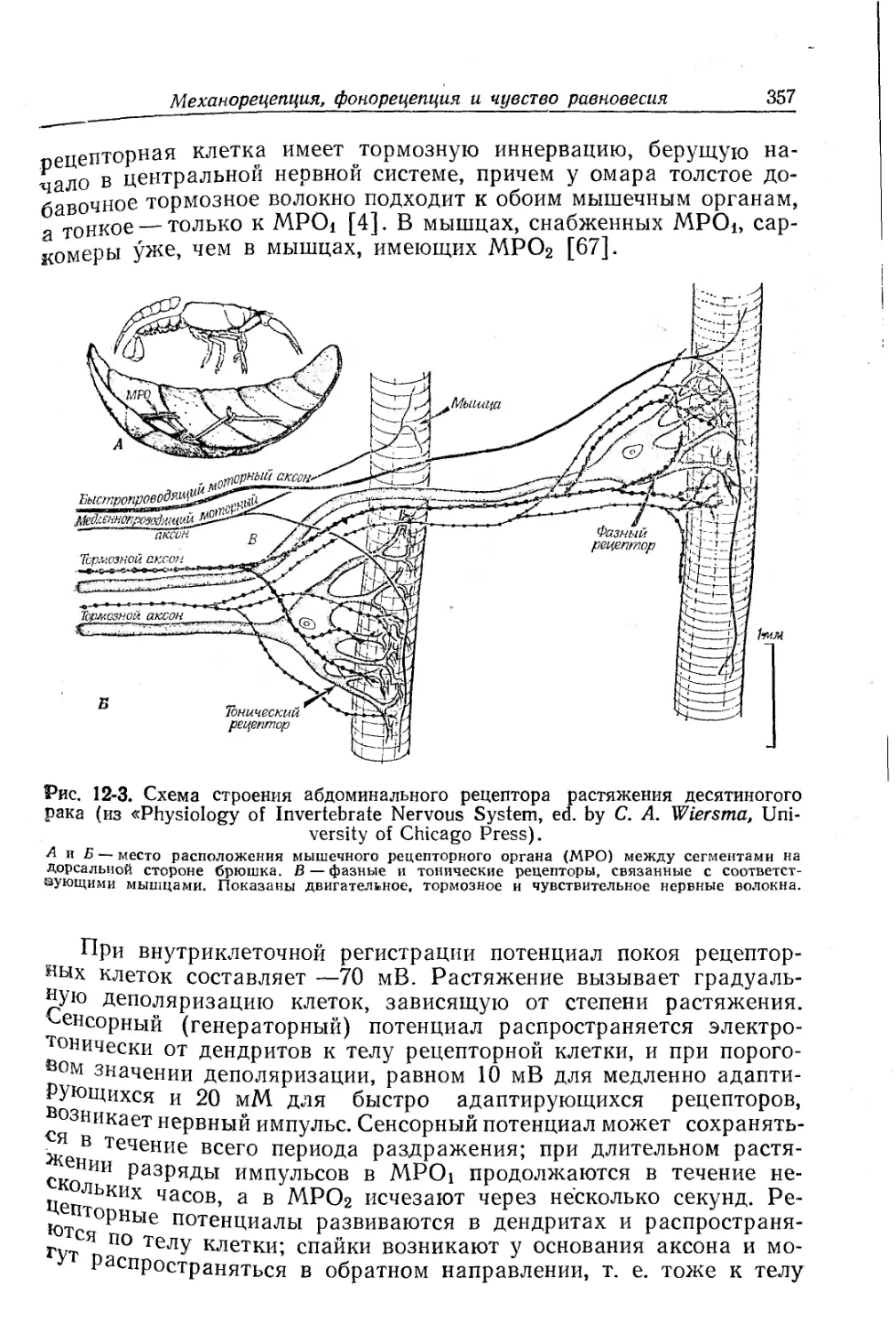
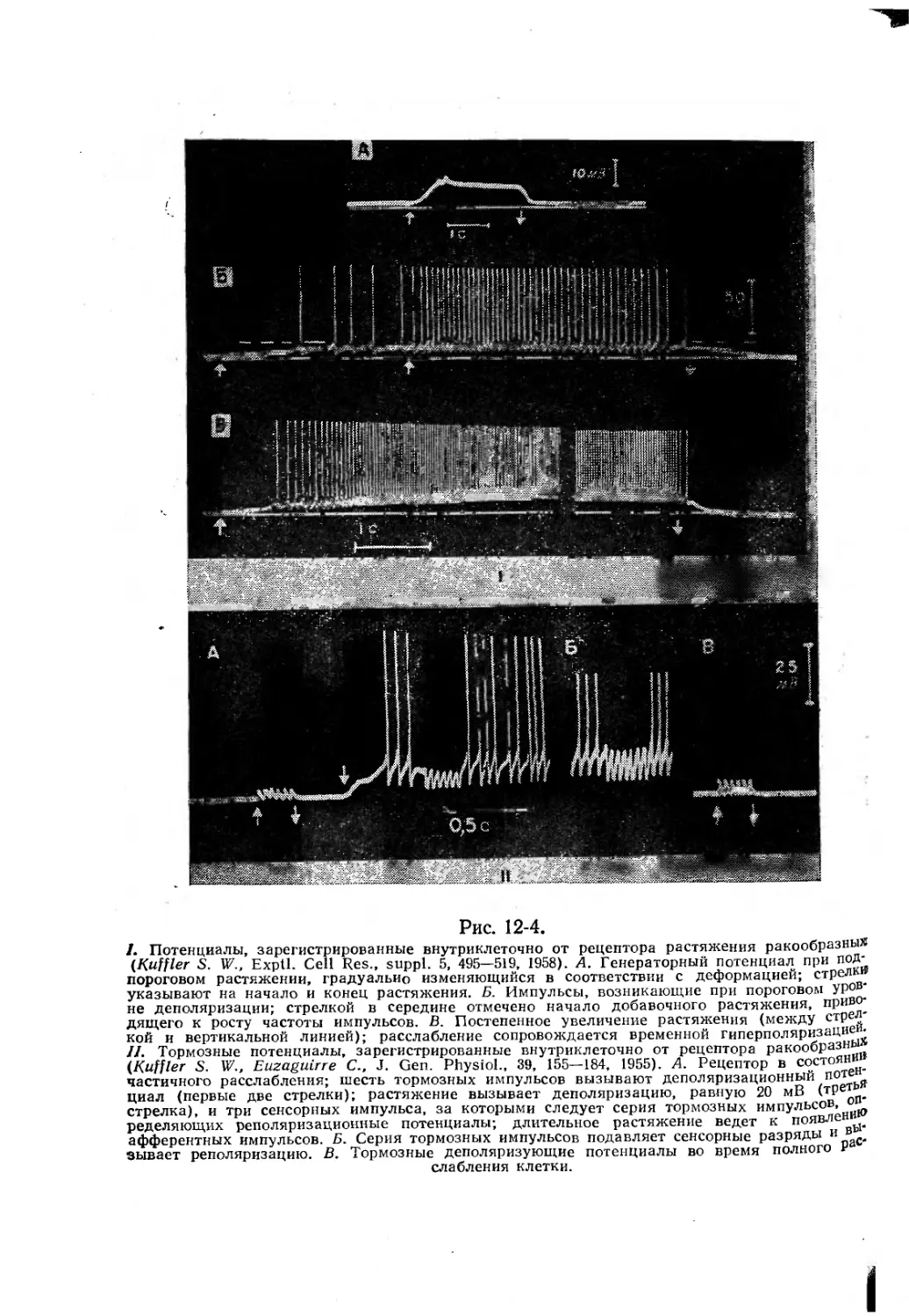
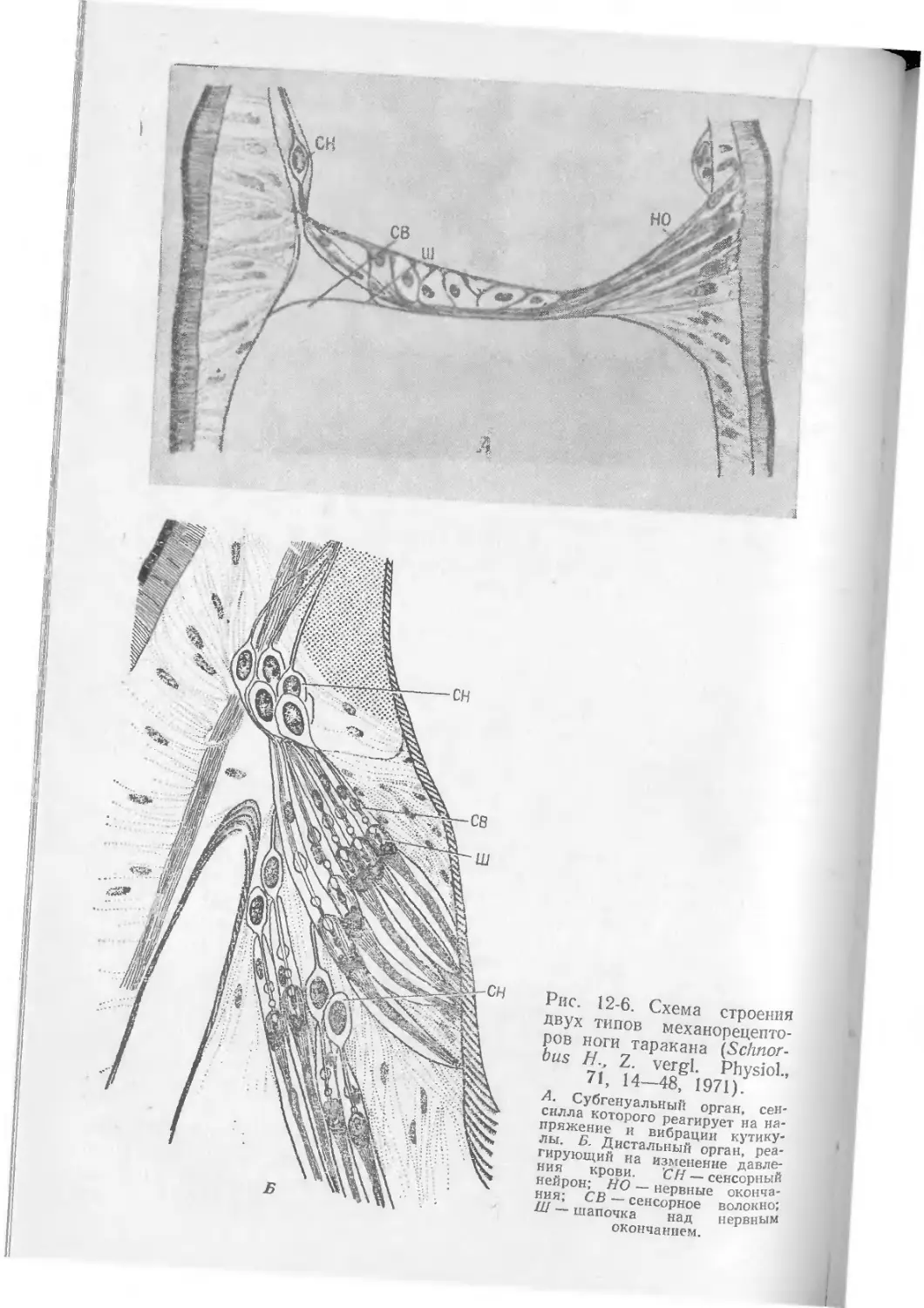
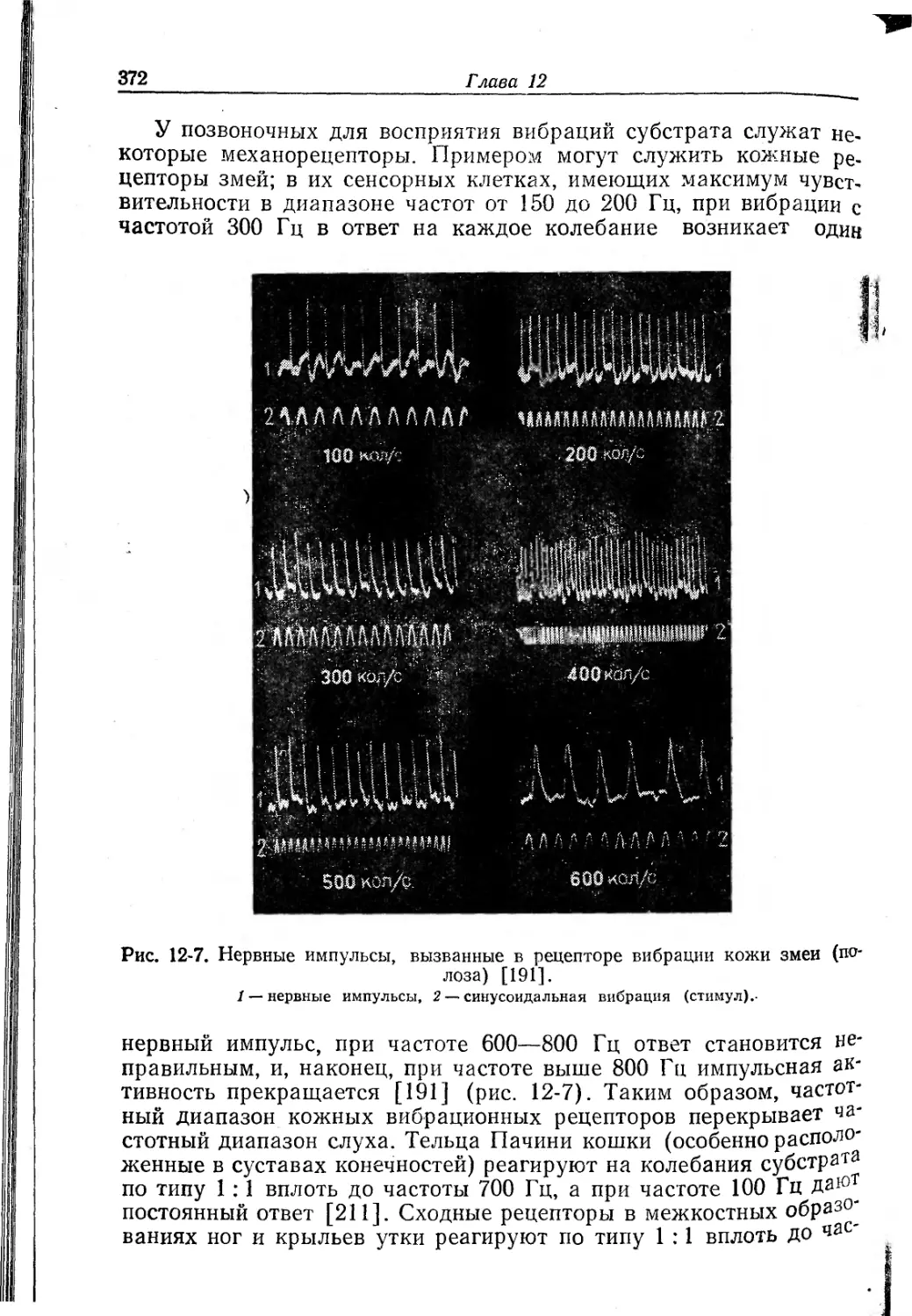
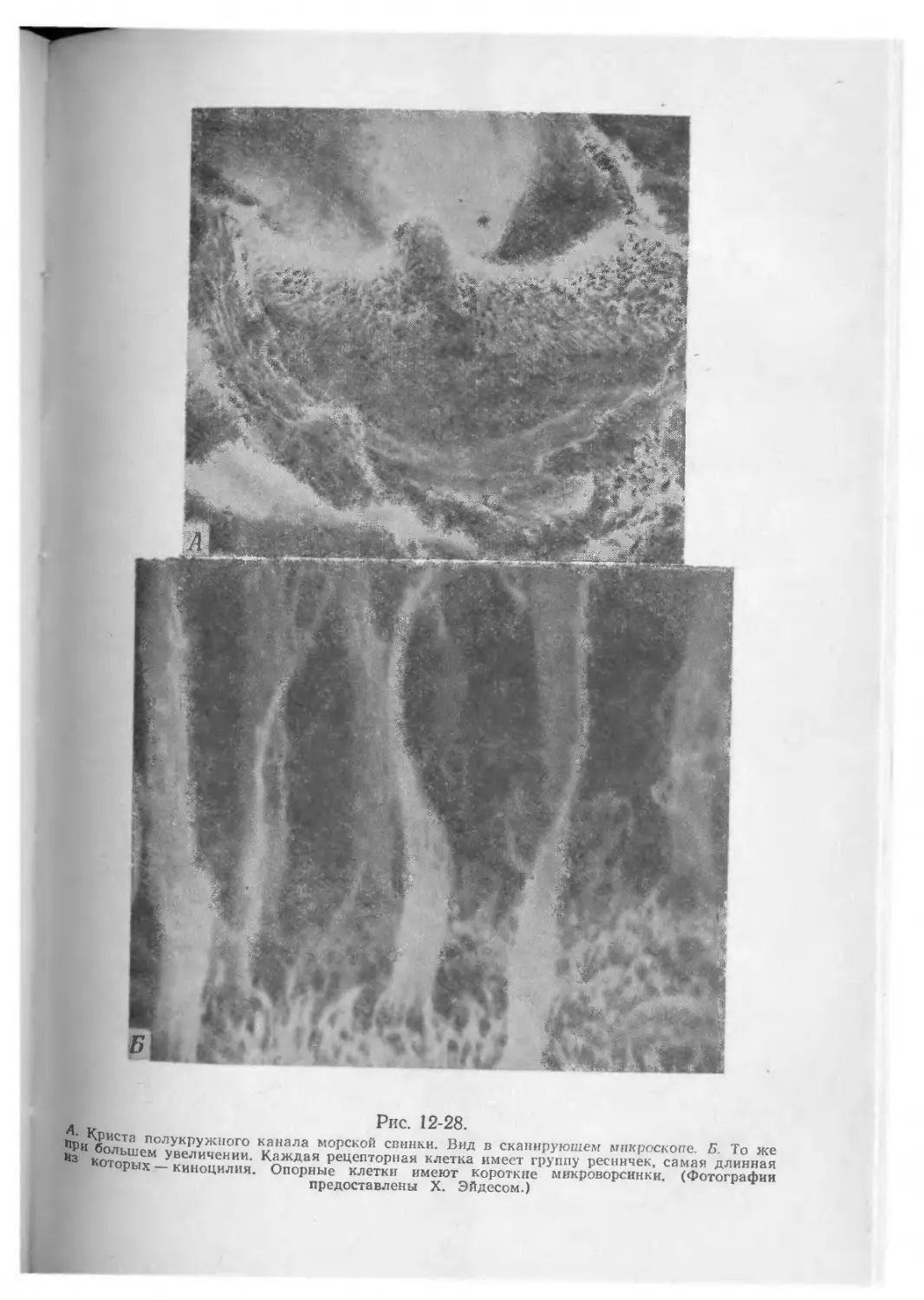
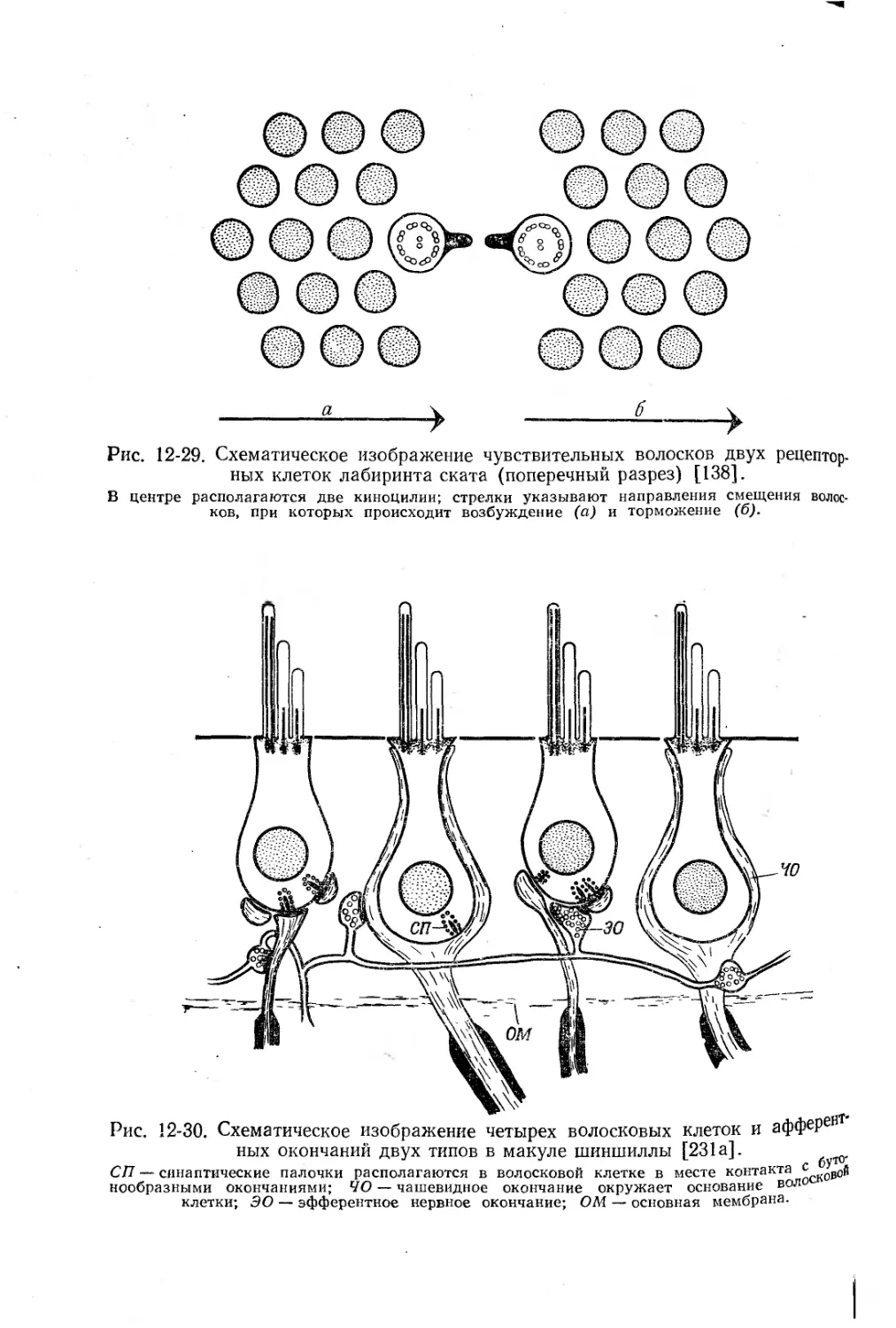
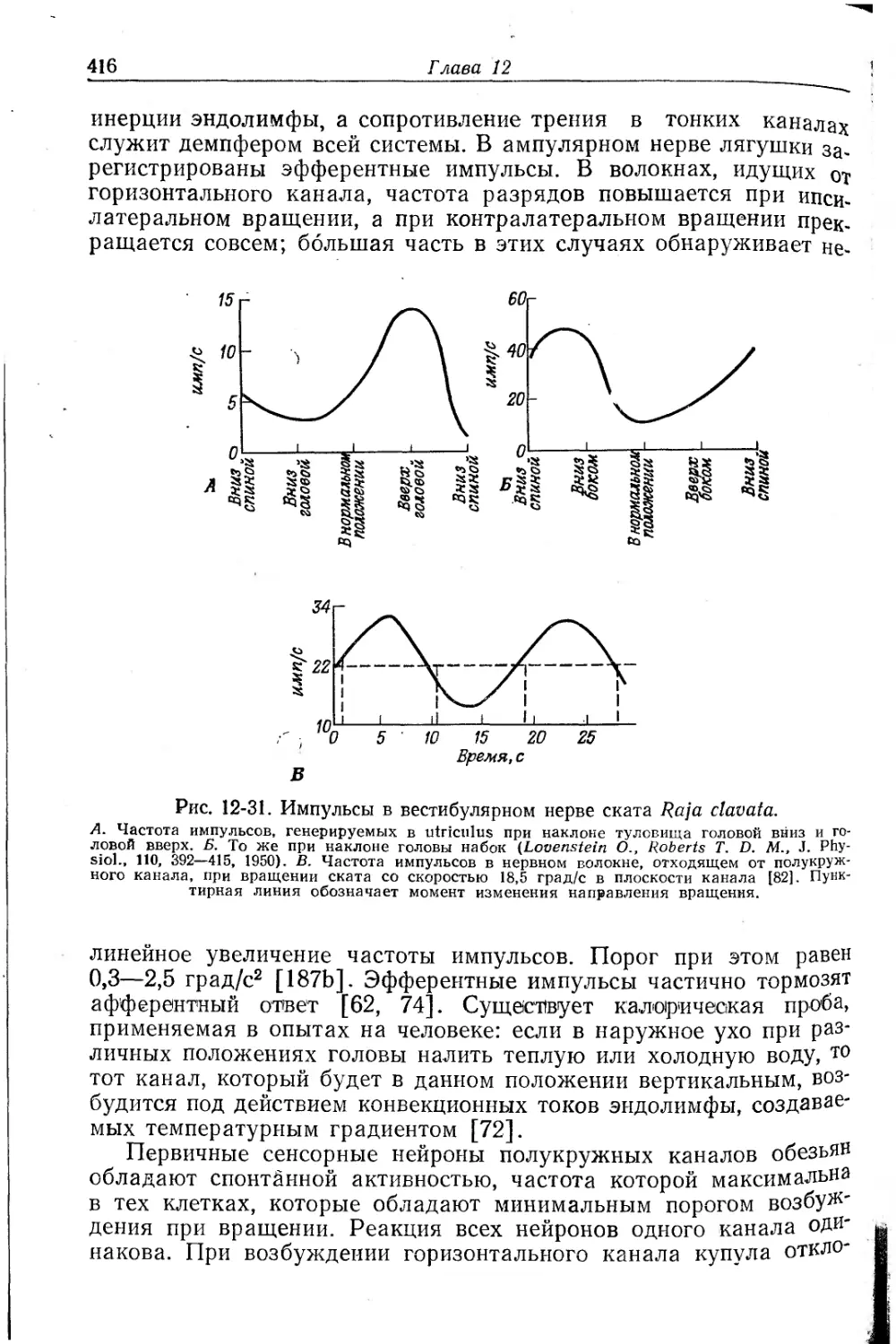
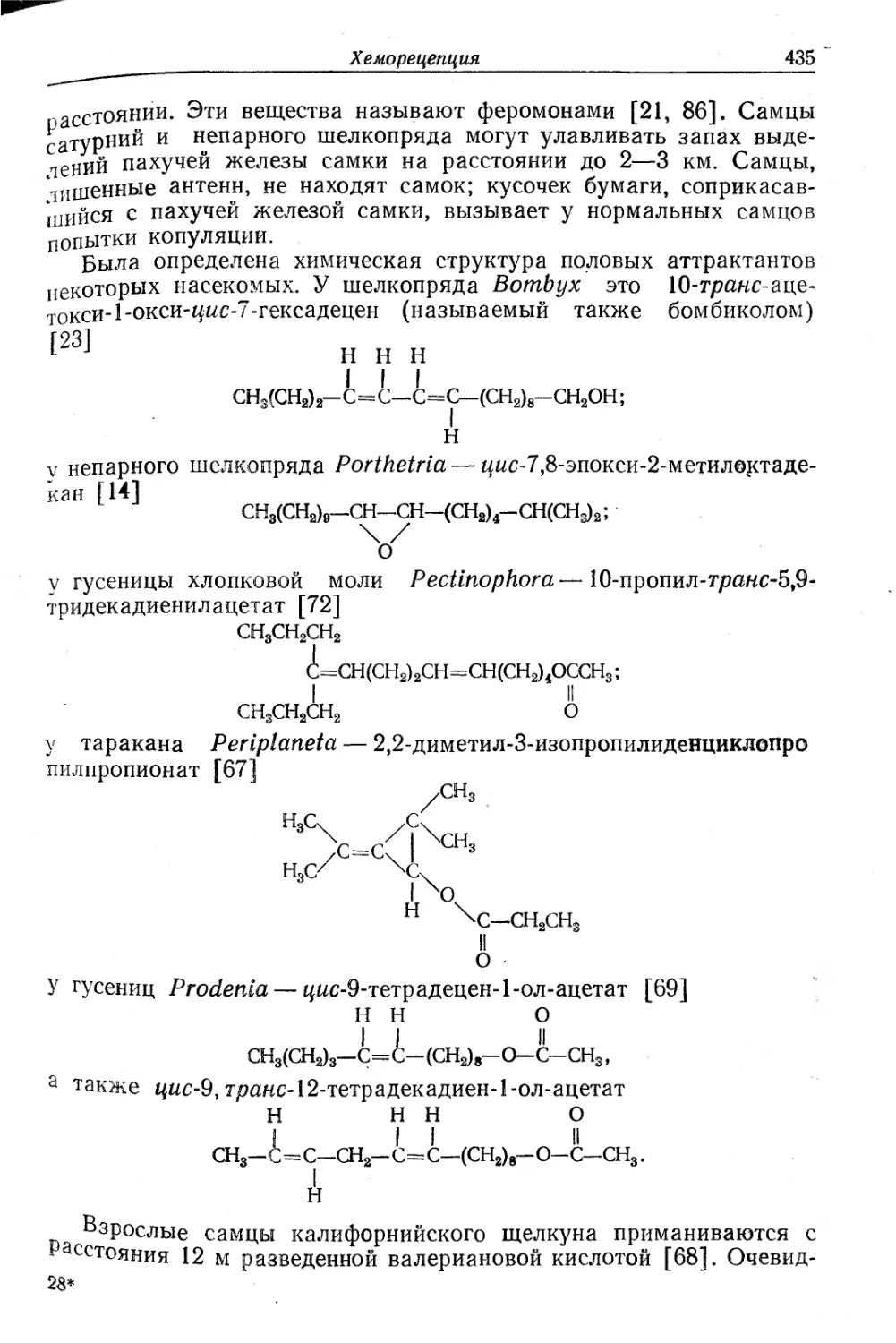
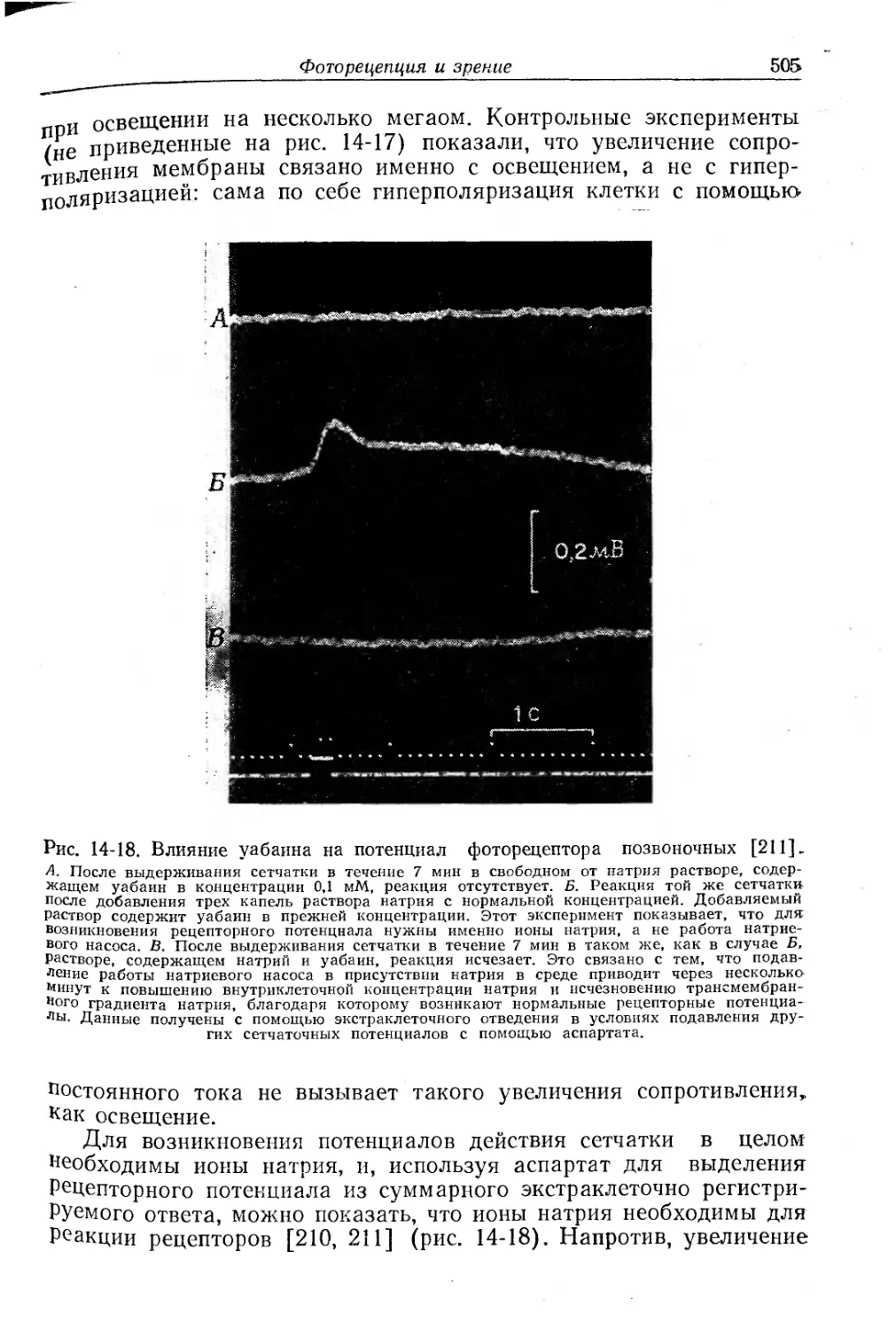
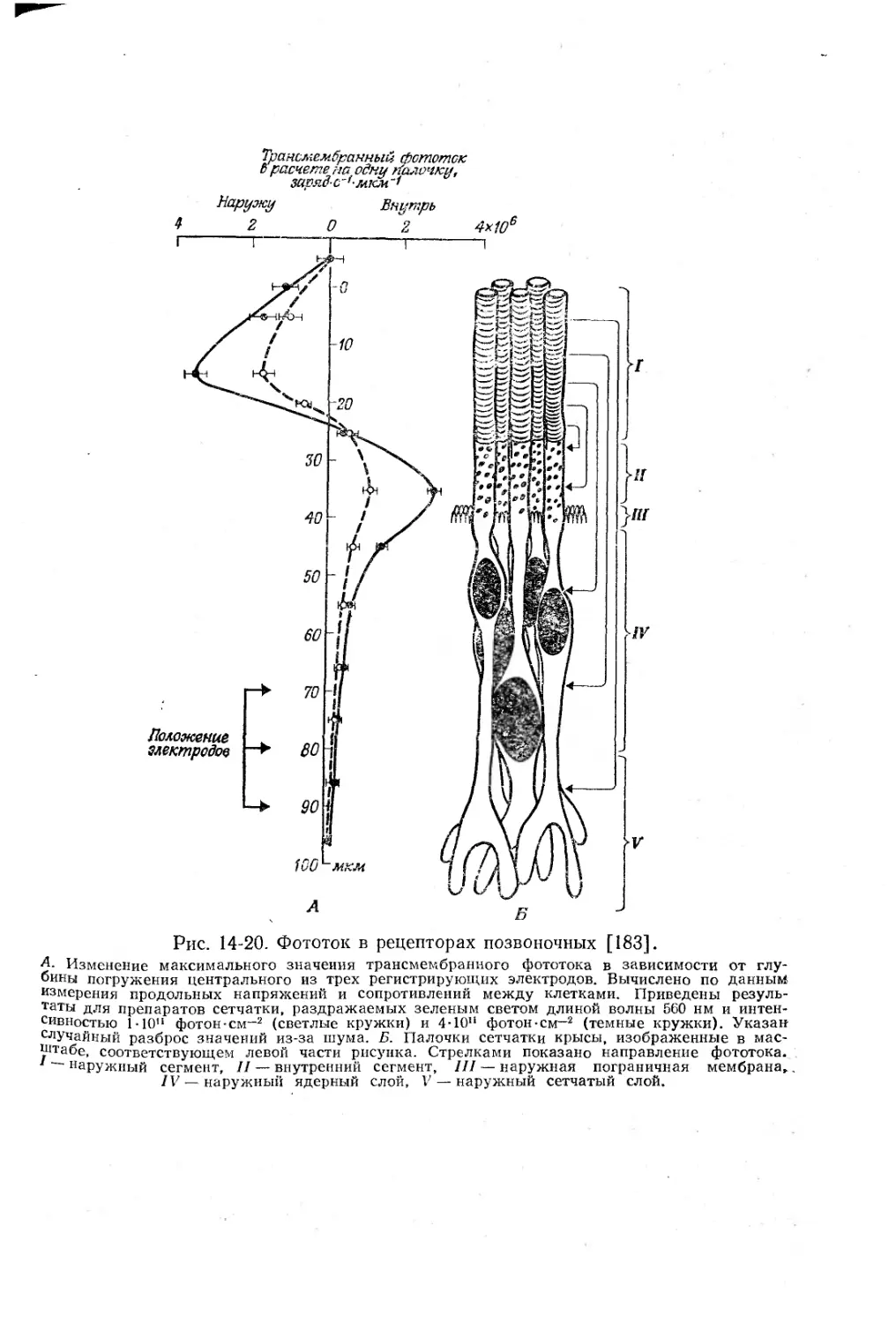
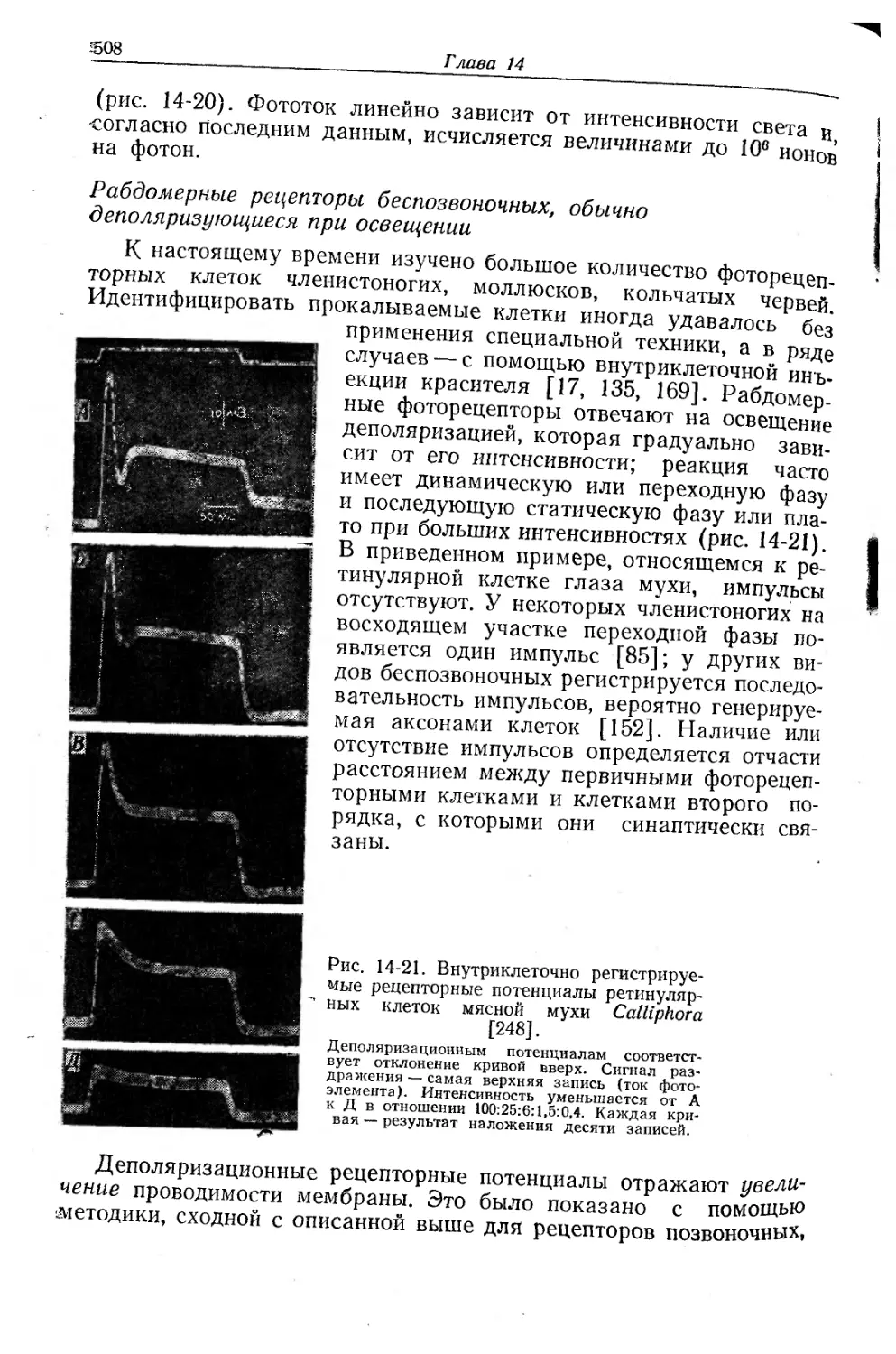
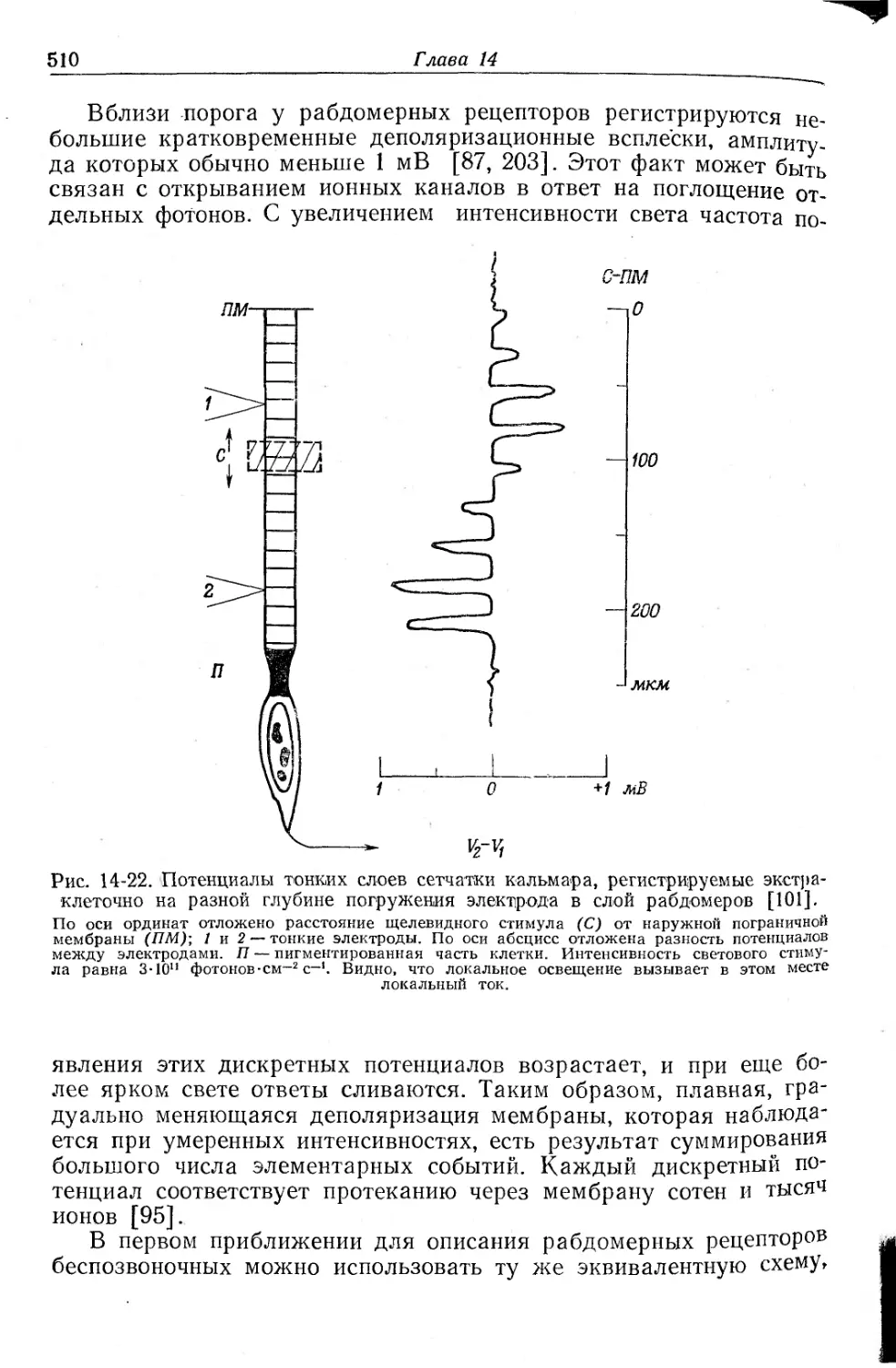
/





































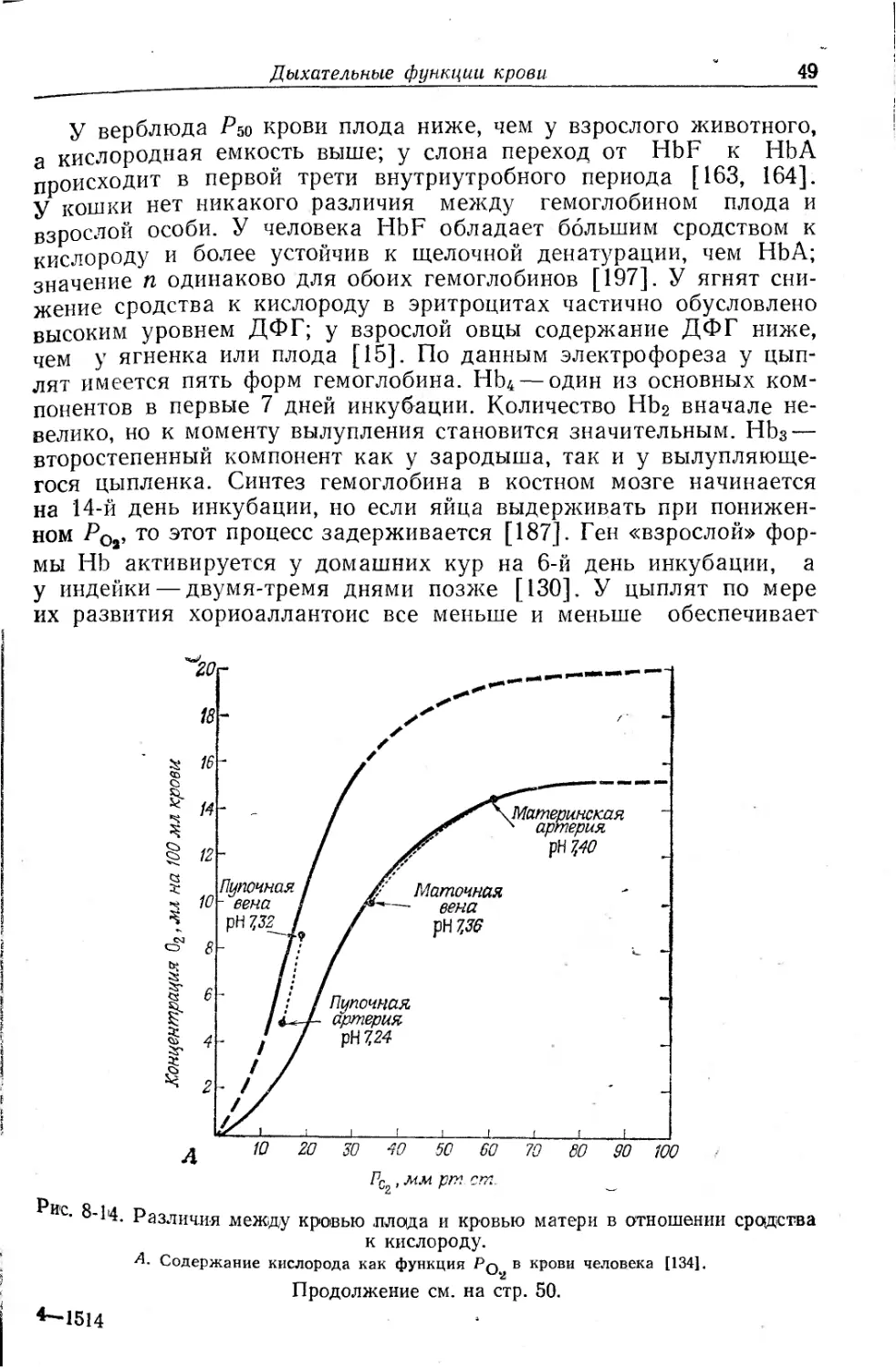
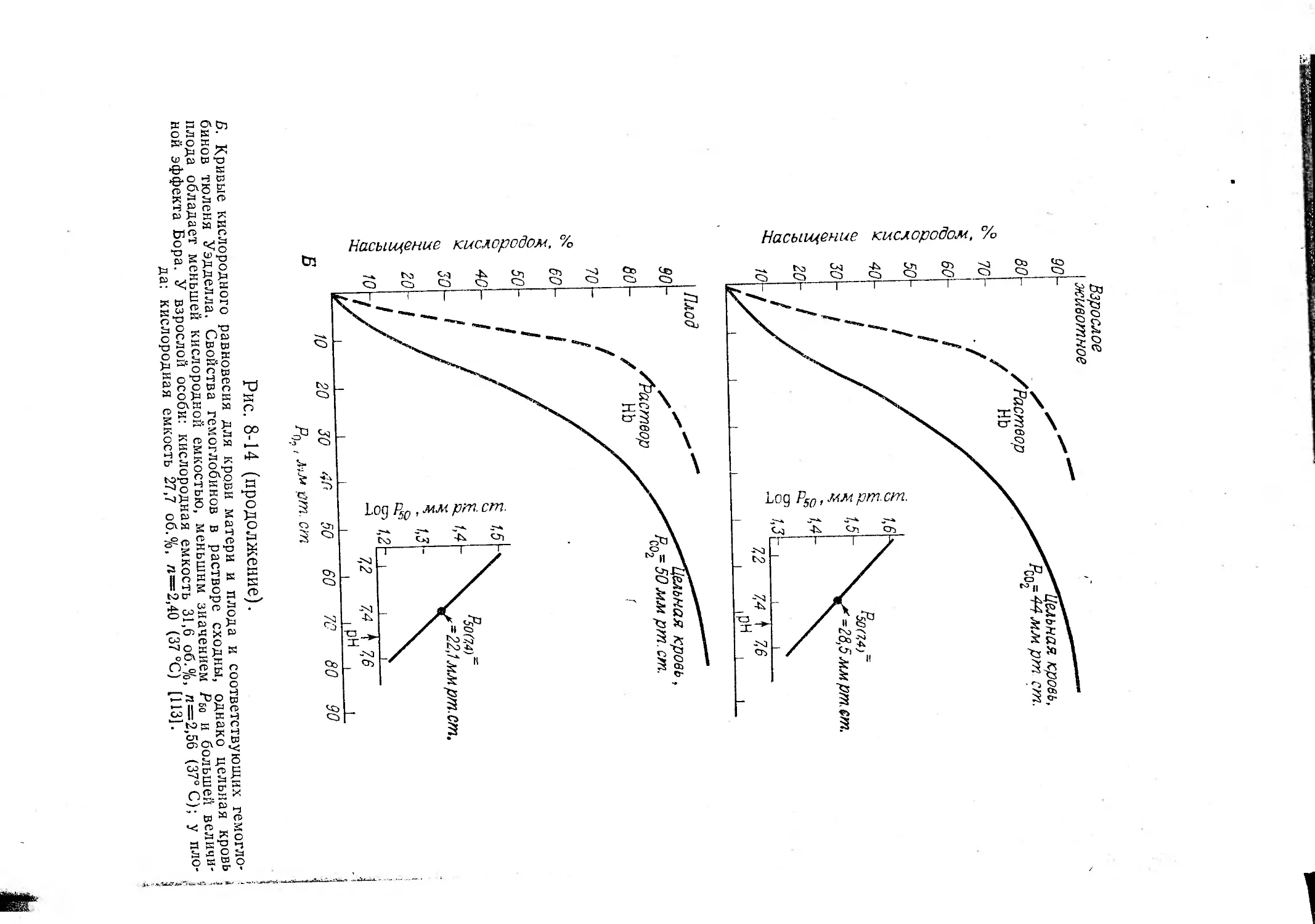
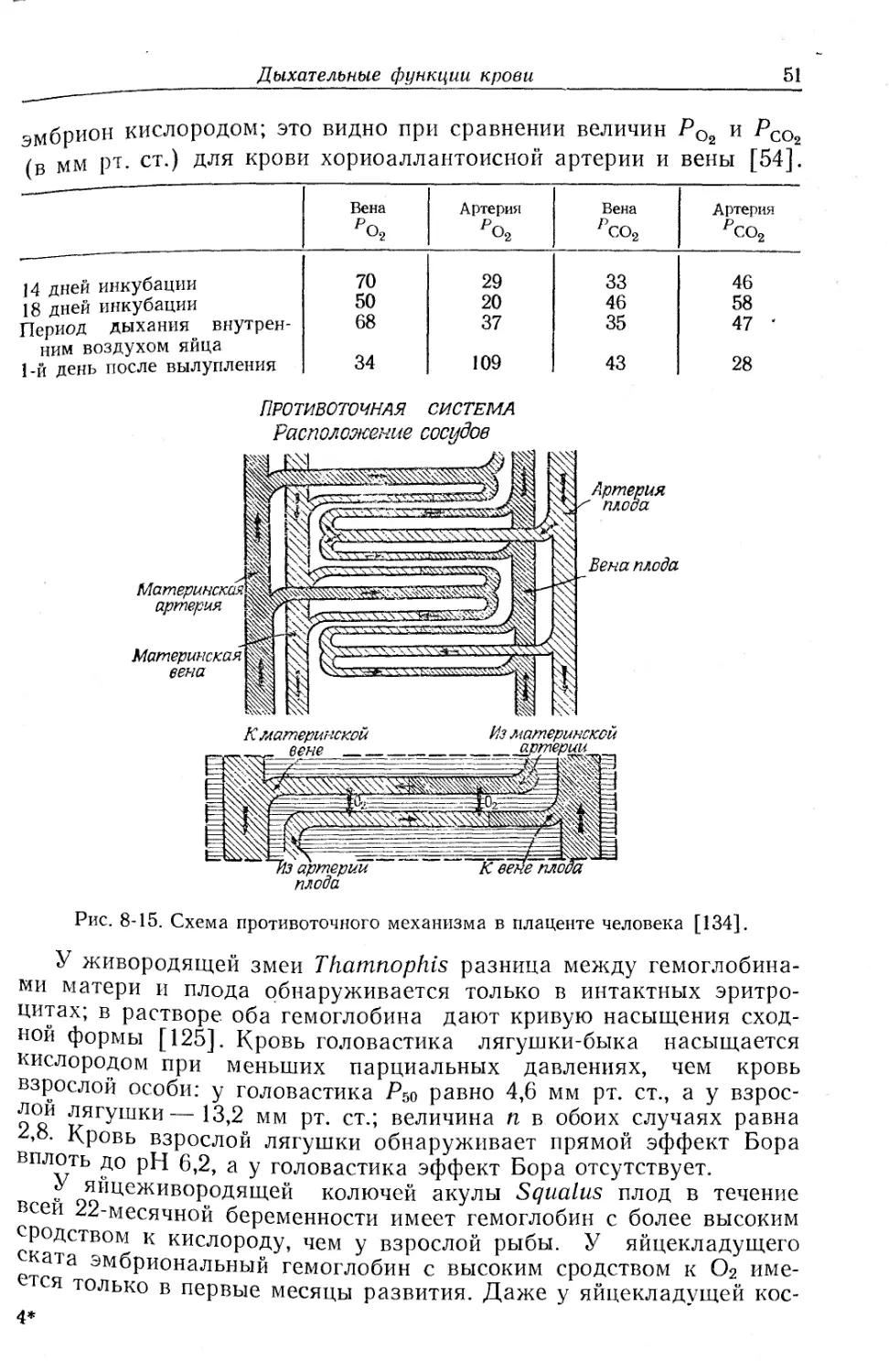

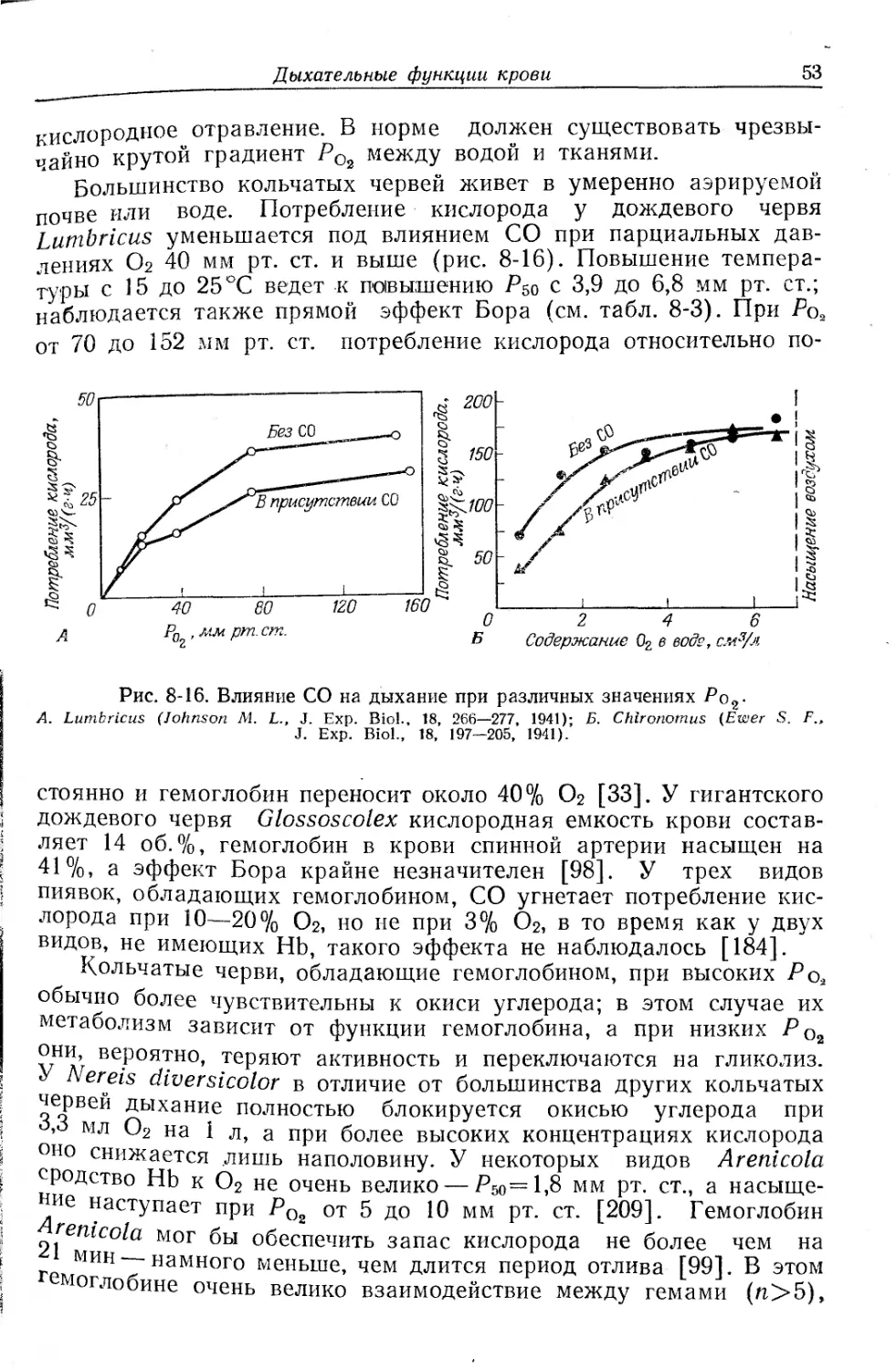

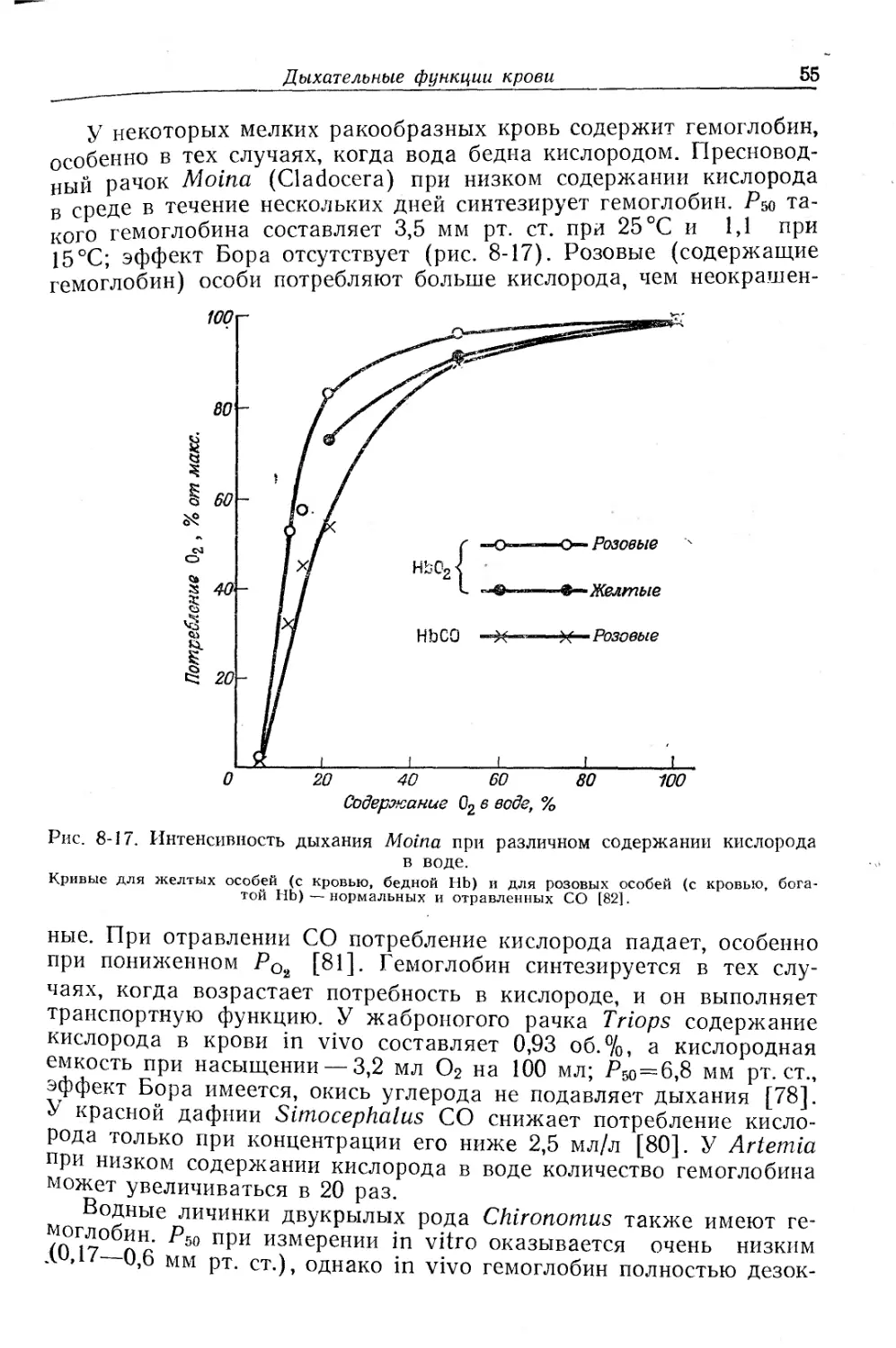









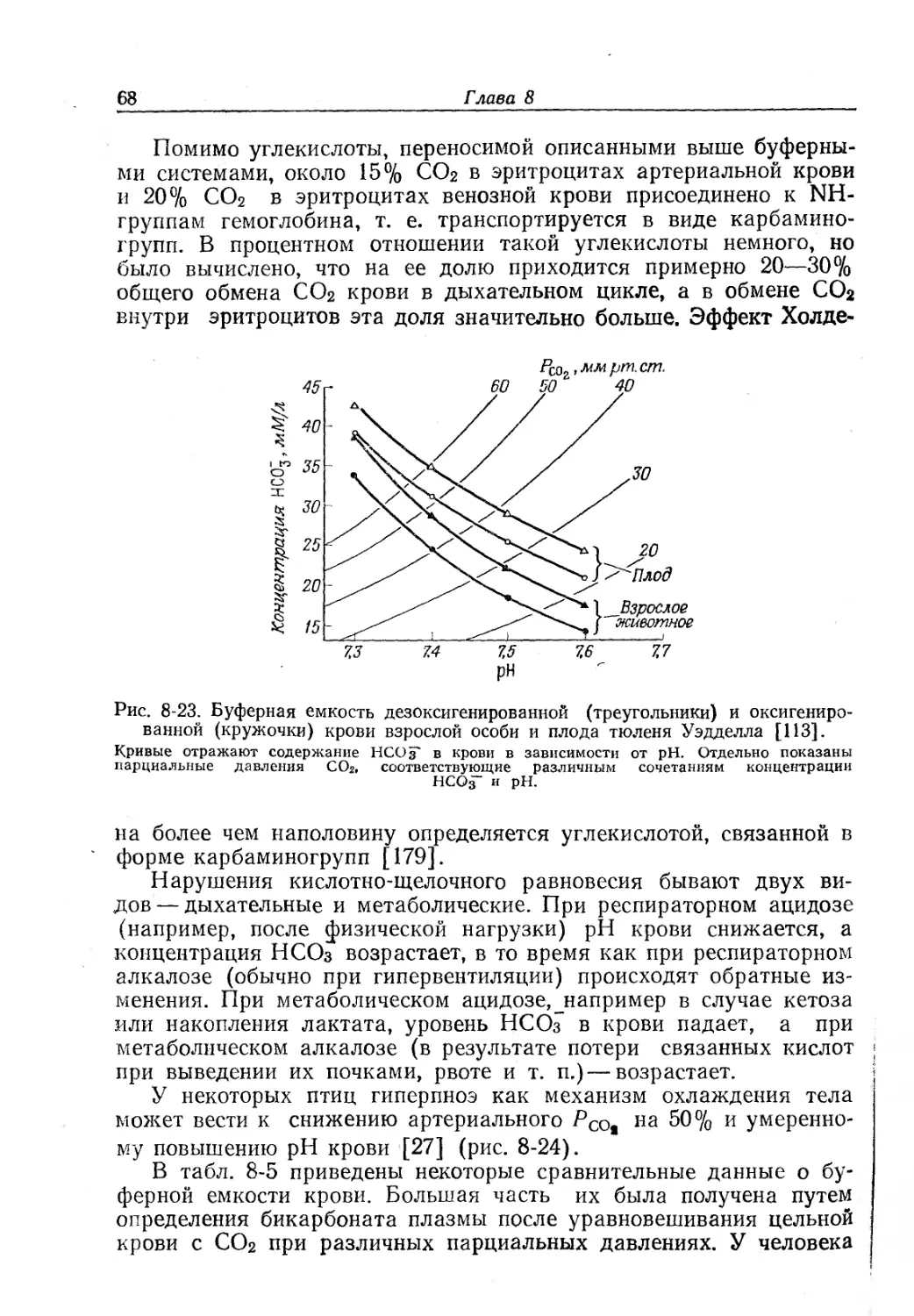
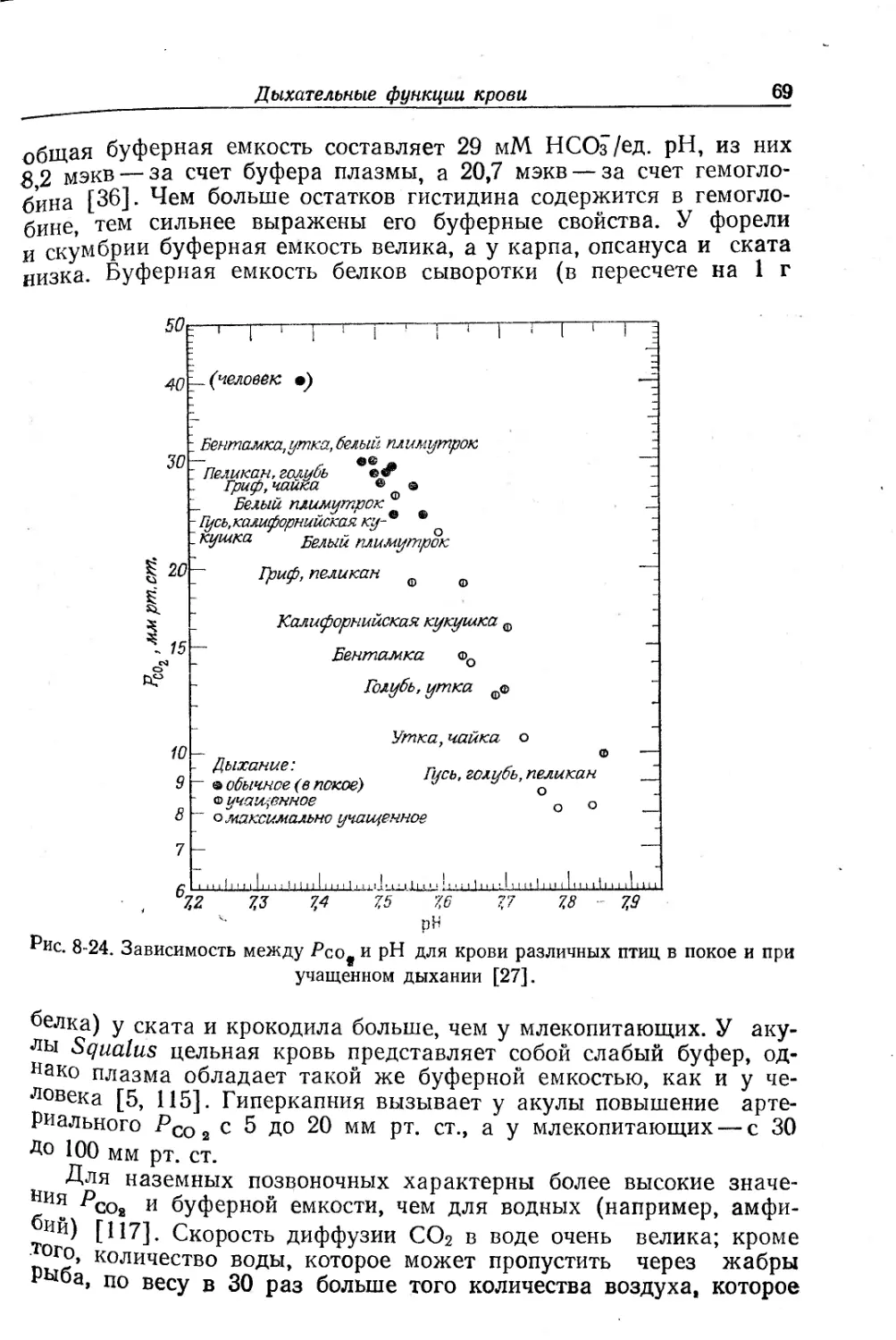

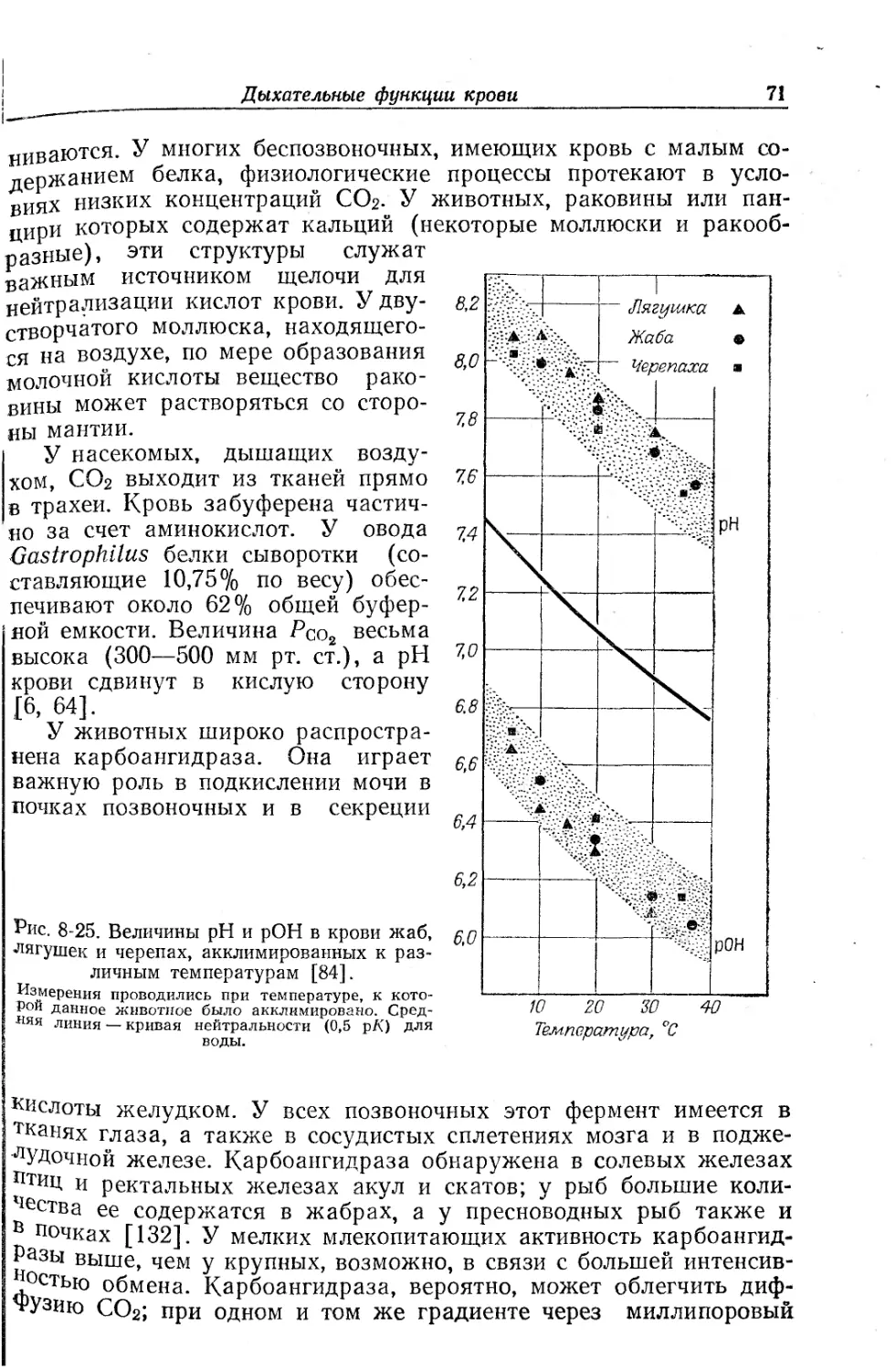



















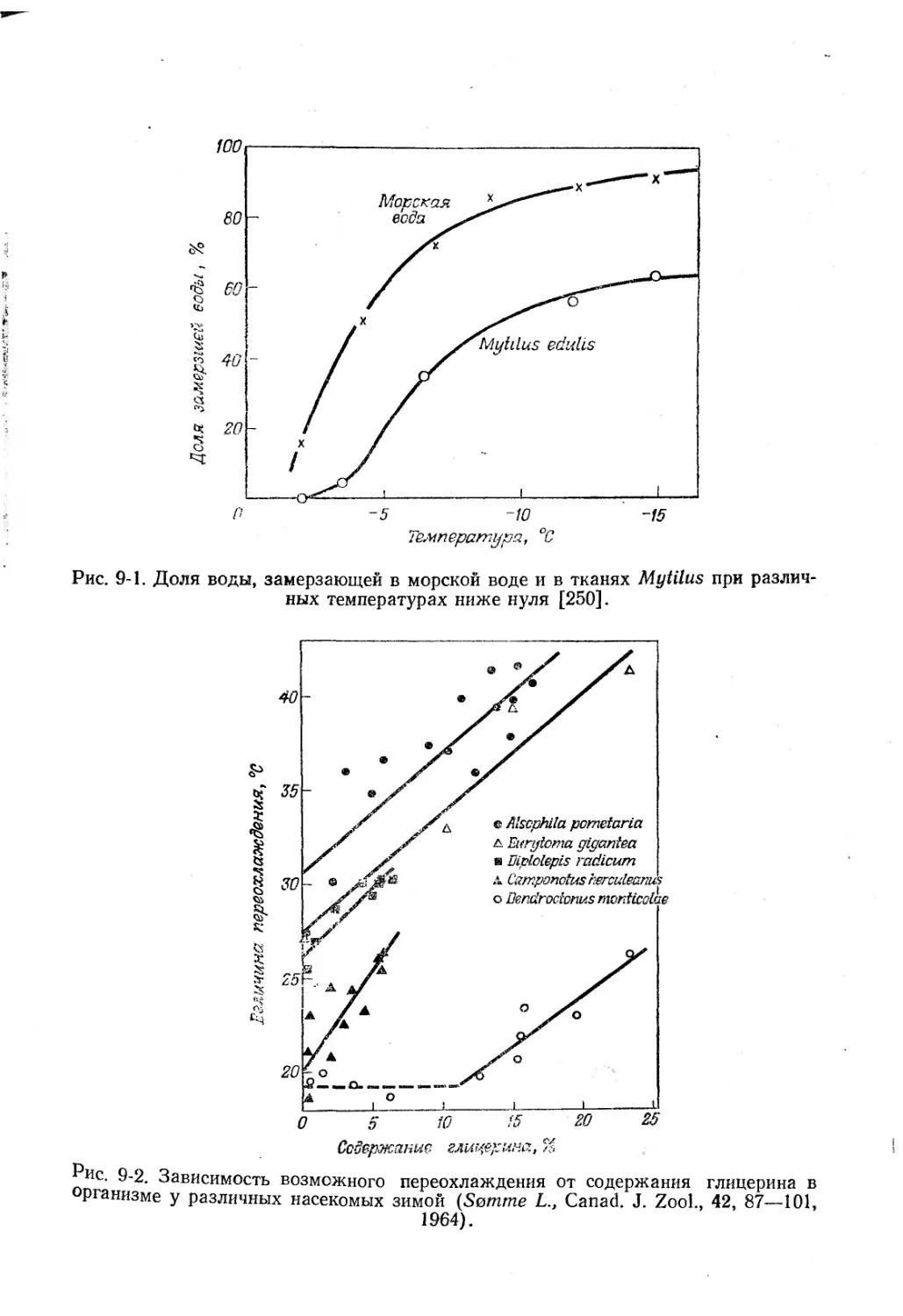













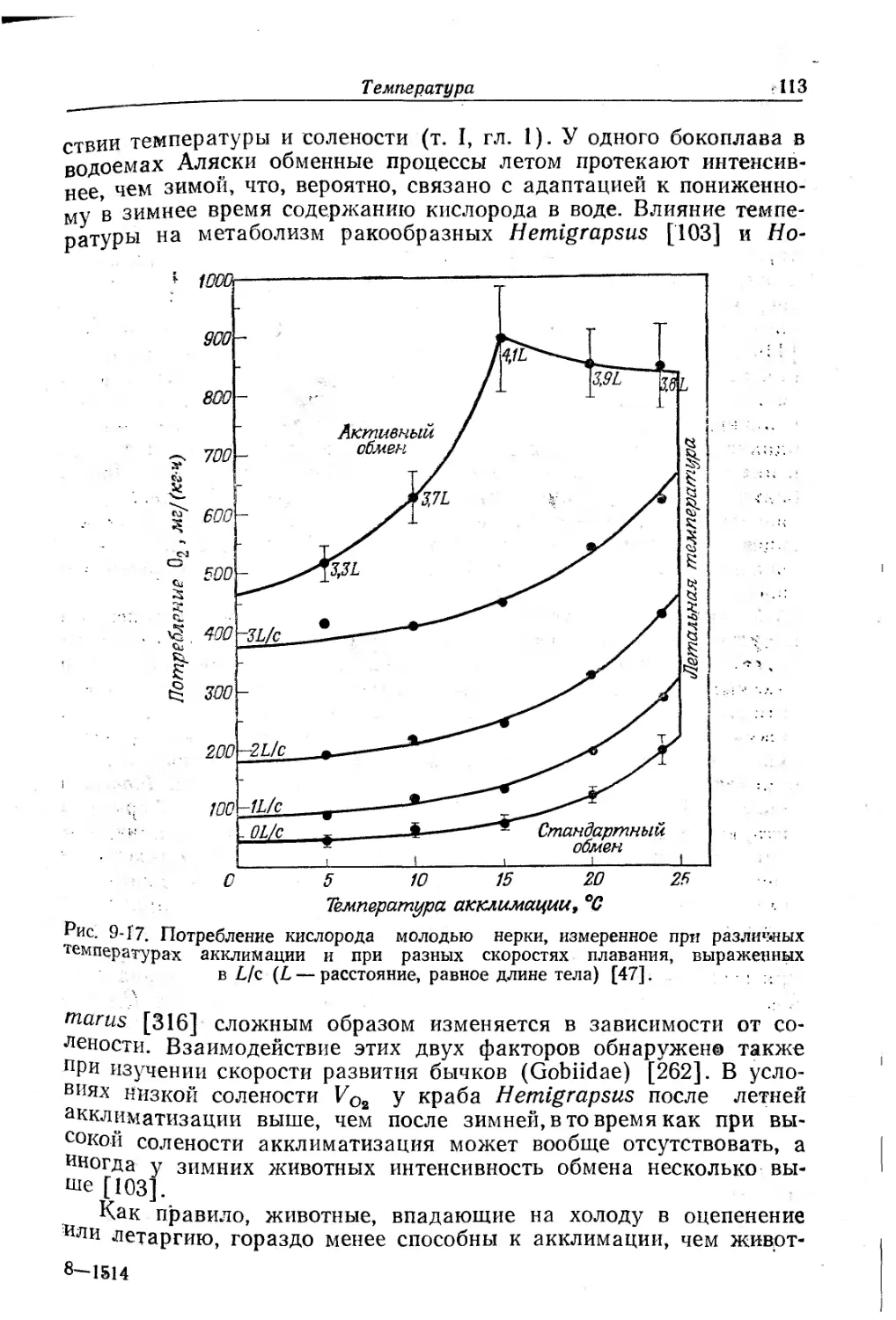





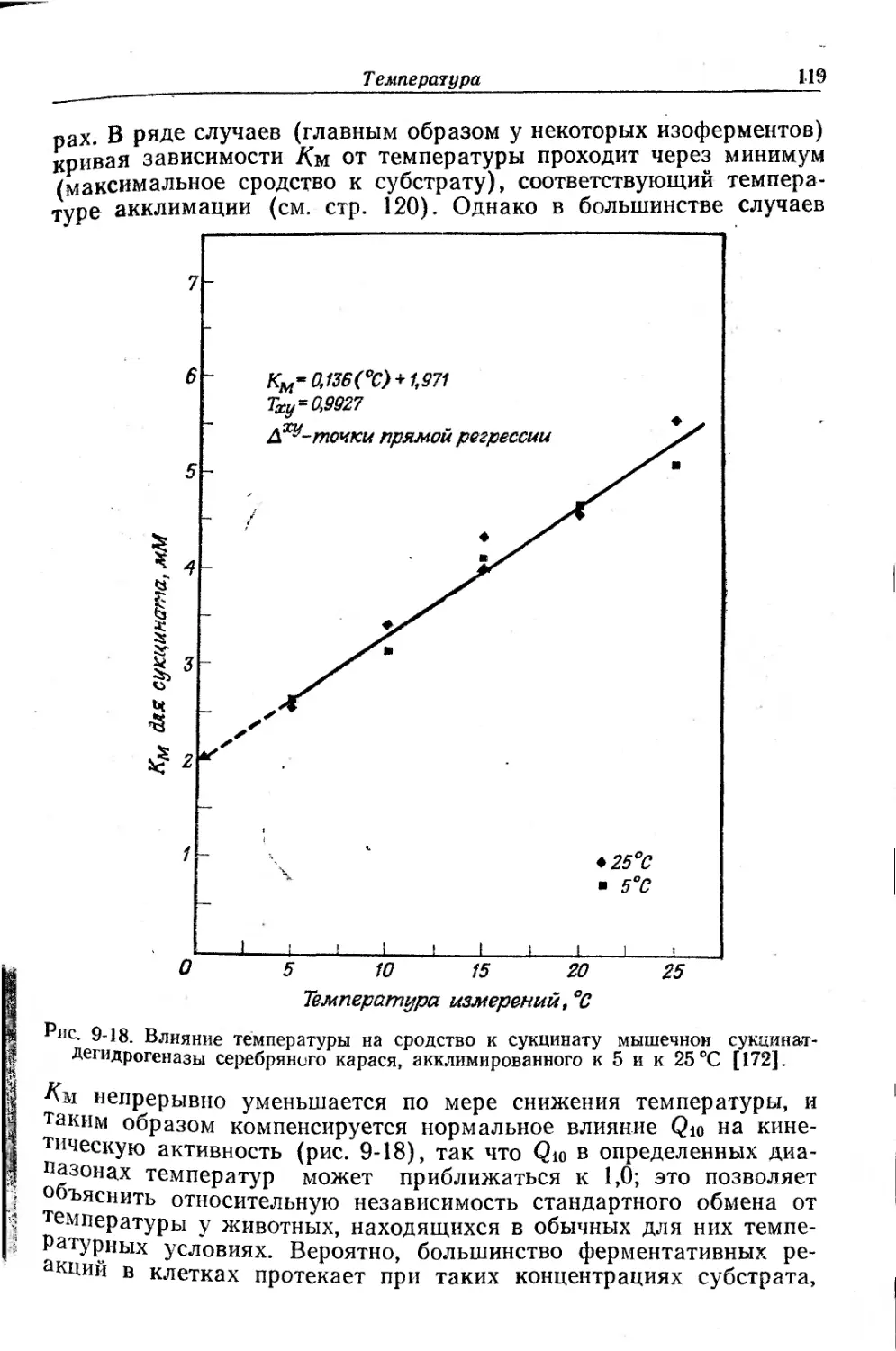

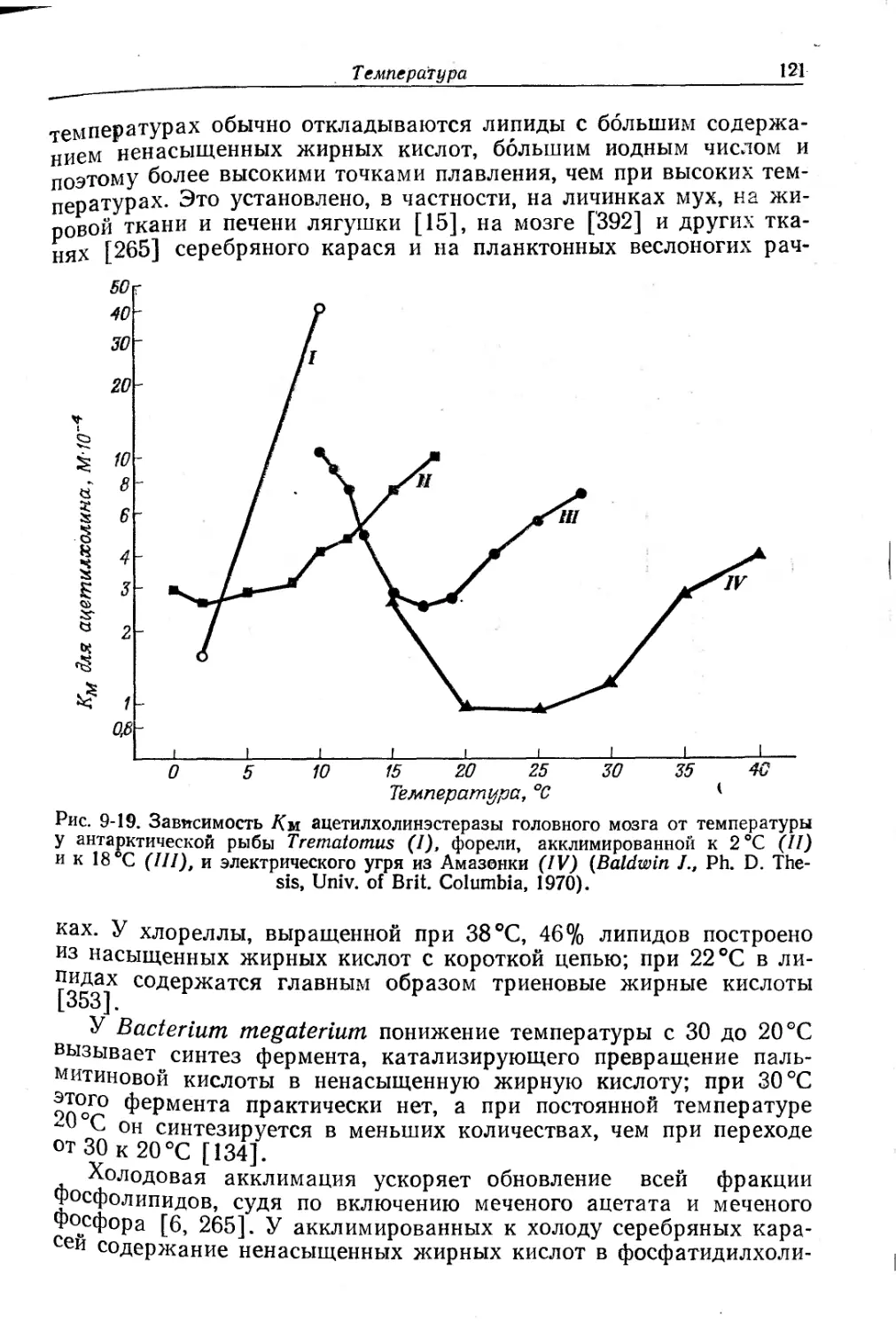
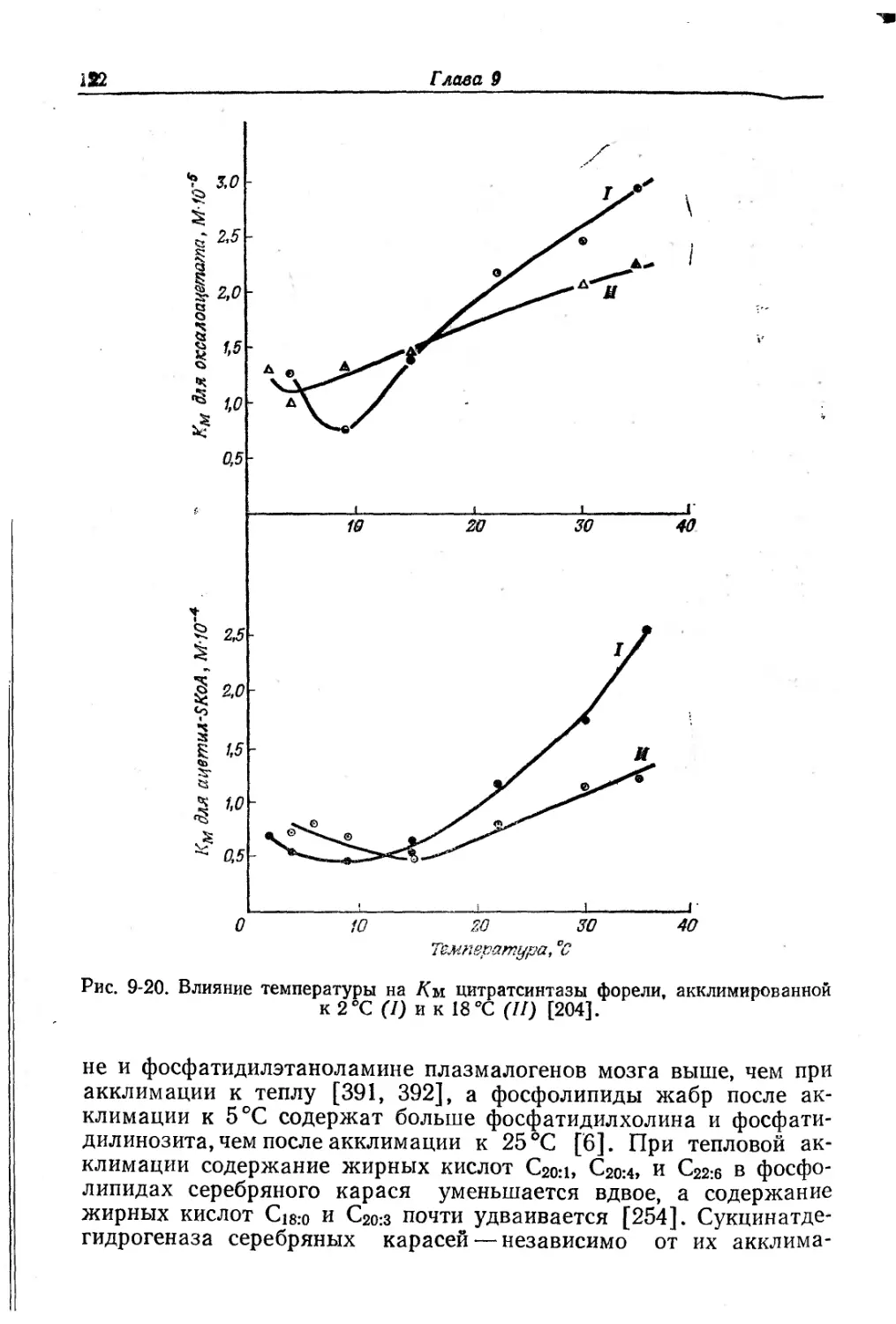
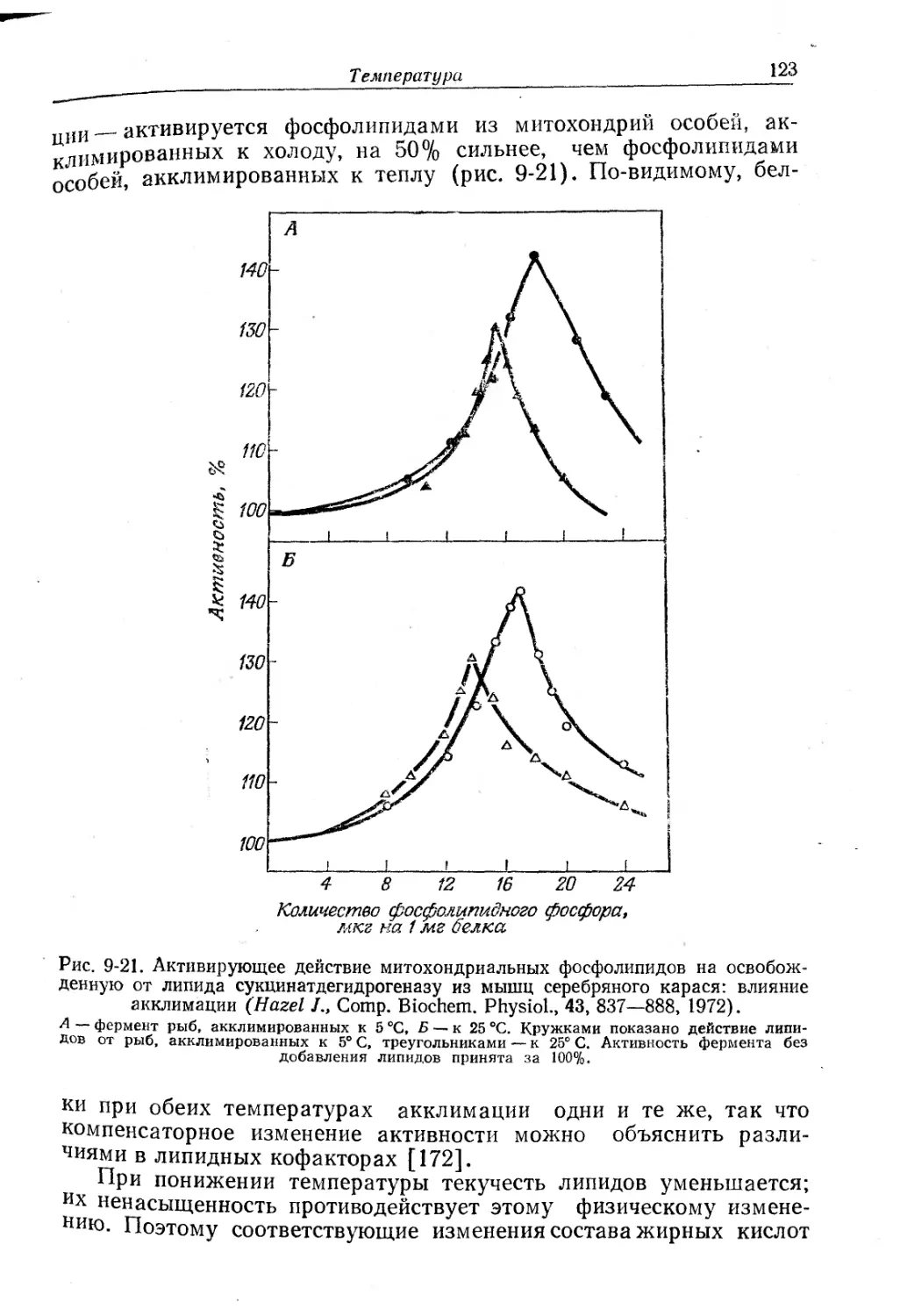
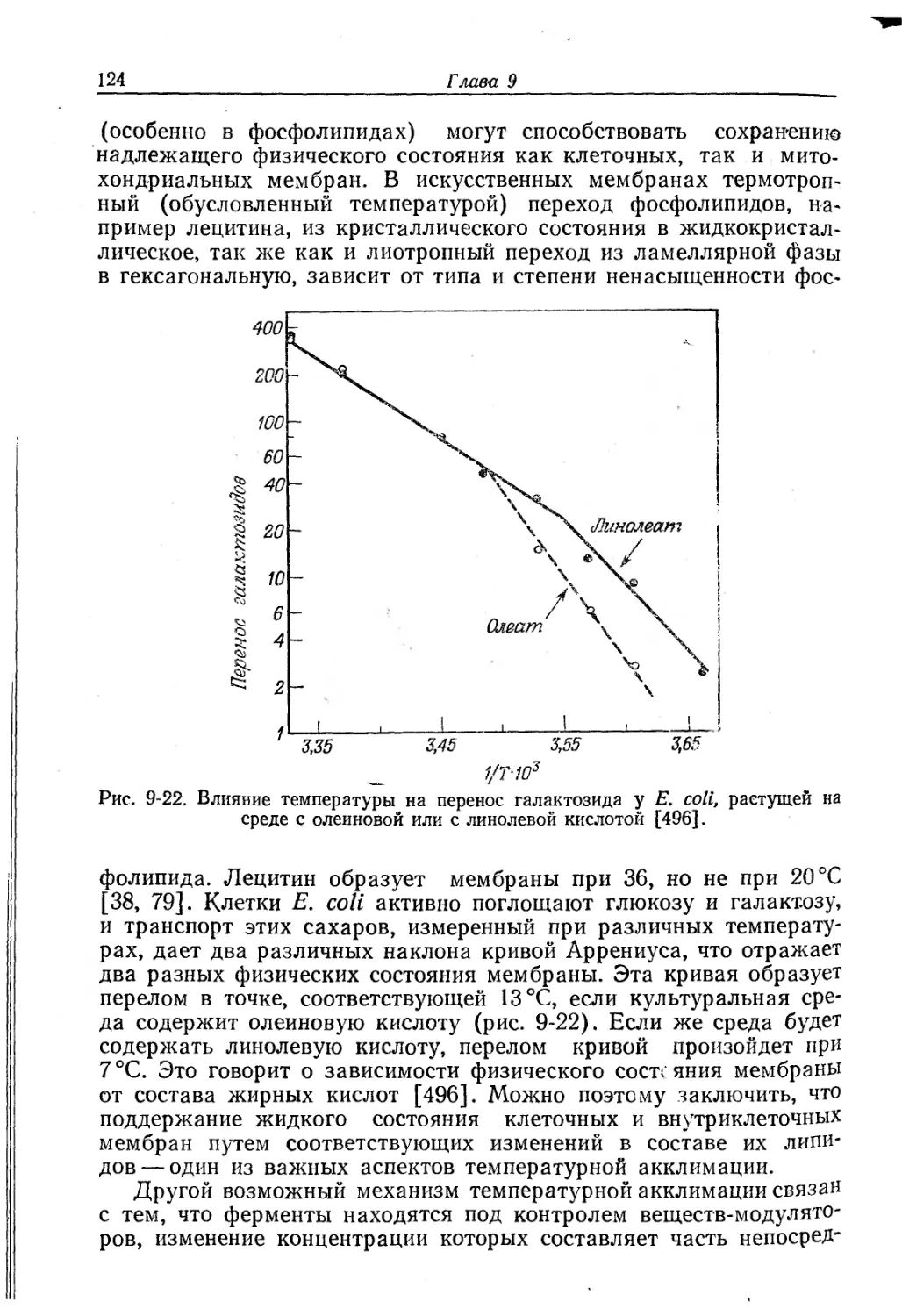





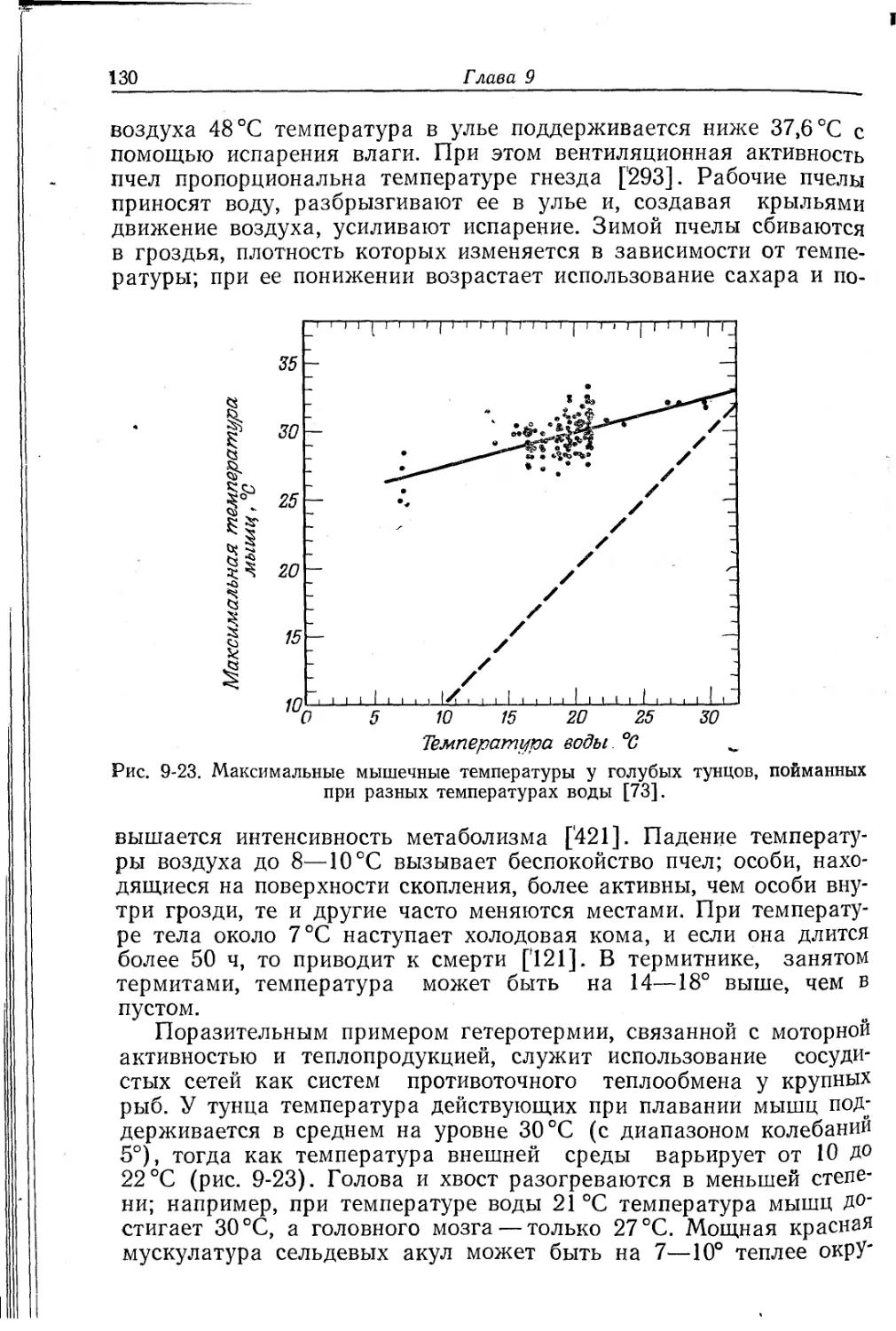
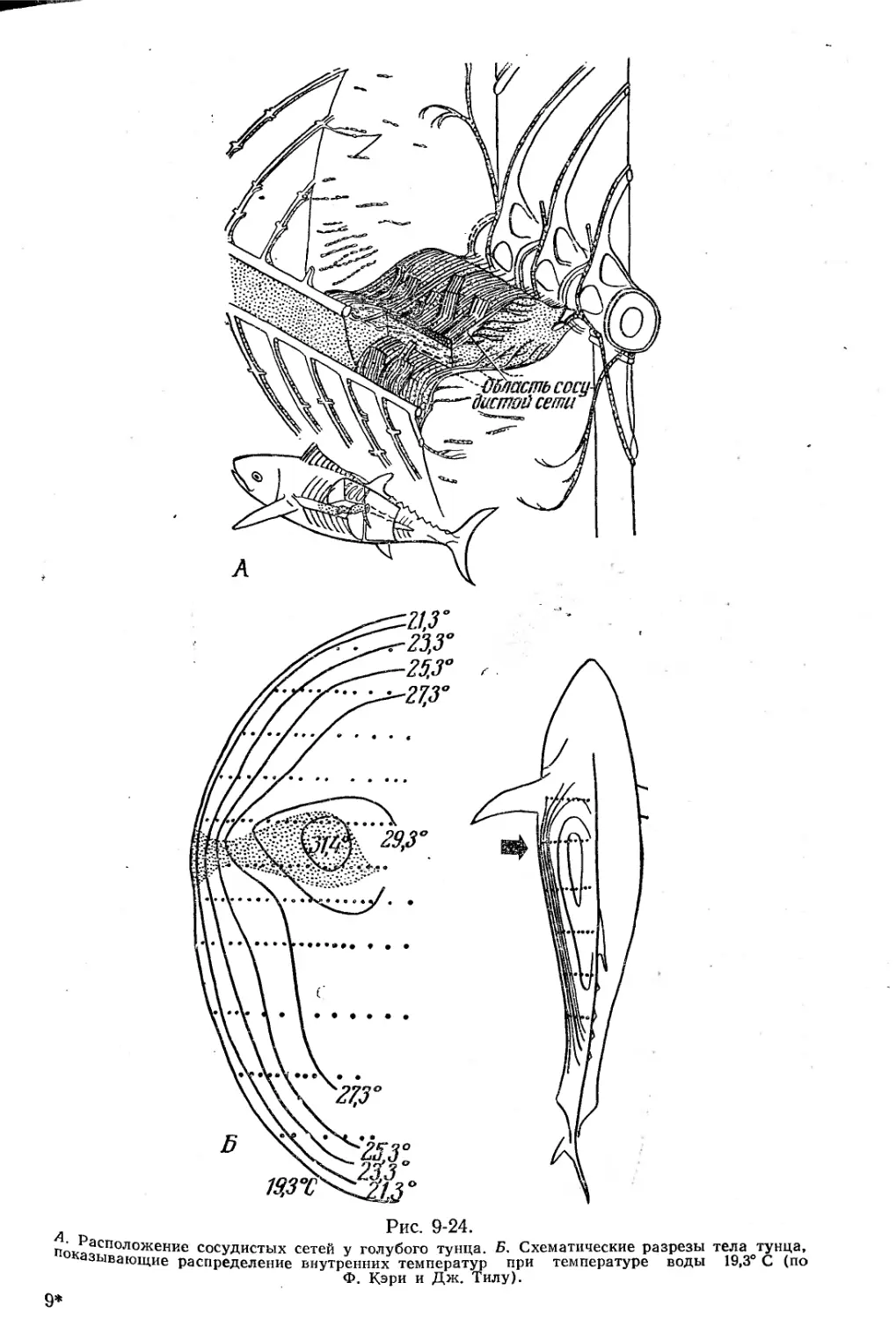






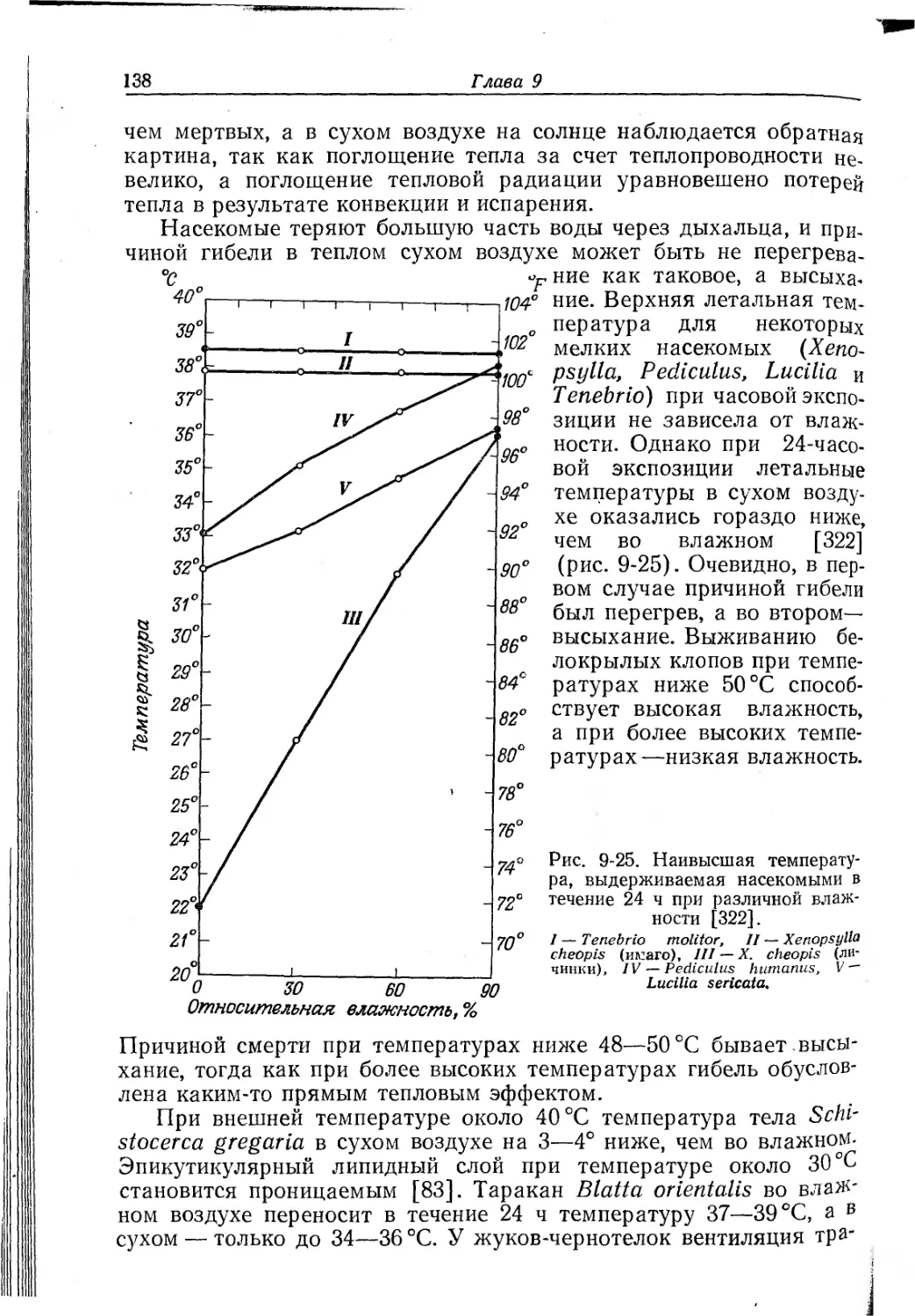



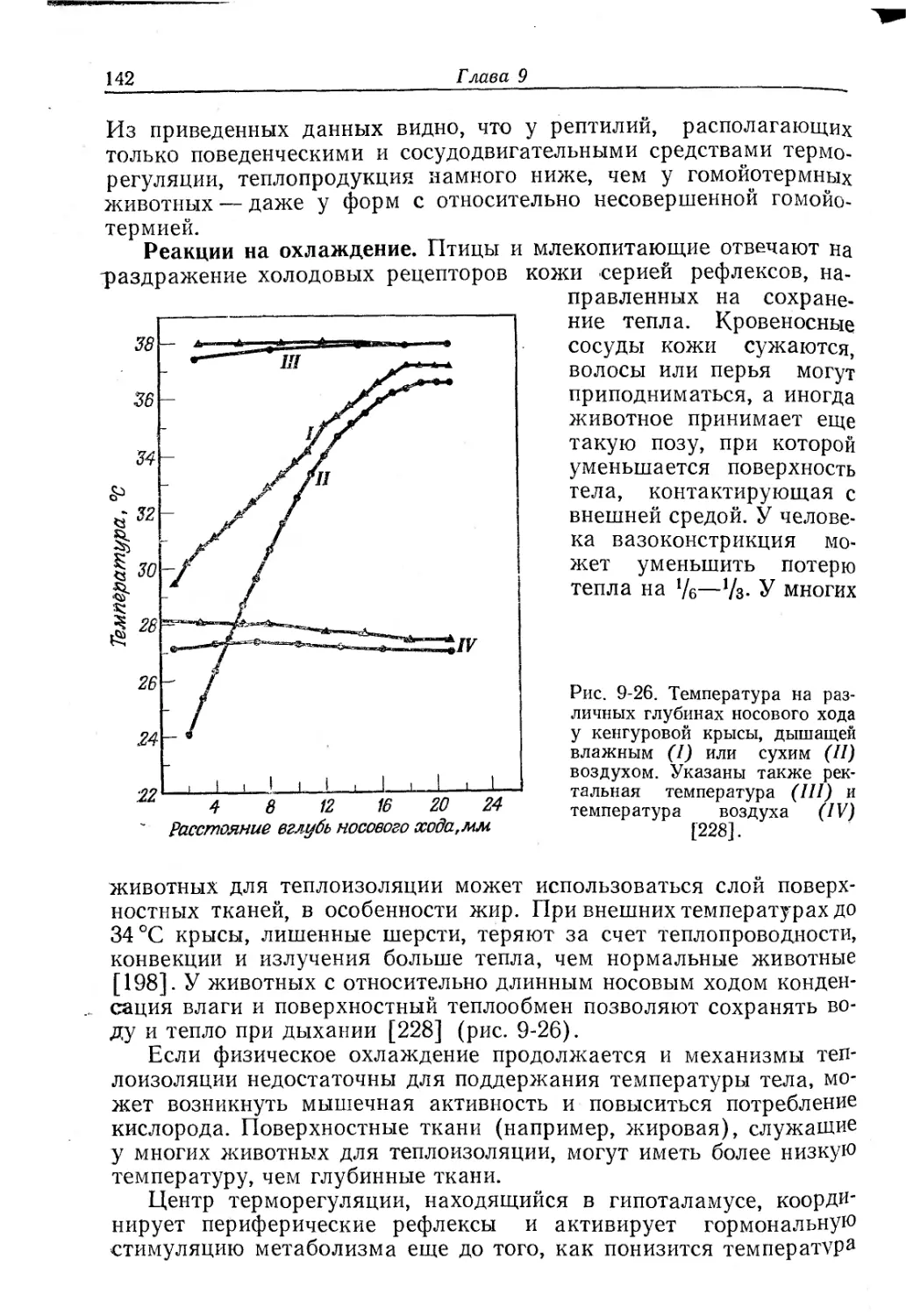

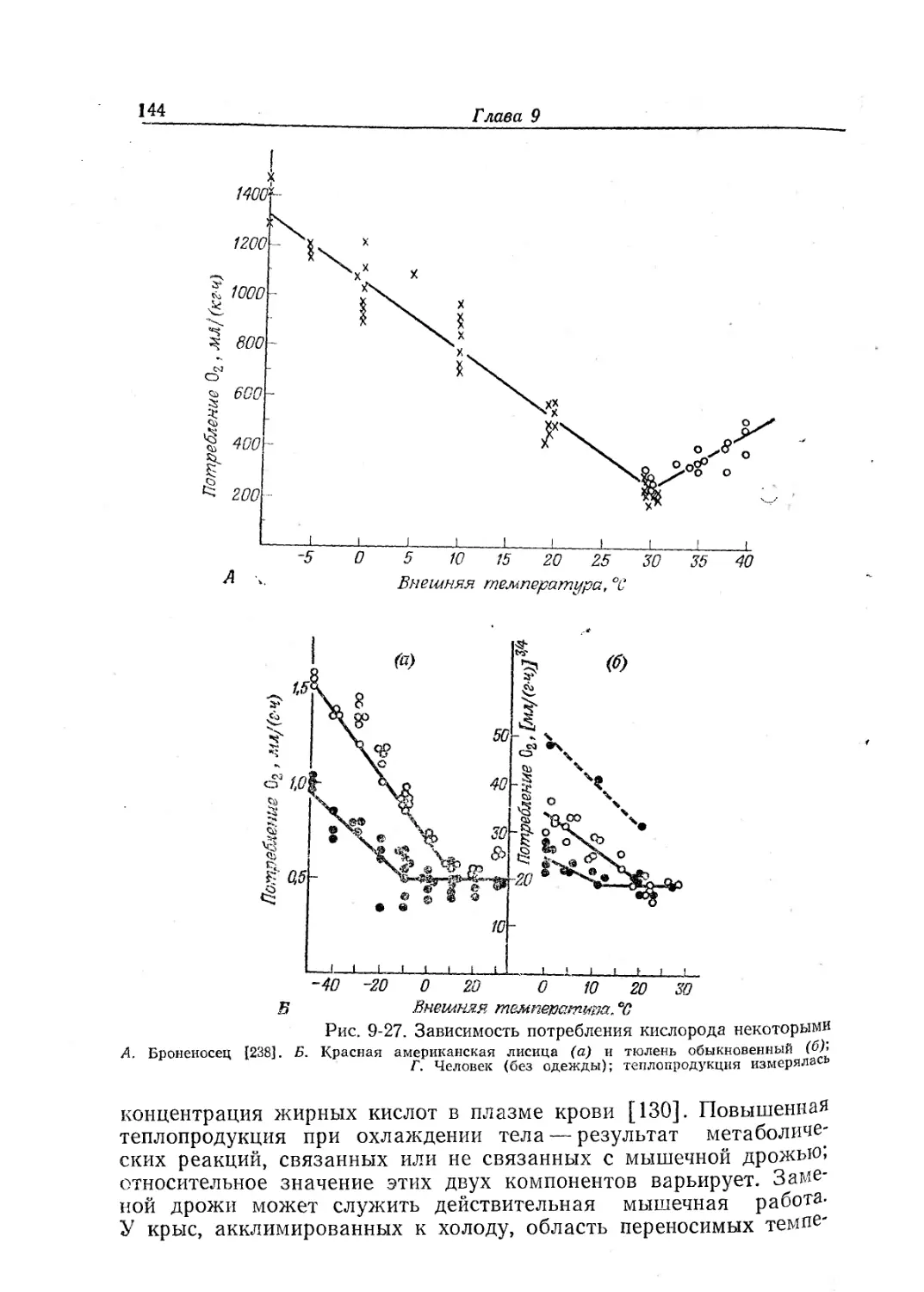
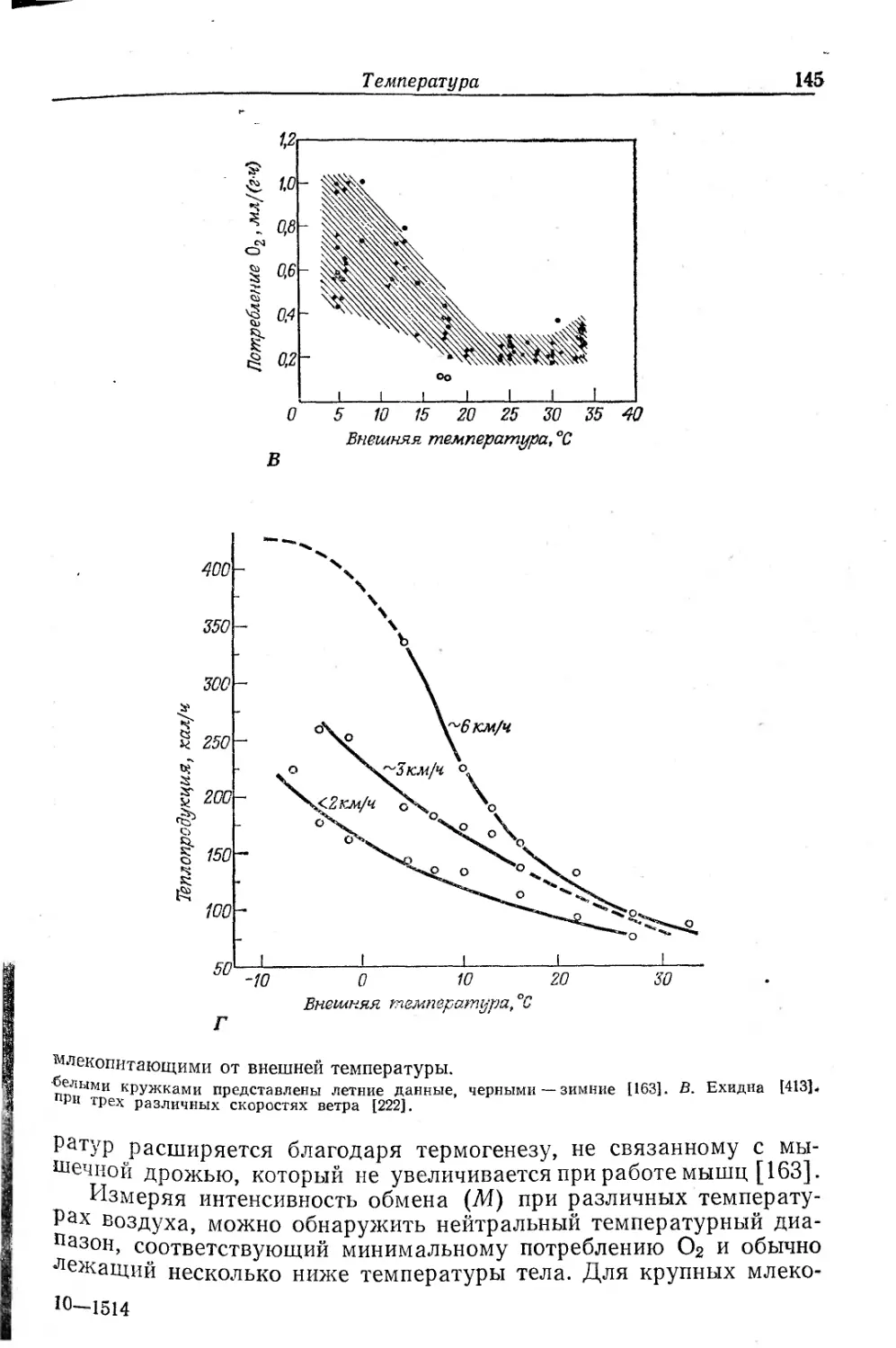






























































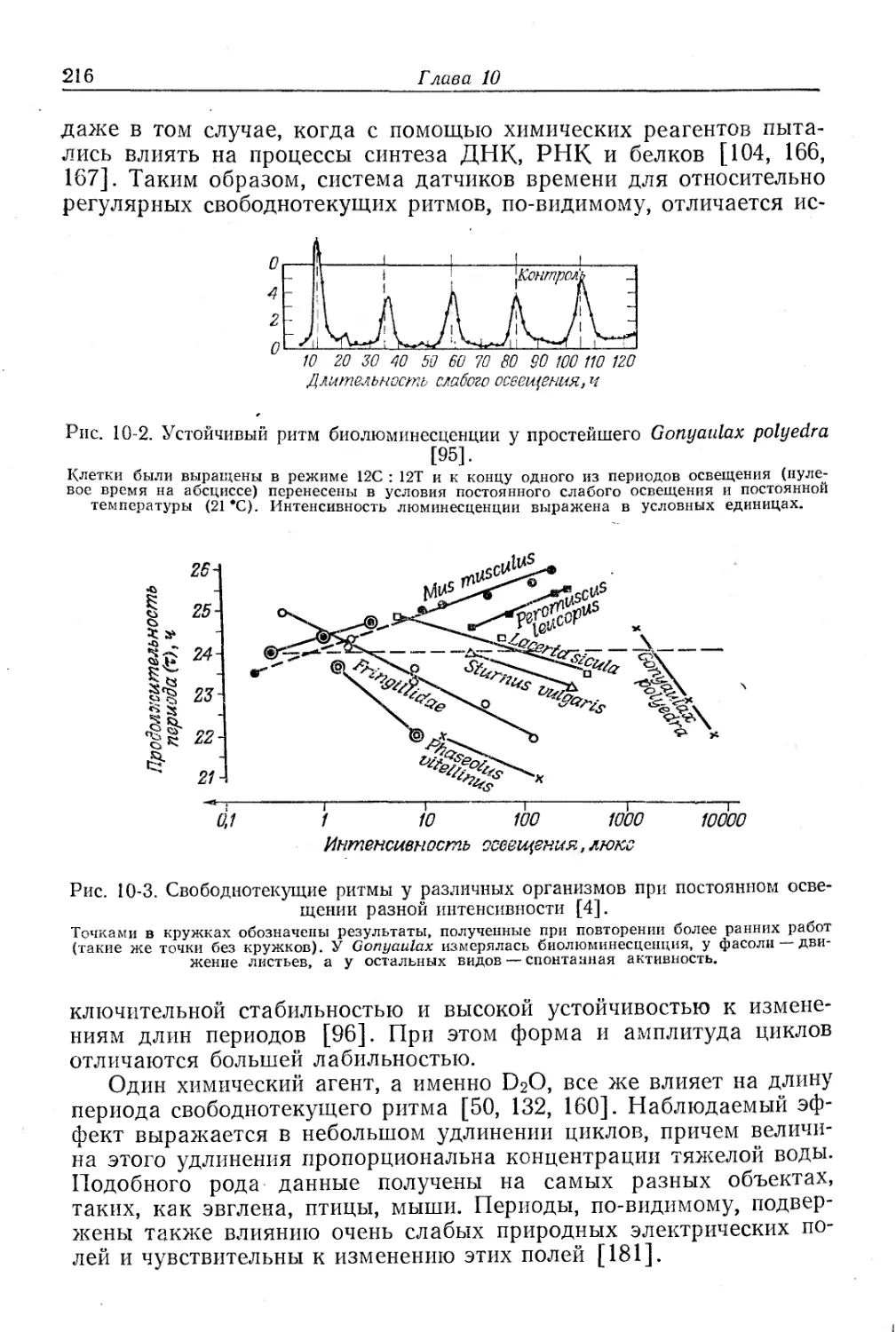

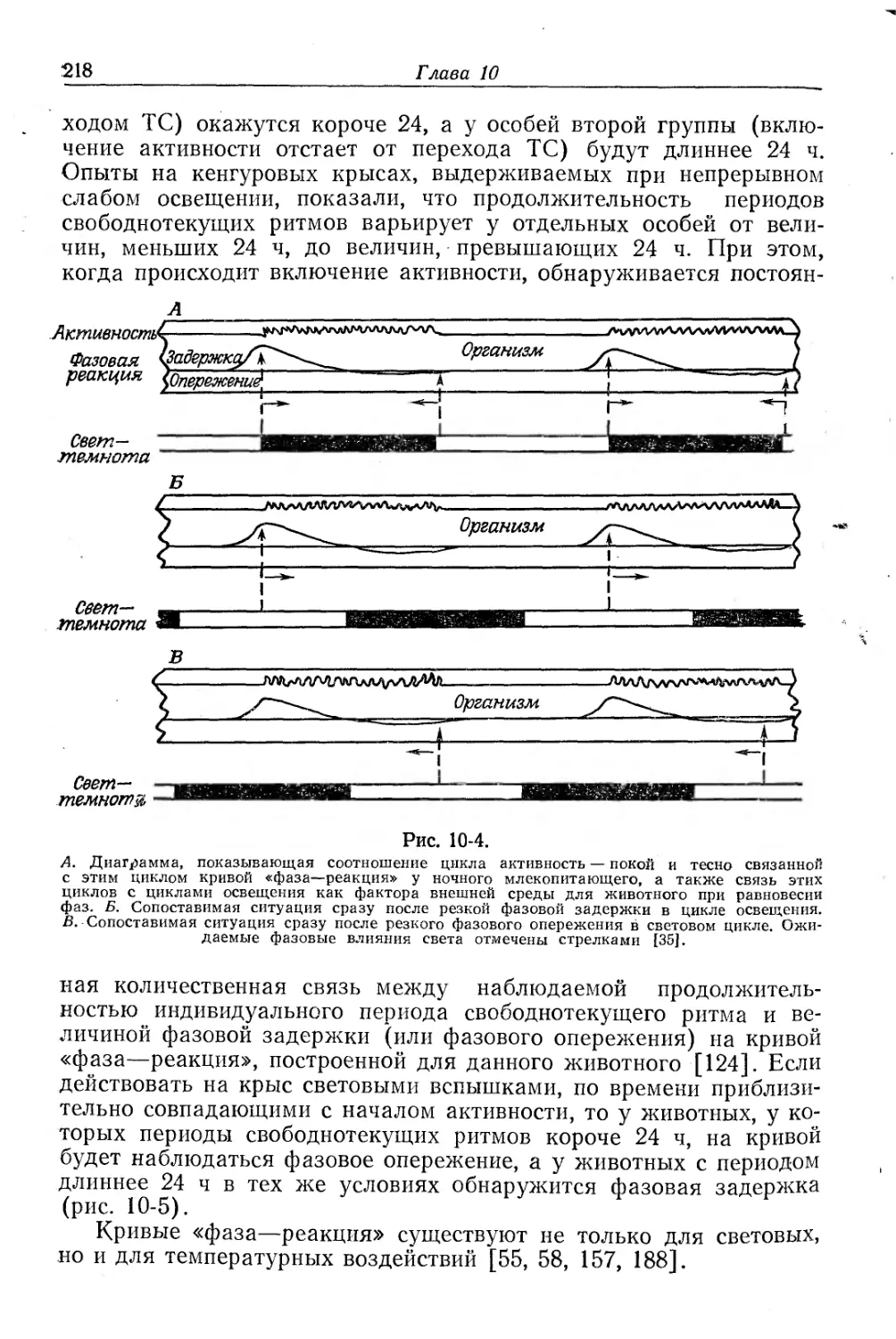
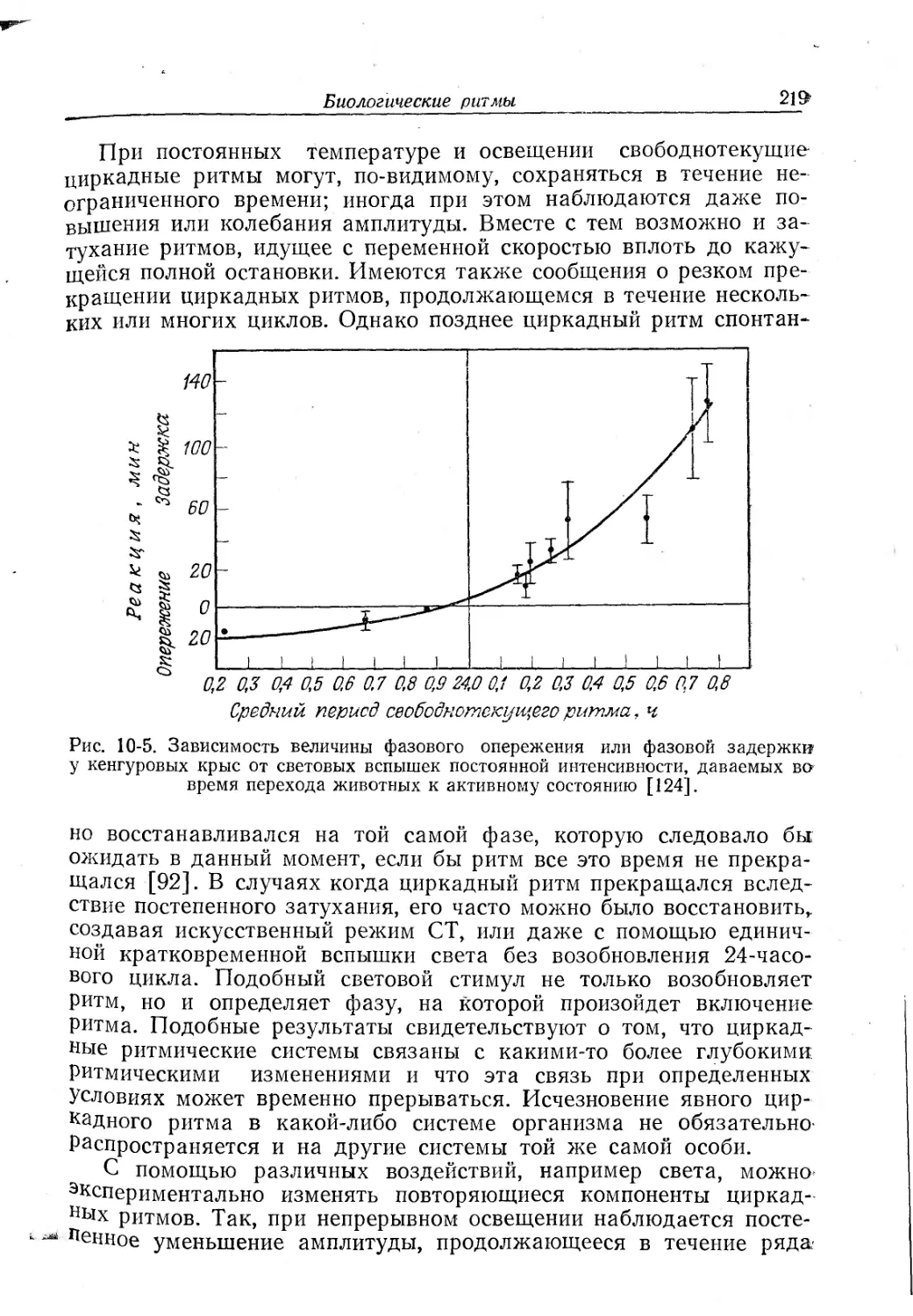





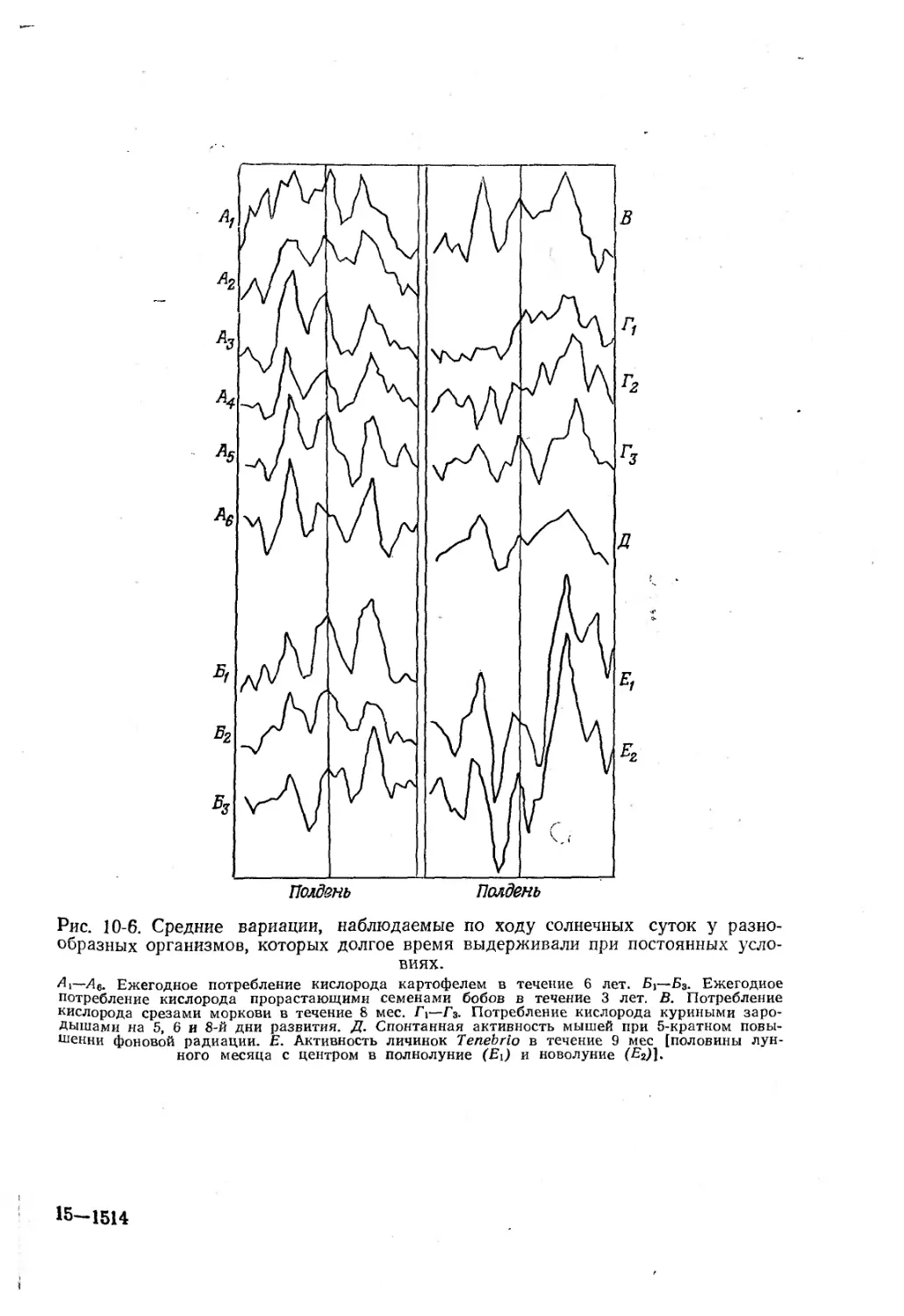






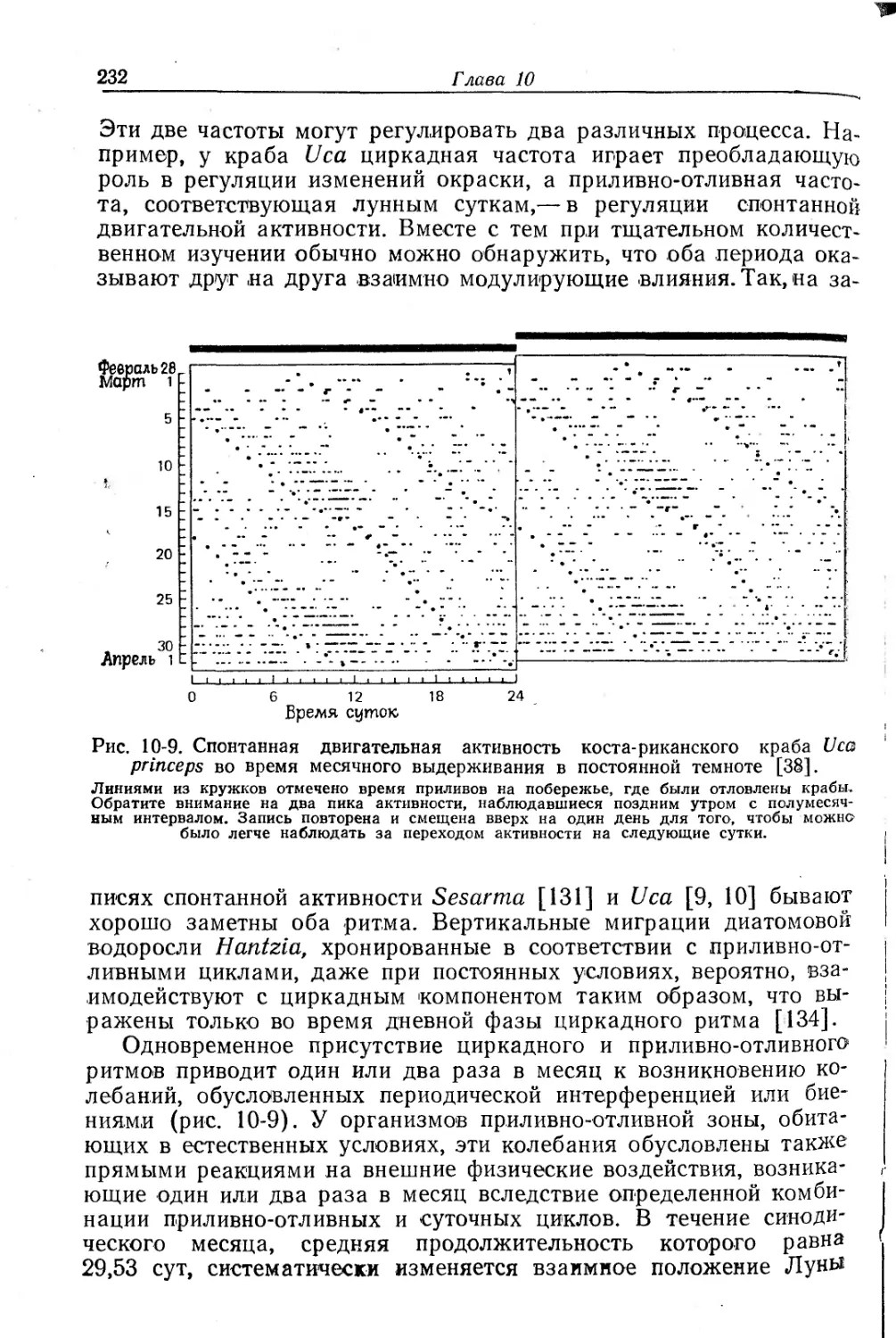










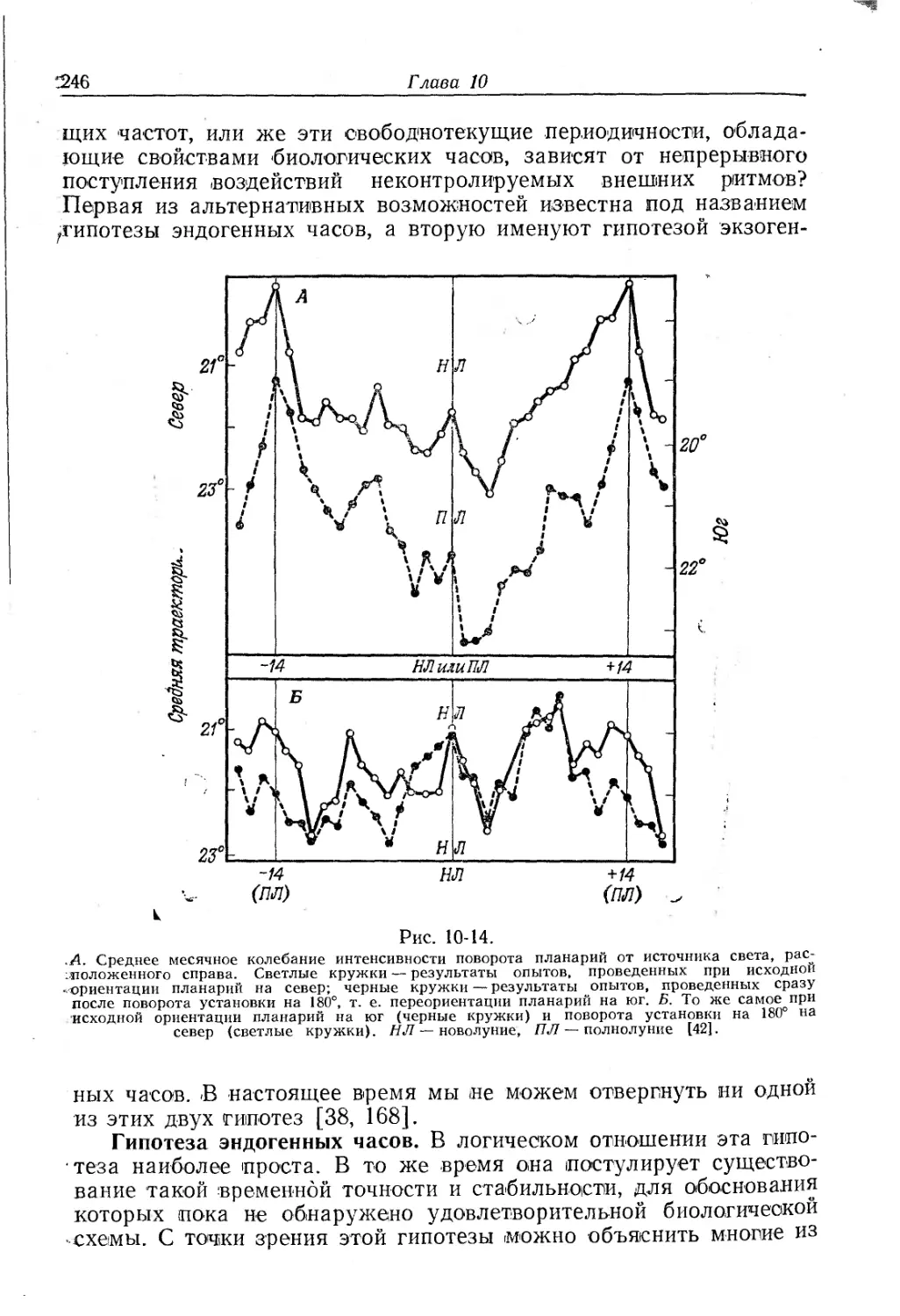

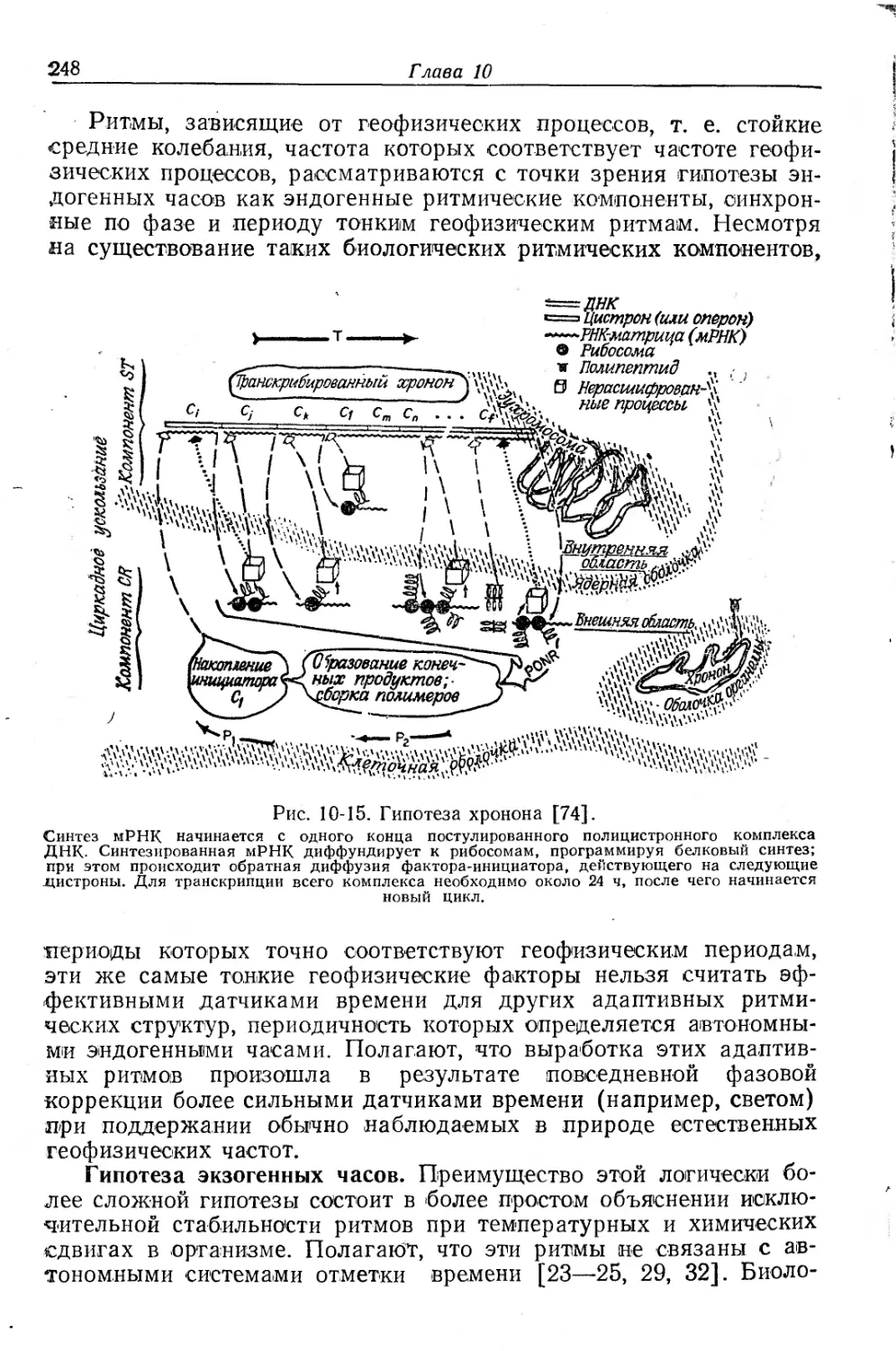

















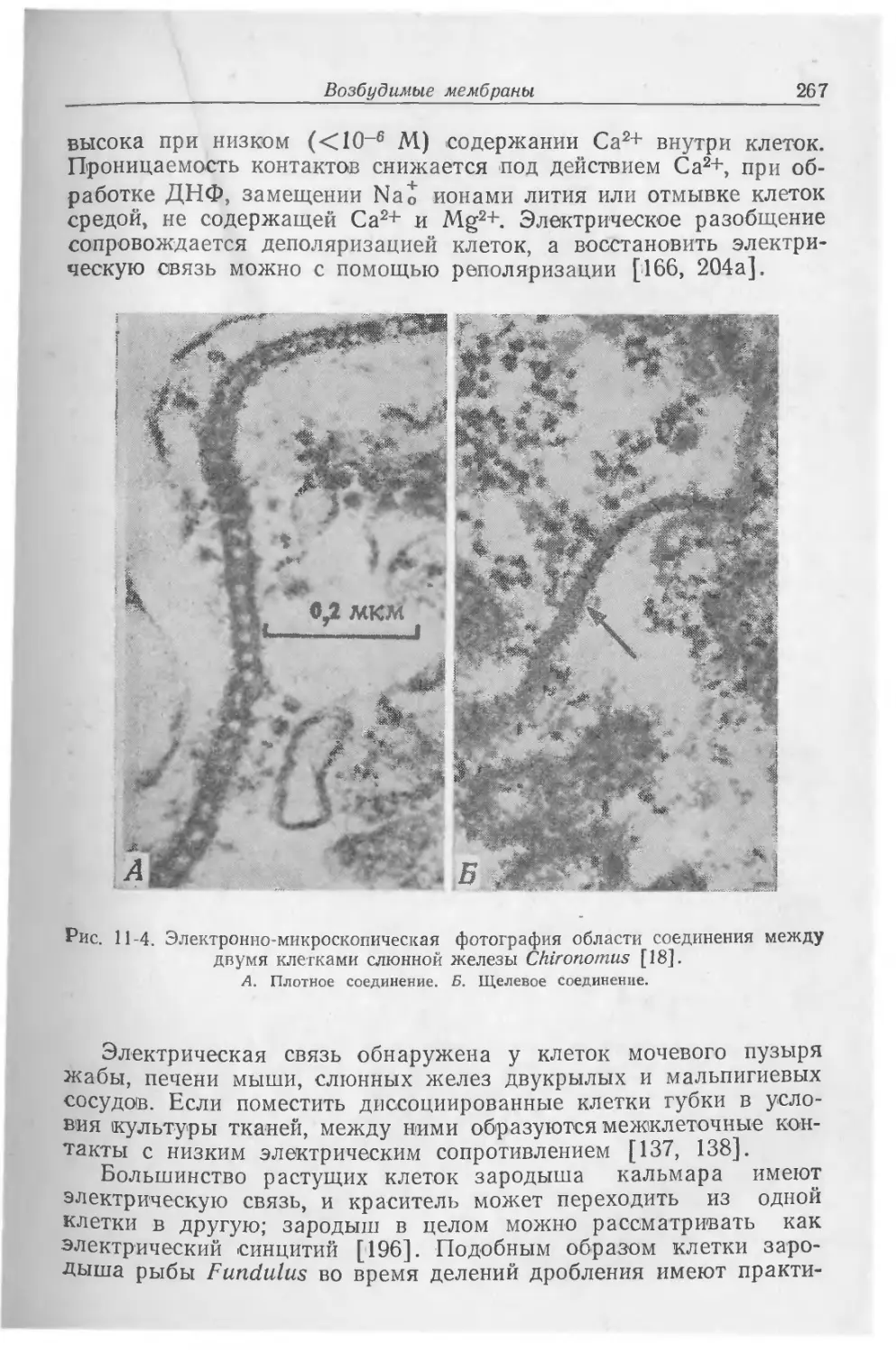











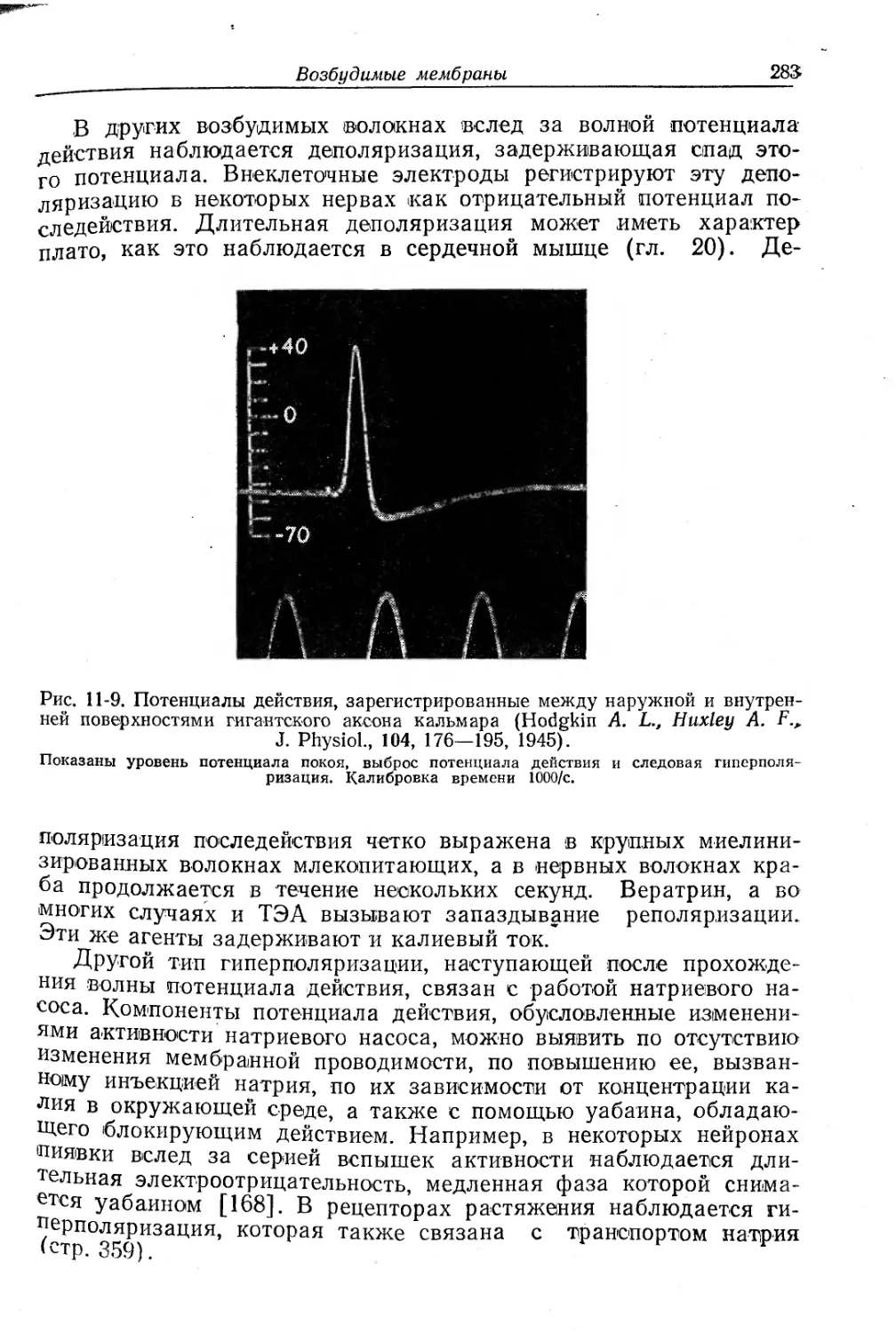








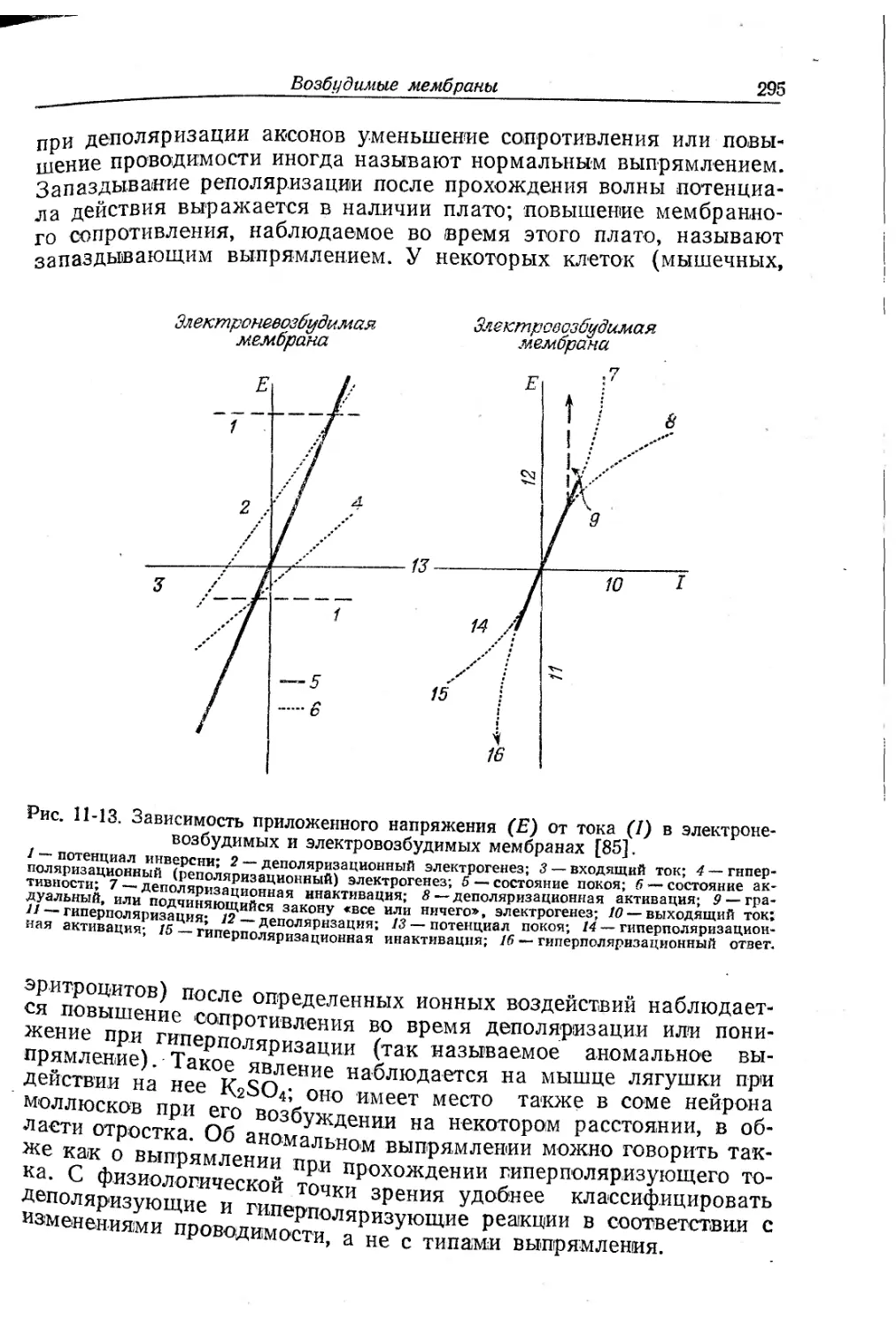












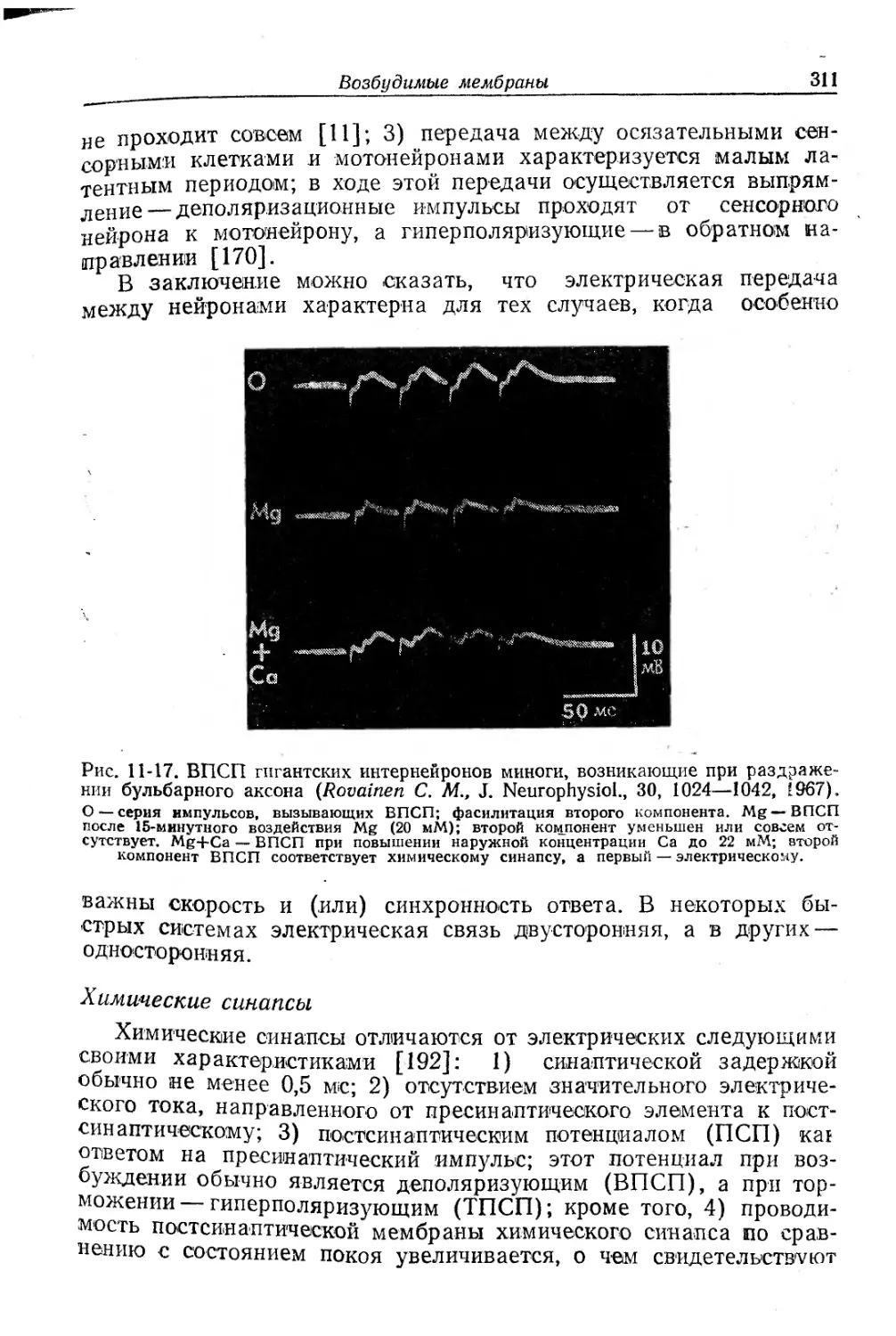



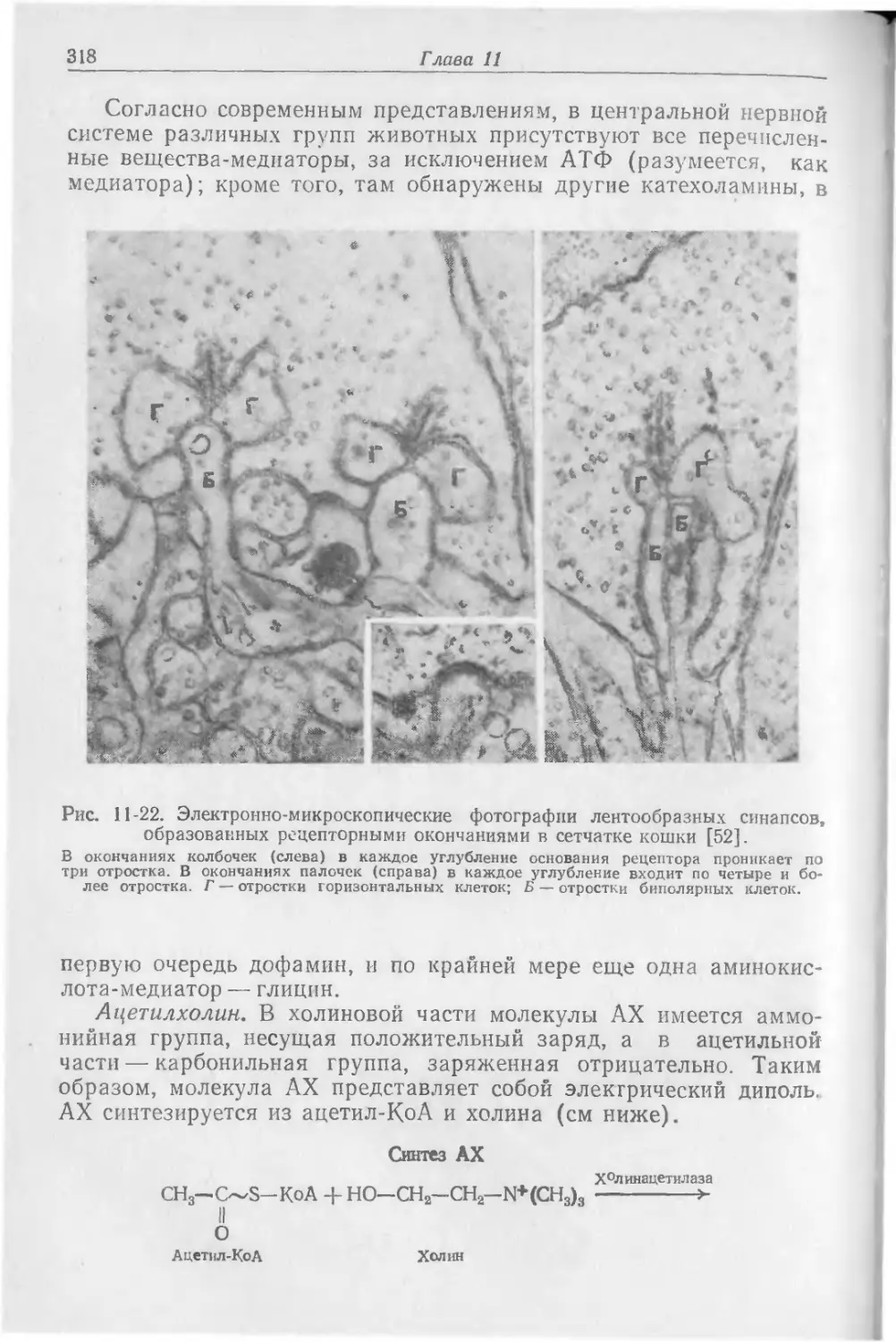






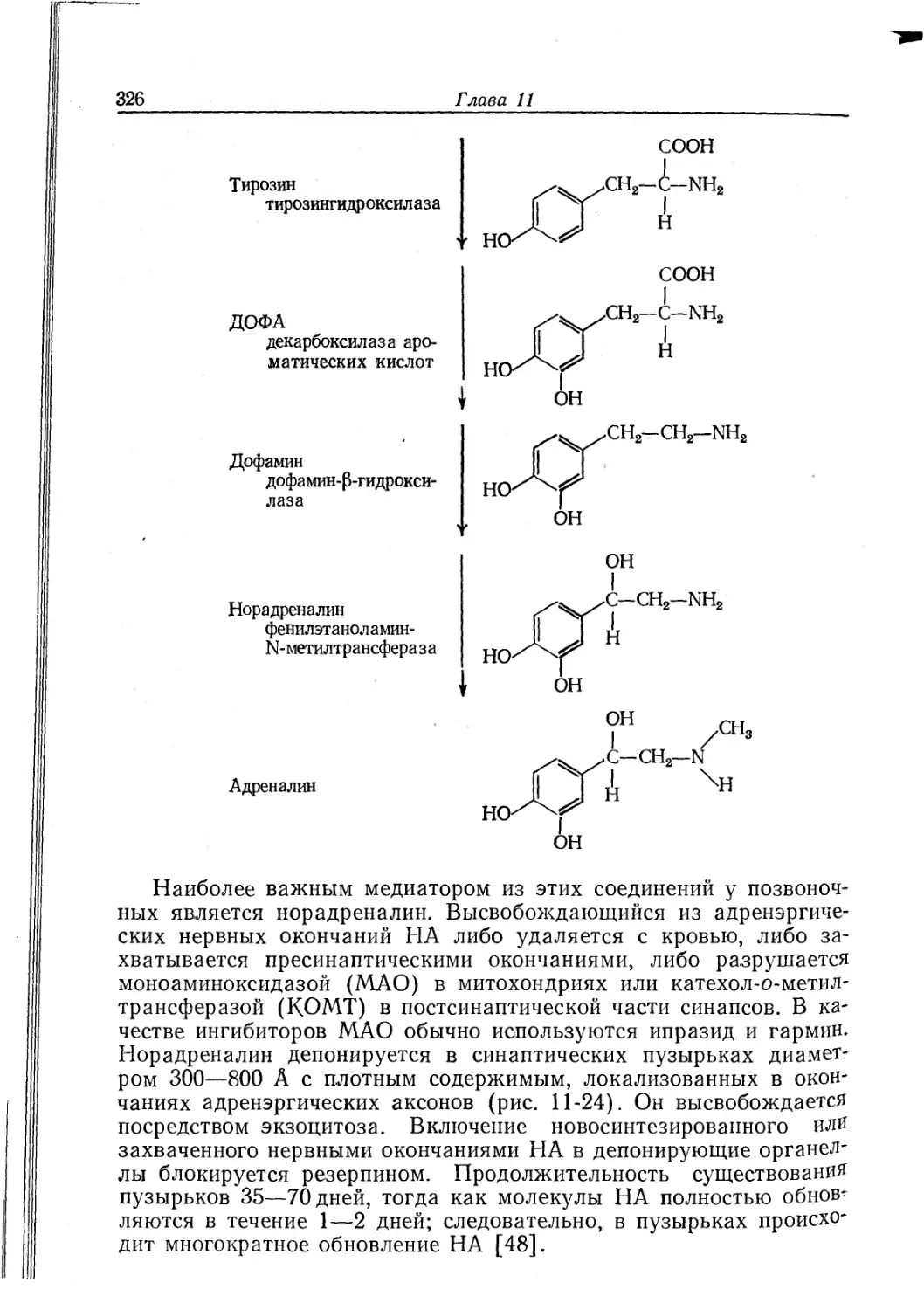
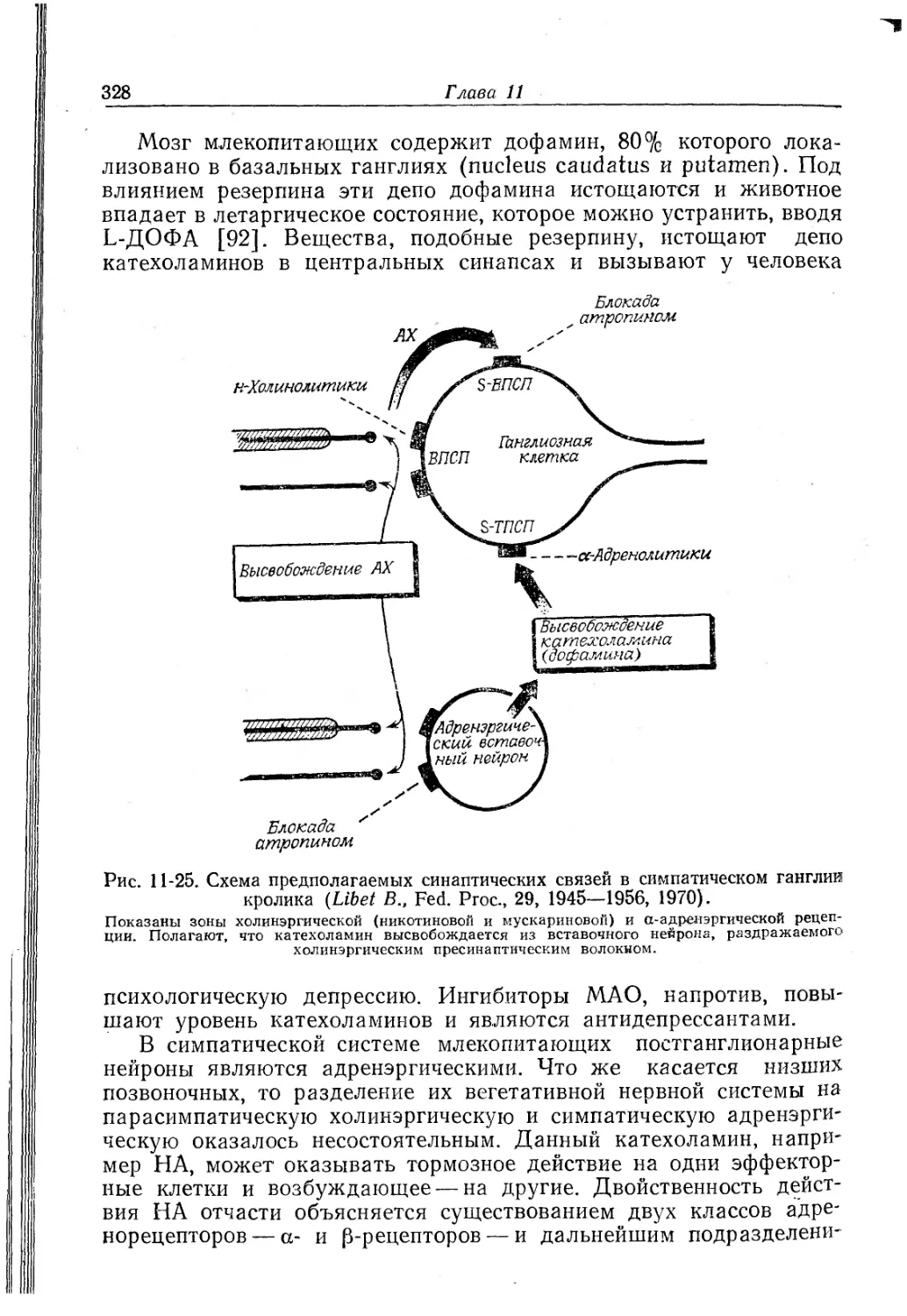






































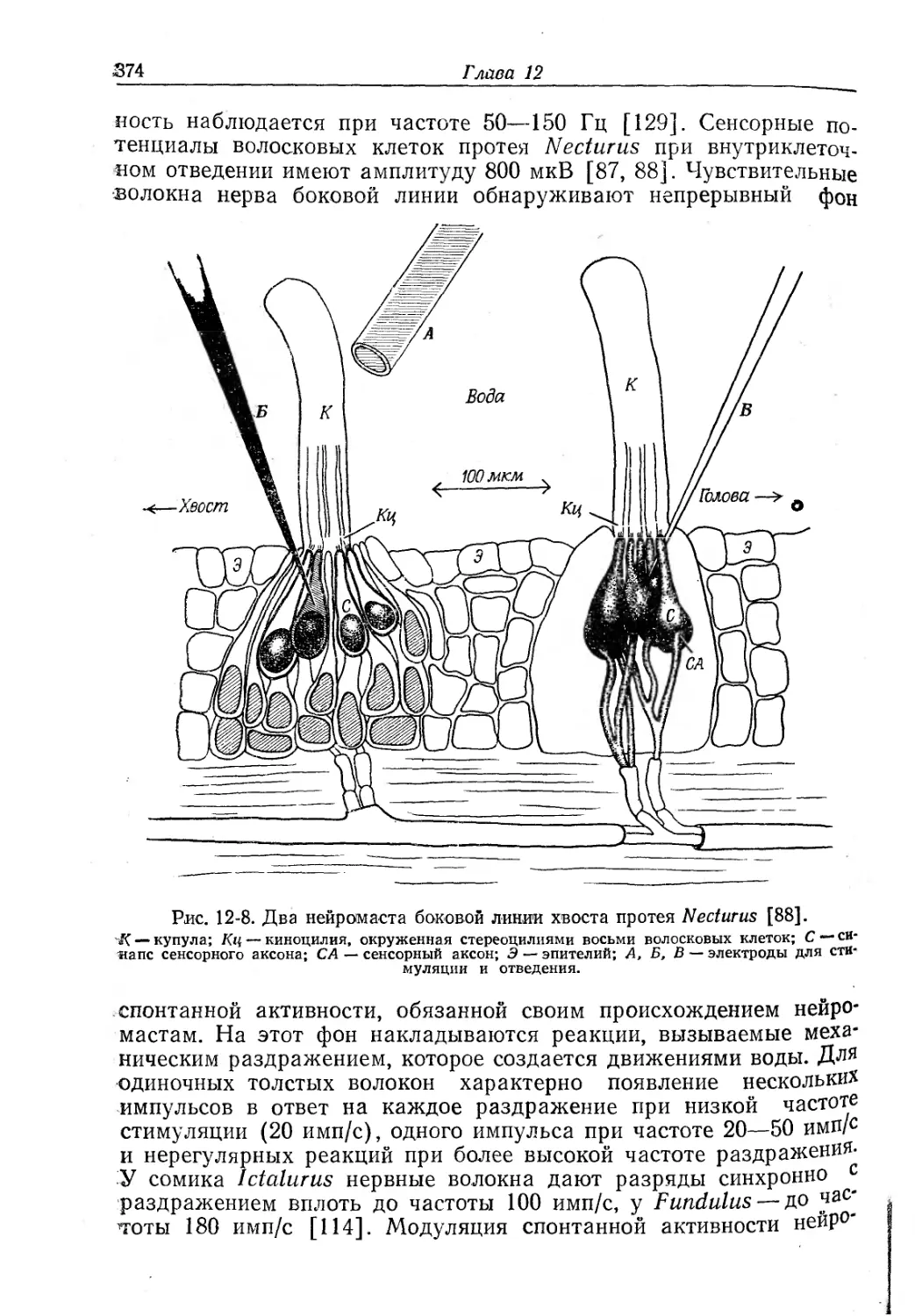
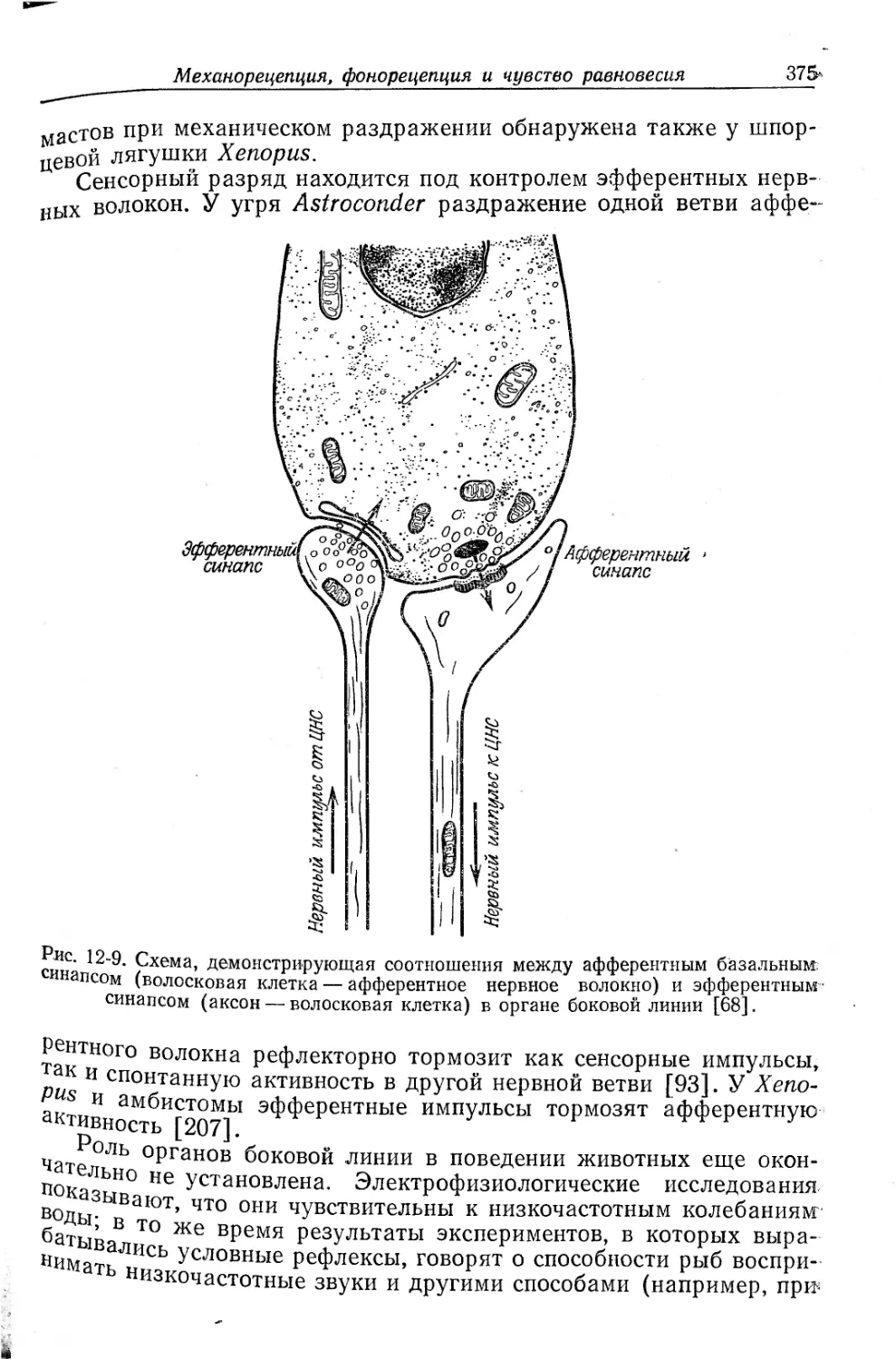

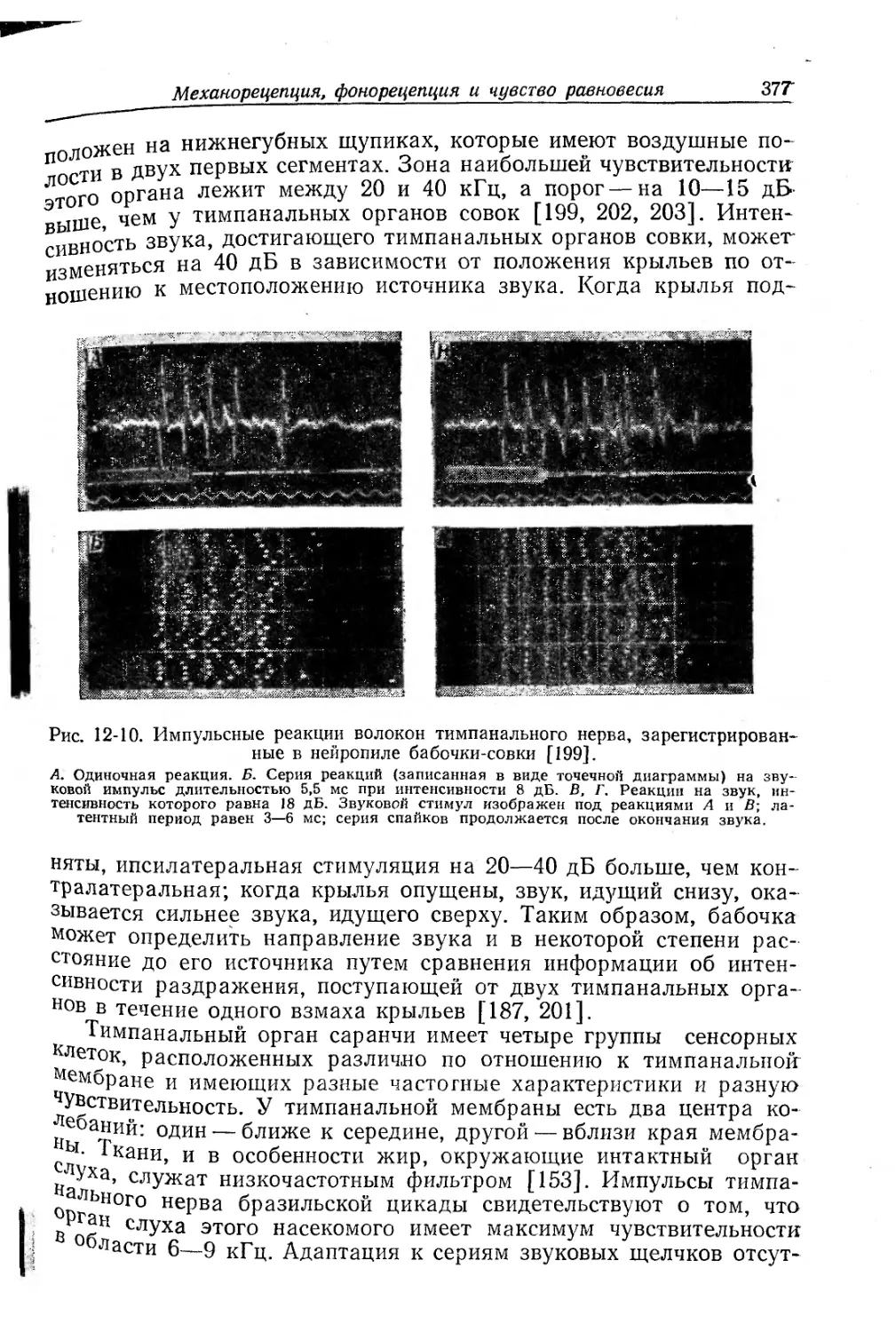

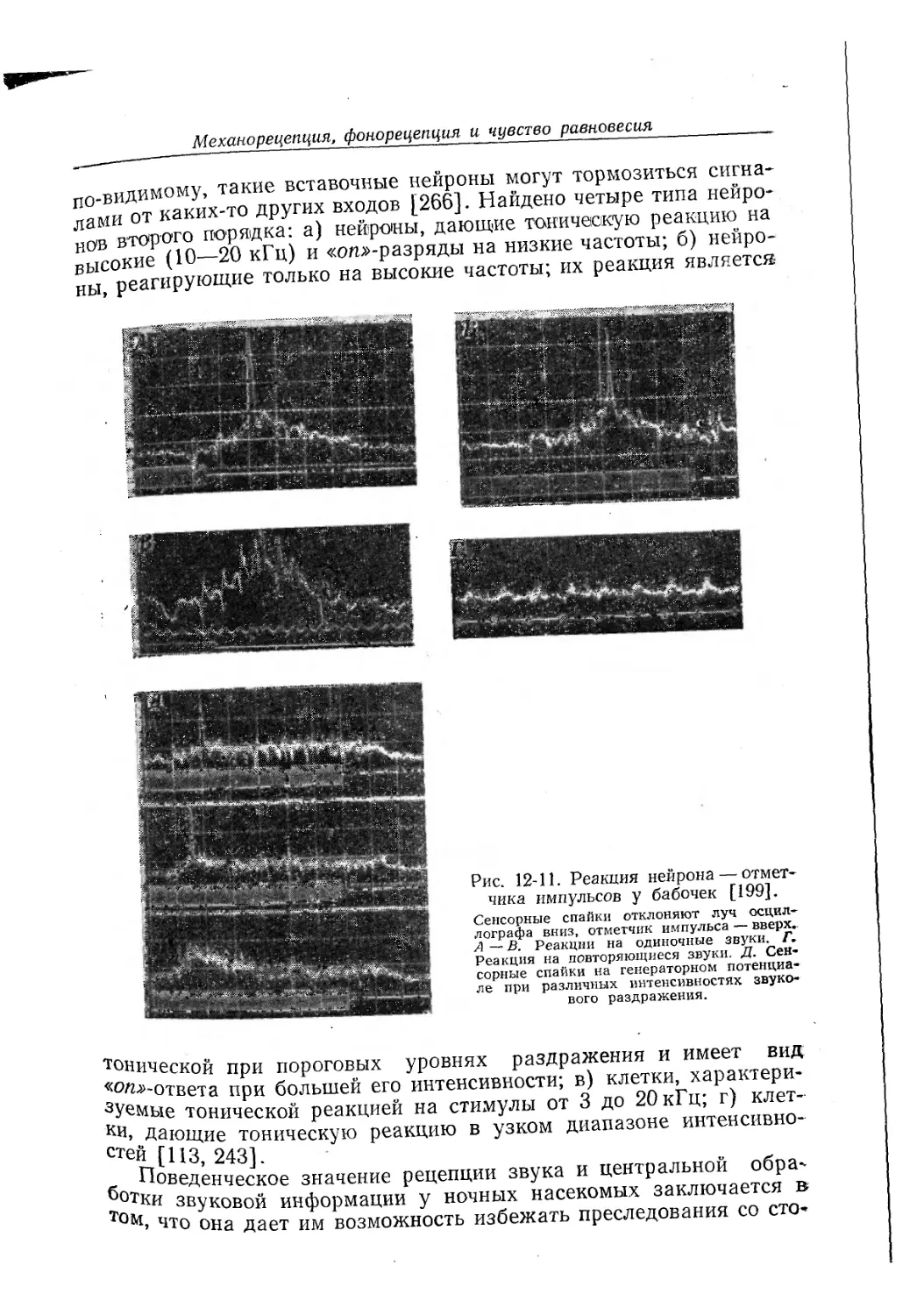









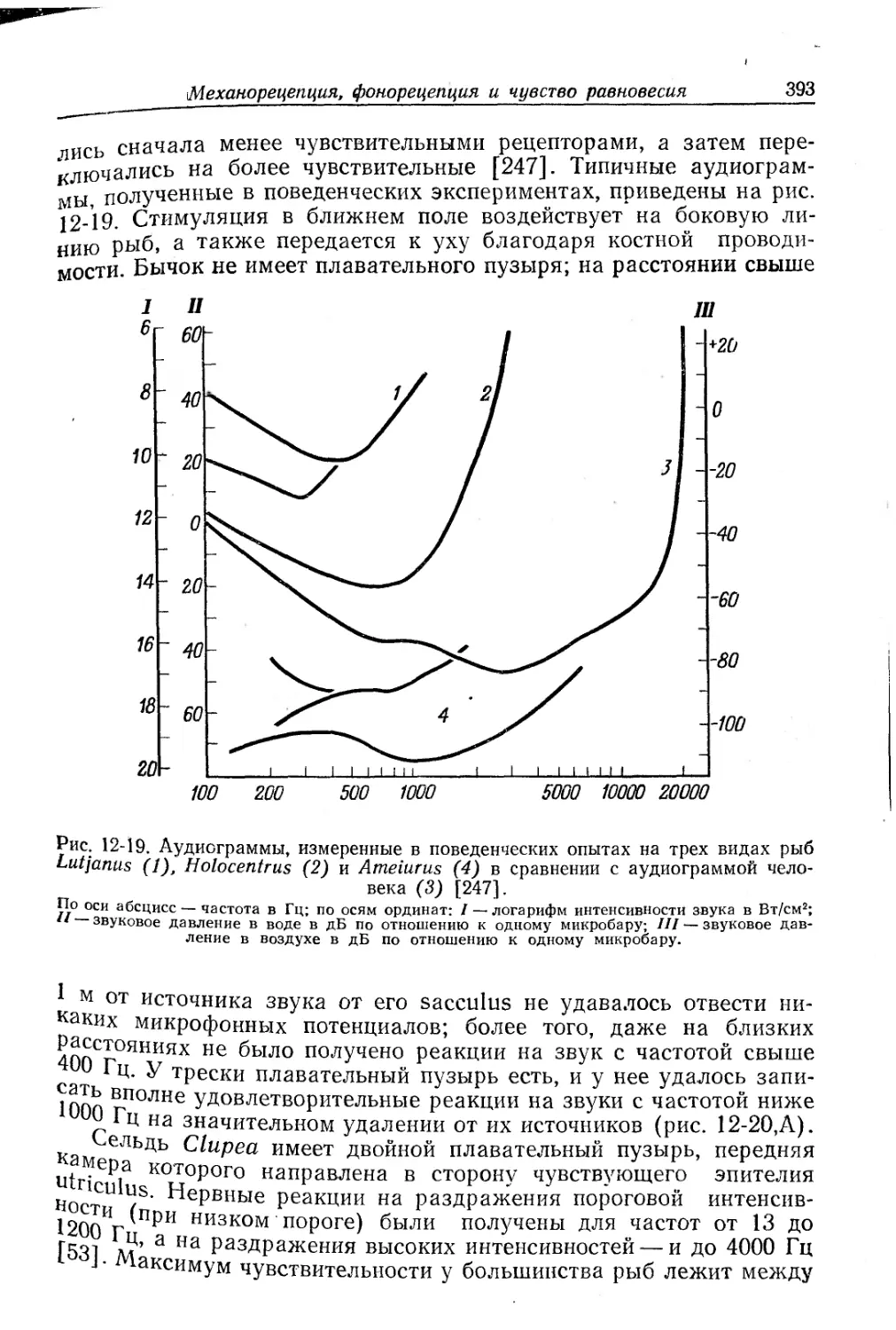
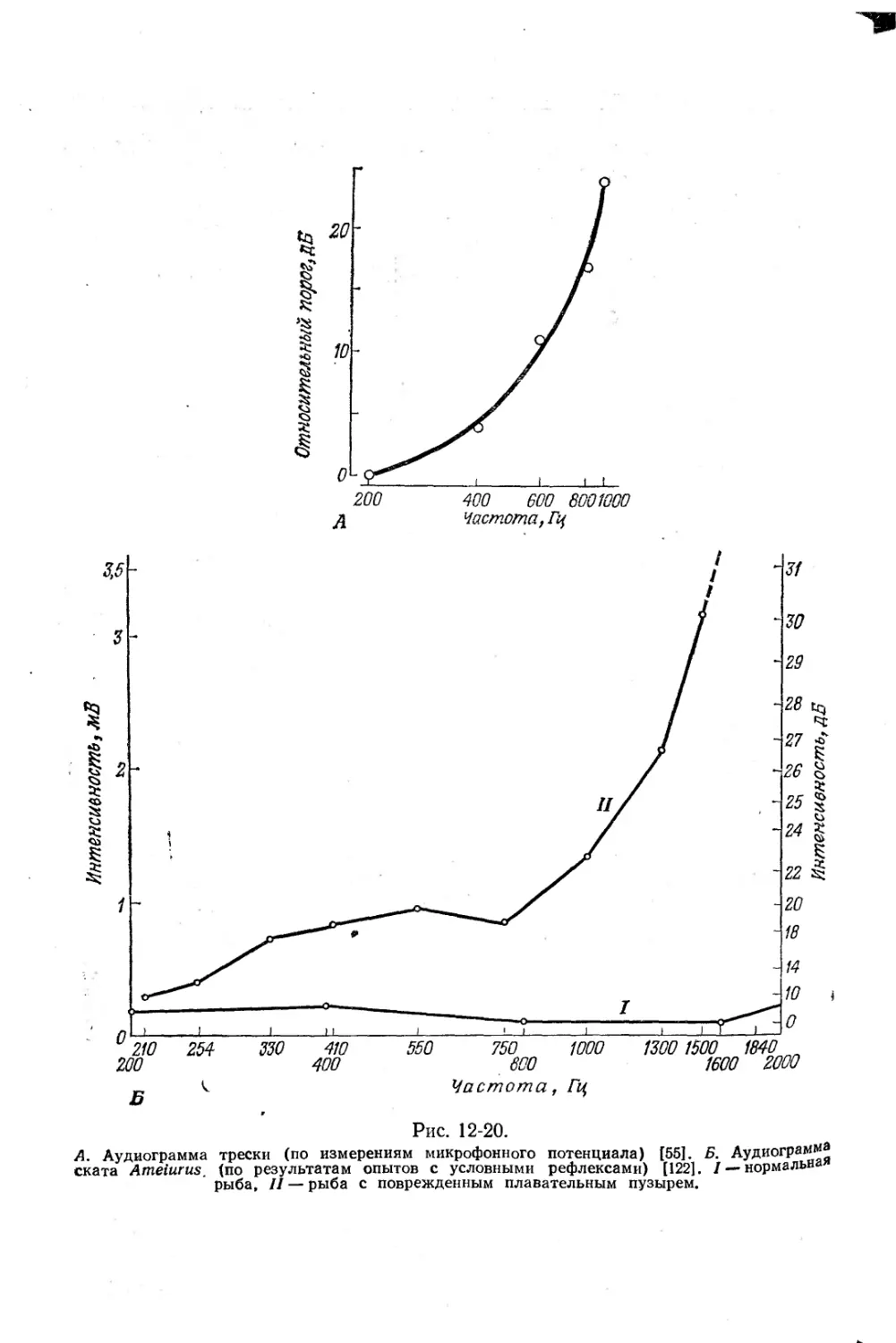

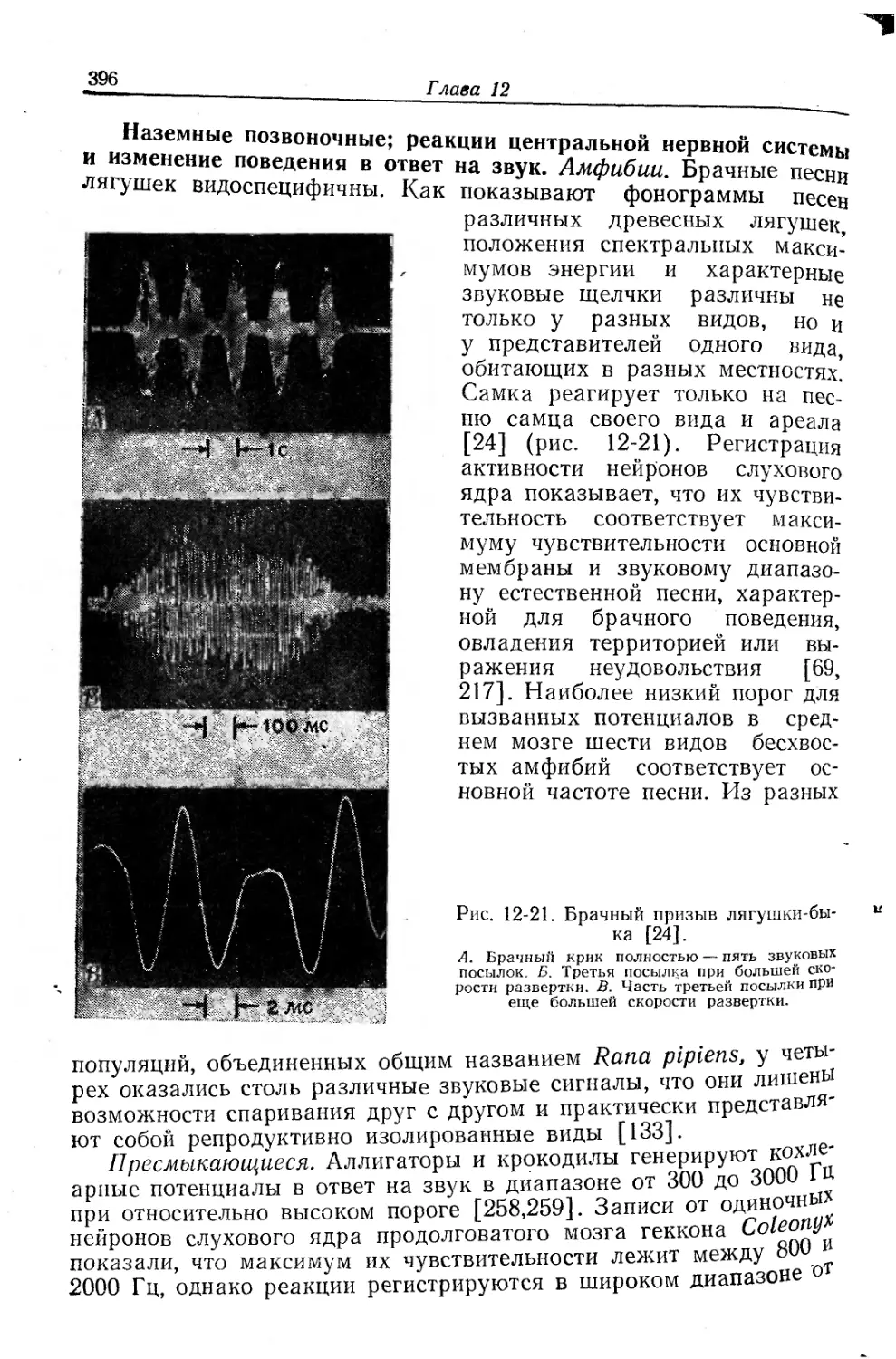

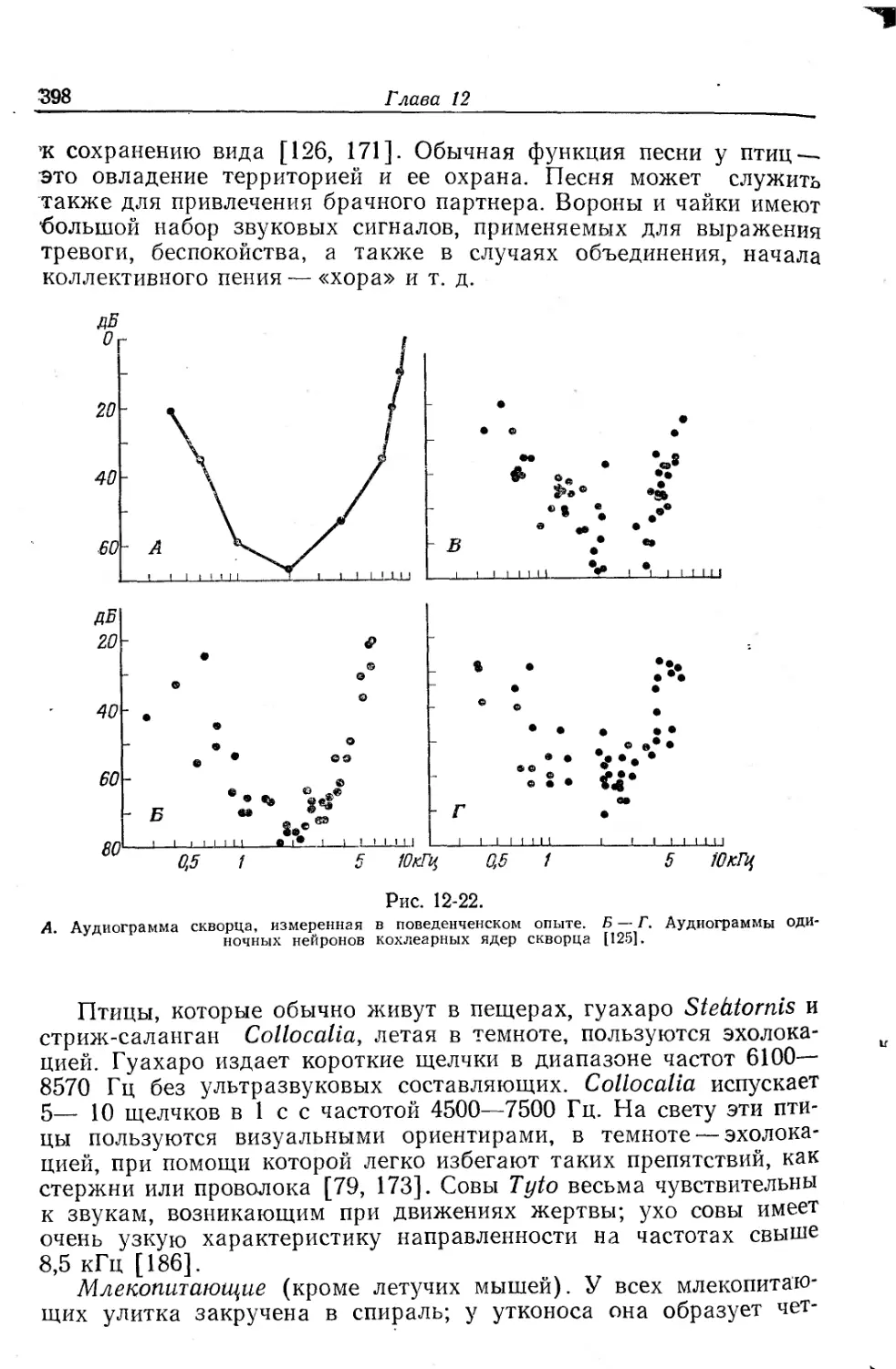
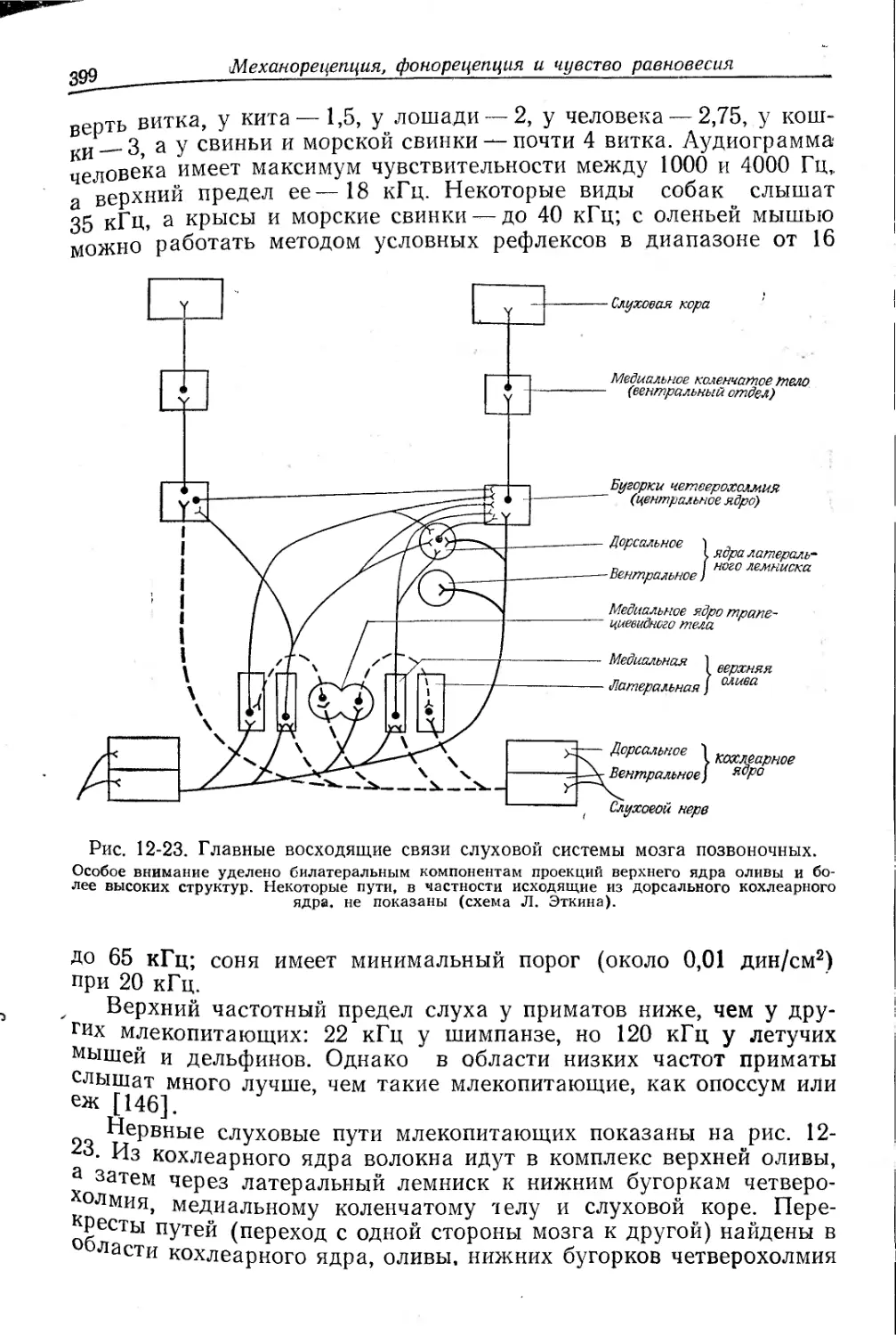




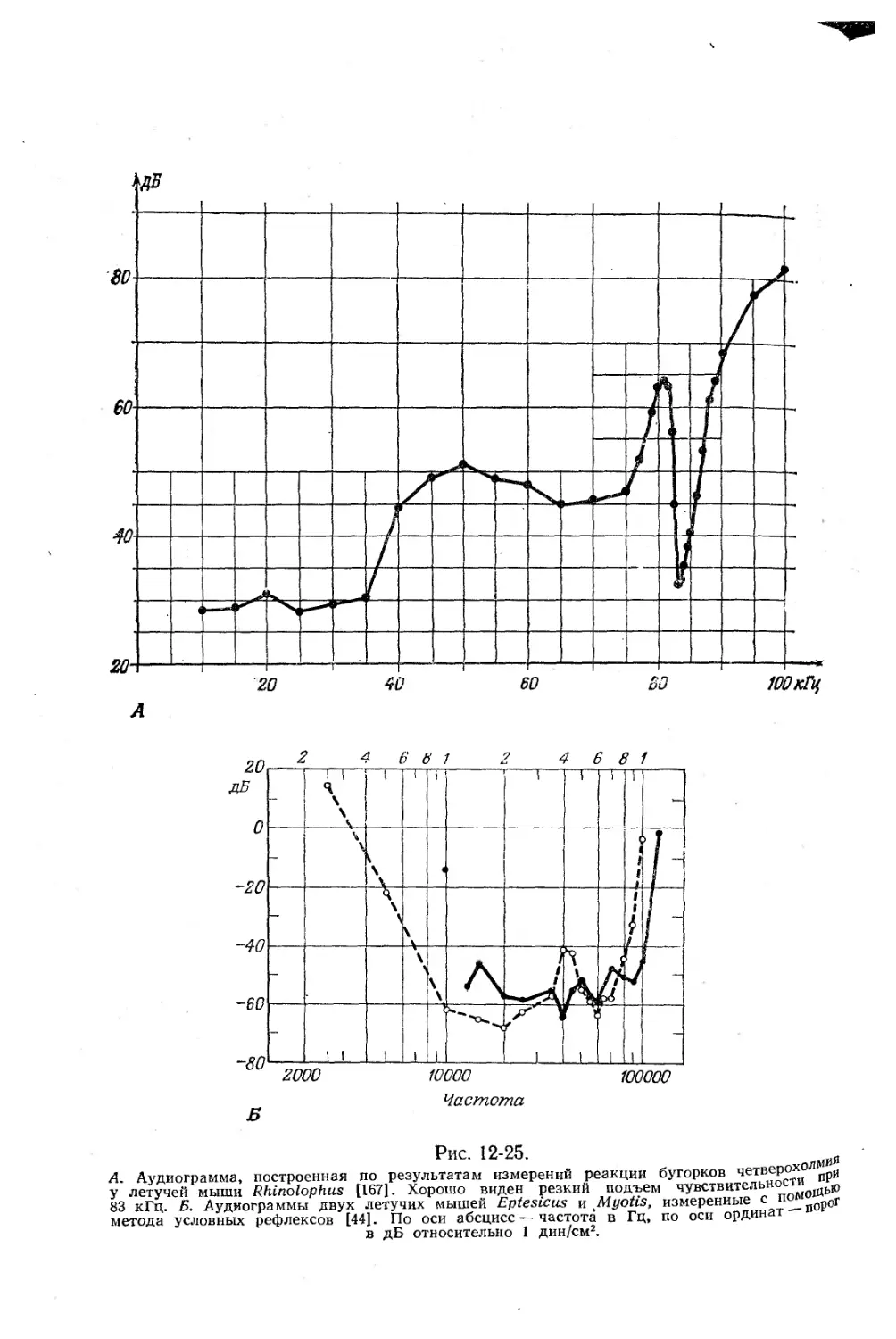

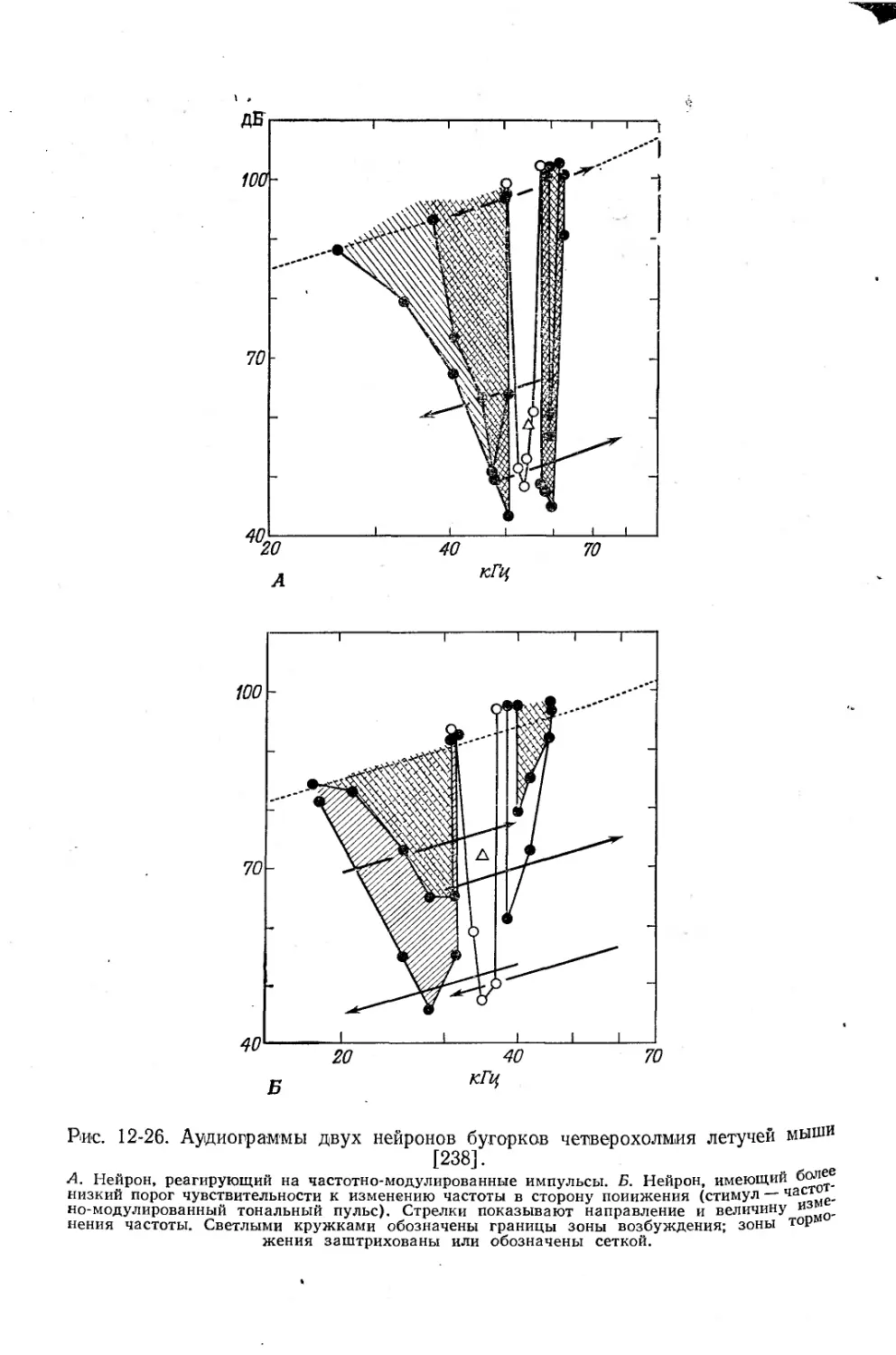




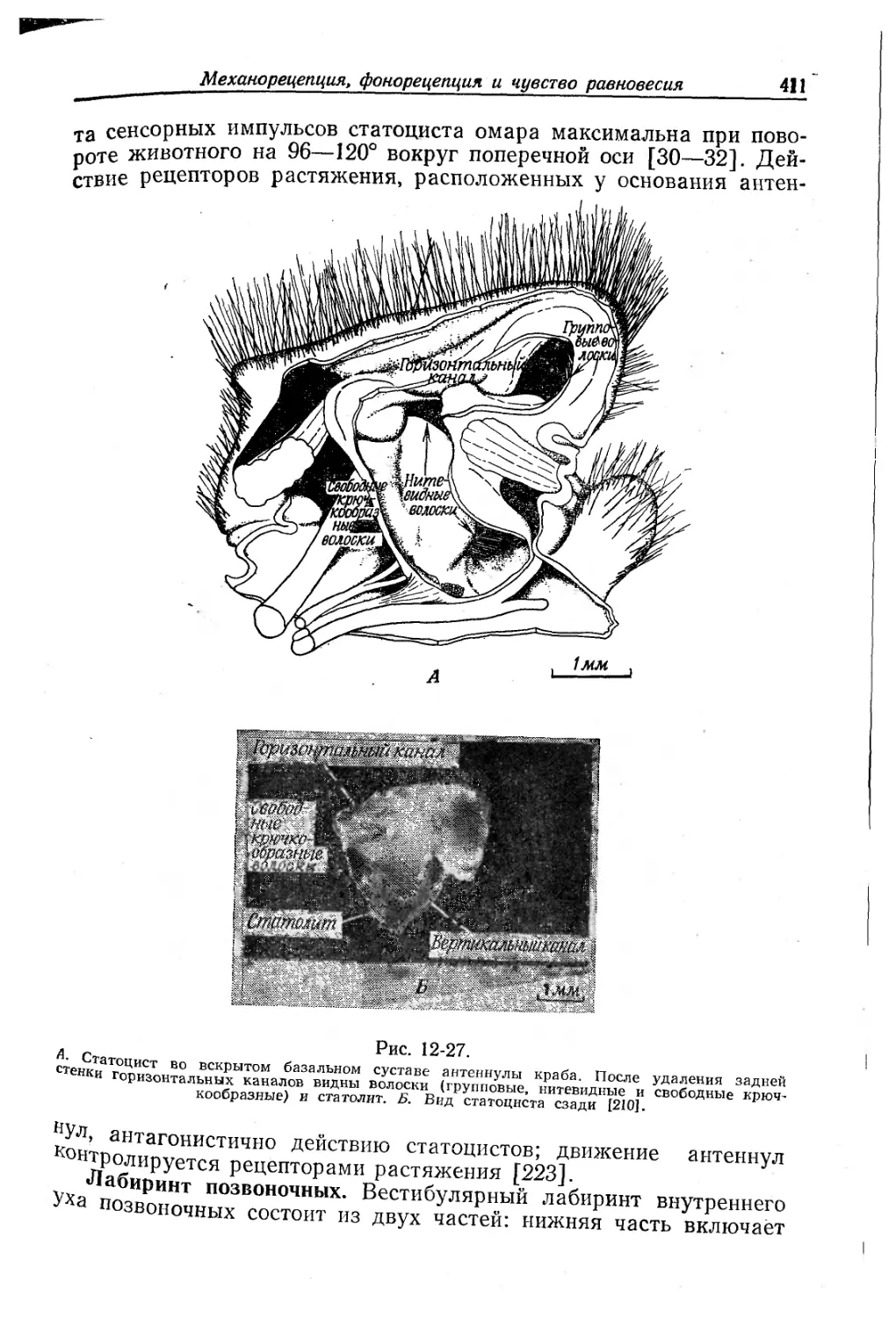
























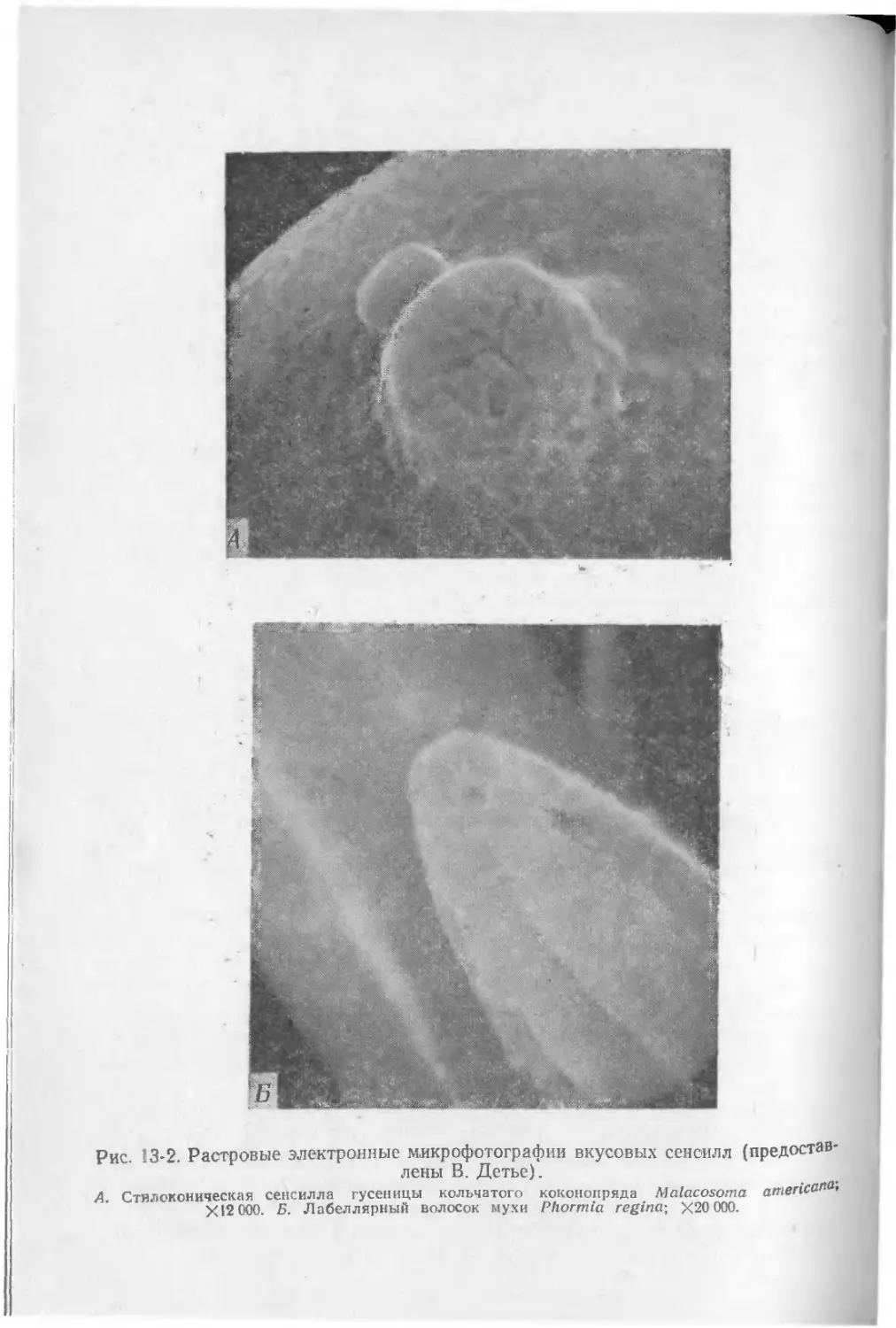

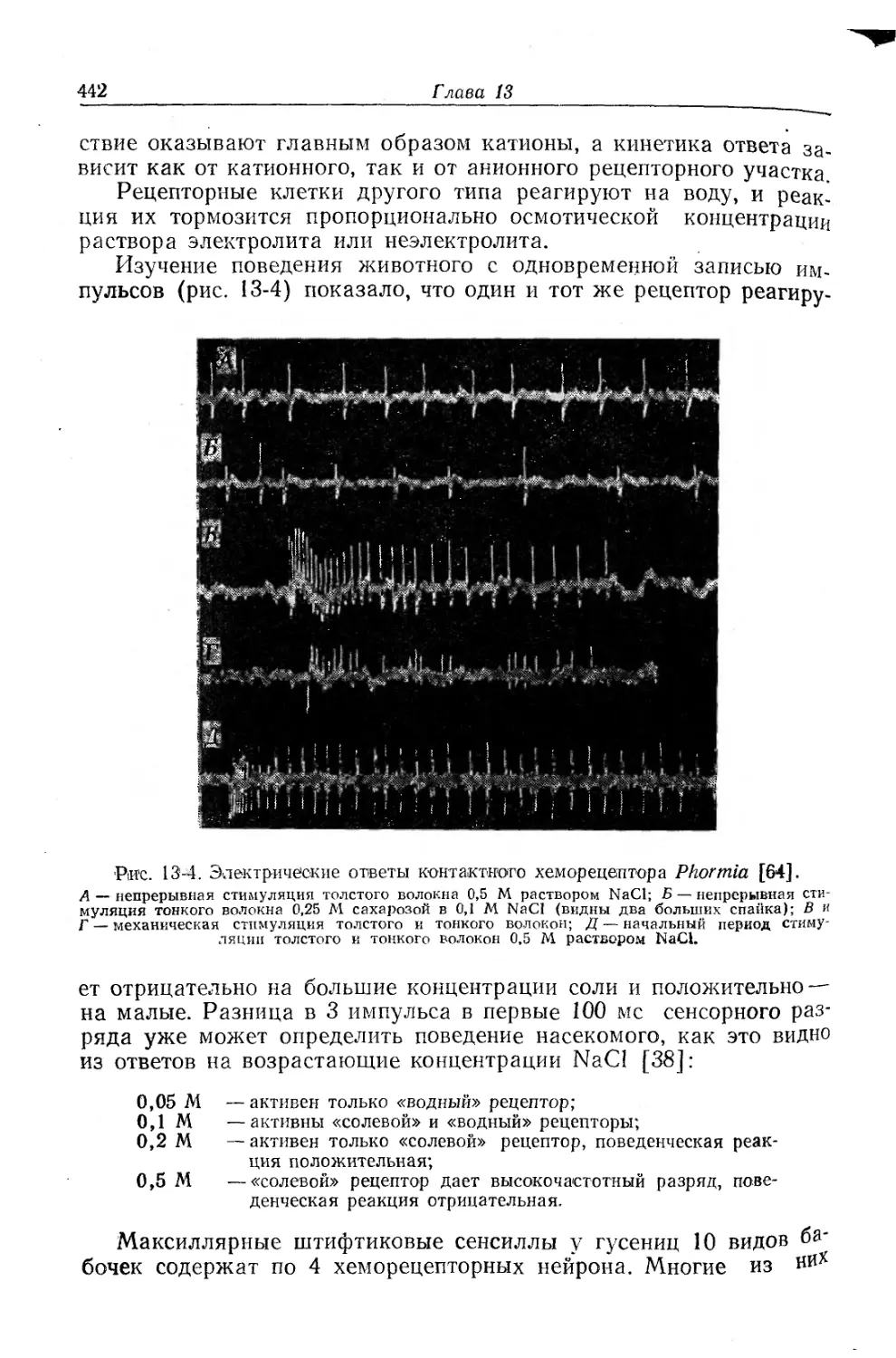

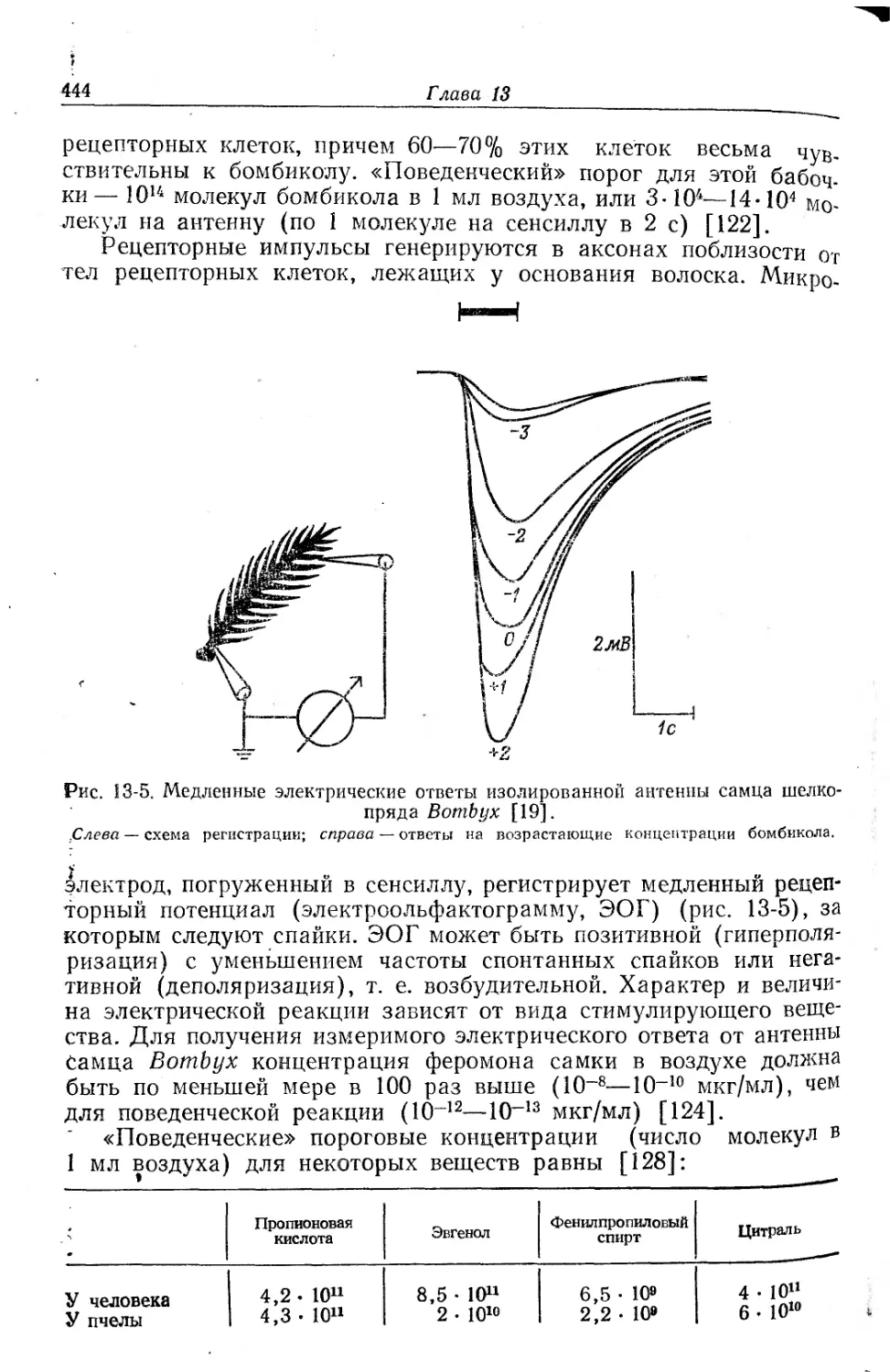

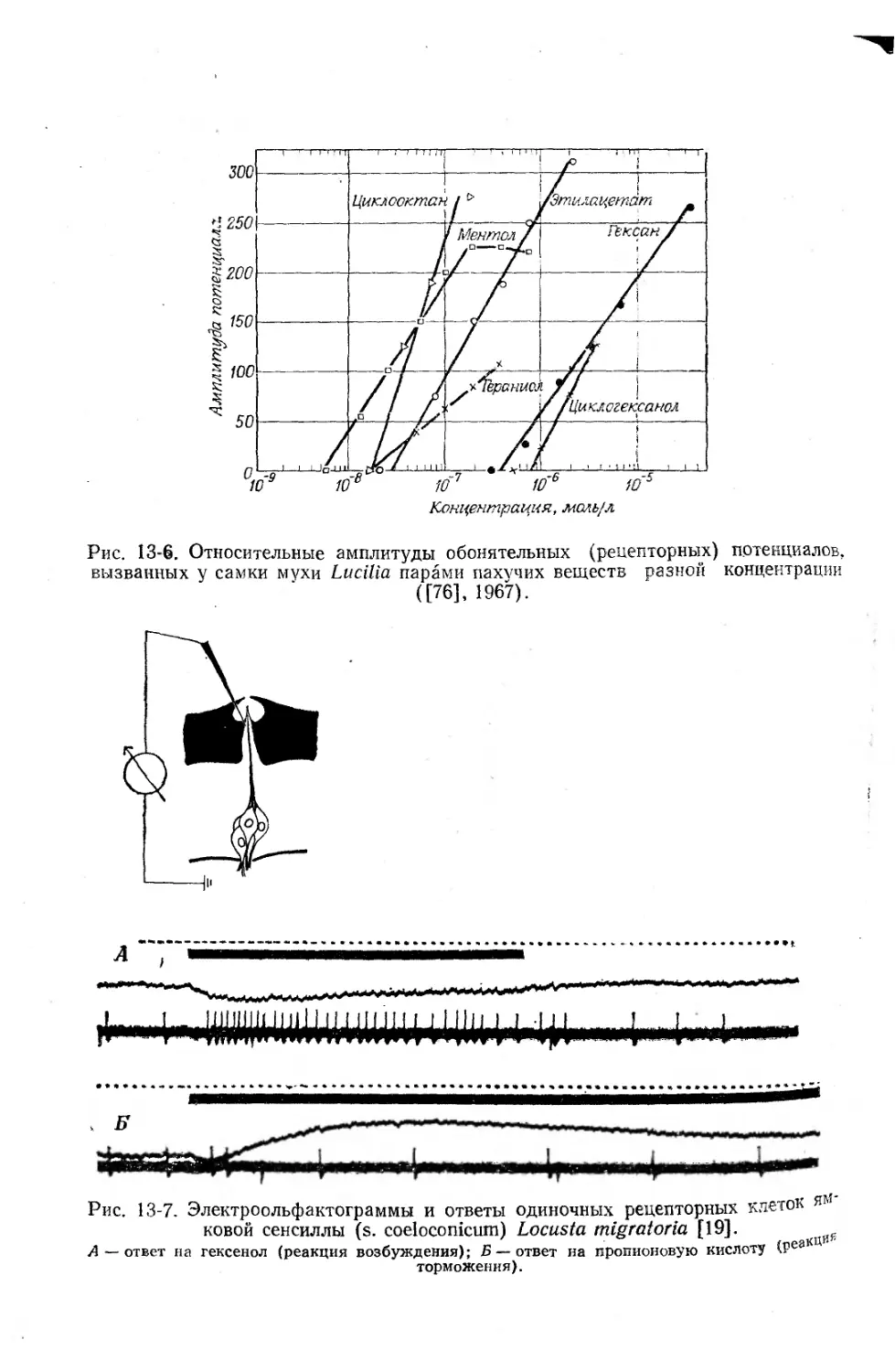


























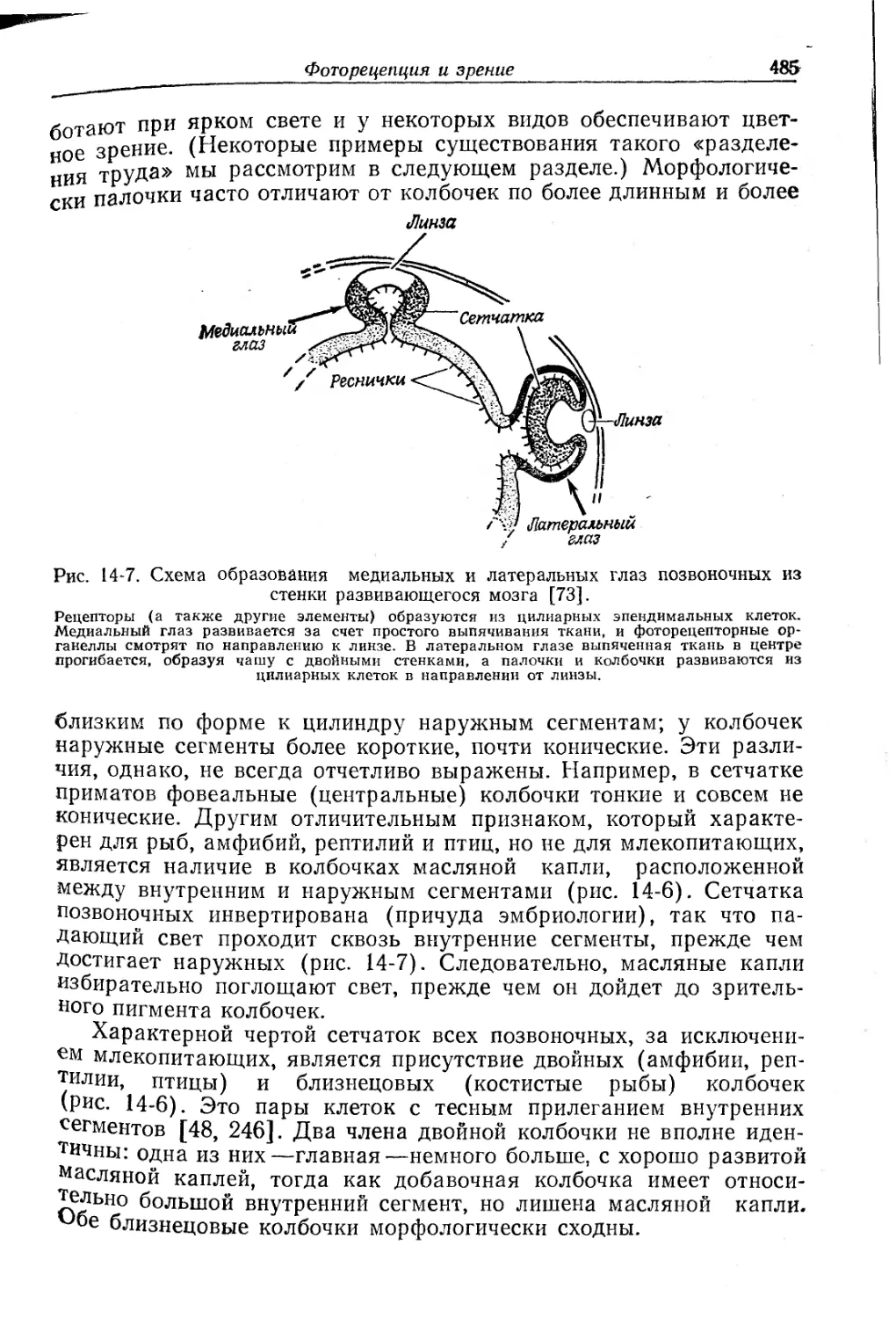


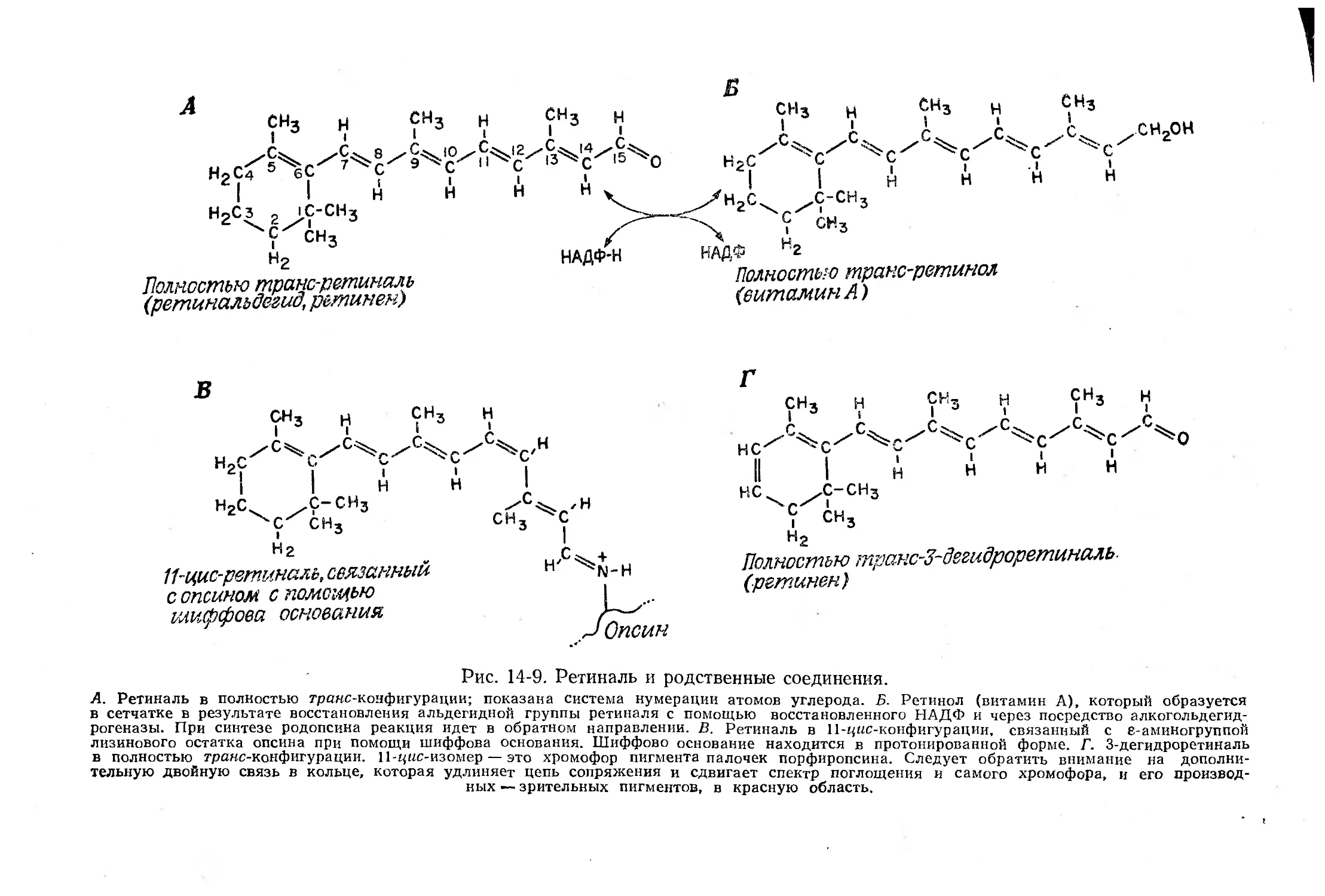


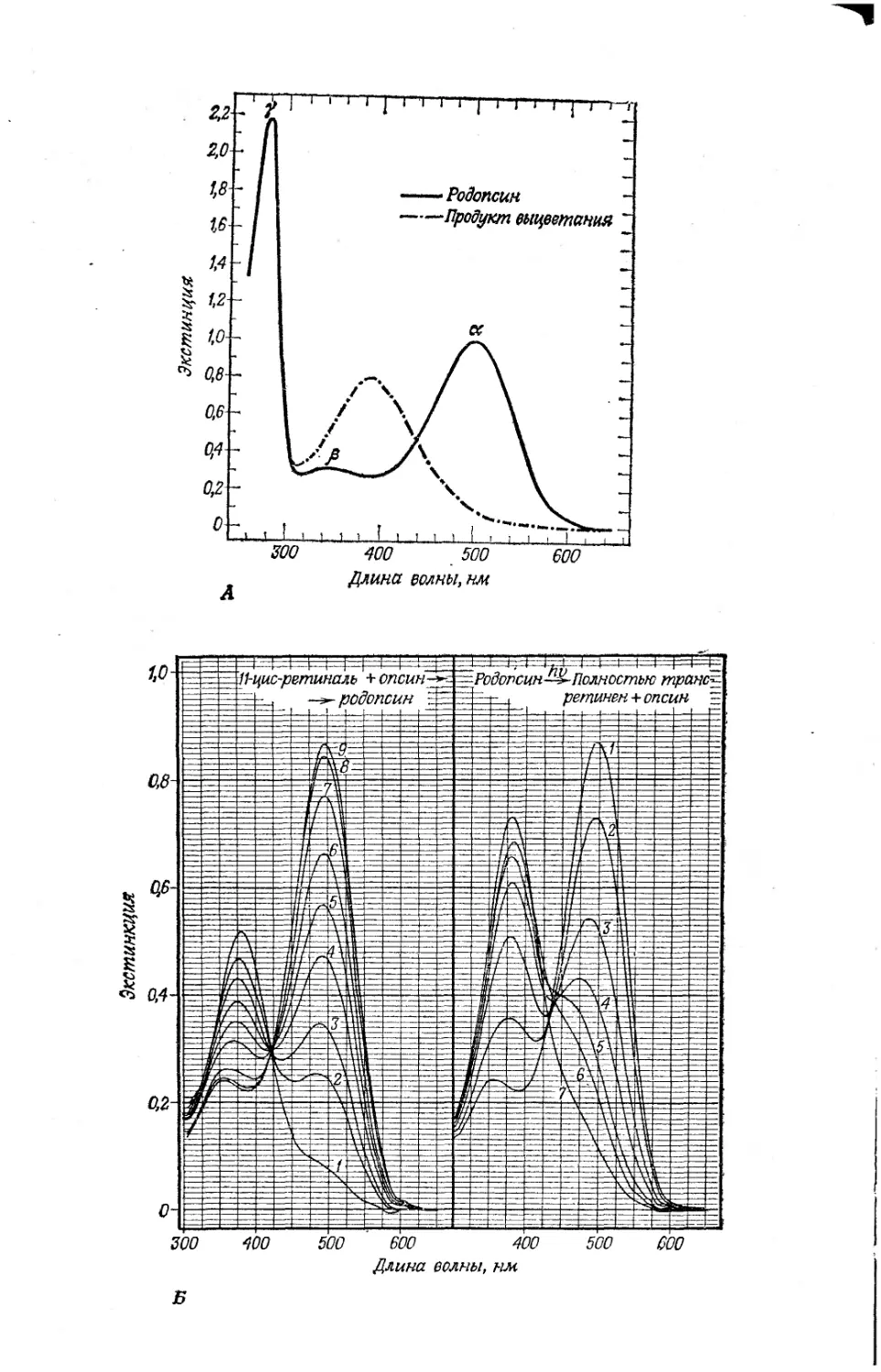










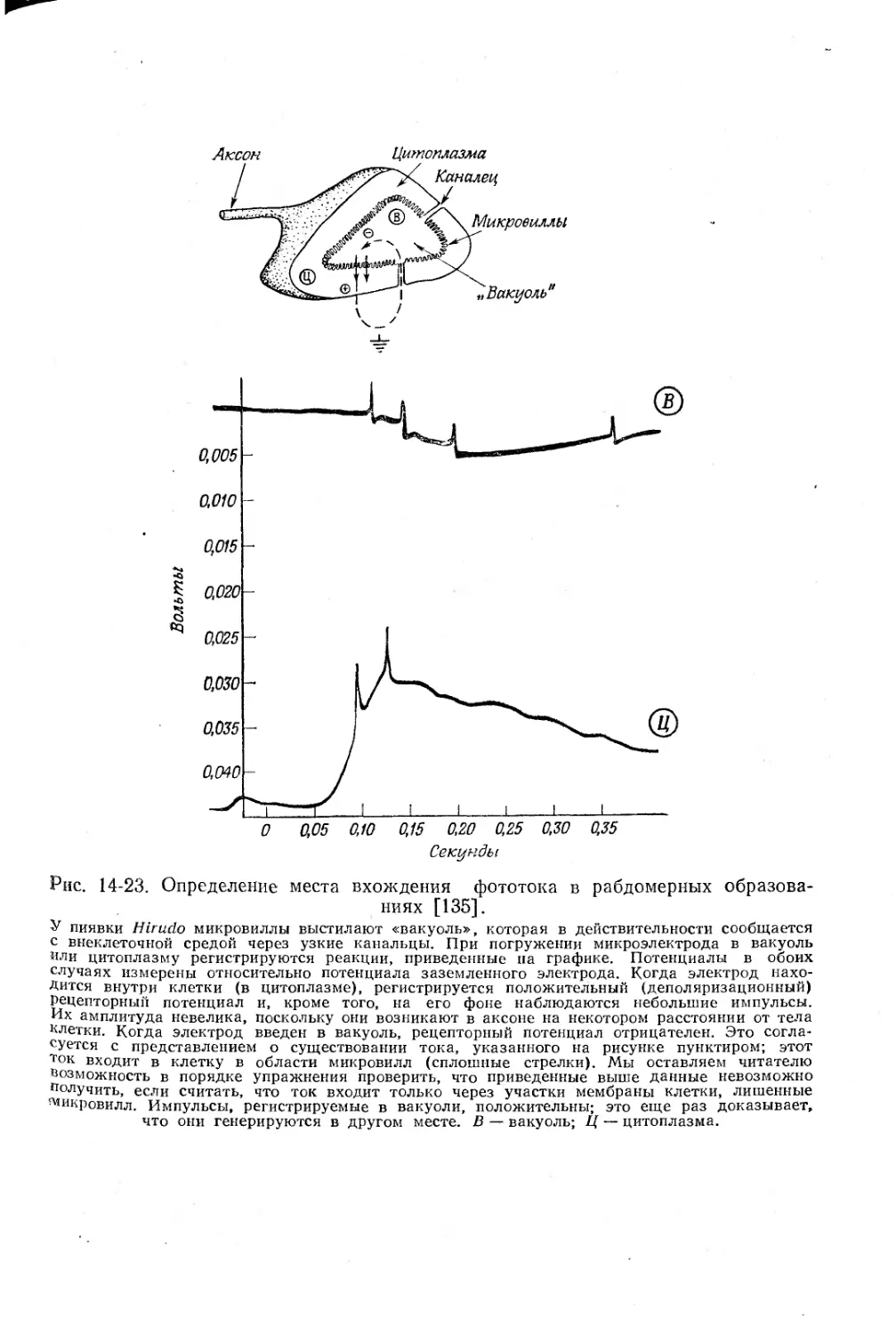







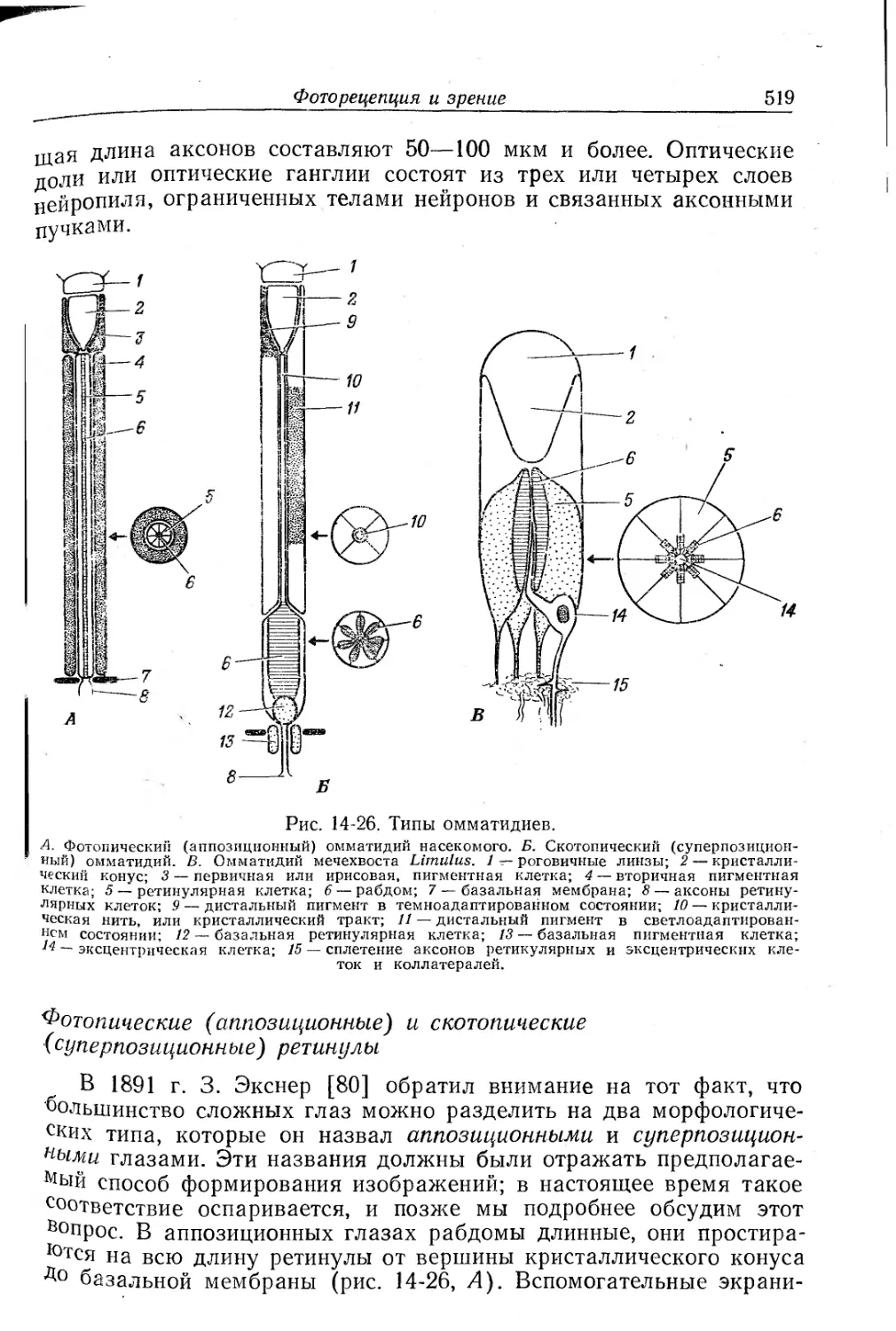


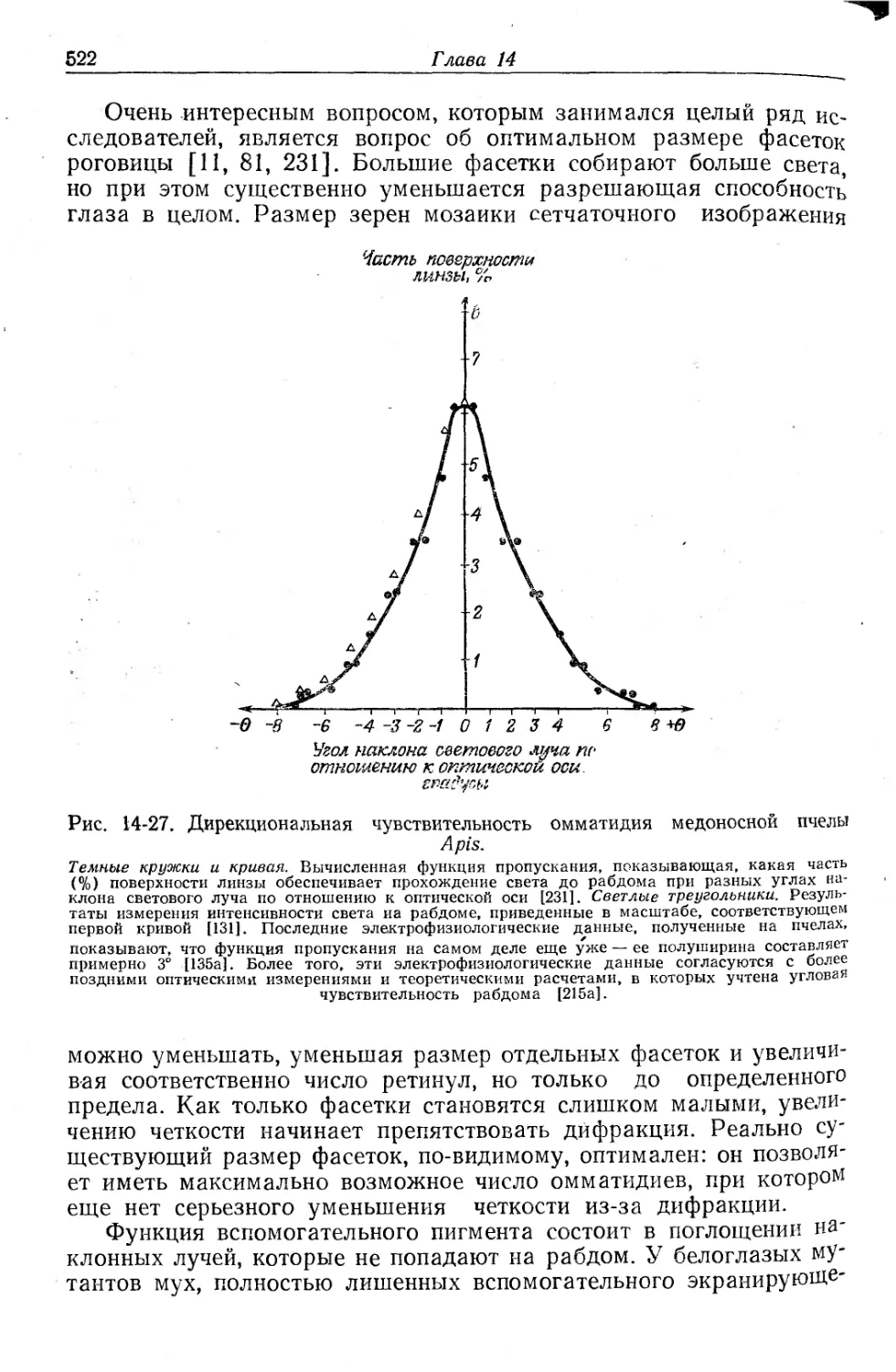


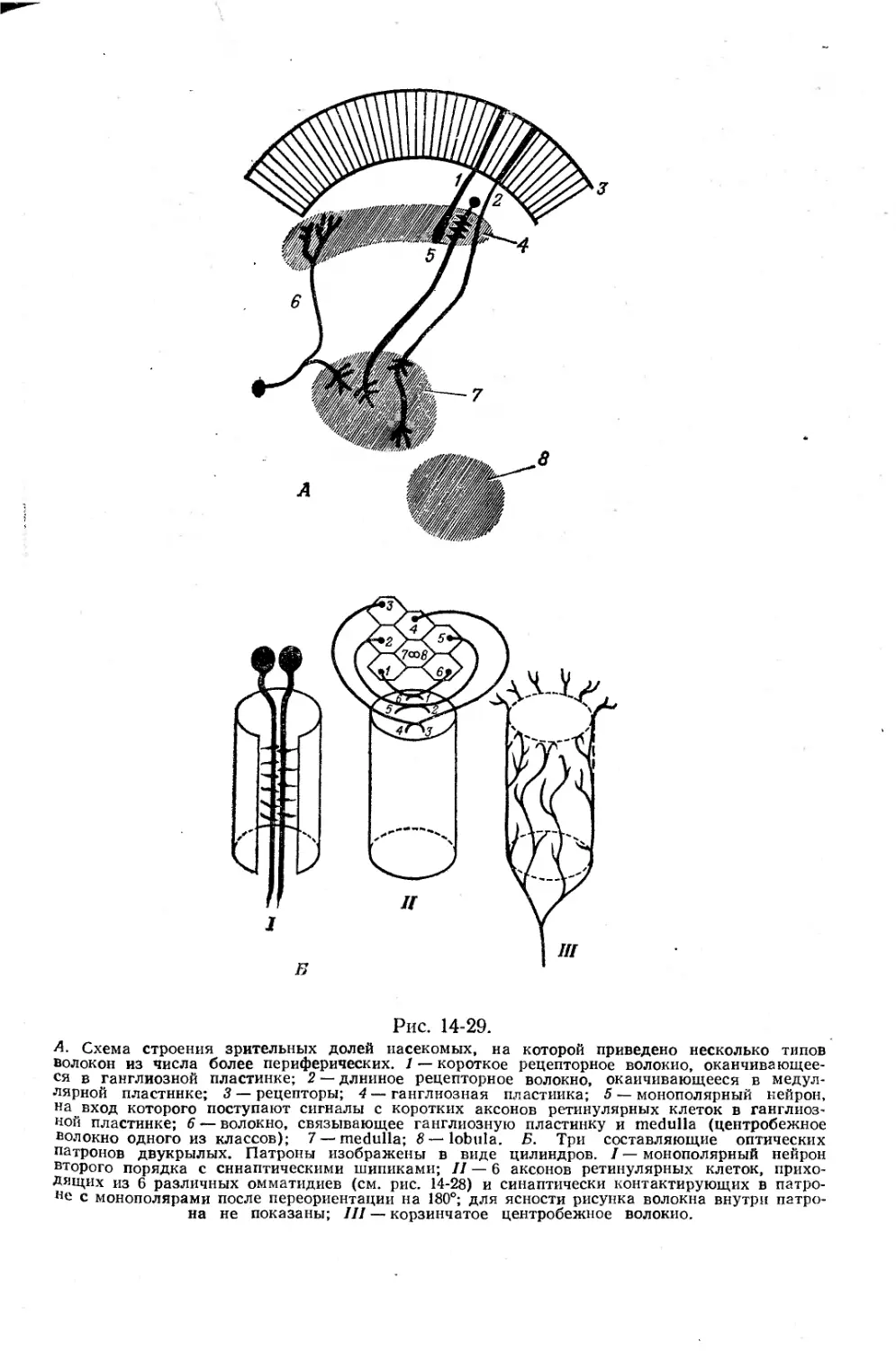



















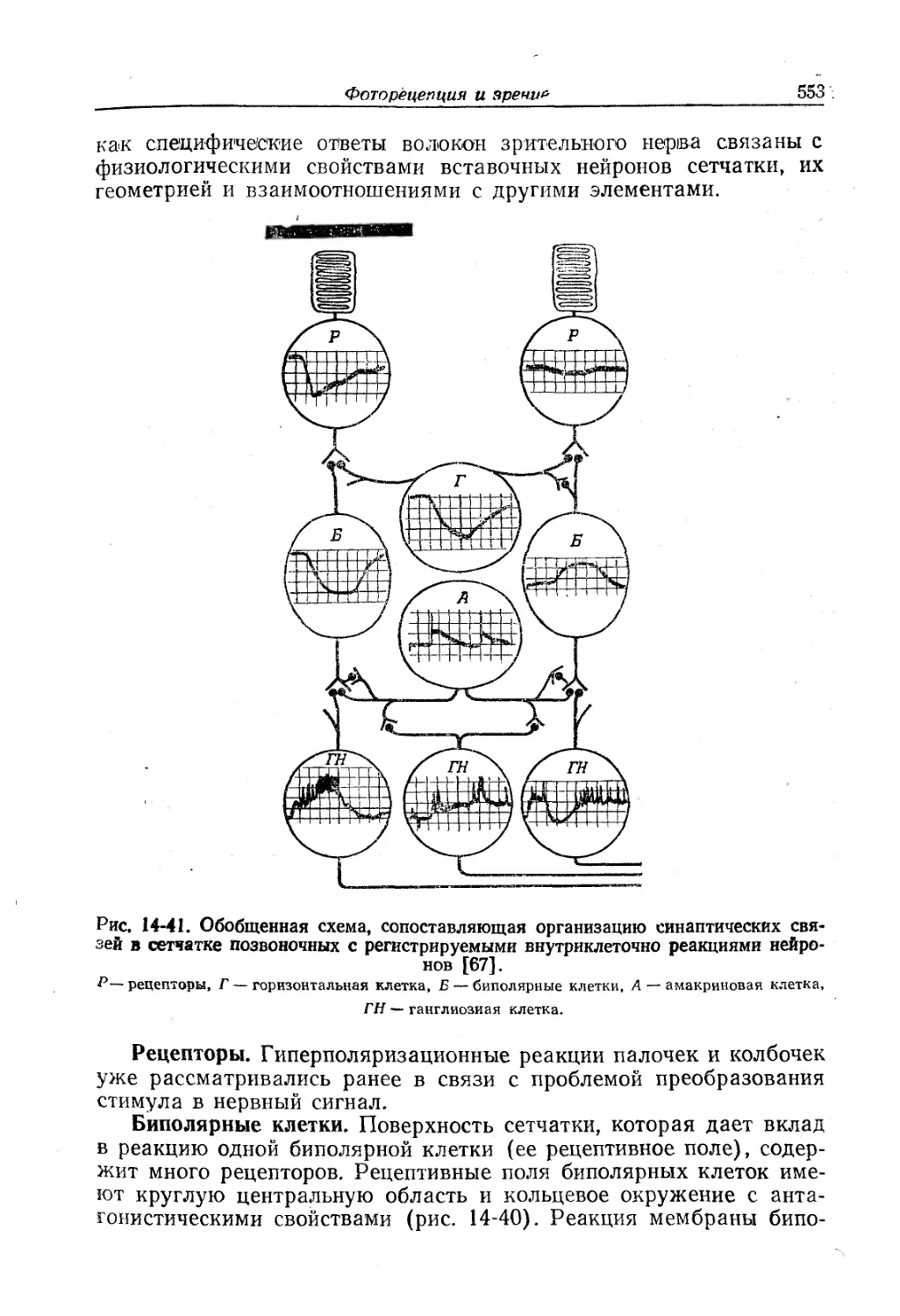


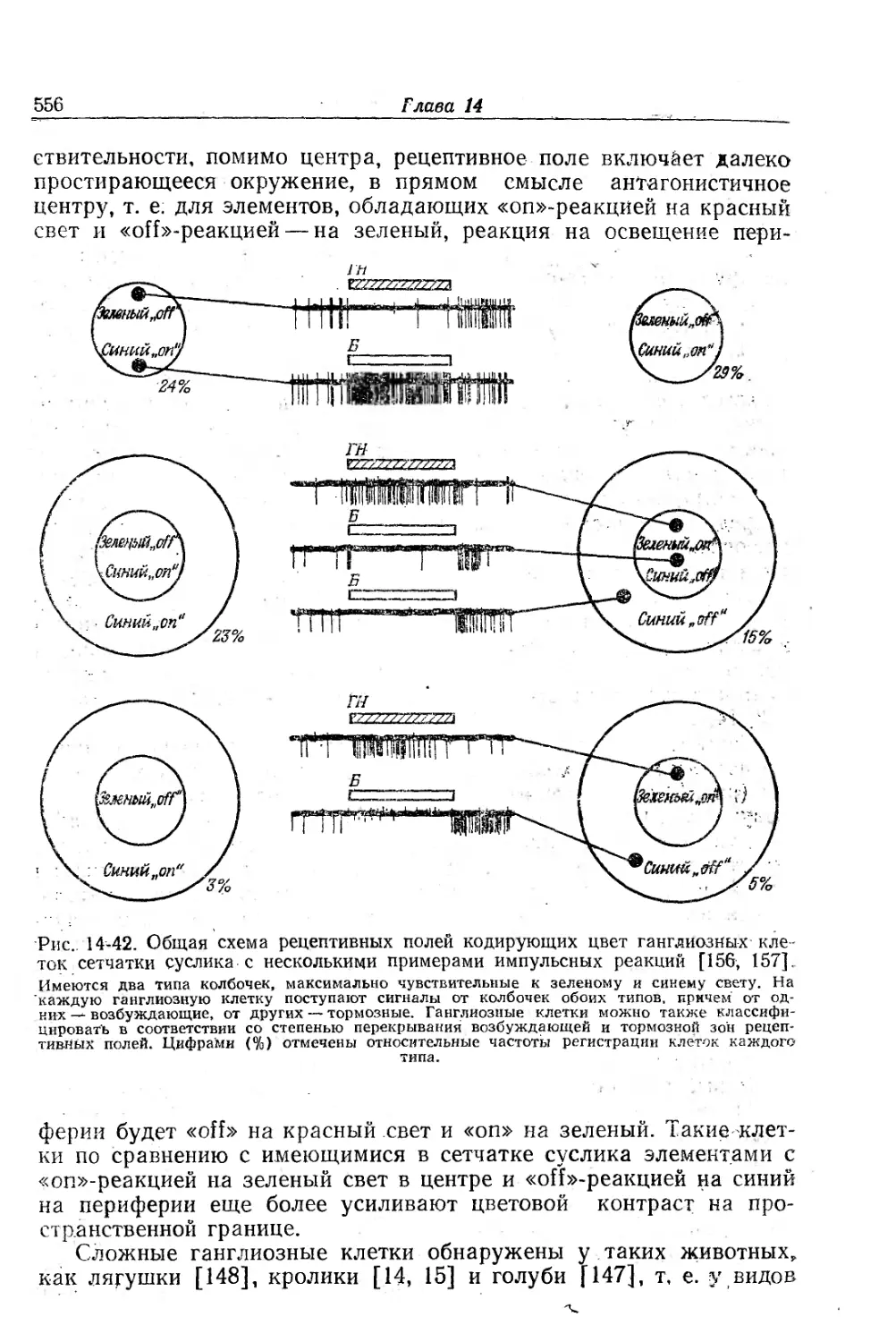
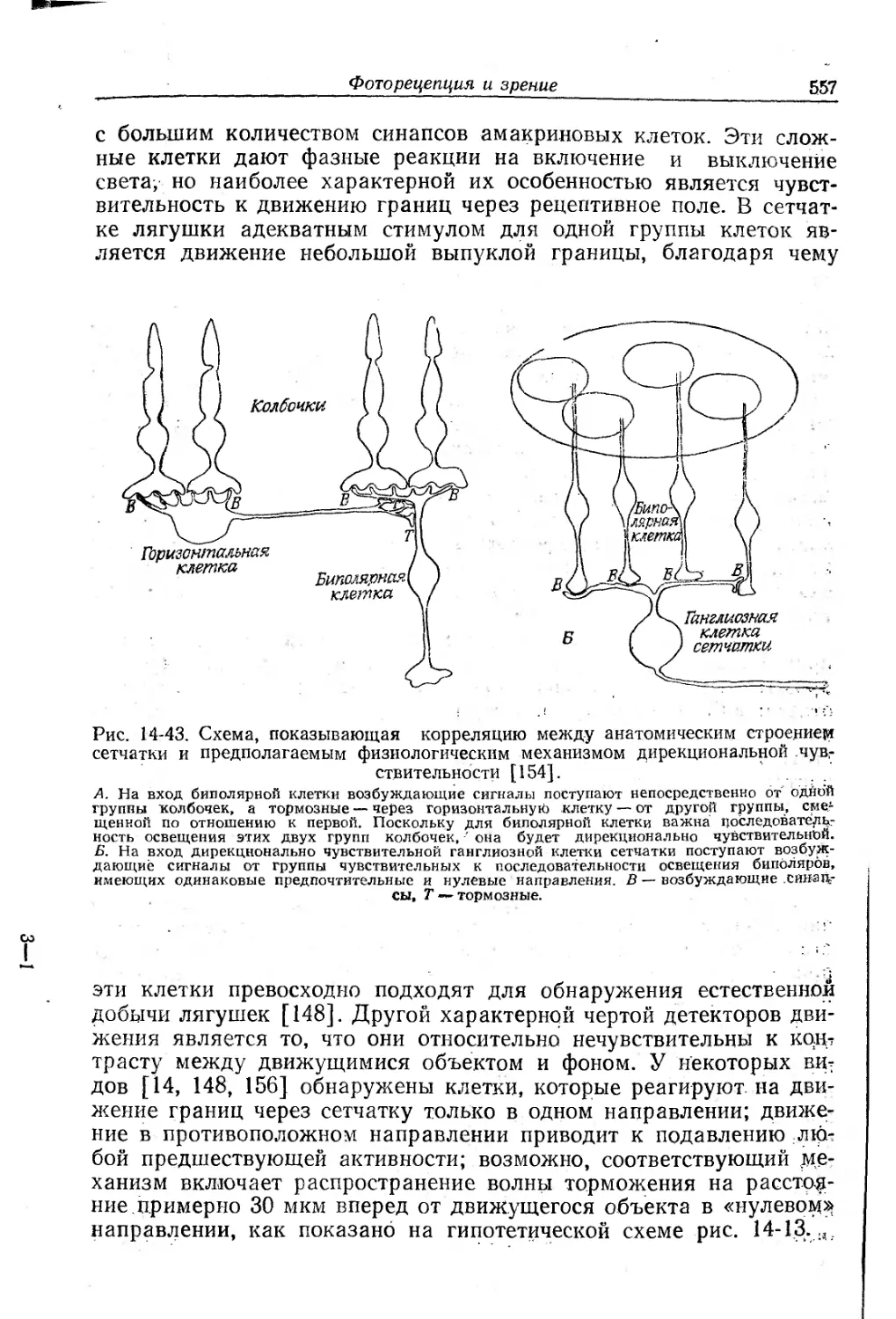























Текст
I
СРАВНИТЕЛЬНАЯ
ФИЗИОЛОГИЯ
животных
Comparative
Animal
Physiology
Edited by C. Ladd Prosser
Professor of Physiology and Zoology
Department of Physiology and Biophysics
University of Illinois at Urbana — Champaign
Third Edition
W. B. SAUNDERS COMPANY
PHILADELPHIA LONDON TORONTO
1973
Сравнительная
физиология
1
животных том II
Перевод с английского
под редакцией
чл.-корр. АН СССР Т. М. Турпаева
ИЗДАТЕЛЬСТВО «МИР»
МОСКВА
1977
УДК 591.1
Фундаментальное руководство по сравнительной физиологии животных; вы-
ходит на русском языке в трех томах. В первый том вошли материалы по регуля-
ции водного обмена, транспорту и обмену неорганических ионов, физиологии и
биохимии пищеварения, основным путям обмена веществ, детоксикации некото-
рых метаболитов.
В настоящий, второй том входят главы, в которых рассмотрены дыхательная
функция крови, терморегуляция, значение биологических ритмов, различные виды
рецепции — механорецепция, фонорецепция, хеморецепция, фоторецепция.
Предназначено для физиологов, биохимиков, экологов, зоологов, студентов,
аспирантов и преподавателей соответствующих кафедр университетов и институ-
тов, а также для инженеров, занимающихся проблемами бионики и кибернетики^
Редакция биологической литературы
50300—370
-----------Поди, изд,
041(01)—77
© 1973 by W. В. Saunders Company
© Перевод на русский язык, «Мир», 1977
Глава 8
Дыхательные функции крови
Л. ПРОССЕР
Если у животного имеется система кровообращения, выполняю-
щая функцию переноса кислорода от дыхательной поверхности к
тканям тела, то в крови у него обычно есть и пигмент — перенос-
чик кислорода. Только у немногих животных кровь может перено-
сить достаточное количество кислорода в растворенном состоянии
без участия пигмента. У человека содержание растворенного кис-
лорода в плазме составляет 0,3, а в цельной крови — 0,24 об. %;
артериальная кровь содержит больше 19 об. % О2, так как 98%
всего кислорода крови связано с гемоглобином. У большинства
животных пигмент участвует в переносе кислорода постоянно,
у некоторых — только при низких парциальных давлениях О2;
у части животных пигмент обеспечивает запас кислорода сверх
того, что существует в растворенном виде. Кроме того, пигменты
крови выполняют функцию буфера при переносе СО2 и, как белки,
участвуют в поддержании коллоидно-осмотического давления
крови.
Все пигменты-переносчики представляют собой металлооргани-
ческие комплексы. Большинство пигментов крови содержит железо,
немногие — медь; известны пигменты и другого строения, но их
дыхательная функция достоверно не установлена.
Распространение пигментов
Гемоглобины. Гемоглобины состоят из железопорфирина (ге-
ма), связанного с белком (глобином). Порфирины широко распро-
странены в природе. Хлорофилл, например, представляет собой
магнийпорфирин. Во всех аэробных клетках содержатся железо-
порфириновые белки — цитохромы. Из пигментов, переносящих
кислород, наиболее полно изучен железопротопорфирин, или гем,
который бывает связан со специфическими белками. Белковая
часть, или глобин, значительно различается у разных животных
по величине, аминокислотному составу, заряду, растворимости и
другим физическим свойствам. Гемоглобины могут быть внекле-
точными — растворенными в жидкостях тела — и внутриклеточны-
6
Глава 8
ми; последние находятся в особых кровяных тельцах или в ткане-
вых клетках, чаще всего в мышечных и нервных. Гемоглобин встре-
чается у представителей далеких друг от друга типов, а в пределах
одного типа он иногда имеется лишь у немногих видов. Сущест-
вует много гемоглобинов, различающихся белковой частью, но
имеющих одинаковый гем.
Ниже приводятся сведения о распространении гемоглобинов и
родственных им пигментов, идентифицированных главным образом
с помощью спектроскопии.
Хордовые
Позвоночные. У всех позвоночных имеется гемоглобин (НЬ) в клетках кро-
ви; в красных мышцах содержится миоглобин (мышечный гемоглобин, или Mb).
Пигмент отсутствует лишь у некоторых рыб — у личинок угря (лептоцефалов)
и у представителей трех родов антарктических рыб.
Эритроциты млекопитающих лишены ядра, имеют форму круглого (за
исключением эритроцитов Camellidae) двояковогнутого диска. У большинства
других позвоночных они двояковыпуклые, эллиптические и содержат ядро.
Безъядерные эритроциты имеют обычно меньшую величину и содержатся в
крови в большем числе, чем эритроциты с ядром. Самые крупные эритроциты —
у амфибий.
Содержание гемоглобина на 100 мл крови составляет у большинства мле-
копитающих и птиц 12—18 г, у амфибий и рептилий 6—10 г, у рыб 6—11 г.
Средний срок жизни эритроцита в крови у человека 113—118 дней, у собаки
90—135, у крысы 50—60, у кролика 50—70, у курицы 28—38 дней, у черепахи —
более 11 месяцев.
Низшие хордовые. У большинства, в том числе у ланцетника (Branchio-
stoma), гемоглобина нет.
Иглокожие
У голотурий (Thyone, Cucumaria, Molpadia, Caudina) гемоглобин имеется в кро-
вяных тельцах.
Аннелиды
У олигохет (Lumbricus, Tubifex) и пиявок (Hirudo, Analastoma) гемоглобин рас-
творен в плазме крови; в мышцах Lumbricus содержится миоглобин.
Полихеты отличаются большим разнообразием пигментов.
А. Полихеты с замкнутой системой кровообращения могут иметь:
1) гемоглобин в клетках целомической жидкости и в плазме крови (Тerebel-
la, Travisia);
2) гемоглобин только в плазме крови (Nereidae, Arenicola, Amphitrite, Cir-
ratulidae, Eunicidae и др.);
3) хлорокруорин в плазме крови (Sabellidae, Serpulidae, Chlorhaemidae,
Ampharetidae). У некоторых представителей Serpulidae в плазме могут
содержаться одновременно гемоглобин и хлорокруорин;
4) гемоэритрин в кровяных тельцах (Magelona);
5) они могут не иметь пигмента ни в крови, ни в целомической жидкости
(Syllidae, Phyllodice, Aphroditidae, Chaetopteridae, Lepidonotus).
Б. Полихеты без замкнутой системы кровообращения могут иметь:
1) гемоглобин в клетках целомической жидкости (Capitellidae, Glyceridae,
Polycirrus hematodes, P. aurantiacus);
2) пеломическую жидкость, не содержащую пигмента (Polycirrus tenuisetis,
Р. arenivorus).
Эхиуриды
Гемоглобин имеется в клетках пеломической жидкости Urechis и Thalassema,
а также в мышцах стенки тела Urechis и Arhynchite.
Дыхательные функции крови
7
Форониды
Гемоглобин содержится в кровяных тельцах Phoronis и Phoronopsis.
Членистоногие
Ракообразные. Гемоглобином обладают многие Entomostraca, но его не
найдено у Malacostraca. Он имеется у Artemia (Anostraca), Daphnia (Cladoce-
ra), Triops и Apus (Notostraca); гемоглобин найден также у одного паразити-
ческого веслоногого (Copepoda), у одного представителя Ostracoda и у одного
из паразитических усоногих (Cirripedia).
Насекомые. Гемоглобин есть у личинок хирономид и у овода Gastrophilus,
у которого пигмент содержится в трахеальных и некоторых других клетках.
Моллюски
Гемоглобин имеется в кровяных тельцах у немногих пластинчатожаберных
(Solen, Area, Pectunculus) и в плазме крови одного брюхоногого моллюска
(Planorbis), миоглобин — в мускулатуре радулы у многих переднежаберных
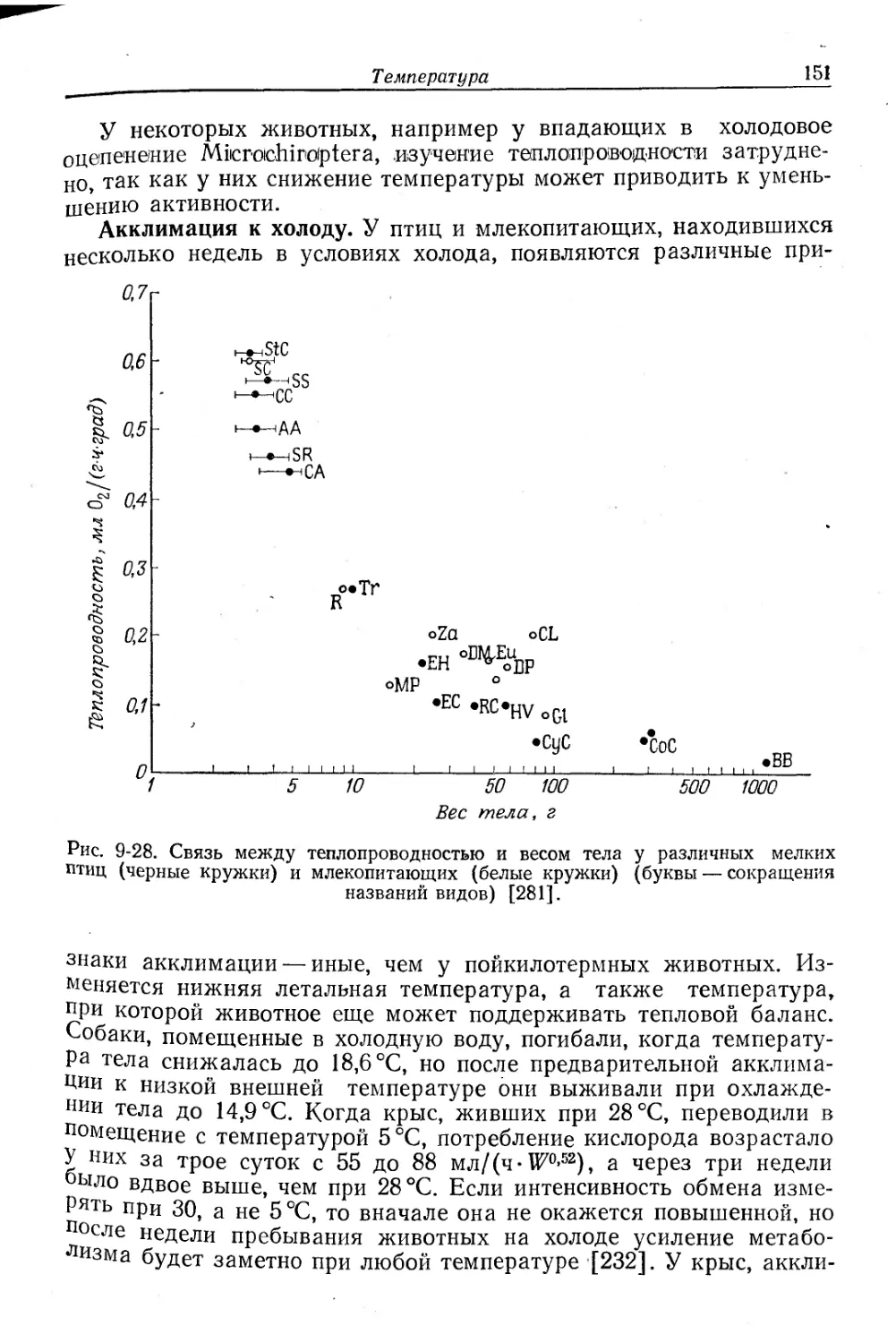
брюхоногих моллюсков и хитонов.
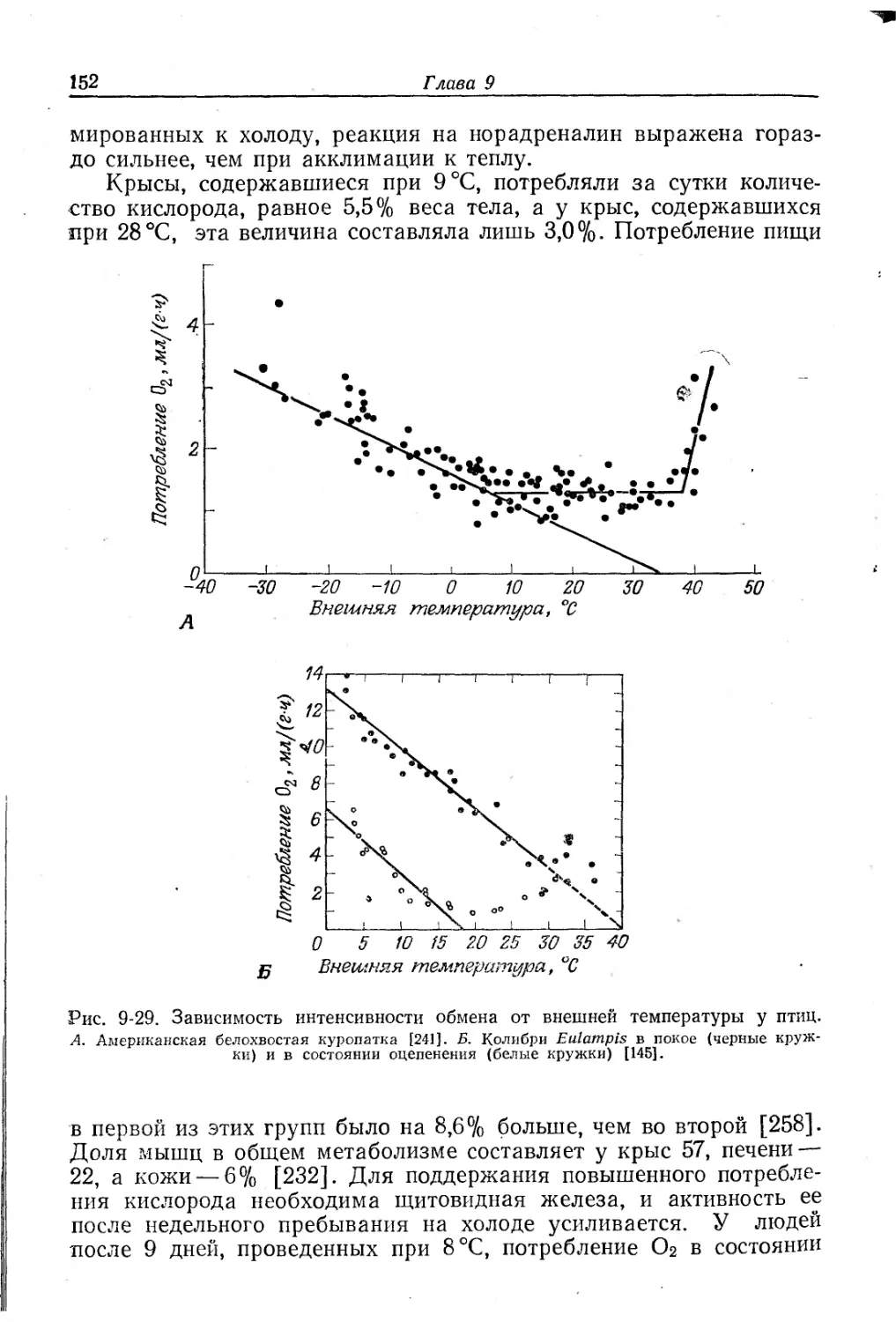
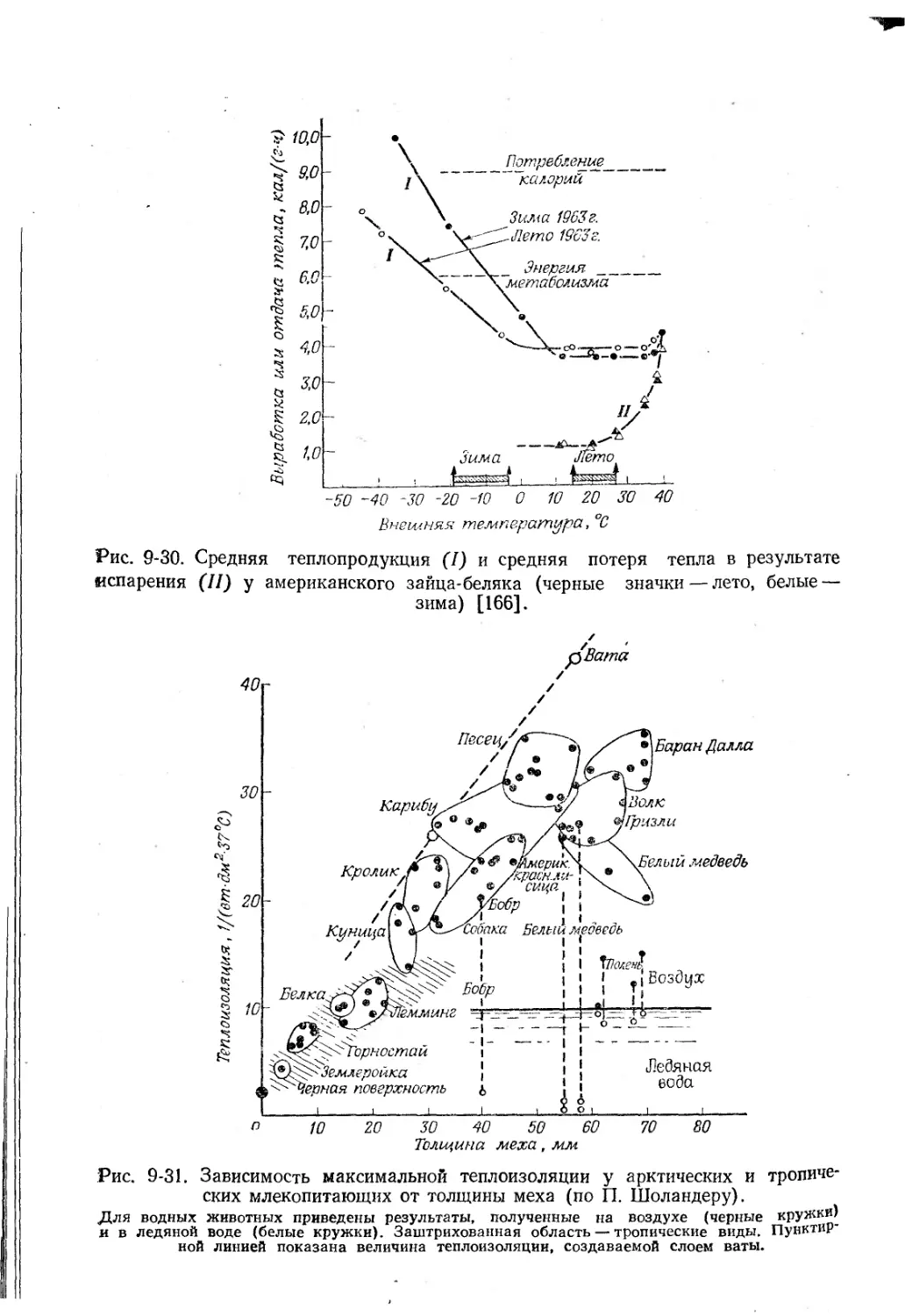
Немертины.
У некоторых немертин гемоглобин содержится в плазме крови, у других — в эри-
троцитах, а у Polia — также в клетках ганглиев.
Плоские черви
Гемоглобин есть у некоторых паразитических сосальщиков и прямокишечных тур-
беллярий (Derostoma, Syndesmis, Telorchis).
Круглые черви
Гемоглобин имеется в псевдоцеломической жидкости и в клетках гиподермы
(Ascaris, Nippostrongylus, Eustrongylides, Camallanus).
Простейшие
Hb найден у Paramecium и Tetrahymena (некоторые линии).
Растения
Корневые клубеньки ряда бобовых содержат гемоглобин, который образуется
только при симбиозе растения с бактериями.
Хлорокруорин. В разбавленном растворе этот пигмент имеет
зеленый цвет; он содержит железопорфирин, отличающийся от ге-
ма. Хлорокруорин найден в плазме представителей по крайней
мере 4 семейств многощетинковых червей, в частности у Sabellidae
и Serpulidae; у 21 вида Serpulimorpha обнаружен хлорокруорин,
а у Potamilla — гемоглобин в мышцах и хлорокруорин в крови.
У Spirorbis borealis имеется хлорокруорин, у S. corrugatus — ге-
моглобин в крови, а у S. militaris нет ни того, ни другого. Кровь
Serpula содержит и гемоглобин, и хлорокруорин [50, 53].
Гемоэритрин. Третий железосодержащий пигмент — гемоэрит-
рин — найден у полихеты Magelona, у сипункулид Sipunculus, Den-
drostomum и Golfingia (Phascolosoma), у плеченогого Lingula и
некоторых приапулид. Этот фиолетовый пигмент находится в кро-
вяных тельцах; железо в его молекуле не входит в порфириновое
кольцо.
Гемоцианин. После гомоглобина наибольшее значение, судя по
его распространенности, имеет медьсодержащий пигмент гемоциа-
нин. Он найден у моллюсков (боконервных, головоногих, некоторых
8
Глава 8
брюхоногих) и у членистоногих — ракообразных (в том числе
у многих высших раков), мечехвостов (Limulus) и некоторых пау-
кообразных. Гемоцианин представляет собой купропротеид без
порфириновой группы; он всегда растворен в плазме. У многих
моллюсков, например у улитки Busycon и боконервного моллюска
Criptochiton, в крови содержится гемоцианин, а в некоторых мыш-
цах — миоглобин.
Химия пигментов — переносчиков кислорода
Гемоглобины
Общее строение. Молекула гемоглобина состоит из нескольких
мономеров, каждый из которых содержит один гем, соединенный
с белком. Молекулярный вес мономера у позвоночных составляет
около 16-103, гемоглобин крови высших позвоночных содержит
4 мономера (т. е. имеет мол. вес от 64 до 65-103). В гемоглобинах
беспозвоночных молекулярное соотношение гем : белок меньше
единицы. Гем представляет собой протопорфирин, состоящий из
4 пиррольных колец с железом в центре (рис. 8-1).
В гемоглобине млекопитающих железо составляет 0,336%,
гем — 4% общей массы. В норме железо находится в закисной
форме. Миоглобин человека содержит 0,318% железа. В основе
функции гемоглобина лежит обратимое присоединение кислорода
к закисному железу (оксигенация) в зависимости от парциального
давления кислорода (Ро2)- При воздействии сильных окислителей
железо может переходить в окисную форму, в результате чего об-
разуется метгемоглобин, который уже не способен обратимо свя-
зывать кислород. Кислород присоединяется к каждому из атомов
железа согласно уравнению равновесия
[Fe][O2] •
Окись углерода также обратимо связывается с гемоглобином, при-
чем сродство последнего к СО обычно намного выше, чем к
Как показывает рентгеноструктурный анализ, гем находится
у поверхности глобулярного белка, который в этом месте образует
«карман» (рис. 8-2). Гем связан с глобином главным образом с
помощью гидрофобных взаимодействий. В p-цепи группа СОО^
одной пропионовой кислоты гема образует связь с серином, а дру-
гой — с лизином; в cz-цепи остаток пропионовой кислоты связан
с гистидином [146].
Молекула дезоксигенированного гемоглобина имеет положи-
тельный магнитный момент, т. е. обладает парамагнитными свой-
ствами. Соединение с кислородом или окисью углерода приводит
к потере магнитного момента, т. е. молекула становится диамаг-
нитной, слабо восприимчивой к магнитному полю, что указывает
Дыхательные функции крови
9
на отсутствие в ней неспаренных электронов. Изменение магнит-
ного момента отражает изменения в составе электронных пар у
атома железа. Изучение оптических спектров и магнитных свойств
молекулы позволяет следить за валентностью железа и природой
лиганда в оксигемоглобине [206]. Окисное и закисное железо мо-
жет существовать в двух спиновых состояниях, отражающих рас-
СН3 CH—СНз
с--——-с
нс=с с=сн
с==с
I I
(СНг)2 СНз
соон
Рис. 8-1. Строение гемоглобина (по Perutz М. F. et al., Nature, 185, 416—422,
1960, видоизменено).
А. Структурная формула гема. Б. Конфигурация а- и P-цепей по данным рентгеноструктур-
ного анализа. Гемы, изображенные в виде дисков, присоединены к белку через гистидин.
Сульфгидрильные (SH) группы располагаются в плоскости, отличной от плоскости гисти-
дина (Гис).
пределение электронов на d-орбитали. В результате сравнения
ряда железопорфириновых соединений установлено, что оксигена-
ция сопровождается частичным переходом электрона от закисного
железа к О2, железо проявляет тенденцию к окислению, а кислород
частично восстанавливается:
НЬ (гем d2) О2 НЬ(гемб|)-ОО .
Таким образом, железо переходит формально из восстановленного
(d6) в окисленное (d5) состояние, а кислород переходит в ОО~.
Молекулы миоглобина и гемоглобина имеют шарообразную
форму и построены из разного числа цепей. Миоглобин состоит
из одной цепи, а гемоглобины большинства позвоночных — из че-
тырех цепей. У человека молекула гемоглобина содержит четыре
гема и состоит из двух одинаковых половин, каждая из которых
включает одну а- и одну p-цепь. <р-Цепь состоит из 8 сегментов
правосторонней а-спирали, а в а-цепи 7 таких сегментов. В мио-
глобине 8 спиральных сегментов, и его цепь состоит примерно из
150 аминокислотных остатков. В НЬА a-цепь содержит 141 оста-
Рис. 8-2. Модель реконструированной молекулы гемоглобина [144].
А. Вид со стороны aiPi-контакта. Б. Вид со стороны сцРг-контакта (в направлении, перпен-
дикулярном предыдущему).
Дыхательные функции крови
11
ток, а р-цепь — 146 остатков. Модели этих молекул разработаны
Перутцем и его сотрудниками [144—146].
Отщепление даже одной или двух концевых аминокислот гемо-
глобина карбоксипептидазой ведет к значительному уменьшению
эффекта Бора и снижению сродства к О2; при этом карбоксипеп-
тидаза легче расщепляет НЬО2, чем НЬ. Таким образом, даже
очень небольшие структурные изменения влияют на дыхательную
функцию пигмента. Антонини [7] показал, что при воздействии на
гемоглобин человека карбоксипептидаза А отщепляет аминокис-
лотные остатки от С-конца p-цепи, а карбоксипептидаза В — такие
же остатки от а-цепи.
Строение гемоглобина некоторых беспозвоночных (иногда на-
зываемого эритрокруорином) было выяснено сравнительно недав-
но. Гемоглобин Paramecium aurelia содержит 100 аминокислотных
остатков [190]. Как показал рентгеноструктурный анализ, а-спи-
ральная структура гемоглобина одинакова у насекомого Chirono-
mus [85], аннелиды Glycera [139] и лошади [146]. У полихеты
Arenicola в отличие от позвоночных гемоглобин содержит один гем
на две белковые цепи, каждая из которых имеет мол. вес около
13000 [201а]. У Ascaris молекула гемоглобина полостной жидко-
сти содержит 8 гемов, и мол. вес ее достигает 328000, а в гемо-
глобине из стенки тела (мол. вес 37 000) только один гем [138а].
Многие животные имеют несколько различных гемоглобинов,
в которых одна из цепей может быть заменена другой. Например,
фетальный гемоглобин человека (HbF) отличается от гемоглобина
«взрослого» типа тем, что p-цепи заменены в нем у-цепями.
У человека описано более 90 аномальных гемоглобинов; за не-
многими исключениями, они отличаются от нормального НЬ всего
лишь одним аминокислотным остатком в одной из полипептидных
цепей. Например, замена валином остатка глутаминовой кислоты
(одного из примерно 290 аминокислотных остатков в каждом ди-
мере тетрамера НЬ) в 100 раз уменьшает растворимость дезоксиге-
нированной формы гемоглобина, и это приводит к так называемой
серповидноклеточной анемии [90, 91].
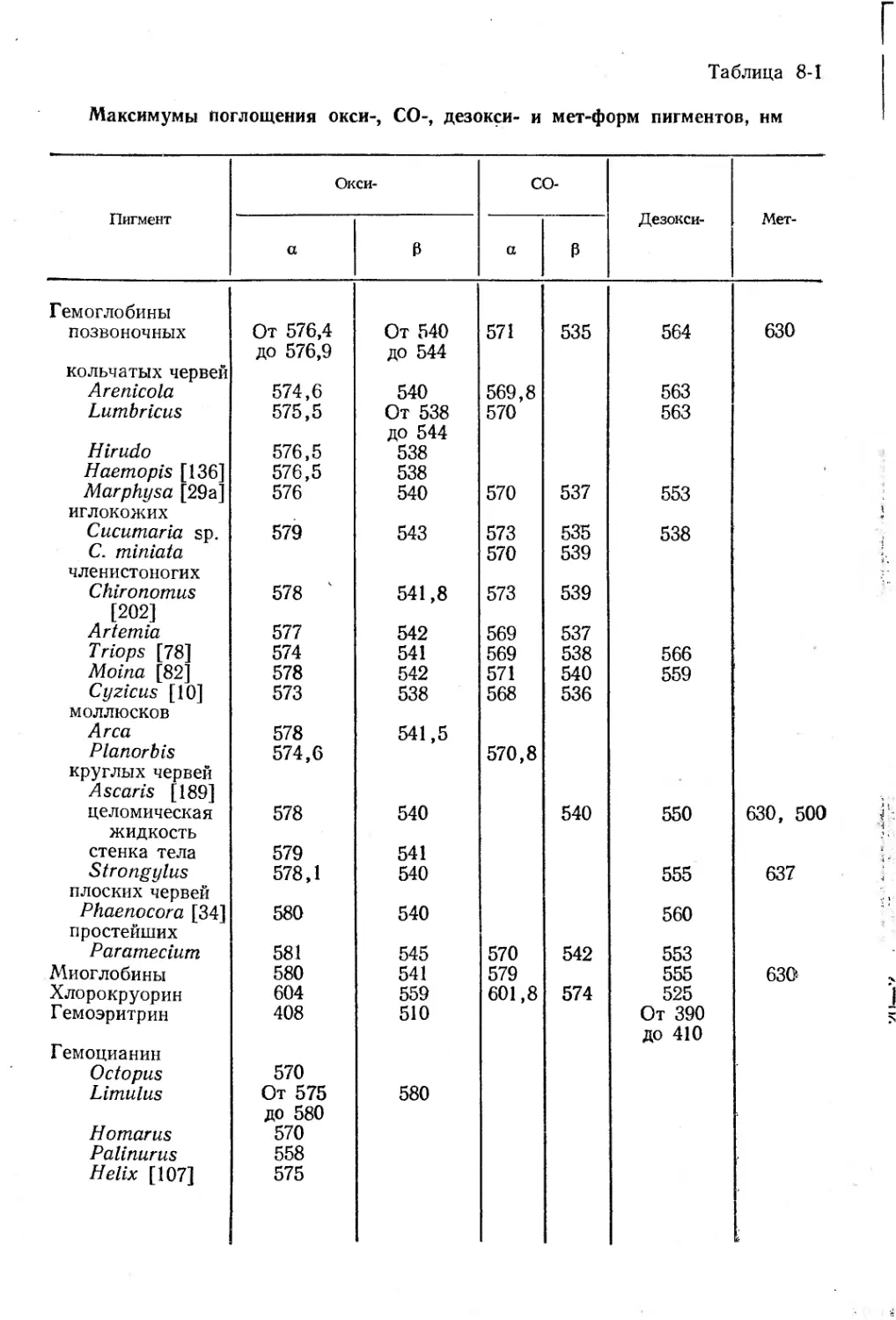
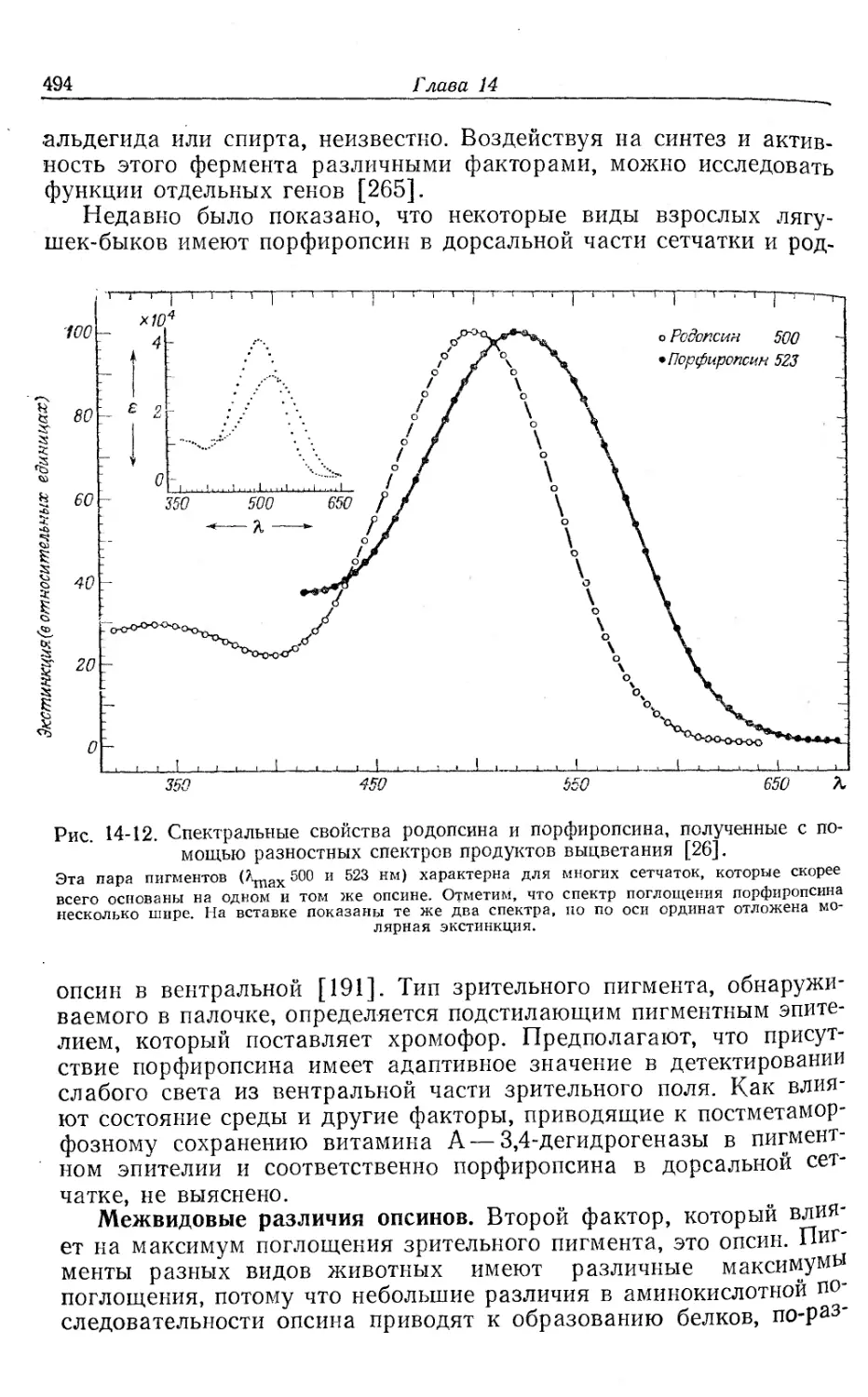
Полосы поглощения. Все соединения, содержащие гем, имеют
характерные спектры поглощения. Значения некоторых максиму-
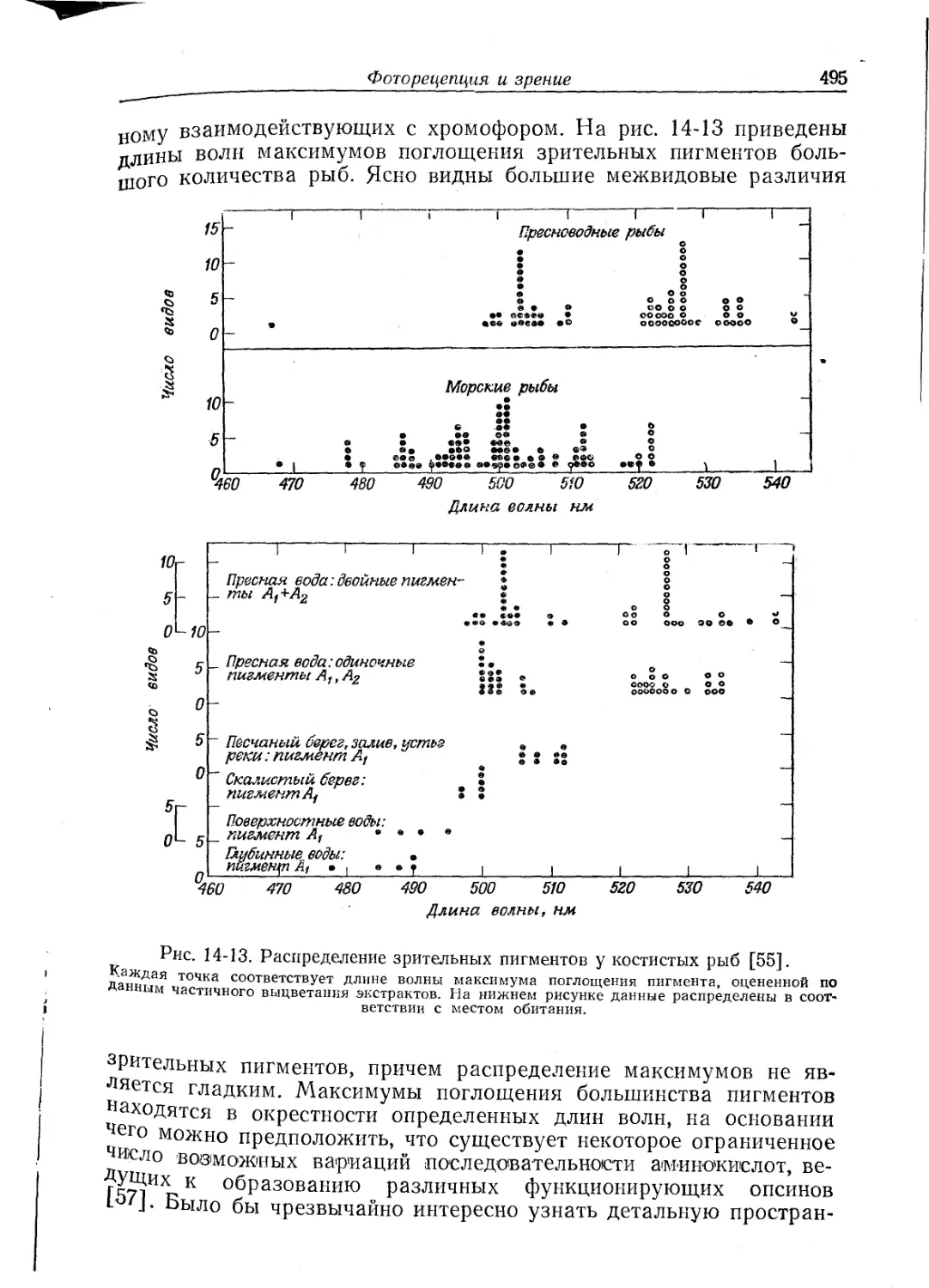
мов поглощения для отдельных видов животных (из огромного
числа изученных) приведены в табл. 8-1. Оксигенированный гемо-
глобин имеет две главные полосы поглощения: a-полосу в желтой
и p-полосу в зеленой части спектра. Карбоксигемоглобин также
имеет две полосы, но при меньших длинах волн, чем оксигемогло-
бин, и расстояние (в нанометрах или ангстремах) между а-поло-
сами окси-НЬ и карбокси-НЬ является характерной величиной для
Данного гемоглобина. Дезоксигенированный гемоглобин имеет
только одну широкую полосу в желто-зеленой части спектра. Для
метгемоглобина характерны четыре полосы поглощения, завися-
щие от pH, из них две очень сильные. Помимо полос поглощения
Таблица 8-1
Максимумы поглощения окси-, СО-, дезокси- и мет-форм пигментов, нм
Пигмент Окси- СО- Дезокси- Мет-
a ₽ a ₽
Гемоглобины
позвоночных От 576,4 От 540 571 535 564 630
до 576,9 до 544
кольчатых червей
Arenicola 574,6 540 569,8 563
Lumbricus 575,5 От 538 570 563
до 544
Hirudo 576,5 538
Haemopis 136] 576,5 538
Marphysa ’29a] 576 540 570 537 553
иглокожих
Cucumaria sp. 579 543 573 535 538
C. miniata 570 539
членистоногих
Chironomus 578 ' 541,8 573 539
[202]
Artemia 577 542 569 537
Triops [78] 574 541 569 538 566
Moina [82] 578 542 571 540 559
Cyzicus [10] 573 538 568 536
моллюсков
Area 578 541,5
Planorbis 574,6 570,8
круглых червей
Ascaris [189]
целомическая 578 540 540 550 630, 500
жидкость
стенка тела 579 541
Strongylus 578,1 540 555 637
плоских червей
Phaenocora [34] 580 540 560
простейших
Paramecium 581 545 570 542 553
Миоглобины 580 541 579 555 630
Хлорокруорин 604 559 601,8 574 525
Гемоэритрин 408 510 От 390
до 410
Гемоцианин
Octopus 570
Limulus От 575 580
до 580
Homarus 570
Palinurus 558
Helix [107] 575 i
Дыхательные функции крови
13
б видимой части спектра, гемоглобин имеет выраженные полосы
в фиолетовой области (полосы Сорэ):
Полосы Сорэ, нм
ньо2 ньсо MetHb НЬ
Гемоглобин 414,5 420 406 425
Миоглобин 418 424 407 435
У различных гемоглобинов полосы поглощения качественно
одинаковы, так как поглощение определяется гемом. Однако раз-
личия в белковой части гемоглобина могут приводить к сдвигу
максимума на несколько ангстрем (в пределах одного класса
животных) или даже на несколько нанометров (у представителей
разных классов или типов). Обычно a-пик выше, чем p-пик, но
в некоторых случаях (например, у Hb Ascaris и корневых клу-
беньков бобовых)1 p-пик выше [102]. Сдвиг a-полосы при переходе;
окси-формы в карбокси-форму для большинства гемоглобинов
составляет 43—56 А, для миоглобина 31—36 А, для гемоглобина
личинки овода (Gastrophilus) —95 А, для гемоглобина корневых,
клубеньков — 100 А. Спектры поглощения гемоглобинов паразита
и хозяина в большинстве случаев слегка различаются между со-
бой (например, у Ascaris и свиньи, у Gastrophilus и лошади). Од-
нако у Nematodirus полосы поглощения НЬ те же, что и у хо-
зяина — овцы. Шесть гемоглобинов морской миноги имеют одина-
ковый молекулярный вес, но слегка различаются в отношении по-
лос Сорэ.
Растворимость, изоэлектрическая точка и щелочная денатура-
ция. В нейтральном фосфатном буфере гемоглобин лошади рас-
творяется лучше, чем гемоглобин человека; у коровы фетальный
гемоглобин в 6 раз (а у овцы в 20 раз) более растворим, чем ге-
моглобин взрослых особей, тогда как у человека эта форма НЬ,
наоборот, менее растворима, чем пигмент «взрослого» типа.
Для гемоглобинов позвоночных характерны изоэлектрические
точки при pH 6,8—7,0, тогда как у большинства беспозвоночных
они лежат в области pH 5,0—6,0. Однако у некоторых рыб изоэлек-
трическая точка НЬ т’оже сдвинута в кислую сторону, например
у карпа 6,5, у опсануса от 5,7 до 6,2 [173]. Гемоглобин круглоро-
тых (Petromyzori) имеет изоэлектрическую точку при pH 5,6 [201],
один из гемоглобинов черепахи Pseudemys — при pH 5,7, а дру-
гой — при 7,2. Изоэлектрические точки гемоглобинов беспозвоноч-
ных различны, например у Gastrophilus 6,2, у Arenicola 5,1 [201а],
у Marphysa 4,6 [29а]. Изоэлектрическая точка гемоглобина клу-
беньковых бактерий лежит при pH 4,5. Ascaris имеет два гемогло-
бина: НЬ стенки тела и НЬ полостной жидкости с изоэлектрически-
ми точками при pH 5 и pH 6,7 соответственно [189]. У угря An-
guilla japonica также два гемоглобина: Ei имеет изоэлектрическую
1 А также у человека и многих позвоночных. — Прим, перев.
14
Глава 8
точку при pH 8,08, а Е2 — при pH 5,96. Кроме того, форма Ei
более устойчива к щелочной денатурации [68].
Скорость денатурации при высоких значениях pH используют
для характеристики гемоглобинов. Для фетального гемоглобина
человека (HbF) эта скорость значительно меньше, чем для гемо-
глобина взрослого (НЬА), тогда как у многих животных — овец,
головастиков, некоторых рыб — эмбриональный гемоглобин дена-
турируется быстрее, чем «взрослый» [101]. Гемоглобин дождевого
червя Lumbricus гетерогенен: он денатурируется в щелочи в три
этапа, и структура его пока неизвестна.
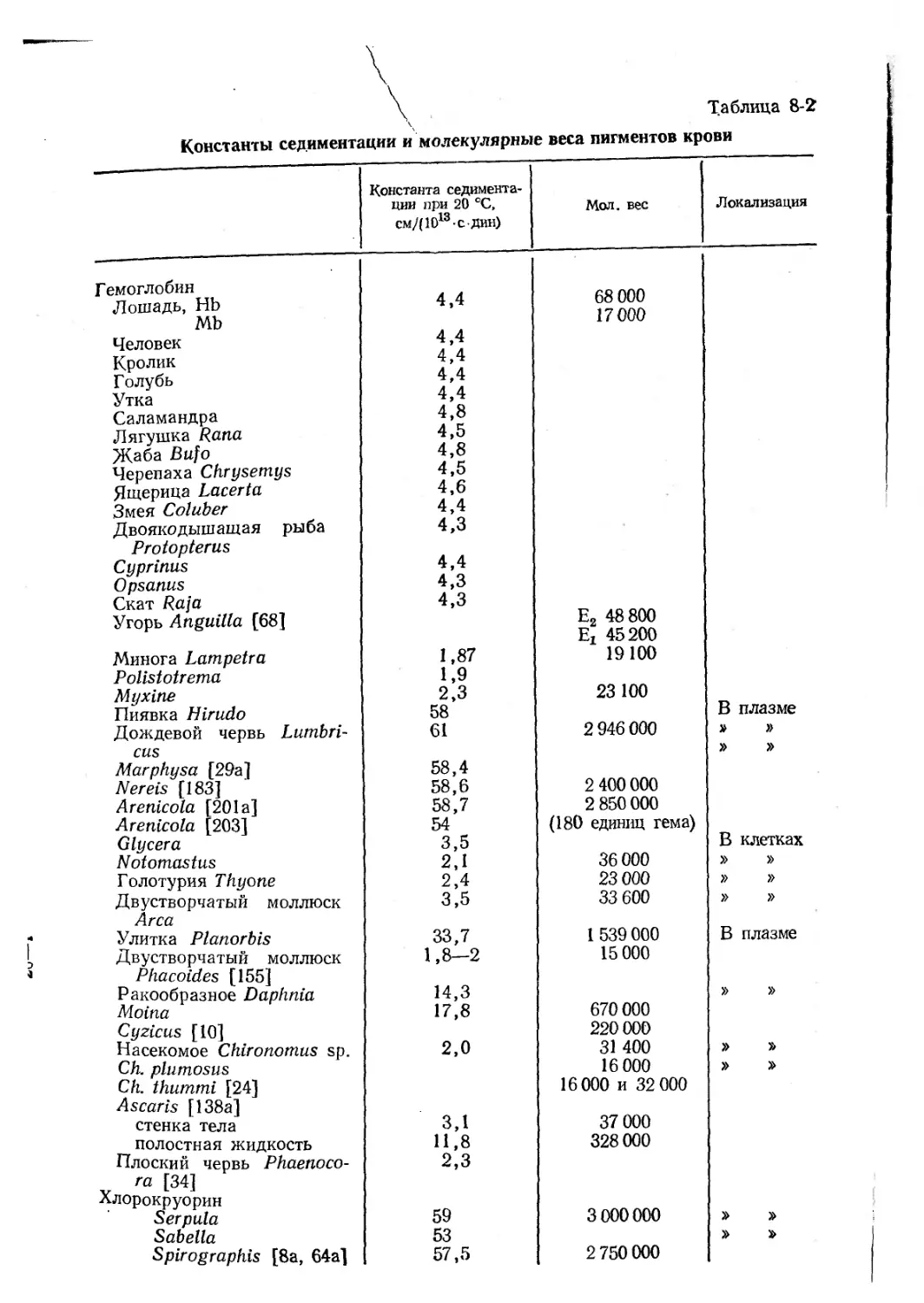
Размеры молекул. Величину молекул многих гемоглобинов оп-
ределяли методом седиментации в ультрацентрифуге. Константа
седиментации s20j w представляет собой скорость осаждения
(в см/с) в стандартном гравитационном поле при 20 °C в среде
с вязкостью, равной вязкости воды [193]. В табл. 8-2 приведены
константы седиментации и рассчитанные по ним молекулярные ве-
са пигментов крови различных животных. Для гемоглобинов поз-
воночных константа седиментации, как правило, составляет 4,3—
4,7-10"13 см/с, а для гемоглобинов беспозвоночных она выше. У уг-
ря Anguilla japonica одна из форм гемоглобина, Еь имеет мол. вес
65200 и состоит из четырех субъединиц с мол. весом 16 300, а дру-
гая форма, Е2, имеет мол. вес 68 800 и состоит ив четырех субъеди-
ниц с мол. весом 17 200 [212]. У амфибий и рептилий молекулы
гемоглобинов обычно несколько крупнее, чем у млекопитающих,
птиц и рыб. У жабы Bufo valliceps гемоглобин состоит из трех
компонентов с константами седиментации 4,8, 7,7 и 12,5S, что,
возможно, указывает на агрегацию субъединиц [193]. При изуче-
нии гемоглобинов у 54 видов черепах в ряде случаев были найдены
величины 4S и 7S, а у остальных видов — более 8S [192]. Гемо-
глобины лягушек и черепах имеют тенденцию полимеризоваться
при гемолизе [165]. Константа седиментации основного компонен-
та гемоглобина из полостной жидкости Ascaris равна 11, но имеют-
ся также компоненты 8S и 2S; для гемоглобина стенки тела этот
показатель равен 1,5S [189, 191]. Отсюда можно предположить,
что каждая молекула гемоглобина из полостной жидкости содер-
жит 8 гемов и состоит из субъединиц с мол. весом 40600. У Para-
mecium гемоглобин имеет мол. вес 13000 и константу седимента-
ции 1,5S (у миоглобина она равна 1,84S [190].
Миоглобин человека имеет константу седиментации 1,81 S и
мол. вес 17 450 (150 аминокислотных остатков); его молекула со-
держит один гем. Гемоглобин крови имеет мол. вес 64 500, и в его
молекуле 4 гема. Гемоглобин крови круглоротых имеет мол. вес
около 17 800, один гем в молекуле и константу седиментации 1,9S.
Гемоглобин Petromyzon marinus представляет собой в оксигениро-
ванном состоянии мономер, а в дезоксигенированной форме об-
разует димеры или тримеры, что связано со ступенчатым характе-
«к-гАтлГлий грмоглобины
\
\ Таблица 8-2
Константы седиментации и молекулярные веса пигментов крови
Константа седимента- ции при 20 °C, см/(1013сдин) Мол. вес Локализация
Г емоглобин Лошадь, НЬ Mb Человек Кролик Голубь Утка Саламандра Лягушка Rana Жаба Bufo Черепаха Chrysemys Ящерица Lacerta Змея Coluber Двоякодышащая рыба Protopterus Cyprinus Opsanus Скат Raja Угорь Anguilla [68] Минога Lampetra Polistotrema Myxine Пиявка Hirudo 4,4 4,4 4,4 4,4 4,4 4,8 4,5 4,8 4,5 4,6 4,4 4,3 4,4 4,3 4,3 1,87 1,9 2,3 58 68 000 17000 Е2 48 800 Ег 45200 19100 23 100 В плазме
Дождевой червь Lumbri- cus Marphysa [29a] Nereis [183] Arenicola [201 a] Arenicola [203] Glycera 61 58,4 58,6 58,7 54 3,5 2 946 000 2 400 000 2 850 000 (180 единиц гема) » » в » » клетках
Notomastus 2,1 36 000 » »
Голотурия Thyone 2,4 23 000 » »
Двустворчатый моллюск Area 3,5 33 600 » »
Улитка Planorbis Двустворчатый моллюск Phacoides [155] Ракообразное Daphnia Moina Cyzicus [10] 33,7 1,8—2 14,3 17,8 1 539 000 15 000 670 000 220 000 в » плазме »
Насекомое Chironomus sp. Ch. plumosus Ch. thummi [24] Ascaris [138a] стенка тела полостная жидкость Плоский червь Phaenoco- га [34] Хлорокруорин 2,0 3,1 11,8 2,3 31 400 16 000 16 000 и 32 000 37 000 328 000 » » »
Serpula Sabella Spirographis [8а, 64а] 59 53 57,5 3 000000 2 750 000 » » »
16
Глава 8
_____________ _ Продолжение
Константа седимента- ции при 20 rC, см/(Ю13 е дин) Мол. вес Локализация
Гемоэритрин Sipunculus Golfingia Гемоцианин Брюхоногие моллюски Вис cinum Littorina Busy con [179a] Helix [107] Helix Головоногие моллюски Loligo [39] Rossia Octopus Eledone Членистоногие Cancer [43] Callianassa [179a] Eriphia [40] Homarus [148] Limulus Pandalus Panulirus Pagurus Astacus Carcinus 102 (132) 99,7 (132) 60 100 130 99 59 (19, 11) 56,2 49,3 49,1 25,4 18 35 24 (16, 5) 16 34,6 (56,6; 16,1; 5,9) 17,4 16,4 16 93 3 23,3 (16,7) 66 Ос 108 000 4 400 000 8 800 000 13 009 000 6 680 000 9 000 OOJ 3 800 000 3 316000 2 785 000 940 000 480 000 1 800 000 950 000 825 000 1 800 000 397 000 В клетках » »
различаются по способности к полимеризации: у аксолотля дезок-
сигемоглобин при pH ниже 6,0 образует октомеры, а оксигемогло-
бин остается в виде тетрамера во всем диапазоне pH; у Rana
esculenta оксигемоглобин не полимеризуется, а дезоксигемоглобин
может образовывать октомеры при pH от 5 до 8; наконец, у Tritu-
rus как окси-, так и дезоксигемоглобин существуют в виде тетраме-
ров при любом значении pH [44]. Миоглобины млекопитающих
способны к полимеризации, особенно при pH между 3,5 и 5,5.
У тех беспозвоночных, у которых гемоглобин растворен в плаз-
ме, молекулярный вес его, как правило, превышает 1 000000; столь
большие размеры препятствуют выходу молекул из системы кро-
вообращения. У беспозвоночных, обладающих эритроцитами, мо-
лекулы гемоглобина имеют меньшую величину; например, у Thyo-
пе и Gastrophilus мол. вес гемоглобина около 34 000 и каждая
молекула, вероятно, состоит из двух субъединиц. Однако у личи-
Дыхательные функции крови 17
нок хирономид гемоглобин с низким молекулярным весом суще-
ствует в свободном виде в плазме. У Chironomus thummi имеются
два типа гемоглобина с мол. весами 16 000 и 32 000 — вероятно,
мономер и димер [24]. У аннелиды Glycera целомический гемо-
глобин имеет мол. вес 18200; число остатков на одну спираль то
же, что и в миоглобине [139]. Гемоглобин Arenicola имеет мол.
вес 2,85-106. Интактная молекула состоит из двух шестичленных
колец, т. е. из 12 субъединиц, каждая из которых обладает мол.
весом 230 000 и содержит 8 гемов. Таким образом, вся молекула
содержит 96 гемов [201а]. У раковинного листоногого рачка Cyzi-
cus гемоглобин крови имеет константу седиментации 11,4S, что
соответствует мол. весу 2,2-105; молекула состоит из 12 или
13 субъединиц [10].
При обработке гемоглобина некоторых млекопитающих такими
агентами, как мочевина, он распадается на частицы с мол. весом
34 000. Дальнейшее расщепление протекает гораздо труднее [211].
Как показывает рентгеноструктурный анализ, молекула гемогло-
бина лошади представляет собой сфероид размером 64X55X60 А.
Миоглобин имеет размеры 40X35X23 А [103, 104]. Кристаллы ге-
моглобина разных животных имеют характерную форму. На элек-
тронных микрофотографиях крупных молекул гемоглобина неко-
торых аннелид, в частности Lumbricus, видно, что они состоят из
двух наложенных друг на друга шестиугольных дисков диаметром
230—265 А. Каждый диск состоит из 6 единиц, содержащих по
3 субъединицы величиной 70 А [173а, 178а].
Электрофоретическая подвижность. Гемоглобины разных жи-
вотных различаются по своей электрофоретической подвижности.
Электрофорез обычно позволяет также обнаружить разные типы
гемоглобина у одного и того же вида животных. Например, мио-
глобин лошади состоит из двух электрофоретических компонентов,
гемоглобин коровы, овцы и буйвола — тоже из двух, а гемоглобин
свиньи оказывается гомогенным [200]. У овец, обитающих на уров-
не моря, преобладает один из двух компонентов, а у живущих на
больших высотах — другой. Разные породы овец можно различать
по соотношению гемоглобинов [45]. В гемоглобине взрослого чело-
века (НЬА) содержится примесь особой формы, HbAs, составляю-
щая в норме 2,5%, а при талассемии — 5,1 % [108]. В пуповинной
крови человека обнаружен фетальный гемоглобин (HbF), у 20-не-
дельного плода имеется гемоглобин с еще меньшей электрофоре-
тической подвижностью — эмбриональный, или «примитивный»
(НЬР). Есть данные о том, что НЬР образуется в мезобластах, в то
время как HbF синтезируется в печени, а НЬА — в костном мозге
[109]. У ряда других позвоночных фетальному гемоглобину тоже
предшествует эмбриональный. У 112 видов из 73 родов хищных
животных гемоглобины имеют электрофоретическую подвижность,
Равную 0,85 подвижности НЬА человека [185].
18
Глава 8
При изучении гемоглобинов птиц оказалось, что у уток и кур
имеются три компонента, у многих диких птиц (20 видов) — два,
у голубей и пингвинов — по одному. Гибрид перепела и дикой ку-
рицы обладает гемоглобинами, характерными для обоих родителей
[130]. Многие виды амфибий и рептилий имеют гемоглобины с не-
сколькими электрофоретическими компонентами. Многокомпонент-
ный гемоглобин обнаружен, например, у лягушки Rana catesbiana,
причем ни одна из электрофоретических полос гемоглобина взрос-
Рис. 8-3. Изменения фракционного состава гемоглобина у североморской сельди
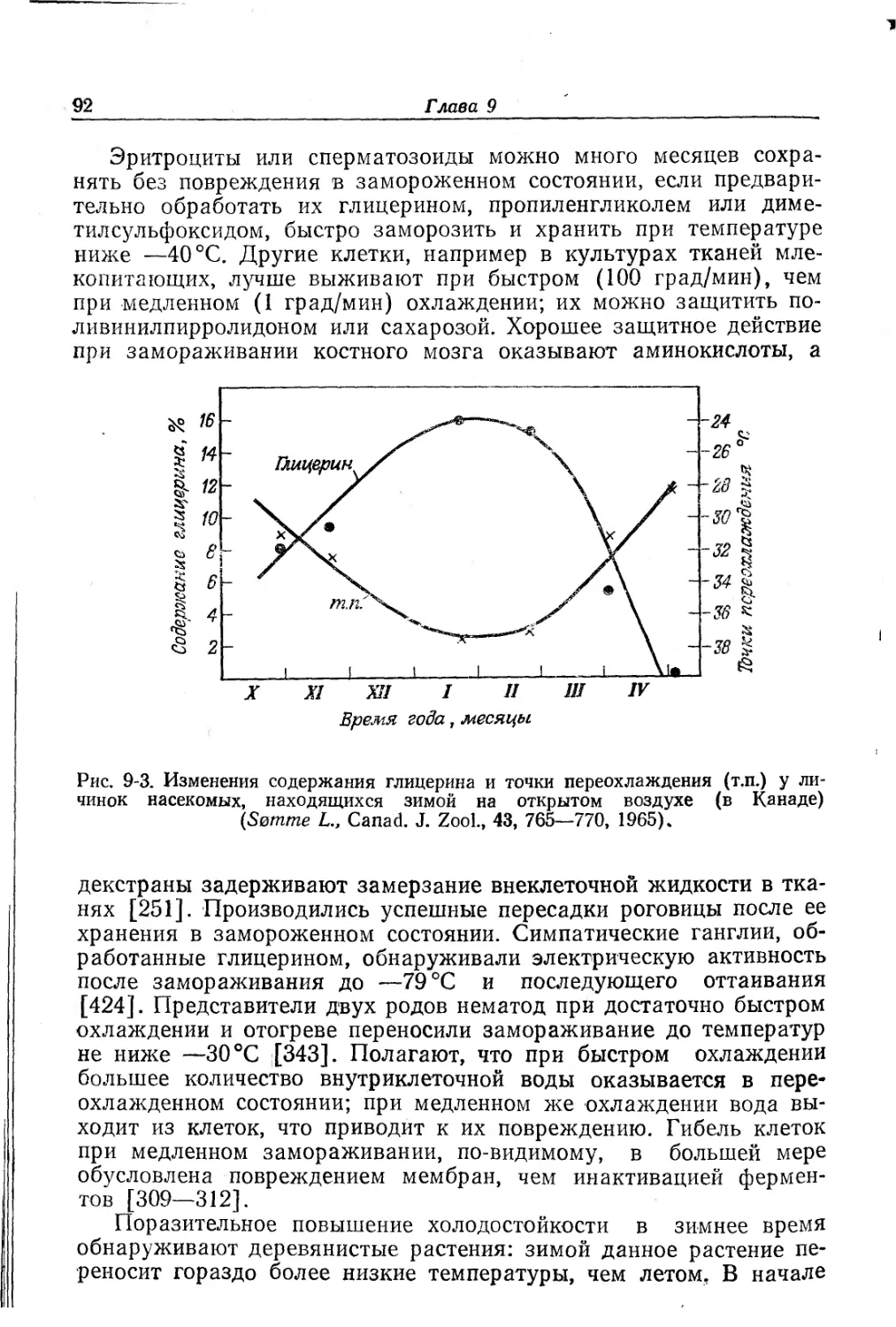
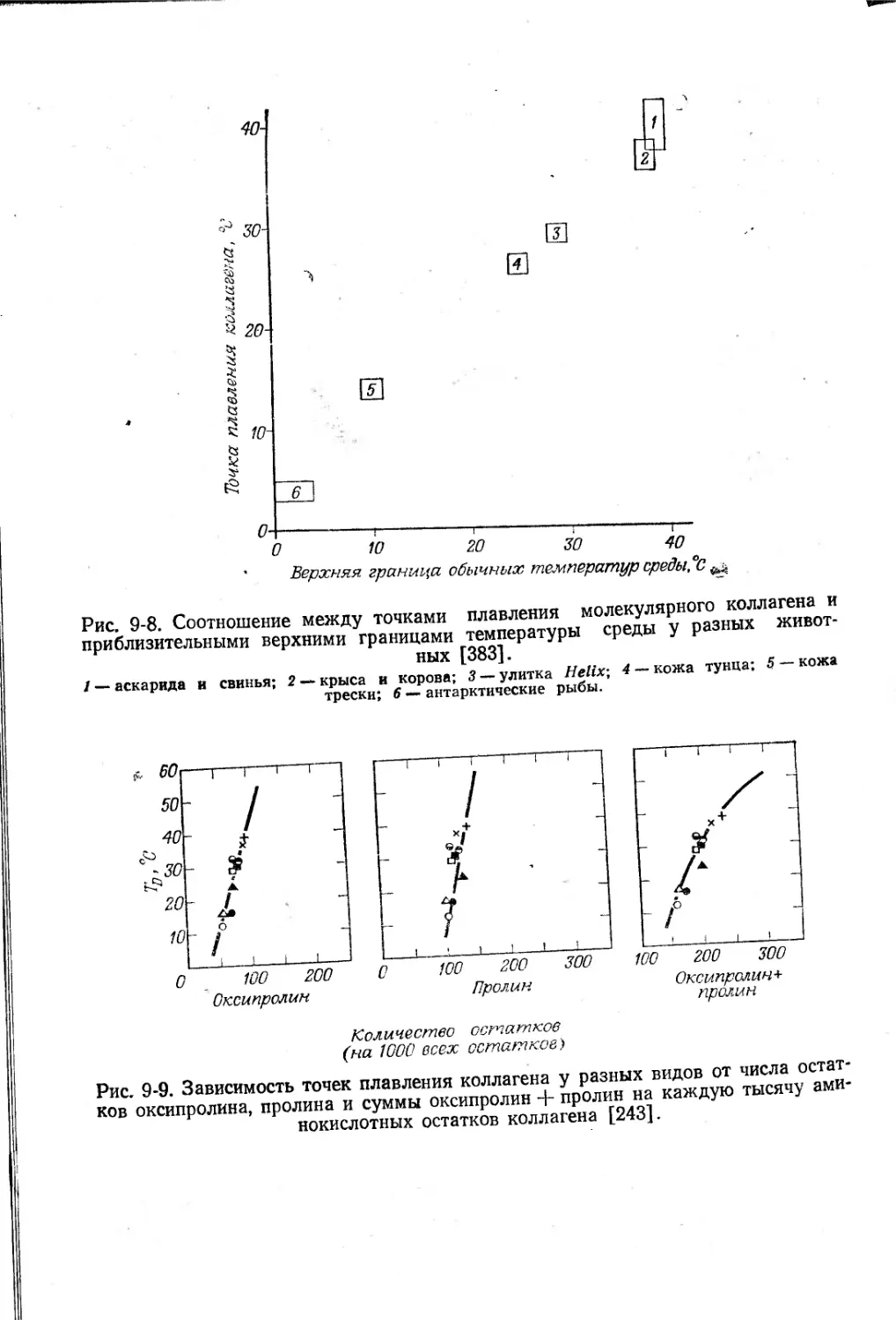
в зависимости от возраста и размеров тела [207].
лого животного не совпадает с полосами, характерными для пиг-
мента головастика [2].
У Tritiirus гемоглобин состоит из двух фракций — «быстрой»
(НЬА) и «медленной» (НЬВ), одна из двух полипептидных цепей
белка является общей для обеих форм [182].
У миноги Petromyzon marinas 6 типов гемоглобина с одинако-'
вым молекулярным весом [180], а у Lampetra fluviatilis гемогло-
бин состоит в основном из двух компонентов. Большинство рыб
имеет несколько циркулирующих в крови гемоглобинов, причем
состав их может быть полезен для целей систематики. Гибриды Fi
между разными видами рыб сем. Centrarchidae (например, уша-
стым окунем и краппи) обычно обладают гемоглобинами обоих
родителей. Однако при скрещивании того же вида ушастого окуня
с хенобриттусом у потомков Fj образуется лишь 25% гибридных
молекул, а другой вид ушастого окуня с хенобриттусом дает по-
томство с 40% гибридных молекул [131].
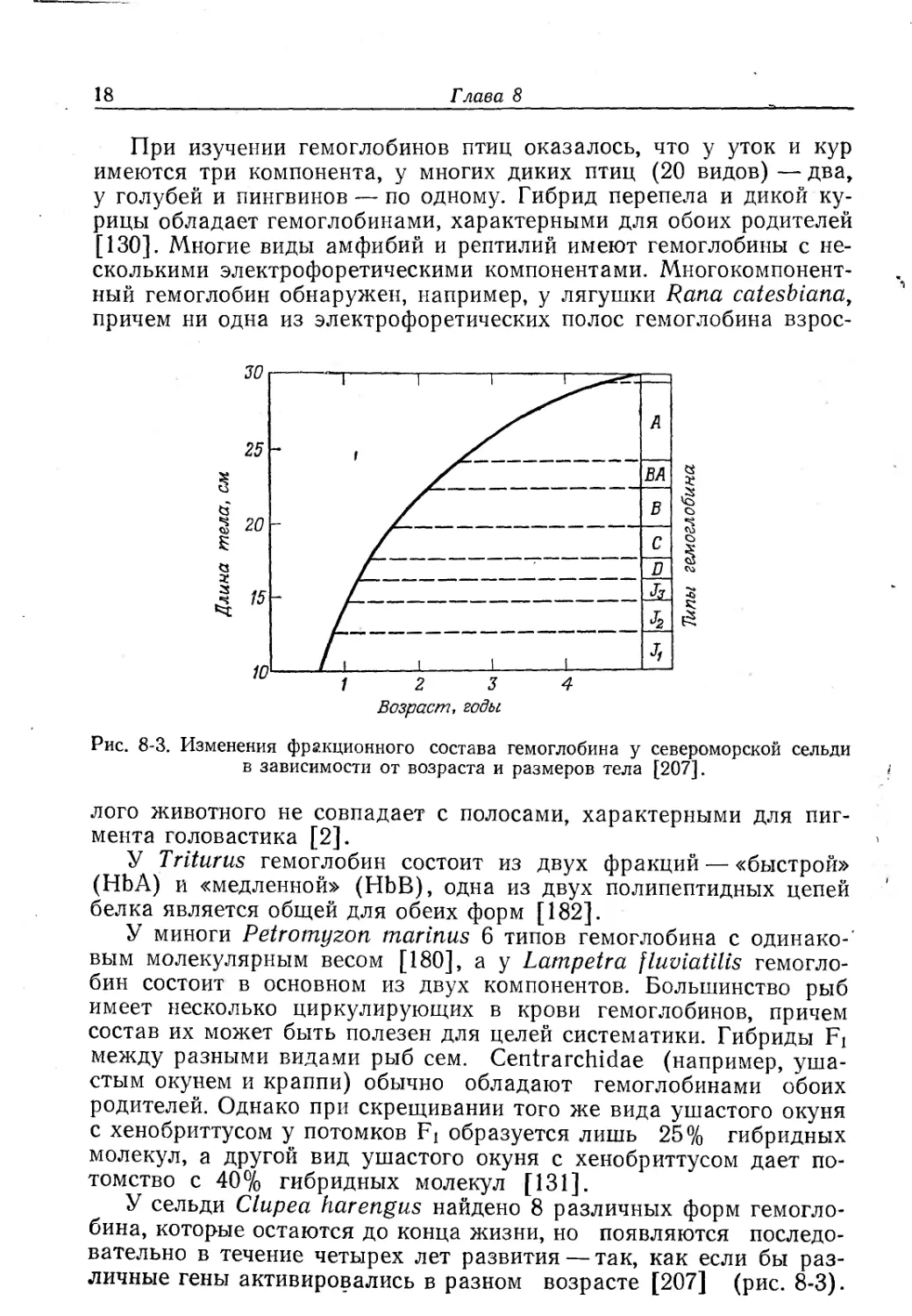
У сельди Clupea harengus найдено 8 различных форм гемогло-
бина, которые остаются до конца жизни, но появляются последо-
вательно в течение четырех лет развития — так, как если бы раз-
личные гены активировались в разном возрасте [207] (рис. 8-3).
Дыхательные функции крови
19
Кижуч (вид лосося) имеет 10 фракций гемоглобина, мигрирующих
к аноду, и 12 фракций, мигрирующих к катоду; у взрослых особей
можно обнаружить еще ряд катодных фракций [199].
У одной голотурии известны два генетических варианта: «тол-
стый» вариант имеет два вида гемоглобиновых цепей, а «тон-
кий» — пять [129].
Очевидно, у большинства животных, обладающих гемоглоби-
ном, этот пигмент одновременно представлен в крови несколькими
формами.
Аминокислотная структура цепей. Разделив гемоглобин на со-
ставляющие его цепи, можно исследовать аминокислотный состав
и структуру полученных полипептидов. Каждый тип цепи кодиру-
ется определенным цистроном, и точечная мутация в нем может
вызвать замену одного аминокислотного остатка в соответствую-
щем пептиде. Гемоглобин взрослого человека (НЬА) состоит из
двух a-цепей и двух p-цепей (с^Рг); фетальный гемоглобин (HbF)
имеет структуру «272- Как показал генетический анализ, гены, оп-
ределяющие синтез а- и p-цепей, находятся в разных хромосомах,
а гены ,р- и у-цепей — в одной хромосоме. р-Цепь больше подвер-
жена мутациям, чем а-цепь. N-концевая последовательность ами-
нокислот в p-цепях четырех мутантных гемоглобинов человека вы-
глядит следующим образом [90]:
НЬА: Вал-Гис-Лей-Тре-Про-Глу-Глу-Лиз
HbS: Вал-Гис-Лей-Тре-Про-Вал-Глу-Лиз
НЬС: Вал-Гис-Лей-Тре-Про-Лиз-Глу-Лиз
HbG: Вал-Гис-Лей-Тре-Про-Лей-Гли-Лиз
Гемоглобин относительно нечувствителен к заменам амино-
кислот на поверхности молекулы, но очень чувствителен к измене-
ниям в области неполярных связей, особенно вблизи гемов. Кроме
того, на функции молекулы сказываются замены аминокислот в
местах соединения а- и р-субъединиц. Некоторые замещения ами-
нокислот не влияют на функцию, в то время как другие изменяют
сродство к кислороду, взаимодействие между гемами или констан-
ту равновесия для агрегации субъединиц [146].
Третичная структура а- и p-цепей в окси- и дезоксигемоглобине
одинакова, однако четвертичная структура может быть различной.
Вообще a-цепи относительно более стабильны, и Ингрэм предпо-
лагает, что у первых позвоночных дыхательный пигмент состоял
из одной-единственной цепи (как миоглобин), а переход к гемо-
глобину современного типа произошел путем удвоения а-цепей;
в дальнейшем по мере эволюционной дивергенции появлялись раз-
личия в аминокислотном составе этих цепей. Согласно этой гипоте-
зе, у- и p-цепи дифференцировались значительно позже [91].
У человека а- и ip-цепи различаются по 85 из 141 аминокислот-
ного остатка; в р- и у-цепях неидентичны всего лишь 7 или 8 остат-
20
Глава 8
ков. у-Цепь (в HbF) содержит больше глутаминовой кислоты и
метионина, но меньше валина и пролина, чем p-цепь (в НЬА).
Гемоглобины человека и человекообразных обезьян почти оди-
наковы; видимо, устойчивые гены, ответственные за синтез а- и
p-цепей, сформировались еще до разделения этих двух ветвей эво-
люции. Сравнение гемоглобинов человека и гориллы показывает,
что они различаются по двум аминокислотным остаткам в а-цепи
и одному в p-цепи, в то время как между гемоглобинами человека
и лошади имеется 15 различий в аминокислотах [214]. У низших
приматов (лемуров) различия в p-цепях больше, чем в а-цепях
[71].
Среди 153 аминокислотных остатков в каждой из четырех по-
липептидных цепей гемоглобина млекопитающих обнаружено
25 различий между лошадью и кенгуру и 31 — между китом и
кенгуру [3]. Для 141 остатка a-цепи гемоглобина число различий
для некоторых видов приведено ниже [17]; видно, что оно тем
больше, чем дальше отстоят друг от друга виды в эволюционном
отношении.
Чело- век Макак- резус Лошадь Коро- ва Кури- ца Гигантский се- рый кенгуру Карп
Человек 0 4 18 17 35 27 71
Макак-резус 4 0 16 16 35 26 71
Лошадь 18 16 0 18 40 29 70
Корова 17 16 18 0 38 26 68
Курица 35 35 40 38 0 41 75
Гигантский серый кен- 27 26 29 26 41 0 74
ГУРУ Карп 71 71 70 68 75 74 0
Овцы полиморфны в отношении гемоглобина : две основные
фракции, А и В, находятся у них в разных соотношениях в зависи-
мости от породы. НЬА имеет большую электрофоретическую по-
движность, чем НЬВ. При анемии НЬА заменяется третьим гемо-
глобином (НЬС). Все три гемоглобина отличаются друг от
друга структурой p-цепей (С от А — по 12 аминокислотным остат-
кам, В от С — по 21 остатку) [22] и функциональными особенно-
стями (см. стр. 43). Высказано предположение о наличии опре-
деленной корреляции между соотношением гемоглобинов А и В и
принадлежностью овец к линиям с высоким или низким содержа-
нием калия в эритроцитах.
Гемоглобин взрослых кур разделяется на две полосы; НЫ со-
ставляет от 75 до 80% общего количества пигмента и содержит
больше лизина, гистидина и аргинина и меньше аспарагина и
глутамина, чем НЬП. У куриного эмбриона имеется третий тип
(НЫП) [88]. У ряда воробьиных птиц электрофоретически обна-
ружено по два и по три типа гемоглобина; каждый гемоглобин
Дыхательные функции крови
21
состоит из четырех цепей, причем а2- и ^-субъединицы в НЫ бо-
лее сходны между собой, чем в НЬП.
Два гемоглобина угря состоят из четырех полипептидных цепей
каждый, как у большинства позвоночных; во всех цепях на N-koh-
це находится валин, а С-концевые последовательности а- и р-цепей
различны [212]. Гемоглобин речной миноги (Lampetra) состоит из.
одной цепи, которая сильно отличается по своей первичной струк-
туре от гемоглобина человека (менее 30% совпадающих последо-
вательностей) [170].
У беспозвоночных гемоглобины содержат относительно меньше
гистидина и лизина, больше аргинина и намного больше цистина„
чем у позвоночных. Гемоглобин миноги, так же как и у млекопи-
тающих, содержит мало аргинина и много лизина, но он сходен
с гемоглобинами некоторых беспозвоночных малым содержанием
гистидина и высоким содержанием цистина [173]. Гемоглобин Аге-
nicola состоит из полипептидных цепей двух типов: в более длин-
ной цепи на N-конце находится аспарагиновая кислота, а на С-кон-
це — аланин [201а]. Гемоглобин аскариды содержит 332 амино-
кислотных остатка, среди которых относительно много изолейцина„
глутамина и аргинина [138а].
У человека при серповидноклеточной анемии эритроциты при-
нимают характерную форму серпа или полумесяца и частично те-
ряют способность переносить кислород. Серповидность проявляет-
ся при низких значениях Ро2 и низких pH. У оленей (по данным
электрофореза) имеется четыре гемоглобина. Один из них ответ-
ствен за феномен серповидности, который проявляется у оленя
при высоких Ро2 и щелочном pH [204].
Насколько часто различные формы гемоглобина встречаются
в одних и тех же эритроцитах, неизвестно. У головастиков крас-
ные кровяные клетки образуются в печени, а у взрослых лягу-
шек — в костном мозге. Иммунофлуоресцентное исследование осо-
бей в период метаморфоза показало, что эмбриональный и «взрос-
лый» гемоглобины находятся в разных эритроцитах [121].
Предполагается, что у млекопитающих в одном эритроците могут
одновременно присутствовать молекулы гемоглобина нескольких,
(мутантных) типов. При исследовании эритробластов взрослого
человека и плода, окрашенных с помощью флуоресцирующих ан-
тител, в тех и других были выявлены как НЬА, так и HbF, причем
соотношение между ними зависело от стадии развития организма;
[34а, 59а].
Хлорокруорин
Хлорокруорин — зеленый железосодержащий пигмент, находя-
щийся в крови сабеллид и серпулид, — содержит порфирин, ко-
торый отличается от гема тем, что в одном из пиррольных колец.
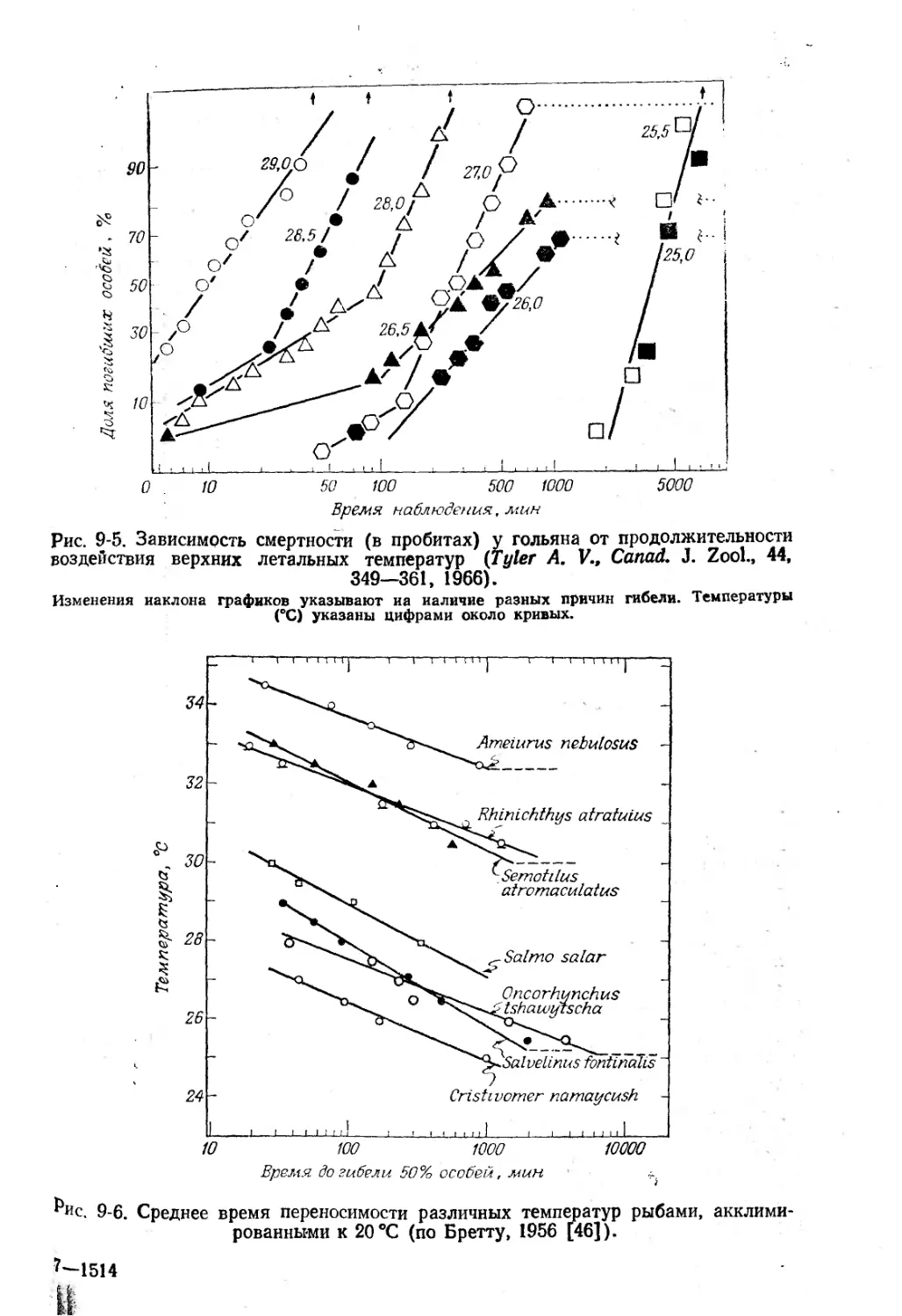
Рис. 8-4. Электронные микрофотографии
Гемоцианин Helix pomatia-, Х184 000. (Фото любезно
Х200 ООО
А Гемоцианин Litnulus; X290 ООО. Б.
Дыхательные функции крови
23
винильный радикал заменен формильным:
СН3— С—С— СНО
N
Хлорокруорин сходен по аминокислотному составу с гемогло-
бинами беспозвоночных. Это близкое сходство с гемоглобином поз-
воляет объяснить тот факт, что в крови некоторых червей содер-
жатся оба пигмента, а иногда в пределах одного рода у одних
видов имеется хлорокруорин, у других — гемоглобин (стр. 59).
Полосы поглощения хлорокруорина, оксихлорокруорина и
СО-хлорокруорина сдвинуты на 20—25 нм в сторону более длин-
ных волн по сравнению с соответствующими полосами гемоглоби-
на (см. табл. 8-1). У Spirorbis хлорокруорин имеет мол. вес около
3000000, а у Spirographis — около 2 750000 [64а]. Изоэлектриче-
ская точка пигмента равна 4,2; он всегда растворен в плазме (см.
табл. 8-2). Как и в гемоглобине, к одному атому железа присоеди-
няются два атома кислорода [50]. У Spirographis молекула хло-
рокруорина содержит 80 гемов [80]. Как показывают электронные
микрофотографии, хлорокруорин Spirographis, подобно гемоглоби-
ну некоторых других аннелид, состоит из двух одинаковых частей,
каждая из которых построена из 6 субъединиц величиной 70 А,
расположенных по углам шестиугольника [64а] (рис. 8-4,5).
Гемоцианин
В гемоцианине медь не входит в состав простетической группы,
а связана непосредственно с белком. Некоторые гемоцианины об-
разуют соединения с углеводами; например, гемоцианин Helix со-
молекул гемоцианина и хлорокруорина.
Предоставлены д-ром
1Ь4а].
Э. Бруггеном.) В. Дезоксихлорокруорин Spirographis при pH 7;
24
Глава 8
держит около 9% углеводов. Если в крови имеется гемоцианин, то
он обычно составляет более 90% всего растворенного белка.
У Loligo концентрация гемоцианина в крови 80 мг/мл, а у Busycon
40 мг/мл. У членистоногих содержание меди в гемоцианине равно
= 0,17%, а у моллюсков — 0,25%. У ряда ракообразных и Limulus
концентрация меди в крови колеблется от 4 до 9 мг%; у головоно-
гих моллюсков она составляет 25 мг%. В дезоксигемоцианине
медь находится в закисной форме; возможность ее присутствия
в окисной форме в настоящее время оспаривается [74].
В дезоксигенированном состоянии гемоцианины бесцветны, но,
как и все белки, поглощают в ультрафиолетовой зоне с максиму-
мом при 280 нм. Оксигенированные гемоцианины имеют синий
:цвет, у них широкий максимум поглощения при 570 нм и четкая
полоса поглощения в области от 340 до 350 нм.
Гемоцианины — очень крупные молекулы: у моллюсков кон-
станты их седиментации соответствуют молекулярному весу в не-
сколько миллионов, у членистоногих — в сотни тысяч. Молекула
гемоцианина может диссоциировать на меньшие компоненты. При
наибольшей степени агрегации молекулярный вес гемоцианина у
членистоногих достигает сотен тысяч. Гемоцианины членистоногих
могут иметь константу седиментации 35S (Lumulus, Callianassa)
и 60S (Lumulus). При высоких значениях pH дезоксигенированный
гемоцианин проявляет тенденцию к диссоциации, а по мере реокси-
генации происходит обратная ассоциация субъединиц. Гемоцианин
омара имеет мол. вес 825 000; в щелочном растворе удаление Са2+
способствует диссоциации на 12 субъединиц с мол. весом около
69 000 каждая. При дальнейшей диссоциации отщепляются актив-
ные центры, связывающие кислород [148]. Гемоцианины членисто-
ногих обычно состоят или из двух медьсодержащих цепей с мол.
весом 37 000, или трех цепей с мол. весом 25 000, в одной из кото-
рых атом меди отсутствует [74]. У Cancer гемоцианин не содер-
жит субъединиц с мол. весом менее 75 000 [179а].
Как показала электронная микроскопия, гемоцианин омара с
константой седиментации 24S имеет форму двойного диска, кото-
рый построен из 168-субъединиц, образующих в проекции прямо-
угольник или шестиугольник величиной до 100—125 А. Гемоцианин
скорпиона в состоянии димера образует большой прямоугольник.
У Limulus молекулы пигмента состоят из цилиндров с гексагональ-
ным расположением осей [39, 74] (рис. 8-4, А).
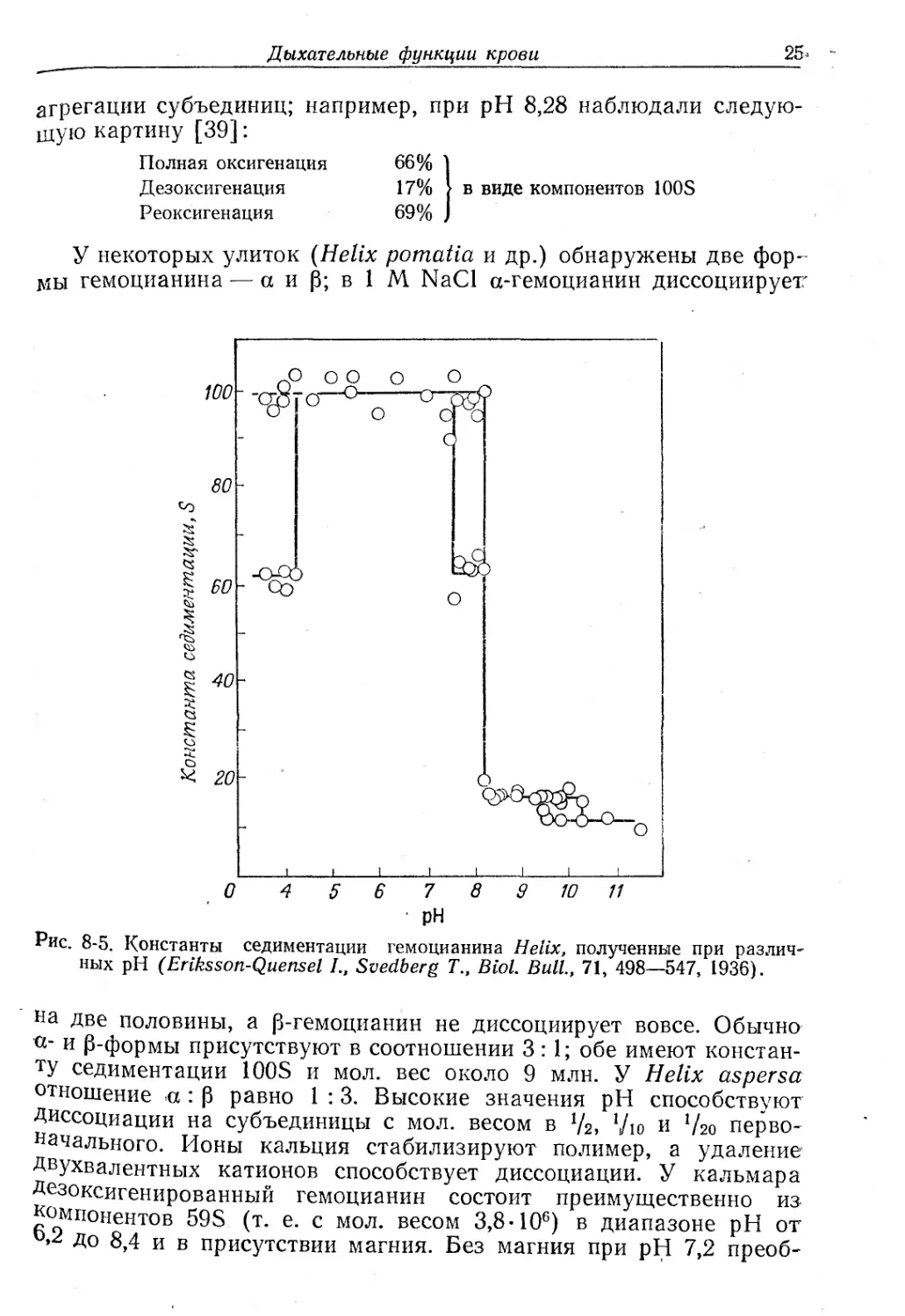
У моллюсков гемоцианины более разнотипны, чем у членисто-
ногих. У брюхоногих молекулы пигмента имеют константу седи-
ментации, равную 100S, и мол. вес от 8,7 до 9 млн. (рис. 8-5). По
данным электронной микроскопии у Busycon молекулы с констан-
той 100S имеют вид полого цилиндра диаметром 350 А и высотой
380 А. Молекулы с константой 60 S — результат поперечного рас-
щепления цилиндра; кроме того, образуются и более мелкие час-
титгы MIS. 15S и 30S). Присоединение кислорода способствует
Дыхательные функции крови
25-
агрегации субъединиц; например, при pH 8,28 наблюдали следую-
щую картину [39]:
Полная оксигенация
Дезоксигенация
Реоксигенация
66%
17%
69%
в виде компонентов 100S
У некоторых улиток (Helix pomatia и др.) обнаружены две фор-
мы гемоцианина — а и Р; в 1 М NaCl а-гемоцианин диссоциирует’
Рис. 8-5. Константы седиментации гемоцианина Helix, полученные при различ-
ных pH (Eriksson-Quensel I., Svedberg Т„ Biol. Bull., 71, 498—547, 1936).
на Две половины, а р-гемоцианин не диссоциирует вовсе. Обычна
°' и P-формы присутствуют в соотношении 3:1; обе имеют констан-
ту седиментации 100S и мол. вес около 9 млн. У Helix aspersa
отношение а : р равно 1 : 3. Высокие значения pH способствуют
Диссоциации на субъединицы с мол. весом в V2, Vio и ’/2о перво-
начального. Ионы кальция стабилизируют полимер, а удаление
Двухвалентных катионов способствует диссоциации. У кальмара
Дезоксигенированный гемоцианин состоит преимущественно из
компонентов 59S (т. е. с мол. весом 3,8-106) в диапазоне pH от
До 8,4 и в присутствии магния. Без магния при pH 7,2 преоб-
26
Глава 8
ладают 195-единицы, а при pH 9,5—HS-единицы [38]. При
80 %-ном насыщении кислородом гемоцианин представлен только
субъединицами; при 100%-ном насыщении происходит ассоциация
их в полимер [39].
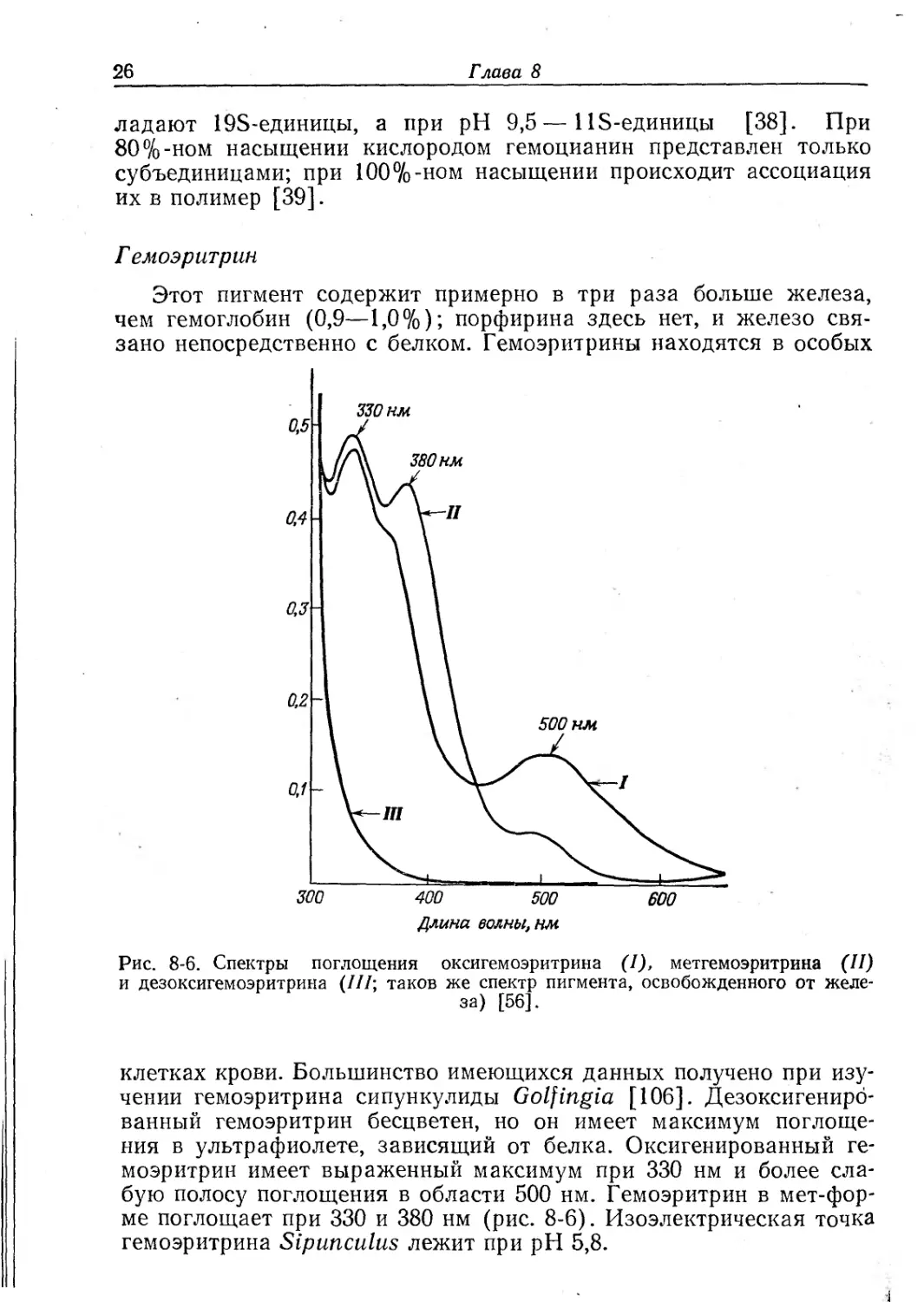
Г емоэритрин
Этот пигмент содержит примерно в три раза больше железа,
чем гемоглобин (0,9—1,0%); порфирина здесь нет, и железо свя-
зано непосредственно с белком. Гемоэритрины находятся в особых
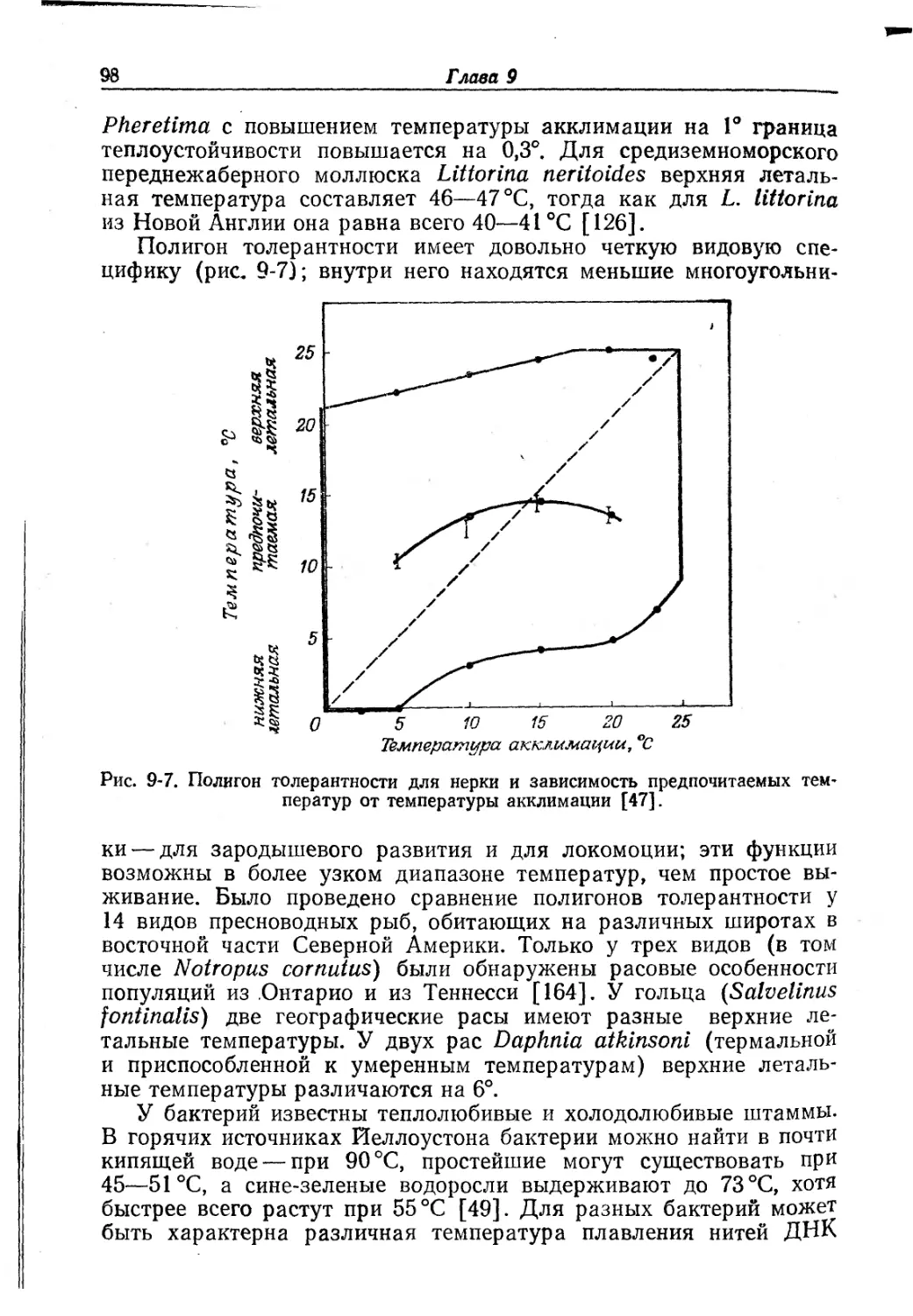
Рис. 8-6. Спектры поглощения оксигемоэритрина (1), метгемоэритрина (II)
и дезоксигемоэритрина (///; таков же спектр пигмента, освобожденного от желе-
за) [56].
клетках крови. Большинство имеющихся данных получено при изу-
чении гемоэритрина сипункулиды Golfingia [106]. Дезоксигениро-
ванный гемоэритрин бесцветен, но он имеет максимум поглоще-
ния в ультрафиолете, зависящий от белка. Оксигенированный ге-
моэритрин имеет выраженный максимум при 330 нм и более сла-
бую полосу поглощения в области 500 нм. Гемоэритрин в мет-фор-
ме поглощает при 330 и 380 нм (рис. 8-6). Изоэлектрическая точка
гемоэритрина Siptmculus лежит при pH 5,8.
Дыхательные функции крови 27
Гемоэритрины имеют константу седиментации 7S и мол. вес
108000. С помощью химических реагентов можно вызвать дис-
социацию молекулы на субъединицы с константой 2S и мол. весом
13 500; каждая субъединица содержит два атома железа, тогда как
в целой молекуле их 16. Есть данные о том, что гемоэритрин су-
ществует в двух формах — в виде мономера и октомера, которые
находятся в равновесии между собой, а димеры и тетрамеры прак-
тически отсутствуют; при обычной высокой концентрации пигмента
в кровяных тельцах большая часть пигмента находится в форме
октомера [56].
По данным пептидного анализа мономер представляет собой
цепь из 113 аминокислот с единственным цистином в 50-м положе-
нии, большая часть цепи (75%) имеет структуру а-спирали. Оба
атома железа двухвалентны и расположены примерно в середине
свернутой цепи [56]. При переходе гемоэритрина в мет-форму же-
лезо может присоединять различные лиганды — О2, С1, цианат,
N з или ОН.
Результаты новейших исследований (химический анализ, изу-
чение эффекта Мёссбауэра, оптических и магнитных свойств) ука-
зывают на то, что при оксигенации происходят значительные кон-
формационные изменения. Предложены следующие структуры
[56]:
/Н2О Н2ОХ
Дезоксигемоэритрин FeH. /FeH
ХН2О Н2СУ
/0\
Метгемоэритрин FeHi^ /FeHi
L
°ч
Оксигемоэритрин FeilL /Fein
of—О........................H—
(L—лиганд)
Соединение гемоглобина с кислородом
Скорости ассоциации и диссоциации
Скорости присоединения и отдачи лигандов пигментами были
измерены с помощью быстрорегистрирующих спектрофотометров.
Процесс связывания и освобождения О2 или СО гемоглобином со-
стоит из четырех этапов, и кинетика его очень сложна [179]. Со-
единение с СО идет на порядок медленнее, чем с О2. Константы
28
Глава 8
скорости суммарной реакции связывания для гемоглобина (моль-1*
-с-1) равны [58]:
Овца
Lumbricus
Arenicola
Кислород
2,6-Ю6
2,3-106
2,3-10®
Окись углерода
1,8-105
2,2-105
2,9-105
Присоединение лиганда идет быстрее, чем диссоциация. С мио-
глобином млекопитающих реакции протекают быстрее, чем с гемо-
глобином; время полунасыщения кислородом составляет для мио-
глобина 0,0004 с, а для гемоглобина 0,038 с. Гемоцианин в этом
отношении сходен с гемоглобином.
При работе с интактными эритроцитами реакции идут медлен-
нее, чем в растворе гемоглобина. Крупные эритроциты (опреде-
ленных животных) больше замедляют реакцию, чем мелкие. Таким
образом, размеры клеток, их проницаемость и скорость диффузии
могут лимитировать скорость присоединения и отдачи лиганда
[77]. Приводим данные о связывании Ог-
Константы скорости оксиге- нации, мМ—1с—1 Объем эритроцита, мкмЗ
эритроциты гемоглобин
Овца 137 .—
Лошадь — 2,4-103
Коза 133 2.7-103 20
Человек 80 2,4-103 90
Собака 65 —
Лягушка 19 2,7-103 680
Структурные изменения при оксигенации
Ряд данных говорит о том, что процессы присоединения и от-
дачи кислорода сопровождаются конформационными изменениями.
Так, расстояние между двумя реактивными остатками цистеина в
р-цепях, составляющее в дезоксигенированном гемоглобине 37,6 А,
уменьшается при оксигенации до 30 А; это связано с изменением
положения p-цепей относительно a-цепей [135а]. При оксигенации
освобождаются протоны гистидина — в среднем 3,8 протона в ге-
моглобине мыши и 1,5 протона в гемоглобине слона. Это означает,'
что оксигемоглобин — более сильная кислота (т. е. он в большей
степени диссоциирует), чем дезоксигемоглобин. При оксигенации
один электрон переходит от атома двухвалентного железа к кисло-
роду, который превращается в отрицательно заряженный ион (об
этом переносе заряда уже упоминалось на стр. 9).
Сродство различных гемоглобинов к кислороду неодинаково.
Например, у Rana catesbiana гемоглобин В наполовину насыщает-
Дыхательные функции крови
29
ся кислородом при Ро2 15,8 мм рт. ст., а гемоглобин С —уже при
7 9 мм рт. ст. [2]. Гемоглобин миноги состоит из одной цепи, но
при дезоксигенации мономеры обнаруживают тенденцию полимери-
зоваться в тетрамеры, которые вновь распадаются при оксигена-
ции. У миноги процесс насыщения НЬ кислородом, по-видимому,
включает три этапа: дезагрегацию, оксигенацию и высвобождение
протонов [25]. У ужа (Natrix) присоединение О2 ведет к диссо-
циации субъединиц гемоглобина [192]. Гемоглобин кошки дает
при электрофорезе две полосы; величины Р50 для обоих компонен-
тов сходны [194].
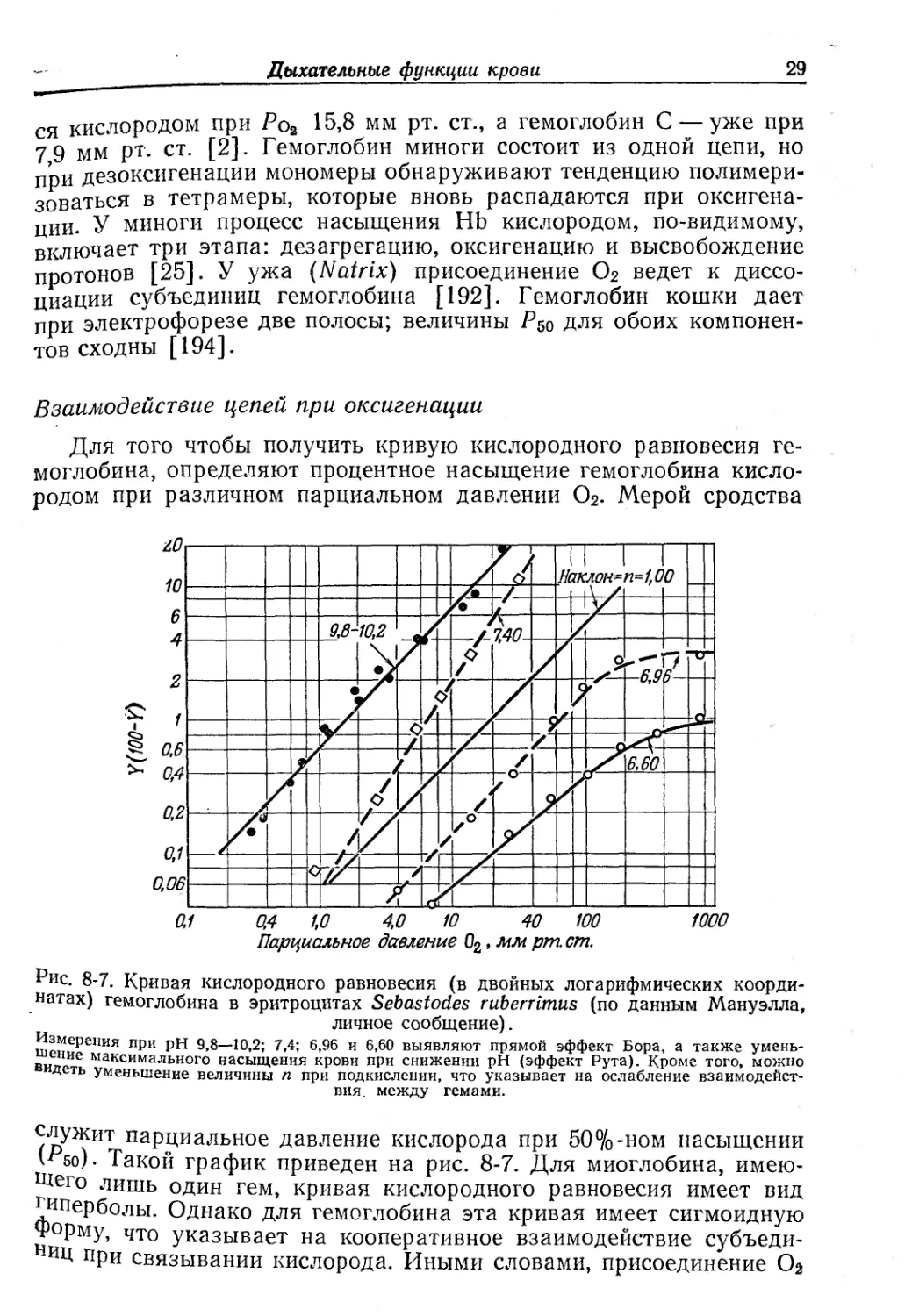
Взаимодействие цепей при оксигенации
Для того чтобы получить кривую кислородного равновесия ге-
моглобина, определяют процентное насыщение гемоглобина кисло-
родом при различном парциальном давлении О2. Мерой сродства
Рис. 8-7. Кривая кислородного равновесия (в двойных логарифмических коорди-
натах) гемоглобина в эритроцитах Sebastodes ruberrimus (по данным Мануэлла,
личное сообщение).
шщерения п₽н 9,8—10,2; 7,4; 6,96 и 6,60 выявляют прямой эффект Бора, а также умень-
в иие максимального насыщения крови при снижении pH (эффект Рута). Кроме того, можно
деть уменьшение величины п при подкислении, что указывает на ослабление взаимодейст-
вия. между гемами.
служит парциальное давление кислорода при 50%-ном насыщении
\”бо). Такой график приведен на рис. 8-7. Для миоглобина, имею-
щего лишь один гем, кривая кислородного равновесия имеет вид
иперболы. Однако для гемоглобина эта кривая имеет сигмоидную
форму, что указывает на кооперативное взаимодействие субъеди-
ИЙ при связывании кислорода. Иными словами, присоединение О2
30
Глава 8
одной субъединицей увеличивает сродство к Ог остальных субъеди-
ниц. Кривую равновесия можно приближенно описать уравнением
У=100х
(Р/Р5о)я
1 + (Р/Р^у ’
где У — процент гемоглобина, связанного с кислородом, или
Ю0(НЬО2)
(НЬ+НЬО2)
, а Р — парциальное давление кислорода в мм рт. ст.
Если У/100—У выразить как функцию logP, то наклон кривой
в точке полунасыщения (где У=50%) будет равен п, т. е. величине,
которую принимают за показатель взаимодействия между гемами
(см. фиг. 8-7). Наклон кривой на обоих ее концах может быть
меньше п. Для миоглобина п=1, для гемоглобинов млекопитаю-
щих эта величина в середине кривой варьирует в пределах от 2,4
до 2,9; для высокомолекулярных гемоглобинов беспозвоночных п
больше 3, например для Hb Arenicola — от 5,0 до 5,7 [201а]. Ге-
моглобин Nereis имеет два значения п [203]. Для хлорокруорина
Spirographis п — 3,2.
Сродство к кислороду одного из гемов в молекуле гемоглобина .
зависит от состояния остальных гемов (от того, связаны ли они
с кислородом). Такое взаимодействие между гемами облегчает
присоединение О2 и является причиной сигмоидной формы кривой
кислородного равновесия; численным выражением этого служит
величина и, которая больше единицы, но меньше общего числа ге-
мов в молекуле. Для гемоглобина миксины Eptatretus п = 1,0, для
НЬ взрослой особи Ichthyomyzon— 1,0, а для личинки— 1,2 [127].
Для личинки Lampetra п = 1,25, а для взрослой особи—1,5; для
разных фракций гемоглобина Petromyzon п колеблется в преде-
лах от 1,1 до 1,55 [9]. Однако при высоких концентрациях гемо-
глобина и 90 %-ном насыщении его кислородом п может возрас-
тать до 3, что указывает на полимеризацию [170]. .
Изучение гемоглобина у 54 видов черепах показало, что спо-
собность его образовывать полимеры весьма различна у предста-
вителей разных семейств. У черепах (в отличие от человека) ве-
личина п зависит от pH; например, у Gopherus при pH 5 она равна
1,3, а при pH 7 — от 2,5 до 2,7 [192].
Дополнительные данные о структурных изменениях при окси-
генации были получены при изучении гибридных молекул гемогло-
бина. Отдельные цепи гемоглобинов мыши (т) и осла (d) смеши-
вали in vitro; значения Р50 для различных комбинаций этих цепей
составляли [171]:
а-цепи p-цепи Р50, мм. рт. ст.
та
^2
^2
т2
т2 12,3
^2 4,7
т2 14,4
d2 4,7
Дыхательные функции крови 31
Сродство гибридной молекулы к кислороду в основном определя-
лось тем гемоглобином, который служил источником р-цепи [171].
О конформационных изменениях при оксигенации свидетельст-
вуют также изменение расстояния между p-цепями (см. стр. 8—11)
и различия в скорости расщепления окси- и дезоксигемоглобина
карбоксипептидазой. В молекуле оксигемоглобина две титруемые
SH-группы, а в дезоксигемоглобине таких групп нет. Величина п
зависит от концентрации солей в окружающем растворе и, кроме
того, может снижаться под влиянием дифосфоглицерата [145].
Сигмоидная форма кривой кислородного равновесия отражает
способность гемоглобина легко присоединять и отдавать кислород,
особенно в случае высокого сродства гемоглобина к О2.
Влияние кислотных групп на связывание кислорода
Эффект Бора. Когда в кровь позвоночного поступает углекис-
лота (как это происходит в тканях), сродство гемоглобина к кис-
лороду падает и кривая равновесия сдвигается вправо (прямой
эффект Бора). При высоких парциальных давлениях СО2 или при
низких pH напряжение кислорода, необходимое для насыщения
гемоглобина, выше, чем при нормальных условиях. В тканях по-
ступление СО2 (или молочной кислоты) облегчает отдачу кисло-
рода; наоборот, в легких или жабрах по мере отдачи СО2 погло-
щение кислорода гемоглобином растет (рис. 8-8). Эффект Бора от-
ражает взаимосвязь между кислородным равновесием и отдачей
протонов (т. е. кислотностью гемоглобина). При низких pH (обыч-
но значительно ниже физиологических) эффект Бора может обра-
щаться, и тогда сродство возрастает; таким образом, при некото-
ром pH Р50 имеет максимальную величину. Количественной мерой
эффекта Бора служит изменение величины Р50 на единицу pH;
выражение дает количество протонов, освобождаемых на
1 моль гема. Для гемоглобина человека изменение P$q составляет
1,1 на единицу pH в диапазоне pH от 6,5 до 9,5 [7]. Величины
эффекта Бора для крови различных животных приведены в
табл. 8-3. Прямой эффект Бора свойствен также хлорокруорину
полихет.
Протоны, освобождающиеся при оксигенации, принадлежат
главным образом имидазольным группам С-концевых гистидинов
p-цепи, а также аминогруппам a-цепей. Если «-аминогруппы
a-цепи блокированы, эффект Бора уменьшается, но не исчезает
совсем [144]. Эксперименты по гибридизации указывают на боль-
шой вклад p-цепей в эффект Бора [171]. Двуокись углерода мо-
жет влиять и на связывание СО [7. Величина эффекта Бора
падает с уменьшением концентрации гемоглобина, с повышением
емпературы и с увеличением ионной силы раствора за счет со-
ей. у мелких млекопитающих эффект Бора выражен в большей
32
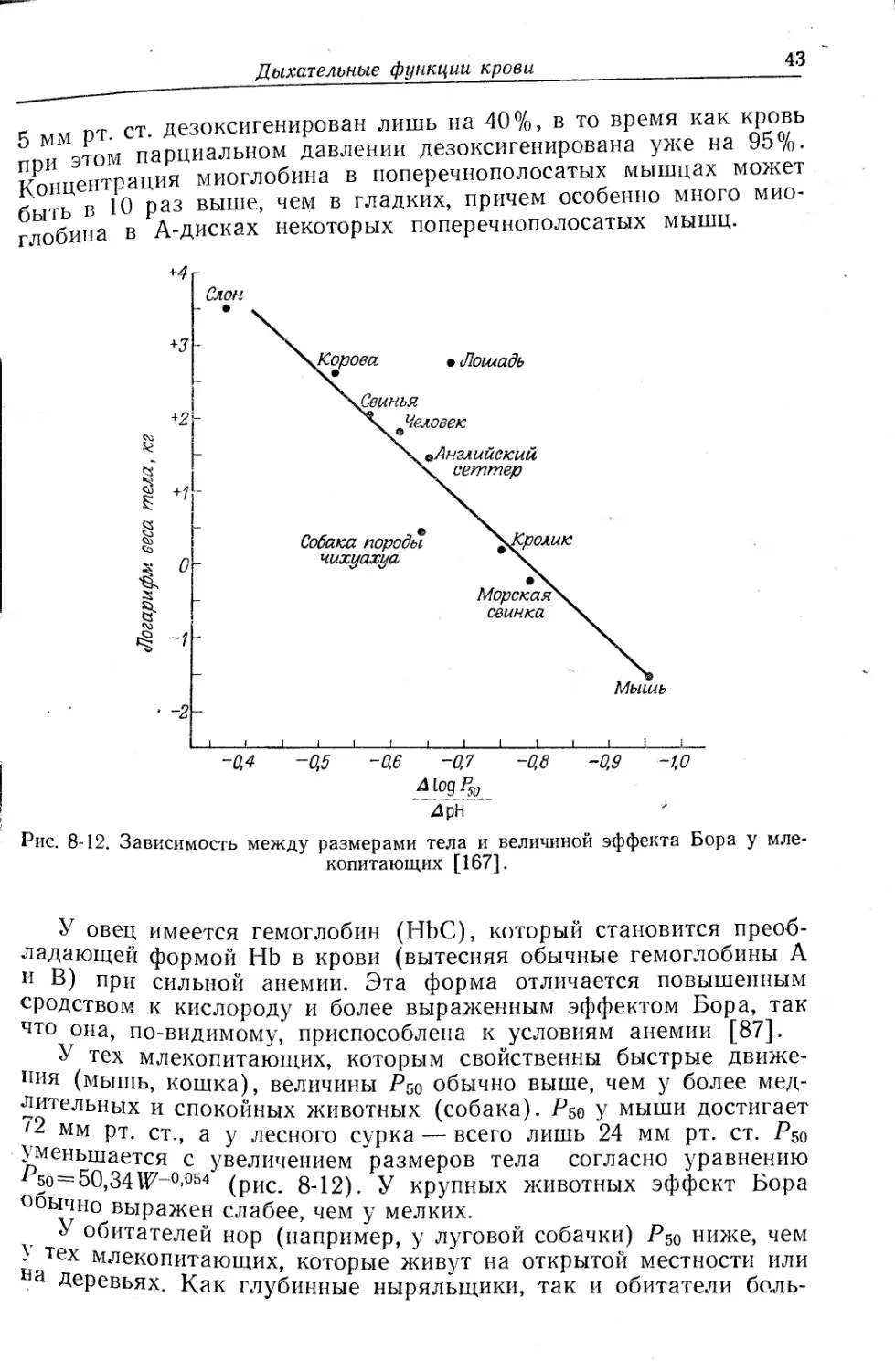
Глава 8
степени, чем у крупных; например, для мыши AlogPso/ApH состав-
ляет 0,9, для человека — 0,65, а для слона — 0,45 [195].
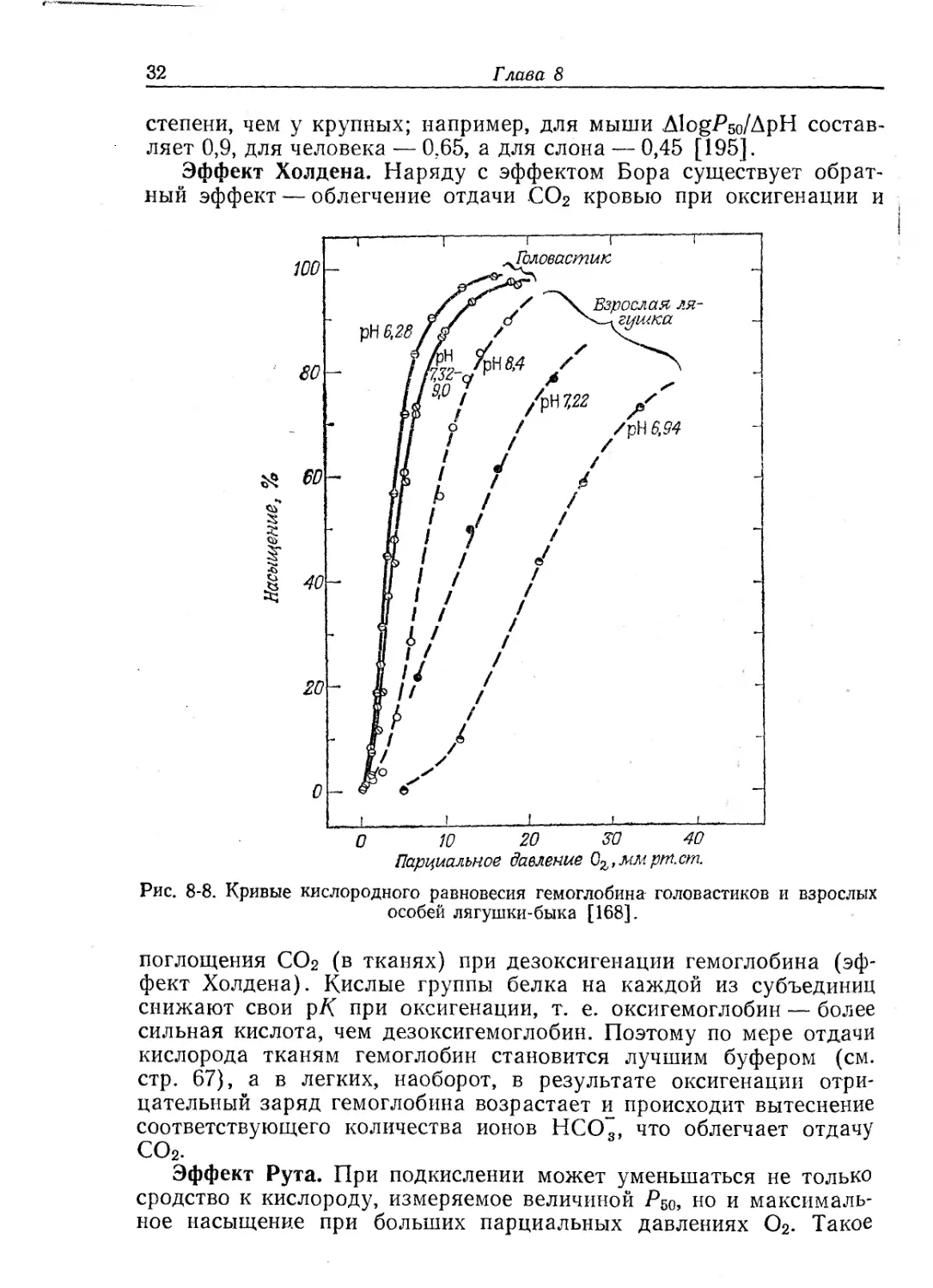
Эффект Холдена. Наряду с эффектом Бора существует обрат-
ный эффект — облегчение отдачи СО2 кровью при оксигенации и
О W 20 20 40
Парциальное давление Оъ,ммрт.ст.
Рис. 8-8. Кривые кислородного равновесия гемоглобина головастиков и взрослых
особей лягушки-быка [168].
поглощения СО2 (в тканях) при дезоксигенации гемоглобина (эф-
фект Холдена). Кислые группы белка на каждой из субъединиц
снижают свои рК при оксигенации, т. е. оксигемоглобин — более
сильная кислота, чем дезоксигемоглобин. Поэтому по мере отдачи
кислорода тканям гемоглобин становится лучшим буфером (см.
стр. 67), а в легких, наоборот, в результате оксигенации отри-
цательный заряд гемоглобина возрастает и происходит вытеснение
соответствующего количества ионов НСО~, что облегчает отдачу
СО2.
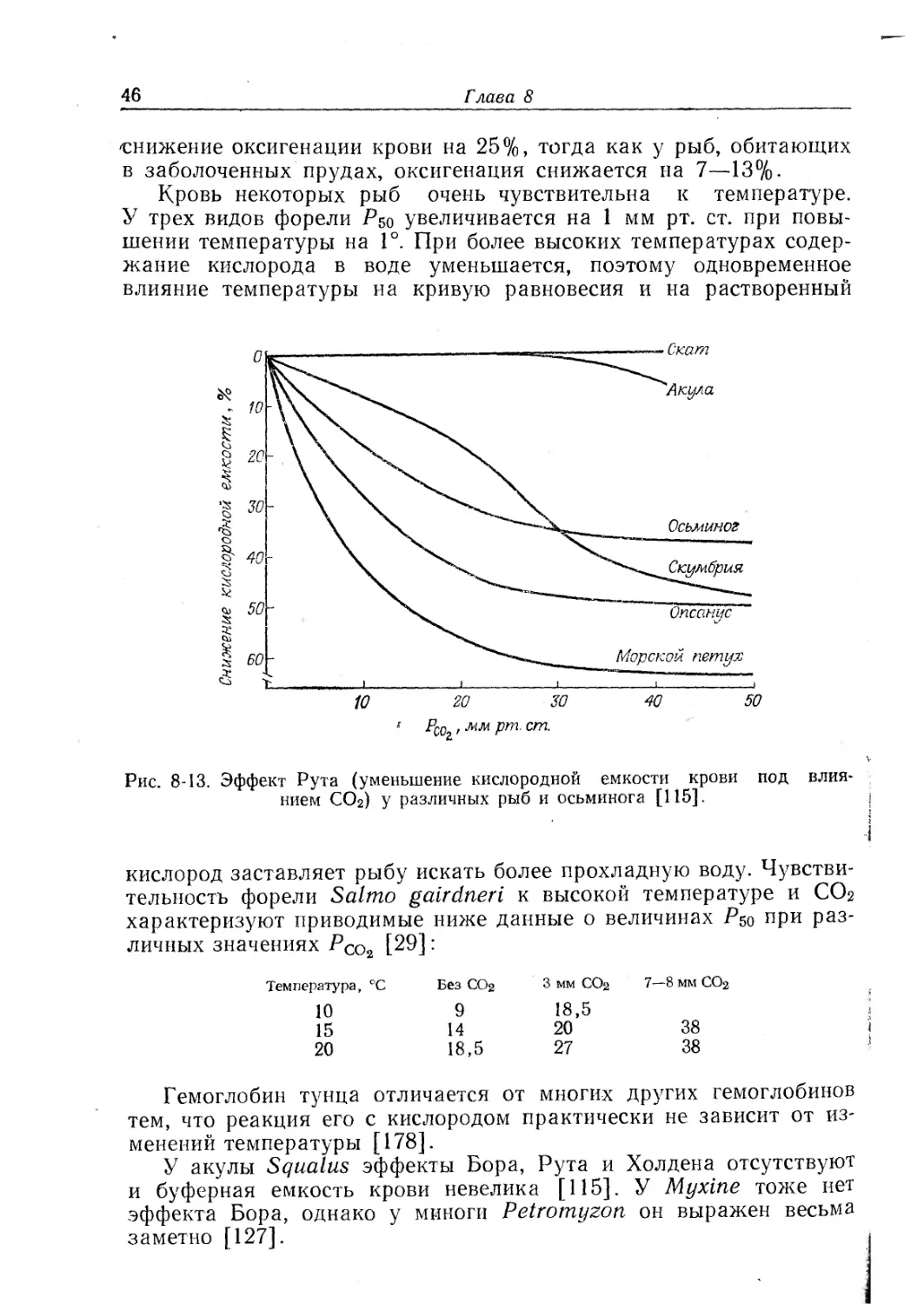
Эффект Рута. При подкислении может уменьшаться не только
сродство к кислороду, измеряемое величиной Р50, но и максималь-
ное насыщение при больших парциальных давлениях О2. Такое
Дыхательные функции крови
33
Таблица 8-3
Величина эффекта Бора в крови различных животных
Животное A log Р50/ДрН Животное A log P50/ApH
/Млекопитающие Тюлень Cystophora —0,66
Человек —0,62 [32а]
цельная кровь [194] Морской котик [118] —0,475
раствор гемоглоби- —0,48 Тюлень обыкновенный —0,534
на [164] —0,49 [ИЗ]
Горилла [163] Тюлень-крылатка [118] —0,48
Шимпанзе [163] —0,59, —0,46 Морж [118] —0,525
Бабуин [112] —0,55 Морской лев [118] —0,696
Лев [142] —0,54 Морская выдра [118] —0,46
Кошка [194] —0,49 Кенгуру [164] —0,54
Собака [194] —0,65 Сумчатый дьявол [164] —0,47
Морская свинка [167] —0,79 Броненосец [38] —0,43
Мышь [167] —0,96 Ехидна [196] —0,54
Кролик [167] —0,75 Птицы
Хомяк [198] —0,41 Пингвин Адели [120] -0,5
Землеройка [198] —0,63 Утка —0,40
Шиншилла [66] —0,53 Рептилии и амфибии
Нутрия [66] —0,56 Iguana [210] —0,52
Лесной сурок [66] —0,63 Necturus [117] —0,131
Луговая собачка [66] —0,51 Amphiuma [117] —0,205
Еж [32а] —0,62 Rana catesbiana [117] —0,288
Полевка -0,54 Рыбы
Лошадь [167] —0,68 Protopterus [116] —0,47
Як [12] —0,68 Neoceratodus [116] —0,42
Лама [12] —0,74 Ictalurus [62] —0,31
Верблюд [12] —0,84 Salmo gairdneri [42a] —0,54
Олень [12] —0,78 Беспозвоночные
Африканский слон [12] —0,58 Lumbricus [33] —0,40
Свинья [167] —0,57 Gastrophilus 0,0
Тюлень Уэдделла [113] Spirographis (хлоро- —0,66
взрослый —0,613 круорин)
плод —0,671 Chironomus [202] От —0,56 до
Бурый дельфин [79] -0,55 —1,3
Бобр [32а] —0,56 Cancer (гемоцианин) —0,27
[96]
снижение верхнего уровня насыщения особенно выражено в крови
некоторых рыб. Это явление — эффект Рута — в присутствии СОг
наступает быстрее, чем в присутствии эквивалентного количества
молочной кислоты.
Описанные выше три вида взаимодействий между связыванием
О2 и СО2 кровью отражают влияние одной части молекулы гемо-
глобина на другую.
Влияние разведения, ионов и фосфатов на сродство к кислороду
Р50 для гемоглобина в интактных эритроцитах, как правило, вы-
чем для того же пигмента, перешедшего в раствор в результа-
те гемолиза. Кроме того, при разведении раствора гемоглобина
3—1514
34
Глава 8
кривая равновесия может сдвигаться в сторону более низких пар-
циальных давлений О2. При диализе раствора гемоглобина потеря
солей приводит к сдвигу кривой влево. Гемоглобин миноги в раз-
бавленном растворе при нейтральном pH представляет собой мо-
номер, если он насыщен кислородом; при дезоксигенации он час-
тично превращается в димер, а при высоких концентрациях пиг-
мента и низких значениях pH может быть и тетрамером.
Увеличение концентрации гемоглобина в 63 раза ведет к 10-крат-
ному снижению его сродства к кислороду [170]. Р50 для цельной
крови овцы при 6 °C составляет 7,2 мм рт. ст., а для раствора ге-
моглобина той же концентрации — 5 мм рт. ст. Разведение раство-
ра в 10 раз снижает эту величину примерно до 2,5 мм рт. ст.
[143]. У миксины Polistotrema величина Р50 равна 3,5 мм рт. ст.
для эритроцитов и 1,8 мм рт. ст. для раствора гемоглобина; соот-
ветствующие величины для голотурии Cucumaria составляют 12,5
и 3,84 мм рт. ст. [123, 124]. Таким образом, микроокружение мо-
лекулы гемоглобина в эритроците влияет на связывание кислоро-
да. Как разведение, так и присутствие солей изменяют Р50. Поэто-
му наблюдения, сделанные на цельной крови, не следует крличест-
венно сравнивать с результатами, полученными при изучении
очищенного белка.
Давно известно, что кривые насыщения для гемоглобина в рас-
творе располагаются левее, чем кривые для цельной крови, и что
цельная кровь более чувствительна к изменениям pH. Важным
компонентом эритроцитов, регулирующим сродство гемоглобина
к кислороду, является дифосфоглицерат (ДФГ). В эритроцитах
человека концентрация ДФГ равна 5-10~3 М, т. е. на каждую мо-
лекулу гемоглобина приходится примерно одна молекула ДФГ.
Это вещество сдвигает кривую кислородного равновесия гемогло-
бина вправо; оно связывается с двумя1 0-цепями тетрамера НЬ
(больше с дезокси-, чем с оксиформой). ДФГ, так же как и NaCl,
увеличивает Р50 и является причиной многих различий в свойствах
изолированного гемоглобина и эритроцитов [19]. У людей, адап-
тированных к большим высотам, эритроциты содержат больше
ДФГ, чем у тех, кто живет на уровне моря [111].
Содержание ДФГ в эритроцитах утробного плода и взрослого
человека практически одинаково, но НЬА чувствительнее к ДФГ,
чем HbF [197]. У ягненка в ранний период жизни содержание
ДФГ возрастает с 8,2 (у плода) до 29,9 мкмоль РО4 на 1 г НЬ,
а сродство НЬ к кислороду и pH внутриклеточной среды падают;
позднее у взрослого животного, когда в клетке уже преобладает
НЬА, содержание ДФГ снижается до 1,1 мкмоль РО4 на 1 г НЬ
[15].
1 ДФГ вклинивается между двумя 0-цепями тетрамера, взаимодействуя при
этом с гистидином, лизином и валином каждой 0-цепи. — Прим, перев.
Дыхательные функции крови
35
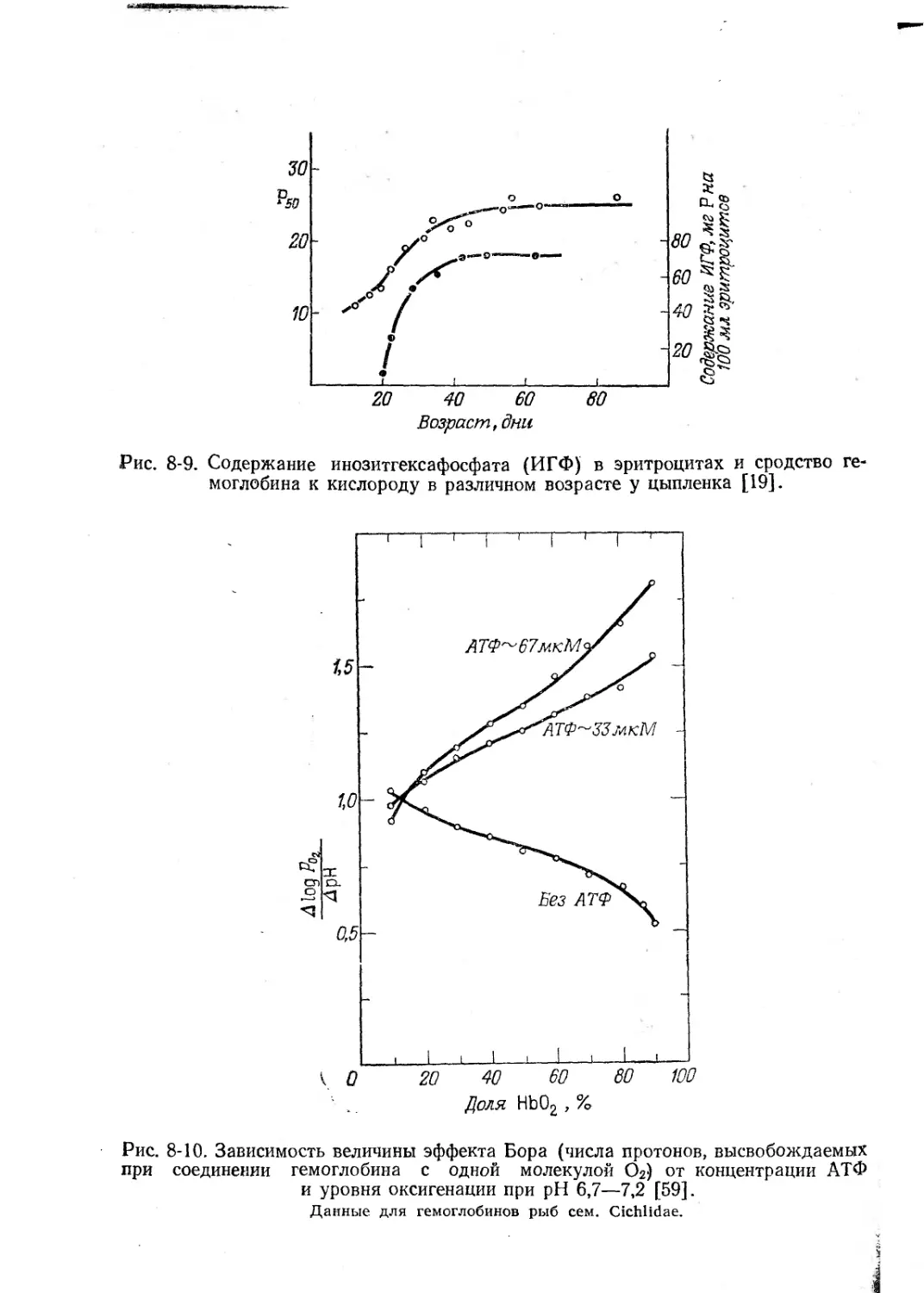
В эритроцитах птиц и черепах вместо ДФГ содержится инозит-
гексафосфат (ИГФ) (рис. 8-9). У рыб аналогичную функцию вы-
полняет АТФ, который может присутствовать в концентрации от
1 до 2-Ю'3 М. На долю этих фосфорилированных соединений при-
ходится 45% всего фосфора эритроцита; они снижают сродство
гемоглобина к кислороду в 30 раз, облегчая таким образом отда-
чу кислорода.
log Р50
для НЬ человека для НЬ курицы
0,01 М NaCl —0,64 -0,59
0,01 М NaCl + ДФГ +0,87 +1,11
0,01 М NaCl + ИГФ + 1,35 + 1,35
У пресноводных рыб из семейства Cichlidae содержание АТФ
соста(вляет 0,76 моля на 1 моль гемоглобина; эффект Бора возрас-
тает с увеличением количества АТФ [59]. У круглоротых органи-
ческих фосфатов в эритроцитах нет, и их гемоглобин к этим ве-
ществам нечувствителен [170].
При добавлении ДФГ к гемоглобину млекопитающих или ИГФ
к гемоглобину птиц эффект Бора увеличивается. Число протонов,
освобождаемых при оксигенации гемоглобинов некоторых рыб,
возрастает на 6О°/о при добавлении АТФ (рис. 8-10). Дифосфогли-
церат снижает сродство гемоглобина к кислороду. Он сильнее свя-
зывается с дезокси-, чем с окси-формой, и прочнее присоединяется
при низких pH и небольшой ионной силе раствора. По-видимому,
молекула ДФГ фиксируется между концевыми ЫН2-группами р-це-
пей; СО2 конкурирует за участок связывания, и частичное освобож-
дение ДФГ при оксигенации сопровождается выходом протонов
Видовая специфичность и экологические аспекты сродства
к кислороду
Кислородная емкость
Пигмент, переносящий кислород, прежде всего должен обла-
дать способностью обратимо присоединять О2 в количестве, доста-
точном для удовлетворения потребностей организма. Кислородная
емкость — это наибольшее возможное содержание кислорода в ок-
сигенированной крови. Содержание О2 обычно выражают в объем-
ных процентах или в молях кислорода на 1 л цельной крови или
эритроцитов. Уравновешивание с воздухом может не дать полного
насыщения, если для насыщения требуется высокое давление кис-
3*
Рис.
8-9. Содержание инозитгексафосфата (ИГФ) в эритроцитах и сродство ге-
моглобина к кислороду в различном возрасте у цыпленка [19].
Рис.
при
8-10. Зависимость величины эффекта Бора (числа протонов, высвобождаемых
соединении гемоглобина с одной молекулой Ог) от концентрации АТФ
и уровня оксигенации при pH 6,7—7,2 [59].
Данные для гемоглобинов рыб сем. Cichlidae.
Дыхательные функции крови 37
порода, особенно в присутствии СО2 и при высокой температуре.
Поэтому для точного определения кислородной емкости лучше
производить насыщение чистым кислородом. Кроме того, нужно
учитывать, что некоторые клетки крови, особенно ядерные эритро-
циты, сами потребляют значительное количество О2. Кислородная
емкость крови (за вычетом кислорода, физически растворенного
в плазме) пропорциональна содержанию в ней гемоглобина или
иного пигмента.
В табл. 8-4 приведены значения кислородной емкости крови
ряда животных. У млекопитающих и птиц кислородная емкость
составляет обычно от 15 до 20 об.%. Некоторые ныряющие млеко-
питающие обладают большей емкостью, а у ламы эта величина
весьма значительна даже на уровне моря. У молодняка кислород-
ная емкость, как правило, меньше, чем у взрослых животных.
У холоднокровных позвоночных она составляет чаще всего от 5
до 12 об. %. У некоторых рыб, способных дышать воздухом, кис-
лородная емкость высока. У таких активных рыб, как, например,
скумбрия, она обычно выше, чем у малоподвижных форм вроде
опсануса. У беспозвоночных весьма значительны индивидуальные
различия. При достаточном количестве гемоглобина (например,
у Arenicola, Urechis и ряда других животных) кислородная ем-
кость может быть в 10 раз больше, чем она была бы без пигмента.
Определение сродства к кислороду по кривым равновесия
Большинство дыхательных пигментов полностью насыщается
при более низком парциальном давлении кислорода, чем РОа в воз-
духе на уровне моря (155 мм рт. ст.). Адаптивные различия меж-
ду гемоглобинами у разных животных проявляются прежде всего
в величинах РОа, при которых происходит отдача и присоединение
кислорода. От этих величин зависит диапазон Р02, в котором дан-
ный пигмент может эффективно функционировать. Так, например,
у человека кровь в легких насыщается при парциальном давлении
около 100 мм рт. ст. Выходя из легких, она содержит 19 об. % кис-
лорода при 80 мм рт. ст., и гемоглобин в ней насыщен на 98%.
В капиллярах кровь проходит через ткани, где напряжение О2
незначительно (5—30 мм рт. ст.), и здесь происходит отдача 25—
<>О/о кислорода. Венозная кровь содержит 14 об. % кислорода при
парциальном давлении около 40 мм рт. ст.
Исходя из кривых кислородного равновесия, сродство лучше
всего выражать величиной Р50 (см. стр. 29 и рис. 8-11). Опре-
деленное значение Р50 имеет смысл лишь в том случае, если указа-
Ь1 состояние пигмента, РСО2 (или pH) и температура. В табл. 8-4
представлены некоторые данные, взятые из очень обширной лите-
нЯТУРЫ П° ЭТОМУ вопросу; они относятся в основном к гемоглобину,
годящемуся в эритроцитах, и получены при условиях, близких
Таблица 8-4
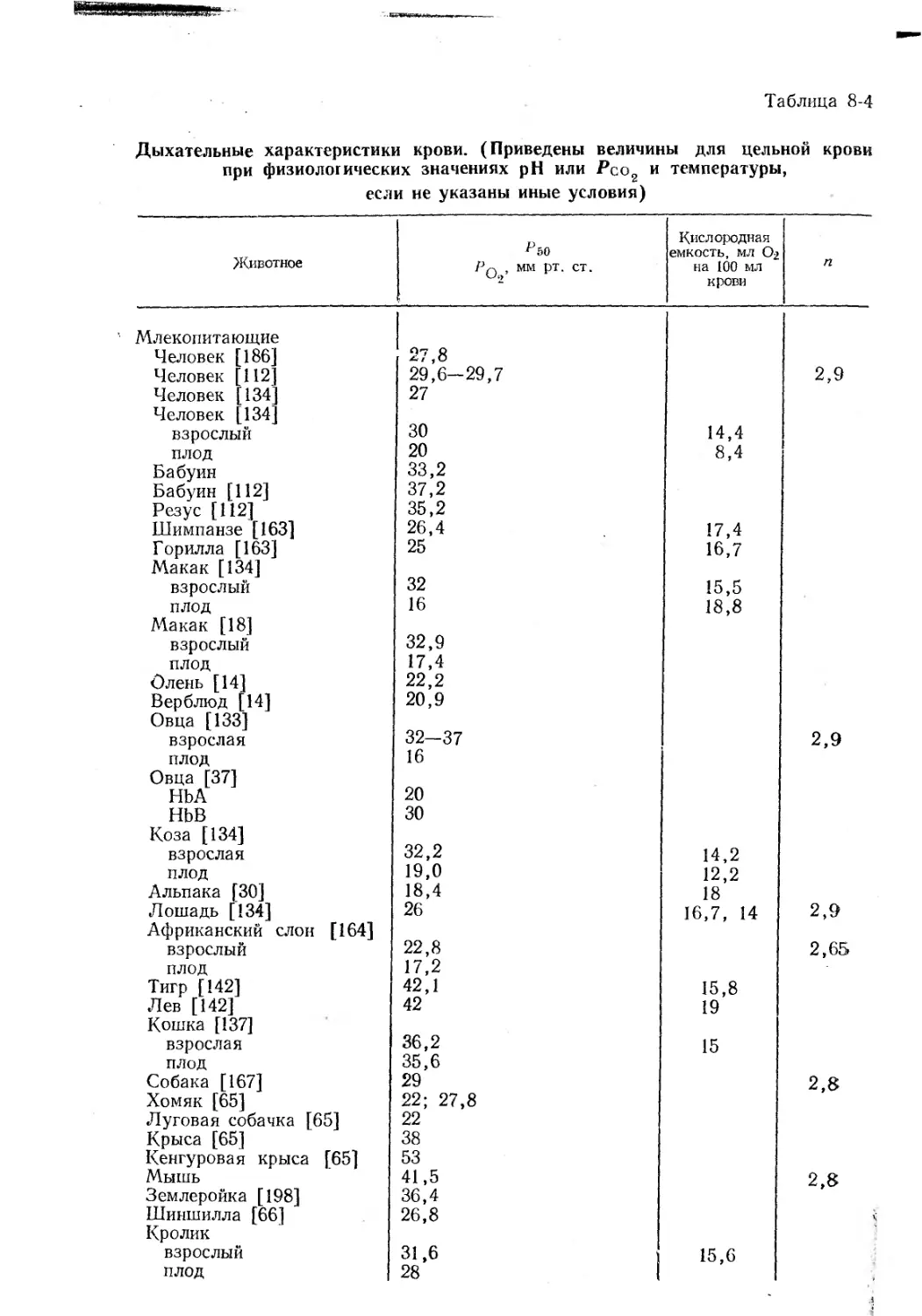
Дыхательные характеристики крови. (Приведены величины для цельной крови
при физиологических значениях pH или Рсо2 и температуры,
если не указаны иные условия)
Животное ^50 Pq , мм рт. ст. Кислородная емкость, мл Ог на 100 мл крови п
Млекопитающие
Человек [186] 27,8
Человек [112] 29,6—29,7 2,9
Человек [134] 27
Человек [134]
взрослый 30 14,4
плод 20 8,4
Бабуин 33,2
Бабуин [112] 37,2
Резус [112] 35,2
Шимпанзе [163] 26,4 17,4
Горилла [163] 25 16,7
Макак [134]
взрослый 32 15,5
плод 16 18,8
Макак [18]
взрослый 32,9
плод 17,4
Олень [14] 22,2
Верблюд [14] 20,9
Овца [133]
взрослая 32—37 2,9
плод 16
Овца [37]
НЬА 20
НЬВ 30
Коза [134]
взрослая 32,2 14,2
плод 19,0 12,2
Альпака [30] 18,4 18
Лошадь [134] 26 16,7, 14 2,9
Африканский слон [164]
взрослый 22,8 2,65
плод 17,2
Тигр [142] 42,1 15,8
Лев [142] 42 19
Кошка [137]
взрослая 36,2 15
плод 35,6
Собака [167] 29 2,8
Хомяк [65] 22; 27,8
Луговая собачка [65] 22
Крыса [65] 38
Кенгуровая крыса [65] 53
Мышь 41,5 2,8
Землеройка [198] 36,4
Шиншилла [66] 26,8
Кролик
взрослый 31,6 15,6
плод 28 1
Продолжение
р50 Кислородная
Животное , мм рт. ст. е>2 мл Ог на 100 мл п
крови
Лесной сурок [65] 25 15,3
Еж [32а] 34 (38 °C) 8,9 (5 °C)
Бобр [32а] 23,3 16,1
Водяная крыса (Arvicola) 38,4 19,4
Тюлень Cystophora [32а] 24 36
Tur slops [79] 24,6 18,20
Phocoenoides [79] 19,1 25,27
Морской лев (Zaophus) 32 17,5 2,6
[Н8] Косатка (Orcinus) [118] 30,7; 37,5 21,5 2,6
Тюлень обыкновенный 31 29,3
[118] Морж [118] 34,2 23,4 2,4
Морской котик [118] 29,7 19,6 2,5
Морская выдра [118] Тюлень Уэдделла [113] 30,8 21,3 2,6
взрослый 28,5 31,6
плод 22,1 27,7
Броненосец [38] 10,1
Ехидна [196] 19,5 19
Утконос [97] 10 (4 мм СО2) 36 (30 мм СО2) 23,5
Птицы
Курица [13]
взрослая 58
в возрасте 10 дней 54,8 10
в возрасте 1 дня 48,4 10,7
17-дневный эмбрион 31,3 10,3
Пингвин Адели [120] 34,4 22,4
Гусь [120] 45
Голубь [120] 42
Утка [120] Рептилии 54
Pseudetnys 19,5 6,6—10,8
Chrysemys 15
Crocodylus 38 8—10
Alligator 28 6,7
Heloderma [152] Eutneces 32 (18—20 °C) 19 10 12 5
Iguana [210] 51 10*5
Agama [152] Sauromalus [152] 34 (37 мм СО2) 24
Амфибии •
Necturus [117] (жаберное 14,5 6,3
дыхание)
Amphiuma [117] (жабер- 27 7,6
ное и легочное дыхание) Рапа catesbiana [117] (кожное и легочное ды- хание) Eana esculenta [168] 39
8,1
взрослая 13,2 9 8
головастик 4,6 (pH 7,2; 20 °C) 7,8
Продолжение
Животное ^50 Pr. , мм рт. CT. U2 Кислородная емкость, мл O2 на 100 мл крови п
Rana esculenta [55], адаптированная к 5 °C 42,5 13,3
адаптированная к 20 °C Bufo marinas 39,1 42 (25 °C) 38 (15 °C) 10,4
Рыбы
Щука 9,0
Серебряный карась [6] Радужная форель [29] 30 (7—8 мм CO2, 15 °C) 10,7
Лосось Голец [20] 19 (1—2 мм CO2, 15 °C) 7 (1 мм CO2, 0 °C)
21 (1 мм CO2, 25 °C)
Электрический угорь Японский угорь [212] 12 (0 мм CO2, 18 °C) 14 (7,4 мм CO2) 19,7 —
НЬЕ, 2,1 (7,0 mm~CO2, 20 °C) 2,4
нье2 14 (7,0 мм CO2, 20 °C) 1
Тrematomus 21,5 (0,1 мм CO2, —1,5 °C) 5,3—7,7
Скумбрия 16 (1 мм CO2, 20 °C) 5 (1—2 мм CO2, 15 °C) 15,7
Карп 12,5
Сом 1,4 (0—1 мм CO2, 15 °C) 13,3
Bagrus (обитает в пресной воде на глубине) bates (обитает в пресной 1,5 (0 мм CO2) 17 (0 мм CO2)
воде, богатой О2)
Neoceratodus [116] 11 (3,5 мм CO2) 7,7
Protopterus [116] 10 (6 мм COo, 25 °C) 6,8
Lepidosiren [95] 10,5 (6 мм CO2, 23 °C) 4,9-6,8
Symbranchus [95] 5—6 14,7
Scyliorhynus [150] 12 (7,01мм CO2, 17 °C) 4,5
Squalus [115] Squalus 17 (0,5Гмм CO2, 11 °C) 4,35
взрослая 16,4 (7,3 мм CO2, 12 °C)
плод 10,6
Lampetra planeri [9]
взрослая 0,77 (Hb|5 мг/мл)
личинка 0,37
Eptatretus [123] Hb 2 4 1
Myxine [123] Hb 8 (7,5 mm’CO2,*25 °C) 1
Petromyzon [123] Ichthyomyzon [123] 14—20 1,2
взрослая 17—19 (25 °C) 1,0
пескоройка 16 1,2
Членистоногие 5,4—11,6 1,1—1,2
Chironomus plumosus 0,39 (12 mm CO2)
[202] 0,21 (0,6 мм CO2)
Moina [10] 3,5 (7,2 мм CO2, 28 °C)
Daphnia [10] 3,1 (7,2 мм CO2, 28 °C)
Cyzicus [10] 0,035 (7,2 мм CO2, 28 °C) 2,3
Triops [78] Черви 6,6 (7,4 мм CO2) 3,2
Urechis 12,3 (8,6 мм CO2, 19 °C) 2,2—6,7
Glycera [72] 7,0 (7,4 мм CO2) 4,1
Glossoscolex [98] 7,0 (7,5 мм CO2, 20 °C) 14
Продолжение
Животное Pso Pfy , мм рт. CT. Кислородная емкость, мл C>2 на 10 мл крови п
Lumbricus [331 Tub if ex Arenicola Arenicdla [201a] Nephthys внутрисосудистая жид- кость целомическая жидкость Иглокожие Thyonella [129] Cucumaria Круглые черви Ascaris [191] полостная жидкость стенка тела Ascaris [138а] полостная жидкость Nematodirus Nippostrongylus Моллюски Phacoides [155] (Hb в жабрах) Cryptochiton Planorbis Простейшие Paramecium [190] Хлорокруорин Spirographis [8] Sabella Гемоэритрин Sipunculus Golfingia Гемоцианин Cancer [96] Речной рак [110] Gecarcinus [160] Cardisoma [157] Скорпион Heterometrus [140] Limulus Panulirus Homarus Helix [107] Cardita Neotia [126] Pusitriton Busycon Cryptochiton Chiton [156] Diodor a [156] Octopus [114] Sepia Loligo Mytilus Asterias 6,8 (7,4 мм CO2, 25 °C) 0,6 (0 мм CO2, 17 °C) 1,8—2 (7,4 мм CO2, 18 °C) 4,0 (pH 7,0) 6 (7,4 мм CO2, 20 °C) 7,4 (7,4 мм CO2, 20 °C) 8 12,5 (7,4 мм CO2, 10 °C) 0,05 0,2 0,001—0,004 0,05 0,2 0,19 (7,4 мм CO2) 18 (7,2 мм CO2, 10 °C) 7 (0 мм CO2, 20 °C) 0,6 1,65 27 (7,7 мм CO2, 20 °C) 8 6,9 (6,3 мм CO2, 20 °C) 19,6 (7,7 мм CO,) 1,6 17 (7,4 мм CO,, 27 °C) 3,5 (25 °C) 16,5 11 (0 мм CO2, 23 °C) 6,5 (7,5 мм CO2, 15 °C) 14 (0 мм CO2, 25 °C) 8; 10 2 7 11 (7,5 мм CO,) 13,5 2 7 (7,8 мм CO2, 25 °C) 15 (0 мм CO2, 23 °C) 18 20—26 5 (10 °C) 15 (0,4 мм CO2, 11 °C) 4 (0,7 мм CO2, 14 °C) 36 (0 мм CO2; 23 °C) 5,1-5,4 5,0 3 1,2 3,44 3,1 2,9 1,8 0,7—2,7 2,0 1,3 1,1—2,2 2,1-3,3 1,3 0,86 3,1 3,8—4,5 0,32 0,46 2,6 2,2—3,0 1,7
42
Глава 8
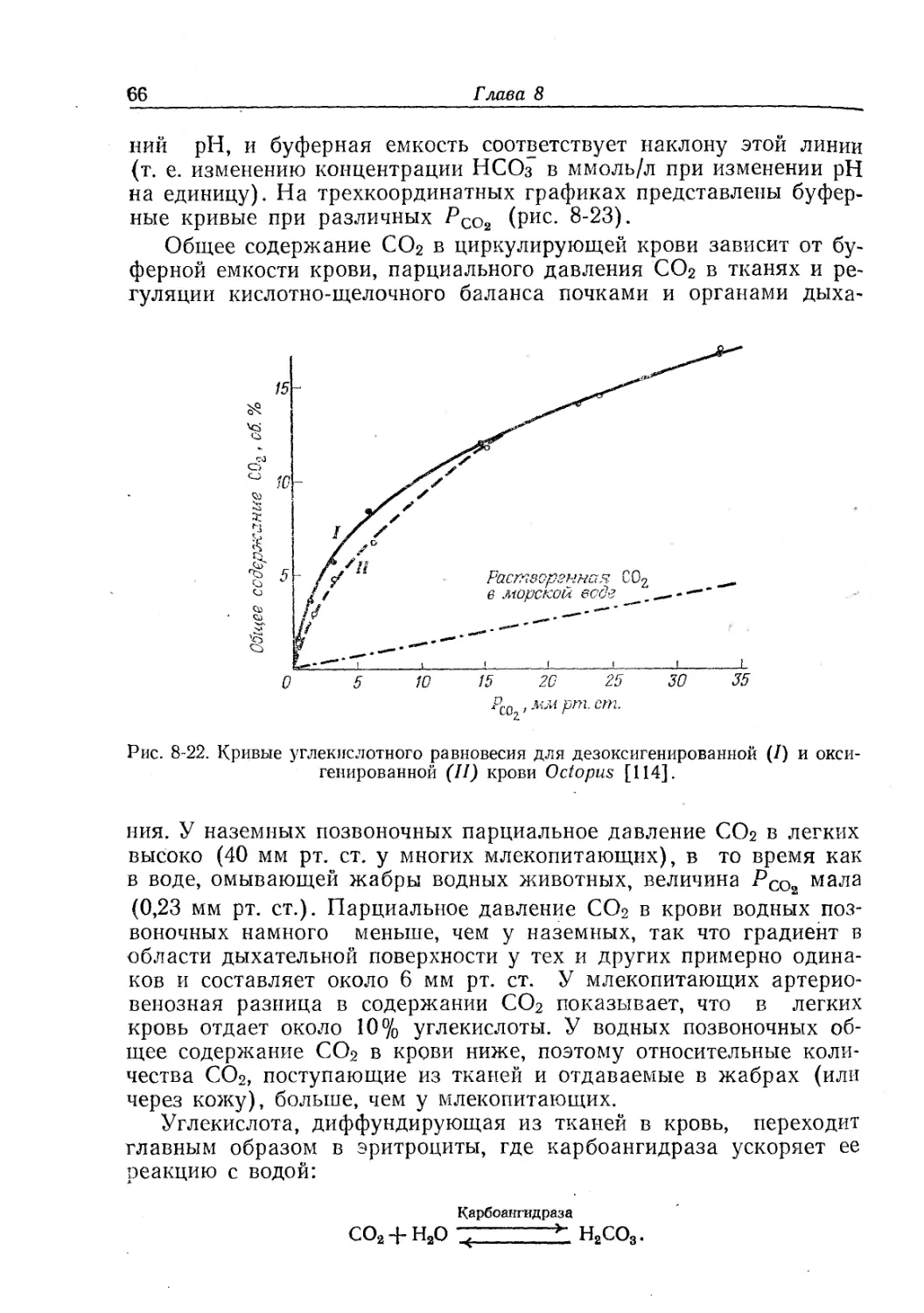
к физиологическим. При наличии у данного вида нескольких ге-
моглобинов приводятся средние величины: функциональные осо-
бенности отдельных фракций гемоглобина известны лишь у немно-
гих животных.
Гемоглобины сильно различаются по своему сродству к кисло-
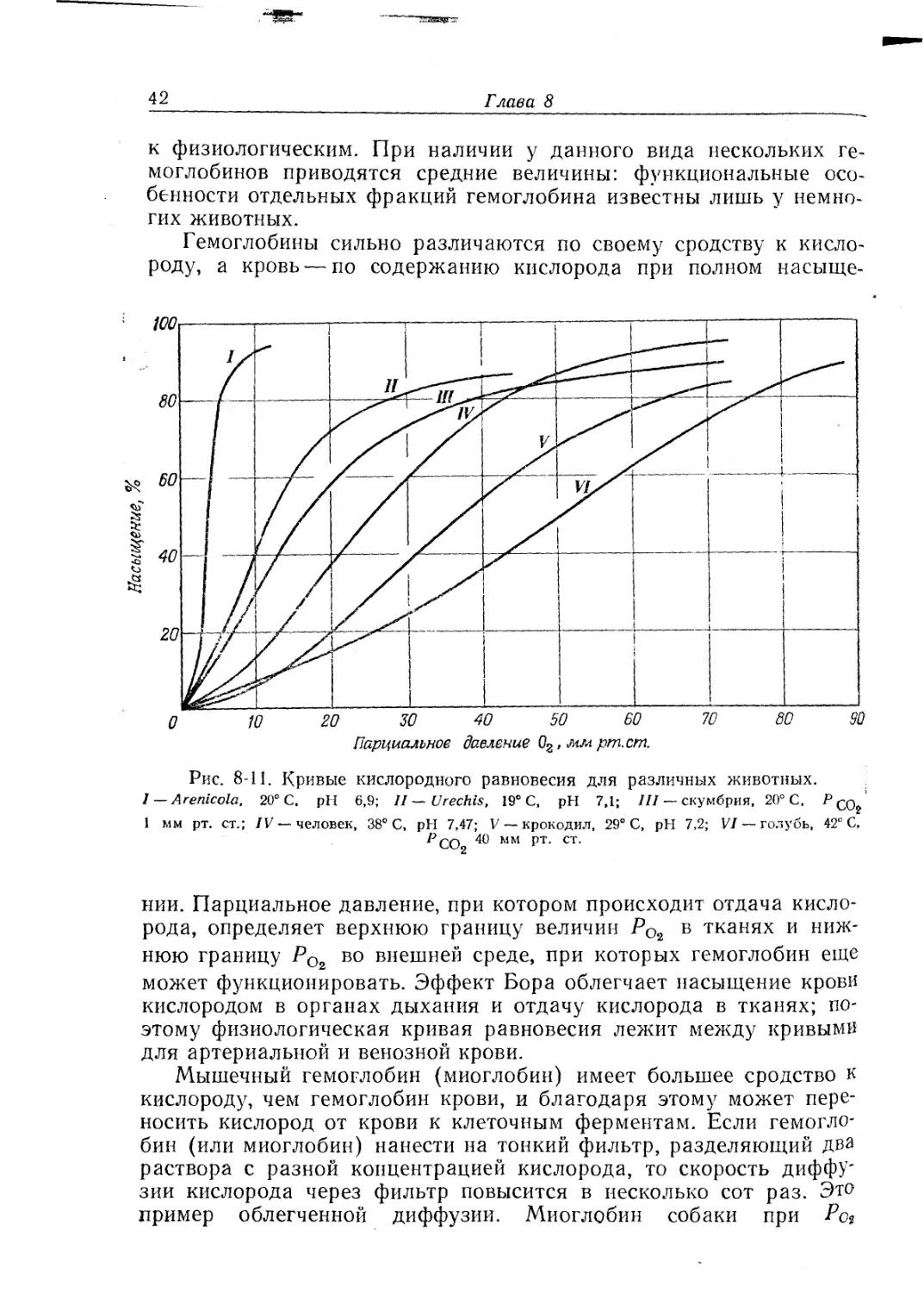
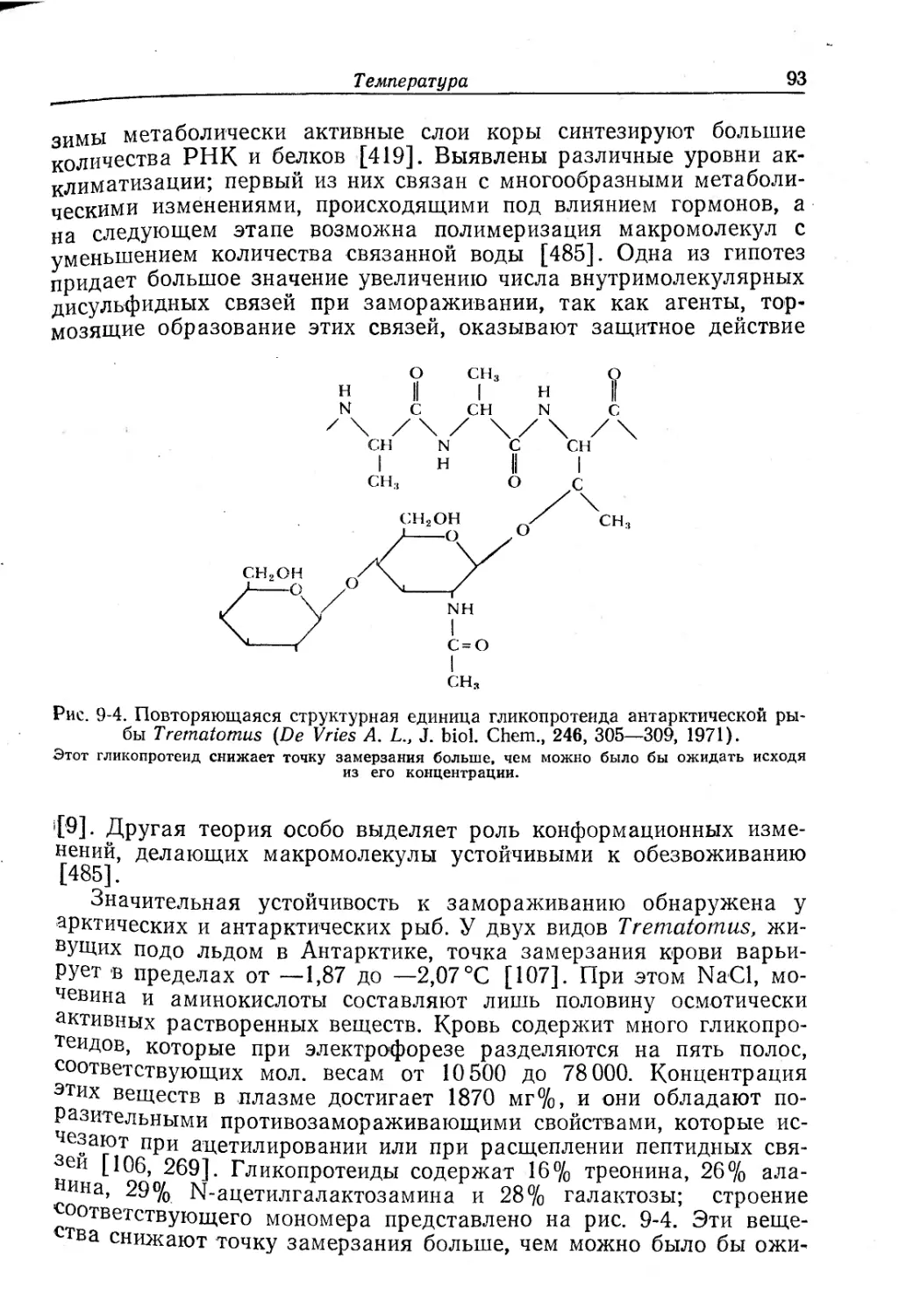
Рис. 8-11. Кривые кислородного равновесия для различных животных.
I — Arenicola, 20° С, pH 6,9; II — Urechis, 19° С, pH 7,1; III — скумбрия, 20° С, Р со?
1 мм рт. ст.; IV — человек, 38° С, pH 7,47; V — крокодил, 29° С, pH 7,2; VI — голубь, 42° С,
r’cOg 40 мм рт. ст.
нии. Парциальное давление, при котором происходит отдача кисло-
рода, определяет верхнюю границу величин РО2 в тканях и ниж-
нюю границу РОг во внешней среде, при которых гемоглобин еще
может функционировать. Эффект Бора облегчает насыщение крови
кислородом в органах дыхания и отдачу кислорода в тканях; по-
этому физиологическая кривая равновесия лежит между кривыми
для артериальной и венозной крови.
Мышечный гемоглобин (миоглобин) имеет большее сродство к
кислороду, чем гемоглобин крови, и благодаря этому может пере-
носить кислород от крови к клеточным ферментам. Если гемогло-
бин (или миоглобин) нанести на тонкий фильтр, разделяющий два
раствора с разной концентрацией кислорода, то скорость диффУ'
зии кислорода через фильтр повысится в несколько сот раз. Это
пример облегченной диффузии. Миоглобин собаки при Рс?
Дыхательные функции крови
43
5 мм рт. ст. дезоксигенирован лишь на 40%, в то время как кровь
при этом парциальном давлении дезоксигенирована уже на 95%.
Концентрация миоглобина в поперечнополосатых мышцах может
быть в 10 раз выше, чем в гладких, причем особенно много мио-
глобина в A-дисках некоторых поперечнополосатых мышц.
J____!___I___I____I___! t I I I I I I i
-0,4 -0,5 -0,6 -0,7 -0,8 -0,9 -1,0
л tog Р50
4рН
Рис. 8-12. Зависимость между размерами тела и величиной эффекта Бора у мле-
копитающих [167].
У овец имеется гемоглобин (НЬС), который становится прео -
ладающей формой НЬ в крови (вытесняя обычные гемоглобины А
и В) при сильной анемии. Эта форма отличается повышенным
сродством к кислороду и более выраженным эффектом Бора, так
что она, по-видимому, приспособлена к условиям анемии [о7].
У тех млекопитающих, которым свойственны быстрые движе-
ния (мышь, кошка), величины Р50 обычно выше, чем у более мед-
лительных и спокойных животных (собака). Р50 У мыши достигает
72 мм рт. ст., а у лесного сурка — всего лишь 24 мм рт. ст. 1 50
уменьшается с увеличением размеров тела согласно уравнению
Л>о= 50,34№~°.°54 (рис 8-12) у крупных животных эффект Бора
обычно выражен слабее, чем у мелких.
У обитателей нор (например, у луговой собачки) Р50 ниже, чем
У тех млекопитающих, которые живут на открытой местности или
на деревьях. Как глубинные ныряльщики, так и обитатели боль-
44
Глава 8
ших высот имеют гемоглобин с высоким сродством к О2 [65].
У зайца-беляка содержание миоглобина в мышцах зимой почти в
два раза больше, чем летом [176]. У овцы гемоглобин А имеет
большее сродство к кислороду, чем гемоглобин В, и те породы
овец, у которых преобладает НЬА, могут страдать от хронической
гипоксии даже на уровне моря из-за низкого РОг венозной крови
[37]. Тюлень Уэдделла способен нырять на глубину 400 м и на-
ходиться под водой до 43 мин. Его гемоглобин отличается сильно
выраженными эффектами _Бора и Холдена и может, связывать
большие количества НСОз [ИЗ]; у утконоса эти два эффекта
также весьма значительны [97]. Кровь тюленя хохлача обладает
высокой кислородной емкостью благодаря высокому содержанию
гемоглобина — 26,4 г на 100 мл [32а]. У зимнеспящих животных,,
например у ежа, насыщение гемоглобина кислородом намного
меньше зависит от температуры, чем у остальных млекопитающих.
У ежа составляет 0,017, а у человека — 0,023 [32а]. Эф-
фект Бора у ежа во время зимней спячки также незначителен.
Как правило, для полного насыщения гемоглобинов птиц нуж-
ны более высокие парциальные давления кислорода, чем для пиг-
мента млекопитающих. У уток и голубей артерио-венозная разни-
ца в насыщении соответствует использованию 60% кислорода
(у человека используется 27%, у черепахи 44%, у ската Raja
66%). У такой хорошо ныряющей птицы, как пингвин Адели, Р50= !
= 34,4 мм рт. ст. [13].
У амфибий, проводящих много времени на суше, кривые кис-
лородного равновесия сдвинуты вправо по сравнению с кривыми |
для чисто водных амфибий [117]. У взрослых особей лягушки-бы-|
ка отмечено увеличение Р$о при снижении pH до 6,2 (этот эффект ]
обратим); у головастиков же Р$о не зависит от pH в диапазоне от
9,0 до 6,2 [168]. В ряду амфибий эффект Бора оказался наимень- '
шим у водной формы (Rana>Amphiuma>Necturus}. У черепах
была найдена обратная зависимость; у водных форм эффект Бора
выражен сильнее, чем у наземных. У сухопутной коробчатой чере-
пахи Р5о составляет 28,5 мм рт. ст., а у ложной каретты, живущей
в воде,— 12 мм рт. ст. У малоподвижной Теггарепе величина Р50
равна 11 мм рт. ст., а у активного пловца Chelydra — 32 мм рт. ст.
У некоторых головастиков, лягушек и черепах содержание гемо-
глобина можно снизить до величины менее 1 % нормы путем введе-
ния фенилгидразина, и они могут выжить, если содержать их в
воде, хорошо насыщенной воздухом: количества О-, растворен-
ного в крови, достаточно для поддержания основного обмена.
Как правило, рыбы, живущие в стоячей воде, имеют низкие
значения Р50, и эффект Бора (который может быть значителен по
относительной величине) не сдвигает у них кривую диссоциации за
физиологические пределы. Однако у рыб с высоким Р50 увеличение
напряжения СО2 с 2 до 10 мм рт. ст. может настолько сдвинуть
кривую вправо, что рыба начинает задыхаться даже при избытке
Дыхательные функции крови
кислорода: углекислота, способствуя отдаче кислорода тканям, в
то же время препятствует насыщению НЬ в жабрах. Например,
когда Ро поддерживали на уровне 160 мм рт. ст., нотропис поги-
бал при 80 мм рт. ст. СО2, в то время как сомик выживал при по-
вышении РСо2 ДО 338 мм рт. ст. [20]. Добавление СО2 в закрытый
сосуд, содержащий кровь форели, может вызвать появление пу-
зырьков кислорода из-за большого эффекта Бора. Незначительное
повышение уровня СО2 может-заставить рыбу с низким сродством
НЬ к кислороду переходить в воды с высоким содержанием О2.
У рыб с высоким сродством НЬ к кислороду эффект СО2 менее
выражен. У некоторых пресноводных рыб (у сома, карпа и ильной
рыбы) при РСо2 Ю мм рт. ст. Р50 не превышает 10 мм рт. ст., в то
время как у чукучана и трех видов форели Р50 в тех же условиях
поднимается выше 35 мм рт. ст. У рыб с низким сродством к О2
и значительным эффектом Бора (таких, как форель) при мышеч-
ной активности в крови появляется больше молочной кислоты, чем
у сома и карпа, которые переносят низкое напряжение кислорода
и обладают НЬ со слабо выраженным эффектом Бора. Среди мор-
ских рыб к СО2 чувствительны опсанус, скумбрия и морской пе-
тух; акулы и скаты отличаются меньшей чувствительностью.
Из крови японского угря были выделены два гемоглобина: Ei
с Р50 2,1 мм рт. ст. и показателем взаимодействия между гемами
(и) 2,4 и Е2, у которого Р50 = 14 мм рт. ст. и п = 1,0. Гемоглобин
Е2 обнаруживает эффект Бора, а у гемоглобина Ei этого эффекта
нет. Экологическое значение обеих форм, которые обычно находят-
ся в определенном количественном соотношении (Ei:E2=3:7),
остается неизвестным [212]. У личинки Lampetra planeri два гемо-
глобина — один с высокой, другой с низкой электрофоретической
подвижностью — имеют сходные значения Pso- Из шести гемогло-
бинов взрослой особи Petromyzon marinas один имеет высокую,
а остальные — низкую величину Р50 [9].
Эффект Рута — уменьшение насыщения крови при подкисле-
нии — наблюдается у мнбгих рыб. У Tautoga и Sebastodes для про-
явления эффекта Pj^ra после гемолиза требуется более сильное
подкисление, чем в цельной крови. Кроме того, для эритроцитов
форма кривой кислородной диссоциации ближе к гиперболической
и значение п снижается по мере подкисления [128]. У озерной
форели (Salvelinus fontinalis) эффект Рута весьма значителен [20]
(рис. 8-13), а у прилипала (Gobiesox) его нет вовсе. В гемоглобине
^corpaenichthys, находящемся в эритроцитах, взаимодействие меж-
ду гемами почти не проявляется, а эффекты Бора и Рута ослабле-
Н(Ъж ДвоякоДышащая рыба Neoceratodus обладает значительньш
эффектом Бора, но не эффектом Рута [116]. Когда такая рыба
аходится на воздухе, дыхательный ацидоз компенсируется мета-
ллическим алкалозом. У одной южноамериканской рыбы, живу-
щеи вблизи водопадов, повышение Рсс>2 до 5 мм рт. ст. вызывает
46
Глава 8
снижение оксигенации крови на 25%, тогда как у рыб, обитающих
в заболоченных прудах, оксигенация снижается на 7—13%.
Кровь некоторых рыб очень чувствительна к температуре.
У трех видов форели Ръ0 увеличивается на 1 мм рт. ст. при повы-
шении температуры на 1°. При более высоких температурах содер-
жание кислорода в воде уменьшается, поэтому одновременное
влияние температуры на кривую равновесия и на растворенный
Рис. 8-13. Эффект Рута (уменьшение кислородной емкости крови под влия
нием СО2) у различных рыб и осьминога [115].
кислород заставляет рыбу искать более прохладную воду. Чувстви-
тельность форели Salmo gairdneri к высокой температуре и СО2
характеризуют приводимые ниже данные о величинах Ры> при раз-
личных значениях Рсо.2 [29]:
Температура, °C Без СО2 3 мм СО2 7—8 мм СО2
10 9 18,5
15 14 20 38
20 18,5 27 38
Гемоглобин тунца отличается от многих других гемоглобинов
тем, что реакция его с кислородом практически не зависит от из-
менений температуры [178].
У акулы Squalus эффекты Бора, Рута и Холдена отсутствуют
и буферная емкость крови невелика [115]. У Myxine тоже нет
эффекта Бора, однако у миноги Petromyzon он выражен весьма
заметно [127].
Дыхательные функции крови
47
О значении гемоглобина для выживания того или иного вида
можно судить по эффекту отравления угарным газом (СО). Пове-
дение серебряных карасей при инактивированном СО гемоглобине'
не изменяется в течение длительного времени [6J, а радужная
форель выживает лишь около 30 мин (при 10—15°C) [76]. У че-
ловека 1,28% СО вызывает токсическое действие уже через 3 мин.
Относительное сродство гемоглобина к СО и к О2 выражается
величиной М в уравнении
СО-НЬ м Рю (СО-НЬ)Ро2
о^нь м РОг' (02-НЬ)(Рсо)
Приводим данные об относительном сродстве различных гемо-
глобинов к СО и О2 [6, 76, 138а]:
м м
Лошадь 550 Карп 60,5
Человек 300 Серебряный карась 63
Плотва 210 Мышцы млекопитающих 21—51
Щука 100 Клубеньковые бактерии 37
Угорь 99—114 Овод Gastrophilus 0,67
Форель 66 Аскарида 0,036
Из различия между форелью и серебряным карасем очевидно,
что высокое сродство гемоглобина к СО — это только один из
факторов, определяющих выживание после инактивации гемогло-
бина.
У антарктических рыб из сем. Chaenichthyidae эритроцитов нет,
а уровень потребления кислорода почти такой же [45 мл/(кг-ч)],
как и у видов, имеющих гемоглобин. Кожа их сильно васкуляризо-
вана [154]. Если путем введения фенилгидразина уменьшить коли-
чество эритроцитов в крови Lagodon до 2,9% нормальной величи-
ны, то потребление кислорода почти не изменится: вероятно, гемо-
гос^ИН здесь слУжит только резервным переносчиком кислорода
0 09 Chaen°cephalus потребление кислорода составляет при 1 °C
0,02 мл/(г-ч), а критическая величина РО2равна 50 мм рт. ст.,
объем крови очень велик, и высока интенсивность кожного дыха-
ния (8 /0 приходится на долю хвоста) [70].
4 J а™™еск°й рыбы Trematomus повышение температуры до
’ С (при норме 1 °C) действует подобно СО2 — уменьшает на-
сыщение крови кислородом, а Р50 зависит от температуры в боль-
ей степени, чем у других рыб; поэтому Trematomus может жить
только в холодных водах [62].
3 всего сказанного видно, что СО2 и температура оказывают
сьма различное влияние на сродство гемоглобинов разных рыб
обхоСЛ°Р°АУ’ ДЛЯ Т0Г0 чтобы сделать какие-то общие выводы, не-
димы дополнительные данные об экологии многих видов.
48
Глава 8
Изменения транспортной функции в процессе эмбрионального
развития
Мы уже говорили о структурных различиях между гемоглоби-
нами плода и взрослого организма (стр. 17). В фетальном гемо-
глобине человека (HbF) на месте p-цепей находятся у-цепи. Р50
для крови взрослого человека составляет 28 мм рт. ст., а для крови
плода — 20 мм рт. ст. [183]. Однако в растворе гемоглобина эта
разница исчезает, и при pH от 6,5 до 7,5 величина Р^о равна 14,3
для фетального и 14,7 для «взрослого» гемоглобинов, а при pH ни-
же 6,5 Р5о для HbF выше, чем для НЬА [9]. Кривые кислородного
равновесия для крови плода лежат намного левее, чем кривые для
крови взрослого (рис. 8-14). У коз Р50 для цельной крови с возра-
стом увеличивается, а кислородная емкость падает [12, 14]. У коз
и овец HbF в растворе обнаруживает большее сродство к Ог, чем
НЬА, хотя это различие не так .велико, как в цельной крови.
У приматов кровеносные сосуды плаценты проходят через мно-
жество ворсинок, и в норме существует градиент РОз от маточной
артерии к венам плода. У овец и коз капилляры плода примыкают
к «резервуарам» с материнской кровью. У кроликов артерии пло-
да идут параллельно материнским артериям, но кровь в них течет
в противоположном направлении, и диффузия кислорода между
этими сосудами приводит к тому, что в венах плода РОг выше, чем
в материнских венах. Поэтому у кролика более важную роль игра-
ет принцип противотока, а у человека, овцы и козы — различие в
сродстве к кислороду [14, 134] (рис. 8-15). Это видно из данных,
приводимых ниже [134] (РО2И Ао даны в мм рт. ст.).
Кровь матери
артериальная венозная р»о Кислородная емкость, %
РО2 PH ро2 PH
Человек 100 7,4 33 7,36 27 15,5
Коза 84 46
Кролик 80 25
Макак 30 32 15,5
Кровь плода
венозная артериальная f’so Кислородная емкость, %
РО2 pH % PH
Человек 29 7,32 17 7,24 19
Коза 33 14
Кролик 46 17 (противоток)
Макак 16 12 19,2 17,8 .
Дыхательные функции крови
49
У верблюда Р5о крови плода ниже, чем у взрослого животного,
а кислородная емкость выше; у слона переход от HbF к НЬА
происходит в первой трети внутриутробного периода [163, 164].
у кошки нет никакого различия между гемоглобином плода и
взрослой особи. У человека HbF обладает большим сродством к
кислороду и более устойчив к щелочной денатурации, чем НЬА;
значение п одинаково для обоих гемоглобинов [197]. У ягнят сни-
жение сродства к кислороду в эритроцитах частично обусловлено
высоким уровнем ДФГ; у взрослой овцы содержание ДФГ ниже,
чем у ягненка или плода [15]. По данным электрофореза у цып-
лят имеется пять форм гемоглобина. НЬ4— один из основных ком-
понентов в первые 7 дней инкубации. Количество НЬ2 вначале не-
велико, но к моменту вылупления становится значительным. НЬз—
второстепенный компонент как у зародыша, так и у вылупляюще-
гося цыпленка. Синтез гемоглобина в костном мозге начинается
на 14-й день инкубации, но если яйца выдерживать при понижен-
ном РОа, то этот процесс задерживается [187]. Ген «взрослой» фор-
мы НЬ активируется у домашних кур на 6-й день инкубации, а
у индейки — двумя-тремя днями позже [130]. У цыплят по мере
их развития хориоаллантоис все меньше и меньше обеспечивает
Пс^, мм рт сгл ~
Рис. 8-14. Различия между кровью .плода и кровью матери в отношении сродства
к кислороду.
А- Содержание кислорода как функция Pq в крови человека [134].
Продолжение см. на стр. 50.
4—1514
Рис. 8-14 (продолжение).
Б. Кривые кислородного равновесия для крови матери и плода и соответствующих гемогло-
бинов тюленя Уэдделла. Свойства гемоглобинов в растворе сходны, однако цельная кровь
плода обладает меньшей кислородной емкостью, меньшим значением Р50 и большей величи-
ной эффекта Бора. У взрослой особи: кислородная емкость 31,6 об.%, л=2,56 (37° С); у пло-
да: кислородная емкость 27,7 об.%, /1=2,40 (37 °C) [113].
Дыхательные функции крови
51
эмбрион кислородом; это видно при сравнении величин РО2 и РСО2
(в мм рт. ст.) для крови хориоаллантоисной артерии и вены [54].
Вена ро2 Артерия Ро2 Вена РСО2 Артерия РСО2
14 дней инкубации 70 29 33 46
18 дней инкубации 50 20 46 58
Период дыхания внутрен- ним воздухом яйца 68 37 35 47 '
1-й день после вылупления 34 109 43 28
Рис. 8-15. Схема противоточного механизма в плаценте человека [134].
У живородящей змеи Thamnophis разница между гемоглобина-
ми матери и плода обнаруживается только в интактных эритро-
цитах; в растворе оба гемоглобина дают кривую насыщения сход-
ной формы [125]. Кровь головастика лягушки-быка насыщается
кислородом при меньших парциальных давлениях, чем кровь
взрослой особи: у головастика Р50 равно 4,6 мм рт. ст., а у взрос-
лягушки—13,2 мм рт. ст.; величина п в обоих случаях равна
Л Кровь взрослой лягушки обнаруживает прямой эффект Бора
вплоть до pH 6,2, а у головастика эффект Бора отсутствует.
„ яйцеживородящей колючей акулы Squalus плод в течение
всей 22-месячной беременности имеет гемоглобин с более высоким
родством к кислороду, чем у взрослой рыбы. У яйцекладущего
етсТЭ змбРИ0Нальный гемоглобин с высоким сродством к О2 име-
я только в первые месяцы развития. Даже у яйцекладущей кос-
4*
52
Глава 8
тистой рыбы Scorpaenichthys происходит изменение НЬ на постли-
чиночной стадии [122].
Разница в сродстве к кислороду между кровью матери и плода
у млекопитающих, некоторых змей и Elasmobranchia служит при-
мером облегчения транспорта кислорода, так как между первичной
кровью с высоким парциальным давлением 0% и эмбриональной
тканью с низким напряжением кислорода функционирует пигмент
с промежуточным сродством к кислороду. У птиц и лягушек это
может быть связано с переходом от эмбриональной жизни в вод-
ной среде к дыханию воздухом. У яйцекладущих рыб значение
этой смены пигментов неясно.
Функция гемоглобина у беспозвоночных
У одних беспозвоночных гемоглобин участвует в переносе кис-
лорода при атмосферном давлении, у других он функционирует
только при низких Pq2. У некоторых форм гемоглобин обеспечивает
запас кислорода на случай гипоксии, а есть виды, у которых он
служит источником гематина для нужд клетки. Для оценки роли
пигмента в транспорте О2 могут быть полезны следующие крите-
рии: 1) положение кривой кислородного равновесия (т. е. способен
ли пигмент присоединять и отдавать кислород при величинах РОа,
свойственных дыхательной поверхности и тканям); 2) наличие эф-
фекта Бора (т. е. облегчает ли присутствие СО2 отдачу кислорода
при тканевом PQz и присоединение его при более высоких РО2)’
3) кислородная емкость крови и содержание в ней связанного и
растворенного кислорода; 4) разница в содержании кислорода
между артериальной и венозной кровью; 5) влияние окиси уг-
лерода, инактивирующей пигмент, на потребление кислорода ор-
ганизмом при различных парциальных давлениях Ог. При экспери-
ментальном отравлении окисью углерода следует контролировать
спектроскопически процесс соединения СО с гемоглобином и не до-
пускать избытка СО, который может повлиять на тканевое ды-
хание.
Некоторые кольчатые черви живут в условиях, близких к ана-
эробным, и все же имеют функционально активный гемоглобин.
Африканская олигохета Alma emini живет в болотах, где уже на
глубине нескольких миллиметров от поверхности практически нет
кислорода. Гемоглобин у этого вида насыщается при Ро2 меньше
2 мм рт. ст., и его функция не зависит от СО2 [16]. У червя
Tubifex, обитающего в застойных дренажных канавах, гемоглобин
полунасыщен при 0,6 мм рт. ст., однако при измерении in vivo по-
лосы поглощения оксигемоглобина исчезают, когда РО2 у поверх-
ности тела снижается до 10 мм рт. ст. [53]. Окись углерода угне-
тает дыхание, особенно при высоких парциальных давлениях кис-
лорода. При повышенных давлениях О2 у этих червей наступает
Дыхательные функции крови
53
кислородное отравление. В норме должен существовать чрезвы-
чайно крутой градиент РО2 между водой и тканями.
Большинство кольчатых червей живет в умеренно аэрируемой
почве или воде. Потребление кислорода у дождевого червя
Lumbricus уменьшается под влиянием СО при парциальных дав-
лениях О2 40 мм рт. ст. и выше (рис. 8-16). Повышение темпера-
туры с 15 до 25°C ведет к повышению Р50 с 3,9 до 6,8 мм рт. ст.;
наблюдается также прямой эффект Бора (см. табл. 8-3). При Р<\
от 70 до 152 мм рт. ст. потребление кислорода относительно по-
Рис. 8-16. Влияние СО на дыхание при различных значениях Ро2-
A. Lumbricus (Johnson М. L., J. Exp. Biol., 18, 266—277, 1941); Б. Chlronomus (Ewer S. F.,
J. Exp. Biol., 18, 197—205, 1941).
стоянно и гемоглобин переносит около 40% О2 [33], У гигантского
дождевого червя Glossoscolex кислородная емкость крови состав-
ляет 14 об.%, гемоглобин в крови спинной артерии насыщен на
41%, а эффект Бора крайне незначителен [98]. У трех видов
пиявок, обладающих гемоглобином, СО угнетает потребление кис-
лорода при 10—20% О2, но не при 3% О2, в то время как у двух
видов, не имеющих НЬ, такого эффекта не наблюдалось [184].
Кольчатые черви, обладающие гемоглобином, при высоких Ро2
обычно более чувствительны к окиси углерода; в этом случае их
метаболизм зависит от функции гемоглобина, а при низких РОг
°н д’ вероятно, теряют активность и переключаются на гликолиз.
У Nereis diversicolor в отличие от большинства других кольчатых
червей дыхание полностью блокируется окисью углерода при
ю мл О2 на 1 л, а при более высоких концентрациях кислорода
оно снижается лишь наполовину. У некоторых видов Arenicola
сродство НЬ к О2 не очень велико — Р50=1,8 мм рт. ст., а насыще-
но наступает при РОг от 5 до 10 мм рт. ст. [209]. Гемоглобин
rerucola мог бы обеспечить запас кислорода не более чем на
ГемМИН намног° меньше, чем длится период отлива [99]. В этом
оглобине очень велико взаимодействие между гемами (п>5),
54
Глава 8
а эффект Бора весьма незначителен [201а]. У Nephthys равно
5,5 мм рт. ст. для гемоглобина крови и 7,5 мм рт. ст. для гемо-
глобина целомической жидкости. В воде, которая остается во время
отлива в песке, окружающем червя, среднее парциальное давление
О2 около 5,7 мм рт. ст., а в свежезаполненных водой норах —
13,7 мм рт. ст.
У многощетинкового червя Glycera гемоглобин содержится в
целомических клетках. Величина Р5о оказалась у него весьма из-
менчивой, но в среднем составляла 7 мм рт. ст. Окись углерода
снижает потребление О2 на 20%, но полностью не подавляет даже
при низких РО2. Гемоглобин из целомических клеток был разделен
на две фракции: легкую с высоким сродством и тяжелую с низким
сродством [72].
Многие брюхоногие и двустворчатые моллюски имеют миогло-
бин в некоторых тканях (например, в красных мышцах радулы).
У одного боконервного моллюска мол. вес миоглобина радулы
33 800 и молекула содержит два гема; миоглобин представлен ди-
мерной и мономерной формами. В жабрах двустворчатого моллюс-
ка Phacoides миоглобин находится, вероятно, в форме тетрамера
[155]. У моллюсков этот пигмент широко распространен, а гемо-
глобин встречается значительно реже. У Cardita гемоглобин крови
растворен в плазме (Г’5о:=11 мм рт. ст.). У мышечного миоглобина
Mercenaria Р50 = 0,55 мм рт. ст. [126].
У пресноводной и болотной улитки Planorbis гемоглобин хоро-
шо приспособлен для переноса кислорода: при 20 °C Р5о =
— 7,4 мм рт. ст., имеется прямой эффект Бора, полосы поглощения
оксигемоглобина исчезают при снижении Pq2 в наружной воде до
25 мм рт. ст. Высокие концентрации СО2, по-видимому, никогда
не встречаются. При РО2 выше 54 мм рт. ст. (насыщение 7,7%)
достаточно бывает кожного дыхания. При насыщении О2 от 7,2 до
3% улитки часть времени проводят у поверхности воды, чтобы
дышать легкими, а при насыщении ниже 3% они находятся у по-
верхности постоянно. Судя по данным о содержании О2 в легких
и артериальной крови, гемоглобин Planorbis активно выполняет
функцию переноса кислорода в диапазоне РО2 от 20 до 60 мм рт. ст.
При больших величинах Pq2 высокое содержание кислорода в ве-
нозной крови препятствует отдаче его тканям. При Pq2 ниже
40 мм рт. ст. артериальная кровь уже не насыщается полностью,
а при давлениях ниже 20 мм рт. ст. гемоглобин теряет способность
служить как переносчиком, так и резервуаром кислорода. Нижний
предел для кожного дыхания выше, чем для легочного. Минималь-
ное Pq2, при котором возможно полное насыщение, составляет
39 мм рт. ст. при РСО2 3,1 мм и 61,5 мм — при РСо2 1,9 мм [100].
Другая улитка, Lymnaea, питается у поверхности воды. У нее нет
гемоглобина и поэтому РО2 в легких не снижается так сильно, как
у Planorbis [100].
Дыхательные функции крови
55
У некоторых мелких ракообразных кровь содержит гемоглобин,
особенно в тех случаях, когда вода бедна кислородом. Пресновод-
ный рачок Moina (Cladocera) при низком содержании кислорода
в среде в течение нескольких дней синтезирует гемоглобин. Р50 та-
кого гемоглобина составляет 3,5 мм рт. ст. при 25 °C и 1,1 при
15°C; эффект Бора отсутствует (рис. 8-17). Розовые (содержащие
гемоглобин) особи потребляют больше кислорода, чем неокрашен-
Рис. 8-17. Интенсивность дыхания Moina при различном содержании кислорода
в воде.
Кривые для желтых особей (с кровью, бедной НЬ) и для розовых особей (с кровью, бога-
той НЬ)—нормальных и отравленных СО [82].
ные. При отравлении СО потребление кислорода падает, особенно
при пониженном Pq2 [81]. Гемоглобин синтезируется в тех слу-
чаях, когда возрастает потребность в кислороде, и он выполняет
транспортную функцию. У жаброногого рачка Triops содержание
кислорода в крови in vivo составляет 0,93 об.%, а кислородная
емкость при насыщении — 3,2 мл О2 на 100 мл; Р50 = 6,8 мм рт. ст.,
эффект Бора имеется, окись углерода не подавляет дыхания [78].
У красной дафнии Simocephalus СО снижает потребление кисло-
рода только при концентрации его ниже 2,5 мл/л [80]. У Artemia
при низком содержании кислорода в воде количество гемоглобина
может увеличиваться в 20 раз.
Водные личинки двукрылых рода Chironotnus также имеют ге-
^50 ПРИ измеРении in vitro оказывается очень низким
. > 0,6 мм рт. ст.), однако in vivo гемоглобин полностью дезок-
56
Глава 8
сигенирован при содержании кислорода в воде, соответствующем
13 мм рт. ст. Повышенное потребление кислорода после анаэробио-
за снимается добавлением СО. При насыщении НЬ выше 50% СО
не влияет на интенсивность дыхания. Гемоглобин может обеспечи-
вать запас кислорода на период остановки дыхания для приема
пищи. Как правило, розовые виды перестают питаться при насы-
щении воды воздухом ниже 10%, а в присутствии СО — при 26%;
неокрашенные виды продолжают активно кормиться при низком
POs и даже в присутствии цианида. У розовых и неокрашенных ви-
дов не обнаружено никаких различий в погашении кислородной
задолженности после анаэробиоза. У личинки Chironomus plumosus
кислородная емкость крови варьирует в пределах от 5,4 до
11,6 об.% в зависимости от количества гемоглобина; Р50 составля-
ет 0,2 мм рт. ст. при pH 8 и 0,6 — при pH 7,4 [202]. Потребление
кислорода равно 250 мл/(кг • ч); вычислено, что кислорода, связанно-
го с гемоглобином, хватило бы всего лишь на 9 мин, что близко к
обычному интервалу между периодами вентиляции. При высокой
температуре сродство к кислороду понижается, а эффект СО2 воз-
растает. Другой вид, Ch. thiimmi, менее чувствителен к снижению
Рое чем Ch. plumosus-. потребление кислорода поддерживается на
постоянном уровне вплоть до РО2 18 мм рт. ст. [202]. Кажется ве-
роятным, что свойства ферментов у различных ракообразных и хи-
рономид могут варьировать в зависимости от того, нуждается ли
данный вид в гемоглобине. Для того чтобы понять биологическую
роль различного содержания гемоглобина, необходимо изучить не
только потребление О2, но и ряд других функций.
У некоторых водных насекомых (например, Notonectidae) име-
ются трахеальные клетки, богатые гемоглобином. Насекомое под-
нимается на поверхность для дыхания каждые 2—5 мин; под влия-
нием СО время пребывания под водой сокращается [134а]. Однако
у некоторых видов гемоглобина нет.
У эндопаразитов функция гемоглобина неясна. У ряда нематод
Ры для гемоглобина в разбавленном растворе меньше 0,05 мм рт. ст.
[174]. Однако in vivo гемоглобин Nippostrongylus дезоксигениро-
ван даже тогда, когда черви находятся в среде с Ро2 13 мм рт. ст.
Обычно концентрация кислорода в кишечнике, где живут паразиты
(4—9 мм рт. ст.), значительно ниже критической точки для их
аэробного обмена. При РОг от 30 до 5 мм рт. ст. потребление кис-
лорода не уменьшается в присутствии СО. Значит, эти черви в
обычных условиях не встречаются с такими концентрациями кис-
лорода, при которых интенсивность обмена и насыщение гемогло-
бина достигали бы у них возможного максимума.
У Ascaris имеются два весьма различных гемоглобина — один
в полостной жидкости, а другой в стенке тела. В атмосфере азота
гемоглобин стенки тела явно дезоксигенируется в отличие от ге-
моглобина полостной жидкости. Время дезоксигенации в присутст-
Дыхательные функции крови 57
вии восстановителя составляет для полостного гемоглобина Ascaris
150 с (для гемоглобина овцы всего лишь 0,008 с) [35]. Темпера-
турный коэффициент (Сю) для дезоксигенации гемоглобина из
стенки тела равен 3, и этот процесс очень чувствителен к pH. Для
полостного гемоглобина Qio = 5, и дезоксигенация нечувствительна
к pH; очевидно, что этот гемоглобин не участвует в транспорте Ог-
Смит [189] предполагает, что этот пигмент может служить источ-
ником гематина, который необходим для созревания яиц.
Пресноводная немертина Prostoma имеет гемоглобин, и потреб-
ление ею кислорода при низком РО2 снижается в присутствии СО
[151]. Давно известно, что гемоглобин есть и у Paramecium', на его
роль в переносе кислорода указывает тот факт, что СО угнетает
дыхание этой инфузории [190].
Таким образом, гемоглобин спорадически встречается у пред-
ставителей нескольких типов беспозвоночных; при этом виды, не
имеющие гемоглобина, выживают, по-видимому, не хуже тех, у
которых он есть. Некоторым беспозвоночным гемоглобин необхо-
дим для транспорта кислорода; у других градиент кислорода меж-
ду средой и тканями может быть очень крутым. У нескольких ви-
дов, имеющих гемоглобин, функция его не выяснена.
Влияние пониженного содержания кислорода в среде
на синтез гемоглобина
Животным, обитающим на больших высотах, обычно свойствен-
но высокое содержание гемоглобина в крови. Если животное под-
нимается с уровня моря в горы, то количество гемоглобина в его
крови возрастает и вместе с ним увеличивается кислородная ем-
кость крови. Когда животное впервые попадает в условия гипо-
ксии, как это происходит на больших высотах, эритроциты выходят
из кровяных депо, например из селезенки, в циркулирующую кровь,
позднее под влиянием гормона эритропоэтина усиливается крове-
творение. У человека при сильной гипоксии синтез гемоглобина
возрастает уже в первые 12 ч, а через три дня он достигает мак-
симума [188]. Объем крови увеличивается — главным образом за
счет увеличения числа эритроцитов. Одновременно с повышением
кислородной емкости кривая кислородного равновесия крови у че-
ловека может сдвигаться вправо, однако изменения величины п
или эффекта Бора не происходит [111]:
"'.'Г" |>|”' Кислородная емкость, об. % Psq, мм рт. ст. Емкость для СОг, %
Уровне моря 18,9 26,8 43,6
ра высоте, у аборигенов 26,4 30,7 36,4
высоте, у вновь прибыв- ших 20,7 30,4 36,6
58
Глава 8
Числа эритроцитов и гематокрит у акклиматизированных овец,
кроликов, собак и людей возрастают; однако у местных живот-
ных— ламы и викуньи (в Южной Америке) — эти величины не
очень заметно меняются в зависимости от нахождения в горах или
низине [65]. Кривые кислородного равновесия у этих животных,
так же как и у других обитателей гор, сдвинуты влево по сравне-
нию с кривыми для родственных видов, живущих на уровне моря.
У собак, выросших на высоте 4500 м, оказалось на 40% больше
гемоглобина в крови и на 67,5% больше миоглобина в мышцах,
чем у собак на равнине [200]. У высокогорных перуанских грызу-
нов диафрагма и мышцы конечностей содержат вдвое больше мио-
глобина, чем у равнинных видов [161, 162].
Увеличение количества гемоглобина при акклиматизации к вы-
соте может оказаться недостаточным для надлежащего снабжения
тканей кислородом. У ягнят обнаружены следующие различия в
зависимости от высоты местности [64]:
1580 м 3870 м
Среднее насыщение, % 84 59
Ро2 в артериальной крови, мм рт. ст. 70 40
Артерио-венозная разность Ро2, мм рт. ст. 28 11
Время жизни эритроцитов в крови человека на большой высоте
и на уровне моря одинаково (111—121 день). У крыс, находивших-
ся в атмосфере с 10% О2 в течение шести месяцев, концентрация
гемоглобина возросла с 14,8 до 22,3 г на 1 мл.
У плода эритроциты образуются в печени, а у взрослого жи-
вотного-—в костном, мозге. Гемоглобин синтезируется в ядерных
проэритроцитах или ретикулоцитах. Из ретикулоцитов, циркулиру-
ющих в крови, выделена система РНК, регулирующая этот синтез.
Потеря крови, а также пониженное парциальное давление кисло-
рода стимулируют образование ретикулоцитов. Активность гормо-
на эритропоэтина оценивают по нарастанию количества ретикуло-
цитов при введении этого гормона мышам с полицитемией; уровень
его в крови возрастает при гипоксии и кровопотере, а также при
введении кобальта. В норме эритропоэтин образуется в почке,
скорее всего в юкстагломерулярных клетках. Возможно, что имен-
но почка является органом, «чувствующим» гипоксию [93]. Тесто-
стерон и пролактин усиливают действие эритропоэтина, а эстроге-
ны ослабляют его. Эритропоэтин обнаружен у млекопитающих,
птиц, лягушек и рыб [177]. Он воздействует главным образом на
стволовые клетки костного мозга, побуждая их поглощать железо
и продуцировать ретикулоциты. Гормон способен также стимули-
ровать синтез гемоглобина в культуре печеночной ткани от 13-
дневного плода мыши; при этом синтез ДНК возрастает через
20 мин, а синтез РНК и самого гемоглобина — через 2 ч [89].
Синтез рибонуклеопротеидных частиц с константой седиментации
150S можно обнаружить через несколько минут. Эритропоэтин
млекопитающих неэффективен в отношении низших позвоночных,
Дыхательные функции крови 59
однако сыворотка частично обескровленной рыбы усиливает эрит-
ропоэз при введении другой рыбе. Следовательно, образование
гормона у рыб, так же как и у млекопитающих, возможно, стиму-
лируется гипоксией [213].
У некоторых ракообразных синтез гемоглобина заметно возрас-
тает при низком парциальном давлении кислорода. Дафнии, со-
бранные или выдержанные в воде, бедной кислородом, имеют крас-
ную окраску — в отличие от тех, которые росли в хорошо оксиге-
нированной воде. Существуют и генетические различия в способ-
ности к образованию гемоглобина. Дафния синтезирует или теря-
ет гемоглобин в течение примерно 10 дней; кроме того, при низ-
ком Ро у нее активируется синтез цитохрома и миоглобина в
мышцах. Artemia может быть ярко-красной от гемоглобина, если
живет в очень соленой воде, бедной кислородом. Концентрация
гемоглобина в крови возрастает при снижении POz не только у
ряда ракообразных (главным образом Cladocera и Phyllopoda),
но и у личинок двукрылых (Chironomus и Anatopynia), а также у
улиток (Planorbis). Однако у многих других беспозвоночных
(кольчатые черви, различные моллюски и др.) подобной стимуля-
ции синтеза НЬ не обнаружено [49, 51]. Участвуют ли в такой
стимуляции гормоны (как у позвоночных), неизвестно.
Функция хлорокруорина
Несмотря на значительно большие размеры молекулы, хлоро-
круорин весьма сходен с гемоглобином по функциональным свой-
ствам. Кривая кислородного равновесия имеет S-образную форму;
константа п Хилла в области полунасыщения равна 4,5. Величина
Ao (30—40 мм рт. ст.), по-видимому, достаточно высока для того,
чтобы пигмент был дезоксигенирован в тканях и насыщен кисло-
родом в пальпах. Кислородная емкость крови, содержащей хлоро-
круорин, весьма велика (9,1 мл Ог на 100 мл) [50]. Сродство пиг-
мента к СО выше, чем у любого гемоглобина, и потребление кис-
лорода червями-сабеллидами, у которых переносчиком его служит
хлорокруорин, подавляется окисью углерода на всем диапазоне
концентраций О2. Кровь Serpula содержит как гемоглобин, так и
хлорокруорин; оба пигмента дезоксигенируются одновременно
[52]. у Potamilla в крови находится хлорокруорин, а в мышцах —
гемоглобин; последний, вероятно, облегчает диффузию кислорода
°т хлорокруорина, обладающего низким сродством к Ог. В целом
можно считать, что хлорокруорин приспособлен для переноса кис-
лорода при высоких уровнях Pq2.
Функция гемоцианина
Как мы уже видели, гемоцианины членистоногих и моллюсков
Различаются по своим физическим свойствам. Однако все гемо-
цианины обладают способностью обратимо связывать кислород—
60
Глава 8
по одной молекуле О2 на два атома меди. У ряда членистоногих
один связывающий центр приходится на 1 частицу с мол. весом
около 75 800, а у моллюсков — на 1 частицу с мол. весом 50800
[74]. В окси-форме гемоцианин имеет голубой цвет и дает интен-
сивные полосы поглощения, а в дезокси-форме бесцветен.
У головоногих моллюсков Loligo и Octopus практически весь
кислород транспортируется с помощью гемоцианина. Артериальная
кровь кальмара насыщена кислородом и содержит 4,27% О2 и
3,82% СО2, а венозная — 0,37% О2 и 8,27% СО2 (по объему). Сле-
довательно, ткани поглощают около 92% кислорода крови, что в
три раза превышает соответствующую величину для человека. Зна-
чение Р50 (36 мм рт. ст. при 23 °C в отсутствие СО2) достаточно
высоко для того, чтобы сделать организм кальмара чувствитель-
ным к колебаниям Pq2 в окружающей воде. Из-за сильно выражен-
ного эффекта Бора кривая кислородного насыщения сдвигается
при добавлении СО2 вправо. Кроме того, взаимодействие между
субъединицами зависит от pH и ионов магния; оно невелико при
низких и высоких pH, но при pH 7,4 п = 3,9 (рис. 8-18). При pH от
6 до 10 дезоксигемоцианин находится в недиссоциированном со-
стоянии (59S), при 80%-ном насыщении кислородом происходит
полная диссоциация на субъединицы 19 S и 11S; однако при
100%-ном насыщении субъединицы реассоциируют [39].
У Octopus кислородная емкость составляет от 3 до 5 об. %
[208]. Присутствие СО2 повышает величину Роа, при которой ге-
моглобин отдает кислород (эффект Бора), и уменьшает общее ко-
личество связанного кислорода при полном насыщении (эффект
Рута); высокая температура также уменьшает связывание О2
[114]. Артериальная кровь при РО2 77,5 мм рт. ст. оказалась на-
сыщенной на 82%, венозная при Pq2 9,7 мм рт. ст. — на 9,6; более
90% кислорода в артериальной крови связано с гемоцианином.
Octopus извлекает из прокачиваемой воды 27% кислорода [94].
Видимо, у головоногих перенос кислорода в основном осуществля-
ется гемоцианином; резерв кислорода в крови крайне мал. Про-
цесс агрегации субъединиц весьма сложным образом зависит от
РОа, pH и двухвалентных катионов.
У улитки Helix pomatia обнаружены две молекулярные разно-
видности гемоцианина, различающиеся по диссоциации в 1 М
NaCl, но сходные по своим функциональным свойствам. Оксигемо-
цианин Busycon в разбавленном растворе представляет собой аг-
регат с константой седиментации 100 S; в дезокси-форме такую
константу имеют всего лишь 17% молекул, остальные имеют кон- ри
станты 60S и 19 S (рис. 8-19) [39]. В отсутствие Са и Mg кривая
кислородного равновесия гиперболическая (ц^1,1); в присутствии
этих катионов кривая становится сигмоидной (п равно 2,3 для
a-формы и 1,4—4,6 для p-формы). В физиологическом диапазоне
pH a-форма обнаруживает небольшой прямой эффект Бора, а
Рис. 8-18. Кривые кислородного равновесия гемоцианина Loligo.
Измерения проводились при pH, указанных цифрами у кривых ([39], 1960).
с- 8-19. Влияние оксигенации на содержание 100S-компонента в гемоцианине
Busycon при pH 8,2 ([39], 1970).
62 1 лава 8
[3-форма — обратный эффект Бора. Возможно, что [3-форма лучше
функционирует при низких концентрациях СО2, а а-форма — при
высоких [107]. У Busycon Р50 ниже, чем у Helix, но, так же как
и у Helix, значение п возрастает в присутствии двухвалентных
катионов; эффект Бора обратный или вообще отсутствует. РС)2 ар-
териальной крови равно 36 мм рт. ст., а венозной — 6 мм рт. ст.
У другой морской улитки, Fusitriton, эффект Бора при физиологи-
ческих значениях pH отрицательный. Высказано предположение,
что из-за очень медленной циркуляции крови накопление СО2 в
каком-либо участке тела вызывает приток кислорода к этому
участку [158, 159]. У Diodor а и Chiton больше половины кисло-
рода транспортируется гемоцианином и величина Р50 мало чувст-
вительна к pH [156, 158]. Некоторые брюхоногие моллюски (Busy-
con, Cryptochiton) используют гемоцианин для переноса кислорода
кровью, но содержат миоглобин в некоторых мышцах, особенно в
мышце радулы; поскольку сродство миоглобина к кислороду выше,
чем у гемоцианина, мышечный пигмент, видимо, облегчает (как и
у позвоночных) диффузию кислорода.
Перенос кислорода гемоцианином изучен также у ракообраз-
ных, Limulus и некоторых скорпионов (рис. 8-20). У омара гемо-
цианин составляет 88,2% белков крови; Р50 равно 6 мм рт. ст. при
РСО2 мм и 25 мм — при РС0.2 9,9 мм. Степень агрегации субъ-
единиц при физиологических значениях pH выше, нежели в щелоч-
ной или кислой зонах; ионы кальция увеличивают значение п.
Удаление кальция ведет к диссоциации крупного агрегата и сдви-
гает кривую кислородного равновесия вправо [148]. В крови, про-
шедшей через жабры, гемоцианин насыщен кислородом на 49%,
а в притекающей к жабрам крови — на 20%. У речного рака при
повышении температуры с 15 до 25 °C Р50 возрастает с 1,57 до
2,79 мм рт. ст., а значение п падает с 3,1 до 2,9. Значение Р5о
максимально при pH 7 и уменьшается при изменении pH как в
кислую, так и в щелочную сторону; в отсутствие Са уменьшение
Р5о при подщелачивании выражено слабее (рис. 8-21) [НО].
В длительном опыте с введенными внутрь катетерами были по-
лучены дыхательные характеристики крови краба Cancer magister,
которые ясно демонстрируют роль гемоцианина в переносе кисло-
рода. У этого краба кислородная емкость крови составляет
3,44 об. % и из воды, пропускаемой через жабры, извлекается 16%
кислорода. Ро,2 в артериальной крови равно 91 мм рт. ст., что
соответствует полному насыщению, а в венозной — 21 мм рт. ст.
(50% насыщения). После пятиминутной физической нагрузки на-
сыщение кислородом падает до 10%, а повышение температуры!
сдвигает кривую насыщения в сторону больших Ро [96].
У двух наземных крабов гемоцианин переносит большую часть
кислорода: у Cardisoma артериальная кровь содержит 1,6—
1,7 об.% О2, а венозная— 0,85—0,99 об.%; у Gecarcinus соответ-
Рис. 8-20. Кривая кислородного равновесия для крови краба Cancer [96].
Рис. 8-21. Зависимость Рьо гемоцианина речного рака от pH.
Кривая имеет характерный максимум, положение которого изменяется в присутствии ионов
кальция [НО].
64
Глава 8
ствующие величины равны 1,45 и 0,61 об.% [157, 160]. У обоих ви-
дов наблюдается прямой эффект Бора; насыщение крови кислоро-
дом в жабрах достигает примерно 85%, а венозная кровь насыще-
на на 20—40%.
У скорпиона Heterometrus кислородная емкость крови состав-
ляет 1,82 об.%, а Ао равно 16,5 мм рт. ст. при 30°С. Эффект Бо-
ра— прямой при pH от 10 до 8 и обратный при pH от 8 до 6; зна-
чение п возрастает по мере оксигенации [140]. У Limulus также
отмечен прямой эффект Бора при pH от 9,1 до 8,2; при более низ-
ких pH он становится обратным. Это означает, что Р$о достигает
максимума при pH выше физиологической границы. Значение об-
ратного эффекта Бора, свойственного почти всем гемоцианинам,
неясно; возможно, он облегчает извлечение кислорода из воды, на-
сыщенной СО2. У многих ракообразных кровь перикарда далека
от насыщения, хотя кислорода, связанного с гемоцианином, намно-
го больше, чем растворенного. При этом надежные измерения по-
казали, что пигмент работает как переносчик. Возможно, что мно-
гие животные с малоподвижным образом жизни (особенно брюхо-
ногие моллюски и Limulus), обладающие гемоцианином, могли бы
обходиться и без него.
Функция гемоэритрина
У Golfingia (а также Sipunculus) кривая диссоциации оксиге-
моэритрина проходит через Р50 при 8 мм рт. ст. Повышение тем-
пературы сдвигает кривую вправо, теплота оксигенации для гемо-
эритрина много выше, чем для гемоглобина. Величина п близка к
1,0 (или чуть больше), т. е. мономеры почти не взаимодействуют
между собой. У Golfingia pH не оказывает заметного влияния на
кривую кислородного равновесия. Эффект Бора не был обнаружен
и при изучении гемоэритринов Dendrostomum и Sipunculus. У пле-
ченогих (Lingula) наблюдается прямой эффект Бора, но гемо-
эритрин у этих животных менее изучен в химическом отношении,
чем у сипункулид. СО не отравляет гемоэритрин. В этом пигмен-
те 1 атом кислорода приходится на 1 атом железа, тогда как в
оксигемоглобине соотношение О: Fe равно 2:1, а в гемоцианине
1 атом О приходится на 1 атом Си.
В сосудистой системе сипункулиды Dendrostomum zosteriocolum
(в сосудах щупалец и главных сократимых сосудах) имеется гемо-
эритрин, электрофоретически отличный от гемоэритрина целомиче-
ской жидкости. А>о Для клеток, содержащих сосудистый гемоэри-
трин, составляет 40—50 мм рт. ст., для целомического гемоэри-
трина— всего лишь 4,5 мм рт. ст. Очевидно, сосудистый гемоэри-
трин переносит кислород от дыхательной поверхности, соприка-
сающейся с хорошо насыщенной О2 морской водой, к целому, а
целомический гемоэритрин доставляет кислород тканям. У зарыва-
Дыхательные функции крови 65
ющейся в ил сипункулиды Siphonosoma ingers), у которой щупаль-
ца не используются для дыхания, сосудистый пигмент обладает
большим сродством к кислороду, чем целомический (так же как
и гемоглобин у Nephthys)-, кислород сначала проходит здесь через
стенку тела в целом, а затем переносится к сосудистому гемоэри-
трину, обладающему высоким сродством к О2 [125].
Транспорт двуокиси углерода
В крови лишь незначительная доля всей СО2 (так же как и
О2) находится в растворенном состоянии. В плазме крови имеет
место равновесие между растворенными СО2, Н2СО3, НСОз и
СО|" Равновесие реакции СО2 с Н2О сильно сдвинуто в сторону
образования Н2СО3, и эту реакцию ускоряет карбоангидраза, по-
этому концентрация растворенной СО2 определяется концентра-
цией Н2СО3. Значение pH, при котором ион НСОзГ диссоциирован,
достаточно высоко, поэтому при физиологических pH концентрация
СО|~ крайне незначительна. Таким образом, общее содержание
СО2 определяется суммой концентраций Н2СО3 и НСОз .
При Рсо2 760 мм рт. ст. и 37,5 °C растворимость СО2 в крови
человека равна 48 об.%. Поскольку парциальное давление СО2 в
альвеолярном воздухе составляет 40 мм рт. ст., количество СО2,
которое могло бы быть растворено в крови, равно 2,5 об. % (ко-
эффициент растворимости, умноженный на парциальное давление).
Однако на самом деле артериальная кровь содержит от 40 до
50 об.% СО2, а венозная — от 55 до 60 об.%. Аналогичный расчет
показывает, что морская вода (коэффициент растворимости в ней
СО2 равен 0,71 при 24°C), находящаяся в равновесии с воздухом,
в котором Рсо2 =0,23 мм рт. ст., должна была бы содержать
0,0215 об. % СО2; однако в действительности она содержит 4,8 об. %
СО2. Это различие обусловлено взаимодействием НСОз с катиона-
ми различных буферных систем. Для любой жидкости, содержащей
буферы, можно построить кривую насыщения СО2, уравновешивая
жидкость с СО2 при известном парциальном давлении и определяя
количество поглощенной СО2 в об.%. В двух приведенных приме-
рах 40—50 об. % СО2 в крови соответствуют РСО2 40 мм рт. ст.,
а 4,8 об. % СО2 в морской воде соответствуют РСОл 0,23 мм рт. ст.
Зависимость между величиной pH и концентрациями НСОз и
Н2СО3 выражается уравнением Гендерсона—Гассельбаха:
₽H=p*+'°gs-
гДе р/(=6,1. Зависимость между РСО2 и концентрацией НСОз (или
Н2СО3) нелинейна, и ее невозможно выразить простым уравнением
(рис. 8-22). Однако зависимость каждой из этих величин (обычно
берут концентрацию НСОз) от pH линейна в средней зоне значе-
5—1514
66
Глава 8
ний pH, и буферная емкость соответствует наклону этой линии
(т. е. изменению концентрации НСОз в ммоль/л при изменении pH
на единицу). На трехкоординатных графиках представлены буфер-
ные кривые при различных РСО2 (рис. 8-23).
Общее содержание СО2 в циркулирующей крови зависит от бу-
ферной емкости крови, парциального давления СО2 в тканях и ре-
гуляции кислотно-щелочного баланса почками и органами дыха-
Рис. 8-22. Кривые углекислотного равновесия для дезоксигенированной (/) и окси-
генированной (II) крови Octopus [114].
ния. У наземных позвоночных парциальное давление СО2 в легких
высоко (40 мм рт. ст. у многих млекопитающих), в то время как
в воде, омывающей жабры водных животных, величина РСО2 мала
(0,23 мм рт. ст.). Парциальное давление СО2 в крови водных поз-
воночных намного меньше, чем у наземных, так что градиент в
области дыхательной поверхности у тех и других примерно одина-
ков и составляет около 6 мм рт. ст. У млекопитающих артерио-
венозная разница в содержании СО2 показывает, что в легких
кровь отдает около 10% углекислоты. У водных позвоночных об-
щее содержание СО2 в крови ниже, поэтому относительные коли-
чества СО2, поступающие из тканей и отдаваемые в жабрах (или
через кожу), больше, чем у млекопитающих.
Углекислота, диффундирующая из тканей в кровь, переходит
главным образом в эритроциты, где карбоангидраза ускоряет ее
реакцию с водой:
Карбоангидраза
СО2 + Н2О — >•. Н2СО3.
Дыхательные функции крови
67
Затем происходит диссоциация:
Н2СО3 НСО3 + Н+
и большая часть НСОз выходит из клеток, обмениваясь на ионы
С1" из плазмы. Эти ионы переходят из плазмы в эритроциты, ког-
да кровь проходит через ткани, и возвращаются обратно в плазму
в легких. Вся реакция протекает по схеме:
Ткань Плазма Эритроциты
СО2
> СО2 --> СО2 —
Н2О ---> Н2О —
Карбоангидраза
нсо;
н2со3
НСОз н+
сг —> сг
Эритро- Плазма Легочные
циты альвеолы
СО2 -----> СО2 --->
Карбоангидраза •—►Н2О -----> Н2О
Н2СО3
t
н+----------нсо- ч--- нсо-
СГ ---> СГ
В артериальной крови около 67% НСОз’ растворено в плазме; для
венозной крови эта величина составляет около 66,5%. В плазме
основным катионом, уравновешивающим НСОз, является Na+.
Внутри эритроцитов буферные свойства среды в основном опреде-
ляются гемоглобином:
ВНЬ + Н+ =₽=* ннь + в+,
где В означает катион, главным образом К+.
Дезоксигенированный гемоглобин — более слабая кислота
(р/(=7,95 для пигмента лошади), чем оксигенированный (р/С=
= 6,68). В связи с этим по мере отдачи кислорода тканям в эритро-
цитах высвобождаются катионы. При отдаче СО2 в легких более
кислый оксигемоглобин связывает большее количество катионов,
кто способствует дополнительному высвобождению СО2. Присоеди-
нение одной молекулы кислорода увеличивает отрицательный за-
ряд молекулы гемоглобина на 0,6 эквивалента, и при этом вытесня-
ется соответствующее количество НСОз. С этим влиянием оксиге-
нации на кислотные свойства гемоглобина связан перенос большей
Пасти СО2.
5*
68
Глава 8
Помимо углекислоты, переносимой описанными выше буферны-
ми системами, около 15% СО2 в эритроцитах артериальной крови
и 20% СО2 в эритроцитах венозной крови присоединено к NH-
группам гемоглобина, т. е. транспортируется в виде карбамино-
групп. В процентном отношении такой углекислоты немного, но
было вычислено, что на ее долю приходится примерно 20—30%
общего обмена СО2 крови в дыхательном цикле, а в обмене СО2
внутри эритроцитов эта доля значительно больше. Эффект Холде-
Рис. 8-23. Буферная емкость дезоксигенированной (треугольники) и оксигениро-
ванной (кружочки) крови взрослой особи и плода тюленя Уэдделла [ИЗ].
Кривые отражают содержание НСОз' в крови в зависимости от pH. Отдельно показаны
парциальные давления СО2, соответствующие различным сочетаниям концентрации
НСОГ и pH.
на более чем наполовину определяется углекислотой, связанной в
форме карбаминогрупп [179].
Нарушения кислотно-щелочного равновесия бывают двух ви-
дов— дыхательные и метаболические. При респираторном ацидозе
(например, после физической нагрузки) pH крови снижается, а
концентрация НСОз возрастает, в то время как при респираторном
алкалозе (обычно при гипервентиляции) происходят обратные из-
менения. При метаболическом ацидозе,_например в случае кетоза
или накопления лактата, уровень НСОз в крови падает, а при
метаболическом алкалозе (в результате потери связанных кислот
при выведении их почками, рвоте и т. п.) — возрастает.
У некоторых птиц гиперпноэ как механизм охлаждения тела
может вести к снижению артериального Pqq на 50% и умеренно-
му повышению pH крови [27] (рис. 8-24).
В табл. 8-5 приведены некоторые сравнительные данные о бу-
ферной емкости крови. Большая часть их была получена путем
определения бикарбоната плазмы после уравновешивания цельной
крови с СО2 при различных парциальных давлениях. У человека
Дыхательные функции крови
69
общая буферная емкость составляет 29 мМ НСОз/ед. pH, из них
g 2 мэкв— за счет буфера плазмы, а 20,7 мэкв — за счет гемогло-
бина [36]. Чем больше остатков гистидина содержится в гемогло-
бине, тем сильнее выражены его буферные свойства. У форели
и скумбрии буферная емкость велика, а у карпа, опсануса и ската
низка. Буферная емкость белков сыворотки (в пересчете на 1 г
50f
я
5=
40
L_ (человек •)
30
: Бентамка,утка, белый плимутрок
' — г в® а,
' Пеликан, голубь
Гриф, чайка ®
Белый плимутрок ®
- Гусь,калмфорнийская ку-9
кушка
20
ф
- - о
Белый плимутрок
Гриф, пеликан ф
15
Калифорнийская кукушка ф
Бентамка ®о
Голубь, утка ф©
10
9
8
Утка, чайка о
— ©
Дыхание. Гись, голубь, пеликан
— ® обычное (в покое) о
ф учащенное о о
— о максимально учащенное
6.
7,2
7,3 7,4 7,5 7,6
7,8 - 7,9
7
Рис. 8-24. Зависимость между Pcof и pH для крови различных птиц в покое и при
учащенном дыхании [27].
белка) у ската и крокодила больше, чем у млекопитающих. У аку-
лы Squalus цельная кровь представляет собой слабый буфер, од-
нако плазма обладает такой же буферной емкостью, как и у че-
ловека [5, 115]. Гиперкапния вызывает у акулы повышение арте-
риального Рсо 2 с 5 до 20 мм рт. ст., а у млекопитающих — с 30
100 мм рт. ст.
Для наземных позвоночных характерны более высокие значе-
ня Лю8 и буферной емкости, чем для водных (например, амфи-
ии) [117]. Скорость диффузии СО2 в воде очень велика; кроме
DbrP’ количество воды, которое может пропустить через жабры
Р оа, по весу в 30 раз больше того количества воздуха, которое
70
Глава 8
Таблица 8-5
Буферная емкость крови (приведены значения для цельной крови,
если не указано иное)
днсоу (мМ) днсоу (мМ) ДрН
ДрН
Человек 30,8 Бобр [32а] 27
плазма 6,5 Водяная крыса [32а] 24
Акула [115] 9 Аллигатор 22,6
плазма 6,5 Крокодил 18,2
Акула [5] 10 Necturus [117] 8,0
плазма 9,8 Amphiuma [117] 9,2
Двоякодышащие рыбы 13 Rana catesbiana [117] 16,4
[П5] Urechis 4,9
Лошадь 25,3 Sipunculus 3,5
пропускают через легкие наземные животные. У животных, дыша-
щих воздухом, выведение электролитов с мочой играет более важ-
ную роль в поддержании кислотно-щелочного равновесия, поэтому
нарушение функции почек может привести к изменению pH крови.
У двоякодышащей рыбы переход к воздушному дыханию сопро-
вождается снижением артериального РО2 и повышением PCOs; ПРН
этом у рыбы развивается метаболический алкалоз, компенсирую-
щий дыхательный ацидоз [116].
При pH 7,0 вода имеет нейтральную реакцию только при 22 °C.
При повышении температуры нейтральное значение pH снижается,
а при понижении температуры возрастает; например, при 35 °C
оно равно 6,84, а при 5 °C — 7,365. У лягушек, жаб и черепах, ак-
климированных к различным температурам, pH крови изменяется
с температурой согласно уравнению ДрН/Д°С =—0,016. Для буфе-
ров крови характерна та же зависимость, что и для воды, поэтому
при различных значениях pH отношение [ОН~] к [Н+] остается
постоянным [83, 84] (рис. 8-25).
У беспозвоночных буферные свойства определяются в основном
белками крови, а главные белки крови — это дыхательные пигмеи- i
ты. У Urechis, например, целомическая жидкость практически ли-
шена буферных свойств, однако буферная сила гемоцитов пример-
но такая же, как у кровяных клеток позвоночных, имеющих сход-
ную кислородную емкость крови. Целомическая жидкость морского,
ежа содержит очень мало белка и является слабым буфером.
Тнтрационные кривые гемоцианинов Limulus и Helix свидетель-
ствуют о наличии нескольких кислотных и основных групп. У Maja,
Octopus и Loligo дезоксигенированный гемоцианин — более слабая
кислота, чем оксигемоцианин, и поэтому связывает больше COs-
Кривые углекислотного равновесия (т. е. зависимости содержания
СО2 от Рсо2 ) вначале круто идут вверх, а затем постепенно вырав*
Дыхательные функции крови
71
Температура, °C
ниваются. У многих беспозвоночных, имеющих кровь с малым со-
держанием белка, физиологические процессы протекают в усло-
виях низких концентраций СО2. У животных, раковины или пан-
цири которых содержат кальций (некоторые моллюски и ракооб-
разные), эти структуры служат
важным источником щелочи для
нейтрализации кислот крови. У дву- в,2
створчатого моллюска, находящего-
ся на воздухе, по мере образования so
молочной кислоты вещество рако-
вины может растворяться со сторо-
ны мантии. 7,8
У насекомых, дышащих возду-
хом, СО2 выходит из тканей прямо 76
в трахеи. Кровь забуферена частич-
но за счет аминокислот. У овода ^4
Gastrophilus белки сыворотки (со-
ставляющие 10,75% по весу) обес- 7
печивают около 62% общей буфер-
ной емкости. Величина Рсо2 весьма
высока (300—500 мм рт. ст.), а pH 70
крови сдвинут в кислую сторону
16, 64]. " 6.8
У животных широко распростра-
нена карбоангидраза. Она играет 66
важную роль в подкислении мочи в
почках позвоночных и в секреции
6,2
Рис. 8-25. Величины pH и рОН в крови жаб, qq
лягушек и черепах, акклимированных к раз-
личным температурам [84].
Измерения проводились при температуре, к кото-
рой данное животное было акклимировано. Сред-
няя линия — кривая нейтральности (0,5 р/() для
воды.
кислоты желудком. У всех позвоночных этот фермент имеется в
тканях глаза, а также в сосудистых сплетениях мозга и в подже-
лудочной железе. Карбоангидраза обнаружена в солевых железах
Птиц и ректальных железах акул и скатов; у рыб большие коли-
чества ее содержатся в жабрах, а у пресноводных рыб также и
в почках [132]. У мелких млекопитающих активность карбоангид-
разы выше, чем у крупных, возможно, в связи с большей интенсив-
остью обмена. Карбоангидраза, вероятно, может облегчить диф-
фузию СО2; при одном и том же градиенте через миллипоровый
I
72 Глава 8
ww
фильтр с карбоангидразой проходит значительно больше СОг, чем
через фильтр, не содержащий фермента.
Карбоангидраза обнаружена у кишечнополостных (например,
в щупальцах Anthozoa); она имеется в дыхательных путях и поло-
вых железах иглокожих, в крови дождевых червей и нереид, в
больших количествах содержится в жабрах полихет, кальмара и
двустворчатых моллюсков, у которых ее много также в мантии.
В жабрах Limulus карбоангидразы больше, чем в жабрах Homarus
и Libinia.
В эритроцитах человека карбоангидраза представлена тремя
изоферментами, которые можно разделить с помощью электрофо-
реза, а в жабрах и мышцах Sepia — двумя формами [1]. У чело-
века все три изофермента содержат 0,2% цинка. У утки аналогич-
ный фермент содержит 0,14% цинка, у черепахи 0,2, а у тунца ;
0,09% [132]. Ингибиторами карбоангидразы являются ненасыщен- j
ные ароматические сульфонамиды. ‘ i
Карбоангидраза ускоряет гидратацию дыхательной СО2 в эрит- 1
роцитах наземных позвоночных и в жабрах многих водных жи-
вотных. Она может облегчать диффузию СО2 подобно тому, как
миоглобин облегчает диффузию кислорода. Карбоангидраза игра-
ет важную роль и в других тканях, где перенос СО2 и образование ,
НСОз не связаны с дыханием. !
Заключение
Дыхательные пигменты, переносящие кислород, представляют
интерес для исследователей, работающих в области физиологиче-
ской экологии, физической биохимии, генетики и эволюции.
Все дыхательные пигменты представляют собой металлопроте-
иды; ферменты, содержащие металл, возникли, вероятно, уже в
самом начале аэробной фазы биохимической эволюции. Для пе-
реноса кислорода в организме животных служат три основных ти-
па пигментов: 1) гемоглобин и сходный с ним хлорокруорин;
2) железосодержащий белок гемоэритрин у сипункулид и плече-
ногих; 3) медьсодержащий гемоцианин у моллюсков и членисто-
ногих.
Гемоглобин встречается спорадически у представителей отда-
ленных филогенетических групп; поэтому можно думать, что ок
возникал неоднократно в процессе эволюции. В пользу этого го-
ворят обнаруженные недавно многочисленные различия между ге-
моглобинами аннелид (особенно Arenicola) и позвоночных. Одна-
ко имеются также факты, которые легче согласовать с противо-
положной точкой зрения: широкая распространенность миоглобина,
разнообразие форм гемоглобина у позвоночных и легкость, с ко-
торой можно индуцировать синтез этого пигмента у многих жИ'
вотных.
Многие специфические свойства пигментов крови можно рас*
сматривать как адаптивные в данных экологических условияХ-Я
Дыхательные функции крови
73
у водных животных, ведущих очень активный образ жизни и оби-
тающих в быстрых реках, значение Р50 пигмента крови выше, чем
у малоподвижных форм, которые часто живут в стоячей воде (на-
пример, гемоцианин кальмара отличается в этом отношении от
гемоцианина некоторых крабов, гемоглобин форели — от гемогло-
бина карпа или сома). Кривая кислородного равновесия для кро-
ви эмбриона или плода у большинства позвоночных сдвинута вле-
во по сравнению с кривой для крови взрослого организма. В ус-
ловиях гипоксии синтез гемоглобина активируется, как, например,
у человека на больших высотах или у Artemia в соленых водое-
мах. Некоторым антарктическим рыбам и многим беспозвоночным
вполне достаточно того количества кислорода, которое растворено
в плазме крови, однако у других близких видов пигмент играет
важную роль, особенно в условиях стресса. Животным, обитаю-
щим в среде с низким содержанием кислорода,— таким, как червь
Tubifex, живущий в иле,— свойственны очень низкие значения Р Ой
в тканях и крутые градиенты кислорода от дыхательной поверх-
ности к тканям.
У животных с высоким уровнем активного обмена добавление
к крови СО2 (или кислот) сдвигает кривую кислородного равно-
весия вправо; напротив, у малоподвижных видов СО2 почти не
влияет на сродство гемоглобина к кислороду. Как правило, СО2
облегчает отдачу О2 тканям, а выход СО2 из крови, наоборот, спо-
собствует ее насыщению кислородом в органах дыхания. Обрат-
ное явление — увеличение буферной емкости пигмента при дезо-
ксигенации — имеет очень большое значение для тех животных,
у которых буферные свойства крови в основном определяются ды-
хательным пигментом. Уменьшение кислородной емкости в присут-
ствии СО2 может играть важную роль в механизме секреции и на-
копления кислорода в плавательных пузырях рыб. Для некоторых
пигментов крови (например, гемоцианина Limulus) характерно
уменьшение сродства к кислороду при подкислении крови до ве-
личин pH несколько ниже физиологических; значение этого эф-
фекта неизвестно. В стенке тела паразитической нематоды Asca-
ris имеется гемоглобин, хотя РО2 здесь никогда не понижается
настолько, чтобы он отдавал кислород; этот факт тоже не получил
еще объяснения.
Применение физических методов при изучении пигментов кро-
пи позволило узнать много нового об их белковых компонентах.
Гемоглобины позвоночных (за исключением круглоротых) состо-
ят из 4 цепей, обычно из двух цепей одного типа и двух — друго-
Го- Для многих гемоглобинов уже установлена последовательность
аминокислот в отдельных полипептидных цепях. С помощью рент-
пеноструктурного анализа и других физических методов уже уда-
лось определить пространственное расположение всех четырех це-
Пеи и групп гема. Однако современные методы еще не позволяют
74
Глава 8
провести такой же анализ структуры гемоцианинов. Валентность
ионов металла и природа связи металла с лигандами еще не вы-
яснена окончательно. Давно установлено, что медь в гемоцианине
и железо в гемоэритрине изменяют свою валентность при связыва-
нии кислорода; судя по новейшим данным, и в гемоглобине воз-
можен перенос электрона с железа на кислород при оксигенации.
Присоединение кислорода сопровождается также конформацион-
ной перестройкой молекулы пигмента. Некоторые гемоцианины
при оксигенации распадаются на субъединицы, и по крайней мере
один из них снова переходит в полимерную форму при полном на-
сыщении кислородом. Ассоциация мономеров характерна и для
гемоглобина круглоротых. Гемоцианины — очень крупные молеку-
лы, нередко состоящие из большого числа субъединиц, которые
агрегируют в определенном диапазоне pH и концентраций Са2+ и
Mg2+. Гемоцианин улиток состоит из двух цепей, аналогичных а-
и p-цепям гемоглобина.
Влияние pH на сродство гемоглобина к О2 обусловлено осво-
бождением одного протона из каждой цепи при оксигенации. Та-
ким образом, оксигемоглобин является более сильной кислотой, а
дезоксигемоглобин — лучшим буфером.
Еще одно загадочное явление, происходящее при оксигена-
ции,— это кооперативное взаимодействие цепей в гемоглобине
или субъединиц в гемоцианине. Присоединение кислорода проис-
ходит постепенно, и одна молекула НЬ связывает в среднем боль-
ше одной, но меньше четырех молекул О2. Сильное влияние на
степень агрегации молекул гемоцианина и на его сродство к кис-
лороду оказывает pH.
Уже давно известно, что гемоглобин, находящийся внутри эри-
троцитов, обладает меньшим сродством к кислороду, чем тот же
гемоглобин в растворе. Судя по новейшим данным, основную роль
в увеличении Р50 и заметную роль в определении величины эф-
фекта Бора играет присутствие в эритроцитах дифосфоглицерата
(у млекопитающих), инозитгексафосфата (у птиц) и аденозин-
трифосфата (у рыб). Соединение пигментов крови с кислородом
происходит очень быстро — период полупревращения составляет
несколько миллисекунд. Реакция гемоглобина с окисью углерода
протекает медленнее, но сродство НЬ к СО выше, чем к О2. Мно-
гие детали столь быстрых и обратимых изменений в молекуле
пигмента при связывании и отдаче кислорода остаются невыяснен-
ными.
Дыхательные пигменты — хороший материал для изучения на-
следственности и эволюции. Полипептидные цепи гемоглобинов
млекопитающих (а- и p-цепи в НЬА и а- и у-цепи в HbF) обла-
дают видовой специфичностью, но это не исключает получения
гибридных молекул. В гомологичных цепях у разных видов отме-
чаются различия в последовательности аминокислот, и делались
даже попытки судить о филогенетической удаленности тех или
Дыхательные функции крови 75
иных форм по числу различий в первичной структуре этих цепей.
Эти попытки вряд ли можно считать достаточно обоснованны-
ми. Дело в том, что различия в первичной структуре цепей зави-
сят не только от видовой принадлежности: обнаружено много му-
тантных цепей и, кроме того, у большинства позвоночных одно-
временно имеется несколько гемоглобинов. У некоторых
животных, например у большей части млекопитающих, в крови
взрослых особей электрофоретически выявляется один или иногда
два главных типа НЬ, однако у других животных (например, у
большинства рыб) обнаруживается несколько электрофоретически
различимых НЬ; у лосося, например, находят около дюжины анод-
ных и столько же катодных фракций. Соотношение различных
форм гемоглобина в крови меняется в процессе развития организ-
ма, некоторые из них различаются по своему сродству к О2; био-
логическое значение такого полиморфизма еще не выяснено.
Две различные формы гемоцианина обнаружены только у од-
ного животного (улитки Helix). Однако для этого пигмента ха-
рактерен полиморфизм иного рода — различной может быть сте-
пень полимеризации, что не свойственно гемоглобину (за возмож-
ным исключением гемоглобина круглоротых).
Так как последовательность аминокислот в цепях многих ге-
моглобинов известна, можно восстановить и генетические коды
для каждой из них. Гемоглобин синтезируется непрерывно, что
обеспечивает его медленное обновление в организме; однако ско-
рость синтеза заметно возрастает при длительной гипоксии или
анемии. Синтез гемоглобина, по крайней мере у позвоночных, ре-
гулируется гормоном эритропоэтином. Вводя этот гормон в куль-
туру клеток, из которых образуются ретикулоциты (а затем и эри-
троциты), можно изучать влияние его на белковый синтез. Появ-
ление новых форм гемоглобина в процессе развития организма,
вероятно, означает, что на разных этапах онтогенеза активиру-
ются различные гены.
Структура и функция пигментов крови — превосходный при-
мер взаимосвязи молекулярного- и физиологического уровней в
биологии, а также пример влияния внешней среды на функцио-
нальную адаптацию специфических макромолекул.
СПИСОК ЛИТЕРАТУРЫ
1- Addink A. D. F., Dissertation, Utrecht University, 1968, 161 рр. Carbonic an-
hydrase of Sepia.
2- Aggarwal S. J., Riggs A., J. Biol. Chem., 244, 2372—2383, 1969. Hemoglobin
in bullfrog.
3- Air G. M., Thompson E. О. P., J. Biol. Sci., 24, 75—95, 1971. Amino acid se-
quence, myoglobin of red kangaroo.
4. Albers C., In: Fish Physiology, Vol. 4, pp. 173—208, edited by W. S. Hoar
Ч ля? D- J- Randall. Academic Press, New York, 1970. Acid-base balance.
Albers C., Pleschka K„ Respir. Physiol., 2, 261—273, 1967. Buffering bv
elasmobranch blood.
76
Глава 8
6. Anthony F. H., J. Exp. Biol., 38, 109—125, 1961. Survival of goldfish in car-
bon monoxide.
7. Antonini E., In: Oxygen in the Animal Organism, edited by F. Dickens and
E. Neil, Pergamon Press, New York, 1965, pp. 121—137; also Physiol. Rev.,
45, 123—170, 1965. Structure and function of hemoglobin and myoglobin.
8. Antonini E. et al., Arch. Biochem. Biophys., 97, 336—342, 343—350, 1962.
Studies on chlorocruorin.
8a. Antonini E., Rossi-Fanelli A., Caputo A., Arch. Biochem. Biophys., 97, 343—
350, 1962. Size and structure of chlorocruorin.
9. Antonini E., Wyman J., Bellelli L., Rumen N., Siniscalco M„ Arch. Biochem.
Biophys., 105, 404—408, 1964. Oxygen equilibrium of lamprey hemoglobins.
10. Ar A., Scheffer A., Comp. Biochem. Physiol., 33, 481—490, 1970. Hemoglobin
in clam shrimp Cyzicus.
11. Atassi M. Z., J. Theor. Biol., 11, 227—241, 1966. Amino acid composition,
hemoglobin.
12. Bartels H., Lancet, 2, 601—604, 1964. Oxygen transport in mammals.
13. Bartels H., Hiller G., Reinhardt W., Respir. Physiol., 1, 345—356, 1966. Oxy-
gen affinity of chicken blood.
14. Bartels H. et al., Amer. J. Physiol., 205, 331—336, 1963; also H. Bartels,
D. E. Yassin, and W. Reinhardt, Respir. Physiol., 2, 149—162, 1967. O2 and
CO2 transport, adult and fetal mammals.
15. Battaglia F. C., McGaughey H., Makowski E. L., Meschia G., Amer. J. Phy-
siol., 219, 219—221, 1970. Effect of diphosphoglyceric acid on O2 affinity of
red cells.
16. Beadle L. C., J. Exp. Biol., 34, 1—10, 1957. Respiration in oligochaete
Alma.
17. Beard J. M., Thompson E. О. P., Austral. J. Biol. Sci., 24, 765—786, 1971.
Amino acid sequences in kangaroo hemoglobin.
18. Behrman R. E., J. Appl. Physiol., 25, 224—229, 1968. O2 dissociation curve,
rhesus monkeys.
19. Benesch R., Benesch R. E., Yu С. I., Proc. Nat. Acad. Sci., 59, 526—532,
1968; also Benesch R. E., Benesch R., Fed. Proc., 29, 1101—1104, 1970.
Reaction between hemoglobin and diphosphoglycerate and inositol hexaphos-
phate.
20. Black E. C., Kirkpatrick D., Tucker H. H., J. Fish. Res. Bd. Canad., 23, 1—13,
1966. O2 dissociation curves of brook trout.
21. Bowen S. T., Lebbenz H. G., Poon M., Chow V. H. S., Grigliatti T. A., Comp.
Biochem. Physiol., 31, 733—747, 1969. Hemoglobins of Artemia.
22. Boyer S. H. et al., Science, 153, 1539—1543, 1966. Hemoglobins in sheep.
23. Bradshaw R. A., Rogers L. A., Hill R. L., Buettner-Janusch J., Arch. Biochem.
Biophys., 109, 571—578, 1965. Amino acid composition of primate hemoglo-
bins.
24. Braun V., Crichton R. R., Braunitzer G., Z. physiol. Chem., 349, 197—210,
1968. Polymers of hemoglobin in Chironomus.
25. Briehl R. W., J. Biol. Chem., 238, 2361—2366, 1963. O2 equilibrium and aggre-
gation of subunits, lamprey hemoglobin.
26. Brunoni M„ Antonini E., Wyman J., Tentori L., Vivaldi G., Carta S., Comp.
Biochem. Physiol., 24, 519—524, 1968; also Biochim. Biophys. Acta, 133,
177—180, 1967. Equilibrium kinetics of frog hemoglobin.
27. Calder W. A., Schmidt-Nielsen K., Amer. J. Physiol., 213, 883—889, 1967; 215,
477—482, 1968. Acid-base balance in birds.
28. Cameron I. N., Comp. Biochem. Physiol., 32, 175—192, 1970. Hematology,
pinfish Lagodon and striped mullet.
29. Cameron J. N„ Comp. Biochem. Physiol., 38A, 699—704, 1971. O2 equilibrium
curve of rainbow trout.
29a. Chew M. У. et al., Biochem. J., 94, 378—383, 1965. Size of Hb molecule in
polychaete.
30. Chiodi H., Acta Physiol. Lat. Amer., 12, 208—209, 1962. O2 affinity in high
altitude animals.
Дыхательные функции крови
77
31. Chiodi Н., Terman J. W., Amer. J. Physiol., 208, 798—800, 1965. Blood gases
32. 'chumley J. H., Holland R. A. B„ Respir. Physiol., 7, 287—294, 1969. Respira-
tory properties of sheep hemoglobin. __
32a. Clausen G., Ersland A., Resp. Physiol., 5, 221—233, 350—359, 1968; 7, 1 6,
1969. Respiratory properties of blood of hedgehog, beaver, water vole, and
bladdernosed seal. w
33. Cosgrove W. B., Schwartz J. B., Physiol. Zool., 38, 206—212, 1965. Properties
of earthworm hemoglobin.
34. Crompton D. W., Smith M. H., Nature, 197, 118—119, 1963. Flatworm he-
moglobin.
34a. Dan M., Hagiwara A., Exp. Cell. Res., 46, 596—598, 1967. HbA and HbF in
same red cell, human.
35. Davenport H. E., Proc. Roy. Soe. bond. B, 136, 255—270, 271—280, 1949.
Hemoglobins of nematodes.
36. Davenport H. W., ABC of Acid-Base Chemistry. University of Chicago Press,
1969, 86 pp.
37. Dawson T. J., Evans J. V., Austral. J. Biol. Sci., 15, 371—378, 1962. Relation
between К levels and O2 transport, sheep red cells.
38. DePhillips H. A., van Hodde К. E., Biol. Bull., 133, 462, 1967. O2 equilibrium
of hemocyanin.
39 DePhillips H. A., Nickerson K. W., Johnson M., van Holde К. E., Biochemistry,
8, 3665—3672, 1960; also J. Molec. Biol., 50, 471—479, 1970. O2 dissociation
of hemocyanin, subunit structure, Busycon, squid.
40. Di Giamberardino L., Arch. Biochem. Biophys., 118, 273—278, 1967. Dis-
sociation of Eriphia hemocyanin.
41. Eaton J. W., Brewer G. J., Grover R. F., J. Lab. Clin. Med., 73, 603—609,
1969. Diphosphoglycerate in man at altitude.
42. Eddy F. B., J. Exp. Biol., 55, 695—711, 1971. Blood gas data for Salmo
gairdneri.
43. Ellerton H. D., Carpenter D. E., van Holde К. E., Biochemistry, 9, 2225—2232,
1970, Subunits of hemocyanin of Cancer.
44. Ellis R., Giuliani A., Tentori A. L., Chiancone E., Antonini E., Comp. Biochem.
Physiol., 36, 163—171, 1970. Hemoglobin of amphibians.
45. Evans J. V. et al., Nature, 178, 849—850, 1956. Genetics of hemoglobin diffe-
rences in sheep.
46. Faura J. et al., Blood, 33, 668—676, 1969. Effect of altitude on erythropoiesis.
47. Flores G., Frieden E., Science, 159, 101—103, 1967. Bullfrogs lacking hemoglo-
bin.
48. Forster R. E., Steen J. B., J. Physiol., 204, 259—282, 1969. The Root shift in
eel blood.
49. Fox H. M., Proc. Roy. Soc. bond B, 135, 192—212, 1948; 136, 388—399, 1949;
138, 514—528, 1951; 141, 179—189, 1953; also Nature, 166, 609—610, 1950;
also Bull. Soc. Zool. France, 80, 288—298, 1955. Hemoglobin synthesis and
function in Daphnia.
50. Fox H. M., Proc. Roy. Soc. bond. B, 136, 378—388, 1949. Comparison of chlo-
rocruorin and hemoglobin.
51. Fox H. M., Proc. Roy. Soc. Lond. B, 143, 203—214, 214—225, 1955. Effect
of O2 on hemoglobin synthesis and function, invertebrates.
52. Fox H. M., Nature, 168, 112, 1951. Blood pigments of Serpula.
53. Fox H. M., J. Exp. Biol., 21, 161—164, 1945. O2 affinities of invertebrate he-
moglobins.
54. Freeman M. В. M., Misson В. H., Comp. Biochem. Physiol., 33, 763—772, 1970.
Respiratory properties of blood of chickens.
55. Gahlenbeck H., Bartels H., Z. vergl. Physiol., 59, 232—240, 1968. O2 affinity
of blood of bullfrog.
56. Garbett K., Darnall D. W., Klotz I. M., Arch. Biochem. Biophys., 142, 455—
470, 1971. Reactivity of residues in hemerythrin.
78
Глава 8
57. Ghosh J., Comp. Biochem. Physiol., 16, 341—360, 1965. Chemical properties
of avain hemoglobins.
58. Gibson Q. H„ Prog. Biophys., 9, 1—53, 1959. Kinetics of reactions between
Hb’s and gases.
59. Gillen R. G., Riggs A., Comp. Biochem. Physiol., 38B, 585—595, 1971. Effect
of ATP on hemoglobin properties in fish.
59a. Gitlin D. et al., Blood, 32, 796—810, 1968. ImmunofIncrescent detection of
two Hb’s in same cell.
60. Gordon A. S„ Physiol. Rev., 39, 1—40, 1959. Hemopoietine.
51. Gray L. H., Steadman J. M., J. Physiol., 175, 161—171, 1964. Dissociation cur-
ves for mouse and rat blood.
62. Grigg G. C., Comp. Biochem. Physiol., 23, 139—148, 1967. Blood of Antarctic
63. Grover R. F„ Ann. N. Y. Acad. Sci., 121, 662—673, 1965. Pulmonary tension
at altitude.
64. Grover R. F. et al., J. Appl. Physiol., 18, 560—566, 567—574, 575—579, 909—•
912, 1963. Oxygen transport at high altitudes.
64a. Guerritore D. et al., J. Molec. Biol., 13, 234—237, 1965. Electron microscopy
of Spirographis chlorocruorin.
65. Hall F. G., J. Biol. Chem., 115, 485—490, 1946; also J. Mammal., 18, 468—
472, 1937. Blood of animals living at high altitudes.
66. Hall F. G„ Proc. Soc. Exp. Biol. Med., 119, 1071—1073, 1965. O2 transport in
chinchilla.
67. Hall F. G., J. Cell. Biol, 68, 69—74, 1966. Hemoglobin properties in toad.
68. Hamada R. et al., J. Biochem. (Tokyo), 55, 154—162, 1964. Eel hemoglo-
bins.
69. Harkness D. R., Ponce J., Grayson V., Comp. Biochem. Physiol., 28, 129—138,
1969. Phosphoglyceric acid cycle in mammalian red cells.
70. Hemmingsen E. A., Douglas E. L., Grigg G. C., Comp. Biochem. Physiol., 29,
467—470, 1969. O2 transport in hemoglobin-free fish.
71. Hill R. L., Buettner-Janusch J., Buettner-Janusch V., Proc. Nat. Acad. Sci.,
50, 885—893, 1963. Evolution of primate hemoglobins.
72. Hoffman R. J., Mangum С. P., Comp. Biochem. Physiol., 36, 211—228, 1970.
Coelomic hemoglobin in Glycera.
73. van Holde R. E., Cohen L. B., Biochemistry, 3, 1803—4808, 1809—1813, 1965;
6, 93—99, 1967. Ultrastructure of hemocyanins.
74. van Holde R. E., van Bruggen E. F. J., in: Biological Macromolecules, Vol. 5,
edited by G. D. Fasman and S. N. Timasheff. Marcel Dekker, Inc. New York,
1971. The hemocyanins.
75. Holeton G. F., Comp. Biochem. Physiol., 34, 457—471, 1970. Antarctic fish
without hemoglobin.
76. Holeton G. F., J. Exp. Biol., 54, 239—254, 1971. O2 uptake in Salmo exposed
to CO.
77. Holland R. A. B., Forster R. E., J. Gen. Physiol., 49, 140—146, 199—220, 1968.
Kinetics of uptake of O2 and CO by red blood cells.
78. Horne F. R„ Beyenbach R. W., Amer. J. Physiol., 220, 1875—1881, 1971. He-
moglobin in branchiopod crustaceans.
79. Horvath S. M., Chiodi H., Ridgway S. H., Azar S., Comp. Biochem. Physiol.,
24, 1027—1033, 1968. Respiratory properties of hemoglobin of porpoise and
sea lion.
80. Hoshi T., Sci. Rep. Tohoku Univ., 23, 35—58, 1957. Hemoglobin in a daphnid.
81. Hoshi T., Robayashi M., Sugano H., Sci. Rep. Niigata Univ. D, 5, 87—98,
1968; also Hoshi T., Sugano H., Sci. Rep. Niigata Univ. D, 2, 13—26, 1965.
Oxygen dissociation curve of hemoglobin in cladocerans.
82. Hoshi T., Shimada T., Sci. Rep. Niigata Univ. D, 2, 1—12, 1965. Hemoglobin
in freshwater cladocerans.
83. Howell B. J., Fed. Proc., 29, 1130—1134, 1970. Acid-base balance in transition
from water to air breathing.
Дыхательные функции крови 79
\-----—---------------------
84 Howell В. J„ Baumgardner F. W., Bondi К., Rahn Н., Amer. J. Physiol., 218,
600—606, 1970. Acid-base balance, cold-blooded vertebrates.
85. Huber R., Formanek H., Epp 0., Naturwissenschaften, 55, 75—77, 1968. Cry-
stal structure analysis of erythrocruorin.
86 Huehns E. R., Dance N., Beaver G. H., Keil J. V., Hecht F., Motulsky A. G„
Nature, 201, 1095—1097, 1964. Human embryonic hemoglobins.
87. Huisman T. H. I., Kitchens J., Amer. J. Physiol., 215, 140—146, 1968. Fun-
ctional properties of Hb A, В and C in sheep.
88. Huisman T. H. J., Van Veen M. S., Biochim. Biophys. Acta, 88, 352—366,
367—374, 1964. Studies in animal hemoglobins.
89. Hunter J. A., Paul J., J. Embryol. Exp. Morph., 21, 361—368, 1969. Adult and
fetal hemoglobin, rat.
90. Ingram V. M., Biochim. Biophys. Acta, 28, 539—545, 1958; 36, 402—411, 1959.
Normal and sickle cell hemoglobins, man.
91. Ingram V. M., The Hemoglobins in Genetics and Evolution. Columbia Uni-
versity Press, New York, 1963, 165 pp.
92. Itazawa У., Bull. Jap. Soc. Sci. Fish., 36, 571—577, 1970. Respiratory chara-
cteristics of fish.
93. Jacobsen L. 0., Krantz S. B., Ann. N. Y. Acad. Sci., 149, 578—583, 1968. Sum-
mary on erythropoietin.
94. Johansen K., Lenfant C., Amer. J. Physiol., 210, 910—918, 1966. Gas exchange
in Octopus.
95. Johansen K., Lenfant C., J. Exp. Biol., 46, 205—218, 1967. Respiratory function
in lungfish, Lepidosiren.
96. Johansen K-, Mecklenburg T. A., Z. vergl . Physiol., 70, 1—19, 1970. O2
transport in Cancer.
97. Johansen K., Lenfant C., Grigg G. C., Comp. Biochem. Physiol., 18, 597—608,
1966. Respiratory properties of blood of platypus.
98. Johansen K., Martin A. W., J. Exp. Biol., 45, 165—172, 1966. O2 transport by
blood of giant earthworm.
99. Jones J. D., J. Exp. Biol., 32, 110—125, 1955. Hemoglobin function in po-
lychaetes Nephthys and Arenicola.
100. Jones J. D., Comp. Biochem. Physiol., 4, 1—29, 1961; 12, 283—295, 1964.
Function of hemoglobin in aquatic snails.
101. Jonxis J. H. P. In: Haemoglobin, edited by F. J. W. Roughton and J. C. Kend-
rew. Interscience Publishers, New York, 1949, pp. 261—267. Properties of fe-
tal hemoglobin.
102. Keilin D., Wang Y. L., Biochem. J., 40, 855—867, 1946. Hemoglobin of Gastro-
philus.
103. Kendrew J. C. et al., Nature, 174, 946—949, 1954; 181, 662—666, 1958; 185,
422—427, 1960. Structure of myoglobin.
104. Kendrew J. C., Science, 139, 1259—1266, 1963. Structure of myoglobin.
105. Kleihauer E., Stoffler G., Molec. Gen. Genet., 101, 59—69, 1968. Embryonic
hemoglobins.
106. Klotz 1. M. et al., Arch. Biochem. Biophys., 68, 284—299, 1957. Active sites in
hemerythrin.
107. Konings W7. N., van Driel R., van Bruggen E. F. J., Gruber M., Biochim. Bio-
phys. Acta, 194, 55—66, 1969. Properties of Helix hemocyanin.
108. Kunkel H. G. et al., J. Clin. Invest., 36, 1615—1625, 1967. Types of human
hemoglobin, especially A2.
109. Kunzer W„ Nature, 179, 477—478, 1957. Human embryo hemoglobin.
HO. Larimer J. L., Riggs A. F., Comp. Biochem. Physiol., 13, 35—46, 1964. Pro-
perties of crayfish hemocyanin.
Hl. Lenfant C. et al., J. Clin. Invest., 47, 2652—2656, 1968. Effect of altitude on
O2 binding by hemoglobin in man.
112. Lenfant C., Aucutt C., Respir. Physiol., 6, 284—291, 1969. Respiratory pro-
perties of blood of monkeys.
113. Lenfant C„ Elsner R., Kooyman G. L., Drabek С. M., Amer. J. Physiol., 216,
1595—1597, 1969. Respiratory properties of blood of seal.
80
Глава 8
114. Lenfant С., Johansen К., Amer. J. Physiol., 209, 991—998, 1965. Oxygen
transport by blood of Octopus.
115. Lenfant C., Johansen K., Respir. Physiol., 1, 13—29, 1966. Respiratory fun-
ction of blood in dogfish, Squalus.
116. Lenfant C., Johansen K., Grigg G. C., Respir. Physiol., 2, 1—21, 1966. Respi-
ratory function of blood of lungfish.
117. Lenfant C., Johansen K., Respir. Physiol., 2, 247—260, 1967. Respiratory
adaptations in amphibians.
118. Lenfant C., Johansen K., Torrance J. D., Respir. Physiol., 9, 277—286, 1970.
Respiratory properties of Hb in sea mammals.
119. Lenfant C., Kenney D. W., Aucutt C„ Amer. J. Physiol., 215, 1506—1511, 1968.
Properties of hemoglobin in whale, Orcinus.
120. Lenfant C. et al., Amer. J. Physiol., 216, 1598—1600, 1969. Respiratory pro-
perties of blood of penguin.
121. Maniatis G. M., Ingram V. M., J. Cell. Biol., 49, 390—404, 1971. Erythro-
poiesis in amphibians.
122. Manwell C., Science, 126, 1175—1176, 1957; also Physiol., Zool., 31, 39—100,
1958. Larval and adult hemoglobins in ovoviviparous dogfish and oviparous
teleost.
123. Manwell C., Biol. Bull., 115, 227—238, 1958. Hemoglobin of hagfish.
124. Manwell C., J. Cell. Comp. Physiol., 53, 75—84, 1959. Absence of Bohr effect
in Cucumaria hemoglobin.
125. Manwell C., Ann. Rev. Physiol., 22, 191—244, 1960. Respiratory pigments.
126. Manwell C., Comp. Biochem. Physiol., 8, 209—218, 1963. Hemoglobin in mari-
ne clams.
127. Manwell C. In: Biology of Myxine, edited by A. Brodal and R. Fange, Oslo
University Press (Universitetsforlaget), Oslo, 1963, pp. 372—455. Blood pro-
teins of cyclostomes.
128. Manwell C., In: Oxygen in the Animal Organism, edited by F. Dickens and
E. Neil, Pergamon Press, New York, 1965, pp. 49—116. Review, comparative
properties of blood pigments.
129. Manwell C., Science, 152, 1393—1395, 1966. Hemoglobins of holothurians.
130. Manwell C., Baker С. M. A., Proc. Nat. Acad. Sci., 49, 496—504, 1963. Survey
of hemoglobins in wild birds.
131. Manwell C„ Baker С. M. A., Childers W., Comp. Biochem. Physiol., 10, 103—
120, 1963. Genetics of hemoglobins in hybrid fish.
132. Maren T. H., Fed. Proc., 26, 1097—1103, 1967. Carbonic anhydrase in the
animal kingdom.
133. Meschia G. et al., Quart. J. Exp. Physiol., 46, 156—160, 1961. Properties of
fetal and adult hemoglobin in sheep at altitude.
134. Metcalfe J., Bartels H., Moll W., Physiol. Rev., 47, 782—838, 1967. Gas ex-
change across placenta.
134a. Miller P. L., J. Exp. Biol., 44, 529—543, 1966. Function of hemoglobin in wa-
ter bug Anisops.
135. Moll W., Pfliiger W., Arch., 299, 247—251, 1968. Facilitated diffusion of oxy-
gen by myoglobin.
135a. Muirhead H. et al., J. Molec. Biol., 28, 117—156, 1957. Structure of hemo-
globin.
136. Needham A. E., Nature, 221, 572, 1969. Absorption spectra of leech hemo-
globin.
137. Novy M. J., Parer J. T., Respir. Physiol., 6, 144—150, 1969. Oxygen affinity
in blood of fetal cat.
138. Novy M. J., Parer J. T., Behrman R. E., J. Appl. Physiol., 26, 339—345, 1969.
O2 dissociation curves, adult and fetal macaque.
138a. Okazaki T. et al., Biochim. Biophys. Acta, 111, 496—502, 503—511, 1965; 140,
258—265, 1967. Properties of hemoglobin from Ascaris perienteric fluid and
body wall.
139. Padlan E. A., Love W. E., Nature, 220, 376—378, 1968. Structure of hemoglo-
bin of Glycera.
Дыхательные функции крови____________81
140. Padmanabhanaidu В., Comp. Biochem. Physiol., 17, 167—181, 1966. Properties
of hemocyanin in scorpion.
141. Parer J. T., Respir. Physiol., 2, 168—172, 1967. O2 dissociation curve of rhesus
monkey. „
142. Parer J. T., Hoversland A. S., Metcalfe J., Respir. Physiol., 10, 30—37, 1970.
Respiratory properties of blood lion and tiger.
143. Paul W., Roughton F. J. W., J. Physiol., 113, 25—35, 1951. O2 equilibrium cur-
ve of sheep hemoglobin.
144. Perutz M. F., Proc. Roy. Soc. bond. B, 173, 113—140, 1969. The hemoglobin
molecule.
145 Perutz M. F., Nature, 228, 726—734, 1970. Stereochemistry of hemoglobin.
146. Perutz M. F. et al., Nature, 219, 29—32, 131—139, 1968; also Perutz M. F.,
Lehmann H., Nature, 219, 902—909, 1968. Crystal analysis and molecular mo-
del of hemoglobin.
147. Perutz M. F. et al., Nature, 222, 1240—1243, 1969. Function of specific resi-
dues in hemoglobin.
148. Pickett S. M., Riggs A. F., Larimer J. L., Science, 151, 1005—1007, 1966. Su-
bunit structure of lobster hemocyanin.
149. Piiper J., Schumann D., Respr. Physiol., 2, 129—134, 1967. O2 exchange in
gills of dogfish.
150. Piiper J., Schumann D., Respir. Physiol., 5, 317—325, 326—337, 1968. Respi-
ratory functions of blood of dogfish.
151. Poluhowich J. L., Comp. Biochem. Physiol., 36, 817—821, 1970. Respiratory
pigment in a nemertean.
152. Pough F. H., Comp. Biochem. Physiol., 31, 885—901, 1969. O2 and CO2 trans-
port by lizard blood.
153. Rahn FL, Yokoyama T., eds., Physiology of Breath-holding; the Ama of Japan.
Publ. 1341, Nat. Acad. Sci., Washington, 1967, 369 pp.
154. Ralph R., Everson I. Comp. Biochem. Physiol., 27, 299—307, 1968. Metabolism
of Hb-less Antarctic fish Chaemocephalus.
155. Read R. R. FL, Biol. Bull., 122, 605—717, 1962; Comp. Biochem. Physiol., 15,
137—138, 1963. Hemoglobin in bivalve mollusc.
156. Redmond J. R., Physiol. Zook, 35, 304—313, 1962. Respiratory function in
Chiton.
157. Redmond J. R., Biol. Bull., 122, 252—262, 1962. O2 transport in crab Cardi-
soma.
158. Redmond J. R., Science, 139, 1294—1295, 1963. Molluscan hemocyanin.
159. Redmond J. R„ Helg. wiss. Meeresunters., 9, 303—311, 1964. O2 transport by
blood of various invertebrates.
160. Redmond J. R., Amer. Zool., 8, 471—479, 1968. Transport of O2 in crab Car-
cinus.
161. Reynafarje C. et al., Proc. Soc. Exp. Biol. Med., 116, 649—650, 1964. Humoral
control of erythropoietic activity in man at altitude.
162. Reynafarje C., Morrison P. R„ J. Biol. Chem., 237, 2861—2864, 1962. Myoglo-
bin in rodents at high altitudes.
163. Riegel R., Bartels FL, Rleihauer E., Lang E. M., Metcalfe J., Respir. Physiol.,
1, 138—144, 145—150, 1966. Respiratory properties of mammalian blood.
164. Riegel R., Bartels FL, Yassin D. E., Oufi J., Rleihauer E., Parer J. T., Metcal-
fe J., Respir. Physiol., 2, 173—181, 182—195, 1967. Respiratory functions in
mammalian blood.
165. Riggs A., Canad. J. Biochem., 42, 763—775, 1964. Structure and function of
hemoglobins.
166. Riggs A., Physiol. Rev., 45, 619—673, 1965. Functional properties of hemoglo-
bins.
167. Riggs A., J. Gen. Physiol., 43, 737—752, 1960. Bohr effect, mammalian hemo-
globins.
168. Riggs A., J. Gen. Physiol., 35, 23—40, 1951. Frog and tadpole hemoglo-
bin.
6—1514
82
Глава 8
169. Riggs A., Proc. Nat. Acad. Sci., 68, 2062—2065, 1971. Enhancement of Bohr
effect in mammalian hemoglobins by diphosphoglycerate.
170. Riggs A., Biology of Lampreys, Academic Press, New York, 1971. Lamprey
hemoglobins.
171. Riggs A., Hemer A. E., Proc. Nat. Acad. Sci., 48, 1664—1670, 1962. Hybridi-
zation of hemoglobins.
172. Riggs A., Rona M., Biochim. Biophys. Acta, 175, 248—259, 1969. Aggregation
of polymerizing mouse hemoglobins.
173. Roche J., Fontaine M., Ann. Inst. Oceanogr. Monaco, 20, 77—87, 1940. Amino
acid composition of various blood pigments.
173a. Roche J. et al., Biochim. Biophys. Acta, 41, 182—194, 1960. Electron micro-
graphs of Hb of annelids.
174. Rogers W. P., Austral. J. Sci. Res. B. Biol. Sci., 2, 287—303, 399—407, 1949.
Hemoglobin function in nematode parasites.
175. Rosenberg M., Proc. Nat. Acad. Sci., 67, 32—36, 1970. Electrophoretic analy-
sis of hemoglobin.
176. Rosenmann M., Morrison P. R., J. Biol. Chem., 240, 3353—3356, 1965. Myo-
globin in snowshoe hare.
177. Rosse W. F., Woldmann T., Hull E., Blood, 22, 66—72, 1963; also Rosse W. F.,
Waldmann T., Blood, 27, 654—661, 1966. Stimulation of erythropoiesis in
frogs and birds.
178. Rossi-Fanelli A., Antonini E., Nature, 186, 895—896, I960; also Rossi-Fanel-
li A., Antonini E., Giuffre R., Nature, 186, 896—897, 1960. Crystalline hemo-
globin from tuna fish.
178a. Rossi-Fanelli M. R. et al., Arch. Biochem. Biophys., 141, 278—283, 1970. Size
and structure of earthworm Hb.
179. Roughton F. J. W., In: Handbook of Physiology, Vol. 1, Sec. 3, pp. 767—825,
edited by W. F. Hamilton. Amer. Physiol. Soc., Washington, D.C., 1964.
Transport of O2 and CO2 (review).
179a. Roxby R., van Holde K., 1972. Molecular size, Hey of Busycon (личное сооб-
щение) .
180. Rumen N. M., Love W. E., Arch. Biochem. Biophys., 103, 24—35, 1963. He-
moglobins of sea lamprey.
181. Saha A., Ghosh J., Comp. Biochem. Physiol., 15, 217—235, 1965. Properties
of avian hemoglobins.
182. Sarcini M., Orlando M., Tentori L., Comp. Biochem. Physiol., 34, 751—753,
1970. Polymorphism of hemoglobin of salamander.
183. Schruefer J. J. P. et al., Nature, 196, 550—552, 1962. Dissociation curves of
hemoglobin and whole blood.
184. Schweer M., Z. vergl. Physiol., 42, 20—42, 1959. Metabolism of leeches with
and without hemoglobin.
185. Seal U. S., Comp. Biochem. Physiol., 31, 799—811, 1969. Hemoglobins of car-
nivores.
185a. Serfaty A. et al., Rev. Canad. Biol., 24. 1—5, 1965. Cardiac reaction to emer-
gence in carp.
186. Severinghaus J. W., J. Appl. Physiol., 21, 1108—1116, 1966. Blood gas calcu-
lator.
187. Simons J. A., J. Exp. Zool., 162, 219—230, 1966. Embryonic development of
hemoglobin in chicken.
188. Siri W. E. et al., J. Appl. Physiol., 21, 73—80, 1966. Erythropoietic responses
of man to hypoxia.
189. Smith M. H., Biochim. Biophys. Acta, 71, 370—376, 1976, 1963. Combination
of Ascaris hemoglobin with O2 and CO.
190. Smith M. H., George P., Preer J. R., Arch. Biochem. Biophys., 99, 313—318,
1962. Hemoglobin from paramecium.
191. Smith M. H., Lee D. L., Proc. Roy. Soc. Lond. B, 157, 234—257, 1963; also
Smith M. H., Morrison M., Biochem. Biophys. Acta, 71, 364—370, 1963. Isola-
tion and metabolism of hemoglobin from Ascaris.
Дыхательные функции крови 83
192. Sullivan В., Riggs A., Comp. Biochem. Physiol., 23, 437—447, 449—458, 459—
474, 1967. Structure, function, and evolution of turtle hemoglobin.
193. Svedberg T., Pederson К. O., The Ultracentrifuge. Oxford University Press,
New York, 1940; Johnson Reprint Corp., 1959, 478 pp.
194. Taketa F., Morell S. A., Biochem. Biophys. Res. Comm., 24, 705—713, 1966.
Oxygen affinity of cat hemoglobin.
194a. Terwilliger R. C., Read K. R., Comp. Biochem. Physiol., 29, 551—560, 1969.
Myoglobin in radular muscle, mollusc.
195. Tomita S., Riggs A., J. Biol. Chem., 246, 547—554, 1971. Effects of body size
on Bohr effect.
196. Tucker V. A., Comp. Biochem. Physiol., 24, 307—310, 1968. O2 dissociation
curve in Echidna.
197. Tyuma I., Shimizu K., Fed. Proc., 29, 1112—1114, 1970. Effect of organic
phosphates on O2 affinity of hemoglobin.
198. Ulrich S., Hilpert P„ Bartels H., Pflug. Arch., 277, 150—165, 1963. Respiratory
properties of rodents.
199. Vanstone W. E. et al., Canad. J. Physiol. Pharmacol., 42, 697—703, 1964. Mul-
tiple hemoglobins in coho salmon.
200. Vaughan В. E., Pace N., Amer. J. Physiol., 185, 549—556, 1956. Myoglobin
changes at high altitudes.
201. Wald G., Riggs A., J. Gen. Physiol., 35, 45—53, 1951. Hemogibbin of (lamprey,
Petromyzon. \
201a. Waxman L., J. Biol. Chem., 246, 7318—7327, 1971. Properties of Arenico-
la Hb.
202. Weber R. E., Konink. Nederl. Akad. van Wetenschappen, 66, 284—295, 1963.
Hemoglobin in larva of Chironomus.
203. Weber R. E., Comp. Biochem. Physiol., 35, 179—189, 1970. Properties of an-
nelid hemoglobins.
204. Weisberger A. S., Proc. Soc. Exp. Biol. Med., 117, 276—280, 1964. Sickling
phenomenon of deer hemoglobin.
205. Whittenberg B. A., Okuzuki T., Whittenberg J. B., Biochem. Biophys. Acta,
111, 485—495, 1965. Hemoglobin of Ascaris.
206. Whittenberg J. B. et al., Proc. Nat. Acad. Sci., 67, 1846—1853, 1970. State of
iron in oxyhemoglobin.
207. Wilkins N. P., Iles T. D., Comp. Biochem. Physiol., 17, 1141—1158, 1966.
Hemoglobin polymorphism in fishes.
208. Wolvekamp H. P., Z. vergl. Physiol., 25, 541—547, 1938. Oxygen transport
and hemocyanin in Octopus.
209. Wolvekamp H. P. et al., Arch. Need. Physiol., 25, 265—267, 1941; 28, 620—
629, 1947. Respiratory function in blood of Arenicola, Helix and Homarus.
210. Wood S. C., Moberly W. R„ Respir. Physiol., 10, 20—29, 1970. Respiratory
properties of blood of Iguana.
211. Wyman J., Adv. Protein Chem., 4, 407—531, 1948. Heme proteins.
212. Yoshioka M. et al., J. Biochem. (Tokyo), 63, 70—76, 1968. Eel hemoglobins.
213. Zanjani E. D., Yu M.-L., Perlmutter A., Gordon A. S„ Blood, 33, 573—581,
1969. Hormonal control of erythropoiesis in fish.
214. Zuckerkandl E., Jones R. T., Pauling L., Proc. Nat. Acad. Sci., 46, 1349—1360,
1960. Evolution of hemoglobins.
6*
Глава 9
Температура
Л. ПРОССЕР
Введение
Экологические и физические соображения
Температура ограничивает распространение животных и опре-
деляет уровень их активности. Диапазон температур на Земле
значительно шире тех границ, в которых возможна активная
жизнь. Температура поверхностных слоев воды в открытом океане
варьирует в пределах от —2 до +30 °C, а в воздухе — от —70 до
+85 °C. Как общее правило, процессы жизнедеятельности возмож-
ны лишь при температурах от 0 до +40 °C; для большинства жи-
вотных температурные границы намного уже. Некоторые организ-
мы выживают в неактивном состоянии и при температурах ниже
0 °C, а иные даже выдерживают замораживание. Ряд животных
обитает в горячих источниках, а отдельные виды бактерий и во-
дорослей живут и размножаются в источниках с температурой во-
ды до 70 °C. Размножение животных возможно в более узком диа-
пазоне температур, чем выживание взрослых особей, хотя для
эмбрионов многих гомойотермных видов пределы переносимых
температур шире, чем для взрослого организма. Температура, как
мера скорости движения молекул, определяет скорость химиче-
ских реакций и является одним из факторов, ограничивающих
рост и метаболизм. Научных работ о температуре очень много,
поскольку ее так легко измерять — ртутным термометром, терми-
стором или термопарой. Теплота — одна из форм энергии, и тем-
пература характеризует содержание тепловой энергии в данном
теле.
У многих животных температура тела изменяется вслед за
температурой внешней среды — это так называемые «холоднокров-
ные», или пойкилотермные, животные. «Пойкилотермия» означает
изменчивость, лабильность температуры. У пойкилотермных жи-
вотных температура тела не обязательно равна температуре окру-
жающей среды. Например, у рыбы весом 40 г измерение мышеч-
ной температуры дало величину на 0,44 °C выше внешней темпе-
ратуры [445].
Относительно меньшая часть животных регулирует температу-
ру своего тела — это так называемые «теплокровные», или гомойо-
термные, животные. Промежуточное место между* пойкилотерм-
ными и гомойотермными формами занимают гетеротермные жи-
Т емпература
85
вотные, способные к частичной регуляции температуры тела. Эта
регуляция ограничивается либо определенными условиями среды,
либо какими-то областями тела.
Температура любой метаболизирующей клетки обязательно
должна быть выше температуры окружающей^ среды, так как при
окислительных процессах и гликолизе происходит выделение теп-
ла. Температура тела животного зависит от соотношения ряда
факторов, влияющих на тепловой баланс организма противопо-
ложным образом. Источником тепла может быть метаболический
термогенез (эндотермия) или же внешняя среда, главным образом
солнечная энергия (эктотермия). Теплоотдача происходит путем
излучения, конвекции, теплопроводности и испарения воды. Поте-
ре тепла способствует циркуляция жидкостей от внутренних ча-
стей организма к поверхности тела, а препятствует-теплоизоляция.
Пойкилотермные животные не совсем лишены способности к тер-
морегуляции. Средствами такой регуляции могут служить пове-
денческие реакции, впадение в спячку, метаболические и нервные
компенсаторные механизмы. Относительное постоянство темпера-
туры тела у гомойотермных животных поддерживается теплопро-
дукцией, а также с помощью защитного поведения и изменений
теплоизоляции, кровообращения и других факторов, изменяющих
перенос тепла. У некоторых «теплокровных» животных периоды ле-
таргии сопровождаются снижением температуры тела, а у других
во время зимней спячки физиологический термостат переключает-
ся на более низкую температуру. Сенсорные механизмы сообща-
ют об изменениях температуры, вызывая соответствующие ком-
пенсаторные и защитные реакции.
Термические свойства воды как главного компонента тела во
многом определяют тепловой баланс организма. Теплопроводность
воды [0,0014 кал/(см-с-град)] ниже, чем, например, у металлов,
но выше, чем у многих жидкостей (у этилового спирта 0,00042, у
оливкового масла 0,000395). Кроме того, вода имеет высокую
удельную теплоемкость— 1,0 кал/(г-град), тогда как, скажем, для
меди соответствующая цифра будет 0,09, а для этилового спир-
та — 0,535. Удельная теплоемкость большинства животных тканей
(за исключением компактного вещества кости) составляет от 0,7
до 0,9 кал/(г-град). Есть еще одна характеристика — коэффици-
ент температуропроводности, который равен теплопроводности, де-
ленной на произведение удельного веса и удельной теплоемкости.
Низкая температуропроводность приводит к медленному прогрева-
нию или охлаждению тканей и к ограниченному распространению
тепла внутри организма; хорошим теплоизолятором оказывается
жир, хотя он и уступает в этом отношении воздуху. Животные с
большой тканевой массой медленно согреваются и медленно ох-
лаждаются; внутреннюю передачу тепла у них обеспечивают глав-
ным образом циркулирующие жидкости тела, так что вялость цир-
куляции обусловливает медленное распространение тепла. Рыба
86
Глава 9
весом 1 г охлаждается со скоростью 1,8 град/мин на разность
температур 1 °C, тогда как у рыбы весом 100 г эта величина со-
ставляет всего 0,4 град/мин [445].
Благодаря высокой теплоемкости и низкой теплопроводности
воды температура естественных водоемов (за исключением горя-
чих источников) обычно не превышает максимальной температу-
ры, при которой может существовать большинство водных живот-
ных. Точка замерзания тканей у водных животных (кроме неко-
торых, в частности морских костистых рыб и ракообразного
артемии) обычно близка к точке замерзания окружающей среды
или ниже этой точки (гл. 1). Поскольку удельный вес льда мень-
ше, чем у воды, водные животные, находящиеся в воде подо
льдом, не замерзают. Наземные животные подвержены действию
гораздо больших колебаний внешней температуры, чем водные,
и температура их тела тесно связана с водным балансом организ-
ма.
Вода имеет высокую теплоту плавления (79,7 кал/г); водные
растворы, особенно в капиллярных пространствах, могут переох-
лаждаться на несколько градусов, а связанная вода устойчива к
замерзанию. Некоторые животные, переохлажденные или частич-
но обезвоженные (а также те, у которых часть воды замещена по-
лигидроксильными органическими растворителями), могут выдер-
живать температуру намного ниже точки замерзания воды, и при
этом их ткани не замерзают. Испарение воды заметно охлаждает
любую влажную поверхность (на испарение 1 г воды при 20 °C за-
трачивается 585 кал), и при высоких температурах воздуха оно
предотвращает перегревание тела у животных.
Температура и скорость химических реакций. Энергия активации
Кинетическая активность, или частота соударения молекул,
пропорциональна абсолютной температуре и возрастает приблизи-
тельно на 3% на каждые 10 °C. Один из методов количественной
оценки влияния температуры на скорости реакций, состоит в оп-
ределении Qio. Qio — коэффициент увеличения скорости реакции
при повышении температуры на 10°С — вычисляется по формуле
10
__/ К1 \ *1—*2
V1° \ К2 /
где Ki и К2— константы скорости реакции при температурах и
t2. Обычно вместо констант скоростей в формулу подставляют ско-
рости Vi и У2. В большинстве случаев при определении темпера-
турной зависимости мы имеем дело не с линейными, а с логариф-
мическими функциями. Величина Qi0 зависит от температурного
диапазона — при низких температурах она больше, чем при высо-
ких. Поэтому необходимо указывать область температур, для ко-
Температура
87
---
торой вычислен Qi0. Физические свойства растворов меньше изме-
няются с температурой, чем скорость катализируемых реакций.
Величина Qio, равная 2,5, соответствует увеличению скорости ре-
акции на 9,6% на один градус; для многих химических реакций
коэффициент Qio близок к этой величине.
Критической энергии активации той или иной реакции соответ-
ствует величина р (иногда обозначаемая как £*) в уравнении
Аррениуса
_1п (\ — _Е_ (—_______L\
Ч Ki / Я \ 7\ /’
—2,37? (log К2 —log КО
Tz Л
где Ki и К2 — константы скорости реакции (пропорциональные из-
меренным скоростям) при абсолютных температурах 7\ и Т2, а
К — газовая постоянная (1,98 кал/моль). Для большинства биоло-
гических и всех простых химических реакций зависимость лога-
рифма скорости реакции от величины, обратной абсолютной тем-
пературе, выражается прямой линией, наклон которой равен
—р/2,ЗД, или —р/4,6. Из этого видно, что при повышении темпера-
туры на 10 °C число молекул, энергия которых превышает крити-
ческую (например, 12 ккал), может удвоиться, хотя прирост ки-
нетической энергии (пропорциональный абсолютной температуре)
будет гораздо меньшим. Специфические катализаторы характери-
зуются определенными величинами р, не зависящими от концент-
рации субстрата, если последняя достаточно высока и скорость ре-
акции лимитирует сам катализатор. Однако при более низких,
более физиологических концентрациях субстрата величины темпе-
ратурных характеристик понижаются. Обычно предполагают, что
для сложной биохимической реакции в целом значение р будет
таким же, как для этапа, лимитирующего ее скорость, но это, ко-
нечно, крайнее упрощение.
В биологическом диапазоне температур величины Qio для мно-
гих метаболических реакций лежат в пределах от 2 до 2,5. В то
же время некоторые сложные изменения скоростей физиологиче-
ских процессов, например циркадные ритмы, относительно неза-
висимы от температуры, а величина Qio для потребления кислоро-
да у некоторых пойкилотермных животных в определенном диапа-
зоне температур лежит между 1 и 2. Сравнивая потребление
кислорода животным, находящимся в активном состоянии и в по-
кое, можно определить характерные для этих состояний уровни
обмена при различных температурных условиях. У моллюска Саг-
dium величины Qi0 для активного состояния и покоя равны соот-
ветственно 1,84 и 1,20; вообще при низких нормальных температу-
рах окружающей воды для многих беспозвоночных в состоянии
покоя характерны невысокие величины Qi0 [344, 345]. У насеко-
88
Глава 9
мых (например, у Calliphora) Qio для мышц и для организма в
целом составляет около 2 [463]. Температурные характеристики
(ц или Qio) ферментативных реакций могут понижаться по мере
снижения концентрации субстрата до нормальных внутриклеточ-
ных уровней, при которых лимитирующим фактором становится
уже концентрация субстрата, а не активность фермента. Если ки-
нетика катализируемой реакции соответствует уравнению Миха-
элиса—Ментен, мерой сродства фермента к субстрату будет Км
(см. гл. 6) или концентрация субстрата, при которой фермент на-
сыщен наполовину (Ks). С понижением температуры у большинст-
ва ферментов кажущееся сродство возрастает (т. е. Лм уменьша-
ется). Это противодействует снижению энергии активации и может
уменьшить кажущуюся величину Qio [206, 436]. В случае слож-
ных реакций — с параллельными и последовательными этапами
и с противоположными влияниями на Км и на Qio для актива-
ции— попытки элементарного анализа температурных эффектов
безнадежны.
Замораживание и холодоустойчивость
Протоплазма, будучи водным раствором, замерзает при темпе-
ратуре на несколько градусов ниже нуля. Медленное заморажива-
ние часто сильнее повреждает организм, чем быстрое. Если ох-
лаждение происходит быстро (около 100 град/с) на протяжении
всего температурного диапазона замерзания, кристаллы льда
обычно не образуются и организм просто затвердевает. Если по-
местить мелких нематод, простейших, мышечные волокна или тка-
невые культуры прямо в жидкий воздух (—197°C), они застыва-
ют, а при быстром отогревании оживают.
Одной из адаптаций, создающих устойчивость к замерзанию,
может быть повышение концентрации растворенных веществ, при-
водящее к понижению точки замерзания. В природе этот меха-
низм действительно имеет некоторое распространение. В крови
холодноводных морских рыб несколько повышена концентрация
солей, а иногда также содержатся значительные концентрации
специфических неэлектролитов (антифризов). Зимующие насеко-
мые и некоторые растения накапливают органические вещества,
повышающие осмотическую концентрацию жидкостей организма.
Более важную роль в механизмах зимовки и адаптации к по-
лярным широтам играет переохлаждение. Если охлаждать воду
или водный раствор, температура падает ниже точки замерзания
еще до появления центров кристаллизации; далее при кристалли-
зации воды и высвобождении теплоты плавления температура по-
вышается. После замерзания возможно дальнейшее охлаждение
льда. При кристаллизации происходит образование водородных
связей между молекулами воды; в случае переохлаждения этот
процесс замедляют различные неполярные органические вещест-»
Температура
89
ва, растворенные в воде. Накопление такого рода защитных ве-
ществ обычно сопровождается умеренной дегидратацией [11].
У многих животных и растений внеклеточная жидкость замер-
зает раньше, чем клетки. Клеточные мембраны каким-то неизвест-
ным образом защищают протоплазму от замораживания. При
медленном замерзании солевого раствора вода кристаллизуется,
а соль накапливается между кристаллами, повышая осмотическую
концентрацию оставшегося раствора. Если это происходит во вне-
клеточной жидкости, то более концентрированный раствор отсасы-
вает воду из клеток и они могут быть чрезмерно обезвожены. Ча-
стичное замещение свободной воды неполярными органическими
молекулами предохраняет от замерзания, изменяя структуру кри-
сталлической решетки и распределение водородных связей (см.
обзор Смита [424]). Скорость роста кристаллов льда обратно
пропорциональна концентрации групп, образующих водородные
связи, в защитных веществах [ПО]. Как показали опыты на аме-
бах, растительных клетках и яйцах морского ежа, замерзание вну-
триклеточной жидкости может приводить к механическому разру-
шению структур протоплазмы [10]. Гидратационная вода или во-
да, связанная с заряженными растворенными веществами,
особенно с белками, замерзает в последнюю очередь, и ее замер-
зание обычно приводит к смерти. Коллаген может связывать боль-
шое количество воды, и с помощью ядерного магнитного резонан-
са было показано, что она, по-видимому, сохраняет жидкое состо-
яние при температурах до —50 °C [102].
Температуры ниже 0°С не только вызывают осмотическую де-
гидратацию, механические разрушения и изменения в распреде-
лении связанной воды, но и резко снижают активность фермен-
тов. Кислород и СО2 диффундируют во льду в 104 раз медленнее,
чем в жидкой воде. При температурах, близких к точке замерза-
ния, может также происходить денатурация некоторых фермент-
ных белков.
Многие клетки выдерживают кратковременное замерзание, но
не могут переносить длительного замораживания, особенно при
очень низких температурах. В портняжной мышце лягушки лед
ооразуется при —0,42 °C, но в переохлажденном состоянии ее
можно сохранять без повреждения в течение двух суток при —4°.
1 пЛо оттаивания этой мышцы, замороженной на неделю при
0,9 °C, на 20 ч при —2 °C или на полчаса при —3,5 °C, ее разд-
ражимость восстанавливается. Основной причиной повреждения
здесь может быть удаление воды из молекулярных структур. Ин-
фузория Tetrahymena после охлаждения в 10%-ном диметилсуль-
со скоростью 4,5 град/мин выжила при температуре
У5 С более 128 дней. У инфузории Paramecium после экстре-
мального замораживания восстанавливалась подвижность, но не
пособность к клеточному делению [483].
90
Глава 9
Морские беспозвоночные, особенно моллюски литоральной
зоны и усоногие раки, часто замерзают зимой. У таких животных
при —15 °C замерзает только 55—65% воды, содержащейся в ор-
ганизме. При дальнейшем понижении температуры процент замер-
зающей воды растет, однако при данной температуре процент
воды, замерзшей в организме, ниже процента замерзшей морской
воды [250] (рис. 9-1). Обитатели литорали Balanus, Littorina и
Mytilus зимой выносят замораживание при температурах от —10
до —20 °C, но летом при таком охлаждении погибают [438—439].
Mytilus повреждается при —10 °C, когда замерзает 64% внутри-
клеточной воды, но животное, предварительно акклимированное к
150%-ной морской воде, выдерживало —15°C и замерзание 80%
воды в организме. При выдерживании животных в 150 %-ной мор-
ской воде количество небелкового азота в жидкостях возрастает
непропорционально их осмотической концентрации [491].
Переохлаждение играет важную роль у холодостойких насеко-
мых. Некоторые насекомые, например пилильщик Cephas, вы-
держивают переохлаждение при температурах от —23 до —30°C,
тогда как другие, менее холодостойкие виды переносят лишь от
—5 до —8 °C [396]. При более быстром снижении температуры те-
ла может быть достигнуто большее переохлаждение [397, 399].
У наездника Вгасоп, паразитирующего на пилильщиках, наблю-
дали переохлаждение до —47,2 °C и рост концентрации глицерина
в гемолимфе до 2,7 М. При исследовании 10 видов зимующих на-
секомых у одного вида был найден высокий уровень сорбита, у
двух других накапливался, вероятно, маннит, а у остальных —
глицерин [438] (рис. 9-2). Увеличение концентрации глицерина
(до 16%) коррелирует со способностью к переохлаждению; с ве-
сенним прекращением диапаузы концентрация глицерина падает.
У гусениц мучной моли Anagasta при холодовой акклимации воз-
растает содержание в организме различных нингидринположи-
тельных веществ, а также глюкозы и глицина; это в большей сте-
пени повышает способность к переохлаждению, чем снижает точку
замерзания [437]:
Температура Время Предел пере- Точка замерзания
акклимации, СС акклимации охлаждения, СС гемолимфы, °C
—6 7 дней —21 — 1,1
+6 7 дней —18,3 —1,0
Яйца кольчатого шелкопряда Malacosoma зимой накапливают
глицерин, концентрация которого в январе достигает 35,2% су-
хого веса, и выдерживают переохлаждение до —40,8 °C, а летом
только до —29,5 °C [155] (рис. 9-3). У аляскинской жужелицы Pte-
rostichus обнаружены следующие сезонные изменения [326]:
Способность Точка замерзания Содержание Содержание
к переохлаждению (оттаивания) глицерина воды
Зима до —35 °C —3,5 °C 25% 54%
Лето до —6,6 °C —0,7 °C 0 65%
100
Рис. 9-1. Доля воды, замерзающей в морской воде и в тканях Mytilus при различ-
ных температурах ниже нуля [250].
Содержание глицерина, %
Рис. 9-2. Зависимость возможного переохлаждения от содержания глицерина в
рганизме у различных насекомых зимой (Somme L., Canad. J. Zool., 42, 87—101,
1964).
92
Глава 9
Эритроциты или сперматозоиды можно много месяцев сохра-
нять без повреждения в замороженном состоянии, если предвари-
тельно обработать их глицерином, пропиленгликолем или диме-
тилсульфоксидом, быстро заморозить и хранить при температуре
ниже —40 °C. Другие клетки, например в культурах тканей мле-
копитающих, лучше выживают при быстром (100 град/мин), чем
при медленном (1 град/мин) охлаждении; их можно защитить по-
ливинилпирролидоном или сахарозой. Хорошее защитное действие
при замораживании костного мозга оказывают аминокислоты, а
Рис. 9-3. Изменения содержания глицерина и точки переохлаждения (т.п.) у ли-
чинок насекомых, находящихся зимой на открытом воздухе (в Канаде)
(Semme L., Canad. J. Zool., 43, 765—770, 1965).
декстраны задерживают замерзание внеклеточной жидкости в тка-
нях [251]. Производились успешные пересадки роговицы после ее
хранения в замороженном состоянии. Симпатические ганглии, об-
работанные глицерином, обнаруживали электрическую активность
после замораживания до —79 °C и последующего оттаивания
[424]. Представители двух родов нематод при достаточно быстром
охлаждении и отогреве переносили замораживание до температур
не ниже —30 °C [343]. Полагают, что при быстром охлаждении
большее количество внутриклеточной воды оказывается в пере-
охлажденном состоянии; при медленном же охлаждении вода вы-
ходит из клеток, что приводит к их повреждению. Гибель клеток
при медленном замораживании, по-видимому, в большей мере
обусловлена повреждением мембран, чем инактивацией фермен-
тов [309—312].
Поразительное повышение холодостойкости в зимнее время
обнаруживают деревянистые растения: зимой данное растение пе-
реносит гораздо более низкие температуры, чем летом, В начале
Температура
93
зимы метаболически активные слои коры синтезируют большие
количества РНК и белков [419]. Выявлены различные уровни ак-
климатизации; первый из них связан с многообразными метаболи-
ческими изменениями, происходящими под влиянием гормонов, а
на следующем этапе возможна полимеризация макромолекул с
уменьшением количества связанной воды [485]. Одна из гипотез
придает большое значение увеличению числа внутримолекулярных
дисульфидных связей при замораживании, так как агенты, тор-
мозящие образование этих связей, оказывают защитное действие
о сн3 о
н || I н ||
N С CH N С
СН3
Рис. 9-4. Повторяющаяся структурная единица гликопротеида антарктической ры-
бы Trematomus (De Vries A. L., J. biol. Chem., 246, 305—309, 1971).
Этот гликопротеид снижает точку замерзания больше, чем можно было бы ожидать исходя
из его концентрации.
'[9]. Другая теория особо выделяет роль конформационных изме-
нений, делаюш‘их макромолекулы устойчивыми к обезвоживанию
Значительная устойчивость к замораживанию обнаружена у
арктических и антарктических рыб. У двух видов Trematomus, жи-
вущих подо льдом в Антарктике, точка замерзания крови варьи-
рует в пределах от —1,87 до —2,07 °C [107]. При этом NaCl, мо-
чевина и аминокислоты составляют лишь половину осмотически
активных растворенных веществ. Кровь содержит много гликопро-
теидов, которые при электрофорезе разделяются на пять полос,
соответствующих мол. весам от 10 500 до 78 000. Концентрация
этих веществ в плазме достигает 1870 мг%, и они обладают по-
разительными противозамораживающими свойствами, которые ис-
чезают при ацетилировании или при расщеплении пептидных свя-
еи [106, 269]. Гликопротеиды содержат 16% треонина, 26% ала-
ина, 29% N-ацетилгалактозамина и 28% галактозы; строение
соответствующего мономера представлено на рис. 9-4. Эти веще-
ва снижают точку замерзания больше, чем можно было бы ожи-
94
Глава 9
дать исходя из их концентрации; каким образом достигается та-
кой эффект, не установлено.
У рыб, живущих в глубоких фиордах Лабрадора с темпера-
турой воды —1,7°C зимой и летом, температура замерзания рав-
на —0,9 °C. Эти рыбы замерзают сразу, если в их теле появляют-
ся кристаллы льда. У рыб, живущих в поверхностных водах с лет-
ней температурой -J-5 и зимней —1,5 °C, точка замерзания крови
зимой —0,8, а летом —1,6 °C. Глубоководные же рыбы все время
находятся в переохлажденном состоянии [417]. Камбала Pseudo-
pleuronectes, выдерживаемая как при —1,0, так и при 4-15 °C,
имеет Ат. з. крови —0,71 °C; в то же время эта камбала благодаря
переохлаждению выживает при —1,8 °C [355, 469].
У карпозубой рыбы Pundulus в Новой Англии описаны следу-
ющие сезонные изменения растворенных веществ в крови [469]:
Лето Зима
Глюкоза, мг на 100 мл 68 348
Холестерин, мг на 100 мл 201 307
Аминокислоты, мг N 11,9 13
Белок, г на 100 мл 4 2,2
Натрий, мМ 183 205
Хлорид, мМ. 145 170
Осмоляльность, моем 394 401
Подъем уровня глюкозы в крови и снижение содержания гликоге-
на в печени, наблюдаемые у этих рыб при холодовой акклимации,
зимой выражены сильнее, чем летом (летняя температура воды
была 4~20, а зимняя —1,5°C). При этом точка замерзания зимой
снижается лишь ненамного больше, чем летом, тогда как устойчи-
вость к переохлаждению очень сильно возрастает. Как предпола-
гают, образованию водородных связей при замерзании воды здесь
препятствуют молекулы, богатые оксигруппами [469].
Хомяков наркотизировали с помощью СОг, перфузировали не-
большим количеством глицерина и охлаждали до ректальной тем-
пературы 15—20 °C, после чего погружали в ледяную воду. Дыха-
ние у них прекращалось при температуре от 2 до 6 °C, а сердеч-
ная деятельность — при 0,8—2,5 °C [424]. Далее животных замо-
раживали в пропиленгликоле при температуре от —3 до —14 °C.
Пробыв более 50 мин при температуре —5 °C, они становились
совершенно твердыми. Но если затем их постепенно отогревали и
делали им искусственное дыхание, то большинство животных ожи-
вало и возвращалось к норме. После замораживания, продол-
жавшегося 1 ч, оживали все животные, у которых замерзло 15%
воды организма, и одна треть животных, у которых замерзло 45%
воды [8, 424].
Подводя итоги, можно сказать, что замерзание тканей ведет
к их повреждению в тех случаях, когда во время образования ле-
дяных кристаллов происходит концентрирование внеклеточной
жидкости, что ведет к обезвоживанию клеток, в которых тоже по-
Температура
95
являются кристаллы льда. Некоторые ткани устойчивы к замора-
живанию, особенно если оно совершается быстро. У некоторых
животных небольшое повышение устойчивости к замерзание в
зимнее время обусловлено повышением осмотической концентра-
ции. Чаще, однако, происходит накопление органических веществ,
что делает возможным более сильное переохлаждение.
Летальные температуры; устойчивость и адаптация
Причины смерти при действии высоких или низких температур
как следует не изучены и, без сомнения, многообразны. Живот-
ные, не приспособленные к зимовке и не переносящие заморажи-
вания, гибнут при температурах значительно выше точки замер-
зания. Снижение температуры тела как у гомойотермных, так и
у пойкилотермных животных ведет к замедлению ритма сердца
и дыхания, в результате чего может наступить гипоксия. Очень
часто клеточные мембраны становятся проницаемыми и не могут
более поддерживать ионные градиенты, а ионные насосы переста-
ют работать. Выработка энергии может стать недостаточной для
осуществления жизненных функций. Зависимость Км от темпера-
туры может быть неодинаковой у разных компонентов сопряжен-
ных ферментативных реакций. Исчезает интеграция, осуществляе-
мая центральной нервной системой, и многие животные при тем-
пературах, значительно превышающих летальные, впадают в со-
стояние холодовой комы. Кривые, отображающие зависимость
смертности от времени воздействия различных низких температур,
показывают, что причины гибели могут быть различными и каж-
дая из них действует в определенном диапазоне продолжительно-
сти п интенсивности охлаждения.
При перегреве смерть также может быть вызвана многими при-
чинами— в зависимости от того или иного сочетания продолжи-
тельности и интенсивности теплового воздействия. При повышении
температуры тела уменьшается перенос кислорода пигментами
крови, а у наземных животных возросшее испарение воды может
привести к обезвоживанию организма. С повышением температу-
ры изменяется состояние липидов и возрастает проницаемость кле-
точных мембран; ферменты становятся более активными, но выше
температурного оптимума инактивация начинает преобладать над
активацией. При еще более высоких, нефизиологических темпера-
турах происходит денатурация белков и из поврежденных клеток
могут высвобождаться токсичные вещества. При очень высоких
температурах становится нестабильной двойная спираль ДНК.
Механизмы гибели оказываются неодинаковыми при изучении
их на уровне организма, ткани и ферментного аппарата. Темпера-
туры, вызывающие при 15-минутной экспозиции тепловую смерть,
составляют для интактных головастиков Rana pipiens 37,5 °C, це-
лых лягушек 38,6, икроножной мышцы 40,2, сердца in situ 42 и
96
Глава 9
седалищного нерва 43 °C [351]. Как общее правило, диапазон пе-
реносимых температур наиболее узок для целых животных, не-
сколько шире для тканей и клеток и значительно шире для изоли-
рованных ферментов. Однако оптимальный температурный диапа-
зон данного фермента, определяемый точкой его максимального
сродства к субстрату, может быть очень узким [433]. Пределы пе-
реносимости высоких и низких температур могут изменяться в
результате акклимации, и в то же время они могут быть ге-
нетически связаны с экологией данных животных. Иногда точки
холодовой и тепловой смерти смещаются при акклимации в одном
и том же направлении, а в других случаях изменяются независи-
мо друг от друга; механизмы этих сдвигов могут быть различ-
ными.
Измерение верхней и нижней летальных температур, проводи-
мое после акклимации к различным температурам без рассмотре-
ния причин гибели, позволяет определить характерную для данно-
го вида «зону толерантности». Можно экспериментально акклими-
ровать к различным температурам большое число особей
какого-либо вида водных животных. Далее группы животных,
акклимированных к различным температурам, помещают в серию
ванн с высокими температурами воды для определения точки теп-
ловой смерти, а другие группы — в ванны с низкими температура-
ми воды для определения точки холодовой смерти. Затем строят
графики, отражающие зависимость смертности (в пробитах) от
времени воздействия данной температуры (рис. 9-5) и зависимость
времени, необходимого для гибели 50% животных данной группы,
от температуры (рис. 9-6) [46]. С увеличением времени экспози-
ции температура, переносимая половиной животных (LD50), сни-
жается, пока не дойдет до уровня, когда даже при длительной
(около 48 ч) экспозиции животные больше не погибают. Этот уро-
вень называют пороговой летальной температурой (верхней); точ-
но так же определяют и соответствующую нижнюю границу. За-
тем вычерчивают две линии, отображающие зависимость пороговых
температур (верхней и нижней) от температуры акклимации (рис.
9-7); эти линии образуют замкнутый «полигон толерантности».
Скорость акклимации у разных видов различна. Тепловая аккли-
мация обычно происходит быстрее, в течение нескольких дней,
тогда как холодовая занимает 10—20 дней. У нерки верхняя ле-
тальная температура изменяется приблизительно на 1° при изме-
нении температуры акклимации на 4° [46]. У серебряного карася
верхняя летальная температура повышается на 1° при повышении
температуры акклимации на каждые 3° вплоть до 36,5 °C; при
этом верхняя летальная температура достигает 41 °C — макси-
мальной возможной величины. Нижняя летальная температура
понижается на 2° при понижении температуры акклимации на
каждые 3° вплоть до 17 °C, когда нижняя летальная температура
доходит до возможного предела (0°С) [131]. У дождевого червя
50 100
500 1000 5000
Время наблюдения, мин
io
Рис. 9-5. Зависимость смертности (в пробитах) у гольяна от продолжительности
воздействия верхних летальных температур (Tyler А. V., Canad. J. Zool., 44,
349—361, 1966).
Изменения наклона графиков указывают иа наличие разных причин гибели. Температуры
(°C) указаны цифрами около кривых.
^ис. 9-6. Среднее время переносимости различных температур рыбами, акклими-
рованными к 20 °C (по Бретту, 1956 [46]).
*—1514
98
Глава 9
Pheretima с повышением температуры акклимации на Г граница
теплоустойчивости повышается на 0,3°. Для средиземноморского
переднежаберного моллюска Littorina neritoid.es верхняя леталь-
ная температура составляет 46—47 °C, тогда как для L. littorina
из Новой Англии она равна всего 40—41 °C [126].
Полигон толерантности имеет довольно четкую видовую спе-
цифику (рис. 9-7); внутри него находятся меньшие многоугольни-
Рис. 9-7. Полигон толерантности для нерки и зависимость предпочитаемых тем
ператур от температуры акклимации [47].
ки — для зародышевого развития и для локомоции; эти функции
возможны в более узком диапазоне температур, чем простое вы-
живание. Было проведено сравнение полигонов толерантности у
14 видов пресноводных рыб, обитающих на различных широтах в
восточной части Северной Америки. Только у трех видов (в том
числе Notropus cornutus) были обнаружены расовые особенности
популяций из Онтарио и из Теннесси [164]. У гольца (Salvelinus
fontinalis) две географические расы имеют разные верхние ле-
тальные температуры. У двух рас Daphnia atkinsoni (термальной
и приспособленной к умеренным температурам) верхние леталь-
ные температуры различаются на 6°.
У бактерий известны теплолюбивые и холодолюбивые штаммы.
В горячих источниках Йеллоустона бактерии можно найти в почти
кипящей воде — при 90 °C, простейшие могут существовать при
45—51 °C, а сине-зеленые водоросли выдерживают до 73 °C, хотя
быстрее всего растут при 55 °C [49]. Для разных бактерий может
быть характерна различная температура плавления нитей ДНК
Температура
99
[2551. Белок флагеллин, выделенный из термофильных бактерий,
содержит больше аминокислот, легко образующих стабильные во-
дородные связи; это указывает на возможную большую стабиль-
ность белка [267]. Величины Qi0 для роста у термофилов ниже,
чем v Е. coli; иными словами, термофильные штаммы менее чувст-
вительны к изменениям температуры [49]. Ферменты этих бакте-
рий (например, фосфофруктокиназа) стабильны вплоть до 95 °C,
хотя их Км зависит от температуры [500]. Известны многие при-
меры генетически обусловленной термостабильности белков [473].
Термостабильность коллагена, измеряемая температурой перехо-
да («точкой плавления», рис. 9-8), пропорциональна содержанию
пролина вместе с оксипролином (пирролидином) [243] (рис. 9-9):
Число остатков Коллаген Ascaris Коллаген Lumbricus
Пролин 291 .7,7
Оксипролин 19 165
Полуцистин 16 0
Температура перехода 52 °C 22 °C
В коллагене тропических рыб этих аминокислот больше, чем
в коллагене холодповодных рыб [144]. Наиболее термостабильны
триплеты аминокислот, соседствующие с пирролидином [243].
Высокая температура может также блокировать определенные пу-
ти биосинтеза: например, некоторые бактерии при 37°C нуждают-
ся в аминокислотах и витаминах, без которых могут обходиться
при 26 °C [279]. Исследование аспартат-глутаматтрансаминазы у
11 родственных видов двустворчатых моллюсков показало, что у
сублиторальных форм она более термостабильна, чем у обитате-
лей верхней литорали [379, 380]; приводим данные об относитель-
ной инактивации этого фермента при 2-минутном нагревании до
56 °C [378].
Инактивация Средняя летальная
трзнсаминазы, % температура, °C
Modiolus (сублитораль) 80—100 25
Mytilus (литораль) 40—50 28
Brachidontes (литораль) -• 0—10 38
Теплоустойчивость мышц, определяемая по их реакции на
электрическое раздражение после нагревания, у самых разнооб-
разных водных животных тесно коррелирует с термостабильностыо
многих мышечных ферментов — альдолазы, холинэстеразы, сукци-
натдегидрогеназы, а также АТФазной активности миозина [473].
При сравнении глубоководных рыб и рыб мелководья обнаружи-
вается параллелизм таких показателей, как термостабильность
мышц, устойчивость к тепловому наркозу и температура, вызы-
47л1Щ'а? 50%-ную инактивацию мышечной холинэстеразы [275,
4'4]. у ряда рептилий термостабильность АТФазной активности
миозина обнаруживает хорошую корреляцию с температурой мак-
симального мышечного напряжения и с предпочитаемой животным
температурой [295].
7*
Верхняя, граница обычных температур среды, °C
Рис. 9-8. Соотношение между точками плавления молекулярного коллагена и
приблизительными верхними границами температуры среды у разных живот-
ных [383].
1 — аскарида и свинья; 2 — крыса и корова; 3 — улитка Helix; 4 — кожа тунца; 5 — кожа
трески; 6 — антарктические рыбы.
о 100 200
Оксипролин
Оксипролин*
пролин
Количество остатков
(на 1000 всех остатков)
Рис. 9-9. Зависимость точек плавления коллагена у разных видов от числа остат-
ков оксипролина, пролина и суммы оксипролин + пролин на каждую тысячу ами-
нокислотных остатков коллагена [243].
Т емпература
101
у некоторых амфибий обнаружены наследственные различия
в нижней летальной температуре; животные, обитающие в Цен-
тральной Америке на больших высотах, сходны с формами из уме-
ренной зоны, а обитатели низких участков — с тропическими ви-
дами [44]. Среди австралийских Anura широко распространен-
ные виды обнаруживают лучшую приспособляемость к температу-
ре, чем виды с узким ареалом [45].
’Температурные границы развития часто хорошо коррелируют
с температурными границами выживания, что можно видеть на
примере двух видов жаб [44]:
Границы развития, °C Границы выживания, °C
Bufo cognatus (Аризона) 19,1—32,0 10—31
Bufo leutkeni (Гватемала) 32,1—34,8 22—39
Rana pipiens из северных штатов США развивается при более
низких температурах по сравнению с южными популяциями (хотя
возможно, что это разные виды) [395]. У клопа Oncopeltus ниж-
няя температурная граница для выживания яиц 5 °C, для эмбрио-
нального развития 14 °C и для формирования жизнеспособных ли-
чинок 18 °C [382]. Верхняя граница теплоустойчивости у Balanus
balanoides из Северной Каролины на 5° выше, чем у того же вида
на побережье Англии; у тепловодных и холодноводных усоногих
сходным образом различаются как верхние, так и нижние темпе-
ратурные границы биения усиков. Проводились эксперименты по
отбору и поддержанию термофильных штаммов бактерий, жгути-
ковых и других организмов. Холодолюбивые нематоды растут при
18°C и погибают при 24,5°C; однако путем постепенного повыше-
ния температуры среды и отбора в течение 100 поколений удалось
получить формы, хорошо растущие при 24,5 °C [53].
Летальная температура зависит не только от температуры ак-
климации и генетических факторов, но также от возраста, разме-
ров тела, гормональных влияний, питания и таких внешних фак-
торов, как длительность фотопериода, аэрация и соленость. У ома-
ров верхняя летальная температура понижается при уменьшении
солености и при низких уровнях Ог [316]. Устойчивость Mytilus
к высоким температурам также понижается при уменьшении со-
лености [409]. Получено много данных о влиянии липидов пищи
на температурную устойчивость пойкилотермных животных. При
кормлении серебряных карасей свиным салом их теплоустойчи-
вость повышается больше, чем при кормлении рыбьим жиром.
Теплоустойчивость личинок мясной мухи возрастает при кормле-
нии насыщенными жирами [210]. У Hemigrapsus oregonensis чув-
ствительность к температуре и к солености зимой и летом неоди-
накова; высокие внешние температуры повышают теплоустойчи-
вость, а низкая соленость понижает ее [461]:
102
Глава 9
Соленость, % от солености MopcKoii воды Летальная температура, °C
после акклима- ции к 20 °C после акклима- ции к 5 °C
35 33,3 30,2
75 34,3 32,6
У речного рака Astacus тканевое дыхание продолжается в течение
часа при высоких температурах, летальных для животного в це-
лом; эти температуры приводят, однако, к понижению уровня Na
и повышению уровня К в крови, и если перфузировать нормальное
животное солевым раствором, в котором содержание Na и К со-
ответствует таковому у перегретых особей, то это ведет к выклю-
чению нервно-мышечных функций. Следовательно, смертность
здесь может быть связана в большей степени с ионными, чем с
ферментативными сдвигами. При температурах от 15 до 20 °C
эвригалинная рыба Cyprinodon растет лучше всего в пресной воде,
а при температурах от 25 до 35 °C — в воде с соленостью 35—55%о
[261, 262]. Высокое парциальное давление кислорода (более
1 атм) способствует выживанию серебряного карася при высоких
температурах [484].
Температуры, при которых наступает тепловая или холодовая
кома, хорошо коррелируют с соответствующими летальными тем-
пературами и изменяются при акклимации. Кома — показатель
обратимого угнетения функций центральной нервной системы.
У гуппи (Lebistes) температура тепловой комы повышается на
0,4° с каждым повышением температуры акклимации на 3°, а тем-
пература холодовой комы при снижении температуры акклимации
на 3° понижается на Г [465].
При изучении ряда моллюсков литоральной зоны обнаружены
видовые различия в температуре тепловой комы, соответствующие
их экологии, а также значительные эффекты акклимации [147].
Вид Местообитание Темпера- тура аккли- мации, °C Средняя (температу- ра тепловой комы, °C
Nerita peleronta Nerita versicolor Nerita tesselata Littorina littorea Температуры тепловой температурами, при кот Верхняя литораль, Би- мини Средняя литораль, Би- мини Ниже среднего уровня воды, Бимини Литораль, Вудс Хоул комы у Littorina npi орых наступает тепло! 15 37 25 37 25 37 5 15 25 шерно зая блок 37 45,8 43 44,6 41 44,2 32,5 35,0 38,8 совпадают с ада спонтан-
Температура
103
ной активности в центральной нервной системе. У моллюсков,
обитающих в жарких пустынях Израиля, средняя летальная тем-
пература при двухчасовой экспозиции равна 55 °C [415а].
Температура, необходимая для возникновения холодовой комы
при 30-минутной экспозиции, коррелирует у насекомых с Qio Для
АТФазной активности миозина [339]:
Температура комы, *С <210
Musca 6,1 3,9
Periplaneta 7,9 2,7
Tenebrio 7,4—10,8 2,2
Galleria 8,8 2,0
У таракана Blattella температура холодовой комы может изме-
няться при акклимации следующим образом [84]:
Температура акклимации, “С Температура комы, *С
35 7
15 —4,4
10 —4,3
Если подвергнуть лосося Salmo действию циклически изменяющей-
ся температуры (с периодом 24 или 48 ч), то температура тепло-
вой комы после этого будет почти такой же, как и после акклима-
ции к постоянной температуре, соответствующей верхней точке
цикла [180]. То же самое для температуры холодовой комы уста-
новлено в опытах на равноногих рачках (но определяющей здесь
оказывается нижняя точка цикла) [116].
Высшие нервные центры функционируют в более узких темпе-
ратурных пределах, чем периферические нервы и мышцы. На рис.
9-10 представлены данные, полученные при изучении холодовой
блокады на пресноводных рыбах [371]. У ската различные реф-
лексы при повышении температуры исчезают в определенной по-
следовательности, причем уже после исчезновения спинальных
рефлексов можно активировать мышцы, раздражая двигательные
нервы [32]. В опытах на изолированной портняжной мышце лягуш-
ки показано, что температура холодовой блокады снижается при
акклимации к холоду [236], однако при этом повышается темпе-
ратура тепловой блокады, что можно рассматривать как неадап-
тивное изменение [365].
Таким образом, гибель от перегревания или охлаждения мо-
жет быть обусловлена различными механизмами, которые еще
плохо изучены. У сложных организмов большое значение могут
иметь вторичные эффекты, например гипоксия. Вообще в гибели
клеток важную роль могут играть такие факторы, как повышение
мембранной проницаемости и блокада ионных насосов, нарушение
равновесия сопряженных ферментативных реакций и кинетиче-
ские ограничения. Выживание при крайних температурах стано-
вится возможным благодаря множеству генетических адаптаций,
апример изменению первичной структуры определенных белков.
104
Глава 9
Как правило, нервная система, по-видимому, более чувствительна
и функционирует в более узком диапазоне температур, чем дру-
гие ткани. Летальные температуры могут изменяться при аккли-
мации; другие внешние факторы также могут влиять на послед-
ствия экстремального перегрева или охлаждения.
Простой рефлекс
J--------1------о-------1---------------1-----,
5 10 15 20 25 30 35
Рис. 9-10. Температура холодовой блокады (*) нервного проведения, спинномоз-
говых рефлексов и условных рефлексов у рыб, акклимированных к различным
температурам [3711.
А — сомик, Б и В — серебряный карась. Цифрами около стрелок обозначено число опытов.
Скорость биологических процессов
Температурные адаптации. У пойкилотермных животных ак-
климация может влиять не только на летальные температуры,
но и на скорости различных биологических процессов, поведение
и биохимическую активность. По временным масштабам следует
различать три группы явлений: 1) изменения температуры, про-
должительность которых измеряется часами, непосредственно
влияют на скорость биологических процессов, причем характер
Температура
105
наблюдаемых реакций определяется внутренними свойствами дан*
ной системы; 2) температурное воздействие, продолжающееся в
течение многих дней или недель, приводит к компенсаторной ак-
климации; 3) на протяжении многих поколений идет отбор гене*
тических вариантов, приспособленных к существующему темпера*
турному режиму. Естественный отбор может использовать те осо*
бенности прямых реакций, которые сводят к минимуму изменение
скорости биологических процессов; он может влиять также на
способность к компенсаторной акклимации.
У большинства пойкилотермных животных резкое повышение
или понижение температуры приводит к избыточной первичной ре-
акции (первичному перерегулированию) или даже к шоковой ре*
акции; этот эффект продолжается секунды или минуты. У Artemia
потребление О2 чрезмерно повышается под действием тепла и
чрезмерно понижается при охлаждении; у серебряного карася из*
быточный подъем интенсивности обмена наблюдается при быстром
изменении температуры в том и в другом направлении. Амплитуда
этого первичного эффекта зависит от величины температурного
сдвига. Эта первичная метаболическая реакция может быть ча*
стично обусловлена повышением двигательной активности в ре-
зультате сенсорной стимуляции, но возможно, что здесь участвует
и какой-то общий клеточный эффект, поскольку первичное пере-
регулирование наблюдается и у дрожжей.
Затем интенсивность обмена стабилизируется в соответствии с
температурой и может оставаться на этом новом уровне многие
часы; именно этот уровень обычно используют при определении
Qio. Стабильный уровень зависит от состояния акклимации, и если
в этот период животное снова окажется в прежних температурных
условиях, то восстановится и первоначальная скорость изучаемых
процессов. Для периода прямой реакции на температурное воздей-
ствие характерно повышение или понижение выработки энергии
и концентраций промежуточных продуктов метаболизма; кроме
того, возможны перераспределения ионов, в особенности натрия и
калия, между клетками и внеклеточной жидкостью и изменения
pH. Может усиливаться или ослабевать спонтанная активность в
центральной нервной системе.
У ряда морских беспозвоночных основной обмен на всем диа-
пазоне «обычных» температур относительно стабилен, в то время
как для активного обмена характерен высокий Qio [344, 345]
(рис. 9-11). Потребление кислорода и частота сокращений сердца
У рептилии Thamnophis из Манитобы (но не из Флоридцу) , умень-
шаются по мере снижения температуры с 30 до 20 °C, затем снова
возрастают, достигая максимума между 15 и 20 °C, и резко пада-
ют при температурах ниже 15 °C [4].
Величину Qlo для ферментативных реакций обычно определяют
ври насыщающих концентрациях субстратов, т. е. при максималь-
ных скоростях реакций. Однако in vivo концентрации субстратов
Рис. 9-11. Минимальное потребление кислорода при различных температурах
(по Ньюэллу и Норткрафту [344]).
А — Actinia, Б — ракообразное Nephros, В — брюхоногий моллюск Littorina, Г — двуствор-
чатый моллюск Cardium,
Рис. 9-12. Влияние концентрации сукцината на Qio для сукцинатдегидрогеназы
мышц из надосевой мышцы серебряного карася, акклимированного к 5 и к 25 °C
[172].
Температура
107
редко бывают насыщающими, так что измерение в условиях полу-
насыщения дает более физиологические результаты. Величина Qio
снижается при уменьшении концентрации субстрата; поэтому мно-
гие данные, полученные при измерении Qio in vitro, практически
не имеют физиологического смысла. При физиологических концен-
трациях субстрата величина Qw может приближаться к 1 (рис.
9-12).
Основные типы акклимации. Если животное в течение мно-
гих дней содержать при измененной температуре, то у него про-
изойдет компенсаторное изменение скорости
биологических процессов, т. е. акклимация.
В случае возвращения в первоначальные
температурные условия скорость этих про-
цессов теперь уже не вернется просто к пер-
воначальному уровню, а будет выше или
ниже в соответствии с направлением аккли-
мации. Прехт [363] предложил удобную
Рис. 9-13. Типы температурной акклимации по
Прехту.
Т2 — исходная температура, Т\ и Те — более низкая и бо-
лее высокая температуры соответственно. Объяснение см.
в тексте.
классификацию типов акклимации (рис. 9-13). Т% означает здесь
промежуточную температуру, из которой животное переносят в ус-
ловия более низкой (Л) или более высокой (Т3) температуры. Пря-
мой реакцией на эту новую температуру будет соответственно по-
нижение или повышение скорости биологических процессов, кото-
рое в дальнейшем может оказаться стабильным (тип 4 по Прехту).
Если же в ходе акклимации эта скорость вернется к исходному уров-
ню, то можно констатировать полную компенсацию (тип 2 по
Прехту). Частичную (неполную) акклимацию относят к типу 3, пе-
ререгулирование (избыточную компенсацию) — к типу 1, а обрат-
НУЮ> или парадоксальную, акклимацию —к типу 5.
Можно получить более полную картину акклимац , если из-
мерять стабилизированные скорости биологических процессов не
только в двух крайних температурных точках, а на всем диапазо-
не температур (рис. 9-14). При полном отсутствии акклимации
ривые, отражающие зависимость скорости от температуры, сов-
*да-т Для всех температур, при которых содержались животные
I )• Часто наблюдается сдвиг кривой без изменения ее наклона,
ак что при всех температурах скорость биологических процессов
ри холодовой акклимации выше (II, А) или ниже (II, Б), чем при
пловой. Возможен также поворот кривой температурной зави-
Температура.
Рис. 9-14. Типы кривых, отображающих зависимость интенсивности биологиче-
ских процессов от температуры у животных, акклимированных к двум различным
температурам.
Сплошные кривые — холодовая акклимации, пунктирные кривые — тепловая акклимации.
Т емпература
109
симости около некоторой средней точки — изменение ее наклона,
т е. Qio (Я^)« Обычно акклимации приводит к некоторому сочета-
нию сдвига и поворота кривой. Если Qio при холодовой акклимации
ниже, чем при тепловой, изменение носит компенсаторный харак-
тер (/Е,Д), тогда как обратная ситуация (Qio при холодовой ак-
клима’цйи выше, чем при тепловой) соответствует некомпенсатор-
ному эффекту (IV, Б). Если же кривые пересекаются при низкой
температуре (V), то высокий Qio при холодовой акклимации
(V,B) будет компенсаторным, а низкий (V,Г)—некомпенса-
торным. :
При' одном из обычных типов акклимации сдвиг кривой соче-
тается с ее поворотом по часовой стрелке (рис. 9-14, IV, А), так
что экстраполяция двух кривых температурной зависимости При-
водит К их пересечению выше нормального диапазона температур
и Qio При холодовой акклимации уменьшается. Примерами такого
эффекта может служить изменение просасывания воды у Мидии
или частоты сокращений сердца у тритона.
Сдвиг кривой без изменения ее наклона (рис. 9-14, II, А) об-
наружен у многих животных — жуков, карпа, Planaria, Pachygrap-
sus и-в жабрах мидии — при изучении потребления кислорода.
Примером почти полной компенсации служит акклимация Chiro-
nomus к 20 и 30°C: при обеих температурах потребление кислоро-
да одинаково [359].
В опытах на язе (Idas} обнаружено, что в случае суточных
температурных циклов акклимационная кривая занимает проме-
жуточное положение между кривыми для непрерывной акклима-
ции к каждой из двух крайних температур цикла [36].
Классификация акклиманионных реакций по Прехту позволяет
установить, существует ли для даной функции компенсация тем-
пературных эффектов. Описанные выше типы акклимации по
Проссеру отражают еще один параметр — Q10. Обе эти классифи-
кации применяли к интактным животным, тканям и ферментам от
различным образом акклимированных особей, а также при
сравнении популяций или видов, различающихся по географиче-
ской Широте обитания, и при анализе сезонных различий. Ни одна
из этих классификаций не позволяет делать обобщения относитель-
но механизмов акклимации. Однако выделение типов акклимации в
сочетании с другими подходами помогает понять долговременные
реакции животных на температуру среды.
Модифицирующие факторы при акклимации. Говоря о типе
акклимации, следует прежде всего учитывать, в каком диапазоне
температур проводились измерения. По последним данным, ком-
пенсаторная акклимация возможна лишь в некоторой средней об-
ласти температур, но не выше и не ниже этой области. У комара
Culex компенсаторные изменения в потреблении О2 хорошо вы-
ражены только между 15 и 25 °C [56]. Потребление кислорода
ушастым окунем, измеренное при различных температурах аккли-
110
Глава 9
мации, обнаруживает эффект компенсации (тип 2 по Прехту)
только в диапазоне 16—17,5 °C (рис. 9-15) [388]. Таким образом,
любые данные об акклимации следует приводить с указанием тем-
пературного диапазона.
Близкий характер имеет усложнение, обусловленное тем, что
кратковременные измерения Qi0, производимые в разных условиях
и в разных температурных диапазонах, приводят к различным ре-
зультатам. В случае ферментативных реакций Qi0 уменьшается с
понижением концентрации субстрата (см. стр. 119). На холоду
кинетическая активность (число столкновений) уменьшается, но
зато возрастает кажущееся сродство фермента к субстрату (стр.
119); поэтому в сложных системах интерпретация температурных
характеристик затруднена, а их измеренные величины сомнитель-
ны. Потребление О2, особенно в покое, может относительно мало
зависеть от температуры, т. е. иметь низкий Qio в ограниченном
диапазоне «обычных» температур. У Littorina и некоторых других
литоральных беспозвоночных Qio для стандартного обмена в
среднем диапазоне температур составляет около 1,0, тогда как
для активного обмена в том же температурном диапазоне Qio зна-
чительно выше [345]. У Balanus balanoides при температурах от
10 до 15 °C Qio всегда, за исключением января, близок к 1,0 [18].
В отличие от этого у мухи Calliphora интенсивность обмена про-
порциональна температуре, причем у особей, акклимированных к
10 °C, наклон кривой (Qio) больше, чем у особей, акклимирован-
ных к 20 или 30 °C [463].
Характер компенсаторной акклимации изменяется в зависимо-
сти от сезона, гормонального баланса и питания. Основной обмен
Salvelinus в период нереста (поздняя осень) вдвое превышает
уровни марта и апреля, что отражается на температурных реакци-
ях [33]. Обмен у речного рака изменяется в зависимости от ста-
дии линочного цикла. В гепатопанкреасе при этом не обнаружено
компенсации температурных эффектов в отношении VO8 , но при
холодовой акклимации возрастает использование гексозомонофос-
фатного шунта в обход гликолиза в межлиночный период (но не
на стадии предлиньки) [321]. Данные об акклимации ферментов
пищеварительного тракта крайне противоречивы — вероятно, по-
тому, что питание животных во время опытов не контролировали.
У Schistocephalus— паразита колюшки — кривые VOa, построен-
ные для зимнего и летнего сезонов, пересекаются в точке около
35 °C, a Qio выше у летних животных. Таким образом, летние жи-
вотные, акклимированные к 15 °C, на всех участках температур-
ного диапазона сходны по величине VO2 с зимними животными,
акклимированными к 10 °C [89].
Еще одна причина различий в характере акклимации — то, что
он неодинаков для разных тканей и функций одного и того же жи-
вотного. Например, из рис. 9-16 видно, что развитие и потребление
кислорода у кобылки Melanoplus по-разному зависят от темпера-
Рис. 9-15. Основной обмен ушастого окуня Lepomis gibbosus, измеренный при
температурах акклимации (данные Дж. Робертса).
Черные кружки — продолжительность фотопериода 9 ч, белые кружки — 15 я.
t/T-104
Рис. 9-16, Кривые Аррениуса для потребления кислорода (белые кружки) и ско-
рости развития (черные кружки) кобылки Melanoplus [382].
112
Глава 9
туры [382]. В опытах на лососях Salmo gairdneri, акклимирован-
ных к 8 и к 16 °C, при этих же температурах измеряли тканевое
дыхание; акклимация организма в целом соответствовала типу 3
по Прехту, жабр — типу 4, мозга — типу 2 и печени — типу 1
[122]. В другой работе [418] угрей помещали в трубку с входны-
ми отверстиями для воды на обоих концах и оттоком в середине;
затем брали мышечную ткань из головного и хвостового концов
тела и измеряли потребление ею кислорода. Если температура во-
ды была везде одинаковой, то величина VO2 для мышц головы
составляла 3Д для хвостовых мышц; после охлаждения голо-
вы, продолжавшегося в течение недели, эта разница не обнару-
живалась, а после охлаждения хвоста она, наоборот, возрастала.
Причина этих региональных различий мышечного метаболизма не-
известна.
Частота сокращений сердца у ящерицы Sauromalus возрастает
по мере повышения температуры от 15 до 45 °C, в то время как
артериальное давление остается постоянным в диапазоне темпе-
ратур от 25 до 45 °C, а пульсовое давление — в диапазоне от 10
до 45 °C; таким образом, регуляция сосудистого русла резко от-
личается от реакции сердца [455]. У улиток при повышении тем-
пературы с 20 до 30 °C гепатопанкреас и гонады обнаруживают
быструю и полную акклимацию, тогда как у мышцы ноги компен-
сация оказывается лишь частичной [368]. Акклимация краба Uca
pugnax из Северной Каролины соответствует типу 3 по Прехту
для животного в целом и типу 2 для цитохромоксидазы мозга; ?<к-
климация U. гарах (как всего организма, так и мозга) соответст-
вует типу 4 [478]. Для цитохромоксидазы жабр Libinia '{no не
Callinectes и Ocypode) описана акклимация типа 3, для мышц
Libinia — обратная акклимация (тип 5) ['479, 480]. Сердце и жабры
речного рака обнаруживают некоторую компенсацию,. в гепато-
пакреасе компенсации не отмечается, а акклимация мышц брюш-
ка соответствует типу 5 (парадоксальная, или обратная, акклима-
ция) [43, 245]. Таким образом, обобщения, касающиеся акклима-
ционных реакций, невозможны даже в случае разных тканей од-
ного и того же животного.
При изучении акклимации в большинстве случаев измеряли
потребление кислорода. Однако кривые зависимости максимально-
го активного обмена от температуры акклимации образуют пик, а
кривые для обмена в покое обнаруживают постепенное повышение
[33] (рис. 9-17); между тем в экспериментах чаще всего не про-
водилось различия между активным состоянием и покоем. У Gam-
marus oceanicus величина VOa в диапазоне от 5 до 10 °C относи-
тельно независима от температуры акклимации, в то время как
интенсивность обмена, в том числе активного, в этом диапазоне
круто возрастает [146].
Наряду с температурой на форму кривых акклимации. могут
влиять и другие внешние факторы. Мы уже говорили о взаимодей-
Температура
113
ствии температуры и солености (т. I, гл. 1). У одного бокоплава в
водоемах Аляски обменные процессы летом протекают интенсив-
нее, чем зимой, что, вероятно, связано с адаптацией к пониженно-
му в зимнее время содержанию кислорода в воде. Влияние темпе-
ратуры на метаболизм ракообразных Hemigrapsus [103] и Но-
Рис. 9-17. Потребление кислорода молодью нерки, измеренное при различных
температурах акклимации и при разных скоростях плавания, выраженных
в £/с (£ — расстояние, равное длине тела) [47].
tnarus [316] сложным образом изменяется в зависимости от со-
лености. Взаимодействие этих двух факторов обнаружен® также
при изучении скорости развития бычков (Gobiidae) [262]. В усло-
виях низкой солености VO2 у краба Hemigrapsus после летней
акклиматизации выше, чем после зимней, в то время как при вы-
сокой солености акклиматизация может вообще отсутствовать, а
иногда у зимних животных интенсивность обмена несколько вы-
ше [ЮЗ].
Как правило, животные, впадающие на холоду в оцепенение
или летаргию, гораздо менее способны к акклимации, чем живрт-
8—1514
114
Глава 9
ные, сохраняющие активность в широком диапазоне температур.
Попытки обнаружить метаболическую акклимацию у речных ра-
ков, лягушек, жаб и черепах не привели к однозначным результа-
там, и обнаруженные изменения оказались незначительными по
сравнению с аналогичными изменениями у активных рыб. У неко-
торых рыб, например у таутоголабруса, нет никаких различий
между результатами зимних и летних измерений. У лягушки-
сверчка Acris при разных температурах практически отсутствуют
какие-либо метаболические различия, связанные с акклимацией
[114]. Улитка Doroceras и насекомое Carausius не обнаруживают
никакой акклимации к низким температурам; обе формы относят-
ся к числу зимнеспящнх. У видов, впадающих в сезонную спячку,
в активной фазе возможна некоторая температурная акклймация,
однако ценность многих работ по этому вопросу снижает то, что
в них не учитывается оцепенение при низких температурах. Мета-
болическая акклимации четко выражена у краба Callinectes, ак-
тивного в течение всего года, за исключением 2—3 холодных ме-
сяцев, в то время как сухопутный краб Ocypode, впадающий в
оцепенение при температуре ниже 15 °C, почти не обнаруживает
способности к акклимации [480].
У животных, обитающих в зонах со значительными годовыми
и суточными колебаниями температуры (например, в умеренном
поясе), генетически закрепленная способность к температурной
акклимации может быть выражена сильнее, чем у родственных жи-
вотных из областей с относительно постоянным температурным
режимом (тропики и полярные области). Потребление О2 акти-
ниями Metridium из Вудс-Хоула обнаруживает акклиманионные
изменения типа 3 по Прехту, тогда как у актиний Haliplanella и
Diadumene из Виргинии подобных явлений нет [401]. Ферменты
антарктических рыб гораздо меньше способны к акклимации, чем
соответствующие ферменты форели ['435]. У одной полихеты из
северной Атлантики Qio составляет около 2 в широком диапазоне
температур, в то время как для южных популяций характерна
большая величина Qio при крайних температурах и малая — при
средних [305]. Популяциям бычков, обитающим в низких широтах,
при всех температурных условиях свойственно невысокое значение
Qio, а популяциям высоких широт — низкий коэффициент Qi0 при
высоких температурах и высокий—при низких температурах [16].
У крабов широко распространенного рода Uca наблюдаются зза-
метные видовые и широтные различия. У Uca pugnax из Северной
Каролины обнаружена температурная компенсация типа 3 (т. е.
частичная), тогда как при лабораторной акклимации U. гарах ни-
какой компенсации не выявлено. Не обнаружено акклимации и у
U. pugnax из Флориды и с Ямайки [478]; бразильские особи так-
же акклимируются очень слабо.
При метаболической акклимации для одних ферментов харак-
терна выраженная компенсация, а для других — нет. В табл. 9-1
Температура
115
Таблица 9-1
Ферменты с выраженной температурной компенсацией
(типы 2 и 3 по Прехту) [171]
Фермент Организм, ткань Источник данных
Ферменты гликолиза Фосфофруктокиназа Серебряный карась,, М [129|
Альдолаза Карась, Ж Орфа, М, П, Ж [1201
Альдолаза [231а]
Лактатдегидрогеназа Серебряный карась, М, П; го- [201]
г лец, М, П; озерная форель
Ферменты гексозомонофосфат- ного шунта 6-фосфоглюконатдегидро- Карась, Ж [1201
геназа Ферменты цикла Кребса и си- стемы переноса электронов Сукцинатдегидрогеназа Rhodeus, П [273]
Малатдегидрогеназа Серебряный карась, М Орфа, М Дождевой червь, М Орфа, М, П, Ж Серебряный карась, М [172] [231а] [400] [231а]
Цитохромоксидаза [128]
Серебряный карась, М, П, Ж, [69]
Сукцинат-цнтохром-с-редук- ГМ Краб Uca, М Орфа, М, П, Ж Серебряный карась, Ж [479] [231а] [69]
таза НАД-цитохром-с-редуктаза То же [69]
Ферменты анаболизма Амнноацилтрансфераза Опсанус, П [1671
Разные ферменты Серебряный карась, К [430]
Протеаза Серебряный карась, Ж Tenebrio, К; Helix, К
М — мышца; П — печень; Ж — жабра; ГМ—головной мозг; К — кишечник.
приведены ферменты, обнаруживающие частичную или полную
акклимации) (2-го и 3-го типа по Прехту) [171]; все они имеют
отношение к выработке энергии — это ферменты гликолиза, пен-
тозного шунта, цикла трикарбоновых кислот, переноса электро-
нов, окисления жирных кислот. К этому можно добавить, что при
акклимации к холоду белковый синтез идет интенсивнее, чем при
акклимации к теплу. Активность аминоацилтрансферазы в печени
Рыбы Opsanus, акклимированнор! к 10°C, на 60% выше, чем при
акклимации к 21 °C [167, 168, 169]. Конечный эффект этих ком-
пенсаций состоит в том, что акклимированные животные могут
поддерживать относительное постоянство активности (она меньше
Понижается на холоду и меньше повышается в тепле).
116
Глава 9
Таблица 9-2
Ферменты с обратной температурной компенсацией или неспособные
к компенсации (типы 4 и 5 по Прехту)
Фермент Организм, ткань Источник данных
Ферменты лизосом и перокси- сом, * • Катййаза Речной рак, М [367]
Угорь, М
» Карп, М, Ж рад
Серебряный карась, Пк [1711
Пероксидаза То же [1711
Дрожжи [363]
Кислая фосфатаза Серебряный карась, Пк [171]
Оксидаза D-аминокислот То же 1171]
Разные ферменты М22+-АТФаза Серебряный карась, Ж, ГМ [431]
Холйнацетилтрансфераза Серебряный карась, К
Серебряный карась, ГМ [182]
Ацётилхолинэстераза То же [171]
Фундулюс, ГМ Ц71!
Щелочная фосфатаза Ящерицы [296]
Аллантоиназа, аллантоиказа, Карп, П [476]
уриказа
Амилаза Tenebrio, К
Липаза Зимующая улитка, К
«Яблочный» фермент Лягушка, П [151
Г-6-Ф-дегидрогеназа Карп, Ж [1ЭД
Лягушка (после голодания), П [15]
Пк—лючка; остальные сокращения те же, что и в табл. 9-1.
В табл. 9-2 перечислены ферменты, которые либо не способны
к акклимации, либо обнаруживают обратную акклимацию. Это
главным образом ферменты катаболизма (пероксидазы, каталаза,
кислая фосфатаза, оксидаза D-аминокислот, ацетилхолинэстераза,
аллантоиказы и аллантоиназа).
Так как ни одна система, участвующая в выработке энергии, не
обнаруживает полной акклимации (типа 2), концентрация про-
дуктов метаболизма может оказаться более высокой в тепле и бо-
лее низкой на холоду. Поэтому понижение активности катаболиче-
ских ферментов в тепле или повышение ее на холоду было бы не-
адаптивным. В отношении пищеварительных ферментов вопрос
неясен: некоторые из них способны к компенсации, другие — нет.
Вероятно, эти различия зависят от условий питания животного.
С транспортными ферментами дело обстоит по-разному. В ки-
шечнике серебряного карася активный транспорт глюкозы сопря-
жен с переносом Na (см. гл. 2). Чувствительная к уабаину
Ыа+К+-АТФаза в кишечнике рыб, акклимированных к холоду,
более активна, чем у рыб, акклимированных к теплу; при аккли-
Температура 117
мации к 30°C ее сродство к Na выше, чем при акклимации к 8°C.
Мд2+_дтФаза изменяется противоположным образом [427, 428,
430, 431]. Перенос аминокислот (особенно нелипофильных) у рыб,
акклимированных к 8 °C, происходит втрое быстрее, чем у рыб,
акклимированных к 30 °C. Ыа+К+-АТФаза в жабрах серебряного
карася также обнаруживает температурную акклимацию.
Подведем итоги. Характер акклимации может быть различным
в зависимости от температурного диапазона — компенсация мо-
жет быть хорошо выражена в области средних температур И от-
сутствовать при экстремальных температурах. На тип акклимации
могут влиять сезон, фаза репродуктивного цикла, гормональный
баланс и питание. Характер метаболических ответов на темпера^-
турное воздействие оказывается различным в зависимости от того,
измеряют ли основной (стандартный) обмен или обмен при обыч-
ной жизнедеятельности и активном состоянии. Разным тканям и
функциям данного животного не обязательно свойствен один и тот
же тип акклимации. Акклимацию к температуре могут видоизме-
нять другие важные параметры среды, например количество кис-
лорода или соленость воды. У животных, впадающих в зимнее
оцепенение, акклимация, как правило, выражена гораздо слабее,
чем у тех, которые остаются активными. Некоторые виды, живу-
жие в относительно постоянных условиях среды, могут быть менее
способными к компенсаторной акклимации, чем виды, подвержен-
ные сильно варьирующим внешним воздействиям. У ферментов,
участвующих в выработке энергии, отмечается большая тенденция
к компенсации, чем у ферментов, расщепляющих промежуточные
и конечные продукты обмена. И компенсаторная акклимация (ти-
пы 1, 2 и 3), и неспособность к акклимации (тип 4), и обратная
акклимация (тип 5) могут иметь приспособительное значение в
определенных физиологических и экологических условиях.
Механизмы акклимации ферментов. Многие ферменты жи-
вотных, наследственно приспособленных к определенным темпе-
ратурам или акклимированных к холоду или теплу, обнаруживают
способность к адаптации. Для оценки происходящих изменений
определяют активность экстрактов, содержащих ферменты, актив-
ность метаболических путей, температурный коэффициент или ки-
нетические характеристики, например Км- Молекулярные меха-
низмы компенсации могут состоять в изменениях синтеза или ки-
нетических свойств данного фермента, в изменениях пропорций
изоферментов, приспособленных к определенным температурам, в
изменениях кофакторов (липидов, коферментов), pH, ионной сре-
ды и т. п.
Изменения первичной структуры (последовательности амино-
кислот) во время акклимации маловероятны, так как они должны
быть закодированы в генетическом материале, а процессы аккли-
мации совершаются в течение коротких периодов времени у от-
дельных особей. Однако некоторые факторы, влияющие на актив-
118
Глава 9
ность ферментов (см. ниже), могут изменять их конформацию или
четвертичную структуру.
При холодовой акклимации усиливается биосинтез белков, из-
меряемый по включению меченых аминокислот. У акклимированно-
го к холоду серебряного карася наиболее интенсивное включение
радиоактивного лейцина в белки наблюдается в печени, мышцах
и жабрах [88]. При одной и той же температуре интенсивность
белкового синтеза в перфузируемой печени рыбы Opsanus состав-
ляет 29 мкг/(г-мин) у особей, акклимированных к 21 °C, и
46 мкг/(г-мин)—у особей, акклимированных к 10°С. Активность
аминоацилтрансферазы в микросомах печени у последних на 60%
выше, чем у рыб, акклимированных к 21 °C [167]. У дождевых чер-
вей и рыб на холоду ускоряется обновление РНК [87]. Возможно,
что одним из первых звеньев биохимической адаптации является
стимуляция переноса РНК, связанной с белковым синтезом. С по-
мощью иммунологических методов было показано, что холодовая
акклимация приводит к избирательному синтезу определенных
белков (цитохромоксидазы) [495а].
Судя по некоторым данным, в мультиферментных системах при
акклимации возможен сдвиг равновесия между взаимосвязанными
путями, координация которых осуществляется путем воздействия
метаболитов на отдельные ферменты. Потребление О2 жабрами
серебряного карася при акклимации к холоду повышается и ста-
новится более чувствительным к цианиду и менее чувствительным
к иодацетату, чем при тепловой акклимации [119]. Клетки Strep-
tococcus cremoris, растущие при высокой температуре, синтезиру-
ют больше ацетоина и меньше бутиленгликоля; культуры, росшие
при низких температурах, продуцируют при одной и той же тем-
пературе больше молочной кислоты, чем «высокотемпературные»
культуры. Антарктическую рыбу Trematomus, обитающую при
—1,9 °C, акклимировали к —2 и к +4 °C; торможение метаболиз-
ма нодацетатом (ингибирующим гликолиз) было сильнее выраже-
но во второй группе рыб, а чувствительность к цианиду — в пер-
вой группе [’435]. Гольца (Salvelinus fontinalis) акклимировали к
15 и к 4 °C; включение ,4СО2 в положения 1. 2, 5 и 6 глюкозы бы-
ло относительно более высоким в тканях рыб, акклимированных
к теплу; отношение C6/Ci при утилизации глюкозы было большим
при более высокой температуре. Активность гексозомонофосфатно-
го шунта у рыб, акклимированных к холоду, была втрое выше, чем
при тепловой акклимации; гликолиз на холоду был ослаблен, а об-
разование жира (включение ацетата в жиры) повышено ['203].
Холодовая акклимация у рыб благоприятствует глюконеогенезу
[34]. Обмен липидов у лосося Salrno при холодовой акклимации
более интенсивен, чем при тепловой [101].
Ферменты, которые в естественных условиях должны функцио-
нировать при низких температурах, приспособлены к своей работе
иначе, чем ферменты, функционирующие при высоких температу-
Температура
119
pax. В ряде случаев (главным образом у некоторых изоферментов)
кривая зависимости /См от температуры проходит через минимум
(максимальное сродство к субстрату), соответствующий темпера-
туре акклимации (см. стр. 120). Однако в большинстве случаев
Температура измерений, °C
Рис. 9-18. Влияние температуры на сродство к сукцинату мышечной сукцинат-
Дегидрогеназы серебряного карася, акклимнрованного к 5 и к 25 °C [172].
^См непрерывно уменьшается по мере снижения температуры, и
таким образом компенсируется нормальное влияние Qw на кине-
тическую активность (рис. 9-18), так что Qio в определенных диа-
пазонах температур может приближаться к 1,0; это позволяет
объяснить относительную независимость стандартного обмена от
температуры у животных, находящихся в обычных для них темпе-
ратурных условиях. Вероятно, большинство ферментативных ре-
кций в клетках протекает при таких концентрациях субстрата,
120
Глава 9
которые весьма далеки от насыщающих. Поэтому происходящие
прд температурной акклимации изменения формы или положения
кривых, отражающих зависимость скорости реакции от концентра-
ции субстрата, позволяют ферментам функционировать при изме-
нившейся температуре и обычных внутриклеточных концентрациях
субстратов, тогда как при максимальных концентрациях субстра-
тов работа ферментов могла бы оказаться невозможной [202,
436]. Вероятно, в процессе эволюции отбор шел больше по при-
знаку сродства фермента к субстрату, чем по признаку „молеку-
лярной активности”, показателем которой служит число оборотов
[433, 436].
Одним из механизмов акклимации ферментного аппарата мо-
жет быть избирательный синтез изоферментов, которые лучше все-
го функционируют (т. е. имеют минимальные Км) в соответствую-
щем диапазоне температур [205, 436]. В головном мозгу радужной
форели <ацетилхолинэстераза представлена двумя изоферментами,
один из которых образуется при акклимации к 17 °C, а другой —
при акклимации к 2 °C; у рыб, акклимированных к 12 °C, имеются
оба изофермента. Величины их Км соответствуют температурам
их синтеза [14] (рис. 9-19). Пируваткиназа в мышцах камчатско-
го краба также представлена двумя формами; одна из них функ-
ционирует при температурах ниже 10 °C и имеет минимальную Км
при 5 °C, а другая действует при температурах выше 9 °C и имеет
минимальную Км при 12°C [434]. В печени форели содержатся
две формы цитратсинтазы — «холодовая» и «тепловая»; при тем-
пературах ниже 15 °C константы Михаэлиса этих форм близки, но
при более высоких температурах Км «холодовой» формы возрас-
тает (т. е. сродство ее к субстрату снижается) гораздо больше,
чем у «тепловой» формы [205] (рис. 9-20). При повышении темпе-
ратуры с 20 до 42 °C Км лактатдегидрогеназы из мышц пред-
плечья ленивца возрастает в 16 раз, а аналогичный фермент мио-
карда менее чувствителен к температуре [204].
У рыб обнаружено несколько генетически детерминированных
изоферментов лактатдегидрогеназы (ЛДГ). Относительные коли-
чества их в тканях серебряного карася и ушастого окуня при тем-
пературной акклимации не изменяются [495а], однако существует
отбор по этому признаку, связанный с географическими условия-
ми. У карпозубой рыбы Fundulus heteroclitus описаны широтные
различия по двум формам ЛДГ печени (и соответствующие раз-
личия в соотношении гомозигот и гетерозигот). Кинетические ис-
следования показали, что одна из этих форм (В'В') имеет пре-
имущество в теплых водах, а другая (ВВ) — при температурах
ниже 12,2 °C [362а]. Таким образом, можно дать кинетическую
интерпретацию географическому полиморфизму в отношении изо-
ферментов ЛДГ.
Еще один важный механизм акклимации состоит в изменении
липидов. У микроорганизмов, растений и животных при низких
Температура
121
температурах обычно откладываются липиды с большим содержа-
нием ненасыщенных жирных кислот, большим иодным числом и
поэтому более высокими точками плавления, чем при высоких тем-
пературах. Это установлено, в частности, на личинках мух, на жи-
ровой ткани и печени лягушки [15], на мозге ['392] и других тка-
нях [265] серебряного карася и на планктонных веслоногих рач-
Рис. 9-19. Зависимость /См ацетилхолинэстеразы головного мозга от температуры
у антарктической рыбы Trematomus (I), форели, акклимированной к 2 °C (II)
и к 18 °C (III), и электрического угря из Амазонки (IV) (Baldwin J., Ph. D. The-
sis, Univ, of Brit. Columbia, 1970).
ках. У хлореллы, выращенной при 38 °C, 46% липидов построено
из насыщенных жирных кислот с короткой цепью; при 22 °C в ли-
пидах содержатся главным образом триеновые жирные кислоты
[353].
У Bacterium megaterium понижение температуры с 30 до 20 °C
вызывает синтез фермента, катализирующего превращение паль-
митиновой кислоты в ненасыщенную жирную кислоту; при 30 °C
оп°оГО ФеРмента практически нет, а при постоянной температуре
С он синтезируется в меньших количествах, чем при переходе
°т 30 к 20 °C [134].
Холодовая акклимация ускоряет обновление всей фракции
фосфолипидов, судя по включению меченого ацетата и меченого
фосфора [6, 265]. У акклимированных к холоду серебряных кара-
еи содержание ненасыщенных жирных кислот в фосфатидилхоли-
152
Глава 9
Рис. 9-20. Влияние температуры на /См цитратсинтазы форели, акклимированной
к 2 °C (7) и к 18 °C (II) [204].
не и фосфатидилэтаноламине плазмалогенов мозга выше, чем при
акклимации к теплу [391, 392], а фосфолипиды жабр после ак-
климации к 5 °C содержат больше фосфатидилхолина и фосфати-
дилинозита, чем после акклимации к 25 °C [6]. При тепловой ак-
климации содержание жирных кислот Сгон, С20:4, и С22:6 в фосфо-
липидах серебряного карася уменьшается вдвое, а содержание
жирных кислот С18.-0 и С20;з почти удваивается [254]. Сукцинатде-
гидрогеназа серебряных карасей — независимо от их акклима-
Температура
123
и__активируется фосфолипидами из митохондрий особей, ак-
климированных к холоду, на 50% сильнее, чем фосфолипидами
особей, акклимированных к теплу (рис. 9-21). По-видимому, бел-
Рис. 9-21. Активирующее действие митохондриальных фосфолипидов на освобож-
денную от липида сукцинатдегидрогеназу из мышц серебряного карася: влияние
акклимации (Hazel J., Comp. Biochem. Physiol., 43, 837—888, 1972).
Л фермент рыб, акклимированных к 5 °C, Б — к 25 °C. Кружками показано действие липи-
дов от рыб, акклимированных к 5° С, треугольниками — к 25° С. Активность фермента без
добавления липидов принята за 100%.
ки при обеих температурах акклимации одни и те же, так что
компенсаторное изменение активности можно объяснить разли-
чиями в липидных кофакторах [172].
При понижении температуры текучесть липидов уменьшается;
их ненасыщенность противодействует этому физическому измене-
нию. Поэтому соответствующие изменения состава жирных кислот
124
Глава 9
(особенно в фосфолипидах) могут способствовать сохранению
надлежащего физического состояния как клеточных, так и мито-
хондриальных мембран. В искусственных мембранах термотроп-
ный (обусловленный температурой) переход фосфолипидов, на-
пример лецитина, из кристаллического состояния в жидкокристал-
лическое, так же как и лиотропный переход из ламеллярной фазы
в гексагональную, зависит от типа и степени ненасыщенности фос-
Рис. 9-22. Влияние температуры на перенос галактозида у Е. coli, растущей на
среде с олеиновой или с линолевой кислотой [496].
фолипида. Лецитин образует мембраны при 36, но не при 20 °C
[38, 79]. Клетки Е. coli активно поглощают глюкозу и галактозу,
и транспорт этих сахаров, измеренный при различных температу-
рах, дает два различных наклона кривой Аррениуса, что отражает
два разных физических состояния мембраны. Эта кривая образует
перелом в точке, соответствующей 13 °C, если культуральная сре-
да содержит олеиновую кислоту (рис. 9-22). Если же среда будет
содержать линолевую кислоту, перелом кривой произойдет при
7°C. Это говорит о зависимости физического состояния мембраны
от состава жирных кислот [496]. Можно поэтому заключить, что
поддержание жидкого состояния клеточных и внутриклеточных
мембран путем соответствующих изменений в составе их липи-
дов— один из важных аспектов температурной акклимации.
Другой возможный механизм температурной акклимации связан
с тем, что ферменты находятся под контролем веществ-модулято-
ров, изменение концентрации которых составляет часть непосред-
Температура
125
ственной реакции организма на изменение температуры. Кроме того,
изменения концентраций промежуточных продуктов обмена могут
репрессировать или индуцировать синтез ферментов. Образо*
вание таких промежуточных метаболитов, как фосфорилированные
сахара и нуклеотиды, повышается под прямым воздействием теп-
ла и понижается на холоду [129]. У лягушек, черепах и некоторых
рыб с повышением температуры понижается pH крови и, вероятно,
тканей, так что ДрН/ДТ=—0,016; в результате сохраняется посто-
янным’отношение ОН/Н [211]. Некоторые важнейшие ферменты,
например фосфофруктокиназа [129], работают в узких диапазонах
pH. При pH 7,5 Лм пирувата для лактатдегидрогеназ форели и
тунца резко возрастает с повышением температуры, а при pH 7,0
этот эффект практически вообще отсутствует [204, 205]. Имеются
данные о небольших изменениях концентрации ионов в крови; у
рыб, находящихся в тепле, уровни Mg2+ и Са2+ повышаются, а
уровни Na+ и С1~ понижаются [375], однако эти ионные сдвиги,
вероятно, слишком малы для того, чтобы изменить тканевый
обмен.
Если какой-либо фермент чувствителен ко многим модулято-
рам— промежуточным продуктам обмена, ионам, pH и т. д., то
его активность может заметно измениться. Например, фосфофрук-
токиназа серебряного карася подвержена влиянию фруктозо-6-
фосфата, АТФ, АМФ, Mg2+ и цитрата и очень чувствительна к pH.
Однако роль прямых изменений концентрации клеточных компо-
нентов в долговременной акклимации ферментных систем еще не
выяснена.
Таким образом, молекулярные механизмы температурной ак-
климации не вполне ясны. Они могут включать стимуляцию син-
теза или обновления белков и фосфолипидов, избирательный син-
тез адаптивных изоферментов, видоизменение мембранных липи-
дов (повышение степени их ненасыщенности на холоду), возмож-
ные эффекты кофакторов и ионов, в том числе Н+. Механизмы
температурной акклимации многообразны, и это не удивительно —
ведь температура является всеохватывающим параметром внешней
среды.
Прямые или косвенные клеточные эффекты. Пока еще не ясно,
каким образом инициируются биохимические изменения при ак-
климации— путем прямого действия температуры на клеточном
Уровне или же при участии нервной или гормональной стимуля-
ции. У микроорганизмов при отсутствии генетического отбора или
в изолированных жабрах и щупальцах моллюсков изменение ак-
тивности ферментов должно быть прямой реакцией [368]. Темпе-
ратурная акклимация карася и ушастого окуня зависит от длины
фотопериода — при 9-часовом фотопериоде эти рыбы акклимиру-
Ются лучше, чем при 15-часовом [388]. Серебряный карась при
Длинном фотопериоде более устойчив к внезапному повышению
температуры, а при коротком — к охлаждению; это указывает на
126
Глава 9
возможный гормональный контроль. В то же время гипофизэкто-
мия не влияет на метаболическую акклимацию ильной рыбы [159],
а тиреоидэктомия или инактивация щитовидной железы — на ме-
таболическую акклимацию серебряного карася [263]. Обработка
тиомочевиной не изменяет верхнюю летальную температуру для
рыбы Tanichthys [81]. Тироксин повышает холодоустойчивость
рыбки-меченосца (Xiphophorus), но не влияет на ее устойчивость
к теплу [457]; экстракт щитовидной железы млекопитающего не-
сколько повышает холодоустойчивость серебряного карася, но
экстракт железы карася не оказывает подобного действия [200].
Таким образом, данные о гормональном контроле температурной
акклимации у рыб весьма противоречивы; получены, в частности,
и отрицательные результаты. Сезонные различия в акклимацион-
ных возможностях могут быть отражением годичных гормональ-
ных циклов, й это затрудняет интерпретацию многих данных, ка-
сающихся действия гормонов.
Поведенческие и локомоторные адаптации
Метаболическая акклимация способствует выживанию при
экстремальных температурах и обеспечивает нормальную жизне-
деятельность пойкилотермных животных в широком диапазоне
температур. Кроме того, у большинства животных наблюдается
адаптивное поведение. Таким образом, нервная система играет
ключевую роль как в устойчивости, так и в способности к адап-
тации.
Выбор градиента и таксисы. При наличии теплового градиента
простая адаптивная реакция сводится к движению животного в
сторону «избираемой», или «предпочитаемой» температуры. Пара-
меции, помещенные в камеру, теплую на одном и холодную на
другом конце, скапливаются в зоне средней температуры; «пред-
почитаемой» здесь может быть температура, соответствующая ми-
нимальному термотаксису [127]. Свободноживущие нематоды
движутся к оптимальной средней температуре, тогда как немато-
ды, паразитирующие на теплокровных животных, перемещаются
в сторону более высоких температур, даже если это приводит к
термическому повреждению [314]. Слизни Umax передвигаются
в сторону предпочитаемой температуры, которая несколько выше
температуры акклимации и изменяется вместе с последней [387].
Избирательное скапливание сельди наблюдается при перепаде
температур воды 0,5°C; однако сельдь способна различать и мень-
шие градиенты. Температура, избираемая рыбой Girella, повыша-
ется с 18 до 24,3°C при повышении температуры'акклимации с Ю
до 30 °C [347]. Аналогичное повышение предпочитаемых темпера-
тур при повышении температуры акклимации наблюдается у нер-
ки и карпа. Чем определяется предпочитаемая температура, неяс-
но; возм1___., это та температура, при которой ионные насосы
Температура
127
одерживают определенную величину мембранных потенциалов.
Серебряных карасей обучили надавливать на рычаг с интервала-
ми 1 мин, чтобы получать пищу; при изменении температуры чув-
ство времени не нарушалось [394].
Насекомые также скапливаются в определенной точке терми-
ческого градиента; положение этой точки у пчел, муравьев, саран-
човых, мучных хрущаков и клещей зависит от температуры ак-
климации. Муравьи, акклимированные к 3—5 °C, скапливаются в
точке 23,5 °C, а акклимированные к 25—27 °C — в точке 32 °C.
В опытах с одной аляскинской жужелицей жуки, помещенные в
термический градиент, летом скапливаются в интервале 7—9 °C,
а зимой идут в сторону минимальной температуры (—25 °C) и за-
мерзают [326]. Избираемые температурные зоны могут быть и от-
носительно узкими, и достаточно широкими; возможно их измене-
ние в зависимости от сезона, питания и стадии жизненного цикла.
Личинки навозных мух во время активного питания избирают
температуры 30—37 °C, соответствующие обычным для них усло-
виям среды, а личинки, готовые к уходу в землю для окукливания,
при помещении их в температурный градиент скапливаются в зоне
15 °C. Питающиеся личинки комнатной мухи предпочитают зону
температур от 15 до 33 °C, а личинки перед окукливанием — от 8 до
20 °C. Кровососущие насекомые — комары, клоп Rhodnius — нахо-
дят свою добычу отчасти по запаху, но также и по исходящему
от нее теплу. Поведение насекомых в тепловом градиенте в боль-
шей степени зависит от температуры воздуха, чем от температуры
субстрата.
У рыб поведение в температурном градиенте коррелирует с об-
щей активностью, таксисами и максимальной скоростью дли-
тельного плавания. В опытах на форели найдены две температу-
ры, которым соответствует максимальная спонтанная двигатель-
ная активность, — это предпочитаемая температура и температура
чуть ниже верхней летальной. В свободных от льда озерах нерка
плавает с максимальной скоростью при 15,5 °C, а кижуч в более
теплых реках при 20 °C. У серебряного карася максимальная ско-
рость наблюдается при предпочитаемой температуре [133]. Мак-
симальное расстояние, проплываемое лососем или форелью в от-
вет на толчок, резко увеличивается при предпочитаемой темпера-
туре, которая варьирует в зависимости от акклимации.
Если вычесть величину потребления Ог в покое (стандартный
обмен), найденную при различных температурах акклимации и
измерения, из величины потребления О2 при максимальной актив-
ности (активный обмен), то полученная разность будет опреде-
лять «цену» активности. У озерной форели эта разность достигает
максимума при температуре, приблизительно соответствующей
температуре максимальной плавательной активности.
Скорость плавания может определяться энергетическими воз-
можностями; в то же время электрическое раздражение может
128
Глава 9
больше повысить потребление Ог, чем максимальная двигательная
активность. Повреждения мозга могут приводить к изменению
скорости плавания и предпочитаемой температуры. Возможно, что
потребление кислорода не ограничивает скорость, а само опреде-
ляется энергетическими нуждами, связанными с активностью.
У мальков нерки интенсивность пищеварения возрастает с повы-
шением температуры до 25 °C и более, но количество поедаемой
пищи оказывается максимальным при 15 °C; таким образом, рост
мальков больше зависит от поведенческих реакций, чем от интен-
сивности пищеварения [48]. При акклимации возможно не только
изменение метаболизма, но и лучшее приспособление контролируе-
мых нервной системой поведенческих реакций к определенным
температурам. Манитобская жаба с усилением зимних морозов
все глубже зарывается в почву, так что всегда оказывается ниже
замерзшего слоя [456].
Существуют также поведенческие адаптации, связанные с вы-
работкой тепла в результате мышечной активности и с получени-
ем тепла от солнца или нагретого субстрата. Таким образом, ог-
раниченная регуляция температуры тела возможна и у пойкило-
термных животных.
Поглощение солнечного тепла играет большую роль у назем-
ных насекомых и рептилий и может быть связано с пигментацией.
Светлоокрашенный жук-долгоносик Compus niveus поглощает
26% падающих на него инфракрасных лучей, а темный жук-мерт-
воед Silpha obscura — 95%. Пустынная саранча (Schistocerca)
совершенно неактивна при температуре ниже 17 °C и начинает
двигаться при 17—20 °C; при этих температурах утром она распо-
лагается на восточных склонах так, чтобы ее тело было перпен-
дикулярно солнечным лучам. При 28 °C саранча начинает мигри-
ровать, а при 40 °C иногда садится на куст, ориентируя тело па-
раллельно солнечным лучам, т. е. так, что их поглощение сводится
к минимуму. При снижении температуры в поздние послеполуден-
ные часы саранча снова располагается перпендикулярно солнеч-
ным лучам. Некоторые бабочки греются на солнце, поглощая сол-
нечные лучи своими крыльями; при высоких температурах они
ищут тень и усиливают охлаждение путем испарения влаги. Дру-
гие бабочки перед полетом разогреваются с помощью движений,
особенно при низких окружающих температурах. Например, Va-
nessa при 11 °C затрачивала на это свыше 6 мин, при 23— 1,5 мин,
при 34— 18 с, а при 37 °C не разогревалась совсем. Danaus повы-
шает температуру грудного отдела со скоростью 1,3 град/мин до
уровня, на 5—8° превышающего внешнюю температуру [247].
Метаболический «разогрев». Многие ночные бабочки, например
бражники, обладают теплоизолирующими волосками и относятся
к эндотермным, активно продуцирующим тепло животным. Поле-
ту у них предшествует некоординированная дрожь летательных
мышц, которая приводит к повышению температуры грудного от-
Температура
129
дела до 32 °C или более. Если согреть грудные ганглии с помощью
термода, полет становится возможным без предварительного мы-
шечного разогрева. Температура груди повышается со скоростью
4 град/мин и доходит до 32—36 °C при внешней температуре 17—
29 °C. Во время предполетного разогрева потребление кислорода
увеличивается примерно в 2,3 раза, главным «топливом» служит
жир [1, 156]. У одного бражника скорость разогрева достигала
4,2 град/мин, а полетная температура варьировала у разных осо-
бей от 34 до 45 °C [111]. Бражник Manduca не начинает полет,
пока температура его груди не повысится до 38 °C; в полете под-
держивается температура 42 °C при окружающей температуре
17—32 °C. Образующееся в груди тепло передается брюшку с
кровью, протекающей по спинному сосуду; пережатие этого сосуда
приводит к перегреву груди и прекращению полета [184].
Сатурния цекропия (Hyalophora) не может лететь при темпе-
ратуре тела ниже 32 °C, и во время полета она поддерживает
температуру 37 °C. Поведенческая регуляция температуры тела
сохраняется и после перерезки связей между сердцем и грудью,
однако повреждение связей в грудных ганглиях нарушает работу
терморегуляторных центров [313]. При местном обогревании
грудного нервного тяжа цекропия переключается с предполетного
разогрева на двигательную активность полетного типа. Значит,
температура нервного тяжа важнее, чем температура мышц
[156]. Как видно из приводимых ниже данных, у цикад и бабочек
температура определяет характер поведения ['177].
Magiclcada, °C Hyalophora, °C
Тепловая смерть 45 46
Тепловое оцепенение 43 44
Полет максимальной продолжительности 37 38
Пение хором 30-34
Минимальная температура для полета 22 34
Холодовое оцепенение 14 15
Холодовая смерть 0 0
Жук Acillius совершает предполетные разогревающие движе-
ния, издавая звуки частотой до 140 Гц [294]. Кузнечик Neocono-
cephalus начинает стрекотать на частотах 150—200 гц при темпе-
ратуре тела 33,5 °C и может в это время разогреваться, поддержи-
вая температуру тела 34,4 °C, т. е. примерно на 15 °C выше окру-
жающей температуры [178].
Колониальные перепончатокрылые и термиты эффективно ис-
пользуют тепло, образующееся при метаболизме. У танцующей
Медоносной пчелы температура тела может быть на 1 °C выше
внешней температуры, а температура грудного отдела может до-
стигать 36 °C [121]. У муравья Formica rufa температурный опти-
мум для развития потомства составляет 23—29 °C, а у пчелы —
^4,5—35 °C. Температура гнезда при выведении потомства доста-
точно хорошо регулируется. В жаркую погоду при температуре
9" 1514
130
Глава 9
воздуха 48 °C температура в улье поддерживается ниже 37,6 °C с
помощью испарения влаги. При этом вентиляционная активность
пчел пропорциональна температуре гнезда [293]. Рабочие пчелы
приносят воду, разбрызгивают ее в улье и, создавая крыльями
движение воздуха, усиливают испарение. Зимой пчелы сбиваются
в гроздья, плотность которых изменяется в зависимости от темпе-
ратуры; при ее понижении возрастает использование сахара и по-
Рис. 9-23. Максимальные мышечные температуры у голубых тунцов, пойманных
при разных температурах воды [73].
вышается интенсивность метаболизма [421]. Падение температу-
ры воздуха до 8—10 °C вызывает беспокойство пчел; особи, нахо-
дящиеся на поверхности скопления, более активны, чем особи вну-
три грозди, те и другие часто меняются местами. При температу-
ре тела около 7 °C наступает холодовая кома, и если она длится
более 50 ч, то приводит к смерти [121]. В термитнике, занятом
термитами, температура может быть на 14—18° выше, чем в
пустом.
Поразительным примером гетеротермии, связанной с моторной
активностью и теплопродукцией, служит использование сосуди-
стых сетей как систем противоточного теплообмена у крупных
рыб. У тунца температура действующих при плавании мышц под-
держивается в среднем на уровне 30 °C (с диапазоном колебаний
5°), тогда как температура внешней среды варьирует от 10 Д°
22 °C (рис. 9-23). Голова и хвост разогреваются в меньшей степе-
ни; например, при температуре воды 21 °C температура мышц Д0'
стигает 30 °C, а головного мозга — только 27 °C. Мощная красная
мускулатура сельдевых акул может быть на 7—10° теплее окрУ*
д Рис. 9-24.
п’ Расположение сосудистых сетей у голубого тунца. Б. Схематические разрезы тела тунца,
зываюгцие распределение внутренних температур при температуре воды 19,3° С (по
Ф. Кэри и Дж. Тилу).
9*
132
Глава 9
жающей воды; большая часть крови у этих рыб проходит через
боковые артерии и вены, между которыми находятся сосудистые
сети. Эта система аналогична описанной у тунца ['71—74] (рис.
9-24).
Терморецепторы пойкилотермных животных. Терморегулятор-
ные поведенческие реакции осуществляются у этих животных при
участии периферических рецепторов и соответствующих отделов
центральной нервной системы. Эти нервные структуры были изу-
чены у насекомых, рыб и змей.
У сверчков и многоножки Lithobius антенны воспринимают
температуру воздуха, а лапки — температуру почвы. Эксперимен-
ты с ампутацией позволили установить наличие антеннальных тер-
морецепторов у клопа Rhodnius, постельного клопа, пчелы и фаз-
мид; у жуков терморецепторы найдены на антеннальных и мак-
силлярных щупиках. Саранча Locusta migratoria имеет чувстви-
тельные к теплу парные сенсорные бляшки на антеннах, груди и
брюшке; после разрушения одной из антеннальных бляшек насе-
комое перестает поворачиваться в соответствующую сторону к ис-
точнику тепла. Спонтанный разряд антеннальных терморецепторов
Periplaneta имеет частоту 45 имп/с; охлаждение ведет к фазиче-
скому повышению частоты импульсов, а обогревание — к ее по-
нижению с максимальной скоростью 200 имп/(с-град) и абсолют-
ной чувствительностью 0,6 имп/(с-град) [299]. На лапках Peri-
planeta имеются термочувствительные подушечки, активность ко-
торых возрастает при охлаждении насекомого ниже 10 °C или со-
гревании до температур выше 30 °C [259]. У сверлящей дерево
златки Melanophila на тазиках ног находятся специфические ре-
цепторы, чувствительные к инфракрасным лучам (с длиной волны
2—4 мкм). Эти рецепторы могут воспринимать тепло на большом
расстоянии — например, тепло лесного пожара, по направлению
к которому насекомые могут затем лететь [123, 124].
У таракана, речного рака и слизней повышение температуры
приводит к временному угнетению спонтанной активности ганглиев,
а понижение — к временному усилению этой активности. Нервный
тяж таракана, акклимированного к 30 °C, реагирует таким обра-
зом при 27 °C. Оптимальная температура, соответствующая наи-
меньшей частоте импульсов, совпадает с оптимальной температу-
рой для поведенческой агрегации животных и с температурой
акклимации (по крайней мере в среднем диапазоне температур)
[257].
У рыб имеются кожные терморецепторы и, кроме того, соко-
вая линия и головной мозг очень чувствительны к температуре-
Скопление рыб в температурном градиенте, рефлекторные ответы
плавников на местное воздействие холодной или теплой воды, тем-
пературные условные рефлексы — все эти реакции, вероятно, опо-
средуются через кожные рецепторы, так как не исчезают после
перерезки боковой линии. У Ictalurus и форели возможна выра*
Температура
133
ботка условных рефлексов на очень малые (менее 0,1°) быстрые
изменения температуры. У рыб рода Leuciscus не найдено рецеп-
торов, чувствительных только к температуре; однако после аккли-
мации рыб к 5 °C механорецепторы оказывались максимально
чувствительными к прикосновению при 18 °C, а после акклимации
к 15 °C —при 22 °C [442].
У акул и скатов на поверхности головы имеются большие сен-
сорные луковицы — ампулы Лоренцини, чувствительные к различ-
ным раздражителям, в том числе к температурным. Частота спон-
танной ритмической активности в афферентных нервных волокнах,
идущих от этих ампул, достигает максимума при 20 °C и снижает-
ся до нуля при температурах ниже 2 и выше 34 °C. Быстрое ох-
лаждение приводит к резкому кратковременному повышению ча-
стоты импульсов, а обогревание — к быстрому ее снижению. Та-
кого рода реакции можно обнаружить даже при изменении тем-
пературы на 0,05° [189]. Ампулы Лоренцини весьма чувствитель-
ны к механическому раздражению и слабым электрическим полям;
функционируют ли они как терморецепторы, не установлено, но
продемонстрирована тоническая и фазическая модуляция их ак-
тивности температурой.
Температурная рецепция хорошо развита у ямкоголовых змей
(Crotalidae), имеющих на лицевой поверхности головы термочув-
ствительные ямки, и у некоторых удавов, у которых такие ямки
расположены на губе. У ямкоголовой змеи перепонка, находящая-
ся на дне ямки, содержит множество свободных нервных оконча-
ний, оплетающих площадь свыше 1500 мкм2. Эта перепонка отде-
лена от подлежащей ткани воздушной полостью, уменьшающей
потери тепла [58]. В идущих к ямке волокнах тройничного нерва
наблюдается спонтанная активность. Этот орган служит специали-
зированным рецептором теплового излучения; он чувствителен к
инфракрасным лучам с длиной волны 0,5—15 мкм, но не реаги-
рует на более коротковолновое инфракрасное излучение и на ви-
димый свет. Змея способна определять, в каком направлении ле-
жит источник тепла, и легко замечает, когда на ямку падает теп-
ловая тень. По изменению фоновой импульсации было вычислено,
что перепонка ямки может сигнализировать об изменении своей
температуры на 0,003°, что соответствует порогу, равному 5-10-4
кал в 0,1 с. При повышении температуры на 0,4° частота нерв-
ных импульсов возрастает с 18 до 68 в 1 с [58]. У другого вида
ямкоголовых змей, чувствительного к инфракрасным лучам с дли-
ной 2,7 мкм и более, пороговая величина оказалась равной
о,1 мВт/см2. Реакция на тепло выражается в фазической вспышке
импульсов с последующим новым тоническим уровнем разряда;
ЭТа реакция не зависит от температуры тела [139]. Приспособи-
тельное значение этих высокочувствительных рецепторов направ-
ленного восприятия состоит в том, что змея может находить в тем-
ноте добычу — теплокровных животных.
134
Глава 9
Действие температуры на возбудимые мембраны
Многие рецепторы, так же как и центральные нервные струк-
туры, обладают спонтанной активностью, и температура модулирует
частоту их разрядов. Кроме того, она влияет на возбудимость ней-
ронов и проведение импульсов. Эти температурные эффекты могли
бы быть связаны с воздействием либо на ферментные системы, от
которых зависит обеспечение мембранных процессов энергией, ли-
бо непосредственно на возбудимые мембраны. Для поведенческих
температурных адаптаций, вероятно, существенны оба механизма..
Потенциалы покоя относительно независимы от температуры
вплоть до верхнего предела переносимости. Амплитуда потенциа-
лов действия в аксонах кальмара с повышением температуры
уменьшается, и максимум ее лежит в области низких температур;
эти потенциалы регистрируются даже при температурах, близких
к точке замерзания [208].
Скорости подъема и спада потенциала действия уменьшаются
при охлаждении. В безмякотных волокнах подкожного нерва кош-
ки холодовая блокада наступает при 2,7 °C, а в миелинизирован-
ных волокнах — при 7,2 °C. В аксонах омара тепловая блокада
наступает при 26, а холодовая — при 3°С. У дождевых червей
Lumbricus, акклимированных к холоду, скорость проведения в ги-
гантских аксонах выше, чем при тепловой акклимации, которая
приводит также к удлинению периода абсолютной рефрактерно-
сти [278]:
Акклимация Акклимация
к 13 °C к 23 °C
Период рефрактерное™ при 6 °C, мс 6,5 12
Период рефрактерности при 13°C, мс 3,1 3,7
При охлаждении Callinectes в аксонах возникают потенциалы дей-
ствия. Если животные находятся в среде, содержащей 10 мМ Саг
этот эффект возникает при снижении температуры на 0,5°; при
более низкой концентрации Са (8 мМ) для стимуляции достаточ-
но и охлаждения на 0,3° [418а]. Генераторный потенциал меха-
норецепторов, например телец Пачини у кошки, с повышением
температуры увеличивается (Qi0 для скорости подъема потенциа-
ла составляет 2,5, а для амплитуды — 2,1) [227]. В рецепторах
растяжения речного рака генераторный потенциал, напротив, воз-
растает при охлаждении, которое приводит и к уменьшению поро-
га для импульсации [59].
Как правило, нервы зимнеспящих животных перестают прово-
дить импульсы при более низких температурах, чем нервы живот-
ных, не впадающих в зимнюю спячку, а безмякотные волокна чув-
ствительнее к холоду, чем миелинизированные [328]. Проведение
в диафрагмальном нерве активного хомяка блокируется при 5 °C,
тогда как у хомяка, находящегося в состоянии зимней спячки, оно
Температура
135
охраняется при температурах вплоть до О °C [441]. Болыпеберцо-
ьш нерв хомяка блокируется при 2,7 °C, а тот же нерв крысы —
в и и—12 °C [327, 328]. Температуры, блокирующие проведение
в хвостовом нерве, коррелируют с обычными условиями жизни жи-
вотного: у бобра —5 °C, ондатры —4,4, аляскинского суслика —3,8,
койота +1,0 и лисицы +3,1 °C [325]. Проведение нервных им-
пульсов с понижением температуры замедляется; это замедление
в расчете на 1 °C сильнее выражено у крыс, живущих в помеще-
нии, чем у крыс, выдерживаемых на открытом воздухе. У лисицы
скорость проведения снижается больше, чем у ондатры.
У серебристых чаек из холодных районов проведение в боль-
шеберцовом нерве блокируется при температурах 11,7—14,4 °C, а
по плюсневому нерву — при 2,8—3,9 °C [80]. Q10 для скорости
проведения в одиночных нервных волокнах млекопитающих со-
ставляет 4,0 в диапазоне 8—18 °C, 2,5 в диапазоне 18—28 °C и 1,6
в диапазоне 27—37 °C [352]. Данные о влиянии холода на прове-
дение импульсов позволяют заключить, что свойства нервных
мембран приспособлены для функционирования в определенных
температурных диапазонах и что эти свойства могут видоизме-
няться.
Центральная нервная система настолько чувствительна к тем-
пературе, что сама может служить терморецептором. В некоторых
мелких нейронах брюшной нервной цепочки Lumbricus спонтанная
активность достигает максимума -при 12 °C [288]. У отдельных
гигантских нейронов в ганглиях Aplysia мембранный потенциал
покоя обнаруживает неодинаковую зависимость от температуры:
У одних нейронов он минимален при 20—25 °C, у других — при
15 °C, тогда как у третьих электроотрицательность прогрессивно
, нарастает по мере снижения температуры [337]. Мотонейроны
|спинного мозга жабы гиперполяризуются под влиянием холода и
|цеполяризуются теплом; возможна индукция импульсного разряда
^повышенными температурами [454]. У серебряного карася темпе-
ратура холодовой блокады для «высших» уровней центральной
нервной интеграции выше, чем для «низших» [371, 393].
У кошки спинномозговые рефлексы усиливаются при охлажде-
нии и достигают максимальной величины при 20—30 °C [50]. Ана-
логичное усиливающее действие холода описано для вызванных
потенциалов в мозжечке серебряного карася при зрительном раз-
дражении. У рыб, акклимированных к 25 °C, максимальная реак-
ция наблюдалась при 15—20 °C [357а].
Как правило, проводящие мембраны аксонов или мышечных
иолокон способны функционировать в более широком диапазоне
емператур, чем синапсы и концевые пластинки. У волокон порт-
яжной мышцы лягушки потенциал покоя относительно незави-
м от температуры; мышечные и нервные мембраны сохраняют
Р активность при температурах до 0 и даже до —3°С, тогда как
нцевые пластинки блокируются при 1 °C (данные получены на
136
Глава 9
лягушках, акклимированных к 10°C). Охлаждение снижает ча-
стоту спонтанных (миниатюрных) потенциалов концевой пластин-
ки; амплитуда их возрастает при охлаждении до 20—15 °C, но при
более сильном охлаждении снижается. Реакция на прямое воздей-
ствие ацетилхолина также повышается в диапазоне 15—20 °C
[236]. Передача возбуждения с диафрагмального нерва крысы на
диафрагму блокируется при 10°C, а проведение по этому нерву —
при 5 °C.
Сокращения сердца обычно возможны в более широком диа-
пазоне температур, чем сокращения скелетной мышцы. Однако у
ската Rafa тепловая блокада сердечной функции наступает при
22—25 °C, а блокада нервно-мышечной передачи — при 26—28 °C
[32].
Различные функции центральной нервной системы более чув-
ствительны к понижению или повышению температуры, чем функ-
ции периферических нервов и мышц. Так, например, у ушастого
окуня и серебряного карася плавательные реакции и условные
рефлексы исчезают при меньшем охлаждении, чем спинальные
рефлексы (рис. 9-10). В мозжечке серебряного карася реакции
клеток Пуркинье на антидромную стимуляцию прекращаются при
6,5 °C, а синаптические ответы на раздражение параллельных во-
локон— при 8—9 °C. Охлаждение рыбы приводит к сужению так-
тильных рецептивных полей для центральных сенсорных нейро-
нов, расположенных в лицевой доле [357а]. Верхняя и нижняя
температурные границы для дыхательных движений и для реак-
ции челюсти на раздражение ствола мозга изменяются при аккли-
мации следующим образом [365]:
Температура акклимации, °C Температурные границы, °C
для дыхательных движений для реакций челюсти
верхняя нижняя верхняя нижняя
31 40 10,6 40,8 5,6
19 38 8,6 39,6 3,1
У Phoxinus верхняя граница различения частоты звука при 16°C
лежит в области 800—1260 Гц, а при 25 °C — в области 1260—
1420 Гц.
В мозгу низших позвоночных найдены чувствительные к темпе-
ратуре нейроны, сходные с некоторыми нейронами в центрах тер-
морегуляции млекопитающих. При исследовании большого числа
нейронов промежуточного мозга форели 12 из них реагировали
на обогревание и 5 — на охлаждение [141]. У ящерицы Tiliq^
некоторые нейроны преоптической области промежуточного мозга
усиливают импульсацию при согревании мозга, а некоторые — при
охлаждении [63, 152].
Температура
137
Время, проводимое оычком-подкаменщиком в теплом участке
аквариума, уменьшалось при обогревании переднего мозга вжив-
ленным термодом; охлаждение мозга давало обратный эффект
Г154]. Более или менее сильное местное обогревание мозга вы-
зывает у ящериц поведенческие терморегуляторные реакции [150,
152, 173]. Ящерица Tiliqua перемещается в теплые или холодные
участки таким образом, чтобы обеспечить «предпочитаемую» тем-
пературу мозга (29,3°C). При повышении внешней температуры
понижается та температура тела, при которой животное перехо-
дит в более прохладное место. Локальное согревание преоптиче-
ской области мозга также понижает эту критическую температу-
ру, охлаждение же дает обратный эффект. По-видимому, терморе-
гуляторное поведение этой рептилии определяется с учетом инфор-
мации как о температуре мозга, так и об общей температуре те-
ла [341].
Роль испарения влаги и поведенческих реакции в регуляции
температуры у наземных пойкилотермных животных
Если воздух не насыщен влагой полностью, температура тела
у наземной улитки или лягушки ниже, чем показывает сухой или
черный термометр, и выше показаний влажного термометра. В во-
де температура тела не отличается от температуры внешней среды.
При отсутствии ветра температура тела лягушки ближе к показа-
ниям сухого термометра, а при движении воздуха приближается
к показаниям влажного термометра. У жабы при внешней темпе-
ратуре 27,6 °C и относительной влажности 82% клоакальная тем-
пература составляла 26,5 °C, а при той же внешней температуре и
относительной влажности 27% температура в глубине тела была
всего лишь 17,5 °C [322]. При 20 °C интенсивность обмена у ля-
гушки соответствует теплопродукции 6 кал/ч, но при этом лягушка
за 1 ч теряет путем испарения 3,2 г воды, на что расходуется
1850 кал [322]. Таким образом, во влажном воздухе потери тепла
при испарении обычно превышают теплопродукцию и температура
тела оказывается ниже, чем у животного, помещенного в воду той
же температуры.
Насекомые. У насекомых и равноногих раков температура тела
может отличаться от температуры воздуха в результате потери
тепла при испарении влаги, выработки метаболического тепла и
поглощения тепловой радиации [83]. У таких крупных насекомых,
как тараканы Blatta и Periplaneta, саранча Schistocerca и кобыл-
ка Gastrimargus, температура тела оказалась на 2,6° ниже темпе-
ОГ)А^Ы В03Духа’ пРичем эти насекомые теряли путем испарения
,06 г воды в 1 ч. Температура тела мокрицы Armadillidium была
О На 0’4° ниже температуры воздуха, а потеря воды составляла
, U2 г/ч. Во влажном воздухе температура живых особей Ligia
хРавноногий рак, обитатель верхней литоральной зоны) выше,
138
Глава 9
чем мертвых, а в сухом воздухе на солнце наблюдается обратная
картина, так как поглощение тепла за счет теплопроводности не-
велико, а поглощение тепловой радиации уравновешено потерей
тепла в результате конвекции и испарения.
Насекомые теряют большую часть воды через дыхальца, и при-
чиной гибели в теплом сухом воздухе может быть не перегрева-
ние как таковое, а высыха-
° ние. Верхняя летальная тем-
, пература для некоторых
мелких насекомых (Хепо-
psylla, Pediculus, Lucilia и
Tenebrio) при часовой экспо-
зиции не зависела от влаж-
ности. Однако при 24-часо-
вой экспозиции летальные
температуры в сухом возду-
хе оказались гораздо ниже,
чем во влажном [322]
(рис. 9-25). Очевидно, в пер-
вом случае причиной гибели
был перегрев, а во втором—
высыхание. Выживанию бе-
локрылых клопов при темпе-
ратурах ниже 50 °C способ-
ствует высокая влажность,
а при более высоких темпе-
ратурах—низкая влажность.
Рис. 9-25. Наивысшая температу-
ра, выдерживаемая насекомыми в
течение 24 ч при различной влаж-
ности [322].
I — Tenebrio molitor, II — Xenopsylla
cheopis (имаго), III — X. cheopis (ли-
чинки), IV — Pediculus humanus, V —
Lucilia sericata.
Причиной смерти при температурах ниже 48—50 °C бывает высы-
хание, тогда как при более высоких температурах гибель обуслов-
лена каким-то прямым тепловым эффектом.
При внешней температуре около 40 °C температура тела Sc hi-
stocerca gregaria в сухом воздухе на 3—4° ниже, чем во влажном-
Эпикутикулярный липидный слой при температуре около 30 °C
становится проницаемым [83]. Таракан Blatta orientalis во влаж-
ном воздухе переносит в течение 24 ч температуру 37—39 °C, а в
сухом — только до 34—36 °C. У жуков-чернотелок вентиляция тра"
Температура
139
хей заметно усиливается при высоких внешних температурах,
у мухи цеце (Glossina) дыхальца начинают открываться и закры-
ваться примерно при 30 °C, а при 39—41 °C они постоянно откры-
ты. Температура тела мухи цеце, находящейся в сухом воздухе,
при 45 °C может быть ниже внешней температуры на 1,66°, а во
влажном воздухе такой разницы нет [117, 196]. Очевидно, испаре-
ние воды через дыхальца у некоторых насекомых служит для ох-
лаждения тела. Потеря воды через кутикулу не контролируется и
возрастает при повреждении воскового слоя высокими температу-
рами; в последнем случае высыхание может свести на нет пользу
от охлаждения.
Рептилии. Поведенческие и метаболические адаптации позво-
ляют рептилиям нормально существовать даже при температурах,
близких к критическому тепловому максимуму. О достижении это-
го максимума будут свидетельствовать дезорганизация движений
и последующая смерть. Доусон [95] разделил семейства ящериц
на две группы — термофильную, или теплоустойчивую (предпочи-
таемые температуры выше 35°C), и менее термофильную, более
чувствительную к теплу (предпочитаемые температуры 30—35°C).
Некоторые змеи отличаются высокой теплоустойчивостью, мало
изменяющейся под влиянием акклимации ['229].
Температура аккли- Температура аккли-
мации 15 °C мации 30 °C
критические температуры, °C
верхняя нижняя верхняя нижняя
Natrix 40 6 41 8,2
Thamnophis 39 4 42 8
используются весьма
и ящерицы греются на
В температурной адаптации рептилий
различные формы поведения. Многие змеи
солнце, поглощая лучистое тепло и получая также тепло от на-
гретых камней и песка. Способность кожи отражать тепловое из-
лучение изменяется в зависимости от пигментации [414]:
Отражение от кожи, % Поглощение, %
Iguana 26 74
Phrynosoma 35 65
Морские игуаны выбирают на берегу такие температурные усло-
вия, при которых температура тела равна 35—37 °C, а в море кор-
мятся при 25 °C. На береговых скалах, где черный термометр по-
казывает свыше 50°C, игуана благодаря принимаемым ею позам
охраняет температуру тела ниже 40 °C [20]. Морские змеи гре-
Тся в солнечных лучах, лежа на поверхности океана.
Жабовидная ящерица Phrynosoma, помещенная в террариум с
вещенными и теневыми участками, обычно перемещается так.
140
Глава 9
чтобы поддерживать температуру тела на уровне 35 °C при мак-
симальной добровольно переносимой температуре 39 и минималь-
ной 28 °C. Она ищет тень при 37,7, зарывается в песок при 40,5 и
тяжело дышит при 43,2 °C [173].
Iguana прячется при 39,2 °C, выходит при 38,3 °C, а при темпе-
ратурах ниже 36 °C становится неактивной [315]. Пустынная яще-
рица Amphibolurus гибнет при 49,3 °C, но может жить при темпе-
ратурах, очень близких к этой летальной границе, сохраняя спо-
собность к координированным движениям вплоть до 48,5 °C [181].
Игуана Dipsosaurus регулирует свою температуру, изменяя позу;
эта ящерица предпочитает температуру тела 38,5 °C, но может про-
извольно выдерживать до 42 °C [108]. К поведенческим реакциям
на температуру относятся также ориентация по отношению к сол-
нечным лучам, способствующая большему или меньшему обогре-
ванию, поиски тени или укрытия, бег «на ходулях», когда тело
приподнято над субстратом. Многие ящерицы и змеи, обитающие
в пустынях, в жаркое время прячутся в норах и ведут ночной об-
раз жизни. Самка питона, высиживая яйца, вырабатывает тепло
с помощью мышечных сокращений. При внешней температуре
33 °C температура ее тела может на 7,3° превышать температуру
субстрата; при 25 °C она потребляет в 9,3 раза больше кислорода,
чем самка, не высиживающая яиц ['221].
У рептилий кожа менее проницаема, чем у амфибий, и поэто-
му они теряют меньше воды путем испарения. При понижении от-
носительной влажности со 100 до 7% (при 20°C) температура те-
ла у жабовидной ящерицы, черепахи и аллигатора снизилась все-
го лишь на 1°, а у лягушки и саламандры — на 8°. Многие репти-
лии, например, сцинки, при приближении температуры тела к
верхней критической границе начинают тяжело дышать, а другие
просто держат рот открытым. В температурном диапазоне 32—
40 °C игуана отдает половину всего метаболического тепла путем
испарения воды, а при 44 °C эта отдача тепла уже на 25% превы-
шает его выработку [414]. Потеря воды у Sauromalus характери-
зуется следующими величинами [85]:
Температура акклимации
26 °C 43,5 °C
Кожное испарение воды, мг/(см2-сут) 2,0 9,5
Потеря воды при дыхании, 1,3 8,0
мг/(см2-сут)
Теплопроводность кожи у этой рептилии втрое выше, чем у мле-
копитающих.
Sphenodon активен ночью и согревается быстрее, чем остывает
[497]. Медленно остывает и ящерица Amphibolurus-, выбор поло-
жения, в котором она греется на солнце, может влиять на скорость
изменения температуры тела [28]. На воздухе морская игуана
охлаждается со скоростью 0,3 град/(кг-мин)—втрое медленнее,
Температура
141
чеМ в воде [25]. Греющийся на солнце варан утром повышает тем-
пературу своего тела со скоростью 0,14 град/мин, а ночью осты-
вает со скоростью 0,02 град/мин [443]. Черепахи Pseudemys в во-
де согреваются быстрее, чем остывают, а на воздухе, наоборот,
остывают быстрее. Во время согревания кровоток в сосудах кожи
возрастает, а во время охлаждения уменьшается. Эти изменения
зависят от местной температуры кожи, а не от общей температу-
ры тела. У рептилий различие скоростей согревания и охлаждения
обусловлено главным образом циркуляторными адаптациями. Ин-
тенсивность обмена у рептилии ниже, а теплопроводность выше»
чем у млекопитающего сравнимых размеров.
Гомойотермные животные
Согласно определению, у пойкилотермных животных темпера-
тура тела непостоянна, у гомойотермных поддерживается на по-
стоянном уровне, а у гетеротермных регулируется не полностью
или резко различна в разных частях организма. Как мы уже ви-
дели, многие так называемые пойкилотермные животные в опре-
деленных условиях (особенно в период активности) могут поддер-
живать относительно постоянную температуру тела. Это может
достигаться за счет поглощения внешнего тепла (эктотермия), на-
пример путем соответствующей ориентации тела по отношению к
его источнику, или за счет собственной теплопродукции (эндо-
термия), как, например, при мышечном разогреве. В то же время
у многих так называемых гомойотермных животных температура
тела не всегда постоянна, и они могут становиться по существу
пойкилотермными. Несмотря на эту нечеткость границ, способ-
ность регулировать температуру тела позволяет все же выделить
среди животных группу «теплокровных», к которым относятся пти-
цы и млекопитающие.
Сопоставим некоторые параметры для эктотермных животных
(ящериц), гетеротермных и гомойотермных млекопитающих и
Двух групп птиц, различающихся по степени активности [92]:
Нормальная темпера- тура тела, °C Метаболическая теплопродукция при обычной темпе- ратуре тела, ккал/(кг3• сут) Теплопродукция, рассчитанная для температуры тела 38СС, ккал/(кг3/а • сут)
Ящерицы 30 7,5 19,5
Однопроходные 30 34 62
Сумчатые 35,5 49 62
Высшие млекопитающие 38 69 62
Птицы (кроме воробьи- 39,5 83 72
пЫд ] Воробьиные 40,5 143 114
142
Глава 9
Из приведенных данных видно, что у рептилий, располагающих
только поведенческими и сосудодвигательными средствами термо-
регуляции, теплопродукция намного ниже, чем у гомойотермных
животных — даже у форм с относительно несовершенной гомойо-
термией.
Реакции на охлаждение. Птицы и млекопитающие отвечают на
'раздражение холодовых рецепторов кожи серией рефлексов, на-
правленных на сохране-
ние тепла. Кровеносные
сосуды кожи сужаются,
волосы или перья могут
приподниматься, а иногда
животное принимает еще
такую позу, при которой
уменьшается поверхность
тела, контактирующая с
внешней средой. У челове-
ка вазоконстрикция мо-
жет уменьшить потерю
тепла на Ve—Уз- У многих
Рис. 9-26. Температура на раз-
личных глубинах носового хода
у кенгуровой крысы, дышащей
влажным (I) или сухим (II)
воздухом. Указаны также рек-
тальная температура (III) и
температура воздуха (IV)
[228].
животных для теплоизоляции может использоваться слой поверх-
ностных тканей, в особенности жир. При внешних температурах до
34 °C крысы, лишенные шерсти, теряют за счет теплопроводности,
конвекции и излучения больше тепла, чем нормальные животные
[198]. У животных с относительно длинным носовым ходом конден-
сация влаги и поверхностный теплообмен позволяют сохранять во-
ду и тепло при дыхании [228] (рис. 9-26).
Если физическое охлаждение продолжается и механизмы теп-
лоизоляции недостаточны для поддержания температуры тела, мо-
жет возникнуть мышечная активность и повыситься потребление
кислорода. Поверхностные ткани (например, жировая), служащие
у многих животных для теплоизоляции, могут иметь более низкую
температуру, чем глубинные ткани.
Центр терморегуляции, находящийся в гипоталамусе, коорди-
нирует периферические рефлексы и активирует гормональную
стимуляцию метаболизма еще до того, как понизится температур3
Температура
143
крови. Часть тепла вырабатывается в результате мышечной дро-
>кИ (у птиц и млекопитающих), а часть — с помощью иных меха-
низмов в мышцах, бурой жировой ткани и других тканях (у мле-
копитающих). Термогенез при охлаждении тела обычно активи-
руется у млекопитающих норадреналином; подъем потребления
О2 пропорционален уровню этого гормона в крови [234]. Инъек-
ция норадреналина оказывает калоригенное действие, которое при.
акклимации к холоду выражено сильнее, чем при акклимации к.
теплу; оно может проявляться у кураризированных животных и у
животных с удаленными брюшными внутренностями, т. е. гормон
непосредственно воздействует на метаболизм мышечной ткани.
Эффект норадреналина наиболее значителен у мелких животных
и пренебрежимо мал у животных весом более 10 кг [187]. В об-
щем повышении интенсивности обмена на холоду участвует также
щитовидная железа; удаление ее приводит к исчезновению реак-
ции на норадреналин. При реакции на холод снижается содержа-
ние ненасыщенных жирных кислот в митохондриях печени, но по-
сле введения тиоурацила этого не происходит. В норме крыса спо-
собна регулировать температуру своего тела при температурах
воздуха до —10 °C, после удаления щитовидной железы или над-
почечников— до —2 °C, а после удаления обеих этих желез —
всего лишь до 4-10 °C. У млекопитающих непосредственным сти-
мулятором термогенеза служит норадреналин, тогда как у птиц
ведущая роль принадлежит щитовидной железе. В опытах на мно-
гих видах птиц инъекции адреналина или норадреналина не ока-
зывали никакого стимулирующего действия [96, 165].
Пребывание в течение нескольких часов на холоде активирует
кору надпочечников — вероятно, через гипофиз (АКТГ); в резуль-
тате могут появляться общие признаки стресса, например умень-
шение числа эозинофилов и лимфоцитов. У человека иногда
уменьшается объем циркулирующей крови и возрастает ее кон-
центрация. У различных животных сохранению тепла способст-
вуют такие поведенческие реакции, как скучивание особей, устрой-
ство гнезд, закапывание в снег или свертывание в клубок, умень-
шающее охлаждаемую поверхность.
Норадреналин вызывает липолиз и ускоряет окисление жир-
ных кислот [214]. Уровень липидов сыворотки у крысы вначале
понижается, а затем медленно восстанавливается. На холоде у
крыс может отмечаться двукратное повышение уровня норадре-
налина в крови и при этом соответственно возрастает потребление
и2 мышцами (но не почками) [233]. Максимальная возможная
теплопродукция у человека — 370 ккал/ч; постоянная температура
ела может поддерживаться при температурах воздуха от —6,7 до
С [222]. У эскимоса при внешней температуре 17 °C интенсив-
ость обмена повышается на 22 кал/(м2-ч) [уровень основного
ПрМе?о— 55 кал/(м2-ч)] [2], У японского перепела (Coturnix)
Ри 4 С снижается содержание гликогена в мышцах и возрастает
144
Глава 9
Рис. 9-27. Зависимость потребления кислорода некоторыми
А. Броненосец [238]. Б. Красная американская лисица (а) и тюлень обыкновенный (б)’
Г. Человек (без одежды); теплопродукция измерялась
концентрация жирных кислот в плазме крови [130]. Повышенная
теплопродукция при охлаждении тела — результат метаболиче-
ских реакций, связанных или не связанных с мышечной дрожью,
относительное значение этих двух компонентов варьирует. Заме-
ной дрожи может служить действительная мышечная работа-
У крыс, акклимированных к холоду, область переносимых темпе-
Температура
145
млекопитающими от внешней температуры. Ехидна 14131-
белыми кружками представлены летние данные, черными зи
при трех различных скоростях ветра L22ZJ.
ратур расширяется благодаря термогенезу, не связанному с ь
шечной дрожью, который не увеличивается при работе мышц [ J-
Измеряя интенсивность обмена (Л4) при различных температу
Pax воздуха, можно обнаружить нейтральный температурный диа
пазон, соответствующий минимальному потреблению Ог и о ычно
•лежащий несколько ниже температуры тела. Для крупных млеко
Ю—1514
146
Глава 9
питающих эта термонейтральная зона может быть широкой, а для
мелких млекопитающих и птиц соответствовать точке минимума
на кривой, отражающей зависимость М от температуры. У неко-
торых птиц М продолжает снижаться по мере приближения к
максимальной переносимой температуре [486]. У большинства
птиц и млекопитающих нижней границе термонейтральной зоны
соответствует критическая температура (нижняя Ткрит), ниже ко-
торой физические механизмы терморегуляции уже недостаточны
и для поддержания температуры тела необходима активация ме-
таболизма. Верхней границе термонейтральной зоны также соот-
ветствует критическая температура (верхняя Гкрит), выше кото-
рой уровень обмена возрастает из-за неэффективности механизмов
охлаждения (рис. 9-27). У домового воробья термонейтральная
зона во время дневной активности имеет характер точки, а во
время ночного сна представляет собой плато [218, 283, 284].
В табл. 9-3 приведены данные о верхней и нижней границах тер-
монейтральной зоны для различных млекопитающих и птиц. Низ-
кие уровни нейтральной температуры (Гн) характерны для видов
с низкими величинами отношения поверхности тела к весу и с хо-
рошей теплоизоляцией, будь то перья или мех. Если теплоизоля-
ция зимой улучшается, то происходит снижение Гн. Понижение
термонейтральной зоны коррелирует с условиями обитания. Так,
например, у живущей в пустыне тауи Аберта Гн соответствует
25—35 °C, а у обитающей на холодных прибрежных склонах корич-
невой тауи — 23—33 °C; Тн для садовой овсянки из южной Кали-
форнии 32—38 °C, а для обыкновенной овсянки из более северных
районов 25—32 °C.
Для крупных тропических животных и для человека нижняя
Ткрит (ниже которой интенсивность обмена возрастает) равна
+26 °C, а для полярной лисицы —30 °C. Температурные кривые
метаболизма для животных с низкой Гкрит имеют обычно меньший
наклон и часто экстраполируются до температуры тела. Малый
наклон кривой свидетельствует о небольшой теплоотдаче (удель-
ной теплопроводности).
Когда все регуляторные механизмы оказываются недостаточ-
ными и температура тела падает, это приводит к замедлению кро-
вотока и может привести к нарушению функций мозга. У крысы
кровоток в сосудах брыжейки слепой кишки прекращается при
20 °C, а у хомяка в сосудах защечного мешка — при 5—10 °C.
Кошка теряет сознание при температуре тела 25—27 °C, но неко-
торые рефлексы сохраняются при снижении температуры пример-
но до 16 °C. У человека некоторое изменение психики отмечается
уже при температуре тела 35 °C; дыхание продолжается при тем-
пературах до 25 °C. У собаки дыхание останавливается при тем-
пературе тела 20—25 °C. У крысы нижняя температурная граница
для локомоции 21, для дыхания 13 и для сердечной деятельности
8 °C [362]. При кормлении крыс пищей, содержавшей 50% жиров,
Таблица 9-3
Температура тела в период активности и термонейтральные зоны
— Термонейтральная зона, °C
Животное Температура тела, °C НИЖНЯЯ граница верхняя граница
Коупные млекопитающие Человек [499] Бабуин [135] Снежный баран [41] Коза [451] Морской котик [225] Верблюд [411] Кит-горбач [330] Грызуны и родственные животные Baiomys [215] Perognathus [468] Р. hispidus [482] Р. longimembris [82] Liomys [219] Microdipodops [51] Peromyscus eremicus [320] P. californicus [320] Tamias [482] a Citellus [300] Даман, Procavia [453] Летучие мыши Macroderma [292] Pteropus polycephalus [31] P. scaphilalus [31] Syconycteris [31] Dobsonia [23] Eutnops [291] Myotis [349] Eptesicus [264] Miniopterus [332] Примитивные млекопитающие Броненосец Dasypus [238] Ехидна [413] Кенгуру Macropus [92] Опоссум Marmosa [333] Сумчатое Cercaertus [24] Крупные птицы Куропатка Lophortyx [217] Lophortyx [54] Голубь инка [316а] Голубь [377] Иволга [386] Красноногая олуша [21] Олуша обыкновенная [21] Малый козодой [283] Козодой Eurostopodius [96] Домашняя курица [130] 37 38,1 37,9 (ночь) 39,8 (день) 37—40 38 38,1 36 35 36—38 36,8—38,7 34,7 38,8 36,6 36,4 36—40,3 35,5—39,5 37—38,5 35—39 35-39 36,4 37 31,4—33,1 35,6 37—39,1 34—36 30,7 33,9—34,5 33,2 (день) 35,7 (ночь) 32—38 40,6 40,6 38,8—42,7 40,0 40,3 (день) 38,0 (ночь) 40,7 (день) 38,3 (ночь) 36—40 39—42 39,8 27 20 (вода) 0—3 (воздух) 29 32 30 31 31 30 30 27 28,5 20 30 18 24 3 3 31 32,5 30 20 30,5 28 31 30 27,3 35 25 34 35 ',5 3 4 ,5 32 36 33,5 34 35 35 34,5 32 30 35 35 35 34,5 30 35 37,5 30
148
Глава 9
Продолжение
Животное Температура тела, °C Термонейтральная зона, °C
нижняя граница верхняя граница
Страус [86] Сова 39,3 38 25 37
Баклан 39—40 40
Лесной аист [246] Буревестник [212] Куропатка [241] 40,7 39,5 (день) 37,7 (ночь) 39,6 4 36
Американская вечерница [486] 41 (день) 16 34
Клёст Loria [98] 38,5 (ночь) 38,5—40 15 28,5
Серая сойка (Perisoreus) [475] 42,3 (день) 36 (лето)
Воробьиные Зебровый ткачик [65] 41 (ночь) 41,5—42,2 7 (зима) 36 42
Зебровый ткачик [67] 39,8—42,4 30 40
Воробей Харриса [385] 41,1 (день) 38,3 (ночь) 2 5
Домовый воробей [218] Колибри Patagona [287] 27
Eugenes [287] 31
Lampornis [285] 31
Синегорлый колибри [285] 38,5—40 31- -33
Eulampia [145] 40 30
Stellula [281] 35-40 27- -30
остановка сердца происходила при температуре тела 2,9 °C, если
это были насыщенные жиры, и при 7,2 °C в случае ненасыщенных
жиров [163].
Для оценки эффективности теплоизоляции использовались раз-
личные показатели. Критические температуры (верхняя и ниж-
няя) и ширина термонейтральной зоны зависят от размеров тела
и от теплоизоляции.
Удельная теплопроводность равна теплопотере с единицы по-
верхности тела при разности температур в 1 °C:
К =кал/(см2 • ч • град).
Площадь поверхности тела можно вычислить с помощью пра-
вила 2/3 (см. гл. 5), но ввиду ненадежности получаемых резуль-
татов лучше относить потерю тепла к весу тела. В этом случае
удельная теплопроводность
С=мл О2/(г-ч-град),
или
Г— м
Тт-Тс ’
1
Температура 149
где М— потребление кислорода, мл/(г-ч), Тт — температура тела
— температура внешней среды. Если С надо выразить в
ккал/(г-ч-град), вводят коэффициент 4,74, соответствующий ка-
лорическому эквиваленту 1 л О2.
Мерой эффективности метаболизма служит отношение всей
метаболической энергопродукции к величине теплопотери, т. е.
М1С.
Испарение с поверхности кожи и дыхательных путей приводит
к значительной потере тепла; поэтому при вычислении теплопро-
водности необходимо ввести соответствующую поправку:
Р___М—(Теплопотеря при испарении)
(Площадь) • (Тт — Тс)
Некоторые величины С приведены в табл. 9-4.
Если животное подчиняется ньютоновскому закону охлаждения
и поддерживает постоянную температуру тела, то рост теплопро-
дукции находится в линейной зависимости от снижения внешней
температуры. Наклон кривой MfTc будет мерой теплопроводности.
В табл. 9-4 приведено также несколько величин теплопровод-
ности, измеренной по потреблению О2 при различных внешних
температурах и постоянной температуре тела. Данные, полученные
при измерении скорости охлаждения мертвых животных, удовле-
творительно согласуются с представлением о том, что мелкие мле-
копитающие остывают быстрее крупных [335] (рис. 9-28).
Вес, г Теплопроводност ь, кал/(см2 ч • град)
Sorex 2,3 0,31 (конечности
выпрямлены)
Clethrionomys 14,3 0,47
Microtus 20,4 0,43
Как правило, наклон кривой метаболизм — внешняя темпера-
тура зимой (а также у арктических животных) меньше, чем летом
(или у тропических животных). Это обусловлено лучшей теплоизо-
ляцией и, следовательно, меньшей теплопроводностью при адап-
тации к холоду (рис. 9-29; см. также стр. 157). О сезонных изме-
нениях теплопроводности свидетельствуют и другие данные [385].
Теплопроводность, мл О2/(г • ч-град)
зима лето
Кардинал Американская вечер- ница Воробей Харриса Зонотрихия белоголо- вая 0,098 0,10 0,125 0,205 0,23
Таблица 9-4
Теплопроводность при температурах ниже термонейтральной зоны
Животное Вес тела, г Удельная тепло- проводность, мл О2/(г • ч • град) Теплопровод- ность, мл О2/ (см2 . г • град)
Мелкие млекопитающие
Perognathus [468] 21 0,19
Р. hispidus [482] 39,5 0,201
Р. longimembris [82] 11,55 0,28
Peromyscus maniculatus [317] 24 0,17
Р. eremicus [320] 21,5 0,18 0,22
Р. californicus [320] 45,5 0,10 0,18
Microtus [335] 47 0,166
Dipodomys [93] 35 0,174
Sorex [333] 3,3 0,54
Marmosa [333] 13 0,26
Летучие мыши
Macroderma [292] 148 0,11
Eumops [291] 56 0,56
Pteropus polycephalus [23] 598 0,026
P. syconycteris [23] 0,66
Другие млекопитающие
Морская свинка 430 0,04
Кролик [166] 1581 0,029
Procavia [453] 2630 0,02
Собака [150] 6666 0,022
Кенгуру Macropus [92] 4960 0,16
Американская красная лисица
[92] 0,14
Птицы
Серая сойка (Perisoreus) [475] 71,2 0,06
Козодой [96] 88 1,7
Американская вечерница [98] 60 1,8
Голубь Scardafella [316а] 42 0,20 (день)
0,14 (ночь)
Кардинал Richmondena [280] 40 0,10
Колибри
Patagona [287] 19,1 0,17
Eugenes [287] 0,30
Archilochus [287] 3,3 0,50
Lampornis [285] 7,9 0,23
Eugenes [285] 6,6 0,30
Calypte [280] 5,4 0,44
Stellula [280] 3 0,61
Selasphorus [280] 3 0,57
Температура
151
У некоторых животных, например у впадающих в холодовое
оцепенение Miicroichiroiptera, изучение теплопроводности затрудне-
но, так как у них снижение температуры может приводить к умень-
шению активности.
Акклимация к холоду. У птиц и млекопитающих, находившихся
несколько недель в условиях холода, появляются различные при-
Рис. 9-28. Связь между теплопроводностью и весом тела у различных мелких
птиц (черные кружки) и млекопитающих (белые кружки) (буквы — сокращения
названий видов) [281].
знаки акклимации — иные, чем у пойкилотермных животных. Из-
меняется нижняя летальная температура, а также температура,
при которой животное еще может поддерживать тепловой баланс,
собаки, помещенные в холодную воду, погибали, когда температу-
ра тела снижалась до 18,6 °C, но после предварительной акклима-
ции к низкой внешней температуре они выживали при охлажде-
нии тела до 14,9 °C. Когда крыс, живших при 28 °C, переводили в
помещение с температурой 5 °C, потребление кислорода возрастало
У них за трое суток с 55 до 88 мл/(ч • Ц7°>52), а через три недели
ыло вдвое выше, чем при 28 °C. Если интенсивность обмена изме-
рять при 30, а не 5 °C, то вначале она не окажется повышенной, но
осле недели пребывания животных на холоде усиление метабо-
изма будет заметно при любой температуре [232]. У крыс, аккли-
152
Глава 9
мированных к холоду, реакция на норадреналин выражена гораз-
до сильнее, чем при акклимации к теплу.
Крысы, содержавшиеся при 9 °C, потребляли за сутки количе-
ство кислорода, равное 5,5% веса тела, а у крыс, содержавшихся
при 28 °C, эта величина составляла лишь 3,0%. Потребление пищи
Внешняя температура, °C
Рис. 9-29. Зависимость интенсивности обмена от внешней температуры у птиц.
А. Американская белохвостая куропатка [241]. Б. Колибри Eulampis в покое (черные круж-
ки) и в состоянии оцепенения (белые кружки) [145].
в первой из этих групп было на 8,6% больше, чем во второй [258].
Доля мышц в общем метаболизме составляет у крыс 57, печени —
22, а кожи — 6% [232]. Для поддержания повышенного потребле-
ния кислорода необходима щитовидная железа, и активность ее
после недельного пребывания на холоде усиливается. У людей
после 9 дней, проведенных при 8 °C, потребление Ог в состоянии
Температура
153
покоя возросло с 0,23 до 0,46 л/мин, а в активном состоянии — с
1,50 до 1,53 л/мин; таким образом, основной обмен повысился в
гораздо большей степени, чем активный обмен [390].
Данные о биохимических изменениях при холодовой акклима-
ции носят противоречивый характер. Это обусловлено тканевыми
и видовыми особенностями, несовпадением результатов, получен-
ных на гомогенатах и на интактных митохондриях, и т. п.; при низ-
ких температурах различия могут также зависеть от того, способно
ли данное животное впадать в зимнюю спячку, а если способно,
то находится ли оно в данный момент в состоянии спячки или в ак-
тивном состоянии. Кроме того, ферменты большей частью иссле-
довали при насыщающих концентрациях субстратов; сведения о
величинах Км и активности ферментов при физиологических кон-
центрациях субстратов практически отсутствуют.
У крыс, акклимированных к холоду (5 °C в течение более
10 дней), потребление кислорода различными тканями при 37°C
повышено. У тиреоидэктомированных крыс метаболизм мышц и
печени повышается в меньшей степени. У леммингов, акклимиро-
ванных к 1 °C, потребление кислорода скелетными мышцами на
40% выше, чем при акклимации к 17°C.
В гомогенатах печени и мышц крыс, акклимированных к холо-
ду, повышена активность сукцинатдегидрогеназы, малатдегидроге-
назы и цитохромоксидазы, отмечается усиленное окисление изо-
цитрата, а-кетоглутарата и глутамата, но понижена активность
ферментов пентозного шунта — 6-фосфоглюконатдегидрогеназы и
глюкозо-6-фосфатдегидрогеназы. Активность лактатдегидрогеназы
и НАД-Н-зависимой цитохром-с-редуктазы снижена или остается
на прежнем уровне [157, 158]. Почечная ткань крыс и хомячков,
содержавшихся две недели при 5 °C, обнаруживает повышенную
способность к глюконеогенезу, т. е. образованию глюкозы из пи-
рувата и из аминокислот [62]. У крыс, адаптированных к 28 °C,
содержание цитохрома с в брюшных мышцах равно 33 мкг/г, а
у адаптированных к 6 °C — 50,5 мкг/г [105]. Акклимированные
к холоду крысы обладают повышенной способностью к окислению
жирных кислот и сильнее реагируют на инъекцию норадреналина
[213, 214]. У крыс и хомячков при холодовой акклимации возрас-
тает степень ненасыщенности белого жира, а у хомячков — также
и ненасыщенность жиров в печени [490]. Кроме того, у хомячков,
снижается содержание линолевой и пальмитиновой кислот в около-
почечном и подкожном жире, тогда как относительные количества
олеиновой и стеариновой кислот, наоборот, возрастают [266]. В пе-
чени крысы заметно уменьшается отношение P/О, особенно при
окислении p-оксибутирата; это говорит о более интенсивном НАД-
зависимом окислении жирных кислот [158]. Многие из этих сдви-
гов могут быть вызваны при более высоких температурах с по-
мощью инъекции тироксина; на холоде заметны признаки повы-
шенной активности щитовидной железы.
154
Глава 9
В отличие от тканевых гомогенатов изолированные митохонд-
рии крыс, акклимированных к холоду, не обнаруживают никаких
изменений в способности окислять сукцинат, глутамат и р-оксибу-
тират и в активности цитохромоксидазы [78, 298]. В митохондриях
не изменяется и отношение P/О. Однако митохондрии из печени
хомячков, содержавшихся на холоде, но не находившихся в состоя-
нии зимней спячки, более активно окисляли сукцинат и р-оксибу-
тират, а величина P/О оказывалась пониженной, особенно в тех
случаях, когда измерения производились при низкой температуре
[298]. У крыс, акклимированных к 5 °C, понижено содержание не-
насыщенных жирных кислот в митохондриях. Этот эффект холо-
довой акклимации можно предотвратить введением тиоурацила и
воспроизвести при более высоких температурах путем инъекции
тироксина [354]. В митохондриях из печени хомячков, акклимиро-
ванных к 35 °C, отмечалось пониженное использование сукцината,
глутамата, изоцитрата и p-оксибутирата по сравнению с мито-
хондриями от животных, акклимированных к 23 °C. В митохондри-
ях из тканей хомячков и сусликов, находившихся в состоянии зим-
ней спячки, при 37 °C можно обнаружить заметно повышенное
окисление сукцината и p-оксибутирата и возросшую активность’
цитохромоксидазы. В митохондриях из бурой жировой ткани таких
животных окисление «-глицерофосфата и изоцитрата повышено в
3—4 раза больше, чем в печени [77, 78].
Таким образом, у животных, не впадающих в зимнюю спячку,
например у крыс, повышение дыхания на холоде обусловлено, по-
видимому, действием внемитохондриальных механизмов, вероятно
находящихся под контролем щитовидной железы, а у зимнеспящих
животных в реакции на холод участвуют и митохондрии. При воз-
действии холода в обеих этих группах животных наблюдается тен-
денция к уменьшению насыщенности жирных кислот, к усиленному
окислению жиров и к уменьшению отношения P/О вследствие воз-
росшего потребления О2. В конечном итоге больше энергии расхо-
дуется на теплопродукцию и меньше — на образование АТФ.
В лабораторных условиях холодовая акклимация крыс и кроли-
ков не приводит к понижению нижней Ткрит, т. е. температуры, ни-
же которой интенсивность обмена возрастает. Между тем при
акклиматизации в природных условиях эта критическая температу-
ра понижается. В лаборатории воздействие холода на крыс и кро-
ликов ведет к усиленной васкуляризации кожи, что, по-видимому,
не имеет приспособительного значения. Частота сокращений серд-
ца in vivo при понижении температуры тела возрастает, a in
vitro наблюдается обратная реакция. Однако после акклимации
к 30—33 °C сердце сокращается in vitro медленнее, чем после ак-
климации к холоду (в опытах при одной и той же температуре)
[215].
Акклиматизация к холоду. Увеличение теплопродукции, наблю-
даемое при холодовом воздействии в лаборатории, в условиях хо-
Температура
155
годного климата может лишь временно разрешить проблему холо-
дового стресса. В зимнее время, а также в Арктике или Антарктике
у животных наблюдаются совсем иные изменения, чем после не-
скольких недель акклимации в холодном помещении. В природе
температура понижается на более долгое время, зимние холода
усиливаются постепенно; к тому же взаимодействие таких факто-
ров, как фотопериод, питание и поведение, тоже существенно из-
меняет ситуацию по сравнению с лабораторными условиями.
Зимой млекопитающие и птицы лучше переносят сильное
охлаждение, чем летом. Peromyscus maniculatus зимой выдержива-
ет в течение 200 мин температуру —35 °C, а летом — только
— 15°C. Уменьшение наклона кривой метаболизм — температура,
наблюдаемое, в частности, у Peromyscus, Zapus и Citellus, свиде-
тельствует о более эффективной теплоизоляции. Возможно, что по-
вышенная устойчивость к холоду связана и с изменениями в цент-
ральной нервной системе. У диких крыс холодовая дрожь слабее,
а не связанный с дрожью термогенез при введении норадреналина
выражен сильнее, чем у лабораторных крыс в холодном помеще-
нии [194]. Кривые метаболизм — температура, построенные для
мелких птиц (домового воробья, дубоноса, голубя), не обнаружи-
вают заметных сезонных особенностей. В зимнее время возраста-
ет потребление пищи, а вызванная холодовым стрессом теплопро-
дукция поддерживается дольше, чем летом [165, 166] (рис. 9-30).
Зимний жир отличается большей ненасыщенностью, чем летний.
У многих птиц, а также у дикобразов, белок, собак и кроликов
теплоизолирующие свойства оперения или меха зимой улучшают-
ся. У английского домового воробья общий вес перьев возрастает
зимой на 29%, а переносимая температура в осенний период сни-
жается со скоростью 6° в месяц [19]. Величина теплоизоляции воз-
растает у серой сойки (Perisoreus) с 0,16 (град-м2-ч)/ккал летом
до 1,0 зимой [475]; нижняя критическая температура летом 36, а
зимой 7 °C. Зимние нижние критические температуры для амери-
канской вечерницы, кардинала и пуночки равны соответственно
16, 18 и 10 °C [98]. У вечерницы большую роль играет интенсив-
ность обмена, а у сойки — теплоизоляция. Нижняя переносимая
температура у домового воробья равна —25 °C в январе и 0°С в
августе; при этом зимой у него больше жира и выше ненасыщен-
ность жирных кислот [19]. У белых крыс, содержавшихся зимой
вне помещения при средней температуре 10 °C, толщина шерстного
________________________________________________ _ ___...........r.i
С, этого не произошло; у животных обеих групп усилилась вас-
Л ' — ------ A XAXV А А ж ж.л и* А 11^ 11 KJ ц 1111 AYjyiyiv, W V/ 1ГХ V/ jp/ KJ t A ХА/А
(например, ушей) наблюдались реже, чем у акклимированных
покрова увеличилась, а у крыс, находившихся в помещении при
куляризация кожи. У акклиматизированных крыс обморожения
[194].
п0 У круглохвостого суслика (Citellus tereticaudus) ТКрИТ летом на
выше, чем зимой [216]. Критическая внешняя температура, при
которой у диких зайцев возникает недостаточность терморегуля-
10,0-
9,0-
8,0-
7,0-
6.0-
5,0-
4,0-
3,0~
2,0~
1,о-
-50
-40 SO -20 -Ю О 10 20 30 40
Внешняя температура, °C
Рис. 9-30. Средняя теплопродукция (I) и средняя потеря тепла в результате
испарения (II) у американского зайца-беляка (черные значки — лето, белые —
зима) [166].
Гч’
х
го
X
£
рВата
40
Песец,
30
Кролик
20
10
ф
о
]Волк
Гризли
* Баран Далла
о®
сица
емминг
Ледяная
вода
. Землеройка
верная поверхность
70 80
10 20 30 40 50
Толщина меха, мм
Куница
Белка 3
1
бобр
Белый медведь
1 *
Белый меовеоь
! I г
I I Тйиеж
’ ‘ TiВоздух
Рис. 9-31. Зависимость максимальной теплоизоляции у арктических и тропиче-
ских млекопитающих от толщины меха (по П. Шоландеру).
Для водных животных приведены результаты, полученные на воздухе (черные кружки)
и в ледяной воде (белые кружки). Заштрихованная область — тропические виды. Пунктир-
ной линией показана величина теплоизоляции, создаваемой слоем ваты.
Температура
157
ции, летом равна +10 °C, а зимой —5 °C; вес меха зимой на 27%
больше, чем летом. Реакция дрожи летом выражена сильнее, чем
зимой, а потребление пищи приблизительно одинаково. Когда
обритых кроликов выдерживали при 6 и при 28 °C, потребление
пищи в первой группе возросло на 41%; введение одной и той же
дозы норадреналина повышало интенсивность обмена в первой
группе на 113, а во второй на 41% [194].
У арктических млекопитающих мех не только толще, но и обла-
дает лучшими теплоизолирующими свойствами. Максимальная теп-
лоизоляция создается (если не считать вакуума) неподвижным
воздухом; она равна 1,9 кло/см. («кло» — это единица, применяе-
мая для измерения теплоизоляционных свойств одежды и харак-
теризующая теплоотдачу при данной разности температур с едини-
цы площади за единицу времени.) Наклон кривой зависимости
теплоотдачи от толщины меха у ряда арктических животных соот-
ветствует 1,5 кло/см (рис. 9-31). Теплоизоляция у арктических
млекопитающих в среднем в 9 раз лучше, чем у тропических. Тю-
лений жир — настолько эффективный теплоизолятор, что темпера-
тура внутренних органов у тюленя поддерживается на постоянном
уровне 37 °C, в то время как кожа лишь ненамного теплее окру-
жающей воды. Зимнее повышение теплоизоляции сильнее выраже-
но у крупных млекопитающих (на 52% у барибала и на 41% у
волка), чем у мелких (на 21% у оленьей мыши и на 16% у зайца).
Теплоизоляция увеличивается в ряду от землеройки до кролика
[163, 166].
Конечности теплокровных животных гораздо устойчивее к
охлаждению, чем животное в целом, и могут продолжать функ-
ционировать при очень низких температурах. Так, температура
ног чайки на плавучей льдине, нижней части ног северного оленя,
ластов тюленя или плавников кита может быть ниже 10 °C.
У свиньи, находящейся на открытом воздухе при 10—12 °C, темпе-
ратура кожи спины равна 10—12 °C, рыла 17—18, нижней части
ног 9, а температура в глубине тела 38,4 °C [224]. Ниже приведе-
ны некоторые свойства подкожного жира свиньи [100]:
Жир Точка плавления, °C Отношение насыщенных жиров к нена- сыщенным
Спинной, наружный слой 26,4 0,66
Спинной, внутренний слой 27,8 0,79
Околопочечный 29,6
У
У
карибу точка плавления жира
арктических' млекопитающих
ноги понижается от бедра книзу,
(песца, карибу, эскимосской со-
158
Глава 9
баки) температура плавления костного жира фаланг может со-
ставлять 10—15 °C, тогда как у жира из области головки бедра
она равна 45°C. Жир в подушечках лап может не затвердевать
даже при охлаждении до 0°С [226].
Ферменты в клетках кожи и различных придатков тела у мно-
гих млекопитающих гетеротермны, т. е. имеют температурный диа-
пазон устойчивости около 35° — шире, чем у многих пойкилотерм-
ных животных. Разные части одной и той же клетки (например,
нервного волокна) химически разнородны. У акклимированной к
холоду серебристой чайки проведение в плюсневом участке нерва
ноги прекращается при 2,8—3,9 °C, а в тибиальном участке того
же нерва — при 11,7—14,4° С [80]. Проводимость большеберцо-
вого нерва у хомячка, не акклимированного к холоду, сохраняется
при понижении температуры до 3,4, а у крысы — до 9 °C.
Существуют многообразные поведенческие адаптации. Если со-
держать крыс вне помещения не поодиночке, а группами, то они
дольше выживают на холоде, обнаруживают большую способность
к теплопродукции, у них менее выражена мышечная дрожь, выше
уровень основного обмена; они более чувствительны к норадрена-
лину, а теплоизолирующие свойства шерстного покрова улучша-
ются у них в меньшей степени. У крыс, которых содержат порознь,
наблюдается большее повышение активности щитовидной железы
[194]. Колонию мышей в течение 10 лет разводили при —3°С, и
они больше ели, имели меньшие выводки, лучше строили гнезда,
обнаруживали меньший исследовательский рефлекс и были мель-
че, чем контрольные мыши, выращенные в тепле; особенно выра-
женной была разница в величине выступающих придатков тела.
В то же время ректальная температура была у животных обеих
групп одинаковой [19а].
Вазомоторные адаптации особенно важны для конечностей.
У некоторых животных артерии и вены проходят близко друг к
другу или переплетаются, образуя сеть. В случае такого тесного
контакта артериальной и венозной крови между ними происходит
передача тепла по принципу противоточного обмена. Переход
тепла из артериальной крови в венозную приводит к тому, что ко-
нечности могут оставаться относительно холодными, получая в то
же время достаточно кислорода. У ленивца, у которого есть такие
сосудистые сети, охлажденная конечность согревается медленно;
конечности носухи или обезьяны, не имеющие подобных сетей, со-
греваются после охлаждения в 5 раз быстрее, чем у ленивца [416].
Сосудистые сети есть и в подмышечных впадинах пингвинов.
Всюду, где имеются такие сосудистые сплетения, теплоотдача уже
не пропорциональна общей поверхности тела. У куропаток и фа-
занов такого рода сеть (сплетение вен вокруг артерии) есть в
верхней части ноги; на холоде температура бедра равна 37,5 °C,
тогда как двумя сантиметрами ниже температура ноги составляет
Температура
159
g 7 °C, а температура пальцев всего лишь 2,7 °C. У чаек и цапель
ноги обеспечивают отведение большей части избыточного тепла,
особенно при высоких внешних температурах. У этих птиц сразу
прекращается характерное «охлаждающее» дыхание, как только
ноги погружаются в воду [444а].
У рыбаков Гаспе, привыкших работать с обледенелыми сетями,
погружение одной руки в ледяную воду вызывает меньшее повы-
шение артериального давления, чем у контрольных индивидуумов;
при этом температура пальцев у рыбаков оказывается более высо-
кой, что свидетельствует о вазомоторной регуляции [290]. Средняя
температура пальцев после их 10-минутного погружения в ледяную
воду (0°С) равна 2,5 °C у представителей кавказской расы и
4,8 °C у эскимосов (в том числе у эскимосов, живущих в теплом
климате) [115]. Одну группу людей выдерживали в течение пяти
недель в тепле, а другую — на холоде, с минимальной защитой от
температурных воздействий; в каждой группе часть людей регуляр-
но занималась физической работой, и у этих людей интенсивность
обмена во время ночного холода была более высокой. Кроме того,
у них отмечали большую теплопроводность кожи, большую частоту
сокращений сердца и некоторые изменения реакции на холод:
относительно уменьшалась роль мышечной дрожи и увеличивался
вклад других видов термогенеза. Только у этих лиц имело место
усиление кровотока в пальцах. Таким образом, физическая нагруз-
ка может повышать устойчивость к холоду за счет большей интен-
сивности обмена и кровообращения [5]. У людей, проживших
12 месяцев в Антарктике, пальцы ног меньше охлаждаются при
погружении их в воду с температурой 5 °C, однако термонейтраль-
ная зона метаболизма остается такой же, как и у обитателей уме-
ренного пояса [498].
Таким образом, акклиматизация к холоду связана с увеличени-
ем толщины и эффективности термоизолирующего покрова, повы-
шением чувствительности к норадреналину, улучшением кожного
кровообращения и адаптивными реакциями со стороны централь-
ной нервной системы.
Реакции на высокую температуру и акклимация к теплу. Вы-
сокая температура окружающей среды, стимулируя тепловые ре-
цепторы кожи, вызывает у животных рефлекторные реакции, повы-
шающие теплоотдачу. Кожные сосуды расширяются, объем крово-
тока в коже возрастает и теплопроводность периферических тканей
увеличивается в 5—6 раз, что приводит к большей теплоотдаче.
Возрастает также незаметное испарение влаги с поверхности те-
Ла- Если этого окажется недостаточно для поддержания теплового
равновесия, то повысится температура кожи; у животных, имею-
щих потовые железы, начнется рефлекторное -потоотделение, а
У Других животных усилится отдача тепла при дыхании.
160
Глава 9
Относительная роль дыхания, потоотделения и незаметного ис-
парения с поверхности кожи в общей потере воды у разных видов
различна. Человек в покое теряет за счет испарения воды около
20% тепла, а при мышечной работе 75—80%. При низких темпе-
ратурах потеря тепла путем излучения больше, чем за счет испа-
рения, а при температурах выше 31 °C главную роль в теплоотдаче
у человека, находящегося в состоянии покоя, играет испарение.
Теплоизоляция уменьшает теплоотдачу, происходящую за счет из-
лучения, конвекции или теплопроводности. У млекопитающих,
имеющих волосяной покров или лишенных его, теплоотдача в воде
вдвое больше, чем на воздухе. Человек в холодном воздухе теряет
за счет испарения около 1 л воды в сутки, а при напряженной фи-
, зической работе в условиях пустыни эта величина может достичь
1,5 л/ч. Пороговая температура кожи, при которой начинается
потоотделение, варьирует для различных участков тела в пределах
от 30 до 32 °C. Потоотделение увеличивается на 20 г/ч при повыше-
нии внешней температуры на Г. У верблюда испарение резко воз-
растает при 35°С, а у осла — при 31 °C [414]. Человек теряет с
потом большое количество солей; содержание хлоридов в поте осла
гораздо ниже. Потеря воды и солей с потом компенсируется умень-
шением мочеотделения.
У человека активность потовых желез в коже ладоней нахо-
дится под эмоциональным контролем; работа других потовых же-
лез контролируется термическими факторами. Потоотделение и
секрецию жидкости эккринными железами стимулируют симпати-
ческие нервные волокна, относящиеся к числу холинэргических.
Апокринные железы разрушаются при выделении секрета и долж-
ны постоянно замещаться новыми; они активируются норадрена-
лином и адреналином. Все высшие млекопитающие, за исключени-
ем грызунов и зайцеобразных, имеют кожные железы, способность
которых секретировать пот сильно варьирует. У барана, собаки
и свиньи секреция пота происходит непрерывно, но на поверхность
кожи он выделяется периодически в результате сокращения мио-
эпителиальных клеток, и это не связано с воздействием тепла или
адреналина [40]. Большинство млекопитающих, не относящихся к
приматам, имеет апокринные железы, которые открываются в во-
лосяные фолликулы или поблизости от них, тогда как эккринные
железы приматов часто обособлены от фолликулов. У осла пото-
отделение носит циклический характер — оно происходит синхронно
на всей поверхности кожи с частотой до 2 циклов в минуту. При
охлаждении кожа выделяет меньше пота, но цикличность сохраня-
ется; потоотделение зависит и от температуры кожи, и от глубин-
ной температуры. У газели и сернобыка это тоже циклический про-
цесс, а у буйвола — непрерывный [58, 389]. У коров, акклимиро-
ванных к различным температурам, теплоотдача распределяется
следующим образом [241]:
Температура
161
Порода Вклад в общую теплоотдачу, %
при аккли- мации к 10 °C при аккли- мации к 27 °C
Коричневая швей- Кожа 16 41
царская Дыхательные 8 12
Брамин пути Кожа 19 46
Шортгорн « 26 53
При повышении температуры среды с 20 до 44 °C легочная вен-
тиляция возрастает у коровы в три раза. Животные с выраженной
тепловой одышкой (например, собаки и коровы) испаряют воду
с поверхности дыхательных путей, что могло бы привести к повы-
шению уровня хлоридов крови и к развитию алкалоза; однако у
собаки основная часть воздуха поступает при этом через нос и сра-
зу же выводится через рот, так что вентиляция легких существенно
не увеличивается [412]. У барана при температуре кожи мошонки
выше 35,3 °C начинается потоотделение и температура тела сни-
жается; таким образом, тепловые рецепторы мошонки могут сти-
мулировать потоотделение независимо от температуры тела. Мест-
ное согревание молочных желез у овцы также активирует потовые
железы [481]. Благодаря потоотделению человек может переносить
внешние температуры до 49,3 °C. Тепловая одышка хуже защища-
ет организм собаки от перегрева: при температуре среды 41,1 °C
температура тела может достичь 42 °C. Бабуин занимает промежу-
точное положение — его ректальная температура может повысить-
ся с 38,1 до 40,6 °C при температуре воздуха 45 °C [135]. У некото-
рых копытных главную роль в охлаждении тела играет тепловая
одышка, а у других — потоотделение. При сильной инсоляции
охлаждение кожи за счет испарения воды с ее поверхности иногда
начинается еще до повышения глубинной температуры тела; зна-
чит, потоотделение может быть отчасти местной реакцией.
Некоторые млекопитающие (например, опоссум и ряд грызу-
нов) при высокой температуре выделяют обильную слюну и обли-
зывают свой мех. Большинство млекопитающих обладает меньшей,
чем у человека, способностью увеличивать отдачу тепла путем ис-
парения воды, и температура тела повышается у них сильнее. Ре-
акция человека на температуру среды отражает его тропическое
происхождение.
Многие млекопитающие ищут тень или спасаются от жары, за-
капываясь в землю; в условиях пустыни они активны по ночам.
Свиней можно обучить включению обогревателя или выключению
°хлаждающего устройства [13].
И-1514
162
Глава 9
Рога козы хорошо снабжаются кровью. Их температура близка
к внешней при 15 °C и ниже и превышает внешнюю температуру,
если последняя лежит в пределах 30—39 °C. При искусственной
теплоизоляции рога наблюдается опасное повышение температуры
мозга. При 30°C с поверхности рогов рассеивается 3% метаболиче-
ского тепла, а при 22 °C — 2%. В теплом воздухе кровеносные со-
суды рогов расширяются, в холодном — сужаются [450, 451].
Когда механизмы охлаждения оказываются недостаточными и
температура тела повышается, потребление Ог возрастает вслед-
ствие прямого действия тепла на клетки и усиленной вентиляции
легких. Возможно, что это повышение интенсивности обмена со-
ставляет часть общего регуляторного механизма, поскольку оно
бывает слабее выражено у крыс после удаления щитовидной же-
лезы или гипофиза. Термонейтральная зона не распространяется
на высокие температуры воздуха, т. е. верхняя Ткрит обычно близка
к температуре тела или чуть ниже ее. У мелких птиц и млекопи-
тающих верхняя и нижняя 7\;р11Т могут совпадать. У крыс после
повышения внешней температуры с 28 до 34 °C уровень обмена воз-
растает и остается повышенным в течение 48—72 ч. То же самое
происходит вначале у коров при повышении внешней температуры
с 21 до 29°C, но через 9 недель уровень обмена снижается на 15%,
а испарение с поверхности кожи возрастает на 85%. Действие го-
рячего воздуха на коров приводит к усилению экскреции натрия
и глюкокортикоидов, уменьшению задержки азота и снижению
экскреции минералокортикоидов. На холоду вначале увеличивает-
ся объем плазмы крови, а затем и объем эритроцитов.
Выше было дано определение теплопроводности (стр. 148) и
приведены ее величины у ряда животных (табл. 9-4). С пониже-
нием внешней температуры теплопроводность у Perognathus умень-
шается следующим образом [82]:
Теплопроводност ь,
мл. Оз/(г • ч град)
Внешняя
температура, °C
2,5
1,3
0,58
0,43
0,42
35
32
25
15
5
При недостаточности терморегуляции и критическом повышении
температуры тела наступает тепловой шок (гиперпирексия). У че-
ловека тепловое истощение может развиться и без повышения тем-
пературы тела — главным образом в результате обезвоживания ор-
ганизма и изменений солевого баланса. В случае потери слишком
большого количества солей тепловое истощение возможно и при
достаточном содержании воды в организме. В результате повреж-
дения мозга происходит так называемый тепловой удар. Смерть от
перегрева наступает в значительной мере вследствие нарушений
Температура
163
кровообращения. При согревании извне переносимая температура
тела ниже, чем при гипертермии эндогенного происхождения. Че-
ловек теряет сознание, если в результате внешнего обогрева его
ректальная температура достигает 38,6 °C, тогда как при лихорад-
ке она может повышаться до 42 °C, а при физической работе — до
40 °C без вреда для организма.
В результате акклиматизации к теплу вырабатывается способ-
ность легче переносить жару и сохранять при этом высокую актив-
ность. Предельная интенсивность потоотделения у человека может
удваиваться, оно начинается при более низкой температуре кожи
и пот содержит меньше солей. У акклиматизированных людей
быстрее снижается температура кожи и повышается эффективность
работы сердечно-сосудистой системы; например, у уроженцев Ни-
герии учащение ритма сердца при повышении температуры возду-
ха выражено слабее, чем у европейцев. При одной и той же высо-
кой температуре воздуха у банту ректальная температура ниже и
потоотделение менее интенсивно, чем у представителей кавказской
расы. Для банту характерны также более низкий температурный
порог для повышения уровня метаболизма (Гкрит) и меньшая теп-
лоизоляция [499].
Многие тропические млекопитающие обладают наследственной
приспособленностью к жаре. Так, например, индийские коровы лег-
че переносят жару, чем европейские; при 27 °C они так же при-
бавляют в весе, как и при 10 °C, в то время как шортгорны на жаре
меньше едят и рост их замедляется [240]. Африканские копытные
используют два способа адаптации к климату жарких пустынь.
Крупные животные (антилопа канна) утром имеют низкую темпе-
ратуру тела (32,8 °C) и медленно согреваются; мелкие животные
(газель) согреваются быстро, и температура тела у них оказывается
выше температуры воздуха (42°C при внешней температуре 40°C);
оба эти способа обеспечивают сохранение воды в организме. По
достижении критической температуры тела в обоих случаях усили-
вается испарение — за счет потоотделения или вследствие усилен-
ного дыхания. У дегидратированного животного тепловая одышка
наступает при более высокой температуре, чем у животного, орга-
низм которого содержит достаточно воды [452]. Шерстный покров
замедляет не только отдачу тепла, но и его поглощение; у остри-
женного верблюда температура тела повышается на солнце быст-
рее, чем у нормального животного. В отсутствие воды ректальная
температура у верблюдов колеблется от 34 до 40 °C; у животных,
получающих воду, эти колебания выражены слабее. Если живот-
ное может выдерживать высокую температуру тела, то темпера-
турный градиент между телом и внешней средой меньше, так что
Меньшим будет и поступление тепла извне [411].
У птиц, обитающих в пустыне, имеются разнообразные адапта-
ции, позволяющие переносить жару, в том числе много поведенче-
ских адаптаций [97]. У птиц нет потоотделения, но существует два
11*
164
Глава 9
метода охлаждения, связанных с дыханием, — усиленное дыхание
и трепетание горла. Частота дыхания у голубя инка при темпера-
туре воздуха ниже 40 °C равна 30 в 1 мин, а при 45 °C повышается
до 650 в 1 мин; при этом, судя по химизму крови, легочная вен-
тиляция возрастает меньше, чем общая вентиляция, так что послед-
няя, очевидно, в значительной части связана с функцией воздуш-
ных мешков. Однако и легочная вентиляция возрастает настолько,
что заметно увеличивается потеря СОя, и это может привести к раз-
витию алкалоза. Кроме того, одновременно с усилением вентиля-
ции при высоких температурах возникает и трепетание горла [68,
316а]. У молодой белой цапли при тепловом стрессе оно может
иметь частоту 965 в 1 мин при частоте дыхания всего лишь 44 в
1 мин. У калифорнийской куропатки (Lophortyx) возникает как
тепловая одышка, так и трепетание горла; кроме того, птица мо-
жет увлажнять свои ноги испражнениями, что приводит к допол-
нительной отдаче тепла путем испарения влаги [54]. При повы-
шении температуры воздуха с 35 до 47 °C испарение воды возрас-
тает у козодоя Phalaenoptilus nuttalli с 2,9 до 23 мг/(г-ч) [282].
У козодоя температура тела 36 °C, и испарение воды резко возрас-
тает у него только тогда, когда температура воздуха поднимается
выше 35 °C: при 1,2 °C потеря воды равна 1,1 мг/(г-ч), при
35,5 °C — всего лишь 2,8 мг/(г-ч), а при 44 °C достигает уже
17,9 мг/(г-ч) [283]. Африканский страус за одно и то же время
на холоде теряет при дыхании 0,4 г воды, а при температуре
воздуха 45 °C — 4,5 г [86]. У аляскинской куропатки Lagopus теп-
ловая одышка начинается при внешней температуре 21 °C [241].
При умеренных температурах потеря воды путем испарения (£)
обнаруживает тесную корреляцию с весом тела птицы (IF); на-
блюдаемая зависимость соответствует эмпирическому уравнению
£=0,432 IF0-585 [85а].
Различные поведенческие реакции птиц уменьшают поглощение
лучистого тепла и усиливают охлаждение за счет испарения влаги.
Африканский страус при температуре воздуха выше 25 °C взъеро-
шивает перья. Терморегуляция у этой птицы настолько эффектив-
на, что позволяет сохранять постоянную интенсивность обмена в
широком диапазоне внешних температур [86]; температура тела
в естественных условиях остается ниже 40 °C даже при температу-
ре воздуха 50 °C. Охлаждение, связанное с испарением, усилива-
ется по мере усиления тепловой одышки, которая благодаря су-
ществованию воздушного шунта в обход легких не приводит к
развитию алкалоза [415]. Мелкие птицы «купаются» в пыли, воз-
можно, увеличивая таким образом теплоотдачу за счет теплопро-
водности. Птицы, обитающие в пустыне, расправляют крылья,
прячутся в тени или парят на значительной высоте. У многих из
них нормальная температура тела лежит в пределах 41—42,6 °C
(см. табл. 9-3, стр. 147), а переносимая внешняя температура не
превышает 46—47 °C [97]. Олуша, нормальная температура тела
Температура
165
которой 38,3—40,7 °C, принимает определенное положение относи-
тельно солнца, держа свои лапы в тени; у этой птицы наблюдается
также трепетание горла. Олуша насиживает яйца при 38—40 °C и
может ощущать температуру яиц и птенцов с помощью терморе-
цепторов лап [21].
У водяных птиц — цапель и чаек — отдача метаболического теп-
ла может происходить через поверхность ног и лап. Аист при пе-
регреве тяжело дышит и увлажняет свои ноги экскретами; после
охлаждения тела оба явления прекращаются [246]. У альбатроса
имеется брюшное наседное пятно, температура которого ниже об-
щей температуры тела; он высиживает птенцов при 38,8 °C. При
более высоких температурах птенцы альбатроса тяжело дышат
и могут балансировать на пятках, не опуская лапы на землю
[212].
Развитие гомойотермии. У новорожденных млекопитающих и
только что вылупившихся птенцов температура тела почти не ре-
гулируется. Двухдневные лабораторные мышата по существу
пойкилотермны; потребление ими О2 достигает максимума при рек-
тальной температуре 32 °C [358]. Десятидневные мышата способны
к терморегуляции при средних температурах воздуха, а двадцати-
дневные— и при экстремальных температурах. У самых мелких
диких мышей (вес взрослой особи 8 г) до 5—8-дневного возраста
охлаждение не вызывает характерной метаболической реакции;
у более крупных видов эта реакция появляется уже в первые сутки.
С момента первого появления мышечной дрожи и пиломоторной
реакции наблюдается устойчивое повышение интенсивности обмена
[82]. В раннем возрасте лабораторные мыши могут переносить
более сильное охлаждение, чем взрослые особи, и отличаются бо-
лее высокой активностью сукцинатдегидрогеназы в клетках бурой
жировой ткани. В течение первых 4 дней мышата не обнаруживают
метаболической реакции на введение адреналина, и только через
3 недели эта реакция достигает уровня, характерного для взрослых
животных: в этот период начинает функционировать щитовидная
железа. Новорожденных мышат также можно сделать чувствитель-
ными к норадреналину, впрыснув им тироксин [276]. Хомяк в
первые 10 дней жизни является пойкилотермным животным: при
повышении внешней температуры с 25 до 36 °C потребление О2 у
него возрастает. В течение последующих 7 дней увеличение VO2
при повышении температуры воздуха снижается до нуля, а между
17-м и 21-м днями жизни достигается гомойотермная стабильность
в термонейтральной зоне (30—36°C); более высокие и более низ-
кие внешние температуры вызывают теперь повышение интенсив-
ности обмена [384].
У шестидневных крысят температура тела всего лишь на 1,3—
выше температуры среды; уровень терморегуляции, характер-
ный для взрослых животных, достигается к 25 дням жизни. Темпе-
ратура тела 10—12-дневных полевок-экономок в гнезде равна
166
Глава 9
35,4 °C, но. вне гнезда при температуре воздуха О °C она варьирует
в пределах от 2,5 до 16 °C. Поросята обладают хорошей терморе-
гуляцией уже в возрасте 1 дня. У опоссума, который рождается
в недоразвитом состоянии, первые признаки терморегуляции появ-
ляются в 2-месячном возрасте.
У птенцовых птиц, например у цапель, неоперившиеся птенцы
требуют большой родительской заботы, и температура их тела
подвержена значительным колебаниям, которые уменьшаются толь-
ко после вылета птенцов из гнезда. Выводковые птицы, например
чайки, уже с момента вылупления обладают хорошей способностью
к терморегуляции. Птенцы домового крапивника в 9-дневнем воз-
расте способны к частичной терморегуляции при внешних тем-
пературах не ниже 26 °C, а в 15-дневном возрасте поддерживают
температуру тела 40 °C даже при 10 °C. Птенцы гнездарей нахо-
дятся в невыгодном положении не только из-за отсутствия перьев,
но и вследствие большой величины отношения поверхности тела
к объему. У вечернего воробья в течение 2-го дня после вылупле-
ния температура тела превышает температуру воздуха менее чем
на 3°, на 4-й день эта разница увеличивается до 10°, а на 7-й—
9-й дни становится возможным поддержание температуры тела на
уровне выше 35°C. Интенсивность обмена в возрасте 1—4 дней
повышается или понижается вместе с температурой, в последую-
щие два дня наблюдается некоторый разброс величин VCv а в воз-
расте 7 дней потребление О2 уже возрастает при охлаждении так
же, как у взрослых особей [94] (рис. 9-32). У птенцов разрисован-
ной куропатки (Excalfactoria) 70% теплоотдачи обусловлено кож-
ным испарением, а у взрослых особей эта величина снижается до
13%. Таким образом, становление гомойотермии связано с умень-
шением кожного испарения и повышением метаболической тепло-
продукции [37].
Отсутствию терморегуляции у птенцов и у новорожденных мле-
копитающих соответствуют более низкие, чем для взрослых живот-
ных, нижние летальные температуры [3]. Становление терморегу-
ляции нельзя связать с каким-либо отдельным фактором — нерв-
ным, эндокринным или метаболическим.
Водные млекопитающие. Термические стрессы могут достичь
для водных млекопитающих большей силы, чем для наземных, по-
скольку теплопроводность воды значительно выше теплопроводно-
сти воздуха. Кроме того, многие водные млекопитающие проводят
часть времени на суше, т. е. могут подвергаться действию сильно-
варьирующих температур. У мелких животных, например у водя-
ных землероек весом 8—18 г, мех содержит прослойку воздуха,
что уменьшает теплоотдачу. Мелким животным свойствен значи-
тельно более высокий, чем у крупных видов, показатель теплопро-
водности [66].
Мех с заключенным в его толще воздухом может обеспечивать
прекрасную теплоизоляцию, пока не намокнет. Шерстный покрои
40\
Рис. 9-32. Развитие гомойотермии.
пй<»^Та^ильная температура тела у мышей разного возраста (в сутках, цифры у кривых) при
различных внешних температурах [276]. Б. Потребление кислорода (слева) и температура
тела (справа) у вечернего воробья в период от вылупления до 7 суток [94].
168 Глава 9
ондатры содержит 21,5% воздуха. У гренландского тюленя первые
полтора месяца жизни проходят на воздухе и теплоизоляция до-
стигается с помощью шерстного покрова, тогда как у взрослых
особей для этой цели служит слой жира [89]. Детеныши морского
котика могут переносить гипотермию, и их мех смачивается (в от-
личие от меха взрослых котиков) [225].
У детенышей тюленя в отличие от взрослых особей обмен ве-
ществ при погружении в воду усиливается: потребление О2 у бель-
ков равно 1048 мл/(кг-ч), а у взрослых тюленей — 444 мл/(кг-ч).
Нижняя критическая температура воздуха для тюленя близка к
0 °C; оценка этой величины в воде дает цифру —20 °C [163]. Та-
ким образом, подкожный жир обеспечивает настолько эффектив-
ную теплоизоляцию, что становится невозможным определить ниж-
нюю Ткрит в воде с помощью кривых метаболизм — температура.
У гренландского тюленя при температуре воды 10 °C температура
поверхности кожи также равна 10 °C, но под кожей на глубине
22 мм регистрируются 22 °C, а на глубине 35 мм 39 °C. Властах
тюленей и плавниках китообразных артерии и вены образуют сеть,
причем всем крупным артериям соответствуют трабекулярные вены
(ср. стр. 131, рис. 9-24).
У белого медведя теплоизоляция достигается с помощью меха,
жира и периферических мышц; при температуре воздуха от —15
до —2 °C температура кожи равна 30—36 °C [350].
В хвостовом и грудных плавниках китов имеется противоточная
система кровеносных сосудов: большая центральная артерия окру-
жена спиральными кольцами мелких вен. Плавники могут функ-
ционировать при более низких температурах, чем остальное тело.
Иногда водные млекопитающие сталкиваются с противополож-
ной трудностью — с проблемой теплоотдачи, борьбы с перегревом.
У китообразных теплоотдача вдвое выше, чем у наземных млекопи-
тающих, но вследствие высокой удельной теплопродукции она ока-
залась бы недостаточной для защиты организма от перегрева на
суше. Температура мертвого кита, даже находившегося частично
в воде, снижалась за 28 ч меньше чем на 1°, а период полуохлаж-
дения составлял около 2 недель [249]. У дельфина слою ворвани
толщиной 2 см соответствует разность температур 30 °C. Дельфин
потребляет всего лишь 380 мл О2 в 1 мин, т. е. приблизительно
вдвое меньше, чем наземное животное сходных размеров [249].
Бобр, находящийся на воздухе, способен успешно регулировать
температуру своего тела, если его хвост опущен в холодную воду;
в противном случае становится возможным перегрев. Таким обра-
зом, хвост используется для регуляции теплоотдачи [444].
Примитивные млекопитающие. Однопроходным, некоторым
сумчатым, броненосцам, ленивцам и муравьедам обычно свойст-
венна невысокая температура тела, изменяющаяся в некоторых
пределах в зависимости от внешней температуры. У ехидны в по-
левых условиях отмечались суточные колебания температуры тела
Температура
169
от 28,7 до 31,5 °C; в лабораторных условиях температура тела со-
ставляла в среднем 30,7 °C при внешних температурах от 5 до
25 °C, причем термонейтральная зона соответствовала 20—30 °C.
При 33 °C у ехидны возможна гипертермия, при этом около трети
метаболического тепла теряется в результате испарения влаги
Г413]. На холоде может наступать оцепенение, особенно при недо-
статке пищи; при температуре воздуха 5 °C температура тела па-
дает до 5,7 °C. Оцепенение может продолжаться 5—10 дней [12].
Утконос {Platypus) на воздухе способен удерживать температуру
тела на уровне 30—31 °C даже при 0°С, но не может сохранять
нормальную температуру тела в холодной (11 °C) воде и поэтому
при добывании пищи ныряет лишь на короткое время. У утконоса
в отличие от ехидны не бывает холодового оцепенения [62а]. Наи-
более совершенной терморегуляцией среди австралийских сумчатых
обладают кенгуру рода Macropus. У фалангеров терморегуляция
менее эффективна, а у сумчатых мышей развита слабо. Централь-
ноамериканский опоссум Metachirus, ведущий ночной образ жиз-
ни, ночью имеет довольно стабильную температуру тела около
36°C при внешних температурах 10—30 °C; однако при 10 °C тем-
пература тела у него может снижаться до 33 °C; а при 30 и 35 °C—
повышаться соответственно до 34 и 40 °C [333].
Активный в ночное время броненосец Dasypus имеет ночью тем-
пературу тела 35 °C, сохраняющуюся при температуре воздуха до
О °C. На холоде он сворачивается в шар, а в жару прибегает к
учащенному дыханию при одновременном расширении кровенос-
ных сосудов, но не способен к потоотделению. К гетеротермным
животным относится и примитивное насекомоядное тенрек, термо-
регуляция у которого носит нестабильный и ограниченный харак-
тер. Интенсивность обмена у тенрека примерно на 50% ниже, чем
у «типичного» млекопитающего той же величины. В жаркое время
температура тела у него может понижаться за счет испарения во-
ды [197].
Большинство сумчатых по своим температурным реакциям сход-
но с высшими млекопитающими. У американского опоссума DI-
delphis интенсивность обмена на холоде возрастает; удаление коры
больших полушарий не ведет к нарушению терморегуляции, тогда
как спинальные животные утрачивают способность сохранять на
колоде постоянную температуру тела. Интенсивность обмена у
сумчатой куницы на 32% ниже, чем у плацентарного млекопитаю-
щего сходных размеров, но особи весом более 100 г поддерживают
температуру тела 37,2 °C [316а]. У тасманийского фалангера тем-
пература тела варьирует в пределах от 32 до 38 °C, составляя в
'Среднем 34,9 °C; термонейтральная зона для этого вида 31—35 °C
1^4].
Очевидно, что у низших млекопитающих используются в общем
Те же механизмы терморегуляции, что и у плацентарных, но они
Поддерживают несколько более низкую температуру тела.
170
Глава 9
Животные, способные впадать в оцепенение. Летучие мышц
(Chiroptera) существенно различаются между собой по характеру
терморегуляции. У большинства Megachiroptera и крупных Micro*
chiroptera температура тела хорошо регулируется. Холод вызыва-
ет у плодоядных летучих мышей реакцию тревоги и не приводит
их в летаргическое состояние. Мелкие Microchiroptera днем впада-
ют в оцепенение; большинство их относится к зимнеспящим видам.
Несколько семейств летучих мышей — Vespertilionidae, Rhinolophi-
dae и Desmodontidae — занимает промежуточное положение, при-
чем у каждого вида терморегуляция имеет свои особенности [301].
Летучая мышь Dobsoma (Megachiroptera) весит 80 г и имеет
постоянную температуру тела 36—38 °C, сохраняющуюся даже при
температуре воздуха 40 °C. У этого вида потеря воды за счет испа-
рения при внешних температурах от 5 до 35 °C составляет
4,5 мг/(г-ч), а при температурах выше 35 °C возрастает [23].
У ложного вампира Macroderma температура тела остается в пре-
делах 35—39 °C при внешней температуре от 0 до 35 °C; при 38 °C
наблюдается усиленная саливация, животное учащенно дышит и
облизывает свои конечности, как это делают и многие грызуны
[292]. При температуре воздуха 10 °C теплопроводность на 29%
меньше, чем при 30 °C. В ряду Megachiroptera весом от 17,5 до
598 г потребление кислорода в термонейтральной зоне снижается
по мере увеличения веса тела (W) в соответствии с уравнением
[23]
VOa мл/(г.ч)=3,8Г-°»27
У ряда Microchiroptera температура тела на 2,5° выше темпе-
ратуры воздуха. При внешней температуре 41,5 °C температура
тела благодаря испарению влаги оказывалась ниже 42,5 °C, но при
43,5 °C животные погибали [297]. Летучая мышь Eptesicus fascus
использует для охлаждения свои крылья. При ректальном обогре-
вании у животного происходит расширение сосудов и температура
крыльев может увеличиваться на 4°. Местное согревание преопти-
ческой области гипоталамуса ведет к повышению температуры
крыльев, а охлаждение — к ее понижению. После повреждения ги-
поталамуса способность к терморегуляции не исчезает, но для
стимуляции регуляторных процессов требуется большее повышение
ректальной температуры, чем у нормальных животных. При вду-
вании воздуха в рот температура гипоталамуса становится ниже
ректальной температуры. Возможно, что малая температурная чув-
ствительность гипоталамуса — это адаптация к полету с открытым
ртом. Крайняя гетеротермия, вероятно связанная с малой термо-
чувствительностью гипоталамуса, свойственна насекомоядным ле-
тучим мышам и птицам [264].
Летом у Myotis наблюдаются суточные колебания температуры
тела или периоды дневной торпидности. Зимой эта летучая мышь
впадает в спячку, и если температура воздуха опускается ниже
Температура
171 -
5°C, то температура тела обычно снижается в меньшей степени,
а отдельные особи пробуждаются [381]. Если Myotis проснется во
время зимней спячки при низкой температуре воздуха, то темпе-
ратура тела может за 40—60 мин подняться с 3 до 35 °C. Если же
охладить животное до 3°С во время летнего оцепенения, то темпе-
ратура тела может повыситься не более чем на 1—2 °C [323]. Зи-
мой летучие мыши способны пробуждаться при более низкой тем-
пературе тела, чем летом. Введение норадреналина может повы-
сить потребление кислорода при пробуждении в 10 раз. Переход
Myotis в негомойотермное состояние происходит в конце сентября,
у летучей мыши Eumops наблюдается дневное оцепенение, но нет
зимней спячки [291]. Легко впадают в оцепенение мелкие насеко-
моядные летучие мыши неотропической области. Эта способность
могла выработаться в условиях тропиков как приспособление, поз-
воляющее в период покоя сохранять энергетические резервы путем
снижения температуры тела [318].
Вес различных колибри варьирует в пределах от 1,7 до 19,1 г.
В термическом отношении колибри весом 3 г напоминает трехграм-
мовую землеройку. В дневное время потребление кислорода у са-
мых мелких колибри в состоянии покоя составляет 11—16, а во
время полета 70—85 мл/(г-ч). Основной обмен у синегорлого ко-
либри соответствует 2,3, а у гигантского колибри — 2,7 мл О2 на
1 г в 1ч. Эти энергетйческие потребности относительно высоки
по сравнению с тем, что мы находим у крупных птиц и млекопи-
тающих. Высок у колибри и индекс теплопроводности С, который
у крупных видов равен 0,17, а у синегорлой разновидности 0,3 мл
О2/(г-ч-град) [145]. Очень высока частота сокращений сердца, со-
ставляющая у синегорлого колибри от 7 до 21 в 1 с. В полете расход
энергии в 6—12 раз больше, чем в состоянии покоя. Таким обра-
зом, для колибри характерны высокие энергетические потребно-
сти, и эти птицы могут выключать свои механизмы гомойотермии,
когда в них нет надобности. Потребление О2 синегорлым колибри
в состоянии оцепенения может составлять всего лишь 0,1 мл/(г-ч)
при температуре воздуха 15 °C и 1,18 — при 27 °C. Переход к тор-
пидности быстр и резок (его Qio равен 3,7—4,8), при этом он не
Bee тела, г Потребление Ог, мл/(г • ч)
оцепенение покой
Колибри Calypte costae 3,2 0,17 10,1
Calypte anna 6,8 0,17 'фку. 9,8
Eugenes 6,6 0,12 7,0
Latnpornis 7,9 0,12 6,5 (2,3
Козодои Phalaenoptilus 40,0 0,15 при 31 °C) 2,7
Chordeiles 75,0 0,18 2,4
172
Глава 9
сопровождается «пробными» снижениями температуры, как у мле-
копитающих при переходе в состояние зимней спячки [145, 285].
Температура тела ниже 8 °C для колибри гибельна. Метаболиче-
скую экономичность торпидности можно проиллюстрировать дан-
ными о потреблении Ог некоторыми птицами, находящимися в оце-
пенении или просто в покое при 15—16 °C (см. стр. 171) [287].
Выход из состояния оцепенения происходит очень быстро. Ко-
либри весом 4 г, повышая температуру тела с 25 до 37,5 °C, расхо-
дует 0,057 кал, тогда как козодою Phalaenoptilus весом 40 г для
этого необходимо 0,57 кал. Для охлаждения тела очень мелким
птицам достаточно нескольких секунд, а у крупной птицы время
охлаждения измеряется часами. Поэтому оцепенение (обычно
дневное) может быть полезной адаптацией только у мелких птиц
и млекопитающих.
Кроме колибри, в торпидное состояние могут впадать лишь не-
многие птицы. Козодои Phalaenoptilus и Caprimulgus могут в те-
чение нескольких часов переносить понижение температуры тела
до 5—8 °C. Эти птицы в отличие от зимнеспящих животных не про-
буждаются при дальнейшем охлаждении тела. Выход из оцепене-
ния происходит только при повышении температуры тела до 13 °C
(колибри) или 15 °C (Phalaenoptilus) [282, 283] (рис. 9-33).
В оцепенение могут также впадать некоторые грызуны, обитаю-
щие в пустыне. При этом их метаболизм оказывается в прямой
зависимости от изменений внешней температуры, тогда как у ак-
тивного животного наблюдается обратная зависимость. У карлико-
вой мыши Baiomys при отсутствии пищи температура тела может
опускаться до 23—25°C (при норме 32—36°C), но не ниже. Эта
мышь выживает при искусственном охлаждении до 15 °C в тече-
ние 4 дней и до 6 °C в течение 5 ч [215]. Карманчиковая мышь
(Perognathus) впадает в оцепенение при падении температуры те-
ла ниже 30 °C; потребление кислорода при этом снижается с 0,9
до 0,19 мл/(г-ч) [468]. Голодающая карманчиковая мышь оцепе-
невает при внешних температурах от 0 до 23 °C; температура тела
остается у нее значительно выше температуры воздуха. Оцепенение
длится не более полусуток, и ему не предшествуют «пробные» по-
нижения температуры тела; у голодных животных оно продолжи-
тельнее, чем у животных, получающих достаточное количество пи-
щи [306]. Бразильский карликовый опоссум Marmosa впадает в
оцепенение ежедневно с 9 до 18 часов. У кенгуровой мыши Micro-
dipodops продолжительность оцепенения может измеряться часами
или днями в зависимости от внешней температуры и условий пи-
тания; это позволяет более экономно использовать пищу [51].
Таким образом, способность впадать в оцепенение встречается
у ряда неродственных видов мелких птиц и млекопитающих; по-
видимому, она неоднократно возникала в процессе эволюции. Меж-
ду торпидностью и зимней спячкой нельзя провести резкой грани-
цы; однако торпидность в отличие от спячки не носит сезонного ха-
Температура
173
пактера и часто бывает ежедневной, животное выходит из нее быс-
трее, а температура тела снижается не так сильно. У некоторых
летучих мышей наблюдаются оба эти феномена. Торпидность —
специфическое приспособление для сохранения энергии, возможное
только у мелких животных, которые способны быстро согреваться
или охлаждаться. Интенсивность обмена при оцепенении сильно
понижается; к сожалению, мы ничего не знаем ни об отдельных
Рис. 9-33. Зависимость частоты сокращений сердца у колибри Patagona gigas
от температуры тела в начале оцепенения (белые кружки) и при выходе из него
(черные кружки) [287].
ферментах, ни о функциях центральной нервной системы в период
торпидности.
Бурая жировая ткань. У детенышей большинства млекопитаю-
щих и у всех зимнеспящих форм имеется особая активная ткань—
так называемая бурая жировая ткань, гистологически весьма не-
сходная с белой жировой тканью и обладающая поразительной
способностью к теплопродукции. У новорожденного крольченка на
бУРую жировую ткань приходится 4,3% веса всего тела, с возрас-
том эта величина быстро уменьшается [220]. Уменьшение количе-
ства бурой жировой ткани идет параллельно с развитием го-
Моиотермии. Термогенная реакция на норадреналин у крольчат
Ропорциональна количеству бурого жира. Бурая жировая ткань
стощается при выдерживании кроликов на холоде; однако пере-
174
Глава 9
резка шейного симпатического нерва приводит к замедлению этого
процесса, а раздражение симпатических нервов усиливает выра-
ботку тепла в бурой жировой ткани. Следовательно, окисление жи-
ра в ней активируется симпатической нервной системой [220].
У новорожденных хомячков и у взрослых хомячков, акклимирован-
ных к холоду, шейная бурая жировая ткань прогревает шейные
позвонки и подавляет холодовую дрожь [52]. У крыс в первые три
дня жизни весь термогенез, не связанный с дрожью, осуществля-
ется в бурой жировой ткани, а позднее на ее долю приходится до
трех четвертей такого термогенеза [17]. У многих млекопитающих
при холодовой акклимации количество этой ткани возрастает; у
акклимированных к холоду землероек и у сурков и сусликов во
время зимней спячки оно приближается к 3% от веса тела. Одно-
временное введение тироксина и норадреналина животному, нахо-
дящемуся в тепле, может привести к такому же увеличению коли-
чества бурого жира [289]. При холодовой акклимации в бурой жи-
ровой ткани повышается активность сс-глицерофосфатдегидрогена-
зы и лактатдегидрогеназы, а также содержание кофермента А.
Особенность метаболических путей этой ткани состоит в том,
что выработка тепла сопровождается лишь незначительным син-
тезом АТФ, т. е. отношение P/О имеет малую величину. Процессы
фосфорилирования и окисления в митохондриях разобщены [143].
У крыс, акклимированных к 6 °C, бурая жировая ткань окисляет
сс-кетоглутарат вдвое быстрее, чем при акклимации к 26 °C, но ве-
личины P/О в обоих случаях остаются сходными [432]. Основным
источником тепловой энергии служит окисление жирных кислот.
Норадреналин, действуя через циклический АМФ, стимулирует
высвобождение жирных кислот из триглицеридов, составляющих
здесь 99% всех липидов. Адреналин, норадреналин и циклический
АМФ (а также олеат) усиливают дыхание в 30—40 раз; разобща-
ющие агенты стимулируют дыхание и уменьшают реакцию на нор-
адреналин [492]. В бурой жировой ткани летучей мыши энергия
активации (температурный коэффициент) и скорость окисления
пальмитиновой, олеиновой и янтарной кислот ниже, чем у крысы
[113]. Жирные кислоты, освобождающиеся в этой ткани под влия-
нием норадреналина, не только дают, окисляясь, тепловую энер-
гию, но регулируют также перенос энергии, действуя, по-видимо-
му, еще до включения неорганического фосфата (подобно
агентам, разобщающим окисление и фосфорилирование) [374].
Двухнедельная обработка крыс ингибитором белкового синте-
за окситетрациклином предотвращает метаболическую реакцию
бурой жировой ткани на холод и на норадреналин. Важную роль
в холодовой акклимации играет синтез специфических окислитель-
ных ферментов в митохондриях бурой жировой ткани (но не в ми-
тохондриях печени и мышц) [200].
Температура
175
Нервные механизмы терморегуляции
Действие холода на нервы было рассмотрено ранее (см.
стр. 134).
Терморецепция. Механизмы терморегуляции активируются при
раздражении терморецепторов кожи, внутренних органов и различ-
ных отделов центральной нервной системы.
Периферическая термо рецепция. Тепловые и холодовые рецеп-
торы распределены в коже определенным образом; тепловые обыч-
но лежат глубже холодовых; холодовые рецепторы более много-
численны. Некоторые терморецепторы инкапсулированы, но в
большинстве случаев это голые нервные окончания. Значительно
выражен эффект суммации, благодаря которой порог ощущения
при раздражении больших участков ниже, чем при стимуляции
одиночных сенсорных окончаний. Для восприятия может быть до-
статочно изменения температуры на несколько тысячных долей
градуса в секунду. Тепловые (инфракрасные) лучи могут вызы-
вать также ощущение боли. Как тепловые, так и болевые сенсор-
ные сигналы передаются по тонким волокнам, но возникают они,
вероятно, в различных нервных окончаниях. Некоторые холодовые
рецепторы чувствительны также к давлению.
У разнообразных млекопитающих зарегистрированы нервные
импульсы, идущие в нервных стволах и отдельных нервных волок-
нах от холодовых и тепловых рецепторов [192]. Рецепторы обоих
типов обладают постоянной (спонтанной) активностью. Волокна,
идущие от тепловых рецепторов, активны в диапазоне 20—47 °C,
причем максимальная частота импульсов наблюдается при 38—
43 °C; волокна от холодовых рецепторов активны в диапазоне 10—
40°C, а наибольшую частоту импульсов дают температуры от 20
до 34°C (рис. 9-34).
Максимальный постоянный разряд холодовых рецепторов имеет
частоту около 10 имп/с, а тепловых рецепторов — 3,7 имп/с; раз-
ряд тепловых рецепторов менее регулярен.
Внезапное повышение температуры кожи приводит к резкому
кратковременному усилению разряда тепловых рецепторов, про-
должающемуся 1—2 с, а затем частота импульсов снижается до
Уровня, характерного для данной температуры. Устранение тепло-
вого стимула приводит к временному урежению или прекращению
разряда в волокнах от тепловых рецепторов. Подобным же обра-
зом снижение температуры приводит к короткому экстраразряду
холодовых рецепторов (до 140 имп/с), после чего устанавливается
новая частота импульсации, более высокая, чем при первоначаль-
ной температуре. После прекращения холодовой стимуляции ак-
тивность холодовых рецепторов временно затормаживается. При
температурах от 35 до 45°C волокна от холодовых рецепторов
неактивны, но при 45—50 °C в них снова возникает импульсация;
это парадоксальный ответ холодовых рецепторов на тепло. Опыты
176
Глава 9
с измерением температуры кожи и с обогреванием поверхности
кожи и подкожного слоя показали, что раздражение терморецеп-
торов зависит от абсолютной температуры нервных окончаний, а
не от температурного градиента [192].
У кошки терморецепторы носовой полости посылают сигналы
по подглазничному нерву с максимальной частотой 45—47 имп/с
для тепловых и 27 имп/с для холодовых рецепторов [190, 191].
Скорость изменения импульсации при изменении температуры
Рис. 9-34. Частота импульсов в афферентных нервных волокнах от кожных тер-
морецепторов приматов [223].
У — группа волокон; // — одиночное волокно.
варьирует от 20 до 80 имп/(с*град). Тепловые рецепторы у мака-
ка-резуса имеют устойчивую частоту разряда при 32 °C; при повы-
шении температуры со скоростью 20 град/с частота импульсации
возрастает, достигая максимума при 40—44 °C, а при дальнейшем
повышении температуры опять снижается [191]. Импульсация не-
которых кожных терморецепторов носит характер вспышек, тогда
как другие дают непрерывный разряд; у собаки максимальная
чувствительность холодовых волокон отмечается при 31—37, а теп-
ловых — при 40 °C [223].
Терморецепторы есть и в глубине тела; раздражение их может
вызвать реакцию дрожи даже тогда, когда температура кожи и
головного мозга остается постоянной. Некоторые из этих рецепто-
ров находятся в венах. Обогревание бедренной вены у собаки при-
водит к повышению кровяного давления [460].
Центральные терморецепторы и центр терморегуляции. Охлаж-
дение спинного мозга вызывает у собаки мышечную дрожь и су-
жение кровеносных сосудов, тогда как обогревание его останав-
ливает дрожь, а также приводит к расширению сосудов и тепло-
Температура
177
вой одышке. Эффект изменения температуры спинного мозга сум-
мируется с эффектом изменения температуры гипоталамуса; при
этом температурные эффекты, возникшие на уровне гипоталамуса,
можно нейтрализовать соответствующим температурным воздейст-
вием на спинной мозг [237]. Не исключено, что холодовой тремор
может начинаться на уровне спинальных мотонейронов. Данные о
чувствительности спинного мозга к охлаждению и согреванию уда-
лось получить не только на собаке [237], но также на кошке [270]
и голубе [377]. При согревании или охлаждении спинного мозга
или гипоталамуса изменяется частота разряда некоторых нейро-
нов гипоталамуса, непосредственно не подвергавшихся темпера-
турному воздействию [142].
Импульсы от периферических рецепторов идут вверх по лате-
ральным спиноталамическим путям и проходят через таламус к
гипоталамусу, где активируют центры различных автономных реф-
лексов. В гипоталамусе имеются также центры, которые сами чув-
ствительны к температуре. Гипоталамические нейроны, чувстви-
тельные к холоду и к теплу, по-видимому, пространственно не обо-
соблены, а перемешаны и могут в некоторой степени тормозить
друг друга. После разрушения заднего отдела гипоталамуса соба-
ки утрачивают способность регулировать температуру тела на хо-
лоде, а кошки с повреждениями в заднелатеральном отделе гипо-
таламуса становятся в известной мере пойкилотермными. Местный
обогрев передней части гипоталамуса вызывает у кошки расшире-
ние периферических сосудов и учащение дыхания. При внешней
температуре 25 °C местное охлаждение преоптической области
гипоталамуса приводит к гипертермии, тогда как при 35°C такое
же охлаждение не дает эффекта. У летучих мышей местный обо-
грев преоптической области гипоталамуса вызывает расширение
сосудов в крыльях, что увеличивает отдачу тепла; охлаждение ги-
поталамуса оказывает обратное действие. Крысы, обученные при
охлаждении нажимать на рычаг, делают то же самое и при
охлаждении гипоталамуса термодом [75].
У бабуина местное охлаждение преоптической области гипота-
ламуса вызывает мышечную дрожь, сужение сосудов, высвобожде-
ние норадреналина и адреналина уменьшается и уровень 17-кето-
Роидов плазмы; когда охлаждение прекращают, происходит рас-
ширение сосудов. При обогревании того же участка высвобожде-
ние норадреналина и адреналина уменьшается и уровень 17-кето-
стероидов в плазме падает [136, 462]. У козы местное охлаждение
переднего отдела гипоталамуса с помощью термодов активирует
китовидную железу и симпатическую нервную систему; при этом
Уисе через несколько минут возрастает содержание в плазме иода,
связанного с белком. Перерезка ножки гипофиза предотвращает
Развитие этих эффектов и блокирует действие норадреналина [7].
У кошек была исследована электрическая активность примерно
Ысячи нейронов переднего отдела гипоталамуса; из них 60% не
12—1514
178 Глава 9
реагировали на местное изменение температуры, а из остальных
нейронов 4/5 усиливали импульсацию при обогревании и Vs — при
охлаждении [162]. У кролика 10% исследованных нейронов, на-
ходившихся в 1—2 мм от средней плоскости гипоталамуса, реаги-
ровали на температуру; из них 2/з отвечали повышением частоты
разряда на обогрев и снижением — на охлаждение, а 7з давала
обратную реакцию; было найдено несколько нейронов, у которых
в узком диапазоне температур (не более чем на 2° выше или ниже
температуры тела) частота разряда была пропорциональна темпе-
ратуре [64, 188]. Локальное охлаждение преоптической области
гипоталамуса вызывает дрожь или поведенческую реакцию, на-
правленную на использование внешнего источника тепла (если он
доступен для животного). У крыс с повреждениями гипоталамуса
нарушено физиологическое повышение теплопродукции, но сохра-
нены соответствующие поведенческие реакции [403, 404].
Очевидно, что термочувствительные нейроны широко распро-
странены; при этом в координированных процессах терморегуля-
ции могут участвовать как периферические рецепторы, так и ре-
цепторы внутренних органов, спинного мозга и гипоталамуса. Ги-
поталамус служит регуляторным центром, действующим через ве-
гетативную и нейроэндокринную системы.
Согласно одной из гипотез о механизме действия гипоталамуса
[125], его реакции зависят от баланса между выделением норадре-
налина и серотонина в промежуточном мозгу. У кошки, собаки или
обезьяны введение в гипоталамус норадреналина вначале приво-
дит к расширению кровеносных сосудов и падению температуры
тела; серотонин оказывает противоположное действие — вызывает
мышечную дрожь и повышение температуры тела. При тепловом
воздействии содержание норадреналина в гипоталамусе возра-
стает. В опытах на кроликах, овцах, козах, коровах и крысах по-
лучена обратная картина: норадреналин вызывает гипертермию,
а серотонин — снижение температуры тела [125]. Эффекты серо-
тонина подробнее рассмотрены в другом месте (гл. 11).
Согласно другой гипотезе, в критических участках мозга изме-
няется соотношение между натрием и кальцием; в опытах с пер-
фузией мозговых желудочков обезьяны увеличение концентрации
Na приводило к повышению температуры тела, а увеличение кон-
центрации Са — к ее понижению [340].
Гипотеза «точки включения», выдвинутая Хэммелом, состоит в
том, что каждая данная регуляторная реакция (например, суже-
ние сосудов) возникает при определенной пороговой температуре:
R — Rq—Ar (^гт ^вкл.д)»
где R— регуляторная реакция, выраженная в кал/(кг-ч); Rg— ис-
ходный уровень; AR — коэффициент, положительный в случае теп-
лоотдачи и отрицательный в случае теплопродукции, 7,ГТ’— темпе-
Температура
179
аТура гипоталамуса, Твкл. л— пороговая температура (которая
служит точкой отсчета).
реакция пропорциональна разности Тщ—TBKJ1.R. TBK3i.r может
зависеть как от температуры кожи, так и от температуры внутрен-
них органов. Например, 7ВКЛ для сужения сосудов может пони-
жаться при высокой температуре тела или высокой внешней тем-
пературе и повышаться при низких температурах тела или возду-
ха. Таким образом, потеря тепла при испарении зависит и от Тгт,
и от внешней температуры [153]. Для каждого регуляторного от-
Вь1сокий Q/e
Низким Qio
Таламус
Холодовые реце-
пторы 1
Тепловые рецед
пторы
Афферентные волокна
от кожи, идущие через
таламус
Дыхательная область
' (тепловая одышка) *
Прессорно-депрессорная область,
связанная с симпатической це-
почкой (кожная вазоконстрикция)
Скелетные мышцы
(дрожь) '
Рис. 9-35. Схема возможной организации нейронных связей в терморегуляторном
центре гипоталамуса [149].
Афферентные волокна от кожных рецепторов идут к нейронам, которые сами обладают раз-
личной термочувствительностью. Эти нейроны в свою очередь воздействуют на центры, уп-
равляющие дыханием, сосудистыми реакциями и мышечной дрожью. Каждый из этих цент-
ров имеет свою собственную «установку» на поддержание определенного баланса между
входными сигналами от терморецепторов.
вета — тепловой одышки, дрожи или вазомоторных реакций —
имеются свои точки включения (пороговые температуры). Порого-
вые температуры преоптической области гипоталамуса могут изме-
няться в зависимости от температуры кожи [230].
Предложены многочисленные модели терморегуляции в виде
блок-схем [153, 160, 161, 446]. В каждой из этих моделей имеется
регулируемая система (температура определенной области тела),
от которой поступает сигнал обратной связи в регулятор, находя-
щийся в гипоталамусе, и здесь происходит сравнение его с «точ-
кой включения». После этого регулятор посылает сигналы, вызы-
вающие такие реакции, которые переводят регулируемую систему
в точку с минимальным сигналом обратной связи [148] (рис. 9-35).
В технике используется четыре основных типа терморегули-
РУющих устройств — двухпозиционные (on-off) регуляторы, систе-
мь! пропорционального регулирования, регуляторы по скорости и
интегральные регуляторы; все эти системы сравнивали с биологи-
чкой терморегуляцией [160, 446]. Работа этих регуляторов опи-
рается следующими уравнениями:
12»
180
Глава 9
а) двухпозиционный
П>7’нач, Г=тах;
т < т У=0-
1 т 2 нач’ *
б) пропорциональный
^0 ~ ® (Тт ^нач)»
в) регулятор по скорости
г) интегральный
h
где Гт— регулируемая температура, 711ач— начальная, или задан-
ная, температура, У — эффекторная реакция, (У1—Уо)—дифферен-
циальная эффекторная реакция, а, [3, у — коэффициенты пропор-
циональности.
В качестве приближенных моделей биологической терморегуля-
ции наиболее полезны системы двухпозиционного и пропорцио-
нального регулирования [161]. Двухпозиционные модели могут
быть использованы при описании поведенческих и физиологиче-
ских терморегуляторных реакций типа «все или ничего» (напри-
мер, когда ящерица переходит с освещенного солнцем участка в
тень) [177]. Система пропорционального регулирования пригодна
как модель управления теплопродукцией и потерей тепла при ис-
парении [150]. Пропорциональный регулятор может обеспечить
постоянную температуру тела, если Гнап изменяется в зависимости
от температуры среды, внутренней температуры тела и состояния
активности [150]. Изменение Тнач обратно пропорционально раз-
мерам тела [179].
Зимняя спячка
Существует целая гамма различных типов гипотермии — от
ежесуточного оцепенения, при котором температура тела обычно
снижается незначительно, до зимней или иной сезонной спячки, ко-
торая может быть глубокой или неглубокой. Зимняя спячка отли-
чается от кратковременного оцепенения тем, что ей обычно пред-
шествуют «пробные» понижения температуры тела; кроме того,
температура тела во время спячки может понижаться значительно
больше, чем при оцепенении. Выход из обоих состояний осуществ-
ляется путем выработки тепла в самом организме. Спячка обычно
бывает один раз в году и наступает после ряда подготовительных
процессов. Оцепенение может наступать ежедневно или при всЯ'
Температура
181'
ком умеренном охлаждении и нехватке пищи. Биохимия и нейро-
физиология обоих состояний пока не расшифрованы и, возможно,
имеют разный характер.
Во время зимней спячки терморегуляторная система организма
переключается на более низкий уровень (часто около 2°C), а ра-
бота большинства систем органов перестраивается так, что даже в
относительно неактивном состоянии поддерживается гомеокинез.
В настоящую спячку впадают лишь немногие млекопитающие —
представители однопроходных, насекомоядных, грызунов и лету-
чих мышей. Некоторые крупные млекопитающие, например медве-
ди, длительное время обходятся без пищи, но продолжают поддер-
живать температуру тела около 31 °C. Согласно одной из рассмот-
ренных выше теорий терморегуляции, точки включения реакций,,
служащих для сохранения тепла, могли бы в таких случаях пони-
жаться под действием сигналов от ретикулярной активирующей
системы мозга. При выходе из спячки периферическая теплопро-
водность уменьшается, резко возрастает теплопродукция (особен-
но в грудной клетке) и температура тела быстро повышается.
Получены убедительные данные о существовании годичного
цикла, лежащего в основе спячки. У сусликов (Citellus), которых,
в течение двух лет содержали при постоянной температуре (0, 21
или 36°C) и 12-часовом фотопериоде, наблюдался одногодичный
цикл изменений веса и потребления пищи. Животные, выдержи-
ваемые при 0 или 21 °C, впадали в спячку, особенно при ограни-
ченном количестве пищи. У других животных, содержавшихся в те-
чение четырех лет при 12 или 3°С, также были обнаружены годич-
ные циклы температуры тела и спячки. При постоянных внешних,
условиях эти циклы были примерно на 15% короче календарного
года [356, 357].
Имеются данные о том, что сыворотка крови сусликов, находя-
щихся в состоянии зимней спячки, даже летом может вызвать
спячку у животных-реципиентов [91].
Подготовка к спячке. У многих зимнеспящих животных, напри-
мер у сусликов, осенью в организме накапливается жир, дыхатель-
ный коэффициент становится больше 1,0 и идет интенсивный син-
тез жира из углеводов. Другие животные, например хомячки Ме~
socricetus, не накапливают жира, но, помещенные на холод, начи-
нают запасать корм и строить гнездо. На холоду (5 °C) у хомяков
откладываются ненасыщенные жиры с более низкой точкой плав-
ления. У ежей во время зимней спячки повышается содержание
инсулина в крови, а щитовидная железа подвергается инволюции;
инъекции тироксина могут задерживать наступление спячки. Вво-
Дя хомяку питуитрин, можно вывести его из состояния зимней,
спячки. Во время зимней спячки наблюдается истощение коры над-
почечников, а животные с удаленными надпочечниками в спячку
е впадают. Введение норадреналина во время зимней спячки не
взывает реакции термогенеза. Зимняя спячка не связана с функ-
82
Глава 9
цией какой-либо отдельной эндокринной железы, но многие из
этих желез имеют к ней какое-то отношение.
У некоторых животных, например у хомяков, холод задержи-
вает переход в состояние зимней спячки, который затем совер-
шается постепенно; у других животных, таких, как карманчиковая
мышь (Рего gnat has), подобной задержки нет; наконец, у третьих
спячке ’предшествуют периодически возникающая мышечная
дрожь, подъемы и падения температуры тела. Обычно температу-
ра понижается медленно, со скоростью 2—4 град/ч, спады тем-
пературы связаны с расширением периферических сосудов и
уменьшением мышечного тонуса. При температуре тела 33—34 °C
частота сокращений сердца резко снижается. Дыхание замедляет-
ся постепенно, и дыхательная система все еще сохраняет чувстви-
тельность к стимулирующему действию гипоксии.
Состояние зимней спячки. При понижении температуры тела
частота сердечных сокращений становится минимальной, а при
температуре чуть ниже 3°С у большинства видов возникает арит-
мия. У суслика в состоянии активности частота сокращений сердца
равна 200—400 ударов в 1 мин, а у спящего — всего лишь 7—10.
Интервал Р—Т электрокардиограммы удлиняется; атриовентрику-
лярная блокада может приводить к десинхронизации сокращений
предсердий и желудочков. Изолированные сердца животных, впа-
дающих в зимнюю спячку, способны сокращаться при более низ-
кой температуре (у суслика ниже 1 °C), чем сердца животных, не
впадающих в спячку (у белки 13—16 °C) [299а]. Сокращения сер-
дец ежа и хомяка прекращаются соответственно при 1,5 и 6,0 °C,
.а крысы и кролика — при 16—18 °C.
Если предсердия суслика охладить до 6 °C, потенциал покоя
почти не изменится, а потенциал действия даже повысится,
тогда как у кролика потенциал покоя уменьшается уже при
температурах ниже 25 °C, а потенциал действия исчезает при тем-
пературе ниже 17 °C [308]. Минутный объем сердца у суслика во
время зимней спячки в 65 раз меньше, чем у активного животно-
го [361]. Остановка сердца происходит у морской свинки при
13,4 °C, у крысы при 6, у бодрствующего хомяка при 10,2, а у хо-
мяка в состоянии спячки при 1 °C [426]. У ежа (Erinaceus) при
температурах ниже 6,4 °C блуждающий нерв почти не влияет на
работу сердца и кровяное давление, а при согревании животного
сердце не отвечает на раздражение блуждающего нерва, пока не
будет достигнута температура 20 °C. В то же время при низкой
температуре сохраняются некоторые реакции на ацетилхолин и
адреналин [239]. Такая же нечувствительность сердца к раздра-
жению блуждающего нерва во время спячки обнаружена у сус-
.лика [300]. В изолированном препарате диафрагмальный нерв —
диафрагма блокада нервно-мышечной передачи наступает при
10 (крыса), 5 (бодрствующий хомяк) и ниже 5°C (хомяк в со-
стоянии спячки ) [441].
Температура
183
При охлаждении тела функции нервной системы не выключают-
ся полностью. Активность коры у хомяка, судя по волнам ЭЭГ,
прекращается при температуре около 19 °C, но некоторые корти-
кальные реакции на раздражение периферических нервов могут
наблюдаться даже при 9,1 °C. У суслика электрическая активность
мозга сохраняется при снижении температуры тела до 12 °C,
а у североамериканского лесного сурка — до 11 °C. У суслика при
5 °C еще можно обнаружить низкоамплитудные волны в коре,,
а при температуре мозга 6,1 °C он способен издавать и восприни-
мать звуки, настораживать уши и двигаться [447].
Хомяки при повреждении заднего отдела гипоталамуса утрачи-
вают способность впадать в спячку. Суслики с повреждениями
переднего гипоталамуса сохраняют эту способность, но не могут
пробуждаться [404, 406]. Вдувание СО2 во время спячки повышает
частоту дыхания, а затем и сокращений сердца; у хомяка такая
стимуляция дыхания не приводит, однако, к пробуждению.
У млекопитающих температура тела во время зимней спячки
почти совпадает с температурой окружающего воздуха и иногда
изменяется в соответствии с колебаниями последней. Температура
толстых кишок и пищевода может быть на 2—3°С выше температу-
ры воздуха. У суслика при внешней температуре 2 и 5,5° С температу-
ра мозга равна соответственно 6 и 8,7 °C. Если температура возду-
ха приближается к нулю, некоторые зимнеспящие животные (хо-
мяк, лесной сурок, соня) имеют тенденцию удерживать темпера-
туру тела около 2 °C, а потребление ими кислорода может
увеличиться. Таким образом, они сохраняют некоторую способ-
ность противостоять чрезмерному охлаждению. Многие зимнеспящие
животные периодически пробуждаются; некоторые при этом едят
и пьют. У меченых летучих мышей было обнаружено много
индивидуальных различий в зимней двигательной активности.
У сусликов периоды пробуждения составляют 7% времени зимней
спячки; они повторяются в среднем 1 раз в 11 суток и длятся по
нескольку часов. За один такой период животное расходует столь-
ко же резервов организма, сколько за 10 суток спячки. Хомяки
пробуждаются через каждые несколько дней.
Интенсивность обмена во время спячки уменьшается в 20—
100 раз. У сурка она обычно составляет 2,8 ккал/(кг-ч), а в период
спячки — всего лишь 0,09 ккал/(кг-ч). При этом дыхательный ко-
эффициент имеет величину, характерную для обмена жиров. Не-
смотря на низкую интенсивность метаболизма, животные во время
спячки сильно теряют в весе. Например, у бурого кожана потеря
Веса за 180 дней составляет 33%, а содержание жира уменьшается
с 28 до 10% веса тела.
У ежей, сусликов и сурков во время зимней спячки обнаружено
пьтсокое содержание магния в сыворотке крови; то же самое най-
дено у летучих мышей, находящихся в спячке при 13, но не при
20 °C, когда содержание магния оказывается нормальным..
384
Глава 9
Высокий уровень Mg не играет причинной роли в поддержании
спячки, поскольку он сам по себе не препятствует пробуждению.
У некоторых зимнеспящих видов сахар.крови удерживается на
низком уровне, тогда как у других концентрация сахара вначале
падает, а затем вновь поднимается до уровня, близкого к нормаль-
ному. Число лейкоцитов может быть меньше обычного, гемато-
крит слегка понижен. Свертывание крови замедлено — вероятно,
вследствие уменьшения количества протромбина. Суслики и лету-
чие мыши во время зимней спячки обнаруживают временную
устойчивость к рентгеновскому облучению. Вначале эффект облу-
чения почти незаметен, но спустя несколько дней, после пробуж-
дения, происходит обычное снижение количества клеток крови.
В срезах различных тканей суслика (за исключением печени)
уровень эндогенного дыхания осенью выше, чем весной; ниже все-
го он оказывается в тканях животных, находившихся в состоянии
зимней спячки [158]. Срезы почек, взятых во время спячки, обна-
руживают повышенную способность к глюконеогенезу и гликолизу,
<с чем, возможно, связана наблюдаемая в этот период устойчивость
к гипоксии [60, 61]. У суслика во время зимней спячки найдена
также большая величина Qi0 для дыхания митохондрий печени,
т. е. чувствительность их к температуре повышена [298]. Однако
у хомяков измерения Qio для окислительного фосфорилирования
в митохондриях сердца и для дыхания и гликолиза в ткани мозга
не выявили существенных различий между периодами спячки и
активности [440]. В то же время были отмечены различия в дей-
ствии внемитохондриальных факторов между зимнеспящими и не-
зимнеспящими видами. Дыхание митохондрий, выделенных из пе-
чени крыс после 5—7 недель содержания животных при 5 °C,
оставалось на обычном уровне, тогда как потребление кислорода
гомогенатами (с добавкой сукцината) возрастало, а отношение
P/О уменьшалось. В отличие от этого дыхание митохондрий из пе-
чени бодрствующих хомяков, находившихся на холоде, оказыва-
лось повышенным, а отношение Р/О — пониженным, тогда как во
время зимней спячки наблюдалась обратная картина [298]. У вы-
держиваемых на холоде крыс митохондрии бурой жировой ткани
обнаруживают усиленное дыхание (опыты с добавкой сукцината),
а чувствительность к стимулирующему действию АДФ при 5-у
15 °C уменьшается. В то же время митохондрии из бурой жировой
ткани хомяков, выдерживаемых на холоде, или из печени сусли-
ков, находившихся в состоянии спячки, обнаруживают при 5—
15 °C повышенную чувствительность к АДФ [298]. Приходится за-
ключить, что не существует какой-то общей схемы метаболических
различий между животными, впадающими и не впадающими
в зимнюю спячку.
Способность печени к липогенезу у хомяков вчетверо выше»
чем у крыс. Когда хомяк пробуждается от спячки, интенсивность
липогенеза у него быстро возрастает [104].
Температура
185
Очень существенно то, что зимнеспящие животные могут под-
держивать нормальные ионные градиенты при низких температу-
рах. Накапливается все больше данных о необычных свойствах
клеточных мембран у этих животных. Способность сохранять нор-
мальное распределение ионов ведет к тому, что клетки на холоду
не набухают, проведение импульсов не блокируется, продолжается
белковый синтез, по-прежнему происходит поглощение сахара и
аминокислот. В поддержании ионных градиентов участвуют как
пассивные перемещения ионов, так и механизмы активного пере-
носа. Достоверно установлены различия в функции этих механиз-
мов у животных, впадающих и не впадающих в зимнюю спячку,
у зимнеспящих форм холод меньше подавляет перенос К+. Напри-
мер, срезы почек суслика могут несколько дней сохранять нормаль-
ную концентрацию калия, если хранить их при 3°С, тогда как
срезы почек крысы в этих условиях теряют калий [493]. Срезы
сердца и диафрагмы крысы при 6 °C набухают сильнее, чем срезы
тех же органов хомяка или суслика [494]. Активное (т. е. чувстви-
тельное к уабаину) поглощение калия эритроцитами при 5 °C со-
храняется у суслика, но падает до нуля у морской свинки или че-
ловека [259]. На+К+-АТФаза почек хомяка или головного мозга
ежа, находящегося в состоянии зимней спячки, устойчива к холо-
ду, тогда как у животных, не впадающих в спячку, этот фермент
чувствителен к охлаждению. Ыа+К+-АТФаза почек хомяка во вре-
мя спячки вдвое активнее, чем у бодрствующего хомяка (рис. 9-36),
это говорит о компенсаторной акклимации вроде той, которая об-
наружена у пойкилотермных животных (см. стр. 107). Вполне
возможно, что свойства мембран гораздо более важны для выжи-
вания зимнеспящих животных при низких температурах, чем осо-
бенности энергетического обмена [495].
Пробуждение. Выход из состояния зимней спячки носит харак-
тер быстрого пробуждения при самосогревании, начинающемся
в области грудной клетки. Летучие мыши, находившиеся без пищи
в холодильнике в течение 144 дней, были способны к полету уже
через 15 мин после переноса в обычные условия. У суслика темпе-
ратура тела может за 4 ч повысится с 4 до 35 °C, а лесная мы-
чювка согревается со скоростью 1 град/мин. Пробуждение млеко-
питающего протекает как процесс самосогревания, не требующий
притока тепла извне. Сначала повышается температура грудной
клетки, а затем — благодаря распределению крови — и темпера-
тура передней половины тела. В задней половине тела сосуды
сжимаются, и приток крови к грудным мышцам возрастает
в 16 раз [239]. Быстро согревается бурая жировая ткань, находя-
щаяся в грудной клетке; активируется окисление триглицеридов —
од воздействием адренэргических симпатических нервов, а может
Ь1ть, и косвенно, при участии АКГТ [422]. За первый час после
Р°оуждения образование СО2 возрастает в 15 раз; дыхательный
Оэффициент достигает 0,7, что говорит об использовании жиров.
Рис. 9-36. Влияние температуры на Na+K+'АТФазу животных, впадающих и не
впадающих в зимнюю спячку (по С. Голдмэну и Дж. Уиллису).
А. Активность фермента в срезах почки крысы (I), бодрствующего хомяка (II) и хомяка,
находившегося в состоянии спячки (III) (в логарифмическом масштабе). Б. Отношение ак-
тивности в мозгу хомяка во время зимней спячки к активности у бодрствующего животного.
Т емпература
187
[449]. Действительно, у суслика сразу после пробуждения обнару-
жен повышенный обмен триглицеридов [449]. У летучей мыши
температура бурой жировой ткани повышается быстрее, чем темпе-
ратура сердца; в данном случае термогенез не связан с мышечной
дрожью: пробуждение возможно у кураризированных животных,
хотя и протекает замедленно. Однако у суслика и сони Glis бло-
када мышечной дрожи сильно задерживает согревание. У летучей
мыши Myotis уровень глицерина в крови повышается в этот период
с 1,3 до 25 мг %• Блокада |3-адренорецепторов пропранололом
вдвое увеличивает время, необходимое для пробуждения; на долю
термогенеза, не связанного с дрожью, приходится 55% общей теп-
лопродукции при пробуждении [186].
Частота сокращений сердца быстро возрастает; например, у су-
слика во время спячки (при температуре тела 5 °C) она составля-
ет 3 в 1 мин, а в период пробуждения увеличивается до 20 (при
8°C), затем до 200 (при 14°C) и 300 в 1 мин (при 20°C). Дыха-
ние уже в самом начале становится регулярным, а затем учащает-
ся. При выходе из спячки наблюдается метаболическое «перерегу-
лирование»; например, потребление кислорода у хомяка возраста-
ет с 0,5 до 8000 мл/(кг-ч), а позднее, после стабилизации темпе-
ратуры, снижается до уровня около 5000 мл/(кг-ч). Электрическая
активность коры больших полушарий заметно повышается при
температуре около 20 °C. Сигнал к окончанию спячки поступает
из вегетативной нервной системы, поэтому важно было бы иметь
данные об электрической активности базальных ганглиев и ретику-
лярной системы.
Таким образом, животные, впадающие в зимнюю спячку, во
многом отличаются от остальных млекопитающих. Для зимнеспя-
щих животных характерны, например, отложение больших коли-
честв бурого жира, способность клеточных мембран сохранять
активность при низких температурах, малая чувствительность нер-
вов к холоду, уникальные свойства многих ферментов и процессов
белкового синтеза. При погружении в спячку наибольшее значе-
ние имеет предполагаемая перенастройка центра терморегуляции,.
а при пробуждении — активация симпатической нервной системы.
Зимнюю спячку нельзя связать с функцией какой-либо одной си-
стемы: по-видимому, существенно то, что зимнеспящие животные
и на низких уровнях активности сохраняют интеграцию всех си-
стем организма.
Заключение
Температура является мерой количества тепловой энергии
Физическом теле. Температура отражает кинетическую энергию
олекул в растворах и, таким образом, определяет скорость хими-
еских реакций. Измерив скорость реакции при различных темпе-
.188
Глава 9
р атурах, можно вычислить энергию активации для данной реак-
ции. При изучении таких сложных систем, как живые клетки,
ценным показателем служит Qlo, хотя его величина зависит от
температурного диапазона, концентрации субстрата и модулирую-
щих факторов. Кроме того, с понижением температуры возрастает
кажущееся сродство фермента к субстрату (Км) — может быть,
вследствие большей стабильности фермент-субстратного комплек-
са. Величины Км генетически детерминированы и имеют приспо-
собительное значение в определенных температурных диапазонах,
соответствующих условиям жизни данного организма.
Температурные границы биологической активности тесно связа-
ны со свойствами воды, денатурацией белков и физическим состоя-
нием липидов. Внутриклеточное образование твердых кристаллов
льда несовместимо с жизнью; замерзание внеклеточной воды при-
водит к дегидратации клеток. Устойчивость животных к заморажи-
ванию может быть обусловлена: 1) замещением части воды каким-
либо органическим растворителем, например глицерином; 2) пони-
жением точки замерзания вследствие повышения количества
растворенных веществ (в этом отношении особенно эффективны
некоторые гликопротеиды); 3) переохлаждением, которое стано-
вится возможным благодаря органическим молекулам, связываю-
щим водород; 4) устойчивостью к образованию льда, обусловлен-
ной какими-то неизвестными свойствами клеточных мембран. В то
же время гибель от холода наступает при температурах значитель-
но выше точки замерзания: причиной ее может быть замедление
процессов, доставляющих энергию, рассогласование различных ме-
таболических реакций, центральные нервные нарушения или недо-
статочное снабжение клеток кислородом, а также, несомненно, из-
менение клеточных мембран, которые начинают «протекать» —
отчасти в результате замедления работы ионных насосов.
Высокие температуры могут быть неблагоприятными для жиз-
ни или летальными вследствие денатурации ферментов (в экстре-
мальных случаях), относительного ускорения одних реакций по
сравнению с другими, создания неоптимальных температурных
условий для отдельных реакций и «расплавления» липидов, осо-
бенно в клеточных и митохондриальных мембранах, которые ста-
новятся при этом свободно проницаемыми.
Изолированные клетки и ткани выживают в более широком
диапазоне температур, чем интактные животные, а для экстраги-
рованных из тканей ферментов температурные границы еще шире.
Как правило, узкий температурный диапазон активности интакт-
ных животных зависит от центральной нервной системы. Однако
причины нарушений нервной деятельности при высоких и низких
температурах пока не установлены. Как для целых животных, так
и для некоторых тканей температурные границы выживания могут
изменяться в результате акклимации, которая, вероятно, ускоряет
синтез специфических липидов и белков. Выработка холодоустои-
Температура
189
чивости у растений связана с целым рядом биохимических изме-
нений. Кроме того, многие ферменты животных и микроорганизмов
представлены генетически детерминированными вариантами, при-
способленными к определенным температурным условиям.
Нормальный ход биологических процессов в пределах «обыч-
ного» диапазона внешних температур возможен благодаря много-
образным компенсаторным механизмам. У пойкилотермных живот-
ных температура тела изменяется вместе с температурой окружаю-
щей среды, но «метаболические компенсации» позволяют им под-
держивать относительно постоянный уровень активности в опре-
деленных температурных границах. У гомойотермных животных
сама температура тела относительно постоянна; в случае охлаж-
дения интенсивность обмена у них повышается и доставляет орга-
низму добавочное тепло. Кроме того, у некоторых представителей
обеих этих групп встречаются компенсаторные реакции третьего
типа: на холоде у них может наступать состояние диапаузы, оце-
пенения или спячки. Приспособления, компенсирующие перегрев,
носят более ограниченный характер и более специфичны для от-
дельных групп животных, чем адаптации, компенсирующие охлаж-
дение.
Реакции организма на изменение внешней температуры можно
разделить по их временному масштабу на три категории. К первой
группе относятся прямые ответы, длительность которых может из-
меряться минутами или часами; они состоят в непосредственном
адаптивном ускорении или замедлении определенных процессов.
Амплитуда таких прямых реакций зависит от величины и скорости
изменения температуры, а характер ответа — от особенностей дан-
ного животного. Вторую группу составляют более медленные реак-
ции — процессы акклимации и акклиматизации, которые компен-
сируют изменение внешней среды. И наконец, к третьей группе
относятся генетические изменения: на протяжении многих поколе-
ний может происходить отбор мутантов, более приспособленных
к определенным температурным условиям.
У пойкилотермных животных способность к акклимации может
быть различной в зависимости от температурного диапазона, обыч-
ного для данного вида, от гормонального баланса, от уровня пита-
ния и от того, впадает ли данное животное на холоде в оцепенение
или остается активным. Животные, обитающие в относительно по-
стоянной среде, могут обладать меньшим «акклимационным по-
тенциалом», чем виды, живущие в изменчивых условиях. Компен-
саторные изменения характерны для ферментов, связанных с вы-
работкой энергии; ферменты, участвующие в деградации метабо-
литов, часто не обнаруживают компенсаторных изменений, а иног-
да их активность изменяется в противоположном направлении.
Пекулярные механизмы компенсации включают ускоренное об-
овление белков (и, вероятно, РНК), избирательный синтез изо-
ферментов с подходящими кинетическими свойствами и изменение
И
190 Глава 9
модулирующих факторов, особенно фосфолипидов (повышение сте-
пени их ненасыщенности на холоде). Предполагается, что как для
устойчивости, так и для адаптации к температуре решающее зна-
чение могут иметь мембранные липиды.
У гомойотермных животных процесс акклимации включает:
1) изменения теплоизоляции (вазомоторные механизмы, подкож-
ный жир, шерстный или перьевой покров); 2) изменение чувстви-
тельности к гормональным регуляторам, особенно к гормонам над-
почечников и щитовидной железы; 3) изменения активности неко-
торых ферментов, пока еще мало изученные; 4) изменение общей
чувствительности к экстремальным температурам. Холодовая ак-
климация более эффективна, чем тепловая.
Между пойкилотермными и гомойотермными животными нет
резкой границы. Пойкилотермные животные не находятся в полной
зависимости от Qi0. Многие из них способны к долговременной ме-
таболической компенсации в условиях холода или тепла, некото-
рые теряют активность при определенном снижении температуры,
а некоторые (в особенности насекомые и рептилии) благодаря
своим поведенческим реакциям могут поглощать тепло извне или
рассеивать избыток тепла; у ряда пойкилотермных животных
(главным образом ночных бабочек и крупных рыб) выработка
тепловой энергии в мышцах делает возможным эндотермное повы-
шение температуры тела. Иногда такого рода поведенческие или
физиологические реакции запускаются благодаря наличию пери-
ферических терморецепторов или способности нервной системы
к непосредственному восприятию температуры. При этом некого- ,
рые структуры мозга могут обладать повышенной температурной
чувствительностью. Подобным же образом многие гомойотермные
животные могут «перенастраивать» свои терморегуляторы и со-
хранять энергию на холоде, впадая в летаргическое состояние или
зимнюю спячку. Терморецепторы у гомойотермных животных
имеются и в коже, и в глубинных сосудах, и в центральной нерв-
ной системе — в спинном мозгу и особенно в гипоталамусе. Иерар-
хия терморегуляторных механизмов хорошо выявляется при тем-
пературном стрессе. Широко распространенным способом пониже-
ния температуры тела служит испарение влаги; однако у ряда
животных возможно повышение температуры тела ради экономии ;
воды.
Необходимо подчеркнуть, что у всех животных в определенном
диапазоне температур сохраняется координация функций различ-
ных систем организма. У зимнеспящих животных головной мозг и
сердечно-сосудистая система способны к упорядоченной работе и
при 37, и при 5 °C. Стоит отказать одной системе (как это проис-
ходит с мозгом животных, не впадающих в зимнюю спячку, при
температуре ниже 25 °C) — и жизнь оказывается в опасности. При
температурной компенсации у пойкилотермных животных проиС'
ходит адаптивное изменение многих ферментных систем. Темпе- l
Температура
191
сатура — это такой физический параметр, который влияет на все
биологические функции, и поэтому реакции на ее изменения и
приспособления к этим изменениям носят многосторонний харак-
тер. В процессе эволюции было отобрано огромное множество
мутаций, способствующих выживанию в широком диапазоне внеш-
них температур и расширению ареала того или иного вида. Фер-
ментные системы «теплолюбивых» или «холодолюбивых» организ-
мов приспособлены к работе соответственно при высоких или низ-
ких температурах.
СПИСОК ЛИТЕРАТУРЫ
1. Adams Р. A., Heath J. Е., Nature, 201, 20—21, 1964. Temperature regulation
in sphinx moth Celerio.
2. Adams T., Covino B. G., J. Appl. Physiol., 12, 9—12, 1958. Racial variations
in response to cold stress.
3. Adolph E., Origins of Physiological Adaptations. Academic Press, New York,
1968, 144 pp. Development of thermal tolerance.
4. Aleksiuk M., Comp. Biochem. Physiol., 39A, 495—504, 1971. Temperature-de-
pendent metabolism of reptile Thamnophis.
5. Andersen K. L., Wilson O., Acta Univ. Lundensis, Sec. II, 1966. Nos. 11—20.
Tolerance of men to cold in field conditions.
6. Anderson T. R., Comp. Biochem. Physiol., 33, 663—687, 1970. Temperature
adaptation, membrane phospholipids in goldfish.
7. Andersson B. et al., Acta physiol, scand., 59, 12—33, 1963; 61, 182—191, 1963.
Control of secretion of thyrotropic hormone and of catecholamines by heat loss
center.
8. Andjus R. Symp. Soc. exp. Biol., 23, 351—394, 1969. Mammalian tolerance
of low body temperature.
9. Andrews S., Levitt J., Cryobiology, 4, 85—89, 1967. Cryoprotective agents in
protection against freezing.
10. Asahina E. In: Cryobiology, edited by H. T. Meryman, Academic Press, New
York, 1966, pp. 451—486. Freezing and frost resistance in insects.
11. Ashwood-Smith M. J. In: Current Trends in Cryobiology, edited by
A. U. Smith, Plenum, Press, New York, 1970, pp. 5—42. Resistance to freezing
in microorganisms.
12. Augee M. L. et al., J. Mammal., 49, 446—454, 1968; 51, 561—570, 1970. Body
temperature and torpor in echidna.
13. Baldwin B. A., Ingram D. L„ Physiol, and Behav., 2, 15—21, 1967. Behavioral
thermal regulation in pigs.
14. Baldwin J., Hochachka P. W., Biochem. J., 116, 883—887, 1970. Interpretation
of isozymes in thermal acclimation.
15. Baranska J., Wlodawer P., Comp. Biochem. Physiol., 28, 553—570, 1969. Effect
of temperature on fatty acids and lipogenesis in frog tissues.
16. Barlow G. W., Biol. Bull., 121, 209—229, 1961. Metabolic differences in nor-
them and southern populations of gobiids.
17. Barnard T., Skala J., Lindberg O., Comp. Biochem. Physiol., 33, 499—508,
509—528, 1970. Brown fat in developing rats.
Barnes H., Barnes M., J. Exp. Marine Biol. Ecol., 4, 36—50, 1969. Effects of
temperature on Ог consumption of barnacles.
19. Barnett L. B., Comp. Biochem. Physiol., 33, 559—578, 1970. Seasonal tempe-
10 rature acclimatization of house sparrow.
’ z,arnett Biol. Rev., 40, 5—51, 1965. Adaptations of mice to cold.
u. Bartholomew G. A., Copeia, 241—250, 1966. Temperature relations of marine
iguanas.
192 Глава 9 ___________________________
21. Bartholomew G. A., Condor, 68, 523—536, 1966. Behavioral regulation of tem-
perature in booby. „ „ ,
22. Bartholomew G. A., Symp. Soc. Exp. Biol., 18, 7—29, 1964. Behavioral ther-
mal regulation in desert animals.
23. Bartholomew G. A., Dawson W. R., Lasiewski R. C„ 1. vergl. Physiol., 70,
196—209, 1970. Thermal regulation and heterothermy in small flying
24. Bartholomew G. A., Hudson J. W., Physiol. ZooL, 35, 94—107, 1962. Tempera-
ture relations of pygmy possums, Cercaertus.
25. Bartholomew G. A., Lasiewski R. C., Comp. Biochem. Physiol., 16, 573—582,
1965. Heating and cooling rates in marine iguana.
26. Bartholomew G. A., MacMillen R. E., Physiol. ZooL, 34, 177—183, 1961. Tem-
perature regulation in kangaroo mouse.
27. Bartholomew G. A., Rainy M., J. Mammal., 52, 81—95, 1971. Regulation of
body temperature in hyrax.
28. Bartholomew G. A., Tucker V. A., Physiol. ZooL, 36, 199—218, 1963. Tempera-
ture control in agamid lizard.
29. Bartholomew G. A., Tucker V. A., Physiol. ZooL, 37, 341—354, 1964. Tempe-
rature relations in varanid lizards.
30. Bartholomew G. A., Tucker V. A., Lee A. K., Copeia, 169—173, 1965. Thermal
relations in Australian skink Tiliqua.
31. Bartholomew G. A. et al., Physiol. ZooL, 37, 179—198, 1964. Temperature re-
lations of bats.
32. Battle H. L, Trans. Roy. Soc. Canad., 20, 127—143, 1926; also Contrib. Canad.
Biol, and Fisheries, N. S., 4, 497—526, 1929. Lethal temperatures in relation
reflexes of skste
33. Beamish F. W. H., Mookherji P. S„ Canad. J. ZooL, 42, 161—175, 1964. Routi-
ne and standard metabolism in trout at different seasons.
34. Behrisch H. W., Hochachka P. W„ Biochem. J., 111, 287—295, 1969. Fructose
diphosphatase in poikilotherms.
35. Berg K., Hydrobiologia, 5, 331—350, 1953. Temperature and metabolism of
freshwater limpets.
36. Berkholz G., "L. wiss. ZooL, 174, 377—399, 1966. Temperature adaptation in '
fish Idas.
37. Bernstein M. H., Comp. Biochem. PhysioL, 38A, 611—617, 1971. Cutaneous
and respiratory regulation of temperature, quail.
38. Bishop D. G., Still J. L., Lipid Res., 4, 87—90, 1963. Temperature and fatty
acid metabolism in bacteria.
39. Blazka P., PhysioL ZooL, 36, 117—128, 1958. Metabolic depression in winter,
crucian carp.
40. Bligh J., Envir. Res., 1, 28—45, 1967. Mechanism of sweat secretion.
41. Bligh J. et al., J. PhysioL, 176, 136—144, 1965. Telemetered temperatures in
ungulates.
42. Bollinger R. E., McKinney С. O., J. exp. ZooL, 161, 21—28, 1966. Development
of temperature tolerance in anuraris.
43. Bowler K., J. Cell. Comp. PhysioL, 62, 119—132, 133—146, 1963. Mechanisms
of acclimation and heat death in Astacus.
44. Brattstrom В. H., Comp. Biochem. PhysioL, 24, 93—111, 1968. Thermal accli-
mation of anurans at different latitudes and altitudes.
45. Brattstrom В. H„ Comp. Biochem. PhysioL, 35, 69—103, 1970. Thermal accli-
mation in Australian amphibians.
46. Brett J. R„ Publ. Ontario Fish Res. Lab., 63, 1—49, 1944; J. Fish. Res. Bd.
Canad., 9, 265—323, 1952; 21, 1183—1226, 1964; 22, 1491—1501, 1965; Quart.
Rev. Biol., 31, 75—87, 1956. Lethal temperatures of freshwater fish.
47. Brett J. R., Amer. ZooL, 11, 99—113, 1971. Energetic responses of salmon to
temperature.
48. Brett J. R., Higgs D. A., J. Fish. Res. Bd. Canad., 27, 1767—1779, 1970. Effect
of temperature on gastric digestion in young salmon.
Температура
193 ~
,о n-ock Т D„ Science, 158, 1012—1019, 1967. Life at high temperature.
50 ‘Brooks С. M., Koiaumi K., Malcolm J. L., J. Neurophysiol., 18, 205—216, 1955.
Fffects of temperature on cat spinal cord.
5j Brown J. H., Bartholomew 0. A., Ecology, 50, 705—709, 1969. Energetics of
toroor in kangaroo mouse.
52 Bruck K., Wunnenberg W., Pflug. Arch., 290, 167—183, 1966. Role of brown
fat in thermal regulation of guinea pigs.
53 Brun J. L., Ann. Biol. Anim. Biochem. Biophys., 6, 127—158, 267—300, 439—
466, 1966. Adaptation to high temperatures in a nematode.
54. Brush A., Comp. Biochem. Physiol., 15, 399—421, 1965. Temperature relations
of California quail.
55. Buetow D. E., Exp. Cell. Res., 27, 137—142, 1962. Effects of temperature on
growth of Euglena.
56. Buffington J. D„ Comp. Biochem, Physiol., 30, 865—878, 1969. Temperature
acclimation in Culex.
57. Bullard R. W., Dill D. B., Yousef M. K-, J. Appl. Physiol., 29, 159—167, 1970.
Responses of burro to desert heat.
58. Bullock T. H., Fed. Proc., 12, 666—672, 1953. Pit viper perception of heat.
59. Burckhardt D., Biol. Zentralbl., 78, 22—62, 1959. Effect of temperature on
crayfish stretch receptors.
60. Burlington R., Comp. Biochem. Physiol., 17, 1049—1052, 1966. Biochemistry
of kidney from cold-exposed rats and hamsters.
61. Burlington R., Klein J., Comp. Biochem. Physiol., 22, 701—708, 1967. Gluco-
neogenesis during hibernation and arousal.
62. Burlington R., Wiebers J. E., Comp. Biochem. Physiol., 17, 183—189, 1966.
Glycolysis in heart of hibernators and nonhibernators.
62a. Bui wick R. E., personal communication.
63. Cabanac AL, Hammel T„ Hardy J. D., Science, 158, 1050—1051, 1967. Tempe-
rature-sensitive units in lizard brain.
64. Cabanac AL, Stolwijk J. A., Hardy J. D., J. Appl. Physiol., 24, 645—652, 1968.
Single unit responses in temperature center of rabbit brain.
65. Cade T. J. et al., Physiol. Zool., 38, 9—33, 1965. Temperature relations of
finch.
66. Calder W. A., Comp. Biochem. Physiol., 30, 1075—1082, 1969. Temperature
relations of water shrew. ,
67. Calder IF. A., King J. R., Experientia, 19, 603—604, 1963. Evaporative cooling
in zebra finch.
68. Calder IF. A., Schmidt-Nielsen K., Proc. Nat. Acad. Sci., 55, 750—756, 1966.
Evaporative cooling and respiratory alkalosis in pigeon.
69. Caldwell R. S., Comp. Biochem. Physiol., 31, 79—93, 1969. Thermal compen-
sation of respiratory enzymes, goldfish.
70. Caldwell R. S., Vernberg J. F., Comp. Biochem. Physiol., 34, 179—191, 1970.
Temperature effects on lipid composition of fish mitochondria.
71. Carey F G„ Teal J. M„ Proc. Nat. Acad. Sci., 56, 1464—1469, 1966. Heat con-
servation in tuna fish muscle.
72. Carey F. G., Teal J. M., Comp. Biochem. Physiol., 28, 199—204, 1969. Warm
bodied sharks.
73. Carey F. G., Teal J. AL. Comp. Biochem. Physiol., 28, 205—213, 1969. Regula-
tion of body temperature in tuna.
74. Carey F. G., Teal J. M„ Kanwisher J. IF., Lawson K. D„ Amer. ZooL, 11, 137—
145, 1971. Temperature regulation in tuna.
5. Carlisle H. J., Nature, 209, 1324—1325, 1966. Hypothalamic temperature-sen-
sitive cells in rat.
76. Carpenter D. O., Comp. Biochem. Physiol., 35, 371—385, 1970. Temperature
effects on membrane Na pump in Aplysia neurones.
‘i. Cassuto Y., Chaffee R. R. J., Amer. J. Physiol., 210, 423—426, 1966. Effect of
heat on cell metabolism of hamster.
°- Chaffee R. R. J. et al., Lipids, 5, 23—29, 1970. Temperature effects on enzymes
of brown fat.
13—1514
194
Глава 9
79. Chapman. D. In: Thermobiology, edited by A. H. Rose, Academic Press, New
York, 1967, pp. 123—146. Effect of heat on cell membranes.
80. Chatfield P. O., Lyman С. P„ Irving L., Amer. J. Physiol., 172, 639—644, 1953.
Temperature adaptations of nerves in leg of gull.
81. Cheverie J. C., Lynn W. G., Biol. Bull., 124, 153—162, 1963. High temperature
tolerance, thyroid activity of teleosts.
82. Chew R. M., Lindberg R. G., Hayden P„ Comp. Biochem. Physiol., 21, 487—
505, 1967. Temperature regulation in mouse Perognathus.
83. Cloudsley-Thompson J. L., Ann. Rev. EntomoL, 7, 199—222, 1970. Thermal
relations of insects.
84. . Colhoun E. H., Entomol. Exp. AppL, 3, 27—37, 1960. Acclimation to cold in
85. Crawford E. C„ Kampe G„ Amer. J. Physiol., 220, 1256—1260, 1971. Respon-
ses of lizard to temperature changes.
85a. Crawford E. C., Lasiewski R. C., Condor, 70, 333—339, 1968. Water loss and
body temperature of large birds.
86. Crawford E. C., Schmidt-Nielsen K., Amer. J. Physiol., 212, 347—353, 1967.
Temperature regulation and evaporative cooling in ostrich.
87. Das A. B., Comp. Biochem. Physiol., 21, 469—485, 1967. RNA and protein
turnover in cold acclimation.
88. Das A. B., Prosser C. L., Comp. Biochem. Physiol., 21, 444—467, 1967. Protein
synthesis.
89. Davies P. S., Walkey M., Comp. Biochem. Physiol., 18, 415—425, 1966. Tempe-
rature effects on metabolism of cestodes.
90. Davydoff A. F„ Makarova A. R., Fed. Proc. Trans. Suppl., 24, T563—T566,
1964. Newborn seals entering water.
91. Dawe A. R., Spurrier W. A., Science, 163, 298—299, 1969. Hibernation indu-
cing factor in blood of ground squirrel.
92. Dawson T. J., Denny M. S., Hulbert A. J., Comp. Biochem. Physiol., 31, 645—
653, 1969. Thermal balance in marsupial Macropus.
93. Dawson T. J., Hulbert A. J., Amer. J. Physiol., 218, 1233—1238, 1970. Meta-
bolism, body temperature, and surface area of Australian marsupials.
94. Dawson W. R., Evans F. C., Condor, 62, 329—340, 403—405, 1960. Deve-
lopment of temperature regulation in young vesper sparrow.
95. Dawson W. R. In: Lizard Ecology: A Symposium, edited by W. W. Milstead,
University of Missouri Press, Columbia, Mo., 1967, pp. 230—257. Temperature
relations of lizards.
96. Dawson W. R., Fisher C. D„ Condor, 71, 49—53, 1969. Responses to tempera-
ture changes by nightjar Eurostopodius.
97. Dawson W. R., Schmidt-Nielsen K. In: Handbook of Physiology, Sec. 4, edited
by D. B. Dill et al. Amer. Physiol. Soc., Washington, D. C., 1964, pp. 481—492.
Terrestrial animals in dry heat.
98. Dawson W. R., Tordoff H. B„ Condor, 61, 388—395, 1959; Auk, 81, 26—35,
1964. Metabolism in relation to temperature in evening grosbeak and cross-
bills.
99. Dawson W. R., Templeton J. R„ Physiol. Zool., 36, 219—236, 1963; Ecology,
47, 759—765, 1966. Physiological responce to temperature in lizards.
100. Dean H. K., Hilditch T. P., Biochem. J., 27, 1950—1956, 1933. Body fat in
Pig-
101. Dean J. M., Comp. Biochem. Physiol., 29, 185—196, 1969. Metabolism of ther-
mally acclimated trout.
102. Dehl R. E., Science, 170, 738—739, 1970. Water content of collagen fibers.
103. Dehnel P. A., Biol. Bull., 118, 215—249, 1960. Temperature and salinity effects
on metabolism of crabs.
104. Denyes A., Baumberg J., Ann. Acad. Sci. Fenn. A. IV 71/9, 131—139, 1964.
Lipogenesis of cold and hibernating hamsters.
105. Depocas F., Canad. J. Physiol. Pharmacol., 44, 875—880, 1966. Cytochrome C
in rat muscles.
Температура
195
infi DeVries A. L., Science, 172, 1152—1155, 1971; also DeVries A L S. К Ko-
matsu and R. E. Feeney, J. Biol. Chem., 245, 2901—2908, 1970. Chemistry of'
anti-freeze glycoproteins from Antarctic fishes.
106a DeVries A. L. In: Fish Physiology, Vol. 6, edited by W. S. Hoar and
D J- Randall, Academic Press, New York, 1971, pp. 157—190.
107. DeVries A. L„ Wohlschlag D. E„ Science, 163, 1073—1075, 1969. Freezing
resistance of Antarctic fishes. , , .
108. DeWitt С. B., Physiol. Zool., 40, 49—66, 1967. Thermoregulation in desert
iguana Dipsosaurus.
109. Dingley F., Smith J. M., J. Insect. Physiol., 14, 1185—1194, 1968. Temperature
acclimation in Drosophila.
110 Doebber G. F., Cryobiology, 3, 2—11, 1966. Cryoprotective compounds.
Ill’ Dorsett D. A., J. Exp. Biol., 39, 579—588, 1962. Preparation for flight by hawk
moths.
112. Drury D. E., Eales J. G„ Canad. J. Zool., 46, 1—9, 1968. Temperature influence
on thyroid activity in brook trout.
113. Dryner R. L., Paulsund J. R., Brown D. J., Mavis Lipids, 5, 15—22, 1970.
Oxidation of fatty acids by brown fat.
114. Dunlap D. G., Comp. Biochem. Physiol., 38A, 1—16, 1971. Metabolism-tempe-
rature curves in frog Acris.
115. Eagan C. J., Int. J. Biometeor., 10, 293—304, 1966. Effect of cold on circula-
tion in fingers of Eskimos and whites.
116. Edney E. B., Physiol. Zool., 37, 364—394, 1964. Acclimation to temperature
in terrestrial isopods.
117. Edney E. B., Barrass R., J. Insect Physiol., 8, 469—481, 1962. Body tempera-
ture of tsetse fly.
118. Edwards G. A., Nutting W. L., Psyche, 57, 33—44, 1950. Metabolism of fi-
rebrat and snow cricket.
119. Ekberg D. R., Biol. Bull., 114, 308—316, 1958. Temperature acclimation and
tissue metabolism, goldfish.
120. Ekberg D. R., Comp. Biochem. Physiol., 5, 123—128, 1962. Anaerobic and me-
tabolism of carp gills in relation to temperature.
121. Esch H., Z. vergl. Physiol., 43, 305—335, 1960. Temperature relations of ho-
neybee.
122. Evans R. M., Purdie F. C., Hickman С. P., Canad. J. Zool., 40, 107—118, 1962.
Temperature effects on metabolism of rainbow trout.
123. Evans W. G., Nature, 202, 211, 1964. Infrared receptors in buprestid beetles.
124. Evans W. G., Ann. Entomol. Soc. Amer., 59, 873—876, 1966. Infrared receptors
in beetles.
125. Feldberg W. et al., Nature, 200, 1325, 1963; also J. Physiol., 191, 501—515,
1967; 197, 221—231, 1968. Regulation of body temperature by monoamines in
hypothalamus.
126. Fraenkel G., Ecology, 42, 604—616, 1961. Resistance to high temperatures in
anail Littorina.
127. Fraenkel G., Gunn D. L., Orientation of Animals. Dover Press, New York,
1961.
128. Freed J. M., Comp. Biochem. Physiol., 14, 651—659, 1965. Temperature accli-
mation of cytochrome oxidase in goldfish.
129. Freed J. M., Comp. Biochem. Physiol., 39B, 747—764, 765—774, 1971.
Phosphofructokinase in temperature adaptation.
130. Freeman В. M., Comp. Biochem. Physiol., 33, 219—230, 1970; 34, 871—881,
1970. Thermoregulation in quail and other fowl.
131. Fry F. E. J., Publ. Ontario Fish. Res. Lab., 66, 1—35, 1946; 68, 1—52, 1947.
Environmental effects on activity of fish.
!32. Fry F. E. J., Brett J. R., Clawson G. H„ Rev. Canad. Biol, 1, 50—56 1942
Lethal temperatures vor young goldfish.
3. Fry F. E. J Hart J. S., J. Fish. Res. Bd. Canad, 7, 169—175, 1949. Swimming
speed of goldfish at different temperatures.
13»
1
196 Глава 9
134. Fulco A. J., Biochem. Biophys. Acta, 218, 558—560, 1970. Induction of fatty
acid desaturation by temperature in bacteria.
135. Funkhouser G. E., Higgins E. A., Adams T., Snow С. C., Life Sci., 6, 1615—
1620, 1967. Response of savanna baboon to heat.
136. Gale С. C„ Jobin M., Proppe D. W., Hotter D„ Fox H„ Amer. J. Physiol., 219,
193—201, 1970. Endocrine responses to hypothalamic cooling in baboons.
137. Galster VF. A., Morrison P., Amer. J. PhysioL, 218, 1228—1232, 1970. Carbo-
hydrate changes during hibernation in ground squirrel.
138. Gatt S„ Science, 164, 1422—1423, 1969. Thermal lability of galactosidase from
salmon liver.
139. Goris R. C., Nomoto M., Comp. Biochem. Physiol., 23, 879—892, 1967. Infra-
red reception by facial pits of crotaline snakes.
140. Grainger J. N. R., Comp. Biochem. PhysioL, 29, 665—670, 1969. Heat death
in terrestrial snail Arianta.
141. Greer G. L., Gardner D. R., Science, 169, 120—122, 1970. Temperature-sensi-
tive neurones in brain of brook trout.
142. Guieu J. D., Hardy J. D., J. Appl. PhysioL, 29, 675—683, 1970. Effect of tem-
perature change of spinal cord on hypothalamic neurones in rabbit.
143. Guillory R. J., Racker E., Biochim. Biophys. Acta, 153, 490—493, 1968. Brown
fat oxidation.
144. Gustavson К. H., Chemistry and Reactivity of Collagen. Academic Press, New
York, 1956, 342 pp.
145. Hainsworth F. R., Wolf L. L-, Science, 168, 368—369, 1970. Metabolism and
body temperature in torpor, hummingbird.
146. Halcrow K., Boyd M., Comp. Biochem. PhysioL, 23, 233—242, 1967. Metabo-
lism of Gammarus.
147. Hamby R., Ph. D. Thesis, Univ. Chicago, 1969. Heat coma in intertidal
snails.
148. Hammel H. T., Spec. Rep. Univ. Missouri Agric. Exp. Station, 73, 1—34, 1966.
Theory of temperature regulation in mammals.
149. Hammel H. T. In: International Symposium on Natural Mammalian Hiberna-
tion 1965; Mammalian Hibernation III, edited by К. C. Fisher, American Else-
vier Publ. Co., New York, 1968, pp. 86—96. Theory of temperature regula-
tion.
150. Hammel H. T., Ann. Rev. PhysioL, 30, 641—710, 1968. Regulation of internal
body temperature.
151. Hammel H. T. In: Physiological Controls and Regulations, edited by
W. S. Yamamoto and J. R. Brobeck, W. B. Saunders Company, Philadelphia,
1965, pp. 71—97. Neurons and temperature regulation.
152. Hammel H. T„ Caldwell F. T„ Abrams R. M., Science, 156, 1260—1262, 1967.
Regulation of body temperature in lizard Tiliqua.
153. Hammel H. T., Jackson D. C., Stolwyk J. A. J., Hardy J. D., Stromme S. B.,
J. Appl. PhysioL, 18, 1146—1154, 1963. Temperature regulation by hypothala-
mic control center.
154. Hammel H. T., Stromme S. B., Myhre R., Science, 165, 83—85, 1969. Beha-
vioral thermal regulation in Arctic sculpins.
155. Hanec W., J. Insect PhysioL, 12, 1443—1449, 1966. Cold hardiness in tent ca-
terpillar Malacosoma.
156, Hanegan J. L., Heath J. E., J. Exp. Biol., 53, 349—362, 1970. Control ol body
temperature in moth Hyalophora.
157. Hannon J. P., Fed. Proc., 19, 100—105, 1960. Intermediary metabolism in cold-
acclimated rat.
158. Hannon J. P„ Larson A. M., Amer. J. PhysioL, 203, 1055—1061, 1962. Fatty
acid metabolism in thermogenesis of cold-acclimated rat.
159. Hanson R. C„ Stanley J. G„ Comp. Biochem. PhysioL, 33, 871—879, 1970.
Hypophysectomy and thermal acclimation in mud minnow.
160. Hardy J. D., PhysioL Rev., 41, 521—605, 1961. Thermoregulating center in
mammalian brain.
Температура
197
iri Hardy J- D. In: Physiological Controls and Regulations, edited by W. S. Yama-
101 ’ moto and J. R. Brobeck, W. B. Saunders Comnapy, Philadelphia, 1965,
DD 98—Ц6. The «set-point» concept in temperature regulation.
162 Hardy J. D., Helion R. F., Sutherland R., J. Physiol., 175, 242—253, 1964. Tem-
perature-sensitive neurones in dog hypothalamus.
163 Hart I- S" sYmP- Soc- Exp. I8> 31—48, 1964. Insulative and metabolic
adaptation to cold in vertebrates.
164^ Hart J. S., Publ. Ontario Fish. Res. Lab., 72, 1—79, 1952. Lethal temperatures
of fish from different latitudes.
165. Hart J. S., Physiol. Zool., 35, 224—236, 1962. Seasonal acclimation in small
birds.
166. Hart J. S., Pohl H., Tener J. S., Canad. J. Zool., 43, 731—744, 1965. Seasonal
acclimatization in hare.
167. Haschmeyer A. E. V., Proc. Nat.'Acad. Sci., 62, 128—135, 1969. Synthesis of
polypeptide chains in temperature acclimation of Opsanus.
168. Haschmeyer A. E. V., Biol. Bull., 135, 130—140, 1968; Comp. Biochem. Phy-
siol., 28, 535—552, 1969. Protein synthesis by liver in temperature acclimation
of toadfish.
169. Haschmeyer A. E. V., Biol. Bull., 136, 28—32, 1969. Oxygen consumption in
temperature acclimation of toadfish.
170. Hayward J. S., Lyman С. P. In: International Symposium on Natural Mam-
malian Hibernation 1965; Mammalian Hibernation III, edited by К. C. Fisher,
American Elsevier Publ. Co., New York, 1968, pp. 346—355. Nonshivering
heat production in arousal from hibernation.
171. Hazel J., Prosser C. L., Z. vergl. Physiol., 67, 217—228, 1970. Interpretation
of inverse acclimation to temperature.
172. Hazel J., Ph. D. Thesis, University of Illinois, 1971. Effects of mitochondrial
lipids on succinic dehydrogenase.
173. Heath J. E., Univ. Calif. Publ. Zool., 64, 97—134, 1965. Temperature regula-
tion by behavior in horned lizards.
174. Heath J. E. In: Evolution and Environment, edited by E. T. Drake, Yale Uni-
versity Press, New Haven, Conn., 1968, pp. 259—278. Evolution of thermal
regulation.
175. Heath J. E., The Physiologist, 13, 399—410, 1970. Theory of behavioral regu-
lation of temperature in poikilotherms.
176. Heath J. E., Adams P. A., J. Exp. BioL, 47, 21-—33, 1967. Regulation of heat
production by large moths.
177. Heath J. E. et al., Amer. Zool., 11, 147—158, 1971. Behavioral thermoregula-
tion in insects.
178. Heath J. E., Josephson R. R., Biol. Bull., 138, 272—285, 1970. Body tempera-
ture and singing in katydids.
179. Heath J. E., Williams A., Mills S., Int. J. Biometeor., 15, 254—257, 1971. Hy-
pothalamic thermosensitivity.
180. Heath W. G„ Science, 142, 486—488, 1963. Thermoperiodism in trout.
181. Heatwole H., Ecol. Monogr., 40, 425—457, 1970. Thermal regulation in desert
dragon Amphibolurus.
182. Hebb C., Morris D., Smith M. W., Comp. Biochem. Physiol., 28, 29—36, 1969.
Acetyl transferase in brain of goldfish from different temperatures.
183. van Heel W. H. D., Experientia, 12, 75—77, 1956. Temperature effect on pitch
discrimination in minnow, Phoxinus.
184. Heinrich B., Science, 168, 580—582, 1970. Thoracic temperature in free flying
moths.
185. Heinrich B., J. Exp. Biol., 54, 141—152, 1971. Temperature regulation of sphinx
moth Manduca.
186. Heldmaier G., Z. vergl. Physiol., 63, 59—84, 1969. Heat production in bat
Myotis.
187. Heldmaier G., Z. vergl. Physiol., 73, 222—240, 1971. Calorigenic responses to
noradrenaline.
198
Глава 9
188. Helion R. F., J. Physiol., 193, 381—395, 1970. Thermal stimulation of hypo
thalamic neurones in rabbit.
189. Hensel H., Z. vergl. Physiol., 37, 509—526, 1955; Pfliiger. Arch., 263, 48—53,
1956. Temperature sensitivity of ampullae of Lorenzini.
190. Hensel H., Pfliiger. Arch., 313, 150—152, 1969. Cutaneous heat receptors in
primates.
191. Hensel H., Kenshalo D. R., J. Physiol., 204, 99—112, 1969. Warm receptors in
nasal region of cat.
192. Hensel H., Zotterman У., Acta Physiol. Scand., 22, 96—105, 106—113, 1951;
23, 291—319, 1951. Heat and cold receptors.
193. Henshaw R. E., Folk G. E., Physiol. Zool., 39, 223—236, 1966. Thermal regu-
lation in seasonally changing climate, bats.
194. Heroux O., Fed. Proc., 22, 789—794, 1963. General patterns of temperature
adaptation in mammals.
195. Herreid C. F., Kessel B., Comp. Biochem. Physiol., 21, 405—414, 1967. Ther-
mal conductance in birds and mammals.
196. Heusner A., Stussi T., Insectes Sociaux, 11, 239—265, 1964. Energy metabolism
of bee.
197. Hildwein G., Arch. Sci. Physiol., 24, 55—71, 1970. Seasonal thermoregulation
in tenrec.
198. Hildwein G., Champigny O., Arch. Sci. Physiol., 21, 45—58, 1967. Thermore-
gulation of hairless rats.
199. Himms-Hagen J., Canad. J. Physiol. Pharmacol., 49, 545—553, 1971. Effect of
tetracycline on noradrenaline action on brown fat.
200. Hoar W. S., Eales J. G., Canad. J. Zool., 41, 653—669, 1963. Thyroid and tem-
perature resistance in goldfish.
201. Hochachka P. W., Arch. Biochim. Biophys., Ill, 96—103, 1965. LDH isozymes
in temperature adaptation of goldfish.
202. Hochachka P. W., Comp. Biochem. Physiol., 25, 107—118, 1968. Glucose and
acetate metabolism in temperature adaptation.
203. Hochachka P. Hayes F. R., Canad. J. Zool., 40, 261—270, 1962. Tempera-
ture acclimation and pathways of glucose metabolism in trout.
204. Hochachka P. II7., Lewis J. K-, J. Biol. Chem., 245, 6567—6573, 1970. Enzyme
variance in thermal acclimation.
205. Hochachka P. W., Lewis J. K., Comp. Biochem. Physiol., 39B, 925—934, 1971.
Effects of temperature and pH on fish LDH.
206. Hochachka P. II7., Somero G. N., Comp. Biochem. Physiol., 27, 659—668, 1968.
Adaptation of enzymes to temperature.
207. Hochachka P. W. In: Fish Physiology, Vol. I, edited by W. S. Hoar and
D. J. Randall, Academic Press, New York, 1970, pp. 351—389. Adaptation of
enzymes to temperature.
208. Hodgkin A. L., Katz B., J. Physiol., 109, 240—249, 1949. Effect of temperatu-
re on action potentials in squid giant axon.
209. Horowitz B. A., Nelson L., Comp. Biochem. Physiol., 24, 385—394, 1968. Mi-
tochondrial respiration in hibernator and nonhibernator.
210. House H. L. et al., Canad. J. Zool., 36, 629—632, 1958. Effect of diet on tem-
perature resistance, insects.
211. Howell B. J., Baumgartner F. W., Bondy K., Rahn H„ Amer. J. Physiol., 218,
600—606, 1970. Acid-base balance as a function of body temperature.
212. Howell T. R., Bartholomew G. A., Auk, 78, 343—354, 1961; Condor, 63, 185—
197, 1961. Temperature regulation in petrels and shearwaters: incubation tem-
perature of albatross.
213. Hsieh A. C. L., J. Physiol., 169, 851—861, 1964. Metabolism of cold-adapted
rats.
214. Hsieh A. C. L. et al., Fed. Proc., 25, 1205—1212, 1966. Metabolic effects of
noradrenaline in cold-adapted rats.
215. Hudson J. W. In: International Symposium on Natural Mammalian Hiberna-
tion 1965; Mammalian Hibernation III, edited by К. C. Fisher, American Else-
Температура
199
216.
217-
218.
219.
220.
221.
222.
223.
224.
225.
226.
227.
1964.
vier Publ. Co., New York, 1968, pp. 30—40. Patterns of torpidity in small
Sn/. W., Ann. Acad. Sci. Fenn. A. IV, 71/15, 217-233, 1964. Tempera-
ture regulation in ground squirrel.
Hudson J. W., Brush A. H., Comp. Biochem. Physiol., 12, 157—170,
Comparative metabolic study of quail and dove.
Hudson J. IF., Kjmsey S. L., Comp. Biochem. Physiol., 17, 203—217,
Metabolism and temperature regulation in house sparrow.
Hudson J. IF., Rummel J. A., Ecology, 47, 345—354, 1966. Temperature
1966.
regu-
lation of primitive rodent, Liomys.
Hull P., Segall M. M., J. Physiol., 181, 449—457, 468—477, 1965. Brown fat
in heat production of newborn rabbit.
Hutchinson V. M., Dowling H. G., Vinegar A., Science, 151, 694—696, 1966.
Thermal regulation in brooding Indian pythons.
lampietro P. F. et al., J. Appl. Physiol., 15, 632—634, 1960. Heat production
from shivering.
Iggo A., J. Physiol., 200, 403—430, 1969. Cutaneous thermal receptors in
mammals.
Irving L., J. Appl. Physiol., 9, 414—420, 1956. Temperature of skin in pig.
Irving L., Peyton L. J. Bahn С. H., Peterson R. S., Physiol. ZooL, 35, 275—
284, 1962. Temperature regulation in fur seal.
Irving L., Schmidt-Nielsen K., Abrahamson N. S., Physiol. ZooL, 30, 93—105,
1957. Melting points of animal fats in cold climates.
Ishiko N., Loewenstein W. R., J. Gen. PhysioL, 45, 105—124, 1961. Effects of
temperature on pacinian corpuscles.
228. Jackson D. C., Schmidt-Nielsen K., Proc. Nat. Acad. Sci., 51, 1192—1197, 1964.
Temperature relations of kangaroo rat.
229. Jacobson E. R., Whitford W. G„ Comp. Biochem. PhysioL, 35, 439—449, 1970.
Effect of acclimatization on responses to temperature in snakes.
230. Jacobson F. H., Squires R. D., Amer. J. PhysioL, 218, 1575—1582, 1970. Ther-
mal regulation center in brain of cat.
231. Jankowsky H. D., Helg. wiss. Meeresunters., 13, 402—407, 1966. Metabolism
and temperature adaptation of eel.
231a. Jankowsky H. D., Personal communication. Metabolic acclimation in golden
orfe.
232.
233.
234.
235.
236.
237.
238.
239.
240.
241.
242.
243.
Jansky L., Canad. J. Biochem., 41, 1847—1854, 1963. Cytochrome oxidase in
cold- and warm-acclimated rats.
Jansky L., Hart J. S., Canad. J. Biochem., 41, 953—964, 1963. Nonshivering
thermogenesis in cold-acclimated rats.
Jansky L. et al., PhysioL Bohemoslov., 16, 366—371, 1967. Noradrenaline
thermogenesis in rats.
Javaid M. У., Anderson J. M., J. Fish. Res. Bd. Canad., 24, 1507—1513, 1967.
Thermal acclimation and temperature selection in salmon trout.
Jensen D., Comp. Biochem. PhysioL, 41A, 685—695, 1972. Effect of cooling on
frog endplate.
Jessen C„ Simon E., Rullman R., Experientia, 24, 694—695, 1968. Interaction
of spinal and hypothalamic temperature detectors in dog.
Johansen K., PhysioL ZooL, 34, 126—144, 1961. Temperature regulation in
armadillo, Dasypus.
Johansen R., Frog J., Reite O., Ann. Acad. Sci. Fenn. A. IV, 71/17, 245—253,
1964. Nervous control of heart in hypothermic regulator.
Johnson H. D. et al., Univ. Missouri Coll. Agric. Res. Bull., 683, 1—31, 1958.
Temperature tolerance of cattle.
Johnson R. E„ Comp. Biochem. PhysioL, 24, 1003—1014, 1968. Temperature
regulation in ptarmigan.
Johnston P. V., Roots B. J., Comp. Biochem. PhysioL, 11, 303—310, 1964.
brain lipid fatty acids and temperature acclimation.
Josse J., Harrington W. F., J. Molec. Biol., 9, 269—287, 1964. Composition of
collagen as a function of temperature.
00
Глава 9
244. Jungreis А. М., Comp. Biochem. Physiol., 24, 1—6, 1968. Glycogen and long-
term temperature acclimation in crayfish.
245. Jungreis A. M., Hooper A. B., Comp. Biochem. Physiol., 26, 91—100, 1968.
Cold resistance adaptation in crayfish.
246. Kahl M. P., Physiol. Zool., 36, 141—151, 1963. Thermal regulation in wood
stork.
247. Kammer A. E., Z. vergl. Physiol., 68, 334—344, 1970. Thoracic temperature
and flight in monarch butterfly.
248. Kanungo M., Prosser C. L„ J. Cell. Comp. Physiol., 54, 259—263, 265—274,
1960. Biochemical changes in temperature acclimation, goldfish.
249. Kanwisher J., Topics in the Study of Life in the Sea. Harper and Row, New
York, 1970, pp. 209—214. Temperature regulation in marine mammals.
250. Kanwisher J. In: Cryobiology, edited by H. T. Meryman, Academic Press,
New York, 1966, pp. 487—494. Freezing in intertidal animals.
251. Karow A. M„ Webb W. A., Cryobiology, 2, 99—108, 1965. Tissue freezing.
252. Kayser C., Rev. Canad. Biol., 16, 303—389, 1957; Ann. Rev. Physiol., 19,
83—120, 1957. Hibernation.
253. Keister M„ Back J., J. Insect Physiol., 7, 51—72, 1961. Temperature and respi-
ration in Phornrm
254. Kemp P., Smith M. W., Biochem. J., 117, 9—15, 1970. Effect of temperature
on fatty acid composition in goldfish.
255. Kempner C. L., Science, 142, 1318—1319, 1963. Upper temperature limits of
life.
256. Kendeigh S. C., Wilson Bull., 81, 441—449, 1969. Energy responses of birds
to thermal environment.
257. Kerkut G. A., Taylor B. J. R., Nature, 178, 426, 1956; J. Exp. Biol., 34, 486—
493, 1957; Behaviour, 13; 259—279, 1958. Effects of temperature on nervous
system of cockroach, slug, and crayfish.
258. Kibler H. H., Silsby H. D., Johnson H. D., J. Geront., 18, 235—239, 1963. Cold
acclimation in rats.
259. Kimzey S. L„ Willis J. S., J. Gen. Physiol., 58, 620—633, 634—649, 1971. Re-
sistance of red blood cells to loss in hibernation and in cold storage.
260. King J. R., Comp. Biochem. Physiol., 12, 13—24, 1964. Oxygen consumption
and body temperature in whitecrowned sparrow.
261. Kinne O., Oceanogr. Marine Biol. Ann. Rev., 1, 301—340, 1963. Effects of
temperature and salinity on invertebrates.
262. Kinne O., Kinne E. M„ Canad. J. Zool., 40, 231—253, 1962. Development of
fish embryos at different temperatures.
263. KHcka J. K., Physiol. Zool., 38, 177—189, 1965. Physiological effects of ACTH,
TSH, and thyroid hormones in goldfish.
264. Kluger M. J., Heath J. E., Comp. Biochem. Physiol., 32, 219—226, 1970. Ther-
mal regulation by bat wing.
265. Knipprath W. G., Mead J. F., Lipids, 3, 121—128, 1968. Temperature effects
on fatty acids in goldfish.
266. Kodama A. M., Pace N., Fed. Proc., 22, 761—765, 1963. Cold-dependent chan-
ges in tissue fat.
267. Koffler H., Mallett G. E., Adye J., Proc. Nat. Acad. Sci., 43, 464—477, 1957.
Molecular basis of heat stability of bacteria.
268. Kohler H., Amarine Biol., 5, 315—324, 1970. Acclimation to temperature and
salinity in an oligochaete.
269. Komatsu S. K., DeVries A. L., Feeney R. E., J. Biol. Chem., 245, 2909—2913,
1970; also Komatsu S. К., H. T. Miller, A. L. DeVries, D. T. ’tisuga, and
R. E. Feeney, Comp. Biochem. Physiol., 32, 519—527, 1970. Glycoproteins and
lipoproteins in blood of Antarctic fish.
270. Kosaka M., Simon E„ Pflugers Arch., 302, 357—373, 1968. Spinal receptors
for cold.
271. Kosaka M„ Simon E., Thauer R., Walther О. E., Amer. J. Physiol. 217 858—
863, 1969. Thermal stimulation of spinal cord.
Температура
201
979 Krog J., Biol. Bull., 107, 397—410, 1954. Seasonal effects on metabolism and
lethal temperature in Alaskan gammarid.
973 Kruger G., Z. wiss. Zool., 167, 87—104, 1962. Temperature adaptation in bit-
terling Rhodeus.
274 Kulzer E„ Z. vergl. Physiol., 50, 1—34, 1965. Temperature regulation bats
from different climates.
975 Kusakina A. A., Tsitologiya, 4, 68, 1962; transl. in Fed. Proc. Transl. Suppl.,
22, T123 —T126, 1963. Thermal stability of muscle enzymes in fishes.
276. Lagerspetz K. Y. H., Helg. wiss. Meeresunters., 14, 559—571, 1966. Deve-
lopment of thermal regulation in laboratory mice.
277. Lagerspetz K. Y. H., Dubitscher D., Comp. Biochem. Physiol., 17, 665—671,
1966. Temperature acclimation of cilia in gill of Anodonta.
278. Lagerspetz K. Y. H., Talo A., J. Exp. Biol., 47, 471—480, 1967. Temperature
acclimation and conduction in nerve fibers, Lumbricus.
279. Langridge J., Ann. Rev. Plant Physiol., 14, 441—462, 1963. Biochemical aspects
of temperature response.
280. Lasiewski R. C„ Proc. XIII Int. Ornith. Cong., 1095—1103, 1963. Energetic
cost of small size in hummingbirds.
281. Lasiewski R. C., Physiol. Zool., 36, 122—140, 1963. O2 consumption of torpid,
resting and active hummingbirds.
282. Lasiewski R. C., Amer. J. Physiol., 217, 1504—1509, 1969. Physiological
response to heat stress in the poorwill.
283. Lasiewski R. C., Dawson W. R., Condor, 66, 477—490, 1964; 72, 332—338,
1970. Temperature responses in night hawk and frogmouth Podargus.
284. Lasiewski R. C., Condor, 66, 212—220, 1964. Energetic relationships of very
small passerine birds to temperature.
285. Lasiewski R. C., Lasiewski J. J., Auk, 84, 34—48, 1967. Responses of humming-
birds to temperature changes.
286. Lasiewski R. C., Snyder G. K., Auk, 86, 529—540, 1969. Temperature in
nesting cormorants.
287. Lasiewski R. C., Weathers W. W., Bernstein M., Comp. Biochem. Physiol., 23,
797—813, 1967. Metabolic responses of hummingbirds to temperature.
288. Laverack M. S., Comp. Biochem. Physiol., 3, 136—140, 1961. Effect of tem-
perature on activity of nerve cord of earthworm.
289. Leblanc J., Villemaire A., Amer. J. Physiol., 218, 1742—1745, 1970. Thyroid
and adrenalin in cold resistance, brown fat.
290. Leblanc J. et al., J. Appl. Physiol., 15, 1031—1034, 1960; 17, 950—952, 1962;
Canad. J. Physiol. Pharmacol., 44, 287—293, 1966. Habituation of Gaspe fisher-
men to cold extremities; laboratory acclimation.
291. Leitner P., Comp. Biochem. Physiol., 19, 431—443, 1966. Metabolism and tem-
perature regulation in mastiff bat Eumops.
292. Leitner P., Nelson J. E., Comp. Biochem. Physiol., 21, 65—74, 1967. Tempera-
ture, metabolism, and heart rate of vampire bat.
293. Lensky Y., J. Insect Physiol., 10, 1—12, 1964. Bee colony, temperature extre-
mes.
.294. Leston D., Pringle J. W. S„ White D. C. S., J. Exp. Biol., 42, 409—414, 1965.
Insect warmup by muscle contraction.
295. Licht P., Comp. Biochem. Physiol., 13, 27—34, 1964. Thermal dependence of
contractility in reptilian muscles.
296. Licht P„ Copeia, 428—36, 1965; Amer. Midi. Nat., 79, 149—158, 1968. Prefer-
o red body temperatures in lizards.
297. Licht P„ Leitner P., Comp. Biochem. Physiol., 22, 371—387, 1967. Responses
_ to high temperatures in bats.
298. Liu C„ Frehn J. L„ LaPorta A. D., J. Appl. Physiol., 27, 83—89, 1969. Liver
oon апс* brown fat responses to cold.
299. Loftus R„ Z. vergl. Physiol., 59, 413—455, 1968; 63, 415—433, 1969. Antennal
9on COfd receptors in Periplaneta.
299a. Lyman С. P., Blinks D. C„ J. Cell. Comp. Physiol., 54, 53—63, 1959. Tempe-
rature effects on hearts of hibernators.
202
Глава 9
300. Lyman С. Р., O’Brien R. С., Ann. Acad. Sci. Fenn., A IV 71/22, 213—320,
1964. Effects of autonomic drugs on hibernating ground squirrel.
301. Lyman С. P., Wimsatt W. A., Physiol. ZooL, 39, 101—109, 1966. Temperature
regulation in vampire bat Desmodus.
302. MacMillen R. E., Trost С. H., Comp. Biochem. PhysioL, 23, 243—253, 1967.
Nocturnal hypothermia in Inca dove.
303. MacMillen R. E., Nelson J. E., Amer. J. PhysioL, 217, 1246—1251, 1969.
Bioenergetics in dasyurid marsupials.
304. Malan A., Arch. Sci. PhysioL, 23, 47—87, 1969. Hypothalamic control of ther-
mal regulation and hibernation in hamster.
305. Mangum С. P., Sassaman C., Comp. Biochem. PhysioL, 30, 111—116, 1969.
Temperature sensitivity and metabolism in polychaete annelid.
306. Markardt J. E., Comp. Biochem. PhysioL, 33, 423—439, 1970. Daily torpor
in mouse Peromyscus.
307. Markert C. L., Faulhaber I., J. Exp. ZooL, 159, 319—332, 1965. LDH isozymes
in fish.
308. Marshall J. M., Willis J. S., J. PhysioL, 164, 64—76, 1962. Effects of tempera-
ture on membrane potentials in isolated atria of ground squirrel Citellus.
309. Mazur P. In: Cryobiology, edited by H. T. Meryman, Academic Press, New
York, 1966, pp. 213—315. Basis of cellular injury in freezing and thawing.
310. Mazur P., Ann. Rev. Plant PhysioL, 20, 419—448, 1969. Freezing injury in
plants.
311. Mazur P., Science, 168, 939—949, 1970. The freezing of biological systems.
312. Mazur P. et al., Cryobiology, 6, 1—9, 1969. Survival of tissue culture cells
after freezing and thawing.
313. McCrea M., Heath J. E., J. Exp. Biol., 54, 415—435, 1971. Dependence of flight
on temperature regulation in moth Manduca.
314. McCue J. F., Thorson R. E., J. ParasitoL, 50, 67—71, 1964. Behavior of para-
sitic helminth in thermal gradient.
315. McGinnis S. M., Dickson L. L., Science, 156, 1757—1759, 1967. Iguana tempe-
rature.
316. McLeese D. W„ J. Fish. Res. Bd. Canad., 13, 247—272, 1956. Effects of tem-
perature, salinity, and oxygen on survival of lobster.
316a. McMillen R. E., Trost С. H., Comp. Biochem. PhysioL, 23, 243—253, 1967.
Thermal regulation in Inca dove.
317. McNab В. K., Ecology, 44, 521—532, 1963. Energy balance in wild mice.
318. McNab В. K., Comp. Biochem. PhysioL, 31, 227—268, 1969. Economics of tem-
perature regulation, neotropical bats.
319. McNab В. K., J. Exp. Biol., 53, 329—348, 1970. Body weight and energetics
of temperature regulation.
320. McNab В. K., Morrison P., Ecol. Monogr., 33, 63—82, 1963. Body temperature
and metabolism in Peromyscus.
321. McWhinnie M. A., O’Connor J. D., Comp. Biochem. PhysioL, 20, 131—145,
1967. Aletabolism and temperature acclimation in crayfish.
322. Mellanby K., J. Exp. Biol., 18, 55—61, 1942; Nature, 181, 1403, 1958. Body
temperature, frogs and insects.
323. Menaker M., J. Cell. Comp. PhysioL, 59, 163—174, 1962. Hibernation in bat
Myotis.
324. Meryman H. T. In: Cryobiology, edited by H. T. Meryman, Academic Press,
New York, 1966, pp. 1—114. Review of biological freezing.
325. Miller L. K., Comp. Biochem. PhysioL, 21, 679—686, 1967. Temperature and
conduction in mammalian nerve.
326. Miller L. K., Science, 166, 105—106, 1969. Freezing tolerance in carabid
beetle.
327. Miller L. K., Dehlinger P. J., Comp. Biochem. PhysioL, 28, 915—921, 1969.
Neuromuscular function in cold.
328. Miller L. K., Irving L., Amer. J. PhysioL, 204, 359—362, 1963. Sensitivity of
rat nerve to cold.
Температура
203 ~
329.
330.
331.
332.
333.
334.
335.
336.
337.
338.
339.
340.
341.
342.
343.
344.
345.
346.
347.
348.
349.
350.
351.
352.
353.
354.
355.
356.
Moore J. A., Evolution, 3, 1—21, 1949. Geographic variation of temperature
adaptation in Rana pipiens.
Morrison G R., Brook F. E., Sobral D. T., Shank R. E., Arch. Biochim. Bio-
nhvs П4, 494—501, 1966. Rat liver enzymes after cold acclimation.
Morrison P., Biol. Bull., 123, 154—169, 1962. Body temperature of humpback
^Morrison P., Biol. Bull., 116, 484—497, 1959. Heat loss in small mammals.
Morrison P., McNab В. K., Comp. Biochem. Physiol., 6, 57—68, 1962. Daily
torpor in opossum.
Morrison P., Ryser R. A., Physiol. Zool., 37, 90—103, 1959. Body temperature
in Peromyscus leucopus.
Morrison P., Tietz W. J., J. Mammal., 38, 78—86, 1957. Insulation in small
mammals.
Mount L. E., J. Physiol., 217, 315—326, 1971. Metabolic responses of hairless
mice to cold.
Murray R. W., Comp. Biochem. Physiol., 18, 291—303, 1966. Temperature
effects on membrane properties of neurones in Aplysia.
Murrish D. E„ Comp. Biochem. Physiol., 34, 853—858, 859—869, 1970. Respon-
ses to diving and temperature in the dipper Cinclus.
Mutchmor J. A., Richards A. G., J. Insect Physiol., 7, 141—158, 1961. Low
temperature and muscle apyrase of insects.
Myers R. D„ Veale W. L., Science, 170, 95—97, 1970; J. Physiol., 217, 381—
392, 1971. Ionic mechanisms of temperature control in hypothalamus.
Myhre K., Hammel H. T., Amer. J. Physiol., 217, 1490—1495, 1969. Behavioral
regulation of temperature in lizard Tiliqua.
Nakayama T., Hammel H. T., Hardy J. D., Eisenman J. S., Amer. J. Physiol.,
204, 1122—1126, 1963. Thermal stimulation of neurones in preoptic hypotha-
lamus.
Наматов T., ДАН СССР, 185, 1382—1385, 1969. Устойчивость нематод
Rhabitis sp. и Anguina tritici к воздействию низких и сверхнизких темпе-
ратур.
Newell R. С., Nature, 212, 426—428, 1966; J. Zool. (London), 151, 299—311,
1967; also Newell R. C. and H. R. Northcraft, J. Zool. (London), 151, 277—
298, 1967. Metabolic independence of temperature over limited ranges in
poikilotherms.
Newell R. C., Pye V. I., Comp. Biochem. Physiol., 38B, 635—650, 1971. Re-
lation between metabolism and temperature in Littorina.
Nopp H., Farahat A. Z., Z. vergl. Physiol., 55, 103—118, 1967. Temperature
and metabolism in snails.
Norris K. S., Ecol. Monogr., 33, 23—62, 1963. Temperature in the ecology
of percoid fish Girella.
Nowell N. W., White D. C„ Life Sci., 8, 239—245, 1969. Noradrenaline in
cold acclimation of rat heart.
O’Farrell M. J., Studier E. H., Comp. Biochem. Physiol., 35, 697—703, 1970.
Metabolism in relation to temperature of Myotis.
Oritsland N. A., Comp. Biochem. Physiol., 37, 225—233, 1970. Temperature
regulation of polar bear.
Orr P. R., Physiol. Zool., 28, 290—302, 1955. Heat death, whole animal and
tissues.
Paintai A. S., J. Physiol., 180, 1—35, 1965. Cold block of mammalian
nerve.
Patterson G. W., Lipids, 5, 597—600, 1970. Effect of culture temperature on
fatty acids of Chlorella.
Patton J. F., Plainer W. S., Amer. J. Physiol., 218, 1417—1422, 1970. Cold
acclimation and thyroid effects on liver fatty acids.
Pearcy W. G., Science, 134, 493—194, 1961. Seasonal changes in osmotic con-
centration of flounder.
E: Asmundson S. M., Comp. Biochem. Physiol., 30, 177—183
1УЬ9. Circadian rhythms in ground squirrel Citellus.
204
Глава 9
357. Pengelley Е. Т., Fisher К. С., Canad. J. Zool., 41, 1103—1120, 1963. Annual
cycles in ground squirrel.
357a. Peterson R. H„ Prosser C. L., Comp. Biochem. Physiol., 42A, 1019—1038,
1972. Cold block of central nervous responses in goldfish.
358. Pichotka J., Helg. wiss. Meeresunters., 9, 274—284, 1964. Temperature regu-
lation in newborn mice.
359. Platzer I., Z. Vergl. Physiol., 54, 58—74, 1967. Temperature adaptations in
tropical chironomids.
360. Pocrnjic Z., Arch. Biol. Nauka, 17, 139—148, 1965. Thermal adaptation of oxy-
gen consumption in the newt.
361. Popovic V., Amer. J. Physiol., 207, 1345—1348, 1964. Cardiac output in hi-
bernating ground squirrels.
362. Popovic P. et al., Biometeorology, 2, 276—282, 1966. Rats, adaptation to hy-
pothermia.
362a. Powers D. A., Personal communication. Polymorphism of LDH in Fundulus.
363. Precht H. In: Physiological Adaptation, edited by C. L. Prosser, Amer. Phy-
siol. Soc., Washington, D. C., 1958, pp. 50—78. Theory of temperature adapta-
tion in cold-blooded animals.
364. Precht H., Z. vergl. Physiol., 42, 365—382, 1959. Resistance adaptation in tro-
pical fish.
365. Precht H., Z. wiss. Zool., 164, 336—354, 354—363, 1960. Thermal resistance
of muscles and muscle endplates.
366. Precht H., Christopherson J., Hensel H., Temperatur und Leben, Springer-
Verlag, Hamburg, 1955, 514 pp.
367. Precht H., Z. vergl. Physiol., 44, 451—462, 1961. Temperature adaptation of
metabolism of eel.
368. Precht H., Christopherson J., Z. wiss. Zool., 171, 197—209, 1965. Temperature
adaptation of ciliated epithelium of clam gills.
369. Prosser C. L. In: Physiology Adaptation, edited by C. L. Prosser, Amer. Phy-
siol. Soc., Washington, D. C., 1958, pp. 167—180.
370. Prosser C. L. In: Handbook of Physiology, Sec. 4., edited by D. B. Dill et al.,
Amer. Physiol. Soc., Washington, D. C., 1964, pp. 11—25. Theory of adapta-
tion.
371. Prosser C. L., Nagai T. In: The Central Nervous System and Fish Behavior,
edited by D. Ingle, University of Chicago Press, Chicago, 1968, pp. 171—180.
372. Prosser C. L., Farhi E., Z. vergl. Physiol., 50, 91—101, 1965. Effects of tempe-
rature on conditioned reflexes.
373. Prosser C. L. In: Molecular Mechanisms of Temperature Adaptation, edited
by C. L. Prosser, Amer. Assoc. Adv. Sci., Washington, D. C., 1967, pp. 351—
376.
374. Prusiner S., J. Biol. Chem., 245, 382—389, 1970. Oxidative phosphorylation in
hamster brown fat.
375. Rao К. P., In: Molecular Mechanism of Temperature Adaptation, edited by
C. L. Prosser, Amer. Assoc. Adv. Sci., Washington, D. C., 1967.
376. Rapatz G., Luyet B., Cryobiology, 4, 215—222, 1968. Cryoprotective agents in
preservation of blood.
377. Rautenberg W., Z. vergl. Physiol., 62, 235—266, 1969. Spinal cord in relation
to thermal regulation in pigeons.
378. Read K. R. H., Comp. Biochem. Physiol., 9, 161—180, 1963. Thermal inactiva-
tion of aspartic-glutamic transaminase from molluscs.
379. Read K. R. H., Proc. Malac. Soc. Lond., 37, 233—241, 1967. Thermal tolerance
of Puerto Rican bivalves.
380. Read K. R. H. In: Molecular Mechanisms of Temperature Adaptation, edited
by C. L. Prosser, Amer. Assoc. Adv. Sci., Washington, D. C., 1967, pp. 93—
106. Thermal stability of proteins in invertebrates.
381. Reiter О. B„ Davis W. H., Proc. Soc. Exp. Biol. Med., 121, 1212—1215, 1966.
Chill in bats.
382. Richards A. G„ Physiol. Zool., 37, 199—211, 1964. Temperature effects on de-
velopment and oxygen consumption in insect eggs.
Температура
205
_____.___t______~~
---------------- х
383. RigbipB. J., Biol. Bull., 135, 223—229, 1968. Melting temperature of coIla-
384 *Rink R. D., J. Exp. ZooL, 170, 117—124, 1969. Metabolism of brown fat in
newborn, hamsters.
385 Rising J. D., Comp. Biochem. Physiol., 25, 327—333, 1968. Temperature effect
on metabolism of a sparrow.
386. Rising J. D., Comp. Biochem. PhysioL, 31, 915—925, 1969. Metabolism and
evaporative water loss in orioles.
387. Rising T. L., Armitage К. B., Comp. Biochem. Physiol., 30, 1091—1114, 1969.
Acclimation to temperature in snails.
388. Roberts J. L., Helg. wiss. Meeresunters., 9, 459—473, 1964. Metabolic respon-
ses of sunfish to photoperiod and temperature.
389. Robertshaw D., Taylor C. R., J. Physiol., 203, 135—143, 1969. Sweat gland
activity in African ungulates.
390. Rodahl K. et al., J. Appl. Physiol., 17, 763—767, 1962. Human metabolism,
cold stress.
391. Roots В. I., Comp. Biochem. PhysioL, 25, 457—466, 1968. Phospholipids of
goldfish brain.
392. Roots В. I., Johnston P. V., Comp. Biochem. PhysioL, 26, 553—560, 1968.
Plasmalogens and environmental temperature.
393. Roots В. I., Prosser C. L., J. Exp. Biol., 39, 617—629, 1962. Temperature accli-
mation and nervous system of fish.
394. Rozin P. N., Science, 149, 561—563, 1965; also Rozin P. N., Mayer J., Scien-
ce, 134, 942—943, 1961. Temperature discrimination in goldfish.
395. Ruibal T., Evolution, 15, 98—111, 1961. Graded temperature preferences of fi-
ve species of Anolis.
396. Salt R. W., Canad. J. ZooL, 34, 1—5, 283—294, 391—403, 1956. Ice formation
and supercooling in insects.
397. Salt R. Canad. J. ZooL. 39. 349—357, 1961. Survival of freezing in larvae
of sawfly.
398. Salt R. W., Nature, 193, 1207—1208, 1962. Intracellular freezing in insects.
399. Salt R. W„ Symp. Soc. Exp. BioL, 23, 331—350, 1969. Survival of insects at
low temperatures.
400. Saroja K., Rao К. P., Z. vergl. PhysioL, 50, 35—54, 1965. Biochemical changes
in invertebrates in temperature acclimation.
401. Sassaman C., Mangum C. P„ J. Marine BioL, 7, 123—130, 1970. Lethal tempe-
ratures in North Atlantic actinians.
402. Sastry A. N., BioL Bull., 138, 56—65, 1970. Seasonal variations in breeding of
Atlantic scallop.
403. Satinoff E., Amer. J. PhysioL, 206, 1389—1394, 1964. Behavioral thermal re-
gulation after brain cooling in rats.
404. Satinoff E., Science, 148, 399—400, 1965. Effect of brain lesions on tempera-
ture regulation in hibernators.
405. Satinoff E., Science, 155, 1031—1033, 1967; Amer. J. PhysioL, 212, 1215—1220,
1967. Disruption of hibernation and thermal regulation by hypothalamic le-
sions in ground squirrel.
406. Satinoff E., Prog. PhysioL Psych., 3, 201—236, 1970. Hibernation and central
nervous system.
407. Satinoff E., Rutstein J., J. Comp PhysioL Psych., 71, 77—82, 1970. Behavioral
. thermal regulation in rats with hypothalamic lesions.
• Scheit H. G., Marine BioL, 6, 158—166, 1970. Effect of temperature adaptation
on denervated muscle of eel.
- Schlieper C. R. et al., Kieler Meeresforsch., 14, 3—10, 1958. Temperature and
дю sanity effects on bivalve molluscs.
4 i ^mid w- D ’ Ecology, 46, 559—560; 1966. Thermal tolerance in toads
E Schmidt-Nielsen K. et al., Amer. J. PhysioL, 188, 103—112, 1957. Temperature
4io Nations in camels.
4. Schmidt-Nielsen K„ Bretz W. L., Taylor C. R., Science, 169, 1102—1104 1970
Panting in dogs.
/
!
206___________________________________Глава 9__________________________/________
/
413. Schmidt-Nielsen К., Dawson Т. J., Crawford E. C., J. Cell. Comp. Physiol., 67,
63—72, 1966. Temperature in echidna. >
414. Schmidt-Nielsen K., Dawson W. R. In: Handbook of Physiology,.Sec. 4, edited
by D. B. Dill, Amer. Physiol. Soc., Washington, D. C., 1963,/pp. 467—480.
Terrestrial animals in dry heat.
415. Schmidt-Nielsen K-, Kanwisher J., Lasiewski R., Cohn J. E„ Bretz W. L., Con-
dor, 71, 341—352, 1969. Temperature regulation of ostrich.
415a. Schmidt-Nielsen K. et al., J. Exp. Biol., 55, 385—398, 1971. Heat tolerance of
416. Scholander P. F. et al., J. Appl. Physiol., 8, 279—282, 1955; 10, 404—411,
1957. Countercurrent heat exchangers.
417. Scholander P. F. et al., J. Cell. Comp. Physiol., 49, 5—24, 1957. Supercooling
and osmoregulation in Arctic fish.
418. Schultze D., Z. wiss. Zool., 172, 104—133, 1965. Temperature adaptation of
muscle of eel.
418a. Shes S., Sigafoos D., Scott D., Comp. Biochem. Physiol., 28, 701—708, 1969.
Thermoexcitability of Callinectes nerve.
419. Siminowitch D. et al. In: Molecular Mechanisms of Temperature Adaptation,
edited by C. L. Prosser, Amer. Assoc. Adv. Sci., Washington, D. C., 1967,
pp. 3—40.
420. Simmonds M. A., Iverson L. L., Science, 163, 473—474, 1969. Effect of tempe-
rature on hypothalamic norepinephrine.
421. Simpson J., Science, 133, 1327—1333, 1961. Honeybee colonies in cold.
422. Smalley R. L., Dryer R. L. In: International Symposium on Natural Mamma-
lian Hibernation, 1963; Mammalian Hibernation III, edited by К. C. Fisher,
American Elsevier Publ. Co., New York, 1968, pp. 325—345. Brown fat in hi-
bernation.
423. Smith H., Comp. Biochem. Physiol., 21, 125—132, 1967. Effect of temperature
on gastric secretion in bullhead Ictalurus.
424. Smith A. U., Monogr. Physiol. Soc. No. 9, Williams and Wilkins, Baltimore,
1961. Biological effects of freezing and supercooling.
425. Smith C. L., J. Exp. Biol., 28, 141—164, 1951. Temperature and pulse rate in
frog.
426. Smith D. E., Katzuns B., Amer. Heart J., 71, 515—521, 1966. Mechanical pro-
perties of heart from hibernating and nonhibernating mammals.
427. Smith M. W., J. Physiol., 182, 559—590, 1966; 183, 649—657, 1966. Effect of
temperature on sodium and glucose transport in goldfish intestine.
428. Smith M. W., Biochem. J., 105, 65—72, 1967. Temperature-dependent intestinal
7\.T P3.se
429. Smith M. W., Comp. Biochem. Physiol., 35, 387—401, 1970. Regulation of ami-
no acid transport by intestine of goldfish.
430. Smith M. W., Colombo V. E., Munn E. A., Biochem. J., 107, 691—698, 1968.
Effect of temperature on ionic activation of ATPase.
431. Smith M. W., Ellory J. C., Comp. Biochem. Physiol., 39A, 209—218, 1971.
Temperature-induced change in Na transport and ATPase in goldfish.
432. Smith M. W., Roberts J. C., Hittelman K. J., Science, 154, 653—654, 1966.
Nonphosphorilating respiration of brown fat mitochondria.
433. Somero G. N., Amer. Nat., 103, 517—530, 1969. Enzymic mechanisms of tem-
perature compensation.
434. Somero G. N., Biochem. J., 114, 237—241, 1969. Pyruvatekinase of Alaskan
king crab.
435. Somero G. N„ Giese A. C., Wohlschlag D. E., Comp. Biochem. Physiol., 26,
223—233, 1968. Cold adaptation of Antarctic fish.
436. Somero G. N., Hochachka P., Amer. Zool., 11, 159—167, 1971. Biochemical
adaptations to temperature in poikilotherms.
437. Somme L., J. Insect Physiol., 12, 1069—1083, 1966. Blood composition and
supercooling in larvae of flour moths.
438. Somme L., J. Lnsect Physiol., 13, 805—814, 1967. Effect of temperature and
anoxia on hemolymph in overwintering insects.
\ Температура_______________________________2и'_
439 Semn\ L„ Nytt Mag. Zool., 13, 52—55, 1967. Seasonal changes in cold tole-
rance oh intertidal animals. .
440 South F\E„ Physiol. Zool., 31, 6—15, 1958. Respiration and glycolysis of heart
and braid from hibernators and nonhibernators.
441. South F.E., Amer. J. Physiol., 200, 565—571, 1961. Nerve-diaphragm m rela-
tion to temperature and hibernation.
442. Spath M„ Z. vergl. Physiol., 56, 431—462, 1967. Action of temperature on
mechanoreceptors of fish Leuciscus.
443. Stebbins R. C., Barwick R. E., Copeia, 541—547, 1968. Telemetry of temperature
from Var anus.
444. Steen I., Steen J. B„ Comp. Biochem. Physiol., 15, 267—270, 1965. Thermal
regulation by beaver’s tail.
444a. Steen I., Steen J. B., Acta Physiol. Scand., 63, 285—291, 1965. Heat loss via
lags of aquatic birds.
445. Stevens E. D., Fry F. E. J., Canad. J. Zool., 48, 221—226, 1970. Cooling cur-
ves of fish.
446. Stolwiyk J. A. J., Hardy J. D., Pflugers Arch., 291, 129—162, 1966. Theory of
temperature regulation in man.
447. Strumwasser F., Amer. J. Physiol., 196, 8—30, 1959. Brain activity in hiber-
nating Citellus.
448. Sumner F. B., Lanham U. N., Biol. Bull., 82, 313—327, 1942. Respiration of
warm cold spring fishes.
449. Tashima L. S., Adelstein S. J., Lyman С. P., Amer. J. Physiol., 218, 303—309,
1970. Glucose utilization by active, hibernating and arousing ground squir-
rels.
450. Taylor C. R., Arctic Aeromed. Lab. Bull. AAL-TR-63-31, 1—103, 1966. Ther-
mal regulatory functions of horns in Bovidae.
451. Taylor C. R., Physiol. Zool., 39, 127—139, 1966. Thermal regulatory functions
of horns in goats.
452. Taylor C. R., Amer. J. Physiol., 219, 1131—1135, 1136—1139, 1970. Tempera-
ture regulation in African ungulates.
453. Taylor C. R., Schmidt-Nielsen K„ Amer. J. Physiol., 220, 823—827, 1971. Tem-
perature relations of African hunting dog.
454. Tebecis A. K., Phillis J. W., Comp. Biochem. Physiol., 25, 1034—1047, 1968.
Effect of temperature on spinal cord of toad.
455. Templeton J. R., Physiol. Zool., 37, 300—306, 1964. Vascular responses to
temperature in Sauromalus.
456. Tester J. R., Breckenridge W. J., Ann. Acad. Sci. Fenn. A IV, 31, 423—430,
1964. Winter behavior in Manitoba toad.
457. Thiede IF., Z. wiss. Biol., 172, 305—346, 1965. Hormonal basis of adaptation
in fish.
458. Thiessen C. L, Mutchmor J. A., J. Insect Physiol., 13, 1837—1842, 1967. Effects
of temperature on muscle apyrase in Periplaneta and Musca.
459. Thoenen H., Nature, 228, 861—863, 1970. Catecholamine metabolism in cold
exposed rats.
460. Thompson F. J., Barnes C. D., Life Sci., 9 (Part I), 309—312, 1970. Thermo-
sensitive elements in femoral vein.
461. Todd M. E.,Dehnel P. A., Biol. Bull., 118, 150—172, 1960. Effects of tempera-
ture and salinity on heat tolerance in grapsoid crabs.
o2. loivola P., Gale С. C„ Neuroendocrinology, 6, 210—219, 1970. Temperature
4R effects on amines in hypothalamus of baboon.
Tribe M. A., Bowler K., Comp. Biochem. Physiol., 25, 427—436, 1968. Tempe-
464 7 U1b • dePendence of standard metabolism in flies.
• iroshin A. S., ed., International Symposium on Cytoecology, Leningrad, 1963.
^er^gamon Press, Oxford, 1967, 462 pp. The cell and environmental tempera-
465. Tsukuda H., Biol. J. Nara Women’s Univ., 10, 11—14, 1960. Temperature to-
lerance in the guppy.
208___________________________________Глава 9________________________/_________
/
466. Tsukuda И., J. Biol. (Osaka City, Japan), 12, 15—45, 1961. Temperature accli-
matization in fish. /
467. Tsuyuki H„ Wold F., Science, 146, 535—537, 1964. Enolase in.iish muscle.
468. Tucker V. A., J. Cell. Comp. Physiol., 65, 405—414, 1965. Relation between
torpor cycle and heat exchange in Perognathus.
468a. Tucker V. A., J. Exp. BioL, 48, 55—66, 1968. High-altitude flight in spar-
rows.
469. Umminger B. L., Biol. Bull., 139, 574—579, 1970. Effects of subzero tempera-
ture on blood of winter flounder.
470. Umminger B. L., J. Exp. ZooL, 172, 283—302, 409—424, 1970. Blood consti-
tuents and supercooling in killifish.
471. Umminger B. L., J. Exp. ZooL, 173, 159—174, 1970. Carbohydrate metabolism
in fish at subzero temperatures.
472. Umminger B. L., Nature, 225, 294—295, 1970. Osmoregulation in fish in fresh
water near freezing.
473, Ushakov В. P., Int. J. Life in Oceans and Coastal Waters, 1, 153—160, 1968;
also pp. 107—130, in: Molecular Mechanisms of Temperature adaptation, edi-
ted by C. L. Prosser, Amer. Assoc. Adv. Sci., Washington, D. C., 1967. Cellular
thermostability of aquatic animals.
474. Ushakov В. P., Lamosova B., Pashkova I., Chernokozheva I., J. Exp. ZooL,
167, 381—390, 1968. Individual variability in heat resistance of cells.
475. Veght J. H., PhysioL ZooL, 37, 316—328, 1964. Metabolic regulation of tem-
perature in gray jay.
476. Vellas F., Ann. LimnoL, 1, 435—442, 1965. Temperature effects on uricolytic
enzymes in carp.
477. Vernberg F. J., Schlieper C., Schneider D. E., Comp. Biochem. PhysioL, 8,
271—285, 1963. Temperature and salinity effects on cilia of molluscan gills.
478. Vernberg F. J., Tashian R. E., Ecology, 40, 585—593, 1959, also Vern-
berg W. B., F. J. Vernberg, Comp. Biochem. PhysioL, 17, 363—374, 1966.
Physiological variation of tropical and temperate zone Uca.
479. Vernberg W. B., Vernberg F. J., Amer. ZooL, 8, 449—458, 1968. Diversity in
metabolism of marine and terrestrial crustaceans.
480. Vernberg W. B., Vernberg F. J., Comp. Biochem. PhysioL, 26, 499—508, 1968.
Effect of temperature on cytochrome C oxidase of Uca.
481. Waites G. M. H„ Quart. J. Exp. PhysioL, 471, 314—323, 1962; Nature, 190,
172—173, 1961; also Waites G. M. H., Voglmayr J. K., Austral. J. Agric. Res.,
14, 839—851, 1963. Temperature regulation by scrotum of ram.
482. Wang L. C., Hudson J. W., Comp. Biochem. PhysioL, 32, 275—293, 1970; 38A,
59—90, 1971. Temperature regulation in chipmunk Tamias and in Perogna-
thus.
483. Wang G. Z., Marquardt W. C., J. ProtozooL, 13, 123—128, 199—202, 1966.
Survival of Tetrahymena and Paramecium following freezing.
484. Weatherley A. H., BioL Bull., 139, 229—238, 1970. Effects of high oxygen on
thermal tolerance of goldfish.
484a. Weathers W. W„ White F. N„ Amer. J. PhysioL, 221, 704—710, 1971. Ther-
moregulation in turtles.
485. Weiser C. J., Science, 169, 1269—1271, 1970. Cold resistance and injury in
woody plants.
486. West G. C., Hart J. S„ PhysioL ZooL, 39, 171—184, 1966. A4etabolic response
of grosbeak to temperature.
487. Wieser W., Kanwisher J., LimnoL Oceanogr., 6, 262—270, 1961. Ecology of
marine nematodes from salt marsh.
488. Williams B. A., Heath J. E., Amer. J. PhysioL, 218, 1654—1660, 1970. Preoptic
heating and cooling in ground squirrel.
489. : Williams D. D., Comp. Biochem. PhysioL, 27, 567—573, 1968. Effect of cold
exposure on norepinephrine heat production in hamster.
490. Williams D. D„ Plainer W. S., Arctic Aeromed. Lab. Bull. AAL-TR-67-5, 1—12.
Cold-induced changes in fatty acids in rat and hamster.
Температура 209
491. Williams R. J., Comp. Biochem. Physiol., 35, 145—161, 1970. Freezing toleran-
ce in Mytilus.
492. Williamson J. R., Prusint S., Olson M. S., Fukami M., Lipids, 5, 1—14, 1970.
Control of metabolism in brown fat.
493. Willis J. S., J. Physiol., 164, 51—63, 64—76, 1962. Resistance of tissues of
hibernators to cold swelling.
494. Willis J. S., Amer. J. Physiol., 214, 923—928, 1968. Cold resistance of kidney
of hibernators.
495. Willis J. S., Li N. M., Amer. J. Physiol., 217, 321—326. Cold resistance of
Na — К ATPase of kidney.
495a. Wilson F. R., Personal communication. Specific protein synthesis in acclima-
tion.
496. Wilson G., Rose S. P., Fox C. F., Biochem. Biophys. Res. Comm., 38, 617—
623, 1970. Lipid unsaturation and sugar transport by E. coli.
497. Wilson K. J., Lee A. K., Comp. Biochem. Physiol., 33, 311—322, 1970. Metabo-
lism and heart rate of Sphenodon at various temperatures.
498. Wyndham С. H., Plotkin R„ Munro A., J. Appl. Physiol., 19, 593—597, 1964.
Reaction of men in Antarctic.
499. Wyndham С. H. et al., J. Appl. Physiol., 19, 583—592, 868—876, 1964. Rea-
ctions of African natives to cold.
500. Yoshida M„ Oshima T„ Imahori K., Biochem. Biophys. Res. Comm., 43, 36—39,
1971. Thermo-stable enzyme PFK from thermophilic bacteria.
501. Zhirmunsky A. V. In: International Symposium on Cytoecology, Leningrad,
1963, edited by A. S. Troshin, Pergamon Press, Oxford, 1967, pp. 209—218.
Cellular thermal stability of marine invertebrates.
14-1514
Глава 10
Биологические ритмы
Ф. БРАУН
В наше время уже никто не сомневается в том, что биологиче-
ская ритмичность — явление чрезвычайно распространенное. Эта
ритмичность связана с основными колебаниями физической среды
и обнаружена у животных, растений и микроорганизмов; по-види-
мому, ее следует считать универсальным атрибутом жизни на Зем-
ле. Биологические ритмы обладают особыми свойствами, отличны-
ми от свойств иных, физиологических ритмов (например, ритмов
сердца и дыхания).
Сходство основных свойств биологических ритмов у всех иссле-
дованных представителей простейших, растений и животных позво-
ляет говорить об этих ритмах как об основополагающих и чрезвы-
чайно древних чертах, присущих всему живому. Поэтому не удиви-
тельно, что наши знания о природе этого явления получены на
организмах, составляющих весь спектр живых существ. Биологи-
ческие ритмы, по-видимому, практически не подвержены ни влия-
нию температуры, ни действию самых разнообразных агентов,
в частности депрессантов, ингибиторов и стимуляторов обменных
процессов. Различия между разными ритмами зависят главным об-
разом от природы видоспецифических биохимических, физиологи-
ческих и поведенческих реакций, ритмические изменения которых
исследуются.
В задачу этой главы не входит описание всего того бесчислен-
ного множества ритмических процессов, которые либо уже доста-
точно хорошо изучены, либо еще только изучаются и которые
вновь и вновь подтверждают, что связь между геофизическими
явлениями и биологической периодичностью следует считать пра-
вилом, а не исключением. Специфические ритмические вариации
уместно описывать при рассмотрении соответствующих конкрет-
ных физиологических функций. Здесь же мы займемся биоритмич-
ностыо как таковой, используя при этом лишь отдельные специ-
ально отобранные примеры. Изобилие подобных примеров можно
найти в многочисленных обзорах [18, 38, 56, 61—63, 92, 120, 147,
155, 161].
Компоненты ритмов. У организмов, находящихся в естественной
для них среде, периодические циклы возникают вследствие одно-
Биологические ритмы
211
временного действия факторов двух типов: а) непрерывной прямой
реакции на периодические изменения внешней среды, как биоти-
ческой, так и абиотической; б) присущей любому организму тен-
денции, повторяя последовательные циклы, создавать на их осно-
ве физиологические и поведенческие стереотипы, которые прояв-
ляются на существующем генетическом или внешнем фоне.
В результате изменение внешних условий вызывает некоторые
изменения рассматриваемых циклических процессов в ходе после-
довательных циклов. Изменяющиеся внешние факторы могут
влиять как на синхронный им цикл, так и на ближайшие после-
дующие циклы.
К наиболее важным изменяющимся факторам внешней среды
следует отнести свет, температуру, влажность, уровни прилива и
отлива, время приема пищи; реакции, вызванные этими фактора-
ми, по-видимому, типичны для адаптивных ответов. Менее значи-
тельные, но тем не менее измеримые воздействия на циклические
процессы оказывают трудноуловимые, но повсеместно распростра-
ненные геофизические факторы, вероятнее всего, электромагнит-
ной природы. Периодические изменения этих факторов постоянно
нарушаются или модифицируются под влиянием изменений пого-
ды, периодических процессов с другими частотами и иных, не иден-
тифицированных факторов.
Организмы, находящиеся в постоянных условиях освещения,
температуры и других очевидных факторов, будут все же непре-
рывно реагировать на эти чрезвычайно трудноуловимые физиче-
ские факторы, от действия которых организм остается незащи-
щенным. Реакции на трудноуловимые факторы могут сохраняться
более или менее долго (в зависимости от исследуемых организма
и системы) на фоне непрерывного возобновления предшествующих
адаптивных ритмических процессов, отражая неотъемлемую тен-
денцию живого к ритмизации. Эта тенденция даже при постоян-
ных внешних условиях продолжает проявляться в возникновении
ритмов, характеризующихся очень высокой устойчивостью к изме-
нениям температуры или к действию факторов, влияющих на ме-
таболизм.
Становится очевидным, что биологические ритмы являются
важной, а 'иногда и решающей временной основой для биологиче-
ской интеграции, осуществляющейся на трех уровнях.
1- Эта основа используется для изменения масштаба времени
тех биологических явлений, которые позволяют организму в рам-
ках своих возможностей поддерживать оптимальное адаптивное
состояние при воздействии таких ритмически меняющихся физи-
ческих внешних факторов, как свет, температура, влажность, океа-
нические приливы и отливы. Подобное адаптивное состояние под-
разумевает не только прямую реакцию на внешнее более или ме-
иее благоприятное воздействие, но также и эффективное «предви-
дение» этого воздействия и успешную подготовку к нему.
14*
212
Глава 10
2. С помощью биологических ритмов успешно осуществляется
координация половой активности, координация различных форм
активности экологически взаимозависимых видов, а порой и коор-
динация динамики экосистемы, которая в норме представляет со-
бой пример замечательной интеграции. В случае экосистем каж-
дый из бесчисленных видов, разнообразие которых важно для
поддержания экологической стабильности, реагирует на опреде-
ленные внешние воздействия лишь в течение отдельных периодов
(периодов оптимальной адаптации), в результате чего достигается
максимальная эффективность и продуктивность.
3. Накапливается все больше данных в пользу гипотезы, со-
гласно которой независимые ритмы индивидуальных органов, тка-
ней, клеток и даже клеточных компонентов отдельного организма
участвуют в создании временной упорядоченности биологических
явлений. Эта упорядоченность делает возможной такую степень
интеграции, которая в противном случае была бы недостижима
для великого множества взаимосвязанных и взаимодействующих
биологических процессов и для компонентов каждого из этих про-
цессов.
Пожалуй, в биологии нет другого столь же универсального
явления, о котором мы бы так мало знали, как о «биологических
часах» и о ритмах (суточных, приливно-отливных, месячных, го-
дичных), определяемых этими часами. Наши знания об основах и
механизмах «биологических часов» столь туманны и умозритель-
ны, что пока еще не представляется возможным разработать соот-
ветствующие экспериментальные подходы.
Суточные ритмы. К числу наиболее изученных биологических
ритмов относятся ритмы, связанные с 24-часовыми солнечными
сутками. В естественных условиях все факторы, участвующие в по-
вторяющихся процессах, обычно имеют 24-часовую периодичность.
Поэтому вклад каждого отдельного фактора в общую совокуп-
ность повторяющихся процессов оценить довольно трудно. Такая
оценка становится отчасти возможной, если проводить наблюдения
в контролируемых лабораторных условиях. Например, можно
L создать для исследуемого организма экспериментальный режим
12С : 12Т (чередование 12 ч на свету и 12 ч в темноте), поддержи-
вая при этом все другие известные факторы внешней среды на по-
стоянном уровне. Подобная схема опыта позволяет выявить суточ-
ные ритмы активности и показать, что у животных, ведущих ноч-
ной образ жизни, активность приурочена к темному периоду, днев-
ные животные активны на свету, а сумеречные проявляют макси-
мум активности при переходах от света к темноте, незадолго до
этих переходов или вскоре после них. Что же касается других жи-
вотных, то для них могут быть характерны очень короткие периоды
максимальной активности, приуроченные к какому-либо опреде-
ленному времени суток. Если, сохраняя формулу 12С : 12Т, сме-
стить на другие часы суток включение и выключение света, то по-
Биологические ритмы
213
допытное животное быстро (обычно в течение нескольких циклов)
перестроит суточный ритм активности, так что она опять будет
соответствующим для данного объекта образом приурочена
к новому времени чередования света и темноты. Поэтому очевид-
но, что животное приспосабливает свою генетически обусловлен-
ную схему поведения, связанную с временем суток, к условиям
освещения. Иными словами, главным эффектом 24-часового ритма
освещения является то, что фазы генетически обусловленного по-
вторяющегося каждые солнечные сутки цикла активности живот-
ных адаптивно связаны с чередованием дня и ночи. Сходным обра-
зом организм, быстро перенесенный по географической долготе,
к востоку или западу, заново приспосабливает свой ритм чередо-
вания активности и покоя к сдвинувшемуся по фазе чередованию
дня и ночи. В реализации этой способности организма свет играет
роль «датчика времени», или фактора совмещения ритма и фазы.
Аналогичную роль фактора фазовой настройки часто могут играть
24-часовые температурные ритмы (особенно в отсутствие циклов
освещения), а иногда также ритмы питания, звуковые ритмы и
даже ритмы социальных взаимодействий.
Наблюдаемые 24-часовые циклы могут быть обусловлены не
только светом или другими фазовыми датчиками. Это явствует
из того, что у животных суточные ритмы, судя по результатам
тщательно проведенных исследований, существенно меняются со
дня на день даже при строгом поддержании постоянного уровня
всех обычных внешних факторов и точном повторении световых
циклов. Кроме того, показано, что если смещать цикл 12С: 12Т
на разное число часов по отношению к местному времени, то эти
смещения могут давать не вполне эквивалентные результаты.
Иногда наблюдаются также полумесячные и месячные модуляции
циклов. Следовательно, если неконтролируемые параметры внеш-
ней среды вызывают какие-либо реакции у животных, то парамет-
ры этих реакций продолжают до некоторой степени отражать
24-часовые пли естественные периодические изменения внешних
условий.
Таким образом, у животных в естественных условиях наблю-
дается 24-часовая периодичность биологических процессов, причем
Достаточно точному выдерживанию этой периодичности не препят-
ствуют сильные колебания среднесуточной температуры, обуслов-
ленные погодой, или колебания других действующих на организм
факторов. В лабораторных условиях при режиме 12С : 12Т отме-
ченная периодичность наблюдается в широком диапазоне темпе-
ратур, и ее можно легко объяснить на основе 24-часового естест-
венного пли искусственного ритма освещения.
Циркадные ритмы. Приуроченные к солнечным суткам биологи-
ческие ритмы могут сохраняться и при полном устранении колеба-
ний света и температуры — двух главных периодически изменяю-
щихся внешних факторов «фазовой настройки». Однако при этом
214
Глава 10
очень часто ритмы теряют свою точную среднюю периодичность,
соответствующую 24 ч. Продолжительность повторяющихся цик-
лов будет теперь составлять немногим больше или немногим мень-
ше 24 ч, а периоды активности — систематически «дрейфовать» ко
ъсе более раннему или все более позднему местному времени
суток.
После того как в указанных экспериментальных условиях обна-
ружили такие измененные ритмы, был введен термин «циркадные
ритмы» (circa — приблизительно, diem — день), подчеркивающий,
что период этих ритмов лишь приблизительно соответствует 24 ч
[87]. Название циркадные ритмы получило широкое распростра-
нение, став синонимом для обозначения дневных, суточных, или
24-часовых, ритмов. Поскольку эти ритмические изменения, конеч-
но, не сохраняют никакой фиксированной связи с местным време-
нем, для нх характеристики стали употреблять эпитет «свободно-
текущий» [67] (рнс. 10-1).
Свойства свободнотекущнх циркадных ритмов, характерных
как для популяций, так и для отдельных особей, интенсивно изу-
чались во многих лабораториях [73, 95]. Было выяснено, что пе-
риоды циркадных ритмов у разных особей одного и того же вида,
выдерживаемых в одинаковых и тщательно контролируемых по-
стоянных внешних условиях, сильно отличаются друг от друга. Эти
периоды варьируют по ходу опыта (по-видимому, спонтанно) даже
у одной н той же особи (рнс. 10-2), причем наблюдаемые колеба-
ния иногда носят градуальный характер, а иногда выражены рез-
ко. Периоды при прочих равных условиях могут зависеть от интен-
сивности постоянного освещения. У дневных позвоночных периоды
свободнотекущих ритмов, как правило, короче 24 ч, а у ночных
обычно больше 24 ч (правило Ашоффа [4]; рнс. 10-3); чем ярче
освещение, тем выраженнее эта тенденция. У пойкилотермных
животных, по-видимому, есть очень слабо выраженная, но сопо-
ставимая зависимость длины периода от температуры [162]. Для
большинства организмов величина Qio лежит в диапазоне от 1,0
до 1,2, но у некоторых видов найдена обратная температурная за-
висимость и Qio составляет от 0,8 до 1,0. При прочих равных усло-
виях продолжительность периода свободнотекущих ритмов опре-
деляется также и генетическими факторами [51, 52, ПО].
Многочисленные попытки использовать факторы, стимулирую-
щие или подавляющие процессы обмена, для изменения периода
свободнотекущих ритмов, приводили, как правило, к отрицатель-
ным, или неопределенным результатам, или к результатам, объяс-
нить которые с наибольшей вероятностью можно было бы тем, что
действующий агент вызывает временный сдвиг фазы, приводящий
к явному изменению продолжительности нескольких циклов, но не
влияет на всю непрерывную периодичность. Ритмическое воздей-
ствие химическими агентами также не вызывало сколько-нибудь
четкого изменения периодов. Отрицательные результаты получали
i I I I I ill I I -I ..4.^1 I J L_J_1 1—I—I till
12 14 16 18 20 22 24 2 4 6 8 10 12
Время суток часы.
Рис. 10-1. Свободнотекущие ритмы летяги, выдерживаемой в полной темноте
(дни 1—9); приучение животного к 24-часовым циклам СТ (дни 10—32, стрелкой
отмечены часы освещения); прекращение циклов СТ и восстановление свободно-
текущих ритмов (дни после 32-го) [67].
216
Глава 10
даже в том случае, когда с помощью химических реагентов пыта-
лись влиять на процессы синтеза ДНК, РНК и белков [104, 166,
167]. Таким образом, система датчиков времени для относительно
регулярных свободнотекущих ритмов, по-видимому, отличается ис-
10 20 30 40 50 60 70 80 90 100 110 120
Длительность слабого освещения, ч
Рис. 10-2. Устойчивый ритм биолюминесценции у простейшего Gonyatilax polyedra
[95].
Клетки были выращены в режиме 12С : 12Т и к концу одного из периодов освещения (нуле-
вое время на абсциссе) перенесены в условия постоянного слабого освещения и постоянной
температуры (21 *С). Интенсивность люминесценции выражена в условных единицах.
Интенсивность освещения,люкс
Рис. 10-3. Свободнотекущие ритмы у различных организмов при постоянном осве-
щении разной интенсивности [4].
Точками в кружках обозначены результаты, полученные при повторении более ранних работ
(такие же точки без кружков). У Gonyatilax измерялась биолюминесценция, у фасоли — дви-
жение листьев, а у остальных видов — спонтанная активность.
ключительной стабильностью и высокой устойчивостью к измене-
ниям длин периодов [96]. При этом форма и амплитуда циклов
отличаются большей лабильностью.
Один химический агент, а именно D2O, все же влияет на длину
периода свободнотекущего ритма [50, 132, 160]. Наблюдаемый эф-
фект выражается в небольшом удлинении циклов, причем величи-
на этого удлинения пропорциональна концентрации тяжелой воды.
Подобного рода данные получены на самых разных объектах,
таких, как эвглена, птицы, мыши. Периоды, по-видимому, подвер-
жены также влиянию очень слабых природных электрических по-
лей и чувствительны к изменению этих полей [181].
Биологические ритмы
217
Изучая влияние освещенности на фазы циркадных ритмов,
определяющие видовые особенности поведения в 24-часовом цикле
с чередованием света (С) и темноты (Т), исследователи обнару-
жили одно интересное обстоятельство. Оказалось, что переходы от
С к Т или от Т к С не вызывают эквивалентных фазовых сдвигов
циркадных ритмов в любое время суток [37, 58, 68, 99, 140, 173].
Величина этих фазовых сдвигов при действии света зависит от
времени суток; это свойство многих организмов может быть изоб-
ражено графически с помощью кривой «фаза—реакция». Так, на-
пример, в опытах на ночном млекопитающем летяге, выдерживае-
мой в условиях постоянной темноты (ТТ), обнаружено, что фазо-
вый сдвиг при 10-минутной световой вспышке зависит от того,
в какой момент циркадного ритма животного эта вспышка произ-
ведена [67]. Продолжительность периодов у летяги в этих экспе-
риментальных условиях обычно слегка отличается от 24-часовой,
и поэтому фазы активности (субъективная ночь) и неактпвности
(субъективный день) систематически смещаются по отношению
к времени суток. Световая вспышка, произведенная в начале пе-
риода активности животного, вызывает максимальную фазовую
задержку (сдвиг на более позднее время), а точно такое же 10-ми-
нутное включение света при завершении периода активности при-
водит к максимальному опережению по фазе; включение света во
время активности животного дает промежуточные величины эф-
фекта вплоть до полного отсутствия фазового сдвига (рис. 10-4).
Полагают, что кривые «фаза—реакция» описывают именно те
свойства циркадной системы, которые ответственны за нормальное
приспособление фазы ритмического процесса к естественному чере-
дованию дня и ночи. Иными словами, постулируется то, что эти
кривые отражают фундаментальное свойство циркадных ритмиче-
ских систем. Можно ожидать, что у ночных животных, например
у летяги, включение циклов фаза—реакция, сопряженных по фазе
с циклами покой—активность, будет задерживаться при вечернем
переходе от С к Т и опережать при утреннем переходе от Т к С.
Однако равновесие фаз настройки при этом не будет нарушено
при условии, что компоненты кривой, соответствующие суточному
опережению или суточной задержке, равны друг другу.
Для кривых «фаза—реакция», по-видимому, характерны неко-
торые индивидуальные особенности, проявляющиеся в деталях
формы и амплитуды. Поэтому можно ожидать, что при одном и
том же цикле СТ будут наблюдаться незначительные индивиду-
альные отличия фазовой регулировки. Обычно это так и есть. На-
пример, при выдерживании вьюрков в режиме 12С : 12Т можно от-
метить, что у некоторых птиц включение активности связано с пе-
реходом от Т к С, тогда как у других — с изменениями освещения
[5]. Если птиц, принадлежащих к обеим этим группам, выдержи-
вать при непрерывном освещении, то свободнотекущие ритмы
У особей первой группы (включение активности совпадает с пере-
218
Глава 10
ходом ТС) окажутся короче 24, а у особей второй группы (вклю-
чение активности отстает от перехода ТС) будут длиннее 24 ч.
Опыты на кенгуровых крысах, выдерживаемых при непрерывном
слабом освещении, показали, что продолжительность периодов
свободнотекущих ритмов варьирует у отдельных особей от вели-
чин, меньших 24 ч, до величин, превышающих 24 ч. При этом,
когда происходит включение активности, обнаруживается постоян-
Рис. 10-4.
А. Диаграмма, показывающая соотношение цикла активность — покой и тесно связанной
с этим циклом кривой «фаза—реакция» у ночного млекопитающего, а также связь этих
циклов с циклами освещения как фактора внешней среды для животного при равновесии
фаз. Б. Сопоставимая ситуация сразу после резкой фазовой задержки в цикле освещения.
В. Сопоставимая ситуация сразу после резкого фазового опережения в световом цикле. Ожи-
даемые фазовые влияния света отмечены стрелками [35].
ная количественная связь между наблюдаемой продолжитель-
ностью индивидуального периода свободнотекущего ритма и ве-
личиной фазовой задержки (или фазового опережения) на кривой
«фаза—реакция», построенной для данного животного [124]. Если
действовать на крыс световыми вспышками, по времени приблизи-
тельно совпадающими с началом активности, то у животных, у ко-
торых периоды свободнотекущих ритмов короче 24 ч, на кривой
будет наблюдаться фазовое опережение, а у животных с периодом
длиннее 24 ч в тех же условиях обнаружится фазовая задержка
(рис. 10-5).
Кривые «фаза—реакция» существуют не только для световых,
но и для температурных воздействий [55, 58, 157, 188].
Биологические ритмы
21Э
При постоянных температуре и освещении свободнотекущие
циркадные ритмы могут, по-видимому, сохраняться в течение не-
ограниченного времени; иногда при этом наблюдаются даже по-
вышения или колебания амплитуды. Вместе с тем возможно и за-
тухание ритмов, идущее с переменной скоростью вплоть до кажу-
щейся полной остановки. Имеются также сообщения о резком пре-
кращении циркадных ритмов, продолжающемся в течение несколь-
ких или многих циклов. Однако позднее циркадный ритм спонтан-
Средний период сеободнотекущего ритма. ч
Рис. 10-5. Зависимость величины фазового опережения или фазовой задержки
у кенгуровых крыс от световых вспышек постоянной интенсивности, даваемых во
время перехода животных к активному состоянию [124].
но восстанавливался на той самой фазе, которую следовало бы
ожидать в данный момент, если бы ритм все это время не прекра-
щался [92]. В случаях когда циркадный ритм прекращался вслед-
ствие постепенного затухания, его часто можно было восстановить,,
создавая искусственный режим СТ, или даже с помощью единич-
ной кратковременной вспышки света без возобновления 24-часо-
вого цикла. Подобный световой стимул не только возобновляет
ритм, но и определяет фазу, на которой произойдет включение
ритма. Подобные результаты свидетельствуют о том, что циркад-
ные ритмические системы связаны с какими-то более глубокими
ритмическими изменениями и что эта связь при определенных
Условиях может временно прерываться. Исчезновение явного цир-
кадного ритма в какой-либо системе организма не обязательно'
Распространяется и на другие системы той же самой особи.
С помощью различных воздействий, например света, можно-
экспериментально изменять повторяющиеся компоненты циркад-
ных ритмов. Так, при непрерывном освещении наблюдается посте-
Пенное уменьшение амплитуды, продолжающееся в течение ряда-
220
Глава 10
последовательных циклов. Если животных перевести затем в усло-
вия постоянной темноты, то достигнутое к моменту переноса сни-
жение амплитуды сохраняется по крайней мере в течение многих
циклов '[39]. А в опытах на проростках бобов, выдерживаемых при
постоянном слабом освещении, показано, что вспышки более ярко-
го света, даваемые в определенный момент цикла установившегося
ритма, вызывают кратковременную поведенческую реакцию. Эта
реакция будет повторяться в соответствующие моменты последую-
щих циклов даже при отсутствии световых стимулов [54]. Сход-
ным образом можно выработать реакцию на время. Так, пчелы,
обученные прилетать за подкормкой в определенное время дня
или циркадного цикла, продолжают прилетать в это же время
на место подкормки в течение нескольких дней после ее прекра-
щения [146]. Следовательно, повторяющиеся компоненты циркад-
ных систем обусловлены не только внешними воздействиями, но
и генетически закрепленными факторами.
Проводились опыты, в которых организмы быстро перемещали
на самолете из одной географической долготы в другую [48, 146].
Это перемещение обычно не оказывало никакого влияния на ход '
свободнотекущнх ритмов. Однако картина становилась иной в тех
случаях, когда исследовались не свободнотекущие ритмы, а рит- '
мы, наблюдаемые в естественных условиях. Вначале сохранялся
выработанный ранее циркадный характер ритмов, но затем датчи- {
ки времени (свет, температура и другие), соответствующие новой
долготе, постепенно в течение нескольких последовательных цик-
лов осуществляли перенастройку фазы циркадного ритма, созда-
вая новое равновесное состояние [99, 158].
Циркадные ритмы обнаружены даже у арктических организмов
во время полярного дня [114, 117] или полярной ночи. Наблюде- i
ния, проводимые на южном географическом полюсе, показали, что I
организмы, выдерживаемые там при тщательно контролируемом I
постоянстве всех известных внешних факторов, сохраняют типич- ;
ные свободнотекущие ритмы [89]. Уникальность и фундаменталь-
ная природа 24-часового периода в циркадных ритмах подтверж-
дается целым рядом наблюдений. Были сделаны попытки удлинить
или укоротить циклы циркадных ритмов, подвергая такие различ-
ные объекты, как цветущие растения или мыши [109, 165], дей-
ствию ритмов СТ широкого диапазона частот. Оказалось, что орга-
низм часто может приспособиться к циклам более коротким (при-
близительно ЮС : ЮТ) или более длинным (14С : 14Т), чем 24-ча-
совой цикл. Однако, если еще более изменить циклы СТ и выйти
за пределы этих относительно узких границ, организм обычно вы-
рывается из навязанного ему ритма и возвращается к характерной
циркадной частоте. В опытах на крабе Uca было обнаружено, что
выдерживание животных в режиме 16С : ЮТ приводит к постепен-
ному угасанию исходного четко выраженного 24-часового ритма
и переходу к 96-часовому ритму, что, по-видимому, является след-
Биологические ритмы
221
ствием биений между непрерывно навязываемым 32-часовым и
упорно удерживаемым 24-часовым ритмами [173]. Возможности
приучения мышей к новым циклам СТ значительно расширяются,
если вместо резкого включения и выключения постепенно изменять
интенсивность освещения [105].
Циркадные ритмы присущи не только целым организмам. Неза-
висимые ритмы можно также выявить у отдельных органов [3],
тканей и даже единичных клеток [7, 148, 153, 163]. Более того,
у одноклеточных организмов и ядро, и цитоплазма могут иметь
самостоятельные циркадные ритмы. Если ядро какой-либо клет-
ки пересадить в энуклеированный цитоплазматический фрагмент,
циркадный ритм которого по фазе не синхронен ритму ядра, то
ядро вызовет постепенный фазовый сдвиг цитоплазматического
ритма вплоть до установления фазовой синхронности [154].
Организм, по-видимому, представляет собой интегрированный
комплекс составляющих его ритмических систем на всех уровнях
организации. 24-часовые ритмические вариации естественных на-
стройщиков фазы, или датчиков времени, приводят в условиях нор-
мального равновесного состояния организма к появлению у этих
организмов ритмических систем, имеющих 24-часовой период.
Фазы этих систем связаны во времени так, чтобы обеспечить опти-
мальное облегчение взаимодействий. Различные ритмические ком-
поненты могут проходить через свои максимальные и минималь-
ные значения в совершенно разное время суток, опережая друг
друга или отставая друг от друга. Подобная картина в значи-
тельной степени зависит как от многочисленных причинно-следст-
венных взаимосвязей, так и от времени, необходимого для завер-
шения последовательных процессов, представляющих различные
звенья общей циркадной цепи. Описывая различные временные
отношения ритмических компонентов, лежащих в основе циркад-
ной ритмичности организма как целого, можно получить циркад-
ную «фазовую карту».
В случае резкого изменения датчика или комплекса датчиков
времени (это изменение может быть естественным, как, например,
при быстром переносе в другой часовой пояс, или эксперименталь-
ным, когда дается эквивалентный резкий сдвиг циклов СТ по фа-
зе) перенастройка организма начинается уже во время первого
Цикла. Однако обычно должно пройти несколько или даже много
Пиклов, прежде чем общая циркадная система организма достиг-
нет нового равновесного состояния, когда все составляющие рит-
мические компоненты займут свое место на «фазовой карте». Раз-
ные компоненты достигают этого нового равновесия с различной
скоростью. Во время переходного периода возможно разобщение
тсх или иных переходных элементов цикла, которое наблюдается
в зависимости от их состояния либо исходно, либо по достижении
нового равновесного состояния. Пониженная работоспособность
Бо время полета на реактивном самолете, вероятно, связана с этим
222
Глава 10
переходным периодом; такое же переходное состояние, возможно,
характерно и для других организмов.
Сходное разобщение было описано для некоторых свободноте-
кущих ритмов при постоянстве всех эффективных датчиков вре-
мени. Например, для человека был описан случай, когда продол-
жительность цикла в ритме сна и бодрствования превышала 33 ч,
а для ритма температуры тела составляла всего 24,8 ч [6].
Переходные циклы циркадных ритмов можно обнаружить,
пользуясь и другими экспериментальными методиками. Одиноч-
ный фазовый стимул может вызвать фазовый сдвиг в ближайшем
последующем цикле свободнотекущего ритма; в ходе нескольких
следующих циклов этот сдвиг постепенно исчезает. Вместе с тем
такой одиночный стимул может вызвать лишь слабый фазовый
сдвиг в ближайшем цикле или вообще на этот цикл не подейство-
вать; однако в течение ряда последующих циклов свободнотеку-
щего ритма возникает серия фазовых сдвигов, или переходных
циклов, продолжающихся вплоть до достижения нового равновес-
ного состояния и до практически полного восстановления исходной
частоты. Предположили, что эти два альтернативных эффекта за-
висят от того, вызвал ли стимул фазовый сдвиг у главного рит-
мического компонента, определяющего фазу.
Мы уже говорили, что при отсутствии любого очевидного дат-
чика времени циркадные ритмы обычно продолжают свободно
протекать о периодичностью, незначительно отличающейся от
24-часовой. Известно также [47, 119, 164], что свободнотекущие
ритмы иногда даже в течение длительного времени сохраняют та-
кую периодичность, не имеющую статистически достоверных отли-
чий от 24-часовой. Это было показано, в частности, для мышей и
крыс, выдерживаемых в постоянной темноте, а также для популя-
ций крабов Uca, кенгуровых крыс и ящериц ядозубов. Сохранение
точного 24-часового цикла встречается гораздо чаще, чем можно
было бы ожидать на основании произвольной выборки циркадных
периодов. Животные могут в течение многих дней и даже недель
сохранять свободнотекущие ритмы, период которых равен точно
24 ч, а затем внезапно переходить на другую частоту. Возможна
и обратная картина, когда животное, в течение некоторого вре-
мени имеющее период, не равный 24 ч, резко и без видимых при-
чин переключается на точный 24-часовой ритм. Поэтому в общей
форме можно сказать, что 24-часовой период, по-видимому, зани-
мает особое место в циркадных ритмах и, кроме того, что жи-
вотные гораздо больше предрасположены к свободнотекущим рит-
мам с точной 24-часовой периодичностью, чем к любым другим
ритмам, периодичность которых значительно отклоняется от 24 ч.
Другой особенностью циркадных систем является их очевид-
ная способность хотя бы в редких случаях адаптивно настраи-
ваться по фазе на местное время. Эта способность обнаруживает-
ся даже при полном отсутствии каких-либо явных внешних дат-
Биологические ритмы
223
чиков времени. В качестве примера приведем опыты, в которых
куриных зародышей* инкубировали при строгом постоянстве всех
известных фазовых стимулов до тех пор, пока у этих зародышей
не завершится формирование сенсорных и моторных компонентов
нервной системы [101]. У выращенных таким способом цыплят
обнаруживался циркадный ритм активности, который к тому же
был адаптивно связан по фазе с местным временем типичным для
данного вида образом. Результаты этих опытов позволяют пред-
положить, что датчиком времени в данном случае служат трудно-
уловимые колебания атмосферы, обусловленные солнечными при-
ливами.
Ритмы, зависящие от геофизических процессов. Существует
еще один периодический компонент, который сохраняется у орга-
низмов даже при постоянстве всех внешних факторов, «задающих»
фазу циркадных ритмов. Этот компонент, по-видимому, воз-
никает как следствие непрерывной реакции организма на те тон-
кие переменные факторы внешней среды, которые все еще не под-
даются контролю в эксперименте. Интенсивность биологической
активности изменяется при прочих равных условиях в зависимости
от местного времени, или часового угла Солнца. Средняя ампли-
туда этих изменений относительно мала, и поэтому они обычно
(маскируются гораздо более выраженными циркадными и други-
ми очевидными ритмическими изменениями. Сверх того, атмосфер-
ное электромагнитное поле, т. е. тот периодический внешний комп-
лекс, от которого, по имеющимся данным, зависят эти слабые
биологические ритмы, сам претерпевает последовательные цикли-
ческие изменения, модулируемые естественными геофизическими
ритмами других частот, перемещениями воздушных масс, а также
изменениями активности Солнца и связанными с ней геомагнит-
ными явлениями. Короче говоря, живая система, по-видимому, по-
стоянно получает информацию о флуктуациях, происходящих в
земной атмосфере. Об этом свидетельствует хотя бы возможность
легко продемонстрировать непрерывные перекрестные корреляции
с трудноуловимыми внешними параметрами как для организмов,
находящихся в естественных условиях чередования дня и ночи
или в поле действия искусственного периодического датчика вре-
мени (например, света), так и для организмов, защищенных от
всех периодических датчиков времени.
В простейшем случае мы можем наблюдать и охарактеризо-
вать средний 24-часовой цикл, определяемый геофизическими фак-
торами, если поместим какой-либо организм, лишенный выражен-
ных циркадных ритмов, в условия, при которых поддерживается
постоянство всех очевидных внешних стимулов; можно воспользо-
ваться и популяцией, состоящей из большого числа особей, так-
поместив их в условия, когда потенциальные синхронизаторы
(датчики времени) отличаются постоянством действия. Для по-
добных популяций предполагается случайное распределение фазо-
224
Глава 10
®ых отношений индивидуальных свободнотекущих ритмов. В тех
случаях, когда наблюдаются четко выраженные свободнотекущие
циркадные ритмы, рассматриваемый 24-часовой цикл накладыва-
ется на них как реакция на неконтролируемые физические пере-
менные. Этот цикл можно выделить, измеряя среднюю величину
активности для каждого часа суток и проводя эти измерения в
течение периода времени, достаточного для того, чтобы получить
суточную развертку циркадной системы целое число раз. Однако
число дней, в течение которых проводятся измерения, должно не
только соответствовать определенным требованиям, которые необ-
ходимо соблюдать для получения развертки циркадной системы
на протяжении солнечных суток, но одновременно охватывать це-
лое число синодических месяцев. Дело в том, что некоторые жи-
вотные обладают четко выраженным ритмическим компонентом,
связанным с лунными приливами; к тому же на все организмы,
вероятно, действуют изменения атмосферы, происходящие в тече-
ние лунных суток и имеющие в общем тот же порядок величин,
что и в случае солнечных суток. Очевидно, описанная процедура
позволяла установить такую величину активности для каждого
часа суток, которая была практически рандомизирована по от-
ношению к фазовым углам как циркадной системы, так и любой
периодической системы, связанной с лунными приливами или лун-
ными сутками.
Для того чтобы проанализировать стойкие циклы, зависящие
от геофизических процессов, чаще всего исследуют изменения ин-
тенсивности окислительного обмена и спонтанной двигательной
активности; оба эти показателя, разумеется, тесно связаны с ин-
тенсивностью стандартного обмена. При изучении первого пока-
зателя применяются респирометры с автоматической записью,
сконструированные так, чтобы осуществлять очень продолжитель-
ный контроль за ритмом при постоянстве всех очевидных внешних
факторов, включая внешнее давление. Для регистрации спонтан-
ной двигательной активности используются актографы различных
систем, связанные с соответствующими системами записи и также
обеспечивающие необходимую продолжительность исследований.
Если циркадные системы, как полагают, обусловливают специ-
фическую и приспособительную связь каждого вида с внешними
изменениями в течение солнечных суток, то изменения, зависящие
от геофизических процессов, менее специфичны и имеют тенден-
цию к тому, чтобы принять ту или иную форму из ограниченного
их числа (рис. 10-6). Наиболее общей является трехвершпнная
кривая с максимумами е 7, 12 и 18 часов и с суточным диапазоном
колебаний обычно около 4—20% среднего суточного уровня. По-
добная кривая описана для самых разнообразных организмов, в
частности для картофеля, бобов, моркови, мучного хрущака, ку-
риных зародышей 4- и 5-дневного возраста в весенние месяцы,
крысы [29, 44, 101]. При действии очень слабых -у-лучей на мы-
Полдень Полдень
Рис. 10-6. Средние вариации, наблюдаемые по ходу солнечных суток у разно-
образных организмов, которых долгое время выдерживали при постоянных усло-
виях.
—Ле. Ежегодное потребление кислорода картофелем в течение 6 лет. £>>—Б3. Ежегодное
потребление кислорода прорастающими семенами бобов в течение 3 лет. В. Потребление
кислорода срезами моркови в течение 8 мес. Л—Г3. Потребление кислорода куриными заро-
дышами на 5, 6 и 8-й дни развития. Д. Спонтанная активность мышей при 5-кратном повы-
шении фоновой радиации. Е. Активность личинок Tenebrio в течение 9 мес [половины лун-
ного месяца с центром в полнолуние (Ei) и новолуние (ЕгЛ-
15-1514
226
Глава 10
шей наблюдалась такая же общая 24-часовая кривая [43]. Дру-
гой общий тип циклических изменений, описанный для крабов
Uca, имеет характер одновершинной кривой с максимумом около
5—6 и минимумом около 17—18 часов [174]. 24-часовая кривая,
наблюдаемая у ранних куриных зародышей осенью и зимой, при-
близительно соответствует зеркальному отображению кривой, опи-
санной для крабов {8].
Различные исследователи пытались идентифицировать те ат-
мосферные параметры, к которым чувствительны организмы. По-
началу было обнаружено, что в определенные моменты суток меж-
ду скоростью биологических процессов и скоростью изменения ба-
рометрического давления существует слабая запаздывающая кор-
реляция. Средние интенсивности метаболизма, наблюдаемые у
картофеля или крабов Uca поздней весной и летом в 5—7 и в 17—
19 часов независимо друг от друга, в высокой степени коррелиру-
ют с изменениями барометрического давления соответственно в
2—6 и в 14—18 часов [24, 25]. Знак этой корреляции у крабов ос-
тается одним и тем же как во время утреннего максимума, так
и во время вечернего минимума. У картофеля утренняя корреля-
ция была положительной, а вечерняя—отрицательной, так что
интенсивность метаболизма в обоих случаях оказывалась макси-
мальной. Аналогичная вечерняя корреляция обнаружена и у цып-
лят [8]. Все эти корреляции сохраняются и тогда, когда организ-
мы выдерживаются в условиях постоянного внешнего давления.
Следовательно, организмы реагируют не на изменения атмосфер-
ного давления как такового, а один или два раза в сутки с ин-
тервалом в 12 или 24 ч отвечают на некоторый атмосферный па-
раметр (или параметры), стимулирующее действие которого в эти
четко определенные интервалы времени коррелирует с происходя-
щими одновременно изменениями атмосферного давления. Таким
образом, в течение солнечных суток хорошо известные атмосфер-
ные приливы могут оказывать непосредственное воздействие на
организм.
Недавно были получены данные, позволяющие сделать предпо-
ложение о природе хотя бы одной из действующих сил. Объектом
исследования служили песчанки, выдерживаемые в режиме 12С:
: 12Т. Оказалось, что суточные особенности средней активности
песчанок, регистрируемые между 15 и 18 часами, находятся в тес-
ной корреляции с наблюдаемыми в те же часы суточными особен-
ностями скорости, с которой изменяется сила горизонтального
вектора земного магнитного поля. В любое другое время суток
и при любой другой временной связи эти два параметра не обна-
руживают корреляции [159]. Известно, что горизонтальный век-
тор земного магнитного поля имеет четко выраженные суточные
колебания.
У растения картофеля, герметически изолированного при посто-
янных внешних условиях (включая температуру), наблюдается
Биологические ритмы Wl
высокая степень корреляции между 24-часовым циклом каждых
суток (в интервале между полуночью и полуднем) и одновременно
регистрируемыми величинами средней суточной температуры. Эта
корреляция положительна для температур ниже 14 °C и отрица-
тельна для более высоких температур, что свидетельствует о роли
какого-то иного атмосферного параметра, влияющего на харак-
терные 24-часовые ритмы, зависящие от геофизических факторов
[24].
Можно допустить, что флуктуации любого атмосферного пара-
метра хотя бы в некоторой степени коррелируют с другими ат-
мосферными параметрами. С этой точки зрения не будет неожи-
данным, если обнаружатся корреляции между интенсивностью
биологических .процессов у организмов, находящихся в постоянных
условиях, и амплитудами суточных изменений общей фоновой ра-
диации [45] или типами вариаций первичных космических лучей
,[49]. Напомним, что в последнем случае, как и в случае ранее
рассмотренных корреляций с атмосферным давлением и темпера-
турой, исключена прямая связь типа «причина — следствие», по-
скольку все организмы защищены атмосферой от действия первич-
ных космических лучей.
Исследования, направленные на выяснение того, к каким имен-
но атмосферным электромагнитным параметрам специфически чу-
вствительны организмы, лишний раз ‘подчеркнули всю сложность
этой проблемы. С помощью простых прямых опытов по изучению
способности живых организмов воспринимать очень слабые маг-
нитные, электрические и радиационные поля, сила которых имеет
тот же порядок, что и соответствующие поля Земли, было пока-
зано, что организмы воспринимают силовые поля всех трех на-
званных типов. Более того, оказалось, что организмы обладают
максимальной разрешающей способностью и улавливают такие
изменения силы и направления векторов, уровни которых близки
к нормальным для данного вида внешним уровням этих парамет-
ров [33, 107, 116, 122, 139, 177, 180, 184].
Лунные ритмы. Ритмические изменения физических внешних
условий могут 'быть связаны и с другим земным периодом, лунны-
ми сутками, 1продолжительность которых равна 24 ч 50 мин; эта
'Периодичность играет особенно важную роль в жизни организмов,
обитающих в приливно-отливной, литоральной зоне. Океанические
приливы и отливы, периоды которых определяются главным обра-
зом положением Луны по отношению к Земле, обычно происходят
по два раза в сутки. Высота обоих ежесуточных приливов может
быть приблизительно одинаковой или неодинаковой, причем во
втором случае различия могут сильно варьировать в разных гео-
графических районах — от незначительных до максимальных, ког-
да в течение суток происходит только один прилив и один отлив.
В течение синодического месяца амплитуда приливов и отливов
каждой данной области претерпевает типичные изменения, явля-
15*
1
228 Глава 10
ющиеся следствием взаимодействия солнечного и лунного притя-
жения. В новолуние и полнолуние (т. е. каждые полмесяца), ког-
да силы притяжений Луны и Солнца складываются, происходят
высокие, или сизигийные, приливы; когда же силы притяжения
Луны и Солнца направлены под прямым углом друг к другу, на-
блюдаются низкие, или квадратурные, приливы. В некоторых рай-
онах наблюдаются также полумесячные или месячные изменения
суточной структуры приливов и отливов.
У береговых организмов временная динамика активности адап-
тивно связана с чередованием приливов и отливов. Если организ-
мы кормятся на литорали во время отлива (диатомеи, плоские
черви Convoluta, крабы Uca, птицы), то максимумы их циклов
активности совпадают с малой водой. Другие организмы, как, на-
пример, моллюски, морские желуди, зеленые крабы, во время от-
лива впадают в неактивное состояние и используют различные
приспособления, препятствующие высыханию и компенсирующие
уменьшившуюся доставку кислорода, а во время прилива прояв-
ляют максимальную активность. Организмы, обитающие в верх-
ней или нижней литорали, подвергаются соответственно затопле-
нию или осушению лишь во время сизигийных приливов или отли-
вов, т. е. один раз в полмесяца или месяц.
Давно известно, что активность плоского червя Convoluta, пе-
ренесенного с литорали в лабораторные условия, сохраняет связь
с приливно-отливным ритмом — червь поднимается к поверхности ,
песка во время отлива и уходит вглубь во время прилива [19, I
82]. Подобная стойкая ритмичность поведения, обусловленная !
приливно-отливным ритмом, описана и для других литоральных
организмов — ракообразных и моллюсков. В последние годы со-
общалось об аналогичных биологических ритмах у простейших и
одноклеточных водорослей, живущих в приливно-отливных водое-
мах, у диатомей приливно-отливной зоны, у крабов Uca и зеленых
крабов, а также у некоторых литоральных рыб [16, 36, 77, 80,
83, 102, 125, 133, 134, 144, 183].
Литоральные организмы в естественных условиях подвержены
действию не только приливно-отливных циклов, но одновременно
и 24-часовых циклов освещения. Фазы приливно-отливных циклов
систематически смещаются по отношению к времени суток со
средней скоростью 50 мин в сутки, так что пики этих циклов мо-
гут происходить в любое время дня или ночи. Активность некото-
рых организмов, например крабов Uca, в естественных условиях
также смещается вместе с приливно-отливными циклами на про-
тяжении всех 24 ч суток. У других организмов, например у днев-
ных птиц, кормящихся на берегу, или у диатомовых водорослей,
у которых от освещения зависят процессы фотосинтеза, на при-
ливно-отливные циклы накладывается значительный дневной
компонент; пики активности соответствуют частоте приливов, но J
происходят только в дневное время.
Биологические ритмы
229
Подобно солнечным, лунные ритмы также характеризуются
видостгецифичностью; благодаря этому осуществляется приспосо-
бительная настройка организмов на приливно-отливные изменения
в данной географической точке. Влияние местных топографиче-
ских условий на прохождение океанической приливной волны мо-
жет приводить к тому, что в точках побережья, находящихся все-
Время суток
8Рис. 10-7. Приливно-ститивный ритм краба Uca pugnax в лаборатории в естествен-
ных условиях освещения [10].
Точками отмечено предсказанное время приливов в том районе, где были отловлены крабы.
а*о в нескольких милях друг от друга, время прилива будет разли-
чаться очень сильно (до 5—6 ч). Поэтому ясно, что между при-
ливно-отливными и суточными колебаниями не может быть зна-
чительной корреляции и что такие хорошо известные регуляторы
>фаз циркадных ритмов, как естественные суточные изменения ос-
вещения и температуры, не участвуют в фазовой настройке био-
•логических ритмов, связанных с приливами и отливами.
Некоторые животные литоральной зоны (крабы Uca [11] и
мидии [144]), перенесенные в лабораторию, т. е. изолированные
от воздействия приливно-отливных циклов, по крайней мере в те-
чение нескольких циклов сохраняют фазовое соответствие с при-
ливно-отливными циклами, характерными для их местообитания
(Рис. 10-7). Показано также, что эти животные, перенесенные в
другую часть побережья, за несколько дней или недель приспо-
230
Глава 10
сабливаются к новому расписанию приливов и отливов. Пока еще
твердо не установлено, какие именно внешние ритмические фак-
торы осуществляют фазовую настройку биологических ритмов,
связанных с приливами и отливами.
Приливно-отливные ритмы устриц и крабов Uca [13], выдер-
живаемых в темноте при постоянной температуре, а также изоли-
рованных от действия приливов и отливов, в течение одной-двух
недель постепенно изменяются по фазе. В итоге максимумы этих
ритмов совпадают уже не с местными фазами океанических при-
ливов и отливов, а со временем верхней и нижней кульминаций
Луны, которое тесно связано с максимумами лунных приливов
в атмосфере. Это позволяет предположить, что тонкие колебания
атмосферы, вызываемые лунными приливами, могут использовать-
ся для фазовой настройки биологических ритмов, связанных с
приливами и отливами, разумеется, если отсутствует обычно доми-
нирующий фазовый регулятор, обусловленный океаническими при-
ливно-отливными изменениями. Крабы Uca, собранные на тех уча-
стках побережья, где низкая вода (и соответственно этому макси-
мальная активность животных) совпадает с временем верхней и
нижней кульминаций Луны, могут свыше 45 сут точно выдержи-
вать приливно-отливные ритмы активности в лабораторных усло-
виях, несмотря на то что они изолированы от прямого действия
океанических приливов и отливов.
Таким образом, одно из главных отличий приливно-отливных
биологических ритмов от циркадных ритмов солнечных суток со-
стоит в том, что первые из них нельзя, а вторые можно синхрони-
зировать с 24-часовыми циклами чередования света и темноты.
Как мы видели, приливно-отливные ритмы сохраняются на фоне
естественных или искусственных циклов СТ. Более того, естест-
венные суточные изменения освещенности в некоторых случаях,,
по-видимому, даже способствуют установлению большей синхрон-
ности этих биологических ритмов и приливных циклов [10, 176].
Однако свет все' же оказывает влияние на приливно-отливный
ритм. Если, изменяя сроки чередования света и темноты, вызвать
фазовый сдвиг циркадного цикла, то это приведет к непосредст-
венному эквивалентному фазовому сдвигу и приливно-отливного’
цикла [36]. Приливно-отливный цикл ведет себя при этом так, как
если бы он был функционально связан с циркадным циклом, но«
лишь во время фазового сдвига; затем, несмотря на продолжаю-
щиеся циклы 'СТ, восстанавливается исходная частота приливно-
отливного ритма, соответствующая лунным суткам. Если выдер-
живать крабов Uca в условиях 24,8-часового цикла СТ, циркад-
ный ритм будет ежедневно отставать по фазе на 50 мин. Видимые
приливно-отливные циклы действительно будут отражать это 50-
минутное запаздывание, а поскольку сверх того они обычно от-
стают ежесуточно еще на 50 мин от циркадных циклов, то в итоге
эти циклы будут продолжаться 25 ч 40 мин [476].
Биологические ритмы
231
Проиллюстрировать только что изложенные данные можно, по-
видимому, следующим наблюдением. У крабов Uca, выдерживае-
мых в темноте (ТТ) и при постоянной температуре, сохраняются
точный в среднем 24-часовой свободнотекущий циркадный ритм
окраски и точный приливно-отливный ритм спонтанной двигатель-
Рис. 10-8. Общая картина активности популяции краба Sesarma при постоянных
внешних условиях [131].
Черными прямоугольниками отмечено время, в течение которого активность была равна
среднему дневному уровню или превышала его.
ной активности. Однако в условиях непрерывного освещения, спо-
собствующих удлинению периодов свободнотекущих циркадных
ритмов до величин, превышающих 24 ч, по-видимому, удлиняются
сходным образом и приливно-отливные циклы, в результате чего
Устанавливается новый свободнотекущий ритм с длиной периода
свыше 24,8 ч .[,10].
У одной и той же особи, находящейся в природных условиях
Или в лаборатории при поддержании постоянного уровня всех
явных внешних факторов, могут одновременно существовать четко
сраженные циркадный и приливно-отливный ритмы, связанные
соответственно с солнечными или лунными сутками (рис. 10-8),,
232
Глава 10
Эти две частоты могут регулировать два различных процесса. На-
пример, у краба Uca циркадная частота играет преобладающую
роль в регуляции изменений окраски, а приливно-отливная часто-
та, соответствующая лунным суткам,— в регуляции спонтанной
двигательной активности. Вместе с тем при тщательном количест-
венном изучении обычно можно обнаружить, что оба периода ока-
зывают друг на друга .взаимно модулирующие влияния. Так, на за-
Время суток
Рис. 10-9. Спонтанная двигательная активность коста-риканского краба Uca
princeps во время месячного выдерживания в постоянной темноте [38].
Линиями из кружков отмечено время приливов на побережье, где были отловлены крабы.
Обратите внимание на два пика активности, наблюдавшиеся поздним утром с полумесяч-
ным интервалом. Запись повторена и смещена вверх на один день для того, чтобы можно
было легче наблюдать за переходом активности на следующие сутки.
писях спонтанной активности Sesarma [131] и Uca [9, 10] бывают
хорошо заметны оба ритма. Вертикальные миграции диатомовой
водоросли Hantzia, хронированные в соответствии с приливно-от-
ливными циклами, даже при постоянных условиях, вероятно, вза-
имодействуют с циркадным компонентом таким образом, что вы-
ражены только во время дневной фазы циркадного ритма [134].
Одновременное присутствие циркадного и приливно-отливного
ритмов приводит один или два раза в месяц к возникновению ко-
лебаний, обусловленных периодической интерференцией или бие-
ниями (рис. 10-9). У организмов приливно-отливной зоны, обита-
ющих в естественных условиях, эти колебания обусловлены также
прямыми реакциями на внешние физические воздействия, возника-
ющие один или два раза в месяц вследствие определенной комби-
нации приливно-отливных и суточных циклов. В течение синоди-
ческого месяца, средняя продолжительность которого равна
29,53 сут, систематически изменяется взаимное положение Луны
Биологические ритмы
233
и Солнца и в соответствии с этим меняется величина и сроки лун-
ного освещения ночью; это можно рассматривать как еще одно
повторяющееся ежемесячно изменение физических внешних ус-
ловий.
Полумесячные и месячные периодичности являются, пожалуй,
наиболее очевидными в случае регуляции генетически обусловлен-
ного типа репродуктивной функции и полового поведения. Имен-
но с помощью этих циклических процессов осуществляется син-
хронизация периода и фазы активности, связанной с размножени-
ем у морских организмов, например у водоросли Dictyota [59],
червя палоло и рыбы леурестес. Точная связь нерестовой активно-
сти с фазами Луны обнаружена также у японских морских лилий
[65]. У определенных видов хирономид и поденок наблюдается
ежемесячное роение [93, 126—128]. Менструация у некоторых
приматов происходит в новолуние, что может свидетельствовать о
связи менструального цикла с фазами Луны. Продолжительность
менструального цикла у человека приблизительно равна месяцу;
в некоторых недавних работах высказываются предположения о
глубокой связи этого цикла с синодическим месяцем [69, 121].
В естественных условиях продолжительность менструального цик-
ла человека может сильно отличаться от синодического месяца
(29,5 сут). Однако ночное освещение, включаемое с 14-х по 17-е
сутки после начала менструации, может индуцировать уменьше-
ние этих естественных колебаний и приблизить продолжитель-
ность индивидуальных менструальных циклов к общему уровню,
приблизительно равному 29,5 сут [69].
У некоторых организмов, подобных Dictyota, ритм размноже-
ния, фаза которого соответствует фазе Луны, по-видимому, опре-
деляется фазовым углом между временем прилива и отлива в
данной точке побережья. Таким образом, связанный с фазой Луны
день высвобождения гамет будет одним и тем же для всех осо-
“бей данного участка побережья, но окажется иным в другом рай-
оне, где приливы и отливы происходят в другие часы. Судя по
имеющимся данным, изменения ночного освещения служат фазо-
вым фактором для связанных с Луной циклов размножения ряда
видов: полихеты Platynereis, некоторых хирономид и, возможно,
некоторых диких приматов [98].
Выраженные биологические ритмы могут быть связаны с вза-
имодействиями Земля—Луна или Земля—Луна—Солнце. Ясно,
что и в том и в другом случае эти ритмы могут сохраняться как
свободнотекущие, если организмы изолированы от всех очевидных
ритмических колебаний соответствующей частоты.
Изучая многие организмы или биологические процессы, иссле-
дователи обнаружили, что при предполагаемом постоянстве внеш-
них условий сохраняются колебания, проявляющиеся раз в полме-
сяца или в месяц. Эти колебания наблюдаются в реакциях гуппи
£112, ИЗ] и планарий на свет, реакциях улиток и планарий на
234
Глава 10
очень 'слабые магнитные поля, реакциях планарий на очень сла-
бые поля у-излучения, в спонтанной активности мышей и хомяков,
выдерживаемых в контролируемом режиме 12С : 12Т, в скорости
обучения планарий, в величине активности и скорости ориентации
насекомых и даже в склонности к кровотечениям после хирургиче-
ских вмешательств у человека [2, 17, 26, 28, 41]. Спонтанная ак-
тивность пресноводных раков и крыс [44] изменяется в зависимо-
сти от часового угла Луны или от лунной фазы; эти изменения
наблюдаются даже при полной изоляции животных от всех свя-
занных с Луной внешних воздействий. Таким образом, существо-
вание лунных ритмов описано для многих организмов, однако зна-
чение этих ритмов остается до сих пор неизвестным.
Мы уже говорили, что изменения внешней среды, происходя-
щие в течение лунных суток и, вероятно, связанные с лунными
приливами и отливами в атмосфере, по-видимому, могут быть гра-
дуальными настройщиками фазы в свободнотекущих приливно-от-
ливных ритмах устриц и крабов. Сопоставимые данные получены
даже на млекопитающих — удалось показать возможность выра-
ботки у них свободнотекущих циркадных ритмов под влиянием
тонких атмосферных процессов, периодичность которых соответст-
вует лунным суткам [30, 46]. Описано приспособление циркадных
ритмов хомячков и крыс к частотам лунных суток; наиболее бла-
гоприятной является такая ситуация, когда эти ритмы по фазе со-
ответствуют циклическим изменениям внешних условий. У чело-
века, длительное время находящегося в постоянных условиях,
наблюдаются циклические изменения температуры тела, период
которых равен 24,8 ч; возможно, что мы имеем здесь дело со слу-
чайным совпадением [6]. Однако другие сообщения о свободноте-
кущих ритмах, наблюдаемых у человека, позволяют предполо-
жить, что периоды, соответствующие лунному приливно-отливному
циклу (24,7—24,9 ч), встречаются чаще, чем можно было бы ожи-
дать при случайных совпадениях.
Таким образом, связанные с лунными сутками биологические
ритмы, по-видимому, распространены достаточно широко; поиски
этих ритмов, проводимые с помощью адекватных методов, в боль-
шинстве случаев оказывались успешными. Возможно, что ритми-
ческие колебания, зависящие от связанных с лунными сутками
геофизических процессов, также распространены не менее широко
и носят универсальный характер подобно биологическим ритмам,
частота которых соответствует солнечным суткам и которые явля-
ются отражением биологических реакций на соответствующие
ритмические компоненты атмосферных процессов. Эти ритмы, как
и циркадные, имеют ограниченное число общих форм или их зер-
кальных отображений. Максимумы и минимумы вершинных цик-
лов, подобных описанным для активности крыс, интенсивности ме-
таболизма улиток, размыкания створок раковины у двустворчатых
моллюсков, имеют тенденцию к совпадению с верхней кульмина-
Биологические ритмы
235
цией Луны (рис. 10-10). В тех случаях, когда связанные с лун-
ными сутками циклы носят по существу двухвершинный характер
(активность крабов Uca, дыхание жука Tenebrio), их максимумы
или минимумы имеют тенденцию совпадать как с верхней, так и
с нижней кульминациями Луны. У некоторых видов, в том числе
мышей, дождевых червей и са-
ламандр, наблюдается слож-
ная картина. Она напоминает
ситуацию у крабов и Tenebrio,
с той только разницей, что
здесь чередующиеся компонен-
ты полусуточных приливно-от-
ливных циклов инвертированы.
Следовательно, все эти циклы,
по-видимому, можно свести к
полусуточным приливно-отлив-
ным колебаниям, причем каж-
дый из этих полусуточных ком-
понентов способен к независи-
мой инверсии.
Рис. 10-10. Формы кривых, соответ-
ствующих средним колебаниям, свя-
занным с лунными сутками [29].
А — Solanum; Б — Daucus; В — Lumbricus;
Г—Triturus; Д— Mus; Е — Fucus; Ж\—
Rattus (1955—1956); Ж2 и Ж — Rattus
(1959); 3 — Venus; И — Nassarius; R — Uca;
Л — Tenebrio. В К — верхняя кульминация
Луны. А, Б, В, Г, Е, И, К, Л — потребле-
ние кислорода; Д, Ж, 3 — спонтанная дви-
гательная активность.
Годичные и сидерические ритмы. Физические условия для все-
го живущего на Земле имеют еще один очень четкий ритмический
период — год с его сезонными изменениями высоты Солнца над
горизонтом, продолжительности дня, а также с соответствующими
колебаниями таких внешних факторов, как температура и освеще-
ние. Сверх того расстояние между Землей и Солнцем изменяется
в течение года приблизительно на 3%, имея минимальную вели-
чину в январе (перигелий) и максимальную в июле (афелий).
I одичные колебания биологических процессов имеют универсаль-
ный характер; они охватывают важные и характерные для вида
совокупности реакций, связанных с размножением и ростом, а так-
же миграции .и различные физиологические и морфологические из-
менения. Годичные колебания описаны даже для тех организмов,
которые населяют экваториальные области; там эти колебания
часто, хотя и не всегда, связаны с ритмическими изменениями та-
236
Глава 10
ких внешних факторов, как выпадение или отсутствие осадков в
течение года.
Показано, что годовые изменения фотопериода для многих ор-
ганизмов могут играть роль фактора, определяющего фазу еже-
годных генетических и адаптивных циклов [20, 53, 64, 78, 79, 88,
90, 143, 187]. Происходящее весной удлинение дня и укорочение
ночи вызывает у видов длинного дня переход к размножению.
У видов короткого дня, размножающихся осенью, аналогичную
роль играет укорочение дня. Существует хорошо обоснованная
гипотеза, согласно которой организмы дифференцируют различ-
ные длины фотопермода с помощью своей циркадной системы,
действующей как механизм отсчета времени. Изменения во внеш-
ней среде различаются здесь благодаря меняющемуся положению
рассвета и сумерек на протяжении циркадного цикла.
Температурные изменения, носящие менее правильный и менее
зависимый от времени года характер, изучены в данном аспекте
хуже, чем световой фактор; по-видимому, они обычно имеют мень-
шее значение в жизни животных и растений.
Годичные ритмы (и даже ритмы, имеющие относительно боль-
шую амплитуду) могут сохраняться при полном постоянстве всех
факторов, передающих очевидную информацию о времени года.
Напомним, что аналогичная устойчивость характерна и для рит-
мов, связанных с солнечными или лунными сутками. У ткачиков,
выдерживаемых в режиме 12С: 12Т при прочих постоянных внеш-
них условиях, было отмечено два более или менее нормальных
ежегодных цикла размножения [118]. У некоторых других видов
птиц, исследуемых при постоянных условиях, по-видимому, сохра-
нялся ритм, приблизительно соответствующий годичному; однако
последовательные циклы, повторяющиеся в последующие годы, на-
ступали на 2—3 мес раньше срока [85, 86]. Помимо этого, были
еще обнаружены неправильные колебания, продолжительность ко-
торых приблизительно соответствовала году.
Описанные устойчивые повторения видоспецифичных поведен-
ческих стереотипов могут отклоняться по продолжительности от
истинного годичного периода. В то же время известны некоторые
другие, вероятно менее специфические, биологические процессы,,
колебания которых точнее совпадают с годичным циклом. Так,
годичный цикл скорости прорастания семян сохраняет свою точ-
ность независимо от того, хранились ли семена при —22 или при
-]-45оС и находились ли они на воздухе или *в атмосфере азота; в
этих условиях представляется крайне маловероятным любое из-
мерение длины цикла, основанное на эндогенных метаболических
процессах [57]. Такие показатели, как скорость восстановления
азота водорослями, реакция на свет у дождевых червей и челове-
ка, реакция планарий на очень слабое у-излучение, интенсивность
потребления О2 прорастающими бобами и картофелем и потребле-
ния пищи североамериканским лесным сурком, систематически из-
Биологические ритмы
237
меняются в зависимости от времени года даже тогда, когда эти
организмы выдерживаются при постоянном уровне всех очевидных
внешних факторов [14, 31, 40, 66, 72, 108].
То, что перечисленные изменения интенсивности биологических
процессов имеют в среднем точную годичную периодичность, го-
ворит об их прямой зависимости от внешних условий, выражаю-
щейся в реакциях на трудноуловимые физические колебания в те-
чение года. В пользу подобного источника годичных ритмов сви-
детельствуют следующие данные: 1) многие годичные циклы име-
ют свой максимум в июле—августе и минимум в октябре—декаб-
ре; 2) изменения периодов освещения и температурных условий
не меняют фазы цикла потребления пищи у североамериканского
лесного сурка [66]; 3) интенсивность дыхания картофеля имеет
выраженный суточный цикл, форма и амплитуда которого отража-
ют действие двух параметров — средней ежесуточной температуры
воздуха, зависящей в свою очередь от погоды, и нарушений фор-,
мы атмосферных приливов и отливов. Показало [24], что интен-
сивность дыхания изменяется вслед за хорошо известными систе-
матическими годичными изменениями обоих названных парамет-
ров.
Еще один естественный период связан с вращением Земли —
это период ее вращения вокруг собственной оси, который состав-
ляет 24 ч 56 мин. Для практических целей можно считать, что
фазовые отношения солнечных и сидерических (звездных) суток
воспроизводятся точно один раз в год. Любое систематическое
ежегодное колебание в цикле солнечных суток (а мы уже гово-
рили о существовании таких колебаний) неотличимо от любого
существующего модулирующего влияния цикла звездных суток,
который отстает от цикла средних солнечных суток примерно на
4 мин в сутки. Тем не менее представляется важным то, что у на-
земных организмов, выдерживаемых при постоянном уровне всех
известных внешних факторов, удалось обнаружить очевидное ко-
лебание, связанное со звездными сутками [22]. Это наблюдение,
сделанное в опытах на картофеле, было подтверждено одиннад-
цатилетними непрерывными измерениями потребления О2 [34].
Средний цикл, диапазон колебаний которого составлял приблизи-
тельно 1%, имел в общем синусоидальную форму с максимумом
около 6-го и минимумом около 18-го часа звездного времени.
Верхняя кульминация 6-го часа звездного времени достигается в
полдень солнечных суток во время летнего солнцестояния, а верх-
няя кульминация 18-го часа звездного времени — в полдень сол-
нечных суток во время зимнего солнцестояния. Возможно, что на
эти биологические циклы влияет солнечная активность. Во вся-
ком случае, крайние величины рассматриваемого «звездного» ко-
лебания, прослеженного в течение одиннадцати лет, по-видимому,
были приурочены ко времени максимума и минимума солнечных
пятен.
238
Глава 10
Циклы продолжительностью в 18 и 18,6 лет. Некоторые другие
ритмы, связанные с геофизическими процессами, вероятно, явля-
ются результатом одновременного протекания рассмотренных вы-
ше периодических процессов. Многие беспозвоночные с внешним
оплодотворением, особенно полихеты, имеют кратковременный и
очень четко ограниченный ежегодный сезон размножения; выме-
тывание половых продуктов происходит синхронно в первую или
третью (или и в первую, и в третью) четверти Луны. Это выме-
тывание обычно приурочено, кроме того, к определенному часу су-
ток. Из всех, подобных случаев наиболее изучены циклы размно-
жеция у тихоокеанского червя палоло Eunice viridis [60] и морской
лилии Cotnanthus japonica [65]. В сезоне размножения этих видов
календарная дата выметывания гамет смещается в соответствии
с фазами Луны. Интервал между ближайшими годами с одинако-
выми календарными датами одноименных фаз Луны (цикл бие-
ния) равен циклу Метона, составляющему 18 лет. Практически
у названных двух видов наблюдается такой же интервал между
годами с совпадающей датой выметывания гамет. Однако невоз-
можно установить, коррелирует ли этот биологический цикл с цик-
лом Метона или с периодом «лунной нутации» (периодом обра-
щения плоскости орбиты Луны). Последний период приводит в
итоге к колебанию, при котором даты максимального лунного
склонения (в 1971 г. приблизительно соответствовавшие первой и
третьей четвертям Луны) разделены периодом в 18,6 лет.
Биологические ритмы, возвращение к «дому» и навигация. Жи-
вотные могут точно определять как время (или фазовый угол в
биологических ритмах — см. выше), так и географическое поло-
жение (широту и долготу), только ориентируясь по небу. Поэто-
му не удивительно, что исследования биологических ритмов, с од-
ной стороны, а также навигации и возвращения животных к «до-
му», с другой, должны были объединиться, сформировав одну
ветвь исследований со своими фундаментальными проблемами,
общими для обеих указанных ветвей [1, 129].
Около двадцати лет назад стало известно, что птицы могут
использовать Солнце как базис отсчета при определении геогра-
фического направления. Это сразу же указало на важность био-
логического «хронометра», определяющего солнечное время [111]-
Вскоре было обнаружено, что для этой цели может использовать-
ся циркадная ритмическая система. Оказалось, что не только пти-
цы, но и рыбы, черепахи, амфибии, насекомые, ракообразные и
пауки могут, используя 24-часовые ритмы, определять время по
Солнцу для того, чтобы получать надежную географическую ин-
формацию, которую приходится постоянно корректировать вслед-
ствие перемещения Земли относительно Солнца [81, 84, 94, 100,
115, 137, 138, 146, 150, 151, 171]. Система отсчета времени отлича-
ется фазовой лабильностью, типичной для циркадного ритма; в
экспериментальных условиях, используя циклы СТ, можно вызвать
Биологические ритмы
239
фазовый сдвиг этой системы, что соответствующим образом на-
рушит у животных, ориентирующихся по Солнцу, способность оп-
ределять направление (рис. 10-11).
Позднее было показано, что бокоплавы — ракообразные, миг-
рирующие по направлению к кромке воды и от нее,— используют
в качестве базиса отсчета при определении географического на-
Рис. 10-11. Повторное включение навигационных «часов» [21].
Верхний ряд — последний опыт перед повторным включением. Рыбы А, Б и В обучены плыть
соответственно на север, юг и запад; ожидаемое направление их движения отмечено тонкой
стрелкой. Нижний ряд — с 7-х по 13-е сутки после повторного включения. Толстой стрелкой
отмечено направление по компасу, в котором рыба должна была бы плыть при условии,
что азимут Солнца изменяется со скоростью 15° в 1 ч и что рыба компенсирует это измене-
ние, смещая направление своего движения на 15° в 1 ч в противоположную сторону. Каж-
дая точка соответствует критическому выбору. Между А, Б, В и А', Б', В' имел место сдвиг
светового периода, равный 14 ч 10 мин.
правления Луну [135, 136]. Естественно, что бокоплавам необхо-
дим биологический «хронометр», позволяющий определять время
лунных суток. При этом используется не только скорость видимо-
го движения Луны по небу в какую-либо отдельную ночь, но и
систематически изменяющееся от ночи к ночи положение Луны,
определяемое )в одно и то же время суток; 'средняя 'продолжитель-
ность полного цикла этих изменений составляет 29,5 сут. Биоло-
гический хронометр необходим также и для того, чтобы измерять
продолжительность синодического месяца.
'Славки во время своих перелетов способны получать всю не-
обходимую информацию о направлении полета и своем местопо-
ложении, ориентируясь по звездам. Этот вывод сделан на основа-
нии опытов, поставленных в планетарии [149]. Положение звезд
на ночном небе (за исключением Полярной звезды в небе Север-
ного полушария) изменяется в зависимости от времени суток.
240
Глава 10
Кроме того, картина звездного неба оказывается разной в зави-
симости от времени года и от географического положения наблю-
дателя. Таким образом, птицы могут ориентироваться по звездам
при возвращении к «дому» или для навигации лишь в том слу-
чае, если они обладают звездным «хронометром» и способны,
кроме того, к точному различению времен года. В планетарии
изучалась также ориентация овсянок [75, 76].
Ясно, что использование небесных тел для выбора направле-
ния и для ориентации на местности становится возможным благо-
даря работе исключительно сложной, точной и надежной системы
определения календарного и суточного времени. Кроме того, для
этого необходима хорошая погода, позволяющая наблюдать за не-
бесными телами. Некоторое усовершенствование этой системы об-
наружено у многих животных, в том числе у рыб и насекомых,
которым для ориентировки бывает достаточно увидеть кусочек го-
лубого неба [172]. Эти животные, наблюдая отдельный участок
неба, способны по структуре поляризованного света установить
точное положение Солнца, недоступного для прямого наблюдения.
С другой стороны, уже давно известно, что хорошая погода не
является обязательным условием возвращения к гнезду или нави-
гации даже для животных, находящихся на чужой территории и
не знакомых с местными ориентирами. Интенсивное изучение пе-
релетных птиц, проведенное недавно методами радарного слеже-
ния и радиотелеметрии, показало, что нормальное направление
перелета может выдерживаться при сплошной облачности и не-
возможности видеть Землю. Почтовые голуби так же легко воз-
вращаются к гнезду, даже если им приходится лететь в облачную
погоду над чужой территорией [106, 170]. Кроме того, было об-
наружено, что, каковы бы ни были в последнем случае факторы,
используемые при ориентации, прямое участие циркадной системы
здесь исключено. С помощью экспериментальных циклов СТ мож-
но изменить фазу циркадной ритмической системы, но это не по-
мешает голубю найти дорогу домой. Птица, несомненно, обладает
какими-то средствами, заменяющими географический компас и
позволяющими получить информацию о направлении к гнезду;
эти средства основаны на использовании тонких геофизических
полей. Следовательно, как биологическая способность к ориента-
ции, навигации и определению географического положения, так и
биологические ритмы с их сопутствующей явной способностью оп-
ределять точки геофизических временных циклов, представляются
зависящими, хотя бы отчасти, от восприятия тонких геофизических
параметров. В земной атмосфере все эти параметры непрерывно
изменяются в четырех измерениях пространства и времени. Ори-
ентация в пространстве — столь же универсальное свойство на-
земных животных, как и их постоянное изменение во времени.
Пространство, время и реакция на геоэлектромагнитные поля.
В течение последних десяти лет появились многочисленные экс-
Биологические ритмы
241
периментальные исследования, 'посвященные как механизмам на-
стройки биологических ритмов во времени, так и средствам, ис-
пользуемым животными при возвращении к «дому» и навигации.
Исследователи, занимающиеся биологическими ритмами, сталки-
вались с тем, что полученные результаты всегда можно было ин-
терпретировать двояким образом и ни одну из этих двух возмож-
ностей нельзя было исключить. Если какое-либо повторяющееся
биологическое явление имеет период, совпадающий с естествен-
ным периодом окружающей среды или близкий к нему, то быва-
ет невозможно определить, зависит ли периодичность этого явле-
ния от ритмически изменяющихся внешних условий или не зави-
сит. Допустим, что, подвергая экспериментальным воздействиям
один из естественных параметров внешней среды, удалось изме-
нить фазу, амплитуду или период биологического ритма. При
этом нельзя будет различить, чем обусловлен полученный эф-
фект— модифицирующим, синхронизирующим или каким-либо
другим влиянием на эндогенную систему, способную давать нор-
мальные ритмические колебания даже при полном отсутствии по-
ля рассматриваемого внешнего параметра, или чем-то совершенно
иным. Очень важен для проблемы ритмов вопрос, способен ли ор-
ганизм определять точки фазовых углов на протяжении временно-
го геофизического цикла. Если это так, то возникает второй серь-
езный вопрос: может ли организм использовать такую организо-
ванную во времени информацию для создания собственной
системы отсчета, позволяющей упорядочить физиологические и по-
веденческие реакции? При этом речь идет либо о генетически обу-
словленных реакциях, либо о реакциях, изменяющихся вследствие
воздействия внешних условий, либо о тех и других вместе.
Очевидно, что изменяющиеся во времени тонкие геофизические
параметры постоянно изменяются и в трех измерениях простран-
ства. При помощи экспериментального исследования пространст-
венных изменений, вероятно, можно было бы решить и вопрос об
изменениях во времени. Проблему можно было бы упростить еще
больше, ограничив изучение пространственных изменений только
одним измерением — в горизонтальной плоскости.
Опыты по подобной упрощенной схеме были впервые проведе-
ны на улитках и планариях [33]. Было показано, что: 1) в любой
Данный момент времени организм способен различать географи-
ческие направления, руководствуясь при этом лишь тонкими гео-
физическими параметрами; 2) в случае любого географического
направления реакция организма изменяется во времени в ответ
на главные геофизические частоты солнечных суток, лунных пе-
риодов и годичных циклов.
Использование этой упрощенной системы позволило обнару-
жить, что организмы необычайно чувствительны к силе и геогра-
фическим изменениям направления горизонтального вектора очень
слабых магнитных полей [26]. Максимальная способность воспри-
16—1514
242
Глава 10
нимать экспериментальные изменения этих параметров соответст-
вует таким силам магнитного поля, которые близки к естествен-
ным величинам. При экспериментальном изменении силы магнит-
ного поля организму необходимо не менее 0,5 часа для видимого
Фаза луны
Рис. 10-12. Связанные с фазой Луны колебания у планарий, первоначально ориен-
тированных к северу (0°) и находящихся в световом поле от двух источников све-
та, расположенных с южной и восточной стороны; продолжительность опытов —
несколько месяцев.
Средние траектории отдельных планарий представлены сплошной линией. Пунктиром изо-
бражены средние траектории этих же особей, повторно определенные после того, как на-
правление горизонтального вектора магнитного поля было изменено на 180° по отношению'
к световому полю. Поворот осуществлен вращением всей экспериментальной установки,
в том числе световых источников, на 180° по отношению к географическим координатам (А)
или приложением обратного магнитного вектора 0,05 гаусс с помощью стержневого магнита,,
расположенного соответствующим образом под установкой (Б). ПЛ — полнолуние, НЛ— но-
волуние. (Canadian Journal of Botany, M, 287—298, 1969).
вектор магнетизма связан с ритмической системой иначе (рис.
10-12). Если планарии ориентированы к северу и источник света
находится справа от них, то величина их отрицательного фототак-
сиса обнаруживает четкие месячные колебания с максимумом в
новолуние и минимумом в полнолуние. Эти месячные колебания
можно мгновенно сдвинуть по фазе на 180°, используя один из
двух следующих методических приемов: 1) повернуть всю экспе-
риментальную установку на 180°; планарии при этом ориентиро-
ваны к югу, а источник света находится с западной стороны;
2) произвести эквивалентное изменение магнетизма путем ревер-
Биологические ритмы
243
«сии магнитного поля слабо намагниченным стержнем; магнитная
индукция обращенного поля может не превышать 0,04 гаусс
[32].
В других опытах обнаружили способность планарий определять
наличие и различать географическое направление горизонтальных
электростатических градиентов, не превышающих 0,1 мкВ/см
«[27]. Реакция планарий на одно и то же направленное поле ме-
няется в зависимости от времени суток.
Планарии определяют силу и даже векторные направления
полей у-излучения, в 2—25 раз превышающего фоновый уровень;
горизонтальные источники этого излучения помещали слева и
справа от ‘воспринимающей системы [28]. При этом также обна-
ружили закономерные циклические изменения чувствительности
.животных, соответствующие естественным геофизическим циклам,
в том числе даже годичному циклу.
В ряде р^бот [122, 123, 184, 184а] было показано, что мали-
новка, птица, проявляющая весной и осенью очень высокую миг-
рационную активность, способна определять нужное направление
миграции. Эта способность сохраняется даже у птиц, находящих-
ся в клетках в лабораторных условиях и лишенных контакта со
всеми видимыми указателями направления (рис. 10-13). Помещая
птиц в обратное геомагнитное поле, созданное с помощью кату-
шек Гельмгольца, можно было вызвать реверсию направления
миграции. Если это экспериментальное поле сильнее или слабее
нормального геомагнитного поля хотя бы приблизительно на 50%,
то птицы могут использовать его для ориентации лишь после 2—
3 дней приспособления к новым величинам магнитного поля.
Очень слабые магнитные поля, по-видимому, могут вызывать ори-
ентационные реакции и у других организмов [12, 15, 116, 130, 152,
177].
Недавно было показано [70, 103], что некоторые пластиножа-
берные рыбы необычайно чувствительны к электрическим полям,
возникающим за счет потенциалов действия, генерируемых в теле
замаскировавшейся потенциальной добычи. Эта чувствительность
пластиножаберных, воспринимающих до 0,01 мкВ/см, имеет непо-
средственное отношение к общей проблеме высокой чувствитель-
ности живых систем к естественным электромагнитным полям и
их изменениям. Согласно имеющимся данным, электрические по-
Дя каким-то образом связаны с периодом и фазой циркадных рит-
мов [71, 180—182] и, может быть, с навигацией угрей в океане
1148а].
Очевидные раздражители и реакция на тонкие электромагнит-
ные поля. Живая система способна связывать свои реакции на
обычные раздражители с одновременно поступающей информаци-
ей, относящейся к тонким электромагнитным полям. Эту способ-
ность демонстрируют наблюдения, согласно которым организмы
знают по опыту о существовании своего «дома», а будучи перене-
16*
Рис. 10-13.
А. Среднее направление, выбранное малиновкой, находящейся в состоянии миграционной
активности (нормальный выбор — ЮЗ). Опыт проводили ночью; птица выдерживалась в маг-
нитном поле 0,41 гаусс и испытывалась в магнитном поле 0,14 гаусс. Стрелкой в центре
обозначена средняя тенденция выбора направления. Б. Птица выдерживалась и испытыва-
лась при 0,14 гаусс. В. Птица выдерживалась и испытывалась в искусственном поле
0,30 гаусс, повернутом в указанном направлении. Г. Птица выдерживалась при 0,14 гаусс
и испытывалась в искусственном поле 0,41 гаусс, повернутом в указанном направлении [123].
Биологические ритмы
245’
сены в новое, совершенно неизвестное для них место, возвраща-
ются домой даже в условиях сплошной облачности, скрывающей*
все обычные небесные ориентиры, причем скорость возвращения
значительно превышает ту, которую можно было бы ожидать
при случайном обнаружении знакомого ландшафта поблизости от
дома. Подобное поведение было обнаружено, например, у почто-
вых голубей.
У голубя удалось выработать условную реакцию на очень сла-
бые магнитные поля; эта реакция выражается в изменении часто-
ты сердцебиений [145]. Показано, что молодые чайки имеют вы-
раженную тенденцию перемещаться в направлении их обычной
осенней миграции; это движение ослабевает, исчезает или даже-
приобретает обратное направление в зависимости от усиления
происходящих одновременно возмущений геомагнитного поля-
увеличения силы поля) [156]. Такие данные весьма убедительно*
свидетельствуют о связи между генетически закрепленными струк-
турами поведения птиц и векторами земного магнитного поля.
Описана способность планарий связывать освещенность окру-
жающей среды с географической сетью, т. е. с естественным элек-
тромагнитным полем Земли в данном районе (рис. 10-14) [42].
При этом световое воздействие не только определяет отчасти ре-
акцию животного, но вызывает также стойкие изменения, кото-
рые до некоторой степени определяют реакцию на другие свето-
вые воздействия, включаемые несколькими минутами позднее.
Гипотезы биологических часов. Таким образом, биологические
ритмы сохраняются даже тогда, когда живые системы полностью-
изолированы от любого внешнего периодического процесса, на ко-
торый они, насколько нам известно, способны реагировать. Отно-
сительно стабильные и надежные биологические периоды сохра-
няются на фоне температурных сдвигов и изменений химического*
состава среды. При рассмотрении этих свойств живой системы
напрашивается аналогия с часами. Организмы, по-видимому,
обладают каким-то высокоспециализированным эквивалентом систе-
мы измерения времени, относительно точно определяющей продол-
жительность естественных геофизических периодов. Была выдви-
нута концепция биологических часов, определяющих периоды раз-
личных биологических ритмов. Биохимические и физиологические-
компоненты биологических ритмов включают в себя множество од-
новременных и последовательных реакций, но ни одна из этих ре-
акций сама по себе, вероятно, не обладает необходимыми свойства-
ми отметчика времени. Однако в совокупности эти реакции, по-ви-
димому, организованы во времени так, что приобретают свойства
сложной системы измерения времени.
Уже с момента первого обнаружения биологических ритмов
закономерно возник следующий основной вопрос: генерируются ли
эти ритмы в организме как автономная колебательная система,
независимая от всех внешних физических ритмов соответствую-
246
Глава 10
щих частот, или же эти свободнотекущие периодичности, облада-
ющие свойствами биологических часов, зависят от непрерывного
поступления воздействий неконтролируемых внешних ритмов?
Первая из альтернативных возможностей известна под названием
^гипотезы эндогенных часов, а вторую именуют гипотезой экзоген-
Рис. 10-14.
.А. Среднее месячное колебание интенсивности поворота планарий от источника света, рас-
положенного справа. Светлые кружки — результаты опытов, проведенных при исходной
• ориентации планарий на север; черные кружки — результаты опытов, проведенных сразу
после поворота установки на 180°, т. е. переориентации планарий на юг. Б. То же самое при
исходной ориентации планарий на юг (черные кружки) и поворота установки на 180° на
север (светлые кружки). НЛ — новолуние, ПЛ — полнолуние [42].
ных часов. В настоящее в<ремя мы не можем отвергнуть ни одной
из этих двух гипотез [38, 168].
Гипотеза эндогенных часов. В логическом отношении эта гипо-
теза наиболее проста. В то же время она постулирует существо-
вание такой временной точности и стабильности, для обоснования
которых пока не обнаружено удовлетворительной биологической
схемы. С точки зрения этой гипотезы можно объяснить многие из
Биологические ритмы
247
установленных свойств свободнотекущих ритмов, а большую часть
других свойств можно отнести за счет способности реагировать на
неконтролируемые тонкие геофизические ритмы, способные при
определенных обстоятельствах действовать как фазовые раздра-
жители, определяющие фазовую настройку ритмической системы,
периоды которой генерируются эндогенными часами. Доводами в
пользу гипотезы эндогенных часов считают следующие свойства
биологических ритмических систем: 1) ритм сохраняется даже тог-
да, когда максимально возможная часть внешних параметров ос-
тается на постоянном уровне; 2) часто оказывается возможным
экспериментально сдвинуть фазу биологического ритма, который
после этого приобретает новую фазу, но продолжается с прежней
периодичностью; 3) периодичность наблюдаемого биологического
ритма может отклоняться от периодичности соответствующего гео-
физического ритма; 4) иногда возможен запуск ритма отдельным
кратковременным стимулом; 5) вещества, подавляющие или тор-
мозящие метаболизм, иногда вызывают фазовую задержку ритма;
6) период свободнотекущего ритма может изменяться в зависимо-
сти от величин температуры или освещения или от концентрации
D2O; 7) ритм может остаться неизменным после быстрого перено-
са организма к востоку или западу и даже на южный географиче-
ский полюс.
Полагают, что периоды биологических часов и наблюдаемые
периоды биологических ритмов совпадают и что любые наблюдае-
мые изменения фазы или периода биологического ритма отража-
ют соответствующие изменения работы биологических часов. Ве-
роятно, в организме существует множество биологических часов,
и это объясняет все случаи диссоциации периодов или фаз био-
логических ритмов, а также сохранение ритмов в изолированных
частях организма.
Были предприняты многие попытки, основанные на рабочей ги-
потезе эндогенных часов, для обнаружения клеточного или биохи-
мического механизма, который имел бы все необходимые харак-
теристики часового механизма, включая независимость от темпе-
ратуры. В некоторых работах была предложена конкретная схема
эндогенных часов, но лишь в одном случае (концепция хронона)
эта схема основывалась на представлениях молекулярной биоло-
гии (рис. 10-15) [74]. Предполагают, что существуют очень длин-
ные фрагменты ДНК, на протяжении которых скорость генетиче-
ской транскрипции регулируется целым комплексом соответству-
ющих клеточных факторов так, что вся система в целом может \
играть роль часов. Однако в большинстве случаев исследования
природы предполагаемых эндогенных часов ограничены описани-
ем ритмических систем как теоретических колебательных моделей
и выяснением степени соответствия между результатами экспери-
ментального нарушения биоритмов и предсказаниями, сделанны-
ми на основе этих моделей [141, 142, 178, 179, 185, 186].
248
Глава 10
Ритмы, зависящие от геофизических процессов, т. е. стойкие
средние колебания, частота которых соответствует частоте геофи-
зических процессов, рассматриваются с точки зрения гипотезы эн-
догенных часов как эндогенные ритмические компоненты, синхрон-
ные по фазе и периоду тонким геофизическим ритмам. Несмотря
на существование таких биологических ритмических компонентов,
^==ДНК
<= Цистрон (или оперон)
е
'РНК-матрица (мРНК)
Рибосома
Полипептид
Нерасшифрован
ные процессы
0
§
I
'г'
Рис. 10-15. Гипотеза хронона [74].
Синтез мРНК начинается с одного конца постулированного полицистронного комплекса
ДНК. Синтезированная мРНК диффундирует к рибосомам, программируя белковый синтез;
при этом происходит обратная диффузия фактора-инициатора, действующего на следующие
цистроны. Для транскрипции всего комплекса необходимо около 24 ч, после чего начинается
новый цикл.
§
I
I
3
^йсутренняя
J область
Внешняя область
Образование конеч-
ных продуктов;-
борка полимеров
периоды которых точно соответствуют геофизическим периодам,
эти же самые тонкие геофизические факторы нельзя считать эф-
фективными датчиками времени для других адаптивных ритми-
ческих структур, периодичность которых определяется автономны-
ми эндогенными часами. Полагают, что выработка этих адаптив-
ных ритмов произошла в результате повседневной фазовой
коррекции более сильными датчиками времени (например, светом)
при поддержании обычно наблюдаемых в природе естественных
геофизических частот.
Гипотеза экзогенных часов. Преимущество этой логически бо-
лее сложной гипотезы состоит в более простом объяснении исклю-
чительной стабильности ритмов при температурных и химических
сдвигах в организме. Полагают, что эти ритмы не связаны с ав-
тономными системами отметки времени [23—25, 29, 32]. Биоло-
Биологические ритмы 249
гические часы зависят от непрерывного «ввода» ритмов извне; с
этой точки зрения объясняют все известные свойства свободноте-
кущих ритмов. Главное отличие этой гипотезы от гипотезы эндо-
генных часов состоит в том, что в случае экзогенного обусловлива-
ния во времени ритмов период наблюдаемых ритмов может не-
сколько отличаться от периода часов, определяющих время
ритмов. Часы, период и фаза которых, согласно рассматриваемой
гипотезе, определяются извне, точно выдерживают среднюю гео-
Сеет,
температура,
пища ит.в^
Внешняя среда
(солнечные, лунные,
годичные ритмы)
/Регулиру-^
ъмые^~^--физ1
логические.
Ритмы,
зависящие
от геофи-
зических
процессов
Магнитное поле,
электростатическое
поле, поле f-излучения
и т.д.
Ритмически
ИЗМЕНЯЮЩИЙСЯ
ОРГАНИЗМ
Рис. 10-16. Гипотеза экзогенных часов (Brown F. A., Jr., Adv. Astronaut. Sci., 17,
29—39, 1964).
Постулируется, что организм имеет скрытый ритмический комплекс, зависящий от геофизи-
ческих процессов и возникающий вследствие прямой реакции на геофизические ритмы тон-
ких параметров. Полагают, что именно этот скрытый комплекс служит носителем главных
свойств часов, в частности относительной стабильности периодов у соответствующих биоло-
гических циклов, обусловленных генетически или вызванных внешними воздействиями (при-
способительных физиологических) и имеющих частоты, равные частотам геофизических рит-
мов или близкие к ним. Наблюдаемые частоты этих регулируемых биологических ритмов
при постоянных условиях могут несколько отклоняться от частот геофизических ритмов,
что выражается в длительных фазовых сдвигах (задержке или опережении).
физическую периодичность; стабильность их периода поэтому на-
поминает стабильность периодических процессов, определяемых
геофизическими факторами (рис. 10-16). Считают, что в естест-
венных условиях периоды биологических ритмов, генетически за-
крепленных или вызванных внешними воздействиями (т. е. цир-
кадных ритмов или ритмов, приблизительно соответствующих лун-
ному или годичному циклам), синхронизированы с соответствую-
щими биологическими часами и с ритмами таких важных внешних
факторов, как свет или температура.
Рассмотрим случай циркадного ритма, когда все силы, могу-
щие быть эффективными датчиками времени для организма, под-
держиваются на постоянном уровне. Согласно гипотезе экзоген-
ных часов, нормальная система фазовой настройки организма,
обычно работающая в ответ на периодический датчик времени, в'
этом случае вызывает слабый фазовый сдвиг, автофазировку, воз-
никновение последовательных циклов в результате взаимодейст-
вия между постоянным параметром отметчика времени и цикли-
250
Глава 10
чески изменяющейся чувствительностью организма к данному дат-
чику [23, 35]. Такой периодический фазовый сдвиг приводит к
'возникновению нового циклического периода, неотличимого от
правильных колебаний с той же периодичностью. Полагают, что
у разных особей одного вида существуют индивидуальные особен-
ности периода колебаний, заключающиеся в различиях формы и
амплитуды кривых реакций; такие отличия были действительно
обнаружены в недавних исследованиях. Эти индивидуальные раз-
личия могут быть результатом как генетических модификаций,
так и колебаний внешних условий. Постулируемый метод возник-
новения периодов, отклоняющихся от естественных геофизических
периодов, позволил бы в то же время ожидать, что величина этих
отклонений может несколько варьировать в зависимости от изме-
нений уровня любого постоянного параметра датчика времени,
взаимодействующего с рассматриваемой системой фазовой реак-
ции. Действительно, изменения уровня двух наиболее известных
датчиков времени — света и температуры — сопровождаются не-
значительными изменениями периода свободнотекущего ритма.
„Действие этих двух датчиков сходно и в количественном, и в ка-
чественном отношениях. Поэтому можно предположить, что сла-
бое влияние температуры на период свободнотекущего ритма в
некоторой степени осуществляется благодаря ее способности ре-
гулировать фазу, а не благодаря неспецифическому кинетическо-
му тепловому эффекту. Аналогичным образом и влияние D2O как
фактора экзогенного хронирования опосредовано, вероятно, через
механизм фазового сдвига.
Полагают также, что циркадные ритмы практически безгра-
нично подвижны в своей зависимости от лежащих в их основе ча-
сов (подобно тому как ведут себя стрелки обычных часов по от-
ношению к часовому механизму) и что в нормальных условиях
датчик времени и другие очевидные факторы-раздражители уп-
равляют фазой и структурой ритма. Другими словами, ритмиче-
ские структуры не имеют явно предпочитаемой фазовой зависимо-
сти от лежащих в их основе часов и обычно не отражают влия-
ния какого-либо важного датчика времени из числа тонких внеш-
них параметров.
Гипотеза экзогенных часов не встречает трудностей при рас-
смотрении проблемы одновременного наличия периодичностей, со-
ответствующих солнечным и лунным суткам и не взаимодейст-
вующих между собой; не противоречит этой гипотезе и стойкость
месячных и годичных свободнотекущих ритмов, для которых по-
стулируется количественно увеличивающаяся степень прямой свя-
зи между внешними факторами и явлениями, происходящими в
организме.
Гипотеза эндогенно-экзогенных часов. Две только что рассмот-
ренные гипотезы оказались очень плодотворными в том отноше-
нии, что они стимулировали постановку экспериментов, расширив-
Биологические ритмы
251’
ших наши представления о различных характеристиках ритмиче-
ских процессов и о зависимости последних от двух групп внеш-
них стимулов. К первой группе относятся очевидные стимулы^
которые отражают важные для выживания организма внешние воз-
действия, 'ко второй — тонкие геофизические поля, влияние кото-
рых труднее распознать и труднее расшифровать. В то время как
крайности обеих гипотез, т. е. гипотез исключительно внутренне-
го или исключительно .внешнего хронирования, представлялись со-
вершенно несовместимыми, в них имелось нечто общее, лежащее-
за пределами простого описания ритмов и поведения этих ритмов.
Прогресс, достигнутый при разработке обеих этих точек зрения,,
оказался, однако, настолько существенным, что стало ясно: при
объяснении природы биологических часов и ритмов невозможно
исключить ни одну из двух рассматриваемых гипотез.
Представляется наиболее вероятным, что неуловимый меха-
низм вездесущих биологических ритмов свойствен тем системам
организма, которые сами по себе обладают генетически закреп-
ленной потенциально ритмической природой. Эти системы имеют:
унаследованные поведенческие программы, позволяющие каждо-
му организму приспособиться к общим и специфическим особен-
ностям ритмических характеристик их физической и биологиче-
ской среды [35]. Используя такие программы, представляющие
собой упорядоченные последовательности биохимических и физио-
логических явлений, организм имеет эндогенные возможности для
возникновения биологических ритмов. Эти ритмы могут обладать,
любым периодом или комбинацией периодов и иметь в основе раз-
личные структуры, приспособленные к различным циклическим
временным разверткам и к систематически изменяющимся фазо-
вым отношениям на протяжении таких сходных периодов, как
солнечные или лунные сутки. На основании современных данных
биохимии и физиологии можно считать, что эта циклизация мо-
жет происходить автономно, но иметь периоды, которые должны
в значительной степени отражать различия температурных уров-
ней и влияние различных химических и физических изменений
внутренней и внешней среды. Если бы такие системы работали
сами по себе, то можно было бы ожидать нерегулярных циклов с
колеблющимися длинами периодов. Биологические часы с их пра-
вильной периодичностью и независимостью от температуры и дру-
гих внешних факторов должны были бы отсутствовать полностью*
или частично, в зависимости от степени развития у организма об-
щей регуляции температуры или других физиологических или по-
веденческих механизмов гомеостаза.
Полагают, что наблюдаемые свойства биологических ритми-
ческих систем обусловлены действием автономных эндогенных ме-
ханизмов и ритмическими изменениями тонких физических полей
внешней среды. Хорошо известна высокоспециализированная чув-
ствительность организмов к этим внешним ритмам. Считают, что.
•252
Глава 10
эти ритмы ведут к выработке генетически закрепленных цикличе-
ских метаболических процессов — суточных, лунных и годичных;
на этих процессах основана работа биологических часов, опреде-
ляющих биологические ритмы и обеспечивающих регулярность и
•стабильность обмена веществ при действии разнообразных воз-
мущающих раздражителей.
Наряду с этим экзогенные факторы также могут снабжать ор-
ганизмы информацией о фазовых углах как во времени, так и в
пространстве, поскольку тонкие электромагнитные поля изменя-
ются в четырех измерениях. Возможна биологическая идентифи-
кация точек и векторных направлений во времени и пространстве;
возможно также выявить жизненно важные адаптивные реакции,
поскольку эта информация нужна для фазовой настройки некото-
рых систем организма или для узнавания направления к дому (а
может быть, даже и самого дома). Птицы способны находить до-
рогу домой либо по небесным световым ориентирам с использо-
ванием биологических часов, либо посредством восприятия тонких
полей; можно рассматривать эту способность как простую утили-
зацию различных участков в диапазоне континуума «время—про-
странство». Организм со всеми его биологическими «часами»,
«компасами» и «секстантами» должен с максимальной эффектив-
ностью использовать внешнюю информацию от всех потенциаль-
ных источников.
Заключение. Для всех живых существ — растений, животных
и микроорганизмов,— по-видимому, характерны периодические ко-
лебания, частоты которых соответствуют геофизическим ритмам.
В естественных условиях эти колебания обычно синхронны суточ-
ным, лунным приливно-отливным или месячным и годичным коле-
баниям физической внешней среды. Эти биологические флуктуа-
ции 1) отражают реакцию организмов на изменения очевидных
внешних стимулов, 2) включают в себя генетически обусловлен-
ные и изменяющиеся в зависимости от внешних воздействий по-
вторяющиеся поведенческие схемы, оптимально настроенные на
внешние циклы, и 3) отражают изменения в организмах, завися-
щие от непрерывных реакций на периодически изменяющиеся
внешние тонкие поля, как, например, электромагнитные.
Организм можно защитить от колебаний всех очевидных внеш-
них раздражителей, в норме вызывающих непосредственные реак-
ции, и от всех ритмических вариаций, действующих как определи-
тели фазы или датчики времени, т. е. настраивающих фазы адап-
тивных повторяющихся структур в соответствии с их характерной
зависимостью от внешних циклов. В этом случае биологиче-
ские ритмы могут стать свободнотекущими и приобрести перио-
дичность, слабо отличающуюся от таковой у естественных геофи-
зических ритмов. В то же время периодичности, являющиеся от-
ветами на тонкие геофизические поля, сохраняют свои точные
средние частоты.
Биологические ритмы
253
Биологические ритмические изменения, по-видимому, могут
служить хронометрами для измерения относительной продолжи-
тельности светлого и темного периода суток и тем самым для
адаптивной фазовой настройки годичных биологических циклов в
соответствии с временем года. Иногда эти изменения могут ис-
пользоваться как часы, работающие в связи с Солнцем, Луной и,
возможно, даже со звездами, для возвращения домой и для нави-
гации.
Недавно было показано, что организмы, выдерживаемые в поч-
ти постоянных условиях, продолжают отвечать на флуктуирующие
параметры неконтролируемых тонких геофизических полей, даю-
щие информацию о вариациях фазовых углов во всех естествен-
ных геофизических циклах. Более того, оказалось, что некоторые
организмы способны получать информацию о географическом на-
правлении непосредственно с помощью самих тонких геофизиче-
ских полей. Изучение природы тонких временных и пространст-
венных сигналов, передающих организму эту информацию, пока-
зало, что живые существа обладают чрезвычайно специализиро-
ванной чувствительностью к очень слабым электрическим,
магнитным и электромагнитным полям Земли и что эта чувстви-
тельность связана с механизмами, посредством которых организ-
мы нормально ориентируются в пространстве и вырабатывают
геофизически обусловленные ритмы. Существуют доказательства
того, что организмы могут наследовать специфичность ответной
реакции на геофизические поля подобно тому, как наследуется
характер ответа на такие факторы, как свет. Кроме того, организ-
мы могут «обучаться» и сохранять на какое-то время связи между
внешними стимулами и пространственно-временными отношения-
ми тонких геофизических полей.
Генетически закрепленные и модифицируемые внешними воз-
действиями схемы поведения, особенно те из них, которые в ес-
тественных условиях синхронны внешним циклам, но при постоян-
ном уровне всех датчиков времени могут стать «свободнотекущи-
ми», до некоторой степени хронируются вполне автономными,
температурно-компенсированными колебаниями в организме. Пе-
риоды этих колебаний лишь приблизительно соответствуют геофи-
зическим периодам и в естественных условиях подвергаются не-
прерывной фазовой коррекции под влиянием датчиков времени
(гипотеза эндогенных часов). Ряд авторов считают, что схемы по-
ведения хронируются неконтролируемыми и всепроникающими
ритмическими геофизическими колебаниями (гипотеза экзогенных
часов). Различные «свободнотекущие» частоты возникают при
этом в результате взаимодействия присущего организмам естест-
венного механизма фазового сдвига с неизменяющимися фактора-
ми датчика времени; это взаимодействие приводит к внутриорга-
низменному фазовому сдвигу (автофазировке или повторному са-
мовключению).
254
Глава 10
Предложена гипотеза, согласно которой в основе большинства
обычных и экстраординарных свойств биологических ритмов ле-
жит взаимодействие эндогенных и экзогенных компонентов. Счи-
тают, что эндогенные ча-сы участвуют в работе генетически закре-
пленных и подвергающихся действию внешних факторов цикличе-
ских систем; эти системы зависят также от последовательностей
биохимических и физиологических процессов и от присущей орга-
низму способности синхронизироваться с внешними физическими
ритмами. Экзогенные часы обеспечивают правильность ритмов и
необходимую стабильность периодов, несмотря на температурные
изменения и на большинство метаболических нарушений. Экзо-
генные часы дают также постоянно сохраняющуюся и весьма на-
дежную точку отсчета, связанную с небесными ориентирами и ис-
пользуемую для разнообразных способов ориентации организма
во времени и пространстве.
СПИСОК ЛИТЕРАТУРЫ
1. Adler Н. Е„ Animal Behavior, И, 566—577, 1963. General problems in animal]
migration.
2. Andrews E. J., J. Florida Med. Assoc., 46, 1362—1366, 1960. Monthly va-
riations in postoperative hemorrhaging.
3. Andrews R. V., Folk G. E., Comp. Biochem. Physiol., 11, 393—490, 1964.
Circadian rhythm in cultured hamster adrenals.
4. Aschoff J., Cold Spring Harbor Symp. Quant. BioL, 25, 11—27, 1960. Circa-
dian period and light intensity.
5. Aschoff J., Comp. Biochem. Physiol., 18, 397—404, 1966. Phase-angle differen-
ces and circadian periods.
6. Aschoff J. In: Life Science and Space Research, edited by A. H. Brown and!
F. G. Favorite, North-Holland Publishing Co., Amsterdam, 1967, pp. 160—173.
Circadian rhythms.
7. Barnett A., J. Cell. Physiol., 67, 239—270, 1966. Circadian rhythm of Parame-
cium mating type.
8. Barnwell F. H., Proc. Soc. Exp. Biol. Med., 105, 312—315, 1960. Geophysically
dependent periodisms in chick.
9. Barnwell F. H., Biol. Bull., 125, 399—415, 1963. Tidal rhythms in Uca.
10. Barnwell F. H., Biol. Bull., 130, 1—7, 1966. Uca from Woods Hole region.
11. Barnwell F. H., Amer. ZooL, 8, 569—583, 1968. Tidal rhythms in Uca.
12. Becker G., Naturwissenschaften, 50, 664, 1963. Insect orientation to magnetic
fields.
13. Bennett M. F., Z. vergl. PhysioL, 47, 431—437, 1963. Rephasing of Uca tidal
cycles.
14. Bennett M. F., Z. vergl. PhysioL, 60, 34—40, 1968. Annual rhythm in earth-
worm light response.
15. Bennett M. F., Huguenin J., Z. vergl. PhysioL, 61, 440—445, 1969. Earthworm
response to geomagnetism.
16. Bennett M. F., Shriner J., Brown R. A., BioL Bull., 112, 267—275, 1957. Tidal
rhythm in Uca.
17. Best J. B., Animal Behavior, 14 (suppl. 1), 69—75, 1966. Monthly rhythm ire
planarian «learning».
18. Biological Clocks, edited by Chovnick A, Cold Spring Harbor Symp. QuanL
BioL, 25, 524 pp., 1960. General account of rhythms.
19. Bohn G., C. R. Acad. Sci. Paris, 137, 576—578, 1903. Tidal rhythm in Con-
voluta.
Биологические ритмы
255
20. Borthwick Н. A., Hendricks S. В., Science, 132, 1223—1228, 1960. Photoperio-
dism and plants.
'21. Braemer W, Cold Spring Harbor Symp. Quant. Biol., 25, 413—427, 1960. Sun
orientation by fish.
22 Brown F. A., Jr., Biol. Bull., 115, 81—100, 1958. Mean sideridal-day cycle.
23. Brown F. A., Jr., Science, 130, 1534—1544, 1959. Autophasing, genesis of cir-
cadian period.
24. Brown F. A., Jr. In Cold Spring Harbor Symp. Quant. Biol., edited by
A. Chovnick, 25, 57—71, 1960. Geophysically dependent rhythms.
25. Brown F. A., Jr., N. Y. Acad. Sci., 98, 775—787, 1962. Geophysically depen-
dent rhythms.
26. Brown F. A., Jr., Biol Bull., 123, 264—281, 1962. Magnetism and Dugesia and
Paramecium.
27. Brown F. A., Jr., Biol. Bull., 123, 282—294, 1962. Planarian response to ele-
ctrostatic fields.
28. Brown F. A., Jr., Biol. Bull., 125, 206—225, 1963. Dugesia response to weak
gamma radiation.
29. Brown F. A., Jr. In: Circadian Clocks, edited by J. Aschoff, North-Holland
Publishing Co., Amsterdam, 1965, pp. 231—261. Geophysically dependent pe-
riodisms.
30. Brown F. A., Jr., Proc. Soc. Exp. Biol. Med., 120, 792—797, 1965. Lunar-day
propensity of hamster.
31. Brown F. A., Jr., Scientia, 1968. Geophysically-dependent rhythms.
32. Brown F. A., Jr., Canad. J. Bot., 47, 287—298, 1969. Extrinsic timing of cir-
cadian rhythms.
33. Brown F. A., Jr., Ann. N. Y. Acad. Sci., 188, 224—241, 1971. Responses to weak
electromagnetic fields.
34. Brown F. A., Jr., Siderial Variation in Potato Metabolism (Unpublished ma-
nuscript) .
35. Brown F. A., Jr., Amer. Sci., 60, 756—766, 1972. The «clocks» timing biologi-
cal rhythms.
36. Brown F. A., Jr., Fingerman M., Sandeen M. I., Webb H. W., J. Exp. Zool.,
123, 29—60, 1953. Tidal rhythm in Uca.
37. Brown F. A., Jr., Fingerman M., Hines M. N., Biol. Bull., 106, 308—317, 1954.
Diurnal variation in phase shifting response in Uca.
38. Brown F. A., Jr., Hastings J. W., Palmer J. D., The Biological Clock: Two
Views. Academic Press, New York, 1970, 94 pp. Review of clock theories.
39. Brown F. A., Jr., Hines M. N., Physiol. Zool., 25, 56—70, 1952. Persistence of
experimentally modified circadian cycle forms.
40, Brown F. A., Jr., Park Y. H., Nature, 202, 469—471, 1964. Gammataxis, seaso-
nal variations, planarians.
41. Brown F. A., Jr., Park Y. H., Proc. Soc. Exp. Biol. Med., 125, 712—715, 1967.
Monthly rhythm in hamster.
42. Brown F. A., Jr., Park Y. H., Biol. Bull., 132, 311—319, 1967. Association for-
mation between light and subtle geophysical fields.
43. Brown F. A., Jr., Park Y. H., Zene J. R., Nature, 211, 830—833, 1966. Geo-
physically dependent periodism in mice.
44. Brown F. A., Jr., Shriner J., Ralph C. L., Amer. J. Physiol., 184, 491—496,
1956. Geophysically dependent periodisms in rat.
45. Brown F. A., Jr., Shriner J., Webb H. M., Biol. Bull., 113, 103—111, 1957.
Organismic activity and background radiation.
46. Brown F. A., Jr., Terracini E. D., Proc. Soc. Exp. Biol. Med., 101, 457—460,
1959. Lunar-day propensity in rat.
47. Brown F. A., Jr., Webb H. M., Physiol. Zool., 21, 371—381, 1948, 24-hour pre-
cision in constant conditions in Uca.
48. Brown F. A., Jr., Bennett M. F., H. M., Proc. Nat. Acad. Sci., 41, 93, 1955.
Geographic translocation experiment.
•49. Brown F. A., Jr., Webb H. M., Bennett M. F., Amer. J. Physiol., 195, 237—243,
1958. Organismic correlations with cosmic radiation.
256
Глава 10
50. Bruce V. G., Pittendrigh C. S., J. Cell. Comp. Physiol., 56, 25—31, 1960.
D2O and Euglena circadian period.
51. Banning E., Jahrbuch. Bot, 77, 283—320, 1932. Inheritance of free-running
period in Phaseolus.
52. Biinning E„ Jahrb. Bot., 81, 411—418, 1935. Inheritance of free-running period
in Phaseolus.
53. Banning E., Ber. Deutsch. Bot. Ges., 54, 590—607, 1937. Circadian rhythms
in photoperiodism.
54. Banning E., Naturwiss. Rundschau, 9, 351—357, 1956. Persistence of expe-
rimentally modified circadian cycle forms.
55. Bunning E., Cold Spring Harbor Symp. Quant. Biol., 25, 1—9, 1960. Phase
response curve for temperature in Phaseolus.
56. Bunning E., The Physiological Clock, Springer-Verlag, New York, 1967,
167 pp. General account of rhythms.
57. Bunning E., Bauer E. W., Zeitschr. Bot., 40, 67—76, 1952. Annual rhythm in
seed germination.
58. Bunning E„ Moser I., Planta, 69, 101—110, 1966. Phase response curve in
Phaseolus.
59. Bunning E., Mailer D., Z. Naturforsch., 16b, 391—395, 1962. Menthly rhythm
in Dictyota.
60. Caspers H., Int. Rev. Ges. Hydrobiol., 46, 175—183, 1961. Swarming rhythm
in palolo worm.
61. Circadian Clocks, edited by J. Aschoff, North Holland Publishing Co., Amster-
dam, 1965, 479 pp. International symposium.
62. Cloudsley-Thompson J. L., Rhythmic Activity in Animal Physiology and Be-
havior, Academic Press, New York, 1961, 236 pp. General account of rhythms.
63. Conroy R. T. W. L., Mills J. N., Human Circadian Rhythms, J. and A. Chur-
chill, London, 1970. 236 pp.
64. Cumming B. G., Canad. J. Bot., 47, 309—324, 1969. Circadian rhythm in pho-
toperiodism.
65. Dan K, Kubota H., Embryologia, 5, 21—37, 1960. Monthly, annual, and
18-year rhythms in Comanthus spawning.
66. Davis D. E., Physiol. Zool., 40, 391—402, 1967. Annual rhythm in wood-
chuck.
67. DeCoursey P., Z. vergl. Physiol., 44, 331—354, 1961. Phase-response curves in
Glaucomys.
68. DeCoursey P„ J. Cell. Comp. Physiol., 63, 189—196, 1964. Phase-response cur-
ve of hamsters.
69. Dewan E. M., Science and Technology, Jan. 20—28, 1969. Monthly rhythm in
man.
70. Dijkgraaf S., Kalmijn A. J., Z. vergl. Physiol., 53, 187—194, 1966. Electroper-
ception.
71. Dowse H. B., Palmer J. D„ Nature, 222, 564—566, 1969. Circadian entrain-
ment by electrostatic fields.
72. Dresler A., Licht, 10, 79—82, 1940. Annual variations of man to spectral
colors.
73. Edmunds L. N., Jr., Funch R. R., Science, 165, 500—503, 1969. Free-running
rhythm in Euglena population.
74. Ehret C. F., Trucco E., J. Theoret. Biol., 15, 240—262, 1967. Chronon concept
for circadian clock.
75. Emlen S. T., Auk, 84, 309—342, 1967. Star navigation in birds.
76. Emlen S. T„ Auk, 84, 463—489, 1967. Star navigation in birds.
77. Enright J. T., T. vergl. Physiol., 46, 276—313, 1962. Tidal rhythm in Amphi-
pods.
78. Farner D. S., Proc. XIV International Ornithological Congress, 107—133, 1967.
Photoperiodism and bird reproductive cycles.
79. Farner D. S., Envir. Res., 3, 119—131, 1970. Photoperiodic control of annual
cycles.
Биологические ритмы
257
80. Faure-Fremier Е., Biol. Bull., 100, 173—177, 1951. Tidal rhythms in micro-
organisms.
81. Ferguson D. E., Landreth H. F., McKeown, Animal Behavior, 15, 45—53, 1967.
Sun navigation in frogs.
82. Gamble F. W., Keeble F., Quart. J. Mier. Sci., 47, 363—431, 1904. Tidal rhythm
in Convoluta.
83. Gompel M., C. R. Acad. Sci. Paris, 205, 816—818, 1937. Tidal rhythms in
fish.
84. Gould E., Biol. Bull., 112, 336—348, 1957. Sun orientation in turtles.
85. Gwinner E., Naturwissenschaften, 54, 447, 1967. Circannual rhythms in mice.
86. Gwinner E., J. Ornithol., 109, 70—95, 1968. Circannual rhythms in migratory
birds.
87. Halberg F., Z. Vitamin-, Hormon and Fermentforschung, 10, 225—296, 1959.
Coining of term «circadian».
88. Hamner К. C., Cold Spring Harbor Symp. Quant. Biol., 25, 269—277, 1960.
Circadian rhythms in photoperiodism.
89. Hamner К. C., Finn J. C., Siroli G. S., Hoshizaki T., Carpenter В. H., Nature,
195, 476—480, 1962. Circadian rhythms at south pole.
90. Hamner W. M., Science, 142, 1294—1295, 1963. Circadian rhythms in photo-
periodism.
91. Harker J. E., J. Exp. Biol., 35, 251, 1958. Phase dissociation of tissues and tu-
mor production.
92. Harker J. E., The Physiology of Diurnal Rhythms. Cambridge University
Press, 1964, 114 pp. General account of rhythms.
93. Hartland-Rove R., Rev. Zool. Bot. Africaine, 58, 185—202, 1958. Monthly
rhythm in mayfly emergence.
94. Hasler A. D., Horrell R. M., Wisby W. J,., Braemer Ж, Limnol. Oceanogr., 3,
353—361, 1958. Sun and fish homing.
95. Hastings J. W., New Engl. J. Med., 282, 435—441, 1970. Review.
96. Hastings J. W., Keynan A. In: Circadian Clocks, edited by J. Aschoff, North
Holland Publishing Co., Amsterdam, 1965, pp. 167—182. Metabolic rhythms
in Gonyaulax.
97. Hastings J. W„ Sweeney B. M„ Biol. Bull., 115, 440—458, 1958. Phase respon-
se curve in Gonyaulax.
98. Hauenschild C., Cold Spring Harbor Symp. Quant. BioL, 25, 491—497, 1960. .
Lunar periodicity in Platynereis.
99. Hauty G. T., Adams T„ Aerospace Med., 37, 1257—1262, 1966. Human circa-
dian rhythms during geographic translocations.
100. Hoffman K., Z. TierpsychoL, 11, 453—475, 1954. Sun navigation and «clock»-
shifting.
101. Johnson L. G., Biol. Bull., 131, 308—322, 1966. Circadian-rhythm self-setting
to local time.
102. Jones D. A., Naylor E., J. Exp. Mar. Biol. Ecol., 4, 188—199, 1970. Tidal
rhythms in isopods.
103. Kalmijn A. J., Nature, 212, 1232, 1966. Electroperception in sharks and
rays.
104. Karakashian M. W., Hastings J. W., J. Gen. PhysioL, 47, 1—12, 1963. Inhibi-
tors of macromolecule synthesis and rhythms.
105. Kavanau J. L., Nature, 194, 1293—1295, 1962. LD transitions and extended
limits for circadian entrainment.
106. Keeton W. T., Science, 165, 922—928, 1969. Pigeon homing without sun.
107. Keeton W. T., Proc. Nat. Acad. Sci., 68, 102—106, 1971. Magnets and pigeon
homing.
108. Kessler E., Czygan F. G., Experientia, 19, 89, 1962. Annual variations in
Ankistrodesmus.
109. Kleinhoonte A., Arch. Need. Sci. Exp. Nat., 5, 1—110, 1929. Limits of circa-
dian entrainment in Phaseolus.
HO. Konopka R. J., Benzer S„ Proc. Nat. Acad. Sci., 68, 2112—2116, 1971. Inhe-
ritance of free-running period in Drosophila.
17-1514
258
Глава 10
111. Kramer G., Ibis, 101, 399—416, 1959. Sun navigation in birds.
112. Lang H. L, Z. vergl. Physiol., 56, 296—340, 1967. Monthly rhythms in guppy
light response.
113. Lang H. J., Verhandl. Deutsch. Zool. Gesellsch., Innsbruck (1968), 291—298,
1968. Monthly rhythm in guppy light response.
114. Lewis P. R., Lobban M. C., J. Physiol., 133, 670—680, 1956. Human circadian
rhythms in Arctic.
115. Lindauer M„ Cold Spring Harbor Symp. Quant. Biol., 25, 371—377, 1960.
Clocks and bee sun-compass.
116. Lindauer M., Martin H., Z. vergl. Physiol., 60, 219—243, 1968. Magnetic
responsiveness of bees.
117. Lobban M. C., Quart. J. Exp. Physiol., 52, 401—410, 1967. Human circadian
rhythms in Arctic.
118. Lofts B., Nature, 201, 523—524, 1964. Annual rhythm in Quelea.
119. Lowe С. H., Hinds D. S., Lardner P. J., Justice К. E., Science, 156, 531—534,
1967. 24-hour precision in circadian rhythms in kangaroo rats and gila mon-
sters.
120. Luce G. G., Biological Rhythms in Psychiatry and Medicine. Public Health
Service Publ. 2088, 183 pp., 1970. General review.
121. Menaker W., Menaker A., Amer. J. Obstet. Gynecol., 77, 905—914, 1959.
Monthly period in human reproduction.
122. Merkel F. W., Ann. N. Y. Acad. Sci., 188, 283—294, 1971. Magnetic fields in
bird navigation.
123. Merkel F. W., Wiltschko W„ Die Vogelwarte, 23, 71—77, 1965. Magnetic
fields and bird orientation.
124. Natalini J. J., Physiol. Zool., 45, 153—166, 1972. Phase response curve of kan-
garoo rat.
125. Naylor E., J. Exp. Biol., 35, 602—610, 1958. Tidal rhythm in Carcinus.
126. Neumann D„ Z. vergl. Physiol., 53, 1—61, 1966. Monthly rhythms in Clunio.
127. Neumann D., Z. vergl. Physiol., 60, 63—78, 1968. Tidal-cycle entrainment of
semimonthly rhythm in Clunio.
128. Neumann D., Oecologia, 3, 166—183, 1969. Timing by combinations of rhythms
periods.
129. Orientation: Sensory Bases, edited by H. E. Adler, Ann. N. Y. Acad. Sci., 188,
1—408, 1971.
130. Palmer J. D., Nature, 198, 1061—1062, 1963. Responses of Volvox to weak
magnetic fields.
131. Palmer J. D., Nature, 215, 64—66, 1967. Diurnal and tidal rhythms in Se-
sarma.
132. Palmer J. D., Dowse H. B., Biol. Bull., 137, 388, 1969. D2O and mouse and
bird circadian period.
133. Palmer J. D., Round F. E., J. Marine Biol. Assoc. U.K., 45, 567—582, 1965.
Tidal rhythm in Euglena.
134. Palmer J. D., Round F. E., Biol. Bull., 132, 44—55, 1967. Tidal rhythm in
diatoms.
135. Papi F., Cold Spring Harbor Symp. Quant. Biol., 25, 475—480, 1960. Amphi-
pod orientation by moon.
136. Papi E., Pardi L., Biol. Bull., 124, 97—105, 1963. Lunar orientation in amphi-
pods.
137. Papi E., Tongiorgi P., Ergeb. Biol., 26, 259—280, 1963. Celestial orientation
by wolf-spiders.
138. Pardi L., Cold Spring Harbor Symp. Quant. Biol., 25, 395—401, 1960. Sun
orientation by amphipods.
139. Picton H. D., Nature, 211, 303—304, 1966. Drosophila response to weak
electromagnetic fields.
140. Pittendrigh C. S., Cold Spring Harbor Symp. Quant. Biol., 25, 159—182, 1960.
Phase-response curve in Drosophila.
141. Pittendrigh C. S„ Harvey Lectures, Ser. 56, 93—125, 1961, Academic Press,
New York. Circadian rhythms as endogenous complex oscillations.
Биологические ритмы
259
142. Pittendrigh С. S„ Bruce V. G., Photoperiodism and Related Phenomena in
Plants and Animals. Amer. Assoc. Adv. Sci., Washington, D.C., 1959,
pp 475—505. Daily rhythms as complex oscillator systems.
143. Pittendrigh C. S., Minis D. H., Amer. Nat., 98, 261—294, 1964. Circadian
rhythms in photoperiodism.
144. Rao К. P., Biol. Bull., 106, 353—359, 1954. Tidal Rhythm in Mytilus.
145. Reille A., J. Physiol. Paris, 30, 85—92, 1968. Pigeon conditioning to magnetic
fields.
146. Renner M. Naturwiss. Rundschau, 14, 296—305, 1961. Persistence of experi-
mentally modified circadian cycle forms: geographic translocation expe-
riment.
147. Rhythmic Functions in the Living System, edited by W. Wolf, Ann. N. Y. Acad.
Sci., 98, 753—1326, 1962. General account.
148. Richter G. Z. Z. Naturforsch., 18B, 1085, 1963. Circadian rhythms in enucleated
Acetabularia.
148a. Rommel S. A., Jr., McCleave J. D., Science, 176, 1233—1235, 1972. Electric
field perception by eels.
149. Sauer F., Z. Tierpsychol., 14, 29—70, 1957. Star navigation by birds.
150. Schmidt-Koenig K-, Z. Tierpsychol., 15, 301—331, 1958. Sun orientation and
«clock» shifting.
151. Schmidt-Koenig K, Z. Tierpsychol., 18, 221—224, 1961. Sun orientation and
«clock» shifting.
152. Schneider F., Ergebn. Biol., 26, 147, 1963. Insect orientation to magnetic
fields.
153. Schweiger E., Walraff H. G., Schweiger H. G., Z. Naturforsch., 19B, 499—505,
1964. Circadian rhythms in enucleated Acetabularia.
154. Schweiger E., Walraff H. G., Schweiger H. G., Science, 146, 658, 1964.
Nuclear determination of circadian phase in Acetabularia.
155. Sollberger A., Biological Rhythm Research, Elsevier Publ. Co., Amsterdam,
1965, 461 pp. General review.
156. Southern IF. E., Ann. N. Y. Acad. Sci., 188, 295—311, 1971. Geomagnetic
strength and gull orientation.
157. Stephens G. C., Physiol., Zool., 30, 55—69, 1957. Phase response for tempe-
rature in Uca.
158. Strughold H., Ann. N. Y. Acad. Sci., 134, 413—422, 1965. Human circadian
rhythms during space flight.
159. Stutz A., Ann. N. Y. Acad. Sci., 188, 312—323, 1971. Gerbil response to geo-
magnetism.
160. Suter R. B., Rawson К S., Science, 160, 1011—1015, 1968. D2O and mouse
circadian period.
161. Sweeney В. M., Rhythmic Phenomena in Plants. Academic Press, New York,
1969, 147 pp. General review.
162. Sweeney В. M., Hastings J. W., Cold Spring Harbor Symp. Quant. Biol., 25,
87—104, 1960. Temperature and circadian rhythms.
163. Sweeney В. M., Haxo F. T., Science, 134, 1361, 1961. Circadian rhythms in
enucleated Acetabularia.
164. Terracini E. D., Brown F. A. Jr., Physiol. Zool., 35, 27, 1962. Propensity for
24-hour periods in mice.
165. Tribukait B., Z. vergl. Physiol., 38, 479—490, 1956. Limits of circadian entrain-
ment in mice.
166. Van den Driessche T„ Biochim. Biophys. Acta, 126, 456—470, 1966. Nucleus
and circadian rhythm in Acetabularia.
167. Van den Driessche T., Bonotto S., Biochim. Biophys. Acta, 179, 58—66, 1969.
Circadian rhythm in RNA synthesis in Acetabularia.
168. Van Laar W., Acta Biotheoretica, 19, 95—139, 1970. The concept of «Biologi-
cal Clock». &
169' Dictyota” V" Zeitschr' Bot’ 51’ 156~173> 1963- Monthly rhythm in
17*
260
Глава 10
170. Walcott С., Michener М. С., J. Exp. Biol., 54, 291—316, 1971. Sun navigation
in pigeons.
171. Wallraff H. G., Proc. XIV International Ornithological Congress, 1967,
pp. 331, 358. Review of bird homing problems.
172. Waterman T. H., Nature, 228, 85—87, 1970. Polarized light and fish orienta-
tion.
173. Webb H. M„ Physiol. Zool., 23, 316—337, 1950. Diurnal variations in phasing
shifting response in Uca.
174. Webb H. M., Brown F. A., Jr., Biol. Bull., 115, 303—318, 1958. Metabolic
periodism in fiddler crabs.
175. Webb H. M., Brown F. A., Jr., Physiol., Rev., 39, 127—161, 1959. RMI hypo-
thesis for biological rhythms.
176. Webb H. W., Brown F. A., Jr., Biol. Bull., 129, 582—591, 1965. Daily and ti-
dal rhythms in fiddler crabs.
177. Wehner R., Labhart T., Experientia, 26, 967—968, 1970. Drosophila response to
geomagnetism.
178. Wever R., Kybernetik, 2, 127—144, 1964. Model of 24-hour clock.
179. Wever R., Z. angewandte Math. Meeh., 46, 148—157, 1966. Mathematical mo-
del for biological clock.
180. Wever R., Z. vergl. Physiol., 56, 111—128, 1967. Electromagnetic fields and
human circadian rhythms.
181. Wever R., Naturwissenschaften, 55, 29—32, 1968. Influence of weak electro-
magnetic fields on circadian period.
182. Wever R., Life Sciences and Space Research VIII, North Holland Publ. Co.,
Amsterdam, 1970, pp. 177—187. Electric fields and human circadian rhythms.
183. Williams B. G., Naylor E., J. Exp. Biol., 51, 715—725, 1969. Tidal rhythm in
Carcinus.
184. Wiltschko W., Z. TierpsychoL, 25, 537, 1968. Magnetic-field orientation in
birds.
184a. Wiltschko W., Wiltschko R., Science, 176, 62—65, 1972. Magnetic compass of
European robins.
185. Winfree A. T., J. Theoret. BioL, 16, 15—42, 1967. Biological rhythms and po-
pulations of coupled oscillators.
186. Winfree A. T., J. Theoret. BioL, 28, 327—374, 1970. Integrated view of reset-
ting a circadian clock.
187. Wolfson A. In: Photophysiology, Vol. 2, edited by A. C. Giese., 1965, pp. 1—49.
Photoperiodism.
188. Zimmerman W. F., Pittendrigh C. S., Pavlidis T., Insect PhysioL, 14, 669—684,
1968. Phase response curve for temperature in Drosophila.
Глава 1 1
Возбудимые мембраны
Л. ПРОССЕР
В основе поведения животных, если рассматривать этот вопрос
па клеточном уровне, лежит проведение сигналов 'Клеточными
мембранами и передача их от клетки к клетке. Сенсорные струк-
туры дают информацию об окружающей среде, центральная нерв-
ная система сопоставляет эту сенсорную информацию с уже су-
ществующими в ней «образами», мышцы и другие эффекторы осу-
ществляют поведенческие реакции. Этим трем компонентам, от-
ветственным за поведение животных, присущи общие клеточные
особенности. Функция мембран в нервной и мышечной тканях ба-
зируется как на электрических свойствах, характерных для возбу-
димых клеток вообще, так и на функциональном взаимодействии
между невозбудимыми клетками.
Для всех нервных и многих других (ненервных) тканей следу-
ет различать проведение сигнала в клетке и передачу его от клет-
ки в клетке. Электрическое проведение бывает: 1) градуальным,
затухающим, т. е. протекающим с декрементом или 2) регенера-
тивным, самораспространяющимся, протекающим по принципу
«все или ничего». Межклеточная передача бывает электрической
и химической. Существуют следующие виды электрической пере-
дачи: а) градуальная, электротоническая и двусторонняя, б) об-
ладающая свойством выпрямления (тока) и (реже) в) тормоз-
ная, под влиянием внешних отрицательных полей. Химическая пе-
редача основана на высвобождении из пресинаптических пузырь-
ков специфических агентов, для чего необходима деполяризация
пресинаптической мембраны; это высвобождение блокируется низ-
ким содержанием Са2+ и (или) высоким содержанием Mg2+ в
среде; выделившиеся специфические агенты химически возбужда-
ют постсинатттическую мембрану.
Ненервное проведение; реакции отдельных клеток
Все клетки окружены поверхностным слоем — плазматической
мембраной, лежащей непосредственно под наружной оболочкой.
Эта мембрана электрически поляризована — в норме она элект-
, роотрицательна со стороны протоплазматического содержимого.
262
Глава 11
Возбуждение обычно состоит в деполяризации — уменьшении или
инверсии нормальной поляризации. Возможна местная и распро-
страняющаяся деполяризация. Проведение — это распространение
по плазматической (мембране волны деполяризации. Ранее полага-
ли, что у инфузорий для проведения сигнала от одной части одно-
клеточного организма к другой и координирования таким образом
биения ресничек служат внутриклеточные фибриллы. Недавние
опыты, проведенные на Euplotes, показали, однако, что если пере-
резать эти фибриллы, то координированные изменения направле-
ния и частоты биения ресничек трех типов сохраняются, к тому
же координация биения ресничек есть и у клеточных фрагментов
[178]. Следовательно, у простейших, как вообще у возбудимых
клеток, проведение осуществляется плазматической мембраной.
Мембранный потенциал инфузории Opalina равен —27 мВ; ес-
ли при электрическом раздражении происходит инверсия биения
ресничек, то внутриклеточная электроотрицательность уменьшает-
ся [233]. У инфузории Paramecium потенциал покоя, измеренный
с помощью внутриклеточных электродов, составляет от —30 до
—40 мВ; тактильное, химическое или электрическое раздражение
вызывает активную электрическую реакцию, изменяющую направ-
ление биения ресничек. Тактильное раздражение переднего конца
клетки вызывает быструю деполяризацию, распространяющуюся
к заднему концу; ее амплитуда зависит от концентрации Са в
окружающей среде. Раздражение заднего конца клетки вызывает
местную гиперполяризацию, которая уменьшается при повышении
концентрации К в окружающей среде и, следовательно, обуслов-
лена оттоком к из цитоплазмы. Мембранный потенциал, связан-
ный с инверсией биения ресничек, обусловлен повышением про-
ницаемости мембраны для Са; градиент [Са2+] направлен внутрь
клетки. При электрическом раздражении или под влиянием ионов
бария в мембране возникают периодически повторяющиеся им-
пульсы потенциала действия; эти потенциалы по величине могут
превосходить нулевой потенциал. Высказывалось предположение,
что у инфузорий генерируются как местные, так и регенеративные
мембранные потенциалы, а у примитивных организмов потенциал
действия определяется изменением проницаемости мембраны для
Са2+, а не Na+ [57—60, 164, 165].
У губок вокруг пор расположены веретеновидные сократитель-
ные клетки. Механическое раздражение приводит к их местному
сокращению, не передающемуся от клетки к клетке, нервные эле-
менты здесь отсутствуют. Для сократительных реакций на при-
косновение необходимо присутствие и одно-, и двухвалентных ка-
тионов; эти реакции протекают и при полном замещении натрия
калием (500 мМ). Сократительная система губок, вероятно, реа-
гирует на прямое раздражение. Некоторые деполяризуемые кали-
ем гладкие мышцы позвоночных могут сокращаться также под
действием медиаторных агентов, например ацетилхолина.
Возбудимые мембраны
263
Передача сигнала от клетки к клетке в ненервных тканях
В различных тканях, например в пластах эпителия или у ран-
них зародышей, передача сигнала может осуществляться без уча-
стия нервных волокон. В таких тканях между клетками существу-
ет электрическая связь — толчки тока, приложенные к одной клет-
ке, регистрируются и в соседних клетках, лишь незначительно
уменьшаясь по амплитуде. Коэффициент электрической связи К
равен отношению потенциала, регистрируемого в данной клетке
(Vi), к потенциалу, приложенному к соседней клетке (Кг):
К—Jx
Л— у2 .
Следует иметь в виду, что в классификации межклеточных свя-
зей для обозначения одной и той же структуры применяется не-
сколько терминов; кроме того, в одной межклеточной связи мо-
жет быть несколько типов контактов. Существует две основные
группы межклеточных контактов: а) контакты с обособленными
мембранами, разделенными пространством до 100—150 А, и
б) прямые контакты или даже слияние наружных слоев мембран.
Первая из двух названных групп включает в себя: 1) десмосомы,
или плотные палочки, расположенные по одну или обе стороны
области, где обособленные мембраны прилежат друг к другу;
2) химические синапсы с синаптическими пузырьками, локализо-
ванными у пресинаптической мембраны, и иногда с десмосомопо-
добными структурами постсинаптической мембраны; 3) разделен-
ные перегородками (септированные) контакты, например между
эпителиальными клетками некоторых беспозвоночных.
При электронно-микроскопическом исследовании прямых кон-
тактных соединений видно 3, 5 или 7 (в зависимости от методики
окрашивания) чередующихся светлых и темных полос. Некоторые
из этих соединений называются плотными (tight junctions); они
существуют в виде так называемых «zonula occludens», или колец,
окружающих один из концов клетки, в виде «facia occludens», или
полосок, и в виде «macula occludens», или пятен. При рассмотре-
нии каждого из этих контактов с лицевой стороны в плоскости
мембраны видны фибриллярные структуры. Тесные контактные
соединения встречаются главным образом в эпителии позвоноч-
ных. Другим типом прямого контактного соединения является ще-
.левое соединение (nexus или gap junction), которое слагается из
субъединиц, имеющих в проекции размер 60—70 А и образующих
межклеточные связи в области наружных пластинок мембраны. Эти
контактирующие участки разделены щелью в 20 А, являющейся,
по-видимому, частью межклеточного пространства. С лицевой сто-
роны субъединицы щелевого соединения имеют вид шестиуголь-
ников, расстояние между центрами которых равно 90—100 А
{151, 200, 238] (рис. 11-1, Ц-2). Всюду, где между контактирую-
264
Глава 11
щими клетками имеется низкое электрическое сопротивление — в
септированных гигантских нервных волокнах, между волокнами
гладких мышц или миокарда, между нейронами с электрической
связью, между некоторыми эпителиальными клетками у позвоноч-
ных или беспозвоночных,—.имеются контакты этого типа [23, 49]
Рис. 11-1. Щелевое соединение в сердечной мышце [151].
Замораживание со скалыванием, травление и напыление платиной (фронтальный срез). На
о
поверхности сети (1) имеются регулярно расположенные углубления (2) диаметром 35—50 А.
С внутренней стороны поверхности (3) выступают многочисленные контактные цилиндры
(4); в правом верхнем углу они показаны при большем увеличении.
(рис. 11-3). Они часто встречаются среди областей перегородча-
тых, или плотных (occludens), соединений [18]. Электрическая
связь может осуществляться через щелевые соединения как при
участии zonula occludens, так и без них.
Слюнные железы личинок двукрылых имеют ряды крупных
Возбудимые мембраны
265
эпителиальных клеток. В норме коэффициент электрической свя-
зи между смежными клетками равен 80—90%, а электрическое
сопротивление между ними может составлять всего лишь 0,01 со-
противления между клеткой и внешней средой [136—139]. Если в
одну из клеток 'ввести краситель (например, флуоресцеин с мол.
весом 330 или процион с мол. весом 550), он быстро появится в
Рис. 11-2. Модель щелевого соединения [151].
Al, и М2 — две плазматические мембраны. Каждая мембрана состоит из двух слоев А и В.
смежных клетках; более крупные молекулы — полипептиды — из
клетки в клетку не переходят. В эпителии слюнной железы дву-
крылых чередуются области тесных и щелевых соединений [18]
(рис. 11-4). Межклеточная связь зависит от количества ионов
кальция, связанных с клеточной мембраной. При высоком содер-
жании кальция в среде (10~4 М для слюнной железы личинок
Chironomus, 10-3 М для печени мыши) межклеточные контакты
становятся непроницаемыми; при полном удалении кальция клет-
ки оказываются разобщенными. Межклеточная проницаемость
Рис. 11-3. Области соединения на примере латерального гигантского аксона реч-
ного рака при большом увеличении (Pappas С. D., Asada Y., Bennet М V L
J. Cell Biol., 49, 173—188, 1971). •
О
Л. Тонкий срез; общая толщина контакта 180 А. Б. Тангенциальный срез через область
соединения после фиксации в присутствии лантана, выявляющего систему шестиугольников.
Г
1
Возбудимые мембраны
267
высока при низком (<106 М) содержании Са2+ внутри клеток.
Проницаемость контактов снижается под действием Са2+, при об-
работке ДНФ, замещении Na о ионами лития или отмывке клеток
средой, не содержащей Са2+ и Mg2+ Электрическое разобщение
сопровождается деполяризацией клеток, а восстановить электри-
ческую связь можно с помощью реполяризации [466, 204а].
Рис. 11-4. Электронно-микроскопическая фотография области соединения между
двумя клетками слюнной железы Chironomus [18].
А. Плотное соединение. Б. Щелевое соединение.
Электрическая связь обнаружена у клеток мочевого пузыря
жабы, печени мыши, слюнных желез двукрылых и мальпигиевых
сосудо®. Если поместить диссоциированные клетки губки в усло-
вия культуры тканей, между ними образуются межклеточные кон-
такты с низким электрическим сопротивлением [137, 138].
Большинство растущих клеток зародыша кальмара имеют
электрическую связь, и краситель может переходить из одной
клетки в другую; зародыш в целом можно рассматривать как
электрический синцитий [196]. Подобным образом клетки заро-
дыша рыбы Fundulus во время делений дробления имеют практи-
268
Глава 11
чески полную электрическую связь, между этими клетками есть
как щелевые, так и тесные контакты. На стадии гаструлы эта
связь выражена слабее [16]. Клетки зародыша морокой звезды
электрически связаны между собой на стадии двух бластомеров;
после второго, третьего и четвертого делений дробления эта связь
исчезает, но на стадии 32 бластомеров снова появляются контакты
с низким сопротивлением [232].
Распространение сигнала в ненервной ткани обнаружено у
многих кишечнополостных. В плавательном колоколе сифонофор
участки эпителия лишены нервных и мышечных волокон, тем не
менее в эпителии наблюдается незатухающее идущее во все сто-
роны распространение электрического потенциала со скоростью
20—25 см/с и с рефрактерным периодом 2—3 мс [141—143]. У ко-
лониальных гидроидов проведение от полипа к полипу осуществ-
ляется частично через ненервную ткань. В эпителиальных клетках
гидромедуз наблюдаются двухфазные импульсы продолжитель-
ностью от 1 до 15 мс, распространяющиеся со скоростью 15—
35 см/с; эти импульсы не блокируются ионами магния, подавля-
ющими работу нервной сети [144]. Вероятно, проведение по мед-
ленной системе у гидроидов, которое происходит со скоростью
6 см/с, имеет высокий порог и непосредственно не отражается на
поведении, также является ненервным [98, 99]. У Hydra сущест-
вует несколько типов электрической активности: ритмические
спонтанные волны потенциала действия, возникающие в обла-
стях— водителях ритма (пейсмекерах), малые потенциалы, свя-
занные с движением щупалец, и большие потенциалы, распростра-
няющиеся со скоростью 5 см/с. Полость тела гидры имеет поло-
жительный потенциал в 15—40 мВ по отношению к наружной
поверхности тела, что обусловлено пространственной асимметрией
потенциалов; распространяющиеся большие потенциалы действия
представляют собой инверсию потенциала через эпителиальный
слой. Щелевые соединения позволяют току протекать от клетки к
клетке и препятствуют его прохождению из клеток через облада-
ющие высоким сопротивлением внешние мембраны [100]. Наруж-
ные поверхности эктодермальных клеток у Hydra связаны между
собой посредством плотных контактов; ниже расположены щеле-
вые контакты дисков, содержащих множество шестиугольных
структур с расстоянием между центрами 95—ПО А [87].
У поздних зародышей и ранних головастиков шпорцевой ля-
гушки Xenopus клетки наружного слоя кожи еще до развития
нервной ткани имеют оба вида прямых контактных соединений..
При раздражении поверхностных клеток генерируются импульсы,,
распространяющиеся по принципу «все или ничего» со скоростью
7,7 см/с. Проведение прекращается при замене натрия инкубаци-
онного раствора трис-ионами или при отравлении тетродотоксином
(стр. 279). Клетки кожи имеют потенциал покоя — 75 мВ и гене-
рируют деполяризующие потенциалы действия продолжительно-
269
Возбудимые мембраны
---------г----------—
стью 100 wfe И амплитудой ПО мВ. Такие кожные потенциалы у
поздних зародышей могут вызывать возбуждение нервных окон-
чаний [201].''
Таким образом, в основе самого простого типа передачи сиг-
нала лежит электрическая связь между клетками с плотными или
щелевыми соединениями. Подобная связь у эмбрионов обеспечива-
ет функциональную непрерывность, а в эпителиальных тканях у
некоторых организмов (например, у кишечнополостных и у заро-
дышей амфибий) опосредует поведенческие реакции. Ниже (стр.
305—308) мы рассмотрим примеры подобной электрической связи
между нейронами.
Проведение в возбудимых тканях, строение мембран
(в основном мембран нейронов)
Каждая возбудимая клетка ограничена плазматической мем-
браной толщиной 50—100 А; к мембране прилегают окружающие-
клетку оболочки. Некоторые крупные немиелинизированные нерв-
ные волокна (как, например, аксоны некоторых ракообразных и
головоногих) окружены глиальной оболочкой. Оболочка пучка
мелких немиелинизированных волокон представлена одной много-
ядерной клеткой. Нервные волокна позвоночных покрыты слоями
миелина, образующими спиральную структуру; тело клетки, обра-
зующей эту оболочку, расположено с ее наружной стороны. У не-
которых беспозвоночных, например у десятиногих раков, ядро та-
кой клетки миелинизированного нервного волокна лежит внутри
одиночного рыхлого липидного или миелинового слоя; в других
случаях миелиновая оболочка состоит из нескольких спиральных
слоев. Оболочка миелинизированных волокон позвоночных преры-
вается перехватами Ранвье; многочисленные боковые ответвления
волокон некоторых беспозвоночных функционируют почти так же,
как перехваты. Обычно в начальной и конечной частях аксона,
т. е. у сомы нейрона и в нервном окончании, оболочки утончают-
ся, а миелин исчезает.
Возбудимая клеточная мембрана состоит из липидов и белков;
она имеет высокое сопротивление и высокую электростатическую
емкость (рис. 11-5). Клетка цилиндрической формы, например мы-
шечное волокно или, скажем, аксон, обладает свойствами электри-
ческого кабеля, характеристиками которого являются временная
константа т и константа длины А. Эти константы обозначают со-
ответственно время затухания и расстояние, на которое распрост-
ранился электрический импульс за время уменьшения его ампли-
туды до величины, составляющей 1/е (приблизительно 37%) от
приложенного напряжения (табл. 11-1).
Временная константа в секундах равна См7?м, где См — емкость
мембраны в Ф/см2, а 7?м — удельное сопротивление мембраны (в
?Ю___________________________Глава 11_________ '
— —- у----------
Ом-см2)1. Пассивное распространение сигнала описывается урав-
нением Г
* х * 0е »
где Vo приложенное напряжение, a Vx — напряжение, измерен-
ное на расстоянии х от точки приложения. В таком случае, если
графически изобразить зависимость In V0/Vx от х, то X будет рав-
на тангенсу угла наклона получившейся прямой; Л определяется
.также из следующего выражения:
' ri+ го
где гм — поперечное сопротивление мембраны, выраженное в Ом*
•см, гг—внутриклеточное сопротивление на единицу длины в
Рис. 11-5. Схема элемента возбудимой мембраны нервного волокна (Hodkin A. L.,
Proc. Roy. Soc. bond. В, 148, 1—37, 1958).
С?м — постоянная емкость мембраны; Rj^a и Кд — каналы переменной проницаемости, по ко-
торым проходят Na+ и К+; Rq — канал, по которому проходят другие ионы, в основном
£j_. ум — мембранный потенциал; Ро—наружное сопротивление; R,—внутреннее сопротив-
ление. У гигантского аксона кальмара эти величины имеют следующие значения /?м=400—
1100 Ом-см2; Ro— 20 Ом-см; R ~30 Ом-см; См —1,1 мкФ/см2; + м— от —60 до —70 мВ;
от —85 до —90 мВ; Vq — от —55 до —60 мВ; Г^а —от +45 до +50 мВ.
Ом*см-1 и г0 — внеклеточное сопротивление на единицу длины в
Ом-см-1.
Поскольку толщина клеточной мембраны не более 100 А, ее
удельное сопротивление имеет порядок 109 Ом-см, т. е. очень ве-
лико. Удельное сопротивление аксоплазмы близко к удельному
сопротивлению плазмы или (морской воды и, напротив, невелико
(ri=200 Ом-см). Проводимость — это величина, обратная сопро-
1 Удельное сопротивление г соответствует сопротивлению кубического санти-
метра материала и измеряется в Ом-см2/см=Ом-см. При измерении поперечного
сопротивления клеточной мембраны толщина мембраны неизвестна, поэтому
удельное сопротивление мембраны /?м — это сопротивление 1 см2 ее поверхности
безотносительно к толщине I, т. е. R№=rl. Измеренное сопротивление R~~r , сле-
довательно, RM=rl—RA и измеряется в Ом-ем2. Электростатическая емкость про-
порциональна диэлектрической постоянной D и площади А при данной толщи-
не /, т. е. С= , следовательно, удельная емкость мембраны См=—=С/Л
/ •
и измеряется в Ф/см2.
\ Возбудимые мембраны 271
1 Таблица 11-1
Характеристики мембран некоторых нейронов (аналогичные данные )иля мышечных мембран приведены в табл. 16-4)
\ \ Животные Сопротивление мембраны (входное сопротивление измеряется в Ом, а удельное— в Ом - см2) Электростатическая емкость мембраны (входная емкость измеряется в Ф, а удельная—в Ф/см2) Временная константа, мс Скорость распростра- нения импульса, м/с
•а 1 Myxicola [19], гигантское волокно Eudistyla [85а], гигантское волокно Пиявка [128], крупные нейроны Таракан [246], гигантские волокна Омар [130а], гигантские волокна Шримс [130а], гигантские волокна Серебряный карась [67а], маутнеровские клет- ки мотонейроны Onchidium [178а], гигантские нейроны Helix [146], нейроны Aplysia [93а], Н-клетки Таракан [32], сенсорные волокна Лягушка [90], перехват Ранвье Минога [150], осязательные клетки барорецепторы болевые рецепторы маутнеровские клет- ки [204b] 1,2-103 Ом-см2 1,5—4,5-Ю3 Ом-см2 1,7-103 Ом-см2 0,8- Ю3 Ом-см2 65 Ом-см2 600 Ом-см2 6,8 МОм 0,98-103 Ом-см2 4-103—10-103 Ом-см2 10—20 Ом-см2 5,4 МОм 6,3 МОм 8,3 МОм 2 МОм (16 000 Ом-см2) 2—3 мкФ/см2 6,3 мкФ/см2 6,5 мкФ/см2 5,0 мкФ/см2 0,27 мкФ 29 мкФ/см2 3—7 мкФ/см2 или 0,6-1,5 пФ 271 пФ 375 пФ 479 пФ 2,8 мкФ/см2 4,2 0,39 3,1 30 100—200 1,2 2,3 3,9 1,5—3,5 8 90 3,3'
тивлению: Удельное сопротивление многих возбудимых
клеточных мембран равно примерно 1000 Ом-см2. Для клеток не-
установленной Формы или неизвестной площади поверхности удоб-
ной количественной характеристикой является входное сопротив-
ление, т. е. сопротивление, измеренное микроэлектродом по отно-
шению к окружающей среде.
1 Е(П/0С2Ь/ мембРан многих возбудимых клеток близка к
мкФ/см (табл. 11-1). Емкость поперечнополосатых мышечных
___________________________Глава И /
волокон и волокон Пуркинье в миокарде существенно выше; ее
можно разделить на два компонента, соответствующих емкости
наружной мембраны и емкости поперечных трубочед. Эти компо-
ненты в волокнах портняжной мышцы лягушки равны соответст-
венно 2,6 и 4,1 мкФ/см2, в мышечных волокнах речного рака —
3,9? и 17 мкФ/см2 [64] и в волокнах Пуркинье — 2,4 и 7 мкФ/см2
Потенциалы покоя. Потенциал покоя (ПП) — это .мембранный
’Потенциал клетки, на которую не действуют никаким раздражите-
лем. Большинство клеток симметричны. Однако некоторые упло-
щенные клетки асимметричны, что выражается в различных элек-
трических свойствах двух поверхностей.
Симметричный потенциал покоя. Потенциал покоя можно из-
мерить, вводя микроэлектроды в крупные волокна или внутрь
клеток или подводя электроды к хорошо изолированным (напри-
мер, с обеих сторон отделенным воздушными прослойками) пере-
хватам Ранвье. У многих нейронов потенциал покоя равен при-
мерно —70 мВ, у волокон многих скелетных и гладких мышц —
соответственно —90 и —40 мВ. Значения потенциала покоя для
клеток разных типов лежат в диапазоне от —4 до —100 мВ. По
сравнению с внеклеточной жидкостью цитоплазма содержит го-
раздо больше К+, меньше Na+ и С1~ и гораздо меньше Са2+. По-
тенциал покоя можно формально описать уравнением Гольдмана
(уравнением постоянного поля), в котором концентрация каждого
иона умножается на соответствующую константу проницаемости
Р, т. е. принята во внимание реальная подвижность ионов в мем-
бране:
F __ RT_ < / Рк [К.1 + PNalNa,] + Pg [С1О]
nF РК [Ко] + Рш [Na0] + РС1 [Ch] ) ’
Где Е — равновесный потенциал, — —множитель, в который вхо-
дят газовая постоянная 7?, абсолютная температура Т, валентность
п и число Фарадея F; индексы i и о относятся соответственно к
содержимому нейрона и окружающей среде. Очень многое о нерв-
ных импульсах удалось узнать из опытов на гигантских аксонах
кальмара, которые достаточно велики (500—1000 мкм), чтобы в
них можно было ввести два или более внутриклеточных электро-
да. PNa равна здесь 0,008Рк в состоянии покоя и ЗОРк в началь-
ной фазе активности.
В нейронах Aplysia относительные проницаемости для К, Na
и С1 равны соответственно 1, 0,01 и 0,45 [207]. Ра в нервном во-
локне краба выше, чем в нерве позвоночных или кальмара.
В мышце лягушки Ра относительно высока, Рк * Риа >Рс\ = 1 :0,9 :
: 1,90 и ПП близок к Есь В миокарде Риа также высока. Вследст-
вие низкой Рма потенциал покоя в большинстве возбудимых тка-
ней не зависит от концентрации Na+ в окружающей среде и чув-
\Возбудимые мембраны 273
ствителен к ионам К; иными словами, мембрана невозбужденной
клетки ведет себя как калиевый электрод. Тем не менее при фи-
зиологических концентрациях К натрий может сильно влиять на
ПП, так что нарушается линейная зависимость ПП от In Ко [1].
Как мы уже говорили, внутриклеточный отрицательный заряд
в основном создается анионами, не проникающими через мембра-
ну. В аксонах кальмара таким анионом является изетионат, у ра-
кообразных это аминокислоты, в некоторых нейронах млекопита-
ющих— N-ацетиласпартат. Потенциалы некоторых нейронов близ-
ки к равновесному потенциалу хлора или несколько меньше его.
В то же время в аксонах кальмара внутриклеточная концентра-
ция С1 в 2—3 раза выше той, которая должна была бы быть при
равновесном состоянии, т. е. ПП отрицателен по отношению к
£сь Потенциал покоя существенно отличается от равновесного по-
тенциала для Na, величина которого — от +20 до +50 мВ. Гра-
диент концентрации Na поддерживается за счет работы натрие-
вого насоса, и при этом выведение Na, в малых количествах про-
никающего в невозбужденную клетку, связано с удержанием
калия. Первые данные о связи мембранной АТФазы с натрий-ка-
лиевым насосом были получены из опытов, поставленных на нерв-
ных волокнах ракообразных. Концентрация ионов кальция в аксо-
плазме кальмара составляет всего 0,0001% их концентрации в
окружающей среде, так что £са приближается к +100 мВ. Такая
низкая концентрация Са2+ обусловлена отчасти внутриклеточным
связыванием его, а отчасти активным вытеснением, зависящим от
[Nao ]. Mg2+ также вытесняется из клетки с помощью механизма,
зависящего от Na [90]. В среде, не содержащей К, транспорт
Na ослабевает; он уменьшается при большом значении Na0 и уси-
ливается при увеличении Nai.
Потенциал покоя обусловлен следующими факторами: присут-
ствием внутри волокна недиффундирующих органических анионов
(что позволяет малым анионам приблизиться к равновесию Дон-
нана); различной проницаемостью разных ионов в состоянии по-
коя; активным вытеснением Na, связанным с удержанием К; свя-
занным с Na вытеснением Са. Возбудимость определяется уров-
нем потенциала покоя. При снижении ПП (т. е. при деполяриза-
ции мембраны, вызванной, например, аноксией) возбудимость
можно восстановить, используя катодную реполяризацию.
Как видно из рис. 11-5, реакция клеточной мембраны может
быть обусловлена изменением трансмембранного сопротивления
(или проводимости 1/7?) или проницаемости для специфических
ионов, а также изменением в ионных батареях. Емкость, как пра-
вило, меняется мало. Иногда сопротивление изменяется несиммет-
рично, и ток в одном направлении идет легче, чем в другом; ины-
ми словами, мембрана может работать как выпрямитель.
Асимметричный потенциал. Асимметричный потенциал образу-
ется в уплощенных клетках, наружная и внутренняя поверхности
18—1514
274 Глава 11
которых различаются по своим электрическим свойствам. Прони-
цаемость этих двух клеточных поверхностей неодинакова, и через
одну из них идет активный перенос ионов (обычно натрия). Хоро-
шо известны асимметричные потенциалы эпителия; впервые их
обнаружили на коже лягушки (стр. 206), в мочевом пузыре жабы
(стр. 208), жабрах ракообразных и рыб и в эпителии желудочно-
кишечного тракта. Жидкости, омывающие две поверхности упло-
щенных клеток, при физиологических условиях часто существенно
различаются по составу. Таким образом, асимметричный клеточ-
ный потенциал возникает вследствие активной перекачки ионов
из одной жидкости в другую и играет важную роль в ионной ре-
гуляции. В гл. 17 даны примеры асимметричных потенциалов дей-
ствия в некоторых электрических органах.
Потенциалы, связанные с активностью. Существует несколько ти-
пов электрической активности возбудимых клеток, причем они
совсем не исключают друг друга. Рассмотрим некоторые из них,
представляющие наибольший интерес с практической точки зре-
ния: 1) местный, или нераспространяющийся, потенциал отдель-
ной клетки; этот потенциал может быть пассивным (зарядка .мем-
бранной электростатической емкости) или активным (изменение
проводимости мембраны); 2) распространяющийся, регенератив-
ный потенциал действия, подчиняющийся закону «все или ниче-
го», и 3) контактный потенциал, возбудительный или тормозный
постсинаптический потенциал (ВПСП или ТПСП). Ранее мы
рассмотрели явление, которое тоже может привести к возникно-
вению потенциала действия,— изменение в работе электрогенных
ионных насосов. В этой главе рассматриваются главным образом
потенциалы действия в нервной ткани, а о мышцах и электриче-
ских органах мы будем говорить в последующих главах.
Местный потенциал и электротоническое проведение. Аксон
или волокно поперечнополосатой мышцы можно представить как
кабель — цилиндрический проводник низкого сопротивления, окру-
женный мембраной с высоким сопротивлением, которая отделяет
его от среды с низким сопротивлением. Местные потенциалы из
любого источника градуальны по своей амплитуде и могут пас-
сивно распространяться с затуханием по 7?С-контуру этого кабе-
ля. Распространение по нервному или мышечному волокну под-
порогового тока определяется временной и пространственной кон-
стантами мембраны. Эта поляризация, или электротонические по-
тенциалы на катоде и аноде одинаковы по величине, но противопо-
ложны по знаку. При увеличении силы раздражения до величи-
ны, составляющей приблизительно половину реобазы (порога
при очень длительном раздражающем токе), местный ответ (пре-
потенциал) появляется на катоде. Это нераспространяющийся
градуальный препотенциал, но увеличивается он не пропорцио-
нально силе раздражителя, т. е. представляет собой активное
уменьшение сопротивления для входящего тока. По достижении
"WH
\Возбудимые мембраны 275
критической амплитуды этот локальный потенциал может приве-
сти к возникновению распространяющегося потенциала действия
(спайка).
При местной блокаде нервного импульса под действием холо-
да или анастетика можно зарегистрировать электротонический по-
тенциал сразу же за пределами блокированного участка. Ток, рас-
пространяющийся в виде .импульсов, пассивно протекает в локаль-
ной цепи от деполяризованного участка волокна к той его ча-
сти, где деполяризация отсутствует. При этом ток течет по
цитоплазматическому сердечнику, затем наружу через недеполя-
ризованную мембрану и в обратном направлении через окружа-
ющую среду. Ток, направленный наружу, деполяризует тот уча-
сток мембраны, через который он протекает. Скорость проведения
снижается при повышении сопротивления сердечника или окру-
жающей среды (например, когда волокно находится в масле или
на воздухе).
В последующих главах приведены многочисленные примеры
возникновения градуальных или местных потенциалов. В каждом
из этих случаев повышается проницаемость мембраны для одного
или нескольких типов ионов. Потенциалы водителей ритма (в ча-
стности, пейсмекера сердца, гл. 20) обусловлены градуальны-
ми ритмическими изменениями потенциала покоя или специфиче-
скими изменениями проводимости, зависящими от временных и
пространственных параметров. Сенсорные потенциалы в чувстви-
тельных к механическому или химическому воздействию нервных
окончаниях и генераторные потенциалы в эпителиальных клетках
приводят к возникновению нервных импульсов; в соответствую-
щих разделах приведены примеры хеморецепторов, рецепторов ра-
стяжения, фонорецепторов и фоторецепторов. Контактные, или
синаптические, потенциалы возникают вследствие деполяризации
или гиперполяризации постсинаптических мембран (синаптиче-
ские потенциалы рассматриваются в гл. 11, а потенциалы нервно-
мышечной передачи — в гл. 16).
При электротоническом распространении местного потенциала
происходит перенос заряда в соответствии с пространственной
константой мембраны и в нейронах с короткими отростками ин-
формация на соседние нейроны не всегда передается с помощью
потенциала действия. Так, в коротких проприоцептивных нейро-
нах мелких ракообразных [27, 185] (рис. 11-6) и в зрительных
волокнах второго порядка в глазах мух [248] информация пере-
дается от рецепторов в центральную нервную систему электрото-
нически, без потенциалов действия. Электротонический потенциал,
вероятно, вызывает в каждой из этих тканей высвобождение како-
го-то химического передатчика. Некоторое количество клеток в
сетчатке позвоночных функционирует последовательно, передавая
информацию лишь с помощью градуальных медленных потенциа-
лов, которые бывают гиперполяризующими или деполяризующи-
18*
276
Глава 11
Рис. 11-6. Вызванные механическим раздражением различной амплитуды (А) или
растяжением различной интенсивности (Б) (нижняя запись) реакции, зарегистри-
рованные внутриклеточными (верхняя запись) или внеклеточными (средняя за-
пись) электродами в сенсорных волокнах мышечного рецептора краба
(Ripley S. М., Bush М. М, Roberts A., Nature, 218, 1170—1171, 1968).
Эти реакции являются генераторными потенциалами, которые распространяются электрото-
нически и активируют интернейроны центрального ганглия.
ми. Такой тип передачи информации, вероятно, встречается в тех
случаях, когда нейрон активируется на одном из полюсов, а дли-
на аксона ненамного больше его пространственной константы.
Электротоническое распространение играет большую роль в
нейронах, не имеющих аксонов, а также при передаче сигнала с
дендритов одного нейрона на дендриты другого (например, в сер-
Возбудимые мембраны
217
печных ганглиях ракообразных). Дендриты некоторых нейронов,,
по-видимому, не способны генерировать потенциалы действия, но
передают информацию, например, между клетками-зернами и ми-
тральными клетками обонятельной луковицы (гл. 15) [198]
или между сильно разветвленными дендритами клеток вентраль-
ного таламуса [199]. У некоторых видов дендриты клеток Пур-
кинье мозжечка не генерируют потенциал действия, а передают
синантические сигналы на сому клеток [56]. В пирамидальных
клетках коры больших полушарий наблюдается градуальная де-
поляризация дендритов. Эта деполяризация, ослабевая в соме’
клеток, оказывается тем не менее достаточной, чтобы модулиро-
вать частоту их разрядов ',[96].
Таким образом, электротоническое распространение граду-
альной деполяризации или гиперполяризации широко использует-
ся для передачи информации там, где нет унитарных импульсов.
Сумма всех положительных и отрицательных электротонических
потенциалов, достигающих места генерирования потенциалов дей-
ствия, определяет частоту разрядов клетки.
Распространяющиеся потенциалы действия (регенеративные
спайки). Если на электр©возбудимую клетку действует раздража-
ющий сигнал пороговой величины, местный потенциал достигает
критической амплитуды и возникает распространяющийся импульс
(рис. 11-7). В нервном волокне ракообразных критическая ампли-
туда местного потенциала равна приблизительно 20% амплитуды
потенциала действия. Отношение амплитуды потенциала действия;
к пороговому уровню деполяризации называется коэффициентом;
(или фактором) надежности. Спайк (нервный или мышечный им-
пульс) отличается от тока в пассивном проводнике тем, что явля-
ется самораспространяющимся (регенеративным), распространя-
ется от источника без затухания (декремента), т. е. подчиняется
закону «все иди ничего», и завершается рефрактерным периодом.
Информация, переносимая потенциалом действия, кодируется не
амплитудой (как в случае электротонического проведения), а
частотой импульсов и их распределением. В норме возбуждение
проявляется как повышение ионной проницаемости, выражающе-
еся в повышенной проводимости (g). В течение восходящей фа-
зы нервного импульса сопротивление мембраны снижается приб-
лизительно в сорок раз, а емкость не изменяется. Проведение на-
блюдается и в отсутствие кислорода. Ферменты аксоплазмы суще-
ственной роли в проведении не играют. Из аксона кальмара,
можно выдавить, аксоплазму и перфузировать его раствором, со-
держащим калий в соответствующей концентрации и какой-либо
непроникающий через мембрану анион (например, сульфат или:
изетионат); в этих условиях мембрана будет проводить нормаль-
ные импульсы [10]. Однако ионные насосы, которые требуют для
своей работы сохранения окислительных механизмов аксоплазмы^
таком волокне функционировать не будут.
278
Глава 11
Д>ля измерения ионных токов, идущих через электрогенную
мембрану, успешно применяется техника фиксации напряжения.
В крупную клетку, например в гигантский аксон кальмара, вво-
дят два электрода — один для подачи тока и другой для регист-
рации напряжения; с помощью
усилителя с обратной связью мож-
но измерить ток, необходимый
для фиксации мембранного по-
тенциала на данном уровне
(деполяризации или гиперпо-
ляризации). Общий мембран-
ный ток /м слагается из
двух компонентов — тока,
Рис. 11-7. Реакции, возникающие в
нервном волокне краба при раздра-
жении электрическими импульсами
возрастающей интенсивности (Hodg-
kin A. L., Proc. Roy., Soc. bond., В,
126, 87—121, 1938).
Кривые, расположенные под осью абсцисс,
и первые 5 кривых над осью абсцисс —
электротонический потенциал на аноде и
катоде соответственно; следующие 4 кри-
вые над осью абсцисс — местная реакция;
последние 3 кривые — возникновение потен-
циала действия; а — другая серия опытов;
электрические изменения на катоде.
необходимого для
сти С, и тока 1ц,
зарядки мембранной электростатической емко-
обусловленного перемещением ионов:
/ —С —4-/ 1
G dt + и’
В условиях фиксации-^- =0 и /м=/и. Ионный ток в нерве яв-
.ляется суммой токов, обусловленных перемещением ионов натрия
и калия, и тока утечки (главным образом утечки хлоридов); каж-
дый из этих токов определяется проводимостью данного иона и
потенциалом, который соответствует мембранному градиенту по-
тенциала инверсии. Потенциал инверсии для каждого иона — это
равновесный потенциал, определяемый концентрационным гради-
ентом иона в соответствии с уравнением Нернста (стр. 32).
Натриевая проводимость grNa==yZ.N^~»
Калиевая проводимость
£к— v _ук •
Возбудимые мембраны
279»
Концентрационный градиент натрия направлен внутрь клетки, а
калия — наружу. Измерения тока, про(водимые в условиях фикса-
ции напряжения одновременно с замещением ионов, показали, что-
при прохождении по аксону волны потенциала действия проницае-
мость мембраны как для натрия, так и для калия повышается,,
хотя этот процесс для разных ионов имеет различные временные
характеристики (рис. 11-8). Во время фазы роста потенциала дей-
ствия проницаемость мембраны для Na увеличивается и Na+ дви-
жется внутрь аксона, поскольку [Na0]>[Nai] и /ца^/ко; выброс
потенциала действия соответствует приближению к равновесному
потенциалу для натрия. Замещение натрия в среде катионом, не-
проходящим сквозь мембрану (холином или трис), уменьшает ам-
плитуду потенциала действия. В некоторых нервах Na+ можно*
заменить литием, аминами и катионами четвертичных аммонийных
соединений азота. Во время нисходящей фазы потенциала дейст-
вия натриевая проводимость падает и . Калиевая прово-
димость повышается гораздо медленнее, чем g’Na, так что к тому-
времени, когда ток натрия начинает уменьшаться, £к еще увели-
чивается и в нисходящей фазе спайка ток, направленный из клет-
ки, обусловливают ионы калия [29]. Таким образом, в основе-
нервного импульса лежит изменение натриевой и калиевой прово-
димости; эти изменения зависят от напряжения и времени. Изме-
рения, проведенные на аксонах кальмара с помощью метки, пока-
зали, что за один импульс в аксон переносится 4пМ Na+/cM2 и
удаляется эквивалентное количество К+; Са2+ проникает в аксон
в количестве 0,006 пМ/см2 за один импульс [116, 117]. Натриевая
проводимость увеличивается в 1000 раз и составляет в гигантском
аксоне омара 0,53 МОм/см2, а в аксоне кальмара — 0,12 МОм/см2'
[158, 159]. После прохождения потенциала действия через макси-
мум проницаемость для ионов К (обусловливающих выходящий
ток) увеличивается, а для ионов Na (обусловливающих входящий
ток) уменьшается.
Каналы, через которые поступает натрий, можно блокировать
тетродотоксином (ТТК) — ядом, полученным из рыбы тетродонта.
Оценки, основанные на определении количества ТТК, необходимо-
го для блокирования импульса, и на допущении, что одна моле-
кула ТТК блокирует один канал, показывают, что на 1 мкм2 мем-
браны приходится 13 чувствительных к ТТК зон. Другой яд, сак-
ситоксин, полученный из некоторых двустворчатых моллюсков,,
также блокирует натриевые каналы, но действует в отличие от
ТТК необратимо [104, 159, 167]. Тетраэтиламмоний (ТЭА) специ-
фически блокирует каналы, через которые выходит К, и поэтому
увеличивает продолжительность потенциала действия; при этом
наблюдается некоторая реполяризация, обусловленная проникно-
вением внутрь клетки хлора. Инсектицид ДДТ поддерживает нат-
риевые каналы открытыми (т. е. препятствует падению натриево-
280
Глава 11
Рис. 11-8.
А. Измерение трансмембранных ионных токов в гигантском аксоне кальмара методом фик-
сации напряжения. Мембранный потенциал внезапно смещают с уровня —56 мВ до нуля.
^Na^K-ток в Уравновешенной ионной среде; /к—калиевый ток при замещении Na хо-
лином; /j\ja вычислен по разности суммарного и калиевого токов.Току, направленному внутрь
соответствует отклонение кривой вниз, а току, направленному наружу, — отклонение кри-
вой вверх. Б. Потенциал действия (кривая V) в гигантском аксоне кальмара; проводимости
Na+ и К+ (gj^) вычислены по величине тока при деполяризации и реполяризации*
'У и — равновесные потенциалы для N+ и К+ (Hodgkin А. Г., Proc. Roy. Soc. Lond. В,
148, 1—37, 1958).
го тока); при этом замедляется также рост и, следовательно,
сохраняется деполяризация мембраны [89]. Многочисленные дан-
ные, в частности полученные при изучении влияния ТТК и ТЭА,
ри" ~
____Возбудимые мембраны____________________281
свидетельствуют о том, что Na+ и К+ проходят по разным кана-
лам. Проводимости различных одновалентных катионов, измерен-
ные в аксоне кальмара ibo .время максимума входящего тока, дают
следующие величины относительной проницаемости [225]:
Li Na К Rb Cs
1,1 1 1/12 1/40 1/60
После прохождения волны потенциала действия натрий-калиевый
насос восстанавливает характерные для состояния покоя ионные
градиенты. В гигантском аксоне, аксоплазма которого заменена
раствором K2SO4, скорость поступления натрия при прохождении
волны потенциала действия в миллион раз выше, чем скорость
его выведения ионным насосом в состоянии покоя [89].
При распространении волны потенциала действия по аксону
увеличивается и кальциевая проводимость, однако это повышение
в 600 раз меньше, чем для натрия. Невозбужденный аксон каль-
мара в норме содержит 0,3 мкМ. ионов Са при общей концентра-
ции Са 400 мкМ; после действия цианида в течение 50—100 мин
внутриклеточная концентрация Са2+ повышается в 500 раз. Если
раздражать аксон с частотой от 50 до 200 имп/с, наблюдается
медленное повышение концентрации Сац восстановление нормаль-
ного уровня после прекращения раздражения происходит с вре-
менной константой 10—30 с [7]. Процесс увеличения кальциевой
проводимости двухфазный; первая его фаза чувствительна к ТТК,
и, возможно, Са2+ в этот период проходит через натриевые кана-
лы. Вторая фаза совпадает по времени с максимумом калиевого
тока, но не чувствительна к ТЭА; эта фаза угнетается Мп2+ [7].
В некоторых клетках кальциевый ток по величине превышает
ток, обусловленный ионами натрия. Например, в гигантских ней-
ронах моллюска Aplysia замещение Na или Са в среде другим
ионом (тцким, как трис) приводит к уменьшению выброса потен-
циала действия, а замещение обоих ионов (и Na, и Са) — к пол-
ному его исчезновению. Натриевый ток блокируется ТТК, а каль-
циевый— кобальтом; в бескальциевой среде весь ток обусловлен
ионами натрия, а в безнатриевой — ионами кальция [73]. Для од-
ного класса нейронов улитки Helix выброс потенциала действия
зависит от Na, для других нейронов величина входного тока опре-
деляется и натрием, и кальцием, а для прочих нейронов — только
кальцием [108, НО].
В некоторых мышечных волокнах (например, в поперечнопо-
лосатых мышцах позвоночных) потенциал действия обусловлен из-
менением натриевой проводимости. В другом случае (например,
в гладких мышцах кишечника позвоночных, в поперечнополосатых
мышцах ракообразных) этот потенциал обусловлен преимущест-
венно изменением кальциевой проводимости. В некоторых мышцах
(вапример, у Branchiostoma, гл. 16) после устранения натрие-
282
Глава 11
вого тока с помощью тетродотоксина или замещения натрия ок-
ружающей среды, а также после устранения калиевого тока про-
каином или ТЭА обнаруживается значительный кальциевый ток
'[856]. У отдельных позвоночных деполяризация миокарда сопро-
вождается появлением входящего кальциевого тока [12]. При
действии адреналина на мышцу предсердия лягушки усиливается
как быстрый входящий ток Na, так и медленный входящий ток
С а [235]. У большинства проводящих мембран имеются, по-види-
мому, ионные каналы нескольких типов. В норме натрий и калий
проходят по разным каналам; кроме того, в большинстве мем-
бран, по-видимому, имеются и кальциевые каналы.
При действии кальция на возбудимые мембраны наблюдаются
и другие явления. Кальций поддерживает нормальную проницае-
мость мембраны или ее сопротивление, т. е. стабилизирует мем-
брану. Изменение концентрации Са в окружающей среде приво-
дит к сдвигу кривых натрий-калиевой проводимости вдоль оси на-
пряжения [118].
Деполяризация клетки может возникнуть еще в одном слу-
чае— при увеличении выходящего тока С1. Электрическое разд-
ражение некоторых растительных клеток, например клеток круп-
ных нитчатых водорослей, приводит к возникновению потенциала
действия, обусловленного повышением gci- В мышцах некоторых
ракообразных имеется обширная система внутриклеточных трубо-
чек, и отток ионов хлора в складки внеклеточного пространства
приводит к деполяризации [81]. Сходным образом у ската при си-
наптической стимуляции электроцита усиливается отток С1~ через
одну из клеточных поверхностей. Если Еа положителен по отно-
шению к ПП, увеличение gci должно привести к оттоку С1~ и де-
поляризации, несмотря на то что химический концентрационный
градиент С1 направлен внутрь клетки. Реполяризация сердечной
мышцы отчасти обусловлена входящим Zci [67]. Адреналин вызы-
вает деполяризацию гладких мышц taenia coli морско^ свинки,
открывая каналы как для С1~, так и для К+; при этом на долю
С1~ приходится 36% прироста проводимости [177].
Последующие процессы и генераторные потенциалы. После
прохождения волны потенциала действия во многих нервных во-
локнах наблюдается гиперполяризация (рис. 11-9). Она выраже-
на очень четко (например, в гигантских аксонах кальмара), а по-
сле тетанического раздражения безмякотных нервных волокон
позвоночных сохраняется в течение 0,1 с или дольше. Эта следо-
вая гиперполяриэация наблюдалась в ранних экспериментах как
положительный потенциал последействия, регистрируемый внекле-
точными электродами. Опыты с мечеными атомами и другие экс-
перименты показали, что гиперполяризация, следующая за воль-
ной потенциала действия, обусловлена сохраняющейся высокой
калиевой проводимостью. В мотонейронах эта гиперполяризация
снимается при уменьшении градиента калия (Ki/K0).
Возбудимые мембраны.
28&
В других возбудимых волокнах вслед за волной потенциала
действия наблюдается деполяризация, задерживающая спад это-
го потенциала. Внеклеточные электроды регистрируют эту депо-
ляризацию в некоторых нервах как отрицательный потенциал по-
следействия. Длительная деполяризация может иметь характер
плато, как это наблюдается в сердечной мышце (гл. 20). Де-
Рис. 11-9. Потенциалы действия, зарегистрированные между наружной и внутрен-
ней поверхностями гигантского аксона кальмара (Hodgkin A. L., Huxley A. F.,
J. Physiol., 104, 176—195, 1945).
Показаны уровень потенциала покоя, выброс потенциала действия и следовая гиперполя-
ризация. Калибровка времени 1000/с.
поляризация последействия четко выражена -в крупных миелини-
зированных волокнах млекопитающих, а в нервных волокнах кра-
ба продолжается в течение нескольких секунд. Вератрин, а во
многих случаях и ТЭА вызывают запаздывание реполяризации.
Эти же агенты задерживают и калиевый ток.
Другой тип гиперполяризации, наступающей после прохожде-
ния волны потенциала действия, связан с работой натриевого на-
соса. Компоненты потенциала действия, обусловленные изменени-
ями активности натриевого насоса, можно выявить по отсутствию
изменения мембранной проводимости, по повышению ее, вызван-
ному инъекцией натрия, по их зависимости от концентрации ка-
лия в окружающей среде, а также с помощью уабаина, обладаю-
щего блокирующим действием. Например, в некоторых нейронах
пиявки вслед за серией вспышек активности наблюдается дли-
тельная электроотрицательность, медленная фаза которой снима-
ется уабаином [168]. В рецепторах растяжения наблюдается ги-
перполяризация, которая также связана с транспортом натрия
84
Глава И
Имеются данные о том, что деполяризация в некоторых тканях
связана не только с изменением проницаемости мембраны для
натрия и кальция, но и с наступающим на ранних этапах измене-
нием калиевой проводимости. При гиперполяризации некоторых
нейронов моллюска Antsodoris и последующей их деполяризации
.возникает выходящий ток калия (1а), который отличается от на-
чального тока при прохождении волны потенциала действия (/^а)
и от тока восстановления (/к). Под влиянием этого возникающего
на ранних этапах тока калия задерживается или замедляется дей-
ствие приложенного напряжения, но в конечном счете входящий
ток Na превышает /д и возникает потенциал действия. Считают,
что в периодически разряжающихся нейронах гиперпол яр из ация,
наступающая после прохождения волны потенциала действия, при-
водит к возникновению раннего калиевого тока, который опреде-
ляет величину интервала до следующей волны возбуждения. Ран-
ний калиевый ток и ток, связанный с реполяризацией при про-
хождении импульса, идут по разным каналам [37].
У находящихся в состоянии покоя клеток ведущего узла серд-
ца позвоночных натриевая проводимость выше, чем проводимость
калия, которая обусловливает устойчивый входящий ток. Эти про-
водимости зависят от времени и от напряжения. Если они соот-
ветствуют пороговой величине, необходимой для генерирования
потенциалов действия, возникают циклически повторяющиеся им-
пульсы [226]. Для некоторых невозбужденных нейронов Aplysia
характерно высокое отношение P^JPk, и возникающие в них цик-
лические импульсы выражены более четко, чем у клеток ведущего
узла сердца позвоночных; в то же время в некоторых нейронах
возникают повторяющиеся разряды и в отсутствие Na+ в среде
[28].
Градуальные реакции, возникающие во многих механорецепто-
рах, обусловлены увеличением натриевой проводимости, даже если
мембрана не генерирует потенциалов действия. Освещение вызы-
вает прирост gNa в фоторецепторах некоторых беспозвоночных
(мечехвост Limulus, морские желуди; стр. 512). В темноте у фо-
торецепторов позвоночных отношение Р^п/Рк высоко; на дисталь-
ном конце рецептора генерируется натриевый ток, который можно
зарегистрировать с помощью внеклеточных электродов. Освеще-
ние уменьшает этот темновой ток и вызывает гиперполяризацию
рецептора (стр. 504). Потенциал покоя в некоторых фоторецепто-
рах гребешка ниже, чем Ек, а отношение Рыа/Т’к велико. Освеще-
ние вызывает гиперполяризацию, обусловленную увеличением gK
[151а].
Синаптические потенциалы в химических синапсах
Постсинаптические мембраны отличаются от мембран проводя-
щих аксонов и мышечных волокон, а также от мембран сенсор-
ных клеток и пейсмекерных клеток. Появлению импульсов в си-
Возбудимые мембраны
285
пайсах предшествуют градуальные по своей природе потенциа-
лы—-это или 1) возбудительные (обычно деполяризующие) си-
наптические или постсинаптические потенциалы (ВПСП), или
2) тормозные (обычно гиперполяризующие) постсинаптические по-
тенциалы (ТПСП). Синаптический потенциал можно зарегистри-
ровать и тогда, когда постсинаптический потенциал действия не
возникает вследствие блокирования его различными препаратами
(ТТК или прокаином) или вследствие утомления (как после пре-
Рис. 11-10. Синаптические потенциалы и потенциалы действия, зарегистрирован'
ные в звездчатом ганглии кальмара (Bullock Т. И., Nature, 158, 500—556, 1946).
Нижняя запись — импульс в преганглионарном волокне. Верхняя запись — постганглионар-
ные потенциалы действия, которые по мере утомления возникают все позже и позже по
отношению к синаптическому потенциалу; в конце концов остается только синаптический
потенциал.
синаптической тетанизации звездчатого ганглия кальмара)
(рис. 11-10), Продолжительность импульса ВПСП больше, чем
длительность рефрактерного периода пресинаптического сигнала,
и поэтому повторяющиеся импульсы могут суммироваться; когда
ВПСП вследствие суммирования возрастает, постсинаптический
импульс возникает раньше, а когда снижается при утомлении, то
позже. Величина синаптического потенциала зависит от количест-
ва выделившегося медиатора. В постсинаптических мембранах
многих синапсов (например, в звездчатом ганглии кальмара и в
концевых пластинках скелетных мышц) возникают спонтанные
миниатюрные синаптические потенциалы (мВПСП) единичной
амплитуды. Считают, что эти потенциалы обусловлены высвобож-
дением некоторой минимальной порции (кванта) медиатора
[153—155], а каждый ВПСП возникает в результате действия оп-
ределенного количества (обычно нескольких сотен) таких квантов.
Во всех известных случаях для высвобождения медиатора не-
обходимы: а) некоторое количество внеклеточного кальция и
286
Глава 11
б) деполяризация пресинаптических окончаний. Высвобождение
медиатора уменьшается при высоком содержании магния и отсут-
ствии кальция. Например, повышение концентрации Са в среде
увеличивает синаптический ток в синапсах звездчатого ганглия
кальмара, образованных гигантскими волокнами, а повышение
концентрации Mg уменьшает этот ток. Импульс ВПСП возникает
при пресинаптичеакой деполяризации, равной 25 мВ, а миниатюр-
ные постсинаптнческие потенциалы появляются даже в присутст-
вии ТТК, блокирующего пресинаптические импульсы. Для сопря-
жения деполяризации с секрецией медиатора необходим Са2+
[129, 155]. В концевых пластинках портняжной мышцы лягушки
кальций можно заменить стронцием, который, однако, значитель-
но менее эффективен [152]. Измерение количества квантов меди-
атора ацетилхолина (АХ), высвобождаемого в нервных окончаниях
портняжной мышцы лягушки при различных концентрациях каль-
ция, показало, что в каждой «критической точке» должно присут-
ствовать четыре иона кальция. Количество квантов высвобождае-
мого АХ пропорционально [СаХ]4, где X — активируемая молеку-
ла. По числу квантов, соответствующих одному ВПСП, можно
судить о числе молекул X, образующих комплекс с Са2+ |[51].
При возникновении ВПСП ,в диафрагме крысы одному кванту ме-
диатора соответствует в среднем 2,7 иона Са, а при миниатюр-
ном ПСП — один ион. В опытах на диафрагме мыши получены
данные, указывающие на кооперативность процесса — высвобож-
дение квантов медиатора ускоряется при повышении концентрации
Са2+, причем при высокой концентрации калия в большей степени,
чем при низкой. Регистрация электрических импульсов в преси-
наптических окончаниях звездчатого ганглия кальмара показала,
что в них возникает регенеративный электрический потенциал,
обусловленный притоком Са2+. В норме этот регенеративный по-
тенциал действия нервного окончания реполяризуется калиевым
током; однако в присутствии ТЭА он выходит на плато, величина
которого зависит от концентрации Са в окружающей среде. Пла-
то уменьшается под влиянием Мп2+, и это сопровождается умень-
шением 7?м [105, 106]. Можно заключить, что деполяризация мем-
браны пресинаттического окончания повышает ее проницаемость
для Са и входящий ток Са2+ приводит к высвобождению медиато-
ра из синаптических пузырьков.
Синаптическая задержка включает в себя время, необходимое
для высвобождения и диффузии медиатора и для его взаимодей-
ствия с рецепторами постсинаптической мембраны. В звездчатом
ганглии кальмара распространение ВПСП начинается с задерж-
кой в 0,5 мс, достигает максимума через 1 мс и затухает с констан-
той 1,2 мс [24]. Время нарастания ВПСП в крупных нейронах
Aplysia достигает 100 мс, а время затухания — 500 мс.
Постсинаптические мембраны не обладают электрической воз-
будимостью, и возникающий в них ток определяется количеством
Возбудимые мембраны
287
химического медиатора. Взаимодействие медиатора с рецептором
мембраны повышает ее проницаемость для определенных ионов.
Если потенциал постсинаптической мембраны меньше, чем равно-
весный потенциал для этих ионов, то ВПСП увеличивается, а ес-
ли больше, то ВПСП меняет знак. Для ТПСП эффект носит об-
ратный характер.
Зависимость между током и напряжением в постсинаптической
хемочувствительной мембране носит линейный характер, т. е. не
происходит выпрямления тока, а мембранное сопротивление явля-
ется омическим, или независимым от мембранного потенциала.
Возбудительные синаптические потенциалы. Нервный импульс,
распространяющийся по электровозбудимой мембране, обусловлен
последовательным повышением натриевой (или кальциевой) и ка-
лиевой проводимости, тогда как ВПСП обычно является следст-
вием повышения одновременно натриевой и калиевой проводимо-
сти. Амплитуда ВПСП в спинальных нейронах понижается, если
мембранный потенциал сдвинут в сторону деполяризации, и рав-
новесный потенциал близок к нулю [44]. В мышечных концевых
пластинках позвоночных равновесный потенциал равен —15 мВ
и ВПСП зависит от одновременного повышения проницаемости
для обоих ионов — Na и К. В синапсе гигантского нервного во-
локна кальмара ВПСП приближается к равновесному потенциа-
лу для Na (ф-45 мВ), а калиевый ток отсутствует [70]. В симпа-
тическом ганглии лягушки потенциал инверсии составляет от —8
до —20 мВ, т. е. является почти таким же, как в концевой плас-
тинке [172, 173].
Использование введенного в сому нейрона электрода для из-
мерения потенциала равновесия ВПСП или ТПСП, или потенциа-
ла инверсии может дать неточные результаты, особенно в том
случае, когда синапс удален от тела нейрона и находится на ден-
дритах или неглавных ветвях аксона. Во всех синапсах и нейро-
эффекторных соединениях участок мембраны с изменяющимся
сопротивлением связан с участками, не изменяющимися под вли-
янием медиатора [80].
Некоторые постсинаптические импульсы могут возникать при
искусственном удлинении ВПСП. Так, ВПСП некоторых интерней-
ронов (клеток Реншоу) в норме длится 50 мс, но под влиянием
ингибитора ацетилхолинэстеразы эзерина его продолжительность
увеличивается до 300 мс, и в течение всего этого времени генери-
руются серии высокочастотных импульсов. Возникновение одиноч-
ных импульсов в постсинаптических клетках других синапсов
обусловлено ^синхронными сложными ВПСП. В двухнейронной
Рефлекторной дуге спинного мозга кошки ВПСП возбуждается
в мотонейроне через 0,5 мс после поступления сенсорного залпа
® спинной мозг; максимум ВПСП достигается через 1—1,5 мс, а
константа затухания равна 4,7 мс [38]. Ток очень быстро возра-
стает и достигает максимума раньше, чем напряжение (рис. 11-
Возбуждающее
синаптическое
Б
Возбудимые мембраны
289
11,Л)- При равной 10 мВ деполяризации мотонейрона, ПП которо-
го’равен —70 мВ, постсинаптический потенциал действия генери-
руется в начальном сегменте нейрона. ВПСП в соме и дендритах
дает деполяризацию, достаточную для возникновения спайка в на-
чальном сегменте аксона; позднее этот спайк может охватить со-
му и дендриты, где пороговая величина деполяризации для гене-
рирования потенциалов действия равна 30 мВ [54, 55]. С другой
стороны, в клетках Пуркинье мозжечка некоторых животных и в
некоторых пирамидальных клетках коры потенциалы действия
могут передаваться через синапсы дендритов. Возможно также
широкое распространение синаптических потенциалов в ветвях
дендритов, как это наблюдается в униполярных нейронах ракооб-
разных.
В шейных симпатических ганглиях кошки, кролика, лягушки
и черепахи наблюдается три типа постсинаптических контактных
потенциалов; относительный вклад их у разных видов неодинаков.
Медиатором возбуждения здесь выступает АХ. У кошки начальный
ВПСП имеет латентный период 4—9 мс и время спада 6—12 мс;
этот потенциал, по-видимому, связан с никотиновыми рецептора-
ми, чувствительными к тубокурарину. Поздний ВПСП кролика
имеет латентный период 200—300 мс и блокируется атропином.
Тормозный ПСП имеет латентный период 35 мс и блокируется
а-адренолитиком дибенамином; по-видимому, этот потенциал воз-
никает при холинэргической активации хромаффинных клеток, ко-
торые в свою очередь выделяют норадреналин, действующий на
клетки симпатического ганглия. Все три типа синаптических по-
тенциалов можно блокировать ядом Bacillus botulinum или пони-
жая отношение Ca/Mg, т е. воздействиями, подавляющими высво-
бождение АХ из преганглионарных волокон [133, 134].
В симпатических ганглиях лягушки (а возможно, и черепахи)
хромаффинных клеток мало или нет вообще и регистрация потен-
циалов в отдельных нейронах свидетельствует об ином механизме
возникновения медленных ПСП, чем те, которые были постулиро-
ваны для кошки и кролика на основании опытов с внеклеточными
электродами. Быстрые ответы у лягушки, как и у млекопитающих,
связаны с никотиновыми рецепторами. В одном из типов клеток
Рис. 11-11.
кпиряо?^ДИтелЬНЫ^ постсинаптический потенциал в мотонейроне (левый график, сплошная
токя в о11 соответствующий постсинаптический ток (пунктирная кривая). В центре —линии
xv гппяояТИВИроваН1ой/^?Напсе’ масштаб по вертикальной шкале увеличен в 10 раз. Ввер-
кпипЛ ^гпг-Яапись впсп в симпатическом ганглии лягушки. Внизу справа — теоретические
Фике к т „ и постсинаптического тока, аналогичные кривым, приведенным на левом гра-
синаптипо1^рлОЗНЬ1Й/ постсинаптический потенциал в мотонейроне (сплошная кривая) и пост-
nucleus ^гСК?? т°к <пУнктиРная кривая) [56]. В. Инверсия ТПСП и ВПСП в мотонейронах
за — „ <rochleans кошки при раздражении вестибулярного нерва на той же стороне. Сле-
и послепг,Н^Иие тпперполяризующей реакции при вливании С1-; справа — уменьшение ВПСП
Дующее изменение его знака по мере увеличения приложенного внутриклеточно де-
поляризующего тока (Lltnas R., Baker R„ J. Neurophys., 35, 484—492, 1972).
19—1514
290
Глава 11
обнаружен медленный ВПСП, по-видимому обусловленный умень-
шением проводимости при возбуждении, т. е. увеличением /?м и
понижением gx. В клетках другого типа обнаружено гиперполяри-
зующее действие АХ, обусловленное снижением проводимости Na,
т. е. сдвигом от [237].
Таким образом, деполяризующие ВПСП являются в том или
ином виде следствием изменения натриевой и калиевой проводи-
мости.
Тормозные постсинаптические потенциалы. Медиатор торможе-
ния обычно вызывает такие изменения проводимости рецепторной
мембраны, которые приводят к возникновению тормозного синап-
тического потенциала и уменьшают реакцию на ПСП возбужде-
ния. ТПСП, как правило, имеют больший латентный период и боль-
шую продолжительность, чем ВПСП. ТПСП обычно гиперполяри-
зующий, но может быть и деполяризующим, если мембранный ПП
отрицателен по отношению к равновесному потенциалу для ионов,
вызывающих ТПСП. Этот переход от гиперполяризующего к депо-
ляризующему ТПСП можно сопоставить с изменением знака
ВПСП, происходящим тогда, когда потенциал покоя положителен
по отношению к равновесному потенциалу соответствующих ионов.
Мышцы ракообразных имеют как возбуждающую, так и тор-
мозную иннервацию. В некоторых мышцах ракообразных тормоз-
ные окончания являются постсинаптическими и тормозные импуль-
сы повышают анионную проводимость мембраны мышечных кле-
ток. В других мышцах наблюдается пресинаптическое торможе-
ние, выражающееся в уменьшении ВПСП и не оказывающее пря-
мого влияния на мышечную мембрану.
На основании данных о ионных концентрациях и величинах на-
пряжения, меняющего знак ПСП, рассчитаны равновесные потен-
циалы спинальных мотонейронов кошки [3, 56]:
Еы (ПП)» мВ —70 ЕСр мВ —80
^Na* мВ +40 £ВПСП, мВ от —10 до 0
мВ —90 ^тпсп, мВ —80
При возникновении ТПСП мотонейронов кошки и маутнеров-
ских клеток серебряного карася основную роль играют ионы хло-
ра, некоторое значение имеют также ионы К [56]. Если с по-
мощью ионтофоретических методов, используя внутриклеточный
электрод, повысить внутриклеточную концентрацию С1_ и умень-
шить входящий ток, ТПСП изменит знак (рис. 11-11,В). При ис-
пытании серии анионов на спинальных мотонейронах показано,
что анионы меньшие, чем СЮГ, могут входить в клетку и таким
образом вызывать гиперполяризацию ТПСП; более крупные анио-
ны такого эффекта не дают [56]. Следовательно, медиатор тормо-
жения открывает мембранные каналы для малых анионов. Ана-
логичные результаты получены на маутнеровских клетках у рыб.
Возбудимые мембраны
291
Постсинаптические потенциалы у моллюсков. Сигналы, веду-
щие к возникновению гиперполяр изующих ТПСП в нейронах ган-
глиев моллюсков, поступают по специфическим нервным волок-
нам; по другим нервным волокнам могут идти сигналы, вызыва-
ющие в том же самом нейроне деполяризующие ВПСП (рис.
11-12). Один и тот же медиатор — ацетилхолин — вызывает в од-
них клетках (D-клетки) ВПСП, а в других (Н-клетки) —ТПСП
[101а]. В некоторых синапсах медиаторами являются нехолинэр-
гические агенты. Так, отдельные нейроны улитки деполяризуются
дофамином низкой (порядка 10~п М) концентрации, а другие —
серотонином (5-ОТ), кроме того, есть неидентифицированный пока
медиатор (передатчик «длительного торможения»), который ока-
зывает гиперполяризующее действие, длящееся много секунд [33,
74—79]. Разнообразие изменений ионной проводимости, свиде-
тельствующее о существовании различных мембранных рецепто-
ров и типов клеток, проиллюстрировано в табл. 11-2.
Действие ацетилхолина на некоторые D-клетки выражается в
деполяризации мембраны, обусловленной повышением натриевой
проводимости; равновесный потенциал близок к нулю [114]. Од-
нако при уменьшении концентрации Na деполяризация изменяется
под влиянием Mg2+, создающего входящий ток [207]. Гиперполя-
ризующее действие ацетилхолина на Н-клетки некоторых ганглиев
обусловлено повышением gci; эта реакция исчезает при замене
С1~ на SOF • В опытах с ионтофоретическим введением в эти ней-
роны Helix различных анионов было обнаружено, что анионы,
меньшие, чем ВгОГ , соответствующие диаметру пор 3 А, создают
входящий гиперполяризующий ток. В норме 90% этого тока, оче-
видно, связано с CI-, а 10% обусловлено выведением К+ [114,
115]. Гиперполяризация одного из нейронов другой улитки, Сгур-
iomphallus, нечувствительна к С1~, но при удалении из среды ка-
лия ТПСП удваивается вследствие более сильного выхода К+ из
клетки [76].
У Helix [ПО] и у Aplysia [207] содержание ионов хлора в Н-
клетках ниже, чем в D-клетках. Это способствует притоку С1~
при гиперполяризации Н-клеток. Потенциал покоя D-клеток Сгур-
tomphallus отрицателен по отношению к Ес\; по-видимому, высо-
кая .концентрация ионов С1 в этих клетках поддерживается за
счет работы С1~-насоса. Если под действием медиатора изменяет-
ся STr Деполяризация может быть вызвана и выходящим током
, причем этот ток может быть направлен даже против концент-
рационного градиента, если созданы соответствующие условия за
счет электростатического градиента [101а].
. Другого типа нейронов Helix гиперполяризация вызывается
мгчЛ*а1хИН0М’ °на нечУвствительна к К+ и С1~ и устраняется уабаи-
паир локирующим работу натриевого насоса [НО]. Сделанное
ня™™ предположение о влиянии тормозных импульсов на работу
натриевого насоса не подтвердилось.
19*
Морская вода
Нет СГ
Морская вода
Морская вода
,,НШ -J11SM
п
Морская вода
Морская вода
^50мВ
Л Б бс
Рис. 11-12. Ионные механизмы синаптического возбуждения (А) и торможе-
ния (Б) в D- и Н-клетках абдоминального ганглия Aplysla (из Kandel Е. R, in:
Electrophysiological studies in Neuropharmacology, ed. by W. Koella, Charles
C Thomas, Springfield, Ill., pp. 385—398, 1968).
Действие возбуждающих импульсов прекращается, если из среды удалить Na, а тормозя-
щих—если удалить С1. При удалении Na (А) происходит инверсия ответа D-клеток, а пР®
удалении С1 (Б) — инверсия ответа Н-клеток,
Таблица 11-2
Постсинаптические явления в нейронах моллюсков (сводная таблица)
Животное и тип клеток Равновесные потенциалы для АХ и постсинаптических ответов, мВ [СЦ], мэкв Потенциал покоя, мВ Механизмы
Aplysia [207]
D-клетка ЕАХ~ ~14 12 -36 t ^Naj
Н-клетка ^АХ= 60 43 От —36 до —48 t^CL
Aplysia [71] I
D-клетка ^ВПСП^ ~14 t^Na}
Н-клетка £ТПСГГ=~'74 —60 t^CIj
Двухфазные клетки Деполяризация до —14, затем f£Na, затем f£cl
Helix [109, НО, 114, 115] гиперполяризация до —63
D-клетка ЕАХ~ —62, 7?впсп= “64 24,7 —50
Н-клетка £Ах=-72 11,2 —55
Клетка, чувствительная к персдат- ^тпсп~ 60 fgK
чику «длительного торможения»»
деполяризуется дофамином до
—80
Cryptomphallus
D-клетка еах~ ~~25,1 —47
Н-клетка —56,5; ^тпсп= 55,8 t^CIj
Клетка с длительной деполяриза- о 1 X t^Naj
цией
Нехолинэргическая клетка с дли- £тпсп~ ““80
тельным торможением
1 Стрелки означают увеличение проводимости.
294
Глава 11
Ранее мы отмечали (стр. 281), что возникновение потенциала
действия у различных нейронов моллюсков обусловлено переносом
Na+, Са2+ или обоих катионов. Теперь к этому можно добавить,
что постсинаптические мембраны обладают избирательной чувст-
вительностью и к различным медиаторам и что реакция мембраны
на раздражение сводится к повышению ее проницаемости для Na,
К, С1, возможно, для Mg, а также к активации натриевого насо-
са.
Изменения мембранной проводимости
Электрически невозбудимые мембраны — это мембраны, обла-
дающие только пассивной реакцией на электрические раздражи-
тели. В нормальных условиях такие мембраны возбуждаются хи-
мическими синаптическими передатчиками (медиаторами) или,
если речь идет о сенсорных мембранах, специфическими химиче-
скими агентами, механически или с помощью света.
Мембраны этого типа функционируют как омическое сопротив-
ление, т. е. зависимость напряжения (Е) от тока (/) является ли-
нейной [83—85] (рис. 11-13). Однако наклон соответствующей
прямой может изменяться при активации, а изменение мембран-
ного потенциала становится обратным, если Ем больше потенциа-
ла инверсии, который определяется из пересечения прямых Е(1)
для пассивного и активного состояний (рис. 11-13). Таким обра-
зом, при деполяризации или гиперполяризации происходит инвер-
сия синаптических потенциалов.
Реакция электровозбудимых мембран является регенеративной,
и кривые Е(1) имеют нелинейные участки, соответствующие депо-
ляризации или гиперполяризации (рис. 11-13). Мембрану можно
назвать активированной, если сила тока увеличивается непропор-
ционально напряжению; при этом возрастает проводимость и ре-
акция оказывается регенеративной. Для большинства электровоз-
будимых мембран характерен деполяризующий электрогенез, ко-
торый бывает градуальным или подчиняется закону «все или
ничего». Реже встречается непропорциональный прирост гиперло-
ляризующего тока (пиперполяризующая активация). Если нели-
нейность выражается в отклонении в сторону непропорционально-
го прироста напряжения, то речь идет об инактивации мембраны.
В большинстве случаев деполяризующие потенциалы действия
обусловлены изменением проводимости Na и Са, которые имеют
большой положительный равновесный потенциал. Инактивация
это понижение проводимости, которое может происходить как при
деполяризующем, так и при гиперполяризующем токе (рис. 11-13)*
Нелинейные участки кривой Е(1) (увеличение или уменьшение
проводимости) соответствуют уменьшению или увеличению меМ'
бранного сопротивления в определенном направлении; говоря тех-
ническим языком, имеет место выпрямление тока. Происходят66
Возбудимые мембраны
295
при деполяризации аксонов уменьшение сопротивления или повы-
шение проводимости иногда называют нормальным выпрямлением.
Запаздывание реполяризации после прохождения волны потенциа-
ла действия выражается в наличии плато; повышение мембранно-
го сопротивления, наблюдаемое во время этого плато, называют
запаздывающим выпрямлением. У некоторых клеток (мышечных,
Рис. 11-13. Зависимость приложенного напряжения (Е) от тока (I) в электроне-
возбудимых и электровозбудимых мембранах [85].
плля»?ТеНЦИал и™е₽сни; 2 — деполяризационный электрогенез; 3 — входящий ток; 4 — гнпер-
тивипсти-Ц170ННЫЙ Н’еполяризационный) электрогенез; 5 — состояние покоя; 6 — состояние ак-
дуальный ' ~ деполяРизационная инактивация; Я—деполяризационная активация; 9— гра-
ц___ГипрпПлпИ П0ДЧИНЯЮ1Иийся закону «все или ничего», электрогенез; 10— выходящий ток;
ная активен яри?яция’ 12—Деполяризация; 13—потенциал покоя; 14 — гиперполяризацион-
ация; it> гиперполяризационная инактивация; 16 — гиперполяризационный ответ.
эритроцитов) после определенных ионных воздеи > пони-
ся повышение сопротивления во время деполярмзац
жение при гиперполяризации (так называемое анома
прямление). Такое явление наблюдается на мышце ля у J
действии на нее K2SO4; оно имеет место также в соме Р
моллюсков при его возбуждении на некотором расстояни ,
ласти отростка. Об аномальном выпрямлении можно говорить
же как о выпрямлении при прохождении гиперполяризующего -
ка. С физиологической точки зрения удобнее классифицирова
деполяризующие и гиперполяризующие реакции в соответствии
изменениями проводимости, а не с типами выпрямления.
296
Глава 11
Сравнительное рассмотрение электрогенеза показывает, что
в различных тканях одного и того же животного существуют раз-
личные электрогенные механизмы и что у разных видов животных
имеется широкий спектр ионных типов электрогенеза. Ниже пере,
числены эти механизмы и приведены соответствующие примеры.
1. Во всех нервах и быстрых мышцах позвоночных деполяри-
зационная реакция на раздражение, почвидимому, сопровождается
повышением натриевой проводимости. Тетродотоксин блокирует
каналы для входящего тока натрия.
2. Калиевая проводимость увеличивается медленнее; в нервах
и во многих мышцах она достигает максимума во время фазы
снижения потенциала действия. Тетраэтил аммоний задерживает
активацию калия, потенциал при этом выходит на плато. Запаз-
дывание этой активации обнаружено в гигантских волокнах дож-
девого червя, миокарде позвоночных и в уретре. Продолжающая-
ся после реполяризации активация К может приводить к выбросу
потенциала действия или к гиперполяризации, как это наблюдает-
ся, например, в аксоне кальмара.
3. В некоторых нейронах моллюсков с ритмической импульса-
цией ранний калиевый ток, проходящий по иным каналам, чем
калиевый ток при спайковой реполяризации, противоположен по
направлению непрерывному входящему натриевому току и опре-
деляет интервалы между потенциалами действия.
4. Повышающаяся при деполяризации кальциевая проводимость
играет важную роль в электрогенезе потенциалов действия мышц
ракообразных и некоторых гладких мышц позвоночных. Значи-
тельное повышение кальциевой проводимости, идущее параллельно
с изменениями натриевой проводимости, обнаружено в некото-
рых нейронах моллюсков [72], мышце ланцетника [85] и в неко-
торых гладких мышцах млекопитающих, например в уретре. Зна-
чительный входной ток кальция наблюдается в сердце лягушки.
5. В основе некоторых колеблющихся пейсмекерных потенциа-
лов (например, в ведущем узле сердца позвоночных) лежит боль-
шая величина отношения Р^а/Рк, а также зависящее от времени и
от напряжения изменение натриевой и калиевой проводимости.
6. Показано, что через электровозбудимые мембраны системы
внутренних трубочек мышц речного рака и омара, через одну из
поверхностей электроцитов ската и, возможно, через мембраны не-
которых гладких мышц млекопитающих интенсивно выводится С1,
и деполяризационный потенциал действия обусловлен здесь повы-
шенной хлорной проводимостью. Этот эффект усиливается при ма-
лых концентрациях С1~ в окружающей среде [81]. Потенциал дей-
ствия у водоросли Chara также обусловлен интенсивным выведе-
нием С1~.
7. По крайней мере в мышце одного насекомого (Tenebrio)
потенциал покоя практически нечувствителен к Ко; в среде, бога-
той калием, активация приводит к повышению калиевой проводи-
Возбудимые мембраны
297
мости и к деполяризации, обусловленной притоком К [13]. Мед-
ленный ВПСП в чувствительной к химическому воздействию мем-
бране (симпатический ганглий лягушки) также обусловлен
снижением £к [237].
8. Деполяризационные реакции многих чувствительных к хими-
ческому раздражению постсинаптических мембран состоят в од-
новременном повышении натриевой и калиевой проводимости. Это
наблюдается, в частности, в концевых пластинках поперечнополо-
сатых мышц позвоночных.
9. Некоторые постсинаптические реакции на специфические ме-
диаторы сводятся к повышению проводимости хлора и притоку
С1-. Эти реакции, обычные для тормозных синапсов, носят харак-
тер гиперполяризации, поскольку Eci часто отрицателен по отно-
шению к ПП.
10. В некоторых нейронах моллюсков Eci менее отрицателен,
чем ПП, и деполяризация при ВПСП может быть следствием воз-
росшей проводимости С1-.
11. В некоторых фоторецепторах гребешка наблюдается гипер-
поляризационная реакция, обусловленная повышением £к- ПП
этих клеток менее отрицателен, чем Ек, a PnJPk в состоянии по-
коя выше, чем при раздражении.
12. Характерный для состояния покоя интенсивный натриевый
ток подавляется при раздражении, как, например, в палочках сет-
чатки позвоночных при переходе от темноты к освещению.
13. Электрические свойства одной поверхности плоских асим-
метричных клеток могут быть иными, чем противоположной; об-
разующиеся при этом асимметричные потенциалы действия могут
последовательно суммироваться. В гл. 17 будут рассмотрены раз-
личные типы электрогенеза у асимметричных электроцитов.
14. Работа электрогенных ионных насосов приводит к воз-
никновению тока, направление которого совпадает с направлением
ионного транспорта. Если насос несимметричен к ионному обме-
ну, возникает разность потенциалов. Работа натриевых насосов в
некоторых случаях обусловливает возникновение потенциалов по-
коя. Ткани, в которых во время хранения на холоду увеличива-
лось содержание Na и уменьшалось содержание К, при повыше-
нии температуры, когда восстанавливается работа натриевого на-
С™Са’ стан,овятся электроотрицательными. Наступающая после
о ветнои реакции гиперполяризация некоторых центральных ней-
р нов (например, в ганглиях пиявки) и некоторых рецепторов
( апример, рецепторов растяжения у речного рака) обусловлена
активным вытеснением Na.
доказано, что колебания активности натриевых насосов
ляются причиной медленных ритмов в некоторых гладких мыш-
х внутренних органов и регулятором ритма вспышек потенциа-
лов действия в некоторых нейронах Aplysia.
электроотрицательными. Наступающая после
ронов (например
298
Глава 11
Неэлектрические физические явления, связанные
с проведением импульсов
При распространении нервного импульса наблюдается ряд не-
электрических явлений. На подъеме потенциала действия в волок-
нах блуждающего нерва кролика высвобождается тепло
(24,5 мккал/г), а на спаде потенциала тепло поглощается [93].
Уменьшается двойное лучепреломление демиелинизированных
аксонов краба или кальмара, измеренное под углом 45° к плоско-
сти поляризации; временной ход этого процесса коррелирует с из-
менением потенциала действия [118]. Уменьшается светорассея-
ние, измеренное под малым углом к оси волокна, и увеличивается
светорассеяние, измеренное под углом 90°. Последний эффект за-
висит от напряжения в меньшей степени, чем светорассеяние, из-
меренное под малым углом, и связан, по-видимому, с увеличением
пери аксонального пространства [35, 36]. Некоторые флуоресци-
рующие красители связываются мембраной аксона; наблюдаемые
при электрической активности изменения флуоресценции красите-
лей, возможно, отражают конформационные изменения в молеку-
лах белков, позволяющие полярным соединениям проходить
сквозь неполярные участки мембраны [221].
Мы надеемся, что использование различных физических мето-
дов позволит выяснить, какие именно изменения происходят в ли-
попротеидах при изменении ионной проводимости.
Адаптивные особенности нервного проведения
Адаптации, увеличивающие скорость проведения. Скорость про-
ведения нервного импульса в разных волокнах и нервных путях
неодинакова. Более быстрое проведение по рефлекторным путям
дает животному преимущества при спасении от хищника или лов-
ле добычи и при стрессовых реакциях. Временные константы воз-
буждения коррелируют со скоростью проведения. Некоторые
обычно используемые константы возбуждения получены на осно-
вании кривых силы тока—длительности или напряжения—вре-
мени; быстро проводящие волокна имеют обычно меньшее время
возбуждения, чем медленно проводящие. Другим параметром воз-
буждения является аккомодация, или увеличение порога возбуж-
дения при прохождении раздражающего тока. В том случае, ког-
да сила тока возрастает медленно, порог возбуждения становится
выше, чем для быстро нарастающего тока, т. е. возможно измене-
ние порога возбуждения в отсутствие реакции, подчиняющейся
закону «все или ничего».
Физические факторы. Скорость проведения импульса nponopj
циональна вызванному деполяризацией повышению натриевой
проводимости; при увеличении скорости нарастания потенциала
действия увеличивается сила местного тока и быстрее возбужда-
Возбудимые мембраны
299
ся прилежащие участки аксона. Скорость проведения пропор-
циональна также отношению величины порогового потенциала к
Цотенциалу покоя, поскольку при более низком пороге возбужде-
п возникает быстрее. Скорость распространения импульса об-
атно пропорциональна емкости мембраны — чем больше емкость,
тем большее время необходимо для ее зарядки, и наоборот. Ско-
рость обратно пропорциональна квадратному корню удельного
внутриклеточного сопротивления, поскольку чем меньше внутри-
клеточное сопротивление, тем больше проходит ионов и тем боль-
ше сила продольного тока при данном напряжении. С повышени-
ем температуры скорость проведения в данном волокне возраста-
ет, но только до определенного предела.
’ Морфологические факторы. В длинных нервных отростках ско-
рость проведения больше, чем, например, в центральных нервных
путях, где проведение замедляется промежуточными синапсами.
Нервная сеть многих кишечнополостных построена из коротких
нейронов, и поэтому скорость проведения здесь невелика. Ско-
рость проведения в нервной сети Calliactis равна 0,04—0,15 м/с,
а в сквозных нервных путях того же животного—1,2 м/с. В ней-
ропиле ганглионарной цепочки аннелид, для которого характерны
сегментарные синапсы, скорость проведения ниже 0,05 м/с, тогда
как в сквозных гигантских волокнах она составляет 10—25 м/с
[224].
Скорость проведения импульса определенным образом связа-
на с диаметром волокна. В крупных миелинизированных волокнах
позвоночных скорость проведения, выраженная в м/с, численно
равна диаметру 'волокна в микронах, умноженному на коэффици-
ент 6—8; для малых немиелинизированных волокон (симпатиче-
ские волокна, волокна обонятельного нерва) коэффициент пропор-
циональности иной. В гигантских аксонах кальмара скорость яв-
ляется степенной функцией диаметра (V=D0-61) [247]; аналогич-
ная зависимость найдена для гигантского волокна Myxicola (V—
— KD -5) [9]. в ходе индивидуального развития животного диа-
метр нервных волокон увеличивается и одновременно с этим
возрастает скорость проведения. Таким образом, зависимость ско-
рости проведения от диаметра волокна неодинакова для различ-
ных типов волокон. У отдельных нейронов во время их роста при
и^учалас индивидУУма эта зависимость адекватным образом не
тоттГИГаНТСКИе волокна —это нервные волокна крайне большой
КОТЩЗИНЫ' СУЩествУет ДБа основных типа гигантских волокон,
живо ь*е В пР°цессе эволюции возникли независимо у разных видов
ный ТНВ1Х‘ Одн0мУ из этих типов соответствует одиночный круп-
клеткНеИР°Н С большим аксоном, как, например, маутнеровская
ные 3 РЬ1б- Гигантские волокна второго типа — это синцитиаль-
отпос“а’ Ф'оРми,РУющиеся следующими двумя способами: а)
и нескольких нейронов сливаются, образуя один аксон (на-
300
Глава 11
пример, гигантский аксон мантии кальмара), или б) сегментарные
нейроны посылают свои отростки в длинные волокна, снабженные
сегментарными перегородками; проходя через многие сегменты
(например, латеральное и медиальное гигантские волокна дожде-
вых червей), эти отростки обеспечивают непрерывную связь меж-
ду ними. Медиальные гигантские волокна ракообразных, например
речного рака, являются отростками отдельных нейронов, тела ко-
торых расположены в мозге, тогда как латеральные гигантские
волокна исходят из сегментарных клеточных тел.
Функция системы гигантских нервных волокон рассмотрена
ниже. Здесь же можно сказать, что эти волокна передают сигна-
лы быстрее, чем интегрированные системы, т. е. имеют большую
скорость проведения, чем построенные из мелких волокон системы
тех же животных. Одноклеточные гигантские волокна обнаруже-
ны у немертин, цестод, некоторых полихет, насекомых и кишечно-
дышащих. У костных рыб, хвостатых амфибий и головастиков бес-
хвостых амфибий в базальной части продолговатого мозга присут-
ствуют крупные маутнеровские клетки, принадлежащие к вести-
булярно-акустической системе. Волокна этих клеток имеют 20—
45 мкм в диаметре и проводят импульс со скоростью 50—80 м/с
при 15 °C. У карпа скорость проведения по волокнам маутнеров-
ских клеток равна 38—63 м/с. Многоклеточный, или функциональ-
но-синцитиальный, тип системы гигантских волокон найден у ан-
нелид, ракообразных и головоногих моллюсков. У всех этих групп
животных гигантские волокна передают сигналы для быстрых дви-
жений и для синхронного сокращения многих мышц. Скорость
проведения по септированному латеральному гигантскому волокну
дождевого червя Lumbricus составляет от 7 до 17 м/с, а по меди-
альному волокну — от 17 до 45 м/с. У кальмара Loligo каждый
гигантский аксон третьего порядка (диаметр 0,1—0,8 мм) образу-
ется в результате слияния отростков от 300—1500 клеток звездча-
того ганглия и обслуживает большую область мантии. Сокраще-
ние мышц выталкивает струю воды из полости мантии через во-
ронку, благодаря чему животное движется вперед или назад в
зависимости от наклона воронки. Скорость распространения им-
пульса в гигантских волокнах лежит в диапазоне от 3 до 25 м/с.
Юнг рассчитал, что благодаря системе гигантских волокон такие
поведенческие реакции, как бегство или пищевая реакция, проис-
ходят вдвое быстрее, чем это было бы возможно за счет проведе-
ния по обычным аксонам [247].
Миелиновая оболочка обеспечивает более быстрое проведение^
снижая См, повышая 7?м и увеличивая константу длины волокна.
Волокна, проводящие импульсы с наибольшей скоростью, у поз-
воночных имеют средние размеры (диаметр от 4 до 15 мкм), но
толстую оболочку, на долю которой приходится 30—50% общего
диаметра волокна. У беспозвоночных эти волокна имеют большой
диаметр (от 50 до нескольких сотен микрон), но тонкую оболочку
301
Возбудимые мембраны
/менее 1% общего диаметра волокна). Волокно диаметром 4 мкм1
из подкожного нерва кошки при 38 °C имеет приблизительно та-
кую же скорость проведения (25 м/с), как гигантское волокно-
кальмара (диаметр 650 мкм) при 20 °C.
Проведение по миелинизированным аксонам позвоночных яв-
ляется сальтаторным— импульс перескакивает от перехвата к пе-
рехвату. Лишь в области перехватов Ранвье местный ток при про-
ведении импульса слагается из входящего и выходящего токов;
участок между перехватами имеет высокое сопротивление и низ-
кую электростатическую емкость. Внешний ток протекает с наруж-
ной стороны миелиновой оболочки. Оптимальное расстояние меж-
ду перехватами Ранвье зависит от фактора надежности и от сопро-
тивления аксоплазмы. Гигантские волокна креветки не имеют пере-
хватов. Однако в точке, где от такого волокна отходит ветвь к дви-
гательному нерву или к нейропилю, регистрируется двухфазный
потенциал действия; в этом месте может произойти и блокада
проведения. Таким образом, точки разветвления способствуют
сальтаторному проведению. Скорость проведения по этому волок-
ну у креветки равна 90 м/с, тогда как в имеющем такой же диа-
метр (100—200 мкм) волокне омара, где проведение не является
сальтаторным, скорость проведения составляет всего 8 м/с [130а].
Итак, можно сделать вывод, что различные группы животных
обладают рядом адаптаций, обеспечивающих различные скорости
нервного проведения. Все они основаны на обычных физических
законах увеличения силы тока, мембранной емкости и мембранно-
го сопротивления, электропроводности аксоплазмы и результирую-
щего коэффициента надежности. У одних групп животных высокая
скорость проведения обеспечивается миелинизированными нерва-
ми с сальтаторным проведением, а у других — нервными волокна-
ми очень большого диаметра, имеющими тонкую оболочку. У не-
которых животных в нервных путях имеются многочисленные ко-
роткие нейроны, а у других используются немиелинизированные
волокна с очень малым диаметром и низкой скоростью проведе-
ния. Скорость проведения нервных сигналов лежит в диапазоне
от 0,1 до 100 м/с, т. е. крайние значения скорости различаются на
три порядка.
ВРввеДение В03буждения у животных с низким содержанием
натрия в крови
Насекомые
калия в гемолимфе некоторых насекомых на-
бы а натРия столь низка, что в такой среде не мог
оапиях ипипп оигналы ни один известный нерв (данные о концент-
наЦ= Т На СТр- 179>’ В нервной системе таракана волок-
РУ ы периневральной оболочкой, потенциал которой ра-
302
Глава II
вен 10—15 мВ (положительный с внутренней стороны). В нор-
мальных покрытых оболочкой гигантских аксонах таракана in
situ потенциал покоя равен —58 мВ, а потенциал действия
—105 мВ; в находящихся в гемолимфе нервных волокнах с уда-
ленной оболочкой потенциал покоя равен —67 мВ, а потенциал
действия —86 мВ. Между нервными, глиальными и периневраль-
ными элементами происходит быстрый обмен натрием; измерение
оттока указывает на существование трех компартментов Na
[231]. Способность К влиять на свойства поверхности аксона ко-
личественно связана с экстранейрональным потенциалом, или по-
тенциалом оболочки [193, 194]. После удаления оболочки нерва
деполяризующее действие калия усиливается и быстрее наступает
вызванная калием блокада проведения импульса [227, 228]. При
высоком уровне К+ временная константа деполяризации для ин-
тактного растянутого гигантского нервного волокна таракана рав-
на 173 с, а после удаления оболочки она понижается до 24 с. Ка-
лий легче проникает в подсушенные или растянутые нервные
стволы.
В гемолимфе палочника Carausius morosus концентрация Na
равна 20 мэкв/л, а в нервной цепочке—102 мэкв/л. Центральная
нервная система окружена нейральной пластинкой, а затем слоем
жирового тела; у периферических нервов этих оболочек нет. Счи-
тают, что клетки слоя жирового тела или глия в периневральных
элементах [227] осуществляют активный перенос натрия и под-
держивают его высокую концентрацию вокруг аксона даже при
низком содержании натрия в гемолимфе. Концентрация Na в ге-
молимфе шелкопряда равна всего 4 мМ, тем не менее нервное
проведение блокируется ТТК, т. е. зависит от Na.
В нервах растительноядных насекомых, как и у других живот-
ных, потенциал покоя обусловлен высоким отношением Ki/Ko, а
потенциал действия — градиентом натрия, направленным внутрь
клетки. Содержание натрия в гемолимфе может быть низким, а
калия — высоким, при этом нервы функционируют благодаря обо-
лочкам, осуществляющим избирательный и (или) активный пере-
нос ионов. Механизм, с помощью которого оболочка поддержива-
ет определенный, отличный от гемолимфы состав экстранейро-
нальной жидкости, не вполне ясен.
Моллюски
Концентрация Na в крови беззубки Anodonta составляет всего
14 мМ, однако при этом идет относительно быстрый обмен натрия
с нервной тканью; временные константы оттока свидетельствуют
о существовании двух компартментов натрия, а внутриклеточная
концентрация Na равна 8,6 мМ. Таким образом, даже если абсо-
лютная концентрация натрия вне и внутри клетки очень низка,
потенциал действия обусловлен направленным внутрь градиен-
том концентрации Na+ [229].
Возбудимые мембраны
303
функции глии
Многие крупные аксоны покрыты клетками оболочки, а мелкие
объединяются в пучки, и каждый такой пучок окружен глиальны-
ми клетками. В центральной нервной системе существует несколь-
ко типов глиальных клеток, разделяющих или окружающих ней-
роны. Свойства глиальных оболочек хорошо изучены в зритель-
ном нерве Necturus и в центральной нервной системе пиявки, где
клетки глии занимают большую часть экстранейронального прост-
ранства. В ганглии пиявки имеется около 10 крупных глиальных
клеток, окружающих нейроны и пучки нервных волокон; эти клет-
ки занимают половину экстранейронального пространства [128].
При [Ко] ниже 20 мМ потенциал покоя глиальных клеток подчи-
няется уравнению Нернста. [Ki], рассчитанное по графику зави-
симости Ew от [Ко], равно 108 мМ для глии и 138 мМ для нейро-
нов [126, 169]. Глиальные клетки, окружающие зрительный нерв
Necturus, связаны .между собой мостиками низкого сопротивления.
Через межклеточные щели могут проходить ионы и сахар, и глия
деполяризуется калием, высвобождающимся при прохождении
импульсов по волокнам зрительного нерва. Эта деполяризация спа-
дает в течение нескольких секунд, когда калий возвращается в
его
аксон.
В нейронах пиявки после распространения волны потенциала
действия наблюдается остаточная деполяризация, медленно исче-
зающая со временем (временная константа этого процесса равна
100 мс). После прохождения по .нейрону серии импульсов происхо-
дит медленная деполяризация глиальных клеток. Согласно прове-
денным расчетам, в щелях размером 150 А, расположенных с на-
ружной стороны аксона, накапливается калий; концентрация
достигает величины 1,1-10~12 М/см2 (0,75 мМ). После серии им-
пульсов потенциал покоя нейрона на протяжении нескольких ми-
нут повышен. Этот потенциал обусловлен работой натрий-калие-
вого насоса, активируемого калием, который накапливается с на-
ружной стороны аксона, и устраняется уабаином. Глия играет
роль некоего барьера, предотвращающего поступление ионов калия
из нервной системы в кровь [11, 127, 168]. Возможно, деполяри-
зация глиальных клеток сетчатки калием, высвобождаемым нерв-
ными элементами, когда они находятся в состоянии возбуждения,
ньщИ[52] °снове волны на электроретинограмме позвоноч-
Функции глии изучены недостаточно. У насекомых она может
выполнять роль насоса, поддерживающего концентрационный гра-
„ионов между экстранейрональным пространством и гемо-
имфои. В нервной системе других групп животных, в частности
озвоночных, глия занимает значительную часть экстранейрональ-
го пространства и служит ионным буфером..
304
Глава 11
Явления, связанные с разными размерами частей одного
и того же нейрона
Один из отростков нейрона может функционально отличаться
от другого, а точки разветвления могут служить частотными филь-
трами. Это обусловлено как зависимостью скорости проведения,
константы длины волокна и порога возбуждения от диаметра во-
локна, так и разными размерами частей одного и того же нейро-
на. У крупных волокон X и См больше, а 7?м меньше, чем у мел-
ких. Если импульс, идущий по тонкому волокну, переходит в об-
ласть большого диаметра, эффективная плотность тока понижа-
ется, для достижения пороговой интенсивности требуется больший
ток, в результате время нарастания потенциала действия удлиня-
ется, поскольку для зарядки большей емкости требуется больше
времени. Напротив, при сужении проводящего пути, например при
переходе из тела крупного мотонейрона в его начальный сегмент,
плотность тока увеличивается; именно в этом месте может воз-
никнуть потенциал действия. В гигантских аксонах таракана име-
ются области сужения, проведние в которых замедляется; при пе-
реходе импульса из области малого диаметра в область расшире-
ния коэффициент надежности снижается, и при повторении им-
пульса может возникнуть блок [184].
В точках разветвления возможен связанный с частотой диффе-
ренциальный 'блок. Например, у речного рака и омара один двига-
тельный аксон иннервирует как медиальный, так и латеральный
пучки мышцы — разгибателя брюшка. Ветвь этого аксона, идущая
к медиальному пучку, имеет более низкий коэффициент надежно-
сти и блокируется при частоте 40 Гц, тогда как ветвь, иннервиру-
ющая латеральный пучок, проводит импульсы с частотой до 80 Гц
[183]. В центральной нервной системе многих животных точки
разветвления нервных волокон служат фильтрами низких частот.
Потенциал действия высокой частоты может приводить к повы-
шению наружной концентрации калия, а результирующая деполя-
ризация уменьшать коэффициент надежности вследствие повыше-
ния порога.
Электрическая и химическая межклеточная передача
Обычно сигналы поступают к нейронам через одни отростки
(дендриты) или через сому, а передаются от нейронов с помощью
другого отростка (аксона), т. е. нейроны поляризованы. Такая си-
туация сохраняется независимо от того, является проведение в
нейроне градуальным или процесс протекает по принципу «все
или ничего», является передача электрической или химической.
Правда, известны примеры неполяризованных электрических кон-
тактов, Во многих нейронах, например в амакриновых клетках и
Возбудимые мембраны
305
в различных звездчатых клетках, между дендритами и аксонами
четкой грани провести нельзя. В некоторых нейронах связь между
отростками (обычно дендритами) осуществляется без участия со-
мы и других отростков. Обратные дендро-дендритные синапсы, су-
дя по электронно-микроскопическим данным, представляют собой
случай, когда один и тот же отросток может и получать, и посы-
лать сигналы. В качестве примера дендр о-дендритных связей мож-
но привести связи в сердечных ганглиях ракообразных, клетках-
зернах и митральных клетках обонятельных луковиц позвоночных
[198], амакриновых клетках сетчатки [52], верхних бугорках чет-
верохолмия и таламуса млекопитающих [199], а также, возмож-
но, зрительной покрышке лягушки.
Электрическая передача
Существует два типа электрической межнейронной передачи—•
двусторонняя и поляризованная.
Электротоническая связь между нейронами. Об электрической
связи между клетками мы упоминали выше, когда рассматрива-
ли эпителиальные и эмбриональныые клетки (стр. 262—269).
В настоящее время известно много примеров существования по-
добной низкоомной связи и между нейронами. Электрическое со-
пряжение в этих межнейронных контактах симметрично: импульс,
приложенный к одной из клеток, регистрируется в соседней клет-
ке; амплитуда его пропорциональна коэффициенту сопряжения
[181, 186, 187]. В каждом ганглии нервной системы пиявки име-
ется пара гигантских нейронов; электрический импульс, передан-
ный с помощью микроэлектрода, введенного в один из этих ней-
ронов, регистрируется в другом нейроне и амплитуда его вследст-
вие затухания составляет от 72 до Vs амплитуды исходного
импульса. Если деполяризация одного из этих нейронов достигает
уровня, при котором возникает потенциал действия, то во втором
нейроне наблюдаются малые электротонически проводимые потен-
циалы действия, соответствующие спайкам первой клетки
(рис. Ц-14) [58]. При перфузии ганглия бескальциевым раство-
ром входное сопротивление повышается с 24 до 70 МОм и функ-
циональная связь между парными нейронами прекращается [188,
Аналогичные низкоомные связи имеются между поперечными
ветвями латеральных гигантских аксонов у речного рака и дож-
девого червя; они обнаружены также в виде межсегментных пе-
регородок в гигантских волокнах дождевого червя. Амплитуда им-
пульса при переходе его от одного латерального волокна к дру-
ому может уменьшаться до 7з первоначального значения, а точки
перехода смещаются от сегмента к сегменту [243].
Медиальные гигантские аксоны речного рака не являются пе-
Р городчатыми и не взаимодействуют между собой, в латераль-
20—1514
306
Глава 11
них же аксонах существует межсегментная электрическая связь
как через перегородку между сегментами, так и от одного -чате’
рального гигантского волокна к другому [103]. Сопротивление
перегородки равно 2-105—4-105 Ом (1,2—3 Ом «см2); септальная
Рис. lil-14. Электрическая связь между нейронами в ганглии пиявки.
/I. Гиперполяризация одного гигантского нейрона (средняя запись) приводит к соответствую-
щей реакции во втором гигантском нейроне (верхняя запись). Б. Вызванные деполяризацией
потенциалы действия в первой клетке (средняя запись); во второй клетке синхронно возни-
кают спайки [58].
задержка при проведении потенциала действия составляет от 50
до 200 мкс. Краситель флуоресцеин может проходить через пере-
городки гигантского аксона речного рака (но не дождевого чер-
вя). Электрический толчок 80 мВ, приложенный к пресептально-
му сегменту латерального гигантского волокна речного рака, вы-
зывает реакцию в постсептальном сегменте; возникающий там сиг-
нал имеет амплитуду 30 мВ [236]. Перегородчатые контакты не
поляризованы. Они проводят импульс без затухания (в отноше-
нии 1:1), но в случае утомления при многократном раздражении
возникает необходимость в облегчении (фасилитации). Серия
Возбудимые мембраны
307
электротонических контактов обусловливает серию линий задерж-
ки; через поперечные связи между двумя латеральными гигантски*
ми аксонами может возникнуть круговая активность [130]. Ток,
обусловливающий проведение импульса вдоль аксона, значительно
больше тока, протекающего по связям между латеральными во-
локнами; в нормальных условиях этого достаточно, чтобы предот-
вратить транссептальную реверберацию [130]. Электронно-мик-
Рис. 11-15. Коэффициент электрической связи V2/Vi между двумя нейронами
буккального ганглия моллюска Navanax при различных частотах поляризации Vi
(Levitan Н., Таис L., Segundo J. Р., J. Gen. Physiol., 55, 484—496, 1970).
роскопичесиие исследования показали, что в перегородках гигант-
ского аксона речного рака имеются щелевые соединения, это
подтверждает наличие электротонического сопряжения последо-
вательных сегментов [86].
В буккальном ганглии моллюска Navanax имеется гроздь из
десяти нейронов, между которыми обнаружена слабая электри-
ческая связь. Она служит скорее для передачи импульсов посто-
ств °ГО ТОКа’ чем для проведения потенциалов действия; в соответ-
ии с этим высокочастотные импульсы гасятся [214] (рис. 11-
и это сопряжение может не иметь функционального значения.
сеРдечных ганглиях ракообразных высокочастотные сигналы
сому^ктеток Че^е3 дендР'ИТЬ1 пейсмекерных нейронов, не входя в
ван^тг^ЛИаЛЬНЬ1Х °боло’1ках в corpora pedunculata мозга пенсиль-
межд Г0 мУРавья„ имеются поры, которые обеспечивают связь
ду телами нейронов. Эта связь осуществляется как посредст-
20*
308
Глава И
вом ряда щелевых контактов, расположенных друг от друга на
расстоянии 50—70 А, так и с помощью химических синапсов (диа-
метр пузырьков от 300 до 600 А) [131].
В мозге рыб также обнаружены многочисленные электротони-
ческие контакты [14, 15]. На дорсальной поверхности продолго-
ватого мозга кузовка расположены два крупных (250 мк) нейро-
на, которые активируют мотонейроны мышц плавательного пузы-
ря. Афферентные входные сигналы, поступающие через спинной
мозг, вызывают синхронные реакции в обоих крупных нейронах, а
электрический импульс, приложенный к одному из нейронов, об-
наруживается с незначительным затуханием и во втором нейроне.
Эти клетки связаны электротонически через аксоны. У некоторых
видов электрических рыб существует электротоническая связь
между нейронами, управляющими разрядом электрического орга-
на (гл. 17). Релейные нейроны, расположенные в продолговатом
мозге мормирид, активируются пейсмекерными нейронами высших
уровней мозга. Каждый медуллярный нейрон генерирует два по-
тенциала действия на одно раздражение; медуллярные нейроны
электротонически связаны между собой с помощью дендро-денд-
ритных мостиков. Соответствующие контакты относятся к числу
щелевых и имеют вид шестиугольников, находящихся друг от
друга на расстоянии 80 А. Медуллярные нейроны оказывают ак-
тивирующее действие на спинальные мотонейроны, связь с кото-
рыми осуществляется через химические синапсы. Однако в гроз-
дях спинальных мотонейронов опять наблюдается электрическая
связь; три импульса, переданных этими нейронами, соответствуют
одному разряду электрического органа [14]. У гимнотид электро-
тоническое сопряжение наблюдается на уровне пейсмекера, где
оно осуществляется через дендро-дендритные контакты. На уров-
нях продолговатого и спинного мозга существуют электротониче-
ские аксо-соматические контакты, а передача с одного уровня на
другой осуществляется посредством химических синапсов. В про-
долговатом мозге электрического сома Malapterurus есть два ги-
гантских электромоторных нейрона, электротоническая связь меж-
ду которыми осуществляется через пресинаптические волокна.
Каждая из таких электротонических связей у электрических рыб
имеет двустороннее проведение, а полярность всей системы обус-
ловлена химическими 'синапсами между ее разными уровнями. Су-
ществующая на каждом уровне электротоническая связь обуслов-
ливает синхронность разряда сопряженных клеток.
Глазодвигательные нейроны некоторых костистых рыб электро-
тонически связаны между собой посредством аксо-соматических
контактов. Кроме того, там имеются и синаптические окончания
с пузырьками. Раздражение глазного нерва приводит к возникно-
вению потенциалов действия поблизости от клеточных тел; эти
разряды синхронизируются с помощью электротонического сопря-
жения [124].
Возбудимые мембраны
309
Односторонние электрические синапсы. Известно несколько
случаев поляризованной электрической передачи. В синапсе меж-
ду латеральным гигантским волокном и мотонейроном у речного
рака потенциал действия пресинаптического волокна является аде-
кватным раздражителем мотонейрона; процесс передачи протекает
без задержки. При ортодромном (нормальном) направлении им-
пульсов в мотонейроне возникают
сильные транссинаптические токи,
регистрируемые внутриклеточно;
при обратном (антидромном) на-
правлении импульсов эти токи от-
сутствуют (рис. 11-16). Таким обра«
зом, синаптическая мембрана дейст-
вует как выпрямитель, пропуская
через контакт с низким сопротивле-
нием только ортодромный ток и не
пропуская ток, идущий в обратном
направлении. Деполяризующий им-
пульс, приложенный к пресинапти-
Рис. 11-16. Ответ в пре- и постсинаптиче-
ском волокнах электрического синапса реч-
ного рака при поступлении антидром-
ных (А) и ортодромных (Б) импульсов
[68].
Отметим, что трансмембранный ток проходит
только ортодромно, т. е. происходит его выпрямле-
ние. пре — пресинаптическое волокно; пост — пост-
синаптическое волокно.
ческому волокну, проходит в постсинаптическое волокно, а гипер-
поляризующий не проходит; с постсинаптического волокна, напро-
тив, передается только гиперполяризующий импульс. Выпрямление
происходит в отношении 50:1. На мотонейронахоканчиваются так-
сш^пНЬ111Ие аксоны третьего корешка; они возбуждают медленные
врте ТИЧеСКИе п<>тенциалы, меняющие знак при —7 мВ, т. е. соот-
твующие синапсы являются химическими [68, 69].
рыбы ।С°И/ поляРиз'ованнь1й электрический синапс обнаружен у
eropelecus> где активация мотонейронов гигантскими
ПУПЫ нроиаходит с задержкой 0,05 мс. Деполяризующий им-
вению в гигантскомУ волокну, приводит к возникно-
пульс не еиР°не сильного тока, а гиперполяризующий им-
УменьшартеРст01ВпДИТСЯ °PtW;0mho. Синаптическое сопротивление
вяртга ггг^т? рИ деполяризации гигантского волокна и увеличи-
ли Деполяризации мотонейрона [5]. Благодаря
ЭТИМ СИ-
310
Глава 11
напсам у Gasteropelecus уменьшается синаптическая задержка и
синхронизируются сокращения мышц грудных плавников.
Физиология маутнеровских нейронов будет рассмотрена позже.
Эти нейроны получают сигналы по нескольким группам путей, в
том числе от контралатерального восьмого нерва через возбужда-
ющие химические синапсы и ипсилатерального восьмого нерва че-
рез возбуждающие электрические синапсы. В свою очередь воз-
вратная коллатераль аксона маутнеровской клетки активирует
вставочный нейрон, аксон которого обвивается вокруг аксонного
холмика маутнеровской клетки. Импульсы вставочного нейрона
приводят к образованию внешнего поля, которое гиперполяризует
аксон маутнеровской клетки и тем самым подавляет возникнове-
ние потенциалов действия. Такое возвратное торможение предот-
вращает возникновение ритмической импульсации после однократ-
ного разряда маутнеровской клетки [69а]. Этот пример показыва-
ет, что электрические синапсы могут быть как возбуждающими,
так и тормозными.
В ресничном ганглии цыпленка в возрасте 3—5 дней осуществ-
ляется и электрическая, и химическая передача. Ответ постгангли-
онарного нерва на преганглионарный раздражитель слагается из
двух компонентов, разделенных интервалом от 1,5 до 5 мс. Тубо-
курарин снимает второй компонент, не действуя на первый, кото-
рый, кроме того, не зависит от поляризации мембраны. Судя по
тому, что антидромное раздражение вызывает небольшой преган-
глионарный ответ, здесь существует двустороннее проведение по
электрически сопряженным путям [148, 149]. В настоящее время
известны многочисленные примеры таких смешанных электриче-
ских и химических синапсов. Например, у морской миноги Petro-
myzon ВПСП, регистрируемый в синапсе, который образуют ак-
соны крупных клеток Мюллера продолговатого мозга и гигантские
вставочные нейроны спинного мозга, состоит из двух компонен-
тов. Один из них является химическим и исчезает при высокой
концентрации Mg2+, а второй компонент электрический [150,
204Ъ] (рис. 11-17).
В ганглии каждого сегмента пиявки существует три типа элек-
трических синапсов: 1) с каждой стороны ганглия имеется группа
приблизительно 14 мотонейронов, соединенных между собой элек-
трическими неполяризованными контактами, так что сигнал от од-
ного нейрона электротонически распространяется на другие; 2) в
каждом полуганглии имеется шесть сенсорных нейронов, реагиру-
ющих на легкое прикосновение к коже; потенциал действия в лю-
бом из этих нейронов приводит к возникновению в остальных ней-
ронах этого сегмента, а также в трех сенсорных нейронах каждого
смежного ганглия потенциала с малой латентностью. Между эти-
ми сенсорными нейронами существует необычное двойное выпрям-
ление, выражающееся в том, что деполяризующий сигнал «может
проходить в любом направлении, тогда как гиперполяризующий
Возбудимые мембраны
311
не проходит совсем [11]; 3) передача между осязательными сен-
сорными клетками и мотонейронами характеризуется малым ла-
тентным периодом; в ходе этой передачи осуществляется выпрям-
ление— деполяризационные импульсы проходят от сенсорного
нейрона к мотонейрону, а гиперполяризующие — в обратном на-
правлении [170].
В заключение можно сказать, что электрическая передача
между нейронами характерна для тех случаев, когда особенно
Рис. 11-17. ВПСП гигантских интернейронов миноги, возникающие при раздраже-
нии бульбарного аксона (Rovainen С. М., J. Neurophysiol., 30, 1024—1042, 1967).
О — серия импульсов, вызывающих ВПСП; фасилитация второго компонента. Mg — ВПСП
после 15-минутного воздействия Mg (20 мМ); второй компонент уменьшен или совсем от-
сутствует. Mg+Ca — ВПСП при повышении наружной концентрации Са до 22 мМ; второй
компонент ВПСП соответствует химическому синапсу, а первый — электрическому.
важны скорость и (или) синхронность ответа. В некоторых бы-
стрых системах электрическая связь двусторонняя, а в других —
односторонняя.
Химические синапсы
Химические синапсы отличаются от электрических следующими
своими хар актер,истиками [192]: 1) синаптической задержкой
обычно не менее 0,5 мс; 2) отсутствием значительного электриче-
ского тока, направленного от пресинаптического элемента к пост-
синаптическому; 3) постсинаптическим потенциалом (ПСП) ка1
ответом на пресинаптический импульс; этот потенциал при воз-
буждении обычно является деполяризующим (ВПСП), а при тор-
можении гиперполяризующим (ТПСП); кроме того, 4) проводи-
мость постсинаптической мембраны химического синапса по срав-
нению с состоянием покоя увеличивается, о чем свидетельствуют
312
Глава 11
изменение величины и полярности ПСП по отношению к мембран-
ному потенциалу и инверсия ПСП по отношению к некоторому рав-
новесному потенциалу; 5) в пресинаптических окончаниях присут-
ствуют синаптические пузырьки, а для постсинаптической мембра-
ны характерно специфическое окрашивание; 6) высвобождение
медиатора блокируется высокими концентрациями Mg2+ и низкими
концентрациями Са2+.
Структура синапса. Синаптические контакты могут находиться
как в области аксональных концевых утолщений, или «пуговок»
(boutons), так и у касательных утолщений по ходу аксона (еп
passage); эти контакты могут быть между аксоном и дендритом,
между аксоном и сомой клетки, а также между аксонами. Денд-
ро-дендритные синапсы обычно бывают у нейронов, не имеющих
аксонов. В области соприкосновения пре- и постсинаптических
мембран элементы оболочек отсутствуют; постсинаптическая мем-
брана на участке напротив скопления синаптических пузырьков
обычно утолщена. Медиатор синтезируется в основном в нервном
окончании, но иногда и в других частях нейрона. В пресинаптиче-
ских окончаниях в большом количестве присутствуют митохонд-
рии, а также многочисленные пузырьки, единообразные или раз-
личающиеся размерами и плотностью (рис. 11-18, 11-19 и 11-20).
При фракционировании нервной ткани можно получить синапто-
сомы — одетые мембраной нервные окончания с синаптическими
пузырьками1; в изолированных синаптосомах содержатся фермен-
ты, участвующие в процессе синтеза медиатора. При многократ-
ном раздражении запасы медиатора, депо.нированно1го в синапти-
ческих пузырьках, истощаются. Например, в двигательных нерв-
ных окончаниях лягушки число и плотность синаптических
пузырьков после длительного раздражения уменьшаются [88].
Катехоламиновые пузырьки оказываются «пустыми» после обра-
ботки резерпином, который блокирует депонирование медиаторов
группы катехоламинов. Медиаторы, например ацетилхолин, лока-
лизованный в пузырьках, находятся в равновесии с медиаторами
цитоплазмы нервного окончания; характер обмена между этими
двумя пулами медиатора неясен. АХ, высвобождаемый через пре-
синаптическую мембрану, по-видимому, весь происходит из синап-
тических пузырьков [242]. В пресинаптическом окончании нервно-
мышечного синапса лягушки содержится до 600 000 пузырьков;
при раздражении, продолжающемся одну минуту, это количество
снижается до 400000. Высвобождение содержимого пузырьков но-
сит характер экзоцитоза, причем мембраны пузырьков, по-видимо-
му, сливаются с плазматической мембраной. Затем эти мембраны
возвращаются в цитоплазму нервного окончания, образуя цистер-
1 Это определение не вполне точное, поскольку компонентами синаптосомЫ
могут быть также синаптическая щель и часть постсинаптической клетки.-*
Прим, перев.
Рис. П-18. Синаптические участки абдоминального ганглия моллюска Aplysia;
фотографии получены с помощью сканирующего электронного микроскопа (Le-
wis Е. R.t Everhart Т. Е., Zeevi Y. У., Science, 165, 1140—1143, 1969).
Рис. 11-19. Пресинаптическое окончание subfornix кошки (фронтальный срез). За-
мораживание-травление (Akert К., Waser Р. G., eds., Mechanism of Synaptic
transmission, Progress in Brain Research, 31, 223—245, 1969, Elsevier, Amster-
dam) .
сннап™ческие пузырьки (1), митохондрия (2), синаптические пузырьки с плотным
ежимым (3), плотная постсинаптическая область (4) и наружная поверхность пресинап-
тической мембраны (5).
314
Глава И
ны, которые заполняются медиатором и образуют новые синапти-
ческие пузырьки (рис. 11-21). Таким образом, происходит повтор-
ное использование белка синаптических пузырьков [88а]. Из моз-
га млекопитающих была выделена фракция синаптических
пузырьков, а из электрического органа ската Torpedo получена
чистая суспензия пузырьков с АХ. Каждый такой пузырек имеет
Рис. 11-20. Синапс, образованный лиановидным волокном на дендрите (D) клет-
ки Пуркинье в мозжечке серебряного карася (предоставлено Kotchabhakdi N.).
1 — синаптические пузырьки, 2 — митохондрия, 3 — пресннаптическая мембрана, 4 — постси-
наптическая мембрана, 5 — синаптическая щель.
диаметр около 840 А и содержит примерно 70 000 молекул АХ
Для идентификации специфических синаптических пузырьков,
содержащих определенный медиатор, сопоставляют присутствие
большого количества пузырьков какого-либо типа с концентрацией
этого медиатора в исследуемой нейральной структуре. В случае
биогенных моноаминов для этой цели используется также флуо-
ресцентная микроскопия. Мелкие агранулярные пузырьки с проз-
рачным содержимым, имеющие диаметр от 300 до 600 А (в сред-
Возбудимые мембраны
315
нем 500А), вероятно, являются холинергическими — они обнару-
жены во всех известных холинергических окончаниях. Мелкие гра-
нулярные пузырьки диаметром 300—600 А с плотной сердцевиной,
вероятно, содержат норадреналин (НА), а крупные (от 900 до
1200 А), с периферическим ободком — другие катехоламины [20,
21]. В висцеральных синапсах обнаружены крупные (1000—
2000 А), не имеющие ободка пузырьки; активным компонентом
их содержимого, возможно, является АТФ [25, 26]. При изучении
мозжечка млекопитающих и нервно-мышечных синапсов ракооб-
разных получены данные, согласно которым мелкие уплощенные
пузырьки локализованы преимущественно в тормозных окончани-
ях, а крупные сферические — в окончаниях возбуждающих воло-
кон [176]. В других системах подобная корреляция, по-видимому,
отсутствует.
В некоторых пресинаптических окончаниях имеются интенсивно
окрашивающиеся столбики с синаптическими пузырьками на обо-
их концах; эти столбики ориентированы перпендикулярно плоско-
сти пресинаптической мембраны. Подобные структуры часто встре-
чаются в синапсах, образованных тремя отростками (так называ-
емые синаптические ленты). Например, в сетчатке позвоночных
синаптические ленты расположены напротив двух или трех пост-
синаптических рецепторов, находясь между рецепторными и би-
полярными или горизонтальными клетками или между биполяр-
ными и амакриновыми или ганглиозными клетками [52, 53]
(рис. 11-22).
Постсинаптические мембраны весьма разнообразны по строе-
нию. У некоторых из них есть десмосомоподобные плотные струк-
туры, в ряде случаев расположенные точно напротив пресинапти-
ческих окончаний. Иногда под мембранами находятся плотные
скопления гранул. Постсинаптические мембраны некоторых холин-
эргических синапсов, например нервно-мышечных синапсов позво-
ночных, дают положительную гистохимическую реакцию на аце-
тилхолинэстеразу (АХЭ). В других холинэргических синапсах этот
фермент имеется и в пре-, и в постсинаптических мембранах.
Прекращение действия медиатора обусловлено следующими
Факторами: 1) расщеплением медиатора ферментами постсинапти-
ческой мембраны; 2) диффузией медиатора, сопровождающейся
его поступлением в систему кровообращения; 3) обратным захва-
том медиатора пресинаптическими окончаниями с последующим
повторным использованием этого вещества; 4) захватом медиато-
ра с его последующим расщеплением в специфических участках
пресинаптических окончанмй.
Медиаторы нервной системы. Для идентификации медиатора
используются следующие критерии: 1) способность малых коли-
честв предполагаемого медиатора воспроизводить эффект преси-
наптической импульсации; 2) присутствие в пресинаптическом ней-
роне как предполагаемого медиатора, так и его предшественни-
Возбудимые мембраны
317 -
ков и ферментов, принимающих участие в его синтезе; 3) высво-
бождение медиатора в перфузат при раздражении пресинаптиче-
ского нерва; 4) наличие механизмов инактивации предполагемого
медиатора; это или специфическое ферментативное расщепление,
или активный обратный захват в пресинаптические волокна;
5) способность специфических фармакологических препаратов по-
тенцировать или блокировать реакции как на пресинаттическую
импульсацию, так и на введение предполагемого медиатора [65,
240]. Число предполагаемых медиаторов невелико, и их молекуляр-
ный вес лежит в пределах от 100 до 3001. Реакции, обусловленные
процессами синаптической передачи, разнообразнее, чем можно
было бы ожидать на основании количества предполагаемых меди-
аторов. Возможно, это связано с большим разнообразием рецепто-
ров [95]. В соответствующих главах мы рассмотрим химическую
передачу в следующих мышцах:
Скелетные мышцы позвоночных и элек-
троциты
Мышцы ракообразных и насекомых; воз-
буждение
Мышцы ракообразных и насекомых; тор-
можение
Сердечная мышца позвоночных; возбуж-
дение
Сердечная мышца позвоночных; тормо-
жение
Мышцы желудочно-кишечного тракта
позвоночных; возбуждение
Мышцы желудочно-кишечного тракта
позвоночных; расслабление
Сердечная мышца моллюсков; возбужде-
ние
Сердечная мышца моллюсков; торможе-
ние
Запирательная мышца двустворчатых
моллюсков; возбуждение
Запирательная мышца двустворчатых
моллюсков; торможение
ацетилхолин
глутамат
у-аминомасляная кисло-
та
норадреналин
ацетилхолин
ацетилхолж
норадреналин, АТФ
серотонин
ацетилхолин
ацетилхолин
серотонин
В свете современных данных это положение
ПО крайней мере несколькими десятками и среди
вещества. — Прим, перев.
неточно — их число измеряется
них есть высокомолекулярные
синапса в портняжной мышце
Рис. 11-21. Ультраструктура нервно-мышечного
лягушки [88а].
нёппн>Пе^ечнь1^ срез; видны толстые и тонкие иити мышечного волокна и клеточное ядро;
вепхн°е 0К01Гчание окружено шванновской оболочкой со всех сторон, за исключением по-
мнтохлСТИ’ °°Рзи^енной к мышце; в окончании видны синаптические пузырьки и плотные
ны мнп ₽ИИ' пР°Д°льный сРез через окончание, не подвергавшееся раздражению; вид-
синаптип°ЧНСЛеННЬ1е синаптические пузырьки и плотные митохондрии; стрелкой отмечено пре-
OTDr>CTvJeCKOe Уплотнение, противостоящее складке на поверхности мышцы. Видны также
чание чяЖ1ВаИНОВСКИХ клеток (г) между нервом и мышцей. В. Продольный срез через окон-
точисленны ксиР°ванное после 15-минутного раздражения электрическим током; видны мно-
пнстерны (2) и расширенные отростки шванновских клеток (1); стрелкой от-
мены пузырьки с неровной оболочкой; митохондрии (3) светлые и набухшие.
318
Глава 11
Согласно современным представлениям, в центральной нервной
системе различных групп животных присутствуют все перечислен-
ные вещества-медиаторы, за исключением АТФ (разумеется, как
медиатора); кроме того, там обнаружены другие катехоламины, в
Рис. 11-22. Электронно-микроскопические фотографии лентообразных синапсов,
образованных рецепторными окончаниями в сетчатке кошки [52].
В окончаниях колбочек (слева) в каждое углубление основания рецептора проникает по
три отростка. В окончаниях палочек (справа) в каждое углубление входит по четыре и бо-
лее отростка. Г — отростки горизонтальных клеток; Б — отростки биполярных клеток.
первую очередь дофамин, и по крайней мере еще одна аминокис-
лота-медиатор — глицин.
Ацетилхолин. В холиновой части молекулы АХ имеется аммо-
нийная группа, несущая положительный заряд, а в ацетильной
части — карбонильная группа, заряженная отрицательно. Таким
образом, молекула АХ представляет собой электрический диполь.
АХ синтезируется из ацетил-КоА и холина (см ниже).
Синтез АХ
Х°линацетилаза
СН3—C~S-KoA +НО—СН2—СН2—N+(CH3)3 ----------------->
II
О
Ацетил-КоА
Холин
Возбудимые мембраны
319
> СН3— С— О—СН2— СН2—N+(CH3)34- Ко A— SH
О
Ацетилхолин
Гидролиз АХ
Кофермент А
АХЭ
СН3— С-О-СН2-СН2—N+(CH3)3 + н2о —>
II
о
---> НО—СН2—СН2—N+(CH3)34-CH3COOH
Холин Уксусная кислота
Ацетилхолин синтезируется во многих нервных тканях. Он де-
понируется в холинэргических нервных окончаниях в синаптиче-
ских пузырьках с прозрачным содержимым диаметром от 300 до
600 А. Гидролитический фермент ацетилхолинэстеразу (АХЭ) мож-
но выявить гистохимическими методами; этот фермент обнаружен
в постсинаптических мембранах и на поверхности некоторых пре-
синаптических окончаний (рис. 11-23). Число оборотов АХЭ в
электрическом органе составляет 300 000 молекул в 1 с, т. е. одна
молекула АХ гидролизуется за 3—4 мкс [163]. Скорость синтеза
АХ в электрическом органе составляет 5—8 мг на 1 г ткани в 1 ч
[160]. Действие АХ обычно состоит в повышении проницаемости
постсинаптических мембран для Na+ и К+, причем для разных
ионов проницаемость меняется в разной степени. В некоторых ней-
ронах моллюсков АХ действует и на проницаемость для С1~ (стр.
293). Иногда происходит спонтанное высвобождение некоторой
порции АХ и возникают миниатюрные ПСП; эти потенциалы по-
стоянны по величине или кратны ей. В моторной концевой пластин-
ке ВПСП возникает при высвобождении около 300 квантов АХ (стр.
285) [121]. Каждый такой квант содержит 103—104 молекул.
Ацетилхолинэстеразу по определению можно рассматривать
как некий рецептор, поскольку гидролиз АХ происходит лишь пос-
ле образования комплекса АХ—АХЭ. Блокада АХЭ любым инги-
битором этого фермента [наиболее известны среди них эзерин, или
физостигмин, и ДФФ (Диизопропилфторфосфат) ] усиливает фи-
зиологическое действие АХ. В молекуле АХЭ, выделенной из элек-
трического органа в виде чистого белка, центр, по которому связы-
вается ингибитор рецептора, не совпадает с каталитическим цент-
ром молекулы фермента. Концентрационные кривые АХЭ сдвига-
ется при действии как рецепторных активаторов, так и рецептор-
ЙЬ1Х„ ингибиторов. Эти данные говорят в пользу аллостерических
свойств АХЭ [30]. С другой стороны, можно блокировать физиоло-
гический холинорецептор, не повлияв на активность эстеразы. Сус-
пензии, полученные из мембран электрического органа Torpedo,
образуют комплексы с негидролизуемыми аналогами АХ (декаме-
д°у5И’ мУскаРон); АХ остается связанным даже после инактивации
образование комплекса блокируется J-тубокурарином [175].
320
Глава 11
Змеиный а-токсин является специфическим блокатором, необра-
тимо соединяющимся с холинорецептором; ковалентно связав этот
комплекс с сефарозой и разделив образовавшиеся частицы с по-
мощью центрифугирования, можно удалить из исходного экстрак-
та 75—100% рецептора; одновременно с этим удаляется только
12% АХЭ и 6% белка [29а, 30].
Синап ти ческие
пузырьки
Митохондрия
, гемиэсолиний
Холин
ацетат
~ Токсин
Вас, botulinum
Постсинаптическая
мембрана
Расширение
пресинаптического
окончания
никотин,
пилокарпин ~
АХ
АХ
АХЭ
гексаметоний,
кураре,
атропин
Ацетил-КоА
холин .
Рис. 11-23. Схема холинэргического синапса, а также пресинаптического синте-
за АХ и постсинаптического гидролиза АХ ацетилхолинэстеразой; показано поло-
жение холинэргических рецепторов (никотиновых или мускариновых) (Rech R. И.,
Moore К. Е., Introduction to Jpsychopharmacology, Raven Pres, New York, 1971)«
физостигмин
Исходя из данных по равновесному диализу солюбилизирован-
ного электрического органа Torpedo, можно сделать вывод, что хо-
линорецептор является простым белком или липопротеидом и, воз-
можно, имеет несколько активных центров. Молекулярный вес это-
го белка по оценкам лежит в пределах от 50 000 до 250000.
Липопротеиды, выделенные из электрического органа электриче-
ского угря Electrophorus, использовали для приготовления искусст-
венных мембран; добавление АХ вызвало изменение ионной прово-
димости мембран, а d-тубокурарин снимал этот эффект [182].
Каждая концевая пластинка диафрагмы крысы содержит до 13 000
рецепторных молекул на квадратный микрон, что соответствует
расстоянию между молекулами 100 А. С помощью меченого бун-
Возбудимые мембраны
321
гаротоксина удалось установить распределение холинорецептора
в скелетной мышце; выявленные места соответствуют точкам мак-
симальной чувствительности к АХ. Эти опыты показали также, что
происходящее после дегенерации нерва распространение чувстви-
тельности к АХ на экстрасинаптическую мышечную мембрану
обусловлено синтезом рецептора в новых местах. Из всех этих
данных сделан общий вывод, что рецептор, ответственный за
изменение ионной проницаемости, является белком, отличным
от АХЭ.
На основании опытов с холинолитиками и холиномиметиками
было высказано предположение о существовании двух типов хо-
линорецепторов — никотиновых (н-холинорецепторов), активируе-
мых низкими концентрациями никотина (рецепторы симпатических
ганглиев или нервно-мышечных синапсов позвоночных), и муска-
риновых (м-холинорецепторов), активируемых мускарином и бло-
кируемых атропином (рецепторы сердечной мышцы позвоночных).
Сравнительное изучение блокаторов н-холинорецепторов показыва-
ет, что они бывают двух типов. Одни из них оказывают деполяри-
зующее действие (как, например, гексаметоний на симпатические
ганглии), а другие блокируют действие АХ путем конкурентного
торможения (как, например, d-ту бокур арин в нервно-мышечных
синапсах позвоночных). Размеры молекул соединений, блокирую-
щих передачу через «никотиновые» и «мускариновые» синапсы, бо-
лее или менее соответствуют структуре ацетилхолина [62].
Связывание н-холинолитиков зависит от акцептора водородных
связей, расположенного приблизительно в 5,9 А от центра поло-
жительного заряда; у м-холинолитиков это расстояние равно
4,4 А [13а].
Роль АХ как межнейронного передатчика впервые четко уда-
лось показать для симпатических ганглиев позвоночных. В симпа-
тическом ганглии лягушки-быка ответ на преганглионарную серию
импульсов слагается из двух компонентов — быстрого ВПСП, бло-
кируемого d-тубокурарином, и отстающего от него приблизительно
на 100 мс второго ВПСП, блокируемого атропином. Таким обра-
зом, в одном и том же нейроне есть оба типа холинорецепторов
(никотиновые и мускариновые) [120]. Если перфузировать верх-
ний шейный ганглий кошки через ветвь сонной артерии и раздра-
жать преганглионарный нервный ствол, то количество АХ, высво-
ождаемого в перфузат, будет пропорционально числу импульсов,
уступающих в ганглий (при условии, что ганглий был предвари-
тельно обработан ингибитором АХЭ эзерином). Инъекция АХ вы-
аает РазРяД ганглиозных клеток. Антидромные импульсы не вы-
Га а1у освобождения значительных количеств АХ, но при пре-
бло лионаРном раздражении, когда постганглионарный эффект
ЬепхИ?^ВаН ” ТУ?ОКУРарином, АХ продолжает высвобождаться,
скол НИИ шейный и звездчатый ганглии кошки синтезируют АХ со
Р стью 3 мг/г сухого веса в 1 ч; перерезка преганглионарных
21-1514
322
Глава 11
волокон за 90 ч снижает эту величину на 80%. АХ синтезируется
и в изолированных нервных окончаниях. Количество АХ, высво-
бождаемого верхним шейным ганглием кошки при 60-минутном раз-
дражении с частотой 20 имп/с, в пять раз превышает исходное со-
держание АХ в ганглии; следовательно, во время периода актив-
ности должно происходить быстрое замещение израсходованного
АХ. Постганглионарные нейроны, кроме двух типов деполяриза-
ционного ответа на холинэргические импульсы, дают гиперполяри-
зационный ответ на адренэргические импульсы пресинаптических
волокон [134]. Таким образом, в симпатических нейронах имеется
по меньшей мере три типа медиаторных рецепторов.
Другим примером холинэргических синапсов являются синап-
сы между коллатералями спинальных мотонейронов и вставочны-
ми нейронами Реншоу; через эти синапсы по принципу обратной
связи происходит торможение мотонейронов. Ответы клеток Рен-
шоу на раздражение двигательного нерва (антидромные импуль-
сы) усиливаются эзерином и блокируются d-тубокурарином, а
клетки стимулируются АХ и никотином. Окончания двигательных
волокон в мышцах также являются холинэргическими. Здесь мы
видим пример так называемого правила Дэйла, согласно которому
все окончания данного нейрона высвобождают один и тот же ме-
диатор.
Предполагают, что АХ играет роль медиатора и в других от-
делах нервной системы позвоночных, однако четких данных по
этому поводу нет. В коре больших полушарий млекопитающих к
АХ чувствительно менее 30% нейронов; некоторые из них возбуж-
даются под действием АХ, а другие тормозятся. АХ высвобожда-
ется корой больших полушарий человека во время глубокого сна
(быстрое движение глаз). Еще одним примером холинэргических
синапсов являются синапсы между параллельными волокнами и
клетками Пуркинье в мозжечке [42].
Под действием ацетилхолина 35% клеток моста и продолгова-
того мозга возбуждается, а 22% —тормозится [22]. АХ и антихо-
линэстеразный препарат ДФФ усиливают, а d-тубокурарин блоки-
рует ответы зрительной коры на импульсы, поступающие по интер-
каллозальным волокнам. Нервные окончания эфферентного оливо-
кохлеарного пучка в улитке дают положительную гистохимиче-
скую реакцию на АХЭ; введение АХ уменьшает ответ восьмого
нерва, обусловленный раздражением эфферентного пучка. Возмож-
но, АХ является медиатором в синапсах между эфферентными
окончаниями и волосковыми клетками кортиева органа [1Ь].
Ацетилхолинэстераза в большой концентрации содержится в
спинном мозге, симпатических ганглиях, таламусе и хвостатом яД-
ре. Неспецифическая холинэстераза встречается также в некото-
рых ненервных тканях — эритроцитах, подчелюстной железе, гли-
альных клетках и плаценте. Содержание АХ в вентральных кореШ"
ках в 300—500 раз выше, чем в дорсальных; особенно много Аа
Возбудимые мембраны
323
содержится в спинальных мотонейронах. Ацетилхолин обнаружен
не только в возбудимых тканях, но также в селезенке и плаценте.
В большинстве сенсорных волокон (дорсальные корешки спинного
мозга, зрительный нерв) активность холинацетилазы сравнительно
низка; однако в сенсорных нейронах второго порядка она выше,
а в хвостатом ядре очень высока. Активность этого фермента об-
наружена в моторной зоне коры, а также в незначительной степе-
ни в пирамидных путях [145]. Приведенные примеры позволяют
предположить, что роль ацетилхолина в центральной нервной сис-
теме не исчерпывается участием его в синаптической передаче и,
таким образом, выяснена далеко не полностью.
Распределение холинэргических ферментов сходно у всех групп
позвоночных. Для мозга рыб характерна очень высокая активность
АХЭ: при гидролизе 0,027 молей АХ у форели за 30 мин высвобож-
I дается 193 мм3 СО2 на 100 мг сырого веса, у линя—123 мм3, по
I сравнению с 67 мм3 у крысы и 59 мм3 у собаки [34].
1АХЭ и связанные с ней ферменты распространены в животном
мире очень широко. Среди простейших АХЭ обнаружена у инфузо-
рий и трипаносом, но отсутствует у плазмодиев. Инфузория Tetra-
। hymena содержит значительные количества АХЭ, а у Trypanosoma
? найдена и холинацетилаза. АХЭ есть у плоских червей (плана-
рий); по гистохимическим данным, этот фермент локализован пре-
имущественно в нервной системе [132]. Высокой чувствитель-
1 костью к АХ обладают мышцы некоторых сипункулид, аннелид и
иглокожих. АХ в большой концентрации (60 мкг/г сырого веса)
> содержится в радиальных нервах морской звезды [190]. Холинэр-
I гическими, вероятно, являются двигательные нервы некоторых си-
| пункулид, аннелид и иглокожих. ВПСП в мышцах дождевого чер-
* вя блокируется тубокурарином, а АХ повышает проводимость
। кальция [94].
Тормозные экстракардиальные нервы некоторых двустворчатых
и брюхоногих моллюсков также относятся к числу холинэргичес-
ких. В опытах с перфузией слюнных желез осьминога Octopus об-
наружено, что раздражение идущего к ним нерва сопровождается
высвобождением АХ [6]. Высоким содержанием АХ отличаются
ганглии мозга головоногих моллюсков.
Содержание ЛХ,нМ/г
Зрительная доля кальмара 2090
Кора больших полушарий 13
морской свинки
Хвостатое ядро собаки 17
Очень высокой активностью отличаются АХЭ и холинацетилаза
пыоГа кальмаРа- Обмен АХ в нервной ткани головоногих характе-
ризуют следующие данные [135]:
21*
324
Глава 11
Содержание АХ, мг/г сырого веса Синтез АХ, мкМ/мг в 1 Ч Гидролиз АХ, мг/100 мг сырого веса в 1 ч
Мозг кальмара Ганглии осьминога 40—80 200—400
зрительный 236 97,9
звездчатый 33 6,2
надпищеводный 225 20
Нерв мантии осьминога 17 3
Несмотря на то что АХ в этих ганглиях содержится в большом
количестве, его функциональная роль в нервной системе головоно-
гих моллюсков неясна. Совершенно точно установлено, что медиа-
тор гигантских синапсов звездчатого ганглия не идентичен ацетил-
холину. Холинацетилазу (ацетилхолинтрансферазу) содержат три
идентифицированных нейрона брюшного ганглия Aplysia, а в соме
этих нейронов возможен синтез АХ.
У асцидии Ciona ганглии содержат 120 мкг АХ/г, а иннервация
продольных мышц, по-видимому, является холинэргической [66].
Данные, полученные на членистоногих, говорят как за, так и
против роли АХ в синаптической передаче у этих животных. Цент-
ральная нервная система речного рака, омаров и крабов содержит
значительные количества АХ и имеет высокую активность АХЭ
и холинацетилазы. Однако АХ даже в очень высоких концентраци-
ях (10~3 г/мл) и даже после блокады АХЭ не действует на синап-
сы ракообразных [230]. Возбуждающим агентом в периферической
нервной системе ракообразных, по-видимому, является глутамат,
а не АХ.
Большая концентрация АХ и высокая активность АХЭ обнару-
жены в нервной системе некоторых насекомых [36а]. Подсчитано,
Насекомое Часть тела Содержание АХ, мкг/г сырого веса Синтез АХ, мг/г сырого веса в 1 ч Гидролиз АХ, мг/г сырого веса в 1 ч
Calliphora Голова Мозг 37,2 500 (по оценкам) 900 2500
Periplaneta Мозг и подпищеводные ганглии 135 53 153
Торакальные ганглии 95 20 331
6-й абдоминальный ганглий 63 18 314
что если бы весь АХ головы синей мухи Calliphora был ло-
кализован в мозге, его концентрация должна была бы равняться
Возбудимые мембраны
325
500 мкг/г, а у мухи Musca —170 мкг/г. После обработки некото-
рыми инсектицидами (ДДТ, тетраэтилпирофосфат) количество
свободного АХ в нервной системе насекомых повышается; наблю-
даемая после такой обработки усиленная активность потенциалов
действия, по-видимому, ведет к высвобождению АХ. Высокая ак-
тивность АХЭ, возрастающая приблизительно в 6 раз в ходе про-
цессов роста и морфогенеза, наблюдается в нейропиле гусеницы
тутового шелкопряда [208]. Ганглии таракана в нормальных усло-
виях нечувствительны к АХ, но приобретают эту чувствительность
после удаления оболочки. Прямое ионтофоретическое введение АХ
(1,3-10~13 М) в некоторые нейроны ганглиев таракана вызывает
возбуждение вследствие повышения натриевой проводимости
[111]. Потенциал инверсии для АХ равен —40,3 мВ, а для ВПСП
составляет —45 мВ [195а]. Таким образом, ганглии насекомых со-
держат ферменты синтеза и деградации АХ, имеют депо АХ, а неко-
торые нейроны этих ганглиев очень чувствительны к локальному
действию АХ [195].
Резюмируя все эти данные, можно с уверенностью сказать, что
АХ является медиатором в нервно-мышечных синапсах позвоноч-
ных, в парасимпатических окончаниях сердца и внутренних орга-
нов позвоночных, в симпатических и парасимпатических ганглиях
млекопитающих и в синапсах между мотонейронами и клетками
Реншоу, а также может функционировать в некоторых проводящих
путях мозга. Возможно, АХ является медиатором в периферичес-
ких нервах некоторых моллюсков, сипункулид и, вероятно, анне-
лид. АХ — медиатор торможения в сердце большинства моллюсков,
а также, возможно, медиатор возбуждения в некоторых централь-
ных синапсах насекомых. Отсутствие корреляции между распреде-
лением АХ, АХЭ и холинацетилазы, с одной стороны, и действием
экзогенного АХ, его литиков и миметиков, с другой, говорит о
том, что, возможно, холинэргическая система выполняет и другие
функции (по-видимому, «трофические»), кроме участия в процес-
сах синаптической передачи.
Моноамины. Вторая группа медиаторов нервной системы — это
моноамины, а именно: 1) катехоламины, производные катехола
(1,2-дио|К,с|ибензола) с алифатической цепью, включающей амино-
группу, и 2) индолилалкиламины. Наиболее важными из кате-
холаминов являются дофамин, норадреналин (НА) и адреналин
(А), а из индолилалкиламинов—5-окситриптамин (5-ОТ), или се-
ротонин. Моноамины в малых количествах можно обнаружить в
канях по яркой флуоресценции, зеленой в случае катехоламинов
желтой для серотонина. Обе группы моноаминов обнаружены в
ервной системе большинства групп животных.
катехоламины. Катехоламины образуются из тирозина по
следуЮЩей схеме [157]:
326
Глава И
Адреналин
Тирозин
тирозингидроксилаза
СООН
ДОФА
декарбоксилаза аро-
матических кислот
Дофамин
дофамин-|3-гидрокси-
лаза
Норадреналин
фенилэтаноламин-
N-метилтрансфера за
ОН
ОН
Наиболее важным медиатором из этих соединений у позвоноч-
ных является норадреналин. Высвобождающийся из адренергиче-
ских нервных окончаний НА либо удаляется с кровью, либо за-
хватывается пресинаптическими окончаниями, либо разрушается
моноаминоксидазой (МАО) в митохондриях или катехол-о-метил-
трансферазой (КОМТ) в постсинаптической части синапсов. В ка-
честве ингибиторов МАО обычно используются ипразид и гармин.
Норадреналин депонируется в синаптических пузырьках диамет-
ром 300—800 А с плотным содержимым, локализованных в окон-
чаниях адренергических аксонов (рис. 11-24). Он высвобождается
посредством экзоцитоза. Включение новосинтезированного или
захваченного нервными окончаниями НА в депонирующие органел-
лы блокируется резерпином. Продолжительность существования
пузырьков 35—70 дней, тогда как молекулы НА полностью обнови
ляются в течение 1—2 дней; следовательно, в пузырьках происхо-
дит многократное обновление НА [48].
Возбудимые мембраны
327
Адреналин и норадреналин содержатся в мозговом веществе
надпочечников. Относительное количество этих двух катехолами-
нов неодинаково у разных позвоночных. У акулы на долю НА при-
ходится 73%, у кита —83, у мыши — 25, у кролика и морской
свинки — 2% [241]. Симпатическая цепочка собаки содержит
7,3 мкг НА/г; у крупного рогатого скота эта величина равна
4 мкг/г [63]. Постганглионарные симпатические нейроны являют-
Рис. 11-24. Схема норадренэргического синапса, образованного расширением аксо-
на {Cooper J. R.r Bloom F. Е., Roth R. H., Biochemical Basis of Neuropharmacolo-
gy. Oxford University Press, Fair Lawn, N. J., 1970).
Показаны пресинаптический синтез и депонирование НА, высвобождение и обратный захват
НА и расщепление НА моноаминоксидазой (МАО).
ся^адренэргическими и оказывают возбуждающее или тормозное
действие в зависимости от характера ткани-рецептора. Медиато-
ром преганглионарных симпатических волокон служит АХ, но не-
которые вставочные нейроны ганглиев являются адренэргическими
и высвобождают, по-видимому, дофамин [133, 134] (рис. 11-25).
НА в больших концентрациях содержится в гипоталамусе, продол-
говатом мозге и симпатических ганглиях, а в коре больших полу-
шарий и базальных ганглиях его мало. В молекулярном слое моз-
жечка крысы есть флуоресцирующие аксоны; содержащие НА
нервные окончания образуют синапсы на клетках Пуркинье, спон-
танная активность которых уменьшается под влиянием НА. Бло-
када обратного захвата НА повышает этот тормозный эффект.
УДя по этим результатам, некоторые тормозные аксоны моз-
ечка высвобождают НА, который через аденилциклазу действует
на клетки Пуркинье [21, 210].
328
Глава 11
Мозг млекопитающих содержит дофамин, 80% которого лока-
лизовано в базальных ганглиях (nucleus caudatus и putamen). Под
влиянием резерпина эти депо дофамина истощаются и животное
впадает в летаргическое состояние, которое можно устранить, вводя
L-ДОФА [92]. Вещества, подобные резерпину, истощают депо
катехоламинов в центральных синапсах и вызывают у человека
Блокада
атропином
АХ
н-Холинолитики
S-ЕПСП
впсп
БТПСП
- -сс-Адренолитики
Блокада
Ганглиозная
клетка
атропином
Рис. 11-25. Схема предполагаемых синаптических связей в симпатическом ганглии
кролика (Libet В., Fed. Proc., 29, 1945—1956, 1970).
Показаны зоны холинэргической (никотиновой и мускариновой) и а-адронэргической рецеп-
ции. Полагают, что катехоламин высвобождается из вставочного нейрона, раздражаемого
холинэргическим пресинаптическим волокном.
Высвобождение Ал
Высвобождение
катехоламина
(дофамина)
психологическую депрессию. Ингибиторы МАО, напротив, повы-
шают уровень катехоламинов и являются антидепрессантами.
В симпатической системе млекопитающих постганглионарные
нейроны являются адренэргическими. Что же касается низших
позвоночных, то разделение их вегетативной нервной системы на
парасимпатическую холинэргическую и симпатическую адренэрги-
ческую оказалось несостоятельным. Данный катехоламин, напри-
мер НА, может оказывать тормозное действие на одни эффектор-
ные клетки и возбуждающее — на другие. Двойственность дейст-
вия НА отчасти объясняется существованием двух классов адре-
норецепторов — а- и |3-рецепторов — и дальнейшим подразделени-
Возбудимые мембраны
329
Таблица 11-3
Адренэргические рецепторы [1а, 203, 204]
а P
Относительная чувствитель- ность Адреналин > норадреналин > > изадрин Изадрин > адреналин > нор- адреналин
Блокирующие агенты Алкалоиды спорыньи Феноксибензамин Фентоламин Дибензилин Дихлоризопропилнорадрена- лин Пропранолол Пронеталол MJ 1999 (соталол) _ _
Место действия и эффект Vas deferens Кишечник (нервное сплетение) Р-клетки поджелудочной желе- зы Жировые клетки (липолиз) Некоторые сосуды (кожа); су- жение Зрачок; расширение Селезенка; сокращение Мигательная перепонка; сокра- щение Матка; сокращение1 Musculus expansor secundario- rum цыпленка Сердце; возбуждение (₽i) Желудочно-кишечный тракт (мускулатура); расслабление (Р2) Некоторые сосуды (кожа); расширение (р2) Бронхи и легкие; расслабление гладкой мускулатуры Матка; расслабление Печень (гликогенолиз); стиму- ляция (а и Р)
1 В матке виргильной кошки стимуляция Р-рецепторов ведет к расслаблению; в обра-
ботанной прогестероном матке кошки стимуляция а-.рецепторов повышает активность.
ем этих классов. Для идентификации адренорецепторов использу-
ют различные адренолитические препараты или определяют отно-
сительную активность различных катехоламинов. Разные органы
одного и того же животного или гомологичные органы разных ви-
дов животных могут иметь «-адренорецепторы, блокируемые фе-
ноксибензамином или фентоламином, или p-адренорецепторы, бло-
кируемые пропранололом [1Ь]. Норадреналин вызывает расслаб-
ление матки крысы, сокращение матки кролика и человека, а на
матку кошки оказывает смешанное действие, характер которого
зависит от того, беременно животное или нет [156]. Раздражение
симпатических волокон, идущих к толстой кишке, тормозит ее мо-
торику; эффект блокируется пропранололом. Свойства а- и р-рецеп-
торов приведены в табл. 11-3. Высвобождение норадреналина бло-
ируется бретилием и гуанетидином (октадином) независимо от
ого, какой тип адренорецепторов свойствен данному синапсу.
Как и в случае многих других гормонов, посредником действия
техоламинов на клетку служит внутриклеточный аденозинмоно-
330
Глава II
фосфат — циклический З^б'-АМФ. В клеточной мембране чувстви-
тельных клеток присутствует фермент аденилатциклаза, он катали-
зирует превращение АТФ в циклический АМ.Ф, который изменяет
активность различных клеточных ферментов, что и ведет к прояв-
лению специфического физиологического действия катехоламинов.
Далее циклический АМФ расщепляется фосфодиэстеразой, превра-
щаясь в АМФ. Во всех случаях, когда катехоламины действуют
через p-адренорецепторы, происходит повышение активности аде-
нилциклазы, приводящее к увеличению количества циклического
АМФ. Так, повышение амплитуды сокращений сердца млекопита-
ющих при адренэргической стимуляции сопровождается повышени-
ем уровня циклического АМФ. Точно так же адренэргическое рас-
слабление матки сопровождается повышением аденилатциклазной
активности. Увеличение количества циклического АМФ предшест-
вует механическим реакциям на катехоламины; препараты, блоки-
рующие эти реакции, предотвращают и подъем уровня цикличес-
кого АМФ [203, 204]. Последствия активации «-рецепторов, по-
видимому, также связаны с изменениями уровня циклического
АМФ. Различия между двумя типами адренорецепторов, возмож-
но, обусловлены наличием двух активных центров аденилатцикла-
зы клеточной мембраны или двумя формами этого фермента,
встречающимися в разных тканях или одноименных тканях разных
видов.
У некоторых беспозвоночных катехоламины идентифицировали
по флуоресценции нервных окончаний или прямым анализом.
В нервной системе дождевого червя содержится 1,4 мкг адре-
налина и 0,32 мкг норадреналина в расчете на 1 г сырого веса.
В нервной системе некоторых насекомых, например личинок муч-
ного червя Tenebrio, также есть НА (1—2 мкг/г сырого веса). Воз-
можно, НА является нейромедиатором, действующим на светя-
щийся орган жука-светляка [211]. Нервная система морских ежей
и звезд содержит дофамин в концентрациях от 3 до 8 мкг/г и нор-
адреналин в более низких концентрациях [41].
Катехоламины, особенно дофамин, являются важными медиато-
рами нервной системы у моллюсков. В нервной системе головоно-
гих моллюсков Eledone, Octopus и Sepia имеются и дофамин, и нор-
адреналин; концентрация этих моноаминов максимальна в зри-
тельных долях и передней буккальной доле [101]. В ганглиях
двустворчатого моллюска Spisula содержится 40—50 мкг/г дофами-
на и 5—6 мкг/г НА [39]. У пресноводных двустворчатых моллюс-
ков в висцеральном ганглии в большом количестве содержится
дофамин, а в педальном ганглии — 5-ОТ; в церебральном ганглии
присутствуют оба эти амина [217]. В статоцисте беззубки Anodon-
ta у трех нейронов наблюдается значительная флуоресценция,
обусловленная катехоламинами. У Aplysia дофамин в концентра-
ции 10~и М возбуждает (деполяризует) Н-клетки, т. е. клетки, ги-
перполяризуемые АХ [41, 78]. В нейронах длительного торможе-
Возбудимые мембраны
331
ния улитки Helix ответ на раздражение мантийного нерва тормо-
зится дофамином [109]. Дофамином тормозятся и другие нейроны
Helix, возбуждаемые 5-ОТ. В силу всего сказанного роль дофами-
на как естественного медиатора торможения у моллюсков вызы-
вает некоторые сомнения.
(серотонин). Серотонин образуется
5-окситриптамин
по следующей схеме:
Серотонин
Триптофан
триптофангидрокси-
лаза
5-окситриптофан (5-ОТФ)
5-ОТ Ф-декарбокси-
лаза
5-ОТ инактивируется моноаминоксидазой, а ингибитором этого
фермента является ипразид (рис. 11-26). Резерпин приводит к ис-
тощению запасов серотонина и блокирует его синтез аналогично
тому, как это было при его действии на адренэргическую систему.
5-ОТ имеется в различных отделах мозга млекопитающих, в част-
ности в ретикулярной формации, и особенно в области шва.
Высокое содержание серотонина характерно для таламуса, об-
ласти шва, гипоталамуса, лимбической системы и эпифиза, а низ-
кое— для коры больших полушарий и мозжечка. Торможение син-
теза 5-ОТ в мозге вызывает бессонницу; если после этого инъеци-
ровать 5-ОТФ, то восстановится нормальный сон.
Функционирование серотонина как медиатора наиболее четко
выяснено для моллюсков. Нервная система многих моллюсков со-
держит 5-ОТ, а также ферменты, катализирующие его образование
из 5-ОТФ, и моноаминоксидазу для его инактивации. 5-ОТ высво-
ождается при раздражении стимулирующих экстракардиальных
ервов у двустворчатых моллюсков, а кроме того, по-видимому,
®Ляе1ся меДиатором, вызывающим расслабление мышцы-ретрак-
ра биссуса и других подобных мышц.
332
Глава 11
В различных тканях беззубки Anodonta содержатся следующие
количества 5-ОТ (в мкг/г сырого веса):
Церебральный ганглий 72,5
Висцеральный ганглий 43,5
Ножной ганглий 66,5
Мышца - аддуктор 0,3
Сердце 0,0
Концентрация серотонина в ганглиях изменяется в зависимости от
локомоторных ритмов [206].
Триптофан
ипразид
резерпин -
Постсинаптическая
мембрана
Расширение пресинапти-
ческого окончания
Триптофан \
Митохондрия
ДЛЮ
МАО
Рис. 11-26. Схема серотонинэргического синапса с указанием метаболических пу-
тей синтеза и распада серотонина (Rech R. И., Moore К. Е„ Introduction to Psy-
chopharmacology, Raven Press, New York, 1971).
Названия веществ-блокаторов взяты в рамку. 5-ОИУК — оксипндолилуксусная кислота.
Синаптические
пузырьки
Б-ОИУК
5-ОТ
।______
пробенецид
У двустворчатого моллюска Tapes иннервация прямой кишки
носит сложный характер и влияние нервной системы осуществля-
ется через периферическое сплетение; вероятно здесь имеют место
возбуждающий и тормозной эффекты ацетилхолина и возбужда-
ющий эффект серотонина [191]. По-видимому, серотонин является
возбуждающим, а АХ — тормозным медиатором в сердце двуствор-
чатого моллюска Mercenaria [140].
5-ОТ широко распространен у различных головоногих; особен-
но много его содержится в нижней буккальной и оптической долях
£101]. У двустворчатого моллюска Sphaerium 5-ОТ содержат все
Возбудимые мембраны
333
флуоресцирующие клетки ножного ганглия [217]. Два симметрич-
но расположенных гигантских нейрона церебрального ганглия Не-
цх и Ытах содержат 5-ОТ и иннервируют группу нейронов бук-
кальных ганглиев; оба этих рецепторных нейрона стимулируются
5-ОТ при концентрации его 10-7 г/мл. Диэтиламид лизерговой
кислоты блокирует ответ этих нейронов как на импульсы, идущие
от указанных 5-ОТ-содержащих гигантских нейронов, так и на
5-ОТ. Следовательно, рассматриваемые гигантские нейроны, по-
видимому, являются серотонинэргическими [40, 179].
В одном из изученных нейронов Aplysia возникает торможение
под действием дофамина (10-10М) и возбуждение под действием
5-ОТ (Ю~8 М) [78]. У Helix окологлоточное кольцо ганглиев со-
держит 5,5 мкг/г дофамина, причем дофамин низкой концентрации
вызывает в клетках этих ганглиев торможение [112, 113]. 5-ОТ
повышает gci в одних и в других нейронах Helix', в обоих слу-
чаях возникает гиперполяризация.
У пиявки есть вставочные нейроны и мотонейроны, содержащие
5-ОТ; они сосредоточены в подглоточном и сегментных ганглиях,
тогда как нейроны, содержащие катехоламины, локализованы пре-
имущественно в головных ганглиях [61, 147]. У дождевого червя
Lumbricus обнаружено большое количество нейронов, обладающих
флуоресценцией, характерной для 5-ОТ, которые, по-видимому, яв-
ляются моторными клетками нервной цепочки. В мозге мечехвоста
Limulus есть нейроны, содержащие 5-ОТ, и нейроны, содержащие
НА; концентрация 5-ОТ в нервной цепочке равна 10 мкг/г [162].
Имеются довольно четкие данные о том, что адреналин являет-
ся медиатором некоторых автономных нервов низших позвоночных,
дофамин и норадреналин — медиаторами в специфических обла-
стях мозга позвоночных и дофамин — медиатором у моллюсков.
Серотонин широко распространен у различных групп животных,
но его роль как медиатора наиболее четко выявлена в случае ней-
ропиля позвоночных и у моллюсков; возможно, что серотонин иг-
рает аналогичную роль и у аннелид.
Аминокислоты. Роль медиаторов, как предполагают, играют
следующие три аминокислоты:
Н
Глутаминовая кислота НООС—СН2—СН2—С—СООН
NH2
> СО2
у-Аминомасляная кис- НООС—СН2—СН2—СН2—NH2
лота (ГАМК)
Н
Глицин Н—С—СООН
nh2
334
Глава И
Имеются веские доводы в пользу того, что ГАМК является тормоз-
ным медиатором в нервно-мышечных синапсах ракообразных и на-
секомых (гл. 16) [122]. Пикротоксин снимает как тормозное
влияние нервов, так и тормозный эффект ГАМК у этих животных.
Показано также, что ГАМК высвобождается при раздражении этих
тормозных нервов [180]. Медиатором возбуждения мышц у этих
животных, по-видимому, является глутамат [121].
У млекопитающих ГАМК синтезируется в специфических нерв-
ных окончаниях (синаптосомах), а также захватывается ими из
^межклеточной среды. Такой захват осуществляется 25—32% си-
наптосом, выделенных из спинного мозга и коры больших полу-
шарий [20]. У млекопитающих передачу возбужения с обонятель-
ных рецепторных нейронов на клетки-зерна, по-видимому, осуще-
ствляет норадреналин, а передачу торможения с этих клеток на
митральные клетки обонятельных луковиц — ГАМК [43]. ГАМК
оказывает тормозное действие на нейроны коры больших полуша-
рий и, возможно, является медиатором торможения в коре, гиппо-
кампе, мозжечке и стволе мозга [46, 47]. Клетки Пуркинье моз-
жечка оказывают тормозное действие на нейроны ядра Дейтерса,
а ГАМК воспроизводит это действие. Оба эти эффекта блокируют-
ся пикротоксином [174]. Возможно, что ГАМК осуществляет пере-
дачу торможения от корзинообразных и звездчатых клеток к клет-
кам Пуркинье в мозжечке лягушки [244].
Тормозное действие на многие центральные нейроны позвоноч-
ных оказывает также глицин, который вполне может являться
главным медиатором торможения в спинном мозге. Глицин гипер-
поляризует мотонейроны; потенциал инверсии равен при этом
—3,8 мВ по отношению к потенциалу покоя, а кривая «доза—эф-
фект» имеет S-образную форму, что свидетельствует о том, что
для начала реакции необходимо наличие нескольких молекул гли-
цина [239, 240]. Концентрация глицина в спинном и продолгова-
том мозге в 2—5 раз выше, чем в остальных отделах мозга [2].
Торможение, обусловленное глицином, подавляется низкими кон-
центрациями стрихнина, в то время как тормозное действие ГАМК
блокируется бикукуллином или пикротоксином [43, 45]. В спинном
мозге стрихнин подавляет преимущественно постсинаптическое
торможение, а пикротоксин — пресинаптическое [44].
ГАМК оказывает тормозное действие на маутнеровские нейро-
ны у серебряного карася; при этом максимальной чувствитель-
ностью к ГАМК обладают коллатеральные окончания, но некото-
рый эффект наблюдается и на периферии латерального дендрита.
Следовательно, тормозное действие ГАМК может здесь быть как
пресинаптическим, хак и постсинаптическим [50].
Концентрация ГАМК в брюшной нервной цепочке омара равна
480 мкг/ г сухого веса, в сенсорных нервах — от 9 до 14, в волок-
нах двигательных нервов—6150 и в тормозящих нервах —
32 800 мкг/г сухого веса [122, 123].
Возбудимые мембраны 335
ГАМК является весьма эффективным ингибитором мышц рако-
образных; ее тормозное действие, как и нервное торможение, бло-
кируется пикротоксином. Действие ГАМК очень четко локализо-
вано [218, 219]. У членистоногих его действие на клетки как при
пресинаптическом, так и при постсинаптическом торможении со-
стоит в повышении проводимости для С1~ [220]. В центральной
нервной системе кобылки ГАМК тормозит ответ крупного вставоч-
ного нейрона (Т-клетки) на импульсы, поступающие от слуховых
органов; на фоне пикротоксина эти клетки реагируют на бинау-
ральное раздражение, тогда как в норме наблюдается только мо-
уральная реакция [216]. В ганглии таракана местная апплика-
ция ГАМК (Ю-13 М) вызывает исчезновение потенциалов дейст-
вия, гиперполяризацию, повышение проводимости СГ~ и в мень-
шей степени К+ [Ш]. В шестом абдоминальном ганглии потен-
циал инверсии для ТПСП равен —79 мВ, а для ответа на местную
аппликацию ГАМК —75 мВ; оба эти ответа блокируются пикро-
токсином [195а].
L-глутамат (но не его D-изомер) оказывает возбуждающее дей-
ствие на некоторые нейроны, расположенные в самых разных отде-
лах нервной системы, в частности в коре головного мозга и в моз-
жечке [46]. Глутаминовая кислота высвобождается корой при
электрическом раздражении и во время пробуждения (но не во
время сна) и оказывает сильное возбуждающее действие на кору
(особенно на гиппокамп) и на некоторые базальные ганглии [97].
Глутамат возбуждает также и нервные клетки ракообразных; в
большой концентрации (0,01—0,1 мг/г) он содержится в нервных
тканях Carcinus. Глутамат оказывает очень сильное возбуждающее
действие и на мышцы ракообразных. Мышцы насекомых также
возбуждаются глутаматом, причем эффективность L-изомера в 10
раз выше, чем D-изомера [11, 234]. Местная аппликация глутама-
та в разведении 1 :108 вызывает сокращение мышцы саранчи
[17]. Он оказывает возбуждающее действие в синапсах гигантско-
го аксона кальмара [154]. В ганглиях улитки глутаматом
(5-10~7 М) возбуждаются нейроны, отличные от тех, которые воз-
буждаются АХ [74, 75]. В нервно-мышечном препарате Helix при
раздражении нерва происходит высвобождение глутамата [107].
Возникает вопрос, почему аминокислоты, являющиеся обычны-
ми компонентами плазмы, служат специфическими медиаторами
нервной системы. Один из возможных ответов на него состоит в
том, что постсинаптическая мембрана в обычных условиях не омы-
вается плазмой крови и концентрация аминокислот в синаптичес-
кой щели во время передачи нервного импульса может быть зна-
чительно выше, чем в плазме. Роль ГАМК как медиатора тормо-
ения (особенно у ракообразных, насекомых и в мозжечке позво-
чных) и роль глицина как медиатора торможения в спинном
та ЗГе ДОстаточно хорошо обоснованы. Что же касается роли глу-
мата как медиатора возбуждения, то для нервно-мышечной пе-
336
Глава 11
редачи у ракообразных и насекомых она показана совершенно оп-
ределенно, а для позвоночных эти данные менее убедительны.
Недавние исследования показали, что некоторые автономные
нейроны нельзя отнести ни к числу холинэргических, ни к числу
адренэргических; они могут использовать в качестве медиатора
АТФ и поэтому именуются пуринэргическими. Такие нейроны обна-
ружены в желудочно-кишечном тракте и мочевом пузыре млеко-
питающих, а также в нервном аппарате кишечника рыб и амфи-
бий [25] (гл. 16).
Вопрос о том, какое именно вещество является медиатором в
каждом конкретном синапсе, выяснен лишь в немногих случаях.
Однако представляется маловероятным, что в дальнейшем будет
обнаружено большое количество новых медиаторов. Реальные раз-
личия, состоящие в разной чувствительности иннервированных кле-
ток и в различных молекулярных механизмах действия данного
медиатора, обусловлены свойствами молекул-рецепторов. Возмож-
но, что для каждого медиатора будет открыто много новых специ-
фических рецепторов.
Заключение
В сравнительной физиологии клеточных мембран в последнее
время получено очень много новых данных. Клеточные мембраны
выполняют большое количество специфических функций, и то, что
все эти функции выполняются слоем липопротеида толщиной всего
100 А, вызывает изумление. Опыты по измерению проницаемости
мембран показали, что в них имеются специальные каналы диа-
метром от 5 до 15 А. Разнообразные процессы активного перено-
са были рассмотрены выше (вопросы ионной регуляции в гл. 2 и
транспорт продуктов пищеварения в гл. 4).
Основная особенность мембран сенсорных клеток, нейронов и
клеток-эффектов состоит в том, что в них есть области, обладаю-
щие электровозбудимостью, и области, чувствительные к химиче-
ским агентам или механическому воздействию. Электровозбудимые
мембраны обычно характеризуются пороговой величиной потен-
циала, после превышения которой возникают регенеративные им-
пульсы. К структурам, реагирующим на химическое воздействие,
относятся: 1) сенсорные окончания, возбуждающиеся при воздей-
ствии на них определенного химического вещества или продуктов
фотохимической реакции, и 2) постсинаптические мембраны, в ко-
торых имеются рецепторы для специфических медиаторов. Многие
нейроны возбуждаются химическими раздражителями на одном
своем полюсе, поддерживают самораспространяющиеся импульсы
в аксонах и выделяют определенный медиатор в своих окончаниях.
Другие нейроны не имеют электровозбудимых проводящих участ-
ков. Если размер нейрона значительно меньше, чем его константа
длины (например, лишенные аксонов амакриновые клетки сетчат-
Возбудимые мембраны
337
ки) то передача сигнала происходит без генерирования потенциа-
ла действия. Электротонический потенциал распространяется без
возникновения спайка от постсинаптической зоны дендритов к со-
ме нейрона и области генерирования потенциалов действия в ак-
соне.
Наиболее изученным типом электрогенеза является потенциал
действия, распространяющийся по принципу «все или ничего» в
электровозбудимой мембране. В большинстве случаев причиной
возникновения такого потенциала в нервах и поперечнополосатых
мышцах является последовательное повышение проницаемости кле-
точной мембраны для натрия и калия. Продолжительность потен-
циала действия изменяется в зависимости от продолжительности
К-активации. К явлениям, наблюдаемым в электровозбудимой
мембране после прохождения волны потенциала действия, отно-
сится задержка реполяризации, обусловленная задержанной К-ак-
тивацией, и остаточная гиперполяризация, обусловленная устой-
чивым оттоком калия. Во многих мышцах (особенно в мышцах
членистоногих и гладких мышцах позвоночных), а также в некото-
рых нейронах моллюсков и у инфузорий возникновение потенциала
действия обусловлено входящим током ионов кальция. Возможно,
что кальциевые спайки характерны для более низкоорганизован-
ных форм, чем натриевые. У всех электровозбудимых мембран,
по-видимому, имеются и натриевые, и кальциевые каналы, причем
у разных клеток или в разных частях мембраны может преобла-
дать какой-то один тип каналов. У немногих мембран деполяриза-
ция обусловлена выходящим током хлора. Отдельные нейроны,
для которых характерна ритмическая импульсация, обладают дву-
кратной калиевой активацией, а колебание активности в некото-
рых пейсмекерных нейронах обусловлено изменением во времени
сопряжения натриевой и калиевой проводимости.
В мембранах, обладающих химической чувствительностью, так-
же обнаружены различные градуальные изменения проводимости.
Обычно деполяризация постсинаптической мембраны обусловлена
повышением натриевой и калиевой проводимости, в то время как
в наиболее изученных гиперполяризующихся постсинаптических
мембранах гиперполяризация обусловлена направленным внутрь
током хлора. Амплитуда возникающего импульса зависит от от-
ношения между потенциалом покоя и потенциалом ионного равно-
весия, а в некоторых случаях играет роль и изменение мембран-
ного сопротивления. Так, гиперполяризация фоторецепторов гре-
ешка обусловлена повышением проводимости калия. Если потен-
Иал равновесия положителен по отношению к ПП, то возникаю-
щая Деполяризация может быть обусловлена повышением прово-
Мости С1 (даже если концентрационный градиент С1 направлен
н УТРЬ Клетки). Третий тип градуального электрического ответа,
х ЛЮДаемый в фоторецепторах позвоночных, обусловлен проис-
Дящим на свету ослаблением асимметричного натриевого тока
22" 1514
338
Глава 11
(темнового тока) между двумя концами рецептора. В нейронах
моллюсков медиаторы вызывают самые разнообразные изменения
проводимости. У многих клеток потенциал покоя обусловлен ра-
ботой натриевого насоса, который в некоторых случаях, например
в рецепторах растяжения ракообразных, ответствен и за остаточ-
ную гиперполяризацию. Существующие в норме колебания работы
насоса могут лежать в основе медленных электрических ритмов в
гладких мышцах и некоторых нейронах.
Во многих тканях — эмбриональных, эпителиальных, а также
в некоторых нервных центрах между клетками существует элект-
рическая связь, которая зависит от условий и от типа клеток, а
также от площади щелевых соединений. Электрическая передача
может быть симметричной и несимметричной (поляризованной),
как в электрических синапсах. Обычно электрические контакты
обеспечивают быстрые синхронные ответы групп нейронов. В не-
которых синапсах сосуществуют зоны химической и электрической
передачи. Известен случай электрического торможения, осущест-
вляемого с помощью экстрааксональной гиперполяризации.
В химических синапсах, в пресинаптических нервных оконча-
ниях или расширениях аксонов происходит синтез специфических
медиаторных веществ (синаптических передатчиков), которые депо-
нируются в особых пузырьках. Высвобождение медиатора происхо-
дит при пресинаптической деполяризации, в результате притока
ионов кальция. Синаптическая задержка отчасти обусловлена тем,
что медиатор должен диффундировать через синаптическую щель
к рецепторам постсинаптической мембраны. Инактивация медиа-
тора может произойти в результате постсинаптического гидролиза,
диффузии его в направлении от рецепторов или возвращения в
пресинаптические окончания. Химические синапсы обнаружены во
всех типах нервной системы, в том числе в нервной сети кишечно-
полостных.
Размер молекул всех известных к настоящему времени медиа-
торов относительно мал. Наиболее широко распространен среди
них ацетилхолин, который, возможно, несет мембранные функции
и во многих ненервных тканях. Установлено, что ацетилхолин яв-
ляется медиатором в синапсах вегетативной нервной системы поз-
воночных, в некоторых центральных синапсах и в окончаниях ре-
гуляторных нервов позвоночных, в сердце моллюсков, а также
участвует в нервно-мышечной передаче многих групп животных.
Кроме того, ацетилхолин является медиатором в некоторых цент-
ральных синапсах насекомых.
Широко распространены также катехоламины, о чем свидетель-
ствуют данные, полученные с помощью флуоресцентной микроско-
пии. Основным медиатором адренэргических нейронов позвоноч-
ных является норадреналин, тогда как у моллюсков главную роль
играет дофамин. В нервной системе различных групп животных
широко распространен также серотонин; у моллюсков он может вы-
Возбудимые мембраны
339
ступать как функциональный антагонист ацетилхолина и (или)
дофамина.
Роль медиаторов выполняют три аминокислоты — глутаминовая
кислота, оказывающая возбуждающее действие, глицин и у-ами-
номасляная кислота (ГАМК), оказывающие тормозное действие,
у членистоногих глутамат и ГАМК являются антагонистами; этот
антагонизм особенно выражен в нервно-мышечных синапсах.
Данный медиатор у одного и того же вида животных может
работать по-разному в зависимости от свойств рецепторных моле-
кул у различных постсинаптических клеток. Например, у позвоноч-
ных холинорецепторы могут быть двух видов — мускаринового и
никотинового. В ганглиях моллюсков АХ деполяризует одни и ги-
перполяризует другие нейроны. Никотиновый холинорецептор уда-
лось изолировать (главным образом из электрического органа
электрических рыб), и оказалось, что этот белок отличен от аце-
тилхолинэстеразы. Норадреналин у позвоночных функционирует
через посредство рецепторов двух типов —а- и р-адренорецепто-
ров. В обоих случаях инициируется цепь внутриклеточных реак-
ций, одним из звеньев которой — своего рода посредником — слу-
жит циклический АМФ; эти реакции в конце концов приводят к от-
вету клетки на норадреналин. Предполагают, что характер измене-
ния ионной проводимости под действием медиатора зависит от
природы рецепторных молекул.
Показано, что свойства мембран возбудимых клеток в разных
ее частях различны. Изменение проницаемости для специфических
ионов происходит в определенных участках мембраны. Раздражи-
телями для липопротеидных молекул, ответственных за возбуди-
мость мембраны, являются электрический ток, специфические хи-
мические вещества или механическая деформация. Амплитуда воз-
никающих импульсов зависит от равновесных потенциалов для
ионов, перемещающихся через мембрану в клетку или из нее. Кро-
ме того, изменение сопротивления может изменять внеклеточные
поля, обусловленные асимметрией клеточных батарей. Механиз-
мы биоэлектрогенеза бывают самыми разными. Одни из них
встречаются чаще, другие реже.
СПИСОК ЛИТЕРАТУРЫ
Г Adrian R. И., Freygang W. Н., J. Physiol., 163, 61—103, 1962. К and Cl con-
1 ^etance, frog muscle.
a- Ahlquist ft. P., Ann. Rev. Pharmacol., 8, 259—272, 1968. Agents blocking adre-
^erg*c receptors.
Amaro J. et al., Brit. J. Pharmacol. Chemother., 28, 207—211, 1966. Neuro-
ransmitter in efferent fibers of auditory system.
prison H. et al., Comp. Biochem. Physiol., 28, 1345—1355, 1969. Distri-
3 zi glycine in brain and spinal cord of vertebrates.
A, Ito M., Oscarsson O., J. PhysioL, 159, 410—435, 1961. Anion per-
4 Дл1 ity m°torneuron synapses.
’ So Ул Bennett M. V. L., J. Cell BioL, 49, 159—172, 1971. Coupling resi-
nce at septa of lateral giant axons, crayfish.
22*
340
Глава 11
5. Auerbach A. A., Bennett М. V. L., J. Gen. Physiol., 53, 183—210, 211—239,
1969. Giant fiber synapses in fish.
6. Bacq Z. M„ Ghiretti F„ Arch. Int. Physiol., 49, 165—171, 1952; also Pubbl.
Staz. Zool. Napoli, 24, 267—277, 1953. Acetylcholine in Octopus.
7. Baker P. F., Proc. XXV Internal. Physiol. Congr., 8, 17—19, 1971. Transport
of Ca and Mg across nerve cell membranes.
8. Baker P. F. et al., J. Physiol., 200, 459—496, 1969. Ouabain-sensitive fluxes
of Na and К in squid giant axons.
9. Baker P. F., Hodgkin A. L., Ridgway E. B., J. Physiol., 218, 709—755, 1971.
Calcium flux in squid axon.
10. Baker P. F. et al., Nature, 190, 885—887, 1961; also J. Physiol., 170, 541—560,
1964. Internally perfused squid axon.
11. Baylor D. A., Nicholls J. G„ J. Physiol., 203, 555—569, 571—589, 1969. Ionic
factors in activity of leech central nervous system.
12. Beeler G. W., Reuter H., J. Physiol., 207, 165—190. 191—209, 1970. Voltage
clamp experiments on mammalian ventricular fibers.
13. Belton P., Grund fest H., Amer. J. Physiol., 203, 588—594, 1962. К spikes in
Tenebrio larva muscle.
13a. Beers W. H., Reich E., Nature, 228, 917—922, 1970. Structure and activity of
acetylcholine.
14. Bennett M. V. et al., J. Neurophysiol., 30, 131—179, 180—208, 209—235, 236—
300, 1967. Physiology and ultrastructure of electrotonic junctions.
15. Bennett M. V. L. In: Central Nervous System and Fish Behavior, edited by
D. Ingle, Chicago University Press, 1968, pp. 147—169. Electrotonic synapses.
16. Bennett M. V. L., Trinkaus J. P., J. Cell Biol., 44, 592—610, 1970. Electrical
coupling between cells in Fundulus embryos.
17. Beranek R., Miller P. L., J. Exp. Biol., 49, 83—93, 1968. Glutamate stimulation
of insect muscle.
18. Berger IF. X., Uhrik B., Z. Zellforsch., 127, 116—126, 1972. Nexuses in dipteran
salivary glands.
19. Binstock L., Goldman L., J. Gen. Physiol., 54, 730—740, 741—754, 755—764,
1969. Conduction in Myxicola giant axon.
20. Bloom F. E. In: The Neurosciences, Second Study Program, edited by
F. O. Schmitt, Rockefeller University Press, New York, 1970, pp. 729—747.
Structure-function correlation in synapses.
21. Bloom F. E., Hoffer B. J., Siggins G. R., Brain Res., 25, 501—521, 1971. Che-
mical transmitters in rat cerebellum.
22. Bradley P. B., Int. Rev. Neurobiol., 11, 1—55, 1968. Chemical transmitters in
brain.
23. Brightman M. W., Reese T. S., J. Cell Biol., 40, 648—677, 1969. Gap junctions,
tight junctions, Mauthner cells.
24. Bullock T. H., J. Neurophysiol., 11, 343—364, 1948. Synaptic transmission,
squid mantle ganglion.
25. Burnstock G., Pharmacol. Rev., 21, 247—324, 1969; 52, 129—197, 1972. Auto-
nomic innervation of visceral and vascular muscles in vertebrates.
26. Burnstock G., Iwayama T., Progr. Brain Res., 34, 389—404, 1971. Autonomic
synapses.
27. Bush В. M. H., Roberts A., Nature, 218, 1171—1173, 1968. Reflexes with-
out impulses from a crab muscle receptor.
28. Carpenter D., Gunn R„ J. Cell. Physiol., 75, 121—127, 1970. Ionic currents in
Aplysia neurons.
29. Chandler W. K„ Meves H„ J. Physiol., 180, 788—820, 1965. Voltage clamp
experiments on internally perfused giant axons.
29a. Changeux J. P., Meunier J., Huchat M„ Molec. Pharmacol., 7, 548—553, 1971-
Properties of acetylcholine receptor.
30. Changeux J. P., Podleski T., Meunier J., J. Gen. Physiol., 54, 225s — 244s,
1969. Acetylcholine receptor, electric organ.
31. Chapman R. A., J. Exp. Biol., 45L, 475—478, 1966. Repetitive firing, crab fi-
bers.
Возбудимые мембраны 341
on Chapman R. A., Parkhurst J. Н., J. Exp. Biol., 46, 63—84, 1967. Conduction
6 ' velocity, cockroach nerve.
33 Chiarandini D. J., Gershenfeld H. M., Science, 156, 1595—1596, 1967. Ionic
3 ‘ mechanisms, molluscan synapses.
34 Close F., Serfaty A., Bull. Soc. Histoire Nat. Toulouse, 92, 205—217, 1957.
Cholinesterase in fish.
35 Cohen L. B. et al., J. Physiol., 211, 495—515, 1970; 218, 205—237, 1971. Opti-
cal changes in nerve during conduction.
36 Cohen L. B., Landowne D. In: Biophysics and Physiology of Excitable Mem-
branes, edited by W. J. Adelman, Van Nostrand Reinhold, New York, 1971,
np. 247—263. Birefringence and other optical changes in nerves.
36a Colhoun E. H., J. Insect Physiol., 2, 108—116, 1958; also Can. J. Biochem.
' Phys., 37, 1127—1134, 1959. Content of ACh, AChE, ACh acetylase, insect
nervous systems.
37 . Connor J. A., Stevens C. F., J. Physiol., 213, 1—19, 21—30, 31—53, 1971.
Early potassium current and repetitive firing.
38 . Coombs J. S. et al., J. Physiol., 137, 326—373, 1955; 145, 505—538, 1959.
Electrical constants of motorneuron membranes.
39 . Cottrell G. A., Brit. J. Pharmacol., 29, 63—69, 1967. Adrenergic synapses.
40’ Cottrell G. A., Nature, 225, 1060—1062, 1970. Serotonin synapses.
41. Cottrell G. A., Laverack M. S., Ann. Rev. Pharmacol., 8, 273—298, 1968. In-
vertebrate pharmacology.
42. Crawford J. M. et al., Nature, 200, 579—580, 1963. Excitation of cerebellar
neurons by acetylcholine.
43. Curtis D. R., Duggan A. W., Johnston G. A. R., Exp. Brain Res., 12, 547—565,
1971. Evidence for glycine as a transmitter in spinal cord.
44. Curtis D. R., Eccles J., J. Physiol., 145, 529—546, 1959; 150, 374—398, 1960.
Time course of synaptic actions in spinal cord.
45. Curtis D. R., Watkins J. C., J. Neurochem., 6, 117—141, 1960. The excitation
and depression of spinal motorneurons by amino acids.
46. Curtis D. R., Watkins J. C., Pharmacol. Rev., 17, 347—391, 1965. Transmitters
in central nervous system.
47. Curtis D. R. et al., Nature, 226, 1222—1224, 1971; also Brain Res., 32, 69—96,
1971. Evidence for GABA as transmitter in spinal cord.
48. Dahlstrom A., Haggendal J., Acta Physiol. Scand., 67, 278—288, 1966. Half-
life of adrenal vesicles.
49. Dewey M. M., Barr L., J. Cell Biol., 23, 553—585, 1964. Structure and distri-
bution of nexus.
50. Diamond J., J. Physiol., 194, 669—723, 1968. GABA and glutamate sensitivity,
Mauthner cells.
51. Dodge F. A., Rahamimoff R„ J. Physiol., 193, 419—432, 1967. Role of calcium
cn 7? neiJromuscular transmission.
Dowling J. E., J. Invest. Ophthal., 9, 655—680, 1970. Retinal synapses.
w. Dowling J. E., Werblin F. S., J. Neurophysiol., 32, 315—338, 1969. Retinal
c. synapses in Necturus.
’ J- C-> The Physiology of Nerve Cells, Johns Hopkins Press, Baltimore,
1957, 270 pp.
• Eccles J. C. In: Handbook of Physiology, Sec. 1, Vol. 1, edited by H. W. Ma-
goun. Amer. Physiol. Soc., Washington, D. C., 1959, pp. 59—74. Central ner-
56 f°US synaPses-
J’ The Physiology of Synapses, Springer-Verlag, Heidelberg, 1964,
57 F h PP'
58 Science, 176, 473—481, 1972. Bioelectric control of ciliary activity.
o-q J’ ^еп. Physiol., 46, 573—587, 1963. Electrical interaction of paired
59 l’a^110n cells in the leech.
rZT- Naitoh y- «I- Gen. Physiol., 55, 467—483, 1970. Bioelectrics of Pa-
60 pniectun.
Naitoh y- Friedman R., J. Exp. Biol., 56, 683—694, 1972. Sensory
chanisms in Paramecium.
342
Глава 11
61. Ehinger В., Falck В., Myhrberg Н. S., Histochemie, 15, 140—149, 1968. Hirudo
catecholamines and indolamines.
62. Ehrenpreis S., Fleisch J. H., Mittag T. W., Pharmacol. Rev., 21, 131—181, 1969.
Molecular nature of pharmacological receptors.
63. Von Euler U. S., Noradrenaline, Chemistry, Physiology, Pharmacology, Clini-
cal Aspects, Charles C. Thomas, Springfield, Ill., 1958, 382 pp.
€4. Falk G., Fatt P., Proc. Roy. Soc. Lond. B, 160, 69—123, 1964. Linear electrical
properties of striated muscle fibers observed with intracellular electrodes.
65. Florey E., Fed. Proc., 26, 1164—1178, 1967. Neurotransmitters and modulators
in the animal kingdom.
66. Florey E., Comp. Biochem. PhysioL, 22, 617—627, 1967. Cholinergic neurons in
tunicates, an appraisal of the evidence.
-67. Fozzard H., J. Physiol., 182, 255—267, 1966. Capacity of heart membranes.
Personal communication, 1973.
67a. Furshpan E. J., Furukawa T., J. Neurophysiol., 25, 732—771, 1962. Conduction
in Mauthner cells.
68. Furshpan E. J., Potter D. D., J. Physiol., 145, 289—325, 1959. Transmission
at giant motor synapses of crayfish.
69. Furshpan E. J., Potter D. D., J. PhysioL, 145, 326—330, 1959. Slow post-
synaptic potentials recorded from giant motor fiber of crayfish.
69a. Furukawa T., Progr. Brain Res., 21A, 44—70, 1966. Electrical inhibition in the
Mauthner cell.
70. Gage P. IF., Moore J. W., Science, 166, 510—512, 1969. Synaptic current at
squid giant synapse.
71. Gardner D., Randel E. R., Science, 176, 675—678, 1972. Diphasic postsynaptic
potential, Aplysia.
12. Geduldig G., Gruener R., J. PhysioL, 211, 217—244, 1970. Aplysia neurons:
1SI3 з n d СЗэ currents
73. Geduldig G., Junge D., J. PhysioL, 199, 347—365, 1968. Sodium and calcium
components of action potentials in Aplysia giant neurons.
74. Gerschenfeld H. M., Nature, 203, 415—416, 1964. Non-cholinergic synaptic in-
hibition in the central nervous system of a mollusc.
75. Gerschenfeld H. M., Science, 171, 1252—1254, 1971. Serotonin, snail neurons.
76. Gerschenfeld H. M., Chiarandini D. J., J. NeurophysioL, 28, 710—723, 1965.
Synaptic potentials and ion conductance changes, snail neurons.
77. Gerschenfeld H. M., Lasansky A., Int. ,J. Neuropharm., 3, 301—315, 1964.
Action of glutamic acid and amino acids on snail central neurons.
78. Gerschenfeld H. M., Stefani E., J. PhysioL, 185, 684—700, 1966. Electro-phy-
siological study of 5-HT receptors of neurons in molluscan nervous systems.
79. Gerschenfeld H. M„ Tauc L., J. PhysioL (Paris), 56, 360—361, 1964. Dopamine
effects on Aplysia neurons.
80. Ginsborg B. L., Pharmacol. Rev., 19, 289—316, 1967. Biophysics of synaptic
and conducting membranes.
81. Girardier L. et al., J. Gen. PhysioL, 47, 189—214, 1963. Anion permeability of
crayfish muscle fibers.
82. Grinnell A. D., J. PhysioL, 182, 612—648, 1966. Interaction between motor-
neurons in frog spinal cord.
83. Grundfest H. In: Properties of Membranes and Diseases of the Nervous
System, edited by D. B. Tower, Springer-Verlag, Heidelberg, 1962, pp. 71—99.
Ionic transport across neural and non-neural membranes.
84. Grundfest H. In: Advances in Comparative Physiology and Biochemistry,
Vol. 2, edited by О. E. Lowenstein, Academic Press, New York, 1966, pp. I"
116. Comparative electrobiology of excitable membranes.
85. Grundfest H. In: The Neurosciences: A Study Program, edited by G. C. Quar-
ton et al., Rockefeller University Press, New York, 1967, pp. 353—371. Sy-
naptic and ephaptic transmission.
«85а. Hagiwara S. et al., Comp. Biochem. PhysioL, 13, 453—460, 1964. Conduction
in giant fibers of Eudistyla.
Возбудимые мембраны
343
.. Hagiwara S., Kidokoro Y., J. Physiol., 219, 217—232, 1971. Ionic components
of action potentials in amphioxus muscle.
86 Hama K., Anat. Rec., 141, 275—294, 1961. Fine structure of giant fibers of
87 WzcMeSR., Gobel S., J. Cell Biol., 52, 397—408, 1972. Septate and gap jun-
ctions in Hudra.
88 Heuser J. E., Miledi R., Proc. Roy. Soc. bond. B, 179, 247-260, 1971. Frog-
neuromuscular ending; La++ stimulation.
88a Heuser J. E., Reese T. S., J. Cell Biol., 57, 315—344, 1973. Turnover of synaptic
' vesicles at neuromuscular junctions.
89 Hille B., J. Gen. Physiol., 50, 1287—1301, 1967; 51, 199—219, 1968. Pharma-
cological analysis of sodium and potassium channels in frog nerve.
90. Hodgkin A. L., Conduction of the Nervous Impulse, Charles C. Thomas,.
Springfield, Ill., 1964, 108 pp.
91. Hoffer B. J. et al., Brain Res., 25, 523—534, 1971.
92. ' Hornykiewicz O., Pharmacol. Rev., 18, 925—964, 1966. Dopamine (3-hydroxy-
triptamine) and brain function.
93. Howarth J. V., Keynes R. D., Ritchie J. M., J. Physiol., 194, 745—793, 1968.
Heat production by nerve.
93a. Hughes G. M„ J. Exp. Biol., 46, 169—193, 1967. Abdominal ganglion of
Aplysia.
94. Ho Y., Kuriyama H., Tashiro N., J. Exp. Biol., 50, 107—118, 1969. Miniature
excitatory junction potentials in somatic muscle of earthworm.
95. Iversen L. L. In: The Neurosciences: Second Study Program, edited by
F. O. Schmitt, Rockefeller University Press, New York, 1970, pp. 768—782.
Neurotransmitters.
96. Jacobson S., Pollen D. A., Science, 161, 1351—1353, 1968. Conduction in api-
cal dendrites.
97. Johnson J. L., Brain Res., 37, 1—19, 1972. Glutamic acid as a synaptic trans-
mitter.
98. Josephson R. K-, J. Exp. Biol., 42, 139—152, 1965. Conduction systems in stalk
of a hydroid.
99. Josephson R. K-, J. Exp. Biol., 47, 172—190, 1967. Conduction and contraction
in the column of Hydra.
100. Josephson R. K-, Machlin M., J. Gen. Physiol., 53, 638—665, 1969. Potentials
across body wall of Hydra.
101. Juorio A. V., J. Physiol., 216, 213—226, 1971. Catecholamines and 5-HT in ner-
vous tissue of cephalopods.
101a. Kandel E. R., Gardner D., In: Neurotransmitters: Proceedings, Vol. 50, Asso-
ciation for Research in Nervous and Mental Diseases, Williams and Wilkins,.
Baltimore, 1972, pp. 91—144. Synaptic actions in molluscan ganglia.
Ю2. Kanno Y. Y., Loewenstein W. R., Nature, 201, 194—195, 1964. Low-resistance
coupling between gland cells.
Rkk Kao C. Y„ J. Neurophysiol., 23, 618—635, 1960. Postsynaptic electrogenesis in’
septate giant axons.
4- Kao C. Y., Pharmacol. Rev., 18, 997—1049, 1966. Tetrodotoxin, saxitoxin, and'
]n_ the study of excitation phenomena.
Katz B., Miledi R„ J. Physiol., 203, 459—487, 1969. TTX-resistant electrical!
Шк activity in presynaptic terminals.
1Ub- Katz B., Miledi R., J. Physiol., 216, 503—512, 1971. Effect of depolarization on-
107 ^YnaPtic transfer in stellate ganglion of squid.
• Kerkut G. A. et al., Comp. Biochem. Physiol., 15, 485—502, 1965. Glutamate as
10R fransmitter in Helix.
p,er,kut G- &., Gardner D. R., Comp. Biochem. Physiol., 20, 147—162, 1967.
109 e calcium ions in action potentials of Helix.
6' H°rn N-> Walker R. J., Comp. Biochem. Physiol., 30, 1061—1074,
110 /г , Long-lasting synaptic inhibition in the snail Helix.
Kerkut G. A., Meech R. W., Comp. Biochem. Physiol., 19, 819—832, 1966. Ionic
conductances in Helix neurons.
344 1 лава 11
111. Kerkut G. A., Pitman R. Al., Walker R. J. Comp. Biochem. Physiol., 31, 611_
633, 1969. lontophoretic application of acetylcholine and GABA onto insect
central neurons.
112. Kerkut G. A., Sedden С. B., Walker R. J., Comp. Biochem. Physiol., 18, 921-.
930, 1966. Deopamine content of snail Helix.
113. Kerkut G. A., Sedden С. B., Walker R. J., Comp. Biochem. Physiol., 21, 687-.
690, 1967. Cellular localization of monoamines.
114. Kerkut G. A., Thomas R. C„ Comp. Biochem. Physiol., 8, 39—45, 1963. Ace-
tylcholine potentials in snail neurons.
115. Kerkut G. A., Thomas R. C., Comp. Biochem. Physiol., 11, 199—213, 1964.
Effect of anion injection on several potentials of IPSP and ACh in Helix.
116. Keynes R. D., J. Physiol., 114, 119—150, 152—182, 1951. Na and К content
and exchange in crab and squid nerve.
117. Keynes R. D., J. Physiol., 169, 690—705, 1963. Chloride in the squid giant
axon.
118. Keynes R. D. In: The Neurosciences, Second Study Program, edited by
F. O. Schmitt, Rockefeller University Press, New York, 1970, pp. 707—715.
Optical changes during nerve conduction.
119. Koketsu K., Nishi S., J. Gen. Physiol., 53, 608—623, 1969. Calcium and action
potentials of bullfrog sympathetic ganglion cells.
120. Koketsu K-, Nishi S., Solda H., Life Sci., 7, 955—963, 1968. Calcium and ACh
potential of bullfrog sympathetic ganglion.
121. Kravitz E. A., In: The Neurosciences; Second Study Program, edited by
F. O. Schmitt, Rockefeller University Press, New York, 1970, pp. 433—443.
Gamma-aminobutyric acid and glutamic acid as neurotransmitters.
122. Kravitz E. A., Potter D. D., J. Neurochem., 12, 323—328, 1965. GABA as in-
hibitor in lobster.
123. Kravitz E. A., Potter D. D., van Gelder N. Al., Biochem. Biophys. Res. Comm.,
7, 231—236, 1962. Gamma-aminobutyric acid distribution in lobster nervous
system.
124. Kriebel M. et al., Science, 166, 520—524, 1969. Electrotonic coupling between
neurons in fish oculomotor center.
125. Krnjevic K-, Phillis J. W., J. Physiol., 165, 274—304, 1963. lontophoretic stu-
dies of neurons in mammalian cerebral cortex.
126. Kuffler S. W., Proc. Roy. Soc. Lond. B, 168, 1—21, 1967. Neuroglial cells.
127. Kuffler S. W., Nicholls J. G., Ergebn. Physiol., 57, 1—90, 1966. Neuroglia fun-
ction in leech nervous system.
128. Kuffler S. W., Potter D. D., J. Neurophysiol., 27, 290—320, 1964. Glia in leech
central nervous system.
129. Kusano K-, J. Gen. Physiol., 52, 326—345, 1969. Relationship between pre- and
postsynaptic potentials in squid giant synapse.
130. Kusano K-, Grundfest H., J. Cell. Comp. Physiol., 65, 325—336, 1965. Circus
reexcitation as a cause of repetitive activity in crayfish lateral giant axons.
130a. Kusano K., LaVail M. M., J. Comp. Neurol., 142, 481—494, 1971. Conduction in
lobster and shrimp giant fibers.
131. Landolt A. Al., Ris H., J. Cell Biol., 28, 391—405, 1966; also Z. Zellforsch., 69
246—259, 1966. Soma-somatic interneuronal junctions in corpus pedunculatum
of wood ant.
132. Lentz T. L., Comp. Biochem. Physiol., 27, 715—718, 1968. Acetylcholinesterase
activity in a planarian.
133. Libet B., J. Neurophysiol., 30, 494—515, 1967. Rabbit sympathetic ganglia.
134. Libet B., Kobayashi H„ Science, 164, 1530—1532, 1969. Adrenergic and choli-
nergic potentials in sympathetic ganglion cells.
135. Loe P. R., Florey E., Comp. Biochem. Physiol., 17, 509—522, 4966. ACh and
ChEs in Octopus.
136. Loewenstein W. R., Ann. N. Y. Acad. Sci., 137, 441—472, 1966. Permeability of
membrane junctions.
137. Loewenstein W. R., Kanno Y„ J. Gen. Physiol., 6, 1123—1140, 1963. Electrical
properties of nuclear membrane.
Возбудимые мембраны
345
138.
139.
140.
141.
142.
143.
144.
145.
146.
147.
148.
149.
150.
151.
151а.
152.
153.
154.
155.
156.
157.
158.
159.
160.
161.
162.
163.
164.
165.
Loewenstein W. R., Каппо У., J. Cell Biol., 33, 225—234, 1967. Electrotonic
coupling, epithelial cells.
Loewenstein W. R., Nakas M., Socolar S. J., J. Gen. PhysioL, 50, 1865—1891,
1967. Junctional membrane uncoupling.
Loveland R. E., Comp. Biochem. PhysioL, 9, 95—104, 1963. 5-Hydroxytrypta-
mine, probable mediator of excitation in heart of Mercenaria.
Mackie G. 0., Amer. Zool., 5, 439—453, 1965. Conduction in nerve-free epithe-
lia of siphonophores.
Mackie G. O., J. Exp. BioL, 49, 387—400, 1968. Electrical activity in hydroid
Cordylophora.
Mackie G. O., Quart. Rev. BioL, 45, 319—332, 1970. Neuroid conduction and
evolution of conducting tissues.
Mackie G. 0., Passano L. M„ J. Gen. PhysioL, 52, 600—621, 1968. Epithelial
conduction in hydromedusae.
McLennan H., Synaptic Transmission, 2nd edition, W. B. Saunders Co., Phi-
ladelphia, 1970, 178 pp.
Майский В. A., Fed. Proc. Transl. SuppL, 23, T1173 — T1176, 1964. Electrical
characteristics of membrane of giant nerve cells of Helix.
Marsden C. A., Kerkut G. A., Comp. Biochem. PhysioL, 31, 851—862, 1969.
Fluorescent microscopy of central nervous system of leech.
Martin A. R., Pilar G., J. PhysioL, 168, 443—463, 1963. Dual mode of synaptic
transmission in avian ciliary ganglion.
Martin A. R., Pilar G., J. PhysioL, 168, 464—475, 1963. Transmission through
ciliary ganglion of chick.
Martin A. R., Wickelgren W. O., J. PhysioL, 212, 65—83, 1971. Sensory cells in
spinal cord of sea lamprey.
McNutt N. S., Weinstein R. S., J. Cell BioL, 47, 666—688, 1970. Ultrastructure
of the nexus.
McReynolds Gorman A., J. Gen. PhysioL, 56, 376—391, 1970. Hyperpola-
rizing responses of Pecten photoreceptors.
Meiri U., Rahamimoff R., J. PhysioL, 215, 701—726, 1971. Activation of trans-
mitter release by strontium and calcium ions at neuromuscular junctions.
Miledi R., Nature, 212, 1240—1242, 1966. Miniature synaptic potentials in
squid nerve cells.
Miledi R., J. PhysioL, 192, 379—406, 1967. Glutamate as possible transmitter
in squid.
Miledi R., Molinoff P., Potter L. T., Nature, 229, 554—557, 1971. Isolation of
the cholinergic receptor protein of Torpedo electric tissue.
Miller M. D., Marshall J. M., Amer. J. PhysioL, 209, 859—865, 1965. Catecho-
lamine receptors in uterus.
Molinojf P. B., Axelrod J., Ann. Rev. Biochem., 465—500, 1971. Biochemistry
of catecholamines.
Moore J. W., J. Gen. PhysioL, 48, SuppL, 11—17, 1965. Voltage clamp studies
on internally perfused axons.
Moore J. W„ Narahashi T., Fed. Proc., 26, 1655—-1663, 1967. Sodium channels
in squid axon.
Morris D., Bull G., Hebb С. 0., Nature, 207, 1265, 1965. Acetylcholine in the
electric organ of Torpedo.
Murdock L. L., Comp. Gen. Pharmacol., 2, 254—274, 1971. Catecholamines in
arthropods.
Myhrberg H. E., Z. Zellforsch., 81, 311—343, 1967. Monoaminoergic neurons in
Limulus.
Nachmansohn D., Chemical and Molecular Basis of Nerve Activity, Academic
press, New York, 1959, 235 pp.
^aiioh У., Eckert R., Z. vergl. PhysioL, 61, 427—452, 453—472, 1968. Para-
ecium, reversal of cilia, electrical responses.
Saitoh Y., Eckert R., Friedman K., J. Exp. BioL, 56, 667—681, 1972. Regene-
ative calcium spikes in Paramecium.
346
Глава 11
]66. Nakas М., Higashino S., Loewenstein W. R., Science, 151, 89—91, 1966. Un-
coupling of an epithelial cell membrane junction by calcium ion withdra-
wal.
167. Narahashi T. In: Electrical Activity of Single Cells, Igakushoin, Hongo, Toky0>
1960, pp. 119—131. Excitation and electrical properties of giant axons of
cockroaches
168. Nicholls J. G., Baylor D. A., Science, 162, 279—281, 1968. Hyperpolarization
after activity of neurons in leech.
169. Nicholls J. G., Kuffler S. IF., J. Neurophysiol., 27, 645—671, 1964. Ionic com-
position of glial cells and neurons of leech.
170. Nicholls J. G., Purves D., J. Physiol., 209, 647—668, 1970. Monosynaptic che-
mical and electrical connections between sensory and motor cells in CNS of
leech.
171. Nicholls J. G„ Wolfe D. E., J. Neurophysiol., 30, 1574—1592, 1967. Extracel-
lular spaces in cells of leech CNS.
172. Nishi S., Koketsu K., J. Cell. Comp., Physiol., 55, 15—30, 1960. Synaptic po-
tentials in frog sympathetic ganglion.
173. Nishi S., Koketsu K., Life Sci., 6, 2049—2055, 1967. Origin of ganglionic in-
hibitory postsynaptic potential, bullfrog sympathetic ganglion.
174. Obata K. at al., Exp. Brain Res., 4, 43—57, 1967. Norepinephrine and cyclic
AMP excitation of cerebellar nuclei.
174a. O’Brien R. D., Eldefrawi M. E., Eldefrawi A. T., Ann. Rev. Pharmacol., 12,
19—34, 1972. Isolation of acetylcholine receptors.
175. O’Brien R. D., Gilmour L. P., Proc. Nat. Acad. Sci., 63, 496—503, 1969; 65,
438—445, 1970. Muscarone-binding material in electroplax.
176. Ochizono K., Nature, 214, 833—834, 1967. Structure of synaptic vesicles.
177. Ohashi H., J. Physiol., 212, 561—575, 1971. Cl and К currents in taenia
coli.
178. Okajima A., Kinosita H., Comp. Biochem. Physiol., 19, 115—131, 1966. Ciliary,
activity and coordination in Euplotes.
178a. Oomura У. et al., Seital no Kagaku, Japan, 13, 31—38, 1962. Membrane con-
stants
179. Osborne N. N., Cottrell G. A., Z. Zellforsch., 112, 15—30, 1971. Biogenic ami--
ncs in Limax.
180. Otsika M. et al., Proc. Nat. Acad. Sci., 56, 1110—1115, 1966. Lobster claw,
GABA inhibition.
181. Pappas G. D., Bennett M. V. L., Ann. N. Y. Acad. Sci., 137, 495—508, 1966.
Electrical transmission between neurons.
182. Parisi M., Rivas E., de Robertis E., Science, 172, 56—57, 1971. ACh receptors
from electric organ.
183. Parnas I., J. Neurophysiol, 1972 (in press). Differential presynaptic block at
high frequency of branches of a single axon innervating two muscles.
184. Parnas I. et al., J. Exp. Biol., 50, 635—649, 1969. Non-homogeneous condu-
ction in giant axons of nerve cord of Periplaneta americana.
185. Paul D. H., Science, 176, 680—682, 1972. Decremental conduction over «giant»
afferent processes in an arthropod.
186. Pay ton B. W., Bennet M. V. L., Pappas G. D., Science, 165, 594—597, 1969.
Electrotonic synapse.
187. Payton B. W., Bennett M. V. L., Pappas G. D., Science, 166, 1641—1643, 1969.
Junctional membranes at an electrotonic synapse.
188. Payton B. W., Loewenstein W. R., Biochim. Biophys. Acta, 150, 156—158, 1968.
Electrical coupling in leech giant nerve cells.
189. Penn R. D., Loewenstein W. R., Science, 151, 88—89, 1966. Uncoupling
nerve cell membrane junction by calcium-ion removal.
190. Pentreath V. W., Cottrell G. A., Comp. Biochem. Physiol., 27, 775—785, 1968-
Acetylcholine and cholinesterase in radial nerve of Asterias rubens.
191. Phillis J. W., Comp. Biochem. Physiol., 17, 909—928, 1966. Neurotransmitter5
in mollusc intestine.
Возбудимые мембраны
347
Q9 Phillis J. W., The Pharmacology of Synapses, Pergamon Press, Oxford, 1970,
_ Pichon Y., P. B., Moreton, J. E. Treherne J. E„ J. Exp. Biol., 54, 757—798,
1971. Ionic basis of extraneuronal potential changes in central nervous system
of cockroach.
194 Pichon Y., Boistel J., J. Exp. Biol., 47, 343—355, 1967. Current-voltage rela-
19 tions in the isolated giant axonn of cockroach.
195 Pitman P. M., Comp. Gen. Pharmacol., 2, 347—371, 1971. Transmitter sub-
stances in insects: a review.
195a Pitman P. M., Kerkut G. A., Comp. Gen. Pharmacol., 1, 221—230, 1970. ACh
' and GABA in cockroach neurons.
196 Potter D. D., Furshpan E. J., Lennox E. S., Proc. Nat. Acad. Sci., 55, 328—336,
1966. Connections between cells of developing squid.
197. Prince IF. T., Berridge M. J., J. Exp. Biol., 56, 323—333, 1972. Salivary gland
potentials in insects.
198. Pall IF., Shepherd G. M., J. Neurophysiol., 30, 1138—1168, 1967; 31, 884—915,
1968. Reconstruction of fields potentials and dendro-dendritic synaptic intera-
ctions in olfactory bulb.
199. Palston H. J., Ill, Nature, 230, 585—587, 1971. Activity in presynaptic dendri-
200. Pevel J. P., Kamov sky M. J., J. Cell Biol., 33, C7 — Cl 2, 1967. Electrical sy-
napses of Mauthner cells and liver.
201. Poberts A., Stirling С. A., T. vergl. Physiol., 71, 295—310, 1971. Propagation
in skin of young tadpoles.
202. Pobertson J. D. In: The Neurosciences: Second Study Program, edited by
F. O. Schmitt, Rockefeller University Press, New York, 1970, pp. 715—728.
Ultrasrtucture of synapses.
203. Pobison G. A., Butcher P. IF., Sutherland E. IF., Ann. N. Y. Acad. Sci., 139,
703—723, 1966. Adenyl cyclase as an adrenergic receptor.
204. Pobison G. A., Sutherland E. IF., Circ. Res., 26, Suppl., 147—161, 1970. Role
of cyclic AMP in adrenergic responses.
204a. Pose B., Loewenstein IF. P., J. Membrane Biol., 5, 20—50, 1971. Effects of
calcium on electrical coupling between epithelial cells.
204b. Povainen С. M., J. Neurophysiol., 30, 1000—1023, 1024—1042, 1967. Electri-
cal and chemical synapses in lamprey giant neurons.
205. Pushforth N. B., Biol. Bull., 140, 255—273, 1971. Contraction pulses in
Hydra.
206. Salanki J., Recent Developments in Neurobiology, Hungary, 3, 67—39, 1972.
Serotonin in neural regulation of bivale.
207. Sato N. et al., J. Gen. Physiol., 51, 321—345, 1968. Ionic permeability changes
during ACh-induced responses of Aplysia ganglion cells.
208. Shappirio D. P., Eichenbaum D. M., Locke В. P., Biol. Bull., 132, 108—124,
1967. Cholinesterase in brain of cecropia silmoth during metamorphosis and
0 Pupal diapause.
oiA S., J. Physiol., 220, 145—175, 1972. Electrotonic spread in barnacle eye.
410. Siggins G. P., Oliver A. P., Hoffer B. J., Bloom F. E., Science, 171, 192—194,
1971. Cyclic AMP and norepinephrine effects on transmembrane properties of
cerebellar Purkinje cells.
!• Smalley K. N., Comp. Biochem. Physiol., 16, 467—477, 1971. Catecholamines
9io *n Photinus.
2- Smith T. G., Stell IF. K., Brown J. E., Freeman J. A., Murray G. C., Science,
*62, 454—458, 1968. Conductance changes associated with receptor poten-
214 ",a^s *n Limulus photoreceptors.
• Сахаров Д. A., Ann. Rev. Pharmacol., 10, 335—352, 1970. Cellular aspects of
214 lnycrtebrate neuropharmacology.
• bptra M. E., Bennett M. V. L., Brain Res., 37, 294—300, 1972. Molluscan sy-
214 haptic control of electrotonic coupling between neurons.
^trumwasser F., J. Psychiat. Res., 8, 237—257, 1971. Spontaneously active
neurons of Aplysia.
348
Глава JI
216. Suga N., Katsukl Y., J. Exp. Biol., 38, 759—770, 1961. Pharmacological stu-
dies in auditory synapses in a grasshopper.
217. Sweeney D. C„ Comp. Biochem. Physiol., 25, 601—613, 1968. Distribution
of monoamines in a freshwater bivalve mollusc.
218. Takeuchi A., Takeuchi N., J. Gen. Physiol., 45, 1181—1193, 1962. Electrical
changes in pre- and postsynaptic axons of giant synapse of Loligo.
219. Takeuchi A., Takeuchi N., J. Physiol., 170, 296—317, 1964; 177, 225—238,
1965. Localized action of gamma-aminobutyric acid in the crayfish muscle.
220. Takeuchi A., Takeuchi N., J. Physiol., 183, 443—449, 1966. Permeability of pre-
synaptic terminal in crayfish neuromuscular junction during synaptic inhibi-
tion by GABA.
221. Tasaki J., Watanabe A., Hallett M., J. Membrane Biol., 8, 109—132, 1972.
Extrinsic fluorescence changes in squid axon.
222. Tauc L., Gerschenfeld H. M., J. Neurophysiol., 25, 236—282, 1962. Cholinergic
mechanism (Inhibitory synaptic transmission) in a molluscan nervous
system.
223. Tauc L., Physiol. Rev., 47, 521—593, 1967. Transmission in invertebrate and
vertebrate ganglia.
224. Taylor G. W„ J. Cell. Comp. PhysioL, 18, 233—242, 1941; 20, 359—372, 1942.
Relation between fiber size, birefringence and conduction velocity.
225. Taylor R. E. In: The Neurosciences; A Study Program, edited by G. C. Quarton
et al., Rockefeller University Press, New York, 1967, pp. 305—312. Initiation
of nerve impulses.
226. Trautwein W., Kassebaum D. G., J. Gen. Physiol., 45, 317—330, 1961. Pace-
maker activity of heart.
227. Treheme J. E., Neurochemistry of Arthropods, Cambridge University Press,
1966, 156 pp.
228. Treheme J. E., Lane N. J., Moreton R. B., Pichon Y., J. Exp. BioL, 53, 109—
136, 1970. Permeability of sheaths of Periplaneta nervous system.
229. Treheme J. E., Mellon DeQ, Carlson A. D„ J. Exp. BioL, 50, 711—722, 1969.
Ionic basis of axonal conduction in central nervous system of Anodonta cyg-
nea.
230. Treheme J. E., Smith D. S., J. Exp. BioL, 43, 13—21, 1965. Penetration of
acetylcholine into central nervous tissues of an insect.
231. Tucker L. E„ Pichon Y., J. Exp. BioL, 56, 441—457, 1972. Na efflux from CNS
of cockroach.
232. Tupper J. et al, J. Cell BioL, 46, 187—191, 1970. Electrical coupling between
cells of starfish embryo.
233. Ueda K., Ann. Zool. Japan, 34, 99—110, 161—179, 1961. Electrical properties
of Opalina.
234. Usherwood P. N. R„ Machili P., Leaf G., Nature, 219, 1169—1172, 1968. L-Glu-
tamate at insect excitation nerve-muscle synapses.
235. Vassort G., et al., Pfliig. Arch., 309, 70—81, 1969. Effects of epinephrine on
inward Ca current in frog atrium.
236. Watanabe A., Grundfest H„ J. Gen. PhysioL, 45, 267—308, 1961. Impulse pro-
pagation at the septal and commissural junctions of crayfish lateral giant
axons.
237. Weight F. F., Votava J., Science, 170, 755—758, 1970. Late synaptic potentials
in frog symapthetic ganglia.
238. Weinstein R. S., McNutt N. S., New Engl. J. Med., 286, 521—524, 1972; also.
J. Cell BioL, 47, 666—688, 1970. Membrane to membrane contacts.
239. Werman R., Davidoff R. A., Aprison M. H., J. NeurophysioL, 31, 81—95, 1968.
Glycine as inhibitory transmitter in spinal cord.
240. Werman R„ XXV Internat. PhysioL Cong., 8, 188—189, 1971. Spinal cord
transmitters.
241. West С. B., Quart. Rev. BioL, 30, 116—137, 1955. Comparative pharmacology
of adrenal gland.
Возбудимые мембраны
349
942 Whittaker V. Р., Ann. N. Y. Acad. Sci., 137, 982—998, 1966; also pp. 761—767
in: The Neurosciences; Second Study Program, edited by F. O. Schmitt, Rocke-
feller University Press, New York, 1970. Properties of synaptosomes.
243 Wilson D. M., Comp. Biochem. Physiol., 3, 274—284, 1961. Connections bet-
ween lateral giant fibers of earthworms.
244. Woodward D. J. et al., Brain Res., 33, 91—100, 1971. Frog cerebellum; sy-
naptic transmission.
245. Woodward D. J., Hoffer B. J., Siggins G. R., Bloom F. E., Brain Res., 34,
73—97, 1970. Development of synaptic activation in rat cerebellar Purkinje
cells.
246. Yamasaki T., Narahashi T., J. Insect Physiol., 3, 230—242, 1959. Electrical pro-
perties of cockroach giant axon.
247. Young J. Z., Proc. Roy. Soc. Lond B, 162, 49—79, 1965. Diameters of fibers
of peripheral nerves of Octopus.
248. Zettler F., Jdrvilehto M., Z. vergl. Physiol., 76, 233—244, 1972. Electrotonic
spread in an insect eye.
Глава 12
Механорецепция, фонорецепция
и чувство равновесия
Л. ПРОССЕР
Клеточные механизмы механорецепции
Основной функцией органов чувств является преобразование
внешнего воздействия в нервные импульсы. Органы чувств можно
возбудить тремя основными способами: химическим, механическим
или электрическим. Рецепторы вкуса и обоняния чувствительны к
воздействию определенными химическими веществами, фоторе-
цепторы возбуждаются в ходе фотохимической реакции. Реакция
терморецепторов заключается в модуляции химически регулируе-
мой спонтанной активности. Статические механорецепторы (неко-
торые рецепторы растяжения, барорецепторы кровеносных сосу-
дов) возбуждаются в течение всего периода деформации их мем-
браны; динамические механорецепторы возбуждаются только в на-
чальный период деформации как при низких частотах механиче-
ского раздражения (тактильные рецепторы), так и при высоких
частотах (фонорецепторы). Механорецепторы играют важную роль
в поддержании позы и положения конечностей; они служат также
для ориентации относительно направления силы тяжести и уско-
рения, для восприятия низкочастотных и высокочастотных вибра-
ций, для опознания поверхности объекта, для оценки скорости
ветра и движения воды и для определения глубины погружения
под воду. Проприоцепторы дают информацию о положении раз-
личных частей тела животного относительно друг друга, тогда как
экстероцепторы сообщают о внешних воздействиях на организм.
У некоторых видов рыб, кроме того, имеются электрорецепторы
(ом. гл. 17).
В процессе эволюции превращение определенных эпителиаль-
ных клеток в рецепторные произошло, вероятно, еще до диффе-
ренцировки нервных клеток. Простейшие механорецепторы пред-
ставляют собой свободные нервные окончания, лежащие либо меж-
ду эпителиальными клетками, либо под ними. К настоящему вре-
мени описано много типов специализированных концевых луковиц,
дисков, телец и спиралей. Нервное окончание часто заключено и
капсулу либо связано с чувствительным шипиком или волоском-
Вспомогательные структуры играют роль «механического фильт-
ра» или усилителя внешнего воздействия, а также могут способ-
Механорецепция, фонорецепция ,и чувство равновесия 351
♦
твовать возбуждению рецептора при его стимуляции только в оп-
еделенной плоскости или направлении. Членистоногие обладают
множеством чувствительных волосков и щетинок. У насекомых
имеются разнообразные специализированные рецепторные клетки,
которые, связываясь с полужесткими участками кутикулы, обра-
зуют чувствительные шипнки, колоколовидные сенсиллы и хордо-
тональные органы. В колоколовидной сенсилле чувствительное
терминальное волокно оканчивается под тонким кутикулярным ку-
полом, увеличивающим поверхностное напряжение. В хордотональ-
ной сенсилле чувствительное волокно связано с эластичным тя-
жем, натянутым между двумя точками наружного скелета. Мно-
гие волосковые пластинки сенсилл насекомых имеют цилиарную
структуру, при этом чувствительное нервное окончание входит в
кутикулярный волосок. Мышцы, сухожилия и суставы содержат
проприоцептивные образования, часто связанные со специализи-
рованными мышечными волокнами.
В локализации ядер чувствительных нейронов наблюдается
большое разнообразие. У дождевого червя эпидермальные чувст-
вительные клетки образуют синаптические контакты в перифери-
ческом нервном сплетении; при этом в сегментарных нервах на
одно афферентное волокно приходится много таких клеток. Во
многих рецепторах ракообразных ядро нейрона лежит у основа-
ния волоска или в рецепторе растяжения. У позвоночных тела
нервных клеток, от которых отходит большинство тактильных и
проприоцептивных волокон, располагаются в ганглиях задних ко-
решков спинного мозга или в стволе мозга.
В некоторых механорецепторах стимул действует непосредст-
венно на нервное окончание, в других его действие передается на
нервное окончание через специализированный эпителий. Для всех
механорецепторов характерно возникновение градуальных рецеп-
торных (генераторных) потенциалов, которые, суммируясь, приво-
дят к генерации распространяющихся нервных импульсов. Меха-
норецепторы, которые отвечают только одним или несколькими
нервными импульсами на каждое раздражение, называются быст-
ро адаптирующимися, другие, дающие длительную серию импуль-
сов в ответ на внешнее воздействие, называются медленно адап-
тирующимися. Быстро адаптирующиеся рецепторы различаются в
зависимости от скорости стимуляции, вызывающей их возбужде-
ние. Чувствительность многих механорецепторов модулируется эф-
ферентными импульсами из центральной нервной системы; неко-
торые механорецепторы обладают спонтанной активностью, а
внешнее воздействие лишь увеличивает или уменьшает частоту их
фоновой активности.
Возбуждение многих нервов и мышц может быть вызвано при
помощи механической деформации. Непонятно, однако, почему од-
ни клеточные мембраны более чувствительны к деформации, чем
другие. Некоторые мышцы при растяжении деполяризуются. Ра-
352
Глава 12
стяжение сердечной мышцы вызывает возрастание частоты ее со-
кращений; многие одиночные гладкие мышцы также активно реа-
гируют на быстрое растяжение. Растягивание поперечнополосатой
мышцы может усилить нервно-мышечную передачу. Амеба прекра-
щает движение при прикосновении к ней. Механическое раздра-
жение нервного волокна вызывает градуальное увеличение прово-
димости его мембраны, при некотором критическом значении ко-
торой возникают спайки. В миелинизированных волокнах нерва
лягушки спайки возникают, если мембрана волокна смещена на
2—5 мкм, в то время как в гигантских аксонах омара для их гене-
рации необходимо смещение на 10—15 мкм. В отсутствие натрия
увеличение проницаемости мембраны не происходит, однако оно
может иметь место в случае, когда фиксация потенциала препятст-
вует деполяризации мембраны [112].
Волосковые клетки внутреннего уха позвоночных возбуждаются
при смещении их всего на 1 А (см. ниже). По-видимому, неболь-
шая деформация изменяет заряд клеточной мембраны и тем са-
мым увеличивает ее проницаемость для натрия.
Тактильные рецепторы, рецепторы движения и растяжения
Тельца Пачина
Тельца Пачини брыжейки, сухожилий и подушечек лап млеко-
питающих можно рассматривать как обобщенную модель механо-
чувствительного органа. Каждое тельце (рис. 12-1) состоит из се-
рии концентрических оболочек, окружающих немиелинизирован-
ное нервное окончание. Начальный участок миелиновой оболочки
и первый перехват Ранвье располагаются внутри капсулы, а вто-
рой перехват Ранвье — уже вне ее. Рецептор возбуждается при
кратковременном смещении на 0,5 мкм (в течение 0,1 мс) или
при смещении на 0,2—0,4 мкм по всей длине нервного окончания
'[134]. Электрическая реакция телец Пачини представляет собой
сенсорный потенциал, амплитуда и латентный период которого за-
висят от силы механического раздражения; когда амплитуда ге-
нераторного потенциала достигает порогового уровня, в первом
перехвате Ранвье возникает нервный импульс. Сенсорный потен-
циал, возникающий в ответ на короткий толчок, длится в течение
4—8 мс и соответствует константе длины 0,8 мм [39, 135]. При
подавлении импульсной активности тетродотоксином сенсорный по-
тенциал продолжает существовать и исчезает только в безнатрие-
вой среде. Сенсорный потенциал увеличивается при гиперполяри-
зации и уменьшается при деполяризации мембраны нервного окон-
чания. Во время длительного раздражения сенсорный потенциал
дает обычно колебания по типу «оп» и «off», а при удалении кап-
сулы он сохраняется в течение всего периода деформации (рис.
12-2). Следовательно, капсула играет роль высокочастотного меха-
А. С
(Quilliam Т.кА„ Sato"'AL, J Phvsiol “¥2Я~~1К7"'17й'* iowTT" “=р“««™в гаиим <«, о, »у
ническое раздражение _1 _ > ,955)^Б. Электрические реакции на меха-
£ОМ, ’.."
Схема строения тельца
ническое ^здражение^ зарегистрированный
Рис. 12-1.
ПД?ИН.И’, показывающая локализацию перехватов Ранвье (а, б, в)
PhVCIol 1О<1 IC7 t'TZ? <nr+-\ г-« r-х ' ' * *
с поляпичяниёа а -----------*" в ™льЦе Пачини [39]: / — без поляризации то-
И N,+, — fbatw пеа^ током. Рп. — рецепторный потенциал; Nb N2, Ns
1+2 фазы реакции, возникающие в соответствующих узлах.
Рецепторные потенциалы телец Пачини, возникающие при механической
л- Декагте, стимуляции [136].
| У»»Р»ва„„ые ,„,|Я. Б-Шотое рецепторный потенциал;
'l^lSM
354
Глава 12
нического фильтра. Температурный коэффициент для сенсорного
потенциала вдвое выше, чем для спайка. Обычно каждый сенсор-
ный потенциал вызывает не более 1—3 импульсов, т. е. тельце
Пачини является быстро адаптирующимся рецептором [136]. Им-
пульсы возникают при частоте раздражения до 150 имп/с при сла-
бых ударах и до 280 имп/с при сильных; частота генераторных
потенциалов хорошо соответствует частоте раздражения вплоть
до 650 имп/с. Быстрая адаптация связана частично со скоростью
возникновения спайков, а частично с механическими свойствами
капсулы [136].
Тактильные окончания
Тактильное нервное окончание в коже лягушки представляет
собой быстро адаптирующийся чувствительный прибор; в ответ
на кратковременное раздражение (менее чем 0,1 с) оно дает один
импульс и в ответ на длительную деформацию — 4—12 импуль-
сов. При повторных кратковременных раздражениях импульсная
активность уменьшается в соответствии с частотой раздражения.
Латентный период механического раздражения составляет 2,8—
4,8 мс, а электрического — 2,3—2,8 мс, тогда как при непосредст-
венной стимуляции нерва он равен всего 0,8 мс. При возрастании
скорости механической деформации латентный период уменьша-
ется; при одном и том же латентном периоде эффективная ско-
рость раздражения увеличивается после обработки кожи колла-
геназой. Существует критическая скорость нарастания раздражаю-
щего стимула. Из всего этого следует, что кожа механически свя-
зана с нервными окончаниями. Эта механическая связь обладает
как упругими, так и вязкими свойствами, причем при низкой ско-
рости механической деформации более важное значение имеет уп-
ругость. При повторных раздражениях по мере возрастания часто-
ты стимуляции происходит возрастание порога; пороговый сти-
мул может поднять порог импульсной активности [25—27]. Спо-
собность следовать за высокой частотой стимуляции ограничена
возможностями механизма возникновения спайков.
Рецепторы, связанные с чувствительными волосками, как пра-
вило, быстро адаптируются к действию раздражителя. У речного
рака каждый чувствительный волосок, расположенный по краю
тельсона и уроподов, дает при сгибании только один импульс.
У птиц и млекопитающих обнаружено несколько типов рецепторов,
связанных с перьями и волосами и различающихся чувствитель-
ностью и скоростью адаптации {27]. У кошки и кролика рецепто-
ры волос подшерстка имеют низкие пороги возбуждения и быстр0
адаптируются, в то время как рецепторы остевых волос имеЮт
высокие пороги возбуждения и адаптируются медленно [15]. Ча-
стота сенсорных импульсов, пропорциональна скорости движения
волоска (в мкм/с), возведенной в некоторую степень. ЧувствителЬ'
Механорецепция, фонорецепция и чувство равновесия
355
ные окончания в коже, покрытой волосами, обладают низким уров-
нем спонтанной активности. Ответ (7?) и стимул (S) связаны меж-
собой степенной зависимостью R = KSn, где rz<Cl для началь-
ной стадии ответа ип^1 для более поздних стадий [256]. Вибрис-
сы кошки связаны с несколькими нервными окончаниями, из кото-
рых одни реагируют только на скорость, а другие — на скорость,
и на амплитуду отклонения [83]. У утки быстро адаптирующиеся
рецепторы пуховых перьев образуют большие рецептивные поля и
имеют низкие пороги возбуждения, а тактильные рецепторы ко-
тки имеют ограниченные рецептивные поля, более высокий порог
возбуждения и определенный диапазон адаптации. Некоторые ре-
цепторы (по-видимому, тельца Гербста) возбуждаются при стиму-
ляции с частотой 400—800 имп/с [46]. В коже аллигатора быстро
адаптирующиеся нервные окончания дают «он»- и off»- ответы и
генерируют импульсную активность при частоте стимуляции, рав-
ной 200 имп/с, в то время как для медленно адаптирующихся ре-
цепторов эта частота пропорциональна смещению [230]. У одних
рецепторов частота прямо пропорциональна смещению, у других
между ними существует логарифмическая зависимость. Медленно
адаптирующиеся нервные окончания в коже рептилий делятся на
две группы, различающиеся мгновенной частотой раздражения,
спонтанной активностью, чувствительностью и скоростью проведе-
ния импульса [118]. Первичные чувствительные нейроны в спинном
мозгу морской миноги напоминают по своим электрическим свойст-
вам кожные нервные окончания [145]:
Типы рецепторов Потенциал покоя чувст- вительного нейрона, мВ Постоянная времени, мс
Тактильные (быстро адапти- рующиеся) 70 1,4
Давления (медленно адап- тирующиеся) 68 2,3
Сильное раздражение 56 3,9
На различных уровнях сенсорного пути обезьян измерялась ча-
стота импульсов, возникающих при надавливании на кожу, в за-
висимости от силы воздействия. Зависимость между ними оказа-
лась нелинейной. В участках кожи, покрытых волосами, частота
импульсов очень резко возрастала в начальный момент стимуля-
ции, а при каждом следующем раздражении увеличивалась значи-
тельно медленнее. Для безволосых участков кожи, напротив, ха-
рактерно медленное нарастание частоты импульсов в начальный
°мент стимуляции, с последующим увеличением, пропорциональ-
ным силе надавливания на кожу. Такая же зависимость была об-
РУЖсна в нейронах таламуса и в коре головного мозга. Следо-
ельно, интенсивность внешнего воздействия кодируется именно
2з*
356
Глава 12
в чувствительных окончаниях, и код в неизмененном виде переда-
ется в центральные отделы нервной системы [159].
В подушечках лап кошки обнаружены три типа механорецеп-
торов: 1) тельца Пачини — быстро адаптирующиеся и наиболее
чувствительные при частоте раздражения 200 Гц; 2) рецепторы,
разряд которых длится 500 мс, и 3) спонтанно активные рецепторы,
дающие частоту импульсов F=K(S—S0)n, где So порог раздраже-
ния, S— раздражающее воздействие, а крутизна п=0,5 [106].
Тактильные окончания, расположенные на голове рыбы Leucis-
cus, максимальный ответ на прикосновение дают при определенной
температуре; этот ответ изменяется в зависимости от температу-
ры адаптации [234].
В ноге моллюска фазные рецепторы генерируют один спайк в
начале смещения и один в конце; при этом их порог равен 60—
100 мкм при скорости 100 мм/с, а при ритмическом (синусоидаль-
ном) смещении один спайк возникает на гребне каждой волны
[177]. Размеры некоторых тактильных'рецепторов речного рака
позволяют вводить микроэлектрод в тело чувствительного нейрона
или в его аксон; регистрируемые при этом рецепторные потенциа-
лы оказываются небольшими и редкими, а спайки возникают, по-
видимому, в дендритах [151].
В щетинках полихеты Наг mot/юё встречаются высокочувстви-
тельные быстро адаптирующиеся механорецепторы; направленные
к центру отростки их нейронов разветвляются таким образом, что
в сегментарном нерве содержится как афферентное волокно из
данного сегмента, так и эфферентный отросток чувствительной
клетки из соседнего сегмента [98].
Колоколовидные сенсиллы насекомых по скорости адаптации
занимают промежуточное положение. Разряд рецепторов суставов
ног насекомых возникает с высокой частотой (от 100 до 300 имп/с),
а через 1—2 с опускается на более низкий уровень [188]'.
Межсегментные рецепторы растяжения ракообразных
У речного рака, омара и некоторых других десятиногих и ро-
тоногих раков имеются крупные рецепторы растяжения, располо-
женные в дорсальной области между сегментами брюшка и груди.
Каждый сегмент содержит две пары мышечных рецепторных орга-
нов— медленно адаптирующихся (MPOi) и быстро адаптирую-
щихся (МРО2). Каждый рецептор состоит из тонкой мышцы, охва-
ченной пальцевидными дендритами большого сенсорного нейрона,
от которого к нервным центрам отходит аксон (рис. 12-3). К мы-
шечным волокнам подходят отростки тех же двигательных аксо-
нов, которые иннервируют разгибательные мышцы; волокна, ин-
нервирующие поверхностные разгибатели, идут к МРОЬ а иннер-
вирующие глубже расположенные разгибатели — к МРОг. У Squtl'
la с каждым сухожилием связаны две рецепторные мышцы. Любая
Механорецепция, фонорецепция и чувство равновесия
357
ецепторная клетка имеет тормозную иннервацию, берущую на-
чало в центральной нервной системе, причем у омара толстое до-
бавочное тормозное волокно подходит к обоим мышечным органам,
а тонкое — только к MPOi [4]. В мышцах, снабженных МРОЬ сар-
комеры уже, чем в мышцах, имеющих МРО2 [67].
Рис. 12-3. Схема строения абдоминального рецептора растяжения десятиногого
рака (из «Physiology of Invertebrate Nervous System, ed. by C. A. Wiersma, Uni-
versity of Chicago Press).
A m Б — место расположения мышечного рецепторного органа (МРО) между сегментами на
Дорсальной стороне брюшка. В—фазные и тонические рецепторы, связанные с соответст-
вующими мышцами. Показаны двигательное, тормозное и чувствительное нервные волокна.
При внутриклеточной регистрации потенциал покоя рецептор-
ных клеток составляет —70 мВ. Растяжение вызывает градуаль-
ную деполяризацию клеток, зависящую от степени растяжения.
Сенсорный (генераторный) потенциал распространяется электро-
тонически от дендритов к телу рецепторной клетки, и при порого-
вом значении деполяризации, равном 10 мВ для медленно адапти-
рующихся и 20 мМ для быстро адаптирующихся рецепторов,
^озникает нервный импульс. Сенсорный потенциал может сохранять-
в течение всего периода раздражения; при длительном растя-
ско ИИ РазРяды импульсов в MPOi продолжаются в течение не-
1тепЛЬКИх часов> а в МРО2 исчезают через несколько секунд. Ре-
юТсТ°РНЬ1е потенциалы развиваются в дендритах и распространя-
Гут по телу клетки; спайки возникают у основания аксона и мо-
Распространяться в обратном направлении, т. е. тоже к телу
Рис. 12-4.
/. Потенциалы, зарегистрированные внутриклеточно от рецептора растяжения ракообразных
(Ruffler S. W., Exptl. Cell Res., suppl. 5, 495—519, 1958). А. Генераторный потенциал при поД-
пороговом растяжении, градуально изменяющийся в соответствии с деформацией; стрелки
указывают на начало и конец растяжения. Б. Импульсы, возникающие при пороговом УР°В'
не деполяризации; стрелкой в середине отмечено начало добавочного растяжения, приво-
дящего к росту частоты импульсов. В. Постепенное увеличение растяжения (между стрел-
кой и вертикальной линией); расслабление сопровождается временной гиперполяризациеи.
7/. Тормозные потенциалы, зарегистрированные внутриклеточно от рецептора ракообразны
(Huffler S. W., Euzaguirre С., J. Gen. Physiol., 39, 155—184, 1955). А. Рецептор в состояни
частичного расслабления; шесть тормозных импульсов вызывают деполяризационный поте
циал (первые две стрелки); растяжение вызывает деполяризацию, равную 20 мВ (JPe?L
стрелка), и три сенсорных импульса, за которыми следует серия тормозных импульсов, о
ределяющих реполяризационные потенциалы; длительное растяжение ведет к появлен
афферентных импульсов. Б. Серия тормозных импульсов подавляет сенсорные разряды и в
зывает реполяризацию. В. Тормозные деполяризующие потенциалы во время полного Р
слабления клетки.
Механорецепция, фонорецепция и чувство равновесия
359
пепторной клетки. Растяжение мышцы и сокращение, вызван-
Рее нервным импульсом, действуют синергично, частота разрядов
51 MPOi (при Данной величине растяжения) увеличивается при
Симуляции двигательных аксонов и достигает 50 имп/с при раз-
дражении последних с частотой 50—80 имп/с [16]. Импульсы,
приходящие по эфферентному (добавочному) волокну или волок-
нам, тормозят сенсорный разряд за счет того, что не дают сенсор-
ному потенциалу достигнуть порогового уровня, при котором воз-
никает разряд. Если рецептор подвергнут растяжению и слегка де-
поляризован, то тормозные волокна усиливают деполяризацию его
мембраны, тогда как при расслабленном состоянии рецептора тор-
мозные импульсы вызывают его реполяризацию. Тормозные им-
пульсы могут приводить мембрану в состояние равновесия на
уровне примерно 6 мВ позитивности по отношению к потенциалу
покоя. Если мембранный потенциал достигнет того же уровня под
действием внешнего тока, то торможение сможет предотвратить
генерацию импульсов без изменения мембранного потенциала [64]
(рис. 12-4). Используя микроэлектрод, внутри клетки можно заре-
гистрировать миниатюрные тормозные синаптические потенциалы
(мТПСП), которые, как и тормозный потенциал, меняют знак
при растяжении мембраны рецептора. Введение ионов С1~ .усили-
вает эти потенциалы. С помощью электрода, заполненного КО,
можно обнаружить мТПСП по наличию деполяризации мембраны
рецептора, а с помощью электрода с цитратом калия — по наличию
ее гиперполяризации. Следовательно, действие тормозного медиато-
ра (по-видимому, ГАМК) сводится к увеличению проводимости
мембраны для ионов С1_ [105].
Различия в скорости адаптации между MPOi и МРОг прояв-
ляется как в случае растяжения, так и в случае внутриклеточного
тока. При подавлении импульсной активности тетродотоксином ге-
нераторные потенциалы не исчезают и характер их в рецепторах
обоих типов одинаков. Сопротивления рецепторных мембран и по-
тенциалы покоя также близки по своим значениям. Вероятно, раз-
личный характер адаптации рецепторов примерно на 30% зависит
от свойств мембраны, генерирующей спайки, а на 70% определя-
ется упругими свойствами мышц [162, 163]. Если при помощи
внутриклеточного электрода через клетку пропустить постоянный
ток, то частота возникающих при этом спайков будет снижаться,
как это происходит в случае нормальной адаптации. Вслед за се-
рией спайков возникает посттетаническая гиперполяризация, тор-
мозятцая генерацию импульсов. Этот гиперполяризующий компо-
нент адаптации связан с натриевым насосом [164, 233]. В обоих
Рецепторах адаптация происходит быстрее при +24, чем при
' & С; внезапное нагревание уменьшает частоту сенсорных им-
Ульсов, а внезапное охлаждение увеличивает ее [20].
Рефлекторная функция и регуляция деятельности рецепторов
стяжения ракообразных еще недостаточно ясны. Некоторые дан-
360
Глава 12
ные говорят о том, что они рефлекторно оказывают тормозное
влияние на разгибательные мышцы [50]. При разгибании брюш-
ка один и тот же мотонейрон возбуждает обе работающие при
этом мышцы — «быструю» мышцу-разгибатель и «медленную»
мышцу рецептора растяжения; затем афферентные волокна из
медленно адаптирующего рецептора возбуждают второй мотоней-
рон, который в свою очередь приводит в действие тонические мыш-
цы-разгибатели. При сгибании брюшка мышцы-разгибатели испы-
тывают центральное торможение, а афферентные волокна из мед-
ленно адаптирующегося рецептора рефлекторно тормозят
рецепторы растяжения в ближайшем переднем сегменте. Пока нет
данных о тормозящем влиянии рецепторов растяжения на мышцы-
сгибатели, а также о функции быстро адаптирующихся рецепто-
ров растяжения [67]. У плавающих ракообразных обнаружены
межсегментные рецепторы растяжения, но они, по-видимому, от-
сутствуют у брахиур, например у Cancer.
Мышечные веретена позвоночных
Многие скелетные мышцы позвоночных содержат чувствитель-
ные веретена и сухожильные рецепторы. У млекопитающих экстра-
фузальные мышечные волокна иннервируются крупными а-мото-
нейронами {147]. Мышечные веретена содержат от 2 до 12 интра-
фузальных (фузимоторных) мышечных волокон, иннервируемых
мелкими у-мотонейронами; однако у некоторых видов животных
к интрафузальным волокнам могут подходить также ветви «-мото-
нейронов [9а]. В центре веретена имеются чувствительные оконча-
ния двух типов — первичные, или аннулоспиральные, и вторичные,
или гроздевидные. Первичные окончания связаны с быстро прово-
дящими (70—120 м/с) нервными волокнами (имеющими диаметр
12—20 мкм), а вторичные — с медленно проводящими (24—72м/с)
волокнами (имеющими диаметр 4—12 мкм). У многих млекопита-
ющих веретена имеют как первичные, так и вторичные окончания,
у некоторых же обнаружены только первичные окончания. На мы-
шечных сухожилиях располагаются окончания Гольджи рецепто-
ров растяжения, связанные с толстыми афферентными волокнами.
Мышечные веретена располагаются параллельно экстрафузальным
волокнам, о чем говорит прекращение их импульсной активности
в момент мышечного сокращения; сухожильные рецепторы распо-
лагаются с экстрафузальными волокнами последовательно, свиде-
тельством чему служит усиление частоты их разрядов в момент со-
кращения. При сокращении экстр афузальных волокон растяжение
веретена уменьшается и сенсорный разряд мгновенно прекращает-
ся (рис. 12-5). При изометрическом сокращении в момент большой
нагрузки для возбуждения мышечного веретена, вероятно, доста-
точно напряжения экстрафузальных волокон [35]. |
Фузимоторные волокна можно разделить на две группы соот-|
ветственно их воздействию на первичные окончания мышечный!
—A
Рис. 12-5.
/ — Двигательный аксон (А) в полусухожильной
мышце ящерицы Tiliquia nigrolutea, ветвящийся
и иннервирующий экстрафузальное (ЭВ) и интра-
фузальное (ИВ) мышечные волокна с помощью
гроздевидного двигательного окончания. (Препа-
рат У. Проске.) II Реакция мышечного веретена
в гл. ibiofubularis той же ящерицы. Потенциалы
Действия, зарегистрированные от тонких нитей
рассеченного седалищного нерва в условиях, когда
Двигательный нерв стимулировался одиночными
Ударами тока возрастающей силы. 1 — разряды
в нерве, 2 — напряжение мышцы. (Обратите вни-
мание на изменение чувствительности миографа
от В к Г; артефакт стимула на записях Б и В
маскируется прямым спайком, вызванным стиму-
лом.) А — подпороговый стимул; Б — порог для
слабого сокращения; остаточное сокращение вере-
тена уменьшается в ходе сокращения и увеличи-
вается при расслаблении (параллельное соедине-
ние двух видов волокон); В — более сильное со-
кращение; веретено растягивается (последова-
тельное соединение двух видов волокон); Г—
очень сильное сокращение экстрафузальных воло-
кон снимает растяжение веретена (осциллограм-
мы У. Проске).
362
Глава 12
веретен при растяжении мышцы с постоянной скоростью [148].
Динамические фузимоторные волокна вызывают реакцию первич-
ных окончаний на скорость растяжения; эта реакция увеличивает-
ся в процессе растяжения. Напротив, статические фузимоторные
волокна вызывают уменьшающуюся реакцию на скорость растяже-
ния, несмотря на значительное возбуждающее действие, которое
они оказывают на веретено при постоянной длине мышцы.
Центральная нервная система может осуществлять независи-
мый контроль за деятельностью статических и динамических фузи-
моторных волокон. Функцией динамических волокон, вероятно, яв-
ляется регуляция реакции веретен на скорость растяжения и, сле-
довательно, регуляция чувствительности рефлекторной петли, свя-
зывающей веретена с экстрафузальными волокнами. Статические
волокна, оказывающие сильное возбуждающее действие на вере-
тена, могут иметь большое значение для предотвращения «замол-
кания» веретена в процессе рефлекторного сокращения мышцы,
что позволяет значительно расширить диапазон работы этого ре-
цептора.
Интрафузальные волокна тонически возбуждаются центрами,
лежащими на более высоких уровнях центральной нервной систе-
мы, и даже после спинномозговой деафферентации разряды мото-
нейронов следуют с частотой 20—60 имп/с [17, 51]. Интрафузаль-
ные волокна сокращаются только на концах, но при этом натяги-
ваются их центральные участки, содержащие чувствительные окон-
чания. У млекопитающих активность у-волокон поддерживает то-
ническую активность мышечных веретен; для возбуждения верете-
на во время тонической активности интрафузальных волокон тре-
буется меньшее натяжение, чем в отсутствие такой активности [78,.
101]. Сухожильные рецепторы генерируют импульсную активность
только при мышечном сокращении и прекращают ее при расслаб-
лении мышцы. Порог возбуждения сухожильных рецепторов при
пассивном растяжении выше, чем при активном сокращении. Каж-
дый сухожильный рецептор возбуждается при сокращении неболь-
шой группы мышечных волокон, с которой он связан, однако при
ответе на пассивное растяжение импульсная активность рецептора
может прекратиться в результате сокращения тех волокон, кото-
рые не располагаются последовательно с рецептором (при таком
сокращении с сухожильного рецептора снимается нагрузка) [99].
При резком растяжении мышцы веретена возбуждаются и их аф*
ферентные импульсы вызывают моносинаптическое рефлекторное
сокращение экстрафузальных волокон. У кошки первичные окон-
чания воспринимают вибрацию, в то время как вторичные оконча-
ния к ней нечувствительны (см. ниже) [10].
В мышечных веретенах млекопитающих и лягушки при прямом
отведении были зарегистрированы градуальные сенсорные потен-
циалы, возникающие при растяжении мышцы и дающие начало
афферентным импульсам. Рецепторный потенциал мышечного вере*
I
Механорецепция, фонорецепция и чувство равновесия
363 "
лягушки чувствителен к Na+ и Са2+ [178]. Период ранней
1б штрой) адаптации сокращается при уменьшении скорости растя-
ния* это отражает изменение тонических свойств сенсорной мем-
^паны* зависящих от длины веретена [179]. Подсчитано, что адап-
тация только на 15—20% определяется механическими свойствами
пецептора, остальное приходится на изменение свойства мембраны
[103]. ’ , .
Мышечные веретена птиц (например, утки) аналогичны вере-
тенам млекопитающих. Разряд веретена прекращается при сокра-
щении моторных волокон; следовательно, веретена параллельно
.связаны с экстрафузальными волокнами и имеют собственную фу-
зимоторную иннервацию [46]. У ящерицы Tiliqua импульсная ак-
'<Si?i flft • а •%' >У»л4 »£<S&- ".'»; IX-Л eY'-tfifJHK.iS» •«'"лй-вг
тивность веретена прекращается при одиночном сокращении мыш-
цы, хотя у некоторых веретен, наоборот, частота импульсов во вре-
мя ’ восходящей фазы сокращения увеличивается. Отсюда следует,
что у ящерицы веретена связаны с экстрафузальными волокнами
последовательно. Из наблюдений над мышечным сокращением и
разрядом веретена при раздражении различных нервных ветвей
был сделан вывод о том, что некоторые нервные волокна иннерви-
руют как фузимоторные, так и экстрафузальные волокна [190].
У лягушки, как и у рептилий, к мышечным веретенам подходят
ветви тех же аксонов, которые иннервируют экстрафузальные во-
локна; иными словами, у лягушки система у-мотонейронов отсут-
ствует. Однако экстрафузальная иннервация у лягушки может
быть двух типов: к «быстрым» мышцам подходят а-волокна, а к
«медленным» (градуальным)—у-волокна [148] (гл. 15). В мыш-
цах лягушки содержатся только первичные чувствительные окон-
чания. У рыб вообще нет мышечных веретен, но в их плавниках
имеются проприоцепторы.
Подводя итог, можно сказать, что рецепторы растяжения и так-
тильные рецепторы чрезвычайно разнообразны, особенно это ка-
сается их вспомогательных структур. Чувствительность к направ-
лению раздражения, постоянство электрического ответа (фазного
или тонического), адаптация и многие другие их особенности за-
висят как от механических свойств самого рецептора, так и от
свойств рецепторной мембраны. По-видимому, в чувствительных
скончаниях всех этих рецепторов под влиянием механической де-
формации возникают местные, или генераторные, потенциалы, ко-
торые, достигнув порогового уровня, вызывают распространяю-
щиеся сенсорные импульсы.
характеристика рецепторов растяжения и связанных с ними
Рефлексов
Че РецептоРЫ растяжения, как правило, адаптируются медленнее,
ць^ Тактильные рецепторы. Импульсы от одиночных веретен мыш-
пальца лягушки в момент начала нагрузки возникают с ча-
364
Глава 12
стотой 120—260 имп/с; затем их частота понижается примерно до
20 имп/с и в дальнейшем в течение многих минут остается на этом
уровне [137]. В барорецепторах каротидного синуса млекопитаю-
щих адаптации фактически не наблюдается. Некоторые рецептор-
ные окончания находятся в состоянии тонической активности, и
частота их разрядов повышается при каждой пульсовой волне;
другие дают четыре или пять импульсов на каждую пульсовую»
волну. У кошки при постоянном давлении в сонной артерии баро-
рецепторы каротидного синуса возбуждаются очень слабо до тех
пор, пока давление не превысит 40—50 мм рт. ст.; после этого ча-
стота разрядов непрерывно возрастает вплоть до давления
200 мм рт. ст. [131].
Рецепторы, воспринимающие растяжение или движение скелет-
ных элементов, встречаются во многих конечностях. У акулы нет
мышечных веретен, но в плавниках у нее имеются медленно адап-
тирующиеся подкожные тельца, реагирующие на синусоидальное
механическое раздражение; частота разрядов этих телец пропор-
циональна угловой скорости и максимальна при максимальной
скорости, когда на каждый локомоторный цикл приходятся две
вспышки активности рецептора. У омара три рецептора ротового
отдела, состоящие из нескольких чувствительных клеток, распола-
гающихся на эластичных тяжах, дают фазные или тонические от-
веты на движения мандибул и пищевода. Третья ходная нога мече-
хвоста Limulus содержит 350 проприцептивных нейронов, большая
часть которых собрана в девять суставных мембранных рецепто-
ров [94]. Миохордотональные органы крабов реагируют как на по-
ложение тела, так и на смещения в суставе, причем некоторые из
них невосприимчивы к направлению воздействия [29, 91, 260].
В зависимости от ориентации рецепторов на поперечной связке од-
ни чувствительные клетки сигнализируют сгибание сегмента, дру-
гие — его разгибание [23, 33]. Когда дактилсшодит краба Carcinus
пассивно открыт, открывающая и закрывающая его мышцы реф-
лекторно тормозятся; точно так же тормозятся при пассивном рас-
тяжении сгибающая и растягивающая мышцы. Оба процесса яв-
ляются следствием механического раздражения хордотональных
органов, расположенных в области сустава [23]. Рецепторы кок-
сальных мышц некоторых крабов генерируют высокие рецепторные
потенциалы, которые пассивно (без возникновения спайков) пере-
даются в расположенные рядом центральные ганглии [197]. Один
сенсорный аксон передает сигналы от статических нервных окон-
чаний, расположенных на эластичных тяжах параллельно мышцам
и чувствительных к изменениям длины мышц, другой собирает
информацию от быстро адаптирующихся динамических окончании,
связанных с мышцами последовательно и реагирующих на ско-
рость их растяжения [23а]. У речного рака дендриты нейронов
первого и второго ганглиозных корешков располагаются в гипо-
дерме вблизи места прикрепления поверхностного сгибателя и ос-
Механорецепция, фонорецепция и чувство равновесия
365
ований плавательных конечностей; при помощи аксонного реф-
лекса эти нейроны подавляют моторные разряды, идущие к сги-
бательным мышцам и брюшным конечностям [181].
у членистоногих известны два типа механорецепторов: рецеп-
торы, располагающиеся между двумя элементами наружного ске-
лета и реагирующие на вибрацию в кутикуле, и рецепторы, связан-
ные с сухожилиями и чувствительные как к движению сухожилий,
так и к изменениям давления (рис. 12-6).
В переднем отделе кишечника мухи Phormia имеются рецепто-
ры растяжения, состоящие из двух биполярных нейронов; раздра-
жение этих рецепторов при растяжении кишечника рефлекторно
тормозит положительную реакцию мухи на хемостимуляцию и
пищу [71]- Педипальпы скорпиона Euscorpius несут на своей по-
верхности по 65 чувствительных волосков, каждый из которых воз-
буждается при его сгибании в сторону дендрита рецепторной клет-
ки. Импульсы возникают при смещении волоска со скоростью 6—
8 град/с и при максимальном отклонении в 2—3°; если волосок
находится вблизи дендрита рецепторной клетки, то рецептор
адаптируется; если же он отклоняется в противоположном направ-
лении — происходит деадаптация [96].
Рецепторные потенциалы одиночной чувствительной клетки из
шейных групп волосков пчелы убывают с двумя постоянными вре-
мени, что говорит о наличии в рецепторном потенциале двух про-
цессов [250]. Возбуждение сенсилл насекомых наступает при сжа-
тии сенсиллы на 30 А или на 0,5% ее диаметра; максимальный
ответ регистрируется при смещении сенсиллы на 0,1 мкм или ее
сжатии на 15% [250]. Рецепторы, расположенные в суставах ног
насекомых, являются детекторами гравитации [195]. Фазные ре-
цепторы у основания крыльев возбуждаются при взмахе крыльев,
тогда как тонические рецепторы реагируют на их положение [182].
На антеннах насекомых располагаются различные механоре-
Цепторы; некоторые из них имеют большое значение для восприя-
тия ветра и волн. У мухи жгутик вращается вокруг своей оси пер-
пендикулярно аристе, при этом резонансная частота его вращения
близка к таковой для взмахов крыла. Если жгутик обездвижить,
то контроль за скоростью взмахов крыльев и их положением со
стороны антенн прекращается [22].
Многие жуки ориентируются по ветру с помощью рецепторов
пторого сегмента антенны, которые измеряют угол между направ-
лением полета и направлением ветра [132]. Жук-вертячка Gyri-
воспринимает движение воды с помощью резонансной (250 Гц)
ибрационной системы, состоящей из второго сегмента антенны и
ЖгУтика [206].
ta К/ЖДЫЙ чУвствительный волосок на церке таракана Periplane-
образован тРихогенн°й (опорной) и рецепторной клетками; сен-
к Рные потенциалы, возникающие в рецепторной клетке, приводят
снерации спайков [251]. При сгибании волоска в одном направ-
Рис. 12-6. Схема строения
двух типов механорецепто-
ров ноги таракана (Schnor-
bus Н., Z. vergl. Physiol.,
71, 14—48, 1971).
А. Субгенуальный орган, сен-
силла которого реагирует на на-
пряжение и вибрации кутику-
лы. Б. Дистальный орган, реа-
гирующий на изменение давле-
ния крови. СИ — сенсорный
нейрон; НО — нервные оконча-
ния; СВ — сенсорное волокно;
Ш — шапочка над нервным
окончанием.
' Механорецепция, фонорецепция и чувство равновесия 367
нии рецепторная клетка деполяризуется, а при сгибании в про-
ЛиВоположном гиперполяризуется, причем амплитуда сенсорного
отенциала пропорциональна степени сгибания [168]. В аналогич-
ных рецепторах саранчи Locusta амплитуда рецепторных потен-
циалов и спайков также пропорциональна сгибанию, однако чув-
ствительности рецептора к направлению воздействия, аналогичной
имеющейся у таракана, обнаружено не было [232].
Поведенческие реакции на прикосновение и давление
Выше уже говорилось о некоторых рефлекторных реакциях на
сигналы, поступающие от кожных и мышечных рецепторов. Много
других примеров известно из поведенческих опытов. Кошка, обу-
ченная подниманию лапы в ответ на тактильное раздражение, мо-
жет воспринимать сигналы от одиночной подушечки лапы [245].
Большинство рыб при одностороннем раздражении области хвоста
поворачивает хвостовой плавник в ту сторону, на которой произво-
дят раздражение. У многих малоподвижных и сидячих организмов
при механическом раздражении переднего (орального) конца со-
кращается все тело. Многие ползающие животные обладают тиг-
мотаксисом, или положительной «реакцией на край». Например,
дождевой червь, ползущий вдоль одной стороны какого-либо пред-
мета, поворачивает в том направлении, в котором он скорее до-
стигнет края раздражающей поверхности. Если после того, как
червь проявил положительную «реакцию на край», раздражающий
предмет удаляют, то животное выпрямляет переднюю часть тела
так, чтобы она составила прямую линию с хвостовой частью. По-
добные контактные реакции, вызванные неодинаковым раздраже-
нием разных сторон тела, обнаружены у кольчатых червей, дву-
парноногих многоножек, личинок насекомых и молодняка млекопи-
тающих.
Другим примером контактной ориентации может служить рео-
таксис, т. е. ориентация в потоке воды. Положительная реакция на
ток жидкости наблюдается у планарий, у которых соответствую-
щие рецепторы рассеяны по всей поверхности тела. Положитель-
ный реотаксис свойствен также инфузории Paramecium. Ослеплен-
ные рыбы не ориентируются в потоке воды до тех пор, пока не кос-
нутся дна, после чего они используют для ориентирования направ-
ление потока у дна.
Анемотаксис, или ориентация в воздушных потоках, наблюда-
ется у большинства летающих животных. У мух рецепторные ор-
ганы, расположенные на антеннах, чувствительны к ветру и вызы-
вают изменение положения тела насекомого в соответствии с на-
правлением и скоростью воздушного потока. На голове саранчи и
ешуекрылых насекомых располагаются ветрочувствительные во-
°ски; в полете эти насекомые оценивают направление по давле-
ию ветра на их крылья, благодаря которому возбуждаются сен-
ллы, расположенные у основания крыльев.
368
Глава 12
Отсутствие раздражения вентральной поверхности тела вызы-
вает реакции перевертывания или выпрямления у улиток, морских
звезд и некоторых насекомых. Если пиявку приподнять за ее рото-
вую часть, она начинает «плавать». Взлет многих насекомых мож-
но вызвать прекращением контактного раздражения лапок. Поло-
жение головы пчелы по отношению к направлению силы тяжести
регулируется рефлексами, вызываемыми раздражением сенсорных
щетинок на эпистерните.
Многие животные выбирают места для своего обитания путем
механического ощупывания поверхности. Например, рак Orconectes
fodiens обитает в водоемах со стоячей водой и илистым дном, а от-
носящийся к тому же роду О. propinquus предпочитает песчаный
или скалистый грунт и проточную воду [13]. Личинки усоногих ра-
ков оседают на неровных поверхностях. Рачок-бокоплав Corophiurn
употребляет для строительства трубчатых домиков в 9 случаях из
10 мелкие песчинки, если строит на освещенном участке, и реже,
если участок затемнен [150].
При погружении в воду гидростатическое давление возрастает
приблизительно на 1 атм на каждые 10 м глубины. Две трети зем-
ной поверхности покрыты водой, 90% океана, или 56% всей био-
сферы, находятся под давлением от 100 до 1100 атм. На глубине
10 км морская вода сжата на 4% под давлением 100 атм. При дав-
лении в несколько сотен атмосфер уменьшается вязкость прото-
плазмы, прекращается деление клеток и погибают обитатели по-
верхностных слоев воды. Некоторые барофильные бактерии океан-
ских глубин, где давление достигает 1000 атм, не развиваются при
давлении в 1 атм. Ферментные системы глубоководных рыб отли-
чаются своей чувствительностью к гидростатическому давлению от
соответствующих систем организмов, живущих на поверхности.
Так как современная техника не позволяет выращивать глубоко-
водные организмы на поверхности (даже под давлением), мы еще
мало знаем о биохимических и физиологических приспособлениях,
позволяющих организмам жить на больших глубинах.
Многие планктонные организмы предпочитают селиться на оп-
ределенной глубине, а некоторые из них ежедневно мигрируют из
области с одним давлением в область с другим давлением [213].
Двигательная активность некоторых ракообразных может менять-
ся в зависимости от перепадов гидростатического давления. Рав-
ноногие раки реагируют на изменение давления всего в 0,01 атм,
хотя ни образований, заполненных газом, ни каких-либо чувстви-
тельных структур у них не обнаружено [58]. Повышение гидроста-
тического давления стимулирует двигательную активность полихе-
ты Nephthys, а понижение давления тормозит ее; при этом перепад
давления должен длиться по крайней мере 5 с, а латентный перй'
од составляет 10 с [158].
Плавательный пузырь рыбы имеет более важное значение для
гидростатической ориентации, чем для дыхания (см. гл. 5). Умень-
Механо рецепция, фонорецепция и чувство равновесия
369
НИ"---
ние гидростатического давления приводит к увеличению количе-
шея газа в плавательном пузыре. Выделение газа способствует
Удержанию плотности тела на определенном уровне, необходимом
Пдя удержания рыбы на постоянной глубине. При повышении гид-
ростатического давления количество газа в плавательном пузыре
уменьшается. Закрытопузырные заглатывают воздух или освобож-
даются от него в зависимости от степени своей плавучести. Phoxi-
ms реагирует на уменьшение давления воды на 5 см вод. ст.;
электрофизиологически (на нерве плавательного пузыря) было ус-
тановлено, что рецепторы, принимающие участие в рефлекторном
выделении газа, располагаются у него в стенке плавательного пу-
зыря [194]. У закрытопузырных рыб, которых принудительно со-
держали вблизи поверхности, после удаления газа из пузыря вос-
станавливалась только половина того количества газа, которое
имелось у них при содержании их на глубине. Резкое изменение
давления в плавательном пузыре вызывает изменение сердечной
деятельности, дыхания и локомоции. После перерезки нервов, иду-
щих к пузырю, все эти реакции исчезают. Очевидно, чувствитель-
ные окончания плавательного пузыря, так же как и внутреннего
уха, участвуют в реакции рыб на гидростатическое давление.
Вибрационная чувствительность
Разделение механорецепторов на рецепторы растяжения и ре-
цепторы вибрации довольно условно. Рецепторы вибрации пред-
ставляют собой быстро адаптирующиеся экстероцепторы, воспри-
нимающие низкочастотные механические раздражения (менее
.100 Гц). Они должны быть быстро адаптирующимися, поскольку
должны передавать информацию о частоте вибраций. Как и все
механорецепторы, они могут быть динамическими или статически-
ми, т. е. реагировать на ускорение или на амплитуду смещения.
К ним относятся рецепторы, контролирующие положение тела, а
также фонорецепторы. На суше вибрации передаются главным об-
разом через субстрат, а звук распространяется по воздуху, причем
скорость распространения вибраций меньше, чем скорость звука;
скорость звука обратно пропорциональна его частоте. Для живот-
ных, обитающих в воде, различия между рецепцией вибраций и
|®Ука менее выражены; гидроидный полип, воспринимая вибрации,
и^ается в направлении локального колебания воды [111].
к Крайне чувствительны к вибрациям твердого субстрата, жид-
и среды и воздуха членистоногие. Их сенсорные элементы обыч-
сенПредставляют с°бой чувствительные волоски и колоколовидные
Кл СИЛлы- Сенсиллы могут состоять из чувствительных нервных
Ны Т°К’ Размещающихся на соединительной ткани, или из мышеч-
Тяжей, связанных в области суставов с хитином. Вибрация
СИл ет предаваться жестким наружным скелетом, а движение сен-
ы может амортизироваться окружающей ее гемолимфой.
1514
370
Глава 12
У многих ракообразных и насекомых описаны чувствительные к
вибрации волоски и сенсиллы суставов. Например, церкальные во-
лоски таракана реагируют на порывы ветра и колебания субстрата
[168]. На голени таракана вблизи ее сочленения с бедром распола-
гаются три рецептора: один из них, воспринимающий изменения в
натяжении кутикулы, имеет порог 10~6 см и частотный диапазон от-
вета 30—500 Гц; второй чувствителен к перепадам давления гемо-
лимфы; третий является вибрационным рецептором с максимумом
чувствительности в диапазоне воспринимаемых частот 1000—
5000 Гц и порогом 10~7—10~10 см [219]. На бедрах третьей пары
ног у саранчи и кузнечика имеются хордотональные органы, содер-
жащие как фазные, так и тонические рецепторы; тонические ре-
цепторы регистрируют угол между бедром и голенью, а фазные
реагируют на скорость движения в суставе [253]. У саранчи ре-
цепторы растяжения, расположенные у основания крыльев, раз-
ряжаются с частотой 10—20 имп/с; частота может изменяться в за-
висимости от скорости синусоидального движения крыльев [180].
На антеннах мух Calliphora в месте сочленения второго сегмен-
та и жгутика располагаются колоколовидные сенсиллы, анало-
гичные джонстоновым органам; во время полета насекомого при
боковом закручивании жгутика под действием потоков воздуха ре-
цепторные клетки сенсилл генерируют фазный ответ. Этот ответ
возникает при закручивании на 2—5°, пропорционален логарифму
скорости движения животного и достигает предела при 100 град/с;
при этом латеральное смещение жгутика возбуждает рецептор, а
медленное — тормозит его [212]. Такие же фазные рецепторы у
падальной мухи реагируют на движение со скоростью до 500 об/с
[21], а у комаров воспринимают направление звука [198]. Антен-
нальный джонстонов орган жуков позволяет им ориентироваться по
направлению ветра, при этом каждая антенна измеряет свое
отклонение от какого-то исходного положения (по ветру или про-
тив него) [132]. Соответствующие рецепторы пчел вызывают реф-
лекторное сближение антенн при усилении встречного ветра, а в
мозгу насекомого при движении жгутика возникают эфферентные
разряды. Афферентные импульсы возникают с частотой от 15 Д°
150 имп/с, по два импульса на каждое колебание. При скорости
ветра 7,4 м/с афферентные импульсы возникают при вибраций
жгутика с частотой 37 Гц. Как показывают поведенческие эксперт
менты, рецепторные органы, расположенные в месте сочленения
жгутика со вторым сегментом антенны, чувствительны не только
к направлению и силе ветра, но и к частоте движений в месте соч*
ленения [95]. Сенсорные волоски саранчи Locusta возбуждаю1'
ся при их смещении под действием ветра и могут разряжаться 0
начальный момент с частотой 245 имп/с, а при длительной стимУ
ляции — с частотой 50—70 имп/с.
Клоп-гладыш Notonecta обнаруживает добычу по колебания
поверхности воды; он особенно чувствителен к колебаниям, имеК)
Механорецепция, фонорецепция и чувство равновесия
371
частоту 100—150 Гц. Рецепторы, расположенные в дисталь-
шИ участках первой и второй ног, воспринимают колебания воды
НЬдМПлитудой в 1 мкм [144]. У водомерки вибрационные рецепто-
С расположены в тибиально-тарзальных суставах. С их помощью
РЬмки этих насекомых находят самцов (по колебаниям воды, ко-
СОг)ые те производят с частотой 20 Гц). У муравьев-листорезов
рецепторы, расположенные в суставах передних ног, при восприя-
тии смещений имеют порог возбуждения 1,3-10~7 см (частота 1—
3 кГц), а при восприятии ускорений — 2,5 см/с (частота 0,1—
2 кГц). Спайки следуют за стимулами вплоть до 100 Гц, а при
учащении стимулов ответ становится менее регулярным [143].
У ракообразных также были обнаружены рецепторы вибраций,
расположенные на антеннах и ходных ногах. На базальном сегмен-
те антеннального жгутика рака-отшельника Petrochirus имеется
хордотональный орган, содержащий биполярные рецепторные
клетки; сенсорные импульсы синхронизуются с вибрациями воды
до частоты от 200 имп/с и могут давать синхронный ответ на виб-
рации до 1000 имп/с [248]. У речного рака в каждой окологлоточ-
ной коннективе обнаружено по одному интернейрону, который ин-
тегрирует нервные импульсы, приходящие из чувствительных к
вибрации воды сенсорных клеток статоциста, а также из рецепто-
ров антенн и антеннальных жгутиков; информация, идущая от
этих рецепторов, антагонистична информации, поставляемой ре-
цепторами ног [249]. Миохордотональный орган краба Ocypode,
связанный с небольшим отверстием в наружном скелете меропо-
дита ходильной ноги, реагирует на распространяющиеся по возду-
ху звуки (максимум чувствительности при 1,5—2 кГц) и на виб-
рацию субстрата (максимум чувствительности при 1—2 кГц) [97].
Самцы крабов Uca подают самкам сигналы вибрациями ног и
постукиванием по субстрату, а некоторые из них даже издают
звуки с помощью стридуляционных органов. Восприятие этих сти-
мулов осуществляется рецепторами ног; ответ на колебания суб-
страта максимален при низких частотах и резко убывает при ча-
стоте 1 кГц, в то время как пороговая кривая для ускорения в
нервном ответе имеет два пика — 600 и 1500 Гц [209]. Омар в
опытах с условнорефлекторными изменениями частоты сердечных
сокращений обнаруживал максимальную чувствительность при ча-
йоте 37 Гц [176].
м ^аУКИ, плетущие паутину, могут отличить живую жертву от
ртвой по колебаниям паутины; этот сигнал имеет неправильную
Г1841^\’Н° максимУм его энергии приходится на частоту 50 Гц
силл ъ паУка Achaearanea имеются лировидные органы (или сен-
НЬ1 Ы ’ лежаш>ие П°Д тонким хитином вблизи тарзо-метатарзаль-
строгСУСТавов’ Каждый такой орган содержит 10 рецепторов,
Анапа5 НастРоенных на определенные частоты главным образом в
^Реда3ваРе °Т 8° Д° 800 Гц и более чувствительных к вибрациям,
ваемым по воздуху, чем к колебаниям паутины [254].
372
Глава 12
У позвоночных для восприятия вибраций субстрата служат не-
которые механорецепторы. Примером могут служить кожные ре-
цепторы змей; в их сенсорных клетках, имеющих максимум чувст-
вительности в диапазоне частот от 150 до 200 Гц, при вибрации с
частотой 300 Гц в ответ на каждое колебание возникает один
Рис. 12-7. Нервные импульсы, вызванные в рецепторе вибрации кожи змеи (по-
лоза) [191].
/ — нервные импульсы, 2 — синусоидальная вибрация (стимул).-
нервный импульс, при частоте 600—800 Гц ответ становится не-
правильным, и, наконец, при частоте выше 800 Гц импульсная ак-
тивность прекращается [191] (рис. 12-7). Таким образом, частот-
ный диапазон кожных вибрационных рецепторов перекрывает ча-
стотный диапазон слуха. Тельца Пачини кошки (особенно располо-
женные в суставах конечностей) реагируют на колебания субстрата
по типу 1:1 вплоть до частоты 700 Гц, а при частоте 100 Гц да-ю^
постоянный ответ [211]. Сходные рецепторы в межкостных образо-
ваниях ног и крыльев утки реагируют по типу 1 :1 вплоть до час
Механорецепция, фонорецепция и чувство равновесия 37$
оты SO®—Ю00 Гц, а затем дают регулярные ответы при частотах
1600 до 2000 Гц и при пороговом смещении, равным 0,5 мкм [45,
471 Первичные мышечные веретена и сухожильные рецепторы
кошки также чувствительны к вибрации и вызывают рефлекторное
сокращение мышцы, подвергшейся вибрационному воздействию
[73]' х
В заключение можно сказать, что рецепторы вибрации являют-
ся высокочувствительными фазными механорецепторами, которые-
расположены преимущественно в суставах и коже и могут воспри-
нимать относительно высокие частоты. Диапазон чувствительности
этих рецепторов в низкочастотной области перекрывается с диапа-
зоном некоторых тактильных рецепторов и рецепторов растяжения,
а в высокочастотной области — с диапазоном рецепторов слуха.
Способность животных воспринимать вибрацию позволяет им как
бы «осязать на расстоянии» субстрат и тем самым выявлять его?
дефекты.
Органы боковой линии
Костистые и пластиножаберные рыбы, а также некоторые ам-
фибии обладают сложной системой сенсорных каналов, которые
располагаются по бокам тела и часто заходят даже на голову.
Каждый канал может представлять собой открытый желобок, а
может быть закрытым по всей длине и иметь только несколько1
отдельных выходов (чаще всего на концах). В стенках канала
расположены опорные эпителиальные клетки и специализирован-
ные волосковые клетки, называемые нейромастами. Нейромасты:
имеют пучок из 40—50 мелких стереоцилий и одну или несколько^
крупных киноцилий (см. гл. 19). Клетки в совокупности могут об-
разовывать чувствительные бугорки, а их волоски вдаются1
в желеобразную купулу (рис. 12-8). Нейромасты иннервируются1
разветвляющимися волокнами нерва боковой линии, который вхо-
дит в головной мозг главным образом в составе X, а частично
в составе VII и IX нервов. Пузырьки в основании волосковых кле-
ток указывают на возможность существования химической пере-
дачи нервных импульсов с клетки на нервное окончание (рис.
12-9). Органы боковой линии воспринимают скорее перемещения
воды вблизи источника колебаний, чем давление; при этом движе-
ния купулы создают срезывающее усилие, под действием которого
возбуждаются чувствительные волоски нейромастов [68]. Некото-
рые нервные окончания, обнаруживаемые на волосковых клетках,
содержат пузырьки; возможно, что они относятся к эфферентным'
олокнам, идущим к рецепторным клеткам [84].
При регистрации активности купулы рыбы Acerina обнаружи-
те ТСЯ генеРатоРНЬ1е (микрофонные) потенциалы, амплитуда ко-
ра?Ь1х зависит от амплитуды механической волны, а частота в два
а превышает частоту раздражения; максимальная чувствитель-
374
Глава 12
ность наблюдается при частоте 50—150 Гц [129]. Сенсорные по-
тенциалы волосковых клеток протея Necturus при внутриклеточ-
ном отведении имеют амплитуду 800 мкВ [87, 88]. Чувствительные
волокна нерва боковой линии обнаруживают непрерывный фон
Рис. 12-8. Два нейромаста боковой линии хвоста протея Necturus [88].
'К — купула; Кц — киноцилия, окруженная стереоцилиями восьми волосковых клеток; С — си-
напс сенсорного аксона; С А — сенсорный аксон; Э — эпителий; А, Б, В — электроды для сти-
муляции и отведения.
спонтанной активности, обязанной своим происхождением нейро-
мастам. На этот фон накладываются реакции, вызываемые меха-
ническим раздражением, которое создается движениями воды. Для
одиночных толстых волокон характерно появление нескольких
импульсов в ответ на каждое раздражение при низкой частоте
стимуляции (20 имп/с), одного импульса при частоте 20—50 имп/с
и нерегулярных реакций при более высокой частоте раздражения.
У сомика Ictalurus нервные волокна дают разряды синхронно с
раздражением вплоть до частоты 100 имп/с, у Fundulus — до уаС'
тоты 180 имп/с [114]. Модуляция спонтанной активности неир0
Механорецепция, фонорецепция и чувство равновесия
375*
мастов при механическом раздражении обнаружена также у шпор-
цевой лягушки Xenopus.
Сенсорный разряд находится под контролем эфферентных нерв-
ных волокон. У угря Astroconder раздражение одной ветви аффе-
Рис. 12-9. Схема, демонстрирующая соотношения между афферентным базальным,
синапсом (волосковая клетка — афферентное нервное волокно) и эфферентным
синапсом (аксон — волосковая клетка) в органе боковой линии [оо].
Рентного волокна рефлекторно тормозит как сенсорные импульсы,
так и спонтанную активность в другой нервной ветви [93]. У Хепо-
Pus и амбистомы эфферентные импульсы тормозят афферентную
активность [207].
чат^°ЛЬ °Рганов боковой линии в поведении животных еще окон-
покеЛЬНО Не Установлена- Электрофизиологические исследования
водьЗЬ1ВаЮТ’ что они чУвствительны к низкочастотным колебаниям
баты’ В Т° Же вРемя результаты экспериментов, в которых выра-
ниматЙЛИСЬ УСЛ0ВНЬ1е рефлексы, говорят о способности рыб воспри-
ь низкочастотные звуки и другими способами (например, при-
376 Глава 12
помощи уха и кожных рецепторов). Реотаксис, или способность
ориентироваться в потоках воды, связан в первую очередь со зре-
нием и с вентральными контактными рецепторами [41, 42]. Уда-
ление нескольких купул из органов боковой линии рыбы Aplochei-
lus (особенно тех, которые располагаются в каналах головы) при-
водит к изменениям поведения, свидетельствующим о потере
-способности воспринимать колебания воды с узкой характеристи-
кой направленности [224]. По-видимому, информация, идущая от
органов боковой линии, в сочетании с информацией, поставляемой
.другими рецепторами, нужна животным для ориентации, поисков
пищи и, возможно, для обучения.
Звуковая рецепция
Фонорецепция и генерация звуков у насекомых
Давно известно, что некоторые насекомые реагируют на звуки
в частотном диапазоне, доступном слуху человека, но лишь недав-
но удалось показать, что насекомые воспринимают ультравысокие
частоты, излучаемые, например, летучими мышами. Орган слуха
ночной бабочки-совки состоит из расположенной на ее грудной
.стороне тимпанальной мембраны, за которой находится воздушная
'полость, пересекаемая связками. В одной из связок находится
нерв, состоящий из трех чувствительных волокон. Два из них идут
к тимпанальному органу от А-клеток, или фонорецепторов; тре-
тий— от механорецептора (В-клетка) [200]. В-клетка проявляет
спонтанную активность с частотой около 10—20 имп/с; при раздра-
жении прикосновением ее частота может возрастать до 200 имп/с;
эта клетка работает как проприоцептор, измеряющий внутренние
смещения. А-клетки тоже проявляют спонтанную активность. Они
отвечают на звук, причем порог одной клетки ниже, чем порог вто-
рой. Активность А-клеток не синхронизируется звуковой волной:
при раздражении чистым тоном они дают вспышку импульсной ак-
тивности с уменьшающейся частотой следования импульсов, проис-
ходящей по мере адаптации к раздражению (рис. 12-10). Они сиг-
нализируют о колебаниях уровня интенсивности звука изменения-
ми частоты следования спайков, укорочением латентного периода
при большей интенсивности, длительностью импульсов последейст-
вия, возникающих в ответ на действие звукового стимула; разница
между порогами обеих клеток составляет около 20 дБ. При интен-
сивности раздражения выше 40 дБ частота спайков достигает
верхнего предела. А-клетки реагируют на звуки, частота которых
превосходит частотный предел для человека. У бражника Celerio
фонорецепторы нижнегубных щупиков наиболее чувствительны
в диапазоне 15—170 кГц. Ухо бабочки представляет собой импуль-
сный детектор, а не частотный дискриминатор; нервные импульсы
не соответствуют звуковой частоте. У бражников орган слуха рас
Механорецепция, фонорецепция и чувство равновесия 377м
пожен на нижнегубных щупиках, которые имеют воздушные по-
П° ти в двух первых сегментах. Зона наибольшей чувствительности
Лт°оСго Органа лежит между 20 и 40 кГц, а порог —на 10—15 дБ-
Э ппе чем у тимпанальных органов совок [199, 202, 203]. Интен-
В вность звука, достигающего тимпанальных органов совки, может-
^меняться на 40 дБ в зависимости от положения крыльев по от-
ношению к местоположению источника звука. Когда крылья под-
Рис. 12-10. Импульсные реакции волокон тимпанального нерва, зарегистрирован-
ные в нейропиле бабочки-совки [199].
А. Одиночная реакция. Б. Серия реакций (записанная в виде точечной диаграммы) на зву-
ковой импульс длительностью 5,5 мс при интенсивности 8 дБ. В, Г. Реакции на звук, ин-
тенсивность которого равна 18 дБ. Звуковой стимул изображен под реакциями А и В; ла-
тентный период равен 3—6 мс; серия спайков продолжается после окончания звука.
няты, ипсилатеральная стимуляция на 20—40 дБ больше, чем кон-
тралатеральная; когда крылья опущены, звук, идущий снизу, ока-
зывается сильнее звука, идущего сверху. Таким образом, бабочка
может определить направление звука и в некоторой степени рас-
стояние до его источника путем сравнения информации об интен-
сивности раздражения, поступающей от двух тимпанальных орга-
нов в течение одного взмаха крыльев [187, 201].
Тимпанальный орган саранчи имеет четыре группы сенсорных
клеток, расположенных различно по отношению к тимпанальной
Мембране и имеющих разные частотные характеристики и разную
Увствительность. У тимпанальной мембраны есть два центра ко-
^соаний: один — ближе к середине, другой — вблизи края мембра-
L *кани, и в особенности жир, окружающие интактный орган
н уха» СЛужат низк°частотным фильтром [153]. Импульсы тимпа-
ОГ)ЛЬного иерва бразильской цикады свидетельствуют о том, что
, е 0^н слУха этого насекомого имеет максимум чувствительности
} ласти 6—9 кГц. Адаптация к сериям звуковых щелчков отсут-
378
Глава 12
ствует, но быстро появляется в ответе на чистый тон [57]. Злато-
глазка Chrysopa имеет тимпанальный орган на жилке вблизи мес-
та прикрепления крыла. Сенсорные разряды имеют латентный пе-
риод 4—6 мс в ответ на звуковые стимулы от 15 до 150 кГц, а ча-
стота повторения импульсов достигает 150 имп/с [154]. Измерение
амплитуды колебаний тимпанальной мембраны у сверчка с по-
мощью эффекта Мёссбауэра показало, что ее максимум соответст-
вует звуку с частотой 4—6 кГц; эта же частота является оптималь-
ной и для ответа нервного волокна. Таким образом, тимпанальная
мембрана механически настроена на узкую полосу частот. Порого-
вая амплитуда оценивается приблизительно в 0,0005 мкм [109].
Ниже приведены величины чувствительности тимпанальных ор-
ганов в области высоких частот в сравнении с чувствительностью
хордотональных джонстоновых (антеннальных) органов и волоско-
вых жлеток [7].
Оптимум, кГц
Anopheles 0,34 Джонстонов орган
Calliphora 0,15—0,25 То же
Locusta 5-12 Тимпанальный орган
Prodenia 15-60 То же
Parametra (паук) 0,3—0,7 Щелевой орган
Gryllus 0,5 Волосковые клетки
Oxya 0,4—0,5 То же
У совок информация от тимпанальных рецепторов поступает
в грудные ганглии [199, 201]. Один интернейрон на каждой сторо-
не является отметчиком спайков (т. е. он отмечает каждый спайк
в серии, идущей от А-рецептора; рис. 12-11). Другой интернейрон
является отметчиком звуковых толчков и отвечает одним (или дву-
мя) импульсом на серию, отмечая таким обрашм интервал меж-
ду сериями. Третий интернейрон реагирует на каждую серию пач-
кой импульсов с интервалами, не зависящими от частоты на входе,
регистрируя длительность серии. Нейроны, расположенные около
средней линии, суммируют сигналы, поступающие от обоих ушей;
один из них стимулируется сигналом от контралатерального орга-
на слуха и тормозится ипсилатеральным сигналом. У бражников,
имеющих орган слуха на нижнегубных щупиках, интернейроны кон-
вергируют сильнее, чем у совок [203].
Одиночные нейроны в надглоточном ганглии Locusta различают-
ся картиной их реакции: одни реагируют на высокие частоты, дрУ'
гие — на низкие; обнаружены также и широкополосные вставочные
нейроны. Возбуждение вставочных нейронов может быть пре-
рвано тормозящим сигналом с частотой противоположной обла-
сти частотного диапазона [1а, 2]. В мезаторакалыюм ганглий
Locusta существуют вставочные нейроны, имеющие резкий
подъем частотной характеристики, в особенности в области
низких частот (3—4 кГц). При перерезке нервных волокон (кроме
идущих к тимпанальной мембране) этот резкий подъем исчезает,
Механорецепция, фонорецепция и чувство равновесия
по-видимому, такие вставочные нейроны могут тормозиться сигна-
лами от каких-то других входов [266]. Найдено четыре типа нейро-
нов второго порядка: а) нейроны, дающие тоническую реакцию на
высокие (Ю—20 кГц) и «ош>-разряды на низкие частоты; б) нейро-
ны, реагирующие только на высокие частоты; их реакция является
Рис. 12-11. Реакция нейрона — отмет-
чика импульсов у бабочек [199].
Сенсорные спайки отклоняют луч осцил-
лографа вниз, отметчик импульса — вверх.
А — В. Реакции на одиночные звуки. А
Реакция на повторяющиеся звуки. Д. Сен-
сорные спайки на генераторном потенциа-
ле при различных интенсивностях звуко-
вого раздражения.
'Тонической при пороговых уровнях раздражения и имеет вид
«ош>-ответа при большей его интенсивности; в) клетки, характери-
зуемые тонической реакцией на стимулы от 3 до 20 кГц; г) клет-
ки, дающие тоническую реакцию в узком диапазоне интенсивно
стей [ИЗ, 243]. „
Поведенческое значение рецепции звука и центральной оор
ботки звуковой информации у ночных насекомых заключается в
том, что она дает им возможность избежать преследования со сто»
'380
Глава 12
роны летучей мыши (рис. 12-12). Чувствительность органа слуха
бабочки в диапазоне частот локационного сигнала летучей мыши
составляет 0,01—0,03 дин/см2, что позволяет бабочке обнаружи-
вать преследователя на расстоянии около 30 м. Бабочка может оп-
ределять направление звука с достаточно высокой точностью, что
видно из меркаторских диаграмм чувствительности органа слуха
для различного положения крыльев (рис. 12-13). В нормальном
полете бабочка нерегулярно порхает вверх и вниз, при прибли-
Рис. 12-12. Реакция тимпанального нерва бабочки Prodenia (!) и запись крика ле-
тучей мыши вблизи бабочки (П) (Roeder К. D., Tread А. Е., J. Exp. Zool., 134,
127—157, 1957).
А. Крик летучей мыши в звуковом диапазоне (частота 10—15 кГц). Б. Более короткий крик
.летучей мыши (частота свыше 20 кГц). Реакция тимпанального органа бабочки продолжается
после окончания стимула.
женин же летучей мыши она бросается круто вниз (возможно,
благодаря активации А2-рецептора органа слуха). Летучая мышь
испускает высокочастотные звуки с частотой порядка 10 имп/с.
Она может обнаружить бабочку по эхо-сигналу примерно в трех
метрах, и тогда частота посылок у нее возрастает до 100 в 1 с. Хо-
тя летучая мышь летает быстрее, чем бабочка, но и она, нападая
на жертву, может промахнуться^ Таким образом, оба животных хо-
рошо приспособились друг к другу: бабочка иногда спасается, а
иногда становится жертвой [200]. Некоторые бабочки из сем. мед-
ведиц обладают цимбальными органами, расположенными в ме-
татораксе; эти органы генерируют звуковые сигналы в частотном
диапазоне, характерном и для сигнала летучей мыши. Есть пред-
положение, что таким образом бабочки искажают сигнал летучей
•мыши [12]. Возможно, однако, что эти бабочки просто не соответ-
ствуют вкусам летучих мышей и последние пренебрегают видами,
которые шлют им предупреждающий ответный сигнал [48].
Многие прямокрылые издают звуки при помощи стридуляцион-
ных органов. Кузнечик трет бедра задних ног, имеющие ряд высту-
пов, о наружную поверхность крыльев. Самцы сверчков и кузнечи-
ков теттигонид трут передние крылья друг о друга. Одно крыло
у них имеет «терку», другое — «пилу». Передние крылья теттиго-
w
Градусы no горизонтали
m % -о го ieo
Градусы no горизонтали
b
Ис. 12-13. Меркаторская проекция интенсивности звука, возбуждающего реакцию
левого органа слуха бабочки, в зависимости от положения крыльев [187].
Крылья подняты. Б. Крылья находятся в горизонтальном положении. В. Крылья опущены.
382
Глава 12
нид могут двигаться с частотой 35—45 движений в 1 с. Зубчатые
звуковые импульсы следуют с частотой 650—800 в 1 с.
при этом несущая частота составляет 18—42 кГц при открыва-
нии и 40—66 кГц при закрывании крыльев. Максимальная чувст-
вительность тимпанального органа лежит в том же диапазоне, что
и издаваемые высокочастотные звуки [240]. «Песни» ухаживания
и соперничества взрослых сверчков можно определить как видо-
специфичные, причем в основе каждой из них лежит упорядочен-
Полегн
Дыхание
Рис. 12-14. Схема, отражающая некоторые ритмичные движения у сверчков: по-
лет, призыв, ухаживание, ходьба, дыхание [130].
Ритм можно объяснить работой двух центральных осцилляторов; х дает период быстрого
ритма (30 Гц), а у — период медленного (3—4 Гц) (Kutsch W., Z. vergl. Physiol. 63, 335—378,
1969).
ная последовательность сокращения мышц [130] (рис. 12-14). |
Самцы пауков-ликозидов издают звуки ухаживания и угрозы при |
помощи пальп; частота первых из них возрастает, когда самка
производит ответные волнообразные движения ногами [205].
«Песня» кузнечика-самца содержит последовательность звуковых i
импульсов, возникающую в результате серии мышечных сокраще- \
ний, эта серия должна быть строго запрограммирована в нервной J
системе [52]. Сравнение звуковых сигналов, испускаемых насеко-
мыми, с пороговыми характеристиками их слуховых органов обна-
руживает четкое соответствие обеих систем друг другу [116]-
Видоспецифичная «песня» цикады имеет сложный рисунок, сос-
тавленный из щелчков и скрежетаний. У Magicicada каждое подер*
гивание мышцы цимбала влечет за собой изгиб серии жестких ре'
бер на мембране цимбала; каждый изгиб вызывает затухаюш116
колебания в воздушном мешке; поэтому сокращение мышцы с час-
тотой 50 раз в 1 с вызывает резонансные звуковые колебания по-
рядка 1000 в 1 с [196]. У саранчи Schistocerca в ответ на звук, ча-
Механорецепция, фонорецепция и чувство равновесия
383
ота которого соответствует наиболее низкому частотному порогу
Симпанального нерва, включается механизм полета [266а].
Т Некоторые виды пчел сообщают о расположении источников
пиши с помощью звуковых сигналов, издаваемых ими в улье (а не
с помощью танца); те пчелы, которые сигнализируют о наличии
пищи специфическими танцами, тоже издают звуки, необходимые
для коммуникации между фуражирами и другими рабочими осо-
бями. Звуки издаются благодаря деформации кутикулярного
скелета, вызываемой крыловыми мышцами с частотой около
250 Гц; обнаружены также высокочастотные обертоны, создавае-
мые, вероятно, другими вибрациями экзоскелета пчелы. Длитель-
ность каждого звукового периода коррелирует с расстоянием до ис-
точника пищи: 0,4 с, если источник пищи находится вблизи улья,
и 1,5 с, если он удален на расстояние 700 м. Предъявляя пчелам
такие звуки, записанные на магнитофон, можно вынудить их по-
кинуть улей. Основную роль в восприятии звука у пчел играет
джонстонов орган, расположенный на антеннах (см. выше) [59,
<60, 61].
Колебания твердого субстрата, воды и воздуха играют в мире
насекомых важную предупреждающую роль, а также способствуют
управлению движением и позой; для некоторых видов они служат
средством коммуникации. Рецепторы, имеющиеся в определенных
сочленениях скелета, весьма чувствительны к колебаниям. Звуко-
вые сигналы воспринимают и ночные бабочки, которые спасаются
•от летучих мышей, едва услышав, что их «засек» сонар преследо-
вателя. У многих прямокрылых звук служит средством коммуни-
кации; он издается скелетными структурами и воспринимается
тимпанальными органами. Кодирование звука у насекомых осу-
ществляется в значительно большей степени с помощью амплитуд-
но-временных характеристик звуковых импульсов, чем с помощью
частоты и гармоник, что более характерно для позвоночных.
Фонорецепция у позвоночных
Свойства звука как стимула. Звук представляет собой регуляр-
ные механические волны, которые в случае чистого тона могут
быть описаны частотой и амплитудой. При прохождении звуковой
волны через среду частицы среды смещаются вперед и назад в
зависимости от величины давления звуковых волн. Скорость распро-
странения и затухания звука зависит от среды. Интенсивность зву-
ка выражается в виде уровня звукового давления в динах на квад-
ратный сантиметр. Наиболее слабый звук, еще слышимый челове-
ком, имеет уровень около 0,0001 дин/см2 (при частоте 1 кГц). Уро- .
вьНь звУка при разговоре составляет примерно I дин/см2, а звук,
Дающий болевое ощущение, соответствует давлению около
цах ^ин/см2. Звуковое давление измеряется в абсолютных едини-
(Дин/см2), но на практике в аудиологии используется относи-
384
Глава 12
Тельная шкала интенсивностей, где единицей служит децибел
равный логарифму отношения двух интенсивностей. Уровень звука*
в децибелах определяется как
7 (дБ) = 10 log—,
70
где 70— стандартная интенсивность (обычно 10-16 В т/см2). В тер-
минах звукового давления, которое пропорционально квадрату ин-
тенсивности, уровень звука в децибелах равен
I (дБ)=20 log
В качестве стандартного уровня давления Ро части принимают ве-
личину, близкую к слуховому порогу человека 2-10~4 дин/см2 (в
особенности для звука, распространяющегося в воздухе). Иногда
(для звуков в другой среде, особенно в воде) Ро выбирают равным
1 дин/см2. Следует заметить, что 1 дин/см2 на 74 дБ больше, чем
2-10~4 дин/см2 (т. е. чем слуховой порог) [119].
Интенсивность звука дается соотношением 7=Р2/рС, где 7 —
интенсивность, выраженная в эрг/см2, Р — давление в дин/см2, р —
плотность среды в г/см3, а С — скорость распространения звука в
см/с. Член рС носит название акустического сопротивления, а его
величина для разных сред при биологических температурах имеет
приблизительно следующие значения:
р, КГ/мЗ С, м/с
Воздух 1,2 344
Вода 1000,0 1481
Морская вода 1026,0 1500
Таким образом, в воде акустическое сопротивление гораздо
больше, чем в воздухе, и скорость звука также значительно выше.
Интенсивность для данного звукового давления в воде ниже, чем
в воздухе.
Фонорецепторы — это органы чувств, в которых звуковые вол-
ны, поступающие из воздушной или водной среды, проходят через
ткани и преобразуются в нервные импульсы. На низкочастотном
конце аудиограммы (около 50 Гц) фонорецепция и вибрационная
чувствительность перекрываются. Звук с частотой выше 20 кГц
(предел, воспринимаемый человеком) иногда определяют как ульт-
развук. Свойство рецептора воспринимать данную частоту еще не
означает, что этот звук будет в той же степени «услышан» нервной
системой. Следовательно, изучение свойств фонорецепторов непре-
менно должно сопровождаться поведенческими опытами и реги-
страцией активности мозга.
Морфология и стимуляция уха позвоночных. Наружное Ух°
(ушная раковина) способствует локализации источника звука 11
концентрирует высокочастотные звуки за счет отражения. СлухО’
Механорецепция, фонорецепция и чувство равновесия
385
бой проход отделен от среднего уха барабанной перепонкой. Сред-
нее ухо содержит слуховые косточки (молоточек, наковальню и
стремечко), которые, действуя по принципу рычага, совершают слож-
ные движения. Основание стремечка контактирует с овальным окном
во внутреннем ухе. Мышцы, сокращающие барабанную перепонку
(tensor tympani), связаны с молоточком, а стремечковая мышца
прикреплена к стремечку. Эти мышцы ограничивают вибрацию ко-
сточек при звуках большой интенсивности. Евстахиева труба сое-
диняет полость среднего уха с глоткой и уравнивает давление в
среднем ухе с давлением во внешней среде. Ниже овального окна
расположено круглое окно, закрытое эластичной мембраной, бла-
годаря которой жидкость внутреннего уха может смещаться.
Функция слуховых косточек состоит в том, чтобы передать звуко-
вые волны из воздуха к жидкости внутреннего уха с минимальны-
ми потерями на отражение (т. е. согласовать импедансы двух
сред). У человека увеличение давления при переходе от барабан-
ной перепонки к овальному окну составляет около 13 дБ. У игуа-
ны среднее ухо имеет коэффициент преобразования около 35 дБ
[258].
У китообразных внешний слуховой проход редуцирован, и звук,
вероятно, проникает в среднее ухо через ткани, особенно жиро-
вые и костные [149, 169, 170]. У животных, не относящихся к мле-
копитающим, среднее ухо содержит только одну слуховую косточ-
ку — колумеллу.
Карпообразные рыбы имеют набор косточек (веберов аппарат),
связывающий плавательный пузырь с внутренним ухом. Соответст-
венно у них слуховой порог значительно ниже, чем у рыб, внутрен-
нее ухо которых не имеет костной связи с плавательным пузырем.
Измерение амплитуды и скорости движения слуховых косточек
и основной мембраны проводилось с помощью миниатюрного ис-
точника гамма-лучей и изменения эффекта Мёссбауэра [3]. На
рис. 12-15 отражена картина смещения стремечка в ухе морской
свинки, ехидны и ящерицы в. широком диапазоне частот. У летучей
мыши скорость смещения барабанной перепонки в пределах от
Ю до 70 кГц постоянна. У рептилий и амфибий движение стре-
мечка с понижением частоты уменьшается. Механические свойст-
ва среднего уха соответствуют диапазону оптимального слуха дан-
ного Животного.
Внутреннее ухо представляет собой систему полостей, запол*
енных жидкостью. Одна часть этой системы известна как лаби-
ринтный орган равновесия, другая (улитка) — как орган слуха,
духовая часть внутреннего уха появилась впервые в виде лаге-
У рыб и амфибий. У млекопитающих это образование удлини-
гелЬ И ПРИНЯЛО форму спирали. Расстояние от овального окна до
у ^КотРема в вершине улитки составляет 55 мм у слона и 7 мм
СНе Ыщи„- Спиральная трубка улитки разделена продольно рей-
Р°вой и основной мембранами на три канала. Два наружных
25~-1514
386
Глава t2
канала — лестница преддверия (scala vestibuli) и барабанная
лестница (scala tympani), так же как и туннель Корти, заполнены
перилимфой. Средний канал (средняя лестница— scala media),
расположенный между рейснеровой и основной мембранами, со-
держит эндолимфу. По сравнению с перилимфой эндолимфа со-
держит в 30 раз больше калия и в 10 раз меньше натрия. В эндо-
лимфе цыпленка содержание калия превышает содержание натрия
Рис. 12-15.
А- Сравнение скорости движения стремечка у морской свинки и ехидны и колумеллы у яще-
рицы Amphibolurus в зависимости. от частоты звука при постоянной его интенсивности. Из-
мерения проводились с помощью эффекта Мёссбауэра. Б. Сравнение амплитуды смещения
барабанной перепонки и стремечка у ехидны, а также отношение амплитуды стремечка
к амплитуде барабанной перепонки у ехидны, кошки и геккона [3].
в 3 раза [156]. Эндолимфа, по всей вероятности, секретируется со-
судистой полоской (stria vascularis) [36, 37]; предполагается также
существование калиевого насоса. При стимуляции звуком концент-
рация натрия в эндолимфе растет, а концентрация калия падает
[244].
Кортиев орган, расположенный на основной мембране, содер-
жит сенсорные клетки: три ряда наружных волосковых клеток и
один ряд внутренних (рис. 12-16). Волоски контактируют с волок-
нистой покровной мембраной, имеющей неклеточное происхожде-
ние. Волосковые клетки иннервируются дендритами нейронов, те-
ла которых лежат в кохлеарном (слуховом) ганглии, а аксоны со-
ставляют большую часть волокон VIII черепно-мозгового нерва.
Каждый нейрон иннервирует одну или несколько волосковых кле-
ток. На электронных микрофотографиях у оснований волоско-
вых клеток видны синаптические пузырьки; к волосковым клеткам
подходят нервные окончания, также содержащие синаптические
пузырьки и представляющие собой, по-видимому, эфферентные ак-
соны, идущие от мозга. Закономерности распространения звуково
волны вдоль основной мембраны установил Бекеши [8, 9] путе
Рис. 12-16.
ей ^ЛитК.а морской свинки — примерно Ч& завитка (вид в сканирующем электронном микро-
к‘оп?2; Кортнев орган виден на радиальном срезе [57а]. ВВК— внутренние волосковые клет-
, ’ 1~3 ~ тРи ряда наружных волосковых клеток; ОМ — основная мембрана; ПМ — покров-
уд- —Мембрана; СГ — спиральный ганглий; КГ— клетки Гензена; КК— клетки Клаудиуса,
кпос/УННЛЛЬ ^°РТИ- -Б- Вил поверхности кортиева органа в сканирующем электронном ми-
Р /_?^/Ф°тогРаФия X. Эйдса). ВВК — внутренние волосковые клетки; КС — клетки-столбы,
J наружные волосковые клетки; КД — фалангеальные отростки клеток Дейтерса.
25*
388
Глава 12
непосредственного наблюдения под микроскопом ее колебаний в
различных зонах, а также с помощью эффекта Мёссбауера, когда
система внутреннего уха возбуждалась синусодиальными колеба-
ниями стремечка. Оба метода показали, что наружная мембрана
волосковых клеток может сдвигаться всего на 0,005 мкм при ча-
стоте, соответствующей максимуму чувствительности [108]. Зона
максимальной амплитуды колебаний перемещается по длине улит-
ки в зависимости от частоты звука. Нормальное ухо человека вос-
принимает частоты от 15—20 до 16 000—20 000 Гц; его чувстви-
тельность к высоким частотам с возрастом ухудшается. Минималь-
ный порог слышимости лежит между 1500 и 2000 Гц. Волокна, иду-
щие к базальному завитку улитки, реагируют на любой тон во
всем диапазоне слышимости, тогда как волокна, иннервирующие
верхний завиток, дают ответ только на звуки низкой частоты. Ло-
кальные повреждения кортиева органа, вызванные продолжитель-
ным действием громкого тонального звука, выражаются в дегенера-
ции групп волосковых клеток и потере чувствительности в соответ-
ствующем интервале частот. Низкие частоты вызывают более зна-
чительные повреждения в дистальном, нежели в проксимальном
ряду клеток [236]. Запись активности одиночных слуховых воло-
кон морской свинки обнаруживает весьма резкую границу слыши-
мости в области высоких частот.
Некоторые электрические параметры улитки могут быть изме-
рены. Эндолимфа средней лестницы электроположительна по от-
ношению к окружающей ткани и перилимфе (рис. 12-17). Этот
постоянный потенциал носит название эндокохлеарного потенциа-
ла (ЭП). Электрод, введенный в кортиев орган (скорее всего вне-
клеточно), регистрирует потенциал от —80 до —90 мВ по отноше-
нию к барабанной лестнице (перилимфа). Таким образом, между
жидкостью над волосковыми клетками и жидкостью, окружающей
их, существует разность потенциалов порядка 160—180 мВ. ЭП
максимален вблизи сосудистой полоски. Он чувствителен к анок-
сии и, вероятно, возникает в результате активного транспорта ио-
нов. Крайние клетки сосудистой полоски образуют множество ла-
теральных и базальных складок, между которыми располагаются
вытянутые митохондрии и микроворсинки, обращенные в просвет.
Такая структура типична для клеток, вовлеченных в ионный тран-
спорт (гл. 6) [49]. ЭП возрастает при инъекции натрия в среднюю
лестницу и слегка снижается при введении ингибитора NaK-АТФа-
зы—уабаина [128]. Он очень чувствителен к динитрофенолу и к
недостатку кислорода. Очевидно, постоянный потенциал генериру-
ется активным насосом, который накачивает в эндолимфу калии
в обмен на натрий. Хлор в эндолимфе находится в той же концен-
трации, что и в перилимфе, и поскольку он не находится в электро-
химическом равновесии, то предполагается существование хлорно-
го насоса [Ю7]. У птиц ЭП меньше (около 20 мВ) и чувствителен
к аноксии. У пресмыкающихся значение ЭП еще ниже — от 2 Д°
Механорецепция, фонорецепция и чувство равновесия
389
7 мВ причем ЭП у них к аноксии нечувствителен [214]. Не все
пуховые отделы с высоким содержанием К+ и низким Na+ име-
т большой положительный потенциал:
Улитка черепахи
Улитка ящерицы
мМ Na'*'»
114 2,7
140 10
Потенциал, мВ
+6
4-9
дополняют актив-
Таким образом, различия в проницаемости
ный транспорт, и оба фактора вместе определяют потенциал эн-
долимфы [Ю7].
При воздействии звука на ухо в улитке генерируется микро-
фонный потенциал (МП). Он легко регистрируется при отведении
от круглого окна (рис. 12-18). Амплитуда МП зависит от интен-
сивности звука, порога МП не имеет. По сути дела, МП — это пе-
ременное напряжение, генерируемое волосковыми клетками и пов-
торяющее форму стимулирующей звуковой волны во всем диапа-
зоне слышимости. МП представляет собой генераторный потен-
циал, уменьшающий уровень ЭП и возникающий в волосковых
клетках, по-видимому, за счет срезывающего усилия, образующе-
гося вследствие того, что покровная и основная мембраны «подве-
шены» в различных местах, МП чувствителен к парциальному
давлению кислорода, воздействию цианида и температуры [165,
246].
Дополнительной реакцией на чистый тон является суммацион-
ный потенциал (СП) [37]. Он также возникает в волосковых клет-
ках; обычно средняя лестница приобретает больший отрицатель-
ный потенциал, чем барабанная лестница. Изгибание волосков
при помощи микроиглы в различных направлениях может вызвать
потенциалы, очень напоминающие микрофонный и суммационный.
Слуховые нервные импульсы, или потенциалы действия (ПД),
представляют собой спайки типа «все или ничего» с рефрактерным
периодом около 1 мс. Одиночные волокна имеют относительно уз-
кую область реакции, или область частот, где чувствительность
максимальна, что предположительно коррелирует с точкой гене-
рации сигнала в кортиевом органе. В волокне можно зарегистри-
ровать от одного до трех импульсов в ответ на одиночную низко-
частотную волну. При низкой частоте и умеренной интенсивности
стимула многие волокна реагируют синхронно, включая даже во-
°кна, идущие от базального завитка [204]. На средних частотах
мпульсы следуют с периодом, большим, чем период звукового сти-
Чай3’ ? Не обРазУют регулярной картины, появляясь как бы слу-
сы Н5*-246]. На высокой частоте первичные волокна дают импуль-
Дани “ а3уЮЩИе хаРактеРные временные последовательности. При
Увел°И частоте звука частота следования спайков возрастает с
ственЧеНИеМ его интенсивности- Многим слуховым волокнам свой-
а спонтанная активность, частота которой не превышает
Кровеносные сосуды
(нулевой потенциал)
Рис. 12-17. «Электрохимия» улитки: ионный состав и потенциалы в вестибуляр-
ной лестнице (перилимфа), средней лестнице (эндолимфа) и барабанной лестнице
(перилимфа) [107].
2С:мКК
Рис. 12-18. Кохлеарные микрофонные потенциалы (МП) и суммационные п°те0.
циалы (СП), отводимые от уха голубя при раздражении звуком разной част
ты [165].
1 — МП, СП-, 2—частота в Гц, уровень в дБ.
Механорецепция, фонорецепция и чувство равновесия
391
ЮО имп/с. Она может быть связана либо с движением волоско-
1 IX клеток, либо с некоторым шумом в окружающей среде. Когда
%йрон возбуждается тональными импульсами оптимальной часто-
ты второй тон, время включения которого подбирается специаль-
но’ может затормозить реакцию по обе стороны от зоны возбуж-
дения; это явление предполагает существование некоторого пери-
ферического торможения в улитке [5].
Возбуждение окончаний слухового нейрона, согласно одной из
теорий, имеет электрическую природу. Эндокохлеарный и внутри-
клеточный потенциалы в совокупности образуют батарею, э. д. с.
которой равна приблизительно 160 мВ; напряжение этой батареи
приложено к волосковым клеткам. Можно думать, что под воздей-
ствием срезывающего усилия изменяется электрическое сопротив-
ление волосковых клеток, что ведет к возникновению МП и СП, а
возникающие при этом изменения тока прямо или косвенно стиму-
лируют нервные окончания [257]. Другая гипотеза полагает, что
нервные окончания возбуждаются химическим медиатором, выде-
ляющимся из волосковых клеток. Волосковые клетки содержат в
базальной области много пузырьков, и эта зона напоминает собой
пресинаптические окончания [57а, 115]. От волокон слухового нер-
ва отводятся синаптические потенциалы, задержанные относитель-
но МП, с флуктуирующей амплитудой, что указывает на квантовый
характер выделения медиатора; кроме них регистрируются спон-
танные миниатюрные потенциалы [70, 104]. Поскольку в хорошо
изученных синапсах (например, в концевой пластинке нервно-мы-
шечного контакта) медиатор высвобождается под воздействием
деполяризации, можно думать, что в органе слуха эту роль игра-
ет микрофонный потенциал, под влиянием которого волосковые
клетки выделяют медиатор.
Птицы. Лагена внутреннего уха птиц вытянута и имеет цент-
ральную основную мембрану. Диапазон чувствительности и разли-
чение высоты тона у них лучше, чем у низших позвоночных. У го-
лубя МП следует за частотой звука вплоть до 25 кГц, наиболее
низкий порог отмечается около 3200 Гц. После удаления лагены
хорошие МП в диапазоне от 100 до 3000 Гц отводятся от ампул
полукружных каналов [226].
Пресмыкающиеся. У этих животных дно лагены напоминает
основную мембрану млекопитающих. У пресмыкающихся, за исклю-
чешгем змей, среднее ухо имеет костную колумеллу. Колумелла
3Мтт пРикРеплена к квадратной кости. У черепах регистрируется
50лл °Н Имеет высокий порог и максимальную частоту около
OU00 Гц [259], у сцинка МП максимален в диапазоне частот от 750
До 3000 Гц [НО].
Амфибии. Лагена амфибий несколько длиннее лагены рыб, и
нут5Нее Ухо бесхвостых амфибий (но не хвостатых) имеет кост-
Ним КолУМеллУ, соединяющую барабанную перепонку с внутрен-
Ухом. Удалось записать МП уха лягушки в диапазоне 50—>
392
Глава 12
3500 Гц с максимумом чувствительности между 400 и
1500 Гц [237].
Сравнение максимумов чувствительности и частотных диапазо-
нов слуха рептилий, птиц и млекопитающих показывает, что пти-
цы обладают гораздо большей чувствительностью, чем пресмыкаю-
щиеся, а диапазон слуха млекопитающих перекрывает диапазо-
ны слуха как тех, так и других. Оптимальные для слуха рептилий
частоты располагаются в нижней части звукового диапазона; мле-
копитающим доступны наиболее высокие звуковые частоты. Одним
из направлений эволюции было удлинение слуховой мембраны, ко-
торое связано с улучшением пространственного кодирования ча-
стоты; слуховая улитка позвоночных позволяет различать наибо-
лее высокие частоты [140].
Водные позвоночные; реакции улитки и центральной нервной
системы, а также изменение поведения в ответ на звук. Звуковая
стимуляция в воде имеет две компоненты — ближнего и дальнего
поля. В первом случае векторные компоненты — скорость движе-
ния частицы и ее смещение — сильно зависят от скалярной компо-
ненты— давления. В дальнем поле векторные и скалярные компо-
ненты имеют постоянные отношения (иными словами, в ближнем
поле фазовые соотношения между давлением и смещением части-
цы меняются, тогда как в дальнем поле они постоянны). В ближ-
нем поле скорость уменьшается обратно пропорционально квадра-
ту расстояния от источника звука. В дальнем поле скорость и дав-
ление убывают с расстоянием линейно. Следовательно, волна
давления в дальнем поле отстает по фазе на 90° от волны в ближ-
нем поле [86]. Условились считать, что ближнее поле простира-
ется от источника звука на расстояние, меньшее чем Х/2л (где
Z — длина звуковой волны), тогда как дальнее поле характеризу-
ется расстояниями, значительно превышающими Х/2л [228]). В во-
де, как и в воздухе, интенсивность равна 1=Р2!рС (см. выше).
Интенсивность звука вблизи источника относительно независима
от частоты, однако низкие частоты с расстоянием сильно затухают
[54, 247].
У пластиножаберных нет плавательного пузыря, однако они
реагируют на низкочастотные звуки даже после перерезки нерва
боковой линии (эта реакция исчезает после перерезки VIII нерва).
Акулы стремятся приблизиться к источнику звука, частота которо-
го лежит между 55 и 500 Гц [161]. У ската Raja в ответ на низко-
частотные звуки (ниже 120 Гц) возникают импульсы в нервных
веточках двух макул sacculuns и одной макулы utriculus, однако
в нерве лагены такие реакции не обнаружены [137].
Рыбы воспринимают звуковые раздражения с помощью кожных
рецепторов, рецепторов боковой линии и трех камер лабиринта;
использование тех или иных зависит, по-видимому, от типа
стимуляции. Первые два вида служат для рецепции смещения
и низких частот. В опытах с обучением некоторые рыбы пользова-
Механорецепция, фонорецепция и чувство равновесия
393
пись сначала менее чувствительными рецепторами, а затем пере-
ключались на более чувствительные [247]. Типичные аудиограм-
мы полученные в поведенческих экспериментах, приведены на рис.
12-19. Стимуляция в ближнем поле воздействует на боковую ли-
нию рыб, а также передается к уху благодаря костной проводи-
мости. Бычок не имеет плавательного пузыря; на расстоянии свыше
Рис. 12-19. Аудиограммы, измеренные в поведенческих опытах на трех видах рыб
Lutjanus (1), Holocentrus (2) и Ameiurus (4) в сравнении с аудиограммой чело-
века (3) [247].
Псуоси абсцисс — частота в Гц; по осям ординат: I — логарифм интенсивности звука в Вт/см2;
1 звуковое давление в воде в дБ по отношению к одному микробару; III — звуковое дав-
ление в воздухе в дБ по отношению к одному микробару.
utri^i3 К0Т°Р°Г0 направлена в сторону чувствующего
HocTtU Нервные реакции на раздражения пороговой
12Ппг ПРИ низком пороге) были получены для частот
1 ц, а н
153]. Макси
м от источника звука от его sacculus не удавалось отвести ни-
каких микрофонных потенциалов; более того, даже на близких
40ПСг°ЯНИЯХ Не бЬ1Л0 получено реакции на звук с частотой свыше
1 Ц. У трески плавательный пузырь есть, и у нее удалось запи-
100П рПолне Удовлетворительные реакции на звуки с частотой ниже
1 Ц на значительном удалении от их источников (рис. 12-20,А).
” <₽поЬД,Ь С1иРеа имеет двойной плавательный пузырь, передняя
эпителия
интенсив-
от 13 до
на раздражения высоких интенсивностей — и до 4000 Гц
лмум чувствительности у большинства рыб лежит между
Интенсивность, дБ
Рис. 12-20.
А. Аудиограмма трески (по измерениям микрофонного потенциала) [55]. Б. Аудиограмма
ската Ameiurus, (по результатам опытов с условными рефлексами) [122]. / — нормальна
рыба, II — рыба с поврежденным плавательным пузырем.
Механорецепция, фонорецепция и чувство равновесия
.395
-РО ц 800 Гц. Рыбы, имеющие веберов аппарат, способны воспри-
ать более высокие частоты (до 6—8 кГц), чем рыбы, лишенные
Нг0 (их верхний предел — 800—3000 Гц) [120, 247]. Таким обра-
зом высокая чувствительность слуховой системы хорошо коррели-
ет с наличием и сложностью плавательного пузыря [55, 56]. Ес-
ли удалить воздух из плавательного пузыря Ameiurus, чувстви-
тельность его слуховой системы на низких частотах (200—300 Гц)
почти не изменится, однако на 1500 Гц она упадет на 30 дБ. Пере-
оезка нерва боковой линии ведет к исчезновению реакций на звуки
низкой частоты [122] (рис. 12-20,В).
Удаление лагены или края sacculus не оказывает почти ника-
кого влияния на микрофонные потенциалы уха у Porychtys, однако
при полном выжигании саккулярной макулы они исчезают [34].
Многие рыбы издают звуки, используемые ими для коммуника-
ции. Звуки генерируются в основном при сокращении мышц, ле-
жащих вдоль плавательного пузыря или между грудным поясом и
плавательным пузырем. При этом плавательный пузырь служит
резонатором. Porychtys издает «хрюканье» с максимальной ампли-
тудой при частоте от 85 до 600 Гц и «жужжание» — на частотах от
85 до 1000 Гц [34]. Therapon издает звуки угрозы и нападения.
Сигнал тревоги морского петуха напоминает хрюканье, а его при-
зывный крик звучит стаккато. Максимум энергии в «хрюкающем»
сигнале морской собачки лежит между 80 и 180 Гц; крик фахана
имеет максимум около 250 Гц. Локальные удары током в область
среднего мозга либо облегчают, либо подавляют реакцию на зву-
ковые щелчки, регистрируемую в слуховом ядре, в зависимости от
интервала времени между импульсом тока и звуковым сигналом
[187а].
В общем пороги поведенческих реакций ниже, чем пороги нерв-
ных реакций. Записи от одиночных волокон в слуховой области
продолговатого мозга сельди Ыиреа обнаруживают большой раз-
брос значений верхнего предела воспринимаемых частот и частоты,
при которой реакция имеет максимум. Зона наилучшей чувстви-
тельности лежит между 100 и 1200 Гц [55]. В мозге серебряного
карася пределы частотно-пороговых кривых для нейронов слухо-
пого ядра шире, чем для нейронов torus semicircularis, и особенно—
Чем для нейронов тегментума; в этой последней зоне некоторые
РеагиРУют как на звуковой, так и на зрительный стимулы
U83]. Импульсные акустические сигналы, не чисто тональные, эф-
фективно используются на практике для отвлечения хищников (на-
Ример, акул) от мест купания человека.
Серебряный карась может слышать тон до 3500 Гц; поврежде-
е utriculus почти не влияет на эту его способность, но после пов-
ни Дения sacculus и лагены у него сохраняется реакция только на
До 6ПпоЧаСТ0ТЬ1 (250 Гц). Гольян Phoxinus слышит частоты вплоть
УДа Гц; способность различать звуки у него также исчезает с
396
Глава 12
Наземные позвоночные; реакции центральной нервной системы
и изменение поведения в ответ на звук. Амфибии. Брачные песни
лягушек видоспецифичны. Как показывают фонограммы песен
различных древесных лягушек
положения спектральных макси-
мумов энергии и характерные
звуковые щелчки различны не
только у разных видов, но и
у представителей одного вида
обитающих в разных местностях’
Самка реагирует только на пес-
ню самца своего вида и ареала
[24] (рис. 12-21). Регистрация
активности нейронов слухового
ядра показывает, что их чувстви-
тельность соответствует макси-
муму чувствительности основной
мембраны и звуковому диапазо-
ну естественной песни, характер-
ной для брачного поведения,
овладения территорией или вы-
ражения неудовольствия [69,
217]. Наиболее низкий порог для
вызванных потенциалов в сред-
нем мозге шести видов бесхвос-
тых амфибий соответствует ос-
новной частоте песни. Из разных
Рис. 12-21. Брачный призыв лягушки-бы-
ка [24].
А. Брачный крик полностью — пять звуковых
посылок. Б. Третья посылка при большей ско-
рости развертки. В. Часть третьей посылки при
еще большей скорости развертки.
популяций, объединенных общим названием Rana pipiens, у четь\
рех оказались столь различные звуковые сигналы, что они лишен
возможности спаривания друг с другом и практически представл
ют собой репродуктивно изолированные виды [133].
Пресмыкающиеся. Аллигаторы и крокодилы генерируют кох
арные потенциалы в ответ на звук в диапазоне от 300 до 3U
при относительно высоком пороге [258,259]. Записи от
нейронов слухового ядра продолговатого мозга геккона Со
показали, что максимум их чувствительности лежит между
2000 Гц, однако реакции регистрируются в широком диапазоне
Механорецепция, фонорецепция и чувство равновесия.
397
100 до 17 000 Гц; отдельные нейроны сильно отличаются друг от
друга и могут тормозиться в узких областях частот [242].
Некоторые первичные слуховые волокна у Trachysatirus прояв-
ляют спонтанную активность; они реагируют на тон начальным
увеличением скорости разрядов, постстимульным торможением и
восстановлением. Оптимальная частота меняется от волокна к во-
локну, и ее максимум был обнаружен при 700 Гц и 3 кГц [ПО].
В тектуме различных видов змей наблюдаются как медленные
реакции, так и спайки. Тональные сигналы, передаваемые воздуш-
ной средой, в диапазоне от 50 до 1000 Гц вызывают реакции с от-
носительно высоким порогом. Когда звук направлен локально,
реакция возникает при стимуляции почти любого участка поверх-
ности тела. Такая чувствительность сохраняется при перерезке
спинного мозга, но уничтожается при удалении внутреннего уха.
Следовательно, звуки, пришедшие из воздуха, должны быть пере-
даны через ткани к уху [89, 90]. Змеи обладают также соматиче-
ской (спинальной) системой, которая воспринимает вибрации суб-
страта в широком диапазоне частот (см. выше).
Регистрация активности кохлеарного ядра некоторых пресмы-
кающихся показала, что максимальный частотный диапазон свой-
ствен тем видам, основная мембрана которых имеет наибольшее
отношение длины к ширине. Так, у каймана диапазон слуха лежит
между 70 и 2900 Гц: в его torus sernicircularis клетки сгруппирова-
ны в колонки; клетки с наилучшей реакцией на низкие частоты
располагаются дорсально, а наиболее чувствительные к высоким
частотам расположены в глубине [141].
_ Птицы. Нейроны латерального мезэнцефального ядра (colliculi
inferioris) голубя обладают спонтанной активностью и дают в от-
вет на звук реакции следующих типов: торможение спонтанной ак-
тивности; возбуждение в центральной области частотного диапазо-
на с торможением на более низких и более высоких частотах; тор-
можение с узкими зонами возбуждения [11]. Пороги нейронов кох-
леарного ядра соответствуют наивысшей чувствительности,
измеренной в поведенческом опыте: канарейка — 2,5, домовый во-
робей— 6,5, юнко — 8,6 кГц (рис. 12-22) [124, 125]. Певчие птицы
не издают звуков с частотой ниже 1 кГц, но могут слышать их. У
скворца некоторые нейроны слухового ядра реагируют в широком
Диапазоне, а некоторые — в узком. Большинство из них имеет мак-
симум чувствительности между 250 и 600 Гц [226].
Опыты, в которых у птиц вырабатывались условные рефлексы,
показали, что попугаи и клесты различают тона с частотой от 40
До 1400 Гц. Фазаны слышат частоты вплоть до 10 500 Гц [235], а
32Пп1гИНСТВО певчих птиц имеет максимум чувствительности около
но ° *Характер песни данного вида предопределен генетически,
ю ’ кР°ме того, он имеет местные вариации, которые запечатлева-
* молодых птиц еще до того, как они начнут петь сами. Этот
гент, возможно, является важным фактором изоляции, ведущим
398
Глава 12
5к сохранению вида [126, 171]. Обычная функция песни у птиц —
это овладение территорией и ее охрана. Песня может служить
также для привлечения брачного партнера. Вороны и чайки имеют
большой набор звуковых сигналов, применяемых для выражения
тревоги, беспокойства, а также в случаях объединения, начала
коллективного пения -— «хора» и т. д.
Рис. 12-22.
А. Аудиограмма скворца, измеренная в поведенченском опыте. Б — Г. Аудиограммы оди-
ночных нейронов кохлеарных ядер скворца [125].
Птицы, которые обычно живут в пещерах, гуахаро Stehtornis и
стриж-саланган Collocalia, летая в темноте, пользуются эхолока-
цией. Гуахаро издает короткие щелчки в диапазоне частот 6100—
8570 Гц без ультразвуковых составляющих. Collocalia испускает
5— 10 щелчков в 1 с с частотой 4500—7500 Гц. На свету эти пти-
цы пользуются визуальными ориентирами, в темноте — эхолока-
цией, при помощи которой легко избегают таких препятствий, как
стержни или проволока [79, 173]. Совы Tyto весьма чувствительны
к звукам, возникающим при движениях жертвы; ухо совы имеет
очень узкую характеристику направленности на частотах свыше
8,5 кГц [186].
Млекопитающие (кроме летучих мышей). У всех млекопитаю-
щих улитка закручена в спираль; у утконоса она образует чет-
399
Механорецепция, фонорецепция и чувство равновесия
рТЬ витка, у кита —1,5, у лошади —2, у человека — 2,75, у кош-
и—-3 а у свиньи и морской свинки —почти 4 витка. Аудиограмма
человека имеет максимум чувствительности между 1000 и 4000 Гц,
а верхний предел ее—18 кГц. Некоторые виды собак слышат
35 кГц, а крысы и морские свинки —до 40 кГц; с оленьей мышью
можно’работать методом условных рефлексов в диапазоне от 16
Слуховая кора
Медиальное коленчатое тело
(вентральный отдел)
— Дорсальное
Вентральное
__Бугорки четверохолмия
(центральное ядро)
Дорсальное
Вентральное
ядра латераль-
ного лемниска
Медиальное ядро трапе-
циевидного тела
- Медиальная
— Латеральная
верхняя
олива
кохлеарное
ядро
Слуховой нерв
Рис. 12-23. Главные восходящие связи слуховой системы мозга позвоночных.
Особое внимание уделено билатеральным компонентам проекций верхнего ядра оливы и бо-
лее высоких структур. Некоторые пути, в частности исходящие из дорсального кохлеарного
ядра, не показаны (схема Л. Эткина).
до 65 кГц; соня имеет минимальный порог (около 0,01 дин/см2)
при 20 кГц.
Верхний частотный предел слуха у приматов ниже, чем у дру-
гих млекопитающих: 22 кГц у шимпанзе, но 120 кГц у летучих
мышей и дельфинов. Однако в области низких частот приматы
|ЛЬ1шат ^много лучше, чем такие млекопитающие, как опоссум или
Нервные слуховые пути млекопитающих показаны на рис. 12-
Из кохлеарного ядра волокна идут в комплекс верхней оливы,
* затем через латеральный лемниск к нижним бугоркам четверо-
°лмия, медиальному коленчатому телу и слуховой коре. Пере-
Ресты путей (переход с одной стороны мозга к другой) найдены в
Ласти кохлеарного ядра, оливы, нижних бугорков четверохолмия
400
Глава 12
и в коре. Ни в одном классе позвоночных, за исключением млеко-
питающих, передний мозг не принимает участия в работе слухо-
вой системы. Взаимодействие сигналов, поступающих от обоих ор-
ганов слуха, происходит прежде всего в оливе, но локализация ис-
точника звука реализуется только в коре [166, 261]. Оливо-кох-
леарные эфферентные волокна, которые идут медиально, иннерви-
руют наружные волосковые клетки кортиева органа. Электриче-
ская стимуляция перекрещивающегося оливо-кохлеарного пучка
уменьшает афферентные реакции одиночных нервных волокон на
тональный сигнал; микрофонный потенциал, однако, может при
этом увеличиваться, а спонтанная активность — оставаться неиз-
менной [262]. Эфферентные волокна вызывают появление тор-
мозных потенциалов и, по всем данным, действуют в базальном
завитке улитки пресинаптически, а в верхней части улитки — пост-
синаптически [127]. Оливо-кохлеарные эфференты активируются
звуком рефлекторно: они могут обострять различение и уменьшать
маскирующий эффект фонового шума [65, 66].
Нисходящие волокна оканчиваются также на клетках кохлеар-
ного ядра, где они могут быть как тормозными (особенно на высо-
ких частотах) [117], так и облегчающими [261].
Стимуляция коры действует тормозяще на одни нейроны меди-
ального коленчатого тела и облегчающе на другие [255]. Нижние
бугорки четверохолмия выступают как центры слуховых рефлек-
сов; они играют также роль релейного блока для коры, куда сиг-
налы восходят через медиальное коленчатое тело. Тонотопическая
локализация сохраняется вдоль всего слухового тракта. Дорсаль-
ные и вентральные латеральные лемнисковые ядра обеспечивают
одно переключение между кохлеарным ядром и бугорками четве-
рохолмия. При классификации нейронов нижних бугорков четверо-
холмия кошки в вертикальном направлении выяснилось, что суще-
ствует четкая последовательность оптимальных частот звуковой
стимуляции — низкие частоты (420 Гц) стимулируют нейроны дор-
сальной области, а высокие частоты (2180 Гц)—нейроны вентраль-
ной области [204]. Сходное тонотопическое распределение обнару-
жено в обоих лемнисковых ядрах: дорсальное имеет бинаураль-
ный вход, тогда как вентральное — только контралатеральный
вход.
Одиночные нейроны слуховой коры способны к различению вы-
соты тона; некоторые из них слабо реагируют на чистый тон, но
дают хороший ответ на «плавающий» тон (частотная модуляция).
Тонотопическая организация в коре выражена нечетко, однако об-
ласть низких частот лучше представлена в задней части коры, а
верхние частоты — в передней. В колонках коры нейронов с ана-
логичными максимумами восприятия частот не обнаружено [1, 75,
76]. Большинство нейронов коры реагирует на сигналы обоих
органов слуха, однако реакция на сигнал с контралатерального
уха выражена у них зачастую больше [75].
Механорецепция, фонорецепция и чувство равновесия
401
Китообразные способны определять направление звука под во-
ой Если дельфину-афалине Tursiops закрыть глаза, он будет ис-
пускать звуковые импульсы длительностью 1—1,5 мс с частотой
повторения около 16 имп/с при свободном плавании; эта частота
возрастает до 190 имп/с, когда он приближается к отражающему
звук предмету, например куску рыбы [170]. При раздражении
очень короткими звуками (0,1 мс) у дельфина были зарегистриро-
ваны вызванные ответы в нижних бугорках четверохолмия, продол-
говатом мозге, а также медиальном коленчатом теле. Максимум
чувствительности этих реакций отмечается в зоне около 60 кГц,
порог круто возрастает при частотах ниже 20 и выше 70 кГц, а
наивысшая обнаруживамая дельфином частота лежит между 120
и 140 кГц. Используя узкую полосу частот, можно получить эф-
фект маскирования, однако реакции на частотно-модулированные
импульсы остаются при этом достаточно большими. Дельфин раз-
личает повторные щелчки вплоть до частоты следования порядка
2000 имп/с. Оптимальная зона рецепции звука лежит в передней
части головы и ниже, и звук, вероятно, достигает улитки через
жировые массы и нижнюю челюсть. Когда дельфин приближается
к цели, он делает движения головой вверх-вниз и в стороны. Веро-
ятно, это нужно ему для того, чтобы знать то положение головы,
при котором эхо-сигнал имеет наибольшую громкость для каждого
уха. Испускаемые звуковые импульсы имеют максимальную гром-
кость в направлении под углом 30—40° вправо и влево от ростру-
ма и чуть-чуть выше горизонтали [19]. Способ издавания звуко-
вых сигналов китообразными изучен мало.
Записи звуковых сигналов горбатых китов показывают, что не-
которые особи излучают последовательности звуков, которые мо-
гут составить «песню», длящуюся несколько минут и повторяющу-
юся много раз. Каждый кит может быть опознан по его индиви-
дуальной «песне». «Песни» китов имеют большую интенсивность,
и в слое воды, предпочитаемом китами, их звуки могут распрост-
раняться на много миль [185]. Некоторые землеройки Blarina ис-
пользуют эхолокацию, издавая звуки длительностью от 4 до 33 мс
и частотой 30—60 кГц [77].
Слух и эхолокация у летучих мышей. Насекомоядные летучие
мыши (Microchiroptera) издают звуки при помощи гортани, пло-
доядные (Megachiroptera) — при помощи щелчков языком. Насеко-
моядные мыши питаются в основном летающими насекомыми и в
лабораторных условиях успешно избегают столкновений с мелки-
ми объектами, такими, например, как подвешенные в темной ком-
ате тонкие проволочки. В полете ночница Myotis непрерывно ис-
Ускает крики, состоящие из периодических «щелчков» длитель-
70^ЬЮ около 2,3 мс. Каждый импульс начинается на частоте
э 80 кГц и оканчивается на частоте 35—45 кГц с максимумом
ергии при 50 кГц [192] (рис. 12-24). Частота повторения им-
Ульсов в свободном полете равна приблизительно 30 имп/с. С при-
2б"1514
Rhlnolophus ferrum-equinum
'•—♦—♦—♦
---НШКгИ---0^
0,2 0,4 0,6 0,8 1,0 c 1,2
Б
Механорецепция, фонорецепция и чувство равновесия
403
ижением к объекту частота повторения возрастает до 50 или
бЛже до 150 имп/с. Myotis издает также звуки с частотой около
?акГц и слышимые человеком щелчки. Звуки производятся путем
атяжения ларингеальных мембран крикотиреоидными мышцами,
едящимися под контролем блуждающего нерва [174, 175].
у Chilonycteris локационные посылки длятся 4 мс при частоте пов-
торения 18 имп/с. При обнаружении отраженного сигнала с рас-
стояния 4—7 м длительность посылки возрастает, так что посылка
и эхо перекрываются примерно на 18—21 мс. Приближаясь к
объекту, мышь укорачивает длительность посылки до минимально
возможного значения — 0,8 мс, однако в процессе сближения с
объектом перекрытие сигнала и эха примерно на 1,2 мс остается
неизменным. Возможно, это перекрытие используется для оценки
расстояния.
Rhinolophus, летящий со скоростью 4 м/с и посылающий сигна-
лы с частотой 81,1 кГц, получает эхо-сигнал спереди с частотой
83,3 кГц (благодаря эффекту Допплера), а с направления, лежа-
щего под углом 90° к курсу, — с частотой 81,4 кГц. Записи вызван-
ных потенциалов в бугорках четверохолмия показывают резкое по-
нижение порога на частоте 83,3 кГц (рис. 12-25,Л), которое увели-
чивает отношение сигнал/шум. Зона максимальной чувствитель-
ности расположена на 30° выше горизонта. За счет попеременных
движений обоих ушей вперед-назад направленность этой реакции
возрастает на 80%. При подлете к объекту импульсы укорачивают-
ся до 10 мс, частота падает до 13—16 кГц, компенсируя, таким об-
разом, допплеровское смещение. За счет этой компенсации, а так-
же контра- и ипсилатерального контраста летучая мышь может
летать с точностью, достаточной для того, чтобы избежать натя-
нутой проволоки, имеющей всего 0,08 мм в диаметре [167, 218].
Megaderma испускает звуки с максимальной энергией в диапа-
зоне от 58,5 до 78,5 кГц и длительностью импульсов от 1,2 мс (в
свободном полете) до 0,7 мс (при приближении к объекту). При
этом в темноте она «замечает» проволоку диаметром всего 60 мкм
[155]. Vampyrum обучали различению мишени разной формы с
помощью эхолокации. Затем мишень заменялась точно такой же,
но изготовленной из другого материала. При этом сохранялась ам-
плитуда, но должны были измениться частотные свойства мишени;
однако только одна из испытуемых особей отличила новую мишень
т старой по изменению частоты сигнала [14].
Зв Рис. 12-24.
пУльс,У сТЫе РМПУльсы, испускаемые Myotis [175]. 1 — ультразвуковой ориентационный им-
ВеТ8и' веохР°СТЬ Раз8еРТки 5 мс; 2 — ориентационный импульс после перерезки мышечной
тРическая НеГ° глоточн°го нерва; заметно понижение частоты; 3 — верхняя запись — элек-
пУльса (покКТИВНость кРпкотиреоидной мышцы во время испускания ориентировочного им-
Ки летучИхКазан внизу); 4 — электрическая активность правой тиреохиодной мышцы. Б. Кри-
а мышей двух видов [218]. 1 — летящая без цели; 2— наметившая цель; 3 — ориен-
тирующаяся на пищу (мучной хрущак) в воздухе.
26*
Рис. 12-25. иЯ
А. Аудиограмма, построенная по результатам измерений реакции бугорков 4eJBJP°*0 пР»
у летучей мыши Rhinolophus [167]. Хорошо виден резкий подъем чувствител ппмо1Иь1°
83 кГц. Б. Аудиограммы двух летучих мышей Eptesicus к Myotis, измеренные с * поРоГ
метода условных рефлексов [44]. По оси абсцисс — частота в Гц, по оси орд
в дБ относительно 1 дин/см2.
Механорецепция, фонорецепция и чувство равновесия 405
Myotis, как это следует из опытов с выработкой условных реф-
лексов, слышит лучше всего при частоте 40 кГц, хотя диапазон
сЛуха простирается у нее от 10 до 120 кГц [44] (рис. 12-25,5).
Базальный завиток ее улитки имеет обычно большой размер, а
мозг строится вокруг центральной слуховой зоны. Нижние бугор-
ки четверохолмия у летучих мышей гипертрофированы, а меди-
альное коленчатое тело и кора нормальные. Записи вызванных ре-
акций и реакций одиночных нейронов нижних бугорков четверо-
холмия показывают, что максимальная чувствительность слуха
данного вида лежит в диапазоне частот излучаемых импульсов,
т. е. обычно между 42 и 80 кГц [81]. Реакции нейронов четверо-
холмия характеризуются дирекциональностью и бинауральным вза-
имодействием; их максимум соответствует контралатеральному по-
ложению источника звука в зоне 15—45° и ипсилатеральному в
зоне 60—90°. Контралатеральная реакция может тормозить ипси-
латеральную на величину, достигающую 20 дБ. Обнаружено так-
же ипсилатеральное торможение контралатеральной реакции. Myo-
tis может снижать маскирующее дйствие шума за счет ориентации
наружного уха под углом максимальной чувствительности. Ней-
роны четверохолмия обладают узкой частотной характеристикой,
по обе стороны от максимума их чувствительность сильно падает
(примерно на 35 дБ/кГц) [80, 81]. Реакции отдельных нейронов
могут повторяться с интервалом 4 мс и возникать при воздействии
звукового импульса длительностью 1 мс. В кохлеарном ядре зона
ответов нейронов шире, чем у большинства нейронов четверохол-
мия. У «широких» нейронов четверохолмия реакция на частотную
модуляцию не зависит от направления изменения частоты, тогда
как у «узких» она зависит от направления изменения частоты и
области звукового диапазона. Тон, частота которого лежит вне об-
ласти реакции нейрона, может оказывать тормозящее воздейст-
вие; некоторые нейроны тормозятся в середине их зоны реакции,
но большинство тормозится на крайних частотах. Нейроны четверо-
холмия не реагируют на чистый тон постоянной частотой повторе-
ния; для них важна не только несущая частота, но и амплитуда
сигнала. Когда время нарастания тонального сигнала увеличива-
ется, порог может возрастать; при этом может измениться диапа-
зон возбуждения и торможения нейронов [239, 241]. Торможение
в отдельных частотных диапазонах может наблюдаться и у нейро-
нов кохлеарного ядра [238] (рис. 12-26).
Билатеральное удаление дорсо-медиальной части четверохол-
мия не сказывается на способности летучих мышей к избеганию
препятствия; удаление же вентральной области оказывает влияние
?п,этУ способность, пропорциональное размеру удаленной зоны
6^41]. у кошек для локализации источника звука необходимо на-
пчие слуховой коры. У летучих мышей двустороннее удаление
пуховой коры привело к исчезновению этой функции лишь у трети
псрированных животных; половина таких животных полностью
Рис. 12-26. Аудиограммы двух нейронов бугорков четверохолмия летучей мыши
[238].
А. Нейрон, реагирующий на частотно-модулированные импульсы. Б. Нейрон, имеющий бол^е
низкий порог чувствительности к изменению частоты в сторону понижения (стимул 4 „Зме-
но-модулированный тональный пульс). Стрелки показывают направление и величину и
нения частоты. Светлыми кружками обозначены границы зоны возбуждения; зоны тор
жения заштрихованы или обозначены сеткой.
Механорецепция, фонорецепция и чувство равновесия
407
охранила возможность ориентироваться с помощью эхолокации.
Следовательно, в осуществлении эхолокации принимает участие
вентральная область четверохолмия, а слуховая кора в этой реак-
ции не участвует [241]. Нейроны слуховой коры имеют узкие час-
тотные зоны ответа; некоторые клетки могут отвечать потенциацией
или торможением также на тон, лежащий за пределами этой зоны.
Чувствительность к частотной модуляции, а не к чистому тону
встречается в нейронах коры чаще, чем в нейронах четверохолмия;
кроме того, в коре обнаружено и больше тонических нейронов. Вы-
сокочастотные нейроны чаще располагаются в передней части коры,
низкочастотные — в задней.
Активация мышцы стремечка среднего уха связана у летучих
мышей с генерацией звука. Эта мышца сокращается как раз перед
возникновением микрофонного потенциала, что ослабляет реакцию
на собственный крик и восстанавливает чувствительность к момен-
ту прихода эхо-сигнала. Мышца стремечка остается в состоянии
сокращения и при возрастании частоты следования импульсов не-
посредственно перед поимкой добычи [193]. Плодоядные летучие
мыши Старого Света и вампиры ориентируются главным образом
с помощью зрения. Одна из них (Rousettus) использует для ориен-
тирования либо зрение, либо слух. Способ ориентации зависит от
условий освещения [172]. Эта мышь излучает в течение 4 мс им-
пульс с затухающей амплитудой с частотой 10—18 кГц.
Подводя итоги, можно сказать следующее. В органах слуха и
органах боковой линии позвоночных при некотором смещении во-
лосковых клеток изменяется электрическое сопротивление мембра-
ны, что, вероятно, через посредство химических синапсов ведет к
стимуляции первичных нервных волокон, подходящих к основа-
ниям этих клеток. Различные дополнительные образования
согласуют импедансы среды (воздух, вода) и жидкости, омываю-
щей волосковые клетки. У рыб фонорецептор находится в одной из
камер лабиринта, и звук передается к нему у некоторых видов че-
рез плавательный пузырь и специальные слуховые косточки. Рыбы
различают тона, но редко выше нескольких килогерц. У наземных
позвоночных рецепторная камера удлиняется и у млекопитающих
превращается в улитку. Это позволяет локализовать восприятие
частот вдоль основной мембраны. Согласование осуществляется с
помощью одной или трех косточек среднего уха. Некоторые мле-
копитающие могут воспринимать частоты, превышающие 80 кГц.
Отдельные виды рыб издают звуки, используя в качестве резона-
т°ра плавательный пузырь. Сигналы лягушек и птиц важны для
них с точки зрения изоляции вида. Некоторые птицы, большинство
^етучих мышей и ряд китообразных для добывания пищи исполь-
•ют эхолокацию. Центральная нервная система кодирует частоту
интенсивность сигнала, при этом одиночные нейроны более высо-
ко порядка реагируют на звук в узкой полосе частот и часто
408
Глава 12
имеют в звуковом спектре как возбуждающие зоны, так и тормо-
зящие. К рецепторным клеткам поступают эфферентные импуль-
сы, которые модулируют входную активность.
Органы равновесия
Любое животное сохраняет положение равновесия в поле силы
тяжести, причем в осуществлении определенной ориентации тела
принимают участие многие органы чувств — зрительные рецепто-
ры, проприоцепторы, тактильные рецепторы и, наконец, специаль-
ные органы равновесия.
Ориентация в поле силы тяжести; геотаксис. Многие организ-
мы ориентируются по отношению к направлению силы тяжести с
помощью каких-то еще не известных механизмов. При прораста-
нии семян ростки проявляют отрицательный геотропизм, а корни—
положительный. Если проростки Avena переворачивать таким об-
разом, чтобы они находились в течение одинакового времени как
вверх, так и вниз корнями, то их рост нарушается; с помощью
соответствующих отклонений во вращении можно показать, что
проростки чувствительны к действию однонаправленных сил вели-
чиной 10~3—10"2g\ а корни 10-4—10-3g. В данном случае рецеп-
торной системой, по-видимому, является аппарат Гольджи клеток
растущих верхушек [227].
Простейшие, такие, как Paramecia, обладают отрицательным
геотаксисом. Многим беспозвоночным, роющим норы, — полихетам,
улиткам, жукам — свойстствен положительный геотаксис.
Другие наземные животные, например легочные моллюски
(улитки), крабы, некоторые жуки, крысята, обладают отрицатель-
ным геотаксисом, благодаря наличию статических рецепторов ра-
стяжения. Если этих животных поместить на наклонную плоскость,
они выбирают такое направление подъема, которое отклоняется от
горизонтали на угол 0, зависящий от угла наклона плоскости а.
Угол ориентации 0 пропорционален логарифму sin а, тогда как си-
ла, под действием которой тело скатывается с плоскости, пропор-
циональна sin а. При увеличении веса брюшка жука с помощью
грузика увеличивается и угол ориентации 0, в то время как увели-
чение веса передней части тела вызывает обратный эффект, т. е.
уменьшение угла 0. У самцов краба Uca наблюдается увеличение
угла 0, когда плоскость наклонена в ту сторону, на которой рас-
положена большая из двух его клешней. Крысята скатываются
вниз, когда мал угол ориентации 0 или велик угол наклона плос-
кости а. Увеличение угла а компенсируется у них увеличением Уг'
ла 0. Геотаксис этих животных зависит от серии позных рефлек-
сов, вызываемых сигналами от статоцистов, проприоцепторов
и рецепторов кожи.
Органы равновесия беспозвоночных. Органы равновесия быва-
ют двух типов: 1) статические, или позные, рецепторы, через ко-
Механорецепция, фонорецепция и чувство равновесия
409
ые поступает информация о воздействии силы тяжести на орга-
Т<изм, и 2) динамические рецепторы (или рецепторы ускорения),
носпринимающие линейные и угловые ускорения. Сила тяжести
БакЖе оказывает действие на динамические рецепторы, однако их
пеакция на ускорения не связана с влиянием силы тяжести. Мно-
гие виды животных имеют оба типа рецепторов равновесия, а не-
которые— либо один из них, либо другой. Деятельность всех ре-
цепторов равновесия основана на возникновении срезывающего
усилия, под действием которого возбуждаются волоски сенсорных
клеток; известно по крайней мере три типа таких волосков.
В статоцистах многих моллюсков первичные нейроны имеют
длинные киноцилии, которые располагаются в заполненной жид-
костью камере, содержащей известковый статолит. У некоторых
видов головоногих моллюсков помимо шести чувствительных к дей-
ствию силы тяжести статоцистов имеются парные кристы, или ре-
цепторы ускорений [6]. Статоцисты сцифоидных и гидроидных
кишечнополостных располагаются по краю колокола. Каждый из
них содержит статолиты из сульфата кальция; отростки первичных
сенсорных нейронов выдаются в полость статоциста. Сенсорные
нейроны в статоцистах гребневиков имеют цилиарные отростки. У
десятиногих раков статоцисты представляют собой мешочки
сферической формы, расположенные у основания антеннул. Они
содержат статолиты — инородные тела (например, песчинки), ко-
торые заменяются при каждой линьке. Волоски рецепторных кле-
ток имеют кутикулярное происхождение и характеризуются раз-
личными размерами и формой. Несколько волосков образуют хор-
ду, которая связана с отростком сенсорного нейрона; нервный
отросток имеет цилиарное строение [221].
Функции статоцистов. При удалении одного или нескольких
статоцистов у сцифоидной медузы она теряет равновесие при пла-
вании; следовательно, статоцист является у нее рецептором гра-
витации. Многие моллюски также имеют статоцисты, расположен-
ные вблизи педального ганглия. Каждый из двух статоцистов мол-
люска Aplysia представляет собой пузырек диаметром 200—
250 мкм, содержащий 13 чувствительных волосковых клеток и ста-
толит, образованный частицами углекислого кальция. При враще-
нии моллюска налево его голова рефлекторно поворачивается на-
право, и наоборот; эта реакция утрачивается после перерезки
^еРва статоциста [43]. У нескольких видов наземных легочных
оллюсков компенсаторная реакция на вращение заключается в
движении щупалец; если улитку перевернуть, частота афферентных
в 1пУЛьсов в нерве статоциста увеличится. Эфферентные импульсы
нерве, перерезанном между ганглием и одним из статоцистов,
гласуются с положением другого статоциста [263, 264].
сое °сьминога имеется пара статоцистов, каждый из которых
орИр°ИТ из тРех рецепторов гравитации (макул со статолитами),
нтированных в разных плоскостях, и рецепторов угловых ус-
410
Глава 12
корений (крист). Удаление одного такого статоциста не оказывает
выраженного действия, удаление же обоих статоцистов влечет за
собой дезориентацию при плавании, которая еще больше усилива-
ется при ослеплении животного [267]. Компенсаторная реакция на
раздражение статоцистов при вращении состоит в движении глаз
Действие шести статоцистов суммируется [18]. Компенсаторные
движения глаз зависят не столько от величины, сколько от направ-
ления срезывающего усилия, возникающего при смещении статоли-
та [18а]. При вращении животного на вращающемся столике у
него возбуждаются кристы, что приводит к возникновению ком-
пенсаторных рефлексов [40]. Регистрация нервных импульсов от
рецепторов угловых ускорений обнаруживает реакцию в системе
продольной кристы при вращении животного вокруг вертикальной
осп, а в системе вертикальной кристы — при вращении его вокруг
вертикальной оси в положении головой вверх и головой вниз.
У некоторых водяных клопов вблизи поверхности тела волос-
ками удерживается тонкий слой воздуха ( см. гл. 5). При повороте
тела пузырек воздуха, перемещаясь, возбуждает прилежащие чув-
ствительные волоски и играет, таким образом, роль своеобразного
газового статолита.
Статоцисты ракообразных (например, омара) представляют со-
бой мешочки сферической формы, расположенные у основания ан-
теннул. Они содержат статолиты (песчинки), которые обновляются
при каждой линьке (рис. 12-27). Если в момент линьки статолит
заменить кусочком железа, то магнит, подносимый сверху, заста-
вит омара перевернуться на спину. Зазубренные стенки статоциста
краба образуют два полых тороида — вертикальный и горизон-
тальный каналы. Вращение тела можно имитировать пропуска-
нием через каналы струи воды. Свободные крючкообразные волос-
ки располагаются в заднем вертикальном канале, «групповые»
волоски — в боковой стенке горизонтального канала, нитевидные
волоски — над центральной чувствительной зоной, а статолитовые
волоски образуют «пятно» в основании вертикального канала. Ни-
тевидные волоски чувствительны к вращению животного вокруг
вертикальной оси, а свободные крючкообразные волоски — к вра-
щению вокруг горизонтальной оси; статолитовые волоски являют-
ся статическими рецепторами, нейроны которых не адаптируются
к возбуждающему воздействию.
Пороговое смещение вершины волоска составляет 0,5 мкм, при
этом для каждого типа волосков имеется свое «оптимальное» со-
стояние, при котором происходит разряд рецептора. Максималь-
ное возбуждение статоцистов возникает при подъеме рострума и
рефлекторно вызывает компенсаторное движение глазных стебель-
ков. Нитевидные волоски являются рецепторами ускорений, ско'
рость разряда которых зависит от направления смещения волоска,
когда волосок находится в каком-то определенном положении,
происходит адаптация сенсорного нейрона рецептора [32]. Часто-
Механорецепция, фонорецепция и чувство равновесия
411
та сенсорных импульсов статоциста омара максимальна при пово-
роте животного на 96—120° вокруг поперечной оси [30—32]. Дей-
ствие рецепторов растяжения, расположенных у основания антен-
Рис. 12-27.
Статоцист во вскрытом базальном суставе антеннулы краба. После удаления задней
стенки горизонтальных каналов видны волоски (групповые, нитевидные и свободные крюч-
кообразные) и статолит. Б. Вид статоциста сзади [210].
НУЛ> антагонистично действию статоцистов; движение антеннул
контролируется рецепторами растяжения [223].
Лабиринт позвоночных. Вестибулярный лабиринт внутреннего
Ука позвоночных состоит из двух частей: нижняя часть включает
412
Глава 12
в себя sacculus и лагену, или улитку, а верхняя—utriculus и полу-
кружные каналы. У всех позвоночных обычно имеется три полу-
кружных канала (за исключением некоторых круглоротых, у ко-
торых нет горизонтального канала). В utriculus, sacculus и лагене
рыб волосковые клетки собраны в группы — так называемые слу-
ховые пятнышки, или макулы, причем нередко одно отделение лаби-
ринта может иметь несколько макул. Перепончатая часть лабирин-
та заполнена жидкой эндолимфой. Как utriculus, так и sacculus
имеют один или несколько статолитов. Статолиты рыб состоят из
арагонита, имеющего удельный вес 2,9, а статолиты млекопитаю-
щих — из кальцита с удельным весом 2,7 [252]. Волосковые клет-
ки полукружных каналов сосредоточены в ампулах, которые рас-
положены у перехода канала в utriculus. В каждой ампуле волос-
ковые клетки сгруппированы в кристу, над которой расположена
желеобразная купула (рис. 12-28). Волосковые клетки синаптиче-
ским путем передают сигналы разветвленным отросткам нейронов
вестибулярного ганглия.
Возбуждение волосковых клеток осуществляется под действием
тангенциального перемещения волосков, т. е. при срезывающем
усилии, направленном параллельно поверхности сенсорного эпите-
лия. Каждая рецепторная клетка имеет от 50 до 60 коротких
стереоцилий и одну длинную киноцилию, расположенную сбоку от
стереоцилий (рис. 12-29). При срезывающем усилии, направленном
от стереоцилий к киноцилии, клетка возбуждается, а при противо-
положном направлении воздействия тормозится [138]. В саккуляр-
ной макуле серебряного карася киноцилия располагается под сте-
реоцилиями в клетках вентральной половины макулы и над ними—
в кетках дорсальной половины макулы [85]. Вблизи рецепторных
клеток вестибулярного аппарата птиц и млекопитающих описаны
два типа чувствительных нервных окончаний — чашевидные и бу-
тонообразные, расположенные у основания волосковых клеток
(рис. 12-30). У рыб и лягушек имеются только сенсорные бляшки.
Наличие пузырьков в волосковых клетках вблизи контакта с нерв-
ными окончаниями указывает на возможность химической переда-
чи возбуждения. Кроме того, к волосковым клеткам, а также к аф-
ферентным нервным окончаниям подходят эфферентные волокна,
окончания которых заполнены пузырьками. Как и в других волос-
ковых клетках (нейромастах, клетках кортиевого органа), в слое
волосковых клеток макулы регистрируется постоянный потенци-
ал— деполяризующий или гиперполяризующий, в зависимости от
направления смещения волосков [252]. Одиночные волокна вести-
булярного нерва ската Raja clavata обнаруживают фон спонтанной
активности. При подъеме рострума ската частота спонтанных раз-
рядов .волокон от utriculus понижается. Частота повышается,
когда ту половину тела, от которой отводят потенциалы, припод-
нимают, и падает, когда эту половину тела опускают. Волокна,
иннервирующие лагену, проявляют максимальную активность при
А Рис. 12-28.
ИриКб'1'СТа пслУкРУЖного канала морской свинки. Вид в сканирующем микроскопе Б. То же
113 котЛЬШеМ Увеличении- Каждая рецепторная клетка имеет группу ресничек, самая длинная
торых — киноцилия. Опорные клетки имеют короткие микроворсинки. (Фотографии
предоставлены X. Эйдесом.)
a
Рис. 12-29. Схематическое изображение чувствительных волосков двух рецептор-
ных клеток лабиринта ската (поперечный разрез) [138].
В центре располагаются две киноцилии; стрелки указывают направления смещения волос-
ков, при которых происходит возбуждение (а) и торможение (б).
Рис. 12-30. Схематическое изображение четырех волосковых клеток и аффеРе
ных окончаний двух типов в макуле шиншиллы [231а].
СП — синаптические палочки располагаются в волосковой клетке в месте контакта С,ковой
нообразными окончаниями; ЧО — чашевидное окончание окружает основание воло
клетки; ЭО — эфферентное нервное окончание; ОМ — основная мембрана.
Механорецепция, фонорецепция и чувство равновесия 415
пмальном положении животного, а наклон тела в любую
«Хону вызывает понижение частоты разрядов. Разряды в
utriculus имеют как статическую, так и динамическую компонен-
та У ската все три отолита участвуют в реакции на изменение по-
ложения животного по отношению к направлению силы тяжести,
причем величина ответа пропорциональна синусу угла, под кото-
рым направлено срезывающее усилие [137, 138]. Сходные резуль-
таты получены при отведении от вестибулярных ядер продолгова-
того мозга костистых рыб (Tinea, Esox, Ameiurus) [220].
Раздражающим воздействием для любого полукружного кана-
ла служит угловое ускорение, действующее в плоскости этого ка-
нала, который работает как крутильный маятник и в котором при
ускорении движение эндолимфы запаздывает по сравнению с дви-
жением стенки канала. У рыб при отведении от эндолимфы реги-
стрируется постоянный потенциал с амплитудой 80 мВ. В эндо-
лимфе содержится 44,4 мМ калия и 15,8 мМ натрия, а в перилим-
фе-— 4,8 мМ калия. Уменьшение амплитуды постоянного потенциа-
ла пропорционально смещению купулы [252]. При ипсилатераль-
ном вращении (ампулой назад) ската Raja с угловой скоростью
3 град/с частота разрядов в нервном волокне, отходящем от ам-
пулы горизонтального канала, повышается, а при контралате-
ральном вращении (ампулой вперед) понижается. Частота разря-
дов в нервном волокне зависит от линейного или вращательного
ускорения. Вертикальные каналы реагируют на вращение относи-
тельно любой из трех осей — каждый канал в своей плоскости.
После прекращения действия раздражителя (вращения) наблюда-
ются эффекты последействия, выражающиеся в подавлении или
усилении разряда в нервных волокнах. При ритмическом покачи-
вании ската изменение частоты импульсов в его ампулярном нерве
либо запаздывает по отношению к моменту перемены направле-
ния движения на обратное, либо опережает его. Понижение часто-
ты после изменения скорости вращения соответствует коэффици-
енту демпфирования (моменту трения) и длится примерно 40 с,
т. е. в течение удвоенного времени возврата купулы [82].
Лабиринт миксины, в котором нет четко выраженного sacculus,
Не дает реакции на вибрацию; однако импульсы, проходящие по
вестибулярному нерву, говорят о наличии четкой реакции на
положение тела и на ускорение, причем реакция выше при движе-
нии головой вверх и головой вниз, чем при боковых перемещениях
(Рис. 12-31) [139].
При отведении от одиночных волокон вестибулярного нерва ля-
[УШки обнаружены реакции нескольких типов: 1) реакция на вра-
щение, которая выражается в появлении спонтанной импульсной
активности или ее подавлении при вращении в двух противополож-
на направлениях; 2) реакция только на наклон; 3) реакция как
Плп1а1Цение’ так и на наклон; 4) отсутствие реакции на ускорение
*• Смещение купулы при ускорении происходит вследствие
416
Глава 12
инерции эндолимфы, а сопротивление трения в тонких каналах
служит демпфером всей системы. В ампулярном нерве лягушки за-
регистрированы эфферентные импульсы. В волокнах, идущих от
горизонтального канала, частота разрядов повышается при ипси-
латеральном вращении, а при контралатеральном вращении прек-
ращается совсем; большая часть в этих случаях обнаруживает не-
Рис. 12-31. Импульсы в вестибулярном нерве ската Raja clavata.
Л. Частота импульсов, генерируемых в utriculus при наклоне туловища головой вниз и го-
ловой вверх. Б. То же при наклоне головы набок (Lovenstein О., Roberts Т. D. М., J. Phy-
siol., ПО, 392—415, 1950). В. Частота импульсов в нервном волокне, отходящем от полукруж-
ного канала, при вращении ската со скоростью 18,5 град/с в плоскости канала [82]. Пунк-
тирная линия обозначает момент изменения направления вращения.
линейное увеличение частоты импульсов. Порог при этом равен
0,3—2,5 град/с2 [187b]. Эфферентные импульсы частично тормозят
афферентный ответ [62, 74]. Существует калорическая проба,
применяемая в опытах на человеке: если в наружное ухо при раз-
личных положениях головы налить теплую или холодную воду, т0
тот канал, который будет в данном положении вертикальным, воз-
будится под действием конвекционных токов эндолимфы, создавае-
мых температурным градиентом [72].
Первичные сенсорные нейроны полукружных каналов обезьян
обладают спонтйнной активностью, частота которой максимальна
в тех клетках, которые обладают минимальным порогом возбуЖ'
дения при вращении. Реакция всех нейронов одного канала оди-
накова. При возбуждении горизонтального канала купула откло-
Механорецепция, фонорецепция и чувство равновесия
417
Ся по направлению к utriculus, а при возбуждении вертикаль-»
Ного канала — в противоположном направлении от него [74а].
Н Ориентация в пространстве. Осуществление нормальной ориен-
тации по отношению к направлению силы тяжести возможно бла-
годаря интеграции сенсорных сигналов, поступающих от лабирин-
тов глаз, проприоцепторов и тактильных рецепторов. У позвоноч-
ных utriculus играет ведущую роль в восприятии положения тела,
тогда как полукружные каналы более важны для ощущения враще-
ния. У человека зрительная стимуляция может компенсировать от-
сутствие стимуляции лабиринта. При рассогласовании вестибуляр-
ного и зрительного входов у человека возникает синдром «укачива-
ния», а у людей, лишенных нормального лабиринта, этого не
Происходит. Имеются данные о том, что в условиях невесомости
чувствительность к движениям головы может возрастать. Компен-
саторные движения глаз достигают максимума при повороте голо-
вы на 60° [222, 231]. Воспринимаемое отклонение головы от нор-
мального положения больше при ускорении, чем под действием си-
лы тяжести (1 g). У человека макула отклонена назад на 30°;
поэтому, если голову наклонить на 30° вперед, утрикулярная маку-
ла окажется в горизонтальном положении и не будет восприни-
мать гравитационных перегрузок. Такой компенсаторный механизм
существует у цыплят и мышей, которых длительное время содер-
жат во вращающихся камерах при высоких значениях g [265].
Удаление одного utriculus у рыб приводит к длительному отклоне-
нию глаз и плавников; эффект от повреждения sacculus или лаге-
ны значительно меньше [221]. После разрушения обоих utriculus
лягушка перестает реагировать на наклон тела.
У ракообразных о раздражении статоциста можно судить по
углу наклона глазных стебельков по отношению к оси тела и по
выпрямлению конечностей. Во время вращения крабов их глаза
совершают компенсаторные движения, а при резкой остановке вра-
щения возникают реакции последействия (нистагм). Ослепление не
ликвидирует этих реакций, удаление же статоцистов ведет к ис-
чезновению реакций последействия и нарушению равновесия.
Крючкообразные волоски, которые поддерживают статолиты, по-
видимому, служат рецепторами положения, а нитевидные волоски,
очевидно, реагируют на угловые ускорения или вращения вокруг
всех трех осей тела. Положение глазных стебельков может в ответ
На Движение полос в зрительном поле измениться (гл 15). Сигналы
От тактильных рецепторов, расположенных на карапаксе (особен-
но вблизи глаз), инициируют колебательные движения глаз; эти
Движения можно вызвать также раздражением статоциста или
Рительной стимуляцией [210].
Некоторые насекомые, например пчелы, способны транспониро-
вать угол меЖду направлением своего движения и направлением
/ Уточник света из горизонтальной плоскости в вертикальную
t TbI), причем направление на источник света заменяется в вер-
1 27—1514
418
Глава 12
тикальной плоскости направлением по отношению к линии дейст-
вия силы тяжести.
Полет насекомого можно вызвать, нарушая контакт его лапок
с субстратом, обдувая его голову или нанося ему сильное неспеци-
фическое раздражение. Полет поддерживается за счет раздраже-
ния встречным потоком воздуха сенсорных органов, расположен-
ных у основания крыльев и на голове насекомого. Входными сен-
сорными устройствами системы равновесия служат органы зрения,
рецепторные органы крыльев и видоизмененные задние крылья —
жужжальца (у Diptera). Во время полета жужжальца колеблются
с частотой 100—200 Гц синхронно с крыльями. На каждый полу-
период колебаний приходится вспышка сенсорных импульсов, иду-
щих от жужжальца, причем разряд сильнее при вращении насе-
комого вокруг вертикальной оси, чем при вращении вокруг гори-
зонтальной. Удаление одного жужжальца не очень сильно сказы-
вается на характере полета, но если насекомое вдобавок еще и
ослеплено, то ориентация значительно ухудшается. При удалении
обоих жужжалец муха оказывается совершенно неспособной сох-
ранять в полете нормальное положение тела [189].
Если лягушку, кролика или обезьяну вращать в подвешенном
состоянии в горизонтальной плоскости вокруг продольной оси, то
конечности, обращенные в сторону вращения, согнутся, а конечнос-
ти противоположной стороны распрямятся. Если четвероногое жи-
вотное наклонить набок, то конечности на наклоняемой стороне
распрямятся, а на противоположной согнутся. Если поднять голову
животного, то тонус разгибателей передних ног возрастет, а зад-
них понизится. Опускание головы понижает тонус разгибателей
передних ног. Нормальная кошка падает на все четыре лапы, но
кошка, подвергнутая двусторонней лабиринтэктомии, не способна
выполнить такую задачу. Зрительные ощущения играют очень
большую роль в восстановлении позы у собак, кошек, человека и
гораздо меньшую у кроликов и морских свинок. Если кролика с
удаленными лабиринтами положить на стол боком, он приводит
голову в нормальное положение; но если к другому боку животно-
го приложить дощечку, нормальное положение головы уже не
восстанавливается. Следовательно, для восстановления позы важ-
но асимметричное раздражение кожных рецепторов.
Общая модель пространственной ориентации выглядит следую*
щим образом. Центральная нервная система в результате воздей- ;
ствия многих сенсорных сигналов инициирует моторные реакции, !
определяющие необходимое направление перемещения. После это- s
го возбуждение статоцистов, достигающее центральной нервной
системы, сравнивается с некоторым «стандартным уровнем», и при |
наличии разности животное выполняет поворот, который длится i
до тех пор, пока сигнал от статоциста не придет в соответствие с |
этим уровнем. I
Механорецепция, фонорецепция и чувство равновесия
419
Заключение
Раздражители, которые воздействуют на механо- и фонорецеп-
•горы и рецепторы равновесия, занимают на шкале частот диапа-
зон от постоянного давления до кратковременных деформаций или
от низкочастотных колебаний до ультразвуковых. Механорецепто-
ры подразделяются в основном на тактильные рецепторы и про-
приоцепторы. Рецепторы равновесия реагируют на гравитационные
воздействия и на ускорения. Фонорецепторы и вибрационные ре-
цепторы стимулируются синусоидальными колебаниями различной
частоты.
Большинство возбудимых клеток чувствительно к деформации
поверхности. Однако неясно, почему одни клеточные мембраны при
этом деполяризуются, а другие — нет. Для механорецепторов ха-
рактерно возникновение генераторных потенциалов, амплитуда ко-
торых возрастает пропорционально величине деформации; это ука-
зывает на увеличение проницаемости мембраны для ионов (глав-
ным образом натрия) в процессе возбуждения. Как показано на
тельцах Пачини, мышечных веретенах и рецепторах растяжения
ракообразных, генераторный потенциал, достигнув порогового
уровня, приводит к возникновению нервного импульса.
Существует два типа механорецепторов: 1) рецепторы, у кото-
рых при механической деформации раздражается часть сенсорного
нейрона; 2) рецепторы, у которых внешний стимул воспринимает-
ся специализированными эпителиальными клетками, а возбужде-
ние передается затем на нервные окончания, связанные с этими
клетками. Примером первого типа механорецепторов могут слу-
жить разветвленные нервные окончания статоцистов моллюсков,
примером второго — волосковые клетки вестибулярного аппарата
позвоночных. В статоцистах десятиногих раков кутикулярные во-
лоски связаны с цилиарными отростками сенсорных нейронов.
В специализированных рецепторах равновесия в процессе эволю-
ции образовалось три типа «волосковых» структур: отростки пер-
вичных нейронов (у моллюсков и кишечнополостных), кутикуляр-
ные волоски (у десятиногих раков) и эпителиальные волосковые
клетки, передающие возбуждение на нервные окончания (у поз-
воночных). Эти структуры являют собой прекрасный пример кон-
вергентной эволюции.
Во всех механорецепторах нервное окончание или рецепторная
клетка связаны со вспомогательными структурами, часто образую-
щими сложные сопрягающие устройства. В качестве примера мож-
° привести тонкие мембраны наружного скелета в суставах ко-
сностей членистоногих, капсулы вокруг нервного окончания тель-
еа Начини, чувствительные образования, реагирующие на гради-
кап давления жидкости в некоторых суставах членистоногих и в
зыГИДН0М синУсе’ передаточное устройство в плавательном пу-
Ре рыб или среднем ухе млекопитающих, статолиты и т. д. В
27*
420
Глава 12
межсегментных рецепторах растяжения ракообразных и мышеч-
ных веретенах имеются сократимые мышечные волокна, связанные
с рецепторами параллельно или последовательно.
Вспомогательные структуры механорецепторов могут обеспечи-
вать их чувствительность к направлению воздействия, приводить
внешний стимул в соответствие с воспринимающим субстратом,
регулировать чувствительность и скорость адаптации, а также ча-
стоту ответа. Адаптация, или уменьшение ответа при длительной
стимуляции, связана как с изменениями в механических структу-
рах рецептора, так и с аккомодацией возбудимой мембраны.
В некоторых рецепторах, обладающих спонтанной активностью,
ответ выражается в увеличении или уменьшении предшествующего
уровня активности. Происхождение спонтанной активности в нер-
вах боковой линии или в волокнах слухового нерва неясно. Ответы
многих рецепторов модулируются эфферентными импульсами. Эф-
ферентные волокна могут либо оказывать тормозящее действие на
мембрану дендрита рецепторной клетки (в рецепторах растяжения
ракообразных), либо каким-то образом тормозить синаптическую
передачу (в органах боковой линии или в волосковых клетках
улитки). Реакция некоторых рецепторов растяжения связана с эф-
фективным воздействием на мышечные волокна.
Раздражение свободных или инкапсулированных нервных окон-
чаний осуществляется путем деформации их мембраны, что приво-
дит к ее деполяризации. В том месте, где возбуждение передается
с волосковой клетки на нервное окончание, обнаруживаются
синаптические пузырьки и другие признаки химических синапсов
(например, в рецепторах органов боковой линии, в кохлеарных или
вестибулярных рецепторах). Остается невыясненным значение
большой разности потенциалов, существующей между эндолимфой
и основаниями волосковых клеток в улитке млекопитающих.
Развитие рецепторов, воспринимающих звуковые колебания
воздуха, шло у членистоногих и позвоночных различными путями.
Ракообразные способны воспринимать звуковые частоты с по-
мощью статоцистов, насекомые—с помощью тимпанальных органов.
У позвоночных развитие сенсорных элементов — органа, с кото-
рым связаны поддержание равновесия и реакция на ускорения
(лабиринта), а также чувствительность к высокочастотным колеба-
ниям (улитки), — шло по пути видоизменения основной структуры
нейромаста органов боковой линии рыб. У рыб чувствительностью
к звуку обладают некоторые части лабиринта (у отдельных ви-
дов— utriculus, а у большинства — система sacculus — лагена),
у амфибий и рептилий — лагена. С преобразованием лагены в
улитку (у млекопитающих) различные участки основной мембраны
оказались способными реагировать на звуковые колебания раз-
личной частоты.
Многим животным важны определенные параметры звуковой
стимуляции. Центральные нейроны прямокрылых насекомых йе
Механорецепция, фонорецепция и чувство равновесия
421
пособны кодировать частоту звука; для них оказался существен-
ным характер звука — форма импульсов и частота модуляции,
у большинства позвоночных отдельные нейроны имеют свои опти-
мальные входные частоты, перекрывающие друг друга. У летучих
мышей, которые слышат в области высоких частот, некоторые
центральные нейроны обладают частотной модуляцией. Звук ис-
пользуется прямокрылыми, лягушками и птицами для идентифика-
ции видов; многие животные используют его для обнаружения вра-
га а хищники (летучие мыши) — для поисков жертвы (с помощью
эхолокации).
Ориентация в пространстве и сохранение нормального положе-
ния тела в покое и при движении возможны благодаря постоянно-
му рефлекторному контролю со стороны органов зрения и равно-
весия и проприоцепторов. Некоторая избыточность сенсорной ин-
формации компенсируется мощными центральными механизмами,
обеспечивающими сбалансированность всех входов.
Механорецепторы по сравнению с другими сенсорными органа-
ми обладают наибольшим разнообразием. Их механизм основан на
способности мембраны к деполяризации под влиянием деформации,
и в этом смысле механорецепция представляет собой наиболее при-
митивную из всех сенсорных функций. Специализация механоре-
цепторов шла главным образом по пути усложнения вспомогатель-
ных элементов, изменения чувствительности и частотного диапазо-
на ответа.
СПИСОК ЛИТЕРАТУРЫ
1.
1а.
2.
2а.
3.
4.
5.
6.
7.
8.
9.
9а.
10.
Abeles М., Goldstein М. FL, Neurophysiol., 33, 172—197, 1970. Functional
architecture of auditory cortex.
Adam L.-J., Z. vergl. Physiol., 63, 227—289. Bioacoustics in Locusta.
Adam L.-J., Schwartzkopff J., Z. vergl. Physiol., 54, 246—255, 1967. Central
nervous responses to sound in Locusta.
Aitkin L. M., Anderson D. J., Brugge J. F., J. Neurophysiol., 33, 421—440,
1970. Tonotopic organization of neurons in inferior colliculus and lateral lem-
niscus.
Aitkin L. M., Johnstone В. M., J. Exp. Zool., 180, 245—250, 1972. Properties of
middle ear of echidna.
Alexandrowicz J. S., Biol. Rev., 42, 288—326, 1967. Stretch receptor organ in
Crustacea.
Arthur R. N., Pfeiffer R. R„ Suga N., J. Physiol., 212, 593—610, 1971. Central
nervous responses to sound.
Barber V. C., Symp. Zool. Soc. bond., 23, 37—62, 1968. Structure of statocysts
in cephalopods.
G’’ Vergl- Physiol., 55, 407—449, 1967. Auditory responses of spi-
G. von, Nature, 169, 241—242, 1952. Mechanics of hearing.
^.,esy G- von, Rosenblith W. A. In: Handbook of Experimental Psychology,
waited by S. Stevens, John Wiley and Sons, New York, 1951, pp. 1075—1115.
^echanical properties of the ear.
errou P et al., J. Physiol., 180, 644—672, 1965. Innervation patterns of mo-
tor muscle fibers.
gmftconi R„ van der Meulen J. P., J. Neurophysiol., 26, 177—190, 1963.
ponses to vibration in mammalian muscle spindles.
422
Глава 12
11. Biederman-Thorson М., J. Physiol., 193, 695—705, 1967. Auditory responses of
neurons in midbrain of dove.
12. Blest A. D„ Collett T. S., Pye J. D., Proc. Roy. Soc. Lond. B, 158, 191—207,
1963. Auditory behavior and sound production by moths.
13. Bovbjerg R. V., Physiol. Zool., 25, 34—56, 1952. Substrate selection by
crayfish.
14. Bradbury J. W., J. Exp. Zool., 173, 23—46, 1970. Echolocation by bat Vam-
pyrum.
15. Brown A. G„ Iggo A., J. Physiol., 193, 707—733, 1967. Responses of cuta-
neous receptors to touch in cat and rabbit.
16. Brown M. C„ J. Exp. Biol., 46, 445—458, 1967. Tonic responses of stretch re-
ceptors in crayfish.
17. Brown M. C., Engberg I., Matthews P. B., J. Physiol., 192, 773—800, 1967.
Sensitivity to vibration in muscle receptors of cat.
18. Budelman B.-U., Z. vergl. Physiol., 70, 278—312, 1970. Statolith organs of
octopus.
18a. Budelman B.-U., Personal communication. Nerve impulses from octopus sta-
tocysts.
19. Bullock T. H. et al., Z. vergl. PhysioL, 59, 117—156, 1968. Electrical respon-
ses in central auditory centers of cetaceans.
20. Burkhardt D., Biol. ZentralbL, 78, 22—62, 1959. Effects of temperature on
stretch receptors of crayfish.
21. Burkhardt D., J. Insect Physiol., 4, 138—145, 1960. Responses to mechanical
stimulation in antenna of Calliphora.
22. Burkhardt D„ Gewecke M., Cold Spring Harbor Symp. Quant. BioL, 30, 601 —
614, 1965. Mechanoreception in arthropods.
23. Bush B. M„ J. Exp. BioL, 42, 285—297, 1965. Proprioreception by chordotonal
organs in leg of Carcinus.
23a. Bush M. В. H., Roberts A., J. Exp. BioL, 55, 813—832, 1971. Coxal stretch
receptors of crab.
24. Capranica R. R., Vocal Response of the Bullfrog: A Study of Communication
by Sound. MIT Press, Cambridge, Mass., 1965, 110 pp.
25. Catton W. T., J. PhysioL, 158, 333—365, 1961. Threshold recovery and fatigue
of tactile receptors in frog.
26. Catton W. T., J. PhysioL, 187, 23—33, 1966. Responses of skin receptors to
mechanical and electrical stimulation.
27. Catton W. T., PhysioL Rev., 50, 297—318, 1970. Mechanoreceptor function.
28. Chadwick L. E., In: Insect Physiology, edited by K. D. Roeder, John Wiley and
Sons, New York, 1953, pp. 577—655. Aerodynamics of flight.
29. Clarac F., Z. vergl. PhysioL, 61, 203—245, 1968. Proprioreception in legs of
Carcinus.
30. Clarac F., Z. vergl. PhysioL, 68, 1—24, 1970. Proprioreceptive function in legs
of crayfish.
31. Cohen M. J., J. PhysioL, 130, 9—34, 1955; also Physiology of Crustacea,
Vol. II, edited by T. Waterman, Academic Press, New York, 1961, pp. 65—108,
Statocysts.
32. Cohen M. J., Proc. Roy. Soc. Lond. B, 152, 30—49, 1960. Neural responses of
crustacean statocysts.
33. Cohen M. J., Comp. Biochem. PhysioL, 8, 233—243, 1963. Crustacean myo-
chordotonal organs.
34. Cohen M. J., Winn H. E., J. Exp. Zool., 165, 355—370, 1967. Sound produ-
ction and reception in fish Porichthys.
35. Crowe A., Matthews P. B., J. PhysioL, 174, 109—131, 1964. Muscle spindle
sensitivity.
36. Davis H„ PhysioL Rev., 37, 1—47, 1957; 41, 391—416, 1961. Mechanisms of
cochlear excitation.
37. Davis H. et al., Amer. J. PhysioL, 195, 251—261, 1958. Summating potential
of the cochlea.
Мёханорецепция, фонорецепция и чувство равновесия 423
Desmedt J. Е., Delwaide Р. J. Exp. Neurol., 11, 1—26, 1965. Function of the
afferent cochlear bundle of pigeon.
„ Diamond J. J-, Gray A. B., Sato M., J. Physiol., 133, 54—67, 1956. Site of
ЗУ’ initiation of impulses in pacinian corpuscle.
40 Dijkgraaf Publ- Statz. Zool. Napoli, 32, 64—86, 1961. Statocysts of
octopus as rotation receptor.
41 Dijkgraaf S., Biol. Rev., 38, 51—105, 1963. Function of lateral line detectors.
42 Dijkgraaf S. In: Lateral Line Detectors, edited by P. Cahn, Indiana Univer-
sity Press, 1967, pp. 83—95. Significance of lateral line organs.
43 Dijkgraaf S., Hessels H. G. A., Z. vergl. Physiol., 62, 38—60, 1969. Structure
and function of statocysts in Aplysia.
44. Dollard J. I., Science, 150, 1185—1186, 1965. Hearing thresholds of bats.
45^ Dorwand P. K-, J. Physiol., 211, 1—17, 1970. Responses of muscle receptors
in ducks.
46. Dorward P. K-, Comp. Biochem. Physiol., 35, 729—735, 1970. Responses of
cutaneous mechanoreceptors in ducks.
47. Dorward P. K., McIntyre A. K., J. Physiol., 219, 77—87, 1971. Vibration-sensi-
tive receptors in duck’s leg.
48. Dunning D. C., Roeder K. D., Science, 147, 173—174, 1965. Moth sounds and
insect capture by bats.
49. Echandia E. L. R., Burgos M. H„ Z. Zellforsch., 67, 600—619, 1965. Fine stru-
cture of stria vascularis, guinea pig.
50. Eckert R. O., J. Cell. Comp. Physiol., 57, 149—174, 1961. Reflex function of
abdominal stretch receptors of crayfish.
51. Eldred E. et al., J. Physiol., 122, 498—523, 1953. Supraspinal control of muscle
spindles.
52. Elsner N., Z. vergl. Physiol., 60, 308—350, 1968. Patterns of muscle activity
in sound production in crickets.
53. Enger P. S., Acta Physiol. Scand. (SuppL), 210, 1—48, 1963. Neural respon-
ses in auditory system of fishes.
54. Enger P. S„ Comp. Biochem. Physiol., 18, 859—868, 1966. Acoustic thresholds
in goldfish.
55. Enger P. S., Comp. Biochem. Physiol., 22, 517—525, 1967. Hearing in herring,
cod, and sculpin.
56. Enger P. S. In: Ciba Foundations Symposium on Hearing Mechanisms in
Vertebrates, edited by A. De Reuck and J. Knight, Williams and Wilkins, Bal-
timore, 1968, pp. 4—11. Hearing in fish.
67. Enger P. S., Aidley D. J., Szabo T., J. Exp. Biol., 51, 339—345, 1969. Sound
reception in cicada.
57a . Engstrom H., Ades H. W., Bredberg G. In: Ciba Foundation Symposium on
Hearing Loss, edited by G. Wolstenholme and J. Knight, Churchill, London,
1970, pp. 127—156. Ultra-structure of cochlea.
58. Enright J. T., Science, 133, 758—760, 1961. Pressure sensitivity of an amphi-
pod.
59. Esch H., 2. vergl. Physiol., 56, 199—220, 1967. Importance of sound for com-
munication in bees.
0. Esch H., Esch I., Kerr W. E., Science, 147, 320—321, 1965. Sound in commu-
nication of bees.
L Esch H., Wilson D., Z. vergl. Physiol., 54, 256—267, 1967. Sounds produced
by flies and bees.
Eyck M. van., Arch. Int. Physiol., 57, 102—105, 231—236, 1949. Electrical
6ч ^?sPonses from semicircular canals in pigeon.
• Eyck M. van, Arch. Int. Physiol., 59, 236—238, 1951. Microphonic poten-
64 tla s’
' ia^gUirr.e C:r Kuffler S. W., J. Gen. Physiol., 39, 87—119, 121—153, 155—184,
*955. Excitation, inhibition, and impulse initiation in stretch receptor of lobster
65 lnd crayfish,
tsex J., j. Acoustical Soc. Amer., 41, 666—675, 1967. Efferent inhibition in
cochlea.
424
Глава 12
66. Fex J. In: Ciba Foundation Symposium on Hearing Mechanisms in Vertebra-
tes, edited by A. De Reuck and J. Knight, Williams and Wilkins, Baltimore,
1968, pp. 169—180. Efferent inhibition in cochlea.
67. Fields H. L., J. Exp. Biol., 44, 455—468, 1966. Proprioceptive control of
posture in crayfish abdomen.
68. Flock A. In: Lateral Line Detectors, edited by P. Cahn, Indiana University
Press, 1967, Ultrastructure of lateral line organ.
69. Frishkopf L. S., Capranica R. R., J. Acoustical Soc. Amer., 35, 1219—1228,
1963; 40, 1262—1263, 1966. Auditory responses in brain of bullfrog.
70. Furukawa T., Ishii У., J. Neurophysiol., 30, 1377—1403, 1967. Auditory respon-
ses in goldfish.
71. Gelperin A., Science, 157, 208—210, 1967. A stretch receptor in foregut of
blowfly.
72. Gernandt B., J. Neurophysiol., 12, 173—184, 1949. Vestibular responses to ro-
tation and thermal stimulation.
73. Gillies J. D., Burke D. J., Lance J. W., J. Neurophysiol., 34, 252—262, 197L
Tonic vibration reflex in cat.
74. Gleisner L., Henriksson N. L, Acta Otolaryngol. (Suppl.), 192, 90—103, 1963.
Effect of efferent impulses on responses in vestibular nerve of frog.
74a. Goldberg J. M., Fernandez C., J. Neurophysiol., 34, 635—684, 1971. Impulses
in neurons of semicircular canals of monkey.
75. Goldstein M. H. In: Physiological and Biochemical Aspects of Nervous In-
tegration, edited by F. D. Carlson. Prentice-Hall, Englewood Cliffs, N. J.,
1968, pp. 131—151. Cortical responses to acoustic stimuli.
76. Goldstein M. H. In: Medical Physiology, edited by V. B. Mountcastle,
С. V. Mosby, St. Louis, 1968, pp. 1465—1498. Auditory periphery.
77. Gould E., Negus N. C., Novich A., J. Exp. Zool., 156, 19—38, 1964. Echolo-
cation in shrews.
78. Granit R., Receptors and Sensory Perception, Yale University Press, New Ha-
ven, Conn., 1955, 369 pp.
79. Griffin D. R., Sathers R. A., Biol. Bull., 139, 495—501, 1970. Echolocation in
cave swiftless.
80. Grinnell A. D., Z. vergl. Physiol., 68, 117—153, 1970. Central auditory respon-
se in bats.
81. Grinnell A. D., J. Physiol., 167, 38—127, 1963. Neurophysiology of audition
in bats.
82. Groen J. J. et al., J. Physiol., 117, 329—346, 1952. Response of horizontal se-
micircular canal in elasmobranch labyrinth.
83. Hahn J. F., J. Physiol., 213, 215—226, 1971. Sensory responses from cat
vibrissae.
84. Hama N, J. Cell Biol., 24, 193—210, 1965. Ultrastructure of lateral line organ
of eel.
85. Hama K„ Zellforsch. Mikr. Anat. 94, 155—171, 1969. Fine structure of saccu-
lar macula of goldfish.
86. Harris G. G. In: Symposium on Marine Hio-Acoustics. American Museum of
Natural History, edited by W. N. Tavolga, Pergamon Press, New York, 1964,
pp. 283—247. Considerations in the physics of sound production by fishes.
87. Harris G. G., Flak A. In: Lateral Line Detectors, edited by P. Cahn, Indiana
University Press, 1967, pp. 135—161. Electrical activity of lateral line in
Xenopus.
88. Harris G. G., Frishkopf L. S., Flak A., Science, 167, 76—79, 1970. Receptor
potentials, lateral line.
89. Hartline P. H., Campbell H. W., Science, 163, 1221—1223, 1969. Auditory and
vibration responses in midbrain of snakes.
90. Hartline P. H., J. Exp. Biol., 54, 349—371, 1971. Sound detection by sna-
kes.
91. Hartman H. B., Boettiger E. G., Comp. Biochem. Physiol., 22, 651—663, 1967.
Proprioceptive organs in crab Cancer.
Механорецепция, фонорецепция и чувство равновесия________________425
—-—-*•"
Harvey R- Matthews Р. В., J. Physiol., 157, 370—381, 1961. Muscle spindles
iгл mammals.
Hashimoto F„ Katsuki N„ Vanagisawa K-, Comp. Biochem. Physiol., 33, 405—
421 1970. Efferent control of lateral line of fish.
94 Hayes W. J., Barber S. B., J. Exp. Zool., 165, 195—210, 1967. Proprioceptors in
Limulus leg.
95 Heran H„ Z. vergl. Physiol., 42, 103—163, 1959. Reflex control of flight in ho-
neybees. , ,
9g Hoffman C„ Z. vergl. Physiol., 54, 290—352, 1967. Structure and function of
mechanoreceptors of scorpion.
97 Horch K. W„ Z. vergl. Physiol., 73, 1—21, 1971. Hearing and vibration sense
in Ocypode.
98 Horridge G. A., Proc. Roy. Soc. Lond. B, 157, 199—222, 1963. Proprioceptors
and tactile receptors in polychaete Harmothoe.
99. Houk J., Henneman E., J. Neurophysiol., 30, 466—481, 1967. Responses of
Golgi tendon organs.
100. Huertas J., Carpenter R. S. In: Fourth Symposium on the Role of Vestibular
Organs in Space Exploration (NASA SP — 187), 1970, pp. 137-—144. Single
unit responses in vestibular nerve of frog.
101. Hunt С. C., Kuffler S. W., J. Physiol., 113, 298—315, 1951. Muscle stretch
receptors and their reflex function.
102. Hunt С. C., J. Physiol., 155, 175—186, 1961. Vibration receptors in cat leg.
103. Husmark I., Ottoson D., J. Physiol., 212, 577—592, 1971. Adaptation of muscle
spindles.
104. Ishii V., Matsaura S., Furukawa T., Japan. J. Physiol., 21, 79—98, 1971. Post-
synaptic responses in auditory nerve fibers of goldfish.
105. Iwasaki S., Florey E., J. Gen. Physiol., 53, 666—682, 1969. Inhibitory poten-
tials in stretch receptors of crayfish.
106. Janig 117., Schmidt R. F., Zimmerman M., Exp. Brain Res., 6, 100—115, 1968.
Single unit responses and tactile efferent outflow, tactile endings of cat.
107. Johnstone В. M. In: Membranes and Ion Transport, Vol. 1, edited by E. E. Bit-
tar, John Wiley and Sons, New York, 1970, pp. 167—184. Ion fluxes in the
cochlea.
108. Johnstone В. M., J. Acoustical Soc. Amer., 47, 504—509, 1970. Movements in
middle ear of guinea pig and bat.
109. Johnstone В. M. et al., Nature, 227, 625—626, 1970. Mechanical responses in
cricket ear.
110. Johnstone J. R„ Johnstone В. M., Exp. Neurol., 24, 528—537, 1969. Responses
in primary auditory fibers of skink.
111. Josephson R. K-, J. Exp. Biol., 38, 17—27, 1961. Response of hydroid to water
disturbance.
H2. ' Julian F. J„ Goldman D. E„ J. Gen. Physiol., 46, 297—313, 1962. Response of
no nerves to mechanical stimulation.
13- Kalmring K., Z. vergl. Physiol., 72, 95—110, 1971. Acoustic responses of ner-
vous system, Locusta.
4. Katsuki У., Japan. J. Physiol., 1, 87—99, 264—268, 1951. Electrical responses
.. _ *rom lateral line organ of fish.
• Katsuki Y„ Prog. Brain Res., 21, 71—97, 1966. Neural mechanisms appearing
Hr l5 ca^s and monkeys.
Katsuki У., Suga N., J. Exp. Biol., 37, 279—290, 1960. Neural mechanisms of
117 v?ar^nS in insects.
• Katsuki Y., Watanabe T., Maruyama N., J. Neurophysiol., 22, 343—359, 1959.
118 v/nSie unit response from auditory tract of cat.
°- Renton B., Kruger L„ Woo M„ J. Physiol., 212, 21—44, 1971. Adaptation in
119 ^echanoreceptors of reptiles.
Ktnsler L. E., Frey A. R„ Fundamentals of Acoustics, 2nd ed., John Wiley and
120 fcvns’ ^ew York, 1962, 524 pp.
• Rieerekoper H„ Chagnon E. C., J. Fish Res. Bd. Canad., 11, 130—152, 1954.
Hearing in fish.
426 Глава 12__________________________________________________________________________
121. Kleerekoper Н., Malar Т. In: Ciba Foundation Symposium on Hearing Mecha-
nisms in Vertebrates, edited by A. De Reuck and J. Knight, Williams and Wil-
kins, Baltimore, 1968,-pp. 188—206. Orientation through sound fishes.
122. Kleerekoper H., Roggenkamp P. A., Canad. J. Zool., 37, 1—8, 1959. Effect of
swim-bladder on hearing in Ameiurus.
123. Kleerekoper H., Sibabin K., Z. vergl. Physiol., 41, 490—499, 1959. Hearing in
frogs.
124. Konishi M., Science, 166, 1178—1181, 1969. Single unit analysis of hearing in
songbirds.
125. Konishi M., Z. vergl. Physiol., 66, 257—272, 1970. Hearing and vocalization
in songbirds.
126. Konishi M., Nottebohm F. In: Bird Vocalization, edited by R. A. Hinde,
Cambridge University Press, 1969, pp. 29—48. Development of bird songs.
127. Konishi M., Slepian J. Z„ J. Acoustical Soc. Amer., 49, 1762—1769, 1971.
Efferent control of cochlear potentials.
128. Kuijpers W„ Houben N. M., Bonting S. L., Comp. Biochem. Physiol., 36, 669—
676, 1970. ATPase activity in cochlea of chicken.
129. Kuiper J. W. In: Lateral Line Detectors, edited by P. Cahn, Indiana LIniversity
Press, 1967, Frequency characteristics of lateral line organs.
130. Kutsch W., Z. vergl. Physiol., 63, 335—378, 1969. Species-specific songs of
crickets.
131. Landgren S„ Acta Physiol. Scand., 26, 1—56, 1952. Excitation of carotid pres-
sure receptors.
132. Linsentnair К. E., Z. vergl. Physiol., 64, 154—211, 1969; 70, 247—277, 1970.
Detection of air currents by beetles.
133. Littlejohn M. J., Oldham R. S., Science, 162, 1003—1005, 1968. Mating calls
in Rana pipiens species complex.
134. Loewenstein W. R„ J. Gen. Physiol., 41, 847—856, 1958. Facilitation and im-
pulse origin in pacinian corpuscles.
135. Loewenstein W. R., Ann. N. Y. Acad. Sci., 81, 367—387, 1959. Receptor po-
tentials and spike initiation in pacinian corpuscles.
136. Loewenstein W. R., Mendelson M., J. Physiol., 177, 377—397, 1965. Receptor
adaptation in pacinian corpuscles.
137. Lowenstein O., Proc. Roy. Soc. Med., 45, 133—134, 1952; also Lowenstein 0.,
Sand A., Proc. Roy. Soc. Lond. B, 129, 256—275, 1940. Equilibrium function
of otolith organs and semicircular canals.
138. Lowenstein O., Osborne M. D., Wer sail J., Proc. Roy. Soc. Lond. B, 160, 1—12,
1964. Innervation of sensory epithelium of labyrinth in ray.
139. Lowenstein O., Thornhill R. A., Proc. Roy. Soc. Lond. B, 174, 419—434, 1970;
176, 21—42, 1970. Responses of labyrinth of hagfish.
140. Manley G. A., Z. vergl. Physiol., 69, 363—383, 1970; also Nature, 230, 506—
509, 1971. Evolution of hearing in vertebrates.
141. Manley J. A., Z. vergl. Physiol., 71, 255—261, 1971. Responses in auditory
units in brain of caiman.
142. Markl FL, Z. vergl. Physiol., 45, 475—569, 1962. Proprioception and tactile
reception in hymenopterans.
143. Markl H., Z. vergl. Physiol., 69, 6—37, 1970. Stridulation, leaf-cutting ants.
144. Markl H., Wiese K., Z. vergl. Physiol., 62, 413—420, 1969. Wave detection by
water beetles.
145. Martin A. R„ Wickelgren W. O., J. Physiol., 212, 65—83, 1971. Sensory fibers
in spinal cord of sea lamprey.
146. Masterton B., Heffner H„ Ravizzo R„ J. Acoustical Soc. Amer., 45, 966—985,
1969. Evolution of mammalian hearing.
147. Matthews В. H. C., J. Physiol., 71, 64—110, 1931; 72, 153—174, 1931; 78,
1—53, 1933. Receptors in mammalian and frog muscle.
148. Matthews P. B., Quart. J. Exp. Physiol., 47, 324—333, 1962; also J. Physiol.,
168, 660 677, 1963. Response of muscle spindle receptors to stretching
different velocities.
Механорецепция, фонорецепция и чувство равновесия_________________427
McCormick J. G., Wever Е. G., Palin J., Ridgway S. H., J. Acoustical Soc.
I49- Amer 48, 1418—1428, 1970. Sound conduction in dolphin ear.
-Ci Meadows’ P. S., J. Exp. Biol., 47, 553—559, 1967. Discrimination of substrate
bv amphipod Corophium.
«-I Mellon D., Jr., Kennedy D., J. Gen. Physiol., 47, 487—499, 1964. Impulse on-
° Pin and propagation in.crayfish tactile receptors.
i-2 Mendelson M., J. Exp. Biol., 45, 411—420, 1966. Origin of impulses in stretch
° receptors of Callinectes.
iw Michelsen A., Z. vergl. Physiol., 71, 49—101, 1971. Physiology of locust ear.
154 Miller L. E., MacLeod E. E., Science, 134, 891—893, 1966. Ultrasonic sensiti-
vity in lacewing Chrysopa.
155 Mohres F. P., Neuweiler G., Z. vergl. Physiol., 53, 195—227, 1966. Detection
of ultrasound by bats.
156. Money К. E. et al., Amer. J. Physiol., 220, 140—147, 1971. Physical properties
of fluids and membranes in vestibular apparatus, pigeon.
157. Money К. E., Sokoloff M., Weaver R. S„ NASA SP-115. pp. 91—97, 1966. Phy-
sical properties of endolymph and perilymph.
158. Morgan E„ J. Exp. Biol., 50, 501—513, 1969; 51, 171—179, 1969. Responses of
polychaete to hydrostatic pressure.
159. Mountcastle V. B. In: The Neurosciences, edited by F. O. Schmitt, Rockefeller
Press, New York, 1967, pp. 393—408. Neural coding of sensory events.
160. Murphey R. K-, Z. vergl. Physiol., 72, 150—167, 1971. Vibration sense in wa-
ter-striders.
161. Myrberg A. A., Banner A., Richard J. D., Marine Biol., 2, 264—276, 1969.
Acoustic stimulation of sharks.
162. Nakajima S., Science, 146, 1168—1170, 1970. Adaptation of stretch receptors
in crayfish.
163. Nakajima S., Onodera K., J. Physiol., 200, 161—185, 1969. Membrane pro-
perties of crayfish stretch receptors.
164. Nakajima S., Takahashi K., J. Physiol., 187, 105—127, 1966. Electrogenic so-
dium pump in stretch receptors of crayfish.
165. Necker R., Z. vergl. Physiol., 69, 367—425, 1970. Cochlear potentials in birds.
166. Neff W. W. I n: Ciba Foundation Symposium on Hearing Mechanisms in Ver-
tebrates, edited by A. De Reuck and J. Knight, Williams and Wilkins, Balti-
more, 1968, pp. 207—233. Localization and lateralization of sound in space.
167. Neuweiler G., Z. vergl. Physiol., 67, 273—306, 1970. Electrical responses in
auditory system of bat.
168. Nicklaus R., Z. vergl. Physiol., 50, 331—362, 1965. Generator potentials and
spikes from cereal nerves of cockroach.
169. Norris K. S. In: Evolution and Environment, edited by E. T. Drake, Yale Uni-
versity Press, New Haven, Conn., 1968, pp. 297—324. Evolution of acoustic
mechanisms in cetaceans.
1*70 . Norris K. S. et al., Biol. Bull., 120, 163—176, 1961. Echolocation in por-
poises.
i7o* Nottebohm F., Science, 167, 950—956, 1970. Ontogeny of bird songs.
Z°°L 137, 443—461, 1958. Orientation in paleotropical
173. Novick A., Biol. Bull., 117, 497—503, 1959. Acoustic orientation in cave
swiftlets.
Novick A., Biol. Bull., 128, 297—314, 1965; also Novick A., Vaisnys J. R.,
17K л? • 127, 478—488, 1964. Echolocation of insects by bat Chilonycteris.
ovlck A., Griffin D. R., J. Exp. oZol., 148, 125—141, 1961. Sound production
176 Ж bats’
G. C., Experientia, 26, 1276—1278, 1971. Acoustic stimulation of lob
j ster.
• Olivo R. F., Comp. Biochem. Physiol., 35, 761—786, 1970. Mechanoreceptors
178 Жfazor clam- foot withdrawal.
i”°son-D\’ J- Physiol., 171, 109—118, 1964; 178, 68—79, 1965. Effects of
ions on isolated muscle spindles.
428
Глава 12
179 Ottoson D„ McReynolds J. S., Shepherd G. M., J. Neurophysiol., 32, 24—34,
1969. Sensitivity of isolated muscle spindle during and after stretch.
180. Pabst H., Z. vergl. Physiol., 50, 498—591, 1965. Electrophysiology of stretch
receptors in Locusta.
181. Pabst FL, Kennedy D., Z. vergl. Physiol., 57, 190—208, 1967. Cutaneous
mechanoreceptors influencing motor output in crayfish.
182. Pabst H., Schwartzkopff J., Z. vergl. Physiol., 45, 396—404, 1962. Phasic and
tonic responses of joint receptors in locusts.
183. Page C„ J. Neurophysiol., 33, 116—128, 1970; also Page C. and Sutterlin A.M.,
J. Neurophysiol., 33, 129—136, 1970. Unit responses incentral auditory
system of goldfish.
184. Parry D. A., J. Exp. Biol., 43, 185—192, 1965. Vibration signals in spider
web.
185. Payne R. S., Science, 173, 585—597, 1971. Humpback whales’songs.
186. Payne R. S., J. Exp. Biol., 54, 535—574, 1971. Directional hearing, barn
owls.
187. Payne R. S„ Roeder K. D., Wallman J., J. Exp. Biol., 44, 17—31, 1966. Di-
rectional sensitivity, ears of noctuid moths.
187a. Piddington R. W., J. Exp. Biol., 55, 569—610, 1971. Central control of audi-
tory responses, goldfish.
187b. Precht W., Llinas R„ Clarke M., Exp. Brain Res., 13, 387—407, 1971. Respon-
ses of frog vestibular nerve to rotation.
188. Pringle J. W. S., J. Exp. Biol., 15, 101—131, 1938. Proprioception in in-
sects.
189. Pringle J. W. C., Phil. Trans. Roy. Soc. Soc. Lond. B, 233, 347—384, 1948; also
Insect Flight, Cambridge University Press, 1957, 133 pp. Gyroscopic mecha-
nism of halteres of Diptera.
190. Proske U., J. Physiol., 205, 289—304, 1969. Responses of muscle spindles in
lizard.
191. Proske U., Exp. Neurol., 23, 187—194, 1969. Vibration receptors in skin of
snake.
192. Pye J. D., Ergebn. Biol., 26, 12—20, 1963. Echolocation in bats.
193. Pye J. D. In: Ciba Foundation Symposium on Hearing Mechanisms in Ver-
tebrates, edited by A. De Reuck and J. Knight, Williams and Wilkins, Baltimo-
re, 1968, pp. 66—84. Hearing in bats.
194. Outob Z., Arch. Neerl. Zool., 15, 1—67, 1962. Swim-bladders of fish as pres-
sure receptors.
195. Rathmayer W., Z. vergl. Physiol., 54, 438—454, 1967. Electrophysiology of
proprioceptors in legs of spiders.
196. Reid К. H., Science, 172, 949—951, 1971. Sound production in cicadas.
197. Ripley S. FL, Bush В. M., Roberts A., Nature, 218, 1170—1171, 1968. Muscle
receptor without nerve impulses.
198. Risler H„ Schmidt K., Z. Naturforsch., 22b, 759—762, 1967. Structure 6f
mechanoreceptors in antenna of Aedes.
199. Roeder' K. D., Science, 154, 1515—1521, 1966; also J. Insect Physiol., 12,
1227—1244, 1966. Auditory system of noctuid moths.
200. Roeder K. D., Nerve Cells and Insect Behavior, Harvard University Press,
Cambridge, Mass., 1967, 238 pp.
201. Roeder K. D., J. Insect. Physiol., 13, 873—888, 1967. Orientation of moths in
an ultrasound field.
202. Roeder K. D., Treat A. E„ J. Insect. Physiol., 16, 1069—1086, 1970. Acoustic
sense in hawkmoths.
203. Roeder K. D., Treat A. E., Vandeberg J. S., Science, 159, 331—333, 1968. Audi-
tory sense in sphingid moths.
204. Rose J. F. et al., J. Neurophysiol., 26, 294—341, 1963; 29, 288—314, 1966;
30, 769—793, 1967. Frequency response and synchrony in auditory fibers.
205. Rovner J. S., Animal Behavior, 15, 273—281, 1967. Acoustic communication in
lycosid spider.
Механорецепция, фонорецепция и чувство равновесия
429
206.
207.
208.
209.
210.
211-
212.
213.
214.
215.
216.
217.
218.
219.
Rudolph Р., Z. vergl. Physiol., 56, 341—375, 1967. Vibration sense in aquatic
/fasseW I. J., J. Exp. BioL. 54, 621—658, 1971. Function of lateral line system
in Xenopus.
Salmon M., Animal Behavior, 15, 449—459, 1967. Visual display and sound
production in sex behavior, fiddler crab Uca.
Salmon M., Horch K. W. In: Behavior of Marine Animals. I. Invertebrates,
edited by H. E. Wynn and B. Olla, Plenum Press, New York, 1972, pp. 60—96.
Acoustic signalling in land crabs.
Sandeman D. C., Okajima A., J. Exp. Biol., 57, 187—204, 1972. Structure and
function, crustacean statocyst.
Salto M., J. PhysioL, 159, 391—409, 1961. Response of pacinian corpuscles to
vibration.
Schlegel P„ Z. vergl. PhysioL, 66, 45—77, 1970. Spikes and receptor poten-
tials from joint receptors in Calliphora.
Schlieper C., Marine BioL, 2, 5—12, 1968. Effects of hydrostatic pressure on
marine animals.
Schmidt R. S., Comp. Biochem. PhysioL, 10, 83—87, 1963. Types of endo-
lymphatic potentials.
Schneider G., Z. vergl. PhysioL, 35, 416—458, 1953. Halteres and equilibrium
in blowflies.
Schneider H., Z. vergl. PhysioL, 47, 493—558, 1964. Bioacoustics in tiger fish
Therapon.
Schneider H., Z. vergl. PhysioL, 61, 369—385, 1968. Bioacoustic studies on
tree frogs.
Schnitzler H., Z. vergl. PhysioL, 57, 376—408, 1968. Ultrasonic detection by
horseshoe bats.
Schnorbus H., Z. vergl. PhysioL, 71, 14—48, 1971. Detectors of pressure
and vibrations.
220. Schoen L., Z. vergl. PhysioL, 39, 399—417, 1957. Reflexes and electrical
responses of vestibular system, fish.
221. Schdne H., Ergebn. BioL, 21, 163—209, 1959. Role of statolith organs and
eyes in space orientation.
222. Schdne H., Z. vergl. PhysioL, 46, 57—87, 1962. Responses to gravity end ro-
tation in man.
223. Schdne H. In: Gravity and the Organism, edited by S. A. Gordon and M. J. Co-
hen, University of Chicago Press, Chicago, 1971, pp. 223—235. Equilibrium
reception in crustaceans.
224. Schwartz E., Z. vergl. PhysioL, 50, 55—87, 1965. Structure and function of
lateral line.
225. Schwartz E., Hasler A. D., Z. vergl. PhysioL, 53, 317—327, 1966. Superficial
lateral line of mud minnow Umbra.
226. Schwartzkopff J., Experientia, 5, 159—161, 1949; also Z. vergl. PhysioL, 31,
527—608, 1949; 34, 46—68, 1952; also Proc. XIII Int. Ornith. Cong., 1059—
1068, 1963. Hearing in various birds; cochlear potentials; conditioned respon-
ses.
227. Shen-Miller J., Hinchman R., Gordon S. A., Plant PhysioL, 43, 338—344, 1968;
a iso Shen-Miller J., Planta, 92, 152—163, 1970. Geotropic responses in Avena
seedlings.
228. Siler W„ J. Accoustical Soc. Amer., 46, 483—484, 1969. Near and far fields in
3 mar*ne environment.
9- Siminoff R_, Exp. Neurol., 21, 290—306, 1968. Slowly adapting mechanore-
230 pt?rs In alligator skin.
' R-’ Kruger L., Exp. Neurol., 20, 403—414, 1968. Cutaneous and
231 Stre^c" receptors of reptiles.
In: Fourth Symposium on the Role of Vestibular Organs in Space
sicknp«V10n Sp-187), 1970, pp. 7—28. Experimental studies of motion
430
Глава 12
231а. Smith С. A., Rasmussen G. L. In: Third Symposium on the Role of Vestibular
Organ in Space Exploration (NASA SP-152), 1968, p. 193. Labyrinth hair
ceils
232. Smola V., Z. vergl. Physiol., 70, 335—348, 1970. Electrical responses from
hair sensilla of locusts.
233. Sokolove P. G., Cooke I. M., J. Gen. Physiol., 57, 125—163, 1971. Inhibition
in a sensory neuron by an electrogenic pump.
234. Spath M., Z. vergl. Physiol., 56, 431—462, 1967. Effect of temperature on
mechanoreceptors in bony fish Leuciscus.
235. Stewart P. A., Ohio J. Sci., 55, 122—125, 1965. Audiogram of pheasant.
236. Stockwell C. W, Ades H. W„ Engstrom H., Ann. Otol. Rhin. Laryngol., 78,
1144—1169, 1969. Damage of hair cells in cochlea due to intense sound.
237. Strother W. F., J Comp. Physiol. Psylchol., 52, 157—162, 1959. Electrical
responses from auditory system of frog.
238. Suga N., J. Physiol., 179, 26—53, 1965. Echolocation by bats.
239. Suga N., J. Physiol., 172, 449—474, 1964; 181, 671—700, 1966. Single unit
activity in cochela nucleus, inferior colliculus and auditory cortex of echo-
locating bats.
240. Suga N., J. Insect. Physiol., 12, 1039—1050, 1966. Ultrasonic production and
reception in neotropical Tettigoniidae.
241. Suga N., J. Jhysiol., 203, 707—739, 1969; 217, 159—177, 1971. Evoked poten-
tials in response to sound in bats.
242. Suga N., Campbell H. W., Science, 157, 88—90, 1967. Auditory responses in
single neurons in gecko.
243. Suga N„ Katsukl У., J. Exp. Biol., 38, 545—558, 1961. Central mechanisms of
hearing in insects.
244. Suga N., Nakashima T., Snow J. B., Life Sci., 9, 163—168, 1970. Na and К in
cochlear endolymph of guinea pig.
244a. Свидерский В. Л., ДАН СССР, 172, 1230—1233. Электрическая активность
рецепторов, связанных с поддержанием памяти у саранчи.
245. Tapper D. N., Exp. Neurol., 26, 447—459t 1970. Behavioral properties of tactile
pad receptors in cat.
246. Tasaki I., Fernandez C., J. Neurophysiol., 15, 497—512, 1952. Cochlear micro-
phonics, impulses in cochlear nucleus, guinea pig.
247. Tavolga W. N., Wodinsky J„ Bull. Amer. Mus. Nat. Hist., 126, 179—239, 1963.
Auditory capacity in fishes.
248. Taylor R. C., Comp. Biochem. Physiol., 20, 709—717, 1967. Anatomy and sti-
mulation of chordotonal organs in antenna of hermit crab.
249. Taylor R. C., Comp. Biochem. Physiol., 27, 795—805, 1968. Water vibration
reception in unrestrained crayfish.
250. Thur tn U., Science, 145, 1063—1065, 1964; also 2. vergl. Physiol., 48, 131—
156, 1964. Mechanoreceptors in cuticle of honeybee.
251. Thurm U., Cold Spring Harbor Symp. Quant. BioL, 30, 75—94, 1965. Theory of
mechanoreception.
252. Trincker D., Symp. Soc. Exp. Biol., 16, 289—317, 1962. Transduction of
mechanical stimulus to nerve impulses by labyrinth receptors.
253. Usherwood P. N., Runion H. I., Campbell J. L, J. Exp. Biol., 48, 305—323,
1968. Structure and physiology of chordotonal organ in locust leg.
254. Walcott C., Amer. Zool., 9, 133—144, 1969; also J. Exp. Biol., 40, 595—611,
1963. Vibration reception by spiders.
255. Watanabe T., Yanagisawa K.., Kanzaki J., Katsuki Y., Exp. Brain Res., 2, 302—
317, 1966. Cortical influence on unit responses of medial geniculate to sound.
256. Werner G., Mountcastle V. B., J. Neurophysiol., 28, 359—397, 1965. Activity
in cutaneous afferent nerves.
257. Wever E. G., Physiol. Rev., 46, 102—127, 1966. Electrical potentials of the
cochlea.
258. Wever E. G., J. Exp. Zool., 175, 327—342, 1970; also Proc. Nat. Acad. Sci.,
68, 1498—1500, 1971. Function of middle ear, reptiles.
s
9
Механорецепция, фонорецепция и чувство равновесия
431
оед Wever Е. G., Vernon J. A., Proc. Nat. Acad. Sci., 42, 213—220, 1956. Audio-
1 ' grams of turtles.
260 Whitear M., Phil. Trans. Roy. Soc. Lond. B, 245, 291—325, 1962. Structure of
2 chordotonal organs in leg of Carcinus.
261 Whifield L. C. In: Ciba Foundation Symposium on Hearing Mechanisms in
Vertebrates, edited by A. De Reuck and J. Knight, Williams and Wilkins, Bal-
timore, 1968, pp. 246—254. Centrifugal control of auditory pathway.
262. Wiederhold M. L., J.. Acoustical Soc. Amer., 48, 966—977, 1970; also Wieder-
hold M. L., Kiang N. Y. S„ J. Acoustical Soc. Amer., 48, 950—965, 1970. Fun-
ction of efferent fibers in mammalian auditory system.
263. Wolff H. G., Z. vergl. Physiol., 69, 326—366, 1970. Statocyst function, pul-
monates.
264. Wolff H. G., Z. vergl. Physiol., 70, 401—409, 1970. Efferent control in stato-
cyst nerves.
265. Wunder С. C. In: Gravity and the Organism, edited by S. A. Gordon and
M. J. Cohen, University of Chicago Press, Chicago, 1971, pp. 389—410. Ef-
fects of chronic acceleration of animals.
266. Yanagisawa K-, Hashimoto T., Katsuki Y., J. Insect. Physiol., 13, 635—643,
1967. Frequency discrimination in central nervous system of locust.
266a. Yinon U., Shulov A., Svilich A. T., J. Exp. BioL, 55, 713—725, 1971. Hearing
in desert locust.
267. Young J. Z., Proc. Soc. Lond., B, 152, 3—29, 78—87, 1960. Statocysts of octo-
pus.
Глава 13
Хеморецепция
Л. ПРОССЕР
Хеморецепция имеет для животных большое значение при поис-
ках пищи, при избегании вредных факторов или хищников, при на-
хождении особи другого пола и узнавании представителей того же
вида, при идентификации хозяина и мест откладки яиц. Животные
отвечают на «аттрактанты» положительным, а на «репелленты»
отрицательным хемотаксисом или реакцией отвергания. Хеморецеп-
торы, обладающие очень высокой чувствительностью и специфич-
ностью,— «дистантные хеморецепторы» — обычно называют обоня-
тельными. Рецепторы средней чувствительности, возбуждаемые
слабыми растворами и функционирующие, как правило, в связи с
приемом пищи, называют вкусовыми или «контактными» хеморе-
цепторами. Наконец, есть еще относительно малочувствительные и
малоспецифичные рецепторные окончания, раздражение которых
вызывает защитную реакцию, — рецепторы «общего химического
чувства». Специфичность хеморецепторов зависит от молекулярной
организации их рецептивных мембран, природа которой еще не
выяснена.
Хеморецепция у беспозвоночных (за исключением насекомых)
У жгутиковых бактерий, таких, как Escherichia coli, мы уже
имеем возможность распознать специализированные хеморецептор-
ные молекулы. Известны мутанты с ферментами, способными (или
неспособными) гидролизовать те или иные субстраты; у других
мутантов имеются пермеазы, необходимые для переноса опреде-
ленных веществ через мембрану; есть также мутанты, способные
«ощущать» некоторые вещества в очень малых концентрациях
(10~7 М), что проявляется в положительном хемотаксисе. У бакте-
рий существуют, например, мутанты с рецепторами для серина,
очень чувствительными также к цистеину, аланину и глицину; му-
танты, реагирующие на аспартат и глутамат; мутанты, чувстви-
тельные к D-галактозе и одновременно к D-глюкозе и D-фруктозе.
Возможно, что из клеточных мембран этих мутантов вскоре удаст-
ся выделить специфические молекулы-рецепторы [1].
Простейшим свойственна общая химическая чувствительность,
Хеморецепция
433
поскольку они избегают щелочей, кислот и солей. Многие катионы
вызывают реверсию активности ресничек и жгутиков; таковы, на-
пример, К+, Li+, Na+ и NH4+ (перечислены в порядке убывающего
действия). Хемотаксисом обусловлена агрегация миксамеб; лейко-
циты приближаются к органическим частицам благодаря положи-
тельному хемотаксису.
Кишечнополостные способны хорошо различать химические ве-
щества. Актинии подносят кусочки пищи ко рту и отбрасывают не-
пищевые частицы в сторону. У гидры пищевая реакция вызывается
очень низкими концентрациями глутатиона; максимальный ответ
зависит от pH, и кривая «титрования» указывает на присутствие
в рецепторном комплексе 4 ионных групп [87]. У других кишечно-
полостных отмечены пищевые реакции на пролин, тирозин и глут-
амин.
Явления симбиоза, комменсализма и паразитизма часто быва-
ют связаны с хеморецепцией. Полихета АгМопоё — комменсал мор-
ской звезды — направляется к последней в результате хемотакси-
са; если этому червю, помещенному в Y-образную трубку, пред-
ложить выбор между чистой морской водой и водой, в которой жи-
ла морская звезда, то он выбирает последнюю, в то время как
вода из аквариума с поврежденной морской звездой отталкивает
его [30]. Полихета Podarke — тоже комменсал морской звезды —
обнаруживает положительный хемотаксис, ортокинез и клиноки-
нез, которые приводят к образованию «физиологической ловушки»
около морской звезды [30]. Мирацидии дигенетических сосальщи-
ков прикрепляются только к тем видам улиток, которые служат
для них хозяевами (даже в присутствии других видов), а сидячие
коловратки Collotheca— только к свежим молодым листьям Utri-
cularia, несмотря на присутствие рядом более старых листьев, а
также листьев других растений. Пресноводные клещи Unionicola
обычно живут как паразиты на жабрах двустворчатого моллюска
Anodonta-, клещи, находящиеся в воде, где нет этого моллюска,
обнаруживают положительный фототаксис, но если добавить не-
много воды из мантийной полости хозяина, у них появляется отри-
цательный фототаксис.
Рыба Amphiprion обычно плавает среди щупалец актинии без
вреда для себя, тогда как другие рыбы, приблизившиеся к акти-
вин, вызывают разрядку ее стрекательных капсул. Если с поверх-
ности тела Amphiprion удалить слизь, эта рыба тоже становится
мишенью для стрекательных капсул. Палочка, покрытая слизью
Различных других рыб, тоже в большинстве случаев вызывает раз-
ряд этих капсул, но слизь Amphiprion тормозит реакцию актинии
иа механическое раздражение палочкой [31].
дождевых червей вся поверхность тела обладает чувствитель-
востью к кислотам. При местном воздействии растворами с
4,2 и ниже в нервах Lumbricus были зарегистрированы им-
вульсные разряды [85]. Дарвин нашел, что дождевые черви с по-
28-1514
434
Глава 13
мощью вкуса различают зеленую и красную капусту, листья морко-
ви и сельдерея.
Улитки используют химическое чувство при поисках пищи, а
двустворчатые моллюски — при регулировании тока воды через
мантийную полость. Морской гребешок (Pecten) особенно чувстви-
телен к какому-то веществу, выделяемому его естественным вра-
гом— морской звездой. Вода, взятая из аквариума с морскими
звездами, заставляет его уплывать прочь. У улиток Buccinium и
Aplysia осфрадии мантийной полости являются «вкусовыми» ре-
цепторами. При стимуляции их разведенными экстрактами мор-
ских водорослей, устриц, креветок или глутаминовой кислотой воз-
никают нервные импульсы [7]. Воздействуя на осфрадии Aplysia
экстрактом морских водорослей или разведенной морской водой,
можно вызвать торможение спонтанной активности одного из
центральных нейронов. На улитку Nassarius стимулирующее дей-
ствие оказывают глицин и лактат, в меньшей степени — бетаин;
особенно сильным стимулятором оказалось термостабильное неле-
тучее вещество, выделенное из мертвых креветок [24]. Осьминог
способен обнаружить с помощью рецепторов на присосках хинин
в концентрации, которая в 100 раз меньше ощущаемой человеком
с помощью вкуса [146].
Ракообразные обладают развитой системой вкусовых рецепто-
ров, которые находятся на ротовых частях, антеннулах и внутрен-
ней поверхности клешней. Рецепторы на клешнях краба Carcinides
чувствительны к глутаминовой кислоте в концентрации 5-Ю-5 М
и могут различать оба изомера глутамата, аспарат и лейцин [26].
Cancer реагирует на DL-аминомасляную кислоту, таурин, L-глут-
аминовую кислоту и серин (эти вещества перечислены в порядке
убывающей эффективности). L-аспарагиновая кислота более эф-
фективна в недиссоциированном виде, а L-аргинин одинаково дей-
ствует как в щелочном, так и в нейтральном растворе [25]. У усо-
ногих раков усики реагируют на разведенные растворы аминокис-
лот, особенно L-глутаминовой и L-пролина, а также на такие ами-
ны, как бетаин и таурин [29]. Омар обнаруживает чрезвычайно
малые концентрации органических кислот — глутаминовой, янтар-
ной и яблочной, а также аланина и пролина; при этом можно на-
блюдать явления синергизма и антагонизма между ними [91]. Та-
ким образом, Crustacea имеют множество рецепторов для амино-
кислот и аминов, но еще нет данных о какой-либо специфичности
отдельных нервных окончаний по отношению к определенным ве-
ществам.
Хеморецепция у насекомых
Обоняние и вкус имеют для насекомых большое значение —в
первую очередь при спаривании, откладке яиц и выборе пиши.
Самцы очень многих насекомых, особенно бабочек и тараканов,
привлекаются видоспецифичными веществами, действующими на
Хеморецепция
435
аттрактантов
10-трсшс-аце-
бомбиколом)
расстоянии. Эти вещества называют феромонами [21, 86]. Самцы
сатурний и непарного шелкопряда могут улавливать запах выде-
лений пахучей железы самки на расстоянии до 2—3 км. Самцы,
лишенные антенн, не находят самок; кусочек бумаги, соприкасав-
шийся с пахучей железой самки, вызывает у нормальных самцов
попытки копуляции.
Была определена химическая структура половых
некоторых насекомых. У шелкопряда Bombyx это
токси-1-окси-цмс-7-гекса децен (называемый также
t23-* Н Н Н
I I I
СН3(СН2)2—С=с—С=С— (СН2)8—СН2ОН;
Н
v непарного шелкопряда Porthetria — ^цс-7,8-эпокси-2-метилоктаде-
кан [14]
СН3(СН2)9—CH—CH—(СН2)4—CH(CHg)2;
у гусеницы хлопковой моли Pectinophora—10-пропил-тр«нс-5,9-
тридекадиенилацетат [72]
СН3СН2СН2
(UcH(CH2)2CH=CH(CH2)4OCCH3 ;
I II
СН3СН2СН2 о
у таракана Periplaneta — 2,2-диметил-З-изопропилиденциклопро
пилпропионат [67]
/СНз
Н3С\ /С\
у. г I ХСН3
Н3(У хСх
|\)
Н \с—сн2сн3
У гусениц Prodenia — ^нс-9-тетрадецен-1-ол-ацетат [69]
нн О
I I II
СН3(СН2)3-С=С-(СН2)8-О-С-СН3,
а также цис-9, транс- 12-тетрадекадиен-1-ол-ацетат
Н НН о
1 II 11
СН3-С=С—СН2-С=С—(СН2)8-О-С—сн3.
I
н
Взрослые самцы калифорнийского щелкуна приманиваются с
Расстояния 12 м разведенной валериановой кислотой [68]. Очевид-
28*
436
Глава 13
но, что половые аттрактанты не принадлежат к какому-то одному
типу органических молекул.
Синтетические феромоны можно было бы использовать для при-
влечения самцов вредителей. У самцов Bombyx были зарегистри-
рованы поведенческие реакции- на обдувание струей воздуха, со-
державшего ничтожное количество бомбикола—10~12 г/мл (т. е.
2-Ю2 молекул в 1 мл) [123]. Подсчитано, что самец этой бабочки
способен реагировать даже тогда, когда всего лишь 40 из 40 000
его рецепторов получают по 1 молекуле бомбикола в секунду
[121].
Насекомые и двупарноногие используют специфические веще-
ства для защиты от врагов. Многие насекомые (например, кома-
ры, пчелы, осы) вырабатывают яды, а некоторые выбрасывают
токсичное вещество в воздух. Среди жужелиц, например, есть ви-
ды, обрызгивающие врагов муравьиной кислотой; полидермиды
вырабатывают цианид, а некоторые двупарноногие — п-бензохинон.
Представитель двупарноногих Amplelorin синтезирует ядовитый
цианоген, а жук-бомбардир выделяет горячий хинон, образующий-
ся в результате экзергонической реакции Н2Ог с производным фе-
нола.
Комары находят теплокровных животных с помощью хеморе-
цепции, причем они чувствительны к ряду различных веществ. Их
привлекает, например, двуокись углерода, они летят по градиенту
в сторону ее высоких концентраций. Они обнаруживают также по-
ложительную реакцию на аминокислоты (лизин в концентрации
1:10000000) [22] и на разнообразные выделения тела млекопи-
тающих. Aedes aegypti реагирует на фильтровальную бумагу, слег-
ка смазанную половыми гормонами млекопитающих [ИЗ].
Пчелы передают некоторую информацию об источниках пищи
с помощью запахов. Муравьи выделяют феромоны тревоги (окта-
нон и нонанон), образующиеся в нижнечелюстных железах [28],
Муравьи из подсем. Dorylinae оставляют свои феромоны на пути
к источнику пищи или к муравейнику. У общественных насекомых
химические средства используются для передачи сигналов трево-
ги, привлечения, пополнения касты, при обмене веществами через
рот или анальное отверстие, для узнавания самцов своего мура-
вейника или особей той или иной касты. Вещества, выделяемые
пчелиной маткой (например, 9-ОН-деценовая кислота), вызывают
роение пчел и стабилизацию роя. 9-кетодеценовая кислота подав-
ляет выращивание маток; трамс-9-кето-2-деценовая кислота служит
половым аттрактантом во время брачного полета [147].
Значительная часть насекомых-фитофагов питается лишь не-
многими видами растений, но таких, которые кормились бы только
одним-единственным видом, вероятно, не существует. У насекомых-
олигофагов круг кормовых растений ограничен определенным р0'
дом, семейством или порядком. Есть, например, насекомые, питаю-
щиеся только на крестоцветных. Насекомые-полифаги используют
Хеморецепция
437
иирокий круг растении, но отвергают те, в которых содержатся
репелленты. Специфичность выбора кормовых растений зависит
де от основных (питательных) веществ, а от вторичных органиче-
ских компонентов, которые делают данное растение либо весьма
приемлемым для определенных насекомых, либо неприемлемым
для всех, за исключением некоторых видов. Колорадский жук и
бражник Protoparce sexta питаются только пасленовыми. Алкало-
иды табака — сильные репелленты для колорадских жуков, в то
время как алкалоиды помидора действуют слабее, а к алкалоидам
картофеля эти жуки относятся безразлично. Колорадские жуки
поедают растения помидора, но не развиваются на них. Бражник
развивается на любом представителе семейства пасленовых, вклю-
чая табак [144а].
Божьи коровки Epilachna varivestis поедают растения рода
Phaseolus и некоторые сорта сои, но не питаются другими Legu-
minosae; их привлекают фазеолунатин или родственные ему циано-
генные гликозиды [98, 99]. Одни виды Papilio кормятся только на
растениях семейства Rutaceae, а другие — на Umbelliferae и иног-
да также на Rutaceae; гусеницы, вскормленные на моркови, при-
влекаются метилгваяколом, а вскормленные на руте — метилнонил-
кетоном [33, 34]. Шелкопряды едят главным образом растения
семейства Могасеае, но их привлекает не какой-то один специфи-
ческий аттрактант, а несколько неспецифических [50].
Активными веществами, привлекающими бабочку-капустницу
(Pieris) на растения семейства крестоцветных, являются горчичные
глюкозиды — синигрин и синальбин [140]. Гусеница Pieris будет
поедать и другие растения, если смазать их глюкозидами горчич-
ного масла. Можно заставить капустную тлю поедать листья вики
(Vicia), если они будут содержать синигрин, всосавшийся из вод-
ного раствора через черешок.
Источники пахучих веществ привлекают насекомых издали.
• Жуки-короеды направляются к местам, удобным для выращивания
потомства, реагируя на различные терпены, выделяемые корой де-
ревьев. Короед Blastophagus привлекается а-терпинеолом, но не
пиненом или лимоненом.
Условия жизни личинки могут изменить хемотактическое пове-
ление взрослого насекомого. Если личинки Drosophila были
вскормлены на среде, содержавшей перечную мяту, то взрослые
мухи обнаруживают положительный хемотаксис по отношению'к
этому растению [139]. Если гусениц бабочки-бражника и Heliothis
выращивать на различных растениях, пригодных для их питания,,
то позднее они будут предпочитать те виды, на которых кормились
раньше, и этот выбор сохраняется после их содержания на искус-
Ственной среде в течение по меньшей мере двух линек; следователь-
Р°> мы имеем здесь дело с выработкой условных реакций [71].
ели кормить личинок колорадского жука и бабочки-бражника
интетической пищей в течение периода, охватывающего несколь-
438
Глава 13
ко линек, а затем предоставить им на выбор экстракты многих
растений, то они выбирают пасленовые. В данном случае пред,
почтение имеет наследственную основу. Однако при выращивании
личинок на одном виде растения они позднее выбирают только его
так что в пределах наследственного предпочтения может действо-
вать импринтинг [65, 150]. Возвращение взрослых насекомых на
определенное растение для откладки яиц, вероятно, зависит глав-
ным образом от «привыкания»
к нему на стадии личинки.
Эволюция цветковых расте-
ний в течение мелового перио-
да протекала в тесной связи с
эволюцией насекомых. Привле-
кательные для некоторых на-
секомых вещества, содержа-
щиеся в цветках, способствуют
их перекрестному опылению.
Как правило, репелленты за-
щищают растения от поедания
насекомыми, но одно и то же
вещество может быть безраз-
личным или даже отталкиваю-
щим для одних насекомых и
привлекающим для других
[50].
Рис. 13-4. Контактный хеморецептор
из длинного базиконического штиф-
тикового органа прямокрылого (Sli-
fer Е., et al., J. Morphol., 101, 359—381,
1957).
Привлечение насекомого специфическими веществами растений
зависит от его физиологического состояния; например, обезвожен-
ная нимфа Dysdercus в меньшей степени привлекается соком хлоп-
чатника, чем такая же нимфа при достаточном количестве воды
в организме. Питание могут стимулировать одни вещества, а от-
кладку яиц — другие [116, 118]. Для защиты культурного растения
от насекомых-вредителей можно выделить из него аттрактант и
обработать им дикорастущие растения на небольшом участке.
Идентификация хеморецепторов по поведенческим реакциям.
Хеморецепторы у насекомых встречаются на различных придатках
тела — антеннах, ротовых частях и лапках ног. Контактными, или
вкусовыми, хеморецепторами часто служат длинные базикониче-
ские штифтиковые органы, а дистантными, или обонятельными,-"
Хеморецепция
439
кроткие целоконические органы [133] (рис. 13-1 и 13-2). Кутикула
над обонятельным рецептором пронизана порами, в которые вхо-
дят отростки сенсорных нейронов, расположенных у основания ор-
гана (рис. 13-3). К дистантным хеморецепторам относятся, в част-
ности, гигрорецепторы.
Определенные вещества, приемлемые для насекомого, при воз-
действии на вкусовые рецепторы вызывают реакцию вытягивания
хоботка, особенно у мух и чешуекрылых. Другие, неприемлемые
вещества вызывают втягивание хоботка. С помощью метода мест-
ного контактного раздражения были найдены вкусовые рецепторы
на сосательных лопастях (например, у слепней), на лапках ног
(у многих двукрылых и чешуекрылых), на максиллярных и лаби-
альных щупиках и на антеннах (у жужелиц).
Пороги вкусовых рецепторов зависят от физиологического со-
стояния организма. Большое значение имеет, например, предва-
рительное кормление. У голодных насекомых вкусовые пороги ни-
же, чем у сытых. Можно продемонстрировать центральную сум-
мацию и торможение, одновременно раздражая две группы рецеп-
торов (например, на двух лапках). Если на одну лапку воздейст-
вовать сахаром (приемлемый раздражитель), а на другую — спир-
том (отвергаемый раздражитель), хоботок втягивается [36].
Реакция отвергания становится более выраженной, если эти два
противоположных раздражителя воздействуют на одну и ту же
лапку, а не на разные. Порог рецепторов лапки наиболее низок при
средних температурах.
У мясных мух раздражение вкусовых рецепторов лапок или
сосательных лопастей водой вызывает вытягивание хоботка. После
инъекции воды этот рефлекс исчезает, а после выпускания гемо-
лимфы (обезвоживания) усиливается. При перерезке у мухи воз-
вратного нерва, идущего от передней кишки, насекомое может
продолжать пить до тех пор, пока не лопнет [34]. Вытягивание
хоботка может быть вызвано дистантным (обонятельным) раздра-
жением антенн, контактным раздражением лапок и, вероятно, так-
же внутренними факторами [33]. Для того чтобы насекомое про-
должало кормиться при открытых ротовых частях, требуется раз-
дражение хеморецепторов, находящихся на межпсевдотрахеальных
сосочках между сосательными лопастями [37]. Во время сосания
сахарного раствора вкусовой порог в результате адаптации рецеп-
торов возрастает; после снижения чувствительности до определен-
ного уровня сосание прекращается, а затем в результате дезадап-
тации (снижения порога) возобновляется вновь. Кормление закан-
чивается под влиянием сенсорных импульсов из передней кишки.
У одного вида комаров пороговая концентрация сахарозы для
Рецепторов лабеллярных щетинок, вызывающая вытягивание хо-
°тка, — 0,011 М, для рецепторов лапок — 0,135 М, а пороговая
онцентрация глюкозы в 10 раз больше [48]. Воду и сахара вос-
Ринимают различные рецепторы. У некоторых гусениц манноза
Рис. 13-2. Растровые электронные микрофотографии вкусовых сенсилл (предостав-
лены В. Детье).
А Стилоконическая сенсилла гусеницы кольчатого коконопряда Malacosoma ameriCO*111’
Х12 000. Б. Лабеллярный волосок мухи Phormia regina; Х20 ООО.
Хеморецепция
441
вызывает только «водный» ответ и может тормозить реакцию на
фруктозу, но не на глюкозу [39].
Если удалить у личинки Bombyx максиллы, она теряет способ-
ность выбирать кормовые растения. При стимуляции максилляр-
ных сенсилл Protoparce соком подходящих кормовых растений в
некоторых рецепторных клетках возникает разряд большей часто-
ты, чем под действием сока непригодных растений. Импульсацию
этих клеток можно усилить
или ослабить, причем каждая
клетка имеет свой собствен-
ный спектр чувствительности
[126]. Каждая пятая клетка
во вкусовых рецепторах на
лапках колорадского жука
стимулируется неприемлемыми
для него алкалоидами [126].
Одним из компонентов «при-
учения» личинки к определен-
ному растению могло бы быть
развитие специфически чувст-
Рис. 13-3. Схема строения штифтико-
вой обонятельной сенсиллы насекомо-
го в продольном разрезе [125].
1 — вакуоль; 2 — футляр; 3 — ресничка; 4 —
трихогенная клетка; 5 — рецепторная клет-
ка; 6 — базальная мембрана; 7 — пора;
8 — дендрит; 9 — кутикула; 10 — эпидер-
мальная клетка; 11—тормогенная клетка;
^-—глиальная клетка; 13—аксон.
вительных рецепторов; другим компонентом является, вероятно,
создание соответствующей модели в центральной нервной си-
стеме.
Электрические реакции. Электрический ответ отдельного вкусо-
вого рецептора можно записать, поместив на одиночную сенсиллу
стеклянный капилляр; этот капилляр может содержать испытуе-
мые вещества и служить, таким образом, и стимулятором, и элек-
тродом. Во вкусовых волосках сосательной лопасти мясной мухи
пять рецепторных клеток: две для соли, одна для сахара, одна для
в°ды и одна механорецепторная. Обычно «сахарные» волокна дают
меиьщие по величине спайки, чем «солевые». Частота импульсов
зависит от концентрации применяемых растворов; сенсорная адап-
ация отмечается спустя 1—13 с [64, 94, 95]. Если принять чувстви-
Льность «сахарных» рецепторов мясной мухи к сахарозе за 1, то
v я глюкозы она будет равна 0,71, а для фруктозы — 0,46 [131].
к показали опыты с различными солями, стимулирующее дей-
442
Глава 13
ствие оказывают главным образом катионы, а кинетика ответа за-
висит как от катионного, так и от анионного рецепторного участка
Рецепторные клетки другого типа реагируют на воду, и реак-
ция их тормозится пропорционально осмотической концентрации
раствора электролита или неэлектролита.
Изучение поведения животного с одновременной записью им-
пульсов (рис. 13-4) показало, что один и тот же рецептор реагиру-
Рис. 13-4. Электрические ответы контактного хеморецептора Phormia [64].
А — непрерывная стимуляция толстого волокна 0,5 М раствором NaCl; Б — непрерывная сти-
муляция тонкого волокна 0,25 М сахарозой в 0,1 М NaCl (видны два больших спайка); В и
Г — механическая стимуляция толстого и тонкого волокон; Д — начальный период стиму-
ляции толстого и тонкого волокон 0,5 М раствором NaCl.
ет отрицательно на большие концентрации соли и положительно —
на малые. Разница в 3 импульса в первые 100 мс сенсорного раз-
ряда уже может определить поведение насекомого, как это видно
из ответов на возрастающие концентрации NaCl [38]:
0,05 М —активен только «водный» рецептор;
0,1 М —активны «солевой» и «водный» рецепторы;
0,2 М — активен только «солевой» рецептор, поведенческая реак-
ция положительная;
0,5 М —«солевой» рецептор дает высокочастотный разряд, пове-
денческая реакция отрицательная.
Максиллярные штифтиковые сенсиллы у гусениц 10 видов ба-
бочек содержат по 4 хеморецепторных нейрона. Многие из нйХ
Хеморецепция
443
еагируют на разведенный раствор NaCl, а одна клетка обычно
Реагирует и на соль, и на некоторые аминокислоты. Определенные
Аминокислоты оказывают возбуждающее действие на одни нейро-
ны и тормозящее — на другие. Возможен также и синергизм, так
что в центральную нервную систему могут поступать разнообраз-
ные по форме вкусовые сигналы [39].
Некоторые аминокислоты не действуют на лабеллярные волос-
ки (табл. 13-1), но эти волоски реагируют неспецифическим обра-
зом на высокие концентрации других аминокислот; при этом «со-
левые» рецепторы чувствительны к одним аминокислотам, а «са-
харные»— к другим.
Таблица 53-1
Реакции вкусовых рецепторов мухи на аминокислоты [132]
Аминокислота Ответ сенсорных клеток мухи Вкус для человека
Глицин Отсутствует Сладкий
Аланин » »
Серин » »
Треонин » »
Цистин »
Тирозин » —-
Пролин Реагирует солевая клетка Сладкий
Оксипролин То же »
Лизин Неспецифический »
Цитруллин .— »
Глутамин — »
Аспарагиновая кислота Неспецифический Кислый
Глутаминовая кислота » »
Гистидин » Кисло-сладкий?
Аргинин » Горький
Валин Реагирует сахарная клетка »
Лейцин То же
Изолейцин » » »
Метионин » » »
Фенилаланин » » »
Гриптофан » » »
Описано несколько морфологических типов обонятельных сен-
силл. Каждая сенсилла имеет или кутикулярный вырост (воло-
сок), или специализированную тонкую пластинку, или ямку. Выра-
женной корреляции между структурой и чувствительностью рецеп-
т°ра нет, но должна существовать специализация на молекулярном
Уровне. Например, рецепторы самцов (но не самок) должны быть
иециализированы по отношению к специфическим половым фе-
fc Р.2Э- Каждый чувствительный волосок имеет около
и пор, к которым от двух или трех рецепторных клеток подходят
сатВеТВЛеНИЯ дендРитов толщиной 100—200 А [15, 19]. У самца
Урнии-полифема насчитывается более 60000 сенсилл с 150000
444
Глава 13
рецепторных клеток, причем 60—70% этих клеток весьма чув-
ствительны к бомбиколу. «Поведенческий» порог для этой бабоч-
ки— 1014 молекул бомбикола в 1 мл воздуха, или 3-104—14-104 мо-
лекул на антенну (по 1 молекуле на сенсиллу в 2 с) [122].
Рецепторные импульсы генерируются в аксонах поблизости от
тел рецепторных клеток, лежащих у основания волоска. Микро-
Рис. 13-5. Медленные электрические ответы изолированной антенны самца шелко-
пряда ВотЬух [19].
Слева — схема регистрации; справа — ответы на возрастающие концентрации бомбикола.
у
электрод, погруженный в сенсиллу, регистрирует медленный рецеп-
торный потенциал (электроольфактограмму, ЭОГ) (рис. 13-5), за
которым следуют спайки. ЭОГ может быть позитивной (гиперполя-
ризация) с уменьшением частоты спонтанных спайков или нега-
тивной (деполяризация), т. е. возбудительной. Характер и величи-
на электрической реакции зависят от вида стимулирующего веще-
ства. Для получения измеримого электрического ответа от антенны
Самца ВотЬух концентрация феромона самки в воздухе должна
быть по меньшей мере в 100 раз выше (10~8—10~10 мкг/мл), чем
для поведенческой реакции (10~12—10-13 мкг/мл) [124].
«Поведенческие» пороговые концентрации (число молекул в
1 мл воздуха) для некоторых веществ равны [128]:
Пропионовая кислота Эвгенол Фенилпропиловый спирт Цитраль
У человека 4,2 • 10й 8,5 • 1011 6,5 • 109 4 • Ю11
У пчелы 4,3 • 1011 2 - 1010 2,2 • 10» 6 • Ю10
Хеморецепция
445
На антеннах жуков-могильщиков имеются сенсиллы двух ти-
пов длинные и короткие, от каждой из которых отходит по одному
аксону. В сенсиллах генерируются рецепторные потенциалы, в ре-
зультате которых возникают спайки с частотой, пропорциональной
эффективности обонятельных стимулов. Жирные кислоты с длин-
ной цепью (более 6 углеродных атомов) обладают деполяризую-
щим действием, а кислоты с короткой цепью вызывают гиперпо-
ляризацию [15]. У жука-могильщика запах падали вызывает не-
гативный генераторный потенциал, а пропионовая кислота — пози-
тивный [16, 17, 18]. У мухи Lucilia пороги возникновения обоня-
тельных генераторных потенциалов в базиконических сенсиллах
сравнимы с порогами поведенческих реакций [76]:
Пороговая концентрация, М
для ЭОГ для поведен- ческих реакций
Изовалериановый альдегид ю-’ IO'6
Этанол 10-7 ю-5
Ментол 5,7 • 10-9
Бутантиол 7,5 • 10-9
Камфора 3,2 • IO'8
У других рецепторов Lucilia порог наиболее низок для спиртов,
цепь которых содержит 7 углеродных атомов, и для углеводов с
цепью из 8 углеродных атомов. Ответы отдельных рецепторов об-
наруживают большое разнообразие [76] (рис. 13-6).
У Locusta 2 или 3 клетки, имеющиеся в отдельной сенсилле, от-
личаются друг от друга по чувствительности к различным запахам.
Одна из них реагирует на запах зеленой травы и имеет пороги 108
Для гексенала, 1010 для капроновой кислоты и 1015 для каприловой
кислоты (в молекулах на 1 см3 воздуха) (рис. 13-7). Наиболее эф-
фективными раздражителями для Locusta служат неразветвленные
кетокислоты и ненасыщенные жирные кислоты с 6 атомами угле-
рода; кислоты с разветвленной цепью, галогениды и ненасыщен-
ные углеводороды относительно неэффективны, а амины с нераз-
ветвленной цепью из 4 углеродных атомов оказывают тормозящее
Действие [74].
У бабочки-сатурнии были изучены реакции 50 обонятельных
клеток антенны на 13 пахучих веществ, и не нашлось ни одной па-
РЫ клеток с совершенно одинаковым спектром чувствительности
1РИС. 13-8). У пчел при исследовании реакций 47 рецепторов на
вещества не было обнаружено четкой корреляции между хими-
ГЯ/пИМн св°йствами этих веществ и их стимулирующим действием
Рис. 13-6. Относительные амплитуды обонятельных (рецепторных) потенциалов,
вызванных у самки мухи Lucilia парами пахучих веществ разной концентрации
([76], 1967).
Л >
Рис. 13-7. Электроольфактограммы и ответы одиночных рецепторных клеток
ковой сенсиллы (s. coeloconicum) Locusta migratoria [19].
A — ответ на гексенол (реакция возбуждения); Б — ответ на пропионовую кислоту (Реа
торможения).
Терпинеол
Изосафрол
Фенилэтиловый спи/.
Коричный альдегид
Фенилэтилаиртат
Гераниол
Вензилацетат
Нитробензол
Эвгенол
Коричный спирт
о г
о т
о
Повышение частоты:
среднее ф сильное
Торможение:
среднее о сильное
{Отсутствие
^реакции
{Вещество не
| испытывалось
Рис. 13-8. Спектры ответов отдельных базиконических сенсилл из чувствительных
штифтиков бабочки Anther ага, [19]. .
На каждой вертикальной линии указаны реакции одной клетки на различные вещества.
448
Глава 13
Хеморецепция у позвоночных
Вкусовые рецепторы позвоночных обычно состоят из сенсорных
клеток, образующих во вкусовом эпителии группы — вкусовые со-
сочки. Рецепторные клетки окружены множеством нервных окон-
чаний. У лягушки и сомика [32а] (и менее четко у крысы) в ос-
новании сенсорных клеток выявляются синаптические пузырьки-
таким образом, стимуляция нервных окончаний во вкусовых ре-
цепторах осуществляется, по-видимому, с помощью химических ме-
диаторов (рис. 13-9).
У млекопитающих и, вероятно, у большинства других позвоноч-
ных вкусовые клетки замещаются каждые несколько дней, в то
время как подходящие к ним сенсорные нервные волокна остаются
постоянными. При изучении развития вкусовых луковиц получены
данные о том, что нервные волокна могут вызывать превращение
эпителиальных клеток в специфические рецепторные клетки.
Обонятельные рецепторы позвоночных представлены первичны-
ми сенсорными нейронами, дендриты которых в виде микроворси-
нок или ресничек обычно проникают в слой слизи (рис. 13-10).
Рецепторы окружены опорными клетками (рис. 13-11) [134]. Ак-
соны первичных обонятельных клеток идут к обонятельной лукови-
це, где образуют синаптические контакты с вторичными нейронами
(например, с митральными клетками), аксоны которых направля-
ются в составе обонятельного тракта в передний мозг.
Рыбы и амфибии
У рыб вкусовые луковицы встречаются не только в полости, рта
(и глотке), но также на жабрах, коже и усиках. Они иннервиру-
ются VII, IX и X черепно-мозговыми нервами. Обонятельные меш-
ки рыб имеют различное строение. У акуловых рыб обонятельные
ямки находятся обычно на вентральной стороне рыла, и вода вхо-
дит в них во время плавания или при дыхательных движениях.
У большинства костистых рыб обонятельная ямка располагается
высоко на дорсальной поверхности головы, и вода поступает в нее
при плавании, при движениях рта или в результате биения рес-
ничек.
Хеморецепция и поведение. О большом значении вкусовой ре-
цепции для рыб свидетельствуют пищевые реакции, возникающие
в ответ на орошение мясным соком боковых поверхностей тела или
усиков. Даже плавниковые лучи чувствительны к жирам и кисло-
там, выделенным из рыб, а коже сомика присуща как общая
химическая чувствительность к кислотам, щелочным растворам и
солям, так и специф-ичеюкая вкусовая чувствительность. У гольянов
можно выработать вкусовые условные рефлексы на слабые PacT®°J
ры сахарозы (2-10~5 М), NaCl (4-10~5 М) и хинина (0,0025%)
[81].
Хеморецепция
449
Значение обоняния для разных рыб различно, и это различие
коррелирует с их анатомией и экологией. Для живущих стаями
нехищных рыб (Phoxinus, Gobio) важны как глаза, так и органы
обоняния. Для одиночных хищников (Anguilla, Lota) обоняние
важнее зрения. Как было показано методом условных рефлексов,
Phoxinus способен узнавать с помощью обоняния различные виды
пыб. Это существенно для отличения представителей собственного
вида от других нехищных рыб и от хищников [56]. Bathygobius
распознает пол другой особи с помощью обоняния [77, 78]. Таким
же образом Ictalurus обучается различать разных особей своего
же вида и защищать свою территорию [72]. Орган обоняния у рыб
образует множество складок, что значительно увеличивает пло-
щадь рецепторной поверхности. На 1 мм2 обонятельного эпителия
Phoxinus имеется 95000 рецепторных клеток. У угря Anguilla дли-
ной 12 см на 1 мм2 приходится 40000 обонятельных клеток, а об-
щее их число около 800000. Для восприятия запаха необходимо,
чтобы вода входила в передние ноздри и выходила через задние
ноздри. Экстракты из кожи особей того же вида или родственных
видов вызывают реакцию тревоги. У миноги Petromyzon плохое
зрение, но хорошо развита обонятельная система, и с помощью
обоняния молодые миноги находят рыбу-жертву, к которой они
присасываются. Эта реакция обнаруживается даже у особей, вы-
ращенных в изоляции, но отсутствует у миног с выключенным обо-
нянием [78]. Гольяны обладают очень высокой обонятельной чув-
ствительностью по отношению к растительной пище и к воде из
различных водоемов, окаймленных неодинаковой растительностью.
Гольяны могут различать смывы родственных видов растений при
большом их разведении. Имеются данные о том, что мигрирующие
рыбы, особенно лососи, находят места, в которых они вывелись,
по специфическим запахам [60, 63]. Если заткнуть у лосося обо-
нятельные мешки, то он будет выбирать реку для икрометания
случайным образом. У угрей Anguilla vulgaris, мигрирующих в
голландские воды, положительный таксис по отношению к воде
каналов определяется ее запахом, а не степенью солености [27].
Опыты, проведенные в многокамерных бассейнах с проточной во-
дой, показали, что у некоторых видов рыб определенные химиче-
ские вещества могут вызывать положительный реотаксис.
Чувствительность к химическим веществам у рыб очень высока.
Phoxinus может обнаружить эвгенол в воде в разведении до
° • Ю14, а фенилэтиловый спирт — до 4,3:1014 [100]. Угри реаги-
руют на р-фенилэтиловый спирт в разведении до 2,8: 1018, а фо-
рель— до 9,9: 109 [138].
Тритоны возвращаются по суше к родному водоему, даже бу-
дучи ослеплены, но после перерезки обонятельных нервов теряют
3тУ способность; значит, они скорее всего находят путь по комби-
g Дацин запахов растений в воздухе [57]. Обонятельные пороги для
29-1514
Рис. 13-9. Микрсфототрафии вкусовых рецепторов (представлены П. Грапиадеи)-
А. Рецептор лягушки Rana pipiens (световая микроскопия; Х3600). Стрелками Укаа^ый
тонкие отростки рецепторных клеток, лежащие между телами опорных клеток; н — нерББ н.
пучок, проходящий к сенсорному эпителию. Б. Рецептор лягушки (сканирующая электр
ная микроскопия, Х12 000). Кольцо из ресничек окружает сенсорную область, которая
стоит из верхушек опорных клеток н находящихся между ними кончиков тонких отрос
вторичных сенсорных клеток.
Рис. 13-9. (продолжение).
микроскопия, Х5000). Сенсорная клетка (ск) посылает
СтРелкамМаТНЧеский отРосток по направлению к базальной мембране. В области, указанной
Ный синап’ Э7?Т отросток соприкасается с нервным волокном (н), образуя с ним афферент-
пузырьки П<г та ФРагменте верху (Х35 ООО) в области этого контакта видны синаптические
мечены я/' Вкусовая луковица крысы (электронная микроскопия, Х2500). Звездочками от-
дра трех вкусовых клеток; н — нервные волокна, п — вкусовая пора, м — митоти-
чески делящаяся клетка перед дифференцировкой во вкусовую.
29*
Рис. 13-10. Строение обонятельного эпителия кролика [88].
Схематически изображены три рецепторные клетки, соединенные аксонами, в окружении
опорных клеток. 1— реснички; 2—пузырьки; 3 — обонятельная булава; 4— замыкатель
пластинка; 5 —дендрит (периферический отросток); 6 — рецепторная клетка; 7 “7 0П.°₽:а).
клетка; 8 — эндоплазматическая сеть; 9 — базальная мембрана; 10 — аксоны (fila olfacto
Б
Схемы
Рис. 13-11. Строение обонятельных рецепторов [134].
«летка"' ?1 стР°ения обонятельного эпителия у позвоночных. 1 — ресничка; 2 — базальная
а; J—слизь; 4 — рецепторная клетка; 5 — опорная клетка; 6—базальная мембрана;
Е. q а, б, в — различные типы рецепторных клеток у гольяна.
<эа. g доставление структуры обонятельных рецепторов у позвоночных и насекомых. 7 — по-
отрос НарУ™й сегмент; 9 — ресничный сегмент; 10 — внутренний сегмент (периферический
и ток рецепторной клетки); //—аксон (центральный отросток рецепторной клетки).
454
Глава 13
масляной кислоты (число молекул на 1 мл воздуха) для различ-
ных видов следующие:
Тритон 4• 109
Человек 7 • 109
Собака 9 • 103
Пчела 1,1 • 103
Электрофизиология. Записи, полученные от сенсорных ветвей
небного нерва карпа, показали, что один вкусовой сосочек содер-
жит рецепторы нескольких типов: на соль, кислоту, хинин, сахар
и человеческую слюну отвечают разные волокна [79, 80]. У Salmo
solar вкусовые рецепторы нечувствительны к аминокислотам, но
реагируют на сахара и органические кислоты, стимулирующее дей-
ствие которых возрастает с увеличением длины углеродной цепи;
была отмечена также высокая чувствительность этих рецепторов к
слюне. В волокнах небного (но не лицевого) нерва регистрирова-
лась реакция на воду [135]. Обонятельный эпителий Salmo salar
чувствителен к очень низким концентрациям аминокислот (напри-
мер, к аланину в концентрации 3-10_7 М). Обонятельные рецепто-
ры в отличие от вкусовых не реагируют на сахара, спирты и али-
фатические кислоты [135]. Реакция на аминокислоты блокирует-
ся солями ртути.
У амфибий вкусовые луковицы расположены на языке. Волок-
на языкоглоточного нерва лягушки реагируют на воду, причем ре-
акция «водных» рецепторов блокируется изотоническими солевы-
ми растворами, но не раствором сахара. Другие волокна того же
нерва отвечают на гипертонический раствор соли и на кислоту
[151]. Вкусовые ответы у лягушки имеют низкие пороги для ионов
Са2+ (менее 0,001 М). Предварительная обработка комплексооб-
разующим агентом (ЭДТА) усиливает реакцию, a NaCl и КС1
тормозят ее [73]. Порог для NaCl колеблется в пределах от 0,02
до 0,4 М. У сомика Ictalurus как обонятельный эпителий, так и
усики чувствительны к аминокислотам [7а].
Как показали записи электрической активности обонятельного
эпителия у нескольких видов рыб, при его раздражении запахами
возникают медленные рецепторные потенциалы (как on-, так и
on-off-типа). В первичных аксонах регистрировались спонтанные
спайки, а также «вспышки» импульсов на вершине медленных ре-
цепторных потенциалов [130]. В обонятельной луковице лосося
можно обнаружить реакцию на воду природных . водоемов [58,
144].
В обонятельных рецепторах и обонятельной луковице лягушки
и налима отмечается значительная спонтанная активность. Были
получены различные реакции на 28 пахучих веществ — в одних слу-
чаях on-off-ответы, в других — длительные on-ответы, в третьих"'
некоторое торможение спонтанной активности [41]. Эпителий ля-
гушки генерирует рецепторный потенциал (электроольфактограМ*
Хеморецепция
455
ЭОГ) с наложенными на него спайками. Амплитуда рецептор-
п(ЭТенциала пропорциональна силе раздражителя (рис. 13-12).
Г°еди обонятельных волокон нет двух таких, которые давали бы
ентичные ответы, но эти волокна можно подразделить на груп-
наиболее чувствительные соответственно 1) к лимонену, кам-
Ла'це и пинену; 2) к кумарину и мускусу; 3) к масляной, валериа-
новой и тиоуксусной кислотам; 4) к бензальдегиду, нитробензолу
Рис. 13-12. Спайковая активность, накладывающаяся на ольфактограмму (гене-
раторный потенциал), в обонятельной луковице лягушки [155].
А. Ответы на камфору (1), на два вдувания лимонена (2), сероуглерод (3) и этилбутират (4).
Б. Ответы на масляную кислоту (5), пиридин (6) и н-бутанол (7).
и амиловому спирту; 5) к пиридину и бутиловому спирту [53, 55].
Потенциалы обонятельной луковицы флуктуируют в зависимости
от движений рта и дыхательных движений [101, 103]. Многие па-
хучие вещества вызывают негативную ЭОГ, некоторые — позитив-
ную. Позитивная ЭОГ была зарегистрирована после дегенерации
обонятельных рецепторов и при некоторых других условиях, когда
негативная ЭОГ отсутствовала. Поэтому возможно, что позитивные
ответы возникают в опорных и секреторных клетках, в то время
как негативные представляют собой генераторные потенциалы
[136]. Ответы, регистрируемые в латеральной и в медиальной вет-
вях обонятельного нерва лягушки при воздействии различных за-
пахов, неодинаковы, что говорит о возможном существовании в
^д^пятельном эпителии механизма пространственного анализа
В различных веточках обонятельного нерва черепахи были за-
Р гистрированы разнообразные формы электрических ответов; ока-
зе^сь’ что они хорошо коррелируют с растворимостью пахучих
456
Глава 13
У серебряного карася антидромная стимуляция бокового дыха-
тельного тракта тормозит активность многих митральных клеток—.
вероятно, через эфферентные нейроны [59]. В обонятельном трак-
те налима обнаруживаются эфферентные импульсы, как спонтан-
ные, так и вызванные активностью ипсилатерального и контрала-
терального обонятельных трактов; эти импульсы могут исчезать в
результате тактильного раздражения кожи [41, 43]. Вероятная
роль этих эфферентных импульсов состоит в модуляции обонятель-
ной чувствительности.
Млекопитающие и птицы
Поведение. Млекопитающие способны различать сладкое, горь-
кое, соленое и кислое. На основании поведенческих реакций и дан-
ных электрофизиологии был сделан вывод, что способность ощу-
щать вкус воды хорошо развита у кролика и кошки и отсутствует
или слабо развита у крысы и человека. Однако реакция на воду
зависит от предшествующей адаптации. Человеку вода кажется
горькой после воздействия на вкусовые рецепторы соленого и слад-
кой после их адаптации к кислому. У животных, особо чувствитель-
ных к соли, реакция на воду после воздействия NaCl выражена
слабо. Поэтому существование истинных «водных» рецепторов,
представляется сомнительным. Обычно кошки безразличны к саха-
ру, и считается, что они лишены способности ощущать сладкое, но
после NaCl они не отказываются от сахара и предпочитают саха-
розу, растворенную в 0,03 М NaCl, сахарозе, растворенной в во-
де [8]. Вкус сладкого очень слабо ощущают также ягнята, а голу-
би не реагируют на горькое (хинин). Регистрация импульсной
активности показала, что отдельное вкусовое волокно реагирует на
различные вкусовые раздражители, но порог для одного из них
ниже, чем для остальных.
У человека самым низким порогом для сладкого обладает пе-
редняя часть языка, для горького — корень и для кислого — боко-
вые поверхности, причем эти зоны перекрывают друг друга. Ко-
каин раньше всего устраняет ощущение горького, а затем после-
довательно сладкого, соленого и кислого. Гимнемовая кислота по;
давляет ощущение сладкого, но не соленого или кислого. Белковый
экстракт из плода Synsepalum не обладает собственным вкусом,
но подавляет сладкий вкус сахарозы, цикламата, D-аминокислот
и уксуснокислого свинца; в то же время он делает кислое (лимон)
сладким [83]. К соленому адаптация наступает быстрее, чем к
сладкому или горькому. Данное вкусовое ощущение (например,
ощущение сладкого) может вызываться рядом несходных в хими-
ческом отношении веществ. Некоторые люди не способны ощуШаТ1г
вкус таких горьких веществ, как n-этоксифенилтиомочевина, к ко-
торым другие лица весьма чувствительны [49].
Крысы, у которых в результате адреналэктомии развились
солевая недостаточность, в большей степени отдают предпочтени
Хеморецепция
457
левым растворам перед водой, чем животные с нормальным со-
С°вым балансом. Однако вкусовые пороги у тех и других сходны,
л к что больший солевой голод адреналэктомированных животных
Тмеет по-видимому, центральное происхождение. Крыс можно
бучить связывать различные запахи с определенными вещества-
ми которых недоставало в пищевом рационе, а затем регулиро-
вать с помощью этих запахов потребление животными пищи и во-
ды [89, ПЗа].
у большинства млекопитающих обоняние играет ведущую роль
в определении качества пищи и обнаружении добычи или хищни-
ков. Запахи также важны для млекопитающих при маркировке и
идентификации своей и чужой территории, узнавании особей дру-
гого пола и половом поведении [104].
Известен ряд желез, вырабатывающих специфический пахучий
секрет: препуциальные железы кабарги и речного бобра, анальные
и промежностные железы куницы, виверры и хорька, промежност-
ные и сальные железы различных млекопитающих [112]. Плото-
ядные более чувствительны к запахам, чем крысы; хорошо извест-
на способность собак идти по следу и отыскивать объекты, нахо-
дящиеся под землей (например, трюфели). Можно натренировать
мышей (Mas и Peromyscus) в Y-образном лабиринте различать с
помощью обоняния особей разных видов или того же вида, но
разного пола и даже двух самцов одного и того же вида [20].
Фалангеры поддерживают сложные общественные отношения, нахо-
дясь в темноте. Они имеют лобные, грудинные, анальные и сумко-
вые железы, секрет которых обладает запахом. С помощью та-
кого секрета животные маркируют себя и своих партнеров, уста-
навливают границы территорий и главенство в колонии без зри-
тельных сигналов. Запахи «чужих» особей вызывают у них агрес-
сивное поведение [127].
Большинство млекопитающих — макросматики; значительная
часть переднего мозга связана у них с функцией обоняния. Прима-
ты и беззубые киты — микросматики (животные с плохо развитым
обонянием). У кролика общее число обонятельных рецепторных
клеток достигает 50—100 млн.
Электрофизиология. Каждый вкусовой сосочек снабжен мно-
жеством тонких нервных окончаний. Вероятно, один нейрон мо-
жет иннервировать несколько сосочков, так как отдельный сосочек
подвержен усиливающему и тормозящему влиянию со стороны
€г° окружения [92]. В каждой вкусовой клетке в ответ на спе-
цифические химические воздействия возникают рецепторные потен-
Характер импульсов в отдельных волокнах вкусовых нервов
л°Шки может обнаруживать «смешанную» специфичность. Одни во-
^°кна реагируют на кислоты при pH ниже 2,5, другие — на NaCl
х°линхлорид. Встречаются также «водные» волокна, отвеча-
йте на отмывание языка от солей. «Водные» нервные окончания
458
Глава 13
стимулируются солевыми растворами, содержащими менее 0,03 Ду
NaCl, тогда как для возбуждения «солевых» рецепторов нужны
концентрации NaCl выше 0,05 М. У кошки лишь немногие вкусо-
вые волокна реагируют на сахар, в то время как для кролика са-
хар служит более сильным раздражителем, чем хинин. Вкусовое
волокно может отвечать на два или большее число вкусовых сти-
мулов (например, на кислоту и хинин), но пороги для них различ-
ны [52]. У собак сахарин или совсем не вызывает ответа, или ак-
тивирует более тонкие волокна, чем сахар; гимнемовая кислота не
подавляет у них реакции на сахарин, но подавляет ответ на сахар,
у человека же она полностью снимает восприятие сладкого.
У обезьяны волокна барабанной струны отвечают на хлористый
натрий, кислоту, хинин, сахар и воду. Цыплята реагируют на соль,
глицерин, этиленгликоль, хинин, воду и кислоту, но не реагируют
на сахарозу или сахарин. В барабанной струне крысы из 48 иссле-
дованных волокон 19 отвечали на три из четырех видов вкусовых
стимулов, а 12 — на все четыре. У хомячка 5 из 28 исследованных
волокон реагировали на один, 5—на два, 11 —на три и 7 — на все
четыре типа вкусовых стимулов, хотя пороги для них были весьма
различными [Н5]. Очевидно, существует несколько видов вкусо-
вых рецепторов, и часть из них реагирует на многие вещества по-
разному в зависимости от их концентрации, возбуждая два раз-
личных ощущения. Вкусовые пороги в поведенческих эксперимен-
тах часто оказываются более низкими, чем при регистрации нерв-
ных импульсов. В таламусе и в соматосенсорной области коры
большого мозга пространственная проекция рецептивных полей со-
ответствует их расположению на языке [40].
Обонятельные рецепторные клетки в отличие от вкусовых пред-
ставляют собой первичные нейроны, от каждого из которых идет
аксон в обонятельную луковицу. В аксонах обонятельных рецепто-
ров птиц и грызунов были зарегистрированы электрические отве-
ты на запахи [9, 143], разряды импульсов синхронизированы с
дыхательными движениями, при которых пахучие вещества прихо-
дят в соприкосновение с обонятельной слизистой.
Первичные обонятельные нейроны образуют синапсы с дендри-
тами митральных клеток в клубочковом слое обонятельной луко-
вицы. Аксоны митральных клеток в составе бокового обонятель-
ного тракта идут к переднему мозгу. Обонятельная луковица со-
держит также клетки-зерна, отростки которых соединяются с ден-
дритами митральных нейронов, образуя таким образом коллате-
ральные связи в пределах луковицы (рис. 13-13). Разряды им-
пульсов в митральных клетках сопровождают каждое вдыхание
паров пахучих веществ, причем отдельные клетки различаются по
своим ответам на разные запахи; следовательно, они должны полу-
чать входные сигналы от определенных рецепторных клеток.
В обонятельной луковице имеет место спонтанная активность, ко-
торая при действии запахов может сменяться ритмическими ра3'
Хеморецепция
459
оЯдами большей или меньшей частоты. Некоторые митральные
нейроны дают постоянный разряд, другие активируются при каж-
дом^ прохождении воздуха через нос и, наконец, третьи специфи-
чески реагируют только на запахи [145]. У человека пороги рит-
мической активности (вызванных потенциалов) обонятельной луко-
вицы хорошо коррелируют с порогами субъективного восприятия
запахов [66]. В экспериментах с регистрацией разрядов митраль-
ных клеток при антидромном возбуждении их аксонов и при орто-
Рис. 13-13. Схема нейронных взаимоотношений в обонятельной луковице кроли-
ка [110].
1 слой обонятельных волокон, которые образуют синапсы с митральными (2) и пучковы-
ми (3) клетками; 4— клетки-зерна; 5 — перигломерулярные клетки.
Дромной стимуляции обонятельных волокон было показано, что
спайки возникают в них у основания аксонов. Вторичные дендриты
митральных клеток, отходящие значительно ниже клубочкового
Дендрита, активируют клетки-зерна, дендриты которых в свою оче-
редь оказывают тормозящее воздействие на те же самые или дру-
гие митральные нейроны. Клетки-зерна лишены аксонов, и воз-
УЖдение их, вероятно, распространяется электротонически по ден-
дриту; в этом отношении они сходны с амакриновыми клетками
тчатки. Между митральными нейронами и клетками-зернами име-
н7Ся РеДипрокные синапсы, содержащие пузырьки; эти образова-
служат структурной основой процессов самоторможения и ла-
460
Глава 13
терального торможения в обонятельной луковице [108, ПО, 111].
В наружном сетевидном слое обонятельной луковицы оканчива-
ются эфферентные волокна, идущие из переднего мозга; они, не-
видимому, тоже модулируют ответы митральных клеток [109].
Клеточные механизмы хеморецепции
Попытки сформулировать единую теорию химической рецепции
не увенчались успехом; по-видимому, в этом процессе участвует
несколько различных взаимодействий, происходящих у поверхно-
сти клетки. Общее химическое чувство связано с деполяризующим
влиянием разнообразных агентов на возбудимые клетки. Простей-
шие в той или иной степени реагируют на различные неорганиче-
ские ионы и на кислоты. Подобным же образом после длительного
латентного периода могут возбуждаться свободные нервные окон-
чания. Действие сильных кислот, особенно неорганических, опре-
деляется концентрацией водородных ионов. Слабые кислоты, обыч-
но органические, по-видимому, проникают в клетку в виде недис-
социированных молекул, и действие их не связано с изменением
pH. Такие ионы, как К+, могли бы возбуждать клетку в результа-
те местной деполяризации. Анионы типа цитрата и оксалата, свя-
зывающие кальций, могли бы воздействовать, удаляя Са и дестаби-
лизируя таким образом мембрану. Гипертонические растворы, воз-
можно, изменяют свойства мембраны, вызывая ее обезвоживание.
Неспецифичность химических стимулов, длительный латентный пе-
риод (секунды или минуты) и относительно высокие пороги отли-
чают общее химическое чувство от вкуса и обоняния.
Ряд исследователей, занимающихся вкусом и обонянием, видо-
изменили применительно к хеморецепции уравнение Михаэлиса—
Ментен, описывающее кинетику ферментативной реакции. Соглас-
но предложенной формуле, ответ — амплитуда электроольфакто-
граммы, частота рецепторных импульсов или интенсивность пове-
денческой реакции — представляет собой S-образную функцию от
логарифма концентрации стимулирующих молекул [46, 75]. Кон-
станта равновесия К определяется уравнением
р a(s — n)
п ’
где s — общее число рецепторных участков, п — число участков?
занятых возбуждающим веществом, и а — эффективная концентра-
ция возбуждающего вещества [10, 11, 75].
Вкусовые луковицы различных позвоночных реагируют на воз-
действие соли через 20—25 мс. Ответ могут вызывать концентра-
ции NaCl от 0,0005 до 5,0 М, так что возбуждающее действие со-
ли определяется не только ионами. Реакция, вызываемая солью?
зависит как от катиона, так и от аниона. И NaCl, и Na2SO4 имеют
соленый вкус, но этого нельзя сказать о NaNOs и NaHCOs; вКУе
Хеморецепция
461
поридов варьирует в зависимости от катиона. Солевые рецепторы
х лапках мясной мухи наиболее чувствительны к KI и в меньшей
Нтепени к КС1. У некоторых насекомых выраженность реакции от-
вергания последовательно изменяется в ряду NH4>K>Na>Li для
катионов и 1>Вг>5О4>С1>Ацетат —для анионов [64].
Ощущение сладкого вызывают у человека сахара, содержащие
фруктозу или глюкозу, а также ряд других, весьма различных по
своей структуре веществ, например сахарин, уксуснокислый свинец
и некоторые соли бериллия. Правовращающий аспарагин имеет
сладкий вкус, а левовращающий безвкусен. Вкус гликолей варь-
ирует от сладкого (этиленгликоль) до горького (гексаметиленгли-
коль). Из 34 исследованных веществ 30 оказались сладкими для
человека и 9 (судя по поведению) — для пчелы. Сладкие для чело-
века маннит, сорбит, ксилоза, галактоза и мелибиоза отвергаются
пчелами [51]Сахара по чувствительности к ним ротовых частей
пчелы образуют следующий ряд [36]: Сахароза = Мальтоза>
>Трегалоза> Фруктоза. Результаты опытов на мухах с некоторы-
ми производными D-глюкозы указывают на взаимодействие ОН'
групп при атомах С3 и С4 с рецепторным участком. Стерические за-
труднения в области этих групп препятствуют стимуляции рецеп-
торов, а метилирование делает глюкозу безвкусной. Первичный
процесс стимуляции вкусовых рецепторов малочувствителен к тем-
пературе. Относительную эффективность сахаров как вкусовых раз-
дражителей можно выразить следующими величинами: мальтоза—
33, фруктоза — 21,6, сахароза— 13,2, глюкоза— 1 [45]. Для кошки,
судя по ответам барабанной струны, сахара образуют ряд: D-фрук-
тоза>Сахароза>Сорбоза>П-манноза>Г)-глюкоза > Мальтоза >
>В-галактоза>Лактоза. Часть нервных волокон барабанной стру-
ны реагировала на все эти сахара, а часть — только на два из них.
Некоторые вкусовые волокна реагируют и на сахар, и на соль.
По-видимому, один и тот же нейрон может получать информацию
более чем от одной вкусовой клетки. Предварительная обработка
вкусового сосочка солью подавляет его ответ на сахар [6].
Субъективное ощущение горького вызывают у человека такие
весьма несходные вещества, как хинин, стрихнин, некоторые соли
магния и аммония. Наследственные различия в способности ощу-
щать горький вкус н-пропилтиоурацила и близких к нему веществ
коррелируют со способностю воспринимать вкус хлорпромазина.
dT0 говорит о возможной химической специфичности рецепторов
в некоторых вкусовых клетках. У человека существует ряд гено-
типов, проявляющихся в определенных дефектах вкусовой рецеп-
ции. В молекулах многих горьких веществ группа, которая служит
Донором протона, находится на таком близком расстоянии (1 А)
т акцептора, что здесь образуется водородная связь. Образование
Утримолекулярной водородной связи лишает вещество способ-
-и вызывать ощущение сладкого [82]. В веществах со сладким
Усом доноры и акцепторы протонов достаточно удалены друг от
462
Глава 13
друга (3 А), так что образование водородных связей невозможно
Изучение мутантов Е. coli показало, что у этих бактерий рецепторы
для определенных аминокислот генетически детерминированы.
Пока неясно, есть ли у насекомых ощущение, аналогичное горь-
кому вкусу. Многие вещества, например спирты, альдегиды и ке-
тоны, отвергаются насекомыми (вызывают втягивание хоботка).
В гомологичных рядах альдегидов, кетонов, спиртов и гликолей от-
рицательное стимулирующее действие возрастает с удлинением
углеродной цепи (т. е. пороговая концентрация снижается).
У мясной мухи вкусовые пороги для отвергаемых ею спиртов со-
ответствуют следующим молярным концентрациям: метиловый
спирт — 11,3; этиловый — 3,2; w-бутиловый — 0,66; н-амиловый —
OJs; н-гексиловый— 0,012; вторичный н-октиловый спирт — 0,0021
[34]. Разветвление цепи повышает вкусовой порог. Наличие вто-
рой ОН-группы, так же как и замещение галогеном, уменьшает
стимулирующую способность вещества. w-Кетоны более эффектив-
ны, чем изо-кетоны [34], а монокарбоновые кислоты действуют
сильнее, чем дикарбоновые; различные хеморецепторные волоски
по-разному реагируют на одну и ту же кислоту при одинаковом
pH [90]. В гомологичных рядах органических веществ вкусовые
свойства часто лишь незначительно возрастают, пока число угле-
родных атомов не достигнет примерно 5, а затем усиливаются го-
раздо более резко. Возможно, что низкомолекулярные вещества
поступают в водную фазу, а вещества с более длинной цепью —
в липидную.
Водные вкусовые рецепторы возбуждаются в результате отмы-
вания с их поверхности солей или сахара; их можно рассматри-
вать как тонические рецепторы, отвечающие off-реакцией при уда-
лении растворенных веществ; возможно, что они служат осморецеп-
торами для сахарозы, но другие растворенные вещества в очень
малой концентрации не тормозят их.
Во всех контактных (вкусовых) хеморецепторах независимо от
того, представлены ли они просто нервными окончаниями или
вкусовыми рецепторными клетками, обнаруживаются градуальные
электрические ответы. Вероятно, это обусловлено изменением ион-
ной проницаемости клеточных мембран, которые содержат высоко-
специфические рецепторные молекулы.
Обонятельные, или дистантные, хеморецепторы обладают наи-
большей чувствительностью и наиболее многообразны по своей
специфичности. На основе субъективных оценок качества запахов,
их маскировки и перекрестной адаптации психологи предложили
ряд различных классификаций пахучих веществ. Запахи подразде-
ляют, например, на эфирные, камфорные, мускусные, мятные, цве-
точные, острые и гнилостные. Представление о многообразии обо-
нятельных рецепторов вытекает из данных о заметных различиях
пороговых концентраций пахучих веществ (во вдыхаемом воздухе)
для человека: например, 7 • 10—13 М для этилмеркаптана, 5,8
Хеморецепция
463
ттпя тиоамилового эфира и 7,0-10~15 М для этилового эфира
l Наиболее полный обзор весьма обширной литературы по хе-
морецепции принадлежит ЛАонкрифу [93], который сформулировал
62 общих положения относительно связи между химической струк-
турой и запахом. В некоторых гомологичных рядах интенсивность
запаха возрастает с увеличением длины углеродной цепи вплоть до
известного оптимума. В случае летучих жирных кислот этот опти-
мум для человека, пчелы и собаки достигается при 4-углеродной
цепи, а в случае летучих эфиров для человека и пчелы'—при
12-углеродной цепи [128]. Возможно, что ольфакторные свойства
более мелких молекул ограничивает растворимость в жирах, а бо-
лее крупных—сама величина молекулы. Из первичных спиртов
наиболее эффективным раздражителем для лягушки оказался
(судя по электроольфактограмме) спирт с 8-углеродной цепью
[Ю2].
Очевидно, в мембранах обонятельных клеток имеются высоко-
специфические рецепторные молекулы, и клетки различаются меж-
ду собой по относительному числу молекул того или иного рода.
В пользу этого говорит весьма высокая специфическая чувстви-
тельность таких клеток (например, у насекомых к феромонам).
Как показала регистрация электрических реакций отдельных ре-
цепторов на различные пахучие вещества, рецепторные клетки об-
ладают индивидуальными спектрами чувствительности. «Сопостав-
ление» информации от большого числа таких клеток в центральной
нервной системе позволяет различать почти безграничное множест-
во запахов. Рецепторы на антеннах самца Bombyx возбуждаются
всего лишь несколькими молекулами бомбикола, в то время как
антенны самок того же вида намного менее чувствительны к нему.
При изучении структурной специфичности оказалось, что в ряду
изомеров бомбикола эффективность изменяется следующим обра-
зом: 10-транс, 12-цис>10-цис, 12-транс>10-цис, \2-цис> 10-транс,
12-транс [17]. На рис. 13-14 представлены типичные спектры реак-
ции обонятельных рецепторов насекомых на пахучие вещества.
Ни одно из известных свойств молекулы, таких, как насыщенность
углеродных атомов, положение двойных связей, длина цепи, нали-
чие функциональных групп, изомеризация или геометрическая фор-
ма, не коррелирует со специфическими обонятельными порогами
Для данных веществ [119].
Индивидуальность рецепторов в обонятельной системе млекопи-
тающих можно было выявить благодаря тому, что нервные волок-
ла, идущие от разных областей обонятельного эпителия, распре-
деляются по разным отделам обонятельной луковицы. С помощью
ивленных электродов записывали интегрированную спайковую
^тивность в различных участках обонятельной луковицы кроли-
н ’ в 6 участках были обнаружены характерные спектры реакций
пахучих веществ; один и тот же тип ответа сохранялся в
464
Глава 13
Испытанные вещества Calliphora, рецептор трупных запахов (s. baslconicum) Necrophorus, рецептор трупных запахов (s. basiconicum) Locusta, s. coeloconicum Apis, рецептор запаха маточ- ного вещества (s. placodeurn)
.Жирные кислоты с нераз- ветвленной цепью Насыщенные С3 с4 Сч 0 0 0 -j -
С6 С.7 0
о
с8 С9 Сю С12, 14, 16, 18 Ненасыщенные Сю (олеиновая, линолевая) 0 0 0 0 + + 0 0 0 ++ "о
Падаль Амины, меркаптаны Гексенал Гексанол Г ексенол Г ексенилформиат Вещество пчелиной матки (9-оксодеценовая кислота) 9-<жсидеценовая кислота 1 1 1 +++
—I—НН- 0 0 0 0 0 +
Рис. 13-14. Спектры
£) — отсутствие реакции.
электрических ответов специализированных обонятельных
клеток у некоторых насекомых [19].
«+» — возбуждение, «—» — торможение. Число знаков соответст-
вует интенсивности реакции.
этих участках в течение нескольких дней [96] (рис. 13-15). Ана-
логично этому суммарные электрические ответы в двух ветвях
обонятельного нерва, идущих от противоположных областей обоня-
тельного эпителия лягушки, оказались при действии разных ве-
ществ различными [97]. Ответ на смесь пахучих веществ нельзя
предсказать по ответам на отдельные ее компоненты. В первич-
ных обонятельных рецепторах лягушки реакция на данный кон-
кретный запах варьирует в зависимости от предшествующих обо-
нятельных раздражителей и от времени, прошедшего после их воз-
действия. Например, клетка, реагирующая на бутанол единичными
спайками, после действия мускуса (который сам по себе не вызы-
вает ответа) может дать на то же вещество целую вспышку раз-
рядов. Таким образом, в обонятельной системе не только существу-
ет различие между рецепторными клетками по их отношению К
разным запахам, но одна и та же клетка может дать разную реак-
Хеморецепция
465
цИю в зависимости от предшествующих обонятельных стимулов
[55].
В каждом из четырех классов запахов обонятельные пороги
пахучих веществ в общем коррелируют с их адсорбцией на поверх-
ности раздела липид — вода [32] и с величиной и формой моле-
кулы [2—5]. Вещества, обладающие камфорным запахом, состоят
из сферических молекул с диаметром 7 А; молекулы веществ с
Рис. 13-15. Реакции на 12 пахучих веществ в шести различных участках обоня-
тельной луковицы [96].
А — н-пропанол, 10—3; Б — н-амилацетат, 10~2; В — м-октан, 10~2; Г — трет-бутанол, 10-2;
Д—а-ионон, 10~2; Е — 2-ундеканон, 10-2; Ж—м-гептанол, 10~2; 3 — скатол, 10—3; И— н-ва-
лериаковая кислота; Д — эвгенол, 10~2; Л — бутиловый эфир, 10~2; М — D-лимонен, Ю-1-6
(цифры отражают концентрации пахучих веществ по отношению к концентрации их насы-
щенных паров при 22° С).
мускусным запахом больше приближаются по форме к дисковид-
ным. Данные об электрической активности отдельных нейронов в
обонятельной луковице лягушки подтверждают наличие корреля-
ции между стереохимией молекул и их ольфакторными свойствами,
выявленной в психофизических экспериментах [43]. Весьма веро-
ятно, что с запахом коррелирует несколько свойств молекулы и что
Рецепторные клетки снабжены гетерогенными мембранами с высо-
коспециализированными липопротеидными участками, соотношение
которых варьирует от клетки к клетке.
Заключение
Химические чувства имеют первостепенное значение для боль-
инства животных. Известны поразительные примеры поведения,
кованного на специфической хеморецепции; привлечение особей
30" 1514
466
Глава 13
другого пола у многих насекомых, видовое и «семейное» узнавание
у некоторых млекопитающих, идентификация кормовых растений
насекомыми, возвращение в родные водоемы лососей и тритонов,
защитное поведение большинства животных, обнаружение запахов
животных собаками. Умение человека распознавать вина и арома-
ты духов — еще одно свидетельство тонкости химических чувств.
Специфичность растительных аттрактантов и репеллентов может
быть использована при разработке биологических методов борьбы
с насекомыми-вредителями.
Общая химическая чувствительность свойственна всем клеткам.
Плазматические мембраны большинства клеток, и в частности кле-
ток возбудимых тканей, реагируют на кислоты, щелочи и многие
ионы; при их воздействии мембраны деполяризуются, клетки округ-
ляются, цитоплазма их сокращается, биение ресничек останавлива-
ется или изменяет свое направление. Величина гомеостатического
ответа клетки зависит от силы стимула. Большая эффективность
некоторых ионов по сравнению с другими определяется, по-види-
мому, природой поверхностных клеточных мембран.
Значительно труднее понять специфическую чувствительность к
различным органическим веществам и отдельным неорганическим
ионам. Всего лишь несколько молекул полового аттрактанта могут
вызвать у бабочки сложное поведение. С такой же тонкой чувстви-
тельностью связано возвращение рыб в родные водоемы. Специфи-
ческая реакция вкусовых рецепторов на определенные аминокисло-
ты и сахара предполагает наличие во вкусовых клетках целого на-
бора рецепторных молекул. Существование таких молекул под-
тверждается и тем, что в популяциях бактерий, насекомых-фитофа-
гов и человека встречаются мутанты, по-разному воспринимающие
определенные вещества. О рецепторных участках вкусовых клеток
мы можем судить главным образом по их порогам к различным
стимулам. Химическая идентификация рецепторных молекул еще
не осуществлена.
Определенные недавно спектры чувствительности отдельных хе-
морецепторов (в частности, у насекомых) показали, что нет двух
хеморецепторных клеток, вполне сходных между собой, и что каж-
дая из них имеет в своей мембране сложную мозаику хеморецеп-
торных участков. Число разных рецепторных молекул должно быть
очень большим; если же число их невелико, то они должны обла-
дать множеством разных рецептивных участков. Многообразие ре-
цепторов и их неодинаковое соотношение позволяют различать
всевозможные комбинации запахов. Рецепторные молекулы долж-
ны быть способны к изменениям (например, при сенсорной адап-
тации); чувствительность их может, например, зависеть от состоя-
ния гидратации клетки и от предварительной обработки ее опре-
деленными веществами. Примером этому может служить инверсия
вкусового ощущения у человека.
Хеморецепция
467
По-видимому, все хеморецепторы при возбуждении генерируют
ецепторные потенциалы. В некоторых органах чувств, в том чис-
во многих сенсиллах насекомых, дендрит (периферический от-
росток) рецепторной клетки подвергается прямой стимуляции.
В ДРУГИХ °Рганах’ например во вкусовом эпителии позвоночных,
рецепторная клетка передает возбуждение на нервные окончания,
вероятно синаптическим путем. Во всех хемосенсорных системах
имеет место значительная конвергенция возбуждения в централь-
ных отделах. У позвоночных обонятельная луковица представляет
собой часть мозга, в которой хорошо развитый нейропиль «смеши-
вает» информацию, поступающую от очень многих рецепторов.
Прежде многие явления, связанные с органами химического
чувства, представлялись загадочными. Сейчас в результате обна-
ружения феромонов, частичного выяснения природы рецепторных
молекул и некоторого постижения механизмов центрального коди-
рования сенсорных сигналов химические чувства становятся более
понятными.
СПИСОК ЛИТЕРАТУРЫ
1. Adler J., Science, 166, 1588—1597, 1969. Chemoreceptors in bacteria.
2. Amoore J. E., Nature, 198, 271—272, 1963. Stereochemical theory of olfaction.
3. Amoore J. E., Cold Spring Harbor Symp. Quant. Biol., 30, 623—636, 1965.
Psychophysics of odor.
4. Amoore J. E. In: Olfaction and Taste, 3rd Internat. Symp., edited by C. Pfaff-
mann, Rockefeller University Press, New York, 1969, pp. 158—171.
5. Amoore J. E., Molecular Basis of Odor, Charles C Thomas, Springfield, 1970,
200 pp.
6. Andersen H. T., Funakoshi M., Zotterman Y., Acta Physiol. Scand., 56, 362—
375, 1962; also in: Olfaction and Taste: A Symposium, edited by Y. Zotterman,
Pergamon Press, New York, 1963, pp. 177—192. Electrophysiological respon-
ses of taste receptors to sugar.
7. Bailey D. F., Laverack M. S., Nature, 200, 1122—1123, 1963. Central nervous
responses to chemical stimulation of a gastropod receptor.
7a. Bardoch J. et al. In: Olfaction and Taste, edited by H. Hayashi, Pergamon
Press, New York, 1967, pp. 647—666. Chemical stimulation in bullhead
fish.
8. Bartoshuk L. M., Homer M. A., Parks L. H., Science, 171, 699—701, 1971. Wa-
ter taste in cat.
9. Beidler L. M„ J. Neurophysiol., 16, 595—607, 1953; also Beidler L. M. et al.,
Amer. J. Physiol., 181, 235—240, 1955. Analysis of taste responses, mam-
mals.
10. Beidler L. M. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman,
11 Г*е{Еатоп Press, New York, 1963, pp. 133—145. Dynamics of taste cells.
1. Beidler L. M. Cold Spring Harbor Symp. Quant. Biol., 30, 191—200, 1965.
19 pOHParison of gustatory and olfactory receptors.
• Beidler L. M. In: Olfaction and Taste. Vol. 2, edited by T. H. Hayashi, Perga-
13 z?011 Press’ New York, 1967, pp. 509—534.
Bell F. Kdtchell ft. L., J. PhysioL, 183, 145—151, 1966. Taste reception in
j goat, sheep, and calf.
• Fieri B. A., Beroza M., Collier C. W., Science, 170, 87—89, 1970. Sex attra-
15 orants of gypsy moth.
oeckh J., Z. vergl. Physiol., 46, 212—248, 1962. Electrophysiology of odor
eception in carrion beetles.
30*
468
Глава 13
16. Boeckh J. In: Olfaction and Taste, Vol. 2, edited by T. H. Hayashi, Pergamon
Press, New York, 1967, pp. 721—736. Single insect olfactory receptors.
17. Boeckh J., Z. vergl. Physiol., 55, 378—406, 1967. Olfactory receptors in several
insects.
18. Boeckh J. In: Olfaction and Taste, 3d, edited by C. Pfaffmann, Rockefeller
University Press, 1969, pp. 34—51. Electrical activity in olfactory receptors.
19. Boeckh J., Kaissling К. E., Schneider D., Cold Spring Harbor Symp. Quant
Biol., 30, 263—280, 1965. Insect olfactory receptors.
20. Bowers J. M., Alexander В. K., Science, 158, 1208—1210, 1967. Olfactory re-
cognition by mice.
21. Brady U. E„ Tomlinson R. G., Brownlee R. G., Silverstein R. M„ Science, 171,
802—804, 1971. Sex attractants in moths.
22. Brown A. W. A., Carmichael A. G., Nature, 189, 508—509, 1961. Lysine as a
mosquito attractant.
23. Butenand A., Triangle (Basel), 5, 24—27, 1961. Biochemistry of ecdyson and
mating scent of silk moths.
24. Carr W. E. S., Biol. Bull., 133, 90—127, 1967. Chemoreception in snail Nassa-
rius.
25. Case J., Biol. Bull., 127, 428—446, 1964. Chemoreceptors on dactyl of Can-
26. Case J., Guilliam G. F., Biol. Bull., 121, 449—455, 1961. Amino acid sensitivity
of chemoreceptors of Carcinides.
27. Crcutzberg F., Nederl. J. Sea Res., 1, 257—338, 1961. Orientation of migrating
elvers of Anguilla.
28. Crewe R. M., Blum M. S., Z. vergl. Physiol., 70, 363—373, 1970. Alarm phero-
mones in ants.
29. Crisp D., J. Biol. Bull., 133, 128—140. Chemoreception in barnacles.
30. Davenport D. et al., Biol. Bull., 100, 71—83, 1951; Animal Behavior, 8, 3—4,
1960. Commensalism between starfish and polychaetes.
31. Davenport D. et al., Biol. Bull., 115, 397—410, 1958. Symbiosis of sea anemone
and pomacentrid fish.
32. Davies J. T., J. Theoret. Biol., 8, 1—7, 1965. Theory of odors.
32a. Desgranges J.-C., Comp. Rend. Acad. Sci. Paris, 263, 1103—1106, 1966. Ultra-
structure of taste endings on barbels of catfish.
33. Dethier V. G., Biol: Bull., 72, 7—23, 1937; 76, 325—329, 1939; also Amer. Na-
turalist, 75, 61—73, 1941. Chemoreception by lepidopteral larvae in relation to
food plants.
34. Dethier V. G., Amer. J. Physiol., 165, 247—250, 1951; also J. Gen. Phvsiol.,
35, 55—65, 1951; also Biol. Bull., 102, 111—117, 1952; 103, 178—189, '1952.
Stimulation of tarsal chemoreceptors in flies by organic molecules.
35. Dethier V. G., Chadwick L. E., J. Gen. Physiol., 33, 589—599. 1950. Relation
between solubility and stimulating effect of organic and inorganic compounds.
36. Dethier V. G. In: Insect Physiology, edited by K. D. Roeder, John Wiley and
Sons, New York, 1953, pp. 544—576. Review of chemoreception in insects.
37. Dethier V. G. et al., Biol. Bull., Ill, 204—222, 1956; also Dethier V. G.,
Evans D. R., Biol. Bull., 120, 108—116, 1961. Relation between taste and in-
gestion of carbohydrates and water, blowflies.
38. Dethier V. G., Science, 161, 389—391, 1968. Chemosensory input and taste
discrimination in blowfly.
39. Dethier V. G., Kuch J. H., Z. vergl. Physiol., 72, 343—363, 1971. Gustatory
responses of lepidopteran larvae.
40. Doetsch G. S., Ganchron J. J., Nelson L. M., Erickson R. P. In: Olfaction and
Taste, 3d, edited by C. Pfaffmann, Rockefeller University Press, New York,
1969, pp. 492—511. Information processing in taste system.
41. Doeing R. B., Acta Physiol. Scand., 66, 290—299, 1966. Olfactory influences
on secondary neurons in fish Lota.
42. Doving R. B., Acta Physiol. Scand., 68, 404—418, 1966. Responses from olfa-
ctory bulb of frog.
______Хеморецепция________________________ 469
43 Deving К. В., Genne G., J. Neurophysiol., 28, 139—153, 1965; 29, 665—674,
4 ' 1966 Efferent olfactory system in fish.
44 Elsner T., Meinwald J., Science, 153, 1341—1350, 1966. Defensive secretions of
arthropods.
43 Evans D. R. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman;,
pergamon Press, New York, 1963, pp. 165—175. Chemical structure and sti-
mulation by carbohydrates.
46 Evans D. R., Mellon D., J. Gen. Physiol., 45, 487—500, 651—661, 1962. Stimu-
lation of primary receptors by salt and water.
47 Feir D., Beck S. D., Ann. Entom. Soc. Amer., 56, 224—229, 1963. Feeding, On-
copeltus. _
48. Feir D., Lengy J. I., Owen W. B„ J. Insect. Physiol., 6, 13—20, 1961. Contact
chemoreception in mosquito.
49 Fischer R„ Griffin F., Nature, 200, 343—347, 1963. Quinine dimorphism.
49a. Fraenkel G., Science, 129, 1466—1470, 1959. Chemical relations of food plants
and insects.
50. Fraenkel G„ EntomOl. Exp. Appl., 12, 473—486, 1969. Review of function of
secondary plant substances.
51. Frisch K. von, Z. vergl. Physiol., 21, 1—156, 1934. Comparative physiology of
taste, particularly in honeybees.
52. Funakoshi M., Zotterman Y., Acta Physiol. Scand., 57, 193—200, 1963. Effect
of salt on sugar response of taste endings.
53. Gesteland R. C., Ann. N. Y. Acad. Sci., 116, 440—447, 1964. Initial events of
electro-olfactograms.
54. Gesteland R. C., Lettvin J. Y., Pitts W. H., Chung S. H. In: Cybernetic Prob-
lems in Bionics, edited by H. L. Oestreicher and D. R. Moore, Gordon & Breach
Science Publishers, New York, 1968, pp. 313—322.
55. Gesteland R. C., Lettvin J. У., Pitts W. H., Rojas A. In: Olfaction and Taste:
A Symposium, edited by Y. Zotterman, Pergamon Press, New York, 1963,.
pp. 19—34. A code in the nose.
56. Goz H., Z. vergl. Physiol., 29, 1—45, 1941. Olfactory discrimination by fish.
57. Grant D„ Anderson O., Twitty V., Science, 160, 1354—1355, 1968. Homing
orientation by olfaction in newts.
58. Hara T. J., Ueda K„ Gorbman A., Science, 149, 884—885, 1965. Brain respon-
ses to odors in salmon.
59. Hara T. J., Comp. Biochem. Physiol., 22, 199—208, 1967. Electrophysiology oF
olfactory system in goldfish.
60. Hasler A. D., Science, 132, 785—792, 1960. Guideposts of migrating fish.
61. Hasler A. D„ Wisby W. J., J. Amer. Fish Soc., 79, 64—70, 1950. Discrimina-
tion of steam odors and pollutants by fish.
62. Hasler A. D., Wisby W. J., Amer. Nat., 85, 223—238, 1951. Discrimination of
stream odors and pollutants by fish.
63. Hasler A. D., Wisby W. J., Quart. Rev. Biol., 31, 200—209, 1956. Discrimina-
tion of stream odors and pollutants by fish.
64. Hodgson E. S., Roeder R. S., J. Cell. Comp. Physiol., 48, 51—76, 1956. Respon-
ses of chemoreceptor hairs in Diptera.
65. Hsiao T. H„, Fraenkel G., Ann. Entom. Soc. Amer., 61, 44—54, 476—503.
1968. Action of secondary plant substances on Colorado potato beetle.
66. Hughes J. R„ Hedrix D. E„ Wetzel H„ Johnston J. W. In: Olfaction and Taste,
3d, edited by C. Pfaffmann, Rockefeller University Press, New York, 1969,
pp. 172—192. Electrophysiology of olfactory bulb and subjective response to-
odor in man.
6Л Jacobson M., Beroza M., Yamamoto R. T., Science, 139, 48—49, 1963. Sex:
attractants of cockroach.
8- Jacobson M., Lilly С. E., Harding C., Science, 159, 208—209, 1968. Sex attra-
ctants of sugar beet wireworm.
Jacobson M. et al., Science, 170, 542—544, 1970. Sex pheromones of army
worm moth. J
470
Глава 13
70. Jahan-Parwar В., Smith M., Baumgarten R. von, Amer. J. Physiol., 216
1246—1257, 1969. Chemical stimulation in nervous system of Aplysia.
71. J er my T., Hanson E. E., Dethier V. G., Entomol. Exp. Appl., 11, 211—230,
1968. Induction of food preference in lepidopteran larvae.
72. Jones W. A., Jacobson M., Martin D. F., Science, 152, 1516—1517, 1966. Sex
attractans of pink bollworm moth.
73. Junge D., Brodwick M. S„ Comp. Biochem. Physiol., 35, 623—630, 1970. Sti-
mulation of frog gustatory units by calcium.
74. Kafka W. A., Z. vergl. Physiol., 70, 105—143, 1970. Chemostimulation in Lo-
custa.
75. Kaissling К. E. In: Olfaction and Taste, 3d, edited by C. Pfaffman, Rockefel-
ler University Press, New York, 1969, pp. 52—70. Kinetics of olfactory re-
ceptor potentials.
76. Kay R. E., Eichner J. T., Gelvin D. E., Amer. J. Physiol., 213, 1—10, 1967;
220, 1473—1487, 1971. Olfactory receptor potentials of Lucilia.
77. Kleerekoper H. In: Olfaction and Taste, 2d, edited by T. H. Hayashi, Perga-
mon Press, New York, 1967, pp. 625—645. Some effects of olfactory stimula-
tion on locomotory patterns in fish.
78. Kleerekoper H., Olfaction in Fishes, Indiana University Press, 1969, 222 pp.
79. Konishi J., Zotterman Y., Acta Physiol. Scand., 52, 150—161, 1961. Taste fun-
ction in carp.
SO. Konishi J., Zotterman У. In: Olfaction and Taste: A Symposium, edited by
Y. Zotterman, Pergamon Press, New York, 1963, pp. 215—233. Taste functions
in fish.
SI. Kriner M., Z. vergl. Physiol., 21, 317—342, 1934. Chemical discrimination by
minnows.
82. Kubota T., Kubo I., Nature, 223, 97—99, 1969. Bitterness and chemical stru-
cture.
S3. Kurihara K., Kurihara Y., Beidler L. M. In: Olfaction and Taste: A Sympo-
sium, edited by Y. Zotterman, Pergamon Press, New York, 1963. Isolation and
mechanism of taste modifiers.
84. Lacher V., Z. vergl. Physiol., 48, 587—623, 1964. Chemical receptors of ho-
neybee.
85. Laverock M. S., Comp. Biochem. Physiol., 2, 22—34, 1961. Tactile and chemi-
cal perception in earthworms.
86. Law J. H„ Wilson E. O., McClosky J. A., Science, 149, 544—545, 1965. Phero-
mones in termites, bees and ants.
87. Lenhoff H. M., Comp. Biochem. Physiol., 28, 571—586, 1969. pH profile of
peptide receptor.
88. de Lorenzo A. J D. In: Olfaction and Taste: A Symposium, edited by Y. Zot-
terman, Pergamon Press, New York, 1963, pp. 5—18. Ultrastructure of cell
membranes and synapses in chemoreceptors.
89. Maier S. F., Zahorik D. M., Wallin R„ J. Comp. Physiol. Psychol., 74, 254—
262, 1971; Psychon. Sci., 17, 309—310, 1971. Specific vitamin hungers in
rats.
*90. McCutchan M. C„ Z. vergl. Physiol., 65, 131—152, 1969. Responses of chemo-
receptors of blowfly to organic acids.
91. McLeese D. W., J. Fish. Res. Bd. Canad., 27, 1371—1378, 1970. Chemoreception
in lobsters.
92. Miller I. J., J. Gen. PhysioL, 57, 1—25, 1971. Interaction among taste papillae
in rat.
93. Moncrieff R. W., The Chemical Senses, 3rd ed., Chemical Rubber Co., Cle-
veland, 1967, 700 pp.
:94. Morita H. In: Olfaction and Taste, 3d, edited by C. Pfaffmann. Rockefeller
University Press, New York, 1969, pp. 370—381. Electrical signs of taste
receptor activity.
95. Morita H„ Shiraishi A., J. Gen. Physiol., 52, 559—584, 1968. Stimulation of
sugar receptors of flesh fly.
Хеморецепция_____________ _____________________1ZL
ng Moulton D. G., Cold Spring Harbor Symp. Quant. BioL, 30, 201—206, 1965.
У ’ Differential sensitivity to odors.
Q7 Mozell M. M., Science, 143, 1336—1337, 1964; also J. Gen. Physiol., 50, 25—
y 41, 1966; 56, 43—63, 1970. Spatiotemporal analysis of odor by olfactory re-
ceptor sheet.
98 Nayar J. K-, Fraenkel G„ J. Insect Physiol., 8, 505—525, 1962. Host plant
selection in silk moth Bombyx mori.
99 Nayar J. K., Fraenkel G., Ann. Entom. Soc. Amer., 56, 119—122, 174—178,
1963. Host selection by sphinx Ceratomia and Mexican bean beetle Epi-
lachna.
100. Neurath H., Z. vergl. Physiol., 31, 609—626, 1949. Olfactory sense in min-
nows.
101. Ottoson D., Acta Physiol. Scand., 43, 167—181, 1958. Electro-olfactogram of
frog.
102. Ottoson D., Acta Physiol. Scand., 47, 136—172, 1959. Olfactory bulb potentials,
rabbit and frog.
103. Ottoson D. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman,
Pergamon Press, New York, 1963, pp. 35—44. Generation and transmission
of olfactory signals.
104. Parkes A. J. Reprod. Fertil., 1, 312—314, 1960. Odors in mammalian repro-
duction.
105. Pfaffmann C. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman,
Pergamon Press, New York, 1963, pp. 257—273. Taste stimulation and pre-
ference behavior.
106. Pfaffmann C. et al. In: Olfaction and Taste, 2d, edited by T. H. Hayashi, Per-
gamon Press, New York, 1967, pp. 361—381. Sensory and behavioral factors
in taste preferences.
107. Pfaffmann C., editor, Olfaction and Taste, 3d, Rockefeller University Press,
New York, 1969.
108. Phillips C. G., Powell J. P. S., Shepherd G. M. J. Physiol., 156, 26P —27P,
1961; 168, 65—88, 1963. Responses of mitral cells to olfactory stimulation,
rabbit.
109. Price J. L., Brain Res., 7, 483—486, 1968. Termination of centrifugal fibers in
olfactory bulb.
110. Price J. L., Powell T. P. S., J. Cell. Sci., 7, 91—188, 1970. Granule cells of
olfactory bulb.
111. Rail W. D., Shepherd G. M., J. Neurophysiol., 31, 884—915, 1968. Rabbit
mitral cells.
112. Ralls K., Science, 171, 443—449, 1971. Mammalian scent marking.
113. Roessler H. P., Z. vergl. Physiol., 44, 184—231, 1961. Chemical attraction of
Aedes.
113a. Rozin P. In: Handbook of Physiology, Vol. 1, edited by C. F. Code, Amer.
Physiol. Soc., Washington D.C., 1967, pp. 411—431. Thiamine specific hun-
ger.
114. Radinsky J. A., Science, 152, 218—219, 1966. Responses of wood beetles to
terpenes.
15. Sato M., Yamashita S., Ogawa H. In: Olfaction and Taste, 3d, edited by
C. Pfaffmann, Rockefeller University Press, New York, 1969, pp. 470—488.
lift Afferent specificity in taste.
6- Saxena K. N., J. Insect Physiol., 9, 47—71, 1963. Ingestion by heteropterous
. insect Dysdercus.
1Л Saxena K. N. In: Olfaction and Taste, Vol. 2, edited by T. H. Hayashi. Per-
gamon Press, New York, 1967. Some factors governing olfactory and gusta-
Hr еГУ resP°nses of insects, pp. 799—819.
’ Saxena K. N., Entomol. Exp. Appl., 12, 751—766, 1969. Changes in food attra-
11Q cuon with hydration, Dysdercus.
rn^ih^er & ’ J' Insect Physi°l-> 15—30, 1962. Olfactory specificity among
l2°- Schneider D. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman,
*472
Глава 13
Pergamon Press, New York, 1963, pp. 85—103. Electrophysiology of insect
olfaction.
121. Schneider D., Symp. Soc. Exp. Biol., 20, 273—297, 1965. Chemical sense com-
munication in insects.
122. Schneider D., Science, 163, 1031—1037, 1969. Insect olfaction.
123. Schneider D„ Block В. C., Boeckh J., Priesner E., Z. vergl. Physiol., 54, 192—
209, 1967. Reaction of male moths to bombykol.
124. Schneider D., Lacher V., I\ais sling К.. E., Z. vergl. Physiol., 48, 632—662, 1964.
Reaction spectrum of odors in saturnid moths.
125. Schneider D., Steinbrecht R. A., Symp. Zool. Soc. bond., 23, 279—297, 1968.
Check list of insect olfactory sensilla.
126. Schoonhouen L. M., Ann. Rev. Entom., 13, 115—136, 1968; also Schoon-
houen L. M., Dethier V. G., Arch. Need. Zool., 16, 497—530, 1966. Sensory
aspects of discrimination of host plant by insect larvae.
>127. Schultze-W estrum T., Z. verg. Physiol., 50, 151—220, 1965. Olfactory commu-
nication in marsupial phalangers.
128. Schwarz R., Z. vergl. Physiol., 37, 180—210, 1955. Olfactory thresholds of
honeybee.
129. Scott J. W., Pfaffmann C., Science, 158, 1592—1594, 1967. Olfactory input to
hypothalamus.
130. Shibuya T., Japan J. Physiol., 10, 317—326, 1960. Electrical response, olfacto-
ry epithelium of fishes.
<131. Shiraishi A., Morita H., J. Gen. Physiol., 53, 450—470, 1969. Labellar sugar
receptors of flesh fly.
'432. Shiraishi A., Kubara M., J. Gen Physiol., 56, 768—782, 1970. Responses of
chemosensory hairs to amino acids.
133. Slifer E., J. MorphoL, 105, 145—191, 1959. Ultrastructure of chemoreceptors,
grasshoppers.
134. Steinbrecht R. A. In: Olfaction and Taste, 3d, edited by C. Pfaffmann, Rocke-
feller University Press, New York, 1969, pp. 3—21. Comparative morphology
of olfactory receptors.
135. Sutterlin A. M., Sutterlin N., J. Fish. Res. Bd. Canad., 27, 1927—1942, 1970;
28, 565—572, 1971. Olfactory and taste responses of salmon.
136. Takagi S. F„ Aoki K., Hho M., Yajima T. In: Olfaction and Taste 3d edited by
C. Pfaffmann, Rockefeller University Press, New York, 1969, pp. 92—108.
Electropositive potentials in olfactory epithelia.
137. Tateda H., Beidler L. M., J. Gen. Physiol., 47, 479—486, 1964. Receptor poten-
tial in taste cell of rat.
138. Teichman H., Z. vergl. Physiol., 42, 206—254, 1959. Olfaction in fish.
139. Thorpe W. H., Proc. Roy. Soc. Lend. B, 127, 424—433, 1939. Preimaginal
olfactory conditioning in insects.
140. Thorsteinson A. J., Canad. J. Zool., 31, 52—72, 1953. Chemotactic responses
of Lepidoptera.
141. Tucker D., J. Gen. Physiol., 46, 453—489, 1963. Physical variables in olfactory
stimulation.
142. Tucker D., Nature, 207, 34—36, 1965. Olfactory responses of birds.
143. Tucker D., Shibuya T., Cold Spring Harbor Symp. Quant. Biol., 30, 207—215,
1965. Physiology and pharmacology of olfactory receptors, turtle.
144. Ueda К., Hara T. J., Gorbman A., Comp. Biochem. Physiol., 21, 133—143, 1967.
Electro-olfactograms in salmon.
144a. Waldbauer G. P., Entom. Exper. Appl., 7, 253—269, 1964. Utilization of sola-
naceous plants by tobacco hornworm.
145. Walsh R. R., Amer. J. Physiol., 186, 255—257, 1956. Single unit response,
olfactory bulb of rabbit.
146. Wells M. J., J. Exp. Biol., 40, 187—193, 1963. Taste responses in Octopus.
147. Wilson E. O., Science, 149, 1064—1071, 1965. Chemical communication in so-
cisl insects
148. Wolbarsht M. L., Cold Spring Harbor Symp. Quant. Biol., 30, 281—288, 196-5.
Receptor sites in insect chemoreceptors.
Хеморецепция
473-
149. Yamamoto С., Yamamoto Т., Iwama К., J. Neurophysiol., 26, 403—415, 1963.
Inhibitory system in olfactory bulb.
150. Yamamoto R. T., Jenkins R. Y., McClusky R. K., Entom. Exp. Appl., 12, 504—
508, 1969. Selection of plants by tobacco hornworm.
151. Zotterman У., Experientia,- 6, 57—58, 1950; also Acta Physiol. Scand., 37,.
60—70, 1956; also Ann. N. Y. Acad. Sci., 81, 358—366, 1959. Species differen-
ces in water, sweet, and salt tastes.
152. Zotterman Y. In: Olfaction and Taste: A Symposium, edited by Y. Zotterman..
Pergamon Press, New York, 1963, pp. 205—216. Studies in the neural mecha-
nisms of taste.
Глава 14
Фоторецепция и зрение
7. ГОЛДСМИТ
Фотобиология и электромагнитный спектр
Введение
В биологии часто говорят о приспособлении организмов к окру-
жающей среде, но при этом всегда имеется в виду, что у данного
вопроса есть и другая сторона. Как указывал несколько лет назад
Гендерсон в своей книге «Взаимное соответствие в окружающей
среде», многие физические характеристики нашей планеты кажутся
удивительно хорошо согласованными с жизнью. Взаимное соответ-
ствие биологических процессов и физических условий окружающей
среды нигде не проявляется так убедительно, как в сфере фото-
биологии.
Электромагнитный спектр охватывает широкий диапазон длин
волн, простираясь от у-лучей с длиной волны меньше 10-3 А
(10~13 м), до радиоволн, длина которых измеряется километрами
(103 м). Фотобиологические процессы приурочены почти исключи-
тельно к узкой полосе спектра от 300 до 900 нм (рис. 14-1), и это
обусловлено двумя взаимосвязанными причинами [237].
С одной стороны, именно эти излучения преобладают около по-
верхности Земли. Максимум спектра солнечного света лежит в об-
ласти, имеющей наибольшее значение для фотобиологических про-
цессов, и на диапазон длин волн от 300 до 1100 нм приходится три
четверти всей солнечной энергии. На пути к биосфере эта полоса
длин волн сужается вследствие поглощения света в атмосфере и
воде (рис. 14-2). В коротковолновой области спектра поглощение
обусловлено озоном верхних слоев атмосферы, который препятст-
вует проникновению к поверхности Земли большей части солнечно-
го света с длинами волн короче 290 нм. Поглощение длинноволно-
вой части солнечного излучения обусловлено СО2, парами воды и
озоном. Излучение, проникающее сквозь толщу чистой воды озер
или открытого океана, сосредоточено в узкой полосе длин волн с
максимумом в голубой области спектра от 475 до 480 нм. В воде,
содержащей большое количество ила или органических остатков,
свет поглощается еще интенсивнее; кроме того, наличие в среде
различных поглотителей сдвигает спектр в сторону более длинных
волн — в желтую или даже оранжевую область [121, 224].
С другой стороны, именно указанные излучения энергетически
лучше всего соответствуют фотобиологическим процессам. Фото-
Фото рецепция и зрение
475
биологические реакции — это специфические химические реакции,
иля осуществления которых нужна энергия, и в фотобиологии эту
энергию поставляет свет. Молекулы поглощают энергию дискрет-
ными порциями, называемыми фотонами или квантами-, энергию
кванта можно вычислить, исходя из соотношения Е=-%-, где h (по-
Фотохимические реакции
фотосинтез в бактериях
фотосинтез в зеленых растениях
Фотоморфогенные реакции
растений
Фототропизм растений
Фототаксис водорослей,
Фотопериодизм насекомых
Зрение пчелы
Зрение человека
Килокалопии на моль фотоноа
Рис. 14-1. Некоторые наиболее важные фотобиологические процессы и соответ-
ствующие спектральные диапазоны.
Относительно экспериментальных данных и ссылок на них см. [28, 34, 46, 83, 90, 102, 180,
188, 234, 237, 238, 266].
стоянная Планка) = 1,58-10-34 кал-с, с (скорость света) —
=3-1010 см‘С-1 и X — длина волны света в см. На рис. 14-1 и 14-2
приведены также энергетические эквиваленты различных длин
волн света в ккал-эйнштейн-1 (один эйнштейн — это 1 моль кван-
Тов, т. е. 6,02-1023 фотонов). Энергия активации реакций между ор-
ганическими соединениями обычно лежит в диапазоне от 15 до
о ккал -моль-1 (от 1900 до 440 нм). Фотохимическая реакция на-
инается с перехода валентного электрона на более высокий энер-
тический уровень, „г и этот процесс требует затраты более
ккал/моль (что соответствует Z<700 нм). Таким образом, при
реходе в инфракрасную область энергия квантов становится не-
статочной для того, чтобы при их поглощении началась фотохи-
Мическая реакция.
Рис. 14-2. Сужение спектра солнечного света на пути к биосфере, обусловленное
поглощением [237].
В коротковолновой области поглощение связано с озоном верхних слоев атмосферы, а длин-
новолновое излучение поглощается водой. Сплошной линией </тах) показаны длины волн
максимумов интенсивности, а пунктирными (/ц)%) отмечен диапазон длин волн, в котором
сосредоточено 90% солнечной энергии на данной высоте над уровнем моря или на данной
глубине океана. Буквы под шкалой длин волн (УФ, Ф, Г и т. д.) означают ультрафиолете
к'зый, фиолетовый, голубой и т. д. Ниже показано, какое место занимает приведенный выы
спектр на общей шкале электромагнитных волн.
Фоторецепция и зрение
477
При длинах волн короче 300 нм (95 ккал/моль) возникает дру-
гая проблема. Здесь фотоны обладают энергией, достаточной для
того, чтобы перевести в возбужденное состояние и вызвать фотохи-
мические повреждения в молекулах белков и нуклеиновых кислот,
структурная целостность которых является необходимым условием
нормального функционирования всего живого. Отсюда можно за-
ключить, что возникновение земных форм жизни требовало в каче-
стве предварительного условия появления озона в верхних слоях
атмосферы для поглощения той части солнечного излучения, кото-
рая гибельна для жизненно важных макромолекул [238]. Из всего
сказанного ясно, почему фотобиологические процессы протекают
именно в указанном выше спектральном диапазоне.
Некоторые фотобиологические реакции
Какие же фотобиологические процессы известны? На рис. 14-1
перечислены наиболее важные из них с указанием спектральных
диапазонов, в которых они протекают.
В ходе фотосинтеза запасается большое количество энергии
(или отрицательной энтропии), которую могут использовать другие
организмы. Другие процессы, протекающие при участии света, ха-
рактеризуются тем, что в их ходе не происходит превращения энер-
гии солнечного света в потенциальную энергию химических соеди-
нений, а свет используется для управления или получения инфор-
мации об окружающей среде. Например, фотопериодические
реакции растений, осуществляемые с помощью пигмента фитохро-
ма,— это адаптивные реакции, синхронизирующие различные этапы
репродуктивного цикла с наиболее подходящим временем года
,[214]. У животных фотопериодические реакции, опосредуемые раз-
личными пигментными системами, регулируют сезон размножения
[267] или синхронизируют время осуществления различных стадий
развития с наиболее благоприятным временем года или даже вре-
менем суток (у насекомых) [34, 83, 137, 264].
Термины фототаксис и фототропизм относятся к реакциям орга-
низмов, которые управляются светом и создают благоприятные ус-
ловия для удовлетворения потребностей организмов, таких, напри-
мер, как питание [82]. Фототропизм — это изгибание частей рас-
тений или прикрепленных организмов, например сидячих кишечно-
полостных [140], по направлению к свету или от него. Фототаксис
'Существует у незакрепленных организмов.
Фототаксис и фототропизм — это скорее реакции целостного ор-
ганизма, чем какой-то один механизм реакции на свет, поэтому эти
термины объединяют ряд различных явлений (табл. 14-1). Напри-
МеР, фототаксис (клинокинез, по терминологии табл. 14-1) фото-
синтезирующих бактерий имеет тот же спектр действия, что и фо-
тосинтез, протекающий в этих бактериях. Уменьшение интенсивно-
€ти света заставляет бактерии менять направление их движения на
478
Глава 14
Таблица Ц.}
Простые движения животных [82]
Вид движения
Кинез
Ортокинез
Клинокинез
Таксис
Клинотаксис
Тропотаксис
Т елотаксис
Характеристика
Ненаправленные движения; ось тела по отношению к раздра-
жителю не ориентирована
Скорость или частота движений зависит от интенсивности
света
Частота или число поворотов в единицу времени зависит от
интенсивности света
Направленные движения; ось тела определенным образом
ориентирована по отношению к раздражителю
Ориентация возникает косвенным путем; интенсивность дей-
ствующего на рецептор раздражения меняется в связи с ре-
гулярными поворотами тела
Ориентация возникает прямым путем; об интенсивности осве-
щения с обеих сторон в каждый момент времени сигнали-
зирует билатеральная пара рецепторов; ориентация опреде-
ляется уравниванием интенсивностей
Близок к тропотаксису, но ориентация определяется каким-
либо одним источником света даже в присутствии конкури-
рующих источников
обратное. В результате они скапливаются в наиболее освещенных
местах [46, 47]. Адаптивное значение этого явления совершенно
понятно.
Фототаксис зеленых водорослей отличается от вышеописанного
тем, что направление движения определяется положением источ-
ника света: водоросли плывут к нему или от него. Этот процесс
осуществляется с помощью фоторецептора, расположенного в осно-
вании жгутика, но не идентичного «глазному пятну». Во время пла-
вания животное поворачивается вокруг своей продольной оси, и
если его траектория находится под прямым углом по отношению
к направлению на источник света, фоторецептор попеременно то
освещается, то затемняется глазным пятном (у некоторых видов,
возможно, хлоропластом). По-видимому, меняя направление дви-
жения по отношению к источнику света, животное может увеличи-
вать или уменьшать освещенность фоторецептора [47].
В качестве третьего примера укажем насекомых, у которых
фототаксис определяется работой глаз и центральной нервной си-
стемой.
У многих животных есть фотореакции, которые нельзя связать
ни с каким специфическим многоклеточным органом [162]. Напри-
мер, движения Amoeba [145], движения игл морского ежа Diadetna
[163], разряд фоторецепторных нейронов шестого брюшного ганг-
лия речного рака Procambarus [126] и зависимость времени выхо-
да из диапаузы куколок шелковичного червя от долготы ДнЯ
[264] — все это случаи, в которых точная локализация фоторецеп-
Рис. 14-3. Простые глаза.
Развитие дирекциональной чувствительности и способности формировать изображение
идет через образование чаш, выстланных рецепторами и окруженных темным пигментом
(слева), или линз, помещенных перед рецепторами (справа), или комбинацией того и дру-
гого. Линзы позволяют лучше собирать световую энергию [164]. Б. Разрез простого глазка
плоского червя Planaria. Два глаза расположены на дорсальной поверхности животного;
пигментные чаши открыты с латеральных направлений. Фоторецепторные нейроны снабжены
Раодомерными окончаниями, внедряющимися в полость пигментной чаши [164]. В. Двой-
ая сетчатка «простого» глаза гребешка Pecten. Дистальная сетчатка содержит фоторецеп-
D ры> Развившиеся из ресничек; проксимальная сетчатка содержит рабдомерные рецепто-
ру [152]. 1 — стенка тела; 2 — рабдомерное окончание; 3 — пигментная чаша; 4 — аксон;
щР“Клеточное тсло фоторецептора; 6 — роговица; 7 — линза; 8 — перегородка; 9—отражаю-
слой; 10 — пигмент; // — проксимальная сетчатка; 12— зрительный нерв; 13— дисталь-
я ветвь нерва; 14 — проксимальная сетчатка; 15 — дистальная сетчатка; 16 — пигментный
эпителий.
480
Глава 14
торов неизвестна, хотя в последних двух примерах они определен-
но находятся в центральной нервной системе. У дождевых червей
Lumbricus светочувствительные клетки разбросаны по всему телу
[37, 186]. Есть много других примеров, когда небольшие группы
специализированных светочувствительных клеток образуют ма-
ленькие глазные пятна, или оцеллии. Примеры некоторых из таких
морфологически примитивных фоторецепторных систем показаны
на рис. 14-3.
Пигменты фоторецепторов, которые поглощают свет и вызыва-
ют реакцию, для многих из этих относительно неспециализирован-
ных систем неизвестны. Часто пытались установить спектр погло-
щения пигмента фоторецептора, измеряя спектр действия. С этой
целью экспериментатор определял относительное число фотонов со-
ответствующих длин волн, необходимое для того, чтобы вызвать
некоторую постоянную физиологическую реакцию. Соответствую-
щий пример показан на рис. 14-35. Именно на таких данных осно-
вана оценка специфических спектральных диапазонов, указанных
на рис. 14-1. Экспериментов такого рода самих по себе, как прави-
ло, недостаточно для установления химической природы пигмента
рецептора.
Выяснена роль хлорофилла в фотосинтезе [188] и фитохрома
в фотоморфогенетических реакциях растений [214, 266]. Для таких
фотопериодических реакций, как диапаузы насекомых [137], цир-
кадный ритм вылупления Drosophila [270], фототаксис самых низ-
ших организмов [47, 102], фототропические реакции грибов [180]
и высших растений [28], равно как и для целого ряда других ре-
акций низших беспозвоночных [93], рецепторные пигменты неиз-
вестны.
Большая часть информации о фотобиологических процессах у
животных получена из опытов по исследованию зрительных систем
животных с крупными глазами. В последующих разделах главы мы
рассмотрим природу рецепторных пигментов, механизм возбужде-
ния рецепторных клеток, способы обработки информации до уров-
ня поведенческой реакции.
Цитологическая организация фоторецепторных клеток
Обычно фоторецепторы имеют органеллы, образованные разви-
той плазматической мембраной, хотя у некоторых чувствительных к
свету клеток (например, Amoeba) [145] отсутствуют какие-либо
видимые признаки морфологической специализации. Структуры»
увеличивающие площадь поверхности, на которой помещается ф°"
топигмент, делятся на два основных класса в соответствии с тем,
связаны они с базальным тельцем ресничек (цилий) или нет [72,
74]. Разделение это интересно в том отношении, что фоторецепто-
ры, развивающиеся из ресничек, обнаружены в основном у позво-
ночных или у вторичноротых (т. е. в тех филогенетических ветвях
беспозвоночных, которые близки к главному стволу позвоночных)»
Фоторецепция и зрение
481
тогда как нецилиарные (или рабдомерные) образования характер-
ны для первичноротых (кольчатых червей, моллюсков, членисто-
ногих) .
Цилиарные образования у беспозвоночных
Парафлагеллярное тело одноклеточной зеленой водоросли
Euglena, возможно, является фоторецептором, связанным со жгу-
тиком, который не потерял способности двигаться [74, 1361. Па-
пафлагеллярное тело является не специализированной частью кле-
точной мембраны, а скорее вздутием, полностью заключенным
внутри жгутика. Глазное пятно (или стигма) представляет собой
чашу с пигментными гранулами (возможно, каротиноид) [92], ко-
торая лежит на поверхности жгутика и считается пассивным зате-
няющим устройством. Природа фотопигмента, находящегося внут-
ри парафлагеллярного тела, и способ, посредством которого воз-
буждение приводит к ориентированным плавательным движениям,
неизвестны.
Чаще всего формирование фоторецепторных органелл связано
с увеличением площади поверхности клеточной мембраны, а не с
развитием внутриклеточных структур, аналогичных парафлагел-
лярному телу.
Модификация ресничек для формирования сенсорных оконча-
ний не ограничивается только случаем фоторецепторов: некоторые
механорецепторы и ольфакторные окончания также являются про-
изводными ресничек. В общем случае сенсорные реснички теряют
две центральные микротрубочки и «ручки» колец 9 дублетов [74]
(гл. 12).
В дистальной сетчатке морского гребешка Pecten (один из слу-
чаев, когда цилиарные рецепторы обнаружены у первичноротых, и,
следовательно, исключение из отмеченного выше филогенетическо-
го обобщения) фоторецепторные клетки имеют по нескольку меш-
кообразных выростов, каждый из которых представляет собой
сильно уплощенную ресничку [10]. У кишечнополостных, иглоко-
жих и некоторых других животных (рис. 14-4) мембрана каждой
реснички бывает разделена на пальцеобразные выросты, тогда как
в фоторецепторных клетках личинок асцидий цилиарная мембрана
изгибается, образуя ряд параллельных дисков, ориентированных
приблизительно параллельно продольной оси реснички [74, 94].
Этот последний тип организации напоминает палочки и колбочки
сетчатки позвоночных.
Палочки и колбочки сетчатки позвоночных
Фоторецепторные клетки сетчатки позвоночных — наиболее вы-
сокоорганизованные из цилиарных фоторецепторов. Строение па-
лочковой клетки показано на рис. 14-5. Клетка состоит из внутрен-
него и наружного сегментов. Наружный сегмент содержит стопку
i з 5оо—1000 мембранных дисков, заключенных внутрь плазматиче-
l3l'~1514
Членистоногие
Простейшие
Рис. 14-4. Схематическое изображение фоторецепторных органелл отдельны
групп «цилиарной» и «рабдомерной» линий эволюции.
(Evolutionary Biology, vol. 2, edited by Th. Dobzhansky, M. K. Hecht and Wm. C. Steere.
I
Фоторецепция и зрение
483
ской мембраны. Наружный сегмент соединяется с внутренним по-
средством ножки, в которой, если рассматривать поперечный срез
Рис. 14-5.
/1. Схема строения палочек из сетчатки млекопитающих по данным электронно-микроскопи-
ческих исследований [213]. Б. Область соединения внутреннего и наружного сегментов в уве-
личенном виде [193]. / — наружный сегмент; 2 — внутренний сегмент; 3 — волокно палочки,
4 — синаптическое тельце; 5 — соединительная ресничка; 6 — ядро; 7 — дендрит биполярного
нейрона; 8— диски; 9— поверхностная мембрана; 10 — цилиарные нити; 11 центриоли,
12 — эндоплазматическая сеть; 13 — митохондрии.
с помощью электронного микроскопа, обнаруживаются характер-
ные 9 пар микротрубочек. Диски являются выростами плазматиче-
ской мембраны. В колбочковых клетках многие из них сохраняют
связь с поверхностной мембраной, и, следовательно, их содержи-
мое сообщается с экстраклеточной средой. У палочек, напротив, по
имеющимся данным, диски полностью отделяются от плазматиче-
ской мембраны [48, 64]. Осмотические измерения показывают, что
содержимое дисков близко скорее к внутриклеточной, чем к вне-
клеточной среде [49, 64]. Этот момент немаловажен, поскольку,
31*
484
Глава 14
как мы увидим, плазматическая мембрана — это структура, по ко-
торой в процессе возбуждения распространяется сигнал. Видимое
отсутствие связи между дисками наружных сегментов палочек и
Рис. 14-6. Некоторые типы палочек и колбочек позвоночных [246].
А. Обычная, или красная, палочка лягушки-леопарда Rana pipiens; темновая адаптация, ми-
оид сокращен. Б. Колбочка R. pipiens; темповая адаптация, миоид удлинен. В. Палочка че-
ловека из области, примыкающей к темпоральному краю фовеа. Г. Двойная колбочка чере-
пахи Chrysemys picta. Д. Близнецовые колбочки костистой рыбы Lepomis macrachirus; све-
товая адаптация, сросшиеся миоиды сокращены. I — наружный сегмент; 1' — наружный сег-
мент дополнительной колбочки: 2 — эллипсоид (митохондрии); 2'— эллипсоид дополнитель-
ного члена пары; 3 — миоид; 4 — наружная пограничная мембрана сетчатки; 5 — ядро;
6 — масляная капля; 7 — основание, или ножка; 8 — параболоид (гликоген).
плазматической мембраной клетки ставит вопрос о механизмах пе-
редачи возбуждения с диска на эту мембрану, которые до конца
не выяснены. Внутренний сегмент и остальная часть клетки содер-
жат обычный набор органелл [48, 64]. В непосредственной близости
к наружному сегменту, как правило, имеется много митохондрий;,
они образуют компактную массу, которая в световом микроскопе
видна как эллипсоид. Базальный конец палочковой клетки пред-
ставляет собой пресинаптическое окончание с синаптической лен-
той и пресинаптическими пузырьками. Более детальное описание
связи рецепторных клеток с другими нейронами сетчатки можно
найти на стр. 547—549.
Сетчатки обычно содержат два типа фоторецепторов: палочки
и колбочки. Палочки обеспечивают зрение при сумеречном свете
и не способны обеспечить цветное зрение. Колбочки, напротив, ра'
Фоторецепция и зрение
485
ботают при ярком свете и у некоторых видов обеспечивают цвет-
ное зрение. (Некоторые примеры существования такого «разделе-
ния труда» мы рассмотрим в следующем разделе.) Морфологиче-
ски палочки часто отличают от колбочек по более длинным и более
Линза
Рис. 14-7. Схема образования медиальных и латеральных глаз позвоночных из
стенки развивающегося мозга [73].
Рецепторы (а также другие элементы) образуются из цилиарных эпендимальных клеток.
Медиальный глаз развивается за счет простого выпячивания ткани, и фоторецепторные ор-
ганеллы смотрят по направлению к линзе. В латеральном глазе выпяченная ткань в центре
прогибается, образуя чашу с двойными стенками, а палочки и колбочки развиваются из
цилиарных клеток в направлении от линзы.
близким по форме к цилиндру наружным сегментам; у колбочек
наружные сегменты более короткие, почти конические. Эти разли-
чия, однако, не всегда отчетливо выражены. Например, в сетчатке
приматов фовеальные (центральные) колбочки тонкие и совсем не
конические. Другим отличительным признаком, который характе-
рен для рыб, амфибий, рептилий и птиц, но не для млекопитающих,
является наличие в колбочках масляной капли, расположенной
между внутренним и наружным сегментами (рис. 14-6). Сетчатка
позвоночных инвертирована (причуда эмбриологии), так что па-
дающий свет проходит сквозь внутренние сегменты, прежде чем
Достигает наружных (рис. 14-7). Следовательно, масляные капли
избирательно поглощают свет, прежде чем он дойдет до зритель-
ного пигмента колбочек.
Характерной чертой сетчаток всех позвоночных, за исключени-
ем млекопитающих, является присутствие двойных (амфибии, реп-
тилии, птицы) и близнецовых (костистые рыбы) колбочек
(рис. 14-6). Это пары клеток с тесным прилеганием внутренних
сегментов [48, 246]. Два члена двойной колбочки не вполне иден-
тичны: одна из них—главная—немного больше, с хорошо развитой
Масляной каплей, тогда как добавочная колбочка имеет относи-
тельно большой внутренний сегмент, но лишена масляной капли.
'J6e близнецовые колбочки морфологически сходны.
Головоногий моллюск
Octopus
Фоторецепция и зрение
487
Теменные и пинеальные глаза рыб, амфибий и рептилий имеют
фоторецепторные клетки с той же структурой, что и латеральные
глаза. [73]. Однако из-за различий в онтогенетическом развитии
этих глаз сетчатка пинеальных глаз не инвертирована (рис. 14-7).
Рабдомерные окончания
Второй основной класс фоторецепторных клеток характеризу-
ется наличием микровилл, не связанных с базальными телами или
центриолями [72, 74, 166]. Обычно на одном конце клетки содер-
жатся многочисленные микровиллы, а на другом начинается нерв-
ное волокно той или иной длины. В относительно неспециализиро-
ванных клетках микровиллы вырастают из нескольких участков по-
верхности сомы. У кальмара дистальный конец фоторецепторной
клетки вытянут в виде длинного плоского пальца, на обеих сторо-
нах которого имеются микровиллы [271]. У членистоногих фоторе-
цепторные клетки собираются в пучки или гроздья, причем каждая
клетка посылает пучки своих микровилл к центру, так что обра-
зуется аксиальная палочкоподобная структура, называемая рабдо-
мом. Каждая маленькая группа фоторецепторных клеток окружена
заполненными пигментом опорными клетками. Каждая группа ре-
цепторов (ретинула) вместе со своими пигментными клетками со-
ставляет омматидий, и сложные глаза образуются за счет скопле-
ния сотен или тысяч омматидиев. На рис. 14-8 показано несколько
способов образования рабдомов пучками микровилл окружающих
ретикулярных клеток.
Зрительные пигменты
Воздействие света на зрительные пигменты
Чувствительность фоторецепторных клеток к свету обусловлена
присутствием в них пигмента, называемого родопсином. (Термин
Рис. 14-8. Некоторые типы рабдомов.
Л. Муха (Arthropoda). У двукрылых и полужесткокрылых рабдомеры прорастают в цент-
ральную полость и остаются изолированными. Такой рабдом называется «открытым» в отли-
чие от «закрытого», показанного на Б и В. У мух имеется по восемь ретикулярных клеток.
Шесть из них (обозначенные цифрами 2, 3, 4, 5, 6 и 8) имеют рабдомеры, простирающиеся
по всей длине ретинулы. Две другие (обозначенные цифрами I и 7) имеют более короткие
рабдомеры, расположенные один над другим. Соответственно на поперечном срезе оммати-
дия виден только один из этих рабдомеров. На данной схеме црчение проходит через ди-
- альную половину омматидия и пересекает клетку 1 — верхнюю центральную клетку. Клет-
HVJI нижняя центральная клетка; она расположена целиком в базальной половине рети-
МрПы 11 3Десь не видна. На более проксимальном срезе был бы виден центральный рабдо-
ВеохнС°еДИНЯ1О1Ш'и’1СЯ с клеткоя была бы видна и клетка 1, но без своего рабдомера.
остапЯЯ 11 НИЖНяя центральные клетки имеют синаптические связи, отличные от связей
’более НЫХ шести Ретинулярных клеток. Б. Пчела (Arthropoda). Такая слитная организация
«Лин мТИПИЧИа для HaceKOMbIX- Каждая из восьми ретинулярных клеток посылает к центру
назывя КР°ВНУЛ по всей длине клетки. Компактная масса микровилл от всех восьми клеток
бокамаеТСЯ РабДомом. В. Рак (Arthropoda). У десятиногого рака микровиллы простираются по
смыкаРетинУляРПых клеток в виде «языков». Имеется семь клеток, и «языки» микровилл
моллюскСЯ’ обРазУя слои толщиной до 5 мкм. Г. Кальмар (Mollusca). Сетчатка головоногих
тгый «nanv Не „°Рганизована в омматидии. Каждая фоторецепторная клетка имеет вытяну-
аружныи сегмент», который состоит из центральной полоски цитоплазмы и двух ла-
теральных каемок микровилл.
488
Глава 14
родопсин, или зрительный пурпур, имеет также более узкий смысл:
так называют один из подклассов зрительных пигментов, найден-
ный в палочках позвоночных. Что именно имеется в виду в каждом
конкретном случае, обычно ясно из контекста.) Значение родопси-
на для зрительного процесса начали понимать почти сто лет назад,
когда Болл [21] и Кюне [130] показали, что розовый цвет темно-
адаптированной сетчатки быстро исчезает при освещении. В по-
следующие годы было получено много данных о свойствах родопси-
нов, взятых у разных животных, и стало ясно, что все они реагиру-
ют на свет сходным образом [25, 168, 241].
Родопсин представляет собой сложный белок, который служит
основным компонентом мембран фоторецепторных органелл. По-
скольку родопсин связан с липидами, растворить его можно только
с помощью детергентов. Для его молекулярного веса приводятся
значения от 28000 до 40 000; конкретные цифры зависят отчасти
от того, насколько хорошо были удалены липиды при приготовле-
нии экстракта [109, 116]. Диаметр молекул родопсина составляет
40—50 А, и они, по-видимому, располагаются в мембранах рецеп-
торных органелл в один слой [268].
Хромофором родопсина является жирорастворимое вещество
ретиналь, или альдегид витамина А, молекула которого содержит
20 атомов углерода (рис. 14-9, А, Б) [9]. Благодаря сложной си-
стеме одинарных и двойных связей ретиналь может существовать
в виде нескольких стереоизомеров. Один из них, 1 l-tywc-ретиналь
(рис. 14-9, В), обнаружен в родопсине [178, 244]. Ретиналь с по-
мощью шиффова основания связан с е-аминогруппой лизинового ос-
татка опсина (рис. 14-9, В) [2, 23]; правда, другая исследователь-
ская школа недавно выступила с обоснованием того, что шиффово
основание соединяет ретиналь с фосфатидилэтаноламином [185].
Предполагается, что белок взаимодействует с боковой цепью
11-1{«с-ретиналя весьма специфическим образом.
Под действием света хромофор переходит из 11-цис- в полно-
стью транс-конфигурацию [118]. В результате стерическое соответ-
ствие между ним и белком становится невозможным и изомериза-
ция сопровождается цепью конформационных изменений в белко-
вой части молекулы. В родопсине позвоночных эти изменения
приводят через ряд промежуточных состояний, которые можно
идентифицировать спектрофотометрически (рис. 14-10), к гидролизу
хромофора [1, 146, 241]. Под действием алкогольдегидрогеназы и
восстановленного никотинамидадениндинуклеотидфосфата (НАДФ)
[88] ретинальдегид обратимо переходит в витамин А [120]
(рис. 14-9, А, Б) и (по крайней мере у крысы) перемещается в пиг-
ментный эпителий [62]. Однако восстановление не является уни-
версальной реакцией: у многих членистоногих [92] и головоногих
моллюсков [119] цепь реакций заканчивается образованием ста-
бильного промежуточного продукта, аналогичного метародопсину I
(рис. 14-10). Подлинного процесса выцветания, т. е. потери цвета,.
СН3
Н2Сд бС
HgC3 2 <<>СН3
4f CH3
Н2
сн3
I °
НАДФ-Н
СНз
н СНз
н
Б
СНз
Н2с
н
I
/с
(3
сн3
!
с.
I
н
I I н
н?с. _Л-сн3
V сн3
НАДФ Н2
Полностью транс-ретинол
(витамин А)
н
I
Сх
I
н
СНз
,с сн2он
"с
I
н
I I Н
н2с. сн3
XfCH3
н2
11-цис-ретиналь, связанный
сопсином с помощью
шиффова основания
н
'Н
СН3 Л
N-H
Опсин
Полностью транс-З-дегидроретиналь.
(ретинен)
Рис. 14-9. Ретиналь и родственные соединения.
А. Ретиналь в полностью транс-конфигурации; показана система нумерации атомов углерода. Б. Ретинол (витамин А), который образуется
в сетчатке в результате восстановления альдегидной группы ретиналя с помощью восстановленного НАДФ и через посредство алкогольдегид-
рогеназы. При синтезе родопсина реакция идет в обратном направлении. В. Ретиналь в Пщнс-конфигурацин, связанный с е-аминогруппой
лизинового остатка опсина при помощи шиффова основания. Шиффово основание находится в протонированной форме. Г. 3-дегидроретиналь
в полностью транс-конфигурации. 11-цас-изомер— это хромофор пигмента палочек порфиропсина. Следует обратить внимание на дополни-
тельную двойную связь в кольце, которая удлиняет цепь сопряжения и сдвигает спектр поглощения и самого хромофора, и его производ-
ных — зрительных пигментов, в красную область.
490
Глава 14
который столь характерен для сетчатки позвоночных, в зрительных
системах многих беспозвоночных, следовательно, не происходит.
Спектры поглощения родопсина лягушки-быка и конечного продук-
та его выцветания показаны на рис. 14-11, А.
При добавлении в раствор обесцвеченного родопсина ll-i{uc-pe-
тиналя хромофор соединяется с белковым компонентом (опсином'
и происходит спонтанный ресинтез зрительного пигмента [120'
(рис. 14-11, Б). Таким образом, проблема ресинтеза в зрительном
Прелюмиродс-псин
(543нм)
\>-f40°C
гиаопсин Люмиродопсин
(498 нм) (497нм)
Парародопсин
\ (465нм)
Метародопсин1^^Метародопсин1/2д^ Регпинсшдегид (зв7нлл
(478 нм) (380нм) Н20 *опсин
Рис. 14-10. Схема превращений молекулы родопсина при выцветании (взято с не-
которыми изменениями из [118, 241].
Хромофор 11-цас-ретиналь специфическим образом связан с опсином. Под действием света
происходит изомеризация хромофора в полностью транс-конфигурацию, что сопровождается
разворачиванием молекулы опснна с экспонированием SH-групп и одной протон-связываю-
щей группы. Выцветание происходит при образовании метародопснна II. Связь между па-
рародопсином и другими промежуточными продуктами пока не установлена. Отметим, что
первые промежуточные продукты стабильны только при низких температурах. Процесс зри-
тельного возбуждения запускается в период между образованием люми- и метародопсина II.
цикле сводится к выяснению механизма, с помощью которого
1 l-ijizc-ретиналь регенерирует из полностью тушнс-ретиналя. В на-
стоящее время механизм этого процесса не вполне ясен [117, 241].
Промежуточные продукты — прелюми- и люмиродопсин — уда-
ется идентифицировать только при низких температурах (т. е. в
искусственных условиях), поскольку при физиологических темпера-
турах время жизни этих соединений слишком мало. С другой сторо-
ны, более поздние реакции, показанные на рис. 14-10, требуют для
своего завершения нескольких секунд и даже минут. Поэтому вни-
мание исследователей было сосредоточено на метародопсине и его
превращениях как возможной триггерной реакции в процессе зри-
тельного возбуждения [1, 241].
Максимумы поглощения зрительных пигментов
Зрительные пигменты разных видов животных значительно раз-
личаются по длинам волн максимумов поглощения. Эти различия
обусловлены следующими основными причинами: 1) разными хро-
Фоторецепция и зрение 49 1
мофорами; 2) межвидовыми различиями в опсине; 3) внутрисетча-
точными различиями в опсине. Рассмотрим каждый из этих фак-
торов.
Дегидроретиналь и порфиропсин. Сетчатки некоторых живот-
ных, например пресноводных рыб, вместо красного палочкового
пигмента родопсина содержат пурпурный пигмент, известный под
названием порфиропсина [27, 129, 233]. Различие между этими
пигментами обусловлено тем, что они содержат разные хромофоры.
Основой порфиропсина является не 1 l-tft/c-ретиналь, а близкий к
нему 11-1{нс-3-дегидроретиналь (рис. 14-9, Г). Дегидроретиналь
имеет одну лишнюю двойную связь в цепи сопряжения, в связи с
чем максимум поглощения сдвигается с 387 примерно к 400 нм.
Когда 3-дегидроретинальдегид соединяется с опсином, максимум
поглощения образовавшегося пигмента также оказывается сдвину-
тым в красную сторону (рис. 14-12). Например, у родопсина ля-
гушки максимум поглощения приходится на 502 нм, а соответству-
ющий порфиропсин (полученный из того же опсина) имеет макси-
мум поглощения примерно при 522 нм.
Зрительные пигменты, основанные на 3-дегидроретинале, извест-
ны только у позвоночных: ни у одного из беспозвоночных ни 3-де-
гидроретинальдегид, ни 3-дегидроретинол не обнаружены.
Бриджес [27] недавно представил обзор распространения и
свойств порфиропсинов. Обычно считают, что порфиропсины харак-
терны для пресноводных рыб, личинок амфибий и пресноводных
черепах. У мигрирующих рыб, обитающих поочередно в море и
пресноводных бассейнах, обнаруживаются и порфиропсин, и родоп-
син. В разные периоды жизненного цикла основным является тот
или иной пигмент, причем переход с одной пигментной системы на
другую предшествует обычно перемещению рыб в среду, которой
соответствует новый пигмент. Подобным образом дело обстоит и
у амфибий: сетчатки личинок лягушек и саламандр содержат пор-
фиропсин, но по мере метаморфозных превращений, предшествую-
щих переходу к наземной жизни, он заменяется родопсином. У пят-
нистого тритона в период второго метаморфоза из красной назем-
ной формы в полностью водную половозрелую форму наблюдается
обратный переход в сетчатке от родопсина к порфиропсину. (Ис-
ключение из этого правила составляют жабы Bufonidae; у них род-
опсин сохраняется в течение всего жизненного цикла.) Эти и дру-
гие данные Уолд [236] рассматривает как указание на то, что пор-
фиропсин филогенетически более примитивный из этих двух пиг-
ментов, однако другие исследователи относятся к такому утверж-
дению скептически [33].
Большинство полученных в последнее время данных по зри-
тельным пигментам рыб [27] указывает на то, что появление 3-де-
гидроретиналя и порфиропсина отражает адаптацию к водной сре-
де, в которой спектральное распределение света сдвинуто от сине-
зеленой области в сторону больших длин волн. По мере расшире-
Б
Фсторецепция и зрение
493
ния набора исследованных в этом отношении рыб стало ясно, что
примерно половина пресноводных рыб имеет как порфиропсин, так
и родопсин и что это справедливо также и для немигрирующих
рыб, которые всю жизнь проводят в пресной воде. Более того, по-
явились сообщения, что некоторые морские формы (Labridae) так-
же содержат порфиропсин. Хотя и нет сомнений в том, что связан-
ные с метаморфозом переходы от порфиропсина к родопсину и на-
оборот являются генетически запрограммированными и контроли-
руются гормонально, а не представляют собой прямую реакцию на
световое окружение, сейчас появляется все больше доказательств
того, что у некоторых рыб отношение родопсин/порфиропсин нахо-
дится под более непосредственным контролем среды. Например, у
молодого лосося можно получить сдвиг в сторону родопсина, уве-
личив интенсивность окружающего света. У немигрирующих рыб
наблюдаются сезонные флуктуации соотношения этих пигментов,
причем максимум содержания порфиропсина приходится на зимние
месяцы; здесь также видна зависимость от общего уровня осве-
щенности. Эксперименты на рыбах с глазами, прикрываемыми по
отдельности, показали, что контроль этот внутриглазной: на каж-
дый глаз можно влиять независимо от другого и от состава цир-
кулирующих гормонов. Имеются сообщения о сходном влиянии
света на отношение родопсин/порфиропсин у карповых рыб [4].
Микроспектрофотометрические измерения, выполненные на оди-
ночных наружных сегментах палочек форели, показали, что родоп-
син и порфиропсин гомогенно перемешаны в наружном сегменте
каждой палочковой клетки, так что в каждый момент времени
все рецепторы имеют идентичные смеси [139]. Другие исследова-
тели [172], однако, обнаружили, что в разных частях сетчатки зри-
тельные пигменты находятся в разных пропорциях, и для выясне-
ния этого вопроса необходимы дополнительные опыты.
Взаимные превращения ретиналя и 3-дегидроретиналя, по-ви-
димому, осуществляются с помощью фермента («дегидрогеназы
терминального кольца»), но как работает этот фермент, на уровне
Рис. 14-11.
Л. Спектры поглощения родопсина лягушки и продуктов его выцветания в водном растворе
Дигитонина; pH 5,55. Пик а обусловлен поглощением хромофора, ответственного в основном
за ^спектральную чувствительность палочек. При выцветании он исчезает и появляется но-
вый пик, который соответствует конечному продукту выцветания. Пик у обусловлен погло-
аР°матических аминокислотных остатков опсина и не меняется при выцветании
UdbJ. Б. Синтез и выцветание родопсина в растворе; 22° С, pH 7,0. Слева: спектры погло-
щения смеси 11-цпс-ретиналя и опсина быка, снятые после инкубации их в темноте в тече-
ние 0,3 мин (1); 2,5 (2); 5 (3); 10 (4); 18 (5); 30 (6); 60 '(7); 120 (8); 180 (9) мин. Поглощение
-цпс-ретиналя (Хтах 380 нм) падало, а родопсина тх 498 нм) возрастало. Справа: родоп-
полученный в первой части опыта (слева), облучали светом длиной волны >500 нм
обп, ение различных промежутков времени, а затем сразу снимали спектр. Общее время
ецУчения составляло: 5 с (2); 10 с (3); 15 с (4); 30 с (5); 120 с (6). Оставшийся препарат
ЦветяВ теч®ние 45 с облучали светом длиной волны >440 нм (7). Отметим, что продукт вы-
И-пнеИЯ 'Полностью трпнс-ретиналь) имеет более высокую молярную экстинкцию, чем
точки НЗОМе₽’ использованный при синтезе (слева). Отметим, также наличие изобестической
и при 418 нм на спектрах, снятых в процессе синтеза, и ее отсутствие на спектрах, сня-
тых при фотолизе [141].
494
Глава 14
альдегида или спирта, неизвестно. Воздействуя на синтез и актив-
ность этого фермента различными факторами, можно исследовать
функции отдельных генов [265].
Недавно было показано, что некоторые виды взрослых лягу-
шек-быков имеют порфиропсин в дорсальной части сетчатки и род-
Рис. 14-12. Спектральные свойства родопсина и порфиропсина, полученные с по-
мощью разностных спектров продуктов выцветания [26].
Эта пара пигментов Umax 500 и 523 нм) характерна для многих сетчаток, которые скорее
всего основаны на одном и том же опсине. Отметим, что спектр поглощения порфиропсина
несколько шире. На вставке показаны те же два спектра, по по оси ординат отложена мо-
лярная экстинкция.
опсин в вентральной [191]. Тип зрительного пигмента, обнаружи-
ваемого в палочке, определяется подстилающим пигментным эпите-
лием, который поставляет хромофор. Предполагают, что присут-
ствие порфиропсина имеет адаптивное значение в детектировании
слабого света из вентральной части зрительного поля. Как влия-
ют состояние среды и другие факторы, приводящие к постметамор-
фозному сохранению витамина А — 3,4-дегидрогеназы в пигмент-
ном эпителии и соответственно порфиропсина в дорсальной сет-
чатке, не выяснено.
Межвидовые различия опсинов. Второй фактор, который влия-
ет на максимум поглощения зрительного пигмента, это опсин. Пиг-
менты разных видов животных имеют различные максимум^
поглощения, потому что небольшие различия в аминокислотной по-
следовательности опсина приводят к образованию белков, по-ра3'
Фоторецепция и зрение
495
ному взаимодействующих с хромофором. На рис. 14-13 приведены
длины волн максимумов поглощения зрительных пигментов боль-
шого количества рыб. Ясно видны большие межвидовые различия
Пресноводные
Морские рыбы
СОCOG G
OQOOQOCOC
°460
•ft®
в®® о © ®
©Ф® 9
бее©
470
490 500
510 520 530 540
•®е
Юг
5~
Длина волны нм
Пресная вода: двойные пигмен-
_ ты Af+A2
*
©
'-Ю
о «о
«©•
9
С©
ооо
so е©
5
5
о-
5i
Пресная вода: одиночные
пигменты А,, А2
Песчаный берег, залив, устье
реки: пигмент А1
~ Скалистый берег:
пигмент А,
«в
сооо „
O0UOOOО
ООО
Поверхностные воды:
пигмент А, •
Глубинные воды:
пигменр! А, • } •>
480 490
°460 470
М.
500
510
520 530
540
О
0
е
О
О
О
5
Длина волны, нм
лятДая точка соответствует длине волны
а иным частичного выцветания экстрактов,
ветствии с
Рис. 14-13. Распределение зрительных пигментов у костистых рыб [55].
максимума поглощения пигмента, оцененной по
На нижнем рисунке данные распределены в соот-
местом обитания.
рительных пигментов, причем распределение максимумов не яв-
яется гладким. Максимумы поглощения большинства пигментов
аходятся в окрестности определенных длин волн, на основании
го можно предположить, что существует некоторое ограниченное
ело возможных вариаций последовательности аминокислот, ве-
Г57]И?КК обРазованию различных функционирующих опсинов
l J- Ьыло бы чрезвычайно интересно узнать детальную простран-
496
Глава 14
ственную структуру молекул этих родопсинов и порфиропсинов
равно как и аминокислот, которые составляют их основу.
На основании приведенных на рисунке данных можно сделать
также некоторое экологическое обобщение. Виды, у которых мак-
симум поглощения зрительных пигментов сдвинут к голубой части
спектра, обитают в более глубоких океанических водах, в глубь
которых лучше всего проникает солнечный свет с длинами волн
от 470 до 480 нм (рис. 14-2). С другой стороны, рыбы, зрительные
пигменты которых чувствительны к более длинноволновому излуче-
нию, обитают преимущественно в более мутных прибрежных и
пресных водах, где максимум проникающего света находится в
зеленой или желтой областях спектра.
Существование «порфиропсиновой» зрительной системы можно
рассматривать как приспособление к среде, которое накладыва-
ется на межвидовые вариации опсинов и служит для сдвига мак-
симума чувствительности в сторону еще более длинных волн. От-
метим, что порфиропсин с наиболее коротковолновым максимумом
(510 нм) обнаружен у морских форм. Рассматриваемая в таком
экологическом контексте способность к сезонной смене родопсино-
вой и порфиропсиновой систем позволяет сдвигать максимум чув-
ствительности на 20 нм и более, причем в зависимости от соот-
ношения между двумя пигментами в рецепторах максимум может
принимать непрерывный ряд значений. Адаптивное значение этого
механизма остается бесспорным независимо от того, что делает
генетическая программа: осуществляет смену пигментов как ста-
дию метаморфоза или позволяет ей осуществиться в ответ на изме-
нение светового периода.
Внутрисетчаточные различия между опсинами: зрительные пиг-
менты колбочек. Одинаковый набор рецепторов, имеющих один
и тот же пигмент или смесь пигментов, не может служить основой
цветного зрения. Чтобы отличить разницу в длинах волн от раз-
ницы в яркостях, животное должно иметь два или более вида
рецепторов с максимальной чувствительностью в различных об-
ластях спектра. В принципе этого можно достичь, имея два или
более зрительных пигмента, распределенных между различными
рецепторными клетками. Существует и другая возможность: если
фоторецепторы индивидуально снабдить цветными фильтрами, по-
добными масляным каплям колбочек, то может быть достаточно
одного пигмента.
В 1807 г. Томас Юнг высказал следующее положение: «Три
простых ощущения, по-разному комбинируясь, позволяют полу-
чить некоторое примитивное различение цветов, однако, посколь-
ку они могут комбинироваться в самых разных пропорциях, полу-
чается бесчисленное множество оттенков». Число «три» оказалось
выбранным очень удачно, поскольку проводимые в течение столе-
тия психофизические эксперименты по смешиванию цветов пока-
зали, что минимальное число приемников с различными функпия-
Фоторецегщия и зрение
497
Таблица 14-2
Пигменты цветного зрения приматов
Сине-чувстви- тельные колбочки нм) Зелено-чув- ствительные колбочки <\пах> нм) Желто- чув- ствительные колбочки ^тах- нм> Техника измерения Источник данных
445, 455*^ 535 570, 570 Торцевая микроспектрофото- метрия [144]
447* 540* 577* То же [142]
450 525 555 » » [33[
440* — -— » » [139]
-— 535* 575—580* Латеральная микроспектрофо- тометрия [139]
440? 527, 535 565, 565 Измерение спектра света, про- шедшего через кусочек сет- чатки [32]
430—440 530—540 565—595 Измерение спектра света, отра- женного от глазного дна [192, 199]
440 550 580—585 Психофизические измерения, [200, 257]
430 540 575 скорректированные с учетом поглощения в оптических сре- дах и в желтом пятне [239]
*) Значения Л, выделенные курсивом, относятся к человеку, остальные — к обезьяне.
Звездочкой помечены общие данные по приматам или данные, происхождение которых
не указано.
ми спектральной чувствительности, необходимое для объяснения
цветного зрения человека, равно именно трем. Однако эксперимен-
тально три типа рецепторов Юнга удалось обнаружить лишь в
последнее десятилетие: было показано, что существует три зри-
тельных пигмента, содержащихся в трех различных колбочковых
клетках (табл. 14-2). Различие между этими пигментами обуслов-
лено скорее опсином, чем хромофорами.
Данные табл. 14-2 не вполне точно соответствуют друг другу,
поскольку они основаны на сравнительно небольшом числе изме-
рений, выполненных в условиях огромных технических трудностей,
некоторые из которых заслуживают упоминания. Микроспектро-
фотометрические измерения затруднены из-за малых размеров на-
ружных сегментов и большой светочувствительности зрительных
пигментов, что не позволяет увеличить интенсивность тестового
луча для улучшения отношения сигнал/шум. Из-за малого диамет-
ра наружного сегмента (~1 мкм) спектр, регистрируемый с тор-
Ца («end on»), по-видимому, искажается светом, который рассеи-
вается латерально внутрь соседних рецепторов [5, 139, 192]. Ла-
терально входящие лучи, фокусируемые на изолированных наруж-
ных сегментах, должны быть свободны от этого недостатка, но в
Данном случае эффективная длина пути очень мала. Использова-
ние большего количества рецепторов облегчает измерение погло-
32-^1514
498 Глава 14
щения, но при этом труднее анализировать спектр, в который да-
ют вклад несколько пигментов, и невозможно сказать, что пигмен-
ты не находятся в одной колбочке. Техника рефлектометрии глаз-
ного дна имеет свои трудности [192, 200], и исследование «крас-
ных рецепторов» этим методом оказывается особенно ненадежным.
400 450 500 550 600
ж 1 Лягушка п.432| л,к502| к575(
Саламандра п432| п,к502| к575|
Геккон п467. п518(
Черепаха к442, п,к502| к562(
Голубь п500| к562|
Цыпленок п 500| к562(
Чайка п508| К562,
Обезьяна к440| П498| к535| к575|
Человек к440| п498| к535. к575|
Серебряный . карась К 45 52 п5222 к5352 к62О2
Протей п5272 к5752
Головастик п4382 п,к5272 к6202
Черепаха к4502 I п,к5182 1 1 к 620g 1 —
400 450 500 550 600
ДЛИНА волны, нм
Рис. 14-14. Сводные данные по одиночным наружным сегментам, выявляющие
внутрисетчаточное разнообразие зрительных пигментов [139].
Числа означают длины волн максимумов поглощения пигментов; буквы (п, к) указывают,
где находится пигмент — в палочке или в колбочке; индексы (1, 2) соответствуют разным
хромофорам — ретиналю и дегидроретиналю.
Таблица 14-2 включает данные, полученные на сетчатках как че-
ловека, так и обезьяны; межвидовые различия если и существуют,
то они меньше, чем ошибки измерения, обусловленные техниче-
скими трудностями.
Последний горизонтальный ряд цифр в табл. 14-2 представляет
данные психофизических измерений спектральной чувствительно-
сти, полученные в условиях адаптации испытуемых к цветному
свету. Эта техника основана на избирательном подавлении выхо-
да двух из трех типов колбочек; третий при этом можно исследо-
вать относительно независимо. Полученные результаты указыва-
Фоторецепция и зрение
499
ют на то, что спектры поглощения, регистрируемые оптическими
методами, соответствуют функциям спектральной чувствительности
трех физиологических процессов (обсуждение трудностей, которы-
ми чревата эта интерпретация, можно найти у Раштона [201а]).
Рис. 14-14 суммирует данные Либмана [139] по спектрофото-
метрическому исследованию рецепторов сетчатки; в нем собрана
большая часть того, что известно о внутрисетчаточном разнообра-
зии зрительных пигментов других позвоночных.
Зрительная система серебряного карася [143] очень похожа
на зрительные системы человека и обезьяны в том отношении, что
в ней также имеется один палочковый пигмент и три колбочковых.
Разница состоит в том, что хромофором у серебряного карася слу-
жит дегидроретиналь и соответственно поглощение пигментов
сдвинуто в длинноволновую область.
Сделаем здесь небольшое отступление в область терминологии.
Уолд [241] говорит о четырех основных пигментах в зрении позво-
ночных (см. ниже). Большинство других исследователей находят
это разделение в большей или меньшей степени неудовлетвори-
тельным. Во-первых, оно неприменимо к пигментам беспозвоноч-
ных. Во-вторых, оно не допускает существования более чем одно-
го типа палочкового или колбочкового пигмента для каждого хро-
мофора. Например, колбочковый пигмент сетчатки серебряного
карася с Zmax = 620 нм — это классический цианопсин, но тогда что
такое пигмент той же сетчатки с Zmax = 535 нм? Такая негибкость
привела к изобретению других наборов терминов, например «эрит-
ролаб», «хлоролаб» и «цианолаб» для колбочковых пигментов че-
ловека. Заметим, что цианолаб, который разлагается при голубом
свете, поглощает в противоположном конце спектра по сравнению
с цианопсином, который имеет голубую окраску.
Ретиналь + Палочковый опсин—^Родопсин (Хтах = 500 нм)
Ретиналь+ Колбочковый опсин—^Йодопсин (Хтах = 562 нм)
3-дегидроретиналь + Палочковый опсин—^Порфиропсин (Хтах =
= 522 нм)
3-дегидроретиналь + Колбочковый опсин—>Цианопсин (Хтах =
= 620 нм)
Подобное обилие наименований не может привести ни к чему,
кроме путаницы. Обычно удобно использовать сокращенные обо-
значения, из которых чаще всего употребляется латинская буква
«Р» — «пигмент», с последующим указанием Zmax (в нм) и с индек-
сами 1 или 2 для обозначения того, что является хромофором: ре-
тиналь или дегидроретиналь. Так, например, Р620г — это вышеупо-
мянутый пигмент сетчатки серебряного карася. Одно дополнитель-
Ное слово указывает, где он находится: в палочках или в колбоч-
ках. В рис. 14-14 с небольшими изменениями использована именно
эта система обозначений.
Возвращаясь снова к рис. 14-14, мы видим, что у птиц имеет-
ся только один колбочковый пигмент и один палочковый. Однако
32*
500
Глава 14
благодаря присутствию в каждом наружном сегменте красных
оранжевых и желтых масляных капель, содержащих каротиноиды’
образуются два или три типа рецепторов с Хтах (вычислено для
голубя) 625, 585 и 562 нм [139]. Дальнейшие исследования в этой
области покажут, является это единственной основой цветного
зрения птиц [127а] или здесь имеются добавочные, пока неизвест-
ные колбочковые пигменты.
Взрослые лягушки имеют два палочковых и два колбочковых
пигмента. Наиболее распространен палочковый пигмент P502i. '
Палочковый пигмент Р4321 находится в так называемых зеленых
палочках. Одиночные колбочки и основные клетки двойных колбо-
чек содержат пигмент Р502ь который спектроскопически неотли-
чим от преобладающего пигмента палочек. У головастиков имеет-
ся соответствующая дегидроретинальная система.
Черепахи Pseudemys также имеют «палочковый пигмент» в сво-
их добавочных колбочках и, кроме того, два других колбочковых
пигмента, поглощающих при больших и меньших длинах волн.
Вдобавок у них имеется четыре типа масляных капель, распреде-
ленных таким образом, что предположительно образуется пять
спектральных классов колбочек [139].
Имеются достаточно веские основания в пользу того, что в гла- ”
зах многих беспозвоночных присутствует несколько зрительных
пигментов. Однако для однозначного ответа на этот вопрос необ-
ходимо иметь значительно большее количество спектров поглоще-
ния. Измерения спектральной чувствительности рецепторов более
многочисленны. Обзор данных по этому вопросу можно найти, на-
пример, в работе [92].
Расположение молекул зрительных пигментов в мембранах
рецепторов
В наружных сегментах палочек и колбочек хромофоры ориен-
тированы приблизительно параллельно плоскостям мембранных
дисков и перпендикулярно продольным осям палочек и направле-
нию падающего света (рис. 14-15, Л). Вследствие того что сопря-
женная система одинарных и двойных связей располагается под
прямым углом к падающему свету, вероятность поглощения долж-
на быть в 1,5 раза выше, чем для тех же молекул, ориентирован-
ных в растворе случайно I Реально хромофоры, как правило, не
1 Хромофор (см. рис. 14-9) поглощает наиболее сильно в том случае, когда
свет поляризован параллельно оси цепочки. В растворе молекулы ориентированы
случайным образом. Если молекулу хромофора считать прямолинейной, то вектор
поглощения такой случайно ориентированной системы можно разложить на
3 взаимно перпендикулярные составляющие: две перпендикулярные и одну па-
раллельную направлению распространения падающего света. Ясно, что поглоще-
ние в этом случае не будет меняться при изменении плоскости поляризации света,
причем независимо от направления луча один из векторов поглощения всегда
Фоторецепция и зрение
50f
совсем параллельны дискам (среднее отклонение составляет не-
сколько градусов), и поэтому измеряемое экспериментально уве-
личение поглощения составляет примерно 1,37 [243].
Такая ориентация молекул выявляется в виде дихроичного по-
глощения палочек при их освещении сбоку (рис. 14-15, Л) [61,.
138, 243]. Когда падающий сбоку свет поляризован перпендику-
лярно оси палочек (параллельно плоскостям дисков), поглощение
велико, а когда плоскость поляризации поворачивается на 90° и
становится параллельной оси палочек, поглощение составляет при-
мерно 22% предыдущего значения, что соответствует отношению»
дихроичности примерно 4,5. По результатам таких измерений мож-
но оценить степень ориентированности хромофоров.
Был измерен и дихроизм рабдомов ракообразных [107, 253].
Рабдомы, отслоенные от ретикулярных клеток, рассматривали
сбоку, а тестирующий луч «загоняли» внутрь одиночного пучка
микровилл (рис. 14-8, В и 14-15, Б). Выбирали такие рабдомы, в-
которых микровиллы различных слоев параллельны или перпен-
дикулярны измеряющему лучу. Когда направление распростране-
ния света совпадало с осью микровилл, поглощение не зависело
от плоскости поляризации, чего и следовало ожидать, исходя из
цилиндрической симметрии микровилл. Когда луч был направлен
перпендикулярно оси микровилл, наблюдался дихроизм. Отноше-
ние дихроичности равнялось примерно 2, а максимальное погло-
щение имело место, когда электрический вектор был параллелен
осям микровилл. Этот результат можно объяснить в предположе-
нии, что хромофоры лежат в поверхностных мембранах микро-
вилл, как это, по-видимому, имеет место у позвоночных. Большее
отношение дихроичности можно получить только в том случае, ес-
ли имеется некоторая предпочтительная ориентация хромофоров
вдоль осей микровилл, что, по-видимому, наблюдается у кальмара
[97, 167].
Из-за перекрещивания пучков микровилл в рабдомах десяти-
ногого рака (рис. 14-8, В) измерения, выполненные на одиночных
Дихроичных пучках, могут дать информацию о поглощении акси-
ально проходящего света в отдельных рабдомерах (рис. 14-15, Б).
Таким образом, измерение дихроизма рабдомов рака может по-
мочь объяснить чувствительность членистоногих к поляризации
света; об этом мы будем говорить позже.
бУДет ориентирован параллельно направлению распространения света и, следова-
тельно, не будет давать вклада в поглощение. Если молекулы имеют определен-
ную ориентацию, как, например, в кристалле, поглощение существенно зависит
от плоскости поляризации падающего света. Это свойство называют дихроизмом..
хромофоров поглощение максимально тогда, когда вектор Е параллелен их оси.
Дисках палочек молекулы частично ориентированы, а именно они, по-видимому^
°гут относительно свободно поворачиваться лишь в пределах плоскости дисков,
то приводит к появлению дихроизма, описанного в тексте и на рис. 14-15.
Наружный сегмент палочки позвоночных
Продольный пучок
света
Сильное
поглощение
Поперечный
пучок света
Слабое
| J поглощение
Б у Рабдомер десятиногого рака
Сильное
поглощение
Ретикулярная
клетка —
Микровилла
Фоторецепция и зрение
503-
реакции мембран рецепторных клеток на возбуждение
£иперполяризация палочек и колбочек позвоночных
при освещении
Как указывалось на стр. 481, фоторецепторные клетки сетчат-
ки позвоночных являются производными ресничек и в этом отно-
шении отличаются от большинства фоторецепторных клеток бес-
позвоночных. Осуществить внутриклеточные отведения потенциа-
лов палочек и колбочек с помощью микроэлектродов удалось
совсем недавно [226]. Рецепторы обычно очень малы и прокалыва-
ются с трудом. К моменту написания этой книги были зарегистри-
рованы реакции колбочек рыбы (карп) [225], колбочек и, возмож-
но, палочек протея Necturus [22, 227, 258], палочек лягушки [226],.
палочек (трансмутированных колбочек?) геккона [227] и колбочек
черепахи Pseudemus [16]. В каждом случае тип проколотой клетки
определяли с помощью инъекции красителя через отводящей элек-
трод. У геккона и лягушки были проколоты наружные сегменты,
в остальных случаях электрод проникал во внутренние сегменты.
Палочковые и колбочковые клетки имеют отрицательный потен-
циал покоя. При освещении клетка гиперполяризуется (т. е. внут-
риклеточная среда становится еще более электроотрицательной по
отношению к наружной). Реакции градуально меняются с измене-
нием интенсивности света, причем никаких импульсов не возникает
(рис. 14-16). Гиперполяризационный ответ клеточной мембраны
весьма интересен тем, что в противоположность реакциям других
возбудимых клеток он связан с уменьшением проводимости мем-
бран (т. е. увеличением сопротивления). На рис. 14-17 приведены
данные, иллюстрирующие изменение проводимости мембраны при
освещении. Реакция мембраны на короткие толчки постоянного то-
ка, пропускаемого через клетку, на свету больше, чем в темноте.
Это объясняется тем, что сопротивление мембраны увеличивается
Рис.
л. в
14-15. Ориентация молекул зрительного пигмента в мембранах фоторецепто-
ров (согласно данным по дихроизму поглощения) [167, 253].
наружных сегментах палочек и колбочек позвоночных хромофоры лежат в плоскости
ембран дисков и ориентированы случайным образом. Следовательно, поглощение акси-
льно входящего света не зависит от плоскости его поляризации. Однако при освещении
лиК^ С1ггуация меняется: свет, плоскость поляризации которого параллельна плоскостям
Дисков (а значит, и осям многих хромофоров), поглощается сильно, а поляризованный
„перпендикулярном направлении — слабо. Б. Результаты измерений дихроизма рабдомеров-
вам Т11НОГих ракообразных согласуются с представлением о том, что хромофоры ориентиро-
ре"Ь1 в поверхностных мембранах фоторецепторов случайным образом. Здесь показана одна
ваю кУля₽ная клетка с несколькими «языками» микровилл. В рабдоме эти языки переслаи-
сбокТСя С Рабдомерами других рецепторов (рис. 14-8, В), однако с помощью направленного
мИкпу микролуча можно освещать отдельный пучок микровилл. Когда луч параллелен осям
Чай °Вилл’ поглощение изотропно, а когда перпендикулярен — дихроично. Последний слу-
Но эквивалентен аксиальному освещению целого глаза. Поглощение при этом максималь-
2, ’ Когда свет поляризован параллельно осям микровилл. Отношение дихроичности равно
х’роЛ°лсогласУется с представлением о случайной ориентации, отмеченной выше; если бы
оформы по отношению к осям микровилл были расположены упорядоченно, отношение
было бы больше.
Рис. 14-16. Градуальные гиперполяризационные реакции колбочек черепахи в от-
вет на вспышки (длительностью 10 мс) нарастающей интенсивности [16].
При самой яркой вспышке на одну колбочку попадает 8,5-106 фотонов. Отклонение кривых
вниз означает увеличение отрицательного потенциала на внутриклеточном микроэлектроде.
Цифры у кривых — интенсивности вспышек в относительных логарифмических единицах."
Рис. 14-17. Падение напряжения, возникающее при пропускании тока через
мембрану фоторецептора позвоночных во время реакции на свет [16].
Наложено 6 записей реакций на вспышки длительностью 10 мс, даваемые в нулевой момент
времени. Во время каждой записи через микроэлектрод пропускался короткий импульс де-
поляризующего тока силой 6,1-10~10 А, дающий кратковременное отклонение потенциала.
Максимальное увеличение высоты отклонения по сравнению с темновым имеет место в мо-
мент максимума реакции на свет и составляет около 5 мВ, что соответствует увеличению
сопротивления мембраны на 8 МОм. Отметим, что вызванное током отклонение включае
и эффекты, связанные с изменениями в цепи электрода, поскольку при таких токах
ные артефакты видны и тогда, когда электрод находится вне клетки. Однако изменени
падения напряжения после вспышки не являются артефактом, а отражают изменение вход
лого сопротивления клетки, так как освещение электрода, не находящегося внутри фотор '
цептора, такого эффекта не вызывает.
Фоторецепция и зрение
505
пои освещении на несколько мегаом. Контрольные эксперименты
(не приведенные на рис. 14-17) показали, что увеличение сопро-
тИВЛеНия мембраны связано именно с освещением, а не с гипер-
поляризацией: сама по себе гиперполяризация клетки с помощью
Рис. 14-18. Влияние уабаина на потенциал фоторецептора позвоночных [211].
Л. После выдерживания сетчатки в течение 7 мин в свободном от натрия растворе, содер-
жащем уабаин в концентрации 0,1 мМ, реакция отсутствует. Б. Реакция той же сетчатки
после добавления трех капель раствора натрия с нормальной концентрацией. Добавляемый
раствор содержит уабаин в прежней концентрации. Этот эксперимент показывает, что для
возникновения рецепторного потенциала нужны именно ионы натрия, а не работа натрие-
вого насоса. В. После выдерживания сетчатки в течение 7 мин в таком же, как в случае Б,
растворе, содержащем натрий и уабаин, реакция исчезает. Это связано с тем, что подав-
ление работы натриевого насоса в присутствии натрия в среде приводит через несколько
Минут к повышению внутриклеточной концентрации натрия и исчезновению трансмембран-
ного градиента натрия, благодаря которому возникают нормальные рецепторные потенциа-
лы. Данные получены с помощью экстраклеточного отведения в условиях подавления дру-
гих сетчаточных потенциалов с помощью аспартата.
постоянного тока не вызывает такого увеличения сопротивления,
Как освещение.
Для возникновения потенциалов действия сетчатки в целом
необходимы ионы натрия, и, используя аспартат для выделения
Рецепторного потенциала из суммарного экстраклеточно регистри-
руемого ответа, можно показать, что ионы натрия необходимы для
Реакции рецепторов [210, 211] (рис. 14-18). Напротив, увеличение
506
Глава 14
концентрации калия в окружающей среде снижает рецепторный
потенциал. Свет не изменяет активность электрогенного натриево-
го насоса, а непосредственно влияет на проницаемость мембраны
.для натрия. В пользу этого говорят описанные выше опыты по из-
менению сопротивления мембраны на свету, а также тот факт, что
рецепторные потенциалы возбуждаются и в таких сетчатках, в ко-
торых катионный насос подавляется с помощью сердечного глю-
Рис. 14-19. Схема, моделирующая реакции мембран фоторецепторных клеток.
Обсуждение смотри в тексте [211].
козида уабаина в течение всего времени, пока не исчезнет транс-
мембранный градиент натрия [211] (рис. 14-18).
Эквивалентная схема мембраны наружного сегмента фоторе-
цептора позвоночных показана на рис. 14-19. Мембранный потен-
циал обусловлен градиентами концентраций натрия и калия и от-
ношением трони’цаем'остей мембраны для этих ионов. На схеме
это представлено в виде натриевой и калиевой батарей с последо-
вательно соединенными сопротивлениями. Сопротивление по нат-
рию регулируется светом и возрастает при освещении, что приво-
дит к сдвигу мембранного потенциала в сторону калиевого потен-
циала равновесия (£к). Ниже мы сравним эту модель с фоторе-
цепторами беспозвоночных, когда будем их рассматривать.
Данные, полученные с помощью внутриклеточных электродов,
очень полезно сопоставлять с данными экстраклеточной регистра-
ции потенциалов в слоях интактной сетчатки. Из такого сопостав-
ления можно вычислить как продольные экстраклсточные токи
между рецепторами, так и токи через мембрану в цитоплазму и
из нее [99, 183]. В темноте существует ток, выходящий из цито-
плазмы через внутренние сегменты и входящий через мембраны
наружных сегментов. При освещении на него накладывается мень-
ший по величине фототок противоположного направления, который
можно рассматривать как результат уменьшения темнового тока
Трансмембранный фототек
б расчете на одну гбалочки,
зарядс-'-мкм ~1
Наружу Внутрь
4 2 0 2 4x10е
Рис. 14-20. Фототок в рецепторах позвоночных [183].
Л. Изменение максимального значения трансмембранного фототока в зависимости от глу-
бины погружения центрального из трех регистрирующих электродов. Вычислено по данным
измерения продольных напряжений и сопротивлений между клетками. Приведены резуль-
таты для препаратов сетчатки, раздражаемых зеленым светом длиной волны 560 нм и интен-
сивностью 1 -10” фотон-см-2 (светлые кружки) и 4-10" фотон-см-2 (темные кружки). Указан
случайный разброс значений из-за шума. Б. Палочки сетчатки крысы, изображенные в мас-
штабе, соответствующем левой части рисунка. Стрелками показано направление фототока.
1 наружный сегмент, // — внутренний сегмент, III — наружная пограничная мембрана,..
IV — наружный ядерный слой, V — наружный сетчатый слой.
S08
Глава 14
(рис. 14-20). Фототок линейно зависит от интенсивности света и
согласно последним данным, исчисляется величинами до 106 ионов
на фотон.
Рабдомерные рецепторы беспозвоночных, обычно
деполяризующиеся при освещении
К настоящему времени изучено большое количество фоторецеп-
торных клеток членистоногих, моллюсков, кольчатых червей
Идентифицировать прокалываемые клетки иногда удавалось без
применения специальной техники, а в ряде
случаев — с помощью внутриклеточной инъ-
екции красителя [17, 135, 169]. Рабдомер-
ные фоторецепторы отвечают на освещение
деполяризацией, которая градуально зави-
сит от его интенсивности; реакция часто
имеет динамическую или переходную фазу
и последующую статическую фазу или пла-
то при больших интенсивностях (рис. 14-21).
В приведенном примере, относящемся к ре-
тикулярной клетке глаза мухи, импульсы
отсутствуют. У некоторых членистоногих на
восходящем участке переходной фазы по-
является один импульс [85]; у других ви-
дов беспозвоночных регистрируется последо-
вательность импульсов, вероятно генерируе-
мая аксонами клеток [152]. Наличие или
отсутствие импульсов определяется отчасти
расстоянием между первичными фоторецеп-
торными клетками и клетками второго по-
рядка, с которыми они синаптически свя-
заны.
Рис. 14-21. Внутриклеточно регистрируе-
мые рецепторные потенциалы ретинуляр-
” ных клеток мясной мухи Calliphora
[248].
Деполяризационным потенциалам соответст-
вует отклонение кривой вверх. Сигнал раз-
дражения — самая верхняя запись (ток фото-
элемента). Интенсивность уменьшается от А
к Д в отношении 100:25:6:1,5:0,4. Каждая кри-
вая — результат наложения десяти записей.
Деполяризационные рецепторные потенциалы отражают увели-
чение проводимости мембраны. Это было показано с помощью
.методики, сходной с описанной выше для рецепторов позвоночных,
Фото рецепция и зрение
509
использующей толчки тока [86], или более изящно, методом фик-
сации напряжения [30, 158].
Экстраклеточные измерения потенциалов вдоль тонких «на-
ружных сегментов» фоторецепторов кальмара, выполненные на
кусочках сетчатки, вырезанных параллельно продольным осям ре-
цепторных клеток, дают дополнительные свидетельства в пользу
этого [Ю1]. На рис. 14-22 показаны разности потенциалов, реги-
стрируемые между двумя фиксированными электродами при дви-
жении узкой полоски света вдоль длинной оси рецептора. Этот
эксперимент показывает, что участок рабдома, находящийся не-
посредственно под световым пятном, заряжен отрицательно по
отношению к остальным частям рабдома, чего и следует ожидать,
если предположить существование локального входа тока под осве-
щенной точкой.
Существуют указания на то, что местом, где меняется проводи-
мость, является сам рабдом. Наиболее прямые аргументы в пользу
такого заключения получены на фоторецепторных клетках пиявки,
у которых микровиллы фоторецепторных мембран простираются в
центральную «вакуоль» внутри клетки [135] (рис. 14-23). В дей-
ствительности вакуоль является экстраклеточным пространством,
которое с помощью узких канальцев сообщается с жидкостью,
омывающей наружную поверхность клетки. При освещении микро-
электрод, введенный в цитоплазму клетки, регистрирует отклоне-
ние потенциала в положительную сторону (деполяризацию). С дру-
гой стороны, микроэлектрод, введенный в вакуоль (что устанавли-
вается инъекцией красителя), при освещении клетки регистрирует
увеличение отрицательного потенциала. Этот результат можно
объяснить, если принять, что во время освещения ток входит в
клетку также и через мембраны микровилл; если бы единственным
местом вхождения фототока являлась часть клеточного тела, сво-
бодная от микровилл, подобных явлений не наблюдалось бы.
Ионные процессы, лежащие в основе рецепторного потенциала,
не вполне ясны. Наиболее важным из ионов является по-видимо-
му, натрий; уменьшение его наружной концентрации ведет к умень-
шению рецепторного потенциала [85, 218]. В ряде случаев играет
роль и калий, однако является ли он носителем заряда или кон-
тролирует натриевую проводимость, в настоящее время не выясне-
но. Рецепторные потенциалы могут сохраняться после многих ча-
сов вымачивания препарата в свободных от натрия растворах —
факт, объяснения которому до сих пор не найдено. В ряде случаев
временной ход изменений проводимости в период развития рецеп-
торного потенциала указывает на то, что в реакцию дает свой
вклад не один тип ионов. Потенциалы реверсии могут лежать вбли-
зи 0 мВ [85, 153] (что указывает на общее закорачивание потен-
циала покоя) или быть явно положительными и более сильно за-
висящими от наружной концентрации натрия [30].
510
Глава 14
Вблизи порога у рабдомерных рецепторов регистрируются не-
большие кратковременные деполяризационные всплески, амплиту-
да которых обычно меньше 1 мВ [87, 203]. Этот факт может быть
связан с открыванием ионных каналов в ответ на поглощение от-
дельных фотонов. С увеличением интенсивности света частота по-
Рис. 14-22. Потенциалы тонких слоев сетчатки кальмара, регистрируемые экстра-
клеточно на разной глубине погружения электрода в слой рабдомеров [101].
По оси ординат отложено расстояние щелевидного стимула (С) от наружной пограничной
мембраны (ПМ)\ 1 и 2— тонкие электроды. По оси абсцисс отложена разность потенциалов
между электродами. П — пигментированная часть клетки. Интенсивность светового стиму-
ла равна 3-1011 фотонов-см-2 с—1. Видно, что локальное освещение вызывает в этом месте
локальный ток.
явления этих дискретных потенциалов возрастает, и при еще бо-
лее ярком свете ответы сливаются. Таким образом, плавная, гра-
дуально меняющаяся деполяризация мембраны, которая наблюда-
ется при умеренных интенсивностях, есть результат суммирования
большого числа элементарных событий. Каждый дискретный по-
тенциал соответствует протеканию через мембрану сотен и тысяч
ионов [95].
В первом приближении для описания рабдомерных рецепторов
беспозвоночных можно использовать ту же эквивалентную схему,
Секунды
Рис. 14-23. Определение места вхождения фототока в рабдомерных образова-
ниях [135].
У пиявки Hirudo микровиллы выстилают «вакуоль», которая в действительности сообщается
с внеклеточной средой через узкие канальцы. При погружении микроэлектрода в вакуоль
или цитоплазму регистрируются реакции, приведенные па графике. Потенциалы в обоих
случаях измерены относительно потенциала заземленного электрода. Когда электрод нахо-
дится внутри клетки (в цитоплазме), регистрируется положительный (деполяризационный)
рецепторный потенциал и, кроме того, на его фоне наблюдаются небольшие импульсы.
Их амплитуда невелика, поскольку они возникают в аксоне на некотором расстоянии от тела
клетки. Когда электрод введен в вакуоль, рецепторный потенциал отрицателен. Это согла-
суется с представлением о существовании тока, указанного на рисунке пунктиром; этот
ток входит в клетку в области микровилл (сплошные стрелки). Мы оставляем читателю
возможность в порядке упражнения проверить, что приведенные выше данные невозможно
Получить, если считать, что ток входит только через участки мембраны клетки, лишенные
^икровилл. Импульсы, регистрируемые в вакуоли, положительны; это еще раз доказывает,
что они генерируются в другом месте. В — вакуоль; Ц — цитоплазма.
512
Глава 14
что и для моделирования реакций палочек и колбочек позвоноч-
ных. Существенная разница состоит в том, что в схеме для бес-
позвоночных свет уменьшает натриевое сопротивление.
На рис. 14-24 показан другой способ сравнения реакций фото-
рецепторов беспозвоночных и позвоночных. В клетках беспозвоноч-
ных реакция на свет увеличивается, если мембрану гиперполяри-
зовать внешним током, уменьшается, если мембрану деполяризо-
Рис. 14-24. Схематическое изображение реакций фоторецепторов беспозвоночных
и позвоночных [227].
Гиперполяризацию и деполяризацию осуществляли с помощью пропускания тока между
внутриклеточным электродом и наружным раствором, как показано на рис. 14-13. Осталь-
ные объяснения даны в тексте.
Световое Световое
раздражение раздражение
вать, и меняет знак при деполяризации за потенциал равновесия.
Эта ситуация обычна при исследовании ионных процессов в воз-
буждающих синапсах. Влияние внешнего тока на фотореакции ре-
цепторов позвоночных показано во второй колонке рис. 14-24. Все
реакции здесь имеют обратный знак по сравнению с реакциями
беспозвоночных. Гиперполяризация увеличивает реакцию, деполя-
ризация ее уменьшает, и по другую сторону от потенциала равно-
весия наблюдается реверсия. Чтобы понять, как возникает ревер-
сия при прохождении через потенциал равновесия, вспомним, что
для отклонения потенциала, обозначаемого термином «деполяри-
зация», через микроэлектрод нужно пропускать ток, вытекающий
через мембрану. Поскольку действие света состоит в увеличении
сопротивления мембраны, во время освещения пропускаемый из-
вне через микроэлектрод ток еще больше деполяризует мембрану-
Фоторецепция и зрение
513
Некоторые фоторецепторы беспозвоночных генерируют
гиперполяризационные реакции
Среди исключений из общего правила, согласно которому фо-
торецепторные органеллы беспозвоночных не развиваются из рес-
ничек, можно назвать фоторецепторные клетки моллюсков. Наи-
лучший из изученных примеров — мантийные глаза гребешка
Pecten. Эти «простые» глаза, располагающиеся в ряд по краям
мантии, замечательны тем, что имеют двойную сетчатку, состоя-
щую из двух отдельных слоев фоторецепторных клеток (рис. 14-3,В).
Каждый слой имеет свою собственную ветвь оптического нерва,
и считается, что между ними нет синаптических связей на пери-
ферии [152, 153].
Проксимальная сетчатка такая же, как обычно. Фоторецептор-
ные клетки имеют выросты в виде микровилл, типичные для бес-
позвоночных, и мембраны реагируют на свет обычным деполяриза-
ционным рецепторным потенциалом.
Фоторецепторные клетки дистальной сетчатки устроены иначе;
они имеют серию слоистых мембран, развивающихся в связи с ци-
лиарными микротрубочками. Оба типа фоторецепторных клеток
содержат зрительный пигмент с максимумом поглощения около
500 нм, но чувствительность дистальных клеток на 2—3 логариф-
мические единицы меньше. Более важно то, что дистальные клет-
ки при освещении гиперполяризуются. Эта гиперполяризация су-
щественным образом отличается от реакций цилиарных фоторе-
цепторов сетчатки позвоночных: она обусловлена увеличением, а
не уменьшением проводимости мембраны. Таким образом, если фо-
тоны сравнивать с молекулами синаптического передатчика, гипер-
поляризация приобретает формальное сходство с тормозными
постсинаптическими потенциалами в центральной нервной системе.
Хотя в сетчатке позвоночных реакцией рецепторов является гипер-
поляризация, совсем не ясно, какое влияние — возбуждающее или
тормозное —она оказывает на высвобождение синаптических пу-
зырьков. Однако гипергюляризация дистальных фоторецепторных
клеток гребешка Pecten является, по-видимому, первичным тормо-
жением. Аксоны этих клеток генерируют импульсы, и действие
света состоит в уменьшении частоты или подавлении генерации.
С точки зрения поведения функцией фоторецепторных клеток
моллюсков, реагирующих первичным торможением, является обес-
печение реакции избегания опасности. Так, если на глаз падает
тень, возникает сильный «ofb-разряд, поскольку аксоны фоторе-
цепторов освобождаются от торможения, и раковина быстро за-
крывается [133]. Этому сенсорному механизму можно противопо-
ставить дорсальные глазки взрослых насекомых, которые также
обеспечивают реакцию на внезапное уменьшение освещения. В этом
случае рецепторы отвечают на освещение деполяризацией и акти-
вируют тормозные синапсы на дистальном конце оцеллярного нер-
33—1514
514
Глава 14
ва [105]. В результате оцеллярный нерв, как и аксоны дистальной
сетчатки Pecten, должен реагировать на затенение залпом им-
пульсов.
Другой случай предположительно первичного торможения об-
наружен в нейронах, клеточные тела которых лежат вдоль ман-
тийного нерва морского моллюска Spisula [126]. Изолированные
участки нерва чувствительны к освещению, причем спектр действия
для торможения имеет Хтах около 540 нм. Этот препарат исключи-
тельно интересен тем, что торможению противопоставляется имею-
щий более длинный латентный период процесс возбуждения с мак-
симумом спектральной чувствительности в области еще больших
длин волн. Имеющиеся в настоящее время данные указывают на
то, что оба фотопигмента находятся в одной и той же клетке, од-
нако полной уверенности в этом нет.
Для подтверждения предположения о том, что одна фоторецеп-
торная клетка может содержать два пигмента, один из которых
вызывает возбуждение, а другой —торможение, необходимы даль-
нейшие исследования. Известно, что в сетчатках позвоночных и,
по-видимому, в глазах всех членистоногих каждая фоторецепторная
клетка содержит единственный зрительный пигмент. Когда наблю-
даются тормозные взаимодействия (а обычно так и бывает), они
осуществляются с помощью синапсов между нейронами второго и
более высоких порядков (см. ниже). Против существования воз-
будительно-тормозных взаимодействий между двумя фотопигмен-
тами одного и того же фоторецепторного нейрона «простого» гла-
за серьезных аргументов нет, хотя это может иметь значение для
сетчатки, на которой формируется изображение, поскольку там
цветовой контраст должен быть соответствующим образом увязан
с пространственными контурами. В дополнение к рассмотренному
выше случаю Spisula можно назвать имеющиеся в медиальных
глазках мечехвоста Limulus клетки, которые деполяризуются при
освещении близким ультрафиолетом и гиперполяризуются под дей-
ствием видимого света, а также другие клетки с реципрокными
взаимоотношениями [176]. Вопрос о том, является ли этот случай
примером первичного торможения, накладывающегося на возбуж-
дение, или тут дело в латеральных тормозных синапсах, пока не
выяснен.
Ранний рецепторный потенциал
Реакции мембран, которые мы только что рассмотрели, иногда
называют поздними рецепторными потенциалами, чтобы отличить
их от раннего рецепторного потенциала (РРП). РРП— это низко,-
амплитудный сигнал без заметного латентного периода, имеющий
другое происхождение. Впервые он был зарегистрирован Броуном
и Мураками [31]. Они заметили, что когда электроретинограмма
(ЭРГ — суммарная реакция всей сетчатки, регистрируемая с по-
Фоторецепция и зрение
515
мощью больших экстр аклеточных электродов) вызывается корот-
кими вспышками света большой интенсивности, ей предшествует
небольшой потенциал, никем ранее не зарегистрированный. РРП
изучали затем в нескольких лабораториях [29, 51, 53, 98, 182]; бы-
,, , max,
Хромофоры нм
Сигнал
раздра-
жения,
нм
7,8 Родопсин д-90
\М
$560нм
Родопсин
7,8 + л •
мета А 490
|-н*
„ „ Родопсин
9,8 +
метав 380
I м
ч560нм
9,8 г метав
7,8 3
500-650 9,8
Форма сигнала
раздражения
рн*
мета А
р’’
Родопсин
+
мета А
Изородопсин ^480
' родопсин
Рис. 14-25. Ранние рецепторные потенциалы в сетчатке кальмара, зафиксирован-
ной с помощью глутаральдегида, при состояниях хромофоров родопсина, указан-
ных на рисунке справа [98].
Температура 0° С. Раствор: 500 мМ трис — НС1; pH указаны на рисунке. Приведены реакции
на одиночные, вспышки света, обладающие энергией (в фотонах на квадратный сантиметр):
(а) 3-1014, (б и в) 2-Ю16, (г) 6-Ю14; (д и е) 8-Ю16, (ж) 3-1016. Хромофоры переводились в ука-
занные состояния с помощью таких адаптирующих вспышек, что в каждом случае поглоща-
лось по крайней мере два фотона на хромофор.
ло показано, что он имеет сложную форму и не меняется под дей-
ствием холода и метаболических ингибиторов, подавляющих позд-
ний рецепторный потенциал. РРП не связан с изменением проводи-
мости мембраны, а генерируется за счет смещения электрических
зарядов при конформационных изменениях в молекуле пигмента.
РРП возникает лишь при упорядоченном расположении молекул
пигмента в мембранах рецепторов и исчезает при нарушении ори-
Зз*
516
Глава 14
ентации (например, при нагревании) [6, 52]. Ранний рецепторный
потенциал наблюдался в рецепторах как позвоночных, так и бес-
позвоночных. Его форма меняется в процессе выцветания зритель-
ного пигмента и зависит от того, что содержит рецептор — родоп-
син или различные промежуточные продукты, которые появляют-
ся при освещении рецептора. На рис. 14-25 показаны эти потенци-
алы, зарегистрированные на рецепторах кальмара.
Хотя величина раннего рецепторного потенциала линейно зави-
сит от интенсивности света, он примерно в миллион раз менее
чувствителен, чем поздний рецепторный потенциал. Действительно,
при внутриклеточных отведениях видно, что световые вспышки, ин-
тенсивность которых соответствует практически полному насыще-
нию позднего рецепторного потенциала, не способны возбудить
сколько-нибудь заметный ранний рецепторный потенциал [226].
В связи с этим ранний рецепторный потенциал не рассматривается
как прямое причинное звено в цепи активации рецептора (см., од-
нако, работу Хагинса и Рюппеля [100]). Тем не менее он является
очень удобным инструментом, позволяющим практически мгновен-
но определить состояние фотопигментов в рецепторах, что невоз-
можно осуществить спектрофотометрически.
Связующее звено между поглощением фотонов и изменением
проводимости
Фотохимические исследования родопсина указывают на то, что
наиболее вероятным процессом, обусловливающим зрительное
возбуждение, являются конформационные изменения, претерпевае-
мые опсином в процессе изомеризации хромофора. С другой сто-
роны, физиологические исследования реакции мембран отдельных
клеток показывают, что изменение мембранной проводимости —
это первое измеримое проявление передачи информации по рецеп-
торам. Выяснение вопроса о том, как структурные изменения мо-
лекул родопсина управляют движением ионов натрия через рецеп-
торную мембрану, в настоящее время является центральной биофи-
зической проблемой зрительного возбуждения.
Поскольку для возбуждения полностью темноадаптированной
палочки достаточно одного кванта [108], было высказано предпо-
ложение, что процесс активации включает в себя значительное уси-
ление. Хагинс [95] подсчитал, что для образования вблизи ножки
палочки (благодаря электротоническому распространению) потен-
циала, превышающего уровень шума, необходимо, чтобы поглоще-
ние одного фотона вызывало переход через мембрану наружного
сегмента около 3000 элементарных электрических зарядов. Более
поздние измерения показали, что константа длины палочковой
клетки составляет всего 25 мкм, так что приведенная выше вели-
чина, вероятно, является оценкой снизу. Подобные расчеты пред-
ставляют собой попытку оценить количественно, в какой мере одна
Фото рецепция и зрение
517
молекула родопсина способна регулировать движение ионов. Не-
давние эксперименты показали, что число ионов, проходящих через
мембрану при поглощении одного фотона, больше 106, что позво-
ляет адекватно регистрировать отдельные фотоны [99].
Уолд [240] обсуждает проблему возбуждения в зрительной си-
стеме скорее в биохимическом плане, уподобляя родопсин профер-
менту, который под действием света переходит в фермент, осуще-
ствляющий усиление благодаря своей способности воздействовать
на многие молекулы субстрата в короткий промежуток времени.
Хотя эта модель возникла как предположение, некоторые недавние
открытия Битенского и др. [20] указывают на то, что родопсин и
в самом деле управляет активностью ферментов.
Авторы указанной работы предполагают, что в процессе зри-
тельного возбуждения участвует циклический АМФ. Оказалось, что
в препаратах мембран наружных сегментов темноадаптированных
палочек активность аденилциклазы в десятки раз больше, чем во
всех известных тканях. При экспонировании препаратов наружных
сегментов активность аденилциклазы падала до одной седьмой ак-
тивности при темновой адаптации. Таким образом, свет действует
как переключатель, инактивирующий аденилциклазу и уменьшаю-
щий образование циклического АМФ. Здесь гипотеза опять опира-
ется на экспериментальные факты. Авторы предполагают, что
регуляция активности аденилциклазы зависит от конформацион-
ных изменений смежных молекул родопсина. Далее, уменьшение
образования АМФ может вести (непосредственно?) к уменьшению
натриевой проводимости. Авторы далее предсказывают, что в раб-
домерных фоторецепторах беспозвоночных действие света должно
выражаться в активации аденилциклазы. Из этих экспериментов
создается впечатление, что свет управляет выходом фоторецепто-
ров аналогично тому, как пептидные гормоны или катехоламины
действуют на соответствующие клетки.
Здесь уместно еще раз напомнить, что диски палочек непосред-
ственно не связаны с поверхностной мембраной наружного сегмен-
та. Поскольку любая из 109 молекул родопсина потенциально спо-
собна активировать палочку [108], возникает вопрос, как возбуж-
дение от центра диска доходит до поверхностной мембраны. (Та-
кого вопроса не возникает в случае рабдомерных образований, где
вся поверхность микровилл находится в непосредственном контакте
с экстраклеточной средой.) Для объяснения механизма передачи
возбуждения большинство исследователей не видят оснований при-
влекать различные процессы миграции энергии в твердом теле
[1, 96]. Иошиками и Хагинс [269] предположили, что высвобож-
дающиеся из дисков ионы Са2+ диффундируют к поверхности мем-
браны и контролируют проводимость по Na+. Поглощение одного
фотона уменьшает темновой ток [128а] примерно на 1%, или при-
близительно на 107 ионов в 1 с, и в принципе резонно предполо-
жить, что какой-нибудь внутренний носитель, например Са2+, тем
518
Глава 14
или иным образом контролирует каналы, через которые проходит
Na+, и обеспечивает это усиление. Однако если поглощение одного
фотона инактивирует только одну из 109 частей активности аденил-
циклазы, не ясно, как это приводит к уменьшению темнового тока
на 1%. Это несоответствие является доводом против того, что аде-
нилциклаза принимает непосредственное участие в изменении нат-
риевой проводимости. Исследования в этой области в настоящее
время ведутся столь интенсивно, что, без сомнения, к моменту вы-
хода этого обзора из печати роль аденилциклазы и ионов кальция
будет много яснее, чем сейчас.
Сложные глаза членистоногих
Общий план строения
Сложные глаза состоят из отдельных единиц, называемых ом-
матидиями. Типичный сложный глаз имеет несколько тысяч омма-
тидиев, однако у некоторых видов с редуцированными глазами их
может быть всего несколько штук. Омматидий — это длинная, по-
хожая на карандаш структура, ориентированная перпендикулярно
поверхности роговицы. Обычно он состоит из восьми фоторецеп-
торных клеток, окружающих рабдом (рис. 14-8, 14-26). На дисталь-
ном конце омматидий имеет маленькую роговицу и вспомогатель-
ное диоптрическое приспособление, называемое кристаллическим
конусом. Рассматриваемые снаружи роговичные линзочки прида-
ют глазу его фасеточный вид. Вокруг каждого пучка фоторецеп-
торных клеток (ретинулы, или маленькой сетчатки) имеется обо-
лочка из пигментных клеток, которая служит для оптической изо-
ляции омматидиев. В качестве обзора по этому материалу можно
рекомендовать работы Уотермана [250], Голдсмита [91], Голдсми-
та и Бернарда [93], Буллока и Хорриджа [37].
Хотя обычно омматидий состоит из восьми ретинулярных кле-
ток, у некоторых видов их только семь (десятиногие ракообразные)
[75, 76, 202], а у других форм — много больше (у Limulus, напри-
мер, двадцать) [160]. Одна или две из ретинулярных клеток могут
быть несколько меньшего размера, иметь дополнительную пигмен-
тацию или занимать особое положение в ретинуле. В качестве при-
мера можно привести базальные ретикулярные клетки омматидиев
ночной бабочки [91] или верхнюю и нижнюю центральные клетки
глаз мухи [228, 229]. В сложном глазе Limulus каждый омматидий
содержит одну или две эксцентрические клетки, которые являются
скорее нейронами второго порядка, имеющими очень тесные связи
с фоторецепторами, чем ретикулярными клетками [160]
(рис. 14-26, Б).
У проксимального конца ретинулы располагается базальная
мембрана, которую пересекают аксоны ретинулярных клеток. Как
правило, расстояние до оптических долей мозга и соответствую'
Фоторецепция и зрение
519
щая длина аксонов составляют 50—100 мкм и более. Оптические
доли или оптические ганглии состоят из трех или четырех слоев
нейропиля, ограниченных телами нейронов и связанных аксонными
пучками.
Рис. 14-26. Типы омматидиев.
4. Фотопический (аппозиционный) омматидий насекомого. Б. Скотопический (суперпозицион-
ный) омматидий. В. Омматидий мечехвоста Limulus. 1 — роговичные линзы; 2 — кристалли-
ческий конус; 3 — первичная или ирисовая, пигментная клетка; 4 — вторичная пигментная
клетка; 5 — ретикулярная клетка; 6 — рабдом; 7 — базальная мембрана; 8 — аксоны ретику-
лярных клеток; 9 — дистальный пигмент в темноадаптированном состоянии; 10 — кристалли-
ческая нить, или кристаллический тракт; 11 — дистальный пигмент в светлоадаптирован-
нсм состоянии; 12 — базальная ретикулярная клетка; 13 — базальная пигментная клетка;
— эксцентрическая клетка; 15 — сплетение аксонов ретикулярных и эксцентрических кле-
ток и коллатералей.
Фотопические (аппозиционные) и скотопические
(суперпозиционные) ретинулы
В 1891 г. 3. Экснер [80] обратил внимание на тот факт, что
большинство сложных глаз можно разделить на два морфологиче-
ских типа, которые он назвал аппозиционными и суперпозицион-
ными глазами. Эти названия должны были отражать предполагае-
мый способ формирования изображений; в настоящее время такое
соответствие оспаривается, и позже мы подробнее обсудим этот
вопрос. В аппозиционных глазах рабдомы длинные, они простира-
ется на всю длину ретинулы от вершины кристаллического конуса
базальной мембраны (рис. 14-26, Л). Вспомогательные экрани-
520
Глава 14
рующие пигменты иногда способны перемещаться, но даже в этом
случае перемещение выражено слабо. Экснер отметил, что этот
тип глаз характерен для животных, которые активны днем и при
сильном освещении; распространяя на этот случай общую термино-
логию палочкового и колбочкового зрения сетчатки позвоночных,
их следует называть фотопическими глазами.
Суперпозиционные глаза отличаются тем, что имеют относи-
тельно короткие, толстые рабдомы, расположенные у базальных
концов ретинул. Между дистальными концами рабдомов и прокси-
мальными частями кристаллических конусов находится кристалли-
ческий тракт — преломляющая среда, образованная либо продол-
жением конусов, либо дистальными, не несущими рабдомеров окон-
чаниями ретикулярных клеток [131, 161]. Перемещения вспомога-
тельного пигмента четко выражены. В состоянии световой адап-
тации он распределен вдоль кристаллического тракта, а когда жи-
вотное адаптировано к темноте, пигмент отходит в дистальную
область, образуя компактную массу между кристаллическими ко-
нусами (у некоторых насекомых), или расщепляется на две части,
одна из которых перемещается дистально, а вторая — к базальной
мембране (у некоторых ракообразных) (рис. 14-26, Б). Такой тин
омматидиев обычно встречается у животных, которые активны
ночью или живут в условиях сумеречного света. Удаление пигмен-
та из области кристаллического тракта — это приспособление, обес-
печивающее увеличение абсолютной чувствительности глаза при-
мерно на 2 логарифмических единицы [111]. Этот механизм более
детально будет рассмотрен в разделе о формировании изображе-
ний. По функциональному сходству с палочковым зрением позво-
ночных глаза с такой морфологией называют скотопическими.
Некоторые сложные глаза нельзя отнести ни к одной из этих
двух категорий. Бабочка-адмирал — это дневное животное, однако
рабдомы у нее короткие, имеется кристаллический тракт и отсут-
ствует перемещающийся экранирующий пигмент [161]. Сетчатка
Dytiscus делится на два яруса. В дистальную сетчатку входит по
одному рабдомеру от каждой ретинулы, и она соединяется с вер-
шиной конуса с помощью короткого кристаллического тракта.
Проксимальная сетчатка лежит глубже, и рабдомы имеют вид, ти-
пичный для скотопического глаза, но они не связываются с диоп-
трическим аппаратом с помощью кристаллического тракта. Счи-
тается, что проксимальная сетчатка не способна разрешать изо-
бражения [Н5].
Формирование изображений в сложных глазах
В 1826 г. Иоганн Мюллер выдвинул гипотезу, согласно которой
каждый омматидий сложного глаза способен воспринимать лишь
узкие пучки света, распространяющиеся параллельно оси оммати-
дия [171]. Экснер считал, что так устроены аппозиционные глаза»
а в адаптированных к темноте суперпозиционных (скотопических)
Фоторецепция и зрение
521
глазах в фокусировании света на уровне рабдомов участвует вся
роговичная поверхность, и, таким образом, изображение создается
в результате суперпозиции лучей, проходящих через многие фасет-
ки. Эта теория многими исследователями подвергалась сомнению,
поскольку не удалось обнаружить ни суперпозиционных образов
там, где, по предположению Экснера, они должны были быть, ни
концентрических слоев с переменным показателем преломления,
которые предположительно должны были располагаться в рогови-
це и кристаллическом конусе. Тем не менее недавние исследования
подтвердили теорию суперпозиции [132]. Эта проблема обсужда-
ется далее на стр. 526 и критически разобрана в работе [93].
Точка зрения Мюллера в своей основе верна, по крайней мере
для фотопических глаз. В качестве примера можно рассмотреть
детально исследованный в последнее время ход лучей через диоп-
трический аппарат пчелы [231]. Падающие на роговицу параллель-
ные лучи света фокусируются примерно на расстоянии 2/з пути от
кристаллического конуса; далее лучи начинают расходиться. По-
скольку апикальный конец кристаллического конуса окружен ци-
топлазмой пигментных клеток с несколько меньшим показателем
преломления, латеральные части конуса света теряются, а цент-
ральные лучи входят в рабдом. Рабдом имеет больший показатель
преломления, чем окружающая цитоплазма, и, таким образом,
служит волноводом. В пользу такого его свойства имеется ряд сви-
детельств. Во-первых, на свежих препаратах глаз, освещаемых че-
рез роговицу, рабдомы вспыхивают, как яркие пятнышки, когда на-
правление на источник света совпадает с их осью. Во-вторых, у
пчелы удается зарегистрировать несколько обычных типов распре-
деления интенсивностей, которые можно объяснить на основании
теории распространения электромагнитной энергии по волноводу.
Благодаря таким свойствам каждый омматидий собирает свет из
узкого угла, и энергия, которая достигает основания кристалличе-
ского конуса, затем направляется в рабдом и тонкую оболочку ци-
топлазмы вокруг него.
Поле зрения каждого омматидия ненамного больше, чем угол
расхождения осей соседних омматидиев. На рис. 14-27 показана
вычисленная зависимость попадающего на рабдом пчелы количе-
ства света от угла наклона луча по отношению к оси омматидия.
Светлые треугольники — это данные измерений интенсивности све-
та у срезанных концов отдельных рабдомов на препаратах глаз,
освещаемых точечным источником, положение которого по отноше-
нию к оси омматидия фиксируется с большой точностью [131].
Как указано в подписи к рисунку, самые последние измерения по-
казали, что функция дирекционалы-юй чувствительности у пчелы
на самом деле еще уже, чем это изображено на рис. 14-27. Изме-
рения дирекциональной чувствительности, выполненные с помощью
внутриклеточных электродов на одиночных ретинулярных клетках
^ух, также дают более узкие распределения [204, 249].
522
Глава 14
Очень интересным вопросом, которым занимался целый ряд ис-
следователей, является вопрос об оптимальном размере фасеток
роговицы [11, 81, 231]. Большие фасетки собирают больше света
но при этом существенно уменьшается разрешающая способность
глаза в целом. Размер зерен мозаики сетчаточного изображения
Угол наклона светового луча по
отношению к оптической оси
enafyw
Рис. 14-27. Дирекциональная чувствительность омматидия медоносной пчелы
Apis.
Темные кружки и кривая. Вычисленная функция пропускания, показывающая, какая часть
(%) поверхности линзы обеспечивает прохождение света до рабдома при разных углах на-
клона светового луча по отношению к оптической оси (231]. Светлые треугольники. Резуль-
таты измерения интенсивности света иа рабдоме, приведенные в масштабе, соответствующем
первой кривой [131]. Последние электрофизиологические данные, полученные на пчелах,
показывают, что функция пропускания на самом деле еще уже — ее полуширина составляет
примерно 3° [135а]. Более того, эти электрофизиологические данные согласуются с более
поздними оптическими измерениями и теоретическими расчетами, в которых учтена угловая
чувствительность рабдома [215а].
можно уменьшать, уменьшая размер отдельных фасеток и увеличи-
вая соответственно число ретинул, но только до определенного
предела. Как только фасетки становятся слишком малыми, увели-
чению четкости начинает препятствовать дифракция. Реально су-
ществующий размер фасеток, по-видимому, оптимален: он позволя-
ет иметь максимально возможное число омматидиев, при котором
еще нет серьезного уменьшения четкости из-за дифракции.
Функция вспомогательного пигмента состоит в поглощении на-
клонных лучей, которые не попадают на рабдом. У белоглазых му-
тантов мух, полностью лишенных вспомогательного экранируют6'
Фоторецепция и зрение
523
Го пигмента, амплитуда деполяризационного ответа чувствительных
клеток никогда не падает ниже 40% максимума при отклонении
источника света даже на 20° от оси омматидия [249].
Альтернативой суперпозиционной теории является другая точка
зрения, согласно которой в скотопических глазах кристаллический
тракт работает как волновод [36]. При световой адаптации, когда
вспомогательный пигмент располагается проксимально, гранулы
Рис. 14-28. Схема строения роговичных фасеток и лежащих под ними рабдомеров,
имеющих одно и то же поле зрения; глаз двукрылых (взято с некоторыми изме-
нениями из работы [12'8]).
Рабдомеры, имеющие одинаково ориентированные оптические оси, обозначены цифрами
I — 6 и обведены кружочками. Аксоны этих шести клеток входят в один и тот же оптиче-
ский патрон в ганглиозной пластинке, как показано на рис. 14-29. Оставшиеся два рабдо-
мера расположены концом к концу, и в любом поперечном сечении виден только один
из них. Это верхняя и нижняя центральные клетки (здесь обозначенные цифрами 7 и 8,
а на рис. 14-8, А — цифрами 1 и 7), и их аксоны представляют собой не что иное, как длин-
ные зрительные волокна, которые проходят между патронами и синаптически переключа-.
ются в медуллярной пластинке.
пигмента окружают кристаллический тракт и увеличивают пока-
затель преломления «обкладки». Эффективность рабдома как вол-
новода становится меньше, и соответственно часть энергии теряется
за счет поглощения в окружающих пигментных гранулах. Вспо-
могательный пигмент, таким образом, играет роль «продольного
зрачка» [131], уменьшая чувствительность рецепторов в состоянии
световой адаптации. При темновой адаптации образное зрение в
глазах с кристаллическим трактом зависит от эффективности ди-
оптрических структур, фокусирующих свет на волноводной систе-
ме, и от того, существуют ли потери в кристаллическом тракте и
в Рабдоме. В зависимости от вида животного работает тот или иной
524
Глава 14
механизм или оба вместе. Для определения относительного вклада
каждого, как и для оценки качества суперпозиционных образов
необходимы тщательные эксперименты. У бабочки-адмирала функ-
ция дирекциональной чувствительности отдельных ретикулярных
клеток достаточно узкая [161], однако это еще не означает, что в
данном случае отсутствует суперпозиция образов [93]. У рака по-
ловина света, попадающего на данную ретикулярную клетку, про-
ходит через соседние фасетки [209], а у жука Dytiscus [115], пред-
ставляющего крайний случай, между дистальной и проксимальной
сетчатками, вероятно, нет эффективного волновода.
Фотопические глаза двукрылых представляют собой особый слу-
чай другого рода. У видов со слитными рабдомами все объединен-
ные ретикулярные клетки, по-видимому, «смотрят» в одну и ту же
точку в пространстве и одинаково возбуждаются при аксиальном
освещении. У двукрылых и по крайней мере у некоторых полужест-
кокрылых рабдомеры отдельных ретинулярных клеток остаются
разделенными (рис. 14-8, А). У мух две из восьми клеток — верх-
няя и нижняя центральные клетки (клетки 1 и 7 на рис. 14-8, Л) —
имеют укороченные рабдомеры, которые лежат один над другим,
как два карандаша, приставленные концом к концу [228]. Таким
образом, на срезе омматидия мухи, сделанном на дистальном кон-
це, видно семь ретинулярных клеток и семь рабдомеров, а на сре-
зе, сделанном на проксимальном конце, — восемь ретинулярных
клеток и семь рабдомеров. Ориентация оптических осей всех семи
рабдомеров несколько различается; оптическая ось каждого из
шести периферических рабдомеров параллельна оси одного из раб-
домеров в каждом из пяти соседних омматидиев [128], как это
показано на рис. 14-28. Хотя все шесть ретинулярных клеток,
имеющих одинаковые оптические оси, лежат в различных оммати-
диях, они посылают свои аксоны к одной и той же паре нейронов
второго порядка [24, 229]. Такой механизм называют «нейронной
суперпозицией»; по-видимому, таким способом увеличивается све-
тосила глаза без увеличения размеров фасеток и снижения остро-
ты зрения (см. ниже).
Синаптические связи в оптических ганглиях
Общее строение оптических долей. В оптических долях обычно
имеется три или четыре фиброзные массы (рис. 14-29, Л). У насе-
комых три соответствующие области от периферии во внутрь назы-
ваются lamina ganglionaris, medulla и lobula. Для ракообразных
терминология несколько другая: lamina ganglionaris, medulla
externa, medulla interna и medulla terminalis [37]. Аксоны, выхо-
дящие из самой проксимальной массы, направляются в мозг. Да-
лее мы будем рассматривать только более дистальные области.
Ганглиозная пластинка содержит два основных типа вставоч-
ных нейронов. Монополярныё клетки получают сигналы от чувст-
J
Рис. 14-29.
Л. Схема строения зрительных долей насекомых, на которой приведено несколько типов
волокон из числа более периферических. 1 — короткое рецепторное волокно, оканчивающее-
ся в ганглиозной пластинке; 2 — длинное рецепторное волокно, оканчивающееся в медул-
лярной пластинке; 3 — рецепторы; 4—ганглиозная пластинка; 5 — монополярный нейрон,
навход которого поступают сигналы с коротких аксонов ретикулярных клеток в ганглиоз-
ной пластинке; 6 — волокно, связывающее ганглиозную пластинку и medulla (центробежное
волокно одного из классов); 7 — medulla; 8—lobula. Б. Три составляющие оптических
патронов двукрылых. Патроны изображены в виде цилиндров. I — монополярный нейрон
второго порядка с синаптическими шипиками; II — 6 аксонов ретикулярных клеток, прихо-
дящих из 6 различных омматидиев (см. рис. 14-28) и синаптически контактирующих в патро-
не с монополярами после переориентации на 180°; для ясности рисунка волокна внутри патро-
на не показаны; III — корзинчатое центробежное волокно.
S26
Глава 14
вительных клеток. Их тела лежат проксимально по отношению к
базальным концам омматидиев, а аксоны направляются далее в
центр к медуллярной пластинке. В месте прохождения аксонов че-
рез нейропиль ганглиозной пластинки — где они являются постси-
наптическими по отношению к аксонам ретикулярных клеток — на
их латеральных поверхностях видны пуговки и шипики. Вставоч-
ные нейроны второго типа — центробежные-, тела их лежат цент-
рально по отношению к пластинке, а отростки распространяются
на периферию в ганглиозную пластинку (или на периферию в ган-
глиозную пластинку и в центр в медуллярную пластинку). Назва-
ние «центробежные» отражает анатомическое положение тел этих
нейронов по отношению к ганглиозной пластинке; функция этих
клеток неизвестна.
У многих видов отчетливо видны связи пучков аксонов ретику-
лярных клеток с монополярными нейронами ганглиозной пластин-
ки [43]. У тех видов, у которых такие «оптические патроны» хоро-
шо выражены, имеется по одному патрону на каждый омматидий.
Синаптические связи у двукрылых. Детальное строение пласти-
нок наиболее полно изучено у мух. При рассмотрении формирова-
ния изображения мы отмечали, что у мух шесть периферических
ретикулярных клеток из шести омматидиев имеют одно и то же
поле зрения. Более того, аксоны этих шести ретикулярных клеток
собираются в одном оптическом патроне (рис. 14-29, Б), где они
переключаются на одну и ту же пару монополярных нейронов.
Каждый из двух монополярных нейронов связан со всеми шестью
пресинаптическими элементами. Таким образом, оптический патрон
суммирует свет, идущий из одной и той же точки пространства че-
рез шесть различных фасеток.
Монополярные нейроны второго типа (не показанные на
рис. 14-29) посылают свои главные отростки между патронами, где
они с помощью коротких коллатералей устанавливают латераль-
ные связи с клетками второго порядка в трех, а иногда и четырех
смежных патронах [222]. Затем аксоны направляются к медул-
лярной пластинке. Помимо этого, описано несколько типов центрон
бежных клеток [220]. Все они в той или иной степени ветвятся
внутри ганглиозной пластинки; некоторые из них имеют соедини-
тельные волокна, которые связывают ганглиозную пластинку с
медуллярной, тогда как другие (амакриновые клетки) имеют вет-
вления только в ганглиозной пластинке. Об их синаптических свя-
зях известно очень мало.
Аксоны двух центральных ретикулярных клеток не оканчива-
ются в ганглиозной пластинке, а проходят эту область, не вклю-
чаясь в патроны. На выходе из ганглиозной пластинки они объ-
единяются с аксонами пары монополярных клеток из патрона, на-
ходящегося непосредственно под их омматидием [220, 228]. Эти
четыре волокна, относящиеся к одной и той же точке поля зрения,
остаются сгруппированными при прохождении через антерио-по-
Фоторецепция и зрение
527
стериорную хиазму между ганглиозной и медуллярной пластинка-
ми и оканчиваются в общих колонках в нейропиле второго зри-
тельного ганглия. Имеются данные, что к этой четверке волокон
присоединяется пятое волокно, связывающее ганглиозную и медул-
лярную пластинки и идентифицируемое в ганглиозной пластинке
как одно из центробежных [ИЗ]. Главное при этом, однако, состо-
ит в том, что топографическая проекция реального мира на гангли-
озную пластинку сохраняется и в медуллярной, хотя она и повер-
нута относительно переднезадней оси.
Другие членистоногие. Хотя сетчатки двукрылых имеют неко-
торые особенности, у видов со слитными рабдо1ма1ми зрительные
ганглии сходны по строению. В ретинуле со слитным рабдомом
все рабдомеры имеют одно и то же поле зрения (кроме Limu-
lus) [189]. Соответственно Хорридж и Майнертцхаген [113] сооб-
щили, что за исключением длинных зрительных волокон, все рети-
нулярные клетки омматидиев со слитными рабдомами (у Schisto-
cerca, Apis, Notonecta и Aeshna) конвергируют на одну и ту же
пару (или группу) монополярных нейронов (т. е. на один патрон,
хотя у этих видов патроны не очень хорошо выражены). Варела
[230] пришел к такому же заключению относительно пчел, однако
для других животных это не столь очевидно [221], а Хамори и
Хорридж [104] даже установили, что у омара аксоны из одной
ретинулы идут в разные патроны.
У ночных бабочек Sphinx, которые имеют скотопические глаза,
приспособленные для того, чтобы видеть в сумерках, дендритные
поля монополярных нейронов второго порядка простираются ла-
терально примерно на 20 мкм [221], указывая на связь со многими
омматидиями, что напоминает высокую конвергенцию палочек на
ганглиозные клетки в сетчатке позвоночных (стр. 548). Таким об-
разом, по имеющимся в настоящее время данным, заключение о
том, что ганглиозная пластинка содержит тонкозернистую топогра-
фическую карту зрительного поля, встречает ряд возражений. Тем
не менее схема проекции волокон из ганглиозной пластинки в ме-
дуллярную, которая была описана для мух, обнаружена также у
некоторых других насекомых с фотопическими (аппозиционными)
глазами [113|.
На основании распределения синаптических пузырьков в окон-
чаниях волокон можно сделать некоторые предварительные за-
ключения о характере нейронной активности в ганглиозной пла-
стинке. Все исследователи единодушны в том, что рецепторы си-
наптически связаны с монополярными нейронами. У омара
Центробежные волокна считаются пресинаптическими по отноше-
нию к монополярным нейронам и постсинаптическими по отноше-
нию к аксонам рецепторов [105]. У пчелы описаны аналогичные
связи, а также переключения с центробежных волокон на рецеп-
торы и контакты между центробежными волокнами [230].
528
Глава 14
Без сомнения, метод окрашивания по Гольджи позволяет со-
ставить лишь весьма смутное представление о некоторых тонких
морфологических деталях. Как уже упоминалось, имеется несколь-
ко типов монополярных и центробежных нейронов. Кроме того,
существует три морфологических вида длинных зрительных воло-
кон, выделяемых по признаку наличия и распределения латераль-
ных отростков в ганглиозной пластинке и по типу окончаний в ме-
дуллярной. У некоторых видов (например, Pieris) имеется также
три типа коротких ретикулярных аксонных окончаний в ганглиоз-
ной пластинке [221]. Разнообразие форм предполагает специали-
зацию функций, так что на основании анатомических данных мож-
но ожидать, что ретикулярные клетки одного омматидия выпол-
няют различные функции. Этому вопросу посвящен следующий
раздел.
Дифференциация функций внутри одной ретинулы
Как мы отмечали, у видов со слитными рабдомами волновод-
ные свойства рабдома обусловливают оптическую связь ретику-
лярных клеток, и даже у двукрылых с разделенными рабдомерами
те единицы, которые имеют одинаковые оптические оси, физиоло-
гически связаны благодаря конвергенции на первом синапсе. Од-
нако наличие разных морфологических типов аксонов ретикуляр-
ных клеток позволяет предположить, что одиночная ретинула мо-
жет представлять собой целый комплекс разных функциональных
элементов.
Электротоническая связь между ретикулярными клетками.
Если различные ретикулярные клетки одного омматидия способны
реагировать независимо, можно ожидать, что они электрически
изолированы. Вводя электроды внутрь разных клеток одного ом-
матидия и пропуская ток, можно обнаружить связи с малым со-
противлением. Такие эксперименты проводились на нескольких
видах животных, причем результаты оказались совершенно различ-
ными. У Limulus между всеми клетками одного омматидия суще-
ствуют разносторонние связи [215]. Это не удивительно, если учесть
характер возбуждения клетки второго порядка (эксцентрической).
Дендрит эксцентрической клетки располагается вдоль оси омма-
тидия между рабдомерами ретикулярных клеток (рис. 14-26, В).
Из-за наличия контактов с малым сопротивлением между всеми
клетками омматидия деполяризация мембран рабдомеров, вызы-
ваемая светом, обусловливает появление тока, выходящего из экс-
центрической клетки. Деполяризация дендрита эксцентрической
клетки электротонически распространяется к основанию аксона,
где возникают импульсы. Электротоническая связь не ограничива-
ется только омматидиями Limulus\ она была обнаружена также в
ретинулах трутней медоносной пчелы [207].
Реакции на поляризованный свет. У некоторых видов членисто-
ногих со слитными рабдомами имеются ретикулярные клетки, ко-
Рис. 14-30. Чувствительность фоторецепторных клеток беспозвоночных к поляризации света [252].
Внутриклеточные отведения (запись при медленной развертке во времени) рецепторных потенциалов, возбуждаемых в ретикулярных клетках
рака Procambarus вспышками монохроматического света длительностью 0,4 с при 18 значениях длин волн — от 400 до 700 им. Вспышки оди-
наковы по количеству квантов и следуют друг за другом с интервалами в 2 с. Плоскости поляризации света в случаях А и Б взаимно пер-
пендикулярны.
530
Глава 14
Таблица 14-3
Спектральная чувствительность сложных глаз насекомых1)
Животное \nax- «и Метод2) Ссылка
Libellula (стрекоза) Peri planet а (таракан) Locusta (саранча) <380 365 420 410 430(515) 520 450—5503) 510 Селективная адаптация Анализ отдель- ных элементов То же » » [1961 [H2J [1691 [181
Notonecta (водный клоп) 350 420 464? 567 » » [351
Calliphora (муха) Apis $ (медоносная 345(470) 345 490(345) 520(345) 535 » » Селективная адаптация [381 [89]
пчела) 340 430 460 530 Анализ отдель- ных элементов [7]
Apis <5 345 340 440 450 535 530 Селективная адаптация Анализ отдель- ных элементов [90] [71
Ч Взято из работы [92]. Цифры в скобках относятся ко второму максимуму чувстви-
тельности той же самой клетки.
2) Избирательная адаптация при экстраклеточном отведении сетчаточных потенциалов
действия или исследование отдельных ретикулярных клеток с помощью внутриклеточных
микроэлектродов.
3) Результаты варьируют. См. также [78].
торые не связаны между собой. Например, у саранчи можно заре-
гистрировать ответы двух ретикулярных клеток, которые имеют
идентичные зрительные поля (из чего можно заключить, что они
лежат в одном омматидии), но чувствительность которых по-раз-
ному зависит от плоскости поляризации света [205, 207, 208].
У ракообразных ретикулярные клетки можно разделить по этому
признаку на две группы (рис. 14-30). Показано, что плоскость по-
ляризации света, к которому клетки наиболее чувствительны, соот-
ветствует ориентации осей’'микровилл в рабдомерах [77]. Отноше-
ние чувствительности отдельных клеток к свету, поляризованному
в оптимальном направлении и под прямым углом, доходит до 10:1
[208, 252]. Эти эксперименты показывают, что возбуждение одной
ретикулярной клетки не обязательно затрагивает другие клетки
омматидия, и определенно свидетельствуют о том, что дифференци-
альная чувствительность к плоскости поляризации света, свойст-
венная отдельным ретикулярным клеткам омматидия, обусловлена
дихроичным поглощением света в микровиллах (рис. 14-15, Б).
Фоторецепция ц зрение
531
Эти эксперименты очень важны и с точки зрения поведения
животных, поскольку многие членистоногие [122,251] и головоногие
моллюски [165] реагируют на свет по-разному в зависимости от
плоскости поляризации. По-видимому, самым известным примером
здесь является медоносная пчела, которая способна анализировать
поляризованный свет неба при ориентации по своему солнечному
компасу [84]. Безусловно, для выяснения с точки зрения нейрофи-
зиологии хотя бы сенсорной стороны такого поведения необходима
дополнительная информация о синаптических связях ретинуляр-
ных клеток, происходящих из одного омматидия.
Периферическая основа цветного зрения. Как было показано с
помощью поведенческих методов и используя тренировки, пчела
способна различать цвета [58, 150]. Функции спектральной чув-
ствительности одиночных рецепторных клеток глаз нескольких ви-
дов членистоногих выявили наличие различных типов клеток с
максимальной чувствительностью в разных областях спектра.
Часть этих данных приведена в табл. 14-3 (см. также Голдсмит и
Бернард [193]). Как эта сенсорная информация используется в
поведении, как правило, неизвестно.
Благодаря выполненным в последнее время исследованиям об-
наружилось, что различные цветовые рецепторы могут существо-
вать в одном омматидии. Изучение микроспектрофотометрическим
методом отдельных рецепторов глаз мухи Calliphora показало, что
шесть периферических ретинулярных клеток имеют зрительный пиг-
мент с максимумом поглощения 510 нм, тогда как две центральные
клетки с длинными аксонами, которые обходят ганглиозную пла-
стинку, содержат другой зрительный пигмент, миисимум погло-
щения которого равен 470 нм [134]. Таракан Periplaneta имеет
слитные рабдомы и два четко выраженных класса рецепторов с
максимальной чувствительностью к ультрафиолетовому (360 нм)
и зеленому (510 нм) свету [169, 247]. Используя микропипетки с
различными красителями [170] или наблюдая специфическое пе-
ремещение экранирующего пигмента при адаптации к цветному
освещению [42], можно идентифицировать оба типа сенсорных
клеток в одном омматидии. Было бы весьма интересно с помощью
аналогичной техники исследовать глаза других видов животных.
Заслуживает также внимания вопрос о различиях между оммати-
диями из разных частей глаза.
Реакция клеток второго порядка
В настоящее время достаточно четко показано, что реакция на
свет монополярных нейронов, с которыми аксоны ретинулярных
клеток синаптически контактируют в ганглиозной пластинке [8,
206], а также терминалей аксонов оцеллярного нерва, с которыми
связаны оцеллярные фоторецепторные клетки [195], состоит в по-
явлении гиперполяризационного потенциала. В ганглиозной пла-
34*
532
Глава 14
стинке монополярные клетки, по-видимому, не генерируют импуль-
сов, а дают медленные гиперполяризационные потенциалы, которые
повторяют с некоторой фильтрацией [122а] рецепторные потенци-
алы и, как предполагают, электротонически распространяются в
более глубокие синаптические области.
Как отмечалось выше, омматидии глаза Limulus анатомически
отличаются от омматидиев глаз насекомых и ракообразных. Здесь
элемент второго порядка — эксцентрическая-клетка, тело которой
лежит внутри омматидия, электротонически связана с ретикуляр-
ными клетками и, следовательно, возбуждается при деполяризации
ретикулярных клеток. Аксоны эксцентрических клеток считаются
единственными волокнами оптического нерва, которые проводят
импульсы [254]. В области, смежной с базальной мембраной и
располагающейся проксимально по отношению к ней, нет больше
никаких нейронных тел, хотя там имеются обширные сплетения
нервных волокон, образованные частично коллатералями аксонов
эксцентрических клеток (рис. 14-26, В). В сплетении осуществля-
ются сериальные тормозные связи между аксонами эксцентриче-
ских клеток соседних омматидиев, в результате функционирования
которых появление импульсной активности в одном омматидии
(эксцентрической клетке) ведет к уменьшению вероятности появ-
ления разряда в близлежащих омматидиях [224а]. Кроме того,
предполагается наличие возвратных коллатералей, образующих
цепи обратной связи с собственными эксцентрическими клетками
и лежащих в основе системы «самоторможения» [106, 187, 190].
Функция аксонов ретикулярных клеток у Limulus неизвестна.
Переработка информации 5
Limulus. Наличие тормозных взаимосвязей в сплетении под ба-
зальной мембраной означает, что данный омматидий может реаги-
ровать на свет независимо от своих соседей только тогда, когда он
является единственным освещенным элементом. Количественное
исследование этой системы Хартлайном и его коллегами [106]
выявило многие ее свойства, которые оказались сходными с функ-
циональными характеристиками сетчатки позвоночных, где имеется
больше различных типов нейронов.
Взаимодействие двух омматидиев в стабильном состоянии мож-
но описать системой двух линейных уравнений [190]:
Г1=в1—Л1,2(г2—г?>2),
Ч1(г1~г2,1)»
где Г1 и Г2 — реакции (число импульсов в единицу времени) каждо-
го из. омматидиев; е± и е%— реакции тех же омматидиев в усло-
виях, когда каждый из них освещается по отдельности; k\t 2 —к0'
эффициент тормозного воздействия элемента 2 на элемент 1; г 12--
Фоторецепция и зрение
533-
пороговая частота разряда, которую элемент 2 должен превысить,
чтобы оказать тормозное воздействие на элемент 1. Из этих урав-
нений видно, что реакция элемента 1 определяется алгебраиче-
ской суммой собственного возбуждения и торможения, поступаю-
щего из другого омматидия. Это торможение в свою очередь пря-
мо пропорционально частоте разряда второго омматидия, если
только она превышает некоторое пороговое значение. Торможение
0,5 мм поверхности
глаза
Рис. 14-31. Частота разряда омматидиев (аксонов эксцентрических клеток), рас-
положенных по линии, перпендикулярной границе светлое — темное при ступен-
чатом распределении интенсивности, показанном вверху справа [190].
Треугольники. Когда каждый из омматидиев освещается отдельно, пространственное распре-
деление частоты разряда отражает объективное распределение интенсивности возбуждающе-
го света. Кружки. Когда изображение проецируется сразу на все рецепторы, частоты раз-
ряда омматидиев ложатся на нижнюю кривую. Усиление и подавление реакций по обе сто-
роны от границы светлое—темное обусловлено латеральным торможением.
между омматидиями взаимное, но не симметричное, т. е. соответ-
ствующие коэффициенты торможения и пороговые значения часто-
ты, вообще говоря, не равны между собой. Величина торможения
уменьшается с увеличением расстояния между омматидиями. Дей-
ствия нескольких омматидиев аддитивны.
Одно из важных следствий такой организации — усиление
контраста изображения на границах. На рис. 14-31 (в рамке) по-
казано ступенчатое распределение интенсивности света, проецируе-
мого на глаз. Верхняя кривая (светлые треугольники) соответст-
вует реакциям отдельных омматидиев в условиях, когда каждый
омматидий освещается по отдельности, и отражает объективное
распределение интенсивности света. Когда вся картина проециру-
ется сразу, выходные сигналы омматидиев ложатся на нижнюю
кривую (светлые кружки). Омматидии, расположенные по светлую
сторону от границы, возбуждаются максимально, поскольку они
534
Глава 14
слабо тормозятся близлежащими элементами из темной зоны. На-
против, элементы, прилегающие к границе с темной стороны, име-
ют самый низкий выход, так как они близки к клеткам с макси-
мальным выходом и тормозятся сильнее, чем все другие элемен-
ты. В результате разница в частоте разряда омматидиев по раз-
ные стороны от границы светлое — темное будет еще больше.
Возможны также другие обусловленные геометрическим распо-
ложением возбужденных омматидиев эффекты, включая снятие
торможения. Оно может осуществляться в ряду из трех (не смеж-
ных) омматидиев, когда два элемента на концах этого ряда на-
столько далеки, что их тормозным воздействием друг на друга
можно пренебречь. Возбуждение одного краевого (тестируемого)
омматидия заставляет его разряжаться. Одновременное возбуж-
дение среднего омматидия уменьшает частоту разряда тестируе-
мого. Если теперь возбудить и третий омматидий, он снизит часто-
ту разряда среднего омматидия и тем самым уменьшит торможе-
ние, поступающее на тестовый.
Динамические аспекты торможения более сложны. Наличие са-
моторможения, т. е. своего рода отрицательной обратной связи,
приводит к некоторому временному обострению реакций и частич-
но объясняет адаптацию, наблюдаемую в аксонах эксцентрических
клеток. Взаимосвязь самоторможения и латерального торможения,
которые по-разному меняются во времени, может также привести
к возникновению переходных эффектов. Например, повышенная ре-
акция клетки после снятия латерального торможения может объ-
ясняться задержкой в развитии процесса самоторможения [106,
187].
Ракообразные и насекомые. Функциональные свойства зри-
тельных ганглиев исследовались электрофизиологически и с по-
мощью поведенческих методов. У ракообразных зрительные ганг-
лии обычно отделены от мозга оптическим трактом или оптическим
стебельком, так что можно выделять активность отдельных эле-
ментов, вводя игольчатые электроды между волокнами. У насеко-
мых аналогичные отведения делаются путем погружения электро-
дов в оптические доли и мозг. Эти эксперименты позволяют клас-
сифицировать волокна по их положению, видам эффективных
стимулов и рецептивным полям (т. е. площади, на которой распо-
ложены рецепторы, оказывающие влияние на данный вставочный
нейрон). Такого рода подход имеет свои внутренние ограничения,
о которых полезно помнить. Отбор волокон не случаен, и актив-
ность многих мелких элементов, возможно имеющих другие функ-
циональные свойства, не регистрируется. Кроме того, по мере про-
движения информации к высшим центрам она становится все бо-
лее абстрактной и интегрированной. Поэтому нужна некоторая
интуиция при определении того, что может служить адекватным
сетчаточным стимулом для зрительных вставочных нейронов
высшего порядка. Так, например, клетки могут быть чувствитель-
Фоторецепция и зрение
535
ны к однонаправленному движению в зрительном поле, но намного
хуже реагировать на простое включение и выключение света. На-
конец, при современной технике часто невозможно соотнести ре-
гистрируемые ответы с определенным морфологическим типом во-
локон, особенно в областях нейропиля.
Нейроны оптического тракта ракообразных имеют большие
зрительные поля: от 30 до 180°. У Podophthalmus, например, это
означает, что число связанных с одним нейроном омматидиев со-
ставляет 300—10 000 [256]. Было описано несколько типов зри-
тельных вставочных нейронов. Тонические волокна — слабо адапти-
рующиеся волокна, которые сигнализируют об освещении больших
участков. Они могут иметь небольшое тормозное окружение, и их
чувствительность больше, когда животное находится в состоянии
возбуждения. Волокна, реагирующие на затемнение, функциональ-
но полностью противоположны тоническим: они тормозятся светом.
Имеется несколько типов волокон, чувствительных к движению..
Один тип — это быстро адаптирующиеся элементы, которые стиму-
лируются внезапным движением в любом месте поля зрения. Раз-
личные части поля зрения адаптируются независимо, так что при-
выкание можно снять перемещением тест-объекта в область поля
зрения, которая перед этим не стимулировалась. Эти клетки мож-
но назвать «клетками новизны». Чем больше ракообразное живот-
ное в своей моторной активности полагается на зрение (сравните,
например, краба и рака), тем больше у него в оптическом тракте
волокон, чувствительных к движению [255, 259, 261—263]. У Po-
dophthalmus имеются волокна, чувствительные к направлению дви-
жения объекта; их можно разделить на подклассы по скорости
движения, которая вызывает максимальную реакцию [256].
Другой важной чертой зрительных трактов ракообразных явля-
ется наличие большого количества эфферентных сенсорных входов
от механорецепторов, статоциста и контралатерального глаза [41,
206]. В действительности большинство реагирующих на движение
волокон получает сигналы не только от фоторецепторов, но и от
чувствительных волосков. Вероятно, самые удивительные из этих
мультимодальных клеток — это константные в пространстве волок-
на, которые одновременно являются тоническими и чувствительны-
ми к движению. В примере, приведенном на рис. 14-32, рецептив-
ное поле занимает часть верхней половины глаза около центра, и,
кроме того, имеются связи с рецепторами статоциста. Когда жи-
вотное поворачивается вокруг своей поперечной оси (наклоняется),
рецептивное поле движется в обратную сторону, и волокно продол-
жает «смотреть» в ту же самую часть неба (рис. 14-32, нижний
ряд). Таким образом, омматидии, которые потенциально являются
частью рецептивного поля, занимают большую центральную об-
ласть глаза, но те, которые оказываются смотрящими ниже линии
горизонта, не способны дать свой вклад из-за торможения, идущего
от статоциста. Когда животное поворачивается вокруг своей про-
536
Глава 14
дольной оси (вращается), размер рецептивного поля меняется. При
повороте глаз вниз все омматидии, потенциально способные вызы-
вать возбуждение вставочного нейрона, со временем оказываются
ориентированными ниже линии горизонта, и волокно «слепнет».
Если животное переворачивается на другую сторону и его глаза
смотрят вверх, рецептивное поле увеличивается до максимума
(рис. 14-32, верхний ряд).
Рис. 14-32. Изменения возбуждающей части поля зрения «константного в про-
странстве» тонического волокна, воспринимающего сигналы только из определен-
ной области пространства.
Оптический тракт рака Procambarus [261]. / — Вид на левый глаз животного спереди. Нор-
мальное положение глаза показано на третьей диаграмме справа. При вращении животного
вокруг своей антеро-постериорной оси рецептивное поле (зачерненная область) меняется.
Л. Вид на левый глаз сбоку. Когда животное поворачивается вокруг дорсо-вентральиой оси,
размер поля зрения и его абсолютные координаты в пространстве остаются неизменными.
Остальные объяснения даны в тексте.
Из насекомых исследовались саранча [39, 40, 114] и кузнечик
[117]. Были обнаружены тонические элементы, волокна, реагирую-
щие на затемнение, фазные «оп»- или «on-ofb-элементы, а также
быстро адаптирующиеся волокна, чувствительные к движению без-
относительно к его направлению. Как и у ракообразных, многие
волокна связаны с механорецепторами, но относительная незави-
симость адаптации по зрительной и механорецепторной модально-
стям может означать, что они связаны с различными частями ден-
дритного поля. Все эти типы нейронов обнаруживаются в медул-
лярной пластинке. По сравнению с ракообразными у насекомых
волокна с малыми рецептивными полями встречаются чаще [151],
но локализованы они в более периферических областях оптических
ганглиев, которые у ракообразных не исследовались. В процессе
многочисленных исследований оптомоторных реакций у насекомых
проводились интенсивные поиски элементов, чувствительных к на-
правлению движения. Такие клетки были найдены в lobula пчел
[123], мух [19] и ночной бабочки Sphinx [50], а также в медул-
лярной пластинке кузнечика и саранчи — видов с менее дифферен1
цированной lobula. По-видимому, там имеется набор из четырех
Фоторецепция и зрение
537
нейронов, реагирующих на движение протяженных границ в гори-
зонтальном и вертикальном направлениях. У перепончатокрылых
и двукрылых клетки имеют монокулярные поля, а у Sphinx — би-
нокулярные. Волокна медленно адаптируются и могут успешно
участвовать в оптомоторных реакциях.
Глаза позвоночных
Строение глаза человека
Глаз человека, схематически изображенный на рис. 14-33, яв-
ляется типичным примером глаза позвоночных. Он имеет почти
сферическую форму и обтянут волокнистой тканью — склерой, пе-
реходящей спереди в прозрачную роговицу. Хрусталик делит глаз-
ное яблоко на переднюю камеру, заполненную водянистой влагой,
и камеру большего размера, расположенную сзади и заполненную
стекловидным телом. Радужка, которая определяет цвет глаза, рас-
полагаясь непосредственно перед хрусталиком, играет роль диа-
фрагмы. Апертура этой диафрагмы называется зрачком.
Изнутри к склере прилегает вторая оболочка, называемая сосу-
дистой, а между ней и стекловидным телом лежит третья — сетчат-
ка. Сосудистая оболочка пронизана сосудами, снабжающими сет-
чатку питательными веществами. Ее внутренняя поверхность кон-
тактирует со слоем сильно пигментированных эпителиальных кле-
ток, содержащих меланин. Сетчатка, о которой более подробно
будет идти речь ниже, представляет собой светочувствительный
слой и состоит из рецепторов (палочек и колбочек) и нескольких
типов нейронов. Наружные сегменты рецепторов обращены к пиг-
ментному эпителию, так что свет, достигающий зрительного пиг-
мента со стороны стекловидного тела, проходит перед этим через
два слоя нервных клеток и внутренние сегменты палочек и колбо-
чек (рис. 14-7). Нервные клетки, граничащие со стекловидным
телом, называются ганглиозными клетками; их аксоны по поверх-
ности сетчатки направляются к оптическому диску, или слепому
пятну, где они собираются вместе, проходят сквозь склеру и обра-
зуют зрительный нерв.
Вблизи того места, где оптическая ось глаза пересекает сет-
чатку, на ней имеется небольшая область — желтое пятно диамет-
ром около 1,5 мм; желтая окраска его обусловлена присутствием
каротиноидов. В центре желтого пятна имеется углубление диа-
метром примерно 0,3 мм, называемое центральной ямкой (фовеа).
Центральная ямка содержит только колбочки; это область сетчат-
ки с наивысшей остротой зрения, и на нее проецируются объекты,
на которые в данный момент направлено внимание наблюдателя.
Сетчатка глаза человека содержит приблизительно 6,5-106 кол-
бочек и 110-Ю6—125-106 палочек [179]. Как показано на рис. 14-34,
рецепторы распределены по поверхности сетчатки неравномерно.
Рис. 14-33. Разрез правого глаза человека [246].
1 — решетчатая пластинка, 2 — сосудистая оболочка, 3 — склера, 4 — сетчатка, 5 — канал
Клоке, 6 — оптическая ось, 7 — заднее хрусталиковое пространство, 8 — отросток цилиарного
тела, 9 — цилиарное тело, 10 — лимбическая зона, 11 — задняя камера, 12 — передняя каме-
ра, 13 — хрусталик, 14 —роговица, 15— радужка, 16— конъюнктива, 17 — шлеммов канал,
18 — цилиарная мышца, 19 — сухожилие прямой мышцы, 20 — волокна цинновой связки,
21 — цилиарный эпителий, 22 — граница сетчатки, 23 — зрительная ось, 24 — стекловидное
тело, 25 — центральная ямка, 26 — желтое пятно, 27 — диск, 28 — оболочка нерва.
Фоторецепция и зрение
539
Плотность колбочек очень высока в области центральной ямки и
существенно ниже на периферии. Плотность палочек, напротив, в
области центральной ямки равна нулю, достигает максимума при-
мерно в 20° от нее и спадает к периферии. Отметим, что на
рис. 14-34 показан разрыв на месте оптического диска, где рецеп-
торы отсутствуют. В зрительном нерве имеется всего около 10е во-
локон; следовательно, к одиночным ганглиозным клеткам должно
сходиться большое число палочек и колбочек. Поскольку палочек
Рис. 14-34. Распределение палочек и колбочек в сетчатке человека [184].
Плотность колбочек максимальна в области
центральной ямки. (Распределение палочек и
нее с носовой стороны не показано, но оно
нию в височной
центральной ямки (0°), а палочек — в 20° от
колбочек в области центральной ямки и около
аналогично изображенному здесь распределе-
части сетчатки.)
примерно в двадцать раз больше, чем колбочек, число палочек, схо-
дящихся к одному волокну зрительного нерва, больше, чем колбо-
чек. В области центральной ямки соотношение между числом кол-
бочек и аксонов ганглиозных клеток равно 1:1. Физиологические
следствия такого устройства мы рассмотрим позже.
Формирование изображений
Глаз позвоночных, как и глаз головоногих моллюсков, часто
сравнивают с фотокамерой. В обоих случаях имеется система линз;
у глаза — это роговица и хрусталик, которые дают перевернутое
изображение объекта на поверхности светочувствительного слоя—
сетчатки. Рассеяние света уменьшается благодаря тому, что за
сетчаткой камера выстлана темным пигментным эпителием; коли-
чество проходящего через хрусталик света регулируется перемен-
ной диафрагмой — радужкой. Изменяя свою форму, хрусталик спо-
'540
Глава 14
собен фокусировать более близкие или более удаленные объекты
(аккомодация).
В отличие от фотокамеры внутренность глаза заполнена желе-
образной средой с коэффициентом преломления (1,33) значитель-
но большим, чем у воздуха. У наземных животных основное пре-
ломление света происходит на границе воздух—роговица. Хруста-
лик имеет коэффициент преломления 1,42 и служит для тонкого
регулирования фокусного расстояния системы. Он подвешен таким
образом, что сокращение цилиарных мышц ослабляет натяжение
волокон, растягивающих хрусталик, благодаря чему он принимает
более выпуклую форму за счет действия упругих сил. Это ведет к
уменьшению фокусного расстояния и позволяет формировать изо-
бражения близко расположенных от глаза объектов. Напротив,
когда цилиарные мышцы расслабляются, хрусталик становится ме-
нее выпуклым и его фокусное расстояние увеличивается [246].
Как и все оптические системы с линзами, система глаза позво-
ночных страдает разного рода аберрациями. Хроматическая абер-
рация связана с тем, что красный свет фокусируется дальше, чем
свет меньшей длины волны. В результате в белом свете вообще
невозможно резко сфокусировать объекты. Стеклянные линзы из-
бавляют от этого недостатка, соединяя части линз с разными по-
казателями преломления, эволюция же нашла другое решение
проблемы. Зависимость показателя преломления от длины волны
наиболее сильно выражена для коротковолновой части спектра, и
позвоночные приспособились ее отфильтровывать. Так, хрусталик
у позвоночных, особенно у дневных видов, часто имеет желтова-
тую окраску, в результате чего отфильтровываются близкий уль-
трафиолет и фиолетовая часть спектра. Желтое пятно — второе
приспособление для поглощения синего света в области централь-
ной ямки. Существует мнение, что той же цели служат масляные
капли в колбочках многих низших позвоночных [12, 246].
Свет, проходящий через краевые области зрачка, возбуждает
колбочки гораздо менее эффективно, чем тот, который проходит
через центр хрусталика, даже когда оба пучка фокусируются на
одном и том же рецепторе. Такая дирекциональная чувствитель-
ность колбочек называется эффектом Стайлса—Крауфорда и об-
условлена волноводными свойствами наружных сегментов колбо-
чек [78, 79, 219]. У колбочек этот эффект выражен гораздо силь-
нее, чем у палочек.
Несмотря на то что оптическая система сконструирована из не-
совершенных материалов, разрешающая способность человеческо-
го глаза ограничена аберрациями хрусталика, по-видимому, толь-
ко при относительно сильно расширенном зрачке. Человек спосо-
бен разрешать решетку с угловым расстоянием между темными и
светлыми полосами в 0,5'. В этом случае изображение решетки на
поверхности сетчатки искажено вследствие дифракции и интенсив-
ность изображения меняется по синусоиде, различаясь в максимуме
Фоторецепция и зрение
541
и минимуме на 25% • Проводились эксперименты, в которых сину-
соидальное распределение интенсивности с различным периодом
н высоким контрастом создавалось на сетчатке с помощью интер-
ференции двух когерентных пучков лазера, а не за счет свойств
оптической системы. При сравнении видимости этих образцов с
видимостью тонких решеток, воспринимаемых в естественных усло-
виях, было обнаружено, что при диаметре зрачка 2 мм качество
сетчаточного изображения эквивалентно качеству изображения в
системе, разрешающая способность которой ограничена дифрак-
цией [44, 45]. Интересно, что зернистость сетчаточной мозаики на-
ходится в соответствии с этими оптическими факторами. Угол зре-
ния в 0,57 отвечает примерно 2 мкм на сетчатке, что приблизитель-
но соответствует расстоянию между центрами наружных сегмен-
тов соседних колбочек [184].
Оценки предела остроты зрения зависят от вида тест-объекта.
Одиночную черную линию на однородном светлом фоне можно за-
метить даже в том случае, когда ее толщина составляет всего 0,5".
Эта величина в 60 раз меньше, чем предел для решеток. На сет-
чатке наблюдателя в этом случае имеется расплывчатая дифрак-
ционная картина линии, причем ряды колбочек, находящихся в
местах затемнения, получают света примерно на 1 % меньше, чем
соседние рецепторы [184].
Теория двойственности
В 1825 г. Пуркинье опубликовал результаты своих наблюдений,
что красные цветы, выглядевшие при сильном дневном свете ярче
голубых, в сумерках кажутся менее яркими и по мере наступления
темноты становятся невидимыми раньше, чем голубые. Этот сдвиг
Пуркинье — смещение максимума чувствительности глаза в сторо-
ну меньших длин волн при уменьшении уровня освещенности —
является одним из свидетельств переключения зрения с колбочек
на палочки.
Различные характеристики палочкового и колбочкового зрения
были суммированы в 1866 г. М. Шульцем в теории двойственности.
Палочки обеспечивают скотопическое зрение, или зрение при низ-
ком уровне освещенности. Палочковое зрение нецветное и не очень
отчетливое. Колбочки обеспечивают фотопическое зрение и работа-
ют при более ярком освещении. Колбочковое зрение цветное и от-
четливое [184].
Палочки очень чувствительны и в состоянии темновой адаптации
могут возбуждаться при поглощении единственного фотона [108,
184]. Благодаря тому что с одним нервным волокном в конечном
счете связано большое количество палочек, эти палочки играют
роль антенны и повышают чувствительность ганглиозных клеток.
Абсолютный порог зрения соответствует одновременному поглоще-
нию шести или семи фотонов на площади сетчатки, содержащей
542
Глава 14
около 500 палочек. Палочковое зрение нецветное, поскольку все
палочки содержат один зрительный пигмент — родопсин, и, следо-
вательно, различия в длине волны воспринимаются только как
различия в интенсивности. Палочковое зрение неотчетливое пото-
му, что конвергенция большого числа палочковых клеток на одно
нервное волокно делает эффективную мозаику сетчатки довольно
грубой.
Рис. 14-35. Спектральная чувствительность (величина, обратная пороговой энер-
гии) палочек и колбочек темноадаптированного глаза человека (по отношению
к максимальной чувствительности в фовеа) [234].
Темные кружки, сплошная линия — палочки, 8° выше центральной ямки; светлые кружки^
сплошная линия — центральные колбочки; пунктирная линия — периферические колбочки, 8°
выше центральной ямки. Диаметр тестового поля 1°, длительность вспышки 0,04 с. Колбоч-
ки несколько более чувствительны к далекому красному свету, чем палочки. Прогиб на кри-
вой фовеальной чувствительности в области 450 нм — это артефакт, связанный с наличием
в желтом пятне каротиноидов.
Колбочковое зрение преобладает при более высокой освещенно-
сти. Вследствие того что по своей природе колбочки менее чувст-
вительны и, кроме того, к одной ганглиозной клетке их сходится
меньше, чем палочек, абсолютная чувствительность колбочковой
системы ниже, но зато уменьшение конвергенции приводит к по-
вышению остроты зрения. Кроме того, у видов, имеющих несколько
типов колбочек с разными зрительными пигментами, поглощающи-
ми в различных областях спектра, возможно цветное зрение.
В этих случаях в результате взаимодействия сигналов колбочек
на последующих этапах зрительной системы интенсивность и дли-
на волны воспринимаются как независимые параметры.
Фоторецепция и зрение
543
Скотопическая кривая чувствительности (зависимость видимой
яркости от длины волны при околопороговых интенсивностях све-
та) определяется спектром поглощения родопсина и имеет макси-
мум вблизи 500 нм. Фотопическая кривая чувствительности (сни-
маемая при больших интенсивностях) определяется поглощением
всех трех типов колбочек и имеет максимум около 560 нм. Обе эти
Рис. 14-36. Темновая адаптация глаза человека, измеряемая в периферической
области (12° выше фовеа), содержащей и палочки, и колбочки [245].
Темновая адаптация колбочек завершается примерно в течение 5 мин, палочек — 45 мин.
Светлые кружки — левый глаз, темные — правый.
кривые приведены на рис. 14-35. Абсолютная чувствительность
колбочек больше, чем чувствительность палочек, только для дале-
кого красного света. В голубой области палочки примерно на 3 ло-
гарифмические единицы чувствительнее, чем колбочки. Рис. 14-35
объясняет явление, обнаруженное Пуркинье, касающееся относи-
тельной видимой яркости цветов. Отметим также влияние погло-
щающего голубой свет макулярного пигмента на чувствительность
фовеальных колбочек.
На рис. 14-36 показано восстановление чувствительности глаза
человека после пребывания на ярком свету, вызывающем обесцве-
чивание зрительных пигментов. Это восстановление называют адап-
тацией к темноте, и, как ясно видно из графика, процесс распада-
ется на две стадии. Первая, более крутая часть кривой, отражает
увеличение чувствительности колбочек, вторая, более пологая, —
544
Глава 14
восстановление палочек. Ход этих частей кривой отражает отно-
сительные скорости синтеза колбочковых и палочковых пигментов.
Другие факторы, влияющие на темновую адаптацию, будут обсуж-
даться в последующих разделах.
Ретиномоторные процессы
Для регулирования количества света, поступающего на рецеп-
торы, в глазах позвоночных имеется несколько вспомогательных
механизмов. Это позволяет животным быть активными в условиях
меняющегося освещения [3, 246].
Перемещение пигмента. У многих рыб, бесхвостых амфибий,
птиц и в меньшей степени у хвостатых амфибий и многих репти-
лий пигментные гранулы пигментного эпителия способны прони-
кать между наружными сегментами рецепторов при ярком осве-
щении и отходить назад, когда освещение падает.
Перемещение колбочек. У тех же групп животных, у которых
имеет место перемещение пигмента, процесс дополняется движе-
нием наружных сегментов колбочек. Как только меланин начинает
приближаться к рецепторам, наружные сегменты колбочек отодви-
гаются от пигментного эпителия вследствие сокращения части
внутреннего сегмента, известной под названием миоида
(рис. 14-6, Б). Следовательно, при фотопических условиях колбоч-
ки остаются на свету.
Перемещение палочек. У многих рыб и бесхвостых амфибий
миоиды палочек перемещают наружные сегменты палочек в на-
правлении, противоположном колбочкам (рис. 14-6, А). Таким об-
разом, когда наружные сегменты колбочек отходят от пигментного
эпителия, наружные сегменты палочек погружаются в него и на-
оборот.
Адаптация с помощью зрачка. Количество попадающего на сет-
чатку света может также регулироваться изменением размера
зрачка. У рыб, за исключением небольшого числа групп, таких,
как угри и плоские рыбы, этот процесс развит плохо. С помощью
зрачка хорошо регулируется чувствительность у некоторых репти-
лий (ночные змеи, крокодилы), птиц и млекопитающих. Таким об-
разом, за исключением птиц, зрачковый рефлекс хорошо развит
в тех группах позвоночных, где не наблюдается перемещений пиг-
мента и колбочек.
У
Световая и темновая адаптация
Глаз человека способен регулировать свою чувствительность в
диапазоне интенсивностей, различающихся примерно в 1010 раз.
Поскольку перемещение пигмента в нем отсутствует, а изменение
диаметра зрачка обеспечивает изменение чувствительности лишь
на 1,2 логарифмических единицы, объяснение такой способности
Фоторецепция и зрение
Б45
надо искать в природе процессов, протекающих в рецепторных
клетках и связанных с ними нейронах.
Восстановительные процессы в палочках удобно изучать на гла-
зах тех видов животных или отдельных особей, которые лишены
колбочек; в противном случае начальную ча^гь процесса темновой
адаптации нельзя измерить, поскольку чувствительность глаза
Рис. 14-37. Зрительная адаптация глаза крысы, определяемая по чувствительно-
сти b-волны электроретинограммы [63].
Во время световой адаптации (светлые кружки, пороговые добавки) увеличение логарифма
порога прямо пропорционально логарифму освещенности фона, исключая область самых
малых значений освещенности. Темновая адаптация (крестики, нижняя шкала) развивается
очень быстро до тех значений освещенности фона, при которых в течение пятиминутного
периода адаптации обесцвечивается значительное количество родопсина (темные кружки,
пунктирная линия). При больших значениях фоновой освещенности виден медленный ком-
понент процесса темновой адаптации, выраженный в разной степени в зависимости от ко-
личества обесцвеченного родопсина.
определяется в это время фотопическим механизмом. В палочковой
сетчатке крысы восстановление чувствительности после умеренно-
го освещения — относительно быстрый процесс, завершающийся в
течение секунд или самое большое 1—2 мин [63, 65]. Если освеще-
ние сильное и выцвело значительное количество родопсина, быст-
рая «нейронная» адаптация сопровождается более медленной, ко-
торую можно соотнести с количеством зрительного пигмента в па-
лочках. На протяжении этой медленной фазы восстановления ло-
гарифм чувствительности меняется линейно с изменением количеств
на ресинтезированного родопсина. Эти соотношения показаны на
рис. 14-37. У живого глаза человека, как показал Раштон, исполь-
35—1514
546
Глава 14
зуя метод отражательной денситометрии, т. е. измеряя свет, вы-
ходящий из зрачка после двойного прохождения через рецепторы,
такое соотношение выполняется для пигмента и палочек, и кол-
бочек. Оказалось, кроме того, что колбочки восстанавливаются
быстрее (рис. 14-36) *[197—199, 201].
Экологические адаптации глаз позвоночных
Глаз человека является наиболее типичным глазом позвоноч-
ных, поскольку он содержит как палочки, так и колбочки и не
обладает ярко выраженной специализацией для дневного или ноч-
ного образа жизни. У животных, которые наиболее активны в
дневные или ночные часы, глаза имеют соответствующие приспо-
собления [246].
Адаптации к дневному образу жизни. У животных, которые ак-
тивны только днем, острота зрения, как правило, увеличивается
ценой уменьшения абсолютной чувствительности. Глаза их обычно
имеют большие размеры (предел зависит от размеров головы) и
на изображение за счет этого приходится большее количество кле-
ток сетчатки. Число колбочек превышает число палочек, а глаза
большинства ящериц, змей, многих птиц, сусликов и черепах во-
обще имеют либо полностью, либо практически полностью колбоч-
ковые сетчатки. Сетчатка глаз птиц и ящериц имеет область, где
нет палочек (area centralis), а колбочки очень длинные и тонкие.
Как и центральная ямка глаза человека, это область минимальной
конвергенции на нейроны высших порядков и максимальной остро-
ты зрения. Центральная ямка птиц, вероятно, наиболее высокораз-
вита. Сетчатка в этом месте значительно тоньше, так что рецепто-
ры лежат в небольшом углублении. Уоллс [246] предположил, что
назначение углубления состоит в некотором увеличении изобра-
жения, попадающего на это место, за счет того, что показатель
преломления сетчатки больше, чем стекловидного тела.
Адаптации к ночному образу жизни. Животные, которые актив-
ны в ночные часы, жертвуют остротой зрения ради чувствительно-
сти. Так, у них увеличено число палочек и нет колбочек. У многих
видов в пигментном эпителии имеется слой отражающего матери-
ала (tapetum lucidum), так что свет, который прошел через сет-
чатку, не поглотившись, отражается назад к наружным сегмен-
там. Этот отражающий слой ответствен за «свечение» сетчатки, или
«горение» глаз животных, смотрящих в направлении на источник
света.
Виды, которые активны ночью, но любят погреться на солнце,
должны изменять размер зрачка в очень широких пределах. Круг-
лый, широко открытый ночью зрачок нельзя достаточно сильно
уменьшить в дневное время с помощью мышц-сфинктеров, и в про-
цессе эволюции был «изобретен» щелевидный зрачок, который да-
ет большое преимущество, например, змеям.
Фоторецепция и зрение
547
Строение сетчатки позвоночных
Пять типов нейронов. Являясь «аванпостом» мозга, сетчатка
представляет собой весьма сложно устроенную нервную ткань. По-
мимо рецепторов, она содержит четыре типа нервных клеток более
высокого порядка. Волокна зрительного нерва отделены от палочек
и колбочек по крайней мере двумя синаптическими переключе-
ниями.
Рис. 14-38. Основные типы клеток и слои сетчатки позвоночных [67].
Препарат сетчатки Nectu.ru.s-, окрашивание по Гольджи. Р — рецептор, Г — горизонтальная
клетка, Б — биполярная клетка, А — амакриновая клетка, ГН — ганглиозная клетка, М —
мюллеровская (глиальная) клетка.
I — наружный ядерный слой, Л — наружный сетчатый слой, 111 — внутренний ядерный слой,
IV— внутренний сетчатый слой, V — слой ганглиозных клеток.
Как показано на рис. 14-38, сетчатка состоит из трех отчетливо
разделяющихся клеточных слоев, характеризующихся скоплением
клеточных ядер. Наружный клеточный слой, ближайший к пигмент-
ному эпителию, состоит из клеточных тел рецепторов. Внутренний
ядерный слой содержит клеточные тела вставочных нейронов трех
типов: биполярных клеток, оси которых вертикальны (радиаль-
ны), а также горизонтальных и амакриновых клеток, которые
расположены в сетчатке горизонтально (тангенциально). Слой
ганглиозных клеток примыкает к стекловидному телу, и, как ска-
зано выше, аксоны этих клеток сходятся в области оптического
диска и образуют зрительный нерв.
Между тремя слоями ядер располагаются две области синап-
тических контактов: в наружном сетчатом слое рецепторные клет-
ки синаптически контактируют с биполярами и дендритами гори-
зонтальных клеток; во внутреннем сетчатом слое биполярные клет-
ки образуют связи с дендритами амакриновых и ганглиозных
клеток.
Таким образом, можно сказать, что наиболее короткий путь
прохождения сигналов через сетчатки — это рецептор — биполяр —
35*
548 Глава 14
ганглиозная клетка. Имеются, однако, дополнительные вставочные
нейроны — горизонтальные и амакриновые клетки, которые осуще-
ствляют латеральные взаимодействия.
Другие клетки. Помимо описанных выше типов нейронов, по
крайней мере у некоторых видов (птицы) имеется небольшое чис-
ло центробежных волокон с неизвестными функциями, оканчиваю-
щихся на отростках амакриновых клеток во внутреннем сетчатом
слое [54, 69].
Глиальные элементы, называемые клетками Мюллера, имеют
вид колонн, поддерживающих слои сетчатки (рис. 14-38). Мюлле-
ровские клетки имеют латерально простирающиеся отростки, ко-
торые вклиниваются между нервными элементами. Мембранные
потенциалы этих клеток меняются при повышении концентрации
калия во внеклеточной среде, обусловленном нейронной активно-
стью. Интересно, что эти пассивные глиальные потенциалы дают
существенный вклад в электроретинограмму (ЭРГ)—суммарный
электрический ответ глаза, который регистрируется макроэлектро-
дами [159].
Виды синаптических контактов. На препарате сетчатки, окра-
шенной по Гольджи, видно, что четыре типа вставочных нейронов
можно разделить на подсистемы по форме их дендритных отрост-
ков. Электронно-микроскопическое исследование синаптических
контактов в сетчатке показало, что их организация в основном
одинакова у многих видов. Однако имеется и ряд интересных
межвидовых различий, например в строении внутреннего сетчато-
го слоя, которые, по всей видимости, находят отражение в физио-
логических свойствах сетчатки (рис. 14-39) [66, 67, 71]. Обо всех
деталях, которые невозможно отметить в таком общем обзоре,
можно прочесть в оригинальных работах.
Большинство синапсов между рецепторами и вставочными ней-
ронами сетчатки имеет характерное строение, выражающееся в на-
личии триад постсинаптических элементов [68, 70, 212, 217].
К расположенному центрально окончанию биполяра с боков при-
мыкают два дендрита горизонтальных клеток, и эти три окончания
входят в углубление (карман) в основании палочки или колбочки.
Рецептор содержит синаптические пузырьки и характерную синап-
тическую ленту — электроноплотную пластинку, ориентированную
под прямым углом к пресинаптической мембране. В сетчатке встре-
чаются и синапсы обычного типа без синаптических лент, связы-
вающие лишь две клетки: рецепторы и биполяры, горизонтальные
клетки и биполяры, две горизонтальные клетки. В наружном слое
рецепторы, таким образом, являются явно пресинаптическими эле-
ментами по отношению как к биполярам, так и к горизонтальным
клеткам, а биполяры являются постсинаптическими элементами по
отношению как к рецепторам, так и к горизонтальным клеткам.
Горизонтальные клетки, напротив, даже на малом участке ден-
дритного дерева нужно рассматривать и как пре-, и как постси-
Рис. 14-39. Общая схема организации синаптических связей в сетчатках позво-
ночных [67].
В наружном сетчатом слое отростки биполярных (Б) и горизонтальных (Г) клеток прони-
кают внутрь углублений в терминалях рецепторов (ТР) и оканчиваются там вблизи синап-
тических лент. Отростки плоских биполярных (ПБ) клеток образуют поверхностные кон-
такты на основаниях некоторых рецепторных терминалей. Горизонтальные клетки с по-
мощью обычных синапсов контактируют с дендритами биполяров и отростками других го-
ризонтальных клеток (здесь это не показано). Поскольку горизонтальные клетки обычно
простираются латерально в наружном сетчатом слое далее, чем дендриты биполяров, можно
предполагать, что далеко отстоящие рецепторы способны воздействовать иа биполяры че-
рез горизонтальные клетки. Предполагают, что во внутреннем сетчатом слое существует
два основных типа синаптических переключений. Окончания биполяров могут контактирог
вать с одним дендритом ганглиозной клетки и одним отростком амакриновой в ленточном
синапсе (на схеме слева) или двумя отростками амакриновых клеток (А) (на схеме справа).
Когда в сетчатке преобладает последняя организация, там обнаруживаются многочисленные
синапсы обычного типа между отростками амакриновых клеток (сериальные синапсы) и
ганглиозные (ГН) клетки контактируют преимущественно с отростками амакриновых кле-
ток (правая часть схемы). Во всех сетчатках отростки амакриновых клеток с помощью
синапсов вбычного типа переключаются обратно на окончания биполяров (возвратные си-
напсы).
550
Глава 14
наптические элементы. Все эти взаимоотношения показаны на
рис. 14-39 и 14-41.
Во внутреннем сетчатом слое также имеются и ленточные си-
напсы, и синапсы обычного типа [67, 68, 70, 71]. Окончания би-
поляров образуют ленточные синапсы с парами (диадами) пост-
синаптических отростков, обычно или дендритами ганглиозной и
амакриновой клеток, или двумя дендритами амакриновой клетки
(рис. 14-39), и изредка с парой дендритов ганглиозной клетки.
У разных видов преобладают те или иные виды пар постсинапти-
ческих элементов. У кошек и приматов 80% пар — ганглиозно-
амакриновые, тогда как у лягушек 75% приходится на долю
амакрино-амакриновых диад. Necturus и кролик занимают проме-
жуточное положение: у них имеется примерно равное количество
тех и других пар. К этим сравнительным данным мы вернемся
ниже.
Между амакриновыми и биполярными клетками, амакриновы-
ми и ганглиозными клетками, различными амакриновыми клетка-
ми, а также биполярными и ганглиозными клетками обнаружива-
ются и обычные синапсы (моносинаптические, без ленты).
Аналогично горизонтальным клеткам, образующим многочис-
ленные латеральные связи и создающим дополнительные парал-
лельные пути от рецепторов к биполярам, амакриновые клетки
также разветвляются латерально и создают дополнительные ли-
нии передачи между биполярными и ганглиозными клетками.
Морфологические данные о наличии обратной связи с горизонталь-
ных клеток на рецепторы не вполне четкие. Неясен также меха-
низм и степень влияния активности горизонтальных клеток на вы-
ходные сигналы рецепторов. В то же время для амакриновых кле-
ток четко установлено наличие возвратных синапсов на оконча-
ния биполяров, и, таким образом, дендриты амакриновых клеток
в пределах участка всего в несколько микрон могут быть по отно-
шению к биполярам и пре-, и постсинаптическими.
Обращает на себя внимание наличие больших видовых вариа-
ций синапсов амакриновых клеток. Как было отмечено выше и
показано на рис. 14-39, у большинства ганглиозных клеток лягу-
шек (в отличие от кошек и обезьян) на вход поступают сигналы
скорее амакриновых клеток, чем биполяров. Соответственно ган-
глиозные клетки являются скорее нейронами четвертого порядка,
чем третьего. Параллельно включению между биполярными и
ганглиозными клетками амакриновых клеток происходит увеличе-
ние общего числа сериальных контактов, в которых амакриновые
клетки являются пресинаптическими элементами.
Соответствующие данные для нескольких видов приведены в
табл. 14-4.
Ленточные синапсы обнаруживаются в окончаниях биполярных
клеток; в большинстве обычных синапсов пресинаптическими эле-
ментами являются амакриновые клетки. Следовательно, увеличе-
Фоторецепция и зрение
551
Таблица 14-4
Относительное количество амакриновых клеток
в сетчатках позвоночных
Организм Отношение числа обычных синапсов к числу ленточных во внутреннем сетчатом слое
Человек (парафовеа) Обезьяна (фовеа) (парафовеа) (периферия) Кошка Крыса Кролик Суслик Лягушка (2 вида) Голубь 1,3 2,0 1,7 2,2 2,5—2,7 2,8 3,7—3,8 5,5 6,3—9,5 10,8
ние отношения свидетельствует об увеличении числа контактов с
амакриновыми клетками [71].
В следующем разделе мы покажем, что различия в характере
связей между элементами сетчатки имеют определенное физиологи-
ческое значение — они проявляются в характере обработки инфор-
мации в сетчатке. По сравнению с кошкой лягушка имеет большее
число синаптических переключений во внутреннем сетчатом слое,
и реакции ганглиозных клеток у нее более сложные. Как правило,
чем более развиты зрительные центры в мозгу, тем меньшая ней-
ронная интеграция имеет место в сетчатке [157].
Переработка информации в сетчатке позвоночных
В течение нескольких последних лет удалось осуществить
внутриклеточные отведения от всех типов нейронов сетчатки поз-
воночных с последующей однозначной идентификацией клеток с
помощью красителя [124, 216, 258]. Дистальные элементы сетчат-
ки реагируют градуальным образом или тонически, в проксималь-
ных клетках появляются также и фазные ответы (рис. 14-40, 14-41).
В зрительном нерве обнаружены волокна, которые чувствительны
не к специфическому виду раздражителя, а к контрасту независи-
мо от того, является ли этот контраст цветовым, пространственным
(граница между областями с разной освещенностью) или времен-
ным (резкие изменения освещенности во времени). У тех живот-
ных, у которых степень переработки информации в сетчатке вы-
сока (например, у лягушек), пространственно-временные взаимо-
действия хорошо развиты, так что в зрительном нерве обнаружи-
ваются волокна, чувствительные, например, к направлению дви-
жения объектов. На данном этапе мы только начинаем понимать,
Пятно
Рис. 14-40. Записи основных типов реакций клеток сетчатки Necturus на стимулы
в виде пятна и кольца радиусом 250 и 500 мкм [258].
Рецепторы имеют сравнительно небольшие рецептивные поля, так что кольцевые стимулы
вызывают слабые реакции. Небольшие потенциалы, регистрируемые в условиях стимуляции
кольцом, возможно, вызваны рассеянным светом. У горизонтальных клеток рецептивные
поля намного больше, поэтому при раздражении кольцом с той же суммарной энергией, что
н у пятна, реакция (правые колонки) ненамного меньше, чем в случае пятна (левая колонка).
Если освещать центр рецептивного поля биполярных клеток, то они дают гиперполяриза-
ционные реакции (левая колонка). Если освещение в центре оставить (правая запись;
обратите внимание на снижение нулевой линии реакции и на подъем линии стимула), то
кольцевой стимул оказывает противоположное действие и наблюдается реакция обратного
знака. В примере, приведенном в средней колонке, кольцо было столь малым, что оно
действовало сразу и на центр, н на периферию. Амакриновые клетки раздражали в тех же
условиях, что и биполярные, онн давали кратковременные реакции в моменты включения
и выключения освещения. Организация их рецептивных полей была близка к концентриче-
ской, у них регистрировалась более сильная «оп»-реакция на пятно и более сильная «off»-
реакция на кольцо радиусом 500 мкм. В случае кольца радиусом 250 мкм клетка одинаково
сильно реагировала и на включение, и на выключение. Ганглиозная клетка, представленная
в верхнем ряду, была фазного типа и реагировала пачками импульсов и на включение, и на
выключение. Организация ее рецептивного поля была аналогична описанной выше для
амакриновой клетки. Ганглиозная клетка, представленная в нижнем ряду, была тонического
типа. При раздражении пятном она давала длительный импульсный разряд. При сохра-
нении раздражения пятном стимуляция большим кольцом (правая колонка) тормозила им-
пульсный разряд в течение всего времени действия. Кольцо меньшего диаметра подавляло
импульсацию в момент выключения (средняя колонка).
Фоторецепция и зрение
553 ;
как специфические ответы волокон зрительного нерва связаны с
физиологическими свойствами вставочных нейронов сетчатки, их
геометрией и взаимоотношениями с другими элементами.
Рис. 14-41. Обобщенная схема, сопоставляющая организацию синаптических свя-
зей в сетчатке позвоночных с регистрируемыми внутриклеточно реакциями нейро-
нов [67].
Р~ рецептары, Г — горизонтальная клетка, Б — биполярные клетки, А — амакриновая клетка,
ГН — ганглиозная клетка.
Рецепторы. Гиперполяризационные реакции палочек и колбочек
уже рассматривались ранее в связи с проблемой преобразования
стимула в нервный сигнал.
Биполярные клетки. Поверхность сетчатки, которая дает вклад
в реакцию одной биполярной клетки (ее рецептивное поле), содер-
жит много рецепторов. Рецептивные поля биполярных клеток име-
ют круглую центральную область и кольцевое окружение с анта-
гонистическими свойствами (рис. 14-40). Реакция мембраны бипо-
554
Глава 14
лярной клетки градуально меняется с изменением интенсивности
света, а импульсов в биполярных клетках не возникает. Реакция
может быть любой полярности: гиперполяризационная и деполяри-
зационная. Если раздражение центра рецептивного поля вызыва-
ет гиперполяризацию, то освещение кольцевого окружения — де-
поляризацию. Поскольку рецептивные поля по площади превышают
область ветвлений дендритного дерева биполярной клетки, пред-
полагают, что на периферии рецептивного поля связь с рецептора-
ми осуществляется через горизонтальные клетки.
Горизонтальные клетки и S-потенциалы. На идентифицирован-
ных горизонтальных клетках было показано, что они реагируют на
освещение медленной градуально меняющейся гиперполяризацией
или же гиперполяризацией в ответ на коротковолновое излучение
и деполяризацией на длинноволновое. Таким образом, эти реакции
относятся к классу S-потенциалов, открытых Светихиным в 1953 г.
и наиболее интенсивно изучавшихся на сетчатке рыб [141, 174,
175, 223]. S-потенциалы представляют собой либо простую гипер-
поляризацию в ответ на освещение (L-тип) (рис. 14-40), либо двух-
фазные цветовые реакции (С-тип). Последние бывают двух типов:
гиперполяризация при освещении зеленым и деполяризация —
красным светом или гиперполяризация при освещении голубым и
деполяризация — желтым светом. У «желто-голубых» элементов
иногда наблюдается дополнительная гиперполяризация в красной
области спектра. Избирательная адаптация сетчатки к свету, при-
ходящемуся на один конец спектра, может усилить реакцию эле-
ментов на излучение, относящееся к другому его концу, что ука-
зывает на существование независимых цветовых входов для за-
пуска механизма генерации S-потенциалов. S-потенциалы облада-
ют рядом свойств, которые трудно объяснить, исходя из свойств
отдельных нейронов [225]. Так, амплитуда реакции не зависит от
потенциала покоя мембраны и не меняется при пропускании тока
через микропипетку. Далее, S-потенциал сопровождается медлен-
ным изменением проводимости, которое отстает от потенциала, а
не предшествует ему. S-потенциалы L- и С-типов иногда претер-
певают взаимные превращения; этот процесс, по-видимому, проис-
ходит спонтанно. И наконец, большие области сетчатки, которые
дают вклад в S-потенциал (миллиметр и более), превышают по
площади область ветвлений латерально распространяющихся ден-
дритов горизонтальных клеток. Большие размеры рецептивных по-
лей можно объяснить наличием электрической связи между гори-
зонтальными клетками, которая была показана для сетчатки рыб
[125].
Амакриновые клетки. Первые фазные реакции на афферентном
пути регистрируются в амакриновых клетках (рис. 14-40, 14-41).
При изменении освещенности амакриновые клетки деполяризуются
и генерируют несколько импульсов. В зависимости от вида стимула
возникают «on»-, «off»- или «on-ofb-реакции. Высказывалось пред-
Фоторецепция и зрение
555
положение, что фазный характер ответа связан с наличием цепей
локальной отрицательной обратной связи, основанных на синапсах
амакриновая клетка — биполяр в непосредственной близости от
синаптических контактов биполяр — амакриновая клетка [67].
Ганглиозные клетки. Ганглиозные клетки деполяризуются и ге-
нерируют серию импульсов, распространяющихся далее по зритель-
ному нерву. Деполяризация, по-видимому, представляет собой ге-
нераторный потенциал в прямом смысле этого слова, поскольку ее
амплитуда определяет частоту разряда аксона.
Рецептивные поля ганглиозных клеток подразделяются на
простые и сложные. Простые рецептивные поля имеют концентри-
ческую структуру, аналогичную рецептивным полям биполярных
клеток, описанным выше. Размер рецептивного поля может пре-
вышать область дендритных ветвлений ганглиозной клетки, что, по-
видимому, связано с наличием латеральных взаимодействий через
посредство амакриновых клеток. Клетки могут реагировать на
включение пятна света («оп»-реакция) в центре рецептивного по-
ля и на его выключение («оН»-реакция) на периферии; другие
ганглиозные клетки имеют «оИ»-центр и «оп»-периферию. Размер
рецептивного поля не постоянен. Степень проявления торможения
в сетчатке, по-видимому, возрастает при световой адаптации, а
при развитии процесса темновой адаптации тормозное окружение
клеток с «оп»-центром в конце концов может вообще исчезнуть
[13, 14, 154].
Ганглиозные клетки с так называемыми простыми рецептивны-
ми полями могут кодировать цвет, если они соединены с колбочка-
ми (рис. 14-42). У суслика, глав которого имеет полностью колбоч-
ковую сетчатку, состоящую ив зелено- и синечувствительных колбо-
чек, те ганглиозные клетки, которые имеют хроматические входы,
могут давать «оп»-реакцию на зеленый свет и «оН»-реакцию на
синий, и наоборот. У некоторых ганглиозных клеток чувствитель-
ная к зеленому свету часть рецептивного поля совпадает с чув-
ствительной к синему. У других чувствительная к зеленому свету
область находится в центре, а к синему — и в центре, и на перифе-
рии. Имеются также клетки, у которых два вида колбочек полно-
стью разделяются: зеленочувствительные находятся в центре, а си-
нечувствительные— на периферии. Такие клетки, по-видимому,
способствуют усилению цветового контраста [155].
У рыб описаны кодирующие цвет ганглиозные клетки с рецеп-
тивными полями размером до 5 мм (40—60°) [60]. Реакции этих
клеток несколько сложнее, чем у суслика. Центр реагирует на
красный цвет «оп»-, на зеленый — «оН»-реакцией (или наоборот).
Области сетчатки, содержащие два типа колбочек, не вполне сов-
падают между собой: зеленочувствительные колбочки занимают
несколько большую площадь. В первых сообщениях об этих ган-
глиозных клетках центр рассматривался как все рецептивное поле
[232]. Использование стимулов в виде колец показало, что в дей-
556
Глава 14
ствительности, помимо центра, рецептивное поле включйет далеко
простирающееся окружение, в прямом смысле антагонистичное
центру, т. е. для элементов, обладающих «оп»-реакцией на красный
свет и «ofb-реакцией — на зеленый, реакция на освещение пери-
1Т1
Рис.. 14-42. Общая схема рецептивных полей кодирующих цвет ганглиозных кле-
ток сетчатки суслика с несколькими примерами импульсных реакций [156, 157].
Имеются два типа колбочек, максимально чувствительные к зеленому и синему свету. На
каждую ганглиозную клетку поступают сигналы от колбочек обоих типов, причем от од-
них — возбуждающие, от других — тормозные. Ганглиозные клетки можно также классифи-
цировать в соответствии со степенью перекрывания возбуждающей и тормозной зон рецеп-
тивных полей. Цифрами (%) отмечены относительные частоты регистрации клеток каждого
типа.
ферии будет «off» на красный свет и «оп» на зеленый. Такие клет-
ки по сравнению с имеющимися в сетчатке суслика элементами с
«оп»-реакцией на зеленый свет в центре и «ofb-реакцией на синий
на периферии еще более усиливают цветовой контраст на про-
странственной границе.
Сложные ганглиозные клетки обнаружены у таких животных»
как лягушки [148], кролики [14, 15] и голуби [147], т, е. у видов
Фото рецепция и зрение
557
с большим количеством синапсов амакриновых клеток. Эти слож-
ные клетки дают фазные реакции на включение и выключение
света, но наиболее характерной их особенностью является чувст-
вительность к движению границ через рецептивное поле. В сетчат-
ке лягушки адекватным стимулом для одной группы клеток яв-
ляется движение небольшой выпуклой границы, благодаря чему
Рис. 14-43. Схема, показывающая корреляцию между анатомическим строением
сетчатки и предполагаемым физиологическим механизмом дирекциональной чувг
ствительности [154].
Л. На вход биполярной клетки возбуждающие сигналы поступают непосредственно от одной
группы колбочек, а тормозные — через горизонтальную клетку — от другой группы, сме?
гценной по отношению к первой. Поскольку для биполярной клетки важна последователе
ность освещения этих двух групп колбочек, ' она будет дирекционально чувствительной.
Б. На вход дирекционально чувствительной ганглиозной клетки сетчатки поступают возбуж-
дающие сигналы от группы чувствительных к последовательности освещения биполяров,
имеющих одинаковые предпочтительные и нулевые направления. В — возбуждающие .синап-
сы, Т — тормозные.
„ "Л
эти клетки превосходно подходят для обнаружения естественной
добычи лягушек [148]. Другой характерной чертой детекторов дви-
жения является то, что они относительно нечувствительны к кон-
трасту между движущимися объектом и фоном. У некоторых ви-
дов [14, 148, 156] обнаружены клетки, которые реагируют, на дви-
жение границ через сетчатку только в одном направлении; движе-
ние в противоположном направлении приводит к подавлению лю-
бой предшествующей активности; возможно, соответствующий ме-
ханизм включает распространение волны торможения на расстоя-
ние примерно 30 мкм вперед от движущегося объекта в «нулевому
направлении, как показано на гипотетической схеме рис. 14-13. ,3 .
558
Глава 14
Сравнение глаз позвоночных и сложных глаз беспозвоночных
Глаз позвоночных, состоящий из большого числа фоточувстви-
тельных клеток и расположенной перед ними линзы, имеет одно
очевидное преимущество перед сложным глазом, который можно
рассматривать как совокупность простых глаз с несколько разли-
чающейся дирекциональной чувствительностью. Камерный глаз
превосходит сложный по остроте зрения или угловой разрешаю-
щей способности. С первого взгляда может показаться, что приро-
да просто нашла два решения проблемы создания глаза, способ-
ного формировать изображение, и только по чистой случайности
сложный глаз является характерным для низших. Однако у глаза
позвоночных есть одно свойство, которое делает его принципиаль-
но неприменимым для очень мелких животных: это его неспособ-
ность фокусировать объекты, находящиеся вблизи роговицы. По-
скольку у сложного глаза нет механизма аккомодации, такой проб-
лемы у него не возникает. На самом деле разрешающая способ-
ность сложного глаза тем больше, чем ближе объект, а в жизни
многих членистоногих способность ясно видеть объекты на расстоя-
нии нескольких миллиметров может иметь большее значение, чем
корошее угловое разрешение.
Для глаз позвоночных коротковолновая граница спектра вос-
принимаемого света определяется необходимостью минимизировать
хроматическую аберрацию. Поэтому глазные фильтры постоянно
отсекают близкий ультрафиолет, а иногда и фиолетовый свет. Гла-
за насекомых, не имеющие аналогичной системы линз, не нужда-
ются в таком ограничении, и у многих наземных видов развились
зрительные пигменты, чувствительные преимущественно к близко-
му ультрафиолетовому излучению. Более того, отражаемый цве-
тами ультрафиолетовый свет и характер поляризации света неба
(главным образом близкого ультрафиолетового излучения) опреде-
ляют поведение некоторых видов [95, 150].
Поразительно, что, несмотря на огромные анатомические разли-
чия, глаза позвоночных и членистоногих имеют определенное сход-
ство. В обеих группах протекают одни и те же фотохимические
процессы, хотя возникающие при этом мембранные реакции могут
несколько различаться.
Морфология нейронов сетчатки позвоночных существенно отли-
чается от структуры вставочных нейронов беспозвоночных. Соот-
ветственно и характер синаптических связей в двух группах совер-
шенно разный. Тем не менее и у позвоночных, и у членистоногих
зрительными вставочными нейронами высшего порядка извлека-
ется сходная информация. Ни разу не были обнаружены нейроны,
чувствительные к специфическому виду стимула и в то же время
сильно чувствительные к контрасту. Детекторы «резких движений»,
которые были обнаружены в оптическом тракте ракообразных и
оптических долях насекомых, имеют много общего с детекторами
Фоторецепция и зрение
559
добычи в сетчатке лягушки. Сходными свойствами обладают и во-
локна обеих систем, чувствительные к направлению движения гра-
ниц в зрительном поле. И хотя для полного выяснения механизма
зрения нам необходимо еще очень многое выяснить как об упомя-
нутых выше, так и о других элементах, уже сейчас можно сказать,
что представление об окружающем мире создается нервными систе-
мами обоих типов сходным образом.
СПИСОК ЛИТЕРАТУРЫ
1. Abrahamson Е. W., Ostroy S. Е., Photochemical and macromolecular aspects
of vision, Prog. Biophys. Molec. Biol., 17, 181—215, 1967.
2. Akhtar M., Blosse P. T., Dewhurst P. B„ The nature of retinal-opsin linkage,
Biochem. J., 110, 693—702, 1968.
3. All M. A., Retinomotor responses: characteristics and mechanisms, Vision Res.,
11, 1225—1288, 1971.
4. Allen D. M., Photic control of the proportions of two visual pigments in a fish,
Vision Res., 11, 1077—1112, 1971.
5. Alpern M., Distal mechanisms of color vision, Ann. Rev. Physiol., 30, 279—
318, 1968.
6. Arden G. B., Ikeda H., Effects of hereditary degeneration of the retina on
early receptor potential and corneo-fundal potential of rat eye, Vision Res.,
6, 171—184, 1966.
7. Autrum H., Physiological basis of color vision in honeybees, In: Ciba Founda-
tion Symposium on Physiology and Experimental Psychology of Colour Vi-
sion. Williams and Wilkins, Baltimore, 1965.
7a. Autrum H., Kolb G., Spectral sensitivity of single retinula cells of Aeschna,
Z. vergl. Physiol., 60, 450—477, 1968.
8. Autrum H., Zettler F., Jarvilehto M., Postsynaptic potentials from single mo-
nopolar neuron of ganglion opticum I of blowfly Calliphora, Z. vergl. Phy-
siol., 70, 414—424, 1970.
9. Ball S., Goodwin T. W., Morton R. A., Preparation of retinenei — vitamin A
aldehyde, Biochem. J., 42, 516—523, 1948.
10. Barber V. C., Evans E. M., Land M. F., Fine structure of the eye of mollusc
Pecten maximus, Z. Zellforsch., 76, 295—312, 1967.
11. Barlow H. B., Size of ommatidia in apposition eyes, J. Exp. Biol., 29, 667—
674, 1952.
12. Barlow H. B., The physical limits of visual discrimination. In: Photephysiolo-
gy, Ed. A. C. Giese, v. 2, Academic Press, 1964, pp. 163—202.
13. Barlow И. B., FitzHugh R., Kuffler S. W., Change of organisation in receptive
fields of cat retina during dark adaptation, J. Physiol., 137, 338—354, 1957.
14. Barlow И. B., Hill R. M., Levick W. R„ Retinal ganglion cells responding se-
lectively to direction and speed of image motion in rabbit, J. Physiol., 173,
377—407, 1964.
15. Barlow H. B., Levick W. R., Mechanism of directionally selective units in rab-
bit retina, J. Physiol., 178, 477—504, 1965.
16. Baylor D. A., Fuortes M. G. F., Electrical responses of single cones in turtle
retina, J. Physiol., 207, 77—92, 1970.
17. Behrens M. E., Wullf V. J., Light-initiated responses of retinula and eccentric
cells in Limulus lateral eye, J. Gen. Physiol., 48, 1081—1093, 1965.
18. Bennet R. R., Tunstall J., H or ridge G. A., Spectral sensitivity of single reti-
nula cells of the locust, Z. vergl. Physiol., 55, 195—206, 1967.
19. Bishop L. G„ Keehn D. G., McCann G. D., Motion detection by interneurons of
optic lobes and brains of flies Calliphora phaenicia and Musca domestic^
J. NeurophysioL, 31, 509—525, 1968.
560
Глава 14
20. Bitensky М. W., Gorman R. E., Miller W. H. Adenyl cyclase as a link between
photon capture and changes in membrane perme — ability of frog photore-
ceptors, Proc. Nat. Acad. Sci., 68, 561—562, 1971.
21. Boll F., Anatomy and physiology of the retina, Mber. Berl. Acad. Wiss., 12,
783—788, 1876.
22. Bortoff A., Norton A. L., Simultaneous recording . of photoreceptor potentials
and PHI component of the ERG, Vision Res., 5, 527—533, 1965.
23. Bownds D., Site of attachment of retinal in rhodopsin, Nature, 216, 1178—
1181,1967.
24. Braitenberg V., Pattern of projection in visual system of fly: retina-lamina
projections, Exp. Brain Res., 3, 271—298, 1967.
25. Bridges C. D. B. In: Comprehensive Biochemistry, Vol. 27, edited by M. Flor-
kin and E. H. Stotz, Elsevier Publishing Co., Amsterdam, 1967, pp. 31, 37.
26. Bridges C. D. B., Spetroscopic properties of porphyropsins, Vision Res., 7,
349—369, 1967.
27. Bridges C. D. B„ The rhodopsin-porphyropsin visual system. In: Handbook of
sensory Physiology, Vol. 7, Photochemistry of Vision, edited by H. J. A. Dart-
nail. Springer-Verlag, Berlin, 1972.
28. Briggs W. R., Phototropism in higer plants. In: Photophysiology, Vol. 1, edi-
ted by A. C. Giese. Academic Press, New York, 1964, pp. 223—271.
29. Brindley G. S., Gardner-Medwin A. R., Origin of early receptor potential of
the retina, J. Physiol., 182, 185—194, 1966.
30. Brown H. M., Hagiwara S., Koike H., Meech R. W., Membrane properties of
barnacle photoreceptor examined by voltage clamp technique, J. Physiol., 208,
385—413, 1970.
31. Brown К. T., Murakami M., A new receptor potential of the monkey retina
with no detectable latency, Nature, 201, 626—628, 1964.
32. Brown P. K., Wald G., Visual pigments in human and monkey retinas, Nature,
200, 37—43, 1963.
33. Brown P. K., Wald G., Visual pigments in single rods and cones of human re-
tina, Science, 144, 45—52, 1964.
34. Bruce V. G., Minis D. H., Circadian clock action spectrum in a photoperiodic
moth, Science, 163, 583—585, 1969.
35. Bruckmoser P., Spectral sensitivity of single sight cells of backswimmer No-
tonecta glauca L. (Heteroptera), Z. vergl. Physiol., 59, 187—204, 1968.
36. deBruin G. H. P., Crisp D. J., Influence of pigment migration on vision of
higher crustaceans, J. Exp. Biol., 34, 447—463, 1957.
37. Bullock T. H., Horridge G. A., Structure and Function in the nervous Systems
of Invertebrates. W. H. Freeman and Co., San Francisco, 1965.
38. Burkhardt D., Spectral sensitivity and other response characteristics of single
visual cells in the arthropod eye, Symp. Soc. Exp. Biol., 16, 86—109, 1962.
39. Burtt E. T., Catton W. T., Electrical responses to visual stimulation in the
optic lobes of the locust and certain other insects. J. Physiol., 133, 68—88,
1954.
40. Burtt E. T., Catton W. T., Properties of single-unit discharges in optic lobe
ol locust, J. Physiol., 154, 479—490, 1960.
41* . Bush В. M. H., Wiersmac A. G., Waterman T. H., Efferent mechanoreceptive
responses in optic nerve of crab Podophthalmus, J. Cell. Comp. Physiol, 64
327—346, 1964.
42. Butler R. Z., Identification and mapping of spectral cell types in retina of
Periplaneta americana, Z. vergl. Physiol., 72, 67—80, 1971.
43. Cajal S. R., Sanchez D„ Contribution al eonocimiento de los centres ner-
viosos de los insectos, Trab. Lab. Invest. Biol. Madrid, 13, 1—164.
44. Cambell F. W., Green D. G., Optical and retinal factors affecting visual re-
solution, J. Physiol., 181, 576—593, 1965.
45. Cambell F. W., Gubisch R. W., Optical quality of human eye, J. Physiol., 186,
558—578, 1966.
Фоторецепция и зрение
561
46. Clayton R. К., Studies in phototaxis of Rhodospirillutn rubrum action spe-
ctrum, growth in green light and Weber Law adherence, Arch. Mikrobiol., 19,
107—124, 1953.
47. Clayton R. K., Phototaxis in microorganisms. Ln: Photophysiology, Vol. 2,
edited by A. S. Giese, Academic Press, New York, 1964, pp. 51—77.
48. Cohen A. 1., Vertebrate retinal cells and their organization, Biol. Rev., 38,
427—459, 1963.
49. Cohen A. I., Linkage to extracellular space of outer segment saccules of frog
cones, J. Cell. Biol., 37, 424—444, 1968.
50. Collet T. S., Blest A. D., Binocular directionally sensitive selective neurons
possibly involved in optomotor response of insects, Nature, 212, 1330—1333,
1966.
51. Cone R. A., Early receptor potential: photoreversible charge displacement in
rhodopsin, Science, 155, 1128—1131, 1967.
52. Cone R. A., Brown P. K., Dependence of early receptor potential on orienta-
tion of rhodopsin, Science, 156, 536, 1967.
53. Cone R. A., Cobbs W. H. Ill, Rhodopsin cycle in liying eye of rat, Nature,
221, 820—822, 1969.
54. Cowan W. M., Powell T. P. S., Centrifugal fibers in avian visual system. Proc.
Roy. Soc. Lond. B, 158, 232—252, 1963.
55. Crescitelli F., Visual cells and visual pigments of vertebrate eye. In: Hand-
book of Sensory Physiology, Vol. 7, Photochemistry of Vision, edited by
H. J. A. Dartnall, Springer-Verlag, Berlin, 1972.
56. Dartnall H. J. A., Visual pigment 467, a photosensitive pigment present in
tench retinae, J. Physiol., 116, 257—289, 1952.
57. Dartnall H. J. A., Lythgoe J. N., Spectral clustering of visual pigments, Vi-
sion Res., 5, 81—100, 1965.
58. Daumer K., Stimulation measurement investigation of color vision in bees,
Z. vergl. Physiol., 38, 413—478, 1956.
59. Daumer K., Flower colors: how bees see them, Z. vergl. Physiol., 41, 49—110,
1958.
60. Daw N. W., Color-coded ganglion cells in goldfish retina: extension of their
receptive fields by means of new stimuli, J. PhysioL, 197, 567—592, 1968.
61. Denton E. J., Contributions of oriented photosensitive and other molecules to
the absorption of the whole retina, Proc. Roy. Soc. Lond. B, 150, 78—94,
1959.
62. Dowling J. E., Chemistry of visual adaptation in rat, Nature, 188, 114—118,
1960.
63. Dowling J. E., Neural and photochemical mechanisms of visual adaptation in
rat; J. Gen. Physiol., 46, 1287—1301, 1963.
64. Dowling J. E.^The organization of vertebrate visual receptors. In: Molecular
Organization and Biological Function, edited by J. M. Allen. Harper and Row,
New York, 1967.
65. Dowling J. E., The site of visual adaptation, Science, 155, 273—279, 1967.
66. Dowling J. E„ Synaptic organization of frog retina: electron microscopic ana-
lysis comparing retinas of frogs and primates, Proc. Roy. Soc. Lond. B, 170,
205—228, 1968.
67. Dowling J. E., Organization of vertebrate retinas, Invest. Ophthal., 9, 655—
680, 1970.
68. Dowling J. E., Boycott В. B., Organization of the primate retina: electron
microscopy, Proc. Roy. Soc. Lond. B, 166, 80—111, 1966.
69. Dowling J. E., Cowan W. M., Electron microscopic study of normal and dege-
nerating centrifugal fiber terminals in pigeon retina, Zellforsch. Mikr. Anat.,
71, 14—28, 1966.
70. Dowling' J. E., Werblin F. S., Organization of retina of mudpuppy Necturus
maculosis: synaptic structure, J. Neurophysiol., 32, 315—338, 1969.
71. Dubin M. W., Inner plexiform layer of the vertebrate retina, J. Comp. Neurol.,
140,479—505,1970.
36—1514
562
Глава 14
72. Eakin R. M. Evolution of photoreceptors. In: Evolutionary Biology, Vol. 2, edi-
ted by T. Dobzhansky, M. K. Hecht, and W. C. Steere. Appleton-Century-
Crofts, New York, 1968.
73. Eakin R. M., A third eye, Amer. Sci., 58, 73—79, 1970.
74. Eakin R. M., Structure of invertebrate photoreceptors. In: Handbook of Sen-
sory Physiology, Vol. 7, Photochemistry of Vision, edited by H. J. A. Dartnall,
Springer-Verlag, Berlin, 1972.
75. Eguchi E., Rhabdom structure and receptor potentials in single crayfish reti-
nular cells, J. Cell. Comp. Physiol., 66, 411—430, 1965.
76. Eguchi E., Waterman T. H., Fine structure patterns in crustacean rhabdoms.
In: The Functional Organization of the Compound Eye, edited by C. G. Bern-
hard, Pergamon Press Ltd., Oxford, 1966.
77. Eguchi E., Waterman T. H., Cellular basis for polarized light perception in
spider crab Libinia, Z. Zellforsch., 84, 87—101, 1968.
77a. Eheim W. P„ Wehner R., Visual field of central ommatidia in apposition eyes
of Apis and Cataglyphis (Apidae, Formicidae, Hymenoptera), Kybernetik, 10,
168—179, 1972..
78. Enoch J. M., Wave guide modes: are they present and what is their role in the
visual mechanism? J. Opt. Soc. Amer., 50, 1025—1026, 1960.
79. Enoch J. M., Nature of the transmission of energy in retinal receptors, J. Opt.
Soc. Amer., 51, 1122—1126, 1961.
80. Exner S„ Die Physiologie der facettieren Augen von Krebsen und Insekten
(Physiology of facetted eyes of crabs and insects), Franz Deuticke, Vienna,
1891.
81. Reynman R. P„ Leighton R. B., Sands M., The Feynman Lectures on Physics,
Vol. 1, Chap. 36, Addision— Wesley Publishing Co., Reading, Mass., 1964.
82. Fraenkel G. S., Gunn D. L., The Orientation of Animals, Clarendon Press,
Oxford, 1940.
83. Frank K. D., Zimmerman W. F„ Action spectra for phase shifts of a circadian
rhythm in Drosophila, Science, 163, 688—689, 1969.
84. Von Frisch K., The Dance Language and Orientation of Bees, Harvard Uni-
versity Press, Cambridge, 1967.
85. Fulpius B., Baumann F., Effects of sodium, potassium, and calcium ions on
slow and spike potentials in single photoreceptors, J. Gen. Physiol., 53, 541—
561, 1969.
86. Fuortes M. G. F„ Initiation of impulses in visual cells of Limulus, J. Phy-
siol., 148, 14—28, 1959.
87. Fuortes M. G. F., Yeandle S., Probability of occurence of discrete potential
waves in eye of Limulus, J. Gen. Physiol., 47, 443—463, 1964.
88. Futterman S., The role of reduced triphosphopyridine nocleotide in the visual
cycle, J. Biol. Chem., 238, 1145—1150, 1963.
89. Goldsmith T. H., Nature of the retinal action potential, and spectral sensitivi-
ties of ultraviolet and green receptor systems in compound eye of worker
honeybee, J. Gen. Physiol., 43, 775—799, 1960.
90. Goldsmith T. H„ Physiological basis of wavelength discrimination in eye of
honeybee. In: Sensory Communication, edited by W. A. Rosenblith, MIT Press,
Cambridge, 1961.
91. Goldsmith T. H., The visual system of insects. In: The Physiology of Insecta,
edited by M. Rockstein, Academic Press, New York, 1964.
92. Goldsmith T. H., Natural history invertebrate visual pigments. In: Handbook
of Sensory Physiology, Vol. 7, Photochemistry of Vision, edited by
H. J. A. Dartnail, Springer-Verlag, Berlin, 1972.
93. Goldsmith T. H., Bernard G. D. In: The Physiology of Insecta, 2nd edition,
edited by M. Rockstein, Academic Press, New York, 1973 (in press).
94. Gorman A. L. F., McReynolds J. S., Barnes S. N., Photoreceptors in primitive
chordates: fine structure, hyperpolarizing receptor potentials and evolution,
Science, 172, 1052—1054, 1971.
95. Hagins W. A., Electrical sings of information flow in photoreceptors. Cold
Spring Harbor Symp, Quant. Biol., 30, 403—418, 1965.
Фоторецепция и зрение 563
96. Hagins W. A., Jennings W. Н., Radiationless migration of electronic exitation
in retinal rods, Disc. Faraday Soc., 27, 180—190, 1959.
97. Hagins W. A., Liebman P. A., Relationship between photochemical and electri-
cal processes in living squid photoreceptors, Abstr. Biophys. Soc. 7th Annual
Meeting, New York, N.Y.M.E., 6, 1963.
98. Hagins W. A., McGaughy R. E., Molecular and termal origins of fast pho-
toelectric effects in squid retina, Science, 157, 813—816, 1967.
99. Hagins W. A., Penn R. D., Yoshikami S., Dark curernt and photocurrent in
retinal rods, Biophys. J., 10, 380—412, 1970.
100. Hagins W. A., Rappel H., Fast photoelectric effects and properties of ver-
tebrate photoreceptors as cables, Fed. Proc., 30, 64—68, 1971.
101. Hagins W. A., Zonana H. V., Adams R. G., Local membrane current in the
outer segments of squid photoreceptors, Nature, 194, 844—847, 1962.
102. Halldal P., Action spectra of phototaxis and related problems in Volvocales,
Ulva-Gametes, and Dinophyceae, Physiol. Plantarum, 11, 118—153, 1958.
103. Hamasaki D. I., Effect of sodium ion concentration on electroretinogram of
isolated retina of frog, J. Physiol., 167, 156—168, 1963.
104. Hamory J., Horrindge G. A., The lobster optic lamina. 1. General organisation,
J. Cell. Sci., 1, 249—256, 1966.
105. Hamory J., Horridge G. A., The lobster optic lamina. II. Types of synapse,
J. Cell. Sci., 1, 257—270, 1966.
106. Hartline H. K., Visual receptors and retinal interaction, Science, 164, 270—
278, 1969.
107. Hays D., Goldsmith T. H., Microspectrophotometry of the visual pigment of
the spider crab Libinia emarginata, Z. vergl. Physiol., 65, 218—232, 1969.
108. Hecht S., Shlaer S., Pirenne M. H., Energy, quanta, and vision, J. Gen. Phy-
siol., 25, 819—840, 1942.
109. Heller J., Comparative stury of a membrane protein, Characterisation of bovi-
ne, rat, and frog visual pigments, Biochemistry, 8, 675—679, 1969.
110. Henderson L. G., The Fitness of the Environment, Macmillan, New York,
1913.
111. Hoglund G., Pigment migration, light screening and receptor sensitivity in
compound eye of nocturnal Lepidoptera, Acta Physiol. Scand., 69, Suppl. 282,
1—56, 1966.
112. Horridge G. A., Unit studies on retina of dragonflies, Z. vergl. Physiol., 62,
1—37, 1969.
113. Horridge G. A., Meinertzhagen I. A., The exact neural projection of the visual
fields upon the first and second ganglia of the insect eye, Z. vergl. Physiol.,
66, 369—378, 1970.
114. Horridge G. A., Scholes J. H., Shaw S., Tunstall J., Extracellular recordings
from single neurons in optic lobe and brain of locust. In: The Physiology of
the Insect Central Nervous System, edited by J. E. Treherne and J. W. L. Bea-
ment, Academic Press, New York, 1965, pp. 165—202.
115. Horridge G. A., Walcott B., loannides A. C., Tiered retina of Dytiscus: a new
type of compound eye, Proc. Roy. Soc. Lond. B, 175, 83—94, 1970.
116. Hubbard R., Molecular weight of rhodopsin and nature of the rhodopsin-di-
gitonin complex, J. Gen. Physiol., 37, 381—399, 1954.
117. Hubbard R., Retinene isomerase, J. Gen. Physiol., 39, 935—962, 1956.
118. Hubbard R., Kropf A., The action of light on rhodopsin, Proc. Nat. Acad. Sci.,
44, 130—139, 1958.
119. Hubbard R., George R. С. C. St., The rhodopsin system of the squid, J. Gen.
Physiol., 41, 501—528, 1958.
120. Hubbard R., Wald G., Mechanism of rhodopsin synthesis, Proc. Nat. Acad.
Sci., 37, 69—79, 1951.
121. Hutchinson G. E„ Treatise on Limnology, Vol. 1, Geography, Physics, and
Chemistry, John Wiley and Sons, New York, 1957.
122. Jander R., Waterman T. H., Sensory discrimination between polarized light
and light intensity patterns by arthropods, J. Cell. Сорт. Physiol., 56, 137—
160, 1960.
36*
564
Глава 14
122а. Jarvilehto М., Zettler F., Localized potentials from pre- and postsyaaptic ele-
ments in external plexiform layer of insect retina, Z. vergl. Physiol., 75, 422—
440, 1971.
123. Kaiser W., Bishop L. G., Directionally selective motion detecting units in
optic lobe of honeybee, Z. vergl. Physiol., 7, 403—413, 1970.
124. Kaneko A., Physiological and morphological identification of hori&ontal, bi-
polar, and amacrine cells in goldfish retina, J. Physiol., 207, 623—633, 1970.
125; Kaneko A. J., Electrical connections between Horisontal cells in dogfish re-
tina, J. PhysioL, 213, 95—105, 1971.
126. Kennedy D., Neural photoreception in a lamellibranch mollusc, J. Gen. Phy-
siol., 44, 277—299, 1960.
127. Kennedy D., Milkman R. D., Selective light absorbtion by lensen of lower ver-
tebrates and its influence on spectral sensitivity, Biol. Bull., Ill, 375—386,
1956.
127a. King-Smith P. E., Absorbtion spectra and function of colored oil drops in pi-
• geon retina, Vision Res., 9, 1391—1399, 1969.
128. Kirshfeld K., Projection of the optic environment on the screen of the rhabdo-
mere in complex eye of Musca, Exp. Brain Res., 3, 248—270, 1967.
128a. Korenbrot J. I., Cone R. A., Dark ionic flux and effects of light on rod outer
segments, J. Gen. Physiol., 60, 20—45, 1972.
129. Kbttgen E., Abelsdorff G., Absorption und Zersetzung des Sehpurpurs bei den
• Wirbeltieren (Absorption and decomposition of visual purples by vertebrates),
Z. Psychol. Physiol. Sinnesorg., 12, 161—184, 1896.
130. Kuhne W., Uber den Sehpurpur. Untersuchungen aus dem Physlologishen In-
stitut der Univ. Heidelberg, 1, 15—103, 1877.
131. Kuiper J. W., Optics of the compound eye, Symp. Soc. Exp. Biol., 16, 58—71,
1962.
132. Kunze P., Behavioral physiology and optical experiments on the Superposition
theory of image location in the compound eyes, eVrh. Zbol. Ges. Koln., 64,
234—238, 1970.
133. Land M., Functional aspects of optical and retinal organisation of mollusc
eye, Symp. Zool. Soc. Lond., 23, 75—96, 1968.
134. Langer H., Thorell B„ Microspectrophotometry of single rhabdomeres in in-
sect eye, Exp. Cell. Res., 41, 673—676, 1966.
135. Lasansky A., Fuortes M. G. F., Site of origin of electrical responses in visual
cells of leech, J. Cell. Biol., 42, 241—252, 1969.
135a. Laughlin S. B., Horridge G. A., Angular sensitivity of retinular cells of dark-
adapted worker bee, Z. vergl. Physiol., 74, 329—339, 1971.
136. Leedale G., Meeuse B. J. D., Pringsheim E. G., Structure and physiology of
Euglena spirogyra, Arch. Mikrobiol., 50, 68—102, 1965.
137. Lees A. D., Photoperiodism in insects. In: Photophysiology, Vol. 4, edited by
A. C. Giese, Academic Press, New York, 1968, pp. 47—137.
138. Liebman P. A., In situ microspectrophotometric studies on pigments of single
retinal rods, Biophys. J., 2, 161—178, 1962.
139. Liebman P. A., in: Handbook of Sensory Physiology, Vol. 7, Photochemistry
; of Vision, edited by H. J. A. Dartnall, Springer-Verlag, Berlin, 1972. Micro-
spectrophotometry of photoreceptors.
140. Lpeb J., Wasteneys H., Relative efficiency of various parts of the spectrum
for heliotropic reactions of animals and plants, Z. Exp. Zool., 19, 23—35,
1915.
141; MacNichol E. F., Jr., Retinal mechanisms of color vision, Vision Res., 4,
119—133, 1964.
142/ MacNichol E. F., Jr., Three pigment color vision, Sci. Amer., 211, 48—56,
1964.
143. - Marks W. B., Visual pigments of single goldfish cones, J. PhysioL, 178, 14—
.32, 1965.
144. . Marks W. B., Dobelle W. H., MacNichol E. F., Jr., Visual pigments of single
primate cones, Science, 143, 1181—>1183, 1964.
Фото рецепция и зрение
565
145. Mast S. О., Stabler N., Relation between luminous intensity, adaptation to
light and rate of locomotion in Amoeba proteus (Leidy), Biol. Bull., 73, 126—
133, 1937.
146. Mathews R. G., Hubbard R., Brown P. K.., Wald G„ Tautomeric forms of
metarhodopsin, J. Gen. Physiol., 47, 215—240, 1963.
147. Maturana H. R., Frenk S., Directional movement and horisontal edge detectors
in pigeon retina, Science, 142, 977—979, 1963.
148. Maturana H. R., Lettvin J. Y., McCulloch W. S„ Pitts W. H., Anatomy and
physiology of vision in frog (Rana pipiens), J. Gen. Physiol., 43 (suppl.),
129—175, 1960.
149. Mazokhin-Porshnyakov G. A., Recognition of colored objects by insects. In:
The Functional Organization of the Compound Eye, edited by C. G. Bernhard,
Pergamon Press Ltd., Oxford, 1966.
150. Mazokhin-Porshnyakov G. A., Insect Vision (translated from Russian), Ple-
num Press, New York, 1969.
151. McCann G. D., Dill J. C., Properties of intensity, form and motion perception
in visual nervous systems of Calliphora phaenicia and Musca domestica, J. Gen.
Physiol., 53, 385—413, 1969.
152. McReynolds J. S., Gorman A. L. F., Membrane conductances and spectral sen-
sitivities of Pecten photoreceptors, J. Gen. Physiol., 56, 392—406, 1970.
153. McReynolds J. S., Gorman A. L. F., Photoreceptor potentials of opposite
polarity in eye of scallop Pecten irradians, J. Gen. Physiol., 56, 376—391,
1970.
154. Michael C. R„ Receptive fields of single optic nerve fibers in a mammal with
an all-cone retina. I. Contrast sensitive units, J. Neurophysiol., 31, 249—256,
.1968.
155. Michel C. R., Receptive fields of single optic nerve fibers in a mammal with
’.о aft all-cone retina, II. Directionally selective units, J. Neurophysiol., 31, 257—
267, 1968.
156. Michael C. R., Receptive fields of single optic nerve fibers in a mammal with
an all-cone retina. III. Opponent color fibers, J. Neurophysiol., 31, 268—282,
: .1968.
157. Michael C. R., Retinal processing of visual images, Sci. Amer., 220, 104—114,
1969.
158. Millecchia R., Mauro A., Ventral photoreceptor cells of Limulus, J. Gen. Phy-
siol., 54, 331—351, 1969.
159. Miller R. F., Dowling J. E., Intracellular responses of the Muller (glial) cells
of mudpuppy retina: their relation to b-wave of the electroretinogramr
< J. Neurophysiol., 33, 323—341, 1970.
160. Miller W. H., Morphology of ommatidia of compound eye of Limulus, J. Bio-
phys. Biochem. Cytol., 3, 421—428, 1957.
161. Miller W. H„ Bernard G. D., Allen J. L., Optics of insect compound eyes. Sci-
ence, 162, 759—771, 1968.
162^ Millot t N., Animal photosensitivity, with special reference to eyeless forms,.
Endeavour, 16, 19—28, 1957.
163. Millott N., Yoshida M., Spectral sensitivity of echinoid Diadema antillarum
Philippi, J. Exp. Biol., 34, 394—401, 1957.
164. Milne L. J.. Milne M., Photosensitivity in invertebrates. In: Handbook of Phy-
siology, Sec. 1,. Vol. 1, Neurophysiology, Amer. Physiol. Soc., Washington,
D.C., 1959, pp. 621—645.
165. Moody M. F., Evidence for intraocular discrimination of vertically and hori-
zontally polarized light by Octopus, J. Exp. Biol., 39, 21—30, 1962.
166. . Moody M. F., Photoreceptor organelles in animals, Biol. Rev., 39, 43—86,
1964.
167. Moody M. F., and Parriss J. R., Discrimination of polarized light by Octopus,.
Z. vergl. Physiol., 44, 268—291, 1961.
168. Morton R. A., Pitt G. A. J., Aspects of visual pigment research. In: Advances-
in Enzimology, Vol. 32, edited by F. F. Nord, John Wiley and Sons, New York...
1969, pp. 97—171.
566
Глава 14
169. Mote М. I., Goldsmith T. H., Spectral sensitivities of color receptors in com-
pound eye of cockroach Periplaneta, J. Exp. Zool., 173, 137—146, 1970.
170. Mote M. I., Goldsmith T. H„ Compound eyes: localization of two color re-
ceptors in same ommatidium, Science, 171, 1254—1255, 1971.
171. Muller I., Zur vergleichenden Physiologie des Gesichtsinnes des Menschen
und Tiere (On the Comparative Physiology of the Visual Senses of Men and
Animals), Chobloch, Leipzig, 1826.
172. Muntz W. R. A., Northmore D. P. M„ Visual pigments from different parts of
the retina in rudd and trout. Vision Res., 11, 551—562, 1971.
173. Munz F. W., The photosensitive pigments from the retinae of certain deep sea
fishes, J. Physiol., 140, 220—235, 1958.
174. Naka К. I., Rushton W. A. H., S-potentials from color units in the retina of
fish (Cyprinidae), J. Physiol., 185, 536—555, 1966.
175. Naka К. I., Rushton W. A. H., Generation and spread of S-potentials in fish
(Cyprinidae), J. Physiol., 192, 437—461, 1967.
176. Nolte J., Brown J. E., Smith T. G., Jr., Hyperpolarizing component of receptor
potential in median ocellus of Limulus, Science, 162, 677—679, 1968.
177. Northrup R. B., Guignon E. F., Information processing in optic lobes of the
lubber grasshopper, J. Insect Physiol., 16, 691—713, 1970.
178. Oroshnik W., Synthesis and configuration of neo-b vitamin A and neoretine-
ne b, J. Amer. Chem. Soc., 78, 2651—2652, 1956.
179. Qsterberg G., Topography of the layer of rods and cones in the human retina,
Acta Ophthal., Suppl. 6, 1935.
180. Page R. M., Photo tropism in fungi. In: Photophysiology, Vol. 3, edited by
A. C. Giese, Academic Press, New York, 1968, pp. 65—90.
181. Pak W. L., Properties of early electrical response in vertebrate retina, Cold
Spring Harbor Symp. Quant. Biol., 30, 493—499, 1965.
182. Pak W. L., Cone R. A., Isolation and identification of the initial peak of the
early receptor potential, Nature, 204, 836—838, 1964.
183. Penn R. D., Hagins W. A., Signal transmission along retinal rods and the ori-
gin of the electroretinographic a-wave, Nature, 223, 201—205, 1969.
184. Pirenne M. H., Vision and the Eye, 2nd edition, Chapman and Hall, ltd., Lon-
don, 1967.
185. Poincelot R. P., Millar P. G., Fimbel R. L. Jr., Abrahamson E. W. Determina-
tion of the chromophoric binding site in native bovine rhodopsin, Biochemi-
stry, 9, 1809—1816, 1970.
186. Prosser C. L., Impulses in the segmental nerves of the earthworm, J. Exp.
BioL, 12, 95—104, 1935.
187. Purple R. L., Dodge F., Self inhibition in eye of Limulus. In: The Functional
Organization of the Compound Eye, edited by C. G. Bernhard, Pergamon Press
Ltd., Oxford, 1966.
188. Rabinowitch E., Govindjee, Photosynthesis, John Wiley and Sons, New York,
1969.
189. Ratliff F., Selective adaptation of local regions in the rhabdom in an ommati-
dium of compound eye of Limulus, in: The Functional Organization of the
Compound Eye, edited by C. G. Bernhard, Pergamon Press Ltd., Oxford,
1966.
190. Ratliff F., Hartline H. F-, Responses of Limulus optic nerve fibers to patterns
of illumination on receptor mosaic, J. Gen. Physiol., 42, 1241—1255, 1959.
191. Reuter T. E., White R. H., Wald G., Rhodopsin and porphiropsin fields in adult
bullfrog retina, J. Gen. Physiol., 58, 351—371, 1971.
192. Ripps H„ Weale R. A., Photophysiology of vertebrate color vision. In: Photo-
physiology, Vol. 5, edited by A. C. Giese, Academic Press, New York, 1970,
pp. 127—168.
193. de Robertis E., Ultrastructure and morphogenesis of photoreceptors, J. Gen.
Physiol., 43 (Suppl. 2), 1—6, 1960.
194. Robinson N., editor, Solar Radiation, Elsevier Publishing Co., Amsterdam,
1966.
Фоторецепция и зрение
567
195 Ruck Р., Electrophysiology of the insect dorsal ocellus, J. Gen. Physiol., 44,
605—657, 1961.
196 Ruck P., Components of visual system of dragonfly, J. Gen. Physiol., 49,
289—307, 1965.
197 Rushton W. A. H., Dark-adaptation of regeneration of rhodopsin, J. Physiol.,
156, 166—178, 1961.
198. Rushton W. A. H., The intensity factor in vision. In: Light and Life, edited
by W. D. McElroy and B. Glass, Johns Hopkins Press, Baltimore, 1961,
pp. 706—729.
199 Rushton W. A. H., A cone pigment in the protanope, J. Physiol., 168, 345—359,.
1963.
200. Rushton W. A. H., A foveal pigment in the deuteranope, J. Physiol., 176, 24—
37, 1965.
201. Rushton W. A. H., Cone pigment kinetics in the deuteranope, J. Physiol., 176,
38—45, 1965.
201a. Rushton W. A. H., Visual pigments in man, in: Handbook of Sensory Physio-
logy, Vol. 7, Photochemistry of Vision, edited by H. J. A. Dartnall, Springer-
Verlag, Berlin, 1972.
202. Rutherford D. J., and Horridge G. A., Rhabdom of lobster eye, Quart. J. Mier.
Sci., 106, 119—130, 1965.
203. Scholes J., Discontinuity of excitation process in locust visual cells, Cold
Spring Harbor. Symp. Quant. Biol., 30, 517—527, 1965.
204. Scholes J., Electrical responses of retinal receptors and lamina in visual
system of Musca, Kybernetik, 6, 149—162, 1969.
205. Shaw S. R., Simultaneous recording from two cells in locust eye, Z. vergL
Physiol., 55, 183—194, 1967.
206. Shaw S. R., Organization of locust retina, Symp. Zool. Soc. Lond., 23, 135—
163, 1968.
207. Shaw S. R., Interreceptor coupling in ommatidia of drone honeybee and locust
compound eyes, Vision Res., 9, 999—1029, 1969.
208. Shaw S. R., Optics of arthropod compound eye, Science, 164, 88—90, 1969.
209. Shaw S. R„ Sense-cell structure and interspecies comparisons of polarized-
light absorbtion in arthropod compound eyes, Vision Res., 9, 1031—1040,
1969.
210. Sillman A. J., Ito H., Tomita T., Studies on the mass receptor potential of the
isolated frog retina. I. General properties of the response, Vision Res., 9,
1435—1442, 1969.
211. Sillman A. J., Ito H., Tomita T., Studies on the mass receptor potential of the
isolated frog retina. II. On the basis of the ionic mechanism, Vision Res., 9,.
1443—1451, 1969.
212. Sfostrand F. S., Ultrastructure of retinal rod synapses of guinea pig eye,.
J. Ultrastruct. Res., 2, 122—170, 1958.
213. Sfostrand F. S., Ultrastructure of retinal receptors of vertebrate eye, Ergebn..
Biol., 21, 128—160, 1959.
214. Smith H„ Phytochrome and photomorphogenesis in plants, Nature, 227, 665—
668, 1970.
215. Smith T. G., Baumann F., Functional organization within ommatidium of la-
teral eye of Limulus, Prog. Brain Res., 31, 313—349, 1969.
215a. Snyder A. W., Angular sensitivity of bee ommatidium, Z. vergl. Physiol., 76,
438—445, 1972.
216. Steinberg R. H., Schmidt R., Identification of horisontal cells as S-potential
generators in cat retina, Vision Res., 10, 817—820, 1970.
217. Stell W. K., Structure and relationship of horizontal cells and photoreceptor-
bipolar synaptic complexes in goldfish retina, Amer. J. Anat., 121, 401—424,
1967.
218. Stieve H., Interpretation of generator potential in terms of ionic processes.
Cold Spring Harbor Symp. Quant. Biol., 30, 451—456, 1965.
219. Stiles W. S., Crawford В. H., Luminous efficiency of rays entering eye pupil
at different points, Proc. Roy. Soc. Lond. B, 112, 428—450, 1933.
568
Глава 14
220. Strausfeld N. J., Golgi studies on insects. II. Optic lobes of Diptera, Phil.
Trans. Roy. Soc. Lond. B, 258, 135—223, 1970.
221. Strausfeld N. J., Blest A. D., Golgi studies of insects. I. Optic lobes of Lepi-
doptera, Phil. Trans. Roy. Soc. Lond. B, 258, 81—134, 1970.
222. Strausfeld N. J., Braitenberg V., The compound eye of the fly (Musca domesti-
ca): connections between the cartridges of the lamina ganglionaris, Z. vergl.
Physiol., 70, 95—104, 1970.
223. Svaetichin G., MacNichol E. F„ Retinal mechanisms for chromatic and achro-
matic vision, Ann. N. Y. Acad. Sci., 74, 385—404, 1958.
224. Sverdrup H. U., Johnson M. W„ Fleming R. H., The Oceans, Their Physics,
Chemistry and General Biology. Prentice-Hall, Inc., New York, 1942.
224a. Tomita T., Lateral inhibition in eye of Limulus, J. Neurophysiol., 21, 419—429,
1958.
225. Tomita T., Electrophysiological study of mechanisms subserving color coding
in fish retina, Cold Spring Harbor Symp. Quant. Biol., 30, 559—566, 1965.
226. Tomita T., Electrical activity of vertebrate photoreceptors, Quart. Rev. Bio-
phys., 3, 179—222, 1970.
227. Toyoda J., Nosaki H., Tomita T., Light-induced resistence changes in single
photoreceptors of Necturus and Gekko, Vision Res., 9, 453—463, 1969.
228. Trujillo-Cenoz O., Melamed J., Compound eye of Diptera: anatomical basis for
integration, J. Ultrastruct. Res., 16, 395—398, 1966.
229. Trujillo-Cenoz O., Melamed J., Electron microscope observations on the pe-
ripheral and intermediate retinas of dipterans, in: The functional Organization
of the Compound Eye, ed. C. G. Bernhard, Pergamon Press Ltd., Oxford, 1966.
230. Varela F. G., Fine structure of the visual system of the honeybee (Apismelli-
fera). II. The lamina, J. Ultrastruct. Res., 31, 178—194, 1970.
231. Varela F. G., Wiitanen W., Optics of compound eye of honeybee (Apis melli-
fera), J. General. Physiol., 55, 336—358, 1970.
232. Wagner H. G., MacNichol E. F., Wolbarsht M. L., Response properties of
single ganglion cells in goldfish retina, J. Gen. Physiol., 43 (Suppl), 45—62,
1960.
233. Wald G., Porphyropsin visual system, J. Gen. Physiol., 22, 775—794, 1939.
234. Wald G., Human vision and the spectrum. Science, 101, 653—658. 1945.
235. Wald G., Photochemistry of vision. Docum. OphthaL, 3, 94—134, 1949.
236. Wald G., Significance of vertebrate metamorphosis, Science, 128, 1481—1490,
1958.
237. Wald G., Life and light, Sci. Amer., 201 (4), 92—108, 1959.
238. Wald G., Origins of life, Proc. Nat. Acad. Sci., 52, 595—611, 1964.
239. Wald G., Receptors of human color vision, Science, 145, 1007—1017, 1964.
240. Wald G., Visual excitation and blood clotting, Science, 150, 1028—1030,
1965.
241. Wald G., Molecular basis of visual excitation, Nature, 219, 800—807, 1968.
242. Wald G., Brown P. N., Synthesis and bleaching of rhodopsin, Nature, 177,
174—176, 1956.
243. Wald G., Brown P. K., Gibbons I. R., The problem of visual excitatin, J. Opt
Soc. Amer., 53, 20—35, 1963.
244. Wald G., Brown P. K., Hubbard R., Oroshnik W., Hindered cis isomers of vi-
tamin A and retinene: the structure of the neo-Ь isomer, Proc. Nat. Acad. Sci.,
41, 438—451, 1955.
245. Wald G., Brown P. K., Smith P. H., Iodopsin, J. Gen. Physiol., 38, 623—681,
1955.
246. Walls G. L., The Vertebrate Eye and its Adaptive Radiation, Bulletin 19,
Cranbrook Institute of Science, Bloomfield Hills, Michigan, 1942.
247. Walther J. B., Changes induced in spectral sensitivity and form of retinal
action potential of cockroach eye by selective adaptation, J. Insect Physiologv,
2, 142—151, 1958.
248. Washizu Y., Electrical activity of single retinular cells in compound eye of the
blowfly Calliphora erythrocephala, Comp. Biochem. Physiol., 12, 369—387,
1964. .
Фоторецепция и зрение
569
249. Washizu У., Burkhardt D„ Streck Р., Visual field of single retinula cells and
interommatidial inclination in compound eye of blowfly Calliphora erythro-
cephala, Z. vergl. Physiol., 48, 413—428, 1964.
250 Waterman T. H., Light sensitivity and vision. In: The Physiology of Crusta-
cea, Vol. 2, edited by T. H. Waterman, Academic Press, New York, 1961,
pp. 1—64.
251 Waterman T. H„ Responses to polarized light: animals. In: Experimental Bio-
logy, edited by P. L. Altman and D. S. Dittmer, Fed. Amer. Soc. Exp. Biol.,
Bethesda, 1966. ,
252. Waterman T. H., Fernandez H. R., E-vector and wavelength discrimination by
retinular cells of crayfish Procambarus, Z. vergl. Physiol., 68, 154—174,
1970.
253. Waterman T. H„ Fernandez H. R., Goldsmith T. H., Dichroism of photosensi-
tive pigment in rhabdoms of crayfish Orconectes, J. Gen. Physiol., 54, 415—
432, 1969.
254. Waterman T. H„ Wiersma C. A. G„ Functional relation between retinular cell
and optic nerve in Limulus, J. Exp. Zool., 126, 59—85, 1954.
255. Waterman T. H„ Wiersma C. A. G., Electrical responses in decapod crustacean,
visual systems, J. Cell. Comp. Physiol., 61, 1—16, 1963. •
256. Waterman T. H., Wiersma C. A. G., Bush В. M. H., Afferent visual responses
in optic nerve of crab Podophthalmus, J. Cell. Comp. Physiol., 63, 135—155,
J964.
257. Wedle R. A., Photochemistry of human central fovea, Nature, 218, 238—240,
1968.
258. Werblin F. S„ Dowling J. E., Organization of retina of mudpuppy Nectus tna-
culosus, J. NeurophysioL, 32, 339—355, 1969.
259. Wiersma C. A. G., Neuronal components of optic nerve of crab Carcinus
maenas, Proc. Konikl. Nederl. Akad. Wetenschapperi (Amsterdam), series C,
73, 25—34, 1970.
260. Wiersma C. A. G., Bush В. M. FL, Waterman T. H., Efferent visual responses
of contralateral origin in optic nerve of crab Podophthalmus, J. Cell. Comp.
Physiol., 64, 309—326, 1964.
261. Wiersma C. A. G., Yamaguchi Y., Neuronal components of optic nerve of
crayfish as studied by single unit analysis, J. Comp. Neurol., 128. 333—358,
1966.
262. Wiersma C. A. G., Yamaguchi T., Integration of visual stimuli by crayfish
central nervous system, J. Exp. Biol., 47, 409—431, 1967.
263. Wiersma C. A. G., Yamaguchi T., Integration of visual stimuli in rock lobster,
Vision Res., 7, 197—204, 1967.
264. Williams С. M., Photoperiodism and endocrine aspects of insect diapause,
Symp. Soc. Exp. Biol., 23, 285—300, 1969.
265. Wilt F. H., Differentiation of visual pigments in metamorphosing larvae of
Rana catesbeiana, Develop. Biol., 1, 199—233, 1959.
266. Withrow R. B., Klein W. FL, Elstad V., Action spectra of photomorphogenic
induction and its photoinactivation, Plant Physiol., 32, 453—462, 1957.
267. Wolfson A., Animal photoperiodism. In: Photophysiology, Vol. 2, edited by
A. C. Giese, Academic Press, New York, 1964,-pp. 1—49.
268. Worthington C. R., Structure of photoreceptor membranes, Fed. Proc., 30,
57—63, 1971.
269. Yoshikami S., Hagins W. A., Light, calcium, and the photocurrent of rods and
cones, Abstr. Biophys. Soc. 15th Annual Meeting, New Orleans, TPM-EI6
1971.
270. Zimmerman W. F., Goldsmith T. H., Photosensitivity of circadian rhvthm and
of visual receptors in carotenoid-depleted Drosophila, Science, 171, 1167—1169
1971.
271. Zonana H. V., Fine structure of squid retina, Johns Hopkins Hosp. Bull., 109.
185—205, 1961. "
ОГЛАВЛЕНИЕ
Глава 8. Дыхательные функции крови. Л. Проссер. (Перевод Л. Н. Грин-
берга.) ................................................................. 5
Распространение пигментов................................................ 5
Химия пигментов — переносчиков кислорода................................. 8
Соединение гемоглобина с кислородом......................................27
Влияние разведения, ионов и фосфатов на сродство к кислороду ... 33
Видовая специфичность и экологические аспекты сродства к кислороду 35
Изменения транспортной функции в процессе эмбрионального развития . . 48
Функция гемоглобина у беспозвоночных.................................52
Влияние пониженного содержания кислорода в среде на синтез гемоглобина 57
Функция хлорокруорина .................................................. 59
Функция гемоцианина..................................................59
Функция гемоэритрина..................................... 64
Транспорт двуокиси углерода ............................................ 65
Заключение..................................................... 72
Список литературы....................................................75
Глава 9. Температура. Л. Проссер. (Перевод Г. А. Бузникова.) .... 84
Введение.............................................................84
Замораживание и холодоустойчивость...................................88
Летальные температуры; устойчивость и адаптация......................95
Скорость биологических процесов.....................................104
Поведенческие и локомоторные адаптации..............................126
Действие температуры на возбудимые мембраны.........................134
Роль испарения влаги и поведенческих реакций в регуляции температуры
у наземных пойкилотермных животных......................................137
Гомойотермные животные..............................................141
Нервные механизмы терморегуляции....................................175
Зимняя спячка.......................................................180
Заключение . 187
Список литературы . ....................................................191
Глава 10. Биологические ритмы. Ф. Браун. (Перевод Г. А. Бузникова.) . . 210
Список литературы * 254
Глава 11. Возбудимые мембраны. Л. Проссер. (Перевод Г. А. Бузникова.) 261
Ненервное проведение; реакции отдельных клеток ....... 261
Передача сигнала от клетки к клетке в ненервных тканях...................262
Проведение в возбудимых тканях, строение мембран (в основном мембран
нейронов) 269
Оглавление
571
Синаптические потенциалы в химических синапсах........................284
Изменения мембранной проводимости.....................................294
Неэлектрические физические явления, связанные с проведением импульсов 298
Адаптивные особенности нервного проведения............................298
Проведение возбуждения у животных с низким содержанием натрия в
крови.................................................................301
функции глии..........................................................303
Явления, связанные с разными размерами частей одного и того же нейрона 304
Электрическая и химическая межклеточная передача......................304
Заключение............................................................336
Список литературы ................................................... 339
Глава 12. Механорецепция, фонорецепция и чувство равновесия. Л. Прос-
сер. (Перевод Ф. Г. Грибакина и А. Д. Поляновского.)..................350
Клеточные механизмы механорецепции....................................350
Тактильные рецепторы, рецепторы движения и растяжения.................352
Звуковая рецепция.................................................... 376
Органы равновесия.....................................................408
Заключение.......................................................... 419
Список литературы ................................................... 421
Глава 13. Хеморецепция. Л. Проссер. (Перевод Р. А. Певзнер.) .... 432
Хеморецепция у беспозвоночных (за исключением насекомых) .... 432
Хеморенепция у насекомых..............................................432
Хеморецепция у позвоночных............................................434
Клеточные механизмы хеморецепции......................................460
Заключение............................................................465
Список литературы ................................................... 467
Глава 14. Фоторецепция и зрение. Т. Голдсмит. (Перевод Г. И. Рожковой.) 474
Фотобиология и электромагнитный спектр................................474
Цитологическая организация фоторецепторных клеток.....................480
Зрительные пигменты................................................. 487
Реакции мембран рецепторных клеток на возбуждение.....................503
Сложные глаза членистоногих...........................................518
Глаза позвоночных.....................................................537
Список литературы.....................................................559
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении, качестве
перевода и другие просим присылать по адресу:
129820, Москва, И-ПО, ГСП,
1-й Рижский пер., д. 2,
издательство «Мир»
ИБ 698
СРАВНИТЕЛЬНАЯ ФИЗИОЛОГИЯ
животных
Редактор Ю. И. Лашкевич
Художник В. Медников
Художественный редактор А. С. Смеляков
Технический редактор Г. Б. Алюлина
Корректор Е. К- Монякова
Сдано в набор 23/П 1977 г.
Подписано к печати 16/VI 1977 г.
Бумага кн. журн. 60X901/i6==18 бум. л.
Печ. л. 36. Уч.-изд. л. 41,56. Изд. № 4/8749.
Цена 3 р. 10 к. Зак. 1514.
ИЗДАТЕЛЬСТВО «МИР»
Москва, 1-й Рижский пер., 2
Московская типография № 11 Союзполиграфпрома
при Государственном комитете Совета Министров
СССР по делам издательств, полиграфии и книж-
ной торговли. Москва, 113105, Нагатинская ул., д. 1.
ИЗДАТЕЛЬСТВО «МИР»
Вышла в свет в 1977 году
Лайтфут Э. Явления переноса в живых системах, перевод с ан-
глийского, 34 а. л.
В последние годы все более заметным становится проникнове-
ние точных методов исследования в различные области биологии.
Полученные при помощи этих методов сведения о химических и
электрохимических процессах в живых организмах позволили ре-
шить важные практические задачи, в результате которых были со-
зданы аппараты типа «сердце—легкие» или «искусственная почка».
В книге известного американского ученого Э. Лайтфута изложены
общие принципы применения математических методов при модели-
ровании ряда физиологических процессов и рассмотрены различ-
ные аспекты биореологии, перенос массы и энергии в многокомпо-
нентных системах, особенности биологических мембран с точки
зрения их моделирования. Дано теоретическое обоснование принци-
пов проектирования медицинской аппаратуры.
Предназначена для физиологов, биофизиков, специалистов, ра-
ботающих в области биоинженерии и биоматематики, для физиков,
химиков и для лиц других специальностей, занимающихся приклад-
ными вопросами массопереноса.
ИЗДАТЕЛЬСТВО «МИР»
Выйдет в свет в 1977 году
Бак 3. Химическая передача нервного импульса, перевод с фран-
цузского, 6 а. л.
Небольшая по объему книга иностранного члена Академии на-
ук СССР известного бельгийского физиолога и радиобиолога 3. Ба-
ка посвящена одному из самых замечательных открытий современ-
ной нейрофизиологии — химическому механизму передачи нервных
импульсов в синаптических структурах. Научная строгость удачно
сочетается с живостью изложения, делающей книгу доступной для
широких кругов читателей.
Предназначена для физиологов, биохимиков, биофизиков, фар-
макологов, врачей, студентов, аспирантов и преподавателей уни-
верситетов, медицинских и педагогических институтов.
Хочачка П., Семеро Дж. Стратегия биохимической адаптации,
перевод с английского, 24 а. л.
Монография посвящена влиянию факторов внешней среды (со-
держание кислорода и углекислоты, влажность, температура, дав-
ление) и изменений состава внутренней среды на процессы обмена
веществ, определяющие адаптацию живых организмов к этим воз-
действиям. В основе биохимической адаптации лежит изменение
типов макромолекул, входящих в состав живых систем, их количе-
ства и строения или регуляции их функций. С этих позиций обсуж-
даются проблемы гомеостаза и физиологической адаптации. При-
ведены обширные данные о межвидовых различиях адаптации на
примере строения и функционирования гемоглобина, а также об
адаптации в онтогенезе.
Предназначена для физиологов, биохимиков, экологов, аспиран-
тов и студентов университетов, педагогических и медицинских ин-
ститутов.
ИЗДАТЕЛЬСТВО «МИР»
Выйдет в свет в 1977 году
Кеннеди К. Экологическая паразитология, перевод с английско-
го, 14 а. л.
Первая .в мировой литературе, краткая сводка, посвященная эко-
логии паразитов животных. Рассмотрены разные типы взаимоот-
ношений между паразитами и хозяевами, проблема специфичности,
регуляция численности и распространения паразитов у разных
групп животных. Книга завершается описанием общих принципов
математического моделирования системы паразит—хозяин.
Предназначена для широкого круга биологов, паразитологов,
эпидемиологов, врачей, ветеринаров, а также студентов и препода-
вателей соответствующих специальностей.
Роллер Э. Открытие основных законов жизни, перевод с англий-
ского, 19 а. л.
Живо написанная история развития молекулярной биологии и
ее основных успехов — от открытия живой клетки до выяснения
механизмов биосинтеза белка, строения и функционирования генов.
Рассмотрены перспективы дальнейшего развития молекулярной
биологии и значение ее достижений для расшифровки механизмов
таких сложных процессов, как дифференцировка, рост, старение
и т. д. В книге помещены интересные иллюстрации научного ха-
рактера, а также портреты выдающихся ученых. Написана она
живо и увлекательно и вместе с тем на достаточно высоком науч-
ном уровне.
Предназначена для широкого круга читателей — биологов, меди-
ков, физиков, химиков, математиков, инженеров, для студентов, а
также и для школьников старших классов.