/


































































































































































































































































































































































































































Текст
К. ЛЛ.ЗА В АДС КИ Й
ВИД
ВИДООБРАЗОВАНИЕ
Академия наук с с с р
ИНСТ И ТУТ
ИСТОРИИ ЕСТЕСТВОЗНАНИЯ И ТЕХНИКИ
К. At. 3 а в адски й
вид
ВИДООБРАЗОВАНИЕ
ИЗДАТЕЛЬСТВО «НАУКА»
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
АЕНИНГРАД • 1968
УДК 576.12 : 675А
Вид и видообразование. Завадский К. М. 1967. Изд-во «Наука», Ле-
нингр. отд., Л. 1—396.
В книге рассматриваются данные о числе существовавших в прошлом
и живущих в настоящее время видов растений и животных, анализи-
руются главные этапы истории изучения вида и видообразования
от древнейших времен до современности, монографически излагается
современное состояние теории вида (критерии вида, определения вида,
проблемы целостности, неравноценности и универсальности вида,
структуры вида и внутривидовых отношений) и учение о типах, спо-
собах и путях видообразования.
Вид рассматривается как одна из основных и первичных форм органи-
зации живого, как поле деятельности естественного отбора и система,
приспособленная к персистированию и к эволюции. Илл. — 22,
табл. — 15, библ. — 790 пазв.
Пр едисловие
Изучение вида и видообразования является одной из фунда-
ментальных проблем биологии. В течение целого столетия эта
проблема сохраняет ключевое положение, и ныне ее значение все
возрастает, так как вид является основой и предметом исследо-
ваний самых различных отраслей. Не только классические науки,
hi кип как морфология, физиология, генетика, биогеография, эко-
логия и палеонтология, опираются на понятие о виде, но в нем
нуждаются и его разрабатывают биохимия, биокибернетика и
другие новые отрасли.
Накоплено множество данных, характеризующих вид с раз-
ных сторон. Рост объема информации и в этой области становится
ней более стремительным. Однако, как правило, знания о виде
до сих пор разрознены. Объясняется это тем, что по традиции
ниучопио вида рассматривается задачей главным образом систе-
матики, которая не в состоянии эти знания синтезировать.
If а к бы некоторые факты и обобщения глубоко не характеризо-
вали вид, если они не способствуют его диагностике, в поле
зрения систематики они не попадали. Так, например, обстояло
цело с, областью, посвященной внутривидовым отношениям и
различным формам и механизмам интеграции надиндивидуальных
живых систем. Давно назрела необходимость создания общебио-
логичоской теории вида и выделения ее в самостоятельную от-
расль пауку о виде, или эйдологию. Одна из попыток создать
ьпнгуры такой теории была предпринята нами недавно в «Уче-
нии о виде».
< Ь нову данной работы составляют идеи, впервые изложенные
в статьях 40—50-х годов, затем обобщенные и систематизирован-
ный (Завадский, 1961) и изложенные бблее сжато в недавней
рабогп (Завадский, 19676). Здесь уместно в двух словах сказать
н । панной идее этих работ.
Пид рассматривается как одна из четырех первичных, универ-
• винных и основных форм организации жизни. Попытки харак-
1*
S
теризовать вид с какой-либо одной стороны неправильны. Спе-
цифичность вида состоит в особом сочетании целого комплекса
показателей (это особый тип организации и множество индиви-
дов, обособленная от других биологическая отдельность и много-
образие форм, целостное и вместе с тем составное образование,
географо-экологическая определенность и т. п.). Являясь надин-
дивпдуальной, самовоспроизводящейся и регулируемой системой,
вид оказывается качественно определенным элементом в экоси-
стемах и специфическим звеном в биологических и биохимиче-
ских миграциях веществ. Поэтому исследование статики вида,
механизмов, поддерживающих его целостность и сообщающих
ему исключительную устойчивость и способность к существова-
нию в течение заранее неопределенного геологического вре-
мени, — одна из важнейших задач современной биологии.
Содержанием жизни вида признаются внутривидовые отноше-
ния, как основные, непосредственно обеспечивающие воспроиз-
ведение вида, так и производные, включающие самые разнооб-
разные противоречия. Совокупность основных внутривидовых
отношений является необходимой предпосылкой самого существо-
вания вида (поддержание численности гамет, зигот, личинок и
всех других возрастных групп, смена поколений и т. п.). Эти от-
ношения непрестанно контролируются и шлифуются естествен-
ным отбором.
Являясь нолем деятельности естественного отбора, вид высту-
пает носителем п узловым пунктом эволюционного процесса.
Способность к самостоятельному эволюционному развитию —
главная черта вида. В центрё современной эволюционной теории
стоят исследования микроэволюционных процессов, т. е. преобра-
зований, протекающих в недрах популяций п завершающихся
видообразованием.
В книге рассматриваются вопросы, связанные только со спо-
собами видообразования и с характером этого процесса. Предла-
гается подразделять видообразование на шесть типов, в пределах
которых рассматриваются многие конкретные способы видообра-
зования. Помимо хорологического принципа выделения типов ви-
дообразования (аллопатрическое и симиатрпческое), автор выде-
ляет типы видообразования по характеру исходных преобразо-
ваний, по времени изоляции молодой формы от материнской
популяции, по числу фаз и по темпу процесса (неоформогенное
и микроаккумулятивное), а также на основании того, лежит ли
в основе процесса объединение двух или нескольких инициаль-
ных систем в одну продуцируемую или же, наоборот, обособле-
ние ранее единой системы на ряд дочерних (синтезогенез. и сег-
регациогенез). Поэтому видообразование в настоящее время сохра-
няет значение главной проблемы эволюционной теории. Однако
такая точка зрения не общепризнана. Некоторые призывают но
придавать слишком большого значения концепции вида, которая
4
нвооы «сужает пути мышления биологов в области эволюционных
проблем» (Эрлих, Холм, 1966 8).
Задача этой книги — обосновать нашу концепцию вида в ста-
тике и эволюции, по возможности на разнообразном материале.
Исторический очерк также призван служить этой цели. Крити-
ческой оценке подвергается материал, который в какой-то сте-
ноп и входит в компетенцию автора. По этой причине об особен-
ное тих проблемы вида и видообразования у таких групп, как
например простейшие или несовершенные грибы, только упо-
минается. ,
До с их пор подавляющее большинство Исследований о виде
осуществлялось систематиками и было ориентировано на обслужи-
вание нужд этой науки. Настоящая же книга написана не систе-
матиком, а биологом, много лет экспериментально и в полевой об-
становке исследовавшим внутривидовые отношения и строение
популяций у некоторых видов высших растений, а та*кже измен-
чивость природных популяций растений по ряду признаков.
Основу ого теоретических представлений составляли прежде всего
именно эти данные. Большое значение сыграло также то, что
в точение многих лет автору приходилось читать курс эволюцион-
ной теории в Ленинградском университете. Это требовало ши-
рокого синтеза знаний о виде и видообразовании, полученных как
в прошлом, так и в наше время.
Проблема вида имеет общебиологическое значение, и разраба-
тывать со следует не только с различных сторон, но и синтетиче-
ски как единое целое. Нами и предпринята попытка создания од-
ного пл вариантов первого контура такого синтетического учения
виде. Оно не призвано быть механической сводкой массы раз-
нообразных сведений о виде, накопленных десятками современных
отраслей биологии. Обобщая различные данные о виде и видооб-
разовании, мы убедились в существовании определенной тенден-
ции к снижению относительного значения знаний, полученных
гиг тематикой, и возрастанию роли знаний, добытых другими от-
раслями биологии, и в первую очередь экологией, этологией, ге-
нетикой популяций и экспериментальными исследованиями мик-
роэаолюции. Этим заключением мы не намерены в какой-либо сте-
пени принижать то действительно огромное значение, которое
имела в этом, вопросе классическая систематика, и ту большую
роль, которую продолжает играть современная систематика.
Состояние всей проблемы вида в целом в настоящее время та-
ково, что число более или менее вероятных гипотез значительно
Польше, чем численность истин, твердо установленных в качестве
незыблемых законов. Некоторые самые коренные вопросы до сих
нор еще не получили своего разрешения. Недостаточная разра-
|»о 1НППОГгь теории вида связана с тем, что до недавнего времени
в этой области часто делались довольно поверхностные обобщения,
oi поив иные па использовании примитивных методов индукции или
5
дедукции и мало опирающиеся на современные методы экспери-
ментального анализа, который включает сочетание разработки
научно обоснованных гипотез с опытами и их сложные взаимодей-
ствия. Надо полагать, что вскоре все разделы проблемы вида
будут разрабатываться именно такими методами, характерными
для передовых областей естествознания. В настоящее время на-
сущной задачей являются попытки синтеза разрозненных знаний
о виде и видообразовании с целью создания единой рабочей ги-
потезы.
Эта книга содержит, разумеется, и спорные утверждения, и
места, сформулированные слишком схематично или же эмпирично.
Трудно избежать таких недостатков, вытекающих из ограничен-
ных возможностей одного автора, из особенностей его подхода и
состояния вопроса, а также из-за Желания более подробно обосно-
вать те части концепции, которые рассматриваются в качестве
главных.
В настоящем издании заново написана глава 1, значительно
переработана и расширена вся историческая часть, в которой
почти целиком обновлены разделы, посвященные изучению вида
и видообразования в новейшее время (главы 4 и 5). Главы, в ко-
торых рассматривается сущность вида, а также пути и способы
видообразования (главы 7 и 10), также сильно расширены за счет
введения новых разделов и пополнены новейшими данными.
В меньшей мере подверглись изменениям остальные главы, допол-
ненные отдельными новыми разделами и данными, опубликован-
ными в последние годы. Кроме того, все разделы книги сущест-
венно пополнены материалами из области зоологии, нехватка
которых была одним из недостатков предыдущих публикаций.
Положения, высказанные нами в ранее опубликованных рабо-
тах, вызвали широкий отклик какв отечественной, так и в за-
рубежной биологической и философской литературе, обсуждались
на ряде совещаний и во время многих личных контактов с био-
логами разных специальностей. «Учение о виде» не раз издава-
лось за рубежом. Это отражает интерес к поднятым вопросам и
к их трактовке.
В заключение считаю своим долгом выразить глубокую
признательность за помощь в подготовке книги Л. Н. Хахиной и
3. М. Рубцовой, а также за ценные замечания Л. Я. Бляхеру и
С, Р. Микулинскому,
Часть I
ИСТОРИЯ ИЗУЧЕНИЯ ВИДА
Глава 1
ЭВОЛЮЦИЯ ЗНАНИЙ И СОВРЕМЕННЫЕ ДАННЫЕ
О ЧИСЛЕ ВИДОВ
Инвентаризация видов уже более двух веков строится на все
более точно определяемой теоретической основе, выражает ту
или иную концепцию вида и является одним из важных итогов
деятельности биологов. Тем не менее сам процесс инвентариза-
ции видов не так уж сильно зависел от прогресса в изучении
сущности вида. Поэтому предпринимаемое нами в этой главе опи-
сание количественной стороны вопроса, отвлекаясь от качествен-
ной, не только удобно, но и принципиально возможно. Дело
в том, что основные этапы первичной инвентаризации видового
состава флоры и фауны Земли только отчасти совпадают с эта-
пами изучения сущности вида. Взаимосвязь между широтой
охвата видов систематикой и глубиной знаний далеко не всегда
несомненна и совсем не проста. Уже начиная с эпохи Линнея
темпы инвентаризации видов более определяются развитием про-
изводительных сил и ростом потребностей в сырье, в защите от
вредителей, числом систематиков и другими факторами социаль-
ной природы, чем углублением познания вида. Это объясняется
тем, что для первичной инвентаризации видов, т. е. для простого
перечня всех видов с поверхностными диагнозами, в общем было
достаточно уровня знаний о виде, достигнутого уже 200 лет на-
зад. Для охвата же всех географических, экологических, ai для
паразитических форм «физиологических» внутривидовых групп
и для учета всех отдельных популяций необходимы уже более
глубокие знания о структуре вида. Для полной инвентаризации
форм, составляющих генетический потенциал хотя бы одного вида,
как это показывает опыт последних десятилетий, накопленный
1лавпым образом по важнейшим культурам (пшеница, кукуруза,
спекла, сахарный тростник, хлопчатник и др.), потребуется иногда
труд, соизмеримый с тем, который был затрачен мировой наукой
на первичную инвентаризацию видов всей флоры и фауны Земли.
7
1. НАКОПЛЕНИЕ ЗНАНИЙ О ЧИСЛЕ ВИДОВ
Число видов в д о л и и и е ев с к и е времена. Если
ио древним источникам мы хотим установить степень изучен-
ности видового состава растений и животных, приходится преодо-
левать затруднения, связанные с неточностью старых описаний
видов (Carns, 1872). Уже у Аристотеля иногда можно встретить
четкое описание видов с применением бинарной номенклатуры
(верблюд дромадер и двугорбый и т. п.) и с указаниями подви-
довых различий. Например, у льва Аристотель различал формы
с очень пышной и со слабо развитой гривой, соответствующие
современным географическим расам (персидской, пли северо-
африканской, и западиоиндпйской). Идентификация древних
описаний и названий с современными определениями позволяет
с уверенностью сказать, что в IV в. до и. э. было известно еще
очень мало видов. Так, Аристотель упоминает приблизительно
500 видов животных (Даппемап, 1932 134), а Теофраст при-
мерно 450 видов растений (Sprengel, 1817). Через 400 лет,
в I в. п. э., судя ио описаниях 11л илня-старшего, число известных
видов животных и растении возросло вдвое — с 1000 до 2000
(Meyer, 1854). В списках Плиния появились не только виды из
Греции, Италии, Малой Азии и сопредельных стран, по и индий-
ские, африканские, а также североевропейские. Знания видов
растений и животных древними учеными Индии, Египта и других
более пли мепее замкнутых цивилизаций iiq были большими по
объему, чем греко-римские. Далее в течение полутора тысячеле-
тий работы по инвентаризации видов либо совсем не проводи-
лись, либр не превосходили античных ио объему и уровню.
В XVI столетии травники и каталоги животных, бывшие чаще
всего комментариями к древним-классикам, содержали описания
форм в алфавитном порядке. Опп включали число видов, не пре-
вышающее уже известное древним авторам (Radi, 1905). Напри-
мер, в многотомных травниках И. Вока (1498—1544 гг.) содер-
жалось описание всего 600 видов растений. В трудах этого ав-
тора, а также К. Гесснера, У Альдрованди и других описания
форм были громоздкими п неопределенными, соответствующими
то роду, виду пли даже внутривидовой форме (преимущественно
в описаниях культурных форм), то гораздо более крупным так-
сонам. Иногда, впрочем, такие расплывчатые описания чередова-
лись с лаконичными диагнозами и бинарными названиями, как
правило заимствованными из греческого или латинского языков
и реже из родпых автору народных местных названий.
Лишь па грани XVII—XVIII в в. впервые среди других сход-
ных явлений был выделен вид и дано определение этого понятия.
В истории познания живой природы это был важный перелом,
так как пришел конец расплывчатым описаниям неопределенных
групп. В XVIII столетии уже была создана возможность прово-
8
iiiil инвентаризацию растений и животных на основе более или
мгнег определенной единицы — эталона, т. е. на единственно
ниучной основе описания видов. Однако работы эти могли полу-
чип, необходимый размах только после внедрения в систематику
днух нововведений. Оставалось применить определение вида как
нинменьшей единицы классификации к практическим нуждам
систематики и создать иерархию таксономических единиц, осно-
iinii которой служил бы вид. Автором этих нововведений был
Г». Линней. После Линнея наступил расцвет инвентаризационных
работ, целью которых было выявление и описание всех видов.
Огромное значение приобрели такие гигантские по размаху опи-
сательные работы, как труд коллектива сотрудников Альфонса
Декандоля. В течение полувека нм было описано 59 000 видов
двудольных растений (De-Candolle, 1880). Было решено считать
условным отправным пунктом работ по инвентаризации видов
растений и животных списки, составленные К. Линнеем в 10-м из-
дании «Системы природы» (1758 г.).
Инвентаризация видов за последние 200 лет.
Период с середины XVIII до середины XIX столетия можно на-
звать эпохой целеустремленной инвентаризации видов и располо-
жения их хотя бы во внешне стройную систему. Велось первона-
чальное накопление чисто описательных знаний о видах.
Таблица 1
Гост изученности видового состава
п о к р ы тосемеплых р а с т е и и й
Год Автор Количество видов Год Автор Количество видов
1753 Линней 5250 1922 Кузнецов 142410
1774 Линней 7000 1936 Энгл ер—Дильс 165600
1808 Вильденов 14460 1938 Бовер 182000
1814 Броун 37000 1943 Смирнов, 1943 250000(300000)
1817 Гумболъд 38000 1951 Lawrence, 1951 —200000 *
1888 Дюран 100220 1956 Spector, 1956 200000
10ОО Вайнз 103000 1963 Воронов, 1963 200000-300000
191/1 Бэсси 132500 1964 Engler, 1964 —220000 **
* При суммировании числа видов, приводимого Дно Лоуренсом по каждому
семейству, оказалось, что в его сводке число видов покрытосеменных несколько
меньше — 175 000—190 000.
** Подсчет видов по Энглеру (Engler. 1954, 1964) дал следующее количество:
"18 .»(к 1—221 900 (с учетом колебаний данных по отдельным семействам, отмеченных
н справочнике).
Прогресс в установлении новых видов на протяжении послед-
них двухсот лет был огромен. Примером этого могут служить
.данные табл. 1.
Еще более велики были приросты числа вновь открытых Bn-
Д'ж животных. В 10-м издании «Системы природы» Линней пе-
9
речисляет около 4200 видов, в том числе 11 видов губок, 74 —
кишечнополостных, 29 — иглокожих, 76 — червей и червеобраз-
ных, 674 — моллюсков, 2038—членистоногих и 1222 вида позво-
ночных. Через полтораста лет было известно уже в 100 раз
больше видов — 413 000, в том числе губок — 1500 видов, кишеч-
нополостных — 3000, иглокожих — 3000, червей и червеобраз-
ных — 9000, моллюсков 50 000, членистоногих — 293 000, по-
звоночных— 33 500 (Mobius, 1898). Двести лет назад было
известно 0.3% числа видов, приводимых в современных справоч-
никах (табл. 3), и примерно 1% числа видов, известных в конце
прошлого века. Как указывает П. В. Терентьев, Линнею было
известно только 215 видов ныне живущих амфибий и рептилий,
Дюмеримо и Биброну в середине прошлого века — 846 видов,
Буланджеру в конце XIX в. — уже 5151 вид, а ныне — около
6950 видов: «Сейчас существуют значительные разногласия, про-
истекающие преимущественно из разного понимания объема
вида» (Терентьев, 1961 8—9).
В течение двух тысячелетий число известных науке видов
млекопитающих не превышало сотни и в 1758 г. Линней назвал
86 видов (183 формы с разновидностями). За последующие
150 лет было открыто много видов млекопитающих, и в конце
XIX в. их число приблизилось к 3500 (Mobius, 1898). Далее за
четверть века оно возросло еще в четыре-шесть раз — до 13 000—
более 20 000 (Hesse, 1929; Spector, 1956:533, прим. 7).
В 40-е годы XX в. многие виды были «разжалованы» в под-
виды и их число упало до 6000—6500, а затем еще сократилось
так, что в настоящее время насчитывается около 3.5 тыс. видов
млекопитающих (Барабаш-Никифоров, Формозов, 1963:11) или
около 4000 видов (Млекопитающие..., 1963:25). Такое сниже-
ние числа видов сопровождалось ростом количества изученных
географических рас или подвидов и явилось следствием реформы
в толковании объема вида.
Казалось бы, что количество видов современных млекопитаю-
щих должно быть подсчитано точпо. Но в действительности и
в этой небольшой п хорошо изученной группе существует немало
неясностей с таксономическим рангом некоторых форм, и есть
основания думать, что в труднодоступных областях живут еще
неизвестные виды.
В заключение рассмотрим рост общего числа известных си-
стематике видов растений и животных за 200-летний период
после К. Липпея и приросты числа видов по полувековым пе-
риодам (табл. 2).
Для табл. 2 взяты усредненные данные из многих сводок по
систематике растений и животных, составленных не более чем
на 10 лет раньше или позднее данной даты. Эти усредненные
цифры были округлены. Важно было получить ясное представле-
ние о порядке величин, характеризующих количественные изме-
10
Таблица 2
Рост числа известных науке видов за 200 лет
Дата Число видов Прирост за полвека
абсолютный относительный
( Середина XVIII в Конец XVIII в Середина XIX в Конец XIX в Середина XX в 10000 30000-35000 100000 500000 1600000 20000—25000 70000 400000 1100000 Трехкратный » Пятикратный Трехкратный
। кмгия в изученности видового состава и приводимые данные до-
статочно точно отражают эти изменения.
Три полувека характеризуются трехкратным приростом числа
Пидов и только в дарвиновскую эпоху (вторая половина XIX сто-
летия) прирост был пятикратным. Однако вряд ли правильно
связывать такой большой рост со стимулирующим действием
эволюционных идей. Скорее он был связан с интенсивным изуче-
нием флоры и фауны колоний, в состав которых входили богатые
видами области влажных тропиков, а также с усилением «дроби-
тольской» тенденции в систематике. Относительный прирост
числа известных науке видов, как это видно из табл. 1 и 2 и из
других приведенных выше данных, шел в общем довольно рав-
номерно и за 200 лет существенно не изменялся. Абсолютные же
темпы прироста вновь описанных видов, естественно, колоссально
возросли (за полвека после Линнея ~ 90 000 видов, а за послед-
ние полвека более 1000 000 видов). Пока нас интересовали, так
сказать, только порядки чисел, и они отражают уровень знаний
числа видов в каждый период. Однако эти округленные данные
по отражают большого варьирования точек зрения разных авто-
ров на число видов в пределах очень многих групп растений и
животных, на которых мы остановимся в следующих разделах.
Причины же этих разногласий будут обсуждены в следующих
главах в связи с вопросом о выборе тех или иных практических
стандартов вида в систематике.
2. ЧИСЛО ВИДОВ ПО СОВРЕМЕННЫМ ДАННЫМ
Общее число видов. Данные о числе ныне живущих
видов представлены в табл. 3.
Нами приведены сведения из наиболее авторитетных справоч-
ников и сводок, дающие понятие о крайних цифрах, называемых
и литературе. Таким образом, общее число видов растений и жи-
вотных, по современным данным, составляет 1200 000—2 000 000.
11
Т а б л и if а
Общее число видов ио с о в р е м е и и ы м [ а и н ы м
Количество видов животных Литературный источник Количество видов растений ЛитературньГ источник
1000000 938000 1120000—1311000 1000000-1500000 1070000 Dobzhansky, 1955 Spector, 1956 Гептнер, 1956 Воронов, 1963 Kaestner, 1965 265000 342730-350000 500000 356800 Dobzhansky, 1955 Spector, 1956 Воронов, 1963 Engler, 1954, 1964
В систематике большинства групп преобладает концепция слож-
ного (полптиппческого) вида крупного объема. Именно эта кон-
цепция в основном и получила свое количественное выражение
в приведенных цифрах. С позиции же концепции мелкого вида,
приравниваемого к одной географическом расе, общее число ви-
дов должно быть увеличегго по крайней мере в четыре-пять раз.
Согласно третьей концепции, вид — это наименьшая из различи-
мых форм, т. е. элементарная единица морфологического, эколо-
гического или генетического анализа. Далее (глава 4) она будет
обсуждена подробно, а здесь отметим лишь, что при таком под-
ходе число «видов» сразу же возрастает на несколько порядков
и будет исчисляться сотнями миллионов пли даже тысячами мил-
лиардов. Эта ультрадробптельская тенденция имела некоторый
успех в начале XX столетия, а йыпе почти всеми оставлена.
Огромные расхождения в подсчетах видов связаны главным
образом с различиями в понимании объема вида. Однако все под-
считываемые с тех пли иных позиции формы реально сущест-
вуют, хотя все они качественно различны. Истинный вид отли-
чен от клона, биотипа, мутанта и т. и. Ошибки в подсчетах
происходят, кроме того, из-за брака в работе систематиков, опи-
сывающих иногда впды-артефакты, и из-за существования раз-
ных названий для одного и того же вида (синонимика).
Различия в подсчетах могут, наконец, проистекать из-за су-
ществования еще недостаточно изученных видов. Например,
в пределах рода подсолнечник (Helianthus) насчитывают 100—
200 пли даже больше видов, из которых только 100 —120 яв-
ляются более или менее строго определенными и общеприня-
тыми, а несколько десятков видов должны быть отнесены к еще
неустоявшимся в науке и к еще пеоиределившимся в природе.
На табл. 4 и 5 представлено число видов основных групп
растений и животных по работам последнего десятилетия, а на
табл. 5 для сравнения, кроме того, приведено число видов живот-
ных по представлениям 20-х годов нашего столетия (Hesse, 1929).
12
Таблица 4
Число видов растений
Илзвание группы Литературный источник
Engler, 1954, 1964 Dobzhan- sky, 1955 Spector, 1956 Воронов, 1963
Виктории 4600 3000-6000
М нкснмицеты 500 — 430 300
Грибы 60650 70000 73500 70000
Водоросли 24800 14000 17535 20000-25000
Лишайники 18000-20000 — 15500 30000
Мхообразные 21600 21000 23820 22000-26500
Папоротникообразные . . 10600 10000 11280 9625—10025
Голосеменные 650 500 665 570-770
Покрытосеменные 218000 150000 200000 200000—300000
однодольные .... 53000 — 34000 —
двудольные 165000 — 166000 —
Хотя разные авторы и приводят разные числа видов одной
и той же группы, разнобой в цифрах не столь уж велик. На-
против, порядок чисел почти для всех групп растений и живот-
ных остается стабильным. Исключение составляют моллюски,
размах данных о числе видов которых очень велик. Причины та-
кой несогласованности цифр сложны. Систематика моллюсков
бурно развивается, разные их группы классифицировались си-
стематиками, различно понимавшими объем вида, некоторые
группы объективно трудно разделять на виды, и т. п. Иногда
один вид моллюска разбивался на многие десятки мелких (на-
пример, у Achatinella).
Асимметрия распределения видов по круп-
ным таксонам. Распределение видов по крупным таксонам,
как это видно из табл. 4 и 5, чрезвычайно неравномерно. Такая
асимметрия наблюдается у всех таксонов от типов и классов
до родов.
Ярко выражена асимметрия при сопоставлении числа видов
одного класса насекомых с числом видов всех остальных 69 клас-
сов животных (Беклемишев, 19646).1 Число видов насекомых
составляет около 80% от общего числа видов животных, т. е.
в четыре раза больше суммы числа всех вместе взятых видов
остальных животных. Такая же асимметрия распределения ви-
дов обнаруживается при сопоставлении количества видов по
разным типам животных.
1 По В. Н. Беклемишеву, из 70 классов животных 50 являются обще-
принятыми, т. е. твердо обоснованными, а таксономический ранг осталь-
ных до сих пор более или менее спорен. Другие авторы выделяют боль-
шое число классов, например по справочнику В. Спектора (Spector,
—79.
13
Таблица &
Число видов животных
Название группы Литературный источник По другим источникам
Hesse, 1929 Spector, 1956 Buchsba- um а. Milne, 1961 Kaesther, 1965
Простейшие — 15000 30000 20000 20000 (Догель и
Губки Кишечнополост- 4500 5000 4500 5000 др.,
ные . . 9000 10100 9100 9000 —
Черви плоские Черви первично- 9000 6000 9000 12400
полостные 5000 10500 12500 25000 (Dob-
Немертины — 500 570 800 zhansky,
Аннелиды Прочие червеоб- 7600 6500 6000 8700 1955)
разные 3300 5150 8600 5800
Моллюски 104000 70000 40000 128000 88000 (Там же)
Членистоногие 802000 750000 700000 — 816000 685000-1070000 (Воронов, 1963)
насекомые 750000 — — 750000 (Dobz- hausky, 1955)*
паукообраз-
ные 28000 15000 • 65000 — 30000 (Там же)
многоножки 8100 7000 — 100000 »
ракообраз-
ные 15500 25000 — 40000 »
Иглокожие Оболочники, по- 4200 50р0 5500 6000 4700 »
лухордовые и др. . . 1600 730 1320 180 1700 »
Позвоночные 70000 60000 — 45000 35000 »
рыбы 20000 40000 — — 18000 »
амфибии 2860 2770 — — 6950 (Терен-
рептилии 5460 6960 — — тьев, 1961) 8700 (Мауг, 1963)
птицы 28000 8600 — —
млекопита-
ющие 13000 3000-5000 3500 (Бара- баш-Никифо- ров, Формо- зов, 1963)
* Некоторые авторы считают, что число видов насекомых больше [например,
Б. Б. Родендорф (1965) определяет его в 1 000 000].
Число типов животных нс является еще твердо установлен-
ным. По В. Н. Беклемишеву (19646), существует 14 типов, но
только семь из них не возбуждают в настоящее время никаких
серьезных сомнений ни в отношении их самостоятельности, нп
объема. Другие авторы принимают значительно большее число
14
пикт: 24 (Kaestner, 1965), 29 ^(Buchsbaum a. Milne, 1961) или
даже 33 (Spector, 1956). Асимметрия распределения видов по
чинам животных видна из следующего сопоставления.
'Тринадцать древних типов включают менее чем по 100 видов
каждый: Mesozoa — семь видов (Buchsbaum a. Milne, 1961), 50 ви-
ч,<>1» (Kaestner, 1965); 2 Ctenophora ~80; Priapulida — четыре или
шесть; Phoronida — 15; Ghaetognatha — 30—50; Hemichordata —
100; Pogonophora — 50—80 до 100; Gastrotricha — 100; Echiuroi-
doa — 60 (Buchsbaum a. Milne, 1961), 150 (Kaestner,> 1965);
Kinorhyncha ~30; Nematomorpha ~80; Entoprocta ~60; Branchio-
Iromata ~80 видов. Эти 13 типов вместе взятые включают всего
700—900 видов. Еще четыре типа включает менее чем по 1000 ви-
дов: Nemertini — 500—800, Acanthocephala ~400, Brachiopoda —
120—260, Sipunculida — 250. Вместе они содержат 1270—1700 ви-
дов. Таким образом, 17 вместе взятых типов животных ныне
представлены всего 1770—2600 видами, т. е. меньшим числом,
чем количество видов в одном из многих средних по численности
родов. Далее, следующие семь типов включают менее 10000 ви-
дов каждый: Porifera — 4500—5000, Coelenterata — 9000—10 000,
Platyhelminthes — 6000—9000—12 400, Rotifera — 1200, Bryozoa —
3000—6000, Annelida — 6000—8700, Echinodermata — 5000—
6000. Эти семь типов вместе имеют 34 700—49 300 видов.
Итак, 24 перечисленных типа3 животных включают всего
36 470—51 900 видов, в то время как, например, только один
отряд двукрылых насекомых — 75 000—120 000 ныне живущих
видов (Родендорф, 1959). Только пять типов животных имеют
более чем по 10 000 видов каждый (Protozoa, Nemathelmintes,
Chordata, Mollusca и Arthropoda) и только два типа — моллюски
и членистоногие — имеют более чем по 100 000 видов.
Число видов в типе простейших не больше 2% числа видов
насекомых. Число видов моллюсков раз в 10—30 больше числа
видов млекопитающих, а число видов паразитических червей
(12 000) — в три-четыре раза.
Асимметрия распределения видов существует не только среди
типов и классов, но и среди более мелких таксонов. Например,
отряд грызунов имеет до 2500 видов, а все остальные вместе взя-
тые отряды млекопитающих — около 1000 видов. При этом такие
отряды, как хоботные, включают всего два ныне живущих вида.
То же можно наблюдать и при сопоставлении числа видов
в родах. Одни роды высших растений (среди орхидных, сложно-
цветных, бобовых и пр.) включают по много тысяч видов каж-
дый, а другие очень малочисленны или даже монотипны.
2 Далее в этом перечне ссылки на авторов даются только при значи-
тельных расхождениях в цифрах.
3 В табл. 5 многие из этих типов не упомянуты п объединены
в группу «прочие червеобразные», или присоединены к другим близким
типам, как например гребневики (Ctenophora) — к кишечнополостным.
15
Причины асимметрии распределения видов по высшим так-
сонам сложны. «Виды, относящиеся к различным родам и клас-
сам, изменялись неодинаково быстро и не в одинаковой степени»
(Дарвин, 1937 420). Затем это различия в интенсивности
адаптивной радиации групп и их обособления в повых экологи-
ческих нишах, связанные с особенностями организации, позво-
ляющими иметь больше пли меньше «потенциальных ниш». На-
конец, это различия в масштабах вымирания видов. На слож-
ность причин, лежащих в основе асимметрии распределения
видов, указывает отсутствие корреляций между численностью
видового состава группы и высотой организации, филогене-
тическим возрастом группы и некоторыми другими показате-
лями.
Вопрос о причинах, определяющих число видов крупных так-
сонов, разработан еще мало. Зависимость числа видов группы от
степени биологического прогресса, иепрямолппейна, так как рост
численности и эколого-географическая экспансия групп ведет
у одних к распадению их на изолированные подгруппы, у дру-
гих к сохранению интеграции подгрупп, т. е. не сопровождается
видообразованием. Число видов в группе некоторые ставят в связь
с размерами тела, большей относительной величиной мозга (или
гапглиев) п скоростью метаболизма (Стрельников, 1962). Дей-
ствительно, число видов крупных животных на несколько поряд-
ков величии меньше, чем число видов мелких.
Между числом видов в крупных таксонах, численностью осо-
бей у отделI,пых видов и их географическим распространением
также пет прямой зависимости. Напрпмер, голосеменных насчи-
тывается всего несколько сот видов, по среди них есть немало
многочисленных п очень широко расселенных (многие листвен-
ницы, ели, пихты, сосны н др.).
Число видов сухопутных животных составляет 93% от общего
числа животных, а водных только 7% (Маут, 1963). Эта более
чем десятикратная разница должна быть связана с существова-
нием на суше гораздо большего разнообразия физико-географи-
ческих и топо-экологических условий, которое ведет к возраста-
нию числа потенциальных экологических ниш. Несмотря на го-
раздо большую площадь и больший объем обитаемой толщи вод,
пригодных для жизни, мировой океан и другие водные бассейны,
как оказалось, дают гораздо меньшие возможности для диффе-
ренциации форм жизни, чем суша. Правда, если исключить насе-
комых, то соотношение численности видов животных суши и
моря резко изменится. По нашим приблизительным подсчетам,
общее число видов животных (без насекомых) па суше и в воде
примерно одинаково и составляет цифру порядка 170 000. При
этом некоторые типы и классы, как известно, являются пол-
ностью водными (губки, кишечнополостные, оболочники и низ-
шие хордовые, ракообразные, рыбы), а другие — тяготеющими
16
ими и поде (моллюски), или к суше (многоножки, птицы, млеко-
IHI пнощие и т. п.).
Отношение числа водных видов растений к сухопутным
почти в точности такое же, как и у животных. Число живущих
и воде видов растений составляет приблизительно 26 000, а на
суше (включая грибы) —около 330 000, соответственно около 8
и 92% [составлено на основании данных Энглера (Engler, 1954,
1964)].
3. СТЕПЕНЬ ИЗУЧЕННОСТИ ВИДОВОГО СОСТАВА
ФЛОРЫ И ФАУНЫ
О возможности инвентаризации современных
и ископаемых видов. В каждую геологическую эпоху
число видов определенно. Достаточно определенно и число видов,
существующих в каждый данный период, например в современ-
ных условиях. Однако оно не может быть строго определенным,
так как наряду с установившимися существует известное число
очень молодых видов с еще расплывчатыми границами. Если же
не касаться таких трудно классифицируемых видов, то следует
признать возможность полного учета всего видового состава
флоры и фауны земного шара.
Надо различать два способа установления новых видов: 1) от-
крытие действительно нового, т. е. ранее неизвестного вида и
2) изменение таксономического ранга уже известной группы.
Наиболее яркими примерами открытий действительно новых для
пауки видов является описание новых видов, принадлежащих
к ранее неизвестным крупным таксонам (например, виды погоно-
фор) или к группам, считавшимся давно вымершими (например,
окапи, латимерия, неопеллпна). Одпако находки новых типов или
классов, так же как и находки «живых ископаемых», имея огром-
ное научное значение, мало влияют на число известных видов.
Подавляющая же масса вновь открытых видов относится к давно
известным господствующим на Земле группам. Выборочное об-
следование нескольких семейств растений и отрядов животных
показало, что свыше 85% вновь открытых в последние 20 лет
видов являются новыми видами уже известных родов, падродов
или триб, принадлежащих к наиболее обильным видами группам
насекомых, паукообразных, многоножек, моллюсков и цветковых
растений. Новые виды усиленно описываются и у малоизученных
групп (бактерии, синезеленые водоросли, грибы, простейшие
и т. п.), но их число во много раз меньше. Кроме того, и само
явление вида у таких групп, как бактерии, несовершенные грибы,
синезеленые водоросли и другие своеобразно и обрисовывается
с большим трудом.
Совершенно иначе должен ставиться вопрос о возможности
инвентаризации ископаемых видов. Из-за неполноты геологиие-
2 К. М. Завадский
17
ской летописи и трудностей разграничения видов во времени точ-
ный подсчет видов, живших в минувшие геологические эпохи, —
задача неосуществимая.
Неполнота геологической летописи очень велика. Она связана
с неизученностью недр, а также является результатом плохой
фоссилизации погибших организмов. Многие наземные животные
(птицы, мелкие млекопитающие и пр.), например, погибали в та-
ких условиях, что фоссилизация их остатков является редким
исключением. Вот почему до сих пор найдено так мало ископае-
мых птиц (по Ромеру, только 400—500 видов, т. е. около 5% от
числа современных видов). Размеры пробелов А. А. Чернов
(1959) иллюстрирует таким примером. Уже в девоне найдены
древнейшие бескрылые насекомые, но затем находки этих насе-
комых исчезают до триаса и далее, после триаса, следует пере-
рыв до третичного времени (балтийские янтари). Почти столь же
велики пробелы в находках ископаемых крылатых насекомых.
Общее число найденных ископаемых видов насекомых не превы-
шает 20000 (из них около 3000 дотретичных), т. е. 2% от числа
современных видов. Такое положение не связано с малочислен-
ностью видового состава насекомых прошлых эпох. Напротив,
характер находок свидетельствует о громадном разнообразии ви-
дов насекомых, по крайней мере с карбона и триаса, а особенно
в третичный период. Видовое разнообразие насекомых уже в те-
чение многих десятков миллионов лет, вероятно, столь же велико,
как и в настоящее время.
Многие древнейшие крупные таксоны (инфузории, гидроме-
дузы, плоские черви, гребневики, бесскелетные губки и т. п.)
почти неизвестны в ископаемом состоянии (только отдельные
находки).
Биологи прошлого века пытались отгадать число видов, су-
ществовавших на Земле за все 3.5 или даже 4.6 млрд лет, про-
шедших от истоков жизни (Горский, 1964). Г. Спенсер называл
цифру в 10 млн видов, а Т. Гексли — 50 млн. Но эти догадки
нельзя назвать прогнозами, так как они по существу совершенно
не были обоснованы. В настоящее время единственным методом,
который мог бы дать хотя бы приближенные цифры, является
пересчет числа ныне живущих видов на средний срок жизни
вида, который должен быть эмпирически установлен палеонтоло-
гами для каждой группы и для каждого геологического периода
отдельно. Однако этот путь решения вопроса сталкивается с боль-
шими трудностями. Например, Д. Симпсон (Simpson, 1952) в за-
метке о числе видов приводит следующие расчеты (табл. 6),
использованные недавно с изменениями Н. П. Дубининым
(1966:394).
В таких сугубо приблизительных расчетах имеют значение
только порядки величин. Однако в таблицу Симпсона вкрались
ошибки именно в порядке величин.
Таблица в
Общее число видов, существовавших на Земле
(Приблизительные подсчеты, ведущие к оценке числа видов)
Среднее количество современных видов (а) Средняя продолжительность жиз- ни вида (б) * Время с начала жизни (в) .... Максималь- ное Среднее Минималь- ное
1000000 500000 2000000000 625000 2750000 1500000000 250000 5000000 1000000000
„ ав Сумма видов —g— 4000000000 341000000 50000000
* В таблице Симпсона значения максимальной и минимальной средней продолжи-
тельности жизни вида переставлены местами; при правильной же перестановке этих
величин десятикратно изменяется итоговая сумма видов и порядки этих сумм оказыва
ются одинаковыми. Н. П. Дубинин поставил максимальную и минимальную среднюю
продолжительность жизни на соответствующие места, но не пересчитал при этом общей
суммы видов, которая в таблице его редакции не соответствует сумме видов в таблице
Симпсона.
Во-первых, число современных видов не может быть меньше
1500 000 (табл. 3). Если же учесть еще неизвестные виды, то
это число должно быть увеличено примерно до 3 000 000, т. е.
в 3—12 раз по сравнению с данными Симпсона. Во-вторых,
Симпсон сильно занизил время существования жизни на Земле.
В третьих, приводимая этим автором цифра средней продолжи-
тельности жизни вида не является обоснованной, так как опи-
рается на произвольно взятые крайние значения. Основываясь
па данных по четвертичным отложениям, минимальную продол-
жительность жизни вида, видимо, можно принять в 20 000—
50 000 лет. Известные же сроки существования древних реликтов
позволяют значительно увеличить также и максимальные сроки
жизни вида (как минимум в 10 раз —до 50 млн лет). Что же
касается средней продолжительности жизни вида, то в настоя-
щее время невозможно еще судить даже и о ее порядке, так как
длительность существования видов одной и той же группы очень
различна (видимо, от тысяч до десятков миллионов лет) и по-
этому среднее значение должно быть выведено из очень большой
выборки, которой трудно располагать. Палеонтология, как пра-
вило, не имеет полных серий для установления сроков существо-
вания видов.
Некоторые считают, что инвентаризация ныне живущих видов
близка к завершению, так как скорость открытия и описания
новых видов в последнее время очень замедлилась (Abelson, 1964).
В последние десятилетия наблюдается значительное снижение
интенсивности описания новых видов в некоторых крупных
группах.
2*
19
Число видов цветковых растений за последние полвека су-
щественно возросло, как это видно из выборки по 12 семействам
(табл. 7).
Таблица 7
Изменения в численности видов и е к о т о ры х
семейств покрытосеменных за по л века (1912—1964 гг.)
Семейство Engler Lawrense 1951 Engler 1964
1912 1936
Орхидные 7200 17000 10000—15000 20000
Эуфорбиевые 4500 7200 7300 7500
Осоковые 2G00 3000 3200 3700
Крестоцветные 1900 3000 2500 3000
Вересковые 1350 1500 1900 2500
Маревые 500 1400 1400 1500
Камнеломковые 650 1100 1200 1200
Лавровые 1000 1056 1100 2250
Фиалковые 450 800 850 850
Гераниевые 630 650 850 780
Сложноцветные 13100 18750 20000 19000
Лилейные 2600 3500 4000 3500
Однако если в первую часть этого периода (1912—1936 гг.)
число видов повысилось во всех семействах, а у восьми семейств
па 50—270%, то во вторую половину периода темп описания
новых видов резко упал: только у одного семейства (лавровые)
наблюдается двукратное увеличение числа видов. У всех осталь-
ных семейств, и в том числе у семи очень обильных видами
(орхидные, сложноцветные, лилейные, крестоцветные, маревые,
осоковые и эуфорбиевые), число видов вовсе не возросло или
увеличилось незначительно. Л. А. Смирнов (1943) сопоставил
число видов в 17 семействах цветковых растений, приводимое
Н. И. Кузнецовым (1922), с данными о числе видов в этих же
семействах jio монографиям 30-х годов. Нами было подсчитано
число видов в этих семействах по данным 50—60-х годов
(Lawrence, 1951; Engler, 1964). Оказалось, что за первое двадца-
тилетие прирост числа видов составил 72%, а- за последующее
двадцатилетие только 29%.
Другие авторы считают, что до сих пор остается еще неиз-
вестной основная масса видов (Blackwelder, 1962), что мы далеки
от стадии первичной инвентаризации; даже флора и фауна об-
ластей умеренного климата еще не вполне изучена в отношении
видового состава, а некоторые тропические области изучены еще
очень мало. Такое разноречие объясняется отсутствием объектив-
ных критериев степени изученности видового состава.
20
На какой же стадии в действительности находится сейчас
инвентаризация видов и каковы причины столь разноречивых
мнений специалистов? Эти противоречия отчасти связаны с раз-
личной степенью изученности отдельных групп и с различиями
в толковании вида, отчасти же — с отсутствием точных критериев
для суждения о степени завершения инвентаризации видов в той
или иной группе. Вот примеры прогнозов числа видов у пауков
н клещей. Характеризуя отряд пауков (Aranei), А. В. Иванов
(1965) пишет, что хотя известно около 21000 видов, однако ис-
тинное число видов, вероятно, больше 50 000. Число видов клещей
(отряд Acarina) определяется в 10 000, по другие специалисты
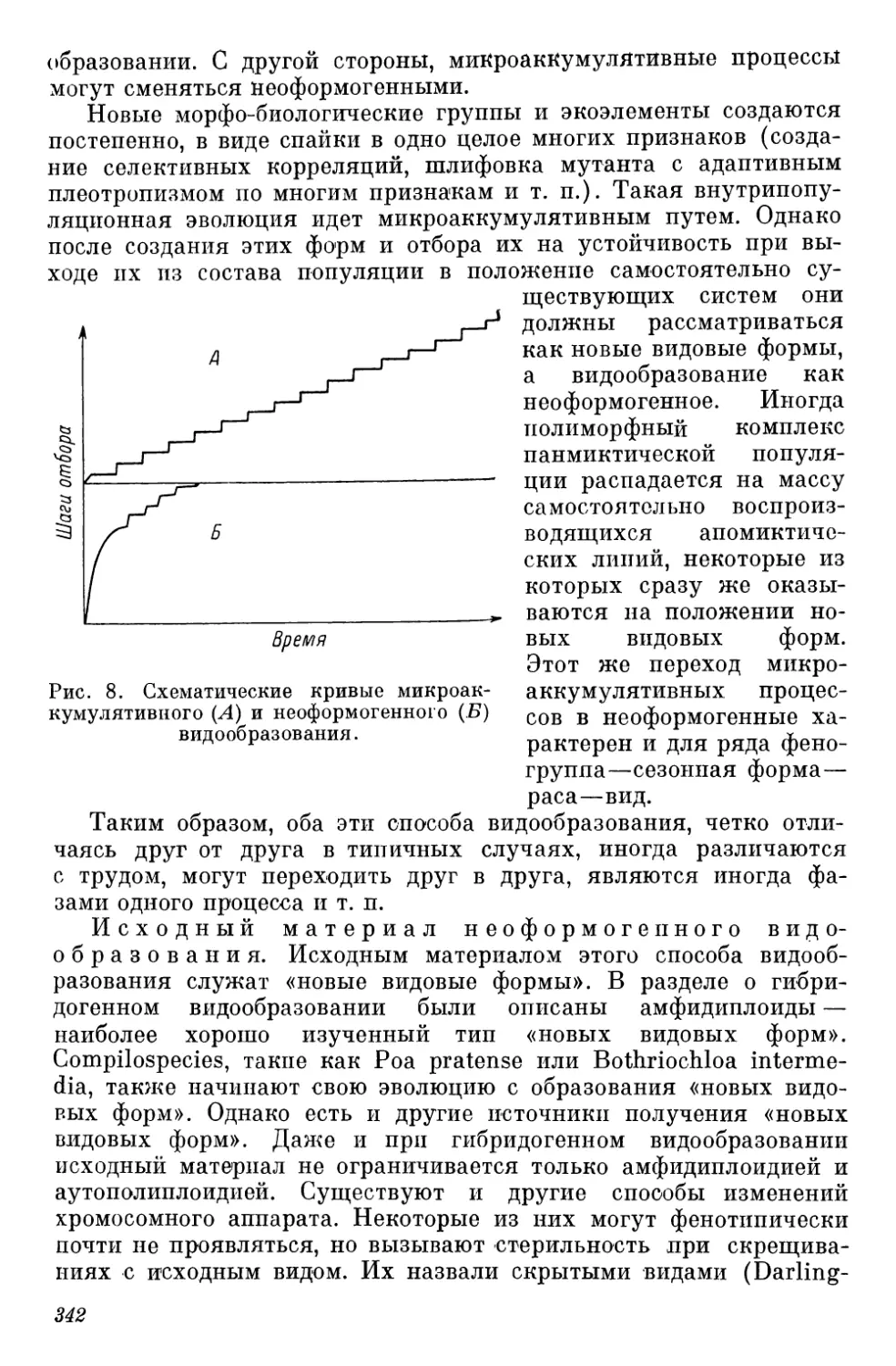
Рис. 1. Кривые скорости описания видов (Steyskal, 1965).
а — птицы (Северная Америка): 1 — виды, 2 — подвиды; б — бабочки
(Rhopalocera; 4 семейства Северной Америки); в — комары (Culicidae).
По оси абсцисс — десятилетия; по оси ординат — число видов.
называют большие цифры (20 000—100 000), а В. Б. Дубинин —
даже 250 000. Широко известно, что об истинном числе видов
насекомых существуют разноречивые прогнозы крупных энтомо-
логов, отличающиеся чуть ли не на полтора миллиона видов (от
1000 000 до 2 500 000). Дж. Гексли в 7-м издании «Эволюции»
(Huxley, 1963 169) пишет, что количество вновь описанных ви-
дов непрерывно увеличивается и только для насекомых состав-
ляет свыше 10 000 в год. Однако число видов животных харак-
теризуется им по данным 40—50-летпей давности (Hesse, 1929).4
Вели верить Гексли, за полвека должно было быть вновь описано
500 000 видов насекомых и их общее число в настоящее время
превышало бы 1 250 000. Между тем данные табл. 5 показывают,
что ничего подобного фактически не произошло. Число известных
пауке видов насекомых в сводках 50—60-х^одов продолжает
сохраняться на уровне числа, приводимого Гессе. Прогнозы эти
4 Гессе пишет, что суммированные им данные отражают состояние
изученности отдельных групп на 1906—1928 гг. Странно, что, многократно
переиздавая свою книгу, Дж. Гексли не обновил этих данных (как, впро-
чем, и многих других, успевших основательно устареть с 1942 г.).
21
Являются скорее лишь интуитивными догадками. Попытка болеб
обоснованно подойти к прогнозированию была недавно сделана
с помощью «кривых скорости описания видов» (Steyskal, 1965).
Примеры таких кривых приведены на рис. 1.
Число видов североамериканских птиц (рис. 1, а) уже к концу
прошлого века полностью стабилизируется (в то время как число
известных подвидов в одном и том же темпе возрастает до наших
дней). Характер кривой видов бабочек (рис. 1, б) указывает на
то, что в пределах четырех семейств на территории Северной
Америки описание видов близко к завершению. На примере роста
числа известных видов комаров (рис. 1, в) обнаруживается роль
социально-экономических стимулов: открытие их значения как
переносчиков инфекции (конец XIX в.) резко ускорило изучение
видов этой группы. Кроме того, в тот же период для диагноза
были использованы половые признаки. Оба эти фактора вызвали
изменение характера кривой, в XX в. вновь круто пошедшей
вверх.
Автор правильно отмечает, что па ход кривых оказывает
влияние количество ученых, работающих над той или иной груп-
пой, что в свою очередь связано с практическим значением
группы. Данные этой работы говорят о незавершенности инвен-
таризации видов животных. Нельзя не упомянуть, что автор об-
ходит вопрос о влиянии на ход кривых изменений в толкованиях
объема вида — процессов дробления “сборных видов или массового
укрупнения видов. Вспомним, что еще в 20—30-х годах насчиты-
вали 32 000—35 000 видов птиц (Дементьев, 1947; Spector, 1956),
но позднее эти виды были «разжалованы» в подвиды и их число
сократилось вчетверо (табл. 5).
Глава 2
ИЗУЧЕНИЕ ВИДА ДО ПОЛОВИНЫ XIX в.
1. ХАРАКТЕРИСТИКА ЭТАПОВ ИЗУЧЕНИЯ ВИДА
На протяжении тысячелетий человечеству не было известно
понятие о виде, но были хорошо знакомы многие конкретные
виды растений и животных. Пусть в еще очень неясной и рас-
плывчатой форме, чрезвычайно медленно и с перерывами, но
задолго до возникновения понятия о виде уже шло накопление
представлений об отдельных его свойствах. Практические знания,
выросшие из разнообразных потребностей общества, позволяли
порой весьма подробно распознавать формы растений и живот-
ных. Вся производственная деятельность охотничьих, пастушеских
и земледельческих племен опиралась на способность отыскивать
нужные формы, избегать ядовитых или опасных и т. п. Навыки
распознавания полезных и вредных форм закреплялись в опыте
племен, фиксируясь в названиях.
Задолго до того как ученые начали писать о виде, во всех
языках фигурировали названия животных и растений, иногда
даже бинарные, с прилагательным-эпитетом. Издавна, например
в европейских языках, к названию птицы прибавлялся эпитет,
определяющий окраску (ворона серая и ворона черная; grey
crow, black crow; Graukrahe, Nebelkrahe, Rabenkrahe). Названия
некоторых растений в русском языке давпо уже фиксировали
различия видового ранга. Таковы, например, названия видов
в родах Populus (осина, тополь, осокорь и др.), Ulmus (вяз, ильм,
карагач, берест) и т. п. В пределах одного рода существуют как
народные бинарные названия, так и односложные (клевер крас-
ный и белый, кашка, лук огородный, чесно^к, шарлот, порей,
мыший чеснок, желтый лук и др.). Однако наиболее распростра-
нены народные названия растений, соответствующие роду, на-
пример солерос (Salicornia), солянка (Salsola), кумарник
(Agriophyllum), шпинат (Spinacia), лебеда (Atriplex) и др. Систе-
матика рода Salix (ива) до сих пор является крайне трудной, но
уже издревле в разговорном языке выделялись отдельные формы,
виды или группы видов (ветла, верба, ракита, ползучая ива,
красная верба, чернотал, тальник, бредина, синяя ива и т. п.).
В ненецком языке словом «хондо» обозначают куропаток и
кур, но ненцы различают «пёро-хопдо», т. е. кустарниковую (La-
gopus lagopus — белая куропатка) и «хало-хопдо», т. е. тундро-
вую (L. mutus). Словом «япту» ненцы обозначают род настоя-
щих гусей (Anser), различая виды (сёнгры-япту и тйник-япту)
(Житков, 1934).
Современные народы, стоящие па низких ступенях развития,
практически и в названиях различают все виды местных позво-
ночных (Мауг, 1963). Племена Тропической Западной Африки
«наряду с прекрасным знанием пищевых растений, а также ра-
стений, дающих древесину для построек, для лодок-пирог, свя-
щенных масок и т. п., хорошо ориентируются в лечебных свой-
ствах растений... в растениях, вырабатывающих ядовитые
соединения (алколопды и глюкозиды). Наряду с нередко встре-
чающимися родовыми названиями почти все виды и даже разно-
видности у каждого племени имеют свое название» (Баранов,
1956 242)'.
Таким образом, основа научной трактовки вида тысячеле-
тиями складывалась в практической деятельности людей.
Почти столь же древни истоки знаний о способности организ-
мов при воспроизведении потомства в ряде поколений сохранять
тип родительских форм. Эти наблюдения были закреплены в по-
нятии породы. Сохранение породой определенного типа строения
и свойств было основой возделывания растений и разведения
животных. В понятии породы были уже заключены трп буду-
щих критерия вида: 1) морфологический — сходство группы осо-
бей, 2) физиологический — способность при внутрипородиых
скрещиваниях воспроизводить плодовитое потомство, 3) генети-
ческий — сохранение потомством типа родителей. Вот почему
понятие породы стало первоосновой для образования более слож.-
ного понятия о виде и длительное время часто употреблялось
в качестве его синонима. Например, в старых русских книгах
словом «порода» пользовались вместо слова «вид» (см. русский
перевод «Философии ботаники» К. Линнея 1800 г.). Э. А. Эвер-
смап в работах 40-х годов XIX в. отождествлял породу с видом
(Колесников, 1951).
Сам же термин «вид» перешел в биологию из логики. Вид
в логическом смысле — выражение сходства пли одинаковой сущ-
ности у группы единичных предметов. Это противоречивая кате-
гория, отражающая объединение индивидов в коллектив и в то
же время разъединение таких коллективов друг от друга в пре-
делах рода (Аристотель, 1934). В логике Аристотеля видом на-
зывается известное множество в том случае, если его удается
24
подчинить, как часть, множеству более высокого ранга — роду.
Вместе с тем Аристотель открыл и другую сторону логического
отношения вида к роду: по содержанию вид шире рода, так как
родовое образует лишь его основу, не исчерпывая специфических
черт, присущих именно данному виду, как одному из членов
рода.
В течение двух тысячелетий после Аристотеля господство-
вала первичная нерасчлепенность представлений о системе со-
подчиненных друг другу форм организмов. Аристотель применил
термин «вид» (греч. для характеристики групп животных
любого ранга, если она оказывалась частью группы большего
объема (рода). Теофраст (371—286 гг. до и. э.), опираясь на
греческие названия растений, использовал терминологию Аристо-
теля в систематике растений (Meyer, 1854). Уже зная, что «жи-
вотные существуют породами», Аристотель еще не обратил вни-
мания на то, что эта группа занимает в системе’ совершенно
особое положение. Применяя к систематизации животных общие
логические основы классификации, Аристотель все внимание об-
ратил на оценку сходств и различий. В дальнейшем систематика
оперировала оценочными суждениями, определяя степени разли-
чий между группами по избранному признаку и «взвешивая» их
достоинства. Между тем уже тогда практически было известно,
что единство породы состоит не только в морфологическом сход-
стве особей, но и в их способности, совместно размножаясь, под-
держивать тип.
В отличие от других групп порода способна к самовоспроиз-
ведению и поэтому является особой единицей. Понадобилось две
тысячи лет, чтобы это уже давно практически известное явление
стало одной из важнейших основ научной классификации.
До конца XVII в. слово «вид» (species) употреблялось
в столь же неопределенном смысле, как п у Аристотеля, пе имея
значения научного термина. Явление вида лишь смутно угады-
валось, еще не было возможности даже ставить вопрос о его
изучении как особой и основной таксономической единицы.
В предлагаемой схеме сохранены ранее выделенные нами
пять этапов изучения вида (Завадский, 1961, 19676) и введен
еще один, I этап (см. стр. 26).
За основу при выделении этапов памп приняты переломы
в накоплении знаний о виде, обусловленные крупными откры-
тиями или обобщениями, углублением и расширением наиболее
существенных характеристик вида.
Существуют п иные подходы к периодизации истории позна-
ния вида. Часто выделяют два основных этапа — креационист-
ский, или метафизический, и эволюционный, а иногда три-два
перечисленных и современность (Radi, 1905, 1909, и др.).
В. Л. Комаров (1940) в истории изучения вида предложил раз-
личать следующие четыре периода: идеалистический, механи-
зм
Этапы изучения вида
Этап Границы и продолжи- тельность этапов Краткая характеристика этапов
I С древнейших вре- мен до начала XVIII в. Первичная нерасчлененность представле- ний о таксонах.
II От Д. Рея до К. Линнея, первая половина XVIII в. Около 70 лет. Первое обоснование понятия «вид».
III От К. Линнея до Ч. Дарвина; с поло- вины XVIII в. до по- ловины XIX в. Око- ло 100 лет. Определение вида как универсальной еди- ницы и неизменной формы. Применение мо- нотипического и политипического морфоло- гических практических стандартов вида. Развитие типологического и суммативнцго понимания вида; начало борьбы по вопросу о реальности вида. Планомерная инвента- ризация видов и расцвет систематики.
IV Вторая половина XIX в. Примерно 40 лет. Утверждение общебиологических идей из- меняемости и неравноценности видов. Нача- ло изучения видов во времени и в простран- стве. Появление мутационной и эволюцион- ной концепций видообразования. Усиление субъективистского понимания вида и кон- цепции «эволюции без видов».
V Первая треть XX в. Обоснование новых критериев вида и введе- ние новых практических стандартов вида в систематике. Обоснова/ние понятия вида как системы в связи с применением экспе- риментального метода и с открытием слож- ной структуры вида. Первые работы по ми- кросистематике и по экспериментальному изучению внутривидовых отношений.
IV Современность, на- чиная с 30-х годов XX в. Развитие популяционной концепции вида, пришедшей на смену типологической. Обо- снование факта целостности вида. Разработ- ка проблемы неравноценности видов. Трак- товка вида как одной из основных и первич- ных форм организации живого.
86
стический, дарвиновский и диалектический. Эта периодизация
полностью не отражает действительности, так как борьба различ-
ных идеалистических толкований вида с материалистическими
проходит через всю историю проблемы и не существовало особых
чисто идеалистического и чисто механистического периодов в позна-
нии вида. Кроме того, следует исходить из конкретных ступеней
познания именно вида, а не из этапов развития общефилософской
мысли. С этой точки зрения диалектический период познания
вида (который, по Комарову, в 1940 г. еще не наступил) некон-
кретен, а поэтому и неприемлем.
Подробное обоснование выделяемых нами этапов составляет
содержание всей первой части книги. Здесь мы ограничимся
только несколькими замечаниями. Подразделение периода от Рея
до Дарвина на два этапа представляется нам бесспорным, так
как работы Линнея сыграли роль настоящего поворотного пункта.
Границы этих этапов обозначены очень четко. Значительно труд-
нее обосновать границы между IV и V этапами, так как на пер-
вый взгляд в начале XX в. концепция вида не претерпела каких-
либо существенных изменений и продолжала разрабатываться
так же, как и в середине XIX в. Выделение же последних двух
этапов и в особенности разграничения между новейшей историей
(первая треть XX в.) и современностью — задача еще более
трудная.
Эти трудности заставили нас излагать вопросы новейшей
истории изучения вида в различном объеме и доводить их разбор
до различных временных границ. Такой границей между исто-
рией и современностью мы избрали 30-е годы. Однако при об-
суждении ряда проблем, таких, например, как критика субъекти-
вистского направления и некоторых других, было удобнее дово-
дить изложение до современного состояния вопроса.
История исследований вида чрезвычайно тесно переплетается
с историей других проблем биологии, и можно было бы расши-
рить рамки исторического очерка путем включения материала,
излагаемого в историях биологии, эволюционного учения, систе-
матики и т. п., развить интересные вопросы о социально-идеоло-
гических корнях тех или иных идей о виде, о связях этих идей
с философскими течениями эпохи и пр. Мы ограничиваемся рас-
смотрением того, как возрастала изученность вида, как возникали
и разрешались основные противоречия в познании вида, пока,
наконец, наши знания о нем не достигли своего современного
состояния.
2. РАЗРАБОТКА ПРОБЛЕМЫ ВИДА ОТ Д. РЕЯ .ДО К. ЛИННЕЯ
Вид по Д. Рею. В течение всего XVII в. теоретические
вопросы систематики включали главным образом принципы рас-
членения многообразия форм на большие таксоны (Меуег, 1854;
27
Osborn, 1913f, л др.). Проблема рода считалась второстепенной,
и никто еще не подозревал, что истинным фундаментом систе-
матики является вид. Некоторые историки пауки считают, что
в XVII в. наблюдался общий застой в разработке теоретических
основ систематики (Radi, 1905; Brewster, 1916). Хотя К. Линней
и считал «первым, истинным систематиком» А. Цезальпина
(1519—1603), это нельзя считать правильным, так как первым
истинным систематиком мог быть ученый, открывший основу
своей науки — вид.
Как особое явление природы вид стал объектом научного по-
знания после работ Д. Рея (John Ray). Заслуга Рея состояла
в том, что он впервые вычленил это явление пз других, дал пер-
вое определение вида, содержавшее описание некоторых его
специфических черт.
В 1686—1704 гг. публиковался основной труд Рея «Historia
generalis plantarum». В этой работе было предложено именовать
видом (species) наиболее мелкие совокупности организмов, прак-
тически тождественных морфологически, совместно размножаю-
щихся и дающих потомство, сохраняющее это сходство. Этим
самым он сблизил термин «вид» с понятием породы.
В «Synapsis methodica animalium» (1693 г.) содержится любо-
пытная аргументация, приводимая Реем в обоснование необходи-
мости введения в биологию понятия о виде. Он писал, что всякая
собака отличается от кошки пли курицы, но разные собаки также
могут сильно отличаться друг от-друга, и возникает вопрос, как
средн массы разных животных выделить группы наиболее сход-
ных индивидов, т. о. виды. Критерием такого объединения Рей
считал способность передавать отличительные признаки всему
своему потомству. Постоянство формы в поколениях, т. е. проис-
хождение подобного от подобного, — вот основа трактовки вида
Реем.
Слово «вид» впервые приобрело значение не чисто логиче-
ского, а естественнонаучного понятия, отражающего относительно
определенный круг явлений органической жизни. Такпм образом,
была создана возможность исследовании одной из основных форм
организации живого.
Такое понимание вида удерживалось в биологип после Рея бо-
лее ста лет, и^например, основа определения вида, предложенного
в 1819 г. Огюстеием Пирамом Декандолем, та же, что и у Рея:
вид — это собрание особей, сходных между собой более, чем
с какими-либо другими организмами, и способных при взаимном
оплодотворении производить па свет плодовитое потомство.
Таким образом, уже на втором этапе изучения вида были
установлены следующие три особенности, которые, как оказалось
впоследствии, действительно являются^ его существенными чер-
тами: 1) вид является множеством организмов, 2) вид объеди-
няет организмы, сходные морфологически и физиологически,
28
3) вид представляет собой самостоятельно воспроизводящуюся
природе единицу. Этим был уже очерчеш относительно опреде-
ленный круг явлений, подлежащих включению в понятие вида,
и создана возможность для дальнейших исследований. Прежде
всего возникла необходимость решить вопрос о распространен-
ности явления вида в органической природе и о значении этой
единицы в классификационных работах. Не менее важно было
уточнить саму характеристику вида в такой мере, чтобы она да-
вала возможность единообразно понимать его границы.
Что касается вопроса о степени устойчивости вида, то в первой
половине XVIII в. он не стал еще основным и чаще всего ре-
шался в духе средневековых принципов наивного трансформизма,
согласно которому яблоня может «порождать» ягпят. Во времена
Роя считалось, что подобное, как правило, порождает подобное,
но иногда могут совершаться и различные превращения, пони-
маемые как игра природы. В течение XVIII—XIX вв. развитие
морфологии, эмбриологии и других дисциплин привело к вытес-
нению наивного трансформизма из науки.
Вид по К. Л и н нею. В течение первой половины
XVIII в. систематика только накапливала данные, необходимые
для следующего этапа в познании вида. Только во второй поло-
вине XVIII в. создалась возможность доказывать универсаль-
ность вида и его основное значение в качестве структурной еди-
ницы живой природы. Для решения этой задачи необходим был
широкий синтез. Этот синтез и был осуществлен Линнеем.
Линней впервые эмпирически выяснил, что явление вида
имеет всеобщее распространение, т. е. что оно универсально для
органической природы. Все роды растений и животных ему уда-
лось расчленить на виды. После этого открытия вид стал основ-
ной классификационной единицей систематики.
Линней пришел к выводу, что вид — это явление природы,
а не продукт деятельности сознания (Linnaei, 1751, гл. VI,
§ 162). В этом состояла материалистическая основа взглядов
Линнея на вид. Многие авторы справедливо подчеркивают, что
огромная заслуга Линнея состояла прежде всего именно в том,
что им доказывалась естественность, конкретность и качествен-
ная определенность вида.
Вид, как явление природы, по Линнею, — это множество род-
ственных, сходных по строению организмов, при размножении не-
прерывно воспроизводящих себе подобных. Такое понимание вида
являлось точным отражением наблюдаемых фактов. Установив
реальность вида, приняв положение об универсальности вида
в качестве основной единицы классификации, Линней пытался
решить вопрос и о сущности вида. Сущность вида Линией искал
в устойчивости формы. Вся концепция вида была основана
’па положении о существовании градации признаков, от очень
изменчивых до самых постоянных из всех известных (Уэвелл,
29
1869). Устойчивость видовой формы в процессе непрерывного
размножения при смене поколений и при расселении в различ-
ные условия среды, устанавливаемая простым наблюдением,
была истолкована Линнеем в «Философии ботаники» как полная
ее неизменность: «Видов насчитываем столько, сколько раз-
личных форм создано в самом начале» (1800, гл. VI, § 157).
Этим, собственно, и было узаконено метафизическое воззрение
на вид.
Однако не следует думать, что творчество Линнея было ли-
шено противоречий, а сам он был безоговорочным сторонником
креационистской концепции вида. Некоторые историки науки
уже давно считают Линнея эволюционистом (Green, 1909).
В «Философии ботаники», в первых 12 изданиях «Системы при-
роды» и в «Видах растений» Линней действительно писал о по-
стоянстве видов и о виде, как об отдельной сотворенной форме.
Но, по-видимому, между 1751 и 1762 гг. мнение его о постоянстве
видов значительно изменилось. В работе, посвященной гибрид-
ным растениям, Линней писал даже, что отдаленная гибридиза-
ция открывает большие перспективы к созданию новых видов
(1759 г.). В «Fundamcntum fructificaliones» (1762 г.), «Systema
vegetabilium» (1774 г.) он подчеркивал, что многие виды имеют
гибридогенное происхождение, а некоторые возникли из-за изме-
нений среды (такое толкование есть и в ранпей работе, посвя-
щенной флоре Лапландии). В 13-м же издании «Системы при-
роды» (1774 г.) Линней допускает изменение не только видов и
родов, но даже семейств. Однако, несмотря на признание изме-
няемости форм в довольно широких пределах, Линней не был
эволюционистом, так как считал неизменными признаки отрядов.
В этой противоречивой трактовке вида как конкретного явления
(самовоспроизводящаяся совокупность особей) и как особой сущ-
ности (неизменная форма, тип) Линней отразил уровень зна-
ний и способ мышления эпохи. Новая постановка проблемы
вида в трудах Линнея (универсальность явления вида и значе-
ние вида как основного элемента системы) выдвинула перед си-
стематикой задачу описания всех видов, которая, как это было
показано в главе 1, еще далеко не исчерпана и в настоящее
время. Однако результатом деятельности Линнея явилась и идея
вида как строго постоянной, неизменной формы.
Итак, именн<\ Линнею принадлежит заслуга установления
того факта, что вид есть основная форма существования живой
природы, реальная и элементарная ее единица, и что из таких
единиц и складывается структура органического мира.
Благодаря этому открытию вид приобрел значение основной
категории в работах по систематизации многообразия форм и
стал главной классификационной единицей систематики. Во вто-
рой половине XVIII столетия систематика выдвинулась на поло-
жение господствующей отрасли биологии. Начались работы по
30
ni.iявлению и описанию новых видов с целью составления их пол-
кип описи.
( )дпако далеко не всегда рост числа вновь описанных видов
характеризовал увеличение объема знаний. Уже в послелиннеев-
( кие времена оно свидетельствовало иногда лишь об изменении
понимания объема вида: уже известный вид просто разделяется
на несколько более мелких. С другой стороны, затруднения, воз-
ки павшие при определении объема и границ видов, служили
как бы стимулом к уточнению характеристики самого вида как
явления природы.
3. ПРОБЛЕМА УСТОЙЧИВОСТИ И ДИСКРЕТНОСТИ ВИДА
Устойчивость вида. Особенность рассматриваемого пе-
риода заключалась в накоплении аргументов в пользу неизмен-
ности видовых признаков, в обосновании и утверждении идеи
постоянства реального вида. К концу XVIII—началу XIX в. к ха-
рактеристике вида прибавились еще две новые черты: устойчи-
вость и дискретность.
Устойчивость вида доказывалась прямыми наблюдениями.
Было установлено, что вид сохраняет свои морфологические осо-
бенности как при смене поколений, т. е. во времени, так и при
изменении условий внешней среды, т. е. в пространстве. Факт
устойчивости вида был чрезвычайно важен для доказательства
его реальности. Действительно, если бы виды не обладали устой-
чивостью, то нельзя было бы констатировать само явление вида
как качественную определенность. Основная трудность проблемы
заключалась в характеристике той степени устойчивости формы,
которая в действительности присуща видам и в установлении
причин, делающих вид устойчивым. Поэтому вполне естественно*,
что вопрос о соотношении между устойчивостью и изменчивостью
вида стал центральной проблемой биологии. Большая пластич-
ность организмов, их изменяемость факторами среды, скрещи-
ваниями, культурой, легко доказывалась такими же прямыми на-
блюдениями, как и устойчивость видовой формы. Создалось про-
тиворечие, которое биологи того времени пытались разрешить,
допустив существование некоей абсолютно постоянной формы как
неизменной сущности вида. В пределах же границ этой постоян-
ной формы изменения организмов признавались возможными.
Такие изменения считали обратимыми, флюктуирующими вокруг
какого-то неизменного среднего значения. Так понималась в то
время внутривидовая изменчивость, выражавшаяся в наличии раз-
новидностей (varietes). Этим допущением хдтели устранить про-
тиворечие между наблюдаемыми фактами устойчивости форм и
изменчивости их признаков. Вид был признан единственно зако-
номерным явлением, а внутривидовые различия рассматривались
как результат незаслуживающих внимания случайностей. Такой
31
позиции придерживался еще Линней: «Разновидность — это расте-
ние, измененное случайной причиной: климатом, почвой, теплом,
ветром и пр. После устранения причин ее признаки естественно
устраняются с восстановлением исходного состояния» (Linnaei,
1751, гл. VI, прим, к § 158). По Палласу, главные особенности
varietes — обратимость и случайность, а вида — постоянство и за-
кономерность. И. И. Мечников писал, что Паллас «больше самого
основателя (т. е. Линнея, — К. 3.) настаивает на постоянстве
вида и все случаи изменчивости относит к вариететам» (1950 : 27).
В итоге разновидность была противопоставлена виду как нечто
случайное — необходимому, как нетипичное п затемняющее
тип — типу, как колеблющееся — постоянному.
Эти обстоятельства и привели к тому, что идея постоянства
вида заняла господствующее положение в биологии и утвердила
креационистскую концепцию вида. Вид стал рассматриваться как
потомство сотворенной особи (пары), сохраняющее при непрерыв-
ном размножении все существенные особенности формы прароди-
теля. Подобных воззрений держались виднейшие биологи того
времени (Pallas, 1780, и др.), по наиболее четко они были сфор-
мулированы Ж. Кювье. Видом он называл «совокупность суще-
ств, принадлежащих к одной из тех форм, которые стойко сохра-
нились с самого начала вещей, не выходя за своп границы»
(Cuvier, 1829 16). «Нот никаких доказательств,—писал
Кювье, — что все различия, наблюдаемые между организмами,
могли быть созданы одними условиями среды. Все, что высказы-
валось в защиту этого мнения, гипотетично. Опыт приводит, по-
видимому, к противоположному заключению: при современном
состоянии Земли разновидности заключены в определенных, до-
вольно тесных границах, и, как бы далеко не проникали мы
в глубину древности, мы видим эти границы теми же, что и ныне»
(там же 9). Это суждение интересно осторожностью формулиро-
вок. Но далее следует фраза, могущая служить как бы девизом
всего метафизического периода. «Итак, обязательно признавать,
что известные формы существуют с самого начала вещей, не вы-
ходя за определенные границы, и все существа, принадлежащие
к одной из этих форм, составляют вид. Разновидности же — это
случайные подразделения вида» (там же).
Следует подчеркнуть, что Кювье (как и Линией), с одной
стороны, рассматривал вид как особый тип организации
( = форму), а с другой — как совокупность сходных организмов.
Таким образом, объективно существующая устойчивость вида
была абсолютизирована и превращена в идею о его полной неиз-
менности не во времена Линнея, а позднее, получив свое наи-
более законченное выражение в работах Кювье. Некоторые исто-
рики биологии подчеркивают также связь между идеей постоян-
ства вида и концепцией преформизма Ш. Боннэ и других
в эмбриологии (Radi, 1905, 1909; Brewster, 1927; Osborn, 1929;
32
К. Линней
Cole, 1930, и др.). Принцип неуничтожимое!'!! зародышей был
гее но связан с идеей неизменной формы.
Главная особенность этого мировоззрения заключалась в при-
знании неизменности всех существенных форм. Ф. Энгельс (1961)
рассматривал его как закономерный этап познания природы, на
котором главная задача состояла в первичном и более простом
описании явлений. Так сформировалась метафизическая концеп-
ция вида, кратко выраженная Уэвеллом следующими словами:
«Виды имеют реальное существование в природе и изменения
одного в другой не происходят» (1869 755).
Дискретность вида. Одной из существенных черт вида,
сформулированных в рассматриваемый период, была дискрет-
ность. Вид оказался образованием, представляющим собой как бы
биологическую отдельность. Эта отграниченность вида от других,
близких ему видов констатировалась как при морфологическом
изучении, так и в опытах по скрещиваниям. Между видами об-
наружились «разрывы сплошности» (хиатусы, зияния) как по
морфологическим признакам, так и в физиологическом отношении
(пескрещиваемость или бесплодие потомства). Этот факт в свою
очередь приводил к выводу, что вид не является произвольно
выхваченным участком непрерывного ряда, а существует в при-
роде реально, обладая более или менее четкими естественными
границами. Считалось, что задача выделения видов была бы не-
осуществимой, если бы границы вида расплывались и исчезали
в переходных состояниях. Подобные суждения казались в то
время полностью обоснованными, хотя в действительности они
были вполне справедливы только по отношению к известной части
хорошо отграниченных видов. Как это будет объяснено в даль-
нейшем, реальное существование вида не обязательно должно
быть связано с наличием у него резко обозначенных границ.
Факт дискретности вида имел решающее значение в практи-
ческой работе систематиков, он дал возможность обосновать ос-
новные критерии вида: морфологический и физиологический.
Сходство между всеми особями, относящимися к одному виду, в их
основных морфологических признаках и неспособность организ-
мов разных видов скрещиваться между собой или давать бес-
плодное потомство являлись хорошим инструментом для клас-
сификационных работ.
Практически в те времена виды определялись только с по-
мощью одного морфологического критерия. Их впоследствии
называли морфологическими видами. Вплоть до XX в. виды, опи-
сываемые систематиками, п являлись фактически только морфо-
логическими видами. Хотя «морфологический Вид — это выраже-
ние нашего невежества» (Кэйн, 1958:155), но, как это пока-
зывает практика, вполне сформированные виды, значительно
обособленные от других (например, древние реликты), отлично
выделяются на основании одного морфологического критерия.
3 К. М. Завадский
33
Что касается физиологического критерия, то его можно было
установить лишь с помощью трудоемких экспериментальных ра-
бот по гибридизации. Такие работы были начаты еще в середине
XVIII в. (в 1761 г.) И. Кельрейтером и успешно развивались
многими учеными (О. Сажрэ, III. Ноденом и др.). Ж. Бюффон
считал физиологический критерий единственно надежным и
объективным критерием вида. Физиологическая несовместимость
видов вплоть до работ Дарвина рассматривалась в качестве наи-
более существенного доказательства постоянства видов. Теорети-
ческое положение о плодовитости внутривидовых гибридов и бес-
плодии межвидовых, как сформулировал М. А. Мензбир, «при
известном взгляде на постоянство видов было возведено на ос-
новании поверхностных наблюдений в закон и- заставило натура-
листов вращаться в круге: если два вида при скрещивании давали
потомство и помеси, происшедшие таким путем, оказывались пло-
довиты между собой, то говорили, что этот факт доказывает, что
взятые для опыта виды были не виды, а разновидности. Напротив,
если две разновидности при скрещивании не давали потомства или
давали потомство неплодное, то считали доказанным, что они не
разновидности, а виды. Никаких исключений из этого мнимого
закона не допускалось» (1927 44). Это высказывание применимо
и к нашим современникам, абсолютизирующим значение «кри-
терия скрещиваемости». В практике же систематиков того вре-
мени физиблогический критерий вида , совсем не применялся.
Таким образом, уже в те времена создался известный разрыв
между теоретической концепцией вида, включавшей мысль о зам-
кнутости вида в сексуальном отношении, и практическим видом
систематики, выделявшимся лишь морфологическим методом.
Кроме морфологического и физиологического критериев вида,
отдельными учеными делались попытки выдвинуть уже и эколо-
гический, а также этологический критерий. Именно так подходил
к виду К. Ф. Рулье. Э. А. Эверсман в работах 40—50-х годов рас-
сматривал вид как реально существующее единство животных,
связанное функцией произведения потомства. Если же виды не
поддаются определению с морфологических позиций, надо «при-
звать на помощь сведения об их отечестве и образе жизни» (цит.
по: Колесников, 1951 139). По Э. А. Эверсману, внутри вида сле-
дует различать типичных особей, уродов (monstra), а также не-
сколько сортов разновидностей — постоянные и непостоянные,
морфологические, физиологические, географические.
Наблюдавшаяся отграниченность видов друг от друга исполь-
зовалась в качестве довода в пользу постоянства вида. В то
время казалось, что гипотеза изменяемости видов не находит себе
фактических подтверждений: если виды изменяются, то вся
структура органической природы должна была бы быть иной,
чем она есть в действительности. Так, например, Кювье утвер-
ждал, что если виды изменяются, то не могут существовать пре-
34
pi.uiiK -I I,io ряды видов, состоящие из обособленных друг от друга
фирм, и границы между видами должны стать расплывчатыми
ник даже вовсе исчезнуть. И наоборот, раз виды обособлены друг
друга, то, следовательно, они не изменяются. Факт обособлен-
.. г и видов друг от друга был, таким образом, абсолютизирован
ннс же, как и факт их устойчивости. Но само явление известной
hi мкпутости вида было настолько очевидно, что оказалось силь-
ным доводом, устоявшим против всех форм эволюционизма, кроме
। нории Дарвина.
Нами рассматривалось представление о виде как о реально су-
ществующем самовоспроизводящемся и обособленном от других
множестве организмов схожего строения, способном при размно-
жении устойчиво сохранять свои специфические особенности.
II этом представлении уже были правильно отражены некоторые
। ущественные черты, характеризующие вид, но две из них тол-
ковались еще слишком грубо: устойчивость — как неизменность,
обособленность — как полная замкнутость.
4. ВЗГЛЯДЫ Ж. Б. ЛАМАРКА НА ПРОБЛЕМУ ВИДА
Рассмотрим теперь взгляды раннего эволюционизма, который
противопоставлял господствующей идее «реального неизменного
вида» концепцию «развития без видов». Классическим вырази-
телем таких взглядов был Ж. Б. Ламарк.
Будучи крупнейшим систематиком растений и начиная об-
ширные работы по классификации беспозвоночных животных
в возрасте 50 лет, Ламарк еще не сомневался в реальности и
постоянстве видов. В 1792 г. в статье «L’espece» он писал: «Вид
должен состоять из совокупности сходных между собой особей,
которые остаются такими же при размножении... нельзя оспари-
вать, что виды действительно существуют в природе... Авторы,
которые сомневались в самом существовании видов в природе. . .
называли видами простые разновидности и. вследствие этого
они имели случай наблюдать постепенное исчезновение большей
части установленных ими разграничений» (Ламарк, 1955 : 779—
780).
В 1799—1800 гг. у Ламарка созревает идея об эволюции и
• разу эту идею он противопоставляет факту реальности вида и
ищет опору в номиналистической доктрине: «Я долго думал, что
н природе есть постоянные виды... теперь же я убедился, что
ото было ошибкой и что в природе есть только особи» (Lamarck,
1802: 141).
В «Философии зоологии» (1809 г.) позиция Ламарка в вопросе
о соотношении между эволюцией и реальностью вида была весьма
противоречивой. Не раз он утверждает, что виды изменяются:
«. . вид... сложился... незаметно и последовательно, имеет ис-
ключительно относительное постоянство и не может быть так
35
3*
стар, как природа» (Ламарк, 1935 65). В чисто дарвиновском
духе он толкует процесс видообразования как постепенное форми-
рование разновидностей, «которые затем станут породами, а со
временем — видами» (там же : 64).
Но наряду с правильными формулировками в «Философии зо-
ологии» встречаются и такие, где полностью повторяется ошибоч-
ное противопоставление изменяемости организмов реальности
вида, приведенное выше. Так, например, Ламарк писал: «Можно
утверждать, что в действительности природа не образовывала,
пи константных классов. ни родов, ни видов, но исключительно
особей, последовательно сменяющих друг друга» (там же 33).
Непоследовательность Ламарка видна также и из следующего его
высказывания: «Итак, природа... дает нам, строго говоря, только
особей, происходящих одни от других; что же касается видов, их
постоянство относительно и неизменяемость их носит исключи-
тельно временный характер. Тем не менее с целью облегчить
изучение и познание огромного числа разных тел небесполезно
прибегнуть к названию вида, как к обозначению всякой группы
сходных особей, сохраняющих из поколения в поколение неиз-
менным свое состояние, пока условия их положения не изме-
нятся настолько, чтобы вызвать перемену в их привычках, харак-
тере и форме» (там же : 71—72). В этой выдержке номинализм
и субъективизм (вид — лишь удобное для классификации услов-
ное обозначение) мирно уживаются с Правильной идеей о том,
что устойчивость вида не абсолютна, а временна и относительна.
Чем же объяснить такую непоследовательность в высказыва-
ниях Ламарка? Ее основа связана с ошибочной трактовкой при-
чин наблюдаемой дискретности видов. Как известно, хиатусы
между близкими видами Ламарк считал не объективным фактом,
а результатом слабой изученности, выделения видов по «малым
выборкам», иногда даже по одному или по нескольким экземпля-
рам. Все выделенные систематиками виды представляют собой,
по Ламарку, лишь случайно выхваченные куски непрерывных
рядов: «По мере того как. обогащаются наши коллекции, почти
все пробелы... заполняются и границы стираются» (там же
59). Виды лишь кажутся нам обособленными, а в природе суще-
ствуют лишь непрерывные ряды индивидов. Подобное толкование
является односторонним, так как абсолютизирует непрерывность
и отрицает объективный характер прерывности.
Такая концепция может быть названа наивным эволюциониз-
мом. Именно эта теоретическая слабость и заставила Ламарка за-
нять непоследовательную позицию в трактовке отношения между
изменяемостью и реальностью видов.
Помимо термина «вид» (I’especes) Ламарк часто употреблял
и термин «раса». Утверждая, что реально существует лишь от-
дельная особь, Ламарк (1935 : 33, 71) в противоречии с этим пи-
сал и о реальном существовании рас (les races). Понятие о расе
36
। и ilk местах Ламарк отождествляет с понятием вида: «Если бы
расы (то, что именуют видами), которые принадлежат живой
природе, были бы хорошо изучены...» [«Si toutes les races (се
>pi'on nomme les especes) qui appartiennent a un regne des corps
\ minis etaient parfaitement conmies. ..»] (там же : 40—50, см.
иноке 51 и др.), а в других толкует по существу как подвид:
Разновидности, которые впоследствии становятся расами и ко-
трые через долгое время составят то, что мы называем видами»
(«I >es varietes qui deviennent ensuit des races, et qui, avec les
I «'in ps, constituent ce que nous nommons des especes») (там же : 3).
В русском издании термин «раса» везде переведен как «порода»
(там же 39, 40, 57, 64 и др.) и это, возможно, наиболее точно
отражает суть дела.1 Во всяком случае Ламарк считал реальное
существование рас (пород) само собой разумеющейся истиной,
так как речь шла о группах особей, тесно связанных между co-
in >й совместным размножением и способных к воспроизведению
себе подобных.
Подводя итоги, можно сказать, что Ламарк, убежденный в ис-
тинности эволюционной идеи, но не умея совместить эту идею
фактом обособленности видов, предпочел отказаться от положе-
ния о реальном существовании видов.
Так в процессе познания вида возникла альтернатива: или
«реальный неизменный вид», или «развитие без видов». Оба ут-
верждения были ложны, но чтобы это доказать, необходимо было
глубже разобраться в проблеме вида, что и было сделано Дарви-
ном.
Вполне справедливо замечание С. Р. Микулинского, что без
«более или менее ясно представляемых понятий „разновидность"
II „вид" нельзя было обсуждать вопрос об изменяемости видов»
(1961 293). Однако Ламарк создал теорию, основанную на да-
леко не ясном и внутренне противоречивом представлении о виде
и разновидности. Его пример показывает извилистость пути по-
знания. Ранние эволюционисты сталкивались с непреодолимыми
трудностями, связанными с неизученностью вида, а изучение ста-
рики вида не могло быть сколько-нибудь глубоким без представ-
ления о виде как узловом пункте эволюционного процесса.
5. УТВЕРЖДЕНИЕ ИДЕИ ПОСТОЯНСТВА ВИДОВ
В ПЕРВОЙ ПОЛОВИНЕ XIX В.
Попытки объединения идей реальности и из-
меняемости видов. Не все биологи того «времени разделя-
лись на сторонников концепции «реального неизменного вида» и
1 Раса в смысле «потомки общего предка» идентична понятиям «по-
рода» и «род» также и в английском языке.
37
сторонников концепции «развития без видов». В следующем раз-
деле будет показано, что наивно-эволюционистский номинализм
был не единственной формой субъективистской оценки понятия
«вид». Сейчас же необходимо дать характеристику позиции сто-
ронников объединения идеи реальности вида с утверждением
о его изменяемости.
В первой половине XIX столетия неоднократно в еще крайне
схематичной форме высказывалась правильная мысль об изменяе-
мости реально существующих видов. Э. Ждффруа Септ-Илер при-
дал этой мысли форму афоризма: «виды изменяются»,
а К. Ф. Рулье избрал его эпиграфом одной из своих работ.
Взгляды этих авторов широко известны.
Некоторые передовые систематики того времени также при-
держивались эволюционистских взглядов и пытались совместить
их с признанием реальности вида. Чаще всего это делалось
в весьма противоречивых формулировках. Например, М. А. Мак-
симович (1831) был эволюционистом и рассматривал виды как
давно возникшие и ставшие посто>янными разновидности. Послед-
ние же он считал не только случайными отклонениями и уродст-
вами, но и наследственно постоянными формами, уклонившимися
от родоначалыюго вида под воздействием среды. Однако рядом
с этими взглядами, близкими идеям Дарвина, у М. А. Максимо-
вича можно найти утверждения о непрерывной цепи изменений,
о принципиальной искусственности категории вида и условности
вообще всех объединений организмов в группы. Если исключить
устаревшую терминологию, то идеи М. А. Максимовича о видооб-
разовании через формирование и упрочение разновидностей в про-
цессе их расселения получили развитие в ранних работах
В. Л. Комарова (1901—1908 гг.).
Интересны взгляды па вид Н. А. Северцова, сложившиеся во
время путешествий и зоогеографических наблюдений. Северцов
был эволюционистом и считал, что причина изменений видов за-
ключается в прямом влиянии среды (в духе идей Э. Ж. Сент-
Илера). В работах 50-х годов этот автбр сформулировал ориги-
нальное представление о целостности вида. Он рассматривал вид
«органической совокупностью», столь же реальной, как и особь.
Вид, по Северцову, может быть уподоблен «коллективно органи-
зованному существу». Как справедливо отмечает Б. Е. Райков
(1959), эти идеи близки ко взглядам на вид, развитым впоследст-
вии В. Л. Комаровым. Основная слабость таких концепций за-
ключалась в отсутствии доказательств. В ее защиту обычно при-
водились только общие впечатления, полученные во время экспе-
диций или при камеральной обработке материала. Такими путями
гипотеза эволюции видов не могла быть разработана и доказана.
В течение десятилетий эти идеи оставались на положении непро-
веренной гипотезы. Иногда они больше напоминали построения
натурфилософии, чем биологическое исследование.
38
Наряду с такими интересными взглядами в первой половине
XIX в. существовали концепции, объединяющие идею изменяе-
мое ги и реальности видов на совершенно иной основе. Речь идет
натурфилософских спекуляциях по поводу всеобщей взаимо-
нрсвращаемости форм, в том числе и видов. Это направление
можно показать на примере взглядов Э. И. Эйхвальда, который
в 1824 г. писал, что «легко произвести наблюдения над превра-
щением класса в класс, порядка в порядок, рода в род, вида в вид,
I. е. одних сходных форм в другие сходные» (цит. по: Никулин-
ский, 1961:310). Такие трансмутации Эйхвальд считал обрати-
мыми, внезапно происходящими и не связанными между собой
преемственностью. Иначе говоря, помысли этого автора, трансму-
тации видов возникают внеисторически. Наивный натурфилософ-
ский трансмутационизм существовал давно, опираясь на предрас-
судки, на незнание морфологии и отсутствие сравнительного ме-
тода. Такое соединение концепций реальности и изменяемости
вида, разумеется, не имело будущего, так как было антиисто-
рично и грубо ошибочно.
Многие предшественники Дарвина защищали тезис об из-
меняемости видов, но никто из них не показал, почему и как
именно совершается этот процесс. Их работы носили характер
плохо обоснованных гипотез или, напротив, являлись выводами
сугубо описательных флористических или фаунистических тру-
дов, в которых высказывалась лишь догадка об изменяемости
видов. Можно сказать, что натуралисты убедились в существова-
нии эволюции только тогда, когда Дарвин показал, почему она
осуществляется в форме закономерной изменяемости видов (Ти-
мирязев, 1939, 7 : 247).
Почему же в течение первых 60 лет XIX в. господствующее
положение сохраняло учение о покоящемся реальном виде? Это
объясняется рядом причин, которым будет посвящен следующий
раздел.
Доводы в пользу постоянства видов. Реальность
вида была очевидной истиной для большинства натуралистов, зна-
комых с ними не только по музейным образцам, но и в природ-
ной обстановке. И поэтому крайняя формула о реальном суще-
ствовании лишь индивидов многих из них просто отпугивала,
заставляя придерживаться традиционной доктрины: раз виды
реальны, значит они постоянны. Такая позиция была выраже-
нием стихийного материализма «среднего» натуралиста того
времени.
Господство метафизической теории вида объясняется, таким
образом, во-первых, тем, что еще несовершенная эволюционная
идея слишком полно отрицала старые знания и отбрасывала не
только содержащиеся в них ошибочные преувеличения, но и са-
мую основу всякой закономерности в биологии — понятие о реаль-
ности вида.
39
Во-вторых, в течение рассматриваемого периода все возра-
стало число доводов в пользу теории постоянства видов, и к
середине XIX в. она была лучше обоснована, чем во времена
Линнея.
В эпоху Линнея в пользу неизменности видов привлекалось
лишь два аргумента — факт устойчивого и непрерывного воспро-
изведения организмами себе подобных в течение многих поколе-
ний и сексуальный барьер между видами. А в 20-х годах XIX в.
усилиями Кювье были специально собраны доводы, как казалось,
прямо опровергающие эволюцию. Из тождества видовых призна-
ков современных животных с найденными в Египетских пира-
мидах Кювье сделал вывод, что если за 5000 лет ни один вид не
изменился, то нет оснований считать их способными измениться
в течение большего времени. Против эволюции видо*в, по Кювье,
свидетельствовали также факты несходства видового состава
фаун и флор в климатически аналогичных территориях, будь они
географическп столь удалены друг от друга, как Новая Зеландия
и Британия, или же, напротив, так близки, как о-ва Ломбок и
Бали (австрало-азиатское биогеографическое разъединение).
В качестве прямых свидетельств в пользу постоянства видов им
рассматривался тот факт, что наряду со многими признаками,
которые легко изменяются при перемене климата, пищи, освеще-
ния и других факторов среды, такие признаки животных, как
строение зубов, число и особенности строения. костей и др., не об-
наруживают пи малейших изменений. Кювье проводил обширные
сравнительно-анатомические работы и всегда сталкивался с суще-
ствованием постоянных признаков, противостоящих любым пере-
менам. Хотя волк и лисица обитают на огромных территориях,
сильно различающихся климатом, но многие признаки их черепов
остаются неизменными. Например, черепа лисиц из Северной
Европы, из Франции и из Египта не имели никаких различий
кроме «индивидуальных вариаций».
Не менее сильным доводом против изменяемости видов было
и существование морфологических и физиологических хиатусов
между всеми наиболее хорошо изученными видами. Связь теории
неизменности видов Кювье с идеей предустановленной гармонии
природы Лейбница была хорошо комментирована Н. П. Вагнером:
«Всю природу Кювье принимал за одно громадное гармоническое
целое. Та же гармония, та же связь существует и в каждом ор-
ганизме, и здесь мы не можем отнять ни одного члена, не нару-
шая или не изменяя этой гармонии. Вот почему. . никто, кроме
творца, не может изменить основного, видового, типа этих суще-
ств» (1860 53—55). Принцип нерушимости гармонии «с необхо-
димостью вел к признанию строгих рамок, за которые ни при
каких обстоятельствах не может выходить изменчивость» (Russel,
1916 39). Многие биологи того времени разделяли взгляды
Кювье об абсолютной целесообразности и были убеждены, что
40
л кое существенное изменение организмов может привести
разрушению идеальной гармонии и к вымиранию вида.
Нельзя, однако, считать, что абсолютизация органической це-
лесообразности логически неизбежно влечет принятие тезиса
неизменности видов. Ламарк, например, по существу также ис-
ходил из признания абсолютного характера органической целесо-
образности: виды не могут вымирать, так как при изменении ус-
ловий они всегда приспособительно изменяются. Иначе говоря,
ио Ламарку, организмам изначально присуща неограниченная
•иособность к целесообразно направленной изменчивости. Таким
образом, как Кювье, так и Ламарк исходили из абсолютизации
органической целесообразности. Противоположными были их вы-
воды: Кювье переоценил устойчивость формы, а Ламарк — ее ла-
бильность.
Все доводы против изменяемости видов, как казалось, свиде-
тел ьствовали о том, что эволюции не только нет фактически, но
что опа и невозможна, так как сама структура органической
природы исключает такую возможность. Эти доводы вынуждали
отклонять идею изменяемости видов даже таких ученых, как
Ч. Ляйелль.
Следует отметить, что до Дарвина биология еще не распола-
гала столь обширными и точными данными о дифференциации
форм внутри видов, чтобы они могли поколебать противопоставле-
ние вида всем формам внутривидовых различий. Здесь был создан
как бы заколдованный круг: господствующая теория тормозила ис-
следование внутривидовой изменчивости (как несущественного*,
случайного), а отсутствие точных сведений в этой области делало
невозможным опровергнуть устарелую теорию.
В год выхода в свет «Происхождения видов» во Франции по-
явилась книга Годрона (Godron, 1859), посвященная доказатель-
ствам постоянства видов. Основная идея этой работы заключалась
в противопоставлении диких видов животных породам и устойчи-
вым расам домашних животных. Видовые признаки не могут
быть приобретены или утрачены, пишет Годрон (там же 51).
Если бы видовые признаки изменялись в течение геологического
времени, то четкие границы видов во времени должны были бы от-
сутствовать, типичных форм должно было быть меньше, чем про-
межуточных. На самом же деле палеонтологи, как правило, встре-
чают формы, отвечающие хорбшо различимым видам, но не пе-
реходные. Палеонтология свидетельствует в пользу постоянства
видов: виды могут погибать, мигрировать, но не изменяться
(там же 144 и др.). По Годрону, дикие виды всегда строго моно-
типны, и все уклонения от типа случайны, единичны и легко об-
ратимы. Он подчеркивает, что в результате одомашнивания возни-
кает совершенно небывалое в диком состоянии разнообразие
форм. Долговременное и глубокое воздействие измененных усло-
вий существования (питания, климата, тренировок), а также дли-
41
тельная и разнообразная метизация привели к образованию пород
и наследственно стойких рас домашних животных. Некоторые счи-
тают, отмечает Годрон, такие породы и устойчивые расы «домаш-
ними видами», но они ясно отличаются от видов способностью
при скрещиваниях производить метисов с нормальной плодови-
тостью, тогда как межвидовая гибридизация неосуществима
или же дает мулов. Годрон рассуждал примерно так: бог сотво-
рил виды, а человеческое искусство — расы и постоянные разно-
видности животных.
Можно было бы привести и другие примеры, свидетельствую-
щие о кажущейся прочности доктрины неизменного вида. В се-
редине XIX в. идея постоянства вида была многосторонне обосно-
вана, а гипотеза эволюции видов рассматривалась чуть ли но
как научная фантастика. В то время еще не осбзнавали, что ре-
шение, казалось бы, частного вопроса об изменяемости видов
может вызвать такую революцию во всем естествознании, которая
совершилась после выхода «Происхождения видов».
6. О СУБЪЕКТИВИСТСКИХ ТОЛКОВАНИЯХ ВИДА
Номинализм и проблема непрерывности. Как
уже отмечалось, некоторые ранние эволюционисты отрицали
реальное существование видов. Причиной этого была их пози-
ция, которую можно назвать наивно.-эволюционистским номина-
лизмом. Основоположником данного направления некоторые счи-
тают Ж. Л. Бюффона. В его объемистых книгах можно обнару-
жить немало противоречий и неясностей, но в полемике
с Линнеем им было четко сформулировано утверждение о том,
что в природе нет видов, а существуют только отдельные особи.
В противовес Линнею, Бюффон считал вид лишь продуктом согла-
шений между систематиками, т. е. субъективной категорией. Дру-
гой современник Линнея — Ш. Боннэ также возражал против
понимания вида как реального явления природы. Однако если воз-
ражения Бюффона скорее следует рассматривать как номинализм
наивного эволюциониста, то возражения Боннэ строились на иной
теоретической основе. Если существует преформация всех заро-
дышей, то, как утверждал Боннэ, все живое было сотворено раз
и навсегда как совокупность индивидуальностей (Brewster,
1927 17).
Виды же являются только nomina, т. е. лишь удобными сло-
весными выражениями. В природе существует идеальная не-
прерывность рядов неизменных индивидов, и их подразделения
всегда искусственны. Только наш разум усматривает в непрерыв-
ных рядах индивидуальностей какие-то объединения — виды, роды
и т. п. Всякие классификации, по Боннэ, являются условными
(Radi, 1909; Brewster, 1927; Osborn, 1929). Таким образом, как
эволюционисты Бюффон и Ламарк, так и антиэволюционист
42
1>оннэ, опираясь на принцип непрерывности, высказывались про-
тив реального существования видов.
Сложность связей между общефилософскими позициями ав-
торов и их точкой зрения на реальность вида можно иллюстриро-
вать и на примере Линнея, который также принимал учение
Лойбница о непрерывности рядов индивидуальностей («природа
ио делает скачков»), но в то же время был убежден в реальном
существовании видов. Все эти идеи были изложены им в форме
канонов (Linnaei, 1751).
Номинализм формальных систематиков. Су-
ществовало еще несколько направлений, отрицавших реальность
вида по совершенно иным мотивам, чем наивно-эволюционный
или преформистский номинализм. Источником этой формы номи-
нализма являются трудности гносеологического порядка, которые
испытывают систематики при списании видов.
Техника работы систематика заключалась в том, чтобы, сопо-
ставляя между собой музейные образцы (особи), вычленить неко-
торые признаки в качестве общих, видовых. При этом необходимо
было не только оценить значение таких признаков именно как
видовых, но и установить, какие степени различий между образ-
цами являются достаточными для разграничения видов. В каче-
стве же критериев при выборе признаков и установлении видо-
вых степеней различия, как правило, применялись показатели
вроде опытности, интуиции или вкуса ученого. Эти-то недостатки
музейно-морфологической характеристики видов и породили у не-
которых систематиков номиналистическое убеждение. Единствен-
ной подлинной реальностью для таких систематиков являлись
отдельные индивиды — образцы. Что же касается видов, то каза-
лось, что они «фабрикуются» самим ученым для того, чтобы
удобнее сгруппировать коллекции в шкафах. Надо сказать, что
большое методическое несо>вершенство описания видов приводило
к ошибкам и многие виды именно «фабриковались» в музее.
В природе такие «виды» отсутствовали. Нередко один автор учи-
нял ревизию в материалах, обработанных предшественниками, и
в результате одни музейные виды упразднялись, другие объединя-
лись или дробились, третьи вновь устанавливались. Наблюдая за
техникой такого описания видов, многие систематики впадали
в скептицизм и переставали верить в реальное существование
вида.
Номинализм формальных систематиков иногда переходил
в субъективно-идеалистическое толкование вида. К такому ис-
кажению понятия вида приводили ошибки в толковании сравни-
тельно-морфологической характеристики вида, как особого типа
организации.
Еще в XVIII в. возникла концепция, согласно которой вид —
не реально существующее множество организмов, имеющих оди-
наковую форму, а сама эта форма как таковая. Вид начали
43
трактовать как особый тип организации, как чистую форму без-
относительно к ее носителям. В основе этого представления
лежала правильная мысль о наличии у вида единого типа орга-
низации. Единство вида связано с тем, что все особи вида обла-
дают однотипной наследственной основой. Это генотипическое
единство вида проявляется как в общности морфологических
структур, так и в общности физиологических отправлений.
Однако трактовка вида как особого типа организации скоро
выродилась в идеалистическую, субъективистскую концепцию.
Это было вполне закономерно по следующим причинам. Прежде
всего тезис о виде как типе организации был противопоставлен
пониманию вида как множества особей. При правильном подходе
к вопросу следовало не противопоставлять эти положения как
исключающие друг друга, а рассматривать их как существенные
черты, характеризующие вид с различных сторон. Понятие «мор-
фологический тин» — результат операции извлечения общего
(«видового») из множества отдельных его носителей (индивидов).
Всю совокупность этих индивидов — «носителей общего», непра-
вильно было бы отождествлять с «типом», т. е. с тем, что их
объединяет. Тип обобщает множество лишь в известном отноше-
нии, и само это множество не может быть сведено только к типу.
Поэтому полная характеристика вида должна включать в себя
и указание на специфичность морфологического типа, как одного
из проявлений общности (генотипа), и указания на черты, при-
сущие ему как множеству индивидов.
Далее, понимание морфологического типа как реального осно-
вания, объединяющего мнржество особей, было подменено пони-
манием типа как отвлеченной идеи. Если толкование вида как
типа организации имело под собой фактическую основу, то тол-
кование вида как чистой идеальной формы было уже отходом от
его научной интерпретации. Корни этой ошибки лежали в непра-
вильном истолковании смысла операций, проводимых системати-
ками при сравнптелыю-морфологическом исследовании. Работа
систематика включала абстрагирование общих черт, присущих
всему множеству образцов, т. е. как_бы извлекала из него «идею
типа». При этом многим казалось, что извлекаемая идея типа —
это и есть особая сущность — вид; так возникло субъективно-
идеалистическое положение о том, что вид это и есть идея общ-
ности, а в природе живут только отдельные особи, носители этой
идеи.
Логическим концом субъективистского истолкования вида было
ошибочное понимание происхождения самой идеи типа. Была от-
брошена мысль о том, что идея типа извлекается из материала.
Утверждалось, что эта идея создается (придумывается) система-
тиками в качестве «организующего начала» работы. В этом
субъективистском смысле вид как тип — это идея, создаваемая
самим ученым с целью технически удобной организации мате-
44
риала и создания методически наиболее простых форм классифи-
кации.
Итак, в додарвиновской биологии сложились и боролись между
собой две концепции вида: как реального явления природы и как
отвлеченной идеи, абстракции, чисто условного понятия.
7. О ПРАКТИЧЕСКИХ СТАНДАРТАХ ВИДА
В ДОДАРВИНОВСКИЙ ПЕРИОД
Определение понятий. Необходимо строго разграни-
чить понятия «теория вида» и «практический стандарт вида в си-
стематике». Теорией вида мы будем называть представления о сущ-
ности и всех проявлениях вида, независимо от того, приме-
няются ли они в качестве критериев в практической работе по
классификации. Под практическим же стандартом вида мы бу-
дем понимать краткую инструкцию-характеристику вида как
классификационной единицы, т. е. принятый систематиками
шаблон, по которому устанавливаются границы и объем описы-
ваемого вида.
Такое подразделение понятий необходимо, так как до сих пор
в литературе еще живет устаревшая традиция смешения этих
понятий. Под теорией вида многие подразумевают совокупность
критериев, на основании которых практически производится выде-
ление вида.
В додарвиновской биологии, когда вид изучался исключи-
тельно систематиками, разработка теории вида действительно
преследовала лишь узкие цели создания удобного стандарта. Это
было закономерно и оправдано для того времени. С победой эво-
люционной идеи вид перестал рассматриваться лишь как единица
классификации, познание вида впервые вышло за рамки система-
тики. Была заложена основа общебиологической теории вида.
Однако еще и во второй половине XIX в. эта теория была срав-
нительно слабым ростком, с трудом пробивающим корку рутинной
музейно-гербарной «теории» вида.
Даже в работах, вышедших в последнее тридцатилетие, обще-
биологическая теория вида подчас недостаточно разграничивается
пли даже смешивается с вопросом о стандарте практического вида
в систематике. Иногда теория вида рассматривается с такой
оглядкой на практический стандарт, что это делает ее односторон-
ней. Современное состояние этого вопроса и будет рассмотрено
дальше, а сейчас перейдем к характеристике практических стан-
дартов вида, господствовавших в систематике с еередины XVIII
до второй половины XIX в.
За всю историю систематики было предложено шесть основ-
ных практических стандартов вида. В той или иной мере все они
используются систематиками и в настоящее время.
45
Первые практические стандарты были разработаны на чисто
морфологической основе. Это были монотипический стандарт вида,
обоснованный Линнеем, и морфологический политипический
стандарт, вошедший в практику в связи с трудностями примене-
ния монотипического стандарта. Они господствовали в система-
тике почти 150 лет (с середины XVIII до конца XIX в.). Четыре
других стандарта вида возникли уже в XX в. и будут рассмотрены
в следующих главах.
Морфологический монотипический вид. Моно-
типическими называются стандарты, основанные на признании
вида далее неразложимым, элементарным образованием, включаю-
щим в себя только особей, тождественных по организации. Основ-
ное требование этой концепции — чистота типа. Монотипические
концепции вида могут быть основаны на различных крите-
риях (морфологическом, морфолого-географическом, генетиче-
ском). Усовершенствование методов расчленения материала уси-
ливало различительную способность систематики, и то, что ранее
принималось за далее неразложимое тождество, впоследствии
удавалось разложить на более элементарные отдельности. При
этом приходилось констатировать сборный характер образований,
ранее принимавшихся за строгий монотип (т. е. за далее нераз-
ложимую единицу). Таким образом, сама идея монотипического
вида включала требование расчленять множество при обнаруже-
нии внутри него существенных различий.
Усовершенствование систематики приводило к выявлению все
более мелких различий «внутри тождества». Однако стандарт
строгого монотипа требовал, чтобы при обнаружении различий
они не оставались внутри тождества, а выносились вовне и ста-
новились бы основой нового типа (т. е. вида).
Таким образом, стремление к чисто монотипическому виду не-
избежно приводило (и приводит) к ультрадробительству (вид =
особи или вид = клону до появления в нем мутантных и других
измененных форм). Абсурдность такого стремления к идеаль-
ному монотипу была доказана всем дальнейшим развитием науки.
Отцы систематики (особенно Линней) ясно понимали, что
монотипический вид должен служить не только единицей для
расчленения рода, но он должен и объединять множество особей
и известное число подгрупп (именно в духе различий внутри тож-
дества, по Энгельсу). Линней понимал, что необходима мера для
характеристики степеней различия, которые должны оставаться
внутри тождества (в данном случае вида) или же служить ос-
нованием для расчленения материала и выделения новых типо*в.
Такой мерой Линнею служили понятия «существенное различие»
(differentia essentialis) и «степень различия».
С. В. Юзепчук (1958 : 138—139) отмечает, что подразделение
признаков по «степени важности» производилось Альфонсом Де-
кандолем, М. Вилькоммом и другими ботаниками-политипистами,
46
по не упоминает автора этой идеи — Линнея. Вопрос о «видовой
степени различия» и о подразделении признаков на существенные
(видовые) и несущественные был в одинаковой степени камнем
преткновения для сторонников монотипического и политипиче-
ского стандартов вида. По С. В. Юзепчуку же, это ошибочное
воззрение неправильно приписывается только политипистам.
Понятия о степени важности различия или существенного
различия несомненно позволяли уточнить принцип монотипа.
Вместе с тем эти понятия сами нуждались в определении. Что
следует считать существенным различием? Какая степень раз-
личия, или степень важности, является достаточной для выделе-
ния вида?
В процессе решения этих вопросов возникали все новые труд-
ности. Некоторые считали эти трудности непреодолимыми и
переходили на позиции субъективизма или же отказывались от
монотипической концепции в пользу политипической. Другие
пытались сохранить этот стандарт вида путем усовершенствова-
ния его критериев.
Морфологический монотипический стандарт вида в настоящее
время применяется вынужденно при предварительном описании
видов малоисследованных групп (а также если число образцов
в коллекции очень мало). По этому стандарту требуется, чтобы
экземпляры, не отвечающие типу вида, выносились за его рамки
и служили основанием для образования нового типа или не при-
нимались во внимание. Во многих случаях число таких экземпля-
ров угрожающе возрастало и невозможно было ни отнести их
к другим уже известным типам, ни признать за новый самостоя-
тельный тип. Первоначально понятие разновидности было приме-
нено именно к таким ситуациям, т. е. объединяло формы, остав-
шиеся как бы за бортом типа (т. е. вида). Накопление материала
вело к накоплению подобных трудно классифицируемых разно-
видностей. Они как бы роились вокруг центра, принятого за тип
вида. В результате создавались противоречия, которые требовали
отказа от мысли об универсалу пости вида («все живое органи-
зовано в виды») или же реформ л самого стандарта вида и пере-
хода к политипической концепции. Наконец, логически было воз-
можно также и сохранение монотиг ического стандарта, но при
использовании не только одного м рфологического, но и других
критериев (например, эколого-ret графического, генетического,
физиологического).
Вид по А. Жордану. Рассмотрим еще одну попытку усо-
вершенствования монотипического стандарта. В середине XIX в.
работа по раздроблению вида обычного объема *на более вырав-
ненные единицы была предпринята ботаником антиэволюциони-
стом А. Жорданом (Jordan, 1873). Объектом его исследований
была крупка весенняя (Erophila verna), размножающаяся само-
опылением и отчасти апомиктически. Опытным путем Жордан
47
установил, что потомство многих растений, имеющих какую-либо
морфологическую особенность, сохраняет ее во многих поколе-
ниях. Им было получено около 300 семей, оказавшихся вполне
постоянными при многолетней культуре. Вид Е. verna был, та-
ким образом, разложен Жорданом на множество наследственно
постоянных форм.
Основной вывод Жордана заключался в признании неизмен-
ности настоящих видов, которыми он считал каждую из выделен-
ных им форм. Вид Е. verna в обычном объеме был признан смесью
множества неизменных чистых (идеально монотипических) мел-
ких видов. Вид — это совокупность особей, практически тождест-
венных по внешнему облику и наследственной основе. Если най-
дутся экземпляры, отличные от остальных хотя бы по одному
регистрируемому признаку, который оказывается наследуемым,
то, по Жордану, необходимо разбить эту совокупность на две.
Концепция жордановского вида — идеального монотипа —
логически более последовательна, чем различные компромиссные
монотипические стандарты вида, включая и классический стан-
дарт вида — монотипа Линнея. В самом деле, компромиссные
стандарты основаны па признании допустимости каких-то степе-
ней различий достаточными или недостаточными для разделения
совокупности. Границы же между этими степенями различий
устанавливаются, как правило, произвольно, субъективно. Основ-
ной дефект жордановского стандарта состоит в "том, что при его
применении природные популяции раздробляются на отдельные
элементы, фактически не существующие и не способные сущест-
вовать самостоятельно, т. е. в изолированном от популяции со-
стоянии.
Тенденции к дробительству, свойственные некоторым систе-
матикам XIX в., не были связаны с идеями Жордана, и благо-
склонное отношение к этим идеям получило некоторое развитие
уже в XX в. Вопрос о дальнейшей судьбе взглядов Жордана будет
рассмотрен в главе 4.
Морфологический п о л и т и п и ч е с к и й вид. В конце
XVIII в. исподволь происходил переход к политипическому стан-
дарту морфологического вида, а к середине XIX в. он стал обще-
употребительным (Radi, 1909; Юзепчук, 1958). Этот переход был
обусловлен слабостями монотипической концепции, о которых
было сказано выше. Конечно, если бы большинство видов имели
структуру древнереликтовых видов, таких как гинкго, летимерия,
секвойя, саговники, опоссум и т. п, то потребность в политипиче-
ском стандарте была бы не столь остра. Но с увеличением числа
морфологически изученных образцов, с улучшением техники диаг-
ностики угрожающе возрастало количество трудно классифицируе-
мых форм. Степень морфологической специфичности таких форм
одним авторитетным систематикам казалась достаточной, чтобы
основать новый «существенный тип» (т. е. вид), а другим — столь
48
ничтожной, что не заслуживала внимания таксономиста или же
выделялась за рамки типа в качестве разновидности. Именно
такое состояние диагностики и заставило Дарвина констатиро-
вать, что при решении вопроса, следует ли известную форму при-
знать за вид, «руководящим началом является мнение натура-
листов» (1937 143) и вопрос решается только большинством
голосов.
Политипический стандарт позволял рассматривать вид как
многообразие форм, как группу подтипов, объединенных по
какому-либо признаку (или признакам).
Такие подтипы теперь уже не выбрасывались за борт ради
очищения монотипа от несущественных примесей. Разновидности,
подвиды, формы и прочие морфологические подразделения по-
литипического вида впервые стали признаваться достойными
изучения. Преимущество политипической концепции вида со-
стояло, таким образом, в разумном расширении понятия видового
типа. Теперь «тип» перестал быть подобным математической
точке и начал пониматься как некоторая ограниченная площадь,
в пределах которой может существовать целый рой разбросанных
точек. Элементарными, далее неподдающимися анализу система-
тика теперь стали низшие рангом единицы (разновидности, под-
виды, формы). Масса конкретных форм, пребывавших за бортом
пауки, теперь была включена в рамки вида. Именно политипиче-
ская концепция вида позволила понять вид как многообразие
форм.
Как это показал Дарвин (1937, гл. II), тупик, в который за-
шла систематика в проблеме вида, был связан вовсе не с разви-
тием политипической концепции вида, а с господством учения
о его неизменности. Кроме того, уже в середине XIX в. все силь-
нее сказывались отрицательные стороны односторонне-морфоло-
гической трактовки вида. Именно эти-то причины и затормозили
дальнейшее изучение вида, а отнюдь не господство политипиче-
ского стандарта, как это доказывал G. В. Юзепчук (1958:139).
Дискуссия между сторонниками монотипического и политипи-
ческого стандартов вида обычно принимала форму споров между
«дробителями» и «укрупнителями».
Необходимо отметить, что существование двух стандартов
вида позволяло более точно отразить многообразие видов, чем это
было бы возможно при наличии одного стандарта (безразлично
какого именно). Миллионы видов не вылеплены по какому-то
одному шаблону, который соответствует какому-либо стандарту
практического вида в систематике. Далее будет показано, что
существует много различных неравноценных типов видов. Так,
например, древние реликтовые виды неравноценны молодым энде-
микам; размножающиеся бесполым путем — перекрестнооплодо-
творяющимся; существующие в закрытых горных долинах с резко
колеблющимися экологическими условиями — широко расселен-
4 К. М. Завадский
49
ним на огромных равнинных пространствах. Каждый из таких
типов видов обладает своеобразной структурой, объемом, степенью
резкости границ и другими особенностями. Неправильно защи-
щать необходимость сохранения какого-нибудь одного или двух
стандартов вида в систематике. Систематика может точнее отра-
жать действительность, если стандарты практического вида будут
исходить из идеи неравноценности (неэквивалентности) видов
в природе.
К середине XIX в. два рассмотренных нами стандарта вида
нередко оказывались прокрустовым ложем для природных обра-
зований и вопрос об их таксономическом ранге решался субъек-
тивно. Как отмечал Дарвин, одни самые высококомпетентные
систематики считали данную форму «хорошим» видом, а другие
столь же авторитетные систематики — бесспорной разновид-
ностью более широкого вида. Такие затруднения возникали
отнюдь не только у сторонников монотипического вида, но столь
же часто и у приверженцев политипического вида.
8. ЗАКЛЮЧЕНИЕ
В течение 150-летнего периода (XVIII и первая половина
XIX в.) вид как явление описывался еще в грубых чертах и
весьма поверхностно. Он изучался в состоянии покоя и рассма-
тривался преимущественно с морфологической точки зрения. Его
исследования были целиком сосредоточены в руках системати-
ков и имели весьма ограниченные цели. Задача состояла в пере-
числении лишь внешних особенностей вида.
Было установлено несколько существенных черт, характери-
зующих вид (вид как множество особей; вид как тип организа-
ции; способность к воспроизведению; устойчивость; дискрет-
ность) .
Практически было показано, что вид — образование универ-
сальное для живой природы, что он является ее основной фор-
мой существования и выступает в качестве особой единицы ее
структуры.
Между признаками, характеризующими вид, было констатиро-
вано наличие противоречий (между устойчивостью и изменчиво-
стью, между видом как множеством особей и видом как морфо-
логическим типом). Первые попытки понять сущность этих объ-
ективных противоречий, присущих виду, не увенчались успехом.
Для их устранения была создана ложная концепция постоянства
вида и столь же ошибочное представление о развитии без ви-
дов; в силу тех же причин возникли две концепции вида (вид
как реальное множество особей и вид как абстрактный тип
строения), ошибочно противопоставленные друг другу.
В течение всего 150-летнего периода теория вида развивалась
в недрах систематики и была подчинена ее нуждам. Созданный
50
вначале монотипический стандарт вида вскоре не смог служить
надежным инструментом в работе: возросшее число отклонений
от типа вынуждало ставить под сомнение правильность определе-
ния самого типа. Эти трудности заставили многих систематиков
перейти к политипическому стандарту вида. Оба указанных стан-
дарта вида были чисто морфологическими. Трудности определе-
ния типичной формы (или границ группы подтипов) вызывали
ожесточенные споры «дробителей» с «укрупнителями» и стиму-
лировали субъективистские воззрения систематиков на природу
основной единицы их работы.
В 50-х годах XIX в. познание вида оказалось на распутье; сло-
жились три противоположные и одинаково ошибочные системы
взглядов, которые можно выразить следующими краткими фор-
мулами: «реальный вид без развития», «развитие без видов»,
«реальна особь, а вид — лишь идея, инструмент для классифика-
ции». Концепция реального развивающегося вида существовала
лишь в форме гипотез и натурфилософских догадок. Однако воз-
росший уровень знаний и необходимость разрешить указанные
выше противоречия подвели биологию к необходимости нового
синтеза, сердцевиной которого и явилось обоснование положения
о «реальном развивающемся виде».
Глава 3
ПРОБЛЕМА ВИДА И ВИДООБРАЗОВАНИЯ
ВО ВТОРОЙ ПОЛОВИНЕ XIX В.
1. ДАРВИН О ВИДЕ И ВИДООБРАЗОВАНИИ
Несмотря на то что теория Дарвина общеизвестна и уже более
столетия существуют ее прекрасные изложения, вопрос о толко-
вании Дарвином вида еще подавно бурно и односторонне диску-
тировался в отечественной бполо! лческой литературе. Наша
цель — сжато охарактеризовать взгляды Дарвина, непосредст-
венно связанные с трактовкой вида и с пониманием видообразо-
вания.
Великий переворот в естествознании был вызван тем, что Дар-
вину удалось убедить подавляющее большинство современников
в факте изменяемости видов. На одних при этом большее впечат-
ление произвели доводы в пользу изменяемости видов, связан-
ные с анализом всего мирового опыта селекции растений и
животных. Среди них особенно выделялись доказательства изме-
няемости признаков, признаваемых за видовые. Другие же стано-
вились эволюционистами под напором массы систематизиро-
ванных в «Происхождении видов» разноообразных косвенных
доводов в пользу изменяемости видов.
Однако сущность учения Дарвина состояла не в простом
обобщении опыта практики п не в сборе косвенных доводов,
а в открытии законов изменения видов. Взаимодействие неопре-
деленной изменчивости, борьбы за существование и естествен-
ного отбора — вот перечень тех общеизвестных законов, которые
были положены Дарвином в основу его теории. Однако только
меньшинство биологов приняло этп доказательства изменяемости
видов. Почти три четверти века теория естественного отбора под-
вергалась экспериментальной проверке, критически оценивались
ее следствия, исследовались факторы элиминации и формы от-
бора, одна за другой отвергались иные гипотезы причин эволю-
52
п,и ir, прежде чем было окончательно доказано, что эволюционный
процесс представляет собою селективное преобразование популя-
ций (Шмальгаузен, 1946а, 1961; Завадский, 1957в, 1965; Huxley,
1963, и др.).
Казавшаяся незыблемой теория постоянства видов была
оставлена, и впоследствии только отдельные биологи пытались
оо восстановить на совершенно повой основе, опираясь на оши-
бочно понятые факты устойчивости гена и заменяя понятие вида
в общепринятом (линнеевском) объеме на принципиально иное
(см. об этом следующую главу). Немало критиков Дарвина сето-
вало на неразработанность в его учении вопросов, непосредст-
венно относящихся к определению вида. Займись он прямыми
изысканиями этого вопроса — и единственно возможный путь до-
казательства эволюции оказался бы утерянным. Обходный же
путь, как известно, состоял в анализе причин приспособленности
организмов и объяснении механизмов, осуществляющих смену
приспособлений. Гениальность открытия Дарвина и логики его
доказательств заключалась именно в том, что, разрешая проблему
адаптациогенеза, ему удалось одновременно найти ключ и к при-
чинному анализу направленных морфо-физиологических преоб-
разований, могущих далеко выходить за рамки видовых.
Сосредоточив свои усилия на доказательствах изменяемости
видов, Дарвин, естественно, меньше касался вопроса q виде как
состоянии. Тем не менее отмечая, что в современной ему систе-
матике термин «вид» часто становится произвольным, придуман-
ным ради удобства для обозначения группы особей, близко между
собой схожих и существенно не отличающимся от термина раз-
новидность, Дарвин опирался на собственные многолетние труды
но систематике усоногих раков. В автобиографии Дарвин вспо-
минает, что работа над систематикой этой сильно варьирующей и
«трудно поддающейся классификации группы видов» оказала ему
пользу при работе над «Происхождением видов». Трудности, ко-
торые испытывал систематик-морфолог, пытаясь разместить
в границах видов циклы форм, по одним признакам налегающих
друг на друга, а по другим более или менее разъединенных,
практически испытал и Дарвин. Ни один из практических стан-
дартов вида не мог уменьшить дозу субъективности в окончатель-
ных суждениях систематиков того времени. Ни монотипический,
ни политипический морфологические стандарты, опиравшиеся на
идею неизменности вида, не обеспечивали достаточно точного
отражения действительности. Констатируя это положение в сов-
ременной ему систематике, Дарвин и писал, что «размеры раз-
личий, признаваемые необходимыми для возведения двух форм
ira степень видов, не поддаются определению» (Дарвин,
1937 154). Изучив состояние дел в систематике, Дарвин выяс-
нил, что «единственным руководящим началом является мнение
натуралистов» и «был крайне поражен, как неопределенно и про-
53
извольйо различие йежду видом и разновидностью» (там же : 143—
144). В итоге Дарвин пришел к выводу, что господствующее пони-
мание превращает вид «в бесполезное отвлеченное понятие, обо-
значающее или подразумевающее отдельный творческий акт»
(там же : 146), и что обсуждать вопрос, следует ли те или иные
формы «называть видами или разновидностями, пока не сущест-
вует общепринятого определения этих терминов, значило бы
болтать на ветер» (там же).
Нельзя не удивляться прозорливости Дарвина: и через столе-
тие еще не создано общепринятого определения вида, хотя эво-
люционное учение за это время гигантски прогрессировало
вглубь и вширь. Гениальность Дарвина состояла прежде всего
в удивительно точном выборе общего направления исследований.
Биология переживала такую эпоху, когда была необходима тео-
рия эволюции видов и внедрение исторического метода.
Видообразование. Основные доказательства Дарвина
в пользу изменяемости видов великолепно выражены в следую-
щих словах: «Дарвин в своем составившем эпоху произведении
исходит из самой широкой, покоящейся па случайности, факти-
ческой основы. Именно бесконечные случайные различия инди-
видов внутри отдельных видов, различия, которые могут усили-
ваться до выхода за пределы видового признака... именно они
заставляют его подвергнуть сомнению прежнюю основу всякой за-
кономерности в биологии — понятие вида в его прежней метафи-
зической окостенелости и неизменности» (Энгельс, 1961 535).
В то время как метафизическая теория вида была основана
на разобщении признававшегося случайным (индивидуальные
изменения и разновидности) от необходимого (вид), усилия Дар-
вина были направлены на доказательства неразрывной связи
между ними. Оказалось, что эта связь осуществляется путем по-
стоянного перехода случайного в необходимое, т. е. перехода
единичных наследственных изменений путем естественного от-
бора в приспособления, закрепленные у целой группы. Если дав-
ление естественного отбора длительное время направлено в одну
сторону, это вызывает направленный в ту же сторону процесс
накопления приспособлений. Однако в пределах вида, расселен-
ного по различным местообитаниям, естественный отбор, как
правило, оказывался направленным не в одну, а в разные сто-
роны. Это вызывает адаптивную радиацию (дивергенцию), т. е.
процесс расхождения групп, в котором каждая из них приобре-
тает специфические приспособления и все более изолируется и
отграничивается от других групп. При таком течении событий
материнский вид преобразуется не в один дочерний (ортоселек-
ция), а в несколько, т. е. не только преобразуется, но при этом и
расщепляется. Терминология Дарвина — «расхождение призна-
ков» и «дивергенция» — менее точна, чем понятия, введенные
впоследствии — «адаптивная радиация» (В. О. Ковалевский и
54
Г Осборн) и «расщепление» (splitting), предложенное Д. Симп-
соном (1948) в связи с необходимостью разграничить явление
преобразования вида во времени («филетическая эволюция») и
дифференциации его в пространстве. Однако все это вопросы усо-
вершенствования терминологии, а сущность представления о рас-
щеплении ранее единого русла развития, происходящего в про-
цессе адаптивного расхождения групп, была в общей форме ясно
сформулирована Дарвином.
Дарвин писал, что под влиянием естественного отбора «в те-
чение длинного ряда изменений, слабые различия, характеризую-
щие разновидности того же вида, возрастают в более резкие
различия, свойственные видам одного и того же рода. Новые и
усовершенствованные разновидности будут неуклонно заменять и
истреблять старые, менее совершенные и промежуточные разно-
видности; и, таким образом, виды становятся определенными и
разъединенными между собой объектами» (1937:552).
Важно подчеркнуть, что Дарвин не считал вымирание проме-
жуточных звеньев и образование хиатусов причинами образова-
ния видов: «Вымирание только очертило группы, но никак не
создало их, потому что если бы все прежде жившие на Земле
формы вдруг ожили, было бы совершенно невозможно указать
границы отдельных групп, но естественная классификация или
но крайней мере естественная группировка была бы возможна»
(там же : 516).
Вместе со многими авторами ранее мы неправильно оценивали
позицию Дарвина в вопросе о реальности вида и приписывали
ему релятивистскую ошибку (Завадский, 1949). Основанием для
такой оценки послужило то, что Дарвин исходил из отсутствия
«ясной пограничной линии между видами и подвидами. . . или
между подвидами и резкими разновидностями и индивидуаль-
ными различиями» (Дарвин, 1937:148) и писал, что термин
«вид» считает условным, ничем существенно не отличимым от тер-
мина «разновидность». Позднее нами было показано, что на са-
мом деле в этих высказываниях Дарвином подчеркивалась ре-
лятивность вида, но он не впадал в релятивизм (Завадский,
1954а и др.).
В 6-й главе «Происхождения видов» подробно разъясняется,
почему «виды делаются чем-то обладающим вполне определен-
ными границами и вообще никогда не представляют неразреши-
мого хаоса нечувствительно изменявшихся и сплетающихся
звеньев» (Дарвин, 1937 252). Каковы же, по Дарвину, эти
причины?
Это медленность образования новых разновидностей * и труд-
ность формирования изменений, могущих дать преимущества но-
вым формам перед видами, уже освоившими данную местность.
Поэтому, по Дарвину, в данной местности в данное время всегда
большинство видов устойчиво и лишь небольшое число видов из-
55
меняется. Это также физико-географическая изоляция, ведущая
к разрыву сплошных ареалов, к обособлению групп, к формиро-
ванию «замещающих» видов. Причем, как отмечал Дарвин,
наиболее процветающие разновидности «будут еще более совер-
шенствоваться отбором и приобретать еще новые преимущества»,
в то время как разновидности, живущие в промежуточной по-
лосе, как малочисленные, будут менее долговечны и «более под-
вержены случайному истреблению» (там же: 253). И, наконец,
«промежуточные разновидности, связывающие близкими звеньями
все виды одной группы, когда-то несомненно существовали, но..
самый процесс естественного отбора постоянно клонится к тому,
чтобы. . истреблять родоначальные формы и связующие их
звенья» (там же).
Как видно, Дарвин четко разделял два явления: 1) существо-
вание установившихся, закончивших свое формирование видов,
имеющих «вполне определенные границы», и 2) зарождение и
развитие молодых видов, при котором нередко (но не всегда)
границы изменяющейся группы становятся расплывчатыми.
Неоднократно подчеркивалась приверженность Дарвина к кон-
цепции медленной, постепенной эволюции, идущей равномерным
темпом. Эта гипотеза развивалась тогда в геологии школой
Ч. Ляйэля. Однако Дарвин понимал эволюцию глубже, чем сто-
ронники этой упрощенной схемы. Исходя из теории отбора, он
настаивал на том, что продолжительность времени сама по себе
не способствует и не препятствует эволюции.
Он неоднократно писал о неравномерности темпов видообра-
зования и характеристика его учения как «постепеновщины»
является необъективной. Так, например, по Дарвину, виды вы-
сокоорганизованные изменяются быстрее, чем низшие, сухопут-
ные — быстрее морских, и т. п. Все зависит от интенсивности
и длительности отбора. Неравномерность темпов видообразования
рассматривалась Дарвином и в пределах одной филетической
линии, где, как правило, короткие периоды относительно быстрых
преобразований сменяются длительными периодами, в течение
которых вид «оставался неизменным». Этот тезис Дарвина не
имеет ничего общего с «постепеновщиной». Напротив, эти мысли
Дарвина близки марксистским иДеям о том, что развитие вклю-
чает как медленную эволюцию, так и быстрые скачки, перерывы
непрерывности. Если в некоторых случаях Дарвин и подчерки-
вал постепенный характер эволюции, то это всегда было свя-
зано с полемикой против «катастрофизма» Кювье. Таким обра-
зом, Дарвин не был последователем вульгарной формы эволю-
ционизма, которую ему приписывали некоторые наши критики.
Он считал, что процесс эволюции не непрерывен: «.. . гораздо
более вероятно, что каждая форма в течение долгих периодов
остается неизменной, а затем вновь подвергается изменению»
(там же : 202).
56
Ч, Дарвин,
Вид и разновидность. Одним из устоев концепции нё-
пзмеиного вида было рассмотренное нами в главе 2 убеждение
в случайном и обратимом характере разновидностей. Вот почему
гак настойчиво Дарвин противопоставлял этому воззрению на
разновидность свое — противоположное. Главная идея Дарвина
пыла сформулирована так: «Разновидности — только виды, за-
х каченные в процессе образования, или.. . зачинающиеся виды»
(гам же: 197). Однако, по Дарвину, далеко не каждая разновид-
ность со временем будет видом, так как он понимал, что многие
разновидности будут отметены отбором, другие окажутся недоста-
точно адаптивными для прогресса до видового ранга. Смысл
итого тезиса заключался в признании преемственности между
видами, а также в идее о закономерном зарождении и развитии
новых видов из недр старых.
Будущие виды длительно развиваются в рамках исходных
видов, являясь еще их частями, и лишь в конце этого историче-
ского преобразования «становятся определенными и разъединен-
ными между собою объектами». Согласно Дарвину, степени раз-
личий не могут быть подразделены на резко различные ранги —
разновидности и вида. Напротив, необходимо ожидать сущест-
вования всех теоретически возможных степеней различий — от
самых малых, создающих впечатление непрерывного ряда, до очень
больших и резко обозначенных. Для проверки этого вывода Дар-
вин обратился к сбору данных о характере неоднородности осо-
бой в пределах видов.
Анализ массового и разнообразного материала (от усоногих
раков до высших растений) показал, что все морфологически
различимые группы в пределах вида действительно образуют
ряд, начинающийся с индивидуальных различий. Далее следуют
слабые групповые различия, не дающие основания даже для опи-
сания разновидностей, затем различия степени слабо выражен-
ных разновидностей и т. д., до более резко выраженных разно-
видностей и, наконец, до подвидов. Наряду с этим Дарвин сопо-
ставил степени морфологических разрывов между видами
в пределах многих родов и показал, что никакой определенной
«видовой степени различия» нет; одпи виды отделены от других
большими разрывами (хиатусами), вторые — гораздо меньшими,
а третьи стоят на грани между видами и подвидами, так как
«до настоящего времени не удалось провести ясной погра-
ничной черты между видами и подвидами, т. е. формами, ко-
торые, по мнению некоторых натуралистов, приближаются к ви-
дам, по не вполне достигают этой степени» (там же: 148). Су-
ществует множество форм, делает вывод ДарвиЗц «которые
одни опытные натуралисты считают разновидностями, дру-
гие — географическими расами или подвидами, а третьи — са-
мостоятельными, хотя и близкими между собой видами» (там
же : 551).
57
Таким образом, наблюдаемые факты оказались хорошо согла-
сованными с ожидаемыми на основании теории видообразования
путем отбора. Однако все это касалось лишь морфологической
стороны вопроса, а в определениях вида уже давно в качестве
основного фигурировал и физиологический критерий.
Сопоставляя различия по морфологическим признакам между
породами голубей, кроликов, собак, кур и многих других групп,
Дарвин показал, что иногда они значительно превышают те, ко-
торые считались достаточными для разграничения «хороших»
видов. Отсюда Дарвин сделал заключение об ограниченности
одного только морфологического критерия вида и о необходи-
мости применения также и физиологического. Ведь только сво-
бодная межпородная скрещиваемость голубей или кроликов за-
ставляет систематиков относить морфологически очень различные
породы к одному виду.
Поэтому особенно важно было показать, что существуют все
степени различий и обособленности видов и в физиологическом
отношении. Вот почему Дарвин, Ноден и другие ученые дока-
зывали существование всех переходов от видов, совершенно не-
способных скрещиваться с ближайшими видами, к видам, легко
скрещивающимся и дающим плодовитое потомство, способное во
многих поколениях сохранять свои особенности. Важно было
показать, что существуют виды хотя и скрещивающиеся между
собой, но дающие совершенно бесплодное потомство; виды, гиб-
ридное потомство которых бесплодно при скрещивании между
собой, но плодовито при скрещиваниях с одной или обеими ро-
дительскими формами и, наконец, виды, гибридное потомство
которых способно воспроизводиться в нескольких поколениях, но
затем или гибнет из-за возрастающего бесплодия, или же возвра-
щается к одной из родительских форм. Этим доказывалось, что
в природе существуют виды, находящиеся на любых из теорети-
чески возможных ступенях перехода от состояния «резко очер-
ченной» разновидности к состоянию «хорошего» вида, вполне
отвечающего физиологическому критерию (нескрещиваемости).
Ныне же известно, что у многих групп морских беспозвоноч-
ных животных и у растений не существует барьера нескрещивае-
мости между «хорошими» видами и даже между видами разных
родов.
Такой была аргументация, подорвавшая абсолютное значение
как морфологического, так и физиологического критериев вида.
Различия между видами по степени физиологической и морфоло-
гической дискретности служили важным доводом в пользу по-
степенного характера их эволюции: разные виды находятся на
различных ступенях обособления.
Теорией естественного отбора Дарвин объяснил причины, по
которым образование вида имеет характер процесса приспособле-
ния целых населений к изменяющимся внешним условиям, со-
58
нровождающегося нарастанием дискретности нового вида (уве-
личением размеров хиатусов). В качестве необходимого след-
ствия теории естественного отбора следовало ожидать именно
такую структуру органической природы, которая и существует
и действительности: наряду с резко обособленными видами, имею-
щими уже вполне четкие границы, обнаруживалось множество
«сомнительных» видов, еще не вполне сформированных, с рас-
плывчатыми границами. Естественное объяснение получил тот
факт, что виды не вылеплены по какому-то одному стандарту,
не равны друг другу по степени дискретности, объему и другим
признакам.
Так был обоснован вывод Дарвина о том, что ясно выражен-
ная разновидность может быть названа зарождающимся видом,
а вид — более резко выраженной и определившейся разновид-
ностью. Эта формулировка выражала основную идею эволюцион-
ной теории Дарвина: виды возникают исторически из зачаточных
состояний и существуют временно, так как рано или поздно вы-
мирают или преобразуются в новые виды. При этом следует еще
раз подчеркнуть, что, по Дарвину, не все зачаточные виды до-
стигают степени видов; многие из них вымирают или же неопре-
деленно долго сохраняются в качестве подразделений исходного
вида. Среди групп, обладающих реальной возможностью в буду-
щем стать новым видом, лишь ничтожное меньшинство стано-
вится таковым в действительности. С современной точки зрения
эти формулировки Дарвина должны быть уточнены, так как
понятие разновидности очень неопределенно и включает разно-
родные формы внутривидовой дифференциации, часто искусст-
венно объединяя материал, весьма разнородный по генезису и
значению для видообразования (см. главу 8).
Дифференциация вида, как доказал Дарвин, является зако-
номерным следствием эволюции, результатом приспособления
к различным условиям среды и в то же время состоянием,
обусловливающим возможность дальнейшей эволюции впда. След-
ствием этого явилось постепенное отмирание господствовавшего
ранее взгляда на неоднородность особей в пределах вида как на
результат случайных отклонений от типичной формы вида
(forma genuina), хотя в практике формальных систематиков,
особенно в палеонтологии, и до сих пор varietes выносится иногда
за рамки «типа» вида.
Дарвину принадлежит первая научная классификация форм
неоднородности особей в пределах вида. Он установил существо-
вание индивидуальных и групповых различий, обратимых и не-
обратимых, имеющих селективное значение и нейтральных, зави-
сящих от пола, возраста, сезона, Лазы (в случае смены поколе-
ний) , местообитания, скрещиваний и не зависящих от этих
факторов, воспроизводящихся видом в ряду поколений в одном и
том же числовом соотношении и непрерывно изменяющихся
59
с течением времени. Наибольший интерес у Дарвина вызывали
микропроцессы в недрах вида, постепенно изменяющие всю
его внутреннюю структуру, т. е. обусловливающие эволюцию
вида. В каждый данный момент эти микропроцессы проявляются
в массе внутривидовых различий.
Вместе с тем Дарвин уделял некоторое внимание и внутри-
видовым различиям иного типа — различиям, обусловливающим
не изменение вида, а, напротив, поддержание status quo. В ка-
честве примеров таких различий, воспроизводящихся со строгим
постоянством, Дарвин приводил гетероморфность растений по
листьям, цветкам, семенам, плодам, ритмам развития и т. н. За-
крепление различий этого типа основано, по Дарвину, на адап-
тивном значении самой разновидности. В ряде случаев и в на-
стоящее время трудно отличать типы дифференциации вида,
представляющие первые шаги адаптивной радиации, от разно-
родности, закрепленной отбором в качестве особого адаптивного
аппарата. Кроме того, столетие назад ничего еще не было из-
вестно о механизмах, поддерживающих эти удивительные
внутривидовые формы.
Обсудим теперь известную мысль Дарвина, вызвавшую
столько споров в наше время. Дарвин писал: «Термин „вид“
я считаю совершенно произвольным, придуманным ради удобства,
для обозначения группы особей, близко между собой схожих, и
существенно не отличающимся от термина „разновидность41, обо-
значающего формы, менее резко различающиеся и колеблющиеся
в своих признаках» (там же: 149). На первый взгляд Дарвин
здесь присоединяется к субъективистскому толкованию вида.
Однако в этой формулировке только констатируются практически
непреодолимые трудности, возникающие при разграничении вида
от разновидности, так как никто еще не предложил какой-то оп-
ределенной степени различия, точно отграничивающей их друг от
друга. Такое состояние систематики имеет объективную под-
основу: виды стоят на разных ступенях своего формирования и
обособления и, расположенные в ряд, образуют цепь, первые
звенья которой состоят из переходных состояний, т. е. таких мо-
лодых видов, которые можно еще отнести и к предшествующему
состоянию (к разновидности).
Конечно, досадно, что в основном труде Дарвина сохранилась
неточная формулировка, давшая повод многим нападкам на его
позицию в вопросе реальности вида. Однако в письмах Дарвина
есть фраза, предельно точно выразившая его взгляды: «Как
будто кто-нибудь сомневается в их временном существовании»
(Дарвин, 1950: 134). Эта мысль и отражает подлинные воззрения
Дарвина на реальность вида. Сформировавшийся вид рассматри-
вался Дарвином как качественно определенное образование, спо-
собное самовоспроизводиться до тех пор, пока отбор не произве-
дет тех или иных преобразований.
60
Естественно, что главное внимание Дарвин уделял факторам,
преобразующим вид. Ибо мог ли он думать, что сразу же вслед
за победой эволюционизма многие биологи столь прямолинейно
уверуют в непрестанную изменяемость вида, что перестанут
воспринимать факты высокой устойчивости видов. Тем не менее
Дарвин все же разобрал ряд причин, вызывающих стабилизацию
вида (сохранение прежних условий существования и отсутствие
(•ильных конкурентов). Он указывал на возможность длительного
сохранения видов, т. е. на возможность, по терминологии
Т Гексли, персистирования видов. Дарвин предполагал суще-
ствование собственно ортоселекции, т. е. преобразования вида
в одном направлении, и отбор на персистирование, т. е. на сохра-
нение ранее сложившейся адаптивной организации. Этот важный
тип действия отбора подвергся анализу только в наше время.
Исторически сложившуюся организацию особи, полиморфизма
популяции и внутривидовых отношений мы будем Далее на-
зывать «адаптивная норма вида». Если в течение предшествую-
щей истории сложившаяся адаптивная норма вида, будучи испы-
танной в различных условиях, обеспечивала высокие коэффи-
циенты выживаемости, то и в будущем отбор может иногда брать
под охрану именно эту адаптивную норму вида, элиминируя те
фенотипы, у которых недостаточно полно обеспечено доминиро-
вание адаптивной нормы над мутантами.
В те времена казалось, что с крушением теории постоянства
вида сразу же откроются просторы для изучения его изменяе-
мости. На самом же деле случилось иное. Дело в том, что пред-
ставления о внутренней структуре вида, высказанные Дарвином
и другими биологами той эпохи, были еще неопределенны и при-
митивны; сложнейшая внутренняя структура вида оставалась
неизученной, что в свою очередь делало невозможным анализ
процессов видообразования. Для конкретного изучения видообра-
зования потребовалось гораздо более глубокое знание статики
вида. Эти знания в сущности накапливаются только в самое по-
следнее время. Вместе с тем следует подчеркнуть, что именно
теория естественного отбора и явилась той отправной точкой,
без которой было бы невозможно изучение статики вида. Замет-
ное продвижение вперед в этой области знаний произошло уже
в XX в., и это стало возможным только благодаря историческому
подходу к виду, впервые обоснованному Дарвином, ина основе его
учения об отборе. Итак, вид оказался явлением развивающимся.
В противовес старой концепции абсолютно устойчивого и абсолютно
обособленного, неизменного морфологического вида была выдвинута
новая эволюционная концепция вида. Это новое понимание вида
опиралось на принципы изменяемости (историчности), экологиче-
ской определенности (т. е. приспособленность к условиям среды
и наличие особого места в экономике природы) и географической
определенности (расселение на особой территории).
61
Вид стал пониматься как временно существующее в природе
образование. Его становление, расцвет, дальнейшая дивергенция
или вымирание оказались этапами единого постепенно идущего
процесса приспособительной эволюции.
На основе теории естественного отбора эволюционная идея
впервые объединилась с учением о реальности вида. Дилемма,
перед которой в течение столетия стояла биология, — либо «ре-
альный неизменный вид», либо «развитие без видов» — была,
наконец, разрешена. Дарвин осуществил высший синтез знаний.
Началась новая эпоха изучения «реального развивающегося
вида». Однако эта концепция вида и видообразования завоевала
господствующее положение после длительной проверки — лишь
в нынешнем столетии. Почти три четверти века понадобилось
для того, чтобы выработать новые методики, применить их для
детальной проверки правильности взглядов Дарвина на вид и
видообразование и подтвердить их, а также установить ошибоч-
ность других представлений (Завадский, 1965).
2. ОБЩАЯ ХАРАКТЕРИСТИКА СОСТОЯНИЯ ПРОБЛЕМЫ ВИДА
Во второй половине XIX в. интересы биологов обратились
к рассмотрению общих законов эволюционной динамики форм,
к процессам, снимающим их стабильное состояние. На базе эво-
люционного учения сложилась новая общебиологическая теория
вида; проблема вида впервые вышла за рамки интересов систе-
матики. Следует, однако, подчеркнуть, что статика вида, как та-
ковая, в этот период еще не изучалась. Вследствие этого создался
определенный разрыв между трактовкой вида в эволюционной
теории и пониманием его в практической работе систематиков.
Вид в систематике по-прежнему оставался формальным, чисто
морфологическим построением.
После победы эволюционной идеи развитие биологии приняло
несколько неожиданный характер. Казалось бы, что интересы
биологов должны были сосредоточиться вокруг проблемы вида.
Однако этого не произошло. В 60—90-х годах XIX в. интересы
биологов переместились от проблемы вида к проблемам, не имею-
щим с нею непосредственной связи. Во-первых, к\общим вопро-
сам эволюции, к изучению «большой филогении». Эти работы
осуществлялись с широким размахом, порой носившим натур-
философский характер, что было особенно характерно для
Э. Геккеля. Об этом периоде развития биологии хорошо написал
В. Гентнер: «В широких очерках и схемах развития происхожде-
ния типов, классов и отрядов, развития органов и систем органов,
построении родословных древес, в поисках групповых „предков14
и „предковых44 групп и т. п. был в сущности потерян вид — пер-
вый и конкретный объект и „носитель44 эволюции» (цит. по Майр,
1947:9). Одновременно в последарвиновское время начался
62
расцвет микроскопической техники и соответствующих отраслей
знания, изучающих организменный, суборганизменный и клеточ-
ный уровни. Таким образом, в биологии образовалось как бы два
полюса, к которым притягивались основные интересы современ-
ников, а проблема вида временно оказалась отодвинутой на вто-
рой план.
Была и другая причина, из-за которой проблема вида в ту
эпоху оказалась почти забытой: утверждение эволюционной кон-
цепции требовало накопления данных о переходных состояниях,
при которых между видами отсутствуют четкие разграничитель-
ные линии. Если виды возникают постепенно, шаг за шагом
обособляясь в процессе нарастающей дивергенции как от пред-
кового (родительского) вида, так и друг от друга, то должны су-
ществовать все градации переходов. Именно с помощью таких
данных и опровергались старые формулировки о полной устой-
чивости и дискретности видов. Вот почему поиски постепенных
переходов превратились в основную задачу. Они отыскивались
палеонтологами среди вымерших видов, систематиками, сравни-
вавшими ныне живущие виды, а также сравнительными эмбрио-
логами.
Уже в ноябре 1861 г. Р. Оуэн обнаружил в юрских отложе-
ниях Золенгофена знаменитую ископаемую птицу Archaeopteryx.
Этим было положено начало целой серии находок переходных
форм, закончившейся в конце века обнаружением Дюбуа яван-
ского обезьяночеловека.
В 1866 г. Ф. Гильгендорф опубликовал работу по миоценовым
Planorbis multiformis (названная Ф. Готшиком в 1920 г.
Gyraulus brachiformis) из неподвергавшихся смещениям слоев
озерных отложений в Вюртемберге. Гильгендорф был первым
палеонтологом, показавшим, как постепенными переходами во
времени образуется филогенетический ряд форм раковин. Сам
термин «ряд форм» (Formenreiche) был введен в 1867 г. Бейри-
хом. В 1875—1888 гг. М. Неймайр опубликовал свои известные
работы по рядам форм раковин паллюдин (Vivipara) из пресно-
водных отложений Словении. Эти работы известны защитой дар-
виновских положений о постепенном характере видообразования
и причин этого процесса. В 60—80-х годах вышли многие работы
по рядам форм у аммонитов (В. Вааген, Л. Вюртенбергер и др.).
Любопытно, что, наблюдая прерывистые ряды, авторы иногда
толковали о внезапном образовании видов (Вааген), а, наблюдая
постепенные переходы, некоторые писали об исчезновении видов,
теряющих свои очертания среди переходных форм.
В 1873—1877 гг. вышли хорошо известные у ндр исследования
В. О. Ковалевского об ископаемых копытных, заложившие основы
дарвинистской экологической палеонтологии. Серия работ
А. О. Ковалевского в области эволюционной эмбриологии, нача-
тых с 70-х годов, в своей основе также имела идею единства
63
организации и поиска переходных форм (развитие ланцетника,
асцидий, открытие Coeloplana и др.).
Важный новый элемент в изучение вида был внесен исследо-
ванием географического распространения форм. В XIX в. уже
многие занимались ареалами видов, но следует выделить А. Гум-
больдта, Огюстена Пирама и Альфонса Декандолей, Ф. Умера,
Е. Форбса, Д. Гукера, А. Уоллеса, М. Вагнера, работами которых
была подготовлена почва для реформы систематики на основе
морфолого-географического метода. Накопление данных по иско-
паемым остаткам позволило начать палеогеографические иссле-
дования изменений ареалов (Engler, 1879; Andersson, 1897,
и др.). Мы не будем увеличивать число этих широко известных
примеров. Надо только подчеркнуть, что подобные работы, часть
из которых была выполнена с большим блеском, отвлекала мно-
гих биологов от изучения статики вида. Увлечение эволюцион-
ной динамикой форм, построением деревьев и рядов форм — это
было знамение эпохи. Поиски переходных состояний, собира-
тельных типов, накопление данных в пользу того, что каждое
состояние может быть истолковано как переход от предыдущего
состояния к будущему, привели к развитию вульгарно-эволюци-
онных идей, к отрицанию реальности вида и к возврату на пози-
ции Ламарка («развитие без видов»). Неоламаркисты охотно
подчеркивали искусственность вида, рассматривая его как произ-
вольно выхваченный кусок непрерывной линии филетического
развития.
Понимание вида в систематике. Период упадка
систематики — так характеризуют конец XIX в. ученые, принад-
лежащие к самым различным течениям (Romanes, 1895; Кома-
ров, 1908, 1912, 1927; Radi, 1909; Юзепчук, 1958, и др.). Как
отметил Р. Э. Регель, систематика, бывшая до этого «строго науч-
ной дисциплиной», начиная с 80-х годов под влиянием неоламар-
кизма «перестала быть таковой так как описывала в качестве
видов или разновидностей все, что представлялось глазу наблюда-
теля, не считаясь с тем, наблюдаются ли наследственные или
ненаследственные признаки» (1917 : 176). В качестве ^хорошей
иллюстрации упадка систематики» Р. Э. Регель приводит сводку
«Synopsis der Mitteleuropeischen Flora» (Ascherson u. Grebner,
1896). Находившиеся под влиянием неоламаркистского толко-
вания формообразования П. Ашерсон и П. Гребнер видели за-
дачу систематики в том, чтобы давать латинское название каждой
морфологически различимой форме. При этом большинство си-
стематиков придерживалось морфологического политипического
стандарта вида, используя многоэтажную систему латинских слов
для обозначения конкретной формы (острую критику такой
«ашерсоновщины» осуществил С. В. Юзепчук (1958).
Нельзя сказать, что в это время большинство систематиков
было сторонниками сборных видов с многоэтаж»м подчинением
64 Ш
разновидностей и рас. Так, например, Э. Ёуассье выделял боль-
шинство морфологически различных форм в виды так, что число
политипических видов у него очень мало (Boissier, 1867—1888).
Но и этот автор нередко возводил в ранг вида ненаследственные
формы, как и сторонники «ашерсоновского» политипического
вида. Упадок систематики на фоне победы эволюционных идей
был закономерным следствием отставания методов работы этой
науки от поставленных проблем.
Для многих систематиков того времени не было еще ясно, что
вид — совершенно особая таксономическая единица. Неправиль-
ное понимание вида лишь как одного из многих таксонов было
характерно, например, для Альфонса Декандоля, мечтавшего
разложить виды на такие же элементы, какими являются виды
внутри рода, роды внутри семейства и т. д. (De-Candolle,
1880:80). Значительная часть систематиков пришла к отрицанию
реальности вида и к утверждению концепции вида как удобной
абстракции.
При вульгарном понимании процесса развития здоровый мо-
мент релятивности, внесенный Дарвином в концепцию вида,
легко переходил в грубый релятивизм, в отрицание его реаль-
ности. Например, палеонтолог Л. Вюртембергер (Wurtemberger,
1880) писал, что раз нельзя указать, где вид начинается и где он
прекращает существование, то понятие о йиде теряет всякую
естественнонаучную почву; палеонтолог С. Никитин (1881) срав-
нивал вид с произвольно вырванным участком непрерывной
ленты, и т. п. Приняв эволюционную идею, многие биологи по-
няли положение о том, что вид — явление текучее, неустанно
меняющееся, слишком буквально, абсолютизировали «текучесть»
вида и упустили из поля зрения его качественную определен-
ность, относительную, но объективную устойчивость.
На почве вульгарно-эволюционного понимания развития воз-
никла новая форма субъективизма в трактовке вида. В резуль-
тате победа эволюционной идеи, как подчеркнул В. Л. Комаров
(1901а: 250), «нанесла сильный удар понятию о виде» и привела
многих систематиков к признанию реального существования
лишь за особью. Многие эволюционисты еще не могли отличить
явление относительности граней, имеющее объективный харак-
тер, от понятия о «субъективности граней», т. е. от представле-
ния, что филогенез — это только непрерывная лента изменений.
Вновь, как и столетие назад, встала проблема соотношения из-
меняемости и устойчивости вида. Однако на этот раз многие
ученые (особенно неоламаркисты) впали в другую крайность,
переоценив степень лабильности вида.
Игнорирование качественной определенности вида было ха-
рактерно не для всех систематиков-эволюционистов 70—90-х го-
дов XIX в. Так, например, механоламаркист Теодор Эймер
(Eimer, 1888—1901), хотя и считал филогенез простым «филе-
5 К. М. Завадский
65
тическим ростом», все же подчеркивал объективную реальность
вида, толкуя вид как остановку роста (генэпистаз). Состояние
устойчивости вида, т. е. генэпистаз может длиться геологическое
время, сменяясь периодами лабильных состояний, в течение ко-
торых собственно вида нет, а есть какое-то «переходное
состояние».
Некоторые систематики дарвинистского направления рас-
сматривали вид как реальное качественно определенное явле-
ние, существующее в течение известного времени. Так именно j
трактовал вид Н. А. Северцов в 1873 г. при описании горных j
баранов (архаров). Таковы характеристики вида и некоторых)
способов видообразования в работах М. Н. Богданова 70— ’
80-х годов, посвященных основам классификации млекопитаю-
щих, систематике сорокопутов русской фауны и т. п.
Все же в целом вопрос о реальности вида в те времена был
изрядно запутан. К тем причинам субъективизма, которые
кроются в технике работы систематиков и связаны с ошибками
в понимании связи между единичным и общим, а также с вуль-
гарно-эволюционным пониманием развития (абсолютизация не-
прерывности) прибавились новые. Приведем некоторые примеры.
В 1867 г. С. А. Усов (1888) выступил с работой, провозглашав-
шей, что истину о виде можно постигнуть только в свете фило-
софии Канта. Подходя к вопросу с кантиантских позиций,
С. А. Усов утверждал, что вид — это ноумен, а особи — реальные
феномены. По Усову, вид — «вовсе не есть сумма, собрание осо-
бей», а некий «абстракт.. . идеальный образ», отвлеченная идея.
К такой идее типа как бы приписываются особи, по мнению
систематика, наиболее ему отвечающие. Вид как абстрактная
идея некоей непознаваемой сущности (ноумен) может сущест-
вовать совсем без особей, проявляться в одной особи или в мил-
лионе особей. В идеалистических построениях Усова отразились
трудности понимания реальности вида систематиками, о которых
уже говорилось в главе 2. Усилению субъективизма способство-
вали также и ошибочные утверждения о том, что «все таксономи-
ческие единицы от подвида и вида... образованы путе^ вымира-
ния.. связующих звеньев» (Бианки, 1907:558), хотя еще
Дарвин показал, что вымирание переходных форм лишь более
резко очеркивает границы возникшего вида, но совсем не со-
здает его.
Таким образом, во второй половине XIX в. возник разрыв
между общебиологпческой эволюционной концепцией вида, раз-
виваемой в теоретических работах, и пониманием вида в прак-
тической систематике. В теоретических работах по эволюцион-
ному учению уже учитывалось, что явление вида многосторонне
и сложно, что природные виды стоят на различных стадиях раз-
вития (зарождающиеся, процветающие, распадающиеся на до-
черние и угасающие) и т. п. В то же время концепция вида
66
в систематике не изменялась в соответствии с требованиями эво-
люционной теории.
Это не значит, что систематики не были эволюционистами;
но, являясь эволюционистами в теории, они тем не менее продол-
жали выделять виды старыми приемами. Вид в систематике
фактически оставался отвлеченным типом, построенным на осно-
вании сравнения морфологических признаков и небольшого числа
образцов, хранившихся в музее (гербарии). Таким образом,
создалась своеобразная ситуация: виды описывались на основании
устарелых приемов, а затем уже проводилось обсуждение фило-
генетического родства и путей эволюции этих чисто «музейных»,
нередко мифических единиц.
Разрыв между общебиологической теорией вида, созданной на
основе эволюционного учения Дарвина, и практическим стандар-
том вида (т. е. формальной единицы музейной работы морфоло-
гов-систематиков) в 60—90-е годы XIX в. пе только сохранялся,
по все более нарастал.
3. ВОПРОСЫ ВИДООБРАЗОВАНИЯ
Поглощающая роль скрещиваний. Одним из су-
щественных, а по мнению многих даже непреоборимых затруд-
нений при объяснении видообразования на основе естественного
отбора явилась возможность нивелирующего действия скрещива-
ний, вследствие которого происходит не накопление адаптивных
различий, а их сглаживание (Уоллес, 1911 157).
Действительно, если в результате скрещивания немногих осо-
бей более приспособительного типа с окружающими их в массе
особями исходного типа получалось потомство, у которого из
поколения в поколение утрачивались бы приспособительные
преимущества, то отбор был бы неэффективен. Его накапливаю-
щая роль ставилась под сомнение. Сам Дарвин понимал серьез-
ность этого затруднения.
Важнейшим открытием, подкрепившим теорию видообразова-
ния на основе отбора, были законы, установленные Г. Менделем.
Однако и до сих пор немало эволюционистов сомневается в воз-
можности или в широком распространении видообразования пу-
тем отбора и дивергенции групп, происходящего в недрах насе-
ления, первоначально образующего единый скрещивающийся
комплекс. Во второй же половине XIX в. многие искали выхода
из заколдованного круга, каким казалась коллизия отбора и по-
глощающего действия скрещиваний. Одни считали, что пережив-
шие формы должны уединяться, мигрируя в другие местности.
В миграционной теории видообразования (Wagner, 1868) прида-
валось исключительное значение изоляции па основе миграций и
была сделана попытка трактовать именно ее, а не отбор в ка-
честве причины видообразования. Другие видели выход в при-
5*
6?
знании одновременных массовых направленных изменений орга-
низмов, при которых исходный тип исчезает фронтально (сто-
ронники прямого приспособления). Дж. Ромене предлагал выход
в гипотезе «физиологической изоляции». Он считал, что учение
Дарвина является не столько теорией видообразования, сколько
гипотезой эволюции приспособлений, что расчленение видов не
связано с процессами адаптации, так как идет по неприспосо-
бительным признакам, которые не могут затрагиваться отбором
(Romanes, 1895).
В XX в. эта проблема вновь была поставлена на генетической
основе в связи с вопросами частоты мутирования, эволюции до-
минантности, относительного значения различных типов изоля-
ции и, наконец, возможности симпатрического видообразования.
Гипотезы мутационного образования видов.
Различные гипотезы внезапного порождения видов возникли еще
в 60-х годах XIX в. в качестве реакции на дарвиновскую теорию
и удерживаются в биологии до сих пор. Датой их рождения
можно считать 1864 г., когда была опубликована работа немецкого
гистолога Кёлликера «Гетерогенное размножение». В этой работе
отрицалось видообразование на основе естественного отбора, мо-
нофилия и дивергенция. Пользуясь аналогиями с метаморфозами
в цикле индивидуального развития медуз и других беспозвоноч-
ных животных, Кёлликер утверждал, что особи одних видов вне-
запно порождают особей других, даже далеких, видов. Видообра-
зование идет путем «гетерогенного размножения» и имеет поли-
филитический характер: «Возможно появление одного и того же
вида из различных филогенетических корней.. . так что перво-
начально различное может стремиться к одинаковой конечной
фазе» (КбШкег, 1872:21). Он считал причины видообразования
внутренними, заложенными в живое изначально (там же: 44
и др.): это — «законы, которые побуждают организмы к эволю-
ции в совершенно определенном направлении, т. е. в сторону
постоянного совершенствования» (там же 38 и др.). Ст. Майварт
также был сторонником внезапного видообразования путем «ино-
рождения» без всяких переходных форм. Разбирая аргументы
Майварта, Дарвин отрицал всеобщность и даже распространен-
ность такого способа видообразования и не допускал такой сте-
пени внезапности (одно поколение), о которой писали мутацио-
писты. Однако, по Дарвину, «можно допустить, что. .переход
мог совершаться иным, более внезапным способом, но такое пред-
положение не может быть принято без представления явной оче-
видности» (1937 : 323).
Дарвин считал внезапное видообразование маловероятным со-
бытием и приводил следующие доводы в пользу такого мнения.
У культурных форм резкие изменения возникают единично, редко
и, как правило, такие особи мало жизнеспособны. Можно
было бы допустить одновременное появление в одном местооби-
68
танин многих особей с одинаковыми изменениями, но, подчерки-
вал Дарвин, опыт практики говорит о невероятности такого
события. О постепенном характере видообразования говорит тот
факт, что виды крупных родов более сходны между собой и
имеют больше разновидностей, чем виды малых родов. Виды
крупных родов, писал Дарвин, «собраны также в небольшие
группы, подобно тому как разновидности группируются вокруг
типичных видовых форм» (там же). Дарвин отмечал невероят-
ность того, что внезапные порождения видов не нашли отраже-
ния в эмбриональной развитии и что все новообразования воз-
никли внезапно, не нарушив при этом всей слаженности целост-
ного организма и его адаптации к среде. В итоге Дарвин делает
вывод, что «допустить все это... значило бы перейти в область
чудес, покинув область науки» (там же 325).
Однако в XIX столетии не один раз делались попытки под-
твердить эту гипотезу. Так, на палеонтологическом материале
В. Вааген в 1869 г. доказывал внезапность видообразования
у аммонитов из группы subradiatus. В построенных им рядах
форм, найденных в смежных отложениях, наблюдались большие
морфологические хиатусы. Они-то и были истолкованы как след-
ствия образования нового вида одной мутацией (названной
«мутацией Ваагепа»). Мутации Ваагена в тысячи раз превышают
генетические сдвиги, которые ныне именуются малыми мута-
циями. Вааген предложил применять понятие «разновидность»
к пространственным (географическим) вариациям вида, а поня-
тие «мутация» — к изменениям во времени. При этом Вааген сде-
лал произвольный вывод, что новые виды возникают только по-
средством мутаций, а не через разновидности (т. е. без участия
таких географических факторов, как расселение, переработка
популяций в пределах больших территорий и пр.).
Впоследствии палеонтологи показали, что скачки типа мута-
ций Ваагена — это отнюдь не следствия гетерогенеза, а, как пра-
вило, следствия неполноты геологической летописи или измене-
ний экологической обстановки, вызвавшей вымирание или мигра-
цию одних форм и распространение других.
В конце ХТХ в. геолог Э. Зюсс, основываясь на данных
палеозоологии, выдвинул гипотезу трехфазности развития всех
филогенетических стволов. Первая фаза — творчество форм пу-
тем внезапной массовой «перечеканки» типов, т. е. путем тех же
больших мутаций. Вторая фаза — адаптация к среде этих ново-
явленных форм, и третья фаза — вымирание. Идеи Зюсса
сыграли заметную роль в возрождении идей катастрофизма
Кювье на новой основе. И ныне подобные идеи защищаются не-
которыми авторами (Степанов, 19596).
Не останавливаясь на других примерах, упомянем лишь из-
вестную попытку С. И. Коржинского в самом конце века за-
щитить гипотезу внезапного порождения видов. По Коржин-
69
скому, «раз сформировавшись, виды остаются неизменными, но
временами отщепляют от себя новые формы путем инорождения
(гетерогенезиса)» (1899 : 863).
С. И. Коржинский собрал много примеров гетерогенных ва-
риаций у культурных растений. По его мнению, все достоверно
установленные случаи возникновения новых форм (сортов) отно-
сятся не к постепенным переходам, а к однократным скачкам.
При этом Коржинский не упоминает о громадном материале, до-
казывающем, что мелкие и мельчайшие изменения встречаются
у культурных форм гораздо чаще, чем прикопавшие его внимание
резкие изменения. Хотя Дарвин свою книгу о домашних живот-
ных и культурных растениях написал за 30 лет до Коржинского,.
она была -свободна от одностороннего выпячивания только резких
(больших) или же только мелких изменений и до сих пор
является образцом всестороннего подхода к вопросу. Коржинский
излагал также давно известные примеры внезапного формообра-
зования: остролистной формы чистотела Ghelidonium majus воз-
никшей в 1590 г. в саду аптекаря Шпрепгера и получившей
бинарное название Ch. laciniatum, формы monophylla обычной
лесной землянки (Fragaria vesca), которую в 1761 г. Дюшен обо-
значил бинарным названием, и т. п. Кроме того, им были собраны
примеры усиления или ослабления разрезности, рассеченности
или зубчатости листовых пластинок у вяза, клена, ольхи, дубаг
липы и др., изменения размеров растений, частей цветка и т. и.
Таким образом, в пользу гипотезы внезапного порождения
видов в XIX в. приводились аналогии с метаморфозом в онтоге-
нетическом развитии, палеонтологические данные об отсутствии
переходных форм, а также флористические наблюдения и данные?
селекционной практики.
Еще Дарвину было хорошо известны примеры крупных мута-
ций по многим признакам, использованных для создания пород:
черноплечий павлин, бульдожий скот ньята, анконская овца, мел-
кие кролики с о. Порто-Санто и пр.
Однако, как бы велико ни было по своим проявлениям одно-
кратное наследственное изменение, обладатель его, по мнению
Дарвина, не может тотчас же стать новым видом. Во-первых, чем
сильнее изменение, тем меньше вероятность повышения жизне-
способности, слаженности отправлений и приспособленности
к среде. Во-вторых, даже если новая форма и оказалась приспо-
собленной к среде и способной размножаться «внутри себя», то,
чтобы стать разновидностью, она должна еще пройти испытание
на конкурентоспособность. Лишь потеснив исходную форму и
закрепившись в пределах определенной территории, новая форма
может перейти к сосуществованию с исходной или же, рас-
селяясь, вытесняет ее. Процесс же расселения новой формы, ис-
пытания ее на конкурентоспособность — это процесс истори-
ческий.
70
Иначе говоря, по Дарвину, процессы наследственной измен-
чивости и видообразования связаны между собой. Гипотеза му-
тационного видообразования, в основе которой лежит идея
о существовании специфической «видообразовательной изменчи-
вости», оказалась не только аптидарвиновской, но и не соответ-
ствующей действительности.
4. ЗАКЛЮЧЕНИЕ
Виды изменяются! Это положение, бывшее туманной гипоте-
зой еще в середине 50-х годов XIX в., через десять лет, т. е. в се-
редйне 60-х годов, стало неоспоримым фактом. Необходимость
процесса развития видов была выведена Дарвином из открытого
им закона естественного отбора. Однако во второй половине
XIX в. немало биологов отрицало видообразовательное значение
естественного отбора. В качестве движущих сил видообразования
назывались различные причины, подробно разбираемые в книгах
по истории эволюционных идей и по теории эволюции. К таким
гипотезам в первую очередь следует отнести различные варианты
мутационизма и неокатастрофизма.
Привлекает внимание парадоксальное замедление темпов изу-
чения вида и некоторый упадок систематики. Казалось бы, что
после быстрого внедрения в биологию эволюционной идеи даль-
нейшее познание вида должно было получить небывалый про-
стор. Тем не менее из-за отсутствия методик исследования струк-
туры вида и скудости знаний о виде как состоянии произошло
замедление в изучении вида и видообразования.
Таким образом, противоречия, возникшие при изучении ви-
дов в состоянии покоя, обусловили крушение теории постоянства
видов и необходимость изучения вида в развитии. С внедрением
исторического метода отпал целый ряд затруднений и учение
о виде получило простор для дальнейшего развития. Была
создана новая общебиологическая эволюционная концепция вида
и учение о виде впервые вышло за рамки систематики. Однако
возникли новые противоречия, связанные с пониманием харак-
тера развития и вопросом о переходных формах, с разрывом
между концепцией вида в теории и концепцией вида на прак-
тике, и т. п.
Эти противоречия можно было разрешить лишь с помощью
новых методов познания вида. После того как эволюционная кон-
цепция вида, опиравшаяся на описательно-морфологические зна-
ния, исчерпала себя, дальнейшее развитие теории вида было
возможно лишь в результате более глубокого изучения его
статики.
Глава 4
РАЗРАБОТКА ПРОБЛЕМЫ ВИДА
И ВИДООБРАЗОВАНИЯ В НАЧАЛЕ XX В.
Новый этап познания вида в течение первой четверти нашего
века характеризовался ломкой морфологической концепции вида
в систематике, поворотом к изучению внутренней организации и
структуры вида и применением новых подходов к изучению вида
(географического, экологического, генетического). Эти события
оказались не безболезненными и вызвали кризис в теории вида,
возникший после разложения вида генетическими методами и от-
крытия его поразительно сложной внутренней дифференциации.
Этот кризис был обусловлен неумением на первых порах совме-
стить новые данные с укоренившимся представлением о виде как
о простом, относительно однородном образовании. Вид в одно и
то же время оказался и элементарной, далее неразложимой еди-
ницей, и сложнейшей системой, включающей в себя массу мелких
форм. Однако объективное содержание этого противоречия было
понято далеко не сразу.
К началу XX в. выяснилось, что на базе скудных знаний
о внутренней организации вида, которыми располагала в то время
биология, невозможно развивать далее эволюционную теорию.
Внимание ученых вновь оказалось прикованным к учению о виде.
Многими признавалось, что суждения об эволюционном про-
цессе в целом останутся бесплодными до тех пор, пока не бу-
дет лучше изучен сам вид и конкретные способы видообразова-
ния.
Это, конечно, не означает, чго изучение микроэволюции в рас-
сматриваемый период не ознаменовалось какими-либо успехами
принципиального значения. Например, еще в самом конце XIX в.
среди английских биометриков выделился молодой зоолог В. Уэл-
дон, показавший в тщательно проведенной работе с крабами
Carcinus maenas в бухте Плимута, как под влиянием естествен-
ного отбора постепенно происходит преобразование состава попу-
72
Р. Веттштейн.
С. И. Коржинский.
пиции. Другие примеры исследований видообразования будут при-
ведены ниже.
И все же, несмотря на отдельные блестящие попытки исследо-
вания процесса видообразования, можно утверждать, что отсут-
ствие знаний о внутренней организации вида служило препят-
ствием для существенного прогресса в изучении закономерностей
п способов видообразования.
1. МОРФОЛОГО-ГЕОГРАФИЧЕСКИЙ МЕТОД В СИСТЕМАТИКЕ
В начале рассматриваемого периода произошел ряд крупных из-
менений в методах изучения вида, приведших к существенной
перестройке всей концепцид. На этот раз перелом коснулся трак-
товки вида в практической работе систематиков.
В 90-х годах XIX в. началась давно назревшая ломка морфо-
логического метода в систематике и замена его новым, морфо-
лого-географическим методом. Так, например, в систематике выс-
ших растений С. И. Коржинским (1892), Р. Веттштейном (Wet-
tstein, 1898) и В. Л. Комаровым (1901а, 1908) было выдвинуто
учение о новой элементарной единице классификации, реально
существующей в природе,— географической расе. Оно и легло
в основу нового способа выделения видов.
Географическая определенность (т. е. специфичность ареала)
как черта вида хотя и была в общем известна еще в середине
XIX в., но до XX в. не использовалась в качестве критерия при
практическом определении вида, если не считать попыток отдель-
ных систематиков, усилия которых в то время не нашли под-
держки.
Географическая определенность как особая черта вида впервые
была введена в его определение А. Кёрнером в 1869 г. (см.
об этом: Комаров, 1940; Юзепчук, 1958). Кёрнер определил вид
как совокупность всех особей, распространенных в пределах опре-
деленного ареала, сходных между собой и в течение продолжи-
тельного времени выраженных в большом числе единообразных
потомков. Однако попытки Кёрнера заменить морфологические
вйды старой систематики видами — географическими расами в то
время были восприняты как ненужное «дробительство», лишь
усложняющее технику классификации.
В з г л я д ы С. И. К о р ж и н с к о г о. С. И. Коржинский (1892)
предложил отказаться от выделения видов с помощью одного мор-
фологического метода, так как при этом их границы устанавлива-
ются слишком субъективно, а сами такие виды — лишь условные
категории систематики и не отражают реальных отдельностей,
существующих в природе. В противовес морфологическому виду
он выдвинул понятие о географической расе растений: «Самостоя-
тельные расы (proles)., суть истинные систематические и гео-
графические единицы... нечто действительно существующее»
7Ь
(там же: 95). «Хороший» вид (species) и подвид (subspecies) яв-
ляются ступенями формирования расы (proles). Подвиды в обла-
стях их общего распространения способны к «многократной пло-
довитой метизации», в то время как виды уже отделены друг
от друга физиологическими барьерами; при совместном обитании,
как правило, они не смешиваются и могут образовывать гибрид-
ные формы лишь спорадически (там же : 95 и др.). Коржинский
усматривал объективное отличие между видом и подвидом в сте-
пени замкнутости расы, как саморазмножающейся группы, а само
это явление характеризовал не только с морфолого-географиче-
ской стороны, но и с физиологической, как более или менее полно
сексуально обособленную группу.
Изучая состав населения в местностях, пограничных между
двумя расами, систематик-флорист иногда может и без экспери-
мента установить степень физиологической замкнутости их; если
здесь обнаруживаются переходные формы, их можно рассматри-
вать как межрасовые гибриды и тогда каждую из рас следует счи-
тать еще подвидом, а вид будет включать два или несколько под-
видов. При отсутствии же переходных (гибридных) растений
каждую из рас следует рассматривать как вид, т. е. образование,
уже полностью изолированное в половом отношении.
Конечно, далеко не всегда в пограничных местообитаниях
можно обнаружить такие переходные формы или с уверенностью
считать их гибридами. Часто флорист оказывается вынужденным
лишь гадать о степени физиологической изоляции рас. В таких
случаях возведение расы в таксономический ранг подвида или
вида, по Коржиискому, не может быть полноценным определе-
нием.
Однако такое положение лишь временно, так как впоследствии,
как наблюдениями -в природе, так и в особенности эксперимен-
тально, всегда можно выяснить действительную степень физиоло-
гической изоляции рас (конечно, при наличии полового размно-
жения) .
Следовательно, вид, по Коржиискому, может состоять из одной
пли из нескольких еще не вполне сформированных рас (подви-
дов). Этим самым Коржинский избежал опасности крайнего моно-
типизма (вид всегда только одна раса) и крайнего политипизма
(вид как группа рас) и не исказил существа дела ради чисто
технических номенклатурных удобств. Таким образом, именно
Коржинский заложил основы для представления о виде как про-
странственно-племенной определенности.
Концепция В. Л. Комарова. Вслед за Коржинским
против музейно-морфологической трактовки вида выступил
В. Л. Комаров и противопоставил ей свое понимание реальной
географической расы растений: «При флористических исследова-
ниях основной единицей... надо считать не отвлеченное типовое
понятие „вид“, а реальную... расу» (1901а: 81). Реальна раса,
а не вид. В то же время ие совсем последовательно он сближай
понятие расы с подвидом морфологической систематики (там
же 79). Стремясь разрушить концепцию морфологического вида,
он пытался при этом уничтожить и самое понятие вида (см.
об этом более подробно: Завадский, 1952). Эту позицию Комаров
занимал лишь в первые годы своей деятельности.
В дальнейшем, с 1908 г., Комаров отказался от противопо-
ставления вида расе и вслед за Коржинским отождествил реально
существующий вид с вполне сформированной географической
расой. В книге «Ламарк» Комаров (1925 : 101) сделал вывод о же-
лательности замены термина «вид» термином «раса», но вместе
с тем подчеркнул, что это «непрактично» и что поэтому «нельзя
выбросить из науки термин „вид“, надо только правильно его
осмыслить». В то же время он продолжал критику морфологиче-
ского вида. «Чисто морфологическое понятие о виде как модели
или типе,— писал Комаров,— разумеется, является абстракцией
п при попытке перенести его на живую природу легко переходит
в отрицание самого вида» (1912:521). С этого времени Комаров
считал вид «не теоретическим построением, а настоящим явлением
природы, не менее реальным, чем особь», и утверждал, что «в ос-
нове этого понятия лежит понятие о племени, о разросшейся и
твердо установившейся расе» (там же : 517, 522).
Представление о расе как населении, «связанном общей пле-
менной жизнью», было выдвинуто Комаровым еще в 1901 г.
(1901а: 79). Он не раз возвращался к мысли, что в основе суще-
ствования расы растений лежат внутренние связи, цементирующие
ее в биологическую целостность. Вид реально существует «как
особое явление природы» (1940:9—10) именно потому, что яв-
ляется активной единицей племенной жизни (1927:39). Таким
образом, по Комарову, вид — это сформировавшаяся раса, или
племя растений, расселившееся на известном пространстве бла-
годаря хорошей организации внутриплемепных отношений и вме-
сте с тем образование относительно хорошо замкнутое (т. е. раз-
множающееся внутри племени).
Географическая изоляция сама по себе не является, по Кома-
рову, причиной обособления вида. Описывая новый вид Phacel-
lanthus continentalis, он отмечает, что географическая изоляция
вначале приводит лишь «к обособлению племенной жизни» у разъ-
единенных частей ранее единого племени, а уже впоследствии
происходит изменение племенных отношений у всех или некото-
рых фрагментов старого племени (Комаров, 1930 274). При-
мерно так же толковался процесс обособления части старого
племени и формирования в пределах этой части особых племен-
ных отношений и при миграциях рас в роде Caragana (1908).
Таким образом, в своих теоретических построениях вслед
за Коржинским и Комаров отводил физиологическому критерию
важное место в определении ранга расы. Однако эти взгляды на
75
роль половой изоляции при окончательном суждении о степени
сформированное™ географической расы не былп реализованы
в предложенном им географо-морфологическом методе выделения
видов. В качестве технического ключа — инструмента для работы
систематика Комаров выдвинул следующее положение: «Вид —
это морфологическая система, помноженная на географическую
определенность» (1927). Практически же это означало, что видом
следует называть всякое однородное население растений, распро-
страненное в известной области, если оно отличается от сходного
населения, живущего в соседней области, хотя бы одним морфоло-
гическим признаком. Эта установка выражала прогрессивную
тенденцию, так как требовала не только морфологического, но и
географического подхода, но теоретическая концепция вида, раз-
работанная Комаровым, была здесь сильно обеднена. Физиологи-
ческий критерий был отброшен, экологический не упоминался,
не было ясно указано, что в качестве признака должны избираться
только константные образования и т. п.
Из сказанного следует, что, создав новую более глубокую кон-
цепцию вида, Комаров не сумел практически ее реализовать для
нужд систематики, хотя и отметил: «Теоретическая подкладка
этого практического приема та, что я признаю существование
у растений племенной жизни и активной единицей таковой жизни
считаю племя» (1927 : 39).
Морфолого-географический метод в систематике растений,
опиравшийся на так называемый камчатский афоризм, оказался
сильным оружием в борьбе против старого морфологического ме-
тода, но сам он таил в себе опасность раздробления единого в ге-
нетико-физиологическом отношении населения на части, так что
термин «вид» мог быть применен не к обособленному племени,
но и к каждой из многих его частей.
О политипическом стандарте видов. Морфолого-
географический метод получил всеобщее распространение не
только в ботанике, но и в зоологии. Рассматривая географическую
расу так же, как основную и реально существующую единицу, си-
стематики животных обычно определяют ее не как вид, а как под-
вид (вид — группа рас). По А. П. Семенову-Тян-Шанскому (1910),
Б. Реншу (Rensch, 1938), Э. Майру (1947), Л. С. Бергу (1922,
1950) и др., вид — образование политипическое, т. е. включает
в себя подчиненные единицы — географические расы (подвиды).
«Раса есть состояние вида до его полного отщепления от ствола
родоначальной формы» (Семепов-Тян-Шапский, 1910:19). Дар-
вин и многие другие эволюционисты в таких случаях употребляли
выражения «рождающийся вид», «молодой вид» и т. д. «Называя
географические расы предпочтительно подвидами, а не видами,—
пишет Майр, мы не только получали гораздо более верную кар-
тину систематических взаимоотношений, но также значительно
упрощали систему» (1947:202).
76
А. П. Семенов-Тян-Шанский.
Согласно А. П. Семенову-Тяп-Шанскому (1910), политипиче-
ским может быть сложный, полиморфный вид. Политипический
стандарт вида этот автор рисовал себе в форме следующей иерар-
хии единиц: вид (species или, точнее, conspecies); раса, или гео-
графический подвид (subspecies); «особо мелкие локально-геогра-
фические единицы» (там же: 20), или племена (natio); «про-
образы рас» (там же : 22), или морфы (morpha)n, наконец, редкие
уклонения от типа — аберрации (aberratio). Для систематика-мо-
нотиписта такая иерархия единиц совершенно неприемлема. Он
будет обязательно возводить в ранг вида расу, часто племя,
а иногда морфу и даже аберрацию.
Политипический стандарт вида, основанный на морфолого-гео-
графическом методе, завоевал господствующее положение в зооло-
гии и ботанике. При этом представители разных специальностей
и различных направлений одно и то же явление обозначали раз-
личными терминами. Такая практика вела к угрожающему росту
терминологической неразберихи. Уже в начале 20-х годов при-
ходилось заниматься следующими сопоставлениями: «Итак,
proles Коржинского, 1892 = Formenkreis Клейншмидта, 1901 =
= conspecies Бианки, 1916 = сборный, или полиморфный, вид
Р. Регеля, 1912» (Берг, 1922 274). Понятие «сингамеон», по
Лотси (Lotsy, 1917), означает группу рас или биотипов (жорда-
нонов), способных эффективно скрещиваться между собой, или
иногда состоит из одного биотипа (гомозиготный, гомогенный
сингамеон). Критерием сингамеона служит нормальная плодо-
витость скрещивающейся совокупности особей. Термин «Kommic-
sum» (Danser, 1929) включает то же самое содержание (круг
скрещивающихся особей). То же содержание выражено и в тер-
мине «coenospecies» новейших авторов, а также в понятии «био-
логический вид» по Добжанскому (Dobzhansky, 1955), Майру
(1947) и Кэйну (1958). Последняя группа терминов толкует об
одном и том же стандарте вида, основанном на физиологическом
критерии (наличие внутренней фертильности и наружной сте-
рильности) .
Политипический стандарт требует подразделения вида па расы
или подвиды. Однако понятие «подвид» остается недостаточно
уточненным. Некоторые систематики превращали термин «под-
вид» в нечто идентичное неопределенному понятию разновидности
старой систематики. Так, по В. В. Алпатову (1924), вид подраз-
деляется на три сорта иерархически соподчиненных подвидов:
географических, экологических и темпоральных. При этом только
географические подвиды соответствуют современному значению
термина. Экологические же «подвиды» входят в состав геогра-
фических подвидов на правах, так сказать, subsubspecies, а фе-
нологические группы — темпоральные «подвиды» являются sub-
subsubspecies! Такая трактовка подвида по существу являлась
возвратом к «ашерсоновщине», т. е. к немотивированным, чисто
77
морфологическим многоэтажным подразделениям вида. Слабая
мотивированность выделения подвидов обнаруживалась часто при
увеличении числа образцов в коллекции, когда разрывы заполня-
лись и вместо подвидов возникали клины. Плохо обоснованные
подвиды браковались томно так же, как браковались и плохо обос-
нованные виды. Если поспешно выделенные подвиды оказа-
лись артефактами, это, однако, еще не может служить доводом
в пользу принципиальной нереальности подвидов, на которой
настаивает, например, П. В. Терентьев (1957, 1966). Несмотря на
упразднения многих ошибочно выделенных подвидов, их реаль-
ное существование — факт не менее обоснованный, чем факт
реального существования видбв. Этот вопрос будет специально
обсужден в главе 7. Более прав в этом вопросе С. С. Шварц
(1966), подчеркивающий, что несмотря на все трудности выделе-
ния, подвиды — реально существующие подразделения вида.
Подведем итог сказанному о реформе в систематике. Приме-
нение морфолого-географического метода в систематике привело
к необходимости пересмотра всех прежде описанных видов, по-
скольку видом стала признаваться совокупность особей, отличаю-
щаяся от других совокупностей не только по морфологическим
признакам, но и по ареалу обитания. Основной единицей класси-
фикации оказалась географическая раса. Вид был отождествлен
с одной географической расой или же рассматривался как группа
близкородственных рас, а сами расы считались однородными по
составу, неделимыми единицами. Такая реформа давала возмож-
ность более точно описывать виды.
Все концепции вида, основанные на прогрессивном для того
времени морфолого-географическом методе, были направлены про-
тив устаревшей односторонне-морфологической трактовки вида.
Противопоставление же монотипической концепции вида полити-
пической — это не более чем спор о терминах, о том, как назвать
одно и то же явление природы (географическую расу), т. е. во-
прос в основном чисто номенклатурный.
2. КРИЗИС В ИЗУЧЕНИИ ВИДА
Разложение вида генетическими методами.
Кроме морфолого-географического подхода к изучению вида с по-
мощью картирования, в начале века зарождается также и эколо-
гический. Работы < экологического направления явились суще-
ственным новым шагом в изучении внутренней структуры вида.
На растениях они были начаты Б5 А. Келлером и И. К. Пачо-
сжим. Келлер (1907) показал существование обособленных эколо-
гических рас, занимающих в пределах одного географического
ареала разные местообитания. Такие замещающие друг друга и
не встречающиеся в одном местообитании расы были названы им
экологическими видами (например, лесная раса и луговая, сухо-
78
дольная и пойменная, приуроченная к кислым почвам и к щелоч-
ным, и т. п.). Понятие «oecospecies», введенное Г. Турессоном
20 лет спустя, по существу тождественно экологическим видам
Келлера. В 1923 г. Ф. Клементс высказал мнение, что только
методы экспериментальной и количественной экологии смогут
вывести проблему вида на путь, где строгие доказательства яв-
ляются системой. Для паразитических форм было введено поня-
тие о физиологической расе. Однако экологический метод в со-
временном его понимании сформировался в самом конце
рассматриваемого периода.
Настоящий же переворот в общебиологических взглядах па
вид произошел в начале века сразу же после переоткрытия за-
конов Г. Менделя и первых успехов гибридологического анализа.
Именно генетические методы и идеи оказали наиболее сильное и
очень противоречивое, влияние на теорию вида. Особо важное
значение для разработки проблемы вида имело применение ме-
тодов экспериментальной генетики. Путем гибридизации и изуче-
ния расщепления признаков, путем инцухта и разведения изоли-
рованных семей, чистых линий, клонов и т. п. впервые удалось
разложить вид на массу мелких константно различающихся ме-
жду собою форм. В результате разложения вида на отдельные
элементарные формы п открытия огромной сложности его состава
была установлена еще одна черта вида — многообразие форм.
До XX в. состав и внутренняя структура вида оставались
почти неисследованными. Предполагалось, что вид в общем про-
стое образование. Было известно, что он включает в себя не-
большое число морфологически различных форм — подвидов и
разновидностей, природа которых оставалась неясной. С позиций
морфолого-географического метода вид также рассматривался как
единица однородная или простая по составу. Однако эксперимен-
тально-генетический анализ показал нечто прямо противополож-
ное: вид неоднороден и может быть разложен на множество на-
следственно устойчивых мелких форм. Например, в составе
мягкой пшеницы (Triticum vulgare = Т. aestivum) было обнару-
жено наличие наследственно различных форм, все разнообразие
которых, согласно Н. И. Вавилову (19356), «во всяком случае
определяется миллионами». Столь же огромным, поражающим
воображение, оказалось и разнообразие форм у других видов.
Таким образом возникло представление о виде как чрезвы-
чайно сложной системе мелких форм и в то же время как об
однородной по своему составу элементарной единице или как
о группе рас, т. е. появились две противоположных и на первый
взгляд взаимоисключающих концепции вида.
Многим тогда казалось, что возникшие противоречия заводят
учение о виде в тупик. В действительности же, как мы увидим
ниже, изучение географической определенности вида и анализ
его генетического состава обогащали и углубляли теорию вида.
79
Вместе с тем разрушение ранее сложившихся представлений
о простоте вида и замена их новым представлением о сложности
внутренней структуры вида не могли не вызвать крупных по-
трясений. Как в физике разложение атома, так и в биологии
экспериментальное разложение вида обусловило крутую ломку
основных понятий. Старые понятия пришли к столкновению
с новыми фактами, и это породило кризис всего учения о виде.
Н е о ж о р д а н и з м. Этот кризис продолжался примерно всю
первую четверть нашего века. Возникли разногласия в истолко-
вании самого факта разложения вида на составные элементы.
Сформировалось и приобрело большую силу особое направление
в понимании вида, называемое неожорданизмом и представлен-
ное главным образом генетиками и селекционерами (Лотси,
Де Фриз, Иогансен, Г. Нильсен, ранний Турессон и др.). Неожор-
данисты признавали «истинными» видами (euspecies) только да-
лее не расщепляющиеся, мелкие, наследственно устойчивые
формы, получаемые в качестве окончательных продуктов экспе-
риментального разложения природных популяций. В центре вни-
мания оказался один изолированно взятый мепделирующий при-
знак. Для разграничения так называемых истинных видов было
достаточно обнаружить различия по одному такому признаку.
К «истинному» виду, таким образом, приравнивалась каждая
чистая линия и практически гомозиготный биотип. Такие «виды»
были названы Г. Де Фризом жорданонами, пли мелкими видами
(microspecies). Это воззрение хорошо выразил Р. Э. Регель, со-
гласно которому, жорданон — это «низшая константная система-
тическая единица, основанная на отдельном самостоятельном на-
следственном факторе (на единице наследственности, гене),
определяемом прп скрещивании на основании закона Менделя,
путем гибридологического анализа» (1912 : 508).
Обыкновенные же виды, принятые в систематике, названные
липпеоиами, рассматривались как «сборные понятия», как смеси
форм или же как «спнгамеоны» — неустойчивые продукты гибри-
дизации между «истинными» видами. По Де Фризу, лпннеон сле-
дует противопоставлять жорданону как практически удобную
единицу систематики, не существующую реально, — истинному
виду. В монографии о происхождении видов и разновидностей
(De-Vries, 1906) можно встретить выражение «систематический
вид» (или «практический вид систематиков»), т. е. линнеон и
«природный» вид, т. е. жорданон. Таким образом, в основе нео-
жордапизма лежала старая идея о виде как абсолютно однород-
ном образовании — строгом монотипе. Руководствуясь этой идеей,
пеожорданпсты последовательно расчленяли полиморфные попу-
ляции вплоть до состояния, прп котором отдельные биотипы
могли признаваться изогенными образованиями (т. е. однород-
ными в генетическом отношении), или даже на практически го-
мозиготные формы. Назвав «истинным» видом такие мельчайшие
80
Г. де Фриз.
частицы природного вида, неожорданисты односторонне преуве-
личивали значение критерия однородности и совершенно игно-
рировали другой более существенный критерий вида — спо-
собность к самостоятельному существованию и воспроизведению
в природной обстановке. Как было установлено впоследствии, вы-
деляемые из состава вида отдельные генетически выравненные
линии не являются самостоятельными природными образова-
ниями и, как правило, пе обладают способностью к длительному
существованию в естественной обстановке. «Истинные» виды
оказались в действительности лишь зависимыми компонентами
популяций, частями пх сложной генетической структуры. Правда,
это утверждение полностью применимо только к группам с пере-
крестным оплодотворением, а у агамов и полных апогамов жорда-
ноны являются более самостоятельными образованиями.
Ошибки неожорданизма усугублялись тем, что некоторые ав-
торы (Лотси, Г. Нильсон) считали «истинней» вид неизменным,
а эволюцию сводили лишь к гибридизации между такими якобы
неизменными «видами» и к расщеплениям. Гипотеза гибридоге-
пеза (Lotsy, 1916а, 1916b, 1917), так же как и гипотеза мута-
ционного образования видов, выдвинутая Де Фризом, своими
корнями опиралась на ложную концепцию вида.
Эти воззрения были раскритикованы К. А. Тимирязевым в се-
рии статей, В. Л. Комаровым (1940) и рядом других авторов.
Недавно Н. П. Дубинин правильно отметил, что «в начале века
проблема вида разрабатывалась менделистами в метафизическом
плане» (1966 215), хотя она и содержала прогрессивные мо-
менты (требование вычленить внутри вида наследственные
формы), и что попытки распылить виды на множество биотипов
вредили работе тех систематиков, которые стремились изучить
вид как реальную систему, вели к отчуждению систематики от
генетики. Одним из последствий обрисованного нами положения
было усиление субъективистских толкований вида, что будет рас-
смотрено в особом разделе.
Неправильно было бы думать, что учение о виде могло бы
выйти из кризисного состояния на основе одних общеэволюцион-
пых построений. Такой знаток учения Дарвина, как К. А. Тими-
рязев, в «Историческом методе в биология» написал главу о виде:
«Естественноисторический вид — отвлеченное понятие или реаль-
ный факт?». Самое название этой главы и ряд содержащихся
в ней утверждений свидетельствовали о неблагополучии с про-
блемой вида в начале XX в. Разнобой в трактовке видов, труд-
ность их разграничения во времени смутили его и привели к двой-
ственной позиции: «В одном смысле вид, очевидно, только
отвлеченное понятие, в другом он — реальный факт. . . Вид, про-
тивополагаемый разновидности, есть, конечно, отвлеченное поня-
тие, но виды, целый ряд видов, противополагаемых друг другу,
представляют несомненный объективный факт» (Тимирязев,
6 К. М. Завадский $2
1922 72). Такие формулировки не могли способствовать прояс-
нению вопроса о сущности вида.
В заключение нельзя не упомянуть о попытках некоторых
биологов выйти из затруднений путем отказа от термина «вид».
К такому приему прибег, как мы уже отмечали, В. Л. Комаров.
В 1901—1907 гг. он выбросил из своего лексикона слово «вид»
и заменил его словом «раса». В том же 1901 г. зоолог О. Клейн-
шмидт внес сходное по форме предложение. Как и Комаров,
Клейншмидт утверждал, что морфологический вид старой систе-
матики является чаще, чем это терпимо в пауке, простой выдум-
кой. В природе нет видов, продолжал Клейншмидт, а существуют
конкретные наследственно постоянные формы, образующие есте-
ственные группы или «круги форм» (Formenkreis). Вместо того
чтобы сказать: в природе нет морфологических видов старой си-
стематики, Клейншмидт выдвинул ошибочную формулу: видов
(Art) не существует, а реальной единицей является «круг форм».
Комаров уже в 1908 г. преодолел свою ошибку и выдвинул пред-
ставление о географической расе как о реальном виде, а Клейн-
шмидт (Kleinschmidt, 1930) еще долго противопоставлял понятие
«круг форм» понятию «вид», хотя к 30-м годам большинству зоо-
логов стало ясно, что его «круг форм» это именно то явление
природы, которое именуют политипическим видом.
Такие попытки предпринимались и впоследствии. Например,
Дж. Гексли в 1942 г. в книге «Эволюция. Современный синтез»,
прекрасно разобрав вопрос о неравноценности видов, вдруг сде-
лал вывод о необходимости ликвидировать понятие о виде, за-
менив его целым рядом понятий, отражающих отдельные типы
видов. Эта реформа была предложена Гексли в качестве одного
из устоев «новой систематики».
Не надо разъяснять, насколько такие предложения усиливали
субъективистские тенденции в толковании вида.
Субъективизм в понимании вида. Кризис теории
вида проявился и в резком усилении субъективизма. В связи
с большими разногласиями в определении вида вновь усилилась
тенденция отрицания его реальности. Одни утверждали, что реа-
лен только классический вид в линнеевском объеме, другие же
считали, что морфологический вид систематиков — отвлеченное
понятие, а реальным признавали лишь географическую расу, име-
нуя и ее видом, третьи считали реальным лишь жордановский
вид и т. п. В этой обстановке и получили развитие не только
старые источники субъективизма, обсужденные выше, но и новые
формы его, связанные с махистской философией. Вид рассматри-
вался как абстракция, в духе термина «архетип»; как условное
понятие, вносящее порядок в наши ощущения; как идея, экономя-
щая работу мышления; как практический удобный символ, и т. п.
Иллюстрируем сказанное примерами. Общепринятое тогда
толкование вида как суммы особей затрудняло понимание его
82
реальности: по В. М. Шимкевйчу (1907 103) «существует завй-
< пмость между четкостью границ вида и численностью образцов
в музее», т. е. оправдывается «положение Бюффона, что природа
не знает видов, а знает лишь особей». Некоторые авторы пыта-
лись узаконить понятие вида — абстракции для систематики,
в то же в£емя считая, что для биогеографии, экологии и других
наук вид — вполне конкретная совокупность особей (Чупров,
1910).
Продолжая старую номиналистическую линию, некоторые ав-
торы утверждали, что «в действительности вида не существует. . .
вид — понятие отвлеченное, как сумма признаков, которыми от-
личаются особи, происходящие от своего родоначальника» (Ры-
тов, 1914:47). По М. В. Рытову, данное растение, допустим
‘подсолнечник, — безусловная реальность, а образуемое нами об-
щее понятие о подсолнечнике как о виде есть абстрагирование
некоего типа строения — и только. С этих же номиналистических
позиций О. Гертвиг в 1916 г. утверждал, что «понятие вида —
чистая абстракция», а реальны лишь индивиды, относимые си-
стематиками то к одной, то к другой идеальной модели (виду
как типу).
Противоречивую позицию в вопросе о реальности вида зани-
мал специалист по эволюционной теории Л. Плате. Взгляды его
представлены в монографии (Plate, 1908), а также и в сжатом
очерке (Плате, 1928). С одной стороны, он формулирует субъек-
тивистскую номиналистическую идею: «В природе мы встречаем
только отдельных индивидов, понятие же о виде абстрагируется
человеком. Как и все понятия, понятие о виде есть продукт че-
ловеческого мышления и вследствие этого не представляет ничего
реального» (там же: 20). Но вступая в противоречие с самим
собой, Плате утверждает, что только в человеческой голове су-
ществует морфологическое определение вида, а вид с физиологи-
ческой точки зрения отражает «реальную связь, кровное родство
между различными индивидами и тем самым связывает членов
вида в естественную единицу. Эту единицу — сумму индивидов,
способных при соединении друг с другом размножаться, — мы и
называем видом, и из этого следует, что каждый вид есть нечто
реальное, тогда как понятие вида, как и всякое понятие, пред-
ставляет, конечно, нечто абстрактное. Неправильно поэтому счи-
тать, как это часто делают, вид лишь абстракцией, а реальное
существование приписывать только индивидам» (там же: 63).
Так Плате с 63-й страницы опроверг Плате с 20-й страницы: на-
туралист не сомневается в реальности, а теоретик сползает
к субъективному идеализму.
Некоторые биологи толковали вид в духе ортодоксальных
принципов махизма. Так, например, автор книги об уродствах
растений П. Виллемен писал, что «принимает понятие вида как
постулат, как рабочий инструмент, необходимый натуралистам»;
6*
83
«важно только то, что понятие „вид“ удобно исследователям» и
что «не следует интересоваться тем, какое явление природы
отражается в этом понятии» (Vuillemin, 1926 VII). По А. Хич-
коку, «концепция вида должна основываться на суждениях от-
дельных систематиков» (Hitchcock, 1931 :228). В том же 1931 г.
К. (Эстенфельд предложил считать видом группу особей, сходных
по признакам, «которые мы условились считать существенными».
Через 70 лет после иронического замечания Дарвина, что
таксономический ранг формы определяют «большинством голо-
сов», этот странный способ доказательства истины вновь начал
приобретать права гражданства. Усиление подобных взглядов —
одно из свидетельств кризиса теории вида. Кризис усугублялся
утверждениями, что виды образуются путем вымирания проме-
жуточных форм (Бианки, 1907), хотя еще Дарвин доказал, что
этим лишь более резко очерчиваются их границы.
Разброду в понимании вида способствовала п нигилистиче-
ская позиция некоторых биологов, считавших, что все виды, опре-
деленные систематиками, реально не существуют. Рассмотрим
одну разновидность таких взглядов. Справедливо обратив внима-
ние на неразбериху, существующую в характеристике вида си-
стематиками, Р. Гольдшмидт (Goldschmidt, 1932) полагал, что
возведение той или иной формы в таксономический ранг больше
зависит от личной склонности систематика, чем от особенностей
самого объекта. Сделав такой вывод, Гольдшмидт пошел дальше
и заявил, что вообще отрицает реальное существование «таксоно-
мических видов». Он считал, что природные виды не имеют ни-
чего общего с видами, которыми занимаются систематики. Такую
точку зрения нельзя назвать субъективистской (реальное суще-
ствование каких-то природных видов признается). Это скорее
нигилистическая точка зрения. Все, что создано систематиками,
рассматривается как субъективное, ничего реального не отражаю-
щее. Такое отношение к результатам работы систематиков более
чем странно.
Отрицание реальности вида, особенно усилившееся в первую
четверть XX в., опиралось на новые формы субъективизма, свя-
занные с механицизмом или разнобоем в генетическом истолкова-
нии вида, которые слились со старыми формами субъективизма
(вид как идеальный тип организации или вид как произвольно
вырванный участок непрерывной «ленты» эволюции). Просущест-
вовав целых два столетия, субъективизм в учении о виде в усло-
виях кризиса теории вида нашел новую питательную среду и
как бы заново возродился. Особо пышное развитие эти ошибочные
взгляды получили до 30-х годов нашего столетия; но и в последую-
щие десятилетия субъективистские воззрения на вид еще сохра-
няли жизнеспособность, хотя и стали менее распространенными.
В следующей главе будут рассмотрены примеры некоторых
современных субъективистских построений.
84
3. ИЗУЧЕНИЕ ВИДООБРАЗОВАНИЯ
Сезонные расы. Большой интерес представляют исследо-
IUI иия видообразования у растений путем обособления сезонных
рис, проведенные в 1893—1904 гг. Р. Веттштейном и его учени-
1и1ми, а также В. Н. Хитрово и особенно Н. В. Ципгером. Прин-
11,111 шальное значение этих исследований состоит в том, что они
положили начало конкретному изучению различных способов
видообразования, включающего как наблюдения в природе, так и
.1К'снерименты на питомниках.
Было установлено, что многие луговые и полевые виды пред-
тавлены двумя совместно произрастающими формами — летней
и осенней. Такое расчленение вида оказалось характерно для
многих горечавок (Gentiana)—10 пар, погремков (Alectorolo-
plins) —8 пар, очанок (Euphrasia) —5 пар и для видов других
родов. Летние расы таких видов быстро растут в начале вегета-
ции, цветут, как правило, в мае—июне, стебель у них простой
пли мало ветвящийся в верхней части, число удлиненных междо-
узлий мало. Осенние расы в начале вегетации растут замедленно,
цветут после середины лета, стебель у них сильно ветвящийся
от основания, число коротких междоузлий увеличено. Отличаются
>ти расы и по другим признакам (по форме листьев, относитель-
ной длине боковых побегов, опушению и окраске стебля и т. п.).
При испытании этих рас в условиях специальных опытов такие
различия сохранялись в течение нескольких поколений (Wett-
slein, 1901).
Образование таких рас объяснялось Веттштейном приспособ-
лением растений к переживанию неблагоприятного периода се-
редины лета, когда на лугах идет сенокошение (Wettstein, 1896,
1897, 1901 и др.). В этих же работах доказывалось, что сезонные
расы постепенно преобразуются в виды. У близких видов тех же
родов, занимающих территории, на которых не происходит сено-
кошения, сезонный диморфизм отсутствует. Наиболее же выра-
женный диморфизм наблюдается в издавна п регулярно скаши-
ваемых лугах. Время сенокоса совпадает с периодом созревания
семян у ранних рас, а поздние в это время еще так малы, что
коса их не срезает. Поздние расы сезонно диморфных пар не
поедаются животными и поэтому сохраняются па уже скошенных
лугах.
Предковый вид, как полагал Веттштейн, включал много раз-
нообразных форм, слегка отличающихся друг от друга по рит-
мам развития. Далее же «регулярный покос полей и лугов, дей-
ствуя избирательно и закрепляя индивидуальные вариации,
с течением времени путем отбора создал сезонно-диморфные
виды» (Wettstein, 1901:341). Не всегда, однако, Веттштейн да-
вал такие объяснения. В том случае, если исходный вид вклю-
чал только поздние формы, появление ранних форм объяснялось
85
г1рЯмь1м приспособлением. В природе удалось обнаружить разный
стадии расхождения сезонных рас. У одних пар оно достигает
уже степени вида (Gentiana norica, G. sturmiana и пр.), у дру-
гих исходная форма рассматривалась как вид, а производная,
ранняя, как подвид, и т. п.
Н. С. Ростова (1967) обращает внимание на забытые интерес-
ные идеи Веттштейна о возникновении сезонного диморфизма
в результате нарастания межгрупповой изоляции во времени
(разрыва в сроках цветения) у совместно живущих родственных
форм.
В России исследования Веттштейна были продолжены в 1905—
1911 гг. В. Н. Хитрово (1911), который показал на очанках, что
сезонные формы существуют в пределах одного местообитания,
но степень разобщения нарастает при расселении их в разные
местообитания. Однако наиболее глубоко вопрос о происхождении
сезонных рас был изучен Н. В. Цингером (1913, 1928). На при-
мере большого погремка (Alectoroloplius major) он показал, что
путь эволюции, исследованный Веттштейпом (образование
именно двух рас), является лишь частным случаем. Оказалось,
что у большого погремка существует не две расы (определенных
Цингером как подвиды), а несколько, различающихся не только
сроками созревания и связанными с ними признаками (число
междоузлий, форма роста и пр.), но и своеобразными адаптив-
ными чертами, характерными, например, для расы, засоряющей
посевы ржи (исчезновение крыловидного придатка у семян, не-
раскрывающиеся коробочка и чашечка, совпадение созревания со
сроками уборки ржи и др.). Образование всех рас (подвидов)
было блестяще объяснено Цингером разными направлениями от-
бора. Исходный тип — позднецветущпй дикорастущий погремок.
При расселении на луга, которые в прошлом выкашивались
в поздние сроки, образовались популяции луговой расы, более
ранней, чем исходный тип. Там же, где покосы стали проводить
в более ранние сроки, возникли еще более скороспелые, цвету-
щие весной, а также и послеукосные расы.х Таким образом, одни
расы произошли за счет обособления форм, уже существовавших
в исходном виде, а другие (ранневесенняя, сорная) возникли
вновь. Эти работы Цингера явились большим шагом вперед по
сравнению с достижениями Веттштейна. Были выяснены кон-
кретные пути селективного происхождения рас. Узкое понятие
сезонного диморфизма было заменено более широкой концепцией
полиморфизма вида. Полиморфизм вида оказался, с одной сто-
роны, проявлением процессов адаптивной радиации рас, а с дру-
гой — их адаптивной стабилизации, получившей в будущем
наименование сбалансированного полиморфизма (Ford, 1940).
Сезонный диморфизм, как показал Цингер (1928), возникает
в тех случаях, когда под влиянием продолжительных покосов
лугов происходит устранение из популяции большого погремка
86
р. гений со средними сроками цветения, и в итоге единая попу-
ил ция распадается на две расы, живущие бок о бок, как типич-
ные симпатрические формы (по Цингеру — ssp. vernalis и ssp.
pnlycladus). Более ранняя работа Цингера (1909) была посвя-
щена анализу видообразования путем отбора форм в процессе
перехода видов рыжика (Gamelina) и торицы (Spergula) к суще-
твованию в качестве специализированного сорняка льна. Эта ра-
бота давно была признана классическим образцом такого рода
исследований и широко излагалась в отечественной литературе
начиная с трудов К. А. Тимирязева. Исследования видообразо-
вания у пыжиков включали не только сравнительный анализ
форм, в различной степени уподобившихся льну, но и серию опы-
тов. Так, например, на питомнике производился отбор яровых
форм среди растений «озимого» вида рыжика (Gamelina micro-
сагра). После повторного отбора были получены семьи, которые
цвели раньше, чем самые ранние родительские формы. Этим ме-
тодом Цингеру впервые удалось моделировать процесс появления
таких новых форм, которые могли основать сезонную расу и по-
лужить базой формирования нового вида.
Наибольшее значение исследований видообразования путем
расчленения единой популяции на сезонные расы состоит в обна-
ружений принципиально важной возможности формирования и
полной изоляции рас, долгое время существовавших бок о бок
в качестве частей единого населения. Такой симпатрический спо-
соб видообразования отрицался ранее и отрицается некоторыми
современными авторами (см. главу 9). Некоторые систематики,
догматически воспринявшие каноны морфолого-географического
метода, были убеждены, что видообразование должно быть обя-
зательно связано с пространственным обособлением рас. Эти воз-
ражения основывались на общих соображениях о невозможности
иного способа видообразования (Soo, 1926—1927, Юзепчук, 1955;
Васильченко, 1958). На непоследовательность критиков, концеп-
ции видообразования через обособление сезонных рас указала
II. С. Ростова (1967): описывая виды погремка (Rhinanthus =
= Alectorolophus) во «Флоре СССР», И. Т. Васильченко назы-
вает видами многие сезонные расы с совпадающими ареалами,
которые даже Цингер считал только подвидами. В главах 8—9
будет показано, что видообразование через обособление сезонных
рас является одним из совершенно реальных путей эволюции.
Попытки экспериментальных доказательств
гипотезы мутационного видообразования. В 1901 г.
Г. Де Фриз (1932) опубликовал статью о мутационном происхо-
ждении новых видов энотер от сорно-полевого вида Oenothera
lamarckiana, завезенного в Европу в XVII в. из Северной Аме-
рики. В 1901—1903 гг. он выпустил двухтомный труд «Мута-
ционная теория» с подзаголовком «Опыты и наблюдения над
происхождением видов в растительном мире», а затем детализи-
87
ровал свою идею в течение целого тридцатилетия. Опыты были
начаты еще в 1886 г., когда Де Фриз собрал семена с нескольких
экземпляров Ое. lamarckiana. Он размножил эти растения и вы-
растил за 12 лет 53 000 особей. В каждом поколении обнаружи-
вались растения, отклоняющиеся от типичной формы. Этих ано-
мальных растений встречалось от 0.07 до 0.5% (от численности
всех растений). Часть таких семей стойко передавала потомству
свои особенности.
Размножая и отбирая семьи, Де Фриз постепенно выделял из
посевов типичной Ое. lamarckiana все больше и больше мутант-
ных форм, стойко передающих свои специфические признаки.
В отличие от типичной энотеры Ламарка каждая такая форма
была обозначена Де Фризом латинским бинарным названием:
•Ое. lata, Ое. gigas, Ое. scintillis, Ое. nanella, Ое. albida, Ое. оЬ-
longa, Ое. rubrinervis.
Этим многолетним исследованием, как отмечает II. И. Вави-
лов, «впервые в экспериментальной обстановке начинается изу-
чение эволюционного процесса» (1936:9). К сожалению, оно
послужило основой для построения ошибочной гипотезы видооб-
разования.
Ошибки Де Фриза состояли в том, что каждый из мутантов
был назван новым элементарным видом и на этой основе было
сформулировано понятие об особой «видообразовательпой измен-
чивости».
Новые виды, согласно Де Фризу, возникнув внезапно, тот-
час же становятся неизменными. В течение геологических «пре-
мутационных периодов» виды постоянны, а в краткие периоды
мутирования порождают сразу много новых элементарных видов.
Для видообразования не надо ряда поколений. Порождение вида
происходит внезапно, и для этого не требуется ни борьбы за су-
ществование, ни естественного отбора. Отбор не приводит к воз-
никновению видов, он лишь устраняет неудачников среди массы
уже готовых новых видов. Естественный отбор — агент, не увели-
чивающий многообразия видов, как полагал Дарвин, а, напротив,
лишь сокращающий его. В противоположность Дарвину, по
Де Фризу, разновидности не являются видами в возможности,
а представляют собой результат регрессивных мутаций, в то
время как видообразование является следствием «прогрессивного
мутирования». Все эти высказывания носили чисто умозритель-
ный характер.
Де Фриз полагал, что виды порождаются не только внезапно,
но и независимо от влияний внешней среды (автогенетически).
Однако, если даже предположить, что внезапное порождение ви-
дов имеет не автогенетическую основу, а осуществляется взаимо-
действием со средой, мутационная концепция видообразования
не перестала бы быть в корне ошибочной. Ошибочность идей
Де Фриза заключалась в отождествлении процессов наследствен-
88
ной изменчивости и видообразования. Эта ошибка лежит в основе
нгех мутационных гипотез видообразования.
Высоко оценивая деятельность Де Фриза по исследованию му-
Н1ЦИОННОГО процесса, Н. И. Вавилов писал, что «антидарвинисти-
'Н‘ские тенденции использовать мутационную теорию для дискре-
дитирования положения Дарвина о роли мелких изменений
и эволюции и о роли естественного отбора опровергаются огром-
ным количеством экспериментальных данных на дрозофиле и
и.вином зеве, показывающих, что мутации, как правило, яв-
ляются в подавляющем большинстве мелкими, охватывающими
не только морфологические, но чаще также физиологические от-
личия, выявляющиеся в большей или меньшей жизненности
форм» (1936 13). В дальнейшем энотера стала предметом мно-
гочисленных цитогенетических исследований (Бэтсона, Реннера,
Клиленда, Дарлингтона, Беллинга, Стертеванта и др.). Было об-
наружено, что значительная часть пыльцы энотер бесплодна и
что все виды имеют сложную гибридогенную природу, являясь
комплексными гетерозиготами. В частности, энотера Ламарка
включает комбинацию двух геномов — velutina и candens, хромо-
сомы которых образуют кольца, благодаря которым при скрещи-
ваниях расщепление идет с большими отклонениями от менде-
левских схем. Де Фриз полагал, что энотера доказывает
мутационную природу видообразования в противовес гибридоген-
ным гипотезам, ио оказалось; что сама она — сложный гибрид.
Мутант, названный Ое. gigas, оказался тетраплоидом гибридоген-
пой природы, некоторые крупные мутанты энотеры были амфи-
диплоидами.
По справедливой оценке Н. И. Вавилова, суровая критика му-
тационной теории видообразования Де Фриза была совершенно
правильна, но так же правильно и то, что экспериментальные
его работы над мутациями и гибридизацией составили эпоху
в ранний период развития генетики; «его труды поучительны
своими великими достижениями и в то же время крупными
ошибками» (там же : 14).
Остановимся вкратце на дальнейшей судьбе мутационной ги-
потезы видообразования. Де Фриз допускал внезапное порожде-
ние не обычных «липнеевских», а лишь «элементарных» видов.
Другие же авторы допускали скачкообразное видообразование
в гораздо больших масштабах. Именно такие взгляды были раз-
виты Л. С. Бергом: эволюция идет «скачками, пароксизмами. ..
захватывающими громадные массы особей на обширной терри-
тории. Виды в силу своего мутационного происхождения резко
разграничены один от другого» (1922 280—281). Это была ре-
ставрация взглядов Кёлликера—Ваагена и других авторов XIX в.
Приняв мутационную гипотезу видообразования, Л. С. Берг
вполне логично подкрепил ее телеологической гипотезой о внут-
реннем стремлении живого к совершенствованию («автономиче-
89
СКпй ортогенез»). Попытки другйх авторов объединять гипотезу
мутационного видообразования с дарвиновскими причинами эво-
люции были эклектичны; единственно логичное причинное объяс-
нение разового порождения видов — телеологический ортогенез.
При этом такой ортогенез не обязательно должен быть основан
па автогенетических факторах, столь же просто «обосновать» его
и онтогенетическими факторами («темной телеологией среды», по
выражению К. А. Тимирязева).
К мутационной гипотезе видообразования склонялись тогда
многие генетики. Так, например, Г. Д. Карпеченко (1927), по-
лучив межродовой гибрид Raphanus X Brassica, нерасщепляю-
щийся, самофертильный, но стерильный при скрещиваниях
с обоими родительскими видами, морфологически своеобразный,
сформулировал результат этой работы как «экспериментальный
синтез нового вида». К таким же мутационистским формулиров-
кам приходили и другие генетики, занимавшиеся гибридным син-
тезом форм (Miintzing, 1930; Huskins, 1931, и др.). Многие
прогрессивные эволюционисты-генетики в начале 30-х годов счи-
тали, «что не все виды образуются путем отбора, а некоторые
возникают через гибридизацию и мутационно» (Haldane,
1932 139).
Анализируя географические различия и мутационную измен-
чивость у тутового шелкопряда, Р. Гольдшмидт (Goldschmidt,
1932), как и Де Фриз, отрывает причины образования новых ви-
дов от причин образования географических вариаций и подвидов.
Он подвергает сомнению приспособительный характер происхо-
ждения признаков, по которым различаются «таксономические
виды». В итоге этот автор склоняется к мысли о мутационном
возникновении новых генетических форм, т. е. «природных» ви-
дов. Т. Морган (1936) хотя и не в отчетливой форме, но все же
склонялся к мутационному возникновению видов. Эти примеры
показывают живучесть гипотезы мутационного видообразования;
поэтому нельзя согласиться с мнением А. Е. Гайсиновича, что
теория Де Фриза «не пользовалась признанием и вскоре была
забыта» (1967 137).
Основная ошибка генетиков того времени состояла в пере-
оценке значения наследственной изменчивости и в недооценке
накапливающегося действия естественного отбора. Отсюда стрем-
ление, обсуждая вопрос о видообразовании, миновать проблему
адаптаций. Можно утверждать, что одним из недостатков гене-
тики раннего периода являлась недооценка проблемы эволюции
адаптаций, сведение эволюционного процесса к простому формо-
образованию и склонность рассматривать вид как новую наслед-
ственно закрепленную форму, безотносительно к степени ее адап-
тивности.
В течение второй четверти века изучение генетики популяций
и эколого-генетические работы позволили в значительной мере
90
устранить из генетики пережитки мутационной гипотезы видо-
образования.
В настоящее время нет данных в пользу мутационного обра-
зования видов. Гипотезы мутационного видообразования неприем-
лемы и в методологическом отношении. Авторы мутационных
гипотез видообразования расходятся в толковании причин скач-
ков (автогенез, эктогенез и пр.). Однако в основе всех- этих ги-
потез лежит признание особой «видообразовательной изменчи-
вости», качественно отличной от обычной наследственной измен-
чивости, а также убеждение, что отбор лишь бракует готовые
виды, т. е. только сокращает многообразие форм и не является
законом их постепенного формирования. К обсуждению этого
вопроса мы еще вернемся в главе о видообразовании.
Примеры палеонтологических работ в обла-
сти видообразования. В самом конце ХТХ и в начале
КХ в. были проведены выдающиеся палеозоологические исследо-
вания видообразования, показавшие историчность и постепен-
ность преобразования видов. Великолепную иллюстрацию такого
способа видообразования можно найти в исследованиях англий-
ского палеозоолога А. Роу (A. Rowe). Еще в 1899 г. он тщательно
изучил постепенный переход верхнемеловых морских ежей Micras-
ter corbovis к формам типа М. cor-anguinum. В отличие от других
подобных работ работа Роу. была построена на анализе массового
материала из серии смежных и несмещенных морских отложений.
Роу показал, как исподволь в популяции, нацело состоящей из
форм типа cor-bovis, возрастает примесь измененных форм. Ока-
залось, что одновременно происходило два процесса: изменение
повой формы до типичной cor-anguinum и возрастание ее числен-
ности в популяции морских ежей.
Таким образом, впервые был изучен не ряд форм (старомод-
ная Formenreiche), а серия изменяющихся во времени популя-
ций. Причем постепенность процесса изменения имела нагляд-
ность кинокадров. И, несмотря на всю медленность формирова-
ния нового типа (cor-anguinum) и замещения им в популяции
ежей исходного типа (cor-bovis), все же четкое качественно
определенное различие исходного и конечного моментов было
столь же очевидным фактом, как и постепенный характер всего
перехода. Такое изучение вида во времени в высшей степени
ценно в познавательном отношении. Существует немало более
поздних работ со столь же полными популяционными сериями.
Например, работа 1929 г. Бринкмана с описанием непрерывного
перехода Юрского аммонита Kosmoceros castor в более молодые
виды К. aculeatum и К. pollux (прослежена дивергенция популя-
ции на два ствола).
Изучая историю видов двустворчатых моллюсков из Dreissen-
siidae верхнетретичных отложений ПонтотКаспия, Н. И. Андру-
сов еще в 1897 г. подчеркивает различную скорость, с которой
91
идет дифференциация видов. Такие виды, как Congeriae mytili-
formes, существуют очень долго, а многие другие быстро расчле-
няются на дочерние, подвергаются частичному вымиранию или
преобразованию. Все это связывается с изменениями солености
бассейна. Например, после опреснения, ведущего к вымиранию
стеногалинных форм, к запустению больших пространств, ранее
занятых конкурентами, происходит внедрение в них видов, ютив-
шихся в устьях рек и в прибрежных озерах, а также эвригалин-
ных морских видов. При относительно неизменной гидрологи-
ческой обстановке видообразование, по Андрусову, происходит
на основе борьбы за существование и конкурентных отно-
шений. Эта работа Андрусова, так же как и более поздняя, по-
священная эволюции Gardiidae, была основана на массовом ма-
териале, иногда собранном из последовательно отложившихся
слоев.
Нами приведены лишь краткие справки о важном, хотя и не
получившем в то время широкого развития, палеопопуляционном
методе изучения видообразования.
Миграционная гипотеза видообразования.
Рассмотрим ее па примере взглядов В. Л. Комарова, развитых
в его ранних работах (1901 —1908 гг.). Видообразование мысли-
лось Комаровым как процесс переработки целых населений в те-
чение многих поколений под влиянием изменений климата или
почвы. Как флорист и ботанико-географ, Комаров отчетливее
всего представлял себе такую переработку именно в процессе
расселения растений. В этих работах Комаров не присоединялся
к дарвиновскому объяснению: «Новые расы (т. е. виды) лишь
в очень редких случаях образуются на почве индивидуальных
отклонений, так как перекрестное опыление и гибридизация
быстро сглаживают возникшие таким образом особенности. .
Для возникновения новой расы (т. е. вида) необходимо, чтобы
характерные ее свойства появились сразу. . у всех неделимых,
населяющих данную географическую область» (1901а : 84). Каза-
лось, что трудности, связанные с нивелирующим действием скре-
щивания, толкнули Комарова к ламаркистской гипотезе видооб-
разования путем «прямого приспособления». Однако рядом
с только что цитированной мыслью расположилось и утвержде-
ние, что «медленные, но строго определенные изменения
климата. отражаются на всех обитателях данной местности,
убивают одних, перерабатывают других» (там же 83). Значит,
признается и фронтальное изменение всех организмов и вместе
с тем отбор. Такая позиция, являясь промежуточной между дар-
винизмом п ламаркизмом, была направлена против мутационизма.
Мутационное возникновение вида из одного резкого индивидуаль-
ного отклонения, по Комарову, «подобно тому, как Минерва яви-
лась из головы Юпитера не только совершеннолетняя, но и
в блеске бранных доспехов» (там же : 82).
92
Миграционная гипотеза видообразования была разработана
Комаровым (1908) на примере рода Garagana. Одна из филоге-
нетических ветвей этого рода, по Комарову, образовалась во
время миграции предкового вида через Среднюю Азию п Джун-
гарию в Центральную Азию и, наконец, в Китай (ряд hamlagu).
11-о мере завоевания новых территорий мигрирующее население
перерабатывалось так, что в новом комплексе физико-географиче-
ских условий возникал новый вид. В результате получается ряд
видов (series), непосредственно примыкающих друг к другу гео-
графически и вместе с тем отражающих филетическпй ряд от
предков к потомкам. Идея видообразования в процессе миграций
была положена в основу объяснения филогении многих групп
растений, изучавшихся Комаровым. Миграционная гипотеза соот-
ветствовала морфолого-географическому подходу систематиков,
хотя Комаров и другие биологи принимали также и гипотезу
видообразования путем повсеместного преобразования вида в пре-
делах старого ареала (Пачоский, 1925) или путем раздробления
первичного ареала на части (Комаров, 1930).
Работы В. И. Та лиев а. «Опыт исследования видообразо-
вания в живой природе», предпринятый В. И. Талиевым (1915),
вызвал очень противоречивые оценки. Если не считать класси-
ческих исследований Н. В. Цингера, то это была единственная
крупная работа по видообразованию в России (Лебедев, 1957).
Фактическая ее основа состояла из наблюдений за изменчивостью
окраски цветков (полихроизм) и формы листьев, сравнительного
исследования видов рода Holosteum с филогенетическими выво-
дами и изучения двух близких видов рода Dentaria. Однако
центральная идея Талиева состояла в резкой критике того экс-
периментального изучения видообразования, которое было рас-
пространено тогда в генетике и в противопоставлении ему метода
наблюдений в самом природе. Как справедливо отмечает Д. В. Ле-
бедев, «диссертация Талиева была безусловно „еретической44
с точки зрения тогдашней генетики, которая не подошла еще при
последовании эволюционных процессов к комбинированию экс-
периментального метода с точными наблюдениями в природе, на
чем основана современная генетика популяций и к чему, соб-
ственно, призывал Талиев» (1965 732). В те времена мнения
авторитетов резко разошлись, и если Комаров отозвался о ней
хвалебно, то Р. Э. Регель написал разгромную рецензию, которая
заканчивалась так: «Основываться на одних голых наблюдениях,
не считающихся с экспериментальными данными, как то делал
Талиев, нельзя. Эго приводит только к наслаиванию догадок на
догадках и исключает возможность прогресса точных наук»
(1917 181).
Проблема видообразования, по Талиеву, — это прежде всего
вопрос о возникновении новых приспособлений. Однако позиция
Талиева была противоречивой и основной его вывод из конкрет-
93
ных исследований сводился к утверждению, что видообразова-
ние — это процесс расщепления более сложных (полиморфных)
комплексов на более простые. Так, например, изучение изменчи-
вости формы листьев и полихроизма цветков привело его к мысли,
что видообразование возможно только в том случае, если отдель-
ные расы, выравненные по форме листьев или по окраске цвет-
ков, так пли иначе обособятся географически. Казалось бы, эта
позиция была близка представлениям ранних генетиков, в кото-
рых видообразование сводилось к распадению первичных слож-
ных гетерозиготных комплексов. Однако Талиев не соглашался
с генетическими толкованиями и, в частности, возражал Де Фризу:
«Элементарные виды представляют собой не зачатки новых видов,
а лишь потухшие очаги деятельности видообразовательного про-
цесса» (1915:259).
Критикуя Талиева, Р. Э. Регель (1917 178) рисовал следую-
щую картину видообразования, «основанную на эксперименталь-
ных данных». Там, где в природе вместе растут и удерживаются
в борьбе за существование близкие наследственно постоянные
формы, дающие при скрещивании плодовитое потомство, полу-
чаются всевозможные промежуточные комбинации, множество из
которых вытесняется с жизненной арены, а немногие основывают
константные расы. Видообразование, таким образом, прежде всего
является следствием возникновения различных наследственных
комбинаций п их просеивания средой и конкуренцией. Однако
возможно, что существует также и процесс настоящего возникно-
вения нового (мутации или прогрессивный гетерогенезис). Про-
блема видообразования, по Регелю, — это прежде всего вопрос
о причинах того, почему число существующих в природе наслед-
ственно различных форм так мало, а не близко к теоретически
ожидаемой цифре порядка многих биллионов. Дискуссия вокруг
диссертации Талиева и характер возражений Регеля могут слу-
жить еще одной иллюстрацией кризисного состояния проблемы
вида.
Как это часто бывает, в пылу дискуссии все проходили мимо
действительно важных данных. Большой интерес представляют,
например, наблюдения Талиева за взаимоотношениями между
близкими видами — Dentaria bulbifera и D. quinquefolia. Оказа-
лось, что в Харьковской и Полтавской областях встречаются
главным образом промежуточные формы: «В действительности
не существует ни D. quinquefolia, ни D. bulbifera как обособлен-
ных систематических единиц, но имеется только некоторый
крайне полиморфный вид с чрезвычайно широкой амплитудой
колебаний признаков между крайними пределами — типом
D. quinquefolia и D. bulbifera» (Талиев, 1915 169). На Кавказе
и в Крыму, где эти виды растут совместно, встречаются как ти-
пичные особи, так и промежуточные. В США виды Dentaria ча-
сто растут совместно и образуют такие серии переходов, что
94
некоторые авторы предлагают объединять эти группы видов
в один (например, D. laciniata, по Культеру). Этот анализ Та-
лиева, проделанный полвека назад, во многом опередил время.
Подобные ситуации приковывают внимание и современных иссле-
дователей. Их можно трактовать как результат вторичной гибри-
дизации между ранее уже сформированными видами (по совре-
менной терминологии, вторичная симпатрия с интрогрессией;
см. главу 9) или же как первичную гибридизацию внутри поли-
морфного видового комплекса с пространственным обособлением
отдельных форм (в духе толкования самого Талиева).
Следует упомянуть о гипотезе гибридогенного видообразова-
ния, которая приобрела в те времена большую силу; на ее
основе пытались объяснить не только формирование целых флор
(Попов, 1927), но и все видообразование в целом (Бэтсон,
Лотси и др.).
4. ПРОБЛЕМА ЦЕЛОСТНОСТИ ВИДА
В эпоху кризиса учения о виде далеко не все биологи отка-
зывались признавать вид реальным явлением природы. Напротив,
многие ученые далее развивали и более глубоко обосновывали
учение о реальном исторически развивающемся виде и высту-
пали с критикой субъективистских идей.
Некоторые авторы рассматривали вид как реально существую-
щее явление, но трактовали его суммативно, как простую сово-
купность особей или как сумму сходных биотипов. Это банальное
и ошибочное толкование вида сохраняется и в современной лите-
ратуре. Между тем уже в начале XX в. зародилось направление,
согласно которому сложность генетической основы вида свиде-
тельствует не о сборном (составном) характере вида, а, на-
оборот, о его целостности. Первые шаги в этом направлении
были сделаны В. Л. Комаровым, Н. И. Вавиловым и И. К. Па-
чоским.
В. Л. Комаров в 1901—1912 гг. поставил вопрос о целостности
видаа но в последующих работах не развил это представление, не
поставил идею целостности вида в связь с данными по разложе-
нию вида генетическими методами, говорящими как будто не
о целостности, а о составном характере вида. Кроме того, разра-
ботка идеи целостности вида требовала экспериментирования,
разложения вида на элементы и работ по изучению интегрирую-
щих механизмов. Поглощенный систематико-флористической ра-
ботой, Комаров реже вспоминал свою замечательную формули-
ровку о виде как о сложно организованной «племенной общности
организмов».
Одним из первых генетиков, поставивших вопрос о целост-
ности вида, был Н. И. Вавилов. Именно ему принадлежала
95
заслуга открытия поразительно сложного генетического состава
многих видов культурных растений. Генетический анализ видов
путем их разложения на изолированные элементы производился
Вавиловым и его сотрудниками в масштабах, позволявших до-
вольно полно описывать «генетический потенциал» этих видов.
Но, трактуя линнеевский вид как систему, Вавилов указывал, что
вид «состоит из связанных друг с другом частей, в котором целое
и части взаимно проникают друг друга» (1931), что отдельные
генетические элементы вида «не представляют собой механически
обособленных частей» (там же). Однако в рассматриваемый пе-
риод взгляды Вавилова на целостность вида наряду с только что
цитированными правильными общими положениями включали
также и односторонне-аналитические формулировки, характерные
для всей генетики первой четверти нашего века. Идеи Вавилова
будут рассмотрены нами в следующей главе.
И. К. Пачоский о целостности вида. Первым био-
логом, во всяком случае в России, действительно начавшим раз-
работку вопроса о целостности вида, был ботаник И. К. Пачоский.
В работах 1904—1925 гг. он пытался синтезировать представле-
ния о виде как племенной общности растений с данными генети-
ческого анализа популяций. Пачоский с самого начала века, как
и Комаров, вел борьбу против музейно-морфологического пони-
мания вида, противопоставляя ему учение о географической расе
растений. Географическую расу Пачоский считал «основной са-
мостоятельно существующей единицей растительной жизни, т. е.
видом» (1907 71). Как и Комаров, он характеризовал такую
расу-вид как географическую и экологическую определенность и
выдвинул мысль о ее целостности.
Положение о целостности вида Пачоский старался развить и
обосновать. Вид-раса, по Пачоскому, — «коллективная единица
растительного бытия», она «живет своей собственной жизнью и
имеет свой собственный округ распространения» (1910а 4;
19106:5), это «известное целое, состоящее из органически свя-
занных частей» (1914:11), являющееся «фактически единицей
самостоятельной» (там же: 35).
Пачоский не считал вид простым по составу и далее не раз-
ложимым, а, напротив, полагал, что это сложное образование,
включающее в себя ряд более мелких форм пли биотипов [в его
терминологии — «элементарные расы» (Пачоский, 1904, 1914)].
Эти «элементарные расы» Пачоский сближал с жорданонами, но,
с одной стороны, в отличие от генетиков рассматривал их в ка-
честве элементов, органически включенных в состав целостного
вида, и с другой — в отличие от Комарова (1940 72) признавал
их формами многообразия вида, а не простым проявлением его
изменчивости.
Пачоского не испугал «огромный переворот, вводимый гене-
тикой в наши понятия о мелких таксономических единицах»
96
Н. В. Цингер.
(1914 34). Он не игнорировал новых данных, как многие систе-
матики, но в то же время вопреки генетикам-пеожорданистам
первичной и основной единицей считал вид — географическую
расу в целом, а не отдельную элементарную расу (биотип):
«Элементарная раса — это только один из составных элементов
более сложного целого, которое уже и является фактически еди-
ницей самостоятельной, хотя построенной сложно и не имеющей
всегда и везде вполне тождественного состава. Такой единицей
является то, что понимают под словом „впд“ (включая и широко
понятый подвид)» (там же: 35). Впоследствии эту мысль выска-
зывали и другие авторы. Например, критикуя Лотси за отожде-
ствление биотипа с видом, В. II. Малеев (1933 27) писал:
«В природе самостоятельное существование имеют не биотипы,
как таковые, а состоящая из них популяция», однако па Пачо-
ского при этом он не сослался.
Далее, Пачоский еще более ясно подчеркивает свою основ-
ную идею, указывая, что элементарная раса (т. е. биотип),
обычно не живет самостоятельной жизнью, входя в состав слож-
ного целого — вида, и именно вид «фактически является дей-
ствительной жизненной единицей» (1914 39). Принципиальная
разница между отношениями близких видов в системе рода и от-
ношениями биотипов внутри вида состоит, по Пачоскому, в том,
что живой связи между видами пет, она была лишь «в прошлом»
(т. е. род не представляет собой целостности) и, «наоборот, наи-
более мелкие расы в пределах вида — это элементы, еще пе по-
терявшие живой связи между собой, элементы, необходимые друг
для друга, могущие пока жить только сообща, но не порознь»
(там же 35).
Несомненно, что Пачоскому удалось правильно поставить дан-
ный вопрос и значительно опередить в этом отношении свою
эпоху. Важно подчеркнуть, что настоящие представления были
развиты им в период кризиса, когда в теории вида господствовал
разброд п видное место занимали идеи иеожорданистов п субъек-
тивистов.
Обсуждая вопрос о целостности вида, Пачоский имел в виду
только перекрестноопыляющиеся формы. Виды самоопылителей,
апогамов и агамов, по Пачоскому, представляют собой не органи-
ческое целое, а «простые конгломераты» биотипов, т. е. «изоли-
рованных элементов» (там же: 11). Считая непрерывное пере-
опыление единственной причиной интегрирования вида, Пачоский
(как п многие специалисты в настоящее время), не ставил во-
проса о существовании иных связей, способных цементировать
растительное население ib целостное образование (например, че-
рез химические взаимодействия и т. п.).
Пачоский не обошел и вопроса о биологическом значении
внутривидового полиморфизма. Не ограничиваясь обычными для
эволюционистов того времени указаниями, что многообразие форм
7 К. М. Завадский
97
внутри популяции есть следствие эволюции и приспособления
к -различным средам, он ставит и обратный вопрос — о многооб-
разии форм как причине устойчивости и процветания вида, как
об особом аппарате приспособления вида. Несмотря па то что
такая мысль в общей форме была высказана еще Дарвином, эта
постановка вопроса была настолько неожиданна в начале XX в.,
что не нашла тогда отклика. Ныне же именно эта постановка
вопроса наиболее актуальна, так как изучение дифференциации
вида с этих позиций подводит к пониманию его сложной внут-
ренней организации. Еще в ранней работе Пачоский (1904) вы-
сказал мысль, что у полиморфного вида, так сказать, на всякий
случай имеются подходящие неделимые, не позволяющие виду
угасать, и что в полиморфности заключается вся сила и жизнен-
ность молодых и новых форм.
Мелкие генетические единицы, по Пачоскому (1914), отли-
чаются различной способностью реагировать на условия среды.
Там, где не может удержаться одна из них, удерживается какая-
либо другая из того же цикла и т. д. Таким образом, приспособ-
ленность, необходимую для существования вида, природа здесь
распределяет между разными неделимыми, образующими все-таки
одпу до известной степени цельную группу.
Эти положения Пачоского представляют выдающийся интерес.
Экологическое мышление было еще чуждо генетикам той эпохи,
всецело поглощенных изучением константности признаков отдель-
ных биотипов п их поведением при скрещивании. Первые генети-
ко-экологические работы велись с позиций идеи о первичности
п самостоятельности биотипа, а вид толковался как явление вто-
ричное и составное, как случайный результат перекомбипацпи
биотипов и отсеивающего действия местообитания (Turesson,
1922а, 1922b, 1929 и др.; Розанова, 1928а, 1932 и др.). Не воз-
никало и мысли, что состав биотипов закономерен, что биотип —
это компонент особого адаптационного аппарата.
Не ограничиваясь положением о роли полиморфизма как спе-
циального аппарата, поддерживающего устойчивость вида, Пачо-
ский (1904, 1914) указал также и па иную его функцию: поддер-
жание высокой жизнеспособности всех особей путем образования
системы переопылений, при которой гетерозисное состояние по-
стоянно поддерживается популяцией.
Понятие о биоэкологпческом потенциале вида было предло-
жено Пачоским (1925) в более поздней работе. «Биоэкологиче-
скпй потенциал вида» — это его большая или меньшая способ-
ность к географическому расселению, к заселению новых
местообитаний и к дальнейшей эволюции и в то же время спо-
собность устоять при резких переменах в условиях. При сравни-
тельной оценке видов следует придавать большое значение
выяснению степени его перспективности в эволюционном
отношении, т. е. биоэкологпческому потенциалу. Полиморфизм
98
вида, по Пачоскому, и является одним из могучих средств,
обеспечивающих повышение его бпоэкологического потен-
циала.
Все эти мысли Пачоского до сих пор пе утратили значения
для понимания целостности вида, по особенно важен его вывод
том, что целостность вида пе противоречит его сложности,
а предполагает ее. Генетики начала века, обнаружив сложность
генетического состава вида, слишком поспешно истолковали вид
как составное явление, а Комаров п систематики-монотиписты
встали по существу на путь отрицания внутренней структуры
вида. Пачоскому же удалось наметить вехи нового воззрения иа
вид, в котором сложность его состава и целостность объясняются
как важнейшие приспособительные черты, обеспечивающие виду
устойчивость в меняющихся условиях и способность к дальней-
шему расселению и развитию. Многообразие форм внутри вида
и взаимодействия между этими формами, сообщающие виду це-
лостность, оказалось особым приспособлением, которое пе сво-
димо ни к отдельной особи, ни к отдельному биотипу и функцио-
нирует лишь па уровне популяции.
Аргументация Пачоского принуждает признать вид объек-
тивно противоречивым, т. е. и единицей жизни, и сложной си-
стемой.
Эти идеи Пачоского получили свое подтверждение и развитие
только в самое последнее время.
* * *
Подведем итог рассмотрению этого этапа изучения вида.
В первую четверть XX в. познание вида было углублено иссле-
дованием трех новых черт — географической и экологической
определенности и полиморфности (вид как многообразие форм),
было начато изучение генетической основы вида; зарождалось
изучение целостности вида. По многим руслам началось исследо-
вание путей п механизмов видообразования. Но одновременно
ломка морфологической концепции лиипеопа и замена ее концеп-
цией расы, разложение вида генетическими методами и другие
факторы привели к развитию неожорданпзма, мутациоиизма,
к усилению субъективизма.
Продолжало господствовать односторонне-аналитическое на-
правление, стремившееся разложить виды на элементы (биотипы)
и описывать их порознь. Вырванные из системы вида отдельные
биотипы рассматривались в качестве основных и первичных еди-
ниц, а вид принимался за образование вторичное, возникшее
путем скрещивания биотипов. Именно таким представлялось со-
отношение между видом и биотипом Иогансену, Лотси (Lotsy,
7*
99
1916а, 1917), Турессону (Tiiresson, 1922b) и многим другим.
Признавалось, что не биотипы возникают внутри вида в процессе
его дифференциации, а именно виды образуются путем гибри-
дизации между биотипами.
В течение первой четверти нашего века в учении о виде и
в представлениях о видообразовании произошло много противо-
речивых событий, вызвавших кризисное состояние проблемы.
Рассматривая эти 25 лет в качестве особого этапа познания вида,
мы наметили лишь его приблизительные границы, не настаивая,
разумеется, на проведении ее точно в определенном году. В сле-
дующей главе будут приведены данные, показывающие, что новый
перелом начался именно в 20-х годах.
Глава 5
НОВЕЙШАЯ ИСТОРИЯ ПРОБЛЕМЫ ВИДА
1. ЭКСПЕРИМЕНТАЛЬНОЕ ИЗУЧЕНИЕ ВИДА
Самой характерной чертой исследования вида в последние
десятилетия является разностороннее применение эксперимен-
тальных методик в сочетании с широким сравнительным изуче-
нием видов в природной обстановке. Экспериментальные направ-
ления изучения вида в современном их понимании начали
складываться в 20-е годы.1 Исследовались структуры видов, кон-
стантность и адаптивность признаков, используемых в качестве
«видовых», способы и механизмы видообразования, закономер-
ности внутривидовых отношений.
В начале 20-х годов стала формироваться экспериментальная
систематика (= дифференциальная систематика, или микроси-
стематика). Ее особенностью являлся комплексный подход к изу-
чению структуры вида и к оценке таксономического значения
выделяемых внутривидовых форм и групп. В процессе эколого-
географического изучения популяций в природе с помощью срав-
нительно-морфологических наблюдений производился сбор исход-
ного материала, так что момент, когда в прошлом изучение вида
в сущности заканчивалось, теперь становился исходным для мно-
голетних комбинированных опытных работ. Исходный материал
клонировался, выращивались генетически однородные линии,
или же исходный материал изучался в нескольких поколениях
в условиях свободных или принудительных скрещиваний. Опыты
велись в питомниках, различающихся географически (климати-
чески) или экологически (например, высотой над уровнем моря)
По мнению некоторых ученых (Clausen, 1958; Мауг, 1963), экспери-
ментальный метод в систематике впервые был применен Линнеем
в 1760 г. В действительности пионерами экспериментального изучения
вида были ученые второй половины XIX в. (Жордан, Кражан и др.).
101
и т. п. Формы, взятые из популяций, различающихся морфоло-
гически или эколого-географически, выращивались в одном пи-
томнике при максимально однородных условиях. Для изучения
генетического потенциала вида проводились диаллельпые скре-
щивания с гибридологическим посемейным анализом потомства.
Для широко расселенных видов, включающих многие тысячи по-
пуляций, такой трудоемкий метод требовал большого числа со-
трудников и усилий в течение целых десятилетий. Однако он
вполне себя оправдывал прп изучении не только таких важней-
ших видов, как кукуруза, пшеница, свекла пли сахарный трост-
ник, по и таких методически удобных, как дрозофила, львиный
зев, божья коровка и т. п. Для экспериментального изучения вида
большую роль сыграла генетика природных популяций. Важное
место заняла также сравнительная и экспериментальная кариоло-
гия и кариосистематика, экспериментально-экологические и физио-
логические исследования с воздействием разливными факторами
для выяснения амплитуды адаптационных реакции у предста-
вителей разных популяций одного вида или близких видов,
сравнительно-биохимический анализ форм, и т. и. Большой вес
приобретали работы в области отдаленной гибридизации и по-
липлоидии, которые также проводились комплексно с примене-
нием цитогенетических, эмбриолого-физиологических и других
методик.
Экспериментально-экологическое направле-
ние микросистематики. Основателями этого направления
были многие ученые. В Швеции Г. Турессон (Turesson, 1922а,
1922b, 1925, 1929, 1931) изучал растения одного вида, взятые из
различных местообитаний (леса, дюны, теневые и осветленные
места и т. п.). Экспериментально расчленив признаки на модифи-
цирующие п фиксированные, отражающие особенности местооби-
таний, Турессон обосновал понятие об экотипе. Экотипом он на-
звал генотипически закрепленную форму, соответствующую опре-
деленному местообитанию. Подчеркивая, что проблема вида «все
больше становится экологической», Турессон настаивал на негео-
графическом (локально-экологическом) характере экотипов.
Однако позднее он применяет понятие «экотип» к широко рассе-
ленным расам (например, южный и северный, сибирский и ев-
ропейский). Это вызвало путаницу понятий. Е. Н. Спнская (1931)
предложила подразделить экотипы на климатические, эдафичс-
ские и цепотические, а М. А. Розанова (19286) ввела понятие
«климатип» и приняла «следующие таксономические единицы
в пределах conspecies: climatipus, oecotipus и isoreagent» (Роза-
нова, 1930 95). Климатип здесь трактуется как единица высшего
ранга, охватывающая группу экотипов и совпадающая с геогра-
фической расой( т. е. с подвидом или видом). Г. И. Поплавская
(1930) на примере Aster amellus из Крыма показала, что явно
климатические экотипы (горный и альпийский) не являются гео-
102
графическими расами. Г. Турессон (Turesson, 1931) предложил
понятие «эковпд» (oecospecies). Эковид — это группа экотипов и
вместе с тем сформированная экологическая раса, входящая в со-
став политипического вида (coenospecies).
С 1922 г. М. А. Розанова (1932) начала анализ трудно рас-
членяемого комплекса лютиков — conspecies Ranunculus aurico-
mus. Это было комбинированное экспериментальное изучение,
проводившееся «на генетической и эколого-географической ос-
нове». Книга Розановой (1930) о современных методах система-
тики растений была, видимо, первой в мире попыткой обобщить
успехи экспериментальной систематики. В работах, посвященных
«экспериментально-генетическому методу в систематике» и ка-
сающихся понимания низших (внутривидовых) единиц, Роза-
нова (1928а, 1935) доказывала необходимость широкого разви-
тия дифференциальной систематики и на экспериментальных об-
разцах показывала, как это надо осуществлять. Экологический
аспект в ее экспериментальных работах был, однако, менее выра-
жен, чем в исследованиях, например, Турессона. Розанова (1940)
чаще интересовалась степенью константности признаков, чем их
адаптивным значением, хотя в обобщениях уделяла много вни-
мания экологическим факторам возникновения рас и видов. Итоги
ее деятельности были подведены в монографии «Эксперименталь-
ные основы систематики растений» (1946).
В том же 1922 г. появилась первая работа Е. Н. Синской,
посвященная экспериментальному изучению изменчивости внутри
видов. Этим было положено начало более чем сорокалетней мно-
гообразной деятельности Синской. Наиболее широко она изучила
проблемы экологической дифференциации видов. Ею было обос-
новано понятие об экоэлементе как внутрипопуляционном зачатке
экотипа и об экотипе как этапе видообразования. В 20-х годах она
занималась межвидовыми скрещиваниями культурных форм рода
Brassica, полиморфизмом популяций Vaccinium, экспериментально
изучала экологическую изменчивость многих видов крестоцвет-
ных, комбинируя эту работу со сравнительно эколого-географи-
ческими наблюдениями популяций в природе, уточнила данные
Н. В. Ципгера по видообразованию рыжика Camelina sativa в ус-
ловиях сорного образа жизни, уподобления основной культуре
(ржи) и климатической дифференциации п др. В обширных опы-
тах она изучала экотипы и популяции многих видов растений
(1938, 1939 и др.). Синская осуществила классические исследо-
вания и иного профиля. Они касались не аутоэкологии внутриви-
довых форм, а познания видов в их взаимоотношениях с расти-
тельным покровом (1931) и истории видов в связи с эволюцией
биоценозов (1933). В этих работах было показано, что вид не-
обходимо исследовать в теснейшей связи с теми сообществами,
элементами которых являются его популяции. Изучение лесных
формаций Кавказа в связи с филогенетической историей видов
103
не было отходом Синской от основного экспериментально-эколо-
гического направления. Для широкого обобщения закономерностей
эволюции видов, которое было сделано ею позднее (1948, 1958,
1961 и др.), было необходимо объединение экспериментально-
генетических и экологических методик со сравнительно-географи-
ческими и эволюционно-филогенетическими.
В США основы генэкологии были заложены Дж. Клаузеном
(Clausen, 1921, 1922). Изучая «коллективный вид» Viola tricolor
генетико-аналитическими методами, Клаузен выделил среди массы
модификаций 19 константных признаков. Далее скрещиваниями
была установлена возможность существования 5 млн различных
фенотипов, образованных сочетаниями этих 19 признаков. Ока-
залось, что существующий в природе полиморфизм V. tricolor
является очень небольшой частью возможных комбинаций. «Поли-
гон скрещиваний в пределах группы V. tricolor огромен», но
большая часть комбинаций устраняется отбором. Сравнение форм,
полученных в эксперименте, с формами из природы, различаю-
щихся экологически или географически, позволило Клаузену рас-
шифровать генетическую основу внутривидовой дифференциации
этого вида. Затем Клаузен с сотрудниками (Clausen a. Hiesey,
1939; Clausen et al., 1940; Hiesey et al., 1942, и др.) исследовали
много других видов. Убедившись, что описательные приемы ра-
боты старой систематики имеют «интуитивную подкладку», Клау-
зен доказывает, что необходимо экспериментально проверять каж-
дый их вывод и что проще работать, не оглядываясь на границы
видов и более мелких единиц, установленных старой системати-
кой (Clausen, 1951).
В Англии Дж. Грэгор, Ф. Сансом, У. Туррилл и др. (Gregor
a. Sansome, 1927, 1930; Gregor, 1930, 1939; Turrill, 1935, 1938)
на пастбищных злаках, подорожниках и других растениях изу-
чали признаки экотипов, степень их константности, изменения
признаков по территории и т. п.
Экологический и экспериментальный подход к изучению вида
осуществлялся и в зоологии. Еще Н. А. Холодковский (1910) об-
наружил у хермесов расы, отличающиеся по эколого-физиологи-
ческим признакам, и назвал их биологическими видами. Впослед-
ствии экологическая дифференциация была обнаружена у многих
видов. Например, у малярийного комара были найдены расы, от-
личающиеся местообитанием и питающими хозяевами, у кукуруз-
ного мотылька — три «пищевых» расы — хмелевая, конопляная й
кукурузная, у американского сверчка (Oecanthus) — расы кустар-
никовая и древесная, а также различающиеся по способам яйце-
кладки и особенностям стрекотания, у рыжего муравья —
экологические расы (лесная и луговая) и т. п. (Арнольдп,
1941).
Существование экотипов у животных было экспериментально
показано Т. Сэмнером п другими авторами па американских гры-
104
лунах Peromyscus polionothus. На выходах лавы живут темно-
окрашенные зверьки, а на песках — светлые, особенности этих
форм оказались наследуемыми.
Обобщая мировой опыт экологического изучения видов жи-
вотных, Д. Н. Кашкаров подчеркивал, что зоологи отстали от бо-
таников, так как значительно меньше опираются на изучение
экологии п ограничиваются «вопросами чистой систематики и
географического распространения, пользуясь к тому же линнеев-
гким понятием вида» (1933 191). Далее Кашкаров одобрительно
отзывается о выводах Турессона, для которого «линнеевский вид
есть абстракция, популяция биотипов» (там же). Интересны дан-
ные Кашкарова по экологической дифференциации видов из
фауны пустынь. Таковы экотипы «поведения», способные, кроме
обычного местообитания, занимать еще и другие. Формы с такими
реакциями рассматривались им в качестве начальной фазы видо-
образования, вслед за которой возникают морфологические адап-
тации. Примером могут служить подвиды ящериц гекконов, внут-
ривидовые формы горной куропатки (кэклика) и т. п. Далее нами
будет показано, что, сетуя на зоологов, Кашкаров был не совсем
прав, так как в начале 30-х годов уже широко проводилось эк-
спериментальное изучение природных популяций у методически
удобных видов животных, подобных дрозофиле.
Рассмотрим подробнее цикл генетико-микросистематических
работ на божьих коровках из группы Epilachna chrysomelina, вы-
полненных в 20—30-е годы Н. В. Тимофеевым-Рессо1вским с сот-
рудниками (1965). Изученная группа форм божьих коровок
включает пять морфолого-географических подвидов. Из них че-
тыре широко расселены в Южной Африке (ssp. capensis) и Эк-
ваториальной Африке (ssp. reticulata), Восточной Африке, Ара-
вии, Юго-Западной и Средней Азии (ssp. orientalis), в Северо-
Западной Африке и Южной Европе (ssp. chrysomelina). Один
подвид имеет небольшой ареал в Восточной Африке (ssp. lim-
bricollis), в пределы которого на севере внедряется ssp. orienta-
lis, а на юге — сразу два подвида (capensis и reticulata). Реаль-
ность всех подвидов была доказана не только географо-морфоло-
гическим методом, но и многолетним биометрическим анализом
сорока количественных признаков. Было обнаружено, что наи-
меньшая степень дивергенции наблюдается между популяциями
одного и того же подвида, а между подвидами chrysomelina и
orientalis, а также capensis и reticulata дивергенция более сильна.
Наибольшая же дивергенция была между подвидами евразиато-
североафриканскими, с одной стороны, и центрально-южноафри-
канскими — с другой. Изучались модификационные возможности
жуков из разных популяций различных подвидов. Разные подвиды
по-разному реагировали на одну и ту же температуру (например,
различно менялась интенсивность пигментации элитр, общая
площадь пятен и т. п.).
105
Генетические исследования сводились к подвидовым скрещи-
ваниям п анализу ряда признакдв. Они подтвердили выводы
о степени дивергенции подвидов, опиравшиеся ранее лишь па
морфологические данные. При скрещиваниях между евразиато-
североафриканскими и центрально-южноафриканскими подвидами
оплодотворенные яйца погибали сразу либо па ранних стадиях
развития, хотя копуляция и была нормальной. Прп скрещиваниях
между географически смежными и морфологически близкими
подвидами (например, между orientalis и chrysomelina и др.)
потомство получалось всегда нормально жизнеспособным и пло-
довитым. Эти скрещивания подтвердили необходимость разде-
ления группы Е. chrysomelina на два вида: Е. chrysomelina
(включая оба средиземноморские подвиды) и Е. capensis (вклю-
чая южно- и центральноафриканские подвиды). Судьба же ssp.
limbicollis, как отмечают авторы, еще неясна. Морфологически
эта раса близка к жукам типа orientalis, но в Экваториальной
Африке в популяциях ssp. reticulata встречаются особи с темным
пронотумом, типичным для ssp. limbicollis, и, наоборот, в попу-
ляциях последнего встречаются особи с более светлым пропоту-
мом, который характерен для восточноафриканского эпдемично'го
подвида. Однако, сознавая неудобство сохранения этого подвида
во «впевидовом» положении, авторы включают его в состав ев-
разиатско-североафрпканского вида.
Большой раздел работы был посвящен изучению наследования
отдельных признаков. Скрещиваниями и селекцией анализирова-
лись многие признаки, по> которым наблюдались различия между
популяциями и между подвидами (форма тела в профиль, густота
волосков на элитрах, площадь черной пигментации в пятнах
и т. п.). Ряд признаков оказался полимерным, а некоторые под-
виды, хотя и расщеплялись моногибридно, поддавались отбору
в разных направлениях (т. е. их проявление частично зависело
от многих генов-модификаторов). Эти работы велись 18 лет, и
общие выводы заключались в доказательстве существования внут-
ривидовых различий по массе признаков и наличия ясной града-
ции степеней дивергенции.
Авторы делают вывод о двух путях образования подвидов:
1) путем гибридизации в зоне контакта уже сформированных
рас и 2) путем постепенного распространения признака из одной
популяции по соседним.
Значение таких работ состояло в применении более точный
определений варьирования признаков, в определении степени ди-
вергенции и судьбы многих признаков в поколениях. Однако на
растительном материале в те годы микросистематики разработали
уже экологический подход и начинали исследовать внутрипопу-
ляционные группы. В рассматриваемой же работе, изучая харак-
тер наследования отдельных признаков, авторы подвергали по-
пуляции травмирующим воздействиям (цапример, длительньщ
Юв
ппцухт и др.). Для чисто генетпческйх целей такая методика
хороша, но для сравнительного изучения природных популяций
непригодна, так как разрушает их. Несмотря на большой мате-
риал, побывавшим в руках авторов, они не располагали доста-
точно представительными выборками природных популяций.
Поэтому исходный материал был подходящим только для изуче-
ния наследования отдельно взятых признаков.
О значении генетики природных популяций.
Для исследований видообразования большое значение имели ра-
боты в области эволюционной генетики и в особенности генетики
природных популяций, начавшиеся в 20-х годах. Важнейшую
роль в создании генетики популяций и изучении процесса видо-
образования сыграла работа С. С. Четверикова (1926). На дро-
зофиле, собранной из природной популяции с Кавказа, Четвери-
ков показал, что однородность касается только фенотипических
признаков, а генотипически материал оказался гетерозиготным и
насыщенным многими мутациями. Оказалось, что в природной
популяции в скрытой форме существует огромный резерв разно-
образных мутаций. Эти мутации служат сырым материалом для
отбора п являются основой, с помощью которой отбор взламывает
установившееся в популяции равновесие и вызывает эволюцион-
ный сдвиг. Если нет мутаций или же нет отбора, вид застынет.
Только противоречивое взаимодействие между мутационным про-
цессом и отбором делает популяции не только мобильными,
но и способными к направленным преобразованиям. Для воз-
никновения нового вида, по Четверикову, помимо благоприят-
ного взаимодействия мутагенеза и отбора, необходима также
п изоляция части старого вида, которая бы не допустила сво-
бодных скрещиваний между дифференцирующимися частями
вида.
На истинно экспериментальную почву было поставлено изу-
чение видообразования. Впервые видообразование рассматрива-
лось генетиком не как формообразование, мутация или комбина-
ция отдельных признаков у одиночных организмов, а как процесс
переработки целых популяций, управляемый взаимодействием
мутагенеза и отбора.
Основываясь на первых работах по генетике популяций,
С. Райт, Дж. Холден и Р. Фишер в начале 30-х годов разрабо-
тали математические модели естественного отбора и предложили
гипотезы эволюции доминантности. Значение генетики популяций
для теории вида, особенно на примере отечественных работ, на-
столько полно обсуждается в книге Н. П. Дубинина (1966), что
это позволяет нам подробно не останавливаться на данном во-
просе.
Отметим только, что важные и разнообразные исследования
по генетике природных популяций дрозофилы в 30—40-х годах
были осуществлены Дубининым с сотрудниками, Ю. М. Олено-
107
вым, Р. Л. Берг п др. Особое значение имело подтверждение
данных Четверикова о насыщенности популяций массой мутант-
ных генов, накопленных исторически благодаря отбору гетерози-
гот на повышение уровня доминирования адаптивной нормы.
Были обоснованы идеи о «мобилизационном резерве наследствен-
ной изменчивости» (С. М. Гершензон), об интеграции генетиче-
ских систем, поддерживающих гетерозисность на основе исполь-
зования гетерозиготности, и др.
Обширные исследования по генетике природных популяций
различных видов дрозофилы в 30—60-е годы были выполнены
Т. Добжанским совместно с Б. и Н. Спасскими, О. Павловской
и др. Изучались направления отбора, роль изоляции, генети-
ческого гомеостазиса, генетического груза (load), полимор-
физма, вопросы о видах-близнецах, о биологической роли ре-
цессивов, сравнивалась хромосомная изменчивость разных ви-
дов и островных популяций с континентальными и многое
Другое.
Для теории вида принципиальное значение имела идея об
уровне мутабилыюсти как адаптивном признаке вида, регулируе-
мым отбором (Дубинин, 1934; Шапиро и Игнатьев, 1945, и др.).
Был поставлен вопрос о существовании коренного противоречия
между тенденцией к поддержанию эволюционной пластичности
вида, основанной на накоплении большого фонда мутаций, и к за-
щите адаптивной нормы вида от дезорганизующего дейст-
вия мутационного процесса. С этой точки зрения мутабильность
вида является не только причиной, но и следствием эволюции.
Наиболее адаптивным, видимо, должен быть такой уровень мута-
бильности, при котором не происходит разрушения уже испытан-
ной на эффективность организации, но обеспечивается накопле-
ние фонда мутаций, достаточного для поддержания эволюционной
пластичности вида.
Что касается предложенной Райтом и Дубининым гипотезы
генетического дрейфа, или генетико-автоматических процессов, то
их роль в видообразовании до сих пор остается неясной. Дуби-
нин (1931) упрекал Четверикова за переоценку роли отбора.
В настоящее время можно с уверенностью сказать, что Четвери-
ков отвел отбору как раз подобающее места. Что же касается
роли генетико-автоматических процессов, связанных со случай-
ными изменениями в концентрации мутаций из-за колебаний
численности популяций, то до сих пор о них нельзя говорить
как об одном из постоянно действующих и всеобщих факторах
видообразования, существующих помимо отбора, т. е. как о само-
стоятельной причине эволюции (Оленов, 1961).
Внутривидовые отношения. Изучение внутривидовых
отношений, не выходившее до 20-х годов за рамки отрывочных
наблюдений, благодаря усилиям целой группы ученых, связавших
его с проблемой борьбы за существование, становится теперь на
108
экспериментальную почву, а также снабжается математической
основой. Именно в этот период к исследованию чрезвычайно за-
путанных явлений внутривидовых отношений впервые приме-
няется принцип моделирования, упрощающий связи в опыте пли
в формулах.
А. П. Шенников (1921) выдвинул задачи изучения проблем
фитоценологии в условиях опытных питомников. Вышли первые
экспериментальные работы Н. Н. Кулешова (1922) о борьбе рас
яровой пшеницы и А. А. Сапегпна (1922) о взаимоотношениях
форм внутри сортосмеси. Несколько позднее подобная работа
появилась и в США (Clements a. Weaver, 1924). В. Н. Любименко
с сотрудниками (1925) изучали соревнование за место в условиях
различных плотностей посева, включая сильное перенаселение,
у ряда видов однолетних растений. Впервые в мире в опытах
по изучению межбиотипной борьбы за существование и отбору
исследуется влияние различных загущений и состава популя-
ции па ее численность и дифференциацию и широко применяется
статистический анализ материала. Это были классические
работы В. Н. Сукачева (1927, 1935, 1941) по изучению сорев-
нования между клонами одуванчика Taraxacum officinale, ов-
сянницы Festuca sulcata и некоторых других видав растений,
а также работы, выполненные его учениками (Сочава, 1926; Смир-
нова, 1928; Успенская, 1929, п др.) на других объектах.
Н. В. Красовская (1931) экспериментально изучала сорев-
нование главным образом между корневыми системами растений.
Ссылаясь именно на эти работы, Дж. Холден (Haldane, 1932) пи-
сал, что дарвинизм в Советском Союзе «поднят на большую
высоту».
Зоологи также начали экспериментальное изучение борьбы за
существование. А. Ильинский (1929) показал зависимость между
плотностью п динамикой численности популяции малого; сосно-
вого лубоеда. В. Полежаев (1934) анализировал такие же зависи-
мости у тополевой моли. В. В. Алпатов экспериментирует с плот-
ностями популяций ряда видов животных и пишет интересный
критический обзор литературы (1934).
Возникает математическое направление в изучении борьбы
за существование, и в течение 20-х годов по этому вопросу пуб-
ликуется ряд работ (Вольтерра, Лотка, Томсон, Перль).
Оригинальные экспериментально-математические работы
Г. Ф. Гаузе (1934, 1935, 1939, Gause, 1934) по борьбе за сущест-
вование у простейших вошли в большинство сводок по теории
эволюции и до сих пор цитируются в качестве классических (см.:
Grebecki et al., 1961 —1962; Huxley, 1963; Механизмы биологиче-
ской конкуренции, 1964:406, и др.). Обобщая обширные наблю-
дения в заповедниках, С. А. Северцов (1936, 1941) строил мате-
матические модели борьбы за существование в популяциях птиц
и млекопитающих.
109
Вместе с тем в ряде экспериментов над животными было ус-
тановлено существование оптимальной плотности, при которой
лучше всего осуществляются все основные процессы жизнедея-
тельности (скорость обмена, рост, активность поведения и, в ча-
стности, пищевая активность, половое созревание и половая ак-
тивность). Во многих случаях оказалось, что при малых плот-
ностях, и в особенности при одиночном существовании, резко
падают не только перечисленные показатели, но снижается и
жизнеспособность, падает иммунитет и т. п. В США Робертсон на
инфузориях показал явление взапмостпмуляцип размножения
(«аллелокатализ»), Р. Чэпмен на мучном жуке выявил наличие
оптимальной и угнетающей размножение плотностей популяции
и открыл, что каннибализм может быть использован в качестве
механизма регуляции плотности (Allee, 1931; Алпатов, 1934;
Парк, 1939, и др.). Таким образом, в 20—30-х годах развернулось
экспериментальное изучение внутривидовых отношений, включав-
шее исследования как борьбы за существование, так и механиз-
мов, поддерживающих адаптивную норму вида.
К а р и о с и с т е м а т и к а и экспериментальная гиб-
ридизация. По этим вопросам сильные группы образовались
в США, СССР, Японии и в Скандинавских странах. Р. Клеланд,
А. Блексли, Д. Бергнегр и другие цитогенетики в работах, продол-
жавшихся около 15 лет, тщательно изучили кариогеографию дур-
мана (Datura stramonium и другие виды), ослинника Oenothera
onagra и других видов, включающих в свой состав ряд типов
структурных гибридов (различающихся транслокациями, числом
колец и т. п.). У дурмана было обнаружено 19 хромосомных типов,
приуроченных к девяти липнеевскпм видам и 680 морфологически
различным формам (микрорасам). Одни из кариотипов дурмана
оказались очень широко расселенными чуть ли не по всем конти-
нентам, а другие узкоэндемпчпымп (например, только па Гавай-
ских островах и в Южной Африке).
Важным направлением было изучение полиплоидных рядов
форм и установление отношений между плопдностью и система-
тическим рангом формы. Делается попытка установить зависи-
мость степени плоидности рас от их эколого-географических осо-
бенностей. В начале 20-х годов Г. Текхольм показал на розах, что
число хромосом у северных рас больше, чем у южных. В 1927 —
1941 гг. О. Хагеруп проверял это заключение па расах Empetrum,
Bicornes, Vaccinium и других родов: повышение плоидности он
связывал с суровостью условий вообще (не только север, но, на-
пример, и Сахара). В 30-х годах появилась серия работ, в кото-
рых приводились факты, опровергающие зависимость между
числом хромосом, ареалом и стойкостью вида. К такому выводу,
в частности, пришли Клаузен, Кэк и Хизи, показавшие, что рас-
пространение специфичных по числу хромосом рас (видов и
экотипов) Viola, Achilea, Potentilla и других групп не соответ-
110
т.вует «правилу крайних условий». В это же время появились
аботы, показывающие, что мпогоплоидпые расы и виды филоге-
етически более молоды, чем диплоидные (в СССР работы
L. П. Соколовской и О. С. Стрелковой). Г. Стеббинс (Stebbins,
950), работая с тремя расами Eriogonum в Калифорнии, также
бнаружил, что тетраплоидпая раса приурочена к флористически
олее молодым формациям, чем диплоидные.
Работы по кариосисте'матпке в Советском Союзе в 20—30-х
одах были теснейшим образом связаны с экспериментальной
пстематпкой и с эволюционными проблемами вообще. Под руко-
одством Н. И. Вавилова во Всесоюзном институте растениевод-
тва сложилась группа цитологов-кариоспстематиков, выполнив-
ши фундаментальные исследования как по отдельным группам
например, по злакам — Авдулов, 1931), так и по общим пробле-
ем карпоспстематикп и ее отношению к эволюционной теорий
Левптскип, 1931, 1939 и др.), а также появились работы
I. Н. Делоне, М. С. Навашина и т. д. В 20—30-х годах в Японии
о цитогенетике и кариосистематике работают многие ученые
Кихара, Кувада, Кагава, Матузима и др.). X. Матсуура (Matsu-
ra, 1935) установил существование карио-экотппов. У Fritillaria
amschatgensis триплоидные расы оказались более влаголюбивыми,
дпплопды — горными (и менее конкурентными).
В заключение необходимо упомянуть, что в этот же период
чепь усилился интерес к работам по межвидовой и межродовой
пбридизацип. Данные работы проводились не только ради созда-
ия новых видовых форм, хотя это направление всегда было са-
ым важным. Экспериментальная гибридизация проводилась
акже и для воссоздания форм видов, которые рассматривались
ак гибридные по происхождению.
Отдаленная гибридизация была использована для синтеза
овых форм И. В. Мичуриным еще в начале века (подробнее
б этом см.: Завадский, 1948а, 1956). Межродовая гибридизация
синтезом новой видовой формы на цитогенетической основе
первые была осуществлена Г. Д. Карпеченко (1927). В 1935 г.
тот же автор написал «Теорию отдаленной гибридизации».
l. Мюнтцингу (Muntzing, 1930) также удалось получить гибрид-
мфидпплоид, сходный с природным видом (см. главу 9). Вскоре
же много экспериментаторов шло по пути экспериментального
есинтеза видовых форм (Розанова, 1938а и 19386 — на малине
землянике; Рыбин, 1935, 1951 — синтез культурной сливы на ос-
ове скрещиваний терна с алычей, и многие другие).
Кроме работ по ресинтезу и синтезу новых форм с помощью
тдалепной гибридизации, изучение видообразования осуществля-
ось и иными методами. Ряд работ, например, был посвящеп ана-
изу генетической природы различий между близкими видами
Клаузена на Viola, Баура на Antirrhinum, Блексли па Datura,
др)-
J11
2. РАБОТЫ Н. И. ВАВИЛОВА
Крупнейший вклад в экспериментальную систематику расте-
нии был сделан в 20—30-х годах Н. И. Вавиловым и его много-
численными сотрудниками. Главной особенностью этих работ
было сочетание генетических испытаний материала в питомниках
с «географическими посевами» образцов на многочисленных стан-
циях с ботанико-географическими и экологическими наблюде-
ниями в природе. Невиданной была широта охвата материала:
если во времена гербарно-экскурсионной систематики каждый вид
был представлен небольшим числом образцов, то при системе,
организованной Вавиловым, он изучался на тысячах образцов.
Исследования внутривидового разнообразия форм охватывали
массу видов растений от хлебных злаков и корнеплодов до мас-
личных и бахчевых культур. Внутри каждого вида изучалось мно-
жество популяций, по возможности из всех частей ареала и из
разных местообитаний. В питомниках проводились скрещивания
между массой образцов, устанавливались генетические особен-
ности каждой географической и экологической формы. Если до ра-
бот Н. И. Вавилова о внутривидовом разнообразии культурных
растений имелись лишь отрывочные данные, то теперь «генети-
ческий потенциал» многих видов удавалось изучать с ранее неви-
данной полнотой.
В итоге Вавиловым и его школой были сделаны многие важ-
ные обобщения.
Центры происхождения видов и центры разно-
образия форм. Учение о центрах происхождения было осно-
вано па изучении огромного гено-географического материала
по массе видов культурных (Вавилов, 1926) и отчасти диких
(Cooper, 1926) видов растений. Установленные Н. И. Вавиловым
центры — это сосредоточие гибридного разнообразия форм на от-
носительно небольших территориях, так что «максимум различных
генов располагался на минимальной территории». Основные^
положения этого представления базируются на данных о приуроА
ченности генетического и фенотипического внутривидового и ви-
дового разнообразия к областям распространения древних цивили-
заций и ко многим горным странам. Как полагал Вавилов, эти
районы мира, наиболее богатые «генофондом», и являются ареной
видообразования. Иначе говоря, признавалось, что видообразова-
ние приурочено к центрам происхождения, а формирование
окончательных ареалов происходит уже после видообразова-
ния. Видообразование трактовалось главным образом как процесс
рекомбинации богатого генного материала при гибридизационных
процессах. В духе генэкологическпх идей 20-х годов рассматри-
валась и дальнейшая судьба возникших форм: местообитания
отсеивают большинство комбинаций и сохраняют подходящие.
Возникшие в этих центрах виды являются вначале не географи-
112
Н. И. Вавилов.
ческими расами, а типичными эковидами (oecospecies) в смысле
Турессона. Формирование географического ареала — это результат
миграции уже сложившихся видов, т. е. не первичный по от-
ношению к видообразованию, а вторичный процесс. Согласно
идеям Вавилова (1926), молодые виды характеризуются первич-
ным эндемизмом. Формирование окончательного ареала представ-
ляет собой в основном процесс реализации уже ранее образован-
ных эволюционных возможностей, а не приобретение таких воз-
можностей в процессе миграции. Так же решался вопрос и
о развитии экоареала видов: число осваиваемых популяциями
вида различных местообитании в большей мере определяется его
бпоэкологическим потенциалом (Пачоский), т. е. генетико-адапта-
ционной гибкостью, приобретенной в прошлом, чем взаимодей-
ствиями со средой в настоящее время. Для Вавилова был харак-
терен исторический подход: настоящее вида в основном
определяется его прошлым.
Фактическая основа учения Вавилова о центрах происхожде-
ния покоилась па огромном числе данных о сосредоточии разно-
образия современных форм в довольно узколокалытых районах.
Понятие о центрах концентрации разнообразия в отличие от по-
нятия о центрах происхождения не включало в себя ничего гипо-
тетического. Это была только констатация фактов. В течение 20-х
годов сам Вавилов и многие его сотрудники посетили десятки
районов концентрации разнообразия форм во всех частях света.
Во время экспедиций Всесоюзного Института Растениеводства
были собраны свыше 150000 образцов более чем по 200 культурам.
В течение 30-х годов «коллекции ВИРа» выросли чуть ли не втрое
и превосходили все подобные собрания, в том числе и американ-
ские. Были обследованы многие районы Азии, Северной и Южной
Америки, Африки, тщательно изучены все районы древних куль-
турных центров Средиземноморья, Закавказья, Средней Азии
и т. п.
Такие центры разнообразия форм были установлены для пше-
ниц и многих других злаков, Горохов и других бобовых, миндаля,
абрикоса и других плодовых, многих корнеплодов, табака, карто-
феля, кукурузы, подсолнечника, хлопчатников и др. Вавилов счи-
тал, что районы максимального разнообразия форм являются
близкими к экологическому оптимуму вида п иногда совпадают
с центрами происхождения вида (если к ним приурочено много
доминирующих признаков), а у климатических границ ареалов
вид часто представлен генетически сильно обедненными популя-
циями. Кроме понятия о центрах разнообразия форм, было вве-
дено также понятие центра распространения вида. В качестве
критериев центра были предложены такие признаки, как макси-
мальная эколого-генетическая дифференциация (полиморфизм)
популяций и наибольшее разнообразие самих шшуляций (по вы-
ражению Вавилова, максимальное формовое разнообразие),
8 К. М. Завадский
113
а также максимальное обилие, максимальное развитие особи
(размеры, продолжительность жизни), максимальная плодови-
тость («коэффициент жизненности» — Cooper, 1926), наибольшее
доминирование вида в ценозах и участие его популяций в наи-
большем количестве ассоциаций. Таким образом, понятие центра
распространения оказалось сложным и весьма емким как с эколо-
гической, так п с генетической точек зрения.
Чем же объясняется тот парадоксальный факт, что очаги
разнообразия культурных растений оказались, как правило,
приуроченными к горным районам, где никогда, разумеется,
не велось планомерной селекции? Объясняется это, возможно,
тем, что древние земледельцы культивировали взятые из природы
целые популяции и мало изменили их искусственным отбором
(Вавилов, 1926, 1935а; Вульф, 1944). Ныне, зная об огромном
генетическом потенциале популяций многих диких видов растений
и животных, не следует удивляться самому факту такого разнооб-
разия форм в горных очагах земледелия. Генетический потенциал
их диких родичей был огромен, по жесткий отбор в условиях
конкуренции в естественных биоценозах, поддерживал один или
несколько (по немного) наиболее адаптивных фенотипов. Гено-
фонд же таких видов состоял из массы рецессивных аллелей.
Фенотипическое (формовое) разнообразие могло возникнуть
только в культуре, при ослаблении отбора, разрушении естествен-
ных ценозов и т. п.
Учение Вавилова о центрах было подкреплено целой серией
монографий (К. А. Фляксбергера и Е. Ф. Пальмовой по пшени-
цам, А. И. Мальцева по овсам, Е. И. Барулиной по чечевице,
Л. И. Говорова по гороху, С. В. Юзепчука и С. М. Букасова
по картофелю, К. И. Пангало по бахчевым и др.). Дальнейшая
разработка проблемы показала, что центры происхождения видов
часто не совпадают с современными центрами разнообразия и
распространения видов. Простейшие случаи таких несовпадений —
массовое размножение в Австралии видов европейского и амери-
канского происхождения или в Европе многих видов американ-
ских вредителей и сорняков. Многие виды и в центре распростра-
нения не доминируют, встречаются одиночно или редко, а их
популяции по составу не отличаются от периферических. Было
введено понятие «центр экологического оптцмума вида» (Cooper,
1926) и показано, что к области экологического оптимума при-
урочен и центр разнообразия видов. Только в области экологи
ческого оптимума популяции вида способны запять максимальное
число местообитаний (стаций), быть наиболее ценотически доми-
нирующими, обильными и т. п. (Синская, 1933, 1948; Odum, 1959;
Andrewartha, 1961, и др.).
Остановимся теперь кратко на критике, которой подверглись
представления Вавилова о центрах происхождения. Некоторые
отвергали идею локализации видообразования в очагах с последу-
Ш
1ощим «распаковыванием» и обеднением первичного генного
клубка за ее близость идеям Бэтсона. При этом суждения о гене-
тической основе эволюции основывались не на фактических дан-
ных, которыми располагал Вавилов, а на критических соображе-
ниях, высказанных в начале века К. А. Тимирязевым и т. п. Такая
критика, хотя она и была очень резкой (обвинения в бэтсопиап-
стве, лотсиапстве и т. nJ, по существу пе затрагивала основ уче-
ния Вавилова и была направлена против отдельных неудачных
формулировок.
Ныне же ясно, что основа учения Вавилова о центрах базиро-
валась на строгом материале и пе была связана с идеями Бэтсона
об эволюции как процессе обеднения генотипов путем выпадения
доминантных генов. Вместе с тем надо напомнить, что и Бэтсон и
Вавилов знали об исследованиях, проведенных еще в конце прош-
лого века и показавших, что приспособительную эволюцию можно
толковать как выбор образца из потенциально возможного много-
образия форм. Например, из множества гибридных комбинаций
ячменей подавляющее большинство оказалось слабо жизнеспособ-
ными или неприспособленными и из сотен в культуре сохранилось
только две-три формы (Rimpau, 1891).
В 20—30-е годы очень мало были изучены темпы мутагенеза,
по был твердо установлен тот факт, что генетически различных
устойчивых форм возникает на несколько порядков величин
больше, чем сохраняется. Современные знания по генетике попу-
ляций подтверждают необходимость различать теоретически воз-
можные комбинации, фактически осуществляющиеся, по погиба-
ющие из-за летальности, существующие короткое время, но пе
закрепляющиеся из-за слабой адаптивности или же вытесняемые
в силу недостаточной конкурентоспособности и, наконец, устой-
чиво воспроизводящиеся формы. В пределах видовых комплексов
может существовать 108—1016 теоретически возможных комбина-
ций, но осуществляется только 105—108, из которых устойчиво
адаптивными бывают только единичные формы или у прогресси-
рующих полиморфных видов 102 —103 форм. Очень отчетливо изме-
нения порядков этих величин было показано не только па культур-
ных, но и на диких видах (например, Viola tricolor—Clausen, 1921,
1922, 1951), а также у декоративных (у гиацинта наблюдались,
например, десятки тысяч морфологически различных форм,
из которых образовалось 5000 сортов, но только несколько десят-
ков из них удержалось в современном ассортименте — Матвеева,
1965). Теперь известно, что видообразование происходит на основе
использования уже ранее накопленного резерва мутаций и может
сопровождаться как обеднением, так и обогащением исходного
генофонда. Возможно, что большинство вновь возникших видов
характеризуются именно обеднением генетического резерва.
Такие виды эволюционно пе перспективны, но они возникают
чаще, чем виды — носители прогресса с высокой концентрацией
8*
115
генетических возможностей, сообщающих им большую эволю-
ционную лабильность и неугасающую приспособительную ак-
тивность.
В итоге можно сказать, что идеи Вавилова, развитые в учении
о центрах происхождения, гораздо ближе современным научным
представлениям, чем ламаркистская концепция адаптивной пере-
делки видов в процессе миграции или при изменении среды после
сформирования ареала, а также чем гипотеза массовой фронталь-
ной перечеканки видов по всему ареалу. Теперь известно, что
далеко не обязательно ареной видообразования должен являться
весь ареал и формирование ареала должно происходить одновре-
менно с видообразованием (Хохлов, 1946). Но несомненно, что
видообразовательный процесс нередко оказывается приуроченным
не к центрам, а как раз к периферии, на которой наиболее ве-
роятно длительное действие преобразующего отбора.
«Линнеевский вид как система». Работа Н. И. Ва-
вилова (1931) под этим названием обобщала знания о виде с пози-
ций генетики, кариологии, ботанической географии и эксперимен-
тальной экологии. Неправильно думать, что в этой концепции
нашло отражение изучение только вида у культурных растений
(Базилевская, 1959) и что опа неприменима к характеристике
вида у других групп растений и животных. Представления о виде
как системе были далее развиты в последующих работах Вави-
лова.
Главная идея этой концепции лучше всего была, пожалуй,
сформулирована так: «Для того чтобы охватить генетически вид,
чтобы подойти к проблемам эволюции и формообразования в свете
новейших знаний, мы должны исходить из видов как целостных
комплексов — систем, стремиться по возможности к исчерпанию
разнообразия наличного состава вида» (Вавилов, 1932:25). Это
действительно исчерпывающее знание генетического состава вида,
по Вавилову, возможно «только путем детального изучения жор-
данонов и генотипов» (1935а: 7).
Н. И. Вавилов писал, что история познания вида характерна
стремлением уложить в стройную систему различные внутривидо-
вые подразделения, «число которых растет по мере углубления
методов распознавания наследственных форм, изучения новых
образцов... Линнеевские виды приходится разбивать на подвиды,
разновидности; разновидности в свою очередь — на расы. Генети-
ческие исследования последних лет посягнули, однако, и на неде-
лимость расы, т. е. мельчайшей морфо-физиологической единицы
систематики (элементарных видов в смысле Де Фриза) и выяс-
нили, что за внешней однородностью могут скрываться различные
генотипы» (там же : 3).
Итак, главная проблема в теории вида — познание его гене-
тического состава, т. е. аналитическое изучение фенотипов из всех
точек ареала и генетическое исследование их потенций.
116
Вот, например, как описывал Вавилов успехи в аналитическом
изучении мягкой пшеницы: «Если 10 лет тому назад мы опреде-
ляли число жорданонов мягкой пшеницы Triticum vulgare Vill.
приблизительно в 3000, то ныне. . . эта цифра должна быть уве-
личена во много раз, и мы не можем определить ее даже прибли-
зительно. При этом дело идет не об искусственных гибридах,
а главным образом о местных формах пшеницы, возделываемых
в различных районах Азии и Европы» (там же 4). Тогда учи-
тывалось свыше 400 генетически значимых признаков, по которым
различаются формы мягких пшениц. При свободном комбиниро-
вании их между собой возможно образование многих мил-
лионов генетически различных внутривидовых форм. Однако
фактически существует, как сказал Н. И. Вавилов в 1939 г., при-
мерно 50 000 различных форм, а остальные комбинации элимини-
рованы из-за летальности, плохой приспособленности или малой
хозяйственной ценности. Оказалось, что столь же велико разно-
образие форм не только у всех видов культурных растений, по и
у любого дикорастущего вида, если он глубоко изучен в прпроде и
в эксперименте. По Н. И. Вавилову, почти каждое дерево дикой
алычи по каким-либо особенностям наследственно отличается
от других, число форм дикой яблони, груши, айвы, граната, мин-
даля «с трудом поддается учету», так же как и внутривидовое
разнообразие сорно-полевых растений, диких клеверов, донника,
желтой люцерны и других кормовых злаков и бобовых. Многие
тысячи генетически различных форм были обнаружены среди
видов тыквенных (работы К. И. Пангало и др.). Упомянув об от-
крытии большого полиморфизма у ели Picea excelsa (работа
Впттрока), у анютиных глазок и т. п., Н. И. Вавилов делает
следующий общий вывод: «Монотипные виды существуют обычно
только до того времени, пока они изучаются в гербарии. Изучение
их в культуре на большом числе образцов неизбежно вскрывает
полиморфную природу видов» (там же 6.)
Таким образом, вид представляет собой систему подчиненных
внутривидовых единиц (1931, 1935а, 19356). Однако наряду с та-
ким пониманием слов «вид как система» у Вавилова встречается
и иное, в котором вид трактуется как обособленная, сложная,
подвижная морфо-фпзиологическая система, связанная в своем
генезисе с определенной средой и ареалом (1931). Слово «система»
в этом случае приобретало иной смысл (не система подчиненных
внутривидовых единиц, а система признаков). Если формулировка
«вид как сложная система внутривидовых единиц» обобщала
успехи аналитической работы, то представление о виде как си-
стеме признаков заслуживает критики (см. следующий раздел).
Следует напомнить, что, рассматривая вид как систему подчинен-
ных единиц, Вавилов вместе с тем писал и о целостности вида
(см. главу 4), т. е. он уже сталкивался с противоречивостью
вида — сочетанием составного характера и целостности. В пони-
727
мание вида как системы входила так>ке и меч^га о создании си-
стемы признаков, схожей, например, с системой Менделеева.
Н. И. Вавилов рисует увлекательную картину будущей система-
тики, которая от примитивного «запоминания бесчисленного мно-
жества форм, называемых по месту нахождения пли в честь лиц»,
перейдет к построению стройных систем форм, как это в свое
время с системами веществ было выполнено химией (1935а: 51).
Такие системы должны быть построены на основании точного
сравнительного анализа форм по отдельным признакам и по ско-
реллированным группам признаков, которые, по мнению Вави-
лова, подчиняются сформулированному им в 1920 г. закону гомо-
логических рядов в наследственной изменчивости. Этот закон
обобщает данные по параллельной изменчивости признаков мно-
гих впдов культурных и отчасти диких растений. Например,
у пырея (Agropyrum repens) и у житняка (A. cristatum) был
обнаружен одинаковый спектр различий (полиморфизм) по 13
признакам (остистые и безостые формы, с чешуямп гладкими и
опушенными, с лежачим, полуразвалистым и прямым кустом,
желто-, красно- и черноколосые. с тонкой и толстой соломиной,
узко- и широколистные, плотно- п рыхлоколоспые, с восковым
налетом и без него, высокорослые и низкие, с опушенной и глад-
кой листвой, раннеспелые и позднеспелые, ксероморфные и гидро-
морфные, с желтыми и фиолетовыми пыльниками). Н. И. Вавилов
полагал, что этот закон поможет упорядочить описание и класси-
фикацию множества внутривидовых форм и тем самым станет
основой изучения структуры вида.
3. ВИД —ЕДИНИЦА ИЛИ СИСТЕМА?
«Учение о виде у растений». Монография В. Л. Ко-
марова (1940) под таким названием представляла собой первый
па русском языке обобщающий труд по проблеме вида. Его автор
подводпл итог почти полувековой работе над изучением ви^а.
Основные идеи этой работы были сформулированы следующим
образом: «Реальность видов. совершенно очевидна и не нуж-
дается в доказательствах» (там же : 30); «вид есть этап эволюци-
онного процесса, а не вещь, замкнутая сама в себе» (там же : 5);
«истина о виде уже высказана Ч. Дарвином» (там же 6). Если
до 30-х годов Комаров в общем склонялся к концепции эволюции,
представляющей собой компромисс между ламаркизмом и дарви-
низмом, то в последней книге он занял дарвинистские позиции,
учитывающие достижения генетики и цитологии. Он подчеркивал,
что вид — основа исследований не только систематики, но и эво-
люционной теории п селекционной работы. Комаров предложил
определять его следующим образом: «Вид есть совокупность поко-
лений, происходящих от общего предка и под влиянием среды и
борьбы за существование обособленных отбором от остального
118
шра живых существ; вместе с тем вид есть определенный этап
процессе эволюции» (там же: 212). Особый интерес представ-
яет последняя (18-я) глава о диалектике вида. Она содержит
еликолеппые по точности и новые формулировки о виде как од-
ом из «способов осуществления жизни», об отношении к вопросу
реальности вида: «Совершенно неправы те авторы, которые
ытались изображать вид как прием классификации, как нечто
словпое... вид есть объективный факт» (там же : 202).
Вместе с тем нельзя не отметить, что эта книга содержала ряд
порных положений. Так, например, развивая на протяжении
сей книги пдею об универсальности явления вида и о необходи-
ости понимать вид как совокупность поколений, связанных
динством происхождения и бытия, в конце автор говорит, что
нет никакой надобности каждый индивид относить непременно
определенному виду, он может относиться пе к виду, а к гиб-
иду или к мутации, или к категории мепделяптов... под словом
вид“ мы подразумеваем совокупность поколений, происходящих
ибо вегетативным путем, либо через автогамию, гейтоногамию
ли ксеногамию, но без скрещивания с соседними видами, а тем
олее, не путем так называемого отдаленного скрещивания»
там же : 208—209).
Всю первую четверть века Комаров характеризовал вид как
леменную общность, а в монографии 1940 г. эта прогрессивная
юрмулировка была заменена значительно менее точной, в кото-
ой вид назывался организмом. Согласно В. Л. Комарову, обра-
овапие каждого отдельного вида надо рассматривать как «движе-
пе по восходящей линии» (там же: 207). Такое отождествление
роцесса видообразования с прогрессивной эволюцией непосред-
твенно вытекает из известной философской формулировки,
тождествляющей всякий процесс развития с переходом от пиз-
iero к высшему. Соответствующая цитата и приведена В. Л. Ко-
маровым перед утверждением, что образование любого нового
ида является процессом восходящей эволюции. Выражение «вид
сть этап на пути развития» (там же) на первый взгляд идеп-
ично трактовке вида как «узлового пункта» в процессе эволюции,
'акое понимание вида пе учитывает того, что многие впды,
озпикнув, оказались способными лишь к персистированию, да и
о иногда существуют геологически недолгое время. Вряд ли такие
иды можно называть этапом эволюции, так как они пе являются
сиователямп новых филогенетических ветвей.
Игнорируя данные экспериментальной систематики, Комаров
трицал существование внутривидовых единиц и сводил все к изу-
ению индивидуальной изменчивости. Такая позиция была след-
твпем концепции таксономической неделимости вида.
В. Л. Комаров раскритиковал предложение Вавилова опреде-
ять вид как особую живую систему: «Вид как морфо-фпзполо-
ическад система пе подходит, цотому что и индивид, и род, и
119
семейство также морфо-физиологическая система. Вид как система
подвидов, разновидностей, рас, форм и пр. не подходит, так как
мы уже видели, что вполне сложившийся вид по существу такой
системой не является, да и вообще вид есть факт, а система
(не совокупность, а система) как будто логическое построение»
(там же: 206). Правда, несколькими страницами ниже можно
найти и иное по духу утверждение, включающее не только само-
критику, по и признающее пригодность слова система: «Если
считать вид системой, то, конечно, это не морфолого-биогеографи-
ческая система (Комаров, 1927) и не морфо-физиологическая си-
стема (Вавилов. 1931), а система поколений (происходящих
от общего предка)» (там же 209). Это критическое высказыва-
ние Комарова нельзя признать четким и последовательным.
Так п оставалось неясным, что же такое вид — единица или
система?
Столкновение двух концепций вида. Две крупней-
шие в СССР школы ботапиииков Комарова и Вавилова почти два
десятилетия вели дискуссию по вопросу об объеме впда. Одна
из них была представлена макросистематиками, морфологами и
флористами, работавшими над «Флорой СССР». Другая — микро-
систематиками, растениеводами и селекционерами-генетиками,
работавшими над сортами и коллекциями ВИРа. Первые занима-
лись видом в системе рода и считали свою работу законченной,
дойдя до вида. Вторые ставили своей целью изучение внутренней
структуры вида; они начинали с вида и стремились дойти до наи-
более элементарных его частей (генетических и географо-экологи-
ческих). Для макросистематиков естественным концом работы
считалось установление «вида как тождества», в то время как
микросистематики с этого момента только начинали свои изыска-
ния, стремясь найти как можно больше «различий внутри тожде-
ства». Одно и то же явление изучалось как бы при разных увели-
чениях: при малом увеличении можно было распознать только
отдельные виды и каждый из них казался далее неразложимым
и элементарным явлением; при большем увеличении тот же вид
оказывался очень сложным образованием, целым «микрокосмо-
Sfcl 1 *
СОМ». *'•
К тому же авторы «Флоры СССР» условились понимать вид
в малом объеме (вид — одна географическая раса), а микро-
систематики толковали его в объеме conspecies, пользуясь полити-
пическпм стандартом.
По Комарову, «вид не система, а обособлении! в процессе эво-
люции отдельный организм. Вид может включать в себя систему
подчиненных ему форм, но сам он не система, а единица. Если
вид „система44, то истинной единицей... придется признать био-
тип» (там же: 204). Из этой выдержки следует, что Комаров
возражал против понимания вида как составного образования,
но не настаивал на абсолютной его однородности.
120
В противоположность этому школа Вавилова рассматривала
вид как явление чрезвычайно сложного состава. По Вавилову, мо-
нотипными виды кажутся обычно до того времени, пока они рас-
сматриваются в гербарии. Экспериментальное же изучение их не-
избежно вскрывает полиморфную природу видов. В результате
расчленения популяций генетическими методами, обобщения
опыта акклиматизации, селекции и широких географических посе-
вов в школе ВИРа сложилось представление, что вид — комплекс
биотипов (Розанова, 1928а, 19286, 1930), система экотипов, эко-
элементов и изореагентов (Спнская, 1938), что «вид является да-
леко не последней неразложимой систематической единицей наи-
меньшего объема, а представляет собой сложный комплекс мелких
константных единиц» (Малеев, 1938 19) и т. д. По Г К. Крейеру,
линнеевские виды следует рассматривать «системами эколого-гео-
графических типов. Мы тем самым не подвергаем их такому
суждению, которое низводило бы эти виды до объема географи-
ческих рас и тем более экотипов путем извлечения из липноев-
скпх видов чего-то нетипичного и оставления за ними чего-то ти-
пичного» (1941 43).
В общем для микросистематиков вид — система, состоящая
из множества иерархически и сетчато связанных между собой
единиц различного объема п различного таксономического до-
стоинства.
Вместе с тем следует помнить, что обе школы содействовали
критическому пересмотру старого стандарта морфологического
вида — линнеона. Действуя разными методами (географо-морфо-
логическим и дифференциальным географо-эколого-генетическпм),
они способствовали разрушению старой концепции вида и разло-
жению музейно-морфологического линнеона. Правда, результаты
этого разложения различны. Авторы «Флоры СССР» разложили
липнеон на географические виды (расы). В. Л. Комаров писал,
что «ряд (series) как бы заменяет собой линнеевский вид, распа-
дающийся в процессе эволюции на современные географически
локализированные реальные виды» (1934:8). Дифференциально-
генетический метод раздроблял линнеон «на огромное число хо-
рошо различимых наследственных форм. Линнеевский вид, счи-
тавшийся в XIX в. основной конечной единицей, в XX столетии
разбит селекционерами и систематиками на большое число жорда-
нонов» (Вавилов, 19356 4).
Сравнение «конечных продуктов» разложения линнеона на-
глядно показывает глубокое различие целей в работах этих школ.
Школа Комарова не хотела идти дальше уточнения границы вида-
расы и как бы закрывала глаза на то, что .«атом разложцм»,
а школа Вавилова разлагала вид на элементы. Увлечение морфо-
лого-географическим методом чаще всего приводило к возведению
в ранг вида региональных экотипов или подвидов, т. е. форм,
все же завоевавших известное географическое пространство.
121
Пульверизация вйдой Генетическими методами Гайт в себе опас-
ность возврата к толкованиям неожорданпстов, отождествивших
«истинный» вид с биотипом, клопом пли элементарным видом и
microspecies (в смысле Де Фриза и Лотси). Такое дробление
представляет собой произвольное перенесение понятия «вид»
на область явлений принципиально иной, более нпзкой «размер-
ности».
Дискуссия 30-х годов отражала известную односторонность
взглядов, явившуюся следствием разделения труда специалистов.
Обе школы оперировали положениями, содержащими часть
истины, и полагали, что они исключают друг друга. В действи-
тельности же они дополняли друг друга, так как вид — и единица,
и система.
Вид, как расселившееся, сформировавшееся и обособившееся
племя растений, в эволюционном процессе и во взаимоотношениях
со средой выступает как качественно определенная единица.
Вместе с тем вид более или менее сложен по составу. Внутри
себя вид однороден только в той мере, в какой все его элементы
обладают одной и той же (видовой) специфичностью. В рамках
видовой специфичности наблюдается дифференциация. Вид высту-
пает как многообразие форм. Наличие внутренней струкутуры,
т. е. определенным образом связанных между собой элементов и
их групп, дает право рассматривать вид как систему.
Об объеме вида у пшениц. Ценность тех пли иных
толкований легче сопоставлять па примерах наиболее изученных
групп. Возьмем, например, пшеницы. Вначале рассмотрим, ка-
ковы были взгляды обеих школ па пшеницы двузернянки (эм-
меры, пли полбы — Triticum dicoccum). С. А. Невский (1934),
руководствуясь монотипическим стандартом вида В. Л. Комарова,
возвел в ранг видов целый ряд сортовых групп двузернянок, рас-
сматриваемых представителями школы Н. И. Вавилова (например,
К. А. Фляксбергером) только в качестве форм. Т. dicoccum var.
ferrum f. armeniacum и f. volgense были обозначены Невским
соответственно как Т. armeniacum и Т. volgense. Под названием
Т. dicoccum, ранее охватывающим все географические расы,
формы и сорта двузернянок, Невский оставил лишь формы
из Германии. В противоположность этому, по Вавилову (19356),
вид Т. dicoccum должен трактоваться широко, включая в себя
три географических подвида: абиссинский, возделываемый также
в Индии п в Йемене; азиатский, распространенный в Грузии,
Армении, Северном Иране н Малой Азии; н европейский, возде-
лываемый в горных областях Западной Европы.
Оба автора (Невский и Вавилов) пользовались морфолого-
географическим методом, но тем пе менее пришли к противопо-
ложным результатам. Очевидно, что сам по себе морфолого-гео-
графический метод не ведет к дроблению или укрупнению стан-
дарта. Эти тенденции связаны непосредственно с узким (мопотп-
122
ппчоским) или широким (политиппческим) толкованием объема
вида.
Описание новых мелких видов полбы и ликвидацию Невским
вида Т. dicoccum в его общепринятом объеме Вавилов считает
«очень необоснованным... и неприемлемым ни для ботанических,
пи для селекционных целей» (там же 9). «Если следовать Нев-
скому, — замечает Вавилов, — то мягкую пшеницу (Т. vulgare или,
но Невскому, Т. aestivum) надо разбить на десятки видов»
(там же). Понимая вид как монотипную географическую расу,
Невский выделил в пределах Т. dicoccum три вида, образующих
один ряд (series) в смысле Комарова, а Вавилов рассматривал
Т dicoccum как один политипический вид. Из этого примера
видно, что обе школы признавали реальное существование эколо-
го-географических рас и необходимость объединения группы род-
ственных рас в высшие таксоны. Расхождения между этими шко-
лами по существу сводились лишь к тому, как именовать геогра-
фическую расу и группу близких рас: видом и рядом (series) пли
же подвидом и видом.
Итак, эта дискуссия в основном носила номенклатурный ха-
рактер. Не замечая этого, С. В. Юзепчук (1958) пытается дока-
зать, что дискуссия комаровцев со сторонниками политипического
вида затрагивала принципиальные основы учения о виде. Между
тем аргументы, выдвинутые Юзепчуком в пользу «Комаровской
концепции вида», также носят номенклатурный характер. Политп-
ппческпп стандарт вида, по Юзепчуку, следует отвергнуть по мо-
тивам его неудобства: удобнее обозначить одним словом «вид»
молодые экологические и географические расы и сформированные,
древние, «хорошие» виды; удобнее не упоминать в названиях низ-
ших таксонов и при помощи такого умолчания «превращать» вид
в конечную таксономическую единицу; удобнее, наконец, но от-
ражать в названиях филогенетические связи групп, а фиксировать
сам факт существования данной расы, и т. п. Нельзя, разумеется,
такую аргументацию рассматривать в качестве раздела теории
вида.
Посмотрим теперь, как будет выглядеть вся система 28-хромо-
сомпых пшениц в целом, если объем вида будет установлен иа ос-
новании одного физиологического критерия, как па этом наста-
ивают Добжанский (Dobzhansky, 1955, 1958), Майр (1947), и
многие другие.
Если вид понимать как группу популяций, хотя бы потенци-
ально способных к внутригрупповым скрещиваниям, то шесть
больших видов Вавилова (Т. durum, Т. polonicum, Т. turgidum,
Т. dicoccum, Т. persicum, Т. orientalis) и еще несколько мелких
видов Невского должны быть объединены в один вид.1 Этот об-
1 По современным данным (Жуковский, 1964), в эту группу входит
И видов, часть которых имеет сложное строение. Например, эммер
123
ширный вид будет сложно-политипическим. Вид в таком объеме
это и есть coenospesies в понимании Клаузена (Clausen, 1951),
т. е. видовой комплекс или superspecies (надвид), по Майру.
Coenospecies может охватывать целую группу видовых рядов
(series) в смысле Комарова и целую группу видов в смысле Ва-
вилова. Виды в объеме школы Комарова соответствуют подви-
дам (subspecies), а иногда и экологическим расам в понимании
Вавилова, Клаузена п зоологов (Л. С. Берг, Майр, Кэйн и др.).
Из этого сопоставления видно, что представители всех точек
зрения на объем вида исходят из признания эколого-географиче-
ской расы в качестве реально существующей в природе и ка-
чественно специфической единицы.
Вернемся, однако, к нашему объекту. В видовой комплекс
или надвид, согласно требованиям определений Добжанского и
Майра, нами были введены пока только шесть видов 28-хромо-
сомных пшениц из восьми стоящих в списке Вавилова (19356 8—
9), так как 2 расы этих пшениц, а именно Т. dicoccoides (дикая
двузернянка с Ближнего Востока в объеме Кёрпике) и Т. ti-
mopheevii (из Западной Грузии в объеме Жуковского) при скре-
щиваниях с другими расами 28-хромосомиых пшениц, обнаружи-
вающих частичную пли значительную стерильность, заслуживают
возведения в ранг видов.
Встает вопрос о ранге этих рас с точки зрения сторонников
применения чисто физиологического критерия. Решение этого
вопроса будет зависеть от толкования выражения «потенциаль-
ная способность к скрещиваниям». Допустим, что этому тезису
придан абсолютный характер. Тогда линяю разъединения надо
проводить там, где генетико-физиологические системы разошлись
столь далеко, что ни при каких условиях и экспериментальных
воздействиях нельзя получить хотя бы единичные экземпляры
фертильных потомков. При такой трактовке в наш падвид пше-
ницы следует включать не только все виды 28-хромосомных пше-
ниц, но и все 14- и 42-хромосомпые виды пшеницы, а также
все те виды пырея и эгилопса, с которыми осуществлена гибри-
дизация пшениц (см. например: Bowden, 1959). Но такой «над-
вид» явно бессмыслен. Он не будет отражать фактически су-
ществующих в природе отношений.
Таким образом, попытка создать «стандарт практического
вида» в форме одного шаблона универсального значения не увен-
чалась успехом.
Концепция «Комаровского» монотипического вида подверглась
критике со стороны ряда зоологов (см. например: С. Чернов,
(Т. dicoccum) включает пять географических подвидов и многие десятки
экотипов. Систематика некоторых из них до сих пор очень запутана, и
объем, границы и состав их еще спорен. Поэтому естественно, что до сих
пор описываются новые виды (Т. ispaghanicum, Т. georgicum, Т. monta-
num).
124
1941; Огнев, 1944; Берг, 1950), однако она мало опиралась на
теорию вида, больше основывалась на технико-номенклатурных
доводах (удобство работы с определителями, преимущества три-
парной номенклатуры и т. п.). В критических работах зоологов
па первый план были выдвинуты мотивы практического удобства
пользования крупными таксонами и неудобств чисто приклад-
ного характера при работе с мелкими классификационными еди-
ницами.
В этой дискуссии, к сожалению, обе стороны совершенно не-
достаточно опирались на положение об объективной неравноцен-
ности видов, которое по существу надо было бы положить
в основу практической концепции вида. Одни пытались нивели-
ровать различия, наблюдающиеся в объеме видов, стремясь под-
вести все виды под крупномерный стандарт, другие — под мало-
мерные стандарты.
Неравнозначность видов — основное явление, и попытки игно-
рировать его могут только усилить крайне нежелательный разрыв
между теорией вида как синтезом всех современных знаний и
«стандартом практического вида» как инструментом работы си-
стематиков. К сожалению, в систематике больше исходили из
технических удобств того пли иного стандарта вида, нежели из
основ современной общебиологической теории вида. Выход
один — отбросить саму идею вида как единицы музейной работы
и опираться на современную общебпологическую концепцию
вида как па единственную основу работы систематиков.
Признание вида единицей не исключает возможности призна-
ния его в то же время и сложным образованием, включающим
разнородные элементы, т. е. системой. Здесь нет альтернативы.
Вид — единица, отдельность и в то же время он система, много-
образие. Несмотря на то что это положение вряд ли можно
оспаривать, до последнего времени оно продолжает быть дис-
куссионным.
4. ТРУДНОСТИ В ПРИМЕНЕНИИ ПРАКТИЧЕСКИХ
СТАНДАРТОВ ВИДА
После внедрения в практику систематики морфолого-геогра-
фического и морфолого-экологического методов трудности в обос-
новании границ вида в ряде случаев нисколько не уменьшились.
В 30-х годах ведущее положение занял морфолого-географиче-
ский стандарт политипического вида (conspecies), которым очень
широко пользовались в зоологии, а за рубежом и ботаники.
Среди ботаников Советского Союза в те годы переживал расцвет
морфолого-географический стандарт монотипического вида. Среди
части ботаников, микробиологов и в меньшей мере зоологов стал
применяться морфолого-генетический стандарт монотипического
мелкого вида (жорданона, пли microspecies). В конце 30-х годов
125
начала пропагандироваться концепция так называемого биологи-
ческого вида, представляющая собой политипический морфолого-
географический стандарт вида, подкрепленный применением кри-
терия скрещиваемости (Huxley, 1938а; Майр, 1947; Dobzhansky,
1955, и др.). После выхода в 1940 г. сборника «Новая систематика»
(«The New-Systematics») и введения понятия о непрерывной
изменчивости признаков (clines, по Huxley, 1938b) делались
попытки ввести в стандарт биологического вида также и приемы
количественной (математической) оценки таксономического зна-
чения признаков. В последнее время они вылились в направле-
ние «числовой таксономии», предлагающей обрабатывать коэф-
фициенты различий по десяткам признаков на электронно-счет-
ных машинах. Однако точность такого метода диагностики
только кажущаяся (Терентьев, 1966), так как признаки имеют
совершенно различное таксономическое значение и математиче-
ская обработка необоснованно избранных признаков — занятие
бесперспективное.
В поисках объективных основ диагностики видов систематики
обратились не только к цитологии, генетике и математике, но
п к серодиагностике, иммунологии, энзимологии и другим раз-
делам биохимии, цитофизиологии и к другим наукам и методи-
кам. В систематике различных царств и групп при различной
степени изученности материала в зависимости от различных по-
зиций систематиков сложилось большое число теоретических
моделей вида, которые выступают в качестве стандартов при их
описании. Систематики-специалисты по отдельным группам —
ихтиологи и альгологи, энтомологи и микологи, бактериологи,
гельминтологи и т. п. — ввели в обиход массу своих стандартов,
при сопоставлении которых обнаруживаются целые цепи сино-
нимов (пли полусиноппмов). Эти терминологические ухищрения
создают видимость специфичности объекта, и через эти словес-
ные преграды подчас с трудом пробиваются общебиологические
принципы теории вида.
Трудности возникали еще и по той причине, что один и тот же
термин употреблялся во многих смыслах.
Не надо забывать, что эти осложнения с разработкой новых
стандартов практического вида в систематике проходили в усло-
виях кризиса учения о виде. Несмотря па пестроту точек зре-
ния, все же можно сказать, что главные направления развива-
лись по двум старым руслам: дробительства и укрупнительства
вида. Новой была только аргументация, которая стала гораздо
более разнообразной.
Тяга к увеличению числа стандартов вида и к новой терми-
нологии имела и серьезные объективные основания, так как на-
копился большой материал о неравноценности видов. Она обна-
руживалась в самых различных планах. Трудно было
сопоставлять объемы, сложность внутренней структуры, харак-
126
гор выражеппостй границ видов микроорганизмов, простейший,
например, с темп же показателями у видов высших растений и
животных; видов с агамным размножением — с «зиготными»
видами, имеющими половое размножение, и т. п. Трудно было
сопоставлять молодые виды, у которых многие признаки еще
трансгрессируют, с хорошо отграниченными, сопоставлять объемы
и границы видов, образующих обширные гибридные циклы, с ви-
дами, размножающимися бесполосеменным путем. Неэквива-
лентны и виды, широко расселенные па обширных равнинах,
узкоэндемичпым видам изолированных горных долин и т. и.
Следует исходить из того, что в идеале число стандартов
должно соответствовать числу неравноценных друг другу типов
видов. Только при глубоком понимании особенности каждого из
этих типов видов можно будет создать такие стандарты, которые
были бы вполне адекватны природному типу вида и не служили
бы для многих из них прокрустовым ложем.
Таким образом, осложнения с пониманием вида в новейшее
время представляют собой результат сплетения объективных и
субъективных причин. Приведем теперь некоторые примеры по-
нимания практического стандарта вида в палеонтологии, микро-
биологии и в систематике групп, трудно поддающихся класси-
фикации.
О стандарте вида в палеоспстематике. Нами
будут обсуждены лишь те аспекты проблемы вида в палеонто-
логии, которые представляют общебиологический интерес. Совре-
менная палеосистематика служит двум хозяевам, потребности
которых несовместимы. Во-первых, опа является отраслью палео-
биологии, изучающей конкретные филогенезы групп. Вид для
палеобиологии — основной предмет исследования. Во-вторых, она
обслуживает биостратпграфию, для которой вид «руководящее
ископаемое», т. е. маркировочный знак, используемый для рас-
членения горизонтов, их синхронизации и для классификации
осадочных пород. Вид для геологов-биостратиграфов — фунда-
ментальное, ио чисто служебное понятие.
С вопросом об объеме и границах вида палеонтологи испы-
тывают столь же большие затруднения, как и неонтологп. В пале-
онтологии накапливалось все больше данных в пользу того, что
«вид во времени» [выражение Д. М. Федотова (1940) и В. Л. Ко-
марова (1940)], или «фратрия» (Кэйн, 1958), изменяется посте-
пенно, т. е. так, как это было констатировано в случаях
непрерывной (клинальной) географической и экологической из-
менчивости. Изменение ископаемых популяций с крайне мелкими
ступенями и постепенными переходами в ряду, поколений может
быть названо «хроноклином». В этих случаях полностью оправ-
дывались слова Ф. Энгельса о том, что развитие в природе ино-
гда осуществляется столь мелкими скачками, что они становятся
незаметными.
127
По мере накопления материалов, относящихся к одной и
той же серин, палеонтологи получили возможность наблюдать не
только отдельные куски вида во времени, представляющие об-
рывки изменяющихся стволов, но целые крупные отрезки радип-
рующих фратрий. Эти материалы доказывали, что адаптивная
радиация является основным путем формирования многообразия
видов-фратрий.
Только палеонтологии доступно прямое исследование вида
как совокупности множества поколений и видообразования как
исторического процесса. Если бы не огромная неполнота ее дан-
ных (Чернов, 1959; Ротай, 1962), можно было бы шаг за шагохм
проследить в последовательных отложениях процесс преобразо-
вания популяций и картировать ход видообразования одновре-
менно во времени и в пространстве. Однако очень часто плохая
фоссилизация создает гигантские пробелы. Из многих миллиар-
дов особей в течение сотен тысяч лет часто пи одна не фоссили-
зируется или же сохраняются* только некоторые части органов.
Такпе пробелы до сих нор создают почву для поддержания мута-
ционистских и пеокатастрофических идей (Степанов, 19596).
Палеонтологи оказались участниками развития кризиса тео-
рии вида и неразберихи со стандартом практического вида в си-
стематике. Эти противоречивые тенденции привели к большому
разнобою в понимании вида в палеонтологии. Помимо борьбы
«дробителей» с «укрупнителямн», среди палеосистематиков все
еще бытует идея о субъективности понятия о виде и о принци-
пиальном различии между понятием вида в палеонтологии и
в систематике современных групп. Такая мысль высказывалась,
например, А. А. Борисяком (1947), А. Кэйном (1958). В палео-
систематике концепция вида иногда сохраняет еще черты старо-
модного морфологического вида и некоторые специалисты
указывают, что именно такой тип вида, как бы изъятый из му-
зеев XVIII в., все же является самым удобным инструментом
для работы. Однако в палеонтологии, по-видимому, верх одержи-
вает современная политиппческая концепция широкого вида
(Степанов, 1959а), чаще всего эквивалентного первым надвидо-
вым таксонам систематиков-пеонтологов. Степень изученности
материала — важный фактор при определении объема стандарта
практического вида. С этой точки зрения правы те палеонтологи,
которые пользуются крупномерным стандартом вида. Надо только
помнить, что такой «вид» — отражение действительности в пер-
вом приближении и значительная часть таких видов будет в даль-
нейшем разукрупнена. В последние годы появились бпостратп-
графы, выступающие за дробительство видов. Стремление
к выделению мелкого стратиграфического вида имеет следую-
щую подоснову.
Определение относительного возраста пород на основании
ископаемых остатков — такова задача биостратиграфии. Но раз-
128
. । мчать горизонты iio ископаемой фауне и флоре возможно .ришь
при условии достоверного изменения хотя бы одного вида или же
целого комплекса видов. Таким образом, вид является основной
единицей стратиграфического исследования только в том случае,
если продолжительность его существования в данных отложе-
ниях невелика. Вид теряет значение руководящего ископаемого,
если существует в течение геологически долгого времени. Страти-
графическое значение вида возрастает с уменьшением сроков его»
существования. Это обстоятельство и явилось поводом для тен-
денций некоторых палеосистематиков к неоправданному дроби-
тельству и к утилитарному обоснованию понятия «стратиграфи-
ческий вид».
Обратимся к примерам. Некоторые авторы считают непра-
вильным объединять в один вид самые близкие формы из раз-
ных горизонтов (Ананьев, 1956; Наливкии, 1964, и др.), так как
боятся, что вид потеряет стратиграфическое значение. Такое
чисто утилитарное «установление вида для стратиграфических
нужд» привело Д. В. Наливкина к смешению понятия о наи-
меньшем различии, улавливаемом палеонтологическими мето-
дами, с понятием вида в палеонтологии. «Выделение видов, ха-
рактерных для определенных горизонтов, полезно, а раз оно
полезно, значит, и правильно в теоретическом отношении» (На-
ливкин, 1964:34). Далее предлагается виды, не имеющие стра-
тиграфического значения, оставлять в большом объеме, а страти-
графические виды предельно дробить и утверждается, что
«выделение стратиграфических видов, свойственных лишь одному
горизонту, представляет сейчас одну из важнейших задач па-
леонтологии» и что возражение против такого чрезмерного дроб-
ления «ошибочно и вредно» (там же).
Вводится «правило», согласно которому один вид соответ-
ствует одному горизонту. Это произвольное использование тер-
мина «вид», возведенное в «правило», должно повести к без-
удержному дробительству. Исходя из этого «правила»,
подвергаются сомнению основы общебиологической теории вида,
утверждается, что теория Дарвина сыграла отрицательную роль,
так как «связь видов между собой послужила причиной ошибоч-
ного методического приема, несколько задержавшего развитие
стратиграфии» (там же 32). Этот «ошибочный прием», оказы-
вается, заключался в объединении близких форм из разных
горизонтов в один вид. Вредность объединения близких форм
в один вид иллюстрируется примером с Atrypa reticularis. Ранее
считалось, что этот вид существовал в течение всего силура и
девона. Ныне же его существование ограничено только силуром,
а в девоне признается наличие уже других видов и при этом,
в нижнем, среднем и верхнем — специфических.
Стратиграфы такого направления считают, что расчленение
разрезов и дробление видов следует продолжать вплоть до мо-
9 К. М. Завадский
129
Мейта, «когда размерь! раковйн руководящих форм йрёвЫсят
мощность выделяемых пластов» (там же). Из этой трактовки
стратиграфического вида вытекает и утверждение, что «видо-
образование происходит фактически одновременно» на громадных
территориях (там же 35), что нет примеров нахождения одного
вида в отложениях разного возраста и что после проверки всегда
оказывалось необходимым дробить «слишком широкие» виды.
Другие же биостратиграфы считают механистичным «ошибочное
допущение, что продолжительность существования вида является
одинаковой или почти одинаковой во всех местах его обитания,
что повсюду он появляется и вымирает одновременно» (Ротай,
1962:19). Не входя в обсуждение проблем стратиграфии и ее
нужд как отрасли, обслуживающей разведку недр, отметим
только всю несообразность утилитаристского подхода к объему
стратиграфического вида. Право авторов — искать удобные ру-
ководящие ископаемые. Слово «вид» также употребляют не
только в биологии, но и в логике, кристаллографии, химии и т. п.
Не следовало бы только смешивать стратиграфический вид как
удобный маркер с видом как явлением живой природы и вносить
поправки в понимание вида биологами.
Неонтологи хорошо знают, что чем хуже изучена группа, тем
крупнее объем выделяемых в ее пределах видов. В палеонтоло-
гии же, как правило, изученность видов невелика, и в соответ-
ствии с нею и объем видов должен быть большим. Ошибочно
изобретать стандарт вида, не соответствующий ни природе са-
мого явления, ни степени его изученности, но отвечающий лишь
иллюзорному стратиграфическому «утилитаризму».
О применении монотипического стандарта
«мелкого» вида. Сторонники этого стандарта стремились тол-
ковать вид как строгий (идеальный) монотип. Однако на базе
этого стандарта сложился весьма пестрый конгломерат толкова-
ний и терминов: клональный вид, вид-штамм, элементарный вид,
microspecies, жорданон, вид — чистая линия, вид — практически
гомозиготная инцухт-линия, вид — биотип, вид — одна узкоэнде-
мичная популяция. Монотипическая концепция мелкого вида
имела известный успех в систематике агамных групп, например
синезеленых водорослей. Пытаясь расчленить морфологически
разнообразную популяцию на отдельные части, систематики при-
менили методику культур на искусственных средах. Выделяя из
такой культуры морфологически отличимые особи и проверяя их
в изолированных линиях па константность, удавалось расчле-
нить обычный политипическпй вид (или, по терминологии аль-
гологов, поливалентный вид) на множество клональных линий.
Каждая такая линия систематиками-дробителями именовалась
элементарным видом, клональным видом или даже просто видом.
Так, species, в толковании Шода (Chodat, 1909, 1914), —это ка-
ждая отдельная клональная линия (клональный вид), отличаю-
130
щаяся от другой хотя бы одним константным признаком- В про-
тивоположность этому крайнему дробительству А. А. Еленкин
(1936), Н. Н. Воронихин (1946, 1951) и В. И. Полянский
(1956, 1958) толковали вид синезеленых водорослей как
совокупность многих клональных линий, т. е. гораздо более
крупно.
Как видно из микробиологической и бактериологической ли-
тературы, для наведения порядка с объемом вида в этих обла-
стях предстоит сделать значительно больше, чем у более высоко-
организованных групп. В качестве свидетельства этих трудностей
приведем мнение К. Ван-Ниля, высказанное им в 1956 г.: «Было
сделано много серьезных попыток привести в упорядоченную
систему огромное число видов микробов. Хорошо служить
своей цели может лишь такая классификация, которая основана
па устойчивых признаках. К сожалению, в прошлом па это тре-
бование обращали, по-видимому, очень мало внимания, особенно
в области классификации бактерий. В новые «виды» и даже
«роды» возводились клоны, отличавшиеся от сходных клонов
всего лишь одним признаком, и вполне возможно, что во многих
случаях отличительные особенности представляли собой проявле-
ние различия в одном гене» (Клюйвер и Ван-Ниль, 1959 167).
Надо сказать, что использование неустойчивых, модифицирую-
щих признаков — это ошибка иного плана, чем использование
лишь одного признака. Ван-Ниль как-то сблизил эти различные
вопросы. .
Далее Ван-Ниль описывает затруднения систематиков, не
знающих, толковать ли вид узко, как монотип, или же как
группу монотипов: «Когда бактериолог выделяет большую группу
штаммов, скажем Pseudomonas fluorescens или аэробных споро-
образующих бактерий, из природного материала обычными ме-
тодами, он обнаруживает, что фактически каждый клон отли-
чается от всех остальных одним или несколькими свойствами. . .
Он может прибегнуть к удобному способу — предложить особое
видовое название для каждого штамма». Эта практика, однако,
имеет тот недостаток, что все вновь выделенные штаммы «прихо-
дится рассматривать как новые виды. Поэтому ее и критиковали
любители объединения, которые считают, что методологически
правильнее применять видовое название только для обозначения
комбинации нескольких признаков. Хотя у такого подхода есть
большие достоинства, он, однако, несвободен от упрека в произ-
вольности» (там же : 168).
Ясно, что Ван-Ниль не решается определенно высказаться
в пользу дробления или объединения видов «микроорганизмов.
Основной упор сделан на нужды коллекционера, начинающего
изнемогать под грузом новых образцов, т. е. на поиск удобных
методик распознавания и идентификации штаммов. Во всяком
случае, по свидетельству Ван-Ниля, мельчайший вид (вид = од-
9*
131
ному клону) до сих пор является одним из полноправных стан-
дартов в систематике микроорганизмов.
Одно время мельчайший монотипический стандарт использо-
вался ботаниками, работавшими с полиморфными апомиктиче-
скими группами. Многие систематики (Бузер, Далыптедт,
Юзепчук и др.) считали видом каждую отдельную бесполосемен-
ную форму, сохраняющую в культуре специфические признаки.
Иначе говоря, это было выделение настоящих жорданонов (по
образцу Erophylla verna). Целые рои таких форм могут суще-
ствовать бок о бок, составляя единое население. Сторонники
крайне мелкого стандарта каждую морфологически различимую
апомиктическую форму или каждый компонент полиморфной
системы именовали видом. В действительности каждый апомик-
тический вид, выделенный на основании обсуждаемого стандарта,
есть не что иное, как один из мелких компонентов исходной по-
пуляции перекрестноопылителя, после нарушения полового раз-
множения оказавшегося на положении самостоятельно воспроиз-
водящейся группы. Систематика многих апомиктических групп
(Hieracium, Alchemilla, Taraxacum, Роа, Potentilla, Crataegus
и др.) разрабатывалась сторонниками мелких видов (жордано-
нов). Поэтому трудно сопоставлять численность видов, размно-
жающихся половым путем, с численностью апомиктических
«видов» (microspecies). Иногда один хороший амфимиктический
вид по объему равен нескольким сотням бесполосеменных «ви-
дов», описанных «дробителями». Приводимые С. С. Хохловым
(1959) интересные количественные данные о распространенности
бесполосеменного размножения (около 20% «видов»), по-види-
мому, несколько завышены.
В начале главы 4 уже отмечалось, что многие генетики и
некоторые систематики в первые десятилетия нашего века стре-
мились к абсолютной монотипичности вида. Они именовали
истинным видом (euspecies) каждую чистую линию, выделенную
из популяции самоопылителей, каждую переставшую расщеп-
ляться (т. е. практически гомозиготную) инцухт-лпнию, каждый
биотип (найденный в природе или полученный при разложении
популяции).
Иной характер носят многочисленные узкоэндемичные изо-
лированные популяции — мелкие виды, выделенные Гуликом
(Gulick, 1905) среди наземных моллюсков из Achatinellae на
Гавайских островах. На одном маленьком островке (Оаху) Гу-
лик обнаружил массу мелких популяций, живущих в лесах гор-
ных долин и полностью изолированных одна от другой. Каждая
маленькая долина имеет морфологически отличимую популяцию,
возведенную Гуликом в ранг вида. Генетическая однородность
материала у ахатинелл, разумеется, отсутствовала.
Как видно из этих примеров, стремление к максимальному
дроблению вида и к соблюдению наибольшей чистоты монотипа
132
привело к смешению различных образований (клоп, штамм, апо-
миктическая форма, чистая линия, инцухт-линия, биотип, изо-
лированная популяция) и к возведению в ранг видов форм как
неспособных к длительному самостоятельному существованию
в природных условиях, являющихся зависимыми частями более
сложного целого (например инцухт-линий), так и образований,
стоящих на видовом уровне (Achatinellae).
5. ОШИБОЧНЫЕ ТОЛКОВАНИЯ ВИДА И ВИДООБРАЗОВАНИЯ
Отрицание реальности вида. Реальное существова-
ние вида подвергается сомнению и некоторыми современными
биологами. По-прежнему эта позиция объясняется несколькими
причинами. По-видимому, наиболее распространено отрицание
реальности вида из-за наивно-номиналистических взглядов: об-
щее существует только через единичное п вне единичного суще-
ствует только в форме идеи, каковой п является понятие вида.
К этой позиции близки и взгляды некоторых морфологов-систе-
матиков, рассматривающих вид прежде всего как таксономиче-
скую категорию — шаблон, с помощью которого удобнее всего
сортировать организмы. Недавно нами (Завадский, 1961, 19676)
была сделана попытка доказать, что\вид не менее реален, чем
особь.
В основе нашей аргументации лежит положение, что вид —
это не сумма особей, а целостная система, основная единица
надорганизменной формы существования живого. Одним из
источников субъективистских ошибок в проблеме вида является
пессимизм, связанный с тем, что в последние десятилетия вместо
консолидации точек зрения возросло число различных подходов
к толкованию вида и разнобой в понимании его сущности (Сие-
not, 1936). Помимо коллизии между видом как единицей эволю-
ции и видом как шаблоном в работе систематиков, возникли
новые коллизии между видом как генетической проблемой и ви-
дом как экологической или историко-географической проблемой,
между видом как проблемой биостратиграфов и видом с точки
зрения биохимика и т. п. С умножением числа подходов к иссле-
дованию вида стена непонимания иногда становилась выше и
казалось, что вид — это действительно лишь понятие-оператор.
Далее на нескольких примерах будут показаны современные
субъективистские толкования вида.
Питательной средой для этой концепции продолжает служить
механоламаркистская трактовка эволюции. СлеДуя за основопо-
ложником, некоторые механоламаркисты до сих пор выступают
против реального существования видов. Э. Рабо (Rabaud, 1953)
основой эволюции считает прямую «физиологическую адапта-
цию», следствием которой являются непрерывные ряды измен-
133
живости. Он отвергает естественный отбор и борьбу за существо-
вание, так же как реальность видов и наличие особого вещества
наследственности. К субъективизму ведет также и переоценка
трудностей с определением границ вида во времени, с которыми
сталкиваются палеонтологи в наши дни. Однако эти затруднения,
разумеется, не свидетельствуют об условности вида, хотя к та-
кому выводу и приходят некоторые авторы (Померанцева, 1964,
и др.). Наличие хроноклинов доказывает расплывчатый (гра-
дуальный), а не четкий (пороговый) характер границы. Но су-
ществование хроноклинов не является свидетельством условности
(т. е. нереальности) самого вида. В тех случаях, когда объект
изучен хорошо, но трудно классифицируем, расплывчатость очер-
таний вида следует рассматривать как неопределенность границ,
а не как результат неточности знаний или нереальности самого
изучаемого объекта.
В сборнике «Новая систематика» наряду с работами, углуб-
ляющими наше понимание вида как реального явления природы,
была опубликована работа Д. Гильмура (Gilmour, 1940), в кото-
рой понятие «вид» истолковывалось с позиций логического пози-
тивизма. Вот каково основное содержание этой статьи: чувственно
воспринимать вид мы не в состоянии; видеть, ощупывать и т. д.
мы можем только отдельные предметы, да и они представляют
смесь чувственных восприятий с идеями. В начале статьи Гиль-
мур характеризует вид как идею, связующую множество чув-
ственных образов, а далее — как смесь чувственного с идеальным.
По-видимому, в начале статьи Гильмур писал о виде как о типе
организации, а затем как о множестве особей. Исходя из таких
взглядов, вид можно определять как совокупность особей, степень
сходства которых «принято считать» достаточной для объедине-
ния в одну группу. Окончательно же степень сходства устанавли-
вается «более или менее произвольными взглядами системати-
ков». Гильмур в принципе по соглашается с теми, кто рассмат-
ривает вид как «биологическую реальность» в отличие от групп,
объединяемых учеными по какому-либо признаку, но в самой
природе не являющихся «естественными единствами». Для Гиль-
мура все классификации основаны на организующем начале со-
знания классификаторов и вид не составляет исключения.
Существование «взаимодополнительных» систем низших таксоно-
мических единиц, основанных на различных подходах (морфоло-
гическом, эколого-генетическом, основанном на степени интер-
фертильностп, па методе клипов), по Гпльмуру, служит
иллюстрацией произвольности выбора критериев и равноценности
этих подходов. Единственное, что может выдвинуть ту или иную
систему на первое место — это согласование вопроса между спе-
циалистами и заключение конвенции.
Разберем кратко интересную попытку ликвидации понятия
о виде, предпринятую с позиций логического позитивизма в США
134
Ё. Верма.1 Этот автор считает бесспорным лишь «логйчесКое су-
ществование вида». В этом смысле вид является «определенным
классом некоторого сорта и как таковой существует, можно ска-
зать, в точности в том же смысле, как и класс единорогов. .. Вид
как класс стоит вне реального существования. .. Никто никогда
не видел мыслительную конструкцию, гуляющую "по улице»
(Burma, 1954 194). Но единичные члены этого класса суще-
ствуют реально (в отличие от единорогов!). Эволюция осущест-
вляется неизмеримо мельчайшими этапами, образуя настоящую
биологическую непрерывность. Исключения касаются лишь не-
которых растений и не мешают «общей линии доказательства».
Раз процесс эволюции — это настоящая непрерывность и, «оче-
видно, не существует никакой естественно различимой единицы
внутри непрерывности», то вид не может быть реальностью и
имеет лишь логическое существование (там же 196).
По Бёрма, все систематики стоят на почве «неизменной ста-
тичной Платоно-Аристотельской философии», так как виды опи-
сываются ими как «дискретные объективные реальные единицы».
Итак, Бёрма, видимо, вполне самостоятельно доходит до старой
мысли, что вид в систематике — «это произвольно вырезанный
отрезок непрерывной фплетической линии». Его призыв оставить
«статическую концепцию вида» и встать на позиции эволюцион-
ной биологии убеждает нас в том, что айтор вульгарный эволюцио-
нист, не понимающий диалектики прерывности и непрерывности.
Позитивная часть работы Бёрма начинается фразой, проник-
нутой верой в чудесную силу слова: «Нужно. . . отбросить тер-
мин „вид“ с его подавляющим грузом нежелательных значений
и употреблять пли создать новое слово для обозначения тех от-
резков филетической линии, которые до сих пор назывались
видами» (там же 199). Вместе с тем Бёрма не является номи-
налистом, так как он признает существование реальных надин-
дивидуальных единиц. Это — элементарная размножающаяся
популяция, весьма непостоянная, потому что каждая особь мо-
жет быть сочленом множества таких первичных популяций, раз-
множающихся половым путем. Вторичная размножающаяся
популяция — совокупность многих первичных популяций. Попу-
ляцией третьего порядка Бёрма называет комплекс вторичных
популяций, в котором скрещивания ограничены не только физи-
ческими барьерами, но уже отчасти и биологическими. Популя-
ции третьего порядка развиваются уже почти независимо от
других популяций и, по Бёрма, являются последней реальной
(естественной) единицей биологической иерархии. Вся иерархия
единиц над названной, по Бёрма, — целиком рассудочна. Под-
водя итог, Бёрма пишет: «Такая высокоабстрактная выдумка,
1 Более подробную критику взглядов Бёрма см.: Завадский 1961 75—
77).
135
как вид. . . способна лишь затемнить понимание эволюционного
процесса, в основу которого надо положить идею реальности
эволюирующих популяций» (там же : 208).
Любопытно, что «размножающаяся популяция третьего по-
рядка», призванная, по Бёрма, упразднить вид, как абстрактную
выдумку, именно и называется видом большинством биологов.
Стоило ли спорить о слове «вид», чтобы признать реальным по-
нятие о виде, выраженное четырьмя другими словами — «размно-
жающаяся популяция третьего порядка».
Номиналистический образ мышления заставил выступить
с попыткой упразднения понятия «вид» и другого американского
ученого Д. Давидсона. Заглавие его статьи очень яркое: «Дефло-
гистонизация проблемы вида». По Давидсону, «с термином „вид“
сейчас происходит то же, что происходило с теорией флогистона
двести лет назад. Реальность вида. . . вероятно, настолько же
действительна, насколько действительна реальность флогистона.. .
Настало время, когда мы, подобно химикам в прошлом, должны
пересмотреть фактические основы проблемы» (Davidson, 1954 :
:246). По его мнению, единственной общей особенностью всех
определений вида является утверждение, что вид состоит из
множества особей, которые обладают известными общими при-
знаками. В отличие от Бёрма Давидсон отмечает, что ряды форм
были бы идеально непрерывны («полный континуум»), если бы
не действовал отбор. Отбор создает перерывы всевозможных раз-
меров п остается найти индикаторы для объективного определе-
ния их размеров. Признав степень стерильности или фертиль-
ности при скрещиваниях неподходящим критерием (почему он
плох, автор умолчал), Давидсон утверждает, что степень пре-
рывности может определяться только субъективно («только
вследствие нашего умственного процесса»).
По Давидсону, «тенденция говорить и думать терминами, от-
носящимися к группам особей», является примером «чисто мыс-
лительного объединения», так как в природе существуют лишь
особи, а любые их группы создаются в мозгу систематиков.
Если имеется случай такого гигантского хиатуса, как например
у Ginkgo biloba, то выделение вида — простая задача. Но, с дру-
гой стороны, есть много трудных случаев (например, с родом
Salix): «Если мы хотим доказать, что виды имеют объективную
реальность, мы можем выбрать такие примеры, как гинкго.
Если мы хотим доказать, что виды вообще не существуют в при-
роде, мы можем выбрать такие примеры, как ивы» (там же : 251).
В итоге автор приходит к выводу, что единственно реальной био-
логической единицей является особь, а виды «скорее являются
мыслительными воображаемыми единицами».
Оба рассмотренных нами автора решительно атаковали тер-
мин «вид». Однако Бёрма доказал, что явление природы, обычно
называемое видом, действительно реально существует, но что он
136
предпочитает его именовать другими словами. Давидсон же, дав
оригинальную заявку на «дефлогистопизацию» проблемы вида,
оказался рядовым номиналистом.
Оба автора по существу направляли свои стрелы против
формально-систематических трактовок вида. В этом состоит по-
ложительное значение их намерений. Однако они не разграни-
чили современную общебиологическую теорию вида и практиче-
ский стандарт вида систематиков, иногда еще сохраняющий
музейно-морфологический облик. Надо было не отбрасывать по-
нятие о виде, а критиковать недостатки, присущие его трактовке
в систематике. Но философское вооружение авторов оказалось
мало приспособленным для этого, и стрелы полетели не по
адресу.
Подведем итог всему сказанному о субъективизме в толкова-
нии вида (главы 2—5).
Самой общей причиной, поддерживающей субъективистские
толкования вида во все времена является недостаточная глубина
познания этого явления и гносеологические трудности, стоящие
перед исследователями. Конечно, здесь речь идет о тех биологах,
чьи субъективистские ошибки им^ют только эмпирическую ос-
нову и не связаны с намеренным’ применением ложных фило-
софских концепций (махизма, логического позитивизма, семан-
тики и т. п.).
Следует заметить, что новые формы субъективизма, связан-
ные с махизмом и другими формами субъективного идеализма,
не сосуществовали, а слились со старыми формами субъек-
тивизма, такими как номинализм систематиков, существую-
щий с додарвиновских времен, примитивный эволюционизм
и т. п.
В заключение необходимо еще раз подчеркнуть, что, по на-
шему мнению, действительно смертельный удар по субъекти-
вистскому пониманию вида может быть нанесен разработкой
теории вида как одной из основных и первичных форм органи-
зации живого и изучением механизмов, интегрирующих вид
в целостное образование.
Псевдодарвинизм. В 40—50-х годах именем «творче-
ского», или «мичуринского», дарвинизма назывались взгляды
Т. Д. Лысенко (1946, 1950, 1956) и его сотрудников. Они осно-
вывались на признании адекватной, или приспособительной из-
менчивости, первичной и единственно значимой для эволюции.
Наследование приобретаемых признаков рассматривалось осно-
вой материализма в биологии и главным законом эволюции.
Данные генетики не только игнорировались, ‘но и были объяв-
лены «лженаучным идеалистическим менделизмом-морганизмом».
Биология отбрасывалась ко временам примитивного отождест-
вления генотипа с фенотипом. Теория естественного отбора под-
менялась «направленным воспитанием» и в то же время отвер-
757
галась, как построенная на случайностях («ультраселекционизм»).
Под творческой ролью отбора понималось прямое приспособление
с помощью «ассимиляции условий» и т. п. Нет нужды останав-
ливаться на доказательствах ошибочности этих ламаркистских
воззрений. Но это направление проявляло большой интерес
к проблеме вида и видообразования.
Т. Д. Лысенко писал, что «в биологической науке, именно
в науке, а не в практике, виды перестали считать реальными,
отдельными качественными состояниями живой природы»
(1950:16). После этого неправильного утверждения автор счел
возможным приписать себе открытие того, что вид — реальное
явление. Разъясняя биологам «новое в науке о биологическом
виде», Лысенко писал: «Согласно нашему представлению, Жучка
это не уклонение от собаки, а одна из форм настоящей собаки.
А мои оппоненты, исходя из неверного теоретического представ-
ления, говорят, что Жучка не настоящая собака, а уклонение от
нее» (1956:25). Так как фамилии оппонентов не назывались,
очевидно, это высказывание было направлено также в целом
против биологии и до сих пор не понимающей того, что Жучка —
это собака.
Реальность вида, в толковании Лысенко, тесно связана с ги-
потезой порождения видов, согласно которой в теле организма
одного вида под влиянием условий жизни зарождаются кру-
пинки тела другого вида и этот процесс идет не постепенно,
а скачкообразно. Сообщалось о порождении в короткие сроки
(2—3 года) сосной — ели, пшеницей — ячменя, ржи, костра и
овсюга, грабом — лещины, пеночкой — кукушки и многое дру-
гое. В печати эта гипотеза оценивалась как «единственная со-
временная научная теория вида и видообразования» («Журнал
общей биологии», № 6 за 1952 г.), как «окончательное разреше-
ние загадки видообразования» и т. п. Утверждалось, что сорные
растения порождаются культурными и что можно «предотвра-
щать порождение нежелательных видов и, наоборот, вызвать
порождение одними видами других, желательных нам» (Лысенко,
1956:5). Это представление о видообразовании называлось диа-
лектико-материалистическим и было противопоставлено взглядам
Дарвина. Последний обвинялся в «постепеновщине», в непони-
мании различий между количественными и качественными изме-
нениями, т. е. в «плоском эволюционизме».
Представление о противоречивости внутривидовых отношений,
факты внутривидового соревнования, пли конкуренции, отверга-
лись и назывались реакционным мальтузианским положением,
вейсманистским лжеучением о внутривидовой борьбе и другими
подобными эпитетами. Главный аргумент состоял в том, что при
межвидовой борьбе виды, внутри которых существуют противо-
речия, будут вытеснены видами, внутри которых существуют
такие же гармоничные регулируемые отношения, как между ор-
13;8
ганами индивида. Утверждалось, что перенаселенности, как пра-
вило, не было и пе может быть, что гибель особей в густом на-
саждении является «самоизреживанием», происходящим «пе
потому, что деревцам уже тесно, а для того, чтобы им в бли-
жайшем будущем пе стало тесно» (там же 17).
Все эти воззрения Лысенко на вид и видообразование, дол-
гое время включавшиеся в учебники для средней и высшей
школы и в другие массовые публикации в качестве непрелож-
ных истин, оказались ошибочными. Проверка показала, что при-
меры порождения видов были получены в опытах, поставлен-
ных с грубыми методическими ошибками, или являются произ-
вольным толкованием хорошо известных явлений (гибридпость
и др.), или же, наконец, оказались простой мистификацией (см.
многочисленные статьи в «Ботаническом журнале», «Бюллетене
Московского общества испытателей природы» и «Вестнике ЛГУ»
за 50-е годы). П. А. Баранов справедливо указывал, что гипо-
теза порождения видов является грубой формой мутацпопизма,
основанной на невероятных допущениях (зарождение крупинок
нового вида и т. п.) и ненаучных способах их защиты, что «при-
нижение и даже полное игнорирование Т. Д. Лысенко роли от-
бора в видообразовании с неумолимой логикой ставит его на
путь, ведущий в другую сторону от материалистической основы
дарвинизма» (1953:683).
Несостоятельность взглядов Лысенко на характер внутриви-
довых отношений была показана многими работами (Сабинин,
1947; Формозов, 1947; Сукачев, 1953, 1956, и др.). На конферен-
ции по дарвинизму в МГУ в феврале 1948 г. (Конференция. .,
1948) А. А. Парамонов, С. Я. Соколов, Е. И. Лукин, Н. П. Нау-
мов, В. С. Ивлев, А. Л. Зеликман, И. М. Поляков и другие по-
казали закономерность и эволюционное значение внутривидовой
конкуренции, неизбежность антидарвинистического разрешения
основных вопросов эволюционной теории при ее отрицании. Было
экспериментально проверено и опровергнуто утверждение Лы-
сенко, что в гнездовых загущенных посевах кок-сагыза отсут-
ствуют конкурентные отношения (Завадский, 1947, 1954г,
1957а). Оказалось, что в этих условиях происходит жест-
кий отбор быстрорастущих форм и вывод, что в густых гнез-
дах нет угнетения одних растений другими и они растут
лучше, чем в редких посевах, был сделан на основании наблю-
дений за небольшой группой переживших растений — победи-
телей.
Вся эта система взглядов может быть названа псевдодарви-
низмом (Завадский, 1965; Завадский, Бермай, 1966), так как
хотя она и являлась эклектическим сочетанием ламаркизма, му-
тационизма и телеологии, но в отличие от всех других форм
антидарвинизма провозглашала себя самым передовым дарви-
низмом.
139
6. ЗАКЛЮЧЕНИЕ
Количество работ по проблеме вида и видообразования, вы-
шедших после 20-х годов, во много раз превышает число работ
по этой проблеме, опубликованных за два предыдущие века.
Однако новейшая история проблемы еще не написана. Отсут-
ствие такого анализа не могло не отразиться на содержании
глав 4 и 5. Эти главы не претендуют на полные историко-науч-
ные обзоры и содержат лишь тот материал, который, с нашей
точки зрения, характеризовал основные этапы в познании вида,
перечисленные в главе 2. Зная, что трудности будут велики, что
возможны ошибки в освещении некоторых вопросов или одно-
стороннее их толкование п сознавая, что эти главы написаны
слишком фрагментарно, я все же решился их опубликовать, так
как надеюсь, что они могут стимулировать более основательное
изучение новейшей истории проблемы, без которой очень затруд-
нена и разработка многих общих вопросов теории вида.
Обилие работ в новейшее время не означает того, что про-
блема вида и видообразования разрабатывалась особенно высо-
кими темпами. Например, из обзора исследований советских
биологов за 30 лет (Камшилов, 1947) видно, что по проблеме
вида и видообразования в нашей стране тогда было сделано
меньше, чем по другим вопросам. Этот невысокий по сравнению
с другими проблемами темп исследований по проблеме вида ха-
рактерен и для других стран.
В ряде случаев вопросы, которые следовало бы осветить
в исторической части, целиком перенесены во вторую часть
книги. К ним, например, относится проблема неравноценности
(типов) видов. Во всех таких случаях мы руководствовались
удобством изложения и желанием избежать излишних повто-
рений.
Научное познание вида, если его истоки датировались от
Д. Рея, продолжается более 250 лет. Вначале стояла задача пер-
вичного описания массы еще неизвестных видов. Надо было
хотя бы просто констатировать их наличие. Были развернуты
чисто описательные работы, дававшие весьма приближенные
представления об отдельно взятом виде. Возникли два стандарта
вида в систематике: морфологический монотипический и, позд-
нее, морфологический политипический. Были поставлены две
тесно связанные между собой задачи: точнее определить само
явление вида, очертить его границы и глубже понять внутрен-
нее строение, организацию вида, его сущность.
Когда само явление вида очерчивалось лишь грубо и поверх-
ностно, вопрос о соотношении покоя и движения еще не выдви-
гался в качестве заслуживающего специального внимания. Вид
изучался как стабильное состояние; оформилось представление
о его абсолютном постоянстве; учение о виде было противопо-
140
ставлено учению о развитии в качестве альтернативы. Дарвин
определил вид точнее, как явление развивающееся; после этого
внимание было приковано к переходным моментам, -к процессам
постоянно снимающим стабильное состояние вида. Однако выяс-
нилось, что без более глубокого изучения статики вида невоз-
можно изучать его эволюцию. Тогда обратились к познанию
внутренней структуры вида. Снова встал вопрос о стабильности
вида; но теперь уже вскрывались ее исторические причины; вид
понимается как узловой пункт эволюции, т. е. как ее момент.
Вид истолковывается как одна из первичных основных форм,
в которой осуществляется жизнь и ее развитие.
Однако познание вида включало и противоречия. Противо-
речивость вида обнаруживалась уже в первых попытках его ха-
рактеристики по отдельным внешним показателям. Факт прак-
тической универсальности вида (всеобщего распространения)
все время сталкивался с идеей вида как частной формы органи-
зации живой природы. Вид делается основной единицей класси-
фикации и основным понятием систематики, по одновременно
возникает и субъективистское его понимание. Представление
о реальности вида в «первой редакции» неразрывно связывается
его постоянством и неизменностью. Число известных видов
возрастает, но систематика занимает господствующее положение
в биологии и познание вида осуществляется только для ее нужд
и не идет вглубь. «Теория вида» отождествляется систематикой
с разработкой удобных практических стандартов вида на основе
признаков, наиболее пригодных для распознавания и каталоги-
зации. Ограничивает познание вида и господство морфологиче-
ской концепции вида. Выдвигается физиологически критерий, но
сам он абсолютизируется. Ложная теория реального неизменного
вида укрепляется, так как ей противостоит наивная концепция
«развития без видов». Возникает субъективистское представле-
ние о виде.
Утверждение теории «реального развивающегося вида» и
исторического метода в истолковании вида, сняв одни противо-
речия, стимулировало новые. Общебиологическая эволюцион-
ная концепция вида возникла за рамками систематики и пришла
в столкновение с чисто морфологической концепцией вида,
утвердившейся в систематике. Разрыв между ними все нарастал
и нарастал. Развиваются субъективистские идеи на базе вуль-
гарного эволюционизма. Гипотезы мутационного видообразова-
ния были противопоставлены дарвинизму.
Более глубокая и всесторонняя характеристика вида как гео-
графической, исторической и экологической определенности
также вызвала к жизни новые трудности. Введение географо-
морфологического ц экологического методов привело к формиро-
ванию на его основе противоположных трактовок объема вида —
монотипической и политипической. Применение генетических
141
методов, разложение вида на элементарные формы и открытие
сложнейшей внутренней структуры вида еще более усложнили
его понимание, так как вид теперь приравнивался то к мелкому
«истинному» виду (жорданону), то к объему линнеона. В этих
условиях возник кризис теоретических основ учения о виде, вы-
разившийся в усилении субъективистских тенденций и в попыт-
ках восстановить представление о неизменности вида.
Последнее сорокалетие характеризуется развитием экспери-
ментальных методов изучения видообразования, исследованием
различных способов видообразования, началом работ по даль-
нейшему изучению статики вида, исходя из знания законов его
эволюции, дальнейшим развитием комплексных генетико-геогра-
фо-экологических исследований внутренней организации вида,
оформлением учения о популяциях, исследований вопроса о не-
равноценности видов (о типах видов), началом эксперименталь-
ной разработки вопроса о целостности вида и о внутривидовых
отношениях. Выяснилось, что без более глубокого исследования
статики вида и познания его внутренней структуры невозможно
изучать его эволюцию. Снова встал вопрос о стабильности вида,
но теперь уже вскрываются ее исторические причины и вид по-
нимается как момент эволюции, как ее статика. Вид истолковы-
вается как необходимая форма, в которой осуществляется жизнь
и ее развитие. Все эти исследования обусловили отход от одно-
сторонне-генетического разложения вида и вывели всю проблему
из состояния кризиса.
В исторической части книги мы намеренно не рассматривали
подробно некоторые из перечисленных только что направлений
исследований, обсуждаемые во второй части.
* * *
В итоге 250-летнего изучения вида было выявлено несколько
его существенных и противоречивых черт. Оказалось, что вид
является не только морфологическим типом, но и специфическим
типом организации ДНК, белков и метаболизма; множеством
сходных особей; общностью особей в физиологическом отноше-
нии, способной к самовоспроизведению в ряду поколений;
дискретной единицей жизни и звеном в ряду близких форм; гео-
графической и экологической определенностью; неделимым эле-
ментом и вместе с тем сложной системой; образованием одно-
родным внутри себя, многообразием форм с центробежными
тенденциями и в то же время целостностью; образованием, все-
цело зависящим от окружающей среды и в то же время способ-
ным к относительно независимой эволюции; стабильным состоя-
нием п в то же время носителем и единицей эволюционного
процесса.
Часть II
ОЧЕРК ТЕОРИИ ВИДА
Глава 6
КРИТЕРИИ И ОБЩИЕ ПРИЗНАКИ ВИДА
1. ПРОБЛЕМА ВИДА И СИСТЕМАТИКА
Эйдология и вид в с и с т е м а т и к е. Общая теория вида,
или эйдология, должна рассматривать вид всесторонне, в том
числе и такие его черты, которые недоступны или не нужны для
систематики. Согласно развиваемой в этой книге концепции, со-
держание жизни вида — это совокупность основных и производ-
ных внутривидовых отношений. Ни старая, ни новая систематика
не занимались и не призваны заниматься изучением внутривидо-
вых отношений. Внутривидовыми отношениями занимаются эко-
логия и этология (Шовен, 1965), генетика популяций, синфизио-
логия и другие отрасли науки. С оформлением эйдологии ее
важнейшим разделом станет исследование содержания жизни
вида, т. е. внутривидовых отношений. Систематика же привлекают
черты, применимые в практике классификации, так как вид для
него — прежде всего единица таксономии. Такой «диагностиче-
ский утилитаризм» естествен, но при этом игнорируются все
черты п критерии, не используемые для классификационной ра-
боты. Можно сказать, что систематика хочет знать пе столько
сущность вида, сколько его характеристику как единицы класси-
фикации. Разрыв между формальной трактовкой вида и теорией
вида до конца не устранен и до сих пор. Некоторые авторы рас-
сматривают систематику как ведущую отрасль в разработке
теории вида (Берг, 1950; Юзепчук, 1958; Cain, 1959, и др.). Еще
встречаются утверждения-анахронизмы, что «систематика, осно-
ванная на филогенетических принципах, является венцом биоло-
гии» (Тихомиров, 1965 : 175). Не принижая значения систематики,
следует все же напомнить, что подлинный венец биологии, разу-
меется, не в классификации, а в открытии глубоких законов,
управляющих функционированием и развитием живых систем,
143
и в практическом использовании этих законов. Задачей исследова-
ний вида является не определение его места в филогенетической
системе, а изучение законов, управляющих его существованием и
эволюцией.
Познание вида как особой формы организации живого нельзя
смешивать с составлением каталогов видов. Для познания вида
вглубь нет необходимости изучать сотни тысяч видов и выяснять
существующие между ними филогенетические отношения. Более
того, как это показывает вековая история изучения организма,
привлечение для этой цели слишком большого числа объектов вы-
нудит исследователей отказаться от углубленного познания сущ-
ности явления и переключиться на поверхностную классификацию
множества объектов на основе нескольких наиболее изученных
признаков. Теория вида не должна сбиваться на экстенсивный
путь работы. На немногих специально подобранных модельных ви-
дах следует вначале углубленно изучать законы существования и
эволюции самых основных типов видов.
Современные систематики нередко сетуют на то, что проблемой
вида начали заниматься «неспециалисты» и требуют признать
решающими именно их воззрения на вид. Например, С. В. Юзеп-
чук, подчеркнул, что систематикам «все время приходится наблю-
дать вид непосредственно в природе, ибо даже гербарные образцы,
с которыми им так часто приходится оперировать, представляют
собой подлинные документы... Поэтому только видам флориста
может быть придано значение конкретных „единиц" эволюции
в той форме, как она действительно протекает в природе»
(1958:131). Далее он утверждал, что «как бы ни подходили
к изучению видовой категории другие дисциплины, как бы они
не понимали и не определяли вид, их точка зрения, если только
она игнорирует задачи и требования флориста, для последнего
не только неприемлема, по и малоинтересна» (там же). Эти фор-
мулировки выражают точку зрения многих современных система-
тиков. Некоторые систематики до сих пор смешивают общебиоло-
гическую теорию вида с практическим стандартом, а сущность
вида с международным кодексом правил номенклатуры (Проха-
нов, 1965а). Эта путаница сильно тормозит познание вида с обще-
биологических позиций, т. е. формирование эйдологии.
Есть ли у систематики объективные права на занятие таких
ключевых позиций в теории вида, на которые, как полагают не-
которые, она должна претендовать? В состоянии ли она взять на
себя труд изучения основных аспектов теории вида?
Истинное положение с трактовкой вида в систематике хорошо
показано Э. Майром: «Что такое вид? Среди систематиков нет
единой точки зрения... Это относится не только к разногласиям
между систематиками растений и систематиками животных или
между паразитологами и зоологами... Разногласия обнаружи-
ваются даже среди специалистов по отдельным группам, напри-
144
В. Н. Сукачев.
В. Л. Комаров.
мер, среди систематиков двукрылых, моллюсков и среди орнитоло-
гов. Это положение ясно указывает, что ни один систематик
не может еще сделать из систематического материала обобщаю-
щих выводов, которые были бы приемлемы для всех его коллег-
систематиков» (1947 : 28—29).
Э. Майр нарисовал картину, соответствующую действитель-
ности и по сей день. Некоторые систематики считают, что «проб-
лема вида менее всего разработана в современной... систематике
и что причина столь явной отсталости систематики в целом
состоит... в несовершенстве методов работы» (Хржановский,
1961 : 255). Высказываются и такие пессимистические точки зре-
ния, удивительные своим консерватизмом: что такое вид, в настоя-
щее время определить нельзя, так как для этого еще не хватает
фактов; одни слишком дробят, другие чересчур объединяют,
третьи экспериментируют, но опереться на все эти данные
нельзя в силу их ненадежности (Васильков, 1965). В конечном
счете автор призывает систематиков по существу вернуться
к старому «гербарному» методу (там же: 39).
Систематика перестала быть «владычицей наук» и сделалась
лишь одной из отраслей, необходимых для изучения вида, при
этом не призванной исследовать внутренние закономерности вида.
Для изучения сущности вида в настоящее время требуются иные,
не используемые систематиками приемы работы. Изучение суще-
ственных особенностей вида должно проводиться па специально
подобранных объектах с применением всех приемов эксперимен-
тального анализа. Вид должен изучаться с позиций молекулярной
биологии, генетики и биохимии, с позиций этологии, экологии и
т. п. Если систематики по-прежнему не будут опираться на новые
данные о виде, получепныё^другимп дисциплинами, это приведет
к тому, что она будет тормозить дальнейшее более глубокое изуче-
ние вида. Существенно новые открытия в области теории вида
в течение последних 150 лет подготовлялись не в недрах система-
тики, а в эволюционной теории, биогеографии, экологии, генетике
и т. и. Учение о виде продвигалось вперед лишь теми системати-
ками, которые пытались всесторонне обобщить опыт изучения
вида, используя данные разных отраслей биологии.
Надо четко разграничить операцию первичного выделения
вида и описания его границ (иногда почему-то именуемую опреде-
лением вида), для выполнения которой необязательно знать
внутреннюю организацию вида и гораздо более трудную задачу
глубокого определения вида. Многие внутренние стороны вида,
не попадавшие в поле зрения систематиков, еще только начинают
исследоваться при участии экспериментальных* отраслей биоло-
гии. Особенно важно изучение внутривидовых отношений (между
гаметами, молодью и взрослыми особями), составляющих содержа-
ние жизни вида. Это далекие от систематики вопросы; поэтому
она и не может быть лидером в разработке теории вида. Вместе
Ю К. М. Завадский
145
с тем следует подчеркнуть большое значение систематики в изуче-
нии положения вида среди рода и высших таксонов. Эксперимен-
таторы много знают о немногих видах, а систематики — по-
немногу о множестве видов. Ошибочно было бы не учитывать и
прогресса самой систематики — внедрения математических (Те-
рентьев, 1960, 1966) и экспериментальных методик. «Новая
систематика» (Huxley, 1938b, 1940, 1956, 1963; Rensch, 1938, 1954,
1958; Clausen, 1951, 1958; Turesson a. Turesson, 1960, и др.) не-
мало сделала для совершенствования ее. Большой прогресс
осуществлен в методах палеобиологов (см. например: Zimmerman,
1959; Muller, 1963, и др.), где практикуется уже популяционно-
экологический подход со статистической обработкой данных.
И среди «обычных» систематиков, разумеется, есть ученые
с широким кругозором, активно работавшие над общебиологиче-
ской теорией вида (например, Семенов-Тян-Шанский, Пачоский,
Комаров, Майр, В. Полянский и др.).
Недавно была предложена идеальная модель описания вида
(«der ideale Typus der Rassen- und Artbeschreibung».— Rensch,
1958:92). Кроме признаков географического Rassenkreise
(Rensch, 1929), в эту модель включаются «физиологические и это-
логические характеристики», генетический анализ популяций
с окраин и из центра ареала (гибридизация, кариология и пр.),
индексы роста органов, данные об индивидуальной изменчивости
и т. п. Из опыта изучения видов культурных растений или дрозо-
филы известно, что для исследований даже одного вида по такой
программе было бы необходимо несколько комплексных институ-
тов. Существует более эффективная стратегия: на немногих ви-
дах (с учетом их неравноценности) глубоко разрабатывать
теорию вида и применять ее для нужд систематики. Симпозиум
1958 г. в г. Упсале и другие выступления «новых систематиков»
показывают их стремление поставить на службу систематики всю
биологию. Систематика должна опираться па общебиологическую
теорию вида, а не подменять ее. Разрабатывать теорию вида
должна новая комплексная отрасль биологии — эйдология (Завад-
ский, 1961).
Практические стандарты вида в современной
систематике. В современной систематике вымерших и ныне
живущих групп применяется шесть основных стандартов вида.
1. Морфологический монотипический стандарт. Главное требо-
вание— чистота «типа», т. е. отсутствие подразделений (varietas
и пр.) и неделимость. Им пользовались на заре систематики,
а впоследствии «дробители»,1 жорданисты, считавшие видом каж-
1 Общую критику «дробптельства» см.: Майр, 1947; Rensch, 1954; Кэйн,
1958; Завадский, 1961 и др., а критику «дробительства» в ботанике
см.: Воронихин, 1946, 1951; Бочанцев и Липшиц, 1955; Грубов, 1955;
В. Полянский, 1956, 1958, и др.
146
о;ую морфологически отличную популяцию, внутрипопУляпионную
форму (биотип) или даже особь (=клон).
2. Морфологический политипический стандарт. Его именно и
называют линнеоном, сборным видом (conspecies), полиморфным
видом. Согласно этому стандарту, «вид является далеко не послед-
ней неразложимой -единицей наименьшего объема, а представляет
собой сложный комплекс мелких константных единиц» (Малеев,
1933:19). Этим стандартом в чистой форме ныне также поль-
зуются лишь вынужденно в систематике малоизученных групп.
По Д. Л. Степанову (1959а), политипическая концепция вида
является наиболее прогрессивной для палеозоологии.
3. Морфолого-географический (= экологический) монотипи-
ческий стандарт Кернера—Веттштейна—Комарова. Вид = геогра-
фической, или экологической, и в том числе сезонной, расе. По
В. Л. Комарову (1934), политипический вид — линпеоп— прооб-
раз более высокого-таксона — ряда (series), или сборного вида
(conspecies), а раса — группа особей, отличающаяся хотя бы по од-
ному константному признаку и имеющая особый ареал.2 Этому
стандарту соответствует эковид (oecospecies) Турессона, климатип
М. А. Розановой и subspecies большинства зоологов (Rensch, 1954,
1965; Шварц, 1959, 1963; Мауг, 1963; Калабухов, 1964, и др.).
Среди зоологов этот стандарт вида, по словам С. А. Чернова
(1941 : 12), был распространен в США: Мэрием только в Северной
Америке выделил 78 видов бурых медведей, а Миллер в каталоге
млекопитающих — 13 видов северных оленей, 14 видов лисиц,
10 видов волков и др. \
4. Морфолого-генетический ^стандарт монотипического вида
отличается от чисто морфологического абсолютизацией принципа
монотипизма на основе генетического эксперимента. Как уже
указывалось, «microspecies» = гомозиготному биотипу = одной
апомиктической линии (клону) = чистой линии = инцухт-линии,
переставшей расщепляться, и т. п. Вид приравнивается к одному
мутанту и у высокогетерозиготных форм, так же как у агамов и
апогамов — к индивиду. Применение этого стандарта ведет
к столь крайнему дробительству, что в результате единое насе-
ление разделяется на много частей, каждая из которых возводится
в ранг вида.
5. Морфолого-географический политипический стандарт. Часто
используется в зоологии и ботанике (Van-Steenis, 1957). Вид рас-
2 Эти взгляды критиковали многие зоологи (Чернов, 1941; Огнев,
1944; Берг, 1950, и др.). Лучшую защиту этого стандарта можно найти
в работе С. В. Юзепчука (1958). Е. М. Лавренко (1964) справедливо кри-
тикует С. В. Юзепчука, идущего дальше В. Л. Комарова в сужении моно-
типического стандарта (см. также: Завадский, 1961). Ошибочное отож-
дествление реального вида с монотипической расой и противопоставление
его политипическому виду как нереальному встречается и в работах по-
следних лет.
10*
147
сматривается как группа географических (экологических) рас —
подвидов, установлено существование тесных роев рас, примыкаю-
щих, перекрывающих, полностью налегающих или иерархически
соподчиненных, а также и рас-видов, стоящих особняком (ре-
ликты типа Ginkgo biloba и Sequoia или Latimeria chalumnae).
При этом, однако, сохраняется извечное для систематики затрудне-
ние — уточнение «достаточной степени различия» (где проводить
границы группы рас и к какой группе относить данную расу?).
6. «Биологический вид» — этот стандарт политипического вида,
основан на трех показателях: специфичности морфологии,
географо-экологической определенности и обособленности в поло-
вом отношении. Роль ведущего критерия отводится физиологиче-
скому критерию (скрещиваемость внутри группы и межгрупповая
изоляция). Майр (1947 192) считает, что этот стандарт введен
в 1937 г. Добжанским, а Кэйн (1958) отдает приоритет Майру.
На самом же деле старые понятия «сингамеон» (Lotsy, 1917),
«kommiesum» (Danser, 1929) и другие имеют то же содержание:
группы форм, способных нормально скрещиваться и воспроиз-
водиться. Таков же и термин «coenospecies» (Clausen, 1951) —
совокупность фактически и потенциально скрещивающихся попу-
ляций. Следует подчеркнуть, что степень стерильности — фертиль-
ности — проверяется экспериментально, как это сделано очень
тщательно с видами дрозофилы (Dobzhansky, 1955, 1958 и др.),
о растениями (Вавилов, 1931, 1935; Clausen et al., 1940; Синская,
1948, 1963; Clausen, 1951, 1958, и др.). Систематики же делают
заключения о величине полового барьера по морфологической
характеристике образцов, собранных в зонах контакта или пере-
крытия ареалов (Rensch, 1954; Кэйн, 1958; Bowden, 1959; Мауг,
1963, и др.).
Такой описательный подход делает принципы биологического
вида в значительной мере декларативными, и по существу под
этим названием большинство зарубежных авторов использует
стандарт морфолого-географического политипического вида (Ren-
sch, 1958, 1965; Мауг, 1963, 1965).
Таковы шесть основных стандартов вида, используемые си-
стематикой. Модификаций же этих стандартов существует много.
При знакомстве с работами систематиков разных групп может
показаться что число стандартов очень велико. Однако терми-
нология, употребляемая специалистами по отдельным груп-
пам, не всегда отражает особенности видов и, возможно, чаще
является лишь сложившейся традицией у микологов, альгологов,
бриологов и т. д. (Cifem, 1932; Голлербах, 1941; Полянский, 1958;
Савич-Любицкая и Смирнова, 1958, п многие другие). То же са-
мое наблюдается и у зоологов.
Особенности объектов микробиологии — слабая изученность
или неразвитость многих черт вида (молекулярной морфологии,
географо-экологической определенности, способов интеграции по-
148
пуляций в связи со способами размножения и обмена генетиче-
ской информацией, дискретности в связи с явлениями трансдук-
ции и трансформации, сложности циклов и др.). Это заставляет
прибегать к поискам новых признаков для построения стандарта,
например физиологических и биохимических, включая серологиче-
ские, иммунпологические,3 токсикологические, паразитологические,
метод фагии, антагонистических культур, анализ специфичности
ДНК и др. Так ставится вопрос микробиологами разных специаль-
ностей (Курсанов, 1945; Красильников, 1947, 1962; Купревич,
1949, 1950; Кудрявцев, 1951; Клюйвер и Ван-Ниль, 1959; Имше-
пецкий, 1961; Белозерский, 1962). По-видимому, определение ос-
новных стандартов вида следует модифицировать для групп, в ко-
торых обычная морфология явно отступает на задний план
(например, риккетсии и некоторые группы бактерий).
Дискуссии о стандарте вида часто носили в большей мере
номенклатурный характер (см. главу 5), хотя их участникам и
казалось, что решается вопрос о сущности вида (Огпев, 1944;
Берг, 1950; Юзепчук, 1958).
Еще недавно в палеонтологии стандарт вида был морфологиче-
ским (Федотов, 1940; Борисяк, 1947). По Д. М. Федотову, «вид
в палеонтологии... отвечает лиинеевскому виду в современной
биологии. К одному виду относятся ископаемые объекты... об-
ладающие достаточно резкими морфологическими различиями,
даже если они связаны переходами в вертикальном направления»
(1940 8). Кэйн (1958 153) ошибается, считая этот стандарт
морфологическим на вечные времена. При возможности эколого-
географического анализа массы образцов вид во времени, т. е.
фратрия, может определяться как популяция пе хуже многих
современных видов. Палеозоолог А. Мюллер (Muller, 1963 : 137—
140 и др.) выступает за выбор разных стандартов; вид может быть
монотипическим и состоять из одной популяции, или политипиче-
ским, включая ряд формотипов (typusform) или ряд географиче-
ских рас (subspecies), или же состоять из непрерывной серии по-
пуляций, нечувствительно переходящих одна в другую. Предложе-
ние Мюллера выгодно отличается от позиций биостратиграфов —
сторонников унифицированного мелкого вида (см. главу 5).
2. КРИТЕРИИ ВИДА
Морфологический критерий. В предыдущих разделах
было показано, какие затруднения возникали в систематике при
попытках применить отдельно взятые критерии вида. Оказалось,
3 Недоверие к морфологическим признакам вызвало увлечение сероло-
гическими (Мед с сотрудниками в 20—30-х годах) и иммунологическими
признаками высших организмов, однако вскоре стало ясно, что эти
признаки в принципе не более весомы, чем структурные.
149
что среди признаков вида нет ни одного, который можно было бы
использовать как единственный, абсолютный видовой критерий.
Морфологический критерий вида, например, не содержит в себе
никаких возможностей определить, какая именно «степень раз-
личия» структур должна быть принята за видовую. Поиск «суще-
ственного различия», проводившийся со времен Линнея, окон-
чился безрезультатно: не существует какого-то одного специфиче-
ского «видового» морфологического признака. Один и тот же
признак у разных групп имеет совершенно различное таксономи-
ческое значение. Например, сросшиеся или открытые створки
плодов, одиночное расположение цветков или по три вместе,
супротивное расположение листьев или попеременное у одних
групп характерно для целого рода или даже семейства, у других
отличает виды, а у третьих все эти признаки встречаются не
только в пределах одного вида, но и одной популяции и т. и.
В одних случаях окраска маркирует виды (куропатки, медведи и
пр.), а в других является типичным «индивидуальным различием»
(домашняя кошка). Большие морфологические различия по це-
лым корелляцпонпым системам признаков являются у домашних
голубей или собак лишь породными, а вдесятеро меньшие у дру-
гих групп — видовыми.
Итак, один и тот же признак в разных группах может иметь
совершенно различное таксономическое значение пли даже не
иметь его вовсе, выражая лишь вариацию непрерывного ряда
изменчивости (возрастной, сезонной, географической и т. п.).
Чтобы наглядно представить значение изолированно исполь-
зуемого морфологического критерия, рассмотрим хотя бы один
пример. Вид шляпочного гриба — осиновика (Boletus versipellis =
Kromholzia aurantiaca) когда-то считали разновидностью березо-
вика (Boletus scaber), а затем он был выделен в особый вид. Было
описано несколько форм осиновика (с оранжевой шляпкой и чер-
ными чешуйками ножки, с красной шляпкой и белыми чешуй-
ками ножки, и т. п.). Кроме того, была описана тундровая форма
осиновика. Вскоре каждая из этих форм была возведена в ранг
нового вида (В. duriuscula, В. rufescens, В. aurantiacus,
В. percandidas и пр.), а тундровую форму, единственную, имев-
шую особый ареал, назвали почему-то разновидностью
(В. versipellis var. arcticus). Ни один автор при этом не изучал
наследственной обусловленности признаков, их экологического
значения и т. п. Такая безосновательная фабрикация новых видов,
к сожалению, имеет место не только в систематике шляпочных
грибов. Б. П. Васильков вначале развенчал все эти виды и разно-
видности в ранг «микоризных форм» одного вида осиновика в ста-
ром объеме на основании приуроченности их к отдельным древес-
ным породам: осине, березе, дубу, сосне, карликовой березе и пр.
Однако затем вновь квалифицировал их как разновидности и, на-
конец, пришел к выводу, что «указанные выше виды, они же —
150
микоризные формы, они же — разновидности», должны квалифи-
цироваться как простые модификанты (Васильков, 1954 : 683), так
как все признаки, по которым их пытались возвести в ранг видов
или разновидностей, оказались обратимыми и заходящими (тол-
щина гиф поверхности шляпки, строение, форма и размеры спор,
базидий и цистид, форма и размеры плодового тела, особенности
чешуек ножки, мякоти и окраски шляпки). «Микоризными фор-
мами,— пишет Васильков,— квалифицировать подразделения
осиновика тоже нельзя, так как. имеются случаи, когда в лесах,
состоящих из одной и той же породы, произрастают различные
осиновики (в сосняках, например, желто-бурый и серый) и,
наоборот, в лесах, состоящих из различных пород, — одинаковые
(тот же желто-бурый осиновик и в сосняках, и в березняках)»
(там же : 689).
Казалось бы, все уже ясно, так как установлено, что система-
тики называли видами и разновидностями осиновика всего лишь
отдельных модификантов. Но Васильков предпочел все же воз-
вести каждого из них в таксономический ранг формы (forma) и
дать ему латинское название. Так вновь появились семь латин-
ских названий форм осиновика. Следовало ли этим заниматься?
«Несомненно да,— отвечает автор,— раз они существуют в" при-
роде, хорошо различимы и достаточно уклоняются от видовой
формы» (там же: 692). В- потомстве одной особи могут суще-
ствовать растения, пораженные хлорозом, вирусными болез-
нями, недоразвитые или жирующие, этиолированные или полу-
чившие избыток света и т. п., но зачем! же каждую такую форму
именовать особым латинским названием? Этот случай типичен
в том отношении, что опираясь в основном на морфологию, систе-
матик часто оказывается в тупике. Вид не имеет не только внешне
морфологических, но и анатомических, гистологических, цитоло-
гических, кариотипических и ультрамикроскопических «видовых»
признаков, которые позволяли бы маркировать ими именно его,
а не иные группы (внутривидовые или надвидовые). Хотя и дока-
зана видовая специфичность хромосомного аппарата 4 (генетиче-
скими и химическими методами), пластид (при реципрокных
межвидовых скрещиваниях), цитоплазмы (например, работами
Лампрехта на фасоли, Махаэлиса па эпилобиумах и пр.), однако
никакой стандартной «видовой степени различия» не сущест-
вует.
Состав ДНК имеет таксономическое значение, существенно
различаясь у близких видов. Однако многие переоценивают роль
«молекулярного» критерия вида, возлагая на него непосильные
задачи (см. например: Анфинсен, 1962, п др.). Один и тот же
4 С. Г. Навашин считал число и форму хромосом «наиболее ранним»
морфологическим признаком, по которому можно различать виды уже
при первом делении зиготы.
151
признак — молекулярный ли он, микроскопический или макроско-
пический — может в одних группах служить критерием вида,
в других — рода, семейства или подвида и даже не иметь ни-
какого систематического значения (Майр, 1947 186—187; Мауг,
1958, 1963; Юзепчук 1958 : 138, и др.).
В молекулярном кодирующем аппарате не существует опреде-
ленных цистронов, оперонов, репликонов или их систем, ответст-
венных именно за видовую специфичность генотипа. Нет никаких
особых молекулярных «видовых признаков». В одних генетиче-
ских системах видовое молекулярное различие может быть след-
ствием внутрицистронных рекомбинаций кодонов, в других —
рекомбинаций цистронов внутри оперона, в третьих — геномных
перестроек, проходящих на уровне целых, хромосом или целых
наборов хромосом. Сравнительный анализ генетических причин
видовых различий, осуществленный на дрозофиле и на других
комплексах близких видов (Dobzhansky, 1955; Дубинин, 1966),
показал, что одна пара «хороших» видов может различаться по
нескольким мелким мутациям, другая — по одной крупной мута-
ции, сопровождающейся мутациями генов-модификаторов, и т. и.
С рассматриваемой точки зрения молекулярная морфология гене-
тических систем хотя и является более глубоким признаком вида,
чем макроморфология фенотипа, тем не менее сама по себе она
не способна стать тем «существенным различием», поисками
которого безуспешно занималась систематика.
Биохимический критерий. Все что сказано о морфоло-
гическом, можно сказать и о биохимическом критерии вида.
Не существует никаких специфических «видовых веществ»,
а также иммунологических и других реакций. Справедливо пи-
сала М. А. Розанова (1946:32), что «иммунитет является та-
ким же относительным признаком, как и другие систематические
признаки». Такие субстанциональные признаки, как последо-
вательность нуклеотидов и их особенности в составе частицы
ДНК, в одних генетических системах обусловливают лишь малую
мутацию, с узким спектором действия, в других это отличие
оказывается комплексным и проявляется в качестве особенности
популяции, в третьих они имеют видовое или даже надвидовое
значение. В этом отношении биохимические признаки ничем
не отличаются от морфологических. Если они используются в ка-
честве критериев вида изолированно от других, возникает прин-
ципиально перешаемый вопрос о «достаточной степени различия»,
сущность вида становится «неуловимой» и границы вида можно
устанавливать лишь произвольно.
Биохимические признаки имеют такой же таксономический
вес, как морфологические, географические и др. Например, тюль-
панное дерево (Liriodendron tulipifera) толковалось как один вид,
включающий и популяции из Китая. Некоторые же (Благовещен-
ский, 19656) выделяют особый вид L. chinensis. Оказалось, что
152
биохимически, по составу белковых комплексов, эти популяций
резко различаются (в %):
Вид Глютелины Альбумины Глубулины
L. tulipifera L. 12.30 75.56 12.14
£. chinensis Sargent 6.36 6.03 87.61
Однако таксономический ранг этих различий сам по себе не-
определен, и правильный вывод может быть сделан при использо-
вании целого комплекса признаков. На растениях показано, что
вид специфичен каким-либо основным протеиновым компонентом,
а отдельные расы и сортотипы вида, кроме того, содержат специ-
фичные для них протеины (Tsuyoshi, 1964). В таких работах
остается еще неясным, насколько подобные соотношения специ-
фичны или универсальны. Существуют ли такие биохимические
показатели, которые могли бы вскрыть глубокую специфичность
вида и в будущем выполнять функцию истинных «существенных
различий», о которых мечтали систематики со времен Линнея?
Такими показателями считают специфические особенности состава
нуклеиновых кислот. Исследование видовой специфичности ну-
клеиновых кислот проводилось на бактериях (Белозерский, 1962,
1964; Спирин, 1962), на грибах и па высших растениях (Ваню-
шин, 1965). У близких видов бактерий ^(в пределах одного рода)
состав ДНК, т. е. отношение А одинаково пли почти одина-
ково, а у далеких видов различается от 0.45 до 2.6. У грибов же
„ г + ц + мц
и особенно у высших растении отношение —А _^_-р— варьи-
рует гораздо меньше, чем у бактерий, и достоверные различия
пока удается обнаружить только у далеко отстоящих друг от друга
видов. Поэтому Б. Ф. Ванюшин (1965) констатирует, что значе-
ние состава ДНК для диагностики видов грибов и высших расте-
ний очень невелико. Этот несколько обескураживающий вывод не
должен давать повода для свертывания работ, так как несомненно,
что видовая специфичность представлена в генетическом коде.
Не следует впадать в крайности и требовать возврата систематики
в гербарий (Васильков, 1965).
В по-следние годы совершенно по-новому поставлены вопросы
о связи между структурой ядерной ДНК, матричной и рибосомаль-
ной РНК, управляющими синтезом белка и фенотипическими при-
знаками. В работах последнего времени (Крика, Ниренберга и
Маттей, Бензера, Очоа, Белозерского, Спирина и «др.) была пока-
зана необычайно заманчивая перспектива перейти от общего
утверждения о видовой специфичности белков и ДНК—РНК
аппарата к изучению конкретных связей между молекулярными
изменениями и их проявлениями. Надо осознать огромное значе-
ние этих работ и их перспективность. Однако нельзя забывать
153
при этом, что вид не сйодйМ к £енетическо;му коду и что Сам КОД
преобразовывается в ходе отбора, идущего по отдельным феноти-
пам или по их группам. Велико желание найти сакраментальную
сущность вида и один «главный» критерий, который состоял бы
в характеристике специфических молекулярных аппаратов, управ-
ляющих синтезом белка, становлением организации клетки, инди-
вида и генезисом всех видовых адаптаций. Этот наивно-оптимп-
стический взгляд основан на неправильных посылках о чисто ге-
нетической основе эволюции и об аддитивном характере всех
взаимодействий на молекулярном, клеточном и более высоких
уровнях организации (Уоддингтон, 1964). Хотя выработка точно
действующих кодирующих систем и явилась фундаментом самой
эволюции, все же ошибочно сводить характеристику вида к харак-
теристике кодирующего аппарата. Фенотипическая «надстройка»
вида входит необходимым и существенным компонентом в комп-
лекс систем, образующих организацию вида. В последних работах
(Федоров и Пименов, 1967) подчеркивается значение химических
признаков, отмечается, что, изолированно взятые, они пе могут
определять таксономического ранга формы и что роль химиче-
ского критерия вида еще не совсем определена.
Географический критерий. Этот критерий, взятый
изолированно, позволяет возводить в ранг расы или вида каждую
пространственно обособленную популяцию. Как показал опыт его
применения, он не гарантирует от механического раздробления на
части населения, единого по всем другим показателям. С другой
стороны, при переоценке роли географического критерия могут
быть объединены заведомо различные виды, если они сосуществуют
на одной территории. Вообще географический метод сам по себе
бессилен разграничить морфологически хорошо обособленные
виды, если они обитают вместе, по может объединить простран-
ственно разобщенное население, даже тождественное по всем дру-
гим показателям. Когда морфолого-географический стандарт вида
применяется со все большим упором на географо-экологические
моменты, возрастает опасность произвольного дробления единого
населения.
Географическая определенность вида может быть признана
существенной его чертой только в том случае, если ареалы обла-
дают значительной устойчивостью. В настоящее время хорошо
изучены случаи быстрого расселения видов и обратные процессы,
когда под влиянием истребления, вырубок, распашки, пожаров
некоторые виды в течение короткого времени сильно сократили
ареалы. Несомненно, что воздействие человека значительно изме-
нило течение естественпоисторических процессов расселения видов.
И тем не менее географические ареалы многих видов достаточно
устойчивы и изменяются лишь в течение геологического времени.
Географический ареал является одной из обязательных черт
вида, и полная характеристика вида невозможна без указаний на
154
специфические особенности его ареала. Во многих случаях по осо-
бенностям географического распространения вида удается восста-
новить его происхождение, родственные связи и фазу эволюции.
Для этого сопоставляются ареалы близких (видов и видов, принад-
лежащих к одной флоре или фауне, а также современное расселе-
ние вида с его палеогеографией. Так, например, на Кавказе рас-
пространен один вид орешниковых — Pterocarya fraxinifolia. Он
встречается вместе с многими формами третичного происхожде-
ния (в Западном Закавказье). Встречаются еще два вида этого
рода: Р. rhoifolia в Японии и Р. stenoptera в Китае. Оба живут
также совместно с формами третичного происхождения. Следова-
тельно, род птерокария имеет разорванный ареал, а каждый из
трех видов имеет эндемичный ареал, приуроченный к местооби-
таниям, насыщенным представителями реликтовой флоры. Кроме
того, в третичных отложениях па Алтае, в Альпах и других ме-
стах найдены остатки птерокарий, сходных с современными ви-
дами, что указывает на широкое распространение этих форм
в прошлом. На основании этих данных можно рассматривать,
ареалы видов птерокарии как остаточные и утверждать, что эн-
демизм данных видов носит реликтовый характер. Этими же прие-
мами удалось доказать реликтовый характер эндемизма группы
американского болотного кипариса (Taxodium).^ состоящего из
трех видов и близкой ему восточноазиатской группы глиптостро-
бус (всего два вида), и т. п.
Совершенно! иной характер имеет эндемизм таких видов, как
например европейские малины (Rubus), американские боярыш-
ники (Crataegus), среднеазиатские одуванчики (Taraxacum), ас-
трагалы (Astragalus) и многих других. В этих группах ареалы
отдельных видов хотя и являются эндемичными или полуэнде-
мичными, но не разъединены, а налегают или примыкают друг
к другу и образуют как бы рои форм. Такие молодые эндемичные
виды характерны для равнинных территорий, недавно освободив-
шихся от ледникового покрова, и в особенности для горпых
стран (Вульф, 1944; Толмачев, 1962, и др.).
Размер ареала, его границы и характер очертаний объяс-
няются различными причинами, которые можно свести к двум
группам: наличие границ пригодных для жизни условий и взаи-
модействие с видами-конкурентами.
Даже границы ареала одного вида могут определяться не-
сколькими причинами, например, на севере — климатическими
или пищевыми факторами, на востоке или западе — физико-гео-
графическими (море, горный хребет, пустыня), а на юге — кон-
куренцией.
Эндемизм флоры и фауны на островах чаще всего объясняется
наличием физических преград для распространения. Более близ-
кие к материку Канарские острова имеют 45% эндемичных видов
высших растений, Мадагаскар — 56%, Корсика —58%, а более
155
удаленные — Св. Елены —85%, Гавайи —82%. В Австралии
свыше 9000 видов растений из 12 000, т. е. более 75% являются
эндемичными для этого континента (по Аудасу). Физико-геогра-
фические причины ограничивают расселение и многих горных ви-
дов: ареал кок-сагыза — несколько долин в горах Тянь-Шаня,
ареал белой акации (Robinia pseudoacacea) — в южной части Ал-
легановых гор в Северной Америке, а ареал конского каштана —
небольшая площадь в горах Балкан. Препятствием для их рассе-
ления является невозможность переброски семян через горные
хребты. В культуре эти виды успешно расселяются на огромных
территориях, но их широкие возможности приспособления к рас-
пространению остаются нереализованными в природной обста-
новке.
Обширный ареал кедрового стланика (Pinus pumila), охваты-
вающий весь северо-восток Азии, по Б. А. Тихомирову (1946),
почти совпадает с ареалами ряда других видов (Betula midden-
dorfii, Larix dahurica, Populus suaveolens и др.). Все эти виды
входят в состав одних и тех же сообществ. Сходство ареалов и
биоценотическая связь данных видов указывает па общность исто-
рических условий, в которых происходило их формирование.
Естественно, что для видов-космополитов «географическая
определенность» теряет значение различающего критерия, как и
для симпатрических видов, например видов-«двойников», а также
для некоторых быстро расселяющихся видов. Все это говорит
о том, что «географическая определенность», как и все другие
признаки, не может служить абсолютным и единственным крите-
рием вида.
Другие критерии. Ни один из других критериев также
не в состоянии безотказно различать виды от других образований.
Экологический критерий (см. главу 5) сам по себе не позво-
ляет отчленить от вида экологические формы внутри вида (и
даже внутри одной местной популяции). Не позволяет он отчле-
нить также и обратимые различия (адаптивные модификации,
морфозы и др.) от необратимых, наследственно фиксированных
различий.
Эколого-цитофизиологический критерий вида, предложенный
Б. П. Ушаковым, интересен, но и его переоценка нежелательна.
Теплоустойчивость гамет и тканевых клеток у многих пойкило-
термных животных является видовым признаком, так как она
одинакова у разных популяций одного вида и различна у близ-
ких видов. Однако этот признак не имеет исключительности,
так как все структурные и функциональные особенности, входя-
щие в изогенный видовой радикал, ведут себя таким же обра-
зом. Ранее Ушаков писал, что теплоустойчивость — это «истин-
ный цитофизиологический критерий вида», подступ к выработке
общебиологического критерия вида (1959:556), но в последних
работах (1963, 1964) правильно снял эти формулировки.
156
Генетический критерий (см. главы 4 и 5), взятый изолиро-
ванно, также бессилен отграничить видовые признаки не только
от генетически значимых признаков внутривидовых форм, имею-
щих географическое или экологическое значение, но и от призна-
ков внутрипопуляционных форм. Ф. Шванитц (Schwanitz, 1963)
справедливо доказывает, что генетика не может быть единствен-
ной основой новой систематики, так как ее влияние противоре-
чиво: она не только помогает определять виды, но и толкает
к безудержному их раздроблению на массу форм и групп.
Наконец, физиологический критерий, т. е. степень половой изо-
ляции, также не имеет абсолютного значения и, кроме того, не-
применим к группам, лишенным полового процесса, и к оценке
ранга пространственно изолированных и давно сформированных
рас. Абсолютизация физиологического критерия наиболее полно
реализована в стандарте биологического вида. Однако многие ав-
торы справедливо предупреждают о недопустимости абсолютиза-
ции репродуктивных барьеров в качестве критериев вида (Beaudry,
1960, и др.), так как при полной географо-экологической изоляции
«расы, разошедшиеся до уровня вида, могут свободно скрещи-
ваться между собой» (Дубинин, 1966 : 308)
Надо опираться только на весь комплекс известных критериев
вида или пользоваться хотя бы многими из них. Иной подход
в большинстве случаев делает проблематичным таксономический
ранг изучаемых объектов. Так, например, близкие одуванчики —
кок-сагыз (Taraxacum kok-saghys) и таджикский одуванчик
(Т. tadshikorum) — отличаются ареалом и экологией (кок-са-
гыз — восточная часть Центрального Тянь-Шаня, более высоко-
горный, холодостоек; таджикский — южные склоны Гиссарского
хребта с сухим и жарким климатом), морфологическими при-
знаками (строение листочков обертки семяпок и форма края ли-
стовой пластинки), по каучуконосности, по ритмам вегетации
(таджикский с эфемероидным развитием). Систематики (Овчин-
ников и Овчаров, 1947; Кудряшева, 1957) выделили таджикские
популяции в особый вид, основываясь на критериях морфолого-
географического метода п на физиологических отличиях. Следо-
вало бы сравнить эти формы также и в условиях опыта. Не ис-
ключена возможность, что тянь-шаньская и гиссарская популя-
ции генетически почти идентичны и в этом случае видовой их
ранг не отвечает действительности.
3. УНИВЕРСАЛЬНОСТЬ И НЕРАВНОЦЕННОСТЬ ВИДА
Универсальность вида. Следует обсудить два вопроса:
1) древен ли вид, как жизнь, или возник позднее; 2) сущест-
вует ли он у всех групп в современных условиях (независимо
от высоты организации, способа размножения и трудностей
классификации).
157
Нет прямых данных о времени появления вида в процессе
растекания живого вещества и формирования биосферы (Вернад-
ский, 1926). Обнаружение видов у риккетсий п подобных форм
как будто позволяет отодвинуть время появления вида к периоду
формирования у протобионтов способности к точной саморепро-
дукции.
Многие авторы, например Ледерберг (1961), Флоркен, Брауп-
штейн, Пасынский, Стенли, Бернал, Пири, Мирский, Шрамм
и др. (Происхождение жизни..., 1959), Бернал, Рич и др. (Го-
ризонты биохимии, 1964), отмечают глубокое единообразие и ог-
ромный консерватизм химических процессов и структур в жи-
вой природе, что свидетельствует об их большой древности.
Можно допустить, что вначале существовали неспецифические по-
лимеры нуклеиновых кислот, редупликация которых была еще
неточной и происходила вне связи с белками. Далее возникает
топохимическая взаимосвязь между ДНК-РНК и белками, мо-
лекулярный вес нуклеиновых кислот растет (по В. Стенли) до
300 000 (около 1000 нуклеотид в пити). Появляется новая функ-
ция точной редупликации — передача наследственной информа-
ции. Эобиопт (по Н. Пири) — еще не организм, так какие обладал
внутренне обусловленным устройством точного самовоспроизве-
дения. До появления такого устройства отсутствовала возмож-
ность устойчивой передачи потомкам тех или иных достоинств
родительской частицы. По Дж. Берналу, раньше существовало
очень неточное копирование, приводящее лишь к черновому сход-
ству потомков с родителями. Без точной редупликации нет устой-
чивости, так же как нет п основ для мутационной изменчивости
и отбора. Надо думать, что число возникших на этом этапе типов
открытых систем было па несколько порядков величин больше,
чем число систем, способных к устойчивому самовоспроизведе-
нию (по А. Пасынскому и Н. Пири).
Вид необходимо связан с устойчивостью аппарата редуплика-
ции и не может существовать без отбора, накапливающего цолез-
ные мутации, число которых должно быть оптимальным. Исходя
из этих посылок, можно предположить, что вид возник при пере-
ходе от эобионтов к устойчиво самовоспроизводящимся прото-
бионтам. Такой первичный тип вида можно назвать eoagamospe-
cies.
Первые фактические данные об ископаемых видах известны
из протерозойских отложений кварцитов Южной Австралии
(Glaessner, 1960), Чехословакии, Норвегии (Горский, 1964)
и т. п. Древность этих отложений исчисляется в 1 —1.5 млрд лет.
В этих слоях, помимо бактерий и синезеленых водорослей, была
обнаружена богатая и разнообразная фауна медуз, гидроидов, ан-
нелид, а также неизвестных животных с трехскладчатой симмет-
рией: Tribrachidium и Parvancornia. Изучение докембрия пока-
зало, что и 1.5 млрд лет тому назад явление вида у растений и
755
животных йе отличалось от того, что мы знаем о нем в настоящее
время.
Характеризуя восходящее развитие живого, Б. Рснш (Rensch,
1959) перечисляет следующие «основные изобретения»: возникно-
вение молекул нуклеопротеидов, способных к саморепродукции;
возникновение мутационного процесса, давшего необходимой число
вариантов молекул или групп молекул для отбора; возникновение
генов, поддерживающих динамическое равновесие в живых си-
стемах (т. е. целостность молекулярных комплексов); объедине-
ние генов в хромосомах (возникновение хромосомного аппарата
наследственности),5 возникновение полового размножения, гран-
диозно умножившего число вариантов путем комбинирования ге-
номов, хромосо(м, их частей и отдельных генов и создавшего но-
вые возможности для накопления мутаций в гетерозиготном
состоянии (рецессивов); возникновение клеточной организации,
оказавшейся замечательным элементом для построения всех бо-
лее сложных живых систем. Эти шесть «изобретений» лежат в ос-
нове эволюции как растительной, так и животной организации.
В этом перечислении не упомянуто «изобретение» вида. Можно
предполагать, что Ренш, являясь сторонником концепции биоло-
гического вида, не считает его первичным для жизни образова-
нием.
Существование и эволюция видов возможны только при на-
личии дискретной структуры наследственной основы. Квантован-
пость генов (кодонов, цистронов и пр.) опирается на известную
самостоятельность нуклеотидов в цепочках РНК и ДНК.
А. А. Малиновский высказал идею, что при «слитной» наслед-
ственности виды не могли бы эволюционировать путем естествен-
ного отбора и что «некорпускулярные виды (если они существо-
вали на заре жизни) совершенно не могли бы соревноваться с ви-
дами, которые обладают корпускулярной наследственностью»
(1941 172). Эволюция на основе отбора могла осуществляться
только при большой свободе комбинативных возможностей при-
знаков, а это неизбежно вызывало сохранение и совершенствова-
ние генотипов с дискретностью генов. «Линейное расположение
генов оказывается чрезвычайно ценным для вида. . Всякое от-
клонение от линейного расположения должно отметаться естест-
венным отбором, как невыгодное, поэтому линейное расположение
неизбежно является формой расположения генов, общей всем
видам» (там же 174).
По вопросу о распространенности явления вида в современ-
ную эпоху не существует единомыслия. Одни считают, что все
5 Этим гарантировалась точная одновременность и равномерность рас-
пределения генов при размножении первичных живых существ, а также
возможность более надежно регулировать посредством отбора сам мута-
ционный процесс.
159
Живое вещество биосферы в современную эпоху организовано
в виды, но при этом типы видов сильно отличаются один от дру-
гого (Завадский 1961, 19676; Дубинин, 1966). Другие допускают
существование «довидовых форм жизни», каковыми признаются
вирусы (Купревич, 1950). Третьи считают вид частной формой
существования жизни, возникшей вместе с половым размноже-
нием и исчезающей в тех случаях, когда этот способ размножения
нарушается пли выпадает вовсе (Комаров, 1934, 1940; Stebbins,
1941 и др., а также все сторонники стандарта биологического
вида). С. С. Хохлов (1950) полагал, что с появлением бесполосе-
менного размножения наступила эра вневидовой эволюции. По
Кэйну (1958 123), «биологическое определение вида приложимо
только к животным и растениям, размножающимся половым^ пу-
тем». Применение термина «вид» к апомиктам некоторые счи-
тают принципиально неверным, так как в ходе эволюции обра-
зуются не виды, а особые таксоны — agamospecies (Тихомиров,
1967 54).
С общебиологических позиций, по Б. М. Козо-Полянскому,
вид универсален и апомиктические группы, «конечно, суть виды
в этом толковании» (1948:127), которые можно выделить
в группу «апомиктантов», отвечающих «множеству старых и но-
вых биологических определений вида... тем более что никакого
общепринятого определения вида в биологии нет» (там же).
Р. Н. Шляков доказывает, что отказ от категории вида у апомик-
тов «не соответствует действительной структуре рода» (1967 : 60)
и неправилен принципиально. По Н. П. Дубинину, «для форм
с закрепленным апомиксисом понятие вида хотя и сохраняет свое
значение, но принимает другой характер» (1966:321). У скаль-
ных ящериц из группы Lacerta saxicola, размножающихся парте-
ногенетически и не имеющих самцов, доказано наличие «хоро-
ших» видов, отвечающих всем критериям, кроме биологического
(Даревский, 1967). Занимаясь систематикой тлей, Г. X. Шапош-
ников (1966, 1967) пришел к выводам о правильности нашей кон-
цепции универсальности вида. Отрицание этой концепции осно-
вано, по его мнению, на переоценке значения генетической сто-
роны проблемы вида и недооценки ее экологической стороны:
«Критерий вида должен включать два момента: возможность
объединения популяций, основанную на их родстве. . и ^на-
правленность или разную направленность их экологической спе-
циализации» (Шапошников, 1967 27).
Опыт систематики агампых микроорганизмов, например сине-
зеленых водорослей и других групп, доказывает, что при описа-
нии видов здесь возникает не больше трудностей, чем в группах
с половым размножением той же примерно степени изученности,
а возникающие трудности принципиально сходны (Еленкин,
1936; Клейн, 1945; Воронихин, 1946, 1951; Кудрявцев, 1951; По-
лянский, 1956, 1958, и др.). Нашу аргументацию в пользу суще-
160
ствование видов у а га иных групп поддержал О. Л. Рудаков
(1964), работающий с несовершенными грибами. В пользу нали-
чия вида у агамных групп свидетельствуют и данные протозо-
ологии. У агамных пресноводных корненожек виды разграничены
не хуже, чем у сексуальных форм той же группы. У жгутиконос-
цев агамные виды Hypermastigida, паразитирующие в термитах,
разграничиваются столь же четко, как и виды группы, паразити-
рующей в тараканах и размножающейся половым путем (Догель
п др., 1962:406).
Поэтому существование видов не должно связываться с появ-
лением полового процесса. По существу отрицание видов у агам-
пых и апотамных групп объясняется только тем, что частное
определение вида у наиболее изученных групп, размножаю-
щихся половым путем, было произвольно превращено в универ-
сальное определение вида. Если видом признать только сексуаль-
ный вид, то все агамные и апогампые з^руииы должны быть при-
знаны стоящими «до вида», «вне вида», «между видами» или
«вышедшими за рамки вида». В действительности же нет основа-
ний отрицать у этих групп существование вида, но тип вида
у них иной, чем при половом размножении. В последнее время
у групп, признававшихся агамными, обнаружены различные
механизмы, обеспечивающие обмен генетической информацией.
Показано, например,, что и у синезелеиых водорослей существует
процесс взаимообмена между хромосомоподобными телами, яв-
ляющийся, видимо, прообразом полового размножения. У бак-
териофагов профаг соединяется с геном (или даже замещает его)
и передается другому хозяину механизмом трансдукции.
О. Л. Рудаков (1964) подчеркивает большое значение образова-
ния у несовершенных грибов устойчивых колоний со многими
генетически различными ядрами — гетерокарионов. Вид при этом
оказывается сложной биологической системой, включающей
в одну популяцию-колонию филогенетически разные и даже дале
кие формы. Мы называем такой тип вида синтезогенетпческим
(см. главу 10). Между ядрами гетерокарионов возникают взаи-
модействия, сходные с половым процессом. Это — парасексуаль-
пый цикл, состоящий в слиянии гаплоидных ядер в одно диплоид-
ное. Эволюционно-генетическое значение парасексуалыюсти со-
стоит в возрастании комбпиативпых возможностей генотипа,
а значит, и новообразований.
Если мы примем тезис об универсальности полового размно-
жения в этом расширенном толковании, то тем более следует при-
знать и положение об универсальности вида.
Прямой опыт показывает, что все ныне существующие группы
организованы в виды, т. е. что в этом смысле явление вида уни-
версально для живой природы.
Каковы же причины, по которым немало биологов считают
вид частным явлением? Отчасти они носят терминологический
П К. М. Завадский
161
характер (тип вида при половом размножении признается един-
ственным заслуживающим наименование «вид»). В основном же
они основаны на игнорировании факта неравноценности видов и
стремлением подогнать виды под единый стандарт наиболее
изученного и привычного типа амфимиктического вида у выс-
ших групп растении и животных.
Примерами чисто семантических решений служат мнение
Дж. Гексли (Huxley, 1963) о замене термина «вид» рядом но-
вых, отражающих отдельные частные его проявления (для ага-
мов, апомиктов, амфимиктов, гибридных циклов и др.) и предло-
жение О. Клейншмидта (Kleinschmidt, 1930) о замене термина
«вид» термином «Formenkreise» и т. п. Большие различия типов
видов заставляют некоторых ученых говорить о принципиальной
невозможности дать общее определение вида (Turesson, 1931).
Все сказанное делает необходимым уже сейчас рассмотреть
вопрос о неравноценности явления вида.
Неравноценность видов. Мысль о существовании раз -
личных типов организации видов является прямым следствием
эволюционной теории. Эти различия объясняются разными фазами
эволюции, па которых они находятся (молодые, процветающие и
древние; широко расселенные п эндемичные; полиморфные и
мономорфные).
Известная неэквивалентность, неравноценность видов может
быть выявлена и по другим показателям. А. П. Семенов-Тян-Шан-
ский предложил несколько типов видов: древние, резко очерчен-
ные, мономорфные виды (species firmae), виды «со! значительной
изменчивостью, зависящей от молодости вида и быстроты его
эволюции (species praecoces) или от вторично возникшего по-
лиморфизма (species labefactae), виды, представляющие комплекс
нескольких рас с преобладанием одной основной формы или без
нее (species polymorphae, или conspecies)» (1910 10). Эта клас-
сификация была для своего времени очень ценной, так как наряду
с уже банальными признаками (величина хиатуса, возраст вида
и др.) выдвигала новые (зависимость от темпа видообразования
и др.).
Е. Дю-Рие (Du Rietz, 1930) предложил классификацию видов
высших растений, основанную на различиях по способам размно-
жения, по структуре и расселенности. Виды с неполовым размно-
жением (асексуальные): а) простые, состоящие из одного биотипа
и имеющие небольшие ареалы, б) сложные, состоящие из несколь-
ких биотипов и имеющие большие ареалы. Виды с половым раз-
множением (сексуальные): а) гомофацпальные, пе дифференци-
рованные географически (состоящие из одного подвида), б) гете-
рофациальные, дифференцированные географически и состоящие
из двух или нескольких подвидов.
Эта классификация заслуживает большого внимания, но тип
«простого вида, состоящего из одного биотипа», является под-
162
разделением спорным, так как Дю-Рие говорит здесь о виде
в жордановском смысле.
Были сделаны попытки разделить виды на £ипы, руковод-
ствуясь не эволюционной, а сравнительной точкой зрения. Так,
например, Дарлингтон (Darlington, 1940) предложил делить все
виды растений на шесть типов, основываясь на различиях в меха-
низмах их размножения: 1) с преобладанием самооплодотворения
(самофертильные диплоиды); 2) перекрестнооплодотворяюгциеся
диплоиды; 3) плодовитые полиплоиды; 4) смешанные виды, вклю-
чающие диплоидные и полиплоидные формы; 5) сложные гетеро-
зиготные виды типа Oenothera lamarckiana; 6) клональные виды,
вообще не размножающиеся половым путем. Клональные виды
в свою очередь могут быть подразделены па субполовые, подобные
Rosa canina, цартепогенетическпе беспорядочно расщепляющиеся,
виды с диплоидными самками и гаплоидными самцами, такие как
у Hymenoptera, и др.
Делались попытки объединить в одной классификации эволю-
ционный принцип со сравнительным. Так, например, С. А. Нев-
ский6 предложил Подразделять виды по способам Происхождения
(гибридогенные, климатогенпые, ценогенные, эдаф1огенные и др.),
а в пределах этих групп различать подгруппы, руководствуясь
структурными и прочими показателями. Подобное же предложе-
ние позднее внес Дж. Гексли (Huxley, 1963), считающий, что
виды должны подразделяться в соответствии со способами видо-
образования, так как при географической изоляции не возникает
таких «хороших» видов, как при изоляции в условиях спмпатрии
и т. п. Недостатком подобных подразделений является то, что в их
основу кладутся не точно известные факты, а толкования, вклю-
чающие гипотетический момент (какой фактор является ведущим
при видообразовании: почвенный, климатический пли биоценоти-
ческий?). Конечно, гибридогенное происхождение вида иногда
может быть констатировано достаточно точно.
Можно подразделять виды по их значению в экосистемах на
массовые, редкие и т. п. Было предложено различать понятие
«редкие виды» и «малочисленные виды» (Dobrowolski, 1963).
Редкий вид может быть малочисленным и многочисленным: его
признак — большая дисперсия по территории. Малочисленный вид
может быть очень обильным на небольшой территории или же
редким на большой. Неравноценность видов по численности свя-
зана не только с фазами их эволюции, но и с местом в биоценозе.
Неравноценность видов проявляется даже в пределах одного рода:
у наземных моллюсков Partula одни виды включают массу рас,
эндемичных для отдельных островов п местообитаний, а другие
представлены одной формой на всем архипелаге.
6 Доклад, прочитанный в 1935 г., оставшийся неопубликованным.
И*
163
Д. Гуд (Good, 1931, 1953) подчеркивает большую неравноцен-
ность видов по толерантности (жизнеспособности-выносливостп),
которая определяет возможности занятия видом «потенциального
ареала». Кэйн (1958) выделяет четыре типа видов: морфологи-
ческий, т. е. вид, установленный лишь по морфологическим при-
Рис. 2. Близнецовые виды группы Drosophila willistoni.
Ареалы пяти морфологически почти неразличимых форм,
полностью изолированных друг от друга в половом
отношении (по Добжанскому).
1 — insularis; 2 — willistoni; з — paulistorum; 4 — tropicalis;
5 — equinoctialis.
шакам; палеонтологический, т. е. тот же морфологический вид,
шисанный на ископаемом материале (такие виды автор считает
ароизвольными отрезками какой-либо филогенетической ветви —
фратрии); биологический вид (рис. 2) содержит формы «с истин-
тым половым размножением, генетически изолированные от всех
других видов, с которыми они сосуществуют» (там же: 154);
1гамный вид у групп, размножающихся исключительно бесполым
тутем; такие виды «можно классифицировать только на основании
эбщего сходства и различия как морфологические виды» (там
ке:155).
W4
До сих пор рассматривались попытки установить типы 'видов,
существующие в природе. В типологии Кэйна один тип выделен
па основании способов распознавания вида и отражает стадию его
познания. Автор сам утверждает, что «морфологический вид —
это выражение нашего невежества» (там же). Палеонтологи-
ческий тип вида также не является особым явлением природы,
а особым способом рассмотрения (вид во времени). Агамные виды
созданы лишь для удобства классификации и возведение их в ранг
вида «всегда произвольно». Абсолютизируя значение физиологи-
ческого критерия, Кэйн утверждает, что биологический вид — «это
единственный сорт вида, который можно определить как таковой»
(там же: 163). Таким образом, классификация, предложенная
Кэйном, неудовлетворительна, так как включает всего лишь раз-
ные точки зрен’ия, которые складывались в зависимости от изучен-
ности материала и подхода к нему.
Как полагает Г. Смит (Smith, 1958), существует три* принци-
пиально различных типа видов: менделевские, клональные, фпле-
тические. При этом виды одного типа независимы й неидентичны
видам других типов. Но критерии, установленные для сексуаль-
ных (менделевских) видов, являются универсальными. Нельзя
согласиться ни с этими критериями выделения типов видов (три
типа — три разных критерия), ни с универсализацией менде-
левского типа (это и приводит к отрицанию видов у агамных
групп).
Руководящим принципом разделения видов на типы должен
быть избран показатель, отражающий эволюцию вида как особой
формы организации живого от eoagamospecies к высокоцелостным
амфимиктпческим видам высших групп. Виды неравноценны
именно потому, что стоят на различных ступенях развития видо-
вой формы организации жизни или же, находясь на одной сту-
пени организации, имеют какие-либо принципиальные конструк-
тивные особенности.
Основным критерием для выделения типов видов должна стать
степень целостности популяции (Завадский, 1957а, 1961, 19676).
С появлением полового размножения возникла совершенно новая
возможность интеграции систем видового уровня. Образование
спсте<м скрещивания, поддержание гетерозиготпостп прп сбаланси-
рованном полиморфизме, гомеостатические аппараты популяцион-
ного уровня, регулирующие доминирование и другие процессы,—
этп и другие механизмы интегрируют популяцию (Шмальгаузен,
1958а, 19586, 1961; Grant, 1963, и др.). Помимо первичноагамного
вида (eoagamospecies), следует выделить несколько типов агам-
ных (клональных видов), далее виды с прообразом полового про-
цесса, способные к обмену наследственной информацией (транс-
формация, трансдукция, как у сальмонелл, и т. п.). Ждет своей
разработки классификация типов сексуального (амфимиктиче-
ского, зиготного) вида. Кроме того, необходимо выделять вто-
165
ричноагамные и апогамные типы видов, возникающие у высших
растений. В пределах каждого из таких типов виды должны под-
разделяться как по фазам развития (молодые, стабилизированные,
угасающие), по способам образования, если они известны, так и
по другим показателям.
Принцип неравноценности видов должен стать основой для
представлений об объеме вида и его границах. В разных филоге-
нетических стволах и даже в одной ветви виды могут быть неэк-
вивалентны друг другу. М. К. Хохряков подчеркивает, что «рав-
ноценных видов грибов может не быть даже в пределах одного
рода» (1964:161), так как виды отличаются не только характе-
ром признаков и объемом, но и стадией развития. Е. Н. Синская
показала, что в пределах рода люцерна существует многоплано-
вая неравноценность видов (по типу организации популяций,
по способам видообразования, по возрасту и др.). Тенденция
унифицировать виды связана со стремлением систематиков упро-
стить выделение нового вида и стандартизировать его ради номен-
клатурных удобств. В предисловии к «Флоре СССР» (Комаров,
1934), например, предлагается унифицировать виды по одному
мелкому стандарту, но наряду с этим отмечается, что в природе
виды неравноценны: в теории вид признается неравноценным,
а на практике рекомендуется один шаблон. Поскольку в этот
стандарт вида не вмещались апогамные виды и гибридные циклы
(как у эвкалиптов, тополей, ив и т. п.), указывалось, что к ним
«концепция вида... применяться не может».
Задача заключается в том, чтобы положение о неравноценности
видов превратить из теоретической идеи в руководящий принцип
систематики.
Однако, вместо того чтобы, опираясь на положение о нерав-
ноценности видов, протестовать против универсализации какого-
нибудь одного практического стандарта вида, некоторые авторы,
справедливо возражая против универсализации мелкого стан-
дарта вида, противопоставляют ему политипический вид в качестве
универсального же стандарта (Бочанцев и Липшиц, 1955; Гру-
бов, 1955; Тахтаджян, 1955).
Одним из особых типов видов являются виды культурных
форм. Специфические черты видов домашних животных и ра-
стений столь очевидны, что некоторые авторы давно предлагали
объединить их в особый тип вида (С. В. Юзепчук, П. М. Жуков-
ский и др.). Однако другие авторы считают, что не существует
принципиальных различий между видом одомашненным п диким
(см., например: Проханов, 1965а).
Итак, можно утверждать, что вид как явление природы
в современную эпоху представляет универсальную форму
организации живого, по в зависимости от различных при-
чин проявляется весьма многообразно, т. е. что виды неравно-
ценны.
166
Ь. ОБЩИЕ ПРИЗНАКИ ВИДА
Характеристика признаков. Если вид — явление уни-
версальное, правомочен и вопрос об определении с7мых общих
признаков вида, а так как различные виды неравноценны друг
другу, надо различать общие признаки, присущие всем видам, и
частные признаки, характерные лишь для отдельных его типов.
Ранее нами были перечислены десять общих признаков вида
(Завадский, 1954а, 19576, 1961, 1967а), и ниже они приводятся
с небольшими изменениями.
1. Численность. Вид включает множество особей, т. с. является
надиндивидуальным образованием. Уровень численности вида вхо-
дит в его качественную характеристику.
2. Тип организации. Вид обладает единой наследственной
основой. Генотипическое единство вида проявляется в специ-
фичности системы ДНК—РНК—белки, в сходстве типа обмен-
ных реакций, процессов морфогенеза, внутреннего и внешнего
строения особей, а также в сходстве внутривидовых отношений
в популяции. Вид — относительно однородное (изоморфное) об-
разование и как тип организации /выступает в качестве особой
единицы.
3. Воспроизведение. Вид — самостоятельно воспроизводящееся
в природе образование, способное в процессе размножения сохра-
нять свою качественную определенность.
4. Дискретность. Вид — биологическая отдельность, существую-
щая и эволюирующая как более или менее обособленное образо-
вание.
5. Экологическая определенность. Вид приспособлен к усло-
виям существования и конкурентоспособен. Он занимает опре-
деленное особое место в экологических системах и выступает
как отдельное звено в круговороте веществ и во взаимосвязях
живого с живым.
6. Г еографическая определенность. Вид расселен в природе
на определенной территории. Ареал, как правило, является
обязательной чертой, входящей в характеристику вида.
7. Многообразие форм. Вид дифференцирован и обладает вну-
тренней структурой. Включая много разнородных форм, вид вы-
ступает как система, основной единицей которой является мест-
ная популяция (система клонов или же дём).
8. Историчность. Вид — система, способная к эволюционному
развитию. Историчность вида проявляется во временном суще-
ствовании его как особой филогенетической ветви (вид во вре-
мени, или фратрия).
9. Устойчивость. Вид не имеет заранее заданного, внутренне
определенного срока существования в природе. Способность со-
хранять качественную определенность в течение известного гео-
логического времени — специфическая черта вида.
167
10. Целостность. Вид является племенной общностью, объеди-
ненной внутренними связями, и пе представляет собой суммы
особей. Вид — пе аддитивное образование, и связями, консоли-
дирующими его в целостную систему, являются видовые адап-
тации, полезные виду как целому, а также особый строй внутриви-
довых отношений. Целостность вида наиболее полно проявляется
в отдельных популяциях. Вид представляет собой надиндивиду-
альную форму организации живой материи.
Видом является только такое образование, которому присущи
все указанные признаки. Все эти признаки входят в характери-
стику любого вида, независимо от способа размножения, высоты
организации, способа питания и т. п.
Наша характеристика позволяет выделить вид из других био-
логических образований, сходных с ним по одному или даже по
нескольким отдельным признакам, но в целом качественно отлич-
ных от него. Так, например, одна особь, как бы она не выделя-
лась генетически или фенотипически из массы особей, не
является видом: типологическое понимание вида окончательно
изжило себя и заменяется статистико-популяционной концеп-
цией (Завадский, 1961, 1965; Мауг, 1963, 1965). Значение чис-
ленности как специфического признака вида и механизмов ее
регуляции у птиц и других животных прекрасно рассмотрено
в книге Д. Лэка (1957). Г. А. Викторов (1965) подчеркивает,
что каждый вид насекомых имеет специфические колебания п
средний уровень численности, которые регулируются главным
образом внутрипопуляционными и биоценотическими отноше-
ниями. Отдельные клоны или практически гомозиготные линии,
как правило, также не являются особыми видами, хотя у неко-
торых групп (синезеленые водоросли, вегетативно размножаю-
щиеся высшие растения) на определенном этапе эволюции они
могут приобрести видовой статут. Две группы популяций, не
различающиеся между собой по типу организации или эколо-
гически, также не являются видами. Не является видом и мно-
жество особей, не имеющее в природе эколого-географической
определенности, или популяция, не доказавшая еще своей при-
способленности и конкурентоспособности. Множество особей
с генетическими отличиями любого размера не является видом,
если оно не доказало способности устойчиво воспроизводиться
в природе. Любые генетически несбалансированные образова-
ния (смеси форм, расщепляющиеся гибриды, формы, существую-
щие и воспроизводящиеся лишь как компоненты единой популя-
ции, т. е. биотипы и другие впутрипопуляцпонные группы) не
являются видами. Разумеется, не является видом и отдельная
раковая опухоль, хотя, как это считает Дж. Гексли, «возник-
шую опухоль логически можно рассматривать как новый биоло-
гический вид со своим собственным присущим ему типом
самовоспроизведения и способностью к дальнейшей эволюции»
168
(1960 12). В отличие от вида, как правило, исходный тип опухоли
не дивергирует, «и каждый вид опухоли исчезает со смертью хо-
зяина». Несомненно, что каждая популяция опухолевых клеток
в отдельных чертах эквивалентна виду, но это еще не означает,
что каждую отдельную опухоль можно рассматривать как но-
вый биологический вид.
Хотя перечисленные признаки присущи всем видам, но это
не значит, что у каждого вида они проявляются одинаково;
у разных типов видов они различны. Какая-либо группа при-
знаков вида может быть более или менее развита. Например,
у агамных и апогамных видов такие черты, как целостность и
дискретность, еще мало развиты пли же подверглись вторич-
ной редукции. У некоторых низших агамных водорослей па
определенном этапе их развития может слабо/'проявляться и
признак многообразия форм, так как при жестком отборе и
большой генотипической однородности особей вид .по существу
оказывается равным одной клональной линии '(Еленкин, 1936;
Полянский, 1956). У водорослей и мхов ведущую роль играют
цитологические, физиологические, географические признаки,
а также способы размножения (Peterfi, Stefureac, 1965), а у
несовершенных грибов — биохимические (Рудаков, 1964),
и т. п.
Степень проявления общих признаков вида зависит от ста-
дии его развития. У молодых видов при некоторых способах ви-
дообразования может быть слабо проявлена дискретность и неко-
торые другие признаки. По разным причинам у эвкалиптов, ив,
тополей, осок, одуванчиков, ястребинок, хохлаток и в других ро-
дах дискретность видов выражена слабее, чем в других группах,
и более расплывчат у них такой признак, как тип организации.
Трудно классифицируемые группы существуют не только среди
агамных микроорганизмов, гибридных роев растений-апомиктов
или среди молодых видовых комплексов. Трудными для систе-
матики являются и некоторые группы млекопитающих. Напри-
мер, систематика бурых и пещерных медведей «чрезвычайно за-
путана в связи с огромной полиморфиостью и изменчивостью
популяций обоих видов при отсутствии четких и объективных
критериев объема систематических категорий» (Верещагин,
1963:55). У древних реликтовых видов дискретность про-
является исключительно четко, так как величина хиатуса бывает
огромной. Естественно, что многие общие признаки давно пер-
систировавших видов значительно отличаются от признаков ви-
дов, продолжающих эволюционные преобразования.
Многообразие форм у одних древних реликтов перестает быть
практически заметным (хотя всегда может быть обнаружено
экспериментально), а у других этот признак может быть отлично
развит. Тау-сагыз (Scorzonera tau-sagiz) имеет эндемичный
ареал в горах Кара-тау и, по данным М. В. Культиасова и др.,
169
является очень древним реликтом, персистировавшим еще в ме-
зозое. Однако он включает такую массу хорошо различимых
форм, что описание их в терминах систематики — совершенно
бессмысленное занятие. Древним реликтом является и гинкго,
однако, судя по нашим наблюдениям за листовой пластинкой,
оно чрезвычайно полиморфно, что подтверждается вариабель-
ностью формы кроны и некоторых других признаков (по Ota).
В гибридных циклах, а также у молодых видов с вторичным
перекрытием ареалов и местообитаний и с возникающей конвер-
генцией или интрогрессией может быть слабо выражена морфо-
физиологическая или сексуальная дискретность. У таких видов,
а также у видов с особыми формами сбалансированного поли-
морфизма расплывчат также и такой признак, как тип органи-
зации вида. У многих водорослей континентальных водоемов
в связи с их полукосмополитизмом снижается значение геогра-
фической определенности вида. Значение этой черты снижается
также под воздействием человека и у некоторых агрессивных
сорно-полевых и водных высших растений, некоторых грибов,
насекомых, быстро расселяющихся и ныне не имеющих устой-
чивых ареалов (Элтон, 1960). Степень проявления отдельных
признаков вида связана и со способами видообразования. На-
пример, у сложного гибридогенного вида, сформировавшегося
с большой скоростью, многие признаки будут значительно отли-
чаться от вида, медленно преобразовавшегося из географически
изолированной расы. Такими же большими будут различия
между видами, возникшими из внутрипопуляционных экологиче-
ских форм и из целых популяций (см. главу 10). Относительное
значение отдельных признаков вида меняется и в зависимости от
характера условий среды (наземные или водные, замкнутый
пресноводный бассейн или океан, горная долина, остров и т. п.).
Наконец, дискретность, экологическая определенность и целост-
ность вида изменяются из-за особенностей положения вида
в биоценозе (место в цепях питания, способ существования, сте-
пень напряженности межвидовых отношений и т. и.).
Таким образом, можно сказать, что неодинаковое проявление
различных признаков у разных видов является следствием не-
равноценности видов и различий в способах видообразования.
Эти различия — свидетельство того, что не существует какого-
то одного стандарта, по которому были бы организованы все
виды.
В предложенных нами десяти общих признаках вида,
точно ли они сформулированы и равно ли существенны, нам ка-
жется, отражен некоторый итог познания вида. Хотя наша ха-
рактеристика общих признаков вида может быть значительно
усовершенствована, она все же лучше отражает действитель-
ность, чем, например, описание сторон вида, сделанное С. Я. Па-
рамоновым (1943): 1) вид как имя, как наименование; 2) вид
170
как фиксационная единица, т. е. как первичная единица меры;
3) вид как таксономическая единица; 4) вид как биологическое
явление (отражение дискретности и цепи живых существ);
5) вид как биологический факт (биологическая изолированная
сущность, ведущая свое особое существование и несущая свои
особые признаки); 6) вид как процесс и 7) вид как понятие
(как реальность или абстракция). Эти семь сторон вида содер-
жат подходы не только к изучению явления природы, но и изы-
скания различных употреблений слова «вид» в логике и биоло-
гии; к виду как явлению природы непосредственно относятся
лишь два-три пункта. Кроме того, принципиально ошибочно
отрывать обсуждение понятия от явления, .которое в нем
отражается, и тем более относить их к различным сторонам
вида.
Некоторые авторы приняли предложенную нами характери-
стику десяти общих признаков вида полностью. (Полянский,
1956, 1958, 1959; Савич-Любицкая и Смирнова, 1958; Снискан,
1961; Карпов, 1962; Caloianu, 1962; Trojan, 1962; Лавренко, 1963;
Olson, 1963; Афанасьев, 1964; Левина, 1964; Petrusewicz, Trojan,
1964; Чижек, 1965; Волкова, Филюков, 1966; Шляков, 1967,
п др.). М. К. Хохряков (1964) отметил, что принимает десять
наших признаков вида, так как они применимы и к грибам, и
что у него нет с нами разногласий, так как ранее им было пред-
ложено девять сходных критериев вида. В первых публикациях
по этому вопросу (Завадский, 1952, 1954а) я, естественно, еще
не мог сослаться на предложения Хохрякова (1955), но в «Уче-
нии о виде» следовало это сделать.
Критерии вида, предложенные М. К. Хохряковым, таковы:
общность происхождения, сходная морфология, одинаковая или
сходная биология, наследование этих признаков и свойств, ареал,
наличие внутривидового разнообразия, физиологическая неразоб-
щенность форм, отсутствие антагонизма между ними и хиатус.
Наши списки сходны, но между ними есть и принципиальные
расхождения. Хохряков не считает критериями устойчивость и
историчность, «как общие и сами по себе подразумевающиеся»
(1964:162). С нашей точки зрения, ошибочно считать общим
критерием вида отсутствие антагонизма внутри вида: он возни-
кает у всех видов при определенных условиях (перенаселение,
бескормица, авитаминоз и т. п.), у некоторых является звеном
внутривидовых отношений (например, каннибализм у окуня,
«расы убийц» у инфузорий, химические токсические выделения,
агрессивные рефлексы некоторых хищных животных). Целост-
ность не исключает антагонизма, а иногда включает его как тип
отношений подсистемы (или функциональных звеньев и фаз
процесса). Хохряков правильно подчеркивает, что вид грибов
включает многообразие форм, и критикует тенденцию к дроблению
вида. Возведение каждой специализированной формы и физиоло-
171
гической расы в ранг вида (например, П. Н. Головиным) свя-
зано с отсутствием экспериментальной проверки изменчивости
форм, закрепленности их признаков, возникающих на разных
видах растений-хозяев (пли на отмерших субстратах), а также
с неправильным представлением о виде как о микромонотипе,
стоящем на пределе различительной способности система-
тика.7
Некоторые авторы приняли наши десять общих признаков
вида, сделав некоторые оговорки. Часть из них уже были нами
обсуждены ранее (Завадский, 1961). Отмечая, что наша харак-
теристика вида «биологическая и эволюционная», что в ее основу
«положена. идея целостности вида», А. Л. Тахтаджян считает,
что она подходит «больше к географической расе (подвиду)»,
а не к виду (1955 794). Но еще более целостны местная попу-
ляция или стадо, интегрированные различными взаимодей-
ствиями. Политипический вид является гораздо менее целостной
системой, а местные популяции широко расселенных рас, если
они удалены друг от друга, могут эволюировать как практи-
чески независимые образования.
Наша характеристика признаков вида может быть применена
ко всем расам, способным самостоятельно, т. е. практически
обособленно, воспроизводиться в природе и способным к продол-
жительному существованию и эволюции. Иначе говоря, она ох-
ватывает все сформировавшиеся и фактически изолированные
расы, приспособленные к особым комплексам физико-географи-
ческих и биотических факторов среды, все ecospecies в смысле
Турессона и Клаузена, а также и политипические, т. е. гетерофа-
циальные виды (Rensch, 1965). Ранее уже говорилось об опас-
ности навязывания природе больших или малых стандартов
вида. В нашей характеристике делается попытка избежать этой
ошибки.
В работе С. В. Юзепчука не было возражений против нашей
характеристики общих признаков вида, и можно сделать вывод,
что автор согласен с этой характеристикой, хотя и считает, что
ее «можно при желании рассматривать в качестве даваемого раз-
вернутого определения вида», которое «вышло чрезмерно громозд-
ким» (1958:157, 158). Наш перечень общих признаков вида не
призван служить ни кратким определением вида, ни краткой ин-
струкцией для практических целей. Он содержит попытку от-
разить существенные и общие черты вида, в том числе и не-
нужные для систематики (во всяком случае не используе-
мые ею).
7 Экспериментальная гибридизация грибов (А. И. Шифман, А. Джану-
заков) является важным методом определения таксономических различий
форм; у грибов гибридизация закономерна только между внутривидовыми
формами, ио иногда возникает и в пределах рода.
172
Некоторые авторы отмечают полноту и многосторонность на-
шего перечня, но не согласны с тем, что в списке все общие
признаки вида стоят в одном ряду без соподчинения. А. И. Па-
лил ов ссылается на то, что в перечне «мы не находим указа-
ния на основное свойство вида» (1957:4). Вслед за этим автор
отмечает, что в тех «немногих случаях, когда это свойство ука-
зывается, обычно не дается достаточно всесторонней характери-
стики вида п оно вычленяется произвольно» (там же). С послед-
ним можно полностью согласиться. И именно чтобы избежать
такого произвола, мы и предложили свою характеристику об-
щих признаков вида. Далее (глава 7) нами выделяются свойства
вида, как основной и первичной формы организации живрго.
«Виды, — пишет Палилов, — необходимо прежде всего харак-
теризовать по их месту в общебиологическом круговороте, ио их
месту в энергетике природы. Следовательно, основной характери-
стикой вида должна быть его энергетическая характеристика,
а место вида в энергетике природы — основным свойством вида.
Все остальные свойства вида являются производными от этого
главного свойства» (там же : 36). Энергетическая сторона отме-
чена нами при описании экологической определенности вида.
В понятие «экологическая определенность вида» входит и его
роль отдельного звена в круговороте веществ. Вне связей вида
с биоценозом невозможно понять адаптивный строй вида, дина-
мику численности, продолжительность жизни особи и массу дру-
гих признаков. Место вида в «экономике природы» —несомненно
важный признак вида. Но почему именно этот признак вида сле-
дует считать самым основным, а признаки организации самого'
вида, делающие его способным выполнять эту энергетическую
функцию, считать производными? Только благодаря наличию
некоторых основных свойств, присущих его внутренней организа-
ции, вид оказался образованием, способным выполнять свою энер-
гетическую роль. Вопрос о соотношении основных и производ-
ных свойств вида, разумеется, не является еще решенным. Под-
держивая в целом нашу характеристику, Т. Абдылдаев также
считает, что среди десяти черт вида «важнейшей является опре-
деленный обмен веществ и энергии» (1966 262).
Кроме того, он не согласен с том, что численность вида —
это его специфическая качественная черта. Тем не менее осо-
бенности пульсаций численности специфичны для вида так же,
как почерк для человека.
Итак, перечисление десяти признаков вида не является по-
пыткой дать его определение как это полагал С. В. Юзепчук
(1958:154—158). Это лишь материал для определения.
Характеристика сущности вида требует выделения среди этих
признаков самых важных, наиболее глубоко определяющих его
специфичность и позволяющих понять его особое положение
среди других образований живой природы.
173
5. О НЕКОТОРЫХ СОВРЕМЕННЫХ ОПРЕДЕЛЕНИЯХ ВИДА
В первой части книги были обсуждены определения вида,
сделанные в разные времена с различных позиций специали-
стами в различных областях биологии. Большинство определе-
ний вида давалось систематиками. Цель таких определений со-
стояла не в характеристике коренных свойств вида, а в перечи-
слении признаков, практически пригодных для классификации
форм. Подобные определения были названы нами инструктив-
ными (Завадский, 1961). В этом разделе будут обсуждены только
наиболее типичные такие определения.
Все современные определения-инструкции сводятся к пере-
числению тех или иных существенных различий (differentia essen-
tialis) по двум или нескольким признакам.
Давно было принято определять вид прежде всего исходя из
собственного практического опыта или же из опыта коллег. Так
появлялись определения вида, основанные на изучении только
одной группы, но претендующие на широкие обобщения. Частное
определение, обобщающее изучение одних групп, механически
переносилось на другие, например из систематики высших расте-
ний в микологию (Ciferri, 1932) или в альгологию (Воронихин,
1946). Поэтому правильно предложение В. И. Полянского (1958)
о разграничении общей и частных сторон в определениях вида.
Удовлетворительных определений-инструкций, как свидетель-
ствуют систематики, пока не создано. Разногласия среди система-
тиков, по Майру, показывают, что «ни один систематик не может
еще сделать из систематического материала обобщающих выво-
дов, которые были бы приемлемы для всех его коллег системати-
ков» (1947:28—29). Микробиологи также считают, что, «не-
смотря на огромное количество проведенных исследований и на-
личие десятков определенnii вида у различных групп микробов,
этот вопрос пе нашел еще научного разрешения» (Карпов,
1962:23).
Итак, среди современных систематиков нет единого мнения
о том, какое природное образование следует именовать видом.
Суммативные определения вида. Большинство
определений вида основывается на том, казалось бы, очевидном
положении, что вид — это сумма (совокупность) особей или
признаков, поколений, биотипов, популяций, экотипов, подвидов.
Суть всех этих определений в признании сборного, составного
или суммативного характера вида, т. е. в отрицании его целост-
ности.
В определениях, носящих инструктивный характер, упоми-
наются только внешние проявления вида, т. е. признаки, исполь-
зуемые при классификационных работах. Так, например, со-
гласно И. А. Рубцову, «под видом разумеют наименьшую, биоло-
гически и морфологически изолированную, развивающуюся
174
генетически независимо от других форм, качественно отличную
совокупность организмов отдельной филогенетической ветви, воз-
никшую в процессе дивергенции путем естественного отбора и по-
этому приспособленную к конкретным условиям существования»
(1941 198).
В. Ф. Купревич считает видом «совокупность особей ряда по-
колений общего происхождения, приуроченных к определенным
субстратам и территориям и характеризующихся обособленным
морфологическим, функциональным или биологическим\единством,
проявляющимся в определенных жизненных условиях и относи-
тельно стабилизированным в границах естественной ,ипдиви1дуаль-
ной или групповой изменчивости» (1949 31).
В этих определениях вид описывается по нескольким общим
признакам (численность, тип организации, воспроизведение, ди-
скретность, экологическая и географическая определенность, исто-
ричность, многообразие форм). Основным их .недостатком
является то, что вид толкуется как простая совокупность
(сумма) особей.
Если в определении вида игнорируется его целостность, то
оно искажает действительность даже и при перечислении всех
других признаков. Понимание вида как суммы особей или суммы
признаков характерно для многих современных определений вида
(Берг, 1950; Арнольди, 1957; Van-Steenis, 1957; Юзепчук, 1958;
Bremekamp, 1959; Пузанов, 1963; Slastenko, 1963, и др.). Большин-
ство суммативных определении вида дано систематиками, а для
них проблема интеграции вида не так существенна, как возмож-
ность разграничения двух совокупностей.
Но иногда сумматпвные определения вида выдвигаются био-
логами-теоретиками. Н. В. Тимофеев-Рессовский считает, на-
пример, самым «строгим», «теоретически удовлетворяющим всем
требованиям» определение, в котором «вид представляет собой
совокупность особей» (1965 14). Недавно были распространены
определения вида как совокупности биотипов, соседствующих
(тогда вид — смесь биотипов) или образующих гибридную си-
стему (тогда вид — гибридный «круг» биотипов). Так понимала
вид М. А. Розанова (1930, 1935).8 Ошибочно определять вид как
сумму популяций, популяцию, как сумму биотипов, а биотип же
как сумму признаков п т. д. Такая иерархия «сумм» — продукт
чисто аналитического подхода с разрушением существующих
в природе системных отношений. При таком подходе целостная
система описывается очень неточно. Так же определяют вид и
8 Позднее М. А. Розанова исключила из своего определения толкова-
ние вида как суммы биотипов: «Вид — это движущаяся и изменяющаяся
популяция в некотором отрезке времени, имеющая общее происхожде-
ние и характеризующаяся определенными морфолого-физиологическими и
экологическими признаками и географическим ареалом, отделенная от дру-
гой подобной популяции прерывистостью в серии биотипов» (1946:182).
175
популяцию некоторые генетики. А. Мюнтцинг (1963:76—82)
считает их механической смесью биотипов, а Н. В. Тимофеев-
Рессовскпй (1958:320)—смесью генотипов. Е. Н. Синская
определяла вид как совокупность экотипов (1938, 1939, 1948,
1961). По Б. Реншу, политипический вид — это «совокупность гео-
графических рас, особи которых скрещиваются друг с другом»
(Rensch, 1965 : 136). Распространенным стало понимание вида как
совокупности популяций (Красильников, 1962; Шварц, 1963, и
многие другие). Надо подчеркнуть различие между сведением
вида к сумме признаков, особей или биотипов и более близкой
к истине трактовкой его как совокупности популяций и экоти-
пов. Признаки, индивиды и биотипы не являются самостоя-
тельно эволюпрующими единицами. Популяции и экотипы же
могут обладать способностью к самостоятельному существова-
нию и к эволюции. Иногда виды включают группы фактически
изолированных популяций (рас, экотипов). Таким образом, по
отношению к отдельным популяциям и экотипам некоторые
виды могут быть названы составными. Недостатком этих опре-
делений является замена одного трудно определимого явления
другим, не менее трудно определимым.
Вид как тип организации. Важную роль продол-
жают играть определения вида, основанные на морфолого-гео-
графическом критерии (см. главы 4 и 5). Такие определения
эффективны, если морфологические признаки группы обособ-
лены, сама она эндемична и можно восстановить ее палеогеогра-
фию. О многом говорит ареал (и экоареал), если он совпадает
с группой ареалов видов, имеющих общие условия формирова-
ния. Труднее определить ранг групп, если морфологические раз-
личия недостаточно надежды (трансгрессия, большая изменчи-
вость и пр.), а ареалы рас разъединены пли перекрывают друг
друга с экологическим разъединением. В таких случаях недоста-
точно определение вида, основанное на морфолого-эколого-гео-
графических критериях, и возникают споры о ранге рас. Такие
споры важны для таксономии, но не для теории вида, так как
обособившаяся раса, как бы ее ни называть, уже существует и
эволюирует вполне самостоятельно. В определениях-инструк-
циях, даваемых с позиций морфолого-географического метода,
самостоятельным видом признается или группа особей, таксоно-
мически однородная и неотделимая в морфологическом отноше-
нии, или же, напротив, не последняя неразложимая система-
тическая единица наименьшего объема, а сложный комплекс
форм.
Недостатком морфолого-географической концепции вида яв-
ляется одностороннее использование признака «тип организа-
ции».
Затруднения, возникающие при характеристике вида как
особого типа организации, заключается в определении степени
176
единства генотипов и степени однородности особей в морфологи-
ческом, биохимическом и других отношениях, достаточных для
определения границ вида.
Какую именно степень сходства следует считать «видовой»?
Среди специалистов, пытавшихся определить вид как тип орга-
низации, наметилось два противоположных направления. Одно
из них настаивает на строгой монотипностп вида: вид — группа
особей, морфологически (или генетически, биохимически) во
всех существенных отношениях тождественных ^руг другу.
Ошибка монотипической концепции вида вскоре стала совер-
шенно очевидной: скрытые различия при усовершенствовании
методики оказались выше «порога различения» и это вело к без-
удержному раздроблению вида. Если отсутствие различий при-
нимается в качестве единственного критерия вида, то всякое
усовершенствование техники различения материала обязательно
должно сопровождаться дроблением видов.
Хорошим примером такого необоснованного раздробления
видов могут служить работы некоторых систематпков-альголо-
гов, подвергнутые справедливой критике В. И. Полянским
(1956) и др. По В. Чурда, видом следует считать все формы,
которые можно различить хотя бы по одному признаку. Приме-
нительно к агамным водорослям это означает, что при совершен-
ной технике различения особым видом является потомство каж-
дой особи, т. е. клон. Всем этим представлениям присущ один
и тот же недостаток: абсолютизация однородности (гомомор-
физма) вида. В действительности вид характеризуется лишь
определенной мерой однородности, и вся трудность состоит
в определении этой меры. Попытки определить ее, не прибегая
к изучению других признаков вида, являются в принципе неосу-
ществимыми.
Второе направление выдвигает правильное толкование типа
организации как некоторого единства в многообразии. Однако
и при этом возникает старый и коренной вопрос о степени раз-
личия, достаточной для определения границы вида. Затрудне-
ния с поиском «видовой степени различия» пытались разрешать
выбором «хорошего видового признака», а правильность этого
выбора зависит от опыта, подготовленности и суждения специа-
листа; в конечном счете границы видового типа устанавливаются
субъективно.
На самом же деле существует принципиальная и вполне объ-
ективная невозможность точно определить вид, руководствуясь
лишь одной его чертой — типом организации. По каким бы при-
знакам не устанавливался этот тип (морфологическим, физиоло-
гическим, генетическим) и как бы правильно не ставился во-
прос о самом характере этого типа, эта черта не может иметь
самодовлеющего значения при характеристике вида. Именно
в этом смысле и следует понимать утверждение Дарвина, что
12 К. М. Завадский
177
«размеры различий, признаваемые необходимыми для возведе-
ния двух форм на степень видов, не поддаются определению»
(1937 137). И именно эта принципиальная неопределенность
видовой степени различия заставила Дарвина писать о «неуло-
вимой сущности вида». Вместе с тем надо подчеркнуть, что
в комплексе с другими чертами вида «тип организации» сохраняет
все свое существенное значение. Определения вида, основанные
на двух признаках — типе организации и географической опре-
деленности — являются в современной систематике наиболее
распространенными «инструментами» для выделения видов.
Определения биологического вида. Среди совре-
менных определений-инструкций видное место занимают харак-
теристики вида, основанные на физиологическом критерии (т. е.
несовместимости организмов в сексуальном отношении). В 1919г.
зоолог Е. Штреземан предложил такое определение вида:
формы, достигшие уровня вида, физиологически дивергировали
да такой степени, что они могут встречаться не скрещи-
ваясь.
Основываясь па этом определении, необходимо возвести
в ранг вида не только все сезонные формы внутри популяции и
сезонные расы, но п отдельные экземпляры — гибриды, хромо-
сомные абберанты и т. д., которые не способны скрещиваться
с другими особями той же популяции. Это определение было
шагом назад от Дарвина, который показал ошибочность абсо-
лютно понятого физиологического критерия. Тем не менее с ус-
пехами цитогенетических и экспериментальных работ, с введе-
нием понятия репродуктивной изоляции оно было преобразовано
и в паши дни некоторыми котируется как самое глубокое (Cain,
1959; Мауг, 1963; Love, 1965, и др.).
Эколог-ботаник Турессон считает видом «популяцию, эле-
менты которой при взаимных скрещиваниях в природе дают
жизнеспособное и плодовитое потомство. При скрещиваниях же
с особями других популяций потомство оказывается маложизне-
способным и более или менее стерильным» (Turesson/ 1929 :
324). Генетик-зоолог Добжанский (Dobzhansky, 1955) считает,
что вид — стадия эволюционного процесса, на которой группы
форм, когда-то фактически или потенциально скрещивавшиеся
между собой, расщепились на два или более набора форм,
ставшие физиологически неспособными к скрещиванию. Орни-
толог-систематик Э. Майр (Мауг, 1963) писал, что вид — это
группы фактически или потенциально скрещивающихся при-
родных популяций, репродуктивно изолированных от других по-
добных групп.
Такое понимание вида многократно повторяется разными
биологами. Оно позволяет не смешивать с видами породы и
расы, резко отличающиеся комплексами константных морфо-
фпзпологических признаков, а также экологией и географиче-
178
ским распространением. Для групп с половым размножением
репродуктивная изоляция — существенная черта вида.
Однако концепция биологического вида сводит всю проблему
к репродуктивной изоляции. В подобных определениях имеется
ряд недостатков. Во-первых, такая точка зрения автоматически
ведет к отрицанию реальности видов у всех агамных и аногам-
ных форм, существование которых доказано многочисленными
прямыми наблюдениями. Во-вторых, заняв эту позицию^ необ-
ходимо считать видами все группы организмов, возникающие
в популяциях и даже единичные уклоняющиеся экземпляры,
потерявшие способность скрещиваться с другими особями попу-
ляции. Между тем известно, что многие из таких хромосомных
аберрантов и других мутантных форм обладают пониженной
жизнеспособностью и слабо приспособлены к среде, а поэтому
неспособны устойчиво воспроизводиться в пр и родгюн обстановке
и тем более увеличивать свою численность п расселяться. Это
не только не виды, но даже и не родоначальники новых под-
разделений вида: возникнув, они вскоре же и погибают. В-треть-
их, ею предписывается не считать видами викарирующие гео-
графические расы, которые фактически давно и полностью изо-
лированы друг от друга (европейские и североамериканские,
африканские и южноамериканские и т. п.), занимают иногда
различное положение, в экологических отношениях, обитают
в различных сообществах, различаются генотипически и морфо-
логически. Чтобы «возвести» такие уже сформированные расы,
давно идущие своим собственным эволюционным путем, в ранг
видов, почему-то признается необходимым доказать, что в при-
родных или лабораторных условиях они не скрещиваются или
дают стерильные гибриды. В-четвертых, все указанные форму-
лировки являются попытками очертить границы вида, но не за-
трагивают вопроса о специфическом содержании самого этого яв-
ления. Абсолютизация репродуктивной изоляции, кроме того,
умаляет значение экологических связей, логически отвергает
возможность межвидовой гибридизации, делает невозможным
диагностику при филетическом видообразовании (Шапошников,
1966, 1967). Не следует вновь абсолютизировать данный при-
знак. Кроме того, надо еще раз подчеркнуть, что этими опреде-
лениями молчаливо признается отсутствие клональных видов
пли же их неопределимость. Например, Н. В. Тимофеев-Рессов-
ский считает, что «для половых и агамных форм нельзя создать
единого... строгого определения вида» (1965:18). По его мне-
нию, «в настоящее время можно дать достаточно строгое и точ-
ное определение понятия вида у половых пер4екрестнооплодотво-
ряющихся организмов» (там же: 6). Анализ же понятия «вид»
у агамных форм встречает якобы непреодолимые затруднения,
гак как основной критерий вида — скрещиваемость, а он непри-
меним к агамным и облигатно-автогамным формам; отсюда и
12*
179
само понятие «вид» «неприменимо к агамным, облигатно-парте-
ногенетическим и облигатно-самооплодотворяющимся формам»
(там же 14). Далее, правда, категоричность этих суждений
уменьшается: «.. .нельзя отрицать того, что под влиянием более
или менее единообразных условий естественного отбора до из-
вестной степени аналогичные таксоны могут формироваться как
у половых форм... так и у агамных» (там же: 18). Однако, да-
вая определение вида, этот автор односторонне ведущее значение
приписывает одному критерию нескрещиваемостп, подчеркивая
этим еще раз, что вид — явление частное, присущее формам с по-
ловым размножением.
* * *
Все приведенные выше определения вида имеют служебное
значение; их функция заключается не в характеристике внут-
ренней организации вида, его сущности, а в указании на такие
внешние проявления вида, с помощью которых можно практи-
чески отличать один вид от другого.
Определения-инструкции до сих пор еще нередко рассматри-
ваются в качестве определений сущности вида как особого яв-
ления природы. Однако неправильно было бы отождествлять
концепцию практического вида, создаваемую для нужд система-
тиков, с современными общебиологическими представлениями
о сущности вида.
Отсутствие строгого, однозначного для всех групп фундамен-
тального определения вида и постоянное смешивание инструк-
тивных определений с попытками вскрыть его сущность вызы-
вает у некоторых авторов пессимистические нотки. Например,
в определение вида наряду с объективными показателями (мор-
фологическое сходство, скрещиваемость и генетическое единство)
предлагается вводить также и «суждение систематиков» (Beau-
dry, 1960). Поскольку все известные критерии нельзя приме-
нить на практике и до сих пор не дано ясного определения вида,
предлагается заменить это слово другими, точно отражающими
начальные фазы дивергенции (Harvey, 1960).
Чтобы скорее приблизиться к глубокому пониманию вида,
следует строго разграничить служебные определения от обще-
биологических. Необходимо разносторонне изучать вид, покончив
со старым предрассудком, согласно которому вид — это проблема
только одной систематики. Надо глубже изучать его структуру
экспериментальными методами (Valentine, 1962), больше вни-
мания уделять экологическим исследованиям вида (Наумов, 1962,
и др.), изучению внутривидовых отношений, составляющих со-
держание жизни вида (Завадский, 1961), и т. п.
Глава 7
ВИД КАК ОДНА ИЗ ПЕРВИЧНЫХ
И ОСНОВНЫХ ФОРМ ОРГАНИЗАЦИИ ЖИВОГО
1. ОСНОВНЫЕ ФОРМЫ ОРГАНИЗАЦИИ ЖИВОГО
И ИХ ПОДРАЗДЕЛЕНИЯ
Системная организованность живого. Принцип
сложной организованности жизни, несводимой к какой-либо од-
ной форме или системе, становится одним из теоретических фун-
даментов биологии. Важнейшей чертой этого принципа является
отказ от признания организма единственно основной и первич-
ной формой организации живого и признание того, что в составе
живого покрова Земли существует множество подчиненных си-
стем, «обладающих различными степенями организованности..
от рыхлых и случайных скоплений живых существ до самых вы-
сокоинтегрированных и высокоорганизованных особей» (Бекле-
мишев, 19646 25). Все больше ученых убеждается в том, что
«организация действительно является первичной категорией био-
логии, все еще требующей особого определения» (Huxley,
1963 XLII).
Сущность этого воззрения прекрасно выражена Сцепт-Дьёрди:
«Одним из основных принципов жизни является организация.
Мы понимаем под этим, что при объединении двух вещей ро-
ждается нечто новое, качества которого неаддитивны и не могут
быть выражены через качества составляющих его компонентов»
(Горизонты биохимии, 1964 : 11).
Понятие «организация» применительно к живому отражает
факт пространственно-временной упорядоченности однотипных
пли разнотипных элементов и блоков элементов, а также более
или менее устойчивых их объединений, имеющих характер «уз-
ловых пунктов». На основе полимеризации и дифференциации
нуклеотидов и аминокислот возникают линейно-цепочные макро-
181
молекулы РНК, ДНК п белка. При переходе от элементарных к
более сложным биологическим системам фундаментом служит пе
только полимеризация химических структур (Шмальгаузен,
1961), но также полимеризация и дифференциация субклеточных
образований, клеток и всех надклеточных живых систем в пре-
делах организма и всех надоргаппзменных систем (Завадский,
1961, 1966а). Следует подчеркнуть также, что, по данным био-
геохимии и в особенности сравнительной биохимии (Э. Болдуин,
М. Флоркен, Е. Крепе, Дж. Бернал и др.), всеобщее глубокое
сходство веществ и типов биохимических превращений является
следствием добпологического происхождения этого сходства или
возникновения его в процессе становления жизни, частично же —
на первых этапах эволюции. Поразительный консерватизм основ-
ных типов химических соединений и циклов также свидетель-
ствует о важности проблемы организации живого.
Живые системы резко различаются по многим важнейшим
показателям: размерам, длительности и степени самостоятель-
ности существования, распространенности в природе, способам
воспроизведения, механизмам интеграции, степени целостности
(т. е. числом степеней свобод элементов в системе) и т. п. Связи
между системами могут быть иерархическими, функциональ-
ными, сетчатыми.
Однако еще недавно безраздельно господствовал взгляд, что
только организм (особь, индивид) является основной и первич-
ной формой организации живого, а все надорганпзменные си-
стемы, начиная с колонии, популяции, вида п биоценоза и кончая
биосферой, — производными и вторичными формами. Целостность
надорганизменных систем отрицалась, их свойства принимались
за аддитивные (Bonner, 1958). Отсюда тенденция к сведению
таких форм к сумме особей пли даже стремление отрицать реаль-
ность их существования. С другой стороны, тканевая клетка и
ткань приравнивались иногда, по степени самостоятельности,
к организму и виду. Общепризнанными считались классификации
«иерархии соподчиненных индивидуальностей» Э. Геккеля,
К. Нэгелли и их более новые модификации. В рамках этой кон-
цепции велись дискуссии лишь о том, какая именно особь-бпопт
может быть принята за элементарную и какое образование в си-
стеме многоклеточных организмов следует считать элементарной
структурной единицей — эквивалентной элементарной особи-
бпонту.
Вид, например, рассматривался Э. Геккелем, как «генеалоги-
ческая индивидуальность» 2-го порядка (а клон — 1-го порядка).
Биологи разных направлений (Икскюль, Комаров, Роза и др.)
высказывались в пользу того, что вид — это особый индивид,
своеобразный организм. Например, Г. Впльморен считал, что всех
представителей вида в целом можно рассматривать как один
организм (1894 7). Биоценоз также пытались относить к осо-
182
бому классу индивидов (И. К. Пачоский, Ф. Клементс, В. Шел-
форд, А. Тенсли и др.). Эти запутывающие вопрос аналогии тем
не менее опирались на факт интегрированности (целостности)
вида и биоценоза. В качестве реакции на эти преувеличения воз-
никали столь же опрометчивые суждения, отрицавшие какую бы
то ни было интегрированность вида п биоценоза, признававшие
их простыми суммами особей. Стремление приравнять вид и био-
ценоз к индивиду связано и с расплывчатостью самого поня^тя
«индивид», которое стало обозначать не только особь (организм),
но также и все то, что в биологии именуют биологическими си-
стемами.
Строились линейные иерархические ряды, первым звеном
которых была клетка, моиера, мицелла и т. п. При этом повто-
рялась главная ошибка старинных «лестниц существ»: в одну
цепочку выстраивались системы, принадлежащие к принци-
пиально различным природным рядам.
Критика линейных иерархических рядов си-
стем. Наиболее существенные ошибки связаны с построением
иерархических рядов с простой линейной последовательностью,
когда в одну цепь соподчиненных звеньев включают элементы,
относящиеся к разным природным рядам, т. е. строят заведомо
искусственные ряды.
Рассмотрим примеры таких рядов. Л. Берталанффи подразде-
ляет иерархические зависимости на пространственные, генети-
ческие, функциональные и каждую из них сводит к линейно-
лестничной последовательности. Вот как выглядит всеобщая
пространственная иерархия систем: поле, плазма, элементарные
частицы, атомы, молекулы, агрегаты молекул, мицеллы, кол-
лоиды, клетки, ткани, органы, организмы, колонии, сообщества
организмов (Bertalanffy, 1949 11). Если касаться только живых
систем, то недостатком этого ряда является совмещение частных
п вторичных образований, неспособных к самостоятельному су-
ществованию в природе (например, ткани и органы) с универ-
сальными и первичными формами организации (организм и со-
общество). Кроме того, этот ряд совмещает системы, являющиеся
этапами эволюции организма (мицелла, клетка, организм, коло-
ния), со структурными подразделениями организма и с надорга-
низменными образованиями. Берталанффи исключил из обсу-
ждаемого ряда популяцию и вид, так как это системы,
относящиеся к генетической, а не к пространственной иерархии,
не имеющие централизованного управления, не являющиеся
«сверхорганизмом» (Bertalanffy, 1949, 1959). Смешение в одной
линейной последовательности систем, принципиально разнород-
ных по способности к самостоятельному существованию и вос-
произведению в природных условиях, по древности и степени
универсальности, характерно и для других авторов (например,
для А. А. Заварзина).
183
Рассматривая «основные уровни интеграции живого»,
И. И. Шмальгаузен предложил следующий ряд: первичная жи-
вая частица, клетка, многоклеточный организм, популяция, вид.
Этот ряд обрывается на виде, так как «выше вида биологическая
интеграция прекращается» (1961 : 122), а сообщество (биоценоз)
хотя и является целостной живой системой, но имеет своеобраз-
ные (отличные от онтогенетических и популяционно-ъидовых)
механизмы интеграции. Иерархии Берталанффи и Шмальгаузена
сходны: в одной ошибочно исключена популяция и вид, в дру-
гой-сообщество (биоценоз). Но Шмальгаузен не допустил
ошибки Берталанффи п исключил из ряда структурные эле-
менты организма (ткани, органы и т. п.). Однако первые три
звена этого ряда, являясь ступенями эволюции организма (докле-
точпой, одноклеточной, многоклеточной) не эквивалентны по-
следним двум (надорганизменные системы). Те же недостатки
свойственны ряду, предложенному Е. Н. Синской (1963) : клетка
организм, популяция, экотип, вид, фитоценоз, геобиоценоз.
Об ошибочности включения клетки в ряд основных форм орга-
низации живого уже говорилось. Экотип является частью вида,
т. е. не имеет коренных отличий от местной популяции и вида.
Ни популяция, ни экотип не могут поэтому рассматриваться
членами иерархии основных форм организации живого. Геобпо-
цеиоз включает неживые компоненты и является членом ланд-
шафтно-географических иерархий. Он не может быть включен
в ряды живых систем.
Некоторые авторы, строя иерархические ряды, руковод-
ствуются не только сложностью системы, но и степенью их ин-
тегрированности, специфичностью механизмов интеграции и т. п.
Сталкиваясь с нелинейным характером соподчинения систем, эти
авторы исключают из иерархии те или иные звенья (например,
вид). И. И. Пузанов приходит к выводу, что «чисто логически
вид является категорией совершенно иного порядка, чем орга-
низм и биоценоз» (1963 157), и поэтому не может являться зве-
ном единой иерархии живых систем. При этом Пузанов отрицает
реальность и целостность вида, рассматривая его с номинали-
стических позиций как удобное понятие, отражающее сумму
сходных и родственных особей. Вот почему он считает, что орга-
низм и биоценоз, с одной стороны, и вид — с другой — «вещи
абсолютно разнородные» (там же). Как правильно отмечает
Р. Е. Левина (1964), ото толкование вида совершенно неприем-
лемо, как чисто субъективистское. Учитывая эту позицию Пуза-
нова, понятно его желание рассматривать вид абсолютно несопо-
ставимым с другими формами организации живого.
В. Н. Беклемишев справедливо отмечает иерархичность строе-
ния живого, но подчеркивает только линейный характер сопод-
чпненности: «Каждое живое существо входит в состав другого
живого целого, вышестоящего по своему конструктивному рангу
184
it. кроме, может быть, самых элементарных, построено из жи-
вых частей низшего конструктивного ранга» (1964:36). Его ряд
включает клетку, многоклеточный организм, биоценоз п живой
покров Земли (геомериду).
Популяция и вид выпали из перечня, как будто в биоценоз
входят не популяции, а ипдивиды-робинзоны. Вместе с тем пер-
вое звено ряда — клетка, как уже отмечалось, является не основ-
ной формой организации живого, а ступенью эволюции орга-
низма или же его структурным элементом.
К. М. Хайлов (1963) правильно поставил вопрос о необходи-
мости пересмотра взглядов ультраэволюционистов, для которьрс
характерна недооценка системной организации живого (на при-
мере вида и видообразования эта ситуация была показана нами
в книге 1961 г.). Вполне приемлемо и его определение живой
системы как способной к развитию дискретной совокупности
взаимодействующих функционально различных элементов. Спор-
ным же является стремление Хайлова соподчинить живые си-
стемы в едином иерархическом ряду и исключить вид из «единой
иерархии живых систем» из-за отсутствия у него механизмов ин-
теграции, основанных на взаимодействиях функционально раз-
личных элементов. В действительности основные внутривидовые
отношения (см. главу 9) являются ярким примером цементиро-
вания системы на основе взаимодействия функционально разно-
родных элементов (полиморфизм половой, экологический, обес-
печивающий поддержание гетерозиготности, гетерозиса и т. п.).
Иногда строят иерархические ряды руководствуясь только
сложностью систем, и системы, относящиеся к различным при-
родным рядам, располагаются в одной цепи. Стремление
построить единую иерархию приводит к недопустимой схемати-
зации. Ряд «координационных уровней», предложенный
Б. П. Ушаковым (1963), включает следующие звенья: 1) моле-
кулярный, 2) органоидный, 3) клеточный, 4) органо-тканевой,
5) системы органов (корреляты), 6) организменный, 7) семейно-
стадный (репродуктивный), 8) популяционный, 9) видовой,
10) межвидовой (ценотический). Н. П. Наумов (1964) предложил
сходную иерархию из 12 уровней, включающую, кроме ценотпче-
ского, также и более высокие уровни (формация, биосфера).
Если руководствоваться лишь чисто формальным признаком
«конструктивного ранга» системы, то, как это отмечает
Б. П. Ушаков (1963), число звеньев в иерархии может быть уве-
личено во много раз. Действительно, лишь одна поперечнополо-
сатая мышца, по Д. Берналу, включает тысячи волокон-клеток,
состоящих из фибрилл, каждая из которых содержит миомеры,
являющиеся системами из миозиновых фибрилл, и фибрилл акто-
мизина, состоящих из сотен полипептидных цепей, включающих
расположенные в специфическом порядке аминокислоты, каждая
из которых состоит из 10—30 атомов. Однако при соподчинении
185
систем нельзя игнорировать такие существенные показатели, как
первичность или вторичность (пронзводпость) системы по отно-
шению к происхождению жизни, универсальный (всеобщность
для живого), пли частный характер системы, степень самостоя-
тельности существования, самовоспроизведения и преобразования
(изменчивость, эволюция), специфичность механизмов интеграции
и т. п.
Между тем в обсуждаемых иерархиях сосуществуют принци-
пиально разнородные системы — первичные и универсальные
(организм, вид, биоценоз, биосфера) со вторичными и частными
(орган, ткань, семья, стадо и т. п.), относительно самостоятельно
воспроизводящиеся и преобразующиеся в природе с неспособ-
ными к этому, и т. п. Даже крайние приверженцы симбиогенеза
не будут, например, утверждать, что органы многоклеточных
в прошлом существовали в качестве самостоятельных бионтов:
эволюция гистологических структур и органогенез осуществля-
лись внутри систем организменного ранга в качестве подчинен-
ных процессов. Органно-тканевые системы филогенетически мо-
ложе организменных, популяцпоппо-впдовых и биоценотпческпх
систем и являются производными. Семейно-стадные системы
возникли на очень высокой ступени эволюции организма и вида.
Образование Metazoa и Metaphyta, по-видимому, шло через со-
единение одноклеточных организмов в колонии и отбор клеток,
наиболее пригодных к зависимому существованию, с подавле-
нием свойств «организменности» (Захваткин, 1953; Беклемишев,
1964а). Одновременно шел отбор наиболее организованных ко-
лоний (т. е. колоний с оптимально развитым соподчинением
клеток целому). Не может быть исключена и гипотеза И. Хаджи
и Е. Хансона о возникновении Metazoa путем целлюлярпзации
полиэпергпдпых инфузорий, в которой защищается идея эволю-
ции многоклеточпостп путем дифференциации эпергпд внутри
исходного бпопта, т. е. по таким же законам, как и филогенети-
ческий процесс гисто- и органогенеза у Metazoa. Но каким бы
путем ни возникла многоклеточная особь, органно-тканевые си-
стемы являются не первичными, а вторичными образованиями.
Следует указать еще и на то, что нет оснований для выделе-
ния популяции и вида в качестве различных самостоятельных
форм организации живого. Известно, что многие молодые узко-
эндемичные и реликтовые виды состоят из одной локальной по-
пуляции. Таковы, например, пицундская сосна (Pinus pithyusa),
произрастающая только па горе Проклети в Албании Wulfenia
baldaccii, крымская Orchis comperiana и многие другие. Виды
улиток Achatinella на Гаваях имели крохотные ареалы — всего
по 14—20 га. В 1938 г. 86 рас комплексного вида A. mustelina
заселяли площадь всего около 250 га (Мауг, 1963). Гигантская
наземная черепаха на о. Альбадо в Мозамбикском проливе жи-
вет одной популяцией, состоящей из 80 000 экземпляров (Pros-
186
peri, 1955). Особенно много видов, состоящих из одной популя-
ции, па островах и в горных странах, но и на равнинах иногда
формируются суперпопуляции, включающие все население вида.
Аргументация в пользу объединения популяции и вида в одну
основную форму организации живого заключается в том, что
этп образования не имеют каких-либо специфических законов
существования и эволюции (Завадский, 1961, 1967а, б).
Ф. Чижек утверждает, что существует следующий единый
естественный ряд, основанный на «возрастании степени целост-
ности систем» (1965 451): 1) макромолекулы белков или ну-
клеиновых кислот, способные к самовоспроизведению, 2) сверх-
молекулярпые частицы протоплазмы, 3) мопера (доклеточпый
организм), 4) одноклеточный организм, 5) первичный многокле-
точный организм, 6) многоклеточный организм, дифференциро-
ванный па ткани и органы, 7) сверхиидпвид (колония н пр.),
8) локальные популяции, 9) вид, 10) биоценоз, 11) формация,
12) биосфера. Этот ряд такой же сборный, как и все только что
приводившиеся. Утверждение, что он основан на возрастании
степени целостности, неверно, так как, например, системы
№№ 3—6 более целостны, чем №№ 7—9 и тем более №№ 10—
12. Автор также располагает звенья, принадлежащие к различ-
ным рядам живых систем, в один искусственный ряд.
Подведем итог этому разделу. Попытки создавать единые
иерархии живых систем неизменно приводят к линейному со-
подчинению разнородных образований. При этом наблюдается
две тенденции. Одна — к классификации систем на основании
нескольких критериев (сложность или конструктивный ранг, пер-
вичность или вторичность, механизм и степень интеграции
и т. п.). Стремление создать единую иерархию с применением
нескольких критериев ведет к неправильному исключению из
ряда таких систем, как биоценоз (сообщество), популяция или
вид и т. п. Другая тенденция /состоит в использовании только
одного критерия — сложности I (конструктивного ранга) и во
включении в линейную иерархию принципиально различных об-
разований.
Существует несколько иерархических рядов соподчиненных
систем. Надо учесть также, что системы связаны между собой не
только иерархически. Далее будут предложены три главных
иерархических ряда.
Нежелание считаться с нелинейным характером соподчине-
ния систем часто ведет к тому, что исключает из иерархий
именно вид. Делается это па том основании, что по сравнению
с организмом и биоценозом вид специфичен по механизмам ин-
теграции. Но каждая из основных форм организации живого
специфична по степени и по механизмам интеграции. Построе-
ние иерархии с непосредственным переходом от организма к био-
ценозу не отражает действительности. В биоценозы входят не
187
отдельные организмы, а популяции. Конечно, жизненный цикл
многих высших животных приурочен далеко не к одному кон-
кретному биоценозу, а при кочевом образе жизни — к многим
биоценозам. Популяции же оседлых животных из поколения
в поколение приурочены к одному и тому же биоценозу. У остров-
ных, горных и других узкоэндемичных видов в состав одного
биоценоза входит весь вид, а широко расселенные виды распа-
даются па тысячи местных популяций, каждая из которых входит
в состав одного или нескольких биоценозов.
О термине «уровень организации». В последнее
время этот термин стал общеупотребительным. Нами вид рас-
сматривался также как один из уровней организации (Завад-
ский, 1961). Однако содержание этого понятия становилось все
менее определенным. Часто вместо кратких «изучение клетки»
или «изучение вида» писалось/об изучении на клеточном (или
видовом) уровне и т. п.
Вместе с тем существование иерархических рядов живых си-
стем требовало установить понятие, фиксирующее не только спе-
цифичность каждой системы, но и ее положение в ряду. Необхо-
димость сопоставлять живые системы по степени их сложности,
независимо от того, к одному или к различным естественным
рядам они принадлежат, также требовало соответствующего по-
нятия. Во всех этих случаях стали применять термин «уровень
организации». Например, в книге об эволюции на «сверхвидовом
уровне» (Rensch, 1959) говорится об эволюции крупных таксо-
нов. Ряд таксономических единиц — от вида до типа — построен
в строгой линейной иерархии, и можно, конечно, говорить не
о низших и высших таксонах, а вводить еще и слово «уровень».
Смысл же при этом сохраняется старый, т. е. высший таксон =
= таксон высшего уровня.
Термин «уровень» часто употребляется в различных смыслах,
которые недостаточно четко разграничиваются. Во-первых,
имеется в виду уровень организации самих живых систем, во-
вторых, уровни, на которых осуществляется изучение живого.
Легко показать, что уровни изучения живого не всегда совпа-
дают с уровнями организации живого. Живое можно изучать на
уровне элементарных частиц, атомов, ионов, малых молекул
и т. п., т. е. элементов, которые заведомо не являются живыми.
Нижний порог сложности организации самой элементарной жи-
вой системы выше самого высокого из только что упомянутых
уровней изучения. Такой элементарной живой системой называют
нуклеопротеид, еще неспособный к точному самовоспроиз-
ведению, а также к эволюции путем мутагенеза и селекции.
Дж. Ледерберг (1961) считает, что первичное живое образование
должно было включать хотя бы один полинуклеотид и один фер-
мент-катализатор. Элементарная живая система может быть
представлена моделью «голого гена», как это считает К. Дар-
255
лингтон, или вироидной стадией (Smith, 1962) и т. п. Так или
иначе, но ни атом и ион, пи малая молекула не могут рассмат-
риваться в качестве уровней организации живого, хотя живое и
исследуется на атомном и молекулярном уровнях (например,
биофизические исследования характеристик электронных оболо-
чек атомов, входящих в состав биополимеров, излучений генери-
руемых клетками и т. п.). Несовпадение понятий «уровень изуче-
ния» и «уровень организации живого» в таких случаях выступает
совершенно отчетливо. Правда, для надмолекулярных живых си-
стем уровень изучения может практически почти совпадать
с уровнем организации живого. Поэтому некоторые авторы под-
черкивают, что уровни изучения органического мира одновре-
менно являются и уровнями его организации (Лавренко,
1964:32, 34).
Недостаточное разграничение этих понятий приводит к сме-
шиванию гносеологического и онтологического подходов. Так,
Н. В. Тпмофеев-Рессовский (1962) пишет о генетическом, онто-
генетическом, эволюционном и хорологическом уровнях органи-
зации, подразумевая различные «этажи» познания, а также раз-
личные подходы и способы анализа (см. также: Ляпунов, 1962).
Н. П. Наумов (1964) предлагает подразделять уровни организа-
ции живого в зависимости от приемов изучения и исторически
сложившейся дифференциации наук, хотя правильнее было бы
настаивать на реформе обратного порядка.
Увлеченные познанием элементарных закономерностей, неко-
торые авторы признают существенным п качественно своеобраз-
ным лишь один молекулярный уровень организации живого. Все
более сложные системы, по их мнению, «являются сильно увели-
ченным отражением составляющих их молекул» (Уолд, 1962 : 19)
или должны рассматриваться лишь как внешние проявления
(феноменология) активных молекул ДНК, хранящих информа-
цию, определяющих специфическое строение молекул белка
(Анфинсен, 1962; Уоддингтон, 1964). Этим самым молчаливо
отрицается возникновение существенно новых типов живых си-
стем, стоящих на более высоких, чем молекулярный, уровнях,
а свойства этих систем признаются аддитивными.
Стремление к «монополии» молекулярного уровня встречает
возраженпя со стороны авторов, показывающих специфичность
организации, возникающей при взаимодействии мембран и орга-
нелл (Вейсс, 1961), не говоря уже о более сложных системах.
Например, по Е. М. Лавренко, исследования на молекулярном
уровне «пе могут полностью разрешить вопросы., видообразова-
ния» (1964 33). И. И. Шмальгаузен (1961) показал существо-
вание своеобразных интегрирующих механизмов у надмолеку-
лярных систем всех уровней.
Элементарная единица жизни может быть как образованием
организменного уровня, так и структурным элементом орга-
189
низма — клетки или тканевой клетки многоклеточной особи.
Во всех этих случаях говорят о «молекулярном уровне», хотя по
существу следовало бы строго разграничивать элементарную еди-
ницу, существующую в качестве первичной особи, от конструк-
тивно сходных единиц, входящих в состав более сложных систем
в качестве зависимых компонентов.
В итоге можно сделать вывод о желательности ограничить
употребление термина «уровень» только значениями, которые
соответствуют уровням изучения.
Первичные и основные формы организации
ж и в о г о. Идею первичности сразу нескольких форм существо-
вания жизни впервые обосновал В. И. Вернадский (1926, 1928,
1931, 1940). Глубоко укоренившаяся мысль о первичности орга-
низма и о пропзводности всех остальных форм существования
жизни, как доказывал Вернадский, не соответствовала биогео-
химическим закономерностям. В процессе возникновения жизни
«должен был одновременно появиться сложный комплекс живых
форм» (Вернадский, 1931 639). Образование первичных особей
шло одновременно с образованием первичных видов, первичных
биоценозов и первичной биосферы. Надоргапизменные системы
являются, таким образом, столь же древними, как и сама жизнь.
Первичная биосфера изначально была неоднородной сложной
системой, включающей много первичных видов и биоценозов,
так как одиночный вид не «мог бы исполнить. все геохимиче-
ские функции жизни, которые существуют в биосфере изначала»
(там же 645). Можно только удивляться могуществу консер-
вативных традиций в биологии, благодаря которому эти идеи
Вернадского долго заслонялись господствующей концепцией «мо-
нополии организма». Поэтому-то с таким трудом завоевывает
умы концепция об одновременном становлении четырех форм
организации живого (организменной, популяционно-видовой,
биоцеиотпческой и биосферной), признающая все эти формы
первичными и основными носителями жизни. Еще немало био-
логов реальностью считают только особь и входящие в ее состав
структуры, а надиндивидуальные формы организации живого
рассматривают лишь в качестве практически удобных, но услов-
ных группировок или же в качестве «особого рода организмов».
Наивное сведение всех форм существования живого к «орга-
низму» опиралось на факт устойчивости и интегрированности
(целостности) вида и биоценоза. Надо сказать, что по крайней
мере одна из причин отождествления вида и биоценоза с орга-
низмом состояла в необычайной расплывчатости понятия об ин-
дивиде. Путаница терминов вела к тому, что слова «особь»,
«организм» и «индивид» рассматривались то в качестве полных
синонимов или полусипонимов, то в них вкладывалось различ-
ное содержание (например, индивид — не только как отдельный
бионт-организм, но и как любая биологическая система).
190
Еще треть века назад взгляды Вернадского не получали при-
знания. Ныне же они приобрели большое значение. В эволю-
ционной теории этому помогла победа популяционистского под-
хода, согласно которому элементарной единицей является не
особь, а местная популяция (Северцов, 1936, 1951; Завадский,
1957 и др.; Haldane, 1959; Синская, 1961, 1963; Шмальгаузен,
1961; Huxley, 1963; Мауг, 1963, и др.); в биоценологии — призна-
ние биоценоза целостной системой, несводимой к сумме индиви-
дов, вне которой невозможно существование отдельно взятых
видов (Odum, 1959; Сукачев, 1960; Andrewartha, 1961; Арнольди
и Арнольди, 1963; Беклемишев, 1964а, и др.); в теоретической
биологии — разработка общей теории систем (Bertalanffy, 1949,
1959; General system, 1956—1964, п др.) и привлечение внима-
ния к проблеме организованности живого. Чем же отличаются
«основные формы» от всех других многочисленных форм орга-
низации живого?
Основными формами являются первичные, относительно вы-
сокосамостоятельные, универсальные, устойчивые живые си-
стемы, связанные между собой иерархическими и сетчатыми
отношениями, которые обусловливают существование и эволю-
цию органического мира.
Первичность организма, вида, биоценоза и геомериды доказы-
вается данными биогеохимии, палеонтологии докембрия, эволю-
ционной биохимии и других наук. Уже на самых первых этапах
биогенеза живое вещество не только «растекалось», но и «само-
организовывалось», так что с появлением эабнопта и протобпонта
возник eoagamospecies и протоценоз. Укоренившееся представле-
ние о производном характере всех надорганизменных форм су-
ществования живого по существу не имеет под собой научной
основы.
Ранее нами было предложено выделять пять основных уров-
ней организации живого (Завадский, 1961): 1) организменный
(единица — особь, индивид); 2) популяционно-видовой (еди-
ница — отдельная популяция или группа популяций, т. е. вид);
3) биоценотический (единица — конкретный биоценоз); 4) фор-
мационный (единица — фауна и флора относительно замкнутой
биогеографической области); 5) биосферный (геомерпда пли все
живое вещество биосферы). Е. М. Лаврепко (1963) высказался
в пользу объединения двух уровней организации — формацион-
ного п биосферного — в один — биостроматический.1 Это предло-
жение должно быть принято, так как, видимо, не существует
специфических закономерностей, имеющих принципиальное зна-
1 Вскоре Лавренко (1964) высказался за существование шести уров-
ней: молекулярного, клеточного, организменного, популяционно-видового,
биоценотического и биостроматического. Но первые два уровня, как о
нами рассматривалось выше, включены в иерархию по иным мотивам,
чем последние четыре, и являются членами иного ряда систем.
191
О с п о в н ы е ф о р м ы организации живого и главные
ступени их э в о л юц и и
Главные ступени эволюции Высшего организма с по- ведением разумного типа Цетрализованно-диффе- ренцированпая Метамерно-полимеризо- Многокле- ванная точная Первично-двуслойная \ Пластинчатая или одно- / слойная Колониальная Полиэнергидная Однокле- г точная Моноэнергидпая Бактериодная Доклеточ- Вироидная пая Протобионтная Эобионтная Высшая ксено- i гамная ; Генетически сба- j лансированная, полиморфная Истинного дёма Ксеиогамная (зи- готная) Протогамная Агамная (кло- нальная) Eoagamospecies 1
Организменная Популяционпо-ви- довая Биоце- нотиче- ская Биостро- ма тич е- ская
Первичный ряд систем (основные формы организации живого)
чение, отличающих биострому больших областей от всего слоя
живого вещества планеты. По только что приведенным сообра-
жениям отказываемся мы и от термина «уровень организации».
Таким образом, следует принять наличие четырех первичных
форм организации жизни, которые были предложены В. И. Вер-
надским. Эти четыре формы организации живого являются чле-
нами иерархического ряда, который можно назвать основным
рядом первичных систем. Каждая из этих четырех первичных
форм организации жизни в течение миллиардов лот претерпела
огромные эволюционные преобразования. «Главные ступени эво-
люции» двух форм — организменной и популяционно-впдовой —
представлены выше на схеме.2 Кроме этих принципов выделения
2 В состав главных ступеней эволюции входят и растения, и живот-
ные. Так, например, в ступень полиэнергидных одноклеточных наряду
192
иерархических рядов, мы предлагаем еще и третий, основанный
на том, что в ходе развития каждая из главных ступеней эволюции
каждой из основных форм организации живого дифференцируется
и включает сложную систему структурных компонентов.
В процессе эволюции организмов происходило усложнение их
Структурных компонентов. Ими были нуклеотиды и макромоле-
кулы, затем на клеточной ступени — органеллы, достигающие
большого разнообразия (ядро или ядра, цитоплазма с митохонд-
риями, рибосомами и мембранами, фибриллярные скелетные
структуры, мионемы, проводящие фибриллы, жгутики, реснички,
парабазальные тела, аппарат Гольджи, вакуоли и др.)* Диффе-
ренциация одноклеточных происходит и в ином плане: возникают
различные фазы жизненного цикла.
Структурными компонентами примитивных многоклеточных
является клетка, а с усложнением многоклеточной особи возни-
кает сложная система тканей, органов, топографических • и функ-
циональных блоков органов (метамеры и пр.). На новой основе
происходит расчленение фаз жизненного цикла. Процесс кон-
центрации и олигомеризации органов сопровождается увели-
чением числа разноименных тканей и органов и т. п. В процессе
эволюции вида структурными компонентами являются клональ-
ные линии и популяции, а после появления полового процесса —
семьи, биотипы, морфо-биологические группы, стада, местные
популяции, экоэлементы, экотипы, расы и подвиды (см. главу 8).
Более или менее полно изучены главные ступени эволюции
и структурные компоненты только организменной формы. Значи-
тельно хуже исследованы они у популяционно-видовой формы
организации живого, и предлагаемая нами схема представляет
собой лишь черновой набросок. Ступени эволюции биоценотиче-
ской и биостроматической форм организации живого исследованы
еще совсем мало. До сих пор преобладают характеристики эво-
люции типов биоценозов по ведущим типам организмов (напри-
мер, псилофитовые биоценозы девона и т. п.), а не по специфи-
ческим структурно-функциональным закономерностям (конструк-
тивные особенности, особые черты биотических связей, особые
циклы круговоротов веществ).
Живые системы из разных рядов четко разграничиваются
только по типичным образцам, а во многих других случаях гра-
ницы между ними более или менее нечетки или даже исчезают
в переходных состояниях. Так обстоит дело и с границами ме-
жду основными формами организации живого «первичного ряда
систем». Определенные при сопоставлении «типичных образцов»,
с инфузориями входят и водоросли типа Caulerpa, а к метамерно-полиме-
ризованным многоклеточным относятся колонии зооидов или фитонов и
колонии типа сифонофор-дисконант и т. п.
13 К. М. Завадский
193
они могут становиться и совершенно расплывчатыми. У кусто-
вых и ползучекорневищных злаков (мятлик луговой, пырей пол-
зучий, виды камыша) существуют плавные переходы от состоя-
ний, отвечающих понятию типичного индивида (особи),
к агрегациям — колониям, клонам, местным популяциям и фито-
ценозам. В зарослях этих растений практически невозможно
строго их разграничивать (Завадский, 1947). Примерами пере-
хода индивида в заросль-сообщество могут служить виды бам-
бука, у которых один сеянец разрастается на территории в де-
сятки гектаров. У смоковницы (Ficus religiosa) от кроны побеги
опускаются до почвы, укореняются и становятся дополнитель-
ными стволами-опорами, так что дерево разрастается, образуя
целый лес. Между высокоцелостным, централизованным организ-
мом высшего животного и зарослью-клоном пырея располагается
целая серия типов индивидов — простые, слаборазвитые, редуци-
рованные индивиды, а также соподчиненные и частичные инди-
виды, входящие в состав сложных колониальных индивидов 2-го
и 3-го порядков или в состав кустов, зарослей, агрегаций, стоя-
щих на грани между особью и клоном (Левин, 1963, 1964; Бек-
лемишев, 19646). При бесполом размножении или вегетативном
разрастании типа «ведьминых колец», мицелия грибов и т. п.
возникают клональные популяции, иногда определяемые неко-
торыми как отдельные виды [например, апомиктические мелкие
виды —колонии ирисов (Anderson, 1936) и т. п.]. Такие образо-
вания легко переходят от организма-агрегата к клону и элемен-
тарному фитоценозу: после обособления полимеризованных ча-
стей организма и перехода их в ранг отдельных бионтов
отношения между ними являются внутрипопуляционными и
в то же время фитоценотическими. Интересен пример с местной
грузинской пшеницей «Зандури». Это сорт — фитоценоз, состоя-
щий из трех видов (Triticum monococcum, 2n = 14; Т. timophee-
vii, 2n = 28; T. zhukovskii, 2n = 42), представляющих поли-
плоидный ряд, в котором пшеница Жуковского — амфидиплоид,
возникший, как полагают (Синская, 1963:84—85), при скрещи-
вании двух других видов. Хотя это интересное образование изу-
чено недостаточно, все же известно, что между всеми видами
в настоящее время существует высокий (но не полный) фи-
зиологический барьер. Получается, что внутрисортовые формы,
являясь в то же время симпатрическими видами, связанными
генеалогически, выступают как компоненты фитоценоза и как
группы единой полиморфной амфимиктической популяции. Та-
ким образом, сорт «Зандури» как бы включает две или три формы
существования живого.
Характерным свойством живого является расплывчатость
границ между образованиями, дискретность которых по вызывает
сомнений, если сопоставляются крайние точки пли типичные
случаи. Как бы ни были капитальны различия между четырьмя
194
формами «первичного ряда системы», и между ними существуют
переходы. Естественно, это создает трудности при классифика-
ции, в особенности если «уточнять» принадлежность системы
в большей мере, чем это позволяют сами объекты. Существуют
трудности и при уточнении взаимоотношений между системами,
входящими в разные ступени эволюции основных форм или от-
носящихся к различным структурным компонентам какой-либо
ступени.
Вместе с тем эти затруднения не должны сбивать исследо-
вателей на старые позиции исканий единых иерархических
рядов.
2. ОСОБЕННОСТИ ВИДА КАК НЛДИНДИВИДУАЛЬНОЙ СИСТЕМЫ
•
Возможность существования вида в течение
неопределенного времени. Работы, посвященные
вопросу о виде как особой форме организации живого, еще
крайне немногочисленны. Представления В. Л. Комарова (1940),
С. А. Северцова (1941) о виде как об «особом способе осуществле-
ния жизни» или «форме существования жизни» безусловно пра-
вильны и имеют фундаментальное значение. Однако они были
высказаны в самой общей форме и нуждаются в более подроб-
ном обосновании.
Если вид представляет собой одну из форм существования
жизни, то саморепродуцирующаяся макромолекулярная система,
клетка, макроорганизм и биоценоз также являются «способами
осуществления» и «формами существования» жизни. Особен-
ность вида состоит в том, что он не стоит в одном ряду с упомя-
нутыми системами. Одни из этих систем он включает в себя в ка-
честве компонентов подчиненного значения, а в другие сам входит
как компонент. Вид — система, представляющая собой особую и
высокую надиндивидуальную форму организации живого.
В чем же состоит это качественное своеобразие видовой
формы организации живого? Прежде всего в наличии следующих
двух свойств: 1) внутренней возможности к существованию в те-
чение неопределенного времени и 2) способности к относительно
самостоятельному эволюционному развитию.
Отсутствие внутренне обусловленного, определенного срока
существования вида в природе является одним из коренных
отличий вида как от индивида, тканевой клетки и макромоле-
кулы, так и от биоценоза. По этому признаку вид идентичен гео-
мериде в целом.
Индивид в зависимости от ступени развития и генотипически
обусловленной конституции способен существовать от немногих
минут до нескольких тысяч лет. Затем он отмирает. Жизнь тка-
невой клетки также связана с субстанционально обусловленным
старением и определенным сроком существования. Внутренняя
13*
195
же структура вида не заключает в себе никаких моментов, зара-
нее ограничивающих его существование определенными сроками.
В этом смысле вид можно назвать открытой во времени си-
стемой.
Попытки обосновать гипотезу о существовании внутренне за-
данного «срока жизни» вида сводились не к доказательствам
этого положения, а к рассуждению о виде по аналогии с индиви-
дом и к иллюстрациям того несомненного факта, что рано или
поздно вид или вымирает, или же преобразуется.
Идея Дж. Брокки—Роза о существовании заранее предопре-
деленного «срока жизни вида^ приписывает виду несуществую-
щие «филетические» фазы развития: юность, зрелость, старость
и дряхлость. Ту же основу имеет и идея «старения вида» (Веиг-
1еп, 1933). В пользу гипотезы «срока жизни вида» используют
факты быстрого и одновременного вымирания видов. Однако
такие концепции неокатастрофизма встречают возражения как
общие, связанные с трактовкой механизмов эволюционного про-
цесса, так и чисто эмпирические. Катастрофизм имеет под собой
почву, пока неполнота геологической летописи очень велика.
Если же число находок возрастает, то, как правило, обнаружи-
вается, что поредение вида всегда предшествует его вымиранию.
Трилобиты вымирают в конце пермского периода, но последние
из известных трилобитов встречаются в очень больших коли-
чествах и на очень больших территориях. Поэтому вероятно, что
в нижнем триасе (а может быть, и позднее) трилобиты еще
существовали, но их популяции были изрежены, а ареал сокра-
щенным. Последним видом вымирающих граптолитов был Мо-
nograptus hercynicus, но он встречается в массах и на очень
больших территориях, так что при длительном угасании этой
группы можно ожидать находок граптолитов и в нижнем девоне
(Наливкин, 1964). Как показывают палеонтологические дан-
ные и как следует из теории естественного отбора, образования,
достигшие видового уровня, обладают внутренней возможностью
к неопределенному сроку существования.
Но она может быть реализована лишь там, где естественный
отбор будет направлен на сохранение уже существующей адап-
тивной нормы и на поддержание ранее сложившихся внутриви-
довых отношений (погашая постоянное давление мутационного
процесса). В природной обстановке такой теоретически возмож-
ный случай полностью никогда не может реализоваться. Измене-
ния среды, обусловленные климатическими, геохимическими и
в особенности биотическими причинами и последствиями жизне-
деятельности самого вида, рано или поздно, но неизбежно вызы-
вают или вымирание вида, или его эволюционное изменение.
Тем не менее некоторые ныне существующие виды представляют
собой весьма древние образования, возникшие в третичный пе-
риод, в мезозое или даже еще раньше.
196
Исключительная устойчивость некоторых видов доказывается
палеонтологическими наблюдениями; «есть формы жизни, неиз-
мененные с кембрия или даже с альгонгка, т. е. порядка 108—
10 9 лет» (Вернадский, 1932:516). Палеозойские Lingula из пле-
ченогих почти не отличаются от современных. За 500 млн лет
они изменились не более чем на величину «спорного рода или
вида» (Simpson, 1953). По Г. Г. Мартинсону (1951), у моллю-
сков Baicalia из третичных отложений у 9 видов из 10 в течение
примерно 20 млн лет наблюдается только внутривидовая измен-
чивость. Характеризуя высокую устойчивость видов, многие
палеонтологи для морских ежей, крабов и других групп приводят
такие же или даже большие сроки.
С триаса почти не изменились крокодилы (Cuenote, 1936;
Cuenote et Tetry, 1951). Из триасовых отложений Германии опи-
сан, например, щитень (Trips contriformis), не отличающийся от
его современного потомка. В течение 150 млн лет видовая спе-
цифичность формы не изменилась. К. К. Шапоренко (1936) по-
казал, что современная Ginkgo biloba ничем не отличается от
юрской G. inteariuscula, меловых G. primordialis п G. laramien-
sis и т. д. Разбивка фратрии G. biliba на части не имеет иных
оснований, кроме обычая палеосистематиков давать видовые
названия образцам из. разных геологических эпох. Такие же
факты приводятся для видов двоякодышащих рыб (неоцерата-
дус и др.), гаттерии, варанов, многих насекомых и др. О боль-
шой устойчивости некоторых видов свидетельствует также рас-
пространение одного и того же вида на разных континентах,
разъединенных еще в древнейшие времена (например, в Цен-
тральной Африке п в Америке). ,
Многие современные виды высших растений были найдены
в третичных отложениях. С нижнего палеогена, по А. Н. Криш-
тофовичу (1957), известны, например, остатки болотного кипа-
риса, некоторых видов веерных пальм, тополей, орехов, водяного
лотоса, камфарного лавра. В миоценовых отложениях различных
стран были обнаружены обыкновенная ель, сибирская листвен-
ница, некоторые современные виды дуба, тополь, осина, грецкий
орех, кавказская птерокария. Что касается видов еще более
древнего происхождения — палеозойских реликтов, то их су-
ществование в современную эпоху хотя еще полностью и не до-
казано, но теоретически вполне допустимо.
Четко разграниченные группы видов клена (Acer) занимали
огромный ареал уже к началу третичного времени, и, очевидно,
этот род успел пройти длинный путь эволюции еще в меловом
периоде (Васильев, 1961). На примере видов водяного ореха
(Тгара) В. Н. Васильев (1965) показал, что продолжительность
существования вида составляет миллионы лет, но в высоких ши-
ротах она меньше, чем в низких (типы видов, давно исчезнув-
шие на севере, продолжают существовать на юге).
197
Столь большие сроки существования отдельных видов сгу-
щают некоторых палеонтологов. Наличие таких издревле сохра-
няющих постоянство так называемых персистентных видов, по
их мнению, стоит в противоречии с эволюционным учением
(Давиташвили, 1956).
В действительности же тот факт, что ныне существующие
виды имеют различный геологический возраст, лишь подтвер-
ждает отсутствие у вида заранее заданного определенного срока
жизни. Еще Дарвин указал, что «продолжительность времени
сама по себе не содействует и не препятствует естественному
отбору». Отбор может неопределенно долго действовать не на пре-
образование, а на поддержание и на стабилизацию уже сложив-
шейся адаптивной нормы. Поэтому при отсутствии веских мор-
фологических оснований для разграничения видов следует не
фабриковать новые видовые названия (основываясь лишь на
«продолжительности времени»), а объединять материал в одну
фратрию (вид во времени). Впоследствии часть таких видов,
возможно, и окажется сборными, но и другие пополнят списки
персистентных форм.
Вид как образование, способное к самостоя-
тельному эволюционному развитию. Все, что из-
вестно в настоящее время об эволюционном процессе, свидетель-
ствует о приуроченности его исключительно к популяционно-
видовой форме существования живого. Вид является носителем
или основной единицей эволюционного процесса. Эволюционный
процесс фактически протекает и может протекать в пределах
только популяции и вида. Такие системы, как макромолекула,
клетка, высший организм, не способны к самостоятельному эво-
люционному развитию. Макромолекулярные системы способны
к не вполне точной редупликации, т. е. к мутациям. В рамках
клеточных и организменных систем осуществляются процессы
морфогенеза, деления и другие преобразования, но не эволюция.
Переход наследственной изменчивости и всех других типов пре-
образований, свойственных макромолекулам, клеткам и организ-
мам, в эволюционный процесс— это в сущности и есть переход
от динамики, присущей низшим формам организации живого,
к динамике, свойственной только популяции и виду.
Поддержание устойчивости и изменение вида осуществляется
принципиально иным путем, чем у организма, клетки и других
систем. Непрерывно идущий мутационный процесс вызывает
массу разнообразных необратимых изменений молекулярных
структур, при известных условиях изменяющих субклеточные
структуры, клетки и индивиды. Мутационный процесс идет
у всех видов, в том числе и у самых устойчивых, древних. Му-
тации чаще всего дезорганизуют живые системы, являясь поме-
хами, и преобразование последних в информацию происходит
в рамках популяции и осуществляется только естественным отбором.
198
Естественный отбор может поддерживать установившуюся
систему вида прямой элиминацией уклонений от наиболее при-
способительной организации, отбором на понижение мутабиль-
ностп,3 отбором па увеличение силы доминирования генетических
комплексов, осуществляющих адаптивную норму, и т. п. В этих
случаях мутационный процесс не будет влиять на систему вида:
она оказывается заблокированной. Однако эволюция основана на
таких взаимодействиях популяций со средой, при которых отбор
перестает охранять «видовой уровень» от изменений. Этим са-
мым отбор вызывает преобразование вида, т. е. эволюционный
процесс. Отбор определяет направление и темп мутационного
процесса, поэтому последний оказывается одним из важнейших
адаптивных признаков вида. Поэтому-то направление и темп му-
тагенеза «для каждого из видов... специфичны и, возможно, ме-
нялись на различных этапах эволюционного процесса» (Шапиро
и Игнатьев, 1945 343).
Важно подчеркнуть, что из всех форм существования жизни
только популяционно-видовая способна быть полем деятельности
естественного отбора. Макромолекула, клетка, орган, высший
организм представляют собой жестко детерминированные си-
стемы. Их компоненты и блоки настолько спаяны в цельные
образования с централизованным управлением функциями, что
внутри этих систем естественный отбор, как правило, действо-
вать не может. Отдельные компоненты популяции (особи, семьи,
стада, биотипы и т. п.) обычно не столь сильно спаяны между
собой, чтобы препятствовать индивидуальному и групповому от-
бору. В то же время они настолько зависимы друг от друга, что
делают популяцию целостной системой. Популяционно-видовая
форма организации живого оказывается способной к эволюции
именно потому, что, объединяя подчиненные компоненты' низ-
ших форм, она постоянно бывает насыщена мутациями и в то же
время служит полем деятельности естественного отбора.
Эволюция организма не является каким-то самостоятельно
идущим вне вида процессом. По существу происходит лишь эво-
люция видов, но самым наглядным, непосредственно наблюдае-
мым результатом этого исторического процесса являются изме-
нения, происходящие с индивидом, т. е. с нижележащей формой
организации живого.
Подчеркивая качественное своеобразие вида, нельзя в то же
время забывать, что он представляет лишь особую организацию
3 Снижение частоты мутирования до известного оптимума может по-
вышать скорость эволюции, так как создается наилучшее соотношение
Между мутабильностью и отбором. Это было показано еще в работах
30—40-х годов. Поэтому странно сейчас слышать, что «с эволюционной
точки зрения обширная вырожденность генетического кода должна при-
вести к стабилизации видов благодаря снижению частоты мутаций»
(Ochoa, 1964:121).
199
нижележащих систем. Популяционно-видовая форма организа-
ции живого— это специфическая система связей между моле-
кулярными, клеточными, тканевыми, организменными, семей-
ными и другими структурами. Это положение весьма важно, так
как им подчеркивается сложный характер популяционно-видовой
системы.
Необходимо рассмотреть еще одну сторону вопроса о способ-
ности вида к самостоятельному эволюционному развитию. Эта
способность присуща всем видам только в форме возможности.
Это означает, что все популяции, доказавшие свою приспособ-
ленность в прошлом, обладают реальной возможностью эволю-
ции и в будущем. Однако наличие у каждого вида потенциаль-
ной возможности к дальнейшей эволюции еще не гарантирует
ее реализации. Большая или меньшая вероятность реализации
этой возможности в конечном счете зависит от характера взаимо-
действия вида с окружающей средой. При этом ведущей сторо-
ной является внутренняя организация самого вида. Именно от
степени совершенства внутренней организации вида и зависит
его эволюционная перспективность. Это область вопросов, свя-
занных со значением воздействий предшествующих состояний
на последующее развитие (Завадский, 1958). На эту связь обра-
тил внимание еще Дарвин, указавший, что широко расселенные
и полиморфные виды, доказав свои достоинства в прошлом и
настоящем, имеют больше шансов преуспевать и в будущем.
Из миллионов видов лишь некоторые осуществляли дальней-
шее развитие. У многих видов складывается организация, делаю-
щая их хорошо приспособленными только к существующим усло-
виям. При изменении же условий они вымирают или сохра-
няются только в особо благоприятных «нишах резервации».
Конкретные причины вымирания видов многообразны, но
в конечном счете основной является недостаточная эволюцион-
ная пластичность, т. е. невозможность осуществить эволюцион-
ные преобразования столь быстро, как это необходимо в данных
условиях. Такой характер эволюционного процесса подтвер-
ждается и палеонтологическими данными.
Большая эволюционная активность относительно малочислен-
ной группы видов, способных к широкой радиации потомков,
приводит к тому, что общая численность видов следующей геоло-
гической эпохи не становится меньшей.
Особенно широкими возможностями эволюции обладают лишь
немногие виды, накапливающие приспособления общего значе-
ния, эффективные в разнообразных, в том числе в неблагоприят-
ных, условиях (подробнее об этом см. главу 10). Поскольку эво-
люционный процесс зависит не только от качеств самого вида,
но и от окружающей среды, постольку более вероятно, что пер-
спективные для дальнейшей эволюции виды могут сформиро-
ваться в группах, имеющих большее число видов. С этим взгля-
200
дом, однако, не могут быть согласованы данные, рассмотренные
в главе 1. Например, Protozoa включает 15 000—20 000 видов,
т. е. они впятеро обширнее класса млекопитающих. Насекомые
(750 000—1000 000 видов) в 40—50 раз обширнее класса рыб.
Грызуны (около 2500 видов) —наиболее многочисленная группа
среди млекопитающих. По Б. Б. Родендорфу (1957), насекомые
еще в верхнем карбоне были необычайно многообразны, и, ве-
роятно, всегда они были наиболее многочисленным классом жи-
вотных.
Из этих примеров следует, что большая численность видо-
вого состава группы отнюдь не прямолинейно связана с ее эво-
люционной перспективностью. С другой стороны, высшие группы
оказываются далеко не самыми многочисленными по видовому
составу (головоногие среди моллюсков, приматы среди млекопи-
тающих). Вопрос о причинах и условиях формирования видов
с чрезвычайно обширными эволюционными возможностями слу-
жит предметом особого раздела эволюционной теории — учения
о прогрессивной эволюции — и не может обсуждаться здесь.
Является ли вид этапом эволюции? Если мно-
гие виды, возникнув, далее не эволюировали (а персистировали
или вымирали), то возникает вопрос, правильно ли в общей
форме утверждать, что «вид — это стадия (этап) эволюции».
Этим утверждением начинаются многие определения вида (Ко-
маров, 1940; Борисяк, 1947; Dobzhansky, 1955, и др.). С другой
стороны, Майром (1947) было выдвинуто противоположное ут-
верждение: «Вид — не стадия, а результат эволюции». Тезис
«вид есть результат эволюции», выражает непреложную истину
и пе может быть истолкован в различных смыслах. Характери-
стика же вида как стадии (этапа) эволюции может получить
различные толкования.
Вводя в определение вида понятие об этапе эволюции, Доб-
жанский отмечал только, что возникновение вида — заключи-
тельный этап процесса видообразования. Этим подчеркивается
качественная определенность состояния, которое приобретается
группой.
Иное содержание в рассматриваемое понятие вкладывают па-
леонтологи и систематики, изучающие филогению крупных так-
сонов. В филетической эволюции (Simpson, 1951а, 1953) вид
выступает как отрезок ствола (фратрия). Можно ставить вопрос,
через сколько этапов (видов-фратрий) прошла эволюция Pro-
tochordata до человека или от эоценовой Notharctus osbornii до
какого-нибудь современного вида американских обезьян, и т. п.
В синхронных процессах расслаивающей эволюции (Smith, 1956)
вид выступает как единица радиации (дивергенции). Однако
если мы захотим рассматривать каждую из многочисленных вет-
вей меловых аммонитов перед ее вымиранием, то все эти виды
окажутся в буквальном смысле слова заключительными этапами
20}
эволюции. Во всех этих примерах вид можно называть началь-
ным, промежуточным или заключительным этапом эволюции ка-
кой-либо филогенетической ветви. Но речь идет здесь о харак-
теристике конкретных видов, а не об общем определении вида.
Для миллионов видов, которые оказались концевыми звеньями
вымерших ветвей, термин «этап эволюции» не может считаться
удачным.
В применении же к далее эволюционирующим видам фор-
мула «вид — этап эволюции» означает лишь то, что эволюцион-
ный процесс идет от предшествующего вида к последующему
(или последующим), т.е. что вид выступает в качестве узлового
пункта развития живой природы. Таким образом, если в общее
определение вида вводится понятие об «этапе эволюции», то оно
может заключать различное содержание и его нельзя употреблять
без дополнительных разъяснений.
Итак, популяционно-видовая форма существования живого
является надиндивидуальной системой, способной как к неопреде-
ленно долгому самовоспроизведению, так и к эволюционному
развитию. Реализация этих способностей зависит от направления
естественного отбора. Популяция и вид являются статистически
детерминированными системами и этим коренным образом от-
личны от организма и всех внутриорганизменных систем вплоть
до нуклеотидов, которые детерминированы жестко.
Вид обладает достаточно большой однородностью компонен-
тов и блоков (генотипический и фенотипический изоморфизм
индивидов, семей и популяций) при невысокой степени их
взаимосвязи. Именно поэтому популяция и вид оказались систе-
мами, пригодными для деятельности естественного отбора, и
только им присущи специфические преобразования, именуемые
эволюционным процессом.
Сформулированное нами (Завадский, 1961, 19676) представ-
ление о виде как специфическом субстрате действия естествен-
ного отбора поддержали некоторые авторы (Лавренко, 1964; Яб-
локов, 1966; Cizek, Hodanova, 1966, и др.).
3. О ЦЕЛОСТНОСТИ ВИДА
Определение понятий. Из всех десяти черт вида
(см. главу 6) наименее общепризнанной является целостность.
В предыдущих разделах целостность вида в значительной мере
принималась аксиоматически. Приведем теперь доводы в пользу
целостности вида, необходимые для характеристики его сущности.
Одна из трудностей проблемы состоит в отсутствии точно оп-
ределенной терминологии. Поэтому прежде всего уточним неко-
торые общие определения. Термины «целостность», «система» и
«форма организации» чдсто употр0бдо)тся как синонимы. Они
обозначают известную степень спаяннрст^ (взаимозависимости)
202
элементов и частей; их упорядоченность (организованность) й
устойчивость; наличие свойств, присущих только всему объекту
и исчезающих (разрушающихся) при разложении объекта; обя-
зательность изменений всего объекта после прибавления, утраты
или изменения его частей. Можно представить ряд, на одном
конце оканчивающийся чисто суммативной совокупностью, эле-
менты которой независимы друг от друга, а свойства аддитивны,
т. е. равны сумме свойств элементов, а на другом конце которого
находится настолько спаянный комплекс, что его элементы пол-
ностью зависимы друг от друга и утратили даже относительную
автономность, а его свойства ни в какой степени несводимы
к сумме свойств элементов. Но в природе не существует идеально
суммативных совокупностей и абсолютно целостных систем.
Реальный ряд включает почти суммативпые неорганизованные
множества, свойства которых лишь практически аддитивны. Такие!
множества мы называем конгломератами. Далее идут совокуп-
ности-системы со слабо выраженными свойствами целостности,
и обычно их тоже считают не системами, а малоорганизованными
множествами. Кибернетики (Р. Эшби) и авторы «теории си-
стемы» (Л. Берталанффи, К. Болдинг, Д. Аукоф и др., см.: Ge-
neral system, 1956—1964) подчеркивают необходимость разграни-
чения систем на детерминированные статистически и динами-
чески, т. е. жестко. Статистически детерминированные системы
имеют меньшую степень целостности. Динамически, т. е. жестко
детерминированные системы характеризуются возрастанием сте-
пени спаянности, упорядоченности и взаимозависимости частей,
ростом значения свойств, присущих всей системе, и т. п. Таким
образом, природный ряд состоит из систем различной степени
целостности. В случае конгломератов можно пренебречь следами
их целостности, рассматривая их как множества. Высокоцелост-
ные системы во многих существенных отношениях являются
столь прочным сплавом элементов и частей, что рассматриваются
уже не как множества (совокупности), а как «далее неразложи-
мые единицы — индивиды». При исследованиях, не требующих
большой точности, можно пренебречь тем, что они обладают сле-
дами суммативности. Эти ряды отражают тенденции развития,
по выражению А. А. Ухтомского (1962 193), идущему «от
кучи... множества к упорядоченному множеству, к ансамблю».
Случайно перемешанные массы животных, бегущих от по-
жара, — вот пример конгломератцвной совокупности. Организм
высших животных — пример высокоцелостной динамически детер-
минированной системы. Популяция и вид — статистически детер-
минированные системы. Клетки, ткани и органы в организме на-
столько прочно взаимосвязаны, что существенным является пе их
самостоятельность, а впаянность в более обширную систему, за-
висимость друг от друга и от специальных управляющих аппара-
тов. Поэтому, хотя в состав организма могут входить десятки
203
триллионов клеток, организм не множество клеток и тканей,
не составная система. При переходе от механических смесей и
слабо связанных систем к высокоцелостным образованиям мно-
жество, составленное из элементов, превращается в более или
менее неделимую единицу. Давно отжили наивные представле-
ния об организме как сумме клеток. Положение о целостности
клетки, ткани, органа и организма, а также о возрастании в про-
цессе прогрессивной эволюцшгстепени целостности организма
(Завадский, 1958, 1960а) принимается в современной биологии
в качестве безоговорочной истины.
Но далеко не все соглашаются с характеристикой популяции
и вида как целостных живых образований, продолжая толковать
вид как простую совокупность (сумму) особей.
О степени целостности вида. Почему же многие
биологи до сих пор не признают целостность вида? Основной до-
вод состоит в указании на большое пространственное обособле-
ние и разобщенность организмов и столь большую свободу их
перемещения в пространство, что якобы прямым наблюдением
можно установить суммативпый характер вида. Индивид мы зри-
тельно воспринимаем как нечто пространственно непрерывное
и компактное, а в противоположность этому элементы вида могут
быть вкраплены отдельными точками в большие территории, на-
пример по одному на гектаре, могут рассеиваться на миллионы
квадратных километров и т. п. В общем, суммативный характер
популяции и вида устанавливается на основе примитивно-ви-
зуальных «данных». Обычно отмечается также безущербная для
целого делимость вида на части и возможность восстановить по-
пуляцию начиная всего с одного или нескольких экземпляров.
Известно, что массовые колонизации иногда начинались со слу-
чайной или преднамеренной интродукции небольших групп или
даже одиночных особей (в Европе элодея, филлоксера, колорад-
ский жук, фитофтора, ондатра и нутрия, мохноногий краб Erio-
cheir sinensis и др., в Америке европейский скворец, домовый
воробей, жук Popillia japonica, гессенская муха, яблоневая пло-
дожорка и др., в Австралии кролики, кактус Opuntia и мно-
жество других видов).
Клетки, ткани и органы являются компонентами настолько
высокоинтегрированных систем, что способность их к самостоя-
тельному существованию и воспроизведению в природных усло-
виях полностью подавлена. После экспериментального изъятия
из системы изолированные ядра и другие органеллы, безъядер-
ные клетки и их фрагменты, ткани и т. и. даже при наиболее со
вершенной технике культивирования, по выражению Ж. Браше,
становятся лишь «более или менее живыми». Такие сопоставле-
ния как будто свидетельствуют против целостности вида. Но они
основаны на крайне грубом разделении систем на суммативные
и целостные. Эталоном целостности при этом избирается высший
204
организм. Слабое развитие теории системности в биологии приво-
дило к необходимости выбирать одну из двух ошибочных концеп-
ций вида — суммативной или отождествляющей вид с организмом.
Достаточно же представить себе ряд систем с градуально из-
меняющейся степенью целостности от идеально аддитивных до
абсолютно целостных (элементы и блоки исчезают и сливаются),
чтобы и организм и вид разместились на этой шкале. Если при-
менить к виду это общее представление о степенях целостности,
то станет ясно, что вид не суммативен, но и не столь целостен,
как высший организм. Самая интегрированная популяция и вид
относятся к системам с относительно малыми степенями целост-
ности. Вид является упорядоченным образованием, т. е. системой,
но механизмы интеграции, поддерживающие его целостность
(панмиксия, химические взаимодействия, коадаптивпое поведе-
ние и другие «законы племенной жизни»), пе только меиее со-
вершенны, чем у организма, по и в принципе настолько иные,
что нет смысла рассматривать целостность вида, отталкиваясь от
привычных характеристик целостности организма.
Математики также приходят к выводу, что существует два
принципа организации живых систем: с централизованным управ-
лением, при котором обеспечивается строго направленная пере-
дача информации, или статистические, в которых накопление и
передача информации ненаправленны. Высшая ступень органи-
зации— это организм, а надорганизменные образования (семья,
стадо, популяция и т. д.) являются менее организованными си-
стемами (Ляпунов, 1966).
В пределах популяционно-видовой формы организации жи-
вого развитие целостности шло гораздо медленнее, чем у орга-
низменной формы. Так, например, быстро действующее центра-
лизованное управление множеством разнородных и взаимосвязан-
ных функций и элементов, столь поразительно развитое
у индивидов животных, чрезвычайно слабо выражено у популя-
ций .только высших стадных животных. При этом централизован-
ное управление ограничено рамками стада (восприятием зри-
тельных, слуховых и обонятельных раздражителей) и не может
простираться на весь вид. Регуляционные механизмы популя-
ции крайне несовершенны и, как правило, управляются не сигна-
лами, а ошибками. Дифференциация и специализация частей
в стаде гораздо меньшая, чем частей в организме, и т. п.
Тем не менее в процессе эволюции повышалась степень це-
лостности популяции и вида. Переход от первичных агамных
видов к видам с простыми формами обмена нуклеопротеидов и
затем к видам с настоящим половым размножением и перекрест-
ным оплодотворением — это и были основные ступени развития
интеграции репродуктивных систем на популяционно-видовом
уровне. Контактная п дистантная регуляции химическими ве-
ществами, а затем и нейрорегуляция также поддерживали надле-
205
Жащую организацию популяции или внутрипопуляционных групп
(стад, стай и пр.).
Таким образом, преувеличение степени целостности популя-
ции и вида так же ошибочно, как и сведение их к сумме особей.
Сравнительное изучение степеней целостности живых систем
только начинается. Пока же подчеркивается сам факт целост-
ности всех живых систем, начиная с молекулы ДНК и кончая
видом и биогеоценозом (Шмальгаузец, 1961).
Идею целостности популяции и вида поддерживают авторы
различных специальностей и направлений (Малиновский, 1947;
Allee et al., 1949; Wright, 1953, 1964; Завадский, 1952, 1961,
19676; Dobzhansky, 1955; Чижек, 1958, 1965; Шмальгаузен, 1958а,
19586, 1961; Полянский, 1959; Haldane, 1959; Odum, 1959; Бек-
лемишев, 1960; Andrewartha, 1961; Синская, 1963; Wynne-
Edwards, 1962, 1965; Botnariuc, 1963; Grant, 1963, и др.). Но не-
редко еще ошибочно противопоставляется популяция (дём) как
целостная система виду как якобы суммативной совокупности.
Широко расселенный вид, включающий географические расы,
конечно, гораздо менее целостен, чем одна панмиктическая попу-
ляция, отдельные компоненты которой неспособны самостоятельно
существовать в природе. Но если у такого вида существуют меж-
расовые контакты, это означает сохранение хотя бы малых степе-
ней целостности.
Иногда в понятие «целостность» включают не только единство
на основе взаимодействия частей, но и единство на основе сход-
ства состава и реакций на среду (Северцов, 1941, 1951; Васнецов,
1953). Сходство фенотипов создается на основе сходства генети-
ческих программ, и это является лишь фундаментом, на котором
возникают те или иные взаимодействия: «Организованным целым
может быть любое упорядоченное множество, содержащее в себе
определенные взаимодействия» (Ухтомский, 1962:185).
Видовые адаптации. Доказательством целостности по-
пуляции и вида могут служить приспособления, полезные не от-
дельной особи, а группе, популяции и виду, названные нами
«видовые адаптации» (Завадский, 1941, 1952, 1961). Видовые адап-
тации несводимы ни к индивидуальным адаптациям, ни к их сум-
мам. С появлением полового размножения надиндивидуальной
становится уже самая элементарная воспроизводящая единица,
а отдельно взятая особь теряет значение самостоятельной воспро-
изводящейся единицы. С выпадением фазы свободноживущей ли-
чинки, переходом к живородности и удлинением периода обяза-
тельной связи родителей с детенышем такой элементарной еди-
ницей становится семья.
У колониальных насекомых особь в сущности низведена на
положение специализированного и целиком от нее зависимого
органа колонии. В этом случае самой элементарной размножаю-
щейся единицей жизни является уже не пара особей или семья,
206
а колония. Элементарная, размножающаяся единица вида ста-
новится все более сложной и все более целостной системой, а от-
дельная особь оказывается ее зависимым компонентом. В резуль-
тате закрепления механизмов, обеспечивающих неродственные
скрещивания и поддерживание гетерозиготности потомства, фак-
тической элементарной размножающейся единицей оказывается
еще более крупное образование — дём, или местная популяция.
Различные формы сбалансированного полиморфизма популяций
(Ford, 1940, 1963; Sheppard, 1959; Dobzhansky a. Pavlovsky, 1960)
следует рассматривать как особый класс видовых адаптаций.
Удивительными примерами видовых адаптаций являются по-
пуляции, в которых закреплены летальные мутации, в гомози-
готном состоянии умерщвляющие организм. Важным условием
выживания видов бобочек — подражательниц несъедобным насе-
комым является малочисленность. При большей численности не
действовал бы «эффект примеси», так как птицы дифференциро-
вали бы иммитаторов от моделей или же атаковали тех и других.
Численность таких видов регулируется леталями, концентрация
которых в популяции обеспечивает нежизнеспособность 60—80%
зародышей каждого поколения. У немертины Linens ruber была
обнаружена красная раса, в 10 раз более многочисленная, чем зе-
леная (Шмидт, 1941, 1946). Процветание красной расы обуслов-
лено, видимо, тем, что ее популяция насыщена леталями, убиваю-
щими 86—97% яиц в каждой кладке (коконе). Отмершие яйца
служат кормом оставшимся личинкам, которые поэтому дольше
живут под охраной кокона, чем личинки зеленой расы, и вылуп-
ляются более крупными и жизнестойкими. В итоге численность
червей красной расы сильно возросла. Так генетически обуслов-
ленная гибель большинства зародышей стала причиной процве-
тания новой расы.
У многих животных (Dobzhansky, 1955, 1959) и растений
(Mather, 1955, Gotoh, 1957, и др.) обнаружены гены, в гомозигот-
ном состоянии летальные или снижающие жизнеспособность,
а в определенных сочетаниях с другими увеличивающие жизне-
способность, иммунитет, экологическую пластичность и другие
важнейшие биологические показатели. Отбором сохраняются по-
пуляции, имеющие генетические системы, способные постоянно
поддерживать состояние более адаптивной гетерозиготности. Ге-
терозиготность поддерживается и составом генов и избиратель-
ными скрещиваниями, а также элиминацией гомозигот и неадап-
тивных гетерозигот.
Примером видовой адаптации может служить оптимальная
плотность популяции, обеспечивающая лучшее воспроизведение
вида. «С точки зрения популяции как недостаточная, так и избы-
точная плотности одинаково вредны» (Парк, 1939:309). Раньше
подчеркивалось, что оптимальная плотность обеспечивает наи-
лучшее развитие особи, например у животных — скорость обмен-
?07
ных реакций, пищевую активность, рост, половое созревание, пло-
довитость и т. п. (Allee, 1931; Elton, 1933; Алпатов, 1934, и др.),
а у растений — максимальный коэффициент жизненности, т. е.
произведение веса на плодовитость (Cooper, 1926), наивысшую
сохранность (Сочава, 1926; Сукачев, 1927, 1935; Смирнова, 1928;
Успенская, 1929), лучшую перезимовку (Синская, 1939) и т. п.
Для сфагновых мхов и некоторых других растений очень высокое
загущение является необходимым условием жизни.
Позднее накопились данные, показавшие, что плотность, наи-
лучшая для развития отдельной особи (физиологический опти-
мум), часто не совпадает с плотностью, наилучшей для воспроиз-
ведения вида (популяционный оптимум) (Завадский, 1961). Огра-
ничение роста, ведущее к снижению биомассы и плодовитости
среднего растения, которое наблюдается в загущенных насажде-
ниях, свидетельствует о том, что известное угнетение и депрессия
особей сопряжено с максимальной биомассой и урожайностью
всего насаждения. В опытах с многолетниками Trinia henningii
(Сукачев, 1941) и кок-сагызом (Завадский, 1947, 19486, 1954а)
бъ1ло показано, что при загущении посева часть растений затор-
маживает рост и впадает в полуанабиоз. В таком состоянии рас-
тения могут находиться несколько лет, не теряя при этом готов-
ности к выполнению видовой функции. Они образуют резервный
фонд популяций, который мобилизуется после изреживания.
Регуляция на популяционном уровне и формирование опти-
мальной плотности осуществляются не только путем изменения
скорости роста особей вплоть до полного его торможения, но и
с помощью элиминации «избыточных особей», изменений группо-
вого поведения или другими способами (Ильинский, 1929; Allee
et al., 1949; Завадский, 1957а, 1961, 19676; Wynne-Edwards, 1962,
1965; Викторов, 1965; Шовен, 1965, и др.).
Видовыми адаптациями являются не только оптимальная плот-
ность, генетическая система популяции с закреплением полезных
летальных мутаций и надиндивидуальный характер элементарной
размножающейся единицы, но и уровень численности вида, вели-
чина плодовитости и т. п. Видовые адаптации создаются совмест-
ным действием индивидуального внутривидового (например, поло-
вого) и группового отбора, при котором переживают лучше орга-
низованные семьи, стада, колонии или целые местные популяции.
Можно различать две группы видовых адаптаций. Одна
группа — это аппараты, функции и реакции, носителями которых
являются организмы, но значение которых состоит в обслужива-
нии не самих носителей, а популяции и вида (например, первич-
ные и вторичные половые признаки, плодовитость и т. п.). Вторая
группа включает настоящие надиндивидуальные приспособле-
ния, возникающие только при определенном множестве организ-
мов и требующие для своего действия какого-то минимума чис-
ленности или разнообразия огранизмов (плотность популяции,
208
внутрипопуляционный полиморфизм и полиморфизм колониаль-
ных насекомых, «генетическая система популяции» и т. п.).
Своеобразную группу видовых адаптаций выделил С. А. Северцов
(1936, 1951) в учении о конгруэнциях. Конгруэнциями он назвал
соответствия в поведении, в строении и функциях органов, воз-
никающие между особями одного вида. Конгруэнции являются
полезными не для сохранения жизни отдельного индивида, а для
воспроизведения вида. Действуя нередко прямо во вред инди-
виду — ее носителю, конгруэнции обеспечивают поддержание
жизни общины, популяции, т. е. служат виду как целому.
С. А. Северцов подразделил конгруэнции на несколько групп.
Во-первых, это соответствия в строении и функциях эмбриона и
материнского организма (связь через плаценту) между размерами
эмбриона и матки; между строением языка и полости рта дете-
ныша и млечных желез; строением и инстинктами детеныша сум-
чатых и строением и инстинктами самки и т. п. Во-вторых, это
соответствия в строении аппаратов размножения самки и самца,
в ритмичности их функции, в поведении животных разных полов,
обеспечивающим оплодотворение, в образовании органов, способ-
ствующих отысканию особи другого пола (пахучие железы, фото-
форы и пр.). В-третьих, это соответствия в организации особей од-
ного пола, возникающие на основе полового отбора (развитие рогов
и других органов турнирного оружия у самцов и т. п.). В чет-
вертых, это соответствия в строении и поведении особей, связан-
ные со стадным образом жизни (окраска, имеющая сигнальное
значение, звуковая сигнализация в стадах млекопитающих и пр.).
И наконец, в-пятых, это соответствия в организации особей — чле-
нов полиморфных колоний у колониальных впдов перепончато-
крылых (пчел, шмелей, муравьев, ос и термитов). С развитием
конгруэнций возрастает не только взаимозависимость особей, но
и зависимость отдельной особи от колонии, стада, популяции.
Существование видовых адаптаций, и в том числе конгруэнций,
является убедительным доказательством целостности вида.
В последнее время с позиций нашей концепции целостности
вида и общевидовых адаптаций получило истолкование явление
гипертелий (Botnariuc, 1963). По мнению этого автора, не суще-
ствует вредных для вида «гипертелических» признаков. Вредные
для отдельно взятой особи, такие признаки адаптивны для вида.
В таком представлении, возможно, несколько преувеличена сте-
пень целостности вида (щфоль группового отбора). В ряде слу-
чаев противоречия между организмом и видом могли, по-види-
мому, обостряться и приводить даже к вымиранию группы.
Во всяком случае нет оснований переоценивать гармоничность
видовых адаптаций и считать, что диспропорции и ошибки в ходе
эволюции возможны только у индивида.
Все сказанное в этом разделе о целостности популяционно-ви-
довой формы организации живого позволяет сделать следующий
14 К. М. Завадский
209
общий вывод. Вид целостен, по степень целостности никогда не
может быть столь большой, чтобы делаться препятствием для
деятельности отбора. Если степень целостности вида превышает
допустимые пороги, система становится слишком сильно связан-
ной и деятельность естественного отбора затрудняется (кроме
группового отбора целых микропопуляций). Такие виды теряют
эволюционную пластичность и вымирают. Таким образом, суще-
ствует капитальная преграда для роста степени целостности вида
за определенные рамки, которыми является наилучшая приспо-
собленность системы для деятельности отбора.
4. ОПРЕДЕЛЕНИЕ ВИДА
Подводя общие итоги, можно дать следующее развернутое оп-
ределение вида или, вернее, определение популяционно-видовой
формы организации жизни. Вид — одна из основных форм суще-
ствования жизни, особый надиндивидуальный уровень организа-
ции живого; будучи статистически детерминированной системой
и полем деятельности естественного отбора, вид обладает как воз-
можностью длительного самовоспроизведения и существования
в течение неопределенного времени, так и способностью к само-
стоятельному эволюционному развитию, он является носителем и
основной единицей эволюционного процесса. Вид внутренне про-
тиворечив: как результат эволюции он выступает в относительно
стабильном состоянии, качественно определенен, целостен, приспо-
соблен к данной среде, устойчив, обособлен от других групп (дис-
кретен), а как узловой пункт и активный носитель эволюции —
менее определенен, имеет составной характер, неустойчив, лаби-
лен, обладает расплывающимися границами.
Эта характеристика, конечно, громоздка, но мы не стремились
к афористическим лаконичным формулировкам. Задача состояла
в определении сущности явления и его основных противоречий.
В последней книге Майра (Мауг, 1963) вид определяется как вы-
сокоорганизованная единица эволюции, основанная на обобщен-
ном эффекте действия всей совокупности коадаптированных чле-
нов, производящих один или несколько оптимальных фенотипов.
Следует сказать, что образование, достигшее популяционно-ви-
дового уровня, имеет гораздо более расплывчатые очертания в
гораздо более многообразно по проявлениям, чем идеал вида так-
сономистов-каталогизаторов. Видовая форма организации живого
включает многие миллионы популяций и рас, из которых лишь не-
которые, прошедшие испытания и преобразования отбором, со-
храняются в качестве «хороших» видов. Лишь немногие виды из
массы оказываются очень долго существующими. Еще меньшее
число видов оказывается способным к длительной эволюции ц
к основанию мощных пучков адаптивной радиации групп.
Глава 8
СТРУКТУРА ВИДА
1. ВВЕДЕНИЕ
Определения основных понятий. Независимо от
высоты организации, способа размножения, величины ареала,
численности и других особенностей каждый вид включает раз-
личные формы, т. е. дифференцирован. Дифференциация эта
может быть различной по степени и по другим показателям, но
полностью мономорфных видов в природе не существует.
В понятие «дифференциация вида» (т. е. полиморфизм вида
в широком смысле слова) входят все типы, разнородности:
биохимическая, физиологическая и морфологическая; половая, воз-
растная, сезонная, связанная с фазами жизненного цикла, эколо-
гическая, географическая и др.; прерывистая и непрерывная; адап-
тивная и неадаптивиая; модификационная и генотипически ста-
билизированная; проявляющаяся в обычных условиях и более
или менее скрытая, реализующаяся в особых «дифференцирую-
щих» условиях или же при нарушениях сложившейся системы
скрещиваний.
Все внутривидовые различия необходимо подразделить на три
категории: вновь возникающие и еще не успевшие подвергнуться
естественному отбору (новые мутации и морфозы); исторически
обусловленные, уже проверенные и переработанные отбором и
имеющие приспособительное значение; неадаптивные, вызываемые
плеотропизмом, селективными корреляциями или генетико-авто-
матическими процессами.
Различия первой категории, будучи сырьевым материалом для
преобразования вида, вызываются постоянно идущим мутацион-
ным процессом и служат единственным источником возникновения
различий второй категории.
В этой главе вид рассматривается нами как уже установив-
шаяся система. При таком подходе новые мутации и морфозы не
14*
211
входят в описания структуры вида, которые ограничиваются лишь
селекционно отработанными группами. Надо учесть, однако, что
часто трудно отличать различия первой и второй категории. Сово-
купность групп, уже доказавших свое приспособительное значе-
ние, можно назвать «адаптивной нормой вида». Она формируется
под воздействием противоположно направленных процессов инди-
видуального и группового отбора, ведущих к адаптивной инте-
грации и к адаптивной радиации групп внутри вида.
Адаптивная интеграция групп внутри вида — это центростре-
мительная тенденция, опирающаяся на полезность объединения
разнообразия и ведущая к усилению системного единства вида.
Интегрирующая деятельность отбора, как показал И. И. Шмаль-
гаузен (1961), начинается на молекулярном уровне и охватывает
все вышележащие уровни организации. Нас будет интересовать
интеграция надиндивидуальных систем (семья, клон, колония,
стадо, популяция и т. п.). По Э. Форду (Ford, 1940, 1963), в по-
пуляции можно обнаружить два сорта многообразия: 1) сбалан-
сированный полиморфизм, характеризующийся стабильным чис-
ленным соотношением между формами, которое поддерживается
в популяции из поколения в поколение; простыми примерами
такого полиморфизма может служить соотношение полов, соот-
ношение форм в колонии пчел или муравьев; 2) транзитивный
полиморфизм, характеризующийся сдвигами численного соотно-
шения форм в популяции, выражающими процесс ее эволюцион-
ного преобразования; более адаптивная форма завоевывает себе
место за счет снижения численности других компонентов популя-
ции, входящих в ранее сложившуюся адаптивную норму.
Рассматривая процесс эволюции как закономерную смену
адаптивных норм, И. И. Шмальгаузен подчеркивал двоякое зна-
чение внутривидового разнообразия форм (как причины и как
следствия эволюции) и в качестве особого его сорта выделял «рав-
новесный гетероморфизм», понимая под этим те самые явле-
ния, которые Форд назвал «сбалансированным полиморфизмом»,
а много позднее Дж. Гексли (Huxley, 1956) — просто «морфиз-
мом». Тенденция к адаптивной интеграции вида наиболее сильно
проявляется в рамках местной популяции. Именно внутрипопу-
ляционное разнообразие включает случаи функциональной специа-
лизации частей, которые воспроизводятся с помощью различных
механизмов из поколения в поколение в заданных количествен-
ных соотношениях и поддерживают разные стороны адаптивной
структуры популяции.
Адаптивная радиация групп внутри вида — центробежная
тенденция, ведущая к образованию возможностей все более обо-
собленного и самостоятельного существования группы в природе,
т. е. к возникновению зачатков новых видов, стоящих на разных
ступенях преобразования и отщепления от исходного вида.
В итоге адаптивная норма является как бы равнодействующей
212
этих тенденций. Она может включать формы, как интегрирован-
ные в системе единой популяции (балансированный полимор-
физм, например половой или у колониальных насекомых, и т. п.),
так и существующие более или менее изолированно друг от друга
(приспособительные экологические и географические формы).
У стабилизированных (персистентных) видов адаптивная норма
сложилась давно и поддерживается отбором, устраняющим все
тенденции к ее нарушению. Стенобионтные и древнереликтовые
виды имеют очень суженую адаптивную норму. Большинство же
видов, сохранивших эволюционную пластичность, включает раз-
нообразие форм, входящее в адаптивную норму вида и в то же
время являющееся стадиями процесса эволюции, протекающих
еще внутри вида, но имеющих возможность выйти за его пределы.
Не следует думать, что между процессами внутривидовой
адаптивной интеграции и адаптивной радиации форм существуют
какие-то твердые границы. Направление отбора может меняться
в зависимости от соотношения факторов, управляющих этим про-
цессом. При одном соотношении селективных факторов усилится
тенденция к консолидации отдельных групп в единую систему,
при другом — к их обособлению, к выходу из состояния зависи-
мых компонентов целого и к превращению в самостоятельно
воспроизводящиеся единицы. Именно направление отбора и дик-
тует изменения не только характера связей между формами, но и
степени их обособленности друг от друга.
Полиморфизм вида в широком смысле слова, включающий все
формы дифференциации, может поддерживаться не только меха-
низмами адаптивной интеграции и процессами адаптивной радиа-
ции, но также и параллельной изменчивостью. Например, при на-
личии нескольких географических рас у каждой или у некоторых
из них независимо друг от друга могут возникнуть одинаковые
экотипы пли же одинаковые неадаптивные признаки в духе гомо-
логических рядов изменчивости по Н. И. Вавилову.
Внутри вида и внутри отдельной популяции различия могут
быть прерывистыми, и тогда на первый план выходит дискрет-
ность форм (по одному или по комплексу признаков), могут же
быть и почти непрерывными, так что отдельные формы становятся
трудно разграниченными пли же и вовсе исчезают. Вид может
включать формы, находящиеся друг с другом в отношении сопод-
чинения и сосуществования. Виды неравноценны также и по ха-
рактеру разнообразия форм: оно может быть не только большим
пли меньшим, но и отличаться качественно.
Характер дифференциации вида зависит от коренных особен-
ностей приспособления к среде (растения или животные, сидячие,
оседлые, подвижные или кочевые и т. п.), от способа размноже-
ния, а’ также от особенностей самой среды. Животные, способные
в процессе передвижения активно выбирать наиболее оптималь-
ные биотопы или же сочетающие высокую устойчивость одной
213
приспособительной формы с подвйжной системой регуляции
физиологического гомеостазиса, как показывают многие работы
(Lerner, 1954; Mather, 1955; Sheppard, 1959; Grant, 1963, и др.),
фенотипически могут быть довольно однородными (при большом
генотипическом разнообразии). Пассивное передвижение, непо-
движность или недостаточное развитие аппаратов физиологиче-
ского гомеостазиса в среде с меняющимися или пестрыми усло-
виями приводит к развитию модификационной изменчивости или
же балансированного полиморфизма. Такие виды обладают го-
раздо большим фенотипическим разнообразием, чем высшие
животные с совершенным физиологическим гомеостазисом.
Все сказанное в этом разделе относится к хорошо изученным
видам. Иначе обстоит дело с плохо исследованными группами.
Например, после проверки в культуре обнаружилось, что многие
виды и более крупные таксоны мхов — это всего лишь модифи-
канты (Савич-Любицкая и Смирнова, 1958). У некоторых групп
из инфузорий Ophryoscolecidae одни авторы описывают серию
форм в пределах нормы реакции (при культуре клонов), а другие
именуют каждую форму видом, так что серия модификантов воз-
водится в ранг рода (Догель и др., 1962).
Исследования структуры вида проводятся многими отраслями
биологии. Однако до сих пор главное внимание уделялось ана-
литическим аспектам — описывались все новые и новые различия
п формы. Гораздо слабее изучены способы связей между фор-
мами, наличие соподчиненности, функциональных отношений и
другие явления, касающиеся интеграции частей популяции и вида
как целостных систем. Само понятие «структура» часто толкуется
адк синоним понятия «состав», но более точно рассматривать его
кад закономерную систему отношений между элементами и ча-
стями. Нельзя, разумеется, изучать структуру вида, не зная со-
ставу, но нельзя и ограничиваться только изучением состава.
Основой всей системы интеграции вида являются отношения,
возникающие между особями разных полов и поколений, между
членами семей и колоний, стад, стай, популяций и других надин-
дивидуальных образований, в процессе воспроизведения вида.
.Такие внутривидовые отношения составляют содержание жизни
вида (Завадский, 1961).
В заключение следует подчеркнуть, что в сложной и недоста-
точно изученной проблеме вида вопрос о его структуре — самый
малоразработанный. Не только разные авторы, но и разные от-
расли биологии приходят к поразительно противоречивым выво-
дам, касающимся факта существования самых основных подраз-
делений вида, таких как разновидность и подвид, экотип и полу-
вид, не говоря уже о формах дифференциации местной популяции.
Представители разных школ традиционной систематики, экспе-
риментальной систематики, экологи и биоценологи, исследователи
микроэволюции и генетики популяций по-разному подходят и
214
по-разному решают коренные вопросы структуры вида. На данном
этапе исследований полезны как создание независимых друг от
друга представлений о структуре вида, основанных па различных
подходах, так и попытки синтезировать всех их на общебиологи-
ческой основе.
К критике понятия о разновидности. Когда-то
разновидностью называли случайное и обратимое уклонение от'
типа вида. В прошлом веке, когда формы дифференциации вида^
еще не были известны, разновидность была единственной катего-
рией для регистрации морфологической изменчивости в рамках
вида. Одним из первых обратил внимание на неопределенность
понятия разновидность Де Фриз: «Термин разновидность.. . при-
меняется к столь различным явлениям, что не дает никакого яс-
ного представления о вещах» (De-Vries, 1906 21). Но сила инер-
ции в науке велика и, хотя эта категория уже теряла свое зна-
чение, на ботаническом конгрессе 1930 г. в Кембридже (Briquet;.
1935) была принята следующая система внутривидовых единиц/
вид, подвид, разновидность, подразновидность, форма, биологи-
ческая форма, специальная форма. Три последние единицы фик-
сируют находки отдельных аберрантных экземпляров, а три
остальные представляют собой вариации разновидности (морфо-
логический подвид — та же разновидность, кажущаясд система-
тику более крупной, чем обычная). В то же время виднейшие
систематики подчеркивали, что разновидность — устарел'ый тер-
мин, указывающий на чисто морфологические особенности (Кома-
ров, 1927), часто на случайную основу выделения признака (Ро-
занова, 1930), на его искусственность (Делоне, 1945). В. П. Ма-
леев отмечал, что разновидность «устанавливается на чистб
морфологической основе и является в значительной степени не-
определенной» (1933 29). По Л. С. Бергу, «название varietas
мы оставляем для тех случаев, когда остается невыясненным,
имеем ли мы дело с подвидом, морфой, расой или иной низшей
таксономической единицей» (1948:15). Большинство современ-
ных систематиков перестало в своих работах пользоваться поня-
тием разновидности. В палеосистематике, по словам Д. Л. Сте-
панова (1959а), часто различают внутри вида только разновид-
ности и даже противопоставляют их «типу» вида или же
пользуются понятиями экспериментальной систематики лишь де^
кларативно (Родендорф, 1959). Примером чисто морфологической
трактовки внутривидовых единиц является известная класспфи?
кация разновидностей пшениц (Komicke, 1885). Эта система ос-
нована на четырех признаках (остистость, окраска зерна, опу-
шение чешуй и окраска колоса). Одна такая * разновидность
объединяет самые различные группы как в историческом, эколого-
географическом и генетическом отношениях, так и по конститу-
ции. В одну разновидность попадают озимые, полуозпмые, позд-
ние и ранние яровые формы, мезоморфные и ксероморфные, юж-
211
ные и северные и т. п. Единственная причина сохранения этих
искусственных и сборных разновидностей Кёрнике — практиче-
ское удобство. У обыкновенной садовой улитки (Helix striata)
описано около 1500 морфологически различных форм. Сотни
форм обнаружены среди видов пресноводной Unio (Жадин,
1952). Иногда их именуют разновидностями.
Разновидность является формальной единицей и совпадает
с какими-либо формами существования вида в природе только
тогда, когда классификационный признак оказывается связанным
с экологически значимыми реакциями (темп роста, ритм разви-
тия и т. п.) или же входит в корреляционную систему важных
признаков конституции (архитектоника куста, мезо- или ксеро-
морфность, форма роста и др.). Такие совпадения — дело случая.
Часто разновидность остается лишь группировкой изолирован-
ных признаков и не отражает каких-либо определенных форм су-
ществования вида. При дальнейшем же изучении может ока-
заться, что разновидность — определенная форма существования
вида, например географическая раса. Так, мелколистная форма
голубики рассматривалась как varietas вида Vaccinium uliginosum,
пока не было установлено, что в Гренландии (73° с. ш.) и на
Крайнем Севере расселена только эта раса и что голубика лес-
ной зоны имеет 48 хромосом, а мелколистная — 24 (Hagerup,
1933). Более полное изучение показало, что это не какая-то не-
определенная разновидность, а географическая раса, имеющая
обособленный ареал и особую экологию. М. Г. Попов (Popov,
1925—1926), описывая азиатские виды Zygophyllum, указывает,
что раньше различали виды Z. rosowii и Z. latifolium чисто мор-
фологически по форме тычиночной чешуи, длине пестиков и
чашелистиков и по форме придатка черешка, а М. А. Максимович
соединил их в один, не различая даже разновидностей. Сам же
Попов выделил var. latifolium как разновидность Z. rosowii,
так как форма с более крупными листочками и коробочками и
менее сизыми листьями и стеблями к северо-западу от оз. Бал-
хаш имеет обособленный ареал, т. е. является географической
расой. И. П. Савоськин (1959) отмечает, что у морского лука
(Urginea maritima) некоторые авторы выделяют три разновид-
ности (var. typica, var. tadlensis и var. stenophylla), в то время
как все они должны рассматриваться как экологические формы.
Неопределенно характеризуется разновидность и Я. И. Прохано-
вым (1947), выделяющим их у хлопчатника то на чисто морфо-
логическом, то на географическом или экологическом основаниях.
Энтомолог В. Никулеску, защищая политипическую концепцию
вида, неожиданно предлагает расчленять вид не только на гео-
графоэкологические расы п подвиды, а и на разновидности (Nicu-
lescu, 1960). Этим термином он обозначает морфологически раз-
личные формы, живущие симпатрически и синхронически в со-
ставе одной популяции. Некоторые систематики мхов придержи-
316
ваются старинного понимания разновидности как совокупности
особей, отличающихся от типа вида морфологическими призна-
ками (Peterfi, Stefureac, 1965). Любопытно, что эти авторы вы-
деляют также подвиды и формы, по каково их отличие от разно-
видностей — неясно.
Среди биологов, пользующихся в настоящее время понятием
разновидности, существует разброд в ее определении. Вот неко-
торые из таких определений разновидности: морфологическое
отклонение от видового типа; любое подразделение вида, начиная
с единичных мутантов и кончая подвидом; одна из форм поли-
морфной группы; вариация, причины которой неизвестны; край-
ние формы непрерывных рядов изменчивости; наследственные
уклонения, не имеющие ареала; формы, место которых в системе
вида еще не изучено; основная форма существования вида; ста-
дия (ступень) видообразования. Из этого перечня видно, что тер-
мин «разновидность» обозначает столь разнородное содержание,
что стал произвольным и неопределенным. Естественно, что нет
смысла в обсуждении того, является ли разновидность формой
существования вида, или же ступенью видообразования. Едва ли
правильно и сопоставление С. С. Шварца: «основная форма раз-
новидности (подвида) — это разновидность географическая, гео-
графическая раса» (1959 93).
2. СИСТЕМЫ ВНУТРИВИДОВЫХ ЕДИНИЦ
О методах изучения дифференциации вида.
Кроме отживших чисто морфологических, существуют географиче-
ские, экологические, генетические и комплексные генетико-эко-
лого-географические системы внутривидовых единиц.
Морфолого-географический метод обладал слабой различи-
тельной способностью и улавливал лишь географические расы
(подвиды). Вид как раса представлялся далее неразложимым
монотипом лишь потому, что существующие различия этим ме-
тодом просто не улавливались.
Новое в географический метод внес Дж. Гексли (Huxley,
1938а, 1938b), предложивший методику клипов. Он показал воз-
можность количественных исследований географических измене-
ний определенного признака. Клинальные изменения признака —
это градиентные его изменения вдоль определенного разреза
на территории одного местообитания (ecoclin), через весь ареал
или его часть (geoclin). Как показали работы Дж. Гексли, а также
Д. Грегора (Gregor, 1939, 1946 и др.), П. В. Терентьева (1957,
1966) и др., метод клипов позволяет улавливать географические
изменения любых признаков. Однако методика изучения кли-
нальной изменчивости также имеет недостатки, так как сосредо-
точивает внимание лишь на отдельных или немногих признаках.
217
Увлекшись изучением клинальной/изменчивосм, некоторые ав-
торы пришли к выводу, что внутри вида вообще не существует
прерывистых различий, дающих основание выделять какие-либо
группы. Однако были установлены нарушения постепенности, рез-
кие сдвиги (step-cline), и идея чистой непрерывности была подор-
вана, Методика клинов является весьма ценной для выяснения
истинной картины территориального изменения данного признака,
но дает основания для суждений лишь о поведении изученного
признака, а не для более широких экстраполяций. Само собой
разумеется, что географические методы не могут быть единствен-
ными, так как внутри вида существует и многообразие экотипов
и форм, живущих совместно в пределах одной и той же терри-
тории.
Применение изолированно взятых экологических и генетиче-
ских методов изучения дифференциации вида также оказались
неэффективными, так как оставляли в тени или степень наслед-
ственной стабильности форм, или же их приспособительное зна-
чение. Только совместное применение географо-морфологического,
экспериментально-экологического и генетического методов привело
к обоснованию современных представлений о дифференциации
вида (например, у растений: Clausen, 1922, 1951, 1958; Turesson,
1922а, 1922b, 1931, 1961; Розанова, 1930, 1946; Синская, 1938,
1948, 1963, и многие другие). Именно комплексные генетико-эко-
' лого-географические исследования, проводимые в природе, на пи-
томниках и в лаборатории, позволяют наиболее полно вскрыть
структуру вида. Такие работы очень трудоемки и длительны.
Чтобы с достаточной полнотой изучить все формы дифференциа-
ции вида, требуются десятилетия экспериментальной работы.
Так, для выявления генетического потенциала вида необходимо
сосредоточить па питомниках коллекции, более или менее полно
представляющие местные популяции вида из всех географически
и экологически различающихся пунктов сбора. Методом близко-
родственного разведения каждая популяция должна быть разло-
жена на совокупность практически выравненных линий. Этим
путем могут быть выявлены генетический потенциал вида и раз-
личия между отдельными местными популяциями, а также не-
расщепляющиеся генетические комплексы, лежащие в основе
стабилизированных конституций. Для изучения степени генетиче-
ских различий между отдельными популяциями необходимо про-
ведение массы межпопуляционных скрещиваний с изучением
потомств в ряде поколений и сопоставление данных этих опытов
с результатами различных скрещиваний между формами, принад-
лежащими к одной популяции. Требуется т(акже выращивание
всей коллекции в различных пунктах (географические посевы,
питомники, расположенные на разной высоте в горах или в раз-
личных почвенных условиях, и пр.) или при различных условиях
(длина дня, увлажнение, состав почв и т. п.).
218
Чтобы выяснить значение отдельных форм для поддержания
высокой жизнеспособности, приспособленности и конкурентоспо-
собности всей популяции, необходимо специальное изучение от-
дельных линий путем введения или исключения их из состава
популяции, а также исследования аппаратов, с помощью которых
в популяции поддерживается нужный баланс форм (подбор со-
опылителей, системы скрещиваний, оптимальная степень гетеро-
зиготности, соотношение доминирования, модификационные спо-
собности генотипов и пр.). Только в результате такпх многооб-
разных по методикам и длительных исследований можно успешно
изучать дифференциацию видов и при этом избегать односторон-
них выводов, которые столь характерны для чисто генетических
и чисто экологических методов работы.
Внутривидовые категории классической систематики, основан-
ные на морфолого-географических признаках, как полагает
В. Грант (Grant, 1960), должны существовать параллельно с ка-
тегориями экспериментальной систематики, хотя он и подчерки-
вает, что биологическими явлениями следует признать только
последние. Правильнее было бы не увековечивать отрыв морфоло-
гических внутривидовых единиц от единиц экспериментальной
систематики. Внутривидовая систематика, основанная на полевых
наблюдениях, должна рассматриваться как необходимый первый
этап работы. Например, таким полевым эколого-морфологическим
методом можно установить, что полынь Artemisia ciniformis из
Копет-Дага дифференцирована на морфологически отличимые по-
пуляции, приуроченные к разным стациям — от северных пологих
щебнисто-мелкозернистых склонов до сухих крутых каменистых
склонов. К познанию вида ничего, однако, не прибавится, если
эти популяции будут возведены в ранг formae и каждая будет эти-
кетирована латинским словом (viridis, gracilis и т. п.). Действи-
тельный шаг вперед связан с использованием комплексных экспе-
риментальных методов.
Система внутривидовых единиц у растений.
Суммируя результаты комплексного изучения дифференциации
вида, главным образом на растениях, систему основных внутриви-
довых единиц можно представить следующим образом.
1. Полувид (semispecies, или hemispecies) — географическая
или экологическая раса, почти достигшая состояния молодого
вида.
2. Подвид (subspecies) —сформированная географическая или
экологическая раса.
3. Экотип — локальная экологическая раса.
4. Местная популяция — относительно обособленное поселение,
устойчиво занимающее определенную территорию и способное
к самовоспроизведению; основная единица населения и бо-
лее или менее отчетливо отграниченный структурный элемент
вида.
219
5. Экоэлемент — внутрипопуляционная форма, связанная со
спаянным, нерасщепляющимся генетическим комплексом и спо-
собная выходить из состава популяции в положение самостоя-
тельно существующего экотипа.
6. Морфо-биологическая группа, или изореагент, — группа ор-
ганизмов внутри популяции, имеющая одинаковую или различную
наследственную основу, различимая морфологически, имеющая
сходный ритм развития (феногруппа) и одинаково реагирующая
на условия среды.
7. Биотип — элементарная единица генетической структуры
популяции; группа организмов, имеющих почти тождественный
генотип и отличающаяся от всех других таких же групп хотя бы
одной мутацией.
Эта система внутривидовых единиц, опирающаяся на данные
многих современных авторов, была разработана нами (Завадский,
1961) главным образом для высших растений, однако и виды жи-
вотных имеют в принципе такую же внутреннюю организацию.
Основной принцип, па котором построена эта система, — призна-
ние местной популяции главным структурным элементом вида.
Части вида, стоящие выше местной популяции, — экотип, подвид
и полувид, представляют собой аллопатрические или викарирую-
щие, т. е. пространственно взаимоисключающие друг друга
группы (не заселяющие территории, занятой другой группой).
Они воспроизводят только свой тип. Части вида, входящие в со-
став местной популяции, обычно живут совместно, в одном и
том же местообитании, т. е. являются симпатрическими и не
только не вытесняют друг друга с занятой ими территории, но
часто самостоятельно не могут даже и существовать в природе
(не выдерживают конкуренции и т. п.). У видов с перекрестным
оплодотворением симпатрические формы, как правило, воспроиз-
водят не только себя, но и другие формы, входящие в состав
той же местной популяции.
Аллопатрические подразделения отличны от симпатрпческих
(а часто даже противоположны им) по своему функциональному
значению для поддержания целостности вида. Аллопатрические
части вида являются выражением значительно продвинутой адап-
тивной радиации групп. Между такими частями возрастают ге-
нетические различия. Они являются изолированными друг от
друга в половом отношении или вступают в контакты лишь в уз-
ких пограничных полосах. В итоге растет способность групп к са-
мостоятельному существованию и независимому от других групп
эволюционному развитию. Появление аллопатрических групп
внутри вида знаменует собой ослабление целостности вида или
даже фактическое ее исчезновение.
Симпатрические группы часто являются зависимыми компо-
нентами целостной местной популяции. Они отражают интеграцию
популяции на общей генетической основе. Симпатрические
220
группы не только не ограждены друг от друга какими бы то нй
было барьерами, но, напротив, условием их существования яв-
ляется именно непрерывное скрещивание между собой (панмик-
сия или же системы скрещиваний на основе избирательности).
Однако их образование создает возможность частичной или полной
дезинтеграции популяции. При определенных обстоятельствах
(которые будут рассмотрены позднее) любая из стабилизи-
рованных внутрипопуляционных групп может выйти из си-
стемы популяции и стать самостоятельно воспроизводящейся
единицей популяционно-видового уровня. Может произойти так-
же и распад местной популяции (например, после выпадения
полового процесса), после которого отдельные ее компоненты
становятся основателями самостоятельно существующих попу-
ляций.
Внутри вида наблюдается разнообразие форм по фенологии.
Оно широко распространено у споровых и цветковых растений,
у простейших и, по-видимому, у многих пойкилотермных живот-
ных. Фенологический полиморфизм наблюдается как у аллопат-
рических форм (например, южные и северные климатипы, высо-
когорные и долинные экотипы, экотипы террас и пойм и пр.), так
и у симпатрических (внутрипопуляционные феногруппы и фе-
нологические расы). Более подробная характеристика каждой из
внутривидовых единиц этой системы будет дана в последующих
разделах этой главы.
Общепринятых систем до сих пор не существует. Е. Н. Син-
ская (1961, 1963) выделяет экотипы, популяции, стабильные и
зачаточные экоэлемепты и биотипы. А. А. Корчагин (1964, 1966)
критикует нашу систему за комплексный подход, считая, что он
ведет к смешению экологических критериев с морфолого-геогра-
фическими, принимая вместе с тем другие наши определения
(см. схему на стр. 222).
С нашей точки зрения, только комбинированные системы
имеют будущее и ошибочно сохранять обособленные системы,
опирающиеся на отдельно взятые критерии. Как правило, клп-
матип и подвид — это синонимы, и нет оснований отрывать их
друг от друга. Исключение составляют горные климатипы, входя-
щие в состав подвидов на правах локальных экотипов. Состояние
подвида и полувида столь важно с таксономических и эволюцион-
ных позиций, что замена этих понятий понятием «климатип» со-
вершенно неприемлема.
По Н. П. Дубинину (1966 408—410), вид распадается на по-
лувиды, подвиды, расы, микротопографические .расы и линии.
В этой системе недостаточно обосновано понятие расы. По опре-
делению (объединение элементарными генетическими системами
и приспособление к определенным ограниченным условиям среды)
оно соответствует местной популяции. Но термин «раса» уже за-
нят, и стоит ли употреблять его в качестве синонима локальной
221
Системы внутривидовых групп
(по Корчагину 1964, с изменениями)
Э колого-географическая Морфологическая
ненаследственные наследственные и морфолого-географическая
Морфо-биолс Модификанты (эка- ды) Био Экотипы климатические » эдафические » ценотические Ценопопуляции )гические группы | Экоэлементы типы Подвиды, иногда виды Разновидности, реже под- виды и виды Разновидности Разновидности Разновидности, лузусы, абберации
популяции? Специфичность микротопографической расы опреде-
ляется элементами неадаптивной дивергенции из-за генетико-ав-
томатических процессов, но еще не выяснена их распространен-
ность и, возможно, что этот сорт рас — частный случай. Опреде-
ление «линии» соответствует определению «биотипа» в нашей
системе.
Представл енпя о дифференциации вида в зо-
ологии. У нас нет возможности рассматривать всю массу си-
стем внутривидовых единиц и терминов, предложенных в различ-
ных отраслях зоологии. Состояние этого важного вопроса
в настоящее время настолько неудовлетворительно, что нет еди-
нодушия в понимании даже подвида, не говоря уже о других
подразделениях. Из-за разнообразия подходов и точек зрения
высказываются иногда взаимоисключающие предложения. Одни
считают, что вид может включать подвиды, которые состоят из
«многочисленных мелких рас и локальных популяций. Подвид
не является необходимо зарождающимся видом, хотя это может
быть и так, если он изолирован географически» (Маут, 1965 129).
Другие же настаивают па таксономической неделимости вида
(«вид как конечный таксон») и предлагают изучать не его
структуру, а некие «кванты топографической изменчивости» (Те-
рентьев, 1966 15). Как полагает Р. Ппмснтал (Pimental, 1959),
если вид распадается на изолированные популяции, отвечающие
«правилу 84%», они достойны возведения в ранг подвида. Дру-
гие же изолированные популяции с меньшим «уровнем диверген-
ции» следует называть изолятами, а географически не изолиро-
ванные, но морфологически различающиеся популяции можно
называть морфами. Для систематиков такие «договорные начала»
в определении подвида может быть и хороши, но для понима-
222
ййя его биологической природы они Неприемлемы. Нельзя не
посетовать на формальность таких определений внутривидо-
вых единиц, которые опираются на «правило 75%» или «пра-
вило 84%».
По Н. П. Наумову (1962), вид включает три рода независимых
друг от друга группировок: территориальные, функционально-био-
логические и «социальные». Территориальные группировки — это
подвиды, включающие географические популяции, которые со-
стоят из экологических популяций, распадающихся на элементар-
ные популяции. Возможно, что у некоторых грызунов имеется
такая многоэтажная иерархия, но у множества других групп под-
разделения подвида на субподвиды не существует, а экотипы не
соподчинены подвидам, а представляют структурную единицу,
им эквивалентную. Под функционально-биологическими группи-
ровками имеются в виду половые и возрастные группы, а также
«пищевые расы» насекомых и «физиологические расы» парази-
тов. Несмотря на значительную подвижности отдельных половых
и возрастных групп и тенденцию к некоторому перемешиванию,
они, разумеется, не могут рассматриваться независимо от популя-
ций. Что же касается пищевых и физиологических рас, то выделе-
ние их из состава экологических подразделений вида ничем не
оправдано.
«Социальные» группировки, как подчеркивает автор, не яв-
ляются всеобщими, так как часть видов животных ведет одиноч-
ный образ жизни. При одиночно-семейном образе жизни возни-
кают охотничьи, гнездовые и т. п. участки, владение которыми
более или менее устойчиво и поэтому явно зависимо от террито-
риальных группировок. Поселения с сохранением индивидуально-
семейных участков и сообщества стадно-стайных животных, где
определенное пространство закреплено за всей группой в целом,
также не могут рассматриваться как подразделения вида, неза-
висимые от территориальных.
Все это позволяет утверждать, что внутри вида существуют не
независимые друг от друга три рода группировок, а сложносопод-
чиненные внутривидовые группы, основой которых является про-
странственно-экологическая структура вида.
Примером классификации внутривидовых групп может слу-
жить следующее подразделение (Лукин, 1962): 1) сезонные,
2) хорологические (географические, экологические), 3) онтогене-
тические (фотопериодизм и др.), 4) биоценотические и 5) кор-
релятивные изменения. Неясны мотивы исключения групп 1, 3
и 4 из экологической внутривидовой изменчивости. Кроме того,
в этой классификации смешаны групповые и индивидуальные раз-
личия, а также стабилизированные группы, адаптивные модифи-
канты и неадаптивные вариации типа морфозов.
Для палеозоологии Д. Л. Степанов (1959а) предложил следую-
щие подразделения: подвид, морфа, форма, аберрация. Наслед-
223
ственно же закрепленными, как считает автор, являются при-
знаки только подвида, а остальные внутривидовые единицы
являются всего навсего собранием модификантов. С. С. Шварц
(1959, 1963) отрицает и наследственную обусловленность подви-
довых различий. Однако сравнительное изучение генетически
обусловленных различий между местными популяциями и даже
между вцутрипопуляционными формами показало, что колоссаль-
ная наследственная гетерогенность является всеобщим свойством
видов.
Следует сказать также и о том, что неравноценность самих
видов распространяется также и на внутривидовые группы. Так,
например, широкая модификационная пластичность каждой особи
или же способность к физиологическому гомеостазису в большой
амплитуде условий, даже и при резких различиях в местообита-
ниях, может привести к замене экотипов модификантами или же
к относительной внешней монотипиости популяций. Некоторые
считают необходимым разрабатывать особые системы внутриви-
довых единиц для видов, отличающихся типами размножения:
половое кссногамное или автогамное, апомиктическое, вегетатив-
ное (Turesson, 1957).
Можно подразделять виды, принципиально отличающиеся по
структуре, на следующие типы.
1. Сверхвид (superspecies) — комплекс рас, генетически отли-
чающихся друг от друга, как полувиды.
2. Ценовид (coenospecies) — обычный политипический вид,
система географических или экологических рас (подвидов).
3. Географический вид (geospecies) —сформированная раса,
имеющая самостоятельный эволюционный путь.
4. Экологический вид (oecospecies) — преобразованный в вид
локальный экотип (или физиологическая, пищевая, сезонная
раса).
5. Комплексный вид — гибридные циклы, включающие рои
константных, расщепляющихся, а иногда и апогамных форм.
6. Агамный вид (agamospecies) — совокупность клональных
популяций, апогамных линий или партеногенетических рас
(= расщепленный вид).
Эти типы видов не обособлены друг от друга и между ними
существуют переходы. Например, существуют виды, включающие
полувиды и подвиды и т. п.
В последние годы сложная дифференциация вида была обнару-
жена у простейших (Полянский, 1957; Догель и др., 1962). У ин-
фузорий вид включает несколько гоупп, изолированных друг
от друга в половом отношении («сиьгены», по Т. Соннеборну).
Одни протистологи выделяют каждый синген в ранг вида-, а дру-
гие рассматривают их в качестве компонентов единой популяции.
224
Дж. Клаузен.
Е. Н. Синская.
У Paramecium putrinum обнаружено три различных системы
внутривидовых групп: морфотппы, типы спаривания и сингены и
миксотипы (Янковский, 1962а, 19626). Пока же положение
со многими внутривидовыми единицами таково, что остается
неясным, является ли та или иная единица специфичной для
определенного крупного таксона, или же это всего-навсего си-
ноним.
3. АЛЛОПАТРИЧЕСКИЕ ГРУППЫ
Общая характеристика. Полувид, подвид и экотип пред-
ставляют собой наиболее крупные подразделения вида. Они со-
стоят из большего или меньшего числа родственных местных
популяций или, реже, представляют одну большую популяцию.
Это более или менее сформированные географические и экологи-
ческие расы, приспособленные к сходным климатическим, почвен-
ным или биоценотическим условиям. Поэтому-то они подчиняются
правилу викаритета (т. е. пространственно взаимоисключают друг
друга) и на территории, занятой одной расой, не может посе-
литься другая. Пространственное взаимоисключение этих единиц
объясняется селекционными преимуществами экотипа в пределах
его экоареала и подвида — в пределах его географического
ареала. Они смешиваются лишь иногда в узких пограничных зо-
нах. В этих подразделениях находит наиболее яркое и отчетли-
вое выражение тенденция к адаптивной радиации населения
вида. Такие части вида, как уже указывалось, воспроизводятся
в природе почти независимо от других таких же частей и ока-
зываются уже способными к самостоятельному существованию
и эволюционному развитию. Это формы существования вида,
почти достигшие видового уровня организации. Поэтому полное
обсуждение этих групп будет осуществлено в главе о видообра-
зовании.
Экологическая раса (экотип). Экотипом называют
популяцию или группу популяций с наследственно закреплен-
ными приспособлениями к условиям определенного местообитания
(Turesson, 1922а, 1922b, 1931; Синская, 1948). Приспособленность
к одинаковым условиям выражается в наличии у них специфиче-
ских наследуемых морфо-физиологических особенностей. Эти
адаптивные генотипически обусловленные различия создаются
в процессе стабилизирующего отбора. Солончаковый (галоморф-
ный) экотип при выращивании на незасоленой почве сохраняет
свой облик и т. п. Экотипы формируются при заселении новых,
ранее недоступных местообитаний. Так, например, часть популя-
ций горного вида растений образует альпийский (карликовый)
экотип, другая — ксероморфный экотип, приспособленный к жизни
на скалах, третья — теневой экотип, живущий под пологом леса.
У равнинного вида одна группа популяций осваивает заболочен-
15 К. М. Завадский
225
ные местообитания, другая заселяет пески и дюны, третья —
опушки лесов и лесные поляны, и т. п. На пастбищах образуются
характерные экотипы с распластанной розеткой, менее подда-
ющиеся вытаптыванию и сохраняющие свои особенности в каче-
стве стабилизированных признаков. У грызунов известен темно-
окрашенный экотип, освоивший биотопы на выходах лавы,
имеющий наследственно закрепленные особенности поведения.
Примером экотипов, специализированных к условиям особого
биоценоза, могут служить растения-сорняки, которые по ряду
признаков уподобляются основной культуре.
Как уже указывалось выше, экотипы существуют и у видов
животных. Экологическая дифференция вида на примере зяблика
была показана В. И. Аникиным (1966). Наблюдения за гнездами
и количественный учет птиц в различных типах лесонасаждений
(дубовая роща, березовая роща, сосняк травянистый, старый
смешанный лес) показали, что виды, гнездящиеся в различных
условиях, в каждой стации имеют определенную, отличную
от других стаций гнездовую плотность, которая является одним
из критериев микропопуляции, населяющей данную стацию. Дру-
гой особенностью является характер освоения территории: неоди-
наковое распределение гнезд, различные способы расположения
и прикрепления гнезд на деревьях. Кроме того, и материал, из ко-
торого строятся гнезда, оказался специфичным.
Дифференциация вида на экотипы приводит к более полному
использованию территории, так как осваиваются новые, ранее
недоступные местообитания (стации). Она в то же время свиде-
тельствует о значительном развитии процесса адаптивной радиа-
ции (дивергенции). При этом географический ареал вида может
и не изменяться, но в его пределах на основе образования новых
локальных экотипов углубляется дифференциация вида и соответ-
ственно возрастает «насыщение» территории населением данного
вида. Этот процесс более полного использования видом территории
путем увеличения числа занимаемых им экологических ниш носит
название расширения экоареала вида.
У многих видов, географическое распространение которых
ограничено физическими барьерами (горные хребты, водные про-
странства и пр.), дифференциация происходит только указанным
путем. Однако адаптивная радиация популяций вида может осу-
ществляться и благодаря расселению в новые географические
области, т. е. путем освоения новых местообитаний за пределами
границ старого географического ареала. Так происходит формиро-
вание географических pad (подвидов).
Географическая раса (подвид). Как считает Майр,
большинство видов у животных имеет географические расы и при-
мерно у трети видов «эти расы отличаются достаточно, чтобы
рассматриваться как хорошие подвиды. Некоторые весьма распро-
страненные виды птиц и млекопитающих состоят из 20—30 и даже
226
в исключительных случаях из 50 хорошо очерченных рас» (Мауг,
1965:126—127). Подвид определяют как «ансамбль популяций,
имеющий сходную генетическую основу и живущий в определен-
ном географическом ареале. Географические расы нельзя отож-
дествлять с экологическими расами, так как последние обычно
географически нс обособлены и часто несколько экорас живет
в одном географическом районе, например расы степных и лесных
муравьев и т. п.» (Rensch, 1965 137).1 Практически подвиды
чаще всего выделяются на основании морфологических различий,
выраженных географически. Маркирующий признак часто оказы-
вается связанным с целым комплексом изменений. Например,
у американской норки (Mustela vison) известно 14 подвидов и
30 форм, различающихся по окраске. Как показал К. Я. Фатеев
(1966а), различия по окраске связаны с целым комплексом
морфо-физиологических особенностей: весом и длиной тела, об-
хватом груди, длиной кишечника, весом внутренних органов и т. д.
Такие примеры могут служить хорошими моделями для понима-
ния процесса дифференциации видов в природе. Окраска часто
служит лишь опознавательным знаком, а в действительности отбор
шел на адаптивное изменение внутренних органов.
На примере обыкновенной сосны, имеющей обширный ареал,
Л. Ф. Правдин (1965) показал, что формирование подвидов (лап-
ландского, кулундинского, крючковатого, сибирского и обыкновен-
ного) связано с историей расселения вида и с изоляцией отдель-
ных очагов в ледниковый период. Не совсем ясно, почему этот
автор считает, что «объем понятия „географическая раса“ более
обширен, чем понятие „климатическая раса“ или „климатический
экотип“» (там же : 74).
Мигрирующие популяции могут занимать в новых областях
те же типы местообитаний, которые были ими освоены в прошлом.
В этом случае расширяется географический ареал, а экоареал
нового подвида сохраняется таким же, как и у исходных популя-
ций. Так, например, яровой рыжик Camelina sativa образовал,
согласно Е. Н. Синской (1948), три географических подвида:
С. linicola, засоряющий посевы льна па северо-западе СССР,
С. sublinicola, засоряющий лен в более южных областях нечерно-
земной зоны, и С. glabrata — в районах черноземных степей.
Подвид linicola отличается раннеспелостью, крупноплодностью,
крупносемянностью и более гидроморфной конституцией. К югу
расселены переходные популяции от типа linicola к типу sublini-
cola и далее типичные популяции подвида sublinicola, среднеран-
1 По В. Г. Гептнеру, «географическая раса — действительно низкая
систематическая категория, однако ее не следует смешивать с низшей
систематической единицей, которая есть вид» (цит. по: Кэйн, 1958:86—87).
Такие высказывания являются примером сведения теории вопроса к но-
менклатурным изысканиям.
15*
227
ние, с плодами и семенами меньшего размера, чем у linicola.
Еще далее на юг встречаются популяции переходные от типа
sublinicola к типу glabrata и, наконец, типичные популяции под-
вида glabrata (самые мелкие семена и наименьшая гидроморф-
ность конституции растения). Весь ряд популяций ярового ры-
жика образует как бы непрерывную цепь форм с тремя большими
узлами (подвидами), и при этом все популяции подвидов linicola
и sublinicola представляют «экотип уподобления», специализиро-
ванный к посевам льна-долгунца, а южный подвид (glabrata) об-
разует менее специализированный экотип, засоряющий не только
посевы масличного льна, но и других культур (овес, ячмень).
Таким образом, если в процессе расселения вид занимает в но-
вой области такие же экологические ниши, как и раньше, но на-
следственно изменяется в сторону приспособления к новой геогра-
фической среде, формируется однотипная группа популяций или
географическая раса (подвид). Таковы, например, географические
расы ели обыкновенной (Picea excelsa): европейская, сибирская,
финская и другие или липы Tilia cordata: европейская, сибирская,
амурская и т. п.
В процессе расселения в новые области ареал может стать
очень большим, крайние популяции приобретают большое своеоб-
разие и единство вида утрачивается, даже если существуют не-
прерывные ряды переходных популяций. Примером такой далеко
зашедшей адаптивной радиации, связанной с образованием геог-
рафических рас, может служить, по П. В. Серебровскому (1941),
камчатский светлый пестрый дятел (Leuconotopicus immaculatus)
и темный мелкий дятел из северного Ирана (L. quadrifasciatus).
Эти расы сами по себе столь различны, что должны были бы рас-
сматриваться как«хорошие» виды, но существование ряда пере-
ходных популяций заставляет считать их еще подвидами.
В процессе образования повой географической расы (подвида)
популяции могут приспосабливаться не только к новым географи-
ческим особенностям среды, но и к новым типам местообитаний,
образуя новые локальные экотипы. У многих видов растений и
насекомых обнаружено, что все местные популяции одной области
занимают определенные местообитания и одно из них оказывается
самым оптимальным, а в другой области оптимальным становится
уже иное местообитание и т. д. (правило смены стаций вида).
Разделение рас на географические и экологические, разу-
меется, не является безусловным. Во-первых, различное положе-
ние в пространстве всегда связано с какими-то различиями
в условиях существования, так что географическая раса в извест-
ном отношении является и экологической. Во-вторых, особые
местообитания, занятые многими экологическими расами, могут
быть более или менее выражены пространственно, так что обра-
зуется ряд переходов от пеогеографических различий к микро-
географическим и т. д.
228
Необходимо указать на существующую неупорядоченность
в терминологии: экологи называют типичные географические
подвиды геоэкотипами, климатипами или региональными экоти-
пами, биогеографы же в типичном локальном экотипе склонны
видеть микрогеографический подвид. В. В. Алпатов (1924) пред-
лагал ввести три категории соподчиненных подвидов: географи-
ческие, экологические и сезонные (subspecies temporalis).
Насчитывают девять типов географических рас, неравноцен-
ных друг другу (Rensch, 1965). Мы бы выделили следующие
типы: равнинные, стабилизированные и генетически обедненные;
горные, эндемичные, генетически очень сложные; островные,
возникшие от одного или немногих «основателей»; полиплоидные
гетеросексуальные пли апогампые; отличающиеся по неадаптив-
ным признакам, возникшие па основе генетического дрейфа.
Дифференциация вида па экологические п географические
расы представляет собой следствие адаптивной радиации популя-
ций. В результате единство вида нарушается. Ранее монолитный
вид распадается на части. Таким образом, формирование эколо-
гических рас, подвидов и полувидов есть, С одной стороны, способ
приспособления данного вида к новым типам местообитаний и
к новым географическим областям, но, с другой, свидетельствует
о далеко зашедших центробежных тенденциях, ведущих к образо-
ванию и обособлению новых видов.
О реальности подвидов и экотипов. До недавнего
времени никто, кроме монотипистов — «дробителей», не отрицал
существование подвидов и экотипов. Однако и «дробители», требуя
ликвидации всех внутривидовых единиц, не отрицали реального
существования географических и экологических рас, а повышали
их таксономический ранг, называя каждую расу видом. Судя по
данным систематики высших растений («Флора СССР»), млеко-
питающих (Шварц, 1959, 1963), птиц (Кэйн, 1958; Мауг, 1963)
и некоторых отрядов насекомых, до 75% видов включают геогра-
фические подвиды. Можно предположить, что их не имеют только
древние реликты, а также молодые, узкоспециализированные п
эндемичные виды.
У видов, недавно расширивших ареал, как это было показано
па жужелице Diskoptera komarovi (Арнольди, 1939), географиче-
ская изменчивость носит непрерывный характер и дифференциа-
ция на экотипы и подвиды еще не обозначилась.
Поводом для отрицания реальности экотипов п подвидов
явились результаты исследований непрерывной (клинальной)
изменчивости признаков, согласно которым границы между ра-
сами создаются не в природе, а в кабинетах ученых, работающих
с малыми выборками. К отрицанию реальности экотипов впервые
пришел Д. Грегор (Gregor, 1939, 1944), установивший непрерыв-
ный характер изменения некоторых признаков подорожников
(Plantago maritima, Р. lanceolata и др.) на территории Англии.
229
Правда, в последней работе (Gregor a. Watson, 1961) этот автор
уже пе отвергает реальности всех экотипов и утверждает только,
что «экотипические разрывы более редки, чем представлялось
ранее».
Существует и тенденция отрицать реальное существование
подвидов. Возникла она в качестве реакции на формальное
использование этой таксономической категории (морфологические
«подвиды», подвид как разновидность и т. и.). П. В. Терентьев
(1957, 1966) считает, что подвиды были важны в прошлом, когда
систематики работали с малыми выборками, а при высокой
изученности вида разбивка его на подвиды теряет смысл. При-
водятся данные по озерной лягушке (Вана ridibunda), из которых
следует, что ранее произведенная автором разбивка этого вида на
северный (длинноногий) и южный (коротконогий) подвиды была
ошибочной. Вычислив «индекс длинноногости» для многих пунктов
сбора, Терентьев обнаружил непрерывное его изменение с севера
на юг, и границы подвидов оказались стертыми. Но стертыми
оказались только границы между подвидами, по не сами подвиды.
Кроме того, нельзя основываться па градуальном характере изме-
нений одного признака. Необходимо судить только по комплексу
(по корреляционным системам). Изменчивость, непрерывная по
одним признакам, оказывается прерывистой по другим. Кроме
того, истинная высокая изученность вида связана не столько
с большим числом описанных экземпляров и признаков, сколько
с применением комплексных экспериментальных методов иссле-
дования. Если бы при скрещивании северной расы лягушек
с южной можно было проследить наследование «индекса корот-
коногости» и других признаков, это дало бы большие основания
для суждений о структуре этого вида, чем детальное вычисление
геоклипа по одному признаку.
Судя по представленным материалам, географическая измен-
чивость некоторых признаков ушастой круглоголовки также имеет
непрерывный характер (Вельдре, 1964) и не дает возможности
проводить границы подвидов (автор пишет о невозможности «вы-
деления» подвидов). Возможно, что у озерной лягушки и уша-
стой круглоголовки не существует географических рас и их вы-
деление было ошибочно. Однако П. В. Терентьев (1966) делает
широкое обобщение, утверждая, что система низших таксономи-
ческих единиц утрачивает свое значение, заменяясь одной «кли-
нальной изменчивостью».
Отрицание реальности экотипов и подвидов уже критиковалось
рядом авторов (Синская, 1948, 1963; Clausen, 1951, 1958; Завад-
ский, 1957а, 1961, 1967а, 19676; Шварц, 1963; Даревский, 1967,
и др.). Было показано, что существуют резкие сдвиги в самих
клипах (step-cline), что наличие непрерывных изменений по
одному или нескольким признакам не исключает прерывистого
характера различий по другим признакам. Отличия между расами
230
должны устанавливаться на основе изучения целых корреляцион-
ных систем. Утверждение, что с ростом изученности группы все
хиатусы исчезают, оказавшееся ошибочным для видов, является
таким же и для внутривидовых подразделений. Например, изуче-
ние изменчивости грызуна Peromyscus difficilis показало, что
клины иногда не только прерываются, но и меняется само направ-
ление градиента (Hoffmeister a. de la Torre, 1961). Этот пример
принципиален, так как Р. difficilis изучен весьма полно и именно
на таком объекте следовало бы наблюдать исчезновение подвидов.
В действительности же прерывистость клипов позволяет не только
выделять подвиды, но и довольно четко очерчивать границы их
ареалов. Однако не следует обобщать и этот случай. Между неко-
торыми расами граница может и отсутствовать. «При наличии
непрерывного ареала подвиды почти всегда переходят один в дру-
гой незаметно» (Майр, 1947 173). Ныне этот автор пишет, что
географически не обособленные расы «не выражают ни биологиче-
ской, ни эволюционной концепции, являясь просто удобными для
таксономии понятиями» (Мауг, 1965 129). Правильнее говорить,
что иногда расы выделяются недостаточно обосновано и что су-
ществуют расы с расплывчатыми границами. Выпадением проме-
жуточных звеньев подвиды не создаются, а лишь более резко
очерчиваются их границы, например, кавказское и беловежское
стада зубров являются подвидами не потому, что они разделены
огромной, не заселенной зубрами территорией. Даже при наличии
всех переходных популяций подвидовая степень обособления край-
них звеньев ряда осталась бы реальной, так как связана в разной
степени значительной морфологической самобытностью (Толма-
чев, 1962). По Б. Реншу (Rensch, 1954, 1965>, обособление рас
может быть одинаковым по размеру как при большом простран-
ственном разъединении, так и при наличии ряда переходных по-
пуляций. Наличие непрерывного ряда вовсе не упраздняет разли-
чий между далеко отстоящими друг от друга звеньями. Если на
одном краю «клина» популяции имеют белую окраску, на другом
темную, то существование градиентных переходов окраски не
может опровергнуть реального существования популяций с белой
и с темной окраской. У широко расселенных видов с географиче-
ски примыкающими друг к другу подвидами вблизи границ ареа-
лов подвидов появляется все больше переходных особей. У крас-
ной полевки (Clethrionomys rutitus) появление таких переходных
между подвидами особей объясняется действием «переход-
ных» условий (Большаков, 1963), но вполне возможно, что при-
чиной этого явления служит постоянная гибридизация. Задача,
таким образом, сводится лишь к изучению природы переходных
популяций.
Степень обособленности рас необходимо изучать методом скре-
щиваний между различными популяциями, относящимися к од-
ной и той же расе и к предполагаемым разным расам. Прп этом
231
учитываются не только степень скрещиваемости и плодовитости
потомства, но п характер разнообразия потомства (особенно по
признакам, представляющим таксономический интерес). Так
именно поступил Дж. Клаузен, изучая расовый состав видового
комплекса Viola tricolor, так изучался расовый состав видов
Anthyrrinum и т. п. Если бы при скрещивании северных длинно-
ногих лягушек с южными коротконогими в Fi оказались проме-
жуточные формы, а в F2 выявились бы родительские признаки,
наличие подвидов можно было бы легко доказать, так как не-
прерывная изменчивость объяснялась бы межрасовой гибридиза-
цией и гибридностыо переходных форм.
С. С. Шварц (1963) приводит данные в пользу того, что кли-
вальная изменчивость и обособление подвидов — это стороны од-
ного процесса географо-экологической изменчивости. Таким обра-
зом, изучение кливальной изменчивости должно не упразднить
понятие подвида, а напротив, обогатить его. Хорошо выраженные
подвиды недавно описал Н. И. Калабухов (1964) у песчанки
полуденной (Meriones meridiamis). Этот широко расселенный вид
включает несколько хорошо морфологически отличимых подвидов.
Оказалось, что зверьки из нредкавказского подвида почти вдвое
крупнее заволжско-уральских и интенсивность теплообразования
и потребления кислорода у них ниже, они менее теплолюбивы.
Различными у этих форм оказались толщина кожи, содержание
гемоглобина в крови, содержание аскорбиновой кислоты и токо-
ферола, чувствительность к чумному микробу. Несмотря на не-
большую ширину разделяющей пх поймы Волги, каждая из попу-
ляций обладает целым комплексом специфических морфо-физиоло-
гических особенностей.
Справедливо, что «концепция подвида отражает объективные
природные закономерности и при правильном употреблении со-
храняет большое общебиологическое значение» (Шварц, 1963
418). Надо, конечно, иметь в виду, что принцип неравноценности
распространяется и на подвиды и экотипы: они обладают различ-
ной эволюционной перспективностью, генетико-морфофизиологи-
ческой самостоятельностью и дискретностью. Нельзя определять
подвид как реакцию вида только на современные условия и умал-
чивать о его исторической и генетической обусловленности.
4. МЕСТНАЯ ПОПУЛЯЦИЯ
Определение понятия. В генетике со времен В. Иоган-
сена иногда употребляется определение популяции как смеси
форм в противоположность генетически однородным множествам
вроде клонов, гомозиготных линий и т. п. (Haldane, 1932, 1959;
Li, 1955, и др.). А. Мюнтцинг (1963 76—82) характеризует по-
пуляцию как совокупность биотипов. Однако и в генетике попу-
ляций начал утверждаться экологический подход (Дубинин, 1966),
232
и содержание термина «популяция» постепенно изменилось (Ап-
drewartha, 1961; Ford, 1963; Grant, 1963). В экологии уже давно
(Allee, 1931; Elton, 1933) популяция толковалась как часть на-
селения вида, занимающая определенную территорию. Для изу-
чения населения видов — демографии, требовалось установить
единицы населения: популяция означает теперь относительно обо-
собленную единицу населения вида. Многие авторы стали назы-
вать ее локальной, или местной популяцией. Для экологии и де-
мографии местная популяция — это единица населения всех видов
независимо от способа их размножения и других показателей, но
некоторые генетики и систематики противопоставляют «популя-
ционные виды», обладающие половым размножением, — «клональ-
ным видам» (Wright, 1951: Dobzhansky, 1955, 1958, и др.).
По А. Л. Тахтаджяпу (1955), популяция возникает только с по-
явлением полового размножения. Поэтому «амфимпктичсский
вид» он называет «популяционным» и противопоставляет его «кло-
нальному виду». Правильнее точка зрения С. А. Северцова (1951 :
44—45), согласно которой популяция — это любой природный
комплекс организмов данного впдд, населяющий определенную
местность, независимо от того, половым или бесполым способом
происходит размножение. «Популяционная жизнь» понимается
пами широко, как проявление любых внутривидовых отношений,
имеющих адаптивное значение, а не только как процессы скрещи-
ваний. С появлением полового размножения возрастает степень
целостности популяций, а не возникает сама популяция. Экологи
часто определяют популяцию как «совокупность особей одного
вида, населяющих определенную территорию» (Allee et al.,
1949 265). Так же определяют популяцию Ч. Элтон (1960), ав-
торы отечественных руководств по экологии растений и живот-
ных (Шенников, Наумов) и др. В разделе о целостности вида
пами подчеркивалась ошибочность суммативного понимания по-
пуляции.
При определении понятия популяции правильнее обращать
внимание на взаимодействия особей между собой и на известную
обособленность территории (Арнольди, 1957). В дополнение к ука-
занным выше признакам популяции существенными являются
еще устойчивость во времени и способность воспроизводиться
(Завадский, 19576, 1961; Беклемишев, 1960). Некоторые авторы
называют местную популяцию поселением, естественной единицей
экологической структуры вида, образуемой из-за неравномерного,
«пятнистого» расселения вида по ареалу (Завадский, 1952, 19576,
1961; Гиляров, 1954, 1959).
Виды живут популяциями. Иногда вид состоит всего из одной
местной популяции. Многие же виды включают сотни и тысячи
местных популяций. Одни местные популяции более обособлены,
устойчивы и более способны к самовоспроизведению и эволюции,
другие менее, но в отличие от случайных скоплений организмов
233
все они обладают известной степенью организованности, устойчи-
вости и способности к эволюции.
Таким образом, местную популяцию можно определить как
относительно обособленное поселение, способное устойчиво, дли-
тельное время воспроизводиться в пределах освоенной террито-
рии и эволюировать. Местная популяция является основной есте-
ственной единицей существования, приспособления, воспроизве-
дения и эволюции населения вида, т. е. основной демографической
единицей. Именно в рамках местной популяции с наибольшей си-
лой действуют как интегрирующие факторы, так и внутривидовые
противоречия. Местную амфимиктическую популяцию было предло-
жено именовать дёмом (Gilmour a. Gregor, 1939; Gilmour, 1960:
Gregor a. Watson, 1961), но пока этот термин не получил широ-
кого применения. А. А. Корчагин (1964, 1966) предложил термин
«ценопопуляция», подчеркивающий связь местных популяций
с биоценозами. Он должен быть принят только для популяций,
всегда приуроченных к одному биоценозу. В последнее время по-
пуляционный подход проникает и в работы систематиков. На по-
пуляциях кизила (Comus mas) было показано, что размах измен-
чивости большинства признаков значительно превышает то, что
известно в литературе. Типы, получившие в систематике наимено-
вание formae, представляют собой лишь выхваченные из непре-
рывных рядов изменчивости более или менее случайные звенья
(Сагащи et al., 1960). Эти авторы приходят к выводу, что в ос-
нову микросистематических исследований должна быть положена
популяция как целое, а не отдельные особи и, следовательно, мор-
фолого-типологический метод, еще господствующий в систематике,
«не соответствует совремейному пониманию вида» (там же 526).
С другой стороны, существуют виды, у которых характер варьиро-
вания признаков многих местных популяций бывает столь одно-
образным, что не дает основания для подразделения вида на бо-
лее мелкие единицы. Характер изменчивости многих признаков
у Melampyrum pratense из Финляндии в 41 изученной популяции
оказался столь одинаковым, что, по мнению авторов (Jalas, Rikki-
nen, 1962), этот вид не может быть подразделен.
О размерах и границах местной популяции.
Население вида распределяется по территории, как правило, бо-
лее или менее неравномерно, так как между отдельными поселе-
ниями лежат пространства, непригодные для данного вида пли же
занятые более сильными конкурентами. В зависимости от особен-
ностей организмов и характера территории размеры таких посе-
лений могут быть самыми разнообразными как по численности
особей, так и по величине занимаемой территории. Если пригод-
ные для существования и доступные территории вкраплены не-
большими пятнами в массивы непригодных или недоступных тер-
риторий (например, солончаки, такыры, «меловые» почвы и т. п.),
прерывистость поселений бывает выражена резко, если же поселе-
234
ййя занимают большие территории и примыкают одно к другому
(например, виды лесов таежной зоны), пространственные границы
поселений становятся расплывчатыми или вовсе исчезают.
Не следует думать, что существует прямая зависимость между
размерами географического ареала и числом местных популяций.
Так, например, наземные улитки Achatinella на маленьком
острове Оаху (Гавайи) образуют сотни мелких местных популя-
ций, совершенно изолированных друг от друга полосами сухих
местообитаний, с открытой солнечным лучам почвой. С другой
стороны, у некоторых широко расселенных видов каждая местная
популяция занимает обширную территорию и число таких попу-
ляций может быть небольшим. Существуют популяции громадного
размера (суперпопуляции — Беклемишев, I960), как у дождевого
червя Dendrobaena octaedra из хвойных лесов Предуралья, где им
населены почти все типы биоценозов и особи легко мигрируют
из одного в другой. Но в пределах сунерпопуляции есть районы
с редким или с густым населением. У других видов население рас-
пределено по многим отдельным популяциям, разграниченным
более или менее четко в зависимости от степени удаленности их
друг от друга и способностей к перемещению.
Размеры и границы популяции могут резко меняться во время
пульсаций численности. В годы массового размножения возникают
гигантские популяции, как это наблюдается у многих видов мы-
шевидных 1рызунов, насекомых-вредителей, некоторых пушных
зверей, сорно-полевых растений, патогенных микроорганизмов.
Снижение численности сопровождается раздроблением такой
сверхпопуляции на многие островки обособленных мелких попу-
ляций, сохраняющихся в особо благоприятных биотопах — очагах
(убежищах). У таких видов суперпопуляция и микропопуляция —
это фазы, постоянно сменяющие друг друга и создающие своеоб-
разные условия для жизни и эволюции вида. У видов растений и
животных с большой продолжительностью жизни, крупными раз-
мерами особи и относительно малой плодовитостью периодиче-
ские колебания численности не имеют такого большого размаха,
а структура популяции более устойчива. У малочисленных видов,
характерных для древесных пород влажных тропических лесов,
тропических бабочек, некоторых крупных хищников (птиц и мле-
копитающих) и т. п., размеры и границы популяций еще более
стабильны.
На границах ареала размеры местных популяций отличаются
от размеров популяций из центральных областей расселения вида.
Пограничные популяции часто бывают мелкими, малоустойчивыми
и обедненными по составу. Через 12 лет после выпуска 14 бобров
в Лапландском заповеднике численность популяции возросла
в 7—9 раз, а затем снизилась и через 25 лет была в 5—6 раз
больше, чем при выпуске. Выяснилось, что лимитирующим факто-
ром является зимняя кормовая база: плотность популяции не мо-
235
Жет сильно возрастать, так как древесная растительность па се-
вере возобновляется медленно (Герман, 1960).
Чрезвычайно интересны сопоставления пограничных популя-
ций при расширении и при сокращении ареала. Наступление леса
на тундру, заселение новых территорий пришельцами из других
стран осуществляется множеством авангардных популяций, часть
которых вымирает, а остальные сливаются в большие популяции,
прочно заселяющие территории, и па новой границе опять воз-
никают мелкие авангардные популяции, и т. д. Впрочем, при
очень быстрых «завоеваниях» новых территорий расселение но-
сит характер вспышки, так что пе только кролики, колорадский
жук и фитофтора, но и медленно размножающийся кактус Opun-
tia расселялся в Австралии и на Гавайях «со скоростью пожара» и
был назван «пожирателем пастбищ». В случаях таких быстрых
расселений вида исчезает часть основных характеристик местной
популяции (обособленность и устойчивость в пределах определен-
ной территории и т. п.). Однако обычно после таких экспансий
вида наступает период известного равновесия, сопровождающийся
формированием местных популяций. По способности воспроизво-
дить себя в природной обстановке вид однотипен местной популя-
ции. Настоящая местная популяция — это образование, достигшее
нижней границы видовой формы организации живого. Однако
далеко не все местные популяции оказываются способными пере-
ступить эту границу.
Большинство видов состоит из многих местных популяций и
поэтому встает вопрос о верхней границе вида. У групп с поло-
вым размножением она будет проходить между такими популя-
циями, которые совершенно пе скрещиваются друг с другом, т. е.
там, где существует полная фактическая изоляция.
Для характеристики вида важное, а иногда и решающее зна-
чение имеют данные о точных границах местной популяции,
а также границы, за которыми прекращаются всякие скрещивания
между разными популяциями. До сих пор, однако, пе накоплено
еще материала, достаточного как для определения степени половой
изоляции одной местной популяции от другой, так и для установ-
ления закономерностей перемешивания населения видов в рамках
целого ареала. Вопрос о границах местной популяции является
при этом основным.
При определении границ местных популяций необходимо учи-
тывать степень оседлости особи или семьи, площадь питания
особи, пространство, доступное для пассивного пли активного
расселения одного поколения, или область, в которой может осу-
ществляться возможность размножения особей одной популяции
(радиус активности особи и популяции), и другие показатели, ко-
торыми пользуются экологи растений и животных.
В понятие радиуса активности как частные входят такие по-
нятия, как «охотничий участок», «выводковый участок», «участок
236
размножения». У растений радиус активности индивида равен рас-
стоянию, на которое может распространиться корневая система и
надземная часть, а также расстоянию, на котором могут действо-
вать биологически активные вещества, выделяемые этой особью
(корневые выделения и фитонциды). Однако если рассматривать
видовые функции особи, т. е. участие ее в воспроизведении вида,
то ее радиус активности может оказаться большим и менее опре-
деленным. Радиус активности видовых функций особи у растений
складывается из расстояний, на которые разносятся пыльца, се-
мена, споры или вегетативные части, служащие для размножения.
При пассивном разносе пыльцы и семян ветром, животными или
водой очевидно, что величина радиуса активности может быть
очень изменчива. Для большинства видов еще нет точных данных
о радиусе активности средней особи и популяции. Изучение гра-
ниц местных популяций растений и изучение закономерностей рас-
селения потомства (т. е. разлета пыльцы, семян и плодов) пред-
ставляет собой трудную задачу. Есть отдельные данные о заносе
пыльцы уральских сосен на сотни километров к северу, а семян —
водой, ветром и животными — даже на тысячи километров, но зна-
чение таких миграций неясно.
У животных малоподвижных, таких как наземные улитки, на-
блюдается высокая степь оседлости особи. Однако и у весьма мо-
бильных форм, таких как птицы, степень оседлости может быть
все же высокой (например, благодаря инстинктам «гнездового кон-
серватизма»), что сильно ограничивает возможности их к пере-
мещению, заложенные в самой организации.
Нельзя, впрочем, переоценивать эту роль «гнездового консер-
ватизма». Так, например, считалось, что воробьиные весьма оседлы
и размеры их местных популяций малы, однако было показано,
что происходит перемешивание населения птиц на больших пло-
щадях (Мальчевский, 1959). Кольцевание птиц у ряда видов
воробьиных (большая синица, горихвостка и др.) показало, что
молодняк (первогодки) после зимовки редко возвращается на
родину, в то время как взрослые птицы, хотя бы однажды гнез-
дившиеся в данном микрорайоне, как правило, в него же и воз-
вращаются. Потомство из данного микрорайона закономерно рас-
селяется по другим микрорайонам. Поэтому население данной
местности ежегодно обновляется за счет прилета «чужих» перво-
годков, но имеет и «оседлое ядро», состоящее преимущественно
из взрослых птиц. Такое перемешивание населения обеспечивает
поддержание неродственных скрещиваний и широкий гаметный
обмен на больших территориях. Вместе с тем часть популяции
является оседлой, что гарантирует виду использование уже
освоенных и проверенных местообитаний. У изученных видов
воробьиных размеры местной популяции оказались значительно
большими, чем ожидалось, а границы ее — довольно неопределен-
ными.
237
Г. Кэрстер определил с помощью маркировки средние размеры
локальной популяции у игуановой ящерицы Scelopodus olivaceus
(Kerster, 1964). Оказалось, что площадь, в пределах которой осу-
ществляются неограниченные скрещивания (панмиксия), равна
10 га, а число особей — 225—270.
Способность многих животных к выбору наиболее подходящего
местообитания приводит к значительному снижению наблюдаемых
передвижений по сравнению с потенциально возможными. Наиме-
нее оседлыми оказываются не только крупные активно кочующие
виды вроде китообразных, но и микроорганизмы и другие формы,
распространение которых осуществляется пассивно, воздухом или
водой. Споры, цисты или активные фазы таких форм могут
разноситься на сотни и тысячи километров. Например, среди тихо-
ходок и простейших многие виды являются весьма широко рассе-
ленными (экологически же большинство из них строго специали-
зировано). Многие виды планктонных форм океана, распростра-
няющиеся морскими течениями, принадлежат к полукосмополитам.
Таким образом, у видов, воспроизводящихся половым путем,
в зависимости от степени оседлости особи и радиуса ее актив-
ности (или способностей расселительных стадий жизненного
цикла) и особенностей заселяемой территории местные популяции
могут иметь самые различные размеры и степени изолированности
от скрещений с другими популяциями. В природе существуют все
переходы от видов, представленных одной панмиктической популя-
цией, до видов, раздробленных на много совершенно изолирован-
ных популяций.
Типы местных популяций. Популяции, размножаю-
щиеся половым путем, в отличие от популяций с агамным и апо-
гамным размножением (клональных популяций) образуют особый
тип.
При половом размножении наименьшей из возможных единиц,
способных к самовоспроизведению, является при раздельнополости
пара особей, а при обоеполости — одна особь. В природной же
обстановке элементарная размножающаяся единица, как относи-
тельно самостоятельно существующее образование, бывает пред-
ставлена не изолированными парами или отдельными особями,
а более сложной системой. Такой размножающейся единицей
часто оказывается даже не семья или стадо, а именно целая мест-
ная популяция. Она включает много родственных, но генотипи-
чески различных особей, между которыми постоянно происходят
скрещивания. В пределах местной популяции могут осуществ-
ляться избирательные или свободные скрещивания (панмиксия).
Вследствие панмиксии и постоянного обмена гаметами популя-
ция приходит к такому состоянию, что в каждой особи может
сосредоточиться большинство генетических возможностей всей
популяции. Постоянное поддержание гетерозиготности обеспечи-
вает более широкую экологическую пластичность и гетерозисное
238
состояние особей и, кроме того, является важным аппаратом,
страхующим панмиктическую популяцию от обеднения наследст-
венной основы во время периодических колебаний численности.
Вот почему выделение из популяций отдельных пар и размноже-
ние изолированных семей «внутри себя» ведет к понижению жиз-
неспособности потомства и к потере экологической пластичности.
Разложение популяции на отдельные линии, как правило, ведет
к снижению конкурентоспособности. Такие линии обычно не мо-
гут самостоятельно существовать в природе (или существуют
только в особо благоприятных условиях).
Итак, при половом размножении элементарной самовоспроиз-
водящейся и относительно самостоятельно существующей в при-
роде единицей служит не изолированная от популяции семья,
а местная популяция как целое.
В клональных популяциях между особями, клонами и линиями
существуют разнообразные химические взаимодействия, контакт-
ные или действующие через среду (Грюммер, 1957, и др.). Ими
осуществляется регулирование скорости размножения и плодови-
тости, роста, продолжительности жизни и других важнейших по-
казателей. Таким образом, отсутствие у агамных и апогамных попу-
ляций половой интеграции не означает того, что они являются
механическими смесями особей и клонов. Однако при бесполом
размножении самовоспроизводящейся единицей является особь.
Клон представляет собой множество наиболее близкородственных
особей, имеющих сходный генотип, но не является самовоспроизво-
дящейся единицей. Коренное отличие клональной популяции от
панмиктической заключается в том, что клон — сумма самовоспро-
изводящихся единиц — особей, а панмиктическая популяция как
целое сама является самовоспроизводящейся единицей. Именно
это различие и является причиной специфических трудностей, воз-
никающих при определении границ у агамных видов. Одна тен-
денция состоит в отождествлении вида с каждой пространствен-
но обособленной колонией различимой морфологически, как это
наблюдается у ирисов (Anderson, 1936), манжеток и ястребинок
(Stebbins, 1941) и др. Другая — в объединении таких колоний —
микропопуляций — в группы большего объема. Еще сложнее раз-
бираться среди апогамных линий — «видов», когда на одной лу-
жайке бок б бок в одной микропопуляции произрастают десятки
клонов, каждый из которых вполне константен и воспроизводится
независимо от других.
Руководствуясь чисто пространственным критерием, В. Н. Бе-
клемишев (1960) предлагает следующие три типа популяций:
1) вид имеет небольшой ареал, содержащий одну популяцию не-
больших размеров (горные долины, острова и т. п.), 2) ареал
вида обширен, сложно расчленен и занят множеством простран-
ственно разделенных друг от друга популяций и 3) ареал вида
обширен, но по сравнению с возможностями особей однороден и
289
занят одной единственной громадной популяцией — суперпопуля-
цией. Между этими типами существуют все переходы.
В пределах суперпопуляции население бывает то густым, то
редким и, хотя здесь нет пространственной дискретности, районы
с густым населением следует рассматривать как субпопуляции.
Многие виды растений и животных, населяющих равнинные
страны, живут суперпопуляциями (например, на огромных прост-
ранствах европейско-сибирской елово-пихтовой тайги до ее вы-
рубки и распашки). Суперпопуляция населяет столь большую
территорию, что самые ее размеры препятствуют взаимодействиям
удаленных друг от друга ее частей. Например, размножение поле-
вок в Башкирии не сказывается в районе Пскова, так как его
влияние, распространяясь с декрементом, постепенно затухает.
Суперпопуляция едина только топографически, но функцио-
нально она расчленена; пространственное расчленение само по
себе недостаточно для суждения о структуре популяции. С функ-
циональной точки зрения суперпопуляция является комплексом
популяций.
Очень важно подразделение популяций на типы по способности
самовоспроизведения, измеряемой величиной притока особей
извне. Существует шесть таких типов: независимые популяции.
способные самостоятельно самовоспроизводиться, приток особей
извне для них несуществен; полузависимые, способные само-
воспроизводиться самостоятельно, но приток особей извне значи-
тельно повышает их численность; зависимые, собственный при-
плод не покрывает смертности и популяция без притока извне
вымирает; псевдопопуляции, вовсе не способны к воспроизве-
дению и целиком основаны на притоке особей извне; временные
или периодически возникающие популяции, образуются за счет
проникновения особей из постоянных поселений в малоблагопри-
ятные биотопы (например, на границе географического или эко-
ареала, а также при выходе из «убежищ» на биотопы с нестабиль-
ным режимом); гемипопуляции, или поселения отдельных фаз
жизненного цикла гетеротопных животных (например, популяции
всех донных морских животных, имеющие планктонические
личинки, слагаются из донных и пелагических гемипопуляций).
Взрослые особи хищной улитки Nucella lapillus живут на камени-
стой литорали, а молодь входит в состав сублиторальных биоцено-
зов Microporella ciliata. Предложения В. Н. Беклемишева важны
для классификации популяций, этого еще очень молодого направ-
ления исследований.
5. СИМПАТРИЧЕСКИЕ ЕДИНИЦЫ
Понятие о симпатричности. Перейдем к рассмотрению
внутривидовых единиц, являющихся компонентами местной попу-
ляции. В отличие от крупных подразделений вида внутрипопуля-
240
Э. Майр.
ционные единицы в общем обладают меньшей способностью
к самостоятельному существованию и воспроизведению в природ-
ной обстановке. Это утверждение относится только к амфимикти-
ческим популяциям, так как клональные популяции включают
менее прочно взаимосвязанные формы.
В противоположность подвидам и экотипам, воспроизводя-
щимся в природе почти или полностью «внутри себя», внутрипопу-
ляционные формы постоянно скрещиваются пли взаимодействуют
между собой иными способами и составляют единый, совместно
размножающийся комплекс. Внутрипопуляционные формы про-
странственно не исключают друг друга, как аллопатрические, и не
обособлены, как местные популяции, а живут бок о бок в одном и
том же местообитании, т. е. являются симпатрическими.
Понятие о симпатричности пе столь просто, как это может
показаться па первый взгляд. Согласно Майру, «две формы (или
два вида) называются симпатрическими, если они’ встречаются
вместе, т. е. если области их распространения перекрывают друг
друга или совпадают» (1947 235). В других случаях Майр тол-
кует симпатричность гораздо уже — как сосуществование форм
буквально рядом, в одном и том же местообитании — биотопе
(там же 308—320 и др.).
Независимо от степени филогенетической близости форм сим-
патричность следует понимать как факт совместного их существо-
вания на одной и той же территории. Симпатрическими являются,
следовательно, как неродственные виды (принадлежащие к раз-
ным родам, семействам и т. п.), но входящие в один и тот же био-
ценоз или ярус (синузию), так и наиболее близкородственные
формы, принадлежащие к одной и той же местной популяции.
Сейчас нас интересуют именно эти внутрипопуляционные симпат-
рические формы.
Нами уже подчеркивалось (Завадский, 1961, 19676), что поня-
тие симпатричности относительно и следует говорить о большей
или меньшей симпатричности. Можно толковать симпатричность
широко (общность только области распространения) или узко
(общность местообитания), по строго симпатрическими следует
считать формы, живущие в смеси в одном микроместообитании.
Однако даже самое тесное сообитание может быть сопряжено со
значительными различиями по ритмам развития, по предпочитае-
мым факторам среды, сильно изменяющимся в пределах даже
самых малых площадей (освещенность, влажность воздуха в при-
почвенной зоне, иногда состав и химизм почвы и т. п.). Наконец,
надо учитывать различия в конкурентоспособности совместно
живущих форм. В последнее время и другие * авторы отметили
неопределенность понятия симпатричности. Было, например, пред-
ложено различать чисто географические явления, сохранив за
ними терминологию Майра, п экологические, введя понятия син-
топичности и аллотопичности (Rivas, 1964). По нашему мнению,
К. М. Завадский $41
обязательно введение еще и понятий синхронности и аллохрон-
ноСти, так как формы, живущие в одном биотопе, могут разли-
чаться по срокам развития как викарные во времени (хроновика-
ритет). Формы, симпатрические в самом строгом смысле этого
термина, в территориально-экологическом отношении могут сосу-
ществовать в течение различного времени. При длительном сшива-
нии они могут рассматриваться как компоненты устойчиво сбалан-
сированной системы, а при вытеснении одними формами других —
как фаза процесса преобразования популяции. Степень симпатрич-
пости может быть связана с активностью передвижения и переме-
шивания животных. Однако даже при таком кочевом образе
жизни, как у китообразных, разграничение алло- и симпатриче-
ских групп затруднено только из-за незнания демографических
закономерностей.
Симпатрия родственных групп может быть первичной или вто-
ричной, но до тех пор, пока не накопится достаточного материала,
нельзя утверждать, что вторичная симпатрия встречается часто,
а первичная — сравнительно редко (Ларина, 1963 167). Разно-
образие родственных форм в больших озерах Африки одни авторы
считают вторичным, возникшим после формирования каждого
вида в условиях географической изоляции и последующей мигра-
ции (Мауг, 1963), а другие — первичным, результатом симпатри-
ческого видообразования (Myers, 1960а, 19606). Примером вто-
ричной симпатричности, возникшей на наших глазах, может
служить распространение американской норки от Англии до Кам-
чатки. Начало этому процессу было положено животными, удирав-
шими из клеток в зверохозяйствах. В 30-х годах начали возникать
отдельные мелкие популяции американских норок. В дальнейшем
к стихийному размножению добавился специальный выпуск зверь-
ков на волю. Численность американской норки быстро возрастает,
и занимаемая ею площадь расширяется. Европейская же норка
испытывает биологический регресс, который в значительной мере
объясняется конкуренцией симпатричного вида — пришельца из
Америки.
Популяция включает еще два сорта симпатрических ’ форм.
Одни не имеют возможности обособиться от популяции и оста-
ются лишь частями целого. Таковы, например, особи-органы
в общинах колониальных насекомых, половой, возрастной, сезон-
ный полиморфизм. Другие формы также являются частями попу-
ляции, но при определенных обстоятельствах могут стать само-
стоятельными единицами — основательницами новых популяций.
Именно к таким формам относятся рассматриваемые в этом раз-
деле внутрипопуляциоиные симпатрические единицы — экоэле-
мент, морфо-биологическая группа и биотип. Эти единицы не
составляют иерархического ряда и даже не являются звеньями
единой системы. Две из них — экоэлемент и морфо-биологическая
группа — наиболее существенны для изучения состава популяции,
242
внутрипопуляционные группы. Изучение популяций
начинается с установления ее состава. Одно время казалось, что
генетические методики «препарирования» популяций наиболее ре-
зультативны и должны вытеснять все другие. Впоследствии вы-
яснилось, что без анализа приспособительного значения отдель-
ных внутрипопуляционных форм с помощью экспериментально-
экологических методик нельзя понять ни состав популяции, ни
тем более ее структуру. Значительная физиологическая неоднород-
ность организмов одной популяции по уровню обмена веществ, со-
вершенству терморегуляции, составу крови, типам поведения,
чувствительности к агентам среды, вариабельности размеров и
активности эндокринных желез и т. п. показана на многих живот-
ных, например Н. И. Калабуховым (1962) па грызунах. Эти раз-
личия могут быть проявлением генотипической неоднородности
популяции или возникать под воздействием различных условий на
генотипически почти однородную популяцию. Практически же
чаще всего встречается физиологическая неоднородность, вызван-
ная взаимодействием генетических и экологических факторов.
К внутрипопуляционным группам относится и возрастной
спектр популяции. Он является видовой адаптацией к среде и
главным образом к ценотическим отношениям. В разных ценозах
возрастной состав популяций одного вида может быть различным.
Это было показано, например, Т. А. Работновым (1950) на
Anemone fasciulata: чем условия ценоза ближе к экологическому
оптимуму вида, тем больше в составе популяций плодоносящих и
хорошо развитых растений. У многолетних растений это объяс-
няется тем, что с ухудшением условий (загущение и т. п.) затор-
маживается не только накопление биомассы, по и генеративное
развитие (Завадский, 1947, 1954г, 1957а). Особенности возраст-
ного состава популяции следует рассматривать как 'общевидовое
приспособление к удержанию пространства (Уранов, 1960).
Вблизи оптимума растения зацветают быстрее и на фоне ускорен-
ного накопления биомассы, так что «видовая функция» средней
особи осуществляется на высоком уровне. Возрастной состав попу-
ляции и процессы смены поколений чрезвычайно разнообразны и
зависят от средней продолжительности жизни особи, от эколого-
ценотических условий и т. п. Разновозрастные компоненты входят
в целостную популяцию, хотя возможны и антагонистические от-
ношения между ними (гибель проростков и ювениальных расте-
ний под розеткой листьев взрослой особи или заглушение старого
растения быстрорастущим молодым).
В генетической литературе составом популяции называют пере-
чень генов или линий, полученных в результате4 разложения по-
пуляции и выявления рецессивных факторов (см., например: Li,
1955; Dobzhansky, 1958, и др.). Но это не состав, а генетический
потенциал популяции. Обычные амфимиктические популяции
весьма гетерозиготны, включают массу рецессивных и полудоми-
16*
243
найтных мутаций, входящих в «мобилизационный резерв йаслед-
ственной изменчивости» популяции (Шмальгаузен, 1939, 1946а;
Гершензон, 1941). Однако если популяцию понимать не как ком-
бинацию генов, а как единицу населения, то и понятие «состав
популяции» приобретает иное содержание. В состав популяции
прежде всего входят формы, уже проверенные отбором, т. е. со-
ставляющие ее адаптивную норму. В адаптивную норму входят
все доминирующие и условно доминирующие системы реакций и
признаков. Применяя ипцухт, принудительные скрещивания с по-
следующим многократным отбором выщепляющихся форм, клони-
рование и другие средства разложения популяции, исследователь
нарушает ее исторически сложившуюся структуру, а тем самым
искажает или вовсе уничтожает предмет своего исследования.
«Трупоразъятие», конечно, имеет значение и на популяционном
уровне, но и здесь оно не может стать самоцелью.
Экспериментально-экологические методики позволяют выяснить
состав популяции без разложения ее на элементы и без разруше-
ния ее целостности (Синская, 1958, 1963; Завадский, 1957а, 1961;
Завадский и др., 1962, и др.). Этим путем в популяции могут быть
выделены группы особей, сходные не только морфологически, но
и по их реакциям на факторы среды. Такие группы фенотипически
сходных особей могут в генотипическом отношении быть как раз-
нообразными, так и однородными, а их признаки и реакции —
как модифицирующими, так и стабилизированными.
Существует несколько путей приспособленйя популяции
к среде. Одни из них используют возможности отдельно взятых
индивидов (физиологический гомеостазис, включающий адаптив-
ное поведение и адаптивные модификации). Другие опираются
на надиндивидуальные системы (генетический полиморфизм, орга-
низация внутривидовых отношений и ир.). Большинство путей
приспособления популяции лежит через создание разнородности.
Однако пе всегда она достигается этим путем. Эвриадаптивносгь
может достигаться расширением нормы реакции каждого инди-
вида, так что, существуя в разнообразных условиях, популяция
может сохранять фенотипическое однообразие. У высших живот-
ных совершенные аппараты физиологического гомеостазиса и вы-
сокоразвитое адаптивное поведение способствуют гораздо большей
фенотипической выравненности популяции, чем у растений и при-
митивных животных. Однако, «как правило, чем больше разнооб-
разна среда обитания, тем больше ее генетическое разнообразие»
(Шварц, 1963 422).
При половом размножении скрещивания постоянно интегри-
руют популяцию, так что в принципе каждая особь может быть
носителем всех ее генетических возможностей. Процессы расщеп-
ления и доминирования, а также модификационная изменчивость
непрерывно дифференцируют популяцию на отдельные формы,
состав и количественные соотношения которых и образуют ее
244
структуру. Целостность популяции поддерживается пе только
интегрирующей функцией скрещивания и всей совокупностью вну-
тривидовых отношений, но и наличием компонентов, каждый из
которых важен не только как приспособительная форма, но и как
соопылитель, костимулятор и пр.
Популяционные закономерности своеобразны. Например, ред-
кая и невысокожизнеспособная форма может приобрести преиму-
щества перед многочисленной и высокожизнеспособной, так как,
находясь в примеси, не привлекает внимания хищников, которые
совершенствуют навыки охоты за часто встречающейся добычей.
Выгоды существования в виде малочисленной примеси могут
быть так велики, что отбор тем или иным способом постоянно
поддерживает невысокую численность такой формы (Sheppard,
1959, и др.).
О генетическом полиморфизме. Сбалансированный
полиморфизм популяции — очень широко распространенное явле-
ние. Он не менее эффективно страхует вид от гибели, чем
структурно-физиологические адаптации особи. У полиморфных
видов бабочек, имитирующих окраску других видов, выживание
определяется генетическим механизмом, обеспечивающим гибель
части яиц и этим самым сохраняющим малую численность
взрослых особей. Этот механизм основан на поддержании опреде-
ленной концентрации летальных генов в половой хромосоме и
срабатывает при увеличении численности особей выше опти-
мума. Он может служить примером аппарата, обеспечивающего
популяционный гомеостазис (Sheppard, 1959). Возможно, что
чаще популяционный гомеостазис достигается прямым селектив-
ным преимуществом гетерозиготы над гомозиготой (при котором
гомозигота легальна или ослаблена), например, из-за близкого
сцепления рецессивной летальной или полулегальной аллели
с аллелью, ответственной за фенотипическое проявление поли-
морфизма (Ford, 1963; Huxley, 1963). Изучение некоторых ти-
пов полиморфизма показало, что часто в его основе лежит на-
стоящий генетический аппарат в комбинации с различными эко-
логическими (фенотипическими) механизмами. Недавно именно
такая комбинация факторов, обусловливающих полиморфизм,
была всесторонне проанализирована на миметических формах
бабочки Papilio darmanus (Clarke a. Sheppard, 1960). У одного
и того же вида устойчивое соотношение между формами в одном
случае может поддерживаться селекцией, условиями местооби-
тания, хищниками или другими агентами среды, а в другом —
специальным генетическим аппаратом, как это было показано
А. Кэйном и Д. Карреем в 1963 г. на улитке Се^аеа nemoralis
(цит. по: Huxley, 1963) и на многих других моллюсках, на на-
секомых, растениях и т. п. Полиморфизм яиц кукушки зависит
как от генетического аппарата, так и от воздействия птиц тех
видов, в гнезда которых подкинуты яйца. Иногда адаптивно не
245
наличие нескольких определенных форм, а само по себе увели-
чение разнообразия (у актиний, морских звезд, некоторых по-
лихет, кузнечиков и т. п.), так как именно максимальное разно-
образие снижает убыль от хищников.
Наблюдения показывают, что среди полиморфных популяций
можно выделить тип, включающий более или менее четко разли-
чаемые структурные единицы, и тип, в котором структура отсут-
ствует или трудно определима.
Популяции с полиморфизмом, который был удачно назван
Е. Н. Синской «разлитым» (1948, 1961), имеют мало определен-
ную структуру. Они могут быть недавно образованными, еще не
отшлифованными отбором, или же подвергшимися разрушению
из-за выпадения отдельных доминантных комплексов и возник-
шего благодаря этому нарушению отселектированного режима
переопылений (например, благодаря скрещиваниям между дале-
кими формами). В пользу такого взгляда говорят многие факты
по скрещиванию культурных растений. Так, например, в сфор-
мированных популяциях создается определенный режим скре-
щиваний, складываются наиболее приспособительные системы
доминирования, а в результате принудительных скрещиваний
вся отселектированная система дезорганизуется, потомство начи-
нает расщепляться в самых различных направлениях, вырабо-
танные типы доминирования целых конституций исчезают, воз-
никает как бы разлитой полиморфизм. При скрещиваниях резко
различающихся форм Anemone pulsatilla многие признаки на-
чали расщепляться по монофакториальной схеме, чего не на-
блюдалось в скрещивациях между близкими формами. В таких
случаях в популяциях трудно или невозможно установить чет-
кие структурные элементы, в них преобладают случайные ре-
зультаты миграции аллелей. Из множества таких комбинаций
в процессе отбора складываются генетические комплексы, спо-
собные создавать адаптивные модификации или же стабильные
фенотипы, имеющие приспособительное значение и • представ-
ляющие собой уже закономерные структурные элементы попу-
ляции.
Экоэлемент. Наиболее крупной и самостоятельной в био-
логическом отношении среди внутрипопуляционных единиц
является экоэлемент (Синская, 1948), т. е. зачаток экотипа,
складывающийся в недрах местной популяции. Экоэлементный
состав популяций изучался Синской у люцерн, тимофеевок, ежи,
овсяниц, тригонелл, эспарцетов и т. п. Экоэлементы могут отли-
чаться друг от друга не только по морфологическим и габитуаль-
ным (форма куста, тип ветвления, размеры), но и по физиологи-
ческим признакам (ритм роста и развития, сроки размножения,
продолжительность жизни, отношение к затенению, к субстратам,
влажности почвы и т. п.). Особенности экоэлементов наслед-
ственно более или менее закреплены. При скрещиваниях и рас-
246
щеплениях признаки экоэлемента нередко наследуются совместно,
образуя нерасщепляющийся комплекс.
Как показали Е. Н. Синская (1948) на Medicago sativa и
М. glutinosa и другие авторы, генетические комплексы могут
стать нерасщепляющимися, передающими при скрещиваниях по-
томству доминантную возможность целой растительной консти-
туции. При наличии в популяции нескольких таких доминирую-
щих п нерасщепляющихся генетических комплексов ее струк-
тура приобретает полную отчетливость и они представляют
собой экотипический «скелет» ее внутренней структуры.
Принципиальное значение имеют опыты с Potentilla glandu-
losa и Р. gracilis (Clausen et al., 1940). В скрещиваниях между
формами, являющимися по существу экоэлементами, был пока-
зан различный характер расщепления и доминирования. Просле-
живалось наследование ряда признаков (сроки созревания, ритм
развития, размеры и окраска лепестков и др.). Оказалось, что
экоэлемепты Р. glandiilosa передавали потомству целый ком-
плекс признаков как одно целое (или все они оказывались рецес-
сивными, или же, как одна скоррелированная система, домини-
ровали). Изучение скрещиваний’между экоэлементами P.gracilis
показало, что у этого вида те же признаки наследуются неза-
висимо друг от друга.
Таким образом, у Р. glandulosa уже не происходит расщеп-
ления по признакам, характеризующим экоэлементы, а у Р. gra-
cilis все признаки свободно расщепляются, что указывает на от-
сутствие сложившегося генетического комплекса.
Изученные нами экоэлементы популяций мари белой (Cheno-
podium album) воспроизводят себя (материнский тип) на 79—
98.5%, т. е. являются в большой степени самовоспроизводящи-
мися единицами, а группы кок-сагыза (Taraxacum kok-saghis)
только на 4—31% (Завадский, 19576, 1961; Завадский и др.,
1962; Савин, 1957а, 19576, 1959). Столь слабая самовоспроизво-
димость групп кок-сагыза сама по себе уже указывает на отсут-
ствие сложившихся экоэлементов.
Позднее была предложена следующая схема образования и
эволюции экоэлементов (Синская, 1961). Отбор создает в попу-
ляциях селективные корреляции, т. е. все более концентрирует
в одной особи проявление многих полезных признаков. При
устойчивой их полезности селективные корреляции после стаби-
лизации становятся генетическими корреляциями, затем зача-
точными экоэлементами и настоящими экоэлементами, а после
выхода из состава популяции — экотипами и, наконец, видами
(совокупностями экотипов). Было предложено определение эко-
элемента, как «группы биотипов. . . состоящей из растений с не-
сколькими более или менее прочно наследственно скрепленными
между собой приспособительными признаками» (Синская,
1963:54, см. также: Синская, 1961:61—62). Вслед за этим ска-
247
зано, что «экоэлемент — это первичное, низшего порядка группо-
вое приспособление» (Синская, 1963) и что экоэлемент может
вырабатываться не только постепенно в процессе отбора, но и
возникать внезапно в результате одной плейотропной мутации,
удачно меняющей проявление сразу нескольких признаков.
Но если экоэлемент — это группа сцементированных отбором
биотипов, то он не может быть «первичной, низшего порядка»
единицей и не может возникать сразу после одной мутации.
Ниже будут приведены критические замечания, касающиеся по-
нятия о биотипе. Видимо, не следует определять экоэлемент ни
как группу биотипов, ни как первичную единицу.
Экоэлемент в нашем понимании — это отшлифованная отбо-
ром, приспособительная внутрипопуляционная форма амфи-
бионтной природы, достаточно стабильная и конкурентоспособ-
ная, чтобы воспроизводиться и вне популяции. Генетический
комплекс, лежащий в основе экоэлемента, является доминирую-
щим. Объясняется это селекцией на усиление доминирования
наиболее приспособительных внутрипопуляционных форм (Har-
land, 1936). Популяция, включающая сформированные экоэле-
менты, представляет собой систему приспособления к широкому
спектру условий к использованию нескольких, иногда резко раз-
личающихся, экологических ниш. Такие популяции состоят из
нескольких дискретных генетических комплексов, обычно не
расщепляющихся и доминирующих при обычных комбинациях
опыления и обычных составах пыльцы в освоенных местооби-
таниях.
Экоэлементы амфимиктических популяций имеют иную исто-
рию образования, чем экоэлементы автогамных и особенно апо-
гамных и агамных популяций.
Экоэлементы являются формами, уже обладающими извест-
ной эволюционной самостоятельностью, остающейся нереализо-
ванной возможностью или реализуемой после дезинтеграции по-
пуляции и при некоторых других обстоятельствах.
Формы, названные Е. Н. Синской зачатками экоэлементов, как
показывают наши наблюдения, еще не имеют стабилизированной
конституции и при выделении из популяции, как правило, по-
нижают конкурентоспособность и испытывают депрессию. На-
стоящие же экоэлементы и при самостоятельном существовании
сохраняют свои адаптационные свойства.
Экоэлементы могут выходить из состава популяции и перехо-
дить па положение самостоятельно существующих форм — осно-
вателей новой популяции. Однако такое независимое положение
экоэлемента не является стойким. В соседних популяциях он
может оставаться лишь их компонентом. Часто происходит не
выход экоэлемента из популяции, а сохранение его после гибели
других компонентов популяции (например, при освоении небла-
гоприятных условий). Популяции, расположенные па Климати-
248
ческой границе ареала вида, отличаются бедностью состава й
иногда включают лишь один наиболее устойчивый экоэлемент.
Таковы, например, высокогорные, крайне обедненные по составу
популяции Achillea borealis (Clausen et al., 1940; Clausen, 1951).
Ряд таких же процессов обеднения состава популяции рассмотрен
Е. Н. Синской (1948).
Экоэлементы не являются строго симпатрическими формами,
так как занимают особые микроместообптания, отличаются обра-
зом жизни, или, наконец, специфичны по ритму развития и сро-
кам размножения. Их правильнее считать полусимпатрпческимп
формами.
Экоэлементы формируются далеко не во всякой популяции и
не при всех условиях. Первоначально популяция осваивает новые
микроместообитания за счет модификационных реакций отдель-
ных особей. Далее, если эти микроместообитайия устойчиво со-
храняют свои особенности, начинается стабилизирующий (Шмаль-
гаузен, 1946а, 1961; Waddington, 1956; Haldane, 1959) или прямой
(Дубинин, 1966) отбор, которым и создается генетическая основа
экоэлемента. Некоторые особенности экоэлемента фиксируются.
Так, например, на пастбищах при перевыпасе наблюдается обед-
нение состава популяции многих растений и сохранение низко-
рослых, карликовых и распластанных форм. У Lolium perenne
наблюдалась уже далеко 'продвинутая фаза процесса и популя-
ция включала преимущественно наследственно фиксированные
карликовые формы( Gregor a. Sansome, 1927). У Роа pratensis,
Dactylis glomerata, по данным Кемпа, пастбищные популяции
также обеднены, но среди низкорослых растений генетически
фиксированных форм не так много, а у других видов на пастби-
щах карликовые формы целиком или почти целиком состоят из
модификантов. Группа адаптивных модификантов — это не эко-
элемент. Некоторые называют такие группы экадой. Причина
превращения экады в экоэлемент — процесс отбора, в котором
необходимо учитывать три момента: усиление степени наследо-
вания данного признака, усиление корреляций между экологи-
чески важными признаками и увеличение численности растений
с генетически фиксированными признаками и корреляциями.
Какую же ступень этого процесса следует считать возникнове-
нием экоэлемента? Очевидно, такую фиксацию ведущих приспо-
собительных признаков и такую численность растений, при кото-
рых группа становится достаточно обособленной, устойчивой и
приобретает готовность к самостоятельному существованию вне
популяции.
Неправильно представлять себе возникновение экотипов как
процесс, идущий только через обеднение состава популяции и
переход экоэлемента к самостоятельному существованию. Во-пер-
вых, популяции часто преобразуются без обеднения состава и без
выхода из их состава отдельных экоэлементов, а во-вторых,
249
внутри самого экоэлемента совершаются процессы накопления
разнообразия, так что, приобретя самостоятельность, он оказы-
вается генотипически столь же гетерогенным, как и материнская
популяция.
Описывая эволюцию экоэлемента, Е. Н. Синская (1963) под-
черкивает, что наряду с упрочением корреляций (спаянности
приспособительных признаков) усиливается и его толерант-
ность — он приобретает способность жить в более широком спектре
условий. Для превращения экоэлемента в экотип это условие
часто является решающим.
Морфо-биологическая группа. Входящие в популя-
цию группы морфологически сходных организмов, одинаково
реагирующие на воздействие факторов среды, К. Раункиер
(Raunkiaer, 1918) назвал изореагентами. Изореагенты в смысле
Раункиера — это приспособительные группы, практически нераз-
личимые в морфологическом отношении, но специфичные по
ритмам развития, по предпочитаемым почвам, условиям освеще-
ния, влажности и т. п. Впоследствии этот удачный термин упо-
треблялся в разных смыслах, в том числе в противоположном
авторскому (им обозначали группы, отличающиеся неприспосо-
бительнымп признаками). Чтобы избежать путаницы в характе-
ристике приспособительных внутрипопуляционных групп, был
предложен термин «морфо-биологическая группа» (Завадский,
1952, 19576, 1961; Гербильский, 1954, 1957; Завадский и др.,
1962).
Морфо-биологической группой мы называем совокупность
воспроизводящихся популяцией организмов, однородную в обыч-
ных условиях по ритмам роста, развития и по другим реакциям
на среду, а также различимую морфологически.
Изучение популяций ряда видов, проведенное нами и на-
шими сотрудниками, показало, что морфо-биологическая
группа — реально существующее подразделение популяции. Ди-
корастущие полиморфные популяции (марь белая, кок-сагыз,
клевер красный, подорожники и др.) включают несколько таких
групп, а отселектированные сортовые популяции (пшеница,
столовая свекла, морковь, помидоры и др.) состоят как бы из
одной морфо-биологической группы, внутри которой можно об-
наружить групповую неоднородность особей. Оказалось, что
у одних видов морфо-биологические группы дискретны, имеют
четкие границы и отличаются друг от друга целыми комплек-
сами скореллироваиных признаков, а у других трансгрессируют,
имеют менее четкие границы, мало специфических признаков,
коррелятивные связи между которыми менее прочны. Морфо-
биологические группы разных видов сильно отличаются также
и по степени стабильности признаков.
Одни группы образованы адаптивными модификантами
(экады) и воспроизводятся популяцией пока есть индуцирующие
250
условия, другие являются смесями модификантов с константными
формами, третьи целиком состоят из стабильных форм и
должны рассматриваться как экоэлементы. Далеко не все морфо-
биологические группы являются самовоспроизводящимися едини-
цами. Одни из них могут совсем не воспроизводить себя (моди-
фиканты в измененных условиях), другие воспроизводят себя
почти целиком, но в довольно узком спектре условий, третьи
являются полностью самовоспроизводящимися в различных усло-
виях. Морфо-биологическая группа, таким образом, общее назва-
ние, объединяющее и закрепленные отбором экоэлементы, и
модификанты (экады), и промежуточные формы. Будучи выделен-
ными из популяции, многие группы, как показали наши наблю-
дения, не могут сохранить жизнеспособность и конкурентоспособ-
ность, свойственную им при существовании в составе популяции,
и испытывают депрессию. Чтобы стать более самостоятельной
единицей — настоящим экоэлементом, такие группы должны под-
вергаться шлифовке отбором.
Удобно подразделять морфо-биологические группы на типы по
приспособлениям, ведущим к их дифференциации.
Типы различаются предпочтением к определенному составу
почвы или ее pH (эдафогруппы), конкурентоспособностью (цено-
группы), ритмами развития п сроками размножения (фено-
группы) . Внутрипопуляционные феногруппы обнаружены у 98 ди-
корастущих видов, относящихся к 32 семействам покрытосемен-
ных растений (по Н. С. Ростовой). В популяциях моркови,
свеклы, капусты, цикория и других корнеплодов встречаются
растения с однолетним циклом («цветуха»), двулетники и много-
летники («упрямцы»). Феногруппы наблюдаются и у семян, спо-
собных прорастать в разное время после созревания благодаря
различной глубине и продолжительности периода покоя. Диффе-
ренциация на феногруппы известна у многих видов рыб (Берг,
1934, 1948 — на лососевых, карповых, окуневых и др.; Гербиль-
ский, 1954, 1962, 1965а — на осетровых и др.). Разделение групп
на типы довольно условно, так как, например, более теневыно-
сливая группа может быть одновременно и феногруппой (более
позднеспелая) и ценогруппой (более конкурентоспособна в гу-
стых смешанных насаждениях).
В популяциях мари белой из Ленинградской области нами
обнаружено шесть морфо-биологических групп, входящих в со-
став трех феногрупп, различающихся сроками зацветания и
продолжительностью вегетации (эфемерная — вегетационный пе-
риод 45 дней, средняя—54—59 дней и поздняя—72 дня). Эфе-
мерная и поздняя феногруппы морфологически оказались еди-
ными, а средняя распадается на четыре группы. Эти группы
различаются конкурентоспособностью в загущенных посевах,
динамикой роста до и после зацветания, высотой растений, фор-
мой и величиной листьев, формой куста и соцветий, различными
251
морфо-генетическими и физиологическими реакциями на изме-
нение длины дня, удобрения и полив и т. и. (Завадский, 19576,
1961; Савин, 1957а, 19576, 1959; Завадский и др., 1962; Шам-
сиев, 1963, и др.). Численность этих групп в различных райо-
нах значительно меняется по годам и в зависимости от почвен-
ных и ценотических условий.
При весенних посевах границы между этими группамибыли
четкими, по при поздних посевах, при выращивании на корот-
ком дне и в условиях большого загущения различия между груп-
пами по срокам развития, по высоте растений, форме листьев и по
другим признакам сгладились.
По данным Л. Н. Хахиной, популяции мари белой из различ-
ных географических пунктов СССР также полиморфны, по со-
став морфо-биологических групп в них иной.
Многие морфо-биологические группы мари белой описыва-
лись систематиками в качестве разновидностей (см. 6-й том
«Флоры СССР» и др.) или даже особых видов. Так, например,
в определителях П. Ф. Маевского (изд. 1954 г.), а также
С. С. Станкова и В. И. Талиева (изд. 1949 г.) описывается осо-
бый вид мари — Chenopodium viride. На самом же деле этот
«вид» является подробно изученной нами ранней внутрипопуля-
ционной морфо-биологической группой полиморфного вида Ch.
album. Она свободно скрещивается с другими группами и вос-
производит в потомстве растения всех других групп. Кроме того,
на коротком дне и при летних сроках посева растения всех
групп, в том числе и ранней, преобразуются в одну или же
в две новые группы, т. е. ранняя группа («вид» viride) уподоб-
ляется другим группам той же популяции.
Чтобы не выдавать внутрипопуляциопные формы за вторично
симпатрические виды, надо изучать материал эксперименталь-
ными методами. И у таволги Filipendula ulmaria мы обнаружили
полиморфные популяции, растения которых различаются сро-
ками зацветания и морфологическими особенностями (высота ра-
стений от 31 до 142 см, длина соцветия от 5 до 25 см, опушен-
ность нижней стороны листьев от голых до беловойлочных, число
боковых листьев от 4 до 10, окраска стебля, листьев и др.).
Биометрически были установлены четыре морфо-биологиче-
ские группы. Коэффициент корреляции между сроками зацвета-
ния и морфологическими признаками г = 0.4—0.9. Все они встре-
чались совместно как к югу от Ленинграда, так и на северном
и южном побережьях Финского залива, на болотах и лугах,
в сырых лесах и по оврагам. Но много растений (до 28%) ока-
зались переходными, а раноцветущая малорослая группа (31 —
76 см) является редкой (2.6%). Простые опыты с притенением,
осветлением и с изменениями влажности почвы дикорастущих
экземпляров показали большую модификационную пластичность
многих признаков (например, первые листья были сильно опу-
252
шенными, а последующие менее опушенными и, наконец, го-
лыми, зелеными, или наоборот). Когда мы обратились к рабо-
там систематиков, обнаружилось, что одни авторы подразделяют
этот вид на ряд разновидностей (var. tomentosa, var. denudata
и др.), а другие разделяют его на два отдельных вида: F. ulma-
ria и F. denudata. Последний «вид» был выделен на основании
одного признака: отсутствия опушения нижней стороны листьев
или же слабого опушения (только вдоль жилок).
Изучение популяции таволги еще раз подтвердило, что мор-
фологические виды систематиков могут резъединять единое
целое на части.
В популяциях кок-сагыза обнаружено несколько групп цве-
тущих в первом году жизни или зацветающих только после зи-
мовки (Королева, 1948). Было показано, что внутрипопуляцион-
ные формы кок-сагыза вполне отвечают нашему .определению
морфо-биологической группы (Завадский, 1957а, 19576, 1961;
Савин, 1957а, 1959; Ростова, 1960, 1963, 1965; Завадский и др.,
1962, и др.). Но в отличие от четко различимых морфо-биологи-
ческих групп мари белой у кок-сагыза границы между ними ока-
зались расплывчатыми. Н. С. Ростова (1967) показала, что из-
менчивость ряда признаков кок-сагыза имеет клинальиый харак-
тер (число листьев на одном растении, размеры и форма листьев
и др.), а по другим признакам обнаруживается двувершинность
кривых (срок начала цветения, число цветоносов и дополнитель-
ных розеток к началу цветения, число листьев в центральной
розетке и др.). Следовательно, растения, цветущие в первый год
жизни и нецветущие, образуют достоверно разграниченные
группы. Морфо-биологические группы кок-сагыза различаются
как феногруппы, однако особенности ритма развития связаны
с габитуальными и морфологическими признаками.
Изменяя многие условия среды, можно в значительных раз-
мерах менять количественные отношения между группами в по-
пуляции. В популяциях кок-сагыза количественные соотноше-
ния между группами сильно изменяются под влиянием почвен-
ного питания и густоты посева (Завадский, 1947, 1954а). Так,
в густом посеве возрастает численность ранних групп, а на пло-
дородной почве уменьшается число растений, не цветущих
в первый год жизни. Сходные данные приводят В. А. Королева
(1957) и Н. С. Ростова (1960, 1963, 1965).
В то же время в состав каждой морфо-биологической группы
кок-сагыза несомненно входит и стабилизированное «ядро». Каж-
дая группа воспроизводит всю популяцию и прд этом воспроиз-
водит себя в относительно скромных размерах (4—35%). В гу-
стых гнездовых посевах преимущественно выживают растения
ранних групп, наиболее быстро растущих в течение первого ме-
сяца вегетации. Пятилетним опытом было доказано, что состав
популяции, выращивавшейся в густых гнездах, сильно сдви-
253
нулся в сторону преобладания ранних морфо-биологических
групп (Завадский, 1954а, 1957а, 19576, 1961). Многократным от-
бором В. А. Королева получила популяции, резко сдвинутые
в сторону преобладания «озимых» форм. По данным Н. С. Ро-
стовой, испытавшей эти популяции, оказалось, что 87—91% ра-
стений не зацветает в первый год. О наличии в каждой группе
кок-сагыза стабилизированного «ядра» свидетельствует также и
сравнение потомства от внутригрупповых скрещиваний с потом-
ством от межгрупповых скрещиваний.
Морфо-биологические группы кок-сагыза формируются, таким
образом, в каждом поколении за счет генотипически различных
форм. Одни из них в порядке онтогенетической адаптации в за-
висимости от условий формируют несколько фенотипов — адап-
тивных модификантов, а другие способны в разнообразных усло-
виях осуществлять один и тот же фенотип группы. Таким обра-
зом, популяция включает генотипически стабильное ядро и
группу лабильных генотипов, дающих модификантов. В природ-
ных популяциях кок-сагыза пе произошло ни стабилизации
групп, ни перехода к чисто модификационному типу воспроиз-
ведения, так как он обитает в изменчивых условиях высокогор-
ных долин Тянь-Шаня, его особь малостойка в конкурентной
борьбе с травостоем и ее плодовитость невелика. Местообитания,
им занимаемые, характеризуются большой пестротой почв,
микрорельефа, микроклимата, растительного покрова. Отдельные
куртины очень невелики по площади (иногда доли гектара), но
расположены близко друг около друга, так что популяция каж-
дой долины расчленена ца много колоний, занимающих разно-
образные местообитания. В то же время между колониями от-
сутствуют барьеры для гаметного и семенного обмена. Если бы
в популяции кок-сагыза сложилось несколько спаянных генети-
ческих комплексов (как у свеклы и мари), то при небольшой
плодовитости растений, анемохории семян и пестроте местооби-
таний не могло быть обеспечено воспроизводство численности
вида.
Стабилизация признаков и анемохория могут быть совмещены
лишь при высокой плодовитости, обеспечивающей большую ве-
роятность заноса семян «нужного» типа в каждое из освоенных
местообитаний. Уже одно наличие летучих семян при малой пло-
довитости и большом разнообразии местообитаний делает более
выгодным иной тип воспроизведения, регулируемый той средой,
в которой сеянцы начинают свое развитие. Отсутствие же чисто
модификационного воспроизведения объясняется, по-видимому,
невозможностью формировать столь широкую норму реагирова-
ния. Наличие в популяциях кок-сагыза нескольких стабилизиро-
ванных «ядер», широкого модифицирования и многих свободно
комбинирующихся признаков увеличивает их пластичность и
приспособляемость. Гибкое сочетание стабильных и модифици-
254
рующих форм обеспечивает и такое важное приспособление, как
растянутость общих сроков цветения популяции и возможность
в наиболее благоприятных условиях плодоносить большинству
растений.
У дикой свеклы Beta maritima две морфо-биологические
группы. Некорнеплодно-мелколистная группа содержит в два
с лишним раза больше растений, стрелкующихся в первый год
(83.2%), чем корнеплодно-крупнолистная (40.6%) (Савин,
1959). Оказалось, что эти группы являются самовоспроизводя-
щимися с такой же полнотой, как и морфо-биологические группы
у мари белой (в обычных условиях выращивания). Тин разви-
тия групп, как у мари белой и дикой свеклы, обусловливает
более легкую их отделимость и значительную степень самосто-
ятельности. Такие группы являются* экоэлементами. В частно-
сти, именно это свойство группы позволило выделить из популя-
ции дикой свеклы корнеплодно-крупнолистную столовую и
кормовую формы. Искусственному отбору оставалось лишь пога-
сить (или снизить) способность этой группы к воспроизведению
некорнеплодных форм и однолетности.
Изучая популяции апомиктических мелких видов манжеток
(Alchemilla) Турессон (Turesson, 1956, 1957) в условиях питом-
ника обнаружил групповые различия по ритмам развития, форме
куста, высоте, по величине, форме и окраске листьев, по имму-
нитету и т. п. Они вполне соответствуют нашим морфо-биологи-
ческим группам. Сохранение признаков в двух-трех поколениях
ири размножении клонами, например, североскандинавских до-
линных и горно-тундровых форм, позволяет рассматривать их
как экоэлемент, а сам автор возводит такие клоны в ранг aga-
mospecies.
Работы с беленой (Козлова, 1956) показали, что биологиче-
ские группы у этого вида воспроизводятся в основном модифи-
кационно через разновременность созревания семян, а это свой-
ство является фиксированным. При таком воспроизведении
групп ведущим дифференцирующим фактором являются разли-
чия в местообитаниях или в сроках развития. Попав в любые из
освоенных популяцией условия, зачаток способен развить соот-
ветствующую им приспособительную форму.
Было показано, что даже самая выравненная отбором попу-
ляция, например чистолинейные сорта пшеницы, представляет
(•обой мономорфную группу только в нормальных для культуры
условиях (Агаев, 1958), а при изменении сроков посева,
длины дня, густоты посева и других условий вскрывается неод-
нородность материала. Даже в пределах чистых линий яровой
пшеницы обнаруживаются как бы зачатки морфо-биологических
групп, имеющие как модификационную, так и стабилизирован-
ную природу. В последнем случае оказывается эффективным от-
бор в дифференцирующих условиях. Наличие обычно скрытой
255
Неоднородности объясняется нами тем, что отбор на выравни-
вание систематически проводился по конечным фазам развития
(Завадский, 19576, 1961). Генетически различающиеся линии
оказались уподобленными друг другу только в тех условиях,
в которых отбирался сорт, и только по конечным фазам. Не обя-
зательно,конечно, такое уподобление возникает на завершающих
фазах. Если отбор на выравнивание проводился на первом году
жизни двух- или многолетних растений, то уподобление возник-
нет на первом году жизни. У столовой плоской египетской
свеклы синхронность развития и однородность по морфологиче-
ским признакам наблюдается только на первом году жизни и
в оптимальных условиях.
Таким образом, одна морфо-биологическая группа может быть
генотипически однородной или разнородной, состоять из моди-
фикантов или сочетать модификантов со стабилизированными
формами, в свою очередь относящимися к одному или к несколь-
ким генотипам, каждый из которых сохранил способность вос-
производить лишь одну адаптивную форму (т. е. имеет лишь
один «канал развития»).
Понятие морфо-биологической группы стало входить в прак-
тику исследований (Синская, 1963:57; Корчагин, 1964, и др.).
Их изучением занялись ученики Е. Н. Синской и, например,
С. Д. Киселева (1961, 1963) у популяций шести видов вик обна-
ружила феногруппы, установила степень корреляции между
сроками цветения и другими признаками, изучила особенности
воспроизведения морфо-биологических групп. Значение таких
исследований подчеркивает С. Л. Мамаев (1965), изучающий
внутривидовые группы у древесных (сосна обыкновенная).
Биотип. В. Иогансен и другие авторы определяли биотип
как группу генетически тождественных (изогенных) организмов.
Биотипы могут состоять из практически гомозиготных или из ге-
терозиготных организмов. По Дж. Клаузену (Clausen, 1951), — это
«основные элементы генетической структуры популяции». Са-
мыми элементарными едницамп, ио Иогансену, являются гомо-
зиготные биотипы у «идеального самоопылителя». В руковод-
ствах по генетике 30—40-х годов писалось, что в популяции
агамных и апогамных видов — это механические смеси биоти-
пов-клонов, которые имеют четкие границы и просто выделяются.
У растений с автогамным размножением — классическом объ-
екте для выделения биотипов — их границы, степень дискретно-
сти и отделимость могут быть не менее четкими, чем у агамов
и апогамов.
Одно время считали, что виды — это комбинации биотипов
и что из биотипов как из первичных элементов составляются во-
обще все таксоны более высокого ранга (Розанова, 1930, 1946,
и др.). Что же такое биотип в популяциях с перекрестным опло-
дотворением? Ясно, что такая популяция не является простой
256
смесью легко отделимых биотипов и не содержит таких биоти-
пов, как клональные и автогамные популяции. В полиморфных
популяциях морфо-биологические группы состоят из генетиче-
ски разношерстного материала. После изоляции от популяции
они расщепляются. Способность таких форм поддерживать тип
только в условиях системных отношений резко отличает их от
биотинов клональных и автогамных популяций. Своеобразие изо-
генных групп в популяциях с перекрестным оплодотворением
оправдывает предложение А. А. Парамонова (1945) именовать
такие единицы «миксобиотипами». Их можно прямо наблюдать
в таких популяциях, где существуют дискретные, достаточно
(‘паянные и нерасщепляющиеся генетические комплексы, под-
вергшиеся отбору на усиление доминирования. Но такие миксо-
биотипы ранее были определены нами как экоэлемепты. Что ка-
сается популяции с нарушенным режимом скрещиваний, то они
не имеют выраженной внутренней генетической структуры, и
явление миксобпотппа в связи с этим здесь расплывается или
исчезает вовсе.
Когда давалось определение биотипа, еще не была известна
интенсивность мутационного процесса в природных популяциях
и большая их гетерозиготпость, существование под покровом
одного или нескольких доминирующих фенотипов («адаптивной
нормы») массы генотипов, различающихся между собой рецес-
сивными факторами, и т. п. В панмиктических популяциях на-
следственными особенностями обладает большинство особей. Еще
больше генетическое разнообразие в популяциях, живущих
в экологически оптимальных условиях: почти каждая особь
должна считаться особым биотипом. Нечто подобное можно ска-
зать и о живущих в оптимальных условиях популяциях само-
опылителей. Более или менее строго изогенными (если прене-
бречь соматическими мутациями) могут быть лишь клоны.
В опытах В. Н. Сукачева (1927, 1935) по межбиотипной борьбе
каждый биотип получился от одной особи черенкованием корней
(Taraxacum officinale) или делением куста (Festuca sulcata).
Являются ли биотипы действительно структурными элемен-
тами популяции? Генетики раннего периода считали структуру
популяции следствием случайного комбинирования массы изоли-
рованных аллелей и механически отсеивающего действия среды.
На самом же деле структура популяции — образование законо-
мерное, исторически формирующееся отбором, т. е. адаптивное.
Лишь при ее разрушении в природе или в эксперименте возни-
кает беспорядочное комбинирование аллелей и случайное рас-
щепление признаков. Селекционно отработанной группой может
быть не только экоэлемент пли морфо-биологическая группа, но
и семья, стадо, колония, заросль и т. п. Между тем понятие
о биотипе позволяет представить себе в качестве структурного
элемента популяции и одну особь, что является абсурдом. В при-
17 К. М. Завадский
257
родной популяции кок-сагыза из урочища Ойгаин, руководству-
ясь лишь морфологическими отличиями листовой пластинки
(длина, ширина, форма, очертания края, окраска и др.), нами
было выделено более 500 клонов. После их размножения корне-
выми черенками на плантации можно было видеть всю массу
этих четко различающихся между собой биотипов-клонов. Если
учесть большую изменчивость кок-сагыза, каждое растение ко-
торого имеет морфологические особенности, то число возможных
биотипов должно быть необозримо большим. Такие биотипы-
клоны не составляют структурных элементов популяции. То же
самое следует сказать и о биотипах инцухт-линиях. Большин-
ство из них в неразложенной популяции находилось в рецессив-
ном состоянии. Инцухт-линии позволяют установить богатство
генетического потенциала популяции, но не могут служить еди-
ницами изучения ее адаптивной структуры. Такие наблюдения
и заставили нас в свое время при анализе структуры популяции
отказаться от понятия биотипа, заменив его понятием морфо-
биологической группы. Биотипы в амфимиктических популя-
циях — это или формы, селектированные на гомозиготность ге-
нетического ядра, что возможно у некоторых экоэлементов, или
чаще всего — это вовсе не структурные элементы популяции,
а продукты ее распада (особи-клоны, инцухт-линии и др.).
У апогамных и агаманых популяций биотип имеет более
четкие границы и выделяется гораздо легче. Структуры таких
популяций, видимо, складываются более просто. Чем менее це-
лостна популяция, тем более четок и легко отделим биотип.
В последнее время понятие о биотипе обросло нежелатель-
ными наслоениями и почти утратило свой первоначально четкий
смысл. Биотипом часто называют группу, выделенную из попу-
ляции на любых основаниях. Многие авторы словом «биотип»
обозначают фенотипически однородные, т. е., по нашей терми-
нологии, морфо-биологические группы. Но эти группы, как уже
подчеркивалось, вовсе не должны обладать генетической однород-
ностью, этим основным признаком биотипа. В некоторых новей-
ших руководствах по генетике элементарной единицей населе-
ния вида признается не местная популяция, а биотип и экотип
рассматривается как сумма биотипов (Мюнтцинг, 1963:467).
Некоторые авторы считают биотип «мельчайшей таксономи-
ческой единицей» (Розанова, 1930, 1946; Вавилов, 1931, 1935а;
Делоне, 1945, и др.), но против этого взгляда было сделано
много возражений (Комаров, 1940; Clausen, 1951; Завадский,
1961; Huxley, 1963, и др.). Таксономической единицей может
стать лишь форма, способная к относительно самостоятельному
и устойчивому воспроизведению в природе. Биотип в обычных
условиях не является самостоятельно существующим образова-
нием, а зависимым элементом, входящим в структуру популя-
ции как системы более высокого уровня. Лишь в особых, напри-
258
мер в крайне суровых, условиях или после перехода к бесполо-
семянности, а также к вегетативному размножению отдельный
биотип может на известное время стать самостоятельной формой
существования вида. Однако и в этом случае еще преждевре-
менно считать биотип «мельчайшей таксономической единицей».
Дело в том, что регистрация фактов свободного существования
биотипов, как бы часто они не встречались, еще не говорит
в пользу их способности к исторически длительному самовос-
произведению в природной обстановке и тем более к эволюцион-
ным изменениям. Нет фактов, доказывающих эволюционную
перспективность отдельных биотипов, выделившихся в природе
из состава популяции.
6. ЗАКЛЮЧЕНИЕ
Как показывают комплексные исследования последних деся-
тилетий, любой вид, как бы он ни был мопоморфеи по внешним
признакам, включает большее или меньшее число различных по
природе и значению подразделений и всегда выступает как мно-
гообразие. Это многообразие может быть генотипическим и скры-
тым или же явным и включать в адаптивную норму несколько ко-
личественно сбалансированных фенотипов; оно может возникать
из-за широкой модификационной способности индивидов. Чаще
всего многообразие фор(м внутри вида представляет собой ре-
зультат взаимодействия фиксированных и модифицирующих при-
знаков и реакций. В этом отношении нет никаких различий
между признаками подвидов и местных популяций или экоэле-
ментов и морфо-биологических групп.
Все сказанное в настоящей главе о многообразии внутриви-
довых форм было основано главным образом на изучении этого
вопроса у высших растений и высших животных. Пока трудно
делать широкие обобщения и обсуждать, например, вопрос
о структуре видов всех типов растений и животных. Структура
вида у бактерий, синезеленых водорослей, несовершенных гри-
бов и у многих других групп почти не изучена, а то, что стано-
вится известным в последние годы о структуре вида у простей-
ших, свидетельствует о большом ее своеобразии и сложности.
Можно сказать, что виды неравноценны прежде всего именно
по своему составу и структуре. Иначе говоря, проблема неравно-
ценности видов должна разрабатываться в основном путем со-
поставления внутривидовых структур. Так, например, многие
древние реликтовые виды не имеют аллопатрических подразде-
лений и их ареалы могут быть сужены до одной или немногих
локальных популяций (крымские Orchis comperiana или Lepi-
dium turczaninowii и пр.). Популяции таких видов могут вклю-
чать всего один морфо-биологический тип, но генетические груп-
повые различия существуют и у них.
17*
259
Виды молодых эндемиков также могут состоять всего из од-
ной или немногих локальных популяций. Йо как поразительно
различен состав популяций первичных (молодых) и вторичных
(угасающих) эндемов! На примере кок-сагыза было показано,
как в изолированных горных долинах складываются высокополи-
морфные популяции. Многие из них столь богаты внутренними
возможностями для расселения, что напоминают сжатую пру-
жину. Именно о таких исключительно полиморфных популяциях
(или группах популяций) и писал Н. И. Вавилов, развивая свое
представление о центрах происхождения культурных растений.
С другой стороны, популяции, расположенные на климатической
границе ареала вида, могут отличаться крайней бедностью со-
ства и включать лишь одну наиболее устойчивую морфо-биоло-
гическую группу. Ряд таких же процессов обеднения состава
популяции подробно проанализирован в монографии Е. Н. Син-
ской (1948). У Lolium perenne на пастбищах при постоянном
перевыпасе происходит обеднение популяции и сохранение кар-
ликовых форм (Gregor a. Sansome, 1927). Популяции Роа рга-
tensis, Dactylis glomerata и некоторых других видов на пастби-
щах также сильно обедняются и включают генетически фикси-
рованные низкорослые формы.
Даже один клон, чистая линия или инцухт-линия включают
в себя многообразие форм. В природе не существует и не может
существовать видов и популяций — идеальных монотипов.
Изучение структуры вида имеет и огромное практическое
значение. Прежде всего надо исчерпывающе знать генетический
и фенотипический потенциал вида, но еще более важно иссле-
довать те генетико-экологические механизмы, которые поддер-
живают оптимальную гетерозиготность и высокую гетерозис-
ность.
Еще недавно массовое производство сортов основывалось на
линейной селекции. При этом, как правило, разрушался (разла-
гался на отдельные линии) исторически сложившийся и прове-
ренный естественным отбором внутрипопуляционный полимор-
физм. Полученные таким образом двойные межлинейные гиб-
риды кукурузы, оказавшись столь эффективными, дают
скоропроходящий эффект, основанный на гетерозисе, исчезаю-
щем уже во втором поколении. Высокожизнеспособный, пла-
стичный, стойкий и постоянно гетерозисный урожайный сорт
должен синтезироваться из многих линий и представлять собой
сложную популяцию. Эффективность каждой линии должна
быть испытана не порознь, а в популяции. Это длинный путь.
Его можно сократить, беря за основу и улучшая природные
популяции. Они обладают уже сбалансированным полиморфиз-
мом и способны ежегодно поддерживать состояние гетерозиса
у массы особей. Анализ таких популяций должен показать, ка-
кие именно генотипы необходимо удалять и заменять другими,
260
более эффективными (в смысле воздействия этих генотипов как
компонентов популяции на все хозяйственно-биологические свой-
ства) .
Прав К. Гото (Gotoh, 1957), указывая, что важнейшая и
труднейшая задача селекции — управление составом популяции
о создание многолинейных сортов. Такие сорта, разумеется,
должны быть константными. Но эту константность следует по-
нимать не как поддержание типа, а как воспроизведение всего
многообразия форм (всей сбалансированной полиморфной гене-
тической системы). Создание подобных сортов позволит капи-
тально решить такие проблемы, как повышение конкурентности
и иммунности культурных форм до уровня наиболее высокожизпе-
способных диких родичей.
Однако основой всей проблемы дифференциации вида яв-
ляется вопрос о структуре популяции как целостной системы и
об аппаратах, обеспечивающих ее поддержание в оптимальном
состоянии. Можно выделить три аппарата, тесно взаимодейству-
ющих и переплетающихся друг с другом, а иногда и действую-
щих параллельно по принципу множественного обеспечения жиз-
ненно-важных функций (Баркрофт).
Популяционно-генетический аппарат создает адаптивный
«остов» популяции за счет формирования комплекса фиксиро-
ванных реакций и признаков. Он служит приспособлением к со-
вокупности устойчивых во времени и мало изменяющихся в про-
странстве компонентов среды; управляет составом и числовыми
соотношениями между миксобиотипами и другими формами ге-
нетической гетерогенности; обеспечивает наиболее выгодный
режим скрещиваний и наиболее адаптивный характер процесса
доминирования. Этот аппарат, как правило, функционирует лишь
при сохранении целостности популяций.
Индивидуально-физиологический аппарат представляет сово-
купность существующих у особей данной популяции онтогенети-
ческих адаптаций (модификаций); обеспечивает более точную
пришлифовку к тем компонентам среды, которые могут быстро
и обратимо изменяться во времени и в разных участках освоен-
ного местообитания.
Индукционно-следовой аппарат основан на явлении «после-
действия» или «следовых реакциях», передающихся через семена
и сохраняющихся в потомстве в течение одного—нескольких
поколений; этот аппарат обеспечивает возможность лучшего при-
способления к тем условиям, изменения которых не столь часты
и вместе с тем имеют закономерно периодический характер.
Глава 9
ВНУТРИВИДОВЫЕ ОТНОШЕНИЯ
1. КЛАССИФИКАЦИЯ внутривидовых отношений
Определение понятия. Внутривидовые 'отношения
обнимают всю совокупность самых разнообразных по содержа-
нию, характеру и значению связей и зависимостей между орга-
низмами и группами организмов одного вида. Внутривидовые
отношения складываются не только из взаимоотношений между
расами, местными популяциями и внутрипопуляционными еди-
ницами; не в меньшей мере они зависят от таких первичных
объединений индивидов, как семья, выводок, колония, стадо,
стая и т. п. Такие первичные объединения особей могут совпа-
дать с одной локальной популяцией (дёмом), могут и входить
в нее как части. Например, может существовать трехступенчатая
иерархическая зависимость: семья — стадо — местная популяция,
но при отсутствии семьи и при больших размерах стада первичное
объединение это и есть местная популяция.
Внутривидовые отношения могут быть постоянными или но-
сить периодический и эпизодический характер. Они могут быть
весьма стабильными и зафиксированными в особых приспособле-
ниях (структурных, функциональных, стереотипах поведения)
или, напротив, лабильными, возникающими на короткое время
и исчезающими. Внутривидовые зависимости охватывают все
возрастные фазы и имеют место между гаметами, зиготами, за-
родышами, личинками и взрослыми формами.
Отдельные особи и группы особей одного и того же вида мо-
гут воздействовать друг на друга прямо и активно (что более
характерно для животных) или же косвенно, через изменение
окружающей среды. Результаты таких отношений для отдельно
взятой особи или же группы особей могут быть различными.
Ими может обеспечиваться или стимулироваться протекание жиз-
ненно важных процессов (рост, развитие, размножение),
262
а также и более или менее ослабляться протекание этих процес-
сов по сравнению с наследственно обусловленной возможностью
их максимального уровня. Наконец, они приводят организм
к бесплодию или к гибели.
На некоторые формы внутривидовых отношений большое
влияние оказывает численность участвующих в них организмов
и степень их скучивания (плотность), но другие формы отноше-
ний от этих факторов не зависят.
Внутривидовые отношения включают как зависимости, воз-
никающие в результате действия отбора, так и процессы, служа-
щие причиной естественного отбора. В* первом случае эти отно-
шения приобретают характер видовых адаптаций и образуют
систему скоррелированных внутренних связей, цементирующих
популяцию в целостное образование (см. главу 7). Во втором
случае внутривидовые отношения сами могут оказаться агентами
отбора и вызывать процесс отбора. Тем самым они становятся
источником эволюционных изменений вида, вызывают сдвиги во
временно установившемся балансе видовых корреляций. Являясь
и предметом, и результатом отбора, внутривидовые отношения
иногда совершенствуются в такой степени, что приобретают ха-
рактер в высшей степени отрегулированных процессов. Взаимная
пригнанность таких процессов может быть очень тонкой. Су-
ществование каждого вида поддерживается благодаря сбалансиро-
ванности и гармоничности внутривидовых отношений, а так как
этот баланс не идеален, то возникают коллизии, ведущие через
отбор к изменению сложившегося типа внутривидовых отношений,
т. е. к изменению вида. Изменения строя внутривидовых отно-
шений могут вызываться путем отбора самыми различными
абиотическими агентами, видами-конкурентами и врагами,
а также и непосредственно внутривидовым отбором (например,
половым). Если любой агент отбора сдвигает сложившийся ба-
ланс внутривидовых отношений, то происходит перестройка этих
отношений, в результате которой возникает новый, более адап-
тивный строй видовой жизни. Таким образом, в зависимости от
характера внутривидовых отношений они могут обусловить как
устойчивость, так и изменяемость вида.
Классификация внутривидовых отношений осуществляется
исходя из различных принципов.
Выделяют отношения, в которые вступают между собой от-
дельные индивиды, и отношения между естественными группами
особей (семьями, стадами, местными популяциями, физиологиче-
скими или экологическими расами и пр.). В первом случар сле-
дует говорить о внутригрупповых индивидуальных отношениях,
во втором — о межгрупповых. В силу соподчинения одних групп
другим необходимо различать и внутригрупповые отношения
между группами разного ранга, например: внутрипопуляционные
(межбиотипные) и внутрирасовые (межпопуляциопные). Меж-
263
групповые отношения могут осуществляться через индивидуаль-
ные, но нередко являются взаимодействиями двух или несколь-
ких коллективов Кроме того это разграничение важно потому,
что отношения между отдельными организмами и между различ-
ными группами по-разному влияют на существование и эволю-
цию вида.
Широко известно подразделение внутривидовых отношений,
основанное на их характере и результатах. Обычно различают
прямую внутривидовую борьбу, внутривидовую конкуренцию или
жизненное состязание и взаимопомощь (называемую иногда бо-
лее нейтральными терминами: взаимоблагоприятствованием или
внутривидовой кооперацией) J
В результате многих и разносторонних исследований в при-
роде и в экспериментальных условиях было установлено, что все
три перечисденные выше формы внутривидовых отношений (пря-
мая борьба, конкуренция и кооперация) не только встречаются
в природе, но и являются закономерными для существования и
развития всех видов растений и животных.
Однако эта классификация имеет и недостатки, так как ею
не охватываются все формы внутривидовых отношений. Так, на-
пример, само содержание таких отношений, как спаривание, вы-
кармливание детеныша, насиживание яиц, изгнание трутней из
улья пчелосемьи, конъюгация инфузорий, не может быть отне-
сено ни к одной из перечисленных форм.
Основные и производные внутривидовые от-
ношения. По значению для воспроизведения вида внутриви-
довые отношения подразделяются на основные и производные
(Завадский,1961).
Основными внутривидовыми отношениями являются все не-
посредственно обеспечивающие воспроизведение вида, например
спаривание, конъюгация, насиживание яиц, выкармливание дете-
ныша, обучение молодняка и др. Это совокупность отношений,
являющихся необходимой предпосылкой самого существования
вида. Ими обеспечивается смена поколений вида п поддержание
определенного уровня численности всех возрастных групп попу-
ляции. Без этих отношений или при недостаточном их совершен-
стве невозможно само существование вида.
Основные отношения включают отношения между особями и
группами особей разных полов, обеспечивающие определенную
численность и необходимый качественный состав зигот (режимы
1 Лучше не употреблять метафорические термины, такие как «взаимо-
помощь», «конкуренция», «кооперация» и пр. (Завадский, 1954 и др.;
Birch, 1957). Нейтральные термины «взаимодействие» и «взаимовлияние»
были бы более точными. Однако пока нет установившейся терминологии
для обозначения отдельных форм таких взаимодействий, старые метафоры
еще могут служить при их точном определении: об этом свидетельствуют
последние работы (Мазер, 1964 332—335; Мильн, 1964:80, и др.).
264
скрещиваний, подбор соопылителей, избирательность оплодотво-
рения, поиски особи другого пола, турнирные бои самцов у по-
лигамных видов, токование и т. п.); отношения родительского
поколения к потомству, обеспечивающие развитие зародышей и
молодняка и снижение их смертности (все формы пассивной и
активной заботы о потомстве: гнездо- и норостроительные ин-
стинкты, инкубация, вынашивание, выкармливание, охрана и
тренировка молодняка и т. п.); наконец, отношения между осо-
бями или группами особей одного пола и возраста или обоих
полов и нескольких возрастов, обеспечивающие сохранение чис-
ленности популяции (постоянные, сезонные или возрастные агре-
гации, скучиванпя, стада, кулиги, стаи, образуемые для лучшей
добычи корма, защиты от врагов, миграций, для брачных игр,
для создания оптимальных экологических условий и т. и.).
Совершенствование основных внутривидовых отношений яв-
ляется одной из фундаментальных закономерностей 'эволюции.
Так, например, постепенно развиваются приспособления, гаран-
тирующие зародышам и молоди стабильные и оптимальные усло-
вия питания и развития, а также защиту от неблагоприятных
факторов среды.
В эволюции позвоночных происходила замена мелкой икринки
крупным яйцом, удлинение периода инкубации, развитие ин-
стинктов оберегания яиц, увеличение пищевых запасов в яйце и
редукция личиночного типа развития, переход к питанию моло-
ком и развитие инстинктов защиты молодняка, переход от яйце-
кладки к живорождению, совершенствование адаптаций, обеспе-
чивающих внутриутробное развитие зародыша, развитие реакций,
поведения, способствующих тренировке молодняка, и, наконец,
совершенствование механизмов высшей нервной деятельности,
благодаря которым новым поколениям передается опыт, при-
обретенный каждым поколением, а не унаследованный от
предков.
Если примитивные группы кольчатых червей имеют еще сво-
бодноживущую и самостоятельно питающуюся личинку (трохо-
фору), то у прибрежных мпогощетинковых кольчецов все раз-
витие личинки уже проходит внутри больших, богатых желтком
яиц, а у малощетинковых и пиявок возникает хорошо защищен-
ный крупный кокон, наполненный пищевым материалом, внутри
Которого и происходит развитие личинок. У акуловых наблю-
дается переход ко все более крупным, богатым желтком яйцам,
к живородности и даже к образованию специальных структур,
осуществляющих функцию кровоснабжения зародыша, подобных
плаценте.
У растений аналогичные процессы можно наблюдать в пере-
ходе от размножения спорами к размножению семенами, в разви-
тии приспособлений, защищающих зародыш и семена, вплоть до
иокрытосемянности, в развитии связей между материнским расте-
265
нием и зародышем через снабжение его пищевыми резервами
(эндосперм) и т. п.
Все это свидетельствует об огромном значении основных
внутривидовых отношений для эволюции. Виды, у которых сло-
жились совершенные и эволюционно перспективные системы
основных внутривидовых отношений, как правило, были способны
к широкой адаптивной радиации. При этом системы внутривидо-
вых отношений, доказавшие свою практическую универсальность
и высокую эффективность, имели тенденцию закрепляться у всех
потомков, приобретая значение крупных усовершенствований,
ведущих к образованию групп высокого таксономического ранга.
Вот почему определенный тип основных внутривидовых отноше-
ний оказывается иногда хорошим классификационным признаком
(сумчатость, плацентарность, птенцовость, покрытосемянность
и т. п.).
В процессе совершенствования основных внутривидовых от-
ношений происходило формирование целого класса взаимопри-
способлений между организмами одного вида (конгруэнциями —
по С. А. Северцову; см. главу 7). Приведем дополнительно при-
меры конгруэнций. Описано конвергентное возникновение аппа-
ратов для впрыскивания молока в рот детенышу у китообразных,
сумчатых и у ехидны; ускоренное развитие передних конечностей
у эмбрионов некоторых грызунов объясняется тем, что детеныш
добывает молоко, массируя млечную железу (Матвеев, 1943).
Величина кладки яиц у птиц и величина семьи у млекопитающих
приводится отбором в соответствии со средним наибольшим числом
детенышей, которых могут прокормить родители (Лэк, 1957).
У колониальных насекомых, выкармливающих свое потомство
(как млекопитающие и птицы), число откладываемых яиц также
соответствует количеству корма. У термитов при первом сезоне
размножения, когда еще мало рабочих, царица откладывает только
15—20 яиц, а после образования большой колонии — до 7000
в день. Та же закономерность существует и у муравьев.
К производным внутривидовым отношениям относятся все за-
висимости, которые непосредственно не связаны с обеспечением
воспроизведения вида и подержанием его численности. Они воз-
никают на базе уже сложившихся основных отношений. У от-
дельных видов или на некоторых фазах их развития производные
отношения могут отсутствовать или, возникнув, исчезают.
Производные внутривидовые отношения можно подразделять
на возникающие между особями в результате роста плотности
популяции (гибель личинок соснового лубоеда при массовых раз-
множениях вследствие попадания в уже ранее проделанный ими
ход, мину — Ильинский, 1929; переход к каннибализму мучного
жука — Chapman, 1931; откочевки животных; усиление диффе-
ренциации и увеличение смертности в насаждениях растений
и т. п.) и на возникающие между особями одного вида на базе
266
оборонительных и пищедобывательных рефлексов или на базе ин-
стинктов самосохранения и защиты корма, непосредственно не
связанные с плотностью популяции (охрана индивидуального уча-
стка или участка семьи, реакция агрессии, борьба за корм, неко-
торые случаи каннибализма и т. п.).
Предложенное нами подразделение внутривидовых отношений
на основные и производные поддержано рядом авторов (Гудош-
ников, 1962; Козлов, 1962; Ларина, 1962; Федюшин, 1962; Ша-
киров, 1962; Лавренко, 1963; Olson, 1963, и др.).
2. ФОРМЫ ИНТЕГРАЦИИ ИНДИВИДОВ
Формы интеграции индивидов зависят от степени развитости
и дискретности самого индивида. В главе 7 уже отмечалось су-
ществование нечувствительных переходов между индивидом, за-
рослью-колонией, местной популяцией и фитоценозом у корне-
вищных и других вегетативно размножающихся растений.
У примитивных агамных групп средства интеграции надин-
дивидуальных образований, разумеется, крайне бедны в сравне-
нии с видами, размножающимися половым путем и обладающими
аппаратами нейроинтеграции; они ограничены химическими
взаимодействиями. Вместе с тем роль отдельно взятого индивида
здесь очень мала (при астрономически огромной численности)
и у многих групп бактерий обнаруживается более или менее
продвинутая тенденция к объединению в колонии (индивиды
2-го порядка). Этот путь интеграции одноклеточных особей, no-
видимому, многократно приводил к образованию колоний, неко-
торые из которых подвергались дальнейшей интеграции и преоб-
разовывались в особи Metazoa и Metaphyta. Интеграция, ведущая
к столь большой степени зависимости элементов и частей от це-
лого, как в многоклеточном индивиде, далеко не во всех случаях
оказывается эволюционно очень перспективной. Так, например,
глубокая специализация на основе разделения функций и пре-
вращение особи в орган колонии у колониальных насекомых
хотя и создала высокоинтегрированные системы вроде роя пчел
или муравейника, обеспечивших известные преимущества по-
томкам, но не привела к прогрессивной эволюции большого мас-
штаба. Замечательные подтверждения этому — пути эволюции
надиндивидуальных систем у термитов с разделением труда, под-
держанием постоянства условий в гнезде, социальным поведе-
нием (Emerson, 1961).
Превращение особи в неполноценный индиеГид, надежно обес-
печивающий только одну из функций колонии, оказалось менее
перспективным, чем путь эволюции с сохранением полноценности
индивида. Но воспроизведение вида на основе высокой специали-
зации особей в колонии (матка, воины, рабочие, особи — храни-
267
тельйицы запасов Меда й т. п.) требовало создания отточеййЬтх й
автоматизированных систем внутривидовых отношений. Р. Эшби
(1959) пришел к выводу, что наилучшие условия функциониро-
вания системы совпадают не с максимумом, а с определенным
оптимумом взаимозависимости частей и что слишком большое
обилие сильных связей лишает систему пластичности и мобиль-
ности. Этот принцип, надо полагать, имеет всеобщее применение
и очень важен при изучении различных форм интеграции особей.
Стадная жизнь многих высших млекопитающих, особенно обезьян
(Шовен, 1965), является великолепным примером высокой инте-
грированности этих надиндивидуальных систем, основой которой
является не обесценивание индивида, а, напротив, максимально
широкий его прогресс. Надиндивидуальные объединения чрезвы-
чайно разнообразны по самым различным показателям: по чис-
ленности и составу объединяемых элементов, по поддерживаю-
щим механизмам, по времени существования, прочности связей
и т. п. Это могут быть семьи с несколькими особями и суперпо-
пуляции со многими миллиардами особей. Объединение может
включать только взрослых особей одного или обоих полов, только
молодняк пли же быть разнополым и разновозрастным в составе
двух и более поколений (семья бурых медведей, прайд львов,
стадо обезьян и т. п.). Образование колонии из одного клона
у бактерий, синезеленых водорослей и низших грибов приводит
к изогенным и почти гомозиготным объединениям, а стада боль-
шинства позвоночных животных генетически гетерогенны и очень
гетерозиготны. Интеграция может быть мимолетной и легко раз-
рушаемой или случайной агрегацией особей, но может быть гео-
логически долговременной, как у многих местных популяций.
Примером примитивной интеграции могут служить скопления
мериносных овец, которые не допускают перегрева тела в жару
(у одиночных температура тела на 0.4—0.8° выше, чем у овец
из скоплений). Животные выстраиваются в две плотные подвиж-
ные шеренги из 80—120 голов. Каждая овца прячет голову под
туловище стоящей напротив и прижимается к соседней (Фляк,
1962).
Существуют многообразные временные объединения, возни-
кающие для лучшего, более эффективного выполнения какой-
либо одной или нескольких биологически важных задач. Таковы
миграционная стая, стадо, кулига и т. п.; коллективная добыча
пищи в трудное время, например зимняя волчья стая; различные
брачные сборища; терморегуляционпое агрегирование (уменьше-
ние теплоотдачи и взаимный обогрев насекомых, как у «солнеч-
ных» кулижек саранчовых). К такого же рода объединениям
относятся агрегации и стаи у простейших, рыб, насекомых, птиц
и других животных, стимулирующие нормальное течение всех
жизненно важных процессов (скорость обмена, рост, морфогенез,
половое созревание, двигательная и пищевая активность, плодо-
268
витость и т. п.). При жизни в одиночку, как показали экспери-
менты (Allee, 1931; Алпатов, 1934; Парк, 1939; Odum, 1959,
и др.), наступает более или менее резкое снижение жизненно
важных показателей по сравнению с показателями при жизни
в сообществах.2 * * * * Действующими7 агентами здесь выступают хими-
ческие вещества, стимулирующие в оптимальной концентрации
скорости тех или иных процессов, как при аллелокатализе у про-
стейших, действии хлореллпна на скорость делений хлореллы
(Pratt et al., 1945), корневых выделениях растений (Грюммер,
1957, и др.) или же рефлекторные взаимодействия, например
тигмотаксис или комплекс раздражений, идущих через зритель-
ный, обонятельный и слуховой рецепторы, вызывающие опти-
мальный для течения физиологических и морфогенетических
процессов уровень возбуждения и т. и. Например, при выращи-
вании молоди сома в одиночку рост ее задерживается и нару-
шается нормальное развитие. Напротив, при выращивании их
в стайках рост и развитие протекают нормально. Объясняется
это тем, что, находясь в движении и часто соприкасаясь, рыбки
раздражают друг друга и находятся в возбужденном состоянии.
Взаимные раздражения через соприкосновения поддерживают
высокий уровень газообмена, обусловливают большую двигатель-
ную активность рыбок. В большом объеме воды рыбы не рас-
сеиваются, а обязательно образуют тесно скученную стайку. Оче-
видно, что такая плотность населения в стайках хотя и высока,
по является оптимальной.
Интеграция особей может быть не временной (скажем, на
время роста до половозрелостп), а существовать в течение жизни
всего поколения. Филогенетическая преемственность форм инте-
грирования может осуществиться различными средствами. Однако
любые объединения формируются и поддерживаются групповым
и индивидуальным отбором. В результате сочетания их действий
переживают лучше организованные объединения (семьи, стада,
популяции), совершенствуются оправдавшие себя механизмы ин-
теграции, преимущественно сохраняются особи с более разви-
тыми «социальными» реакциями. При этом используются разно-
образные механизмы от регуляций с помощью химической
взаимоиндукции, от половых инстинктов, инстинктов заботы
о потомстве, тигморефлексов, различных стадных условных реф-
лексов вплоть до многообразной звуковой сигнализации в стадах
млекопитающих и «речи» дельфинов, выделении вожаков с бо-
лею или менее широкими функциями (охрана, регулирование
отношений, поддержание необходимой организации стада
2 Показано, что плотность популяции может кардинально менять те-
чение физиологических процессов от основного обмена и роста до пове-
дения, полового созревания и плодовитости. На этой основе начинает
разрабатываться синфизиология, т. е/физиология сообществ, а не индиви-
дов (Harper, 1964).
269
й т. п.). У многих видов обезьян стадо организуется совмест-
ным действием врожденных и многих приобретенных рефлексов,
формирующихся в процессе воспитания молодняка (Тих, 1950).
Прекрасные описания надорганизменных образований у жи-
вотных недавно опубликовал Р. Шовен (1965). Сложные струк-
туры семей, стад и стай с иерархическими отношениями, особая
и высокая интеграция особей в этих структурах, приводящая
к их неделимости, примеры поведения особей и групп особей по-
лезных колонии — все эти данные Шовена доказывают, что вну-
тривидовые отношения действительно составляют содержание
жизни вида.
При высокоразвитых формах стадности наблюдаются не
только разделение функций, но и хорошо организованные формы
кооперации, например общественное выкармливание молодняка
(у обезьян, птиц и других животных), совместная оборона мо-
лодняка и беременных самок у стадных копытных, приматов и
др., гнездовий у колониальных птиц («птичьи базары»), очище-
ние от накожных паразитов и т. п. У птиц с развитой стад-
ностью имеются вожаки, осуществляется не только совместное
выкармливание, но и высиживание птенцов, совместная добыча
корма (пеликаны и др.), защита от врагов, специфический строй
в полете, создаются наилучшие условия для скрещиваний с по-
стоянным поддержанием гетерозисности и оптимальной гетеро-
зиготности и т. п.; весь уклад жизни стаи устойчив и специфи-
чен для вида.
Устойчивые сообщества муравьев имеют довольно постоян-
ную общую численность особей гнезда и колонии, а также чис-
ленность отдельных групп (половых, рабочих и т. д.). Гнезда
и колонии занимают определенную охраняемую территорию,
размеры которой у каждого вида определяются пищевыми ре-
сурсами, численностью сообщества и его конкурентоспособ-
ностью (Длусский, 1965).
Полезность повышения порядка интеграции можно иллю-
стрировать на примере образования союзов муравейников — над-
колоний у рыжего лесного муравья (Formica rufa). По наблю-
дениям П. И. Мариковского, между населениями одиночных му-
равейников существуют враждебные отношения, объясняемые
«перенаселенностью территории» (1962:85). В некоторых слу-
чаях от муравейника отделяются колонии-почки и возникает
надколония. Внутри таких надколоний между муравейниками
складываются дружественные отношения. Встречаются гигант-
ские надколонии с несколькими тысячами близко расположен-
ных муравейников. В Западной Сибири наблюдаются надколонии
с плотностью 1 муравейник на 56 м2 (в 111 муравейниках на
площади в 6100 м2 жило 140 млн особей). Автор отмечает, что
столь высокая плотность «является следствием более совершен-
ной формы общественной жизни» (там же: 87), характерной
270
для надколоний. Между муравейниками надколонии поддержи-
ваются связи: взаимное посещение, перенос личинок, куколок
и иногда самок, взаимообмен рабочими особями. Значение этих
взаимоотношений, как полагает ав^ор, состоит в том, чтобы пре-
пятствовать развитию враждебности и вырождению колонии: от-
сутствие обмена особями ведет к появлению враждебности и
взаимной охоты. В данном случае более высокая интеграция
увеличивает экологический потенциал вида за счет снижения
нерациональной убыли численности.
Для понимания степени интегрированности тех или иных
частей вида необходимо знать интенсивность обмена мигрантами
между ними и характер этого обмена (постоянный, периодиче-
ский или эпизодический; только взрослые самцы или все воз-
растные группы обоих полов и т. п.; взаимный или односторон-
ний; отдельные экоэлементы или биотипы или вся популяция
в целом). Например, у млекопитающих и птиц такое перемеши-
вание бывает связано с откочевками, перераспределением кор-
мовых участков и мест гнездования, расселением молодняка за
пределы уже занятых участков, а между популяциями рыб на-
блюдаются различные способы перемешивания — активные не-
рестовые или нагульные миграции, скат молодняка и т. п. Из-
вестно чрезвычайное разнообразие первичных объединений ин-
дивидов. Одни виды животных всегда ведут «общественный»
(стадный) образ жизни, другие — лишь в определенные периоды
жизни или сезона, третьи вовсе не образуют стад. У обыкновен-
ной колюшки развита забота о потомстве, а у других видов ко-
лючеперых строй внутривидовой жизни совсем иной. Стая мо-
жет быть организована на основе безусловных рефлексов (ку-
лига саранчи) или на основе сложных сочетаний безусловных
рефлексов с условными (гаремы у морских котиков).
Для понимания структуры вида недостаточно знать состав
его аллопатрических групп, местных популяций и симпатриче-
ских групп. Надо учитывать также и первичные объединения,
все формы интеграции индивидов. Содержание жизни вида —
это и есть различные отношения, возникающие внутри таких
первичных объединений и между отдельными объединениями.
С. А. Северцов справедливо подчеркивал, что внутривидовые от-
ношения связывают вид в «некоторое целостное единство»
(1951:44).
3. О ХАРАКТЕРЕ ВНУТРИВИДОВЫХ ОТНОШЕНИЙ
Противоречивость основных внутривидовых
отношений. Все формы основных и пройзводпых внутриви-
довых отношений, как бы хорошо они не были отрегулированы,
какими бы важными видовыми адаптациями они не являлись,
осуществляются весьма противоречиво. Все объединения орга-
271
низмов закреплялись через отбор, т. е. противоречиво. Совершен-
ствование внутривидовых отношений в форме прогресса простой
семьи, семьи-колонии, различных агрегаций, стад и т. п. осуще-
ствлялось преимущественно через групповой отбор. Прямая ин-
дивидуальная и групповая борьба, конкуренция и взаимопомощь
не только легко переходят друг в друга, но иногда и существуют
в рамках одного и того же явления, образуя то его разные сто-
роны, то фазы. Многочисленные примеры перехода отношений
взаимопомощи в конкуренцию и обратно дает изучение стадных
и колониальных животных. Выше было показано жизненно важ-
ное значение существования молоди некоторых видов рыб (сомов
и др.) в скученных стайках. В то же время пищевая активность
молоди весьма высока и рыбки вступают друг с другом в драки
за пищу, даже если ее достаточно. Значительные индивидуаль-
ные различия в размерах и силе рыбок позволяют некоторым из
них скорее овладеть пищей и без прямой борьбы. В результате
возникает дифференциация молоди па фракции сильных, круп-
ных, активных особей и слабых, мелких. Из этого примера ясно
видно, что жизнь в стайке осповапа на взаимодействии противо-
положных отношений — кооперации, жизненного состязания и
прямой борьбы, которые буквально проникают друг в друга
(рис. 3).
Самые ответственные акты внутривидовых отношений, на-
пример видовые адаптации, обеспечивающие сохранение гамет,
яиц, личинок и взрослых особей, осуществляются противоре-
чиво. Внутренне противоречив, например, процесс эволюции ко-
лониальное™ у перепончатокрылых насекомых, так как совер-
шенствование и усложнение размножающейся единицы вида
(колонии) сопровождается увеличением зависимости индивидов
и ростом их неполноценности.
Нельзя признать гармоничной организацию полигамных се-
мей у морских котиков, хотя она и обеспечивает поддержание
численности вида. При рождении соотношение полов примерно
1 1, а у взрослых — один самец на 30—50 самок. В первые
6 лет самцы и самки гибнут с одинаковой интенсивностью.
Убыль самцов резко возрастает в возрасте 6—7 лет, когда они
вступают в бои за гаремы. Каждый год гибнет около трети сам-
цов, часть из них — во время боев, а другие — в море, будучи
ослабленными от ран или от истощения в течение двухмесячной
голодовки во время охраны самок на лежбище. По данным
С. А. Северцова (1951), во время боев самцы придавливают не-
которых детенышей, особенно на песчаных лежбищах, где нет
укрытий. Отбор наиболее сильных самцов привел к резко выра-
женному половому диморфизму: взрослый самец (секач) весит
до 170 кг, а самка — всего 32 кг. Такой способ воспроизведения
вида является малосовершенным, так как значительная часть
поголовья гибнет, не принося существенной пользы виду. Возра-
272
стание размеров самцов вследствие внутривидового (полового)
отбора не является экологически необходимым, так как самки,
ведя в океане сходный с самщами образ жизни, не отличаются
от них по выживаемости. Возможна гораздо более экономная
организация внутривидовых отношений при полигамии. Проти-
Рис. 3. Схема взаимодействия между особями (Мазер, 1964).
Между особями, обладающими сходными потребностями, могут существо-
вать конкуренция, нейтральные отношения или сотрудничество (левая
часть схемы). Между особями, отличающимися по своим потребностям,
могут существовать только нейтральные отношения или сотрудничество
(правая часть схемы.) Взаимодействие по нескольким потребностям при-
водит к взаимоотношениям полной конкуренции, полного сотрудниче-
ства или же конкуренции по одним и сотрудничества по другим факто-
рам (средняя часть схемы). Этот тип взаимодействия лежит в основе
биологической организации на всех уровнях и обусловливает необходи-
мость регуляции, без которой конкуренция погубит взаимоотношения
сотрудничества, необходимые на высших уровнях организации.
воречивость внутривидовых отношений может выражаться в из-
бытке самцов, занимающих территорию, но не участвующих
в воспроизводстве вида. У некоторых птиц существуют бигам-
ные семьи при наличии самцов-холостяков (Verner, 1964).
Внутривидовые противоречия могут иметь разную напряжен-
ность; иногда существуют самые острые противоречия антагони-
стического характера. Таков «самопаразитизм» у насекомых
(пчел и многих других). Например, у первичного паразита ка-
273
18 К. М. Завадский
лифорнийской щитовки, перепончатокрылого насекомого из афи-
линид — Prospaltella perniciosi, иногда наблюдается паразитиче-
ское развитие самцов из неоплодотворенных яиц, отложенных
внутри личинок самок своего же вида. Такой вторичный па-
разитизм самцов на самках авторы (Чумакова и Горюнова,
1963) рассматривают как приспособление, обеспечивающее
воспроизведение вида в периоды низкой численности хозяина
(щитовки).
Постоянный и периодический каннибализм различной интен-
сивности, свойственный многим хищным и многоядным видам
рыб (поедание икры своего вида, мальков и взрослых рыб мень-
шего размера), хищным птицам и другим группам животных,
также может служить примером антагонистически противоречи-
вых внутривидовых отношений. Самец американской саламандры
Cryptobranchus alleganiensis обладает инстинктом защиты потом-
ства и охраняет кладку яиц, никуда не отлучаясь в течение пе-
риода инкубации. При этом, однако, он утоляет голод частью
охраняемых им яиц. Здесь в одном акте сталкиваются канниба-
лизм и забота о потомстве. Инстинкт заботы о потомстве у сам-
цов американского скрытожаберника развился, видимо, из
инстинкта охраны корма. Такое переключение инстинкта оказа-
лось возможным, так как снижение числа яиц в кладке компен-
сируется большей плодовитостью самок (ими обеспечивается не
только воспроизведение вида, но и корм самца — защитника
кладки). Конечно, эволюционная перспективность таких неэконо-
мичных систем заботы о потомстве очень мала.
Наличие антагонистических отношений внутри вида подтвер-
ждается, например, данными Т. Соннеборна, Р. Зигеля и других
на инфузории Paramaecium aurelia. У этой инфузории были
обнаружены генетически фиксированные линии, присутствие ко-
торых приводит к гибели остальных членов популяции. Оказа-
лось, что инфузории таких линий выделяют в среду особый анти-
биотик парамеции, безвредный для них, но токсичный для дру-
гих линий. Присутствие в популяции значительного числа
«убийц» делает невозможным сосуществование с ними «чувстви-
тельных» линий. Некоторые линии «убийц» действуют губи-
тельно только в момент конъюгации с «чувствительными» осо-
бями (Полянский, 1957). Трудно ответить на вопрос, является ли
этот случай примером сбалансированного диморфизма единой
популяции, или же, напротив, проявлением начавшейся дивер-
генции вида.
Такой же характер могут носить некоторые конгруэнции.
Цикл развития гамет не всегда координирован с интересами ор-
ганизма, и, например, у аннелиды Polygordius epicotus овогонии
фагоцитируют кишечник матери. Съев кишечник, нефридии и
мускулатуру, они разрывают кутикулу и выходят наружу, унич-
тожая часть сегментов материнского организма,
274
Следует учитывать, что формы внутривидовых отношений
иногда оказываются очень подвижными и могут легко перехо-
дить одна в другую.
Иногда, казалось бы, самые незначительные обстоятельства
могут менять знак отношений. Забота о потомстве может пере-
ходить в каннибализм. У млекопитающих самка иногда поедает
детенышей из-за нарушений в ее кормовом рационе, во время
резкого возбуждения, а самка одного паучка заботится о дете-
ныше, пока он связан с нею паутиной, но при разрыве нити
тотчас съедает его. И напротив, каннибализм, как у американ-
ского скрытожаберника, может быть использован в качестве
звена, впаянного в цепь реакций самца, направленных на за-
щиту кладки яиц.
У луговых злаков дочерние побеги после ослабления связей
с материнским растением выступают как их конкуренты (За-
вадский, 1947; Смелов, 1947).
Поэтому нет и не может быть никаких безусловных границ
между основными и производными отношениями, кооперация и
сотрудничество существуют бок о бок с жизненным состязанием
и прямой борьбой, образуя то фазы, то стороны единого процесса
(Механизмы..., 1964). «Организация живых систем во всей ее
сложности и на всех уровнях зависит от согласованности про-
тивоположных тенденций конкуренции и сотрудничества» (Мазер,
1964 : 354). Например, организмы, остро конкурирующие из-за
одних элементов экологической ниши, вступают между собой и
в отношения временного или даже постоянного сотрудничества.
Так, например, личинки дрозофил в культуре, поглощая дрожжи,
при известной плотности конкурируют друг с другом за пищу.
Разрыхляя же поверхность дрожжей, они, возможно, препят-
ствуют росту численности вредителей (бактерий и грибов) и
в этом случае помогают друг другу.
Модельные опыты на растениях. Противоречивый
характер внутривидовых отношений был обнаружен в опытах
с гнездовыми посевами кок-сагыза. В загущенных посевах де-
сятки всходов совместно пробивают почвенную корку и содей-
ствуют этим друг другу, в то время как одиночные проростки
погибают, не сумев преодолеть сопротивление корки. Однако,
после того как совместно взошедшие растения подрастают,
между ними возникают острые противоречия, связанные с ис-
пользованием ограниченного объема почвы (Завадский, 1947,
1954г, 1957а, 1961). В каждом гнезде выделяется группа расте-
ний с наибольшей скоростью роста корневые систем. Эти расте-
ния успевают перехватить питательные вещества и воду. Уже
через 20—30 дней эти процессы завершаются гибелью отставших
в росте растений. В менее густых гнездах возникают и менее
острые взаимодействия между всходами. Переживание растений
с большей скоростью роста на ранних фазах жизни, как это
18*
275
было выяснение экспериментально, представляет случай меЖ-
грушювого жизненного состязания внутри популяции. Оказа-
лось, что выжившие растения обладают наследственными осо-
бенностями «раннеяровой группы» и состав популяции оказался
сдвинутым в сторону преобладания именно этой группы.
При равномерном размещении растений на делянках
(рис. 4), в различных густотах известное загущение вначале
Рис. 4. Изменение числен-
ности растений кок-сагыза
на делянках с равномер-
ным размещением растений
(Завадский, 1947).
Цифры при кривых — число
семян, высеянных на 1 дм2;
1 —середина делянки, 2— край
делянки.
создает оптимальную освещенность для
всходов (некоторое притенение), но
затем в силу неодинакового роста розе-
ток листьев одни растения начинают
затенять соседей и часто вовсе подав-
ляют их. В этом случае повторяется та
же смена форм отношений между рас-
тениями, которая была описана выше
для корневых систем: вначале взаимо-
благоприятствование, впоследствии бо-
лее или менее острое жизненное со-
стязание и в итоге отбор внутри попу-
ляции наследственно различающихся
групп. В опытах с кок-сагызом наблю-
дались и иные противоречия. Семена
кок-сагыза различаются по периоду по-
коя и всходы бывают очень растяну-
тыми. При значительном загущении
создаются условия, при которых все
растения из поздних всходов погибают,
так как попадают под уже сформиро-
ванные розетки ранее взошедших рас-
тений. Таким образом, выживают орга-
низмы, первыми захватившие местооби-
тания. При этом происходит отбор
линий, дающих большой процент скоро-
прорастающих семян.
Альбиносные растения подсолнечника (неспособные в силу
генетических дефектов к образованию хлорофилла) обычно гиб-
нут после израсходования запасов семядольных листочков. Од-
нако если альбинос выращивается в окружении нормальных
зеленых растений, то у него начинается образование хлорофилла
и иногда такие растения даже заканчивают цикл развития.
Единственным объяснением этого факта является предположение
о передаче соседними зелеными растениями альбиносу каких-
то ему недостающих, ио необходимых для синтеза хлорофилла
веществ через корневые выделения (Лященко и Лященко, 1957).
С другой стороны, &то же тесное соседство, явившееся единст-
венным шансом выживания альбиноса, ведет и к его ослаблению,
так как более мощно развитые корни соседей отнимают пита-
276
пие, a irx кроны затеняют. Йа других объектах подобные Же
перемены знака отношений и направлений отбора былй уста-
новлены рядом авторов (Sakai, 1951, 1953, 1956; Сакай, 1964;
Wynne-Edwards, 1962; Дональд, 1964; Мазер, 1964, и др.).
Внутривидовые противоречия могут приводить к самым раз-
личным результатам. Их в этом отношении можно подразделить
па две большие группы: а) противоречия, поддерживающие сло-
жившиеся основные внутривидовые отношения и ведущие в ко-
нечном счете к преимущественному переживанию особей, отве-
чающих адаптивной норме вида, и б) противоречия, приводящие
к изменению состава популяции и к перестройке внутривидо-
вых отношений.
Противоречия первой группы могут быть закреплены отбо-
ром и превратиться в видовое приспособление, а противоречия
второй группы вызывают через отбор эволюционные изменения
вида, в результате которых они сами могут быть устранены.
Давно было замечено (Haldane, 1932, а‘ затем и многие другие
авторы), что мутантная более конкурентоспособная форма эффек-
тивна лишь в окружении массы особей дикого типа. По мере
размножения мутантного типа и вытеснения особей исходного
дикого типа преимущество более конкурентоспособной формы
все уменьшается. Этот факт был экспериментально показан нами
на кок-сагызе. Вытесняя особей исходного типа, мутантные особи
сами погашают свои преимущества, так как они все чаще взаи-
модействуют друг с другом и все реже с менее конкурентной
формой. Это простая истина, но ее необходимо всегда учиты-
вать: конкурентоспособность — свойство относительное.
4. ЧИСЛЕННОСТЬ КАК ВИДОВОЕ ПРИСПОСОБЛЕНИЕ
Определенный уровень численности особей и характер его ко-
лебаний входят в качественную характеристику вида. У прими-
тивных видов уровень численности особей чрезвычайно высок
(многие и многие миллиарды особей), а у высокоорганизован-
ных крупных и долгоживущих растений и животных гораздо
более низок (порядка 103—104 * * 7 особей).
Снижение численности вида в течение продолжительного пе-
риода свидетельствует о его недостаточной приспособленности и,
если оно не прекращается, ведет к его вымиранию. Напротив,
периодическое снижение численности вида, сменяющееся ее ро-
стом, т. е. пульсация численности («волны жизни»), представ-
ляет собой нормальное явление. Каждый вид может быть оха-
рактеризован специфическим характером этих пульсаций чис-
ленности.
Длительное возрастание численности вида есть показатель
('го процветания. Оно может явиться результатом приобретения
видом нового приспособления или же изменением среды в благо-
277
приятную сторону, Ё обоих случаях рост численности вида мо-
жет привести или к его расселению, или же к повышению плот-
ности населения. В последнем случае естественный отбор может
вызвать снижение уровня численности вида, о чем будет под-
робнее сказано ниже. При снижении уровня численности вида
вследствие отбора в условиях повышенных плотностей населения
вскоре устанавливается новый более низкий средний уровень ее,
который может стабилизироваться. Если же численность вида
падает из-за недостатков его общей приспособленности, то этот
процесс может совершаться непрерывно.
Численность является важным фактором, обеспечивающим
надежность существования вида. Иногда численность вида рас-
сматривают как случайное следствие сложного баланса сил —
потенциальной способности вида к безграничному размножению
и совокупного действия сдерживающих сил природы (стихий,
а также врагов, конкурентов и прочих факторов так называе-
мого биоценотического равновесия). Эта точка зрения не может
быть признана справедливой.
Процветающие виды — как раз те, которые ломают этот ба-
ланс сил. Эволюционный процесс в сущности и состоит в посто-
янном преодолении временно устанавливающихся «балансов
сил», т. е. в преодолении равновесных состояний. Угасающие
виды также не относятся к поддерживающим этот «баланс».
Что же касается видов, численность особей которых в дан-
ный период стабилизирована (а таких видов, очевидно, очень
много), то относительное постоянство уровня их численности не
может быть объяснено лишь случайным соотношением этих сил.
Являясь следствием сочетания очень сложных процессов, чис-
ленность вида, сбалансированная на определенном уровне, пред-
ставляет вместе с тем и причину его процветания. Поэтому идет
отбор таких популяций, которые тем или иным путем оказы-
ваются способными поддерживать среднюю численность вида
на уровне, обеспечивающим его воспроизведение. Определенный,
специфический для вида уровень численности следует рассмат-
ривать как важнейшее видовое приспособление; как всякое при-
способление, он создается естественным отбором.
Для того чтобы понять, почему, например, уровень числен-
ности видов микроорганизмов во много раз выше, чем у высших
групп, необходимо разграничить два показателя: выживаемость
вида и выживаемость средней особи. Выживаемость вида харак-
теризуется изменением его численности, а выживаемость сред-
ней особи определяется отношением числа взрослых (размно-
жающихся) особей к числу продуцируемых зачатков (в среднем
для каждого данного поколения при обычных внешних усло-
виях).
У видов примитивных групп выживаемость средней особи
ничтожно мала (порядка мелких долей процента), а у видов
278
крупных млекопитающих и птиц она достигает величин порядка
10—30%. У примитивных видов с очень низкой выживаемостью
средней особи (например, одна на миллион продуцируемых за-
чатков) очень мала вероятность выполнения именно данной
особью функций размножения (т. е. основной видовой функции
особи).
У бактерий, простейших, низших водорослей и грибов зна-
чение одной отдельно взятой особи в жизни вида ничтожно
мало. Выживаемость же вида тем не менее может быть весьма
высокой. Высокая выживаемость таких видов, у которых сте-
пень выживаемости средней особи мала, обеспечивается их
очень большой плодовитостью и численностью. Уровень числен-
ности вида должен быть достаточным для погашения всяких
постоянно и случайно действующих неблагоприятных факторов,
могущих в известный момент снижать численность вида до угро-
жающе малых величин. Существование таких видов гаранти-
руется в первую очередь не индивидуальными качествами от-
дельной особи и не соответствующими внутривидовыми отноше-
ниями (защита потомства, обучение потомства и т. п.), а именно
численностью, которая и выступает поэтому как одно из важ-
нейших видовых приспособлений.
У видов с совершенной организацией особи и внутривидовых
отношений действие неблагоприятных случайностей преодоле-
вается уже не столько «числом», сколько «уменьем». Гаранти-
рующая существование вида роль статистических процессов
(связанная с большой численностью) дополняется все возра-
стающим значением другого принципа, основанного на совер-
шенстве адаптационных аппаратов каждой особи. Значение особи
в жизни вида соответственно возрастает, а уровень численности
вида снижается, как это мы и наблюдаем у видов высших групп
животных и, в меньшей степени, у видов высших растений.
Из сказанного не следует, конечно, делать вывода о том, что
с ростом выживаемости особи вообще снижается значение чис-
ленности в качестве видового приспособления. Снижается лишь
относительное значение численности среди других приспособле-
ний вида.
Выживаемость вида всегда представляет собой результат со-
пряженного действия трех взаимосвязанных групп приспособле-
ний, гарантирующих его существование: 1) приспособлений ин-
дивида, 2) определенной организации основных внутривидовых
отношений и 3) численности. Если две первые системы приспо-
соблений развиты еще недостаточно, то выживаемость вида мо-
жет быть тем не менее высокой за счет-компенсаторного разви-
тия третьей. Напротив, совершенное развитие двух первых си-
стем приспособлений делает возможной поддержание достаточно
устойчивой выживаемости вида также и при невысоком уровне
численности.
279
То, что уровень численности вида может регулироваться спе-
циальными аппаратами, известно на примере ряда видов бабо-
чек, подражающих различным несъедобным насекомым (Ford,
1940).
Известно, что численность видов древесных растений во
влажных тропических лесах исключительно низка по сравнению
с численностью видов древесных умеренного пояса. По-види-
мому, малочисленность древесных пород в тропических лесах
объясняется сложными межвидовыми отношениями, т. е. сдер-
живающим фактором здесь является именно «биоценотическое
равновесие» (полог леса образуется не одним или двумя-тремя
видами, как в лесах умеренного пояса, а многими десятками ви-
дов). Таким образом, уровень численности вида представляет
собой не результат случайных соотношений между стремлением
вида к безграничному размножению и совокупным действиям
сдерживающих внешних факторов, а более или менее совер-
шенно селекционно отработанную специфическую его особен-
ность. Являясь важнейшим видовым приспособлением, уровень
численности вида входит в его характеристику как один из су-
щественных признаков.
5. РЕГУЛИРОВАНИЕ ПЛОТНОСТИ ПОПУЛЯЦИИ
Оптимальная плотность популяции. У всех ви-
дов происходят колебания численности, закономерно приводя-
щие то к возрастанию плотности популяции, то к ее снижению.
У мышевидных грызунов выгоды массовых размножений плот-
ность популяции иногда достигает 20—30 тыс. норок на гектар,
а в годы депрессии численности — только 1—2. Чрезвычайно
велики колебания плотности популяции у некоторых видов на-
секомых — массовых вредителей (непарный шелкопряд, тополе-
вая и яблоневая моли, совка-гамма, луговой мотылек, златогузка,
боярышница и др.). В то же время у многих других видов плот-
ность популяции изменяется весьма незначительно.
Густота стояния растений одного вида может колебаться
в широких пределах от центра ареала к его краям, от одного
местообитания к другому. Лишь определенное насыщение терри-
тории, специфическое для каждой группы, обеспечивает наи-
большую устойчивость численности вида.
Под определенным оптимумом насыщения мы понимаем не
строго фиксированную плотность, а известную зону плотностей,
в которых обеспечивается лучшее воспроизведение вида и наи-
большая его устойчивость. Существование такого оптимума на-
сыщения установлено для многих видов животных рядом иссле-
дователей (Allee, 1931; Elton, 1933; Алпатов, 1934, и др.).
Т. Парк (1939:309) отметил: «С точки зрения популяции как
недостаточная плотность, так и избыточная плотность одина-
28Q
Ково вредны». Для одних видов оптимальной насыщенностью
будет уже редкое стояние (например, древесные породы в са-
ваннах) , для других, наоборот, чрезвычайная загущенность
(сфагновые мхи, камыши, тростники). Оптимальная густота сто-
яния может, таким образом, рассматриваться как исторически
обусловленный адаптивный признак вида.
У прогрессирующих и многих относительно стабилизирован
ных видов фактическое насыщение территории имеет тенден-
цию смещаться в область густоты выше оптимума. Разумеется,
что у границ своих географических и экологических ареалов та-
кие виды представлены популяциями, не насыщающими пло-
щади до оптимальной густоты.
У многих видов фактическое насыщение территории колеб-
лется вокруг оптимальной густоты. У некоторых же намечается
тенденция к сдвигу в область густоты ниже оптимума. Условием
стабильности вида является наличие популяций, насыщающих
пространство до оптимальной густоты.
Разумеется, оптимальная для вида плотность не является
признаком, постоянным во всех биоценотических и почвенно-
климатических условиях. У растений величина этого оптимума
меняется также с возрастом насаждений. Большую сложность
приобретает этот вопрос для разновозрастных насаждений.
Наконец, оптимум загущения зависит и от площади элемен-
тарного насаждения (гнездо-куртина или заросль большей пло-
щади).
Возможны два подхода к выбору критериев оптимальной гу-
стоты. Во-первых, можно основываться на состоянии средней
особи и воспользоваться величиной произведения веса зеленой
массы на плодовитость, т. е. коэффициентом жизненности (Co-
oper, 1926). Оптимальная густота будет определяться макси-
мальным значением этого коэффициента. Во-вторых, к опреде-
лению оптимальной густоты можно подходить от характеристики
насаждения в целом. В этом случае оптимальной густотой при-
знается та, при которой наблюдается наименьшая смертность ра-
стений и максимальный урожай семян (численность биологиче-
ски полноценных семян с единицы площади).
С нашей точки зрения, густота стояния — это явление попу-
ляционной жизни п за элементарную единицу анализа следует
принять не особь, а отдельное насаждение. Густота стояния ра-
стений, при которой будет достигнуто наивысшее развитие ви-
довой функции насаждения, и должна быть признана оптималь-
ной.
Выживаемость и урожайность семян — это показатели, объ-
ективно отражающие степень развития видовой функции насаж-
дения только в том случае, если они будут рассматриваться сов-
местно. Если упустить, например, показатель выживаемости, то
в разряд оптимальных густот можно отнести явно перенаселен-
ия!
ные в молодом возрасте насаждения, ставшие вторично малочис-
ленными из-за большого изреживания. Оптимально густое на-
саждение должно давать максимальный урожай при наилучшей
сохранности растений.
Остановимся теперь на вопросе о степени развития особи
в условиях оптимальной густоты.
Какую биомассу и плодовитость особи следует признать адап-
тивной нормой, наиболее соответствующей ее видовой функции?
Таким критерием часто признают показатели, достигаемые
особью в условиях экологического оптимума при свободном сто-
янии. Всякое же ограничение роста, ведущее к снижению био-
массы и плодовитости, наблюдающееся в загущенных посевах,
рассматривается как угнетение. Против такого подхода высказы-
вались В. Б. Сочава (1926), Л. И. Успенская (1929) и мы (За-
вадский, 1947 и др.). В самом деле, пластичность роста — одно
из коренных приспособлений растений. Отбор на увеличение
пластичности роста, как и всякий отбор на усиление приспособ-
ляемости (на расширение нормы реагирования), вследствие фи-
зиологических корреляций создает известный «запас адаптиро-
ванности». Максимальные показатели, полученные в условиях
свободного роста, часто представляют собой именно эту, обычно
нереализуемую в естественной обстановке, возможность.
Оптимальными размерами растения являются такие, при
которых лучше всего реализуется видовая функция средней
особи. Максимальный размер особи может осуществляться при
одиночном стоянии, но именно в этих условиях выживаемость
иногда бывает низкой. Например, 95% особей гибнет, а 5%
оставшихся, несмотря на полное развитие всех потенций, все же
не компенсирует высокой смертности и не обеспечивает воспро-
изведения вида. Напротив, при известном загущении потенциалы
роста и воспроизведения каждой средней особи оказы-
ваются реализованными далеко не полностью, но выживаемость
в этих условиях может быть максимально высокой. В итоге ви-
довая функция каждой средней особи реализуется здесь гораздо
полнее. Максимальное развитие особи совпадает с оптимумом
видовой функции особи только в тех случаях, когда достигается
лучшая выживаемость в условиях свободного стояния. У многих
видов максимум выживаемости сдвинут в область более или
менее значительных загущений, и поэтому оптимум видовой
функции средних особей сопряжен с постоянным ограничением
их роста и плодовитости. Именно эти размеры и плодовитость
растения (нередко сильно сниженные по сравнению с макси-
мально возможными) и следует принять за адаптивную норму
для вида.
Итак, отождествление максимально возможного развития
отдельно взятой особи с ее оптимальным развитием является
ошибочным, так как основано на произвольном отрыве особи
282
от вида. Показанное нами противоречие носит объективный ха-
рактер.
Сам способ существования растений делает невозможным
строгую фиксированность границ роста, которая присуща, на-
пример, высшим животным. Именно широкая приспособляемость
в онтогенезе к изменениям размеров площади питания — одна из
коренных адаптивных особенностей наземных высших растений.
Конечно, и в эволюции растений происходит сужение широких
приспособительных изменений роста (например, ограничение
роста при переходе от древесной формы роста к травянистой,
у однолетних монокарпических видов и пр.). Но это сужение
никогда не бывает столь полным, как у высших животных.
Указанная широта приспособляемости растений к количеству
пищевых ресурсов вызывает трудности при определении состоя-
ния перенаселения. При поверхностном подходе к вопросу боль-
шие густоты определялись как перенаселение. Какие же крите-
рии избирались при этом для определения нормальной густоты?
Таким критерием являлось то самое максимальное развитие ра-
стения в свободном состоянии, о котором речь шла выше. При-
меняя такие критерии, можно прийти к субъективному опреде-
лению перенаселения, называя им всякое большое загущение,
при котором естественно ограничивается рост особей.
Объективный анализ таких загущений предполагает изучение
реакции самих растений в связи с выполнением ими видовых
функций. Если эти реакции носят адаптивный характер, выжи-
ваемость велика, видовая функция насаждения осуществляется
на высоком уровне, то такую густоту, как бы она количественно
ни была высока, нельзя относить к вредно-избыточной, т. е. к пе-
ренаселению.
Одна и та же урожайность насаждения может быть достиг-
нута различными сочетаниями следующих сопряженно изменя-
ющихся показателей: численности растений на площади, веге-
тативного роста (веса, биомассы) одного растения, плодовитости
одного растения, размаха изменчивости растений по весу и по
плодовитости. Она может быть достигнута, например, малым чис-
лом крупных высокоплодовитых особей или же большим числом
мелких низкоплодовитых особей, а также средним числом осо-
бей, очень различающихся по весу и плодовитости, и т. д.
В природе известны самые различные способы осуществле-
ния рассматриваемых соотношений. Одни популяции воспроиз-
водятся по принципу «мало крупных», другие же — по принципу
«много мелких» и т. п. Максимальная же урожайность вида
в данных экологических условиях осуществляется всегда при
определенном сопряжении показателей густоты насаждения, веса
п плодовитости растений. Именно эту-то густоту мы и называем
оптимальной. У тех видов, которые обладают высокой приспо-
собляемостью к загущению, корреляция между численностью ра-
283
стений на площади, их весом и плодовитостью будет близка
к единице не в одной строго определенной густоте стояния,
а в целой зоне густоты, в пределах которой проявляется высокая
отрегулированность всей системы.
При любых колебаниях численности можно установить опти-
мальные плотности, при которых достигается самая высокая вы-
живаемость и воспроизводймость вида. Абсолютная величина
оптимальной плотности может быть очень различна у разных
видов. Меняется она и у одного и того же вида в зависимости
от возраста и условий существования.
Для многих видов высших растений было установлено (Су-
качев, 1927, 1935, 1941, 1953; Успенская, 1929; Смирнова, 1928;
Завадский, 1947, 1954а, 1957а, и др.), что наивысшая сохран-
ность растений и максимальная продуктивность насаждения до-
стигаются не в очень редких и очень густых посевах, а в опре-
деленной зоне густот, специфической для каждого вида.
В результате обширных исследований, проведенных на жи-
вотных у различных видов инфузории, иглокожих, ракообраз-
ных, насекомых, рыб, была установлена та же закономерность.
В зависимости от плотности изменяются такие важные показа-
тели, как рост особи, интенсивность обмена, продолжительность
периода созревания, иммунность и т. и. При этом в ряде слу-
чаев оказалось, что наилучшее морфологическое и физиологиче-
ское развитие особи достигается не при одиночном содер-
жании, а при определенных и иногда весьма значительных
плотностях.
Плотность, наилучшая для развития отдельной особи (физио-
логический оптимум), нё всегда совпадает с оптимальной плот-
ностью для воспроизведения вида (популяционный оптимум), и
эти показатели могут быть даже очень далеки друг от друга.
Могут сложиться такие условия межвидовой борьбы или такие
сочетания абиотических факторов, при которых лучшая сохран-
ность вида достигается в плотностях, иногда значительно огра-
ничивающих развитие особи. Так, например, в опытах с рядом
растений (гречиха, просо, морковь, пшеница, лен и др.) было
показано, что при малых плотностях гибель растений оказы-
вается такой большой, что не компенсируется преимуществами
в росте п плодовитости немногих сохранившихся растений. При
больших же плотностях сохранность растений значительно воз-
растает и, хотя при этом ограничиваются рост и плодовитость
средней особи, воспроизводимость вида все же оказывается са-
мой высокой.
Видовую функцию средняя особь популяции осуществляет не
изолированно, а будучи связана системными отношениями. Эти
системные отношения (в данном случае плотность) определен-
ным образом ограничивают особь и дают ей возможность проя-
вить лишь такие реакции, которые адаптивны для системы в це-
284.
лом. Поэтому уменьшение размеров особи и снижение ее
плодовитости в условиях оптимальной плотности (по сравнению
с возможными для данного генотипа в иных условиях) необхо-
димо рассматривать не как угнетение особи, а как адаптивную
(для вида) модификацию.
Поддержание определенной плотности популяции имеет боль-
шое значение для существования вида и постоянно служит
объектом деятельности отбора.
Способы регулирования численности. На плот-
ность популяции воздействует много факторов. Одни действуют
односторонне, изменяя численность, но не регулируя ее. Другие
действуют по принципу отрицательной обратной связи, изменяя
свою силу в зависимости от плотности популяции, и поэтому
являются регулирующими. Справедливо указывается на несо-
вершенство действия этих регуляторов, реагирующих не па само
воздействие, а на его следствие, т. е. па отклонение плотности
(Pimental, 1961; Milne, 1962, и др.). Известно, что такие регу-
ляции осуществляются с запозданием, вызывая более или ме-
нее значительные флюктуации (Эшби, 1959). Регулировать плот-
ность популяции насекомых могут только некоторые биотические
факторы, а все абиотические факторы действуют односторонне
(Викторов, 1965). С нашей точки зрения, обоснованной опытами
на модельных популяциях растений, такое подразделение искус-
ственно, так как любые факторы могут способствовать повыше-
нию плотности популяции, а верхний ее предел определяется
в конечном счете «автоматическим контролем» (Элтон, 1960),
т. е. авторегуляторно (Завадский, 1947, и др.). И у насекомых
повышение плотности популяций после достижения известного
порога вызывает разнообразные авторегуляторные реакции.
Д. Лэк выступает в защиту идеи саморегуляции численности и
утверждает, что «регулирующие факторы действуют сильнее при
высокой, чем при низкой, плотности населения» (1957:334). До-
казано, что у одних видов снижается плодовитость, у других
происходит резорбция яиц, у третьих возникает или усиливается
каннибализм, у четвертых начинается миграция гусениц или
взрослых форм, у пятых наступает диапауза, у шестых тем или
иным путем возрастает смертность, и т. и.
По новым данным Р Морриса и других канадских энтомо-
логов (Викторов, 1965), у листоядных и стволовых вредителей
наблюдаются две формы регуляции численности, сменяющиеся
в зависимости от плотности: при низкой численности она удер-
живается только биоценотическими факторами — естественными
врагами, а при высокой — лимитируются запасы пищи, за обла-
дание которыми возникает острая внутривидовая конкуренция.
В тех случаях, когда главным фактором, ограничивающим чис-
ленность вида у животных, является пища, следует учитывать,
что сильную задержку роста популяции может вызвать даже и
285
кратковремнное голодание в критические периоды, например
у птиц в конце зимы и в конце лета (Лэк, 1957).
У некоторых видов, постоянно испытывающих вспышки мас-
сового размножения (некоторые стадные саранчовые, совки
и др.), возникают адаптации, позволяющие избегать перенаселе-
ния с помощью миграций. Когда плотность популяции достигает
определенного порога, частота контактов особей возрастает так,
что служит сигналом для формирования миграционной фазы, или
фазы активной гусеницы.
У колониальных видов регулирование численности особей
в колонии осуществляется, как уже отмечалось, генетическими
механизмами, а также кормлением, особым воспитанием, изгна-
нием лишних индивидов, их умерщвлением и т. п.
Волны жизни являются неизбежным следствием существова-
ния такой регуляции. Совокупность признаков этих волн (длина,
высота, вариабельность самих волн й пр.) специфичны для
вида, являясь выражением взаимодействия генетической и фено-
типической систем вида с регулирующими и односторонне изме-
няющимися факторами. Например, у видов, приспособленных
к интенсивной элиминации хищниками или паразитами, спо-
собных безущербно и быстро восстанавливать численность с
помощью высокой плодовитости, быстрой смены поколений
(ранняя половая зрелость, низкая средняя продолжительность
жизни, малые размеры), волны жизни имеют совершенно иные
параметры, чем у крупных долгоживущих млекопитающих,
и т. п.
При возрастании, плотности популяции высших животных,
помимо голодания, эпизоотий, нехватки мест для размножения и
убежищ, высокая плотность выступает как самостоятельный
фактор, вызывающий снижение размножаемости и элиминацию
особей с наибольшей реактивностью (ранимостью) на избыток
контактов с другими особями по сравнению с определенной адап-
тивной нормой. Это явление было названо стрессом (Селье).
«Подавление интенсивности размножения животных при высо-
кой их плотности доказано для млекопитающих и, очевидно, свя-
зано с реакцией их центральной нервной системы на избыток
„посторонних" особей и последующим нарушением нормальной
деятельности ряда эндокринных желез» (Калабухов, 1962:31).
Речь идет об увеличении продукции адренокортикотропинов из-
за гиперфункции надпочечников. Многие авторы считают стресс
важным фактором регуляции плотности популяций. В полевых
опытах с сурками было показано, что при отлове 30% взрослых
особей снизилась эмбриональная смертность и возросло число
беременных и рожающих самок (Snyder, 1961). В эксперименте
(морские свинки, обезьяны, грызуны) увеличение плотности мо-
дельных популяций вызывало эффект стресса, т. е. понижение
плодовитости, иммунитета и рост смертности (Christian, 1961).
286
Однако К. Мандэй в обстоятельном анализе доказывает, что роль
стресса в регуляции численности до сих пор не может считаться
вполне доказанной, так как перенос выводов лабораторных опы-
тов на природные популяции «не представляется возможным»
(1964:239).
Эти данные интересны тем, что показывают один из возмож-
ных механизмов регулирования плотности популяции с помощью
«ошибок», т. е. перенаселения, или «вредной избыточности».
Регулирование плотности популяции при ее отклонениях от
нормы у примитивных форм осуществляется путем выделения
в окружающую среду биологически активных веществ, действу-
ющих на темп размножения. У водоросли Chlorella vulgaris ве-
ществом, регулирующим плотность популяции, является хлорел-
лин. Как установили Р. Пратт с соавторами (Pratt a. al., 1945),
хлореллин выделяется в окружающую среду самой водорослью
и в малых концентрациях стимулирует ее размножение, а в боль-
ших, наоборот, тормозит его. При возрастании численности осо-
бей в определенном объеме воды увеличивается и концентрация
хлореллина, а этим самым задерживается дальнейший рост плот-
ности популяции.
У высших растений регулирование плотности популяции осу-
ществляется как с помощью конкуренции, приводящей к гибели
известное количество особей, так и путем изменений скоростей
ростовых процессов в зависимости от густоты стояния. Эта спо-
собность растений, которую можно назвать аккомодацией роста
(Завадский, 1947, и др.), представляет собой приспособительную
реакцию, с помощью которой сохраняется необходимое соответ-
ствие между площадью питания и размерами (а значит, и по-
требностями) особи. В опытах с Trinia henningii В. Н. Сукачев
(1941) впервые отметил адаптивный характер такой реакции на
загущение: в течение ряда лет существовали особи, которые
почти не росли и не плодоносили, но не потеряли способности
к росту и плодоношению при увеличении площади питания. Та-
кие растения образуют как бы резервный фонд популяции, так
как их видовая функция оказалась только временно заторможен-
ной. Эта своеобразная видовая адаптация многолетников к усло-
виям высоких загущений была обнаружена и специально изу-
чена нами у кок-сагыза.
В больших густотах (18 000 всходов на 1 м2) растения кок-
сагыза надолго как бы замораживались в ювенильном состоянии.
При этом выживаемость оказалась весьма высокой: через 3.5 года
от начала опыта — более 40%. Интересно, что в контроле (35
растений на 1 м2) в живых осталось лишь* 18% растений. Го-
товность к цветению таких мелких растеньиц была подтверждена
опытами с пересадкой «на свободу», с прореживанием густых
посевов и с подкормкой. Если судить по площади питания, не-
обходимой для обычного среднего растения кок-сагыза (300 см2),
287
То густота стояния растений в разбираемом опыте была чрезвы-
чайно высокой (площадь питания растений была 0.6—1.8 см2,
т. е. в 600—150 раз меньше обычной). Вместе с тем ростовые
взаимодействия в этих густотах почти отсутствовали, так как
растения находились в состоянии полуанабиоза.
Эта обратимая инактивация многолетников в больших загу-
щениях является адаптивной реакцией на ухудшение условий
питания и на отсутствие необходимого пространства. Однако до-
статочно активизировать ростовые процессы, чтобы возникли
самые острые взаимодействия между растениями (Завадский,
1954:532).
Многие однолетние растения способны к аккомодации роста
в чрезвычайно широких пределах (Polygonum aviculare, Setaria
erigeron и др.). При выращивании мари белой (Chenopodium
album) по два растения на 1 м2 вес отдельных особей достигает
нескольких килограммов, а плодовитость 100—250 тыс. семян,
в то время как в очень больших густотах (10 000 на м2) вес
растений надает до 0.7—7.0 г, а плодовитость — до 1 — 63 (сред-
няя 57) штук семян (Завадский, 1954г, 1957а). Общее же число
семян, собранных с площади (урожайность), и выживаемость
растений как в редких, так и в густых посевах оказываются
примерно одинаковыми. Таким образом, уменьшение размеров
особи в тысячи раз не сопровождалось какими-либо болезнен-
ными явлениями, которые привели бы к нарушению ее видовой
функции.
Аккомодация роста изучалась нами у садового мака. Расте-
ния, выросшие без полива на гальке с песком, плодоносили, имея
вес 0.2 г и высоту 1.5 см. Такую же карликовость вызывало и
очень большое загущение. При редком посеве с поливом и под-
кормками плодоносящие растения в среднем весили более 400 г.
С помощью аккомодации роста в популяции может поддер-
живаться оптимальная плотность и при этом воспроизводитель-
ная способность всей группы растений остается постоянной;
у некоторых видов при этом увеличивается продолжительность
жизни особи и т. п. Подобная реакция присуща и некоторым
древесным (например, «замирание» на многие годы молодых
дубков под материнским пологом).
В работе А. Н. Папонова (1959) показано, что салат и то-
маты реагируют на загущение замедлением развития. Эти факты
на первый взгляд противоречат положению, названному мной
ранее (Завадский, 1947, 1954г) «правилом Сукачева». Это пра-
вило состоит в том, что однолетним растениям с тенденцией
к эфемерности в ответ на загущение свойственно ускорять раз-
витие, а многолетним растениям — замедлять. Такая способность
является важной адаптацией, позволяющей приурочивать сроки
размножения к временам года и биоценотической обстановке,
прп которых наиболее вероятно сохранение потомства.
288
Данные А. Н. Папонова интересны, но, как нам представ-
ляется, они не противоречат «правилу Сукачева». Вот что, на-
пример, пишет о томате настоящем (Lycopersicum esculentum)
11. М. Жуковский (1964:670): «Разводят томат как однолетнее
растение, но по природе своей он является многолетником».
Примерно то же можно сказать и о салате. Эти объекты не
имеют тенденции к эфемеризации развития.
6. ПЕРЕНАСЕЛЕНИЕ И ЕГО СЛЕДСТВИЯ
Определение понятий. Вопрос о перенаселении яв-
ляется частью более широкой проблемы динамики численности
популяций. Перенаселение — это противоречивое по своей основе
следствие процветания вида, роста его численности. Такие виды
ломают сложившиеся межвидовые отношения, выводят экологи-
ческие системы из равновесия и не только расселяются, по и на-
сыщают уже ранее освоенную территорию (Connell, Orias, 1965;
Lack, 1965; Zarzycki, 1965, и др.). Именно для таких процветаю-
щих, а не для стабилизированных или тем более не для угасаю-
щих видов и закономерно явление перенаселения (Завадский,
1947, 1957а).
В вопросе о распространенности явления перенаселения
в природе и о ее значении для эволюции были высказаны раз-
личные точки зрения. Одни считают, что перенаселение имеет
всеобщее распространение и является необходимым фактором
естественного отбора (Уоллес, 1911 27—40; Тимирязев, 1939,
6 154; Васнецов, 1946:194—195, и др.) или же имеет большое
значение для эволюции, но не является всеобщим и единствен-
ным фактором, вызывающим процесс естественного отбора (Дар-
вин, 1937). Другие полагают, что перенаселение — частный слу-
чай динамики численности вида; его следствием является узкая
специализация, сокращение плодовитости, снижение темпа эво-
люции, закрытие «возможности развития в направлении широ-
кой прогрессивной эволюции» (Шмальгаузен, 19466:254).
По Ф. Энгельсу (1961 621), перенаселение имеет место на из-
вестных ступенях развития растительного царства и на низших
ступенях развития животного царства, но основной путь эволю-
ции осуществляется без его участия. Соглашаясь с тем, что пере-
население — довольно распространенное явление, некоторые ут-
верждали, что оно представляет собой фактор, скорее не обус-
ловливающий, а сдерживающий эволюцию (Мечников, 1950: 146—
186). Существует и убеждение, что перенаселение возникает
спорадически и для эволюции имеет лишь отрицательное зна-
чение (Берг, 1922), так как вызывает угнетение всех организ-
мов или их гибель (теоретические соображения в защиту этой
идеи можно найти у К. Ф. Кесслера в работе 1880 г.,
у П. А. Крапоткина в работе 1890 г., а попытки эксперимен-
19 К. М. Завадский
289
тальных доказательств этого положения см.: Любименко и др.,
1925; Самофал, 1931 126, и др.). Возможность перенаселения
отрицал еще Ламарк, считавший, что «род необходимого равно-
весия» всегда сохраняется благодаря неодолимым границам
размножения, предусмотрительно поставленным природой
(1935:88-89).
Из перечисленных определений некоторые полностью несосто-
ятельны, другие построены на абсолютизации частных случаев
и также ошибочны. Точки зрения Дарвина, Энгельса и Шмаль-
гаузена в целом согласуются с данными современной биологии,
однако они в известной мере схематичны, так как основаны
в большей степени на общих логических предпосылках, нежели
на обобщении точных фактов. Основным недостатком является
то, что само-то явление перенаселения не уточнено, оно прини-
малось как сравнительно простое, всегда одинаковое по содер-
жанию и следствиям.
Очень возможно, что разные авторы под перенаселением
имели в виду различные конкретные явления, например различ-
ные степени плотности, и поэтому приходили к разным выводам
относительно его значения.
Вся проблема в настоящее время упирается в неразработан-
ность самого понятия о перенаселении. Не установлен круг
явлений, который должен быть отнесен к перенаселению. Прежде
всего надо найти показатели (критерии), позволяющие точно
выявлять состояние перенаселения. Оно возникает при таком со-
четании плотности популяции с условиями (пища, площадь
и т. п.), которое вызывает острые взаимодействия между особями.
С биологической точки ‘ зрения только эти показатели являются
критериями перенаселения.
Основываясь на опытах с растениями, нами было предложено
ввести понятия «абсолютное перенаселение», «относительное»,
«возрастное» и «условное», а также понятия «степень пере-
населения» и «локальное перенаселение» (Завадский, 1957а,
1961).
При абсолютном перенаселении неизбежно возникает массовая
гибель, имеющая избирательный или неизбирательный характер.
Примером абсолютного перенаселения с избирательной гибелью
являются густые посевы кок-сагыза в гнездах малой площади (см.
выше), в которых переживают немногие «победители» в смысле
Дарвина, т. е. происходит жесткий отбор. Тотальная элиминация
при перенаселении обнаружена у личинок мухи Lucilia, живущих
в трупах овец, если начальная плотность (яиц) очень велика и
личинки успевают съесть все запасы пищи задолго до окончания
развития (Nicholson, 1950).
Под относительным перенаселением понимаются плотности,
вызывающие некоторое увеличение гибели по сравнению с ги-
белью при оптимальной плотности [т. е. выше «пороговой
290
густоты» в смысле Де Вита (1964)]. Как правило, гибель расте-
ний в этих случаях имеет избирательный характер, но приводит
к более мягким формам отбора, чем при абсолютном перенаселении.
Возрастное перенаселение возникает прп обострении взаимо-
действий между индивидами, которое приурочено к определенным,
иногда небольшим отрезкам времени, названным нами критиче-
скими периодами в жизни популяции. В результате популяция,
бывшая перенаселенной, быстро изреживается (Закарьян, 1930).
Условное перенаселение — такие плотности, в которых времен-
ная и обратимая приостановка роста растений снижает остроту
взаимодействий между ними пли же, прп полной инактивации,
прекращает их. Достаточно вынудить растения к активному росту,
и возникает уже настоящее перенаселение.
Степень перенаселения может быть различной в зависимости
от сочетания многих факторов.
Первичная плотность, т. е. численность, зачатков (Семян, всхо-
дов, яиц, личинок) па единице площади определяет только воз-
можность перенаселения. Если первичная плотность мала, воз-
можность перенаселения исключена, по даже и огромная
первичная густота сама по себе не может вызвать перенаселения.
Важным фактором является соответствие условий обитания эко-
логическому оптимуму вида и размеры запасов доступных пита-
тельных ресурсов. У растений этот фактор влияет на скорость
роста и при данной первичной густоте может обусловить тот пли
иной срок начала взаимодействий и их интенсивность. Следова-
тельно, существует прямая зависимость между степенью перена-
селения и совокупностью условий, стимулирующих рост. Один
из факторов — гибель растений из-за действия биотических и
абиотических агентов (поломки, болезни, поедания, вымерзание,
вымочки и пр.). Этот фактор находится в обратной зависимости
от степени перенаселения. Фактором, влияющим на степень пере-
населения, выступает и состав популяции: чем разнороднее попу-
ляция по признакам, биологически важным в условиях загущения,
тем степень перенаселения при равенстве остальных условий
будет меньше (закон Дарвина: максимум жизни достигается
при наибольшем разнообразии компонентов). Площадь элементар-
ной популяции как фактор стоит в обратной зависимости к нали-
чию доступных запасов питания и к скорости роста и с уменьше-
нием ее размеров (при достаточно высокой первичной плотности)
степень перенаселения увеличивается. Наконец, фактором, обус-
ловливающим степень перенаселения, является и адаптивное
реагирование на плотность (откочевка, изменение группового по-
ведения, аккомодация роста, уход в покой и т.* п.). Чем выше
развита эта реакция, тем совершеннее популяция нейтрализует
и родные последствия недостатков в питании, в местах размноже-
ния, в пространстве и т. п. Она способна уменьшить степень пе-
ренаселенности или вовсе снять это состояние.
19*
291
Локальными перенаселениямп мы называем скопления расте-
ний на столь малой территории, что переживание той или иной
особи в основном объясняется не местоположением ее в насажде-
нии (краевое или серединное), а индивидуальными качествами.
Если вокруг такого элементарного насаждения есть свободная
территория, то это только способствует более энергичному росту
всходов, а следовательно, более интенсивным взаимодействиям и
ведет к более энергичному вытеснению одних особей другими.
Наиболее яркие примеры действия естественного отбора получены
нами именно при локальном перенаселении.
О последствиях перенаселения. В наших модельных
опытах (Завадский, 1947, и др.) установлено, что перенаселение
может вызвать естественный отбор, либо адаптивное реагирование
путем торможения ростовых процессов и генеративного развития,
либо, наконец, общую гибель растений (образование «кольцевого»
насаждения).
Было подтверждено существование внутривидового отбора и
доказано, что его направление может быть весьма разнообразным
в зависимости от конкретного сочетания напряженности отбираю-
щего агента, особенностей популяции и характера условий, в ко-
торых протекает этот процесс. Одно из изученных нами направ-
лений — отбор на скорость роста корневой системы. У кок-сагыза
он приводит к переживанию более быстроцветущих групп;
в 5-летнем опыте (1949—1954 гг.) одни и те же условия перена-
селения были созданы для пятп поколений, в результате чего
состав популяции оказался сдвинутым в сторону групп, зацве-
тающих в первый год жизни. Другое направление — отбор на ско-
рость образования и большую мощность розетки листьев — у кок-
сагыза возникает в насаждениях менее загущенных, чем в преды-
дущем случае, когда взаимодействия начинаются позднее.
Наблюдался отбор на скорость прорастания семян: растения,
взошедшие первыми, занимают пространство, а поздние всходы,
попадая под розетки соседей, подавляются ими и элиминируются.
Интересен случай отбора на увеличение продолжительности
периода покоя семян: при очень больших первичных густотах
растения, взошедшие первыми, все погибают, а переживают «пе-
реждавшие» в семенах. В этом случае, как и в предыдущем,
большое значение для определения направления отбора имеет
количественное соотношение фракций семян по длине периода
покоя.
Наконец, нами был изучен отбор на более совершенную
аккомодацию роста или на способность к уходу в летний покой;
такое направление отбор приобретает в том случае, если менее
совершенная регуляция роста пли более низкий порог активации
ростовых процессов ставят растение в невыгодное положение
по отношению к соседям. Так, например, в густых насаждениях
кок-сагыза из растений, ушедших в покой, гибнут те, которые
292
провоцируются к выходу из покоя даже малыми дозами полива;
начав рост, такие растения испытывают трудности в водоснабже-
нии, ио, утратив способность вновь уйти в покои, погибают; пере-
живали же те формы, у которых покои более глубок и которые
не провоцировались на активацию роста. Кроме того, были уста-
новлены преимущества большей теневыносливости сеянцев и
наследственно обусловленной неветвистости и некустистости
(а при иных загущениях, наоборот, ветвистости п кустистости),
т. е. прямостоячих форм куста над раскидистым и распластанным
(а при иных условиях — наоборот), и т. п.
При известных степенях перенаселения вполне сохраняется
селективное значение различий между особями внутри популя-
ции и между морфо-бпологическими группами и др. Наличие
естественного отбора в условиях перенаселения было по-
казано А. Л. Зеликманом и А. К. Гейнрих (1959) на циклопах
п т. п.
Имея все это в виду, мы не можем согласиться с мнением
И. И. Шмальгаузена (1946а, 19466), что в результате перенасе-
ления всегда переживают узкоспециализированные формы. Надо
учесть, что направление отбора меняется в зависимости от сте-
пени перенаселения. При определенных условиях перенаселения
происходит отбор таких форм, которые вполне заслуживают
наименование победителей в смысле Дарвина. Одно пз следствий
отбора в условиях перенаселения — развитие адаптивных реакций
па загущение (аккомодация роста, уход в покой и пр.). Этот
процесс эволюционного расширения зоны оптимальных густот
приводит к тому, что у многих прогрессирующих видов густоты,
ранее бывшие излишними, впоследствии становятся нормальными.
Такие густоты, войдя в норму адаптивного реагирования, пере-
стают быть перенаселением.
Разобранный путь эволюции характерен для биологически
прогрессирующих видов, живущих в условиях резко изменяю-
щихся густот. Если же повышение густоты оказывается более
постоянным (часто встречающимся) условием расселения, то от-
бор идет не па увеличение пластичности роста, а па генетически
фиксированное уменьшение скорости и периода роста. В резуль-
тате сокращаются максимально возможный размер растения и его
плодовитость.
У видов, приспособившихся к условиям повышенных загуще-
ний, происходит смещение экологического оптимума в сторону
показателей, обычных для таких густот (освещенность, темпера-
тура, влажность, движение и газовый состав воздуха, водоснаб-
жение и минеральное питание, и т. п.). Естественно, что для таких
форм изреживание насаждений и тем более «свободное стояние»
могут вызвать повышенную смертность (Успенская, 1929), общую
дспрессию или даже общую гибель (как у некоторых сфагновых
м хов).
293
У животных установлено много разнообразных механизмов,
противодействующих перенаселению. Эти механизмы могли воз-
никнуть путем отбора в условиях перенаселения.
Естественно ожидать, что у процветающих видов с высокой
плодовитостью и с большими волнами жизни значительные сте-
пени перенаселения возникают часто, а иногда и периодически
почти при каждом массовом размножении (пике численности).
О некоторых следствиях эволюции в таких условиях уже упомп-
0 0.5 1.0 1.5 20 2.5 3.0 3.5 k.O Ь.5 5.0 5.5 6.0
Расстояние между особями,мм
Рис. 5. Распределение расстояний между со-
седними особями Spirorbis в углублениях,
окаймляющих главные жилки вайи Fucus
serratus (Найт-Джонс и Мойз, 1964).
1 — большая плотность, 2 — малая плотность попу-
ляций
налось в предыдущем
разделе. Сейчас же по-
дробнее рассмотрим не-
которые примеры.
У видов, имеющих
активно движущихся
личинок и прикреплен-
ных взрослых особей,
возникают специальные
поведенческие адапта-
ции прикрепления.
Примером может слу-
жить выбор места для
посадки на субстрат
у личинок устриц, мор-
ских желудей и червей
(Spirorbis из полихет,
рис. 5). Этот выбор,
как было доказано
(Найт-Джонс и Мойз,
1964), сопровождается
сложными движениями
личинки, которая долго
кружится над дном и всегда садится не на случайное место,
а в центр участка, свободного от других личинок. С увеличением
плотности заселения будущей устричной банки поведение личи-
нок не меняется. Такой «механизм распределения в пространстве
составляет часть специфического поведения личинок, связанного
с прикреплением» (там же: 98).
Виды, сверлящие субстрат, например личинки, минирующие
ткани листа, как у тополевой моли, или древесину, как у лубоедов
или у «корабельного червя» (Toredo), при высоких плотностях
все же избегают встречи друг с другом. Такие личинки, вероятно,
имеют какие-то устройства, предупреждающие о близости пустот,
а в некоторых случаях способные даже отличать на расстоянии
ходы своего вида от чужих ходов. При сверх гереиаселении, од-
нако, может произойти «самоудушение популяции», т. е. общая
гпбель личинок, как это наблюдали многие (Ильинский, 1929;
Полежаев, 1934, и др.). В гельминтологической литературе при-
294
водится Много данных об огромных изменениях размеров парази-
тических червей в зависимости от плотности популяции. Как и
у растений, эти изменения размеров можно рассматривать как
приспособительную «аккомодацию роста». Все эти приспособления
могли возникнуть только в процессе внутривидового отбора в ус-
ловиях перенаселения. Именно под влиянием перенаселения
могли элиминироваться личинки с ошибочными реакциями, веду-
щими к вредной скученности взрослых организмов и к ухудшению
существования всей популяции,.
Телеономический характер реакций и структур присущ и
падорганизменпым системам. Регуляционные процессы, останав-
ливающие тем или иным путем рост плотности популяции, отно-
сятся именно к таким телеономическим реакциям. Причиной телсо-
номичности надоргапизменпых систем, в частности регулирую-
щих. плотность, численность популяций и плодовитость* является
комбинированное действие индивидуального и группового отбора.
7. ВИДОВЫЕ КОРРЕЛЯЦИИ
Различные связи и зависимости между различными популя-
ционно-видовыми и организменными адаптациями можно назвать
видовыми корреляциями. Так, например, это зависимости между
численностью вида, оптимальной плотностью популяции, спосо-
бами и степенью совершенства первичных надиндивидуальных
объединений, заботой о потомстве и других основных внутриви-
довых отношений, средней выживаемостью особей, плодовитостью,
размерами тела, долголетием, сроками созревания, ритмами раз-
множения и т. п.
Структура вида и строй внутривидовых отношений находятся
в тесной зависимости от соотношения между отдельными
звеньями видовых корреляций. Вид с населением в сотни трил-
лионов субмикроскопических агамных особей, живущих только
десятки минут, с выживаемостью 0.0001%, коренным образом
отличается от вида с населением в сотни тысяч многоклеточных
крупных долголетних особей с выживаемостью 20% при низкой
плодовитости (например, одподетпость), длительной и совершенно
организованной заботе о молодняке в стаде и т. п. Значение
отдельной особи в жизни вида у таких форм резко различно.
У малочисленных, малоплодовитых видов с высокоразвитым долго-
живущим индивидом и совершенной интеграцией стад или других
объединений роль отдельно взятой особи в жизни вида весьма
велика по сравнению с примитивными видами, нечувствительными
к гибели десятков миллиардов особей. Вместе с тем совершенство
индивида и надиндпвпдуальной интеграции в миллионы раз повы-
шает вероятность выполнения каждой особью ее видовой функ-
ции, может компенсироваться огромной размножаемостью. Можно
представить себе два вида, которые не отличаются по суммарной
295
биомассе, но в то время как у одного вида она концентрируется
в небольшом числе огромных индивидов, у другого распылена
в мириадах мельчайших индивидов. Поэтому-то виды, одинаково
успешно воспроизводящиеся в природе, могут резко различаться
по соотношению между отдельными показателями видовых кор-
реляций. 1
Хорошая приспособленность видов примитивных групп (бак-
терии, сппезелепые водоросли, простейшие) достигается на основе
совершенно иных видовых корреляций, чем у видов высших групп
(например, млекопитающих), как это видно из следующей схемы:
Группа
видов
Численность
вида
Средняя Размеры
выживаемость Плодовитость тела 0Сдби
особи
Долголетие
Примитив- Очень боль- Очень низ-
ные. шая. кая.
Высшие. Малая. Высокая.
Очень вы- Микроско- Очень ма-
сокая. пические. лое.
Низкая. Большие. Большое.
Не следует, однако, сводить различия видовых корреляций
только к различиям в высоте организации. Велико бывает и воз-
действие биоценотпческих отношений.
Изменение всей системы видовых корреляций начинается
со сдвига в каком-нибудь одном звене. Например, появление
охраны потомства у рыб и амфибий обычно ведет к многократ-
ному повышению средней выживаемости икры и мальков и
к снижению плодовитости по сравнению с таковыми у видов, ли-
шенных заботы о потомстве. Колюшка в нерест мечет около 100
икринок, а близкие ей виды — тысячи. Пипа и жаба-повитуха
в год откладывают от 40 до 120 ипкринок, а некоторые лягушки
по многу десятков тысяч. Меняется и строй внутривидовых отно-
шений, например, в связи с тем, что функцию заботы о потом-
стве выполняет сцмец (колюшка, повитуха) или самка (пипа).
Рост пеизбирательпой смертности ведет к повышению плодови-
тости, что в свою очередь часто сопряжено с уменьшением разме-
ров особи, сокращением продолжительности жизни, увеличением
амплитуды пульсаций численности и т. п., как это наблюдается,
например, у некоторых грызунов (полевки, белки и т. п.).
Многие показатели в системе видовых корреляций бывают
более или менее строго фиксированными и изменяются только
в процесе отбора, а другие сильно модифицируют. Так, у многих
растений в зависимости от площади питания, богатства почвы и дру-
гих условий плодовитость, скорость роста и размеры особи могут
модифицировать в очень широких рамках. Аккомодация роста —
это видовая адаптация растений, позволяющая изменять видовые
корреляции в порядке онтогенетической регуляции (см. стр. 287).
Изменения видовых корреляций. Изменения видо-
вых корреляций у немертины Linens ruber были изучены
296
Г А. Шмидтом (1941 и др.). Он обнаружил, что резкое возраста-
ние численности взрослых немертин красной расы по сравнению
с зеленой объясняется увеличением продолжительности периода
инкубации личинок и большим снижением их смертности. Повы-
шение выживаемости личинок красной расы пе только компен-
сировало снижение плодовитости, по и привело к возрастанию
численности этой расы.
Нельзя не обратить внимания на обстоятельства, связанные
с причинами этого изменения видовой корреляции.
Во-первых, оставшиеся в живых личинки питаются трупами
своих сестер и братьев. Они используют их в качестве дополни-
тельного питания и поэтому дольше живут под охраной кокона,
выходят на свободу в более взрослом состоянии и гибнут в мень-
шем числе, что способствует процветанию вида, которое дости-
гается особой формой каннибализма (некрофагией).
Во-вторых, гибель большей части зародышей, ло-видимому,
объясняется накоплением в популяции мутаций с летальным
проявлением на ранних фазах развития. Насыщенность ими по-
пуляции весьма высока (отмирает 86 —97%. яиц в каждом коконе).
Такое насыщение популяции деталями поддерживается отбором
и является сбалансированным.
Далее было выяснено, что в пределах красной расы популяции
из Бретани (Франция) имеют или меньше япц в кладке, или
меньше нормально развивающихся личинок, чем популяции
из-под Мурманска п из Белого моря. Кроме того, поздние личинки
немертин пз Бретани оказались способными к дополнительному
заглатыванию отмерших яиц. Таким образом, красная раса сама
оказалась дифференцированной, а бретанская популяция — более
совершенной.
Рассмотрим еще один пример изменений видовых корреляций.
С. А. Северцов (1936, 1941, 1951) показал пути, которыми у видов
млекопитающих и птиц достигается связь между повышением
нормы выживания особи и понижением плодовитости. Этот про-
цесс протекает в несколько фаз. Отбор на приобретение новых
приспособлений приводит к повышению выживаемости средней
особи, в результате чего численность популяции возрастает. Если
новое приспособление позволяет виду выйти за пределы освоенной
нм территории (стации), то начинается расселение вида. Если же
возникает приспособление, дающее преимущества лишь в преде-
лах старой стации, то рост численности вызывает увеличение
плотности популяции. Возрастает конкуренция между особями
за пищу и т. и., и их условия существования ухудшаются.
У большинства видов .млекопитающих и птиц родители затра-
чивают много энергии на продуцирование потомства и его воспи-
тание. Величина кладки яиц у птиц изменяется отбором в соот-
ветствии со всей цепью видовых корреляций, главным звеном
которых является среднемаксимальное число птенцов, которые
297
могут быть выкормлены родителями (Лэк, 1957). В условиях
перенаселения самки ослабляются, наблюдаются случаи частичной
резорбции эмбрионов, сроки наступления половой зрелости затя-
гиваются, увеличивается число яловых самок и т. п. Наиболее
плодовитые самки оказываются более ослабленными ,п наименее
переживающими. Они будут погибать вместе с потомством чаще,
чем менее плодовитые самки (Шмальгаузен, 19466). Преимуще-
ство получают «наследственно менее плодовитые особи». Отбор
па понижение плодовитости приводит к уменьшению потенциала
воспроизведения, а это в свою очередь при постоянной выжива-
емости особей приводит к снижению численности популяции
до уровня, при котором устраняется ее слишком высокая плот-
ность. Уменьшение плотности популяции автоматически устра-
няет давление отбора в сторону понижения плодовитости. Таким
путем организуется новая структура видовых корреляций: выжи-
ваемость особей возрастает, плодовитость уменьшается, а числен-
ность в итоге может сохраниться па старом (исходном) уровне.
Для того чтобы был достигнут такой эффект, плотность популяции
продолжительное время должна быть высокой.
Следует обратить внимание еще и па обстоятельства, при кото-
рых новый уровень численности может оказаться более низким,
чем исходный. Это очень важный случай для понимания эволюции
видовых приспособлений. Такое понижение уровня численности
вида может произойти в том случае, если новым приспособлением
является, например, увеличение размеров тела животного или
повышение его пищевой активности. При этом во время перена-
селения давление отбора в сторону понижения плодовитости будет
более сильным, а уровень численности, при котором отбор в дан-
ном направлении прекратится, будет меньшим, чем исходный. Та-
кой результат является неизбежным следствием изменения вели-
чины оптимальной плотности популяции в связи с увеличением
индивидуальной площади питания (охотничьего участка и т. п.).
Оптимальная плотность популяции достигается теперь прп мень-
шей фактической плотности. Таким образом, увеличение размеров
тела животного или усиление его пищевой активности может
привести не только к повышению выживаемости и понижению
плодовитости, но и к снижению уровня численности вида.
На первый взгляд представляется, что этот пример отражает
не типичный для эволюции, а какой-то исключительный случай.
Казалось бы, что типичным результатом приобретения новых
приспособлений является расселение популяции за пределы ста-
рой стации, ведущее к совершенно иным эволюционным послед-
ствиям, чем только что разобранные. Напомним, что А. Н. Север-
цов (1939) считал этот последний случай основным и даже
единственным результатом приобретения видом нового приспо-
собления. «Биологический прогресс вида», по Северцову, всегда
проявляется в увеличении численности, расселении и дифферен-
298
Циации на подчиненные группы. Анализ современных экологиче-
ских данных показывает, что биологический прогресс вида
в понимании Северцова не является общим случаем. Бесспорно,
что приобретение популяцией нового приспособления обусловли-
вает увеличение ее численности. Однако при этом далеко пе всегда
происходит расселение популяции, т. е. занятие новых стаций.
Этому могут помешать чисто внешние преграды (если границы
ареала определяются физико-географическими факторами), основ-
ная экологическая специализация группы (степная, лесная,
живущая па каменистых россыпях и т. п.) или же занятость со-
седних и потенциально доступных стаций близкими расами (эво-
люция которых протекает параллельно), имеющими более высо-
кую конкурентоспособность. В случае островной, горной и т. п.
изоляции, так же как при узкой специализации к определенному
типу территории, возможность расселения вида часто практически
совершенно исключена. В подобных случаях увеличение числен-
ности приводит не к расселению, а к возрастанию плотности попу-
ляции.
При наличии потенциально доступных, но уже занятых стаций
тот или иной результат будет зависеть от относительной, конку-
рентоспособности видов (рас) при межгрупповом столкновении.
При этом новые приспособления, оказавшиеся эффективными
в условиях уже освоенной стации, могут и не повысить конкурен-
тоспособности вида в борьбе за новую стацию. Для захвата новых
стаций и вытеснения конкурентов вид должен приобрести приспо-
собления, дающие ему крупные преимущества, как это, например,
имело место при расселении серой крысы (и вытеснении черной)
или при расселении рака Astachus leptydactylus и вытеснения его
конкурента — рака A. astachus. Чаще, однако, возникают приспо-
собления, эффективные лишь в пределах данной стации. Таким
образом, более обычно возрастание численности без немедленного
расселения, т. е. биологический прогресс в пределах уже освоен-
ной стации, ведущий к повышению плотности популяции. При
этом следует учитывать степень оседлости и подвижности вида,
наличие пли отсутствие миграционных инстинктов, скорость,
с которой возрастает плотность популяции, и т. п.
Наконец, нельзя обойти и вопроса о действии на вид, увеличи-
вающих его численность факторов «биоценотического равнове-
сия». Допустим, что новое приспособление состоит в изменении
терморегуляции и численность вида начнет возрастать из-за сни-
жения гибели части особей во время зимовок. В этом случае воз-
можно быстрое увеличение силы действия факторов «биоценоти-
ческого равновесия» (например, хищников), и численность вида —
объекта, поднявшись, вновь упадет почти до старого уровня
(Гаузе, 1934; Gause, 1934, и др.).
Совсем иной результат будет достигнут в том случае, когда
повое приспособление позволяет сломать сложившееся биоцепо-
299
тическое равновесие, например, если возрастет пассивная или
активная оборона вида от хищников, иммунитет к паразитам и т. и.
Если возникшее приспособление способно на продолжитель-
ное время снизить смертность, вызываемую хищниками или пара-
зитами, то в течение известного периода будут беспрепятственно
осуществляться процессы роста численности, повышения плот-
ности популяции, отбора на понижение плодовитости и т. д.
Кроме того, не следует представлять себе дело так, что в усло-
виях роста численности и перенаселения действует лишь одна
форма отбора (т. е. внутривидовой отбор на снижение плодови-
тости). Отбор в природе одновременно происходит в нескольких
направлениях. Так, например, среди менее плодовитых самок
могут переживать и более защищенные от хищников пли более
иммунные. При ослаблении голодом животные гибнут чаще всего
под воздействием биотических агентов. Поэтому совсем не обя-
зательно, чтобы некоторый «отрыв» вида от хищников или пара-
зитов носил лишь временный характер. Разрыв биологических
связей жертвы и хищника может закрепиться п сохраниться
не только в силу приобретения видом жертвы новых приспособ-
лений (папример, сила животного и организованное стадо у бизо-
нов) , но также и потому, что не всегда у хищника возникают
приспособления, достаточные для компенсации возросшей защи-
щенности жертвы.
8. ЗАКЛЮЧЕНИЕ
Внутривидовые отношения не только сложное и многообразное,
по и крайне противоречивое биологическое явление. Будучи
одним из главных объектов отбора, они представляют собой отно-
сительно стройную систему регуляций и зависимостей между осо-
бями п группами особей. В результате деятельности отбора
формируются основные видовые отношения, выступающие в форме
видовых адаптаций. Однако внутри вида непрерывно возникает
масса различных противоречий. У процветающих видов одним
из ведущих противоречий является рост численности, могущий
привести к перенаселению. Эти противоречия одна из основных
причин внутривидового отбора. Подвергаясь отбору, популяции
реорганизуются, некоторые из внутривидовых противоречий полу-
чают разрешение, формируются новые видовые адаптации и про-
изводные внутривидовые отношения переходят в основные.
Чрезвычайно важно не упускать из виду того, что, хотя основ-
ные внутривидовые отношения и являются адаптациями, они
сами внутренне противоречивы. На многих примерах (конгруэн-
ция, сбалансированный полиморфизм популяций насекомых,
некрофагия у немертины Linens ruber и др.) можно убедиться
в далеко не гармонической основе их образования. Популяционно-
видовая адаптация организуется за счет снижения физиологиче-
ских показателей у организма или же гибели части зигот (личи-
300
нок) из-за накопления деталей и т. п. Изменение строя внутриви-
довых отношений, как бы оно ни было полезно виду, никогда не
может устранить этих противоречий. Также невозможно устранить
несовпадение физиологического оптимума с популяционно-видо-
вым оптимумом и главенство последнего над первым.
Нельзя отождествлять противоречия между видовой и организ-
менной формами организации живого, существующие в рамках
уже селекционно отшлифованных и сложившихся видовых адап-
таций, с противоречиями, возникающими прп перенаселении у мо-
лодых, прогрессирующих видов и при других формах производных
внутривидовых отношений. Противоречия в сложившихся видовых
адаптациях — это свидетельство недостатков их организации, или,
на языке кибернетики,— это помехи в* системе (Шмальгаузен,
1958а и 19586). Противоречия производных внутривидовых отно-
шений— это как бы магма, из которой путем отбора формируются
новые видовые адаптации; такие противоречия возникают у видов
с хорошо отшлифованным строем основных внутривидовых отно-
шений п являются пе «помехами системы», а причиной и движу-
щей силой их совершенствования.
Целостность образований на популяционно-видовом уровне
объясняется существованием сложных связей и зависимостей
между особями и их группами, консолидирующая роль которых
прежде всего проявляется в пределах местной популяции. Вместе
с тем в результате группового отбора могут быть закреплены и
некоторые межпопуляционные связи (становящиеся после селек-
ционной отработки видовыми адаптациями). Видовыми адапта-
циями такого сорта может стать закономерный обмен гаметами,
семенами, а также обмен молодняком п взрослыми особями между
соседними популяциями (или между целой серией популяций).
Эти внутривидовые межпопуляционные отношения не сводятся
к поддержанию гетерозиса, оптимальной гетерозпготпости и
к другим наиболее изученным примерам. Существует много еще
очень слабо изученных форм связей (у простейших и растений —
химических и биофизических, действующих через среду или
контактно, у животных — нейрофизиологических и т. п.).
Крайне недостаточная разработанность этой проблемы объ-
ясняется в первую очередь тем, что до спх пор изучение внутри-
видовых отношений осуществляется слабыми силами и без доста-
точной их координации. До самого последнего времени исследова-
ния внутривидовых отношений пе имели тесной связи
с проблемой вида, являясь разделами экологии, биоценологии
пли эволюционной теории. Существовало противоестественное
положение: внутривидовые отношения представляют собой содер-
жание жизни вида, но изучались они однобоко — лишь как один
из факторов эволюции. Надо полагать, что в ближайшем будущем
внутривидовые отношения займут место основного раздела в тео-
рии вида.
Глава 10
ВИДООБРАЗОВАНИЕ
1. ВИДООБРАЗОВАНИЕ И ФОРМООБРАЗОВАНИЕ
Микроэволюция и видообразование. Сущностью
эволюционного процесса является адаптациогенез, идущий на ос-
нове естественного отбора. Эволюция любых масштабов пред-
ставляет собой адаптивное преобразование популяций или целых
совокупностей популяций.
По классической схеме видообразование является процессом
адаптивной радиации групп. С этой точки зрения сам вопрос,
адаптивно ли видообразование, является неправомочным. Однако
не раз делались попытки разъединить проблемы адаптациогенеза
и видообразования. Одним из главных доводов для этого служил
тот факт, что многие признаки, по которым систематики распо-
знавали виды, не имели ясного адаптивного значения, и можно
было думать, что некоторые пз них вовсе пе адаптивны. Так по-
явилась проблема пеадаптивпых видовых признаков. Не углуб-
ляясь в этот вопрос, укажем лишь па то, что в качестве «видовых»
избираются признаки-маркеры, среди которых могут встретиться
и неадаптивные признаки, связанные с адаптациями лишь кор-
релятивно. Следует учитывать также и то, что случаи, когда
признаку ошибочно приписывалось неадаптивное значение, до сих
пор довольно часты.
Если следовать классической схеме, то адаптивными являются
не только преобразования исходного вида в производный (при
ортоселекции) или в несколько производных (с исчезновением
или же с сохранением самого исходного вида), по и различия
между самими дочерними видами. В настоящее время нет ни-
каких оснований отказываться от этих представлений. Напротив,
все новые данные подтверждают их.
Видообразование может рассматриваться как заключительный
акт предшествующих ему процессов адаптивных преобразований,
302
протекающих внутри вида. Как было показано в главе 8, все эти
адаптивные преобразования внутри вида в каждый данный мо-
мент воплощаются в структурных подразделениях вида.
Микроэволюция рассматривается нами как совокупность
прежде всего таких адаптивных преобразований внутри вида.
Материалом микроэволюции служат как отдельные мутации, це-
лые генотипы и генетические системы, так и адаптивные фено-
типы с малой степенью наследственного закрепления. Микро-
эволюция осуществляется естественным отбором. Элементар-
ными микроэволюционными изменениями являются включение
в популяцию мутации с адаптивным фенотипическим выраже-
нием или образование новой адаптивной реакции (модификации).
Все направленные отбором адаптивные изменения — сдвиг коли-
чественного баланса форм в полиморфной популяции, образование
новой морфо-биологической группы и экоэлемепта, изменение или
возникновение новых популяций, локальных экотипов, физиоло-
гических рас, географических подвидов и т. п. (см. главу 8), если
они протекают в пределах вида, рассматриваются как микроэво-
люция.
Процессы микроэволюции чаще всего не завершаются видооб-
разованием. Некоторые из этих процессов (например, увеличение
полиморфизма популяции), как правило, не ведут к видообразо-
ванию, а лишь несколько изменяют ранее существующий вид.
Такое определение микроэволюции отличается от приводимого
Д. Симпсоном (1948:154) и Н. В. Тимофеевым-Рессовским
(1958, 1965), так как в нем подчеркивается адаптивный характер
даже самых мелких микроэволюциоипых преобразований.
А. Мюнтцинг (1963:391) под микроэволюцией понимает «наслед-
ственное изменение в одном признаке», т. е. фенотипическое про-
явление мутации. Это определение неприемлемо, так как, игнори-
руя адаптивный характер микроэволюцип, автор, как и генетики
начала века, сводит эволюцию к мутации.
Как отмечалось в предыдущих разделах, многие адаптивные
преобразования популяций при определенных условиях могут
выйти за рамки вида, т. е. возникает процесс видообразования.
Иерархия внутривидовых единиц, как часто считается, отра-
жает шаги процесса микроэволюцип, и погтепепное нарастапие
величины различий между этими единицами должно рассматри-
ваться в качестве ступеней или стадий видообразования (см.,
например: Rensch, 1965 — определение географической расы, и
др.). Существует много попыток линейного расположения подраз-
делений вида в иерархическую цепочку, которое одновременно
должно служить моделью процесса видообразования. Рассмотрим
подробнее только один пример такой иерархии (Niculescu, 1960).
Основываясь на данных по систематике чешуекрылых насекомых,
В. Никулеску намечает семь случаев расчленения вида, которые
он называет «стадиями видообразования»: 1) одна популяция
303
состоит из двух подпопуляций, различающихся сроками лёта (но
тождественных генетически и морфологически); 2) такие подпо-
пуляции различаются п морфологически; 3) образуется две попу-
ляции, живущие па различных видах растений (такие популяции
иногда возводятся в ранг видов); 4) существование в популяции
симпатрическпх и синхронических форм, четко различающихся
морфологически; автор применяет к этому случаю термин «раз-
новидность», хотя из приводимых им примеров вытекает, что
такие формы могут быть и двойниковыми видами (Мауг, 1963),
и компонентами единой полиморфной популяции; 5) наличие
аллопатрических форм, различающихся и морфологически, — слу-
чай, описываемый большинством систематиков как подвиды;
6) в зонах контакта аллопатрические популяции хотя и скрещи-
ваются, но образуют неустойчивые гибриды промежуточного
характера — случай полувида; 7) наличие столь больших генети-
ческих различий между популяциями, что возникает барьер не-
скрещиваемости, который п позволяет говорить о завершении
видообразования и появлении новых «хороших» видов. Подоб-
ная же идея лежит и в основе структуры вида растений, предло-
женной Дж. Клаузеном (Clausen, 1951).
Взаимоотношения между микроэволюцией и видообразованием
сложны, и если при многих типах видообразования иерархия
внутривидовых единиц схематично, но отражает стадии видообра-
зования, то некоторым типам этого процесса свойственно меньшее
количество стадий или качественно иные стадии.
Видообразование — основное явление эволюции. Образование
нового вида является фундаментом процесса эволюции и ее
узловым пунктом, в том смысле, что вся эвлюция может быть
представлена как совокупность видообразовательных процессов.
Когда говорят об эволюции рода, семейства или отряда, под-
разумевается совокупность npoi 1,ессов видообразования, идущих
в нескольких пли во многих «горячих» точках исходной радии-
рующей системы видов. Не существует никаких особых специфи-
ческих закономерностей образования высших таксонов, которые
нельзя было бы свести к видообразованию. Родообразованпе и
даже отрядообразование являются целиком аддитивными процес-
сами (т. е. это суммы результатов конкретных видообразований).
Однако некоторые признают, что типогепез (Schindewolf, 1954)
принципиально отличен от обычного видообразования и что новые
сложные адаптивные механизмы, вроде новых систем органов или
фаз жизненного цикла и т. и., связаны со внезапными «пере-
чеканками», а пе с видообразованием. Большинство современных
эволюционистов, к которому принадлежит и автор этой книги,
считает, что между видообразованием и мегаэволюцией нет
принципиальных различий Если образуются длинные стволы
ортоселективной эволюции, то следует признать, что они состоят
из множества переходящих один в другой видов. Эти стволы от-
304
ражают множество последовательно шедших процессов видообра-
зования.
Видообразование включает как приобретение и накопление
новых приспособлений, т. е. трансформацию вида (фратрии) во
времени, так и пространственное^ расщепление или расслоение
ранее единой популяции на несколько (schistic evolution — Smith,
1956). Иначе говоря, это процесс, совершающийся во времени и
в пространстве одновременно.
Поэтому характеристика видообразования’ только во времени
или только в пространстве недостаточна. Однако сведения о точ-
ных границах видов во времени, как правило, отсутствуют. Желая
рассматривать видообразование как смену изменяющихся поколе-
ний, мы постоянно наталкиваемся на несколько капитальных
затруднений, нередко действующих совместно. Эти затруднения
состоят в неполноте геологической летописи, в малочисленности
предковых популяций или тем более предковых инициальных
внутрипопуляциопных форм, в высокой скорости или же, напро-
тив, в большой постепенности процесса видообразования, в пере-
мещениях популяций, так что первые фазы процесса проходят на
одной территории, а остальные — на другой или же на других, и
т. п. Каждое из этих затруднений может быть столь большим, что
во многих случаях делает практически невозможным палеонтоло-
гическое исследование видообразования. Даже в таких богатых
массовыми остатками группах, как моллюски, и в таких важных,
как гоминиды, восстановление пути эволюции современных видов
носит весьма приблизительный характер. Разбивка филогенетиче-
ской ветви, идущей к современному виду, на отрезки-фрат-
рии, т. е. на виды во времени, как правило, является очень пе-
сочной.
Видообразование характеризуется неравномерностью темпов.
(Существует различие как в темпах видообразования разных видов,
сак и у разных географических или экологических рас одного и
того же вида при различных способах видообразования (о чем
будет сказано ниже). Неравномерность темпов эволюции отдель-
ных популяций и даже отдельных внутрипопуляциопных форм
одного вида может привести к тому, что одновременно будет
существовать предковая популяция вида и разной степени эволю-
ционной продвпнутости популяции — потомки.
Исследование образования новых видов является центральным
моментом всех эволюционных теорий. Поэтому естественно, что
к этому процессу давно приковано внимание биологов. Мы не бу-
дем здесь возвращаться к рассмотрению причин видообразования.
В этой главе анализируются вопросы, связанные только со спосо-
бами видообразования и с характером этого процесса.
Критика понятия о видообразовательной и з -
м е п ч и в о с т п. Выше уже объяснялась необходимость разграни-
чения эволюционного процесса как исторических изменений попу-
20 К- М. Зава деки*'
305
ляций и видов и мутационного процесса, как физиологического и
даже биофизического процесса, осуществляющегося на уровне
других более низких форм организации живого. Мутационная
изменчивость постоянно поставляет материал для эволюционного
процесса, однако не может быть к нему приравнена. Современное
эволюционное учение полностью подтвердило необходимость раз-
граничения эволюционного и мутационного процессов (Шмальгау-
зен, 1946а, 19466, 1961; Wright, 1951, 1964; Huxley, 1956, 1963;
Waddington, 1956; Завадский, 1957в, 1961, и др.). Формообразова-
нием в широком смысле мы называем образование новых структур
п реакций независимо от степени их наследования и адаптивного
значения.
Формообразование служит материалом для видообразования, но
это принципиально разные процессы: формообразование возникает
на молекулярном и клеточном уровнях, п в конечном итоге при-
суще организменной форме существования живого, а видообра-
зование возникает на уровне дёма (местной популяции) и
присуще популяционпо-видовой форме организации живого.
Понятие видообразовательиой изменчивости (Де Фриз),
игравшее решающую роль в мутационной гипотезе видообразова-
ния (каждая мутация — новый вид), впоследствии было поддер-
жано разными учеными (Морган, 1936, и др.). Ранее уже отмеча-
лась ошибочность этого понятия (Завадский, 1954а, 1954в, 1961,
19676). Видообразовательиой изменчивости нет и не может суще-
ствовать, так как изменчивость не непосредственно переходит
в видообразовательный процесс. Обоснование мутационной теории
видообразования в сущности состояло только в том, что словом
«вид» была названа каждая новая мутантная форма. Неясности
в определении понятия «вид», разнобой в его критериях, нераз-
работанность типологии видов — все это не может не тормозить
разработку проблемы видообразования. И правильно, что некото-
рые авторы еще раз обращают внимание на этот вопрос (Harvey,
1960).
Видообразование осуществляется не через лапласовский тип
детерминизма, а статистически, через вероятностный механизм
селекции. Подтвердилась правильность основного тезиса Дарвина:
от «неопределенной» изменчивости через элиминацию неадаптив-
пых вариантов к накапливающему и направленному процессу
преобразования популяции — таков путь от изменчивости к видо-
образованию.
Мутационизм не подтвержден фактами, однако уже столетие
в разных формах (гетерогенезис, перечеканка, катаклизмогенез
и др.) эта гипотеза противостоит теории исторического развития
видов.
Некоторые вирусологи и микробиологи присоединялись к фан-
тастической гипотезе порождения видов. Например, работая
с одной из форм вируса табачной мозаики, К. С. Сухов пишет,
306
что пассировал ее через растения различных семейств и затем
после инокуляции табака «была получена измененная форма ви-
руса, несущая видовые отличия от исходного вида» (1953:8).
Новая форма вируса вызывала на листьях табака некрозы более
крупных размеров. Если бы была доказана ее константность,
то это был бы мутант. В работе нет и попытки обосновать воз-
ведение этой формы в ранг вида. Столь же необоснованно К. С. Су-
хов применяет слова «вид» и «видообразование» и к другим
формам вирусов (переход форм, образующих гексагональные
кристаллы Ивановского в формы, образующие игловидные пара-
кристаллы под влиянием выращивания проростков томатов
в темноте и т. п.). Чем менее ясны границы видов, тем легче
подменять изучение видообразования изучением изменчивости,
при этом не различая даже ее формы (мутации, модификации
и т. п.).
Н. Н. Жуков-Вережпиков п А. П. Пехов (1963 251) пишут,
что «видообразующая изменчивость бактерий» доказана и поэтому
ее отрицание — это «слишком категорическое» суждение К. М. За-
вадского. При этом авторы справедливо отмечают, что теперь
общебиологические основы учения о виде и видообразовании
«в значительной степени применимы и к микроорганизмам»
(там же: 250). Если это так, то необходимо сделать другие вы-
воды. В самом деле, границы и объемы видов у микроорганизмов
изучены гораздо слабее, чем у высших форм, надежные критерии
для «возведения в ранг» вида часто отсутствуют (Имшенецкип,
1961; Красильников, 1962; Карпов, 1962). Такое положение
иногда лишает микробиологов возможности уверенно обсуждать
вопрос о характере того или иного изменения штамма (Клюй-
вер и Ван-Ниль, 1959). Многие микробиологи не рискуют сле-
довать мутационизму и признавать новыми видами каждую но-
вую наследственную форму (мутанта), клон и даже модифи-
канта.
Вид представляет собой такую целостную и дифференцирован-
ную племенную общность, доказавшую способность к само-
стоятельному воспроизведению и эволюции, которая может изме-
няться только исторически в процессе естественного отбора, не-
зависимо от способов этих изменений, от скорости процесса и от
качества исходного материала (едва уловимые сдвиги состава
популяции или же новая форма, резко отличающаяся от роди-
тельского типа).
В разных группах органического мира видообразование совер-
шается по-разному, но всюду на основе естественного отбора, т. е.
имеет характер постепенного процесса. Именно* так идет видо-
образование у низших водорослей (Полянский, 1956, 1958) и
у высших растений (Clausen, 1951, 1961; Синская, 1961, 1963;
Stebbins a. Daly, 1961, и др.). У позвоночных животных «исходная
форма должна пройти длительный путь относительно само-
20*
307
стоятельного развития, прежде чем ее дифференциация достигнет
уровня, характерного для вида» (Шварц, 1963:431).
Вся совокупность данных современной биологпии свидетель-
ствует о том, что видообразование — процесс, совершающийся
среди масс особей. Образование нового вида — это эволюционное
преобразование популяции или группы популяций. Видообразо-
вание — это приспособительная трансформация популяций, осуще-
ствляемая естественным отбором. Сущность видообразования со-
стоит в том, что преобразуемые популяции приобретают все боль-
шую способность к самостоятельному (отдельному) существованию
и развитию в природе, становятся устойчивыми и конкуренто-
способными. Это проявляется в увеличении численности и проч-
ном захвате места в биоценозах и экосистемах.
2. ТИПЫ ВИДООБРАЗОВАНИЯ
Принципы выделения типов. Лучшей, несомненно,
была бы классификация, опирающаяся па различия в способах
действия, путях, темпах и других особенностях естественного от-
бора. Индивидуальный отбор приводит к иным формам видообра-
зования, чем групповой или же чем индивидуально-групповой
отбор. И. И. Шмальгаузен (19466) показал, что отбор мелких
групп (таких как кладка и семья) приводит к иным результатам,
чем отбор крупных групп (стад, локальных популяций и т. п.).
Дж. Холден и Дж. Гексли еще в 30-х годах доказывали, что
внутривидовой отбор (агент — конкуренция индивидов и групп
своего же вида) приводит к иным эволюционным последствиям,
чем межвидовой или же отбор абиотическими агентами. Известно,
что совершенно различны последствия действия одного и того же
агента (например, голода, морозов, засухи и пр.), если в одном
случае он вызывает прямую элиминацию или же стерильность,
а в другом — градацию случаев физиологической ослаблснности
(дифференциальная плодовитость переживших, различно ослаб-
ленная имунность и пр.). Интегрирующий отбор будет поддер-
живать сложившуюся адаптивную норму в то время, как дисруп-
тивный отбор (Smith, 1962) формирует сбалансированно
полиморфную популяцию, а изолирующий отбор способен
преобразовывать отдельные внутрипопуляционные группы в само-
стоятельно воспроизводящиеся и самостоятельно эволюционирую-
щие расы — молодые виды. Давно была показана зависимость
видообразования от интенсивности отбора (давление отбора) и т. п.
В настоящее время, одпако, еще нет достаточного материала
для обоснованного выделения типов видообразования по формам
естественного отбора. Объясняется это, по-впдимому, тем, что
проблема отбора исследовалась до сих пор главным образом
в связи с теорией адаптациогенеза, а учение о видообразовании
оставалось в тени. Великолепный анализ отбора па уподобление
308
сорняков основной культуре, проведенный Н. В. Ципгером (см.
главу 4) и продолженный Е. Н. Синской, был редким образцом
совместного изучения отбора и видообразования. Лишь в послед-
ние годы началось сближение исследований по отбору с теорией
видообразования.
Наибольшее число современных авторов следует хорологиче-
скому принципу подразделения и вслед за 0. Майром различает
аллопатрическое и симпатрическое видообразование. Однако нали-
чие или отсутствие территориальной разобщенности эволюциони-
рующих групп пе сводится только к наличию викарирующих от-
ношений между ними или же к сосуществованию форм в смешан-
ных популяциях. За пространственными отношениями стоят
коренные биологические отношения. Поэтому эти типы видообра-
зования различаются не только природой групп начинающих
видообразование и характеристикой стадий процесса, по и внутри-
видовыми отношениями между этими группами, значением раз-
личных форм изоляции, темпов процесса и пр.
Хорологический принцип выделения типов видообразования
хотя и стал общепризнанным, все же далеко не исчерпывает всего
многообразия типов видообразования. Многие известные случаи
видообразования существенны другими чертами. Поэтому нами
намечаются еще четыре типа видообразования, выделяемые на
основании ряда существенных признаков.
По характеру исходных преобразований, по времени изоляции
молодой формы от материнской популяции, по числу фаз и по
темпу процесса видообразования мы предложили различать нео-
формогенное и микроаккумулятивное видообразование (Завад-
ский, 1966а, 1967). Чертой неоформогенного видообразования
является такое мутационное изменение инициальной формы, кото-
рое адаптивно и сразу приводит к несовместимости ее с особями
материнской популяции, а также к ускорению темпа видообразова-
ния. Такая форма по существу является не простым сырьем для
отбора, а полуфабрикатом, требующим только некоторой шли-
фовки и «доводки». Эту инициальную форму мы предложили
именовать «новая видовая форма» (Завадский, 1961, 1966а, 1967).
Часто новая видовая форма является высокоадаптивной в сома-
тической сфере, но фертильность ее понижена. В этих условиях
успех процесса зависит от эффективности отбора на повышение
фертильности этой формы при внутригрупповых скрещиваниях и
па создание устойчивой и достаточно высокой плодовитости.
Микроаккумулятивное видообразование — следствие накопле-
ния в популяции мелких адаптивных особенностей и объедине-
ния их в комплексы, которые частично подвергаются стабили-
зирующему отбору. Это медленно идущий процесс, требующий
длительного, геологического времени. Он опирается на отбор, иду-
щий по мелким мутациям со слабым фенотипическим проявле-
нием.
309
Наконец, мы предлагаем различать еще два самых общих типа
видообразования: сиптезогенез и сегрегациогенсз. Принципом их
выделения является способ преобразования, связанный или с объ-
единением нескольких инициальных систем в одну продуцируе-
мую или же, наоборот, с обособлением ранее единой системы на
ряд дочерних. Логически возможны и фактически наблюдаются
только два способа эволюционной дифференциации систем: синте-
зогенез, т. е. объединение нескольких систем в одно целое, и
сегрегациогепез — подразделение единого целого па части, кото-
рые в дальнейшем обособляются как друг от друга, так и от мате-
ринской системы. Таким образом, общим принципом выделения
этих типов видообразования является соотношение между инте-
грацией и дифференциацией систем у инициальных и производ-
ных видов.
Каждый из этих шести типов видообразования включает много
конкретных способов видообразования. Аллопатрическое видообра-
зование включает географическое и собственно экологическое, ко-
торые в свою очередь могут быть подразделены на более мелкие
группы (например, миграционное, партпкуляционное, топографи-
ческое, бпоцепотическое п т. п.). Симпатрическое видообразование
также может быть подразделено па соответствующие группы.
Если в популяции формируется несколько дискретных групп,
а затем некоторые из них выходят из популяции и становятся
родоначальниками повой популяции и вида — это дпсруптивно-
изолирующее видообразование. Если соседствующие внутрппопу-
ляционные группы различаются по скорости или ритмам развития
и в конце концов возникает изоляция из-за различий в сроках
созревания половых продуктов — это хроносимпатрическое видо-
образование. Неоформогенное видообразование включает такие
способы видообразования, как гибрпдогепез и полиплоидия или
анеуплоидия, а также апомиксис, педоморфоз, тератогенез и др.
Микроаккумулятпвное видообразование включает все способы,
связанные с накоплением мелких адаптивных признаков и с по-
степенным преобразованием популяций и рас. К этому типу видо-
образования относятся, по-видимому, часть случаев географиче-
ского и экологического видообразования, дисруптивно-изолирую-
щее видообразование и др.
О способах, которыми происходит синтезо- и сегрегациогене-
тическое видообразование, будет сказано ниже.
Из этого перечисления видно, что одни и те же способы видо-
образования входят в разные типы видообразования. Это означает,
что предлагаемые типы не являются членами дихотомически
выдержанной классификации.
Синтезогенез и с е гр е г а ц и о г е н е з как типы
видообразования. Самым изученным путем видообразования
с помощью синтезогенеза является гибридогенез, т. е. отдаленная
гибридизация, которой посвящен специальный раздел этой главы.
310
Здесь же мы остановимся, может быть, па самом ярком проявле-
нии синтезогенеза — видообразовании путем многократных и
сложных межвидовых скрещиваний с объединением в одном
генотипе отдельных фрагментов, частей или даже целых геномов
нескольких видов. Такие гибридизационные процессы происходят
в пределах обширных родов или даже между родами. У эвкалип-
тов, роз, хлопчатников, пшениц, тыкв, анютиных глазок и других
групп высших растений видообразование путем такой повторной
отдаленной гибридизации распространено очень широко. Недаром
трансгрессия признаков в пределах таких родов (и даже в преде-
лах триб) очень велика, а сами они целиком или частично входят
в особо трудно классифицируемые группы. Видообразование пу-
тем сложных повторных межвидовых скрещиваний с последую-
щим отбором лучших семей характерно для таких видов мятли-
ков, как Роа pratensis, Р. arctica и др. (Akkerberg, 1942). Недав-
ними работами экспериментально (цитогенетически) доказано, что
Р. pratensis — синтезогенетический вид, включающий признаки
нескольких видов Роа, с которыми его предки вступали в кон-
такты (Clausen, 1961; Harlan a. Wet, 1963). Этими авторами
в комплексных исследованиях некоторых злаков обоснована кон-
цепция «compilospecies». Особенно интересен Bothriochloa inter-
media (рис. 6) Оказалось, что этот вид вступает в половые кон-
такты (и сложно интерградирует) с различными другими видами
(в Северной Австралии с Capillipedium parviflorum и с Bothrio-
cliloa ewartiana, на северо-западе Пакистана и в Индии с Dichan-
thium annulatum, с Bothriochloa grahamii, с обычным евразиат-
ским В. ischaemum и т. д.). Вид В. grahamii сам оказался
продуктом гибридизации Dichanthium annulatum с расой Bothrio-
chloa intermedia из долины Ганга (раса gangetica). Многие другие
виды этого сложного комплекса, как было экспериментально вос-
произведено, также содержали большую или меньшую часть
генотипа В. intermedia. Стало очевидно, что В. intermedia сам
является синтезированным по крайне мере из пяти видов, принад-
лежащих к трцм родам. В коллекции содержалось свыше 200 об-
разцов этого вида из Южной и Восточной Африки, Южной и
Юго-Восточной Азии, Индонезии, Новой Гвинеи и Австралии, и
пн один из них с уверенностью нс может быть назван основной
формой вида. Бесспорно, что первооснова этого вида тетраплоид-
ная, хотя встречаются популяции с 5—8 п.
Рассматриваемый вид может служить моделью «compilospe-
cies», т. е. вида агрессивного в генетико-эволюционном отноше-
нии, захватывающего у родственных видов при скрещиваниях
ценные признаки и даже способного полностью* ассимилировать
некоторые виды. При этом поглощение может приводить к исчез-
новению ассимилированного вида и в то же время укреплять
агрессивный вид. Полиморфность, широта адаптивной нормы,
высокая пластичность и приспособляемость compilospecies, по-
311
Число колосков Число соцветии
020.30.40.50.6 0.8 ' 1.0 1.2 1.4 1.6 1.8 20 2.2 2.4 2.6 ^3
Длина основной оси / Длина соцветия
1.4 1.61.8 2.0 2.2 2.6 3.0 3.4 3.8 4.2 4.6 5.0 6.0 8.0 1.0 >10
Длина основной оси / Длина соцветия
Рис. 6. Интрогрессия между compilospecies Bothriochloa
intermedia и Dichanthium annulatum из Южной Азии, долины
Ганга и Пенджаба (Л) и видами Capillipedium в северо-во-
сточной Австралии (Б). Растения типа В. glabra морфоло-
гически идентичны растениям расы gangetica (Harlan a. Wet,
1963).
1 — D. intermedia; 2 — paca gangetica В. intermedia; з — D. annula-
tum; 4 — В. glabra; 5 — G. spicigerum; 6‘ — C. parviflorum.
видимому, делает этот способ видообразования весьма перспектив-
ным. Мягкая пшеница, сахарный тростник, мятлик Р. pratense
могут служить классическими образцами генетически агрессивных
compilospecies (Harlan a. Wet, 1963). Не следует, однако, отож-
дествлять синтезогенез с гибридогенезом. Видообразование путем
сиптезогенеза может опираться и на иные пути синтеза. Упомя-
нем, например, гетерокариоз, открытый Понтекорво у несовершен-
ных грибов и являющийся источником видообразования. К эво-
люции путем сиптезогенеза относится и явление трансдук-
ции микроорганизмов, т. е. присоединения к организму одного
таксона частей организмов других таксонов.
Эволюция путем закрепления симбиотических отношений
также может служить примером сиптезогенеза. Таким путем
совершается видообразование пе только у низших (лишайники)
и высших (микориза) растений, но и у животных. Ассоциация
макрооргаиизма с некоторыми вирусами, как это давно предпола-
гал К. Дарлингтон, может привести к такому включению вирус-
ной ДНК в генетическую систему макроорганизма, что чужерод-
ная ДНК будет воспринята как обычное мутационное изменение.
Как полагает М. М. Камшилов, видообразование, ведущее
к прогрессивному усложнению биосинтеза, «произошло не как
следствие серии случайных мутаций у одного вида, а в результате
обмена различными веществами между членами биоценоза/)
(1961 101), т. е. что один вид мог объединять генетический
материал многих видов.
Итак, можно считать, что как у животных, так и у растений
видообразование путем синтезогепеза играло известную пли даже
важную роль в эволюции, принимая различные формы (трансдук-
ция, гетерокариоз, симбиогенез, гибридизация и явление compi-
lospecies и пр.).
Видообразование же путем сегрегациогенеза является давно
известным и очевидным путем эволюции, о котором подробно бу-
дет сказано в разделах, посвященных аллопатрическому и сим-
патрическому видообразованию. Подчеркнем только, что этот тип
видообразования включает все преобразования исходного вида,
сопровождающиеся расчленением целого на части (партикуляция,
фрагментация) или же выходом части из состава целого (морфо-
биологическая группа -> экоэлемент -> экотип вид) и т. п.
3. АЛЛОПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
Географическое видообразование. Видообразова-
ние путем формирования и обособления географических рас пред-
ставляет собой распространенный и лучше других изученный
случай. Многие виды на равнинных территориях тундры, лесной
зоны, степей и пустынь имеют большие ареалы, во много раз пре-
восходящие ареалы видов горных стран. Некоторые из них имеют
313
географические подразделения (подвиды). Очень Много видов,
имевших в третичный период большие ареалы, изменялись путем
образования и обособления географических рас. Этот процесс свя-
зан с расселением и приспособлением популяции к почвенно-
климатическим и биотическим особенностям окружающей среды.
Происходило образование популяций, размножающихся «внутри
себя» вследствие удаленности других популяций или же из-за
изолирующего влияния достаточно широких полос непригодных
местообитаний. Приспособление таких популяций к специфиче-
ским условиям среды и фактическая их полупзоляция от других
популяций приводят к постепенному формированию географиче-
ских рас. В дальнейшем географическое обособление рас может
усилиться (например, вымирание популяций на промежуточных
территориях в ледниковый период пли вытеснение одних сооб-
ществ новыми и пр.). Возникает фактическая изоляция геогра-
фических рас. Каждая из них образует практически замкнутую
племенную общность, размножающуюся только внутри себя. На-
конец, между такими расами иногда может возникнуть и столь
большое генетико-физиологическое несоответствие, что если
впоследствии они вновь поселяются па одной территории, то ока-
зываются уже неспособными к скрещиваниям или дают стериль-
ное потомство.
Образованию новых видов путем обособления географических
рас посвящено много работ. По В. Л. Комарову (1940), ландыш
Convallaria majalis представляет видовой комплекс, состоящий
из европейской расы (собственно С. majalis); закавказской расы
(С. transcaucasica), более крупной, с тычиночными нитями без
треугольного расширения к основанию, с венчиком более ци-
линдрической формы; дальневосточной расы (С. manchurica),
также более крупной, с более широким венчиком, имеющей крас-
ные черешки и более сильный восковой налет па листьях, а также
сахалшю-японской расы (С. keiskei) и североамериканской
(С. majauscula). По Комарову, ландыш сформировался в миоцене
и расселился в широколиственных лесах. В четвертичный период
он был частично уничтожен, частично же сдвинут па юг в гор-
ные области и по мере продвижения лесов на север вновь распро-
странился по Европе. Распадение общего ареала ландыша на
пять самостоятельных, удаленных друг от друга площадей яви-
лось причиной образования самостоятельных рас. Это «дало воз-
можность проявиться молодому видообразованию, не сделав,
однако, этих молодых видов ландыша общепризнанными»
(там же : 158).
Сравнение восточноевропейской и дальневосточной рас лан-
дыша (С. majalis и С. keiskei) показали (Зоз и Черных, 1961),
что на Дальнем Востоке существуют не две расы (С. keiskei и
С. manshurica), а одна и что С. keiskei отличается от С. majalis
по целой группе признаков (многожилковые прицветники, форма
314
зубцов околоцветника, крылатые оси соцветия, более плотные и
толстые верхние листья и пр.).
Таков же, по Б. К. Шишкину (1954), способ видообразования
у лесного купыря Anthriscus silvestris. Единый в третичное
время вид распался на три ясно выраженные географические
расы — европейскую, сибирскую (A. aemula) и кавказскую
(A. nemorosa).
Видообразование, связанное с обособлением географических
рас, описано и у сорного растения спорыша (Polygonum
aviculare). По Комарову (1937), оно было связано не с распадом
третичного широкого ареала на обособленные части, а с недавним
расселением популяций и с заносами семян из Европы в восточ-
ном и южном направлениях.
Географическое видообразование — это процесс приспособле-
ния к новым климатическим и биоценотическим условиям. При
образовании одного из северных видов мака (Papaver radicatum)
возникла группа признаков адаптивных в условиях Крайнего
Севера: уплотнение дерновинки за счет укорочения и уменьше-
ния поверхности листа, увеличение опушенности листьев, быст-
рое развитие коротких цветоносов (чем обеспечивается раннее
цветение и расположение цветков ближе к почве) п т. п. (Тол-
мачев, 1953).
Классическим примером географического видообразования мо-
жет служить видовой комплекс большой синицы (Parus major).
Зта синица широко расселена. Ее географические расы отли-
чаются по окраске спины и брюшка, толщине клюва, длине
хвоста п по другим признакам, а также экологически. При этом
одпи расы сосуществуют в зонах контакта не скрещиваясь, а дру-
гие образуют на этих территориях гибридные популяции. Гра-
ницы между одними расами по морфологическим признакам обо-
значены четко, а между другими имеются плавные переходы;
одни расы занимают громадные ареалы (например, обыкновенная
европейская большая синица), другие локализованы в отдельных
горных областях или па островах. В результате у большой си-
ницы образовалась сложная система популяций, стоящих па раз-
ных этапах обособления. Так, например, восточноазиатскпе
популяции европейской расы (ssp. major) в Приамурье сосуще-
твуют с северными популяциями китайской расы (ssp. minor)
и не скрещиваются между собой. В то же время китайская раса
связана переходами с индийской (ssp. cinereus), а последняя
в свою очередь в Северном Иране образует переходные, гибрид-
ные популяции с обыкновенной европейской расой.
Итак, существует кольцо популяций и по крайней мере в од-
ном пункте звенья этого кольца между собой не спаяны, а лишь
налегают друг на друга. Приамурский разрыв позволяет рассмат-
ривать европейскую и китайские расы как вполне сложившиеся
виды. Налицо четкий морфологический хиатус, различие в ареа-
315
лах (при небольшом налегании) и половая изоляция. Однако,
по-видимому, во всех других звеньях кольца отдельные популя-
ции генетически более близки к соседним и в конце концов по-
лучается, что китайская раса в южной части кольца выступает
по отношению к обыкновенной большой синице еще как подвид,
а в Приамурье — уже как вид. Существование таких кольцевых
видов свидетельствует о том, что в пределах одной географиче-
ской расы (подвида) существуют популяции, сильно различаю-
щиеся между собой в генетическом отношении.
Широко расселенные формы чайки — клуша (Larus fuscus) и
серебристая чайка (L. argentatus) — в Европе живут совместно,
по не скрещиваются. Однако их географические расы образуют
полное циркумполярное кольцо и при этом между североамери-
канской расой серебристой чайки через восточноазиатские и за-
падносибирские расы устанавливается цепь переходов к типичной
европейской клуше (Stresemann u. Timopheef-Ressovsky, 1947
и др.).
Возможны следующие два крайних случая географического
видообразования. Во-первых, это фрагментация ареала материн-
ского вида. Евриадаптивпый вид вначале широко расселяется, но
впоследствии под воздействием различных климатических и био-
ценотических факторов формируются географические расы, они
обособляются друг от друга, и в конечном итоге возникают моло-
дые виды. При этом большой ареал материнского вида раз-
дробляется па несколько дочерних рас или видов, имеющих менее
широкое распространение. Конечно, среди молодых видов неко-
торые могут выделиться как более прогрессивные и вторично
завоевать большой ареал. Процесс видообразования путем фраг-
ментации ареала материнского вида осуществляется преимуще-
ственно стабилизирующим отбором и приводит чаще к большей
специализации потомков. Как считает В. Н. Васильев, «формиро-
вание новых видов происходит в результате распада древнего
(материнского) вида и его ареала на молодые (дочерние) виды
и пх ареалы вследствие дифференциации физико-географических
условий» (1961 1588). Итак, видообразование происходит пе
в процессе самой миграции, а после ее окончания, вследствие из-
менений среды, возникающих после образования обширного
ареала.
Во-вторых, это видообразование в процессе миграции, когда
переработка популяций происходит по мере их расселения. Шаг
за шагом идет расширение ареала, и именно в этот период про-
исходит интенсивное преобразование и формирование новых по-
пуляций, некоторые из которых могут стать родоначальниками
новых видов. Этот процесс внешне может проходить именно так,
как его описал В. Л. Комаров (1908) в монографии рода
Caragana. Очень интересен особый случай видообразования на
архипелагах («archipelago speciation» — Мауг, 1963), т. е. адап-
316
тивпая радиация (одновременная или разновременная), идущая
не от одной предковой популяции, а от нескольких близких
предковых популяций (Гавайи, Галапагосы, Соломоновы острова
II т. д.).
Важное значение для видообразования в процессе миграции
имеют преобразования популяций, расположенных на границах
ареала вида. Установлено, что на некоторых пограничных участ-
ках экспансия вида проходит путем образования за пределами
сплошного ареала отдельных мелких поселений, являющихся
как бы форпостными укреплениями наступающего вида. Такие
форпосты остаются и после спада очередной волны жизни.
По соседству могут происходить и временные отступления вида,
в результате которых в приграничной полосе еще недавно сплош-
ной ареал становится как бы продырявленным, площади, запя-
тые отдельными популяциями, сокращаются, а некоторые попу-
ляции полностью исчезают.
В некоторых случаях такие форпостные популяции выживают
п укрепляются за пределами старого ареала. Немецкие авторы
(Luers u. Ulrich, 1954 : 620) именуют их словами «Bruckenkopfe»
и «Jgelstellungeh», т. е. «предмостное укрепление» и «круговая
оборона» («еж»).
Этот способ расширения ареала иллюстрируется примерами за-
летов и внедрения в Англию черной горихвостки (Phoenicurus
ochurus) (Лэк, 1957) и др. В отличие от такого расселения мел-
кими группами могут иметь место быстропротекающие миграции
с массовым размножением (колорадский жук в Европе, кролики
в Австралии и т. п.). Однако, по-видимому, всегда имеют место
колебания численности — популяционные волны. У одних видов
они огромны, у других очень пологи и почти незаметны. Но во
всех случаях волны жизни усиливают- давление отбора и, воз-
можно, иногда приводят к случайным генетическим эффектам
(Оленов, 1961).
Постоянные колебания численности пограничных популяций,
расселение одних популяций и гибель других благоприятствуют
усиленному давлению естественного отбора. В результате все
время происходит преобразование состава популяций, а также
преимущественное расселение некоторых наиболее адаптивно
организованных популяций.
Этим путем может осуществляться пе только расселение при
«географическом видообразовании», но и заселение новых стаций
при «экологическом видообразовании».
Выдвигая тезис о единстве расселения и эволюции, С. С. Хох-
лов (1955) предлагает различать три типа расселений: гомоэко-
логическое, при котором расселение происходит в пределах из-
давна освоенных местообитаний; гетероэкологическое, при кото-
ром осваиваются новые местообитания в пределах географического
ареала, и географическое расселение, приводящее к расширению
317
этого ареала. Мысль о единстве расселения и эволюции в такой
общей формулировке требует уточнения. Гомоэкологическое рас-
селение происходит непрерывно и не обязательно связано
с микроэволюцией. Географическое расселение, как это пока-
зывают факты быстрых массовых миграций, отнюдь не
всегда сопровождается эволюционными сдвигами. Только гете-
роэкологпческое расселение необходимо связано с микроэволю-
цией.
Австралийские зоологи недавно дискутировали вопрос о при-
уроченности видообразования к тому или иному району мате-,
ринского ареала. Одни настаивали на том, что видообразование
животных преимущественно происходит в краевых популяциях,
где типичные особи вида оказываются менее приспособленными
(М. White, 1959), а другие, — что оно происходит в центре ареала
и развивается центробежно, так как именно там сосредоточен мак-
симум генетического и фенотипического полиморфизма. Как будет
показано дальше, действительность сложнее таких односторонних
схем и при известных условиях видообразование протекает и
в центре, и па краю ареала, с изменением эколого-географических
границ обитания материнского вида или без их изменения, и т. п.
Не следует упрощать вопроса, полагая, что любая окраинная попу-
ляция может служить материалом видообразования. Окраинные
популяции генетически отличаются от популяций из центра ареала
и, как это видно из опытов с дрозофилой (Drosophila robusta),
менее генетически полиморфны и менее адаптивны (Carson,
1961). Обеднение генотипа па окраине ареала, которое было уста-
новлено Н. И. Вавиловым на растениях, было открыто и на дро-
зофиле. Высокогетерозпготные центральные популяции спо-
собны приспособляться к разнообразным условиям, а более
гомозиготные окраинные являются узкоспециализирован-
ными.
Справедливо, видимо, что видообразование в смысле расщеп-
ления одного вида па два чаще происходит путем дивергенции
изолированных популяций, в которых под давлением отбора про-
исходит достаточно сильная реорганизация старых генетических
шаблонов (Мауг, 1963).
Подвид и полувид как стадии видообразо-
вания. Подвид является не только крупной частью вида, но
и одной из заключительных стадий1 эволюционных изменений,
протекающих в рамках вида. Примерно около трех четвертей
современных видов имеют подвиды, но молодые и древние
эндемы, узкоспециализированные виды и, напротив, виды, особи
которых способны адаптивно перестраиваться при широких коле-
баниях условий, не образуют подвидов.
1 Некоторые рассматривают географический подвид как «начальную
стадию видообразования» (Rensch, 1965).
318
Если сформированные географические пли экологические
расы обособляются настолько, что получают возможность к само-
стоятельному существованию и эволюционному развитию, по эта
возможность реализуется неполностью (наличие переходных
форм или эффективность скрещиваний), то такие расы и име-
нуют подвидами.
Последние десятилетия проводились исследования, показав-
шие, что среди рас наряду с только что начавшими обособляться
и вполне отвечающими понятию подвида, можно обнаружить
также и более обособленные, по существу находящиеся в про-
цессе перехода от состояния подвида к состоянию типичного
вида. Разумеется, далеко пе все расы, достигшие ступени под-
вида, оказываются способными к следующему эволюционному
шагу и развиваются в виды. Многие в течение неопределенного
времени так и остаются на положении подвида и потом выми-
рают. При внутривидовых миграциях в пределы ареала подвида
могут внедриться отдельные экоэлемепты или целые полиморф-
ные популяции других подвидов, что приводит к межгрупповой
гибридизации и к уменьшению степени обособления подвидов.
При этом граница между подвидами, бывшая четкой, расплы-
вается п возникает зона трансгрессии. В таких случаях конста-
тируется вторичное уменьшение (или уничтожение) известной
обособленности рас. Как отмечалось в главе 8, образование гиб-
ридных переходных зон с клипальной изменчивостью, как пра-
вило, не ведет к ликвидации подвидов.
Однако следует сделать существенное дополнение. Дальней-
шая эволюция таких гибридных популяций различна и зависит
от многих факторов. Если подвиды представлены суперпопуля-
циями (Беклемишев, 1960), то гибридные популяции займут
маленькую полоску между массивами подвидов. По мере углуб-
ления в эти массивы численность типичных особей подвидов
будет возрастать, а гибриды исчезать. Однако при небольших
ареалах и значительных селекционных преимуществах гибридов
возможно расширение зоны трансгрессии на весь ареал. Вместо
подвидов образуется единая большая популяция. Этот процесс
может сопровождаться значительным ортоселективным преобра-
зованием новой популяции, дающим право считать ее новым ви-
дом. Такой путь видообразовапия основан на гибридном синтезе
подвидов, адаптивности этих гибридов и связан с гибридогенной
ликвидацией подвидов.
Разберем более подробно вопрос об эволюции подвидов до
состояния, при котором квалификация их как подвидов оказы-
вается уже недостаточной. Степень обособления рас, при которой
возникают состояния не вполне отвечающие подвиду, но и не
являющиеся еще видами, с эволюционной точки зрения пред-
ставляет большой интерес, так как позволяет рассмотреть по-
следние акты видообразования.
319
Среди 755 видов североамериканских птиц около 10% отно-
сятся к группам, промежуточным между типичным подвидом и
настоящим видом (Мауг, 1963).
Примером разбираемого явления могут служить морские ко-
тики, образующие отдельные расы («стада») па Курильских в
Командорских островах и на Аляске. Каждое такое «стадо» одни
считают атдельным видом, а другие — подвидом. Эти расы четко
обособлены друг от друга, имеют особые пути миграций, собст-
венные лежбища для размножения, никогда не смешиваются
между собой, каждая раса имеет специфическую динамику
численности, не влияющую на изменения численности дру-
гих рас (Северцов, 1951). Обособление рас здесь, по-види-
мому, действительно значительно большее, чем обычное у под-
видов.
Степень обособления европейской и сибирской косули, рыже-
ватого и краснощекого суслика, степного и темного хоря, зайца
русака и толая, по Г. П. Дементьеву (1954), превышает харак-
терную для обычных подвидов, по в некоторых отношениях еще
недостаточна для отнесения этих групп к видам: это обособление
объективно является промежуточным, пограничным состоянием
подвида и вида.
Хорошими примерами такого состояния служат расы фазанов
(Rensch, 1929, 1933, 1954), столь обособленные друг от друга, что
они отличаются от типичных подвидов. Как показал П. В. Сереб-
ровский (1941), именно для таких степеней обособления Ренш и
предложил вместо понятия «вид» понятие «круг рас». Таковы же
расы барсуков Meles leptorhynchus и М. meles (Огнев, 1944) и
многие другие.
Пограничные состояния между подвидом и видом всегда явля-
лись камнем преткновения для систематиков: материал вы-
нуждал учитывать переходные стадии, по не существовало поня-
тия, которое отражало бы такое состояние объекта. Трудно «за-
ковать в неподвижную формулу вечно изменчивую сущность
вида» (Комаров, 1934), и систематик был вынужден относить
переходные состояния к одному из двух понятий, принимаемых
за смежные, — к подвиду или к виду. Требовалось ввести особое
понятие, отражающее большую степень обособления расы, чем
у подвида, но меньшую, чем у вида. Дж. Гексли (Huxley, 1938а)
предложил такую стадию называть «полувид» (semispecies). При
скрещиваниях между собой полувидов в потомстве возникают
различные аномалии, свидетельствующие о существенных раз-
личиях генетических систем. Такие последствия скрещивания и
являются критериями полувида. Примером полувида может слу-
жить Hemizonia angustifolia (Clausen, 1951). Ареал этого вида
растения в Калифорнии разделен горами на северную и южную
части. В результате обособились две расы. Северная имеет более
тонкие побеги, более раскидистый куст, меньшего размера и ме-
320
нее скученные головки соцветенпй, чем южная. Эти морфологиче-
ские отличия наследуются. Наличие таких особенностей и особого
ареала давало право некоторым систематикам каждую из этих
рас возводить в ранг вида. Клаузен произвел множество скрещи-
ваний различных популяций северной расы с южной. При этом
обнаружилось, что хотя первое поколение и было вполне плодо-
витым, но во втором часть растений оказалась карликами, почти
в 1000 раз меньшими, чем родительские формы. Такие аномалии
в потомстве, пе наблюдавшиеся при внутрирасовых скрещива-
ниях, и свидетельствуют о значительном расхождении генетиче-
ских систем. Основываясь на всей совокупности данных, Клаузен
отмечает, что обособление этих рас у Hemizonia достигало сте-
пени полувида. С позиций морфолого-географического метода
каждая из рас Н. angustifolia является «хорошим» видом, пол-
ностью удовлетворяя его критериям (стойкая морфологическая
специфичность комплекса признаков и четко обособленный
ареал). Кроме того, через горный хребет (Сапто-Люция) невоз-
можен обмен семенами и пыльцой, т. е. эти расы фактически
изолированы и размножаются внутри себя. С другой стороны,
с позиций стандарта биологического вида эти расы являются
типичными подвидами, так как межрасовое скрещивание эффек-
тивно и гибриды оказываются плодовитыми. Это резкое противо-
речие выводов, полученных при применении различных концеп-
ций вида, полностью разрешается при комплексном подходе и
при изучении особенностей, возникающих у гибридных растений
второго и последующего поколений.
Углубленное изучение таких спорных случаев показывает на-
личие стадий, переходных от подвида к виду, и доказывает прин-
ципиальную важность введения понятия полувида. «Биологиче-
ские единицы на такой переходной ступени развития являются
очень важными, так как их существование отражает промежу-
точные ступени эволюции видов» (Clausen, 1951:86).
С. С. Шварц (1959, 1963) правильно подчеркивает постепен-
ность и историчность видообразования. Однако его заключения
о принципиальных различиях между видовыми и подвидовыми
адаптациями и о том, что отличия подвидов не закреплены от-
бором (Шварц, 1959:25 п др.), являются спорными. Вопрос об
адаптивности видовых и расовых признаков изучается давно
(Кашкаров, 1939, и др.). Некоторые систематики и полевые эко-
логи маркировали подвиды иногда сильно модифицирующими
признаками, экспериментально пе проверяя размах их изменчи-
вости. Ныне доказано, что высококонстантными (с малым значе-
нием С %) бывают не только подвидовые, но и многие меж- и
внутрипопуляционные различия. Существование промежуточных
стадий, таких как полувид, показывает, что нужно не противо-
поставлять видовые различия подвидовым, а считать их стадиями
единого процесса.
21 К. M. Завадский
321
Экологическое видообр азование. Образование
новых видов путем формирования локальных экотипов и их по-
следующего обособления — процесс осуществляющийся в общем
теми же путями, что и географическое видообразование. Нами
уже отмечалось, что разделение рас на географические и эколо-
гические является в известной мере условным, так как различное
положение-популяций в пространстве всегда связано с какими-то
различиями в условиях существования. Это положение и было
зафиксировано в понятии «климатип» (климатический экотип).
Занятие географической расой новых биотопов и биоценозов мо-
жет служить основанием рассматривать ее как особый эдафоэкотпп
или ценоэкотип. С другой стороны, локальные экотипы зани-
мают какое-то пространство и могут толковаться как микрогео-
графические расы. Между широко расселенной географической
расой и узколокальным экотипом могут быть расположены целые
серии популяций переходного типа. По содержанию экологиче-
ским является и географическое видообразование и симпатриче-
ское, так как во всех случаях действует отбор по адаптивным
различиям. В тесном смысле слова экологическим видообразова-
нием мы будем называть случаи обособления рас, приуроченных
к различным биоценозам (биотопам, стациям и т. д.) без измене-
ния географического ареала.
Интересные данные по видообразованию у растений в пойме
Волги сообщил А. Д. Фурсаев (1937). Поймы рек в геологиче-
ском отношении — молодые образования. Заселяющие поймы
популяции морфологически малоотличимы от родственных попу-
ляций из внепойменных местообитаний, но все имеют ряд специ-
фических экологических особенностей. Начало вегетации у них
резко сдвинуто на более поздние сроки (конец мая—начало
июня), что соответствует средним датам окончания паводка. Та-
ким образом, пойменные популяции оказались изолированными
от внепойменных не только пространственно, но и во времени
развития, образуя особые экологические расы. Изоляция этих рас
очень, велика из-за того, что большинство видов не имеет приспо-
соблений к распространению семян ветром. Такие эндемичные
для поймы расы обнаружены Фурсаевым у многих видов ивы и
травянистых растений.
Хорошим примером видообразования путем обособления эко-
логических рас могут служить многие рыбы пресноводных зам-
кнутых бассейнов, например севанская форель. У этой форели
существуют группы, различающиеся сроками нереста, располо-
жением нерестилищ (у одних в озере, но на разных глубинах,
у других в определенных реках, ближе или дальше от их устья),
районом нагула. Существует и ручьевая форма севанской форели,
постоянно живущая и размножающаяся в горных реках. Некото-
рые из этих групп имеют п четкие морфологические отличия.
По Н. Л. Гербильскому (1957), такая дифференцированность
322
севанской форели позволяет ей наиболее полно использовать от-
носительно небольшие площади, пригодные для нереста, и кор-
мовые ресурсы озера.
Сдвиги в сроках нереста в сочетании со специализацией по
местам нереста и районам нагула, по-видимому, гарантируют пол-
ную изоляцию каждой группы. Каждая такая сезонно-экологиче-
ская группа должна быть поэтому признана вполне сформирован-
ной расой. Более того, эти экологические расы превратились уже
в самостоятельные биологические единицы, стоящие па видовом
уровне. Севанская форель представляет собой уже видовой
комплекс (coencspecis). Ихтиологи же обозначают такие единицы
разными терминами (infraspecies и пр.).
Образование видов специализированных сорняков льна —
наиболее разработанный и замечательный случай видообразова-
ния путем уподобления основному виду, доминирующему в фито-
ценозе. На примере рыжика льняного (Gamelina linicola) было
показано, что многие расы тонко приспособлены к особенностям
различных сортов льна. В сторону уподобления эволюируют ритмы
развития, габитус, размеры растения, окраска, величина, вес и
коэффициент парусности семян и другие признаки. Ф. Швапитц
(Schwanitz, 1954:513) отмечает, что Ротмалеру удалось показать
это па ряде видов — специализированных сорняков льна (Lolium
remotum, L. brasilicum, Spergula camara, Silene linophila и Came-
lina alyssum).
Обширные данные об экологическом видообразовании, идущем
по этапам экоэлемент -> экотип -> вид, собраны в работах
Е. Н. Синской. Особенно интересны ее исследования по постепен-
ному нарастанию комплекса различий у экотипов кавказских лю-
церн, тригонелл, овсяниц, тимофеевки и многих других расте-
ний.
Недавно Синская писала, что в довоенных ее работах «не были
достаточно оттенены различия между экотипом и видом. Теперь
становится ясно, что при типичном видообразовании экотипиче-
ские популяции подвергаются сложной переработке» (1963 63).
Эта переработка, по Синской, может осуществляться разными
путями. Вид пырея среднего (Agropyrum intermedium) явился
родоначальником пырея волосоносного (A. trichophorum). Этот
процесс шел путем преобразования наиболее засухоустойчивых
(ксероморфных) экоэлементов пырея среднего и до сих пор еще не
завершен. Отбор объединил и спаял в единую систему особого
экотипа ксероморфные признаки разных экоэлементов родитель-
ского вида, возникшие независимо один от другого в различных
популяциях. Признаки молодого вида пырея волосоносного вна-
чале возникают в популяциях пырея среднего единично и редко.
С севера на юг увеличивается частота встречаемости этих при-
знаков. Далее на юг они спаиваются в комплексы и, наконец,
образуют целую ксерощорфную конституцию. Таким же пред-
21*
323
ставляет себе Синская образование вида анатолийской ржи (Secale
anatolicum) от более влаголюбивого предка.
При взаимодействии генетически еще близких групп могут
возникать сложные процессы. При встрече в одном местообита-
нии сорно-полевого вида Melandrium album с видом, заселяющим
лесные опушки (М. rubrum), возникают как процессы скрещи-
ваний, так й конкуренции. М. album — вид, лучше приспособлен-
ный к суровым зимам (глубокое заложение зимующих почек
и пр.), и на безлесных территориях он имеет преимущество
(так же как и гибриды типа album). Вблизи леса более конку-
рентоспособны М. rubrum и гибриды с доминированием типа
rubrum. Все эти процессы имеют определенную тенденцию: уси-
ление изоляции и территориально-экологического разобщения
форм (Akerlund, 1934).
В последнее время получены доказательства того, что в неко-
торых случаях эколого-географическое видообразование идет не
от зачатка экологической расы к экотипу и к виду, а иначе, что
«у некоторых родов образование экологических рас и видообразо-
вание — по-видимому, совершенно различные явления» (Lewis,
1962:270). На видах Clarkia из США этот автор обосновывает
значение катастрофического отбора для видообразования. Давно
известно существование массовой элиминации, ведущей к гибели
почти всей популяции, как это часто наблюдалось при воздей-
ствиях на насекомых ядохимикатами, или же к гибели целых
популяций. Однако экспериментально значение такого отбора
для видообразования проверялось впервые. Изучались популяции
на границах ряда видов Clarkia юго-запада США. Наступающие
там в отдельные годы сильные засухи вызывают массовую гибель
растений, принимающую катастрофический характер в окраинных
популяциях. В работе анализируются отношения между рядом
видов. Мы же рассмотрим только взаимоотношения близких ви-
дев — С. biloba и С. lingulata из Калифорнии. Последний вид —
специфичный тетраплоид с некоторыми внутрихромосомными из-
менениями. Его ареал прилегает к южным границам распростра-
нения С. biloba, но не образует с этим видом смешанных поселе-
ний. При экспериментальных скрещиваниях оказалось, что
гибриды между ними стерильны (Lewis, 1961). В смешанных
экспериментальных посевах С. lingulata не имела никакого пре-
имущества перед С. biloba, но в природе С. biloba более чувстви-
тельна к засухе.
В годы исключительно сильной засухи популяции С. biloba
изреживаются больше, чем популяции С. lingulata. Одна из при-
чин устойчивости С. lingulata состоит в ее раннеспелости (пики
цветения различаются на две недели), позволяющей упредить
засуху и существовать в более южных районах Сьерра-Невады,
чем С. biloba. Наблюдалось, как на южной границе ареала
С. biloba во время сильной засухи две популяции погибли и их
324
местообитания были заняты С. lingulata. Автор предполагает, что
С. lingulata возникла из одной или нескольких особей С. biloba,
переживших сильные засухи и образовавших исходные популя-
ции. Эти популяции заняли пустующие местообитания по южной
кромке ареала С. biloba. Прекращение катастрофических засух
лишает С. lingulata преимуществ, и она не расширяет своего
ареала до очередной засухи. Однако после следующих сильных
засух новая популяция занимает еще некоторые выморочные
местообитания С. biloba. Поскольку в зонах контакта новой расы
со старой не происходит гибридизации, они сталкиваются уже
как два разных вида. Следует подчеркнуть, что хромосомные из-
менения нового вида множественны и поэтому пельзя объяснить
видообразование внезапным катастрофическим отбором. Пра-
вильнее рассматривать его как агент, выделивший новую видо-
вую форму, которая затем шлифовалась отбором по мелким му-
тациям и доводилась до состояния молодого вида.
Экологическое видообразование иногда называют микрогеогра-
фическим, а симпатрическое — микроэкологическим. Это верно
потому, что всякое видообразование — это селекция на улучше-
ние адаптаций, т. е. по существу процесс экологический. Суще-
ство преобразований рас при географическом видообразовании
состоит в их приспособлении к различным комплексам физико-
географических и биотических условий. Иногда видообразование
рассматривают как особую форму радиации, независимую от эко-
логии (Remane, 1964), но это мнение фактически необосновано,
так как автор оперирует ископаемыми скорпионами, коллемба-
лами и другим совершенно неподходящим (из-за слабой изучен-
ности) материалом.
Название «экологическое», возможно, следует сохранить лишь
за теми случаями, когда вид образуется из локального экотипа.
Этот способ образования видов распространен не менее, чем ти-
пично географический, и широко известен у растений и живот-
ных. Изменения в нишах, в питании и в циклах развития привели
к возникновению новых видов рыб Haplochromis в африканских
озерах, пещерных рыб Anoptichys из Мексики, жуков Plagithysmus
на Гаваях и т. п. (Tjtfnneland, 1964). Анализируя видообразование
у животных оз. Байкал, М. М. Кожов (1960) подчеркивает, что
ведущим фактором была не географическая изоляция рас, а эко-
логическая, связанная с тем, что литоральные популяции вы-
теснялись на большие глубины. Именно этот процесс вызывал
замедление роста и созревания, удлинение срока жизни, редук-
цию глаз, окраски и т. п. Заселение глубинных зон озера
М. М. Кожов рассматривает как симпатрическое видообразова-
ние, хотя приведенные им примеры относятся к экологическому
видообразованию.
Ь. СИМПАТРИЧЕСКОЕ ВИДООБРАЗОВАНИЕ
О возможности симпатрического видообразо-
вания. Специфической чертой симпатрического видообразова-
ния является сосуществование формирующегося вида с исход-
ным в одном местообитании в рамках одной популяции. Более
или менее строгая симпатричность может при этом сохраняться
лишь до поры до времени. В дальнейшем же процессы адаптив-
ной радиации и конкуренции устраняют или по крайней мере
ослабляют симпатричность. Проблема состоит, таким образом,
в возможности осуществления симпатрическими группами пер-
вых фаз видообразования.
Возможность симпатрического видообразования оспаривалась
еще во времена Дарвина. Считалось, что «поглощающее действие
скрещивания» способно растворить малочисленную новую группу
в массе особей «среднего типа». Далее работами Г. Менделя и
генетиков была доказана неосновательность этих опасений.
В наше время, однако, вновь возникли сомнения в возможности
симпатрического видообразования. С наиболее развернутой кри-
тикой этой возможности выступили Э. Майр и А. Кэйн, а также
Д. Лэк (1957). Поэтому уместно рассмотреть возражения именно
этих авторов.
Первое возражение: «Существует очень серьезное генетиче-
ское затруднение, которое необходимо преодолеть каждому сто-
роннику постепенного симпатрического видообразования. Когда
в самом деле возникает новая форма, то, если она действительно
симпатрична нормальной форме, она растворится в ней в резуль-
тате скрещивания между ними» (Кэйн, 1958 225).
Под «действительной симпатричностью» автор подразумевает
жизнь в совершенно однородной среде (там же 210) и ссылается
на модельные опыты Г. Ф. Гаузе (Gause, 1934), проведенные
в пробирках. Настаивая же па реальности растворения мутантных
форм в массе нормальных, автор вступает в противоречие с фак-
тами. Известны многочисленные примеры сбалансированного по-
лиморфизма популяций, основанного не только на простых гене-
тических изменениях, но и на сложных комплексных измене-
ниях.
Если происходит «растворение», то каким же путем формируются
миксобиотипы, экоэлементы и другие типы внутрипопуляцион-
ного симпатрического полиморфизма? Собрано множество приме-
ров существования такого симпатрического полиморфизма (Ford,
1940; Mather, 1947, 1955; Синская, 1948, 1961, 1963; Stebbins, 1950;
Clausen, 1951; Huxley, 1956, 1963; Завадский, 1957a, 1961;
Dobzhansky a. Pavlovsky, 1960; Завадский и др., 1962; Grant,
1963, и др.). Такие полиморфные популяции могут быть устой-
чивыми системами, но могут и преобразовываться и дезинте-
грироваться.
326
Балансированный полиморфизм, как показал Е. Форд и мно-
гие другие, может переходить в «транзитивный» в том случае,
если отбор на поддержание сложившейся адаптивной нормы
(в которую входит несколько симпатрических форм) сменяется
отбором на ее преобразование. При этом изменяется численное
соотношение форм в популяции, может быть элиминирована
одна или несколько форм и популяция становится мономорфной,
но может возникнуть новая форма и т. п. Положительное селек-
тивное значение самого полиморфизма может быть столь боль-
шим, что устойчивость всей системы и каждой из форм охра-
няется очень строго. Вместе с тем происходит элиминация
линий, при скрещиваниях дающих потомство, у которого разру-
шаются (расщепляются) селективно скоррелированные комплексы
признаков. Отбор в этом направлении и создает экоэлементы и
морфобиологические группы, способные устойчиво .воспроизво-
диться в популяции. Спаянность таких селективно созданных
комплексов бывает достаточной для поддержания стабильности
в условиях системных отношений популяций (ее пыльцевого ре-
жима, типов скрещиваний и пр.). Однако нередко такие формы
оказываются константными и при существовании вне популяции,
в качестве самостоятельно воспроизводящихся форм. Если в по-
пуляции возникло несколько «адаптивных пиков» на модифика-
ционной основе и их положительное селективное значение оста-
валось постоянным, то стабилизирующий отбор преобразует их
в более или менее константные внутрипопуляционные формы.
Иногда же такие константные внутрипопуляционные формы воз-
никают на основе одной мутации и отбор только регулирует
проявление плейотропизма. Например, у мягкой пшеницы мута-
ция Spherococcum, кроме шаровидности и мелкости зерен, вызы-
вает усиление побегообразования, укорочение стебля, уменьше-
ние его ломкости, уменьшение колосков, вздутость чешуй и др.
Таким образом, вопреки опасениям А. Кэйна, многие формы
могут не только создаваться и сохраняться в составе панмикти-
ческой популяции, но и становиться все более константными и
дискретными образованиями, не разлагающимися даже и после
выхода из ее состава.
Второе возражение: «Не известно процесса, который допу-
скал бы развитие и усовершенствование биологических изоли-
рующих механизмов в экологических расах, пока они находятся
в широком контакте с соседними популяциями» (Майр,
1947 307). Это утверждение фактически неверно. Эксперимен-
тально доказана эффективность прямого отбора на усиление
половой изоляции в смешанных популяциях, например в опытах
с дрозофилой (Drosophila persimilis и D. pseudoobscura). При
совместном содержании этих рас К. Купман (Кооршап, 1950) на-
бюдал снижение численности гибридных форм в пяти поко-
лениях с 49 до менее ,чем 5%. Это снижение происходило из-за
327
элиминации линий, имевших большую склонность к межрасо-
вым скрещиваниям, и из-за преимущественного сохранения ли-
ний с большей избирательностью к внутрирасовой копуляции.
Причина такого направления отбора — в дефектах межрасовых
гибридов (стерильность самцов, маложизнеспособное потомство
от скрещиваний гибридных самок с самцами обоих видов). Меж-
групповая гибридизация оказалась вредной, снижала потенциал
воспроизведения обоих видов, и в короткий срок отбор резко по-
высил физиологический барьер между этими видами. Такие же
данные на этих видах позднее получил Добжанский.
Прямой отбор на снижение межгрупповых скрещиваний воз-
можен только при отсутствии других препятствий (пространст-
венных, временных), т. е. именно у симпатрических групп. В по-
лиморфных популяциях такой отбор может отгораживать адап-
тивные формы (комбинации) от маложизнеспособных или же
оберегать особо высокоадаптивные формы от скрещиваний, веду-
щих к их разрушению. Отбор на усиление межгрупповой изоля-
ции может привести к образованию групп, уже переставших
быть компонентами единой скрещивающейся системы, но еще
продолжавших жить совместно с другими членами этой попу-
ляции.
Итак, отбор может сохранить полезную форму и в условиях
панмиксии и формировать ее дальше, потом изолировать от менее
адаптивных форм и, наконец, вывести на дорогу самостоятельного
существования.
Третье возражение: «Если полагать, что образование видов
идет, так сказать, через индивидуумы, то признание процесса
симпатрического видообразования неизбежно, так как эти два
понятия очень тесно связаны Теперь все больше выясняется,
что виды, вообще говоря, происходят путем эволюции популяции
в целом» (Майр, 1947 294). Здесь правильно подчеркивается
несостоятельность мутационизма, но при этом ставится знак
равенства между ним и симпатрическим видообразованием. Майр
(как и Кэйн и др.) проходит мимо уже известных возможностей
видообразования путем развития в вид отдельных внутрипопу-
ляционных групп. Симпатричность здесь налицо, но, разумеется,
процесс идет не через отдельных особей, а через группу в целом,
становящуюся обособленной популяцией и молодым видом. В этом
вопросе Майр п Кэйн непоследовательны. В самом деле, они
признают существование «внезапного видообразования» (Майр,
1947 : 294 и др.; Кэйн, 1958: 228), происходящего, по их мнению,
сразу с появлением одной особи, если она* неспособна скрещи-
ваться с родительской формой. Можно сказать, что в этом пункте
концепция биологического вида слилась с мутациоиистской кон-
цепцией вида. Далее будет показано, что симпатрическое видо-
образование протекает на популяционном уровне и что идея
«внезапного видообразования» ошибочна. Симпатрическое видо-
328
образование осуществляется путем преобразования популяций,
даже и в том случае, если оно начинается с образования адап-
тивного аллополиплоида.
Четвертое возражение сводится к тому, что не существует на-
дежных данных в пользу такого процесса: «Все случаи, приво-
дившиеся в качестве примеров симпатрического видообразования,
можно объяснить при помощи географического видообразования»
(Кэйн, 1958 226). Ошибочность этого утверждения А. Кэйна
будет показана в следующих разделах.
Примеры симпатрического видообразования.
Интересны данные Н. И. Лариной (1961, 1962, 1964), изучавшей
взаимоотношения лесной мыши (Apodemus silvaticus) и желто-
горлой (A. tauricus).
Географо-экологическая изменчивость обоих рас настолько
велика, что ни один отдельно взятый признак пе является
надежным и определение возможно только по комплексу призна-
ков (длины тела, черепа, задней ступни и т. п.): с северо-востока
на юг (Кавказ) степень различий снижается. В разных частях
ареала складываются различные межрасовые отношения:
1) расы живут в одиночку; 2) на больших территориях их
ареалы перекрыты, но местообитания различны; 3) во многих
районах они занимают одни и те же биотопы, т. е. симпатричны,
но живут как антагонисты и благодаря психофизиологическому
барьеру размножаются только внутри себя, как «хорошие» био-
логические виды; 4) на юге ареала обе расы строго симпатричны
(не образуют даже отдельных поселений), между ними не только
отсутствует антагонизм, но происходят половые контакты (меж-
расовые и гибридов с чистыми формами); 5) в ряде районов
Кавказа вполне устойчивы все три симпатричные формы (лес-
ная, желтогорлая и гибридная); 6) в Армении гибридные формы
имеют селективное преимущество над желтогорлой мышью, ареал
которой поэтому имеет расплывчатую южную границу, как бы
растворяясь в гибридных популяциях.
Н. И. Ларина справедливо подчеркивает, что эти факты дока-
зывают реальность и закономерность симпатрического видообра-
зования. К этому, однако, опа добавляет: «на основе межвидовой
гибридизации». Между тем взаимоотношения между лесной и
желтогорлой мышами на Кавказе, особенно в Армении, характе-
ризуются настолько большим сближением в морфологическом,
экологическом и генетико-физиологическом отношениях, что пра-
вильнее говорить о формах внутри единой панмиктической попу-
ляции (подобных экоэлементам и морфобиологическим группам
растений). Автором показано, что на юге ареала взаимоотноше-
ния между расами имеют характер внутривидовых (Ларина,
1962 200—201). Таким образом, если на северо-востоке эти мыши
различаются как географические виды, затем как экологические
виды, далее как биологические виды, то на юге несколько изме-
329
ненные формы следует рассматривать не как виды или расы,
а лишь как симпатрические формы — компоненты целостной по-
пуляции. Если внутри таких диморфных популяций возникают
гибриды, которые становятся устойчивыми и конкурентоспособ-
ными, то возникает триморфная популяция с транзитивным по-
лиморфизмом, т. е. происходит строго симпатрическое видообра-
зование. Сходные ситуации обнаружены и у растений. Виды го-
рицвета (Silene vulgaris и S. maritima) не скрещиваются из-за
экологической изоляции, а на северной окраине ареала живут
в одном местообитании, образуя пеструю гибридную попу-
ляцию.
Наглядным примером симпатрического видообразования у рас-
тений на основе межвидовой гибридизации является Spartina
townsendii (2n = 120). В 70-х годах прошлого века на морском
побережье юга Англии было описано новое растение гибридного
облика пз рода Spartina. Как было показано (Huskins, 1931),
этот вид действительно был гибридогепным аллополиплоидом
между S. alternifolia (2п = 70) и S. stricta (2п = 50). Эта новая
популяция «доказывает сейчас свою приспособленность действи-
тельно по-дарвиновски, уничтожая своих родителей» (Haldane,
1932: 106). А. Мюнтцинг приводит этот случай как пример гиб-
ридогенного образования «настоящих видов: S. townsendii
«чрезвычайно успешно соперничает с другими видами, так что
область его распространения расширяется» (1963:392).
Строго симпатрическим является видообразование у насеко-
моопыляемых растений благодаря специализации переносчиков
пыльцы на опылении цветков какой-либо одной внутрипопуляци-
онной группы растений. Оказалось, что энтомофилия в ряде слу-
чаев вызывает полную половую изоляцию одних групп растений
от других, живущих бок о бок. Причина этого явления заключа-
ется в избирательности насекомых, предпочитающих определен-
ные формы растений внутри популяции («видообразование по
типу экобиотической дифференциации»—Huxley, 1963). Расы
львиного зева Antirrhinum majus и A. glutinosum можно без
затруднений скрещивать между собой. Гибриды между ними
вполне плодовиты. В природе же, в местах, где эти расы живут
как симпатрические формы, возникает не более 3% гибридов.
Изолирующим агентом являются пчелы. Пчела никогда не пере-
ходит с облета цветков одной расы на другую расу (Mather, 1947).
Позднее такие же данные были получены на расах Pentstemon,
на видах Aquilegia.
Симпатрическое видообразование возможно также и путем
обособления внутрипопуляционных феноформ, преобразования их
в сезонные расы и в молодые виды (см. главу 5 и рис. 7).
В наших опытах с марью белой была показана возможность пол-
ной селективной изоляции ранних морфо-биологических групп
мари белой от поздней (через выпадение форм с промежуточными
330
сроками цветения). Такое обособление ранее уже сформирован-
ных сезонных форм при благоприятных условиях может стать
существенным этапом видообразования. В исследованных пами
популяциях мари белой с Карельского перешейка почти отсутст-
вуют все четыре среднеранние морфобиологические группы,2 так
что ранняя группа (I) почти обособилась от поздних (VI). Если
Зтап I
Единая популяция
со сбалансированным
соотношением численностей
феногрупп и преобладанием
средней группы (Б)
Сроки осуществления
жизненного цикла (или одной
фазы-цветения и т.д.
Зтап II д
Популяция еще едина I —
и триморфна, но средняя £
феногруппа подвергается
элиминации,и полиморфизм q
стал транзитивным •==
Зтап 111
Образовались две
самостоятельно
воспроизводящиеся
сезонные расы
Соотношение числа
особей в феноеруппах
А
В
t==i
Рис. 7. Видообразованием путем обособления внутрипопуляцион-
ных групп растений (схема).
в Ломоносовском и Гатчинском районах в 1960 и 1961 гг. попу-
ляции содержали 65% растений, относящихся к четырем средне-
ранним группам, то на Карельском перешейке — только 16%.
В экспериментальных условиях элиминация промежуточных
групп приводила уже в третьем поколении к образованию почти
полного разрыва между двумя крайними частями популяции
У многих рыб в одной популяции сосуществуют формы, раз-
личающиеся по скорости роста и размерам, а также по питанию,
срокам нереста, иногда по соотношению полов (сельдь, окунь,
сазан и др.). Сопоставляя размеры таких различий у разных
2 Краткую характеристику морфобиологических групп мари белой
см. в главе 8, а подробнее: Завадский, 1957а, 1961 : 151—155, 1962.
331
видов, можно составить целую серию переходов от единых поли-
морфных популяций к сосуществованию уже значительно обосо-
бившихся групп — сезонных рас — и, наконец, до совместно
живущих близких, но уже самостоятельных видов (например,
большая и малая голомянка из оз. Байкал). По Б. М. Медникову
(1963), подобный путь видообразования у рыб очень распростра-
нен. Он считает, что материалом для симпатрического видообра-
зования у рыб служат внутривидовые группировки. Например,
различие в размерах и неравномерность полового созревания
могут привести к обособлению внутривидовых групп без какой бы
то ни было географической изоляции.
Таким образом, симпатрическое видообразование у рыб может
осуществляться тем же путем, что у растений при образовании
в популяции феногрупп и обособления их в сезонные расы.
Вообще любая адаптация, приводящая к изменению предпочитае-
мого местообитания, будь она модификационной или мутацион-
ной, облегчает видообразование (Завадский, 1961; Huxley, 1963;
Мауг, 1963).
Одним из наиболее интересных и специфических путей сим-
патрического видообразования является отдаленная гибридиза-
ция. В первых поколениях развитие гибридов проходит бок о бок
с родительскими формами, т. е. характеризуется большими или
меньшими степенями симпатричности.
Попытки математического моделирования некоторых ситуа-
ций, возникающих при симпатрическом видообразовании (Базы-
кии, 1965), являются безусловно полезными. Очень интересно за-
ключение о том, что две нескрещивающиеся популяции, совместно
живущие в однородной экологической нише, представляют собой
очень неустойчивую систему и малейшая флюктуация условий
способна привести к вытеснению одной из таких популяций.
С и м п а т р и ч е с к о е видообразование и внутриви-
довая конкуренция. Вопрос о преобразовании внутривидо-
вых отношений в межвидовые существен только при симпатриче-
ском видообразовании. Пространственное (географическое, топо-
графическое или биотопическое) разъединение рас связано
с такими адаптивными преобразованиями, которые могут совсем
не затрагивать особенностей внутривидовых отношений или за-
трагивать их лишь коррелятивно. Строй внутривидовых отноше-
ний дочерних видов может быть полностью или почти полностью
унаследован от материнского вида. При хроноэкологическом видо-
образовании строй внутривидовых отношений должен нарушаться
после выхода феногруппы из состава популяции, так как меняется
режим скрещиваний и складывается новая генетическая струк-
тура. Однако сущность хроноэкологического видообразования со-
стоит не в преобразовании внутривидовых отношений в межвидо-
вые, а в нарастании временной изоляции между преобразован-
ными частями исходной популяции.
332
Только при истинном симпатрическом видообразовании вну-
тривидовые отношения превращаются в межвидовые. Вначале су-
ществует обычная внутривидовая межгрупповая конкуренция, ко-
торая, как известно, может достигать большого напряжения. Опа
может приводить к незначительному нарушению полной панмик-
сии внутри популяции. Если конкурирующие группы становятся
устойчивыми и фенотипы этих групп оказываются селекционно
ценными, автоматически начинается отбор на закрепление этих
фенотипов. Дйсруптивный отбор, т. е. отбор, формирующий поли-
морфизм популяции (Smith, 1962), может поддерживать устойчи-
вое соотношение групп, не увеличивая небольшого нарушения
панкмиксии, но он же способен действовать в сторону элимина-
ции межгрупповых гибридов. Сохранение равновесия между кон-
куренцией и устойчивым полиморфизмом осуществляется тогда,
когда селекционно ценны не только чистые фенотипы внутрипо-
пуляционных групп, по и их гибриды, так что отбор поддержи-
вает не только полиморфизм, но и панмиксию. Совершенно
иной результат получается в популяциях, у которых по разным
причинам селективное значение межгрупповых гибридов стано-
вится отрицательным. «Если приспособленность гетерозигот
низка, то следствием дисруптивиого отбора будет возрастание
репродуктивной изоляции и внутригрупповых спариваний»
(Smith, 1962:62). Приспособительные фенотипы обособляются и
в половом отношении, так что дисруптивный отбор, поддержи-
вавший когда-то сбалансированный полиморфизм сексуально еди-
ной популяции, сменяется на противоположный ему изолирующий
отбор. Если при этом внутрипопуляционные межгрупповые конку-
рентные отношения не приведут к территориальному или хроно-
экологическому разобщению групп, то быстро наступает полная
межгрупповая половая изоляция. У животных одним из распро-
страненных механизмов такой изоляции служит возрастание пси-
хофизиологического барьера на основе перехода острой внутри-
видовой конкуренции за пищу, убежища и т. п. в антагонизм,
("удя по нашим опытам с популяциями растений, среди них также
существует переход дисруптивиого отбора в изолирующий и
внутривидовых отношений в межвидовые, хотя механизмы этих
процессов совершенно иные, чем у высших животных.
Итак, при настоящем симпатрическом видообразовании дей-
ствительно происходит разрушение основных внутривидовых от-
ношений и формирование межвидовых путем усиления конку-
рентных отношений между внутрипопуляционными группами.
После половой изоляции групп эти отношения становятся антаго-
нистическими и по своей природе должны быть отнесены к био-
ценотическим. Говоря о переходе внутривидовой конкуренции
в антагонизм, мы не хотим этим создать мнение, что в процессе
симпатрического видообразования обязательно возрастает интен-
сивность конкуренции. Под антагонизмом мы понимаем конку-
333
ренцию пе в пределах единой популяции, целостность которой
поддерживается скрещиваниями и другими формами основных
внутривидовых отношений, а между обособленно воспроизводящи-
мися системами, использующими одни и те же биотопы и пиши.
Некоторые считают, что зарождение межвидовых отношений
происходит с того момента, когда конкуренция из-за террито-
рии ведет к’вытеснению части населения в менее благоприятные
местообитания (Ларина, 1961). Но, во-первых, в этом случае
в начале процесса исчезает симпатричпость, а во-вторых, оши-
бочно рассматривать сам факт противоречивости внутривидовых
отношений как явление, относящееся к межвидовым отношениям.
Поэтому неправильно рассматривать межвидовые отношения как
механизм симпатрического видообразования. Здесь сказалось
влияние идей о полной гармоничности внутривидовых отношений
и о невозможности внутривидовой конкуренции. В основе симпат-
рического видообразования лежит внутривидовой межгрупповой
отбор (дисруптивный и изолирующий), вначале формирующий
сбалансированный полиморфизм популяции, а в дальнейшем ис-
пользующий селективные преимущества разных форм и обособ-
ляющий их друг от друга.
5. ГИБРИДОГЕННОЕ ВИДООБРАЗОВАНИЕ И ПОНЯТИЕ
О «НОВОЙ ВИДОВОЙ ФОРМЕ»
Гибридогенные виды в природе. Отдаленная
(межвидовая, межродовая) гибридизация играет известную роль
в процессах видообразования растений и животных. Среди цвет-
ковых растений, рыб и других групп давно отмечались виды,
совмещающие признаки двух близких ныне существующих видов
(или родов). В последние десятилетия было обнаружено мно-
жество «гцбридогеппых видов», хотя природа их происхождения
не всегда точно устанавливалась.
О классическом примере со Spartina townsendii уже было ска-
зано (стр. 330).
Хорошим примером молодого вида (расы) гибридного проис-
хождения является рябинокизильник (Sorbocotaneaster), описан-
ный Л. К. Поздняковым (1952) и А. И. Поярковой (1953).
От рябины Sorbus sibirica он унаследовал вкус и окраску плодов,
форму листа, форму почек, характер зубчатости листа, а от ки-
зильника Cotaneaster melanocarpa — строение плода, форму со-
цветия, характер опушения. Растет рябинокизильник по опушкам
хвойных лесов на юге Якутии на берегах Алдана не в одном
месте, а на протяжении 300 км. Цветет и плодоносит нормально.
В данном случае обнаружены не просто единичные экземпляры
межродового гибрида, а целая популяция, освоившая значитель-
ную территорию и занявшая определенное место в биоценозах.
На Украине были обнаружены естественный гибрид Primula
334
officinales X P. acaulis, некоторые экземпляры которого по от-
дельным признакам уклонялись к родительским видам, гибриды
пыреев Agropyrum oristatum X A. intermedium, а также A. re-
pens X A. cristatum (Троицкий, 1960). Этот же автор доказы-
вает гибридогенное происхождение Andropogon caucasicus
(A. ischaemum X Sorghum helepense), крушины Rhamnus
spathulaefolia (R. pallasii X R. cathartica), а также Papavcr de-
sertorum, Galium nupercreatum, Potentilla procumbens, Eremos-
tachys canescens и др.
Иногда удается регистрировать первые шаги завоевания гиб-
ридом места в биоценозе, начальную фазу процесса эволюции
гибридной формы в самостоятельную популяцию. В США целое
десятилетие вели наблюдения за популяцией спонтанного меж-
видового гибрида Helianthus annuus X Н. bolonderi, сумевшего
отвоевать участок среди дикой растительности. Авторы (Stebbins
a. Daly, 1961) установили, что численность популяции за 10 лет
колебалась от 6 тыс. до 75 тыс. растений. Вначале был единый
массив, но потом он был разделен па два островка барьером
в 120 м, возникшим после внедрения другого вида. На каждом
из островков наблюдались своеобразные изменения растений.
На одном из них, с большей численностью, стали преобладать
промежуточные типы, по-видимому, более конкурентоспособные
и плодовитые.
Большой интерес представляют находки популяций отдален-
ных гибридов, если они пе только многочисленны, по и начинают
занимать определенные местообитания, отличные от родитель-
ских. По наблюдениям И. А. Райковой и Н. В. Кузьминой (1958)
над распространением на полях Памирской биостанции межро-
дового спонтанного гибрида между ячменем Hordeum turcestani-
cum и клпнэлимусом Clinelymus nutans, этот гибрид растет не
только в местах контакта местообитаний родительских видов,
но и в местах, отличающихся от типичных для родительских ви-
дов. Наличие гибридов на стыках местообитания родительских
видов в качестве симпатрических форм, еще не свидетельствует
о том, что они приобрели устойчивость и известную эволюцион-
ную самостоятельность (возможна постоянная гибридизация).
Иное дело — наличие гибрида в нетипичных местообитаниях.
Если гибрид населяет их в массе и постоянно, то уже можно го-
ворить об образовании популяций молодого вида, нашедших себе
особое место в экономике природы.
Пример с ячмень-клинэлимусовым гибридом интересен также
и потому, что симпатричность здесь оказалась первой фазой
процесса, а аллопатричность — второй.
В литературе приводятся многие другие данные о гибридных
популяциях в природе. Например, в Молдавии В. А. Рыбиным
(1962) обнаружена популяция межродового гибрида между алы-
чой и абрикосом.
335
Среди полиплоидных видов растений преобладают гибридо-
генные амфидпплоиды, по, возможно, некоторое участие в обра-
зовании полиплоидных видов принимала также и аутополи-
плоидия. Некоторые считают полиплоидию исключительно ред-
ким фактором видообразования или значимым лишь в условиях
Арктики (Исландия — 55% и о. Шпицберген — 80% полиплоид-
ных видов) и в других крайне суровых районах. Однако в Сред-
ней Европе (ФРГ) полиплоидных видов высших растений — 44%.
Экспериментально показано, что удвоение числа хромосом меняет
норму реакции. Поэтому, возникнув рядом с родителями, т. е.
симпатрично, формы, отличные по плоидности, вскоре занимают
особые экоареалы или становятся географически викарирующимп
расами. Образование видов картофелей, дурманов, пшениц, хри-
зантем, мятликов, тополей, капуст и многих других растений было
связано с многократным увеличением плоидности, так что возникли
полиплоидные ряды видов. Наиболее высокоплоидные виды яв-
ляются более молодыми, производными от видов с низкой плоид-
ностью. Численность высокоплоидных видов весьма значительная
для всех суровых местообитаний, например горных областей, пу-
стынь, Крайнего Севера, о. Сахалин (Соколовская, 1960, и др.).
Длинные полиплоидные ряды видов описаны у многих родов,
имеющих декоративное значение (Матвеева, 1965). Например,
у Zephyranthes диплоидные виды имеют 2п = 12, а наиболее вы-
сокоплоидный — 2п = 96, у Gladiolus соответственно 2п = 30 и
180, у Gagea 2п = 24 и 102, у Crocus 2п = 6 и 30, у Scilla
2п = 12 и 58, и т. д. Число видов в таких рядах бывает различ-
ным — от 4—5 до 10—15 и более (включая и анеуплоидные
виды). Интересные превращения при полиплоидии наблюдаются
у видов Heliantlius: диплоидные виды (2п = 34), одно- или мно-
голетники, не имеют даже зачатков клубней, тетраплоидные виды
(2п = 68) образуют ряд от форм с корневищами к формам с клу-
беньками, а у гексаплоидных видов (2п = 120) возникают на-
стоящие клубни, как у топинамбура (Марченко, 1965). Поли-
плоидия является фактором видообразования и у мхов (Peterfi,
Stefureac, 1965). Работая с арахисом, Ю. П. Мирюта (1965) при-
шел к важному обобщению, объясняющему причины поддержа-
ния гомозиготности и самоопыления у полиплоидов. Чистые
линии арахиса сохраняют устойчивую гетерогеномность, которая
осуществляется при самоопылении па основе избирательной
конъюгации сестринских (а не только гомологичных) хромосом.
Гомозиготность при самоопылении рассматривается как вспомо-
гательный механизм, обеспечивающий поддержание гетерогеном-
ности как одной из форм гибридного состояния организмов.
Можно полагать, что явление гетерогеномности лежит в основе
закрепления гетерозиса и является важнейшей генетической при-
чиной видообразования и вообще приспособительной эволюции
самоопылителей путем полиплоидии.
336
У животных также существуют виды бесспорно гибридогеп-
пого происхождения, например среди рыб у осетровых и карпо-
вых и т. д., у птиц, грызунов и некоторых других групп
позвоночных, хотя число их много меньше, чем у растений (Ру-
байлова, 1963). У беспозвоночных животных число гибридоген-
ных видов, по-видимому, очень значительно, но эти группы
пока слабо изучены (Беклемишев, 19646). Б. Л. Астауров
(1965) подчеркивает, что в эволюции животных полиплоидия
играла известную роль, но эта область исследований сильно недо-
оценивалась.
Можно сказать, что в природе существует много видов гиб-
ридогенного происхождения. Вместе с тем не следует смешивать
это утверждение с ошибочной концепцией гибридогенеза. Со-
гласно последней, гибридизация служит единствеипой причиной
видообразования (Lotsy, 1916а, 1916b, 1917), чуть ли пе все
виды покрытосеменных растений имеют гибридное происхожде-
ние (Попов, 1928, 1956) и т. п. Эта концепция содержит большие
преувеличения.
Изменение вида может происходить и путем интрогрессии,
т. е. через частичный обмен генов между уже сформированными
видами. Интрогрессия — процесс редкий у животных и несколько
более частый у растений. Особенность интрогрессии состоит
в том, что вид приобретает отдельные признаки другого вида и
при известных условиях может все более и более насыщаться
ими. В концепции compilospecies зафиксирована эта способность
отдельных видов к ассимиляции других видов через повторные
скрещивания с ними.
Воссоздание форм, близких природным гиб-
рид огенным видам. А. Мюнтцингу (Muntzing, 1930) впер-
пые удалось воссоздать форму, почти идентичную уже извест-
ному в природе виду Galeopsis tetrahit (2n = 32). Это растение
предположительно относилось систематиками к гибридогенпым,
и для проверки гипотезы были скрещены близкие виды: G. рп-
bescens (2п = 16) п G. speciosa (2п = 16). Первое поколение
оказалось почти стерильным, а во втором было получено три-
плоидное растение (2п = 24), скрещенное с родительским видом
G. pubescens. В третьем поколении была получена тетраплоидная
форма (2п = 32), объединившая наборы хромосом родительских
видов. Этот амфидиплопд как морфологически, так и по поведе-
нию при скрещиваниях был трудно отличим от природного вида
G. tetrachit: не скрещивался с родительскими видами, но скре-
щивался с другим видом — G. bifida. Синтезированная форма
оказалась нерасщепляющейся и вполне самофертильной, т. е.
самовоспроизводящейся. Однако это была лишь форма, очень по-
хожая на природный вид. Чтобы стать видом, она должна дока-
зать свою приспособленность к абиотическим условиям и конку-
рентоспособность, образовать устойчивые местные популяции,
22 К. М. Завадский
337
войти в состав биоценозов, расселиться, т. е. приобрести эколо-
гическую и географическую определенность.
Впоследствии методом отдаленной гибридизации было синте-
зировано много амфидиплоидных форм, близких к уже сущест-
вующим в природе видам. Так, Д. Костову (1941 — 1943) и
М. Ф. Терповскому (1960) удалось синтезировать амфидиплоид-
ные формы, весьма близкие к табаку Nicotia-na tabacum, путем
скрещивания других близких видов Nicotiana. Хорошим примером
ресиитеза видовой формы путем аллоплоидии является получе-
ние малины Rubus maximus. М. А. Розанова (19386, 1946) вос-
создала форму этого вида, скрещивая R. idaeus с R. speciosa.
В. А. Рыбину (1935, 1951) удалось синтезировать гибрид, близ-
кий домашней сливе (Prunus domestica), путем скрещиваний
алычи (Р. divaricata) с терном (Р. spinosa). В последние годы
были получены новые факты о происхождении мягкой пшеницы
(Triticum aestivum = Т. vulgare) с гексаплоидным набором хро-
мосом (2п = 42). При скрещивании тетраплоидного эммера
Т. dicoccoides (2и = 28) с Aegilops squarrosa (2n = 14) в США
и Японии были получены синтетические гексаплоидные формы,
подобные спельтам. Они легко скрещиваются с природными
спельтамп и с другими мягкими пшеницами, а их гибриды
вполне плодовиты. Как и предполагалось ранее, 42-хромосомные
пшеницы оказались аллополиплоидами. Однако вопрос о путях
синтеза мягкой пшеницы остается далеко еще не решенным.
Например, одни (Писарев и Жилкина, 1965) считают, что в этом
синтезе участвовало три дикорастущих вида: Triticum boeoticus,
Aegilops speltoides и Ae. squarrosa. Спонтанная гибридизация
между первыми двумя привела к созданию дикой однозернянки
Triticum dicoccoides, а ее гибридизация с Aegilops squarrosa —
гексаплоидной спельты, которая «и явилась родоначальным ви-
дом для обширной группы Triticum aestivum» (там же: 57). Опи-
раясь на тот факт, что спельта не имеет генного комплекса Q,
существующего у мягкой пшеницы, П. М. Жуковский (1965)
склоняется к тому, что последняя возникла через амфидиплоид
от скрещивания Т. carthlicum с Aegilops squarrosa. А. Р. и
Э. А. Жебракам (1965) удалось создать новые видовые формы
пшениц (2п = 70). Эти высокоплоидные формы они получили
как от скрещиваний твердой пшеницы с мягкой, так и мягкой
с Triticum timopheevii. В Швеции была получена синтетическая
брюква. Для этой цели у гибрида от скрещивания турнепса
с капустой с помощью колхицина было удвоено число хро-
мосом. Путем гибридизации Brassica campestris с В. oleracea
был получен синтетический рапс. Вначале у обоих видов уд-
воением числа хромосом получают тетраплоиды. После скре-
щивания их друг с другом сразу (в Fi) возникает форма син-
тетического рапса, вполне константная и плодовитая (Мюнт-
цинг, 1963).
338
Всей совокупностью подобных работ показано, что у высших
растений немало видов возникло в природе путем преобразования
в процессе естественного отбора «новых видовых форм», образо-
вавшихся в результате гибридизации.
Экспериментальный синтез «новых видовых
форм». Более полувека назад И. В. Мичуриным были полу-
чены отдаленные гибриды, резко отличные от родительских ви-
дов: церападусы — от скрещиваний вишни с японской черемухой,
фиалковая лилия — от скрещивания лилии Шовицпапум с ли-
лией Тунберга, и др. При этом Мичурин не считал вновь полу-
ченные гибридные формы новыми видами: «Гибриды вишни и
черемухи... в будущем, очевидно, дадут при селекции совершенно
новые самостоятельные виды» (1948, 2:200). Такая точка зре-
ния является совершенно верной: как бы удачно не был сфор-
мирован новый гибрид, без «доводки» отбором — он всего лишь
форма, имеющая более или менее отдаленную возможность стать
видом.3
На цитогенетической основе синтез новых видовых форм от-
даленных гибридов впервые был осуществлен Г. Д. Карпеченко
(1927, 1929). В результате скрещивания редьки (Raphanus)
с капустой (Brassica) был получен амфидиплоид. Он оказался
не только самовоспроизводящейся и константной (нерасщепляю-
щейся) формой, но и обнаружил неспособность к скрещиваниям
с обоими родительскими видами. Морфологически это растение
имело не только черты обоих родителей, но и ряд новых, отсут-
ствовавших у них признаков. По Г. Д. Карпеченко (1927 386—
387), такие формы следует характеризовать как новые виды
(species novae) или даже роды. Поэтому автор и дал новой форме
родовое название (Raphanobrassica). Такие формы действительно
обладают рядом существенных черт настоящего вида: морфоло-
гическим своеобразием, способностью воспроизводить тип при
размножении, полной фертильностью при скрещиваниях внутри
себя и полной половой изоляцией от родительских видов. В то же
время у них еще отсутствует ряд других существенных черт
природного вида: географический ареал, особая экологическая
ниша, определенный уровень численности, наличие внутренней
структуры и др. Поэтому ошибочно считать такие формы ви-
дами (species). Гибридная форма, если она жизнеспособна, пло-
довита, константна и изолирована в половом отношении, яв-
ляется только «видом в возможности». Развитие этой возмож-
ности (видовой формы) в действительность (природный
вид) — исторический процесс, осуществляемый естественным от-
бором. Поэтому надо согласиться с М. А. Розановой (1946:59),
что «не следует думать, что амфидиплоиды, возникающие в при-
3 О взглядах И. В. Мичурина на видообразование как па историче-
ский процесс подробнее см.: Завадский, 1948а, 1956.
22*
339
роде, все жизнеспособны и могут завоевать себе название
вида».
Смешение еще не апробированных и не отшлифованных от-
бором гибридных форм с новыми видами — ошибка, характерная
для мутациопизма. Эту же ошибку делают и генетики, склонные
абсолютизировать значение межгрупповой стерильности и возво-
дящие в ранг species novae как амфидиплоиды многих растений,
так и автотетраплоиды энотеры (типа gigas), дурмана, ячменя
и многих других растений только потому, что все они не скре-
щиваются с диплоидными родителями. Так, например, поступает
А. Мюнтцинг (1963:335, 392 и др.). Однако в другой работе
он отказался от представления о том, что всякий аллополиплоид
является новым видом и предложил понятие о «цитотипе», соот-
ветствующее, в нашем понимании, одному из случаев образова-
ния новых видовых форм (Miintzing, I960). Дж. Гексли (Huxley,
1963) аллополиплоидию у растений так же истолковывает как
«внезапное видообразование». Некоторые принимают эту мысль
с оговорками и признают, что «полиплоидия является фактором
скачкообразного возникновения новых видов», но если они выде-
ляются по количественным признакам, то даже наличие барьера
нескрещиваемости не дает все же права рассматривать их «как
отдельные виды» (Щербаков, 1965 : 252).
Иную позицию занял Н. П. Дубинин: «Видовое значение
аллополпплоиды приобретают лишь после длительной эволюции
генотипов» (1966 403).
Интересным направлением синтеза новых видовых форм, ко-
торое, возможно, в какой-то мере воспроизводит пути древней
спонтанной отдаленной гибридизации, являются скрещивания
пшениц с рожью и получение амфидиплоидов типа Triticale
(2п = 56). Эти ржано-пшеничные гибриды перспективны как
формы, более зимостойкие, чем пшеницы.
Таким образом, полиплоидия и в особенности амфидиплои-
дия являются источником новых форм, играющих специфическую
роль в видообразовании. Некоторые из них имеют сбалансиро-
ванный генотип, константы, вполне жизнеспособны, при скрещи-
вании между собой стойко воспроизводят свой тип, стерильны
при попытках скрещиваний с родительскими формами. В свою
очередь часть из этих новых форм оказываются приспособлен-
ными и конкурентоспособными. Именно они и начинают рассе-
ляться, формировать популяции, как это только что было рас-
смотрено. При географическом и экологическом видообразовании
генетическая несовместимость является косвенным следствием
длительной радиации, а не стартовым условием процесса обособ-
ления. В противоположность этому у амфидиплоидов такая не-
совместимость — это одна из начальных предпосылок видообра-
зования. Именно в «стартовой стерильности» и следует видеть
материал для образования новых видов.
340
6. НЕОФОРМОГЕННОЕ И МИКРОАККУМУЛЯТИВНОЕ
ВИДООБРАЗОВАНИЕ
Определение понятий. Неоформогенпым предла-
гается называть видообразование на основе отбора наиболее адап-
тивных «новых видовых форм», с самого начала изолированных
от исходного вида, и преобразования их в популяции, способные
к самостоятельной эволюции. Микроаккумулятивньтм можно на-
звать видообразование путем накопления мелких адаптивных
сдвигов. Таким образом, подразделение па неоформогенное и
микроаккумулятивное видообразование основано па различиях
в исходном материале и в сроках обособления новых форм.
Скорости видообразования, как известно, бывают весьма раз-
личны. Неравномерность темпов эволюции очень велика. Иногда
недостаточно и десятков или даже сотен тысяч лет для диверген-
ции до степени «хорошего» вида (папример, расы ландышей,
зубр, бизон и др.). Панамский перешеек образовался около
3 млн лет тому назад, но и такая изоляция не привела атланти-
ческие и тихоокеанские популяции многих беспозвоночных и рыб
к дивергенции до вида, но известны и факты быстрого образова-
ния рас (птиц за 500 лет, ящериц за 9000 лет и др.). Многие
расы растений и животных, сформировавшиеся во время послед-
них оледенений Евразии, существуют не более 5000—10 000 лет,
но гораздо быстрее шло преобразование мелких кроликов
с о. Порто-Санто (за 400 лет), домовой фарерской мыши (за
500 лет). Наконец, гибридогенная Spartina townsendii оформи-
лась в видовую популяцию за 50—70 лет и ныне вытесняет ро-
дительские виды и активно расселяется. Пеоформогенпое видооб-
разование, по-видимому, всегда протекает значительно скорее,
чем микроаккумулятивное (рис. 8). Именно его многие ошибочно
называли «внезапным», «мутационным» и т. п. В книге Э. Майра
(Мауг, 1963, гл. 18) показано, что время видообразования может
варьировать от многих миллионов лет (только пространственная
изоляция популяций) до нескольких тысяч лет или даже меньше
(изоляция с «эффектом Райта», преобразования вследствие от-
даленной гибридизации и т. п.).
Такие различия в темпах видообразования могут объясняться
качеством исходного материала, формами и причинами изоляции,
характером отбора. Микроаккумулятивное видообразование чаще
бывает ’аллопатрическим, а неоформогенное на первых фазах
всегда носит более или менее строго симпатрический характер.
Необходимо подчеркнуть, что по характеру вовлекаемого в про-
цесс материала неоформогенное видообразование только вначале
отличается от микроаккумулятивного. После того как «новая
видовая форма» преобразуется в поселение — мпкропопуляцию,
отбор идет уже по мелким особенностям, т. е. занят такой же
«дошлифовкой» деталей, как и при микроаккумулятивном видо-
341
образовании. С другой стороны, микроаккумулятивные процессы
могут сменяться неоформогенными.
Новые морфо-биологические группы и экоэлементы создаются
постепенно, в виде спайки в одно целое многих признаков (созда-
ние селективных корреляций, шлифовка мутанта с адаптивным
плеотропизмом по многим признакам и т. п.). Такая внутрипопу-
ляционная эволюция идет микроаккумулятивным путем. Однако
после создания этих форм и отбора их на устойчивость при вы-
ходе их из состава популяции в положение самостоятельно су-
Рис. 8. Схематические кривые микроак-
кумулятивного (Л) и неоформогенного (Б)
видообразования.
ществующих систем они
должны рассматриваться
как новые видовые формы,
а видообразование как
неоформогенное. Иногда
полиморфный комплекс
панмиктической популя-
ции распадается на массу
самостоятельно воспроиз-
водящихся апомиктиче-
ских линий, некоторые из
которых сразу же оказы-
ваются на положении но-
вых видовых форм.
Этот же переход микро-
аккумулятивных процес-
сов в неоформогенные ха-
рактерен и для ряда фено-
группа—сезонная форма—
раса—вид.
Таким образом, оба эти способа видообразования, четко отли-
чаясь друг от друга в типичных случаях, иногда различаются
с трудом, могут переходить друг в друга, являются иногда фа-
зами одного процесса и т. п.
Исходный материал неоформогенного видо-
образования. Исходным материалом этого способа видооб-
разования служат «новые видовые формы». В разделе о гибри-
догенном видообразовании были описаны амфидиплоиды —
наиболее хорошо изученный тип «новых видовых форм».
Compilospecies, такие как Роа pratense или Bothriochloa interme-
dia, также начинают свою эволюцию с образования «новых видо-
вых форм». Однако есть и другие источники получения «новых
видовых форм». Даже и прп гибридогенном видообразовании
исходный материал не ограничивается только амфидиплоидией и
аутополиплоидией. Существуют и другие способы изменений
хромосомного аппарата. Некоторые из них могут фенотипически
почти не проявляться, но вызывают стерильность при скрещива-
ниях с исходным видом. Их назвали скрытыми видами (Darling-
342
ton, 1940), но правильнее говорить о скрытых видовых формах.
В популяциях «гибридных циклов» (как у ив, малин и др.) хро-
мосомные аберранты встречаются довольно часто. Размножение
и селективное преобразование отдельных аберрантов может при-
вести в конце концов к возникновению симпатрических и морфо-
логически трудно различимых групп, каждая из которых, однако,
полностью сексуально обособлена. Такие случаи называют «виды-
двойники», или «виды-близнецы» (sibling-species). Надо, разу-
меется, в каждом отдельном случае еще доказать, что симпатрич-
ность таких «сборных» популяций не является вторичной,
а изоляция — полной и связанной с дивергенцией хромосомных
аппаратов.
Одним из интереснейших путей возникновения новых видо-
вых форм является переход к бесполосеменпому размножению.
Апомиктические виды, как и все прочие типы видов, форми-
руются в процессе естественного отбора, опирающегося, видимо,
на все формы мутационной изменчивости (Петров, 1963). Од-
нако момент половой изоляции от исходной популяции насту-
пает пе в конце видообразования, а при переходе к бесполосе-
мепному размножению. Правда, образование новой апогамной
видовой формы (т. е. линии, выпавшей из системы панмиктиче-
ской популяции) обычно не является таким одноактным собы-
тием, как возникновение амфидиплоида. У многих видов обна-
ружены непрерывные ряды от форм с обычным половым раз-
множением к формам полностью апомиктичным. Это может
указать на постепенность перехода к бесполосеменному размно-
жению.
Существование апомиктических особей в качестве «новых ви-
довых форм» служит доказательством того, что этот способ
размножения имеет некоторые преимущества перед ксеногамией
(Хохлов, 1950; Цингер и др., 1965). Однако по таким важным
показателям, как скорость обогащения генотипа путем гибрид-
ного синтеза и интрогрессии, возможность накопления мобили-
зационного резерва рецессивных мутаций и их рекомбинаций под
покровом доминантной адаптивной нормы, поддержание высокой
гетерозпготностп п сбалансированного полиморфизма па панмик-
тической основе в целостной популяции, — апомиктическое размно-
жение уступает ксеногамии (Завадский, 19666). Апомиктическая
особь может стать «новой видовой формой» в тех случаях, когда
выгоднее закрепить существующий гетерозис более простым и
надежным путем, чем с помощью сложного генетического меха-
низма, основанного на расщеплениях, в суровых условиях, где
затруднено переопыление (Мирошниченко, 1963). Однако, с па-
шей точки зрения, видообразование на основе апомиктических
форм носит черты инадаптивности в смысле В. О. Ковалевского.
Адаптивный эффект достигается быстро, но он носит частный,
тактический характер, а в дальнейшем рано или поздно должны
343
обнаружиться такие «стратегические промахи», как разрушение
целостности популяции и резкое сокращение объема наследствен-
ной информации, получаемой каждым потомком (при панмиксии
в конечном счете от всей популяции и расы, а при апомиксисе
только от одной линии).
Важным источником образования новых видовых форм яв-
ляется тератогенез. Еще В. М. Шимкевич (1907) собрал мате-
риал, подчеркивающий значение уродств животных для видооб-
разования. Значение различных аномалий в строении растений
для формообразования подробно рассмотрено Ал. А. Федоровым
(1958). При создании многих сортов были использованы резкие
уклонения от обычного строения цветков, соцветий, стеблей,
плодов и листьев, т. е. тератные формы, махровые сорта декора-
тивных, лучшие сорта землянки и клубники, ветвистоколосые,
пестролистые, вивипарные и многие другие формы. О значении
тератов для видообразования Ал. А. Федоров высказывается осто-
рожно, признавая, что «элементы тератогенеза имеют место в про-
цессе обычного видообразования, хотя, вероятно, закрепление
растениями аномалий в качестве признаков структуры представляет
собой как бы боковой путь их эволюции» (там же 280).
Велико значение тератогенеза для породообразования птиц,
млекопитающих, рыб. Тератогенез соматических органов может
сопровождаться и той или иной степенью половой изоляции.
Эффективность искусственного отбора при сорто- и породообра-
зоваиии на основе тератных форм является моделью, доказываю-
щей возможность подобных процессов и при видообразовании.
При видообразовании могут использоваться также и новые
видовые формы, образуемые путем наследственного фиксирова-
ния неотении (педоморфоз). Ускорение полового созревания мо-
жет приводить к едва заметному омоложению взрослых стадий,
но может вызвать и полный «обрыв» морфогенеза с «выталкива-
нием» из онтогенеза последних фаз и превращением личиночной
(ювенильной) организации во взрослую. Работами В. Гарстанга,
Ж. Де-Бира, И. И. Шмальгаузена, А. Харди и других на жи-
вотных, Б. М. Козо-Полянского, И. Т. Васильченко, А. Л. Тах-
таджяна и других на растениях показана значительная роль
педоморфоза в формо- и видообразовании.
Переход от древесных форм к травянистым покрытосеменным,
как полагают, совершался путем неотении, как приспособление
к горному климату (Тахтаджян, 1966). Сокращение жизненного
цикла является адаптивным в условиях высокогорья. Через
педоморфоз, возможно, шла эволюция предков мшанок, хордовых
и других крупных стволов эволюции животного мира. Отбор на
ускорение созревания домашних животных (свиньи и др.) при-
водит к ювенилизации взрослых форм. По П. П. Сушкину,
верхнепермский стегоцефал Dvinosaums был наследственно фик-
сированным гигантским аксолотлем: во взрослом состоянии он
344
сохранял окостеневший жаберный аппарат, поразительно сход-
ный с таким же аппаратом личинок современных амфибий.
Примером видообразования путем педоморфоза могут служить
ручьевые миноги, возникшие от паразитических через закрепле-
ние преждевременного полового созревания (Hardisty, 1963).
Педоморфоз сопровождается уменьшением плодовитости и сокра-
щением размеров взрослой особи. Непаразитпческие виды воз-
никли параллельно в нескольких группах паразитических миног.
Все они морфологически неразличимы от своих паразитических
предков (кроме размера) и в смешанных популяциях изолиро-
ваны в половом отношении.
Наконец, следует упомянуть об образовании новых видовых
форм также и путем симбиоза.
Итак, можно полагать, что пеоформогеппое видообразование,
начинающееся симпатрической фазой, играло очеш» большую,
возможно ведущую, роль в филетическом видообразовании.
Для неоформогенного видообразования характерна трехфаз-
ность процесса (Завадский, 1949): скачкообразное появление
новой видовой формы, образование поселения (местной популя-
ции) и, наконец, расселение и внедрение в биоценозы на основе
отбора к биотической среде. По В. Б. Дубинину (1953, 1954),
такая трехфазность присуща видообразованию у перьевых
клещей.
При отдаленной гибридизации и других способах спнтезоге-
неза, а также при тератогенезе, педоморфозе и т. и. возможно
многократное возникновение одних и тех же или генетически
почти идентичных форм (в разных местах в разное время).
В однотипных условиях их отбор приведет к одному и тому же
окончательному состоянию. Поэтому надо согласиться с возмож-
ностью не только политопного (Толмачев, 1958), но и полихрон-
ного видообразования. Политопность и полихронность видообра-
зования чрезвычайно маловероятны лишь при настоящем геогра-
фическом видообразовании на микроаккумулятивной основе.
Не исключена возможность политопного и полихронпого
видообразования и при экологическом видообразовании. Один и
тот же экоэлемент, сезонная форма или же локальный экотип
могут сформироваться в популяцию молодого вида независимо
в нескольких районах и в разное время. Вполне реальна воз-
можность их параллельной эволюции и выход па положение
одного и того же сформированного вида. Чем больше признаков
должно быть спаяно в одной системе, тем меньше вероятность
высокой степени идентичности параллельно эволюирующих по-
пуляций.
Возможность политопного и полихропного видообразования
требует признания возможности также и внутривидовой полифи-
лии. Полифплия в обычном широком понимании, т. е. происхо-
ждение одного таксона в объеме вида, рода и т. п. от различных
345
таксонов того же объема, настолько маловероятна, что следует
считать ее практически несуществующей. Умеренная же внутри-
видовая полпфилия, в нашем понимании, является необходимым
следствием политопного или полихронного видообразования. Если
однотипные амфидиплоиды образуются в разных популяциях и,
подвергаясь отбору в одном и том же направлении, образуют
в конце концов единый вид, то это и есть внутривидовая поли-
филпя. В генетически или эколого-географически различных
популяциях одного вида могут создаться условия, при которых
отбор будет протекать в одном и том же направлении. В итоге
такого процесса в разных районах возникнут однотипно пре-
образованные популяции, сохраняющие, кроме того, и некоторые
особенности. Новый вид будет первично полиморфным.
Таким образом, в настоящее время не выдерживает критики
ультрамонофплитическая точка зрения. На наш взгляд, наиболее
точную характеристику принципа монофилии дал А. Н. Север-
цов, понимавший, что монофплия связана с признанием адаптив-
ной радиации (дивергенции) от одного вида, по от различных его
экологических или географических подвидов (1939 116).
7. ИЗОЛЯЦИЯ И ВИДООБРАЗОВАНИЕ
О классификациях форм изоляции. В предыду-
щих разделах было показано значение для видообразования раз-
ных форм изоляции. Задача этого раздела состоит в обобщении
уже рассмотренного материала и его классификации.
Под изоляцией в широком смысле понимают такое обособле-
ние отдельных особей, внутрипопуляционных форм, популяций,
видов или же целых фаун и флор, которое препятствует тем или
иным взаимодействиям (скрещиваниям, трансдукции и транс-
формации, химической индукции, конкуренции и пр.). По при-
чинам, ее вызывающим, изоляцию можно подразделить на про-
странственную (физико-географическую и локально-экологиче-
скую), временную (хроноэкологическую: изменение сроков
размножения) и репродуктивную (половую или физиологиче-
скую). Иногда изоляцию делят на территориальную и биологиче-
скую (?), а последнюю подразделяют на экологическую,
физиологическую и собственно генетическую (Тимофеев-Рессов-
ский, 1958) пли на макротопографическую (географическую),
мезотопографическую (экологическую), микротопографическую
(биологическую) и генетическую (Huxley, 1963).
Некоторые насчитывают большее число форм изоляции, вклю-
чая географическую, экологическую, сезонную, этологическую,
психическую, физиологическую и миксиологическую, при кото-
рых оплодотворение протекает нормально, но потомство стерильно
(Bouillon, 1959).
346
Согласно Тимофееву-Россовскому, все формы изоляции «мо-
гут быть разбиты на две основные группы: территориально-меха-
нической и биологической изоляции» (1965:6). Слово «биоло-
гическая» скрыло такие совершенно различные формы изоляции,
как психофизиологическая, генетическая и временная несовме-
стимости, анатомо-морфологические несоответствия. Возмож-
ность межвидовой гибридизации, обнаруживаемая опытным пу-
тем, в природе реализуется редко из-за экологических, механи-
ческих или, у животных особенно, этологических изолирующих
факторов (Панов, 1965). Особенности поведения самцов одной
группы действуют отпугивающе на самок другой группы. Для
обеспечения спаривания важно сходство в брачном поведении
(специфичные демонстрационные позы и другие опознавательные
признаки поведения).
По степени обособления следует различать полную и непол-
ную изоляцию (полуизоляцию). При полной изоляции прекра-
щаются обмены гаметами, взаимная миграция, заносы спор и
семян и другие формы взаимодействий между группами. Если
число межгрупповых взаимодействий близко к числу внутригруп-
повых, состояние полуизоляции исчезает п группы сливаются
воедино, а при снижении числа межгрупповых взаимодействий
степень изоляции возрастает.
Полуизоляцию следует в свою очередь подразделить на пе-
риодическую и хроническую.
При пульсациях численности происходит чередование фаз
изоляции отдельных популяций (в период депрессии числен-
ности) с фазами слияния их в одну большую популяцию (в пе-
риод пиков численности). Хроническая полуизоляция возникает
при всех сортах барьеров, если длительное время преобладают
межгрупповые взаимодействия (например, 10% скрещиваются
с особями из других популяций, а 90% —внутри группы).
В отличие от суперпопуляций, по-видимому довольно инерт-
ных в эволюционном отношении, большие полуизолированные
комплексы популяций в структурном отношении являются очень
перспективными для эволюции. Такая структура способствует
оптимальному сочетанию адаптаций общего значения, распро-
страняющихся по всей расе с частными адаптациями, локализо-
ванными в отдельных популяциях.
Некоторыми подчеркивается различное значение оседлости и
кочевого образа жизни для расоэбразования: препятствуя изоля-
ции, номадизм этим самым замедляет и видообразование
(С. White, 1959).
Вопрос о значении разных форм изоляции для видообразова-
ния был поставлен давно (Ч. Дарвин, А. Уоллес, М. Вагнер,
Д. Ромене), но до сих пор еще остается дискуссионным. Некото-
рые авторы рассматривают изоляцию как самостоятельный фак-
тор эволюции, действующий наряду или только при вспомога-
347
тельном участии отбора (Gulick, 1905; Wright, 1934, 1953, 1964;
Mayr, 1949, 1963, и др.). При этом одни выпячивают роль мигра-
ций (Майр, Кэйн, Лэк), другие — роль случайных генетических
дифференцировок в мелких популяциях при пульсациях числен-
ности (Райт, Симпсон и др.) Правильнее, однако, считать изоля-
цию не самостоятельным фактором видообразования, а одним из
факторов естественного отбора (Шмальгаузен, 1946а; Завадский,
1961; Оленов, 1961, и др.). Нельзя поэтому согласиться с рас-
пространенным утверждением, что всякая изоляция является
прелюдией к образованию нового вида. Тем не менее некоторая
степень пространственной изоляции, как полагают некоторые,
все же «является необходимой предпосылкой для видообразова-
ния» (Мауг, 1963).
Пространственная (топографическая) изо л я-
ц и я. Эта форма возникает пз-з.а непригодности территорий для
жизни или непреодолимости их для миграций и заносов зачатков,
а также из-за больших расстояний между частями одной широко
расселенной популяции. Физико-географическими барьерами мо-
гут быть горные страны или хребты, большие акватории или
реки, массивы суши или перешейки, пустыни, леса, положение
острова, оазиса или озера и т. п. Локально-экологическими
барьерами могут быть пространственно мало выраженные пре-
пятствия (дюны, пойма, лесок и т. п.), неблагоприятные физико-
химические показатели воздуха, воды, почвы (температура,
влажность, химический состав, осмотическое давление, соленость,
pH, продуваемость и т. п.), непригодные биоценозы (отсутствие
пищи, мест для размножения и т. п.), биологически «непрони-
цаемые» биоценозы (наличие сильных врагов, конкурентов
и т. п.). Следует подчеркнуть, что понятие геоморфологической,
физико-химической или биотической преграды приобретает смысл
только в отношении к конкретным видам. Для высокоподвижпых,
кочевых и эвриадаптивных видов пе является барьером и тысяча
километров, и горный хребет пли пустыня, а стенобионтный пли
малоподвижный и оседлый вид неспособен перейти в соседнюю
долинку или колонизировать ближайший лесок. Например, для
многих наземных птиц даже ничтожный водный рубеж, как это
ни парадоксально, часто является непреодолимой преградой.
Именно в этом видят причину удивительного островного энде-
мизма наземных птиц на архипелагах Лэк (галапагосские
вьюрки), Майр (многие птицы Полинезии) и др. Семена же и
плоды растений с архипелагов Тихого океана преодолевают по
воде во много раз большие расстояния. Но у многих растений
изолирующим фактором может служить и небольшая полоса —
массив растений другого вида, — препятствующая перелету насе-
комых-опылителей или же переносу пыльцы. Особым изолирую-
щим фактором служат сами насекомые-опылители, обладающие
высокой избирательностью. Один вид не способен преодолеть
348
«экологическое сопротивление» биоценоза, который оказывается
непроницаемым барьером, а другой вид легко преодолевает эту
преграду.
Существуют столь подвижные и эвриадаптивные виды, что
изолирующими факторами для них служат лишь океаны и очень
сильные изменения климата. Примером может служить сибир-
ский шелкопряд (Dendrolimus sibiricus), очень широко рассе-
ленный (от Урала до Дальнего Востока), многоядный (гусеницы
питаются хвоей почти всех видов хвойных), политопный (засе-
ляет леса разнообразные в климатическом, почвенном и биоти-
ческом отношениях) и легко преодолевающий различные пре-
грады (Коломиец, 1962). Достаточно помочь таким видам преодо-
леть физико-географические барьеры, как они покажут свои
ранее скрытые возможности. Например австралийский червец
Jcerya purchasi заселил пригодные местообитания всех континен-
тов и островов, так же как и европейская бабочка’ репница Pie-
ris гарае (Элтон, 1960).
Особой формой пространственной изоляции является влияние
расстояния как такового, не связанное с территориальной пре-
рывистостью популяций. Из-за слабой изученности процессов
обмена гаметами в природной обстановке, правда, еще трудно
судить о степени изолирующего воздействия «фактора расстоя-
ния». У многих видов, живущих суперпопуляциями, сильно уда-
ленные друг от друга участки, по-видимому, оказываются практи-
чески полностью изолированными. Однако радо учесть, что обмен
особями п зачатками может проходить исподволь и для распро-
странения влияния, скажем, уральских популяций на забайкаль-
ские необходимы тысячелетия. Изучение изоляции расстоянием
может пролить свет и на значение других форм пространствен-
ной изоляции. Многие авторы сомневаются в перспективности
для видообразования больших нерасчлененных популяций (Clau-
sen, 1951, 1958; Wright, 1953; Беклемишев, 1960, и др.) и счи-
тают «наилучшим исходным материалом для возникновения
новых видов популяции среднего размера, полностью или ча-
стично обособленные от исходной популяции» (Мюптцинг,
1963:389). Однако эти мнения, как правило, больше основы-
ваются на гипотезе Райта, чем па анализе процессов в природ-
ных популяциях (Оленов, 1961).
Для понимания механизма изолирующего воздействия рас-
стояния важны исследования зон налегания ареалов близких рас.
В таких зонах популяции, как правило, имеют сложный состав.
Они могут включать чистые формы рас, гибриды промежуточного
типа, гибриды, близкие по фенотипу к одному из родителей, по
в известной степени «насыщенные» адаптивными признаками
другой расы (Anderson, 1953), а также модификантов. Часто
промежуточные формы не проникают далеко в глубину ареалов
обеих рас. Объясняется это селекционным преимуществом типич-
349
ной формы в центральных районах ареала, а также степенью
оседлости и радиусом активности особей (радиусом разлета се-
мян и пыльцы) и величиной занимаемой ими зоны. В результате,
несмотря на то что в зоне контакта идет гибридизация и поддер-
живаются переходные популяции, основная масса популяций
таких рас скрещивается только «внутри себя» и фактически ока-
зывается изолированной «расстоянием» от соседних рас. Если же
гибриды оказываются более приспособленными не только в погра-
ничной зоне, но и в пределах основного ареала расы (или обеих
рас), происходит расширение площади, занятой переходными
популяциями за счет вытеснения исходных популяций. При
взаимодействии между экологически различными популяциями,
занимающими небольшие площади, особенно если их взаиморас-
положение по территории напоминает «лоскутное одеяло», обмен
гаметами (пли занос семян) может совершаться почти беспре-
пятственно, и пространственная изоляция невозможна (кроме
случаев очень большой оседлости и специализации). Вся эта
скрещивающаяся система становится объектом группового отбора
на организацию структуры размножающейся единицы, включаю-
щей несколько микропопуляций. Создается сложная популяция
со сбалансированным полиморфизмом и с широкой способностью
к модифицированию. Такие полиморфные популяции могут сло-
житься в пределах небольших, сильно экологически расчлененных
территорий. Они представляют собой «сжатую пружину» и при
наличии возможности могут быстро и широко расселяться. Ко-
нечно, при той же расчлененности местообитаний вероятен и
иной исход. При высокой селективной ценности одной экологи-
ческой формы, например особенно эвриадаптивной, все другие
микропопуляции будут вытеснены и вся площадь будет заселена
только одной формой. Возможен и третий исход — усиление изо-
ляции каждой микропопуляции и обособление их в мелкие ло-
кальные экотипы.
Очень распространена точка зрения, согласно которой про-
странственная изоляция — это необходимый фактор видообразо-
вания. Существование симпатрического видообразования опро-
вергает эту точку зрения. Пространственная изоляция сама по
себе не только не является необходимым фактором видообразо-
вания, но до сих пор не вычленено ее подлинное значение.
Не исключена возможность того, что в некоторых или во многих
случаях пространственная изоляция является не действующим,
а лишь сопутствующим фактором. Действующим же фактором
будет отбор к пространственно разграниченным комплексам кли-
матических и биотических условий.
Обилие видов при островном, горном, оазисном и других слу-
чаях эндемизма говорит в пользу большого значения простран-
ственной изоляции. Разнообразие же видов планарий в оз. Бай-
кал, брюхоногих в оз. Охрид, муравьев в некоторых лесах и
350
Другие случай, которых, вероятно, йе меньше, чем фактов геогра-
фического викаритета, говорит о том, что интенсивное видообра-
зование идет и без пространственной изоляции.
Временная и з о л я ц и я. Временная, пли хропо-эколо-
гическая изоляция занимает важное место среди изолирующих
механизмов. В нее входят изменения сроков размножения (цве-
тения, брачного периода, течки), препятствующие скрещиваниям,
и сдвиги всего цикла жизни, снижающие пли устраняющие кон-
курентные отношения. Значение ее для видообразования подчер-
кивалось давно. Однако в связи с недоверием к симпатрпческому
видообразованию роль временной изоляции все же недооцени-
валась. Известно, что различия между близкими, по уже сложив-
шимися видами во многих случаях носят хроноэкологический
характер (у рыб, насекомых, птиц, цветковых растений и т. д.).
У лягушек Rana esculenta период размножения начинается
в конце мая, когда у R. ridibunda он уже завершен. Такие же
различия обнаружены у моллюсков, например у сепии (Cuenote,
1936). Все эти различия обычно истолковывались как вторичные,
возникшие у географических рас, впоследствии мигрировавших
в одни п те же местообитания. Эта распространенная точка зре-
ния не учитывает возможности эволюции зачатков внутрипопу-
ляционных феноформ в сезонные расы и в виды. Она уже рас-
сматривалась нами в разделах о симпатрическом видообразовании
и о структуре популяции. Вряд ли есть основания считать вто-
ричными озимые и яровые формы в популяциях растений и
озимые и яровые расы рыб (Берг, 1948), «темпоральные под-
виды» (Alpatov, 1924) и т. п.
Хроно-экологическая изоляция может возникать тремя пу-
тями. Во-первых, путем обособления крайних феноформ внутри
единой полиморфной популяции. Если популяция включает цепь
феноформ, различающихся друг от друга по средним срокам раз-
множения, то каждая из феноформ лишь отчасти является само-
воспроизводящейся единицей, так как скрещивается с более
ранними и более поздними формами. Барьер между смежными
феногруппами может быть очень небольшим. Между край-
ними же гаметный обмен осуществляется только окольно, через
ряд форм-посредников. Обычные феноформы могут быть модифи-
кантами или смесью модификантов и стабилизированных форм.
Они, как правило, выщепляют все или большинство других фено-
форм, входящих в данную популяцию. После селекции фено-
форма становится стабилизированной и самовоспроизводящейся
единицей. Элиминация близких феноформ или отбор на нескре-
щиваемость одной феноформы с другими приврдят к тому, что
из структурного компонента единой полиморфной популяции опа
становится уже самостоятельной биологической единицей — се-
зонной расой. Такая сезонная раса может широко расселиться
и стать географической расой.
351
Ёо-вторЫх, путем преобразования полиморфной популяций
в мономорфные, развивающиеся из различных фепоформ. Напри-
мер, при заселении высокогорий сохраняется только одна из
феноформ, как это показано на примере калифорнийской
Achillea borealis (Clausen, 1951), а при заселении засушливых
местообитаний .в долине выживает другая феноформа. Когда
такие популяции сравнивают, говорят уже о разных экотипах,
которые вместе с тем являются и сезонными расами.
В-третьих, путем изменения сроков размножения у близких
экологических или географических рас, ранее бывших мономорф-
ными по срокам размножения. Это изменение часто происходит
как приспособление к новому климату или каким-либо другим
особенностям местообитания. Тогда к существующим простран-
ственным барьерам может присоединяться еще и временной
барьер. В зоне контакта такие географические или экологические
расы будут относиться друг к другу и как сезонные расы.
При обособлении сезонной расы из внутрипопуляционной фе-
ноформы временной барьер возникает раньше, чем формируется
раса, и служит одним из факторов ее эволюции. Временные же
барьеры между географическими расами возникают в процессе
приспособления этих рас к различным условиям существования,
являясь следствием их преобразования, и лишь усиливают обо-
собленность рас, уже существующую из-за пространственной
изоляции. Пространственные и временные барьеры обусловли-
вают фактическую нескрещиваемость изолированных групп. Од-
нако их генетико-физиологические системы могут быть столь
близкими, что, при исчезновении или ослаблении указанных
барьеров между ними возникает свободное скрещивание.
Репродуктивная (половая) изоляция. Такая
изоляция возникает из-за несоответствия в анатомо-морфологп-
ческих особенностях половых аппаратов, психофизиологических
различий и несовместимости генетических систем. К репродук-
тивной изоляции следует отнести также и переход к бесполому
размножению (апомиксис у растений и др.). Не существует пря-
мой зависимости между степенью общих морфо-физиологических
различий и степенью репродуктивной изоляции.
На примере популяций Viola tricolor (Clausen, 1951) было
показано что между морфологически неразличимыми популя-
циями может существовать значительная генетическая изоляция,
а легко скрещивающиеся популяции бывают морфологически
несходными. Отсутствие полной корреляции между этими пока-
зателями было подтверждено многими другими работами на жи-
вотных и растительных объектах.
Репродуктивная изоляция может служить как абсолютным, так
и относительным барьером. Обнаружены все степени несовмести-
мости от свободной скрещиваемости при едва заметном понижении
жизнеспособности или плодовитости потомства и случая «полу-
552
вида», как у Hemizonia angustifolia, до полной стерильности по-
томства и, наконец, до полной нескрещиваемости форм.
Полное бесплодие обычно наблюдается у близкородственных
видов, живущих в одном и том же или в территориально смежных
биоценозах, сходных по своей экологии и ритмам размножения.
У столь же родственных видов, изолированных пространственно-
экологическими и временными барьерами, как правило, полного
бесплодия не возникает, а иногда наблюдается даже хорошая пло-
довитость межвидовых гибридов. Примером могут служить гиб-
риды между видом влажных лугов Geum rivale и лесным, более
сухолюбивым, G. urbanum. По-видимому, полное бесплодие у епм-
патрических видов чаще возникает в результате прямого отбора
на избирательные внутригрупповые скрещивания и на подавление
гибридных форм, т. е. так, как это было показано в модельных
экспериментах (Koopman, 1950; Dobzhansky, 1955). У близких
видов животных одна экологическая изоляция часто ле может
служить барьером для межвидовой гибридизации, тем более что
иногда местообитания перекрываются и виды оказываются вре-
менно синтопическими (Rivas, 1964). На американских мышах
Peromyscus leucurus и Р. gossipinus было показано, что в таких
ситуациях важную роль играют реакции поведения. У самок
Р. leucurus и обоих полов Р. gossipinus, взятых из синтопических
популяций, ясно выражено предпочтение особей своего вида,
а мыши из аллотопических популяций такого предпочтения почти
не обнаруживают (Me.Carley, 1964).
Как подчеркивает Р. Хайнд (Hinde, 1959), обязательным усло-
вием отбора на нескрещпваемость у симпатрических популяций
позвоночных животных является несколько пониженная' адаптив-
ность и жизнеспособность межпопуляционных гибридов, а одним
из распространенных механизмов отбора — выработка и закрепле-
ние специфических стереотипов поведения. Изолирующая роль
реакций поведения присуща не только птицам и млекопитающим,
ио и амфибиям. У австралийской квакши Hyla ewingi половая
изоляция симпатрических популяций основана па резких разли-
чиях брачного крика (по числу импульсов, продолжительности
крика, доминантной частоте), который является основным изоли-
рующим механизмом у многих близких видов бесхвостых амфи-
бий (Littlejohn, 1965). При полной половой изоляции близкие
виды могут быть строго симпатрическими, заселяя один и тот же
охотничий участок и ведя идентичный образ жизни, как это обна-
ружено у птиц Юго-Восточной Азии Garrulax pectoralis и G. шо-
niliger и др. (Мауг, 1963).
Известно, что многие межвидовые и межродовые гибриды воз-
никают изредка и не удерживаются в популяциях, так как эли-
минируются или поглощаются при повторных обратных скрещи-
ваниях (заяц беляк с русаком, виды мышей и других грызунов,
волк с шакалом, глухарь с тетеревом и многие другие виды птиц,
’/4 23 К. М. Завадский
353
виды ужей и других змей, тритонов, жаб и других амфибий, мно-
гие виды рыб и т. и.). В таких случаях нежизнеспособность гиб-
ридов прежде всего объясняется тем, что сами они малоадаптивпы.
Неверно, что межвидовая гибридизация «во всех случаях» ведет
не к изоляции, а к стиранию границ видов (Майр, 1947 407) и
«тормозит видообразование» (там же 342). Вспомним злак Spar-
tina townsendii — гибридогенный вид, вытесняющий родительские
виды. Все дело в степени адаптивности гибрида и в его селектив-
ных преимуществах перед чистыми видами. Изолирующей причи-
ной может явиться затрудненность или невозможность самого
спаривания, возникающая из-за несоответствий в строении поло-
вых органов или из-за психофизиологических преград. При скре-
щиваниях на грани межвидовых у млекопитающих репродуктив-
ная изоляция часто сказывается уже с большой силой: если и
происходит спаривание, то может не произойти оплодотворения
или беременная самка гибнет из-за несоответствия между разме-
рами эмбриона и матки, оказываются невозможными роды, рож-
даются мертворожденные или, наконец, погибает молодняк.
У растений репродуктивный барьер может состоять в том, что
пыльца одной формы пе прорастает па рыльце другой или про-
растает, но пыльцевые трубки растут медленно и не происходит
оплодотворения. Если происходит оплодотворение, то яйцеклетка
может погибнуть на первых фазах эмбриогенеза или погибает бо-
лее пли менее сформированный зародыш. Гибель зародыша объяс-
няется или его генетическими дефектами, или же несоответст-
виями между ним и материнским организмом (так как прп изоля-
ции он оказываемся жизнеспособным).
Не существует и прямой зависимости между степенью репро-
дуктивной изоляции и таксономическим различием групп. Напри-
мер, у птиц, рыб, у многих беспозвоночных известна широкая спо-
собность к межродовой гибридизации, а у млекопитающих опа от-
сутствует. При таких сопоставлениях, правда, приходится
сталкиваться с различным толкованием объема вида и рода. На-
пример, род Raphanus включает с десяток близких видов, удов-
летворяющих морфолого-географическому критерию, но настолько
не изолированных друг от друга в репродуктивном отношении и
взаимофертильных, что весь этот род при желании может быть
истолкован как один политипический вид. Отсутствие репродук-
тивной изоляции между родами толкает некоторых (Bowden,
1959) на такие странные новшества, как например объединение
в один вид множества видов злаков разных родов (Triticum, Se-
cale, Aegilops, Agropyrum и пр.).
По своему происхождению репродуктивная несовместимость
может быть трех родов. Во-первых, она может быть косвенным
(коррелятивным) следствием общих различий, возникших в про-
цессе преобразования популяции. При формировании географи-
ческих или экологических рас даже продолжительная фактпче-
354
ская их изоляция может и не приводить к их репродуктивной не-
совместимости. Такая несовместимость не является причиной фор-
мирования и обособления рас, а возникает как одно из побочных
следствий адаптивной радиации. Стерильность многих межвидовых
гибридов объясняется случайными генетическими изменениями
внутри небольших популяций на границе ареала вида, которые
оказались изолированными экологически или географически (Le-
wis, 1962; Мюнтцинг, 1963, и др.).
Во-вторых, несовместимость может быть результатом прямого
отбора. Отбор на нескрещиваемость возможен только в отсутствии
других барьеров (т. е. у симпатрических форм). Действуя внутри
полиморфных популяций, он отгораживает более адаптивные
формы от менее ценных. При этом могут быть использованы раз-
личные механизмы: сдвиг времени созревания половых продук-
тов, изменение рефлекторных реакций (па запах, позу, окраску
и т. п.), изменение копулятивного аппарата и пр. В результате
адаптивная форма оберегается от гибридизации и, перестав быть
составной частью популяции, начинает самостоятельное сущест-
вование. В этом случае несовместимость возникает уже после
образования новой адаптивной внутрипопуляционной формы и не
в результате побочного (косвенного) следствия эволюции, а пу-
тем прямого отбора на охрану уже сложившейся «адаптивной
нормы». Именно такие процессы и являются, по нашему мнению,
основой одного из распространенных путей симпатрического видо-
образования.
Существуют иногда неожиданные причины, из-за которых от-
сутствуют переходные (гибридные) формы между симпатриче-
скими видами, способными свободно скрещиваться между собой.
В опытах с симпатрическими видами мака Papaver dubium и
Р. lecoqii были получены вполне жизнеспособные, хорошо расту-
щие и цветущие гибриды Fi, но в природе они совершенно отсут-
ствуют (Naughton, Harper, 1960). Оказалось, что у гибридных се-
мян исчез период покоя. Поэтому они всходят осенью и зимой на-
цело погибают. Другая причина хиатуса между этими маками
в том, что все растения Fi стерильны и поэтому даже при их вы-
живании переходная группа растений поддерживалась бы только
за счет ежегодно происходящих межвидовых скрещиваний.
И, наконец, в-третьих, репродуктивная изоляция может быть
продуктом геномных преобразований, тератогенеза и т. п. Как
уже указывалось, в этом случае половая изоляция новой видовой
формы является исходным моментом («стартовым условием»),
а не итогом более или менее длительных процессов.
Итак, с помощью репродуктивной изоляции внутрипопуляцион-
пые симпатрические группы приобретают возможность эволюции
самостоятельными путями, т. е. — видообразования. Было бы
однако неправильно преувеличивать значение репродуктивного
барьера для видообразования. Для географического и экологиче-
23 К- М. Завадский
355
ского видообразования указанный барьер вообще не имеет ника-
кого значения. В случае преобразования сезонных рас в эковиды
этот барьер также не необходим.
В заключение рассмотрим особый случай репродуктивной изо-
ляции, обусловленной переходом к бесполому размножению. Этим
способом размножаются, согласно данным С. С. Хохлова (1959),
до 20% всех видов растений. Хотя систематика апомиктов сильно
запутана и для нее характерна «пульверизация» видов, тем не
менее их число все же весьма велико. Спорным следует признать
убеждение в превосходстве такого способа размножения над по-
ловым, а также мысль об особой эре бесполосеменных растений
(Хохлов, 1946, 1950). Апомиктические виды (обыкновенный оду-
ванчик, некоторые виды ястребинки, мятлика, ежевики, манжетки
и т. п.) широко расселены и весьма конкурентоспособны. Поэтому,,
надо полагать, ошибочны и воззрения, согласно которым апогам-
ные формы — быстро отмирающие осколки панмиктических попу-
ляций (Комаров, 1940, и др.). Последние гистохимические иссле-
дования показывают, что, например, у апомиктических одуван-
чиков по сравнению с половыми сильно повышена энергия
метаболизма (Циигер и др., 1965).
Нами высказывался взгляд, согласно которому наиболее про-
грессивным способом размножения является сочетание полового
размножения с бесполосеменным: недостатки каждого из них
«подстраховываются» другим способом размножения, а преимуще-
ства суммируются (Завадский, 19666). Примером могут служить
высокополиморфные виды Роа pratensis, Р. alpina и др., популяции
которых включают, формы с половым размножением, апомиктов и
гибридов между ними, выщепляющих как половые, так и апо-
миктические формы (Мирошниченко, 1963).
Справедливо то, что апомикты — продукт распада панмикти-
ческой популяции (Stebbins, 1941, 1950 и др). Утрата полового
процесса ведет к немедленному репродуктивному обособлению
всех генетически стабильных форм, бывших компонентами еди-
ной популяции, к превращению популяции в фитоценоз, а от-
дельных форм, как это уже отмечалось, — в новые видовые формы.
Новая апогамная форма может стать доброкачественным мате-
риалом для неоформогенного видообразования. Другой вопрос,
какова дальнейшая судьба таких видов, способны ли они к даль-
нейшей эволюции и т. п. Нельзя согласиться с тем, что распад
панмиктической популяции на рой апогамных форм — это только
«имитация эволюционного процесса» (Mather, 1955), а не один
из его путей. Таким образом, репродуктивная изоляция при пе-
реходе к бесполосеменному размножению является таким же
«стартовым условием» видообразования, как и другие случаи об-
разования «новых видовых форм». Апомиксис, по-видимому, яв-
ляется таким же боковым путем эволюции, как и самоопыление
(Stebbins, 1957) или вегетативное размножение. Его значение
356
для филетической эволюции еще не исследовано, но для «рас-
слаивающей» эволюции оно несомненно.
В заключение следует сказать, что изоляция является частой,
но не обязательной предпосылкой видообразования, которое может
совершаться и внутри панмиктических популяций. Существует и
иная распространенная точка зрения, согласно которой новый вид
не может обособиться от материнского чисто селективным путем
и полная изоляция является практически необходимой (Simpson,
1965).
8. ВИДООБРАЗОВАНИЕ И ПРОГРЕССИВНАЯ ЭВОЛЮЦИЯ
Видообразование и неравноценность видов.
Видообразование — это процесс, идущий далеко не у всех, воз-
можно, только у меньшинства видов. Многие популяции, достигнув
видового уровня, вскоре же вымирают, другие вымирают, просу-
ществовав геологически довольно продолжительное время, третьи
персистируют, неопределенно долго подвергаясь отбору на охрану
уже сложившейся адаптивной нормы и претерпевая только микро-
эволюционные изменения. Некоторые авторы считают, что под-
держание устойчивости (персистирование) даже более частое
следствие преуспевания вида, чем преобразование (Simpson, 1953;
Rensch, 1954; Huxley, 1958). Полное же отрицание явления пер-
систирования (Давиташвили, 1956) является неубедительным,
так как при этом недооценивается значение поддерживающего от-
бора и обходятся факты существования «стационарных» групп,
в изобилии установленные палеонтологией. Наконец, известная
часть новых видов трансформируется в дочерние виды. При этом
одни из них существуют кратковременно, так как в процессе адап-
тивной радиации целиком преобразуются в дочерние виды, а дру-
гие, отщепляя от себя дочерние виды, длительно сосуществуют
с ними и даже с внучатыми видами. Неравноценность видов в от-
ношении эволюционных перспектив очень велика. Одни радиируют
слабо, преобразуясь в несколько или в десятки видов, другие силь-
нее, преобразуясь в сотни и тысячи видов, наконец, некоторые под-
вергаются необычайно энергичной адаптивной радиации и в ко-
роткие геологические сроки преобразуются во многие тысячи ви-
дов-потомков.
Эволюционную перспективность вида-предка можно, таким об-
разом, измерять числом видов-потомков. Однако это только одна
сторона вопроса, связанная с «расслаивающей» эволюцией. Если
рассматривать филетическую эволюцию, т. е. изменение фратрий
или таксонов во времени, то их перспективность можно измерять
скоростью морфо-физиологических изменений безотносительно
к численности дочерних таксонов.
Нами было предложено понятие «степень ароморфности вида»
(Завадский, 1958). Ароморфные или эрогенные виды — это виды,
23*
35Т
доказавшие свою эволюционную перспективность и ставшие родо-
начальниками более или менее разнообразных и более или менее
сильно измененных групп. Понятие об эрогенном виде вместе
с тем и очень неопределенно, так как в нем не ограничиваются
масштабы эволюционной перспективности вида. Если судить
post factum, как это делают филогенетики, то эрогенным видом
бесспорно следует назвать родоначальника хордовых или покрыто-
семенных, копытных, приматов или сложноцветных. Но как быть
с видами — родоначальниками отдельных отрядов или семейств?
Очевидно, что такие виды являются менее эрогенными. Следует
различать несколько степеней арогенности видов.
Мегаарогенные виды — это виды, оказавшиеся родоначаль-
никами классов и типов, или, иначе говоря, мегаэволюционных
филумов (Simpson, 1953).
Макроарогенные виды — это виды, оказавшиеся способными
на адаптивную радиацию потомков, давшую роды, трибы, семей-
ства, отряды. И, наконец, микроарогенные виды — это виды,
эволюционная перспективность которых оказалась весьма неболь-
шой и привела к образованию подродов, серий и т. п. Множество
видов не прошло испытаний па эволюционную перспективность и
существует большее или меньшее геологическое время без сущест-
венной адаптивной радиации (брадителлические виды в смысле
Симпсона). При этом часть из них могла достичь состояния над-
видов и больших политипических видов, другие — обычных хоро-
ших видов, третьи — только эковидов или географических рас.
Появившись, ароморфные виды оказываются способными пе
только к более иди менее широкой радиации потомков, но и
к быстрой эволюции. Повышение скоростей эволюции — харак-
терная черта ароморфного видообразования. Сколько можно су-
дить по палеонтологическим данным (Muller, 1963) и по опытам
«со скоростью отбора в модельных популяциях, скорости эволюции
могут временами возрастать в тысячи крат. При оптимальных ус-
ловиях отбора, при оптимальной структуре популяции и организ-
мов на некотором отрезке времени ароморфная фратрия может
преобразоваться с ошеломляющей скоростью, а затем столь же
быстро радиировать на множество видов-потомков. В геологически
короткое время одна фратрия способна преобразоваться так глу-
боко, что дает принципиально новый тип организации, а другая
фратрия за десятки миллионов лет не переходит рамок рода.
По А. В. Благовещенскому, общий ход эволюции цветковых, иду-
щей брадителлически, «прерывается периодами подъема на более
высокий энергетический уровень (ароморфозы и арохимозы) и
энергичным приспособительным видообразованием» (19656:13).
Неравномерность темпов эволюции — это важнейший закон, дей-
ствующий на популяционно-видовом уровне.
Особенности эрогенных видов. Вопрос об орга-
низации популяций и особенностях их взаимоотношения со сре-
3'58
дой, ведущих к ароморфной эволюции — большая и особая
проблема. Здесь уместно лишь краткое упоминание некоторых ее
сторон. Найболее перспективными в эволюционном отношении
являются виды, живущие крупными популяциями, в свою очередь
подразделенными на много мелких популяций (микропопуляций) ,
лишь частично изолированных друг от друга в половом отношении
(Wright, 1934; Li, 1955; Haldane, 1959, и др.). Можно полагать,
что в^ таких популяциях, характерных для горных стран, все
адаптации общего значения, возникшие у одной микропопуляции,
вскоре могут стать достоянием всей большой популяции в целом,
а существующие ограничения в этом обмене создают возможность
накопления адаптаций, имеющих частное значение лишь в рам-
ках одной микропопуляции. Эволюционно перспективный вид
должен быть высокотолерантным, т. е. обладать высокой жизне-
стойкостью к неблагоприятным факторам среды, быть эвриадап-
тивным и конкурентоспособным (Good, 1953) или, по выражению
С. А. Северцова (1951), иметь «запас адаптированности» (одни
виды при возрастании элиминации снижают численность и вы-
мирают, другие же дифференцируются и перестраиваются). Поли-
гонные и мигрирующие виды имеют большие преимущества над
стенотопными и очень оседлыми. Некоторые полагают, что основ-
ным источником возникновения новых таксонов крупного мас-
штаба являются изолированные популяции, живущие на границе
ареала («типострофические популяции» —Schindewolf, 1954).
И. И. Шмальгаузен (1961) доказывает большое значение для эво-
люционных возможностей вида полимеризации единиц всех уров-
ней от молекулярного (полигенность, увеличение числа нуклеотид
в молекулах нуклеиновой кислоты — числа структурных единиц
ядра и цитоплазмы) и до популяционного (возрастание числен-
ности особей в популяции и числа популяций до размеров, обес-
печивающих эффективное сочетание индивидуального и группо-
вого отбора, т. е. сохранения лучших особей в наиболее хорошо
организованных популяциях). Степень ароморфности вида воз-
растает при определенном пути разрешения капитального проти-
воречия между приспособленностью к существованию в данных
условиях и приспособленностью к эволюции (Завадский, 1958).
Процветание вида связано с укреплением проверенной отбо-
ром адаптивной нормы и с повышением силы ее доминирования
по отношению к мутантным аллелям. Эволюционная же перспек-
тивность вида связана не только с богатством наследственных воз-
можностей (большая гетерозиготность, обилие рецессивов), но и
со способностью при нужде мобилизовать эти возможности. Усло-
вием такой мобилизуемое™ скрытых возможностей является не
слишком большой избыток генетических средств защиты адаптив-
ной нормы. Иначе говоря, адаптивная норма эволюционно пер-
спективного вида должна обладать не слишком высоким, уровнем
доминирования над мутантными аллелями. Трудно сказать, суще-
359
ствуют ли в данном случае оптимальные решения противоречия,
или же во всех ароморфных стволах эволюция идет по «компро-
миссным» путям.
Для ароморфных видов характерно не только приобретение
новых приспособлений, имеющих значение более или менее ко-
ренных усовершенствований общего значения, но и сохранение
способности к усовершенствованиям в будущем,
По своему происхождению ароморфные изменения, как и все
приспособления, — результат накапливающего действия естест-
венного отбора. Сначала они служат приспособлениями к тем
частным условиям, в которых они возникли. Впоследствии, при
расселении потомков, ароморфные приспособления оказываются
эффективными для широкого круга новых условий и помогают
расселяться все далее и далее. В итоге после испытаний на при-
годность в самых различных условиях из массы первоначально
возникших вариантов приспособлений сохраняются лишь те не-
многие, которые доказали свое общее значение. Мегаарогенез —
это коренное усовершенствование путем смены организации на
принципиально новую, это приспособление практически универ-
сального значения (например, переход от одноклеточности к мно-
гоклеточное™, от размножения икрой и наружного оплодотворе-
ния к размножению богатыми пищей и хорошо защищенными
крупными яйцами, к внутреннему оплодотворению, к живород-
ности и кормлению молоком и т. п.). Вероятность возникновения
крупных ароморфозов мала, так как они представляют собой наи-
более удачные варианты коренных усовершенствований всей ор-
ганизации. Появление > мегаарогенного вида — редчайшее собы-
тие. Из сотен тысяч или даже из миллионов видов лишь один
оказывается столь удачно организованным, что становится родо-
начальником очень крупного эволюционного ствола. На этом вы-
соком уровне эволюции также действуют статистические законо-
мерности. Мегаарогенпый вид обычно появляется среди крупных
ансамблей близких видов (в мощных пучках адаптивно ради-
ирующих и параллельно развивающихся фратрий). Арогенные
виды более низких степеней — явления не столь редкие.
Переход к апомиктическому размножению, по С. С. Хохлову
(1950), — крупный ароморфоз в эволюции растений. Однако эволю-
ционная перспективность апомиктических видов не является
столь уж большой и может служить примером умеренных, а глав-
ное — кратковременных успехов вида. Бесполосеменное размно-
жение закрепляет генетическую конституцию и особо удачные
гетерозиготы фиксируются и строго воспроизводят материнский
тип, образуя множество вторичноклональных линий. В малоиз-
меняющихся условиях, где успех связан со скоростью «наводне-
ния» местообитания, апомиксис полезен, но в меняющихся усло-
виях более полезным является как раз не стандартизация потом-
ства, а его разнообразие, связанное с половым размножением и
360
перекомбинированием генотипов. Наиболее эволюционно перспек-
тивными, возможно, являются виды, способные постоянно соче-
тать амфимиктическое размножение как с апомиктическим, так и
с вегетативным размножением, как это наблюдается среди мят-
ликов, ежевик и т. п. (Завадский, 19666). Эволюционные возмож-
ности самооплодотворяющихся видов не могут быть признаны
ароморфными, так как они редко эволюировали в новые роды и
«не могут ни при каких обстоятельствах давать новые семейства»
(Stebbins, 1957 352-353).
Связи организмов со средой при мегаарогеиезе, с одной
стороны, усложняются (например, с возникновением органов
чувств, с возрастанием различительной способности рецепторов),
с другой — происходит автономизация организма от некоторых из-
менений среды (теплорегуляция и постоянная температура тела,
изменение порогов реагирования, развития гомеостатических
устройств). Накопление коренных усовершенствований создает
ее носителю решающие преимущества и начинается широкое и
быстрое расселение потомков. Возникают новые господствующие
группы, вытесняющие своих менее совершенных предшественни-
ков и проникающие в ранее недоступные им области (выход на
сушу, освоение воздушного пространства, пустынь и т. п.).
Повышение выживаемости и степени целесо-
образности особи и вида. Важными показателями аро-
генного видообразования являются повышения средней выжи-
ваемости организма, т. е. отношения числа родившихся к числу
доживших до периода размножения, и повышение степени целе-
сообразности особи и вида (Завадский, 1958, 1961).
Повышение средней выживаемости организма в миллионы раз
(от 0.000001 до 10—30%) объясняется накоплением множества
ароморфных приспособлений (возникновение полового процесса
и зиготного размножения; увеличение запасов пищи в яйце и
защищенности эмбриона; переход от икринки к яйцу и к живо-
родности; питание молоком; активация ферментов и более эф-
фективное использование пищи; более энергичное окисление;
совершенствование теплорегуляции; развитие органов высшей
нервной деятельности и совершенствование рефлекторного аппа-
рата; использование высшей нервной деятельности для охраны и
научения молодняка; приспособление к новой среде не только
путем морфо-физиологических изменений, но и через изменение
поведения, и т. д.). Арогенное повышение выживаемости особи
сопровождается развитием адаптаций к неблагоприятным факто-
рам среды не путем инактивации (образование цист, уход в по-
кой, анабиоз, развитие пассивных защитных приспособлений, уход
от действия фактора), а нейтрализацией и “преодолением этих
факторов при сохранении активной жизнедеятельности. Приспо-
собление ко все более усложняющейся и изменчивой среде дости-
гается не «избеганием» агентов, а их все более совершенной амор-
361
тизацией. Благодаря развитию аппаратов, амортизирующих дей-
ствие неблагоприятных факторов среды, организм оказывается
способным сохранять физиологическую норму, несмотря на по-
стоянное действие «потенциально повреждающих» факторов.
Такие гомеостатические аппараты создают организму известную
независимость от воздействий факторов среды и создают возмож-
ность вступать во все более сложные отношения со средой. Число
связей со средой возрастает, сами они становятся более сложными.
Таким образом, увеличение «независимости» организма от изме-
нений среды увеличивает возможность усложнения его условий
существования.
У видов с низкой выживаемостью особи (например, одна на
миллион) вероятность выполнения видовой функции именно дан-
ной особью очень мала. Случайности имеют здесь еще большой
простор, и преодоление их у таких видов осуществляется главным
образом путем повышения рождаемости (плодовитости). Значе-
ние отдельной особи в жизни вида крайне мало. Существование
вида обеспечивается главным образом высокой плодовитостью и
высоким уровнем численности, т. е. статистическими механиз-
мами. Например, у видов морских беспозвоночных существует
резко выраженная обратная зависимость между размерами и за-
щищенностью яиц (и личинок) и плодовитостью. Среди видов
с хорошей защитой яиц и молоди за один раз откладывается 10—
100 крупных яиц, среди видов менее защищенных — 100—
1000 яиц, а у видов, не имеющих защитных приспособлений и
мечущих яйца непосредственно в воду, — 1000—500 000 000 мел-
ких и мельчайших, очень бедных желтком яиц (Thorson, 1950).
При изменениях внешней среды поддержание численности
вида может происходить двумя путями: повышением рождае-
мости (плодовитости) и повышением выживаемости особи. Пер-
вый путь бесперспективен для ароморфной эволюции, а второй
является основным ее направлением.
В первом случае эволюция вида идет несмотря на изъяны
в организации особи, вызывающие большую смертность, а во вто-
ром — благодаря устранению этих изъянов в организации особи, и
ведет к снижению смертности. С повышением выживаемости
особи все возрастает значение отдельной особи в жизни вида.
Преодоление случайностей осуществляется уже не столько чисто
статистическими механизмами, сколько возросшей защищен-
ностью, совершенством организации каждой особи.
Эволюционная перспективность вида, например, значительно
возрастает с появлением адаптаций, увеличивающих вероятность
встречи яйца со сперматозоидом. Отсутствие аппаратов, повы-
шающих вероятность этой встречи, ведет к огромному росту чис-
ленности гамет, статистически обеспечивающих количество опло-
дотворений, необходимое для поддержания численности вида.
Широкое поле деятельности, предоставленное случайности в про-
362
цессе, имеющем коренное значение в жизни вида, свидетельствует
о небольшой перспективности такой организации. Господство
статистических механизмов встречи гамет ведет к уменьшению
размеров яиц и поэтому к ряду фундаментальных ограничений
(например, к долгой личиночной жизни, вызывающей большую
смертность до половой зрелости, и т. п.).
Эволюция мегаарогенных видов характеризовалась законо-
мерным повышением выживаемости особи и снижением плодови-
тости. Если из миллиона особей лишь одна выполняет видовые
функции, нельзя признать вид совершенным: слишком велика
«дань», оплачиваемая видом за несовершенство его организации.
Колоссальные массы продуцируемых гамет и молодняка затра-
чиваются не на воспроизведение вида, а на преодоление случай-
ностей. Такая организация жизни малоэкономична. Если же в вос-
произведении вида принимает участие, например, каждая из
пяти родившихся особей, — это высокоэкоиомическая система,
в качественном отношении стоящая на гораздо более высоком
уровне.
Наибольшей эволюционной перспективностью обладают виды,,
у которых повышение выживаемости организма связано не только
со снижением плодовитости, по и с увеличением его размеров и
продолжительности жизни до определенных оптимальных величин.
Численность же таких видов становится гораздо меньшей, чем
у видов примитивных групп.
Не менее важной чертой ароморфных видов является повыше-
ние степени целесообразности организации индивида и вида. Це-
лесообразность проявляется как в приспособлении особи и вида
к внешней среде, так и в отношениях между элементами самих
живых систем (слаженность организации особи и вида, целост-
ность системы).
Повышение степени целесообразности живого включает
в себя возрастание целостности вида и увеличение степени при-
способленности вида к среде. Организация вида претерпела слож-
ную эволюцию. Прогресс в организации вида тесно связан с про-
грессом в организации особи, но представляет собой особое
явление со своими закономерностями, более высокого уровня,
чем закономерности, действующие в рамках одного организма.
У примитивных агамных групп степень целостности вида
очень мала. Интегрирующие связи (внутривидовые отношения)
представлены лишь химическими воздействиями через среду, спо-
собными регулировать скорость размножения, рост и некоторые
другие процессы. С возникновением полового размножения
появился новый аппарат, интегрирующий популяцию и вид.
Скрещивания цементировали систему вида. С поляризацией по-
лов возникла одна из основных форм внутривидовой дифферен-
циации. В дальнейшем все усложнялись формы внутривидовой
дифференциации и интеграции: вид становился более целостным
36%
образованием. С развитием высшей нервной деятельности воз-
никли новые типы внутривидовых отношений и целостность вида
еще более возросла.
В отличие от особи вид не имеет определенного заданного
в его структуре «срока жизни» и в этом отношении противопо-
ложен индивиду.
Приспособленность многих видов не может обеспечиваться
только простым самовоспроизведением, выживанием на основе
сохранения старой организации в течение длительного времени.
Многие виды представляют собой не только форму осуществле-
ния жизни, но и единицу ее развития. В определение приспособ-
ленности вида необходимо поэтому ввести и показатель его эво-
люционной перспективности (пластичности), способности к из-
менениям в будущем. Таким образом, повышение целесообразности
живого представляет собой процесс возрастания целостности особи
и вида, повышение выживаемости особи, а также поддержание или
увеличение эволюционной пластичности вида. Следует указать на
отдельные, особенно характерные показатели повышения степени
целесообразности живого. Важными показателями этого процесса
служат закономерные изменения обмена веществ организма и обо-
рота веществ внутри вида: обмен становится все более экономич-
ным. Структуры становятся все более работоспособными, более на-
дежными, лучше обеспечивающими функции. Ускоряются и стано-
вятся более точными процессы возврата к «норме» при опасных
сдвигах обмена, отклонениях функций и повреждениях структур
(совершенствование физиологического гомеостазиса).
Более экономичным становится и оборот веществ на уровне
вида, так как по мере увеличения целостности и выживаемости
особи уменьшаются «непроизводительные» потери. Снижение не-
производительных потерь в процессах самовоспроизведения вида
свидетельствует о непрерывном уменьшении относительного зна-
чения вероятностных (статистических) адаптаций (плодовитость,
численность) и о росте значения адаптаций индивида и надинди-
видуальных систем.
В этом отношении прогрессивная эволюция сопоставима со
свойствами цепи Маркова: с удалением от старта возрастает жест-
кость детерминирования и снижается вероятность появления
случайных флюктуаций.
От организации вида, при которой случайности преодоле-
ваются главным образом «числом», постепенно происходит переход
к более высокой организации, где большинство случайностей
преодолевается «не числом, а уменьем», т. е. совершенством от-
дельно взятой особи и надорганизменных систем. Следовательно,
принцип «больший эффект достигается при меньшей затрате ве-
щества и энергии» осуществляется не только в эволюции инди-
вида, но и вида.
ЛИТЕРАТУРА
Абдылдаев Т. 1966. Философские вопросы учения о виде. Фрунзе.
Авдулов Н. П. 1931. Карио-систсматическое исследование семейства
злаков. Тр. по прикл. бот., ген. и сел. Прилож. 44, Л.
Агаев М. Г. 1958. Морфо-биологическая неоднородность внутри сортов
хлебных злаков. Автореф. канд. дисс. Л.
Алпатов В. В. 1924. Изменчивость и низшие систематические катего-
рии. Русск. зоол. журн., 4, 1—2 : 227—244.
Алпатов В. В. 1934. Плотность населения как экологический фактор.
Усп. соврем, биол., 3, 2: 229—251.
Ананьев А. Р. 1956. Вопросы видообразования в палеонтологии. Уч. Зап.
Томск, унив., 27 : 81—94; 113—114.
Аникин В. И. 1966. К вопросу о внутривидовой экологической диффе-
ренциации птиц на примере микропопуляций зяблика. В сб.: Внутри-
видовая изменчивость наземных позвоночных животных и микро-
эволюция, Свердловск.
Анфинсен К. 1962. Молекулярные основы эволюции. М.
Аристотель. 1934. Метафизика. М.
Арнольди К. В. 1939. К вопросу о непрерывной географической измен-
чивости. Зоол. журн., 18 : 685—710.
Арнольди К. В. 1941. К вопросу об экологической дивергенции видо-
вых популяций: дивергенция у жуков Tentyria nomas Pall. Тр. Зоол.
инет. АН СССР, 6, 4: 33—68.
Арнольди К. В. 1957. О теории ареала в связи с экологией и про-
исхождением популяций. Зоол. журн., 36: 1609—1630.
Арнольди К. В. и Арнольди Л. В. 1963. О биоценозе как одном
из основных понятий экологии. Зоол. журн., 42 : 161—183.
Артюшенко 3. Т., Харкевич С. С. 1956. Рапневесснпио декоратив-
ные растения природной флоры Карпат. Бот. журн., 41 : 1604—1616.
Астауров Б. Л. 1965. Экспериментальная полиплоидия у животных.
В сб.: Полиплоидия и селекция, М.—Л.
Афанасьев В. Г. 1964. Проблема целостности в философии и биоло-
гии. М.
Базилевская Н. А. 1959. Проблема вида в трудах советских ботани-
ков. В сб.: Анналы биологии, 1, М.
Б а з ы к и н А. Д. 1965. О возможности симпатрического видообразования
Бюлл. МОИП, 70, 1:161—165.
Барабаш-Никифоров И. И., Формозов At Н. 1963. Териология. М
Баранов П. А. 1953. О видообразовании. Бот. журн., 39:669—695.
Баранов П. А. 1956. В тропической Африке. М.
Беклемишев В. Н. 1960. Пространственная и функциональная струк-
тура популяций. Бюлл. МОИП, 65, 2 :42—48.
365
Беклемишев В. И. 1964а. Об общих признаках организации жизни.
Бюлл. МОИП, 69, 2:22—38.
Беклемишев В. Н. 19646. Сравнительная анатомия беспозвоночных
животных, 1—2. М.
Белозерский А. Н. 1962. Видовая специфичность нуклеиновых кислот.
Тр. V биохим. конгр., Симпозиум 3, М. : 202—214.
Белозерский А. Н. 1964. Состав нуклеиновых кислот и эволюцион-
ная систематика. В сб.: Молекулярная биология. М.
Берг Л. С. 1922. Номогенез. Л.
Б е р г Л. С. 1934. Яровые и озимые расы у проходных рыб. Изв. АН СССР^
отд. матем. и ест. наук, 5 : 711—732.
Берг Л. С. 1948. Рыбы пресных вод, 1. М.—Л.
Берг Л. С. 1950. О ботанической номенклатуре и понятии вида у бота-
ников. Природа, 9 :30—33.
Берг Р. Л. 1944. Вид как эволюирующая система. Рефераты работ. Отд.
биол. наук АН СССР за 1941—1943 гг., М.
Берг Р. Л. 1957. Типы полиморфизма. Вести. ЛГУ, 21 :115—139.
Бианки В. Л. 1907. Формы родов Montifrihgilla Brehm. Ежегодн. Зоол.
муз. Акад, наук, 12 : 555—597.
Бианки В. Л. 1916. Вид и подчиненные ему таксономические формы.
Русск. зоол. журн., 1:287—297.
Благовещенский А. В. 1965а. Биохимические критерии таксономии
цветковых растений. В сб.: Проблемы современной ботаники. 1.
М.-Л.
Благовещенский А. В. 19656. О белковых комплексах семян некото>-
рых древнейших групп цветковых растений. В сб.: Проблемы фило-
гении растений, М.
Богданов М. 1881. Сорокопуты русской фауны и их сородичи. СПб.
Большаков В. Н, 1963, О приспособительном характере природы под-
видов на примере красной полевки. В кн.: Зоогеография суши. Таш-
кент. (Тр. 3-го Всесоюзн. совещ.).
Б о р и с я к А. А. 1947. Основные проблемы эволюционной палеонтологии.
М.-Л.
Б о ч а н ц е в В. П. и Липшиц С. Ю. 1955. К вопросу об объеме вида,
у высших растений. Бот. журн., 40: 542—547.
Вавилов Н. И. 1926. Центры происхождения культурных расте-
ний. Л.
В а в и л о в Н. И. 1931. Линнеевский вид как система. Тр. по прикл. бот.,
ген. и сел., 26, 3 : 109—134.
Вавилов Н. И. 1932. Генетика на службе социалистического земледе-
лия. Соц. растениеводство, 4 : 19—42.
В а в и л о в И. И. 1935а. Закон гомологических рядов в наследственной
изменчивости. М.—Л.
Вавилов Н. И. 19356. Научные основы селекции пшеницы. М.—Л.
Вавилов Н. И. 1936. Гуго Де Фриз. Соц. растениеводство, 17:5—15.
Вагнер Н. 1860. Ж. Кювье и 3. Жоффруа Сент-Илер. Казань.
Вайпагий И. В. 1962. Биология генеративного размножения травяни-
стых растений Украинских Карпат. Автореф. канд. дисс. Киев.
Ванюшин Б. Ф. 1965. Нуклеиновые кислоты и систематика растений.
В сб.: Проблемы филогении растений, М.
Васильев В. Н. 1961. О миграционных гипотезах в видообразовании.
Бот. журн., 46 : 1584—1601.
Васильев В. Н. 1965. О продолжительности существования вида.
В сб.: Проблемы современной ботаники, 1, М.—Л.
Васильев И. В. 1953. Липы СССР. Автореф. докт. дисс. Л.
Васильков Б. П. 1954. Опыт изучения вида у шляпочных грибов на
примере осиновика. Бот. журн., 39:681—693.
Васильков Б. П. 1965. К проблеме вида. В сб.: Споровые растения
Средней Азии и Казахстана, М.
366
Васильченко И. Т. 1958. О сезонно-диморфных видах. В сб.: Проблема
вида в ботанике, 1, М.—Л.
Васнецов В. В. 1946. Дивергенция и адаптация в онтогенезе. Зоол.
журн., 25 : 185—200.
Васнецов В. В. 1953. Целостность экологии вида у рыб. (Экологиче-
ские корреляции). В сб.: Очерки по общим вопросам ихтиоло-
гии, М.
Вейсс П. 1961. Взаимодействия между клетками. В сб.: Современные
проблемы биофизики, 2, М.
Вельдре С. Р. 1964. О реальности подвидов ушастой круглоголовки.
Вести. ЛГУ, сер. биол., 3 : 34—40.
Верещагин И. К. 1963. Географические изменения черепа и размеров
пещерных и бурых медведей голарктики. В кн.: Зоогеография суши.
Ташкент. (Тр. 3-го Всесоюзн. совещ.).
Вернадский В. И. 1926. Биосфера. Л.
Вернадский В. И. 1928. Эволюция видов и живое вещество. Природа,
3 : 227—250.
Вернадский В. И. 1931. Об условиях появления жизни на Земле.
Изв. АН СССР, 5, Б-5 633—653.
Вернадский В. И. 1932. Проблема времени современной науке.
Изв. АН СССР, 7, 4:511—541.
Вернадский В. И. 1940. Биогеохимические очерки. М.
Викторов Г. А. 1965. Колебания численности насекомых как регули-
руемый процесс. Журн. общ. биол., 26, 1: 43—55.
Виль морен Г. 1894. Наследственность у растений. СНб.
Волкова Э. В., Филю ко в А. И. 1966. Философские вопросы теории
вида. Минск.
Ворон и х и н Н. Н. 1946. О полиморфизме Spirulina platensis (Nordst)
Geitl в связи с вопросом о виде у синезеленых водорослей. Сов. бот.,
4: 239—246.
Воронихин Н. Н. 1951. О некоторых водорослях Боровского заповед-
ника в связи с вопросом о виде. Тр. Всесоюзн. гидробиол. общ.,
3 : 217—220.
Воронов А. Г. 1963. Биогеография. М.
Вульф Е. В. 1938. Понятие о реликте в ботанической географии.
В сб.: Проблема реликтов во флоре СССР, М.—Л.
Вульф Е. В. 1944. Историческая география растений. М.—Л.
Г а й с и п о в и ч А. Е. 1967. Зарождение генетики. М.
Г а у з е Г. Ф. 1934. Экспериментальное исследование борьбы за существо-
вание между Paramecium caudatum, Paramecium aurelia и Stylo-
nychia mytilus. Зоол. журн., 13 : 1—17.
Г а у з e Г. Ф. 1935. Исследование над борьбой за существование в сме-
шанных популяциях. Зоол. журн., 14 : 243—270.
Г а у з е Г. Ф. 1939. Исследования по естественному отбору у простейших.
Зоол. журн., 18: 631—641.
Гексли Дж. 1940. Теория полового отбора. Усн. соврем, биол., 13,
2 (5) : 306—323.
Гексли Дж. 1960. Рак как биологическая проблема. М.
Гентнер В. Г. 1956. О числе видов фауны СССР и об отношении ее
к мировой фауне. Зоол. журн., 35: 1780—1790.
Г е р б и л ь с к и й Н. Л. 1954. Сравнительное исследование проявлений
внутривидовой разнокачественности у осетровых. Тез. докл.
3-й эколог, конфер., 2, Киев: 33—36.
Гербильский Н. Л. 1957. Внутривидовая биологическая дифференциа-
ция и ее значение для вида в мире рыб. Вестн. ЛГУ, сер. биол., 21,
4:82-92.
Гербильский И. Л. 1962. Теория биологического прогресса осетровых
и ее применение в практике осетрового хозяйства. В сб.: Воспро-
изводство рыбных запасов в связи с гидростроительством, Л.
367
Г е р б и л ь с к ий Н. Л. 1965а. Теория биологического прогресса вида и ее
использование в рыбном хозяйстве. В сб.: Теоретические основы
рыбоводства, М.
Гербильский Н. Л. 19656. Сложные формы поведения как элемент
видовых адаптаций. В сб.: Сложные формы поведения, М.—Л.
Герман А. Л. 1960. Восстановление бобра в Лапландском заповеднике.
Бюлл. МОИП, 65, 3: 13—19.
Гершензон С. М. 1941. «Мобилизационный резерв» внутривидовой
изменчивости. Журн. общ. биол., 2, 1 : 85—107.
Гиляров М. С. 1954. Вид, популяция и биоценоз. Зоол. журн., 33: 769—
778.
Гиляров М. С. 1959. Проблемы современной экологии и теория есте-
ственного отбора. Усп. соврем, биол., 18, 3 (6) : 267—278.
Г о л л е р б а х М. М. 1941. О понятиях «тип» и «изменчивость» в система-
тике водорослей. Сов. бот., 3 : 161—166.
Горизонты биохимии. 1964. Сборник статей. М.
Горский И. И. 1964. О древнейших ископаемых остатках животных и
растений. В сб.: Вопросы закономерностей и форм развития органи-
ческого мира, М.
Г о р т и н с к и й Г. Б. 1963. Аллелопатия и биогеоценология. Бюлл. МОИП,
68, 6 : 103-110.
Грубов В. И. 1955. К вопросу об объеме вида на примере аптечной ва-
лерианы. Бот. журн., 40 : 815—824.
Грюммер Г. 1957. Взаимное влияние высших растений. М.
Гудошников С. В. 1962. Некоторые аспекты внутривидовых отно-
шений у растений. В сб.: Проблемы внутривидовых отношений,.
Томск.
Давиташвили Л. Ш. 1956. Очерки истории учения об эволюционном
прогрессе. М.
Давиташвили Л. Ш. 1961. Теория полового отбора. М.
Даннеман Ф. 1932. История естествознания, 1. М.
Дарвин Ч. 1937. Происхождение видов. М.—Л.
Дарвин Ч. 1950. Избранные письма. М.
Даревский И. С. 1967. Скальные ящерицы Кавказа. Автореф. докт.
дисс. Л.
Де Вит С. 1964. Пространственные взаимоотношения в популяциях
одного или нескольких видов растений. В сб.: Механизмы биологи-
ческой конкуренции, М.
Делоне Л. И. 1945. Опыт определения основных категорий систематики,
селекции и генетики. Зап. Харьковск. с.-х. инет., 4: 25—36.
Дементьев Г. П. 1947. Число видов птиц земного шара и нашей
фауны. Природа, 2 : 35—37.
Дементьев Г. П. 1954. Замечания о виде и некоторых сторонах видо-
образования в зоологии. Зоол. журн., 33: 525—537.
Дементьев Г. П., Ильичев В. Д., Курочкин Е. Н. 1966. Функцио-
нальный анализ некоторых явлений полиморфизма у птиц и на-
чальные этапы формообразования. В сб.: Внутривидовая изменчи-
вость наземных позвоночных животных и микроэволюция, Сверд-
ловск.
Де Ф р и з Г. 1932. Избранные произведения. М.
Джерард Р. 1966. Концепция информации и биологические системы.
М.: 327-340.
Длусский Г. М. 1965. Охраняемая территория у муравьев. Журн. общ.
биол., 26, 4:479-489.
Догель В. А., Полянский Ю. И. и Хейсин Е. М. 1962. Общая
протозоология. М.—Л.
Дональд С. 1964. Конкуренция за свет у сельскохозяйственных культур
и пастбищных растений. В сб.: Механизмы биологической конку-
ренции, М.
368
Дубинин В. Б. 1953. Понятие о виде в зоологии. Зоол. журн., 32: 1095—
1109.
Дубинин В. Б. 1954. Понятие о виде у паразитических животных.
Тр. пробл. и тематич. совещ. Зоол. инет. АН СССР, 4: 163—185.
Дубинин Н. И. 1931. Генетико-автоматические процессы и их значение
для механизма органической эволюции. Журн. экспср. биол., 7,
5—6.
Дубинин Н. П. 1934. Экспериментальное уменьшение числа пар хромо-
сом у Dr. melanogaster. Биол. журн., 3: 719—734.
Дубинин Н. П. 1966. Эволюция популяций и радиация. М.
Еленкин А. А. 1936. Синезеленые водоросли СССР. Общая часть. Л.
Жадин В. И. 1952. Моллюски пресных и соленых вод СССР. Л.
Жебрак А. Р. и Жебрак Э. А. 1965. Теоретические аспекты использо-
вания полиплоидных форм пшеницы и гречихи. В сб.: Полиплоидия
и селекция, М.—Л.
Житков Б. М. 1934. Предшественники Линнея. Бюлл. МОИП, 43,
2 : 248—255.
Жуйкова И. В. 1959. Эколого-морфологические юбенности некоторых
растений Хибин. Автореф. каид. дисс. Л.
Ж у к о в - В е р с ж н и к о в II. Н. и Пехов А. II. 1963*. Генетика бакте-
рий. М.
Жуковский П. М. 1964. Культурные растения и их сородичи. Л.
Жуковский П. М. 1965. Эволюция культурных растений на основе по-
липлоидии. В сб.: Полиплоидия и селекция, М.—Л.
Завадский К. М. 1941. Конспект курса дарвинизма. Л.
Завадский К. М. 1947. Влияние густоты насаждений на численность
и рост кок-сагыза. Канд. дисс. ЛГУ.
Завадский К. М. 1948а. И. В. Мичурин как преобразователь при-
роды. Л.
Завадский К. М. 19486. Роль перенаселения в эволюции. Тр. конфер.
по дарвинизму, изд. МГУ : 35—52.
Завадский К. М. 1949. Критика эволюционизма и трансформизма.
Вести. ЛГУ, 12 : 28—53.
Завадский К. М. 1952. О прогрессивном значении взглядов В. Л. Ко-
марова на реальность вида. Вести. ЛГУ, 4: 3—25.
Завадский К. М. 1954а. О некоторых вопросах теории вида и видо-
образования. Вести. ЛГУ, 10 : 3—15.
Завадский К. М. 19546. В. И. Ленин о требовании всесторонности
в исследованиях и некоторые нарушения этого требования в био-
логии. Вести. ЛГУ, 1 : 3—20.
Завадский К. М. 1954в. К вопросу о понимании перехода «старой» на-
следственности в «новую». Вести. ЛГУ, 4: 3—16.
Завадский К. М. 1954г. О причинах выпада растений в гнездовых по-
севах. Бот. журн., 39 : 515—544.
Завадский К. М. 1956. Научное наследие И. В. Мичурина и некото-
рые задачи биологии. Бот. журн., 41 : 3—22.
Завадский К. М. 1957а. Перенаселение и его роль в эволюции. Бот.
журн., 42 : 426—449.
Завадский К. М. 19576. К вопросу о дифференциации вида у высших
растений. Вести. ЛГУ, 21, 4: 18—44.
Завадский К. М. 1957в. Предмет и задачи современного дарвинизма.
Бот. журн., 42:583—595.
Завадский К. М. 1958. К пониманию прогресса в органической при-
роде. В сб.: Проблемы развития в природе и обществе, Л.
Завадский К. М. 1960а. Ароморфоз. В кн.: Философская энциклопедия,
1. М.
Завадский К. М. 19606. Биология. В кн.: Философская энциклопедия,
1. М.
Завадский К. М. 1961. Учение о виде. Л.
369>
Завадский К. М. 1965. Теория эволюции и ее преподавание. Журн. общ.
биол., 6 : 731—743.
Завадский К. М. 1966а. Основные формы организации жизни и их
подразделения. В сб.: Философские проблемы современной биоло-
гии, Л.
Завадский К. М. 19666. Проблема вида у апомиктических растений.
Совещание по проблеме апомиксиса, Саратов: 17—21.
Завадский К. М. 1967а. Объем вида и внутривидовые подразделения.
Совещание по объему вида и внутривидовой систематике, Л.: 8—10.
Завадский К. М. 19676. Вид как форма существования жизни. Струк-
тура вида. Видообразование. В кн.: Современные проблемы эволю-
ционной теории. Л.
Завадский К. М., Берман 3. И. 1966. Об одной распространенной
форме антидарвинизма. Вести. ЛГУ, 9, 2: 5—22.
Завадский К. М., Горобец А. М., X о д ь к о в Л. Е. и Хахина Л. Н.
1962. Некоторые результаты изучения популяций высших растений.
Тр. Петергофск. биол. инет., 19: 17—34.
Завадский К. М., Ермоленко М. Т. 1966. О критике неономогенеза.
В сб.: Философские проблемы современной биологии, Л.
3 а к а р ь я и С. Ф. 1930. Борьба за жизнь между всходами солянок. Баку.
3 а х в а т к и и А. А. 1953. Сборник научных работ. Изд. МГУ.
Зеликман А. Л. 1966. Эволюционная теория Ч. Дарвина. В кн.: Исто-
рия эволюционных учений в биологии. Л.
Зеликман А. Л. и Г о й прих А. К. 1959. К вопросу о влиянии плот-
ности популяций на смертность и развитие ее компонентов у цикло-
пов. Бюлл. МОИП, отд. биол., 64, 4 : 125—139.
Зоз И. Г. и Черных И. А. 1961. Сравнительное изучение дальневосточ-
ного и восточноевропейского ландышей. Бот. жури., 46 :562—569.
Иванов А. В. 1960. Погонофоры. Фауна СССР, новая серия, 75. Л.
Иванов А. В. 1965. Пауки. Л.
Ильин М. М. 1938. Реликтовые элементы широколиственных лесов
во флоре Сибири и их возможное происхождение. В сб.: Проблема
реликтов во флоре СССР, М.—Л.
Ильинский А. 1929». Закономерности в размножении малого соснового
лубоеда и теоретическое обоснование мер борьбы с ними в лесу.
Бюлл. Защита раст. от вредит., 5, 5—6 : 523—542.
Имшенецкий А. А. 1961. Теоретические основы селекции полезных
форм микроорганизмов. Тр. Ипст. микробиол. АН СССР, 10:5—24.
К а л а б у х о в II. И. 1962. Физиологическая неоднородность популяции
животных как один из факторов динамики их численности.
В сб.: Вопросы экологии, 4, Киев.
Калабухов Н. И. 1964. Адаптивные особенности двух географических
форм полуденной песчанки. В сб.: Вопросы внутривидовой измен-
чивости позвоночных животных и микроэволюция, Свердловск.
Камшилов М. М. 1947. Исследования советских биологов в области
эволюционной теории за 30 лет. Усп. соврем, биол., 24, 2 (5) : 297—
Камшилов М. М. 1961. Значение взаимных отношений между организ-
мами в эволюции. М.—Л.
Карпеченко Г. Д. 1927. Полиплоидные гибриды Raphanus sativus L. X
X Brassica oleracea L. Тр. по прпкл. бот., ген. и сел., 17, 3 : 305—410.
Карпеченко Г. Д. 1929. К синтезу константного гибрида из 3 ви-
дов. Тр. Всесоюзн. съезда по ген., сел., сем. и плем. жив., 2:
277—294.
Карпеченко Г. Д. и Щавинская С. 1930. О половом обособлении
тетраплоидных Raphanus X Brassica. Тр. Всесоюзн. съезда по ген.,
сел., сем. и плем. жив., 2:267—276.
Карпов С. П. 1962. Проблема вида в микробиологии. В сб/ Проблемы
внутривидовых отношений, Томск.
370
Кашкаров Д. П. 1933. Среда и сообщество. Л.
Кашка ров Д. Н. 1939. Адаптивна ли эволюция и что такое видовые
признаки? Зоол. журн., 18: 612—630.
Ке. лер Б. А. 1-907. Очерки по флоре юга Царицынского уезда. В кп.:
Н. А. Дим о и Б. А. Келлер. 1907. В области полупустыни.
Саратов.
Кирпичников М. М. 1948. К вопросу о количественном анализе в си-
стематике растений. Бот. жури., 33 : 333—344.
Киселева С. Д. 1961. Морфобиологическио группы и распределение их
по экотипам у вики озимой. Сб. трудов аспирантов ВИРа, 2,
6 : 151 — 155.
Киселева С. Д. 1963. Изучение состава популяции Vicia villose и
V. pannonica. Авторсф. канд. дисс. Л.
К л е п п Б. И. 1945. Понятие о виде в общей и медицинской микробиоло-
гии. Микробиология, 14, 1 : 35—44.
Клоков М. В. 1947. Эндемизм украинской флоры. Докт. дисс. Киев.
Клоков М. В. 1967. Рациональные основы биологической таксономии.
Тез. докл. Совещания но объему вида и внутривидовой систематики,
Л. : 40—42.
К л ю и в с р А. и Ban Пиль К. 1959. Вклад микробов в биологию. М.
Ковальский В. В. 1963. Возникновение и эволюция биосферы. Усн.
соврем, биол., 55, 1 : 45—67.
Кожанчиков И. В. 1948. Биологическая специфика видов насекомых.
Усп. соврем, биол., 25. 2 251—263.
Кожов М. М. 1960. О видообразовании в оз. Байкал. Бюлл. МОИП, 65.
6 39-47.
Козлов К. А. 1962. О дифференциации вида у микроорганизмов. В сб.:
Проблемы внутривидовых отношений, Томск.
Козлов а Л. М. 1956. О приспособительном значении периода покоя
семян. Автореф. канд. дисс. Л.
К о з о - П о л я и с к и й Б. 1948. О филогенетическом значении апомиксиса.
Бот. журн., 33: 123—127.
Колесников А. Н. 1951. Борьба за дарвинизм в Казанском универси-
тете в 60—70 годы ХТХ века. Автореф. канд. дисс. ЛГУ.
Коломиец Н. К. 1962. Паразиты и хищники сибирского шелкопряда.
Новосибирск.
Комаров В. Л. 1901а. Флора Маньчжурии, 1. СПб.
Комаров В. Л. 19016. Формы изменяемости Sagittaria. Тр. СПб. общ.
естествоиспыт., 32, 1, Прот. засед. № 7—8:319—323.
Комаров В. Л. 1902. Вид и его подразделения. Дневник XI съезда рус-
ских естествоиспыт. и врачей, 6:250—252.
Комаров В. Л. 1908. Введение к флорам Китая и Монголии, Моно-
графия рода Caragana. СПб.
Комаров В. Л. 1912. Видообразование. В сб.: Итоги пауки, 6, Изд. «Мир».
Комаров В. Л. 1925. Ламарк. М.
Комаров В. Л. 1927. Флора полуострова Камчатки, 1. Л.
Комаров В. Л. 1930. Род Pbacellantbus Sicb. el Zucc. (Orobanchaceae)
на Дальнем Востоке. Изв. АП СССР, 7, 3 : 267—274.
Комаров В. Л. 1934. Предисловие. Флора СССР, 1. Л.
Комаров В. Л. 1937. Род Poligonum. Флора СССР, 5. Л.
Комаров В. Л. 1940. Учение о виде у растений. М.—Л.
Конференция по проблемам дарвинизма. 1948. Тезисы докладов. М.
Концепция информации и биологические системы. 1966. М.
Коржинский С. 1892. Флора востока Европейской России в ее систе-
матическом и географическом отношениях. Томск.
Коржинский С. 1899. Гетерогенезис и эволюция. К теории происхож-
дения видов. СПб.
Королева В. А. 1948. Биологический анализ популяций кок-сагыза. Тр.
по прикл. бот., ген. и сел., 28, 1 163—174.
371
Королева В. Л. 1957. Внутривидовое разнообразие кок-сагыза. Бог.
журн., 42 : 414—425.
Корчагин А. А. 1964. Внутривидовой (популяционный) состав расти-
тельных сообществ и методы его изучения. В сб.: Полевая геобота-
ника, 3, JI.
Корчагин А. А. 1966. Значение выявления внутривидового (популя-
ционного) состава растительных сообществ при стационарных геобо-
танических исследованиях. Бот. журн., 51 : 777—782.
Костов Д. 1941—1943. Цитогенетика рода Nicotiana. София.
Красильников Н. Л. 1947. Понятие о виде у бактерий. Микробиоло-
гия, 16, 5 : 381—393.
Красильников Н. А. 1962. Основные принципы классификации микро-
организмов. Руководство по микробиологии, клинике и эпидемиоло-
гии инфекционных болезней. 1. Общая микробиология. М.
Красовская И. В. 1931. К вопросу о соревновании первичных и вто-
ричных культур в совместных посевах. Тр. по прикл. бот., ген. и
сел., 25, 3:273—323.
Крашенинников И. М. 1946. Опыт филогенетического анализа неко-
торых евроазиатских групп рода Artemisia в связи с особенностями
палеографии Евразии. Материалы по истории флоры и раститель-
ности СССР, 2, Л.
Крей ер Г. К. 1941. Теория стадийного развития систематике расте-
ний. Сов. бот., 1 -2 : 39—50.
К р и штофов и ч А. II. 1957. Палеоботаника. 7.
Кроган П. 1964. Конкуренция и механизмы приспособления к осмо-
тическому давлению. В сб.: Механизмы биологической конкурен-
ции, М.
Крылов Г. В. 1967. Вид и внутривидовые подразделения у древесных
растении. Совещание по объему вида и внутривидовой система-
тике, Л.
К у д р я в ц е в В. И. 1951. К проблеме вида у микроорганизмов. Тр.
Инет, микробиол. АП СССР, 1 :86—107.
Кудряшева О. И. 1957. Некоторые биологические особенности таджик-
ского одуванчика. Изв. отд. естеств. наук АП Тадж. ССР, 20 : 46—53?.
Кузнецов Н. И. 1922. Количество видов растений на земном шаре.
Изв. Главн. бот. сада РСФСР, 21, 2 : 92—108.
Кулешов IT. Н. 1922. Борьба рас яровой пшеницы. Харьков.
Купревич В. Ф. 1949. Проблема вида у гетеротрофных и автотрофных
растений. М.—Л.
Купревич В. Ф. 1950. Вид как этан эволюции гетеротрофных и авто-
трофных растений. В сб.: Проблемы ботаники, 1, Л.
Кур санов Л. И. 1945. Понятие о виде у низших растений. Микробио-
логия, 14, 4 : 210—214.
К э й н А. 1958. Вид и его эволюция. М.
Лавренко Е. М. 1963. К. М Завадский. Учение о виде. [Рецензия].
Бот. журн., 8 : 1217—1223.
Лавренко Е. М. 1964. Об уровнях изучения органического мира. Изв.
АН СССР, сер. биол., 1 32—46.
Ламарк. Ж. Б. 1935. Философия зоологии, 1. М.—Л.
Ламарк Ж. Б. 1955. Избранные прпзведения, 1. М.
Ларина И. И. 1958. К вопросу о диагностике близких видов лесной и
желтогорлой мыши. Зоол. журп., 37 1719—1732.
Ларина И. И. 1961. О постоянстве видовых границ и эволюционном
значении межвидовых отношений. Автореф. докт. дисс. Саратов.
Ларина И. И. 1962. Внутривидовые отношения позвоночных животных
и проблема симпатрического видообразования. В сб.: Проблемы
внутривидовых отношений, Томск.
Ларина Н. И. 1963. Симпатрия близких видов и ее возникновение.
В сб.: Зоогеография суши, Ташкент.
372
Ларина Н. И. 1964. Географическая изменчивость некоторых эколого-
географических признаков лесных и желтогорлых мышей. В сб.: Во-
просы внутривидовой изменчивости позвоночных животных и микро-
эволюция, Свердловск : 67—68.
Лебедев Д. В. 195-7. Эволюционные взгляды В. И. Талиева. Бот. журн.,
42: 1338—1353.
Лебедев Д. В. 1965. Из истории некоторых идей отечественной гене-
тики. Бот. журн., 50: 730—733.
Левин Г. Г. 1963. Жизненные циклы растений, их связь и эволюция.
Бот. журн., 48 : 1039—1059.
Левин Г. Г. 1964. Индивидуальность и жизненные циклы растений. Бот.
журн., 49 : 272—280.
Левина Р. Е. 1964. Проблема вида в биологической науке. (К. М. Завад-
ский. Учение о виде). Вести. ЛГУ, сер. биол., 9, 2:131—134.
Левитский Г. 1931. Морфология хромосом и понятие кариотипа в си-
стематике. Тр. по прикл. бот., ген. и сел., 27, 1 : 187—240.
Левитский Г. 1939. Цитогенетические основы эволюции. Природа,
5 : 33—44.
Ледерберг Дж. 1961. Итоги и проблемы генетики. Журн. Всесоюзн.
хим. общ., 6, 3 298—304.
Линней К. 1800. Философия ботаники. СПб.
Л и ч к о в Б. Л. 1965. К основам современной теории Земли. Л.
Лукин Е. И. 1962. Внутривидовые экологические изменения организмов.
В сб.: Вопросы экологии, 4, Киев.
Лысенко Т. Д. 1946. Естественный отбор и внутривидовая конкурен-
ция. В кн.: Т. Д, Л ы с е н к о. Агробиология. М.
Лысенко Т. Д. 1950. Новое в науке о биологическом виде. Агробиоло-
гия, 6 : 15—25.
Л ы с е н к о Т. Д. 1956. О биологическом виде и видообразовании. Агробио-
логия, 4 : 3—30.
Л э к Д. 1957. Численность животных и ее регуляция в природе. М.
Л ю б и м е и к о В. Н., Щеглова О. и Булгакова 3. 1925. Опыт над
соревнованием за место у растений. Журн. Русск. бот. общ., 10,
3-4 : 293—338.
Ляпунов А. А. 1962. Об управляющих системах живой природы и
общем понимании жизненных явлений. М.
Ляпунов А. А. 1966. Математическая интерпретация биологических за-
кономерностей. В сб.: Математическое моделирование жизненных
процессов, М.
Ляпунов А. А., Стебанев И. В. 1964. О биоценологическом уровне
управления в рамках биосферы. В сб.: Проблемы кибернетики, М.
Лященко И. Ф. и Лященко И. И. 1957. Биологические особенности
растений альбиносов подсолнечника. Уч. зап. Ростовск. унив. 58,
4: 43-45.
Мазер К. 1964. Конкуренция и сотрудничество. В сб.: Механизмы био-
логической конкуренции, М.
Майр Э. 1947. Систематика и происхождение видов. М.
Максимович М. А. 1831. Систематика растений. СПб.
Малеев В. П. 1933. Теоретические основы акклиматизации. Л.
Малиновский А. А. 1941. Закономерности наследственности в свете
учения об отборе. Усп. соврем, биол., 14, 1 : 171—176.
Малиновский А. А. 1947. Вид как эволюционное целое. Рефераты
научно-исслед. работ за 1945 г., М. : 286—287.
Мальчевский А. С. 1959. Гнездовая жизнь певчих птиц. Л.
Мамаев С. А. 1965. Вариации сосны обыкновенной по окраске генера-
тивных органов. В сб.: Внутривидовая изменчивость древесных
растений, Свердловск.
Мандэй К. 1964. Физиологический и экологический аспекты стресса.
В сб.: Механизмы биологической конкуренции, М.
24 Б. М. Завадский
373
М а р и к о в с к и й П. Й. 1962. О внутривидовых отношениях рыжего лес-
ного муравья Formica rufa L. Энтомол. обозр., 41 : 83—93.
Марковский Б. П. 1952. К стратиграфии девонских отложений Лен.
области. В сб.: Палеонтология и стратиграфия, Л.
Мартинсон Г. Г. 1951. Третичная фауна моллюсков Прибайкалья.
Тр. Байкальск, лимыол. станц. АН СССР, 13:5—93.
Марченко И. И. 1965. Полиплоидия в роде Helianthus и ее использова-
ние в селекции подсолнечника и топинамбура. В сб.: Полиплоидия
и селекция, М.—Л.
Матвеев Б. С. 1942. О приспособлениях к лактации у млекопитающих.
Зоол. журн., 21, 3 : 47—68.
Матвеева Т. С. 1965. Спонтанная и экспериментально вызванная поли-
плоидия у некоторых декоративных однодольных растений. В сб.:
Полиплоидия и селекция, М.—Л.
Медников Б. М. 1963. Экологические формы рыб и проблема симпатри-
ческого видообразования. Зоол. журн., 42: 70—77.
Мензбир М. 1927. За Дарвина. М.—Л.
Механизмы биологической конкуренции. 1964. Сборник статей. М.
Мечников И. И. 1950. Очерк вопроса о происхождении видов. Избран-
ные произведения, М.
Микулинекий С. Р. 1961. Развитие общих проблем биологии в Рос-
сии. М.
Мильн А. 1964. Определение понятия «конкуренция» у животных. В со.:
Механизмы биологической конкуренции, М.
Миняев Н. А. 1965. Арктические и аркто-альпийские элементы во флоре
северо-запада СССР. В сб.: Ареалы растений флоры СССР, Л.
Мирошниченко Е. Я. 1963. К вопросу об использовании апомиксиса
для селекции мятлика. В сб.: Апомиксис и некоторые новые методы
селекции растений, Новосибирск.
М и р ю т а 10. П. 1965. Об избирательной конъюгации хромосом у поли-
плоидов. В сб.: Полиплоидия и селекция, М.—Л.
Мичурин И. В. 1929. Итоги полувековых работ, 1. М.
Мичурин И. В. 1948. Собрание сочинений, 1—4, М.
Млекопитающие фауны СССР, 1. 1963. М.—Л.
Морган Т. Г. 1936. Экспериментальные основы эволюции. М.
Мюнтцинг А. 1963. Генетические исследования. М.
Найт-Джонс Е., Мойз Дж. 1964. Внутривидовая конкуренция у при-
крепленных форм морских животных. В сб.: Механизмы биологиче-
ской конкуренции, М.
Наливкин Д. В. 1964. Установление вида в палеонтологии. В сб.: Во-
просы закономерностей и форм развития органического мира, М.
Наумов Н. П. 1962. Популяционная экология и ее основные проблемы.
В сб.: Вопросы экологии, 4, Киев.
Наумов Н. П. 1964. О методологических проблемах биологии. Философ-
ские науки, 1: 136—145.
Невский С. А. 1934. Род Triticum. Флора СССР, 2. Л.
Невский С. А. 1935. К вопросу о системе рода Triticum. Сов. бот.,
6 : 120—128.
Неймайр М. 1898. Корни животного царства. М.
Никитин С. 1881. Дарвинизм и вопросы о виде в палеонтологии.
«Мысль». 8—9 : 229—245.
Овчинников П. И. и Овчаров К. Е. 1947. Новый каучуконосный
одуванчик из Таджикистана. Сообщ. Таджикск. фил. АН СССР,
1 : 3-5.
Огнев С. И. 1944. Проблема систематики. Вопрос о виде. Зоол. журн.,
23 : 1—16.
Огнев С. И. 1947. Вид, низшие категории вида, насущные задачи систе-
матики. Бюлл. МОИП, отд. биол., 52, 1 : 3—21.
О леиов 10. М. 1961. Некоторые вопросы генетики и дарвинизма. Л.
374
П а а в е р К. Л. 1966. О вековой изменчивости как форме групповой из-
менчивости животных во времени. В сб.: Внутривидовая измен-
чивость наземных позвоночных животных п микроэволюция, Сверд-
ловск. в
Палилов А. И. 1957. К вопросу о философских основах проблемы вида.
Уч. зап. Белорусск. унив., сер. биол., 37 : 3—40.
П а л ь м о в а Е. Ф. 1935. Введение в экологию пшениц. М,—Л.
Панов Е. Н. 1965. Этологические изолирующие механизмы. Бюлл. МОИП,
отд. биол., 6 : 163.
Папонов А. Н. 1959. Зависимость развития растений от площадей пи-
тания на разных по плодородию почвах. Изв. Тимирязевск. с.-х.
акад., 5 : 197—210.
Папонов А. Н. 1962. Влияние площади питания и плодородия почвы па
урожай п развитие растений. Автореф. капд. дисс. Л.
Парамонов А. А. 1945. Курс дарвинизма. М.
Парамонов С. Я. 1943. Что такое вид в биологии, Сов. бот., 2:
3-18.
Парк Т. 1939. Аналитическое изучение популяции животных и его отно-
шение к общей экологии. Усп. соврем, биол., 10, 2 : 304—314.
Пачоский И. К. 1904. Очерк растительности Днепровского уезда Таври-
ческой губернии. Одесса.
Пачоский И. К. 1907. Цветные расы растений. Тр. Бот. сада Юрьевск.
унив., 8, 2 : 67—72.
Пачоский И. К. 1910а. Современные задачи изучения растительного
покрова. Одесса.
Пачоский И. К. 19106. Основные черты развития флоры юго-западной
России. Херсон.
Пачоский И. К. 1914. Херсонская флора. Введение. Херсон.
Пачоский И. К. 1925. Ареал и его возникновение. Журн. Русск. бот.
общ., 10, 1—2 : 135-138.
Петров Д. Ф. 1963. Генетически управляемый апомиксис и его значение
для эволюции и селекции. В сб.: Апомиксис и новые методы селек-
ции растений, Новосибирск.
Петровская-Баранова Т. П. и Цингер Н. В. 1965. Атавистиче-
ские приспособления к половому процессу в семяпочках партеноге-
нетических видов Taraxacum. В сб.: Проблемы филогении расте-
ний, М.
Петрусевич К. 1960. О внутривидовых и межвидовых отношениях.
Зоол. журн., 39 : 1591—1596.
Писарев В. Е. и Жилкина М. Д. 1965. Использование полиплоидии
в перестройке геномного состава мягкой пшеницы. В сб.: Полиплои-
дия и селекция, М.—Л.
Плате Л. 1928. Эволюционная теория. М.—Л.
Поздняков Л. К. 1952.. Межродовой гибрид из семейства розоцветных.
ДАН СССР, 85, 5:1161—1164.
Полежаев В. 1934. Борьба за существование у тополевой моли. Зоол.
журн., 13 : 485—506.
Полянский В. И. 1956. О виде у низших водорослей. М.—Л.
Полянский В. И. 1958. Понятие о виде в альгологии. В сб.: Проблема
вида в ботанике, 1, Л.
Полянский В. И. 1959. О целостности вида и ее эволюции. Бюлл. МОИП,
отд. биол., 64, 4 : 5—13.
Полянский Ю. И. 1957. О внутривидовой дифференциации и структуре
вида у простейших. Вести. ЛГУ, сер. биол., 21,<4: 45—64.
Померанцева А. А. 1964. О виде и видообразовании в связи с дан-
ными палеоботаники. Вопросы закономерностей и форм развития
органического мира, М.
Поплавская Г. И. 1930. Об экотипах некоторых растений в Крыму.
Журп. Русск. бот. общ., 15, 4 339—349.
24*
375
Попов М. Г. 1927. Географо-морфологический метод систематики и гибри-
дизационные процессы в природе. Тр. прикл. бот. и сел., 17,
1 : 221—290.
Попов М. Г. 1928. Гибридизационные процессы в природе и значение
их для эволюции. Дневник Всесоюзн. съезда ботаников в Ленин-
граде : 92—93.
Попов М. Г. 1948. Род Erigeron в горах Ср. Азии. Тр. Бот. инет. АН СССР,
сер. I, 7 : 6—44.
Попов М. Г. 1956. К вопросу о происхождении покрытосеменных. Бот.
журн., 41: 768—769.
Пояркова А. И. 1953. Sorbocotoneaster Pojark. — новый естественный
межродовой гибрид. Бот. матер, герб. Бот. инет. АН СССР, 15: 92—
108.
Правдин Л. Ф. 1965. Внутривидовая систематика и ее значение для
селекции. В сб.: Проблемы современной ботаники, 1, Л.
Происхождение жизни на Земле. 1959. Тр. Междуиародн. симпоз.
в Москве, М.
Проханов Я. И. 1947. Конспект новой системы хлопчатника (род Gos-
sypium L.). Бот. журн., 32:61—78.
Проханов Я. И. 1965а. Вид у культурных и у дикорастущих растений.
В сб.: Проблемы современной ботаники, 1, Л.
Проханов Я. И. 19656. Возникновение двудольных многолетних трав.
В сб.: Проблемы филогении растений, М.
Пузанов И. И. 1963. Книга по общим вопросам таксономии (К. М. За-
вадский. Учение о виде). Вести. ЛГУ, 3, 1 : 155—160.
Работнов Т. А. 1946. Возрастной состав популяций некоторых пустын-
ных растений. Усп. соврем, биол., 22, 3 (6) : 435—438.
Работнов Т. А. 1950. Вопросы изучения состава популяций для целей
фитоценологии. В сб.: Проблемы ботаники, 1, Л.
Райков Б. Е. 1952—1959. Русские биологи-эволюционисты до Дарвина,
1—4. М.-Л.
Райкова И. А. и Кузьмина Н. В. 1958. Спонтанные памирские меж-
родовые гибриды злаков. Тр. Среднеазиатск. унив., 136:111—127.
Р е г е л ь Р. Э. 1912. Селекция с научной точки зрения. Тр. Бюро по прикл.
бот, 5 : 425^623.
Р е г е л ь Р. Э. 1917. К вопросу о видообразовании. Тр. Бюро по прикл.
бот, 1 : 157—181.
Р и ч А. 1964. Передача биохимической информации и проблема эволю-
ции. В сб.: Горизонты биохимии, М.
Родендорф Б. Б. 1957. Палеоэнтомологические исследования в СССР.
Тр. Палеонтол. инет. АН СССР, 66 : 5—99.
Родендорф Б. Б. 1959. Вопросы палеозоологической систематики. Па-
леонтол. журн, 3 : 15—26.
Родендорф Б. Б. 1965. Многообразие органического мира и его исто-
рическое развитие. Журп. общ. биол, 26, 4 : 40'5—415.
Розанова М. А. 1928а. Экспериментально-генетический метод в систе-
матике. Журн. Русск. бот. общ, 13, 3—4: 245—270.
Розанова М. А. 19286. О низших таксономических единицах. Журн.
Русск. бот. общ, 13, 3—4: 323—342.
Розанова М. А. 1930. Современные методы систематики растений. Л.
Розанова М. А. 1932. Опыт аналитической монографии consp. Ranuncu-
lus auricomus Korsh. Тр. Петергофск. биол. инет, 8: 19—148.
Розанова М. А. 1935. Экспериментальная систематика растений и ее
задачи. Тр. Петергофск. биол. инет, 15:135—148.
Розанова М. А. 1938а. О полиплоидном возникновении видов.
ДАН СССР, 18: 677-680.
Розанова М. А. 19386. Межвидовая гибридизация в пределе родов
Rubus и Fragaria в связи с вопросами формообразования. Изв.
АН СССР, 3:667—679.
376
Розанова М. А. 1940. Пути изменения и возникновения рас и видов
у растений. Усп. соврем, биол., 12, 2: 203—234.
Розанова М. А. 1946. Экспериментальные основы систематики расте-
ний. М.—Л.
Ростова Н. С. 1960. О природе так называемого «озимого биотипа» кок-
сагыза. Тр. Петергофск. биол. инет., 18: 84—101.
Ростова Н. С. 1963. Структура популяции кок-сагыза. 2. Яровая часть
популяции. Вести. ЛГУ, 9, 2:42—54.
Ростова Н. С. 1965. Структура популяции кок-сагыза. 3. Вести. ЛГУ,
15, 3 : 138—152.
Ростова Н. С. 1967. Концепция сезонных рас Р. Веттштейна—Н. В. Цин-
гера в связи с проблемой внутривидового фенологического полимор-
физма. Бот. жури., 5 704—715.
Ротай А. П. 1962. Палеонтологический метод и проблема вида в страти-
графии. Киев.
Ру б ай лов а П. Г. 1963. Проблема отдаленной гибридизации животных
в ее историческом развитии. Автореф. канд. дпсс. М.
Рубцов И. А. 1941. Теория круга форм. Тр. Зоол. инет. АП СССР, G,
4 191-206.
Рудаков О. Л. 1964. Эволюционные предпосылки образования несовер-
шенных грибов и биологическая роль гетерокариоза. Тр. Всосоюзп.
инет, защиты раст., 23 175—183.
Рыбин В. 1935. Опыт синтеза культурной сливы из родственных ей ди-
ких видов. Соц. растениеводство, 15 87—100.
Рыбин В. А. 1951. Экспериментальные данные по вопросу гибридного
происхождения садовой сливы Prunus domestica. Тр. Крымск, фи л.
АН СССР, 1:45—61.
Р ы б п и В. А. 1962. Естественный процесс гибридизации между алычой и
абрикосом. Изв. АН Молд. ССР, 12 : 18—31.
Рыт о в М. В. 1914. Овощное семеноводство. М.
Сабинин Д. А. 1947. О внутривидовой борьбе в насаждениях растений.
В сб.: Внутривидовая борьба у животных и растений, М.
Савин В. Н. 1957а. К характеристике морфобиологических групп в по-
пуляциях некоторых перекрестноопыляющпхся растений. Вести. ЛГУ,
сер: биол., 3, 1 44—51.
Савин В. Н. 19576. Характеристика морфобиологических групп мари бе-
лой. Вести. ЛГУ, сер. биол., 21, 4 : 148—152.
Савин В. И. 1959. Сравнительное изучение состава популяций некото-
рых видов перекрестноопыляющпхся растений. Автореф. канд. дисс.
ЛГУ.
С а в и ч - Л ю б и ц к а я Л. И. и Смирнова 3. II. 1958. Понятие о виде
в бриологии. В сб.: Проблема вида в ботанике, 1, Л.
Савоськин И. П. 1959. Биологические основы введения в культуру
морского лука и подснежника Воронова. Автореф. канд. дисс. Томск.
С а к ай Кан-Ихи. 1964. Конкурентоспособность растений, ее наследуемость
и некоторые связанные с ней проблемы. В сб.: Механизмы биологи-
ческой конкуренции, М.
Сал ганский А. А. 1962. К проблеме одомашнивания. В сб.: Вопросы
экологии, 4, Киев.
С а м о ф а л С. А. 1931. Дарвинизм в лесоводстве. Тр. по леей. хоз. и лесн.
пром., 18: 107—141.
Сапегин А. А. 1922. Наблюдения над «перерождением» искусственного
сорта-смеси. Тр. Одесск. с.-х. сел. станц., 6 : 5—23.
Северцов А. Н. 1939. Морфологические закономерности эволюции. М.—Л.
Северцов С. А. 1936. Морфологический прогресс и борьба за существо-
вание. Изв. АН СССР, 5: 895—944. w
Северцов С. А. 1940а. Хищник и жертва. Тр. Инет, эволюц. морфол.,
2, 1: 5—67.
Северцов С. А. 19406. О конгруэнциях. Бюлл. МОИП, 9 (5—6) 97—103.
377
Северцов С. А. 1941. Динамика населения и приспособительная эво-
люция животных. М,—Л.
Северцов С. А. 1951. О конгруэнциях и понятии целостности вида.
В сб.: Проблемы экологии животных, 1, М.
С е м е н о в - Тян-Ш а н с кий А. С. 1910. Таксономические границы вида
и его подразделений. Зап. Акад, наук, 25, 1 : 1—29.
Серебровский П. В. 1941. Критика теории «кругов форм». Тр. Зоол.
инет. АН СССР, 6, 4: 158—190.
Симпсоп Д. 1948. Темпы и формы эволюции. М.
Син ска я Е. Н. 1931. К познанию видов в их динамике и взаимоотно-
шениях с растительным покровом. Тр. по прикл. бот., ген. и сел.,
25, 2:1-97.
Синская Е. Н. 1933. Основные черты эволюции лесной растительности
Кавказа в связи с историей видов. Бот. журн., 18: 370—406, 487—515.
Синская Е. Н. 1938. Учение об экотипах в свете филогенеза высших
растений. Усп. соврем, биол., 9, 1 : 1—45.
Синская Е. Н. 1939. Проблема популяций у высших растений. Усп.
соврем, биол., 10, 3: 446—470.
Синская Е. Н. 1946. К вопросу о растительной конституции и выра-
жающих ее признаках. Журн. общ. биол., 7, 2 : 147—160.
Синская Е. Н. 1948. Динамика вида. М.—Л.
Синская Е. Н. 1958. Проблема популяций у высших растений. Вестп.
ЛГУ, 9, 2:5-13.
Синская Е. II. 1961. Об уровнях группового приспособления в расти-
тельных популяциях. В сб.: Проблема популяций у высших расте-
ний, 1, Л.
Синская Е. Н. 1963. О категориях и закономерностях изменчивости
в популяциях высших растений. В сб.: Проблема популяций у выс-
ших растений, 2, Л.
Синская Е. Н. 1964а. Об общих закономерностях эколого-географиче-
ской изменчивости дикорастущих и культурных растений. Тр. по
прикл. бот., ген. и сел., 36, 2 : 3—13.
Синская Е. Н. 19646. Рядовая изменчивость. Бюлл. МОИП, отд. биол.,
69, 6:120-129.
Смелов С. П. 1947. Основные этапы жизни побегов лугового злака при
вегетативном возобновлении и динамика связей между их смеж-
ными поколениями. Бот. журн., 32 : 79—88.
Смирнов Л. А., 1943. Современное состояние вопроса о численности
видов высших растений. Сов. бот., 5: 39—49.
Смирнов Ю. 1963. Размещение волка в Казахстане. В кн.: Зоогеогра-
фия суши. Ташкент. (Тр. 3-го Всесоюзн. совещ.).
Смирнова Е. А. 1928. О влиянии фито-социальных условий на ход
борьбы за существование между посевными и сорными растениями.
Изв. Главп. бот. сада, 2: 161—187.
Соколовская А. П. 1960. Географическое распространение полиплоид-
ных видов растений (Исследование флоры о. Сахалин). Вести. ЛГУХ
сер. биол., 21, 14 42—58.
Сорокина О. 1937. Плодовитый и константный 42-хромосомный гибрид
A. ventricosa X Tr. durum. Тр. по прикл. бот., ген. и сел., 2, 7 5—12.
С о ч а в а В. Б. 1926. Этюды по экспериментальной фитосоциологии. Зап.
Ленингр. с.-х. ипст., 3 160—215.
Спирин А. С. 1962. Современные представления о строении нуклеино-
вых кислот и нуклеопротеидов. В сб.: Усп. биол. химии, 4, М.
Степанов Д. Л. 1959а. Политппическая концепция вида в палеонтоло-
гии. Палеонтол. журн., 3 : 3—14.
Степанов Д. Л. 19596-. Неокатастрофизм в палеонтологии наших дней.
Палеонтол. журн., 4 : 11—16.
Стрельников И. Д. 1962. Значение нервной системы в экологии жи-
вотных. В сб.: Вопросы экологпи, 4, Киев.
378
Сукачев В. Н. 1927. К вопросу о борьбе за существование между био-
типами одного п того же вида. В сб.: Памяти акад. Бородина, Л.
Сукачев В. II. 1935. Опыт экспериментального изучения межбиотипной
борьбы за существование у растений. Тр. Петергофск. биол. инет.,
'15 : 69—88.
Сукачев В. Н. 1941. О влиянии интенсивности борьбы за существова-
ние на развитие. ДАН СССР, 30, 8: 752—755.
Сукачёв В. Н. 1953. О внутривидовых и межвидовых взаимоотноше-
ниях среди растений. Сообщ. Инет, леса АН СССР, 1: 5—44.
Сукачев В. Н. 1956. О внутривидовых отношениях в растительном
мире. Бюлл МОИП, отд. биол., 61,2: 5—20.
Сукачев В. II. 1960. Соотношение понятий биогеоценоз, экосистема и
фация. Почвоведение, 6 : 1—10.
Сухов К. С. 1953. Видообразование у вирусов. Изв. АН СССР, сер. биол.,
3 : 3—15.
Талиев В. И. 1915. Опыт исследования процесса видообразования в жи-
вой природе, 1. Харьков.
Тахтаджяп А. Л. 1955. Некоторые вопросы теории вида в систематике.
Бот. журн., 6 : 789—796.
Тахтаджян А. Л. 1966. Система и филогения цветковых растений. Л.
Терентьев П. В. 1957. О применимости понятия «подвид» в изу-
чении внутривидовой изменчивости. Вести. ЛГУ, сер. биол., 21,
4: 73—81.
Терентьев П. В. 1960. Дальнейшее развитие метода корреляционных
плеяд. В сб.: Применение математических методов в биологии, Л.
Терентьев П. В. 1961. Герпетология. М.
Терентьев П. В. 1966. Методические соображения по изучению внутри-
видовой географической изменчивости. В сб.: Внутривидовая измен-
чивость позвоночных животных и микроэволюция, Свердловск.
Т е р н о в с к и й М. Ф. 1960. Итоги и перспективы работ по межвидовой
гибридизации в роде Nicotiana. В сб.: Отдаленная гибридизация
растений, М.
Тимирязев К. А. 1922. Исторический метод в биологии. М.
Тимирязев К. А. 1939. Собрание сочинений, 6. М.—Л.
Тим о ф е е в - Р е с с о в с кий Н. В. 1958. Микроэволюция. Элементарные
явления, материал и факторы микроэволюционного процесса. Бот.
журн., 43 : 317—336.
Тимофеев-Рессовский Н. В. 1962. Некоторые проблемы радиацион-
ной биогеоценологии. Свердловск.
Тимофеев-Рессовский Н. В. 1965. О микро- и макрофилогонии
у половых перекрестнооплодотворяющихся организмов. В сб.: Ра-
диационная цитогенетика и эволюция, Свердловск.
Тимофеев-Рессовский II. В., Т и м о ф о о в а - Р с с с о в с к а я Е. А.,
Циммерман К. 1965. Экспоримептально-систематичоский анализ
географической изменчивости и формообразования у Epilachna chry-
somelina (Coleoplera, Coccinellidao). В сб.: Радиационная цитогене-
тика и эволюция, Свердловск.
Тих Н. А. 1950. Стадная жизнь обезьян и средства их общения в свете
проблемы антропогенеза. Автореф. докт. дисс. Л.
Тихомиров Б. А. 1946. К происхождению ассоциаций кедрового стла-
ника. В кн.: Материалы по истории флоры и растительности СССР,
2. Л.
Тихомиров В. Н. 1965. О некоторых новых взглядах на происхождение
цветковых растений. В сб.: Проблемы филогении растений, М.
Тихомиров В. Н. 1967. Некоторые особенности систематики облигат-
ных апомиктов. Совещание по объему вида и внутривидовой систе-
матики, Л.: 53—55.
Толмачев А. И. 1953. О некоторых вопросах теории видообразования.
Бот. журн., 38: 530—555.
379
Толмачев А. И. 1956. К изучению арктической флоры СССР. Бот.
журн., 41 : 783—796.
Толмачев А. И. 1958. Ареал вида и его развитие. В сб.: Проблема вида
в ботанике, 1, Л.
Толмачев А. И. 1962. Основы учения об ареалах. Л.
Троицкий Н. А. 1960. Исторический обзор и современное состояние
учения о гибридогенном возникновении видов у растений. Тр.
1-й научн. копфер. преп. биол., с.-х., хим. дисциплин Пед. ипст.
УССР, Киев : 9—32.
Уоддингтон К. 1964. Морфогенез и генетика. М.
Уолд Дж. 1962. Филогения и онтогения на молекулярном уровне. Тр.
V Международн. биохим. конгр. Симпозиум 3, М.: 19—58.
Уоллес А. 1911. Дарвинизм. Изд. Сабашниковых. М.
У р а н о в А. А. 1960. Жизненное состояние вида в растительном сооб-
ществе. Бюлл. МОИП, отд. биол., 65, 3: 77—92.
Усов С. А. 1888. Таксономические единицы и группы. Сочинения, 1, М.
Успенская Л. И. 1929. К вопросу о влиянии интенсивности жизнен-
ного состязания между растениями на их развитие. Зап. Ленингр.
с.-х. инет., 5, 4 : 37—54.
Ухтомский А. А. 1962. Собрание сочинений, 6, Л.
Ушаков Б. П. 1959. Физиология клетки и проблема вида в зоологии.
Цитология, 1, 5 : 541—565.
Ушаков Б. П. 1963. Изменение теплоустойчивости клетки в онтогенезе
и проблема консервативности клеток высших холоднокровных жи-
вотных. В сб.: Проблемы цитоэкологии животных, Л.
Ушаков Б. П. 1964. Анализ теплоустойчивости клеток и белков пойки-
лотермных животных в связи с проблемой вида. Автореф. докт.
дисс. Л.
У э в е л л В. 1869. История индуктивных наук, 3. СПб.
Фатеев К. Я. 1966а. Морфо-физиологическая изменчивость американ-
ской норки (Mustela vison Briss.). В сб.: Внутривидовая изменчи-
вость наземных позвоночных животных и микроэволюция, Сверд-
ловск.
Фатеев К. Я. 19666. Особенности внутривидовой изменчивости европей-
ского крота Talpa curopaea. В сб.: Внутривидовая изменчивость
наземных позвоночных животных и микроэволюция, Свердловск.
Федоров А. А. и Пименов М. Г. 1967. Хемосистематика, ее проблемы
и практическое значение. Растительные ресурсы, 3, 1 : 3—16.
Федоров Ал. А. 1958. Тератогенез и его значение для формо- и видо-
образования у растений. В сб.: Проблема вида в ботанике, Л.
Федотов Д. М. 1940. О проблеме вида в палеонтологии. Палеонтол.
обзор, 2 : 2—9.
Федюшин А. В. 1962. О качественных отличиях внутри- и межвидовых
отношений. В сб.: Проблемы внутривидовых отношений, Томск.
Фляк В. 1962. Скопления мериносовых овец как средство защиты
от интенсивной инсоляции. В сб.: Вопросы экологии, 4, Киев.
Формозов А. Н. 1947. Наблюдения над внутривидовой борьбой у позво-
ночных. В кн.: Внутривидовая борьба у животных и растений. М.
Фурсаев А. Д. 1937. К вопросу видообразования в условиях пойм рек.
Сов. бот., 3 : 33—40.
Хайлов К. М. 1963. Проблема системной организованности теоретической
биологии. Журн. общ. биол., 5 : 324—331.
Хитрово В. Н. 1911. Сезонные расы, их морфологические признаки и
значение для видообразования. Протоколы общ. естествоиспыт.,
Киев : 43—45.
X о л о д к о в с к и й Н. А. 1910. О биологических видах. Изв. Акад, наук,
4 : 751—771.
Хохлов С. С. 1946. Бесполосеменные растения. Уч. зап. Саратовск. унив.,
380
Хохлов С. С. 1950. Перспективы эволюции высших растений. Са-
ратов.
Хохлов С. С. 1955. К вопросу о видообразовании у растений. Научи,
ежегодн. Саратовск. унив. за 1954 г. 251—253.
Хохлов С. С. 1959. О количестве видов растений, размножающихся бес-
полосеменным путем. Уч. зап. Саратовск. унив., 64 : 117—123.
Хохлов С. С. 1965. Полиплоидия и апомиксис у покрытосеменных расте-
ний. В сб.: Полиплоидия и селекция, М.—Л.
Хохряков М. К. 1955. О виде у грибов. Бот. журн., 40 : 34—45.
Хохряков М. К. 1964. Проблема вида и видообразования у грибов.
Тр. Всесоюзн. инет, защиты раст., 23: 159—166.
Хржановский В. Г. 1961. К проблеме биологического вида у покрыто-
семенных. Докл. Тимирязевск. с.-х. акад., 72 : 255—261.
Ц инг ер Н. В. 1905. К вопросу о влиянии климата па форму и жизнь
растений. Зап. Киевск. общ. естествоиспыт., 19 : 5—46.
Ц ингер Н. В. 1909. О засоряющих посевы льна видах Camelina и Ярег-
gula и их происхождении. Тр. Бот. музея Акад, наук, 6.
Ц инг ер Н. В. 1913. Подвиды Alectorolophns major Rcliub., живущие
в местах, подвергающихся влиянию с.-х. культуры, и их происхожде-
ние путем естественного отбора. Тифлпс.
Ц и н г е р II. В. 1928. О подвидах большого погремка (Alectorolophns
major). Вологда.
Цингер Н. В., Поддубная-Арнольд и В. А., Петровская Т. П.,
Полунина Н. Н. 1965. К вопросу о причинах апомиксиса.
В сб.: Проблемы филогении растений, М.
Чернов А. А. 1959. О неполноте геологической летописи. Палеонтол.
жури., 4 : 17—29.
Чернов С. А. 1941. Проблема вида. Тр. Зоол. инет. АН СССР, 6, 4:5—15.
Четвериков С. С. 1926. О некоторых моментах эволюционного про-
цесса с точки зрения современной генетики. Журн. экспер. биол.,
2, 1 : 1—54.
Ч и ж е к Ф. 1956. Проблема реальности вида. Автореф. канд. дисс. ЛГУ.
Ч и ж е к Ф. 1958. Проблема целостности вида. Уч. зап. ЛГУ, 248, сер.
философск. наук, 13:65—78.
Ч и ж е к Ф. 1965. Философские вопросы современной биологии. В сб.: Фи-
лософия и естествознание, М.
Чумакова Б. М. и Горюнова 3. С. 1963. Развитие самцов Prospaltella
perniciosi Tow. (Hymenoptera, Aphelinidae) — паразита калифорний-
ской щитовки. Энтомол. обозр., 42, 2 : 320—328.
Чупров А. А. 1910. Очерки по теории статистики. СПб.
Ш а киров Ф. X. 1962. Взаимоотношения среди растений и практиче-
ские вопросы лесоразведения. В сб.: Проблемы внутривидовых отно-
шений, Томск.
Ш а м с и е в Г. Ш. 1963. Влияние различных условий выращивания на
развитие морфобиологических групп мари белой. Вести. ЛГУ, 9,
2 : 55—63.
Шапиро Н. И., Игнатьев М. В. 1945. Эволюция мутабильности. Усп.
соврем, биол., 20, 3 : 325—344.
Шапоренко К. К. 1936. Ближайшие предки Ginkgo biloba L. Тр. Бот.
инет. АН СССР, сер. I, 2 : 5—32.
Шапоренко К. К. 1937. Liriodendron — тюльпанное дерево. Тр. Бот.
инет. АН СССР, сер. I, 4 : 93—170.
Шапошников Г. X. 1966. Возникновение и утрата репродуктивной
изоляции и критерий вида. Энтомол. обозр., 45 : 3—33.
Шапошников Г. X. 1967. Эволюция тлей в связи со специализацией
и сменой хозяев. Автореф. докт. дисс. Л.
Шварц С. С. 1959. Некоторые вопросы проблемы вида у наземных позво-
ночных животных. Тр. Инет. биол. У ФАН СССР, И, Свердловск : 3—
130.
381
Шварц С. С. 1963. Внутривидовая изменчивость млекопитающих и ме-
тоды се изучения. Зоол. жури., 42:417—433.
Шварц С. С. 1966. Экспериментальные методы исследования начальных
стадий микроэволюционного процесса (постановка проблемы). В сб.:’
Внутривидовая изменчивость наземных позвоночных животных и
микроэволюция, Свердловск.
Шенников А. П. 1921. Фитосоциология и опытные питомники. Журн.
Петрогр. агроном, общ., 3—4 : 4—19.
Ш и м к е в и ч В. М. 1907. Уродства и происхождение видов. СПб.
Шишкин Б. К 1954. Выступление на дискуссии о виде. Вестн. ЛГУ,
10, 4:43—45.
Шишкин Б. К. и Юзепчук С. В. 1937. О правилах ботанической
номенклатуры. Сов. бот., 4 : 6—16.
Шляков Р. Н. 1967. О виде у ястребинок. Совещание по объему вида
и внутривидовой систематике, Л.: 60—61.
Шмальгаузен И. И. 1940. Пути и закономерности эволюционного
процесса. М.—Л.
Шмальгаузен И. И. 1946а. Факторы эволюции. М.
Шмальгаузен И. И. 19466. Проблемы дарвинизма. М.
Шмальгаузен И. И. 1958а. Регулирующие механизмы эволюции. Зоол.
журн., 37 1291—1306.
Шмальгаузен И. И. 19586. Контроль и регуляция в эволюции. Бюлл.
МОИП, отд. биол., 63, 5 : 93—121.
Шмальгаузен И. И. 1961. Интеграция биологических систем и их
саморегуляция. Бюлл. МОИП, отд. биол., 66, 2 : 104—135.
Шмидт Г. А. 1941. Эмбриональный диморфизм у немертин Lineus
gesserensis ruber. Тр. Инет, цитол., гистол., эмбриол. АН СССР,
1 : 67—144.
Шмидт Г. А. 1946. Родственные и экологические особенности литораль-
ных немертин рода Lineus. ДАН СССР, 51, 5 : 401—403.
Шмидт Г. А. 1958. О различиях эмбриональных приспособлений геогра-
фических рас немертины Lineus ruber. Folia Biol., 6, 4:265—285.
Шмидт Г. А. 1962. Изменение экологических отношений у взрослых
особей и эволюция эмбриогенеза. Зоол. журн., 41 168—192.
Шмидт Г. А., Янковская Л. А. 1937. Биология размножения Lineus
gesserensis subsp. ruber из Роскова и Кольского залива. Биол. журн.,
6, 2 : 365—384.
Ш о в о н Р. 1965. От пчелы до гориллы. М.
Шумаков Е. М. 1940. Причины стадности у сарапчовых. Докл. ВАСХНИЛ,
21 10-15.
Щербаков В. К. 1965. Роль полиплоидии и анеуплоидии в эволюции
покрытосеменных растений. В сб.: Проблемы филогении растений, М.
Эйтинген Г. 1934. Рубки ухода за лесом в новом освещении. М.
Элтон Ч. 1960. Экология нашествий животных и растений. М.
Энгельс Ф. 1961. Диалектика природы. В кн.: К. Маркс и Ф. Энгельс,
Сочинения, 20.
Эрлих П. и Холм Р. 1966. Процесс эволюции. М.
Эшби У. Р. 1959. Введение в кибернетику. М.
Юзепчук С. В. 1939. Проблема вида в свете учения Дарвина. Сов. бот.,
6-7 12-34.
Юзепчук С. В. 1955. Род Очанка — Euphrasia L. Флора СССР, 22, М.—Л.
Юзепчук С. В. 1958. Комаровская концепция вида. В сб.: Проблема вида
в ботанике, 1, Л.
Яблоков А. В. 1966. Актуальные проблемы эволюционной теории. М.
Янковский А. В. 1962а. Процессы конъюгации Paramecium putrinum
Clap, et Lachm. III. Множественная система типов спаривания
у Р. putrinum. Журн. общ. биол., 23, 4: 276—282.
Янковский А. В., 19626. Процессы конъюгации Paramecium putrinum
Clap et Lachm. Цитология, 4, 4:434—444,
383
Abelson P. 1964. ftapid evolution of the frontiers is a hazard for scien-
tists young and old. Science, 143, 3603 : 218—223.
Akerlund E. 1934. Ein Fall von Naturselection in einer Kreuzungspopu-
lation. Journ. Herod., 18 : 16—33.
Akker be r g E. 1942. Cytogenetic studies in Poa pratensis and its hybrid
with P. alpina. Journ. Hcred., 28 : 1—126.
Allee W. C. 1931. Animal aggregation. Chicago.
Allee W. C., Emerson A., Park 0. and Park T. 1949. Principles
of animal ecology. Philadelphia—London.
Alpatov V. V. 1924. Die Definition der untersten systematischen Katego-
rien vom Standpunkto des Studiums der Variabilitat der Ameisen
und Crustaceen. Zool. Anz., 60, 5—6 : 161—168.
Anderson E. 1936. The species problem in Iris. Ann. Missourri Bot. Gard.,
23, 3:457-509.
Anderson E. 1953. Introgressive hybridization. Biol. Rev., 28, 3:280—307.
Anderson E. and Stebbins G. 1954. Hybridization as an evolutionary
stimulus. Journ. Evolution, 8, 4 : 378—388.
Andersson G. 1897. Die Geschichte der Vegetation, Schwedens. Engler’s
Bot. Jahrb., 22 : 433—548.
Andre warth a H. 1961. Introduction of animal populations. London.
Ascherson P. und G r e b n e r P. 1896. Synopsis der Mitteleuropeischcn
Flora, 1. Leipzig.
Beaudry I. R. 1960. The species concept: its evolution and present status.
Rev. Canad. Biol., 19, 3 : 219—240.
Bertalanffy L. 1949. Das biologische Weltbild, 1. Bern.
Bertalanffy L. 1959. Modern concepts on biological adapt. The historical
development of physiological thought. A Symposium Held at the state
Univ, of New York : 265—288.
В e u r 1 e n K. 1933. Vom Aussterben der Tiere. Natur u. Museum, 63, 1—3 :
1-8.
Birch L. C. 1957. The meanings of competition. Journ. Amer. Nat.» 91,
856 : 5—18.
Black welder R. 1962. Animal taxonomy and the new systematics. In: Sur-
vey of biological progress, 4. N. Y.—London : 1—57.
Blakeslee A., Berger A., Avery A. 1937. Geographical distribution
of chromosomal prime types in Datura stramonium. Journ. CytoL,
2 : 1070—1093.
Boissier E. 1867—1888. Flora Orientalis. Paris.
Bonner J. 1958. The evolution of development. Cambridge Univ. Press.
Botnariuc N. 1963. La Question des pretendues hypertelies. Tr. Museum
d’histoire nat. «Grigore Autipa», 4 : 47—71.
Bouillon J. 1959. Speciation et adaptation. Naturalist. Belg., 40, 8:
249—261.
Bowden W. 1959. The taxonomy and nomenclature of the wheats barleys,
and ryes and their wild relatives. Canad. Journ. Bot., 37, 4 : 657—684.
Braun W. 1953. Bacterial genetics. Philadelphia—London.
Bremekamp С. E. B. 1959. Specific and infraspecific delimitation, 1. The
practical aspect of the problem. Proc. Koninke. nederl. akad. wet.,
62, 2 : 91—99.
Brewster E. T. 1916. Creation. A History of non-evolutionary theories.
Indianopolis.
Brien P. 1962. Reflexions sur la classification, la speciation et 1’evolution
en biologie. Bull. Soc. roy. Sci. Liege, 31, 3—4: 233—246.
Briquet J. 1935. International rules of botanical nomenclature. Jena.
Buchsbaum R. and Milne L. 1961. The lower animals. N. Y.
Burma B. 1954. Reality, existence and classification: a discussion of the
species problem. Madrono, 12, 7 : 193—210.
Cain A. 1959. The post-Linnaean development of taxonomy. Proc. Linn. Soc.
London, 170, 3 : 234—244.
383
Caloianu M. 1962. К. М. Zavadskii. Studies about the species. [Рецензия].
Stud, si cercet. biol. 3 : 480—481.
Carau^u D., TudoseJ., Zi lb er man S., Cerchez N. 1960. Variabi-
litatea citorva caractere ?i Ensu^iri la specia Cornus mas L. An.
stuntifice ale univ. «А1. J. Cusa» din Ja$i, 6, 3 : 513—528.
Carson H. 1961. Relative fitness of genetically open and closed experimen-
tal population of Dr. robusta, Genetics, 46, 5 : 553—567.
C a r u s V. F. 1872. Geschichte der Zoologie. Leipzig.
Chapman R. 1931. Animal ecology. N. Y.
C h о d a t R. 1909. Etude critique et experimentale sur le polymorphisme
des algues. Geneve.
Chodat R. 1914. La notion scientifique de Гехрёсе. Bull. Soc. Bot. Geneve,
2, 6 : 83—226.
Christian J. 1961. Phenomena associated with population density. Proc.
Nat. Acad. Sci. USA, 47, 4 : 428—449.
C i f e r r i R. 1932. The criteria for definicion of species in mycology. Ann.
MycoL, 30, 1—2 : 122—136.
C i z e к F. 1960. Universalnost a neravnocennost druhii. Biologia Casopis
sloven, akad. vied. Bratislava, 15, 9 : 641—652.
Cizek F., Hodanova D. 1966. Prirodni vyber jako hlavni faktor evoluce.
Praha.
Clarke C. and Sheppard P. 1960. The genetics of Papilio dardanus.
Genetics, 45, 4 : 439—457.
Clausen J. 1921, 1922. Studies on the collective species Viola tricolor L.
Bot. Tidskr., 37, 1 : 205—221; 2 : 363—416.
Clausen J. 1951. Stages of the evolution of planta species. N. Y.
Clausen J. 1958. The function and evolution of ecotypes, ecospecies.
In: Systematics of to-day. Uppsala—Wiesbaden.
Clausen J. 1961. Introgression facilitated by apomixis in polyploid poas.
Journ. Euphytica, 10, 1 : 87—94.
Clausen J. and H i e s e у W. 1939. The concept on species based on expe-
riment. Amer. Journ. Bot., 26, 2: 103—106.
Clausen J., Keck D. and H i e s e у W. 1940. Experimental studies in the
nature of species, 1. Washington.
Clausen J., Keck D. and H i e s e у W. 1945. Experimental studies on
the nature of species, 2. Washington.
Clausen J., Keck D. and H i e s e у W. 1947. Heredity of geographically
and ecologically isolated races. Journ. Amer. Nat., 81, 797 : 114—133.
Clements F. 1929. Experimental methods in adaptation and morphogeny.
Journ. Ecology, 17, 2 : 356—379.
Clements F. and Weaver J. 1924. Experimental vegetation. Washington.
Cole F. 1930. Early theories of sexual generation. Oxford.
Connell J., E. О r i a s. 1965. The ecological regulation of species diver-
sity. Journ. Amer. Nat., 98, 903 : 399—414.
Cooper W. 1926;. The fundamentals of vegetational change. Journ. Eco-
logy, 7, 4: 391—413.
Cuenote L. 1936. L’espece. Paris.
Cuenote L. et A. Tetry. 1951. L’evolution biologique. Les faits. Les in-
certitudes. Paris.
Cuvier G. 1800. Lemons d’anatomic comparee, 1. Paris.
Cuvier G. 1829. La regne animate, 1. Paris.
D a n s e r B. 1929. Uber die Begriffe Komparium, Kommiskuum und Kon-
vivien. Genetica, 11 : 399—450.
Darlington C. 1940. Taxonomic species and genetics systems. In: The
New-Systematics. Oxford : 137—170.
Davidson J. F. 1954. A dephlogisticated species problem. Madrono, 12,
8: 246-254.
De-Candolle A. 1855. Geographic botanique raisonnee. Paris.
De-Candolle Alph. 1880. La Phytographie. Paris.
384
De-Candolle Aug. P. 1819. Tbeorio olemoulairo do la bolaniquo. Paris.
De-Vries H. 1901, 1903. Dio Mutationsllicorie, I 2. Leipzig.
De-Vries H. 1906. Arion und Varivtalen und ill re Ihislohuiig (lurch
Mutation. Berlin.
Diels L. 1924. Die Mellioden dor Phylographio und dor Sysleuialik dor
Pflanzen. In: Handbucli dor biologiscbon ArboilMinollmdeii, Abt. 11, I,
Berlin.
Dobrowolski K. 1963. Proba analizy pojecia «rzadki galunok», Ekol.
polska, 9, 3 : 207—212.
Dobzhansky Th. 1955. Goiielic. and the origin of specie. 3 od. N. Y.
Dobzhansky Th. 1958. Species after Darwin. In: Л century of Darwin.
London—Toronto.
Dobzhansky Th. 1959. Variation and evolution. Amor. Philos. Soo. 103,
2 : 252-263.
Dobzhansky Th., Pavlovsky (). 1960. How stable is balansed poly
morphism? Proc. Mat. Acad. Sci., 46, 1 :41—47.
Dodson E. O. 1957. Textbook of evolution. Philadelphia- London.
Dowdeswell W. II. 1960. The mechanism of evolution. N. Y.
Du Rietz E. 1930. The fundamental units of biological taxonomy. Sv. But.
Tidskr., 24, 3 : 333-428.
Eimer E. 1887. Die Entstehung der Arlen auf Grund von vererben orwor
bener Eigenschaften. Jena.
Eimer G. H. T. 1888—1901. Die Entstehung der Arten auf Grund von er-
worbener Eigenschaften nach den Gesetzen organischen Wachsens,
1—3. Jena.
Elias M. K. 1946. Taxonomy of tertiary flowers and herbaceous Seed.
Amer. Midland. Nat., 36 : 373—380.
Elton C. 1933. The ecology of animals. Oxford.
Emerson A. 1961. Vestigial characters of termites and processes of re-
gressive evolution. Evolution, 15, 2 : 115—131.
Engler A. 1879. Versuch einer Entwicklungsgeschichte der Pflanzenwelt
selt der Tertiarperiode, 1. Berlin.
Engler A. 1954, 1964. Syllabus der Pflanzenfamilien, 1, 2. Berlin.
Ford E. 1940. Polymorphism and Taxonomy. In: The New-Systematics.
Oxford.
Ford E. 1963. Ecological Genetics. London.
Fryxell P. A. 1962. The «Relict species» concept. Acta Biotheoret., 15,
1—3 : 105—118.
Gause G. 1934. The struggle for the existence. Baltimore.
General system. 1956—1964. Yearbook of the society for general systems
research. Michigan.
Gilmour J. 1940. Taxonomy and philosophy. In: The New-Systematics.
Oxford.
Gilmour J. 1960. The deme terminology. Rep. Scottish Pb. Breed. Stat.
99-105.
Gilmour J. and Gregor J. 1939. Demes: a suggested new terminology.
Nature, 144, 3642 : 333.
Glaessner M. 1960. Precambrian fossilis from South Australia. Intern.
Geol. Gongr., 21, Session, Melburn.
G о d г о n D. 1859. De 1’espece et des races chez les etres organises. 1. Paris.
Goldschmidt R. 1932. Untersuchungen zur Genetik der geographischen
Variation. Arch. Enlwickl.-Mech. Org., 126 : 227—324.
Goldschmidt R. 1940. The material basis. of evolution. New Haven—
London—Oxford.
Good D. 1931. A theory of plant geography. The New Phytologist, 30, 3 :
99—108.
Good D. 1953. The geography of the flowering plants. London.
Gotoh K. 1957. Genetic analisis of varietal differentiation in Cereals. VII
Japan. Journ. Bot., 16, 1 :46—60.
385
G r a n t V. 1963. The origin adaptation. N. Y.
Grant W. F. 1960. The categories of classical and experimental taxonomy
and the species concept. Rev. Canad. Biol., 19, 3 : 241—262.
Grebecki A., Kinastowski W., Kuznicki L. 1961, 1962. Ewolu-
cjonizm, 1—2. Warszawa.
Green E. 1909. Linnaeus as an evolutionist. Washington Acad, sci., 31—
27—39.
Gregor J. 1930. Experiments on the genetics of wild populations. Journ.
Genetics, 22, 1: 15—25.
Gregor J. 1939. Experimental taxonomy. The New Phytologist, 38:
293________322.
Gregor J. 1944. The ecotype. Biol. Rev., 19, 1 : 20—30.
Gregor J. 1946. Ecotypic differentiation. The New Phytologist, 45, 2:
254—270.
Gregor J. and Sansome F. 1927. Experiments on the genetics of wild
populations. Journ. Genetics, 17, 3:349—364.
Gregor J. and Sansome F. 1930. Experiments on the genetics of wild
populations. Journ. Genetics, 22, 3 : 373—387.
Gregor J. and Watson P. 1961. Ecotypic differentiation: observations
and reflections. Evolution, 15, 2 : 166—173.
Grene M. 1958. Two evolutionary theories. Brit. Journ. Philos., 9, 34:
110—127.
G u 1 i с к J. T. 1905. Evolution, racial and habitudial. Washington.
Gustafsson A. 1946, 1947. Apomixis in higher plants. Lund, gleorup.
Leipzig.
Hagerup O. 1933. Studies on polyploid ecotypes in Vaccinum uligino-
sum L. Journ. Hered., 18: 122—128.
Haldane J. 1932. The causes of evolution. London.
Haldane J. B. S. 1954. The statics of evolutions. In: Evolution as a pro-
cess. London.
Haldane J. B. S. 1959. The theory of natural selection to day. Nature, 183,
4663: 710—713.
Hardisty M. 1963. Fecundity and speciation in lampreys. Evolution, 17,
1:17-22.
Harlan J., Wet J. 1963. The compilospecies concept. Evolution, 17, 4:
497—501.
Harl and S. 1936. The genetical conception of species. Biol. Rev., 9,
1: 83—112.
Harper J. L. 1964. The individual in the population. Brit. Ecol. Soc. Jubilee
sympos. London, 1963. Oxford.
Harper R. 1923. The species concept from the point of view of a mor-
phologist. Amer. Journ. Bot., 10, 5 : 229—233.
Harvey L. 1960. Speciation. Sci. Progress, 48, 191: 535—539.
Harvey W., Hall M. and Clements F. 1923. The phylogenetic method
in taxonomy. Washington.
Heim R. 1958. Les criteres chimiques dans les Macromycetes. In: Syste-
matics of to-day. Uppsala—Wiesbaden.
Hesse R. 1929. Bericht uber das Tierreich. S. B. Preuss. Akad. Wissensch.:
40-44.
H i e s e у W., Clausen J. and Keck D. 1942. Relations between cli-
mate and intraspecific variation in plants. Amer. Nat., 76 (762)
5—22.
Hinde R. A. 1959. Behaviour and speciation in birds and lower vertebra-
tes. Biol. Rews Cambr. Philos. Soc., 34, 1 : 85—128.
Hitchcock A. S. 1931. Discussion the species concept. Fifth intern. Bot.
Congres. Cambridge. Report of proceedings.
Hoffmeister D. and De la Torre. 1961. Geographic variation in the
mouse peromyscus difficilis. Journ. Mammal., 42, 1 : 1—13.
Hooker I. 1847. The botany of the antarctic voyage, 2. London.
386
Huskins C. L. 1931. The origin of Sparlina Townsendii, Nailin', 127, 3212
781.
Huxley J. 1938a. Species formation and geographical isolalion. I’ror Linn.
Soc. London, 150, 4 : 253—264.
Huxley J. 1938b. Clines an auxiliary taxonomic principle. Nullin', 142,
3587; 219—220.
Huxley J. 1940. Introductory: Towards the new-systornalic. In: The Now
Sistematics. Oxford.
Huxley J. 1956. Morphism as a clue in the study of population dynamic*
Proc. Roy. Soc. London, 145, 929:319—322.
Huxley J. 1958. Evolutionary process and taxonomy with special role
rence to grades. In: Systematics of to-day. Uppsala—Wiesbaden.
Huxley J. 1963. Evolution. The modern Synthesis. Sec. ed. London.
J al as J., Rikkinen K. 1962. Populationsstudien an Melampyrum pr<
tense L. in Finnland. Ann. bot. soc. «Vanamo», 32, 4 : 1—24.
Jordan A. 1873. Remarques sur le fait de 1’existence en societo a I’elat
sauvage des especes vegetales affinees et sur d’autres faits rolatifs
a la question de 1’espece. Lyon.
Kaestner A. 1965. Lehrbuch der Speziellen. Zoologie, I. Jena.
Kerster H. W. 1964. Neighborhood size in the’rusty lizard, Scoloporus
olivaceus. Journ. Evolution, 18, 3 : 445—457.
Kinsey A. 1936. The origin of higher categories in Cynips, 4. Bloomington,
Indiana.
Kleinschmidt P. 1930. The formenkreis theory and the progress of orga
nic world. London.
Kloss K. 1964. Beitrag zum Artbergiff in der Biologie. Wiss. Z. E. M. Arndt.
Univ. Greifswald. Math.-naturwiss. Reih. 13, 2—3 : 283—291.
Koopman K. 1950. Natural selection for reproductive isolation between
Drosophila pseudoobscura and Drosophila persimilis. Journ. Evolution,
2 : 135—149.
Koilicker A. 1872. Morphologic und Entwicklungsgeschichte des Ponnatu
lidestammes, nebst allemeinen Betrachungen zur Descendenzlohro.
Frankfurt.
Kornicke F. 1885. Handbuch des Getreidebaues. Arten und Vcriotaton
des Getreides. Berlin.
Lack D. 1965. Evolutionary ecology. Journ. Animal. Ecol. 34, 2:223- 231.
Lam H. 1936. Phylogenetic symbols, past and present. Acta Biotlieoret.
2, 3 : 153—194.
Lamarck J. 1802. Recherclies sur 1’organisation des corps vivans. Paris.
Lamarck J. 1873. Phylosophie zoologique, 1. Paris.
Lawrence G. H. 1951. Taxonomy of vascular plants. N. Y.
Lerner G. M. 1954. Genetic homeostasis. N. Y.
Lewis H. 1961. Experimental sympatric populations of Clarkia. Amor. Nat.,
96, 882 : 155—168.
Lewis H. 1962. Catastrophic selection as a factor in speciation. Evolution,
16,3:257—271.
Li С. C. 1955. The stability of an equilibrium and Ibe average filin'. of
a population. Amer. Nat., 89 : 281—296.
Linnaei C. 1751. Phylosophia botanica. Slockliolmiae.
Linnaeus C. 1762. Species plantarum. Ed. 2. ITolmiae.
Littlejohn M. 1965. Premating isolation in the Hyla owingi complex
(Anura). Evolution, 19, 2 : 234—245.
Lotsy J. P. 1912. Zeitschrift fii.r inductive Abstamung- und Vererbiingslobro.
The Hagne.
Lotsy J. 1916a. Qu’est-ce qu’une espece? Arch. Neorland., ser. 3B, 3:
57—110.
Lotsy J. 1916b. Evolution by means of hybridization. The Hague.
Lotsy J. 1917. La quintessense de la tbeorie du croisement. Arch. Neorland.,
ser, 3B? 3, 2 : 178—238,
387
Love A. 1965. Adress of outgoing president JOPB. Taxon 14 (2) : 43—46.
Liier s H. und Ulrich H. 1954. Genetik und Evolutions — forschung bei
Tieren. In: Die Evolution der Organismen, 3. Stuttgart.
Mather K. 1947. Species crosses in Antirrhinum, 1. Journ. Herod., 1, 2:
175—186.
Mather K. 1955. Polymorphism as an outcome of disruptive selection.
Journ. Evolution, 9, 1 : 52—61.
Matsuura H. 1935. On karyo-ecotypes of Fritilaria camschatcensis. Journ.
Fas. Sci. Hekkaido Univ., 5 : 219—232.
M а у r E. 1949. Speciation and selection. Proc. Amer. Philosoph. Soc., 9316 :
514—519.
Mayr E. 1958. Evolutionary significance of the systematic categories.
In: Systematics of to-day. Uppsala—Wiesbaden.
Mayr E. 1963. Animal species and evolution. Harvard Univ. Press. Cam-
bridge.
Mayr E. 1965. Les races dans 1’evolution animale. Polymorphisme, polyty-
pism et monotypism. Rev. intern, des sci. sociales, 17, 1 : 126—129.
McCarley H. 1964. Ethological isolation in the coenospecies Peromyscus
leucopus. Journ. Evolution, 18, 2 : 331—332.
Meyer E. 1854. Geschichte dor Botanik, 1—4. Berlin.
Miller I. L. 1965. Living systems: basic concepts. Behavioral sciense, Ann.
Arbor., 10, 3 : 193—237.
Milne A. 1962. On a theory of natural control of insect population. Journ.
Theor. Biol., 3, 1 : 19—50.
Mobius K. 1898. Uber den Umfang und die Einrichtung des Zoologischen
Museums zu Berlin. SB. PrenB. Akad. Wiss.: 363—374.
Muller A. 1963. Lehrbuch der Palaozoologie, 1. Jena.
Miintzing A. 1930. Uber Chromosomenvermehrung in Galeopsis-Kreuzun-
gen. Journ. Hered., 14 : 153—172.
Miintzing A. 1932. Apomictic and sexual seed formation in Poa. Journ.
Hered, 17 : 131-154.
Miintzing A. 1939. Chromosomaberrationen bei Pflanzen und ihre gene-
tische Wirkung. Zeitschr. ind Abstam. u. Vererbung, 76, 1—2:
324—350.
Miintzing A. 1960. Chromosomes in relation to species differentiation
and plant breeding. Confer, on Chromosomes. Netherlands: 1—37.
Myers G. S. 1960a. The endemic fish fauna of lake Lanao, and the evolu-
tion of higher taxonomic categories. Evolution, 14, 3: 323—333.
Myers G. S. 1960b. Fish evolution in lake Nyassa. Evolution, 14, 3:
384-390.
Naughton I, Harper I. 1960. The nature of barriers isolating simpatric
populations of Papaver dubium and P. lecogii. The New Phytologist.
59, 2>: 129—137.
Newell N. D. 1956. Catastrophism and the fossil record. Evolution, 10,
1 : 97—101.
Nicholson A. 1950. Population oscillations caused by competition for
food. Nature, 165 : 476—477.
Niculescu V. 1960. Les formes d’existence de I’espece et la speciation
chez les Lepidopteres. Bull. Soc. Ent. Mulhouse april,: 1—6.
Ochoa S. 1964. The genetic code. In: New Perspectives in Biology. Amster-
dam—London—N. Y.
Odum A. 1959. Fundamentals of ecology. Philadelphia.
Olson E. 1963. Russian viewpoint on evolution. Journ. Evolution, 17, 1 :
119—121.
Osborn G. 1913. From the greeks to Darwin. N. Y.—London.
Ownbey M. and W. Weber. 1943. Natural hybridization in the genus
Balsomophora. Amer. Journ. Bot, 3, 30: 179—187.
Pallas P. 1780. Memoire sur la variation des animaux. Acta Acad. Sc.
Petrop, 2 :69—102.
388
Peterfi S., Stefureac T. 1965. Conceptia despre specie la alge §i brio-
fite. Studii si cercetari biol. ser. bot., 17, 1 : 101—117.
Petruse wicz K., Trojan P. 1964. Zrodla wspolczesnej Koncepcji gu-
tuncu. Kosmos (Polska), ser. A, 13, 4 : 283—295.
P i men tai R. 1959. Mendelian infraspecific divergence levels and their
analisis. System Zook, 8, 3 139—159.
Pimental R. 1961. Animals population by the genetics feed-back mecha-
nism. Journ. Amer. Nat., 95, 881 : 65—79.
Plate L. 1908. Selektionsprinzip und Problem der Artbildung. Leipzig-
Berlin.
Popov M. 1925—1926. Generis Zygophylii species asiaticae. Бюлл. Средне-
азиатск. унив., И 105—125.
Pratt R., Oneto I. and Pratt I. 1945. Studies on Chlorella vulgaris.
Amer. Journ. Bot., 32, 7 : 405—408.
Prosperi R. 1955. Gran Comora. Milano.
Rabaud E. 1953. Le hasard et le vie les especcs. Paris.
Radi E. 1905, 1909. Geschichte der biologischen Theorien in der Neuzeit,
1—2. Leipzig.
Raunkiaer C. 1918. Uber die Begriff der Elementart im Lichte der mo-
dernen Erblickheitsforschung. Zeitschr. ind. Abstam. u. Vererbung.,
19 :225-240.
R e m a n e A. 1964. Zum Problem der Radiationen. Zool. Anz., 173, 1
92—95.
Rensch B. 1929. Das Prinzip geographische Rassenkreise und das Problem
der Artbildung. Berlin.
Rensch B. 1938. Some problems of geographical variation and species for-
mation. Proc. Linn. Soc. London, 150, 4 : 275—285.
Rensch B. 1954. Die phylogenetische AJjwandlung der Ontogenese. In: Die
evolution der Organismen, 1. Stuttgart.
Rensch B. 1958. Die ideale Artbeschreibung. In: Systematics of to-day.
Uppsala—Wiesbaden.
Rensch B. 1959. Evolution above the species level. Columbia Univ.
Press. N. Y.
Rensch B. 1965. Les races geographiques en zoologie. Rev. intern, sci.
soc., 17, 1 : 136—139.
Rimpau W. 1891. Krewzingsprodukte ladwirschaftlichen Kulturpflanzen.
Berlin.
Rivas L. R. 1964. A reinterpretation of the concepts «sympatric» and «allo-
patric» with proposal of the additional terms «syntopic» and «alloto-
pic». System. Zool., 13, 1 : 42—43.
Romanes D. 1895. Darwin und nach Darwin, 1, 2. Leipzig.
Russel E. S. 1916. Form and function. London.
Sakai K. 1951. Экспериментальное изучение значения отбора в селекции
растений. (Сообщ. 1). Зап. по растениеводству в Японии, 20,. Токио
(на японск. яз.).
Sakai К. 1953. Studies on competition in plants. Japan. Bot., 14, 1 :
161—168.
Sakai K. 1956. Значение конкуренции в растительных популяциях.
В сб.: Международный симпозиум по генетике популяций, Токио
(на японск. яз.).
Sakai К. 1965. Contributions to the problem of species colonization from
the viewpoint of competition and migration. The genetics of colonizing
species. Acad, press inc.: 215—241.
Sanson A. 1900. L’espece et la race en biologie generale. Paris.
Schindewolf O. 1950. Der Zeitfactor in Geologie und Palaontologie.
Stuttgart.
Schindewolf О. H. 1954. Uber die moglischen Ursachen der grossen erd-
geschichtlichen Faunenschnitte. Neues Jahrb. Geol. u. Palaontol.
Monatsh. Dtsch., 10 : 457—465.
25 К. М. Завадский
389
Sc hull H. 1923. The species concept from the point of view of a geneticist;
Amer. Journ. Bot., 10, 5 : 221—228.
Schwanitz F. 1954. Cenetik und Evolutionsforschung bei Pflanzen.
In: Die Evolution der Organismen, 3; Stuttgart.
Schwanitz F. 1963. Genetik, Evolution und Systematik. Planta med., 11,
3 : 252—267.
S h e p p a r d P. M. 1959. Natural selection and heredity. N. Y.
Simpson G. G. 1951a. The species concept. Evolution, 5, 4: 285—298.
Simpson G. G. 1951b. The meaning of evolution. New Amer. Library, N.Y.
Simpson G. G. 1952. How many species? Evolution, 6, 3 : 342.
Simpson G. G. 1953. The major features of evolution. N. Y.
Simpson G. G. 1961. Principles of animal taxonomy. N. Y.
Simpson G. G. 1964. Organisms and molecules in evolution. Science, 146,
3651 : 1535—1538.
Simpson G. G. 1965. Les races dans revolution animale. Rev. intern. Sci.
soc., 17, 1 : 148—150.
Slastenko E. P. 1963. Species and speciation. Biol. Jahrb., 31:158—
168.
Smith H. M. 1956. Schistic evolution. Journ. Evolution, 10, 2:228—
229.
Smith H. M. 1958. The syntetic natural populational species in biology.
System. Zool., 7, 3: 116—119.
Smith J. 1962. Disruptive selection, polymorphism and sympatric specia-
tion. Nature, 195, 4836 : 60—62.
Smith J. 1964. Group selection and kin selection. Nature, 201:1145—1147.
Snyder R. 1961. Evolution and integration of mechanisms that regulate
population growth. Proc. Nat. Acad. Sci. USA, 47, 4:449—455.
Soo R. 1926—1927. Systematische Monographic der Gattung Melampyrum.
1—3. Fedde’s Repert, 23—24.
Spector W. S. (Ed.) 1956. Handbook of biological data. Philadelphia-
London.
Sprengel K. 1817. Geschichte der Botanik, 1. Leipzig.
Stebbins G. 1941. Apomixis in the Angiosperms. Bot. Rev., 7, 10:
507—542.
Stebbins G. 1950. Variation and evolution in plants. N. Y.
Stebbins G. 1957. Self fertilization and population variability, Amer. Nat.,
91, 861: 337—354.
Stebbins G. and Daly K. 1961. Changes in the variation pattern of
a hibrid population of Helianthus over eight-year period. Journ. Evo-
lution, 15, 1: 60—71.
Stegmann В. K. 1934. Uber die Formen der grossen Moven und ihre ge-
genseitigen Beziehungen. Journ. OrnithoL, 82, 3 : 340—380.
Steyskal G. 1965. Trend curves of the rate of soecies description in
zoology. Science, 149, 3686 : 880—882.
Stresemann E. 1931. Die Zosteropiden der indo-australischen Subregion.
Berlin.
Stresemann E. und Timofeef-Ressovsky N. W. 1947. Artenste-
hung in geographischen Formenkreisen. 1. Der Formenkreis Larus
argentatus—cachinans—fuscus. Biol. Zentrbl., 66 : 5—76.
Thellung A. 1918. Neuere Wege und Zeile der botanischen Sistematik.
Naturwis. Wochenschr.: 32—33.
Thoday L, Gibson. 1962. Isolation by disruptive selection. Nature, 193,
4821 : 60—68.
Thorson G. 1950. Reproductive and larval ecology of marine bottam in-
vertebrates. Biol. Rev., 25 : 1—45.
Timopheef-Ressovsky N. W. 1940. Mutation and geographycal va-
riations. In: The New-Systematics. Oxford.
Tjtfnneland A. 1964. Artsdannelse pa tfkologisk grundlang. Natur. vorben.,
Febr.: 49—61.
390
Trojan P. 1962. Analysis of the species concept in the genus Tabanus L.
(Diptera) as shown by taxonomic practice. Ecologia polska, sor. A. T.
10, 6 : 123—229.
Try x ell P. A. 1962. The relict species concept. Acta biotheorot., 15, 1—3
105—118.
Tsuyoshi M. 1964. On the application of the serological method towards
the studies on the phylogenetic differentiation in cultivated plants.
Journ. Fac. Agric. Kyushu Univ., 13, 1.
Turesson G. 1922a. The genotypical response of the plant species to the
habitat. Journ. Hered., 3 : 211—350.
Turesson G. 1922b. The species and the varieties as ecological units.
Journ. Hered., 3 : 100—113.
Turesson G. 1925. The plant species in relation to habitat and climate.
Journ. Hered., 6 : 147—236.
Turesson G. 1929. Zur Natur und Bergenzung der Arteinheiten. Journ.
Hered., 12 : 323—324.
Turesson G. 1931. The selective effect of climate upon the plant species.
Journ. Hered., 14 : 1—152.
Turesson G. 1956. Variation in the apomictic microspecies of Alchemilla
vulgaris L. II. Progeny tests in agamotypes with regard to morpholo-
gical characters. Bot. Notiser, 109, 4: 400—404.
Turesson G. 1957. Variation in the apomictic microspecies of Alchemilla
vulgaris L. III. Geographical distribution and chromosome number.
Bot. Notiser, 110, 4:413—422.
Turesson G. 1961. Haditat modifications in some widespread plants spe-
cies. Bot. Notiser, 114, 4:435—452.
Turesson G. and Turesson B. 1960. Experimental studies in Hieracium
pilosella L. Journ. Hered., 46 : 717—736.
Tur rill W. 1935. Some aspects of experimental taxonomy. Proc. 6 Intern.
Bot. Congr. Amsterdam, 3 : 57—58.
T u г г i 11 W. 1938. The expansion of taxonomy with special reference to
Spermatophyta. Biol. Rev., 13, 4 : 342—373.
Turrill W. 1951. Some problems of plant range and distribution. Journ.
Ecology, 39, 2 : 205—228.
Turrill W. 1958. Genetic variations within taxonomic species. In: Syste-
matics of to-day. Uppsala—Wiesbaden.
Valentine D. 1962. La taxonomie experimentale. Tendences actuelles.
Rev. cytol. et biol. veget., 25, 3—4 : 255—266.
Van-Steenis C. 1957. Specific and infraspecific delimination, Flora Ma-
lesiana, ser. I, 53: 167—229.
Verner J. 1964. Evolution of polygamy in long-billed Marsh Wren. Evolu-
tion, 18, 2 : 252—261.
Vuillemin P. 1926. Les anomalies vegetales. Paris.
Waddington C. 1956. Genetic assimillation of the «Bithorax phenotype».
Evolution, 10 : 1—13.
Waddington C. 1957. The strategy of the genes. N. Y.
Wagner M. 1868. Die Darwinische Theorie und das Migrationsgesetz der
Organismen. Jena.
Watkins A. 1935. Heredity and evolution. London.
Wettstein R. 1895. Der Saison-Dimorphismus als Ausgengspunkt fur die
Bildung neuer Arten in Pflanzenreiche. Ber. deutsch. Bot. Ges., 13.
Wettstein R. 1896. Monographie der Gattung Euphrasia. Leipzig.
Wettstein R. 1897. Die europaischen Arten Лег Gattung Gentiana.
Wien.
Wettstein R. 1898. Grundziige der geographische-morphologische Methode
der Pflanzen-systematik. Jena.
Wettstein R. 1901. Untersuchungen fiber den Saison-Dimorphismus im
Pflanzenreiche. Wien.
White M. 1959. Speciation in animals. Austral. Journ. Sci., 22, 1 32—39.
25* 391
White С. M. N. 1959. Nomadism, breeding and subspeciation in some Afri-
can larus. Bull. Brit. Ornithol. Club., 79, 4: 53—57.
Wright S. 1934. Physiological and evolutionary theories of dominance.
Amer. Nat., 68 : 24—53.
Wright S. 1953. Gene and organism. Amer. Nat., 87, 832 : 5—18.
Wright S. 1964. Evolution. Encyclop. Brit., 8: 915—929.
Wiirtenberger L. 1880. Studien uber die Stammesgeschichte der Am-
moniten. Ein geologischer Beweis fiir die Darwinische Theorie. Leipzig.
Wynne-Edwards V. 1962. Animal dispersion in relation to social
Behaviour. Edinburgh—London.
Wynne-Edwards V. 1965. Self-regulating systems in populations of ani-
mals. Science, 147, 3665 : 1543—1548.
Zarzycki K. 1965. Obecny stan badan nad konkurencja (wspolzawod-
nictwem) roslin wyzszycb. Konkurencja miedzygatun Kowa. Ekol.
polska, 11, 3 : 195—210.
Z e i 11 e r R. 1900. Elements de paleobotanique. Paris.
Zimmerman W. 1959. Die Phylogenie der Pflanzen. 2 Aufl. Stuttgart.
ИМЕННОЙ УКАЗАТЕЛЬ
Абдылдаев Т. 173.
Авдулов Н. П. 111.
Агаев М. Г. 255.
Алпатов В. В. 77, 109, 110, 208, 229,
269, 280.
Альдрованди У. 8.
Ананьев А. Р. 129.
Андрусов Н. И. 91, 92.
Аникин В. И. 226.
Анфинсен К. 151, 189.
Аристотель 8, 24, 25.
Арнольди К. В. 104, 175, 191, 229,
233.
Арнольди Л. В. 191.
Астауров Б. Л. 337.
Аудас В. 156.
Аукоф Д. 203.
Афанасьев В. Г. 171.
Ашерсон П. 64.
Базилевская Н. А. 116.
Базыкин А. Д. 332.
Барабаш-Никифоров И. И. 10, 14.
Баранов П. А. 24, 139.
Баркрофт Дж. 261.
Барулина Е. И. 114.
Баур Е. 111.
Бейрих К. 63.
Беклемишев В. Н. 13, 14, 181, 184,
186, 191, 194, 206, 233, 235, 239,
240, 319, 337.
Беллинг Дж. 89.
Белозерский А. Н. 149, 153.
Бензер С. 153.
Берг Л. С. 76, 77, 89, 124, 125, 143,
147, 149, 175, 215, 251, 289, 351.
Берг Р. Л. 108.
Бергнер Д. 110.
Берман 3. И. 139.
Бернал Дж. 158, 182, 185.
Берталанффи Л. 183, 184, 203.
Бёрма Б. 135, 136.
Бианки В. Л. 66, 77, 84.
Биброн Ж. 10.
Благовещенский А. В. 152, 358.
Блексли А. 110, 111.
Бовер Ф. 9.
Богданов М. Н. 66.
Бок И. 8.
Болдинг К. 203.
Болдуин Э. 182.
Большаков В. Н. 231.
Бонна Ш. 32, 42, 43.
Борисяк А. А. 128, 149, 201.
Бочанцев В. П. 146, 166.
Браунштейн А. Е. 158.
Браше Ж. 204.
Бринкман Р. 91.
Брокки Дж. 195.
Броун Р. 9.
Буассье Э. 65.
Бузер Дж. 132.
Букасов С. М. 114.
Бэсси Ч. 9.
Бэтсон В. 89, 95, 115.
Бюффон Ж. Л. 34, 42, 83.
Вааген В. 63, 69, 89.
Вавилов Н. И. 79, 88, 89, 95, 96,
111—124, 148, 213, 258, 260, 318.
Вагнер М. 64, 347.
Вагнер Н. П. 40.
Вайнз М. 9.
Ван-Ниль К. 131, 149, 307.
Ванюшин Б. Ф. 153.
Васильев В. Н. 197, 316.
Васильков Б. П. 145, 150, 151, 153.
Васильченко И. Т. 87, 344.
Васнецов В. В. 206, 289.
Вейсс П. 189,
Вельдре С. Р. 230.
Верещагин Н. К. 169.
Вернадский В. И. 158, 190, 192, 197.
Веттштейн Р. 73, 85, 86, 147.
Викторов Г. А. 168, 208, 285.
Виллемен П. 83.
393
Вильденов К. Л. 9.
Вилькомм М. 46.
Вильморен Г. 182.
Виттрок В. 117.
Волкова Э. В. 171.
Вольтерра В. 109.
Воронихин Н. Н. 131, 146, 160, 174.
Воронов А. Г. 9, 12—14.
Вульф Е. В. 114, 155.
Вюртембергер Л. 63, 65.
Гайсинович А. Е. 90.
Гарстанг В. 344.
Гаузе Г. Ф. 109, 299, 326.
Геккель Э. 62, 182.
Гексли Дж. 21, 82, 162, 163, 168,
212, 217, 308, 320, 340.
Гексли Т. 18, 61.
Гейнрих А. К. 293.
Гентнер В. Г. 12, 62, 227.
Гербильский Н. Л. 250, 251, 322.
Герман А. Л. 236.
Гертвиг О. 83.
Гершензон С. Н. 108, 244.
Гессе Р. 21.
Гесснер К. 8.
Гильгендорф Ф. 63.
Гильмур Д. 134.
Гиляров М. С. 233.
Говоров Л. И. 114.
Годрон Д. 41, 42.
Голлербах М. М. 148.
Головин П. Н. 172.
Гольдшмидт Р. 84, 90.
Горский И. И. 18, 158.
Горюнова 3. С. 274.
Гото К. 261.
Готшик Ф. 63.
Грант В. 219.
Гребнер П. 64.
Грегор Дж. 104, 217, 229.
Грубов В. И. 146, 166.
Грюммер Г. 239, 269.
Гуд Д. 164.
Гудошников С. В. 26/.
Гукер Д. 64.
Гулик Дж. 132.
Гумбольдт А. 9, 64.
Давидсон Д. 136.
Давиташвили Л. Ш. 198, 357.
Далынтедт Г. 132.
Данпеман Ф. 8.
Дарвин Ч. 16, 26, 27, 34, 35, 37—39,
41, 50, 52—62, 65—71, 76, 81, 84,
88, 89, 98, 118, 129, 141, 177, 178,
198, 200, 289, 290, 291, 293, 306,
326, 347.
Даревский И. С. 160, 230.
394
Дарлингтон К. 89, 163, 189, 313.
Де-Бир Ж. 344.
Де-Вита С. 290.
Декандоль Альф. 9, 46, 64, 65.
Декандоль О.-П. 28, 64.
Делоне Л. Ы. 111, 215, 258.
Дементьев Г. П. 22, 320.
Де-Фриз Г. 80, 81, 87—90, 94, 116,
122, 215, 306.
Джанузаков А. 172.
Дильс Л. 9.
Добжанский Ф. 77, 108, 123, 124,
148, 164, 178, 201, 328.
Догель В. А. 14, 161, 213, 224.
Длусский Г. М. 270.
Дональд С. 277.
Дубинин В. Б. 21, 345.
Дубинин Н. П. 18, 19, 81, 107, 108,
152, 157, 160, 221, 232, 249, 340.
Дюбуа Е. 63.
Дюмеримо 10.
Дюран Ф. 9.
Дю-Рие Е. 162, 163.
Дюшен 70.
Еленкин А. А. 131, 160, 169.
Жадин В. И. 216.
Жебрак А. Р. 338.
Жебрак Э. А. 338.
Жилкина М. Д. 338.
Житков Б. М. 24.
Жордан А. 47, 48, 101
Жуковский П. М. 123, 124, 166, 194,
289, 338.
Жуков-Вережников Н. Н. 307.
Заварзин А. А. 183.
Закарьян С. Ф. 291.
Захваткин А. А. 186.
Зеликман А. Л. 139, 293.
Зоз И. Г. 314.
Зигель Р. 274.
Зюсс Э. 69.
Иванов А. В. 21.
Ивлев В. С. 139.
Игнатьев М. В. 108, 199.
Икскюль Я. 182.
Ильинский А. 109, 208, 266, 294.
Имшепецкий А. А. 149, 307.
Иогансен В. 80, 99, 232, 256.
Кагава Ф. 111.
Калабухов Н. И. 147, 232, 243, 286.
Камшилов М. М. 140, 313.
Кант Э. 66.
Карпеченко Г. Д. 90, 111, 339.
Карпов С. П. 171, 174, 307.
Каррей Д. 245.
Кашкаров Д. Н. 105, 321.
Келлер Б. А. 78, 79.
Кесслер К. Ф. 289.
Кёлликер А. 68~ 89.
Кёльрейтер И. 34.
Кёрнер А. 73, 147.
Кёрнике Ф. 124, 216.
Киселева С. Д. 256.
Кихара X. 111.
Клаузен Дж. 104, ПО, 111, 124, 172,
218, 232, 256, 304, 321.
Клейн Б. И. 160.
Клейншмидт О. 77, 82, 162.
Клеланд Р. 110.
Клементс Ф. 79, 183.
Клиленд Р. 89.
Клюйвер А. 131, 149, 307.
Ковалевский А. О. 63.
Ковалевский В. О. 54, 63, 343.
Кожов М. М. 325.
Козлов К. А. 267.
Козлова Л. М. 255.
Козо-Полянский Б. М. 160, 344.
Колесников А. Н. 24, 34.
Коломиец Н. К. 349.
Комаров В. Л. 25, 27, 38, 64, 65,
73—76, 81, 82, 92, 93, 95, 96, 99,
118—124, 127, 146, 147, 160, 166,
182, 195, 201, 215, 258, 314—316,
320, 356.
Коржинский С. И. 69, 70, 73—75,
77.
Королева В. А. 253, 254.
Корчагин А. А. 221, 222, 234, 256.
Костов Д. 338.
Кражан Ф. 101.
Крапоткин П. А. 289.
Красильников Н. А. 149, 176, 307.
Красовская И. В. 109.
Крейер Г. К. 121.
Крепе Е. 182.
Крик Ф. 153.
Криштофович А. П. 197.
Кувада И. 111.
Кудрявцев В. И. 149, 160.
Кудряшева О. И. 157.
Кузнецова Н. И. 9, 20.
Кузьмина Н. В. 335.
Кулешов Н. Н. 109.
Культер 95.
Культиасов М. В. 169.
Купман К. 327.
Купревич В. Ф. 149, 160, 175.
Курсанов Л. И. 149.
Кэйн А. 33, 77, 124, 127, 128, 146,
148, 149, 160, 164, 165, 227, 229,
245, 326—329, 348.
Кэк Д. 110.
Кэрстер Г. 238.
Кювье Ж. 32, 34, 40, 41, 56, 69.
Лавренко Е. М. 147, 171, 189, 191,
' 202, 267.
Ламарк Ж. Б. 35—37, 41, 42, 64, 88,
89, 290.
Лампрехт К. 151.
Ларина Н. И. 242, 267, 329, 334.
Лебедев Д. В. 93.
Левин Г. Г. 194.
Левина Р. Е. 171, 184.
Левитский Г. 111.
Ледерберг Дж. 158, 188.
Лейбниц Г. В. 40, 43.
Линней К. 7, 9—11, 24, 26—30, 32,
40, 42, 43, 46—48, 101, 150, 153.
Липшиц С. IO. 146, 166.
Лотка А. 109.
Лотси Дж. 77, 80, 81, 95, 97, 99, 122.
Лоуренс Дж. 9.
Лукип Е. И. 139, 223.
Лысенко Т. Д. 137—139.
Лэк Д. 168, 266, 285, 286, 298, 317,
326, 348.
Любименко В. Н. 109, 290
Ляйель Ч. 41, 56
Ляпунов А. А. 189, 205
Лященко И. Ф. 276
Маевский П. Ф. 252.
Мазер К. 264, 273, 275, 277.
Майворт С. Т. 68
Майр Э. 62, 76, 77, 123, 124, 126, 144—
146, 148, 152. 174, 178, 201, 210,
226, 231, 241, 309, 326—328, 341,
348, 354.
Максимович М. А. 38, 216.
Малеев В. П. 97, 121, 147, 215.
Малиновский А. А. 159, 206.
Мальцев А. И. 114.
Мальчевский А. С. 237.
Мамаев С. А. 256.
Мандей К. 287.
Мариковский П. И. 270.
Мартинсон Г. Г. 197.
Марченко И. И. 336
Матвеев Б. С. 266.
Матвеева Т. С. 115, 336.
Матсуура X. 111.
Маттей И. 153.
Матузима К. 111.
Махаэлис П. 151.
Медников. Б. М. 332.
Менделеев Д. И. 118.
Мендель Г. 67. 79, 80, 326.
Менсбир М. А. 34.
Мец К. 149.
Мечников И. И. 32, 289,
395
Микулинский С. Р. 37, 39.
Миллер С. 147.
Мильн А. 264.
Мирошниченко Е. Я. 343, 356.
Мирский А. 158.
Мирюта Ю. П. 336.
Мичурин И. В. 111, 339.
Мойс Дж. 294.
Морган Т. 90, 306.
Моррис Р. 285.
Мэрием Ч. 147.
Мюллер А. 149.
Мюнцинг А. 111, 176, 232, 258, 303,
330, 337, 338, 340, 349, 355.
Навашин М. С. 111.
Навашин С. Г. 151.
Найт-Джонс Е. 294.
Наливкин Д. В. 129, 196.
Наумов Н. П. 139, 180, 185, 189, 223,
233.
Невский С. А. 122, 123, 163.
Неймайр М. 63.
Никитин С. 65.
Никулеску В. 217, 303.
Нильсен Г. 80, 81.
Ниренберг М. 153.
Ноден Ш. 34, 58.
Нэгелли К. 182.
Овчаров К. Е. 157.
Овчинников П. Н. 157.
Огнев С. И. 125, 147, 149, 320.
Оленов Ю. М. 107, 108, 317, 348, .349.
Олсон Е. 171.
Осборн Г. 55.
Остенфельд К. 84.
Оуэн Р. 63.
Очоа С. 153.
Павловская О. 108.
Палилов А. И. 173.
Паллас П. 32.
Пальмова Е. Ф. 114.
Пангало К. И. 114, 117.
Панов Е. Н. 347.
Папонов А. Н. 288, 289.
Парамонов А. А. 139, 257.
Парамонов С. Я. 170.
Парк Т. 110, 207, 269, 280.
Пасынский А. Г. 158.
Пачоский И. К. 78, 93, 95—99, ИЗ,
146, 183.
Перль Р. 109.
Петров Д. Ф. 343.
Пехов А. П. 307.
Пименов М. Г. 154.
Пиментал Р. 222.
Пири Н. 158.
396
Писарев В. Е. 338.
Плате Л. 83.
Плиний (старший) 8.
Поздняков Л. К. 334.
Полежаев В. 109, 294.
Поляков И. М. 139.
Полянский В. И. 131, 146, 148, 160,
169, 171, 174, 177, 206, 224, 307.
Полянский Ю. И. 274:
Померанцева А. А. 134.
Поплавская Г. И. 102.
Попов М. Г. 95, 216, 337.
Пояркова А. И. 334.
Правдин Л. Ф. 227.
Пратт Р. 287.
Проханов Я. И. 144, 166, 216.
Пузанов И. И. 175, 184.
Рабо Э. 133.
Работнов Т. А. 243.
Райков Б. Е. 38.
Райкова И. А. 335.
Райт С. 107, 108, 348, 349.
Раункиер К. 250.
Регель Р. Э. 64, 77, 80, 93, 94.
Рей Д. 26—28, 140.
Реннер О. 89.
Ренш Б. 76, 159, 176, 231, 303, 320.
Рич А. 158.
Робертсон А. 110.
Родендорф Б. Б. 14, 15, 201, 215.
Роза Д. 182, 196.
Розанова М. А. 98, 102, 103, 111, 121,
147, 152, 175, 215, 218, 256, 258,
338, 339.
Ромене Дж. 68, 347.
Ромер А. 18.
Ростова Н. С. 86, 87, 251, 253, 254.
Ротай А. П. 128, 130.
Ротмалер В. 323.
Роу А. 91. <
Рубайлова И. Г. 337.
Рубцов И. А. 174.
Рудаков О. Л. 161, 169.
Рулье К. Ф. 34, 38.
Рыбин В. А. 111, 335, 338.
Рытов М. В. 83.
Сабинин Д. А. 139.
Савин В. Н. 247, 252, 253, 255.
Савич-Любицкая Л. И. 148, 171,
213.
Савоськин И. П. 216.
Сажрэ О. 34.
Сакай К. 277.
Самофал С. А. 290.
Сансом Ф. 104.
Северцов А. Н. 298, 346.
Северцов Н. А. 38, 66.
Северцов С. А. 109, 191, 195, 206, 209,
233, 266, 271, 272, 297, 320, 359.
Селье Г. 286.
Сапегин А. А. 109.
Семенов-Тян-Шанский А. П. 76, 77,
146, 162.
Сент-Илер Ж. Э. 38.
Серебровский П. В. 228, 320.
Симпсон Д. 19, 55, 303, 348, 358.
Синская Е. Н. 102—104, 114, 121, 148,
166, 171, 176, 184, 191,194, 206, 208.
218, 221, 225,' 227, 230, 244, 246—
250, 256, 260, 307, 309, 323, 326.
Смелов С. П. 275.
Смирнов Л. А. 9, 20.
Смирнова Е. А. 109, 208, 284.
Смирнова 3. Н. 148, 171, 213.
Смит Г. 165.
Соколов С. Я. 139.
Соколовская А. П. 111, 336.
Соннеборн Т. 224, 274.
Сочава В. Б. 109, 208, 282.
Спасский Б. 108.
Спасский Н. 108.
Спектор В. 13.
Спенсер Г. 18.
Спирин А. С. 153.
Станков С. С. 252.
Стеббинс Г. 111.
Стенли В. 158.
Степанов Д. Л. 69, 128, 147, 215, 223.
Стертевант А. 89.
Стрелкова О. С. 111.
Стрельников И. Д. 16.
Сукачев В. Н. 109, 139, 191, 208, 257,
284, 287.
Сухов К. С. 306, 307.
Сушкин П. П. 344.
Сцент-Дьёрди А. 181.
Сэмнер Т. 104.
Талиев В. И. 93—95, 252.
Тахтаджян А. Л. 166, 172, 233, 344.
Текхольм Г. ПО.
Тенсли А. 183.
Теофраст 8, 25.
Терентьев П. В. 10, 14, 78, 126, 146,
217, 222, 230.
Терновский М. Ф. 338.
Тимирязев К. А. 39, 81, 87, 90, 115,
289.
Тимофеев-Рессовский Н. В. 105, 175,
176, 179, 189, 303, 346.
Тих Н. А. 270.
Тихомиров Б. А. 156.
Тихомиров В. Н. 143, 160.
Толмачев А. И. 155, 231, 315, 345.
Томсон С. 109.
Троицкий Н. А. 335.
Турессон Г. 79, 80, 99, 102, 103, 105,
113, 147, 172, 255.
Туррилл У. 104.
Умер Ф. 64.
Уоддингтон К. 154, 189.
Уолд Дж. 189.
Уоллео А. 64, 67, 289, 347.
Уранов А. А. 243.
Усов С. А. 66.
Успенская Л. И. 109, 208, 282, 284,
293.
Ухтомский А. А. 203, 206.
Ушаков Б. П. 156, 185.
Уэвелл В. 29, 33.
Уэлдон В. 72.
Фатеев К. Я. 227.
Федоров А. А. 154.
Федоров Ал. А. 344.
Федотов Д. М. 127, 149.
Федюшин А. В. 267.
Филюков А. И. 171.
Фишер Р. 107.
Флоркен М. 158, 182.
Фляке В. 268.
Фляксбергер К. А. 114, 122.
Форбс Е. 64.
Форд Е. 212, 327.
Формозов А. Н. 10, 14, 139.
Фурсаев А. Д. 322.
Хагеруп О. 110.
Хаджи И. 186.
Хайлов К. М. 185.
Хайнд Р. 353.
Хансен Е. 186.
Харди А. 344.
Хахина Л. Н. 252.
Хизи В. 110.
Хитрово В. Н. 85, 86.
Хичкок А. 84.
Холден Дж. 107, 109, 308.
Холодковский Н. А. 104.
Хохлов С. С. 116, 132, 160, 317, 343,
356, 360.
Хохряков М. К. 166, 171.
Хржановский В. Г. 145.
Цезальпин А. 28.
Цингер Н. В. 85—87, 93, 103, 309, 343,
356.
Чернов А. А. 18, 128.
Чернов С. А. 124, 147.
Черных Н. А. 314.
Четвериков С. С. 107, 108.
Чижек Ф. 171, 187, 206.
Чудра В. 177.
397
Чумакова Ё. М. 274.
Чупров А. А. 83.
Чэмпен Р. 110.
Шакиров Ф. X. 267.
Шамсиев Г. 252.
Шапиро Н. И. 108, 199.
Шапоренко К. К. 197.
Шапошников Г. X. 160, 179.
Шванптц Ф. 157, 323.
Шварц С. С. 78, 147, 176, 217, 224,
229, 230, 232, 244, 308, 321.
Шелфорд В. 183.
Шенников А. П. 109, 233.
Шимкевич В. М. 83, 344.
Шифман А. И. 172.
Шишкин Б. К. 315.
Шляков Р. Н. 160, 171.
Шмальгаузен И. И. 53, 165, 182—184,
189, 191, 206, 212, 244, 249, 289,
290, 293, 298, 301, 306, 308, 344, 348,
359.
Шмидт Г. А. 207, 297.
Шовен Р. 143, 208, 268, 270.
Шода Р. 130.
Шпрингер 70.
Шрамм Г. 158.
Штреземан Е. 178.
Щербаков В. К. 340.
Эверсман Э. А. 24, 34.
Эймер Т. 65.
Эйхвальд Э. И. 39.
Элтон Ч. 170, 233, 285, 349.
Энгельс Ф. 33, 46, 54, 127, 289, 290.
Энглер А. 9, 17.
Эшби Р. 203, 268, 285.
Юзепчук С. В. 46—49, 64, 73, 87, 114,
123, 132, 143, 144, 147, 149, 152,
166, 172, 173, 175.
Яблоков А. В. 202.
Янковский А. В. 225.
Abelson Р. 19.
Akerlund Е. 324.
Akkerberg Е. 311.
Allee W. 110, 206, 208, 233, 269, 280.
Alpatov V. 351.
Anderson Е. 64, 194, 239, 349.
Andrewartha Н. 114, 191, 206, 233.
Ascherson Р. 64.
Beaudry J. 157, 180.
Bertalanffy L. 183, 191.
Beurlen K. 196.
Birch L. 264.
Blackwelder R. 20.
Boissier E|< 65.
Bonner J. 182.
Botnariuc N. 206, 209.
Bouillon J. 346.
Bowden W. 124, 148, 354.
Bremekamp C. 175.
Brewster E. 28, 32, 42.
Briquet J. 215.
Buchsbaum R. 14, 15.
Burma B. 135.
Cain A. 143, 178.
Caloianu M. 171.
Carau§u D. 234.
Carson H. 318.
Carus V. 8.
Chapman R. 266.
Chodat R. 130.
Christian J. 286.
Ciferri R. 148, 174.
Cizek F. 202.
Clarke C 245.
Clausen j. 101, 104, 115, 124, 146, 148,
218, 230, 247, 249, 256, 258, 307,
311, 320, 321, 326, 349, 352.
Clements F. 109.
Cole F. 33.
Connell J;. 289.
Cooper W. 112, 114, 208, 281.
Cuenot L. 133, 197, 351.
Cuvier G. 32.
Daly K. 307.
Danser B. 77, 148.
Darlington C. 163, 342.
Davidson J. 136.
De-Candolle Alph. 9, 65.
De-Vries H. 80, 215.
Dobrowolski K. 163.
Dobzhansky Th. 12-14, 77, 123, 126,
148, 152, 178, 201, 206, 207, 233,
243, 326, 353.
Du Rietz E. 162(.
Eimer E. 65.
Elton C. 208, 233, 280.
Emerson A. 267.
Engler A. 9, 12, 13, 17, 20, 64.
Ford E. 86, 206, 212, 233, 245, 280, 326.
Gause G. 109, 299, 326.
Gilmour J. 134, 234.
Glaessner M. 158.
Godron D. 41.
Goldschmidt R. 84, 90.
Good D. 163, 359.
398
Gotoh К. 207, 261,
Grant V. 165, 206, 213, 233, 326.
Grant W. F. 219.
Grebecki A. 109.
Grebner P. 64
Green E. 30.
Gregor J. 104, 217, 229, 230, 234,249,
260.
Gulick J. 132, 348.
Hagerup 0. 216.
Haldane J. B. S. 90, 109, 191, 206,
232, 249, 277, 330, 359.
Harlan J. 311—313.
Harland S. 248.
Harper I. 355.
Harper J. 269.
Harvey L. 108, 306.
Hesse R. 10, 12, 14, 21.
Hiesey W. 104.
Hinde B. 353.
Hitchcock A. 84.
Hodanova D. 202.
Hoffmeister D. 231.
Huskins C. 90, 330.
Huxley J. 21, 53, 109, 126, 146, 162.
163, 181, 191, 212, 217. 245, 258,
306, 320,326,330,332,340,346, 357.
Jalas J. 234.
Jordan A. 47.
Kaestner A. 12, 14, 15.
Kerster H. 238.
Kleinschmidt O. 162.
Koopman K. 327, 353.
Kolliker A. 68.
Kornicke F. 215.
Lack D. 289.
Lamarck J. 35.
Lawrence G. 9, 20.
Lerner G. 213.
Lewis H. 324, 355.
Li C. 232, 243, 359.
Linnaei C. 29, 32, 43.
Littlejohn M. 353.
Lotsy J. 77, 81, 99,* 147, 337.
Love A. 178.
Liiers IL 317.
Mather K. 207, 213, 326, 330, 356.
Matsuura H. 111.
Mayr E. 14, 16, 24, 101, 147, 148, 152,
168, 178, 186, 191, 210, 222, 227,
229, 231, 242, 304, 316, 318, 320,
332, 341, 348. 353.
McCarley H. 353.
Meyer E. 8, 25, 27.
Milne A. 14, 15, 285.
Mobius K. 10.
Muller A. 146, 149, 358.
Miinzing A. 90, 111, 337, 340.
Myers G. S. 242.
Naughton I. 355.
Niculescu V. 217, 303.
Ochoa S. 199.
Odum A. 114, 191, 206, 269.
Olson E. 267.
Orias E. 289.
Osborn G. 28, 32, 42.
Pallas P. 32.
Pavlovsky O. 207, 326.
Peterfi S. 169, 217, 336.
Petrusewicz K. 171.’
Pimental R. 222, 285.
Plate L. 83.
Popov M. G. 216.
Pratt R. 269, 287.
Prosperi R. 186.
Rabaud E. 133.
Radi E. 8, 25, 28, 32, 42, 48, 64.
Raunkiaer C. 250.
Ray J. 28.
Remane A. 325..
Rensch B. 76, 146—148, 159, 172, 176,
187, 227, 229, 231, 318, 320, 357.
Rikkinen K. 234.
Rimpau W. 115.
Rivas L. R. 241, 353.
Romanes D. 64, 68.
Rowe A. 91.
Russel E. 40.
Sakai K. 277.
Sansome F. 104, 249, 260.
Schindewolf O. 304, 359.
Schwanitz F, 157, 323.
Sheppard P. M. 206, 213, 245.
Simpson G. G. 18, 197, 201, 357, 358.
Slastenko E. P. 175.
Smith H. M. 165, 201, 305.
Smith J. 189, 308, 333.
Snayder R. 286.
Soo R. 87.
Spector W. S. 9, 10, 12—15, 22.
Sprengel K.
Stebbins G. Ill, 160, 239, 307, 326,
356, 361.
Stefureac T. 169, 217, 336.
Steyskal G. 21, 22.
Stresemann E. 316.
399
Tetry A. 197.
Thorson G. 362.
Timopheef-Ressovsky N. W. 316.
Tj0nneland A. 325.
Torre D. 231.
Trojan P. 171.
Tsuyoshi M. 153.
Turesson G. 98, 99, 102, 103, 146, 162,
178, 224, 225, 255.
Turrill W. 104.
Ulrich H. 317.
Valentine D. 180.
Van-Steenis C. 147, 175.
Verner J. 273.
Vuillemin P. 84.
Waddington C. 249, 306.
Wagner M. 67.
Watson P. 230, 234.
Weaver J. 109.
Wet J. 311—313.
Wettstein R. 73, 85.
White M. 318.
White С. M. N. 347.
Wright S. 206, 233, 306, 348, 349,
359.
Wiirtemberger L. 65.
Wynne-Edvards V. 206, 208, 277.
Zarzycki K. 289.
Zimmerman W. 146.
ОГЛАВЛЕНИЕ
Стр.
Предисловие ....................................................... 3
Часть I
ИСТОРИЯ ИЗУЧЕНИЯ ВИДА
Глава 1. Эволюция знаний и современные данные о числе видов 7
1. Накопление знаний о числе видов.............................. 8
Число видов в долиннеевские времена (8). Инвентаризация ви-
дов за последние 200 лет (9).
2. Число видов по современным данным........................... 11
Общее число видов (И). Асимметрия распределения видов по
крупным таксонам (13).
3. Степень изученности видового состава флоры и фауны . ... 17
О возможности инвентаризации современных и ископаемых ви-
дов (17).
Глава 2. Изучение вида до половины XIX в.......................... 23
1. Характеристика этапов изучения вида........................ 23
2. Разработка проблемы вида от Д. Рея до К. Линнея .... 27
Вид по Д. Рею (27). Вид по К. Линнею (29).
3. Проблема устойчивости и дискретности вида................... 31
Устойчивость вида (31). Дискретность вида (33).
4. Взгляды Ж. Б. Ламарка на проблему вида...................... 35
5. Утверждение идеи постоянства видов в первой половине XIX в. 37
Попытка объединения идей реальности и изменяемости видов
(37) . Доводы в пользу постоянства видов (39).
6. О субъективистских толкованиях вида........................ .42
Номинализм и проблема непрерывности (42). Номинализм фор-
мальных систематиков (43).
7. О практических стандартах вида в додарвиновский период 45
Определение понятий (45). Морфологический монотипический
вид (46). Вид по А. Жордану (47). Морфологический политипиче-
ский вид (48).
8. Заключение.................................................. 50
Глава 3. Проблема вида и видообразования во второй половине
XIX в....................................' ...... . 52
1. Дарвин о виде и видообразовании............................. 52
Видообразование (54). Вид и разновидность (57).
2. Общая характеристика состояния проблемы вида................ 62
Понимание вида в систематике (64).
401
Стр.
3. Вопросы видообразования....................................... 67
Поглощающая роль скрещивания (67). Гипотезы мутационного
образования видов (68).
4. Заключение.................................................... 71
Глава 4. Разработка проблемы вида и видообразования в начале
XX в........................................................ 72
1. Морфолого-географический метод в систематике................. 73
Взгляды С. И. Коржинского (73). Концепция В. Л. Комарова
(74) . О политипическом стандарте вида (76).
2. Кризис в изучении вида........................................ 78
Разложение вида генетическими методами (78). Неожорданизм
(80) . Субъективизм в понимании вида (82).
3. Изучение видообразования...................................... 85
Сезонные расы (85). Попытки экспериментальных доказательств
гипотезы мутационного видообразования (87). Примеры палеонто-
логических работ в области видообразования (91). Миграционная
гипотеза видообразования (92). Работы В. И. Талиева (93).
4. Проблема целостности вида .................................... 95
И. К. Пачоский о целостности вида (96).
Глава 5. Новейшая история проблемы вида............................ 101
1. Экспериментальное изучение вида.............................. 101
Экспериментально-экологическое направление микросистематики
(102). О значении генетики природных популяций (107). Внутриви-
довые отношения (108). Кариосистематика и экспериментальная ги-
бридизация (110).
2. Работы Н. И. Вавилова........................................ 112
Центры происхождения видов и центры разнообразия форм (112).
«Линпеевский вид как система» (116).
3. Вид—единица или система?..................................... 118
«Учение о виде у растений» (118). Столкновение двух концепций
вида (120). Об объеме вида у пшениц (122).
4. Трудности в применении практических стандартов вида ... 125
О стандарте вида в палеосистематике (127). О применении моно-
типического стандарта «мелкого» вида (130).
5. Ошибочные толкования вида и видообразования.................. 133
Отрицание реальности вида (133). Псевдодарвинизм (137).
6. Заключение................................................... 140
Часть II
ОЧЕРКИ ТЕОРИИ ВИДА
Глава 6. Критерии и общие признаки вида............................ 143
1. Проблема вида и систематика.................................. 143
Эйдология и вид в систематике (143). Практические стандарты
вида в современной систематике (146).
2. Критерии вида............................................... 149
Морфологический критерий (149). Биохимический критерий (152).
Географический критерий (154). Другие критерии (156).
3. Универсальность и неравноценность вида....................... 157
Универсальность вида (157). Неравноцеии эсть видов (162).
4. Общие признаки вида.......................................... 167
Характеристика признаков (167).
5. О некоторых современных определениях вида.................... 174
Суммативные определения вида (174). Вид как тип организации
(176), Определения биологического вида (178).
402 с
Глава ?. Вид Как одна из первйчпыч и <н фирм к|н uni hi
ции живого.............................
1. Основные формы организации жпного и п\ нндрн ни» »»»пнн
Системная организованность живого (|н|) Kpiiiiixii ч и и нП и ы ч
иерархических рядов систем (183). О тсрмппв «ypoimiiii opi hiiiiiiiihhii •
(188). Первичные и основные формы оргпнпппции iiumhhi) iluni
2. Особенности вида как надиттдивидупльной сиг тми 10!«
Возможность существования вида в точении iH»onpi»B,«»iii>iiiioro при
мени (195). Вид как образование, способной к rnMoc i'oii n"i|1|ni'\i у пни
люционному развитию (198). Является ли вид згапом initoiinipiih' (''иI >
3. О целостности вида.................................................... ‘‘02
Определение понятий (202). О степени целое»щи ти ншн» i 'и »»
Видовые адаптации (206).
4. Определение вида....................................................... ’in
Глава 8. Структура вида........................................... 'II
1. Введение..................................................... II
Оперделение основных понятий (211). IC критике ш hi и hi н о |ны
новидности (215).
2. Системы внутривидовых единиц............................... ’’I/
О методах изучения дифференциации вида (217). Система iniy ipn
видовых единиц у растений (219). Представления о диффергнцпицпн
вида в зоологии (222).
3. Аллопатрические группы.......................................
Общая характеристика (225). Экологическая раса (экотип) 122!и
Географическая раса (подвид) (226). О реальности подвидоп и экотн
пов (229).
4. Местная популяция.......................................... 2:12
Определение понятия (232). О размерах и границах местной по
пуляции (234). Типы местных популяций (238).
5. Симпатрические единицы....................................... М)
Понятие о симпатричности (240). Внутрипопуляциоппые группы
(243). О генетическом полиморфизме (245). Экоэлемепт (24В). Морфи•
биологическая группа (250). Биотип (256).
6. Заключение.................................*................ 259
Глава 9. Внутривидовые отношения................................. 2(>2
1. Классификация внутривидовых отношений...................... 2(»2
Определение понятия (262). Основные и производные внутри-
видовые отношения (264).
2. Формы интеграции индивидов................................. 2(17
3. О характере внутривидовых отношений......................... 271
Противоречивость основных внутривидовых отношений (271) Мо-
дельные опыты на растениях (275).
4. Численность как видовое приспособление...................... 277
5. Регулирование плотности популяции........................... 280
Оптимальная плотность популяции (280). Способы регулировании
численности (285).
6. Перенаселение и его следствия.............................. 289
Определение понятий (289). О последствиях перенаселения (292).
7. Видовые корреляции...........................................295
Изменения видовых корреляций (296).
8. Заключение..........................4....................... 300
Глава 10. Видообразование......................................... 302
1. Видообразование и формообразование.......................... 302
Микроэволюция и видообразование (302). Критика понятия
о видообразовательной изменчивости (305).
403
2. Типы видообразования........................................... 308
Принципы выделения типов (308). Синтезогенез и сегрегациоге-
пез как типы видообразования (310).
3. Аллопатрическое видообразование................................ 313
Географическое видообразование (313). Подвид и полувид как
стадии видообразования (318). Экологическое видообразование (322).
4. Симпатрическое видообразование................................. 326
О возможности симпатрического видообразования (326). Примеры
' симпатрического видообразования (329).
5. Гибридогенное видообразование и понятие о «новой видовой
форме»........................................................... 334
Гибридогенные виды в природе (334). Воссоздание форм, близких
природным гибридогенным видам (337). Экспериментальный синтез
«новых видовых форм» (339).
6. Неоформогенное и микроаккумулятивное видообразование ... 341
Определение понятий (341). Исходный материал неоформоген-
ного видообразования (342).
7. Изоляция и видообразование.................................... 346
О классификациях форм изоляции (346). Пространственная
(топографическая) изоляция (348). Временная изоляция (351). Репро-
дуктивная (половая) изоляция (352).
8. Видообразование и прогрессивная эволюция....................... 357
Видообразование и неравноценность видов (357). Особенности
эрогенных видов (358). Повышение выживаемости и степени
целесообразности особи и вида (361).
Литература............................................................365
Именной указатель.....................................................393
Кирилл Михайлович Завадский
ВИД И ВИДООБРАЗОВАНИЕ
Утверждено к печати
Институтом истории естествознания и техники Академии наук СССР
Редактор издательства Ф. И. Кричевская Художник В. В. Грибакин
Технический редактор М. Н. Кондратьева Корректоры Н. В. Лихарева,
Г. А. Мирошниченко иТ. Г. Эдельман
Сдано в набор 26/Х 1967 г. Подписано к печати 2/II 1968 г. РИСО АН СССР № 54—125В.
Формат бумаги бОхЭО1/^. Бум. л. 131/<. Печ. л. 25’/4 4- 10 вкл. (1‘/4 печ. л.) = 261/2 усл. печ. л.
Уч.-изд. л. 28,40. Изд. № 3424. Тип. зак. № 623. М-22751. Тираж 7000.
Бумага типографская № 1. Цена 2 р. 20 к.
Ленинградское отделение издательства «Наука»
Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. издательства «Наука». Ленинград, В-34, 9 линия, д. 12