T I C L E
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
14.
15.
16.
20.
21.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
5 January 2024
45.
46.
47.
48.
49.
50.
51.
AC KNOWLED GME NTS
The authors thank E. Berg, A. Stern, I. Rozhansky, and N. Avraham
Tayar for useful discussions. Funding: This work was supported by
the European Union (ERC, MoireMultiProbe – 101089714): Views
and opinions expressed are however those of the author(s) only
and do not necessarily reflect those of the European Union or
the European Research Council. Neither the European Union nor
the granting authority can be held responsible for them (to E.Z.); the
Minerva Foundation, with funding from the Federal German
Ministry of Education and Research Grant 140687 (to E.Z.); United
States–Israel Binational Science Foundation (BSF) Grant 2022013
(to E.Z. and L.S.L.); Andre Deloro Prize for Scientific Research
and Leona M. and Harry B. Helmsley Charitable Trust grant
2112-04911 (to E.Z.); Goldfield Family Charitable Trust (to E.Z.);
Science and Technology Center for Integrated Quantum Materials,
NSF grant DMR1231319, and Army Research Office Grant W911NF-181-0116 (to L.S.L.); Sagol Weizmann-MIT Bridge Program (to E.Z.
and L.S.L.); European Research Council (ERC Consolidator Grant
“NonlinearTopo,” 815869) (to B.Y.); Israel Science Foundation ISF
grant 2932/21 (to B.Y.); JSPS KAKENHI grants 21H05233 and
23H02052 (to K.W. and T.T.); World Premier International
Research Center Initiative (WPI), MEXT, Japan (to K.W. and T.T.);
MIT Pappalardo Fellowship (to A.U.); VATAT Outstanding
Postdoctoral Fellowship in Quantum Science and Technology (to
A.U.); and a VATAT Outstanding PhD Fellowship in Quantum
Science and Technology (to M.B.) Author contributions: Local
magnetization measurements: M.B., I.R., and S.G. Sample
fabrication and transport measurements: M.U. SOT fabrication and
tuning fork feedback: I.R., M.L., and Y.M. Development of
experimental setup: A.U. Design and building of the SOT readout
system: M.E.H. BS calculations: J.X., B.Y., and M.B. Development
of theoretical model (CMB): M.B., E.Z., L.S.L., and Z.D. Data
analysis: M.B., I.R., M.U., and S.G. hBN crystals: K.W. and T.T.
Writing of original manuscript: M.B., E.Z., L.S.L., J.X., M.U., I.R., and
Z.D. Editing and review of manuscript: all authors. Competing
interests: The authors declare no competing interests. Data
and materials availability: The data from this study and code for
BS calculations are available at the Weizmann Institute of Science
repository (51). License information: Copyright © 2024 the
authors, some rights reserved; exclusive licensee American
Association for the Advancement of Science. No claim to original
US government works. https://www.science.org/about/sciencelicenses-journal-article-reuse
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adh3499
Materials and Methods
Supplementary Text
Figs. S1 to S9
References (52–72)
Movies S1 to S3
Submitted 26 February 2023; accepted 23 November 2023
10.1126/science.adh3499
7 of 7
,
23.
44.
y
22.
42.
43.
y g
18.
19.
L. A. Ponomarenko et al., Nature 497, 594–597 (2013).
C. R. Dean et al., Nature 497, 598–602 (2013).
B. Hunt et al., Science 340, 1427–1430 (2013).
Y. Cao et al., Nature 556, 80–84 (2018).
H. S. Arora et al., Nature 583, 379–384 (2020).
P. Stepanov et al., Phys. Rev. Lett. 127, 197701 (2021).
J. M. Park et al., Nat. Mater. 21, 877–883 (2022).
A. Alexandradinata, C. Wang, W. Duan, L. Glazman, Phys. Rev. X
8, 011027 (2018).
C. Guo et al., Nat. Commun. 12, 6213 (2021).
F. Guinea, M. I. Katsnelson, A. K. Geim, Nat. Phys. 6, 30–33
(2010).
N. Levy et al., Science 329, 544–547 (2010).
J. Mao et al., Nature 584, 215–220 (2020).
S. A. J. Wiegers et al., Phys. Rev. Lett. 79, 3238–3241
(1997).
I. Meinel et al., Phys. Rev. Lett. 82, 819–822 (1999).
N. Ruhe, J. I. Springborn, C. Heyn, M. A. Wilde, D. Grundler,
Phys. Rev. B Condens. Matter Mater. Phys. 74, 235326
(2006).
T. Windisch et al., Phys. Rev. B Condens. Matter Mater. Phys.
80, 205306 (2009).
J. G. E. Harris et al., Phys. Rev. Lett. 86, 4644–4647
(2001).
T. Iwasaki et al., Nano Lett. 20, 2551–2557 (2020).
E. Arrighi et al., Non-identical moiré twins in bilayer graphene.
arXiv:2205.01760 (2022).
A. S. Alexandrov, A. M. Bratkovsky, Phys. Rev. Lett. 76,
1308–1311 (1996).
R. A. Shepherd et al., Phys. Rev. B Condens. Matter 60,
R11277–R11280 (1999).
J.-Y. Fortin, E. Perez, A. Audouard, Phys. Rev. B Condens.
Matter Mater. Phys. 71, 155101 (2005).
K. Kishigi, Y. Hasegawa, Phys. Rev. B 104, 085412
(2021).
H. Zhou et al., Science 375, 774–778 (2022).
C. E. T. G. da Silva, L. M. Falicov, Phys. Rev., B, Solid State 8,
527–535 (1973).
M. I. Kaganov, A. A. Slutskin, Phys. Rep. 98, 189–271
(1983).
D. Shoenberg, Magnetic Oscillations in Metals (Cambridge Univ.
Press, 1984).
P. Steda, A. H. MacDonald, Phys. Rev. B Condens. Matter 41,
11892–11898 (1990).
P. H. Beton et al., Phys. Rev. B Condens. Matter 43,
9980–9983 (1991).
V. M. Gvozdikov, Phys. Rev. B Condens. Matter Mater. Phys. 75,
115106 (2007).
P. Walmsley et al., Phys. Rev. B 102, 045150 (2020).
J. F. Linnartz et al., Phys. Rev. Res. 4, L012005
(2022).
C.-K. Lu, H. A. Fertig, Phys. Rev. B Condens. Matter Mater. Phys.
89, 085408 (2014).
A. Alexandradinata, L. Glazman, Phys. Rev. Lett. 119, 256601
(2017).
41.
Materials and methods are available as supplementary materials.
R. Krishna Kumar et al., Science 357, 181–184 (2017).
J. Barrier et al., Nat. Commun. 11, 5756 (2020).
D. Vasyukov et al., Nat. Nanotechnol. 8, 639–644 (2013).
Y. Anahory et al., Nanoscale 12, 3174–3182 (2020).
A. Y. Meltzer, E. Levin, E. Zeldov, Phys. Rev. Appl. 8, 064030
(2017).
P. Moon, M. Koshino, Phys. Rev. B Condens. Matter Mater. Phys.
90, 155406 (2014).
M. Yankowitz et al., Nat. Phys. 8, 382–386 (2012).
J. R. Wallbank, A. A. Patel, M. Mucha-Kruczyński, A. K. Geim,
V. I. Fal’ko, Phys. Rev. B Condens. Matter Mater. Phys. 87,
245408 (2013).
P. San-Jose, A. Gutiérrez-Rubio, M. Sturla, F. Guinea,
Phys. Rev. B Condens. Matter Mater. Phys. 90, 075428
(2014).
J. Jung, A. Raoux, Z. Qiao, A. H. MacDonald, Phys. Rev. B
Condens. Matter Mater. Phys. 89, 205414 (2014).
Y. Jiang et al., Nature 573, 91–95 (2019).
P. Lucignano, D. Alfè, V. Cataudella, D. Ninno, G. Cantele,
Phys. Rev. B 99, 195419 (2019).
M. Angeli et al., Phys. Rev. B 98, 235137 (2018).
X. Lu et al., Nature 574, 653–657 (2019).
M. Koshino et al., Phys. Rev. X 8, 031087 (2018).
M. Bocarsly et al., Data and code for: de Haas–van Alphen
spectroscopy and magnetic breakdown in moiré graphene. WIS
Library (2023); https://doi.org/10.34933/e6c6e1ae-7c9c45ec-a607-fbbdb00a9616.
y
11.
12.
13.
17.
Owing to the extreme sensitivity of the QOs
to the BS details, our measurement provides a
distinctive probe of multiband moiré FSs and
their low-energy electronic properties. Crucially, the ability to detect thermodynamic QOs at
low fields allows probing of the BS with high
energy resolution and without perturbing it
with high magnetic fields. In particular, our
results show that the hBN-graphene coupling
is substantially weaker than previously estimated values, giving rNB = tNC/tBC ≅ 0.5 and a
large lattice relaxation with w = tAA/tAB ≅ 0.5,
a direct demonstration of a weak moiré potential with strongly overlapping bands and small
energy gaps. This fine potential modulation is
readily washed out at elevated magnetic fields,
leading to a full breakdown in which the carriers orbit mostly along the original BLG FS
unperturbed by the moiré potential (fig. S9)
(35). The nH measured at Ba = 4.2 T shown in
Fig. 1D shows essentially no signs of the moiré
multi-pocket BS resolved at low fields.
Our findings open up a previously uncharted
regime of exotic CMB physics at unusually low
Bocarsly et al., Science 383, 42–48 (2024)
RE FERENCES AND NOTES
35.
36.
37.
38.
39.
40.
g
Discussion and outlook
Ba, arising from cyclotron orbits delocalized
in k-space and supporting states coherently
entangled among different subbands. This
regime is manifested through QOs that do not
obey Onsager’s FS area sum rule. Instead, we
observed a fractional Onsager quantization
relation that indicates the occurrence of particlehole superposition states shared by adjacent
bands and exhibiting a high degree of interband phase coherence. The real-space cyclotron
radius of the observed CMB
orbits in Fig. 5A
f0
S 1=2
≅
500 nm, a value
is as large as Rc ¼ 2pB
a p
comparable with the characteristic scales of
disorder and sample dimensions (35), which
calls for further theoretical and experimental
studies for understanding the QOs in the presence of BS disorder. The particle-hole coherence induced by CMB is an appealing and
not-yet-explored direction for band engineering in moiré materials.
p
in Fig. 5C is a good fit between the calculated
fMB (Fig. 5C, green) and the experimentally
unaccounted for QO line. The fMB line is derived from the fundamental fi, with no adjustable parameters and over their entire range.
A gap of Dk ≅ 0.01 nm–1 corresponds to tunneling probability P ≈ 0.5, allowing the carriers to orbit along both the closed FPs and
along the CMB trajectories. As a result, both
the fundamental (Fig. 5C, red and pink) and
the CMB frequency lines (Fig. 5C, green) were
observed concurrently. Similar CMB QOs were
observed in device 2 with a larger hBN twist
angle q = 1.15° (fig. S7).
The BS at n = –12.28 is shown in Fig. 5D,
displaying three degenerate hole pockets in
the V4 band and an electron pocket at K in the
V3 band. CMB creates an electron trajectory
that flows along the inner and outer edges of
the V3 and V4 pockets (Fig. 5D, green dashed
line), which explains well the unaccounted QO
line in Fig. 5E as indicated with the calculated
green fMB line. The fundamental fV3 line (Fig.
5E, purple) is essentially invisible in the experiment. This can be understood in view of
the extremely small gap between the V3 and
V4 pockets with Dk ≅ 0.02 nm–1 and very small
radii of curvature R. As a result, the electrons
tunnel between the pockets with P ≅ 1, leaving
essentially no carriers that circulate exclusively in the V3 pocket and hence no detectable
fV3. Moreover, in contrast to the usual CMB
behavior (32) and unlike the fMB line in the
conduction bands (Fig. 5C, green), the fMB line
in Fig. 5E cannot be expressed as either integer
or fractional Onsager combination of the fundamental frequencies because of nontrivial
evolution of the FS with doping.
RES EARCH
ELECTROCATALYSIS
Direct propylene epoxidation via water activation
over Pd-Pt electrocatalysts
Minju Chung1, Joseph H. Maalouf1, Jason S. Adams2, Chenyu Jiang2,
Yuriy Román-Leshkov1, Karthish Manthiram2*
Direct electrochemical propylene epoxidation by means of water-oxidation intermediates presents a
sustainable alternative to existing routes that involve hazardous chlorine or peroxide reagents. We
report an oxidized palladium-platinum alloy catalyst (PdPtOx/C), which reaches a Faradaic efficiency
of 66 ± 5% toward propylene epoxidation at 50 milliamperes per square centimeter at ambient
temperature and pressure. Embedding platinum into the palladium oxide crystal structure stabilized
oxidized platinum species, resulting in improved catalyst performance. The reaction kinetics suggest that
epoxidation on PdPtOx/C proceeds through electrophilic attack by metal-bound peroxo intermediates.
This work demonstrates an effective strategy for selective electrochemical oxygen-atom transfer from water,
without mediators, for diverse oxygenation reactions.
1 of 7
,
5 January 2024
We examined the bulk and surface properties
of our annealed PdPt/C, Pd/C, and Pt/C catalysts
(PdPtOx/C, PdOx/C, and PtOx/C, respectively)
y
Chung et al., Science 383, 49–55 (2024)
Catalyst characterizations
y g
*Corresponding author. Email: karthish@caltech.edu
Previously studied catalysts for electrochemical propylene epoxidation can be clustered
into palladium-derived (Pd) (9–12), platinumderived (Pt) (13–15), and silver-derived (Ag)
(16, 17) groups (Fig. 1B and table S1), and the
best catalytic performance for electrochemical
epoxidation has been observed with oxidized
Pd and Pt catalysts. Studies on Pd catalysts
have shown that epoxidation occurs on the
surface oxide layer at anodic potentials >1.1 V
versus reversible hydrogen electrode (RHE),
whereas allylic oxidation producing acrolein
and acrylic acid occurs on the metallic surface
at lower potentials (9, 12). The performance
enhancement of oxidized Pt catalysts was correlated with an increase in surface presence of
Pt(II) and Pt(IV) species, whereas pure PtO2
was less active (14). Moreover, a theoretical study
has identified weak oxygen-binding metal oxides
as promising catalyst candidates for propylene
epoxidation and cautioned that the metallic
phase of strong-binding metals, such as Pt and
Pd, may facilitate the production of unwanted
side products through dehydrogenation (18).
These findings suggest that the catalytic performance may be further optimized by adjusting the oxidation states of the metal. However,
preparing Pt oxide catalysts with high concentrations of oxidized Pt species is challenging because of their instability. By contrast,
stable Pd oxide catalysts can be obtained through
high-temperature annealing, but they generally exhibit lower rates of epoxidation.
y
Department of Chemical Engineering, Massachusetts
Institute of Technology, Cambridge, MA 02139, USA.
2
Division of Chemistry and Chemical Engineering, California
Institute of Technology, Pasadena, CA 91125, USA.
Catalytic performance of PdPtOx/C
and design principle
g
1
possible to directly transfer reactive intermediates typically seen in water oxidation to
the alkene before the intermediates are fully
oxidized into O2 (Fig. 1A, top) (6–8). This direct electrochemical alkene-epoxidation approach does not involve hazardous oxidizing
agents, such as Cl2, making it an attractive avenue for high-demand propylene epoxidation.
p
P
ropylene oxide (PO) is an important chemical building block used in the production of many commodity chemicals, such
as polyurethanes and polyesters (1). Currently, PO production relies on highly reactive, corrosive, or explosive chemical oxidants
such as molecular chlorine (Cl2) or peroxides (2).
Although direct oxidation of propylene with molecular oxygen (O2) has been explored as an alternative solution, achieving high selectivity
for PO is challenging because of the allylic hydrogen stripping in propylene and consequent
further oxidation (2). In this regard, a selective
direct propylene-epoxidation pathway that uses
water as a cleaner and an easily accessible oxygen source would present a compelling alternative to the current epoxidation methods.
Electrochemistry offers a promising approach for using water as a sustainable source
of oxygen atoms at ambient temperature and
pressure. In water electrolysis, typically oxygen
and hydrogen are generated, but alternatively, commodity chemicals can be synthesized
through oxidative processes at the anode of
an electrochemical cell combined with hydrogen evolution at the cathode. For example,
chloride oxidation can be conducted at the
anode to generate Cl2, which acts as a redox
mediator in alkene epoxidation (Fig. 1A, bottom)
(3–5). This electrified version of the chlorohydrin
process was demonstrated to be effective at
industrially relevant current densities and selectivity (4). However, this method produces
byproducts such as hypochlorite, chlorinated
organic compounds, and brine streams that
pose serious environmental challenges unless
100% selectivity and recycling of the chloride
electrolytes can be achieved. Rather than relying on Cl2 as a redox mediator, it should be
Alloying Pt with Pd can be a strategy that
leverages complementary properties of the
two elements, enabling the attainment of optimal oxidation states and structures for an
epoxidation catalyst (Fig. 1C). Under aerobic
conditions, Pt nanoparticles tend to sinter into
a metallic form when annealed above 400°C
(19) because volatile PtO2 species mediate the
sintering process by facilitating Ostwald ripening (20, 21). By contrast, stable PdO nanoparticles can be produced through annealing of
Pd nanoparticles without substantial sintering (19, 22). PdO can trap mobile PtO2 species, which results in less sintering of Pd-Pt
alloy nanoparticles than of Pt nanoparticles
(20, 23). In addition, Pd and Pt share very
similar crystal structures and atomic sizes, allowing homogeneous distributions of Pd and
Pt in their alloys.
In this study, we designed a Pd-Pt alloy catalyst containing both Pd oxide and Pt oxide by
embedding and stabilizing Pt oxide species in Pd
oxide. A series of alloys with varying ratios of
Pd and Pt were synthesized using a coreduction
method (24) on an amorphous carbon substrate
and subsequently annealed under static air
before being tested for their activity in propylene epoxidation (materials and methods
and figs. S1 to S3). Our catalyst is compatible
with a blended electrolyte composed of acetonitrile and water (Fig. 1D) that can accommodate a broad spectrum of alkene substrates, as
well as aqueous electrolytes (Fig. 1E) that can
minimize overall cell voltage for gaseousalkene epoxidation (fig. S4). The alloy catalyst
with an equimolar composition of Pd and Pt
annealed at 500°C (PdPtOx/C) was most active
toward propylene epoxidation and outperformed previously reported catalysts for direct
anodic epoxidation under ambient conditions
(Fig. 1B and table S1). Furthermore, alloys with
metal compositions of Pd3Pt1 and Pd1Pt3 exhibited epoxidation activities on par with those
of PdPtOx/C; when Pd and Pt are mixed at the
nanometer scale (figs. S2 and S3), Pd-Pt alloy
catalysts can achieve enhanced epoxidation rates
and Faradaic efficiencies (FEs) relative to either
Pd or Pt alone (Fig. 1, D and E, and fig. S5).
In an electrolytic cell equipped with circulating liquid electrolytes (fig. S4), we demonstrated continuous PO production with high
FE averaging 66 ± 5% [propylene glycol (PG);
1.3 ± 0.3%] for 3 hours at a constant current
density of 50 mA/cm2. Additionally, PdPtOx/C
catalyzed cyclooctene epoxidation with an FE
of ~90%, a notable improvement over the results observed in our previous studies, in which
we used Ir-MnOx (~50%) and MnOx (~30%)
catalysts (table S2) (7, 8).
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
Chung et al., Science 383, 49–55 (2024)
5 January 2024
to 40.0° in PdPtOx/C. Furthermore, the broader
peaks in the PdPtOx/C XRD pattern indicate
that the PdPtOx/C has smaller crystallites than
those of PtOx/C (table S3), which is consistent
with our hypothesis that the sintering of PdPt/C
is less severe than that of Pt/C upon annealing.
Extended x-ray absorption fine structure
(EXAFS) measurements revealed that, within
fitting errors, Pd and Pt coordinate with the
same number of oxygens in PdPtOx/C; the Pd-O
and Pt-O coordination numbers were estimated
to be 3.7 ± 0.4 and 2.8 ± 0.6, respectively (Fig. 2,
B and C, and tables S4 to S6). The local coordination environments of Pd in PdPtOx/C
and PdOx/C were similar to that of PdO, whereas PtOx/C resembled the Pt foil standard and
did not show any contribution from the Pt-O
scattering path. By contrast, the local environment of Pt in PdPtOx/C was different from that
in the Pt foil or PtO2 standard, and the first-shell
scattering path of PdPtOx/C was fit with a combination of Pt-O and Pt-Pd/Pt scattering paths.
These EXAFS data suggest that blending Pt in
the PdO structure induced a local coordination
2 of 7
,
to shed light on the enhanced performance of
PdPtOx/C compared with PdOx/C and PtOx/C.
X-ray diffraction (XRD) patterns showed
that although PdOx/C and PtOx/C only exhibited oxide and metallic phases, respectively,
the PdPtOx/C contained both phases (Fig. 2A
and fig. S6). The oxide phase of PdPtOx/C resembles that of PdOx/C because the (101) peak
in PdPtOx/C at 33.7° occurs at a similar angle
as the PdOx/C peak at 33.9°. Similarly, the metallic phase of PdPtOx/C parallels that of PtOx/C,
with the (111) peak shifted from 39.8° in PtOx/C
performance with different compositions of Pd-Pt-on-carbon catalysts in (D)
water-acetonitrile electrolyte and (E) aqueous electrolyte. Potentials in this
figure were 100% iR-compensated (i, current; R, resistance). The vertical error
bars represent standard deviations from the mean of multiple replicates (n = 3)
of the same experiment.
y
Fig. 1. Background and demonstration of direct electrochemical propylene
epoxidation. (A) Comparison of chlorine-mediated and direct electrochemical
propylene epoxidation. (B) Compilation of previously reported direct electrochemical
propylene epoxidation and our work. (C) Effect of annealing on Pt, Pd, and
Pd-Pt nanoparticles. (D and E) Direct electrochemical propylene-epoxidation
RES EARCH | R E S E A R C H A R T I C L E
p
g
Electrochemical kinetic study and proposed
reaction mechanism
Understanding the mechanism of direct propylene epoxidation on PdPtOx/C can help us
develop more efficient catalysts and process
conditions for producing PO. The reaction
mechanism was investigated with a multifaceted
approach, including kinetic rate measurements,
deuterium kinetic isotope effect measurements,
3 of 7
,
5 January 2024
Furthermore, EXAFS fitting of catalysts under operating conditions showed that coordination numbers and scattering-path lengths of
Pd and Pt did not change during electrolysis
(tables S5 and S6). Applying a cathodic potential
followed by an anodic potential demonstrated
that the edge energies and the coordination
environment of Pd and Pt respond to applied
potentials in a partially, albeit not fully, reversible fashion (Fig. 3 and figs. S10 and S11).
The results from XRD, CV, and the elemental analysis collectively suggest that Pd and
Pt are mixed in both the bulk and surface of
the PdPtOx/C catalyst. XANES and XPS results revealed that a larger quantity of oxidized
Pt species is stabilized in PdPtOx/C than in
PtOx/C. XAS data suggest that Pd stabilizes
Pt at a similar oxidation state (around +2) and
crystal structure as those of PdO, and these combined factors improve catalytic performance.
y
Chung et al., Science 383, 49–55 (2024)
S8 and S9), indicating that the higher ratio of
oxidized Pd and Pt species is contributing to
the enhancement in the catalyst performance.
Operando x-ray absorption spectroscopy (XAS)
experiments probing the Pd K-edge and Pt L3edges during epoxidation highlighted again
that achieving more oxidized Pt may have a
critical role in improving catalytic activity
toward propylene epoxidation. Progressively
more oxidative potentials were applied from
0.45 to 1.15 V versus Fc/Fc+, followed by a reductive step to −1.35 V and then two oxidative
steps back to 1.15 V. The average metal valency
at each condition was estimated from linear
combination fitting of x-ray absorption near
edge structure (XANES) spectra by using reference spectra from standard materials. The
results indicate that PdO stabilizes oxidized
Pt species in Pd-Pt alloys, leading to a higher
average oxidation state of Pt in PdPtOx/C (+2)
compared with that in PtOx/C (0). This difference is maintained under anodic potentials (0.45 to 1.15 V versus Fc/Fc+, Fig. 3A).
Unlike the stark contrast in Pt valency between PdPtOx/C and PtOx/C, the Pd valency
stayed the same (+2) in both PdPtOx/C and
PdOx/C (Fig. 3B). These observations suggest
that the more-oxidized Pt species contribute
to the enhancement in the production rate
and selectivity of PO.
y g
environment of Pt resembling that of PtO, which
is unstable alone.
The mixing of Pd and Pt on the surface,
matching the bulk composition, was confirmed
through cyclic voltammetry (CV) and x-ray
photoelectron spectroscopy (XPS) analyses.
A single peak of surface-oxide reduction [at
~0 V versus ferrocene/ferrocenium (Fc/Fc+)]
was observed in the CV of the PdPtOx/C electrode, suggesting that Pd and Pt coexist on the
catalyst surface (Fig. 2D and fig. S7). Furthermore, near-surface compositions derived from
XPS survey spectra also indicate that Pd and Pt
are present in an equimolar ratio near the surface (table S7), matching the bulk composition
determined with inductively coupled plasmaoptical emission spectrometry (table S8).
XPS analysis showed that alloying Pt with Pd
stabilizes oxidized Pt on the catalyst surface.
Pt(0) species were dominant in the PtOx/C,
whereas a higher content of Pt(II) species was
observed in PdPtOx/C (Fig. 2F). The ratio of
Pt(II):Pt(0) increased from 1.0 to 6.4 as the
catalyst treatment temperature was increased
from 400° to 500°C (table S9). A small amount
of Pd(0) was present in PdPtOx/C annealed at
400°C, whereas Pd(II) was exclusively observed
in the catalyst annealed at 500°C (Fig. 2E). The
epoxidation rate and selectivity also improved
with the treatment temperature for PdPt/C (figs.
voltammetry traces of the catalysts. The scans were recorded at 50 mV/sec scan
rate with 85% automatic iR compensation (i, current; R, resistance). (E) Highresolution Pd 3d XPS spectra of annealed Pd/C and PdPt/C catalysts. (F) Highresolution Pt 4f XPS spectra of annealed Pt/C and PdPt/C catalysts.
y
Fig. 2. Ex situ characterizations of annealed PdPt/C, Pd/C, and Pt/C
catalysts. (A) Magnified XRD patterns around the peak associated with the oxide
and metallic phases of annealed catalysts at 500°C. (B and C) EXAFS spectra
of standards and catalysts at (B) Pd K-edge and (C) Pt L3-edge. (D) Cyclic
RES EARCH | R E S E A R C H A R T I C L E
y
,
4 of 7
y g
5 January 2024
0.48 mA/cm2. Diverse derivatives of this mechanism involving the OO* epoxidation intermediate are plausible, depending on whether
a dual or single site is presumed for OO* and
O* intermediates. We selected the model with
the lowest RMSE as our representative mechanism (Fig. 4D), and the model provides a
quantitative explanation of the data illustrated
in Fig. 4, A to C. We also simulated surface coverage with the model (Fig. 4E and supplementary text S1). At higher propylene pressures
where O* and OH* are abundant, the overall
epoxidation rate would be largely determined
by the rate of water oxidation to form OOH*.
However, the comparable prevalence of different surface-oxygen intermediates underscores
the necessity for a comprehensive kinetic model
that does not assume a single dominant reactive
intermediate and a rate-limiting step.
Kinetic isotope labeling and stilbene control experiments further support our hypothesis asserting OO* as the key intermediate in
epoxidation. The use of deuterated water lowered the epoxidation rate (kinetic isotope effect value kH/kD = 1.8 ± 0.5; fig. S15 and table
S10), and the FE remained similar. The difference observed in the epoxidation rate is consistent with the kinetically relevant water-activation
steps involving O–H cleavages in our proposed
mechanism. Moreover, our mechanism identifies OO* as the branching intermediate for both
epoxide and O2 evolution (Fig. 4D), providing
an explanation for the marginal selectivity difference because neither of these pathways from
OO* involves O–H cleavage. In addition, cisstilbene was used as a probe substrate to help
determine whether epoxidation proceeds through
a concerted mechanism with OOH* or a stepwise pathway with OO* (25, 26). Cis-stilbene
(96%) epoxidation with PdPtOx/C produced 23%
cis-stilbene oxide and 77% trans-stilbene oxide,
which suggests that alkene epoxidation over
y
Chung et al., Science 383, 49–55 (2024)
first order at a higher water ratio (Fig. 4B and
fig. S12). At lower propylene pressures, the
water dependence showed more linear approximately first-order dependence in the entire
water concentration range. For propylene, the
rate showed first-order dependence on propylene activity at lower propylene pressures, but the
dependence decreased at elevated propylene
pressures (Fig. 4C and fig. S13).
To address the complexity arising from noninteger reaction orders and variations in orders
with reactant concentrations, we implemented
a multiparameter kinetic model and fitted it
to the kinetic data collected over a wide range
of experimental conditions. We initially hypothesized that surface-adsorbed hydroperoxo
(OOH*), peroxo (OO*), or oxo (O*) species might
serve as the reactive oxygen species for epoxidation. We then proposed various elementary steps and corresponding rates for each
hypothesis (supplementary text S1). Given that
acetonitrile is a less-effective proton acceptor
than water, we suggested elementary steps in
which two distinct water molecules play explicit roles in the reaction, one as a reactant
and the other as a proton acceptor. Then, we
employed kinetic modeling to deduce which
intermediate would most likely react with
propylene. This approach was viable because
the reaction of different species (OOH*, OO*,
and O*) with propylene generates distinct surface intermediates after epoxidation (OH*, O*,
and *, respectively). By contrast, the evolution
of O2 invariably leaves vacant (*) surface sites.
The goodness-of-fit of each model was evaluated
by the root mean square error (RMSE) between
the experimental data and the simulated data
from the model, with the lowest value indicating
the best-fit model.
Our experimental data best matched the
mechanism suggesting OO* as the key intermediate for epoxidation, with an RMSE of
g
probe substrate analysis, and substrate-based
descriptor assessment.
Electrochemical kinetic rate measurements
were conducted to examine electrolyte species
dependences for direct anodic epoxidation on
PdPtOx/C (Fig. 4 and figs. S12 and S13). The
reaction rates, represented by the partial current density toward epoxidation, were measured
with chronoamperometry in conjunction with
product quantification performed with proton
nuclear magnetic resonance (1H-NMR) (materials and methods).
We hypothesized that the direct alkeneepoxidation pathway at the anode may involve
multiple coupled proton-electron transfer steps
from water to the electrode, followed by oxygenatom transfer from intermediates typically
seen in water oxidation to propylene. O2 was
observed to be the major byproduct with the
PdPtOx/C catalyst, although a small amount
of CO2 was also observed (fig. S14). It is worth
noting that the selectivity of PO over other
propylene-derived products (PG, acetone, and
CO2) reaches 92%. Therefore, in our proposed
mechanism, only the competition between epoxidation and oxygen evolution was considered.
In cases in which the surface coverage is primarily determined by a single reactive intermediate and a rate-limiting step is evident, the
measured Tafel slope of 119 ± 5 mV/decade implies that the rate-limiting step involves a single
electron transfer and that no pre-equilibrated
electron transfer steps occur between the abundant reactive intermediate and the rate-limiting
step (Fig. 4A). Nevertheless, noninteger and nonlinear reaction orders were observed in the water
and propylene dependences, signaling the presence of more intricate kinetic control.
At 1 atm of propylene, the observed dependence of rates on water molarity exhibited approximately second-order behavior at lower
water ratio, which decreased to approximately
p
Fig. 3. Operando x-ray absorption spectroscopy. (A) Changes of Pt average valency in PdPtOx/C and PtOx/C in responses to applied potentials. (B) Changes of Pd
average valency in PdPtOx/C and PdOx/C in responses to applied potentials. Potentials in this figure were 100% iR-compensated (i, current; R, resistance).
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
Additional insights into direct anodic epoxidation on PdPtOx/C were obtained from evaluating the correlation between the electrophilicity
of alkene substrates and the epoxidation rate.
Previous studies on thermochemical epoxidation suggest that either electrophilic (27–29)
or nucleophilic (30, 31) epoxidation can be a
reasonable reaction pathway for electrochemical propylene epoxidation on late-transition
metal catalysts (supplementary text S2). By
understanding the electronic nature of the
oxygen transfer during the reaction, we can
enhance our understanding of the mechanism
and guide future catalyst development.
Chung et al., Science 383, 49–55 (2024)
5 January 2024
kenes with various structures and functional
groups were selected, with w− values ranging
from 12 to 44 kJ/mol. To ensure consistent reactant chemical potential across different substrates, the gaseous substrates were fed until
saturation, and the liquid substrates were added
to the electrolyte near the phase-separation
point, corresponding to ~0.2 M substrate for a
0.11 M tetrabutylammonium tetrafluoroborate
(TBABF4), 10 M water in acetonitrile (table S11).
The substrates were tested at the same potential referenced to the ferrocene redox couple
(1.05 V versus Fc/Fc+).
A negative correlation between the logarithm
of rate and the alkene electrophilicity was observed, with more nucleophilic alkenes displaying higher reactivity toward epoxidation
(Fig. 5). The results suggest that the epoxidation may proceed through an electrophilic
5 of 7
,
Correlation between alkene electrophilicity
and epoxidation rate
The global electrophilicity index (w) can be
calculated by taking the square of alkene chemical potential divided by chemical hardness
and is expressed in units of energy (32, 33). A
higher value of w indicates that the molecule
is more electrophilic. Furthermore, the Fukui
functions can be used to determine the local
reactivity of each atom in a molecule by predicting the redistribution of electron density
during electrophilic, nucleophilic, and radical attacks. The local electrophilicity (w−) at
each atom center can be obtained by multiplying the global electrophilicity by the value
of the Fukui function for the removal of an
electron.
The average w− of the vinyl carbons as a
substrate-based descriptor was assessed against
the epoxidation rates of a wide range of alkene
substrates. A total of 11 distinct aliphatic al-
y
PdPtOx/C occurs through a stepwise mechanism (fig. S16) with OO* reactive intermediates.
including the direct anodic-epoxidation and oxygen-evolution pathways. (E) Simulated
propylene partial-pressure dependences of intermediate species coverage at
10 M water, 1.1 V versus Fc/Fc+ condition. Potentials in this figure were 100% iRcompensated (i, current; R, resistance). Dotted lines in (A) through (C) represent best
fit from kinetic model of proposed mechanism in (D) (more details are shown in
supplementary text S1, Case 2d). Vertical error bars represent standard deviations
from the mean of multiple replicates (n = 3) of the same experiment.
y g
Fig. 4. Kinetic data on direct anodic epoxidation with PdPtOx/C catalysts.
(A) Anodic potential dependence of current density at 1 atm propylene in a 10 M
water, 0.4 M tetrabutylammonium tetrafluoroborate (TBABF4) acetonitrile solution.
ipo+pg, partial current density toward PO and PG. (B) Water molar concentration
dependences of propylene-epoxidation current density at different propylene partial
pressures. (C) Propylene partial-pressure dependences of propylene-epoxidation
current density at different water concentrations. (D) A putative reaction mechanism
RES EARCH | R E S E A R C H A R T I C L E
Fig. 5. Correlation between
the alkene electrophilicity
index and the direct anodicepoxidation rate. Pearson
correlation coefficient (r)
value = −0.85.
,
6 of 7
y
5 January 2024
y g
Chung et al., Science 383, 49–55 (2024)
y
The successful operation of direct epoxidation
through water activation hinges on whether
the electrolyte system is compatible with a particular combination of catalyst and alkene substrate. Water-acetonitrile blended electrolytes
were used to investigate the epoxidation of
various alkene substrates because the acetonitrile cosolvent enhances the solubility of hydrophobic alkene substrates in the water-rich
phase. However, the use of an organic solvent
can lead to an increase in the ohmic resistance
throughout the electrolyte (fig. S4). The use of
water alone as the solvent for the electrolyte,
without an organic cosolvent, would be ideal
because water is required as a reactant and can
effectively solvate electrolyte ions, minimizing
the ohmic resistance of the overall electrolyte.
For gaseous substrates that are sparingly soluble in water, such as propylene, gas-diffusion
electrodes (GDEs) could be used to reduce
the distance that reactant gases must diffuse
through the electrolyte to reach the catalyst,
and therefore, aqueous electrolytes would be
sufficient for epoxide production. Indeed, the
PdPtOx/C catalyst showed ~60% FE toward
propylene epoxidation at 1.65 V versus standard hydrogen electrode (SHE) in a sodium
phosphate aqueous electrolyte when incorporated into a carbon-paper GDE with Nafion
binder (Fig. 1E and fig. S4). Adding the ionomer onto the electrode was critical to achieve
epoxidation performance commensurate with
g
Electrolyte choice and the effect of solvent
medium on the reaction kinetics
water-acetonitrile electrolytes because the hydrophobic backbone of the ionomer generates
partially wetted channels that are effective for
transport of the reactant gases to the catalyst.
The microenvironment of electrolytes also
affects the conversion of epoxides into glycols.
A noticeable amount of PG was observed in
the aqueous propylene epoxidation (~10% FE),
higher than that observed in the blended wateracetonitrile electrolyte. Furthermore, using 0.1 M
HClO4 or any acidic electrolyte results in the
exclusive formation of PG, with no detected
PO (table S1). Because epoxides are known to
undergo acid-catalyzed hydrolysis to afford
glycols, it is imperative to find a pH regime in
which hydrolysis is minimized to ensure high
selectivity toward the product of interest in
aqueous electrolytes. The reaction rates of PO
to PG in aqueous electrolytes with pH 4 to 7
were measured with 1H-NMR: The hydrolysis
of PO to generate PG was found to be faster
in the pH 4 buffer solution, with almost 40%
consumption of PO after 7 hours, whereas the
hydrolysis in less-acidic solutions, at pH 5 to 7,
was largely suppressed, with less than around
10% PO to PG conversion (fig. S18). Thus, maintaining a near-neutral pH in the aqueous electrolyte is essential to optimize PO production.
The practical industrial operation of this system will likely require the use of a membraneelectrode assembly (MEA) resembling a water
electrolyzer with humidified propylene gas
fed to the anode. The MEA is advantageous for
minimizing ohmic loss and separating PO from
liquid electrolytes. However, our analysis on the
influence of pH on the PO hydrolysis rate suggests that minimizing the interfacial pH gradient at the anode would be the key challenge
for maximizing PO production over PG in MEAs
operated at high current densities.
As long as the issues of mass-transport limitation and hydrolysis of PO are addressed,
mechanistic insights on propylene epoxidation
p
attack by oxygen species and that the energy
barriers to this attack are lower for more nucleophilic alkenes. Furthermore, the correlation suggests that a frontier-orbital interaction
between the lowest unoccupied molecular orbital (LUMO) of the oxygen species and the
highest occupied molecular orbital (HOMO) of
the alkene modulates the reactivity.
obtained in water-acetonitrile electrolyte may
be extended to aqueous systems. The investigation on water activity in water-acetonitrile
electrolyte suggests that the aqueous electrolyte
may be viewed as an extreme case of the wateracetonitrile electrolyte with the water molar
ratio approaching unity. In the water-acetonitrile
electrolyte system, positive deviation from
Raoult’s law (fig. S17) indicates that the water
in this electrolyte prefers self-interactions over
mixing, meaning that the activity coefficient
of water in this case is greater than unity
(gwater > 1) and that it approaches unity with
increasing water concentration. The comparison between the rate order with respect to water activity and water concentration provides
information about the activity coefficient of
the transition state (g‡) as well (supplementary
text S4). At lower water concentrations (1 to
5 M), the propylene-epoxidation rate showed
approximately fourth-order dependence on
water activity, whereas approximately second-order dependence on water molar ratio
was observed. The second-order dependence
is consistent with our hypothesis that the predominant rate-determining step leading to the
formation of OOH* involves a first water molecule that is a reactant and a second water molecule that is a proton acceptor. The abnormally
high apparent fourth-order dependence on water activity suggests that the stabilization of the
transition state may be achieved by augmenting the water content. However, approximately
second-order dependences in both water activity and concentration were observed at higher
water concentrations (5 to 12 M). This can be
rationalized if g‡ scales with gwater at low water
concentrations but remains relatively unchanged
at higher water concentrations. This hypothesis aligns with our chemical intuition: Once a
sufficient quantity of water is present around
the transition state, adding more water does
not substantially alter the near–transition state
chemical environment. Therefore, once the water concentration exceeds 5 M in acetonitrile,
the chemical environment at the active site
should be akin to that of an aqueous electrolyte. In this case, the transition from wateracetonitrile electrolyte to aqueous electrolyte
may be a simple extrapolation of water molar
ratio to one, without a discontinuous change
in the solvent properties. This interpretation is
supported by the measured reaction rates in
aqueous electrolytes that is consistent with
the predictions from the mechanistic model
derived from kinetic studies in water-acetonitrile
electrolytes (fig. S12).
Collectively, the results from multiple orthogonal analyses provide a comprehensive
understanding of the mechanism of electrochemical epoxidation on PdPtOx/C. Our findings
are consistent with a mechanistic hypothesis
that OO* is the key species involved in the electrophilic epoxidation of propylene. The PdO
RES EARCH | R E S E A R C H A R T I C L E
RE FE RENCES AND N OT ES
33. R. Pal, P. K. Chattaraj, J. Comput. Chem. 44, 278–297 (2023).
34. M. Chung et al., Direct propylene epoxidation via water
activation over Pd-Pt electrocatalysts, Version 3, Dryad
(2024); https://doi.org/10.5061/dryad.s7h44j1dd.
AC KNOWLED GME NTS
We are grateful to S. Ehrlich, L. Ma, E. Stavitski, and D. Leshchev
for their help in operating XAS experiments. We also appreciate
A. Penn for taking high-resolution energy dispersive spectroscopy
mapping of catalysts. Funding: This research was supported
by the US Department of Energy (DOE) Office of Science, Office of
Basic Energy Sciences, Catalysis Science Program, award no.
DE-SC0023207. K.M. gratefully acknowledges the support of the
Sloan Foundation. This research used resources of the National
Synchrotron Light Source II, a US DOE Office of Science User
Facility operated by Brookhaven National Laboratory under
contract no. DE-SC0012704. Author contributions:
Conceptualization: M.C. and K.M. Methodology: M.C. Investigation:
M.C., J.H.M., and J.S.A. Validation: M.C. and J.S.A. Visualization:
M.C. Funding acquisition: K.M. Project administration: M.C.
and K.M. Supervision: K.M. and Y.R.-L. Writing – original draft: M.C.
Writing – review and editing: M.C., J.H.M., J.S.A., C.J., Y.R.-L.,
and K.M. Competing interests: M.C. and K.M. have filed a
provisional patent application (no. 63/544,664) regarding the
design of catalysts and electrodes. Data and materials
availability: There are no restrictions on materials. All data needed
to evaluate the conclusions in the paper are present in the paper
or the supplementary materials. Tabular data underlying the
figures are deposited in the Dryad data repository (34). License
information: Copyright © 2024 the authors, some rights reserved;
exclusive licensee American Association for the Advancement of
Science. No claim to original US government works. https://www.
science.org/about/science-licenses-journal-article-reuse
g
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adh4355
Materials and Methods
Supplementary Text
Figs. S1 to S25
Tables S1 to S12
References (35–39)
y
1. M. W. Allsopp, G. Vianello, Ullmann’s Encyclopedia of Industrial
Chemistry – Wiley Online Library (Wiley, 2012); https://
onlinelibrary.wiley.com/doi/book/10.1002/14356007.
2. S. J. Khatib, S. T. Oyama, Catal. Rev. Sci. Eng. 57, 306–344
(2015).
3. J. A. M. Leduc, “Electrochemical Process for the Production of
Organic Oxides,” US Patent 3,288,692 (1966).
4. W. R. Leow et al., Science 368, 1228–1233 (2020).
5. M. Chung, K. Jin, J. S. Zeng, K. Manthiram, ACS Catal. 10,
14015–14023 (2020).
6. J. A. M. Le Duc, “Electrolytic Production of Olefine Oxides,”
US Patent 3,427,235 (1969).
7. M. Chung, K. Jin, J. S. Zeng, T. N. Ton, K. Manthiram, J. Am.
Chem. Soc. 144, 17416–17422 (2022).
8. K. Jin et al., J. Am. Chem. Soc. 141, 6413–6418 (2019).
9. X. C. Liu et al., J. Am. Chem. Soc. 144, 20895–20902
(2022).
10. A. Winiwarter et al., Energy Environ. Sci. 12, 1055–1067
(2019).
11. R. P. H. Jong, E. Dubbelman, G. Mul, J. Catal. 416, 18–28
(2022).
12. S. Koroidov et al., Catal. Sci. Technol. 11, 3347–3352
(2021).
13. K. Otsuka, T. Ushiyama, I. Yamanaka, K. Ebitani, J. Catal. 157,
450–460 (1995).
14. S. Iguchi, M. Kataoka, R. Hoshino, I. Yamanaka, Catal. Sci. Technol.
12, 469–473 (2022).
15. I. Yamanaka, K. Sato, K. Otsuka, Electrochem. Solid-State Lett.
2, 131–132 (1999).
16. L. L. Holbrook, H. Wise, J. Catal. 38, 294–298 (1975).
17. J. Ke et al., Nat. Commun. 13, 932 (2022).
18. H. Li, C. S. Abraham, M. Anand, A. Cao, J. K. Nørskov, J. Phys.
Chem. Lett. 13, 2057–2063 (2022).
19. M. Chen, L. D. Schmidt, J. Catal. 56, 198–218 (1979).
20. C. Carrillo et al., J. Phys. Chem. Lett. 5, 2089–2093
(2014).
21. P. N. Plessow, F. Abild-Pedersen, ACS Catal. 6, 7098–7108
(2016).
22. E. D. Goodman et al., J. Chem. Phys. 151, 154703
(2019).
23. A. Aitbekova et al., Nat. Mater. 21, 1290–1297 (2022).
24. D. Gao et al., J. Am. Chem. Soc. 137, 4288–4291 (2015).
25. D. T. Bregante, P. Priyadarshini, D. W. Flaherty, J. Catal. 348,
75–89 (2017).
26. C. K. Sams, K. A. Jørgensen, S. V. Lindeman, J. Songstad,
Acta Chem. Scand. 49, 839–847 (1995).
27. C. Kim, T. G. Traylor, C. L. Perrin, J. Am. Chem. Soc. 120,
9513–9516 (1998).
28. X. Yang et al., Angew. Chem. Int. Ed. 54, 11946–11951
(2015).
29. Y. Lei et al., Science 328, 224–228 (2010).
30. A. R. Chianese, S. J. Lee, M. R. Gagné, Angew. Chem. Int. Ed.
46, 4042–4059 (2007).
31. A. Zanardo, F. Pinna, R. A. Michelin, G. Strukul, Inorg. Chem.
27, 1966–1973 (1988).
32. F. De Vleeschouwer, V. Van Speybroeck, M. Waroquier,
P. Geerlings, F. De Proft, Org. Lett. 9, 2721–2724
(2007).
p
crystal structure embedding Pt(II) species was
found to be effective for catalyzing direct anodic epoxidation, suggesting that other materials
with similar structures and stronger inductive
effects on Pt may also produce effective catalysts by promoting more electrophilic epoxidation pathways. Additionally, we discovered
through fundamental kinetic analysis that the
identity of reactive oxygen species may vary
between catalysts. For instance, the proposed
reactive oxygen species for epoxidation on
PdPtOx/C and MnOx-based catalysts (7, 8) are
OO* and O*, respectively. Developing approaches
to stabilize OO* may enhance epoxidation on
noble metals while avoiding oxygen evolution.
Our work contributes toward advancing the
sustainable synthesis of epoxides, which currently have substantial energy and environmental
footprints. The molecular-level understanding
of direct anodic epoxidation obtained from this
study may have implications for other electrocatalytic oxygen-atom transfer reactions driven
by water-activation intermediates.
Submitted 4 March 2023; resubmitted 12 September 2023
Accepted 29 November 2023
10.1126/science.adh4355
y g
y
,
Chung et al., Science 383, 49–55 (2024)
5 January 2024
7 of 7
RES EARCH
NEUROSCIENCE
Balancing risk-return decisions by manipulating the
mesofrontal circuits in primates
Ryo Sasaki1*, Yasumi Ohta2, Hirotaka Onoe3, Reona Yamaguchi4, Takeshi Miyamoto1,5,
Takashi Tokuda6, Yuki Tamaki1†, Kaoru Isa1, Jun Takahashi7, Kenta Kobayashi8,
Jun Ohta2, Tadashi Isa1,3,4*
Decision-making is always coupled with some level of risk, with more pathological forms of risk-taking
decisions manifesting as gambling disorders. In macaque monkeys trained in a high risk–high return
(HH) versus low risk–low return (LL) choice task, we found that the reversible pharmacological
inactivation of ventral Brodmann area 6 (area 6V) impaired the risk dependency of decision-making.
Selective optogenetic activation of the mesofrontal pathway from the ventral tegmental area (VTA) to
the ventral aspect of 6V resulted in stronger preference for HH, whereas activation of the pathway
from the VTA to the dorsal aspect of 6V led to LL preference. Finally, computational decoding captured
the modulations of behavioral preference. Our results suggest that VTA inputs to area 6V determine
the decision balance between HH and LL.
1 of 7
,
5 January 2024
Because reward information is critical for riskdependent decision-making, we next investigated
the role of input from the ventral tegmental
area (VTA)—a core area of the midbrain dopaminergic system (26), which projects to the
superficial and deeper layers of area 6VV (Fig. 3E,
y
Sasaki et al., Science 383, 55–61 (2024)
Optogenetic activation of the mesofrontal
pathway in risk-return decision-making
y g
*Corresponding author. Email: sasaki.ryo.3r@kyoto-u.ac.jp (R.S.);
isa.tadashi.7u@kyoto-u.ac.jp (T.I.)
†Present address: Japanese Red Cross Otsu Hospital, Otsu-shi, Shiga
520-0046, Japan.
Six macaque monkeys were trained to perform a cue-target choice task (Fig. 1A). While a
monkey fixated on a central point, we presented
two colored cues (selected from either 16 or
25 possible colors, respectively) in the left
and right hemifields. We assigned different
probabilities and EVs of water reward delivery
to the different colors (Fig. 1B). The colors corresponded to different reward probabilities
with the same EV along the horizontal axis
and changed in EVs systematically along the
vertical axis [four (n = 2 monkeys) or five
(n = 4 monkeys) levels between 100 and 250 ml].
As an example, when a red cue is selected, the
reward would be given in only 10% of the trials
with a reward size of 1000 ml, corresponding to
HH choice, whereas the blue cue would be rewarded in 90% of the trials with a reward size
of 111.1 ml, representing a LL choice. In both
cases, the resulting net EV of the reward would
be the same (100 ml). Thus, this task allows us
to clarify the decisional balance between HH
and LL preference with fixed net EV.
In a two-dimensional (2D) heatmap representing the proportion chosen, which is computed based on the frequency of choosing a
given option against all other options in the
matrix, all six monkeys (monkeys Y, S, M, J, C, and
H) showed a preference for the left-uppermost
corner of the matrix, corresponding to the HH
choice with the largest EV (Fig. 1C). The line plot
y
Division of Physiology and Neurobiology, Department of
Neuroscience, Graduate School of Medicine, Kyoto
University, Kyoto-shi, Kyoto 606-8501, Japan. 2Division of
Materials Science, Graduate School of Science and
Technology, Nara Institute of Science and Technology,
Ikoma-shi, Nara 630-0192, Japan. 3Human Brain Research
Center, Graduate School of Medicine, Kyoto University,
Kyoto-shi, Kyoto 606-8507, Japan. 4Institute for the
Advanced Study of Human Biology (WPI-ASHBi), Kyoto
University, Kyoto-shi, Kyoto 606-8501, Japan. 5Japan Society
for the Promotion of Science, Chiyoda-Ku, Tokyo 102-0083,
Japan. 6Institute of Innovative Research, Tokyo Institute of
Technology, Meguro-Ku, Tokyo 152-8550, Japan.
7
Department of Clinical Application, Center for iPS Cell
Research and Application, Kyoto University, Kyoto-shi, Kyoto
606-8507, Japan. 8Section of Viral Vector Development,
National Institute for Physiological Sciences, Okazaki-shi,
Aichi 444-8585, Japan.
Identification of frontal brain areas regulating
risk-dependent decision-making
g
1
have also been reported in nonhuman primates
(16, 17), this again could not clearly dissociate
risk levels and EVs. Because decision-making is
usually linked to more variable combinations of
risk levels and EVs, we investigated the brain regions involved in risk-dependent decision-making
using macaque monkeys trained to perform a
HH versus LL decision task with a variable combination of reward probabilities and EVs.
p
T
he real world is full of uncertainty, and
our decisions are always accompanied by
some level of risk. Decision-making between high risk–high return (HH) and low
risk–low return (LL) choices is regulated
by the subject’s utility function (1, 2), which varies
substantially from person to person. Pathological risk-taking decisions are known to underlie problematic behaviors, such as gambling
disorders (3–6). Dysregulation of the reward
circuitry, including the ventral striatum and
the orbitofrontal and medial prefrontal cortices,
is implicated in gambling disorders (7–12);
however, their causal role is still elusive owing
to technical limitations of human studies.
Risky choice behavior has also been investigated in psychopharmacological studies in
rats (13, 14). In humans and rodents, riskdependent decisions are frequently assessed
using variants of the Iowa Gambling Task
(15). However, this task cannot uncouple HH
and LL choices from the negative and positive
expected values (EVs), respectively. Furthermore, although preferences for risky choices
above the matrices indicates the risk-dependent
choices of the monkeys at consistent EVs with
a linear regression, showing that the monkeys
prefer HH options without any EV dependence.
The HH preference was also statistically not significantly different depending on the EV blocks
[paired t test, t(5) = 2.021, P = 0.099]. We also
estimated each monkey’s utility and probability
weighting functions from their choice using
standard parameters described in the literature (18–20), which suggested the risk attitude
in decision-making (fig. S1). This was common
in all of the monkeys, which fits with the framework of utility function in monkeys (2) and prospect theory of decision-making in humans (21).
To identify the frontal brain areas involved
in processing risk information, we injected a
g-aminobutyric acid type A (GABAA) receptor
agonist, muscimol (0.2 to 0.5 ml, 5 mg/ml), into a
variety of bilateral symmetric prefrontal regions, including ventral Brodmann area 6 (area
6V) and area 12 (Fig. 2, A to C), which has previously been suggested to play a role in coding mental operation (22). Neurons in this area
have also been associated with probability of
reward in decision-making (23) [described as
the ventrolateral prefrontal cortex (vlPFC)],
which suggests that area 6V might be a potential candidate for the representation of complex
risk-return computation. Muscimol injection
into the ventral aspect of area 6V (6VV) (Fig. 3C)
eliminated the risk dependency of decisionmaking, which recovered the next day (Fig.
2E) [compare the difference of slope (“post pre”) to HH-LL between muscimol and saline
injection in six experiments in two animals:
monkey S, Wilcoxon rank-sum test, P = 0.026,
and monkey M, Wilcoxon rank-sum test, P =
0.016]. Area 6VV is known for its roles in representing motor actions in many studies. Additionally, a few studies (22, 24, 25) have suggested
its role in coding for human decision-making.
The EV dependency of decision-making was
not affected by area 6VV inactivation (Fig. 2F;
Wilcoxon rank-sum test, P > 0.05). We observed
no effect from other candidate areas—muscimol
injections into the orbitofrontal cortex [OFC
(area 14)] (four sessions) and the dorsal anterior cingulate cortex (dACC) (four sessions)
had no effect (Fig. 2, E and F; Wilcoxon ranksum test, P > 0.05). These results suggest that
the area 6VV plays an important role for regulating HH-LL decisions, whereas the OFC
and dACC play no critical role in such complex
computations.
RES EARCH | R E S E A R C H A R T I C L E
C
A
1.0
Monkey Y
Monkey C
0.5
1.0
0.5
Fixation point
Target
Reward cue
(w/ or wo/)
-1000
-500
0
500
1000
Time from target onset (ms)
LL
HH
Probability x Reward = Expected
size
value
: 90%
111.1 μL
100 μL
: 10% 1000.0 μL
100 μL
0.0
250.0
212.5
212.5
175.0
175.0
137.5
137.5
100.0
1.0
0.5
0.0
100.0
10
Proportion chosen
Monkey H
Reward
0.0
250.0
No-reward
30
50
70
90
1.0
0.5
0.0
1.0
0.0
0.0
250.0
250.0
212.5
212.5
175.0
175.0
137.5
137.5
2500
Monkey M
1500
212.5
30
50
70
90
Probability (%)
1.0
1.0
0.5
0.0
30
50
70
90
Probability (%)
1.0
Monkey S
0.5
0.0
0.0
250.0
250.0
200.0
200.0
150.0
150.0
g
1000
10
Proportion chosen
0.5
2000
175.0
90
100.0
10
Proportion chosen
250.0
70
p
B
0.0
50
0.5
100.0
0.5
30
Probability (%)
1.0
Monkey J
0.5
1.0
10
Proportion chosen
Probability (%)
137.5
500
100.0
10
30
50
70
90
LL
HH-LL
based on Probability (%)
10
30
50
70
90
y
HH
0
Probability (%)
100.0
100.0
1.0
0.5
0.0
Proportion chosen
20
40
60
80
Probability (%)
1.0
0.5
0.0
20
Proportion chosen
0
40
60
80
Probability (%)
0.2
0.4
0.6
0.8
1
Proportion chosen
5 January 2024
P < 0.05; Fig. 3J). We stimulated the VTA-6VV,
6VD, and/or vlPFC pathways optogenetically
with red LEDs right after target onset (Fig. 3B;
three repetitions of 20 ms ON and 80 ms OFF,
625-nm wavelength; intensity: 0.782 mW at
2 mA, 1.824 mW at 4 mA). Saccades were initiated at 200 to 500 ms after target presentation. In these experiments, one of the cues was
always fixed at 50% reward probability and
175 ml EV (Fig. 3A, right), and another cue was
selected from the 25 color possibilities (Fig.
3A, left). During these sessions, we divided the
trials randomly into 50% stimulated (LED-ON)
and 50% unstimulated (LED-OFF) trials. In
monkey J (Fig. 3M, top), photostimulation of the
VTA-6VV pathway, delivered at 2 mA during
epoch II (0 to 220 ms after cue presentation) led
to a shift in decision-making to the HH-preferring
mode, without affecting EV dependency (Fig.
2 of 7
,
Sasaki et al., Science 383, 55–61 (2024)
the injection site in the VTA of two monkeys (J
and H), we observed that tdTomato was expressed in the majority of VTA neurons around
the injection site, with 78% (776/1000) and 66%
(570/860) in monkeys J and H, respectively
(Fig. 3, E to H). These neurons were tyrosine hydroxylase (TH)–positive, which suggests that most
of the activated neurons were dopaminergic.
Next, to determine whether the neural activities in 6VV, 6VD, and/or vlPFC were modulated on the basis of HH or LL choices, we
recorded their activity with the ECoG electrodes
during the HH-LL choice task. Cue-related
a-band (8 to 13 Hz) activity in all channels at
100 to 500 ms after cue presentation (Fig. 3I),
when the monkey presumably made its decision, was higher for HH choices (10% reward
probability) compared with LL choices (90%
reward probability) (Wilcoxon rank-sum test,
y
right) (27). We expressed a red light–activatable
opsin, ChrimsonR (28), in the bilateral VTA using
an adeno-associated viral vector (AAV2.1-SynChrimsonR-tdTomato) (fig. S2) and selectively
stimulated the combination of light-emitting
diode (LED) channels belonging to the VTA6VV pathway, the VTA–dorsal aspect of area
6V (6VD) pathway, or the VTA-vlPFC pathway
coupled with electrocorticogram (ECoG) electrodes (29) that were chronically implanted on
the surface of the bilateral target areas (Fig. 3, C
and D, and fig. S3) in monkeys J, C, Y, and H. We
used a nonselective promotor (Syn) to express
ChrimsonR because we could not see the axon
terminals in the frontal cortex using immunohistochemistry staining in a pilot experiment
using a dopamine neuron–specific promotor,
which likely suggests weak expression with the
specific vector. Upon confirming expression at
computed by all comparison against other options, are shown as a color contour
map for each of the six monkeys. The scatter plots show the proportion of
choices for reward probabilities (top) computed by the consistent EV, and the
proportion of choices for EVs (left) computed by the consistent probability. The
lines in the scatter plots represent their linear regressions.
y g
Fig. 1. Task design and behavior. (A) The monkeys were required to choose
one of two cues by saccade to receive a certain amount of water reward.
(B) Assignment of the reward probabilities and EVs for each cue color (left) and
the relationship between the reward probabilities and the reward sizes (right).
(C) Proportions of choosing each combination of reward probability and EV,
RES EARCH | R E S E A R C H A R T I C L E
A
B
I II III IV
6VV
C
I
II
III
IV
dACC
Area 6VV
D
1.0
OFC
Pre
30 min
60
130
200
24h
0.5
10 30 50 70 90
Probability (%)
E
1.0
Monkey S
Monkey M
0.5
1.0
0.5
Accumulation of stimulus effects
on risk-dependent behavioral choices
0.0
OFC
Muscimol
dACC
Muscimol
-1.0
-0.5
Area 6VV
Muscimol Saline
OFC
Muscimol
dACC
Muscimol
-1.0
5 January 2024
3 of 7
,
Sasaki et al., Science 383, 55–61 (2024)
epochs I, III, and IV): the period before cue
onset (epoch I, −500 to −280 ms), after cue onset
(epoch III, 500 to 720 ms), and at reward delivery (epoch IV, 1000 to 1220 ms). We observed
no change in the risk- and EV dependency of
decision-making when photostimulation was
applied during epochs I, III, and IV (fig. S4, A
to G; Wilcoxon signed-rank test, P > 0.05).
Thus, optogenetic stimulation of VTA terminals in the 6VV resulted in modification of riskdependent decisions (but not EV dependent
decisions) to HH-preferring only during epoch II,
when the monkeys were presumably making decisions to select a cue.
By contrast, photostimulation of area 6VD
relieved the HH-preferring mode (Fig. 3, N and
P, top). The decision changed to LL-preferring
y
3M, bottom). Analysis of the population data
(Fig. 3O, top), whereby the preference for HH
was evaluated by a threshold for HH-preferring
decisions in the psychometric function, showed
that the preference for HH was higher during
the LED-ON trials compared with the LED-OFF
trials (Wilcoxon signed-rank test, z = −3.89,
P = 1.01 × 10−4), whereas EV dependency was
not affected (Fig. 3O, bottom; z = 0.26, P = 0.79).
Furthermore, the cue-related a-band response
in channels located in area 6VV was enhanced
during the LED-ON trials compared with the
LED-OFF trials [Fig. 3K; (LED-ON, 3213 trials;
LED-OFF, 3203 trials) Wilcoxon rank-sum test,
P < 0.05]. To understand the temporal dynamics of this response, we also applied the stimulation during various time epochs (fig. S4A;
y g
Fig. 2. Effects of muscimol injections into frontal cortical regions on risky choice decisions. (A) Surface
view of the macaque cortex showing the rostro-caudal levels of the planes in (C) and the approximate location of
area 6VV (yellow). (B) MRI images of the coronal plane from monkey S, including area 6VV. The arrowheads
indicate the Gd3+ injection site. (C) Effective muscimol injection sites in area 6VV (purple-filled circles).
The results from two animals are superimposed. (D) Proportion chosen for a single session (monkey S)
plotted against the reward probability computed by the consistent EV at each time window from injection
(preinjection, 0 to 30 min, 30 to 60 min, 60 to 130 min, 130 to 200 min, and 24 hours). (E) The difference
of slope to reward probability (HH-LL) between before and after injection at each injection site [OFC (area 14)
and dACC] in two monkeys. (F) The same arrangement as (E) for the difference of slope to the EV. Plots with
an asterisk exhibit a statistically significant effect (Wilcoxon rank-sum test, *P < 0.05).
The photostimulation-dependent effects on
choice behavior accumulated over time. In
monkey J, after six control sessions without
stimulation, we introduced photostimulation
at 6VV or 6VD, randomly interleaved (Fig. 4A).
Thereafter, the HH-preference indices for both
the LED-ON and LED-OFF trials increased
gradually (lower threshold of the psychometric
function for HH preference). During the initial
six photostimulation sessions, HH preference
was higher in the LED-ON trials compared with
the LED-OFF trials, but after the seventh session,
HH preference became saturated, and HH preference in the LED-ON and LED-OFF trials became nearly equal. This result implies that the
HH-preferring mode might accumulate more
easily compared with the LL-preferring mode. To
clearly distinguish the stimulation effect between
6VV and 6VD, we stimulated each area in a block
task design for the second monkey (monkey H;
Fig. 4B). The preference for HH became higher in
the LED-ON trials compared with the LED-OFF
trials after three sessions (sessions 23 to 25).
The effect became saturated at session 40, and
stimulation was stopped for nine sessions (sessions 41 to 49). Upon restarting stimulation, the
HH preference reappeared quickly (sessions 49
to 50) before it became saturated again (Fig.
4B, magenta line). We subsequently began photostimulation at 6VD, whereby HH preference
clearly dissipated over time (Fig. 4B, cyan line).
Similar behavioral effects of 6VD stimulation
were also observed in the third monkey (C),
where HH preference was relieved over time
and saturated (Fig. 4C, cyan line).
y
Area 6VV
Muscimol Saline
g
*
*
0.0
-0.5
F
p
0.0
during the LED-ON trials rather than during
the LED-OFF trials (Wilcoxon signed-rank test,
z = 4.55, P = 5.33 × 10−6) while not affecting the
EV dependency (Fig. 3P, bottom; z = −0.57, P =
0.56). Additionally, the cue-related a-band responses in area 6VD were inhibited during the
LED-ON trials compared with the LED-OFF
trials (Fig. 3L; Wilcoxon rank-sum test, P < 0.05),
hence having an opposite effect compared with
photostimulation at area 6VV. It is worth emphasizing that more-localized photostimulation still convincingly enhanced or relieved
the HH-preferring mode (fig. S5). Photostimulation primarily at vlPFC caused no effect in
both risk dependency (Fig. 3Q, top; z = 0.10,
P = 0.91) and EV dependency (Fig. 3Q, bottom; z = 0.61, P = 0.61). Behavioral threshold
and brain activities in the control animal (monkey Y), who was injected with the control vector
carrying no ChrimsonR sequence (AAV2.1-SyntdTomato), were not affected (fig. S6). Notably,
we could largely exclude that these effects
were a result of side effects of heat generated
from photostimulation because the temperature changes induced by LED stimulation were
<0.5°C (fig. S7).
RES EARCH | R E S E A R C H A R T I C L E
A
B
250.0
250.0
212.5
212.5
175.0
175.0
C
Pulses: 20 ms ON, 80 ms OFF, @10Hz
vlPFC
Fixation point
vs
10 30 50 70 90
10 30 50 70 90
Probability (%)
Probability (%)
HH
HH
LL
D
6VD
LED STIM.
100.0
100.0
PS
Reward cue
(w/ or wo/)
137.5
137.5
AS
Target
-1000
0
-500
500
1000
ECoG+LED
Time from target onset (ms)
LL
E
I
6VV
HH (10%)
LL (90%)
70
70
20
10
5
20
10
5
0.4
0.2
0.0
−0.2
1
1
−0.2 0 0.2 0.4 0.6 0.8 −0.2 0 0.2 0.4 0.6 0.8
ECoG+LED
−0.4
Time from target onset (s)
VTA
J
0.6
0.5
AAV2.1-Syn-ChrimsonR-tdTomato
p
G
F
HH option
LL option
p < 0.05 t-test
0.4
AS
0.3
H
PS
0.2
0.1
0.0
−0.1
0.0
0.2
0.4
0.6
0.8
g
−0.2
−0.2
Time from target onset (s)
K
L
0.3
LED OFF
LED ON
0.2
LED OFF
LED ON
0.2
Monkey H
Moneky J
Monkey C
AS
Q
PS
Monkey H
Monkey J
-0.1
-0.1
-0.1
-0.2
-0.2
-0.2
-0.3
-0.3
-0.4
-0.4
0.1
0.1
0.0
0.0
−0.1
−0.1
−0.2
−0.2
M
PS
6VD
0.4
0.3
AS
P
AS
PS
y
6VV
0.4
O
−0.2 0.0 0.2 0.4 0.6 0.8
−0.2 0.0 0.2 0.4 0.6 0.8
Time from target onset (s)
Time from target onset (s)
1.0
LED OFF
LED ON
N
1.0
0.8
0.6
0.6
0.4
0.4
0.2
LED OFF
LED ON
Monkey H
Moneky J
-0.4
-0.4
-0.3
-0.2
-0.1
Threshold HH-LL, LED OFF
100
-0.4
100
-0.3
-0.2
-0.4
-0.1
Threshold HH-LL, LED OFF
100
-0.3
-0.2
-0.1
y g
0.8
-0.3
Threshold HH-LL, LED OFF
0.2
0.0
-40
-20
0
20
40
0.0
-40
-20
0
20
40
Δ Probability (%)
Δ Probability (%)
1.0
0.8
0.8
0.6
0.6
0.4
0.4
0.2
0.2
80
80
60
60
60
40
40
40
20
20
20
y
1.0
80
-40
0
40
80
0.0
-80
-40
0
40
80
0
0
Δ Expected value ( μ l )
Δ Expected value ( μ l )
20
40
Threshold
60
, LED OFF
EV
Fig. 3. Optogenetic stimulation of the mesocortical pathway. (A) Cues in the
optogenetic experiments; one from 25 choices versus the gray one. (B) Photostimulation timing indicated by vertical color bars. (C) Location of the LED or ECoG
probes on area 6VV, 6VD, and vlPFC. AS, arcuate sulcus; PS, principal sulcus.
(D) Experimental design. (E) (Left) Low-magnification view of the VTA with anti–red
fluorescent protein (RFP) immunohistochemistry. Scale bar, 10 mm. (Right) AntiRFP immunohistochemistry showing positive axons in 6VV. Scale bar, 500 mm. (F to
H) High-magnification view of the VTA. Anti-RFP (F), anti-TH immunostaining (G),
and merged image (H) are shown. Scale bar, 100 mm. (I) Averaged time-frequency
plots of ECoG at 6VV for HH (left) and LL choice (right). (J) a-Band activity
Sasaki et al., Science 383, 55–61 (2024)
5 January 2024
80
0
100 0
20
40
Threshold
60
80
, LED OFF
EV
0
100 0
,
0.0
-80
20
40
Threshold
60
EV
80
100
, LED OFF
for the HH and LL choices in (I). (K and L) The time course of a-band activity for
the LED-ON (colored curve) and LED-OFF (black curve) conditions in each 6VV
(K) and 6VD (L). (M) Psychometric function of reward probability (top) and
EV (bottom) (stimulation given at 6VV). The smooth curve shows cumulative
Gaussian fitting to the data. (N) The same arrangement as (M) (stimulation given
at 6VD). (O to Q) Comparison population behavioral thresholds with LED-OFF and
LED-ON for the HH-LL (top) and EVs (bottom). Stimulation given at VTA-6VV
pathway (n = 33) (O), the VTA-6VD pathway (n = 73) (P), and the VTA-vlPFC
pathway (n = 16) (Q). Photostimulations were delivered subareal sets; either one
or two local lines or subareal spots (see also fig. S5).
4 of 7
RES EARCH | R E S E A R C H A R T I C L E
A
B
Monkey J
2 mA
C
Monkey H
2 mA
4 mA
Monkey C
4 mA
-0.1
-0.1
-0.05
-0.2
-0.10
-0.3
Early period
Late period
-0.5
-0.20
-0.25
LED OFF
LED ON (6VV)
LED ON (6VD)
-0.6
-0.2
-0.15
6VV 6VD
-0.4
-0.3
-0.30
-0.35
-0.7
-0.4
0
10
20
30
60
0
70
40
50
60
70
80
90
100
0
Sessions
Sessions
D
30
E
F
G
H
0.5
0.5
0.4
0.3
0.3
0.4
0.4
0.3
0.3
0.3
0.2
0.2
0.2
0.2
0.2
0.1
0.1
0.1
0.1
0.0
0.0
0.0
0.0
−0.1
−0.1
Time from target onset (s)
−0.2
−0.3
−0.2 0.0 0.2 0.4 0.6 0.8
−0.2
−0.3
−0.2 0.0 0.2 0.4 0.6 0.8
Time from target onset (s)
0.1
0.0
−0.1
−0.2
−0.3
−0.2 0.0 0.2 0.4 0.6 0.8
Early period
Late period (6VV)
Late period (6VD)
P < 0.05 t-test
Time from target onset (s)
sessions, respectively. (B and C) The same arrangement as (A) for monkey H (B)
and monkey C (C). The light gray horizontal line for monkey H (B) indicates the
sessions with 2-mA stimulation, which was ineffective. For monkey C (C), only 6VD
stimulation was completed. (D to H) a-Band ECoG activity in 6VV [(D) and (F)] and
6VD [(E), (G), and (H)] during the LED-OFF trials are compared between the early
(grayish red and blue line traces) and late (light red and blue line traces). The inset
panel on the right represents the channels used for this analysis at 6VV (magenta
circles) and 6VD (cyan circles).
g
Fig. 4. Accumulation of the stimulus effect. (A) Accumulation of the stimulus
effect. Behavioral thresholds are plotted for each experiment session in monkey J. The
magenta trace indicates LED-ON trials stimulated at 6VV, the cyan trace indicates
LED-ON trials stimulated at 6VD, and the black trace indicates LED-OFF trials. The light
orange vertical band indicates the six sessions before the start of photostimulation
for 6VV photostimulation, the grayish magenta band indicates the five early sessions,
and the light magenta band indicates the three late sessions. For 6VD photostimulation, the grayish cyan band and the light cyan band indicate early and late
PS
p
−0.2
−0.2
−0.2 0.0 0.2 0.4 0.6 0.8
−0.2 0.0 0.2 0.4 0.6 0.8
80
AS
0.4
−0.1
60
Sessions
0.4
−0.1
20
y
We first identified the dimension of the power
signals relevant to HH-LL by demixed principal components analysis (dPCA) (30) (Fig. 5A).
The time course of normalized power of ECoG
activities for each axis (HH-LL or EV) were
clearly separated depending on each stimulus
intensity (Fig. 5B). The 2D trajectories of a-band
population activity in HH-LL and EV axes
were also clearly separated under each stimulus condition (Fig. 5C), and the most separated
demixed principal components (dPCs) (Fig. 5D
and Fig. 5C, dots of x-y plane) were roughly
consistent with the stimulus dimension. Thus,
the first dPC should be the most prominent
decoder axis of HH-LL as a linear classifier.
For this purpose, we trained the decoder separately for LED-ON and LED-OFF conditions
(see materials and methods for details). We
then performed linear decoding to classify a
given target as a HH (i.e., 10 and 30%) or LL
(i.e., 70 and 90%) option based on power signals of all channels in each frequency band to
measure the decoder threshold as HH-LL pref-
5 of 7
,
5 January 2024
Linear decoding of lower band activity
captures behavioral preference modulation
by photostimulation
erence. The HH-LL decoder preferred HH
options when area 6VV was photostimulated,
with the decoder performance curve becoming
steeper with photostimulation (Fig. 5E, magenta and black curves). By contrast, decoding
of 6VD responses showed that the performance
curve became shallower with photostimulation
(Fig. 5F, cyan and black curves). Both decoders
revealed a pattern of results quite similar to behavior (Fig. 3, M and N, top). Results for the
HH-LL decoder were similar to behavioral performance for both LED-ON and LED-OFF conditions. The decoder threshold for HH-LL was
extremely sensitive when photostimulation was
applied [Fig. 5G, magenta symbols; t test; monkey H, t(58) = −5.36, P = 1.50 × 10−6; monkey J,
t(58) = −5.07, P = 4.34 × 10−6]. By contrast, the
HH-LL decoder significantly diminished HH
preference when area 6VD was photostimulated
[Fig. 5, F and G, cyan symbols; t test; monkey
C, t(58) = 4.26, P = 7.59 × 10−4; monkey H, t(58) =
3.99, P = 1.84 × 10−4; monkey J, t(58) = 8.31,
P = 1.79 × 10−11], whereas photostimulation of
the vlPFC had no effect. In light of these results, neural population decoding could capture the behavioral preference modulation
induced by optogenetic photostimulation in
area 6V, which suggests that neural dynamics of the mesofrontal pathway are involved
in cognitive computation for modulation of
the risk-return decision balance.
y
Sasaki et al., Science 383, 55–61 (2024)
balancing. To directly address this question, we
examined whether neural population decoding
can capture the modulation of behavioral preference caused by the optogenetic stimulation.
y g
To investigate whether brain activities are
coupled to these behavioral changes, we compared the cue-related a-band ECoG responses
during the early period (Fig. 4A: 6VV, grayish
magenta area; 6VD, grayish cyan area) of photostimulation when the facilitatory effects promoting HH preference occurred and during the
late period (Fig. 4A: 6VV, light magenta; 6VD,
light cyan area) when stimulation-induced HH
preference became saturated. The cue-related
a-band responses were measured from both stimulus locations, namely 6VV (Fig. 4, inset, magenta circles) and 6VD (Fig. 4, inset, cyan circles).
Upon 6VV photostimulation, the cue-related
a-band responses became markedly enhanced
even in the LED-OFF trials because HH preference was enhanced without stimulation during
the late period (2757 trials) compared with the
early period (952 trials) (Fig. 4D; Wilcoxon ranksum test, P < 0.05). Similar results were also observed in monkey H (643 trials in the early and
536 trials in the late period; Fig. 4F). Consistent
with the behavioral responses, upon 6VD photostimulation, we observed a response inhibition
that was opposite to the effect of 6VV stimulation (Fig. 4, E, G, and H; 507 to 958 trials).
Because it remained unclear exactly how neural representation in these regions (activation or
inhibition) was causally linked to behavioral HH
or LL preference mode, we next investigated the
changes in neural dynamics for this decision
RES EARCH | R E S E A R C H A R T I C L E
A
Ch-1
Ch-2
B
Demixed principal
components
HH-LL
10%
0.9
HH-LL axis
E
1.0
90%
LED OFF
LED ON
0.8
−0.9
−0.2 0.0 0.2 0.4 0.6 0.8
Ch-3
EV axis
EV
Ch-4
Others
0.4
250.0 μl
2.5
C
Time from target onset (s)
D
-20
0
20
40
Monk H
Monk J
AS
PS
−0.2
AS
PS
−2.0
−0.2 0.0 0.2 0.4 0.6 0.8
Probability
0.0
-40
Others
0.0
−0.1
0.2
−1.0
−0.2 0.0 0.2 0.4 0.6 0.8
Ch-36
G
0.6
100.0 μl
1.6
Δ Probability (%)
F
1.0
LED OFF
LED ON
0.8
2.2
−0.3
−0.4
−0.5
0.6
Monk C
Monk H
Monk J
0.4
−0.6
0.2
1st dPCEV
1st dPCHH-LL
−1.3
1.4
0.0
-40
1st dPCHH-LL
−0.7
−0.7
−0.6
−0.5
−0.4
−0.3
−0.2
−0.1
0.0
ThresholdHH-LL LED OFF
condition matrix (inset). The dots in the x-y plane indicate data at times when
the trajectories are most separated apart (time at 0.2 s). (D) View of the x-y
plane from (C). (E and F) Psychometric functions constructed by choice of
decoders that use the first dPC of HH-LL as a linear classifier. The proportion
chosen was computed by aggregating over all 20 trial repetitions (assuming
a single session). (G) Summary of decoder behavioral thresholds as a comparison
between LED-ON and LED-OFF trials. Plots and error bars represent the means
and 95% confidence intervals across 20 decoding sessions, respectively.
6 of 7
,
sponses in area 6VV and a suppression in area
6VD (Fig. 4, D to H) in LED-OFF trials, respectively. Such changes in the neural responses are
likely induced by long-term dopamine-dependent
plasticity mechanisms (45). Moreover, the existence of subareas with distinct functions might
open up an interesting new direction for future
studies. One can speculate that the distinct dopamine subcircuits to areas 6VV and 6VD might
separately encode for different risk-return computations. Another possibility might be like the
nucleus accumbens (NAc), where distinct subpopulations of D2R+ cells (46) separately encode the selection of risky options. Thus, the
pathway from areas 6VV or 6VD to these distinct
populations in NAc neurons may be interesting
to investigate in the future studies.
We further extended our optogenetic experiment to the computational framework. We observed that neural population decoding could
capture the photostimulation effect of HH preference mode of subjects. Thus, we directly compared neural and behavioral correlates of
dynamically changing HH-preferring mode,
allowing for a more direct assessment of whether
changes in neural activity with HH-preferring
mode can explain behavior. However, only future studies will reveal whether this state in primates is comparable to that in patients with
gambling disorders. Because the D3 receptor
agonist pramipexol administered for treatment of Parkinson’s disease promotes gambling disorder (47), our findings not only have
broad implications for answering such clinically
y
mosaic-like fashion, whereby risk-seeking hot
spots are adjacent to risk-averse hot spots. More
detailed structural analysis will be necessary
to ascertain the validity of this possibility in
the future.
The dACC and OFC play important roles
for the reward value computation in monkeys
(23, 37–41). Therefore, the role of area 6V, and
not that of dACC and OFC, in risk-dependent
decision-making was highly unexpected, especially because the macaque area 6V has long been
considered to be a motor-related area (42–44).
Nonetheless, because EV dependency might be
more widely prevalent in frontal cortical regions
and in the midbrain (23, 38–41), it is likely that
area 6VV is not the sole center responsible for
this behavior. Evidence from human magnetic
resonance imaging (MRI) [area 6 (22); the inferior frontal gyrus (IFG) (24, 25)] and patient
studies (7, 8, 10, 11) supports the notion of area 6
playing a role in behavioral flexibility and risky
decision-making in humans, thereby suggesting the evolutionarily conserved function of
these neural circuits between nonhuman primates and humans.
We also observed that activation of the VTA–
area 6V pathway induced an accumulation
of the modulatory effects; HH preference in
LED-OFF trials gradually enhanced and saturated after a series of sessions with area 6VV
photostimulation. By contrast, HH preference
was relieved with area 6VD photostimulation.
The behavioral changes were accompanied by
an enhancement of the cue-related a-band re-
y g
5 January 2024
Δ Probability (%)
40
y
Sasaki et al., Science 383, 55–61 (2024)
20
g
Until recently, pathway-selective optogenetic manipulation studies aiming to modulate behavior
were rare in macaques (31, 32), although widely
spread nonselective activation could promote
learning of reward value (33, 34) and produce
plastic changes in cortical blood oxygenation
level–dependent (BOLD) responses (35, 36). We
successfully modulated risk-dependent decisionmaking in macaques by selective manipulation
of the mesofrontal pathway from the VTA to area
6V. Activation of the VTA-6VV pathway enhanced
HH-preferring cue-related responses in area 6VV
and facilitated HH-preferring decisions, whereas activation of the VTA-6VD pathway inhibited HH-preferring cue-related responses in area
6VD and relieved HH-preferring decisions. The
effects observed by activation of the VTA-6VV
pathway may be a result of the short-term modulation of synaptic transmission through the
activation of dopamine receptors because most
vector-infected cells in the VTA were dopaminergic (Fig. 3, F to H). The success of optogenetic
manipulation might be a result of the application of ChrimsonR, which can be activated by
red light that penetrates brain tissue better
than green light to activate channel rhodopsin.
Another reason might be that the rich axonal
projections from the VTA to area 6V terminate
in the most superficial layer of area 6V (27),
which enables better access by the optogenetic
stimulation from outside the brain. Though unlikely, we are unable to exclude the possibility that this cortical area may be organized in a
0
p
Fig. 5. Decoding of risk preference in low-dimensional power signal.
(A) Schematic diagram of dimensionality reduction by dPCA, which decomposes
the population activity of individual channels into dimensions relevant to HH-LL,
EV, and condition-independent axes. (B) The first dPCs for each variable in the a
band for each reward probability (top), EV (middle), and others (bottom) from
monkey C. The solid and dashed vertical lines indicate the timing of target onset and
mean reaction times, respectively. (C) Trajectory of lower-band population
activity (≤13 Hz) in HH-LL and EV axes. The line color corresponds to the
Discussion
-20
−1.2
RES EARCH | R E S E A R C H A R T I C L E
relevant questions but also provide opportunities to better understand the underlying neural
mechanism of gambling disorder in humans.
RE FE RENCES AND N OT ES
48. R. Sasaki, Data – “Balancing Risk-Return Decisions by
Manipulating the Mesofrontal Circuits in Primates,” version 1, data
set Zenodo (2023); https://doi.org/10.5281/zenodo.8371506.
AC KNOWLED GME NTS
We thank W. Vanduffel and S. Goulas for critical reading and
constructive suggestions for our manuscript. We also thank
J. Yamashita, M. Nakamura, and Y. Shinto for technical assistance.
Funding: We acknowledge funding from KAKENHI grants from
JSPS 19K21209, 21K19430, and 21H02803 (R.S.); JST/PRESTO
JPMJPR21S6 (R.S.); the Uehara Memorial Foundation (R.S.);
the Takeda Science Foundation (R.S.); the Naito Foundation (R.S.);
the Brain Science Foundation (R.S.); the Fujiwara Memorial
Foundation (R.S.); JST/CREST JPMJCR1651 (T.I.); AMED
20DM0107151 (T.I.); AMED 20DM0307005 (T.I.); and KAKENHI
grants from JSPS 19H05723 and 22H04992 (T.I.). Author
contributions: Conceptualization: R.S. and T.I. Data curation: R.S.,
T.M., and Y.T. Formal analysis: R.S., T.M., R.Y., K.I., and Y.T.
Funding acquisition: R.S. and T.I. Investigation: R.S., T.M., and
Y.T. Methodology: R.S., T.I., Y.O., T.T., J.O., K.K., H.O., and J.T.
Project administration: R.S. and T.I. Resources: T.I., R.S., J.O., and
K.K. Software: R.S., T.M., and R.Y. Supervision: R.S. and T.I.
Validation: R.S. and T.I. Visualization: R.S. and T.I. Writing – original
draft: R.S. and T.I. Writing – review & editing: R.S. and T.I.
Competing interests: The authors declare that they have no
competing interests. Data and materials availability: All data are
available in the manuscript or the supplementary materials or
are deposited at Zenodo (48). License information: Copyright ©
2024 the authors, some rights reserved; exclusive licensee
American Association for the Advancement of Science. No claim to
original US government works. https://www.science.org/about/
science-licenses-journal-article-reuse
p
SUPPLEMENTARY MATERIALS
g
science.org/doi/10.1126/science.adj6645
Materials and Methods
Figs. S1 to S7
References (49–51)
MDAR Reproducibility Checklist
y
1. J. von Neumann, O. Morgestern, Theory of Games and
Economic Behavior (Princeton Univ. Press, 1944).
2. W. R. Stauffer, A. Lak, W. Schultz, Curr. Biol. 24, 2491–2500
(2014).
3. M. Brand et al., Psychiatry Res. 133, 91–99 (2005).
4. W. S. Slutske, A. Caspi, T. E. Moffitt, R. Poulton, Arch. Gen.
Psychiatry 62, 769–775 (2005).
5. D. Brevers, A. Bechara, A. Cleeremans, X. Noël, Front. Psychol.
4, 665 (2013).
6. A. Fujimoto et al., Transl. Psychiatry 7, e1085 (2017).
7. A. Bechara, H. Damasio, D. Tranel, S. W. Anderson, J. Neurosci.
18, 428–437 (1998).
8. A. Bechara, D. Tranel, H. Damasio, Brain 123, 2189–2202
(2000).
9. P. Cavedini, G. Riboldi, R. Keller, A. D’Annucci, L. Bellodi,
Biol. Psychiatry 51, 334–341 (2002).
10. F. Manes et al., Brain 125, 624–639 (2002).
11. L. K. Fellows, M. J. Farah, Cereb. Cortex 15, 58–63 (2005).
12. W.-S. Yan, R.-R. Zhang, Y. Lan, Y.-H. Li, N. Sui, Sci. Rep. 6,
39233 (2016).
13. K. Wallin-Miller, G. Li, D. Kelishani, R. I. Wood, Behav. Neurosci.
132, 152–160 (2018).
14. J. N. Ferland et al., J. Neurosci. 39, 1842–1854 (2019).
15. A. Bechara, A. R. Damasio, H. Damasio, S. W. Anderson,
Cognition 50, 7–15 (1994).
16. X. Chen, V. Stuphorn, Curr. Biol. 28, 3114–3122.e4 (2018).
17. T. R. Smith, M. J. Beran, Learn. Behav. 48, 301–321
(2020).
18. W. R. Stauffer, A. Lak, P. Bossaerts, W. Schultz, J. Neurosci. 35,
3146–3154 (2015).
19. S. Farashahi, H. Azab, B. Hayden, A. Soltani, J. Neurosci. 38,
4383–4398 (2018).
20. S. Ferrari-Toniolo, P. M. Bujold, W. Schultz, J. Neurosci. 39,
2915–2929 (2019).
21. D. Kahneman, A. Tversky, Econometrica 47, 263–292
(1979).
22. T. Hanakawa et al., Cereb. Cortex 12, 1157–1170 (2002).
23. P. H. Rudebeck, R. C. Saunders, D. A. Lundgren, E. A. Murray,
Neuron 95, 1208–1220.e5 (2017).
24. K. Miyamoto et al., Neuron 109, 1396–1408.e7 (2021).
25. G. I. Christopoulos, P. N. Tobler, P. Bossaerts, R. J. Dolan,
W. Schultz, J. Neurosci. 29, 12574–12583 (2009).
26. W. Schultz, J. Neurophysiol. 80, 1–27 (1998).
27. M. Zubair et al., Cereb. Cortex 31, 2913–2931 (2021).
28. N. C. Klapoetke et al., Nat. Methods 11, 338–346
(2014).
29. Y. Ohta et al., IEEE Access 9, 127937–127949 (2021).
30. D. Kobak et al., eLife 5, e10989 (2016).
31. K.-I. Inoue, M. Takada, M. Matsumoto, Nat. Commun. 6, 8378
(2015).
32. L. Nurminen, S. Merlin, M. Bijanzadeh, F. Federer, A. Angelucci,
Nat. Commun. 9, 2281 (2018).
33. W. R. Stauffer et al., Cell 166, 1564–1571.e6 (2016).
34. J. T. Arsenault, S. Rima, H. Stemmann, W. Vanduffel, Curr. Biol.
24, 1347–1353 (2014).
35. J. T. Arsenault, W. Vanduffel, Nat. Commun. 10, 3591
(2019).
36. S. R. Murris, J. T. Arsenault, R. Raman, R. Vogels, W. Vanduffel,
Neuron 109, 1381–1395.e7 (2021).
37. D. L. Hocker, C. D. Brody, C. Savin, C. M. Constantinople, eLife
10, e70129 (2021).
38. C. Padoa-Schioppa, J. A. Assad, Nature 441, 223–226
(2006).
39. M. O’Neill, W. Schultz, Neuron 68, 789–800 (2010).
40. P. H. Rudebeck, E. A. Murray, Neuron 84, 1143–1156
(2014).
41. T. C. Blanchard, B. Y. Hayden, E. S. Bromberg-Martin, Neuron
85, 602–614 (2015).
42. N. Picard, P. L. Strick, Cereb. Cortex 6, 342–353
(1996).
43. S. P. Wise, D. Boussaoud, P. B. Johnson, R. Caminiti, Annu. Rev.
Neurosci. 20, 25–42 (1997).
44. G. Rizzolatti, C. Sinigaglia, Nat. Rev. Neurosci. 17, 757–765
(2016).
45. J. R. Wickens, J. N. J. Reynolds, B. I. Hyland, Curr. Opin. Neurobiol.
13, 685–690 (2003).
46. K. A. Zalocusky et al., Nature 531, 642–646 (2016).
47. T. J. Moore, J. Glenmullen, D. R. Mattison, JAMA Intern. Med.
174, 1930–1933 (2014).
Submitted 28 July 2023; accepted 6 November 2023
10.1126/science.adj6645
y g
y
,
Sasaki et al., Science 383, 55–61 (2024)
5 January 2024
7 of 7
RES EARCH
IMMUNOLOGY
Microbiota-dependent activation of CD4+ T cells induces
CTLA-4 blockade–associated colitis via Fcg receptors
Bernard C. Lo1, Ilona Kryczek2,3, Jiali Yu2,3, Linda Vatan2,3, Roberta Caruso1, Masanori Matsumoto1,
Yosuke Sato4, Michael H. Shaw4, Naohiro Inohara1, Yuying Xie5, Yu Leo Lei6,
Weiping Zou1,2,3, Gabriel Núñez1*
Immune checkpoint inhibitors can stimulate antitumor immunity but can also induce toxicities
termed immune-related adverse events (irAEs). Colitis is a common and severe irAE that can lead to
treatment discontinuation. Mechanistic understanding of gut irAEs has been hampered because robust
colitis is not observed in laboratory mice treated with checkpoint inhibitors. We report here that this
limitation can be overcome by using mice harboring the microbiota of wild-caught mice, which develop
overt colitis following treatment with anti-CTLA-4 antibodies. Intestinal inflammation is driven by
unrestrained activation of IFNg-producing CD4+ T cells and depletion of peripherally induced regulatory
T cells through Fcg receptor signaling. Accordingly, anti-CTLA-4 nanobodies that lack an Fc domain
can promote antitumor responses without triggering colitis. This work suggests a strategy for mitigating
gut irAEs while preserving antitumor stimulating effects of CTLA-4 blockade.
5 January 2024
1 of 9
,
Lo et al., Science 383, 62–70 (2024)
y
*Corresponding author. Email: gabriel.nunez@umich.edu
To investigate the role of the gut microbiota in
ICB-induced intestinal inflammation, we first
utilized C57BL/6 [wild-type (WT)] mouse lines
with dissimilar gut microbiota composition
from various commercial vendors raised under SPF conditions in addition to mice colonized with the WildR microbiota. In our initial
y g
Immune checkpoint blockade triggers colitis
in mice colonized with wild microbiota
Department of Pathology and Rogel Cancer Center, University
of Michigan, Ann Arbor, MI 48109, USA. 2Department of
Surgery, University of Michigan, Ann Arbor, MI 48109, USA.
3
Center of Excellence for Cancer Immunology and
Immunotherapy, Rogel Cancer Center, University of Michigan,
Ann Arbor, MI 48109, USA. 4Takeda Pharmaceuticals
International Co., Cambridge, MA 02139 USA. 5Department of
Computational Mathematics, Science and Engineering, Michigan
State University, East Lansing, MI 48824, USA. 6Department
of Periodontics and Oral Medicine, Rogel Cancer Center,
University of Michigan, Ann Arbor, MI 48104, USA.
y
1
g
use of gene-targeted mice predisposed to intestinal inflammation (8–11). Concurrently,
there has been increasing evidence that the
gut microbiota composition may play a role
in the susceptibility to colitis induced by ICB
therapy, although microbial alpha diversity
was similar in patients who developed ICBinduced colitis compared with patients undergoing ICB who did not develop colitis (12–14).
However, the immunological mechanisms and
the role of the microbiota underlying irAEs in
the gut remain poorly understood.
We sought to establish a model of ICB-induced
colitis using mice colonized with a referenced
gut microbiota originally derived from wild-caught
mice, referred to as wild mouse microbiomereconstituted (WildR) mice (15). Laboratory
mouse strains colonized with free-living microbiota exhibit immunological features that more
closely resemble humans thus overcoming
some of the limited utility of animal models
of human diseases associated with the artificially hygienic status of conventionally raised
specific pathogen-free (SPF) mice (16–18). Previously, WildR mice were shown to provide a
more reliable model than SPF mice for anticipating the outcome of clinical trials of immunemodulating biologics (18). Therefore, we
hypothesized that mice harboring the WildR
microbiota would be a suitable system to examine host-microbiota interactions relevant
to ICB therapy and irAEs.
p
M
onoclonal antibodies targeting CTLA-4,
PD-1, and PD-L1 are widely used to promote antitumor immune responses in
a range of human cancers, but can also
lead to inflammatory toxicities, collectively referred to as immune-related adverse
events (irAEs) (1–3). Colitis is a common and
severe irAE that can lead to treatment discontinuation, particularly in patients receiving
a CTLA-4 inhibitor alone or in combination
with PD-1 inhibitors; intestinal inflammation
triggered by CTLA-4 blockade typically involves the colon and is characterized by the
accumulation of CD4+ T lymphocytes and neutrophils in the intestinal tissue (3–5). Analyses
of colitis-associated lymphocytes of melanoma
patients receiving CTLA-4 and PD-1 inhibitors
have further implicated the pathogenic contributions of tissue-resident CD8+ T cells with
cytotoxic features (6). Our understanding of
the immunological mechanisms of immune
checkpoint blockade (ICB)–induced colitis has
been impeded by the lack of robust animal
models of colitis caused by immune checkpoint inhibitors. Laboratory mice are highly
resistant to intestinal inflammation following
treatment with antibodies targeting immune
checkpoints (7, 8). Therefore, current models
of ICB-induced colitis require supplemental
interventions to induce colitis such as exposure to dextran sulfate sodium (DSS) or the
experiments, we challenged mice with antiCTLA-4 and anti-PD-1 antibodies because dual
blockade was associated with a higher clinical
risk of incurring gut irAEs compared with
single ICB treatments (3, 4). We also relied on
fecal lipocalin-2 (LCN-2) measurements to track
the kinetics and severity of intestinal inflammation after starting ICB treatment (19). Consistent with previous reports (7, 8), we did not
observe induction of fecal LCN-2 in SPF WT
mice originating from Jackson Laboratories
(JAX), Taconic Biosciences, or Charles River
Laboratories after treatment with anti-CTLA-4
and anti-PD-1 antibodies (fig. S1). By contrast,
fecal LCN-2 was highly induced in WT mice
colonized with WildR microbiota after two injections of dual immune checkpoint inhibitors,
and the intestinal inflammation was sustained
for up to 21 days of the ICB treatment course
(fig. S1).
To directly assess the impact of the microbiota on host susceptibility to ICB-associated
colitis, we colonized germ-free C57BL/6 (WT)
mice with SPF JAX microbiota or WildR microbiota. Germ-free (GF) WT mice colonized with
SPF JAX microbiota did not develop colitis following ICB as indicated by fecal LCN-2 measurements and assessment of gut histology
(Fig. 1, A and B). Although GF WT mice colonized with WildR microbiota treated with isotype antibodies did not develop colitis, GF WT
mice colonized with WildR microbiota injected
with anti-CTLA-4 and anti-PD-1 antibodies lead
to clear and sustained induction of fecal LCN-2
(Fig. 1A). In addition, severe immunopathology
was observed in the large intestinal tissue sections of these animals characterized by epithelial
damage, marked infiltration of inflammatory
cells in the lamina propria and submucosa, crypt
abscesses, and apoptotic cells in the crypt epithelium that are prominent features of the
disease in humans (Fig. 1B) (5). In patients,
immune checkpoint inhibitors can cause irAEs
in multiple organs (3). In the current model,
dual ICB induced immune cell aggregates in
the liver at a higher frequency in GF mice
colonized with WildR microbiota compared
with GF mice colonized with SPF JAX microbiota (fig. S2, A and B). Meanwhile, minimal or
no evidence of inflammation was detected in
the kidneys, heart, or lungs after treatment with
ICB (fig. S2, A and B).
We evaluated the bacterial composition of
the microbiota by performing 16s ribosomal
RNA sequencing of fecal pellets collected from
the GF mice colonized with SPF JAX microbiota or WildR microbiota during treatment
with dual ICB or isotype controls. The alpha
diversity of the bacterial communities of SPF
JAX microbiota and WildR microbiota represented by Shannon indexes were similar
and unaltered by treatment with dual ICB (fig.
S3A). Moreover, a nonmetric multidimensional scaling plot of Bray and Curtis dissimilarity
RES EARCH | R E S E A R C H A R T I C L E
B WildR ISO
3000
****
2000
1000
N.S.
N.S.
N.S.
N.S.
9 12
Days
15
**
40
30
20
10
0
*
600
400
IS
R
SP
*
40
30
20
N.S.
10
0
N.S.
Fig. 1. Immune checkpoint blockade induces colitis in C57BL/6 mice
harboring microbiota of wild-caught mice. (A) GF C57BL/6 (WT) mice were
colonized with SPF JAX microbiota or WildR microbiota followed by treatment
with anti-CTLA-4 antibodies and anti-PD-1 antibodies (ICB) or isotype (ISO)
every 3 days. Colitis was assessed by fecal lipocalin-2. N.S., P > 0.05; ***, P <
0.001; ****, P < 0.0001 versus WildR ISO, Dunnett’s multiple comparisons test.
Data presented as mean ± SEM. (B) Hematoxylin and eosin (H&E)-stained
sections of cecal tissues 15 days after starting ICB treatment, and histology
scores. Scale bars, 200 mm. White arrowheads point to epithelial damage, black
arrowheads point to regenerating epithelia, the gray arrow indicates a crypt
abscess, and open arrows indicate inflammatory infiltrates. Each point represents
Lo et al., Science 383, 62–70 (2024)
5 January 2024
ag
R
BL
/
12
57
6
9
Days
C
0
6
IC
0
1 -/IC
R
B
ag
1 -/IS
O
B
N.S.
Rag1-/- ISO
,
N.S.
ild
R
Rag1-/- ICB
Rag1-/- ISO
*
200
W
ild
W
***
800
Rag1-/- ICB
C57BL/6 ICB
y
ng / g feces
D C57BL/6 ICB
Lipocalin-2
1000
y g
C
6
Histology Score
0
O
0
F
***
N.S.
Histology Score
***
4000
SPF JAX ICB
y
****
WildR ICB
WildR ISO
WildR ICB
SPF ICB
I
JA CB
X
IC
B
We then compared the respective contributions
of antibodies targeting CTLA-4 or PD-1 in the
development of ICB-driven gut inflammation.
WildR microbiota mice treated with dual ICB
or anti-CTLA-4 antibodies alone were similarly
susceptible to intestinal inflammation as evidenced by matching fecal LCN-2 induction and
disease severity in intestinal tissues (fig. S4, A
and B). By contrast, WildR microbiota mice
that received anti-PD-1 antibodies alone did
not develop intestinal inflammation (fig. S4,
A and B). These data indicate that CTLA-4
blockade is the principal driver of colitis in
this microbiota-dependent model, which is
consistent with human studies (3, 4). Furthermore, profiling of cecal immune cells by flow
cytometry revealed the accumulation of IFNg+,
IL-17+, and double positive IFNg+ IL-17+ CD4+
T helper (TH) cells 9 days after commence-
Lipocalin-2
5000
ng / g feces
CTLA-4 blockade elicits intestinal inflammation
driven by CD4+ T cells and IFNg
ment of CTLA-4 blockade coinciding with
high disease severity indicated previously
by fecal LCN-2 levels (Fig. 2A). We also observed the induction of IFNg+ CD8+ T cells
after treatment with anti-CTLA-4 antibodies
(fig. S5). Consistent with the skewed IFNg response, we found robust Tbet but relatively
mild RORgt expression by CD4+ T cells in the
intestinal tissue, further highlighting a biased
TH1 response during ICB-mediated inflammation (Fig. 2B and fig. S6). Moreover, antiCTLA-4 antibody–induced colitis was associated
with an increase in multiple myeloid cell subsets including neutrophils, monocytes, macrophages, and dendritic cells in the gut (fig.
S7, A and B).
Intestinal regulatory T cells (Tregs) expressing Foxp3 are critical for immune homeostasis and can be categorized as thymic-derived
Tregs (tTregs) or peripherally induced Tregs (pTregs)
which are responsive to the gut microbiota
(20, 21). In GF mice, most gut Foxp3+ Tregs
display features suggestive of thymic origin
whereas the colonization with intact microbiota or select bacterial species results in the
induction of pTregs including a subset expressing RORgt (22–28). In assessing Foxp3+ Tregs
g
A
bodies (Fig. 1, C and D). Collectively, these
data indicate that induction of colitis in mice
by ICB requires both the microbiota of freeliving animals and an intact adaptive immune
response.
p
indexes of individual mice indicates that the
bacterial community structures of SPF JAX microbiota and WildR microbiota are dissimilar
(fig. S3B). Alterations in the WildR microbiota
were also observed in response to dual ICB coinciding with intestinal inflammation (fig. S3B).
To compare bacterial taxa of the WildR microbiota and SPF JAX microbiota, we performed
linear discriminant analysis effect size (LEfSe).
We identified 132 OTUs at greater than 97%
nucleotide sequence identity, which were differentially abundant in the WildR microbiota
compared with SPF JAX microbiota (fig. S3C).
These findings suggest that the bacterial communities of the SPF JAX microbiota and freeliving WildR microbiota are similarly diverse
yet highly divergent.
To determine the contributions of adaptive
immunity in the present model of ICB-driven
colitis, we colonized GF WT or GF Rag1−/− mice
lacking lymphocytes with WildR microbiota
and challenged them with dual ICB or isotype
antibodies. As colitis was triggered in GF WT
mice colonized with WildR microbiota by dual
ICB, GF Rag1−/− mice colonized with WildR
microbiota did not develop colitis after treatment with dual ICB or isotype control anti-
an individual mouse. n = 4 to 6 mice per group. Representative data of two
independent experiments. N.S., P > 0.05; **, P < 0.01, Kruskal-Wallis with Dunn’s
test. (C) GF C57BL/6 and GF Rag1−/− mice were colonized with WildR microbiota
and treated with ICB or ISO. Colitis was assessed by fecal lipocalin-2. N.S., P > 0.05;
*, P < 0.05; ***, P < 0.001 versus Rag1−/− ICB, Dunnett’s multiple comparisons
test. Data presented as mean ± SEM. (D) H&E-stained sections of cecal tissues
12 days after starting ICB treatment and histology scores. Scale bars, 200 mm. White
arrowheads point to epithelial damage, black arrowheads point to regenerating
epithelia, and open arrows indicate inflammatory infiltrates. Each point represents an
individual mouse. n = 3 to 5 mice per group. Results were confirmed using SPF mice.
N.S., P > 0.05; *, P < 0.05, Kruskal-Wallis with Dunn’s test.
2 of 9
RES EARCH | R E S E A R C H A R T I C L E
receptors (TCRs) (26, 28, 29). Moreover, RORgt+
pTregs are highly enriched in the large intestine but are mostly absent across systemic sites
and in tumors (28). Consistent with previous
studies, colonization of GF mice with WildR
microbiota similarly induced RORgt+ pTregs
which have higher expression of CLTA-4 compared with a Helios+ subset of tTregs (fig. S9, A
and B). These data indicate that CTLA-4 blockade induces preferentially a TH1 response and a
biased depletion of RORgt+ pTregs in the gut.
The accumulation of CD8+ T cell subsets at
mucosal surfaces is a common feature of lab-
2.81
55
4.84
37.3
25
20
15
10
5
0
IFN
Gated on CD4+ cells
0
0.5
0.0
isotype
anti-CTLA-4
anti-CTLA-4
91.2
7.02
D
1.76
0.06
3
2
1
0
isotype
anti-CTLA-4
20
10
0
**
54.6
30.2
42.3
13.2
***
5
4
3
2
1
0
isotype
anti-CTLA-4
ROR t
E
F
IF
N
ot
an
0
,
is
10
e
0
yp
10
20
ti-
20
**
30
y
5 January 2024
Histology Score
30
40
y g
****
anti-IFN
ot
40
Fig. 2. Anti-CTLA-4 antibodies elicit IFNg and CD4+ T cell-mediated
colitis. (A) FACS analysis of lymphocytes isolated from cecal tissues of WildR
microbiota mice 9 days after starting anti-CTLA-4 antibody or isotype treatment.
Pseudocolor plots depict IFNg and IL-17 expression by CD4+ T cells and numbers of
cytokine-expressing CD4+ T cells. Events displayed in flow plots: 2808, isotype
and 6388, anti-CTLA-4 antibody-treated. (B) Numbers of Foxp3− CD4+ T cells
expressing Tbet and RORgt isolated from cecum. (C) Pseudocolor plots of Foxp3
expression by cecal T cells and percentage expression of Foxp3 by T cells. Events
displayed in flow plots: 5906, isotype and 14803, anti-CTLA-4 antibody-treated.
(D) Pseudocolor plot of Helios and RORgt expression by Foxp3+ T cells and ratio of
Helios+ Treg (tTreg) to RORgt+ Treg (pTreg) numbers. Events displayed in flow plots:
940, isotype and 1039, anti-CTLA-4 antibody-treated. Each dot represents an
individual animal. n = 4 mice per group. Representative data of two independent
experiments. **, P < 0.01; ***, P < 0.001; ****, P < 0.0001; unpaired two-tailed
Lo et al., Science 383, 62–70 (2024)
isotype
n.s.
an
anti-CD8
yp
e
tiC
D
4
an
tiC
D
8
anti-CD4
Histology Score
isotype
****
anti-CTLA-4
isotype
isotype
anti-CTLA-4
Foxp3
25
20
15
10
5
0
Gated on Foxp3+ T cells
**
30
**
is
2.1
15.9
1.0
5
4
3
2
1
0
ROR t+
y
CD3
82
1.5
Foxp3+
% CD4+ T cells
isotype
***
2.0
Tbet+
g
C
****
Helios
94.6
2.9
0
B
IFN +
IL-17A+
IL-17A+
p
IL-17A
2.56
CD4+ T Cells (x104)
isotype
+
IFN
anti-CTLA-4
oratory mouse strains colonized with the microbiota of free-living animals (16, 18). Moreover,
tissue-resident CD8+ T cell subsets with cytotoxic effector features have been implicated
in the pathogenesis of ICB-induced colitis in
the clinic (6, 30), and we noted the induction
of IFNg+ CD8+ T cells in the present mouse
model (fig. S5). To clarify the involvement of
CD4+ and CD8+ T cells in ICB-driven intestinal
inflammation, we selectively depleted each
T cell subset in WildR microbiota mice during
ICB treatment (fig. S10, A and B). Anti-CD8
antibodies delayed LCN-2 induction but had
tTreg : pTreg
A
CD4+ T Cells (x104)
in the gut during ICB, we observed a reduction
in the percentages of Foxp3+ T cells (Fig. 2C)
and notably the selective depletion of RORgt+
pTregs whereas the relative percentages of a
Helios+ subset of tTreg cells remained largely intact (Fig. 2D). The reduction of RORgt+
pTregs was confirmed in response to anti-CTLA-4
antibody treatment preceding peak inflammation (fig. S8, A and B). Previous reports demonstrate that RORgt+ pTregs selectively induced
by the microbiota possess distinct immunosuppressive features including exaggerated expression of CTLA-4, IL-10, and bacteria-reactive T cell
t-test. (E) WildR microbiota mice undergoing CTLA-4 blockade were treated with
isotype control antibodies or cell-depleting antibodies against CD4 or CD8. H&Estained cecal sections 12 days after starting ICB treatment and corresponding
histology scores. Scale bars, 200 mm. White arrowheads point to epithelial damage,
black arrowheads point to regenerating epithelia, and open arrows indicate
inflammatory infiltrates. Each point represents an individual mouse. Data were
combined from three independent experiments. N.S., P > 0.05; ****, P < 0.0001;
Kruskal-Wallis with Dunn’s test. (F) WildR microbiota mice receiving anti-CTLA-4
antibodies were treated with neutralizing anti-IFNg antibodies or isotype control.
H&E-stained cecal sections 12 days after starting ICB treatment, and histology
scores. Scale bars, 200 mm. White arrowheads point to epithelial damage, the black
arrowhead points to regenerating epithelium, and open arrows indicate inflammatory
infiltrates. Each dot represents an individual mouse. Representative data of two
independent experiments. **, P < 0.01; Mann-Whitney test.
3 of 9
RES EARCH | R E S E A R C H A R T I C L E
no impact on reducing subsequent disease severity whereas mice that received antibodies
that ablate CD4+ cells were protected from ICBinduced gut inflammation (Fig. 2E and fig.
S11). Furthermore, animals treated with IFNg-
A
neutralizing antibodies displayed highly diminished inflammation in the gut during the
ICB treatment course (Fig. 2F). We also found
that IL-17A/F KO mice displayed severity of
gut immunopathology similar to that of WT
B
CD4+ T cells
Th2
Tfh
B-cells
supCPM_2
6
Naïve T cells
4
1
+
−2
3
C
Tbet+ PD-1+ Th1
p
−4
Tbet+ PD-1- Th1
TCF-1 T-cells
9 10 11 12 13 14
0
11 Th17
8
Th22
13 5
12
8
9 2 10
7
5
2
7
14
4
6
2
Treg
1
2
3
4
5
6
7
8
9
10
11
12
13
14
3
1
4
mice in this disease model (fig. S12, A and
B). Thus, intestinal inflammation triggered
by CTLA-4 blockade in WildR mice is driven
primarily by CD4+ T cell–mediated responses
and IFNg.
−10
−5
0
supCPM_1
5
isotype
anti-CTLA-4
C5: Treg
g
re
g
g
pTreg
tTre
tT
Tbet+ Treg
pT
1
reg
pT
0
2
y
tTreg
−2.5
Tbet +Treg
Tbet + T re g
supCPM_2
2.5
D
re
3
g
1
2
3
5.0
−5.0
−4
0
2
supCPM_1
C1: Tbet+ PD-1- Th1
isotype
4
C3: Tbet+ PD-1+ Th1
C5: Treg
anti-CTLA-4
C5-1: pTreg
C5-2: tTreg
T cells with
unique
TCR
anti-CTLA-4
Fig. 3. Gene expression and clonotype analyses of CD4+ T cells in
ICB-induced colitis. (A) Sort-purified intestinal CD4+ T cells from WildR mice
treated with isotype or anti-CTLA-4 antibodies underwent scRNA-seq analyses;
each sample contained pooled cells from tissues of 3 mice per treatment
group. CD4+ T cell clusters were visualized by supervised capacity preserving
mapping (supCPM). (B) Circos plot showing relative contribution to each cluster
identity by CD4+ T cells from each treatment. (C) Cluster 5 Treg cells were
Lo et al., Science 383, 62–70 (2024)
5 January 2024
segregated and underwent further clustering and supCPM rendering and
(D) circos plots showing relative abundance of each Treg subcluster by treatment
group. (E) Doughnut plots showing relative abundance of clonotypes in each
cluster classified by three levels of clonal expansion. CD4+ T cells with moderate
clonal expansion include clonotypes with frequencies between two and four,
and CD4+ T cells with marked clonal expansion include clonotypes with
frequencies of five or greater.
4 of 9
,
Marked
clonal
expansion
y
Moderate
clonal
expansion
isotype
y g
E
−2
RES EARCH | R E S E A R C H A R T I C L E
B
WT (ICB)
***
FcR KO (ICB) FcR KO (ISO)
*
N.S.
0
20
10
2.64
93
2.39
39.6
79.1
12
IFN
D
IFN
****
****
****
80
60
20
FcR KO (ISO)
21.4
19.9
WT (ICB)
0
Foxp3
44.7
23.2
43.9
FcR KO (ICB)
57.1
Helios
Helios
22.3
WT (ICB)
30
10.9
ROR t
ROR t
5 January 2024
0
ICB
ISO
ICB
WT
FcR KO
ISO
ICB
10
0
WT
FcR KO
ISO
ICB
****
****
****
4
2
0
WT
FcR KO
ISO
ICB
isotype or anti-CTLA-4 antibodies. Events displayed in flow plots: 2949, WT
(ISO); 2515, FcRg KO (ISO); 8060, WT (ICB); 1165, FcRg KO (ICB). (D) Numbers
of cytokine-expressing T cells. Each point represents an individual mouse.
Combined data from two independent experiments for each treatment group.
**, P < 0.01; ***, P < 0.001; ****, P < 0.0001; Dunnett’s multiple comparisons
test. (E) Pseudocolor plots of Foxp3 expression by CD4+ cells isolated from
cecal tissues, and percentages of Foxp3 expression by T cells. Events displayed
in flow plots: 5949, WT (ISO); 6487, FcRg KO (ISO); 6664, WT (ICB); 4870,
FcRg KO (ICB). (F) Subsetting Helios+ tTregs and RORgt+ pTregs, and the ratios of
tTregs to pTregs. Events displayed in flow plots: 1272, WT (ISO); 1293,
FcRg KO (ISO); 567, WT (ICB); 1024, FcRg KO (ICB). Each point represents
an individual mouse. Representative data from two independent experiments.
***, P < 0.001; ****, P < 0.0001; Dunnett’s multiple comparisons test.
ter visualization (31) and observed diverse
immune cell populations similar to clusters
reported by the original investigators (fig. S13A
and table S1) (6). We noted a singular Treg cluster (cluster 7), defined by high expression levels
of Foxp3, Ctla4, Il10, and Cd4, which were elevated in relative abundance in ICB-treated
patients who developed colitis compared with
ICB-treated patients who did not develop co-
litis and normal control samples, which is
consistent with the previous report (6) (fig. S13,
A and B, and table S1). We further segregated
the Treg cluster (cluster 7) and identified three
Treg subclusters: Treg subcluster 7-1 was marked
by high expression of Helios, encoded by Ikzf2;
Treg subcluster 7-2 was defined by lower expression of Ikzf2; and Treg subcluster 7-3 was
characterized by high expression of Sell and
5 of 9
,
Lo et al., Science 383, 62–70 (2024)
0
y
To assess whether ICB alters Treg subset
abundance in a clinical setting, we examined
a comprehensive single cell RNA sequencing
(scRNA-seq) data set of intestinal immune
cells isolated from ICB-treated melanoma patients who developed colitis, ICB-treated melanoma patients who did not develop colitis, and
control cases (6). We utilized supervised capacity preserving mapping (supCPM) for clus-
42.4
5
y g
Fig. 4. Colitis induced by anti-CTLA-4 antibodies requires Fcg receptors.
(A) WT and Fcer1g−/− (FcRg KO) mice were colonized with WildR microbiota
and treated with anti-CTLA-4 antibodies (ICB) or isotype control antibodies
(ISO). Colitis was assessed by fecal lipocalin-2. Statistical significance between
ICB-treated WT and FcRg KO mice was determined by Sidak’s multiple
comparisons test and indicated by N.S., P > 0.05; *, P < 0.05; ***, P < 0.001.
Data presented as mean ± SEM. (B) H&E-stained sections of cecal tissues 12 days
after starting indicated treatment and histology scores. Scale bars, 200 mm.
White arrowheads point to epithelial damage and open arrows indicate inflammatory
infiltrates. Each point represents an individual mouse. N.S., P > 0.05; *, P < 0.05;
Kruskal-Wallis with Dunn’s test. Representative data from two independent
experiments. (C) Pseudocolor plots of cytokine expressing CD4+ T cells isolated
from ceca of WildR microbiota-colonized WT and FcRg KO mice treated with
29.2
2
20
6
tTreg : pTreg
FcR KO (ISO)
10
**
y
WT (ISO)
4
**
**
****
***
***
40
CD4
CD4
F
21
15
g
Foxp3
8.51
***
***
Foxp3+
FcR KO (ICB)
% of CD4+ T cells
WT (ISO)
6
IFN +
IL-17A+
***
40
ISO
E
IL-17A+
+
p
IFN
51.5
FcR KO (ICB)
0.77
8.15
CD4+ T cells (x104)
93.6
WT (ICB)
5.71
3.19
FcR KO (ISO)
3.74
0.32
IL-17A
IL-17A
WT (ISO)
3.42
0.34
N.S.
0
6 9 12
Days
WT (ICB)
FcR KO (ICB)
FcR KO (ISO)
C
*
30
Histology Score
Lipocalin-2
2500
2000
1500
1000
500
0
Fc WT
R
(I
Fc KO CB)
R
(
KO ICB
(IS )
O
)
ng / g feces
A
RES EARCH | R E S E A R C H A R T I C L E
300
100
0
10
5
0
9 12 15
Days
600
an
anti-CTLA-4
+ anti-PD-1
200
0
0 3 6 9 12 15 18
Days
H11-HLE
+ anti-PD-1
*
** ********
0 6 9 12 15 18
Days
H11-HLE + anti-PD-1
H11-HLE + anti-PD-1
an
isotype
anti-CTLA-4 + anti-PD-1
** *
g
isotype
anti-CTLA-4 + anti-PD-1
25
20
15
10
5
0
p
0
***
** ****
*
Isotype
TL
A
I
11 -4 + sot
yp
-H
a
LE nt e
i-P
+
an D-1
tiPD
-1
**
400
F
C
1000
****
****
***
Lipocalin-2
Histology Score
E
CT26
1500
500
6
15
isotype
anti-CTLA-4
H11-HLE
ng / g feces
2000
**
** *
20
o
ti- typ
e
C
TL
H A-4
11
-H
LE
0 3 6 9 12 15
Days
D
H11-HLE
* **
0
0
isotype
anti-CTLA-4
H11-HLE
Tumor volume (mm3)
*
*
200
anti-CTLA-4
Histology Score
*
500
Isotype
*
Is
***
1000
C
Lipocalin-2
400
H
B
MC38
ti-
1500
ng / g feces
Tumor volume (mm3)
A
Considering the pathogenic contributions
of CD4+ T cells in the current mouse model of
CTLA-4 blockade–associated colitis, we sought
to further define CD4+ T cell responses by single cell transcriptomics. Sort-purified CD4+
T cells from the intestinal tissues of isotype
or anti-CTLA-4 antibody-treated mice were
labeled with hashtag antibodies to minimize
batch effects, and pooled samples underwent
droplet-based scRNA-seq for gene expression
and paired TCR⍺ and TCRb clonotype analyses. After filtering out cells with mitochondrial DNA content over 25% and those with a
unique molecular identifier of <200 or >7500,
6 of 9
,
5 January 2024
CTLA-4 blockade induces oligoclonal TH1 cells
with cytotoxic features
we acquired 9671 high-quality transcriptomes.
We clustered cells by using the top 5000 most
differentially expressed genes and a panel of
markers to stabilize immune lineage segregation (fig. S14 and table S2). To better preserve
the accurate correlation between geographic
distance and biological distance and improve
the visualization of intracluster variance, we
employed supCPM to project the clusters onto
a two-dimensional space (31) (Fig. 3A). We
identified nine CD4+ T cell functional subsets
and a small cluster of B cells (Fig. 3A and fig.
S14). Following anti-CTLA-4 antibody treatment,
we found a selective and marked increase in
IFNg+ Tbet+ TH1 cells defined by clusters 1 and
3 which were mostly absent in isotype-treated
mice (Fig. 3, A and B). One of the TH1 clusters
showed enhanced expression of PD-1 (cluster
3) and was further distinguished by elevated
expression of transcripts related to cytotoxicity
y
Lo et al., Science 383, 62–70 (2024)
These data suggest that nonthymic Treg persistence is selectively associated with intestinal homeostasis during ICB therapy.
y g
Ccr7 (fig. S13, C and D). Although human Treg
subset-defining transcripts remain imperfect,
high Ikzf2 expression may aid in the identification or enrichment of human tTregs (32, 33).
We found that the Ikzf2hi tTreg subcluster 7-1,
the most abundant Treg subset, was similarly
elevated in patients undergoing ICB therapy
who developed colitis as well as ICB-treated
patients who did not develop colitis compared
with control cases (fig. S13E). By contrast, the
relative proportion of the Ikzf2lo nonthymic
Treg subcluster 7-2 was diminished in ICBtreated patients who had developed colitis
compared with ICB-treated patients who did
not develop colitis or control cases (fig. S13E).
Moreover, the percentage of Ikzf2lo Treg subcluster 7-2 is sustained in ICB-treated patients
who did not develop colitis compared with
control cases (fig. S13E). Treg subcluster 7-3
abundance was unaltered across patient groups.
independent experiments. (D to F) BALB/c mice colonized with WildR microbiota
were injected subcutaneously with CT26 tumor cells, then treated with isotype,
anti-CTLA-4 antibodies and anti-PD-1 antibodies, or H11-HLE and anti-PD-1
antibodies. Tumor volumes were tracked (D), and intestinal inflammation was
assessed by fecal lipocalin-2 (E). (F) H&E-stained cecal sections 18 days after
starting ICB treatment and histology scores. Scale bars, 200 mm. White
arrowheads point to epithelial damage and open arrows indicate inflammatory
infiltrates. Each point represents an individual mouse. *, P < 0.05; **, P < 0.01;
***, P < 0.001; ****, P < 0.0001; Dunnett’s multiple comparisons test (D, E),
or Kruskal-Wallis with Dunn’s test (F) versus anti-CTLA-4 and anti-PD-1 group.
Top symbols denote P values of comparisons with isotype treatment group, and
bottom symbols denote P-values of comparisons with H11-HLE and anti-PD-1
treatment group (E). Data presented as mean ± SEM (D and E). Representative
data of two independent experiments.
y
Fig. 5. Anti-CTLA-4 nanobodies stimulate anticancer immunity without
inducing intestinal inflammation. (A to C) WildR microbiota-colonized WT
mice were injected subcutaneously with MC38 tumor cells, then treated with
isotype, anti-CTLA-4 antibodies, or anti-CTLA-4 H11 nanobodies with half-life
extender (H11-HLE). Tumor volumes were tracked (A), and intestinal inflammation was assessed by fecal lipocalin-2 (B). (C) H&E-stained cecal sections
15 days after starting ICB treatment, and histology scores. Scale bars, 200 mm.
White arrowheads point to epithelial damage and open arrows indicate
inflammatory infiltrates. Each point represents an individual mouse. *, P < 0.05;
**, P < 0.01; ***, P < 0.001; Dunnett’s multiple comparisons test (A and B), or
Kruskal-Wallis with Dunn’s test (C) versus anti-CTLA-4 group. Top symbols
denote P-values of comparison with isotype treatment group and bottom
symbols denote P-values of comparison with H11-HLE treatment group (B).
Data presented as mean ± SEM (A and B). Representative data of two
RES EARCH | R E S E A R C H A R T I C L E
,
7 of 9
y
5 January 2024
Given the requirement of Fc-FcgR interactions
in the induction of colitis by anti-CTLA-4 antibodies, we hypothesized that camelid heavy
chain–only antibody fragments (VHHs) or
nanobodies which lack an Fc domain may be
used for CTLA-4 blockade therapy while reducing the risk of gut irAEs. We utilized the
CTLA-4-binding H11 VHH stabilized with a
half-life extender (H11-HLE) that was previously found to exhibit anticancer activity in
some settings (35, 36). We sought to compare
the antitumor activity of H11-HLE and anti-
y g
Lo et al., Science 383, 62–70 (2024)
Anti-CTLA-4 nanobodies stimulate antitumor
immunity without inducing colitis
y
The depletion of intratumoral Tregs by antiCTLA-4 antibodies following Fcg receptor (FcgR)
engagement is thought to be one of the mechanisms involved in ICB-induced tumor rejection, although antitumor effects without Treg
depletion in tumor tissue have been observed
in several studies (34–40). To test whether
analogous Fc-dependent processes occur in the
gut, we employed mice that lack the common
FcRg chain which is required for the assembly and function of FcgR family members (41).
FcRg-deficient mice colonized with WildR microbiota were highly resistant to intestinal
inflammation following treatment with antiCTLA-4 antibodies as demonstrated by minimal fecal LCN-2 induction during the course
of ICB treatment and no evidence of immunopathology in cecal tissues compared with anti-
CTLA-4 antibodies in syngeneic tumor models; mice inoculated with tumors were also
colonized with WildR microbiota to permit
simultaneous tracking of ICB-mediated tumor rejection and colitis. First, to examine
anticancer responses in a monotherapeutic
checkpoint blockade setting, we inoculated
mice subcutaneously with MC38 adenocarcinoma cells, and targeted CTLA-4. Compared
with isotype-treated mice in which tumors
grew as expected and reached the experimental endpoint by day 15 of the treatment course,
we found that mice receiving anti-CTLA-4
antibodies displayed reduced tumor sizes by
days 12 and 15 after starting CTLA-4 blockade
(Fig. 5A and fig. S17, A and B). Furthermore,
the same animals receiving anti-CTLA-4 blockade
developed colitis as indicated by fecal LCN-2
induction during the treatment course and
high histological disease scores in intestinal
tissues compared with animals treated with
isotype antibodies (Fig. 5, B and C). Notably,
tumor-inoculated mice receiving H11-HLE
exhibited similar antitumor activity as mice
receiving antibodies against CTLA-4 but by
contrast did not show evidence of colitis (Fig.
5, A to C, and fig. S17, A and B). While assessing MC38 intratumoral T cells—in comparison to the isotype control—the percentages
of Foxp3+ Tregs in CD4+ T cells were reduced in
animals treated with either anti-CTLA-4
antibodies or H11-HLE, though to a lesser extent (fig. S18, A to C). Moreover, treatment
with anti-CTLA-4 antibodies did not alter the
percentages of Helios+ or RORgt+ intratumoral
Tregs compared with isotype controls thus
indicating the tissue-specific effects of antiCTLA-4 antibodies on large intestinal Treg
subsets (fig. S18, D to F). In addition, the frequencies of CD4+ Foxp3− T cells among T cells
were similarly elevated in tumors of mice
treated with anti-CTLA-4 antibodies and H11HLE compared with isotype controls (fig. S18G).
Collectively, these data suggest that Fc domain–
lacking nanobodies that block CTLA-4 can stimulate antitumor responses without inducing
gut irAEs.
Combination CTLA-4 and PD-1 blockade can
stimulate highly potent antitumor immune
responses but also increase the risk and severity of irAEs (3, 4). We therefore hypothesized
that the use of the Fc-lacking H11-HLE CTLA-4
inhibitor with PD-1 blockade can provide similar therapeutic benefits while overcoming
the limitations of its associated gut toxicities.
To test this, we inoculated WildR microbiota
mice with CT26 tumor cells which require
both CTLA-4 and PD-1 blockade for efficient
tumor rejection (42). We found that as tumors
grew in isotype-treated animals, mice treated
with anti-CTLA-4 antibodies and anti-PD-1
antibodies had similarly efficient antitumor
responses compared with mice treated with
H11-HLE and anti-PD-1 antibodies, as evidenced
g
Colitis induced by anti-CTLA-4
antibodies requires depletion of Tregs
by Fcg receptors
CTLA-4 treated WT mice and isotype-treated
FcRg-deficient mice (Fig. 4, A and B). While we
observed an increase in CD4+ T cells expressing IFNg, IL-17A, and both IFNg and IL-17A in
WT mice treated with anti-CTLA-4 antibodies,
the induction of cytokine-producing CD4+ T cells
was not observed in FcRg-deficient mice similarly receiving the anti-CTLA-4 antibodies nor
mice receiving isotype antibodies (Fig. 4, C and
D). Moreover, under anti-CTLA-4 antibody treatment conditions, the percentages of intestinal
Foxp3+ Tregs in WT mice were reduced compared with FcRg-deficient mice and isotypetreated mice with a notable reduction in the
proportion of RORgt+ pTreg (Fig. 4, E and F).
These data suggest that FcRg-dependent
depletion of Tregs is required for gut inflammation in response to treatment with antiCTLA-4 antibodies.
We further assessed the function of the antiCTLA-4 antibody Fc domain during ICB-driven
intestinal inflammation using a humanized
mouse model of CTLA-4 blockade. We challenged a CTLA-4 transgenic mouse strain in
which the extracellular and transmembrane
domains of the mouse Ctla4 sequence were
replaced by the human ortholog (huCTLA-4
KI) with anti-human CTLA-4 antibodies containing a mouse IgG2a Fc domain (ipilimumab
mIgG2a), or anti-human CTLA-4 antibodies
with three amino acid substitutions (L234A,
L235A, P329G) in the Fc domain (ipilimumab
LALAPG) rendering the Fc domain inert to interact with FcgRs (36). We found that huCTLA-4
KI mice colonized with WildR microbiota and
treated with ipilimumab mIgG2a developed
intestinal inflammation as indicated by fecal
LCN-2 levels and assessment of intestinal tissue sections (fig. S16, A and B). However, there
was minimal or no indication of disease in
huCTLA-4 KI mice colonized with WildR microbiota treated with ipilimumab LALAPG,
and in huCTLA-4 KI mice re-colonized with SPF
microbiota treated with ipilimumab mIgG2a
(fig. S16, A and B). Results from this humanized CTLA-4 mouse model further highlight
the requirement of a functional Fc domain for
microbiota-dependent intestinal inflammation
induced by ipilimumab.
p
(Gzma, Gzmb, Prf1) and chemokines (Ccl3,
Ccl4, Ccl5) involved in myeloid cell responses
(Fig. 3, A and B, and fig. S15A). Anti-CTLA-4
treatment resulted in a decrease in the relative abundance of follicular helper T-cells,
naïve CD4+ T cells, Tregs, Tcf7+ CD4+ T cells,
TH22 cells, TH17 cells, and TH2 cells (Fig. 3, A
and B). To better assess the impact of antiCTLA-4 treatment on distinct Treg subsets,
we separated cluster 5 and performed further
clustering and supCPM rendering. We observed three distinct Treg clusters: pTreg (cluster
5-1), tTreg (cluster 5-2), and an intermediate
Treg subcluster defined by elevated Tbet expression (cluster 5-3) (Fig. 3C and table S3).
Consistent with previous findings, anti-CTLA-4
antibody treatment resulted in the selective
depletion of pTregs (42% reduction compared
with isotype-treated mice) whereas tTregs and
Tbet+ Tregs remained intact (Fig. 3D). Although
most cells from TH1 clusters 1 and 3 of the
isotype-treated mice expressed unique TCRs,
CTLA-4 blockade led to a substantial increase
in the frequency of clonally expanded TH1 cells
whereas a major portion of clonally expanded
Tregs were eliminated (Fig. 3E). Further assessment of clonotype abundance in TH1 clusters 1 and 3 revealed that only five clonotypes
with a frequency of 14 or higher were detected,
all of which were selectively expanded by antiCTLA-4 antibody treatment (fig. S15B). Among
the five enriched clonotypes, a highly dominant
clonotype (33% of C3 cells from anti-CTLA-4–
treated mice) was observed in the cytotoxic
PD-1+ TH1 cluster and was 14 times more abundant than the next most frequent clonotype
within the same cluster (fig. S15B). Collectively, these observations indicate that colitic
TH1 cells are normally restricted under homeostatic conditions but can be selectively induced
by anti-CTLA-4 antibodies in the presence of
microbes of the WildR microbiota.
RES EARCH | R E S E A R C H A R T I C L E
Discussion
AC KNOWLED GME NTS
,
8 of 9
y
The authors thank K. Martin and A. Aboud for animal husbandry,
P. Kuffa for laboratory management, T. Duval, L. Keeton, and
L. Kennedy of the University of Michigan (UM) Germ-Free Animal
Core, T. Tamsen and M. Coon of the UM Advanced Genomics Core,
W. Trim and A.M. Deslauriers of the UM Flow Cytometry Core,
S. McClintock of the UM Pathology Flow Cytometry Core, C. Strayhorn
of the UM Dentistry Histology Core, K. Warren of the UM Tissue
and Molecular Pathology Histology Core, and the UM Host
Microbiome Initiative. Funding: This work was supported by NIH
grants R01 DK121504 and R01 DK095782 and a grant from
Takeda/Millennium Pharmaceuticals (to G.N.). B.C.L. was
supported by a Canadian Institutes of Health Research Fellowship.
Additional funding provided by a Crohn’s and Colitis Foundation
Senior Research Award (to R.C.), NIH grants R01 DE026728
and R01 DE030691 (to Y.L.L.), and NSF grant IOS-2107215 (to
Y.X.). Research reported in this publication was supported by the
National Cancer Institutes of Health under Award Number P30
CA046592 by the use of the following Cancer Center Shared
Resources: Single Cell Spatial Analysis, and Tissue and Molecular
Pathology. Author contributions: B.C.L., I.K., J.Y., L.V., R.C.,
and M.M. performed experiments, and analyzed data. Y.S. and M.H.S.
provided advice, discussion, and critical materials. N.I. performed 16s
rRNA analysis, and Y.X. and Y.L.L. performed single-cell immune
profiling analyses. W.Z. designed experiments and provided advice.
B.C.L. and G.N. conceived the project and designed experiments.
G.N. supervised the study. B.C.L and G.N. wrote the manuscript
with input from all authors. Competing interests: Y.S. and M.H.S.
are employees of Takeda Pharmaceuticals International Co.,
Cambridge, MA, U.S.A. The other authors declare that they have no
competing interests. Data and materials availability: C57BL/6
y g
5 January 2024
1. A. D. Waldman, J. M. Fritz, M. J. Lenardo, Nat. Rev. Immunol.
20, 651–668 (2020).
2. W. Zou, J. D. Wolchok, L. Chen, Sci. Transl. Med. 8, 328rv4
(2016).
3. F. Martins et al., Nat. Rev. Clin. Oncol. 16, 563–580
(2019).
4. E. Soularue et al., Gut 67, 2056–2067 (2018).
5. Y. Wang et al., Inflamm. Bowel Dis. 24, 1695–1705 (2018).
6. A. M. Luoma et al., Cell 182, 655–671.e22 (2020).
7. A. V. Heul, T. Stappenbeck, J. Allergy Clin. Immunol. 141, AB119
(2018).
8. Y. Zhou et al., J. Exp. Med. 220, e20221333 (2023).
9. E. Perez-Ruiz et al., Nature 569, 428–432 (2019).
10. F. Wang, Q. Yin, L. Chen, M. M. Davis, Proc. Natl. Acad. Sci. U.S.A.
115, 157–161 (2018).
11. K. Adam, A. Iuga, A. S. Tocheva, A. Mor, PLOS ONE 16,
e0246168 (2021).
12. M. C. Andrews et al., Nat. Med. 27, 1432–1441 (2021).
13. K. Dubin et al., Nat. Commun. 7, 10391 (2016).
14. R. C. Simpson et al., Nat. Med. 28, 2344–2352 (2022).
15. S. P. Rosshart et al., Cell 171, 1015–1028.e13 (2017).
16. L. K. Beura et al., Nature 532, 512–516 (2016).
17. D. Masopust, C. P. Sivula, S. C. Jameson, J. Immunol. 199,
383–388 (2017).
18. S. P. Rosshart et al., Science 365, 6452 (2019).
y
Lo et al., Science 383, 62–70 (2024)
RE FERENCES AND NOTES
19. B. Chassaing et al., PLOS ONE 7, e44328 (2012).
20. E. V. Russler-Germain, S. Rengarajan, C. S. Hsieh, Mucosal Immunol.
10, 1375–1386 (2017).
21. T. Tanoue, K. Atarashi, K. Honda, Nat. Rev. Immunol. 16,
295–309 (2016).
22. K. Atarashi et al., Science 331, 337–341 (2011).
23. G. J. Britton et al., Immunity 50, 212–224.e4 (2019).
24. M. B. Geuking et al., Immunity 34, 794–806 (2011).
25. S. K. Lathrop et al., Nature 478, 250–254 (2011).
26. C. Ohnmacht et al., Science 349, 989–993 (2015).
27. J. L. Round, S. K. Mazmanian, Proc. Natl. Acad. Sci. U.S.A. 107,
12204–12209 (2010).
28. E. Sefik et al., Science 349, 993–997 (2015).
29. M. Xu et al., Nature 554, 373–377 (2018).
30. S. C. Sasson et al., Gastroenterology 161, 1229–1244.e9
(2021).
31. Z. Zhai, Y. L. Lei, R. Wang, Y. Xie, Bioinformatics 38,
2496–2503 (2022).
32. K. R. James et al., Nat. Immunol. 21, 343–353 (2020).
33. A. M. Thornton et al., J. Immunol. 184, 3433–3441
(2010).
34. F. Arce Vargas et al., Cancer Cell 33, 649–663.e4
(2018).
35. D. Bauché et al., J. Immunother. Cancer 8, e001584
(2020).
36. Y. Sato et al., Cancer Immunol. Immunother. 71, 2421–2431
(2022).
37. A. Sharma et al., Clin. Cancer Res. 25, 1233–1238
(2019).
38. Y. Bulliard et al., J. Exp. Med. 210, 1685–1693
(2013).
39. M. J. Selby et al., Cancer Immunol. Res. 1, 32–42
(2013).
40. T. R. Simpson et al., J. Exp. Med. 210, 1695–1710
(2013).
41. T. Takai, M. Li, D. Sylvestre, R. Clynes, J. V. Ravetch, Cell 76,
519–529 (1994).
42. K. Kim et al., Proc. Natl. Acad. Sci. U.S.A. 111, 11774–11779
(2014).
43. W. Wang et al., Nature 569, 270–274 (2019).
44. Y. Wang et al., Nat. Med. 24, 1804–1808 (2018).
45. H. Xu et al., J. Exp. Med. 218, e20210777 (2021).
46. B. Akagbosu et al., Nature 610, 752–760 (2022).
47. R. Kedmi et al., Nature 610, 737–743 (2022).
48. M. Lyu et al., Nature 610, 744–751 (2022).
49. W. Ise et al., Nat. Immunol. 11, 129–135 (2010).
50. K. S. Peggs, S. A. Quezada, C. A. Chambers, A. J. Korman,
J. P. Allison, J. Exp. Med. 206, 1717–1725 (2009).
51. L. F. Mager et al., Science 369, 1481–1489 (2020).
52. T. Tanoue et al., Nature 565, 600–605 (2019).
53. M. Vétizou et al., Science 350, 1079–1084 (2015).
g
The establishment of a robust animal model
of ICB-driven colitis would be beneficial for
gaining mechanistic insights into irAEs observed in the clinic and for improving current
antitumor ICB therapies with reduced toxicities. In this study, we demonstrate the critical role of the microbiota in colitis induced by
ICB and reveal key contributions of CD4+ TH1
cells and IFNg in the pathogenesis of disease
which is consistent with previously reported
features of the human disease (4, 6) (fig. S20).
Several strategies have been proposed for suppressing colitis without altering the beneficial
antitumor effects of ICB including neutralization of TNF or IL-6 (8, 9). Moreover, the
modification of the gut microbiota by fecal
microbiota transplantation can mitigate intestinal inflammation induced by ICB (44).
Here, we report that CTLA-4 inhibitors that
lack an Fc domain do not induce colitis in mice
which are otherwise susceptible to colitis when
receiving conventional anti-CTLA-4 antibodies.
Importantly, anti-CTLA-4 VHHs can effectively
stimulate antitumor responses in mice without
causing gut irAEs, thus circumventing the necessity for the neutralization of various proinflammatory cytokines or other strategies to
modify the microbiota to remedy intestinal
inflammation. These results also indicate that
many immunological redundancies exist in
the gut to prevent inappropriate T cell activation by the gut microbiota and that although
the microbiota-induced Tregs are present,
CTLA-4 blockade by itself is not sufficient to
induce colitis.
In several genetic models, direct perturbation of gut pTregs renders mice more susceptible
to intestinal immunopathologies (26, 28, 29, 45).
Furthermore, mice lacking a population of
tolerogenic RORgt+ antigen-presenting cells,
which are required for the induction of bacteriareactive pTregs, appear to develop intestinal
inflammation associated with exaggerated proinflammatory T cell responses (46–48). However, CTLA-4 can also restrain T cell activation
independently of Tregs in autoimmunity and
antitumor responses (49, 50). Therefore, further studies are needed to understand the
potential cumulative effects of T cell CTLA-4
blockade and anti-CTLA-4 antibody-mediated
depletion of Tregs during colitis.
We find that a CTLA-4 inhibitor lacking an
Fc domain had similar antitumor immunitypromoting effects as conventional anti-CTLA-4
antibodies in mice harboring WildR microbiota. However, some—but not all—reports indicate that anti-CTLA-4 antibodies containing
an Fc domain with high binding affinity to Fc
receptors and functional Fc receptor activity
is required for optimal tumor clearance through
the selective depletion of intratumoral Tregs
(34–40). This discrepancy may be explained,
at least in part, by the dissimilarities in gut
microbiota composition described in these
animal models, as select gut bacterial strains
or microbial communities have a profound impact on extra-intestinal immunity and responsiveness to CTLA-4 blockade for anticancer
therapy (51–53). Given that the gut microbiota
of free-living mice is highly immunogenic, the
threshold of immune tolerance disruption required for activating optimal antitumor responses may be lower in WildR mice. Moreover,
as recent data indicate that experimental mouse
models that use the microbiota of free-living
animals more accurately recapitulate human
responses to immune-modulating therapies
(18), our work provides evidence for the potential utility of an Fc-null anti-CTLA-4 antibody,
either alone or in combination with PD-1 or
PD-L1 blockade, in effectively stimulating anticancer immune responses without inducing
intestinal inflammation.
p
by reduction in tumor sizes during the treatment course compared with isotype-treated
controls (Fig. 5D and fig. S17, C and D). Moreover, only mice receiving both anti-CTLA-4 and
anti-PD-1 antibodies developed colitis whereas
isotype control mice or mice receiving H11-HLE
and anti-PD-1 antibodies did not display evidence of intestinal inflammation (Fig. 5, E and
F). We further evaluated the effects of H11HLE nanobodies and conventional anti-CTLA-4
antibodies, in combination with PD-L1 blockade,
in a syngeneic B16F0 melanoma implantation
model (43). Treatment with H11-HLE and antiPD-L1 antibodies was similarly effective as antiCTLA-4 antibodies and anti-PD-L1 antibodies
in limiting subcutaneous B16F0 tumor growth
but did not induce overt intestinal inflammation (fig. S19, A to E). These results indicate the
potential benefits of an Fc-lacking CTLA-4
inhibitor in combination with PD-1 or PD-L1
blockade without incurring the complications
of colitis.
RES EARCH | R E S E A R C H A R T I C L E
mice harboring WildR microbiota were obtained from Taconic
Biosciences via a material transfer agreement. Fecal microbiota
16s sequencing data and single CD4+ T cell RNA sequencing data
are available at the NCBI SRA under BioProject number
PRJNA944829. License information: Copyright © 2024 the
authors, some rights reserved; exclusive licensee American
Association for the Advancement of Science. No claim to original
US government works. https://www.sciencemag.org/about/
science-licenses-journal-article-reuse
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adh8342
Materials and Methods
Figs. S1 to S20
Tables S1 to S3
References (54–60)
Submitted 16 March 2023; resubmitted 8 September 2023
Accepted 17 November 2023
10.1126/science.adh8342
p
g
y
y g
y
,
Lo et al., Science 383, 62–70 (2024)
5 January 2024
9 of 9
RES EARCH
POLYMERS
Thermodynamically stable plumber's nightmare
structures in block copolymers
Hojun Lee1, Sangwoo Kwon2, Jaemin Min1, Seon-Mi Jin3, Jun Ho Hwang3, Eunji Lee3,
Won Bo Lee2, Moon Jeong Park1*
Block copolymer self-assembly affords diverse nanostructures, spanning from spheres and cylinders to
networks, offering meticulous control over properties and functionalities at the nanoscale. However,
creating thermodynamically stable network structures with high packing frustration remains a challenge.
In this study, we report a methodology to access diverse network structures such as gyroid, diamond,
and primitive phases from diblock copolymers using end group and linker chemistry. The stability of the
medial packing of polymer chain ends (plumber's nightmare structure) over skeletal aggregation (gyroid)
is attributed to the interplay between the strength of the end-end interactions and the initial shape
of the curvature. Our study establishes an approach to develop tailored network structures from block
copolymers, providing an important platform for using block copolymers in nanotechnology applications.
1 of 7
,
5 January 2024
y
Lee et al., Science 383, 70–76 (2024)
Several experimental and theoretical studies
on diblock copolymers have identified the Ia3d
structure as the most stable network phase,
y g
*Corresponding author. Email: moonpark@postech.ac.kr
Emergence of plumber’s nightmare
for diblock copolymers
y
Department of Chemistry, Pohang University of Science and
Technology (POSTECH), Pohang 37673, Republic of Korea.
School of Chemical and Biological Engineering, Institute of
Chemical Processes, Seoul National University, Seoul 08826,
Republic of Korea. 3School of Materials Science and
Engineering, Gwangju Institute of Science and Technology
(GIST), Gwangju 61005, Republic of Korea.
2
g
1
Beyond the Ia
3d structure, block copolymers have also been theoretically predicted to
andIm3m
space
form cubic lattices withPn3m
groups that have Schwarz diamond (D) and
primitive (P) surfaces, respectively (28). However, these structures have a higher packing
structure because
frustration than the Ia3d
excessive chain stretching is required to access
the constant-mean-curvature structure, thereby substantially limiting their feasibility for
experimental observations (29). In particular,
diblock copolymers have difficulty filling the
space to form minimal P surfaces with six intersecting tubes compared with gyroid (three
tubes) and diamond (four tubes) structures (30).
To explore ways for diversifying the type of accessible network structures from block copolymers, extensive experimental studies (31, 32)
have been conducted. However, all observed
double primitive structures were not in thermodynamic equilibrium.
One current challenge is the creation of thermodynamically stable network structures with
high packing frustration. A recent theoretical
work by Reddy et al. (33) may provide important insights in this regard. According to them,
manipulating the stability of the network structures by controlling the packing of polymer
chain ends could be a prospective method to
realize structures with high pacing frustration
(34). However, there is no known method yet
to control the arrangement and localization of
the polymer chain ends.
Fddd,
In this study, the stabilization of Ia3d,
and Im3m structures was realized using diend-functionalized block copolymers with different linkages. A polystyrene-b-poly(ethylene
oxide) (PS-b-PEO, SEO) diblock copolymer with
a nominally symmetric molecular weight of
7.5-b-7.4 kg mol–1 and a PEO volume fraction
(fPEO) of 0.48 was synthesized. As briefly shown
in Fig. 1A, the hydroxyl terminal of SEO was
first converted to a diol or primary amine group
to form an intermediate, followed by a Michael
p
S
tarting with the theoretical prediction
of the phase behavior of diblock copolymers by Leibler (1), experimental efforts
have established phase diagrams of block
copolymers based on the block volume
fraction (f), degree of polymerization (N), and
Flory-Huggins interaction parameter (c) between two blocks (2, 3). Recent advances in
polymer synthesis have further allowed the exploration of the effects of the molecular architecture (4, 5), interfacial molecular composition
(6, 7), and conformational asymmetry (8–11) on
the self-assembly behavior of block copolymers.
Among the various self-assembled structures
formed by block copolymers, network structures characterized by triply periodic minimal
surfaces are particularly intriguing (12) owing
to their improved mechanical (13, 14) and transport properties (15–17) across inherently bicontinuous nanodomains. Through the controlled
domain sizes of the network structures with
integrated functional moieties, block copolymers can serve as advanced nanomaterials
in soft electronics (18, 19) and energy storage
systems (20, 21). In this context, the structure )
property relationship of the gyroid ( Ia3d
structure (minimal G surface) has been of particular interest (14, 15). However, the gyroid
structure is stable only over a narrow window
of the phase diagram of block copolymers, and
its feasibility relies on tedious composition
screening (22). The manipulation of the polymer chain conformation at the interfacial region (23–26) and compositional nonuniformity
at the interface (27) have been shown to play a
structures.
crucial role in stabilizing the Ia3d
reaction with diethyl vinylphosphonate (dPE).
After the ethyl group was removed by hydrolysis, two types of SEOs with diphosphonic acid
(dPA) end groups were prepared, in which the
termini were connected to PEO by different
linker molecules. Hereafter, the dPA-ended SEOs
are referred to as SEO-N-dPA (amine linker)
and SEO-O-dPA (ether linker). The detailed synthetic procedures and data are provided in figs.
S1 and S2. The end group conversion rates were
higher than 95% and the molecular weight distribution of SEO was unaffected by the terminal modification reactions (fig. S3).
Figure 1B shows representative small-angle
x-ray scattering (SAXS) profiles and transmission electron microscopy (TEM) images of pristine SEO, SEO-amine, SEO-N-dPA, SEO-diol, and
SEO-O-dPA. Prior to the measurements, the
freeze-dried SEOs were annealed at 130°C for
one week and cooled down to 25°C. Pristine
SEO exhibited a lamellar (lam) structure with
a domain spacing (d10) of 15.9 nm. SEO-amine
and SEO-diol also exhibited analogous lam
structures with a d10 of 16.4 nm. Notably, the
inclusion of dPA end groups in the SEO intermediates led to the evolution of peculiar network morphologies. For SEO-N-dPA, we saw a
mixture of orthorhombic networks (Fddd, hkl
(hkl index in black)
index in blue) and Ia3d
structures, with Fddd being dominant. The
unit cell parameters of Fddd were (a:b:c) =
(1:1.97:3.41), with a = 20.0 nm. Bright-field
TEM micrographs were recorded in each area
were observed, as shown
where Fddd andIa3d
in the bottom inset images of Fig. 1B.
Notably, upon changing only one linker molecule from amine to ether, SEO-O-dPA exhib structure with a d110 of 31.4 nm
ited an Im3m
(hkl index is marked in the scattering profile
of Fig. 1B). The top insets of Fig. 1B show the
tomography images and a captured TEM im
age for the direct visualization of the Im3m
structure in SEO-O-dPA. These clearly reveal
the primitive cubic morphology in the [100]
projection with the interconnected PS domains
(bright phases). Unprecedentedly, diverse network structures were observed from a single
SEO by functionalizing the end group, which
accounts for less than 1% of the total number
of repeating units.
Electron energy loss spectroscopy mapping in
TEM was performed to elucidate the type of ele
ments constituting each domain of the Im3m
structure. The bright-field TEM micrograph and
oxygen mapping image of SEO-O-dPA in the
structure are shown
[111] projection of the Im3m
in fig. S4, revealing that the PEO chains form
minor phases (shown dark by RuO4 staining).
RES EARCH | R E S E A R C H A R T I C L E
A
SEO-N-dPA
Michael reaction
SEO
hydrolysis
(211)
B
SEO-O-dPA
SEO-O-dPA
4
100 nm
2
SEO-diol
(315)
(242)
(062)
g
1
10
50 nm
SEO-O-dPA
(131)
(400)
(202)
(422)
(113)
(220)
10
p
Scattering Intensity (a.u.)
Im3m
(310)
(222)
(400)
(330)(411)
(420)
(332)
(422)
(431)
(521)
(440)
(611)(532)
(541)
(631)
(710)(550)
3
10
(110)
(200)
(211), (111)
(220)
10
Fig. 1. Stabilization
of diverse network
structures using di-end–
functionalized block
copolymers. (A) Synthetic
routes for the preparation
of polystyrene-b-poly(ethylene
oxide) (SEO) having diphosphonic acid (dPA) end
groups. (B) SAXS profiles of
end-functionalized SEOs
measured at 25°C. hkl indices
are indicated in parenthesis
(blue, Fddd; black, Ia3d).
q, scattering vector. a.u.,
arbitrary units. (Bottom
insets) Bright-field TEM
images of SEO-N-dPA. (Top
insets) TEM tomography
images and a captured TEM
image are shown for the
visualization of the Im3m
structure in SEO-O-dPA.
Dotted SAXS profiles show
the morphologies of SEO
intermediates, SEO-diol,
and SEO-amine.
SEO-N-dPA
SEO-N-dPA
y
0
10
SEO-amine
SEO
-1
10
0.2
0.4
0.6
0.8
1.0
q (nm-1)
Fddd
(40). The P surface can be bent to form the G
and D surfaces, complying with the aD:aG:aP =
1:1.58:1.28 ratio (41). This was not the case with
our experiments, in which the Im3m-to-Ia
3d
transitions occurred without changes
-to-Im3m
in the domain size. Thus, a transition mech and Ia3d
structures
anism between the Im3m
through the merging and splitting of nodes
was proposed. As schematically depicted in the
upper inset of Fig. 2A, when viewed along the
and
[111] direction, the (211) planes of Im3m
were parallel to the center of the nodes.
Ia3d
two enantiomeric nodes
Upon cooling Ia3d,
with three intersecting channels merged
through PEO crystallization, resulting in a
These
new node with six channels of Im3m.
are illustrated on the right side of each scattering profile.
structure of SEO-O-dPA was viThe Im3m
sualized by electron density reconstruction
using MATLAB and SUPERFLIP software with
structure factors extracted from the SAXS profile in Fig. 2A (42). Subsequently, we performed
Le Bail refinement using JANA2006 (43). On
2 of 7
,
5 January 2024
SEO-O-dPA. As shown by the SAXS profiles in
Fig. 2A, heating the sample from 25° to 50°C
caused the destabilization of Im3m,
as evidenced by the disappearance of the (110) peak.
phase transition,
This led to an Im3m-to-Ia
3d
and the Ia
3d structure remained intact up to
150°C (fig. S5). When the sample with the Ia3d
structure was cooled from 110° to 25°C, an immediate reemergence of the (110) peak of
structure was observed along with
the Im3m
Upon monitoring the
the coexistence of Ia3d.
changes in morphology of the cooled sample
at room temperature, the slow reenactment
from Ia3d
through the coinciding
of Im3m
d211 of two cubic structures was observed. This
transition is
indicates that the Im
3m-to-Ia3d
structure
thermally reversible, and the Im3m
is in equilibrium at room temperature. The SAXS
results obtained for unannealed SEO-O-dPA
during in situ temperature changes are shown
in fig. S6. The results for the annealed sample
during sample rotation are presented in fig. S7.
Minimal D, G, and P surfaces are related
to each other by the Bonnet transformation
y
Lee et al., Science 383, 70–76 (2024)
Ia3d
y g
which exists in the narrow phase window between the hex and lam structures (22). The
structure, called plumber’s nightmare,
Im3m
was identified in block copolymer–templated
mesoporous silica by Finnefrock et al. (35) two
decades ago; however, evidence of primitive
cubic structures in equilibrium does not exist
for any neat block copolymers. In fact, computational studies based on Gaussian statistics have reported that excessive stretching of
the polymer chains in double diamonds and
double primitives is not preferred over gyroids
owing to the entropic penalty (36, 37). For block
copolymer–homopolymer blends, the possibility of the existence of a thermodynamically
stable plumber’s nightmare structure was theoretically predicted (28, 38, 39). However, it is
only applicable over a finite region of the phase
diagrams when the requirements for the composition or conformational asymmetry of the
block chains and the architecture or length of
the homopolymer are met.
This prompted us to investigate the ther structure of
mal stability window of the Im3m
50 nm
50 nm
1.2
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
Lee et al., Science 383, 70–76 (2024)
5 January 2024
sity isosurface and charge-density contour maps
structure of SEO-O-dPA along the
of the Im3m
[100] and [111] directions, confirming the characteristic structural patterns of the plumber’s
nightmare. The lower panels of Fig. 2C display
bright-field TEM images and the related inverse Fast Fourier transformed (FFT) images
from the [111], [211], and [531] projections. Additional sets of TEM tomography images are
shown in fig. S10 and movie S1.
A close correlation between the hexapod
stability and crystallization of dPA-ended PEO
chains was envisaged for SEO-O-dPA. Essen is favored over Ia3d
at low temtially, Im3m
peratures when the dPA end groups are well
arranged at the center of the PEO domains
of SEO-O-dPA. As the temperature increases,
the PEO crystals melt, and the increased PEO
chain mobility disorganizes the end-end arrangement. This results in the splitting of the
bulky hexapod into smaller tripods in the direction of reducing packing frustration. This
was investigated in more detail by simultaneous SAXS and wide-angle x-ray scattering
3 of 7
,
the basis of the goodness-of-fit parameter and
the weighted profile R-factor, we affirmed the
reliability of our structure determination. The
representative Le Bail refinement result in comparison with the experimental SAXS profile is
presented in fig. S8. Rigorous structural validation through the simSAXSLee software package built on the MATLAB platform (fig. S9)
demonstrates the close correlation between
the measured scattering intensity of the hkl
reflection (Ihkl) and calculated Ihkl values (44).
Figure 2C displays the resultant electron den-
dPA-ended PEO arrangements in hexapod (Im3m) and tripod (Ia3d). (C) Electron
density isosurface and charge-density contour maps of the Im3m structure of
SEO-O-dPA in the [100] and [111] directions and the bright field TEM and related
inverse FFT images from the [111], [211], and [531] projections.
y
structures in SEO-O-dPA. (A) SAXS
Fig. 2. A deeper look into the Im3m
profiles of SEO-O-dPA upon heating and cooling. The epitaxial relationship
between the Ia
3d and Im
3m structures and changes in the nodes owing to the
structural transition are schematically illustrated in the insets. (B) Proposed
RES EARCH | R E S E A R C H A R T I C L E
A
mPEO
mPEO-O-dPA
mPEO-N-dPA
-N-dPA:
N-dPA: Zwitterionic
Absorbance
-O-dPA: H-bond
1150
1100
1050
2700 2600 2500 2400
1000
Wavenumber (cm-1)
B
lam
mPEO
twisted lam
mPEO-O-dPA
D
helical
@ 25 oC
mPEO-N-dPA
p
Fig. 3. Factors affecting the
stability of the network
structures of end-functionalized
SEOs. (A) FT-IR spectra,
end-end interactions, and (B) 2D
scattering patterns of mPEO,
mPEO-O-dPA, and mPEO-N-dPA.
(C) SAXS profiles of di-end–
functionalized SEOs (14.5-b-9.5 kg
mol–1) measured at 25°C.
Schemes depicting the medial
versus skeletal chain packing of
PEO are shown in the top right.
(D) Phase diagram of SEOs
with different types of di–end
groups and linker molecules. The
strength of end-end interaction
and the degree of packing frustration were included as variables. (E) Projected phase
diagram of dPA-ended SEOs in a
low-cN regime. Conventional
phase diagram is indicated with
gray dotted lines for
comparison.
C
g
y
Scattering Intensity (a.u.)
vs.
SEO-O-dPE
E
L
L
SEO-O-dPA
0.2
0.4
0.6
0.8
1.0
1.2
N
0.2
-1
q (nm )
analyses with isothermal crystallization of
SEO-O-dPA (fig. S11).
For SEO-N-dPA with the amine linkage, in
the temperature range 25° to 170°C, coexisting
were always observed
phases of Fddd and Ia3d
over Fddd in(fig. S12). The fraction of Ia3d
creased with increasing temperature, and the
Ia
3d -to-Fddd transition occurred reversibly
with cooling, which was relatively rapid comLee et al., Science 383, 70–76 (2024)
5 January 2024
transition. The
pared with the Ia3d-to-Im
3m
results of molecular dynamics (MD) simulations
based on the MARTINI approach are shown
in fig. S13 (45). Contrary to the flat interface
observed for pristine SEO, percolated structures appeared for SEOs with dPA end groups.
Further, despite the small number fraction (<1%)
of dPA terminals relative to the overall chain
length of SEOs, a considerable increase of more
0.3
0.4
,
Ia3d
SEO
y
nteracti
end-end interactions
SEO-N-dPE
SEO-N-dPA
y g
lam
hex
0.5
fPEO
than 20% in the interfacial areas was evident
for the dPA-ended SEOs, leading to the disruption of interfaces.
Mechanisms underlying the emergence of
diverse network structures in dPA-ended SEOs
The effect of the end-end interactions on the
conformation of the PEO chain was examined
by synthesizing a set of w-functionalized poly
4 of 7
RES EARCH | R E S E A R C H A R T I C L E
y g
y
,
5 of 7
y
5 January 2024
the strengthened end-end interactions, resulting in the appearance of hex structures; this
tendency is opposite to that of the symmetric
SEOs. We propose that the medial versus skeletal chain packing of PEO is determined by the
interplay between the strength of the end-end
interactions and the initial shape of the curvature. To validate this supposition, another set
of SEOs with dinitrile (dCN) ends was prepared,
referred to as SEO-O-dCN and SEO-N-dCN. In
these samples, the −C≡N⋅⋅⋅C≡N dipole-dipole
interaction was considered; its strength was
predicted to be weaker than dPA and slightly
stronger than dPE because of the formation of
dCN dimers. As summarized in table S1, the diend moieties exhibited substantially increased
binding energies compared with their monoend counterparts owing to the doubled interaction sites. Overall, the di–end groups connected
by amine linkages exhibited 10 to 30% higher
binding energies than their ether-linked counterparts, with the values increasing in the order
dPA, dCN, and dPE. The highest binding energy of 123.0 kJ mol–1 was predicted for -N-dPA,
whereas the lowest value of 21.0 kJ mol–1 was
obtained from -O-dPE.
With symmetric SEO (7.5-b-7.4 kg mol–1),
both SEO-O-dCN and SEO-N-dCN showed lam
structures (fig. S20A), which were analogous
to the dPE-ended SEOs. Meanwhile, with asymmetric SEO (14.5-b-9.5 kg mol–1), the morphologies differed, with SEO-O-dCN and SEO-N-dCN
and hex structures, respecexhibiting Im3m
tively (fig. S20B). When the PEO composition
was minor and the end-end interaction was
structures emerged in a
modest, the Im3m
more diverse set of samples, namely SEO-OdCN, SEO-O-dPE, and SEO-N-dPE. When the
end-end interactions strengthened inside the
minor PEO domains, the PEO chains no longer
exhibited medial packing and the curvature was
preserved. By contrast, for SEOs with symmetric
compositions, it was more difficult for the PEO
chains forming a flat interface to reach the
constant mean curvature of the P surface, as
excessive chain stretching and high concentration fluctuation were required. This was
therefore achieved only for SEO-O-dPA. The
morphologies of all SEOs end-functionalized
with dPE, dPA, and dCN that use ether (red)
and amine (blue) linkages are summarized in
Fig. 3D.
The morphology of dPA-ended SEOs in the
low-cN regime was further investigated. For
this, an SEO with a low molecular weight of
3.2-b-3.5 kg mol–1 was used as a precursor, which
exhibited a disordered morphology (fig. S21).
Modifying the –OH terminal of SEO with dPE
did not lead to the disorder-to-order transition.
However, hydrolyzing them into SEO-N-dPA
and SEO-O-dPA resulted in the formation of
structures, respectively. We believe
hex andIa3d
structure of SEOs
that the stability of the Im3m
increased in the high-cN regime by enabling
g
Lee et al., Science 383, 70–76 (2024)
The self-assembled structures of diblock copolymers can be radically altered by fine-tuning
the end-end interactions through ingenious
linker chemistry. Next, the effects of the shapes
of the initial block interfaces on the stabilization of the negative Gaussian curvature of
di-end–functionalized SEOs was investigated.
For this, an SEO with a molecular weight of
14.5-b-9.5 kg mol–1 (fPEO = 0.38) was synthesized, representing a hex structure in the temperature range 25° to 170°C. The hydroxyl
terminal of SEO was modified with dPE moieties following the same synthetic routes to
yield SEO-N-dPE and SEO-O-dPE. Notably, a
morphological transition from hex to Im3m
structures was observed in the prehydrolysis
form (Fig. 3C). This is in sharp contrast to the
results obtained from the dPE-ended SEO (7.5b-7.4 kg mol−1) in which the lam structures
were preserved (fig. S18). Therefore, we envisioned that facile access of the hexapod
intersection for the SEO with initially curved
interfaces would be possible by introducing
relatively weak dPE-dPE interactions at the
end of the PEO chains.
was better
The scattering profile of Im3m
resolved for SEO-O-dPE than for SEO-N-dPE
because the ether linkage also worked in favor
of relieving the high packing frustration. The
dPE-ended SEOs also exhibited a thermally
reversible structural transition from Im
3m to
when the temperature was raised above
Ia3d
the melting temperature of PEO (fig. S19). No to Im3m
during
tably, the transition from Ia3d
cooling was substantially faster for asymmetric
SEO (14.5-b-9.5 kg mol–1; within tens of minutes) than for symmetric SEO (7.5-b-7.4 kg mol–1;
several months for the transition to complete).
This indicates that the low mobility of the glassy
PS chains had a negligible impact on the kinetics of the structural transition. Our proposition is that the PEO crystallization kinetics
played a crucial role in determining the rate of
this transition, primarily owing to the anticipated
substantial increase in local modulus with the
merging of enantiomeric PEO intersections
through PEO crystallization. This increase in
local modulus is expected to be substantial
enough to displace the amorphous PS chains.
However, despite this change in local modulus,
directional shear forces are unlikely to be generated, as PEO crystallization occurred within
the three-dimensionally symmetric channel
structures. Consequently, we do not anticipate
gyroid distortion to occur with cooling. This
supposition is supported by the fact that the
isotropic 2D scattering patterns remained consistent, and that there was no noticeable alteration in the 1D scattering profiles when the
-forming samples were rotated during
Im3m
the measurements (fig. S7).
Upon hydrolyzing the polymers into the
acid forms, SEO-N-dPA and SEO-O-dPA, the
structures were destabilized owing to
Im3m
p
(ethylene glycol) methyl ether homopolymers
(mPEO-O-dPA and mPEO-N-dPA, 2 kg mol–1).
Prior to taking the measurements, all mPEOs
were heated to 90°C for 30 min and cooled
down to 25°C at a rate of 1°C min−1. As shown in
Fig. 3A, the Fourier-transform infrared (FT-IR)
spectra exhibited negligible –PO3H2···EO backbone interactions. Meanwhile, the enhanced
vibration of the P–OH bond of mPEO-O-dPA
indicated −PO3H2⋅⋅⋅PO3H2 hydrogen bonding
interactions. The terminal moieties of mPEON-dPA adopted the zwitterionic form, as determined from n(P–O−) at 1033 cm–1 and n(N+–H)
at 2677 and 2491 cm–1. This strengthened the
dPA-dPA interactions, as confirmed by density
functional theory (DFT) calculations (table
S1) (46). The dissimilar end-end interactions of
-O-dPA and -N-dPA are schematically shown in
the right panels of Fig. 3A.
These different end-end interactions altered
the PEO chain structures. At 30°C, contrary to
the well-defined lam structure of pristine mPEO,
two types of coexisting crystals were observed
for mPEO-N-dPA and mPEO-O-dPA, accompanied by an increase in the interlamellar thickness (L) by 10 to 20% (fig. S14). Kratky analysis
revealed that this increase was primarily due
to the widening of the amorphous region, with
Lcrystalline and Lamorphous values of 9.9 and 2.9 nm
for mPEO, 8.8 and 5.1 nm for mPEO-O-dPA,
and 9.5 and 6.2 nm for mPEO-N-dPA, respectively. This suggests that the inclusion of dPA
end groups led to the stretching of PEO chains
within the amorphous regions. Notably, although the PEO crystallinity decreased in all
dPA-ended samples, this reduction was more
pronounced in mPEO-O-dPA and SEO-O-dPA
compared with the samples with amine linkage (fig. S15). This is in good agreement with
the more substantial reduction in Lcrystalline for
mPEO-O-dPA in comparison with that for mPEON-dPA, suggesting that the dense localization
of end groups in mPEO-N-dPA is facilitated
by electrostatic interactions. At 50°C, twisted
lam structures were observed for mPEO-OdPA with different d10 values of 13.4 and 10.6 nm.
This is expected to be closely related to the
of
relieved packing frustration of the Im3m
SEO-O-dPA, as more stretched PEO chains
could approach the medial limit. For mPEON-dPA, the coulomb end-end interactions
resulted in the development of a hex lattice
through the arrangement of the PEO chain
in helical form. The two-dimensional (2D)
scattering patterns of the three samples are
compared in Fig. 3B. The chain-chain correlation of mPEO-O-dPA and mPEO-N-dPA
remained intact even at a high temperature
of 110°C (fig. S16) owing to the robust end-end
interactions over a wide temperature range.
This also improved the viscoelastic properties of
the dPA-ended SEOs, with an increase of more
than 10 times in the plateau moduli compared
with that of the pristine SEO (fig. S17).
RES EARCH | R E S E A R C H A R T I C L E
p
Fig. 4. Unveiling the
universal impact of end
groups and linker molecules on stabilizing
network structures in
diblock copolymers. SAXS
profiles of (A) SMB-O-dPA
(Pm3n), SMB-O-dCN (Ia3d),
neat SMB (lam), (B) SMB-NdPA (Pn3m), and SMB-NdCN (lam), all measured
at 25°C. All morphologies
were thermally stable in
temperature range of 25° to
170°C. (C) (Left) Chemical
structures and proposed
end group arrangements
within the tetrapod of
dPA-ended PMB domains.
(Right) Phase diagram
of SMBs with controlled
end-end interactions and
packing frustration.
g
y
y g
With the objective of demonstrating the overarching role of end groups and linker molecules
in achieving meticulous control over packing
frustration for various types of block copolymers, we synthesized additional sets of block
copolymers, polystyrene-b-polymethylbutylene
(PS-b-PMB, SMB), with symmetric (7-b-7 kg
Lee et al., Science 383, 70–76 (2024)
5 January 2024
asymmetric SEO-O-dCN (Im3m)
and SEO-NdCN (preserved hex structures), which can be
attributed to relatively modest end-end interactions. For SMB-O-dPA and SMB-N-dPA, which
comprised substantially stronger end-end
and Pn3m
structures
interactions, Pm3n
were achieved. This highlights the systematic
modulation of the packing frustration across
diverse network structures from SMB-O-dCN
to SMB-N-dPA, and further to SMB-O-dPA,
through end group and linker chemistry. Notably, unlike their SEO counterparts, the network structures of dPA-ended SMBs displayed
thermal stability in a temperature range of 25°
to 170°C owing to the absence of a melting
transition. Figure 4C presents the morphologies of end-functionalized SMBs, determined by
the strength of end-end interactions and the
degree of packing frustration. The structures
obtained with amine linkage are represented by
6 of 7
,
End group and linker chemistry: a versatile
approach for stabilizing network structures in
block copolymers
mol−1) and asymmetric (9-b-5 kg mol−1) compositions. Both SMBs displayed well-defined lam
structures with d10 values of 16.6 and 17.5 nm,
respectively (fig. S22). SMBs are hydrophobic
and amorphous block copolymers in stark
contrast to the amphiphilic semicrystalline
SEOs. This makes them ideal candidates for
validating the versatility of end group and
linker chemistry in stabilizing network structures in diblock copolymers. To this end, di-end
functional groups were introduced by varying
linker molecules to yield SMB-O-dCN, SMB-NdCN, SMB-O-dPE, SMB-N-dPE, SMB-O-dPA,
and SMB-N-dPA, analogous to the SEOs.
The SAXS profiles of the di-end–functionalized SMBs (9-b-5 kg mol−1) are shown in Fig.
4A (ether linker) and Fig. 4B (amine linker). In
structure
SMB-O-dCN, an equilibrium Ia3d
emerged, whereas the lam structure persisted
in SMB-N-dCN, mirroring the observations for
y
the organized packing of end groups through
PEO chain stretching. The fact that SEO-NdPA showed the hex structure rather than the
lam structure in the vicinity of a critical cN
value and had an fPEO of ~0.5 is also intriguing,
suggesting that the augmented end-end interactions due to the decreased molecular weight
shifted the phase boundary considerably, as
illustrated in Fig. 3E.
RES EARCH | R E S E A R C H A R T I C L E
AC KNOWLED GME NTS
science.org/doi/10.1126/science.adh0483
Materials and Methods
Figs. S1 to S25
Tables S1 and S2
References (50–59)
Movies S1 to S3
Submitted 7 February 2023; resubmitted 12 September 2023
Accepted 8 November 2023
10.1126/science.adh0483
7 of 7
,
SUPPLEMENTARY MATERIALS
y
We express our sincere gratitude to B. Lee for his invaluable
assistance in the structural analysis and validation process.
Funding: This work was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(nos. NRF-2022R1A2C3004667, NRF-2017R1A5A1015365, and
NRF-2018M3D1A1058624). M.J.P. also acknowledges financial
support from the Korea Toray Science Foundation. E.L.
acknowledges financial support from NRF grant no. NRF2022R1A2C2012889. Author contributions: M.J.P. conceived the
idea and designed the project, and H.L. carried out the
experiments and analyzed data. H.L. and J.M. carried out the
structural analysis, and S.K and W.B.L. conducted MD simulations.
S.-M.J., J.H.H, and E.L performed TEM experiments. M.J.P. and H.L.
wrote the paper. Competing interests: The authors declare that
they have no competing interests. Data and materials
availability: All data are available in the main text or the
supplementary materials. License information: Copyright © 2024
the authors, some rights reserved; exclusive licensee American
Association for the Advancement of Science. No claim to original
US government works. https://www.science.org/about/sciencelicenses-journal-article-reuse
y g
1. L. Leibler, Macromolecules 13, 1602–1617 (1980).
2. F. S. Bates, G. H. Fredrickson, Annu. Rev. Phys. Chem. 41,
525–557 (1990).
3. F. S. Bates, Science 251, 898–905 (1991).
4. F. S. Bates et al., Science 336, 434–440 (2012).
5. V. Percec et al., Nature 391, 161–164 (1998).
6. T. Hashimoto, Y. Tsukahara, K. Tachi, H. Kawai, Macromolecules
16, 648–657 (1983).
7. O. A. Scherman, G. B. W. L. Ligthart, H. Ohkawa, R. P. Sijbesma,
E. W. Meijer, Proc. Natl. Acad. Sci. U.S.A. 103, 11850–11855 (2006).
8. M. W. Matsen, F. S. Bates, J. Polym. Sci., B, Polym. Phys. 35,
945–952 (1997).
9. S. Lee, M. J. Bluemle, F. S. Bates, Science 330, 349–353 (2010).
10. K. Kim et al., Science 356, 520–523 (2017).
11. M. W. Bates et al., Proc. Natl. Acad. Sci. U.S.A. 116,
13194–13199 (2019).
12. D. A. Hajduk et al., Macromolecules 27, 4063–4075 (1994).
13. B. J. Dair et al., Macromolecules 32, 8145–8152 (1999).
14. H. Pernot, M. Baumert, F. Court, L. Leibler, Nat. Mater. 1, 54–58
(2002).
15. B. K. Cho, A. Jain, S. M. Gruner, U. Wiesner, Science 305,
1598–1601 (2004).
16. G. Jo, H. Ahn, M. J. Park, ACS Macro Lett. 2, 990–995 (2013).
17. L. Yan, C. Rank, S. Mecking, K. I. Winey, J. Am. Chem. Soc. 142,
857–866 (2020).
18. S. W. Robbins et al., Sci. Adv. 2, e1501119 (2016).
19. J. Min, D. Barpuzary, H. Ham, G. C. Kang, M. J. Park, Acc. Chem.
Res. 54, 4024–4035 (2021).
20. S. Choudhury et al., ACS Nano 9, 6147–6157 (2015).
y
5 January 2024
RE FERENCES AND NOTES
21. C. F. Cheng et al., J. Mater. Chem. A Mater. Energy Sustain. 6,
13676–13684 (2018).
22. E. W. Cochran, C. J. Garcia-Cervera, G. H. Fredrickson,
Macromolecules 39, 2449–2451 (2006).
23. M. W. Matsen, F. S. Bates, J. Chem. Phys. 106, 2436–2448 (1997).
24. T. A. Shefelbine et al., J. Am. Chem. Soc. 121, 8457–8465 (1999).
25. K. Kawamoto et al., J. Am. Chem. Soc. 138, 11501–11504 (2016).
26. B. Yu, R. Li, R. A. Segalman, Macromolecules 54, 5388–5396 (2021).
27. R. Roy et al., Macromolecules 44, 3910–3915 (2011).
28. F. J. Martinez-Veracoechea, F. A. Escobedo, Macromolecules
42, 1775–1784 (2009).
29. H. Hasegawa, T. Hashimoto, S. T. Hyde, Polymer 37, 3825–3833 (1996).
30. M. W. Matsen, F. S. Bates, Macromolecules 29, 7641–7644 (1996).
31. Y. La et al., Nat. Chem. 6, 534–541 (2014).
32. C. Y. Chang et al., Proc. Natl. Acad. Sci. U.S.A. 118, e2022275118 (2021).
33. A. Reddy, M. S. Dimitriyev, G. M. Grason, Nat. Commun. 13,
2629 (2022).
34. H. Y. Jung et al., Macromolecules 50, 3224–3233 (2017).
35. A. C. Finnefrock et al., Angew. Chem. Int. Ed. 40, 1207–1211 (2001).
36. M. S. Dimitriyev, A. Reddy, G. M. Grason, Macromolecules 56,
7184–7202 (2023).
37. A. Reddy, X. Feng, E. L. Thomas, G. M. Grason, Macromolecules
54, 9223–9257 (2021).
38. F. J. Martinez-Veracoechea, F. A. Escobedo, Macromolecules
40, 7354–7365 (2007).
39. P. Padmanabhan, F. Martinez-Veracoechea, F. A. Escobedo,
Macromolecules 49, 5232–5243 (2016).
40. J. F. Sadoc, J. Charvolin, Acta Crystallogr. A 45, 10–20 (1989).
41. S. T. Hyde, S. Andersson, Z. Kristallogr. Cryst. Mater. 168, 213 (1984).
42. Z. Jiang, B. Lee, Appl. Phys. Rev. 8, 011305 (2021).
43. V. Petříček, M. Dušek, L. Palatinus, Z. Kristallogr. Cryst. Mater.
229, 345–352 (2014).
44. A. J. Senesi, B. Lee, J. Appl. Cryst. 48, 1172–1182 (2015).
45. S. J. Marrink, H. J. Risselada, S. Yefimov, D. P. Tieleman,
A. H. de Vries, J. Phys. Chem. B 111, 7812–7824 (2007).
46. W. Kohn, A. D. Becke, R. G. Parr, J. Phys. Chem. 100, 12974–12980
(1996).
47. C. Y. Chu et al., Macromolecules 45, 2471–2477 (2012).
48. V. Schädler, J. Spickermann, H. J. Räder, U. Wiesner,
Macromolecules 29, 4865–4870 (1996).
49. R. H. Zha, B. F. M. de Waal, M. Lutz, A. J. P. Teunissen,
E. W. Meijer, J. Am. Chem. Soc. 138, 5693–5698 (2016).
g
Lee et al., Science 383, 70–76 (2024)
chemistry, wherein the thioether became a
structures
linking molecule and yielded Ia3d
at room temperature (34). However, there are
no known methods to assess other types of network structures with a high packing frustration.
We believe that conventional phase diagrams
of block copolymers should be revised to consider the type of end-end interactions, linker
chemistry, and geometry of terminal packing.
The lam structures do not occupy a large window of the phase diagram for end-functionalized block copolymers if well-defined end-end
interactions exist. Instead, the diverse network
structures can be stabilized in a large f window. The overall shape of the phase boundary between the ordered and disordered states
is unlikely to be symmetric because the strength
of end-end interactions and end group arrangements are substantially influenced by block
curvatures, that is, confinements.
The crucial role of end group and linker chemistry in relieving the packing frustration of the
network structures of diblock copolymers was
established. The first cases of thermodynami structures for neat block
cally stable Im3m
copolymers were disclosed through the precise
control of the end-end interactions and end
group arrangements. The findings were consistent for two distinct types of block copolymers, underscoring the importance of end group
chemistry in precisely manipulating nanoscale
self-assembly in soft materials. This versatile
approach can potentially uncover network structures from a diverse spectrum of soft materials, which can drive advancements for various
nanotechnologies.
p
blue schemes, whereas those with ether linkage are depicted by pink schemes. Proposed
arrangements of dPA end groups within the
tetrapod of PMB domains are also illustrated.
Two points deserve emphasis: First, the existence of a Pm3n (A15-like unicontinuous cubic
network; fig. S23) structure has not been reported for diblock polymers thus far; second,
(double diamond) structure with a
a Pn3m
tetrapod intersection existing in equilibrium
is rare for diblock copolymers [the only equilibrium cases were reported for polypropyleneb-polystyrene achieved through the tacticity
control of polypropylene blocks (47)]. Our findings hold substantial importance, as we enabled the stabilization of a tetrapod structure
solely by modifying the terminal groups, without
altering the polymer backbones. With symmetric SMBs, the presence of the strongest endend interactions led to the emergence of Fddd
(SMB-N-dPA) structures,
(SMB-O-dPA) andIa3d
both characterized by tripod intersections (fig.
S24). The ether linkage again worked in favor of
relieving the high packing frustration, aligning
with the tendency observed in SEO analogs.
The consistency of these findings across two
distinct types of block copolymers highlights
the importance of end group and linker chemistry, irrespective of the chemical details of the
polymer backbones.
To gain insights into the phase behavior of
end-functionalized diblock copolymers, we conducted dissipative particle dynamics (DPD) simulations. In these simulations, we modeled a
diblock copolymer with a symmetric composition of A15B15 and introduced an additional
end group, represented as a C bead. We investigated the phase behavior of the A15B15C
triblock copolymer by varying the end-end
interaction. The interaction between the block
copolymer and end group was systematically
varied, effectively strengthening the interactions between the end-group beads. As illustrated in fig. S25 and visualized in movies S2
and S3, our DPD simulations revealed a facile
morphological transition from lam to bicontinuous networks with increased end-end interactions, which is in good agreement with
our experimental results.
Modulation of the microphase separation
behavior of block copolymers by modifying the
end group was reported by Schädler et al. (48).
Thereafter, the “one is enough” approach was
proposed by Jo et al. (16), and several research
groups conducted various follow-up studies
(49). Nevertheless, the observed morphologies
were limited to conventional lam, hex, and
body-centered cubic phases. Jo et al. used diol
and dicarboxylic acid moieties to modify the
terminal of an SEO through thiol-ene click
RES EARCH
ORGANIC CHEMISTRY
Cobalt-catalyzed synthesis of amides from alkenes
and amines promoted by light
Mason S. Faculak, Alexander M. Veatch, Erik J. Alexanian*
Catalytic methods to couple alkene and amine feedstocks are valuable in synthetic chemistry. The
direct carbonylative coupling of alkenes and amines holds promise as a perfectly atom-economical
approach to amide synthesis, but general methods remain underdeveloped. Herein, we report an alkene
hydroaminocarbonylation catalyzed by unmodified, inexpensive cobalt carbonyl under mild conditions
and low pressure promoted by light. Silane addition after the reaction enables sequential cobalt-catalyzed
amide reduction, constituting a formal alkene hydroaminomethylation. These methods exhibit
exceptional scope across both alkene and amine components with high chemo- and regioselectivity
and proceed efficiently even in the absence of solvent. The formation of a hydridocobalt through
photodissociation of a carbonyl ligand is proposed to enable catalytic activity under mild conditions,
which addresses a long-standing challenge in catalysis.
y g
y
,
Faculak et al., Science 383, 77–81 (2024)
y
*Corresponding author. Email: eja@email.unc.edu
g
Department of Chemistry, University of North Carolina at
Chapel Hill, Chapel Hill, NC 27599, USA.
The Lewis basicity of alkylamines is particularly problematic in current approaches because
of poisoning of metal catalysts and inhibition
of requisite catalytic intermediates (i.e., metal
hydrides) (17, 18). The development of a hydroaminocarbonylation that uses an earth-abundant
metal under mild conditions to efficiently couple amines with diverse alkenes remains a major challenge in catalysis.
p
C
atalytic transformations that efficiently
couple alkenes and amines have broad
utility in the atom-economical production of N-containing small molecules.
The importance of this goal has led to
many breakthroughs in synthetic chemistry
spanning transition metal and photoredox
catalysis (1–4). One notable area of development is the metal-catalyzed carbonylative coupling of alkenes and amines, whereby instead
of direct C–N bond coupling between the two
building blocks (i.e., alkene hydroamination),
a molecule of carbon monoxide (CO) is inserted
between the reacting components (Fig. 1A).
Metal-catalyzed hydroaminomethylation, which
involves an initial alkene hydroformylation using
CO and H2 followed by a reductive coupling
with an amine, is a prototypical example of
such a process and is widely used in the synthesis of alkylamines (5–8).
Despite substantial progress, key limitations
involving the metal-catalyzed carbonylative
couplings of alkenes and amines remain. Catalytic hydroaminomethylations generally require high temperatures and pressures, and
there are virtually no examples with earthabundant catalysts and alkylamines as substrate (9, 10). Moreover, a direct carbonylative
coupling encompassing terminal and multisubstituted alkenes with amine feedstocks,
known as alkene hydroaminocarbonylation, is
unknown (11). Such a catalytic method would
provide a 100% atom-economical approach to
the synthesis of amides, the most frequently
constructed functionality in pharmaceutical
synthesis (12). Current hydroaminocarbonylations are limited in scope across both the
alkene and amine partners and require the
use of precious metal catalysts under high temperatures and pressures to proceed (13–20).
Prior studies in our research group involving the aminocarbonylation of aromatic electrophiles (21, 22) led us to hypothesize that a
classic Co catalyst for alkene hydroformylation
could be repurposed to facilitate a general
synthesis of amides and amines from alkenes.
Roelen’s 1937 discovery of the hydroformylation (oxo) reaction with Co2(CO)8 as a catalyst,
generating HCo(CO)4 in situ upon reaction
with H2, was critical to the development of
industrial hydroformylation (23). It is widely
accepted that the generation of the tricarbonyl species HCo(CO)3 is necessary for alkene
coordination, and the high temperatures and
pressures typical of hydroformylation using
Co2(CO)8 are required to generate and stabilize this intermediate (24). Studies have speculated that ligand dissociation to generate
HCo(CO)3 could be promoted by ultraviolet
(UV) light (25, 26). During our prior studies of
light-promoted aminocarbonylation involving
unmodified Co2(CO)8 as the precatalyst, our calculations were consistent with the generation
of anionic [Co(CO)3]– species (22). We reasoned
that if HCo(CO)3 could likewise be generated
in the presence of alkenes, and if this species
were stable under the reaction conditions and
tolerant of Lewis basic alkylamines [because
HCo(CO)4 is a strongly acidic metal hydride],
Fig. 1. Carbonylative transformations and catalysts. (A) Fundamental carbonylative couplings of alkylamines
and alkenes. (B) Light-promoted dissociation of CO generates the active metal hydride catalyst in the presence
of Lewis basic alkylamines. (C) Co-catalyzed synthesis of amides or amines by hydroaminocarbonylation
and hydroaminomethylation under mild conditions promoted by light.
5 January 2024
1 of 5
RES EARCH | R E S E A R C H A R T I C L E
then unmodified, inexpensive Co2(CO)8 could
enable a general hydroaminocarbonylation
under mild reaction conditions promoted by
light (Fig. 1B). Herein, we report the successful
development of such a process, which also enables a general alkene hydroaminomethylation
upon the addition of an organosilane reductant (Fig. 1C).
Our initial efforts indicated that unmodified
Co2(CO)8 is indeed a highly effective catalyst
for the hydroaminocarbonylation, requiring
only 1.0 mol % catalyst and low CO pressure
when irradiated with 390-nm purple lightemitting diodes (LEDs). Furthermore, these
reactions are efficient using only 1 equiv of
both alkene and alkylamine coupling partners
with no additional reagents and thus 100%
atom economy. In virtually all cases, the only
p
g
y
y g
y
,
Fig. 2. Hydroaminocarbonylations catalyzed by Co2(CO)8. All yields are of isolated product. Percent selectivity is provided in examples involving minor
regioisomers. See the supplementary materials for reaction details. *2 equiv alkene; †5 equiv amine; ‡5 mol % Co2(CO)8.
Faculak et al., Science 383, 77–81 (2024)
5 January 2024
2 of 5
RES EARCH | R E S E A R C H A R T I C L E
material after the reaction is product and
remaining substrate. During reaction optimization, our experiments indicated that whereas the reactions can be performed at ambient
temperature, heating generally increases the
linear/branched (l/b) ratio of the products,
likely through enhanced chain walking (see
below). No reaction occurs in the absence of
either Co2(CO)8 or 390-nm light.
Substrate scope exploration
y
y g
The hydroaminomethylation of alkenes is a
widely applied transformation for the synthesis
of alkylamines, proceeding through sequential
alkene hydroformylation, amine condensation,
and imine reduction (5, 6). This fundamental
catalytic reaction commonly relies on the use
of precious metal catalysts (i.e., Rh) and tuned
phosphine ligands under high pressures of synthesis gas (CO/H2), with no general examples
involving first-row, earth-abundant transition
g
Reduction to amines by silane addition
metal catalysts and alkylamine building blocks.
However, a recent report indicated that less
basic anilines are viable partners when catalyzed by a Co complex featuring a bulky, modified xantphos ligand (28). Previous examples
of Co-catalyzed hydroaminomethylations with
alkylamines require pressures of ~100 atm and
temperatures of ~180°C to obtain low to moderate efficiencies and selectivities, with recent computational investigations questioning
whether a practical Co-catalyzed hydroaminomethylation is even feasible because of its poor
chemoselectivity (10). Standard catalytic hydroaminomethylations suffer from chemoselectivity issues arising from possible hydrogenations
of the alkene or requisite aldehyde intermediates, as well as formation of aldol condensation by-products (29). We postulated that our
catalytic system could unlock a distinct approach to hydroaminomethylation by alkene
hydroaminocarbonylation and sequential catalytic amide reduction with a hydrosilane (30).
We indeed found that upon completion of
the hydroaminocarbonylation, the simple addition of PhSiH3 to the reaction mixture afforded the desired alkylamines in good overall
yields (Fig. 3). Control experiments using isolated amides indicated that the CO atmosphere
and 390-nm LEDs both increased the efficiency
of the amide reduction. Secondary and tertiary
amines are easily accessed with Earth-abundant
Co as a catalyst at low CO pressure, avoiding
the chemoselectivity issues observed in standard hydroaminomethylations involving aldehydes by instead proceeding through amides
as amine precursors.
We next performed transformations to highlight the practicality of the approach. A reaction mixture composed of equal amounts of
all four octene isomers as substrate, which is
representative of industrial octene production, produced amide 2 in similar yield and
selectivity to the reaction of 1-octene, demonstrating the highly regioconvergent nature of
p
A representative substrate scope of the hydroaminocarbonylation demonstrating the successful coupling of major classes of alkenes and
amines is depicted in Fig. 2. Terminal alkenes
including propylene gas are excellent coupling partners, efficiently delivering the linear
products with high regioselectivity (1 to 8).
Reactions involving both cyclic and acyclic 1,1and 1,2-disubstituted alkenes likewise yield
amides in good to excellent yields (9 to 13).
With the 1,2-disubstituted alkenes 4-octene
and 6-dodecene, extensive chain walking of
the catalyst is observed, leading to selective
functionalization at the end of the chain (9
and 10). In these reactions, unreacted alkene
regioisomers are observed after the reaction.
Whereas tri- and tetrasubstituted alkenes are
well known to present reactivity challenges in
catalytic hydrocarbonylations (27), the successful hydroaminocarbonylations of b-citronellol,
1-methyl cyclohexene, and tetramethylethylene (14 to 16) clearly indicate that the current catalytic system has a rare capability to
transform sterically encumbered alkenes. The
functionalization of the trisubstituted alkene
of b-citronellol favors the terminal product as
expected (14), whereas that of 1-methyl cyclohexene produces a mixture of regioisomers
favoring the 4-position of the substrate (15).
We next applied the catalytic amide synthesis to a diverse range of unsaturated natural
products to test the limits of reactivity and
selectivity of the system (17 to 23). Several
terpenes containing 1,1-disubstituted alkenes
react efficiently to deliver amide products in
good yields (17 to 19). The hydroaminocarbonylations of (–)–sclareol, cholesterol, and quinine
are all successful, highlighting the compatibility of the catalytic system with complex alkenes
in molecules containing unprotected polar functionality (20 to 22). As a final demonstration,
we sought to hydroaminocarbonylate the trisubstituted alkene of the complex alkaloid brucine, which presents a formidable challenge in
alkene functionalization. The reaction with
hexylamine notably delivers a single regioisomer
and diastereomer of amide 23 in good yield
and demonstrates the specific capability of this
catalytic system to furnish amides through the
functionalization of even the most challenging alkenes.
The reaction scope with respect to the alkylamine coupling partner is also excellent (Fig.
2). Diverse primary and secondary cyclic and
acyclic amines all reacted efficiently with vinylcyclohexane with high regioselectivity (24 to
34; Fig. 2). In addition, hydroaminocarbonylation with ammonia gas (35) was also successful; for example, with a solution of NH3 in
iPrOH, the primary amide formed in moderate
yield. Reactions of both the drug compound
fluoxetine hydrochloride (36) after the formation of the free base and the tBu ester of the
amino acid tyrosine (37) also delivered amides
in good yields.
y
,
Fig. 3. Hydroaminomethylations catalyzed by Co2(CO)8. All yields are of isolated product. See Fig. 2 for l/b ratios and the supplementary materials for reaction
details. *2 equiv alkene. Rt, room temperature.
Faculak et al., Science 383, 77–81 (2024)
5 January 2024
3 of 5
RES EARCH | R E S E A R C H A R T I C L E
Fig. 4. Highlighting the practicality of the amide synthesis. (A) Hydroaminocarbonylation of a mixture
of octenes converges to the linear product with high regioselectivity. (B) Hydroaminocarbonylation is efficient with
low catalyst loading on decagram scale in the absence of reaction solvent.
p
g
y
This coordination is then followed by hydrocobaltation, leading to regioisomeric alkylcobalt
intermediates that are capable of isomerization through sequential b-hydride elimination
and reinsertion processes, favoring the terminal
alkylcobalt with acyclic substrates. The high
linear selectivity of the reaction indicates rapid
chain walking of intermediate alkylcobalt species (34). We hypothesize that these equilibria
may contribute to the reduced catalytic efficiencies of reactions involving alkenes with
higher substitution, where hydrocobaltation
is slower (27). Migratory insertion of a CO ligand then takes place, furnishing an acylcobalt
tricarbonyl. The amine nucleophile then coordinates to the metal center, followed by a concerted addition and/or reductive elimination
to generate the amide product [Co(CO)3]– (22),
which undergoes proton exchange to regenerate HCo(CO)3. This specific catalytic mode of
unmodified Co2(CO)8 enables a general hydroaminocarbonylation of alkenes, which constitutes a very efficient, 100% atom-economical
approach to amides from fundamental chemical building blocks under mild reaction conditions. The successful transformations of alkenes
ranging from propylene to complex alkaloid
natural products, and amines from ammonia
to drug compounds, highlight the versatility of
this method, which we anticipate will prove
broadly useful in contexts ranging from the
preparation of medicinally relevant, functionalized small molecules to industrial-scale chemical production.
REFERENCES AND NOTES
A catalytic mechanism consistent with our results is depicted in Fig. 5. After disproportionation of the Co catalyst (33), light-promoted
loss of a CO ligand generates [Co(CO)3]–, which
is subsequently protonated, yielding HCo(CO)3.
Previous UV-visible light absorption studies
from our group indicated that [Co(CO)4]– can
absorb light in the range of the 390-nm LEDs
(21). A high quantum yield was observed for
the hydroaminocarbonylation (F = 15.7), consistent with multiple catalytic turnovers after
CO photodissociation and before another
photodissociation is required, potentially due
to the protonation of [Co(CO)3]– outcompeting CO coordination at the low reaction pressures. These conditions favor the formation of
a hydridocobalt that is stable to Lewis basic
amines and primed for alkene coordination.
5 January 2024
4 of 5
,
Faculak et al., Science 383, 77–81 (2024)
Mechanistic considerations
y
the hydroaminocarbonylation (Fig. 4A). To
test the limits of catalytic activity, we next performed the hydroaminocarbonylation of cyclohexene with piperidine on decagram scale at
0.1 mol % catalyst loading and in the absence
of reaction solvent. Under these challenging
conditions, amide 11 was produced in excellent yield, supporting the scalability of the
hydroaminocarbonylation (Fig. 4B). The ability to perform the amide synthesis under neat
conditions in the absence of any by-products
is consistent with many important principles
of green chemistry (31). Indeed, the development of catalytic methods for amide synthesis
and mild reduction of amides, both demonstrated using the present approach, were recently
cited as two of the top 10 most important green
chemistry research areas for drug discovery
and development (32).
y g
Fig. 5. Proposed catalytic cycle for the hydroaminocarbonylation.
1. A. Trowbridge, S. M. Walton, M. J. Gaunt, Chem. Rev. 120,
2613–2692 (2020).
2. J. Escorihuela, A. Lledós, G. Ujaque, Chem. Rev. 123,
9139–9203 (2023).
3. R. C. DiPucchio, S.-C. Rosca, L. L. Schafer, J. Am. Chem. Soc.
144, 11459–11481 (2022).
4. J. M. Ganley, P. R. D. Murray, R. R. Knowles, ACS Catal. 10,
11712–11738 (2020).
5. P. Kalck, M. Urrutigoïty, Chem. Rev. 118, 3833–3861 (2018).
6. C. Chen, X.-Q. Dong, X. Zhang, Org. Chem. Front. 3, 1359–1370
(2016).
7. X.-F. Wu et al., Acc. Chem. Res. 47, 1041–1053 (2014).
8. S. Hanna, J. C. Holder, J. F. Hartwig, Angew. Chem. Int. Ed. 58,
3368–3372 (2019).
9. K. Murata, A. Matsuda, T. Masuda, J. Mol. Catal. 23, 121–132 (1984).
10. H. M. Bruijn, C. F. Guerra, E. Bouwman, F. M. Bickelhaupt,
ChemistrySelect 5, 13981–13994 (2020).
11. S. Cai, H. Zhang, H. Huang, Trends Chem. 3, 218–230 (2021).
12. S. D. Roughley, A. M. Jordan, J. Med. Chem. 54, 3451–3479 (2011).
13. K. Dong et al., J. Am. Chem. Soc. 137, 6053–6058 (2015).
14. X. Fang, R. Jackstell, M. Beller, Angew. Chem. Int. Ed. 52,
14089–14093 (2013).
15. T. Xu, F. Sha, H. Alper, J. Am. Chem. Soc. 138, 6629–6635 (2016).
16. Y.-H. Yao et al., J. Am. Chem. Soc. 143, 85–91 (2021).
17. J. Liu et al., Angew. Chem. Int. Ed. 55, 13544–13548 (2016).
18. G. Zhang, B. Gao, H. Huang, Angew. Chem. Int. Ed. 54,
7657–7661 (2015).
19. B. Gao, G. Zhang, X. Zhou, H. Huang, Chem. Sci. 9, 380–386
(2017).
20. F. Bede, L. Kollár, N. Lambert, P. Huszthy, P. Pongrácz, Eur. J.
Org. Chem. 26, e202300571 (2023).
21. A. M. Veatch, E. J. Alexanian, Chem. Sci. 11, 7210–7213 (2020).
22. A. M. Veatch, S. Liu, E. J. Alexanian, Angew. Chem. Int. Ed. 61,
e202210772 (2022).
23. B. Cornils, W. A. Herrmann, M. Rasch, Angew. Chem. Int. Ed.
33, 2144–2163 (1994).
24. F. Hebrard, P. Kalck, Chem. Rev. 109, 4272–4282 (2009).
RES EARCH | R E S E A R C H A R T I C L E
25. M. J. Mirbach, M. F. Mirbach, A. Saus, N. Topalsavoglou,
T. N. Phu, J. Am. Chem. Soc. 103, 7594–7601 (1981).
26. V. Dragojlovic, D. B. Gao, Y. L. Chow, J. Mol. Catal. Chem. 171,
43–51 (2001).
27. H.-Y. Yang, Y.-H. Yao, M. Chen, Z.-H. Ren, Z.-H. Guan, J. Am.
Chem. Soc. 143, 7298–7305 (2021).
28. J. Yang, F. G. Delolo, A. Spannenberg, R. Jackstell, M. Beller,
Angew. Chem. Int. Ed. 61, e202112597 (2022).
29. D. Crozet, M. Urrutigoïty, P. Kalck, ChemCatChem 3, 1102–1118
(2011).
30. T. Dombray, C. Helleu, C. Darcel, J. Sortais, Adv. Synth. Catal.
355, 3358–3362 (2013).
31. A. D. Kreuder et al., ACS Sustain. Chem. Eng. 5, 2927–2935
(2017).
32. M. C. Bryan et al., Green Chem. 20, 5082–5103 (2018).
33. J. Guo, H. D. Pham, Y.-B. Wu, D. Zhang, X. Wang, ACS Catal.
10, 1520–1527 (2020).
34. H. Sommer, F. Juliá-Hernández, R. Martin, I. Marek, ACS Cent. Sci.
4, 153–165 (2018).
ACKN OWLED GMEN TS
We thank M. Brookhart and S. Meek for helpful discussions and
C. Vanderwal and the Vanderwal group at University of California,
Irvine, for access to their library of alkene substrates. Funding:
This work was supported by the National Institute of General Medical
Sciences (NIGMS) of the National Institutes of Health (NIH grant
R35 GM131708 to E.J.A.). The University of North Carolina (UNC)
Department of Chemistry’s Mass Spectrometry Core Laboratory
provided expertise and instrumentation that enabled this study
with support from the National Science Foundation (NSF grant
CHE-1726291) and the NIGMS (NIH grant R35GM118055). The UNC
Department of Chemistry’s NMR Core Laboratory provided
expertise and instrumentation that enabled this study with support
from the NSF (grants CHE-1828183 and CHE-0922858). Author
contributions: E.J.A. conceived of the work. All authors designed
the experiments. M.S.F. and A.M.V. performed and analyzed the
experiments. E.J.A., M.S.F., and A.M.V. prepared the manuscript.
Competing interests: E.J.A., M.S.F., and A.M.V. are inventors
on US provisional patent application 63/468,573 covering the
Co-catalyzed hydroaminocarbonylations and hydroaminomethylations
of alkenes promoted by light, filed by UNC. Data and materials
availability: Experimental and characterization data are
available in the supplementary materials. License information:
Copyright © 2024 the authors, some rights reserved; exclusive
licensee American Association for the Advancement of Science. No
claim to original US government works. https://www.science.org/
about/science-licenses-journal-article-reuse
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adk2312
Materials and Methods
Figs. S1 to S4
Tables S1 to S3
NMR Spectra
References (35–40)
Submitted 9 August 2023; accepted 20 November 2023
10.1126/science.adk2312
p
g
y
y g
y
,
Faculak et al., Science 383, 77–81 (2024)
5 January 2024
5 of 5
RES EARCH
CARBENE CHEMISTRY
A stable rhodium-coordinated carbene with a s0p2
electronic configuration
Chaopeng Hu, Xin-Feng Wang, Jiancheng Li, Xiao-Yong Chang, Liu Leo Liu*
Isolable singlet carbenes have universally adopted a s2p0 electronic state, making them s-donors
and p-acceptors. We present a rhodium-coordinated, cationic cyclic diphosphinocarbene with a s0p2
ground state configuration. Nuclear magnetic resonance spectroscopy studies show a carbene carbon
chemical shift below −30.0 parts per million. X-ray crystallography reveals a planar RhP2C configuration.
Quantum chemical calculations rationalize how s-electron delocalization/donation and p-electron
negative hyperconjugation together stabilize the formally vacant s orbital and the filled p orbital at the
carbene center. In contrast to traditional carbene counterparts this carbene can undergo synthetic
transformations with both a Lewis base and a silver salt, producing a Lewis acid/base adduct and a
silver p-complex, respectively. Exhibiting ambiphilic reactivity, it can also form a ketenimine through
reaction with an isocyanide.
,
1 of 5
y
5 January 2024
y g
Hu et al., Science 383, 81–85 (2024)
Subsequently, 3 reacted with NaBPh4 or AgNTf2
to produce 4[BPh4] (31P NMR: 79.8 ppm) or
4[NTf2] (31P NMR: 78.6 ppm), respectively.
Probing further, the proton-decoupled 13C NMR
y
*Corresponding author. Email: liuleoliu@sustech.edu.cn
Characterization of the carbene
g
Department of Chemistry, Southern University of Science
and Technology, Shenzhen 518055, P. R. China.
tified (18). In addition, an excited singlet state
with a s1p1 configuration is also conceivable.
Alternatively, singlet carbenes can exhibit two
nonbonding electrons paired within a single
s or p orbital, leading to two possible singlet
states (17). Of these, the s2p0 state is typically
more stable than the s0p2 state, owing to the
low-lying s orbital that benefits from substantial s-orbital contribution. Notably, all isolable
singlet carbenes identified to date display a
s2p0 singlet ground state, achieved through
a well-established approach of incorporating
p-donating heteroatoms onto the carbene carbon (Fig. 1C) (19, 20). These electronic states have
laid the foundation for countless investigations into carbene chemistry, inspiring applications and expanding our understanding of
molecular properties. However, compelling
inquiries have arisen regarding the potential
isolation of a singlet carbene bearing a s0p2
singlet state.
The ground state of the parent carbene (H2C:)
is known to be a triplet s1p1 configuration,
which is predicted to be ~60 kcal/mol lower
in energy than the s0p2 singlet state (21). This
substantial energy gap underscores the considerable challenge of engineering carbenes that
adopt a s0p2 ground state. In 2013, Hoffmann
and Borden computationally investigated the
stabilization of the s0p2 singlet ground state
of carbenes through combinations of in-plane,
lone pair donor, and p acceptor substituents (22).
Nevertheless, carbenes of this type have only
been detected and trapped in low-temperature
matrices by infrared spectroscopy (23, 24). In
fact, as early as 1980, Pauling proposed that
ideal substituents for stabilizing singlet carbenes should preserve the electroneutrality of
the carbene center (25). Following this logic, a
s0p2 electronic configuration could potentially be realized by incorporating two substituents that act concurrently as p-attractors and
s-donors, thus exerting a pull/pull mesomeric
effect along with a push/push inductive in-
p
C
arbon typically abides by the octet rule,
reacting so as to fill its valence shell with
eight electrons. However, there are exceptions. Among these is the carbene (R2C:),
a class of divalent carbon molecules both
highly reactive and versatile. The early notion of
“methylene” proposed by Dumas (1) in 1835 set
the stage for the subsequent characterization
of carbenes (Fig. 1A). This initial concept was
solidified in 1954 when Doering and Hoffmann
offered evidence for the viability of dihalocarbenes as reaction intermediates (2). In parallel
to the revolutionary impact of Gomberg’s persistent trityl radical (3) on free radical chemistry, the field of carbene chemistry was indelibly
altered by the reports of isolable carbenes (4–7).
In 1988, Bertrand reported an isolable singlet carbene, a (phosphino)(silyl)carbene (Fig.
1A) (8). Three years later, Arduengo unveiled
an isolable, crystalline N-heterocyclic carbene
(NHC) (9). These seminal findings revealed that
carbenes—traditionally thought of as transient
and unstable entities—could in fact be tamed,
crystallized, and handled much like any other
persistent chemical reagent. As a result, stable
carbenes have become invaluable tools in various fields, including catalysis (10–12), materials science (13, 14), and medicinal chemistry
(15, 16).
Conventionally, carbenes have been classified into four primary categories according
to their electronic state (Fig. 1B), with their
frontier orbitals systematically referred to as
s (in-plane) and p (out-of-plane) (17). Triplet
carbenes, for instance, feature two nonbonding electrons situated in separate orbitals, both
with parallel spins, appropriately described
by a s1p1 configuration. Several persistent carbenes with a s1p1 triplet state have been iden-
fluence (Fig. 1C). Diborylcarbenes exemplify
this conceptual approach; however, attempts
to isolate these molecules have hitherto remained elusive (26–30). Other attempts to isolate s0p2 carbenes involve exploring postulated
metallodiphosphinocarbenes with high electrophilicity (31–34). Based on chemical trapping experiments, the transient existence of
these carbenes was proposed although not
directly observed.
In our endeavor to isolate a singlet carbene
exhibiting the s0p2 electronic state, we selected
a-(bisphosphino)diazomethane 2, bearing sterically demanding mesityl substituents on both
N-heterocycles, as the precursor (Fig. 2A). This
compound, characterized by a 31P nuclear magnetic resonance (NMR) shift of 108.2 parts per
million (ppm) and a diagnostic infrared absorption at 1997 cm−1 for the N=N stretch,
was synthesized through the desilylation of
(phosphino)(silyl)diazomethane 1, followed
by a salt metathesis reaction (35). Within the
solid-state architecture of 2, the P atoms of
phosphino substituents display a trigonalpyramidal arrangement, with two distinct
P lone pairs of electrons residing within the
P(1)–C(1)–P(2) plane (fig. S49).
Liberation of dinitrogen from phosphinodiazomethane instigates a simultaneous rotation of the P–C bond and a planarization at
the phosphorus atom, converging in a singlet
phosphinocarbene characterized by the s2p0
electronic configuration (20, 36). Such a mechanistic transformation poses an intricate experimental challenge: the retention of the
planar alignment of the lone pair, necessitating a circumvention of the P–C bond rotation.
To impose this geometrical fidelity, our design
hinged on the integration of a d8 transition
metal, typically inclined to form a square-planar
configuration with four coordinating ligands,
into the P–C–P plane. The subsequent formulation of a rigid P–C–P–M four-membered ring
would thus serve as scaffolding to maintain
the in-plane alignment of the dual phosphorus lone pairs.
In pursuit of this structural archetype,
we orchestrated a reaction involving 2 and
[Rh(AdNC)2Cl]2 (Ad, 1-adamantyl), giving rise
to metallocycle 3 (31P NMR: 68.8 ppm). The
expulsion of dinitrogen was confirmed by the
intensity loss of infrared n(CN2) absorption in 3
whereas x-ray diffraction provided definitive
insights into the structural formulation of the
product as [[(CH2)(NMes)]2P]2[Rh(AdNC)2]C(Cl)
(Mes, mesityl) (Fig. 2B). A noteworthy observation was the planar geometry of the Rh–P–
C–P four-membered ring in metallocycles.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
Fig. 1. Background and key concepts. (A) Timeline showing pivotal milestones in the evolution of stable carbene chemistry. iPr, isopropyl; Ad, 1-adamantyl.
(B) Depiction of primary electronic configurations of carbenes. (C) The strategy employed for stabilizing the singlet state of carbenes, with green arrows representing
the inductive effect, red arrows denoting the mesomeric effect, orange arrows showing in-plane s-delocalization/donation, and purple arrows signifying negative
hyperconjugation.
2 of 5
,
5 January 2024
than those found in 3 (101.04(9)°) and the fourmembered NHCs of Cp2Ti(tBuN)2C (122.7(1)°)
(40) and (iPr)2NB(DippN)2C (94.0(1)°) (41) (Dipp,
2,6-diisopropylphenyl). The C(1)–Rh(1) distance of 4[BPh4] (2.316(3) Å) considerably surpass the Pyykkö standard for a C–Rh single
bond (2.0 Å) (42), yet the distance is markedly
shorter relative to that in 3 (2.9551(18) Å). This
may suggest subtle transannular interactions
between C(1) and Rh(1). Additionally, the bond
lengths of C(1)–P(1) in 4[BPh4] (1.66 7(2) Å) are
shorter than those in 3 (1.7316(18) Å). These
reduced C–P bond lengths in 4[BPh4] mirror a
typical C–P double bond (1.67 Å) (42), hinting
at the in-plane donation of the P lone pairs
into the vacant orbital at C(1).
The solid-state structure of 4 is in line with
the findings from our DFT calculations performed at the B3LYP(D3)/def2-SVP level (table
S8). These calculations predict that 4 has a
singlet ground state, which lies 25.2 kcal/mol
below the triplet state (table S9). Analysis of the
frontier molecular orbitals of 4 yielded notable insights (Fig. 3A). The HOMO, situated
at −7.67 eV, predominantly displays a C(1)centered p lone-pair character. This lone pair
y
Hu et al., Science 383, 81–85 (2024)
unoccupied molecular orbital (LUMO) and
HOMO-1→LUMO (fig. S56). Moreover, species
4[BPh4] and 4[NTf2] displayed notable stability, remaining intact in the solid state under a
nitrogen atmosphere for over a month without noticeable decomposition.
From slow evaporation of a concentrated solution of dichloromethane and hexane at room
temperature, we successfully grew single crystals of 4[BPh4] suitable for x-ray diffraction
analysis. This experiment definitively validated
the formation of 4[BPh4] through chloride abstraction of 3. In the solid state, there was no
evident interaction between the tetraphenyl
borate anion and the cation 4, indicating the
ionic nature of the complex. The P–C–P–Rh
four-membered ring preserves its planar conformation, a testament to the constrained planar alignment of the P lone pair. Compared with
precursor 3, complex 4 manifests: (i) a notably
reduced C(1)–Rh(1) distance; (ii) shortened bond
lengths throughout the four-membered ring;
(iii) widened bond angles for P–C–P and P–Rh–P,
but more acute angles for C–P–Rh.
Delving deeper, the P–C–P angle was measured
to be 133.42(17)° for 4[BPh4]. This angle is wider
y g
spectra revealed notable patterns: the central
carbon signals for 4[BPh4] and 4[NTf2] display
peaks at −30.9, and −32.3 ppm, respectively.
These spectral shifts, in alignment with our
theoretical predictions (4: −31.7 ppm; table S10),
are upfield-shifted compared with those observed for 3 (64.2 ppm). These NMR characteristics markedly deviate from the signals seen
for the carbene carbons in P-heterocyclic carbenes
(PHCs) (>180 ppm) (37) and N-heterocyclic
carbenes (NHCs) (typically >185 ppm) (38).
This divergence underscores a distinct electronic structure separating 4 from the realm of
PHCs and NHCs featuring a s2p0 electronic
state. It is worth noting that, without the presence of Rh coordination, attempts to synthesize
a diphosphinocarbene through photolysis of 2
were unsuccessful, instead leading to the generation of a 1,4,5,6-tetrahydro-1,4,2-diazaphosphinine
derivative, 8 (fig. S8) (39).
The UV-visible spectrum of 4[BPh4] in tetrahydrofuran (THF) reveal two broad peaks at 436
and 385 nm (fig. S54). Based on time-dependent
density functional theory calculations, these
peaks arise from electronic transitions of highest occupied molecular orbital (HOMO)→lowest
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
,
Fig. 2. Synthesis, characterization, and reactivity of metallocyclic carbenes. (A) Schematic representation of the synthetic route to the metallocyclic carbenes
4[BPh4] and 4[NTf2]. Mes, mesityl; Ad, 1-adamantyl; Tf, trifluoromethanesulfonyl. (B) Solid-state structures of 3 and 4[BPh4]. Hydrogen atoms and the noninteracting
tetraphenyl borate anion are omitted for clarity. (C) Reactivity of 4[BPh4] or 4[NTf2] toward 4-dimethylaminopyridine, AgNTf2 and AdNC.
is polarized toward the P–N s*-antibonding
orbitals of the 1,3,2-diazaphospholidine framework. Conversely, the LUMO, at −3.69 eV, largely
resembles a s-type sp2-hybridized orbital centered at C(1). This electron distribution is also
illustrated by the electron localization function (43) plot of 4 showing an intensified localized electron density at the C(1) p orientation
(fig. S57). This is in contrast to the patterns calculated for NHCs (fig. S58). This observation
Hu et al., Science 383, 81–85 (2024)
5 January 2024
confirms that 4 exhibits a s0p2 electronic state,
which is markedly different from the states observed in previously isolated singlet carbenes.
Moreover, our calculations suggest that an increase in the P–C–P bond angle does not affect the
s0p2 electronic configuration of 4 (figs. S59 and
60). Based on gauge-independent atomic orbitalDFT NMR analyses, the distinct electronic
configuration of 4 enhances three magnetic
shielding components, leading to a prominent
upfield chemical shift for the carbene carbon
(fig. S62).
Intrinsic bond orbital (IBO) (44) calculations
offer deeper insights into the bonding scenario
of 4 (Fig. 3C and fig. S63). At the C(1) atom, a
s bond is formed with both P(1) and P(2) (fig.
S63, A and B). Simultaneously, an essentially
nonbonding lone pair orbital resides at C(1),
oriented perpendicularly to the CP2Rh plane
(Fig. 3C, IBO1). This lone pair has minor
3 of 5
RES EARCH | R E S E A R C H A R T I C L E
Together, these electron push-pull effects contribute substantially to the stability of the s0p2
electronic state of the carbene 4.
To further understand the impact of d orbital size on the stabilization of the s0p2 electronic
configuration of carbenes, we computationally
examined the Co and Fe analogs of 4, denoted
as 4-Co and 4-Fe (figs. S66 and 67). Although
the bonding framework of 4-Co aligns with
that of 4 suggesting that d orbital size might
play a limited role in stabilizing the s0p2 configuration, 4-Fe presents as a formal Fe(II)
complex. In this case, the carbon appears formally dianionic as a result of intramolecular
charge transfer, highlighting the critical influence of the energy level of d electrons. Their
considerable reducing power can lead to distinctly different bonding scenarios.
Accordingly, the Wiberg bond indices for
C(1)–P(1), Rh(1)–P(1), and C(1)–Rh(1) in 4 are
1.36, 0.41, and 0.26, respectively. The natural
population analysis shows positive charges on
the P atoms [1.61 atomic units (a.u.)] whereas
both Rh(1) (−0.20 a.u.) and C(1) (−1.13 a.u.)
exhibit negative charges. In light of these findings, four plausible resonance forms of 4
are depicted in Fig. 3B, resembling those observed for NHCs (19). Nevertheless, it is imperative to underscore that in the context of
4, ylide structures emerge as a consequence
of s delocalization/donation as opposed to p
donation, characteristic of NHCs.
Reactivity of 4 as a p-donor and s-acceptor
Contrary to PHCs (37) and NHCs (19), which
function as s-donors and p-acceptors, our computational insights point toward carbene 4
operating as a p-donor and s-acceptor. In
support of this notion the interaction of
4[BPh4] with the potent s-donor Lewis base,
4-dimethylaminopyridine (DMAP), results in
the occupancy of the vacant s orbital of 4 by
DMAP, culminating in the formation of 5[BPh4]
in 89% yield (Fig. 2C), as evidenced by the
x-ray structure of 5[BPh4] in fig. S50. When
the small Lewis acid AgNTf2 is employed, the
p
delocalization toward P(1) and P(2), which
aligns with the HOMO of 4. Although the two
lone pairs at P(1) and P(2) predominantly coordinate to Rh(1), they also delocalize to the
unoccupied s orbital of C(1) (IBO2 and IBO3).
Furthermore, one of the filled d orbitals at Rh
(1) provides slight stabilization for the in-plane
unoccupied s orbital of C(1) (IBO4). Theoretically, this in-plane s delocalization/donation resembles the p-donation found in hitherto known
singlet carbenes featuring the s2p0 electronic
state (fig. S64). However, in this instance, the
formally vacant orbital at C(1) manifests as an
in-plane sp2-hybridized s orbital, diverging from
the conventional out-of-plane pp-orbital. This inplane triple donation is instrumental in pushing
the lone pair of electrons into the C(1) p orbital
(Fig. 3D). Conversely, the P–N s*-antibonding
orbitals within the 1,3,2-diazaphospholidine
scaffold play a crucial role in drawing p electron
density away from C(1), as evidenced by the
analysis of second-order perturbation theory
using the natural bond orbital method (fig. S65).
g
y
y g
y
,
Fig. 3. Theoretical analysis of 4. (A) Energy diagram for the frontier Kohn-Sham orbitals. HOMO, highest occupied molecular orbital; LUMO, lowest unoccupied
molecular orbital. (B) Illustration of the four distinct resonance structures. (C) Visual representation of selected intrinsic bond orbitals. (D) Schematic representing the
essence of stabilization: push/push/push s-electron delocalization/donation and pull/pull p-electron negative hyperconjugation.
Hu et al., Science 383, 81–85 (2024)
5 January 2024
4 of 5
RES EARCH | R E S E A R C H A R T I C L E
37. J. D. Masuda et al., Chem. Asian J. 2, 178–187 (2007).
38. D. Tapu, D. A. Dixon, C. Roe, Chem. Rev. 109, 3385–3407
(2009).
39. S. Marrot, T. Kato, F. P. Cossío, H. Gornitzka, A. Baceiredo,
Angew. Chem. Int. Ed. 45, 7447–7450 (2006).
40. E. P. Beaumier et al., J. Am. Chem. Soc. 142, 8006–8018 (2020).
41. Y. Ishida, B. Donnadieu, G. Bertrand, Proc. Natl. Acad. Sci. U.S.A.
103, 13585–13588 (2006).
42. P. Pyykkö, M. Atsumi, Chemistry 15, 12770–12779 (2009).
43. A. Savin, R. Nesper, S. Wengert, T. F. Fässler, Angew. Chem.
Int. Ed. 36, 1808–1832 (1997).
44. G. Knizia, J. E. M. N. Klein, Angew. Chem. Int. Ed. 54,
5518–5522 (2015).
45. M.-M. Gan et al., Chem. Rev. 118, 9587–9641 (2018).
46. D. R. Tolentino et al., J. Am. Chem. Soc. 141, 9823–9826
(2019).
69. L. Falivene, L. Cavallo, Coord. Chem. Rev. 344, 101–114 (2017).
AC KNOWLED GME NTS
Funding: We gratefully acknowledge financial support from the
National Natural Science Foundation of China (22350004 and,
22271132 to L.L.L. and 22301122 to J.L.), Shenzhen Science and
Technology Innovation Program (JCYJ20220530114806015),
Guangdong Innovation and Entrepreneurial Research Team
Program (2021ZT09C278), and Guangdong Provincial Key
Laboratory of Catalysis (2020B121201002). We also acknowledge
the assistance of SUSTech Core Research Facilities. The theoretical
work was supported by the Center for Computational Science
and Engineering at SUSTech. Author contributions: L.L.L. devised
and supervised the project and wrote the manuscript. C.H.
synthesized and isolated all compounds 1-8, determined the X-ray
crystal structures and the NMR analysis. L.L.L. and C.H. performed
the computational work. X.-F.W. was involved in the preparation
of 1 and provided support for synthesizing Li(THF)1.5CN2TMS.
J.L. helped in the synthesis of 5[BPh4] and 7[BPh4] and provided
valuable discussions. X.-Y.C. provided help in the determination
of X-ray crystal structures. Competing interests: The authors
declare no competing financial interests. Data and materials
availability: General information, experimental procedures,
1
H NMR/13C NMR/31P NMR/19F NMR/11B NMR spectra, high
resolution mass spectrometry data, infrared spectrometry data
and computational details are provided in the Supplementary
Materials. Full crystallographic data for structures 2, 3, 4[BPh4],
5[BPh4], 6[NTf2], 7[BPh4] and 8 are available free of charge
from the Cambridge Crystallographic Data Center under reference
numbers 2267592, 2267593, 2267596, 2267599, 2267595,
2267590 and 2302945, respectively. License information:
Copyright © 2024 the authors, some rights reserved; exclusive
licensee American Association for the Advancement of Science. No
claim to original US government works. https://www.sciencemag.
org/about/science-licenses-journal-article-reuse
y g
SUPPLEMENTARY MATERIALS
y
science.org/doi/10.1126/science.adk6533
Materials and Methods
Supplementary Text
Figs. S1 to S73
Tables S1 to S11
References (47–68)
5 of 5
,
Submitted 3 September 2023; accepted 20 November 2023
10.1126/science.adk6533
y
5 January 2024
1. J.-B. Dumas, E. M. Péligot, in Annales de chimie et de physique
(1835); pp. 5-74.
2. W. von E. Doering, A. K. Hoffmann, J. Am. Chem. Soc. 76,
6162–6165 (1954).
3. M. Gomberg, J. Am. Chem. Soc. 22, 757–771 (1900).
4. M. N. Hopkinson, C. Richter, M. Schedler, F. Glorius, Nature
510, 485–496 (2014).
5. J. P. Moerdyk, D. Schilter, C. W. Bielawski, Acc. Chem. Res. 49,
1458–1468 (2016).
6. M. Melaimi, R. Jazzar, M. Soleilhavoup, G. Bertrand,
Angew. Chem. Int. Ed. 56, 10046–10068 (2017).
7. G. Guisado-Barrios, M. Soleilhavoup, G. Bertrand, Acc. Chem. Res.
51, 3236–3244 (2018).
8. A. Igau, H. Grutzmacher, A. Baceiredo, G. Bertrand, J. Am.
Chem. Soc. 110, 6463–6466 (1988).
9. A. J. ArduengoIII, R. L. Harlow, M. Kline, J. Am. Chem. Soc. 113,
361–363 (1991).
10. D. Enders, O. Niemeier, A. Henseler, Chem. Rev. 107,
5606–5655 (2007).
11. E. Peris, Chem. Rev. 118, 9988–10031 (2018).
12. Y. Xia, D. Qiu, J. Wang, Chem. Rev. 117, 13810–13889 (2017).
13. A. V. Zhukhovitskiy, M. J. MacLeod, J. A. Johnson, Chem. Rev.
115, 11503–11532 (2015).
14. C. A. Smith et al., Chem. Rev. 119, 4986–5056 (2019).
15. L. Oehninger, R. Rubbiani, I. Ott, Dalton Trans. 42, 3269–3284
(2013).
16. W. Liu, R. Gust, Coord. Chem. Rev. 329, 191–213 (2016).
17. D. Bourissou, O. Guerret, F. P. Gabbaï, G. Bertrand, Chem. Rev.
100, 39–92 (2000).
18. K. Hirai, T. Itoh, H. Tomioka, Chem. Rev. 109, 3275–3332
(2009).
19. H. V. Huynh, Chem. Rev. 118, 9457–9492 (2018).
20. J. Vignolle, X. Cattoën, D. Bourissou, Chem. Rev. 109,
3333–3384 (2009).
21. A. Kalemos, T. H. Dunning Jr., A. Mavridis, J. F. Harrison,
Can. J. Chem. 82, 684–693 (2004).
22. B. Chen, A. Y. Rogachev, D. A. Hrovat, R. Hoffmann,
W. T. Borden, J. Am. Chem. Soc. 135, 13954–13964 (2013).
23. J. P. Wagner, J. Am. Chem. Soc. 144, 5937–5944 (2022).
24. G. Maier, J. Endres, Chemistry 5, 1590–1597 (1999).
25. L. Pauling, J. Chem. Soc. Chem. Commun. 688–689 (1980).
26. P. H. M. Budzelaar, P. R. Schleyer, K. Krogh-Jespersen,
Angew. Chem. Int. Ed. 23, 825–826 (1984).
27. R. Wehrmann, H. Klusik, A. Berndt, Angew. Chem. Int. Ed. 23,
826–827 (1984).
28. H. Meyer, G. Baum, W. Massa, S. Berger, A. Berndt, Angew. Chem.
Int. Ed. 26, 546–548 (1987).
29. H. Meyer, G. Baum, W. Massa, A. Berndt, Angew. Chem. Int. Ed.
26, 798–799 (1987).
30. Y. Shibutani, S. Kusumoto, K. Nozaki, J. Am. Chem. Soc. 145,
16186–16192 (2023).
31. J. Ruiz, V. Riera, M. Vivanco, M. Lanfranchi, A. Tiripicchio,
Organometallics 17, 3835–3837 (1998).
32. J. Ruiz et al., Angew. Chem. Int. Ed. 42, 4767–4771 (2003).
33. J. Ruiz, M. E. G. Mosquera, G. García, F. Marquínez, V. Riera,
Angew. Chem. Int. Ed. 44, 102–105 (2004).
34. J. Vignolle et al., J. Am. Chem. Soc. 129, 978–985 (2007).
35. M. J. Menu et al., J. Organomet. Chem. 372, 201–206 (1989).
36. T. Kato, H. Gornitzka, A. Baceiredo, A. Savin, G. Bertrand,
J. Am. Chem. Soc. 122, 998–999 (2000).
g
Hu et al., Science 383, 81–85 (2024)
RE FERENCES AND NOTES
p
p-lone pair of 4[NTf2] coordinates to the Ag
center, giving rise to 6[NTf2] with a 63% yield.
X-ray diffraction analysis of single crystals of
6[NTf2] (fig. S51) unequivocally shows Ag(1)
orienting toward the C(1) p lone pair, marked
by a C(1)–Ag(1) bond length of 2.684(15) Å. Moreover, discernible Rh–Ag and arene–Ag interactions manifest, as denoted by the Rh(1)–Ag(1),
C(2)–Ag(1), and C(3)–Ag(1) separations, which
stand at 2.849(6) Å, 2.634(8) Å, and 2.587(8) Å,
respectively. The p-coordination between C(1)
and Ag(1), supported by DFT calculations (figs.
S68 and 69), stands in contrast to the conventional s-coordination seen in NHC silver
complexes (45).
In a further notable development, 4[BPh4]
undergoes a reaction with AdNC, a reagent
known for [1+1] cross-coupling reactions with
ambiphilic or electrophilic singlet carbenes
(20), yielding 7[BPh4] at a 73% yield, as corroborated by the x-ray structure delineated
in fig. S52. Although similar results, such as
the emergence of a ketenimine, were recorded
for cyclic(alkyl)(amino)carbenes (CAACs) (46)
possessing the s2p0 electronic state and 4[BPh4],
our computational exploration reveals distinct
mechanistic scenarios for a CAAC and 4 when
reacting with isocyanide (figs. S70 to 73). This
distinction is driven by the specific electronic
states.
Over three decades have passed since the pioneering work by Bertrand (8) and Arduengo
(9) on the isolation of stable singlet carbenes.
These molecules have since become indispensable tools in chemistry and have even found
applications in medicine and materials science. The molecular design principles and synthetic strategies we present here offer a clear
roadmap for the creation and stabilization of
singlet carbenes with a reversed s0p2 electronic state and the capacity for s-accepting
and p-donating behavior. Given their distinct
electronic properties, we expect that these singlet carbenes can be harnessed more broadly
for the activation of small molecules and for
fine-tuning the physical and chemical properties of transition-metal complexes to suit specific applications.
RES EARCH
PEROVKSITES
Supramolecular assembly of blue and green halide
perovskites with near-unity photoluminescence
Cheng Zhu1,2,3†, Jianbo Jin4†, Zhen Wang1, Zhenpeng Xu1, Maria C. Folgueras1,2,3, Yuxin Jiang2,4,
Can B. Uzundal2,4, Han K. D. Le2,4, Feng Wang2,3,5, Xiaoyu (Rayne) Zheng1,6, Peidong Yang1,2,3,4*
The metal-halide ionic octahedron is the optoelectronic unit for halide perovskites, and a crown
ether–assisted supramolecular assembly approach can pack various ionic octahedra into tunable
symmetries. In this work, we demonstrate near-unity photoluminescence quantum yield (PLQY) blue
and green emission with the supramolecular assembly of hafnium (Hf) and zirconium (Zr) halide
octahedral clusters. (18C6@K)2HfBr6 powders showed blue emission with a near-unity PLQY (96.2%),
and green emission was also achieved with (18C6@K)2ZrCl4Br2 powders at a PLQY of 82.7%. These
highly emissive powders feature facile low-temperature solution-based synthesis conditions and
maintain high PLQY in solution-processable semiconductor inks under ambient conditions, and they were
used in thin-film displays and emissive three-dimensional–printed architectures that exhibited high
spatial resolution.
1 of 8
,
5 January 2024
y
Zhu et al., Science 383, 86–93 (2024)
We explored a supramolecular synthetic route
in which 18C6 greatly increased the solubility
y g
*Corresponding author. Email: p_yang@berkeley.edu
†These authors contributed equally to this work.
Crown ether–assisted supramolecular assembly
y
Department of Materials Science and Engineering, University of
California Berkeley, Berkeley, CA 94720, USA. 2Materials
Sciences Division, Lawrence Berkeley National Laboratory,
Berkeley, CA 94720, USA. 3Kavli Energy NanoScience Institute,
Berkeley, CA 94720, USA. 4Department of Chemistry, University
of California Berkeley, Berkeley, CA 94720, USA. 5Department
of Physics, University of California Berkeley, Berkeley, CA
94720, USA.USA. 6Energy Technologies Area, Lawrence
Berkeley National Laboratory, Berkeley, CA 94720, USA.
g
1
blocks in the crystal structure (19–21), and,
given the ionic nature and high chemical tunability of halide perovskite structures (22), different compositions and packing geometries
of [MX6]n− could be explored for light-emission
applications. The vacancy-ordered double perovskite (A2MX6 phase) has been proposed to incorporate tetravalent metal cation octahedra,
such as [TeX6]2− (23–25), [SnX6]2− (26, 27), and
[PtX6]2− (28, 29). Although the [PbX6]4− octahedra are corner-shared in all three dimensions in the prototypical CsPbX3 structure (30),
the [MX6]2− octahedra in the A2MX6 phase
are isolated because a vacancy occupies every
other M site in the crystal structure (23). A few
[MX6]n− emitters as well as some non-octahedral
emitters (31) with high PLQY (~95%) have
been identified with yellow emission, such as
[SnX6]4− (32). However, emission of high PLQY
with shorter wavelengths is still very rare. The
isolated nature of the octahedra affects their
optoelectronic properties in that the strong
coupling of the exciton with lattice vibrations
greatly lowers the energy level of the exciton
and forces it into transient self-trapped exciton (STE) states with a range of self-trapped
energy levels (33, 34). As a result, the A2MX6
systems generally have broadband emissions
with a large Stokes shift.
Although the A2MX6 phase has been widely
studied in various compositions, octahedra
with Hf4+ or Zr4+ centers, especially [HfBr6]2−
and [ZrBr6]2− octahedra, have rarely been the
subject of research (35, 36), even though they
have interesting optoelectronic properties.
Cs2HfBr6 crystals have a blue emission with the
PL peak at 435 nm (35), and colloidal Cs2ZrBr6
nanocrystals have been demonstrated to have
a green emission with a PLQY of 45% (36).
There are several reasons why they are less
explored. Theoretical (37) and experimental
studies (38) have shown that the Hf4+ and Zr4+
metal centers are extremely air- and moisture-
p
B
lue and green emission with high photoluminescence quantum yield (PLQY) is
at the forefront of solid-state lighting
and color display research. Although Si
and Zn codoped GaN can exhibit a PLQY
of 90% (1), these covalent semiconductors require high purity to prevent rapid nonradiative recombination at crystal structure defects
(2–5) and rely on solid-state synthesis at elevated temperatures near 1000°C (6). As an
alternative to covalent semiconductors, ionic
halide perovskites have received attention
given their high optical absorption coefficient
(7), tunable bandgap (8–10), high defect tolerance (11, 12), and efficient photo- and electroluminescence (13, 14). For example, the blue
and green emissive colloidal CsPbClxBr3-x quantum dots have exhibited PLQY values of ~80%
(15, 16). In addition, low-dimensional halide
perovskites like the n = 1 Ruddlesden-Popper
phase (C6H5CH2NH3)2PbBr4 show blue emission with a PLQY of 79% (17). Despite the notable optoelectronic properties of lead-based
halide perovskites, the toxicity of lead and the
complex colloidal synthesis complicate largescale applications. Moreover, suitable ligands
are still needed to prevent aggregation of
these low-dimensional nanostructures during
use (18).
Recent studies have revealed that the optoelectronic properties of halide perovskites stem
from the [MX6]n− (where M is a metal cation
and X is a halide anion) fundamental building
sensitive in the A2MX6 phase. Their synthesis
requires the vertical Bridgman-Stockbarger
method at ~1000°C in sealed quartz ampoules
(36, 39, 40). Finally, it is difficult to prepare
high-purity samples that do not contain a secondary impurity, such as CsBr (36). Thus, a new
methodology is needed for the synthesis of
more stabilized and purer solid phases containing the [HfBr6]2− or [ZrBr6]2− octahedra.
Recently, we proposed a general crown ether–
assisted supramolecular assembly approach for
tetravalent metal octahedra (41). Two crown
ether@alkali metal complexes can sandwich
a tetravalent metal octahedron into a (crown
ether@A)2MX6 dumbbell structural unit. The
composition of the dumbbell structural unit is
highly tunable, with crown ether = 18-crown-6
(18C6) or 21-crown-7 (21C7); A = Cs+, Rb+, or K+;
M = Te4+, Sn4+, Se4+, Ir4+, Pt4+, Zr4+, Hf4+, or Ce4+;
and X = Cl−, Br−, or I−. In this work, we extended
this general supramolecular assembly approach
to [HfBr6]2− octahedra to achieve a structure
with formula (18C6@K)2HfBr6 that features blue
emission with near-unity (96.2%) PLQY. We also
optimized the synthetic route by replacing the
challenging high-temperature solid-state synthesis with a low-temperature organic solution–
based synthesis. Moreover, an efficient green
emission was also achieved by tuning the composition of the (crown ether@A)2MX6 dumbbell
structural unit. (18C6@K)2ZrCl4Br2 demonstrated green emission with 82.7% PLQY. By
studying the photophysics of the supramolecular assembled samples, we could attribute
the emission to STE states and observed a very
strong electron-phonon coupling constant
(represented by the Huang-Rhys parameter)
of >90 for (18C6@K)2HfBr6. The supramolecular assembled samples had longer PL lifetimes (in the microsecond timescale) compared
with those of other halide perovskite systems that reflected a low rate of nonradiative
recombination.
The structural integrity and impressive optical properties of the supramolecular assembled
solid powders were further maintained by generating a powder suspension in nonpolar organic solvents, such as dichloromethane (DCM),
to create an ink system. Polystyrene (PS) polymer
was dissolved into the ink to further increase the
solution processability. We used these inks to
fabricate thin films through fast solvent evaporation. In combination with a digitally controlled excitation source, the (18C6@K)2HfBr6/PS
composite thin film could be used as a display
with bright color contrast and fast response
time. A solution-processable ink also allowed
three-dimensional (3D) printing of the powders into various blue-, green-, and dual-color–
emitting structures.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
Zhu et al., Science 383, 86–93 (2024)
5 January 2024
ature to 200°C by combining mechanical forces
with heat to facilitate solid-state diffusion. Details
of the synthesis for (18C6@K)2HfBr6 and K2HfBr6
can be found in the supplementary materials.
The crystal structure of (18C6@K)2HfBr6 was
determined from single-crystal x-ray diffraction (SCXRD). (18C6@K)2HfBr6 crystallized in
space group with lattice parameters of
the R3
a = 14.1332 Å and c = 21.0189 Å (Fig. 1A and
table S1). The (18C6@K)2HfBr6 dumbbell structural unit belongs to the S6 point group, where
two K+ cations and the Hf4+ cation sit on the
S6 axis. The sixfold symmetry axis of the 18C6
and the S6 axis of the Oh-symmetric [HfBr6]2−
octahedron were aligned (Fig. 1B). The K2HfBr6
crystals were face-centered cubic (fcc) (Fig. 1C)
(42, 43), in which the [HfBr6]2− ionic octahedra
were charge balanced by the surrounding K+
cations (Fig. 1D).
The purity of the (18C6@K) 2 HfBr 6 and
K2HfBr6 powders was investigated with powder x-ray diffraction (PXRD) (Fig. 1E). The PXRD
pattern of the (18C6@K)2HfBr6 powders matched
with the calculated pattern generated from
the single-crystal structure with no visible diffraction peaks from impurities. The quality of
the PXRD pattern for the K2HfBr6 powders
was much lower because of their extreme air
sensitivity. The measurement had to be collected in 5 min with an inert atmosphere
2 of 8
,
of the KBr and HfBr4 precursors in polar organic solvents for low-temperature solutionbased synthesis. A clear precursor solution
was obtained with acetonitrile (ACN) at 80°C
with the concentration of 4 mM for 18C6 and
KBr and 2 mM for HfBr4. Our previous study
of the supramolecular assembly of tetravalent metal octahedra (41) indicated that a
(18C6@K)2HfBr6 dumbbell structural unit was
formed in ACN. We grew (18C6@K)2HfBr6
powders and single crystals using the antisolvent crystallization method (39). K2HfBr6
powders were also synthesized by using a modified solid-state synthesis method. We increased
the purity and decreased the synthesis temper-
diffraction patterns. K2HfBr6 showed quite poor PXRD quality because
of its poor stability during measurement. a.u., arbitrary unit. (F and G)
Band structure and corresponding total pDOS of (18C6@K)2HfBr6 (F) and
K2HfBr6 (G). When [HfBr6]2− octahedra were assembled in the supramolecular
approach, the dispersion of the bands decreased, and 18C6 contributed
to the VB.
y
Fig. 1. Two assemblies of the [HfBr6]2− ionic octahedron. (A and B) The
rhombohedral unit cell (A) and the dumbbell-shaped structural unit (B)
of (18C6@K)2HfBr6. (C and D) The Fm-3m unit cell [from the Open
Quantum Materials Database (OQMD) (36, 37)] (C) and the isolated [HfBr6]2−
ionic octahedron building block (D) of K2HfBr6. (E) The PXRD patterns
for synthesized (18C6@K)2HfBr6 and K2HfBr6 powders and the calculated
RES EARCH | R E S E A R C H A R T I C L E
(E) The CIE 1931 chromaticity diagram for the emission of (18C6@K)2HfBr6
powders and (18C6@K)2ZrCl4Br2 powders. “B” stands for the blue
emission of (18C6@K)2HfBr6, and “G” stands for the green emission of
(18C6@K)2ZrCl4Br2. The coordinates for the emission colors of (18C6@K)2HfBr6
and (18C6@K)2ZrCl4Br2 were (0.17438, 0.16922) and (0.30597, 0.41533),
respectively.
p
Fig. 2. Blue emission with near-unity PLQY (96.2%) from
(18C6@K)2HfBr6 powders and green emission with a PLQY of 82.7%
from (18C6@K)2ZrCl4Br2 powders. (A) (18C6@K)2HfBr6 powders under white
lamp and 254-nm UV excitation. (B) PL and PLE spectra of (18C6@K)2HfBr6
powders. (C) (18C6@K)2ZrCl4Br2 powders under white lamp and 302-nm UV
excitation. (D) PL and PLE spectra of (18C6@K)2ZrCl4Br2 powders.
g
3 of 8
,
Compared with K2HfBr6, (18C6@K)2HfBr6 had
greatly enhanced emission intensity. Figure 2A
shows the extremely bright blue emission of
(18C6@K)2HfBr6 powders under 254-nm ultraviolet (UV) excitation. The photoluminescence
(PL) spectrum of (18C6@K)2HfBr6 powders was
measured at 275-nm excitation (Fig. 2B). The
powders had a blue emission centered at 445 nm
(2.79 eV), and the full width at half maximum
(FWHM) was 0.73 eV. Photoluminescence excitation (PLE) spectra revealed a large Stokes
shift (1.35 eV).
The emission intensity of the (18C6@K)2HfBr6
powders was quantified with PLQY measurement, and a near-unity value of 96.2 ± 1.2% was
obtained for the (18C6@K)2HfBr6 powders over
six measurements from two batches of samples (fig. S5). The specific value for each mea-
y
5 January 2024
Optical characterization of the blue and
green emitters
surement is shown in table S2. By contrast, the
PLQY of K2HfBr6 powders was 12.8% (fig. S6).
K2HfBr6 also had an even larger Stokes shift
(1.42 eV) and broader emission, with a peak emission wavelength of 457 nm and a FWHM of
0.90 eV (fig. S7). The color purity of the emission
from the [HfBr6]2− octahedra was also enhanced
by the supramolecular approach. Figure S8 shows
the emission color of (18C6@K)2HfBr6 and K2HfBr6
powders on the CIE 1931 chromaticity diagram.
(18C6@K)2HfBr6 had a much purer blue emission color compared with K2HfBr6.
The [ZrBr6]2− units enabled high-PLQY green
emissions.Upon290-nmexcitation,(18C6@K)2ZrBr6
had a PL peak at 547 nm, and the FWHM of the
PL was 0.69 eV (fig. S9). For the same excitation
wavelength, the PL peak of K2ZrBr6 was at 560 nm,
and the FWHM of the PL was 0.70 eV (fig. S10).
The PLQY of (18C6@K)2ZrBr6 was 49.8% (fig.
S11), which was slightly greater than the PLQY
of K2ZrBr6 (46.3%) (fig. S12). Although the peak
position of the PL spectrum was in the green
region, simply analyzing the peak emission
wavelength was insufficient given the broadness of the STE-based emission because this
crystal actually produced a yellow-green emission color (fig. S13).
Given the great chemical tunability of the
dumbbell structural unit, an alloying approach
at the halide site was proposed to achieve a purer
green emission with near-unity PLQY. For CsPbX3
(where X = Cl−, Br−, or I−) nanocrystals, the emission color can be easily controlled by tuning
the halide composition (44); introducing Cl− in
the halide site may generate a shorter wavelength emission color. By carefully tuning the
y g
Zhu et al., Science 383, 86–93 (2024)
The conduction band (CB) of (18C6@K)2HfBr6
was composed of Hf 5d and Br 4p orbitals, as
was the CB of K2HfBr6. However, the valence
band (VB) compositions were quite different
in these two materials. The VB of K2HfBr6 was
mainly composed of the Br 4p orbital, but 18C6
contributed to the VB of (18C6@K)2HfBr6. Thus,
the 18C6 molecules were electronically coupled
to the [HfBr6]2− octahedra,
which indicates that the entire (18C6@K)
2HfBr6 dumbbell building block became a
new electronic unit. DFT calculations of (18C6@
K)2ZrBr6 showed that the contribution from
18C6 to the VB and the band structures were
more discrete than those in K2ZrBr6 (fig. S4).
y
sample holder to prevent the degradation of
the powders and measurement of the degradation product. Although the quality of the
K2HfBr6 PXRD pattern was not ideal, the most
dominant peaks of the fcc K2HfBr6 phase were
still identifiable. Moreover, no HfBr4 or KBr
diffraction peaks were present (Fig. 1E), which
showed that all of the precursor materials transformed into the K2HfBr6 phase. A Raman spectrum of K2HfBr6 further confirmed the presence
of the [HfBr6]2− octahedra in the crystal structure (fig. S1).
The crown ether–assisted supramolecular approach was generalized to produce other emissive centers. For example, (18C6@K)2ZrBr6
single crystals and powders were successfully
synthesized by the same method, and the same
crystal structure as (18C6@K)2HfBr6 was obtained (fig. S2 and table S1). Figure S3 shows
that phase-pure (18C6@K)2ZrBr6 powders could
be obtained with our established solution-based
synthesis. K2ZrBr6 powders were also synthesized
with the same solid-state method as K2HfBr6.
The (18C6@K)2HfBr6 and (18C6@K)2ZrBr6
dumbbell building blocks were also the electronic units of the new crystal. To elucidate
the effect of 18C6 on the electronic structures
of the assembled [HfBr6]2− octahedra, density functional theory (DFT) calculations were
performed on (18C6@K)2HfBr6 (Fig. 1F) and
K2HfBr6 (Fig. 1G) to determine their electronic band structures and partial electronic
density of states (pDOS). The electronic bands
of (18C6@K)2HfBr6 were less dispersive compared with K2HfBr6 because the [HfBr6]2− octahedra were more separated in (18C6@K)2HfBr6.
RES EARCH | R E S E A R C H A R T I C L E
Fig. 3. Photophysical analysis
of (18C6@K)2HfBr6 and
(18C6@K)2ZrCl4Br2. (A) PL
spectra of (18C6@K)2HfBr6
powders at 4, 50, 100, 150, 210,
and 293 K. (B) FWHM of the
PL spectra of (18C6@K)2HfBr6
powders at different temperatures,
with the orange and teal solid
lines denoting the least-square fit
to Eq. 1 at low (4 to 190 K) and
high (190 to 293 K) temperature
ranges, respectively. (C) PLE
spectroscopy of (18C6@K)2HfBr6
powders. (D) Normalized PL
decay curves of (18C6@K)2ZrCl4Br2
and (18C6@K)2ZrBr6 single
crystals.
p
g
y
4 of 8
,
5 January 2024
cluding their large Stokes shift and broadband
nature, are primarily attributed to the electronphonon coupling effect. To unravel the STE
emission mechanism, we performed lowtemperature PL measurements to examine the
electron-phonon coupling in (18C6@K)2HfBr6.
With increasing temperatures, the PL peak
gradually broadened, and the peak was slightly red-shifted, indicating greater phonon participation at higher temperatures (Fig. 3A). A
small shoulder peak at ~550 nm was present
that was especially distinct at lower temperatures, which we attribute to the Zr impurity
in the HfBr4 precursor (35). Inductively coupled
atomic emission spectroscopy revealed an
~0.5 atomic % (at %) ZrBr4 impurity in the asobtained HfBr4 precursor, and that there was
an ~0.6 at % Zr4+ impurity in the synthesized
(18C6@K)2HfBr6 single crystals (table S5).
To deconvolve the emission from
(18C6@K)2ZrBr6 impurities, a two-peak Gaussian fitting was applied to the PL spectrum at
each temperature. Figure S22 shows an example at 4 K. The FWHMs of the (18C6@K)2HfBr6
peaks were obtained from the Gaussian fittings
and are summarized in table S6. The temperature dependence of the FWHM of the emission peak was modeled using the theory of
Toyozawa (45), which applies a configuration
coordinate model to explain the broadening of
the emission originating from electron-phonon
y
Zhu et al., Science 383, 86–93 (2024)
system. The (101) and (110) diffraction peaks of
(18C6@K)2ZrCl4Br2 were slightly shifted to
larger 2q values compared with the corresponding PXRD peaks of (18C6@K)2ZrBr6, which suggests smaller lattice constants (fig. S21).
Figure 2C shows the bright green emission
of (18C6@K)2ZrCl4Br2 powders under 302-nm
UV lamp excitation. The PL spectrum of
(18C6@K)2ZrCl4Br2 powders was measured at
295-nm excitation (Fig. 2D). The green emission
had a similar Stokes shift (1.36 eV versus 1.35 eV)
and FWHM (0.80 eV versus 0.73 eV) compared
to the blue emission of the (18C6@K)2HfBr6
powders, which suggests similar emission properties of the Hf and Zr metal centers in the
supramolecular assembly materials system. The
PLQY of the emission from (18C6@K)2ZrCl4Br2
powders was 82.7 ± 0.9%, which was determined through the measurement of four samples from two batches (table S4). Therefore, we
achieved highly emissive powders with blue and
green emission colors based on the supramolecular assembly approach. The blue and green
colors of the emissions from (18C6@K)2HfBr6
and (18C6@K)2ZrCl4Br2, respectively, are summarized in the CIE 1931 diagram (Fig. 2E).
Next, we conducted a comprehensive photophysics analysis to confirm and gain deeper
insights into the STE emission mechanism
that underlies these blue and green emissions.
The distinctive features of STE emissions, in-
y g
KCl/KBr and ZrCl4/ZrBr4 precursor ratio in
the synthesis, the Cl−/Br− ratio in the obtained
(18C6@K)2ZrX6 dumbbell structural unit can
be precisely controlled. As expected, a larger
Cl − /Br − ratio created a more blue-shifted
PL (figs. S14 and S15). For example, (18C6@K)
2ZrCl3Br3 and (18C6@K)2ZrCl4Br2 had green
emission with PL peaks at 534 and 530 nm,
respectively. Increasing the Cl− content to a
composition of (18C6@K)2ZrCl4.5Br1.5 changed
the PL color to a cyan (bluish green) color. The
established halide site alloying approach not
only generated a purer green emission color
but also boosted the PLQY of the emission to
near-unity. For the Cl−/Br− ratio from 1:1 to 2:1 to
3:1, the PLQYs were 69.1, 82.7, and 87.0%, respectively (figs. S16 and S17).
Because the 2:1 Cl−/Br− ratio composition had
both pure-green emission color and high PLQY,
we selected (18C6@K)2ZrCl4Br2 for detailed
studies of green emission. (18C6@K)2ZrCl4Br2
single crystals were synthesized by controlling
the Cl−/Br− precursor ratio to be 2:1. The formula of (18C6@K)2ZrCl4Br2 was determined
by SCXRD (Cl−:Br− = 4.3:1.7) (fig. S18 and table
S3) and energy-dispersive x-ray spectroscopy
(EDX) elemental mapping (Cl−:Br− = 4.1:1.9)
(fig. S19). The Cl and Br atoms were perfectly
miscible in the crystal structure. PXRD of the
(18C6@K)2ZrCl4Br2 powders (fig. S20) also indicated that this composition was a phase-pure
RES EARCH | R E S E A R C H A R T I C L E
coupling. The FWHM depends on the Boltzmann
constant kB, the effective phonon energy Eph,
the temperature T, and the Huang-Rhys electronphonon coupling parameter S (46)
pffiffiffi
FWHM ¼ 2:36 SEph
A
rffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
Eph
coth
2kB T
ð1Þ
The relation between FWHM and temperature is shown in Fig. 3B. Analyzing the data
according to Eq. 1 yielded a coupling factor
S1 = 92.2 ± 3.6 and an effective phonon energy
Eph1 = 21.4 ± 0.5 meV. This phonon mode
corresponded to the asymmetric stretching
mode (Eg) of the [HfBr6]2− octahedra, which
was observed at 20.4 meV (164.5 cm−1) in the
Raman spectrum (fig. S23). However, this pho-
non mode was only responsible for STE formation up to 190 K. For temperatures >190 K,
a higher energy phonon mode dominated STE
formation.
Shifting the zero temperature of Eq. 1 by
190 K, a second fit could be obtained with a
coupling factor S2 = 108.8 ± 12.4 and an effective phonon energy Eph2 = 25.8 ± 1.6 meV.
This phonon mode corresponded to the sym-
Solution Processibility with Polymers
Facile casting
Single crystals
Thin FIlm
+Polystyrene
+DCM
DMD chip
Powders
Mild stirring
and Sonicating
Programmable Display
p
Bulk Materials
Processible Inks
PLQY: 90.77%
C
PLQY: 75.01%
D
PLQY: 80.29%
E
PLQY: 68.99%
g
B
y
y g
F
G
y
,
Fig. 4. Solution processability and display application of highly emissive blue
and green semiconductor inks. (A) Schematic illustrating the thin-film fabrication
method and the display application. The inks were formed by mixing (18C6@K)2HfBr6
or (18C6@K)2ZrCl4Br2 powders and PS in DCM. Thin films were obtained by drop
casting, and they demonstrated programmable display capability. DMD, digital
micromirror device. (B) (18C6@K)2HfBr6/PS-DCM ink under white light and 254-nm
UV lamp excitation. (C) (18C6@K)2ZrCl4Br2/PS-DCM ink under white light and
Zhu et al., Science 383, 86–93 (2024)
5 January 2024
302-nm lamp excitation. (D) (18C6@K)2HfBr6/PS composite thin film under white
light and 254-nm UV excitation. (E) (18C6@K)2ZrCl4Br2/PS composite thin film under
white light and 302-nm UV excitation. The scale bars for (B) to (E) are 1 cm. The
PLQYs of all samples are shown in the photos. (F) Image of the “Cal” logo blue
emission on the (18C6@K)2HfBr6/PS composite thin film. (G) Snapshots of a video
showing the alphabet, A to Z, with 0.1 s per letter on the (18C6@K)2HfBr6/PS
composite thin film. The scale bars for (F) and (G) are 3 mm and 4 mm, respectively.
5 of 8
RES EARCH | R E S E A R C H A R T I C L E
y
,
6 of 8
y g
The high PLQY of the blue and green emission
colors in the powders were preserved in thin
films, which would enable various optoelectronic device applications (54–56). Because
the powders were stable in nonpolar organic
solvents, they could be evenly dispersed into
solution to form inks. We used DCM because
its low boiling point (39.6°C) leads to high
volatility for drying films, and we added PS to
create inks suitable for drop casting or spin
coating by increasing the viscosity (Fig. 4A)
(57, 58). The image of (18C6@K)2HfBr6/PS
ink under a white lamp (Fig. 4B) shows that
a uniform white suspension was achieved that
exhibited a bright blue emission under 254-nm
excitation. The emission was solely from the
(18C6@K)2HfBr6 powders in the ink because
the shape of the PL spectrum was the same as the
PL shape of the powders (fig. S26). The solution
PLQY was 90.8% (Fig. 4B and fig. S27), which
was only 5.5% less than the powder PLQY. This
reduction was expected because DCM and PS do
not absorb strongly in the blue color wavelength
region (fig. S28), and the suspended powders
in the ink could cause losses through scattering.
The (18C6@K)2ZrCl4Br2/PS ink also preserved
the green emission of the (18C6@K)2ZrCl4Br2
powders with a solution PLQY of 75.0% (Fig.
4C and fig. S29).
y
5 January 2024
Blue-green dual-color display and 3D printing
The inks could be drop casted under ambient
conditions, and, after rapid solvent evaporation,
a uniform thin film forms (Fig. 4, A, D, and E).
PXRD patterns of the (18C6@K)2HfBr6/PS composite (fig. S30) and the (18C6@K)2ZrCl4Br2/PS
composite (fig. S31) showed that the structural
integrity of the powders was preserved in the
PS matrix. Scanning electron microscopy (SEM)
imaging of the (18C6@K)2HfBr6 powders and the
(18C6@K)2HfBr6/PS composite thin-film surface
(fig. S32) indicated that the submicrometersized powders were uniformly dispersed. Crosssectional SEM imaging of the thin film (fig. S33)
proved the presence and uniformity of the powders across the thin film. Under UV irradiation,
(18C6@K)2HfBr6/PS and (18C6@K)2ZrCl4Br2/PS
composite thin films showed bright blue and
green emissions, respectively (Fig. 4, D and E).
The shapes of the PL spectra of the thin films
were the same as those for the powders (figs.
S25 and S34), and the PLQYs were 80.3% (fig.
S35) and 69.0% (fig. S36) for blue- and greenemitting composites, respectively.
The stability of the air-sensitive Hf and Zr
octahedral clusters was further enhanced in
the PS polymer composite. Both Cs2HfBr6 and
Cs2ZrBr6 double-perovskite structures are predicted to be thermodynamically unstable in
the presence of water and oxygen (37), and we
found that K2HfBr6 and K2ZrBr6 powders turn
from a white to a brownish color after a few
minutes of air exposure and became nonemissive. By contrast, the (18C6@K)2HfBr6/PS
and (18C6@K)2ZrCl4Br2/PS composites maintained their blue and green emission colors,
respectively, after 1 month of storage in the air
(fig. S37). The air-stable PS polymers along
with the hydrophobic crown ethers could greatly protect the air-sensitive Hf and Zr metal
emission centers.
We explored display applications of the powderPS composite thin films. A digital mirror device
with a pixel resolution of 2560 by 1440 sequentially patterned 250-nm UV light through projection optics onto the (18C6@K)2HfBr6/PS
composite thin film with a spot size of 6.9 by
3.9 mm at a frame rate of 60 Hz (schematic of
the process is illustrated in Fig. 4A). An emissive blue “Cal” logo was illuminated on the thin
film with dimensions 3.8 mm in height and
4.7 mm in width (Fig. 4F). The logo exhibited
high luminosity characterized by sharply defined edges. To further demonstrate dynamically
changing display luminescence, we illuminated
the alphabet sequence (from A to Z) onto the
thin film with a fast flipping rate (0.1 s per
letter). A video of 2.6 s was recorded (movie S1).
Although the duration of each letter was very
short, the blue emission with the shape of the
letters was sharp and bright, as illustrated in
the snapshot photos (Fig. 4G). The size of the
letters was only 3.1 mm in width and 3.9 mm
in length, but every feature of the letters was
clearly visible with similar emission intensity
g
Zhu et al., Science 383, 86–93 (2024)
emitting diode (OLED) community has used
a xenon lamp to simulate solar irradiation, dissolving Ir complexes in deuterated toluene for
reference measurements of green and blue
emission (51, 52). To ensure a fair comparison,
we applied identical irradiation energy density
(62 mW/cm2) and temperature (35°C), and
deuterated toluene was used to disperse the
(18C6@K)2HfBr6 and (18C6@K)2ZrCl4Br2 powders. Figure S25 shows the PL intensity decay
of the (18C6@K)2HfBr6 and (18C6@K)2ZrCl4Br2
samples under continuous irradiation. Both
decay trends could be accurately described
by the integrated rate law for the first-order
reaction [ln(It/I0) = −kt]. The rate constants of
photodegradation were estimated to be 5.1 ×
10−3 h−1 and 3.0 × 10−3 h−1 for (18C6@K)2HfBr6
and (18C6@K)2ZrCl4Br2, respectively. Notably,
even under stringent irradiation conditions,
the PL intensities of (18C6@K)2HfBr6 and
(18C6@K)2ZrCl4Br2 decreased to 80% after
43 and 73 hours, respectively. These findings
underscore the superior photostability of the
supramolecular assembled samples compared
with most Ir complexes, rivaling the bestreported green-emitting fac-[Ir(ppy)3] reference (k = 2.6 × 10−3 h−1) (51, 52). Previous studies
on the photodegradation of the Ir complexes, such as Ir(ppy)3 and Ir(piq)3, have identified singlet oxygen attack and interaction
of the excited-state molecule with its local
environment as primary degradation pathways (53).
p
metric stretching mode (A1g) of the [HfBr6]2−
octahedra at 25.1 meV (202.5 cm−1). The large
Huang-Rhys factor S in both scenarios indicated a very strong electron-phonon coupling
in this material. For example, S for CsPbX3
(where X = Br− or I−) is <1 (47), and the S for
double perovskite Cs2AgBiBr6 is only ~12 (48).
STE behavior is closely related to the octahedra
packing dimensionality. Through our supramolecular approach, the [HfBr6]2− octahedra
were more isolated by the bulky (18C6@K)+
complexes, which led to stronger self-trapping
with larger S values.
Excitation wavelength–dependent PL mapping of (18C6@K)2HfBr6 (Fig. 3C) showed that
for excitation wavelengths <285 nm, a broad
PL peak at 445 nm emerged. The PL peak
position and shape were independent of the
excitation wavelength <285 nm. Thus, for abovebandgap excitation, the emission originated
from the relaxation of the same excited state.
However, for excitation wavelengths >285 nm,
a much weaker PL peak at ~550 nm replaced
the previous PL peak that arose from the ~0.6
at % (18C6@K)2ZrBr6 impurity, and 2D PLE
mapping of (18C6@K)2ZrBr6 showed a single
PL peak at ~550 nm from 245-nm to 330-nm
excitation (fig. S24).
Time-resolved PL (TRPL) studies on the supramolecular assembled single crystals revealed
that the PL decay of the (18C6@K)2ZrBr6 could
be mostly described by a monoexponential decay profile on the microsecond timescale, with
a PL lifetime of 6.80 ms (Fig. 3D and table S7).
By contrast, Cs2ZrBr6 bulk crystal featured a
triple-exponential PL decay, yielding decay time
constants of 40 ns (8.9%), 0.99 ms (24%), and
4.6 ms (68%) (35). Additionally, Cs2ZrBr6 nanocrystals showed a double-exponential PL decay with time constants at 0.78 and 4.5 ms (49).
The PLQY was related to both the radiative and
nonradiative decay rates [PLQY = krad/(krad +
knonrad)], so a more sluggish radiative decay did
not necessarily correlate to a lower PLQY. Notably, the PLQY of (18C6@K)2ZrBr6 powders
(49.8%) was greater than that of K2ZrBr6 powders
(46.3%, from our measurement) and Cs2ZrBr6
nanocrystals [~44%, from (49)]. This observation suggested that the nonradiative decay rate
of the supramolecular sample was slower than
that of the vacancy-ordered double perovskite
phases and may indicate a lower defect density
in our assembled crystals. The PL decay of the
(18C6@K)2ZrCl4Br2 single crystal could also
be fit with a single exponential function with
an even longer PL lifetime (12.08 ms) (Fig. 3D
and table S8). Cs2ZrCl6 had a slightly longer
PL lifetime (7.5 ms) compared with that of
Cs2ZrBr6 (50). This result suggested that the
supramolecular material system had a longer
PL lifetime and slower nonradiative decay rate.
We also evaluated the photostability of these
highly emissive blue and green emitters. Notably, previous research in the organic light-
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
Zhu et al., Science 383, 86–93 (2024)
5 January 2024
and, consequently, the printing resolution. However, dyes absorb light and color the final printed
parts. To avoid interference with our blue and
green emitters, we developed a photoabsorberfree resin mainly composed of photomonomer
poly(ethylene glycol) diacrylate (PEGDA) but
with a high content of photoinhibitor to control the printing resolution. The polymerized
PEGDA resin exhibited minimal absorption
within the visible spectrum, featuring a modest absorption peak from 355 to 425 nm (fig.
S38A). Also, under 250-nm UV excitation, the
resin exhibited substantially low emission intensity (fig. S38B). Hence, the emission colors
in the blue and green range of our emitters
remained largely unaffected.
Upon stirring and sonication, the powders were
uniformly dispersed into the PEGDA resin. We
exploited a multimaterial digital light-printing
method (59, 60) to achieve a 3D assembly of the
blue and green emitters into complex macroand microarchitectures. Under 405-nm structured UV light illumination, the resin rapidly
converted into solid 3D structures (Fig. 5A).
The PL spectra from the (18C6@K)2HfBr6/PEGDA
and the (18C6@K)2ZrCl4Br2/PEGDA composites
were similar to those of the powders (fig. S38,
C and D; fig. S39; and fig. S40). The printed
7 of 8
,
owing to the high uniformity of the thin film.
Furthermore, the response time of the display
should be fast because the PL decay rate of
the (18C6@K)2HfBr6 powders was ~3 orders
of magnitude faster than the frame rate of
the digital mirror device. The letters switch
extremely fast, with no blurring, ghosting, or
trailing effects.
These emissive powders could also be processed with high-resolution 3D printing technologies after blending them uniformly into
a monomer resin. Conventional resins for 3D
printing typically use dyes as the photoabsorber to control the depth of UV penetration
structures and geometric shapes, including cuboctahedron, tetrakaidecahedron,
and Menger sponge structures, with the blue and green emitters or their
combinations, respectively. These printed architectures were photoexcited at
254 nm. The scale bars for (B) to (G) are 5 mm. The scale bar for the zoom-in
image of (E) is 0.6 mm. The scale bar for (H) is 4 mm.
y
Fig. 5. Implementation of the blue-green dual-color 3D printing. (A) Schematic
illustrating the multimaterial 3D printing process. (B and C) Two 3D-printed
light-emitting Eiffel Towers under white light (B) and 254-nm UV (C)
excitation. (D) A dual-color–emitting Eiffel Tower under 254-nm UV excitation.
(E to H) Conformal and twisted octet trusses with varying hierarchical
RES EARCH | R E S E A R C H A R T I C L E
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adi4196
Materials and Methods
Figs. S1 to S44
Tables S1 and S8
References (61–67)
Movie S1
Submitted 24 April 2023; resubmitted 15 October 2023
Accepted 29 November 2023
10.1126/science.adi4196
8 of 8
,
The authors thank A. M. Oddo (UC Berkeley) for valuable
discussions on the design of the experiments. We thank
N. Settineri (UC Berkeley) for his assistance in SCXRD collection.
Funding: This work was supported by the US Department of
Energy, Office of Science, Office of Basic Energy Sciences,
Materials Sciences and Engineering Division, under contract no.
DE-AC02-05-CH11231 within the Fundamentals of Semiconductor
Nanowire Program (KCPY23). SCXRD studies were performed
at the UC Berkeley College of Chemistry X-ray Crystallography
(CheXray). We thank the MGCF (Molecular Graphics and
Computation Facility, NIH S10OD023532) and the Kavli Energy
NanoScience Institute (KENI) at UC Berkeley for support on
Materials Studio 2020 and the CASTEP package. C.Z. and M.C.F.
acknowledges support from the Kavli ENSI Philomathia Graduate
Student Fellowship. J.J. acknowledges fellowship support from
Suzhou Industrial Park. H.K.D.L. acknowledges support from the
National Science Foundation’s Graduate Research Fellowship
Program (NSF GRFP) under grant DGE 1752814. X.Z. acknowledges
funding support from NSF 2048200, a DARPA Young Faculty
Award (D20AP00001-02), and the Office of Naval Research
(N00014-23-1-2797). The TRPL measurement was supported by
the US Department of Energy, Office of Science, Office of Basic
Energy Sciences, Materials Sciences and Engineering Division
under contract no. DE-AC02-05-CH11231 (van der Waals
heterostructure program KCFW16). Author contributions: All
authors contributed substantially to the work presented in this
manuscript. C.Z., J.J., and P.Y. conceived the study. C.Z. conducted
the synthesis of the powders, single crystals, inks, and thin films.
C.Z. conducted the PXRD, Raman spectroscopy, UV-vis, PL, PLE, 2D
PLE mapping, PLQY, CIE chromaticity diagram, SEM imaging, and
photodegradation tests. J.J. and C.Z. conducted the SCXRD
measurements. J.J. conducted the DFT calculations and designed
the schematics. Z.W., C.Z., and J.J. took the digital camera images.
Z.W. designed the photoabsorber-free resin for 3D printing. Z.W.,
Z.X., X.Z., and C.Z. conducted the 3D printing experiments. Z.X.,
X.Z., and C.Z. conducted the display demonstrations. M.C.F. and
C.Z. conducted the low-temperature PL measurements. C.B.U.,
F.W., and C.Z. conducted the TRPL measurements. Y.J. conducted the
inductively coupled plasma measurements. H.K.D.L., Y.J., and C.Z.
conducted the SEM-EDX measurements. C.Z., J.J., and P.Y. organized
the manuscript. All authors participated in discussing the results and
providing various sections and comments for the paper. Competing
interests: Lawrence Berkeley National Laboratory has filed a patent
application related to this work for which P.Y. and C.Z. are inventors
(US patent application no. 63/509,821). The remaining authors
declare no competing interests. Data and materials availability:
All data needed to evaluate the conclusions in the paper are present
in the paper or the supplementary materials. The crystallographic
information files (CIFs) have also been deposited in the Inorganic
Crystal Structure Database under reference numbers CSD 2225998,
2225999, and 2292758. These data can be obtained free of
charge through https://www.ccdc.cam.ac.uk/structures/ or by
emailing data_request@ccdc.cam.ac.uk. License information:
Copyright © 2024 the authors, some rights reserved; exclusive
licensee American Association for the Advancement of Science. No
claim to original US government works. https://www.science.org/
about/science-licenses-journal-article-reuse
y
5 January 2024
1. M. A. Reshchikov et al., J. Appl. Phys. 111, 073106 (2012).
2. M. A. Reshchikov, R. Y. Korotkov, Phys. Rev. B 64, 115205 (2001).
3. A. K. Viswanath, J. I. Lee, D. Kim, C. R. Lee, J. Y. Leem,
Phys. Rev. B 58, 16333–16339 (1998).
4. W. Shan et al., Phys. Rev. B 54, 16369–16372 (1996).
5. C. Hauswald et al., Phys. Rev. B 90, 165304 (2014).
6. M. Shibata, T. Furuya, H. Sakaguchi, S. Kuma, J. Cryst. Growth
196, 47–52 (1999).
7. W.-J. Yin, T. Shi, Y. Yan, Appl. Phys. Lett. 104, 063903 (2014).
8. R. J. Sutton et al., Adv. Energy Mater. 6, 1502458 (2016).
9. M. R. Filip, G. E. Eperon, H. J. Snaith, F. Giustino, Nat. Commun.
5, 5757 (2014).
10. L. Protesescu et al., Nano Lett. 15, 3692–3696 (2015).
11. J. Kang, L.-W. Wang, J. Phys. Chem. Lett. 8, 489–493 (2017).
12. K. X. Steirer et al., ACS Energy Lett. 1, 360–366 (2016).
13. F. Deschler et al., J. Phys. Chem. Lett. 5, 1421–1426 (2014).
14. F. Di Stasio, S. Christodoulou, N. Huo, G. Konstantatos,
Chem. Mater. 29, 7663–7667 (2017).
15. J. Li, L. Gan, Z. Fang, H. He, Z. Ye, J. Phys. Chem. Lett. 8,
6002–6008 (2017).
16. S. Wang, C. Bi, J. Yuan, L. Zhang, J. Tian, ACS Energy Lett. 3,
245–251 (2018).
17. X. Gong et al., Nat. Mater. 17, 550–556 (2018).
18. H. Luo, Y. Huang, H. Liu, B. Zhang, J. Song, Chem. Eng. J. 430,
132790 (2022).
19. M. C. Folgueras et al., Nano Lett. 21, 8856–8862 (2021).
20. J. Jin et al., Nano Lett. 21, 5415–5421 (2021).
21. M. Gao et al., Matter 4, 3874–3896 (2021).
22. J. S. Manser, M. I. Saidaminov, J. A. Christians, O. M. Bakr,
P. V. Kamat, Acc. Chem. Res. 49, 330–338 (2016).
23. M. C. Folgueras et al., J. Phys. Chem. C 125, 25126–25139 (2021).
24. D. Ju et al., ACS Energy Lett. 4, 228–234 (2019).
25. A. D. Nicholas, B. W. Walusiak, L. C. Garman, M. N. Huda,
C. L. Cahill, J. Mater. Chem. C 9, 3271–3286 (2021).
26. J. Zhou et al., Adv. Opt. Mater. 7, 1900139 (2019).
27. Z. Tan et al., Adv. Mater. 32, 2002443 (2020).
28. H. Yin et al., Angew. Chem. Int. Ed. 60, 22693–22699
(2021).
29. I. N. Douglas, J. V. Nicholas, B. G. Wybourne, J. Chem. Phys.
48, 1415–1416 (1968).
30. C. Li et al., Acta Cryst. B64, 702–707 (2008).
31. Q. He et al., ACS Mater. Lett. 2, 633–638 (2020).
32. C. Zhou et al., Chem. Sci. 9, 586–593 (2017).
33. M. Li, Z. Xia, Chem. Soc. Rev. 50, 2626–2662 (2021).
34. J. A. Steele et al., ACS Nano 12, 8081–8090 (2018).
35. K. Saeki et al., Jpn. J. Appl. Phys. 57, 030310 (2018).
36. A. Abfalterer et al., ACS Mater. Lett. 2, 1644–1652 (2020).
37. C. Kaewmeechai, Y. Laosiritaworn, A. P. Jaroenjittichai,
Results Phys. 30, 104875 (2021).
38. W. Thomas, H. Elias, J. Inorg. Nucl. Chem. 38, 2227–2229
(1976).
39. E. V. van Loef, G. Ciampi, U. Shirwadkar, L. Soundara Pandian,
K. S. Shah, J. Cryst. Growth 532, 125438 (2020).
40. S. Kodama et al., Opt. Mater. 106, 109942 (2020).
41. C. Zhu et al., J. Am. Chem. Soc. 144, 12450–12458 (2022).
42. J. E. Saal, S. Kirklin, M. Aykol, B. Meredig, C. Wolverton,
J. Miner. Met. Mater. Soc. 65, 1501–1509 (2013).
43. S. Kirklin et al., NPJ Comput. Mater. 1, 15010 (2015).
44. Q. A. Akkerman et al., J. Am. Chem. Soc. 137, 10276–10281
(2015).
45. Y. Toyozawa, Prog. Theor. Phys. 27, 89–104 (1962).
46. W. Stadler et al., Phys. Rev. B 51, 10619–10630 (1995).
47. Y. Song et al., Mater. Res. Express 6, 115064 (2019).
48. S. J. Zelewski et al., J. Mater. Chem. C 7, 8350–8356 (2019).
49. V. Vaněček et al., J. Cryst. Growth 573, 126307 (2021).
50. K. Saeki, Y. Fujimoto, M. Koshimizu, T. Yanagida, K. Asai,
Appl. Phys. Express 9, 042602 (2016).
51. H.-H. Kuo et al., Adv. Sci. 5, 1800846 (2018).
y g
Zhu et al., Science 383, 86–93 (2024)
AC KNOWLED GME NTS
RE FERENCES AND NOTES
y
We have demonstrated a supramolecular assembly strategy for achieving halide perovskite
blue and green emitters with ultrahigh PLQYs.
Specifically, (18C6@K)2HfBr6 warranted a blue
emission with a near-unity (96.2%) PLQY, and
(18C6@K)2ZrCl4Br2 showed a green emission
with a PLQY of 82.7%. The emission of the
supramolecular assembled samples originated
from the STE emission, with strong electronphonon coupling and microsecond PL lifetimes. The supramolecular approach is very
promising for solution processability. The
(18C6@K)2HfBr6/PS-DCM ink maintained a
high PLQY of >90%. Uniform thin films were
fabricated from this ink through a drop-casting
technique. The (18C6@K)2HfBr6/PS composite had blue emission with a PLQY of >80%,
making it favorable for patterning, display,
52. H.-H. Kuo et al., J. Mater. Chem. C 6, 10486–10496 (2018).
53. S. Schmidbauer, A. Hohenleutner, B. König, Beilstein J. Org. Chem.
9, 2088–2096 (2013).
54. W. Hui et al., Science 371, 1359–1364 (2021).
55. F. Mathies et al., J. Mater. Chem. A 4, 19207–19213
(2016).
56. S. X. Li et al., ACS Appl. Mater. Interfaces 13, 31919–31927 (2021).
57. M. Kaseem, K. Hamad, Y. G. Ko, Eur. Polym. J. 79, 36–62 (2016).
58. Y. Tu et al., J. Mater. Chem. 20, 1594–1599 (2010).
59. H. Cui et al., Science 376, 1287–1293 (2022).
60. R. Hensleigh et al., Nat. Electron. 3, 216–224 (2020).
g
Discussion
and printing applications. The powders with
blue and green emissions were also highly
compatible with the 3D printing technology.
The supramolecular assembly approach for
halide perovskite building block catalyzes further investigation into the synthesis and characterization of supramolecular assembled
functional materials, laying the foundation
for substantial progress in the field.
p
architectural models of the Eiffel Tower (Fig.
5B), after excitation at 254 nm, showed their
respective blue and green colors (Fig. 5C). The
dimensions of the two Eiffel Towers were within
a few centimeters, with high-resolution spatial
features (Fig. 5B). The submicrometer scale of
these powders and a printing layer thickness
of 40 mm enabled even distribution throughout each layer and ensured a homogeneous
emission color profile across the entire architectural construct.
A single 3D-printed structure could also manifest emissions in both blue and green by alternative resins during the printing procedure. An
Eiffel Tower design characterized by blue emissions at its upper and lower segments with
green emissions in its central region is shown
in Fig. 5D, and a second-order hierarchical
lattice structure (octet truss) was realized with
one half radiating in blue and the other in
green (Fig. 5E). Notably, a close-up view of the
boundary between these blue- and greenemitting regions within the octet truss structure
revealed the high precision in color transition
without any color crossover on either side.
Twisted (Fig. 5F and fig. S41) and conformal
(Fig. 5G and fig. S42) octet truss architectures
with dual emissions were also achieved with
bright emissions and high structural accuracy.
Other complex topologies, such as cuboctahedron, tetrakaidecahedron, octet truss,
and Menger sponge with the blue emitter embedded (Fig. 5H and figs. S43 and S44), were
also obtained to exhibit the variety of structures that could be printed with the lightemitting ink. These demonstrations served
as a proof of concept for integrating emissive
ionic powders with 3D printing technology.
The potential applications of 3D-printed lightemitting structures are extensive and constantly evolving, ranging from intricate interior
ambient-lighting solutions to seamless integration into wearable devices.
RES EARCH
CATALYSIS
Dealuminated Beta zeolite reverses Ostwald ripening
for durable copper nanoparticle catalysts
Lujie Liu1†, Jiaye Lu2†, Yahui Yang3, Wolfgang Ruettinger3, Xinhua Gao4, Ming Wang5, Hao Lou6,
Zhandong Wang6, Yifeng Liu7, Xin Tao8, Lina Li8, Yong Wang9, Hangjie Li1, Hang Zhou1,
Chengtao Wang1, Qingsong Luo1, Huixin Wu1, Kaidi Zhang1, Jiabi Ma5*, Xiaoming Cao2,10*,
Liang Wang1*, Feng-Shou Xiao1,11*
Copper nanoparticle–based catalysts have been extensively applied in industry, but the nanoparticles tend
to sinter into larger ones in the chemical atmospheres, which is detrimental to catalyst performance. In this
work, we used dealuminated Beta zeolite to support copper nanoparticles (Cu/Beta-deAl) and showed that
these particles become smaller in methanol vapor at 200°C, decreasing from ~5.6 to ~2.4 nanometers in
diameter, which is opposite to the general sintering phenomenon. A reverse ripening process was
discovered, whereby migratable copper sites activated by methanol were trapped by silanol nests and
the copper species in the nests acted as new nucleation sites for the formation of small nanoparticles.
This feature reversed the general sintering channel, resulting in robust catalysts for dimethyl oxalate
hydrogenation performed with supported copper nanoparticles for use in industry.
,
1 of 8
y
5 January 2024
y g
Liu et al., Science 383, 94–101 (2024)
y
*Corresponding author. Email: majiabi@bit.edu.cn (J.M.);
xmcao@ecust.edu.cn (X.C.); liangwang@zju.edu.cn (L.W.);
fsxiao@zju.edu.cn (F.-S.X.)
†These authors contributed equally to this work.
We performed a proof-of-concept experiment
by comparing the stability of Beta-deAl (Cu/
Beta-deAl; figs. S1 and S2) and amorphous
silica–supported Cu NPs (Cu/SiO2, Cu loading
content at 3.0 wt %) in methanol vapor. The
Cu NP sizes and distributions were initially
characterized by scanning transmission electron microscopy (STEM). The fresh Cu/SiO2
(as prepared by calcination in air and prereduction in hydrogen) showed that the NPs had
an average size of 3.0 ± 0.8 nm (fig. S3, A and
D). After the methanol treatment at 200°C for
12 hours (methanol feeding rate of 0.03 mlliquid
min−1 in hydrogen with a flow rate of 15 ml
min−1), the Cu NPs on Cu/SiO2 sintered into
ones with an average size of 4.0 ± 1.4 nm (fig.
S3, B and E), and some NPs were larger than
7.0 nm (fig. S3, C and E). Methanol-triggered
Cu sintering is known to occur under similar
conditions and leads to the deactivation of Cu
catalysts in industrial processes (7, 28–30).
However, the Cu NPs on the Cu/Beta-deAl (Cu
content at 3.0 wt %) exhibited an opposite
phenomenon after the equivalent methanol
treatment. The initial average Cu NP size of
5.6 ± 1.6 nm (fresh catalyst; Fig. 1A and fig. S4,
A and B) was reduced to 2.4 ± 0.7 nm, and
we could not detect NPs larger than 6.5 nm
(Fig. 1B and fig. S4, C and D). The Cu content
(3.0 wt %) was unchanged after the methanol
treatment. These results suggest a redispersion of Cu NPs on the Cu/Beta-deAl in methanol vapor.
The redispersion of Cu NPs on Cu/Beta-deAl
was further confirmed by in situ x-ray diffraction (XRD) characterization. Compared with
the bare Beta-deAl zeolite, the fresh Cu/BetadeAl exhibited peaks at 43.3° and 36.5° that
we assigned to metallic Cu and Cu2O phases,
respectively (31, 32) (Fig. 1C and fig. S5). Methanol treatment reduced these diffraction peaks
until they disappeared after 12 hours, which
confirmed the elimination of large Cu or Cu2O
particles. These results were consistent with those
of the TEM characterizations and supported
the reversed Cu sintering on the Cu/Beta-deAl
in methanol vapor (Fig. 1D). By contrast, after
the equivalent methanol vapor treatment, the
diffractions of Cu-related NPs were enhanced
on the Cu/SiO2 because of the formation of
larger Cu NPs (fig. S6). These results suggest
that the Beta-deAl zeolite was crucial for the
redispersion of Cu. The Cu NPs on other zeolites,
g
1
Key Lab of Biomass Chemical Engineering of Ministry of
Education, College of Chemical and Biological Engineering,
Zhejiang University, Hangzhou 310027, China. 2Joint
International Research Laboratory of Precision Chemistry
and Molecular Engineering, Feringa Nobel Prize Scientist
Joint Research Center, School of Chemistry and Molecular
Engineering, East China University of Science and
Technology, Shanghai 200237, China. 3BASF Advanced
Chemicals Co., Ltd., Shanghai 200137, China. 4State Key
Laboratory of High-efficiency Utilization of Coal and Green
Chemical Engineering, School of Chemistry and Chemical
Engineering, Ningxia University, Yinchuan 750021, China.
5
Key Laboratory of Cluster Science of Ministry of Education,
School of Chemistry and Chemical Engineering, Beijing
Institute of Technology, Beijing 102488, China. 6National
Synchrotron Radiation Laboratory, University of Science and
Technology of China, Hefei, Anhui 230029, China.
7
Department of Chemistry, Zhejiang University, Hangzhou
310027, China. 8Shanghai Institute of Applied Physics,
Chinese Academy of Sciences, Shanghai 201204, China.
9
Center of Electron Microscopy and State Key Laboratory of
Silicon Materials, School of Materials Science and
Engineering, Zhejiang University, Hangzhou 310027, China.
10
School of Chemistry and Chemical Engineering, Shanghai
Jiao Tong University, Shanghai 200240, China. 11Beijing
Advanced Innovation Center for Soft Matter Science and
Engineering, Beijing University of Chemical Technology,
Beijing 100029, China.
Redispersion of the sintered metal NPs into
smaller ones on the support surface could
efficiently reverse deactivation. Sintering often
occurs by the loss of mobile metal atoms from
small NPs and diffusion and capture by larger
NPs, a process known as Ostwald ripening
(18–20). Reversing this process would require
activation of the metal sites on large NPs and
the capture of these metal species by the support. However, this process requires hybrid
supports that have distinguishable interactions
with the metal species that cause them to move
from a weak-interaction region to a stronginteraction region (9, 21). For example, Cu
species migrated from silica to the ceria on the
SiO2-CeO2 mixed supports to form smaller
clusters (21), but this movement changed the
intrinsic catalytic properties of the Cu NPs.
The reverse ripening of metal NPs on a
homogeneous support should return the sintered catalysts to ones with identical performances to the original catalysts, but achieving
this is thermodynamically challenging. Treatment with halogens that coordinate with the
metal sites can initiate their emission from
bulky NPs and transmission on the support.
Such postregeneration of the deactivated catalysts (22–24) has been used in industrial processes, such as oxychlorination for redispersing
Pt NPs on alumina support (23), which can
generate environmentally unfriendly wastes
and corrode the reactor. Ideally, in addition
to the postregeneration of deactivated catalysts, the redispersion that occurs under the
reaction processes would realize a catalyst with
superior durability against sintering (25–27).
We report that dealuminated Beta zeolite
(Beta-deAl, Si/Al atomic ratio at ~1250; fig. S1)
is an efficient support for the reverse ripening
of Cu NPs in methanol vapor at 200°C. This
feature maintained small Cu NPs (2 to 3 nm)
and even redispersed large Cu particles (15 to
Redispersion of Cu NPs
p
M
etal nanoparticles (NPs) on the surface of heterogeneous catalysts encounter reactant and product molecules
at high temperatures (1–7), and these
conditions often cause the NPs to
sinter into larger ones. The resulting loss of
surface area and reduced number of active sites
can lead to irreversible deactivation (1–3, 8–12),
and it costs billions of dollars per year to shut
down the reaction process and replace the
catalysts after deactivation (13). For metals
with low melting temperatures and, hence,
higher surface diffusion, such as copper, sintering can lead to deactivation for various hydrogenation (14, 15) and reforming reactions
(16, 17).
250 nm) on siliceous Beta-deAl support. We
focused on dimethyl oxalate (DMO) hydrogenation, a structure-dependent reaction that has
been industrially used for the conversion of
coal to ethylene glycol (EG) (28, 29). As a result, the Cu/Beta-deAl catalyst maintains high
conversion and selectivity in a continuous reaction for long periods, even during ambientpressure DMO hydrogenation.
RES EARCH | R E S E A R C H A R T I C L E
A
B
5 nm
fresh
methanol-treated
2
50 nm
C
Cu2O
4
6 8 10 12 14
Diameter (nm)
50 nm
D
Cu
6
fresh
6h
8h
Si
10 h
Si
12 h
Beta-deAl
30
40
50
60
HO
OH
HO
Si
silanol nest
70
2 (°)
We evaluated Cu/SiO2 and Cu/Beta-deAl in
the hydrogenation of DMO in methanol vapor
at 200°C with a liquid hourly space velocity
(LHSV) of 0.8 hour−1 (mlDMO mlcat−1 hour−1)
based on a feeding rate of DMO per milliliter
of catalyst. The data that characterize the performances are shown in Fig. 2A, where the
initial DMO conversions were maintained at
~40% for evaluating the catalyst durability.
Liu et al., Science 383, 94–101 (2024)
5 January 2024
2 of 8
,
Catalytic evaluation
The Cu/SiO2 catalyst showed continuously decreased DMO conversion with reaction time;
the conversion of 35.2% after 24 hours dropped to 19.8% after 80 hours and to 9.1% after
110 hours. TEM characterization of the spent
Cu/SiO2 catalyst after testing for 80 hours
showed an average Cu NP size of 4.4 ± 1.3 nm
(fig. S13), which was larger than that of the
fresh catalyst (3.0 ± 0.8 nm). In previous studies
of DMO hydrogenation, abundant methanol
in the reaction system, from the solvent and
product, also caused Cu sintering (28–31).
In the DMO hydrogenation under equivalent conditions for Cu/Beta-deAl, we observed
improved DMO conversion over time; conversion increased from 33.4 to 45.0% in the initial
12 hours and then remained at 45.1 to 48.1% in
the test for another 188 hours (Fig. 2A). TEM
y
such as commercial Beta (Si/Al ratio at 13),
ZSM-5 (aluminosilicate MFI zeolite with
Si/Al ratio at 13), S-1 (siliceous MFI zeolite),
and silanol-modified MFI zeolites (ZSM-5-OH
and S-1-OH), also readily sintered into larger
NPs after the methanol treatment at 200°C
(figs. S7 to S12).
y g
Fig. 1. Redispersion of Cu on the Beta-deAl support by methanol treatment. (A and B) High-angle
annular dark-field STEM images of fresh Cu/Beta-deAl (A) and Cu/Beta-deAl (B) after the methanol
treatment at 200°C for 12 hours. The insets in (B) show Cu NP size distributions of the fresh and methanoltreated catalysts (bottom left) and a high-resolution TEM image of the methanol-treated catalyst (top
right). (C) In situ XRD patterns characterizing the change of Cu NPs on the Cu/Beta-deAl during the
methanol treatment at 200°C. (D) Illustration showing the reverse Ostwald ripening of Cu NPs on the BetadeAl support. The silanol nest, which is formed after removing the tetracoordinated Al sites from the
Beta zeolite framework, usually involves four silanols in a local region with hydrogen bonds between them.
y
6
Si
OH
g
Intensity
4h
p
methanol-treated for
2h
characterizations of the spent Cu/Beta-deAl
after reaction for 10 hours showed an average Cu NP size of 2.3 ± 0.4 nm (fig. S14) versus
5.6 ± 1.6 nm for the fresh sample. This result
confirmed a self-optimization process in the
initial reaction. Interestingly, the formed smallsize Cu NPs on the Cu/Beta-deAl were constant
during the reaction period, and the average Cu
NP size was 2.5 ± 0.7 nm on the spent Cu/BetadeAl after testing for 80 hours (fig. S15). In the
reaction with 1,4-dioxane as solvent, the Cu/
Beta-deAl catalyst also exhibited constant performance, with an average Cu NP size of 2.4 ±
0.9 nm after reaction for 120 hours (figs. S16
and S17).
We then adjusted the LHSV of DMO for
evaluating the catalysts with high or even full
conversion of DMO. After pretreatment with
methanol vapor at 200°C for 10 hours, the Cu/
Beta-deAl exhibited full DMO conversion under
given reaction conditions (LHSV at 0.4 hour−1,
200°C) with EG selectivity >98.5% (Fig. 2B).
Increasing the LHSV to 0.8 hour−1 resulted in
a lower DMO conversion at 45.4% with methyl
glycolate (MG) from semihydrogenation as a
dominant product. This incomplete conversion provided a way to evaluate the activity
change of the Cu catalysts. In a continuous
test for 120 hours, we periodically switched
the LHSV between 0.4 and 0.8 hour−1 every
30 hours, and the Cu/Beta-deAl exhibited constant performance in each period. For example, after the test in which the LHSV was at
0.8 hour−1 and was then switched back to
0.4 hour−1, the Cu/Beta-deAl catalyst gave a
DMO conversion and EG selectivity similar to
those in the initial reaction. By contrast, the
Cu/SiO2 showed continuously decreased DMO
conversion with reaction time, suggesting a
deactivation process (Fig. 2B).
The durability of Cu/Beta-deAl was further
evaluated under a constant condition (LHSV
at 0.4 hour−1, 200°C) for 200 hours. After an
activation period of 12 hours, the DMO was
fully converted, with EG selectivity up to 98.7%
(Fig. 2C). In this case, the selectivity to ethanol
and other heavier products was <0.5%. A longer reaction life was reasonably expected because of the undetectable deactivation trend
and unchanged Cu NP size (average size of
2.7 ± 0.7 nm; figs. S18 and S19) after the test.
However, the Cu NPs on the other zeolites,
such as S-1, ZSM-5, and aluminosilicate Beta
zeolites, exhibited EG yields that were much
lower than that of the Cu/Beta-deAl (fig. S20).
A high EG selectivity under full conversion
of DMO at sufficient LHSV is required for potential commercial utilization. By raising the
Cu loading content to 18 wt % on Beta-deAl
support, DMO hydrogenation was efficiently
catalyzed under a rapid feed. For example, at
200°C with a LHSV of 1.8 hour−1, an EG yield
of ~96.2% at full DMO conversion was obtained without any deactivation trend in the
RES EARCH | R E S E A R C H A R T I C L E
90
90
80
80
MG sel.
70
60
70
60
Conv.
50
50
40
40
EG sel.
30
30
20
20
LHSV,
0.8 h-1
10
B
100
10
90
0
20
40
60
80
70
60
50
40
30
60
MG sel.
50
EG sel.
40
30
20
20
LHSV,
0.4 h-1
10
20
O
O
50
OH
(EG), major
+
(DMO)
O
40
OH
30
70
60
50
40
O
(MG), minimal
30
20
10
MG sel.
LHSV,
0.4 h-1
0
10
0
0
20
40
60
80
100 120 140 160 180 200
Time on stream (hours)
methanol, 400°C
40
20
0
Commercial Cu/SiO2-C
Cu/Beta-deAl-18%
in DMO hydrogenation. Reaction conditions were as follows: 200°C, 2.5 MPa,
and LHSV of 0.4 hour−1. (D) Data showing the EG yields in DMO hydrogenation
over fresh and methanol-treated commercial Cu/SiO2-C and Cu/Beta-deAl18% catalysts. Conditions for methanol treatment were as follows: methanol
feeding rate of 0.03 mlliquid min−1, 400°C, and 48 hours. The error bars represent
standard deviation.
whereas the Cu/SiO2-C was obviously deactivated, given the extremely low EG yield at
3.9% (DMO conversion at 45.4%, EG selectivity at 8.6%). Characterizations by XRD and
TEM showed that the sintered Cu NPs had an
average diameter of up to 7.4 ± 3.3 nm on the
Cu/SiO2-C after the methanol treatment at
400°C (fig. S22), whereas the Cu/Beta-deAl18% exhibited Cu NPs with an average size of
3.8 ± 1.0 nm and even had abundant ones that
were smaller than 3.0 nm (fig. S23).
The DMO hydrogenation at ambient pressure poses greater challenges to catalyst stability. A previous study achieved efficient DMO
hydrogenation at ambient pressure on a Cu/
SiO2 catalyst with fullerene as an electron
buffer, but the Cu sintering still occurred, as
suggested by the XRD characterization (28).
The Cu/SiO2 catalyst without an electron buffer
deactivated quickly during the ambient-
pressure reaction (28, 34–36). Interestingly,
the Cu/Beta-deAl-18% could efficiently catalyze the DMO hydrogenation, resulting in an
EG yield of 90.8% at 200°C under an LHSV of
0.6 hour−1 and an EG yield of 90.1% at 230°C
at an LHSV of 3.6 hour−1; both reactions were
performed at ambient pressure (fig. S24 and
table S3). This catalyst was stable in the continuous reaction for 300 hours and maintained
a Cu NP size of 4.0 ± 1.0 nm (figs. S24 and S25).
In addition to the Cu sintering, another issue
that challenges the durability of general CuSiO2–based catalysts is silica leaching, in which
the tetramethoxysilane-related species is formed
in methanol vapor and leached from the catalyst (37, 38). By contrast, this problem was efficiently avoided on the Cu/Beta-deAl catalyst
(fig. S26) because of the superior stability of
crystallized zeolite in methanol vapor (39–41)
relative to the amorphous silica.
3 of 8
,
5 January 2024
60
y
Liu et al., Science 383, 94–101 (2024)
80
y g
Fig. 2. Catalytic tests in DMO hydrogenation. (A) Catalytic performances
of the Cu/SiO2 (open circles) and Cu/Beta-deAl (solid circles) in DMO
hydrogenation. Reaction conditions were as follows: 200°C, 2.5 MPa, LHSV of
0.8 hour−1, and n(H2)/n(DMO) of 45. (B) Effect of the methanol treatment (initial
10 hours) on the performances of the Cu/SiO2 and Cu/Beta-deAl catalysts in
DMO hydrogenation. (C) Data showing the durability of the Cu/Beta-deAl catalyst
test for 200 hours (fig. S21). By contrast, the
commercial silica-supported Cu-based catalyst (Cu/SiO2-C) under the same conditions
showed an EG yield at 90.1 to 93.4% (table
S1). Even after raising the LHSV as high as
3.0 hour−1, the Cu/Beta-deAl-18% still catalyzed the full DMO conversion with an EG
selectivity at 95.8% (table S2).
The DMO hydrogenation is exothermic, so
inadequate heat transfer could form hot regions in the industrial reactor that cause irreversible catalyst deactivation from Cu sintering
(33). To evaluate thermal stability under harsh
conditions, the Cu/Beta-deAl-18% and commercial Cu/SiO2-C were treated in methanol vapor
at 400°C for 48 hours and then tested in the
DMO hydrogenation. As shown in Fig. 2D, the
EG yields over the Cu/Beta-deAl-18% (full DMO
conversion with EG selectivity at 96.5%) were
unchanged before and after the treatment,
120
y
20
100
100
EG yield (%)
Cu/Beta-deAl
80
g
Conversion (%)
60
60
D
Selectivity to MG or EG (%)
O
O
10
Time on stream (hours)
80
HO
LHSV,
0.8 h-1
p
80
70
40
EG sel. 90
Conv.
LHSV,
0.4 h-1
0
0
100
90
LHSV,
0.8 h-1
0
100 120 140 160 180 200
100
80
70
Time on stream (hours)
C
90
Conv.
80
0
0
100
100
Selectivity to MG or EG (%)
Cu/Beta-deAl
methanol treatment
Cu/SiO2
Conversion (%)
100
Selectivity to MG or EG (%)
Conversion (%)
A
RES EARCH | R E S E A R C H A R T I C L E
A
CuO
Cu2O
Cu foil
B
C
methanol-treated
Cu/Beta-deAl
2
Cu
fresh Cu/Beta-deAl
fresh
fresh Cu/SiO2
2
methanol-treated
for 10 h
Intensity
methanol-treated
Cu/SiO2
FT k 2 (k)
XANES intensity
methanol-treated Cu/Beta-deAl
fresh Cu/Beta-deAl
methanol-treated Cu/SiO2
fresh Cu/SiO2
30 h
CuO
70 h
Cu2O
120 h
Cu foil
190 h
8960
9000
9040
9080
0
9120
1
2
3
4
5
6
20
30
R (Å)
Energy (eV)
50
60
p
D
40
2 (°)
F
E
bulky Cu
g
1.2 1.8 2.4 3.0 3.6 4.2
1.2 1.6 2.0 2.4 2.8 3.2
Diameter (nm)
Diameter (nm)
y
50 nm
Beta -deAl
G
bulky Cu
50 nm
50 nm
Cu powder
Cu NP
daverage size
y g
~2.2 nm
Beta-deAl
70 h
190 h
y
Characterizations
In situ XRD characterizations of the Cu/BetadeAl in methanol vapor at different temperatures showed that the redispersion of Cu NPs
was undetected at 100°C (fig. S27A) but detected at 200°C (fig. S5) and accelerated at
300°C (fig. S27B). Further raising the treatment
temperature to 400°C resulted in Cu NPs of
similar size to those obtained with the fresh
Cu/Beta-deAl catalyst (figs. S27C and S28).
Liu et al., Science 383, 94–101 (2024)
5 January 2024
These results suggest that the redispersion of
the Cu NPs was related to the temperature. The
influence of methanol partial pressure was investigated at 200°C, where the higher methanol
partial pressure (e.g., >67 kPa) would benefit
the Cu redispersion (fig. S29). A general view on
the methanol-triggered Cu sintering includes
methanol decomposition to form CO and COinitiated Cu migration (30, 34, 42). However,
CO was unable to change the average diameter
of Cu NPs by exposure of Cu/Beta-deAl to a CO
atmosphere at 200°C (figs. S30 and S31). The
other molecules, such as H2, N2, and water, were
also unable to decrease the Cu NP sizes (figs. S30
and S31). Formaldehyde, which was produced
from the methanol dehydrogenation (fig. S32),
greatly promoted the redispersion of Cu particles, resulting in an average Cu NP size of 2.3 ±
0.9 nm for the formaldehyde-treated Cu/Beta-deAl
sample (figs. S30 and S33).
4 of 8
,
Fig. 3. Methanol-triggered structure change of the Cu particles. (A) Normalized Cu K-edge XANES spectra of fresh and methanol-treated Cu/SiO2 and Cu/BetadeAl catalysts. The methanol treatments were performed at 300°C for 12 hours. (B) Fourier-transformed (FT) magnitudes of the experimental Cu K-edge EXAFS
spectra. The plot lines with open circles show fitting results of samples. R, bond distance. (C to F) XRD patterns (C) and STEM images [(D) to (F)] characterizing the
mixture of Cu powder and Beta-deAl zeolite during the methanol treatment at 200°C for different periods. The insets in (E) and (F) show particle size distribution of
small Cu NPs. (G) Illustration showing the formation of small-size Cu NPs on the Beta-deAl zeolite from the physically mixed commercial Cu powder. d, diameter.
RES EARCH | R E S E A R C H A R T I C L E
,
5 of 8
y
5 January 2024
y g
Liu et al., Science 383, 94–101 (2024)
Previous studies have shown that the Cu on
relatively inert supports such as silica sinters
through particle migration and coalescence
under thermal conditions but tends to proceed
by Ostwald ripening under chemical triggering
conditions (8, 20, 43), such as the hydrogenation in methanol vapor. The ripening route
involves the formation and diffusion of metal
atoms, complex intermediates, or both (20, 44, 45).
A support with strong interactions might trap
the metal atoms or complex intermediates. We
used synchrotron vacuum ultraviolet photoionization mass spectrometry (SVUV-PIMS;
fig. S41) to detect Cu species in methanol vapor.
In these tests, methanol was introduced to the
physical mixture of Cu powder with Beta-deAl,
and the possible intermediates were captured
through a vacuum channel and detected by
mass spectrometry. The signals of methoxyl
[mass/charge ratio (m/z) of 31] and formaldehyde (m/z of 30) species from methanol
conversion (m/z of 32) appeared (Fig. 4A), as
well as other possible species of (CHO)Cu1*
(m/z of ~92 and 94), (CH2O)Cu1* (m/z of ~93
and 95), and (CH3O)Cu1* (m/z of ~94 and 96)
(Fig. 4B). These results suggest the existence of
Cu1-related intermediates during the migration.
In addition, the signal intensities for (CHO)Cu1*
and (CH3O)Cu1* gradually increased with temperature during the methanol treatment (fig.
S42), which is related to the easier formation
and faster escape of these intermediates at
higher temperatures. These insights are in good
agreement with general knowledge about the
Ostwald ripening processes with chemicaltriggered metal sintering (8, 20, 44–47), but here
we provide direct experimental evidence and also
reverse this process with the Beta-deAl support.
We also performed density functional theory (DFT) calculations, in which we first considered the desorption of a Cu atom from the
Cu NP matrix as the primary step that initiates
ripening. The direct detachment of a Cu atom
from the matrix is difficult because of the strong
Cu–Cu bonding, but the methanol-derived species could coordinate with the surface Cu atom
and induce its migration. We considered the
formation process of methanol-derived species
(Fig. 4C and figs. S43 and S44) and showed
y
The redispersion of Cu on the Beta-deAl zeolite
was further confirmed by methanol treatment
of a physical mixture containing Beta-deAl
and commercial Cu powders (15 to 250 nm;
figs. S34 and S35). The XRD patterns recorded
the change of Cu crystals in methanol vapor
for different periods. The diffraction peaks
assigned to Cu crystals decreased in intensity during the treatment period, which suggested a continuous migration of Cu species
from the bulky crystals (Fig. 3C). A STEM
image of the fresh sample showed only bulky
Cu particles with undetectable small Cu NPs
(Fig. 3D). The small-size Cu NPs were observed
on the Beta-deAl support when methanol was
introduced to the mixture at 200°C (an initial
stage of 5 hours; fig. S36). After the methanol
treatment at 200°C for 70 hours, more Cu NPs
appeared with an average particle size of 2.1 ±
0.4 nm (Fig. 3E and fig. S37; bulky ones were
not included in the calculation), whereas the
bulky Cu particles were still observed but their
sizes were smaller than those of the parents.
After the treatment in methanol vapor for
190 hours, the Cu peaks were undetectable in
Mechanism study
that the production of *CH3O, *CHO, and *CO
was exothermic, whereas the production of
other species was endothermic (figs. S43 and
S44 and table S5). Lower energy inputs for the
Cu detachment and migration than for the
ligand desorption for (CHO)Cu1* and (CH3O)
Cu1* suggest that the migration is more favorable in the reverse ripening process (fig. S43).
For example, the *CHO exhibited an energy input of 0.42 eV for Cu detachment and migration from the Cu matrix to the silanol nest (four
silanols in a local region; fig. S45), whereas direct desorption of the *CHO without Cu detachment required 1.03 eV (fig. S43). These
results indicated that the complexes would
preferentially detach the Cu atoms from the
surface of the Cu matrix to form the intermediates that were ready for the migration,
which is consistent with the results of SVUVPIMS. In this case, lower energy inputs for
(CHO)Cu1* migration from the Cu matrix to
the silanol nest than for (CH3O)Cu1* suggest
that (CHO)Cu1* is a preferential intermediate
in the reverse ripening process.
The *CHO could be further dehydrogenated
into *CO, but the *CO was unable to assist
with the detachment of Cu atoms from the Cu
matrix; thus, CO would be directly desorbed at
the reaction temperature (fig. S43), which is in
good agreement with the experimental results
(fig. S30). In addition, the free-energy barrier
for the dehydrogenation of *CHO to *CO was
slightly lower than that of (CHO)Cu1* moving
from the matrix to the Cu/Beta-deAl surface
(0.29 eV). These results indicated that *CHO
decomposition to CO competes with the detachment of (CHO)Cu1* (fig. S44).
We further studied the methanol-triggered
Cu detachment by exploring the reactions between the gas-phase Cux+ clusters and methanol
with time-of-flight mass spectrometry (48, 49).
In our tests, the 63Cu2+ cations were generated
by laser ablation of the Cu matrix surface, and
then methanol was introduced for reaction. As
shown in Fig. 4D, the peak intensity of 63Cu2+
decreased after reaction with methanol and
led to the formation of abundant Cu1 species
[63Cu1(CH3OH)+]. Increasing the partial pressure of methanol further reduced the Cu2+
and enhanced the Cu1+ signal magnitudes. Although the reactions of 63Cu2+ with CH3OH
could not fully represent the types of ligandCu intermediate because of the obvious scale
difference between the atomic 63Cu2+ cluster
and Cu NP surface, these results strongly supported that methanol triggers Cu–Cu bond
cleavage. In sharp contrast, the 63Cu2+ cations
were inert toward CO and H2 under similar
reaction conditions, which confirms the undetectable ability of these gases to trigger
the Cu detachment (Fig. 4D). These results were
also in good agreement with the results obtained
when Cu/Beta-deAl catalyst was treated with CO
and H2 (figs. S30 and S31).
g
Redispersion of bulky Cu particles
the XRD pattern, which confirmed the removal of bulky Cu crystals. A STEM image of
the sample showed an average Cu NP size of
2.2 ± 0.5 nm on the Beta-deAl support, with
undetectable bulky Cu particles (Fig. 3F and
figs. S37 and S38). The equivalent test on the
mixture of commercial Cu powders with amorphous silica resulted in unchanged XRD patterns, and small Cu NPs were absent (figs. S39
and S40). Based on these results, we present a
schematic illustration showing the formation
of small-size Cu NPs on the Beta-deAl zeolite
from the bulky Cu crystals (Fig. 3G).
p
We characterized the Cu/SiO2 and Cu/BetadeAl catalysts before and after the methanol
treatment at 300°C (the temperature required
to amplify sintering or redispersing NPs on
the catalysts) by x-ray absorption fine structure spectroscopy. The x-ray absorption nearedge structure (XANES) spectrum of fresh
Cu/SiO2 fell between those of Cu foil and Cu2O,
which suggested a multivalent Cu0 and Cu+
composition (Fig. 3A) that was consistent with
the general feature of the Cu/SiO2 with Cud+-OSiOx interaction (7, 28–31, 34, 43). The methanoltreated Cu/SiO2 was more metallic because
the spectrum was near that of the Cu foil that
was caused by Cu NPs sintering that destroyed
the interaction with silica. Extended x-ray absorption fine structure (EXAFS) analysis (Fig.
3B) showed an increased coordination number (CN) of the Cu-Cu shell (3.5 to 8.2) and the
absence of Cu-O or Cu-O-Cu signals after the
methanol treatment (table S4).
By contrast, the fresh Cu/Beta-deAl exhibited a XANES spectrum near that of Cu foil.
After the methanol treatment, the Cu species
were partially oxidized, giving a XANES spectrum near that of Cu2O. The EXAFS fitting
results showed the CN of the Cu-Cu shell at
9.0 on the fresh Cu/Beta-deAl, which dropped
to 2.3 with the CN of the Cu-O shell at 1.5 and the
CN of the Cu-O-Cu shell at 1.7 on the methanoltreated sample (Fig. 3B and table S4). Considering that the methanol treatment with hydrogen
carrier provided a reductive environment, the
formation of more-oxidized Cu species could
be attributed to the formation of a Cud+-O-SiOx
interaction during the methanol-triggered redispersion of Cu.
RES EARCH | R E S E A R C H A R T I C L E
A
C
B
20000
Intensity
Free energy (eV)
CH3OH
Intensity
2
CH3O*
*CHO
100
CH3O
(CH3O)Cu1*
(CHO)Cu1*
0.90 eV
1
CH3OH*
0
1.08 eV
1.09 eV 0.81 eV
0.96 eV
CH3OH(g) CH3O*
*CHO
HCHO
0.88 eV
0.70 eV
Formation
Migration
-1
30
31
92
32
m /z
93
94
95
96
D
63
E
Cu2+
Gas -phase migration
(CHO)Cu1*(g)
He
0.68 eV
63
Cu1(CH3OH)+
63
+
Cu2H
Intensity
CH3OH, 0.65 Pa
H
H
H
O
O
O
-0.26 eV
OH
silanol nest
Si
Si
Si
O O
CH3OH, 1.15 Pa
(CHO)Cu1*(g)
CO
60
70
80
90
100
110
120
130
140
Beta-deAl
Surface migration
0.40 eV 0.22 eV
H
H
H
O
O
O
O O
-0.20 eV
OH
silanol nest
Si
Si
Si
O O
O
Beta-deAl
y
m/z
F
O O
O
g
H2
2
0.95
TS1
Si
0
H
CO(g)+H 2(g)
-1.64
-2.10
Cu1-Cu 3
TS5
-3.72
Cu@Beta-deAl+CO(g)+H2(g)
-3.77
(CHO)Cu1*+Cu3
HCu4+CO(g)
(CHO)Cu1*+HCu4
-3.86
CO(g)
Cu5+CO(g)+H2(g)
y g
(CHO)Cu1*+H(Zeolite)
Cu4-Cu 5
-6
Cu 4-Cu 5
Cu 1-Cu 3
TS4
-1.16
-2
C
(CHO)Cu 1*
-0.57
-0.99
-4
O
TS3
TS2
Free energy (eV)
Reaction Coordinate
m /z
p
Fig. 4. Experimental and
theoretical insights of
the reverse ripening
process. (A and B) Data
showing the signals at
different m/z regions in
the SVUV-PIMS tests that
characterize the mixture
of Cu powder and BetadeAl during methanol
treatment. The background
signals were excluded.
The possible species
corresponding to the peaks
are given, where the signal
of m/z 31 should be from
the fragmentation of methanol. (C) Free energies of
(CH3O)Cu1* and (CHO)
Cu1* intermediates, and the
formation and migration
of the Cu1 intermediates
from the Cu particles to
surface hydroxy groups with
the methanol treatment.
(D) Time-of-flight mass
spectra for reactions of the
mass-selected 63Cu2+ cations
with methanol, CO, and
hydrogen. The reaction time
was about 1.1 to 1.2 ms,
and the effective reactant
gas pressures for CO and
H2 were around 1.0 Pa.
(E) Illustration showing
the gas-phase and surface
migration of Cu species
on the catalyst surface.
(F) Energy profile showing
the nucleation process
from Cu1 to Cu5 at the
silanol nests. TS, transition
state; Cux, Cu cluster with
x number of Cu atoms.
-5.04
y
,
-8
-10
TS1
TS2
TS3
TS4
TS5
Reaction Coordinate
With respect to the migration of Cu1 intermediates on the zeolite surface, both surface and gasphase migration might occur during the reverse
Ostwald ripening process (figs. S46 to S49)
(20, 25, 27, 34, 44–46). With the (CHO)Cu1 intermediate as a model, the gas-phase migration
would cost 0.68 eV for desorption from the solid
surface and −0.26 eV for capture by the silanol
nests (Fig. 4E). In the case of surface migration
Liu et al., Science 383, 94–101 (2024)
5 January 2024
(Fig. 4E), the first step is the migration of the
(CHO)Cu1 intermediate from the Cu cluster to
the adjacent terminal silanol on the zeolite surface (0.40 eV). Next, the (CHO)Cu1* migrates on
the terminal silanol (0.22 eV) (20) and finally arrives at the silanol nest (−0.20 eV). Slightly lower
energy input indicates that the surface migration
would be the preferential pathway for (CHO)Cu1*
diffusion during the dynamic change of the
Cu species that occurs with the methanol
treatment.
Figure 4F shows an energy diagram that
characterizes the stepwise nucleation process
by which the silanol nest captures the (CHO)Cu1*
intermediate, which, with a free energy of
−1.16 eV, is obviously exothermic. With respect
to the interaction between (CHO)Cu1* and the
silanols in the nest, a stable Cud+-(OSi)2 linkage
6 of 8
RES EARCH | R E S E A R C H A R T I C L E
18.
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49.
AC KNOWLED GME NTS
We thank G. Zhu and F. Chen for assistance with TEM
characterization and the Center for High Performance Computing
at Shanghai Jiao Tong University for providing the computing
resources of the Siyuan-1 cluster. We appreciate use of the Hard
X-ray Spectroscopy Beamline (BL11B), the User Experiment Assist
System, and the Chinese Academy of Sciences–Shanghai Science
Research Center of the Shanghai Synchrotron Radiation Facility as
well as TILON Group Technology Limited for data collection. We
appreciate being able to obtain the SVUV-PIMS measurements
through use of the Atomic and Molecular Physics Beamline
(BL09U) at the National Synchrotron Radiation Laboratory in Hefei,
China. Funding: This work was supported by the National Key
Research and Development Program of China (2022YFA1503502
and 2019YFA0405602); the National Natural Science Foundation
of China (22288101, 22241801, 22202176, 22022302, and
92045303); the China Postdoctoral Science Foundation
(2021M702802); the Shanghai Municipal Science and Technology
7 of 8
,
1. B. C. Gates, Chem. Rev. 95, 511–522 (1995).
2. P. Munnik, P. E. de Jongh, K. P. de Jong, Chem. Rev. 115,
6687–6718 (2015).
3. C. Mondelli, G. Gözaydın, N. Yan, J. Pérez-Ramírez, Chem. Soc.
Rev. 49, 3764–3782 (2020).
15.
16.
17.
y
RE FERENCES AND NOTES
14.
y g
We have demonstrated the reverse ripening
phenomenon of Cu NPs on the dealuminated
Beta zeolite support, in which the Cu NPs tend
to become smaller under the methanol vapor
treatment. This feature enables the realization
of an ideal catalyst with superior durability for
the hydrogenation of DMO that outperforms
the classic Cu-silica–based catalysts. We believe
that zeolites with abundant silanol nests act
as powerful supports to optimize the dynamic
change of Cu NPs, and the reverse ripening concept on this support should open a productive
route to more-stable catalysts for industrial
processes. By considering the importance of
Cu-silica–based catalysts in multiple industrial reaction processes, our work provides a
step forward in overcoming the disadvantage
of instability and may improve the vitality of
scalable applications.
12.
13.
A. T. Bell, Science 299, 1688–1691 (2003).
X. Li et al., Nature 611, 284–288 (2022).
E. V. Makshina et al., Chem. Soc. Rev. 43, 7917–7953 (2014).
M. B. Gawande et al., Chem. Rev. 116, 3722–3811 (2016).
S. Hu, W.-X. Li, Science 374, 1360–1365 (2021).
J. Jones et al., Science 353, 150–154 (2016).
J. C. Matsubu et al., Nat. Chem. 9, 120–127 (2017).
T. W. van Deelen, C. Hernández Mejía, K. P. de Jong, Nat. Catal.
2, 955–970 (2019).
Z. Li et al., Nat. Catal. 1, 349–355 (2018).
M. D. Argyle, C. H. Bartholomew, Catalysts 5, 145–269
(2015).
X. Jiang, X. Nie, X. Guo, C. Song, J. G. Chen, Chem. Rev. 120,
7984–8034 (2020).
W. Zhou et al., Chem. Soc. Rev. 48, 3193–3228 (2019).
D. Li et al., Nat. Catal. 5, 99–108 (2022).
D. R. Palo, R. A. Dagle, J. D. Holladay, Chem. Rev. 107,
3992–4021 (2007).
P. Wynblatt, N. A. Gjostein, Acta Metall. 24, 1165–1174
(1976).
S. C. Parker, C. T. Campbell, Phys. Rev. B 75, 035430
(2007).
R. van den Berg et al., ACS Catal. 5, 4439–4448 (2015).
D. Yao et al., Nat. Commun. 14, 1123 (2023).
S. Feng et al., Nat. Commun. 10, 5281 (2019).
S. Chen et al., Chem. Soc. Rev. 50, 3315–3354 (2021).
A. Bishara, K. M. Murad, A. Stanislaus, M. Ismial, S. S. Hussian,
Appl. Catal. 7, 351–359 (1983).
Y. Nishihata et al., Nature 418, 164–167 (2002).
N. Felvey et al., J. Am. Chem. Soc. 144, 13874–13887
(2022).
H. Li et al., ACS Catal. 13, 1197–1206 (2023).
J. Zheng et al., Science 376, 288–292 (2022).
J. Gong et al., J. Am. Chem. Soc. 134, 13922–13925 (2012).
J. Lin, X. Zhao, Y. Cui, H. Zhang, D. Liao, Chem. Commun. 48,
1177–1179 (2012).
C. Xu et al., Nat. Commun. 9, 3367 (2018).
J. Sun et al., Sci. Adv. 4, eaau3275 (2018).
H. Yue et al., AIChE J. 58, 2798–2809 (2012).
J. Zheng et al., J. Phys. Chem. C Nanomater. Interfaces 119,
13758–13766 (2015).
C.-S. Chen, W.-H. Cheng, S.-S. Lin, Chem. Commun. 2001,
1770–1771 (2001).
Z. Gao, B. Ma, S. Chen, J. Tian, C. Zhao, Nat. Commun. 13,
3343 (2022).
C. Wen, Y. Cui, W.-L. Dai, S. Xie, K. Fan, Chem. Commun. 49,
5195–5197 (2013).
S. Qing et al., Int. J. Hydrogen Energy 44, 16667–16674
(2019).
C. D. Chang, A. J. Silvestri, J. Catal. 47, 249–259 (1977).
Z. Liu, J. Liang, Curr. Opin. Solid State Mater. Sci. 4, 80–84
(1999).
S. Lin et al., J. Am. Chem. Soc. 143, 12038–12052 (2021).
B. Eren et al., Science 351, 475–478 (2016).
R.-P. Ye et al., ACS Catal. 10, 4465–4490 (2020).
K. Ma et al., Chem. Sci. 10, 2578–2584 (2019).
R. Ouyang, J.-X. Liu, W.-X. Li, J. Am. Chem. Soc. 135,
1760–1771 (2013).
Q. Wan et al., Nanoscale 10, 17893–17901 (2018).
T. W. Hansen, A. T. Delariva, S. R. Challa, A. K. Datye, Acc. Chem.
Res. 46, 1720–1730 (2013).
Y. X. Zhao, Z. Y. Li, Z. Yuan, X. N. Li, S. G. He, Angew. Chem.
Int. Ed. 53, 9482–9486 (2014).
Z. Yuan et al., J. Phys. Chem. C Nanomater. Interfaces 118,
14967–14976 (2014).
y
5 January 2024
Discussion
4.
5.
6.
7.
8.
9.
10.
11.
g
Liu et al., Science 383, 94–101 (2024)
silanol nests, through (CHO)Cu1* intermediates
to bulky Cu particles with the formation of CO
and H2, which is known as the classical Ostwald
ripening route. This process is endothermic,
with an energy of 0.76 eV. These findings indicate that the nucleation of Cu atoms at locations with small Cu clusters at silanol nests is
preferred over ripening on larger Cu particles.
The redispersion of Cu NPs on the Cu/BetadeAl thus appeared to proceed through the
following steps: (i) the activation of an alcohol
molecule on the surface of the Cu matrix to
promote the cleavage of the Cu–Cu bond, (ii) the
formation of the Cu1 intermediate, (iii) the migration of the Cu1 intermediate to silanol nests,
and (iv) the anchoring of the Cu1 and then more
Cu atoms to form clusters. The silanol nest capture and methanol decomposition would cooperatively regulate the (CHO)Cu1-mediated
reverse ripening route. Notably, except for
methanol, the other alcohols such as ethanol,
n-propanol, n-butanol, and n-pentanol failed to
trigger the redispersion of Cu under the equivalent tests (fig. S61). We calculated the free
energy in each step for the Cu NP redispersion
process for different alcohols. As shown in table
S8, the biggest difference in the energy diagram
for the alcohol-triggered Cu redispersion occurred at the step during which the (RCO)Cu1*
intermediate was captured by the silanol nests.
The results show that the migration of (RCO)Cu1*
intermediates with butanol to the silanol nests
is much more difficult than with methanol.
The (RCO)Cu1* with a longer carbon chain
would have a larger steric hindrance (fig. S62),
thus prohibiting the redispersion of Cu species
on Beta-deAl.
p
with adjacent silanol groups formed, which
was supported by a crystal orbital Hamilton
population (COHP) analysis (figs. S50 and S51).
CO and H2 were preferentially produced from
the reaction between *CHO and the hydrogen
of the silanols (fig. S52). As a result, the Cu1 intermediates could be captured, and the silanol
nest of the Beta-deAl zeolite displayed a crucial
role because of its strong interaction with the Cu
sites (fig. S53). This result is further supported by
Fourier transform infrared spectroscopy, which
showed the reduced signals that were associated
with silanol nests on the Cu/Beta-deAl during
the methanol treatment (fig. S54). However, the
Beta zeolites with a different dealumination degree (table S6) and the different MFI zeolites,
which both have a relatively low content of silanol nests, failed to realize the Cu redispersion
(figs. S11, S12, and S55 to S59).
Compared with the silanol nest, the terminal silanol has a much weaker interaction with
the (CHO)Cu1* intermediate, as confirmed by the
COHP values that represent the strength of the
Cu–O bond (fig. S50). This renders the terminal silanols inoperable with respect to capturing the Cu intermediate (fig. S52), where these
(CHO)Cu1* intermediates would be trapped by
another bulky Cu particle, leading to Ostwald
ripening (20, 43). As a result, the general Cu/SiO2
catalyst with only terminal silanols failed to have
a redispersing feature but caused the rapid Cu sintering to form larger particles (29, 33, 34, 36, 43).
To further confirm this hypothesis, we rationally synthesized a siliceous Beta zeolite (Beta-Si)
with abundant terminal silanols rather than
silanol nests (fig. S58) and used it as support
for Cu NPs. Cu sintered rather than redispersed
after the methanol treatment on the Beta-Si
(fig. S60).
After capturing the first Cu atom, the silanol
nest with a Cu atom became a nucleation site,
and the second and third (CHO)Cu1* species
were captured under the assistance of silanol
groups to form Cu3 clusters with a total freeenergy change less than −2.61 eV. The Cu clusters were efficiently stabilized by the Cud+-OSi
linkage, which was in good agreement with
the experimental results on abundant positively
charged Cu sites after the methanol treatment
(Fig. 3, A and B). The Cu3 cluster could efficiently capture more Cu1-CHO intermediates, with
a total free-energy change at −1.27 eV for capturing the fourth and fifth Cu atoms, which
finally led to the formation of small-size Cu
NPs (Fig. 4F). Regarding the single step of Cu1
intermediate capture on the Cu4 cluster to form
a Cu5 cluster at the silanol nests, the adsorption
energy is −2.80 eV, whereas the Cu1 adsorption
on the bulky Cu surface is −2.22 eV (table S7).
These results suggest that it is more energetically favorable for Cu1 to be trapped by the
small Cu clusters at the silanol nest than at the
bulky Cu surface. We also considered the channel of Cu migration from the Cu5 cluster at the
RES EARCH | R E S E A R C H A R T I C L E
Major Project (2018SHZDZX03); Group Research Asia of BASF
Advanced Chemicals Co., Ltd.; and the Qizhen Project of Zhejiang
University. Author contributions: L.Liu performed the catalyst
preparation, characterization, and catalytic tests. J.L. and X.C.
performed the theoretical simulations and wrote the corresponding
sections of the paper. Y.Y. and W.R. provided helpful discussions
on catalytic performances, catalyst structures, and comparisons
with the commercial catalysts. Y.L., X.G., X.T., and L.Li participated
in the XRD and XAFS characterizations. H.Li, H.Z., C.W., Q.L., H.W.,
K.Z., and Y.W. participated in the catalyst synthesis and
characterization. M.W., J.M., H.Lou, and Z.W. performed the mass
spectra characterization and analyzed the data. L.W. and F.-S.X.
designed the study, analyzed the data, and wrote the paper.
Competing interests: The authors declare no competing interests.
Data and materials availability: All data are available in the
manuscript or the supplementary materials. License information:
Copyright © 2024 the authors, some rights reserved; exclusive
licensee American Association for the Advancement of Science. No
claim to original US government works. https://www.science.org/
about/science-licenses-journal-article-reuse
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adj1962
Materials and Methods
Figs. S1 to S62
Tables S1 to S8
References (50–65)
Submitted 12 June 2023; accepted 5 December 2023
Published online 21 December 2023
10.1126/science.adj1962
p
g
y
y g
y
,
Liu et al., Science 383, 94–101 (2024)
5 January 2024
8 of 8
RES EARCH
SIGNALING COMPLEXES
Molecular insights into atypical modes of b-arrestin
interaction with seven transmembrane receptors
Jagannath Maharana1†, Fumiya K. Sano2†, Parishmita Sarma1†, Manish K. Yadav1, Longhan Duan3,
Tomasz M. Stepniewski4, Madhu Chaturvedi1, Ashutosh Ranjan1, Vinay Singh1, Sayantan Saha1,
Gargi Mahajan1, Mohamed Chami5, Wataru Shihoya2, Jana Selent4, Ka Young Chung3,
Ramanuj Banerjee1*, Osamu Nureki2*, Arun K. Shukla1*
1 of 8
,
5 January 2024
y
Maharana et al., Science 383, 101–108 (2024)
To visualize atypical modes of barr recruitment, we focused our efforts on M2R, which
has a short C terminus, with most of the potential phosphorylation sites localized in ICL3,
y g
*Corresponding author. Email: ramanujb@iitk.ac.in (R.B.);
nureki@bs.s.u-tokyo.ac.jp (O.N.); arshukla@iitk.ac.in (A.K.S.)
†These authors contributed equally to this work.
Structures reveal atypical barr binding modes
to GPCRs
y
1
Department of Biological Sciences, Indian Institute of
Technology Kanpur, Kanpur, India. 2Graduate School of
Science, The University of Tokyo, Tokyo, Japan. 3School
of Pharmacy, Sungkyunkwan University, Suwon, Republic of
Korea. 4Research Program on Biomedical Informatics,
Hospital del Mar Research Institute and Pompeu Fabra
University, Barcelona, Spain. 5BioEM Lab, Biozentrum,
University of Basel, Basel, Switzerland.
a P-X-P-P–type phosphorylation motif in the
C terminus of a broad set of GPCRs, where
P is a phosphorylation site, as a critical determinant of barr interaction and activation
(17, 18).
There are several GPCRs, for example the
human muscarinic receptor subtype 2 (M2R),
that contain a short C terminus with few potential phosphorylation sites, but they harbor
phosphorylation sites primarily in their third
intracellular loop (ICL3) (5, 17, 21–23). Sitedirected mutagenesis and biochemical studies
have shown that phosphorylation sites in the
intracellular loops of some of these receptors contribute to barr binding (24, 25). Whether these receptors engage the same binding
interface with barrs and impart similar activation features as GPCRs with phosphorylation
sites on their C terminus remains unexplored
in terms of direct structural visualization. Several seven transmembrane receptors (7TMRs),
such as the human decoy D6 receptor (D6R),
are sometimes classified as nonsignaling or
nonfunctional GPCRs, as they lack functional
G protein coupling. However, these proteins
robustly interact with and signal through barrs
(26–29). The molecular mechanisms used by
these receptors, known as atypical chemokine
receptors (ACKRs) or arrestin-coupled receptors (ACRs), to bind and activate barrs are
also mostly elusive with respect to the binding interface and activation-dependent conformational changes vis-à-vis prototypical
GPCRs (30–33).
g
b
-arrestins (barrs) are multifunctional
proteins that interact with and regulate a large repertoire of G protein–
coupled receptors (GPCRs) at multiple levels (1–4). The interaction of
GPCRs and barrs is typically conceived to be
driven primarily by agonist-induced receptor
phosphorylation and receptor activation, although emerging studies have started to suggest additional contributing factors such as
membrane interaction, catalytic activation,
and the role of specific phospholipids (2–10).
Structures of GPCR-barr1 complexes have
provided the first glimpse of high-resolution
information about this interaction (11–16).
However, considering the divergent sequences
and phosphorylation patterns of GPCRs, the
molecular mechanisms driving the broadly
conserved nature of GPCR-barr interaction
and activation have been more elusive. Recent
studies have shed light on phosphorylationmediated components of GPCR-barr binding
through broadly conserved phosphorylation
motifs identified in a large number of GPCRs
(17–20). For example, structural and biophysical studies have proposed the framework of
phosphorylation codes and modulatory sites
in the GPCR C terminus as a possible mechanism governing phosphorylation-mediated barr
interaction (19, 20). Two independent structural studies identified that the presence of
p
b-arrestins (barrs) are multifunctional proteins involved in signaling and regulation of seven transmembrane
receptors (7TMRs), and their interaction is driven primarily by agonist-induced receptor activation and
phosphorylation. Here, we present seven cryo–electron microscopy structures of barrs either in the
basal state, activated by the muscarinic receptor subtype 2 (M2R) through its third intracellular loop, or
activated by the barr-biased decoy D6 receptor (D6R). Combined with biochemical, cellular, and biophysical
experiments, these structural snapshots allow the visualization of atypical engagement of barrs with
7TMRs and also reveal a structural transition in the carboxyl terminus of barr2 from a b strand to an
a helix upon activation by D6R. Our study provides previously unanticipated molecular insights into the
structural and functional diversity encoded in 7TMR-barr complexes with direct implications for exploring
novel therapeutic avenues.
and D6R, which is an intrinsically barr-biased
receptor with no detectable G protein activation despite robust barr binding and signaling.
We also determined the structure of barr2 in
the basal conformation and in complex with
a phosphopeptide derived from a prototypical
GPCR, the complement C3a receptor (C3aR)
(Fig. 1). For M2R, we used a full-length, wildtype receptor phosphorylated in cellulo through
coexpression of a membrane-tethered G protein–
coupled receptor kinase 2 construct (GRK2CAAX)
and agonist-induced phosphorylation followed
by incubation with purified barr1 and Fab30
to reconstitute the complex (fig. S1, A and B).
Subsequently, we attempted to determine the
structure of this complex using cryo–electron
microscopy (cryo-EM). Although the receptor
component was not resolved at high resolution, presumably because of inherent flexibility, we successfully determined the structure
of receptor-bound barr1 at 3.1-Å resolution
with focused refinement (Fig. 1C and fig. S2).
Attempting to reduce the flexibility of the receptor component in this complex, we crosslinked the preformed M2R-barr1-Fab30 complex
using on-column glutaraldehyde cross-linking
(34) followed by cryo-EM data collection. The
receptor exhibited flexible positioning relative
to barr1, such that we could determine the
structure of only the receptor-bound barr1 at
3.2-Å resolution (Fig. 1C and figs. S1, C and D,
and S3). These structural snapshots nevertheless allowed us to identify the phosphorylated
region of the ICL3 in M2R that forms the key
interaction interface with barr1 and thereby
allowed us to synthesize and validate the corresponding phosphopeptide (M2Rpp) (fig. S4,
A and B) and determine the structure of the
M2Rpp-barr2-Fab30 complex at 2.9-Å resolution (Fig. 1C and figs. S1, E and F, and S5).
For D6R, we have reported previously that
the critical determinants of barr recruitment
are located primarily in its C terminus (27). We
therefore generated a set of phosphopeptides
corresponding to the phosphorylated D6R and
tested their ability to activate barrs in vitro
using Fab30 reactivity or limited proteolysis
as readouts (fig. S4, C to F). On the basis of
these assays, we identified D6Rpp2 (referred
to hereafter as D6Rpp) as activating barrs
most efficiently, and we used it to reconstitute D6Rpp-barr1/2-Fab30 complexes (fig. S1,
G and H) and determined their structures at
3.4- and 3.2-Å resolution, respectively (Fig.
1D and figs. S6 and S7). We also determined
the structures of wild-type barr2 in its basal
conformation stabilized by Fab6 (Fig. 1A and
figs. S1I and S8) and barr1 in complex with a
C terminus phosphopeptide of the complement C3a receptor (C3aR), a prototypical GPCR
(Fig. 1B and figs. S1J and S9), as references
for basal and typical active conformations.
EM densities of the phosphorylated receptor
domains and the key loops in barrs in these
RES EARCH | R E S E A R C H A R T I C L E
A
GPCR phosphorylation
B
C
Prototypical GPCRs
Complement 3a receptor
(C3aR)
D
GPCRs with long ICLs
Muscarinic receptor M2 (M2R)
-arrestin biased 7TMRs
Atypical chemokine receptor 2 (ACKR2 or D6R)
Agonist
Agonist
M2R
Phosphorylation
by GRKs
?
ICL3
GRK
-arrestin
recruitment
-arrestin
i
i
?
arr1
i
G-protein
iv
M2R
C3aR
arr2
Agonist
D6R
Agonist
M2R
C3aR
GPCR
vii
M2R
arr1
ii
iv
Fab30
arr1
ii
arr1
v
arr1
viii
arr2
Fab30
Fab30
arr2
Fab30
arr1
arr1
D6R
D6R
arr1
Fab30
ii
arr2
i
M2R
Fab30
Fab6
-arrestin
arr2
Fab30
ii
v
Fab30
arr1
arr2
D6Rpp
Fab6
3.9
3.5
iii
M2RICL3
Fab30
M2R-ICL3
C3aRpp
Fab30
ix
3.4
3.2
Fab30
iii
Fab30
vi
arr1
arr2
M2Rpp
D6Rpp
arr2
D6Rpp
2 of 8
,
In contrast to prototypical GPCRs, some chemokine receptors, such as CXCR7, D6R, and a complement C5 receptor (C5aR2), lack G protein
coupling but maintain robust barr recruitment and downstream signaling (27, 35–38).
These receptors, referred to as ACKRs or ACRs,
are essentially intrinsically barr-biased and
represent an excellent model system to probe
structural and functional diversity of barrs.
We thus attempted to reconstitute D6R-barr
complexes using coexpression of the receptor, GRK2 or GRK6, and barr1/2, followed by
in cellulo assembly of the complex through
y
barr signaling complexes with atypical
chemokine receptors
y g
mutagenesis. Subsequently, we measured agonistinduced barr1 recruitment to these mutants
vis-à-vis the wild-type receptor using NanoBiT
and coimmunoprecipitation assay. Mutation
of T-V-S-T, but not T-N-T-T, nearly ablates barr
binding (Fig. 2, L and M, and fig. S12). These
observations establish the key contribution of
the T-V-S-T motif in M2R-ICL3 in driving barr
recruitment and underscore the shared mechanism of barr activation by M2R and other
prototypical GPCRs despite distinct receptor
domains engaging barrs.
y
C3aRpp-barr1 (iii). (C) A 3D reconstruction (left) showing a “hanging” mode of
complex organization in M2R. High-resolution structures of M2R-ICL3–bound barr1/2
are shown below. 2D class average (i), overall 3D map (ii), and structure of
M2R-barr1 (iii); M2R-barr1 of cross-linked complex (iv, v, and vi); and M2Rpp-barr2
(vii, viii, and ix). (D) 2D class average (i), overall dimeric 3D map (ii), and structure
(iii) of D6Rpp-barr1, and D6Rpp-barr2 (iv, v, and vi). The estimated resolutions for
all the structures are shown next to each map.
(Fig. 2, I and J). The barr1 and barr2 in these
structures exhibit an interdomain rotation of
~18° and 23°, respectively (Fig. 2, F, H, and J);
disruption of the three-element and polar-core
network (figs. S11, A to D, and S24); and reorientation of the critical loops compared with
the basal conformation (fig. S11E). The phosphate groups in the M2R-ICL3 stretch resolved
in these structures are organized in a P-X-P-P
pattern, where P is a phosphorylation site, and
are engaged in ionic interactions with conserved Lys and Arg residues in barrs organized
in a K-R-K–type pattern involving R7/R8, K10/
K11, K11/K12, R25/R26, K107/K108, and K294/
K295 (Fig. 2K). A comprehensive list of residueresidue contacts between the phosphopeptides
and barrs is provided in data S2.
The sequence analysis of M2R reveals that
there are two plausible P-X-P-P–type motifs in
the ICL3, one represented by T307-V-S309-T310,
which is observed in the structures presented
here, and the other represented by T340-NT342-T343 (Fig. 2L). To validate the contribution of the T-V-S-T stretch in M2R-ICL3 in
barr engagement and activation, we generated
two different mutants of the receptor with the
phosphorylation sites in each of these P-X-P-P
motifs changed to Ala residues by site-directed
5 January 2024
2.9
g
Maharana et al., Science 383, 101–108 (2024)
M2Rpp
arr1
M2R-ICL3
Fig. 1. A structural approach to understand the atypical modes of barr
interaction with 7TMRs. (A) Cryo-EM structure of full-length barr2 sheds light on
its basal-state conformation. 2D class average (i), overall 3D map of barr2 bound
to Fab6 (ii), and structure of barr2 alone (iii). (B) b-arrestins adopt two distinct
modes of interaction with phosphorylated typical GPCRs. The phosphorylation
pattern of complement receptor C3aR was used to delineate the “hanging” mode of
barr interaction. 2D class average (i), overall dimeric 3D map (ii), and structure of
above-mentioned structures are presented in
fig. S10.
M2R-barr1-Fab30 resembles a hanging conformation observed previously for prototypical
GPCRs (11, 34) with a space between the receptor and barr components, presumably
owing to their interaction mediated primarily
through the long ICL3 (~150 residues) in the
M2R (Fig. 2, A to E). This space is observed
in M2R complexes with both isoforms of barrs
and after receptor phosphorylation by either
GRK2 or GRK6 (Fig. 2, A to D), suggesting
that hanging conformations represent a major
M2R-barr population irrespective of barr or
GRK isoforms. Glutaraldehyde cross-linking
appears to stabilize a more closely engaged
complex, as reflected in negative-staining twodimensional (2D) class averages (Fig. 2G), but
did not improve resolution of the receptor
component in cryo-EM. The structure of M2Rbound barr1 revealed a phosphorylated stretch
of ICL3 in the receptor that harbors the residues from E305 to G313 with four phosphorylation sites (Thr307, Ser309, Thr310, and Ser311)
and docks on the N-domain of barr1 (Fig. 2, F
and H). M2Rpp derived from the ICL3 sequence visualized in M2R-bound barr1 structure binds to an analogous interface on barr2
3.2
vi
arr1
arr1
Basal
arr2
Fab30
iii
D6Rpp
arr1
M2RICL3
3.1
p
iii
Fab30
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
,
Fig. 2. Structural insights into ICL3-driven barr interaction with M2R.
(A to D) Negative-staining EM class averages of M2R, endogenously phosphorylated
by GRK2/6 in complex with barr1 or barr2. (E) Cryo-EM 2D classes, 3D
reconstruction of “hanging” M2R-barr1-Fab30 complex. (F) Structure of barr1 bound
to phosphorylated M2R-ICL3. The EM density of ICL3 and surrounding residues
within 4 Å are shown in the inset. barr1 attains an active conformation with a
Maharana et al., Science 383, 101–108 (2024)
5 January 2024
C-domain rotation of 18.4° with respect to the N-domain. (G) Representative
negative-staining EM 2D classes depicting the effect of cross-linking. Yellow
arrows show potential transition of the complex subunits. (H) Structure of the crosslinked M2R-barr1 complex. The EM density of ICL3 and surrounding residues within
4 Å are shown in the inset. C-domain rotation value with respect to N-domain is
18.6°. (I) Sequence of phosphopeptide derived from the ICL3 of M2R. (J) Structure
3 of 8
RES EARCH | R E S E A R C H A R T I C L E
of M2Rpp-barr2 in ribbon representation. M2Rpp is shown in yellow and barr2 in
blue. Density map of phosphopeptide and surrounding residues within 4 Å are
displayed to the right. barr2 attains an active conformation with 23.4° rotation of
C-domain upon activation with M2Rpp. (K) The phosphorylated residues from ICL3
making critical contacts with Lys and Arg residues of barr1 (upper) and barr2
(lower) are highlighted in blue. (L) Cartoon representation illustrating the presence
of possible phosphorylation clusters in the ICL3 of M2R. Mutations of the two
phosphor-motifs: TVST and TNTT were generated to assess the barr recruitment
measured by bystander NanoBiT assay (receptor+SmBiT-barr1+LgBiT-CAAX).
Substitution of phosphosites of TVST to AVAA leads to abrupt reduction in barr
recruitment, whereas TNTT to ANAA substitution maintains barr recruitment, suggesting
a critical role played by TVST on barr recruitment to M2R (mean ± SEM; n = 3 inde-
,
4 of 8
y
5 January 2024
A cryo-EM structure of a chimeric M2R with
engineered vasopressin receptor 2 C terminus
(M2-V2R) with barr1 has been determined previously (11); however, the ICL3 of M2R was not
resolved in the structure. It thus has not been
known how M2R or other similar GPCRs with
a short C terminus but relatively longer ICL3
engage barrs (42). Our structures of M2R-barr1
and M2Rpp-barr2 underscore that the key interaction interface and the activation mechanism remains conserved despite distinct domains
on the receptor being used to engage barrs.
Our work reveals how two barr isoforms are
able to interact with and regulate a broad set
of receptors with a structurally conserved interface and activation mechanism. Comparing
barr structures determined thus far, including
the C3aRpp-barr1 structure presented here, a
higher interdomain rotation is observed in
barr2 than in barr1 (fig. S17). We speculate that
this observation may provide a mechanism for
how class B GPCRs, which have a relatively
stable barr interaction, exhibit apparently higher affinity for barr2 over barr1 (43). A comparison of our M2R-bound barr1 structure with a
previously reported M2R-V2R-barr1 complex
reveals the hanging conformation of barr1 with
respect to the receptor (fig. S18). This observation further underlines the occurrence of the
hanging conformation as a major population in
the context of native M2R-barr interaction and
offers a structural framework to design guided
experiments to probe functional outcomes in
future studies. However, the active conformations of barr1 were similar in terms of the
interacting residues on N-domain, key loops,
and C-domain rotation values (fig. S19). We observe that M2R-barr complexes can also form
engagements that resemble fully engaged conformations of GPCR-barr complexes, as is apparent in cryo-EM and negative-staining data
and more pronounced after cross-linking. Thus,
it is conceivable, although not yet established,
that barr-mediated M2R internalization and
desensitization follows the same paradigm as
other GPCRs, with the hanging conformation
y g
Previous structures of activated barrs, either
in complex with phosphopeptides or full-length
receptors, have used either truncated barrs or
the C terminus is not resolved structurally.
Even in the crystal structure of barr2 in its
basal conformation, only part of the C terminus is structurally resolved (39, 40). We
determined the cryo-EM structure of wildtype, full-length barr2 and resolved a longer
stretch of the C terminus (Fig. 4, F and G, and
fig. S16). A stretch of the barr2 C terminus
adopts a b strand in its basal conformation
and docks to the N-domain to maintain the
barr in an inactive conformation. A previous
structure of the b-appendage domain of adaptin
(AP2) in complex with a peptide corresponding to the C terminus of barr1 also exhibits an
a-helical conformation of the peptide that is
positioned onto a groove in the platform subdomain of the b-appendage (Fig. 4E) (41). The
propensity of the C terminus in barr1 and barr2
Discussion
y
Resolution of an extended barr C-terminal tail
to adopt an a-helical conformation should be
explored further.
g
Maharana et al., Science 383, 101–108 (2024)
the key loop regions compared with the basal
state, and disruption of the three-element and
polar core network (Fig. 3, G and H, and figs.
S11 and S24). A comprehensive list of residueresidue contacts between the phosphopeptides
and barrs are given in data S2.
We observe that the distal C terminus of
barr2 (Tyr391 to Lys408) in the D6Rpp-bound
conformation adopts an a-helical structure,
which is positioned in the central crest of
barr2 (Fig. 4, A and B) through extensive interactions (fig. S15). This a helix in barr2 forms
a key dimerization interface for the two protomers in this structure and is arranged in an
antiparallel coiled-coil fashion with contacts
across the two protomers (Fig. 4C and data
S3). We analyzed this a helix using molecular
dynamics (MD) simulations and observed that
it exhibits robust stability over a 2-ms simulation time frame (Fig. 4D). We also observed
that this stretch of barr2 C terminus has a
propensity to adopt an a-helical conformation even in isolated form, meaning, without
the barr2 core being present. We did not observe this a-helical structure in D6Rpp-bound
barr1, although the corresponding segment is
not resolved in the structure.
p
agonist stimulation and stabilization using
Fab30. Although we observed clear complex
formation and a typical architecture by negative staining that is reminiscent of the hanging conformation (Fig. 3, A and B), attempts
to scale up the complex for cryo-EM analysis
were not successful. Therefore, we focused our
efforts to determine the structures of barrs in
complex with a phosphorylated peptide corresponding to the C terminus of D6R (D6Rpp)
(Fig. 3D). We first confirmed that D6R-barr
interaction depends on receptor phosphorylation by truncating the C terminus of D6R
harboring the phosphorylation sites, which
resulted in near-complete ablation of agonistinduced barr1 recruitment (Fig. 3C). Subsequently, we characterized D6Rpp using in vitro
proteolysis and Fab30 reactivity assays (fig.
S4, C to F) and further validated barr activation by this peptide using hydrogen/deuterium
exchange mass spectrometry (HDX-MS). We
observed that D6Rpp binding resulted in robust activation of barrs, as reflected by conformational changes in multiple b strands
and loop regions in the N-domain (Fig. 3, E
and F, and fig. S13). We also observed notable differences between the HDX-MS pattern of barr1 versus barr2, such as reduced
solvent exposure of b strands XIV and XV in
the C-domain of barr2, which suggests isoformspecific differences between activation of barr1
versus barr2.
We determined the structures of barr1 and
barr2 in complex with D6Rpp, stabilized by
Fab30, at resolution of 3.4 and 3.2 Å, respectively (Figs. 1D and 3, G and H). We observed
a similar interaction interface of D6Rpp on
N-domains of barr1 and barr2, although seven
phosphates were resolved in the barr2 structure compared to three in barr1 (Fig. 3, G and
H). However, we observed that three phosphate groups, on Ser348, Ser350, and Ser351, were
organized in a P-X-P-P pattern and were engaged in interactions with selected Lys and
Arg residues in the N-domain of barrs (Fig. 3I).
As in M2R, there are two putative P-X-P-P
motifs in D6Rpp, but our structural snapshots
only capture barrs associated with one of them
(fig. S14). We also observed interdomain movement in D6Rpp-bound barrs, reorientation of
pendent experiments; normalized with respect to highest ligand concentration signal for
M2RWT as 100%). (M) Role of TVST in barr recruitment is further corroborated by
coimmunoprecipitation assay. On carbachol stimulation, M2RAVAA showed drastic
reduction in barr1 recruitment. A representative blot and densitometry-based
quantification are presented (mean ± SEM; n = 4 independent experiments; normalized
with M2R 30-min stimulation condition signal as 100%; two-way analysis of variance,
Tukey’s multiple comparisons test). The exact P values are as follows: M2RWT 0 min
versus 15 min, P = 0.0006; M2RWT 0 min versus 30 min, P = <0.0001; M2RANAA 0 min
versus 15 min, P = 0.0008; M2RANAA 0 min versus 30 min, P = <0.0001 (***P = 0.0001;
****P < 0.0001; ns, nonsignificant). Single-letter abbreviations for the amino acid
residues are as follows: A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G, Gly; H, His; I, Ile; K, Lys;
L, Leu; M, Met; N, Asn; P, Pro; Q, Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; and Y, Tyr.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
,
Fig. 3. Structural insights into D6R-barr complex interaction and activation.
(A) Negative-staining EM 2D class averages of D6R-barr1/2 complexes endogenously
phosphorylated with GRK2/6. (B) A representative 2D class average highlighting
the “hanging” mode of barr1 interaction with the receptor. (C) Dose response curve
for CCL7-induced barr1 recruitment for the mentioned D6R constructs using
NanoBiT assay (Receptor-SmBiT+LgBiT-barr1) (mean ± SEM; n = 3 independent
experiments; normalized with respect to the lowest ligand concentration signal as 1).
(D) Design of selected phosphopeptide derived from the C terminus of D6R. (E and
F) HDX-MS plots to show the potential of generated phosphopeptides from D6R to
activate barr1 and barr2, respectively. Among regions (a to f) showing significant
Maharana et al., Science 383, 101–108 (2024)
5 January 2024
changes upon deuterium exchange, the fragment at the C terminus (f) has been
demonstrated to show activation of barrs upon D6Rpp binding. (G) Structure of
D6Rpp-barr1 complex in ribbon representation. The density map of D6Rpp and
surrounding residues within 4 Å are shown to the left. C-domain rotation of barr1
bound to D6Rpp is 19.8°. (H) Structure of D6Rpp-barr2 complex in ribbon
representation. The density map of D6Rpp and surrounding residues within 4 Å are
shown in the inset. C-domain rotation of barr2 bound to D6Rpp was calculated to be
22.3°. (I) The phosphorylation pattern from D6Rpp engages with a network of Lys
and Arg residues present on the N-domains of barrs. Residues highlighted with blue
circles show the Lys and Arg residues in barr1 (upper) and barr2 (lower).
5 of 8
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
,
Fig. 4. Discovery of a C-terminal helix in D6R-activated barr2. (A) Cartoon
representation of barr2 bound to D6R phosphopeptide. barr2 and D6Rpp are
presented in gray and yellow, respectively, and the sequence of the C-terminal
helix is shown in the inset. (B) D6Rpp-barr2 structure displayed in surface
representation in two different views to highlight the pose of the helix. The
C-terminal helix (green) and D6Rpp (yellow) are shown as ribbon diagrams.
(C) Dimeric organization of D6Rpp-barr2 structure shown in ribbon representation (top left). Formation of antiparallel coiled-coil by the C-terminal helix of
barr2 at the dimeric interface (top right) shown as cartoon representation.
The antiparallel coiled-coil exhibits mixed ad layers. Helical wheel representation
of the antiparallel coiled-coil shows Asp at position d of one helix, which
forms a salt bridge with Arg at position g in the other helix (bottom left). Heptad
helical representation of the antiparallel coiled-coil residues in the barr2
Maharana et al., Science 383, 101–108 (2024)
5 January 2024
sequence (bottom right). (D) MD simulations confirm stability of the distal
C-terminal helix/barr2 interface. Structural snapshots (one snapshot every
10 ns, 7 × 250 ns of simulation time) presented here are of the position of the
C-tail during simulation. For each residue, frames where it assembles an
a-helical conformation are colored green. Fragments of the C-terminal helix
can spontaneously assemble an a-helical conformation (right corner, blue
cartoon) in three out of four independent MD simulations (each 2 ms) which is
overlayed with the crystallized C-tail for comparison (green cartoon). For each
residue, frames where it assembles a helical conformation are colored green.
Comparison of a spontaneously assembled helical conformation of the barr2
C-tail (blue) with that present in the structure (gray). (E) Structure of AP2
b-appendage protein in complex with barr1 C-terminal peptide (PDB ID 2IV8) is
shown as cartoon representation (left). The barr1 C-terminal peptide can be seen
6 of 8
RES EARCH | R E S E A R C H A R T I C L E
to adopt similar helical conformation as the C-terminal helix in the D6Rppbound barr2 structure (right). The sequence alignment of the C-terminal
stretches of barr1 and barr2 are shown in the inset. (F) Cryo-EM density map of the
isolated C terminus of barr2 and surrounding residues within 4 Å. (G) The peptide
stretch sequence (top) of the C-tail in basal barr2 transforms into a helical
conformation in the D6Rpp-bound state (highlighted in cyan circles). (H) The C-tail
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
45.
46.
47.
49.
50.
51.
52.
53.
54.
55.
56.
AC KNOWLED GME NTS
We thank M. Ganguly for assistance with structural analysis and
S. Mishra, A. Dalal, and N. Zaidi for help with functional assays.
Cryo-EM on basal state barr2, C3aRpp-barr1, and D6Rpp-barr
7 of 8
,
5 January 2024
24.
y
48.
1. D. S. Kang, X. Tian, J. L. Benovic, Curr. Opin. Cell Biol. 27,
63–71 (2014).
2. V. V. Gurevich, E. V. Gurevich, Mol. Cell. Endocrinol. 484, 34–41
(2019).
3. S. Ahn, S. K. Shenoy, L. M. Luttrell, R. J. Lefkowitz, Cell 182,
1362–1362.e1 (2020).
4. J. Maharana, R. Banerjee, M. K. Yadav, P. Sarma, A. K. Shukla,
Curr. Opin. Struct. Biol. 75, 102406 (2022).
5. R. Ranjan, H. Dwivedi, M. Baidya, M. Kumar, A. K. Shukla,
Trends Cell Biol. 27, 851–862 (2017).
6. J. Grimes et al., Cell 186, 2238–2255.e20 (2023).
7. J. Janetzko et al., Cell 185, 4560–4573.e19 (2022).
8. K. Eichel et al., Nature 557, 381–386 (2018).
9. K. Eichel, D. Jullié, M. von Zastrow, Nat. Cell Biol. 18, 303–310
(2016).
10. R. Ranjan, P. Gupta, A. K. Shukla, Curr. Biol. 26, R285–R288
(2016).
23.
y g
RE FERENCES AND NOTES
22.
D. P. Staus et al., Nature 579, 297–302 (2020).
W. Huang et al., Nature 579, 303–308 (2020).
Y. Lee et al., Nature 583, 862–866 (2020).
C. Cao et al., Neuron 110, 3154–3167.e7 (2022).
W. Yin et al., Cell Res. 29, 971–983 (2019).
J. Bous et al., Sci. Adv. 8, eabo7761 (2022).
J. Maharana et al., Mol. Cell 83, 2091–2107.e7 (2023).
P. Isaikina et al., Mol. Cell 83, 2108–2121.e7 (2023).
D. Mayer et al., Nat. Commun. 10, 1261 (2019).
X. E. Zhou et al., Cell 170, 457–469.e13 (2017).
A. B. Tobin, A. J. Butcher, K. C. Kong, Trends Pharmacol. Sci.
29, 413–420 (2008).
A. B. Tobin, Br. J. Pharmacol. 153 (suppl. 1), S167–S176
(2008).
J. L. DeGraff, V. V. Gurevich, J. L. Benovic, J. Biol. Chem. 277,
43247–43252 (2002).
R. Pals-Rylaarsdam et al., J. Biol. Chem. 272, 23682–23689
(1997).
M. M. Hosey et al., Life Sci. 64, 363–368 (1999).
S. Pandey, J. Maharana, X. X. Li, T. M. Woodruff, A. K. Shukla,
Trends Biochem. Sci. 45, 693–705 (2020).
S. Pandey et al., Mol. Cell 81, 4605–4621.e11 (2021).
S. Rajagopal et al., bProc. Natl. Acad. Sci. U.S.A. 107, 628–632
(2010).
E. M. Borroni et al., Sci. Signal. 6, ra30 (2013).
M. H. Ulvmar, E. Hub, A. Rot, Exp. Cell Res. 317, 556–568
(2011).
G. J. Graham, M. Locati, A. Mantovani, A. Rot, M. Thelen,
Immunol. Lett. 145, 30–38 (2012).
C. Cancellieri, A. Vacchini, M. Locati, R. Bonecchi, E. M. Borroni,
Biochem. Soc. Trans. 41, 231–236 (2013).
R. J. Nibbs, G. J. Graham, Nat. Rev. Immunol. 13, 815–829
(2013).
A. K. Shukla et al., Nature 512, 218–222 (2014).
S. Pandey et al., J. Biol. Chem. 294, 9416–9429 (2019).
D. E. Croker et al., Immunol. Cell Biol. 94, 787–795
(2016).
Y. C. Yen et al., Sci. Adv. 8, eabn8063 (2022).
H. T. Nguyen et al., Cell Biosci. 10, 134 (2020).
M. Han, V. V. Gurevich, S. A. Vishnivetskiy, P. B. Sigler,
C. Schubert, Structure 9, 869–880 (2001).
X. Zhan, L. E. Gimenez, V. V. Gurevich, B. W. Spiller, J. Mol.
Biol. 406, 467–478 (2011).
E. M. Schmid et al., PLOS Biol. 4, e262 (2006).
M. Seyedabadi, M. Gharghabi, E. V. Gurevich, V. V. Gurevich,
Biomolecules 11, 218 (2021).
R. H. Oakley, S. A. Laporte, J. A. Holt, M. G. Caron, L. S. Barak,
J. Biol. Chem. 275, 17201–17210 (2000).
L. L. Porter et al., Nat. Commun. 13, 3802 (2022).
Q. Liu et al., Nat. Commun. 11, 4857 (2020).
Q. T. He et al., Nat. Commun. 12, 2396 (2021).
S. Coffa, M. Breitman, B. W. Spiller, V. V. Gurevich,
Biochemistry 50, 6951–6958 (2011).
Y. K. Peterson, L. M. Luttrell, Pharmacol. Rev. 69, 256–297
(2017).
E. Ghosh et al., Cell Rep. 28, 3287–3299.e6 (2019).
N. R. Latorraca et al., Nature 557, 452–456 (2018).
K. N. Nobles et al., Sci. Signal. 4, ra51 (2011).
E. Reiter, S. Ahn, A. K. Shukla, R. J. Lefkowitz, Annu. Rev.
Pharmacol. Toxicol. 52, 179–197 (2012).
A. K. Shukla et al., Proc. Natl. Acad. Sci. U.S.A. 105,
9988–9993 (2008).
Q. Chen et al., J. Mol. Biol. 433, 166790 (2021).
V. V. Gurevich, E. V. Gurevich, Int. J. Mol. Sci. 23, 7253
(2022).
Q. Chen et al., Nat. Commun. 8, 1427 (2017).
y
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
g
Maharana et al., Science 383, 101–108 (2024)
volve isolated phosphopeptides with defined
phosphorylation patterns without the transmembrane core of the receptors. The receptor core imparts additional conformational
changes in barrs (49, 50), and it is likely that
additional mechanisms and/or conformations
of barrs are induced by receptors, especially in
terms of the positioning of the proximal region
of the phosphorylated segment. However, the
conserved principle of “P-X-P-P key” to open
the “K-K-R-K-R-K lock” is likely to be maintained and guide barr activation even in the
context of full receptors (fig. S22). Previous
studies have demonstrated that different GRKs
may phosphorylate distinct residues in GPCRs,
resulting in a phosphorylation barcode that
may impart distinct barr conformations and
functional outcomes (51–53). Precise mapping
of the phosphorylation patterns of the receptors used here by different GRKs and visualizing
the resulting barr conformations is necessary.
Like GPCRs, barrs are conformationally dynamic proteins, and it is plausible that they
may explore multiple active conformations
depending on the phosphorylation patterns.
Stabilizing chaperones such as Fab30 may
preferentially recognize some of these conformations better than the others. barrs have
a strong tendency to form various oligomeric
states including dimers, trimers, tetramers,
and even higher-order oligomers (54, 55), although previous structures of barr2 either
bound to IP6 (56) or GPCR phosphopeptides
have revealed trimers (17). Future work should
explore whether the dimeric arrangement observed here is specific to D6R. We predominantly
observed dimeric arrangement in negative
staining carried out at low protein concentrations that are easily achievable in a cellular
context, although trimeric classes were also
present.
p
being sufficient to drive endocytosis and signaling, whereas the closely engaged conformation
is required for desensitization.
The observation of an a-helical conformation in barr2 upon activation by D6Rpp is
intriguing from multiple perspectives. The
same conformation is not observed in barr1,
and, although this may simply be due to higher
flexibility of the C terminus in barr1, it would
be anticipated that extensive interactions would
allow structural visualization of the a helix if
it was being formed. D6Rpp-bound barr2 exhibits a dimeric assembly that is different from
the previously determined active-like structures, such as those bound to V2Rpp, C5aR1pp,
M2Rpp, and IP6 captured in a trimeric state
(fig. S20 and data S3). In addition, the a helix
observed in the C terminus of barr2 in D6Rppbound state is also absent from the previously
determined barr2 structures. Although it cannot be completely ruled out that these differences may arise from a preferential orientation
of the samples on cryo-EM grids, we speculate
that these differences underscore the conformational signatures in barrs upon interaction
with GPCRs as compared with ACRs, which
should be investigated further. The a helix in
D6Rpp-barr2 also suggests a “chameleon”
nature of the distal C terminus to adopt a
b strand in the basal state while transitioning to
an a helix upon activation (Fig. 4H). Such secondary structure switching is also observed for
several other proteins that exhibit functional
diversity (44). The positioning of the a helix in
the central crest of barr2 may interfere with the
core interaction of barr2 with the receptor,
although it remains to be experimentally visualized. This idea is supported by the overlay
of D6Rpp-bound barr2 with previously determined GPCR-barr structures where either of
the ICLs of the receptors appears to clash with
the a helix in barr2 (Fig. 4I and fig. S21). Previous studies have demonstrated the dynamic
nature of barrs and conformational space that
they explore upon activation by distinct phosphorylation patterns, which are linked to distinct scaffolding outcomes and downstream
functions (45–48).
There are several key questions that remain
to be answered in the context of GPCR-barr
interaction. Several prototypical GPCRs are
likely to engage barrs through their ICL3 but
lack the P-X-P-P motif, and some of the ACRs,
such as CXCR7 and C5aR2, lack this motif in
their C terminus, but they still recruit barrs.
The structural snapshots presented here in-
of barr2 exhibits a chameleon-like property, adopting a helical conformation in the
active state from a b strand in the basal state. (I) Ribbon representation of the
b1AR-barr1 structure superimposed with D6Rpp-barr2 on barrs (left) shows
positioning of the C-terminal helix on the central crest of barrs. Upon structural
superimposition with all reported GPCR-barr1 structures, ICL1/2/3 of various
receptors reside on the central crest as a C-terminal helix on D6Rpp-barr2 (right).
RES EARCH | R E S E A R C H A R T I C L E
complexes was performed at the BioEM lab of the Biozentrum at
the University of Basel, and we thank C. Alampi and D. Kalbermatter
for their excellent technical assistance. Funding: Research in
A.K.S.’s laboratory is supported by the Senior Fellowship of the DBT
Wellcome Trust India Alliance (IA/S/20/1/504916) awarded to
A.K.S., the Science and Engineering Research Board (CRG/2022/
002646, SPR/2020/000408, and IPA/2020/000405), the Council of
Scientific and Industrial Research [37(1730)/19/EMR-II], the Indian
Council of Medical research (F.NO.52/15/2020/BIO/BMS), a Young
Scientist Award from Lady Tata Memorial Trust, and IIT Kanpur. A.K.S.
is an EMBO Young Investigator and Sonu Agrawal Memorial Chair
Professor. This work was supported by grants from the JSPS
KAKENHI, grant numbers 21H05037 (O.N.), 22K19371 and 22H02751
(W.S.), and 23KJ0491 (F.K.S.); the Kao Foundation for Arts and
Sciences (W.S.); the Takeda Science Foundation (W.S.); the Lotte
Foundation (W.S.); and the Platform Project for Supporting Drug
Discovery and Life Science Research [Basis for Supporting Innovative
Drug Discovery and Life Science Research (BINDS)] from the
Japan Agency for Medical Research and Development (AMED), grant
numbers JP22ama121012 (O.N.) and JP22ama121002 (support
number 3272; O.N.). HDX-MS work in K.Y.C.’s laboratory was
supported by grants from the National Research Foundation of Korea
funded by the Korean government (NRF-2021R1A2C3003518 and
NRF-2019R1A5A2027340). T.M.S. acknowledges support from the
National Center of Science, Poland (2017/27/N/NZ2/02571)
and Sara Borrell grant CD22/00007 funded by the Institute of Health
Carlos III (ISCIII). J.S. acknowledges funding from the Instituto de
Salud Carlos III (ISCIII) (AC18/00030) and the resources of grant
2021 SGR 00046 funded by Agència de Gestió d'Ajuts Universitaris i
de Recerca Generalitat de Catalunya (AGAUR). Author
contributions: Conceptualization: J.M., F.K.S., P.S., J.S., K.Y.C., R.B.,
O.N., and A.K.S. Methodology: J.M., F.K.S., P.S., M.K.Y., L.D., T.M.S.,
Ma.C., A.R., V.S., S.S., G.M., Mo.C., W.S., J.S., K.Y.C., R.B., O.N., and
A.K.S. Investigation: J.M., F.K.S., P.S., M.K.Y., L.D., T.M.S., Ma.C.,
A.R., V.S., S.S., G.M., Mo.C., W.S., J.S., K.Y.C., R.B., O.N., and A.K.S.
Visualization: J.M., F.K.S., P.S., M.K.Y., L.D., T.M.S., M.C., A.R., V.S.,
S.S., G.M., Mo.C., W.S., J.S., K.Y.C., R.B., O.N., and A.K.S. Funding
acquisition: J.S., K.Y.C., O.N., and A.K.S. Project administration: J.S.,
K.Y.C., O.N., and A.K.S. Supervision: J.S., K.Y.C., O.N., and A.K.S.
Writing – original draft: J.M., R.B., O.N., and A.K.S. Writing – review &
editing: J.M., F.K.S., P.S., M.K.Y., J.S., K.Y.C., R.B., O.N., and A.K.S.
Competing interests: The authors declare that they have no
competing interests. Data and materials availability: The cryo-EM
structures are deposited in Protein Data Bank (PDB) and Electron
Microscopy Data Bank (EMDB) under the following accession
numbers: Basal barr2, PDB ID 8J9K and EMDB ID EMD-36110;
M2R-barr1-Fab30cross-linked, PDB ID 8J97 and EMDB IDs EMD-36090
and EMD-36091; M2R-barr1-Fab30, PDB ID 8JAF and EMDB IDs EMD-
36126 and EMD-36093; M2Rpp-barr2-Fab30, PDB ID 8J8R and EMDB
ID EMD-36078; C3aRpp-barr1-Fab30, PDB ID 8JA3 and EMDB ID
EMD-36124; D6Rpp-barr1-Fab30, PDB ID 8J8Z and EMDB ID EMD36082; D6Rpp-barr2-Fab30, PDB ID 8GO9 and EMDB ID EMD-34174;
and D6Rpp-barr2-Fab30-local-refined, PDB ID 8J8V and EMDB ID
EMD-36081. All the other data pertaining to the manuscript are
present in the main text and supplemental materials. License
information: Copyright © 2024 the authors, some rights reserved;
exclusive licensee American Association for the Advancement of
Science. No claim to original US government works. https://www.
science.org/about/science-licenses-journal-article-reuse
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adj3347
Materials and Methods
Figs. S1 to S25
Table S1
References (57–80)
MDAR Reproducibility Checklist
Data S1 to S4
Submitted 21 June 2023; accepted 29 November 2023
10.1126/science.adj3347
p
g
y
y g
y
,
Maharana et al., Science 383, 101–108 (2024)
5 January 2024
8 of 8
RES EARCH
EVOLUTION
Convergence in carnivorous pitcher plants reveals
a mechanism for composite trait evolution
Guillaume Chomicki , Gustavo Burin †, Lucas Busta †, Jedrzej Gozdzik †, Reinhard Jetter
Beth Mortimer6†, Ulrike Bauer7*‡
1
2
3
4
†,
4,5
Composite traits involve multiple components that, only when combined, gain a new synergistic function.
Thus, how they evolve remains a puzzle. We combined field experiments, microscopy, chemical analyses,
and laser Doppler vibrometry with comparative phylogenetic analyses to show that two carnivorous
Nepenthes pitcher plant species independently evolved similar adaptations in three distinct traits to
acquire a new, composite trapping mechanism. Comparative analyses suggest that this new trait arose
convergently through “spontaneous coincidence” of the required trait combination, rather than
directional selection in the component traits. Our results indicate a plausible mechanism for composite
trait evolution and highlight the importance of stochastic phenotypic variation as a facilitator of
evolutionary novelty.
1 of 6
,
5 January 2024
y
Chomicki et al., Science 383, 108–113 (2024)
y g
*Corresponding author. Email: u.bauer@exeter.ac.uk
†These authors contributed equally to this work.
‡Present address: Biosciences, Geoffrey Pope Building, University
of Exeter, Stocker Road, Exeter EX4 4QD, UK.
Pitcher plants use slippery surfaces (21) to
trap insect prey in cup-shaped leaves (Fig. 1,
A and G) with a roof-like lid. The lid is not
typically involved in trapping; however, in
Southeast Asian Nepenthes gracilis, it acts as
a rain-actuated “springboard” that catapults
insects into the fluid-filled trap (Fig. 1D and
movie S1) (22). We discovered (23) that Nepenthes
pervillei, endemic to the Seychelles and separated from N. gracilis by 4000 km of ocean,
uses the same springboard mechanism (Fig.
1J and movie S2).
Springboard trapping requires three independent components: (i) a horizontal lid that
launches prey directly into the pitcher; (ii) a
moderately slippery underside that allows insects to access the still lid but not withstand a
y
Department of Bioscience, Durham University, South Road,
Durham DH1 3LE, UK. 2Natural History Museum London,
Cromwell Road, London SW7 5BD, UK. 3Department of
Chemistry and Biochemistry, University of Minnesota Duluth,
308 HCAMS, 1038 University Drive, Duluth, MN 55812, USA.
4
Department of Chemistry, University of British Columbia,
2036 Main Mall, Vancouver, BC V6T 1Z1, Canada.
5
Department of Botany, University of British Columbia,
3200-6270 University Boulevard, Vancouver, BC V6T 1Z4,
Canada. 6Department of Biology, University of Oxford,
Mansfield Road, Oxford OX1 3SZ, UK. 7School of Biological
Sciences, University of Bristol, 24 Tyndall Avenue, Bristol
BS8 1TQ, UK.
Springboard trapping—a composite trait
with three distinct components
To probe whether springboard trapping evolved
convergently in N. gracilis and N. pervillei, we
first tested (23) whether it is underpinned by
the same three crucial adaptations in both
species. The orientation of the pitcher lid—
recorded as deviation from horizontal (Fig. 1, A
and G)—is close to horizontal in both N. gracilis
[8.0° ± 1.2° (mean ± SEM); n = 68 pitchers]
and N. pervillei (8.8° ± 1.8°; n = 37 pitchers).
These lid angles are significantly lower than
the average lid angles of 55 other investigated
Nepenthes species (24.8° ± 5.0°, lower 95%
confidence interval = 15.0°) (23). Thus, both
species fulfill the first requirement for springboard trapping.
In both species, the lid’s lower surface is
covered by epicuticular wax “pillars” (Fig. 1,
B and H), which differ drastically from the
much more slippery wax platelets inside the
pitcher (Fig. 1, C and I) (25). Wax crystal morphology is determined by chemical composition
(26). Gas chromatography–mass spectrometry of cuticular waxes (23) revealed similar
chemical profiles for corresponding surfaces of
N. gracilis and N. pervillei (Fisher-FreemanHalton’s exact test, df = 5, c2lid = 7.3, PBonferroni =
0.8; c2inner wall = 2.9, PBonferroni = 1.0), but strong
differences between the lid and pitcher wall in
both species (c2gracilis = 42.1, PBonferroni < 0.001;
c2pervillei = 46.8, PBonferroni < 0.001; Fig. 2). Lid
waxes of both species were dominated by nalcohols, whereas the pitcher wall waxes consisted mainly of aldehydes, as in other Nepenthes
species (27). Cuticular wax biosynthesis follows a stepwise pathway where C2 building
blocks are added to elongate fatty acid precursors and form very long-chain aliphatic
compounds that can then be enzymatically
modified into alcohols, esters, aldehydes, alkanes, and other wax compounds (28). Owing
to this modular assembly process, shifts between major compound classes can be achieved
relatively easily by redirecting substrate flux
through different enzymatic pathways. Both
species thus fulfill the second requirement for
springboard trapping.
High-speed video analysis (23) confirmed
that lids of both species respond to a drop
impact with fast pivoting oscillations (Fig. 1,
E and K). N. gracilis lids had lower area and
mass (305 ± 27 mm2; 85 ± 11 mg; n = 11 pitchers)
and higher resonant frequencies (88 ± 8 s−1)
than lids of N. pervillei (836 ± 102 mm2,
318 ± 48 mg, 58 ± 5 s−1; n = 5 pitchers). For
direct kinematic comparison using laser Doppler vibrometry, we thus selected relatively
large N. gracilis lids (315 ± 46 mm2, 87 ± 13 mg,
g
1
are mediated by epigenetic modifications which,
at least in plants, may be heritable (16, 17). In
addition to genetic variation, plasticity sensu
lato can contribute substantially to the total
phenotypic variation under selection (18); however, empirical data demonstrating its role in
trait evolution are scarce (19).
The evolution of any trait is the result of a
combination of natural selection, wherein traits
are shaped by adaptive pressures and tend to
change in a specific direction over time (determinism), and random events and processes,
such as mutation or genetic drift (stochasticity), in the context of the evolutionary history
(contingency) of the trait (20). Accordingly, composite traits may result from natural selection
acting on multiple traits at once, random effects, or a combination of both. Here, we report
evidence that a complex, composite trapping
mechanism found in two carnivorous pitcher
plant species likely evolved convergently through
spontaneous coincidence of a new beneficial
trait combination, facilitated by high stochastic phenotypic variation.
Similar adaptations in two species originate
from convergence
p
C
omposite traits consist of multiple independent components that, in combination, gain a new synergistic function. For
example, bird wings combine adaptations
of morphology, bone anatomy, and skin
appendages to convert a forelimb into an aerofoil (1), and independent modifications of shape
(2), coloration (3), and biosynthetic pathways
(4) underpin the insect-trapping leaves of carnivorous plants. Explaining the origin of composite traits has puzzled biologists since Darwin
(5–8), as it requires coordinated evolution of
multiple independent components (9, 10). Thus,
additional factors such as phenotypic plasticity (11) have been proposed to play a role in
composite trait evolution. Levis et al. (12) demonstrated the emergence of a carnivorous tadpole
morph with three independent morphological
adaptations in Scaphiopodidae species, arising
from high phenotypic plasticity and subsequent adaptive refinement of the new composite trait. Plasticity sensu lato includes both
induced phenotypic variation, expressed by the
same genotype in response to different environments (plasticity sensu stricto), and stochastic
phenotypic variation, expressed by the same
genotype under identical conditions (13–15).
Neither type of plasticity is mediated by genetic differences; rather, both types of plasticity
drop impact; and (iii) a stiff, pivoting lid that
acts as a spring (24). Each trait only gains a
trapping function in the presence of the other
two (22).
RES EARCH | R E S E A R C H A R T I C L E
N. gracilis
B
A
lid
inner
wall
G
lid
N. pervillei
H
inner
wall
rim
5 μm
5 μm
rim
C
I
N. bicalcarata, two species that stand in between
N. gracilis and N. pervillei in the phylogeny (30)
but were not included in our analysis owing to
low sample sizes (23), lacked lid wax crystals
(fig. S2). We then used a recent Nepenthes
phylogenomic tree (30) to test if the composite
trait and its components evolved independently in N. gracilis and N. pervillei. Ancestral-state
estimations (fig. S3) and phylogenetic analyses (figs. S4 to S7 and tables S1 and S2) strongly support an independent origin of springboard
trapping and all three component traits in
N. gracilis and N. pervillei.
Two alternative scenarios for the evolution
of springboard trapping
5 μm
t = 0 ms
E
10mm
2
t = 6.5 ms
5 μm
10 mm
K
2
1
0
0
-1
-1
-2
-2
t = 12 ms
t = 20 ms
t = 15 ms
0
F
100
1
200
p1
p2
p3
0
L
200
t = 38 ms
1
t = 36 ms
t = 48 ms
100
0
0
-1
-1
p4
p5
p6
p7
t = 54 ms
t = 68 ms
0 0.2 0.4 0.6 0.8 1.0
10 mm
Chomicki et al., Science 383, 108–113 (2024)
suggests either a single evolutionary origin
(evolutionary contingency) (29) or convergent
evolution. To test whether (i) springboard trapping as a whole and (ii) the three distinct component traits share common ancestry or evolved
independently in N. gracilis and N. pervillei, we
assessed all three traits in 55 Nepenthes species
(23), corresponding to one-third of the genus
and covering all major clades. Twelve species
had horizontal lids and 8 had pivoting lids,
but only 5 species (including N. gracilis and
N. pervillei) presented pillar-shaped crystals
(Fig. 3 and fig. S1). Moreover, N. vieillardii and
5 January 2024
2 of 6
,
92 ± 5 s−1; n = 3 pitchers) and small N. pervillei
lids (252 ± 39 mm2, 80 ± 9 mg, 81 ± 18 s−1; n =
4 pitchers). When comparing similar lid masses
and lever lengths, we found that both species
behaved almost identically. The initial downstroke was faster and ended with a more abrupt
deceleration than the subsequent, damped oscillations (Fig. 1, F and L). Thus, lids of both
species act as torsion springs that generate high
jerk forces (24), thereby fulfilling the third requirement for springboard trapping.
The prominent similarity of springboard trapping and its component traits in both species
y
Fig. 1. Adaptations for springboard trapping in two Nepenthes pitcher plant species. (A to F) Nepenthes
gracilis. (G to L) N. pervillei. [(A) and (G)] Pitchers with near-horizontal lid orientation. [(B) and (H)] Lid wax
“pillars” provide limited grip for insects. [(C) and (I)] Wax platelets render the internal pitcher surface extremely
slippery. [(D) and (J)] A drop impact dislodges an ant from the lid. [(E) and (K)] High-speed video tracking
and [(F) and (L)] laser Doppler vibrometry of three N. gracilis (p1-3) and four N. pervillei (p4-7) lids show that all
lids respond with a rapid initial downstroke, followed by regular damped oscillations.
y g
5 mm
y
t = 24 ms
t = 0 ms
g
1
J
10mm
p
D
10 mm
The convergent origin of springboard trapping
provides an opportunity to probe two alternative scenarios for the evolution of this composite trait: (i) a “directional selection” scenario
wherein all component traits evolve jointly
and are under directional selection toward
the state required for the synergistic combination (31); and (ii) a “spontaneous coincidence”
scenario wherein the synergistic constellation
occurs by chance and the composite trait is
subsequently fixed by selection for the new
function. The pool of possible random character state combinations is maximized when
the phenotypic variation of each component
trait is high and each component evolves
independently.
A directional selection scenario implies that
(i) the components of the composite trait show
correlated evolution and (ii) phenotypic variation is reduced when a component trait approaches the required state for springboard
trapping, irrespective of the other traits. By
contrast, a spontaneous coincidence scenario
posits that (i) each component evolved independently, and (ii) phenotypic variation in the
absence of the composite trait is higher than
in springboard-trapping species, irrespective
of the trait value, as the individual traits are
not under (strong) selection unless they occur
in the beneficial combination. The two scenarios are not mutually exclusive: Under a “mixed
scenario,” some traits are under selection, leading to exaptation (32) for the composite trait,
whereas high phenotypic variation in traits
not under selection increases the odds for a
synergistic coincidence.
First, we used trait evolution modeling to test
for correlated versus independent evolution of
some or all component traits in 55 Nepenthes
species. Next, we investigated the stochastic
phenotypic variation of the lid angle and response to mechanical loading (Fig. 4) in a subset
of 42 species for which a minimum of five replicates could be sampled. For lid surface waxes,
we could not acquire enough samples to quantify intra-individual variation; however, scanning electron micrographs (Fig. 4 and fig. S1)
indicate high variability of presence, density,
RES EARCH | R E S E A R C H A R T I C L E
Proportion of total cuticular wax (%)
30
Lid
20
N. gracilis
N. pervillei
10
0
10
20
24 26 28 30 32
30
Acids
22 24 26 28 30 32 34
Alcohols
27 29 31 33
Esters
Alkanes
24 26 28 30 32 34
Aldehydes
Interior
wall
Bs AaBa Fr
?
Terpenoids
Lack of correlated evolution between the
three component traits supports the
spontaneous coincidence scenario of
composite trait evolution
5 January 2024
3 of 6
,
Next, we measured lid angles of 900 pitchers
from 42 Nepenthes species to test whether the
stochastic phenotypic variation was generally reduced in species with predominantly
horizontal lids (indicating directional selection),
or in springboard-trapping species only (indicating spontaneous coincidence). Although our
data mainly reflect intra-individual variation,
we cannot exclude a minor genetic variability
component (23).
Deviation from horizontal lid angles is detrimental for the springboard function of the
lid. Therefore, stabilizing selection should limit
the variability of lid angles in springboardtrapping species, regardless of the evolutionary
scenario. As expected, the lid angles of N. gracilis
(SD = ± 9.9°, n = 68 pitchers) and N. pervillei
(SD = 11.0°, n = 37 pitchers) showed lower than
average stochastic variation (in the bottom
20% of values; mean SD for 42 species = 17.4°;
Fig. 3 and fig. S8).
In other species, horizontal lids could also
be selected for more effective rain protection
(35), although a larger lid may compensate for
y
Chomicki et al., Science 383, 108–113 (2024)
Stochastic variation of lid angles supports
the spontaneous coincidence scenario
y g
Altogether, we fitted nine models of trait evolution that ranged from traits evolving independently to a model in which the evolution of
each trait is linked to the other two traits (23)
(Fig. 5). To account for a possible mixed scenario, we included models wherein two of the three
traits evolved in a correlated fashion whereas
the third evolved independently. Each of these
models represents a different hypothesis about
how these traits evolved. We used an implementation of Pagel’s model of correlated evolution
(33) in corHMM (23, 34), a modeling environment that expands the original model and allows testing for correlation among all three
traits at once. We independently confirmed our
results by testing for correlation in pairwise
combinations of traits using BayesTraits V3
(23). In both cases, we first excluded marginal
cases (near-horizontal lid orientation, pivot-like
loading response, inconsistent wax crystal presence or shape) when coding trait presence and
then repeated the analysis including them.
The independent models of trait evolution
were consistently found to fit our data best
(Fig. 5 and tables S1 and S2). In some cases, we
found weak support for correlated evolution
between two traits; however, the single bestfitting model was always an independent model
(table S1). Thus, there is strong evidence for independent evolution of the three traits, favoring
the spontaneous coincidence hypothesis for the
evolution of springboard trapping.
To assess variation in lid loading response, we
manually loaded lids and assigned them to
one of six deformation types (pivoting, pivotlike, bending, buckling, mixed bending-buckling,
and other, more complex responses; Fig. 4) (23).
N. gracilis and N. pervillei exhibited close to
100% pivoting—the crucial response for springboard trapping (index of dispersion ID for
N. gracilis = 69, c2 = 5181.3, df = 75, P < 0.01;
and N. pervillei ID = 19, c2 = 447.9, df = 24, P <
0.01; these were the highest and fourth-highest
ID of all species, respectively; ID ≫ 1 indicates
highly uniform response). Other species typically showed a range of loading responses (median ID = 3, 95% confidence interval = 35.1; Fig. 3
and fig. S9). Thus, the loading response was
less variable in springboard-trapping species
than in other species, as expected. However, lid
loading response variability was also reduced in
two out of six other species with a pivoting lid
response (N. tobaica, ID = 35; N. mikei, ID = 34;
Kruskal-Wallis test, H = 9.83, df = 2, P = 0.008;
fig. S9), suggesting that this trait might be developmentally constrained or under selection
for a different function.
Pivoting requires the lid to be mechanically reinforced to resist bending or buckling (24), thus
implying increased construction costs. Indeed,
N. gracilis pitchers have relatively high lignin
content compared to other sympatric Nepenthes
species (38). Structural compounds such as cellulose and lignin are comparatively “cheap” for
carnivorous plants, which are limited by mineral
nutrients, not by water and light (39). Tissue reinforcement is also correlated with trap longevity
(38). Stiffer lids and their pivoting load response
might thus result from selection for more durable traps, with slow life histories (40) potentially favoring springboard trapping.
y
and morphology of lid wax crystals between
species.
Stochastic variation of lid loading responses
points to a spontaneous coincidence
or mixed scenario
g
Fig. 2. Cuticular wax composition (mean ± SD) on the lower lid surface (upper part of the graph)
and pitcher inside (lower part of the graph) of N. gracilis (white bars) and N. pervillei (gray bars).
Both species show similar differences between the lower lid surface (predominantly alcohols) and the pitcher
inside (predominantly aldehydes). Numbers indicate C-chain lengths. Bs, b-sitosterol; Aa, a-amyrin; Ba,
b-amyrin; Fr, Friedelin. For statistics, see main text.
p
40 42 44 46 48 50 52
a steeper angle. Horizontal lids might also be
advantageous for photosynthesis; however,
pitchers contribute little to photosynthesis
(36), lids are small compared to the leaf lamina, and a broad range of lid angles may be
equally effective for light capture (37). If there
was strong selection for a horizontal lid, we
would expect the median of lid angles to be
close to 0°. For our 42 species, however, median lid angles (23°) were clearly above horizontal. Moreover, lid angles of species with
near-horizontal lids (−20° ≤ lid angle ≤ 20°,
median SD = 16.14) and species with steeper
lids (median SD = 18.16) were equally variable
(Mann-Whitney U test, n1 = 20 species, n2 = 22
species, U = 186.0, P = 0.40), and lid angle variation was not correlated with mean lid angle
(Spearman’s test, n = 42 species, r = 0.10, P =
0.53). Our data thus suggest that the lid angle
is not under directional selection and its evolution follows the spontaneous coincidence scenario.
RES EARCH | R E S E A R C H A R T I C L E
p
g
y
y g
y
,
Fig. 3. Mapping the presence of the three component traits of springboard
trapping on a recently published Nepenthes phylogeny (30) reveals their
independent evolution. Branch lengths (solid lines) represent phylogenetic
distance. For clarity, only species for which we collected trait data are shown.
Squares indicate horizontal lid orientation, circles denote pivoting lids, and stars
represent pillar-shaped wax crystals. Lighter shades denote marginal cases
Chomicki et al., Science 383, 108–113 (2024)
5 January 2024
(near-horizontal lids, pivot-like load responses, and sparse or morphologically
variable wax crystals) (23). Trait variability data reveal lower stochastic
phenotypic variation of lid angles and lid loading responses in springboardtrapping N. gracilis and N. pervillei than in the majority of species without this
composite trait. For detailed analysis of trait variability including statistics, see
main text and figs. S8 and S9.
4 of 6
RES EARCH | R E S E A R C H A R T I C L E
changes in individual component traits. Examples include the baleen of whales (43) or the leaf
mimicry of butterfly wings (44). Our study reveals convergence of a composite trait in tropical
pitcher plants that likely arose by spontaneous
coincidence of the component traits, possibly in a
mixed scenario combining directional selection
in one component with high plasticity sensu lato
(15) in one or more others. Trait plasticity,
whether environmentally induced or stochastic
in nature, enhances phenotypic diversity and can
become canalized when chance combinations
Fig. 4. Lid angles, loading responses, and
surface waxes in Nepenthes. (A to E) Lid angles.
(A) Horizontal, N. pervillei. (B) Steeply angled,
N. albomarginata. (C) Steeply angled, N. inermis.
(D) Upright, N. biak. (E) Reflexed, N. ampullaria.
(F to J) Loading responses. (F) Pivoting, N. gracilis.
(G) Pivot-like, N. adnata. (H) Bending, N. andamana.
(I) Buckling, N. burbidgeae. (J) Mixed bendingbuckling, N. vogelii. (K to O) Surface waxes. (K) Wax
pillars, N. pervillei. (L) Wax pillars, N. khasiana.
(M) Wax spikes, N. mira. (N) Scarce platelets,
N. dactylifera. (O) Smooth wax film, N. glabrata.
Scale bars, 2 mm.
A
B
C
D
E
F
G
H
I
J
K
L
M
N
O
Conclusions
The evolution of composite traits has frequently
been traced back to cumulative selection-driven
g
16 species), which further points to the absence of
notable selective pressure for a specific load response. Although pivoting as a predominant
lid loading response was only found in 8 out of
42 investigated species, and intra-individual variation of lid loading response was high in 38 out of
42 species, we cannot completely rule out that there
might be a degree of selection for pivoting lids.
p
Lid loading response could also be under selection for damage prevention (41) and for effective water shedding (42). For both functions, we
would expect any elastic deformation (pivoting,
pivot-like, bending, twisting) to be effective, and
all loading responses except buckling to be favorable. This is corroborated by the distribution
of predominant loading responses across the
42 investigated species, where all responses except buckling (2 species) and bending-buckling
(none) are common (fig. S9). The most common
response is “variable” (no predominant response,
y
,
5 January 2024
y
Chomicki et al., Science 383, 108–113 (2024)
y g
Fig. 5. Testing the predictions for component
trait evolution under directional selection,
spontaneous coincidence, and under a mixed
scenario. We compared one fully correlated model
(consistent with directional selection), three fully
independent models (consistent with spontaneous
coincidence), and three partially correlated models
(consistent with a mixed scenario), implemented
in the R package corHMM (23). (A) Examples of the
evolutionary models are illustrated for the evolutionary
transition from wax crystal absence (top row of trait
cases) to presence (bottom row), and three exemplary
model cases: fully independent (left), wax crystals
and load response correlated (middle), and all three
traits correlated (right). Thicker arrows indicate higher
transition rates. (B) The proportions of 1000 trees
sampled from the posterior distribution of topologies
supporting each scenario are reported for repeated
analyses with marginal cases (light-colored symbols) for
each trait either coded as present or absent. The
model-fitting results strongly support the independent
evolution of all three component traits, and therefore
the spontaneous coincidence scenario of composite
trait evolution (for detailed results, see table S1).
5 of 6
RES EARCH | R E S E A R C H A R T I C L E
produce a synergistic function. Our findings
highlight the importance of stochastic phenotypic variation, not only for selection to act on,
but also for increasing the likelihood of cooption into a new function (32). Notably, this
spontaneous coincidence mechanism for the
evolution of composite traits is applicable to
any type of variation on which selection can act
when beneficial combinations arise by chance.
Our findings emphasize the role of stochasticity
in composite trait evolution and call for further
investigation into the role of plasticity, and stochastic phenotypic variation in particular.
RE FE RENCES AND N OT ES
46. G. Burin, gburin/nepenthes-springboard: Code for corHMM
analyses, Zenodo (2023); https://doi.org/10.5281/zenodo.8277649.
AC KNOWLED GME NTS
This work was made possible by support from the Seychelles
Bureau of Standards, Seychelles National Parks Authority, Island
Conservation Society, Plant Conservation Action Group, Seychelles
National Herbarium, K. Beaver, L. Chong-Seng, C. Morel, T. Padayachy,
F. Fleischer-Dogley, D. Dogley, S. Sanghera, Universiti Brunei
Darussalam, U. Grafe, H. Tinggal, H. Bin Engin, Chester Zoo,
P. Esseen, P. Leach, Kew Gardens, R. Hilgenhof, Zürich Botanic
Garden, M. Scharmann, and S. Hartmeyer. Comments from
S. Renner, N. Levis, T. Caro, and four anonymous reviewers improved
the manuscript. Funding: G.C.: NERC (UK) Research Fellowship
(NE/S014470/1), ERC/UKRI frontier grant (EP/X026868/1); G.B.:
Leverhulme Trust Research Project Grant RPG-2019-323; L.B.: NSF
(US) Research Fellowship IOS 1812037; R.J.: NSERC (Canada)
Discovery Grant 2018-04909; B.M.: Fellowship of the Royal
Commission for the Exhibition of 1851 and Royal Society (UK)
Research Fellowship URF/R1/191033; U.B.: Royal Society (UK)
Research Fellowship UF150138 and UF\R\221028. Author
contributions: Conceptualization: G.C., R.J., U.B.; Methodology: G.C.,
G.B., L.B., B.M., R.J., U.B.; Investigation: G.C., G.B., L.B., B.M., J.G.,
U.B.; Formal analysis: all authors; Project administration: U.B.;
Visualization: G.C., U.B.; Writing – initial draft: G.C., L.B., B.M., R.J.,
U.B.; Writing – review and editing: all authors. Competing interests:
None. Data availability: All raw data and Matlab code are deposited
at Dryad (45). The code used for the CorHMM analysis is available
at Zenodo (46). License information: Copyright © 2024 the
authors, some rights reserved; exclusive licensee American
Association for the Advancement of Science. No claim to original
US government works. https://www.sciencemag.org/about/
science-licenses-journal-article-reuse
p
g
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.ade0529
Materials and Methods
Figs. S1 to S9
Tables S1 and S2
References (47–60)
MDAR Reproducibility Checklist
Movies S1 and S2
y
1. B. W. Tobalske, J. Exp. Biol. 210, 3135–3146 (2007).
2. K. Fukushima et al., Nat. Commun. 6, 6450 (2015).
3. K. J. Gilbert, J. H. Nitta, G. Talavera, N. E. Pierce, Biol. J. Linn.
Soc. Lond. 123, 321–337 (2018).
4. T. Renner, C. D. Specht, Curr. Opin. Plant Biol. 16, 436–442 (2013).
5. F. Jacob, Science 196, 1161–1166 (1977).
6. C. Darwin, The Origin of Species by Means of Natural Selection
(John Murray, ed. 6, 1872).
7. R. Goldschmidt, The Material Basis of Evolution (Yale Univ.
Press, 1940).
8. R. E. Lenski, C. Ofria, R. T. Pennock, C. Adami, Nature 423,
139–144 (2003).
9. E. Mayr, in Evolution after Darwin, S. Tax, Ed. (Univ. of Chicago
Press, 1960), vol. 1, pp. 349–380).
10. G. P. Wagner, V. J. Lynch, Curr. Biol. 20, R48–R52 (2010).
11. M. J. West-Eberhard, Developmental Plasticity and Evolution
(Oxford Univ. Press, 2003).
12. N. A. Levis, A. J. Isdaner, D. W. Pfennig, Nat. Ecol. Evol. 2,
1289–1297 (2018).
13. C. H. Waddington, The Strategy of Genes: A Discussion of Some
Aspects of Theoretical Biology (George Allen and Unwin, 1957).
14. R. C. Lewontin, The Triple Helix: Gene, Organism, and
Environment (Harvard Univ. Press, 2000).
15. G. Vogt, J. Biosci. 40, 159–204 (2015).
16. Y.-Y. Zhang, M. Fischer, V. Colot, O. Bossdorf, New Phytol. 197,
314–322 (2013).
17. S. Cortijo et al., Science 343, 1145–1148 (2014).
18. M. Medrano, C. M. Herrera, P. Bazaga, Mol. Ecol. 23, 4926–4938
(2014).
19. K. Laland et al., Nature 514, 161–164 (2014).
20. Z. D. Blount, R. E. Lenski, J. B. Losos, Science 362, eaam5979
(2018).
21. B. E. Juniper, R. J. Robins, D. M. Joel, in The Carnivorous Plants
(Academic Press, 1989), pp. 47–126.
22. U. Bauer, B. Di Giusto, J. Skepper, T. U. Grafe, W. Federle,
PLOS ONE 7, e38951 (2012).
23. Materials and methods are available as supplementary materials.
24. U. Bauer, M. Paulin, D. Robert, G. P. Sutton, Proc. Natl. Acad.
Sci. U.S.A. 112, 13384–13389 (2015).
25. E. Gorb et al., J. Exp. Biol. 208, 4651–4662 (2005).
26. C. E. Jeffree, E. A. Baker, P. J. Holloway, New Phytol. 75,
539–549 (1975).
27. M. Riedel, A. Eichner, R. Jetter, Planta 218, 87–97 (2003).
28. L. Samuels, L. Kunst, R. Jetter, Annu. Rev. Plant Biol. 59,
683–707 (2008).
29. S. J. Gould, Wonderful life: The Burgess Shale and the Nature of
History (Norton, 1989).
30. B. Murphy et al., Mol. Phylogenet. Evol. 144, 106668 (2020).
31. T. K. Suzuki, J. Exp. Zool. B Mol. Dev. Evol. 328, 304–320
(2017).
32. S. J. Gould, E. S. Vrba, Paleobiology 8, 4–15 (1982).
33. M. Pagel, Proc. Biol. Sci. 255, 37–45 (1994).
34. J. D. Boyko, J. M. Beaulieu, Methods Ecol. Evol. 12, 468–478
(2021).
35. C. Clarke, Nepenthes of Borneo (Natural History Publications,
1997), p. 15.
36. A. Pavlovič, E. Masarovicová, J. Hudák, Ann. Bot. 100, 527–536
(2007).
37. W. G. Duncan, Crop Sci. 11, 482–485 (1971).
38. O. O. Osunkoya, S. D. Daud, F. L. Wimmer, Ann. Bot. 102,
845–853 (2008).
39. T. J. Givnish, E. L. Burkhardt, R. E. Happel, J. D. Weintraub,
Am. Nat. 124, 479–497 (1984).
40. P. B. Adler et al., Proc. Natl. Acad. Sci. U.S.A. 111, 740–745 (2014).
41. K. J. Niklas, New Phytol. 143, 19–31 (1999).
42. R. M. Ginebra-Solanellas, C. D. Holder, L. K. Lauderbaugh,
R. Webb, Agric. For. Meteorol. 285–286, 107924 (2020).
43. T. A. Deméré, M. R. McGowen, A. Berta, J. Gatesy, Syst. Biol.
57, 15–37 (2008).
44. T. K. Suzuki, S. Tomita, H. Sezutsu, BMC Evol. Biol. 14, 229 (2014).
45. G. Chomicki et al., Raw data and Matlab code for: “Convergence
in carnivorous pitcher plants reveals a mechanism for composite
trait evolution,” Dryad (2023); https://doi.org/10.5061/
dryad.v41ns1s2.
Submitted 22 July 2022; resubmitted 9 June 2023
Accepted 7 November 2023
10.1126/science.ade0529
y g
y
,
Chomicki et al., Science 383, 108–113 (2024)
5 January 2024
6 of 6
RES EARCH
EVOLUTION
The genetic basis of a recent transition
to live-bearing in marine snails
Sean Stankowski1,2,3*, Zuzanna B. Zagrodzka1, Martin D. Garlovsky4, Arka Pal2, Daria Shipilina2,5,
Diego Garcia Castillo2, Hila Lifchitz2, Alan Le Moan6,7, Erica Leder7,8, James Reeve7,
Kerstin Johannesson7, Anja M. Westram2,9, Roger K. Butlin1,7
Key innovations are fundamental to biological diversification, but their genetic basis is poorly
understood. A recent transition from egg-laying to live-bearing in marine snails (Littorina spp.) provides
the opportunity to study the genetic architecture of an innovation that has evolved repeatedly across
animals. Individuals do not cluster by reproductive mode in a genome-wide phylogeny, but local
genealogical analysis revealed numerous small genomic regions where all live-bearers carry the same
core haplotype. Candidate regions show evidence for live-bearer–specific positive selection and are
enriched for genes that are differentially expressed between egg-laying and live-bearing reproductive
systems. Ages of selective sweeps suggest that live-bearer–specific alleles accumulated over more than
200,000 generations. Our results suggest that new functions evolve through the recruitment of many
alleles rather than in a single evolutionary step.
1 of 6
,
5 January 2024
y
Stankowski et al., Science 383, 114–119 (2024)
We used whole-genome sequences from 108
individuals to test the existing hypothesis of a
single origin of live-bearing, which was inferred
by parsimony analysis in earlier phylogenetic
studies (Fig. 1D, figs. S3 and S4, and tables S1
and S2) (7, 8). Rather than forming a single
clade, live-bearers formed two separate, wellsupported clades in a genome-wide phylogenetic
tree (Fig. 1E). One clade contains all L. saxatilis
y g
*Corresponding author. Email: s.stankowski@sussex.ac.uk
Live-bearing snails do not form
a monophyletic group
With the above expectation in mind, we used
topology weighting (Fig. 2A) to identify genomic regions associated with reproductive
mode. For each genomic window, topology
weighting calculates the degree of monophyly
toward three possible taxon subtrees (Fig. 2,
B and C, and fig. S7): (i) the background topology (Tb) observed in our genome-wide analysis; (ii) the reproduction topology (Tr), in
which samples cluster by reproductive mode;
and (iii) the control topology (Tc), which is of
no specific interest but provides a control for
distinguishing incomplete lineage sorting
from other processes that cause genealogical
discordance (e.g., gene flow). We used nonoverlapping 100–single nucleotide polymorphism (SNP) windows (mean size, 5.8 kb;
fig. S8) and calculated topology weights (13)
for each window by sampling 10,000 subtrees
(Fig. 2A).
We analyzed the joint distribution of topology weights in a ternary framework. This
approach exploits the geometric properties
of the ternary plot, allowing us to visualize
and quantify various properties of the genomewide distribution of discordance (Fig. 2A).
We used simulations to illustrate how different factors, including the timing of population splits and gene flow between nonsister
lineages, shape the ternary distribution of topology weights for large numbers of loci (Fig.
2B, supplementary text, figs. S9 to S19, and
tables S3 and S4).
We expected the empirical distribution of
weights to be biased toward Tb because this
y
Ecology and Evolutionary Biology, School of Biosciences,
University of Sheffield, Sheffield S10 2TN, UK. 2Institute
of Science and Technology Austria (ISTA), 3400
Klosterneuburg, Austria. 3Department of Ecology and
Evolution, University of Sussex, Brighton BN1 9RH, UK.
4
Department of Applied Zoology, Faculty of Biology,
Technische Universität Dresden, 01069 Dresden, Germany.
5
Department of Ecology and Genetics, Program of
Evolutionary Biology, Uppsala University, SE-752 36 Uppsala,
Sweden. 6CNRS and Sorbonne Université, Station Biologique
de Roscoff, 29680 Roscoff, France. 7Department of Marine
Sciences, Tjärnö Marine Laboratory, University of Gothenburg,
452 96 Strömstad, Sweden. 8Natural History Museum, University
of Oslo, 0562 Oslo, Norway. 9Faculty of Biosciences and
Aquaculture, Nord University, N-8049 Bodø, Norway.
Topology weighting reveals rampant
genealogical discordance and loci associated
with reproductive mode
g
1
brood pouch where embryos develop inside the
mother. Live-bearing is the only taxonomic
character that is diagnostic of L. saxatilis (8),
because no other known trait differs consistently between the live-bearing and egg-laying
individuals (8). In fact, sympatric populations
are so similar that the difference in mode was
long thought to reflect within-species polymorphism (8, 9), and molecular markers are
needed to identify males and juveniles in locations where live-bearing and egg-laying species coexist (10).
Live-bearing is thought to be an adaptation that allows snails to reproduce in areas
where eggs would be exposed to harsh conditions (8). This is reflected in the much broader
ecological and geographic distribution of
L. saxatilis compared with the two closely
related egg-laying species, L. arcana and
L. compressa (8) (Fig. 1, B and C, and fig. S2).
Egg-laying and live-bearing species have
adapted in parallel to contrasting environments across the intertidal zone (8, 11), largely
decoupling reproductive mode from other
axes of phenotypic divergence (Fig. 1B). There
is also evidence for rare gene flow between
egg-layers and live-bearers (10). These features
provide an opportunity to identify and study
the genetic changes underlying the live-bearing
innovation.
p
E
volution is a gradual process but occasionally results in sudden changes in form
and function that allow organisms to exploit new ecological opportunities (1, 2).
These game-changing traits, including
flight, vision, and the bearing of live offspring,
are known as “key innovations” (2–5). Key innovations are all around us and have catalyzed
the diversification of many groups (1). Despite
their significance, we know surprisingly little
about the origins and genetic basis of innovations (1). This is because most of them originated
deep in the past, making it difficult to disentangle causal loci from the countless other
genetic changes that accumulated up to the
present.
A recent transition in female reproductive
mode offers a rare opportunity to study the
genetic basis of an innovation that has evolved
many times across the animal kingdom (6).
We focus on a clade of intertidal gastropods
(genus Littorina) in which the ancestral state
is to lay a large egg mass, but one species gives
birth to live young (Fig. 1A and fig. S1) (7, 8).
Egg-layers have a gland that embeds fertilized
eggs into a protective jelly. In the live-bearer,
L. saxatilis, this structure has evolved into a
from Iberia (hereafter “Iberian saxatilis”) and
another includes all other L. saxatilis (hereafter “northern saxatilis”) that were sister to
a clade of egg-laying L. arcana (the same pattern was observed in an ML tree, phylogenetic
network, and principal components analyses;
figs. S5 and S6).
The discordance between evolutionary relationships and reproductive mode has several
possible explanations. One interpretation of
the genome-wide tree is that there has been
more than one transition between egg-laying
and live-bearing. However, because the tree
represents the aggregate signal from all loci in
the genome, it does not necessarily reflect the
evolutionary history of these groups or of any
single locus or trait (12). This means that it is
also possible that live-bearing may have evolved
once and causal alleles became associated with
two different lineages through the interaction
of gene flow and selection (12). If this were the
case, then we would expect genealogies for loci
that cause live-bearing to be strongly discordant from the genome-wide tree, with samples
grouping by reproductive mode.
RES EARCH | R E S E A R C H A R T I C L E
C Distributions of egg-laying and live-bearing
A Reproductive modes
Egg-laying (oviparity)
Live-bearing (viviparity)
Jelly gland
Crawl-away
offspring
Egg mass
Birth live young
Reproduce year-round
L. saxatilis
wave
ecotype
L. compressa
L. saxatilis
crab
ecotype
E Whole-genome NJ phylogeny
Single origin
1.7 - 0.06 mya
from egg-laying
ancestor
Wave exposed
Live-bearers
England II
England III
Scotland I
Sweden
L. saxatilis
Russia
Norway
England I / Isle of Man
Ireland / Scotland II
France
Wales II
Iceland
USA
Live-bearers
Egg-layers
Wales I & II
France
0.005
England
III & IV
Scotland I
Norway I & II
L. saxatilis
L. arcana
Wales II
y
France
Other
barnacle
ecotype
Northern group
All ecotypes
g
Crab infested
Egg-layers
Lay a large egg mass
Reproduce seasonally
D Hypothesis for the
origin of live-bearing
p
Habitats
B Modes, species, and ecotypes
L. arcana
Live-bearing only
Egg-laying and live-bearing
Human introductions
(live birth only)
Brood pouch
Iberia I
Iberia II
brackish ecotype
France / Norway II /
Wales II / Ireland
Fig. 1. Variation in reproductive mode in Littorina. (A) Anatomical differences
between modes. (B) Egg-layers reproduce during a limited breeding season, whereas
live-bearers release offspring year-round. The two egg-layers share their habitats with
ecotypes of the live-bearer L. saxatilis. (C) Approximate distributions of the modes
L. compressa
highlighting the broader distribution of live-bearing. (D) Existing hypothesis for the
origin of live-bearing inferred by past phylogenetic studies (7, 8). (E) Neighborjoining phylogenetic tree based on whole-genome sequences (108 individuals and
18.5 million variable sites). All nodes have 100% bootstrap support.
y g
5 January 2024
wide level (DLR for regions outside inversions =
–0.007, P = 0.074; fig. S20).
Much stronger asymmetry was observed between the far left and right subtriangles, corresponding to windows that more strongly fit
one of the alternative topologies (Fig. 2D and
fig. S24). However, the asymmetry was in the
opposite direction from the genome-wide pattern, with a large excess of windows strongly
biased toward the reproduction tree compared
with the control tree (Tr > 0.7 = 1151 windows
versus 461 for Tc; DLR = –0.43, P = 1 × 10–5). A
total of 88 windows perfectly fit the reproduction topology (i.e., Tr = 1; table S7) compared with no windows that perfectly fit the
control topology (DLR = 1.00, P = 1 × 10–5; Fig.
2E and fig. S25).
Evidence for live-bearer–specific
positive selection
Although neutral gene flow can generate strong
asymmetry under some circumstances, we
2 of 6
,
Stankowski et al., Science 383, 114–119 (2024)
ogy (Fig. 2B and supplementary materials)
(14). We detected asymmetry using a new statistic, DLR (Fig. 2D and fig. S19). A genome-wide
test performed by calculating DLR between the
two halves of the triangle, revealed a 3.4% excess
of windows shifted toward the control topology
(DLR = 0.034, permutation test P = 1 × 10–5). DLR
calculated between analogous left and right
subtriangles revealed that this asymmetry was
driven by an excess of trees with a small bias
toward Tc (Fig. 2D and table S5). Further exploration showed that this bias was caused by
10 previously identified chromosomal inversions (16), none of which was associated with
reproductive mode (figs. S20 to S23, table S6,
and supplementary materials). For each inversion, one arrangement was more common
in Spanish L. saxatilis and L. arcana, and the
other was more common in L. compressa and
northern L. saxatilis. When the chromosomal
inversions were removed, we found no significant left-right asymmetry at the genome-
y
was the topology observed in the genomewide analysis. However, the observed bias was
only slight (Tb = 0.380, Tc = 0.310, and Tr =
0.308), with just 62 of ~155,000 genomic regions
perfectly fitting Tb (i.e., Tb = 1) (Fig. 2C). Instead, the bulk of the distribution fell close
to the center of the triangle. This indicates that
sequence variation is broadly shared between
groups, a pattern that could be caused by extensive incomplete lineage sorting due to rapid
diversification relative to the effective population size, widespread gene flow during divergence, or both (14, 15). Thus, although well
supported statistically, the genome-wide tree is
a very poor predictor of evolutionary relationships at any given genomic region.
We found substantial left-right asymmetry
in the distribution of topology weights (Fig.
2D). Such a bias is not expected to arise from
incomplete lineage sorting because there is
an equal chance that a given gene tree will
more closely resemble either alternative topol-
RES EARCH | R E S E A R C H A R T I C L E
p
g
3 of 6
,
5 January 2024
(Fig. 3E). The 88 perfectly associated windows
mapped to 50 contigs in our genome assembly (mean 1.7 ± 1.5 windows per contig; table
S9). Associated regions were narrow, mostly
spanning <20 kb (mean 12 ± 14.4 kb). Slidingwindow analysis of each contig generally
revealed clear peaks of allele frequency differentiation (FST) and sequence divergence
(dxy) between the egg-layers and live-bearers,
as well as valleys of p in live-bearers (Fig. 3E).
We also inferred ancestral recombination graphs
(ARGs) for selected contigs to refine candidate
regions (Fig. 3E). Unlike trees inferred from
genomic windows with arbitrary start and end
positions, each tree in an ARG corresponds to
an inferred nonrecombining segment of the genome (19). Thus, by applying topology weighting
to the sequence of marginal trees, we were able
to identify more precisely the segment of genome
retained by all live-bearing samples after the selective sweep. In both cases, the core live-bearing
haplotype spanned <2 kb. Live-bearers showed
much shallower coalescence in these regions than
egg-layers, as expected after a sweep (Fig. 3E).
y
Stankowski et al., Science 383, 114–119 (2024)
(Tb and Tc) were weak and similar for both
groups, confirming that reduced p in livebearers is specific to Tr rather than being a general feature of windows with extreme weights
(fig. S28). The site-frequency spectra (SFS) and
sample-size–corrected estimates of private alleles for perfectly associated regions provide
further evidence for selection (Fig. 3, B to D;
figs. S29 to S31; and tables S9 to S11): the livebearer SFS was strongly skewed toward rare
variants [Tajima’s D = –1.89, 95% confidence
interval (CI) = –1.77 to –2.01; fig. S29], the majority of which (80%) were private to live-bearers.
Both results are expected during the phase
when diversity is recovered by mutation after a
selective sweep (18). By contrast, the SFS for egglayers was much closer to the neutral expectation (Tajima’s D = –0.24, 95% CI = –0.037 to
–0.437), with polymorphic sites being 2.14 times
more abundant in egg-layers after accounting
for the difference in sample size.
We next characterized footprints of selection within contigs to estimate the number
and size of candidate regions more accurately
y g
are unable to explain the observed Tr bias
without invoking natural selection (see the
supplementary materials and table S8). We found
strong additional evidence for live-bearer–
specific positive selection in regions associated
with reproductive mode. First, window-based
estimates of nucleotide diversity (p) in livebearers decreased substantially with increasing Tr weight (Fig. 3A), but we found no such
relationship in egg-layers. Eighty-four (95%) of
the 88 perfectly associated regions showed reduced p in live-bearers (mean plive-bearer = 0.0029
versus pegg-layer = 0.0065; paired Wilcoxon test,
P = 1.313 × 10–15; Fig. 3A and fig. S26). These
results are consistent with selection having
purged diversity from haplotypes associated
with live-bearing (17). Although this result
could in principle result from a live-bearer–
specific demographic bottleneck, we can rule
this out because live-bearers and egg-layers
have similar levels of genome-wide diversity
(mean plive-bearer = 0.0065 versus pegg-layer =
0.0062; Fig. 3A and fig. S27). Further, relationships between p and the other weights
symmetrical between the left and right half-triangles. Additional factors, including
gene flow, create a bias toward one discordant genealogy (iv to vi). (C) Possible
topologies and the empirical distribution of weights for the 154,971 100-SNP
windows (“C,” compressa; “A,” arcana; “I” Iberian saxatilis; “N,” northern saxatilis).
Hexagonal bins are colored by window count. (D) Counts of windows in the left
and right half-triangles, with asymmetry quantified using DLR. Further division
into subtriangles reveals left-right asymmetry throughout the distribution.
Asterisks indicate significant asymmetry between corresponding left- and rightsided subtriangles. (E) Distributions of weights >0.7.
y
Fig. 2. Topology weighting reveals genomic regions associated with
reproductive mode. (A) For each genomic window, we inferred a tree for all
haplotypes and then classified 10,000 “subtrees” by randomly picking one
haplotype per group. Topology weights are the proportions of each topology
among all subtrees. Windows were plotted in a ternary plot based on the weights.
(B) Simulated distributions of weights. A greater opportunity for lineage sorting
(i to iii) biases the distribution toward the topology that matches the
demographic history. Incomplete lineage sorting yields genealogies that are a
better fit to one of the discordant trees, but the distribution is always
RES EARCH | R E S E A R C H A R T I C L E
A Relationship between π and Tr
0.015
Genomewide π
C
A
0.005
IN
CA
I
N
0.000
0.6
0.7
0.8
0.9
0.4
−0.5
Tajima’s D
0.010
0.0
C &A
G-test p < 2.2e-16
B > E (4)
B < E (84)
C Tajima’s D
Live-bearers
Egg-layers
0.5
Proportion of loci
Genetic diversity (π)
B Folded Site Frequency Spectra
Live-bearers π for 88 windows
where Tr = 1
Egg-layers
0.3
0.2
Live-bearers
n = 80
−2.0
0.0
1.0
n = 28
Egg-layers
−1.0
−1.5
0.1
A only
n = 24
Minor allele count
Topology weight (Tr > 0.6)
p
Example
trees Tr = 1
D Trees
Tr = 1
π & dxy
Contig 1808, LG4
fixed differences
0.6
1.0
0.3
0.5
Contig 3201 | un-mapped
0.6
0.5
0.0
2
0.0
0.02
Live-bearers
Egg-layers
dxy Between
0.01
0.00
0.00
Live-bearers
Egg-layers
5.0
2.0
y g
0.5
0.0
4
0.0
0.02
0.01
5.0
2.0
0.3
0.5
0.1
y
TMRCA (Ne)
1.0
g
Tr weight FST
E Example contigs
0.1
8
8
12
16
50
Physical position (kb)
55
60
65
Physical position (kb)
y
Approximate estimates of the timing of each
selective sweep at the mode-associated loci,
determined from the accumulation of private
mutations (T = pprivate/2m), span a broad
range from ~20,000 to 200,000 generations
before present, with a median of 70,000 generations before present (fig. S32). Assuming two
generations per year, this equates to 100,000
Stankowski et al., Science 383, 114–119 (2024)
each region is shown. Top panel: FST between egg-layers and live-bearers in
3-kb sliding windows (30-bp steps). Tr shows the results of topology weighting
applied to marginal trees obtained from inferred ARGs. Purple arrows show
fixed differences between modes. Middle panel: p and dxy in sliding windows.
Bottom panel shows traces of time to the most recent common ancestor
(TMRCA) obtained from ARGs. Bold lines are median estimates. Envelopes are
95% CIs. The red box shows the inferred length of the core haplotype block
associated with live birth.
to 10,000 years before present, with a median
time of 35,000 years.
Mode-associated regions are widespread and
enriched for genes that are differentially
expressed between reproductive systems
The assignment of contigs to a genetic map
revealed that reproductive-mode–associated
5 January 2024
windows are widespread across the genome
rather than colocalizing to one or a few genomic regions (Fig. 4A). As expected for a polygenic trait, the number of mode-associated
windows on each linkage group (LG) was strongly predicted by LG size (Tr > 0.7, r = 0.79, P <
0.0001; Tr > 0.9, r = 0.71, P < 0.005). Associated
regions were also widespread within linkage
4 of 6
,
Fig. 3. Evidence for positive selection on haplotypes associated with live
birth. (A) Relationship between p and Tr for both reproductive modes. Triangles
on right indicate genome-wide p. Violin plots indicate distributions of p for
windows where Tr = 1. (B) Folded SFS for each mode in perfectly associated
regions projected at the same sample size for comparison. (C) Estimates of
Tajima’s D with 95% CIs for perfectly associated regions. (D) Examples of trees
for windows where Tr = 1. (E) Variation across two example contigs that
contain a window where Tr = 1 (span of the orange box). The tree associated with
RES EARCH | R E S E A R C H A R T I C L E
A Distribution of high Tr windows across LGs
Tr weight
Number of windows
80
1
0.9 - 1
0.8 - 0.9
0.7 - 0.8
60
Mapped
64%
36%
Expected
count
20
LG1 2
3
4
5
6
7
8
C Expression by tissue
Clustering by expression
(13,492 genes)
Brood pouch (North L. saxatilis)
Foot
Jelly gland (L. arcana)
9 10 11 12 13 14 15 16 17
5
6
7
10
11
12
15
17
8
13
9
14
Tr weight
1
0.9 - 1
0.8 - 0.9
0.7 - 0.8
20 cM
E Association of DEGs
and Tr weight
Tr bin
0.0 – 0.1
Jelly gland
0.1 – 0.2
Up in jelly gland
4
2
0
log2(CPM + 1)
-2
-4
Up in brood pouch
0.2 – 0.3
0.3 – 0.4
0.4 – 0.5
0.5 – 0.6
0.6 – 0.7
0.7 – 0.8
0.8 – 0.9
Reproductive tissue
Foot tissue
Foot & reproductive
0.9 – 1.0
Proportion of each expression class
Fig. 4. Candidate regions are widespread across the genome and enriched for DEGs between reproductive systems. (A) The number of high Tr windows (Tr > 0.7) assigned to each of the 17 L. saxatilis
LGs. Dots show the expected number of windows given the total assigned to each LG. (B) Distribution of
high-Tr windows across LGs. (C) Clustering of tissues by expression and the number of DEGs in each
expression class. (D) Clustering of reproductive tissues based on patterns of expression. (E) The proportion
of genes in each differential expression class after binning genes according to the Tr weight.
5 of 6
,
5 January 2024
y
Stankowski et al., Science 383, 114–119 (2024)
1044 DEGs and showed much higher rates of
tissue-specific differential expression between
reproductive systems (Fig. 4C and fig. S33).
To increase power to detect DEGs between
the reproductive systems, we sequenced additional pools of reproductive tissue (Fig. 4D and
fig. S34). This analysis detected 1450 DEGs,
66.1% (858) of which showed higher expression in the brood pouch of live-bearers. To
test for the enrichment of DEGs in regions
associated with reproductive mode, we binned
each DEG according to the Tr score of its associated genomic region (Fig. 4E, fig. S35, and
table S12). We found that the proportion of
reproductive mode DEGs strongly increased
with increasing Tr weight (Spearman’s rho =
0.903, P = 9 × 10–4) (table S13). No correlation
was observed between Tr weight and foottissue only DEGs (Spearman’s rho = –0.410,
P = 0.217) (table S13).
Gene ontology (GO) analysis and functional
annotation suggested that the transition to
Our analyses show that live-bearing in Littorina
is associated with selection on many loci, as
was the case in the only comparable analysis
in Zootoca lizards (20). Although our genomewide analysis suggested two independent origins of live-bearing, the high sequence similarity
of live-bearer–specific alleles indicates that
they had a single origin. Given the number of
associated loci, the history of their origin and
spread may be highly complex, potentially
varying among loci. One possible scenario is
that all live-bearer–specific alleles originated in
one location (e.g., in the ancestor to the Iberian
clade), after which the range of live-bearers
expanded until they encountered egg-layers.
Hybridization may have then allowed beneficial live-bearing alleles to introgress onto the
local egg-laying genetic background, or it may
have eroded genome-wide differentiation between egg-layers and live-bearers, whereas selection maintained alternative sets of alleles at
mode-associated loci. It is also possible that
alleles arose in numerous locations, and that
associations built up between them at different
times, perhaps as live-bearing spread. Regardless
of the precise history, which we cannot resolve
at present, the interaction between gene flow
and selection has allowed us to identify loci
associated with reproductive mode.
Live-bearing is the only known trait that
consistently distinguishes L. saxatilis from the
egg-laying species, making associated loci good
candidates for causing the difference in reproductive mode. We found supporting evidence
in our expression analysis, because associated
regions were strongly enriched for DEGs between the reproductive systems. This suggests
that selection has acted on differences in gene
expression, driving the evolution of live-bearing,
including the development of the brood pouch.
Because reproductive mode is a complex trait,
y g
groups, in some cases with strong associations
near opposite ends of the same LG (Fig. 4B).
Candidate regions also showed strong enrichment of genes that are differentially expressed
between female live-bearing and egg-laying
reproductive tissues. To identify differentially
expressed genes (DEGs), we collected reproductively mature female L. arcana and northern
L. saxatilis at peak breeding season from a
single location (to control for environmental
effects) where sympatric egg-layers and livebearers are morphologically cryptic aside from
their reproductive anatomies. We first compared transcriptomes from pools of reproductive systems (brood pouch versus jelly gland)
and foot tissue paired from the same individuals. Clustering analysis based on patterns of
gene expression (13,492 genes) revealed that
pools of reproductive tissue grouped by system type, but egg-laying and live-bearing species did not group based on expression in foot
tissue. Differential expression analysis revealed
Conclusions
y
72%
4
g
13%
16
1,450 DEGs
Percent DEGs
by tissues (n = 1044)
15%
3
live birth involved genes with diverse functions. Separate GO analyses conducted on a
sequence-based gene set (574 genes in regions
where Tr > 0.7) and expression-based gene
set (1450 reproductive mode DEGs) yielded
37 enriched GO terms, including transmembrane transport, calcium-ion binding, and ion
channel activity (fig. S36). We examined the
putative functions of the 27 genes found in
both sets in more detail (table S14). These included genes putatively associated with antibacterial activity (lectin L6-like protein; higher
expression in the brood pouch), the synthesis
of mucin-type oligosaccharides (GALNT10like; higher expression in the brood pouch),
the formation of structural tissue (IFB-like
and CMP-like, both higher expression in the
brood pouch), and two secretory genes involved in egg mass production in another marine snail (both with lower expression in the
brood pouch).
p
Foot
2
D Differential gene expression
between reproductive tissues
(with additional samples)
Brood pouch
Reproductive
LG1
Contigs
Un-mapped
40
0
B Distribution across map positions
RES EARCH | R E S E A R C H A R T I C L E
key innovations are not visible to selection
before they arise, models of saltational evolution invoke large-effect macromutations to
explain their evolution (28). We do not know
which mutation caused the threshold from
egg-laying to live-bearing to be crossed. Some
potentiating mutations may have preceded
live-bearing but were critical to its origin, and
others may have refined live-bearing after it
arose. Nevertheless, our results suggest that
new functions evolved gradually through the
recruitment of alleles at many loci rather than
arising in a single evolutionary step (20, 29, 30).
RE FERENCES AND NOTES
AC KNOWLED GME NTS
y
We thank J. Galindo, M. Montaño-Rendón, N. Mikhailova,
A. Blakeslee, E. Arnason, and P. Kemppainen for providing
samples; R. Turney, G. Sotelo, J. Larsson, T. Broquet, and S. Loisel
for help collecting samples; Science Animated for providing the
snail cartoons shown in Fig. 1; M. Dunning for help in developing
bioinformatic pipelines; R. Faria, H. Morales, and V. Sousa for
advice; and M. Hahn, J. Slate, M. Ravinet, J. Raeymaekers,
A. Comeault, and N. Barton for feedback on a draft manuscript.
Funding: This work was supported by the Natural Environment
Research Council (grant NE/P001610/1 to R.K.B.), the European
Research Council (grant ERC-2015-AdG693030-BARRIERS to
R.K.B.), the Norwegian Research Council (RCN Project 315287 to
A.M.W.), and the Swedish Research Council (grant 2020-05385
to E.L.). Author contributions: Conceptualization: S.S., A.M.W.,
K.J., R.K.B.; Formal analysis: S.S., Z.B.Z., M.D.G., A.P., D.S., D.G.C.,
H.L., E.L., J.R., A.L.M.; Writing – original draft: S.S., R.K.B.; Writing –
review and editing: S.S., Z.B.Z., M.D.G., A.P., D.S., D.G.C.,
A.L.M., E.L., J.R., K.J., A.M.W., R.K.B. Competing interests: The
authors declare no competing interests. Data and materials
availability: Raw sequence data are available on the short-read
archive (SRA) under bioproject PRJNA626520. VCF files have been
archived on Zenodo (31). All analysis scripts have been archived
on Zenodo (31) and are available on Github at https://github.com/
seanstankowski/Littorina_reproductive_mode. License information:
Copyright © 2024 the authors, some rights reserved; exclusive
licensee American Association for the Advancement of Science. No
claim to original US government works. https://www.science.org/
about/science-licenses-journal-article-reuse
g
SUPPLEMENTARY MATERIALS
science.org/doi/10.1126/science.adi2982
Materials and Methods
Supplementary Text
Figs. S1 to S36
Tables S1 to S14
References (32–79)
MDAR Reproducibility Checklist
Submitted 25 April 2023; accepted 25 October 2023
10.1126/science.adi2982
y g
1. A. Wagner, The Origins of Evolutionary Innovations:
A Theory of Transformative Change in Living Systems (Oxford
Univ. Press, 2011).
2. A. H. Miller, J. T. Stroud, J. B. Losos, Trends Ecol. Evol. 38,
122–131 (2023).
3. A. H. Miller, in Ornithologie als Biologische Wissenschaft,
E. Mayr, E. Schüz, Eds. (Carl, Winter, 1949), vol. 84 pp. 84–88.
4. D. A. Baum, A. Larson, Syst. Biol. 40, 1–18 (1991).
5. A. de Queiroz, Syst. Biol. 51, 917–929 (2002).
6. C. M. Whittington et al., Biol. Rev. Camb. Philos. Soc. 97,
1179–1192 (2022).
7. D. G. Reid, P. Dyal, S. T. Williams, Zool. Scr. 41, 125–136
(2012).
8. D. G. Reid, Systematics and Evolution of Littorina (The Ray
Society, 1996).
9. G. Seshappa, Nature 160, 335–336 (1947).
10. S. Stankowski et al., Philos. Trans. R. Soc. London B Biol. Sci.
375, 20190545 (2020).
11. K. Johannesson, J. Sea Res. 49, 107–117 (2003).
12. M. W. Hahn, L. Nakhleh, Evolution 70, 7–17 (2016).
13. S. H. Martin, S. M. Van Belleghem, Genetics 206, 429–438
(2017).
14. W. P. Maddison, Syst. Biol. 46, 523–536 (1997).
15. R. R. Hudson, Oxf. Surv. Evol. Biol. 7, 44 (1990).
16. J. Reeve, R. K. Butlin, E. L. Koch, S. Stankowski, R. F. Faria,
Mol. Ecol. 10.1111/mec.17160 (2023).
17. J. M. Smith, J. Haigh, Genet. Res. 23, 23–35 (1974).
18. J. M. Braverman, R. R. Hudson, N. L. Kaplan, C. H. Langley,
W. Stephan, Genetics 140, 783–796 (1995).
19. D. Shipilina, A. Pal, S. Stankowski, Y. F. Chan, N. H. Barton,
Mol. Ecol. 32, 1441–1457 (2023).
20. H. Recknagel et al., Nat. Ecol. Evol. 5, 1546–1556
(2021).
21. R. Shine, Herpetologica 39, 1–8 (1983).
22. D. B. Lowry, J. H. Willis, PLOS Biol. 8, e1000500 (2010).
23. F. C. Jones et al., Nature 484, 55–61 (2012).
24. M. Wellenreuther, L. Bernatchez, Trends Ecol. Evol. 33,
427–440 (2018).
25. R. Faria et al., Mol. Ecol. 28, 1375–1393 (2019).
26. E. L. Koch et al., Evol. Lett. 5, 196–213 (2021).
27. H. E. Morales et al., Sci. Adv. 5, eaav9963 (2019).
28. G. Theissen, Theory Biosci. 128, 43–51 (2009).
29. Z. D. Blount, J. E. Barrick, C. J. Davidson, R. E. Lenski, Nature
489, 513–518 (2012).
30. J. R. Meyer et al., Science 335, 428–432 (2012).
31. S. Stankowski et al., Data and code for: The genetic
architecture of a recent transition to live-bearing in marine
snails, Version V1, Zenodo (2023); https://doi.org/10.5281/
zenodo.8318995.
p
associated loci may also underpin a diverse range
of biological functions, including differences
in the synchronization of egg production (8),
differences in embryo retention times (21), and
variations in immune function and metabolism
(21). It is, however, important to emphasize that
some loci may not be causally associated with
reproductive mode and may instead underlie
other less conspicuous traits that are functionally linked or associated with the live-bearing
or egg-laying lifestyle.
Polymorphic inversions often underpin local
adaptations (22, 23) and are thought to maintain beneficial sets of alleles by suppressing
recombination (24). This makes a role in the
evolution of a key innovation seem likely, but
we found that known chromosomal inversions
in Littorina are not associated with the difference in reproductive mode. Many large inversions are shared among these Littorina
species (16), and there is evidence that they
play a key role in ecotype formation and reproductive isolation (25–27). For example, in
L. saxatilis, repeated adaptation to contrasting crab-infested and wave-swept environments
usually involves 8 to 12 major inversions, with
the alternative arrangements contributing to
differences in morphology and behavior
(26, 27). However, egg-laying species coexist
with L. saxatilis and likely use the same inversions to adapt to local selection pressures
(16). Thus, the independence of genetic architectures for ecotype formation and reproductive mode may be a major factor permitting the
local coexistence of egg-laying and live-bearing
species.
Our estimates of the timing of sweeps suggest that alleles associated with live-bearing
were recruited gradually over the past 200,000
generations (~100,000 years). This finding
is relevant to long-standing debate about the
genetic basis of evolutionary novelty. Because
y
,
Stankowski et al., Science 383, 114–119 (2024)
5 January 2024
6 of 6
WORKING LIFE
By Andrew Sharo
The call of activism
p
“I
am not presently indicted for or otherwise criminally or civilly charged …” Reading the statement I
needed to affirm on the annual renewal form for my National Science Foundation (NSF) Graduate
Research Fellowship, I suddenly felt at a crossroads. Three months earlier, I had been charged with
felony theft—one of the crimes listed on the form—because of my animal rights activism. I believed
my actions were legal, not theft, and I was hopeful a jury would agree. But in my worst ruminations,
I feared losing my fellowship and ending up in prison. I never doubted I had done the right thing.
Yet I now faced the prospect that my activism would jeopardize my research career.
science.org SCIENCE
,
5 JANUARY 2024 • VOL 383 ISSUE 6678
y
122
Andrew Sharo is an NSF postdoctoral fellow at University of California,
Santa Cruz. Send your career story to SciCareerEditor@aaas.org.
y g
“I now faced the prospect that
my activism would jeopardize
my research career.”
y
ism would make me look less serious about my research.
The possibility of losing my fellowship came on top of the grind
of my qualifying exam and trial
preparation, and the stress became overwhelming. Also, I feared
prison. So, I took a plea deal, even
though I believed my actions had
been justified. In a stroke of legal
creativity, my lawyer negotiated for
me to plead guilty to two counts of
burglary, which, unlike theft, was
absent from NSF’s no-no list. I felt
relieved and ready to get back to
my research. I didn’t abandon my
activism, but pursued actions that
were lower risk, including community building and animal care—
even adopting some chickens from
a factory farm.
Eventually, two of my co-defendants went to trial, where I testified as a witness. The high stakes were nerve-wracking—if
I messed up, friends would go to jail—yet the experience
was also cathartic. The jury found them not guilty, deciding
the piglets were so sick their value to the company was effectively negative. The trial was a watershed case in farmed
animal welfare. To know that I contributed felt just as gratifying as publishing each dissertation chapter.
I am now a postdoc, studying conservation genetics. I
believe my research will have positive impacts in the long
term, but many days it still feels plodding. In contrast, the
appeal of activism is constant, reignited every time I read
the news or friends tell me about their clashes with the legal system. Sometimes it works out, and sometimes, as I
found, you suffer the consequences. But, as in science, the
rare successes make it all worthwhile. j
g
My Ph.D. had been going well
enough. I loved my work analyzing the genomes of children
with severe inherited immunodeficiencies. Yet the research was
often plodding, and I rarely felt I
was making much of an impact.
Compelled to find another way to
make a tangible difference, I joined
a group planning an undercover
investigation of one of the largest
factory pig farms in the country. All
activism comes with risk, but this
work—to determine whether the
farm had made good on its promise to phase out inhumane gestation crates—felt relatively safe. The
group included two lawyers, and
most folks involved in similar actions were never charged or their
charges were quickly dropped.
At the facility, we discovered
gestation crates were still in use.
We also witnessed widespread diarrheal diseases, untreated
wounds, and high piglet mortality. I can still smell the
ammonia-soaked air and hear pigs banging their heads
against metal bars, a typical stress behavior. We documented the investigation online, garnering national media
coverage. We also rescued two sick piglets and brought them
to medical care. When the facility eventually announced it
would significantly reduce operations, I remember hoping
my research would one day be equally impactful.
One year later, my co-investigators and I were charged
with second degree felony theft and burglary for the rescue.
If found guilty, we faced decades in prison.
The other students in my Ph.D. cohort were supportive, some even encouraging. My program coordinator was
unfazed. My adviser offered his help, but he was deeply
concerned; no one wants to lose a student, particularly to
prison. I was reluctant to reach out for support or to bring
up my situation with other faculty out of fear that my activ-
LIFE SCIENCE TECHNOLOGIES
Produced by the Science/AAAS Custom Publishing Office
ne w prod ucts
5 JANUARY 2024 • VOL 383 ISSUE 6678
science.org/custom-publishing
SCIENCE
,
120
y
Newly offered instrumentation, apparatus, and laboratory materials of interest to researchers in all disciplines in academic, industrial, and governmental organizations are featured in this
space. Emphasis is given to purpose, chief characteristics, and availability of products and materials. Endorsement by Science or AAAS of any products or materials mentioned is not
implied. Additional information may be obtained from the manufacturer or supplier.
y g
Electronically submit your new product description or product literature information! Go to www.science.org/about/new-products-section for more information.
y
''
)'
'-
'
&
'
(
'
"
)
&
'
'
"
) (
"),
'
&
3
(()(
(
Real-time PCR system for IVD
Bio-Rad Laboratories have announced the launch of the CFX Opus
Deepwell Dx Real-Time PCR System, a real-time PCR system listed
with the U.S. Food and Drug Administration for in vitro diagnostic
(IVD) testing and developed in accordance with the European Unionís
/0123
&+4'& '5% '6/7
821'7/9
8/4'#*-':,$
'
&921'& '
use in Canada and Singapore has also been acquired, and Bio-Rad
plans to continue seeking regulatory approvals for additional global
markets in the coming months. The product is the latest addition to
3 /'
& 27' & 3&1
&'&' /213
6/' '
+43 6/+34'&)/
+0' /8
4/'
2+3
"823
&+'!
3 '/
1
3'& ' /283
&+4' '3&'$:'(1'
+'2'
!/11'
format for quantitative PCR diagnostic assays. The system is also
292
121/'
+'2'
'2+7'.!/11' /283
&+'1&8'& 623' 11'443/64'
+'
the CFX Opus Dx Real-Time PCR family are open platforms that can
613
1/' '3&'"9/'32 0/34'3&'/+21/'/ 8
/+3'5% '2442'7/9/1& 6/+3'2+7'7
20+&43
8'3/43
+0'2+7'&)/ '7/43& '62+20/6/+3'2+7'
analysis software with integrated security features for electronic
signature, audit, and user control.
Bio-Rad
& '
+&'$'$,.
,
bio-rad.com/cfxopus-dx
g
PCR Barcode Reader
!
"
#%
&
ing Individual Access 2D coded PCR plates from fellow Azenta Life
Sciences subsidiary 4titude. The Individual Access PCR plate is part
of the Framestar range from 4titude, which combines the rigidity
of a polycarbonate frame with the thermal transfer properties of
polypropylene in each thermal cycler well. Individual wells in this
versatile plate can be removed and independently sealed using an
Azenta semi-automated sheet or individual access heat sealer. This
'(
)
#
)
'+-
&
#
'
/0
-((
'
"(
)'
*.
)'!
'
-
12
-
23(
+
'
#
'
against consumable wastage. Each well in an Individual Access PCR
plate has an individual, randomly generated Data Matrix 2D barcode
on the base, giving it a unique ID that cannot be erased or lost. For
lab scientists who only want to read a single Individual Access plate
well, the Ziath Uno single tube is a lower cost solution. The Uno
simply reads the 2D barcode on the Individual Access PCR plate well
and puts the result into any open program on the desktop using
Microsoft Keyboard Wedge software.
Ziath
'&
$
0/
https://ziath.com/products/barcode-scanning
(EV), including exosomes, as well as addressing EV heterogeneity
and high-throughput solutions for biomarker discovery. Traditionally
ultracentrifugation has been the favored method for EV isolation or
separation. Though it is still widely used, ultracentrifugation not only
tends to alter the vesicle shape and functionality but also requires
#
#
#
(
#
# !#/"#
$%
$
#&$
#
range from AMSBIO now allows researchers to select an optimized
&
(
$
#$%
$
#' #$
#!
&#$*
& #
#,$&#
researchers looking for a method that recovers the highest amount
of extracellular material, regardless of its vesicular or non-vesicular
nature, AMSBIO has introduced the ExoPure Kit. This high-recovery
#%$*+
(
#&
$
#-
##$% ! %
#%
$%#$#
separate and isolate exosomes from your sample, requiring a short
incubation time of just one hour. The product range also includes
!&#
0&
#&$
#1$&#
%#$*#(%&
$
#) ,, # #&
#
#
#!$#*
% ##1$&# &
$
#
#&
(
$
#
of biomolecules that has now been successfully applied to isolate
and separate extracellular vesicles. For labs that need a method
that recovers mixed EVs along with some free proteins, AMSBIO has
&$
#!&#
0&
# $%$
#$1# ,,#(%&#
'%#1$&#/"#
&
(
$
#/"#
$
&
$
#
#/"#
+' # &
$
# !#- #
'
(#$1#!# ,,#&$
# &#! #! # &#& '% #* ! '% #
#
'%#1 #
$
&
$
#$1#
%#
# #
%%#
#$&#&
#
prior to EV isolation. They also allow easy removal of small molecules
and ions from the EV preparation and can be used for EV dialysis
#'0&#
!
# 0&
#1 #/"#&
(
$
#1&$# %%#)#.%#
$#% &#$%#)#$##%#$1#
#
#$
% ##
# #
ExoUltra Size Exclusion Chromatography column can be integrated
$# %$#
#% '$& $& #*$&-$*# !# '%#
$%
#
#'#
%
#
#&##$#(#
#
'%
#/"#&
#
&$
#
# %
$
#
%
#/"#
$%
$
#1&$#
%%#
#'
$
#
#%
#&
#&
(
$
#$1#/"#1&$#
$
#
#
and removal of excess dye after EV labelling. Using a combination
of AMSBIO TFF Filters and ExoUltra SEC columns, researchers will be
able to separate subtypes of EVs based on their size, thereby eliminating non-EV components.
AMSBIO
& '
+&'$
$,..'
https://www.amsbio.com/exosomes-isolation
p
INTEGRA Pipettes Used For Antibody
Therapy
INTEGRA Biosciences has awarded 50 EVOLVE
manual pipettes, GRIPTIPS pipette tips, and
various lab accessories to UK-based start-up
Creasallis, supporting its research seeking to
improve the penetration of therapeutic antibodies into tumors. The prize is part of the
ìINTEGRA Supports Start-Upsî competition,
and will allow the company to accurately pipette volumes of 0.2
! "
# ! #
!!
! #
!
!
!!
!! ! ly penetrate tumors, resulting in only surface-level treatment. The
!!
! !
in this area with its patented CreaTap (Creasallis Tumor Antibody
Penetration) plug-and-play technology, which can be incorporated
into any existing antibody. Creasallis currently operates in a multiuse laboratory at the Babraham Research Campus, and previously
only had access to a limited number of communal pipettes, shared
" !
!
#
!!
focus. The team was therefore thrilled to win the competition,
which ran until April 2023. ìIt is a very generous prize, far greater
than I expected, said Dr. Joyce Ratti, head of Protein Sciences at
Creasallis. ìOur 50 brand new EVOLVE manual pipettes will allow
us to designate a set to each task, such as cell culture or protein
"
!
!
!
!
!
!
!
!! !! ent quantities much easier and far more accurate. Most importantly, we now have the resources to expand our capabilities in the
lab and take on more employees in the future when we are ready,
accelerating this novel area of research.î
INTEGRA
integra-biosciences.com