/

































































































































































































































Автор: Пол Д. Крамер Теодор Т. Козловский
Теги: прикладные науки медицина технология физиология растений физиология древесных растений
Год: 1983




Текст
ПОЛ Д. КРАМЕР, ТЕОДОР Т. КОЗЛОВСКИЙ
ФИЗИОЛОГИЯ ДРЕВЕСНЫХ РАСТЕНИЙ
Перевод с английского И.Г. ЗАВАДСКОЙ, Ц.П. ВИКТОРОВА, М.В. РАЙХИНШТЕЙНА
Москва
"Лесная промышленность" 1983
УДК 6.10*161.4 (03.20)
Крамер Пол Д., Козловский Теодор Т. Физиология древесных растений: Пер. с анг. - М.: Лесн. пром-сть, 1983. — 464 с., ил.
Авторы - известные американские физиологи рассматривают все жизненные процессы, происходящие в деревьях: фотосинтез, дыхание, биохимизм, тропизм, рост и развитие растений, в том числе регуляторы роста. Изложены строение деревьев и взаимодействие древостоев со средой. Значительное место отведено описанию экологический физиологии, изучающей влияние экологических факторов на физиологические процессы.
Для научных работников, специалистов лесного хозяйства, охраны природы.
Табл. 95, ил. 195
Рекомендовано к изданию Управлением воспроизводства лесных ресурсов Гослесхоза СССР
Paul J. Kramer
Department of Botany
Duke University
Durham, North Carolina
Theodore T. Kozlowski
Department of Forestry
University of Wisconsin
Madison, Wisconsin
Physiology of Woody Plants 1979
Academic Press New York San Francisco London
28_8J on (01) -u
© 1979, by Academic Press, Inc.
© Перевод на русский язык ’’Лесная промышленность”, 1983
ОТ ИЗДАТЕЛЬСТВА
В целях успешного решения ответственных задач по восстановлению и приумножению лесных запасов, все более повышается значение науки о лесе, использования ее достижений. В этой связи возрастает роль физиологии растений — отрасли биологии, которая имеет своей целью изучение и управление ростом и развитием растительных организмов для повышения их продуктивности.
Этим вопросам посвящена книга известных американских физиологов лесной растительности П. Крамера и Т. Козловского ’’Физиология древесных растений”. Первое издание этой книги вышло в 1963 г. За это время проведены фундаментальные исследования по изучению фотосинтеза, дыхания, водного режима, питания, роста и развития растений и других физиологических процессов, протекающих в дереве. Все это нашло отражение в новой редакции книги ’’Физиология древесных растений”. По сравнению с первым изданием авторами несколько изменена структура книги, введен ряд новых разделов, в том числе и из их работы ’’Рост и развитие древесных растений” (1971 г.).
Монография является обширной сводкой современных знаний по физиологии древесных растений, подготовленной преимущественно по работам зарубежных ученых, и предназначена для широкого круга специалистов лесного хозяйства, садоводов, озеленителей, преподавателей вузов. Она может стать полезным пособием и студентам.
Книга издана в переводе с некоторыми сокращениями. В частности, исключены разделы, которые не представляют большого интереса для советского читателя, или достаточно глубоко проработаны в нашей стране. В связи с тем, что в работе приведены в подавляющем большинстве ссылки на зарубежные издания и публикации, в основном повторяющие первое издание, библиография в книге не приводится.
Несмотря на некоторые сокращения, издательство надеется, что книга окажется полезной для всех специалистов, занимающихся вопросами физиологии древесных растений.
ПРЕДИСЛОВИЕ
Эта книга предназначена для студентов как учебное пособие, как источник информации для исследователей и для желающих лучше понять, как растут и развиваются древесные растения. Она будет также полезна и широкому кругу специалистов, включая лесоводов, физиологов растений, экологов, озеленителей, а также всем, пожелавшим подробнее узнать о деревьях. Трудно писать для такой разнообразной аудитории из-за больших различий в их знаниях и интересах ..Однако мы предполагаем, что читатели владеют некоторыми знаниями элементарной ботаники и физиологии. Понятно, что одни могут найти некоторые части книги слишком теоретическими, а другие те же разделы будут считать чрезмерно упрощенными. Для того чтобы выращивать деревья высокого качества, надо понять как они растут, В связи с этим следует напомнить тем, кто найдет некоторые разделы слишком специальными и теоретическими, что для понимания таких физиологических процессов, как фотосинтез и дыхание, играющих существенную роль в росте дерева, необходимо иметь элементарное представление о биохимии. Тем же, кто сочтет некоторые разделы упрощенными, напоминаем, что эта книга предназначена скорее для лесоводов и садоводов, чем для физиологов растений. Цель книги — объяснить, каким образом физиологические процессы вовлекаются в рост древесных растений и как на них влияют окружающие условия, и, кроме того, дать представление о механизмах этих процессов.
С этой точки зрения книга несколько отличается от большинства учебников по физиологии растений, так как в ней основное внимание уделяется структуре дерева, взаимодействию деревьев и древесных насаждений с окружающей средой, действию факторов окружающей среды на физиологические процессы. Таким образом, наш подход более экологический, чем биохимический, хотя он и дает ясное представление о том, что под влиянием окружающих условий происходят изменения на клеточном и молекулярном уровнях. Например, свет и температура влияют на фотосинтез на клеточном уровне, воздействуя на поведение замыкающих клеток устьиц, а также и на молекулярном уровне в результате воздействия на биохимические и биофизические процессы, происходящие в хлоропластах. Имеются также данные о внутренних контролирующих системах, которые регулируют и координируют ростовые процессы различных тканей и органов, изменяют скорость разных процессов, в результате которых возникают гармоничные взаимодействия структуры и функции, необходимые для эффективного роста растений.
Особое значение придаем тесному взаимоотношению между структурой и функцией, так как нельзя понять процессы растения без понимания его структуры. Более того, структура дерева возникает в результате сложных физиологических процессов, в совокупности обозначаемых общим термином рост. Поэтому и уделяем особое внимание структуре и физиологическим процессам, от активности которых зависит образование различных тканей и органов деревьев. В книге детально рассматри
Предисловие
5
ваются важные вегетативный и половой способы размножения, иллюстрирующие к тому же интересные и важные физиологические процессы.
Эта книга представляет собой пересмотренное, расширенное и дополненное современными данными издание двух ранее опубликованных книг ’’Физиология деревьев” (в 1963 г. издана на русском языке) и ’’Рост и развитие древесных растений” (в 2-х томах, 1971 г.). В книге рассматривается физиология древесных пород, используемых в озеленении, а также лесных и садовых деревьев. Это объясняется тем, что у всех видов древесных и травянистых растений одинаковы физиологические процессы.
В этой книге мы использовали данные мировой литературы, чтобы сделать работу более авторитетной и хорошо документированной. Многие ссылки относятся к современным работам, но цитируются также и старые статьи, исторически интересные или содержащие данные, которых нет в современных работах. В течение двух десятилетий, прошедших с момента выхода в свет ’’Физиологии деревьев”, опубликованы многочисленные статьи. Нами приведена лишь часть литературы, которая непосредственно относится к обсуждаемой проблеме. Несмотря на огромное количество исследований и публикаций, ощущается недостаток информации по некоторым важным темам, многие вопросы остаются нерешенными. Имеются и противоположные точки зрения по одной и той же проблеме. В таких случаях мы даем собственную интерпретацию, исходя из имеющихся в нашем распоряжении данных. Следует отметить, что новые сведения, на основании которых делаются заключения, нуждаются в проверке, поэтому надеемся, что читатели будут готовы изменить свою точку зрения, если в этом возникнет необходимость.
В тексте использованы общепринятые названия для хорошо известных видов и латинские названия для менее распространенных видов.
ЗНАЧЕНИЕ ФИЗИОЛОГИИ РАСТЕНИЙ
В противоположность книгам по лесоводству и садоводству, которые описывают как деревья вырастить, эта книга дает представление о,том, как деревья растут, т.е. в ней рассматривается физиология древесных растений любой величины. Из-за их больших размеров, продолжительной жизни и относительно небольшого отношения фотосинтезирующей ткани к нефотосинтезирующей деревья будут нами наиболее часто использоваться в качестве примеров. Тем не менее основные физиологические процессы сходны у всех семенных растений, поэтому то, что говорится о деревьях, применимо и к кустарникам с учетом их размеров и продолжительности жизни.
На различных этапах развития человеческого общества деревья для людей имели разное значение. Для наших предков они были источником топлива и убежищем, а иногда и предметом поклонения. Для владельцев домов они создавали летом приятную тень, хотя осенью они причиняли им неприятности, осыпая газоны листьями. Лес был единственным источником топлива. В дальнейшем, когда другие виды топлива станут менее доступны, деревья вновь смогут стать важным его источником. Для работников лесопаркового строительства деревья являются декоративными объектами ландшафта. Защищая бассейны рек и дичь, они обеспечивают приятный отдых для любителей природы и спортсменов. Деревья служат источником сырья для бумажной и деревообрабатывающей промышленности. Садоводы изучают деревья как источник плодов. Для физиологов деревья являются сложными биохимическими фабриками, которые вырастают из семян и буквально строят сами себя. Поэтому физиологи интересуются многочисленными метаболическими процессами, которые в совокупности обеспечивают то, что мы называем ростом.
Об интенсивности синтезирующих процессов деревьев дают представления данные по продуктивности: с 1 га в умеренной лесной зоне получается 20 т сухого вещества ежегодно, а с 1 га тропического влажного леса - более 100 т. Читатели возразят, что эти цифры относятся к валовой, а не чистой продуктивности (по данным Кира, 1975, и табл. 6.1, эти цифры значительно меньше). Вся эта масса образуется из относительно простых веществ: воды, углекислого газа, азотистых соединений и минеральных солей. Таким образом, продуктивность деревьев и других зеленых растений зависит от интенсивности синтеза углеводов, азотсодержащих соединений, липидов и от преобразования их в новую растительную ткань. Однако рост зависит не только от процессов синтеза. Для этого необходимы поглощение достаточного количества воды и элементов минерального питания, передвижение органических и неорганических веществ к точкам роста, а также специальные системы, которые координируют многочисленные процессы. В этой книге сделана попытка объяснить, как происходят эти процессы, как на них влияют окружающие условия и агротехнические мероприятия и как все эти взаимодействующие факторы влияют количественно и качественно на рост.
Физиологи растений в первую очередь заинтересованы в изучении того, как деревья растут, а работники лесопаркового хозяйства, лесоводы и садоводы - в том, как вырастить деревья лучшего качества. Эти две цели взаимосвязаны более тесно, чем может казаться, так как для успешного выращивания деревьев необходимо понимание основных физиологических процессов, управляющих ростом, и влияние на них факторов окружающей среды и агротехнических мероприятий. Таким образом, основные физиологические исследования могут содействовать успешной работе лесоводов, садоводов и наоборот. Физиологи, узнав больше о том, как растут деревья, а лесоводы и садоводы, лучше изучив физиологию деревьев, смогут совместными усилиями разрешить проблему эффективного выращивании деревьев.
Значение физиологии растений
7
РОЛЬ ФИЗИОЛОГИИ РАСТЕНИЙ
Основная задача физиологии растений — объяснить, как растут растения и как физиологические процессы и внутренняя среда реагируют на окружающие условия и антропогенные воздействия. Исследования таких процессов, как фотосинтез, передвижение веществ, ассимиляция, дыхание и транспирация, могут показаться далекими от практических задач лесоводства и садоводства. Однако рост — это результат взаимодействия физиологических процессов, и чтобы понять, почему деревья растут неодинаково в разных окружающих условиях и при различных агротехнических воздействиях, необходимо знать природу этих физиологических процессов и как на них влияют окружающие условия.
Рост деревьев, так же как и других организмов, контролируется их наследственными генетическими потенциальными возможностями и окружающими условиями, но данных о механизме этого контроля слишком мало. Известно, что затенение или засуха ослабляют рост, что новая комбинация генов ответственна за быстрый рост гибрида, однако это не объясняет каким образом осуществляется наблюдаемый эффект. Необходимо более глубокое изучение различных причин, способствующих соответствующему эффекту. Представленная ниже диаграмма показывает, как наследственность и окружающие условия влияют на внутренние физиологические процессы и состояние деревьев и таким образом контролируют количество и качество роста. Диаграмма также показывает связи между областями науки, изучающими эти явления.
Наследственные потенциальные возможности
Факторы окружающей среды
Область генетики
Селекция и скрещивание
Потенциальная скорость роста, размеры и продолжительность жизни деревьев
Типы ксилемы, глубина и простирание корневой системы
т
Область экологии, почвоведения, климатологии, метеорологии и т.д.
Радиация, температура, минеральные вещества, конкуренция, вредители, лесовод-ственные приемы и т.д.
Физиологические процессы и условия
Область физиологии растений
Фотосинтез, углеводный и азотный метаболизм
Дыхание, передвижение
Водный баланс растений и его влияние на рост и метаболизм
Регуляторы роста и т.д. •
I-------------------.-------------------
Количество и качество роста
Область ландшафтного лесоводства, лесоводства и садоводства Количество и качество образующейся древесины, плодов и семян ВспггатианыЙ рост в сравнении с репродуктивным Соотношение роста корней и побегов
8 Глава 1
КОНЦЕПЦИЯ КЛЕБСА. Эту схему иногда называют концепцией Клебса, так как немецкий физиолог растений Клебс (1933, 3914) одним из первых указал на то, что факторы окружающей среды могут влиять на процессы в растениях, изменяя внутренние условия. Лан-дегард (3933) также внес важный вклад в развитие этой точки зрения. Концепция Клебса придает особое значение основному биологическому принципу, согласно которому наследственность и окружающая среда могут влиять на рост организма, воздействуя лишь на внутренние процессы или условия роста. Физиологические процессы дерева представляют собой механизмы, посредством которых наследственность и факторы окружающей среды осуществляют контроль за ростом. В связи с этим, чтобы понять, почему на деревья влияет отдельный фактор, то или иное агротехническое воздействие, необходимо знать, каким образом они влияют на физиологические процессы.
ПРИМЕНЕНИЕ КОНЦЕПЦИИ КЛЕБСА. Применение этой концепции можно проиллюстрировать несколькими конкретными примерами. Так. если в одинаковых условиях гибрид растет быстрее родителей, то это происходит благодаря новой комбинации генов, обеспечивающей эффективный баланс физиологических процессов, что создает внутренние условия для интенсивного преобразования питательных веществ в растительные ткани. Это может быть результатом повышения эффективности либо структуры, либо процессов, либо того и другого вместе. Скорость фотосинтеза на единицу поверхности листа'может возрастать при повышении содержания хлорофилла или вследствие структурных изменений, усиливающих поглощение углекислого газа, а также за счет более выгодного положения листьев по отношению к свету. Протяженная (экстенсивная) корневая система или толстый слой кутина могут стать причиной большей тургесцентности клеток у дерева, которая в свою очередь окажет благотворное влияние на фотосинтез и на рост клеток.
Если в одинаковых условиях какое-либо дерево болотной сосны образует больше смолы, чем соседнее дерево, это, вероятно, обусловлено генетической структурой, способствующей ускорению процессов превращения питательных веществ в смолу. Другая возможность — это структурные различия, которые являются причиной ускорения истечения смолы. Если один вид холодно- или засухоустойчивее или более теневынослив, чем другой, это может быть результатом особенностей генетической структуры, обеспечивающей различие в строении и ходе тех физиологических процессов, от которых зависит устойчивость к неблагоприятным условиям среды. Однако даже если изменения скорости роста можно приписать некоторым структурным модификациям, они в основном зависят все-таки от изменений физиологических процессов. Структура является результатом биологических и физиологических процессов, влияющих на ростовые процессы. Изменения в структуре приводят к изменениям в ходе процессов. Таким образом, процессы контролируют структурообразование, хотя рами они модифицируются под влиянием изменений в структуре.
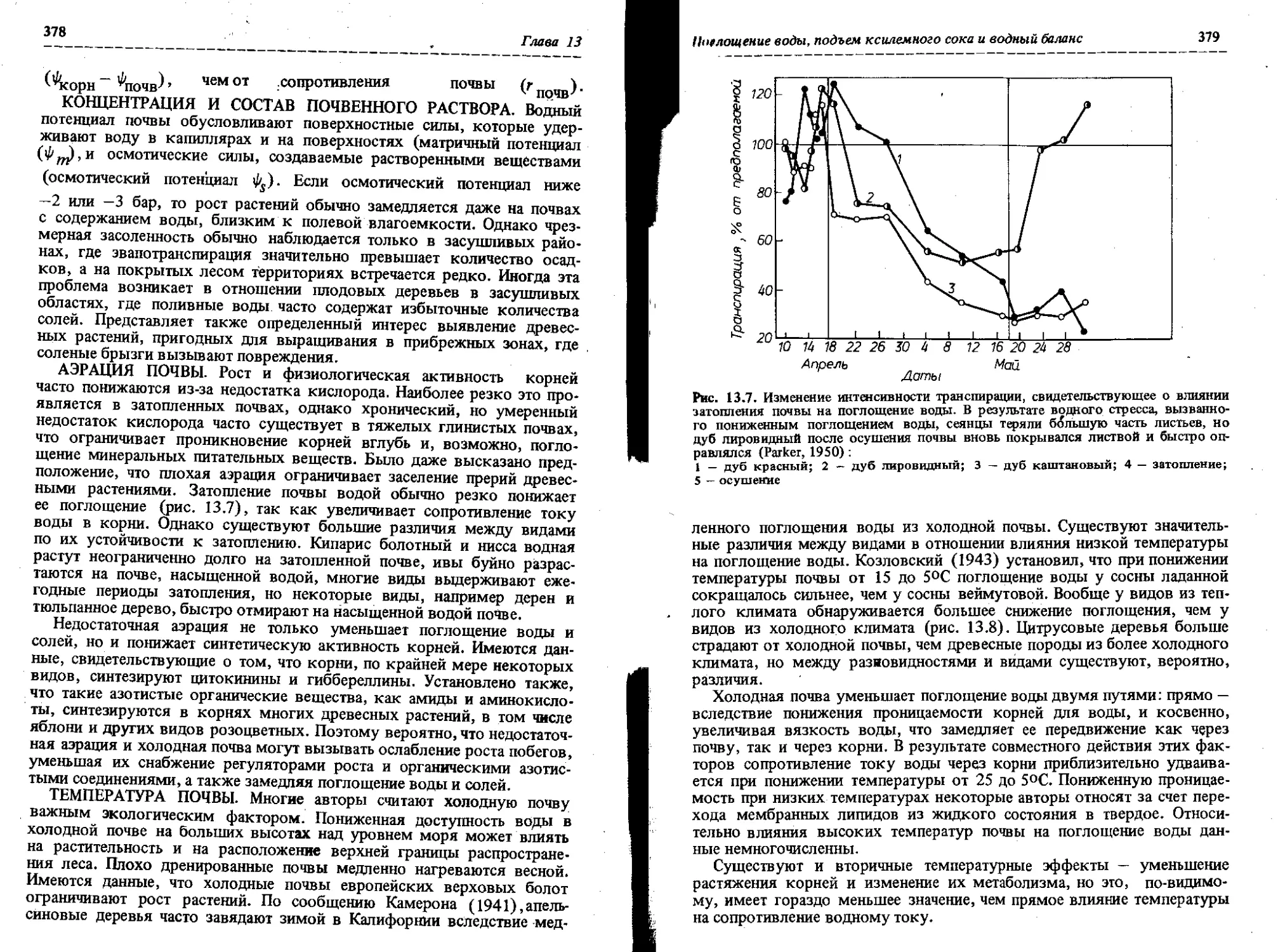
Неблагоприятные окружающие условия, влияющие на существенные физиологические процессы, снижают рост дерева. Например, водный дефицит тормозит рост, так как при этом закрываются устьица,
Значение физиологии растений
9
снижается интенсивность фотосинтеза, уменьшается тургесцентность, прекращается рост клеток и возникают другие неблагоприятные условия внутри дерева. Снижает рост дерева и недостаток азота, который является существенной составной частью белков, необходимых для формирования новой протоплазмы, ферментов и других необходимых веществ. Очень важны для роста дерева фосфор, калий, кальций, сера и другие минеральные элементы, входящие в состав коферментов, буферных и других биохимических систем, необходимых для осуществления различных биохимических процессов.
Насекомые и грибы, поражая дерево, тормозят его рост или вызывают заболевание, если повреждение достаточно глубоко нарушает один или несколько физиологических процессов. Дефолиация дерева непосредственно не снижает ростовые процессы, а косвенно влияет на скорость фотосинтеза и синтез регуляторов роста в кроне. При повреждении флоэмы уменьшается перемещение питательных веществ и регуляторов роста к корням, а при повреждении корневой системы сокращается поглощение воды и питательных веществ из почвы. Фитопатологи, и энтомологи иногда слишком увлекаются описанием и классификацией болезнетворных организмов и не учитывают тот факт, что в действительности они имеют дело с физиологическими проблемами. Устойчивость к поражению насекомыми и грибами в значительной мере является биохимической проблемой, и повреждение является результатом нарушения биохимических и физиологических процессов. Борьбе с заболеваниями человека способствует использование биохимических и физиологических Подходов. Борьба с древесными заболеваниями станет также более эффективной, если энтомологи и фитопатологи станут больше уделять внимания физиологическим аспектам проблемы вместо простого описания болезнетворных организмов и использования химических средств.
Мы повторяем еще раз, что единственный способ, которым генетические различия, внешние факторы, агротехнические воздействия, а также болезни и насекомые могут влиять на рост, — это их влияние ла внутренние условия и физиологические процессы дерева. Усилия лесоводов-озеленителей, лесоводов и садоводов должны быть направлены на создание подходящих генотипов и факторов окружающей среды для того, чтобы управлять физиологическими механизмами, которые действительно регулируют ростовые процессы. Для результативного и умелого подхода они должны понять природу основных физиологических процессов, их роль в ростовых процессах и реакцию на действие различных факторов внешней среды.
КРУГ ВОПРОСОВ ФИЗИОЛОГИИ ДЕРЕВА
Как установил Губер (1937), деревья всегда рассматривались не-Позаниками как представители вершины развития растительного царила. Из-за размеров и красоты, их часто наделяли особыми свойствами и даже поклонялись им. Для деревьев характерны специфические свой-<та. которые, однако, являются скорее количественными, нежели отличными по существу от свойств других растений. Деревья проходят
10
Глава
1
те же стадии роста, им присущи те же процессы, как и другим семенным растениям, но их большие размеры, медленное развитие и продолжи-тельность жизненного цикла выдвигают некоторые особые проблемы, отличные от тех, которые присущи небольшим растениям с более коротким жизненным циклом. Наиболее очевидное отличие деревьев от травянистых растений - это большее расстояние, которое должны преодолевать при передвижении в них вода, минеральные и питательные вещества, а также больший процент нефотосинтезирующих тканей. К тому же вследствие более продолжительной жизни они в большей степени, чем однолетние и двулетние растения, подвергаются действию экстремальных температур и влиянию других климатических и почвенных условий. ' -Таким образом, вследствие крупных размеров деревьев создаются не-которые специфические проблемы, касающиеся физиологических про- \ цессов. <?’
НАИБОЛЕЕ ВАЖНЫЕ ФИЗИОЛОГИЧЕСКИЕ ПРОЦЕССЫ И УСЛО-ВИЯ ИХ ОБЕСПЕЧЕНИЯ. Успешный рост деревьев зависит от взаимо-действия ряда физиологических процессов и условий. Некоторые наи-более важные из них перечислены ниже (в скобках указаны главы, в которых они рассматриваются). Фотосинтез — синтез углеводов из углекислого газа и воды в хлорофиллоносной ткани деревьев. Углеводы — основные питательные материалы, используемые в других процессах (глава 5). Азотный обмен - включение неорганического азота в органические соединения, что делает возможным синтез белка и протоплазмы (глава 9). Липидный, или жировой, обмен — синтез липидов и родственных им соединений (глава 8). Дыхание - окисление питательных веществ в живых клетках, в результате которого высвобождается энергия, используемая при ассимиляции, поглощении минеральных веществ и других процессах, идущих с затратами энергии (глава 6). Ассимиляция — преобразование питательных элементов в новую протоплазму, клеточные оболочки и другие структуры; процесс роста (глава 6). Аккумуляция питательных элементов - запасание питательных веществ в семенах и паренхимных клетках древесины и коры (глава 7). Аккумуляция солей - концентрация солей в клетках и тканях с помощью механизма активного транспорта, протекающего с затратами метаболической энергии (глава 10). Абсорбция - поглощение' воды и минеральных веществ из почвы, кислорода и углекислого газа из воздуха (главы 5, 10 и 13). Транслокация - передвижение воды, минеральных и питательных веществ и гормонов от одной части дерева к другой (глава 11). Транспирация - потеря воды в виде паров (глава 12). Рост — постоянное увеличение размеров в результате взаимодействия физиологических, перечисленных выше процессов (глава 3). Половое размножение — образование в результате взаимодействия многих физиологических процессов стробилов или цветков, плодов и семян (глава 4). Вегетативное размножение играет важную роль у некоторых видов (глава 4). Регуляция роста — сложные взаимодействия гормонов и баланса питательных веществ (главы 15 и 16).
К важным физиологическим условиям, влияющим на рост, отно- I
сятся также следующие: Количество и эффективность хлорофилла •
(глава 5). Состав и количество углеводов и их взаимные превращение, например, переход крахмала в сахара и наоборот (глава 7). Состав и
Чначение физиологии растений
11
количество азотистых соединений и отношение углеводов к азоту (гла-на 9). Состав и количество других компонентов, таких, как жиры (гла-па 6, 8). Свойства протоплазмы, например холодо- и засухоустойчивость, частично обусловлены этими особыми свойствами (глава 16). С )смотическое давление клеточного сока, увеличение осмотического дав-лепия часто связано С наступлением засухи и холода (глава 14). Тургес-цснтность клеток — потеря тургора вызывает прекращение роста и пл инет на скорость различных физиологических процессов (глава 13 и 16).
Задача физиологов заключается в исследовании процессов, изучении их механизмов, наблюдении их реакции на различные окружающие условия и определении их роли в ростовых процессах. От полноты знаний физиологов о механизмах основных физиологических процессов зависит помощь, которую они смогут оказать лесоводам и садоводам в решении их практических задач. При достаточных знанйях физиологических требований деревьев можно предсказать поведение отдельных видов в определенных почвенных и климатических условиях, или их реакцию на те или иные агротехнические воздействия.
('ЛОЖНОСТЬ ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ. Такие физиологические процессы, как фотосинтез, дыхание или транспирация представляют совокупность химических и физических процессов. Чтобы попять механизм физиологического процесса, необходимо выделить его физические и химические компоненты. В связи с этим физиология растений все более нуждается в усовершенствованных методах биохимии. Биохимический подход оказался очень плодотворным в исследованиях таких сложных процессов, как фотосинтез и дыхание.
Основная цель этой книги — объяснить, каким образом растет дерево. Наш подход при этом скорее экологический, чем биохимический, так как больше внимания уделяет способам воздействия факторов окружающей среды на процессы, чем детальному обсуждению их физиологической природы. Однако мы будем, хотя и кратко, рассматривать, как факторы окружающей среды могут влиять на растение на клеточном и молекулярном уровнях. Такие факторы, как температура, вода и спет, влияют на физиологические процессы непосредственно, прямыми iivniMH, которые можно легко объяснить, а также и косвенно, через иоОочные регулирующие системы, которые понять труднее. Например, при понижении температуры снижается скорость дыхания вследствие ммедления скорости молекулярных превращений и биохимических реакций. При этом также уменьшается проницаемость мембран иувели-чппас'тся вязкость протоплазмы и, как следствие, снижается скорость перемещения участников реакции к активным центрам на мембранах. Ьолес того, низкая температура оказывает сложное косвенное воздей-пнис: например, нарушаются покой семян и ’’выход в стрелку” или происходит преждевременное цветение, которое вызывается главным < iti pat ом изменениями в концентрации гормональных регуляторов рос-tti )го должно приводить к активизации или инактивации генов — ре-i у шпоров синтеза специфических белков-ферментов. Водный стресс юрмозит рост клеток и открывание устьиц непосредственно из-за сниже-nmi i ургора клеток, но он также оказывает и важное косвенное влияние на ферментативно опосредствованные процессы, такие, как синтез бел-
12
Глава
1
ка. Свет прямо воздействует на фотосинтез, но он влияет и косвенно на рост и цветение (фотоморфогенез) через генную регуляцию фермен-тативно-регулирующего синтеза ростовых гормонов. Минеральные вещества, являясь составными частями важных клеточных компонентов, \ оказывают прямое действие, а также и косвенное - в качестве кофер- I ментов. ?
ПРОЦЕССЫ, КОНТРОЛИРУЮЩИЕ РАЗЛИЧНЫЕ СТАДИИ РОСТА.
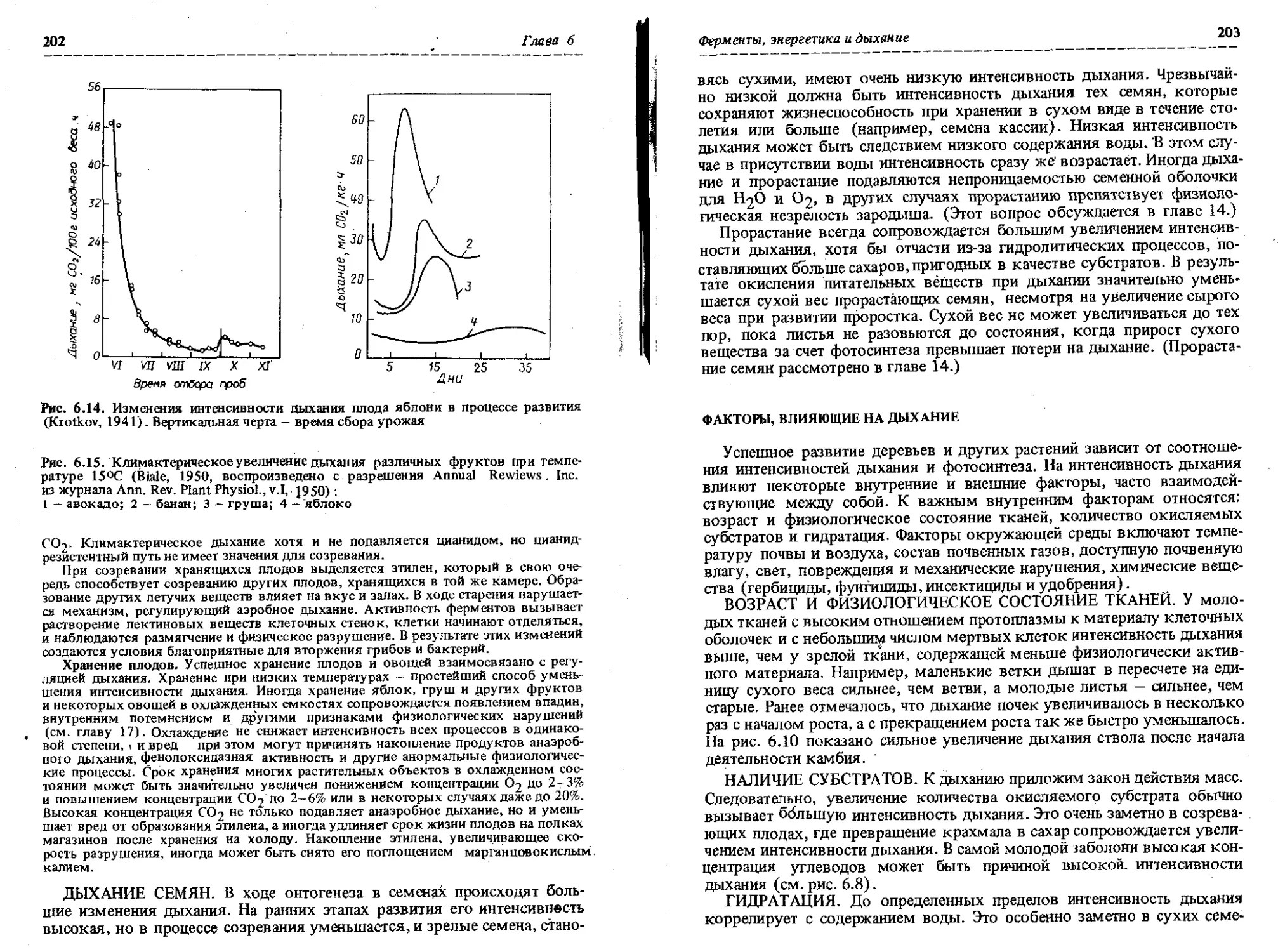
Еще во времена Сакса было установлено, что не все физиологические процессы одинаково важны на разных стадиях роста. Например, в насаждениях сосны условия бывают настолько благоприятными для прорастания семян, что появляются бесчисленные всходы, но эти же условия могут быть неблагоприятны для их укоренения в почве, и они через год-два
1.1. ОСНОВНЫЕ ПРОЦЕССЫ И ФАКТОРЫ ОКРУЖАЮЩЕЙ СРЕДЫ НА РАЗНЫХ СТАДИЯХ РОСТА
Стадия Процессы и условия Наиболее важные окружающие факторы
Прорастание Поглощение воды Температура
семян Усвоение питательных веществ Дыхание Ассимиляция Вода Кислород
Укоренение Фотосинтез Свет
проростков Ассимиляция Водный баланс Вода Температура Питательные вещества
Вегетативный Фотосинтез Свет
рост Дыхание Ассимиляция Передвижение Водный баланс Вода Температура Питательные вещества
Размножение Фотосинтез C/N баланс Готовность к цветению Заложение цветочных зачатков Накопление питательных веществ Свет Питательные вещества Температура
Старение Не выяснены (возможно, водные и гормональные отношения, передвижение и соотношение между фотосинтезом и дыханием) Вода Питательные вещества Насекомые и болезни
погибают. Климатические и почвенные условия в регионе могут быть благоприятны для вегетативного роста, но неблагоприятны для репродукции. Деревья некоторых видов прекрасно растут в искусственных насаждениях, но они не способны к репродукции. В табл. 1.1 указаны факторы окружающей среды и основные процессы, рассматриваемые как доминирующие на каждой стадии роста. У нас пока нет полного понимания этих процессов, и, возможно, что наши представления о роли каждого из них будут уточняться в дальнейших исследованиях.
Значение физиологии растений
13
ПРОБЛЕМЫ И ПЕРСПЕКТИВЫ
РАЗЛИЧНЫЕ ПРОБЛЕМЫ ЛЕСОВОДОВ, САДОВОДОВ И РАБОТНИКОВ САДОВО-ПАРКОВОГО СТРОИТЕЛЬСТВА. Цели садоводов и лесоводов-озеленителей при выращивании деревьев совершенно разные, соответственно их интересуют и разные физиологические проблемы. Лесоводы заботятся о получении максимального количества древесины с единицы площади. При этом они имеют дело с древостоем и вынуждены считаться с факторами, влияющими на конкурентные отношения деревьев в них. Главная цель садоводов - снять больше плодов, поэтому их усилия направлены на то, чтобы добиться цветения и плодоношения деревьев в более раннем возрасте. Из-за высокой ценности фруктовых деревьев садоводы, как и озеленители, часто встречаются с проблемами отдельного дерева, и они особенно озабочены тем, как снизить поражаемость деревьев насекомыми и грибными заболеваниями. Лесоводы-озеленители заинтересованы в том, чтобы вырастить деревья и кустарники хорошей формы независимо от почвенных и других условий среды. Поэтому у лесоводов-озеленителей часто возникают трудности вследствие плохого дренажа, недостаточной аэрации, повреждений корней во время строительства, газовых утечек, загрязнения воздуха и других неблагоприятных факторов окружающей среды. Несмотря на различные цели лесоводов, озеленителей и садоводов, общей задачей для них является глубокое понимание физиологии дерева.
СОВРЕМЕННЫЕ И БУДУЩИЕ ПРОБЛЕМЫ ФИЗИОЛОГИИ. Значительные изменения в методах, применяемых в лесоводстве и садоводстве, уже сейчас создали некоторые проблемы, и насколько позволяют наши знания физиологии дерева, можно предположить, что со временем этих проблем будет еще больше. Возросшая эксплуатация тропических лесов создала проблемы, с которыми не сталкиваются в умеренных зонах, а увеличивающийся интерес к древесине высокого качества требует лучшего понимания факторов, влияющих на плотность древесины и другие ее свойства. Уменьшение возраста рубки вызывает необходимость добиваться ускоренного роста деревьев и, вероятно, улучшения минерального питания при условии более полной утилизации всей массы дерева (см. главу 10). Большинство приемов, используемых в лесоводстве и садоводстве (прореживание, обрезка, удобрение и др.), только тогда бывают эффективными, когда положительно действуют на физиологические функции деревьев.
Некоторые экономически выгодные приемы могут нежелательно воздействовать на физиологические процессы. Например, предполагали, что если листопадные плодовые деревья в центральной долине Калифорнии обработать осенью дефолиантами, то обрезку весной можно будет начинать раньше. Однако в конце вегетационного периода фотосинтез вносит существенный вклад в резервы углеводов, поэтому преждевременное опадение листьев нежелательно. Часто бывает целесообразным выкопать сеянцы из питомника и хранить их упакованными до момента иосадки, но это нежелательно, из-за истощения в них запасов углеводов но время хранения. Также не обязательно, что самые крупные саженцы будут успешнее переносить пересадку. В связи с этим необходимы бо
14
Глава
1
лее полные знания о том, как создавать физиологически полноценные саженцы. Для этого требуется дополнительная информация о накоплении питательных веществ, о сезонных колебаниях корнеобразующей способности саженцев. Возрастающая потребность разведения саженцев в контейнерах повышает интерес к их физиологии. Программы селекции и улучшения деревьев создают необходимость методов, индуцирующих более раннее зацветение, большее образование семян и более успешное укоренение черенков (см. главу 4). Это создает противоречивые требования, так как для цветения и образования семян желательна более ранняя физиологическая зрелость, а более эффективное укоренение черенков происходит в молодом, незрелом состоянии. Настало время интенсифицировать исследования по получению желаемых генотипов с применением методов клеточной и тканевой культуры.
Садоводы достигли больших успехов, чем лесоводы, в познании физиологии деревьев, особенно в области минерального питания. Однако и у них есть свои проблемы, например сокращение времени, необходимого для перехода плодовых деревьев в стадию плодоношения, устранение двухгодичного цикла плодоношения у некоторых сортов и уменьшение интенсивного опадения плодов. Старая проблема, которая становится более серьезной в связи с тем, что неосвоенных земель остается все меньше, - это изыскание возможностей пересадки старых плодовых деревьев, проблема ’’пересадки”. Это важно и для лесоводов при использовании более коротких периодов оборота рубки (см. главу 17). В связи с возрастающим интересом к карликовым деревьям, часто используемым в качестве живой изгороди, возникает необходимость снизить затраты труда на .обрезку, опрыскивание и ощипку плодов. Этот новый прием вызвал большой интерес к корневым подвоям и, вероятно, создаст новые физиологические проблемы.
Лесоводы-озеленители также заинтересованы в маленьких, компактных деревьях для небольших городских участков земли. Как озеленители, так и садоводы сталкиваются с проблемой возраста растений вследствие короткой жизни некоторых ценных фруктовых и декоративных деревьев. К сожалению, практически ничего неизвестно о биохимических и физиологических основах, почему, например, деревья сосны остистой или секвойи живут до 3000—4000 лет, а деревья персика и некоторых других видов всего лишь несколько десятилетий.
Следует также обсудить малые, но измеримые изменения таких факторов окружающей среды, как концентрация СО2 в атмосфере и температура. Концентрация СС>2 постоянно возрастает и к 2000 г. она может достигнуть 0,04%, что повлечет увеличение температуры — это так называемый оранжерейный эффект. С другой стороны, температура в Северном полушарии в течение нескольких десятилетий понижается, но как она будет отклоняться в будущем, трудно предсказать. Было бы желательно знать, в какой мере увеличение содержания СО2 в воздухе и отклонение температуры в ту или иную сторону будет отражаться на физиологических процессах деревьев. Разные виды могут реагировать по-разному, и знание этого было бы полезным при планировании будущих посадок, особенно там, где температура уже сейчас является лимитирующим фактором. Загрязнение атмосферы также будет продолжаться и станет серьезной проблемой в некоторых областях. В связи
Значение физиологии растений
15
с этим необходимо знать причины большего повреждения одних видов по сравнению с другими. Некогда деревья были основным источником топлива, и до сих пор они остаются важным источником химического сырья. Заботы об источниках энергии в будущем — причина возросшего интереса к лесам как возможным существенным источникам топлива и химического сырья, получаемым в настоящее время из угля и нефти. Согласно данным Смита и Джонсона (1977) леса более продуктивны и экономически выгодны по сравнению с годовым урожаем технических культур, если судить по выходу получаемой энергии на единицу затраченной энергии. Так как затраты на посадку и прочистку значительно превосходят стоимость получаемого древесного сырья с единицы площади, то удлинение периода рубок могло бы увеличить эффективность затрат. Возможно, следует пересмотреть применяемые в настоящее время приемы выращивания деревьев, отказаться от краткосрочных оборотов рубок и даже пересмотреть, какой тип деревьев наиболее целесообразен.
БОЛЕЕ ЦЕЛЕСООБРАЗНОЕ ИСПОЛЬЗОВАНИЕ ФИЗИОЛОГИИ. Лесоводы и садоводы иногда сетуют на то, что физиологические исследования в значительной степени объясняют давно известные свойства растений и мало содействуют улучшению образования древесины и плодов. Действительно, данные физиологии скорее объясняют, чем прогнозируют. Например, исследования физиологии корней дали объяснение, почему холодные или переувлажненные почвы повреждают корни и снижают поглощение воды и минеральных веществ. Изучение водного режима растений способствовало пониманию* причин снижения роста во время засухи, а исследования холодоустойчивости показали, почему одни виды лучше, чем другие, выживают при более низких температурах. Результаты физиологических исследований следует шире использовать в работах по выведению и отбору растений, наиболее стойко переносящих различные стрессовые условия. Для этого необходимо определить, какого рода стрессы являются наиболее лимитирующими в различных ситуациях и в результате воздействия на какие физиологические процессы подавляется рост и снижается урожай растений. Селекция обычно направлена на повышение холодо- и засухоустойчивости, а также на увеличение выхода древесины, измеряемого величиной диаметра и высотой растений, или урожая плодов. Многие физиологические процессы регулируют урожайность. Поэтому физиологи должны выяснить, какие именно из них лимитируют получение урожая и к каким факторам внешней среды они более чувствительны. Может быть, это водный стресс? Если так, то нужно знать, влияет ли он на движение устьиц, снижая поглощение СО2, а следовательно, и на скорость фотосинтеза, или только частично уменьшает площадь листа, или повреждает хлоропласты? Можно ли избежать действия водного стресса путем селекции растений с более глубокой корневой системой, с лучшим устьичным и кутикулярным контролем транспирации, или каких-то других свойств растений? После того, как физиологи определят процессы, которые лимитируют рост в определенных окружающих условиях, растениеводы смогут работать более направленно, создавая растения устойчивые в этих условиях.
Медленный прогресс в разрешении проблем ускорения роста деревьев происходит не столько оттого, что нет информации, сколько из-за неудач в использовании имеющейся информации. Это обусловлено отчасти недостатком творческого подхода и частично недостатком обмена идеями между работниками полей и лабораторий. Успешное разрешение проблем лесоводства и садоводства требует совместных исследований ученых разного профиля.
Перспективы для плодотворных исследований физиологии деревьев в настоящее время вполне реальны благодаря лучшему пониманию роли физиологии, усовершенствованию оборудования и техники. К важным технологическим усовершенствованиям относятся хроматография, применение приборов-самописцев, методы исследования с мечеными изотопами, инфракрасные газовые анализаторы, диффузионные порометры и методы измерения водного потенциала. Использование камер со строго контролируемыми условиями для анализа действия факторов среды, использование компьютеров для моделирования роста и прогнозирования действия окружающих факторов, а также применяемых в лесоводстве способов обработки — особенно важные усовершенствования для проведения физиологических исследований.
СТРОЕНИЕ
Для понимания физиологических процессов знание различных форм и строения древесных растений не менее существенно, чем знание химии. Например, свойства кроны воздействуют на многие физиологические процессы, которые в свою очередь влияют на различные ростовые процессы, включая рост ствола, апикальное доминирование, камбиальный рост, рост корня. Особенности кроны играют роль и и конкурентных взаимоотношениях между древесными растениями.
Необходимо знать строение листа, чтобы понимать, каким образом окружающие факторы влияют на фотосинтез и транспирацию. Сведения о стволе дают возможность понять, каким образом происходят ток воды и передвижение питательных веществ, а также камбиальный рост, Изучение строения корня важно для понимания механизма поглощения воды и солей. На любой физиологический процесс в той или иной степени влияет строение тканей или органов, в которых он происходит, поэтому знание анатомии важно для понимания ростовых процессов древесных растений.
Можно считать, что дерево состоит из шести частей. Три части - листья, ствол, корни вегетативные структуры; цветы, плоды и семена - репродуктивные ор-110(1.1, Каждая из этих частей состоит из большого количества тканей. Особенно важные из них - ксилема и флоэма, так как они образуют проводящую систему длн воды, солей и питательных веществ от кончиков очень глубоко расположенных корней до листьев вершины кроны.
лисг
Листья древесных растений играют первостепенную роль в фотосинтезе, с помощью которого энергия поступает в нашу биосферу. Знание структуры листа необходимо для понимания процессов фотосинтеза и транспирации. В процессе транспирации теряется вода из листьев. Для осуществления фотосинтеза, необходимого для поддержания интен-сииного роста, требуется большая площадь листьев.
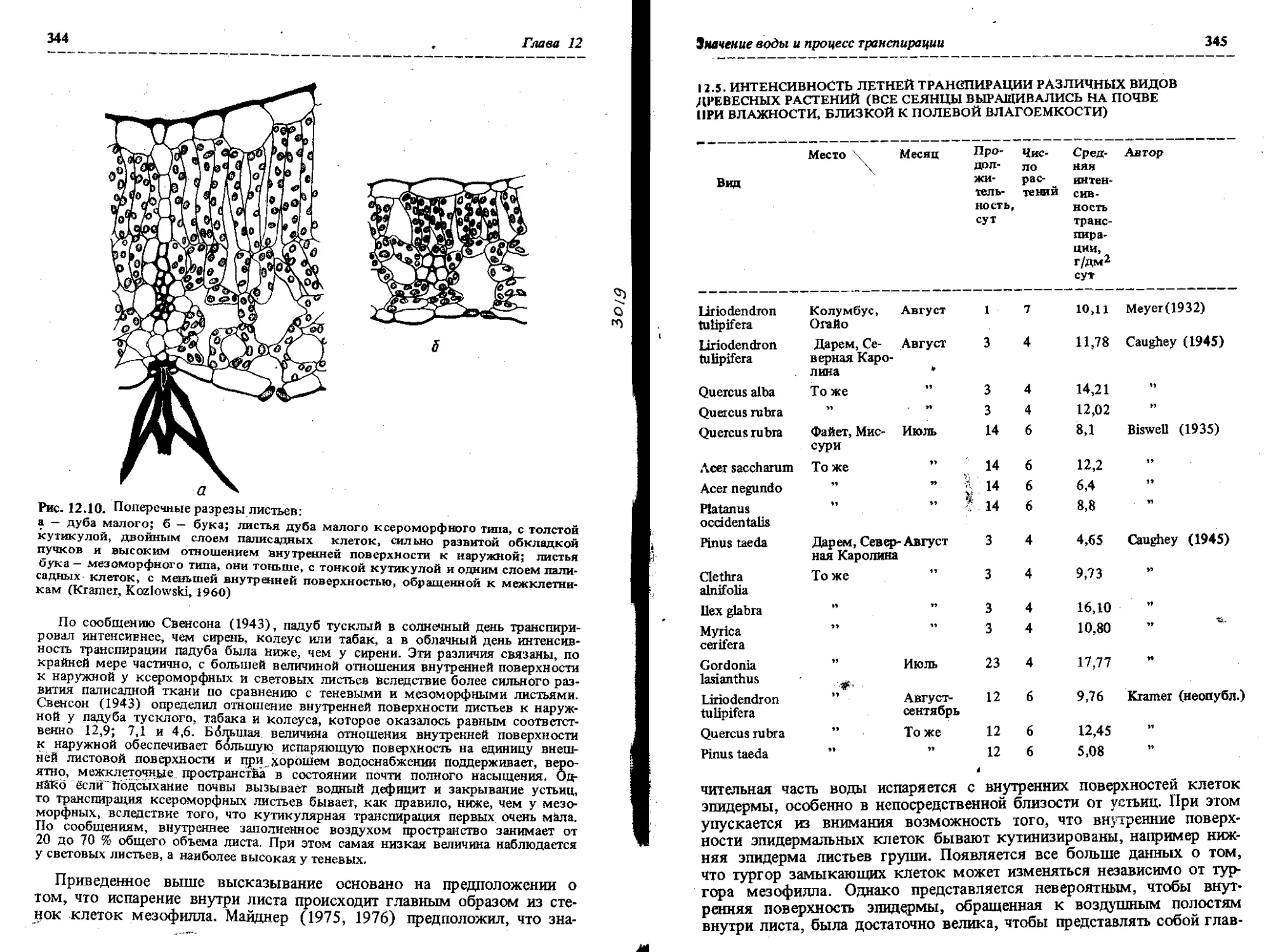
СТРОЕНИЕ ЛИСТА ПОКРЫТОСЕМЕННЫХ. Типичный лист лиственных покрытосеменных состоит преимущественно из первичных тканей. Пластинка листа обычно широкая и плоская, поддерживается черешком, содержит основную ткань, или мезофилл, окруженный сверху и снизу эпидермой. Мезофилл состоит из различного количества палисадной ткани и губчатой паренхимы. Столбчатые, правильной формы палисадные клетки располагаются под верхней эпидермой (а иногда над нижней эпидермой) и легко отличаются от неправильной формы клеток губчатой паренхимы, примыкающих к нижней эпидерме (рис. • 1) Палисадная ткань может состоять из одного слоя клеток, расположенных перпендикулярно верхней эпидерме, или из трех слоев (см. рис 12.10). В паренхиме, содержащей более одного слоя клеток, клетки гйм<н() наружного слоя наиболее удлиненные, а клетки слоя, расположенного ближе к центральной части листа, по форме и размерам могут Пыи. сходными с клетками губчатой паренхимы. При достаточно от-чнлииом различии между клетками столбчатой и губчатой паренхимы можно увидеть, что большая часть хлоропластов локализована в палисадной гкани.
Питательные вещества, вода и минеральные соли перемещаются и нистьях по жилкам, густо пронизывающим мезофилл (рис. 2.2). Жил-
18
Глава 2
Рис. 2.1. Поперечный срез части пластинки листа лиственного дерева: 1 — кутикула; 2 — верхняя эпидерма; 3 — склеренхима; 4 — клетки палисадной ткани; S — ксилема; 6 — клетки губчатого мезофилла; 7 — флоэма; 8 — склеренхима; 9 — замыкающая клетка
Рис. 2.2. Сетчатое жилкование листа березы грабо-листной (Фото D.H. Franck)
кование может быть сетчатое, как у двудольных, или параллельное, как у однодольных. При параллельном жилковании жилки взаимосвязаны латерально многочисленными маленькими пучками. Сосудистые пучки главной жилки пронизывают паренхимные ткани и изолированы от мезофилла. Наоборот, малые жилки обычно расположены в верхней части губчатой паренхимы и делят мезофилл на небольшие участки, называв- ; мые ареолами. '’I
Устьица. Ткань мезофилла содержит многочисленные межклеточ-ные пространства, сообщающиеся с внешней средой многочисленными J отверстиями (устьицами) в эпидерме, ограниченными двумя специализирован- t ными замыкающими клетками (см. рис. 2.2 и 2.3). Устьица играют существенную роль в физиологии растений, так как они служат воротами, через ко- i: торые испаряется вода из листьев и поступает углекислый газ, используемый Л в фотосинтезе. У большинства листвен- [ ных деревьев устьица располагаются только на нижней стороне листа, но у '' некоторых древесных растений, напри- ; мер тополя, они расположены на обеих г, поверхностях листа. При наличии устьиц ; на обеих сторонах листа их обычно болы ше на нижней поверхности (табл. 2.1).
Особое физиологическое значение имеют специфические для вида и генотипически обусловленные размеры и частота устьиц. Например, размер устьиц (длина замыкающих клеток) варьирует у 38 видов деревьев от 17 до 56 мкм, а плотность распределения устьиц - приблизительно от 100 до 600 устьиц на 1 ^м^ поверхности листа (табл. 2.2). Обычно у видов с небольшим числом устьиц на единицу поверхности листа устьица
Строение
19
Рис. 2.3. Устьица древесных покрытосеменных (Esau, 1965):
А ' — В — устьица с прилежащими клетками листа персика, срез сделай в плоскостях, указанных на схеме Г пунктирными линиями аа, бб, вв; Д — Ж — устьица листа бересклета и плюща обыкновенного, срез в плоскости аа; Ж — замыкающая клетка устьица листа плюща обыкновенного, срез в плоскости; 1 — кутикула;
2 - замыкающие клетки; 3 — устьичная щель
крупные. Например, у кленов сахарного и серебристого очень много небольших устьиц, а у ясеня и березы пушистой количество устьиц меньше, но они крупные. Виды дуба в этом отношении - исключение, так как имеют и большие и многочисленные устьица. Размеры устьиц и плотность их распределения широко различаются у таких родов, как боярышник, ясень и дуб (1973). В табл. 2.1 приведены пяя-ные, показывающие изменчивость размеров устьиц и их распределение у видов и клонов тополя.
2.1. РАСПРЕДЕЛЕНИЕ УСТЬИЦДИТ/СМ2 НА НИЖНЕЙ
И ВЕРХНЕЙ ПОВЕРХНОСТИ ЛИСТА ВИДОВ
( Siwecki and Kozlowski, 1973)
Клон
Нижняя поверх- Верхняя поверх-
ность
поста
Pop ulus maximowiczii х Р. nigra
Populus maximowiczii
Populus trichocarpa
Populus deltoides
Populus nigra
45 351 + 1,003 4 216^155
33 521 + 868 7 730+ 242
23 378 + 581 5 013 + 193
22 628 + 408 18 693 + 575
20 450 + 434 5 762 + 313
20
Глава 2
2.2. СРЕДНЯЯ ДЛИНА И ПЛОТНОСТЬ РАСПРЕДЕЛЕНИЯ УСТЬИЦ У ДРЕВЕСНЫХ ПОКРЫТОСЕМЕННЫХ
(Davies et al., 1973)
Вид Длина устьиц, мкм Число устьиц, шт/ мм2 Вид Длина устьиц, мкм Число устьиц, шт/ мм^
Acer saccharinum 17,3 418,8 Malus sp. 23,8 219,5
Acer saccharum 19,3 463,4 Populus deltoides 30,4 163Л
Acer negundo 21,6 233,9 Prunus serotina 30,5 306,3
Betula nigra 39,4 281,3 Prunus virginiana 27,1 244,6
Betula papyrifera 33,2 172,3 Que reus rubra 26,7 532,1
Catalpa bignonioideg 23,2 328,6 Quercus palustris 24,0 575,9
Crataegus sp. 1 22,3 399,1 Quercus palustris 30,9 530,4
Crataegus sp. 11 37,4 221,4 Rhus tyhina 19.4 633,9
Fraxinus americana 24,8 257,1 Robinia pseudoacacia 17.6 282,1
Fraxinus pennsylvanica 29,3 161,6 Salix fragilis 25,5 215,2
Ginkgo biloda 56,3 102,7 Tilia americana 27,2 278,8
Gieditsia triacanthos 36,1 156,3 Ulmus americana 26,3 440^2
Hamamelis mollis 25,3 161,6 Vitis vinifera 29,7 120,5
Juglans nigrs 25,7 242,0
Воски листьев. У листьев многих покрытосеменных и голосеменных растений потеря воды при кутикулярной транспирации снижается благодаря восковым покрытиям. Кутикула — это бесклетйый слой, рас- j
положенный на эпидермальных клетках. Она часто распространяется в устьичные щели, выстилая их тонким слоем. Кутикула представляет собой слой из окисленных жирных кислот, обычно связанных слоем воска (см. главу 8). Количество поверхностного воска значительно t различается у разных видов, у некоторых оно может составлять 15% сухого веса листа. У многих видов устьичные поры закупориваются отложениями воска, что сокращает диффузию воды и углекислого газа.
СТРОЕНИЕ ЛИСТА ГОЛОСЕМЕННЫХ. Листья голосеменных, за ! исключением небольшого числа родов Larix и некоторых видов Тахо-dium, вечнозеленые. У большинства голосеменных листья линейные или ; копьевидные, двусторонне уплощенные, но встречаются листья и другой формы. Например, у Podocarpus, ели и лиственницы листья обычно в поперечном сечении тетрагональные. Чешуеподобные листья характерны для Sequoia, Cupressus, Chamaecyparis, Thuja Libocedrus. Широкие овальные и плоские листья характерны для араукарии. У Abies, Pseu-dotsuga, Dacrydium, Sequoia, Taxus, Torreya, Ginkgo, Araucaria, Podocarpus мезофилл листа дифференцирован на палисадные клетки и губчатую паренхиму (рис. 2.4). У листьев двух последних родов палисадная паренхима расположена с двух сторон. У сосны мезофилл не дифференцирован на палисадные клетки и губчатую паренхиму (рис. 2.5).
Хвоя сосны собрана в полусферические (двухвойные виды) или треугольные (треххвойные виды) пучки. Хвоя однохвойной сосны съедобной на поперечном срезе круглая. Иногда в определенных уело-
Строение
21
Рис. 2.4. Поперечный срез вторичной хвоинки сосны
веймутовой: 1 - смоляной
ход; 2 - трансфузионная ткань; 3 — эндодерма; 4 флоэма; 5 - ксилема; 6 - замыкающая клетка;
7 - устьице; 8 -хлоренхима; 9 -кутикула; 10 -эпидерма; 11 -гиподермальная склеренхима
Рис. 2.5. Устьица голосеменных (Есан, 1965):
А - изображение сверху поверхности эпидермы с погруженным устьицем I’inus merkusisi; Б - Г - устьица сосен; Д и Е - устьица Sequoia; пунктирные линии на схеме А показывают положение в плане срезов, представленных на схемах Б-Е: аа - на Б, Д; бб - на Г; вв - на В, Е; 1 -побочные клетки 2 замыкающие клетки; 3 - эластичные участки клеточных стенок
22
Глава 2
Рис. 2.6. Диаграмма медианного продольного среза дерева, показывающая годичные приросты ксилемы (1-2) в стволе и главных ветвях: а — годичное кольцо; б — кора
виях количество хвои в пучке варьирует. Это часто является реакцией на необычные условия питания, на повреждение или при неправильном развитии, У сосны устьица глубоко погружены в ткань и располагаются рядами (рис. 2.6). Ниже эпидермы и мезофилла имеется толстостенный гиподермаль-ный слой. Паренхимные клетки мезофилла глубоко погружены в ткань. Один или два проводящих пучка в хвое окружены трансфузионной тканью, состоящей из мертвых клеток — трахеид и живых паренхимных клеток. У хвои сосны имеются два или несколько смоляных канальцев. Клетки эндодермы, окружающие трансфузионную ткань, толстостенные. Эпидерма хвои сосны покрыта плотным слоем кутикулы и в подустьичной камере часто обнаруживается значительное количество воска.
СТВОЛ
Ствол древесных растений поддерживает крону, проводит воду и минеральные вещества вверх от корня, передает питательные вещества и гормоны от места их синтеза к участкам, где они используются в ростовых процессах или откладываются в запас.
На рис. 2.7 и рис. 2.8 изображен развитый ствол средней части дерева, обычно состоящий из конусообразных колонок древесины (ксилемы). Эти колонки древе-
сины слагаются из нескольких слоев, или годичных приростов, расположенных друг" над другом, подобно серии нанизывающихся колпачков, покрытых сверху корой. На верхушке ствола и на каждой ветке имеется терминальная точка роста, определяющая увеличение ствола в длину. Между корой и древесиной ствола, ветвей и главного корня располагаются камбий проводящих тканей (далее будет называться просто камбий) и тонкий слой покровной латеральной меристемы (фелоргена).
ЗАБОЛОНЬ И ЯДРО. Молодая древесина (ксилема), или заболонь, проводит пасоку (главным образом воду), укрепляет ствол и служит в некоторой степени для запасания питательных веществ. В живых паренхимных клетках заболони находятся запасные питательные вещества. Заболонь состоит из поперечно ориентированных лучевых клеток, а у многих видов древесных растений и из вертикально ориентированных осевых паренхимных клеток. В среднем лишь 10% клеток заболони живые. С возрастом все живые клетки ксилемы отмирают. Стенки клеток темнеют, образуя центральный цилиндр из темноокра-
1
2
J ’
гнс. 2.7. Схематическое строение ствола дерева, показывающее расположение основных тканей:
1 - наружная кора; 2 - внутренняя кора; 3 — камбий; 4 — заболонь; 5 — ядро
(Фото St. Regis Paper Со.) 1
Рис. 2.8. Изменения в диаметре и распределении сосудов в годичных кольцах прироста:
а - рассеянососудистого вида — клен серебристый; б — кольцесосудистого вида — дуб белый (х50) (Фото Лесной службы США)
24
Глава 2
шейной мертвой ткани, называемый ядром. Ядро обеспечивает механическую прочность. В физиологических процессах оно участвует недолго, что подтверждается примером старых деревьев, долгие годы живущих с разрушенных ядром и поддерживаемых тонким слоем заболони. При образовании ядровой древесины в клетках откладываются разнообразные соединения, включая масла, смолы, камеди и танины (см. главу 16). У деревьев с темным ядром содержится много фенольных соединений, высокотоксичных для грибов. К таким соединениям относятся терпеноиды, трополены, флавоноиды и стильбены. Ядро увеличивается по диаметру в течение всей жизни дерева. Ширина слоя заболони сильно различается у разных видов. В стволе молодых деревьев она широка, у старых деревьев узкая.
ПРИРОСТ КСИЛЕМЫ И ГОДИЧНЫЕ КОЛЬЦА. У деревьев умеренной зоны годичные кольца древесины (ксилемы) четко выделяются по всему сечению ствола и ветвей, так как формирующаяся весной ксилема состоит из клеток большего диаметра, чем ксилема, появляющаяся позднее. Из-за однородности строения ксилемы голосеменных изменение толщины клеточных стенок зависит от изменений плотности древесины. У покрытосеменных плотность древесины зависит не только от толщины клеточных стенок, но и от соотношения различного типа клеток. Это соотношение относительно постоянно в пределах видов и даже в пределах многих родов, хотя оно и изменяется в течение сезона. Расположение и соотношение различного типа клеток существенно варьирует у разных родов покрытосеменных. Вследствие последовательного периклинального (тангентального) деления камбиальных клеток (см. главу 3) молодые, недифференцированные клетки ксилемы и флоэмы организованы в виде радиальных рядов. У голосеменных такое радиальное устройство сохраняется при дифференциации трахеид. Наоборот, у покрытосеменных первоначальное радиальное расположение в ряд элементов ксилемы, образующихся в результате деления камбия, нарушается, так как клетки проводящей ткани могут сильно разрастаться и изменять положение сердцевинных лучей и прилегающих к ним клеток. Вследствие этого не всегда узкий луч огибает крупный сосуд. На достаточно широкие лучи разрастание элементов сосудов не влияет.
2.3. ШИРИНА ЗАБОЛОНИ У ПОКРЫТОСЕМЕННЫХ (Sargent, 1926)
Род,вид Число Род, вид Число годичных годичных колец в колец в заболони, заболони, шт. шт.
Catalpa speciosa Robinia pseudoacacia Juglans cinerea Maclura pomifera Sassafras officinale 1-2 Aesculus glabra 10-12 2-3 Juglans nigra 10-20 5-6 Prunus serotina 10-12 5-10 Gleditsia triacanthos 10-12 7-8
Строение
25
Рис. 219. Переход ранней древесины к поздней у 10-летнего дерева сосны ладанной: а — в верхней; б — нижней части ствола; в нижней части ствола вследствие большего диаметра клеток ранней древесины и более толстостенных клеток поздней древесины переход более резкий, по сравнению с переходом в верхней части (Фото Лесной службы США)
У покрытосеменных имеются два типа расположения просветов сосудов: кольцевое и диффузное (рис. 2.9). У деревьев с кольцевым расположением сосудов (дуба, ясеня, ильма) диаметр сосудов, образованных в начале сезона, значительно крупнее, чем сформированных позднее, У рассеянососудистых деревьев (тополя, клена, березы) диаметр сосудов небольшой, у рано- и позднообразовэнных сосудов он примерно одинаков.
Наружные слои годичных приростов ксилемы могут существенно различаться. В областях с большим количествам осадков и холодной зимой пограничные слои между годичными приростами ксилемы лучше различимы, чем у видов, произрастающих в жарких, сухих районах. В молодой части ствола нормального дерева переход от одного годичного кольца к другому около сердцевины постепенный. В более поздно образованной древесине такой переход более четкий. У старых деревьев граница между приростами ксилемы очень отчетлива. Ширина годичных колец часто уменьшается вследствие засухи. Этот признак часто используют при изучении климатических условий прошлого и даже для того, чтобы датировать древние строения, создаваемые человеком.
Ранняя и поздняя древесина. Древесина малой плотности, формирующаяся обычно (но не всегда) в ранний период вегетации, называется ранней древесиной. Часть годичного прироста ксилемы, которая образуется обычно в поздний период вегетации и плотнее ранней древесины, называется поздней древесиной. От соотношения ранней и поздней древесины зависит качество древесины (см. главу 16).
Термины ранняя древесина и поздняя древесина в литературе используются как синонимы весенней древесины и летней древесины. Это не совсем верно, так как оба типа древесины могут формироваться в разное время вегетационного периода. Чок (1937) считает, что следует отказаться от терминов весенняя древесина и летняя древесина. Однако,
26
Глава 2
несмотря на неточность, эти термины широко используются до сих пор. В литературе имеются возражения против употребления терминов ранняя древесина и поздняя древесина, так как поздняя древесина иногда обнаруживается в начальном периоде роста или лишь в виде фрагментов в годичных приростах. Противники этих терминов предпочитают использовать названия легкая древесина и плотная древесина, акцентируя внимание на структуру ткани, а не на время ее формирования или относительное распределение в годичных слоях прироста.
Граница между ранней и поздней древесиной одного и того же кольца может быть хорошо выраженной, резкой или нечеткой. У твердых сосен, дугласии, лиственницы и можжевельника переход от ранней древесины к поздней четкий. У кольцесосудистых покрытосеменных обнаружен резкий переход от ранней древесины к поздней, у рассеянососудистых видов — постепенный переход. Предлагались различные методы для четкого различия ранней и поздней древесины. Наиболее часто используется метод Морка (1928), согласно которому удвоенная ширина общих стенок между двумя соседними трахеидами поздней древесины должна быть равна ширине просвета или больше его. Если эта ширина меньше ширины просвета, ксилема считается ранней. Все измерения были сделаны в радиальном направлении. Первоначально метод Морка применили для оценки ксилемы ели, но затем его стали использовать и для древесины других голосеменных. По утверждению Доли (1974), это определение оказалось неприемлемым для древесины покрытосемен-
Рис 2 10. Сезонные изменении в формировании ранней древесины, переходнойк Дрезине и поздней древесины на разной высоте ствола сосны смоли-растяжение побега; 2 - активный рост; 3 - замедление роста; 4 - прекращение роста; 5 - удлинение хвоинок; 6 - начало роста; 7 - замедление ррста;
8 - прекращение роста; 9 - ратин древесина; 10 - переход к поздней древесине; 11 - itomiHHH древесина; 12 - начало сезона; 13 - середина сезона, 14 ко нац сезона
Строение
27
Рис- 2.11. Строение древесины голосеменных:
1 - поперечный срез; 2 - радиальный срез; 3 — тангентальный срез; 4 — трахеи* ды; S — срединная пластинка; 6 — ранняя древесина; 7 - поздняя древесина;
8 — годичное кольцо; 9 — луч в древесине; 10 — лучевая трахеида; 11 — веретеновидный луч; 12 — простая пора; 13 — окаймленная пора; 14 — горизонтальный смоляной ход; 15 — вертикальный смоляной ход (Фото Лесной службы США)
ных, у которых часто трудно различить раннюю и позднюю древесину.
В пределах годичного кольца ширина ранней древесины обычно уменьшается, а ширина поздней древесины увеличивается к основанию дерева. У голосеменных трахеиды ранней древесины в пределах одного и того же слоя прироста шире у основания ствола, чем на вершине дерева. Граница между трахеидами ранней древесины, сформированными в конце раннего периода вегетации, и первыми трахеидами поздней древесины также более четкая в нижней части ствола, чем в верхней. Это зависит отчасти от большего диаметра клеток ранней древесины и более толстых стенок клеток поздней древесины в нижней части ствола. Более четкое разделение между слоями поздней и ранней древесины обусловлено также очень толстыми стенками клеток поздней древесины в нижней части ствола. Узкий диаметр клеток ранней древесины верхней части ствола определяет менее четкое, чем в нижней его части, разделение между поздней и ранней древесиной (рис. 2.10).
28
Глава 2
Некоторые трахеиды могут быть отнесены к поздней древесине вследствие уменьшения их радиального диаметра без соответствующего изменения толщины стенок. Другие трахеиды, однако, относятся к поздней древесине из-за увеличения толщины стенок, но без изменения в диаметре. Размеры тех и других трахеид изменяются от вершины к основанию ствола на всем протяжении формирования поздней древесины. В верхней части ствола часто формируется переходная поздняя древесина, которую внешне невозможно уверенно отнести ни к ранней, ни к поздней древесине (рис. 2.11).
Кольца прироста у тропических деревьев. Различный камбиальный рост тропических древесных растений в значительной степени определяется наследственностью. У многих видов прирост ксилемы может продолжаться круглый год. Поэтому у ряда тропических деревьев, особенно у рассеянососудистых видов, годичные кольца отсутствуют или выражены очень слабо. У других видов, наоборот, образуется в год более одного кольца прироста. В субтропической южной части Флориды некоторые вечнозеленые виды, такие, как Swietenia mahagoni,имеют кольца прироста, у других видов, сбрасывающих листья, таких, как Ficus spp., колец прироста нет. Возможна взаимозависимость образования годичных колец с ростом побегов. Например, у молодого деревца фикуса рост побегов происходит периодически, с интервалами 42 дня, появление каждого побега связано с приростом ксилемы и образованием колец прироста. У некоторых тропических сосен отмечается более пяти вспы- ; шек образования новых побегов в год, но при этом формируется только одно кольцо. Специфические анатомические признаки, характерные для колец прироста, сильно варьируют у разных видов тропических древесных растений. У Acacia catechu, например, кольца прироста окружены тонким слоем терминальной паренхимы, иногда толстостенными волокнами самой внешней поздней древесины. Кольца прироста Bomb ах • malabaricum характеризуются радиально сжатыми волокнами и паренхимными клетками во внешней поздней древесине. Древесина Shorea г robusta содержит большое количество неправильной формы полос паренхимы, которые иногда ошибочно принимают за годичные кольца. Они не всегда образуют в древесине кольца и формируются в течение ' года неоднократно, но если и образуют по одному кольцу в год, то это происходит не в определенные месяцы. ‘
ПРИРОСТ ФЛОЭМЫ. Годичный слой зрелой, флоэмы значительно тоньше колец прироста ксилемы. Это объясняется тем, что флоэма хотя и формируется ежегодно, но менее интенсивно, чем ксилема. Увеличение слоя флоэмы ограничено частым разрушением ее старых тканей и отторжением отшелушивающейся наружной нефункционирующей части флоэмы. У многих древесных растений флоэма имеет некоторые отличительные особенности, по которым можно выделить новый прирост. Однако это не всегда заметно так четко, как при формировании годичных приростов ксилемы. Часто структурные различия между ран- ; ней и поздней флоэмой становятся неразличимыми вследствие сплющивания ситовидных трубок и разрастания паренхимных клеток.
У некоторых видов годичный прирост флоэмы различим, так как клетки ранней флоэмы увеличиваются в объеме больше клеток поздней флоэмы. У груши тангентальные тяжи лубяных волокон и клеток.
Строение 29
содержащих кристаллы, разграничивают годичные приросты флоэмы. Прирост ранней и поздней флоэмы иногда можно отличить по некоторым характерным особенностям паренхимы флоэмы. Например, паренхимные клетки флоэмы, образовавшиеся в ранний период вегетации, содержат мало танина, к тому же они бывают сплющенными, если прекратилось функционирование флоэмы (глава 11). Напротив, окрашенные танином паренхимные клетки поздней флоэмы становятся тургорес-центными, и по этому признаку годичные приросты легко различимы. У некоторых видов годичные приросты флоэмы могут быть идентифицированы по числу хорошо заметных различных типов клеток.
Наиболее трудно определить годичные приросты вторичной флоэмы у голосеменных. Диаметр ситовидных клеток ранней и поздней флоэмы хотя и различен, но они часто сплющиваются под давлением разрастающихся клеток паренхимы. У кипарисовиков (Chamaecyparis) волокна годичного прироста, сформированные в начале года,имеют более толстые стенки, чем образующиеся позднее. Согласно данным Губера (1939) и Голдхейда (1951) ранняя флоэма у растений семейства сосновых состоит почти целиком из ситовидных элементов. При сдавливании ситовидные клетки образуют темные тяжи, которые четко ограничивают годичный прирост. Используя такой показатель, Сривастава (1963) пытался идентифицировать годичные приросты флоэмы у разных видов голосеменных. Полученные результаты оказались не однозначными. У некоторых видов, таких, как сосна Жеффрея, ель голубая, ель европейская и лиственница европейская, границы между годичными приростами были четкими. У многих других видов различить годичные приросты было трудно из-за того, что паренхимные клетки флоэмы были расположены диффузно, или за счет того, что четкие тяжи разрушенных ситовидных элементов становились незаметными между последовательно расположенными тяжами паренхимы флоэмы.
Проводящая и непроводящая флоэма. Слой флоэмы, состоящий из проводящих ситовидных трубок, очень узок. Например, слой проводящей системы ясеня равен 0,2 мм; дуба, бука, клена и березы 0,2— 0,3 мм, грецкого ореха и ильма 0,4—0,7 мм, ивы и тополя от 0,8 до 1 мм. Вследствие структурных нарушений элементов непроводящей флоэмы проводящая зона очень узка, и ее легко узнать по характерным признакам. К таким признакам относятся: форма различных элементов флоэмы, наличие утолщенных оболочек с перламутровым блеском, структура ситовидных пластинок и варьирование клеток паренхимы. После того, как ситовидные‘элементы прекращают функционировать, в флоэме происходят существенные изменения, включающие интенсивную склерификацию, отложение кристаллов, сдавливание ситовидных элементов и расширение тканей флоэмы вследствие растяжения и деления клеток осевой и лучевой паренхимы. Степень этих изменений у разных видов различна. Сезонные изменения флоэмы обсуждаются в главе 12 в связи с ее ролью в передвижении веществ.
СТРОЕНИЕ ДРЕВЕСИНЫ ГОЛОСЕМЕННЫХ
У большинства голосеменных продольные элементы ксилемы ствола состоят преимущественно из трахеид и небольшой части паренхимных
30
Глава 2
Рис. 2.12. Различия в переходе от ранней к поздней древесине у голосеменных: а — постепенный переход у сосны Ламберта; б -• резкий переход у сосны австралийской (х27.5) (Фото Леской службы США)
и эпителиальных клеток (рис. 2.12). Осевые (тяжевые) паренхимные клетки содержатся в ксилеме секвойи и кипарисовика, но они не встречаются в ксилеме сосны. Небольшое количество поперечно ориентированных элементов представлено лучевыми трахеидами, клетками лучевой паренхимы и эпителиальными клетками. Имеются также продольно и поперечно ориентированные смоляные ходы, которые являются скорее межклетными пространствами посткамбиального развития, чем клеточными элементами. Смола выделяется в эти канальца примыкающими к ним эгаггелиальными клетками. Смоляные ходы являются нормальным образованием у сосны, ели, лиственницы и дугласии. Кроме того, бывают смоляные ходы травматического происхождения, которые иногда можно обнаружить вместе с нормальными смоляными каналами. В некоторых случаях смоляные ходы травматического происхождения возникают у видов, не имеющих в нормальном состоянии смоляных ходов, например у кедра, тсуги и пихты (коммерческое получение смолы рассматривается в главе 8).
ПРОДОЛЬНЫЕ ЭЛЕМЕНТЫ. Более 90% древесины голосеменных состоит из вертикально ориентированных, примыкающих друг к другу трахеид, расположенных обычно ровными радиальными рядами. Эти клетки в поперечном сечении четырех- или шестигранные, длина их примерно в 100 раз превышает ширину. Длина трахеид от 3 до 7 мм, у большинства видов голосеменных умеренной зоны она равна в среднем 3—5 мм. Трахеиды, сформированные в ранний период вегетации, больше в поперечном сечении, стенки их тоньше, чем у трахеид, образовавшихся в конце вегетационного периода. Переход от крупных клеток ранней древесины к мелким поздней древесины может быть пос-
Строение
31
тененным, как у сосны Ламберта, или резким:, как у сосен ладанной и болотной (рис. 2.13).
Стенки трахеид снабжены разного типа порами, облегчающими передвижение веществ из одной клетки в другую (рис. 2.14). Между соседними трахеидами развиваются крупные окаймленные поры. Между трахеидами и лучевыми трахеидами образуются окаймленные поры меньшего размера. Полу окаймленные поры формируются между трахеидами и клетками лучевой паренхимы. Поры на стенках трахеид образуются главным образом на радиальных поверхностях и концентрируются ближе к концам трахеид. Пары окаймленных пор имею* общую мембрану (пленку поры), состоящую из двух первичных стенок
Рис. 2.13. Пора древесины (Waidiop, 1962):
а — голосеменных; б — покрытосеменных; 1 —
первичная оболочка; 2 — срединная пластинка;
31 — наружный слой вторичной оболочки; З2 — средний слой вторичной оболочки; З3 — внутренний слой вторичной оболочки; 4 — вторичная of. клочка; 5 — мембрана поры; 6 - торус; 7 -инициальное пограничное утолщение
Рис. 2.14. Трахеиды ранней и поздней древесины сосны:
1 — мратряуриттмые окаймленные поры; 2 — окаймленные поры лучевых трахейд;
3 — пол у окаймленные поры на стенках, смежных с лучевой паренхимой (Фото Лесной службы США)
32
Глава 2
и межклетной пластинки. У такой пары пор вторичная стенка каждой из соседних клеток нависает над камерой поры. У многих голосеменных пленка поры имеет выпуклое утолщение, называемое торусом, который окружен тонким участком мембраны — марго (см. рис. 2.14). Строение пленки поры различно. Например, у трахеид ранней древесины сосны todvc плоский и дискообразный, у трахеид поздней древесины он имеет форму выпуклой линзы. У Thuja, Gnetum, Cycas, Welwitshia торуса нет. Пленки окаймленных пор состоят из целлюлозных тяжей, радиально распространяющихся от торуса к краю камеры поры. При расположении торуса посредине жидкость легко проникает через отверстия в марго мембраны. Если же торус прижат к окаймленной поре или мембраны пор инкрустированы аморфными веществами, ток жидкости через поры затрудняется (см. главу 16).
Отверстия в пленках одной и той же окаймленной поры ксилемы голосеменных различны по величине — от 1 нм до нескольких нанометров. Диаметр пор у разных видов также различен. У голосеменных окаймленные поры одного и того же годичного кольца многочисленнее и крупнее в ранней древесине по сравнению с порами в поздней древесине (см. рис. 2.13). В связи с этим транспорт воды в поздней древесине значительно затруднен. У голосеменных осевая паренхима формируется длинными тяжами. Достаточно много осевой паренхимы содержится в древесине секвойи и болотного кипариса, меньше у лиственницы и дугласии, а у сосны ее нет.
ПОПЕРЕЧНЫЕ ЭЛЕМЕНТЫ. Лучи древесины содержат большое количество поперечно ориентированных элементов. Они представляют собой лентообразные скопления клеток, пронизывающих ствол подобно велосипедным спицам. Как уже упоминалось, лучи играют очень важную физиологическую роль в радиальном транспорте воды, минеральных и органических веществ. У голосеменных имеются два типа лучей: 1) узкие, шириной обычно в одну клетку (однорядные), хотя встречаются виды с двурядными лучами; 2) широкие веретеновидные, в которых присутствуют поперечные смоляные ходы. Высота узких лучей большинства голосеменных около 10—15 клеток. У некоторых видов, таких, как болотный кипарис, они могут быть высотой равной 60 клеткам.
Каждый луч у голосеменных может состоять только из клеток лучевой паренхимы, или из клеток лучевой паренхимы и лучевых трахеид, или из одних лучевых трахеид. Лучевые трахеиды образуются у ели, лиственницы и дугласии. Реже их обнаруживают у пихты, болотного кипариса, секвойи, кедра, либоцедруса и можжевельника. Формирующиеся прозенхимные лучевые трахеиды могут располагаться рядами по краям лучей или среди слоев клеток лучевой паренхимы. Клетки лучевой паренхимы тонкостенные и содержат живой протопласт, если они локализованы в части луча, находящейся в заболони. Стенки лучевых трахеид толстые и лигнифицированы. Лучевые трахеиды сосны смолистой и сосны болотной называют зубчатыми, так как их внутренние извилистые стенки образуют зубчики.
Веретеновидные лучи обнаружены у сосны, ели, дугласии и лиственницы. Они состоят из краевых лучевых трахеид, клеток лучевой
Строение
33
паренхимы и эпителиальных клеток вблизи горизонтально ориентированных смоляных ходов. Веретеновидные лучи встречаются редко, они составляют не более 5% общего количества лучей.
СТРОЕНИЕ ДРЕВЕСИНЫ ПОКРЫТОСЕМЕННЫХ
Древесина покрытосеменных состоит из продольных прозенхим-ных клеток (вытянутых с заостренными концами элементов — сосудов, трахеид, волокон), осевой (продольной) паренхимы и эпителиальных клеток (рис. 2.15). У покрытосеменных нет поперечно ориентированных прозенхимных клеток. Их поперечные элементы состоят из лучевой паренхимы и очень редко из эпителиальных клеток. У некоторых тропических видов имеются продольные и поперечные смоляные ходы, у видов умеренной зоны они не встречаются.
Рис. 2.15. Строение древесины покрытосеменных:
1 — поперечный срез; 2 — радиальный срез; 3 — тангентальный срез; 4 — сосуд;
5 — перфорированная пластинка на конце сосуда; 6 - волокна; 7 — пора; 8 — лучевая древесина; 9 — годичное кольцо; 10 — ранняя древесина; 11 или 12 — поздняя древесина; 13 — срединная пластинка (Фото Лесной службы США)
34
Глава 2
ПРОДОЛЬНЫЕ ЭЛЕМЕНТЫ. В ксилеме покрытосеменных содержатся клетки значительно больших типов» чем у голосеменных. Среди них наиболее выделяются сосуды, которые являются главными элементами водопроводящей системы. У сосудов покрытосеменных нет концевой стенки, поэтому сопротивление току воды у них значительно меньше по сравнению с сосудами у голосеменных, трахеиды которых снабжены стенкой на конце. Сосуды состоят из члеников сосудов. Это одиночные клетки с разрушенной стенкой на обоих концах, накладывающиеся одна на другую. Таким образом создается трубчатая система, длина которой может быть от нескольких сантиметров до нескольких метров. В поперечном сечении форма сосудов овальная, но у более примитивных видов сосуды в сечении угловатой формы. Расположение сосудов всегда специфично для каждого вида, что является хорошим признаком для определения древесины. Например, сосуды могут быть одиночными или организованными в группы. У падуба сосуды в виде цепочки, у ильма и каркаса сгруппированы в поздней древесине в форме концентрических волнистых зон.
Трахеиды — это отдельные клетки меньшего, чем сосуды, диаметра. У покрытосеменных имеются трахеиды двух типов: сосудистые и вази-центрические. Сосудистые трахеиды - это неперфорированные клетки, напоминающие по форме и положению маленькие членики сосудов. Вазицентрические трахеиды обычно короткие, неправильной формы, расположены вблизи сосудов, не являются составной частью определенных осевых тяжей.
Основная масса древесины покрытосеменных состоит из волокон, похожих на трахеиды, но с более толстыми оболочками, меньшим количеством пор и просветов. Ксилемные элементы покрытосеменных не образуют упорядоченно радиального порядка, характерного для голосеменных. Беспорядочное расположение элементов ксилемы у покрытосеменных, по-видимому, является результатом экстенсивного роста в диаметре члеников сосудов после дифференциации их из камбия. Вследствие этого у других клеток нарушается нормальное расположение и узкий луч изгибается вокруг больших сосудов. Такое беспорядочное расположение обусловлено отчасти также диспропорцией между менее интенсивным делением камбиальных инициалей и более быстрым разрастанием сосудов по сравнению с инициалами в той зоне, где не формируются крупные сосуды ранней древесины.
У покрытосеменных жидкость перемещается вертикально по перфорированным сосудам. Латеральное передвижение жидкости происходит через окаймленные и полуокаймленные поры. Поры покрытосеменных могут соединять волокна, сосуды с волокнами, волокна с лучевыми клетками и сосуды с клетками луча. Пленкой окаймленных пар пор покрытосеменных является первичная пластинка примыкающих друг к другу клеток. В этой пленке нет отверстий, она является скорее составной частью беспорядочно расположенных микрофибрилл, чем радиально ориентированных микрофибрилл, как у голосеменных.
Масса осевой паренхимы в древесине большинства покрытосеменных значительно больше, чем у голосеменных. У некоторых тропических деревьев более половины объема древесины состоит из осевой паренхимы. У большинства деревьев умеренной зоны осевая паренхима сос
Строение
35
тавляет менее 50 % объема древесины, а у некоторых видов лишь несколько процентов. У тополя масса осевой паренхимы ничтожна. Расположение осевой паренхимы у разных видов деревьев различно, что является хорошим отличительным признаком при определении древесины.
ПОПЕРЕЧНЫЕ ЭЛЕМЕНТЫ. Ширина, высота и протяженность лучей у покрытосеменных варьирует значительнее, чем у голосеменных. Обычно лучи в ширину состоят из одного или многих рядов клеток, например у дуба их может быть до 30 и более. У некоторых видов покрытосеменных лучи по рядности могут быть двух типов, причем более мелкие лучи состоят из одного ряда клеток. У некоторых родов древесных растений (ольхи, граба, лещины) лучи агрегатные, т. е. состоят из груп
Рнс. 2.16. Различия рисунков.*
1 — радиального; 2 — таигентального раскроя (Фото Лесной службы США)
пы узких, тесно расположенных друг у друга лучей, между которыми внедряются трахеальные ткани. Такие агрегатные лучи часто выглядят как простые очень широкие лучи. Высота луча у покрытосеменных также очень различна — от нескольких микрон до 5 см. Лучи покрытосеменных состоят в основном из паренхимных очень разнообразных клеток. Клетки могут быть ориентированы радиально (лежачие клетки) или вертикально (стоячие клетки). Лучи разделяются на гомоцеллюлярные, состоящие из морфологически однородных клеток, и гетероцеллю-лярные, клетки которых по форме и размерам не одинаковы. Структура и размеры пор клеток лучевой паренхимы могут использоваться в качестве диагностических признаков при определении древесины.
Разнообразие рисунка или структуры древесины обусловлено различной структурой лучей покрытосеменных. Например, на радиальной распиловке обнаруживается больше лучевых тканей, чем при танген-тальной (рис. 2.16). Расположение мелких лучей на радиальном срезе может создавать слабозаметный рисунок, как у клена, или красивую текстуру из четких широких лучей, как.у дуба.
КОРА
Кора — значительно более сложная система тканей, чем древесина. У взрослого дерева кора включает все ткани, расположенные снаружи от камбия, включая живую флоэму и мертвые покровные ткани (ри-тидом). Более точно, в тканях, возникших при вторичном утолщении, кора состоит из первичной и вторичной флоэмы, первичной коры и
36
Глава 2
перидермы. В стволе, в котором не происходит вторичного утолщения, кора включает только первичные флоэму и кору.
Флоэма играет существенную роль в передвижении веществ, перидерма снижает потерю воды и обеспечивает защиту от механического повреждения. Волокна флоэмы некоторых деревьев (лубяные волокна) раньше использовали для изготовления веревок и матов, английское название lasrwood, т.е. лубяная древесина, связано с тем, что ее кора служила главным источником лубяных волокон. На тихоокеанских островах очень интенсивно использовали внутреннюю кору Broussonetia papyrifera для изготовления тапи (полинезийской ткани). Из флоэмных волокон изготавливают полотно, пеньку и джут.
Вторичная флоэма, образующаяся в результате клеточных делений в камбии, состоит из вертикально и поперечно ориентированных систем клеток. У голосеменных вторичная флоэма относительно проста, состоит только из вертикально расположенных ситовидных клеток, паренхимных клеток и волокон. Поперечно ориентированные лучи (преимущественно однорядные) содержат лишь паренхиму или паренхиму и альбуминовые клетки. Вертикальная паренхима часто включает смолу, кристаллы и танины. Вторичная флоэма покрытосеменных устроена значительно сложнее и имеет больше морфологических различий у разных видов, чем у голосеменных. Флоэма покрытосеменных состоит из вертикально расположенных ситовидных трубок, обычно окруженных клетками-спутниками, паренхимных клеток и волокон. Лучи бывают однорядные, двурядные и много рядные. В состав осевой и поперечной систем часто входят также склереиды, млечники, секреторные элементы, идиобласты и кристаллы.
ФОРМИРОВАНИЕ КОРЫ. Основной прирост по диаметру древесных растений происходит преимущественно за счет камбиальной активности. Небольшая доля в этом принадлежит и меристематической активности другой латеральной меристемы, феллогена, или пробкового камбия, локализованного снаружи от камбия. Периклинальным делением феллогена, состоящего только из одного типа инициалей, образуются клетки феллемы (пробки) снаружи, а клетки феллодермы—внутри (рис. 2.17). Три слоя (феллема, феллоген и феллодерма) вместе составляют перидерму, покровную вторичную ткань, которая у многих видов уже на ранних этапах развития проростка заменяет эпидерму.
Суберинизированные мертвые клетки пробки располагаются радиальными рядами без межклетных пространств. Вследствие такой клеточной структуры пробка обладает ценными свойствами: устойчивостью к проникновению жидкости, упругостью, сжимаемостью, способностью держаться на воде, низкой теплопроводностью и химической инертностью. Существуют два типа пробки, и оба могут быть у одного и того же дерева. Для одного типа характерны клетки полые, тонкостенные, широкие в радиальном направлении. У пробки другого типа клетки толстостенные и радиально уплощенные. Пробка пихты, дугласии, тополя, ивы, секвойи состоит в основном из толстостенных клеток. У некоторых видов березы кора расслаивается на тонкие полосы из-за наличия тонкостенных и толстостенных клеток по прослойкам, между которыми и происходит разрыв. Клетки феллодермы, отличающиеся от пробковых клеток, но сходные с клетками кортикальной паренхимы, оста-
Строение
37
Рис. 2.17. Поперечный срез части ствола:
а — происхождение феллогена; б — развитие тканей путем деления клеток феллогена: 1 — кутикула; 2 — эпидерма; 3 — первые пробковые клетки; 4 — феллоген;
5 — феллодерма; 6 — первичная кора; 7 — кутикула; 8 — эпидерма; 9 — пробка;
10 — феллоген; 11 — феллодерма; 12 — первичная кора
ются живыми и их стенки не суберинизированы. Иногда в них осуществляется фотосинтез и откладывается крахмал. Клеток пробки образуется больше, чем феллодермы, но слой пробки у разных видов различен. У некоторых видов пробковый слой растрескивается и отшелушивается, поэтому толщина слоя не увеличивается. У других видов, например у пробкового дуба, формируется мощный слой пробки и очень мало клеток феллодермы.
Образование перидермы. Эпидерма ствола разрывается и отшелушивается в первый же год жизни дерева, когда оно начинает увеличиваться в диаметре в результате камбиального роста. У большинства деревьев огшелушцванию эпидермы предшествует формирование цилиндрической перидермы на различной глубине первичной коры снаружи от камбия. Этот прирост относительно постоянен для данного вида. В стволе большинства родов древесных растений исходная перидерма возникает в первичной коре непосредственно под эпидермой. Это характерно для липы, катальпы, ясеня, дуба, тополя, грецкого ореха и вяза. Иное отмечается у груши. У нее в стволе эпидермальные и субэпидермальные клетки дают начало исходной перидерме. У белой акации, гледичии, некоторых видов сосны и лиственницы первичный феллоген формирует второй или третий коровый слой ствола. Особенно глубоко закладывается первый слой перидермы в стволах туи, барбариса и ви-
38
Глава 2
Рис. 2.18. Формирование перидермы:
а — молодой ствол; б — зрелая кора видов, у которых не образуется ритидом;
в — зрелая кора видов, у которых образуется мощный слой ритидома: 1 — эпи-: дерма; 2 — перидерма; 3 — первичная кора; 4 — первичная флоэма; S — вторичная флоэма; 6 — камбий; 7 — вторичная ксилема; 8 — первичная ксилема; 9 — , перидерма; 10 — пробковый слой; 11 — вторичная флоэма; 12 — камбий; 13 — ; ритидом; 14 — вторичная флоэма; 15 — перидерма; 16 — вторичная флоэма; 17 — камбий (Фото Лесной службы США)
Рис. 2.19. Схема, демонстрирующая образование последовательно появляющихся слоев перидермы в коре типичного ствола древесных растений (Eames MacDaniels, 1947):
а — одногодичная ветвь, в которой исходная перидерма образует замкнутый цилиндр внутри эпидермы; б - двухгодичная ветвь, в которой эпидерма и исходная
Строение 39
i[ограда. У этих родов первый феллоген закладывается вблизи от флоэмы или даже в паренхиме флоэмы.
У некоторых деревьев (березы, пихты, граба американского, бука) исходная перидерма остается активной на протяжении многих лет. У большинства же видов деревьев исходная перидерма в течение нескольких лет или даже в первый год замещается слоями перидермы, которые формируются последовательно на все большую глубину в первичной коре (рис. 2.18). У видов, для которых характерна закладка исходной перидермы в очень глубоко расположенных тканях, например у винограда, последовательно формирующиеся слои перидермы, подобно слоям исходной перидермы, представляют собой правильной формы цилиндры. Однако есть виды, у которых первый слой перидермы образует правильный круг, второй и последующие слои имеют вид сводов или лунок, искривленных наружу от оси растения (рис. 2.19). Активность последовательно образующихся слоев перидермы сохраняется различное время. У некоторых видов, таких, как бук, они функционируют всю жизнь, у пихты и березы всего несколько лет. У винограда и некоторых других видов они закладываются в течение всего года, у белой акации - только в вегетационный период. Время образования й развития перидермы зависит также от окружающих условий, от агротехнических мероприятий, особенно от снабжения водой, от интенсивности света, фотопериода, температуры и применения гербицидов. Ткани, расположенные снаружи от перидермы, быстро отмирают, так как они изолированы непроницаемыми слоями пробки в перидерме и не снабжаются питательными веществами и водой.
Развитие основных признаков коры. Кора древесных растений у разных видов может быть гладкой или неровной в зависимости от способа и условий ее формирования (рис. 2.20). Характерный вид коры зависит от радиального положения первой перидермы, характера формирования последующих слоев перидермы и от строения и расположения клеток флоэмы. У деревьев с гладкой корой исходная перидерма существует либо в течение длительного времени, как у некоторых видов березы, либо на протяжении всей жизни дерева, как у тополя осинообразного и бука. У видов с гладкой корой слои перидермы увеличиваются по окружности за счет антиклинальных делений и разрастания клеток феллогена. Прирост флоэмы по окружности приводит к образованию расширяющихся тканей за счет антиклинальных делений и тан-гентального разрастания клеток лучевой флоэмы. Ткани гладкой коры обычно сбрасываются медленно и скорость этого процесса взаимосвязана со скоростью образования пробки.
перидерма разорваны, а новые дугообразные слои образованы более глубоко в первичной коре; в — трехгодичный ствол, в котором наружные ткани отшелушены и начинает развиваться вторичная флоэма; г — развитие этого процесса на примере четырехлетнего ствола; д — наружная часть зрелого ствола, видна узкая полоса живой флоэмы и глубоко расщепленная наружная кора, которая состоит из слоев мертвой флоэмы и перидермы; значительная часть тканей отшелушена: I — флоэма; 2 - первичная кора; 3 — первдерма; 4 — эпидерма; S — ксилема; 6 — эпидерма; 7 — перидерма; 8 — флоэма; 9 — сердцевина; 10 — ксилема; 11 — первичная кора; 12 — перидерма; 13 — ксилема; 14 — мертвая флоэма и перидерма; 15 - живая флоэма
40
Глава 2
Рис. 2.20. Различные виды коры:
а — явор; б - мангровое дерево; в ~ кария овальная; г — секвойя; д — сосна желтая; е — береза белая (Фото St, Regis Paper Со.).
У пробкового дуба первая перидерма, развивающаяся в эпидерме, состоит из большого количества клеток пробки и относительно малого количества клеток феллодермы. При снятии у 12—15-летнего дерева первого слоя пробки феллоген отмирает, а новый, закладывающийся в более глубоких слоях первичной коры, образует пробку быстрее исходного феллогена. После второго и последующих сборов пробки, производимых через каждые 9-10 лет, развиваются другие слои феллогена. Таким образом, процесс формирования пробки происходит на протяжении всей жизни дерева.
У видов с чешуеобразной корой первая перидерма может занимать поверхностное положение в течение очень длительного времени. Постепенно под первой перидермой возникают небольшие участки дополнительной перидермы, которые затем формируются на протяжении всей жизни дерева. В результате образуется толстая наружная кора, которая состоит из чередующихся слоев перидермы и тканей, отделяющихся от ствола в форме чешуек, как у сосны обыкновенной, или пластинок, как у клена белого. Перидерма карии овальной возникает в виде длинных вертикальных ленг, поэтому ткани коры отделяются вертикальными пластинчатыми полосками, отражающими ориентацию слоев перидермы и расположение взаимосвязанных многочисленных волокон флоэмы.
У видов с морщинистой корой флоэма содержит большое количество склеренхимных тканей, особенно волокон. Обычно полукружия перидермы возникают в наружной флоэме, а клетки снаружи от перидермы отмирают, но не отделяются от нее вследствие прочно сцепленной системы волокон. Это приводит к формированию очень глубоко расчлененной, рыхлой и волокнистой коры, как у Sequoia sempervirens. Морщинистая кора ивы, дуба, ильма, ясеня и ореха образуется сочетанием в разных соотношениях клеток пробки и склеренхимных тканей Мягкая кора ильма американского содержит большое количество пробковых клеток и мало склеренхимных тканей, а в твердой коре дуба и ясеня много склеренхимных клеток и относительно мало пробковых. ’’Волокнистая” кора некоторых видов Euoalytus образуется за счет формирования ритидома из волокон. Некоторые эвкалипты с твердой корой, известные под названием iron-bark, содержат много твердых смолистых веществ типа камеди, называемых ’’кино”, препятствующих разрыву ритидома.
У видов с кольцевой корой (виноград, ломонос, жимолость и виды сем. Cupressaceae) первая перидерма закладывается в тканях очень
Строение 41
глубоко. Второй и последующие слои перидермы составляют концентрические цилиндры, что приводит к сбрасыванию полых цилиндров рыхлой коры. Отделение происходит по тонкостенным клеткам пробки, но ритидом остается прикрепленным, что придает наружной коре характерный шероховатый вид. Более подробную информацию по этому вопросу можно получить в работах Боргера (1973).
КОРНИ
Корни служат для закрепления растения в почве, поглощения воды и минеральных солей, а также для запасов питательных веществ. Корневая система деревьев состоит из сети относительно крупных многолетних корней и множества небольших недолговечных боковых корней. Разнообразие в распределении и протяженности корней имеет большое значение для древесных растений. Глубоко проникающая и разветвленная корневая система позволяет поглощать воду и минеральные вещества из большей массы почвы, чем менее развитая корневая система. Корневая система у разных видов проникает на различную глубину и часто не наблюдается соответствия между размерами корней и величиной надземной части растения. Например, корни чайного куста могут проникать глубже корней высоких деревьев. Иногда корни проникают значительно глубже, чем это обычно полагают. Корневая система кал-лиандры и некоторых других деревьев и кустарников сухих местообитаний проникает на глубину 6—10 м, а корни яблони, растущей на хорошо аэрированных лёссовых почвах в Небраске, обнаруживаются на глубине более 10 м. У растений сухих мест обитания часто отмечается высокое отношение корень/побег. По данным Фернандеса и Кол-дуэла (1975), отношение корней к побегам у кустарников холодной полупустыни Юты может быть 9:1, у деревьев яблони или молодых растений сосны ладанной — всего 1:5. Барбор (1973) утверждает, что отношение корней к побегам пустынных растений обычно меньше единицы. Это было подтверждено данными, полученными для кустарников сухих мест обитания.
Колдуэл (1976) считает, что энергетические и материальные затраты растений на рост корней очень значительны. Гибель массы мелких всасывающих корней, потеря активной абсорбирующей поверхности и необходимость постоянного образования новых корней приводят часто к тому, что половина питательных веществ у некоторых растений затрачивается на рост корней. Причины таких на вид непроизводительных затрат заслуживают дальнейших исследований.
Корни деревьев часто распространяются латерально за пределы ширины кроны, однако протяженность распространения корней зависит в большой степени от типа почвы. Например, корни плодовых деревьев на песчаных почвах распространяются латерально в 3 раза дальше пределов границ полога кроны, на суглинистых почвах — в 2 раза, на глинистых - только в 1,5 раза.
У лесных деревьев большая часть корней распространяется в поверхностных горизонтах почвы, вероятно, вследстзие лучшей их аэрации и обогащенности минеральными питательными веществами, чем в более
42
Глава 2
глубоких горизонтах. Кроме того, они лучше оводняются во время летних ливневых дождей. Это особенно типично для тропических лесов, но также относится и к лесам умеренной зоны. С другой стороны, корни иногда проникают и на очень большую глубину. Пробстиг (1943) обнаружил в суглинистых почвах садов Калифорнии максимальную концентрацию корней плодовых деревьев на глубине 0,5—1,5 м, некоторые из них вырастали до 5 м длиной^
КЛАССИФИКАЦИЯ КОРНЕЙ. Корневая система большинства проростков древесных растений наследственно закреплена и разделяется на два типа: 1) главный корень быстро растет вниз и проникает глубоко в почву; 2) первичный корень растет медленно и вглубь не проникает, но разрастаются интенсивно боковые корни. Классифицировать корневые системы взрослых деревьев часто затруднительно, так как почвенные условия могут существенно изменить характер их роста и видовые особенности. Например, у видов эвкалипта на сухих местах развивается мощный главный корень и небольшое количество слаборазвитых боковых. На хороших участках, однако, у них формируется мочковатая корневая система, разрастающаяся в поверхностных слоях почвы. Клен красный на болотах образует слабые боковые корни, а на возвышенностях у него формируется глубоко проникающий стержневой корень. У явора корневая система состоит из сильноразвитого стержневого корня и равномерно распределенных по нему коротких боковых корней. Этот тип корневой системы существенно не изменяется под
1
Рис. 2.21. Схематическое изображение корневой системы взрослого дерева:
1 — периферическая корневая система; 2 — центральная корневая система (сред- . г ний диаметр кроны); 3 — корневая шейка; 4 — гумусовый слой; 5 — горизон-тальные корни; 6 — сердцевинные корни; 7 — стержневой боковой корень; £ — | стержневой корень; 9 — глубина корневой системы; 10 — вертикальные корни; Ji-
ll — наклонные корни; 12 — отвесные корни; 13 - поверхностная корневая система
Строение
43
влиянием окружающих условий. Несмотря на пластичность корней у некоторых видов, созданы различные системы номенклатуры корней. Одна из них, используемая в Европе для обозначения различных частей корневой системы, представлена на рис. 2.21.
Гетероризные корни. Большая часть видов сосны имеет гетерориз-ную корневую систему, включающую длинные и короткие корни. Длинные корни состоят из главного и быстрорастущих боковых корней первого и второго порядка. Они ответственны за развитие корневой системы в целом. Длинные корни функционируют на протяжении всей жизни дерева и увеличиваются в диаметре за счет активности камбия. Эфемерные и медленнорастущие короткие корни, располагающиеся по бокам длинных корней, имеют апикальную меристему и лишены истинного корневого чехлика. Короткие корни лишены вторичного роста и большая их часть исчезает в первый или второй год жизни. Они обычно образуют микоризу. Длинные корни сосны, имеющие большое значение в развитии остова корневой системы, подразделены Ноэлем (1910) и Олдрич-Блэйком (1930) на корни-пионеры и материнские корни. Уилкокс (1964) добавил третью категорию для сосны смолистой — подчиненные материнские корни. Корни-пионеры (так они названы Ноэлем из-за их значения для быстрого распространения корневой системы) — самые крупные по диаметру в корневой системе сосны. Корни-пионеры не встречаются в большом количестве, обычно их можно обнаружить в период наиболее активного роста корней. Материнские корни, названные так из-за их обильного ветвления, меньше в диаметре и короче, чем корни-пионеры. Подчиненные материнские корни сосны смолистой, которые были описаны Уилкоксом (1964), тоньше и меньше в диаметре материнских корней. Их можно сравнить с боковыми корнями второго порядка двухлетних проростков (табл. 2.4). В прежних работах по лесоводству, как подчеркивает Уилкокс (1962), не всегда обращалось внимание на то, что в корневой системе, есть длинные и короткие корни. Это, по-видимому, обусловлено тем, что короткие корни принимали за микоризу. Такие корни развиваются у всех представите-
2.4. КЛАССИФИКАЦИЯ, РАСПРЕДЕЛЕНИЕ И ДИАМЕТРЫ КОРНЕЙ (В СКОБКАХ - СРЕДНЕЕ ЗНАЧЕНИЕ) 2-ЛЕТНИХ СЕЯНЦЕВ И ВЗРОСЛЫХ ДЕРЕВЬЕВ СОСНЫ СМОЛИСТОЙ (Wilcox, 1964)
Диаметр, мм
Класс корней
Число корней . взрослых деревьев сеянцев
Пионеры
Материнские
Материнские подчиненные
Короткие корни: .
первичные
боковые первого порядка
боковые второго порядка
10
26
37
207
1,32-1,81 (1,66)
0,74-1,21 (0,85)
0,36-0,71 (0,54)
0,23-0,50 (0,42)
0,26-0,41 (0,34)
(1,03)
(0,99)
(0,55)
44
Глава 2
лей сем. Pinaceae и у некоторых голосеменных, а также у видов сем. Betulaceae, Fagaceae, но они отсутствуют у многих древесных растений. Например, у кедра ладанного, как и у других представителей сем. Cup-re 5§асеае,гетероризная корневая система не образуется.
Система классификации, в основу которой положена длина корней, не всегда приемлема, так как короткие корни могут превратиться в длинные или в утолщенные опорные, поглощающие воду участками вблизи от кончика корней.
Одревесневающие и неодревесневающие корни. Клоус (1950) и Лифорд и Вильсон (1964) считают, что классификация корней по их длине непригодна для некоторых покрытосеменных. Лифорд и Вильсон предпочитают классифицировать корни клена красного на одревесневающие и неодревесневающие. Они считают, что зрелая корневая система является остовом из одревесневших корней, на которых появляются пучки неодревесневших, недолговечных корней. Одревесневающие корни распространяются от основания ствола в стороны почти горизонтально, быстро образуя вблизи от ствола сетчатые структуры, которые малы в диаметре (до 2,5 мм) и достаточно длинны (до 25 м). Боковые одревесневающие корни ответвляются от главного корня на расстоянии от 1 до 5 м. Первые неодревесневшие корни появляются под прямым углом к одревесневшему корню с интервалами от 1 до 3 см. На одревесневших корнях первого и более высокого порядка образуются сложные перистые ’’пучки корней” длиной 20—40 см (рис. 2.22). У старых и толстых корней таких недолговечных корневых пучков с возрастом становится все меньше. Как видно из табл. 2.5, чем выше порядок неодревесневших корней, тем меньше их диаметр и длина, но у них образуется соответственно больше микоризы и боковых ответвлений.
L Щ t ...
ТК 5
Рис. 222. Отношение одревесневших и неодревесневших корней у клена красного: 1 - ствол; 2 - придаточные корни в зоне крутого конуса; 3 - боковой корень;
4 - неодревесневшие пучки корней; 5 - кончик одревесневшего корня и неодревесневших корней первого порядка; 6 - неодревесневшие корни второго и более высоких порядков, появляющиеся из неодревесневших корней первого порядка; 7 - неинфицированный кончик неодревесневшего корня второго порядка с корневыми волосками; 8 - неодревесневший корень третьего порядка с бисероподобной микоризой; 9 - неодревесневший корень четвертого порядка с микоризой
Строение
45
Суберцнизированные и несуберинизированные корни. Отдельные корни деревьев могут продолжать расти в течение нескольких недель и образовывать боковые корни. Иногда рост Корней прекращается значительно раньше. Наружные кортикальные ткани корней в течение короткого времени остаются неолробковевшими и бесцветными. Этот период у яблонь может длиться от 1 до 4 недель летом и до 3 месяцев зимой. Затем кортикальные ткани становятся бурыми и деградируют. Остающийся центральный цилиндр иногда претерпевает вторичное утолщение. Побурение кортикальных тканей связано с их суберинизацией (структурные изменения во время вторичного роста корней древесных продемонстрированы на рис. 3.20, 13.1, 13.2). С возрастом все большая часть поверхности корней суберинизируется. Не претерпевают суберинизацию лишь самые молодые корни, поверхность которых очень мала по сравнению с поверхностью всей корневой системы. Крамер и Баллок (1966) проследили сезонные изменения растущих и суберинизированных (опробковевших) корней сосны ладанной и липы американской, произрастающих в Северной Каролине. Площадь поверхности растущих, несуберинизированных корней у сосны ладанной составляла менее 1% поверхности всей корневой системы (табл. 2.6). Эта величина превышала 1% лишь в июле после проливных дождей. Большая часть несуберинизированных корней приходилась на микоризу, причем сумма поверхности растущих кончиков корней и микоризы в период активного роста не превосходила 7%. Таким образом, 93-99% поверхности корней суберинизирована, это дает возможность допустить, что поглощение воды и минеральных веществ в большой мере осуществляется суберинизированными корнями (см. главы 10 и 13).
Специализированные корни. Многие древесные растения обладают специализированной, или морфологически измененной, корневой системой, которая часто участвует в важных физиологических процессах во время роста. К таким корневым системам относятся микоризы корней, сросшиеся, воздушные, клубеньковые и опорные корни.
2.5. СВОЙСТВА КОРНЕЙ КЛЕНА-КРАСНОГО (Lyford, Wilson, 1964)
Типы корня Приблизительный диаметр кончика корня, мм Приблизительная конечная длина Частота Частота рас-ра спреде-пределения пения бо- микоризы ковых корней на 1 см длины
Одревесневшие 20 10-25 м 0,3-1 Нет
Неодревесневшие:
первого порядка 0,5-1,0 20-40 см 2-3 Нет
второго порядка 0,5-1,0 10—15 см 3—10 Низкая
третьего порядка 0,3-0,5 1-2 см 3-10 Высокая
выше третьего порядка и ми- 0,2-0,3 1—10 см - Почти сплошь
кориза микориза
46
Глава 2
Микоризные корни. Корневая система большинства древесных растений в значительной степени модифицирована присутствием микоризы (рис. 2.23). Эти структуры, образованные внедрением гиф гриба в молодые корни, являются симбиотической ассоциацией между непатогенными или слабопатогенными грибами и живыми клетками корней. Большая часть микориз делится на две четкие группы — эктотрофные формы, которые существуют и внутри и снаружи корня, и эндотрофные формы, существующие исключительно в клетках хозяина. Эти ассоциации образуются всегда в клетках первичной коры корней хозяина и не возникают в эндодерме или стеле.
Эктотрофные микоризы встречаются у многих экономически цен-
Рис. 2.23. Разновидности микоризы на корнях дугласии:
1 — микоризные пучки; 2 — серая перистая микориза; 3 — оранжевая микориза;
4 — желтая микориза, показывающая ризоморфы грибных симбионтов (Фото Лесной службы США)
Рис. 2.24. Строение микоризных и неинфицированных корней-Fucalyptus (Chil-vers, Pryor, 1965): срединные продольные срезы корня: а — микоризного; б — неинфицированного корня; поперечные срезы полностью дифференцированной зоны корня: в — микоризного; г — неинфицированного; 1 — корневой чехлик; 2 — меристематическая зона; 3 — гифальный чехол, или мантия; 4 — сеть Гартига; 5 - утолщенные оболочки внутренней первичной коры; 6 — эпидерма; 7 — наружная кора первичная; 8 — внутренняя первичная кора; 9 — эндодерма (заштрихованы участки, пропитанные танином) ; 10 — корневые волоски; 11 — одревесневшая протоксилема; 12 — сплющенные остатки клеток чехлика
48
Глава 2
них пород деревьев. Они обнаружены среди видов сем. Pinaceae. Sali-caceae, Betulaceae, Fagaceae, Juglandaceae, Caesalpinioideae, Tiliaceae.
К важным родам голосеменных, которые обладают эктотрофной микоризой, относятся Pinus, Picea, Abies, Pseudotsuga, Cedrus, Larix, а среди покрытосеменных Quercus, Castanea, Fagus, Nothofagus, Betula, Alnus, Salix, Carya, Populus. Эндотрофные микоризы хотя и широко распространены в растительном царстве, но обнаружены они лишь у отдельных видов Liriodendron, Acer, Liquidambar и различных представителей сем. Ericaceae.
При эктотрофной микоризе гифы грибов распространяются по поверхности корней, мицелий может в виде тонкой сетки плотно оплетать корень, либо образовывать компактные псевдопаренхиматозные структуры. Грибы проникают в первичную кору между кортикальными клетками, не входя в собственные клетки хозяина. На поперечных срезах клетки первичной коры оказываются разделенными сетью гифов грибов, называемой сетью Гартига.
Различия в структуре корней Eucalyptus, имеющих микоризу и без микоризы, показаны на рис. 2.24. Количественные различия в структуре состоят в следующем: 1) у микоризных корней не бывает корневых волосков, а у неинфицированных корней корневые волоски развивают
2.6. СЕЗОННЫЕ ИЗМЕНЕНИЯ ПЛОЩАДИ СУБ ЕРИНИЗИРО ВАННОЙ
И НЕСУБЕРИНИЗИРОВАННОЙ ПОВЕРХНОСТИ КОРНЕЙ 34-ЛЕТНИХ ДЕРЕВЬЕВ СОСНЫ ЛАДАННОЙ В ПОСАДКАХ В СЕВЕРНОЙ КАРОЛИНЕ, % (Kramer, Bullock, 1966)
Дата Растущие кончики Микориза Все несубери-низированные Все суберинизи-ро ванные
1.Ш 0,6 0,53 0,59 99,41
8.Ш 0,15 1,20 1,35 98,65
15.Ш 0,13 1,18 1,31 98,69
22.Ш 0,13 1,64 1,77 98,69
29.Ш 0,13 2,06 2,39 97,61
8.1У 0,43 2,27 2,70 97,30
13.1У 0,39 5,09 5,48 94,52
22.1У 0,53 ' 2,77 3,30 96,70
ЗОЛУ 0,34 5,30 5,64 94,36 ;
9 .У 0,72 5,76 6,48 93,52 j
31.У 0,30 3,95 4,25 95,75 :
7.У1 0,25 6,06 5,31 93,69 ‘ !
17.У1 0,48 3,05 3,53 96,47
24.У1 0,38 3,00 3,38 96,62
1.УП 0,22 3,00 3,22 96,78
15-УП 0,54 2,38 2,92 97,08
29.УП 1,36 2,81 4,17 95,83
11.Х1 0,61 2,84 3,55 95,88
.Л
Строение
49
ся; 2) у микоризных корней слабо развиты ткани корневого чехлика, редко развивается более двух слоев клеток между апексом и оберткой из гиф, а для неинфицированных корней характерно образование тканей корневого чехлика; 3) зона дифференциации у микоризных корней расположена значительно ближе к апексу по сравнению с неинфицирован-ными корнями. Морфология микоризных корешков сходна со структурой корней, развивающихся при неблагоприятных условиях. К количественным отличительным признакам микоризных корней относятся также утолщение внутренней первичной коры и радиальное удлинение эпидермальных клеток. Грибы эндотрофной микоризы внедряются в клетки и никогда не соприкасаются, с поверхностью почвы. Они вызывают меньше изменений в морфологии корней хозяина, чем эктотрофные формы.
Микориза играет важную роль в физиологии дерева, увеличивая поглощение минеральных веществ, особенно соединений фосфора. Деревья могут успешно расти без микоризы при некоторых условиях, например при очень высоком плодородие почвы, но они значительно лучше растут с микоризой. Имеется много данных о полезном симбиотическом существовании деревьев и микоризных грибов. Деревья снабжают грибы углеводами и другими необходимыми для них метаболитами. Грибы увеличивают доступность для деревьев азотных, фосфорных и других питательных веществ. Есть также данные о том, что микоризные грибы могут защищать дерево-хозяин от болезней, используя избыток углеводов, действуя как физический барьер, секретируя фунгистатические вещества и. благоприятствуя защитным организмам ризосферы. (Физиологическое значение микоризы в поглощении минеральных веществ обсуждается в главе 10.)
Срастающиеся корни. Срастание корней представляет значительный интерес, так как от одного растения к другому через них передаются вода, метаболиты и различные токсические вещества (см. главу 11). Естественное срастание корней — распространенное явление.
Это наблюдалось более чем у 150 видов древесных растений и несомненно имеется и у других видов. Козловский и Кули (1961) обнаружили внутривидовые срастания корней у представителей покрытосеменных и голосеменных. Корни объединялись как у молодых, так и у старых деревьев, но более мощное срастание корней происходило у старых деревьев.
Часто сообщества деревьев органически объединяются при помощи сложной системы внутривидовых срастаний корней. Основное условие для срастания корней, по-видимому, совместимость и тесное расположение корней. Питающие корни срастаются реже. Угол сближения между корнями не имеет значения, так как они срастаются при самых разных положениях. Обычно срастание происходит при сдавливании корней, когда они растут вплотную к камням и тесно при этом сближаются. Однако срастание обнаруживается и при отсутствии камней в почве. Кроме настоящего срастания, во многих случаях при сдавливании во время роста наблюдается образование каллуса в местах пересечения корней. В насаждениях срастание корней начинается лишь при разрастании деревьев, когда их корни начинают сближаться и вступать в контакты.
50
Глава 2
Срастание корней в посадках ели обыкновенной у молодых деревьев составляло 3-5%, у 40-60-лвтних деревьев оно наблюдалось в 25-35 случаях из 100. По данным Шульца и Вудса (1967), 5-25% деревьев в древостоях 'сосны ладанной в возрасте 25-85 лет были связаны между собой корневыми: срастаниями. Однако и в питомниках наблюдаются срастания корней при очень тесной Иосадке сеянцев. Основными факторами, способствующими срастанию корней, Эйс (1972) считает совместимость тканей, очень близкое расположение деревьев, маломощные каменистые почвы и большую концентрацию корней на единицу площади.
При срастании корней образуется связь между проводящими тканями за счет объединения камбия, флоэмы и ксилемы корней, прежде не связанных между собой. Было показано, что ток воды осуществляется через сросшиеся корни, а выживание на протяжении 10 лет и более окольцованных деревьев указывает на то, что у них имеется общая функционирующая флоэма. Наиболее обычные типы слияния корней -это самосрастание корней одного и того же дерева или разных деревьев одного вида. Близкородственные межвидовые срастания происходят значительно реже.
Эйс (1972) описал процесс срастания корней у хвойных. После установления прочного физического контакта между корнями совмещаемых деревьев лучевые камбиальные клетки быстро разрастались, образуя валик из тканей каллуса. Деление клеток начиналось перед началом формирования поздней древесины. Массы каллуса возникали одновременно у расположенных друг против друга молодых корней. У молодых мелких корней эти массы каллуса прорывали кору в течение второго вегетационного периода и два камбия почти сразу же соединялись. Ста* рые корни с толстой корой срастались значительно медленнее, иногда для этого требовалось не менее 4 лет. На крупных корнях с обширными площадями контакта каллусные массы противоположных корней не всегда возникали друг против друга. В таких случаях каллус одного корня не мог проникнуть через кору другого корня и слияния не было. Плотная кора больших корней - серьезный барьер для срастания.
Воздушные корни. Большинство корней — подземные органы, однако это не абсолютное правило. Часто корни оказываются на поверхности земли, когда семена прорастают в небольшом слое почвы, который позднее смывается и корни оказываются обнаженными.
Козловский и Кули (1961) нередко наблюдали обнажение корней у березы желтой. Это происходило при прорастании семян на старых вырубках или лесозаготовительных участках. Корни проростков вначале развивались под древесными остатками, но после разрушения остатков корни оказывались на поверхности почвы. Подземные корни старых деревьев также могут оказаться незащищенными вследствие эрозии почвы или при близком их произрастании от мест проведения дорожно-строительных работ и других мероприятий.
Воздушные корни характерны для многих тропических древесных растений. Такие корни очень разнообразны по форме. У некоторых древовидных папортников они возникают из воздушных стебелей. У многих лиан корни возникают из наземных органов и становятся наружными ’’обнимающими” корнями. Фикус образует свободно свисающие воздушные корни, которые дают начало боковым корням и подвергаются вторичному утолщению, прежде чем достигнут почвы.
Многие виды образуют корни-ходули, которые возникают из главного ствола дерева, нагибаются вниз и входят в почву. Корни-ходули часто ветвятся на поверхности земли и дают начало вторичным и третичным
Строение
51
Рис. 2.25. Вертикально ориентированные тянущиеся на воздух корни у Avicennia (а), корни-ходули или подпорки у Rhizophora (б). (Scholander et al., 1955)
корням, уходящим в почву. Мангровые (Rhizophora) являются лучшим примером деревьев, образующих корни-ходули (рис. 2.25). Такие же корни формируют и многие другие деревья, растущие на пресноводных болотах и во влажных лесах. Среди родов древесных растений, образующих ’’ходули”, выделяются Pandanus, Clusia, То vomit a, Eleocarpus, Xylo pi a, Dillenia, Eugenia, Musan ga. У другого мангрового растения Avicennia nitida образуются воздушные корни, которые ответвляются от горизонтальных корней и растут вертикально (см. рис. 2.25 и главу 6). Такие же воздушные корни имеются у Атпоога, Сагора и Heritiera.
У некоторых видов образуются вертикальные корни-колена. Происхождение этих корней у разных видов различнее. Корни болотного кипариса развиваются в форме вертикальных колен вследствие локализованной камбиальной активности, которая накапливает мощные слои древесины на верхней поверхности корня. У Bruguiera и ceriops колена возникают в результате роста вверх горизонтального корня до того времени, пока, он появится из земли. У Avicennia marina колена обра-зуются$если боковые ответвления горизонтальных корней в течение некоторого времени растут вверх, а затем вновь направляются вниз, внедряются в почву и там растут горизонтально. Коленообразные корни Mitragyna stipulosa развиваются либо вследствие усиленной местной активности камбия, в результате которой возникают вертикальные выросты на горизонтальных корнях, либо вследствие изгибания корней над поверхностью почвы.
У кипариса колена не развиваются в глубокой воде или в случае очень короткого периода разлива. На периодически затопляемых участках такие корни-колена вырастают величиной равной уровню подъема воды. Вероятно, у этих растений колена развиваются за счет камбиальной активности, локализованной на участке, который лучше аэрируется, чем остальные части корня. (Роль в дыхании колен кипариса и других пневматофоров рассматривается в главе 6.)
Опорные корни. У некоторых видов древесных растений умеренной зоны, произрастающих на болотах, например у ниссы водной, и у многих видов тропических лесов от основания ствола образуются опорные корни. У ниссы водной опорные корни узкие, отходят от раз-
52
Глава 2
бухшего основания ствола, а у тропических видов они различны по фор-ме - от плоских пластин до широких желобов. Эти образования увеличиваются с возрастом дерева, они могут достигать 10 м в высоту, причудливо изгибаясь, подниматься вверх и вновь опускаться. Опорные корни у большинства тропических деревьев состоят из 3—4 лопастей, их может быть и более 10. Образование опорных корней обусловлено наследственно. Обычно они распространены у деревьев влажных тропических лесов из сем. Dipterocarpaceae, Leguminosae, Sterculaceae. Процесс образования этого вида корней зависит также от окружающих условий. Опорные корни развиваются интенсивнее у деревьев, произрастающих на низких местах, во влажных лесах, на маломощных почвах. Смит (1972) считает, что опорные корни обеспечивают большую устойчивость дереву, чем одно цилиндрическое основание ствола того же самого объема. j
Опорные корни формируются только у деревьев с поверхностной корневой системой и хорошо развитыми боковыми корнями. У моло- \ дых деревьев камбиальный рост корней протекает нормально, боковые корни на поперечном срезе концентрические. Однако вскоре камбиальный рост верхней стороны боковых корней и ствола над ними значительно ускоряется, происходит чрезмерное их утолщение, ведущее к образованию опорных корней.
Нижняя часть ствола деревьев с опорными корнями очень оригинальна, на поперечном срезе часто бывает звездообразной. Центральный ствол таких деревьев сужен внизу на уровне восхождения лопастей корней, сужающихся в свою очередь кверху до этого места. У молодых деревьев не бывает конусообразного сужения ствола, но по мере формирования опорных корней у них также начинается сужение ствола, и оно прогрессирует в течение формирования лопастей корней. При этом включается контрольный механизм, который позволяет направлять метаболиты к местам формирования опорных корней.
Клубеньки корней. На корнях многих растений образуются ц, узелки или клубеньки (рис. 2.26). Очи участвуют в фиксации азота,
Рис. 2.26. Клубеньки на корнях Alnusglutinosa fR pricings 1972)
Строение 53
поэтому представляют большой интерес для физиологов растений. Клубеньки образуются на корнях многих древесных бобовых, встречаются также на корнях некоторых небобовых растений. К небобовым видам из голосеменных с клубеньковыми корнями относятся Agathis, Araucaria, Libocedrus, Phyllocladus, Dacrydium, Sciadopitys. К небобовым растениям из покрытосеменных с клубеньковыми корнями принадлежат Alnus, Casuarina, Coriaria, Eleagnus, Hippophae, Shepherdia, Comp-tonia (Myrica), Ceanothus, Purshia. Клубеньки на корнях небобовых многолетние, увеличиваются в размерах в течение каждого вегетационного периода, достигая иногда величины теннисного мяча.
На корнях бобовых клубеньки формируются в результате внедрения бактерий Rhizobium в волоски или через ранки. Описано два типа клубеньков, причем представители наиболее распространенного из них образуются вследствие инфицирования клеток коровой паренхимы, В процессе фиксации азота у древесных бобовых растений участвуют бактерии из рода Rhizobium. В клубеньках небобовых этот процесс осуществляют виды Actinomycetes. У некоторых из этих видов, однако, эндофитами являются также бактерии рода Rhizobium (Фиксация азота более подробно рассматривается в главе 9).
ВЕГЕТАТИВНЫЙ РОСТ
Рост древесных растений может быть охарактеризован с количественной, анатомической, морфологической и физиологической точек зрения. Исследователи количественных показателей часто используют классические технические приемы анализа ростовых процессов, при помощи которых оценивают скорость роста, а не величину растений и урожай. На ранних стадиях роста увеличение размеров растений, сухого веса, органов или тканей происходит приблизительно линейно. Со временем, однако, различные внутренние механизмы, контролирующие рост, вызывают отклонения от линейных отношений, и ростовые процессы на протяжении долгого периода характеризуются сигмоидной кривой. Рост в течение сезона и на протяжении жизни побегов, корней и репродуктивных органов, как правило, подчиняется этой закономерности.
Прирост растений в высоту и по диаметру происходит за счет активности меристематических тканей, которые составляют очень небольшую часть общей массы растений. Различные части растений \ увеличиваются с неодинаковой скоростью, и нередко активный их рост происходит в разное время года. Например, древесные растения умеренной эоны переходят от состояния контролируемого эндогенно глубокого зимнего покоя тканей к меристематической активности в течение вегетационного периода. Однако и в безморозный период происходит торможение роста, за которым следует восстановление ростовых процессов. Считается, что тропические деревья растут непрерывно в течение года, но при более тщательных наблюдениях обнаружилось, что и у них скорость роста не остается постоянной, их рост также цикличен.
В этой главе будут обсуждаться природа и периодичность роста побегов, камбиального роста и роста корней деревьев умеренной зоны и тропиков, Это необходимо для последующего обсуждения роста как конечного результата координированных процессов, таких, как синтез питательных веществ, их превращение в растворимые формы, их передвижение, распределение и ассимиляция новыми тканями (Рост репродуктивных органов будет обсуждаться в главе 4, регулирование вегетативного и репродуктивного роста — в главах 16 и 17.)
РОСТ ПОБЕГОВ
Растущие побеги, которые обычно слагаются из стебля и листьев, состоят также из узлов и междоузлий. Узлы — это участки стебля, к которым прикрепляются листья* Узлами также называют места прикрепления к стеблю удлиненных побегов или мутовок веток. Междоузлия - это части стебля между двумя соседними узлами. Растяжение побегов происходит в результате раскрывания почек и активного деления клеток во многих точках роста (апикальной меристемы), распределенных по стеблю, боковым побегам и ветвям. Раскрывание почек может сопровождаться разрастанием листьев, меристематической активностью в междоузлиях или большей скоростью роста на внутренней поверхности почечных чешуек по сравнению с таким же ростом на наружной поверхности (рис. 3.1). При увеличении размеров побегов продолжительность роста междоузлий и листьев часто бывает разной, что зависит частично от тургора клеток и поступления питательных веществ.
Апекс растущего побега, состоящий из апикальной меристемы и субапикальных тканей, представляет собой гармонично развивающуюся массу клеток. Поверхностный рост побега происходит вследствие деления клеток апикальной меристемы и их последующего растяжения, дифференциации и созревания. Эти этапы роста проходят последов а-
Вегетативный рост
SS
Рис. 3.1. Почки гикори на разных стадиях раскрывания (Фото Лесной службы США)
тельно на участках побега, рас- положенных на разных расстояниях от верхушек боковых побегов. Удлинение почти всех1 побегов является результатом растяжения междоузлий.
СТРОЕНИЕ ПОЧЕК. Зрелая почка - это эмбриональный побег или часть побега, несущая на верхушке апикальную меристему, за счет которой она формируется. Большинство боковых почек закладывается в пазухе листа и возникает в относительно поверхностных тканях. Закладка почки начинается с деления клеток в клеточных слоях в пазухе листа, в результате образуется почечный бугорок и возникает апикальная меристема.
Почки в зависимости от места их локализации, строения-или активности обычно клас-на концевые, боковые, пазушные, спящие, адвентивные, цветочные или смещенные. Любая из этих почек может быть активной и спящей. Вегетативные почки различаются по степени зрелости. Они могут состоять почти целиком из апикальной меристемы, но обычно они содержат небольшое количество меристематических тканей, узлы, междоузлия и маленькие зародышевые листья с почками в их пазухе. Все это покрыто почечными чешуйками. Цветочные почки содержат зародышевые цветки и рудиментарные листья. Смешанные почки имеют и цветки и листья.
Во время формирования вегетативных почек появляются зачатки листьев в последовательности сверху вниз. Поэтому самые большие и старые листовые зачатки локализованы у основания почки, а более мелкие рудиментарные листочки расположены ближе к точкам роста (рис. 3.2). Листья образуются за счет деления поверхностных клеток апикальной меристемы. Появляющиеся листья располагаются очень тесно друг к другу, узлы и междоузлия почти неразличимы. Затем, когда активируется меристема между листьями, начинается интерка-лярный рост и тогда обозначаются междоузлия.
Спящие и адвентивные почки. Не все почки образуют побеги, некоторые из них остаются спящими на протяжении всей жизни дерева. Спящие почки, заложенные в пазухах листьев, соединяются с корой через листовой след. Побегообразование из спящих почек - обычное явление (рис. 3.3). Почки, которые формируются на более старых частях растения, а не на верхушках стебля или в пазухе листьев, называются ад
56
Глава 3
вентивными. Они образуются на частях корня или ствола, у которых нет связи с апикальной меристемой. Они могут возникать как из глубоко ^расположенных, так и из поверхностных тканей. Например, адвентивные почки могут возникнуть из тканей каллуса вокруг раны, из камбия или из зрелых тканей эндодермы и участков перицикла. Адвентивные почки, в противоположность спящим, не имеют связи с сердцевиной.
Многие побеги у покрытосеменных, появляющиеся из корневой шейки и нижней части ствола после обрезки и вырубок, а также отрастающие побеги голосеменных после пожаров и других повреждений,' возникают скорее из спящих почек, а не из адвентивных. Корневые побеги (корневая поросль) образуется только из адвентивных почек/ Размножение корневыми побегами хорошо известно у осины, тополя крупнозубчатого, оно также часто наблюдается у бука, айланта, белой акации, сирени, ниссы лесной и сассафраса.
РОСТ ЛИСТА. Древесные растения обладают несколькими типами листьев, среди которых различают семядоли, катафиллы и настоящие листья. Семядоли, или семенные листья, развиваются в семенах и со-
Рис. 3.2. Верхушка вегетативного побега вин отрада (Fahn, 1967) :
1 — самый молодой листовой примордий; 2 — листовой примордий; 3 — апекс;
4 — прилистник; 5 — главный апекс; 6 — пазушная почка; 7 — примордий нецветоносного побега; 8 — прицветник
Вегетативный рост
57
Рис. 3.3. Образование ветвей из спящих почек сосны ежовой
(Little, Somes,
1956)
держат большое количество запасных питательных веществ (см. главу 14). Они обычно отличаются по размерам и форме от первых настоящих листьев (рис. 3.4). Катафиллы (чешуи) выполняют защитную функцию или содержат запасные питательные вещества, либо осуществляют то и другое вместе. Они представлены в форме почечных чешуек.
Происхождение и развитие листьев. Клеточные деления происходят главным* образом на ранних стадиях развития зачатков листа. Постепенно лист достигает своих конечных размеров и формы вследствие деления и увеличения объема клеток, причем последнее при этом преобладает. Окончательная величина листьев зависит от количества клеток в листовом зачатке, скорости и продолжительности деления клеток и размеров закончивших рост клеток. Наибольшее значение в этом процессе имеет количество клеток в листовом зачатке.
Покрытосеменные. Листья покрытосеменных формируются только на верхушках побегов. Верхушка побега разрастается в бугорок, и появляются листовые зачатки, которые вначале состоят из одинаковых меристематических клеток. После прекращения деления клеток в нижней части апекса происходит дифференциация клеток и формируется основание листа. Верхняя часть листового зачатка продолжает делиться, в результате развивается пластинка. Черешок листа образуется позднее из промежуточной меристематической зоны. Разные части листа, такие, как черешок, пластинка, влагалище и трилистник, закладываются вскоре порле формирования листового зачатка.
На первых этапах рост осуществляется на верхушке листа, но это продолжается недолго, так как начинается интеркалярный рост (за
58
Глава 3
Рис, 3.4. Однонедельный приросток клена ясенелистного. Видны различия в размерах и форме вытянутых цельнокрайних семядолей и зубчатых первых листьев (Фото Лесной службы США)
счет активации меристемы, расположенной между участками более или менее дифференцированных тканей) . Вследствие этого лист увеличивается в длину. Разрастание пластинки в ширину происходит в результате активации деления меристемы, локализованной по краям от листовой оси. Эпидермальная ткань, из которой образуются устьица, дифференцируется рано. В связи с этим большая часть устьиц обнаруживается у листиков в молодых почках, однако их дифференциация возможна и на более поздних стадиях формирования листа. Проводящая система также начинает формироваться рано, в процессе образования пластинки листа. В зависимости от продолжительности деления и роста клеток в эпидерме и мезофилле формируются палисадная и губчатая ткани. У эпидермальных клеток процесс растяжения длительнее, чем у палисадных, вследствие этого возникают межклетники.
Голосеменные. У проростков голосеменных формируются последовательно три отдельных типа листовых придатков. Первый тип — семядоли, формирующиеся в зародыше, второй тип — первичная хвоя у молодых проростков, третий тип — вторичная хвоя, образующая настоящую листву. Рост листа голосеменных начинается с листового зачатка, локализованного по бокам апикальной меристемы. Ось листа образуется благодаря апикальному росту и активности меристемы жилки, но апикальный рост менее продолжителен. У узкой листовой пластинки начинается маргинальный рост.
Особенности роста листьев в течение сезона. Известно несколько типов образования листьев кроны. У некоторых видов максимальная площадь листьев достигается уже в начале вегетационного периода и на протяжении года новые листья не появляются. У других видов новые листья в течение сезона добавляются за счет постоянного образования и разрастания новых листовых зачатков или путем нескольких периодических вспышек роста, способствующих формированию, раскрьгцйо почек и их распусканию. Продолжительность разворачивания отдельных листьев урнзных видов различна, что также зависит от типа побега, его
Иматативный рост
59
положения на дереве, от окружающих условий, особенно температуры и снабжения водой.
Так, у яблоки распускание листьев из спящих почек происходило в течение 2 дней, другие листья распускались на протяжении 15 дней. У березы пушистой и гойоля осинообразного ранние листья, которые содержались в зимних почках, разворачивались в течение 2 недель после раскрывания почек. Отдельные листья ииноградника могут разворачиваться и созревать на протяжении 40 дней.
В противоположность быстрому росту отдельных листьев листопадных видов покрытосеменных, у многих голосеменных листья разрастаются медленно. Например, растяжение хвоинки сосны веймутовой в Англии продолжалось до начала августа. В Соединенных Штатах растяжение междоузлий у сосны веймутовой и сосны смолистой заканчивалось в начале вегетационного периода. Однако xbohip ки удлиняются все лето. У некоторых голосеменных хвоя часто увеличивается в длину более года, например у пихты субальпийской и сосны болотной. Листья вечнозеленых покрытосеменных обычно разворачиваются медленно. Например, листья цитрусовых увеличивались в размере в течение 130 дней. Даже после того, как пластинка листьев вечнозеленых цитрусовых полностью развернется, они могут продолжать утолщаться и у них увеличивается сухой вес. Листья листопадных деревьев могут также утолщаться и увеличивать сухой вес в течение большей части вегетационного периода (см. рис. 13.21).
ИНДЕКС ЛИСТОВОЙ ПОВЕРХНОСТИ. Поверхность листа часто выражают в виде индекса листовой поверхности (ИЛП), т.е. отношения площади листовой поверхности растения или насаждений к площади поверхности почвы под растениями. Поскольку единицы измерения площади листьев и почвы одни и те же (м2), ИЛП не указывает на размеры всего листового покрытия. Величина ИЛП зависит от размеров растения, возраста, пространственного распределения и других факторов, влияющих на размеры листьев. Многие сообщества листопадных древесных растений имеют ИЛП от 3 до 6, широколиственных вечнозеленых растений — до 8. У большинства голосеменных ИЛП значительно более высокий, часто достигающий 16. У сообщества пустынных растений значения ИЛП более низкие, а у некоторых быстрорастущих покрытосеменных ИЛП выше 8. Например, у гибридных тополей при хороших условиях ИЛП необычайно высок и достигает значений от 16 до 45, в за-
висимости от распределения листьев в кроне.
Самая высокая продуктивность леса будет при условии такого расположения листь ев, при котор ом фотосинтетически активная радиация абсорбируется как можно полнее. Обычно это достигается при ИЛП около 4 (рис. 3.5). При ИЛП менее 4 освещенность каждого отдельного растения будет высокой, а следовательно, и эффективность фотосинтеза также будет высокой. Несмотря на это, урожай на единицу поверхности почвы в таком открытом насаждении будет низкий. Максимальная суммарная величина продуктивности достигается при ИЛП от 8 до 10. При очень тесном пространственном
Индекс листовой поверх-
ности
Рис. 3.5. Модель продуктивности леса (Odum, 1971):
1 — валовая первичная продуктивность (ВПП) J 2 — чистая (нетто) первичная продуктивность (ЧПП); 3 -оптимальная пр одуктив-
ность зрелого леса
60 Глава 3
распределении деревьев, листья сильно перекрывают друг друга, интенсивность света в затененных местах недостаточна, чтобы поддержать положительный баланс СОг- Вследствие этого видимый фотосинтез и урожай снижаются из-за больших потерь на дыхание, необходимого для поддержания большого объема листьев и опорных тканей.
ТИПЫ ПОБЕГОВ И СПОСОБЫ ИХ РОСТА
Побеги классифицируются по местоположению, характеру развития или типу почек, из которых они образовались. По местоположению почки различают терминальные, латеральные и базальные побеги. Порослевые побеги - побеги, возникшие из спящих почек, расположенных вблизи от основания ствола древесного растения. Как уже упоминалось, корневая поросль образуется из адвентивных почек на корнях. Другие важные типы побегов - это детерминированные и недетерминированные, развивающиеся из спящих почек, преформированные и гетерофильные побеги, периодически появляющиеся побеги, удлиненные и укороченные побеги, аномальные позднесезонные побеги.
ДЕТЕРМИНИРОВАННЫЕ И НЕДЕТЕРМИНИРОВАННЫЕ ПОБЕГИ. У некоторых древесных растений (сосны, ели, дуба и гикори) побеги образуются из терминальных (верхушечных) почек главной оси и его основных ветвей. После удлинения терминальных побегов наступает период неактивного состояния до сформирования и раскрытия новых терминальных почек. У таких видов растений с детерминированным (моноподиаль-ным) ветвлением ежегодно может раскрываться только одна концевая почка и образовываться один побег. Иногда могут сформироваться и раскрыться последовательно две и более почек в течение 1 года, при недетерминированном (симподиальном) ветвлении побеги образуются не из концевых почек главного ствола, а из почек вторичных осевых ветвей. Симподиальный рост возникает часто в случаях, когда на концах ветвей расположены репродуктивные структуры или недоразвиты кончики веток. У видов с недетерминированным ветвлением противолежащие почки, которые часто ошибочно принимают за концевые, можно рассматривать как боковые, возникшие в результате недоразвитости верхушки побега. Недоразвитые верхушки побегов встречаются часто у таких родов, как Betula, Carpinus, Catalpa, Corylus, Diospyros, Gleditsia, Plat anus, Robinia, Salix, Tilia и Ulmus.
ПОБЕГИ, РАЗВИВАЮЩИЕСЯ ИЗ СПЯЩИХ ПОЧЕК. Спящие почки на главном стебле или на боковых ветвях деревьев часто активируются в результате внезапного изменения интенсивности света и образуют побеги (так называемую жировую поросль). Побеги, возникающие из спящих почек, нежелательны, так как они образуют сучки, снижающие качество лесоматериалов. Такие побеги появляются у покрытосеменных чаще, чем у голосеменных. Их образование зависит от степени воздействия различных факторов среды и от вида растений. У разных видов формируется различное количество спящих почек, поэтому тенденцию к порослеобразованию можно предвидеть. Например, дуб имеет склонность к обильному порослеобразованию из жировых побегов, у ясеня поросль возникает редко (таб. ЗЛ). У молодых и небольших
|Йтгмвный рост
61
1,1. ВИДОВЫЕ РАЗЛИЧИЯ СПОСОБНОСТИ К ОБРАЗОВАНИЮ И ЛЕВЫХ ПОБЕГОВ
(Nunih, 1966; Trimble, Seegirst, 1973)
Ко'Отчество порослевых IIO-вегон Виды Количест- Виды во порослевых побегов
Очень Дуб белый
МНОГО Дуб красный
Много Липа американская Черемуха поздняя Дуб каштановый
Мило Бук
Мало Кария
Тюльпанное дерево Клен красный Клен сахарный Береза вишневая Очень мало Ясень американский
деревьев поросль образуется в большем количестве, чем у крупных и старых экземпляров. Угнетенные деревья по сравнению с мощными экземплярами того же вида жируют реже. На формирование побегов из спящих почек также влияет густота стояния деревьев в лесных массивах. Например, деревья дуба белого в хорошо прореженном древостое формировали из спящих почек более 35 побегов на одно дерево; у сред-нспрореженных - 21, у непрореженных — 7. Блюм (1963) отметил, что склонность к образованию жировых побегов более выражена у деревьев, растущих на границе древостоя, чем у произрастающих в средней его >исти.
ПРЕФОРМИРОВАННЫЕ И ГЕТЕРОФИЛЬНЫЕ ПОБЕГИ. Побеги могут образовываться путем фиксированного роста, неограниченного роста или тем и другим способами одновременно. При так называемом фиксированном росте происходит растяжение преформированных частей побега после периода покоя. Например, зимние почки взрослых деревьев многих видов покрытосеменных и голосеменных содержат примордии (зачатки) всех листьев, которые распустятся весной. У таких видов дифференциация в почках происходит в первый год п, а растяжение всех укороченных частей в побег осуществляется на следующий год п + 1. Такой ход образования побегов обнаружен у некоторых северных видов сосны (сосен смолистой и веймутовой), ели, тсуги и бука. У этих видов растяжение преформированных (фиксированных) побегов полностью заканчивается в короткий срок, иногда в первую половину безморозного периода. У тсуги западной, например, растяжение преформированных побегов происходит примерно в течение 7 недель.
Время формирования листьев в фиксированных почках неодинаково. У многих видов все листья в почках, которые вскроются весной, сформировались поздно осенью предшествующего года п. У некоторых видов все листья могут сформироваться в почках уже в середине лета года л, у других, немногих, .видов они формируются незадолго перед раскрыванием почек года л + 1. Быстрорастущие клоны деревьев образуют больше листьев, чем медленнорастущие, которые продолжают
62
Глава 3
12 г
Рис. 3.6. Развитие зимней почки Populus trichocarpa. Первый лист недоразвит. Хри листовых зачатка (8 -10) изображены в увеличенной шкале вверху справа (Critch field, 1960) 1 — 10 — порядковые номера листьев от основания почки
накапливать листовые зачатки в почках поздней осенью года п. При неограниченном росте побегов в противоположность фиксированному росту растяжение побегов происходит одновременно вследствие заложения и растяжения новых частей побега.
У взрослых деревьев некоторых видов одни побеги полностью пре-формированы в почке (фиксированы), другие побеги не преформиро-ваны. Преформированные побеги образуют листья рано и обычно бывают небольшого размера. Междоузлия побегов гинкго длиной всего 1—2 мм, у клена пенсильванского 1—2 см. Побеги, не полностью преформированные в зимних почках, обычно развиваются длинными и гетерофильными, так как образуют листья двух видов: 1) ранние листья, относительно хорошо развитые в зимних почках; 2) поздние листья, распускающиеся из листовых зачатков, заложенных в почках, или чаще всего из листовых примордиев, которые продолжают формироваться и расти в течение года п +1 во время растяжения побега (рис. 3.6 и 3.7). Такой неограниченный рост напоминает тип роста у травянистых растений. К концу вегетационного периода у гетерофильных побегов листьев оказывается больше, чем было заложено в зимних почках. На этих побегах имеются листья двух типов, различающиеся по таким признакам: размеру, , дольчатости, жилкованию, зубчатости, толщине пластинки и развитию устьиц. Ранние листья обычно крупнее поздних (рис. 3.9). К древесным растениям, у которых обнаружен неограниченный рост, относятся: тополя, яблони, береза пушистая, береза желтая, эвкалипт, жимолость, несколько видов клена, гинкго, лиственница и некоторые тропические сосны.
У немногих видов, которые ранее из-за невнимательных наблюдений относили к видам, образующим преформированные побеги в зимних почках (т.е. проявляющих исключительно фиксированный рост), часто на стадии проростка обнару-
Даты
Рис. 3.7. Развитие гетерофильного побега Populus trichocarna (1958) (Critchfield, 1960)
а - рост в длину ранних листьев (1—6); б - пд^дьих листьев (7—12); в — рост междоузлий (4—12) ; порядковые номера междоузлий те же, что и номера листьев на верхнем конце; стрелкой указано формирование терминальной верхушечной почки
64
Глава 3
живаются как неограниченный, так и фиксированный способы роста. Например, у ели черной неограниченный рост побегов происходил только у молодых проростков. У видов, отнесенных к гетерофильным, склонность к неограниченному росту и образованию удлиненных побегов снижается с возрастом дерева. Например, у 15-летнего дерева гинкго развивались только удлиненные побеги, а у 100-летнего дерева были и укороченные и удлиненные побеги. У 6-летнего дерева тополя осинообразного удлиненные побеги составляли 13% кроны, а у 52-летнего дерева удлиненных побегов не было.
Козловский и Клаузен (1966) обнаружили у взрослого дерева тополя осинообразного лишь предетерминированные побеги, при раскрывании почек весной у них распускались все листья. У кленов пенсильванского и красного в раннем возрасте укороченных побегов было значительно больше, чем удлиненных, а у старых деревьев ежегодно формировалось всего 4—5% удлиненных побегов.
Следует соблюдать осторожность при классификации деревьев на группы по способу образования побегов, если род включает большое количество видов, имеющих различные ареалы. Например, у представителей рода Pinus, насчитывающего более 100 видов, распространенных от тропиков до Арктики, развитие побегов очень разнообразно. У некоторых видов северных сосен побеги развиваются исключительно путем фиксированного роста, у тропических сосен побеги образуются только за счет неограниченного роста.^Строение и способы роста побегов различны у видов Асет. Некоторые виды имеют побеги, полностью префор-мировэнные в зимних почках, другие виды образуют удлиненные гетерофильные побеги. Однако большинство видов рода Acer принадлежит к гетерофильной группе, в которую входят: A. rubrum, A. saccharinum, A. spicatum, A. pseudoplatanus, A. tataricum, A. ginnala, Al caudatum, A. campestre, A. monospessulanum, A. orientale. К видам рода Acer, побеги которых почти полностью преформированы в почках, относят-l/ ся Acer saccharum и Acer platanoides. У большинства гетерофильных видов Acer ранние и поздние листья четко различаются и по размерам
Рис. 3.8. Изменения формы и размеров ранних и поздних листьев нескольких гетерофильных видов Асет (Critchfield, 1971);
1 - A. tataricum; 2 - A. rufinewa; 3 - A. campestre var. leiocarpum; 4 -A. buergerianum;5 - A monspessulanum;6 - A. orientale
Вегетативный рост
65
Рис. 3.9. Изменение типов роста в высоту в течение вегетационного периода северных и южных видов сосен. Две северные сосны имеют пр сформированные побеги (ограниченный рост) - у них только один период побегообразования в год; у южных сосен периодический рост побегов (Kramer, 1943) :
1 — веймутова (Сев: Каролина); 2 — смолистая (Сев. Каролина) ; 3 — смолистая (Нью-Гемпшир) ; 4 — веймутова (Нью-Гемпшир); 5 — среднее для трех южных сосен (Сев. Каролина) ; 6 — безморозный период в Нью-Гемпшире; 7 — в Сев. Каролине
Рис. jl.lO. Сезонный рост в высоту молодых деревьев гетерофильного тополя дельтовидного
и форме (рис. 3.8). У A. sacchannum поздние листья удлиненных побегов лишь слегка отличаются от ранних листьев, расположенных у основания побега.
’ ВОЛНОВОЕ” ОБРАЗОВАНИЕ ПОБЕГОВ. У некоторых сосен умеренной зоны (например, сосен ладанной и замечательной), большинства тропических сосен и у многих тропических и субтропических двудольных рост побегов происходит периодически в виде волн или вспышек в течение вегетационного периода. При этом на побеге распускается несколько терминальных почек в год. После формирования побега из первой почки с ее фиксированными дополнительными листьями сразу же образуется вторая почка на апексе того же самого побега. Эта почка очень скоро распускается, давая удлиненный побег. За второй фазой роста могут следовать дополнительные волны роста из почек, последовательно распускающихся возле верхушки того же осевого побега. У южных сосен США (например, ладанной и длиннохвойной) первая вспышка роста в вегетационном периоде обычно самая продолжительная. Количество последовательно формирующихся и раскрывающихся почек на одном и том же побеге не постоянно у отдельных деревьев и видов и зависит от места положения побега на стволе, а также от климатических условий. Терминальным лидерам и побегам верхних мутовок свойственно обильное формирование почек, и у них бывает больше периодов побегообразования, чем у нижних ветвей. Так, терминальный лидер дерева среднего возраста сосны ладанной не увеличивался за год более чем в 2—3 раза, но у некоторых видов было зарегистрировано до 7 йоследовательных удлинений побега в течение лета.
66
Глава 3
ПРОДОЛЖИТЕЛЬНОСТЬ РОСТА ПОБЕГОВ В ТЕЧЕНИЕ ВЕГЕТАЦИОННОГО ПЕРИОДА. Этот показатель значительно варьирует у разных видов: у одних побег может полностью закончить рост в течение нескольких недель, у других этот процесс длится несколько месяцев (рис. 3.9). Длительность роста побегов зависит также от времени развития компонентов побега в почке, У видов с полностью преформиро-ванными, неразвернутыми побегами и побегами в зимней почке (фиксированный рост) побеги удлиняются очень быстро, например у некоторых дубов через 2-- 3 недели.
Побеги гетерофильных видов, не полностью преформированные в зимних почках, удлиняются значительно дольше, по сравнению с удлинением пр сформированных побегов. Например, у побегов молодых деревьев тополя дельтовидного удлинение продолжалось с конца апреля до конца сентября (рис. 3.10). Аналогично увеличивались в размерах с конца мая до сентября побеги яблони, которым свойствен неограниченный рост. У видов с периодическим побегообразованием удлинение побегов длится обычно очень долго и волнообразно.
АНОМАЛЬНЫЕ ПОБЕГИ, ФОРМИРУЮЩИЕСЯ В КОНЦЕ ВЕГЕТАЦИОННОГО ПЕРИОДА. У некоторых деревьев в конце вегетационного периода наблюдается аномальный взрыв побегообразования из только что сформированных почек, которые должны были бы раскрыться на следующий год. Позднеосенние побеги бывают двух основных типов: Ивановы побеги, появляющиеся из терминальной почки? и пролепти-ческие побеги, .образующиеся за счет пробуждения боковых почек у основания терминальной почки. На одном и том же дереве могут быть либо побеги только одного вида — Ивановы или пролептические, либо появляются те и другие вместе. Они могут быть короче или длиннее побегов, появившихся в первый ранний период побегообразования. Аномальные позднесезонные побеги характерны для многих голосеменных и покрытосеменных. Они обнаружены у Quercus, Fagus, Carya, Alnus, Ulmus, Pinus, Pseudotsuga.
Формированию Ивановых и пролептических побегов часто способствуют обильные осадки в конце вегетационного периода. Но способность к образованию этих побегов находится под строгим генетическим контролем. Установлено, что склонность к формированию аномальных позднесезонных побегов у экземпляров сосны Банкса, выращенных из семян южной репродукции, выражена сильнее по сравнению с удлинением побегов из семян северной репродукции.
Ивановы и пролептические побеги не всегда достаточно зимостойки, поэтому они часто повреждаются зимой. Такие побеги могут стать причиной неправильного формирования ствола. Раздвоение ствола вызывается появлением пролептических побегов. При исследовании обнаружено раздвоение ствола в результате позднеосеннего образования пролептических побегов у 63—94% обследованных деревьев сосны смолистой. При формировании на одной и той же ветви Ивановых и пролептических побегов между ними возникает конкуренция за приоритет в апикальном доминировании.
РАЗЛИЧНЫЕ ТИПЫ РОСТА ПОБЕГОВ ОДНОГО ДЕРЕВА. На одном и том же дереве часто встречаются побеги, обладающие различными типами роста. Например, скорость роста и количество пневых побегов
вегетативный рост
67
1.2. РОСТ ЛИСТЬЕВ И МЕЖДОУЗЛИЙ УДЛИНЕННЫХ ПОБЕГОВ НЕПОДСТРИЖЕННЫХ ДЕРЕВЬЕВ И ОТРАСТАЮЩИХ ПОБЕГОВ ДЕРЕВЬЕВ КЛЕНА КРАСНОГО, ПОДСТРИЖЕННЫХ В РАЗНОЕ ВРЕМЯ ГОДА В 1967 Г.
(Wilson, 1968)
Время подстригания Число пар листьев, образуемых за неделю Длина листьев, мм Длина междоузлий, мм
Максимальная Максимальное удлинение за неделю Максимальный прирост в высоту за неделю, мм
Максимальная Максимальное удлинение за неделю
Неподстриженные 1,о-1;з 87-116 20-30 78-86 25-40 79-126
деревья Осенью 1966 г. 1,3-2,3 180- 208 44-66 107-183 50-88 202-306
31 мая 1967 г. 1,0-1,6* — — — 87-141
14 июня 1967 г. 0,8-1,5 — — — — 70-133
30 июня 1967 г. 1,0-1,3 — — — 114-170
28 июля 1967 г. 1,0-1,5 — — — 10-86
24 августа 1967 г. 1,0=** — — — — 3
* Для отрастающих побегов, подстриженных от 31 мая по 24 августа, взяты только данные по скорости образования пар листьев й скорости роста в высоту.
** Только один отрастающий побег; одна почка Дала одну пару листьев.
иногда значительно превосходит скорость роста побегов другого типа. У клена красного рост листьев и междоузлий пневых побегов у деревьев, обрезка которых производилась зимой, был значительно интенсивнее роста удлиненных побегов контрольных (необрезанных) деревьев (табл. 3.2). Интенсивность роста зависела от времени обрезки деревьев.
Апикальное доминирование. У большинства голосеменных терминальный побег удлиняется значительно больше боковых ветвей, расположенных ниже его. Это приводит к образованию более или менее конической формы дерева, которую часто описывают как сбегающее ветвление. Например, у сосны смолистой терминальный лидер ежегодно вырастает значительно больше любой из боковых ветвей, а годичный прирост ветвей в длину максимальный в самой верхней мутовке (табл. 3.3). Уменьшается он в мутовках, расположенных ниже. Таким образом, очевидно строго коррелятивное ингибирование роста, в результате которого величина ежегодного прироста побегов в длину снижается прогрессивно сверху вниз и внутрь кроны дерева. Фрэзер (1962) также описал апикальное доминирование у ели канадской. У четырех боковых ветвей двух верхних мутовок удлинение в среднем составляло 2/3 от удлинения терминального лидера. Прирост в длину ветвей нижних мутовок был значительно меньше, чем у ветвей верхних мутовок. Апикальное доминирование не наблюдается лишь у немногих голосеменных. Например, у ветвей второго порядка Araucaria excelsa отсутствует наследственная способность к апикальному доминированию, поэтому
68
Глава 3
удаление верхушечного побега не вызывает формирования нового главного побега из какого-либо бокового. Укорененные черенки этого вида могут расти горизонтально в течение длительного времени. У ряда сосен апикальное доминирование сохраняется на протяжении многих лет, у некоторых же видов, таких, как Pinus pinea, эта закономерность утрачивается рано.
Различия в характере роста побегов у одного и того же дерева более свойственны голосеменным, чем покрытосеменным. У некоторых покрытосеменных многие боковые побеги удлиняются с одинаковой скоростью, образуя многочисленные ветви и округлую крону. О таких деревьях говорят, что у них низбегающее или многоствольное ветвление. Такой способ ветвления характерен для ильма, который обычно лишается своей терминальной почки, в результате чего ветвление продолжается до тех пор, пока главный ствол не утратит основного положения в широкой раскидистой вершине. У плодовых деревьев скорость и длительность роста побегов различная. Одни побеги растут в течение всего вегетационного периода, другие распускаются рано, и удлинение прекращается очень быстро. Для некоторых побегов характерен преры-
З.Э ВАРИАЦИИ В УДЛИНЕНИИ ПОБЕГОВ РАЗЛИЧНОЙ ЛОКАЛИЗАЦИИ НА СТВОЛАХ 6-ЛЕТНИХ ХВОЙНЫХ ДЕРЕВЬЕВ, СМ
Хвойные Основная ось (главный побег) Номер осевой мутовки второго Номер осе-
порядка вой мутов- ки третич- 4 иого поряд- ка
1 2 3
2
Сосна смолистая 57,7±6,2 34,6 ±7,7 32,5 ±4,9 27,7+5,4 15,7±8,О 17,1±5,8
Сосна веймутова 444*7,6 25,5 ±8,2 24,8±7,0 19,1±4,1 14,1 ±2,2 12,3+5,1
Ель канадская 33,5 ±5,7 19,8±4,1 17,4±3,5 15,3±2,8 11,8 ±3,0
Ель черная 23,8±6,4 17,3±3,5 16,5 ±2,5 14,1±3,3 10,0 ±3,2
ПРОДОЛЖЕНИЕ ТАБЛ. 3.3
Хвойные Номер осевой мутовки третичного Номер осевой мутовки
порядка четвертичного порядка
3 4 3 4
верхнее нижнее верхнее нижнее положе- положе- положе- положение ние ние ние
Сосна смолистая 15,3±3,4 13,7±4,5 11,5±4,9 8,1 ±4,8 6,2±2,7 4,7±2,5
Сосна веймутова 10,5±3,4 9,5±3,4 7,8±8,4 7,3±3,7 4,3±О,6 3,О±О,5
Ель канадская - — - " - — -
Ель черная _____ _
Негативный рост
69
иистый рост: между двумя вспышками бурного роста у них бывает период замедленного роста. Есть и такие побеги, у которых происходят периодические вспышки роста с незначительными периодами покоя между ними. (Значение апикального доминирования в формировании дерева обсуждается в главе 16.)
Наличие апикального доминирования необходимо учитывать лесоводам. Когда лидирующий побег сосны веймутовой поражался долгоносиком (Pissodes strobi), а один из боковых побегов первой мутовки шиимал главное положение, рост других побегов этой мутовки подавлялся. Конкуренция между побегами длится продолжительное время, пока один из побегов мутовки не окажется доминирующим, а другие будут подавлены. На протяжении этого времени дерево, как потенциальный строевой материал, теряет ценность из-за раздвоения, которое развивается в стволе в результате повреждения главного побега. Подавление апикального доминирования часто оказывается желательным у новогодних елок. Многие хвойные имеют большие междоузлия у 1лавного ствола и ветвей, что придает им веретенообразный вид. Стрижкой и обрезанием отдаленных от главного ствола побегов или удалением почек удается затормозить удлинение побегов новогодних елок и стимулировать раскрывание спящих почек, а также вызвать формирование и последующее раскрывание новых почек. Эти приемы способствуют образованию новых боковых побегов и созданию пушистых, высокого качества новогодних праздничных елок.
Удлиненные и укороченные побеги. Несколько родов голосеменных, таких, как Larix, Ginkgo, и некоторые покрытосеменные, например Betula, образуют как удлиненные побеги с растянутыми междоузлиями, так и укороченные побеги (иногда их называют пшоровыми побегами). У них нет заметных междоузлий, а листья собраны в пучки па концах побегов.
У лиственницы вся хвоя на укороченных побегах при раскрывании почек появляется сразу. Почки быстро разворачиваются и, как правило, хвоинки бывают несколько длиннее, чем ранняя хвоя удлиненных побегов. В этом случае не происходит удлинения междоузлий. На удлиненных побегах образуются и ранние и поздние хвоинки. Вся ранняя хвоя появляется при открывании почки. Поздняя хвоя образуется постоянно в течение большей части лета. На удлиненных побегах лиственницы вся базальная хвоя и около половины стеблевых хвоинок пре-формированы в почках. Заметных морфологических различий между преформированной в почках хвоей и хвоей, которая формировалась путем неограниченного роста после открывания почек, не обнаружено.
МАКСИМАЛЬНЫЙ РОСТ В ВЫСОТУ. Высота взрослых деревьев у разных видов значительно варьирует. Самые высокие деревья видов Sequoia и Eucalyptus. Менее высокие, достигающие 65 м в высоту, деревья дугласии, пихты благородной и сосны веймутовой. Максимальная высота деревьев обычно зависит больше от их долговечности, чем от ежегодной скорости роста побегов, типа образуемых побегов или продолжительности удлинения побегов в вегетационном периоде. Деревья тополей крупнозубчатого и осинообразного часто растут быстро, но никогда не достигают большой высоты, так как они недолговечны.
70 Глава 3
Некоторые относительно медленнорастущие, но долгоживущие деревья, такие, как дуб белый, часто достигают более 30 м в высоту.
Относительно долгоживущая сосна Ламберта, обладающая яреформи-рованными побегами в зимних почках, в зрелом возрасте выше сосны замечательной, для которой характерно периодическое побегообразование. Хотя скорость роста в высоту сосен ладанной и ежовой (у обоих видов периодическое побегообразование) в течение 70 лет выше по сравнению со скоростью роста в высоту у сосны веймутовой, последняя в зрелом возрасте достигает большей высоты. Это обусловлено тем, что сосна веймутова продолжает расти в высоту с большой скоростью около 100 лет. Большая высота некоторых видов, таких, как дугласия и ель ситхинская, обусловлена высокой скоростью роста, сохраняющейся на протяжении многих лет у этих долгоживущих видов.
РОСТ ПОБЕГОВ В ТРОПИКАХ
Характер роста побегов древесных тропических растений очень разнообразен. Рост у них происходит скачкообразно, в течение года бывает до нескольких вспышек побегообразования. Так растут побеги у деревьев какао, кофе, маслин, цитрусовых, каучуконосного фикуса, чая, манго и многих видов лесных деревьев. Интервалы между периодами побегообразования различны, что зависит от вида, способов культивации, климатических режимов, индивидуальных особенностей особи в пределах вида и даже от свойств каждой ветви данного дерева. Классификация типов роста у многих видов затруднительна, так как способы роста их побегов широко варьируют в различных регионах. Например, виды Thespasia и Duabanga в Сингапуре считаются вечнозелеными, а в Индии — листопадными. В то время как у Acrocarpus fraxinifolius, Chlorophora excelsa и Schizolobium excelsum отмечается склонность образовывать побеги 12 раза в год, у деревьев какао может быть до пяти вспышек побегообразования. У манго интервалы между периодами бурного побегообразования иногда бывают более 12 месяцев, у чая -всего несколько дней.
В тропическом климате с характерным чередованием влажных и сухих периодов побегообразование у растений также сезонное. В Бразилии, например, большая часть вспышек роста побегов у деревьев какао отмечается в сентябре-октябре, в ноябре и апреле бывает всего две-три вспышки. У цитрусовых в зависимости от локализации побегов на дереве и климатических условий обычно имеется две мощные и до трех слабых вспышек побегообразования. У молодых деревьев данного вида периоды побегообразования бывают чаще, чем у старых деревьев: например, у молодых деревьев Litchi chinensia и какао отмечено больше периодов побегообразования, чем у старых деревьев. У молодых деревьев манго побеги образуются непрерывно, а у старых деревьев между периодами роста побегов имеются продолжительные интервалы неактивного состояния. У проростков Bombax в Нигерии новые листья появлялись на протяжении 8 месяцев в году, у старых деревьев лишь в течение 1—3 месяцев.
^негативный рост 71
Удлинение междоузлий и разворачивание листьев у многих тропических деревьев может быть очень быстрым. Бамбук способен увеличиваться в высоту за день на 1 м. Примеры быстрого роста в высоту приводят в своей работе Лонгман и Джник (1974): у Terminalia superba ДК м за год, у Musanga cecropioides 3,8 м, у Ochroma lagopus 5,5 м. 'hi кая высокая скорость роста часто определяется произрастанием на открытых пространствах или на границе леса. Внутри лесных сообществ скорость роста деревьев значительно меньше. Она быстро снижается 1акжс с увеличением возраста деревьев. У тропических древесных растений проявляется несколько типов соотношения между заложением листовых примордиев и появлением листьев. Например, у Огеорапах большая часть листовых зачатков формируется незадолго перед разворачиванием листьев. У чая зачатки листьев образуются более или менее постоянно, но листья разворачиваются только во время вспышек роста. Другой тип роста встречается у Rhizophora. У этого вида формирование листовых зачатков и разворачивание листьев идет синхронно, скорость заложения листьев в разные периоды вегетации различна.
В тропических лесах листья опадают постоянно. Однако бывают и периоды более интенсивного сбрасывания листьев, что зависит от вида растения и количества выпадающих осадков. Опадение листьев у растений тропических лесов легко вызывается даже слабым водным стрессом. Лонгман и Джник (1974) различают четыре типа деревьев тропических лесов по способности удерживать и сбрасывать листья (рис. 3.11), но они указывают, что резкой границы между типами провести невозможно: 1. Периодический рост, листопадный тип. Почки раскрываются сразу после опадения листьев. Период жизни листьев 4—11 месяцев. Без листьев все деревья или отдельные ветви бывают от нескольких недель до нескольких месяцев. Между опадением листьев и раскры-
тие. 3.11. Типы отделения и сохранения листьев у тропических лесных деревьев: а — периодический рост, листопадный тип; б — периодический рост, тип, меняющий листья; в — периодический рост, вечнозеленый хип; г — непрерывный рост, вечнозеленый тип (Longman, Jenik, 1974)
72
Глава 3
тием почек видимой связи нет (например, ТегшшаНа ivorensis). 2. Периодический рост, характерна смена листьев. Опадение листьев совпадает с открыванием почек. Период жизни листьев часто равен 6-12 месяцам. Новые листья появляются почти одновременно с опадением старых. У Terminalia catappa это происходит дважды в году, у Dillenia indica, Entand ophragma angolense и Ficus variagata - 1 раз в год. 3. Периодический рост, вечнозеленый тип. Листья опадают через длительный срок после раскрытия почек. Период жизни листьев 7—14 месяцев. Отдельные ветви или деревья целиком вечнозеленые. Примером могут служить Celt is mildbraedii и Mangifera indica. 4. Непрерывный рост, вечнозеленый тип. Закладка и опадение листьев происходят непрерывно. Спящие почки не образуются. Период жизни листьев варьирует, но может быть до 14 месяцев. Образование листьев и опадение их нерегулярно и зависит от окружающих условий. К такому типу относятся: Trema guineensis, Dillenia auffruticosa и молодые сеянцы некоторых других видов.
Рост побегов сосны в тропиках иногда не отличается от роста побегов в умеренной зоне, но может и существенно отличаться. Нормальный рост характеризуется периодическим побегообразованием. При этом ось увеличивается за счет растяжения последовательно формирующихся почек на терминальном лидере главного ствола. После периода растяжения рост ствола прекращается и сразу же формируется пучок новых терминальных почек. Вскоре после этого сформированные почки увеличиваются в объеме, терминальный лидер удлиняется и одновременно формируются мутовки боковых ветвей. Обычно происходит от двух до четырех таких вспышек роста в год. Наоборот, некоторые деревья сосны развиваются аномально и растут непрерывно, так как не способны к заложению почек, из которых должны формироваться боковые ветви. Лойд (1914) описал такой тип роста и назвал его ’’лисохвос-тым”, так как верхняя часть ненормально вытянутого побега напоминает хвост лисы.
Эта удивительная форма возникает в результате чрезмерно усиленного апикального доминирования и часто это проявляется в отсутствии у дерева ветвей на протяжении 6, а иногда и 13 м. Такая реакция на окружающие условия является проблемой при выращивании сосны в тропиках. К образованию ’’лисьих хвостов” способны такие виды сосны; Pinus canariensis, Р. caribaea, Р. cembroide, Р. echinata, Р. elliottii, Р. ke-siya (insularis), P. mercusii, P. оосагра, P. palustris, P. radiata, P. taeda, P. tropicalis. Формирование ’’лисьего хвоста” — наглядный пример неограниченного роста. Недалеко от верхушки побега в форме ’’лисьего хвоста” неразвернувшаяся хвоя плотно окружена цельной оберткой. Несколько ниже этого образования хвоинки проходят сквозь обертку и увеличиваются в размерах, придавая верхней части непрерывно разворачивающегося побега коническую форму. Около апекса побега постоянно формируются новые зачатки хвои, которые последовательно разворачиваются, развиваются и увеличиваются в размерах. В периоды формирования ’’лисьего хвоста” хвоя дольше сохраняется на дереве. По наблюдениям, проведенным в Малой Азии, части ствола, лишенные ветвей, сохраняют хвою в течение нескольких вегетационных периодов,
Вегетативный рост 73
но теряют их сразу, как только дерево возвращается к нормальному способу роста.
Различные степени образования ’’лисьих хвостов” наблюдались в Малайзии на обширных плантациях Р. caribaea var. hondurensis у деревьев В возрасте до 15 лет. Например: 1. У некоторых деревьев после пересаживания в поле непрерывный рост терминального лидера продолжался без пауз. В результате на единственном стебле не было ни одной боковой ветви. 2. Образование ’’лисьего хвоста” начиналось сразу после пересаживания в поле и прекращалось через несколько лет, после чего начиналось нормальное формирование побегов. 3. Деревья некоторое время росли нормально, а затем начиналось формирование ’’лисьих хвостов”. 4. Образование ’’лисьих хвостов” происходило сразу крепе пересадки в поле. Через несколько лет процесс прекращался и затем начиналось формирование главного побега. Таким образом, наблюдались переходы от непрерывного растяжения побегов к периодическому побегообразованию и наоборот.
Сосны, дающие ’’лисьи хвосты”, и сосны с периодическим побегообразованием обеспечивают прекрасный экспериментальный материал для изучения гормональных (Отношений, регулирующих рост побегов, (Апикальное доминирование рассматривается в главе 16.)
В большинстве сообщений процесс образования ’’лисьих хвостов” рассматривается как присущий терминальному побегу, однако боковые побеги также обладают этой способностью. Такие боковые побеги постепенно или внезапно становятся ориентированными вертикально. Апикальные участки ветвей, несущих ’’лисьи хвосты”, морфологически не отличаются от аналогичных частей главного побега. Чередование нормального роста и роста с образованием ’’лисьего хвоста” иногда приводит к формированию у отдельных деревьев гротескных форм.
Большинство лесоводов считают процесс появления ’’лисьих хвостов” нежелательным явлением, так как при этом снижается устойчивость деревьев к ветрам, уменьшается рост по диаметру, отсутствует поздняя древесина, образуется реактивная, снижается продуктивность семян.
КАМБИАЛЬНЫЙ РОСТ
Рост деревьев по диаметру происходит преимущественно за счет меристематической активности камбия, цилиндрической латеральной меристемы, локализованной между ксилемой и флоэмой ствола, ветвей и корней. Камбий состоит из удлиненных веретеновидных клеток (веретеновидных инициалий) и коротких лучевых инициалей. Последние часто описываются как изодиаметрические клетки. Однако показано, что у Pinaceae лучевые инициали часто имеют различные размеры и бывают двух типов: 1) краевые Лучевые инициали, у которых радиальный диаметр равен длине оси или немного меньше ее; 2) радиально вытянутые инициали внутренней части лучей. Веретеновидные инициальные клетки и лучевые инициальные клетки отделены друг от друга.
Камбий обычно делят на ярусный и неярусный. У ярусного камбия (например, у белой акации) веретеновидные инициальные клетки располагаются горизонтальными рядами, что видно на тангентальных срезах. У неярусного камбия (например, у голосеменных и тополей) длинные веретеновидные инициальные клетки имеют перекрывающиеся
74
Глава 3
вторичный, рост
Зрелая флоэма
Дифференцирующаяся флоэма
Камбий
Дифференцирующаяся ксилема
____ Созревающая флоэма
Радиально разрастающаяся флоэма
Делящаяся флоэма (материнские клетки флоэмы) Камбиальная инициальная клетка (делящаяся)
Делящаяся ксилема (материнские клетки ксилемы)
Радиально разрастающаяся ксилема
Созревающая ксилема
Зрелая ксилема
Рис. 3.12. Терминология для описания типов клеток и тканей, связанных с камбиальным ростом (Wilson, et ‘al., 1966)
концы. У ясеня обыкновенного камбий занимает промежуточное положение между двумя типами, у него инициали как в ярусных ,так и в неярусных рядах.
Долгое время имелись разногласия относительно того, считать ли камбием только один слой камбиальных инициальных клеток, или к камбию следует отнести также их меристематические производные, материнские клетки ксилемы и флоэмы. Проблема заключается в том, что трудно индентифицировать единственный (однорядный) слой инициал ей. Установить наличие такого слоя часто полезно при использовании термина ’’камбиальная зона” для всего слоя делящихся клеток (однорядный слой плюс материнские клетки ксилемы и флоэмы). Удобная терминология для различных типов клеток и тканей, связанных с камбиальной активностью, приведена на рис. 3.12. Камбиальная зона у деревьев в состоянии покоя может включать 1 — 10 рядов клеток, у растущих деревьев ширина этой зоны сильно различается. Есть данные, что ширина камбиальной зоны у медленно растущих деревьев состоит из 16—18 рядов клеток, у быстрорастущих — из 12—40 рядов клеток.
ДЕЛЕНИЯ КЛЕТОК КАМБИЯ. В камбии возможно клеточное деление двух типов: аддитивное и мультипликативное. При аддитивном делении веретено видные инициальные клетки делятся перикл йнально (в тангентальном направлении), давая материнские клетки ксилемы и флоэмы, которые в свою очередь создают клетки ксилемы и флоэмы. При мультипликативном делении веретеновидные инициальные клетки делятся антиклинально (радиально), что приводит к увеличению количества периферических клеток камбия.
Формирование ксилемы и флоэмы. После зимнего покоя камбий деревьев умеренной зоны вновь активизируется и откладывает внутренние клетки ксилемы и наружные клетки флоэмы. Эта активность возобновляется под действием гормонов, которые синтезируются в апексе и перемещаются вниз по стволу (см. главу 16). Ежегодный прирост ксилемы и флоэмы обусловливается образованием новых слоев
в&мтативный рост
75
их тканей между ранее образованными, что приводит к утолщению лнола, ветвей и главных корней.
Большинство исследователей согласны с тем, что камбиальная реактивация происходит в два этапа, в результате которых изменяются некоторые внешние признаки камбия (изменяется цвет, клетки становится полупрозрачными и слегка набухают). Эти изменения связаны г митотической активностью, продуцирующей камбиальные производные. При второй фазе первые клеточные деления могут происходить прерывисто и возникать на разных уровнях ствола больших деревьев, обладающих почками на многочисленных боковых ветвях. Однако в начале сезонного роста камбия волна камбиального деления распространя
ется сверху вниз, возникая у основания почек.
Время образования ксилемы и флоэмы. Многие исследователи ранее считали, что ежегодному образованию ксилемы предшествует формирование флоэмы. Однако, как указал Эверт (1963), в ранних исследованиях дифференцирующиеся ситовидные элементы были изучены недостаточно квалифицированно. Кроме того, применялись разные кри-
терии для оценки развития клеток по обе стороны от камбия. Например, дифференциация клеток в сторону флоэмы, которые перезимовали
в камбиальной зоне, обычно интерпретировалась как созревание материнских клеток флоэмы, оставшихся от предшествующего сезона. Считалось, что образование новой флоэмы не начнется до тех пор, пока нс появятся из камбиальных инициален новые материнские клетки флоэмы. Наоборот, дифферен
циация перезимовавших материнских клеток ксилемы воспринималась как доказательство камбиальной активности и образования ксилемы. Следовательно, критерий, на основании которого пришли к выводу, что формированию ксилемы предшествует образование флоэмы, недостаточно аргументирован. У некоторых кольцесосудистых видов покрытосеменных дифференциация флоэмы по всей оси растения начинается несколько раньше или одновременно с дифференциацией ксилемы.
Ж
Рис. 3.13. Сезонные изменения камбиальной активности у деревьев груши (Evert, 1960) :
I—ХП - месяцы; 1 — дифференциация перезимовавших элементов; 2 — ксилемы и склер ендов; 3 — флоэмы; 4 — нефун-хционирующая флоэма; 5 —
камбиальная активность; 6 — прекращение функционирования
Например, у белой акации дифференциация флоэмы происходила почти на неделю раныпе дифференциации ксилемы. У многих видов рассеянососудистых покрытосеменных и у голосеменных сначала дифференцируется флоэма, предшествуя иногда на несколько недель дифференциации ксилемы (рис. 3.13). Например,
76
Глава 3
у груши дифференциация флоэмы происходила на 8 недель раньше, чем у ксилемы, а у тополя осинообразного, сосен Банкса, смолистой и веймутовой формирование флоэмы начиналось раньше на 6 недель. У лиственницы, пихты бальзамической и ели черной прирост флоэмы начинался до начала дифференциации новой ксилемы.
Количество образующейся ксилемы и флоэмы. Порядок формирования камбиальных производных, по-видимому, не имеет прямого отношения к количеству образуемых тканей по обе стороны от камбия. К концу вегетационного периода количество клеток ксилемы, отложенных камбием, значительно превосходит число вновь образованных клеток флоэмы. Аналогичное отмечается даже у видов, у которых формирование флоэмы начинается значительно раньше формирования ксилемы. Например, у дугласии в радиальных рядах ежегодно образуется более 100 рядов трахеид и только 10—12 рядов элементов флоэмы. У пихты одноцветной соотношение клеток ксилемы и флоэмы составляло 14:1.
У некоторых видов прирост ксилемы более чувствителен к стрессам в окружающей среде, чем прирост флоэмы. При неблагоприятных для роста условиях отношение ксилемы к флоэме часто снижается. У туи западной отношение ксилемы к флоэме изменялось от 15:1 до 2:1 соответственно ухудшению состояния деревьев. У пекана годичный прирост небольших 2-летних плодоносящих ветвей происходит в результате увеличения ксилемы и флоэмы в отношении менее 3, а у быстрорастущих ветвей это отношение было более 3. Бэннан (1955) обнаружил, что годичный прирост флоэмы быстрорастущих деревьев туи западной был таким же, как у медленнорастущих. У груши наряду с однообразным приростом флоэмы наблюдались различия в приросте кСилемы. Это дало основание предполагать, что флоэма образуется до развития ксилемы. Эти отношения, вероятно, не сохраняются у тех тропических видов, у которых невозможно различить годичные кольца флоэмы. У Eucaliptus camaldulensis, например, отношение ксилемы к флоэме под влиянием внешних условий изменялось незначительно. Одинаковые отношения между ксилемой и флоэмой (от 1 до 4) были обнаружены у быстрорастущих и медленнорастущих деревьев. Отношение н₽ зависело от ширины камбиальной зоны.
Дифференциация. Дочерние клетки ксилемы и флоэмы после отделения от камбия проходят последовательно следующие фазы дифференциации: растяжение клеток, формирование вторичной оболочки, лиг-нификацию и утрату протопласта (рис. 3.14). Одна фаза переходит к другой, перекрывая друг друга. Например, формирование вторичной оболочки начинается до окончания роста первичной оболочки. Во время дифференциации клеток большая часть производных камбия превращается в специализированные элементы различных тканевых систем.
Производные камбия, которые отделяются от него внутрь, образуют ксилему и могут дифференцироваться в одних из четырех типов элементов: членики сосудов, волокна, трахеиды, паренхимные клетки. Членики сосудов, трахеиды, волокнистые трахеиды и волокна либриформа претерпевают вторичное утолщение оболочек, а оболочки на концах члеников сосудов становятся перфорированными. Производные лучевых инициал ей мало изменяются во время дифференциации. Лучевые
Вегетативный рост
77
Рис. 3.14. Радиальные ряды трахеид сосны смолистой в разное время вегетационного периода (Whitmore, Zahner, 1966) :
а — начало; б — середина; в — конец; 1 — зона первичной стенки; 2 — зона цитоплазмы; 3 — уплощенные клетки поздней древесины; 4 — мертвые клетки поздней древесины; 5 — зрелая ранняя древесина; 6 — флоэма; 7 — поздняя древесина предыдущего года
Рис. 3.15. Изменение формы трахеид голосеменных в ходе дифференциации (Wardrop, 1954) :
А — камбиальная инициапь; Б — зрелая трахеида. Трахеида, увеличиваясь в объеме, растягивается и концы становятся более острыми. Радиальные стенки широкие, тангвитальные стенки узкие. Соответствующие части обозначены арабскими и римскими цифрами и буквами а и б. Длина клетки уменьшена примерно в 50 раз
трахеиды голосеменных изменяются существенно, когда у них развиваются вторичные оболочки и исчезает протопласт. У различного типа производных камбия также очень сильно изменяются размеры клеток.
Полная дифференциаций отдельных производных камбия обычно осуществляется в течение нескольких недель. Время, необходимое для дифференциации, зависит от вида растений, типа клеток, а также от времени вегетационного периода, в течение которого данный тип элементов проводящей системы формировался. Например, радиальное разрастание трахеид ранней древесины сосны замечательной происходило примерно в течение 3 недель. Этот же процесс в поздней древесине проходил примерно в 2 раза быстрее. Формирование вторичной стенки к концу вегетационного периода замедлялось, утолщение оболочек весной занимало 3—4 недели, осенью 8—10 недель. Как уже упоминалось, скорость образования разных типов дериватов камбия различна. У кольцесосудистых покрытосеменных дифференциация сосудов обычно предшествует формированию других элементов.
Увеличение размеров клеток. При поступлении воды в вакуоли производных элементов камбия увеличивается тургор и клетки растя
78 Глава 3
гиваются (рис. 3.15). Камбиальные производные, развивающиеся в членики сосудов, быстро расширяются в радиальном и тангентальном направлениях, но не увеличиваются в длину. Трахеиды и волокна подвергаются некоторому расширению в радиальном направлении и удлинению, степень которых различна у разных растительных групп. У покрытосеменных трахеиды и волокна сильно растягиваются, трахеиды голосеменных удлиняются слабо. Согласно данным Бэйли (1920) производные камбия покрытосеменных удлиняются на 500%, у голосеменных трахеиды удлиняются всего на 20%. Камбиальные инициали у голосеменных значительно длиннее, чем у покрытосеменных. Поэтому, несмотря на относительно ограниченное удлинение производных камбия у голосеменных, полностью сформированные они длиннее, чем у покрытосеменных.
Рост в длину производных камбия осуществляется в концах клеток или в их апикальной зоне. Увеличение в длину происходит за счет вставочного роста при удлинении отдельных клеток. Для этого необходима сочлененность клеток в пределах тканей, расположенных в вертикальном направлении. чПо данным ряда авторов, увеличивающаяся в длину клетка, отчлененная камбием, выделяет фермент, ослабляющий срединную пластинку между нею и двумя соприкасающимися с нею клетками. При этом клетки вследствие увеличения тургорного давления округляются и разъединяются по уголкам. Конец растягивающейся клетки заполняет образовавшееся пространство.
Утолщение клеточных оболочек. Стенки большинства клеток зрелой ксилемы состоят из тонкой первичной и толстой вторичной оболочек. Первичная оболочка формируется при делении клеток камбия и окружает протопласт во время поверхностного роста клеток. Вторичная оболочка формируется после окончания поверхностного роста. Первичная оболочка лишена различных слоев и характеризуется тесной упаковкой микрофибрилл. Вторичная оболочка накладывается тремя слоями (обозначаемыми Др В2 и ^з) на первичную оболочку (рис. 3.16). Эта оболочка состоит из фибриллярных компонентов — целлюлозы, кроме того, содержит инкрустирующие вещества, прежде всего лигнин, а также гемицеллюлозу, пектин и небольшое количество белка. Целлюлозные микрофибриллы укладываются наложением, а лигнин и другие инкрустирующие вещества внедряются в целлюлозную решетку. Степень лигнифйкации, различная в пределах одной и той же клетки, зависит от вида растений, типа клеток. Зрелые трахеиды голосеменных и сосуды покрытосеменных сильно лигнифицированы, а волокна трахеид и волокна либриформа покрытосеменных инкрустированы лигнином слабо. В противоположность клеткам ксилемы, у которых развиваются толстые оболочки, у большинства клеток флоэмы оболочки мягкие, которые впоследствии разрушаются или становятся сильно разрушенными. У волокон флоэмы образуются вторичные оболочки.
Потеря протопласта. Заключительные стадии созревания клеток ксилемы, например члеников сосудов и трахеид, включают разрушение и автолиз протопласта. В начале процесса формирования вторичной оболочки клетки содержат все важные органеллы. В конце созревания клеток мембраны вакуолей, цитоплазма, органеллы и плазмалемма подвергаются автолизу, ядро дезинтегрирует и. таким образом, жизнь
Вегетативный рост
79
Рис. 3.16. Диаграмма строения клеточной стенки зрелой трахеиды (Cote, 1967) "Wood Ultrastructure”, Washington University Press, Seattle, Washington
1 — срединная пластинка; 2 — первичная стенка; 3, 4, S — вторичные стенки; 6 — бородавчатый слой
Рис. 3.17. Тангентальный вид веретенообразных инициальных клеток, показывающий различные типы антиклйгального деления, участвующего в увеличении количества камбиальных клеток (Ваппап, 1967) : а — в — псевдопоперечное деление; г, д — латеральное деление
протопласта трахеид заканчивается. Лизис протопласта дифференцирующихся элементов ксилемы происходит быстро, например у трахеид тсуги канадской примерно в течение 4 дней, сосны обыкновенной на протяжении 2—5 дней.
УВЕЛИЧЕНИЕ КАМБИАЛЬНОГО СЛОЯ. По мере роста дерева увеличивается площадь, занимаемая камбиальной зоной. Это обусловлено появлением новых камбиальных клеток, что происходит двумя путями: 1) удлинением протяженности камбиальной зоны вследствие образования новых клеток, отделяемых от прокамбия, расположенного под апексами корней и стебля; 2) увеличением периферических слоев камбия в результате антиклинального деления веретеновидных камбиальных клеток за счет либо продольного деления (у покрытосеменных с ярусным камбием) , либо псевдо поперечного деления (у покрытосеменных с неярусным камбием и у голосеменных). Кроме того, небольшой процент веретеновидных клеток камбия делится антиклинально, отчленяя новые сегменты рядом с инициальной клеткой (рис. 3.17).
При псевдопоперечном делении оболочка клетки образуется около центра делящейся веретеновидной инициали и может быть короткой, почти поперечной или вытянутой и наклонной. Обычно отделяемая часть составляет около 15% всей длины делящейся клетки, но иногда отделенная часть может составлять одну треть или половину длины исходной клетки. После псевдо поперечного деления каждая из двух новых дочерних клеток-инициалей оказывается несколько длиннее
80
Глава 3
половины длины поделившейся инициали. Затем клетки в результате нескольких периклинальных делений удлиняются еще больше и достигают длины первоначальной инициали. После этого может происходить следующее леев до по перечное деление.
При псевдо по перечном делении может образоваться избыток веретенообразных инициален по окружности, но бывают при этом и потери. В результате этого прирост количества клеток камбия обычно невелик. Размеры клеток и степень контакта с лучами - существенный фактор в выживании веретенообразных инициал ей при внутрикамби-альной конкуренции. Более длинные сестринские инициали псевдо-поперечного деления обладают большей жизнеспособностью, чем короткие. У выживающих длинных инициалей больше контактов с лучевыми клетками. Вновь сформированные короткие инициали, имеющие мало точек соприкосновения с лучами, утрачивают способность к периклинальному делению и превращаются в зрелые элементы ксилемы или уменьшаются в размерах и превращаются в лучевые инициали. В результате роста веретеновидных камбиальных клеток одни клетки могут внедряться между другими. У некоторых видов один конец веретеновидных инициальных клеток удлиняется больше. У Pinaceae, например, рост веретенообразных инициалей после псевдопоперечного деления был направлен вверх, у туи западной вниз, а у груши не наблюдалась полярность в направлении удлинения клеток.
Скорость и распределение псевдо поперечных делений камбиальных инициалей изменяются с возрастом дерева. У быстрорастущих молодых деревьев псевдопоперечное деление происходит часто в течение всего вегетационного периода и степень выживания новых инициалей очень высокая. У таких деревьев и новые веретенообразные инициали и все производные камбия короткие. У взрослого дерева псевдо-поперечные деления происходят реже и преимущественно в конце вегетационного периода. К этому времени основная часть годичных колец уже сформирована. У старых деревьев снижается к тому же и процент выживания вновь сформированных инициалей. У выживших инициалей больше пространственных возможностей для разрастания, поэтому формируются более длинные верстеновидные инициали и производные камбия, так что в конце концов они достигают максимальных размеров, характерных для вида.
РАЗЛИЧНЫЕ ТИНЫ ПРИРОСТА. Камбиальная активность не постоянна: она может наблюдаться у деревьев в определенное время, может быть смещена в результате, например, засухи. Вследствие этого у деревьев формируется слой ксилемы, толщина которого зависит от высоты ствола и размера ветвей. При этом толщина слоя также может существенно различаться на определенной высоте ствола на разных его сторонах. Иногда у деревьев формируется полный прирост ксилемы по всему диаметру в верхней части ствола и не продуцируются годичные кольца ксилемы в нижней его части. Такое явление ’’недостающих колец” особенно характерно для угнетенных и старых деревьев. Кроме того, у нижних ветвей в случае последовательного подавления их новыми, расположенными выше ветвями не формируется ксилема в месте прикрепления к главному стволу (см. главу 17).
Негетативный рост
81
Бывает, что в течение года формируется не одно годичное кольцо (рис. 3.18). Такие ложные или множественные кольца часто возникают вследствие прекращения камбиальной активности под влиянием стрессов, например засухи, а затем возобновления активности. При этом отмечается чередование ранней i и поздней древесины. Ложные кольца могут образоваться также в результате повреждений насекомыми, грибами или пожарами. Возможны отмирание камбия или переход его в состояние покоя на одной стороне дерева, что приводит к формированию частичных или прерывают щихся колец, которые не создают по окружности замкнутого кольца. Прерванные кольца обнаруживаются у старых деревьев, лишенных листьев, у угнетенных деревьев, стареющих ветвей и в стволах деревьев с односторонней кроной. В последней группе годичное кольцо прерывается по радиусу ствола ниже недоразвитой кроны. В корнях часто обнаруживаются прерванные, эксцентричные в поперечном сечении кольца.
ПРОДОЛЖИТЕЛЬНОСТЬ КАМБИАЛЬНОГО РОСТА В ТЕЧЕНИЕ ВЕГЕТАЦИОННОГО
Рис. 3.18. Ложные кольца ветвей кипариса аризонского, сформированные в 1939 и 1940 гг. (Glock et al., 1960)
ПЕРИОДА. Период активности
камбия зависит от климата, вида растений, класса кроны, а также от частей ствола и ветвей. В одной и той же климатической зоне продолжительность активности камбия у вечнозеленых обычно больше, чем у листопадных деревьев. Камбий угнетенных деревьев по сравнению с камбием здоровых экземпляров того же вида может активно делиться на протяжении вегетационного периода непродолжительное время. Сезонный камбиальный рост деревьев начинается почти одновременно с активизацией ростовых процессов побегов, но камбиальный рост часто продолжается еще долгое время после прекращения удлинения побегов. Сезонный камбиальный рост в верхней части ствола длится дольше, чем в нижней. Следует учитывать, что камбиальный рост чутко реагирует на неблагоприятные условия окружающей среды. Поэтому он часто прекращается во время засухи и возобновляется после дождей. (Регули
82
Глава 3
рование камбильного роста рассматривается более подробно в главах 16 и 17.)
АНОМАЛИИ КАМБИАЛЬНОГО РОСТА. Большая часть данных об особенностях камбиального роста относится к деревьям умеренной зоны. Для этих растений вторичное утолщение считается нормальным. У многих видов тропических деревьев и лиан камбиальный рост часто отклоняется от нормального хода. Например, Обатон (1960) сообщает об аномалиях камбиального роста у 108 видов древесных лиан из 21 семейства, распространенных в Западной Африке. Аномальный, или нетипичный, камбиальный рост Может быть обнаружен у некоторых растений, камбии которых занимает нормальное положение. У других растений камбий занимает несвойственное ему положение. Часто аномалии камбиального роста являются результатом неравномерной активности различных камбиальных сегментов, изменений в количестве и положении ксилемы и флоэмы или вследствие отклонений от нормы активности вновь Образованных камбиальных клеток. Разные формы аномалий камбиального роста трудно классифицировать на определенные группы из-за их многообразия и большого количества промежуточных форм камбиального роста.
У некоторых семейств покрытосеменных (например, Amaranthaceae Chenopodiaceae, Menispermaceae и Nyctaginaceae) и у представителей голосеменных (лианы рода Gnetum, Welwischia и у некоторых Cycadal-es) формируются серии последующих функционирующих клеток камбия. Обычно нормальный камбий некоторое время функционирует, а затем отмирает. Новые слои камбия образуются последовательно к наружной поверхности ствола. Каждый последующий слой камбия функционирует нормально, но в течение ограниченного времени. Таким образом, древесина состоит из чередующихся слоев ксилемы и флоэмы. Примером видов, формирующих последовательно кнаружи слой камбия, может служить Avicennia res ini f era. У этого вида первый дополнительный слой камбия возникает путем деления внутренних клеток первичной коры, а последующие образуются в производных предшествующих слоев камбия. На поперечном срезе зрелый ствол Avicennia состоит из ряда участков, каждый из которых является продуктом деления отдельного камбия. В каждом участке кнаружи располагаются последовательно паренхима, вторичная ксилема, вторичная флоэма и склереиды.
У Bougainvillea также формируются последовательно ряды камбия, каждый из которых образуется среди производных предшествующего камбия (рис. 3.19). Каждый ряд камбия образует внутрь ксилему, кнаружи флоэму. Васкулярные ткани формируются в таком порядке: соединительные ткани и волокна ксилемы; флоэма; еще ксилема с сосудами и еще флоэма. Флоэма и ксилема дифференцируются из последовательно, радиально расположенных производных. Однако радиальная последовательность может быть скрыта делением инициалей флоэмы и ростовыми процессами в дифференцирующейся ксилеме.
У некоторых древесных растений тяжи вторичной флоэмы находятся во вторичной ксилеме. Происхождение такой ^’включенной флоэмы” различно. "У Comb return, например, часть камбия может отдежть производные внутрь, которые будут дифференцированы во флоэму.
Рис. 3.19. Поперечный срез корня Bougainvillea, показывающий аномальную активность Камбия:
а — аномальный рост камбия; б — камбий формирует снаружи от ксилемы непрерывный слой; флоэма указана стрелкой; в — новый слон камбия образовался снаружи от флоэмы самого старого пучка и далее продолжается вместе со старым камбием (плотная стрелка) ; новый камбий (светлые стрелки) на более поздней стадии развития, чем на рис. б (Esau, Cheadle, 1969)
84
Глава 3
У Strychnos полоски камбия прекращают деление. Если камбий возле этого участка будет продолжать делиться, то возникшая щель постепенно зарастет, а тяж флоэмы окажется включением в ксилему. У некоторых видов определенные участки камбиального цилиндра образуют больше ксилемы, чем флоэмы, а другие участки, наоборот, отделяют больше флоэмы. У ряда пустынных кустарников, таких, как Peganum harm ala, Zygophyllum dumosum, Zilla spinosa заметна прерывистость камбиальной активности по кругу главной оси, в результате формируется складчатый ствол, который часто растрескивается. Интересный тип аномалий камбиального роста имеется у Bignoniaceae. После сформирования камбиального цилиндра в конце первичного роста четыре участка камбия прекращают образовывать ксилему, но .продолжают формировать клетки флоэмы. Отсюда возникают два типа камбиальной активности, у одного из которых активность двусторонняя, у другого односторонняя.
РАНЕНИЯ И ЗАЖИВАНИЕ РАН. Раны на деревьях служат путями для патогенных организмов. Раны возникают в результате поломки ветвей, вершин или корней, обнажения ксилемы вследствие механических повреждений, ранений животными, пожарами и т.д. Скорость и эффективность реакции растения на райевое воздействие зависит, с одной стороны, от глубины и степени повреждения, с другой стороны, от состояния растения. Повреждения, разрывающие только кору или слегка задевающие камбий, обычно заживают быстро.
Шиго (1975) считает, что-патологические процессы у деревьев, получивших ранения, проходят три последовательные стадии. Первая стадия включает все процессы, связанные с ответными реакциями дерева-хозяина на ранение. На этой стадии происходит некоторое обесцвечивание ксилемы вследствие возникающих химических процессов. В ответ на ранения количество запасных веществ в клетке уменьшается, накапливаются фенольные и хиноновые соединения, изменяется характер активности ферментов. Вторая стадия включает реакции, которые наступают, когда микроорганизмы, преодолев химические защитные барьеры, вторгаются в сферу ксилемы. Прежде всего поражаются лучевые паренхимные клетки. Первыми из проникающих сюда организмов (но не всегда) являются бактерии и негименомицетные грибы. Обесцвечивание древесины усиливается вследствие взаимодействия между живыми клетками ксилемы и внедрившимися микроорганизмами. Когда ткань отмирает и обесцвечивается, усиливается ее влажность и накопление минеральных веществ. У некоторых видов изменения наступают без обесцвечивания, а влажная древесина формируется вследствие деятельности бактерий. На третьей стадии микроорганизмы, особенно Ну шепоту ceres, внедряются и разрушают вещества клеточных оболочек. К этому времени все клетки ксилемы мертвы. После внедрения первых гнилостных микроорганизмов проникают и многие другие микроорганизмы, вступающие в конкурентные отношения с предшественниками за остатки тканей. К таким организмам относятся Phycomycetes, Acty-nomycetes и нематоды. Пройдут ли процессы через все три стадии, зависит во многом от мощности дерева-хозяина и глубины ранения. В небольшой ране на мощном дереве процессы обычно не заходят далее первой стадии.
Вегетативный рост
85
Реакция дерева-хозяина на повреждение. Живая заболонь активно реагирует на ранение: древесина сразу обесцвечивается вокруг места, куда проникли микроорганизмы, заполняется различными экстрактивными веществами. Такая ’’защитная” древесина устойчива к дальнейшему действию пришельцев. Если микроорганизмы преодолевают химические защитные барьеры, дерево-хозяин часто отвечает отторжением пораженных тканей. У некоторых видов защитная реакция против микроорганизмов проявляется в закупорке сосудов, у других в формировании тиллов, или каллуса, в продуцировании камбием толстостенных клеток ксилемы и лучей. Эти образования создают барьер, отделяющий поврежденные ткани от тех, которые формируются после ранения. По тканям, устойчивость которых ослаблена, внедряющиеся микроорганизмы распространяются вертикально через локализованно поврежденные ткани. При повторном ранении дерева вновь формируется барьерная стенка, окружая внутренний участок поврежденной ткани.
Физиологические и биохимические реакции на ранение и инфекцию очень сложны и недостаточно изучены. Как уже упоминалось, при ранении возникают интенсивные изменения в метаболизме белков, увеличивается количество митохондрий и интенсивность дыхания, возрастает активность ферментов, особенно полифенолоксидазы и пероксидазы, усиливается образование этилена в поврежденных и прилегающих к ним неповрежденных тканях. Часто изменяется концентрация ростовых веществ. Все это указывает на то, что какое-то вещество или вещества должны перемещаться из поврежденных тканей и оказывать действие на гены - регуляторы специфических ферментов в прилегающих клетках. Некоторые исследователи приписывают эту роль этилену, синтезирующемуся в поврежденных клетках, но достаточных доказательств пока нет. Более вероятно, что имеются специфические вещества, иногда называемые раневыми гормонами, воздействующие на обменные процессы в прилегающих тканях и вызывающие также изменения в метаболизме. Эти изменения завершаются отторжением пораженных участков и формированием каллуса, способствующего заживлению ран.
Заживление ран. Процесс заживления заключается в затягивании ран и изолировании прилегающих к ранам тканей, пораженных микроорганизмами. Многие обширные раны старых деревьев никогда не затягиваются, но заживляются в результате описанных выше реакций живых тканей.
Камбиальная активность при заживлении ран. Заживление глубоких ран ствола происходит благодаря образованию каллуса и формированию нового камбия путем превращения клеток каллуса в камбиальные клетки. Феллоген также регенерирует в процессе заживления ран. Интенсивное формирование каллуса обычно связано с заживлением продольных морозобоин на стволах деревьев. Такие раны могут открываться и закрываться в ответ на внезапное повышение и понижение температуры. В течение повторного заживления ран вертикально ориентированные выступы, формирующиеся из тканей каллуса, так называемые ’’морозные жилки”, часто развиваются вдоль края раны. Образование каллуса у разных видов происходит по-разному, но у большинства древесных растений большое значение в формировании каллуса имеют
86
Глава 3
сердцевинные лучи. Иногда и другие компоненты камбиальной зоны -участвуют в продуцировании ткани каллуса. Таким образом, раневый каллус может быть образован паренхимой ксилемных и флоэмных лу-, чей, недифференцированными клетками ксилемы и тканями первичной коры.
Общий объем каллуса, формирующегося при заживлении ран ствола, зависит от размеров раны. При неглубоких ранах образование каллуса иногда ограниченно или он не образуется. Количество и скорость продуцирования каллуса после повреждения также зависят от вида растений. Например, при повреждении ствола Populus и Acer каллус образуется быстрее и в большем количестве, чем у Pyru s. Формирование нового камбия не зависит от количества или скорости образования каллуса. Регенерации камбия предшествует образование феллогена. Новый феллоген становится активным после то го,как подушечка каллуса разовьется достаточно хорошо. Заложение нового камбия у раненых деревьев часто связано со старым камбием, расположенным по краям раны, как у Hibiscus и Populus. У некоторых видов, однако, регенерация нового камбия nfe зависит от места локализации или присутствия существующего камбия по обе стороны раны. Например, у поврежденного ствола Trema orientalis новый камбий дифференцируется в средней части каллуса.
Ранняя активность вновь восстановленного камбия может быть нормальной и аномальной. У Hibiscus новый камбий образует нормально расположенные ткани. При заживлении ран ствола Trema orientalis, однако, отмечалось аномальное распределение проводящих тканей. Аналогичное наблюдалось у Acer, Malus, Pyrus. У Populus новые элементы ксилемы и флоэмы отделялись сразу же после формирования камбия (примерно через 9 дней после ранения), но первоначально образованные производные камбия были аномальны и разрушались. Нормальное формирование ксилемы и флоэмы наблюдалось приблизительно через 20 дней после ранения.
Факторы, влияющие на заживление ран. Скорость затягивания раны зависит от активности камбия. У мощных деревьев раны заживают очень быстро. У деревьев умеренной зоны рост за счет камбия происходит преимущественно в мае, июне и июле, поэтому раны, нанесенные до мая, заживают быстрота повреждения, полученные после июля, заживают плохо. Скорость заживления почти не зависит от формы раны.
На протяжении долгих лет раны покрывали материалом типа асфальта, шеллаком, красками, вазелином. Эффективность этих приемов широко обсуждалась. Нили (1970) пришел к выводу, что покрытие ран существенно не влияет на скорость их заживления. Вазелиновое покрытие даже снижает скорость заживления. Шиго и Вильсон (1977) обнаружили незначительное влияние нескольких применяемых для замазывания ран веществ на скорость их затягивания, вертикальное распространение обсцвечивания и загнивания древесины или на проникновение в раны гнилостных грибов. Покрытие не предотвращает инфицирование раны гнилостными грибами. Некоторые авторы признают, что покрытие ран физиологически целесообразно, но пользу для ускорения заживления ран они продемонстрировать не смогли.
Возможно, лучший способ борьбы с повреждением деревьев - предотвращение образования ран путем удаления неполноценных ветвей,
Вегетативный рост
87
коры и древесины. При ранах следует стимулировать камбиальную активность деревьев подкормкой удобрениями и орошением, следует также удалять менее ценные экземпляры, которые могут увеличить процент поврежденных деревьев.
РОСТ КОРНЯ
Семя содержит первичный корешок, или меристему корня, в зародыше, из которого развивается главный корень. Первичный корень ветвится и удлиняется, образуя разветвленную корневую систему, иногда он отмирает. Боковые побеги на стволе образуются из периферических тканей, боковые же корни возникают из глубоко расположенного слоя стелы, называемого перициклом.
При заложении боковых корней несколько клеток перицикла становятся меристематическими и делятся периклинально, образуя клетки, которые затем делятся и периклинально, и антиклинально. При этом формируется выступающий боковой зачаток, который прорастает через эндодерму, первичную кору и эпидерму. Боковой корешок, перед тем как прорваться через поверхностные ткани главного корня, развивает хорошо различимую апикальную меристему и корневой чехлик. В прорастании боковых корешков через первичную кору участвуют как лизис окружающих сг’о тканей, так и механическое сдавливание. Заложение боковых корней регулируется совместным влиянием нескольких гормональных регуляторов роста.
Степень многократного ветвления одревесневших и неодревесневших длинных корней удивительна. Подсчитано, что взрослое дерево дуба красного имеет минимум 500 млн. живых кончиков корней. У очень молодых древесных растений установлена быстрая пролиферация корней.
Обнаружены большие различия в ветвлении корней среди разных видов (табл. 3.4). Так, у цветущего 6-летнего проростка кизила, выращенного в оранжерее, было более 2 тыс. корней общей длиной 5144 см, а у проростков сосны ладан-
3.4. РАЗВИТИЕ КОРНЕЙ СЕЯНЦЕВ ДЕРЕНА ФЛОРИДСКОГО
И СОСНЫ ЛАДАННОЙ (ЧИСЛИТЕЛЬ - ЧИСЛО КОРНЕЙ;
ЗНАМЕНАТЕЛЬ - ОБЩАЯ ДЛИНА КОРНЕЙ, СМ)
Порядок ветвления Дерен флоридский Сосна ладанная
Первый 1/44,1 1/32,2
Второй 93/859,7 71/187,5
Третий 1035/2714,4 496/146,1
Четвертый 1336/1357,0 199/21,1
Пятый 191/168,1 0,0
Шестой 1/0,6 —
Общее число 2657/5143,9 767/387,0
Длина, м - /51,4 - /3,87
Примечание. У 6-месячных сеянцев не было конкурентных отношений.
88
Глава 3
ной того же возраста насчитывалось лишь 760 корней общей длиной 387 см. У обоих видов способность к росту отдельных корней более высоких порядков прогрессивно снижалась. По данным Колесникова (1966), у проростков яблони в течение одного вегетационного периода появлялось около 40 тыс. корней общей длиной 230 м, распределенных в 7 порядках ветвления.
У длинных одревесневших корней возможны боковые корни трех типов. Новое ответвление может стать длинным корнем, который в конце концов вторично утолщается и становится частью постоянной древесной корневой системы. Второй, наиболее распространенный, тип ветвления корней — образование коротких корней. Третий тип ветвления развивается в случаях, когда боковой короткий корень превращается в длинный. Ответвления длинных корней обычно замещают корни, у которых повреждается верхушка. Повреждения неодревесневших боковых корней вызывают формирование замещающих корней и образование развилки.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ КОРНЕЙ. Корневая система древесных растений состоит из относительно долго живущих корней и большого количества недолговечных небольших корней. Неблагоприятные условия окружающей среды, повреждения насекомыми, другими организмами и грибами, старение деревьев приводят к отмиранию корней. У здоровых деревьев многие мелкие корни отмирают вскоре после формирования.
У деревьев яблони, например, мелкие боковые корни живут приблизительно около недели. По данным Колесникова (1966), кончики главных корней проростков плодовых деревьев отмирают к 2-месячному возрасту. У разных видов долговечность мелких корней различна. У ели европейской большинство всасывающих корешков обычно живет 3-4 года, лишь около 10% отмирает в течение первого года и 20% живет более 4 лет. Хэд (1966) наблюдал, что корни черной смородины жили более 1 года. Однако многие более мелкие корни (так называемые питающие корни) плодовых и лесных деревьев живут менее года. Гибель мелких корней зависит от того, какому порядку ветвления они принадлежат. Например, у сосны смолистой быстрее отмирают боковые корни второго порядка, затем -корни первого порядка.
В умеренной зоне наибольшее число корней отмирает в холодные месяцы. У Juglans regia более чем 90% всасывающих корней утрачивалось в течение зимы. По данным Воронкова (1956), сухой вес корней растений чая в феврале был примерно на 12% ниже, чем в декабре. В начале апреля формирование новых корней становилось более интенсивным, количество утраченных в зимнее время корней восстанавливалось.
УДЛИНЕНИЕ КОРНЕЙ. Копчики корней могут быть заостренными у длинных корней и округлыми у коротких. На продольном сечении молодого корня видны четыре зоны, состоящие из различных клеток. На кончике сосредоточена защитная масса клеток, образующих корневой чехлик. За ним расположен конус роста, или меристематическая зона, состоящая из небольших тонкостенных, кубической формы клеток с плотной цитоплазмой. Длина конуса роста около 1 мм. В нем часто видны митотические фигуры. При увеличении количества клеток часть из них отходит к корневому чехлику, а другая часть — к зоне растяжения локализованной выше меристематической зоны. Здесь клетки, продуцированные в конусе роста, быстро увеличиваются в размерах, в основном в продольном направлении. Выше зоны растяжения расположена зона дифференциации и созревания. В конечном счете клетки, пока-
Вегетативный рост
89
лизованные у основания зоны растяжения, теряют способность к дальнейшему росту и дифференцируются на эпидерму, первичную кору и стелу.
Имеется значительное разнообразие в описаниях зон корней, относящихся к разным видам и различным тканям корней. У микоризных корней сосны, например, не бывает корневого чехлика. Зону дифференциации часто бывает трудно определить точно, так как клетки разного типа дифференцируются на разных расстояниях от кончика корня. Кроме того, расстояние зоны дифференциации от апекса зависит от скорости роста. Уилкокс (1954) установил, что у пихты благородной различные элементы медленно растущих корней становились зрелыми в зоне, расположенной ближе к апикальным инициалам.
Растущие кончики корней пробиваются сквозь почву в результате растяжения клеток. При этом корни отклоняются от нормального пути, так как им приходится-обходить препятствия. Тем не менее длинные горизонтальные корни клена красного распространяются радиально от основания ствола дерева на 25 м по прямой линии. Отклонившись из-за какого-то препятствия, обогнув его, корни возвращаются к первоначальному направлению. Подданным Хэда (1965), корням свойственно расти по спирали, но иногда они причудливо закручиваются. По наблюдениям Уилсона (1964), корень клена на протяжении 22 м перекручивался более 4 раз. Другие авторы также наблюдали некоторое закручивание корней у сосны смолистой, но роста по спирали при этом не было.
После прекращения сезонного роста корни часто приобретают бурый цвет в результате процесса, называемого метакутинизацией. При этом происходит лигнификация и суберинизация клеточных стенок первичной коры и находящегося в состоянии покоя корневого чехлика. У многих корней кончик остается белым, хотя и у них есть слой метакутини-зации. Наличие или отсутствие белого кончика корня зависит от количества слоев мертвых клеток, отделенных наружу от метакутинизиро-ванного слоя.
Корневые волоски. Поглощающая поверхность корней многих видов древесных растений увеличивается в значительной степени за счет формирования корневых волосков. Эти выросты, напоминающие трубочки, обычно появляются в виде, выпячиваний из наружных боковых стенок клеток эпидермы, хотя у некоторых видов они возникают из клеток первичной коры на один-два слоя глубже эпидермы. У хвойных корневые волоски коротких корней образуются из клеток поверхностного слоя, а у длинных корней они появляются из второго или третьего слоя клеток первичной коры. Корневые волоски возникают из поверхностного слоя лишь в том случае, если нет устойчивых слоев корневого чехлика. Место формирования корневых волосков обычно расположено в непосредственной близости от наиболее активной меристематической ткани, их длина уменьшается по направлению к апексу. Формирование волосков приурочено ко времени-снижения скорости растяжения клеток эпидермы.
Продолжительность жизни сильно вакуолизированных тонкостенных корневых волосков различна. Большинство из них живет лишь несколько часов, иногда дней или недель и отмирает с началом вторич
90
Глава 3
ного утолщения, суберинизации и лигнификации корня. Зона корневых волосков мигрирует, так как после отмирания старых корневых волосков новые формируются выше точки роста растягивающегося корня. У некоторых деревьев, таких, как белая акация, кофейное дерево в Кентукки и цитрусовые в Валенсии, суберинизированные и лигнифициро-ванные корневые волоски могут сохраняться в течение нескольких месяцев и годов. Такие устойчивые корневые волоски утрачивают способность к поглощению.
Количество корневых волосков зависит от вида растений и от факторов окружающей среды. По данным Козловского и Шольтеса (1948), у 7-недельного сеянца белой акации, выращенного в оранжерее, развилось 11 тыс. корневых волосков (520 см^), а у проростков того же возраста сосны ладанной в тех же условиях было менее 600 корневых волосков ^217 см2, табл. 3.5). Различия в способности формировать корневые волоски у растений клена белого и дуба красного были обнаружены Ричардсоном (1953). У клена белого отмечено нормальное развитие корневых волосков. Зона корневых волосков у него локализована непосредственно за растущим кончиком корня на расстоянии 2—3 мм. По сравнению с этим у дуба красного корневые волоски были только на корнях самого низкого порядка ветвления. Образовывались они лишь после прекращения роста корней.
У многих деревьев (например, авокадо и ореха-пекана) корневые волоски не образуются. Корневых волосков нет также у некоторых голосеменных и на микоризных корнях. У главного корня формирование корневых волосков стимулируется окружающими условиями, которые снижают развитие микоризы и ускоряют удлинение корней. Однако развитие микоризы и корневых волосков не исключают полностью друг друга. Был обнаружен большой диапазон вариаций в развитии корневых волосков под влиянием микоризы: в одних случаях влияние не выявлено, в других развитие корневых волосков полностью подавлялось. У дугласии на микоризных корнях корневые волоски не развивались, но у длинных боковых корней корневые волоски и сеть Гартига хорошо совмещались.
Скорость роста корней. Скорость удлинения корней у древесных растений зависит от вида, генотипа, возраста дерева, времени года, места произрастания и от окружающих условий. Корни во время максимальной активности ростовых процессов могут удлиняться от 1 до 25 мм и более в день.
3.5. РАЗВИТИЕ КОРНЕЙ И КОРНЕВЫХ ВОЛОСКОВ
У 7-НЕДЕЛЬНЫХ СЕЯНЦЕВ БЕЛОЙ АКАЦИИ И СОСНЫ ЛАДАННОЙ (Kozlowski, Scholtes, 1948)
Корни Длина корней, мм Площадь поверхности, см 2 Корневые волоски, шт. Площадь поверхности корневых волосков, см^
Белой акации: первичные вторичные третичные Всего Сосны ладанной: первичные вторичные Всего 16,20 115,62 30,60 162,42 6,45 5,93 12,38 3,45 15,72 3,12 22,28 2,73 0,97 2,70 1,166 3,63 8,321 25,22 2,081 5,18 11,568 34,03 520 корневых волосков/см2 215,0 2,80 371,0 2,080 217 корневых волосков/См2
Вегетативный рост
91
Длинные корни яблони в Ист-Малинге вырастали до 4 6 см в неделю, у вишни 7-8 см в неделю. По данным Гофмана (1966), новые корни белой акации и видов тополя росли с исключительно высокой скоростью,- достигавшей 5 см в день. Имеются данные о том, что скорость роста корней ночью"больше, чем днем. Рид (1939) наблюдал на плантациях южных сосен в период максимальной скорости роста удлинение корней до 2-3 мм в день. По данным Барнея (1951), средняя скорость роста корней сеянцев сосны ладанной в оранжерее при температуре почвы 25°С была 5,2 мм в день, при 5°С только 0,17 мм в день.
Сезонные изменения роста. Годичный рост корней включает два компонента: удлинение уже имеющихся корней, заложение новых боковых корней и последующее их удлинение. В умеренной зоне растяжение корней начинается рано весной и продолжается до конца осени, дольше, чем удлинение побегов того же дерева (рис. 3.20). Период между прекращением удлинения побегов и окончанием прироста корней сильно различается у разных видов.-Растяжение корней может продолжаться в течение многих недель у тех видов, у которых побеги полностью образованы в зимних почках и быстро разворачиваются. Однако у гетерофильных видов, побеги которых растут в течение многих недель, растяжение корней может продолжаться лишь ненамного дольше удлинения побегов. У южных сосен прирост корней в длину происходит ежемесячно в течение всего года. Зимой рост сдерживается низкими температурами, а летом — сухостью почвы.
Рис. 3.20. Различия в сезонном росте побегов и корней семи видов лесных деревьев (заштрихованные участки - рост побегов, зачерненные - рост корней). Начало и окончание роста в сезоне обозначены стрелками (Lvt, Hoffman, 1967)
92
Глава 3
Скорость роста корней в течение вегетационного периода не постоянна. На рис. 3.20 видно, что скорость удлинения растущих корней была максимальной в начале лета, но у некоторых видов интенсивный рост наблюдался и в конце вегетапион-ного периода. Рост корней молодых деревьев Pinus nigra в Южном Онтарио продолжался с апреля до конца ноября. В течение лета ежедневный прирост корней в среднем составлял 6,5 мм, осенью — всего 2,9 мм в день.
Многие Исследователи наблюдали, что сезонный рост корней у лесных и садовых деревьев происходит неравномерно. В ответ на действие окружающих факторов среды часто бывает более двух пиков увеличения интенсивности роста. (Подробнее этот вопрос обсуждается в главе 17.)
КАМБИАЛЬНАЯ АКТИВНОСТЬ КОРНЕЙ. За первичным ростом'у некоторых корней следует вторичный, который включает формирование вторичных проводящих тканей за счет камбия, а феллоген начинает продуцировать перидерму. Вторичное утолщение начинается в течение первого или второго года роста корней (рис. 3.21). Сначала некоторые клетки паренхимы и йерицикла становятся меристематическими и формируют волнистые камбиальные полосы на внутреннем крае тяжей флоэмы и на внешнем крае ксилемы. В конечном счете камбий продуцирует законченный цилиндр ксилемы- Вскоре после сформирования
Рис. 321. Вторичный рост древесных корней, показывающий развитие камбия и образование вторичной ксилемы и флоэмы. Разрастание за счет добавления вторичных тканей разрушает первичную флоэму и эндодерму, отделяет первичную кору (Esau, 1965):
1 _ первичная кора; 2 - эндодерма; 3 — перицикл; 4 — первичная флоэма; 5 -первичная ксилема; 6 — вторичная ксилема; 7 — камбий; 8 — вторичная флоэма;
9 — перицикл; 10 — первичная кора
Рис. 3.22. Годичные приросты ксилемы по всей длине главного бокового корня сосны смолистой (Faille, 1968)
Вегетативный рост
93
Рис. 3.23. Эксцентричность камбиального роста на поперечном срезе горизонтального корня туи восточной (Фото Лесной службы США)
камбия некоторые клетки перицикла начинают делиться и образуют феллоген (пробковый камбий), который внутрь образует ткани фелло-дермы, а наружу — пробковую ткань. С началом формирования пробки первичная кора с ее эндодермой утрачивается и расположение тканей после этого становится сходным с расположением тканей ствола.
Формирование камбием ксилемы начинается в корнях, локализованных вблизи от поверхности почвы, затем процесс образования ксилемы распространяется к корням, расположенным на разной глубине. Это волнообразное распространение камбиальной активности в корнях сверху вниз часто протекает медленнее, чем в стволе.
Так, у деревьев апельсина камбиальная активность в стволе и в боковых ветвях начиналась в апреле и распространялась к главному корню в течение 2 недель. Затем камбиальная активность в корневой системе замедлялась, и ксилема не формировалась в боковых корнях до конца июля, у некоторых мелких корней -до конца сентября.
Камбиальная активность в корнях значительно менее регулярна, чем в стволе. Она заметно варьирует по длине корня и его окружности. Максимальное формирование ксилемы обнаруживается всегда у корней расположенных ближе к поверхности почвы, а по длине корня — в участках, менее удаленных от ствола. Поэтому годичный прирост ксилемы быстро сходит на конус от высоко расположенных в почве корней к глубоко расположенным и постепенно сужается к концам корней боковых разветвлений (рис. 3.22). Возможны и отступления от этой схемы. Например, Хэд (1968) обнаружил нерегулярные утолщения корней яб
94 Глава 3
лони вдоль их длины. Иногда заметное утолщение начиналось в более отдаленных от ствола частях корней, а в некоторые годы камбиальная активность не обнаруживалась вообще.
Молодые корни на поперечном срезе обычно округлые. С возрастом расположение ксилемы по окружности корня становится неравномерным, поэтому старые корни на поперечном срезе обычно эксцентричны. Они содержат в большом количестве ложные и двойные кольца ксилемы (рис. 3.23). У горизонтальных корней многих тропических видов значительно больше образуется ксилемы вдоль верхней стороны по сравнению с нижней стороной, что приводит к формированию опорных корней. Одна из причин образования колен у болотного кипариса заключается, возможно, в очень высокой камбиальной активности на верхней поверхности корней, У одного и того же дерева может быть очень большое разнообразие в образовании ксилемы у разных корней. Обычно более эксцентричный рост бывает у боковых горизонтальных корней, чем у вертикальных?или корней, отклоненных от центральной части корневой системы.
ИЗМЕРЕНИЕ И АНАЛИЗ РОСТА
Рост дерева может быть измерен различными методами в зависимости от целей исследователя. Для лесоводов важно оценить годичный прирост древесины всего древостоя. Оценка производится на основании измерения диаметра и высоты ствола с поправкой на величину конуса ствола. Продуктивность древостоя обычно выражается как объем стволов на единицу площади, занимаемой древостоем (для некоторых целей лучше выражать в тоннах на единицу площади). Высота дерева часто используется как показатель качества участка. Этот индекс определяется высотой среднего доминирующего дерева в возрасте 50 лет. Для оценки медленнорастущих пород определяют высоту 100-летних деревьев. Физиологи заинтересованы в более детальных исследованиях сезонного хода прироста по диаметру и в высоту, а также в изучении влияния окружающих условий и агротехнических воздействий на рост древесных растений. Их также интересует характер ростовых процессов разных частей растения. Такие исследования требуют более частых и точных измерений, чем это необходимо для оценки годичного прироста древесины.
РОСТ В ВЫСОТУ. Трудно точно измерить высокие деревья. Обычно высота таких деревьев измеряется косвенно при помощи гипсометров различных типов. Они основаны на принципе отношения сторон подобных треугольников или на отношении между сторонами и тригонометрическими функциями углов прямоугольников. Геодезический теодолит также может быть использован для более точных измерений высоты деревьев. Высоту молодых деревьев можно измерять метровой линейкой или градуированной палкой. Для измерения деревьев высотой 10—15 м приспосабливают соединенные между собой шесты. Высоту сеянцев измеряют при помощи рулетки, линейки и метровой планки. Для постоянной регистрации суточных изменений роста в высоту сеянцев Крамер (1976) применял самописец, переделанный из гидротермографа.
Вегетативный рост
95
КАМБИАЛЬНЫЙ РОСТ. Измерениям камбиального роста уделяется много внимания вследствие значительного его вклада в увеличение дерева в объеме, из-за его чувствительности к различным внутренним и внешним факторам и легкости его измерения. Для измерений камбиального роста используются разного рода дендрометры, кронциркули и рулетки. Имеются два основных типа дендрометров: 1) прибор для измерения окружности ствола при помощи специального обруча; 2) приборы, при помощи которых измеряют изменения радиуса ствола путем определения расстояния между фиксированной заанкеренной планкой в древесине дерева и точкой поверхности коры. К последней группе относятся дендрометры с градуированным лимбом, с микрометрическим кронциркулем и с регистрирующим устройством (обычно называются дендрографами). Различные оптические дендрометры используются для измерения диаметра верхней части ствола растущего дерева.
Специальным ободом можно измерить изменения окружности ствола, но при этом нельзя определить, за счет коры или древесины увеличился диаметр, или это произошло вследствие изменений в оводненности тканей. Дендрометр с градуированным лимбом (рис. 3.24) дает возможность зарегистрировать начало камбиальной активности и периодическое сокращение ствола более точно, чем измерительный древесный обод. Дендрограф с электрическим управлением измеряет и регистрирует рост в течение длительного времени. На рис. 3.25 изображен пример продолжительного роста в радиальном направлении, который накладывался на суточные сокращения ствола, вызываемые водным стрессом (уменьшения диаметра не отмечалось 13 марта из-за дождя).
К сожалению, одиночное измерение высоты или один замер окружности ствола дерева не дают точной информации о высоте всего дерева. Как указывалось выше в этой главе, в нижней части
О $10 1$ 20 2$ 30
Числа месяца
Рис. 3.24. Дендрометр для измерения радиального роста ствола дерева (Buram, Doolittle, 1950): а—б — расстояние, периодически измеряемое кронциркулем с микрометром; 1 - кора; 2 — винты; 3 - рама; 4 - камбий; 5 — рабочий винт; 6 - заболонь
Рис. 3.25. Дневные измерения радиуса, накладывающиеся на продолжительный радиальный рост дерева Pinus canariensis в Австралии. Отмечено влия”.ие дождя 13 марта (Holmes, Shim, 1968)
96
Глава 3
ствола угнетенных деревьев камбиальная активность часто бывает значительно слабее, чем в верхней части. Она различается и на разных сторонах дерева, поэтому следует устанавливать по окружности ствола несколько дендрометров, чтобы точно зарегистрировать рост в радиальном направлении.
Диаметры стволов и объем древесины отдельных деревьев иногда оцениваются по диаметру кроны. Предлагались методы определения диаметра кроны при помощи фотографической техники. Большинство дендрометров сконструировано для работы с крупными деревьями. Для измерения диаметра сеянцев используются обычно кронциркули с микрометром. Сплинтер (1970) приспособил регистрирующий микрометр для продолжительных измерений. Козловский (1970) проводил постоянные измерения изменений диаметра проростков, установив дендрограф на вспомогательную раму. Ствол опытного сеянца поддерживался против перемещаемой стальной пластинки. Рычаг дендрографа, касающийся противоположной стороны ствола, постоянно регистрировал изменения его диаметра. Эти изменения записывались на карточки, укрепленные на барабане, который приводился в действие часовым механизмом. Малые изменения в диаметре про- с-ростков измерялись приборами с усилителями и датчиками.
АНАЛИЗ РОСТА. По мнению Ледига (1976), одна из важнейших задач физиологии растений заключается в том, чтобы понять, какое ; распределение ростовых процессов между органами растений является > оптимальным. Это требует значительно более детального анализа роста растений, чем измерения диаметра и высоты деревьев. Чтобы понять, J как могут повлиять намост' растения окружающие условия и агротехни-ка, необходима более подробная информация о том, как распределя-ются ростовые процессы между корнями, стволом, ветвями и листьями, £
Накопление сухого вещества в основном зависит от величины фото- 'с синтезирующей поверхности и скорости фиксации углерода (фотосинтеза) на единицу поверхности листа. Для сельскохозяйственных культур и древесных насаждений листовая поверхность выражается как индекс листовой поверхности, как листовая поверхность на единицу поверхности земли, а увеличение веса на единицу листовой поверхности или массы выражает скорость нетто-ассимиляции (Н). Физиологи поэтому заинтересованы в том, чтобы узнать, на какой из компонентов в первую очередь влияют окружающие факторы — на скорость фотосинтеза, выраженную на единицу листовой поверхности, на величину самой листовой поверхности или на распределение продуктов фотосинтеза между корнями, стволом, ветвями и листьями. Уотсон (1952) и другие исследователи в Англии утверждали, что большие колебания в урожае сельскохозяйственных культур зависят скорее от различия в скорости увеличения листовой поверхности, чем от различия в скорости нетто-ассимиляции. Имеются доказательства, что образование биомассы леса положительно коррелирует с биомассой листьев или индексом листовой поверхности до определенного оптимального его значения. За пределами этой величины может наступить снижение скорости фотосинтеза вследствие увеличения затененности нижних листьев. Продолжительность жизни листьев также имеет значение, так как сохранение большой листовой поверхности способствует осуществлению фотосинтеза до поздней осени и вследствие этого накоплению сухого вещества. Азотные удобрения могут затормозить старение листьев и продлить фотосинтетическую активность.
ПРОБЛЕМЫ, ВОЗНИКАЮЩИЕ ПРИ ИЗУЧЕНИИ РАСПРЕДЕЛЕНИЯ ПРОДУКТОВ ФОТОСИНТЕЗА. Распределение сухого вещества или
Вветтивный рост у]
биомассы среди различных частей у древесных растений исследовать труднее, чем у травянистых. Это связано с тем, что большие размеры деревьев ограничивают возможность брать пробы. Оценка образования сухого вещества без повреждения при этом дерева такими методами, как измерение поглощения С©2, не надежна. Иногда определяют распределение сухого вещества в листьях, ветвях, стволе и корнях одного дерева среднего размера и результаты экстраполируют на все деревья древостоя. Это надежно только в том случае, если деревья произрастают на участке сходных размеров, так как распределение общей биомассы среди разных компонентов растения зависит от его возраста и размеров (табл. 3.6).
Более приемлем метод, при котором учитывают накопление массы и измеряют отдельные компоненты ее у нескольких деревьев разных размеров. Затем регрессивным анализом относят все компоненты к какому-либо легко измеряемому параметру, например к диаметру ствола.
ОТНОСИТЕЛЬНЫЕ СКОРОСТИ РОСТА. Установлено, что рост растений подчиняется закону сложных процентных отношений. При этом величина прироста в единицу времени зависит от количества материалов или от размера растений в начальный период роста. Таким образом, сеянцы, развивающиеся из крупных семян, увеличиваются в размерах быстрее сеянцев того.же вида из мелких семян. Это факт привел к развитию концепции об относительной скорости роста (ОСР) или меры прироста сухого вещества в единицу времени на единицу растущего материала, часто выражаемую в граммах на грамм сухого веса в неделю. Это позволяет сравнивать влияние различных факторов окружающей среды на скорость роста независимо от размеров сравниваемых растений.
3.6. ПРИМЕРНЫЕ ОТНОШЕНИЯ ЛИСТЬЕВ К МАССЕ ВЕТВЕЙ И СТВОЛА У БЕРЕЗЫ И СОСНЫ ЛАДАННОЙ РАЗНОГО ВОЗРАСТА (Switzen et al., 1968)
Приблизительный воз- Отношения
раст посадки, лет ----------------------------------------------------
Береза Сосна ладанная
Листья:ветви Листьягствол Листья:ветви Листья:ствол
5 1:1 1:1 1:1 1:10
25 1:6 1:20 1:2 1:20
45 1:12 1:50 1:6 1:50
55 1:12 1:50 1:8 1:70
Для расчета относительной скорости роста используют уравнение
ОСР = 1пВ2- D2 ~Dy ,
где В, и Вл — сухой вес в начале и конце периода взятия проб; и /X. - даты 1 Z 1Z
взятия проб; in - натуральный логарифм числа.
98 Глава 3
Рост зависит от скорости фотосинтеза и площади или веса фотосинтезирующей ткани, поэтому отношение может быть выражено так: ОСР = СНАОПЛ. Здесь СНА (скорость нетто-ассимиляции) — прирост сухого веса на единицу поверхности листа, ОПЛ — отношение поверхности листа к общему сухому весу. Эти выражения отнесены к определенному интервалу времени Т>2 — ^1, равному обычно 1 — 2 неделям.
Развитие и применение этих концепций обсуждается в ряде работ, более детально в книге Эванса (1972). Такой подход позволяет сравнивать скорость роста растений разных размеров и в разных фазах онтогенеза, исследовать относительное значение величины поверхности листа и скорости фотосинтеза, выраженной на единицу поверхности листа. Можно провести аналогию с оборотом капиталовложений, подчиняющихся закону сложных процентных отношений- Капиталовложение фотосинтеза — это поверхность листа: чем больше продуктов фото- ' синтеза превратится в листовую площадь, тем больше будет скорость роста. По утверждению Ледига (1^76), это может привести к нелепому заключению, что наиболее продуктивные растения должны состоять исключительно из листьев. В действительности, ствол и ветви необходимы для наилучшего расположения листьев по отношению к свету, а корни обеспечивают растения водой, минеральными веществами и регуляторами роста. Таким образом, у продуктивных деревьев или других растений продукты фотосинтеза оптимально распределяются между растущими листьями и структурами, которые поддерживают листья. Деревья по сравнению с однолетними растениями находятся в невыгодном положении, так как у них постоянно увеличивающаяся масса су-,хого веса древесины фотосинтетически не продуктивна (см. табл. 3.6). узким образом, относительная скорость роста с увеличением возраста сумеет тенденцию снижаться. Обычно она ниже у многолетних древесных ^растений по сравнению с однолетними травянистыми.
С экономической точки зрения эффективным можно считать рост в том случае, если получается самый высокий урожай желательной про-дукции: древесины, плодов, семян, даже цветов. В связи с этим селекционеры создают такие растения, у которых максимально возможная часть продуктов фотосинтеза превращается в экономически выгодную / продукцию. Для лесоводов такой продукцией является обычно древесина. Способность деревьев формировать древесину зависит от взаимодействия трех основных компонентов: от скорости фотосинтеза на единицу поверхности или массы листа, на единицу хлорофилла; от структуры, > долговечности, экспозиции и площади кроны; от распределения продуктов фотосинтеза между разными органами дерева, что отражено в раз- ! личном содержании сухого вещества в кроне, стволе и корнях. Важнаья £ задача физиологов растений — определить методы воздействия на ком- •_ поненты, от которых зависит формирование древесины, чтобы получить как можно больший ее выход. В отличие от лесоводов садоводы стре- < мятся получить максимально возможный урожай плодов. Это криво- j дит к необходимости создавать карликовые деревья, у которых большая f часть продуктов фотосинтеза превращается в плоды, так как их листья % поддерживаются меньшей частью нефотосинтезирующих тканей, чем листья крупных деревьев. . ’
РЕПРОДУКТИВНЫЙ РОСТ
Одна из наиболее важных задач в лесоводстве и садоводстве - получение доста-Iочного количества высококачественного посадочного материала. Большинство древесных растений размножается в природе семенами, но происходит также и вегетативное размножение отпрысками, корневыми побегами и отводками. Часто удобнее размножать древесные растения вегетативным (черенками, прививкой, отводами или при помощи культуры тканей), чем половым путем. Известно, что плодовые деревья не сохраняют своих свойств при размножении семенами, поэтому их следует размножать вегетативно. Установлено, что большинство деревьев высоко гетерозиготно, так как зародыш семени получает различные участки хромосом от родительских растений, в связи с чем он генетически отличается от обоих родителей. Даже зародыш, образованный в результате самоопыления, не будет идентичен родителю, из семени которого возник. Это невозможно из-за высокой степени гетеро зиготности и случайного характера разделения хромосом во время редукционного деления и рекомбинации при слиянии яйцеклетки и спермия. В отличие от этого вегетативно размноженные растения содержат генетическую информацию только родительского растения.
Имеются и другие причины для вегетативного размножения древесных растений помимо получения отпрысков сходных с родителями. Например, при окули-|х>вке или прививке верхняя часть дерева с определенными желательными признаками может быть скомбинирована с корневой системой другого типа дерева, которое энергично растет или обладает большей устойчивостью к болезням. Так, часто прививают черенки различных сортов винограда на подвои американских сортов, устойчивых к филлоксере. Другой пример вегетативного размножения, но уже с целью получения желательного типа роста, формирование карликовых деревьев яблонь путем прививки на подвои, способные к образованию карликовых деревьев. Так получен сорт яблони Mailing IX (М9). Карликовые груши получают, прививая их на подвой айвы (см. главу 17). Бесполое размножение необходимо и в случаях, когда нужно вырастить растения, не образующие жизнеспособных семян (например, некоторые сорта винограда, инжира, цитрусовых и бананов). Кроме того, вегетативное размножение надежнее полового для растений, семена которых прорастают медленно, или для растений с очень длительной ювенильной стадией, во время которой они не образуют цветов, плодов и семян, а иногда также имеют другие нежелательные свойства (например, склонность к образованию колючек).
Одна из наиболее важных и менее всего разработанных проблем в садоводстве и лесоводстве - это нерегулярное и непредсказуемое образование цветов, плодов и семян. Урожаи плодов и семян варьируют не только у отдельных видов, но и у деревьев того же самого вида. Как подчеркивает Мэтьюз (1963), успех генетиков и селекционеров при выведении пород высшего качества и улучшенных культур плодовых и лесных деревьев зависит от надежной репродукции в раннем возрасте. Результат зависит также от понимания внутренних процессов в репродуктивной фазе роста. К таким процессам относятся: заложение зачатков цветов, цветение, опыление (перенос пыльцы от пыльника к рыльцу) .рост и развитие зародыша, рост плода и семени до зрелости и созревание плодов и шишек. Для получения хорошего урожая плодов и семян, каждая из этих последовательных репродуктивных фаз должна проходить успешно. К сожалению, переход от одной фазы к другой часто тормозится, что препятствует завершению репродуктивного роста.
ПЕРИОДИЧНОСТЬ РОСТА РЕПРОДУКТИВНЫХ ОРГАНОВ
Многие ботаники определяют цветок как детерминированный спо-« рогенный побег, образующий плодолистики, ограничивая применение термина только к покрытосеменным. В данной книге термин цветок
100
Глава 4
будет использован шире, чтобы им могли оперировать и лесоводы и садоводы. Предлагается рассматривать цветок как детерминированный спорогенный побег. Это дает возможность относить его и к молодым шишкам, или стробилам голосеменных, обозначая их как цветки.
Деревья умеренной зоны. Древесные растения на первых этапах жизни проходят ювенильную стадию, в течение которой они не образуют цветов. После завершения этой стадии они становятся способными к цветению, и эта способность сохраняется как периодическое (сезонное) явление. Длительность ювенильного периода значительно варьирует среди видов (табл. 4.1). Некоторые сорта яблонь, как Wealthy, могут зацветать в возрасте 3—4 лет, другие, как No г the гп? часто не зацветают до 10-15 лет. Время первого цветения часто различно в пределах рода, так же как и между родами. У сосны Банкса шишки образуются на 3-й год, у сосны скрученной широко хвойной на 5—6-й год. У сосны Эллиота, однако, шишки появляются редко ранее 10-летнего возраста, до 20-летнего возраста шишек образуется очень мало.
. 4.1. ПРОДОЛЖИТЕЛЬНОСТЬ ЮВЕНИЛЬНОГО ПЕРИОДА У ДЕРЕВЬЕВ, ОПРЕДЕЛЯЕМОГО ВРЕМЕНЕМ ПЕРВОГО ЦВЕТЕНИЯ
Вид Ювенильный период, годы Авторы
Pinus. sylvestris 5-10 Wareing, (1959)
Larix decidua 10-15 Wareing (1959)
Pseudotsuga menziesii 15-20 Wareing (1959)
Picea abies 20-25 Wareing (1959)
Abeis aiba 25-30
Betula pubescens 5-10 Wareing (1959)
Fraxinus excelsior 15-20 Wareing (1959)
Acer pseudoplat anus 15-20 Wareing (1959)
Que reus robur 25-30 Wareing (1959)
Fagus sylvatica 30-40 Wareing (1959)
Чай (Camellia thea) . 5 Wareing (1961)
Яблоня (Malus) 7.5 Wareing (1964)
Груша (Pyrus) 10 Visser (1964)
Мандарин (Citrus reticulata) 5-7 Furr et al. (1947)
Апельсин (Citrus sinensis) 6-7 Furr et al (1947)
Танжело (Citrus paradisi 7-8 Furr et al. (1947)
5-8 Furr et al. (1947)
x C. reticulata)
Длительность ювенильного периода в значительной степени определяется факторами окружающей среды, влияющими на скорость роста. Мощные деревья вступают в стадию цветения раньше, чем угнетенные • экземпляры того же вида. Случайное преждевременное зацветание у некоторых видов не означает, что они достигли стадии зрелости. У цитрусовых, например, цветы могут образовываться у очень молодых
^продуктивный рост 101
сеянцев, а затем на протяжении нескольких лет у них продолжается лишь вегетативный рост, пока полностью' не закончится ювенильная фаза роста. После этого у них начинается нормальное цветение, как у взрослых деревьев.
Древесные растения, достигнув стадии зредости, не цветут ежегодно, гак как физиологические условия и окружающие факторы могут влиять на заложение цветочных зачатков (примордиев). Деревья, растущие на открытых, хорошо освещенных местах и опушках леса, часто зацветают в более раннем возрасте и образуют больше семян, чем деревья, [ыстущие в густых насаждениях и тени (см. главу 17). Древесные растения одного и того же вида, растущие в теплых местах, зацветают раньше, чем в более холодных условиях.
Действие окружающих условий на цветение подтверждается наблюдениями, проведенными Тидеманом (1964) в течение 43 лет. Оказалось, что в Англии полное цветение деревьев яблони сорта Сох orange Pippin происходило от 15 апреля до 23 мая в зависимости от температуры.
Пример периодичности репродуктивного роста хорошо известен у плодовых деревьев с двухгодичным циклом. Некоторые сорта яблонь в один год обильно цветут и дают большой урожай плодов, на следующий год урожай бывает небольшой или яблони вообще не плодоносят. Тенденция к двухгодичному циклу плодоношения больше выражена у сортов York Imperial и Golden Delicious, чем у Rome Besuty или Jonatan. Чередующееся по годам плодоношение -физиологическое свойство отдельных деревьев, так же как и отдельных частей дерева. Отдельные экземпляры плодовых деревьев могут обильно цвести и плодоносить, другие деревья плодоносят очень слабо. На следующий год порядок меняется: деревья, прежде обильно плодоносившие, дают низкий урожай, а те, у которых был плохой урожай, образуют много плодов. Кроме того, одновременно одна ветвь дерева может образовать много цветов и плодов, а другие ветви остаются неплодоносящими. На следующий год, наоборот, прежде плодоносившие ветви остаются бесплодными. Двухгодичное чередование плодоношения деревьев может быть нарушено ранним повреждением органов плодоношения. Сильные заморозки весной могут повредить развивающиеся цветы и обещавший быть урожайным год становится вегетативным. Однако на следующий год вновь завязывается много цветов и плодов, и двухгодичный цикл восстанавливается.
В противоположность более или менее предвидимому ходу репродуктивного роста у плодовых у лесных деревьев цветение и плодоношение очень нерегулярны и непредсказуемы. Количество семян, образуемых лесными деревьями, сильно различается среди видов и по годам. Обычно цветение и плодоношение более регулярно в насаждениях, чем в естественных древостоях. Для некоторых видов, таких, как нисса и южная магнолия, характерно ежегодное образование хороших урожаев семян. Другие виды, такие, как бук, дают хороший урожай семян через несколько лет. Некоторые виды, например клен сахарный, образуют семена с четкими интервалами между годами плодоношения, а у ореха черного и тополя желтого урожаи семян нерегулярны. В промежутках между обильными урожаями семян образуется мало или совсем не бывает. Разные виды одного рода часто очень заметно отличаются по урожаю семян. Например, Populus trichocarpa, Р. balsamifera, Р. delto-ides образуют много семян лишь раз в 4-5 лет. (Нерегулярное плодоношение обсуждается также в главе 16.) Образование семян у различных видов деревьев в одном и том же древостое может значительно различаться. Тсуга западная обильно образует семена, а пихта гигантская и сосна горная из того же сообщества дают небольшие урожаи семян.
102
Глава 4
ДЕРЕВЬЯ ТРОПИКОВ. Тропические леса никогда не бывают лишены цветов из-за большого видового разнообразия и вариаций среди видов по времени цветения. Некоторые тропические деревья цветут круглогодично, но у большинства видов цветение происходит в определенное время или же они цветут нерегулярно.
Некоторые тропические деревья легко классифицируются на основании периодичности их цветения. Многие виды классифицировать по этому признаку трудно, так как периодичность их цветения зависит от окружающих условий, особенно сезонного распределения осадков и режима фотопериода. Например, деревья какао в Коста-Рике цветут в течение всего года, так как там выпадение дождя не приурочено к определенному сезону, а колебания температуры незначительны. В Бразилии, Байе и Эспириту-Санту, для которых характерны четкие дождливые периоды, цветение какао приурочено к влажным месяцам от октября до июня.
Идея о разной периодичности цветения среди видов в тропиках положена в основу классификации деревьев в работе Альвима (1964). Тропические древесные растения распределены им на четыре обширные группы: 1. Вечноцветущие виды. Цветы формируются непрерывно на протяжении всего года. К этой группе относятся виды: Hibiscus spp., Ficus spp. и Carica papaya, а также Trema guineensis в Гане и Dillenia suffruti-cosa в Малайзии. 2. Виды с несезонным цветением. У этих видов имеются вариации периодичности цветения от растения к растению и от ветви к ветви. Их цветение приурочено к определенному сезону при некотором удалении от экватора. Примерами таких видов являются: Spathodea campanulata, ich lia cha paca, Cassia fistula, Cassia splendens и Lagerstroe-mia flosregina. 3. Виды, зацветающие одновременно во всех точках ареала. У этих видов цветочные почки формируются регулярно, но долгое время (недели или месяцы) остаются закрытыми и открываются под влиянием существенных изменений окружающей среды. Они обычно зацветают при выпадении дождей после периода засухи. Примерами являются виды: Coffea spp., Clerodendron inci sum, Pte ro carpus indicus, ? Tabebua serrati folia. Через длительные интервалы цветут Bambusa, / Strobilanthes, Hopea и Schomia. Например, Bambusa впервые зацветает на 7-13 год в южной части Бразилии, а в других районах на 30-й год.
4. Сезонно цветущие виды. Эти виды цветут сезонно в тех местах, где у.' чередуются засушливые и влажные периоды или происходят сезонные изменения длины дня. В некоторых областях они становятся вечноцветущими или цветут несезонно. К сезонноцветущим видам относятся Monodoга tenuifolia и Bosqueia angolensis в Нигерии. В Тринидаде, для которого характерна смена сезонов, около половины листопадных деревьев цветет во время дождливого сезона, а половина — в засушливый период.
ПОЛОВОЕ РАЗМНОЖЕНИЕ У ПОКРЫТОСЕМЕННЫХ
СТРОЕНИЕ И РАСПОЛОЖЕНИЕ ЧАСТЕЙ ЦВЕТКА. Полностью сформированные цветы покрытосеменных состоят из четырех частей, расположенных на цветоложе (рис. 4.1). Самые нижние из них — чашели-
^продуктивный рост
103
Рис. 4.1. Типичный цветок (а — до оплодотворения; б — вскоре после слияния ядер спермин и яйцеклетки). Часть лепестков опала, и тычинки подсыхают ("The Ripening of Fruit”, j. В. Biale. Авторское право с 1954 Sci Atncrikan, Jnc. Все права сохранены) :
I - пыльник; 2 — тычиночная нить; 3 — пыльца; 4 — рыльце; 5 — лепесток; 6 — столбик; 7 — чашелистик; 8 — антиподы; 9 — семяпочка; 10 — полярные ядра; 11 ~ ядро яйцеклетки; 12 — синергиды; 13 — микропиле; 14 — ядро спермин; 15 — пыльцевая трубка
стики, вместе образующие чашечку. Выше чашелистиков располагаются лепестки, образующие венчик. Чашелистики и лепестки вместе составляют околоцветник. Внутри околоцветника находятся продуцирующие пыльцу тычинки, в совокупности называемые андроцеем, и плодолистики, которые составляют гинецей. Цветок может иметь от одного до нескольких плодолистиков, или карпеллей. Они обычно состоят из нижней, фертильной, части — завязи и верхней, стерильной, части - столбика. На кончике столбика расположено рыльце, на которое попадаются пыльцевые зерна, перед тем как оплодотворить семяпочки. Семяпочка важный орган, так как в ней формируются мегаспоры и развивается женский гаметофит, называемый зародышевым мешком.
Модификации цветка. У древесных растений цветок подвергается многочисленным модификациям, причем полностью сформированный он часто утрачивает некоторые части. Например, цветки тополей и черного ореха не имеют венчика, а цветок ивы лишен и чашечки и венчика. Другая модификация цветка — срастание его частей. Например, у винограда и рододендрона срастаются плодолистики, у катальпы - лепестки, у калины — чашелистики. Части околоцветника или плодолистиков могут срастаться либо в начале их образования, либо на более поздней стадии их развития.
Некоторые цветки имеют нектарники — поверхностные участки органов, выделяющих сахаристую жидкость, называемую нектаром, который привлекает пчел и других насекомых. Различают два основных типа нектарников: 1) особые части цветка, секретирующие нектар; 2) нектарники, образованные из органов, которые первоначально вы-
104
Глава 4
подняли другие функции. Нектарники могут развиваться на репродук* тивных и на вегетативных тканях. В цветке нектарники иногда локализуются у основания частей околоцветника, как, например, у гибискуса. Они могут быть также расположены в виде неглубоких или вогнутых колец между тычинками и основаниями завязи (Cercis siliquastrum, Robinia, Prunus, Rubus), между тычинками и столбиками завязи, если завязь нижняя (Eucalyptus), или как четкое кольцо вокруг основания завязи (Citrus). Нектарники часто образуются вне цветка (экстраф-лорально), на зубчиках листьев. Нектар может выделяться из нектарников несколькими способами: диффузно через тонкостенные эпидермальные клетки; диффузно из тонкостенных эпидермальных сосочков; экскрецией из кончиков волосков; через устьица, модифицированные для выполнения этой функции; экскрецией в результате разрыва кутикулы, вызванного набуханием наружных стенок. У магнолии весь цветок может называться нектарным, так как нектар диффундирует через кутикулу лепестков и выделяется через устьичные поры на основания лепестков и на поверхность пестика.
Цветки многих плодовых деревьев очень яркие, у большинства лесных деревьев они незаметные. Цветки покрытосеменных бывают одиночные или чаще собраны в различного типа соцветия. Цветки яблони собраны в зонтиковидные щитки, состоящие из 3-7, чаще из 5 цветков. У груши, как и у яблони, цветочные почки раскрываются терминальным щитком из 5 цветков. У маслины цветки собраны в метельчатые соцветия, каждое из которых состоит из 15 цветков. Соцветия появляются на побегах первого или второго года развития. Цветки развиваются полноценные, с функционирующими тычинками и пестиками. Иногда тычинки развиваются нормально, а пестики остаются недоразвитыми. У магнолии и тополя желтого цветки расположены поодиночке в пазухе листа. У черемухи поздней и клена пенсильванского они собраны в кид-ти, у тополей, берез и ольхи — в сережки, у конского каштана - в метелки, у бузины и калины — в щитки.
Большинство лесных деревьев однодомно, с тычиночными и пестичными цветками на одном и том же дереве (береза и ольха). Другие виды, такие, как хурма виргинская, тополь и ива, двудомные; у них тычиночные и пестичные цветки образуются на разных деревьях. Очевидно, что мужские деревья не могут образовывать семена. При выращивании декоративных двудомных кустарников или деревьев, таких, как остролист, из-за их ягод необходимо позаботиться, чтобы мужские экземпляры оказались рядом с женскими. В этом случае происходит опыление цветков и образуются ягоды. У немногих родов, таких, как Aescuhis на одном и том же дереве появляются обоеполые цветки и однополые, образующие только тычинки и пестики. Встречается и другая комбинация, как у Rhamnus и Fraxinus, у которых цветки обоеполые, или только тычиночные или пестичные.
Заложение и развитие цветка. У многих покрытосеменных деревьев умеренной зоны зачатки цветка начинают формироваться рано. Время заложения органов цветка у разных видов различно, но все-таки для большинства из них оно приходится на начало мая — конец июля. Например, после завершения вегетативного роста к концу июня почти все почки взрослых деревьев клена ложноплатанового вегетативные, но
^продуктивный рост 105
позднее апекс большинства этих почек проходит через переходные стадии и начинается формирование цветков. В результате деления и растяжения клеток цветковый апекс формирует соцветие, которое остается неразвитым в течение зимы. Быстрое его развитие возобновляется весной, и деревья зацветают к концу мая или в начале июня.
Время заложения цветков зависит от погоды, условий местообитания и приемов возделывания. В связи с этим период заложения цветков ежегодно изменяется. Различен он также у разных видов. Степень структурной дифференциации цветка в течение первого сезона сильно варьирует у разных видов. Некоторые вариации во время заложения цветка наблюдались у цветков, расположенных на побегах. У яблони, например, » терминальных почках побегов цветки закладывались позднее, чем в ночках веточек. В терминальных почках веточек двухгодичных побегов они возникали также позднее по сравнению с трехгодичными. В пределах щитка центральный цветок формируется: позднее боковых, но развивается быстрее.
Цветок развивается из апикальной меристемы. Части цветка возникают у верхушки цветоложа, окруженного выступами с меристематической активностью. Обычно развитие происходит акропетально. Первыми появляются чашелистики, за ними следуют по порядку лепестки, тычинки и плодолистики. Ранние стадии формирования отдельных частей цветка сходны с такими стадиями при формировании листьев на вертушке ствола. Части цветка образуются в результате периклинальных делений клеток, находящихся под поверхностными слоями апикальной меристемы. За этими делениями следуют другие, в том числе антиклинальные деления. Части цветка увеличиваются в длину и ширину в результате апикального и маргинального роста,
ФОРМИРОВАНИЕ И РАСПЫЛЕНИЕ ПЫЛЬЦЫ. В год массового цветения иногда возможны низкие урожаи семян и плодов, обусловленные неблагоприятными условиями для опыления. У деревьев образуется огромное количество пыльцы, особенно у ветроопыляемых видов. По данным Фэгри и Иверсена (1975), один пыльник ветроопыляемой березы может образовать 10 тыс. пыльцевых зерен, в то время как пыльник насекомоопыляемого клена формирует только 1 тыс. пыльцевых зерен. Пыльца березы, тополей, дубов, ясеней, ильмов, гикори, сикоморы и голосеменных рассеивается ветром. Пыльца липы, клена, ивы и большинства плодовых деревьев разносится насекомыми, в основном пчелами. При беспорядочном распылении пыльцы ветром большая ее часть оказывается бесполезной по сравнению с более упорядоченным переносом от цветка к цветку насекомыми. Опыление пчелами в значительной степени случайно, происходит при сборе ими нектара, но в немногих случаях имеются особые отношения между насекомыми и растениями, которые они опыляют Примером может служить сложный процесс опыления инжира колхидской осой.
Влияние климатических условий на рассеивание пыльцы. Ежедневные колебания температуры и влажности существенно влияют на освобождение и рассеивание пыльцы. Сарвас (1962) показал, что в Финляндии большая часть пыльцы высыпается в самые теплые дни вегетационного периода. Обычно максимальное рассеивание происходило около полудня, его совсем не было ночью. Очень редко, когда относительная влаж
106
Глава 4
ность воздуха была низкой, большое количество пыльцы рассеивалось также и ночью. Низкая влажность подсушивает клетки пыльника и способствует его раскрыванию. Дождь часто тормозит раскрывание тычинки и рассеивание пыльцы. Холодная, дождливая погода во время опыления снижает также и активность насекомых, что сокращает завязывание плодов. Хорошие погодные условия имеют большое значение, так как цветение и опыление могут закончиться за короткий срок, часто в течение одного-нескольких дней.
Дальность распространения пыльцы. Пыльца большинства древесных видов разносится на большие расстояния, чем семена. Лесные генетики особенно интересуются тем, какое влияние оказывает дальность распространения пыльцы на деревья в семенных плантациях, от чего зависят опыление и развитие локальных популяций. Райт (1953) подчеркивает важность расстояния распространения пыльцы для генетики лесных пород. Например, если мужские и женские экземпляры ясеня американского при посадках или прореживаниях лесных насаждений окажутся удалены друг от друга на 90 м, то всего лишь 1% семяпочек будет оплодотворяться. При сокращении расстояния до 60 м, будет оплодотворяться до 15% семяпочек. Ширина изоляционной полосы вокруг семенного питомника также зависит от дальности распространения пыльцы.
У разных видов дальность распространения пыльцы различна, поэтому мнения о важности распространения пыльцы для лесных пород не совпадают. Райт (1953) представил данные, доказывающие, что даже наиболее далеко переносимая пыльца падает довольно близко от дерева-источника и большая часть перекрестных опылений происходит между деревьями, растущими близко друг от друга. На это указывает и тот факт, что большинство гибридов находится вблизи от родительских форм. Там, где расстояние рассеивания пыльцы относительно короткое, большая часть семян свободноопыляемых видов образуется в результате опыления соседних деревьев. В результате деревья, растущие друг от друга в пределах 100 м, генетически более похожи, чем деревья, разделенные километрами.
Отмечается и очень дальнее распространение пыльцы, особенно у хвойных. Например, Сарвас (1955) наблюдал более мощное выпадение пыльцевого дождя
4.2. РАСПРОСТРАНЕНИЕ ПЫЛЬЦЕВЫХ ЗЕРЕН ЛЕСНЫХ ДЕРЕВЬЕВ (Wright, 1952)
Pinus cembroides var. edulis Pseudotsuga menziesii Cedrus spp.
Picea excelsa Fraxinus spp.
Corylhis avellana
Juglans regia
Ulmus americana
Populus deltoides
Populus nigra var. italica
Примечание. Виды перечислены в порядке увеличивающегося расстояния распространения.
^Продуктивный рост 107
ни пароходе, находившемся в 20 км от берегов Финляндии, чем в прибрежном несу. Андерсон (1963) обнаружил на скалистых островах в 4—8 км от ближайше-IV леса слой пыльцы ели толщиной около 1 см. Ланнер (1966) предположил, что пыльца поднимается вверх током воздуха, переносится на большие расстояния и затем выпадает с дождями.
Пыльцевые зерна ели, дугласии, сосны и ясеня распространяются па меньшие дистанции и быстрее выпадают, чем пыльца тополя, ильма, ореха и лещины (табл. 4.2). По-видимому, нет заметных различий в расстояниях распространения пыльцы насекомыми и ветром. Не обнаружена также четкая корреляция между размерами пыльцы и дальностью ее распространения.
Пыльца деревьев и сенная лихорадка. Широколиственные деревья являются одной из причин ранневесенней лихорадки, совпадающей с периодом распространения пыльцы дуба, ильма, тополя, березы, клена, ореха и ивы. Некоторые виды ивы часто растут рядом, поэтому пыльца такой группой может распространяться в течение нескольких недель. Также могут расти вместе несколько видов березы, и хотя у каждого вида пыльца осыпается менее чем за неделю, время рассеивания пыльцы каждого из них накладывается друг на друга. Таким образом, вся группа видов в целом рассеивает пыльцу в течение 3—4 недель. Наоборот, виды ильма обычно растут одиночно, поэтому у них период осыпания пыльцы короткий. В связи с этим тяжелая сенная лихорадка, которую они вызывают, длится недолго. Голосеменные, образующие очень много пыльцы, не являются существенным фактором, вызывающим сенную лихорадку»
В течение вегетационного периода сенную лихорадку вызывают не только древесные растения. В начале лета причиной сенной лихорадки в основном является пыльца трав. В более поздний период сенную лихорадку в значительной мере вызывают пыльца амброзии и дурнишника.
Хранение пыльцы. Сбор и хранение пыльцы часто необходимы для скрещивания древесных растений. В умеренной зоне период цветения многих видов деревьев очень короток, так что пыльцу необходимо собирать быстро. Лучше всего пыльцу хранить в холодных и сухих условиях. Райт (1976) рекомендует хранить пыльцу в течение короткого периода в пробирках, закрытых ватной пробкой, помещенных в эксикатор с относительной влажностью 25% и ниже, при температуре от 3 д0 - 4°С. или при возможности глубокого замораживания, при минус 10-20°С.
В селекционной работе лучше использовать свежую пыльцу. Пыльцу винограда, сосны и ели можно хранить в течение года при — 18°С и очень низкой влажности. Пыльцу можно запасать и на длительный срок, если предварительно подвергнуть ее обработке. Например, пыльцу дугласии успешно хранили в течение 2 лет, подготовив ее следующим образом; сначала пыльцу подсушили на воздухе в течение 4 ч до остаточной влажности 8%; в течение 30 дней выдержали при 0°С; поместили на 1 ч в вакуум при -77°С, затем пробирки с пыльцой герметически закупорили.
ОПЛОДОТВОРЕНИЕ. Необходимое условие для оплодотворения -восприимчивость рыльца при попадании на него жизнеспособной пыльцы. Такая синхронность встречается на одном и том же дереве у одно
108
Глава 4
домных цветов. В других случаях осыпание пыльцы и готовность рыльца не синхронны. Например, у отдельных растений некоторых однодомных видов, таких, как клен сахарный, пыльца осыпается прежде, чем рыльце становится восприимчивым. У других растений пыльца рассеивается после того, как период готовности рыльца уже миновал. Подобные различия отмечаются и у некоторых двудомных видов растений с однополыми цветами. Такие несовпадения во времени двух процессов предотвращают самоопыление. Это ценное свойство, так как потомство при перекрестном опылении обычно более мощное, чем при самоопылении.
Основные органы семяпочки внутри завязи — это интегументы, микропилярное отверстие на противоположном конце ножки и зародышевый мешок, занимающий большую часть семяпочки. Перед оплодотворением зародышевый мешок обычно содержит восемь ядер. Три из них, локализованные у микропилярного конца, представляют ядро яйцеклетки и две синергиды. Два полярных ядра, локализованных в центре зародышевого -мешка, и три ядра на конце противоположном микропиле являются антиподами (см. рис. 4.1).
Пыльцевые зерна, попав на рыльце, пропитываются водой и прорастают, образуя пыльцевые трубки. Пыльцевая трубка, проникнув через поверхность рыльца, растет между клетками столбика, выделяя ферменты, которые размягчают пектины срединной пластинки. Большое количество дополнительных ферментов, выделяемых пыльцой вовремя прорастания, стимулирует изменения метаболизма тканей столбика и, вероятно, образует субстраты для роста пыльцы. Пыльцевые трубки, прорастая с разной скоростью через столбик, достигают завязи, содержащей семяпочки. Многие пыльцевые трубки могут достигнуть завязи, но лишь одна из них проникает в семяпочку. Ядро пыльцевой трубки и генеративное ядро пыльцевого зерна оба проникают в пыльцевую трубку. Генеративное ядро подвергается раннему митотическому делению и образуются два спермия. Пыльцевая трубка, достигнув семяпочку, проникает в зародышевый мешок и освобождается от двух спермиев. Одно из них сливается с яйцеклеткой, другое с двумя полярными ядрами, образуя ядро эндосперма. После оплодотворения и образования зиготы оставшиеся ядра зародышевого мешка — две синергиды и три антипода разрушаются. Пыльцевые трубки растут очень быстро (иногда на несколько миллиметров в час), поэтому временной интервал между опылением и оплодотворением очень короток.
По данным Магешвари (1950), у большинства покрытосеменных между опылением и оплодотворением проходит лишь 24-48 ч. Этот интервал значительно варьирует среди древесных видов и может быть 12-14 ч у Coffea arabica, 3-4 месяца у Corylus avellana и до 12-14 месяцев у некоторых видов дуба.
РАЗВИТИЕ ПОСЛЕ ОПЛОДОТВОРЕНИЯ. После оплодотворения интенсифицируется клеточная активность семяпочки, характеризующаяся быстрым ростом эндосперма и передвижением питательных веществ к разрастающейся семяпочке. Нормальный рост и дифференциация зародыша зависят от эндосперма. Зародыш, увеличившись в размерах, примыкает к клеткам эндосперма и у некоторых видов может израсходовать почти весь эндосперм. Зрелый эндосперм богат углевода
Репродуктивный рост 109
ми, жирами, белками и ростовыми гормонами. Доказана важность эндосперма как питательной ткани в период роста зародыша. К моменту оплодотворения в зародышевом мешке нет достаточного количества питательных веществ. Во время развития эндосперма в нем накапливаются питательные вещества, необходимые для формирования зародыша. Зигота обычно не начинает делиться до тех пор, пока не начнет развиваться эндосперм. Более того, зародыш развивается нормально только тогда, когда организован эндосперм. Если прерывается формирование эндосперма,сразу же тормозится и развитие зародыша.
ПОЛИЭМБРИОНИЯ. В семенах деревьев покрытосеменных иногда развивается много зародышей. Полиэмбриония бывает ложная и истинная. При ложной полиэмбрионии происходит слияние двух или более ядер или развиваются два и более зародышевых мешка в одном пуцеллусе. При истинной полиэмбрионии в зародышевом мешке возникают дополнительные зародыши вследствие дробления зиготы или из клеток синергид и антипод. Адвентивные (придаточные) зародыши также являются примером истинной полиэмбрионии. Придаточные зародыши возникают из наружных тканей зародышевого мешка (т.е. из клеток нуцеллуса или интегумента). Затем они проникают в зароды-шевый мешок, где и развиваются до зрелого состояния. Придаточные зародыши отличаются от половых эмбрионов боковым расположением, неправильной формой и отсутствием подвеска (суспензора). Они отличаются также и генетически, так как придаточные эмбрионы имеют гены лишь материнского растения. У некоторых видов полиэмбриония взаимосвязана с генетическим фактором и появляется при гибридизации.
Полиэмбриония имеет особенно важное практическое значение при размножении определенных видов деревьев. Например, придаточные эмбрионы, наследующие признаки лишь материнского растения, используются для получения генетически однородных сеянцев манго и цитрусовых. У цитрусовых они используются для формирования подвоя, на который прививается другой сорт. Кроме того, у клонов цитрусовых с ослабленными свойствами в результате повторных вегетативных размножений благодаря нуцелларным эмбрионам может быть восстановлена энергия исходных сеянцев. Такая вегетативная стимуляция, или обновление, может объясняться гормональным влиянием зародышевого мешка. Гофмейер и Обергольцер (1948) вырастили из придаточных зародышей лучшие сеянцы цитрусовых, чем из черенков. Эта разница объясняется тем, что черенки поражались вирусными болезнями, а нуцеллярные зародыши были к ним устойчивы.
АПОМИКСИС. Семена возникают обычно в результате полового размножения, но у немногих видов древесных растений имеется бесполый способ образования семян. Растения, у которых зародыши возникают без оплодотворения семяпочки, называются апомиктами. У многих видов боярышника и ежевики на востоке США встречаются апомиктические дериваты. Предполагается, что апомиксис должен быть и у лесных деревьев, но до сих пор он пока у них не обнаружен. Апомиксис представляет большой интерес для генетиков, так как все сеянцы, полученные из апомиктических семян, имеют однородную генетическую конституцию.
110
Глава 4
ПАРТЕНОКАРПИЯ. Обычно развитие зрелых плодов требует оплодотворения яйцеклетки. У немногих видов, однако, плоды завязываются и созревают без развития семян и без оплодотворения яйцеклетки. Такие плоды, называемые партенокарпическими, хорошо известны у некоторых сортов инжира, груши, персика, яблони, вишни, сливы и цитрусовых. Партенокарпия бывает также у многих лесных деревьев, таких, как клен, ильм, береза и желтый тополь. Некоторые типы партенокарпии требуют опыления, другие не требуют его. Например, плоды у цитрусовых и банана могут развиваться без опыления. У других видов плодовых деревьев, таких, как вишня и персик, возможна бессемян-ность, если зародыши погибают перед созреванием плодов. У некоторых видов опыление стимулирует развитие плода, но пыльцевая трубка не достигает семяпочки и плоды созревают без предварительного процесса оплодотворения.
Физиология партенокарпии недостаточно выяснена. Плоды с несколькими семяпочками часто являются партенокарпическими. Это дает основание предположить, что семяпочки содержат химические вещества, стимулирующие завязывание и формирование плода. Тот факт, что в семяпочках синтезируется ауксин, и что после опрыскивания пестиков синтетическим ауксином у некоторых видов завязываются плоды, дает основание сделать вывод, что ауксин необходим для завязывания плодов. Однако вследствие такого приема партенокарпические плоды не завязываются у большинства листопадных плодовых деревьев (за исключением инжира, груши и некоторых сортов яблони). Поэтому допускается, что для завязывания плодов необходим какой-то дополнительный фактор. Химическое индуцирование партенокарпии наиболее успешно у видов, у которых возможно естественное завязывание парте-нокарпических плодов. У мастин, авокадо и манго применение ауксина может способствовать завязыванию плодов, однако для их дальнейшего развития необходимо опыление. После ауксиновой обработки часто завязываются плоды, развиваются зародыши, которые могут быть оплодотворены, но они часто бывают недоразвитыми. (Роль ростовых веществ в вегетативном и репродуктивном росте обсуждается подробнее в главах 15 и 16.)
РОСТ ПЛОДОВ. Период от цветения до созревания плодов продолжается от 3 недель (у земляники) до 60 недель (у некоторых сортов апельсина). У многих видов этот интервал не превышает 15 недель. Различные плоды растут с разной скоростью и достигают в зрелом состоянии различных размеров. Например, маслины и смородина растут очень медленно (около 0,01—0,02 см^ в день). Следует напомнить, что скорость роста плодов значительно зависит от сезона, окружающих условий, агротехнических приемов. Различна она также у разных плодов одного сбора. Рост плодов происходит за счет клеточных делений и увеличения размеров клеток. В период цветения клеточное деление ограничено, но после завязывания плодов многие их ткани становятся меристематическими, к ним поступает очень много углеводов. У некоторых плодов (например, у смородины, ежевики) деление клеток (за исключением клеток зародыша и эндосперма) заканчивается к моменту опыления. У яблони и цитрусовых клетки делятся в течение короткого времени после опыления. У некоторых других видов деление клеток
Репродуктивный рост
Ш
после опыления продолжается долго. У авокадо клетки продолжают делиться в течение всей жизни плода. У большинства видов увеличение размеров клеток делает основной вклад в общий рост плода. У винограда вследствие увеличения количества клеток размер плода удваивается, а за счет увеличения объема клеток размер плода возрастает в 300 раз.
Кривые роста. Имеются два основных типа кривых роста плодов. Первый из них - простой сигмоидальный, при котором вначале отражается экспоненциальное увеличение размеров плодов, за которым следует спад. В результате кривая; роста приобретает сигмоидальный вид (рис. 4.2). Этот тип кривой рфта характерен для яблони, груши, апельсина, финика, банана, авокадо, земляники, манго и лимона. Определенная форма кривой роста часто несколько нарушается у разных сортов. Кривая роста развивающихся плодов раннеспелых сортов яблони приближается к прямой линии, у сортов позднеспелых кривая выравнивается к концу сезона. Скорость роста плодов также сортовая характеристика: у раннеспелых сортов плоды растут быстрее, чем у позднеспелых.
Второй тип кривой роста характерен для косточковых (инжира, вишни, персика, абрикоса, сливы, маслины) и ряда некосточковых плодов (винограда, смородины). Это двойной сигмоидальный тип, отражающий три стадии роста (рис. 4.3). У косточковых плодов во время 1 стадии завязь, нуцеллус и интегументы семени растут быстро, зародыш и эндосперм растут очень медленно. Во время П стадии зародыш и эндосперм растут быстро, завязь увеличивается в размерах мало. К концу П стадии начинается склерификация ямки, зародыш достигает окончательного размера и сильно увеличивается эндосперм. В Ш стадии возникает новая волна роста завязи, которая не спадает до созревания плода. Длительность трех стадий роста различна: у раннеспелых сортов П стадия проходит в течение нескольких дней, а у поздне-созревающих она может длиться месяц (табл. 4.3).
По данным некоторых авторов, семена и перикарпий активно конкурируют за питательные вещества. Семена являются более слабыми конкурентами из-за их положения в системе передвижения веществ. Тем не менее считается, что семена регулируют рост плода, так как
Рйс, 4.2. Рост в длину у раннеспелых сортов яблони (а - Early harvest, б -Mclutosh, в — Rome) в период от полного цветения до созревания плодов (Tukey, Young, 1942);
1 — целого плода; 2 — плодолистика; 3 — нуцеллуса и интегумента; 4 — зародыша
112
Глава 4
Рис. 4.3. Рост в длину у сортов вишни (а - Farly Richmond. б -English Morello), созревающих в разное время вегетационного периода
(слева - полное цветение, справа -созревание плодов; Tukeu, 1935): 1 — прикарпия; 2 — нуцелпуса и
интегумента; 3 — зародыша
Даты
они дают сигнал для передвижения питательных веществ к развивающемуся плоду. Фактическая стимуляция передвижения, по мнению Чалмерса и ван ден Энда (1977), вызывается этиленом, который синтезируется в перикарпии.
Ни один из двух основных типов кривых роста не относится к какому-то определенному морфологическому типу плодов. Например, рост некоторых ягод, яблок может характеризоваться одним или другим типом кривой роста.
СОЗРЕВАНИЕ ПЛОДОВ. Созревание плодов связано с многочисленными изменениями в метаболизме, который в свою очередь зависит от энергии дыхания. Изменения, связанные с созреванием плодов, включают потерю хлорофилла и демаскирование других пигментов, размягчение мякоти плода, появление аромата, вкуса и уменьшение сухого веса. Эти изменения протекают одновременно с образованием этилена, гидролизом нерастворимых пектинов, превращением крахмала в сахара в одних плодах и исчезновением танина в других. Некоторые из этих изменений являются основными и происходят у многих Плодов. Большинство плодов созревает на дереве, немногие, такие, как авокадо, созревают иначе. Созревание плодов может быть ускорено их удалением с дерева.
Изменения, сопровождающие созревание плодов, взаимосвязаны с критическим увеличением скорости дыхания у многих видов. Например, резкое снижение интенсивности дыхания, за которым следует критический подъем, завершается физиологическим распадом или инвазией грибов. Считается, что климактерический период является критическим, означающим окончание развития, созревания и начало старения. Ряд изменений, связанных с созреванием, происходит во время или сразу прсле критического периода: например, изменение цвета зеленого в желтый у некоторых сортов яблони, груши, банана и папайи, зеленого в красный
Репродуктивный рост
113
4.3. ПРОДОЛЖИТЕЛЬНОСТЬ СТАДИЙ РАЗВИТИЯ ПЛОДОВ
У ВИШНИ, ДНИ (Тикеу, 1935)
Сорт i Стадия От полного — цветения до созревания
П Ш
Farly Ruchmond 22 5 14 41
Montmorency 22 12 23 57
English Morello 21 28 17 66
у сливы и манго, зеленого в темно-коричневый у некоторых сортов авокадо. Изменения цвета могут быть результатом исчезновения хлорофилла (у банана) или образования каротиноидов (у апельсина). Кроме того, плоды размягчаются. У груши столовая спелость создается в критический период. У яблок, бананов и авокадо самые высокие вкусовые качества появляются вскоре после климактерического цикла. (Более подробно дыхание плодов обсуждается в главе 6.)
Вкусовые качества плодов. Вкусовые качества плодов обсуловлены изменениями химического состава и физических внутренних условий во время их развития. Как уже упоминалось, нерастворимые пектины превращаются в растворимые в клеточных оболочках созревающих плодов. Наступает разрыв некоторых клеток, сок освобождается и поступает в межклетники, повышая вкус плодов. Вкусовые качества плодов определяются соотношениемсахаров, кислот и'вяжущих компонентов. Сахароза, глюкоза и фруктоза - основные сахара - у разных плодов находятся в различных соотношениях. Яблоки содержат все три сахара, у персиков и абрикосов преобладает сахароза, у вишни большая доля приходится на глюкозу и фруктозу. Столовые плоды, включая виноград, бананы, апельсины и яблоки, содержат от 9 до 20% сахара. У винограда бывает и еще более высокое содержание сахара. В противоположность семенам у мясистых плодов не накапливается большого количества крахмала или запасных белков.
На вкус развивающихся плодов влияют также органические кислоты Органические кислоты, вероятно, передвигаются в плоды из листьев, так как содержание кислот в плодах снижается, если сокращается площадь листьев у винограда и яблони. Более высокое содержание кислот обнаружено в центральной части растений. Концентрация органических кислот в плодах высокая, достигающая часто 1-2% свежего веса в яблоках и грушах и до 4% у черной смородины. Наиболее важные органические кислоты в плодах - яблочная в яблоках, лимонная у цитрусовых, виннокаменная в винограде, изолимонная в ежевике. В других плодах содержится смесь этих кислот. Вкус плодов связан также с ароматом, который придается амиловыми эфирами муравьиной, уксусной, капроновой и каприловой кислот.
Размягчение плодов. Этот процесс связан с изменениями в пектиновых соединениях и с гидролизом крахмала илт жиров. По мере развития плода нерастворимый протопектин откладывается в первичных стенках клеток, накапливается до высокой концентрации в яблоках., грушах и цитрусовых. Превращения нерастворимых пектинов (протопектинов и кальциевых солей пектиновой кислоты) в растворимые соединения, такие, как пектин и пектиновая кислота, тесно связаны с созреванием. Превращения пектиновых соединений во время созревания, вероятно, происходят благодаря активности протопектиназы. При переходе от зеленой к зрелой стадии в плодах персика количество растворимых пектинов может увеличиться в 2, в яблоках в 12, в грушах в 20 раз. Из перезрелых плодов пектиновые вещества исчезают,и фрукты сильно размягчаются. В созревающих плодах цитрусовых происходят такие же, но в меньшей степени, изменения в пектиновых соединениях. В конечной стадии старения плодов протопласт распадается и клеточные оболочки разрушаются. Действительный распад плодов связан скорее с дезорганизацией и утратой контроля над ферментативными процессами, чем с истощением запасов питательных веществ.
Глава 4
114
ОТДЕЛЕНИЕ ОРГАНОВ РАЗМНОЖЕНИЯ. Репродуктивные органы отделяются в результате процессов, сходных в основном с такими процессами при опадении листьев. Флоральные структуры опадают как части цветка, целые цветки или целые соцветия. Лепестки обычно опадают в результате размягчения срединной пластинки. Исследования Лотта и Симонса (1964) показали одни и те же закономерности в развитии зон отделения чашечки цветка и столбика у большого числа видов Prunus. Были обнаружены и существенные различия среди видов в ско-
4.4. СКОРОСТЬ РАЗВИТИЯ ЦВЕТОЧНОЙ ТРУБКИ И ОПАДЕНИЯ СТОЛБИКА
У РАЗНЫХ ВИДОВ Prunus (Lott, Simons, 1968)
Вид Сорт Число дней от опыления
до опадения лепестков до появления отделит ельн ой зоны цветочной трубки до отделения столбика
Р. avium Starting Harby Giant 3-4 5 -6 3-4
Р. avium Stark Gold 3-4 5-6 5-6
Р. cerasus Monmorency 3-4 5-6 5-6
Р. armeniaca Wilson 3-5 6-8 18- 20
Р. persica Delicius 3-6 7-10 20 - 24
рости формирования зоны отделения. Так у Р. cerasusn Р. avium она развивается быстрее, чем у Р. persica и Р. armeniaca (табл. 4.4).
Иногда плоды отделяются преждевременно, сразу после опыления и завязывания плода или во время роста молодого зародыша. Преждевременное отделение незрелых плодов — наиболее серьезная проблема в отношении яблонь, груш, апельсинов и грейпфрутов. Для яблони характерны три нормальных периода опадания: раннее опадание — меж-
4.5. ОПАДЕНИЕ ЖЕЛУДЕЙ ДУБА БЕЛОГО В РАЗНЫЕ СТАДИИ РАЗВИТИЯ (Williamson, 1966)
Стадия развития
1962 г.
1963 Г.
Исходная дата Про- Исходная дата Процент
цент опадения
опадения
Опыление 30 апреля 55,6 24 апреля 28,4
’азвитие семяпочек 17 мая 10,6 20 мая 37,9
Оплодотворение 7 июня 15,8 8 июня 18,1
Развитие зародыша 6 июля 16,7 3 июля 10,5
Созревание 19 сентября 1,3 20 сентября 5,1
100,0 100,0
Репродуктивный рост
115
ду начальным растяжением завязи и началом развития эндосперма1 июньское опадание молодых плодов; опадание перед сбором урожая.
Известно преждевременное отделение плодов у ветроопыляемых видов лесных деревьев. Например, часто преждевременно опадает болы шая часть потенциального урожая желудей белого дуба (табл. 4,5). Вильямсон (1966) обнаружил, что от начала мая до середины июля в период опыления и развития семяпочки и после оплодотворения у дуба белого теряется до 90 % желудей возможного урожая.
Локализация слоев отделения плодов варьирует у видов. Место отделения яблока расположено у основания плодоножки. Плодики сливы отделяются вместе со стеблем в первые дв₽ из трех волн сезонных опаданий плодов и без стебля в третье опадание. У черешни имеются две зоны отделения плодов, расположенные между плодоножкой и стеблем и между стеблем и Шипом. Плоды сливы, манго, авокадо и апельсина отделяются у основания. Плоды многих тропических видов после сформирования у них плодоножки отделяются от нее за счет второго слоя отделения. Осыпание плодов вместе с их плодоножкой широко распространено в умеренной зоне у ивы, тополя, липы, белой акации, ильма.
ПОЛОВОЕ РАЗМНОЖЕНИЕ У ГОЛОСЕМЕННЫХ
У голосеменных нет чашечки, венчика, тычинки и пестика. Цветы состоят из мужских и женских шишек (шишек, несущих тычинки, и шишек, несущих семяпочки), образующихся у большинства видов на одном и том же дереве. Созревшие мужские шишки часто ярко-желтого, пурпурного или красного цвета, состоят из оси, несущей серию спирально расположенных чешуек. Каждая из чешуек на нижней поверхности имеет два пыльцевых мешка. Женские шишки также иногда ярко окрашены. Они обычно больше и устойчивее мужских шишек, состоят из оси, несущей чешуйки с семяпочками, каждая из которых появляется на оси кроющего листика. Две семяпочки появляются в виде выростов на верхней стороне чешуйки. У конца семяпочки, вблизи от оси шишки, имеется микропиле, через которое могут проникать пыльцевые зерна.
Семена голосеменных обычно развиваются от 1 до 3 лет. Однолетний жизненный цикл характерен для цикадовых, Ginkgo, Ephedra, Gnetum и для большинства представителей Gupressaceae и Taxodiaceae; 1—2-летний — у Podocarpus (в зависимости от вида); 2-летний — у Caphalotaxus й Тоггеуа. У Pinus шишки закладываются осенью или в начале зимы, опыление происходит весной, оплодотворение — на следующий год. Созревание женских шишек и опадение семян завершается в течение следующей осени. Отсюда, жизненный цикл женской шишки сосны охватывает три вегетационных периода.
ЗАЛОЖЕНИЕ И РАЗВИТИЕ ШИШКИ. Для хвойных характерны ив я типа заложения шишек. Почки шишек у кедра, тсуги, лиственницы и ели нормально формируются путем перехода вегетативных побегов в репродуктивные. Для этого перехода у красной сосны требуется 2 недели, у дугласии 11 недель. У сосен, пихты, дугласии и некоторых елей закладывающиеся новые осевые побеги развиваются в генеративные почки быстрее, чем побеги, которые уже функционировали как вегетативные.
116 Глава 4 ;
'p:
Время заложения шишек значительно варьирует у разных видов, а в пределах вида зависит от окружающих условий. При благоприятных условиях шишки закладываются в период, предшествующий опылению. Время это различно: у дугласии шишки закладываются ранней весной, у туи — в начале лета, у некоторых сосен — осенью. После заложения шишки у всех видов развиваются в течение многих месяцев. Порядок морфологических изменений не зависит от режима окружающих условий, но время прохождения каждой фазы развития значительно различается. Как видно из табл. 4.5 и рис. 4.4, мужские и женские шишки дифференцируются в разное время года и с разной скоростью, причем мужские шишки обычно формируются первыми и дифференцируются быстрее. У сосны, дугласии и туи женские шишки в огромном количестве образуются в верхней части кроны, мужские шишки — в нижней части кроны. У пихты женские шишки всегда концентрируются на самых верхних ветвях, мужские шишки расположены ниже, в верхней половине кроны.
4.6. ВРЕМЯ ЗАЛОЖЕНИЯ МУЖСКИХ И ЖЕНСКИХ ШИШЕК
Вид Время заложений шишек Источник
мужских женских
Кипарисовик нуткан- Середина июня - Конец июня - се- Owens, Molder,
ский начало июля редина июля Я9?4?
Криптомерия япон- Конец июня - Середина июля ~ Hashizume
ская конец сентября середина сентяб- (1962)
Сосна Эллиота Конец июня - ря Конец августа Mergen, Koerting
июль (1957) „
Можжевельник за- Начало июня Июль Owens, Pharis
ладный (1971)
Тсуга западная Июнь Июль Molder (1974)
Иесяа Стадии роста
Январь март Апрель Май Июнь Июль Август C&ffUlfyl Ммф ИвяВрь йркаврь Вегетативные почки Почки женской шишки Почки мужской шишки
Спящие вегетативные почки Спящие Вегета -тивные почки Спящие веге- • тативные почки 3
Раннее заложение почечных чешуек Распускание Sese-таяшвных почек позднее заложение почечных чешуек Раннее заложение пачечных чешуек Вскрывание вегетативных почек Позднее заложение почечных чешуек Формирование 'Заложение па-терминпльных [зишных почек пачек “
1 г Заложение почечных чешуек
переход
Раннее (Зыс-трое) заложение листьев Позднее(медлен-ное заложение листьев Спящие веге-татидные почки переход 1 1 Разви- ! Разви- 1 тие mue 1 микро- [микро- 1 споро- спорая- ранние сриллп гия ^стадии i ; мейоза
t i Раз- ! Раз- [Появление витие\вшпие\мееиспср при- че~ ,машерин-цзет-,щуек ских ника \ [клеток
спящие ночки женской шишки Спящие почни мужской шишки
Рис. 4.4. Фенология развития вегетативной почки, почек женской и мужской шишек у тсуги западной, растущей недалеко от Виктории (Британская Колумбия; Owens, Molder, 1974)
Репродуктивный роет 117
ОПЫЛЕНИЕ И ОПЛОДОТВОРЕНИЕ. У разных видов образуется различное количество пыльцы. Процесс формирования пыльцы зависит от состояния древостоя и отдельных ветвей, особенностей отдельных деревьев. Время опыления также различается по годам. В Финляндии у сосны смолистой пыление отмечалось в течение 5-10 дней. В отдельных древостоях пыльца разлеталась у сосны смолистой дольше, чем у ели европейской или березы повислой. В годы, когда пыльцы образовывалось очень много, большая ее часть осыпалась в середйне периода цветения. В годы, когда пыльцы формировалось мало, установить периоды ее максимального разлета не удавалось. Период готовности женских шишек к опылению обычно продолжается не более недели. Исключение составляет дугласия, у которой этот период длится около 20 дней. Длительность периода зависит также от расположения шишек на дереве. На готовность шишек влияют основные факторы окружающей среды, особенно температура, влажность и ветер, поэтому, вероятно, период готовности различен в разные года.
У голосеменных происходит обычно перекрестное опыление, так как женские цветы концентрируются на верхних ветвях, а мужские на нижних. Переносимые ветром пыльцевые зерна застревают между чешуйками шишки и вступают в контакт с семяпочками. У большинства голосеменных, за исключением Abies, Cedrus, Larix, Pseudotsuga и Tsuga, выделяется сахаристая опылительная капля возле микропиле. Во время периода оплодотворения эта жидкость заполняет микропи-лярный канал. Опылительная капля выделяется утром и исчезает днем. Выделение секрета продолжается в течение нескольких дней или до тех пор, пока пыльца не попадет в семяпочку. Пыльцевые зерна попадают в жидкость, и при втягивании капли в канал они вместе с ней всасываются в микропиле и вступают в контакт с нуцеллусом.
Пыльцевые зерна прорастают, образуя пыльцевые трубки, которые растут вниз, в нуцеллус. При растяжении каждой пыльцевой трубки ее генеративные клетки делятся, образуя инициальную антеридиальную и соматическую клетки. Соматическая клетка последовательно делится и образует две клетки спермиев. Более крупный спермин сливается с ядром яйцеклетки в- архегонии, и оплодотворение заканчивается. Другой спермий разрушается. Период между опылением и оплодотворением различен у разных родов голосеменных. Пыльцевые зерна остаются на нуцеллусе неактивными у ели в течение нескольких дней, у дугласии около 3 недель, у Cedrus около 9 месяцев. У сосны оплодотворение происходит примерно через 13 месяцев после опыления.
ПОЛИЭМБРИОНИЯ. Наличие более одного зародыша — распространенная особенность эмбриогении у Pinaceae. У голосеменных встречаются два типа полиэмбрионии: простая полиэмбриония, при которой добавочные зародыши образуются в результате оплодотворения нескольких архегониев гаметофита; полиэмбриония, при которой возникают несколько зародышей в результате расщепления зародышевых клеток одной зиготы. При полиэмбрионии в результате расщепления иногда можно определить на очень раннеей стадии развития, какой из зародышей будет развиваться дальше (детерминированная полиэмбриония в результате расщепления), иногда это трудно предугадать (недетерминированная полиэмбриония в результате расщепления). Все
Глава 4
118
роды Pinaceae имеют простую полиэмбрионию, а полиэмбриония вследствие расщепления встречается у Pinus, Cedrus, Tsuga, Taxodium, CryptO' , meria, Cunninghamia, Sequoia, Podocarpus. <
Несмотря на то, что у женского гаметофита Pinus существует мно- i жественное оплодотворение, большинство зрелых семян содержит лишь один зародыш. Редко в семенах Pinus может быть более одного зародыша. Появление нескольких зародышей чаще встречается у сосен с крупными семенами, чем с мелкими. Берлин (1962) обнаружил по четыре зародыша на зрелое семя приблизительно у одной трети проверен' ных семян сосны Ламберта и пинии и в немногих семенах сосны вейму' товой. В другом исследовании он (1967) обнаружил восемь зародышей в одном семени сосны Ламберта.
ПАРТЕНОКАРПИЯ. Развитие неопыленных шишек с полностью сформированными, но обычно пустыми семенами называется парте* нокарпией. Обнаружена она у многих родов голосеменных. Партенокарпия распространена у родов Abies, Juniperus, Larix, PiceafTaxus и Thuja, отмечалась также у Chamaecyparis, Cryptomeria, Pseudotsuga и Tsuga, редко встречается у Pinus. Например, у сосны обыкновенной лишь 0,4% шишек имели неразвитые семяпочки и только у 1 из 76; развивающихся шишек сосны смолистой были недоразвитые семяпочки.
4.7. ВРЕМЕННОЙ ХОД ПРОЦЕССОВ, ПРОИСХОДЯЩИХ В ПОЧКАХ
И ЖЕНСКИХ ШИШКАХ ДУГЛАСИИ, РАСТУЩЕЙ ВБЛИЗИ
ОТ КОРВЕЛЛА (ОРЕГОН)
(Owens, Smith, 196S)
Процессы, происходящие в почках и женских шишках Дата Время, протекающее от заложения почек,мес
Все почки:. заложены зачатки боковых почек Начало апреля Почки с семяпоч-
становится видимой зональность Середина мая ками вскрываются 1,5
заканчивается заложение катафиллов Середина июля 3,5
(чешуек), происходит апикальное удлинение, начинается заложение листьев, кроющих листочков, или микроспорофиллов Шишки с семяпочками: начинается заложение чешуек Начало сентября 5,0
заложены все кроющие листья Начало октября 6,0
заложены все чешуйки и почки Начало ноября 7,0
с семяпочками в стадии покоя возобновляют рост почки с семя- Начало марта 11,0
почками вскрываются почки с семяпочка^ Начало апреля 12,0
ми, происходит опыление оплодотворение Начало июня 14,0
закончено удлинение шишек Начало июля 15,0
закончено созревание, открыва- Начало сентября 17,0
ются шишки, освобождаются семена
Репродуктивный рост
119
ПРОДОЛЖИТЕЛЬНОСТЬ И ВРЕМЯ РАЗВИТИЯ ШИШЕК. Некоторое представление о сходстве и различиях в развитии шишек разных видов голосеменных умеренной зоны дает сравнение приуроченности ко времени основных событий репродуктивного цикла сосны смолистой и Дугласии. В центральной части Висконсина зачатки шишек сосны смолистой дифференцировались за один сезон, но шишечки не были видны до конца мая — начала июня следующего года. Опыление происходило в начале июня. Растяжение шишек начиналось после опыления, затем чешуйки смыкались. К концу июля шишки удлинялись до 10—12 мм и до конца первого года в длину почти не увеличивались. Пыльцевые зерна прорастали, но пыльцевые трубки прекращали рост и оставались в покое в течение лета и зимы. Мегаспоры формировались приблизительно через 3 недели после появления женской шишки. В результате последовательных клеточных и ядерных делений одной из мегаспор мегагаметофит увеличивался в размерах. Затем наступал период зимнего покоя. Шишки возобновляли рост рано весной второго сезона и достигали наибольшей длины к началу июля. Рост мегагаметофита возобновлялся также рано весной, но пыльцевые трубки не начинали расти до середины июня второго года, когда происходило оплодотворение. После этого зародыш начинал быстро развиваться, и семена в Висконсине созревали к началу сентября. В табл. 4.7 приведены данные о времени и продолжительности основных этапов репродуктивного цикла у дугласии в Орегоне.
УВЕЛИЧЕНИЕ РАЗМЕРОВ И СУХОГО ВЕСА ШИШЕК И СЕМЯН. У сосны шишки формируются в течение 2 лет. Максимальное увеличение размеров и сухого веса шишек приходится на второй год их развития. Например, к концу первого года роста шишки сосны смолистой в Висконсине имели около 1/4 веса, 1/30 части объема и 1/3 длины зрелой шишки второго года развития. Сезонный ход увеличения сухого веса и размеров шишек первого и второго года развития сосны смолистой в Висконсине показан на рис. 4.5 и 4.6. Сухой вес одногодичной шишки возрастал с постоянной скоростью до конца сентября. К этому време-
Рис. 4.5. Сезонные изменения у однолетних шишек сосны смолистой:
а — сухого веса, веса воды (на шишку) и процентного содержания воды; б — длины, ширины и объема (Dickmann, Kozlowski, 1969); 1 - вес воды; 2 -сухой вес; 3 — влажность; 4 — длина; 5 — объем; 6 — ширина
Глава 4
120
Рис. 4.6. Сезонные изменения у двухлетних шишек сосны смолистой:
а — сухого веса, веса воды (на шишку) и процентного содержании виц<л; б -длины, ширины и объема (Dickmann, Kozlowski, 1969); 1 — вес воды; 2 — сухой вес; 3 — влажность; 4 — длина; 5 — объем; 6 — ширина
ни средний вес шишки был немного меньше 0,2 г. Динамика увеличения сухого веса была сходной в течение трех последовательных вегетационных периодов. Длина и ширина одногодичных шишек увеличивались до середины июля, а затем изменялись мало. К концу первого сезона диаметр шишек был в среднем 8 мм и длина 14—15 мм.
На второй год развития шишки сосны смолистой возобновляли рост в середине апреля. Сухой вес медленно увеличивался в течение мая, ускоряясь к началу июня, и до начала августа сухой вес возрастал с большой скоростью. Размеры шишек второго года развития максимально увеличивались в июне. Двухгодичные шишки достигали максимальных размеров за месяц до того, как отмечался максимальный прирост сухо
го веса.
Сезонные изменения сухого и сырого веса и содержания воды оплодотворенных семяпочек или семян сосны смолистой показаны на рис.4.7. Сухой вес семян быстро возрастал с июня до конца августа. После этого прирост прекращался. Содержание воды (в процентах на сухой вес) к концу июня было 600%, к концу августа оводненность резко падала до 50%. Затем содержание воды в семенах постепенно снижалось до 17% к началу октября.
Изменения в диаметре и длине женских шишек дугласии (которые созревают в течение года) показаны на рис. 4.8.
ФИЗИОЛОГИЯ РАЗВИТИЯ ШИШЕК. В этом разделе будут рассматриваться некоторые аспекты углеводных, минеральных и водных отношений во время роста и развития шишек (Роль гормональных регулято-
Рис. 4.7. Изменения в развивающихся семенах от момента оплодотворения до созревания (Dickmann, Kozlowski, 1969): 1 - сырого веса; 2 — сухого веса; 3 - влажности
Репродуктивный рост
121
Даты
Рис. 4.8. Изменения длины и ширины женских шишек у четырех деревьев дугласии вблизи КорвеЙла (Орегон) (Ching, Ching, 1962): 1, 2, 3, 4 номера деревьев; а - ширина; б - длина
ров роста в контролировании ростовых процессов рассматривается в главе 6.)
Углеводы. Быстро развивающиеся шишки голосеменных требуют большого количества углеводов. Фотосинтез зеленых шишек не обеспечивает потребности их в углеводах. Имеются многочисленные противоречивые данные об углеводах, потребляемых растущими щитками. Дикман и Козловский (1968, 1970) показали, что у сосны смолистой при этом мобилизуются углеводы, синтезируемые самими репродуктивными тканями в процессе их формирования. Были также обнаружены закономерности сезонных изменений органов, являющихся источником продуктов фотосинтеза для образования шишек в текущем сезоне. Однолетняя хвоя была главным источником продуктов фотосинтеза текущего года для развития шишек в весенний период. В июне 2 и 3-летняя хвоя также участвовала в образовании продуктов фотосинтеза для развития шишек, но с конца июня ее роль резко снижалась. Старые хвоинки к концу сезона все меньше' вырабатывали фотосинтетических продуктов, и шишки все больше и больше мобилизовывали углеводы, продуцируемые хвоей текущего года.
Дикман и Козловский (1969) изучали изменения в резервными структурных углеводах во время развития 1 и 2-летних шишек сосны смолистой в Висконсине. Их соотношения изменялись в основном только в течение последних 3 месяцев развития. Главные компоненты
а. Даты 6
Рис. 4.10. Сезонные изменения содержания в двухлетних шишках сосны смолистой (Dickmann, Kozlowski, 1969):
а — данные в процентах на сухой вес; б — на вес шишки; 1 — экстрагируемые запасные вещества; 2 — лигнин; 3 — целлюлоза; 4 — гемицеллюлоза
Рис. 4.9. Сезонные изменения содержания в однолетних шишках сосны смолистой (Dickmann, Kozlowski, 1969): а - данные в процентах на сухой вес; б -на вес шишки; 1 — экстрагируемые запасные вещества; 2 — лигнин; 3 - гемицеллюлоза; 4 - целлюлоза
Репродуктивный рост
123
шишек в течение первого и начале второго года включали запасные углеводы и продукты фотосинтеза текущего года. В этот период количество запасных углеводов увеличивалось с такой же скоростью, как и масса молодых шишек. Запасные углеводы составляли около половины сухого веса шишек. Углеводы, накапливаясь со скоростью, соответствующей их включению в запасные и структурные вещества, терялись в процессе дыхания. По этим причинам резервы стабильно сохранялись в течение первого сезона и в апреле-мае второго года развития шишек (рис. 4.9 и 4.10).
В последующем периоде второго года развития шишек произошел сдвиг от высокого уровня резервных и низкого уровня структурных компонентов, особенно целлюлозы, к обратным их отношениям (рис. 4.10). Содержание экстрагируемых резервных веществ быстро увеличивалось от начала мая к концу июня. Позже количество запасных углеводов постепенно возрастало до начала августа, хотя концентрация их
Рис. 4.11. Сезонные изменения содержания в однолетних шишках сосны смолистой (Dickmann, Kozlowski, 1969в):
а — концентрации макроэлементов; б — веса макроэлементов на шишку; в — концентрации микроэлементов; г — концентрации № и Al; 1 — азот; 2 — калий;
3 — фосфор; 4 — кальций; 5 — марганец; 6 — магний; 7 — цинк; 8 — железо; 9 — бор; 10 — натрий; 11 — алюминий
124
Глава 4
резко снижалась к середине июня. В это же время отмечалось резкое увеличение целлюлозы, которое продолжалось до августа. Данные показали, что от середины июня до сентября огромные количества запасных углеводов превращались в целлюлозные компоненты вторичных клеточных оболочек. В конце июня крахмал содержался в больших количествах в чешуйках и сердцевине шишек, к началу сентября он практически исчез. Дальнейшие наблюдения показали, что запасные вещества превращались в структурные материалы.
Минеральные вещества. Дикман и Козловский (1969) изучали также роль минеральных веществ в 1—2-летних женских шишках и развитии семян сосны смолистой. В течение первого года основные макроэлементы постоянно перемещались к развивающимся шишкам. По скорости и величине накопления элементы располагались в следующем порядке: N > К > Р > Са > Mg. Затем концентрация N снижалась, концентрация других макроэлементов была стабильной. В течение первого года микроэлементы также передвигались к шишечкам (рис. 4.11).
Рис. 4.12. Сезонные изменения содержания в двухлетних шишках сосны смолистой (Dickmann, Kozlowski, 1969) :
а - концентрации макроэлементов; б — веса макроэлементов на шишку; в — концентрации микроэлементов; г — концентрации Na и А1 ; 1 азот; 2 - калий; 3 - фосфор; 4 - кальций; 5 - марганец; 6 -магний; 7 — цинк; 8 - железо; 9 — бор; 10 — медь; 11 — натрий; 12 — алюминий
Репродуктивный рост
125
Рис. 4.13. Сезонные изменения содержания в развивающихся семенах сосны смолистой (Dickmann, Kozlowski, 1969):
а — концентрации макроэлементов; б — веса макроэлементов в семенах на одну шишку; в — концентрации микроэлементов; г — концентрации № и Al; 1 — азот; 2 - калий; 3 — фосфор; 4 - марганец; S — кальций; 6 — магний; 7 — цинк;
8 — железо; 9 - медь; 10—бор; 11—натрий; 12 —алюминий
Сезонный ход поглощения и накопления питательных веществ б течение второго года развития шишки отличается от хода в первый год их развития. В течение периода быстрого роста шишек, отмеченного сразу после оплодотворения, поглощение макро- и микроэлементов еще продолжалось. Но к середине июня; когда шишки достигали максимальных размеров, поступление Мп в них резко прекращалось. Начиная с середины июля N, Р и К в созревающих шишках не было. Другие макро- и микроэлементы продолжали накапливаться в шишках в течение всего второго года их роста (рис. 4.12). При старении шишек N, Р, К, Mg, Мп, Zn и Fe активизировались для развития семян. Азот накапливался в больших концентрациях. Стареющие ткани шишек обеспечивались N, Р и К, поступившими в семена. Кальций содержался в семенах в низ
126
Глава 4
ких концентрациях, Na накапливался в больших количествах. После оплодотворения алюминий не поступал в Семена (рис. 4.13).
Водный режим. По исследованиям Дикмана и Козловского (1969), сезонный ход гидратации у шишек первого и второго года развития различен. Однолетние шишки сосны смолистой в начале сезона быстро оводнялись. Этот период длился до середины июня. В дальнейшем количество воды в шишках оставалось постоянным, но процент влажности снижался прежде всего из-за увеличения сухого веса шишек. В шишки второго года вода поступала до конца июня, максимальная оводнен-ность, выраженная в процентах, отмечалась в середине июня. Наибольшее содержание воды определялось ко времени достижения шишками максимальных размеров- Сухой вес при этом продолжал возрастать. Затем шишки теряли воду и по мере созревания уменьшались в размерах (рис. 4.14). Таким образом, стабилизация сухого веса сопровождалась постепенной дегидратацией, характерной для стадии шишек, предшествующей их созреванию.
дйдндндидй дидид
ндндйдндндйдиднд
29 И- 7UL
днДЙДНДНДНДНДНД 13-20-HL
днд н д н д н днд ндн д 20-27-Ц
днднднднднднднд дндндн дндндндндн 261Ш.-4Л 20 ~27 -IX.
Даты, Время суток
Рис. 4.14. Изменение диаметра двухлетних шишек сосны смолистой (Dickmann, Kozlowski, 1969а):
Д — полдень; Н — полночь
Репродуктивный рост
127
Шишки открываются, очевидно, скорее вследствие высыхания, а не из-за процессов, связанных с ростом. Виллис (1917) заметил, что шишки дугласии не открывались до тех пор, пока содержание воды в них не снижалось на 20 %. Все шишки полностью открывались, когда теряли около половины имеющейся воды. Было продемонстрировано, что раскрывание чешуек шишек больше зависело от сокращения вентральных тканей, чем дорзальных. Авторы установили, что дорзаль-
Рис. 4.15. Суточные изменения в разное время вегетационного периода (Chaney, Kozlowski, 1969): 1 — диаметра шишек сосны Банкса; 2 — изменений дефицита давления пара (ДДП)
24
20
16
12
8
4
0
24 -31'^ПЕ
Дефицит давления паров,
^а.ты, время суток
ная зона чешуек шишек сосны состоит из одревесневших волокон, а вентральная - из полосок коротких прямоугольных толстостенных клеток. При высыхании волокна дорзальных тканей мало различаются по длине. Наоборот, вентральные ткани сокращаются в зависимости от вида растений, в длину на 10—13 %.
На некоторых стадиях развития шишек относительное содержание воды в них снижается прежде всего вследствие быстрого увеличения сухого веса, а не из-за поступления или выхода воды из шишек. Например, шишки в целом, их чешуйки и семена у ели канадской и лиственницы в конце мая — начале июня достигали самой высокой оводнен-ности (примерно 400%). Затем они постепенно подсыхали, и в сентябре
128
Глава 4
оводненность снижалась до 50%. Первоначально (в мае-июне) процент оводненности увеличивался в результате истинного увеличения содержания воды, которое превосходило прирост сухого веса. От начала июня до конца июля процентное содержание воды заметно снижалось, хотя истинный вес воды на шишку оставался прежним или становился даже выше. Процентное снижение воды было незначительным по сравнению с увеличением сухого веса. В начале августа сухой вес перестал возрастать, оводненность снизилась в основном вследствие потери воды. Таким образом, процентное содержание воды в развивающихся шишках и семенах может изменяться за счет транслокации воды, изменений сухого веса или того и другого вместе.
Суточные изменения размеров шишек. Помимо изменений размеров шишек вследствие ростовых процессов, происходят и обратимые изменения, отражающие колебания в содержании воды. В'среднем периоде развития шишки часто уменьшаются в размерах днем и увеличиваются ночью. Величина суточных колебаний размеров шишек зависит от стадии развития. Чани и Козловский (1969) показали, что шишки у ели канадской и сосны Банкса на ранних этапах развития увеличивались в диаметре неравномерно. Шишки расширялись вечером и ночью, когда дефицит давления паров воздуха снижался или был очень низок, и не изменялись в диаметре в дневные часы, когда дефицит давления паров был высок. На средних стадиях развития шишек также происходили обратимые изменения в объеме, они тоже были связаны с изменениями влажности воздуха (рис. 4.15), но по мере созревания шишек их диаметр уменьшался постепенно.
БЕСПОЛОЕ РАЗМНОЖЕНИЕ
К способам вегетативного размножения древесных растений относятся: корневая поросль, черенкование, отводки, прививки, использование культуры тканей и клеток.
КОРНЕВАЯ ПОРОСЛЬ. Отрастание - типичная реакция на внезапное отмирание верхней части дерева в результате пожара, рубки, болезней или физиологических нарушений. Способность древесных растений давать поросль имеет огромное значение для их выживания. Побеги, возникающие от корневой шейки и нижней части ствола, обычно формируются из спящих почек (см. главу 3). Такие пневые побеги - наиболее важный тип вегетативного размножения лиственных лесных древостоев. Например, многие лиственные леса Новой Англии и большинство дубовых лесов Пенсильвании возникли от пневых побегов. Корневая поросль развивается из адвентивных почек, образующихся между корой и древесиной пня. Обычно корневая поросль живет недолго и не играет заметной роли в вегетативном размножении.
Способность к образованию поросли распространена среди покрытосеменных и редко встречается у голосеменных. Но среди видов покрытосеменных имеется множество вариаций. Например, по данным Литтла (1938), дубы белый и малый формировали больше побегов от пня, чем дубы бархатный и американский карликовый, но эти побеги были меньших размеров. У пня дуба горного возникало почти столько же побегов, как и у дуба белого, однако поросль дуба горного
Репродуктивный рост
129
к концу вегетационного периода была мощнее, чем у группы бархатного дуба. К 14-летнему возрасту наиболее мощная поросль (по высоте деревьев) развилась у дуба американского шарлахового, несколько слабее у бархатного дуба. По способности к образованию поросли виды дуба можно расположить в следующем порядке: малый, белый, горный, бархатный и американский шарлаховый. На образование корневой поросли влияет также время года и количество углеводов в тканях (см. главу 7),
Не все покрытосеменные образуют одинаковое количество порослевых побегов в разных точках ареалов. Например, клен сахарный, бук, граб виргинский дают мощную поросль на севере ареала и не способны к порослеобразованию в южных его частях. Многие дубы образуют поросль во всех районах их распространения. Возобновление растений путем образования поросли не имеет большого значения для голосеменных, однако некоторые их виды (калифорнийское Мамонтове дерево, сосны жесткая, ежовая, поздняя и кипарис болотный) дают обильную поросль после пожаров. По данным Чиней (1942), в Америке лишь три рода (сосна поздняя, кипарис болотный и калифорнийское Мамонтове дерево) образуют поросль, которая достигает товарных размеров. Часто считают, что после пожаров или вследствие повреждения сеянцев поросль образуется из адвентивных почек. Однако было обнаружено, что у некоторых видов сосен маленькие почки в пазухах первичных хвоинок появлялись как раз над семядолями. В этом месте хвоя располагалась очень тесно и почки после утолщения ствола были собраны в пучки. Эти почки часто давали начало побегам у основания ствола или от корневой шейки. На срезе ствола видно, что проводящие ткани почки имеют строение однолетнего ствола. Это свидетельствует о том, что поросль возникает скорее из спящих, а не из адвентивных почек (см. главу 3).
При повреждении древостоев лиственных деревьев интенсивными вырубками и пожарами некоторые виды деревьев восстанавливаются в значительной мере за счет корневой поросли, образующейся из адвентивных почек на корнях срубленных деревьев. Размножение корневыми побегами наиболее известно у осины, но оно отмечается и у других видов (у ликвидамбра смолоносного, бука, айланта, белой акации, ниссы лесной и сассафраса лекарственного). Значение вырубок для образования корневых отростков в древостоях осины установили Штоколер и Мэзон (1956). Они насчитали более 1000 порослевых побегов осины на 1 га на вырубках и менее 150 на нетронутых участках.
СТЕБЛЕВЫЕ ЧЕРЕНКИ. Укоренение стеблевых черенков — один из лучших методов бесполого размножения древесных .растений. Этот способ заключается в том, что отрезки стеблей с одной или более почками помещают в условия благоприятные для образования адвентивных корней. Термин адвентивный применяется к корням и их ответвлениям, которые образуются не от эмбрионального корня. Адвентивные корни могут возникать от вновь индуцированных или преформирован-ных зачатков, которые развиваются во время ветвления или формирования стебля. Адвентивные корни могут образоваться на побегах, ветвях, листьях, воздушных стеблях, на корневищах и старых корнях.
Индуцированное образование зачатков корней. Хассинг (1974) изло
130
Глава 4
жил результаты работ нескольких исследователей, которые показали, что у многих тканей можно вызвать образование зачатков корней. Почки, эпидерма, первичная кора, перицикл, флоэма, камбий, ксйлема и паренхима сердцевины содержат клетки, способные к заложению зачатков корней. Установлено, что камбий, флоэма и перицикл наиболее часто образуют индуцированные зачатки корней. Первичная- кора, сердцевина и ксилема менее способны к этому. Для образования корней на месте пореза у одних видов должен/образоваться каллус, для других каллус не требуется. У видов, черенки которых трудно укореняются, большая часть корневых зачатков закладывается у основания каллуса.
Время, в течение которого формируются зачатки корней, значительно различается у видов. У легко укореняющихся черенков корневые зачатки образуются в течение нескольких дней- У трудно укореняющихся древесных черенков образование корневых примордиев может длиться несколько месяцев или они вообще не образуются. Протяженность формирования и появление адвентивных корней могут различаться у молодых и взрослых особей одного вида. Адвентивные корни в междоузлиях черенков ювенильной фазы растений английского плюща возникали в паренхимных лучах флоэмы над базальным срезом в течение 6—10 дней. Корни появлялись через 1,5-2 недели после черенкования. У черенков от взрослых растений адвентивные корни закладывались в течение 2-4 недель в паренхимных лучах флоэмы междоузлий и в каллусе вблизи от базального конца черенка. Корни появлялись к концу 3-5 недели.
Преформированные зачатки. У преформированных зачатков меньше вариаций в скорости заложения, чем у индуцированных. Преформированные примордии формируются наиболее часто в узлах из клеток камбия или клеток лучевой паренхимы только что отделенных от камбия. У некоторых деревьев, например у тополей, преформированные зачатки формируются в продольных тяжах между узлами. По данным Карлсона (1938, 1950), преформированные корневые зачатки дифференцируются в течение первого года и развиваются медленно. Дифференциация происходит обычно в течение 2-3 лет, но иногда может продолжаться в течение многих лет. При увеличении ствола по диаметру корневые зачатки удлиняются перпендикулярно оси ствола соответственно камбиальному росту. Некоторые преформированные корневые зачатки не развиваются. Однако при благоприятных условиях окружающей среды даже самые молодые и наименее дифференцированные преформированные зачатки могут стать развитыми корнями.
ЛИСТОВЫЕ ЧЕРЕНКИ. Некоторые растения, например определенные виды рододендронов, размножаются листовыми черенками легче, чем черенками из стеблей. Пучки хвои некоторых сосен также укореняются легче, чем черенки стеблей. Однако почки и побеги при этом обычно не формируются. У сосны Банкса это может быть преодолено за счет использования пучков с преформированными почками.
КОРНЕВЫЕ ЧЕРЕНКИ. Некоторые виды древесных растений, размножающиеся в природе корневыми отпрысками, могут быть размножены частями корней лучше, чем стеблевыми черенками. Размножение корневыми черепками будет более успешным, если черенки корней брать от молодых растений в конце зимы или рано весной, до начала роста побегов. Самый близкий к стволу конец черенка корня должен
Репродуктивный рост
131
быть ориентирован вверх или горизонтально, но ни в коем случае не вниз.
КОНТРОЛЬ ЗА УКОРЕНЕНИЕМ ЧЕРЕНКОВ. Укоренение черенков зависит от многих факторов: вида растения, возраста дерева, от которого взяты черенки, положения отрезанной части растения на дереве, времени года, когда взяты черенки, пола родительского дерева, запасов питательных веществ в черенке, окружающих условий, при которых укореняются черенки. Одна из наиболее четких характерных черт молодых древесных растений — относительно легкое укоренение их черенков по сравнению с укоренением черенков, взятых от старых деревьев того же вида. Это относится к большинству видов и составляет одну из наиболее серьезных проблем для размножения растений, так как древесные растения, проявляющие желаемые свойства в определенном возрасте, не могут быть в это время размножены черенками.
У некоторых видов часто отмечаются сезонные в'ариации в способности корнеобразования. Черенки таких древесных растений, как падуб, легко укореняются в течение недели или месяца, у других видов (бука и сирени обыкновенной) этот период еще короче. Черенкование лучше проводить весной, до того как побеги тронутся в рост, или сразу после начала побегообразования. Черенки, взятые позднее, образуют корни с трудом и не заканчивают рост до конца сезона. Более подробную информацию о технике укоренения стеблевых черенков древесных растений можно получить в специальной литературе. (Внутренние факторы, контролирующие укоренение черенков, обсуждаются в главе 16.)
ОТВОДКИ. При размножении отводками корни формируются на ветвях, контактирующих с влажной средой. Укоренившийся стебель отделяют от растения. Естественное размножение отводками распространено у малины, ежевики, смородины, крыжовника и некоторых видов лес-щдх деревьев, нижние ветви которых касаются земли. Например, ель черная и кипарисники на торфяных вырубках возобновляются отводками. По данным Лутца (1939), отводками размножаются Tsuga, Picea, Abies, Chamаесуреris, Taxus и редко Finns. Литтл (1944) описал интересный случай естественного возобновления сосны отводками после сильного снегопада, пригнувшего ветви к земле. Он наблюдал образование отводков у клена красного, кизила флоридского, Viburnum pubescens и Clethra alnifolia.
При размножении растений используются несколько модификаций искусственного получения отводков. Например, у многих клонов черенки укореняются трудно, а размножение отводками происходит успешно. Метод ограничивается теми видами, для которых естественно образование отводков, или в тех случаях, когда другими способами они не могут размножаться. Метод отводков применяется для размножения подвоев яблони, фундука и мускатного винограда. Вероятно, из методов искусственного размножения отводками наиболее пригодны простые и траншейные отводки. Метод простых отводков заключается в том, что ветвь пригибают к земле и часть ее присыпают почвой до начала роста. Несколько сантиметров конца ветви не прикапывают, на этой части формируются листья. Иногда часть стебля, которую прикапывают, сначала надрезают и обрабатывают ростовыми веществами. У надрезанного отводка развиваются корни и побег. При методе воздуш
132
Глава 4
ных отводков на ветви делают надрез, обрабатывают его ростовым веществом, закрывают это место влажным сфагнумом, а сверху обво-рачивают водоотталкивающей бумагой или пластиком. После образования вокруг ранки корней, часть побега с корнями отделяют и высаживают в почву.
ПРИВИВКИ. При размножении древесных пород издавна применяют различные технические приемы прививки. Например, Гартман и Кестер (1975) напоминают, что прививка использовалась в Китае за 1000 лет до н. э., что Аристотель (384- 322 до н. э.) подробно описал прививки, что прививки были особенно популярны в Древнем Риме.
В ходе прививки камбий отделенной ветви (привой) совмещают с камбием укорененного растения (подвоя) так, чтобы соединились камбии обоих компонентов. Для получения двух прививочных соединений между подвоем и привоем иногда вставляют кусок стебля (промежуточный подвой). Прежде чем произойдет дифференциация соединяющихся проводящих тканей, формируется каллусная ткань, образуемая привоем и подвоем, которая заполняет пустоты между ними. Подвой и привой могут образовать одинаковое количество каллуса, но возможно, что один из членов этого союза образует каллуса значительно больше другого. У прививаемых компонентов сосны, например, подвой продуцирует значительно больше раневых каллусных тканей. У дугласии на привое в первые 7 дней каллуса образуется больше, чем на подвое, но через 10 дней вклад подвоя в образование каллуса становится значительно больше.
Поврежденные во время прививки клетки буреют и отмирают. Клетки каллуса, образуемые подвоем и привоем, разделяются этой бурой пограничной линией, состоящей из мертвых клеток, называемой изоляционным слоем. У некоторых родов, например у Citrus, вдоль изоляционного слоя может появляться раневая камедь. Неповрежденные клетйи вскоре разрастаются по обе стороны от изоляционного слоя и активно делятся, образуя каллус. В течение нескольких недель начинают дифференцироваться изолированные трахеальные элементы в каллусе и через изоляционный слой. Вследствие этого изоляционный слой нарушается и исчезает. Новый камбий образуется через смешанные камбии подвоя и привоя. Он формируется там, где камбий подвоя if привоя находится в контакте с каллусной тканью. В дальнейшем в каллусе происходит деление клеток, ведущее к формированию камбия, который смыкается с вновь образованными камбиальными клетками. После образования новым камбием ксилемы и флоэмы между подвоем и привоем возникает целостность проводящей системы. При прививке глазком смыкающийся камбий дифференцируется в массе каллуса, обычно формирующейся по краю среза коры, затем распространяется по линии раневой поверхности пока не соединится, наконец, с камбием глазка. Вскоре после этого начинается быстрый рост камбия щитка почки, и почка постепенно приподнимается из продольной щели коры. Прививка становится надежной, когда сформируются нормальные проводящие ткани из соединенных камбиев прививаемых компонентов.
Несовместимость прививаемых компонентов. Успех прививки зависит не только от контакта между камбием и другими меристематическими тканями, но также и от генотипической совместимости участ
Репродуктивный роет
133
ников црививки. Несовместимость прививаемых компонентов — распространенное явление, которое на протяжении столетий было бедствием для тех, кто занимался размножением древесных пород. Несовместимость может происходить из-за неспособности прививаемых партнеров прочно соединиться механически, из-за слабой их жизнеспособности или преждевременной гибели прививок, когда неудача обусловлена различиями между подвоем и привоем. Симптомы несовместимости иногда проявляются вскоре после прививки — прививаемые компоненты погибают в первый же год. Но возможны и скрытые признаки неудачной прививки, когда в первый год происходит активный рост, а со временем он прекращается. Иногда симптомы несовместимости не проявляются многие годы.
Причины несовместимости прививаемых партнеров лучше изучены у плодовых деревьев, чем у лесных. Герреро (1956) разделил причины несовместимости прививок на четыре категории; 1) неудачное соединение прививаемых компонентов, в результате чего почка не вырастает; 2) прививаемые компоненты поражаются вирусами; 3) слабое механическое соединение, при котором причиной гибели дерева бывает обычно слабый рост, механическое нарушение и поломка в месте соединения; 4) слабый рост происходит не в результате неудачного соединения компонентов, а в связи с ненормальным распределением крахмала.
Мосс (1962) разделил несовместимые прививочные комбинации плодовых деревьев на два типа: транслокационную и локализованную несовместимость. Ни один из этих типов не является специфичным для вида, оба могут быть обнаружены у одной и той же прививочной комбинации. Транслокационная несовместимость имеет несколько причин: 1) накопление крахмала над местом соединения прививаемых компонентов и отсутствие крахмала ниже места прививки; 2) разрушение флоэмы; 3) различное поведение прививаемых частей; 4) нормальное состояние проводящих сосудов в месте соединения, но при этом возможен интенсивный рост привоя; 5) ранний рост. Локализованная несовместимость может быть обусловлена следующими причинами: 1) нарушением камбиальной целостности; 2) сходным поведением эквивалентных комбинаций;. 3) постепенно наступающим голоданием корневой системы и медленным развитием внешних признаков нарушения.
Коупе (1970) обнаружил два основных внутренних признака несовместимости прививаемых частей деревьев дугласии: 1) заложение суберинизированнойзоны и проникновение ее в кору объединенных тканей; 2) заложение и развитие раневых ксилемных участков. Суберинизированные зоны, заложенные в клетках первичной коры соединяемых компонентов, были едва заметны в течение лета в год прививки, с возрастом интенсифицировались. Участки раневой ксилемы формировались лишь в зоне соединения, где прежде суберин развивался ео внутренней флоэме или в камбии. Раны развивались в начале второго'года и в течение последующих лет. Участки раневой ксилемы были результатом естественной перепрививки тканей подвоя и привоя в начале каждого вегетационного периода.
Раннее определение несовместимости подвойно-привойных комбинаций затруднительно, так как внешние признаки несовместимости (например, клороз и опадение листьев) могут появиться только через несколько лет после прививки. Отсутствие роста корней - ранний признак несовместимости, но его определение затруднительно в полевых условиях. Однако Коупе (1969) заметил, что отсутствие развивающихся вегетативных почек у клонов дугласии в начале второго года после прививки было хорошим признаком для определения неудачной прививки.
134
Глава 4
Рис. 4.16. Восстановление древесного растения из каллуса или культуры суспензии клеток
(Winton, Huhti-
nen,1976): 1 — суспензионная культура; 2 - клетки и агрегаты клеток; 3 — эмбриоид; 4 — эмбрион; 5 — инокулум; 6 — культура каллуса; 7 — побег; 8 — вырезка побега; 9 — корни; 10 — растение; 11 — стерилизация поверхности; 12 -
субкультура; 13 — изоляция каллуса; 14 — образование каллуса; 15 — эксп-лантант; 16 — родительское дерево
Широко распространено мнение, что внутривидовые прививки более успешны, чем межвидовые, межродовые и прививки между представителями разных семейств. Поэтому для удачного совместимого соединения целесообразно близкое генетическое родство между подвоем и привоем. У дугласии случаи несовместимости практически удваивались, когда ее прививали на подвой не близкородственной породы деревьев. Коупе (1974) установил также, что прививка у дугласии контролировалась прежде всего аддитивными генами. Он показал, что улучшить совместимость можно тщательным подбором родительских пар и перекрестным размножением среди лучших родительских форм. Сообщалось также, что несовместимость привитых пар внезапно проявлялась у 14-летних, уже дающих семена деревьев сосны замечательной.
КУЛЬТУРА КЛЕТОК И ТКАНЕЙ. За последние годы повысился интерес к использованию неорганизованных систем, таких, как каллус, или культур клеточных суспензий при размножении древесных растений. Эти культуры могут происходить от одной клетки, способной к делению. Они растут в определенной среде в стерильных условиях. Одна или масса клеток каллуса способны затем делиться и создавать листья, стебель, корни и целое растение (рис. 4.16).
Культивирование вегетативных тканей. Образование каллуса методом клеточных культур и слияния протопластов способствует улучшению древесных растений. Масса каллуса может образовываться от одной клетки камбия, первичной коры или сердцевины стебля. В культуре каллуса клетки тфеимущественно паренхимные. Каллус может поддерживаться в субкультуре неопределенно долгое время при условии правильно подобранной для каждого вида среды. Хорошие результаты получены с культурой каллуса, взятого от сеянцев, корневых отростков, одногодичных ветвей двудольных и одногодичных ветвей однодольных растений.
Ченг и Воки (1977) получали растеньица дугласии при определенных условиях культуры тканей из семядольных эксплантантов от 2—4-недельных проростков.
Репродуктивный рост 135
Они разрезали семядоли на сегменты, что вызывало образование каллуса. Затем стимулировали рост почек и корней при помощи НУК (нафтиленуксусной кислоты) . Выживало 90% пересаженных в почву сеянцев.
Слияние протопластов используется для бесполой межвидовой гибридизации. Протопласты могут быть получены из мезофилла листа, меристемы, микроспор и мегагаметофитов. Для разрушения клеточных стенок обычно применяется смесь ферментов пектиназы и целлюлозы. Лишенные клеточных стенок протопласты могут слиться и образовать новые клетки так же, как и при слиянии половых клеток во время оплодотворения.
Культивирование тканей и клеток гаметофита. Многие проблемы размножения могли бы быть решены культивированием частей гаметофита или репродуктивных органов древесных растений. Возможны два основных источника гаплоидных клеток: репродуктивные ткани (микроспоры) в пыльниках и питательные клетки в мегагаметофитах голосеменных. Культивирование гаплоидных клеток гаметофитов имеет два главных преимущества: 1) гаплоидные клетки могут быть использованы для получения протопластов, при слиянии которых гибридные клетки будут иметь диплоидный набор хромосом соматических клеток; 2) растения, выросшие из гаплоидных клеток, можно стимулировать к удвоению набора хромосом и получить диплоидные растения с гомозиготными аллелями каждого локуса гена. Это обеспечивает получение ценных особенностей самоопыляющихся растений, которые проявятся в следующих поколениях.
Неразрешенные проблемы. Основная трудность применения метода культуры тканей заключается в том, чтобы заставить неорганизованную систему стать организованной, т.е. индуцировать дифференциацию и формирование органов. Оказалось, что специальные обработки, индуцирующие дифференциацию, очень специфичны для каждого растительного материала. Размножение древесных растений при помощи культуры тканей намного продолжительнее размножения таким способом травянистых растений. У большого числа травянистых видов многие генетически однородные растения могут быть получены в течение нескольких недель. По сравнению с ними методом культуры тканей получено очень мало деревьев двудольных и некоторых трудно размножающихся хвойных. По-видимому, основная причина ограниченного успеха в работе с лесными деревьями — недостаточное внимание к этому.
Несмотря на перечисленные трудности, потенциальные возможности улучшения деревьев при помощи культуры клеток и тканей очень велики. Особенно многообещающими представляются методы отсева с целью идентификации лучших генотипов среди клеток гамет для селекции растений. Гаплоидные растеньица, полученные в культуре тканей, могут быть использованы для отбора таких желательных качеств, как мощность развития, устойчивость к болезням и вредителям, засухоустойчивость и др. Например, были получены свободные от вирусов гополя из инфицированных клонов тополя при помощи техники, вызывающей дифференциацию растеньиц из каллуса. Скорость роста различных клонов можно быстро выяснить в широком диапазоне окружающих условий.
ФОТОСИНТЕЗ
Фотосинтез - это процесс, при котором световая энергия поглощается и используется на синтез восстановленных углеродсодержащих соединений из двуокиси углерода и воды. Этот процесс происходит только в освещенной зеленой ткани, потому что хлорофилл играет существенную роль в превращении световой энергии в химическую Термин фотосинтез был предложен Чарльзом Рейдом Бан сом из Чикагского университета в начале этого столетия. В Европе этот процесс часто называют ассимиляцией или ассимиляцией углерода. Большинство американских физиологов растений предпочитают употреблять термин ассимиляция, когда речь идет об образовании новых тканей из углеводов и азотистых соединений (см. главу 6).
Значение фотосинтеза нельзя переоценить, потому что вся энергия:, содержащаяся в нашей пище, накапливается прямо или косвенно благодаря процессу фотосинтеза, а источником большей части используемой энергии, на которой работают заводы, являются горючие ископаемые, где она была запасена посредством фотосинтеза в далеком прошлом. Лит (1972, 1975) подсчитал, что наземные растения образуют за год 100*1 СР т сухого вещества, из которых более 2/3 вырабатывают деревья. Общая энергия, ежегодно накапливаемая в растениях при фотосинтезе, приблизительно в 100 раз превышает энергию, содержащуюся в угле, добываемом в течение года во всех шахтах мира. Хотя леса и занимают только 1/3 поверхности суши земного шара, они вырабатывают около 2/3 сухого вещества, в то время как возделываемые земли занимают приблизительно 9% и дают только 9% сухого вещества.
Пока человек не стал широко использовать уголь, большая часть потребляемой им тепловой энергии получалась от сжигания древесины. В настоящее время древесина шире используется как строительный материал и источник получения бумаги, чем в качестве топлива, хотя интерес к последнему опять возрастает. Независимо от того, как используется древесина, необходимо помнить, что содержащиеся в ней энергия и сухое вещество накапливаются благодаря процессу фотосинтеза. В связи с этим рациональное ведение лесного хозяйства должно быть направлено на повышение количества продуктов фотосинтеза на единицу земной поверхности и эффективности их превращения в растительный материал.
ПИГМЕНТЫ ХЛОРОПЛАСТОВ
Хлорофилл заслуживает особого внимания, потому что в процессе фотосинтеза он является светопоглощающим пигментом, а также и потому, что создает доминирующую окраску земной поверхности. У некоторых декоративных деревьев и кустарников иногда бывают видны желтые пигменты—каротиноиды. Эти пигменты обнаруживаются и вследствие того, что условия становятся неблагоприятными для синтеза хлорофилла или его сохранения. Листья некоторых разновидностей древесных растений, например, темно-пунцовой формы бука европейского, клена дланевидного, имеют красную или пурпурную окраску из-за присутствия в клеточном соке (а не в пластидах) антоцианов. Многие другие деревья вырабатывают антоцианы осенью. (Образование осенней окраски листьев будет рассмотрено в главе 7.)
Хлорофилл и каротиноида/ встречаются, кроме листовых пластинок, во многих тканях, в том числе в черешках, почках, семядолях, в коровой паренхиме молодых побегов и в феллодерме более старых стеблей некоторых видов. Обычно хлоропласты редко встречаются в эпидермальных клетках, за исключением .замыкающих клеток устьиц. Изредка
Фотосинтез
137
Рис. 5.1. Электронная микрофотография хлоропласта (Фото R.F. Evert): 1 — граны; 2 — пластоглобулы; 3 — строма; 4 — межгранные ламеллы
они развиваются в корнях, выставленных на свет, обычно содержатся в частях цветка, в молодых плодах, иногда даже и в зародышах семян. Наибольшее количество хлорофилла и каротиноидов сосредоточено в хлоропластах клеток мезофилла листьев. Хлоропласты представляют собой овальные, дисковидные или блюдцевидные тела диаметром около 5 мкм и толщиной 2—3 мкм, окруженные избирательно проницаемыми мембранами. Внутренняя структура хлоропласта сложная: он состоит из бесцветного белкового матрикса, или стромы, в котором расположены многочисленные более или менее цилиндрические тельца, называемые гранами. Последние состоят из множества дисковидных мембранных структур, называемых ламеллами или малыми тиллакоидами, с которыми связан хлорофилл. Граны соединены друг с другом одинарными межгранными тиллакоидами. Сложная система мембран играет важную роль в разделении электрических зарядов, транспорте электронов и фотофосфорилировании. Пигменты и ферменты, участвующие в фотохимической фазе фотосинтеза, ассоциированы с системой мемб-
138
Глава 5
СН=СНг I Н
✓СХ Xе
HjC-CT т СГ
f ' l' 4
I СПз I 'НС=О/
С~СгНз
НС
С—N
Мд
СН
С С-СНз
снг сн2
НС
СООСНз СООСгоНзд
с=о
Рис. 5.2. Структурная формула хлорофилла а. Хлорофилл b имеет такую же формулу за исключением того, что во II пиррольном кольце вместо группы СН3, отмеченной пунктирным кружком, находится группа НС = 0. Возможно другое расположение лврйных связей. Координационные связи магния показаны пунктирными линиями.
N—С
N
С
С
N—С
С
ран. Ферменты, участвующие в фиксации углерода, находятся в строме (рис. 5.1).
Хлоропласты содержат кроме пигментов (хлорофилла и каротиноидов) мембранные белки, липиды (в том числе триглицериды, фосфолипиды, сульфолипиды и галактолипиды), ферменты, крахмал, растворимые углеводы, аминокислоты, РНК, ДНК и неорганические элементы.
Некоторые авторы считают хлоропласты физиологически и генетически автономными структурами.
ХЛОРОФИЛЛЫ. В царстве растений встречается несколько разновидностей хлорофилла, но в древесных растениях имеют значение только хлорофиллы а и Ь, Хлорофилл а имеет формулу C55H72O5N4Mg; формула хлорофилла b — CssHyoOgNiiMg. Хлорофилла имеет структуру, состоящую из четырех замкнутых пиррольных колец с боковыми цепями. Пиррольные кольца координационно связаны с магнием, расположенным в центре (рис. 5.2). Хлорофилл флуоресцирует, поглощая волны видимого света определенной длины, преимущественно в красной и синей частях спектра, и излучая красный свет с большей длиной волны.
КАРОТИНОИДЫ. Желтая или оранжевая окраска, наблюдаемая иногда у пестрых листьев или у листьев, выдержанных в темноте, вызывается каротинами и ксантофиллами — двумя группами жирорастворимых пигментов, которые также содержатся в хлоропластах. В растениях встречается более 60 различных каротиноидов. Каротины — это углеводороды, имеющие эмпирическую формулу С40Н56 и состоящие из восьми остатков изопрена (см. главу 8). Существует несколько изомеров, из которых 3-каротин, предшественник витамина А, наиболее распространен и может составлять до 0,1% сухой массы листьев. Красный пигмент ликопин, содержащийся в зрелых томатах, плодах шиповника и в других частях растений, также является изомером каротина. Большинство ксантофиллов имеют формулу C4QH56O2 и окраску от желтоватой до коричневатой. Ксантофиллы можно отделить от каротинов, так как они более растворимы в спирте и гораздо менее раство
Фотосинтез
139
римы в петролейном эфире, чем каротины. Из ксантофиллов в листьях наиболее обычен лютеин, который иногда содержится в более высоких концентрациях, чем /3-каротин.
Каротины, в отличие от хлорофилла, образуются в некотором количестве и в темноте, поэтому сеянцы, выращенные в темноте, обычно имеют желтую окраску. Каротиноиды к тому же более устойчивы, чем хлорофилл, к разрушению при неблагоприятных условиях, поэтому листья, выдержанные в течение нескольких дней в темноте или подвергнутые воздействию водного стресса, обычно становятся желтыми, так как распад хлорофилла выявляет желтые пигменты — каротиноиды. Более быстрое разрушение хлорофилла, чем каротиноидов, также вызывает осенью пожелтение листьев некоторых видов (см. главу 7). Каротиноиды выполняют функцию вспомогательных пигментов при фотосинтезе, могут также защищать хлорофилл от необратимого фотоокисления. Кроме того, они являются основной причиной желтой и красной пигментации, встречающейся у животных.
ОТНОСИТЕЛЬНЫЕ КОЛИЧЕСТВА ПИГМЕНТОВ ЛИСТА. Количест ва и соотношения пигментов зависят от вида растений, внешних условий и возраста листьев. Как правило, хлорофилла в листьях приблизительно в 3 раза больше, чем каротиноидов. Общее количество хлорофилла широко варьирует у разных видов, некоторые из которых содержат в 2 или в 3 раза больше хлорофилла на единицу сухой массы листьев, чем другие. Хлорофилла а обычно в 2—3 раза больше, чем хлорофилла Z», а каротины превосходят ксантофиллы в сходной пропорции. Отношение хлорофилла а к хлорофиллу Ъ часто заметно изменяется у одного и того же растения. Например, осенью в листьях, подвергнутых действию водного стресса, хлорофилл а разрушается быстрее хлорофилла Ъ. Пигменты хлоропластов можно легко экстрагировать при помощи таких растворителей, как ацетон или метиловый спирт. Затем разные пигменты, содержащиеся в вытяжке, можно разделить по их различной растворимости или хроматографическими методами.
ФОТОСИНТЕТИЧЕСКАЯ ЕДИНИЦА. Молекулы хлорофилла организованы внутри хлоропластов в фотосинтетические единицы, каждая из которых состоит из нескольких сотен молекул. По мнению Альберти и других исследователей (1976), фотосинтетические единицы хвойных и лиственных деревьев менее многочисленны, но значительно крупнее, чем обнаруженные у травянистых растений. Фотосинтетические единицы теневой хвои сосны также крупнее, чем у световой. Хлорофиллы организованы в комплексы нескольких типов, каждый из которых имеет особую белковую часть и определенные спектральные свойства. Один из них, известный под названием П700 хлорофилл а —белковый комплекс, содержит также 0-каротин я связан t фотосистемой 1. Другой, известный под названием светособирающий хлорофилл а /й—белковый комплекс, содержит хлорофиллы а и Ъ и незначительные количества каротиноидов. Он связан в основном с фотосистемой П. Две фотосистемы осуществляют нециклическое фотофосфорилирование и вырабатывают ’’восстановительную силу” (НАДФ-Н2) и АТФ, необходимые для фиксации СО2 (рис. 5.3), По утверждению Альберти и др. (1976), отношение хлорофилла a lb и другие свойства пигментов сходны у хвойных и лиственных древесных растений и у исследованных
Глава 5
140
Рис. 5.3. Схема, показывающая поток электронов в фотохимической фазе фотосинтеза. Световая энергия, поглощенная пигментами фотосистемы II, вызывает фотолиз воды, высвобождение С>2 и Н+ и возбуждение электронов до высокого энергетического уровня, где они присоединяются к неидентифицированному акцептору Q. Затем электроны передвигаются вниз через ряд переносчиков, в том числе пластохинон, цитохромы и пластоцианин, к хлорофиллу фотосистемы 1. Часть энергии используется в процессе нециклического фотофосфорилирования на образование АТФ (аденозинтрифосфат), вероятно, в двух разных местах. Пигменты фотосистемы 1 также поглощают световую энергию, в результате чего возбужденные электроны передвигаются к акцептору X, а оттуда к ферредоксину. НАДФ восстанавливается до НАДФ • Нз ионами Н\ высвобожденными при фотолизе воды. Иногда происходит также непосредственное ’’перетекание*’ энергии от системы П к системе 1. Фотосистема П поглощает свет с длиной волны около 680 нм, тогда как фотосистема 1 действует преимущественно при больших длинах волн. Наиболее важные продукты фотохимической фазы: АТФ, НАДФ • Нг и 02
Фотосинтез
141
травянистых видов. Однако предполагают, что немногочисленность фотосинтетических единиц в хлоропластах деревьев может служить, по крайней мере частично, объяснением того факта, что интенсивность фотосинтеза листьев древесных растений, как правило, ниже интенсивности фотосинтеза травянистых растений.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ОБРАЗОВАНИЕ ХЛОРОФИЛЛА
Пожелтение, или хлороз, листьев — результат их неспособности увеличивать или сохранять содержание хлорофилла. Всесторонним изучением установлено, что это явление зависит от ряда внутренних и внешних факторов.
ВНУТРЕННИЕ ФАКТОРЫ. Наиболее важный фактор — генетический потенциал растения, так как иногда в результате мутаций, приводящих к полной потере способности образовывать хлорофилл, возникают короткоживущие альбиносные сеянцы. Чаще процесс синтеза хлорофилла нарушается лишь частично, проявляясь в отсутствии хлорофилла в некоторых участках листьев (пестролистно сть) или в общем низком содержании хлорофилла. Вследствие этого листья принимают желтоватый оттенок золотистых разновидностей деревьев и кустарников, часто используемых в качестве декоративных растений. Иногда в результате почковых мутаций возникают альбиносные или пестролистные ветви на нормальных в других отношениях деревьях и кустарниках. В синтезе хлорофилла и сборке его в фотосинтетические единицы участвуют многие гены, поэтому в образовании хлорофилла распространены аномалии. Развитие хлоропластов зависит как от ядерной, так и от пластидной ДНК, а также от цитоплазматических и хлоропластных рибосом.
Достаточное снабжение углеводами необходимо, по-видимому, для образования хлорофилла, и листья, испытывающие недостаток растворимых углеводов, могут не зеленеть, даже если все прочие условия благоприятные. Такие листья, опущенные в раствор сахара, обычно начинают образовывать хлоро’филл. Часто образованию хлорофилла препятствуют вирусы, вызывая ’’желтуху”, характеризующуюся общим хлорозом или бронзовой окраской листовых пластинок, а также просветлением жилок. Крапчатые листья некоторых декоративных растений — результат вирусной инфекции.
ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ. Главными внешними факторами, влияющими на образование и сохранение хлорофилла, являются: свет, температура, минеральное питание, вода и кислород. Синтез хлорофилла очень чувствителен почти к любому фактору, нарушающему метаболические процессы.
Свет. Свет необходим для образования хлорофилла, хотя сеянцы некоторых хвойных и небольшое число других видов растений образуют хлорофилл в темноте. Относительно низкая освещенность эффективна для инициации или ускорения образования хлорофилла. Выращенные в темноте желтые сеянцы содержат протохлорофилл — предшественник хлорофилла а, для восстановления которого до хлорофилла а требуется свет. Очень яркий свет вызывает разложение хлорофилла. Следовательно, хлорофилл всегда одновременно синтезируется и разрушается.
142 Глава 5
На ярком свету равновесие устанавливается при более низкой концентрации хлорофилла, чем при свете малой интенсивности. Теневые листья обычно имеют более высокую концентрацию хлорофилла, чем световые. Кроме того, отношение хлорофилла а к хлорофиллу Ъ у световых листьев выше, чем у теневых.
Температура. Синтез хлорофилла происходит, по-видимому, в широком интервале температур. Вечнозеленые растения умеренной зоны синтезируют хлорофилл от температур близких к температурам замерзания до самых высоких температур в середине лета. Многие хвойные становятся зимой до некоторой степени хлоротичными, вероятно, вследствие того, что распад хлорофилла превышает его синтез при очень низких температурах.
Минеральное питание. Одна из наиболее частых причин хлороза — недостаток какого-либо необходимого элемента. Недостаток азота — обычная причина хлороза древесных растений, особенно у старых листьев. Другая распространенная причина хлороза заключается в недостатке железа, преимущественно у молодых листьев. Достаточное снабжение железом необходимо, очевидно, Для синтеза хлорофилла. В состав хлорофилла железо не входит, 'но оно служит кофактором для предшественника хлорофилла. Магний является составной частью хлорофилла, поэтому его недостаток, естественно, вызывает хлороз. Недостаток большинства макроэлементов и некоторых микроэлементов может быть причиной хлороза. Можно предположить, что почти любое нарушение нормального метаболизма препятствует синтезу хлорофилла. (Хлороз, вызываемый недостатком минеральных элементов, обсуждается более подробно в главе 10.)
Вода. Умеренный водный стресс замедляет образование хлорофилла, а сильное обезвоживание растительных тканей не только нарушает синтез хлорофилла, но и вызывает распад уже имеющихся молекул. В результате листья растений, подвергшихся воздействию засухи, имеют тенденцию к пожелтению. Наиболее сильно действует водный стресс на светособирающий хлорофилл a lb — белковый комплекс. Листья деревьев и кустарников могут также пожелтеть при насыщении водой почвы вокруг их корней. Действие засухи и плохой аэрации почвы является до некоторой степени косвенным: синтез хлорофилла задерживается вследствие общего нарушения метаболизма (см. главы 16 и 17).
Кислород. При отсутствии кислорода сеянцы не образуют хлорофилла даже на свету. Это указывает на то, что аэробное дыхание необходимо для некоторых процессов образования промежуточных соединений, что для синтеза хлорофилла необходим приток метаболической энергии.
МЕХАНИЗМ ФОТОСИНТЕЗА
Фотосинтез заключается в восстановлении атмосферной СО^ до углеводов с использованием световой энергии, сопровождаемой освобождением кислорода из воды. Эту реакцию можно суммировать при помощи общего уравнения
СО, + 2Н,0----(СН,О) + о, + н,о.
2 хлорофилл 4
Фотосинтез
143
Фотосинтез, подобно многим другим физиологическим процессам, состоит из нескольких последовательных этапов. Вследствие их сложности подробное обсуждение этих этапов не входит в задачу данной книги. Однако огромное значение фотосинтеза требует краткого обсуждения основных особенностей механизма данного процесса. Знание природы процесса необходимо для понимания того, как влияют на него различные факторы внешней среды.
Фотосинтез включает несколько последовательных этапов: 1) поглощение световой энергии хлоропластами; сопровождающие хлорофилл пигменты (например, каротиноиды) играют в фотосинтезе вспомогательную роль, перенося энергию на хлорофилл а ; 2) расщепление воды и освобождение богатых энергией электронов и О2; 3) перенос Электронов, приводящий к генерации химической энергии в форме АТФ и восстановительной силы в виде НАДФ-Н2; 4) заключительный этап, включающий потребление энергии АТФ и восстановительной силы НАДФ-Н2 на фиксацию молекул СО2 в фосфоглицериновой кислоте и восстановление ее до фосфоглицеральдегида и, наконец, превращение этого соединения в более сложные углеводы, (сахарозу, крахмал, целлюлозу и гемицеллюлозу).
В процессе фотосинтеза особая роль, естественно, принадлежит углеводам как основным продуктам. Однако большие количества первичных продуктов сразу же превращаются наряду с глюкозой в другие соединения (липиды, органические кислоты и аминокислоты), играющие в равной степени важную роль в метаболизме растений. (Некоторые пути этих превращений показаны на рис. 6.20.)
СВЕТОВЫЕ И ТЕМНОВЫЕ РЕАКЦИИ. Фотосинтез включает две фазы: фотохимическую, для которой требуется свет, и химическую, протекающую в темноте. В течение световой фазы образуются НАДФ-Н2, АТФ и О2, а восстановление СО2 происходит в химической фазе. Хотя для темновых реакций и не нужен свет, но они не происходят при отсутствии света, так как зависят от НАДФ-Н2 и АТФ, которые являются продуктами световой реакции.
Фотохимическая фаза протекает очень быстро (за 0,00001 с). Первичная фотохимическая реакция не зависит от температуры, хотя транспорт электронов — процесс, зависящий от температуры. Темновая фаза происходит более медленно, приблизительно за 0,04 с при 25°С, замедляясь при низких температурах. Интенсивность фотосинтеза ограничивается реакцией, протекающей наиболее медленно. В связи с этим на ярком свету, когда ограничивающей является химическая фаза, процесс фотосинтеза чувствителен к температуре, а при слабой освещенности более вероятно, что ограничивающей становится фотохимическая фаза. При нормальных условиях освещений концентрация СО2 чаще бывает ограничивающим фактором, чем температура, за исключением, возможно, вечнозеленых растений зимой.
Фотохимическая фаза фотосинтеза состоит из двух световых реакций, осуществляемых фотосистемами 1 и II, объединенных переносчиками электронов. Эти системы действуют под воздействием различных пигментов. Способный к окислению и восстановлению хлорофилл в реакционном центре фотосистемы 1 представляет собой один из видов хлорофилла а . Вследствие того, что его максимум поглощения соответ
144
Глава 5
ствует 700 нм, он известен под названием П700. Дополнительные пигменты фотоситемы 1, в том числе остальные формы хлорофилла а и каротиноиды, также участвуют в переносе световой энергии к активным центрам П 700. Пигменты фотосистемы II представлены активной формой хлорофилла а с максимумом поглощения около 680 нм, несколькими дополнительными видами хлорофилла а, хлорофиллом b и каротиноидами. (Постулированная связь фотосистем 1 и II показана на рис. 5.3.)
Действие каждой системы включает поглощение одного кванта световой энергии каждым реакционным центром, о котором в таком случае говорят, что он находится в ’’возбужденном” состоянии. В фотосистеме 1 ’’возбужденный” реакционный центр отдает электрон акцептору электронов, а затем электрон передвигается, понижая свой энергетический уровень, к ферредоксину и участвует в восстановлении НАДФ до НАДФ-Н2. В фотосистеме П поглощенная энергия используется на ’’возбуждение” электронов и фотолиз воды. Фотолиз воды дает Н+, электроны (е—) и О2:
2Н2О -> 4е + 4Н+ +
Возбужденные электроны передвигаются к акцептору Q (см. рис. 5.3) и затем через ряд переносчиков электронов, включая пластохинон, цитохромы и пластоцианин — к возбужденному хлорофиллу П700 фотосистемы 1. Часть энергии расходуется на образование АТФ путем нециклического фотофосфорилирования, вероятно, в двух разных местах. Циклическое фотофосфорилирование показано на лабораторных препаратах и клетках водорослей. НАДФ восстанавливается до НАДФ-Н£ электронами, освобождаемыми при фотолизе воды. Не все этапы в этой системе полностью объяснены. Однако основными результатами фотохимической фазы являются выработка АТФ и НАДФ-Н2, необходимых для восстановления СО2, и выделение О2 в атмосферу. Образование АТФ и НАДФ-Н2 происходит в тиллакоидах или на них, восстановление СО? до углеводов — в строме.
ПУТИ ФИКСАЦИИ УГЛЕРОДА. Фиксация углерода из СО2 в стабильные со.единения включает последовательные сложные реакции, которые могут происходить при отсутствии света, но зависят от снабжения АТФ и НАДФ-Н2, которые вырабатываются в фотохимической фазе. Информация относительно соединений, образующихся при фиксации углерода, была получена при подаче радиоактивной Ct>2 одноклеточным водорослям в течение постепенно укорачивающихся промежутков времени. Затем клетки фиксировались и подвергались анализу. В течение 1 мин после подачи 14СО2 меченый углерод был обнаружен в сахарах, сахарофосфатах, органических кислотах и других соединениях (рис. 5.4). Этот цикл обычно называют циклом Кальвина—Бенсона. Кальвин и его сотрудники разработали большую часть цикла. Существенные особенности данного процесса можно резюмировать следующим образом: 1. Карбоксилирование 5-углеродного С5 сахарофосфата (ри-булозо-1,5-дифосфата) с образованием двух молекул фосфоглицериновой кислоты С3. 2. Превращение фосфоглицериновой кислоты в триозофосфат С3 при помощи НАДФ-Н2 и АТФ, выработанных при
Фотосинтез
аз световых реакций
Кислота (12 молекул)
кислота (12 молекул)
15С)
РуДЧ> карбоксилаза
из световых реакции
6 АДФ+6Ъ
Рибулозодисроссрат (6 молекул )
. из световых реакции
(12 молекул) лх itAV р
Р^улоэо-
моносроссрат (5Ci
(6 молекул)
Глкжоза (6С) и друеие соединения
Рис. 5.4. С3 - путь фиксации углерода, или цикл Кальвина - Бенсона (По Govindjee и Govindjee, 1975) (Другие соединения, образующиеся из первичных продуктов фотосинтеза, приведены на рис. 6.20)
фотохимических реакциях. 3. Взаимопревращение сахаров. На шесть молекул СС>2 образуется одна молекула гексозы, а шесть молекул рибулоз о-1,5-дифосфата регенерируют и продолжают цикл. Кроме того, для превращения рибулозомонофосфата в рибулозо-1,5-дифосфат требуется молекула АТФ. На каждую молекулу глюкозы общее уравнение реакции можно суммировать так:
6СО2 + 12НАДФН2 + 18АТФ->-С6Н12О6+ 12НАДФ+ 18АДФ+ 18ФН + 6Н2О.
Большинство сельскохозяйственных культур и почти все древесные растения называют Сз-растениями, так как после присоединения СС>2 к 5-углеродному сахару (рибулозо дифосфату в цикле восстановления углерода) он превращается в две ъ элекулы 3-углеродного соединения, фосфоглицериновую кислоту (см. рис. 5.4). Рибулозодифосфаткар-боксилаза (РуДФ-карбоксилаза) является карбоксялирующим ферментом. Этот фермент функционирует так же, как оксигеназа. Вследствие этого С>2 является конкурентным ингибитором фиксации СОз-Однако у некоторых видов, таких, как кукуруза и сахарный тростник, первыми обнаруживаемыми продуктами фотосинтеза являются 4-уг-леродные соединения, главным образом аспарагиновая, яблочная и щавелевоуксусная кислоты. В этой системе, известной под названием путь Хетча - Слэка, акцептором СО2 служит фосфоенолпировиноградная кислота Cj, а фосфоенолпируваткарбоксилаза (ФЕП-карбоксилаза) является карбоксилирующим ферментом. Это фермент очень близок
144
Гмм 5
Меэосрилл
реакций
Рис. 5.5. Сд - путь фиксации углерода, путь Хетча-Слэка. (По Govindjee и Govindjee, 1975)
к СО2 и не ингибируется кислородом, в отличие от РуДФ-карбоксилазы цикла Кальвина — Бенсона. В типичных Сд-растениях первоначальная фиксация углерода в Сд-органические кислоты происходит в клетках мезофилла. Затем эти кислоты транспортируются в характерные дня Сд-растений клетки обкладки крупных пучков 1, где происходит декарбоксилирование, освобождая СС>2, который используется в цикле Кальвина, а пировиноградкислота возвращается в систему Хетча — Слэка (рис. 5.5). Благодаря этому транспортному процессу СО2 концентрируется там, где РуДФ-карбоксилаза может легко фиксировать ее в фосфоглицериновую кислоту. Следует подчеркнуть, что путь превращения углерода Хетча — Слэка является не альтернативой циклу Кальвина ~ Бенсона, а дополнительной системой, которая повышает его эффективность при некоторых условиях. Только цикл Кальвина является действительно автокаталитическим в том смысле, что он может генерировать больше акцептора СО2, чем было первоначально, и поэтому способствует увеличению связанного углерода.
Кроме различия в первом продукте фотосинтеза и более высокой компенсационной точки СО2, Сз-растения достигают светового насыщения при более низкой интенсивности освещения, чем С д-р астения, а ин-
1 Наиболее характерными здесь являются крупные хлоропласты этих клеТок, (Нримеч. пер.)
Фотосинтез
147
тенсивность их фотосинтеза увеличивается, если концентрация кислорода понижается. Кроме того, они не имеют характерных для листьев С4-растений клеток обкладки крупных пучков, где может концентрироваться СС>2- Высокую компенсационную точку СО? Cj-растений можно в какой-то степени отнести за счет того, что карбоксилазная активность РуДФ-карбоксилазы конкурентно ингибируется кислородом. Этим объясняются также результаты наблюдений, показывающие, что уменьшение концентрации О2 приводит к значительному повышению фиксации СО2 Сз-растениями. Увеличение концентрации СОп ослабляет оксигеназную активность РуДФ-карбоксилазы, что способствует ускорению наблюдаемой фиксации СО2-
Важной особенностью растений, для которых характерен С^путь, является очень низкая интенсивность фотодыхания по сравнению с фотодыханием Сз-растений. Фотодыханием называют светозависимое образование СО2 фотосинтезирующей тканью (оно не связано с обсуждаемым в главе 6 основным дыханием, в котором участвует цитохромная система). Субстратом фотодыхания служит один из продуктов фотосинтеза, вероятно, глиоксилевая кислота, образующаяся из рибулозодифосфата вследствие .оксигеназной активности РуДФ-карбоксилазы. Интенсивность процесса у С 4-растений незначительна. Это объясняется , вероятно, тем, что почти вся освобождаемая при фотодыхании СО2 вновь вовлекается в оборот более эффективной комбинацией карбоксилаз объединенных циклов Хетча — Слэка и Кальвина, чем в одном цикле Кальвина. Тот факт, что фотодыхание встречается у всех.зеленых растений, наводит на мысль, что оно выполняет какую-то важную функцию, но конкретная его роль неизвестна. Предполагают, что оно способствует синтезу аминокислот, а может быть, и синтезу АТФ в митохондриях, но это могут быть только второстепенные альтернативные метаболические пути.
Вследствие большой потери углеводов в процессе фотодыхания (до половины углерода, связанного при фотосинтезе) возникла дискуссия о возможности повышения нетто-фотосинтеза и урожайности сельскохозяйственных культур путем поиска генетических или химических средств уменьшения фотодыхания. Однако все основные виды древесных растений, за исключением, может быть, некоторых мангровых и немногих других, используют Сз-путь усвоения углерода и осуществляют фотодыхание и тем не менее весьма успешно произрастают в естественных условиях. Некоторые исследователи считают, что ингибирование фотодыхания не сопровождается усиленным фотосинтезом, а наоборот, понижает его. Несмотря на то, что С^путь более эффективен, чем Сз-путь, с точки зрения углеродного баланса на уровне отдельных листьев, он, по-видимому, дает небольшие преимущества на уровне фитоценозов. К тому же еще неизвестно, часто ли рост растений при благоприятных условиях внешней среды ограничивается потенциальной интенсивностью фотосинтеза на единицу листовой поверхности.
Другая разновидность фиксации углерода, известная под названием обмена органических кислот по типу толстянковых — crassulacean acid metabolism (САМ), обнаружена у многих суккулентов и некоторых других растений, способных связывать большие количества СО? в темноте. Реакции карбоксилирования у CAM-растении приводят к о>
148
Глава 5
разованию щавелевоуксусной и яблочной кислот в темноте благодаря действию фосфоенолпируваткарбоксилазы. На свету яблочная кислота декарбоксилируется, давая пировиноградную кислоту и СО2, который используется в процессе фотосинтеза по пути цикла Кальвина — Бенсона. У этих растений ночью кислотность становится очень высокой, а в течение дня снижается, одновременно увеличивается содержание сахара. У многих суккулентов ночью устьица открыты, что способствует поступлению СС>2, а днем они обычно закрыты, предотвращая потерю воды. Такая особенность очень эффективна для растений, произрастающих в сухих местообитаниях, У древесных растений она не встречается.
Как указывалось выше, большой интерес представляет поиск методов повышения эффективности фотосинтеза. Естественный отбор, действовавший в течение тысячелетий, создал согласованность биохимии и биофизики фотосинтеза, которую будет очень трудно улучшить в полевых условиях.
ПОГЛОЩЕНИЕ ДВУОКИСИ УГЛЕРОДА ФОТОСИНТЕЗИРУЮЩИМИ ТКАНЯМИ. СО2 поступает преимущественно через устьица, хотя небольшое количество, по-видимому, диффундирует через кутикулу и эпидермальные клетки. Направленная внутрь диффузия СО2 подчиняется тем же законам, которые управляют направленной наружу диффузией водяного пара, и может быть представлена в виде следующего уравнения:
поток СО2 -Свозд- С мез / гвозд - глист>
где выражение ^В03д — Смез означает разность между концентрацией СО2 в наружном воздухе и в хлоропластах клеток мезофилла; гВозд -г лист - сопротивления диффузии в наружном воздухе и в листе. Величина глист содержит два компонента: г усГ' устьичное совротивле-ние и гмез- сопротивление мезофилла, которое сравнительно велико для водной фазы в клетках мезофилла. Сопротивление воздуха изменяется приблизительно от 0,5 с/см при его движении до 2—3 с/см в спокойном воздухе. Устьичное сопротивление колеблется от 1—7 с/см при открытых устьицах до 50-150 с/см при закрытых. Сопротивление мезофилла для водяного пара незначительно, для СО2 оно составляет около 6—10 с/см.
Таким образом, устьица, закрываясь, увеличивают сопротивление передвижению воды гораздо сильнее, чем сопротивление диффузии СО2, и, очевидно, понижают транспирацию в большей степени, чем фотосинтез. Это создает обоснование для использования метаболических анти-тр ан спирантов, которые вызывают закрывание устьиц, для уменьшения транспирации (см. главу 13).
ТКАНИ, ОСУЩЕСТВЛЯЮЩИЕ ФОТОСИНТЕЗ. Фотосинтез, происходящий главным образом в листьях, отмечается в некоторой степени и в других зеленых тканях, в том числе в семядолях (см. главу 15), почках, стеблях, цветках и плодах. У большинства видов фотосинтетическая деятельность тканей, не входящих в состав семядолей и листьев, незначительна. Например, фотосинтез зеленых плодов белой акации, лимона, апельсина, авокадо, винограда и сливы обычно слишком слаб для того, . чтобы внести заметный вклад в их рост. Фотосинтез быстро разрастаю-’"щихся зеленых шишек сосны смолистой недостаточен для того, чтобы
Фотосинтез
149
уравновесить наблюдающееся у них интенсивное выделение СО2 в процессе дыхания. Большая часть необходимых для роста шишек углево-цов мобилизуется из других источников.
Фотосинтез коры у большинства видов невелик, составляет 5% обще-'о у тополя дельтовидного, но фотосинтез ветвей и коры очень важен щя некоторых растений засушливой зоны. Ряд пустынных кустарников, :брасывающих листья в сухое время года, имеет устойчивые фотосинтетически активные стволы и ветви. Например, Cercidium microphyl-lum находится в безлистном состоянии большую часть года, и его зеленые ветви могут вырабатывать до 40 % всех ежегодных продуктов фотосинтеза.
ИЗМЕНЕНИЯ ИНТЕНСИВНОСТИ ФОТОСИНТЕЗА
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов. К тому же техника измерений бывает иногда неудовлетворительной. Часто бывает трудно сравнить количественно фотосинтез морфологически несходных растений. Интенсивность фотосинтеза растений разных размеров обычно выражают в количестве СО2, поглощенного на единицу сухой массы листьев, поверхности листьев или поверхности, на которой расположены устьица. У некоторых видов с тонкой кутикулой заметное поглощение СО2 происходит непосредственно листовой поверхностью, не имеющей устьиц. У других видов листовая поверхность, лишенная устьиц, относительно непроницаема для СО2. Козловский (1949) установил более высокую интенсивность поглощения СО2 сеянцами дуба, чем сеянцами сосны, при пересчете как на поверхность, так и на сухую массу листьев. Величина разности между полученными значениями изменялась в зависимости от того, на какую физиологическую единицу производился расчет фотосинтеза.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы.
В противоположность этому кратковременные измерения поглощения СО2 обычно проводились чаще в контролируемых, чем в изменяющихся условиях среды. Это может быть благоприятным для всех или только для нескольких сравниваемых видов. Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
150
Глава 5
ВИДОВЫЕ И ГЕНЕТИЧЕСКИЕ РАЗЛИЧИЯ. Фотосинтетическая способность часто значительно различается у разных видов, а также среди разновидностей, клонов и особей одного вида, но разного происхождения. Такое разнообразие обычно связано с основными различиями в метаболизме или в анатомической структуре листьев. Кроме того, разные виды и разновидности одного вида заметно различаются по развитию кроны, поэтому образование большего количества листьев или более продолжительный вегетационный период часто компенсируют низкую интенсивность фотосинтеза на единицу листовой поверхности или сухой массы.
Видовые различия. Некоторые исследователи сообщали о различиях в фотосинтетической активности разных видов. Например, среди лесных деревьев особенно высокая интенсивность фотосинтеза наблюдалась у некоторых покрытосеменных (у тополя, яблони, ясеня и эвкалипта) и у голосеменных (таких, как дугласия, лиственница и метасеквойя). Обнаружено, что поглощение СО2 на единицу листовой поверхности значительно выше у дуба красного и белого, чем у дерена флоридского или сосны ладанной. Наблюдалась высокая интенсивность фотосинтеза у дугласии, средняя у сосны веймутовой и низкая у ели европейской. Определено, что метасеквойя, имеющая очень высокую интенсивность фотосинтеза и слабое дыхание, является продуктивнее бука, дуба, сосны, если или лиственницы. Часто интенсивность фотосинтеза заметно различается у видов одного рода. Например, Eucalyptus globulus имеет гораздо более высокую интенсивность, чем Е. marginata. Имеются также сообщения о различиях среди плодовых деревьев. Поглощение двуокиси углерода на единицу поверхности листьев груши и яблони в 3 раза больше, чем у листьев таких вечнозеленых многолетних растений, как цитрусовые. В других исследованиях фотосинтез изменялся в следующем порядке: яблоня > груша > вишня > слива. У листьев яблони фотосинтез был не только наиболее интенсивным, но и дольше сохранялся осенью, а слива была первой, у которой прекращалось поглощение СО2*
Генетические различия. Имеется много примеров генетических различий в интенсивности фотосинтеза.
Например, обнаружены, большие различия в фотосинтетической способности двух клонов лиственницы европейской. Установлено, что сеянцы дугласии по происхождению с острова Ванкувер имели при определенной температуре более высокую интенсивность фотосинтеза, чем сеянцы, происходящие из Монтаны. Однако иногда варьирование среди сеянцев одного и того же географического происхождения было большим, чем различия между сеянцами из Цвух источников. Люк-канен и Козловский (1972) обнаружили большие различия между клонами тополя по интенсивности фотосинтеза, фотодыхания и химического (темнового) дыхания на единицу листовой поверхности и по компенсационной точке COj- Представители секции Aigeiros имели более низкую интенсивность фотосинтеза и более высокие компенсационные точки, чем представители секции Tacamahaca.
Различия фотосинтетической способности, наблюдаемые среди генетического материала, могут быть результатом анатомических или биохимических особенностей или тех и других одновременно. Часто изменения признаков, характеризующих устьица, являются причиной различий в поглощении СО2.
Фотосинтез
151
Большой интерес лесных генетиков вызывает использование интенсивности фотосинтеза в качестве показателя потенциального роста деревьев. Однако между фотосинтетической способностью и ростом деревьев установлена как высокая, так и низкая и даже отрицательная корреляция. Имеются сообщения о положительной корреляции между интенсивностью фотосинтеза и ростом у тополя, у гибрида осины и тополя, у сеянцев дугласии. Некоторые исследователи обнаружили даже обратные отношения между фотосинтезом и потенциальным ростом. Например, установлено, что интенсивность фотосинтеза и рост сеянцев сосны обыкновенной зависят от происхождения семян. Однако отмечено, что у наиболее хорошо росших сеянцев была самая низкая интенсивность фотосинтеза в расчете на сухую массу листьев. У быстрорастущих сеянцев фотосинтетически активной была меньшая часть общей массы хвои. Это обусловлено более сильным взаимным затенением хвои у быстрорастущих растений или, что более вероятно, анатомическими или биохимическими различиями хвои, связанными с происхождением семян.
По мнению некоторых исследователей, кратковременные измерения фотосинтетической способности не всегда являются надежными для оценки потенциального роста, потому что его детерминируют наряду с интенсивностью фотосинтеза, по меньшей мере, еще три важные физиологические обстоятельства. Сюда входят продолжительность роста f или характер сезонных изменений фотосинтеза, отношение фотосинтеза к дыханию и распределение продуктов фотосинтеза внутри дерева..
Отсутствие положительной связи между фотосинтезом и ростом часто наблюдается вследствие неадекватной выборки результатов кратковременных измерений фотосинтеза. Растение с высокой интенсивностью фотосинтеза на одной стадии его сезонного цикла может иметь на другой стадии низкую интенсивность.
Так, у горного экотипа сосны обыкновенной с апреля по август выявлена более высокая интенсивность фотосинтеза, чем у двух долинных экотипов. Затем интенсивность фотосинтеза горного экотипа быстро уменьшалась и осенью была ниже, чем у обоих долинных экотипов. Таким образом, прогноз прироста сухого вещества по результатам измерения фотосинтеза должен основываться как на интенсивности, так и на продолжительности фотосинтеза. Важный вывод из этого исследования заключается в том, что географические расы с очень высокой интенсивностью фотосинтеза в октябре и ноябре имели и наибольшую скорость роста.
ДНЕВНОЙ ХОД ИЗМЕНЕНИЙ ФОТОСИНТЕЗА. Ранним утром светлого безоблачного теплого дня фотосинтез идет слабо вследствие небольшой интенсивности света и низкой температуры, несмотря на высокую влажность листьев и высокую концентрацию двуокиси углерода в межклетниках листьев. При повышении интенсивности света, нагревании воздуха устьица открываются, нетто-фотосинтез начинает быстро увеличиваться и может достичь максимума до полудня. Часто вслед за максимумом наступает полуденное понижение, которое может быть небольшим или резким. Полуденная депрессия нередко сменяется новым повышением фотосинтеза в более поздние послеполуденные часы, а затем окончательно снижается. Наблюдается это, как правило, ранним
152
Глава 5
£>ремя
Рис. 5.6. Суточные колебания фотосинтеза хвойных в ясный летний день (Hodges, 1967):
1 — свет; 2 - температура; 3 — дефицит давления водяного пара (ДДП) ; 4 — сосна обыкновенная; 5 — пихта благородная; 6 — пихта великая
Полночь Полдень Полночь
время
Рис. 5.7. Суточные колебания фотосинтеза хвойных в пасмурный летний день (Hodges, 1967):
1 — свет; 2 — температура; 3 — дефицит давления водяного пара ( ЛИП) ; 4 — пихта великая; 5 — пихта благородная
вечером вслед за уменьшением интенсивности света и температуры. Вследствие изменений условий внешней среды в разные дни и в пределах одного дня дневной ход фотосинтеза часто значительно отклоняется от описанного выше (рис. 5.6 и 5.7). Большинство дневных изменений фотосинтеза, за исключением полуденного понижения, хорошо взаимодействуют с изменениями интенсивности света.
Например, на открытом пространстве максимум интенсивности фотосинтеза трех видов покрытосеменных наблюдался в полдень и соответствовал максимуму интенсивности общей солнечной радиации. Под пологом леса интенсивность фотосинтеза значительно изменялась в течение дня. При этом наивысшая интенсивность наблюдалась в солнечных бликах. Дневной ход фотосинтеза голосеменных был очень различным в облачные и в солнечные дни. Без затенения в пасмурные или облачные дни типичная картина была такой: интенсивность нетто-фотосинтеза возрастала, достигая максимума около полудня, затем или уменьшалась, или оставалась более или менее постоянной в течение 1-2 ч и потом понижалась. В ясные солнечные дни фотосинтез обычно быстро возрастал, достигал максимума между 9 и 12 ч утра, затем, до поздних послеполуденных часов, понижался. Позднее он вновь усиливался и достигал второго максимума, но значительно более низкого.
Причины дневных изменений фотосинтеза. Регуляция дневного хода фотосинтеза под влиянием разнообразных внешних и внутренних факторов очень сложна. Главными из внешних факторов являются, по-видимому, свет, температура, наличие воды, содержание СС>2 в воздухе и различные взаимодействия между ними. Внутреннюю регуляцию поглощения COj относили за счет разных причин: водного стресса, закрывания устьиц, чрезмерного дыхания, накопления конечных продук
Фотосинтез 153
тов фотосинтеза и фотоокисления ферментов. Значение отдельных факторов, влияющих на поглощение СОэ, нередко изменяется. Например, утром, когда клетки тургесцентны, повышение интенсивности фотосинтеза взаимосвязано с нагреванием воздуха и увеличением интенсивности света. Однако полуденный спад при наивысшей интенсивности света часто происходит, особенно в жаркие дни, вследствие слишком большой потери воды, сопровождаемой закрыванием устьиц.
СЕЗОННЫЕ ИЗМЕНЕНИЯ. Необходимо различать сезонные изменения фотосинтетической способности деревьев, вызываемые развитием листьев и состоянием метаболизма, и наблюдаемую в полевых условиях фактическую интенсивность, которую определяют как фотосинтетическую способность, так и накладывающиеся друг на друга факторы внешней среды. Это разграничение важно в связи с тем, что при исследовании сезонных изменений фотосинтеза растения часто периодически переносили из открытого грунта в лабораторию и измеряли фотосинтез при стандартных и благоприятных внешних условиях. Фактическая интенсивность фотосинтеза в полевых условиях гораздо сильнее изменяется в разные дни вследствие изменений факторов внешней среды, чем интенсивность, измеренная при стандартных условиях.
Сезонные изменения фотосинтетической способности у голосеменных происходят более постепенно, чем у листопадных покрытосеменных. Когда температура весной повышается и ночные заморозки становятся менее частыми, фотосинтетическая способность голосеменных постепенно увеличивается. Осенью интенсивность фотосинтеза также постепенно понижается. У листопадных покрытосеменных фотосинтез быстро ускоряется весной, когда деревья вновь покрываются листвой, остается высоким в течение лета, быстро уменьшается в конце лета, когда листья стареют, и окончательно падает до нуля, когда они опадают. Сезонные изменения фотосинтетической способности различаются у видов с разным характером развития листьев. У видов, побеги которых полностью формируются в зимующих почках, листовая поверхность достигает максимума в начале вегетационного периода. Гетерофильные и периодически отрастающие виды продолжают увеличивать количество листьев постепенно или путем периодического образования новых побегов в течение всего сезона (см. главу 3). Следовательно, сезонные изменения фотосинтетической способности происходят при изменении площади листьев. У голосеменных фотосинтетическая способность также изменяется при появлении новой листвы, а осенью голосеменные дольше, чем покрытосеменные, сохраняют свою способность к заметному фотосинтезу. В районах с теплыми зимами фотосинтез у вечнозеленых голосеменных может происходить в течение всех месяцев года.
Общий фотосинтез дерева и характер его сезонных изменений часто заметно варьируют в разные годы вследствие различий в размерах листовой поверхности и климатических изменений.
Сезонные изменения фотосинтетической способности сеянцев сосны ладанной и сосны веймутовой в Северной Каролине исследовали Грегор и Крамер (1963). Сеянцы выдерживали на открытом воздухе. Периодически их переносили в лабораторию и измеряли поглощение СО2 при 25°С и освещенности 43 000 лк. Начиная с февраля интенсивность фотосинтеза в расчете на один сеянец медленно увеличивалась у обоих видов до апреля, затем быстро возрастала и потом снижалась в течение Осени и зимы (рис. 5.8). Максимальная интенсивность у сосны ладанной
154
Глава 5
Рис. 5.8. Сезонные изменения нетто-фотосмнтеза и дыхания на единицу листовой поверхности сеянцев сосен. Сеянцы содержались на открытом воздухе, фотосинтез измерялся в помещении при 25°С и интенсивности света 43000 як (McGregor, Kramer, 1963):
1 — сосна ладанная; 2 — сосна веймутова
Рис. 5.9. Сезонные изменения нетго-фотосинтеза и дыхания сеянцев сосен в пересчете на одно растение (McGregor, Kramer, 1963):
1 — сосна ладанная; 2 — сосна веймутова
Фотосинтез
155
достигалась в середине сентября, после чего происходило быстрое осеннее пони* жение. Максимум фотосинтеза у сосны веймутовой наблюдался между 15 июля и 15 сентября. Осеннее снижение было более постепенным. Более высокий и более поздний максимум фотосинтеза сосны ладанной был обусловлен В значительной степени тем, что у сеянцев трижды отрастали побеги, йрибавляя новую хвою до конца лета. В го же время сеянцы сосны веймутовой давали новые побеги только 1 раз, в начале вегетационного периода.
Повышение интенсивности фотосинтеза после 9 апреля отчасти могло быть отнесено за счет увеличения ассимилирующей поверхности у обоих видов. Однако значительное увеличение с 14 февраля до 9 апреля нельзя объяснить этой причиной, так как к 9 апреля новая листва еще не распустилась. Это увеличение должно было происходить в результате восстановления фотосинтетической активности уже имевшейся хвои. Уменьшение фотосинтеза после максимума, наблюдавшегося в середине сезона, было вызвано уюбоих видов не потерей хвои, а скорее пониженной фотосинтетической способностью существовавшей хвои.
Сезонные изменения интенсивности фотосинтеза на единицу листвы значительно отличались от сезонного хода интенсивности фотосинтеза в пересчете на один сеянец (рис, 5.9). У сосны ладанной интенсивность фотосинтеза на единицу длины пучка хвои заметно повышалась с февраля по март, несмотря на то, что в этот период новая хвоя не появлялась. Максимальная интенсивность на единицу длины пучка была достигнута в мае, за 4 месяца до максимума интенсивности на один сеянец. Высокая интенсивность сохранялась до сентября, а затем интенсивность фотосинтеза на единицу длины хвои и на один сеянец начала понижаться. У хвои сосны веймутовой фотосинтез также заметно повышался с февраля по март, но максимум не был достигнут до июля. Высокая интенсивность сохранялась в течение сентября, несколько понизилась в ноябре и достигла минимума в январе.
Зимний фотосинтез вечнозеленых растений. В районах с мягкими зимами у вечнозеленых растений фотосинтез осуществляется в течение всего года. В местах с суровыми зимами фотосинтез может быть незначительным несколько недель или месяцев.
Наблюдаемое поглощение СО£ или прирост сухой массы в течение зимы были установлены для сосны ежовой и сосны ладанной на юге Соединенных Штатов, для сосны замечательной и сосны обыкновенной в Аберистуите (Уэльс) и для широколистных вечнозеленых растений в Северной Италии. Деревья дугласии накапливали значительные количества продуктов фотосинтеза зимой в мягких климатических условиях прибрежного района северо-запада Соединенных Штатов. В засушливый год чистый прирост продуктов фотосийтеза’в течение зимы приближался к 1/4 общего прироста за весь год. Вдоль всей прибрежной зоны Норвегии у сосен и елей уменьшалась сухая масса в течение коротких периодов зимой, но за всю зиму у них обнаружился чистый прирост, доказавший превышение фотосинтеза над дыханием. Сухая масса сеянцев ели ситхинской в Южной Шотландии удваивалась за период с конца сентября и до середины апреля. Большая часть прироста приходилась на конец марта - начало апреля, но некоторое количество сухой массы накапливалось в середине зимы.
В районах с холодными зимами интенсивность фотосинтеза у вечнозеленых растений наблюдалась вплоть до точки замерзания или даже немного ниже. Например, около Мюнхена поглощение COj елью европейской происходило в дни, когда температура воздуха была немного ниже нуля. В долине около Инсбрука (Австрия) фотосинтез ели европейской продолжался и зимой до наступления сильных морозов. Однако на границе древесной растительности около горы Патчеркофель морозная погода в ноябре (от -10° до —15°) вызвала прекращение фотосинтеза. Пизек и Випклер (1958) показали, что интенсивность нетто-фотосинтеза ели европейской и сосны кедровой европейской была заметной до поздней осени. Последующие изменения температуры на несколько градусов нижем выше нуля вызывали колебания поглощения СО> Как только температура падала ниже -4° или -5°С, фотосинтез прекращался. Если же после этого морозы повторялись в течение нескольких ночей, то днем фотосинтез был ингибирован, даже если температура поднималась выше нуля. После мороза от -6° до -8°С нетто-фотоснитез прекращался, и требовалось несколько дней с мягкой погодой для восстановления фото
156
Глава 5
синтетической способности. Полностью фотосинтез возобновлялся с повышением температуры весной. Особенно это касалось частей кроны, в которых происходил распад хлорофилла в течение зимы. Весной вместе с колебаниями температуры колебалась и интенсивность фотосинтеза. Следовательно, фотосинтетический аппарат оставался функционально активным только до тех пор, пока зимой не наступали морозы, У верхней границы распространения леса температуры были в течение 4—5 месяцев такими низкими, что фотосинтез по существу был невозможен.
ИЗМЕРЕНИЕ ФОТОСИНТЕЗА. Говоря об интенсивности фотосинтеза, следует вспомнить некоторые проблемы, связанные с измерением этого процесса. В течение длительного периода лучшей мерой нетто-фотосинтеза считался прирост сухого вещества дерева или древостоя. Так, лесовод измеряет урожай по объему древесины или древесной массы, а эколог — по массе сухого вещества, выработанного на единицу земной поверхности. Однако эти методы неприменимы для кратковременных измерений, необходимых в исследованиях, которые в большинстве случаев основаны на определении поглощения С02- Методы определения поглощения СО2 были усовершенствованы после 1930-х годов, когда Хэйпике и другие исследователи улавливали СО2 в заполненных щелочью поглотительных колонках и проводили трудоемкую операцию титрования щелочи. Позже титрование было заменено кондуктометрическим определением изменения концентрации щелочи. После второй мировой войны почти повсеместно стали использоваться инфракрасные газоанализаторы, хотя кратковременное введение радиоактивной метки при помощи Г4сС>2 применяется в качестве полевого метода измерения поглощения СО2 отдельными- листьями. Измерения поглощения СС>2 проводятся при помощи замкнутых приемников, размеры которых колеблются от больших камер с кондиционированным воздухом, вмещающих часть фитоценоза, до маленьких кювет, в которых зажимают часть листа на короткое время (около 1 мин). Таким образом, техника измерения фотосинтеза хорошо разработана.
Достоверность измерений фотосинтеза может быть понижена рядом легко допускаемых ошибок. К числу таких ошибок относятся: движение воздуха через приемник со скоростью, недостаточной для предотвращения чрезмерного истощения запасов СО2: перегрев кювет; невозможность поддерживать постоянный дефицит давления водяного пара в кювете; возникновение незамеченного и неизмеренного водного дефицита; отсутствие оценки существующего физиологического состояния материала. При измерении должна также учитываться история растительного материала (например, где растение выросло — в поле или теплице).
Трудно сравнивать результаты измерений фотосинтеза, выполненных разными исследователями, так как интенсивность фотосинтеза дается в пересчете на различные величины: сырую массу, сухую массу, площадь листьев, поверхность, занятую устьицами, объем листьев и содержание хлорофилла. Наилучшей основой для пересчета является, по-видимому, площадь листьев, однако необходимо указывать, относится ли это к одной поверхности или к обеим. Площадь листьев тоже может вводить в заблуждение, так как толстые листья содержат хлоропластов на единицу поверхности больше, чем тонкие. Вследствие этого они обычно имеют более высокую интенсивность поглощения СО2 на единицу поверхности независимо от того, чем вызвано различие в толщине листьев. Возможны
Фотосинтез
157
также различия в размерах и количестве фотосинтетических единиц. Эти особенности могут иметь значение для объяснения различий в поглощении СС>2 световыми и теневыми листьями, растениями, выращенными в специальных камерах и открытом грунте, а также листьями растений, испытывающих недостаток элементов минерального питания и обеспеченных ими.
Измерение света. В исследованиях фотосинтеза одной из трудных проблем является точное измерение радиации как с точки зрения единиц измерения, так и в отношении места, где она измеряется. Интенсивность света заметно уменьшается внутри кроны дерева и в пологе сомкнутого насаждения, и никакое единичное измерение не может точно охарактеризовать световые условия внутри полога. Как упоминалось выше, отдельные листья достигают светового насыщения при гораздо более низкой интенсивности света, чем фитоценоз. Кроме того, многие измерения величин светового насыщения вводят в заблуждение, так как при этом не принимаются во внимание экспозиция листьев и самозатенение, возраст листьев и условия, при которых находились листья до того, как проводилось измерение.
Было также немало сомнений относительно наиболее подходящих единиц, используемых при измерении радиации. Измеряли освещенность и в футо-свечах, или люксах, и как фотосинтетически активную радиацию (ФАР), т.е. радиацию с длиной волны от 400 до 700 нм, часто выражаемую в мкм EIcn^Ic, или же как общую энергию в таких единицах, как Вт/м2, мВт/cm^ или Ленгли/мин. Физики обычно предпочитают измерение общей энергии. Однако при этом игнорируется тот факт, что такие измерения включают в себя большое количество энергии бесполезной для фотосинтеза и могут привести к неправильным выводам относительно физиологических процессов, особенно при сравнении солнечного света со светом ламп накаливания и ламп дневного света.
Например, в камере фитотрона, где отношение подводимой мощности в ваттах памп дневного света к лампам накаливания составляет 2,5:1, отношение освещенности в люксах от двух источников равно 9,4:1, отношение ФАР в мкм Е/м*/с около 7:1, а отношение обшей энергии в Вт/см^ от этих двух источников - 0,56:1.
По-видимому, в исследованиях по фотосинтезу наиболее подходит измерение световой энергии в диапазоне ФАР в мкм Е/м^/с или других подобных единицах.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ФОТОСИНТЕЗ
Фотосинтез значительно изменяется, так как на него влияет комплекс часто взаимодействующих внешних и внутренних факторов. Некоторые факторы, такие, как свет и СО2? оказывают прямое влияние на фотосинтез, другие (например, вода и минеральные вещества) нередко действуют косвенно. В течение дня ограничивающим может быть сначала один, а затем другой фактор. Например, утром фотосинтез, часто коррелирует с интенсивностью света, а в более позд ние часы дня его ограничивает водный стресс листьев, влияющий на ширину устьичных щелей и поглощение СО2. Наблюдается также сезонная замена одного ограничивающего фактора другим: зимой и весной фотосинтез ограничивает температура, летом — устьичное сопротивление, двуокись углерода и свет.
158
Глава 5
ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ. Основными внешними факторами, влияющими на фотосинтез деревьев, являются: свет, температура, концентрация СО2 в воздухе, вода, плодородие почвы, вещества, загрязняющие атмосферу, и применяемые химические препараты, насекомые и болезни. Вследствие этого фотосинтез деревьев легко реагирует на такие агротехнические мероприятия, как прореживание древостоев, обрезка, ирригация и внесение удобрений.
Интенсивность света. В темноте фотосинтез не происходит, потому что образующаяся при дыхании двуокись углерода выделяется из листьев. С увеличением интенсивности света достигается компенсационная точка, при которой поглощение двуокиси углерода при фотосинтезе и ее освобождение при дыхании уравновешивают друг друга. Вследствие этого газообмен между, листьями и атмосферой отсутствует. Эта световая компенсационная точка зависит от вида растения и генетических особенностей материала, типа листа (у теневых листьев световая компенсационная точка ниже, чем у световых листьев), возраста листа (у молодых листьев эта точка выше, чем у старых), концентрации СО2 в воздухе и от температуры. С повышением температуры дыхание усиливается быстрее, чем фотосинтез, .поэтому световая компенсационная точка также повышается, достигая очень больших значений при температуре выше 35°С.
30/5
Рис. 5.10. Влияние интенсивности света на фотосинтез сеянцев сосны ладанной ? я широколистных деревьев. Измерения проводились при 30°С (Kramer, Decker, 1944): ’
. 1 _ дуб белый; 2 — дуб красный; 3 — кизил; 4 — сосна ладанная
Фотосинтез
159
Рис. 5.11. Изменение интенсивности света внутри густой кроны кипариса (слева) и рыхлой кроны маслины (справа). Интенсивность света, измеренная в полдень в середине лета, выражена в процентах от освещенности на открытом месте (Larcher, 1975):
1 - 100 - SO; 2 -50-25; 3 - 25-10;
4 - 10-5; 5 -5 — 1; 6 — безлистная зона
I--1 1 ES3 2 з КИ 4 Ш 5 Ml б
С увеличением освещенности выше компенсационной точки фотосинтез возрастает пропорционально интенсивности света, пока не наступает световое насыщение, после которого интенсивность фотосинтеза становится более или менее постоянной. У некоторых видов очень высокая интенсивность света может даже вызвать уменьшение фотосинтеза.
Существует мнение, что у большинства растений световое насыщение фотосинтеза достигается при 1/4 или 1/2 полного солнечного освещения. Это справедливо для отдельных листьев и иногда даже для сеянцев, но не для больших деревьев, кустарников или для фитоценозов.
Крамер и Декер (1944) сравнивали фотосинтез сеянцев дубов красного и белого, дерена флоридского и сосны ладанной при интенсивности света от 3200 до 108 000 лк. У всех четырех видов фотосинтез быстро возрастал с увеличением интенсивности света при низкой освещенности. Фотосинтез сосны ладанной повышался до интенсивности света эквивалентной полному солнечному. Однако у трех лиственных видов максимум фотосинтеза достигался при 1 /3 полного освещения или менее. Дальнейшее увеличение интенсивности света не усиливало фотосинтез (рис. 5.10). Сеянцы Других лиственных лесных деревьев, таких, как клен красный и дуб крупноплодный, достигали максимальной интенсивности фотосинтеза при относительно низкой интенсивности света.
Различная реакция фотосинтеза некоторых деревьев на интенсивность света зависит от типа листвы. Сеянцы сосны ладанной с листвой, состоящей только из первичной хвои или с примесью развивающейся вторичной хвои, достигают максимума фотосинтеза при относительно низкой интенсивности света. Иная реакция наблюдается у более старых сеянцев, фотосинтез которых достигает максимума только при интенсивности света, соответствующей полному солнечному освещению. По-видимому, радиальное расположение первичных хвоинок создает лучшую экспозицию и меньшее самозатенение, чем пучки хвои более старых сеянцев.
160
Глава 5
(Woodman 1971^*й1Ие Фотосинтеза 0 Р^ных частях кроны 38-легней дугласии
Рис. 5.13. Зоны относительной эффективности фотосинтеза на северной и южной сторонах кроны 38-летнеЙ дугласии (Woodman^ 1971)
Рис. 5.14. Интенсивность фотосинтеза при семи различных значениях интенсивности света (Kramer, Clark, 1947) :
1 сеянцев дуба красного; 2 — отдельных хвоинок сосны ладанной; 3 — сеянцев сосны ладанной
Эффект взаимозатенения листьев. Интенсивность света в разных частях кроны дерева различна, поэтому с увеличением глубины расположения листьев фотосинтез быстро уменьшается. Однако скорость понижения интенсивности света от периферии кроны внутрь ее сильно различается у разных видов и зависит от формы кроны и густоты листвы. У некоторых деревьев, например у кипариса, крона настолько густая, что свет не доходит до ее внутренней части и листья не развиваются в сильно затененных участках. У деревьев с более рыхлой кроной облиственные ветви простираются до внутренней части кроны, куда проникает рассеянный свет (рис. 5.11).
Эффективность фотосинтеза сильно изменяется также в зависимости от высоты кроны, в значительной степени вследствие различий в количестве листвы и степени затенения. Непроизводительные нижние ветви с их относительно немногочисленными и сильно затененными листьями часто совсем не дают углеводов для роста главного стебля. У 38-летнего дерева дугласии с 18 мутовками ветвей максимальный фотосинтез наблюдался у хвои текущего года около 7-й мутовки от верхушки, в зоне меж’:.у полным освещением и полной тенью. Затем по направлению к основанию кроны интенсивность фотосинтеза постепенно понижалась (рис. 5.12). Фотосинтез на южной стороне постоянно был выше, чем на других сторонах той же мутовки ветвей. Несмотря на дневные и сезонные изменения фотосинтеза по всей высоте кроны, ее поверхность можно разделить на зоны по эффективности фотосинтеза (рис. 5.13). Отношение между зонами было постоянным в течение весны и лета, хотя абсолютные величины интенсивности фотосинтеза значительно варьировали.
фотосинтез
161
Характерные признаки листьев и их расположение очень различны у разных видов, и соответственно различается эффект взаимного затенения листьев. Листья лиан на стенах обычно образуют мозаику, в кото-. рой каждый лист получает максимальное освещение. Такая же ситуация часто отмечается и у листьев, расположенных на поверхности кроны дерева. Листовая мозаика является, по-видимому, результатом изгибания черешков в ответ на фототропический стимул. Однако хвоя сосен располагается в пучках, что вызывает значительное взаимозатенение. Хвоя сосны ладанной, разложенная плашмя в кювете и полностью выс-1 тавленная на свет, достигает светового насыщения приблизительно при такой же интенсивности света, как и листья сеянцев лиственного дерева (рис. 5.14). Однако сеянцам сосны требуется для насыщения в 3 раза более высокая интенсивность света вследствие взаимозатенения.
Приспособления к различной интенсивности света. Тсуга канадская и западная," бук, клен и дерен устойчивы к затенению и прекрасно растут в нижнем ярусе, а такие виды, как осина, тюльпанное дерево, березы и многие сосны, не выносят затенения и могут хорошо расти только на солнце. Различная теневыносливость во многом связана с разной способностью фотосинтетического аппарата приспосабливаться к низкой интенсивности света. Например, интенсивность фотосинтеза теневых листьев теневыносливого бука европейского в 4-5 раз больше, чем у световых листьев, если проводить измерения при очень низкой интенсивности света. Интенсивность фотосинтеза теневых листьев клена сахарного также сравнительно велика. В противоположность этому, Козловский (1949) не обнаружил признаков приспособления к затенению у теневых листьев сосны ладанной и дуба лировидного. Однако исследования некоторых хвойных (Cryptomeria japonica и Chamaecyparis obtusa, сосны смолистой, пихты бальзамической, елей белой и Зигельмана) показали, что фотосинтез у световой хвои выше, чем у теневой, при пересчете как на массу хвои, так и на ее объем или поверхность.
В большинстве случаев у световых и теневых листьев имеются суще- ственные морфологические различия и различия в механизме фотосинтеза. Световые листья, как правило, мельче и толще и имеют больший объем и больше хлорофилла на единицу поверхности, чем теневые листья. Кроме того, у них обычно более низкие сопротивление мезофилла и устьичное сопротивление диффузии СО2- Благодаря этому световые листья имеют более высокую интенсивность фотосинтеза на единицу листовой поверхности и достигают светового насыщения при большей интенсивности света, чем теневые листья. Нобель (1976) объясняет более высокую интенсивность фотосинтеза мелких, толстых световых листьев пустынного кустарника Hyptis emoryi их более развитой тканью мезофилла, большим содержанием хлорофилла и более высоким отношением внутренней поверхности к наружной, чем у теневых листьев. В некоторых экспериментах различия в фотосинтезе световых и теневых листьев, наблюдавшиеся при расчете на листовую поверхность, уменьшались или совсем исчезали при пересчете на вес или объем листа.
На биохимическом уровне более толстые световые листья содержат больше карбоксилирукпцих ферментов и больше переносчиков электронов на единицу листовой поверхности, чем теневые листья. По мнению
162 Глава 5
Эльберта и других исследователей (1976), теневые листья содержат более крупные фотосинтетические единицы и имеют более высокую долю светособирающего хлорофилл а/о-белкового комплекса, чем световые листья. Это увеличивает эффективность использования световой энергии при низкой интенсивности света, но уменьшает ее при высокой интенсивности вследствие понижения уровня, при котором наступает световое насыщение. По-видимому, приспособления, благоприятствующие высокой эффективности при одной экстремальной величине интенсивности света, являются неблагоприятными при другой крайности.
Другая проблема касается степени, до которой может проявиться адаптация листьев, развившихся при одной интенсивности света, а затем оказавшихся в других условиях освещения (например, когда густой древостой подвергается сильному* прореживанию). В этих случаях существенные морфологические и анатомические изменения невозможны. Однако эксперименты с травянистыми растениями показывают, что после недельной экспозиции листья кукурузы и Amaranthus, перенесенные с солнца в тень или наоборот, имели интенсивность фотосинтеза, сравнимую с той, которая была у листьев растений, постоянно находившихся при данной интенсивности света. В значительной степени это относили за счет изменений концентрации карбоксилирующих ферментов (РуДФ-карбоксилазы у Су растений, ФЕП-карбоксилазы у С4-растений).
ТЕМПЕРАТУРА. Нередко температура влияет на фотосинтез так же значительно и сложно, как и действует свет. Вслед за более ранними исследователями Парк (1977) предполагает, что влияние температуры на поглощение СО2 является результатом того, что некоторые зависящие от температуры этапы ограничивают интенсивность фотосинтеза и действуют отдельно или совместно. Можно допустить существование трех главных компонентов: 1) экспоненциально возрастающего процесса, доминирующего при низких температурах; 2) антагонистического процесса или процессов, постепенно ускоряющихся с повышением температуры (вероятно, темновое дыхание); 3) инактивации процесса фиксации СО2 при высоких температурах. Таким образом, наряду с прямым действием на синтез и активность карбоксилирующих и, вероятно, других ферментов существует косвенное влияние температуры через изменение интенсивности темнового дыхания и проводимости устьиц для СО2. Высокая температура часто усиливает транспирацию, вызывая водный дефицит, вследствие чего закрываются устьица и сокращается снабжение хлоропластов углекислотой.
Фотосинтез древесных растений происходит в широком диапазоне -от температур около 0 до 40°С и выше. Этот диапазон зависит от возраста и происхождения растения и от времени года. У большинства видов умеренной зоны интенсивность фотосинтеза увеличивается от температуры около 0°С и достигает максимального значения при 15 25°С. У тропических видов минимальная температура для фотосинтеза обычно составляет несколько градусов выше нуля, оптимум - значительно выше 25°С (рис. 5.15). Действие температуры на-фотосинтез зависйт от интенсивности света, наличия СО2, водоснабжения и предшествующего влияния факторов внешней среды. У деревьев часто наблюдается адаптация фотосинтеза к температурному режиму, при котором они рос-
Фотосинтез
163
Рис. 5.15. Влияние температуры на фотосинтез деревьев умеренного и тропического поясов. По. Larcher (1969, с изменениями) :
1 — Pinus cembra; z — pa<mS (световые листья); 3- Fictfsretusa; 4 -Acacia crLspedocarpa
ли. Например, у сеянцев пихты бальзамической был обнаружен кливальный характер адаптации температурного оптимума фотосинтеза по отношению к градиенту высоты: изменение оптимальной для фотосинтеза температуры составляло 2,7°С на каждые 300 м высоты над уровнем моря.
5.1. ВЛИЯНИЕ ТЕМПЕРАТУРЫ И ВОДНОГО ПОТЕНЦИАЛА ПОЧВЫ НА НЕТТО-ФОТОСИНТЕЗ СЕЯНЦЕВ
Pinus radiata (по Babalola И ДР., 1968)
Температура почвы, °C Водный по- -Наблюдаемое Темпера- Водный поту ра поч- тенциал Наблюдаемо е поглощение COj (10— 5 мл-см_“7- •с"1)
тенциал почвы, бар поглощение СС>2 (10— 5 мл*см~2. •с" х)
вы, °C почвы, бар
10,0 -0,35 2,90 10,0 -2,50 1,66
15,6 -0,35 3,46 15,6 -2,50 1,99
21,1 -0,35 4,06 21,1 -2,50 2,23
26,7 -0,35 4,28 26,7 -2,50 2,36
Температура почвы, так же как и температура воздуха, влияет на фотосинтез: при низких температурах интенсивность поглощения СО2 понижается (табл. 5.1). Этот эффект в значительной степени обусловлен уменьшением поглощения воды при низкой температуре, что часто приводит к закрыванию устьиц.
При хорошем снабжении СО^ и высокой интенсивности света нетто-фотосинтез обычно увеличивается с повышением температуры до не-
К>4
Глава S
КО
S 90
Е -
НН fit?
з§ W cj Э §1» 20
О 4 <3 12 16 20 24 28 32 36 40 44 4850
Дни после морозной мчи*
Рис. 5.16. Влияние температуры на фотосинтез, дыхание и нетто (или наблюдаемый)-фотосинтез сеянцев сосны кедровой европейской. Сплошные части кривых - по фактическим измерениям, штриховые - по вычислениям (TranquiUinii, 1955):
1 — истинный фотосинтез; 2 — нетто-синтез; 3 — дыхание
Рис. 5.17. Влияние на фотосинтез сеянцев сосны желтой одной 16-часовой ночи при отрицательных температурах. Растения выдерживали и измеряли фотосинтез при 3°С (Pharis и др., 1970) :
1 — минус 4°С; 2 — минус 8°С; 3 — минус 6°С; 4 — минус 10°С
которой критической точки, выше которой начинает быстро снижаться. Это часто происходит вследствие того, что дыхание продолжает усиливаться при температуре выше критической) когда фотосинтез начинает уменьшаться (рис. 5.16). У альпийского вида сосны кедровой европейской нетто-фотосинтез сеянцев увеличивался до 10—15°С, а затем уменьшался с повышением температуры. Приблизительно при 35°С листья выделяли СО2, что указывало на превышение дыхания над фотосинтезом. 3 противоположность этому, максимальный фотосинтез у пустынного кустарника Ну pt is emoryi наблюдается при температуре около 30°С.
Иногда снижение нетто-фотосинтеза при высокой температуре больше влияет на фотосинтез, чем на дыхание, так как ингибирование фотосинтеза слишком велико, чтобы его можно было объяснить только Усилением темнового дыхания. При высоких температурах может также иметь значение усиленное фотодыхание. Как правило,- температурный оптимум фотосинтеза тропических видов выше, чем у растений умеренной зоны, за исключением некоторых пустынных растений, которые растут при очень высоких температурах. Эти наблюдения наводят на мысль, что фотосинтез до некоторой степени ограничивается вследствие инактиви-рования ферментов при очень высоких температурах. Привыкание фотосинтетического аппарата к высокой температуре, наблюдаемое у некоторых растений, приписывают большей жаростойкости мембран тиллакоидов, вырабатываемых при высоких температурах.
Последствия воздействий температур. Если растение подвергается действию высокой или низкой температуры, то это влияет впоследствии на интенсивность фотосинтеза при другой температуре. Температуры ниже нуля повреждают фотосинтетический аппарат, однако повреждение обычно бывает обратимым после пребывания растения в течение некоторого времени при температурах выше нуля.
Фотосинтез
165
Так, сеянцы дугласии и сосны желтой подвергались Воздействию температур ниже нуля (от -2В * * 11 до - 12°С) в течение нескольких ночей. В результате фотосинтез понизился на величину, которая зависела от числа ночей с отрицательной температурой и от того, насколько холодной была но>я>. У большинства растений фотосинтез становился нормальным при 3°С. Время восстановления зависело от вида растения и от предварительного воздействия низких температур (рис, 5.17). После одной ночи с температурой -4°С фотосинтез сосны желтой полностью восстанавливался в течение 6 дней, у дугласии - только частично через 60 дней. После воздействия температурой от -6° до -8°С фотосинтез сосны возвращался к норме только частично, а после обработки температурой -10°С ни один из двух видов не восстанавливал фотосинтез при возвращении к температуре 3°С. У растений, оправившихся после воздействия морозом и нагревавшихся медленно, фотосинтез впоследствии был более интенсивным, чем у тех, которые нагревались быстро.
°) Ингибирующее влияние на фотосинтез предшествующей обработкой 5 высокой температурой сможет сохраняться в течение многих дней, причем влияние на фотосинтез оказывается гораздо более сильным, чем . на дыхание. Ответная реакция фотосинтеза на предварительное воздействие жарой часто бывает различной у разных видов.
Так, по данным Бауэра (1972), ингибирующее воздействие на фотосинтез наблюдалось при 20°С у пихты белой после нагревания до 38°С, у явора после выдерживания при 42°С. При увеличении температурного стресса степень ингибирования фотосинтеза была выражена сильнее у явора, чем у пихты. Фотосинтез восстанавливался у обоих видов после обработки высокими температурами, если не происходило отмирания тканей. Скорость оправления зависела от предшествующей температуры и была меньше после обработки более высокой температурой. Как только фотосинтетическая способность восстанавливалась при 20°С после одного периода теплового стресса, растения подвергали действию теплового стресса второго периода. При этом фотосинтез оказывался менее чувствительным. Следовательно, первая обработка высокой температурой, по-видимому, жароустойчивость. Результаты воздействия необходимо учитывать при планировании и оценке измерений фотосинтеза.
ДВУОКИСЬ УГЛЕРОДА. Фотосинтез хорошо освещенных деревьев ограничивается низкой концентрацией двуокиси углерода в воздухе, которая повышается приблизительно на 0,0001% в год.
В 1976 г. концентрация СО2 была около 330 частей на миллион, или'0,033% по объему, т.е. значительно выше, чем концентрация 300 миллионных долей, приводившаяся в учебниках несколько десятилетий назад. По-видимому, образование двуокиси углерода при сжигании ископаемого топлива превышает скорость, с которой она может быть использована растениями и поглощена океаном. Концентрация, вероятно, будет и дальше увеличиваться, пока будет продолжаться сжигание больших количеств газа, нефти и угля. Это, по-видимому, может вызвать увеличение интенсивности фотосинтеза, на что указывают эксперименты с повышенной концентрацией двуокиси углерода.
В некоторых районах концентрация СО2 может подниматься значительно выше глобальной средней (330 частей на миллион) вследствие индустриальной деятельности или падать ниже средней из-за потребления и в процессе фотосинтеза. В безветренные дни обычно наблюдаются
суточные колебания содержания двуокиси углерода в воздухе внутри
растительности и над ней с минимумом в послеполуденные часы. Около
поверхности почвы концентрация СО2 часто бывает высокой вследствие ее выделения при гниении, а концентрация в растительном покрове иногда несколько понижается, так как СО2 используется на фотосинтез. В полдень концентрация двуокиси углерода в лесных древостоях может
166
Глава 5
уменьшаться на одну четверть или более, по-видимому, вследствие поглощения двуокиси углерода при фотосинтезе.
В туманные дни, если свет не является ограничивающим фактором, возможен усиленный фотосинтез, так как в такие дни содержание дву -окиси углерода в воздухе может быть выше, чем в ясные дни. Кроме того, туман часто считают благоприятным для роста растений благодаря его положительному влиянию на поддержание листьев в тургесцентном состоянии. Туман способствует, вероятно, более равномерному освещению внутри растительного покрова.
Объемный процент двуокиси углерода в воздухе одинаков на разных высотах над уровнем моря, поэтому часто высказывалось предположение об отсутствии влияния СОп на распределение видов по высоте. Однако, как указывает Декер (1947), диффузия двуокиси углерода в лист является скорее функцией парциального давления этого газа, чем его концентрации. Парциальное давление СО2 в воздухе изменяется прямо пропорционально общему атмосферному давлению. Нормальное давление атмосферной двуокиси углерода составляет на уровне моря около 0,228 мм ртутного столба, а на высоте 4500 м только 0,130 мм. Этот градиент давления СО2 Вполне может быть одним из комплекса факторов, определяющих вертикальную зональность видов.
Независимо от внешней концентрации доступность двуокиси углерода для фотосинтезирующей ткани сильно ограничивают сопротивления на пути ее диффузии внутрь. Эти сопротивления подобны тем, которые влияют на диффузию водяного пара наружу, в том числе сопротивление пограничного слоя воздуха, кутикулы, устьиц и мезофилла. Сопротивление пограничного слоя у хвои большинства хвойных невелико, хотя оно может увеличиваться по направлению к основанию сосновой хвои в плотных пучках. Сопротивление повышается с увеличением размеров листа и уменьшается при возрастании скорости ветра. Типичные значе-
5.2. СОПРОТИВЛЕНИЕ ПЕРЕДВИЖЕНИЮ ВОДЯНОГО ПАРА И СО2 ( Holmgren и др., 1965)
Растение Нетто фото синтез, мг СО'^/дм2/ /ч Сопротивление передвижению, с*см’“1
водяного пара rS со2 г'т
Га rS г с
Populus tremula 12,2 0,57 2,33 3,96 8,2
Betula verrucosa 14,0 0,67 1,36 56 2,31 6,3
Quercus robur 7,5 0,88 7,5 150 12,8 10,6
Acer platanoides 8,8 0,69 4,7 85 8.0 7,3
Helianthus annuus 35,7 0,55 0,38 — 0,65 2,4
Примечания: 1. Опыты проводились при 22°С и интенсивности света 10 или 11-104 эрг-см-2^-1, за исключением Helianthus annuus. опыты с которым проводились при 22°С и интенсивности света 18-Ю4 эог-см-2.с-1.'
2. Буквами обозначены сопротивления: га — пограничного слоя; г $ -устьичное, г с — кутикулярное; г т~ мезофилла.
Фотосинтез
167
ния для листа шириной 1 см составляют 1 и ОД с/см при скорости ветра 10 см/с и 10 м/с, для листа шириной 10 см они соответственно равны 3 и 0,3 с/см. Поверхность кутикулы большинства листьев малопроницаема для двуокиси углерода, вследствие чего устьичное сопротивление становится доминирующим фактором. Отметим, что фотосинтез часто ограничивается закрыванием устьиц, вызываемым водным дефицитом листьев. Сопротивление мезофилла включает ограничения интенсивности фотосинтеза, связанные с диффузией двуокиси углерода в клетки и с биохимическими процессами фиксации углерода. Поэтому сопротивление мезофилла обычно является довольно высоким по сравнению с сопротивлением открытых устьиц (табл. 5.2).
ВОДОСНАБЖЕНИЕ. Наличие воды — важный фактор для фотосинтеза, интенсивность которого уменьшается под влиянием водного дефицита листьев. Как недостаток, так и избыток почвенной влаги могут вызвать водный дефицит листьев, который замедляет фотосинтез в результате сокращения листовой поверхности, закрывания устьиц и уменьшения степени гидратации протоплазмы. По мнению некоторых исследователей, наиболее серьезное последствие засухи — сокращение фотосинтезирующей поверхности из-за ухудшения процесса растяжения листьев и их преждевременного старения, что приводит к уменьшению прироста сухой массы. Имеет также значение и снижение интенсивности фотосинтеза на единицу листовой поверхности. Большое уменьшение интенсивности фотосинтеза на единицу площади листьев у растений, подвергшихся водному стрессу, обычно бывает связано с закрыванием устьиц. Это доказывается одинаковым сокращением как транспирации, так и фотосинтеза.
Влажность почвы и фотосинтез. В течение ряда лет продолжалась полемика относительно влияния влажности почвы на фотосинтез. Некоторые исследователи обнаруживали значительное торможение фотосинтеза уже при небольшом подсыхании орошаемой почвы, другие сообща-
Рис. 5.18. Негго-фотосинтез при высокой интенсивности света и различной влажности почвы;
1 - амбровое дерево (Bormann, 19S3) ; 2 - сосна ладанная (Kozlowski, 1949)
Рис. 5.19. Изменение фотосинтеза листа апельсина при различной температуре листа (Kriedemann, 1968):
1 - во влажном; 2 — в сухом воздухе
Рис. 5.20. Влияние водного потенциала побегов на фотосинтез деревьев дугласии (Brix, 1972)
168
Глава 5
ли об отсутствии заметного торможения фотосинтеза до тех пор, пока не была исчерпана большая часть доступной почвенной влаги. Например, фотосинтез сосны ладанной и дуба белого понижался задолго до наступления влажности устойчивого завядания (рис. 5.18), а подавление фотосинтеза сосны густоцветной происходило при небольшом уменьшении влажности почвы ниже полевой влагоемкости.
Некоторое расхождение во мнениях относительно ответной реакции фотосинтеза на условия водоснабжения объясняется тем, что исследователи часто допускают существование прямой пропорциональности между водным дефицитом растений и водным дефицитом почвы. Однако у деревьев, растущих на сухой почве, может не обнаруживаться высокий внутренний водный дефицит, если атмосферные условия способствуют слабой транспирации. Наоборот, в периоды с высокой температурой и низкой относительной влажностью воздуха у листьев деревьев даже на влажной почве может развиваться сильный водный стресс. Водный стресс растений зависит не только от поглощения, но и от относительной скорости поглощения воды и транспирации (см. главу 13). Следовательно, хотя умеренный водный дефицит листьев часто и понижает интенсивность фотосинтеза, небольшое уменьшение содержания почвенной влаги не всегда вызывает такой же эффект. Значение для фотосинтеза изменений влажности воздуха показано на рис. 5.19. Эти наблюдения подчеркивают важность установления связи ответных реакций фотосинтеза скорее с внутренним водным дефицитом растений, чем с содержанием почвенной влаги.
Водный дефицит растений и фотосинтез. У многих мезофитных видов умеренной зоны интенсивность фотосинтеза начинает уменьшаться, когда листья лишь слегка обезвоживаются. Например, когда водный потенциал листьев (см. главу 12) понижается до минус нескольких бар, фотосинтез прекращается или становится незначительным при нулевом тургоре. Однако между разными видами и генотипами существуют некоторые различия в отношении величины критического водного де-фицита листьев, при котором начинается подавление фотосинтеза.
Интенсивность фотосинтеза сеянцев сосны ладанной снижалась, когда i ' водный потенциал падал ниже —4 бар, а когда он достигал —11 бар, фотосинтез был незначительным. На скорость восстановления поглощения СС>2 после водного стресса сильно влияет корневая система. Вероятно, понижение в одо поглощаю щей или проводящей способности корневой системы во время завядания являлось причиной медленного оправления от водного стресса листьев при возобновлении полива почвы. Фотосинтез винограда очень чувствителен к обезвоживанию листьев, он уменьшается при водном потенциале листьев ниже —5 бар и падает до нуля приблизительно в интеревале от —12 до —15 бар. У 23-летних деревьев дугласии нетто-фотосинтез начинал понижаться при снижении водного потенциала побегов примерно до —10 бар (рис. 5.20); у сосны ладанной при —5 или —6 бар.
Бойер (1976) и другие авторы указывают, что недостаточно выяснено, в какой мере снижение фотосинтеза при водном стрессе растений вызывается закрытием устьиц, а в какой мере уменьшением фотосинтетической способности. Брике (1962) обнаружил, что фотосинтез и транспирация сосны ладанной понижались в одинаковой степени при усилении
Фотосинтез
169
Рис. 5.21. Влияние высыхания почвы на нет-то-фотосинтез сеянцев дугласии (Zavitkovski, l errelL, 1970):
• — первый цикл засухи; б — второй цикл засухи; 1 — полив
влажность почвы, %
а 5
водного стресса, следовательно, оба процесса сокращались вследствие закрытия устьиц.
По данным некоторых исследователей, поглощение СС>2 и устьичной проводимости у сеянцев ели ситхинской быстро снижалось при водном стрессе побегов ниже -12 бар, а при -23 или.-24 бар поглощение СС>2 прекращалось. При этом поглощение СС>2 уменьшалось сильнее проводимости устьиц. Следовательно, можно сделать вывод, что увеличивалось сопротивление мезофилла. Наблюдалось небольшое понижение активности карбоксилазы или активности фотосистем 1 или II, но это могло быть связано с методикой измерения активности. Уменьшалось также содержание хлорофилла а, но не хлорофилла Ъ.
Механизм, при помощи которого растения приспосабливаются к водному стрессу, точно не установлен, но он, вероятно, включает в себя изменения структуры мембран. Если растения, перенесшие засуху, снова полить, то фотосинтез может возвратиться к норме или может не восстановиться в зависимости от вида растения, силы и продолжительности засухи и от сухости воздуха. Так, фотосинтез сосны густоцветной, перенесшей засуху, восстанавливал свою интенсивность до уровня контрольных растений через день после полива. Для* сравнения: фотосинтез сеянцев дугласии, подвергнутых действию засухи, не восстанавливался после полива до значений, наблюдавшихся перед засухой (рис. 5.21). Растениям, перенесшим стресс, обычно требуется больше времени для восстановления фотосинтетической способности после возобновления полива, чем для восстановления интенсивности транспирации. Это указывает на то, что нужно длительное время для того, чтобы протоплазма после восстановления оводненности вновь достигла полной эффективности фотосинтеза. Неспособность растений, перенесших водный стресс, полностью восстанавливать фотосинтетическую активность после полива может также служить признаком длительного повреждения, в том числе нарушения структуры хлоропластов, опадения листьев, повреждения устьиц, отмирания кончиков корней. Часто засушливый период вызывает такое повреждение устьиц, которое подавляет их способность открываться, несмотря на восстановление тургора листьев.
Избыток почвенной влаги, В затопленных почвах избыток гравитационной воды вытесняет воздух из пор. Плохая аэрация затрудняет поглощение воды корнями, вызывая подсыхание листьев (см. главу 13), что приводит к уменьшению фотосинтеза. Обнаружено, что листья затопленных яблонь содержали меньше воды, чем листья контрольных деревьев, росших на незатопленных почвах. Фотосинтез обычно понижался через 2-*7 дней после затопления. Имеются сообщения; что призатоп-
170
Глава 5
лении корней пекана в течение нескольких дней значительно сокращался фотосинтез листьев. После полудня фотосинтез уменьшался сильнее,, чем утром, что, вероятно, было вызвано водным дефицитом. Фотосинтез затопленных деревьев, растущих на песке, понижался до 11% нормы, на более тяжелых почвах при затоплении он прекращался. При удалении избытка воды фотосинтез увеличивался, но в течение нескольких дней не восстанавливался до нормального уровня.
ПЛОДОРОДИЕ ПОЧВЫ. Недостаток необходимых питательных веществ как прямо, так и косвенно влияет на фотосинтез. Непосредственное действие, понижающее фотосинтез, оказывает ослабление синтеза хлорофилла, которое часто наблюдается при недостатке минеральных веществ. Фотосинтез сокращается также вследствие уменьшения листовой поверхности, а возможно, и изменений структуры листьев и понижения активности устьиц. Несомненно влияние плодородия почвы на ферменты и на внутренние процессы в целом. Доказательством служит то, что фотосинтез иногда понижается вследствие недостатка питательных веществ даже при отсутствии таких видимых симптомов, как хлороз или некроз.
Макроэлементы. Полное удобрение NPK значительно усиливает фотосинтез листьев винограда (Коденко, Ерыгина, 1953) и гибридов тополя. Действие удобрений наиболее сильно выражено в случае, если другие факторы (свет, температура) наиболее благоприятны для фотосинтеза. Азот влияет на фотосинтез, действуя на синтез хлорофилла и белков, на размеры листьев и на устьичные реакции. Содержание хлорофилла пропорционально уровню азотного питания, поэтому недостаток азота задерживает фотосинтез.
Келлер и Кох (1962) установили, что поглощение СО2 листьями тополя, содержащими мало (1,8%) или много (3,4%) азота, было почти одинаковым при интенсивности света до 5000 лк. Однако листья, испытывавшие недостаток азота, достигали светового насыщения при 20 000 лк, а при 40 000 лк интенсивность нетто-фотосинтеза этих листьев составляла только 60% соответствующего показателя листьев с высо- ? ким содержанием азота. Размеры листьев и содержание хлорофилла ? также коррелировали с содержанием азота. Подобная взаимозависимость между содержанием азота, хлорофилла и "интенсивностью фотосинтеза наблюдалась у сеянцев сосны и ели, выращенных на различных j питательных растворах. Брике (1971) обнаружил, что причиной влияния Т азота на фотосинтез дугласии частично было увеличение листовой поверхности и частично - повышение его интенсивности. Реакция фотосинтеза на азотные удобрения нередко изменяется в зависимости от формы вносимого азота. Например, интенсивность фотосинтеза молодых сеянцев сосны обыкновенной больше повышалась под влиянием, азотных удобрений в форме хлористого аммония, чем нитрата аммония.
Недостаток фосфора может несколько задерживать фотосинтез вследствие нарушения переноса энергии в системе АДФ - АТФ (аденозиндифосфат — аденозинтрифосфат). Внесение фосфорного удобрения способствовало формированию у яблони крупных листьев с высоким содержанием (%) палисадной ткани. Количество устьиц- на единицу лис- ' товой поверхности уменьшалось, но их регуляция была более эффективной. Минеральное питание влияет на структуру листьев, поэтому имеет
фотосинтез
171
шачение величина, на которую производят пересчет интенсивности фотосинтеза. Недостаток калия так же, как и фосфора, может затрудни и. перенос энергии при фотосинтезе и усиливать дыхание, понижая интенсивность нетто-фотосинтеза. Кроме того, поток ионов калия святи и с устьичными реакциями (см. главу 13). Несмотря на то, что магний входит в состав молекулы хлорофилла и играет роль активатора мно-1их ферментных систем, умеренный недостаток этого элемента сущест-нспно не влиял ни на фотосинтез, ни на рост тополя.
Микроэлементы. Недостаток железа тормозит фотосинтез, вызы-ппч хлороз и влияя на активность ферментов. Железо содержится в ферредоксине и в цитохромах — важных компонентах электротранспортных систем фотосинтеза и дыхания. Содержание хлорофилла в листьях удобренных растений. тополя взаимосвязано с содержанием железа. Фотосинтез при всех интенсивностях света постоянно был выше у удобренных растений по сравнению с неудобренными. У растений, испытывающих недостаток железа, уменьшалось поглощение СС>2 на единицу листовой поверхности. Общее поглощение СО? также понижалось вследствие сокращения размера листьев (табл. 5.3).
S.3. ВЛИЯНИЕ УДОБРЕНИЯ ХЕЛАТОМ ЖЕЛЕЗА НА ФОТОСИНТЕЗ Populus euramericana marilandica* (Keller, Koch, 1964)
Интенсивность света, лк
Фотосинтез, мг СС^/дм^/ч
Контроль Удобрено
Увеличение, %
500 0,96 1,26 31,0
2500 1,98 . 2,94 48,0
5000 3,60 5,03 40,0
10000 5,86 9,34 59,0
20000 7,92 13,57 71,0
40000 10,09 14,72 45,9
* Выращенные в сосудах растения были политы в конце июня 0,2%-ным раствором Хелата железа. Фотосинтез ^измеряли в конце августа — начале сентября.
Марганец необходим для освобождения кислорода, он может также служить активатором других ферментных систем, влиять на фотосинтез, если содержится в слишком малых или избыточных количествах. Однако установлено, что недостаток марганца сокращает поглощение СО2 листьями тунга в большей степени вследствие уменьшения площади листьев, чем из-за понижения эффективности фотосинтеза на единицу листовой поверхности. Листья, у которых наблюдался частичный хлороз, имели несколько меньшую продуктивность, чем листья, восстановившие нормальную зеленую окраску после применения сульфата марганца. Резкий недостаток меди и цинка понижал фотосинтез тунга соответственно на 30 и 53%, не вызывая таких видимых симптомов, как хлороз или некроз. Медь входит в состав пластоцианина. Кроме то-10, цинк и медь являются кофакторами ферментов, участвующих в носстановлении нитратов.
172
Глава 5
ВЕЩЕСТВА, ЗАГРЯЗНЯЮЩИЕ ВОЗДУХ, И ПРИМЕНЯЕМЫЕ ХИМИЧЕСКИЕ ПРЕПАРАТЫ. Вещества, загрязняющие атмосферу, и разнооб- j разные химические препараты, используемые для борьбы с грибными j
болезнями, вредными насекомыми и сорняками и для уменьшения £
транспирации, часто заметно понижают фотосинтез. Действие этих соеди- J
нений на фотосинтез проявляется: 1) в засорении устьиц и задержке |
поглощения СО2; 2) в изменении оптических свойств листьев вследст- Г вие того, что изменяется отражение и уменьшается проникновение све- •
та; 3) в изменении теплового баланса листьев; 4) в изменении мета- £
болизма листьев; 5) в изменении анатомического строения листьев; |
6) в различных комбинациях перечисленных явлений. Кроме того, фотосинтез понижается под влиянием химических препаратов, вызываю- „ щих повреждение, хлороз, побурение листьев или листопад.
Вещества, загрязняющие воздух. Действие загрязняющих воздух ; веществ на лесные деревья и плодовые сады особенно проявляется вблизи крупных заводов, так же как и их влияние на теневыносливые и декоративные деревья в больших городах <см. главу 17). Целый ряд веществ, загрязняющих воздух, подавляет фотосинтез. У загрязняющих веществ часто наблюдается синергизм.. При этом даже переносимый растениями уровень отдельного вещества может повреждать их в присутствии даже небольших количеств другого загрязняющего, агента. Наибольшее значение для фотосинтеза имеют такие загрязняющие воздух вещества: окислы серы, озон, фториды, пероксиацилнитраты, окис-лы азота и взвешенные частицы.
Приводим несколько примеров действия загрязняющих воздух веществ на фотосинтез, полученных различными исследователями. Двуокись серы. После окуривания еловых веток SO2 (концентрации 1 часть на миллион) у них через несколько часов понизился фотосинтез. Фотосинтез восстанавливался после прекращения окуривания, но если окуривание возобновлялось, то поглощение СО2 падало до еще более низкого уровня. Окуривание сеянцев сосны SO2 той же концентрации также уменьшало фотосинтез. При невидимых повреждениях листьев фотосинтез быстро восстанавливался. При отсутствии видимых повреждений хвои ’ у ели европейской и сосны швейцарской горной, подвергнутых одночасовому окуриванию SO2 (концентрации 10-20 частей на миллион), фотосинтез понижался, а при явных признаках сильного повреждения фотосинтез уменьшался до очень низкого уровня или прекращался. Кратковременное окуривание сеянцев ясеня американского и корневых отпрысков тополя осинообразного SO2 (концентрации 1 часть на миллион или выше) в течение 2 ч понижало у обоих видов фотосинтез, дыхание не нарушалось. Повреждения, вызываемые SO2, обычны в лесах, расположенных даже за много километров от электростанций, сжигающих большие количества угля с высоким содержанием серы.
Озон. Концентрация озбна в воздухе городских районов обычно поднимается выше нормы в солнечные дни вследствие фотоокисления веществ автомобильных выхлопных газов и летучих промышленных отходов. В городских районах, где концентрация озона возрастает выше 0,1 части на миллион, встречаются широко распространенные повреждения травянистой и древесной растительности. У сеянцев сосен Эллиота, поздней, веймутовой и ладанной, подвергнутых воздей-сгвию озона (от 0,05 до 0,16 миллионных долей) в течение нескольких недель, фотосинтез понижался, а дыхание увеличивалось. Интенсивность фотосинтеза сок-ращалась под влиянием концентраций слишком низких для того, чтобы вызывать % видимое повреждение хвои. В других экспериментах обнаруживалось, что у ветвей 5-летнеЙ сосны веймутовой, последовательно подвергнутых воздействию доз 0,5-0,8 части на миллион в течение 4 ч и 0,8-0,9 части на миллион в течение 3 ч, фотосинтез понижался незначительно, а повторная обработка дозой 0,9-1 мил- к лионную долю в течение 5 и 3 ч уменьшала фотосинтез пнчти на 80%. Установлено £
значительное колебание чувствительности среди отдельных молодых деревьев ?
Фотосинтез
173
сосны веймутовой. С учетом уменьшения фотосинтеза выше порога 0,5 частей на миллион деревья были разделены на три группы по их чувствительности к озону. У чувствительных деревьев после обработки их дозой 0,9-1 миллионную долю в течение 10 ч прекращался фотосинтез. У особей со средней чувствительностью зга концентрация понижала фотосинтез примерно наполовину. У устойчивых деревьев фотосинтез не изменялся.
Фториды. Растения повреждаются фторидами обычно вблизи рудоплавильных и алюминиевых предприятий и заводов, производящих удобрения, кирпич и керамику. Растения очень чувствительны к фтору. Так при воздействии в течение нескольких часов концентрацией 0,02-0,05 части на миллион у плодовых деревьев уменьшается фотосинтез. Замедление фотосинтеза связано с пониженным синтезом флорофилла, ослаблением реакции Хилла и повреждением тканей листа.
Взвешенные частицы. Такие вещества, как пыль цементных печей, некоторые фториды, сажа, окись магния, окись железа, литейная пыль и аэрозоли серной кислоты, закупоривая устьичные щели, затеняя листья или влияя на метаболизм клеток листа, могут задерживать фотосинтез. Чистые листья Ulmus pumila, Tilia cordata увеличивали сухую массу за 6 дней на 40% больше, чем листья, покрытые сажей (Ершов, 1957). Пылевидные фтористые соединения (Na; F, Са, г<2 и криолит) сильно ингибировали фотосинтез сосны обыкновенной и березы бородавчатой, нередко без видимых повреждений.
Ядерное излучение. Радиоактивные осадки от ядерных взрывов и отходы ядерных реакторов излучают ионизирующую радиацию, которая оказывает значительное .подавляющее влияние на фотосинтез. Особенно чувствительны древесные растения. Хроническое облучение часто не снижает фотосинтез в течение длительного времени после первоначальной экспозиции, тогда как сильная радиация действует немедленно. При хроническом облучении сосны жесткой ее фотосинтез понижался в течение зимы я восстанавливался весной. Ингибирующий эффект не был заметен после месячной экспозиции по 1300 рентген/сут или после 5 месяцев воздействия 30 рентген/сут. Для сравнения: сильное облучение деревьев сосны ладанной дозой всего лишь 1250 рентген, которая была дана по 8000 рентген/ч, быстро понижало поглощение СО2- Через 21 день степень депрессии была пропорциональна дозе облучения, причем наивысшая доза (равная в сумме 13 200 рентген) сокращала интенсивность фотосинтеза на 80%. Обнаружено, что водный стресс во .время облучения повышал степень повреждения сосны ладанной.
При использовании радиоактивных изотопов в физиологических исследованиях на растения воздействует потенциальный внутренний источник радиации, которая при критических дозах может быть токсичной для растений. Это было продемонстрировано Урсино (1973), подвергавшего сеянцы сосны веймутовой действию в течение 1 ч разной активности. Через 2,5 месяца после обработки сеянцы, которые содержали ПО микрокюри 14СС>2 или больше, имели соответственно более низкую интенсивность фотосинтеза и более короткую хвою, чем контрольные растения.
Применяемые химические препараты. В прошлом для обработки деревьев применяли большие количества фунгицидов и инсектицидов без учета их физиологического действия. Так поступали, пока у растений не возникали явные признаки повреждений. В последние годы стало ясно, что некоторые пестициды оказывают физиологическое действие, не обнаруживаемое невооруженным глазом.
Инсектициды могут понижать эффективность фотосинтеза. При опрыскивании нижней поверхности листьев он уменьшается зна
174
Глава 5
чительнее, чем при опрыскивании верхней поверхности. Причиной снижения фотосинтеза может быть засорение устьиц, нарушения метаболизма или одновременно и то и другое. Маслянистые инсектициды подавляют фотосинтез на очень длительный срок. Например, по данным Уед-динга (1952), фотосинтез цитрусовых был подавлен в течение 2 месяцев после применения бензиномасляных эмульсий. Некоторые инсектициды не на масляной основе, например метасистокс (0,1%), паратион (0,02%) и др., подавляли фотосинтез винограда только в течение 1-2 дней. Длительное действие диметоата несколько ослабляло фотосинтез тополя в течение 2—4 дней. Линдан за 1 ч понижал фотосинтез на 60%, но ингибирование сохранялось только в течение 1 дня. Фосфатные инсектициды диазинон, гутион и этион уменьшали фотосинтез! листьев яблони Рэд Дилишес. Максимальное ингибирование наблюдалось сразу после обработки, а затем понижалось, но оставалось еще значительным и через 14 дней.
Фунгициды могут задерживать фотосинтез, уменьшать интенсивность света, падающего на лист, закупоривать устьица или влиять на метаболизм. Опрыскивание нормальными дозами фунгицидов может понизить освещенность листьев* винограда на 200-500 лк, а 1,5%-ная известково-серная эмульсия или чрезмерно высокие дозы других фунгицидов могут уменьшить интенсивность света на 1000 лк. Бордоская жидкость действует скорее физиологически, чем механически, причем уменьшение фотосинтеза вызывает фракция растворимой меди. Серные фунгициды заметно снижают фотосинтез. Опрыскивание яблонь сорта Болдуин известково-серной эмульсией уменьшало фотосинтез приблизительно наполовину в течение первых 5 дней после применения. Не все фунгициды уменьшают фотосинтез. По данным Бэрнера (1961), органические фунгициды (зирам, зинеб, каптан и фалтан) слегка повышали фотосинтез тополя, , тогда как неорганический фунгицид оксихлорид меди понижал его.
Гербициды. Многие из них тормозят фотосинтез, нередко вследствие ингибирования реакции Хилла. Наблюдаемый эффект во многом зависит от вида и разновидности растения, от химической природы гербицида и его дозы, от срока применения, возраста растения, времени года, фазы роста растения, типа почвы, погоды и других факторов. Сеянцы в фазе семядольных листьев особенно чувствительны к гербицидам (см. главу 14). Ингибирующее действие различно даже среди химических препаратов с родственной структурой. Например, фотосинтез молодых сеянцев подавляли некоторые триазиновые гербициды, но симазин и атразин действовали гораздо сильнее, чем пропазин или ипазин.
Имеет значение и способ применения гербицида, так как его влияние на фотосинтез зависит от количества гербицида, поглощенного растениями. Фактическую токсичность гербицидов для растений нельзя определить путем их внесения в почву, так как они часто теряются в различной степени вследствие испарения, выщелачивания, микробиологического или химического разложения и необратимой адсорбции почвой. Ингибирующее влияние на фотосинтез поступающих в продажу гербицидов может вызываться активными или неактивными ингредиентами, а также синергистическим действием тех и других.
Фотосинтез
175
Фрелих (1961) установил, что опрыскивание деревьев ели и березы в течение 30 мин 3%-ным 2,4,5-Т полностью подавляло фотосинтез и вызывало резкое усиление дыхания, которое продолжалось, несколько дней или до тех пор, пока растение не отмирало. Уединнг и др. (1954) пришли к заключению, что ингибирование фотосинтеза деревьев апельсина 2,4-Д связано с накоплением недиссоциированных молекул 2,4-Д кислоты.
А н тит р ан спир ант ы. Имеется много данных о положительных результатах применения антитран спирантов. Например, плеяЬчные антитранспиранты улучшали водный баланс саженцев цитрусовых, усиливали образование плодов у укоренившихся деревьев маслины и персика, понижали транспирацию декоративных растений.
Необходимо учитывать, что пленочные антитранспиранты более проницаемы для водяного пара, чем для СО2> поэтому полное покрытие листьев пленкой антитранспиранта сокращает поглощение СС>2 сильнее, чем транспирационную потерю воды. Влияние антитранспирантов на. фотосинтез зависит от их химического состава и используемой дозы, от вида растения, способа применения и от условий внешней среды.
Дэвис и Козловский (1975) установили, что некоторые пленочные антитранспиранты сильно понижали фотосинтез сеянцев ясеня американского и сосны смолистой в течение длительного времени. Наблюдались значительные различия в продолжительности ингибирующего эффекта, когда разные соединения переставали действовать или расщеплялись, но некоторые антитранспиранты понижали поглощение СО^ по крайней мере в течение 32 дней (длительности эксперимента). Нередко после прекращения прямого физического действия соединения выявлялась его физиологическая токсичность. Ингибирование фотосинтеза выражалось гораздо сильнее, если листья погружали в растворы антитранспирантов, а не опрыскивали их. Строение листьев и особенно степень закупорки устьиц зернами кутикулярного воска у некоторых видов влияет на эффективность антитранспирантов. Например, фотосинтез и транспирация понижались под влиянием пленочных антитранспирантов в большей степени у клена сахарного, имеющего мелкие устьица, закупоренные зернами воска, чем у ясеня американского с крупными устьицами, не закрытыми воском. Антитранспиранты уменьшали фотосинтез сосны смолистой на 90%, образуя в сочетании с воском в устьичных отверстиях непроницаемые пробки. Закупорка устьиц у покрытосеменных и голосеменных сопровождалась изменением метаболизма, хлорозом, побурением и уменьшением роста листьев. Фитотоксичность иногда обнаруживалась вскоре после обработки, в других случаях — в более поздние сроки. Установлено, что некоторые пленочные антитранспиранты не только понижали фотосинтез 3-летних растений сосны смолистой в лесном питомнике, но и уменьшали скорость превращения первичных продуктов фотосинтеза в нерастворимые в этаноле соединения, после того как обработанные сеянцы были помещены в атмосферу с 14сО2-
Эксперименты с метаболическими антитранспирантами давали различные результаты. Атразин, фенилмеркурацетат и янтарная кислота оказались фитотоксичными, абсцизовая кислота (АБК) этим свой
176
Глава 5
ством не обладала. АБК, вызывая закрывание устьиц, быстро понижала фотосинтез. При восстановлении тургора устьица постепенно вновь открывались и поглощение СС>2 обработанными растениями становилось интенсивнее, чем у контрольных растений.
БОЛЕЗНИ. У больных деревьев фотосинтез подавляется вследствие утраты фотосинтезирующих тканей, повреждения хлоропластов, ингибирования синтеза хлорофилла, изменения метаболизма или совместного действия этих факторов. Слабый рост листьев, некротические повреждения, пятнистость листьев, потеря листвы — все эти симптомы болезней уменьшают синтез углеводов в результате сокращения листовой поверхности, а также понижения эффективности действия остающихся фотосинтезирующих тканей. Фотосинтез листьев карии, пораженной ложной пятнистостью, понижался сильнее, чем это можно было ожидать на основании одного только уменьшения поверхности фотосинтезирующих тканей. Вирусные болезни часто вызывают хлороз и разрушение хлоропластов. Деккер и Тио (1958) установили, что интенсивность фотосинтеза листьев дынного дерева с симптомами мозаичной болезни, вызываемой вирусом, составляла только 36% интенсивности фотосинтеза здоровых листьев. Некоторые вирусные болезни, например ”фони” персика, ингибируют фотосинтез, не влияя непосредственно на содержание хлорофилла.
Некоторые болезни сильно понижают фотосинтез из-за нарушения водообмена. Например, вследствие заболевания корней ослабляется поглощение воды, что приводит к закрыванию устьиц и уменьшению поглощения СО2 листьями. У деревьев, зараженных сосудистыми патогенами, фотосинтез тормозится из-за уменьшения содержания хлорофилла, высыхания листьев, опадения листьев и закрывания устьиц. Интенсивность фотосинтеза часто уменьшается до того, как происходит завя-дание. Фотосинтез быстро понижался у сеянцев ильма американского, инокулиров энных возбудителем голландской болезни (Ceratocysris ulmi) или подвергнутых действию засухи.
ВНУТРЕННИЕ ФАКТОРЫ. Важнейшими внутренними факторами, влияющими на фотосинтез, являются возраст и структура листьев, размер, количество и реакция устьиц, содержание хлорофилла, внутренний водный дефицит, использование и накопление углеводов. У очень молодых листьев интенсивность фотосинтеза невелика, затем она увеличивается до определенного критического уровня зрелости (обычно до полного развертывания листьев), в дальнейшем с увеличением возраста понижается. Изменения интенсивности фотосинтеза листьев голосеменных и вечнозеленых покрытосеменных сходны. Интенсивность фотосин-
Рис. 5.22. Влияние возраста хвои голосеменных на нетто-фотосинтез (Freeland, 1952): 1 — сосна черная австрийская; 2 — сосна обыкновенная; 3 — сосна веймутова; 4 — сосна желтая; 5 — пихта одноцветная; б — ель колючая
фотосинтез
177
теза хвои сосны обычно возрастает до достижения ею максимальных размеров, а затем уменьшается, Интенсивность фотосинтеза однолетней или более старой хвои постепенно понижается с увеличением ее возраста (рис. 5.22).
Возрастные изменения интенсивности фотосинтеза связаны с анатомическими и физиологическими изменениями листьев. Увеличение листьев в течение фазы растяжения связано с развитием внутренней структуры листьев и устьиц, синтезом хлорофилла, понижением диффузионного сопротивления, повышением интенсивности фотофосфорилирования и синтеза белков, увеличением активности РуДФ-карбоксилазы и резким уменьшением митохондриального дыхания. Постепенное понижение фотосинтеза после окончания развертывания листьев связано с увеличением диффузионного сопротивления, уменьшением синтетической активности фосфорилирующей системы хлоропластов, содержания белков, РНК и ДНК и активности РуДФ-карбоксилазы. Кроме того, наблюдается увеличение фотодыхания до сравнительно высокого уровня и заметное понижение митохондриального дыхания.
Изменения фотосинтетической способности листьев разного возраста у голосеменных имеют существенное значение для соотношения между оттоком метаболитов и их потреблением в растущих тканях. Например, в одном из экспериментов, проведенных Дикманном и Козловским (1968), быстрорастущая хвоя сосны смолистой имела такую высокую потребность в углеводах, что не могла обеспечить достаточным количеством продуктов фотосинтеза свой собственный рост. В начале вегетационного периода она получала большие количества ассимилятов от однолетней хвои, у которой интенсивность фотосинтеза была выше, чем у 2-3-летней хвои. Снабжение новых побегов продуктами текущего фотосинтеза преимущественно из старой хвои уменьшалось в более поздние сроки, после достижения новой хвоей максимальных размеров. К этому времени у новой хвои была наивысшая интенсивность фотосинтеза и она заменила старую хвою в качестве главного поставщика углеводов. По данным Кларка (1961), вскоре после распускания почек новая хвоя пихты бальзамической выделяла больше СО^ при дыхании, чем поглощала при фотосинтезе. К середине июня у новой хвои не наблюдалось ни поглощения, ни выделения СО2, но до конца августа фотосинтез новой хвои не был таким высоким, как у однолетней хвои.
Устьица. Сопротивление, оказываемое устьицами поглощению СС>2 листьями, часто является главным ограничивающим фактором фотосинтеза. Поэтому различиями в размерах устьиц, в густоте их расположения или в регуляции степени открытости устьиц можно объяснить различия устьичного диффузионного сопротивления, а следовательно, и фотосинтеза.
Значение устьичной регуляции проявляется в одновременном сокращении степени открытости устьиц и фотосинтеза при усиливающейся засухе и в полуденной депрессии фотосинтеза в течение периодов временного закрывания устьиц. Кридеман (1971) обнаружил тесную связь между диффузионным сопротивлением листьев (мерой степени открытости устьиц) и интенсивностью поглощения СО2 листьями цитрусовых на высыхающей почве (рис. 5.23). Было показано также, что большие различия в интенсивности фотосинтеза у разных видов связаны с изме
178
Глава 5
нениями сопротивления устьиц диффузии СО2- Одновременное уменьшение интенсивности транспирации и фотосинтеза с усилением водного стресса также указывает на то, что фотосинтез ограничивается диффузионным сопротивлением при поступлении СО2 в значительной степени, в результате закрывания устьиц. По мере развития водного стресса, вызываемого засухой, фотосинтез сначала понижается вследствие закрывания устьиц, а прямое ингибирующее действие на фотосинтетический аппарат не проявляется, пока обезвоживание не станет гораздо более сильным.
Содержание хлорофилла. Фотосинтетическая способность листьев с ненормальным бледно-зеленым цветом обычно ниже, чем у листьев со здоровой, темно-зеленой окраской. О значении содержания хлорофилла' для фотосинтеза свидетельствует уменьшение количества хлорофилла и фотосинтеза у листьев, испытывающих недостаток минеральных веществ, а также у стареющих листьев. В контролируемых условиях часто обнаруживается четкая корреляция между содержанием хлорофилла и поглощением СО2 (рис. 5.24). Однако в полевых условиях интенсивность фотосинтеза может не подвергаться большим изменениям при значительных колебаниях окраски листьев. По-видимому, содержание хлорофилла часто имеет меньшее значение, чем другие факторы, влияющие на интенсивность фотосинтеза. Организация хлорофилла с точки зрения числа и размеров фотосинтетических единиц может быть такой же важной, как и его количество. Сильный хлороз, какой бы причиной он ни вызывался, неизменно сопровождается пониженным фотосинтезом.
Зимнее снижение фотосинтеза часто бывает связано с дезорганизацией хлоропластов и распадом хлорофилла. Однако существуют, по-видимому, значительные различия между деревьями, даже принадлежащими
Рис. 5.23. Влияние диффузионного сопротивления листа (степени открытости устьиц) на фотосинтез листьев апельсина (Kriedemann, 1971)
Рис. 5.24. Связь между содержанием хлорофилла и фотосинтезом (КеПеги
Wehrmann, 1963):
Ь — сосны обыкновенной; 2 — ели европейской
Фотосинтез
179
к одному виду, в отношении степени дезорганизации фотосинтетического аппарата в течение зимы. Как содержание хлорофилла, так и интенсивность фотосинтеза сеянцев сосны кедровой европейской в альпийском местообитании уменьшались в течение зимы и увеличивались весной.
Скорость роста. Связь между ростом и фотосинтезом сложна. Скорост роста растений обычно зависит от притока продуктов фотосинтеза. Однако не следует думать, что повышение интенсивности фотосинтеза автоматически будет приводить к усилению роста. Часто предполагают, что растения с высокой интенсивностью фотосинтеза на единицу листовой поверхности будут вырабатывать больше сухого вещества и расти быстрее, чем растения с низкой интенсивностью фотосинтеза. Это иногда верно, но не всегда. Например, Ледиг и Перри (1967) измеряли интенсивность фотосинтеза 1 & семей сосны ладанной и обнаружили, что хотя наиболее быстрорастущие семьи имели наивысшую интенсивность фотосинтеза, корреляция между поглощением СОп и ростом была несколько отрицательной для всех 18 семей, картер (1972) ссылается на ряд других исследователей,, которые обнаружили слабую корреляцию или ее отсутствие между поглощением СО£ на единицу листовой поверхности и ростом. Хелмс (1976) также анализирует трудности, возникающие при определении соотношения между ростом и интенсивностью фотосинтеза. Эти затруднения встречаются также и в отношении травянистых растений. Слабую корреляцию можно отчасти объяснить тем, что измерение фотосинтеза обычно проводят только несколько раз за вегетационный период, тогда как накопление сухого вещества (рост) представляет собой произведение интенсивности фотосинтеза, площади листьев и продолжительности фотосинтеза, причем все три показателя изменяются в течение вегетационного периода.
Вент (1958) высказал предположение, что при определенных условиях скорость роста может непосредственно влиять на интенсивность фотосинтеза, изменяя скорость передвижения ассимилятов из клеток листа. В пользу этой точки зрения имеется немало оснований. Таким образом, можно задать вопрос, в какой степени интенсивность фотосинтеза определяет скорость роста и в какой мере скорость роста регулирует интенсивность фотосинтеза? Существуют доказательства того, что имеются обе ситуации, причем последняя встречается наиболее часто при благоприятных для фотосинтеза условиях. Для ряда видов древесных и травянистых растений было показано, что даже при оптимальных условиях интенсивность фотосинтеза обычно оказывается гораздо ниже максимально возможной, и это происходит за счет того, чтб потребность в продуктах фотосинтеза меньше максимальной. Из этого еле дует, что механизм фотосинтеза у растений часто несет нагрузку значительно ниже его максимальной способности.
Интенсивность фотосинтеза экспериментально регулировали, изменяя отношение между потреблением и оттоком углеводов. При удалении части листовой поверхности яблони изменялся характер транспорта углеводов, фотосинтез оставшихся листьев усиливался, так как они снабжали большее количество потребителей притекающих углеводов. Частичная дефолиация сосны замечательной сопровождалась заметным повышением фотосинтеза оставшихся листьев даже при насыщающей интенсивности света. Интенсивность фотосинтеза увеличивалась также при удалении части корневой системы. Этот прием стимулирует образование многочисленных новых корней, что повышает потребность в продуктах фотосинтеза.
180
Глава 5
Удаление половины корневой системы у молодых яблонь увеличивало нетто-ас-симилицию на 17%. Уменьшение количества потребителей притекающих ассимиля-тов путем удаления цветков или плодов также приводит к понижению фотосинтеза яблони, а плодоношение часто сопровождается увеличением интенсивности фотосинтеза. Если предотвратить быстрый отток продуктов фотосинтеза из листьев, то это также понижает фотосинтез, Нетто-фотосинтез листочков карии значительно сокращался через 1—2 дня после кольцевания ветвей, вследствие чего предотвращалось передвижение углеводов. Пониженный фотосинтез все еще наблюдался через 50 дней после окольцовывания ветвей. Кроме того, при кольцевании ветвей усиливалось дыхание листьев, так что наблюдаемое уменьшение нетто-фото синтез а отражало, no-видимому, не только торможение фотосинтеза, но и более интенсивное дыхание.
Уоринг и другие исследователи (1968) изучали механизм, изменяющий фотосинтетическую активность листьев при изменении отношения между оттоком и потреблением органических веществ. При частичной дефолиации деревьев оставшиеся листья приобретали более интенсивную зеленую окраску и становились крупнее. Кроме того, повышалось общее содержание белков (табл. 5.4), а также уровень карбоксилирующих ферментов. Авторы сделали вывод, что усиление фотосинтеза после частичной дефолиации являлось следствием увеличения количества карбоксилаз. Последнее происходило за счет усиления синтеза белков, вызванного увеличением поступления цитокининов из корней в оставшиеся листья. Это вывод подкреплялся тем, что удаление части корней предотвращало усиление фотосинтеза листьев, оставшихся после частичной дефолиации. Данный вывод основан на предположении, что фотосинтез обычно ограничивается количеством РуДФ-карбоксила-зы. Это предположение некоторые исследователи подвергают сомнению.
5.4. ВЛИЯНИЕ ЧАСТИЧНОЙ ДЕФОЛИАЦИИ НА СОДЕРЖАНИЕ БЕЛКОВ В ЛИСТЬЯХ
Salix viminalis
Вариант опыта Содержание белков, мг/г сырой массы
0 Дни после обработки
5 10 15
Контроль 0,49 0,55 0,72 0,88
Частичная дефолиация 0,49 0,75 0,98 1,15
Результаты этих и других экспериментов показывают, что между ростом и фотосинтезом существуют сложные взаимодействия и что интенсивность фотосинтеза может повышаться или понижаться при изменении скорости роста. На фотосинтез могут влиять изменения отношения между и оттоком и потреблением углеводов, или гормональные взаимодействия, или и го и другое. Торн и Келлер (1974) считают, что углеводы как регуляторы фотосинтеза играют менее важную роль, чем гормоны. Эту точку зрения поддерживают Уоринг и другие исследователи (1968).
ФЕРМЕНТЫ, ЭНЕРГЕТИКА И ДЫХАНИЕ
К числу важных процессов, происходящих в живых организмах, относятся освобождение при дыхании запасенной в питательных веществах химической энергии и ее использование для превращения углеводов, жиров и белков в новую протоплазму и новые ткани, т.е. ассимиляция. Понимание того, как происходят эти сложные процессы, требует, по меньшей мере, элементарных представлений об активности ферментов и переносе энергии.
ФЕРМЕНТЫ И ЭНЕРГЕТИКА
ФЕРМЕНТЫ. Одна из наиболее важных особенностей живых клеток -высокая скорость, с которой протекают в них химические реакции при температурах от 5 до 40°С. Эти же реакции в лабораторных условиях при тех же температурах если и происходят, то очень медленно. Например, древесина, уголь и другие виды топлива не загораются, пока их не нагреют до критической температуры, после чего они горят самопроизвольно. Даже глюкоза должна быть нагрета до высокой температуры, чтобы гореть (окисляться) на воздухе, а в живых клетках она легко окисляется при 5- 10°С. Это происходит потому, что большинство химических реакций, даже приводящих к освобождению энергии, не идет самопроизвольно, для их начала требуется введение дополнительной порции энергии — энергии активации. Ферменты — это органические катализаторы, понижающие энергию активации до значения, при котором реакция может протекать при обычных температурах. Это достигается благодаря кратковременному связыванию молекул субстратов на поверхности молекул ферментов, что увеличивает вероятность протекания реакции. Большинство ферментов очень специфично и катализирует только одну реакцию или один тип реакции. Специфичность обусловлена тем, что ферменты являются белковыми молекулами со специфической структурной конфигурацией, которая допускает связывание молекул субстрата с определенной структурой. Пока молекулы субстрата временно связаны на поверхности фермента, происходит перегруппировка атомов и химических связей, приводящая к образованию иных молекул, часто с другой свободной энергией. Многие ферменты катализируют обратимые реакции, которые идут в ту или иную сторону в зависимости от концентрации реагирующих веществ, pH и других факторов. Иногда ферменты существуют в нескольких молекулярных формах, действующих на один и тот же субстрат и известных под названием изоэнзимов.
В живых клетках имеются сотни разнообразных ферментов, многие из которых строго локализованы в различных органеллах. Например, ферменты, участвующие в цикле Кребса, содержатся в митохондриях, участвующие в переносе электронов — в митохондриях и в хлоропластах. Ферменты, участвующие в гликолизе и пентбзо фосфатном цикле (шунте), отмечаются преимущественно в цитоплазме. Некоторые экстрацеллюлярные ферменты встречаются даже на внешней поверхности клеток или диффундируют в окружающую среду.
182
Глава 6
Основу структуры фермента составляет молекула белка. Ферменты различаются по разной последовательности аминокислот в этих белках. Некоторые ферменты, такие, как уреаза и папаин, состоят только из одной молекулы белка, но многие нуждаются в небелковом компоненте, часто именуемом кофактором или коферментом, который тесно ассоциирован или связан с белковой молекулой. Как неотъемлемая часть фермента кофакторы, такие, как медь в тирозиназе и аскорбаток-сидазе, или железо в каталазе, называются простетическими группами. Многие -ферменты активны только в присутствии иолов, например, Mg2+, Мп2+, Са2+ и К+. Эти ионы известны как металлы-активаторы. Значение микроэлементов для растений и животных заключается прежде всего в том, что они служат простетическими группами или металлами-актираторами ферментов. Некоторые ферменты активны только в присутствии сложных органических молекул, из которых прочно связанные называются простетическими группами, а связанные слабо — кофакторами или коферментами.
Отдельные витамины, особенно группы В, играют важную роль в ка- > честве кофакторов^ ферментов. Так, пиридиновые нуклеотиды очень I важны как кофакторы ферментов, участвующих в метаболических пе- 1 реносах энергии. НАД (ник отинамидаденин динуклеотид) и НАДФ f (никотинамидадениндинуклеотидфосфат) — необходимые коферменты s ферментов окислительно-восстановительных систем живых клеток — !
дыхания и фотосинтеза. В своей восстановленной форме, НАД-Н2 или НАДФ Н2, они являются высокоэнергетическими соединениями, служащими всстановителями в таких процессах, как транспорт электронов « и восстановление углерода при фотосинтезе. Коферментами двух других , важных ферментов, участвующих в окислительно-восстановительных реакциях, являются флавиновые нуклеотиды ФМН (флавинмононуклеотид) и ФАД (флавинадениндинуклеотид) — производные витамина В2, рибофлавина.
Тиаминпирофосфат образуется из витамина В| (тиамина) и служит коферментом различных декарбоксилаз, оксидаз и транскетолаз. Пиридоксаль, пиридоксин и пиридоксамин составляют комплекс витамина Bg, от которого происходит пиридоксальфосфат - важный участник синтеза аминокислот. Другой витамин, пантотеновая кислота, является предшественником кофермента А, играющего важную роль в метаболизме (см. рис. 6.1 и 6.20). Таким образом, некоторые витамины необходимы вследствие их роли как коферментов в важных реакциях метаболизма.
Набор ферментов, вырабатываемых растениями, определяется прежде всего его генотипом. Иногда нарушения обмена веществ вызываются мутациями генов, элиминирующими специфические ферменты. Вне ядра также существует некоторый генетический контроль, особенно в хлоропластах. К наиболее важным в растениях относятся мутации, из-за которых возникают нарушения образования хлорофилла, но имеются и многие другие. Часто они приводят к гибели проростков и потому проходят незамеченными. Изучение мутаций дает обширную информацию о роли ферментов в метаболизме. Регуляция активности ферментов сложна и не до конца понята, она включает обратную связь посред-
Ферменты, энергетика и дыхание
183
Крахмал tl глюкоза - 1-фоСфат
6 - сросфоглюконат^. глюкозо-б-фосфаггг
.глюкоза
#
СО?
)ЗО- 6-фоссрат
пентоэофосфаты /дфуктозо~1,6-дисросфат
Промежуточные ^З-фосфоглицерино&ый соединения альдегид
Фотосинтез
2>
АТФ
АДФ
+ 2НАД Н2
1,3 - дифосфоглиирриновая кислота _
|| х------АТФ
3~ фосфо глицериновая кислота
У
2 ~ фосфо глицериновая кислота
2~ сроссроенолпировиноградная -----------------------АДФ
||'-------*АТФ
пировиноградная кислота
.фосфодиокси. -ацетон
~^С02 +2Н аиртилкоф&рмент А жирные кислоты
Н2О
щавелево-уксусная кислота
лимонная кислота
С02*2Н
яблочная кислота
Н20.___А
Цикл Кребса
фумаровая
кислота «
изолимонная / кислота /
оС - кето глутаровая^ф кислота
-2Н
/-----нго
'нтарная
кислота'
“Л/Wj
:гл/таминовая
кислота
’ЛОК
2>
другие аминокислоты
Рис. 6Л. Схема основных этапов гликолиза и цикла Кребса. Проходя по терминальной электротранспортной цепи, показанной на рис. 6.2, каждая отделившаяся пара атомов водорода участвует в образовании трех молекул АТФ и молекулы воды. На рис. 6.20 показаны взаимоотношения между различными продуктами этих реакций и другими важными соединениями, встречающимися в растениях. Приведенные в линейной части рисунка процессы, названные гликолизом, происходят в цитоплазме, тогда как цикл Кребса функционирует в митохондриях
184
Глава 6
ством накопления продуктов, активацию метаболитами и регуляцию запасов энергии.
Помимо генетического контроля, активность ферментов зависит от таких факторов, как температура, концентрация ионов водорода, концентрация фермента, субстрата и конечных продуктов, гидратация и различные регуляторы роста. У микроорганизмов образование ферментов часто индуцируется в присутствии субстрата. Некоторые примеры индукции встречаются и у семенных растений. Так, образование нитратредуктазы, по-видимому, индуцируется в присутствии нитрата. Активность ферментов может быть также подавлена различными соединениями. В результате использования селективных ингибиторов стало многое известно о процессах метаболизма. Имеются два основных класса ингибиторов: конкурентные и неконкурентные. Конкурентными ингибиторами являются вещества, которые так похожи на молекулу субстрата,что частично замещают его в активных центрах ферментов и препятствуют нормальному действию фермента. Примером может быть блокирование превращения янтарной кислоты в фумаровую кислоту при добавлении малоновой кислоты, которая напоминает по строению янтарную кислоту. Неконкурентные ингибиторы, такие, как фторид, цианид, азид, медь и ртуть, образуют прочные соединения с молекулами ферментов, лишая их активности.
Классификация ферментов. Ферменты классифицируют по катализируемым ими реакциями. 1. Оксидоредуктазы, катализирующие реакции окисления-восстановления (например, оксидазы и дегидрогеназы). 2. Трансферазы, катализирующие перенос химических групп от донора к акцептору (например, трансаминаза). 3. Гидролазы, катализирующие гидролитическое расщепление связей С—О, С—N, С—С и некоторых других (например, сахараза). 4. Лиазы, катализирующие отделение химических групп от субстратов негидролитическим путем. Эти ферменты расщепляют С—С, С -О, С—N и другие связи удалением групп с образованием двойных связей или присоединением групп по двойным связям (например, декарбоксилазы). 5. Изомеразы, катализирующие превращение соединения в какие-то его изомеры (например, триозофосфатизомераза). 6. Лигазы (синтетазы), катализирующие соединение двух молекул с гидролизом пирофосфатной связи в АТФ (например, тиокиназы).
ЭНЕРГЕТИКА. Существование живых организмов зависит от постоянного притока энергии для использования при синтезе новой протоплазмы, для поддержания структуры органелл и мембран, для активного переноса ионов и молекул через мембраны и для выполнения механической работы, такой, как движение протоплазмы. У подвижных организмов много энергии расходуется на передвижение, у теплокровных — на поддержание температуры тела. Непосредственным источником этой энергии является пища, т.е. углеводы, жиры и белки, накопленные организмом, но первичные источники различаются. Автотрофные организмы сами вырабатывают пищу при фотосинтезе (как описано в главе 5) 4или при хемосинтезе. Примеры хемосинтеза встречаются среди бактерий, получающих энергию для синтеза углеводов, путем окисления NH3 до NO 2 ’ №*2 Д°* NO з , H?S до SO^_ или иона двухвалентного железа в ион трехвалентного. Гетеротрофные организмы, включаю-
Ффммгы, энергетика и дыхание
185
тис животных и незеленые растения, зависят от зеленых растений в отношении пищи, служащей для них источником энергии. Можно сказать, •ио успешное произрастание растений зависит от их способности добы-нить, сохранять и освобождать энергию по мере надобности. (Приобре-гспис и хранение энергии у зеленых растений в процессе фотосинтеза описаны в главе 5.)
Необходимо кратко обсудить энергетику окислительно-воссганови-1С/П.ПЫХ реакций. Известно, что вещество окисляется, когда оно теряет шектроны или атомы водорода, и восстанавливается, приобретая электроны или атомы водорода. Очевидно, что при восстановлении одного соединения другое окисляется. В следующем уравнении окисляется соединение А, а восстанавливается соединение В:
АН2 + В дегидрогеназа А +
«
Окисление глюкозы протекает так:
С6Н12О6+ 6О2—^6СО2 + 6Н2О+ 686 ккал.
Углерод глюкозы окисляется до СО2 путем отщепления водорода и соединения его с кислородом с образованием воды. В результате преобразований межатомных связей освобождается большое количество энергии. Когда этот процесс протекает в лаборатории как обычное горение, энергия выделяется в виде тепла. В живых клетках около 2/3
устойчивое к цианиду_______
дыхание ^H2fJ
восстановленный суб- НАД страт S’
АТФ
окцеленмьзй juum. в " убихцнзн X /*^ ре 2* '
ленный НАДИ; субстрат — —
АДФ'Фь
‘цит.в
мовленный убихинон
| АДФ+Ф»
I
I
I
I амитал
кис лота кислота
АТ'-Р
цит. с, ' Ре3*'
кРе* цит. с “1
АТФ
восстанов- ? ценная щито-храмоксиОаэак/
антцмицин
/\Нг0 окисленная* | цитохром- [ оксидаза
CN'CO, Nj,
АД Ф+Фц
Рис. 6.2. Схема митохондриальной системы транспорта электронов. Убихинон известен также как KoQ. Фн означает неорганический фосфат, в отличие от фосфорилирования в гликолизе, где большая часть фосфата для образования АТФ поступает от субстрата. Наличие альтернативной оксидазной системы показывает, что н некоторых тканях, обладающих цианидрезистентныи дыханием, минуется обычкам цитохромная система. Указаны точки действия некоторых распространенных ингибиторов. Амитал является мощным ингибитором в растительных и животных тканях, ротенон - только в животных (Icuma, 1972)
186
Глава 6
энергии удерживается и сохраняется в АТФ, откуда позже она может быть извлечена для совершения химической работы. В живых клетках этот процесс требует около 25 ферментов и происходит многостадийно (рис. 6.1 и 6.2). В противоположном направлении протекает восстановление углерода углекислоты до глюкозы. Для него необходим приток свыше 686 ккал/моль световой энергии, используемой на расщепление Н2О с освобождением Н+. Ион водорода соединяется с НАДФ с образованием восстановленного соединения НАДФ-Н2, которое служит источником водорода, необходимого для восстановления углерода. Энергия света используется также для образования АТФ, снабжающего энергией другие этапы цикла углерода. Как описано в главе 5, углерод восстанавливается, НАДФ-Н2 окисляется, а энергия запасается в продуктах фотосинтеза:
со2 + 2НАДФ-Н2 —---СН2О + 2НАДФ + Н2О.
Эта реакция также осуществляется многоэтапно и для ее протекания нужно около 20 ферментов (см. рис. 5.2 и 5.3). Таким образом, восстановление сопровождается увеличением свободной энергии, окисление — освобождением энергии. К примеру, восстановленные формы НАД и НАДФ (НАД-Н2 и НАДФ-Н2) имеют значительно большую свободную энергию, чем окисленные формы. Их называют "восстановительной способностью”, имея в виду их способность служить источником атомов водорода. То же можно сказать о ФМН и ФАД и их восстановленных формах (ФМН-Н2 и ФАД-^). В качестве примера роли НАД может служить превращение яблочной кислоты в щавелевоуксусную, при котором яблочная кислота теряет 2 атома водорода, а НАД восстанавливается до НАД-Н2:
~ СООН + ндд малатдегидрогеназа ^2 + идд.^
НО - СН - СООН------------------------О--С СООН 2
Окисление и восстановление цитохромоксидазы сопровождается изменением валентности железа, переходящего из ЬеЗ+ (окисленная форма) в Fe2+ (восстановленная форма).
ДЫХАНИЕ
При дыхании освобождается энергия, необходимая не только для поддержания структурной целостности и существования протоплазмы, но также и для синтеза многочисленных соединений, необходимых для образования новой протоплазмы и структуры растений (ассимиляции). Количество энергии, необходимой для синтеза различных соединений, приведено на основе данных потребления О2 и образования СО2 (см. табл. 6.3). Часть образовавшихся при фотосинтезе углеводов, не окисленная при дыхании или не использованная в ассимиляции, накапливается в плодах, семенах и вегетативных органах в виде крахмала, белка или жиров. (Ассимиляция рассмотрена в конце этой главы, а накопление углеводов — в главе 7.)
Ферменты, энергетика и дыхание
187
Возраст насаждений, лет
возраст насаждений, лет
Рис. 6.3. Процент образовавшихся питательных веществ, используемых на дыхание насаждениями бука разного возрасти (Moller et al., 1954)
Рис. 6.4. Количество сухого вещества, использованного при различных процессах в насаждениях бука европейского разного возраста, растущих' на хороших участках в Дании (MoUer et al., 1954) :
1 — общая, или валовая, продукция фотосинтеза; 2 — наблюдаемый фотосинтез; 3 — количество сухого вещества, прибавлявшегося при ежегодном росте; А — потери сухого вещества при дыхании листьев; Б — потери вследствие опадания листьев; В — потери сухого вещества при дыхании корней, стволов и веток; Г — потери при отмирании корней и веток; Д — прирост насаждения
Дыхание может быть определено как окисление пищи (субстрата) в живых клетках, приводящее к освобождению энергии. Это энергия, запасенная в виде химической энергии молекул субстрата. Освободившаяся энергия используется на поддержание структуры протоплазмы (поддерживающее дыхание), на синтетические процессы (ростовое дыхание), на транспорт веществ, на механические процессы (такие, как движение протоплазмы), часть энергии приходится на электрическую энергию, часть рассеивается в виде тепла.
ОСНОВНЫЕ ПОНЯТИЯ. Во всех живых клетках растений дыхание происходит постоянно. В физиологически неактивных структурах, таких, как покоящиеся семена, его интенсивность чрезвычайно низка. Наиболее интенсивно дышат меристематические участки, например, камбий, корни, кончики стебля и очень молодые ткани. Иногда дыхание интенсивно протекает также в созревающих плодах, где много энергии освобождается в виде тепла, не играющего, вероятно, никакой полезной для растения роли. Срок хранения плодов и семян может быть значительно продлен при хранении их в условиях, поддерживающих низкий уровень дыхания. Однако снижение дыхательной активности в растущих тканях при пониженной температуре или низкой концентрации кислорода нежелательно, поскольку оно уменьшает скорость роста.
Для выживания необходима некоторая минимальная интенсивность дыхания, а для роста более высокая интенсивность, но часто интенсивность дыхания поднимается даже выше необходимого уровня. Это приводит к непродуктивному потреблению питательных веществ, которые могли бы быть использованы при ассимиляции для образования новых тканей или аккумулированы в запасающих органах. Большая доля питательных веществ, образованных деревьями, используется на дыхание нефотосинтезирующих тканей (рис. 6.3, 6.4, табл. 6.1). Поль-стер (1950) предполагает, что продуктивность лесов могла бы быть
188
Глава 6
6.1. ПРОДУКТИВНОСТЬ И ПОТЕРИ СУХОГО ВЕЩЕСТВА
РАЗЛИЧНЫМИ ОРГАНАМИ ДЕРЕВЬЕВ В ЛЕСАХ ДВУХ ТИПОВ, Т/ГАТОД (Larcher, 1975)
Наименование показателей Влажный тропический Буковый лес в Да-
лес НИИ
Увеличение биомассы:
листья 0,03 0,0
стволы 2,90 5,3
корни всего 0,20 3,13* (2%) 1,6 6,9 (35%)
Потери в результате гибели и разло-
жения:
листья 12,0 2,7
стволы 13,3 1,0
корни 0,2 0,2 .
всего Использовано на дыхание: 25,5 (20%) 3,9 (20%)
листья 60,1 4,6
стволы 32,9 3,5
корни 5,9 0,7
всего 98,9** (78%) 8,8 (45%)
Согласно Sanchez (1973) восстановление леса в Гватемале достигает приблизительно 10 т/гатод в течение первого десятилетия после рубки, что намного больше величины, приведенной в таблице для более старого тропического леса.
Недавняя работа Kira (1975) показывает, что общие потери сухого вещества на дыхание влажных тропических лесов должны равняться 55 т/гатод, или 65% истинного фотосинтеза.
увеличена скорее снижающими дыхание воздействиями (такими, как удаление паразитических ветвей), чем воздействиями, усиливающими фотосинтез.
КЛЕТОЧНОЕ ДЫХАНИЕ. Глюкоза является обычным дыхательным субстратом, и ее полное окисление в идеальных условиях может быть представлено следующим уравнением:
^6^12^6 + —* ^€>2 + 6Н2О+ 686 ккал/моль.
Разумеется, этот процесс намного сложнее и включает намного больше этапов, чем показано этим суммарным уравнением, не объясняющим, как используется кислород, как образуются двуокись углерода и вода или как выделяется в доступной форме энергия. Попытаемся разъяснить, как все это происходит.
Биологическое окисление. Как уже упоминалось, окисление обычно включает отнятие атомов водорода или перенос электронов от субстратов к акцептору. Атомы водорода (протон плюс электрон) в клетках отщепляются от субстратов ферментами, известными как дегидрогеназы, и переносятся на акцептор или окисляющий агент. Этим акцептором обычно являются НАД (никотинамидадениндинуклеотид) или НАДФ (никотинамидадениндинуклеотидфосфат), которые в восстановленной
Ферменты, энергетика и дыхание
форме становятся НАД Н2 или НАДФ-Нт. Затем атомы водорода переносятся -от восстановленных НАД и НАДФ по цепи акцепторов терминальной оксидазной системы, содержащей цитохромы, и соединяются с кислородом с образованием воды (см. рис. 6.2). В это же время высвобождается энергия, и часть ее используется на образование высокоэнергетического соединения АТФ (аденозинтрифосфата) путем присоединения фосфата к АДФ (аденозиндифосфату).
АТФ - самое важное высокоэнергетическое соединение растений. Аденозин образуется из пурина-аденина и сахара-рибозы. Когда к аденозину добавляется через обычную эфирную связь фосфатная группа, получается АМФ (аденозинмонофосфат). При добавлении через пирофосфатную связь второго фосфата получается АДФ, а добавление третьей фосфатной группы через другую пирофосфатную связь дает АТФ. Это соединение называется высокоэнергетическим потому, что при разрыве концевой пирофосфатной связи путем гидролиза при pH 7,0 и 25°С освобождается около 7,3 ккал/моль энергии по сравнению с всего лишь 2,2 ккал/моль при разрыве эфирной связи в АМФ. Следует подчеркнуть, что высокая энергия заключена не в связи, но относится к разности уровней энергии между исходным соединением и его продуктами. Энергия высвобождается потому, что после отщепления концевого фосфата получается новое распределение электронов с гораздо более низкой общей энергией, чем имевшаяся у АТФ. Фактическое количество энергии, освобождающейся при реакции: АТФ—АДФ + Фн, несколько изменяется в зависимости от pH и других факторов, но чаще всего приводится 7,3-7,6 ккал/моль, несмотря на то, что в существующих внутри клеток условиях оно может достигать 12 ккал/моль.
Метаболически активные ткани содержат большие количества аденозинфосфатов. Например, зародыши прорастающих семян сосны желтой содержали в 80 раз больше аденозин фосфатов, чем непроросшие семена. Отношение АТФ и АДФ к общему содержанию аденозинфосфатов называется аденилатным энергетическим зарядом, и оно в растущих тканях значительно выше, чем в нерастущих.
Другие макроэнергетические соединения. Существуют другие фосфат-содержащие соединения, например фосфоенолпируват, 1,3-дифосфо -глицерат и ацетилфосфат-, имеющие даже более высокие стандартные свободные энергии гидролиза, чем АТФ. Высокой свободной энергией обладает ацетилкофермент А, а пиридиновые нуклеотиды в восстановленной форме (НАД-Н2, НАДФ-!^) также имеют высокую свободную энергию. Однако АТФ занимает исключительное положение, находясь посередине энергетической шкалы и обладая способностью переносить фосфатную группу от высокоэнергетических соединений к тем, у которых свободная энергия ниже, напрймер к таким, как глюкоза и глицерин, и образовывать глюкозо-6-фосфат и 3-глицерофосфат. Таким образом, АТФ и АДФ участвуют почти во всех ферментативных реакциях переноса фосфата в клетках, контролируя поток энергии.
ГЛИКОЛИЗ И ЦИКЛ КРЕБСА. Окисление глюкозы осуществляется в ряде этапов, распадающихся на две группы: линейную последовательность преобразований (гликолиз), в которых глюкоза превращается в пировиноградную кислоту, и следующий за ними цикл Кребса, или цикл трикарбоновых кислот, в котором пировиноградная кислота окис
190
Глава 6
ляется до углекислого газа и воды (см. рис. 6.1). Гликолиз осуществляется в цитоплазме, а реакции цикла Кребса и окислительное фосфорилирование происходят в митохондриях.
При полном аэробном окислении 1 моля глюкозы образуется 38 молей высокоэнергетического АТФ. В ходе гликолиза образуется всего 10 молей, но 2 моля расходуются на образование фосфорилированной гексозы, в результаты чистый выход составляет 8 молей. Три макро-эргические фосфатные связи образуются от каждой из пяти пар атомов водорода, выделенных при окислении молекулы пировиноградной кислоты в цикле Кребса, что дает в сумме 30 молей на 2 моля пировиноградной кислоты, образовавшиеся из каждого моля глюкозы. Если принимать запасенную при образовании концевых пирофосфатных связей в I моле АТФ энергию равной 7,3 ккал, то в АТФ запасается 277,4 ккал (38-7,3), или 40% всей энергии, аккумулированной в глюкозе. Если же свободную энергию принять равной 12 ккал/моль, что вероятнее для клетки, то запасается 66%. Остальная энергия рассеивается в виде тепла. Гликолиз и цикл Кребса можно представить в таком виде:
Гликолиз: С6Н12О6 + О2 + 8АТФ+8ФН-------2СН3СОСООН +
(глюкоза) (пировиноградная кислота)
2Н2О + 8АТФ + 58,4 ккал.
Цикл Кребса: 2СН3СОСООН + 5О2 + 30АДФ + 30Фн *-6СО2 + 4Н2О + ЗОАТФ +219 ккал.
Таким образом, каждый моль глюкозы дает 2 моля пировиноградной кислоты, 2 моля воды и 8 молей АТФ, а каждый моль пировиноградной кислоты, поступающей в цикл Кребса, дает 3 моля двуокиси углерода, 2 моля воды и 15 молей АТФ, или в сумме во всем процессе: 6 молей СС>2, 6 молей Н2О и 38 молей АТФ. Вода образуется в сложной терминальной электронтранспортной системе при соединении водорода, отщепленного дегидрогеназами, с кислородом.
Этапы сложной цепи превращений (см. рис. 6.1) начинаются фосфорилированием глюкозы путем переноса фосфата от АТФ. В результате образуются глюкозо-6-фосфат и АДФ. Запасы питательных веществ в виде крахмала могут быть превращены в глюкозо-1-фосфат фосфорилазами, а затем в глюкозо-6-фосфат. Другая перегруппировка и добавление второго фосфата дают фруктозо-1,6-дифосфат, а более поздние превращения приводят к образованию АТФ и НАД-Но. Окисляясь при участии цитохромной системы, последний дает 3 АТФ. Конечный продукт гликолиза — пировиноградная кислота. В аэробных условиях от пировиноградной кислоты отщепляются молекула СО2, 2 атома водорода и образуется ацетилкофермент А. Он соединяется со щавелевоуксусной кислотой в цикле Кребса с образованием шестиуглеродной лимонной кислоты. По мере отщепления молекул СО2 и атомов водорода лимонная кислота превращается в пяти- и четырехуглеродные кислоты, и в конце концов цикл снова приводит к щавелевоуксусной кислоте. Атомы углерода отщепляются декарбоксилазами при образовании ацетилко-фермента А; а-кетоглютаровой и янтарной кислот, образуя 3 молекулы СО2 на молекулу окисленной пировиноградной кислоты. Пары атомов
Г/or
Ферменты, энергетика и дыхание 191
водорода отщепляются дегидрогеназами при образовании ацетилкофер-мента А и а-кетоглютаровой, янтарной, фумаровой и щавелевоуксусной кислот. Эти пары проходят через цепь акцепторов терминальной электронтранспортной цепи (см. рис. 6.2) и соединяются с кислородом, образуя воду- При этом на каждые 2 моля отщепленного водорода образуются 3 моля АТФ. Хотя при окислении молекулы пировиноградной кислоты отщепляются 5 пар атомов водорода, но 3 молекулы воды используются в цикле, так что чистый выход составляет 2 моля воды на 1 моль пировиноградной кислоты.
ПЕРЕНОС ЭЛЕКТРОНОВ И ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ. Превращение водорода в воду — сложный и очень важный процесс, поскольку он сопровождается образованием АТФ. Этот процесс называется окислительным фосфорилированием. Общая схема приведена на рйс. 6.2, хотя некоторые детали процесса до сих пор не объяснены. По этой схеме, НАД принимает водород, отщепленный в цикле Кребса, и восстанавливается в НАД-Hj. Водород переносится на ФМН,. затем ФМН-Н2 передает ионы водорода и соответствующие электроны убихинону и потом цитохрому. В этой системе может быть от 3 до 7 цитохромов, и в конце концов один из них переносит два Н+ и два электрона на атом кислорода с образованием воды. Согласно современным представлениям, электронтранспортная система прокачивает протоны через мембрану, к которой прикреплены ферменты, создавая градиент концентрации Н+, обеспечивающий энергией процесс образования АТФ. Эта схема известна как хемиосмотическая теория Митчелла. На каждую пару атомов Н образуется одна молекула АТФ (см. рис. 6.2). Так восстанавливается кислород с образованием воды на последнем этапе дыхания и образуется большая часть АТФ. В противоположность такому окислительному фосфорилированию, образование АТФ при гликолизе называют субстратным фосфорилированием. Это обусловлено тем, что фосфат, присоединяющийся к АДФ для образования АТФ, поступает преимущественно от субстрата, а не от неорганического фосфата, как при окислительном фосфорилировании и фотефосфорилировании (связанном с фотосинтезом).
Из сказанного видно, что цикл Кребса выполняет две важные функции - Одна из них — образование промежуточных соединений, важных для синтеза таких веществ, как аминокислоты и жирные кислоты. Другая — образование больших количеств АТФ, поставляющего энергию для различных биосинтезов. Многочисленные ферменты, участвующие в цикле Кребса и в окислительном фосфорилировании, находятся в митохондриях, а участвующие в гликолизе ферменты — в цитоплазме.
Другие оксидазы. Помимо цитохрома, в растениях имеется ряд других оксидазных систем. Значительно представлен содержащий железо фермент каталаза. Он расщепляет перекись водорода на воду и молекулярный кислород, в то время как пероксидаза переносит водород от донора на перекись с образованием двух молекул воды. Пероксидаза окисляет также НАД-Н^ и НАДФ-Н2, что может регулирующе воздействовать на метаболиз клетки. Пероксидаза и полифенолоксидаза способны участвовать в синтезе лигнина. Оксидазы фенолов — это содержащие медь ферменты, вызывающие потемнение поверхностей срезов растительных тканей (например, у яблока или картофеля). Вероятно,
192
Глава 6
в интактных тканях они строго изолированы или отделены, но при повреждении клеток освобождаются и вызывают изменение окраски. Они также препятствуют выделению растительных ферментов и митохондрий. Другая важная оксидазная система — гликолатоксидаза — присутствует в стеблях и листьях (в корнях ее нет) и осуществляет важную для фотодыхания реакцию превращения гликолевой кислоты в глиок-силевую. Эти оксидазы называют растворимыми, потому что они находятся в цитоплазме в отличие от цитохромов, которые встречаются в мембранах митохондрий.
ПЕНТОЗОФОСФАТНЫЙ ПУТЬ (ШУНТ). В растениях обнаружен альтернативный путь окисления глюкозы — пентоз офосфатный, или гексозомонофосфатный. Глюкозо-6-фосфат окисляется в 6-фосфоглю-конат, а НАДФ восстанавливается в НАДФ-Г^. Это сопровождается другим превращением, приводящим к образованию пентозы рибозо-5-фос-фата, углекислого газа и еще одной молекулы НАДФ-Н^. Таким образом, в пентозофосфатном шунте образуется значительная восстанавливающая способность в форме НАДФ-Нз, используемого в таких реакциях, как синтез жирных кислот. Пентоза подвергается ряду превращений, сходных с превращениями в цикле Кальвина — Бенсона и является источником соединений, необходимых для синтеза нуклеиновых кислот, пиридиновых коферментов и других веществ. Дальнейшая перегруппировка приводит к 3-фосфогпипериновому альдегиду и пировиноградной кислоте или в обратном направлении к глюкозо-6-фосфату. Хотя на этом пути образуется меньше АТФ, чем в гликолизе и цикле Кребса, он имеет важное значение как источник восстановительной способности и веществ, необходимых для процессов биосинтеза. По мнению некоторых исследователей, пентозофосфатный шунт является главным путем метаболизма гексоз в прорастающих семенах.
АЦЕТИПКОФЕРМЕНТ А. Это соединение образуется из пировиноградной кислоты и кофермента А. Стандартная свободная энергия ацетильной группы намного выше, чем у АТФ. Ацетипкофермент А — исходное соединение метаболизма, так как углеводы, жирные кислоты и аминокислоты образуют его в качестве основного продукта их распада, вступающего в цикл Кребса. Большая часть углерода всех соединений поступает в дыхательный цикл через ацетипкофермент А. Например, при окислении жирных кислот пары атомов углерода одна за другой отщепляются от кислоты и переносятся при участии ацетилкофер-мента А в цикл Кребса. Он участвует также в синтезе жирных кислот.
АНАЭРОБНОЕ ДЫХАНИЕ. Терминальная электротранспортная система не может функционировать без кислорода. При этом нет и окислительного фосфорилирования, поэтому единственным источником энергии является гликолиз, что видно на схеме:
п Пировино-
Промежуточные * соединения —
кислота
аэробно
-------
анаэробно
СО, + Н2О 68о ккал Органические кислоты, альдегиды, спирты, СО2 15,2 ккал
Таким образом, анаэробное дыхание очень неэффективно и не дает достаточно энергии для поддержания быстрого роста. Более того, на-
Ферменты, энергетика и дыхание
193
копление не полностью окисленных соединений, таких, как спирты и альдегиды, часто причиняет вред растению.
ДЫХАТЕЛЬНЫЙ КОЭФФИЦИЕНТ. Природа субстрата заметно влияет на дыхательный коэффициент - отношение образовавшегося СО2 к использованному О2, что иллюстрируется следующими суммарными уравнениями.
При полном окислении глюкозы дыхательный коэффициент равен 1:
С6Н12°6 + 6О2--*-6СО2 + 6Н2О; 6СО2/6О2=1.
Полное окисление сильно восстановленных соединений, таких, как белки и жиры, дает дыхательный коэффициент менее 1. Например, при окислении обычного жира трипальмитина дыхательный коэффициент равен 0,7:
С51Н98О6 + 72.5О2->51СО2 + 49Н2О; 51СО2/72,5О2 = 0,7.
В окисляющих органические кислоты растениях, таких, как суккуленты, дыхательный коэффициент может быть значительно выше 1. Для щавелевой кислоты он будет равен 4, для яблочной — 1,33:
С4Н6°5 + ЗО2—^.4СО2+ ЗН2О; 4СО2/ЗО2 = 1,33.
Когда углеводы превращаются в органические кислоты, кислород может расходоваться без образования СО2- В растительных тканях окисляться могут сразу несколько различных субстратов и одновременно могут происходить несколько реакций, поэтому дыхательный коэффициент не всегда является хорошим показателем преобладающего типа реакций.
ФОТОДЫХАНИЕ. Много лет физиологи растений спорили о том, больше ли, меньше или одинаково дыхание фотосинтезирующих тканей на свету и в темноте. В конце концов опыты Декера (1955, 1958, 1959), Вина (1958) и многие другие более поздние исследования показали, что у растений с С3 путем фотосинтеза образование СО2 на свету намного выше, чем в темноте. Процесс фотодыхания окончательно не выяснен, но известно, что он отличается от темнового дыхания, так как иначе реагирует на инигибиторы и кислород. Он тесно связан с фотосинтезом, поскольку субстратом для фотодыхания служит, видимо, первичный продукт фотосинтеза, вероятно, глиоксилевая кислота. Эта кислота происходит от гликолевой кислоты, образующейся из рибулозодифосфата. Сз-растения используют на фотодыхание от 20 до 50% С, связанного при фотосинтезе. Поэтому значительный интерес представляет возможность увеличения урожая культурных растений в результате снижения фотодыхания либо путем селекции, либо некоторыми биохимическими средствами. (Связь фотодыхания с ростом рассмотрена в главе 5.)
ДЫХАНИЕ РАСТЕНИЙ И ИХ ЧАСТЕЙ
КОЛИЧЕСТВО ПИТАТЕЛЬНЫХ ВЕЩЕСТВ, ИСПОЛЬЗУЕМЫХ НА ДЫХАНИЕ. Общее количество питательных веществ, расходуемых на
194
Глава б
дыхание растений, представляет интерес ввиду его связи с количеством веществ, которые могут быть использованы для процессов ассимиляции, связанных с ростом, и на количество веществ, накопленных в плодах и семенах. Все питательные вещества образуются в листьях, но при дыхании потребляются всеми живыми клетками. В результате общее их количество, использованное листьями, побегами и живыми тканями стволов и корней, составляет значительную долю продуктов фотосинтеза.
ЦЕЛЫЕ ДЕРЕВЬЯ. Известны попытки измерения баланса питательных веществ растений, иногда называемого балансом углерода.
По оценке Кинерсона (1975), на дыхание 14-летних деревьев сосны ладанной использовалось 58% ассимилятов, а Лар кер (1975) утверждает, что во влажных тропиках леса могут использовать на дыхание свыше 70% продуктов фотосинтеза (см. табл. 6.1). В примечании к более поздней работе Кира (1976) сообщается о переоценке дыхания влажного тропического леса, которое должно составлять около 55 т/га, или 65% суммарной продукции фотосинтеза.
Мюллер с соавторами (1954) провели глубокое исследование количества питательных веществ, используемых на различные процессы насаждениями бука, растущими на хороших участках в Дании. Как видно на рис. 6.3, доля использованных на дыхание питательных веществ возрастала с 40% продуктов фотосинтеза у 25-летних деревьев и приблизительно до 50% у 85-летних деревьев. Это объясняется тем, что отношение дышащих тканей к фотосинтезирующим увеличивается с возрастом. Согласно приведенным на рис. 6.4 данным ежегодный прирост сухого вещества в насаждении бука уменьшается с возрастом частично за счет увеличившейся потери корней, веток и побегов, а частично за счет небольшого уменьшения фотосинтеза и увеличения дыхания. ^Вероятно, по мере старения деревьев отношение фотосинтезирующих тканей к дышащим снижается, что уменьшает снабжение питательными веществами и замедляет скорость роста. У деревьев сосны ладанной с возрастом отношение массы листвы к массе ствола уменьшается от 1:2 в 10 лет до 1:6 в 50 лет. Другие данные по соотношениям листьев, стволов и ветвей приведены в табл. 3.6, Одной из причин медленного роста переуплотненных насаждений может быть чрезмерное использование питательных веществ на дыхание, поскольку множество малых стволов имеет большую поверхность дышащего камбия, чем несколько больших стволов на той же площади.
Например, 100 деревьев с диаметром 5 см располагаются на той площади, что и одно дерево с диаметром 50 см, но имеющие в 10 раз большую окружность и в 10 раз большую поверхность камбия. Можно было
7
О 0J 0,2 0,3 0Л 0,5 0,6 0,7 Относительное расстояние от основания вепрей во вершины кроны
Рис. 6.5. Участие ветвей на разной высоте в ежегодном приросте главного ствола 15-м стр ошлх 30-летних деревьев сосны ладанной. Максимальный объем ствола достигается при отношении высоты кроны к ширине равном 0,4, при отношении равном 0,3 размеры ствола составляют лишь 89 % максимально возможных
(Labyak, Schumacher, 1954)
Ферменты, энергетика и дыхание
195
бы ожидать, что растущие на открытом участке деревья с большими кронами должны иметь более высокие отношения листовой поверхности к древесине и, следовательно, более высокие отношения фотосинтезирующей поверхности к дышащим тканям, чем деревья, растущие в сомкнутых насаждениях, и поэтому должны расти быстрее. Вообще это верно, но нижние ветви часто вносят малый вклад (или вообще никакого) в рост ствола и даже могут паразитировать, т.е. тратить на дыхание больше, чем накоплять в процессе фотосинтеза. Было показано, что с нижних двух третей ствола сосны ладанной ветви можно удалять без существенного снижения роста (рис. 6.5). По всей вероятности, большая часть углеводов, синтезируемых листьями нижних ветвей, используется в них на рост и дыханием не достигает ствола.
ДЫХАНИЕ РАЗЛИЧНЫХ ЧАСТЕЙ РАСТЕНИЙ. Интенсивность дыхания различных частей дерева широко варьирует в основном из-за различий в соотношениях физиологически активных тканей.
Листья составляют наименьшую часть массы дерева, но они обладают наибольшей интенсивностью дыхания среди всех частей дерева, так как содержат больший процент живого материала, чем одревесневшие части. На их долю приходится 50% общего дыхания 60-летнего букового леса, 60% влажного тропического леса и 32% молодого насаждения сосны ладанной. В дневное время интенсивность дыхания листьев лимонных и апельсиновых деревьев составляла от 15 до 20% интенсивности фотосинтеза. Если бы такая интенсивность сохранялась в течение всех 24 ч, это привело бы к потреблению 30—40% всех синтезированных углеводов. Более низкие температуры в ночное время должны существенно
• Рис. 6.6. Сезонная динамика дыхания почек грушевого дерева (Thom, 1951):
.1 - поглощение кислорода; 2 — выделение СО?; 3 - окончание периода покоя фруктового сада; 4 - начало периода покоя (треугольник л- 10 почек, образовавшихся в 1948 г.; кружок — 110 почек, образовавшихся в 1949 г.)
196
Глава 6
Рис. 6.7. Влияние удаления почечных чешуй (сплошная линия — без чешуи; штриховая - с чешуей) на дыхание почек .сосны веймутовой на разных этапах развития (Kozlowski, gentile, 1958):
1 — в пересчете на сухой вес; 2 — в пересчете на сырой вес
Рис. 6.8. Интенсивность дыхания, рассчитанная по поглощению кислорода, различных частей ствола ясеня черного до и после раскрытия почек (Goodwin, Goddard, 1940):
1 — потребление кислорода; 2 — в мае; 3 — в апреле
снижать ночную интенсивность дыхания по сравнению с дневной. Было подсчитано, что листья влажного тропического Леса используют 47% всех ассимилятов, букового леса - 28%, а молодого насаждения сосны ладанной — 32%. Более высокий процент ассимилятов, используемых на дыхание листьев влажных тропических лесов, по-видимому, связан с тем, что их индекс листовой поверхности равен 11,4 (т.е. вдвое выше, чем у букового леса), многие листья затенены и имеют низкую интенсивность фотосинтеза, а высокие ночные температуры вызывают повышенное дыхание.
Почки. Эти органы хотя и составляют очень малую часть массы дерева, в период роста имеют высокую физиологическую активность. Естественно, существует заметная сезонная цикличность их активности (рис. 6.6). На рисунке видно, что с началом роста покоящихся почек интенсивность дыхания возрастала в 5 раз, затем резко уменьшалась в августе, когда прекращалось удлинение ствола. При этом дыхание побегов долго оставалось интенсивным и после перехода почек в состояние покоя и не достигало минимума до января.
Работы ряда исследователей показали, что почечная чешуя препятствует проникновению кислорода. Измеренное по поглощению кислорода дыхание интактных почек клена остролистного составляет лишь половину дыхания тех почек, у которых чешуи были_удалены (рис. 6.7). Имеются также доказательства анаэробного дыхания у них, которое может быть причиной накопления ингибиторов и вызывать состояние покоя или продлевать его.
Ферменты, энергетика и дыхание
197
Стебли, В стволах и больших ветвях дыхание наблюдается в основном в новой флоэме и ксилеме, примыкающих к камбию. Гудвин иГод-дард (1940), изучавшие дыхание древесных стволов, наиболее интенсивное дыхание нашли у клена красного в камбиальной зоне. У ясеня черного оно было наибольшим во вновь дифференцирующейся ксилеме непосредственно внутри камбия (рис. 6.8). Клетки живых сердцевинных лучей и осевой паренхимы поглощают кислород, но их количество так мало, что суммарная интенсивность дыхания невелика. В опытах этих исследователей ядровая древесина поглощала небольшое количество кислорода, но это происходило, вероятно, в результате скорее окисления органических веществ в мертвых тканях, чем дыхания, поскольку прокипяченные блоки из ядра поглощали почти столько же кислорода, что и некипяченая древесина.
Мюллер (1946) также отмечал самую низкую интенсивность дыхания в старых частях древесных стволов. Он установил, что дыхание концентрировалось возле камбия у дуба в большей степени, чем у бука. В буке интенсивность дыхания уменьшалась от камбия внутрь постепенно. Дыхание сосредоточено в камбиальной зоне ствола, поэтому можно ожидать, что в лесном насаждении, состоящем из небольшого числа деревьев большого диаметра, отношение фотосинтеза к дыханию будет более значительным, чем в насаждении на той же площади, состоящем из большего числа деревьев меньшего диаметра.
Рис. 6.9. Сезонные изменения дыхания различных органов сосны ладанной в 14-летнем насаждении (по Kinerson, 1975). Интенсивность дыхания дана в пересчете на 1 поверхности почвы. Для листьев шкала в 20 раз больше, чем для других органов:
1 — воздух; 2 — почва; 3 — ствол; 4 - листва; 5 — ветвь; 6 — корень
Рис. 6.10. Дыхание древесных стволов при различных температурах до (светлые кружки) и после (темные кружки) начала активности камбия. Рассчитанная средняя величина Qjq равна 2,9 (Kinerson, 1975)
198
Глава б
тайный температурный коэффициент был равен 2,9. Зависимость включает как прямое действие на интенсивность дыхания, так и влияние потепления и охлаждения на ростовой пик л. При высокой температуре дыхание стебля сильнее возрастает после того, как камбий становится активным, чем до того (рис. 6.10). Иохансон (1933) наблюдал строгую сезонную цик-личность дыхания стволов и ветвей хвойных и лиственных деревьев в Дании. Масимум дыхания приходился на время наиболее быстрого роста, а поздним летом интенсивность дыхания у лиственных понижалась быстрее, чем у хвойных деревьев. Существуют суточные колебания .интенсивности дыхания древесных стволов, и, как сообщают, интенсивность выше на солнечной стороне стволов, чем на затененной.
ГАЗЫ В ДРЕВЕСНЫХ СТВОЛАХ. Двуокись углерода образуется живыми клетками древесины и накапливается в сосудах и трахеидах, поскольку камбий относительно непроницаем для диффузии газов. В результате в ксилеме накапливается значительное количество газа, особенно в больших сосудах ранней древесины, где при сильной транспирации, приводящей к высокому натяжению в соке, водные нити стремятся разорваться. Считают, что у сосны и секвойи большая часть ствола, за исключением наружных трех или четырех годичных колец, заполнена газом.
Цилиндр камбия и заболони настолько непроницаем для газа, что в ядре концентрация С>2 обычно намного ниже, a CCh выше, чем в окружающем воздухе, по крайней мере в период роста (рис. 6.11). Состав газа также подвержен изменениям, связанным с изменениями метаболизма: в период роста СО2 больше, а О2 меньше, чем зимой (рис. 6.12). Сообщалось, что в стволах быстрорастущих деревьев, таких, как топойь, СО2 накапливается больше, чем в стволах медленнее растущих видов, таких, как крупноплодный и красный дубы. Кажется правомерным предположение, что недостаток 62 в старой древесине играет важную роль, вызывая развитие ядровой древесины. В коре газ накапливается редко, потому что ее межклеточные пространства обычно связаны с атмосферой чечевичками. Кроме того, расстояние, через которое газ должен диффундировать, чтобы достичь камбия, достаточно мало, так что камбий .вряд ли подвержен дефициту кислорода.
Приводились данные о ненормально высоком давлении газа в стволах ряда видов. Иногда давление бывает настолько высоким, что ло-
Рис. 6.11. Сезонные изменения концентрации кислорода и двуокиси углерода в ядре и заболони ствола тополя. Пунктирными линиями обозначены периоды, когда нельзя было взять пробы (Chase, 1934): 1 - заболонь; 2 — ядро
Месяцы
Ферменты, энергетика и дыхание
199
Рис. 6.12. Сезонные изменения концентрации кислорода и двуокиси углерода в стволах деревьев трех видов (Chase, 1934) : а - дуб крупноплодный; ’ б — вяз американский; в — сосна веймутова; 1 — О2; 2 — СО2
паются внутренности насекомых, протачивающих ходы в дереве. Иногда газ горючий. Высокое давление газа отмечается в деревьях, страдающих от болезни, известной как ’’водослой”. По-видимому, такое давление вызвано газом, образуемым разлагающими древесину организмами, .и не имеет ничего общего с нормальным дыханием. Это доказывает, что внешняя древесина относительно непроницаема для газа даже под давлением.
КОРНИ И ПНЕВМАТОФОРЫ. Дыхание корней и других подземных структур представляет особый интерес, поскольку они часто попадают в условия низких концентраций О2- Дыхание корней и почвенных организмов уменьшает содержание О2 и увеличивает содержание СО2- Затопление осложняет ситуацию.
Корни. "Меристематические участки корней имеют высокую интенсивность дыхания. Маленькие корешки содержат большое число живых клеток, в результате они часто страдают от недостаточной аэрации. Известно, что в корнях затопленных растений накапливаются спирт, молочная кислота и другие не полностью окисленные соединения. Обычно растущие на влажной почве виды (например, кипарис, нисса водная, ива и мангровые деревья) намного выносливее в отношении недостаточной аэрации и затопления, чем виды, обычно растущие на хорошо дренированной почве (например, дерен, тюльпанное и амбровое деревья). Распространение многих видов ограничено их высокой потребностью в кислороде, что препятствует произрастанию их в районах, где почва часто бывает насыщена водой. Вероятно, растения некоторых видов устойчивы к затоплению по двум причинам: или структура их стебля и корня допускает диффузию О2 от побегов к корням, или корни при-
200
Глава 6
,8.10.62
8 17.62 18,0/2,5 k,0/>2,00
1013-62 1$0/l,5 4,0/>2,00
11.1662 19,0/1,0 4p/>20,0
3.31 63 - 1,0/>20,0
8,0/12,5
4,5/15,5
6,0/15,0
__________________I-----------------I—
0 6 12
24 (руты
Рис. 6.13. Изменения концентрации кислорода и двуокиси углерода под слоем песка и глины (Yelenosky, 1964) :
1 - глинистая почва; 2 — песок и сланец; 3 — измерительное устройство 1; 4 -дорожка из гравия; S — измерительное устройство 2; 6 — насыпка глины; 7 — два погибающих дерновых дерева (высота 14 футов, толщина ствола 4 дюйма);
8 — измерительное устройство 3; дробными числами обозначены показатели введенных в почву анализаторов воздуха, %; числитель — О2; знаменатель — СО2
способились к анаэробному дыханию. В корнях некоторых древес-ных видов, таких, как мангровые деревья, наблюдаются оба способа адаптации. Распространенная причина гибели деревьев - насыпка почвы над корнями. Ее влияние на состав почвенных газов показано на рис. 6.13. В плохо аэрируемой почве накапливаются ядовитые вещества: сульфиды, метан, двухвалентное железо и другие восстановленные соединения. Сообщалось, что в корнях затопленных растений накапливается этилен и диффундирует вверх к побегам, где он вызывает эпинастию и хлороз характерные для затопленных растений. Аноксия более вредна, чем накопление ядовитых веществ. Поэтому меньшую устойчивость к затоплению ели ситхинской по сравнению с сосной скрученной объясняют худшим внутренним поступлением Оэ от побегов к корням.
Согласно данным, суммированным Лархером (1975), дыхание корней составляет около 8% общего дыхания букового леса и 6% дыхания тропического влажного леса. Данные Киннерсона (1975) показывают, что на долю корней приходится только около 3% дыхания 14-летнего насаждения сосны ладанной, хотя в корнях содержится 18—20% сухого вещества. Возможно, дыхание корней было подавлено низкой концентрацией О2 в почве.
Пневматофоры. У деревьев, произрастающих на болотах или подверженных. приливному затоплению (таких, как мангровые деревья), часто образуются, специализированные корневые системы, именуемые пневматофорами, которые участвуют в газообмене. Растения типа мангровых, представителем которых является Avicennia nitida, могут образовывать тысячи воздушных корней, или пневматофоров, торчащих из ила вокруг основания дерева. Как сообщают Шоландер с соавторами (1955), воздух засасывается через чечевички вертикальных пневматофоров Avicennia во время отлива и выталкивается во время прилива.
Ферменты, энергетика и дыхание 201
Столбовидные корни Rhizohora mangle имеют на поверхности чечевички, которые связаны воздушными полостями с корнями, зарывшимися в ил. Вследствие закупоривания чечевичек жиром понижается содержание С>2 в корнях, погруженных в ил. Это означает, что столбовидные корни служат аэрирующими приспособлениями для погруженных корней.
У таксодиевых деревьев, растущих на часто затопляемой почве, развиваются конические вертикальные образования — корневые выросты, образующиеся благодаря активной, локализованной деятельности камбия, добавляющего мощные спои древесины на верхней поверхности корней. Хотя неоднократно и утверждалось, что корневые выросты болотных кипарисов, подобно другим пневматофорам, служат органами аэрации, снабжающими кислородом погруженные корни, доказательства такой их роли не убедительны. Если перенос О2 от выростов в корневой системе существует в норме, тогда при отделении выростов количество поглощаемого ими О2 должно сразу же уменьшаться. Однако Крамер с соавторами (1952) нашли, что у отделенных выростов в течение 2 дней дыхание было сильнее, чем у прикрепленных, но постепенно уменьшалось. Учитывая высокое содержание в выростах активной камбиальной ткани, можно полагать, что большая часть кислорода утилизируется на месте, и что корневые выросты кипарисов не имеют значения как органы аэрации. (Рост пневматофоров рассмотрен в главе 2.)
ДЫХАНИЕ ПЛОДОВ, Дыхание плодов представляет научный интерес ввиду Климатических* изменений во время созревания. Оно имеет и практическое значение, поскольку срок хранения плодов зависит от контроля дыхательной активности.
Дыхание растущих плодов. Дыхание развивающихся плодов значительно изменяется в ходе развития. Исследования показывают, что дыхание яблок наиболее интенсивно сразу после их завязывания, а затем уменьшается быстро в начале лета и медленно в конце, с подъемом, получившим название климактерического, после сбора плодов (рис, 6.14), В яблоках климактерическая фаза наблюдается иногда еще на деревьях, но в прикрепленных плодах авокадо - никогда. Эпидерма яблока покрыта относительно непроницаемым слоем кутина, препятствующего газообмену. Вследствие этого дыхание внутренних тканей больших плодов, вероятно, лимитировано низким содержанием Иногда анаэробное дыхание вызывает обесцвечивание внутренних тканей плодов, особенно яблок, а также латука и капусты, если их хранят или перевозят при недостаточном аэрировании. (Более подробно рост плодов рассмотрен в главе 4.)
Дыхание собранных плодов. Биохимии и физиологии собранных плодов уделялось большое внимание с целью найти пути продления срока их хранения. Заключительный этап созревания многих или, возможно, большинства плодов характеризуется сложным набором изменений. Этот период высокой активности ферментов и усиленного дыхания, как уже сказано, назван климактерическим. В итоге он завершается размягчением и порчей плодов, становящихся все более восприимчивыми к разрушению микроорганизмами. Быстрая порча плодов в процессе старе ния указывает на потерю контроля,активности ферментов, возможно, из-за нарушения компартментации, когда пространственно разделенные в норме ферменты освобождаются. Одно время считали, что усиление дыхания необходимо для снабжения энергией процесса созревания, но это сомнительно. Более вероятно, что дыхание усиливается благодаря присущим созреванию* биохимическим изменениям, которые обеспечивают повышенные количества дыхательных субстратов. Дыхание плодов происходит, видимо, и по циклу Кребса и через пентозо фосфатный шунт. Климактерический подъем образования СО2 наблюдается у многих плодов, за исключением цитрусовых. Подъем отмечается у плодов авокадо после сбора, но если их оставить на деревьях, подъема не будет. Из рис. 6.15 видны различия в количестве, образовавшегося во время климактерического усиления дыхания
202
Глава 6
Рис. 6.14. Изменения интенсивности дыхания плода яблони в процессе развития (Kiotkov, 1941). Вертикальная черта - время сбора урожая
Рис. 6.15. Климактерическое увеличение дыхания различных фруктов при температуре 15°С (Biale, 1950, воспроизведено с разрешения Annual Rewiews. Inc.
из журнала Ann. Rev. Plant Physiol., v.I, J950) :
1 — авокадо; 2 — банан; 3 - груша; 4 - яблоко
СОэ. Климактерическое дыхание хотя и не подавляется цианидом, но цианид-резистентный путь не имеет значения для созревания.
При созревании хранящихся плодов выделяется этилен, который в свою очередь способствует созреванию других плодов, хранящихся в той же камере. Образование других летучих веществ влияет на вкус и запах. В ходе старения нарушается механизм, регулирующий аэробное дыхание. Активность ферментов вызывает растворение пектиновых веществ клеточных стенок, клетки начинают отделяться, и наблюдаются размягчение и физическое разрушение. В результате этих изменений создаются условия благоприятные для вторжения грибов и бактерий.
Хранение плодов. Успешное хранение плодов и овощей взаимосвязано с регуляцией дыхания. Хранение при низких температурах ~ простейший способ уменьшения интенсивности дыхания. Иногда хранение яблок, груш и других фруктов и некоторых овощей в охлажденных емкостях сопровождается появлением впадин, внутренним потемнением и другими признаками физиологических нарушений , (см. главу 17). Охлаждение не снижает интенсивность всех процессов в одинаковой степени, । и вред при этом могут причинять накопление продуктов анаэробного дыхания, фенол оксидазная активность и другие анормальные физиологические процессы. Срок хранения многих растительных объектов в охлажденном состоянии может быть значительно увеличен понижением концентрации до 2-3% и повышением концентрации СС>2 до 2-6% или в некоторых случаях даже до 20%. Высокая концентрация СО2 не только подавляет анаэробное дыхание, но и уменьшает вред от образования этилена, а иногда удлиняет срок жизни плодов на полках магазинов после хранения на холоду. Накопление этилена, увеличивающее скорость разрушения, иногда может быть снято его поглощением марганцовокислым калием.
ДЫХАНИЕ СЕМЯН. В ходе онтогенеза в семенах происходят большие изменения дыхания. На ранних этапах развития его интенсивность высокая, но в процессе созревания уменьшается, и зрелые семена, стало-
Ферменты, энергетика и дыхание
203
вясь сухими, имеют очень низкую интенсивность дыхания. Чрезвычайно низкой должна быть интенсивность дыхания тех семян, которые сохраняют жизнеспособность при хранении в сухом виде в течение столетия или больше (например, семена кассии). Низкая интенсивность дыхания может быть следствием низкого содержания воды. В этом случае в присутствии воды интенсивность сразу же’ возрастает. Иногда дыхание и прорастание подавляются непроницаемостью семенной оболочки для Н2О и Ch’ в других случаях прорастанию препятствует физиологическая незрелость зародыша. (Этот вопрос обсуждается в главе 14.)
Прорастание всегда сопровождается большим увеличением интенсивности дыхания, хотя бы отчасти из-за гидролитических процессов, поставляющих больше сахаров, пригодных в качестве субстратов. В результате окисления питательных веществ при дыхании значительно уменьшается сухой вес прорастающих семян, несмотря на увеличение сырого веса при развитии проростка. Сухой вес не может увеличиваться до тех пор, пока листья не разовьются до состояния, когда прирост сухого вещества за счет фотосинтеза превышает потери на дыхание. (Прорастание семян рассмотрено в главе 14.)
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ДЫХАНИЕ
Успешное развитие деревьев и других растений зависит от соотношения интенсивностей дыхания и фотосинтеза. На интенсивность дыхания влияют некоторые внутренние и внешние факторы, часто взаимодействующие между собой. К важным внутренним факторам относятся: возраст и физиологическое состояние тканей, количество окисляемых субстратов и гидратация. Факторы окружающей среды включают температуру почвы и воздуха, состав почвенных газов, доступную почвенную влагу, свет, повреждения и механические нарушения, химические вещества (гербициды, фунгициды, инсектициды и удобрения).
ВОЗРАСТ И ФИЗИОЛОГИЧЕСКОЕ СОСТОЯНИЕ ТКАНЕЙ. У молодых тканей с высоким отношением протоплазмы к материалу клеточных оболочек и с небольшим числом мертвых клеток интенсивность дыхания выше, чем у зрелой ткани, содержащей меньше физиологически активного материала. Например, маленькие ветки дышат в пересчете на единицу сухого веса сильнее, чем ветви, а молодые листья — сильнее, чем старые. Ранее отмечалось, что дыхание почек увеличивалось в несколько раз с началом роста, а с прекращением роста так же быстро уменьшалось. На рис. 6.10 показано сильное увеличение дыхания ствола после начала деятельности камбия.
НАЛИЧИЕ СУБСТРАТОВ. К дыханию приложим закон действия масс. Следовательно, увеличение количества окисляемого субстрата обычно вызывает большую интенсивность дыхания. Это очень заметно в созревающих плодах, где превращение крахмала в сахар сопровождается увеличением интенсивности дыхания. В самой молодой заболони высокая концентрация углеводов может быть причиной высокой, интенсивности дыхания (см. рис. 6.8).
ГИДРАТАЦИЯ. До определенных пределов интенсивность дыхания коррелирует с содержанием воды. Это особенно заметно в сухих семе-
204
Глава 6
150 mo 130 120 Я ип
| 70 80 50 40 30\
-4-B-12-16-202420323MIHHS водный потенциал, дары
Рис. 6.16. Влияние обезвоживания на интенсивность дыхания сосны ладанной. Недостаток воды выражен через водный потенциал листьев, интенсивность дыхания - в процентах от интенсивности' при полевой влагоем-кости (Brix, 1962)
нах, где интенсивность дыхания уменьшается по мере того, как семена созревают и становятся сухими, но увеличивается при увлажнении семян. Дыхание до некоторой степени подавляется недостатком воды, но имеются и некото-
рые исключения. Паркер (1952) указывал, что когда побеги и хвоя хвойных деревьев резко обезвоживаются, происходит временное усиление дыхания, а затем ослабление его. Брике (1962) сообщил о сходном явлении у сосны ладанной (рис. 6.16). Согласно Бансу и Миллеру (1976) недостаток воды усиливал дыхание на свету у нескольких видов деревьев сухих мест, но уменьшал у видов, растущих во влажных местах.
ВЛАЖНОСТЬ ПОЧВЫ. Неблагоприятное действие могут оказывать как дефицит, так и избыток почвенной влаги. Влияние избытка влаги на дыхание корней уже обсуждалось. Побеги страдают от недостатка почвенной влаги намного раньше корней. Так, по данным Шнайдера и Чилдерса (1941), при снижении влажности почвы намного ниже полевой влагоемкости дыхание листьев яблони возрастало, хотя фотосинтез и транспирация уменьшались. Наиболее интенсивное, дыхание наблюдалось при наиболее низких транспирации и фотосинтезе, когда листья начинали вянуть. Возможно, дыхание усиливалось потому, что обезвоживание способствовало превращению крахмала в сахар, обеспечивая больше субстрата для дыхания. При повторном поливе почвы до полевой влагоемкости дыхание снижалось до нормального. Затопление яблонь также вызывало водный дефицит в листьях и усиливало дыхание.
ТЕМПЕРАТУРА ПОЧВЫ И ВОЗДУХА. Интенсивность дыхания сильно зависит от температуры и потому, как было показано на рис. 6.9 и 6.10, варьирует с изменением температуры почвы и воздуха. Влияние температуры на процессы часто выражают величиной Qi о. являющейся отношением скорости процесса при температуре Т к скорости при температуре Т = 10° С. Если скорость удваивается, Q10 равно 2. Кинерсон (1975) высчитал, что для стволов сосны ладанной Qio ~ 2,9.
Определено достоверное дыхание стволов тополя осинообразного при — 11°С, самой низкой из использованных температур. Фотосинтеза при температуре ниже -3°С не было (рис. 6.17). Эти результаты согласуются с полученными для хвойных деревьев, у которых выделение СО2 начиналось при прогреве стволов до 3°С, вероятно, потому, что при этой температуре оттаивали коровые клетки.
При высоких температурах может наблюдаться вредное усиление дыхания, потому что оптимальная для фотосинтеза температура обычно ниже оптимума для дыхания. Например, у сосен веймутовой и смолис-той повышение температуры с 20° до 40°С значительно уменьшало
Ферменты, энергетика и дыхание
205
а — ночная температура стебля; б — интенсивность падающего на стебель излучения; в — интенсивность метаболизма стебля: 1 — листопад; 2 — раскрывание ^чек; 3 — темновое дыхание; 4 — истинный фотосинтез
наблюдаемый фотосинтез и снижало способность деревьев этих видов накапливать углеводы. Низкая температура удлиняет срок хранения плодов и овощей, потому что она снижает интенсивность дыхания и других физиологических процессов. Как уже упоминалось, эффективность действия низкой температуры при хранении некоторых плодов и овощей может быть узеличена созданием газовой среды с низким содержанием кислорода и высоким содержанием двуокиси углерода.
СОСТАВ АТМОСФЕРЫ. Значительных изменений концентрации О2 в атмосфере не бывает, но концентрация О2 часто становится лимитирующим фактором в стволах деревьев, внутри почек, плодов и семян. Внутренняя флоэма, по-видимому, хорошо снабжается кислородом путем диффузии внутрь через чечевички и трещины в коре. Диффузия О2 резко уменьшается в растворе, и дыхание тканей внутри камбия должно часто понижаться из-за низкой концентрации О2 Опыты Поллока (1953) показали, что концентрации С>2 выше нормальных не изменяли дыхания покоящихся почек, но усиливали его в растущих почках (рис. 6.18). Влияние недостаточной аэрации обсуждалось в разделе, посвященном дыханию корней. Дыхание корней и почвенных организмов приводит к уменьшению концентрации О2 и повышению концентрации СС>2. Отклонение от нормы обычно увеличивается с глубиной почвенного слоя. Летом оно больше, чем зимой- Поэтому затопление почвы зимой менее вредно, по сравнению с затоплением летом.
По некоторым исследованиям, интенсивность дыхания отделенных кончиков косней нескольких видов хвойных было при 5% От наполовину меньше, чем при Й) % О2, а скорость роста корней снижалась до 20 % от контроля. Корни хвойных погибали за 2 дня в неаэрируемой среде, но корни ивы оставились живыми и имели 50% нормальной скорости роста. Сделаны предположения, что корни ивы получают кислород от стеблей.
Насыпка грунта над принявшейся корневой системой часто повреждает или убивает деревья и кустарники из-за худшего снабжения корней
Глава 6
206
Рис. 6.18. Влияние времени года и концентрации кислорода на поглощение его почками клена при 25°С (Pollock, 1953):
а — клен остролистный; б — клен сахарный; 1—10 апреля; 2 — 4 августа; 3 — 28 февраля; 4 — 23 ноября; 5—1 марта; 6 — 2 августа; 7 — 27 октября
кислородом (см. рис. 6.13). При выращивании деревьев иногда для усиления газообмена устраивают скважины, кладут черепицу и насыпают гравий. Некоторые виды легче переносят насыпку грунта, предположительно потому, что их корневые системы устойчивее к пониженным концентрациям О2. Недостаточная аэрация приводит к ряду нарушений метаболизма, обычно снижаются рост корней и поглощение минеральных веществ и воды. Из-за пониженного поглощения воды в стеблях часто происходит водный стресс, уменьшающий фотосинтез и усиливающий дыхание. Понижение интенсивности нитрификации уменьшает снабжение азотом. В результате таких нарушений ослабляются рост и мощность растения или даже оно погибает.
ПОВРЕЖДЕНИЯ И МЕХАНИЧЕСКИЕ РАЗДРАЖИТЕЛИ. В листьях значительно увеличивается интенсивность дыхания, если до них дотронуться, потереть или согнуть (рис. 6.19). Следовательно, при измерении интенсивности дыхания (а может быть, и других процессов) необходима осторожность во избежание грубых прикосновений к растительной ткани. Ранения, такие, как срезание плодов, вырезание куска коры или древесины, обычно сопровождаются усилением дыхания. Это можно объяснить различно. Удаление куска ткани усиливает снабжение данной поверхности кислородом. Оно также нарушает существующую в норме компартментацию в клетках и освобождает ферменты и субстраты. Например, потемнение поверхностей разреза растительных тканей происходит в результате окисления фенольных соединений под действием фенолоксидаз, освободившихся при повреждении клеток. Иногда интенсивность дыхания увеличивается при грибных заболеваниях растения, но о механизме этого явления известно мало.
ХИМИЧЕСКИЕ ВОЗДЕЙСТВИЯ. Дыхание чувствительно к ряду химических реагентов, подавляющих различные этапы всего процесс? Например, превращение фосфоглицериновой кислоты в фосфоеноль
207
Ферменты, энергетика и дыхание
Рис. 6.19. Влияние механических повреждений на дыхание листьев лавровишни лекарственной. Контрольную группу старались не беспокоить, опытную теребили руками (Godwin, 1935). Условия исследований:
1 — 14—20°С, свет, сухой воздух, теребили 5 мин; 2 — 14—21°С, свет, сухой воздух, теребили 3 мин; 3 - 13,5-21°С, без света, без сухого воздуха, не дотрагивались 10 мин; 4 — температура без изменений, свет, без сухого воздуха, не дотрагивались 10 мин; 5 — 16—21°, без света, сухой воздух, теребили 7—8 мин ; 6 — температура без изменений, без света, без сухого воздуха, теребили 7 мин ; 7 — контроль
ровиноградную ингибируется фторидом, а специфические этапы цикла Кребса блокируются фторацетатом и маловатом. Антимицин А действует между цитохромами Ъ и с, а такие ингибиторы, как цианид, азид и окись углерода, подавляют конечный этап транспорта электронов (см. рис. 6.2). В некоторых растительных тканях значительная доля дыхания нечувствительна к цианиду и другим ингибиторам терминального участка, что указывает на существование альтернативного пути, по которому электроны переносятся, минуя цитохромы, или, по крайней мере, цито-хромоксидазу (см. рис. 6.2). Цианидрезистентное дыхание также усиливается в некоторых запасающих тканях при хранении в аэробных условиях. О циаиидрезистентном дыхании в одревесневших тканях данных нет.
Дыхательные пути в растительных и животных тканях в общих чертах сходны, они одинаково реагируют на одни и те же ингибиторы, но имеются и некоторые различия. Например, ротенон — мощный ингибитор транспорта электронов в митохондриях животных относительно неэффективен в растениях. Имеющиеся ограниченные данные указывают на отсутствие различий между древесными и травянистыми видами в их отношении к ингибиторам дыхания. Между дыхательными путями проростков и более старых тканей возможны некоторые различия, вызывающие разные реакции на ингибиторы, но по этому вопросу нет достаточной информации. Некоторые гербициды сильно увеличивают интенсивность дыхания. На нее влияют также различные инсектициды, фунгициды и другие химикаты, загрязняющие воздух вещества вроде SO? и составные части смога (см, главу 17).
208
Глава 6
АССИМИЛЯЦИЯ
Термин ассимиляция используется для обозначения превращения питательных веществ, т.е. углеводов, жиров и белков, в новую ткань. Это требует не только больших количеств энергии, поставляемой дыханием, но также и материалов, синтезированных в разных частях дыхательного никла. Нуклеотиды и нуклеиновые кислоты, аминокислоты, жирные кислоты и другие важные для метаболизма растений вещест-дыхательного цикла (рис. 6.20) крахмал ы мальтаза
ва возникают в различных частях
нуклеиновые кислоты
глюкоза - 1 - фосфат глюкоза
АДФ, АТФ НАД, НАД<Т> ФИН, Ко А цитокинины
ДНК РНК
•нуклеотиды-— пентаэо-фосфаты
глюкоза - о-фоа
лромежуточнь^ с
глццеральЗегцаи
фосфодижси-аиртон
целлюлоза
индолцл-уксусная кислота
Фотосинтетический цикл
громе соединения
оС- глицерофосфат
триптофан тирозин , jX фенилаланин алкалоиды, флавоноиды, Лигнин
5елкиУ^"
оруеие
аминокислоты
щикимовая
фосфоёналпиро-бцногредмц» кислота
соединения
кислоты
липиды и родствен-
аланин
кислота
ацетц
лимонная^ кислота
кислота кислота
блочная кислота
изопреноиды, терпены, гиббереллины, каротиноиды, стероиды , абсцизовая кислота
глиоксалевая
кислота
-------
фумаровая кислота
цикл Кребса
изо^имомная кислота.
глутаминовая
{глицин , серин
янтарная
К. -кетоглу кислота
кислота
0г*2Н Нг0-
белки
другие, аминокислоты
порфирины J хлорофиллы , фикоцианины, фитохром, цитохромы , каталаза полифенолоксидазы
Рис. 6.20. Некоторые метаболические пути в растениях
Ферменты, энергетика и дыхание
209
Многие из метаболических путей синтеза и распада этих веществ проходят через ацетилкофермент А, являющийся узловым для ряда важнейших реакций метаболизма.
Ассимиляция — неотъемлемая часть роста, поэтому она наиболее заметна в меристематических участках, таких, как камбий и кончики корней и стеблей. Простые углеводы, передвигаясь в эти меристематические участки, превращаются в целлюлозу, пектиновые вещества и лигнин клеточных стенок. Аминокислоты и амиды превращаются в структурные белки и ферменты новой протоплазмы. Протоплазма клетки образует не только новую протоплазму и новые клеточные стенки, но и обширный ряд других веществ: органические азотсодержащие соединения, хлорофилл, каротиноиды и другие пигменты, липиды и производные изопрена (эфирные масла, смолы и каучук), стероиды, таннины, алкалоиды, гормоны и многочисленные другие соединения. Большинство их играет важную роль в метаболизме растений, но некоторые, такие, как алкалоиды и каучук, не имеют известных существенных для растений функций.
6.2. ХИМИЧЕСКИЙ СОСТАВ ПОБЕГОВ СОСНЙ1 ЛАДАННОЙ (Chung, Barnes, 1977) *
Фракция
Группа веществ
Содержание, %
от сухого от веса
веса фракции
Азотсодержащие 8,4
соединения Аминокислоты 7,2
Белки 90,7
Нуклеиновые кислоты 2,1
Углеводы 38,0
Редуцирующие сахара 5,1
Сахароза 8,2
Целлюлоза 56,0
Гемицеллюлозы 25,7
Пектин 5,0
Липиды 5,3
''7 Жирные и смоляные кислоты 74,8
Глицериды 6,0
Неомыляющиеся липиды 19,2
Лигнин 23,3
Органические 3,5
кислоты Шикимовая кислота 48,7
Хинная кислота 51,3
Фенольные соединения 20,0
Неорганические ве- 1,5
щества
* Таблица несколько перестроена переводчиком Д.П. Викторовым.
В табл. 6.2 приведен примерный химический состав побегов (хвои и прилегающих веток) сосны ладанной. Состав близок к тому, который можно было ожи-
210
Глава 6
дать, за исключением высокого содержания фенолов, что удивительно ввиду высоких энергетических затрат на их синтез и неопределенности в отношении их использования растением. Относительное количество израсходованного субстрата и СО^, образовавшегося в процессе синтеза 1 г каждой из главных составных частей, показано в табл. 6.3. Расход субстрата и выделение COj возрастают вместе со степенью, до которой восстанавливается вещество, так как для образования сильновосстановленных соединений, таких, как липиды и фенолы, нужно больше энергии. Например, синтез 1 г углевода требует около 1,2 г глюкозы и 0,11 г О2 с выделением 0,149 г СОу, тогда как синтез 1 г более восстановленного липида требует 3 г глюкозы и 0,3г О2. При этом выделяется 1,5 г СО2. При синтезе лигнина также расходуется большое количество углеводов.
6.3. КОЛИЧЕСТВО ИСПОЛЬЗОВАННОГО СУБСТРАТА
И ОБРАЗОВАВШЕЙСЯ ДВУОКИСИ" УГЛЕРОДА В ХОДЕ СИНТЕЗА ОСНОВНЫХ ХИМИЧЕСКИХ КОМПОНЕНТОВ СОСНЫ ЛАДАННОЙ, Г/Г ФРАКЦИИ
(Chung, Barnes, 1977)
Фракция
Использован- Побочные прений субстрат дукты
Глюкоза °2 с°2 н2о
Азотсодержащие соединения 1,58 0,28 0,40 0,65
Углеводы 1,18 0,11 0,13 0,16
Липиды 3,02 0,30 1,50 0,82
Лигнин 1,90 0,04 0,27 0,66
Органические кислоты 1,48 0,35 0,48 0,35
Фенольные соединения 1,92 0,37 0,56 0,73
Ход ассимиляции и состав образуемых веществ определяются присутствием определенных ферментов. Состав ферментов определяется наследственными факторами, хотя количество и набор присутствующих ферментов могут изменяться под влиянием окружающей среды. Различные семейства растений образуют характерные химические соединения. Например, каждый вид сосны образует свои собственные специфические смолы, а иногда различия имеются и между географическими расами. Делались попытки увязать классификацию растений с химическим составом.
УГЛЕВОДЫ
Углеводы имеют особое значение в жизни растений, так как ‘они являются прямыми продуктами фотосинтеза и, следовательно, играют роль первичных запасающих энергию соединений и исходных органических веществ, из которых-синтезируется большинство других соединений, обнаруживаемых в растениях, Углеводы -это также и основной компонент клеточных оболочек. Они представляют собой отправную точку в синтезе жиров и белков. Большие количества углеводов окисляются при дыхании (см. главу 6), много их накапливается в виде запасных питательных веществ, а часть их различными путями теряется растением. Растворимые углеводы увеличивают осмотическое давление клеточного сока, а такие углеводы, как пентозаны, пектиновые вещества, камеди и слизи, увеличивают водоудерживающую способность тканей. В количественном отношении углеводы являются наиболее важной составной частью древесных растений, заключая в себе около 3/4 их сухого веса.
КЛАССЫ УГЛЕВОДОВ
Углеводы состоят из углерода, водорода и кислорода в полном или приблизительном соответствии с эмпирической формулой (Cf^O)^. Многие углеводы содержат также другие элементы, например фосфор или азот. Углеводы можно подразделить на три основные группы: моносахариды, олигосахариды и полисахариды (рис. 7.1).
МОНОСАХАРИДЫ. - Эта группа включает простые сахара и их производные. Это основные углеводные единицы, из которых образуются более сложные соединения. Моносахариды состоят из атомов углерода, к которым присоединяются водородные атомы, гидроксильные п ?
группы и либо альдегидная (—С—Н), либо кето иная (—С—R) группа. Количество атомов углерода в моносахаридах варьирует от 3 до 8, но чаще всего их 5 (пентозы, C5H1QO5) или 6 (гексозы, С^Н^Об)- Моносахариды не дают гидролитическим путем сахара с меньшим молекулярным весом.
В древесных растениях встречается много простых сахаров, обычно в очень небольших количествах, вероятно, из-за быстрого включения их в полисахариды. Исключение составляют шестиуглеродные сахара глюкоза и фруктоза. Глюкоза присутствует в больших количествах, особенно в некоторых плодах, имеется, по-видимому, в каждой живой клетке. Фруктоза также встречается обычно в больших количествах, но в более низких концентрациях по сравнению с глюкозой. Глюкозу и фруктозу находят не только в живых клетках, но также и в ксилемном соке некоторых деревьев, например кленов и берез (см. главу 11). Фосфорилированные производные (т.е. с присоединенными фосфатными группами) глюкозы и фруктозы являются исходными элементами многих метаболических превращений углеводов (см. рис. 6.20). Многие гексозы встречаются в конденсированной форме, например галактоза и манноза — в виде галактанов и маннанов. Свободные пентозы найдены в растениях только в следовых количествах, но продукты их конденсации, пентозаны, являются важными составными частями клеточных оболочек. Пентозы арабиноза и ксилоза редко встречаются в свободном виде, но часто присутствуют в качестве компонентов поли-
212
Глава 7
Моносахариды Олигосахариды: Полисахариды Другие
Дисахариды Трисахариды Тетрасахариды соединения
Пентозы(CsH1Q05):________ ______________________
щадйуадза, ксилоза, ридаза
Гексозы(С6нпо5); — ....——---------——
глюкозп целлобиоза мальтоза фруктоза—— сахароза 2 глюкозы "I ________
1 фруктоза J
1 глюкоза "I • фруктоза?-* ..—...
1 галактозу
мелецитоза
рафиноза
Пентозаны (С5нв0ч)п: 4
пептоновые вещества, гемицеллюлозы, камеди, слизи
Гексозаны fCsHJ0^:
целлюлоза, крахмал
!ликозиды, антоцианы, антоксантины, тонн ины, нуклеиновые кислоты
1 глюкоза ] 1 фруктоза.
2 галактозы]
стахиоза
Рис. 7.1. Взаимоотношения между некоторыми важными углеводами и продуктами их гидролиза
меров клеточных оболочек: арабанов и ксиланов. Другая пентоза, рибоза, также встречается в связанном виде как составная часть нуклеотидных коферментов: АТФ, НАД, НАДФ, ФАД и КоА. Рибоза входит также в состав рибонуклеиновой кислоты (РНК). Дезоксирибоза встречается в составе нуклеотидов, составляющих дезоксирибонуклеиновую кислоту (ДНК).
Многие моносахариды участвуют в цикле КаЛьвина — Бенсона, темновой фазы фотосинтеза и в пентозофосфатном цикле дыхания. Главным соединением, образующимся в первом и используемым во втором цикле, является глюкоза. Пентоза рибулозо-1,5-дифосфат служит местом связывания СС>2 при фотосинтезе. Оба цикла, особенно пентозо-фосфатный шунт, служат источником пентоз и многих других моносахаридов, входящих в состав более сложных молекул.
ОЛИГОСАХАРИДЫ. Группа состоит из соединенных двух или более молекул моносахаридов. К основным олигосахаридам относятся: дисахариды (сахароза и мальтоза), трисахариды (рафиноза и мелецитоза) и тетрасахариды (стахиоза). Дисахарид сахароза считается основным олигосахаридом растений благодаря высоким концентрациям * в клетках, широкому распространению и значению для метаболизма. Вместе с крахмалом сахароза является главным резервным углеводом. Во многих растениях сахарозой представлено свыше 95% (по сухому весу) материалов, перемещаемых по ситовидным трубкам флоэмы. Мальтоза встречается в растениях в меньших концентрациях, чем сахароза.
Помимо сахарозы и мальтозы часто обнаруживаются в различных количествах и другие сахара. Например, в ситовидных трубках некого-
Углеводы
213
----------Сахароза----------
---------------Рафиноза ——---------------
---------------Стахиоза----------------------------------- --------------- Вербаскоза -------------------------------
Рис. 72. Олигосахариды семейства рафинозы (Zimmerman, Brown, 1971)
рых растений найдены небольшие количества высших олигосахаридов семейства рафинозы: рафиноза, стахиоза и вербаскоза. Все эти сахара сходны, они состоят из сахарозы с разным числом присоединенных к ней галактозных единиц (рис. 7.2). Сахара семейства рафинозы имеют относительно малое значение для большинства растений, но они важны для нескольких семейств, в том числе Bignoniaceae, Celastraceae, Сэт-bretaceae, Myrtaceae, Oleaceae, Verbenaceac.
Состав углеводов в разных тканях различен. Например, по сообщению Дитриха и Шаха (1965), у бука углеводы включали сахарозу, глюкозу, фруктозу, мальтозу (или целлобиозу), рафинозу, стахиозу и пять не-идентифицированных кетоз. Сахароза, глюкоза и фруктоза были основными сахарами по всему дереву. Рафиноза и иногда стахиоза встречались в почках и молодых листьях. Сок флоэмы содержал преимущественно сахарозу с некоторым количеством рафинозы и следами фруктозы.
ПОЛИСАХАРИДЫ. Самые важные полисахариды деревьев — целлюлоза и крахмал. Целлюлоза — самое распространенное органическое вещество. Приблизительно из 30 млрд, т углерода, превращаемых растениями в органические соединения, около 1 /3 приходится на целлюлозу. Целлюлоза — главная составная часть клеточных оболочек, образующих каркас древесных растений. Каждая молекула целлюлозы состоит не менее чем из 3000 остатков глюкозы, связанных кислородными мостиками между 1-ми 4-м атомами углерода смежных молекул с образованием длинных, прямых, неразветвленных цепочек (рис. 7.3). Цепочки уложены вместе в мицеллы, организованные в микро фибриллы. В чистых целлюлозных клеточных стенках (например, в волокнах хлопка) свободные пространства заняты водой. В тканях древесины они частично заполняются лигнином и пектиновыми веществами, кутином или суберином в тех тканях, где эти вещества имеются. Особенность целлюлозы — ее нерастворимость в воде и органических растворителях и высокая устойчивость к химическому и ферментативному
214
Глава 7
Молекула целлюлозы
Модель поперечносвязанных параллельных цепей
Рис. 7.3. Строение целлюлозы, состоящей из чередующихся (1и /Зединиц глюкозы. Гидроксильные группы, выступающие с обеих сторон цепи, образуют водородные связи с соседними гидроксильными группами, в результате образуется пучок поперечно-связанных параллельных цепей
разрушению. Распад- целлюлозы до глюкозы требует двух ферментов: целлюлазы, катализирующей образование целлобиозы, и целлобиазы, доводящей расщепление до глюкозы.
Крахмал - самый распространенный резервный углевод древесных растений. Крахмальные зерна не могут передвигаться из клетки в клетку, следовательно, крахмал должен синтезироваться в тех тканях, где он обнаруживается. Крахмал образуется путем конденсации нескольких сотен молекул глюкозы в длинную спиральную цепь. В крахмале, как и в молекуле целлюлозы, остатки глюкозы связаны друг с другом кислородными мостиками между 1-ми 4-м атомами углерода, но у крахмала это* a-связь, а у целлюлозы 3-связь. В отличие от целлюлозы крахмал легко расщепляется ферментами и потому участвует в метаболических процессах. Крахмал состоит из двух компонентов, различающихся некоторыми физическими свойствами. В крахмале преобладает амилопектин, состоящий из очень длинных молекул с многочисленными боковыми цепями. Другой компонент, амилоза, состоит из неразветв-ленных цепей, содержащих 300-1000 остатков (рис. 7.4). По сравнению с амилопектином амилоза дает более темную синюю окраску с йодом, лучше растворяется в воде и она более вязкая. Крахмал накапливается в особых зернах, состоящих из многих слоев, что придает им слоистый вид. Крахмальные зерна часто встречаются в живых клетках осевой и лучевой паренхимы заболони древесных растений, а также в живых клетках флоэмы внутренней части коры. Количество крахмала в древесине деревьев изменяется по сезонам. В больших количествах крахмальные зерна встречаются в живых клетках почти всех листьев.
Гемицеллюлозы представляют собой плохо очерченную группу полисахаридов, встречающихся во всех древесных тканях. Эта группа включает арабаны, ксиланы, галактаны и маннаны. В отличие от целлюлозы гемицеллюлозы растворимы в разбавленных кислотах и щелочах. Иногда они используются в качестве запасных питательных веществ. Гемицеллюлозы обнаруживаются в некоторых семенах (в том числе у хурмы и некоторых пальм). Во время прорастания семян они расщепляются и используются.
215
Углеводы
Рис. 7.4. Строение крахмала: а — амилоза; б — амилопектин
Пектиновые вещества — гидрофильные соединения, встречающиеся в срединной пластинке и первичных оболочках клеток, особенно в плодах. В одревесневших тканях в больших количествах не присутствуют. Камеди и слизи — это углеводы, напоминающие пектиновые вещества. Известный пример камеди — еловая камедь. Примеры слизей: слизистое вещество внутренней части коры вяза ржавого и клейкие вещества, найденные в бобах рожкового дерева и гледичии обыкновенной. Хорошо известный гуммиарабик (аравийская камедь) получают из акании аравийской. Из ран на стволах вишневых, сливовых и персиковых деревьев также выделяются камеди. Усиленное образование камеди в результате ранения у сливовых, персиковых, цитрусовых и некоторых других деревьев называется гуммозом. Это явление вызывается гидролизом составных частей клеточных стенок. В химическом отношении эта камедь сильно отличается от смолы, выделяемой ранами хвойных деревьев.
ПРЕВРАЩЕНИЯ УГЛЕВОДОВ
Многие углеводы растений постоянно подвергаются превращениям из одной формы в другую или превращаются в соединения, используемые на дыхание и синтез жиров, белков и других веществ (см. рис.6.20). Взаимопревращения крахмала и сахарозы характерны для вегетативных и репродуктивных органов. У развивающихся семян, например, имеется период, в течение которого сахара (в основном сахароза) превращаются в крахмал. В ряде созревающих плодов крах
216
Глава 7
мал превращается в сахара. При инкубации растений с ^СО2 радиоактивный углерод быстро включается во множество сахаров, органических кислот и других соединений.
Так, после 1-минутной экспозиции побегов пихты в атмосфере около 50% метки было в сахарах и крахмале, а 30-40% в фрсфоглицериновой кислоте, У проростков сосны веймутовой после воздействия в течение 8 ч около 98% растворимого в этаноле 14с было во фракции сахаров, а остальные 2% поровну распределялись между аминокислотами и органическими кислотами, Сахароза составляла около 75% фракции сахаров, глюкоза и фруктоза вместе еще 20%. Рафиноза обнаруживалась только в небольших количествах. Нельсон (1964) подвергал 3-летние проростки сосны веймутовой воздействию *4(702 в разное время года в течение 1 ч с последующей ассимиляцией в течение 8 ч на воздухе, Основная масса 14с, выделенного затем из корней, обнаруживалась в сахарах. Главным сахаром во все сезоны была сахароза, составлявшая 79% сахаров в апреле, 89% в мае, 94% в июле, 86% в августе и 88% в октябре. Рафиноза составляла 16% сахаров в апреле, 6% в октябре, в июле не обнаруживалась, Глюкоза и фруктоза присутствовали в небольших количествах в течение всего года: обычно по 2-5% каждой, Трип и другие исследователи (1963) освещали отделенные листья 20 видов растений в атмосфере с 14сО2 и нашли радиоактивный углерод в следующих соединениях: маннит в 4 видах, вербаскоза -3, стахиоза — 9, рафиноза - 16, мелибиоза - 2, мальтоза - 15, сахароза — 20, галактоза - 8, глюкоза - 17, фруктоза - в 12 видах,
ФОСФОРИЛИРОВАНИЕ. Первым этапом многих превращений углеводов является фосфорилирование — процесс, в котором моносахариды взаимодействуют с АТФ с образованием фосфорных эфиров сахаров, а АТФ превращается в АДФ. Например, один сахар может превращаться в другой по общей схеме: сахар А + АТФ Фермент А фосфат сахара А + АДФ;
фосфат сахара А фермент £ фосфат сахара В
Примером может служить превращение глюкозы в глюкозо-6-фосфат и затем во фруктозо-6-фосфат:
Н
Н- С=О
1
Н - С -ОН
I
но-с-н
ц огт гексокин^а
Н - С - ОН а1'Ф а/ф
I
н - с - он
11
Глюкоза
Н- С=О I
Н - С - ОН HO-L-H н - А - он
н-А-он о
Гон * он
Н - С - ОН
СГ° фосфоглюкоизомераза^ НО - Н
Н - С - ОН
Глюкозо-6-фосфат
н-^-он О
Н-С-О-р-ОН
Н ОН
Фрукто зо-6-фосфат
Н С I н
Фосфорилированные моносахариды встречаются среди первичных продуктов фотосинтеза. Они либо прямо включаются в химические реакции с образованием других фосфорных эфиров, либо превращаются в передвигающиеся по растению и накапливающиеся формы. Крахмал при высоком содержании сахаров накапливаете)!, при низком — превращается в сахара. При пониженных температурах равновесие сдвигается в сторону образования сахаров. Фосфорные эфиры сахаров играют особо
Углеводы
217
важную роль, так как являются промежуточными веществами в синтезе и распаде крахмала и сахарозы, а также субстратами гликолиза, брожения, фотосинтетической фиксации СО2 и ряда окислительных процессов. Кроме того, фосфорные эфиры входят в качестве составных частей в нуклеиновые кислоты и коферменты. Превращения углеводов не ограничиваются только сахарами. Например, сахара могут превращаться в спирты, как в случаях образования сорбита и маннита из глюкозы и маннозы.
САХАРОЗА. Образование сахарозы из глюкозы и фруктозы возможно в случае, когда один из этих сахаров связан в комплекс с нуклеотидом. Например, глюкоза в форме уридиндифосфатглюкозы (УДФГ) может взаимодействовать с фруктозой с образованием сахарозы:
УДФГ + фруктоза ду 1 ___сахароза + УДФ.
УДФГ может также реагировать с фруктозо-6-фосфатом с образованием сахарозофосфата, который в свою очередь гидролизуется фосфатазой, в результате чего образуется свободная сахароза:
УДФГ + фрукто зо-6-фосфат*ь=^ сахарозо фосфат + УДФ.
При гидролизе сахароза дает глюкозу и фруктозу. Эта реакция, катализируемая сахаразой, необратима:
С12Н22ОЦ + Н2О С6Н12О6 + С6Н12О6.
сахароза глюкоза фруктоза
КРАХМАЛ. Синтез крахмала происходит несколькими путями. Самые важные из них, по-видимому, следующие:
1. Фосфорилазная реакция, при которой молекулы глюкозо-1-фосфата объединяются с образованием крахмала по такой схеме:
глюкозо-1-фосфат + цепочка остатков глюкозы (и единиц) фосфорилаза цепочка остатков глюкозы (п + 1 единиц) + фосфат
2. Путь при участии уридиндифосфатглюкозы:
гпюкоэо-1 -фосфат + УТФ Дироф^форилаза удфг + пирофосфат;
УДФГ + цепочка остатков глюкозы (и единиц) УРансг;1йк°удф + цепочка остатков глюкозы (и+1 единиц)
3. Путь при участии аденозиндифосфатглюкозы (АДФГ): t
глюкоэо-1-фосфат + АТФ пи рофо сфорил^а ддфг + пирофосфат;
АДФГ + цепочка остатков глюкозы («единиц) трансглико зил аза адф + цепочка остатков глюкозы (« + 1 единиц)
Крахмал распадается двумя раздельными путями — с участием фосфорилаз или гидролаз. Реакция при участии фосфорилазы преобладает в тканях, не играющих главной роли в хранении питательных веществ. Гидролазы обнаруживаются в наибольших концентрациях в тканях, выполняющих функцию основных хранилищ питательных веществ.
218
Глава 7
Гидролазная реакция заключается в гидролизе крахмала путем разрыва связей с одновременным присоединением воды. Мальтазная реакция состоит в гидролизе крахмала до глюкозы через промежуточные продукты:
крахмал ——>- декстрины----мальтоза------глюкоза
Крахмал распадается до мальтозы под действием а-амилазы и /3-амилазы, каждая из которых действует на амилозу и амилопектин. Окончательное превращение мальтозы, в глюкозу осуществляется ферментом мальтазой.
ИСПОЛЬЗОВАНИЕ УГЛЕВОДОВ
Углеводы, образующиеся при фотосинтезе, выполняют несколько функций. Большая часть используется на рост после их передвижения в кончики стеблей и корней, в камбий и репродуктивные структуры, где они превращаются в новую протоплазму, клеточные оболочки и различные пррдукты метаболизма. Другая часть накапливается в виде запасных питательных веществ и затем используется на рост. Значительное количество углеводов окисляется при дыхании с освобождением энергии, необходимой для связанных с ростом синтетических процессов (см. главу 6). Углеводы могут через сросшиеся корни передвигаться к соседним растениям, а иногда к полупаразитам и облигатным паразитам (см. главу 11). Часть углеводов вымывается и улетучивается из листьев и теряется с корневыми выделениями. Часть углеводов отводится к симбионтам, таким, как микоризы и азотфик-сирующие бактерии. Микоризы получают сахарозу, глюкозу и фруктозу от растения-хозяина и превращают их в такие формы углеводов (маннит, трегалозу и гликоген), которые не могут быть снова поглощены хозяином. По подсчетам Харли (1971), до 10% потенциальной годовой продукции древесины елового леса уходило на образование плодовых тел микоризных грибов. Однако следует учитывать и значительные выгоды, извлекаемые микоризными деревьями благодаря усиленному поглощению питательных веществ.
Пропорции, в которых фонд углеводов используется на разные цели, сильно различаются у разных видов и в различных условиях. По некоторым данным, около 35% углеводов идет в плоды, 45% на рост вегетативных частей и 18% на дыхание. По оценке Лархера <1975), около 35% ассимилятов бука в Дании используется на рост, 45% на дыхание, а 20% теряется при сбрасывании листьев, ветвей и других частей растений, Как отмечалось в главе 6, на дыхание деревьев в тропиках теряется большая доля ассимилятов, чем у деревьев умеренной зоны.
Соотношение углеводов, используемых на рост кроны, ствола и корневой системы, изменяется с возрастом. В старых деревьях больше \ сухого вещества сосредоточено в главном стволе и соответственно меньше в кроне и корневой системе. На корни молодых деревьев сосны обыкновенной приходится почти половина общей массы растений, а в старых деревьях намного меньше.
219
Углеводы
Рис. 7.5. Влияние плодоношения на содержание крахмала у вечнозеленых деревьев авокадо (Cameron, Borst, 1938) :
1 — не плодоносящие; 2 — плодоносящие; 3 — плоды собраны
РОСТ. Как запасные, так и только что синтезированные углеводы используются на ростовые процессы. Когда весной начинается рост, резервы углеводов в корнях и стеблях истощаются из-за их передвижения в растущие ткани. Различные потребители углеводов в первую очередь обычно истощают те резервы, которые расположены поблизости от места утилизации. Вследствие того, что различные части деревьев растут с разной скоростью и в разное время года, интенсивность утилизации углеводов значительно варьирует по оси дерева. Например, на растяжение побегов большинства видов углеводы используются в течение более, короткого периода, чем на камбиальный рост у тех же деревьев (см. главу 3), а продолжительность сезонного удлинения корней больше продолжительности активности камбия и растяжения побегов. Следовательно, удлинение корней истощает запасы углеводов особенно длительное время. Интенсивность и продолжительность использования углеводов на камбиальный рост в стеблях, ветвях и главных корнях также значительно варьируют.
Процент доступных углеводов, используемых на образование новых Тканей, зависит от вида и года. Большие годовые изменения количества питательных веществ, используемых на вегетативный рост деревьев данного вида, часто бывают связаны с нерегулярностью репродуктивного роста.
В годы обильного плодоношения или образования семян большая доля углеводного запаса переключается с вегетативного над>епродуктивный рост (рис.7,5). В деревьях, у которых плодоношение не строго двухлетнее (например, персиковое
220
Глава 7
дерево), распределение ассимилятов на вегетативный и репродуктивный рост*с возрастом постепенно меняется. Вегетативный рост персиковых деревьев был быстрым до первого года плодоношения, т.е. до 6-летнего Возраста. С этого времени вегетативный рост снижался с возрастающей скоростью, а плодоношение увеличивалось. Ассимиляты отводились на рост плодов в основном из корней и главного ствола, а рост листьев и побегов снижался незначительно. Следовательно, как только деревья начинали плодоносить, сразу устанавливались и впоследствии усиливались строго конкурентные отношения между отдачей и потреблением.
НАКОПЛЕНИЕ УГЛЕВОДОВ
В каждом вегетационном периоде накопление резервов питательных веществ необходимо для выживания многолетних растений тех мест, где зимы слишком холодны для фотосинтеза. Сказанное верно, в частности, в отношении лиственных деревьев, у которых поддержание жизни зимой и возобновление роста весной зависят от наличия питательных веществ, накопленных за предшествующий вегетационный период. Хотя главным запасным продуктом в древесных растениях является крахмал, но запасаются и другие вещества, в том числе жиры, азотистые соединения, сахароза, рафиноза, фруктозаны и гемицеллюлозы.
Баланс углеводов древесного растения можно выразить так:
Приход= углеводы, образовавшиеся при фотосинтезе.
Расход= углеводы, использованные на ассимиляцию и дыхание.
Сальдо= углеводы, накопленные- растением.
РАСПРЕДЕЛЕНИЕ УГЛЕВОДОВ. Запасные углеводы накапливаются преимущественно в клетках паренхимы. Гибель.клеток сопровождается утилизацией или потерей запасов. Много исследований посвящено накоплению крахмала и сахаров в различных тканях и органах. Количество углеводов в разных частях древесных растений заметно различается. Наблюдаются также сезонные колебания количества и форм углеводов. Имеются также различия в характере сезонных колебаний накопления углеводов между листопадными и вечнозелеными видами тропиков и умеренной зоны.
Необходимо различать общее содержание и концентрацию углеводов в разных частях растений. Распределение углеводов часто выражают в процентах от сухого веса различных тканей. Это может ввести в заблуждение, так как высокие концентрации углеводов часто наблюдаются в тканях, составляющих небольшую долю сухого веса всего растения. Например, в молодых и старых деревьях концентрация углеводов в корнях обычно выше, чем в надземных частях. Тем не менее у взрослых деревьев именно надземные части являются основными вместилищами углеводов, поскольку ствол, ветви и листья имеют больший сухой вес, чем корни.
У изученных Марником (1933, 1942) яблонь концентрация углеводов в корнях была выше, чем в стволах, но тем не менее надземные части, которые приблизительно в 3 раза тяжелее корней, содержали больше углеводов (табл. 7.1). Распре-
Углеводы
221
7.1. РАСПРЕДЕЛЕНИЕ КРАХМАЛА И САХАРОВ В ЯБЛОНЯХ СОРТА ГРАЙМС В СЕРЕДИНЕ ОКТЯБРЯ (ПО Muineek, 1942) (ЧИСЛИТЕЛЬ - ФУНТЫ; ЗНАМЕНАТЕЛЬ - % ОТ СУХОГО ВЕСА)
Наименование показателей Сухой вес, фунты Крахмал и сахар Крахмал Сахар
Листья 32,82 3,09/9,41 0,92/2,80 2,17/6,61
Укороченные побеги Древесина, лет: 6,33 0,69/10,90 0,37/5,85 0,34/5,05
1 5,94 0,65/10,94 0,35/5,89 0,30/5,05
2 7,12 0,73/10,25 0,45/6,32 0,28/3,93
3 8,74 0,85/9,73 0,51/5,84 0,34/3,89
4-6 46,27 2,48/5,36 1,32/2,85 1,16/2,51
7-10 124,00 7,20/5,81 4,46/3,60 2,74/2,21
11-18 101,20 5,95/5,88 4,01/3,96 1,94/1,92
Главный ствол 68,19 7,57/11,10 6,14/9,00 1,43/2,10
В с е г о в надземных частях 400,61 29,21/7,29 18,53/4,63 10,68/2,67
Осевой корень Корни, лет: 62,80 9,94/15,83 6,91/11,00 3,03/4,83
18-14 46,90 8,22/17,53 6,34/13,52 1,88/4,01
13-7 22,52 5,52/24,51 3,97/17,63 1,55/6,88
6-1 5,40 1,10/20,37 0,86/15,93 0,24/4,44
В с е г о в подземных частях 137,62 24,78/18,01 18,08/13,14 6,70/4,87
В с е г о в растении, 538,23 53,99/10,03 36,61/6,75 17,38/3,23
среднее
деление углеводов в надземных и подземных частях также является функцией возраста растений, поскольку отношение масс подземной и надземной Частей с возрастом постепенно уменьшается. В 1-летних яблонях запасы углеводов распределялись между подземными и надземными частями почти поровну.
В листьях концентрация углеводов обычно высокая, но они составляют относительно небольшую долю всех углеводов древесного растения. Хвоя сосны виргинской, исследованной Гентингом (1945), содержала 16,6% углеводов, что почти в 7 раз превышает их концентрацию в древесине ствола. Содержание углеводов (крахмал плюс сахара) в листьях яблони доходило до 9%, что намного больше величин, приводимых для других тканей. Общее содержание углеводов в листьях составляло лишь примерно 5% запасов углеводов всего дерева.
В стволах, ветвях и корнях клетки осевой и лучевой паренхимы и клетки сердцевины служат основными местами накопления крахмала. При определении количества накопленных питательных веществ в стволах древесных растений желательно делать различия между заболонью и ядром, поскольку последнее не имеет живых клеток и обычно не накапливает питательных веществ. В коре крахмал откладывается в паренхиме и альбуминовых- клетках,разбросанных по ряду структур. Концентрация углеводов в коре часто очень высока,
222 Глава 7
но у многих видов общее количество их в коре меньше, чем в древесине. Во внутренней живой флоэме ветвей, главных стволов и корней , взрослых буковых деревьев зимой концентрация запасных углево-1 дов часто в 2—3 раза выше, чем в древесине, хотя общее их количество' в древесине больше, чем в коре. Венджер (1953) также нашел в ;• стволе амбрового дерева более высокие концентрации углеводов в коре, чем в древесине, но общее содержание углеводов было намного больше в древесине. По данным Хептинга (1945), в коре корней сосны виргинской концентрация углеводов была намного выше, чем в древесине корня.
Роль, лучей в хранении и передвижении углеводов. Клетки лучевой паренхимы выполняют важную роль в хранении запасных веществ. Количество и тип запасаемых в лучах веществ меняются с удалением от камбия. Количество крахмальных зерен, многочисленных в лучевых клетках, находящихся вблизи камбия, уменьшается по направлению к внутренней части заболони, В лучевых клетках ядра содержание крахмала незначительно или его вообще нет. В противоположность радиальному распределению крахмала в клетках лучей ксилемы содержание жиров в лучевых клетках внутренней части заболони выше, чем в клетках, расположенных близко к камбию.
Доказательство участия лучей в латеральном передвижении основывается на нескольких источниках. Радиоактивная сахароза, нанесенная на участок камбия, передвигается вдоль лучей в глубь заболони, а жизнеспособность веретеновидных инициальных клеток; камбия зависит от контакта с клетками лучей. Циглер (1965), обработав листья древесных растений радиоактивной серой, обнаружил впоследствии радиоактивность во флоэмных и ксилемных лучах, что еще раз подчеркивает важную роль лучевых клеток в латеральном транспорте. Количество запасенных в лучевых клетках углеводов подвержено сезонным изменениям.
Лучи, а точнее их специализированные клетки (называемые контактными клетками), связанные с сосудистыми элементами большими порами, кроме транспорта углеводов от флоэмы к ксилеме, выделяют сахара в сосуды- Саутер (1972) обнаружил, что крахмал начинал пропадать весной из клеток ксилемной паренхимы клена сахарного одновременно с появлением сахара в пасоке. Эти изменения совпадали с набуханием почек. Скорость выделения сахаров в сосуды во много раз больше скорости, которая может быть объяснена диффузией. Выделение сахаров в сосуды контактными клетками зависит от метаболической активности клеток и от температуры. Ингибитор ПХМБ (парахлормеркурибензоат) резко снижает выход сахаров из клеток паренхимы в сосуды. В связи с тем, что ПХМБ подавляет большинство участвующих в дыхании дегидрогеназ, создается впечатление, что дыхательная активность требуется для выхода сахарозы в сосуды.
ИСПОЛЬЗОВАНИЕ ЗАПАСНЫХ УГЛЕВОДОВ. Наиболее очевидно использование запасных углеводов для поддержания дыхания и роста в периоды, когда питательные вещества не поступают непосредственно от фотосинтеза. Вся метаболическая активность в ночное время зависит от питательных веществ, накопленных за день. У листопадных деревьев
Углеводы
223
дыхание и другие процессы поддерживаются зимой благодаря использованию запасов питательных веществ. По исследованиям, у молодых яблонь менее четверти углеводного запаса использовалось весной на рост новых тканей, а большая часть оставшихся углеводов была использована на дыхание.
Запасные углеводы, помимо их роли в поддержании дыхания в ночное время и зимой, важны и в начале вегетационного периода, что подчеркивается фактом роста корней многих видов листопадных деревьев до распускания листьев. Образование поросли и корневых отпрысков также зависит от мобилизации углеводных запасов.
Клэрк и Лиминг (1953) обнаружили, что отрастание дуба после кольцевания происходило в первый и второй сезону пока не исчерпались запасы углеводов. У деревьев, окольцованных на более позднем этапе вегетационного периода, отрастание в следующем году замедлялось. По Наблюдениям Тью (1970), продолжительность периода, в течение которого у осины образовывались корневые отпрыски, зависела от количества запасных углеводов. Шир и Засада (1973) нашли положительную корреляцию между массой корневых отпрысков осины и количеством запасных углеводов. Эти наблюдения показывают, что отрастание уменьшается по мере исчерпания запаса углеводов. С другой стороны, Венджер (1953) сообщил о втором минимуме отрастания у амбрового дерева в конце лета, когда содержание углеводов в корнях было относительно высоким. Он сделал вывод о том, что регуляторы роста играли более важную роль, чем
запасы питательных веществ,
При рубке деревьев в начале лета, когда их листва только что распустилась и запасы углеводов малы, пневая поросль развивается слабо по сравнению с обильным отрастанием пней деревьев, срубленных в период покоя (рис. 7.6).
Вследствие отсутствия у лиственных деревьев умеренной зоны фотосинтезирующих листьев во время
Рис. 7.6. Влияние срока кольцевания на количество и размеры побегов, образованных деревьями дуба мерилендского в течение шести сезонов после кольцевания (Clark, Liming, 1953):
а — число деревьев, образующих побеги после кольцевания в различные месяцы: 1 — произведен надрез; 2 — снята кора; б — высота и толщина этих побегов; контуры сплошной линией - кольцевание надрезом; контуры штриховой линией — кольцевание снятием коры
224 Глава 7
набухания почек и увеличения их сухого веса начальные этапы роста побегов зависят от углеводных запасов.
Случай особой важности запасных углеводов для роста побегов отметил Шимпер (1903), сообщавший, что почки тропического вида Brownia несколько дней разрастались со скоростью 2,6 см в день, в то время как листья оставались свернутыми и фотосинтеза не было. Следовательно, рост побегов поддерживался за счет запасов углеводов, передвигавшихся из других частей дерева. Другой пример значения запасов демонстрирует появление второй партии листвы из покоящихся почек у деревьев умеренной зоны весной, после гибели первой партии молодых листьев от заморозков.
Важность запасов для листопадных деревьев еще раз подчеркивают: во-первых, быстрое исчезновение запасных углеводов после распускания почек и развития побегов, и, во-вторых, мобилизация меченых радиоактивным углеродом запасов. Согласно подсчетам Хенсена (1971), от 1/2 до 2/3 углеводов, необходимых для роста цветков и побегов яблони в самом начале вегетационного периода, поступали скорее из запасов, чем от продуктов текущего фотосинтеза. Это относилось только к периоду появления 5—6 первых листьев, после которого рост плодов и побегов был связан с утилизацией непосредственных продуктов фотосинтеза.
В отличие от листопадных вечнозеленые деревья к началу ежегодного роста побегов уже располагают функционирующей фотосинтетической системой. Например, у сосен синтезированные старой хвоей углеводы используются уже на самых ранних этапах удлинения побегов. Как и многие листопадные деревья, вечнозеленые не переносят потери кроны. Такие наблюдения привели к сомнениям относительно значения запасных углеводов для роста побегов у вечнозеленых. Однако имеются веские доказательства того, что запасные углеводы все-таки играют важную роль в росте вечнозеленых растений. Так, в рано начинающемся росте корней утилизируется значительная доля запасов углеводов.
Рис. 7.7. Изменение содержания углеводов в однолетних и новых побегах дугласии (Krueger, 1967):
а — концентрация; б — общее количество; 1 — однолетние побеги; 2 — новые побеги
Углеводы
225
Рис. 7.8. Сезонные изменения содержания гексоз, сахарозы, крахмала и общего содержания углеводов во внутренней и внешней древесине (внешней части заболони) клена сахарного на севере штата Вермонт (Jones, Bradlee, 1933):
а — внутренняя древесина; б — внешняя древесина; 1 — общее содержание; 2 — гексоза; 3 — крахмал; 4 — сахароза
Кроме того, накопление и использование запасов — важная особенность роста надземных тканей.
Часть ассимилятов, образующихся в вечнозеленых деревьях умеренной зоны в конце сезона, сохраняется и используется на рост в следующем вегетационном сезоне. Когда хвою проростков сосны смолистой выдерживали в атмосфере С02 в конце августа, после прекращения сезонного роста побегов, меченые фотосинтаты передвигались и запасались в корнях, стволах и ветвях. Весной запасенные в ветвях углеводы использовались на рост побегов. Использование запасов на рост побегов у голосеменных подтверждают наблюдения за быстрым исчезновением углеводов из Ветвей при развитии побегов.
Например, от апреля до начала июня, когда новые побеги дугласии бурно развивались, количество углеводов в 1-летних побегах быстро уменьшалось. Концентрация и количество углеводов в новых, развивающихся побегах увеличивались от раскрытия почек до середины июня (рис. 7,7), Когда старую хвою сосны смолистой выдерживали в 14СО2 в конце вегетационного периода, некоторая часть 14<Г закреплялась в запасах. В течение следующего вегетационного периода часть 14с мобилизовывалась побегами и использовалась на их рост. По сообщению Шира (1970), основная часть ^С-ассимилятов, запасенных осенью 2-летней хвоей сосны смолистой, использовалась на рост тканей следующей весной. Значительное уменьшение сухого веса старой хвои сосны смолистой и пихты бальзамической с началом развития новых побегов также свидетельствует о передвижении углеводов из старой хвои в дополнение к использованию продуктов текущего фотосинтеза в начале вегетационного периода. У молодых растений тиса большая часть углеводов, использованных на рост побегов, хранилась до утилизации в старых листьях.
226
Глава 7
СЕЗОННАЯ ЦИКЛИЧНОСТЬ СОДЕРЖАНИЯ УГЛЕВОДОВ
Древесные растения накапливают запасы углеводов в периоды их избыточного образования, а затем расходуют их, когда скорость утилизации превышает скорость новообразования. В частности, во время наиболее быстрого роста запасные углеводы исчезают из ветвей, стволов и корней в порядке, зависящем от видовых особенностей роста. Сезонные циклы содержания углеводов наиболее выражены у многих листопадных деревьев умеренной зоны. Общее содержание углеводов в
стволах и ветвях достигает максимума осенью, незадолго до начала листопада. В конце зимы оно начинает уменьшаться и быстро уменьшается ранней весной, когда углеводы используются на усиленное дыхание и рост новых тканей (рис. 7.8 и 7.9).
У многих видов обнаружены два сезонных максимума содержания крахмала: весной и в конце лета или начале осени. Вслед за весенним
максимумом содержание крахмала
падает в результате утилизации резервов на рост побегов и камбия. Содержание крахмала зимой снижается вследствие его превращения при низких температурах в сахарозу (рис. 7.10).
Годичные циклы углеводов у видов с периодическим ростом характеризуются исчезновением углеводов с каждой вспышкой роста, после чего новые углеводы занимают их место. Годичные циклы углеводов у вечнозеленых деревьев отличаются от таковых у листопадных деревьев. Вечнозеленые виды накапливают углеводы зимой намного позже. Сезонные колебания запасов углеводов у них намного меньше, чем у листопадных деревьев.
Рис. 7.9. Сезонные изменения (данные приведены на 1-е число месяца) концентраций растворимого сорбита и сахарозы в побегах, коре и древесине ветвей и ствола молодых яблонь (Hansen, Grauslund, 1973): а побеги; б — кора; в — древесина; 1 — 1^С; 2 — cgp-бит; 3 — сахароза
Углеводы
227
Рис. 7.10. Сезонные изменения содержания различных фракций углеводов во внутренней коре деревьев белой акации, растущих под Оттавой, Канада
(Siminovitch et al., 1953):
1 — общее содержание; 2 - сахароза;
3 — крахмал; 4 — редуцирующие сахара
Месяцы
Так, в ветвях каштана калифорнийского, у которого листва опадает при засухе, содержание углеводов в ветвях в течение года варьировало гораздо больше, чем у вечнозеленого дуба траволистного. У каштана калифорнийского осенью, в период образования плодов, стебли были без листьев, а содержание углеводов снижалось с 17 до 10% (рис. 7.11), Когда в феврале развивались новые листья, запасы углеводов расходовались еще быстрее, пока не уменьшались до самого низкого в сезоне уровня - 5%. В марте и апреле, когда деревья были полностью покрыты листвой, содержание углеводов в ветвях восстанавливалось до самых больших за год значений. По сравнению с этим содержание углеводов в облиственных стеблях дуба траволистного было постоянным, варьируя приблизительно от 3,5 до 5%.
Даже у деревьев, растущих в тропическом климате, отмечаются годичные циклы содержания углеводов, хотя в общем диапазон этих циклов ограничен по сравнению с такими же циклами у деревьев более холодных климатов. У периодически растущих тропических деревьев с каждым усилением роста наблюдается некоторое истощение углеводов.
ОСЕННЯЯ ОКРАСКА
Антоциановые пигменты, вызывающие розовую, красную и пурпурную осеннюю окраску листьев, связаны с углеводами и накопление углеводов способствует их образованию. Антоцианы — это гликозиды, возникающие при соединении различных сахаров с циклическими
Рис. 7.11. Сезонные изменения суммы неструктурных углеводов в ветвях каштана калифорнийского и дуба траволистного (Mooney, Haus, 1973) ;
1 - каштан калифорнийский; 2 - дуб траволистный; 3 - с листьями, заливкой обозначен период плодоношения, штриховкой — рост стебля
жи х лт: л л тг г и иг
Меаць!
228
Глава 7
соединениями, называемыми антоцианидинами. Антоцианы растворимы в воде и обычно встречаются в клеточном соке. & кислых растворах они имеют красный цвет, а с увеличением pH могут становиться фиолетово-синими. Количество антоциановых пигментов зависит прежде всего от наличия определенных наследственных факторов их образования, но окружающая среда также оказывает влияние.
С понижением температуры осенью листья перестают образовывать хлорофилл. В это время некотрые виды, содержащие много углеводов и обладающие наследственной способностью, начинают образовывать в листьях антоцианы. После прекращения синтеза хлорофилла имеющийся хлорофилл начинает разрушаться, и только что образовавшиеся антоцианы становятся заметными. У деревьев, не образующих антоциановых пигментов, осенний распад хлорофилла обнаруживает относительно более стабильные желто-оранжевые каротины и ксантофиллы, что придает листь’ям светло-желтую окраску, или же примесь красного антоциана в желтом каротине, что дает ярко-оранжевый цвет (как у некоторых видов клена). У другцх видов хлорофилл и каротиноида! распадаются одновременно, и синтезируются новые каротиноиды. Так путем разрушения зеленых пигментов, выявления желтых, образования красных пигментов или всеми тремя способами сразу листья могут приобретать различные оттенки желтого, оранжевого, малинового, пурпурного и красного цветов.
У таких деревьев, как ольха и белая акация, изменения окраски выражены слабо. В противоположность им, листья большой группы растений, включающей орех черный, катальпу, вяз, карию, липу и платан становятся ржаво-зелеными и желтыми. Окраска листьев тополей, гледичии трехколючковой, гинкго, бука и большинства видов березы изменяется в различные оттенки желтого цвета. Но, несомненно, наибольшее впечатление оставляют красные тона клена красного и сахарного, сассафраса лекарственного, сумаха, дуба белого и американского шарлахового, ирги и бересклета крылатого — растений, образующих большие количества антоциановых пигментов. Разные виды и экземпляры клена демонстрируют постепенный переход окраски от желтой до темно-красной.
Деревья одного и того же вида, растущие вместе, часто сильно различаются по окраске благодаря индивидуальным различиям в количестве растворимых углеводов. Некоторые окрашиваются позже других. Дубы окаршиваются поздней осенью, обычно после того, как наиболее выраженная окраска кленов успевает поблекнуть. Желто-коричневая окраска бука и некоторых видов дуба вызывается и присутствием в листьях одновременно и желтых каротиноидов и, танинов.
Различия в скорости изменения осенней окраски у разных видов свидетельствуют о глубоких различиях интенсивности распада хлорофилла. По сообщению Вицковского (1958), в то время как у одного из видов магнолии наблюдалось быстрое разрушение хлорофилла (за 35 дней), у шелковицы белой распад происходил медленно (более 60 дней). Перед опадением листья явора и бука теряли весь хлорофилл, тогда как листья сирени только 40%. Вольф (1956) показал широкое варьирование содержания хлорофилла в листьях и скорости его распада осенью (табл. 7.2). У многих видов хлорофилл а рячрутттялся быстрее хлорофилла Ь.
Углеводы
229
7.2. СОДЕРЖАНИЕ ХЛОРОФИЛЛА В ЗЕЛЕНЫХ (ЧИСЛИТЕЛЬ) И ЖЕЛТЫХ (ЗНАМЕНАТЕЛЬ) ОСЕННИХ ЛИСТЬЯХ ЛЕСНЫХ ДЕРЕВЬЕВ (Wolf, 195 6)
Вид
Содержание хлорофилла
общее, мг/г
й, % от об- уменыпе-
щего ние, %
Liriodendron tulipifera 2,19/0,29 67,5/40,1 86,8
Populus nigra var. italica 1,79/0,27 73,6/63,4 85,2
Magnolia grandiflora 1,74/0,14 75,4/47,9 91,9
Cercis canadensis 1,55/0,12 71,8/43,0 92,2
Acer saccharum 1,38/0,19 69,4/56,5 86,5
Liquidambar styraciflua 1,23/0,05 70,1/57,4 96,2
Acer saccharinum 1,19/0,26 62,5/42,7 78,1
Juglans nigra 1,10/0,26 65,4/49,4 76,4
Celtis occidentalis 1,06/0,32 71,5/65,1 69,7
Cornu'S florida 0,97/0,18 64,9/53,6 81,4
Uhnus amencana 0,93/0,15 70,4/57,9 83,6
Quercus macrocarpa 0,92/0,11 68,0/47,4 87,6
Fagus grandifclia 0,90/0,22 64,4/57,4 75,3
Quercus palustris 0,87/0,17 71,3/54,2 80,6
Carya sp. 0,76/0,16 70,7/67,1 78,9
Гудвин (1958) проследил изменения содержания хлорофилла и каротиноидных пигментов с июня по ноябрь в сливе черной, дубе чер0щ-чатом и яворе. У дуба и явора содержание хлорофилла и каротиноидов уменьшалось почти до нуля. У дуба они расходовались одновременно, тогда как у явора падение содержания хлорофилла предшествовало снижению содержания каротиноидов. У сливы каротиноиды первыми проявляли тенденцию к уменьшению, но содержание их и хлорофилла снижалось только наполовину. По другим данным, при изменении окраски листьями клена образовывался каротиноидный пигмент, отличавшийся от имевшихся петом каротиноидов, а общее содержание каротиноидов уменьшалось.
Любой фактор, влияющий на синтез углеводов или на превращение нерастворимых углеводов в растворимые, способствует образованию антоциана и появлению ярких осенних окрасок. К наиболее важным факторам окружающей среды, определяющим осеннюю окраску, относятся температура, свет и водоснабжение. Понижение температуры ниже нуля способствует образованию антоциана. Ранние суровые морозы делают красные осенние цвета менее яркими, чем они были бы без них. Яркий свет также способствует появлению красной окраски, так как антоциановые пигменты обычно образуются в листьях, находящихся на свету. Если во время образования красных пигментов один лист был накрыт другим, в нижнем листе красный пигмент обычно не образуется. Снабжение водой также влияет на образование антоцианов: засуха способствует появлению ярко-красной окраски. Дождливые дни с недостатком света незадолго до периода наивысшего развития окраски существенно уменьшают яркость красок листопада. Следовательно, самые лучшие осенние цвета наблюдаются при ясной, сухой и прохладной (но не морозной) погоде.
ЛИПИДЫ, ТЕРПЕНЫ И РОДСТВЕННЫЕ ИМ ВЕЩЕСТВА
Рассматриваемые в этой главе соединения составляют разнородную группу, имеющую мало общего, за исключением их плохой растворимости в воде и хорошей растворимости в органических растворителях (ацетоне, бензоле и эфире) -Сюда входят простые липиды, кутин, суберин, воски и соединения различных веществ с глицеридами (например, фосфолипиды и гликолипиды). Другую большую группу соединений составляют изопреноиды, или терпеноиды - производные изопрена. Эта группа включает эфирные масла, смолы, каротиноиды и каучук. Взаимоотношения между главными группами видны из рис. 8.1 (терпеноиды, или изопреноиды, брлее подробно показаны на рис. 8.3).
Липиды - важные компоненты протоплазмы, особенно протоплазматических мембран. В виде кутина, воска и суберина они образуют защитные покровы на внешней поверхности листьев, плодов и стеблей. В некоторых тканях встречают-ся внутренние отложения кутина или близких к нему липидов. Липиды также играют важную роль как форма запасания питательных веществ особенно в семенах и плодах, а некоторые имеют большую практическую ценность, например, пальмовое, оливковое и тунговое масла. Так называемые эфирные масла, экстрагируемые из многих деревьев, используются в качестве ароматизирующих веществ. Терпены, добываемые из сосен, важны как источник технической канифоли и скипидара. Латекс^ из которого, изготавливается натуральный каучук, получают в основном из Hevea brasiliensis, а изомер каучука - гуттаперчу -из Palaquium gutta и других деревьев.
Простые липиде/
Неочищенные липиды или зерирный экстракт (Все ве- ---*-
щества экстрагируемые Эфиром,бензолам, хлороформом и т.р.)
Эфиры мирных кислот
Эфиры жирных кислот и глицерина. Жиры и масла. Эфиры жирных кислот с длинной цепыа и спиртами, отличны -ми от глицерина.
Кутин,суаерин и воска
и спиртов
Сложные' липиды
Эфиры жирных кислот и спиртов, содержащих добабоч-ч*~ные группы.
Фосфолипиды, гликолипиды, ецльфолипи-ды
Соединения, не являющиеся
эфирами жир-
Производные липидов
ных кислот, изопреноиды
Продукты деградации липидов. Жирные кислоты
Эфирные масла.
Терпены,экстракционные эфирные масла,иос-цизовая кислота, гиддеоеллины. Коротины. Каучук
Рис. 8.1. Взаимоотношения основных групп вещества, обнаруженных в эфирных экстрактах из тканей растений
Липиды, терпены и родственные им вещества
231
J ЛИПИДЫ
(Термин липиды включает простые триглицериды, обнаруживаемые в обычных маслах и жирах, различные сложные липиды, такие, как фосфо- и гликолипиды, и другие соединения (кутин, суберин и воска).
'* ПРОСТЫЕ ЛИПИДЫ. Самые простые и наиболее распространенные
I липиды — триглицериды. Это эфиры глицерина и различных жирных кислот, составляющие природные масла и жиры. Если при обычной температуре эфир представляет собой жидкость, его называют маслом, если он твердый — жиром. Общая формула для масел и жиров:
Rj-CO-O—СН2 r2 - со - о - £н r3-co-o—сн2,
где R1, К-2»и R3 — углеводородные цепочки одной и той же или разных жирных кислот.
!' В самом простом виде синтез масел и жиров включает взаимодей-
! ствие между тремя гидроксильными группами глицерина и тремя j молекулами жирных кислот. Связанные с данной молекулой глицерина жирные кислоты часто бывают разными, образуя смешанные глицери-К ды. Сущность реакции может быть показана следующим образом:
| н2 ~/с -он с15н31соон с15н31соосн2
Г Н - С - ОН + C15H3 jCOOH липа^ с15н31соосн + зн2о
н2-с-он C25H3JCOOH —- с15н31соо6н2
Глицерин Пальмитиновая Трипальмитин
кислота
Исходными соединениями при синтезе жиров являются, вероятно, «-глицерофосфат и жирные кислоты, построенные из остатков ацетата, происходящих от ацетил кофермента А. (Синтез жиров в общей схеме метаболизма показан на рис. 6.20.) Относительно места синтеза жиров существует некоторая неопределенность: и митохондрии, и названные глиоксисомами органеллы, и эндоплазматический ретикулум причастны к метаболизму жиров. В листьях синтез липидов осуществляется, по-видимому, также, и в хлоропластах. Липидные вещества составляют свыше 50% ламеллярных мембран. Синтез липидов, очевидно, происходит в тех клетках, в которых они обнаруживаются, поскольку их нерастворимость в воде делает невозможным передвижение липидов от клетки к клетке.
ЖИРНЫЕ КИСЛОТЫ. Эти кислоты подразделяют на насыщенные, т.е. не имеющие двойных связей, и ненасыщенные — с двойными связями между атомами углерода. Важнейшие из найденных в древесных растениях жирных кислот приведены в табл. 8.1. В древесных растениях наиболее широко из жирных кислот распространена пальмити-
232
Глава 8
8.1- ВАЖНЕЙШИЕ ЖИРНЫЕ КИСЛОТЫ, ОБНАРУЖЕННЫЕ В РАСТИТЕЛЬНЫХ ЖИРАХ
Насыщенные кислоты
Лауриновая С12Н24О2 СН3 (СН2) юСООН
Миристиновая Пальмитиновая С14Н28°2 С1бН32°2 СНз (CH2)i2COOH СН3 (СН2) 14СООН
Стеариновая С18Н36°2 СНз (СН2) igCOOH
Ненасыщенные кислоты
Олеиновая C1RH34O2 СН3 (СН?) ?СН = CH (СН2) 7СООН
Линолевая С18Н32О2 СН3 (СН2)4СН =СН - СН - СНу= СН(СН2) 7СООН
Линоленовая С18Н30О2 СН3 - СН2 - СН ~СН - СН2 - CH =СН - СН2 - СН —
- СН (СН2) 7- СООН
новая кислота, но большинство жирных кислот деревьев ненасыщенные, чаще всего олеиновая и линолевая. Масла с высоким содержанием ненасыщенных жирных кислот на воздухе присоединяют кислород и образуют твердую пленку, характерную для высыхающих масел, таких, как масла, добываемые из тунга и льна. Необычную способность масла какао оставаться твердым почти до температуры тела и. затем вдруг растапливаться относят за счет особого расположения жирных кислот в его триглицеридах.
Большинство кислот, найденных в жидких триглицеридах (маслах) — ненасыщенные (олеиновая, линолевая и линоленовая), в то время как большинство жирных кислот твердых триглицеридов (жиров) — насыщенные (пальмитиновая и стеариновая). Растительное царство наиболее богато жирными кислотами — олеиновой и пальмитиновой Ткани листьев большинства растений содержат сходные жирные кислоты, в основном пальмитиновую, линолевую и линоленовую. В хлоропластах присутствуют значительные количества линоленовой кислоты. В отношении жирных кислот, обнаруженных в семенах, наблюдаются гораздо большие различия между видами по сравнению с кислотами в листьях. Жирные кислоты семян могут отличаться от найденных в вегетативных структурах того же растения.
По некоторым исследованиям, растущие в холодном климате растения обычно образуют больше ненасыщенных жирных кислот, таких, как линолевая и линоленовая, чем растения, произрастающие в теплом климате. Вероятно, виды с широким климатическим ареалом содержат больше ненасыщенных жирных кислот в более холодных областях своего распространения, хотя Миров (1967) указал на некоторые исключения у сосен. По-видимому, температура, при которой в клеточных мембранах происходит фазовый переход из жидкого в твердое состояние, ниже в случаях, когда преобладают ненасыщенные жирные кислоты. Это изменение состояния существенно влияет на физиологические процессы. Например, устойчивость к повреждающему действую холода может быть связана с низкой температурой фазового перехода. Разли
Липиды, терпены и родственные им вещества
233
чия в сопротивлении корней поглощению воды между растениями теплого и холодного климата в холодной почве также можно объяснить разными температурами фазового перехода, связанными с видом содержащихся в корнях жирных кислот.
Подробно метаболизм липидов изучал Штумпф (1976) и другие исследователи. Мы еще раз отметим, что жирные кислоты с длинной цепью построены из остатков ацетата, и что ацетилкофермент А играет важную роль и в синтезе и в распаде жирных кислот (см. рис. 6.20). Например, при усвоении масла и жира жирные кислоты распадаются до ацетилкофермента А, который затем может либо попадать в цикл Кребса и там окисляться с освобождением энергии, либо вступать в клиоксилатный цикл и превращаться в сахара.
ВОСКИ, КУТИН И СУБЕРИН
КУТИКУЛА. Внешняя поверхность травянистых стеблей, листьев, плодов и даже цветочных лепестков обычно покрыта относительно непроницаемым для воды слоем, кутикулой. Кутикула состоит из воска и кутина и прикреплена к клеткам эпидермы слоем пектина (рис. 8.2). Типичная кутикула на затененных растениях тонкая и намного толще на деревьях, находящихся на ярком свету. Утверждают, что между различными видами и сортами растений существуют генетические различия в строении кутикулы. Помимо роли в уменьшении потерь воды, по мнению некоторых исследователей, кутин служит преградой для вторжения патогенных организмов, хотя другие в этом сомневаются.
Шенхер (1976) изучал проницаемость для воды слоев кутина, снятых с листьев цитрусовых, груши и чешуи лука. Он не обнаружил больших различий в проницаемости целого эпидермиса и эпидермиса со
Рис. 8.2. Схема наружной клеточной стенки верхней эпидермы листа груши, демонстрирующая строение кутикулы и воска (по Norris, Bukovac, 1968) :
1 - матрикс из кутина; 2 - воск этикутикулы; 3-кутикула; 4 - пектиновые вещества; 5 — клеточная стенка из целлюлозы; 6 — двоя копр еломляющий слой воска, внедренный в кутин; 7 — антиклинальная стенка
Глава 8
234
снятым с него слоем кутикулы. Проницаемость для воды зависит скорее от содержания в кутикуле экстрагируемого воска, чем от толщины матрикса. Экстракция воска гидрофобными растворителями увеличивала проницаемость кутикулярных мембран в 300—500 раз, что указывает на роль воска в регуляции проницаемости.
ВОСКИ. Это эфиры одноатомных спиртов с длинной цепью и жирных кислот с более длинной цепью, чем у кислот в простых липидах, т.е. с углеводородной цепью, состоящей более чем из 20 атомов углерода. Воски содержат также алканы с нечетным числом атомов углерода, в основном спирты, и свободные жирные кислоты с очень длинной цепью. Обнаружено, что в яблоках и листьях некоторых деревьев синтез воска происходит в клетках эпидермы. Этот процесс должен протекать обязательно поблизости от места отложения ввиду трудности транспортировки такого нерастворимого вещества. По всей вероятности, воски синтезируются в клетках эпидермы в виде капель, проходят через клеточную оболочку по до сих пор не обнаруженным каналам и образуют слои на внешней поверхности. Часть воска выдавливается через слой кутина с воском, образуя отложения воска в виде пластинок и палочек на кутикуле - ’’пушок”, характерный для поверхности некоторых листьев и плодов. Согласно Чемберсу и другим исследователям (1976), у эвкалипта молекулы воска, видимо, достигают через ламеллярные каналы в кутикуле ее наружной поверхности, передвигаются в масляном растворе и откладываются в различной конфигурации по мере испарения растворителя. Воск, очевидно, накапливается на внешних поверхностях растений, в противоположность суберину, накапливающемуся в клеточных оболочках, и кутину, который иногда накапливается как на внутренних, так и на внешних поверхностях. Исключение составляет накопление жидкого воска в семенах Simmondsia chinensis. Количество воска на поверхности листьев в зависимости от вида и условий окружающей среды значительно различается: от следов до 15% сухого веса листьев. Воск, распределяясь неравномерными массами, затрудняет смачивание поверхности листьев. Вследствие этого при опрыскивании для полного покрытия всей поверхности необходимо добавлять смачивающий агент.
Некоторые воски имеют важное промышленное значение. Среди самых известных — карнаубский воск, добываемый из листьев пальмы коперниция восконосная (Copemicia cerifera), растущей в Бразилии. Он содержит около 80% простых алкиловых эфиров жирных кислот с длинной цепью и 10% свободных одноатомных спиртов. Пальмовый воск встречается в виде слоев толщиной 2—3 см на стволе андийской восковой пальмы (Ceroxylon andicola), около одной трети его составляет настоящий воск, остальное—смола. Другие технические пальмовые Ъоски получают из Attalea excelsa и из высушенных листьев Raphia pedunculata. Воск дают также растущий в Тасмании Eucalyptus gunni, var. acervula и листья белого санталового дерева (Santalum album). Листья Му пса carolinensis дают ароматный воск, используемый для изготовления свечей. Интересен жидкий воск, который можно экстрагировать из пустынного кустарника симмонд-зии — этот воск может служить заменителем спермацетового масла.
Липиды, терпены и родственные им вещества
235
КУТИН. Колатакиди (1975) дал описание кутина как полимера жирных оксикислот. О его синтезе известно немного, но, по-видимому, этот полиэфир самопроизвольно образуется из оксикислот. Подобно воску он должен синтезироваться недалеко от места отложения, предположительно в клетках эпидермы листьев. Иногда кутин или кутиноподобные вещества накапливаются также на поверхностях клеток, обращенных к межклеточным пространствам.
СУБЕРИН. Это компонент водонепроницаемых слоев в растениях, преимущественно в коре. В старых одревесневших стеблях и корнях стенки наружных слоев клеток флоэмы, входящих в состав коры, пропитываются суберином и становятся относительно непроницаемыми для воды. Суберин содержит килоты с очень длинной цепью, спирты и фенольные соединения, которые в кутине либо отсутствуют, либо их там меньше.
ВНУТРЕННИЕ ЛИПИДНЫЕ СЛОИ
Не так давно было показано, что поверхности клеточных оболочек, обращенные к межклетникам, часто бывают покрыты гидрофобным липидным слоем. Скотт (1964) считает, "что отложение этих кутино-или субериноподобных слоев происходит всякий раз, когда клетки в процессе роста отрываются друг от друга и их поверхности соприкасаются с воздухом. Это значительно увеличивает сопротивление испарению воды с поверхности клеток мезофилла и снижает давление пара в межклеточных пространствах. Тот же исследователь (1963) утверждает, что кутиноподобный слой образуется на внешней поверхности молодых клеток корня, но он, по-видимому, часто покрыт слоем пектиновых веществ. Отложения суберина образуют пояски Каспари клеток эндодермы. Имеются сообщения и о других примерах внутренней кутини-зации и суберинизации. В связи с этим следует напомнить, что хотя слои кутина и воска в сухом виде относительно непроницаемы для воды, но они более проницаемы во влажном состоянии, как это отмечается внутри листьев и в корнях в сырой почве. Кроме того, при смачивании поверхности листьев растворенные вещества легко проникают через кутикулу.
СОСТАВ липидов
*
Липиды образуют сложные соединения с фосфором (фосфолипиды), углеводами (гликолипиды) и серой (сульфолипиды).
ФОСФОЛИПИДЫ. К данной группе относятся эфиры фосфорной кислоты и различных спиртов, в том числе глицерина, инозита и фитосфингозина, которые встречаются во всех живых клетках. В растениях наиболее широко распространены глицерофосфатиды. Фосфолипиды содержат как нерастворимую — гидрофобную — в воде часть (жирную кислоту), так и водорастворимую - гидрофильную - часть (глицерин, инозит или холин). Этим, видимо, объясняется их значение для мембран.
236
Глава 8
ГЛИКОЛИПИДЫ. В гликолипидах концевая гидроксильная группа соединена с сахаром — галактозой или глюкозой. Содержащие галактозил и дигалактозил диглицериды обнаружены в хлоропластах, где они этерифицированы линоленовой и линолевой кислотами. Гликолипид, содержащий глюкозу и серу, называется сульфолипидом. Его также особенно много в хлоропластах, но он встречается и в нефотосинтезирующих тканях.
РАСПРОСТРАНЕНИЕ ЛИПИДОВ В ДРЕВЕСНЫХ РАСТЕНИЯХ
Липиды встречаются во всех живых клетках растений и животных, но концентрация их в вегетативных структурах растений обычно довольно мала: часто менее 1% общего сухого веса. Однако даже и в этих малых количествах липиды очень важны благодаря их роли как обязательных компонентов клеточных мембран. Большинство данных о содержании липидов относится к эфирным экстрактам, содержащим, кроме простых липидов, значительные количества иных веществ. Например, суммарный эфирный экстракт из листьев обычно составляет менее 5% сухого веса, но он включает воск, кутин и другие вещества, поэтому содержание собственно липидов довольно низко. Содержание липидов в древесине также невелико, редко превышает 2—3%, при этом в заболони концентрация обычно ниже, чем в ядре. В коре концентрация липидов выше, чем в древесине, по-видимому, благодаря содержащемуся в коре суберину.
Фишер (1891) и другие исследователи того времени пытались разделить деревья на группы ’’маслянистых” и ’’крахмалистых” на основе доминирующей в древесине в зимнее время формы запасных питательных веществ. Синнот (1918) в США и Аррениус (1942) в Швеции относили хвойные деревья и липу к группе богатых липидами, а ясень, дуб, вяз и клен - к деревьям, содержащим мало липидов. Работавший в Японии Исибе (1935) также сделал вывод о существовании значительных различий между видами в содержании липидов, Так, у сосны густоцветной липидов намного больше, чем у дуба. Как и другие исследователи, он обнаружил низкое содержание липидов в корнях.
Имеются некоторые доказательства сезонных изменений содержания липидов: тенденции к накоплению зимой или весной у некоторых видов и уменьшению летом у всех Изученных видов. В 20-е годы появились сообщения о значительном превращении углеводов в липиды, происходящем в листьях хвойных деревьев зимой, но они основывались на микрохимических тестах. Позже наблюдения Доуля (1938) показали, что примененные тесты не специфичны в отношении липидов, и что нет надежных доказательств накопления липидов в листьях хвойных деревьев зимой. Имеющиеся данные позволяют предположить, что сезонная цикличность содержания липидов в большей степени регулируется общими закономерностями метаболизма, чем сезонными изменениями температуры.
В плодах и семенах концентрация липидов намного выше, чем в вегетативных структурах. Например, в клетках паренхимы плодов (околоплодника) авокадо и маслин и в семенах масличной пальмы накапливается масло. Плоды оливкового дерева содержат 20—^0% масла, семена американского ореха, кокосовой пальмы, сосны однохвойной и карии пекан 60—75%, желуди дубов красного и американского шарлахового 20—30%. Масло может откладываться в эндосперме, как у кокосовой пальмы, или в зародыше, как в желудях. В семени сим-
Липиды, терпены и родственные им вещества
237
мондзии воск составляет 50—60% сырого веса семядолей. При прорастании он превращается в углеводы, подобно жирам, содержащимся в желудях и некоторых других маслянистых семенах.
Синтез масла и жира, очевидно, происходит в тех органах, где они запасаются, поскольку нерастворимость в воде делает невозможным их передвижение. Тип накапливаемых масел может быть различным в таких смежных тканях, как плодовые оболочки и семена пальмы или листья и плоды других видов. Синтез и накопление масел и жиров протекают очень быстро в течение короткого периода развития плодов и семян.
Уже говорилось о значений липидов как составных частей клеточных мембран и защитных покрытий на поверхностях. Липиды также являются очень важной формой запасания питательных веществ, особенно в семенах, где, как отмечалось ранее, они часто служат основной формой запасных питательных веществ. Липиды содержат в пересчете на углерод и водород намного меньше кислорода, чем углеводы, поэтому при их распаде и окислении освобождается больше энергии. В процессе прорастания семян запасной жир превращается в жирные кислоты и глицерин, которые распадаются далее. Глицерин через «-глицерофосфат превращается в фосфодиоксиацетон, который может распадаться гликолитически и окисляться в реакциях цикла Кребса или прообразовываться в сахар. Жирные кислоты превращаются в аце-тилкофермент А, а затем или окисляются в цикле Кребса с образованием АТФ и НАД-Н2 или вступают в глиоксилатный цикл и превращаются в сахара.
ИЗОПРЕНОИДЫ, ИЛИ ТЕРПЕНОИДЫ
Вещества этой группы интересны с точки зрения биохимии и экономики. Это углеводороды, построенные из разного числа изопреновых единиц. К ним относятся: эфирные масла, смолы, каротиноиды и каучук (рис. 8.3). Все растения могут синтезировать каротиноиды и стероиды, но способность синтезировать другие терпены у представителей растительного царства крайне неравномерна. Атомы углерода поступают обычно от ацетата, который иногда называют перекрестком путей метаболизма растений ввиду того, что он является исходной точкой для синтеза многих соединений.
ЭФИРНЫЕ МАСЛА. Эти масла представляют собой ациклические или циклические соединения: моно-, сескви- и дитерпены, разнообразные свойства которых определяются природой связанных с ними химических групп. Эфирные масла — источник большинства запахов цветов, плодов и древесины многих растений. Наиболее часто они встречаются в видах семейств Pinaceae, Umbelliferae, Myrtaceae, Lauraceae, Rutaceae, Labiatae и Compositae. Содержать эфирные масла могут все органы растений, в том числе кора, древесина и листья. Часто эфирные масяа образуются в группах железистых клеток или в железистых волосках цветков, листьев и стеблей. Иногда они выделяются в специализированные вместилища (ходы) листьев и стеблей. Эфирные масла не выполняют насколько известно, каких-либо существенных функций в метаболизме
238
Глава 8
Класс Монотерпены ^еск&ллерпены Д/итерпены Тритерпены Тетратерпены Политер-терпенов пены
Типы дстре- Эфирные Эфирные масла, Эфирные Смолы, Каротины Каучук >
чающихся в масла смолы, абсцц - масла, латекс гуттаперча
природе зоба я кислота смолы,
соединений : гиббере
ллины
Рис. 8.3. Взаимоотношения различных групп изопреноидов (Bonner, 1950)
растений, хотя они могут быть полезными для привлечения насекомых-опылителей и для отпугивания вредителей. Многие из них летучи и испаряются, особенно в теплые дни, распространяя запахи, характерные для различных цветов и хвойных лесов.
Много летучих веществ выделяется из засыхающей растительности, которая служит источником запаха свежескошенной травы, сена и запаха опавших осенних листьев.
В промышленности эфирные масла экстрагируются в небольшом объеме путем перегонки с водяным паром из хвои сосны, туи западной, ели черной, пихты бальзамической и тсуги канадской, древесины кедра, коры березы граболистной, из корней и почек сассафраса лекарственного. Самое важное эфирное масло, добываемое из деревьев, — скипидар. Приятные запахи хвои объясняются присутствием в ней борнеола и борниловых эфиров. Большие количества эфирных масел в листьях некоторых кустарников делают их очень горючими и намного увеличивают скорость, с которой распространяется огонь.
СМОЛЫ. Это разнородная смесь смоляных кислот (С20Н30О2),жирных кислот, эфиров этих кислот, стеролов, спиртов, восков и резе-нов. Хвойные и лиственные деревья синтезируют смолы, но у хвойных их обычно намного больше. Как сообщали Уайз и Джейн (1952), выход смолы у хвойных деревьев составляет от 0,8 до 25% по сравнению с 0,7—3% у лиственных деревьев. Большую часть используемой в промышленности смолы получают из деревьев семейств Pinaceae, Leguminosae и Dipterocarpaceae. Копалы — это известные твердостью и высокой точкой плавления смолы, экстрагируемые из деревьев семейства Leguminosae. Деревья семейства Dipterocarpaceae образуют
Липиды, терпены и родственные им вещества 239
смолу о торговым названием даммар. Другую важную для промышленности смолу, каури, получают из дерева агатис (Agathis australis), растущего в Новой Зеландии. Балтийскйй янтарь — это ископаемая смола видов сосен, произраставших 40—50 млн. лет назад.
Большая часть смол выделяется в специальные ходы или каналы (смоляные ходы) окружающим их слоем паренхимных клеток. Часто каналы сильно разветвлены, так что при прокалывании или надрезании одного из ответвлений смолы начинают перетекать к поврежденному участку издалека. Смолы иногда обнаруживаются внутри клеток и в клеточных оболочках. В качестве запасных питательных веществ они не используются, их роль в метаболизме древесных растений не известна.
Хэглунд (1951) разделил смолы на физиологические и патологические. Первые встречаются внутри древесных тканей в норме, вторые образуются в результате повреждения коры или древесины. Бюсген и Мюнх (1931) подчеркивают защитную природу смол, указав на тот факт, что смолистая древесина сосны более устойчива к гниению, чем древесина ели, содержащая мало смолы. Присутствие смолы замедляет развитие грибов, вызывающих гниение, о чем свидетельствует долговечность пней и сучьев сосны и смолистого ядра сосновой древесины.
Роль смол в устойчивости хвойных деревьев к нападениям короедов была предметом серьезной дискуссии. Обильный ток смолы, часто сопровождающий деятельность точильщиков, иногда оказывается для них смертельным и предотвращает дальнейшие нападения. Иногда же он привлекает других насекомых. Утверждалось, что специфичность паразитов в отношении сосен связана с типом вырабатываемой смолы. Согласно Каллхэму и Смиту (1966) каждый вид точильщиков устойчив к терпенам, образуемым его хозяином, но не выносит тех, которые образуются другими видами. Есть также некоторые доказательства образования терпенов хвойными деревьями в ответ на грибную инфекцию, а также терпенов, отпугивающих или угнетающих короедов. По данным ряда исследователей, при уменьшении выделения смолы вследствие воздействия окружающей среды (например, водного стресса) сосны становятся гораздо более восприимчивыми к нападению короедов. Роль терпенов и других растительных продуктов как защитных агентов заслуживает дальнейшего изучения.
Чрезмерное истечение смолы может происходить при ранениях или при заражении грибами. Такое истечение характерно для стволов хвойных деревьев, пораженных серянкой сосны веймутовой, раком лиственницы и красной кольцевой гнилью ели. Резко выраженное истечение может наблюдаться у корневых шеек сосны при армилляриеллезной гнили, вызванной Armillaria mellea.
ЖИВИЦА. Это наиболее важная в промышленном отношении смола, добываемая из различных сосен. Термин живица означает вязкую жидкость, которую выделяют клетки, окаймляющие смоляные ходы хвойных деревьев. Она состоит примерно из 66% смоляных кислот, 25% скипидара (эфирного масла), 7% нелетучих нейтральных веществ и 2% воды. Живицу дают паренхимные ткани корней, листьев, древесины и внутренней коры. Она образуется в живых так называемых эпителиальных клетках, которые делятся, формируя смоляные каналы или
240
Глава 8
ходы, особенно активные во внешней части заболони. При надрезе древесного ствола смоляные ходы обнажаются и живица начинает сочиться из них. В промышленных масштабах живицу добывают из сосен подсочкой или разрезом коры с обнажением поверхности заболони. Количество и продолжительность вытекания смолы можно увеличить опрыскиванием раневой поверхности серной кислотой. Выход живицы варьирует не только по видам, но также и у разных деревьев одного и того же вида. Отмечено, что интенсивность истечения у надрезанной сосны Эллиота зависит от количества и размеров смоляных ходов, давления смолы и вязкости эксудата. Давление смолы подвержено суточным колебаниям: самое высокое давление наблюдается на рассвете, а наименьшее—в послеполуденные часы, когда содержание воды в стволе минимально. По некоторым сообщениям, суточные колебания давления, под которым вытекает живица у сосны ладанной, связаны с влажностью почвы и атмосферы.
Живица откладывается в заболони, образуя пропитанную смолой древесину (просмоленную древесину) — важный источник используемых материалов, таких, как канифоль и скипидар. Большая часть этих веществ добывается перегонкой с паром смолистой древесины срубленных деревьев или подсочкой живых деревьев. Если у молодых деревьев удается вызвать образование достаточного количества просмоленной древесины, то эти деревья можно рубить при достижении ими размеров, пригодных для получения балансовой древесины, и экстрагировать из нее смолы, а щепу использовать как древесную массу.
Показано, что инъекция гербицида параквата (1,Г -диметил-4, 4-дипиридий-хлорида) в стволы сосен увеличивает выделение живицы участками древесины, нередко простирающимися на несколько метров выше места инъекции. Инъекция вызывает также типичные раневые реакции, например усиление дыхания и образование этилена. По сообщениям Воягера (1977), инъекция этрела (2-хлор-этилфосфоновой кислоты - соединения, выделяющего этилен) стимулирует образование смолы в хвойных. Это позволяет предположить, что усиленное образование смолы Досле ранения может быть вызвано выделяющимся из поврежденной ткани этиленом. Содержание живицы после воздействия на ткань увеличивается, как полагают, скорее благодаря превращению уже присутствующих в этой ткани предшественников, чем за счет передвижения предшественников с некоторого расстояния.
К сожалению, паракват™ убивает клетки камбия на значительном расстоянии выше места обработки, в результате образуется полоса мертвой древесины и коры. Это ограничивает протяженность дуги по окружности ствола, которую можно обрабатывать. Обработанные деревья к тому же более восприимчивы к нападению короедов, и в целом в результате обработки увеличивается количество погибших деревьев. Тем не менее возможность усиления образования пропитанной смолой древесины путем химического воздействия представляет большой интерес, поскольку этим методом, возможно, удастся в течение ряда лет добывать больше смолы и скипидара, чем общепринятыми в настоящее время способами.
После удаления перегонкой из живицы летучего скипидара остается твердая смола различного цвета — от янтарного до почти черного, именуемая канифолью. Ее основной компонент — абиетиновая кислота.
Коммерческое значение имеют канадский бальзам, добываемый из пихты бальзамической, и орегонский бальзам из дугласии. Первый из них выделяется в смоляные ходы, образованные отделением клеток коры, и встречается в виде маленьких пузырьков под корой. Второй
Липиды, терпены и родственные им вещества
241
содержится в полостях, образованных под действием ветра. Венецианский терпентин, используемый в живописи, добывается из лиственницы европейской.
КАРОТИНОИДЫ. Единственными природными тетратерпенами являются каротиноиды с формулой С40Н56- Каротины—это группа углеводородов, в которую входят встречающиеся во всех органах растений красные, оранжевые и желтые пигменты. Они участвуют в поглощении света и, вероятно, защищают хлорофилл от фотоокисления. Ксантофиллы - желтые или коричневые пигменты, встречающиеся в листьях и водорослях. Они содержат несколько атомов кислорода и имеют формулу С40Н56О2 (см. главу 5).
КАУЧУК. Это политерпен, состоящий из соединенных линейно 500—5000 изопреновых единиц
СН3 СН3
I I
- СНа -с=сн- сн9 сн, -с=сн -сн9 -
В латексе каучук находится в форме более или менее сферических глобул с диаметром 5—6 мкм, стабилизированных тонкой пленкой адсорбированного белка и фосфолипида.
Каучук образуют около 2000 видов растений, в том числе травы, кустарники, деревья и лианы. Он синтезируется только двудольными покрытосеменными растениями и не образуется у однодольных, голосеменных или низших растений. Особенно широко представлены каучуконосные виды в семействах Euphorbiaceae, Могасеае, Аросупасеае, Asclepiadaceae и Compositae. Большинство древесных каучуконосов произрастает в тропиках. Единственное древесное растение умеренной зоны, образующее достаточно каучука для промышленного получения, — гваюла (Parthenium argentatum). Основной источник натурального каучука — тропическое дерево Hevea brasiliensis из семейства Euphorbiaceae. Транс-изомер каучука — гуттаперчу — добывают в основном из гуттаперчевого дерева (Palaquium gutta), принадлежащего к семейству Sapotaceae.
Иногда каучук находится в паренхимных клетках, например, у гваюлы, у которой для его получения надо срезать верхушку растения. Но чаще он присутствует в виде глобул, взвешенных в латексе — сложной жидкой системе, содержащей множество растворенных или суспендированных веществ. Среди них: производные терпенов, сахара, крахмальные зерна, органические кислоты, стеролы и ферменты. Точный состав сильно различается у разных видов и даже у отдельных растений одного и того же вида. Крахмальные зерна встречаются в латексе молочая, но их нет в гевее. В латексе фикуса много белка, в латексе мака снотворного много алкалоидов опиума. Из латекса дынного дерева получают фермент папаин. Сырье для изготовления жевательной резинки добывают из латекса тропического дерева Achras zapota, растущего в Мексике, Центральной Америке и Венесуэле. По данным Боннера (1965), у гевеи каучук содержится в виде частиц, взвешенных в высокоспециализированной цитоплазме млечников, имеющих ядра, митохондрии, рибосомы и ферменты.
242 Глава 8
Каучук обнаруживается в млечном соке не всех растений, а если присутствует, то обычно в очень низких концентрациях. Только у немногих видов каучук содержится в экономически выгодных для добычи количествах, особенноу гевеи, где на его долю приходится 20—60% латекса в пересчете на сухой вес, и у гваюлы, где иногда его содержание превышает 10% сухого веса растения. Ни латекс, ни каучук не используются растениями в качестве запасных питательных веществ. Латекс редко образуется в паренхимных клетках (как у гваюлы), но синтезируется в млечных сосудах — каналах, образованных удлинением одиночных клеток или объединением многих специализированных клеток в сложные системы, У древесных растений (например, у гевеи) млечники находятся в коре в виде вертикальных трубок длиной до нескольких метров.
Каучуконосные деревья обычно надрезают, делая в коре спиральный желобок длиной до половины или более окружности ствола под углом 25-30° к поверхности земли. Млечный сок стекает из вскрытых млечников и собирается в нижней части надреза. Сначала ток происходит благодаря упругому сокращению млечников, но затем । осмотическое передвижение воды в млечники уменьшает вязкость и концентрацию каучука в латексе. Через несколько часов ток прекращается, так как на воздухе латекс коагулирует, и регулярно через день тонкий слой удаляют с нижней части надре:1а, вызывая возобновление тока. Ранение стимулирует метаболическую активность флоэмы, и потерянные при истечении млечного сока рибосомы, митохондрии, ферменты и частицы каучука быстро регенерируются. На стволах гевеи подсочку делают не глубоко, чтобы не повредить камбий, поэтому в течение нескольких лет кора регенерируется и процесс можно повторять снова.
Давление и интенсивность вытекания латекса зависят, по крайней мере частично, от внутреннего водного баланса и мощности дерева, потому что варьируют по сезонам, времени дня и в связи с условиями обитания. Между деревьями имеются также генетически обусловленные различия. Иногда стебли с высокой потенциальной способностью к образованию каучука прививают на корневые подвои, устойчивые к болезням. Ток млечного сока обычно сильнее утром, чем днем. Он уменьшается и в сухую погоду. Делались попытки усилить ток латекса различными химикатами: широко применялись 2,4-Д, 2,4,5-Т и этрел. В некоторых случаях 2,4,5-Т усиливал выход латекса на одну треть в течение нескольких лет без повреждения деревьев. По некоторым сообщениям, применение ауксина 2-(3,4-дихлорфенокси)-триэтиламина увеличивало синтез каучука у гваюлы в 2-3 раза.
РОДСТВЕННЫЕ СОЕДИНЕНИЯ. Производными терпенов являются некоторые важные соединения.
Абсцизовая кислота — важный регулятор роста растений, относящийся к сесквитерпенам (см. главу 15).
Гиббереллины — другая группа важных регуляторов роста растений — производных дитерпенов (см. главу 15).
Стероиды — важная группа стероидов (стеролов), производных изопреновых соединений, содержащихся в растениях и животных. Тритерпен сквален — предшественник холестерина, который в свою очередь является предшественником других стероидов. Те стеролы, которые образуются в растениях, часто называют фитостеролами, например стигмастерол и эргостерол — предшественник витамина D.
Липиды, терпены и родственные им вещества 243
Функции стеролов в растениях в основном не известны, но они играют важную роль в физиологии животных, а некоторые растительные стеролы используются в качестве исходных веществ для синтеза гормонов животных.
Терпеноидные гликозиды. Некоторые терпеноиды, особенно стеролы, существуют в форме гликозидов. К ним относятся сапонины и сердечные гликозиды, в том числе получаемые из Digitalis.
Фитол - спирт, производное дитерпена, является компонентом хлорофилла.
ФУНКЦИИ ВЕЩЕСТВ ВТОРИЧНОГО ПРОИСХОЖДЕНИЯ В РАСТЕНИЯХ
Считают, что группа терпенов, или изопреноидов, состоит из более разнообразных соединений, чем любая другая группа веществ растительного происхождения, хотя алкалоиды, видимо, почти так же разнообразны. Каротиноиды, абсцизовая и гибберелловая кислоты выполняют важные физиологические функции в растениях, а некоторые эфирные масла и компоненты живицы, по-видимому, отпугивают вредителей или привлекают опылителей. О функциях многих из синтезируемых растениями соединений, особенно терпенов и алкалоидов, известно мало. Иногда утверждают, что все соединения, входящие в состав организма, должны иметь свои функции. Однако представляется неправдоподобным, чтобы все аспекты метаболизма растений имели одинаковое значение в процессе естественного отбора. Более вероятно, что многие из этих веществ являются просто продуктами синтетических процессов, которые случайно возникли у древних предков и сохранились лишь потому, что не оказывали настолько отрицательного влияния на выживание, чтобы быть удаленными естественным отбором. С другой стороны, большое внимание уделялось возможным защитным функциям фенольных и других соединений против патогенных организмов и насекомых-вредителей.
Небольшая доля питательных веществ, превращающаяся у дикорастущих растений в каучук, не снижает их конкурентоспособности, так что синтез каучука сохранился, даже несмотря на то, что он представляется бесполезным для образующего его растения. Человек недавно выделил каучуконосы из условий естественной конкуренции и вывел формы, которые превращают питательные вещества в каучук, по-видимому, за счет вегетативного роста. Маловероятно, что такие растения смогли бы выдержать конкуренцию в природных условиях.
Делались попытки использовать жирные кислоты, эфирные масла и терпены в качестве признака для классификации растений. Жирные кислоты в этом отношении бесполезны, поскольку одни и те же встречаются в семенах неродственных друг другу растений, и к тому же существуют генетические различия в составе жирных кислот внутри родов и даже между отдельными особями одного и того же вида.
Миров (1961) изучал различия в составе терпенов с целью подобрать ключ к таксономическим взаимоотношениям сосен. Сквилэйс (1971, 1976) обсуждал наследование состава терпенов у сосны Эллиота, а также и различные методы
244 Глава 8
анализа. Флэйк с сотрудниками (1969) и другие авторы использовали состав терпенов для изучения распределения популяций можжевельника виргинского. Использование веществ вторичного происхождения в систематике растений рассмотрел Эрдтман (1973).
Все большее внимание уделяется применению биохимических методов в дополнение к морфологическим при описании и идентификации новых форм растений, для составления диагнозов и для других целей. Общепринятые описания недостаточны, поскольку размеры, форма и окраска варьируют с изменением условий среды. Однако некоторые биохимические характеристики должны быть относительно стабильными. Ларсен (1969) предложил использовать для определения различий между сортами изоферменты, а Кюн и Фретц (1978) разделили их у сортов розы с помощью электрофореза. Эйсен (1977) сообщил, что использование жидкостной хроматографии высокого давления позволяет быстро охарактеризовать набор флавоноидов у растений. Тилджес (1972) обнаружил различия в составе полифенолов хвои сосен разного происхождения, а Фретц (1977) обнаружил различия между многими разновидностями можжевельника распростертого, измеряя методом газовой хроматографии различия в концентрациях и соотношениях ряда терпенов у растений, выросших в сходных условиях. В этой области много работают, и представляется, что биохимические методы смогут существенно дополнить морфологические описания, используемые в настоящее время для идентификации сортов и рас растений.
МЕТАБОЛИЗМ АЗОТА И АЗОТНОЕ ПИТАНИЕ
Содержащие азот соединения составляют лишь небольшую часть общей сухой массы древесных растений, но физиологически они чрезвычайно важны. Содержание азота в листве яблони и сосны достигает 1-1,2% сухого веса, тогда как в древесине оно намного меньше. Низкая концентрация соединений азота контрастирует с содержанием целлюлозы, которая является структурным компонентом клеточных оболочек и составляет свыше половины сухого вещества древесных растений, но физиологически и биохимически она неактивна. В живых клетках, где азотсодержащие соединения играют существенную роль в биохимических и Физиологических шюцессах, их содержание относительно невелико. Первое место по значению среди, этих веществ занимают белки, образующие структуру протоплазмы, и ферменты, катализирующие биохимические процессы в растениях. Большие количества белка накапливаются также в семенах некоторых растений. Много азота входит в состав таких веществ, как амиды, аминокислоты, нуклеиновые кислоты, нуклеотиды и другие азотистые основания, гормоны, витамины и алкалоиды (см. рис. 6.20). Большинство этих веществ имеет огромное физиологическое значение, но алкалоиды, хотя и хозяйственно важные вещества, являются, по-видимому, просто побочными продуктами метаболизма, в лучшем случае предоставляющими растению некоторую защиту от нападения водителей.
Азот необходим на всех этапах роста — от проростков до взрослых деревьев. Недостаток его является лимитирующим рост фактором, наиболее распространенным после водного дефицита. Наибольшее количество азота требуется для образования белков, используемых при формировании протоплазмы новых клеток. Азот используется на синтез хлорофилла и цитохромов. Значительная часть азота листьев находится в ферментах хлоропластов и митохондрий. Один из наиболее общих признаков дефицита азота - бледная или пятнистая окраска листьев в результате недостаточного синтеза хлорофилла. Недостаток хлорофилла снижает интенсивность фотосинтеза и тем'самым косвенно уменьшает рост.
Потребность в азоте тесно связана со скоростью роста. Деревья, у которых большая часть годового прироста происходит в начале вегетационного периода, используют в это время много азота. Об источнике этого азота данных мало, но ясно, что большая его часть должна поступать из запасов дерева, так как типичная лесная почва бедна доступными формами азота, в результате чего поглощение его должно быть медленным. Садоводы обычно подкармливают плодовые деревья соединениями азота, чтобы обеспечить достаточное снабжение в этот критический период роста. Большинство лесов в определенной степени страдает от недостатка азота. В связи с этим лесоводы считают целесообразным применение удобрений, по крайней мере, при некоторых условиях. Однако дефицит азота — менее серьезная проблема для деревьев с их длинным вегетационным периодом (в течение которого длительное время может происходить поглощение веществ из почвы) по сравнению с полевыми культурами, которые в основном растут и нуждаются в азоте в течение нескольких недель. Азот в растениях очень мобилен, он обычно передвигается из неактивных тканей в активные, что делает дефицит его не таким очевидным по сравнению с менее мобильными элементами.
РАСПРЕДЕЛЕНИЕ И СЕЗОННЫЕ КОЛЕБАНИЯ СОДЕРЖАНИЯ АЗОТА
Колебаниям содержания азота в различных частях древесных растений ввиду физиологической важности его посвящено множество работ. Количество имеющегося азота варьирует в разных тканях, с возрастом или по стадиям роста и временам года. Большая часть азота находится в протоплазме в связанном виде, в составе физиологически активных веществ. Поэтому самые высокие концентрации азота обнаруживаются в тканях, состоящих в основном из физиологически активных клеток:
246 • Глава 9
листьях и меристематических тканях, таких, как камбий и кончики корней и стеблей. Семена также часто богаты азотом, но там он присутствует в форме запасных питательных веществ и физиологически относительно неактивен. (Данные о содержании белка в семенах приведены в главе 14.)
НАКОПЛЕНИЕ В РАЗЛИЧНЫХ ТКАНЯХ, В табл. 9.1 показано распределение азота в пробах из различных частей 18-летней яблони, взятых в середине октября. В листьях концентрация азота была выше, чем в любой другой части дерева, возможно, за исключением лишь самых молодых корней, и она уменьшалась от молодой к самой старой древесине. Из общего содержания азота в яблоне 75% приходилось на надземные части и около 20% на листья. В ходе вегетационного периода содержание азота в листьях (% от общего) было, вероятно, даже больше, поскольку отток, происходящий перед листопадом, по-видимому, должен был в середине октября уже начаться. В укороченных побегах концентрация азота также была относительно высокой. Та же самая ситуация, очевидно, имеет место в лиственных вечнозеленых деревьях, поскольку, как обнаружили Кеймерон и Комптон (1.945), приблизительно 50% имевшегося у плодоносящих апельсиновых деревьев азота находилось в листьях, около 10% в побегах и 25% в ветвях и стволе. Менее 20% общего азота было в корнях. В другой работе Камерон и Эплеман
9.1. РАСПРЕДЕЛЕНИЕ АЗОТА В ЯБЛОНЕ В СЕРЕДИНЕ ОКТЯБРЯ (СРЕДНИЕ ДАННЫЕ ПО ТРЕМ СОРТАМ: ГРАЙМС, ДЖОНАТАН И ДЕЛИШЕС) (Murneek, 1942)
Наименование Сухой вес, Азот
показателей кг --------------------------
кг % от сухого
веса
Листья 13,43 0,166 1,23
Укороченные побеги 2,57 0,027 1,04
Древесина, лет:
1 4,56 0,043 0,93
2 5,55 0,037 0,67
3 5,38 0,029 0,54
4-6 - 19,88 0,070 0,35
7-10 65,48 0,177 0,27
11-18 62,93 0,102 0,16
Главный ствол 30,75 0,044 0,14
Всего в надземных частях 210,52 0,694 0,33
Осевой корень 22,75 0,059 0,26
Корни,лет:
1-6 2,45 0,030 1,24
7-13 13,30 0,080 0,60
14-18 20,96 0,067 0,32
Всего в подземных частях 59,45 0,236 0,40
Итого в дереве 269,97 0,930 0,34
Метаболизм азота и азотное питание
247
Рис. 9.1. Содержание азота в заболони и ядре дуба красного. Сходное распределение наблюдается у ясеня американского, ели ситхинской и сосны веймутовой (Merrill, Gowingl1966): 1 - ядро; 2 - включает сердцевину
Рис. 9*2. Относительное накопление сухого вещества и азота в листве, коре и древесине в течение пятого года развития насаждения сосны ладанной (Nelson et al., 1970) :
а — листва; & — кора; в — древесина; 1 — сухой вес; 2 — азот
(1933) показали, что у 3,5-летних апельсиновых деревьев 60% общего азота находилось в листьях, а у 10-летних деревьев листья содержали лишь 40% общего азота. В корнях концентрация хотя и была довольно высокой, но в них никогда не находилось более 21% всего азота деревьев. Как у яблонь и апельсиновых деревьев, так и у сосны ладанной, около 25% азота содержалось в листьях (табл. 9.2). Создается впечатление, что вообще концентрация азота в листьях выше, чем в любой другой части деревьев.
Эта высокая концентрация азота легла в основу предложения, согласно которому при спиливании деревьев листву следует сохранять и перерабатывать в содержащие азот добавки к пище для крупного рогатого скота и даже для людей. Уже ряд лет белок листьев рекомендуют в качестве добавки в рационы с недостатком белков для людей. Концентрации азота, углеводов и пигментов в листьях тополя близки к таким концентрациям у кормовых культур (например, люцерны). В связи с этим листва лесных деревьев может служить хорошим потенциальным источником дополнительного питания для скота, а также и для людей. Пищевая ценность листьев и веток деревьев и кустарников подтверж
248 . Глава 9
дается животными (оленями, жирафами и слонами), которые охотно их ощипывают.
Содержание азота в ядровой древесине обычно намного ниже, чем в заболони (рис. 9.1). Это уменьшение содержания азота от заболони к ядру связано с отмиранием клеток паренхимы и с передвижением их азота к зонам роста. Сердцевина и примыкающее к ней годичное кольцо содержат больше азота, чем масса ядра, а иногда и больше, чем заболонь. Меррил и Каулинг (1966) обнаружили сходное распределение в стволах хвойных и лиственных деревьев. Они сообщали, что высокое содержание азота способствует развитию грибов, вызывающих гниение древесины. Значительные количества растворимых соединений азота, в основном амидов и аминокислот, находятся в ксилемном соке.
Флоэма (кора) содержит значительно больше азота, чем древесина (рис. 9.2), и является важным источником азота для роста. У апельсиновых деревьев каждый подъем роста сопровождается уменьшением содержания азота в коре соседних ветвей.
Семена часто содержат много азота, поскольку у некоторых видов основным запасным веществом является белок. В желудях нескольких видов в среднем содержится около 1,1% азота, в семенах сосны однохвойной — 2%, сосны Сабина — 5%, сосны болотной — 6% азота.
9 2 СОДЕРЖАНИЕ АЗОТА (ЧИСЛИТЕЛЬ - КГ/ГА; ЗНАМЕНАТЕЛЬ - %) В РАЗЛИЧНЫХ ЧАСТЯХ ДЕРЕВЬЕВ 16-ЛЕТНЕГО НАСАЖДЕНИЯ
СОСНЫ ЛАДАННОЙ (Wells et al., 1975)
Часть дерева
Содержание азота
Хвоя текущего года 55/17,1
Вся хвоя 82/25,5
Живые ветви 34/10,6
Мертвые ветви 26/8,0
Древесина ствола 79/24,6
Кора ствола 36/11,2
Сумма надземных частей 257/80,0
Корни 64/20,0
Всего 321/100,0
СЕЗОННЫЕ ИЗМЕНЕНИЯ СОДЕРЖАНИЯ АЗОТА. Изменения содержания азота и других компонентов в листьях и тканях древесины многолетних растений вызывают большой интерес, поскольку эти части поставляют материал, необходимый для образования побегов весной. Интересно также выяснить, когда эти материалы накапливаются, так как такая информация может указать наилучшее время для внесения удобрений и помочь определить глубину повреждений, наносимых потерей листьев в результате нападения насекомых, развития патогенных организмов или из-за бурь. Изучению накопления азота и других минеральных веществ посвящены многочисленные работы (выполненные в основном на фруктовых деревьях и в лесных насаждениях).
Метаболизм азота и азотное питание
249
У листопадных и вечнозеленых деревьев концентрация азота в тканях древесины имеет тенденцию к увеличению осенью и зимой, уменьшению с началом роста и затем снова к увеличению при замедлении и остановке роста. Тромп (1970) резюмировал данные по сезонным изменениям содержания общего азота, белкового азота, растворимого азота и аргинина в древесине и коре молодых неплодоносящих хорошо удобренных яблонь. Содержание всех форм азота было высоким в течение зимы, уменьшалось весной и оставалось низким до прекращения роста. Осенью снова наблюдалось накопление, достигавшее максимума в декабре—январе. Возможно, накопление азота начинается раньше, но при расчете на процентной основе оно незаметно из-за увеличения содержания сухого вещества, наблюдающегося в течение вегетационного периода. Содержание азота в древесине и коре снижается весной в результате его передвижения в развивающиеся почки и новые побеги. ДлЯ яблонь, персиков и винограда показано, что азот, запасенный в коре и древесине, легче утилизируется при росте, чем вводимый извне. Такая точка зрения основана на наблюдениях, согласно которым с началом роста из запасающих тканей удобренных деревьев уходит почти столько же азота, сколько и из неудобренных. Имеются и другие доказательства. Сообщалось, что содержание азота в коре (живой флоэме) груши сорта Бартлетт уменьшалось с началом роста, но начинало увеличиваться, когда рост замедлялся, и достигало максимума зимой. Леви и Ковлинг (1968) обнаружили, что по мере развития листьев заметно уменьшается содержание азота в заболони дуба серповидного. Аналогично каждое усиление роста апельсиновых деревьев приводит к уменьшению содержания азота в прилегающих одревесневших тканях. Вероятно, основным источником азота является флоэма, так как в древесине содержание азота изменяется меньше, чем в коре.
Несколько ограниченные данные о сезонных изменениях содержания азота у хвойных показывают их сходство в этом отношении с лиственными деревьями. Получены данные, доказывающие передвижение азота из коры и древесины во время быстрого удлинения стеблей, а также увеличение его содержания в конце лета и осенью до наступления зимнего максимума, сопровождавшегося уменьшением азота в конце зимы. Отток азота из древесины и коры в конце зимы начинается до возобновления процесса удлинения стеблей, но азот используется, вероятно, в ходе первого периода роста.
Имеются публикации, в которых сообщается о влиянии времени года и суток, освещенности и минерального питания на количество и состав азотистых веществ у хвойных. Сделаны выводы о необходимости большего внимания к изменения^ состава аминокислот в связи с теневыносливостью и другими факторами окружающей среды.
Тромп (1970) сделал обзор множества работ по сезонным изменениям состава фракции растворимого азота в древесине и коре фруктовых деревьев. Создается впечатление, что аргинин является важным компонентом этой фракции в период покоя, но его содержание в древесине и коре уменьшается весной, видимо, вследствие передвижения в растущие побеги и включения в новый белок, а может быть, в результате превращения в другие соединения. Высказано предположение, что аргинин синтезируется осенью и зимой из амидов, но пока это не дока-
250
Глава 9
9.3. СООТНОШЕНИЕ РАЗЛИЧНЫХ АМИНОКИСЛОТ В КСИЛЕМНОМ СОКЕ В ТЕЧЕНИЕ ВЕГЕТАЦИОННОГО ПЕРИОДА У ЯБЛОНЬ, РАСТУЩИХ В НОВОЙ ЗЕЛАНДИИ, % ОТ СУММЫ АМИНОКИСЛОТ (В ПЕРЕСЧЕТЕ НА АЗОТ) (Bollard, 1958 )
Соединение Дата
12.Х 19.Х 26.Х 29.Х1 29.ХП 26.1
Аспарагиновая кислота 12 10 22 31 58 41
Аспарагин 65 59 52 65 34 48
Глутаминовая кислота + + + 1 2 3
Глутамин 20 27 24 4 6 7
Серин + 1 + + 2
Треонин 2 1 1 + + +
Метионин + валин 1 1 1 + + 1
Лейцин 1 1 + + + +
Общий азот, мг/л 41 117 131 48 22 12
Примечание. Знак + указывает, что содержание данной аминокислоты менее 1 %,
зано. Применение азотных удобрений, по видимому, увеличивает содержание аргинина больше, чем других растворимых соединений азота. Сообщают также, что концентрация аргинина в ксилемном соке яблонь довольно высока осенью и зимой, но уменьшается в течение вегетационного периода, когда основными соединениями азота сока становятся аспарагин и аспарагиновая кислота. Содержание двух последних соединений в значительной степени снижается в период покоя (табл.9.3). Как обнаружили Симинович и Бригс (1949), действие низких температур на деревья сопровождалось увеличением содержания водорастворимых белков. Паркер (1958) сообщал, что общее содержание белков во внутренней части коры деревьев нескольких видов увеличивалось с августа по декабрь. Концентрация нескольких свободных аминокислот в это время уменьшалась, но количество пролина возрастало. Изменения азотного питания, водный стресс и другие факторы окружающей Чреды часто значительно изменяют состав азотистых соединений в деревьях.
ОСЕННЕЕ ПЕРЕДВИЖЕНИЕ ИЗ ЛИСТЬЕВ. Как уже отмечалось, в листьях может находиться свыше 40% общего содержания азота в деревьях. К счастью для растения, значительная часть азота и других компонентов возвращается в побеги и ветви перед опадением листьев. Это передвижение имеет важное значение, так как в противном случае большая доля азота растений терялась бы, по крайней мере, на время, при листопаде. Оланд (1963) тщательно исследовал изменения содержания основных компонентов в ходе старения и опадения листьев 12-летних яблонь в Норвегии. По его подсчетам, в течение 3—4 недель старения сухое вещество уменьшилось до 16%, азота до 52%, фосфора до 27%, калия до 36%. Содержание магния за этот период не изменялось, а содержание кальция увеличивалось на 18%. Потери путем вымывания
Метаболизм азота и азотное питание
251
Рис. 9.3. Изменение с возрастом содержания сухого вещества и азота в листовой пластинке листьев яблони (Oland, 1963)
за время опыта считались незначительными. Большая часть материалов передвигалась в структурные элементы многолетней древесины яблонь. Динамика осеннего уменьшения содержания сухого вещества и азота показана на рис. 9.3.
По данным некоторых исследователей, осенний отток азота из листьев наблюдается у многих других видов листопадных деревьев, в том числе у бука, вяза, конского каштана, вишни, груши, сливы и тиса. Отток из листьев колеблется в пределах от одной до двух третей содержания в них азота. Однако при некоторых измерениях не учитывались потери путем вымывания в то время, когда листья еще находились на деревьях. Для получения удовлетворительных данных необходимо измерять одновременно и прирост содержания азота в одревесневших частях растений и потери их из листьев, но это очень трудно осуществить. В противоположность данным по листопадным видам, Камерон и Апплеман (1933) сообщали, что содержание азота в листьях апельсиновых деревьев перед их опадением не снижалось. Для хвойных деревьев пока накоплено мало данных, кроме факта уменьшения концентрации азота с возрастом.
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ РАСПРЕДЕЛЕНИЯ АЗОТА. Изменения содержания азота, связанные с возрастом, накладываются на сезонные изменения, особенно в листьях листопадных деревьев. Например, по мере старения листьев возрастает доля материала клеточных стенок, что вызывает кажущееся снижение содержания азота и других компонентов, когда результаты выражают в процентах от сухого веса. Как сообщил Мадгвик (1970), содержание азота в хвое сосны виргинской уменьшалось с 1,2% в однолетней хвое до 1% в трехлетней, но часть этого уменьшения была вызвана увеличением сухого веса. Другие исследователи считают, что 30—50% азота сосновой хвои возвращается в дерево перед ее опадением, но авторы не указали, как рассчитывались результаты их работы. Большая часть азота у дубов Гэмбела и Кел-лога в Калифорнии передвигается в листьях в начале их развития, затем растяжение листа продолжается, в результате содержание азота в пересчете на единицу площади листа в течение лета постепенно падает.
Глава 9
9.4. ВЛИЯНИЕ ВОЗРАСТА ЯБЛОНЬ НА СОДЕРЖАНИЕ АЗОТА (ЧИСЛИТЕЛЬ - % ОТ СУХОГО ВЕСА; ЗНАМЕНАТЕЛЬ - Г) В ЛИСТЬЯХ, НОВЫХ ПОБЕГАХ. СТВОЛАХ С ВЕТВЯМИ, КОРНЯХ И ПЛОДАХ (Gardner et al,, 1952, С РАЗРЕШЕНИЯ MC-Graw-Hill Book Company)
Возраст, лет Листья Новые побеги Стволы и ветви Корни Плоды
1 1,71/0,44 0,30/0,29 0,39/0,20
2 2,09/1,51 — 0,57/1,36 0,88/1,14 —
5 1,76/7,84 0,89/1,93 0,48/17,20 0,64/9,85 —
9 1,70/61,50 0,82/9,08 0,35/85,50 0,58/81,00 0,31/10^55
30 2,08/394,00 0,95/13,60 — 0,31/258,00
100 1,04/435,00 1,04/390,00 0,27/2863,00 0,22/417,00
9.S. ИЗМЕНЕНИЯ СОДЕРЖАНИЯ АЗОТА С ВОЗРАСТОМ В РАЗЛИЧНЫХ ТКАНЯХ СОСНЫ ЛАДАННОЙ, РАСТУЩЕЙ НА ХОРОШИХ УЧАСТКАХ, % (Switzer et al., 1968)
Возраст дерева, лет Листва текущего года Старые ветви Кора ствола Древесина ствола
4 1,00 0,41 0,42 0,16
8 0,95 0,24 0,24 0,06
18 1,08 0,23 0,23 0,06
30 1,22 0,22 0,19 0,04
56 1,16 0,21 0,17 0,03
Осенью, по мере старения листьев, содержание азота резко уменьшается, поскольку азотистые соединения перед опадением листьев передвигаются обратно в древесину.
Уменьшение содержания азота с возрастом деревьев особенно заметно в одревесневших частях. По концентрации азота в новых побегах, листьях или плодах яблонь разного возраста различия невелики (табл. 9.4), но в древесине концентрация азота у более старых деревьев уменьшается. У сосны ладанной в одревесневших частях концентрация азота уменьшается с возрастом, но в листве текущего года более старых деревьев она остается высокой (табл. 9.5). Таким образом, у хвойных и лиственных деревьев тенденция, видимо, одинакова. Ранее указывалось, что заболонь содержит больше азота, чем ядро, и что самую высокую концентрацию азота имеет самая молодая заболонь. Это объясняется, вероятно, тем, что в молодой заболони больше всего живых паренхимных клеток. Следовательно, азот покидает стареющие клетки и концентрация азота в старых тканях обычно ниже, чем в молодых.
Метаболизм азота и азотное питание
253
ОСНОВНЫЕ СОЕДИНЕНИЯ АЗОТА
К наиболее важным соединениям, в которых содержится азот, относятся: аминокислоты, амиды, нуклеийовые кислоты, нуклеотиды и нуклеозиды, белки и алкалоиды (см. рис. 6.20). Большая часть наших знаний о метаболизме азота вытекает из исследований, проведенных на микроорганизмах. С белками растений часто трудно работать, так как при размельчении растительной ткани освобождаются оксидазы и протеолитические ферменты, и, кроме того, белки нередко осаждаются таннинами, которыми богаты некоторые растения.
АМИНОКИСЛОТЫ, Эти соединения являются основными ’’кирпичиками”, из которых строятся белки протоплазмы. Формула большинства аминокислот R-CHNHj -СООН. Поскольку все аминокислоты имеют аминогруппу (NH2) и карбоксильную группу СООН, они проявляют свойства как оснований, так и кислот. В простейшей .аминокислоте, глицине, радикал R представлен атомом водорода CH2NH2COOH. У других радикал может быть очень сложным и содержать дополнительные амино- и карбоксильные группы. Обычными компонентами растительных белков считаются около 20 аминокислот, и все они, по-видимому, встречаются в древесных растениях.
Синтез аминокислот. Аминокислоты могут образовываться несколькими путями, в том числе восстановительным аминированием, пере-аминированием, химическим превращением амидов кислот или других азотистых соединений и ферментативным гидролизом белков. По-види-мому, наиболее важными являются первые два способа.
Восстановление нитратов. Нитраты — самый обычный источник азота для растений, а восстановление нитратов является важным этапом метаболизма азота. Нитраты легко поглощаются деревьями и обычно быстро восстанавливаются, хотя при плохом снабжении углеводами и низком уровне метаболической активности они могут накапливаться. Активность нитратредуктазы также может варьировать, что приводит к колебаниям содержания нитратов в растениях. Вероятный путь восстановления нитратов выглядит так:
- 2—
NO3 —►- NO2 —— N2O2 -------------NH2OH ------------— NHg
Нитрат Нигрит Гипснигриг Гидроксиламин Аммгик
По другому пути, существование которого склонны признавать некоторые исследователи, азот более прямо восстанавливается до аммиака. Энергию для восстановления нитрата поставляет окисление углеводов. Образующийся аммиак в конце концов реагирует с органическими кислотами с образованием аминокислот. Некоторые промежуточные продукты процесса восстановления нитрата токсичны в небольших концентрациях, но они обычно не накапливаются до существенных количеств, в которых могли бы причинить вред.
Принято считать, что у хорошо удобренных растений полевых культур восстановление нитратов происходит главным образом в листьях, куда нитраты передвигаются из корней с ксилемным соком.
254
Глава 9
Однако у ряда изученных древесных растений нитраты, по-видимому, восстанавливаются в корнях, так как в их ксилемном соке преобладают органические формы азота. Болард (1958) обнаружил следы или небольшое количество нитратов в ксилемном соке лишь у одной трети из 131 вида изученных деревьев и трав, но в пасоке всех видов присутствовали многие аминокислоты и амиды. Весь азот сока яблок и плодов нескольких других представителей семейства Rosaceae находился в форме аминокислот и амидов, причем в течение всего вегетационного периода наиболее часто встречались аспарагин, аспарагиновая кислота и глутамин. Барнес (1963) обнаружил 17 аминокислот и уреидов в ксилемном соке 60 видов растений в июне-июле (Северная Каролина, США). От 73 до 88% органического азота в 7 изученных видах сосны составляли цитруллин и глутамин, а Болард (1957) нашел, что те же самые соединения преобладали в ксилемном соке сосны замечательной, растущей в Новой Зеландии. Учитывая высокое содержание органического азота в ксилеме, кажется вероятным предположение о том, что восстановление нитратов происходит в корнях данных растений.
Восстановительное аминирование. Образующийся при восстановлении нитрата аммиак может взаимодействовать с органической кислотой по пути восстановительного аминирования:
соон
(J-0 __
2 +^з + НАД - Н2 дегидрогеназа
СН2 глутаминовой
СООН кислоты
a-к етог лутаровая кислота
СООН
СН - NH2
£н2 + Н2О + НАД
СООН
Глутаминовая кислота
Эту реакцию катализирует фермент глутаматдегидрогеназа с НАД или НАДФ в качестве кофермента. а-Кетоглутаровую кислоту поставляет цикл Кребса (цикл трикарбоновых кислот). Менее важный в количественном отношении путь начинается от пировиноградной кислоты и дает аланин и щавелевоуксусную кислоту, от которой происходит аспарагиновая кислота.
Переаминирование заключается в переносе аминогруппы от одной молекулы на другую. Примером может служить реакция между глутаминовой и щавелевоуксусной кислотами с образованием а-кето-глутаровой и аспарагиновой кислот при участии фермента трансаминазы;
СООН СН -nh2 СООН СООН £-0 £00Н
К + f=° трансаминаза ) 2 (pl - nh2
сн9 toOH fh2 соон сн2 соон fH2 СООН
Г луг амино- Щав влево- а-кетоглутаровая Аспарагин сж ая
вая кислота уксусная кислота кислота вая кислота
Метаболизм азота и азотное питание
255
Другая группа аминокислот берет начало от шикимовой кислоты, из которой образуются тирозин, фенилаланин и триптофан (см. рис. 6.20). Фенилаланин и тирозин могут либо прямо включаться в белки, либо служить предшественниками алкалоидов или компонентов лигнина. Из триптофана также могут синтезироваться алкалоиды, но он более известен как предшественник растительного гормона индолилуксусной кислоты.
ПЕПТИДЫ. Как и белки, пептиды состоят их аминокислот, соединенных пептидными связями. Это означает, что карбоксильная группа одной молекулы аминокислоты соединяется с аминогруппой другой молекулы. Если молекулярный вес образующегося соединения ниже 6000, его условно принято считать пептидом. С точки зрения патологии человека, некоторые из небольших пептидов представляют значительный интерес. Трипептидом является пенициллин. Аллергены пыльцы — также пептиды. Другие пептиды обладают фармакологической активностью.
АМИДЫ. Среди распространенных соединений азота, обнаруженных в растениях, важную роль играют амиды. глутамин и аспарагин. Глутамин образуется из глутаминовой кислоты путем присоединения аммиака:
СООН
CHNH9
?н2 + NHj + АТФ
СН2 синтетаза
СООН
I
Глутамин овая
кислота
СООН
chnh2
^Н2 +АДФ+Н3РО4
сн2
£onh2
Глутамин
Считается, что синтез этих амидов предотвращает накопление в растениях вредных концентраций аммиака. Они также появляются в проростках при использовании запасных белков на рост. При остром дефиците углеводов амиды могут окисляться, но это сопровождается выделением вредных количеств аммиака. Глутамин имеет важное значение для метаболизма, вероятно, он служит донором групп NH2 при синтезе карбамоилфосфата и, возможно, для аспарагиновой кислоты при образовании аспарагина.
НУКЛЕИНОВЫЕ КИСЛОТЫ И РОДСТВЕННЫЕ СОЕДИНЕНИЯ. Производные пуриновых и пиримидиновых оснований являются компонентами многих чрезвычайно важных соединений. К ним относятся: нуклеиновые кислоты (РНК и ДНК), нуклеотиды (АМФ, АДФ и АТФ), никотинамидные нуклеотиды (НАД и НАДФ), тиамин, кофермент А и цитокинины (см. рис. 6.20). Нуклеотиды— это фосфорные эфиры нуклеозидов, нуклеиновые кислоты’ — высокомолекулярные полимеры, образованные длинными цепями из четырех типов нуклеотидных единиц, являющихся производными аденина, гуанина, цитозина, тимина или урацила. Генетический материал ядра представлен дезокси-
256
Глава 9
рибонуклеиновой кислотой, каждая молекула которой состоит из двух полинуклеотидных цепей, расположенных в виде двойной спирали. Небольшие количества ДНК обнаруживаются вне ядра — в митохондриях и хлоропластах. Молекулы РНК состоят из простых цепочек. В состав РНК входит сахар рибоза, а в ДНК входит дезоксирибоза. В молекулах РНК вместо тимина содержится урацил. Существуют 3 типа РНК, присутствующих в рибосомах, ядре и других органеллах: матричная (м-РНК), транспортная (т-РНК) и рибосомная (р-РНК).
БЕЛКИ, Это главные органические компоненты протоплазмы. Белки — чрезвычайно сложные азотсодержащие высокомолекулярные вещества, различающиеся по форме, размерам, поверхностным свойствам и функции. Тем не менее у всех белков есть общее: они состоят из аминокислот, амфотерны и имеют свойства коллоидов. Амфотерное соединение выступает или как кислота, или как основание. Белки содержат основные группы NH2 и кислые группы СООН и заряжены положительно при pH ниже критической ’’нейтральной” величины, известной как изоэлектрическая точка, отрицательно — при pH выше изоэлектрической точки. Помимо того, что белки составляют структурную основу протоплазмы, они также являются ферментами и запасными питательными веществами (последнее особенно характерно для семян).
Белки всегда имеют высокий молекулярный вес, но он может значительно варьировать у разных типов белков. Некоторые из белков семян (запасных белков) имеют молекулярный вес от 200 000 до 400 000: у амандина, белка миндаля, молекулярный вес 329 000, а обнаруженный в семенах конского кащтана белок имеет молекулярный вес 430 000. Молекулы некоторых ферментов также очень велики: молекулярный вес уреазы 400 000, каталазы — 500 000. С другой стороны, некоторые молекулы белков довольно малы и имеют молекулярный вес 10 000— 50 000. В безводном состоянии белки содержат обычно 50—55% углерода, 6—7% водорода, 20—23% кислорода и 12—19% азота. Все растительные белки содержат небольшое количество серы, а некоторые также и фосфор.
Традиционный способ классификации белков основывался на их растворимости, но разделение по этому признаку все меньше удовлетворяет исследователей. Полезней могла бы быть классификация на функциональной основе. Например, их можно разделить на структурные (протоплазматические.), запасные белки и ферменты. К структурным относятся белки протоплазмы и таких ее компонентов, как хлоропласты. Запасные белки, которыми особенно богаты семена, служат важным источником пищи для человека и животных, а также и для проростков. Некоторые ферменты, такие как уреаза и папаин, представляют собой простые белковые молекулы, и их каталитические свойства определяются только расположением аминокислотных остатков, из которых они состоят. Другим требуется связанная с белковой молекулой простетическая группа, или кофактор. В некоторых случаях кофактор представлен ионом металла, например меди или железа. Другие ферменты нуждаются в присутствии небелковых простетических групп или менее прочно связанных кофакторов, таких, как пиридиновые нуклеотиды НАД и НАДФ.
Метаболизм азота и азотное питание
257
Распадаясь под действием кислот, щелочей или ферментов, белки дают смесь аминокислот. При этом в процессе гидролиза образуется ряд промежуточных продуктов по такой схеме:
Белки —протеозы —пептоны—полипептиды дипептиды—-»- аминокислоты.
Ферментативный распад более специфичен: ферменты обычно атакуют специфические химические связи и отщепляют определенные аминокислоты или группы аминокислот в отличие от больших неопределенных групп, отщепляемых при кислотном или щелочном гидролизе.
Молекула белка состоит из длинной цепи аминокислот, соединенных пептидными связями, в которых карбоксильная группа одной аминокислоты присоединяется к аминогруппе другой аминокислоты с отщеплением воды. Примером образования пептидной связи может быть соединение двух молекул глицина:
™тт СНо-СООН + нон
___ СН2 - СООН । 2
СЩ-СО » OH+lT-N-=>CH2-CO-NH
)2 С-’"| I2
nh2 н nh2
Как видно из формулы образующегося при этой реакции дипептида глицилглипина, у него имеются свободная карбоксильная группа и свободная аминогруппа, которые могут соединяться с другими аминокислотами. Независимо от того, сколько еще аминокислот присоединятся к такому дипептиду, в образующейся сложной молекуле всегда будут свободные амино- и карбоксильные группы. Таким путем при соединении нескольких сотен аминокислот при помощи пептидных связей образуется белок. Структурную основу молекулы белка можно изобразить следующим образом:
Радикалы в этой цепи представляют собой остатки различных аминокислот. В случае глицина R = Н, в аланине (CH3CHNH2COOH) R=CHj.
В живых растениях белки находятся в состоянии динамического равновесия: они постоянно разрушаются и образуются снова, но общее содержание белка может оставаться постоянным в течение длительного периода, поскольку их распад сбалансирован с синтезом. Разные белки различаются по стабильности и имеют характерные скорости оборота. Биосинтез белка наиболее энергично протекает в клетках меристемати
258
Глава 9
ческих участков, в которых происходит деление клеток, и в запасающих органах, где. накапливается белок (например, в эндосперме семян). Биосинтез белка происходит на рибосомах в цитоплазме. Рибосомы — это органеллы, состоящие из белка и РНК. Последовательность аминокислот, а тем самым и вид образующегося белка определяются генетически матричной РНК. Молекулы м-РНК образуются в ядре под контролем ДНК и мигрируют к рибосомам, где они функционируют в качестве шаблона (модели) для образования молекул белка. Транспортные РНК участвуют в переносе аминокислот на рибосомы.
АЛКАЛОИДЫ. Они составляют большую и сложную группу циклических соединений, содержащих азот. Уже выделено около 2000 различных соединений этой группы, некоторые из которых представляют интерес для фармакологии, например морфин, стрихнин, атропин, колхицин, эфедрин, хинин и никотин. Наиболее часто они. встречаются в травянистых растениях, некоторые обнаруживаются и в деревьях, в основном тропических видов.
Алкалоиды накапливаются в отдельных органах: листьях, коре или корнях. Например, никотин синтезируется в корнях табака, но 85% его находится в листьях. Алкалоиды хинного дерева добывают из коры. Иногда алкалоиды встречаются в древесине. Древесина некоторых видов из семейств Anacardiaceae, Apocynaceae, Euphorbiaceae, Legu— minosae, Rutaceae, Rubiaceae содержит так много алкалоидов, что вызывает дерматиты. Среди алкалоидов, добываемых из деревьев, наиболее известны алкалоиды хинного дерева, используемые для борьбы с малярией. Они встречаются у растущих в Андах представителей родов Chinchona и Remijia семейства Rubiaceae.
Несмотря на широкое распространение алкалоидов в растениях, не обнаружена их существенная физиологическая роль. Возможно, что в некоторых случаях они препятствуют повреждению растений грибами, бактериями или насекомыми. Алкалоиды представляются побочными продуктами метаболизма азота, которые обычно не приносят ни вреда, ни пользы вырабатывающему их растению. Количество попадающего в них азота слишком мало для того, чтобы играть роль в естественном отборе.
ФИТОТОКСИНЫ И ПАТОГЕННОСТЬ. К наиболее экзотическим областям химии азота относятся изучение фитотоксинов и биохимия патогенеза. Ряд продуцируемых грибами фитотоксинов образуется из аминокислот. К тому же считается, что специфичность взаимоотношений хозяина и паразита в значительной степени зависит от азотистых соединений. Альберсайт и Андерсон-Прот (1975) считают, что восприимчивость к патогенным организмам часто зависит от наличия в плазматических мембранах растительных клеток белков, которые являются специфическими рецепторами для антигеноподобных углеводов патогенных организмов. Кроме того, разрушающие полисахариды ферменты, образуемые патогенными для растений организмами, играют важную роль в инфекции. Эта область только начинает изучаться, и, несомненно, ей в будущем будут уделять больше внимания.
Метаболизм азота и азотное питание
259
ПОТРЕБНОСТЬ В АЗОТЕ
Потребность лесных и плодовых деревьев в азоте определялась в многочисленных работах (табл, 9.6). Почти треть всего азота, необходимого за год молодым деревьям, и около одной пятой годовой потребности более старых деревьев поступает в плоды и уносится при сборе урожая. По мере старения деревьев относительно меньше азота накапливается в плодах и большая доля от общего азота используется на рост корней и надземных частей. От 5 до 6% азота временно теряется вследствие сбрасывания весной цветков и молодых плодов, а 30—40% теряется при опадении листьев осенью.
9.6. РАССЧИТАННАЯ ГОДОВАЯ ПОТРЕБНОСТЬ ЯБЛОНЬ В АЗОТЕ ЧИСЛИТЕЛЬ - Г; ЗНАМЕНАТЕЛЬ - %)
Расход азота 20-летние деревья (Murneek,1942) 25-летние деревья f Magness. Regei-rhbal, 193&)
На урожай плодов 180/30,53 150/21,57
Временные потери при опадении цветков и плодов 30/4,59 40/5,88
Временные потери при опадении листьев 180/30,53 270/39,22
На поддержание роста корней и стеблей 160/26,72 230/33,33
Удаляется при обрезке ветвей 50/7,63
Всего 590/100,00 690/100,00
По сообщениям, в годы обильного плодоношения у деревьев запасается мало азота, а уже имеющиеся запасы значительно или полностью истощаются. В результате уменьшается вегетативный рост и подавляется образование цветочных почек в течение данного и даже следующего года. Иногда для полного возвращения к норме требуется более года. Такие последствия обильного плодоношения напоминают его действие на содержание запасных углеводов (см. главу 7).
Подсчитано, что в Европе насаждения лиственных деревьев используют 30-60 кг азота на 1 га и возвращают около 80% с опавшими листьями. По расчетам Бакера (1950), вынос составлял 40—50 кг/га, из которых почти 80% возвращаются в землю с опавшими листьями. Однако по приведенным ранее в этой главе данным, у некоторых видов перед листопадом одна треть азота листьев передвигается обратно в ветви, и потери его с листьями намного меньше 80%.
По данным Швитцера с соавторами (1968), 20-летнее насаждение сосны ладанной содержит свыше 300 кг/га азота, а на образование новых тканей ежегодно использует около 70 кг/га (табл. 9.7). Из них 38 кг поступают из почвы, а остальное — из самого дерева, преимущественно из листьев. (Распределение азота в молодых соснах показано в табл. 9.2.) < 1бщие потери азота в лесной подстилке (листья, побеги, ветви) состав-
260
Глава 9
9.7. СРАВНИТЕЛЬНОЕ РАСПРЕДЕЛЕНИЕ АЗОТА В ЭКОСИСТЕМАХ ИСКУССТВЕННЫХ НАСАЖДЕНИЙ, КГ/ГА (Switzer et al., 1968)
Компоненты экосистемы
37-летняя сосна черная калабрийская (Шотландия)
20-летняя сосна ладанная (штат Миссисипи, США)
Деревья:
кроны 83 89
стволы 60 85
корни 113 138
всего 256 312
Живой напочвенный покров 8 0
Лесная подстилка 322 124
Сумма органического вещества 586 436
Почва 1376 1900
Всего 1962 2336
ляют 30 кг/га за год. Эти потери, разумеется, временные, так как большая часть азота возвращается в деревья. Следовательно, необходимо лишь около 8 кг/га нового азота. Круговорот азота будет рассмотрен ниже, отметим, что много азота может поступать с осадками.
Борман с соавторами (1977) установили, что содержание азота в 55-летнем буковом, кленовом и березовом лесу в Новой Англии (северо-восток США) составляет 351 кг/га в надземной биомассе и 180 кг/га в подземной. На рост используется около 120 кг, из которых 1/3 извлекается из запасов в растительных тканях. Каждый год в систему поступает около 20 кг/га, из них 14 кг добавляется путем азотфиксации в почве, а 6 кг - с осадками. Эти леса теряют очень мало азота.
Эти данные показывают, что для роста деревьев нужно значительно меньше азота, чем для выращивания сельскохозяйственных культур. Кукуруза, например, поглощает свыше 175 кг/га азота за вегетационный период, люцерна может использовать более 200 кг/га. При этом большая часть должна поглощаться в течение короткого периода. Соя использует около 300 кг азота на 1 га, из которых приблизительно 25% фиксируется в корневых клубеньках. Плодовые деревья обычно имеют высокую потребность в азоте, - 60-175 кг/га, в зависимости от числа и размеров деревьев. Лесные деревья получают большую часть нужного им азота благодаря разложению подстилки и атмосферной фиксации, но плодовые деревья необходимо удобрять.
Имеются свидетельства, что древесные породы различаются по потребности в азоте, так как некоторые виды встречаются только на плодородной почве, а другие могут расти и на бедной. После широкого изучения минерального питания 12 распространенных видов лиственных деревьев Митчел и Хандлер (1939) разделили их по потребности в азоте на три группы. Красный, белый и каштановый дубы, тополь осинообразный и клен красный наиболее выносливы в отношении низких концентраций азота. Кария голая, клен сахарный, бук и нисса лесная занимают промежуточное положение. Ясень американский, тюльпанное дерево и липа имеют высокую потребность в азоте. Сообщалось о значительном варьировании реакции представителей разных форм сосны ладанной на внесение азотного удобрения.
Метаболизм азота и азотное питание 261
ИСТОЧНИКИ АЗОТА
Деревья способны усваивать азот в форме нитратов, нитритов, солей аммония и органических соединений азота (таких, как мочевина). Независимо от исходной формы большая часть азота поглощается, вероятно, в виде нитрата или аммония. В корнях азот превращается в органические соединения, в основном в амиды и аминокислоты, которые затем передвигаются в побеги с пасокой (ксилемным соком). Ранние вегетационные опыты Адомса (1937) показали, что сосна ладанная может поглощать как нитратный, так и аммонийный азот, предпочитая при низких значениях pH нитратный, а при высоких — аммонийный азот. По некоторым данным, европейские породы хвойных деревьев поглощают большую часть необходимого им азота в виде аммиака, тогда как лиственные породы, особенно ясень, бук и дуб, получают его в основном в виде нитрата. Фарис с соавторами (1964) сообщали о сложных взаимодействиях между снабжением различными формами азота и кальция в связи с ростом сеянцев сосны ладанной, а Хаук (1968) рассмотрел взаимодействия почвы и удобрений, включая pH, содержание воды, аэрацию и другие факторы, затрудняющие обобщение знаний о наилучшем источнике азота.
Основными источниками азота для лесных деревьев являются: связанный почвенными микроорганизмами, поступающий из атмосферы с дождем и снегом и освобождаемый благодаря разложению лесной подстилки. Выпускаемые промышленностью удобрения — наиболее важный источник азота для плодовых деревьев и декоративных кустарников. Удобрения начинают использовать и для лесных деревьев. Некоторая часть азота теряется, попадая в атмосферу в результате процессов аммонификации и денитрификации, а часть вымывается при сильных ливнях.
АЗОТФИКСАЦИЯ. Атмосфера приблизительно на 80% состоит из азота, но атмосферный азот весьма инертен. Этот потенциальный источник азота может использоваться деревьями только после соединения азота с другими элементами, т.е. после его фиксации. Фиксация азота микроорганизмами, а также во время атмосферных разрядов возмещает потери азота почвой вследствие вымывания, пожаров и поглощения растениями и предотвращает полное истощение его запасов.
Фиксация азота происходит одинаково и у свободноживущих, и у симбиотических организмов. Ферментный комплекс, называемый нитрогеназой, восстанавливает N2 в несколько этапов путем связывания с водородом с образованием NH3, который при соединении с органическими кислотами дает аминокислоты (см. рис. 6.20).-Для этого необходимо наличие АТФ, и весь процесс может быть представлен следующим уравнением:
N2 + 6 е' + 6Н+ + пАТФ—2NH3 + пАДФ + пфн-
Гемоглобин, найденный в корневых клубеньках (единственное место в растительном царстве, где он присутствует), не участвует прямо в азотфиксации, но, по-видимому, играет косвенную роль, контролируя концентрацию кислорода. Нитрогеназа способна восстанавливать ряд
262
Глава 9
других субстратов, в том числе Н+. Ацетилен (С2Н2) легко восстанавливается нитрогеназой до этилена (С2Н4). Эта реакция широко используется для определения нитрогеназной активности. Свободножив у щие бактерии связывают меньше азота на единицу массы протоплазмы по, сравнению с клубеньковыми бактериями, поскольку у них больше энергии используется на рост.
НЕСИМБИОТИЧЕСКАЯ ФИКСАЦИЯ АЗОТА. Связывание азота в почве происходит в значительной мере в результате деятельности сапрофитных бактерий родов Azotobacter и Clostridium. Эти бактерии относятся к свободноживущим, но найдено несколько видов, чье распространение ограничено ризосферой отдельных растений. Некоторые синезеленые водоросли также фиксируют азот и успешно заселяют слаборазвитые почвы и другие экстремальные местообитания. Считается, что анаэробные формы чаще встречаются в лесных почвах, чем аэробные, возможно, потому, что высокая кислотность, обычная для лесных почв, неблагоприятна для аэробных азотфиксирующих бактерий. По мнению Русела (1973), некоторые азотфиксирующие бактерии настолько редки во многих почвах, что вызывает сомнение их способность связывать большие количества азота.
СИМБИОТИЧЕСКАЯ АЗОТФИКСАЦИЯ У БОБОВЫХ. Как упоминалось в главе 2, бактерии рода Rhizobium проникают в корни многих видов бобовых растений, образуя корневые клубеньки, в которых фиксируется азот. Симбиотическая азотфиксация имеет наибольшее значение для лесоводства там, где деревья тесно ассоциированы с многочисленными дикими травами семейства бобовых (например, на юго-востоке США). Хапман (1935) сообщал об усиленном росте в высоту и толщину у нескольких видов лиственных деревьев, посаженных рядом с белой акацией. Он также обнаружил самые высокие величины общего содержания азота в почве возле деревьев белой акации и сделал вывод, что рост улучшался в результате фиксации азота акацией.
На фиксацию азота клубеньками может различными путями влиять водный стресс. При потере воды клубеньками в сухой почве ослабляются инфицирование корней бактериями и образование клубеньков. Не исключена возможность, что водный стресс у растений уменьшает транспорт соединений азота из корней к побегам. Высказывалось также предположение, что пониженное снабжение углеводами побегов, попавших в условия водного дефицита, уменьшает фиксацию азота клубеньками. Создается впечатление, что наряду с прямым действием водного \ стресса на клубеньки имеется и косвенное влияние водного стресса в побегах. На интенсивность азотфиксации часто (но не всегда) влияет интенсивность фотосинтеза, поскольку клубеньки являются крупными потребителями ассимилятов. Увеличение концентрации СО2 в воздухе с 0,08 до 0,12% вызывает у сои увеличение сухого веса и азотфиксации.
АЗОТФИКСАЦИЯ У РАСТЕНИЙ, НЕ ОТНОСЯЩИХСЯ К БОБОВЫМ. Азотфиксирующие клубеньки на корнях обычно связывают с семейством Leguminosae. Однако клубеньки встречаются и на корнях ряда небобовых двудольных растений, в том числе многих видов деревьев и кустарников различных семейств: Betulaceae, Eleagnaceae, Myricaceae, Rhamnaceae, Casuarinaceae, Coriariaceae, Zygophyllaceae и Rubiaceae, Все растения этих семейств приспособлены к росту на бедных почвах.
263
Метаболизм азота и азотное питание
Бонд и его сотрудники (1967, 1976) показали наличие заметной азот-фиксации у нескольких небобовых древесных растений, имеющих клубеньки: Alnus, Myrica, Hippophae, Shepherdia и Casurina. Выращенные из семян в оранжерее растения ольхи, у которых вызывали развитие клубеньков путем инокуляции суспензией разрушенных клубеньков от растущих в поле сеянцев, хорошо росли на субстрате, лишенном связанного азота. Это указывает на связь фиксации молекулярного азота с клубеньками. В другом опыте Бонд (1956) показал, что клубеньки ольхи в полевых условиях, как и в оранжерее, нормально фиксируют азот. Автор отметил, что количество фиксированного азота было значительным. По сообщению Бонда и Гарднера (1957), в условиях эксперимента фиксация азота у ольхи черной и восковницы обыкновенной в течение первого года роста превышала азотфиксацию у однолетних бобовых и белой акации.
Фиксация азота у небобовых растений, имеющих клубеньки, по-видимому, в определенных районах имеет большое экологическое значение. Например, Крокер и Майер (1955) отмечали, что в Глэсье Бей (Аляска) под зарослями ольхи накапливалось азота в среднем 61,6 кг/га, что создавало благоприятные условия для. роста ели ситхинской, сменявшей ольху. Возрастает интерес к азотфиксирую-щей способности ольхи красной в лесах на Тихоокеанском побережье северо-запада США. Ценность метода введения ольхи в плантации хвойных деревьев для улучшения роста последних уже давно признана европейцами, практикуется также и в Японии. Благоприятное влияние в этом случае оказывает, несомненно, увеличение содержания доступного азота. Виртанен (1957) показал, что ель, растущая возле ольхи, получает азот, фиксированный клубеньками на корнях ольхи. Он подсчитал, что в ольховой роще при плотности 10 тыс. деревьев на 1 га и гысоте деревьев около 2,5 М опавшие листья и остающиеся в земле корни должны прибавлять около 200 кг/га азота. Потери азота в этих расчетах не принимались во внимание. Для сравнения, Харди и Хавелка (1976) установили, что соя потребляет около 300 кг азота на 1 га, из которых около 25% фиксируется клубеньками. Содержащие актиномицеты клубеньки на корнях растений калифорнийского чапарреля также связывают значительные количества азота.
Клубеньки небобовых состоят, по-видимому, из сильно разветвленных боковых корней, а клубеньки бобовых обычно развиваются из клеток коры. Кроме того, почти во всех клубеньках небобовых растений типа Alnus фиксацию азота осуществляют актиномицеты рода P'rankia. Исключение составляет присутствие бактерий Rhizobium в клубеньках на корнях деревьев рода Trema из семейства Ulmaceae, растущих в Новой Гвинее. Следует отметить, что утолщения на корнях могут быть вызваны нематодами и другими агентами, поэтому без тщательной проверки наличие клубеньков не должна приниматься в качестве доказательства присутствия азотфиксирующих организмов. Сипе зеленые водоросли, как известно, вступают в симбиотические оиюшения и снабжают азотом мхи, лишайники и некоторые семенные растения, но только у рода травянистых растений Gunnera они действи-...... внедряются в клетки. Сообщалось, что некоторые бактерии оЬразуют азотфиксирующие клубеньки на листьях определенных растении, в том числе из родов Psychotria, Pavetta, Ardisia и Dioscorea. (Ьщако Ван Гов (1976) на основании ацетиленового теста и изучения роста растений сделал вывод, что восстановление азота если и происходит в этих листовых клубеньках, то слишком незначительно для того, чтобы играть роль в азотном балансе растений. Возможно, что
Глава 9
264
эти бактерии вырабатывают регуляторы роста (например, цитокинин) или другие вещества полезные для растений. Утверждают также, что бактерии, обитающие на поверхностях листьев (филлосфере), могут фиксировать азот. Например, Ионес (1970) считает, что живущие на поверхности хвои дугласии бактерии фиксируют измеримые количества азота.
Весьма интересно было бы отыскать способы увеличения количества азота, связываемого растительностью. В эту задачу входят поиски методов усиления азотфиксации у тех растений, где она уже имеется, и, может быть, попытки вызвать микробиологическую фиксацию азота в ризосфере не только тропических видов трав (где она уже обнаружена), но и других растений. Более оригинальный подход включает генетические манипуляции с использованием рекомбинантных молекул ДНК для внедрения генов азотфиксации в те виды растений, у которых их нет. Другая возможность заключается в слиянии протопластов для переноса в другие растения гена или генов, делающих бобовые растения хорошими хозяевами для Rhizobium. Однако для получения желаемых типов растений этими методами предстоит преодолеть громадные трудности.
СВЯЗЫВАНИЕ АЗОТА В АТМОСФЕРЕ. Заметные количества азота попадают в почву с дождем и снегом в форме аммиака и окислов азота, образующихся при электрических разрядах, выделяющихся вследствие вулканической активности и производственной деятельности. Небольшое количество соединений азота вымывается из лесного полога. По подсчетам Бакера (1950), ежегодно из атмосферы в почву возвращается около 5 кг/га азота. Ларк ер (1975) приводит другую максимальную величину — 10 кг/га за год. Он считает, что это лишь 1/7 часть того количества азота, которое связывается в результате деятельности микробов. Хотя это количество выглядит небольшим, но Веле с соавторами (1975) утверждают, что дождь приносит лесам юго-востока США примерно столько же азота, сколько из них выносится при рубке 16-летних сосновых насаждений для переработки в бумажную массу: около 6,5 кг/га в год.
ВЫДЕЛЕНИЕ АЗОТА ЛЕСНОЙ ПОДСТИЛКОЙ. Некоторая часть поглощенного деревьями азота возвращается в почву из лесной подстилки. Поддержание плодородия лесных почв частично зависит от возврата азота и минеральных питательных веществ при разложении подстилки. Листья и ветви, ежегодно пополняющие лесную подстилку, могут приносить до нескольких тысяч фунтов органического материала, содержащего приблизительно 1% азотистых веществ.
Вакер (1950) приводит цифру 3400 кг/га для предельно уплотненных лесов на хороших участках в США в качестве приемлемой средней величины ежегодного опада листьев и ветвей. Наблюдаются и большие отклонения. Например, отмечались такие низкие величины, как 500 кг/га для бука на плохих участках. В то же время в самых лучших насаждениях бука в Европе до 6700 кг/га возвращается в виде листьев и обломков ветвей.
Количество азота в подстилке сильно варьирует в зависимости от видов растений. Согласно Хандлеру (1941) содержание азота в подстилке лиственных лесов штата Нью-Йорк1 колебалось от 0,43 до 1,04%, в среднем 0,65%. Коль (1937) приводит величины от 0,50 до 1,25% для хвойных и лиственных лесов Пьемонта (штат Южная Каролина). Листва лиственных пород обычно содержит больше
Метаболизм азота и азотное питание
265
азота, чем листва хвойных. Опавшая хвоя содержит около 0,6—1% азота, тогда как опавшие листья содержат от 0,8 до 2% азота. Для некоторых лиственных деревьев приводились и величины значительно ниже 0,8%.
Если принять среднюю величину прибавления лесной подстилки за год равной 3400 кг/га, а среднее содержание азота 0,6-2%, то возврат азота в почву составит приблизительно 20-70 кг/га. Лархер (1975) сообщает, что деревья смешанного лиственного леса в Бельгии теряют с опавшими листьями за год 61 кг/га азота, а древостой вечнозеленого дуба на юге Франции - 33 кг/га. Это составляет примерно 70% поглощенного азота.
Скорость разложения лесной подстилки зависит от вида, условий почвенного плодородия, влажности, аэрации и температуры. Хвоя хвойных пород разлагается медленнее, чем листья у лиственных. Разложение протекает медленнее в северных широтах, быстрее — в тропических районах. В северных лесах для освобождения всего азота подстилкой из однолетних ветвей и листьев может понадобиться до 50 лет и вследствие малой скорости распада у деревьев может развиться серьезный дефицит минеральных веществ. Повышению интенсивности роста после прореживания переуплотненных молодых насаждений частично способствует освобождение азота и других питательных веществ при гниении валежника. В тропических лесах разложение протекает очень быстро и соответственно быстро совершается круговорот. Считается, что в неглубоких кислых неплодородных почвах тропиков грибковые пленки, часто обнаруживаемые в поверхностном слое, играют важную роль в ускорении оборота азота и других питательных веществ.
АЗОТНЫЕ УДОБРЕНИЯ. Удобрение плодовых и декоративных деревьев уже давно стало обычной практикой. Расширяется и удобрение лесных насаждений. Плодовым и декоративным деревьям часто недостает микроэлементов. В Австралии, Новой Зеландии и в некоторых других регионах, таких, как сырые прибрежные равнины юго-востока США, приносит пользу внесение фосфора. Лесным породам наибольшую выгоду дает применение азотных удобрений.
Существуют различные азотные удобрения. Сравнению достоинств аммонийного и нитратного азота, мочевины и медленно выделяющих азот составов посвящены многие исследования. Хаук (1968) указывал на отсутствие явных доказательств того, что одна форма азота намного лучше другой, поэтому определяющим фактором при выборе форм азота для внесения может быть стоимость удобрений и их применения; Реакция растения на удобрение азотом зависит от других факторов окружающей среды, таких, как снабжение водой, аэрация почвы и достаточное снабжение другими элементами минерального питания. Например, по подсчетам Веллса с соавторами (1976) одобрение жердняка сосны ладанной в течение 5 лет на почве с недостатком азота увеличивало образование древесины на 14 м3/га, а на почве с высоким содержанием азота — только на 2,1 м3/га.
КРУГОВОРОТ АЗОТА
Рост деревьев и других растений ограничивается чаще недостатком азотного питания, чем дифицитом какого-либо другого вещества, за исключением воды. Изучению скорости, с которой азот поглощается и теряется в ходе круговорота в различных фитоценозах и экосистемах,
266
Глава 9
Отток из болееи старых тканей (32 кг/га)
\ Использу- г ется на рост s (70кг/га)
[ Годовая потеря
I , при опадении
Поглощается де- листьев и деток
к возвращается 0почву
Рис. 9.4. Упрощенная схема основных компонентов круговорота азота у 20-летнего насаждения сосны ладанной (составлена по данным Switzer et al., 1968) :
1 — отток азота из более старых тканей — 32 кг/га; 2 — азот, используемый на рост — 70 кг/га (общий азот деревьев 312 кг/га) ; 3 — годовая потеря при опадении листьев и веток — 30 кг/га; 4 — возвращается в почву при распаде; 5 — поступает из атмосферы с осадками 5-10 кг/га;
6 — азот почвы — 1900 кг/га; 7 — поглощается деревьями из почвы 38 кг /га
Азот ^/поступает почвы цз атмосферы (1300 кг/га) с осадками
(5-10 кг/га)
посвящены многие исследования. Лархер (1975) и Курлин (1970) подвели итог обширной информации по этому вопросу. Согласно Швитцеру и другим исследователям (1968) изученное ими 20-летнее сосновое насаждение содержало 2300 кг/га азота, в том числе около 1900 кг/га находилось в поверхностном слое почвы. За год использовалось лишь 70 кг/га, из которых 38 кг/га поступало из почвы, а остальное — из хвои и ветвей перед их опадением. Опавшая хвоя и другие растительные части возвращали около 30 кг/га в почву, а на долю других источников азотного снабжения оставалось 8 кг/га, вероятно, за счет связывания азота в атмосфере. Упрощенная схема круговорота азота в этом сосновом насаждении приведена на рис. 9.4.
Митчел с соавторами (1975) рассчитали ориентировочный баланс азота для зрелого лиственного леса в юго-западной части Северной Каролины (США). Ниже приведены сокращенные данные.
Компоненты экосистемы Азот, кг/га
Репродуктивные органы...........у........................ 2,7
Листья...................................................95,0
Стволы и ветви........................................... . 310,0
Корни....................................................150,0
Почвенная подстилка......................................137,0
Всего азота в деревьях.............................. 694,7
Органические вещества почвы........................... 3873,0
Почвенные организмы......................................175,0
Всего азота в почве...................................86,5
Итого азота в системе.............................. 4829,2
Создается впечатление, что эта экосистема содержит значительно больше азота, чем в каждом из сосновых насаждений табл. 9.7, но ведь она и значительно старше. Как и в случае сосновых насаждений, около 80% азота находится в почве и нсмно-
Метаболизм азота и азотное питание
267
гим более 10% в деревьях. Смена листьев происходит быстрее, чем смена хвои, но разница в скорости круговорота азота в других частях системы в сосновых и лиственных лесах, по-видимому, невелика. Согласно Борману и другим исследователям (1977), в лиственном лесу Новой Англии (США) около 90% азота находится в органическом веществе почвы, 9,5% - в растительности и 0,5% - в форме доступного для растений азота почвы. В противоположность этому приблизительно 30% азота тропической лесной экосистемы находится в Деревьях.
Обобщенная схема круговорота азота на глобальной основе показана на рис. 9.5. Прирост азота состоит из фиксации симбиотическими и свободноживущими бактериями и другими организмами, атмосферной фиксации при электрических разрядах, азота, выделяющегося вулканами и в промышленных процессах, и того, который прибавляется за счет удобрений и распада растительных и животных остатков. Последнее на самом деле не прирост, а возврат азота, потерянного в ходе разложения. (Фиксация азота более подробно рассматривалась ранее и не требует дальнейшего обсуждейия.)
Потери азота вызываются деятельностью денитрифицирующих бактерий, восстанавливающих нитраты до молекулярного азота, потерями аммиака при гниении растительных и животных остатков, пожарами и вымыванием. Лесные пожары, вызываемые молниями или человеком, происходят с незапамятных времен и существенно влияют на типы древостоев. В последние годы планируемое выжигание стало общепринятым лесохозяйственным методом контроля болезней, снижения скорости возобновления леса и уменьшения ущерба от низовых пожаров, В этой связи большой интерес представляет изучение влияния пожара на минеральное питание деревьев. При сгорании подстилки и органического вещества нижнего яруса леса теряется много азота, но освобождаются другие элементы, становясь непосредственно доступными. Вследствие этого рост может усиливаться. Виро (1974) сделал вывод, что в Северной Европе польза от повышенной минерализации
Рис. 9.5. Общая схема круговорота азота
Растительные
Атмос:
Смерть
Аммиак
Растительные и животные остатки
Аминокислоты
Восстановление 6 растениях
Животные белки
/ Потери. - Бактерии \беншп^1 Фиксация сим-тиотическими и не-ХАимоиотическими^. \\организмам
Бактериальная нитрификация
Нитриты
Бактериальная ^рис^шауЯ'''
Потери при пожаре.
Распад
Аммонификация
268 Глава 9
азота после пожара намного превышают потери во время пожара. Этот вывод может быть верным и для зарослей калифонийского чапарреля, где минерализация протекает медленно, а после пожара содержание нитратного азота возрастает. На юго-востоке США сосновые насаждения также неоднократно горели в течение многих лет без заметного уменьшения скорости роста. (Влияние огня на качество местообитания обсуждается в главе 17.)
Потери азота и других питательных веществ вследствие вымывания в ненарушенных лесах незначительны, и в результате планируемого выжигания обычно не наблюдается их увеличения. Если уничтожается вся растительность, потери могут быть тяжелыми. Однако обычно растительность быстро отрастает после сплошной рубки или выжигания, и это возвращает большую часть питательных веществ, освобождаемых при разложении валежника или остающихся в золе после пожара.
и®
МИНЕРАЛЬНОЕ ПИТАНИЕ И ПОГЛОЩЕНИЕ СОЛЕЙ
МИНЕРАЛЬНОЕ ПИТАНИЕ
Минеральное питание имеет большое значение для физиологии дерева, поскольку для нормального роста необходимо достаточное снабжение определенными минеральными элементами. Растениям требуются кислород, вода, двуокись углерода, азот и более 10 минеральных элементов в качестве реагентов или ’’сырья” для различных синтетических процессов и для других целей. Люди, не знакомые с физиологией растений, часто называют минеральные питательные вещества ’’пищей” растений, а в лесоводстве используют термин ’’подкормка”, когда вносят удобрения. Физиологи растений не одобряют применения термина ’’пища” в этих случаях, так как они обычно ограничивают его значение углеводами, жирами и белками, которые могут быть непосредственно использованы растением в качестве источника энергии, окисляясь при дыхании, или как строительный материал при образовании новых тканей (см. главу 6). Минеральные вещества и двуокись углерода обычно рассматриваются физиологами растений скорее как сырье, реагенты или вспомогательные вещества, необходимые для синтеза питательных веществ, чем в качестве самой пищи.
ОСНОВНЫЕ ФУНКЦИИ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. У минеральных питательных веществ в растениях много важных функций. Они могут играть роль структурных компонентов растительных тканей, катализаторов различных реакций, регуляторов осмотического давления, компонентов буферных систем и регуляторов проницаемости мембран. Примерами значения минеральных веществ как составных частей растительных тканей могут быть кальций в клеточных стенках, магний в молекулах хлорофилла, сера в определенных белках, фосфор в фосфолипидах и нуклеопротеидах. Азот, хотя и не относится к минеральным элементам, часто включают в их число, поэтому следует еще раз отметить его значение как компонента белков. Некоторые элементы, в том числе железо, медь и цинк, требуются в очень небольших количествах, но они необходимы, поскольку входят в состав простетических групп или коферментов определенных ферментных систем. Другие элементы, такие, как марганец и магний, функционируют в качестве активаторов или ингибиторов ферментных систем. Некоторые элементы, например бор, медь и цинк, необходимые для функционирования ферментов в незначительных количествах, в более высоких концентрациях очень ядовиты. Токсичность этих и других ионов, таких, как серебро и ртуть, вероятно, связана с их повреждающим действием на ферментные системы.
Осмотическое давление клеточного сока в основном относят за счет растворимых углеводов, но ощутимую его долю дает присутствие минеральных солей, а главной причиной высокого осмотического давления у галофитов часто являются соли. Фосфаты образуют одну из важных буферных систем растений, а такие элементы, как кальций, магний и калий, входят в качестве катионов в буферные системы органических
270
Глава iq
кислот. Состав имеющихся ионов часто влияет на гидратацию протоплазмы и на проницаемость клеточных мембран. При этом двух- и трехвалентные катионы обычно уменьшают, а одновалентные увеличивают проницаемость. Определенные ионы способны уравновешивать действие других, что получило название антагонизма ионов. Например, для уравновешивания иона натрия и предотвращения нарушения, наблюдающегося в растворе одного только хлористого натрия, требуется небольшая концентрация иона кальция.
Значение достаточного снабжения растений минеральным питанием для хорошего роста уже давно оценено в полеводстве и садоводстве, но до последнего времени им пренебрегали лесоводы. Даже сейчас большинство исследований проводится на сеянцах в питомниках, но растущая стоимость земли и более высокие цены на лесоматериалы меняют это положение. Использование новых программ улучшения деревьев способствует получению саженцев с более высокой способностью к росту, поэтому для реализации ростового потенциала саженцев часто оказывается необходимым адекватное снабжение минеральными питательными веществами. Более того, укороченные обороты и использование помимо ствола также й других частей деревьев резко увличивают скорость извлечения минеральных веществ из почвы. По этим причинам удобрение искусственных насаждении лесных пород может приобрести такое же значение, какое имеет в настоящее время удобрение сельскохозяйственных культур. Дальнейшее изучение специфических потребностей деревьев в минеральных веществах необходимо в качестве основы для практического лесоводства и обеспечения лучшей адаптации деревьев к условиям местообитания.
НЕОБХОДИМЫЕ ЭЛЕМЕНТЫ. В растениях найдено более половины элементов периодической системы, и вполне вероятно, что корнями поглощается любой элемент, находящийся в окружающей их среде. По крайней мере 27 элементов были идентифицированы в некоторых образцах древесины сосны веймутовой, а другие безусловно присутствуют в небольших количествах. Не все найденные в деревьях элементы необходимы. Например, Паркер (1956) обнаружил в листьях сосны желтой платину, олово и серебро. В растениях находятся значительные количества алюминия, кремния и натрия, но ни один из этих элементов не считается необходимым.
Минеральный элемент считается необходимым, если, во-первых, без него растения не могут завершить жизненный цикл, и, во-вторых, если он входит в состав молекулы какого-либо необходимого компонента растений. Необходимость элемента можно определить только в тщательно контролируемых условиях, исключающих возможность загрязнения этим элементом в ходе исследований через соли, воду, сосуд, в котором выращивают растения, и даже пыль из воздуха. Минимальные количества различных элементов, необходимые для успешного роста, легче всего определить путем использования почвенных, песчаных или водных культур или в полевых опытах с удобрениями. Уровень обеспеченности различными элементами в полевых условиях можно определять по анализам почвы и растительных тканей (листовая диагностика), а также по наблюдениям за результатами внесения различных элементов в почву или непосредственно в листву.
Минеральное питание и поглощение солей
271
10.1, ОТНОСИТЕЛЬНЫЕ КОЛИЧЕСТВА РАЗЛИЧНЫХ ЭЛЕМЕНТОВ В ЛИСТЬЯХ ЗДОРОВОГО РАСТЕНИЯ
(Bollard, 1955)
Элемент Содержание, мкг/г Элемент Содержание, мкг/г сухого веса сухого веса
Азот 20 000 Железо 100
Калий 15 000 Бор 40
Кальций 15 000 Марганец 40
Магний 3 000 Цинк 40
Фосфор 2 500 Медь 25
Сера 2 000 Молибден 1
10.2. СОДЕРЖАНИЕ N, Р, К, Са И Mg В РАЗЛИЧНЫХ ЧАСТЯХ ДЕРЕВЬЕВ 16-ЛЕТНЕГО НАСАЖДЕНИЯ СОСНЫ ЛАДАННОЙ, КГ/ГА (WellsetaL, 1975)
Часть дерева N Р К Са Mg
Хвоя текущего года 55 6,3 32 8 4,8
Вся хвоя 82 10,3 48 17 7,9
Живые ветви 34 4,5 24 28 6,1
Мертвые ветви 26 1,5 4 30 3,0
Древесина ствола 79 10,7 65 74 22,7
Кора ствола 36 4,2 24 38 6,5
Всего в надземных частях 257 30,9 165 187 46,2
Корни 64 16,9 61 52 21,9
Итого 321 47,8 226 239 68,1
ю.з. содержание Mn,Zn, Fe, Al, Na и Си в надземных ЧАСТЯХ ДЕРЕВЬЕВ 16-ЛЕТНЕГО НАСАЖДЕНИЯ СОСНЫ ЛАДАННОЙ, Г/ГА (Wellsetal., 1975)
Часть дерева Мп Zn Fe Al Na Си
Хвоя текущего года 1 222 166 334 2 178 258 22
Вся хвоя 2 544 327 650 4 116 356 32
Живые ветви 1 716 345 915 2 519 1 384 64
Мертвые ветзи — 289 1 281 2 902 314 70
Древесина ствола 8 445 1 086 1 830 1 790 3 640 275
Кора ствола 951 336 1 126 9 705 590 59
Всего 13 656 2 383 5 802 21032 6 284 499
В больших количествах (по крайней мере, 1 мг на 1г сухого веса растений) необходимы азот, фосфор, калий, кальций, магний и сера, которые называют макроэлементами. В число элементов, необходимых в меньших количествах, входят железо, марганец, цинк, медь, бор,
272
Глава if)
10.4. РАСПРЕДЕЛЕНИЕ МИНЕРАЛЬНЫХ ВЕЩЕСТВ (ЧИСЛИТЕЛЬ -ФОСФОР; ЗНАМЕНАТЕЛЬ - КАЛЬЦИЙ) В РАЗЛИЧНЫХ ЧАСТЯХ ДЕРЕВЬЕВ СОМКНУТЫХ НАСАЖДЕНИЙ ЕВРОПЕЙСКИХ ПОРОД, РАСТУЩИХ НА ТИПИЧНЫХ УЧАСТКАХ (Baker, 1950, С РАЗРЕШЕНИЯ McGraw-Hill Book Company)
Вид Всего используется, кг/га в древесине % от общего количества
в коре в хворосте в деревьях, срубленных при прореживании в подстилке
Ель 4,5/30,8 1/3 3/1 7/2 17/8 72/86
Пихта 6,3/35,4 3/1 3/1 5/2 17/4 72/92
Сосна 2/7/11,8 5/10 3/2 6/4 17/16 69/68
Бук 6,3/45,8 13/3 2/1 6/2 6/4 73/90
Дуб 5,0/35,4 8/3 2/1 5/3 15/9 70/84
Сухое вещество в каждой части, % — 50 5 7 32 6
молибден и хлор. Возможно, со временем этот список будет расширен. s Например, некоторые виды рода Atriplex нуждаются в натрии, а селен, видимо, необходим некоторым видам Astragalus. Те элементы, которые требуются в небольших количествах, обычно называют микроэлементами (табл. 10.1—10.4, рис. 10.1).
Специфические потребности в минеральном питании лесных пород изучены мало, но опыты с плодовыми деревьями показывают, что их потребности в общем те же, что и у травянистых растений, хотя тре- >' буемые количества некоторых элементов могут быть ниже. Необхо- •• димость элемента должна быть подтверждена для растений нескольких далеких друг от друга видов. Тот факт, что элемент способствует улучшению роста, еще не доказывает его необходимость. Например, натрий,
Рис. 10.1. Средние концентрации азота и фосфора в различных частях деревьев четырех видов (Ralston, Prince, 1965) :
1 — сосна ладанная; 2 — дуб белый; 3 — дуб красный; 4 — кария; а — ствол; б — большие ветви; в — малые ветви; г — листья
Минеральное питание и поглощение солей
273
видимо, улучшает рост сахарной свеклы и ряда других видов, но он не считается для них необходимым, поскольку они могут расти и без него.
ФУНКЦИИ РАЗЛИЧНЫХ ЭЛЕМЕНТОВ. Большинство исследований роли различных элементов проведено на травянистых растениях, так как их небольшой жизненный никл позволяет изучать их в течение короткого времени. Некоторые опыты поставлены на плодовых деревьях и сеянцах лесных пород. Обсуждаемые здесь материалы основываются на приемлемом предположении, что различные элементы как в травянистых, так и в древесных растениях осуществляют одни и те же функции.
Азот. Хорошо известна роль азота как составной части аминокислот — строительных блоков, из которых состоят белки. Азот входит также во множество других соединений: в пурины, алкалоиды, ферменты, регуляторы роста, хлорофилл и клеточные мембраны. При недостатке азота нарушается синтез нормального количества хлорофилла, вследствие чего при крайнем дефиците развивается хлороз и более старых листьев и молодой листвы. (Роль азота обсуждалась подробнее в главе 9.)
Фосфор. Этот элемент является компонентом нуклеопротеидов и фосфолипидов. Макроэнергетические связи между фосфатными группами служат основным посредником в переносе энергии в растениях. Фосфор встречается в органической и неорганической формах. Он легко перемещается по растению, по-видимому, в обеих формах. При недостатке фосфора часто отмечается резкое отставание в росте молодых деревьев без каких-либо видимых симптомов.
Калий. Растениям необходимо большое количество калия, но его органические формы не известны. По-видимому, он необходим для активности ферментов. Его недостаток затрудняет передвижение углеводов и метаболизм азота, но это действие может быть скорее опосредованным, чем прямым. Интересно отметить, что растительные клетки различают калий и натрий, и последний не может быть полностью замещен первым. Считается также, что калий играет роль осмотического агента в открывании и закрывании устьиц. Калий в растениях очень мобилен.
Сера. Этот элемент является компонентом цистина, цистеина, других аминокислот, биотина, тиамина, кофермента А и многих других соединений, чаще всего в виде сульфгидрильных групп. Недостаток серы вызывает хлороз и нарушение биосинтеза белков, вследствие чего накапливаются аминокислоты. По сравнению с азотом, фосфором и калием сера менее мобильна.
Кальций. В значительных количествах кальций обнаруживается в клеточных стенках в виде пектата кальция, влияющего, вероятно, на эластичность клеточных стенок. Он участвует также каким-то образом в метаболизме азота. Кальций активирует несколько ферментов, в том числе амилазу. Он относительно мало подвижен. При недостатке кальция повреждаются меристематические участки, особенно кончики корней. Излишек кальция часто накапливается в виде кристаллов оксалата кальция в листьях и одревесневших тканях.
Магний. Этот элемент входит в молекулу хлорофилла и участвует в работе ряда ферментных систем. При недостатке магния обычно
274
Глава 10
наблюдается хлороз. Магний участвует также в поддержании целостности рибосом: при его отсутствии рибосомы распадаются. У большинства растений он легко передвигается.
Железо. Дефицит железа - один из наиболее распространенных и заметных видов недостаточности питания деревьев микроэлементами. Недостаток железа наблюдается преимущественно на щелочных и известковых почвах, где высокие величины pH препятствуют его поглощению. Большая часть железа листьев находится в хлоропластах, где оно участвует в синтезе пластидных белков. Железо входит также в ряд дыхательных ферментов, таких, как пероксидаза, каталаза, ферредоксин и цитохромоксидаза. Железо относительно неподвижно, поэтому в молодых тканях часто развивается дефицит из-за того, что оно не передвигается из более старых тканей.
Марганец. Этот элемент также необходим для синтеза хлорофилла. Основная его функция, очевидно, заключается в активации ферментных систем. Кроме того, он, вероятно, влияет на доступность железа. Недостаток марганца часто вызывает деформацию листьев и образование хлоротичных или мертвых участков. Марганец относительно неподвижен. Он ядовит (за исключением небольших концентраций) и в листьях деревьев его концентрации часто приближаются к токсичному уровню. У деревьев, растущих на известковых почвах, встречается дефицит марганца.
Цинк. У деревьев нескольких видов недостаток цинка приводит к деформациям листьев, напоминающим вирусные болезни, возможно, потому, что он участвует в синтезе триптофана — предшественника индолилуксусной кислоты. Цинк входит также в состав карбоангидразы. Описаны ярко выраженные симптомы недостатка цинка, в том числе развитие плоской кроны у сосны замечательной в Австралии. У плодовых культур широко встречается недостаточность цинка. В отношении лесных пород имеются сообщения о дефиците цинка только в Австралии. Цинк очень токсичен даже в относительно низких концентрациях, и не будет неожиданностью, если по мере развития лесоводства обнаружатся примеры ущерба от избытка цинка.
Медь. Этот элемент также является компонентом некоторых ферментов, в том числе аскорбинатоксидазы и тирозиназы. Растениям необходимы очень небольшие количества меди, высокие концентрации токсичны. Доказана необходимость меди для сеянцев ели и для сосны замечательной. Недостаток меди вызывает суховершинность у цитрусовых, у лесных пород, по-видимому, не бывает заметного дефицита меди. Симптомы недостатка меди довольно неопределенны.
Бор. Это еще один элемент, требующийся в очень малых количествах. В зависимости от вида растения потребность в боре колеблется от 5 до 15 частей на миллион. К сожалению, оптимальная для роста концентрация бора у некоторых видов очень близка к токсичной концентрации. Хвойные, по-видимому, выдерживают меньшие концентрации бора по сравнению с покрытосеменными растениями. Недостаток бора вызывает серьезные повреждения и отмирание апикальных меристем. Бор, вероятно, необходим для передвижения сахаров. Растения, которым не хватает бора, содержат больше jJaciso-римых сахаров и пентозанов, поглощение воды и транспирация у них
Минеральное питание и поглощение солей 275
проходят медленнее, чем у нормальных растений. Дефицит бора часто встречается у плодовых деревьев. По сообщению Стона (1968) это также один из наиболее распространенных случаев дефицита микроэлементов в лесных насаждениях на всех континентах.
Молибден. Среди всех необходимых элементов молибден нужен растениям в самой низкой концентрации: большинству достаточно менее 1 части на миллион. Молибден входит в состав нитратредук-тазной ферментной системы. Он выполняет , по-видимому, и другие функции. Недостаток молибдена редко встречается у плодовых деревьев и лесных пород, но при недостатке его может снижаться интенсивность азотфиксации у деревьев семейства бобовых и у ольхи.
Хлор. Этот элемент, по-йидимому, необходим растениям и может участвовать в расщеплении воды при фотосинтезе. Большого дефицита хлора у древесных растений, вероятно, не бывает.
Другие элементы. В некоторых растениях в больших количествах содержатся алюминий, натрий и кремний, но хотя эти элементы иногда и усиливают рост, их обычно не считают необходимыми. Избыток алюминия очень ядовит, и способность выдерживать высокие его концентрации очень важна для успешного роста некоторых зерновых культур на кислых почвах, На деревьях эта проблема не изучалась. Между различными элементами существуют многочисленные и сложные взаимодействия: один элемент влияет на поглощение и утилизацию другого. Следует упомянуть, однако, что различия в росте конкурирующих видов могут частично зависеть от разной способности выдерживать ненормально высокие или низкие концентрации определенных элементов.
КОНЦЕНТРАЦИЯ И РАСПРЕДЕЛЕНИЕ МИНЕРАЛЬНЫХ ЭЛЕМЕНТОВ В РАЗЛИЧНЫХ ОРГАНАХ. В общей форме соотношение концентраций минеральных веществ, выраженных в процентах от сухого веса, выглядит так: листья > небольшие ветви > большие ветви > стволы. (Графически это показано на рис. 10.1 для сосны ладанной и трех видов лиственных деревьев.) Относительные количества минеральных веществ в различных частях нескольких видов деревьев были приведены в табл. 10.2, 10.3 и 10.4. Иногда концентрация кальция и магния в коре выше, чем в листьях. Согласно сообщению Ралстона и Принка (1965) насаждение дуба белого в Пьемонте (Северная Каролина, США) содержит в 2 раза больше азота, фосфора и калия и в 15 раз больше кальция, чем насаждение сосны ладанной на такой же площади. На основании этих данных авторы сделали вывод, что если лиственные деревья будут расти с той же скоростью, что и сосны, им понадобится значительно больше минеральных питательных веществ.
Концентрация минеральных веществ в листьях намного выше по сравнению с таковой в одревесневших частях, но она значительно варьирует в зависимости от возраста и положения листа на дереве. Некоторые данные, иллюстрирующие сезонные изменения концентрации минеральных веществ в листьях апельсиновых деревьев, приведены на рис. 10.2. Сезонные изменения содержания кальция и калия в листьях листопадных видов показаны на рис. 10.3. Этот рисунок показывает также влияние увеличения сухого веса листьев на кажущееся содержание минеральных веществ.
276
Глава 10
I Ж Г
1948&. 7949г.
т > жжj т т
/9483. '/9493.
Рис. 10,2. Сезонные колебания концентрации минеральных веществ в листьях апельсиновых деревьев сорта Валенсия (Jones, Parker, 1951) :
1 — кальций; 2 — азот; 3 — калий; 4 — магний; 5 — фосфор; 6 — натрий
Рис. 10.3. Сезонные изменения (данные приведены на 1-е число месяца) содержания калия и кальция в листьях дубов и кленов (по Mitchell, 1936): а — калий; б - кальций; 1 — дуб; 2 - клен
СИМПТОМЫ НЕДОСТАТОЧНОСТИ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Недостаток минеральных веществ вызывает изменения биохимических и физиологических процессов, в результате чего часто наблюдаются морфологические изменения, или видимые симптомы. Иногда вследствие дефицита рост подавляется до появления других симптомов.
Видимые симптомы дефицита. Наиболее существенный результат недостатка минеральных веществ — снижение роста. Однако наиболее заметный эффект - это пожелтение листьев, вызванное уменьшением биосинтеза хлорофилла. Листья, по-видимому, особенно чувствительны к дефициту. При недостатке минеральных веществ у них уменьшаются размеры, изменяется форма или структура, бледнеет окраска,
Минеральное питание и поглощение солей
277
а иногда даже образуются мертвые участки на кончиках, краях или между главными жилками. В некоторых случаях листья собираются в пучки или розетки, а сосновые иглы иногда не могут разделиться и образуется ’’слившаяся хвоя”. Общий признак определенного типа недостаточности минеральных веществ в травянистых растениях -подавление роста стебля и снижение роста листовых пластинок, что приводит к образованию розеток небольших листьев, часто с сетью хлоротичных участков. Видимые симптомы дефицита различных элементов настолько характерны, что опытные наблюдатели могут идентифицировать дефицит по внешнему виду листьев.
Иногда при недостатке минеральных веществ деревья образуют избыточные количества камеди. Это явление получило название гоммоза. Выделение смолы вокруг почек характерно для страдающих от недостатка цинка деревьев сосны замечательной в Австралии.. Камедь обнаруживается также на коре плодовых деревьев, страдающих от суховершинности, вызванной недостатком меди. Значительный дефицит часто вызывает гибель листьев, побегов и других частей, т.е. развиваются симптомы, описанные как суховершинность. Отмирание побегов, вызванное недостатком меди, наблюдалось у многих лесных и плодовых деревьев. При отмирании верхушечных побегов яблони, страдающие от дефицита меди, приобретают кустообразный, чахлый вид. Недостаток бора вызывает засыхание верхушечных точек роста и в конце концов гибель камбия у цитрусовых и у сосен, отмирание флоэмы и физиологический распад плодов у других видов. Недостаток одного элемента иногда способствует появлению нескольких различных симптомов, например дефицит бора у яблонь вызывает деформацию и хрупкость листьев, некроз флоэмы, повреждения коры и плодов.
Хлороз. Наиболее распространенный симптом, наблюдаемый при недостатке самых различных элементов, — хлороз, возникающий в результате нарушения биосинтеза хлорофилла. Характер, степень и тяжесть хлороза у молодых и старых листьев зависят от вида растения, от элемента и степени дефицита. Чаще всего хлороз бывает связан с недостатком азота, но он может быть вызван и дефицитом железа, марганца, магния, калия и других элементов. Более того, хлороз может быть вызван не только дефицитом минеральных веществ, но и множеством других неблагоприятных факторов окружающей среды, в том числе избытком или недостатком воды, неблагоприятными температурами, ядовитыми веществами (например, двуокисью серы) и избытком минеральных веществ. Причиной хлороза могут быть и генетические факторы, вызывающие появление различно окрашенных растений: от альбиносов, полностью лишенных хлорофилла, до зеленоватых проростков или проростков с различной полосатостью и пятнистостью листьев.
На основе многочисленных факторов, вызывающих хлороз, можно заключить, что он происходит в результате как общего нарушения метаболизма, так и специфического влияния отдельных элементов. Ильин (1951) сделал вывод, что вызванный известью хлороз некоторых видов древесных и травянистых растений, растущих на известковых почвах, происходит в результате ненормального метаболизма азота и органических кислот. В хлоротичных листьях он обнаружил в 5—15
278
Глава 10
раз больше растворимого азота и значительно больше органических кислот,чем в нехлоротичных, и предположил, что аномальные концентрации названных веществ препятствуют биосинтезу хлорофилла.
Эту точку зрения поддержал и Стайнберг (1951), обнаруживший повышенную концентрацию аминокислот и пониженное содержание белка в растениях табака, страдающих от недостатка ряда минеральных элементов. Он показал, что избыток изолейцина вызывал типичное заболевание табака и что другие свободные аминокислоты вызывали типичный хлороз, некроз и деформации листьев. Автор выдвинул предположение, согласно которому вызванное дефицитом любого элемента нарушение метаболизма азота приводит к нарушению равновесия между различными азотистыми соединениями. В результате появляются такие видимые признаки, как хлороз и повреждение листьев.
Одним из самых распространенных и причиняющих наибольшие нарушения развития растений типов хлороза является тот, который обнаруживается у большого числа плодовых, декоративных и лесных деревьев, растущих на щелочных и известковых почвах. Его обычно вызывает недоступность же де за при высоких значениях pH, но иногда причиной бывает дефицит марганца. Наиболее тяжелый хлороз, вызванный известью, наблюдается у растений на плохо аэрируемых, холодных почвах с мелким механическим составом, где условия для поглощения минеральных веществ неблагоприятны. Вызванный дефицитом железа хлороз может усиливаться при засухе, что часто бывает у азалий.
При хлорозе у покрытосеменных средняя и более мелкие жилки листьев остаются зелеными, а участки между жилками становятся бледно-зелеными, желтыми или даже белыми. Обычно самые молодые листья страдают от хлороза наиболее сильно. У хвойных деревьев молодая хвоя делается бледно-зеленой или желтой, а при большом дефиците иглы могут становиться коричневыми и опадать.
Интерпретация хлороза в случае недостатка железа сильно осложнилась утверждениями о том, что в хлоротичных тканях были найдены такие же или более высокие концентрации железа, что и в здоровых тканях. Однако Стон (1968) ссылается на исследования, показавшие, что большая доля обнаруживаемого ранее железа принадлежала поверхностным загрязнениям, и что в отмытых перед анализом листьях содержание железа было намного меньше. Хлороз, вызванный недостатком железа, можно частично или полностью устранить, понизив pH почвы. С этой целью растения рекомендуется опрыскать раствором, содержащим железо, или ввести железо непосредственно в деревья. Понизить pH почвы можно добавлением серы или сульфата алюминия. Раньше при недостатке железа часто в почву вносили сульфат железа или опрыскивали им листву, но сейчас обычно применяют опрыскивание хелатами железа. Положительные результаты для некоторых, но не для всех, видов плодовых и декоративных деревьев были получены при введении солей железа в отверстия, высверленные в стволах деревьев, или даже при забивании гвоздей в ствол, но доступность хелатов железа делает такие меры сегодня излишними.
Анатомические и цитологические признаки. При недостатке минеральных веществ наряду с явными внешними симптомами происходят изменения, видимые только в микроскоп. Дэвис (1949) обнаружил, что при недостатке кальция у сеянцев сосны ладанной были меньше не
Минеральное питание и поглощение солей
279
только почки и кончики стеблей, но и листья (с меньшим числом более мелких клеток и с менее развитой ксилемой и флоэмой). При этом в стеблях было меньше первичных и больше вторичных тканей, у корней, по сравнению с корнями нормальных сеянцев, были ненормально тупые кончики с реже встречающимися митотическими фигурами у клеток. Деление клеток при недостатке кальция замедлялось сильнее, чем их растяжение или дифференциация, В результате наблюдалось одревеснение ксилемы очень близко к меристематической эоне.
Дефицит минеральных веществ часто вызывает заметные изменения в структуре клеток. Рид и Дуфрени (1935) обнаружили, что клетки столбчатой паренхимы страдающих от дефицита цинка листьев цитрусовых были шире, чем в нормальных листьях, ромбовидной формы вместо столбчатой и часто разделенные поперек. Хлоропластов было меньше, и они отличались ненормально высоким содержанием жиров, тогда как крахмальные зерна были небольшими и вытянутыми. Микрохимические тесты выявили присутствие фитостерола или лецитина в клеточных вакуолях листьев при недостатке цинка, чего не было в листьях нормальных растений. Позже Рид (1938) наблюдал при недостатке цинка задержку дифференциации в листьях, приводящую к образованию очень компактного мезофилла с уменьшенными межклетниками. Для объяснения действия минеральной недостаточности на клеточном уровне желательны дополнительные анатомические и цитологические исследования.
Связь симптомов с подвижностью ионов. Локализация симптомов недостаточности различных элементов, вероятно, в такой же степени зависит от их относительной подвижности, как и от их функций. Например, у более старых листьев появляются признаки дефицита азота, фосфора, калия и магния, в то же время молодые листья остаются здоровыми, поскольку эти элементы легко передвигаются из старых тканей в молодые. В противоположность этому симптомы дефицита бора и кальция проявляются в растущих верхушках стеблей, а симптомы дефицита железа, марганца и серы — у молодых листьев, так как все эти элементы слабо передвигаются от старых тканей в молодые.
ФИЗИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ НЕДОСТАТКА МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Видимое морфологическое действие или симптомы дефицита минеральных веществ являются результатом изменений различных внутренних биохимических или физиологических процессов. Однако из-за сложных взаимоотношений между ними бывает трудно определить, каким образом недостаток отдельного элемента вызывает наблюдаемые эффекты. Например, недостаток азота может тормозить рост вследствие худшего снабжения азотом процессов биосинтеза новой протоплазмы. Но одновременно снижается скорость синтеза ферментов и хлорофилла и уменьшается фотосинтезирующая поверхность. Это вызывает ослабление фотосинтеза, ухудшающее снабжение ростовых процессов углеводами. В результате возможно дальнейшее снижение скорости поглощения азота и минеральных веществ. Один элемент часто выполняет в растении несколько функций, поэтому непросто определить, нарушение какой именно функции или сочетания функций вызывает появление видимых симптомов. Марганец, например, помимо
280
Глава 10
активирования определенных ферментных систем, также необходим для синтеза хлорофилла. Его недостаток вызывает некоторые функциональные расстройства.
Интересные исследования проведены на яблоне и тунге по выяснению связи недостаточности минеральных веществ с главными физиологическими процессами: дыханием, фотосинтезом, метаболизмом азота, биосинтезом жиров и водным режимом. На лесных породах таких работ мало, поэтому необходимы дальнейшие исследования, особенно по влиянию дефицита минеральных веществ и внесения удобрений на фотосинтез и водный режим. Некоторые результаты такого влияния будут обсуждены в других главах, при рассмотрении главных физиологических процессов. Недостаток азота обычно вызывает заметное ослабление фотосинтеза, но влияние недостатка других элементов не так определенно.
Недостаток минеральных веществ снижает и биосинтез углеводов и их передвижение к растущим тканям. Часто дефицит по-разному влияет на фотосинтез и дыхание (см. главу 5). Например, существенный дефицит калия замедляет фотосинтез и усиливает дыхание, уменьшая тем самым количество углеводов, которые могут использоваться на рост. Иногда также подавляется передвижение углеводов. Этот эффект резко выражен у бор-дефицитных деревьев с некрозом флоэмы. В результате уменьшения количества доступных углеводов снижается скорость роста тканей в одной части дерева, но в то же время может происходить накопление углеводов в другой части. Иногда из-за низкого содержания запасных углеводов уменьшается образование семян. Обильное внесение азотного удобрения приводило к значительному усилению процесса образования семян у деревьев бука и клена сахарного, увеличивались процент здоровых семян и сухой вес семян клена. Образование шишек и семян у молодой сосны ладанной также резко усиливалось после внесения удобрения. Если же деревья не испытывают дефицита минеральных веществ, внесение больших количеств азотных удобрений может снижать образование плодов и семян вследствие стимуляции вегетативного роста,
БОЛЕЗНИ. Очень трудно предвидеть влияние удобрений на восприимчивость к болезням. Фостер (1968) утверждал, что наиболее важные заболевания корней прогрессируют по мере увеличения количества вносимых удобрений. Однако признаки мелколистности, возникающей при поражении корней грибами Phytophthora cinnamomi, можно устранить обильным удобрением азотом. Удобрение уменьшает тяжесть ряда других болезней, связанных с усыханием точек роста. По данным Паркера с соавторами (1947), у молодых деревьев голландская болезнь вяза протекает легче на плодородных почвах, чем у деревьев, растущих на почвах с недостатком минеральных питательных веществ. Имеются свидетельства, что применение удобрений препятствует нападению точильщиков и объедающих листья насекомых, но увеличивает вред от сосущих насекомых. Очень сложно также действие удобрения на почвенные организмы, которые могут быть полезными или вредными.
ВИДОВЫЕ И ИНДИВИДУАЛЬНЫЕ РАЗЛИЧИЯ РЕАКЦИИ НА НЕДОСТАТОК МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Известно, что виды растений
Минеральное питание и поглощение солей
281
различаются по способности накапливать разные элементы. Например, листья кизила флоридского и дуба белого содержат в 2 раза больше кальция по сравнению с листьями дуба малого и сосны ладанной, растущих на той же почве. Апельсин трехлистный чувствительней к недостатку цинка и железа, чем некоторые другие корневые подвои (такие, как цитрус джамбири), особенно на известковых почвах, по-видимому, вследствие ограниченного транспорта веществ от корней к побегам. Он также чувствительней к дефициту магния в присутствии высоких концентраций калия.
Вследствие различной способности к поглощению и передвижению минеральных веществ деревья разных видов и даже отдельные экземпляры одного и того же вида по-разному реагируют на их дефицит. Так, по данным Найберга и Вита, на плоских песчаных равнинах штата Нью-Йорк, непригодных для сельского хозяйства вследствие низкого содержания калия и общего неплодородия, у деревьев сосны смолистой наблюдается общая задержка роста с небольшими различиями признаков дефицита между отдельными экземплярами. Сосна веймутова, однако, демонстрировала большие различия между отдельными деревьями: рядом с хорошо растущими деревьями встречались мертвые. У елей обыкновенной и канадской наблюдались даже еще более выраженные симптомы недостаточности с множеством мертвых деревьев, но на небольших живых деревьях был хороший урожай шишек. В противоположность этому у кедрового стланика признаки дефицита не наблюдались, а у сосны обыкновенной и сосны Банкса проявлялся только слабый хлороз.
Хобс (1944) обнаружил, что у сеянцев четырех видов сосны, растущих в культурах при недостатке магния, вслед за верхушечным хлорозом развивался некроз. Выраженность симптомов варьировала по видам, причем тяжелее страдали более быстро растущие сосны ладанная и смолистая, чем медленнее растущие сосны жесткая и ежовая. Пессин (1937) обнаружил несколько иные симптомы дефицита магния у сосны болотной, у которой некроз развивался без предшествующего пожелтения. По наблюдениям Стона (1953), наиболее заметным признаком недостатка магния у нескольких видов хвойных деревьев в штате Нью-Йорк было изменение цвета хвои текущего года на ярко-желтый. Хлороз проявлялся осенью и затрагивал в основном верхнюю часть деревьев. Всякий раз при развитии острой недостаточности после пожелтения хвоя отмирала или преждевременно опадала. У сосны веймутовой хлороз был гораздо менее заметным, чем у сосны смолистой. Ель обыкновенная реагировала на дефицит магния скорее общим пожелтением, чем апикальным пожелтением новых игл, наблюдающимся у сосен.
Отмечено, что выращенные из семян в песчаных культурах ильмовик, айлант высочайший и катальпа прекрасная в условиях дефицита различных элементов вели себя по-разному. Например, при недостатке калия у катальпы листья были темно-зелеными, у аиланга имели промежуточную окраску, а у ильмовика были желтоватыми. Поэтому необходимо вести наблюдения за внешним видом деревьев, о которых известно, что они страдают от недостатка определенных элементов, чтобы выявить соответствующие реакции. Но даже в этом случае не
282
Глава 10
исключена возможность, что различия в климатических условиях могут изменить эти реакции.
ПРИЗНАКИ ИЗБЫТКА МИНЕРАЛЬНЫХ ВЕЩЕСТВ. В лесных почвах редко наблюдается избыток минеральных питательных веществ, но обильное удобрение садов и питомников иногда приводит к концентрации солей, вполне достаточной для того, чтобы нанести вред. Встречаются также большие площади засушливых земель, где большинство видов растений не может существовать из-за высокого содержания солей. Орошение водой, содержащей много солей, также наносит ущерб. Это происходит вследствие увеличения осмотического давления, неблагоприятных для растений сдвигов pH, нарушения баланса различных ионов или в результате комбинации этих факторов.
Повышенное осмотическое давление почвенного раствора уменьшает поглощение воды, усиливает дефицит воды в листьях и в результате приводит к повреждению тканей от подсыхания в те дни, когда ветер и высокие температуры вызывают сильную транспирацию. При более продолжительном и глубоком обезвоживании наблюдается также закрывание устьиц, препятствующее фотосинтезу. Высокие концентрации солей в почве могут вызвать повреждение корней путем плазмолиза, особенно в песчаных почвах, что мешает синтетической деятельности корней. Иногда листья повреждаются в результате нанесения на них жидких удобрений в высоких концентрациях.
Вредное Действие избыточного удобрения зависит от вида растения, типа использованного удобрения и времени внесения. По сообщениям, тутовые деревья выдерживают без ущерба лишь в 5 раз менее концентрированные почвенные растворы, чем яблони. Различные виды и сорта цитрусовых различаются по солевынослив ости. Бакстер (1943) установил, что можжевельник и туя чрезвычайно подвержены повреждениям от избытка удобрений.
Возникли серьезные разногласия об относительной важности осмотического давления почвенного раствора, его состава и pH как факторов, причиняющих вред растениям в результате избытка солей в почве. Исследователи лаборатории по изучению засоленности в Риверсайде (Калифорния) считали влияние осмотического давления наиболее важным фактором, снижающим скорость поглощения воды, но признавали также некотоое действие ионов. Рост корней персиков сильнее подавлялся сульфатами, чем хлоридами, при одинаковом осмотическом давлении. По некоторым данным, для деревьев косточковых пород, растворы хлорида кальция более токсичны чем изоосмотические растворы хлорида натрия. По неопубликованным данным, хвойные более чувствительны к повреждающему действию избытка солей, чем лиственные.
Возможно, при наличии всех необходимых минеральных элементов хороший рост корней может происходить в значительном диапазоне pH, хотя Лейтон (1952) наблюдал наилучший рост корней ели ситхинской в пределах pH от 4 до 5. Вредное действие очень кислых почв обусловлено, по-видимому, повышенной растворимостью алюминия, тогда как при более высоких величинах pH наиболее часто растения страдают от пониженной доступности таких элементов, как железо и марганец. Ховел (1932) нашел, что сеянцы сосны желтой выживали пои pH от 2,7 до 11, но хорошо росли только при pH от 3 до 6. Гюсти Капман (1944) не обнаружили прямого повреждающего действия на корни сеянцев апельсина сладкого pH в пределах 4-9, хотя косвенное влияние наблюдалось в обеих крайних точках шкалы pH. Один из способов косвенного действия ненормальных значений pH или химического состава может заключаться в усилении повреждения патогенными организмами.
Минеральное питание и поглощение солей
233
Позднее избыточное удобрение плодовых и декоративных деревьев иногда продлевает вегетационный период до такой степени, что у деревьев и кустарников не остается времени для приобретения холодостойкости до заморозков. Слишком поздно сформировавшиеся ветви очень чувствительны к повреждению морозом. Избыточное удобрение иногда стимулирует образование большого числа ветвей, цветков и плодов на старых деревьях. Сосны могут реагировать на него образованием гроздьев шишек. Среди других типов реакции растений на избыточное удобрение — фасциация, или уплощение стеблей, и внутренний некроз коры. На сеянцах нежелательное действие избытка удобрений проявляется в виде чрезмерного верхушечного роста, приводящего к низким величинам соотношения подземной и надземной частей, в результате чего растения часто плохо приживаются после пересадки.
Применение избыточных количеств удобрений расточительно с экономической точки зрения. Оно нежелательно также и для окружающей среды, так как избыток может вымываться и попадать в водоемы или грунтовые воды. Особенно большое значение имеет вымывание избытка азота, обычно в форме нитрата, но проблема загрязнения среды может возникнуть и при внесении в избыточных количествах любого элемента.
ПОЛЕВАЯ ДИАГНОСТИКА ДЕФИЦИТА МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Для сельского хозяйства, лесоводства и садоводства очень важно предвидеть или, по крайней мере, обнаружить дефицит минеральных веществ до того, как он причинит серьезный ущерб. Большие плантации часто располагаются на участках с несколькими типами почв, которые могут сильно различаться по плодородию, в результате чего в разных частях плантации могут возникать разные проблемы минерального питания. В предсказании возможности дефицита часто полезны анализы почвы, хотя существует некоторая неопределенность относительно наилучшего метода экстракции прочно связанных элементов, например фосфора. Доказана также полезность анализа листвы после установления уровней содержания элементов, необходимых для хорошего роста. Этот метод широко используется в садоводстве и овощеводстве для определения потребности в удобрениях. Некоторые данные по концентрациям фосфора и калия в листьях апельсиновых деревьев как показателям состояния минерального питания приведены в табл. 10.5. Некоторые исследователи предлагают использовать анализ листвы у цитрусовых и
10.5. ОПРЕДЕЛЕНИЕ УРОВНЯ СНАБЖЕНИЯ МИНЕРАЛЬНЫМИ ЭЛЕМЕНТАМИ ПО РЕЗУЛЬТАТАМ АНАЛИЗА ЛИСТЬЕВ АПЕЛЬСИНОВЫЕ ДЕРЕВЬЕВ СОРТОВ ВАЛЕНСИЯ И НАВЕЛЬ (Embleton et al., 1975)
Элемент
Снабжение, % от сухого веса листьев
Дефицит Мало
Оптимум Много Избыток
Азот Фосфор Калий
0,09
0,40
2,2-2,3 2,4-2,6
0,09-0,11 0,12-0,16
0,40-0,69 0,70-1,09
2,7-2,8 0,17-0,29 1,10-2,00
2,8 0,30
2,30
284
Глава 10
указывают на необходимость стандартизации методов с учетом возраста и расположения листьев, отбираемых для определений.
Опытные наблюдатели часто могут определить тип дефицита по внешнему виду листвы. По мере накопления опыта методы листовой диагностики лесных пород могут быть так же надежны, как в сельском хозяйстве и садоводстве.
МЕРЫ БОРЬБЫ С ДЕФИЦИТОМ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Совершенствование способов диагностики дефицита минеральных элементов и распознавания его причин в практике лесоводства и садоводства способствовало разработке приемов предотвращения дефицита. Эти попытки предпринимались в нескольких направлениях, в числе которых: внесение удобрений, селекция форм, наиболее эффективно утилизирующих имеющиеся элементы, а иногда использование азотфйксирующих видов (например, ольхи) в качестве подлеска для улучшения снабжения деревьев азотом.
Удобрения. Применение удобрений долгое время было общепринятым способом количественного и качественного улучшения роста плодовых и декоративных деревьев, а также кустарников. Внесение удобрений многие годы практиковалось в лесных питомниках, но до недавнего времени в лесном хозяйстве США использовалось редко. В Европе удобрения применяются в течение более длительного периода, поскольку высокая стоимость земли и ее обработки,, ухудшение участков, вызванное удалением подстилки, и высокие цены на лесную продукцию заставили европейских лесоводов признать удобрения рентабельными задолго до того, как этот метод всерьез приняли во внимание в США- В настоящее время, с увеличением стоимости земли, при более высоких ценах на лесную продукцию и с высадкой специально выведенных саженцев с высоким потенциалом роста, применение удобрений становится все более целесообразным для североамериканских лесоводов. Вследствие этого были проведены обширные исследования по удобрению лесов в основный лесных районах Северной Америки.
Удобрение больших участков леса и даже садов часто производят с самолетов и вертолетов. В некоторых случаях вносили шлам от переработки сточных вод в жидком виде. Иногда мочевиной и другими питательными веществами опрыскивают листву и ветви плодовых и декоративных деревьев. Стоат (1950) сообщал, что страдающие от недостатка цинка деревья сосны замечательной в Южной Австралии удалось вылечить опрыскиванием плантаций. Внесение основных пи-тате льных веществ путем опрыскивания листвы обычно рассматривается скорее как дополнение, а не заменитель почвенных подкормок. Однако опыт обработки цитрусовых деревьев в Калифорнии и Флориде показывает, что внесение азота и калия в почву и через листву часто оказывается одинаково эффективным и выбор метода должен определяться экономическими соображениями. Питательные вещества, попавшие при опрыскивании на кору деревьев, впитываются через имеющиеся трещины и щели, раны от обрезки. Опрыскивание коры — современный вариант старого обычая обмазывать навозом стволы деревьев. Дефицит цинка и железа удавалось ликвидировать даже простым забиванием в кору деревьев кнопок, покрытых цинком, или железных гвоздей.
Минеральное питание и поглощение солей
285
Следует подчеркнуть, что в садоводстве удобрения могут оказывать многообразное влияние как на качество, так и на количество продукции, будь то цветы, плоды или декоративные кустарники. Обильное внесение азота увеличивает урожай, но часто ухудшает окраску яблок и задерживает их созревание.
У плодов некоторых лиственных деревьев, как сообщают, внесение удобрений также действует на аромат илежкость. Наиболее глубокие исследования влияния удобрений на качество плодов были выполнены на цитрусовых. По-видимому, вносить удобрения следует так, чтобы соблюдалось оптимальное соотношение между качеством плодов и их урожаем.
Дефицит макроэлементов. В лесных почвах широко распространен недостаток азота, а в некоторых районах наблюдается значительный дефицит фосфора и калия. Эти элементы являются наиболее важными и для минерального питания плодовых деревьев. В Австралии и Новой Зеландии широко распространен дефицит фосфора. Благоприятное действие оказывает фосфор на рост сосен на почвах сырых прибрежных равнин юго-востока США. Вероятно, не очень важно, в какой форме вносятся азот и фосфор, хотя, может быть, выгодно вносить часть азота в плохорастворимой форме. Реакция нескольких лесных пород на удобрение показана в табл. 10.6.
10.6. ВЛИЯНИЕ ВНЕСЕНИЯ 335 КГ/ГА АЗОТА ВМЕСТЕ С ФОСФОРОМ
ИЛИ БЕЗ НЕГО (ЧИСЛИТЕЛЬ - ЧИСЛО ДЕРЕВЬЕВ, ЗНАМЕНАТЕЛЬ -ПЛОЩАДЬ ОСНОВАНИЯ ОДНОГО ДЕРЕВА, СМ2) НА РОСТ ДЕВЯТИ ВИДОВ ДЕРЕВЬЕВ В 38 ЛЕСАХ ДОЛИНЫ ТЕННЕССИ, США
(Jones, Gurlin, 1968)
Порода Контроль Азот Азот + фосфор
Дуб северный 118/19,5 85/52,0 (167) 73/39,0 (100)
Дуб белый 310/56,7 260/77,1 (36) 321/64,1 (13)
Дуб каштановый 78/21,4 76/55,7 (61) 53/57,6 (70)
Дуб серповидный 45/49,2 68/61,3 (25) 55/99,4 (102)
Тюльпанное дерево 139/56,7 173/83,6 (48) 136/95,7 (69)
Черемуха поздняя 8/19,5 6/29,7 (52) 2/33,4 (71)
Кария 149/16,7 174/60,4 (261) 215/45,5 (172)
Огуречное дерево 13/13,9 30/28,8 (107) 20/42,7 (207)
Дерен 86/10,2 107/13,9 (36) 70/27,9 (173)
Примечание. В скобках приведены данные об усилении роста, %.
Дефицит микроэлементов. Большинство работ по применению удобрений в лесах выполнено на макроэлементах: азоте, фосфоре и калии. Однако у плодовых и декоративных деревьев часто наблюдается дефицит микроэлементов (таких, как железо, цинк, медь и бор), особенно на известковых или песчаных почвах. Дефицит микроэлементов начинает обнаруживаться в лесных насаждениях во всем мире. Без сомнения, будут появляться новые примеры недостаточности микроэлементов по мере того, как экзотические виды станут выращиваться в климатических условиях и на почвах, к которым они мало приспо
286 , Глава 10
соблены. Опыты с цитрусовыми показывают, что наилучшим способом ликвидации дефицита марганца, цинка и молибдена является опрыскивание листвы. Однако цитрусовые деревья не стоит опрыскивать соединениями железа. В известковые почвы железо следует вносить в виде хелатов.
Подлесок. В сельском хозяйстве для увеличения содержания азота в почве используют азотфиксирующие бобовые культуры, а для увеличения содержания органического вещества выращивают покровные культуры. В хвойных насаждениях Европы и Японии в качестве подлеска встречается ольха, которая может благодаря азотфиксирующим организмам на ее корнях снабжать хвойные деревья достаточным количеством азота для улучшения их роста. Для усиления роста ели на бедных азотом почвах в Великобритании использовали ракитник (Cytisus sp.), в Германии — люпин (Lupinus sp.), но опыты не дали желаемых результатов. По сообщению Хапмана и Лана (1951), на заброшенных землях лиственные породы хорошо выращивать вместе с белой акацией, отчасти потому, что азот, фиксированный клубеньками на ее корнях, становится доступным для соседних деревьев. Ранее упоминалось, что богатая кальцием подстилка лиственных пород облегчает разложение хвойной подстилки. Поскольку цены на удобрения растут, можно ожидать, что все больше внимания будет уделяться определению сочетания видов, наиболее эффективно поддерживающих плодородие почв с минимальными добавками удобрений.
Растительность нижнего яруса иногда конкурирует с деревьями за минеральные вещества. Боллард (1957) и другие авторы наблюдали, как иногда травяной покров снижает урожай яблок. По сообщению Рихардсона (1953), трава в результате конкуренции за азот уменьшала рост молодых деревьев явора в специальных вегетационных сосудах, а Лейтон (1954) сообщал, что при ослаблении роста вереска при его затенении усиливался рост молодых елей.
Видовые и клоповые различия. Существуют большие различия между деревьями одного вида и между разными видами по их способности, измеренной по образованию древесины, поглощать и использовать минеральные вещества. Бенгтсон (1968) привел несколько таких примеров в отношении макроэлементов у лесных пород, Ральстон и Принк (1965) отметили различия между соснами и лиственными деревьями, а Герлоф (1963) также рассмотрел ряд примеров, в основном для травянистых растений. Енес и Курлин (1968) сообщали о различиях между клонами тополя (рис. 10.4). Стон (1968) указывал, что при исследовании на садовых культурах выявлено много различий их чувствительности к недостатку микроэлементов, причем некоторые различия несомненно генетические. У сосен разная реакция на удобрение наблюдается в отношении образования не только древесины, но и семян: у некоторых клонов образование семян сильно увеличивается, в то время как у других оно не изменяется. По-видимому, следует уделять больше внимания селекции генотипов с благоприятными физиологическими признаками, в частности с эффективным использованием минеральных питательных веществ.
Вызывает опасения, не будет ли усиление роста, чато происходящее после внесения удобрения, сопровождаться ухудшением качества дре-
Минеральное питание и поглощение солей
287
Рис. 10.4. Рост клонов тополя в зависимости от внесения в почву 330 кг/га азота. Заштрихованная полоса внутри каждого столбика соответствует росту без удобрения. Клон А-8 отличался наилучшим ростом и с удобрением, и без него. Клон А-16 плохо рос без удобрения, после внесения удобрения рост многократно усиливался. Клон К-1 со средней скоростью роста хуже других реагировал на внесение удобрения (Jones, Curlin, 1968)
Азот, фунт/акр
Рис. 10.5. Влияние увеличивающихся доз азота на рост трех видов деревьев с различной потребностью в азоте (Mitchell, Chandler, 1939) : 1 — высокой; 2 — средней; 3 — низкой
весины. Шмидтлинг (1973) ссылается на работу, показывающую, что в некоторых случаях уменьшались длина и плотность волокон. Однако в своих экспериментах он обнаружил, что скашивание травы для уменьшения конкуренции с ее стороны и внесение удобрений в течение 9 лет сильно увеличивали образование древесины у сосен болотной, Эллиота и ладанной без какого-либо уменьшения длины или плотности волокон. Связь между удобрением и качеством древесины заслуживает дальнейшего изучения.
Факторы, ограничивающие реакцию на удобрение. Максимальные результаты могут быть получены от внесения удобрений только при отсутствии других существенно ограничивающих факторов. Летние засухи могут настолько серьезно ограничивать скорость роста, что внесение удобрений лишь немного усилит рост или вообще не повлияет на него. Заболачивание почвы и нападение нематод и патогенных грибов могут повреждать корневую систему и уменьшать поглощение минеральных веществ. Потеря листвы, вызванная насекомыми или грибами, может снижать фотосинтез до той степени, когда рост ограничивается скорее недостатком углеводов, чем йедостатком минеральных веществ. Вредной может быть также и конкуренция с травянистой растительностью. При интерпретации результатов опытов с удобрениями следует учитывать погодные условия и другие факторы внешней среды. Очевидно, нельзя ожидать хороших результатов в условиях, когда неблагоприятные факторы окружающей среды снижают интенсивность основных физиологических процессов до уровня, при котором эти процессы уже не изменяются при улучшении минерального питания. Даже при благоприятных условиях закон убывающего плодородия остается в силе
288
Глава 10
в отношении удобрения (рис. 10.5). Как сильно, так и слабо нуждающиеся в азоте виды одинаково хорошо отзываются на внесение азота при его низком содержании, но с увеличением количества азота прибавка роста уменьшается даже у видов с высокой потребностью в азоте.
КРУГОВОРОТ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Понятие круговорота (цикла) связано с тем, что большая часть поглощенных растительностью минеральных веществ возвращается в почву при разложении опавших листьев и других растительных тканей. Круговорот минеральных веществ — важное свойство любых экосистем, но особенно в природных лесах. Круговорот минеральных веществ в лесу состоит из нескольких подциклов различной длины. Самый короткий цикл характерен для элементов, вымываемых дождем из листьев и из экскрементов насекомых, поедающих листья, поскольку эти элементы могут сразу же использоваться. Разложение листвы и ветвей освобождает минеральные вещества за время от нескольких недель или месяцев в темплом климате до нескольких лет в холодном. Входящие в состав крупных ветвей и стволов вещества не вступают в оборот в течение десятилетий или даже веков, что зависит от долговечности дерева и скорости разложения древесины.
В лесах умеренных зон большая доля азота и минеральных веществ, находящихся в обороте, содержится в почве. Согласно Швитцеру и Нельсону (1972) около 20% общего азота и минеральных веществ лесной подстилки и биомассы 20-петнего насаждения сосны ладанной вступают в круговорот. Из них около 80% идет на образование листвы, а 15% — стволов и ветвей. По данным Занхеца (1973), в тропических лесах биомассе принадлежит значительно большая доля минеральных веществ, находящихся в круговороте, по сравнению с лесами умеренных зон. К примеру, в Гане свыше 40%'Na, Р, К, Са и Mg, содержащихся в растительности в верхних 30 см поЧвы леса, находились в растениях. Растительность содержала 30% азота, 50% обменного кальция и магния, 58% обменного фонда калия и свыше 90% фосфора. По данным Бормана с соавторами (1977), в северном лиственном лесу лишь 9,5% азота находится в растительности, около 90% — в органическом веществе почвы.
В связи с большим интересом, вызываемым круговоротом минеральных веществ, опубликовано много работ по изучению количества азота и различных минеральных элементов в деревьях и лесной подстилке, главным образом в лесах северной умеренной зоны. По подсчетам Бакера (1950), средняя масса подстилки в лесах США составляет около 3400 кг/га. Овингтон (1956) привел сходные величины для уплотненных лесов Англии, но для хвойных лесов умеренно теплой зоны Брэй и Горам (1964) дали величину 5425 кг/га* год, Стон (1975) отметил затруднения, с которыми приходится сталкиваться при попытках объяснения различий в относительном количестве минеральных веществ, поставляемых опадом от деревьев разных видов, Образование подстилки в тропических лесах значительнее по сравнению с образованием ее в лесах умеренных зон и составляет от 5 до 15 т/га. Лиственные и хвойные леса дают примерно одну и ту же массу подстилки на единицу площади. Однако подстилка лиственных лесов часто содержит более высокие концентрации минеральных веществ (табл, 10.7) и разлагается быстрее. Отсюда и круговорот минеральных веществ в лиственных лесах обычно происходит быстрее, чем в хвойных. Очень быстро происходит он в тропических лесах, где половина сухого вещества может минерализоваться за 8—Ю недель, но в холодных северных хвойных лесах оборот часто происходит очень медленно.
289
Минеральное питание и поглощение солей
10.7. КОЛИЧЕСТВО МИНЕРАЛЬНЫХ ЭЛЕМЕНТОВ, возвращаемых в почву при листопаде, кг/га
Штат и насаждение N Р К Са Mg Опад за
год
Штат Нью-Йорк: лиственный лес 17,8 3,7 15,1 73,5 98,9 3042
(Chandler, 1941) хвойный лес, среднее по 7 видам. (Chandler, 1944) (Штат Юж. Каролина) Metz, 1952): лиственный лес 25,4 2,0 7,3 29,7 5,0 2759
24,8 99,1 23,4 4142
смешанный лес 28,6 — — 57,7 12,5 4285
сосновый лес 14,2 — — 19,4 6,25 3292
В дополнение к минеральным веществам, освобождающимся при разложении подстилки, заметное количество их вымывается осадками из листвы и возвращается h почву. Ощутимые количества азота и минеральных веществ вводятся из атмосферы в почву с дождем. Некоторое количество азота добавляется благодаря нитрификации в почве, а содержание минеральных веществ увеличивается в результате разложения горных пород. Происходят потери и при вымывании с дренажными водами, но в ненарушенных экосистемах эти потери невелики. Например, через 9 лет после удобрения сосны смолистой калием 60% обнаруженного в листве калия приходилось из удобрения, а через 23 года — 40%. Значительные количества внесенного калия оставались также в подстилке и в поверхностном слое почвы, что свидетельствует о небольших потерях калия в этом типе почвы. Согласно Рихардсону и Лунду (1975) потери минеральных веществ от сплошной рубки деревьев часто невелики, но при полном уничтожении всей растительности потери могут быть большими.
10.8. СРЕДНИЕ величины прибавки минеральных веществ ПОЧВЫ ЗА СЧЕТ ОСАДКОВ И ПОТЕРИ ИХ С ДОЖДЕВЫМИ ПОТОКАМИ ЗА ГОД НА ПЯТИ ПОРОСШИХ СОСНОЙ ВОДОРАЗДЕЛАХ НА СЕВЕРЕ ШТАТА МИССИСИПИ, КГ/ГА (Schreiber et al., 1976, С РАЗРЕШЕНИЯ American Society of Agronomy, Crop Science Society of America and Soil Science Society of America)
Вещества Прибавка t Потери Прибыль или убыль
NO3“ 3,12 0,32 + 2,80
^4 з 5,73 3,35 + 2,38
0,07 0,04 + 0,03
г 2+ 4,98 3,31 + 1,67
2+ 7,72 6,21 + 1,51
Mg 3,03 3,05 -0,02
290
Глава 10
Количество некоторых элементов, особенно серы, вносимых в почву с дождем, увеличилось за последние годы из-за усиливающегося загрязнения атмосферы в результате деятельности людей. Веллс с соавторами (1975) установили, что добыча древесины общепринятыми способами, при которых валежник остается гнить в подстилке, приводит к выносу азота примерно с той же скоростью, с какой он вносится в почву из атмосферы. Данные Шрайберга и других исследователей (1976) показывают, что прибавление некоторых минеральных элементов, приносимых дождем из атмосферы, часто превышает их потери вследствие вымывания потоками воды (табл. 10.8). Все большее значение приобретает растущая кислотность дождевой воды в Скандинавии, Западной Европе и на северо-востоке США, вызванная превращением в воздухе двуокиси серы в серную кислоту. Образующийся при этом ’’кислый дождь” вызывает нежелательное изменение pH озер и, продолжаясь в течение длительного периода, может привести к снижению pH почв, имеющих слабую буферность.
В тех садах и лесах, где собирают урожай, картина сильно отличается от описан- г ной выше, С урожаем уносятся большие количества минеральных веществ и азота, накопленных в плодах и семенах. Согласно Кромарку и Монку (1975) свыше 10% азота и минеральных веществ опада дубового насаждения содержалось в желудях. Сообщают, что с урожаем яблок уносится более 80 кг/га мине-ральных веществ, а хороший урожай апельсинов уносит более 200 кг/га азота и ; минеральных веществ. Для сравнения: уборка кукурузы на зерно уносит около t 160 кг/га азота и минеральных веществ, а уборка зерна и стеблей - более 1
300 кг/га. Согласно Веллсу и другим исследователям (1975) практикуемое для 1
получения древесной Пульпы прореживание 16-летнего насаждения сосны ладан- / ной уносит около 61 т/га древесины, содержащей 54 кг/га азота, т.с. 21% общего азота экосистемы, и 120,5 кг/га Р, К, Са и Mg, т. е. 28% их общего содержания. Однако полный вывоз целых деревьев при прореживании удаляет 74 т/га биомассы и пропорционально большие количества азота и минеральных веществ. Полный вывоз всех надземных частей пепевьев насаждения унес бы 257 кг/га азота и 429 кг/га Р, К, Са и Mg. Уборка . корней унесла бы еще 64 кг/га азота и более 150 кг/га Р, К, Са и Mg. При полной уборке также были бы унесены большие количества микроэлементов. Но поскольку Мп,2пиСи имеют тенденцию к накоплению в древесине, то при общепринятом сйособе прореживания удаляются большие доли этих элементов, чем азота и фосфора.
Среди лесоводов все чаще обсуждается возможность вывоза целиком надземных частей деревьев и даже корней. Приведенный выше расчет и данные табл. 10.9 показывают, что такие методы сильно увеличат количество уносимых из экосистемы минеральных веществ и обострят проблемы минерального питания. Боль (1976) рассмотрел способы учета влияния заготовки целых деревьев на качество участка и отметил, что это действие наиболее ярко проявится на бедных минеральными веществами почвах. Все более широкое использование быстрорастущих видов с короткими периодами ротации, без сомнения, создаст дополнительные проблемы с минеральным питанием.
Пожары иногда вызывают потерю азота, но увеличивают доступность других элементов. Некоторые сосновые леса на юго-востоке США многократно горели в течение десятилетий без потери плодородия почвы или ее физических свойств. Старк (1977) сделал вывод, что на западе штата Монтана предписанное выжигание можно неограниченно применять без снижения плодородия почв. Круговорот азота рассмотрен в главе 9, а предписанное выжигание — в главе 17.
Минеральное питание и поглощение солей
291
10-9. ВЕЛИЧИНЫ ПОТЕРЬ БИОМАССЫ (ЧИСЛИТЕЛЬ - ОБЩАЯ; ЗНАМЕНАТЕЛЬ - ГОДОВАЯ) И ЭЛЕМЕНТОВ МИНЕРАЛЬНОГО ПИТАНИЯ ПРИ СПЛОШНОЙ РУБКЕ И ДРУГИХ СПОСОБАХ
УБОРКИ 16-ЛЕТНИХ ДЕРЕВЬЕВ (Wells et al., 1975)
Способ уборки Биомасса, т/га Элементы, К г/га ‘Год к Са Mg
N Р
Полная уборка надземных 185/11,6 17,6 2,3 12,6 12,8 3,6
частей и корней более 4 см в диаметре Полная уборка надземных 156/9,8 16,1 1,9 10,3 11,7 2,9
частей Балансовая древесина до 116/7,2 6,5 0,9 5,0 6,4 1,6
высоты 8 см Балансовая древесина до 102/6,4 4,6 0,8 '3,8 4,3 1,3
высоты 8 см без коры
ПОГЛОЩЕНИЕ СОЛЕЙ
Поглощение минеральных питательных веществ так же важно для успешного роста растений, как и поглощение воды. Однако поглощение солей изучено пока недостаточно, поскольку оно значительно сложнее. Поглощение солей целыми растениями состоит из нескольких этапов: 1) движения ионов из почвы к поверхности корней; 2) накопления ионов в клетках корней; 3) радиального перемещения ионов от поверхности корней в ксилему; 4) передвижения ионов из корней в побеги.
ТЕРМИНОЛОГИЯ. Перед рассмотрением вопроса о поглощении солей обсудим некоторые термины. Проникновение веществ в клетки или ткани независимо от механизма этого процесса обычно называют поглощением или абсорбцией. Термин накопление относится к концентрированию данного вещества внутри клеток, происходящему против градиента электрохимического потенциала или концентрации, и требующему затраты метаболической энергии. Передвижение веществ против градиента концентрации или электрохимического потенциала, происходящее в результате потребления метаболической энергии, называется активным транспортом в отличие от пассивного движения путем диффузии по градиенту концентрации - массового перетекания, вызываемого градиентом давления. Примером массового перетекания может служить ток воды в корни и далее, вверх по ксилеме у транспирирую-щих растений.
Мембраны. Накопление ионов можно обнаружить только за относительно непроницаемыми мембранами, поскольку через проницаемые мембраны вещества будут просачиваться обратно путем диффузии с той же скоростью, с какой они передвигаются за счет активного транспорта. Мембрану можно определить как пограничный слой, отличающийся от разделяемых им фаз по проницаемости. Мембраны, позволяющие одним веществам проходить быстрее других, называют избирательно проницаемыми или, менее точно, полупроницаемыми мембранами.
292
Глава 10
В клеточные мембраны входят моном ©пекулярные слои, мембраны, окружающие органеллы (например, ядра и пластиды), наружные и внутренние границы цитоплазмы: вакуолярная мембрана, или тонопласт, и плазмолемма. Кроме того, большое значение для растений имеют многоклеточные мембраны, такие, как эпидерма, эндодерма и кора. Некоторые авторы рассматривают также всю кору молодых корней как многоклеточную мембрану, но обычно решающей для проникновения воды и солей мембраной корней считается эндодерма, так как пояски Каспари на ее радиальных стенках делают корни относительно непроницаемыми для воды и растворенных в ней веществ. Как будет показано ниже, значение эндодермы, по-видимому, переоценивалось.
Свободное пространство. Важное понятие в проблеме передвижения ионов в корни — кажущееся свободное пространство, или внешнее пространство. Это та часть клетки или ткани, куда ионы или другие растворенные вещества свободно проходят за счет диффузии, в отличие от недоступных для диффузии участков, в которых вещества могут накапливаться активным транспортом. Имеются доказательства того, что свободное пространство состоит только из заполненных. водой частей клеточных стенок, хотя некоторые авторы утверждали, что оно включает и цитоплазму. В молодых корнях свободное пространство состоит из воды в стенках клеток эпидермы и коровой паренхимы. Растворенные вещества могут двигаться вследствие диффузии и массового перетекания вдоль этих клеточных стенок вплоть до эндодермы. Конечно, некоторые катионы связываются с отрицательно заряженными веществами стенок.
После прохождения через клетки эндодермы в симпласт ионы в конце концов освобождаются в ксилемный сок и передвигаются вверх к листьям. Там они переходят из ксилемы жилок листа в клеточные стенки, составляющие свободное пространство Из этого раствора вещества избирательно извлекаются клетками листа. Наличиё значительных количеств ионов в свободном пространстве листьев объясняет факт вымывания солей во время дождя, а также поглощение веществ, нанесенных на поверхность листьев. Следуя терминологии Мюнха (1930), Крафт (1961) называет свободное пространство апопластом, апетальное пространство — симпластом. Последний термин относится к протопластам соседних клеток, соединенных плазмодесмами.
Активный транспорт. По свободному пространству ионы движутся неизбирательно, обратимо и независимо от метаболизма. В то же время поглощение солей растительными клетками избирательно, необратимо и зависит от метаболической активности. Например, некоторые ионы накапливаются в клетках до концентраций, намного более высоких, чем другие (относительно концентрации вне клетки). Еще много лет назад Хогленд и другие исследователи показали, что солевой состав клеточного сока очень отличается от состава среды, в которой растут растения. Таким образом, следует признать, что поглощение ионов клетками во многом зависит от избирательно действующего механизма активного транспорта. С другой стороны, практически все ионы, содержащиеся в окружающей среде, обнаруживаются в различных концентрациях в надземных частях растений, что свидетельствует о преодолимое™ ионных барьеров в корнях.
Минеральное питание и поглощение солей
293
Для объяснения активного транспорта через клеточные мембраны и накопления ионов в вакуолях предложено несколько теорий, но наиболее широкое признание получила теория переносчиков. Согласно этой теории ионы образуют на наружной поверхности мембран соединения с особыми молекулами, получившими название переносчиков, почти так же, как ферменты вступают во временное соединение с молекулами субстратов. Образующийся комплекс может перемещаться через мембраны, непроницаемые для ионов. На внутренней стороне мембран ионы отделяются от переносчиков и передвигаются в вакуоль или некоторые другие части внутреннего пространства клетки. Молекулы переносчика снова оказываются способными переносить другие ионы. Скорость оборота должна быть чрезвычайно высокой, а концентрация молекул переносчика, по-видимому, довольно низкая. Поэтому скорость активного поглощения часто лимитируется наличием переносчиков. Подлинная природа переносчиков неясна, но это могут быть ферменты или фосфорилированные соединения. Метаболическая энергия используется на синтез молекул переносчиков и, возможно, на образование комплекса переносчика с ионом, или на расщепление этого комплекса. Энергия необходима также для поддержания мембранной структуры. Избирательность транспорта ионов контролируется, по-видимому, различной способностью переносчиков образовывать комплексы с разными ионами.
Следует отметить, что некоторые исследователи подвергают сомнению общепринятое объяснение поглощения ионов при участии переносчиков и отрицают существование ионных насосов, поскольку для паботы последних потребовалось бы слишком много энергии. Они рассматривают клетки как ионообменные системы, имеющие отрицательно заряженные участки, с которыми связываются катионы. В таких системах пространство занято структурированной водой, играющей роль в определении типа поглощаемых ионов.
ПОГЛОЩЕНИЕ СОЛЕЙ КОРНЯМИ. Поглощение минеральных питательных веществ через корни включает: передвижение ионов из почвы к поверхности корней, накопление в клетках корней, движение из них в ксилему и выделение в ксилемный сок, с которым они переносятся в листья с транспирационным током.
Передвижение ионов в почве. Движение ионов в почве к корням происходит путем диффузии и массового перетекания в направлении к поверхности корней интенсивно транспирирующих растений. Относительное значение диффузии и массового перетекания зависит от вида и концентрации ионов в почвенном растворе, скорости, с которой ионы накапливаются корнями, и скорости тока воды к корням, регулируемой интенсивностью транспирации. Концентрация ионов в почвенном растворе лесных и окультуренных почв обычно низка. Некоторые анионы (например, фосфаты) обычно связаны с частицами почвы и органическими веществами, тогда как другие (например, нитрагы) относительно мобильны. Некоторые авторы проводят различия между поглощающей зоной корневой системы, общей массой почвы, занятой корнями, и поглощающей зоной поверхности корня, за которую они принимают слой почвы толщиной 1 см от корня (рис. 10.6). Рассчитанный таким образом объем поглощающей зоны корня молодой сосны
294
Глава 10
Рис. 10.6. Схема, показывающая, что общий объем почвы, занятый корневой системой, больше той части объема, которая находится в тесном контакте с корнями, т. е. собственно ризосферы (по Bray, 1У54):
1 — поглощающая зона для мобильных веществ; 2 — поглощающая зона для малоподвижных веществ; 3 — участок конкуренции корней за мобильные вещества
составляет лишь около 10% общего объема почвы, занятого ее корневой системой. Авторы тем не менее делают вывод, что количество доступного кальция и калия в поглощающей зоне корневой системы было достаточным для удовлетворения годовых потребностей изученных ими молодых деревьев сосны жесткой-
Корни лесных деревьев способны образовывать плотный ковер в поверхностном слое почвы, который перехватывает минеральные вещества, выделяющиеся при разложении подстилки. Однако корни, глубже проникающие в почву, должны поставлять значительное количество минеральных веществ и большую часть воды. Объем занятой корнями почвы — важный фактор при определении количества минеральных веществ и воды, доступных для деревьев и кустарников. Протяженность корней становится особенно важной, когда речь идет о поглощении медленно диффундирующих минеральных веществ, например фосфатов. Болезнь мелколистности у сосны ежовой наблюдается, когда сочетание неблагоприятных для роста корней почвенных условий и поражение грибом Phytophthora cinnamomi снижают поглощающую поверхность до такой степени, что развиваются симптомы тяжелой азотной недостаточности. Фрид и Бросарт (1967) опубликовали данные о сложных взаимоотношениях между почвой и корнями в связи с поглощением ионов. Эти связи еще больше усложняются присутствием бактерий, грибов, нематод и простейших, которые часто обнаруживаются в ризосфере или слое почвы, окружающем корни.
Передвижение ионов внутрь корней. Часто допускают, что радиальное движение воды и солей внутрь корней происходит через протопласты клеток эпидермы и коры. Однако теперь признано, что движение происходит в основном в клеточных стенках, где сопротивление потоку гораздо меньше, чем в протопластах, Часть солей аккумулируется при прохождении раствора через стенки клеток коровой паренхимы, а большая часть солей, по-видимому, переходит в симпласт путем активного транспорта у эндодермы. Вероятно, затем соли выделяются в пасоку клетками ксилемной паренхимы, прилегающими к элементам ксилемы.
У слабо транспирирующих растений корни проявляют тенденцию функционировать в качестве осмометра и вода движется к проводящему цилиндру в основном путем диффузии. У быстро транспирирующих растений вода движется путем массового перетекания, неся с собой соли по меньшей мере до эндодермы. Эффективность эндодермы как
Минеральное питание и поглощение солей
295
ионного барьера, возможно, переоценивают. Прорывы ветвления, образующиеся при развитии боковых корней, позволяют хотя бы временно пройти какой-то части солей с массовый потоком у транспирирующего растения. В молодых корнях эндодерма функционирует, вероятно, как основной барьер для ионов или избирательно проницаемая мембрана, регулирующая проникновение ионов, но ее нет в более старых корнях, в которых вторичный рост вызывает потерю коровой паренхимы и эндодермы. Местонахождение ионного барьера в старых корнях неизвестно, но им может быть камбий. О существовании ионного барьера в старых корнях свидетельствует развитие корневого давления.
Поглощающая зона. Принято считать, что поглощение солей происходит в основном на участке непосредственно за кончиком корня. Однако опыты Виба и Крамера (1954), Канинга и Крамера (1958), Руселла и Сандерсона (1967), Кларксона и Сандерсона (1974) и других исследователей показывают, что соли легко проходят в участки корней, расположенные в нескольких сантиметрах от кончиков. Опробковению подвергается 95% или больше поверхности корней лесных пород (см. главу 13). Во время вторичного роста эпидерма, корневая паренхима и эндодерма исчезают, а наружная флоэма обычно подвергается сильному опробковению. Хотя такие корни обычно считаются непроницаемыми для воды, неоднократно показано, что через них могут проходить значительные количества воды и солей, по-видимому, через чечевички, борозды между пластинами крры и щели, образованные отмиранием небольших боковых корней. Соли передвигаются, вероятно, путем диффузии и массовым перетеканием. Ионный барьер опробковевших корней еще не идентифицирован, но отчасти таким барьером может служить камбий, потому что из-за его тонких клеточных стенок большая часть воды и солей на пути к ксилеме вынуждена пересекать протопласты. Роль опробковевших корней в поглощении солей и воды заслуживает дальнейшего изучения.
Микориза. Эффективная поглощающая зона корней у многих древесных и травянистых растений видоизменена из-за наличия микоризы. Мицелий образует вокруг корней чехол (эктотрофная микориза) или врастает внутрь ткани корня (эндотрофная микориза). Тяжи гиф прорастают в почву, во много раз увеличивая поверхность соприкосновения с частицами' почвы. Эксперименты с использованием радиоактивных изотопов показали, что поглощенные гифами ионы быстро передаются сеянцам, на которых растут грибы. Бовен (1973) отмечает, что корни древесных видов занимают почву не так плотно, как корни многих травянистых видов, но это компенсируется тяжами мицелия и гифами, тянущимися на несколько сантиметров от микоризных корней. К тому же продолжительность поглотительной деятельности микоризных корней намного превышает время существования неопробковевших участков любых немикоризных корней. Согласно Марксу (1973) микоризные корни также менее подвержены поражению такими патогенными организмами, как Phytophthora, Pythiun и Fusarium, чем корни, не имеющие микоризы.
Передвижение ионов из корней в побеги. Различные минеральные элементы, попав в ксилемный сок корней, переносятся путем массового перетекания с транспирационным током в побеги, в основном в неорга-
296
Глава 10
Рис. 10.7. Сезонные колебания концентраций азота, фосфора, калия и магния в ксилемном соке яблони в Новой Зеландии. Азот передвигается в органической форме, другие элементы - в основном или полностью в неорганической (BoDard, 1958) :
а — период цветения; б — время сбора плодов; 1 ~ азот; 2 — калий; 3 — фосфор ; 4 — магний
нической форме. Как отмечалось в главе 9, у древесных растений большая часть или весь азот передвигается в форме органических соединений, в основном аминокислот и амидов. Некоторая часть серы и фосфора также передвигается в органической форме, но большая часть этих двух элементов и все остальные передвигаются в виде неорганических ионов. Некоторые ионы, особенно цинк и железо, иногда осаждаются в ксилеме и не могут достичь побегов. Солевынослив ость некоторых видов и форм растений (например, винограда) частично относят за счет необычной способности их корневых систем удерживать соли и не давать им достичь побегов.
Сезонные изменения состава и концентрации ксилемного сока хорошо известны (рис. 10.7). Самые высокие концентрации наблюдаются обычно во время цветения плодовых деревьев, самые низкие — в период покоя. Концентрация подвержена также суточной цикличности: наименьшие концентрации наблюдаются в полдень, когда в результате сильной транспирации вода поступает в корни быстрее солей, разбавляя ксилемный сок. По мере того, как соли поднимаются вверх по ксилеме, некоторые ионы передвигаются из ксилемного сока радиально во флоэму и там используются или перемещаются вниз, к корням. Иногда из ксилемного сока выводится так много солей, что их концентрация значительно уменьшается.
Большая часть достигающих листья солей (но не все) накапливается в клетках листьев активным транспортом. Значительные количества мобильных элементов, таких, как азот, фосфор и калий, передвигаются из старых в более молодые листья. Кальций и магний относительно неподвижны и по мере старения листьев проявляют тенденцию к накоплению (см. рис. 10.3). Когда в листья вводят меченые
Минеральное питание и поглощение солей
291
соединения (например, радиоактивный фосфор), они обычно вскоре обнаруживаются в корнях, а также и в более молодых листьях, что свидетельствует о передвижении минеральных питательных веществ вниз так же, как и вверх.
Влияние транспирации, Выше уже говорилось о том, что ток воды к поверхности корней транспирирующих растений усиливает передвижение солей к корням. Транспирация может увеличивать и скорость их передвижения в корнях. Согласно классической точке зрения передвижение солей в корнях происходит путем активного транспорта и мало зависит от скорости поглощения воды. Однако быстрый ток воды через ксилему корней может увлекать с собой соли, а уменьшение концентрации должно усиливать активный транспорт, К тому же у старых корней, где чечевички, отверстия, вызванные отмиранием боковых корней, и другие щели делают возможным некоторое массовое перетекание, заметное количество солей переносится в центральный цилиндр, по-видимому, с транспирационным током. Передвижение солей вверх по ксилеме ствола, из нее в жилки листьев и в конце концов в стенки клеток листьев также можно считать массовым перетеканием с транспирационным током.
Избирательное или неизбирательное поглощение. Существование более чем одного пути передвижения солей внутрь корней объясняет довольно противоречивый факт, что, несмотря на высокую степень избирательности в поглощении ионов клеткамц растений, они содержат почти все элементы, обнаруживаемые в окружающей корни среде. Причина этого парадокса заключается в том, что, хотя большая доля минеральных веществ растений избирательно накапливается в протопластах живых клеток, другая значительная их часть находится в свободном пространстве клеточных стенок и в мертвых тканях. Эти две фракции невозможно различить химическим анализом растительных тканей.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОГЛОЩЕНИЕ СОЛЕЙ. Количество и сотав поглощаемых деревьями ионов широко варьируют в зависимости от вида дерева и условий окружающей среды. Ниже кратко рассматриваются факторы, влияющие на этот процесс.
ВИДОВЫЕ РАЗЛИЧИЯ. В отношении содержания солей между видами существуют некоторые, четкие различия. Например, Кель (1937) установил, что в листьях цветущих дуба белого, кизила и амбрового дерева зольность примерно в 2 раза выше (7—7,2%), чем у сосен ладанной и ежовой (3—3,5%), растущих на тех же почвах. Он также обнаружил, что листья цветущих деревьев кизила, церциса канадского, тюльпанного дерева и дуба белого содержали более 2% кальция, в то время как листья дубов шарлахового, малого и,сосны ладанной содержали менее 1% кальция. Согласно данным Бесона с соавторами (1955) нисса лесная накапливает во много раз больше кобальта, чем большинство других видов, растущих на той же почве, шишкоягода накапливает цинк, а клетра накапливает большие количества и цинка и кобальта. О значительных различиях между видами по содержанию минеральных веществ сообщали и другие авторы.
Лесоводы Европы и США утверждали, что присутствие в лесах деревьев с высоким содержанием кальция и фосфора в листьях желательно, поскольку они усиливают распад подстилки и увеличивают
298
Глава 10
10.10. СПОСОБНОСТЬ ВИДОВ ДЕРЕНА ОГРАНИЧИВАТЬ НАКОПЛЕНИЕ МАРГАНЦА (Gerloff et al., 1966)
Вид Содержание марганца, частей на млн. Содержание ^Мп у других видов на том же участке, частей на млн. pH почвы Дата отбора проб
Comus canadensis 149 936 4,0 12.07.59
Comus racemosa 71 905 5,1 18.06.59
Comus racemosa 28 122 5,8 13.07.60
Comus rugosa 64 210 5,6 29.08.59
Comus stolonifera 37 52 7,4 14.07.60
плодородие почвы, особенно под насаждениями хвойных деревьев. Однако Стон (1975) ставит под сомнение способность отдельных видов или сочетаний видов оказывать заметное влияние на плодородие почвы в течение нескольких десятилетий.
Некоторые виды мангровых, растущих в морской воде, не пропускают соли, но другие виды поглощают 10-20% солей и выделяют их через особые железы, имеющиеся в листьях. Некоторые галофиты, например Atriplex, также отличаются способностью выделять соли, тогда как другие, по-видимому, не пропускают их. Во многих случаях различия в поглощении ионов определяются корнями, возможно, благодаря их способности связывать определенные ионы и предотвращать их поступление в побеги. Сообщают, что апельсины, растущие на подвое апельсина трехлистного, более чувствительны к недостатку цинка и железа по сравнению с теми, которые растут на других подвоях, например цитруса джамбири. Известно, что в некоторых случаях различия в поглощении ионов между травянистыми видами и разновидностями контролируются генетически, но в отношении древесных растений информации мало. Пример того, как несколько видов Comus не допускают накопления марганца, приведен в табл. 10.10.
Состав и концентрация почвенного раствора. Лесоводы и фермеры знают, что рост растений сильно зависит от концентрации необходимых минеральных питательных веществ в почве. Кроме того, рост древесной растительности, так же как и травянистых растений, осложняют: наличие высоких концентраций ядовитых элементов, таких, как свинец, цинк и медь, pH почвы и избыток соли на засоленных почвах. Между ионами существуют сложные взаимодействия, влияющие на их доступность, причем особо важную роль играет кальций. Некоторые виды растений, известных как кальцефилы, встречаются или исключительно на известковых, щелочных почвах, а другие — кальцефобы — встречаются на бедных кальцием, кислых почвах. В некоторых случаях кальцефилы, по-видимому, нуждаются в больших количествах кальция, чем родственные виды растений, относящиеся к кальцефобам. При этом важное значение имеют и другие факторы, например изменения величины pH. Растения можно выращивать в водных культурах при
Минеральное питание и поглощение солей 299
широком диапазоне pH, если принять меры для поддержания необходимых элементов в растворенном состоянии. Однако в природе pH играет важную роль, влияя на растворимость некоторых элементов. На щелочных почвах наблюдается дефицит железа и фосфора, а на кислых растениям часто недостает кальция и магния, причем повышенная растворимость алюминия, марганца и других ионов на кислых почвах может привести к концентрациям, токсичным для растений.
Для серпентинизированных почв, простирающихся вдоль Тихоокеанского побережья Северной Америки, характерны низкие концентрации кальция, высокие концентрации магния и плохой рост растительности. Следует отметить, чтр некоторые растения успешно растут на почвах с высоким содержанием токсичных элементов, не пропуская эти элементы, другие же растения без всякого ущерба поглощают их в больших количествах. По крайней мере, некоторые растения, видимо, образуют хелатные соединения, связывающие различные элементы в клеточных стенках и делающие их безвредными. Хорошим примером растений, растущих в неблагоприятных почвенных условиях, могут служить галофиты, но и они не все выживают по одной и той же причине. Как уже упоминалось, некоторые виды мангровых и Atriplex не пропускают соли, однако другие поглощают много солей и избавляются от них, по крайней мере отчасти, выделяя через специальные железы.
Биогеохимические изыскания. Существуют стойкие различия между видами по способности накапливать как необходимые элементы (например, кальций и калий), так и элементы, не являющиеся необходимыми (например, селен и кремний). Твердо также установлено, что на состав и концентрацию обнаруживаемых в растениях минеральных элементов влияют состав и концентрация элементов, присутствующих в почве, на которой они растут. Этот факт составляет основу листовой диагностики как показателя потребности в удобрениях. Он является также основой для биогеохимических изысканий, в которых состав минеральных веществ растительных тканей используется для выявления наличия или отсутствия определенных минеральных элементов в почве. Этот метод широко применяется во всем мире с использованием в качестве индикаторов древесных и травянистых растений.
Влажность почвы. Чрезмерно низкое или высокое содержание воды в почве оказывает настолько сильное влияние на рост корней, что трудно отличить прямое действие на поглощение минеральных веществ от косвенного, вызванного изменениями скорости роста и дифференциации корней. В насыщенных водой почвах происходит денитрификация, а в сухих почвах активность микробов понижена. Иногда в насыщенных водой почвах концентрация восстановленных форм таких элементов, как железо и марганец, достигает токсичного для растений уровня. На сухих почвах растениям может недоставать фосфора и калия, так как эти элементы связываются почвой, и к тому же рост корней понижен и их поглощающая поверхность уменьшается. Симптомы дефицита железа и марганца также наиболее часто проявляются у растений, растущих на сухой почве.
Обмен веществ в корнях. Поглощение минеральных веществ включает активный транспорт, зависящий от затрат метаболической энергии. Поэтому на поглощение солей влияют такие факторы окружающей
300
Глава J0
Рис. 10.8. Влияние концентрации кислорода на поглощение калия отделенными микоризными корнями бука (Harley, 1956)
Рис. 10.9. Влияние температуры на передвижение радиоактивного фосфора из грибов в сердцевину микоризных корней (Harley, 1956) :
1 - 15°С; 2 - 5°С
10.11. ДЕЙСТВИЕ ИНГИБИТОРОВ ДЫХАНИЯ НА НАКОПЛЕНИЕ РАДИОАКТИВНОГО ФОСФОРА МИКОРИЗНЫМИ И НЕМИКОРИЗНЫМИ КОРНЯМИ СОСНЫ ЛАДАННОЙ (ЧИСЛИТЕЛЬ - РАДИОАКТИВНОСТЬ, ИМП/МИН; ЗНАМЕНАТЕЛЬ - % ОТ КОНТРОЛЯ) (Kramer, 1951)
Воздействие при pH Коичики корней Микоризные
4,7—4,8 ' участки
Контроль
25 мМ малоновая кислота
1 мМ азид натрия
5 мМ фторид натрия
81 713/100,0 77 330/94,6 308/0,4 989/1,2
77 306/100,0
67 408/87,2
965/1,2
1 620/2,1
среды, как аэрация и температура. Большая часть исследований по этому вопросу проведена на корнях травянистых растений, но имеющиеся немногочисленные данные свидетельствуют о том, что аналогично ведут себя и корни древесных видов. Влияние кислорода на поглощение калия микоризными корнями показано на рис. 10.8, а на рис. 10.9 -влияние температуры на передвижение фосфора. Приведенные в табл. 10.11 данные показывают, что ингибиторы дыхания снижают поглощение фосфора микоризными и немикоризными корнями примерно в равной степени. Установлено, что недостаточная аэрация часто ограничивает скорость поглощения питательных веществ как травянистыми культурами, так и большинством древесных видов. Однако, как и в отношении почвенной влаги, трудно отличить прямое действие недостаточной аэрации и низкой температуры на поглощение солей от косвенного влияния на рост корней, что также уменьшает поглощение солей. Примером сложных последствий недостаточной аэрации и заражения грибами служит болезнь мелколистное™ у южных видов сосны (ладанной, болотаой, жесткой, голой, ежовой, карибской и Эллиота).
к
ПЕРЕДВИЖЕНИЕ ВЕЩЕСТВ
Успешное произрастание растений высотой более нескольких сантиметров зависит от передвижения воды, минеральных веществ и некоторых органических соединений от корней к побегам и от передвижения углеводов и других органических соединений от побегов к корням. Продолжительный рост требует снабжения различных меристематических зон достаточными количествами воды, минеральными питательными веществами, органическими соединениями азота и определенными регуляторами роста. Таким образом, транспорт этих веществ к участкам, где они используются в качестве строительного материала и субстратов для дахания, так же важен для роста, как и синтетические процессы, при которых они образуются.
У небольших, относительно недифференцированных организмов в каждой клетке могут осуществляться все необходимые процессы, так как ни одна клетка не удалена от источника сырья. У таких низкорослых растений, как мхи и печеночники, для необходимого перемещения веществ на короткие расстояния достаточно диффузии, которую возможно, облегчает движение цитоплазмы. Для существования крупных наземных растений, таких, как деревья, необходимо быстрое перемещение веществ на большие расстояния в обоих направлежях (вверх и вниз), так как рост корней зависит от поступления из листьев сахаров и, вероятно, регуляторов роста., В свою очередь листья зависят от корней, откуда поступают вода, минеральные питательные вещества, органические соединения азота и некоторые регуляторы роста. Корни и листья часто находятся на расстоянии 30-100 м друг от друга, и большие количества веществ должны одновременно передвигаться в противоположных направлениях, поэтому проблема становится достаточно сложной.
Эволюция сосудистой системы, делающей возможным быстрый транспорт на большие расстояния, была решающим моментом для появления высоких растений. Ее механическая прочность выполняет существенную функцию поддержания большой массы листьев, блаюдаря чему растение получает много солнечной энергии. Таким образом, развитие современной сосудистой системы обусловлено изменением морфологии и физиологии, сделавшим возможным существование деревьев, Для развития растений необходимо правильное распределение метаболитов по различным органам и тканям, в том числе к кончикам корней и стеблей, камбию и развивающимся плодам и семенам. Следовательно, надо учитывать факторы, регулирующие передвижение веществ. Знание того, какие ткани участвуют в передвижении, и какие силы влияют на него, нужно не только для понимания физиологии деревьев, но также и для понимания того, какой вред причиняют, деревьям патогенные организмы, насекомые и механические повреждения (например, кольцевание).
Проросткам, в которых транспорт происходит в основном от клетки к клетке, вполне достаточно диффузии, но по мере их подрастания диффузии недостаточно. Если при расстояниях, измеряемых в нанометрах или даже в миллиметрах, диффузия молекул происходит быстро, то при расстояниях, измеряемых метрами,-за длительный период. Например, подсчитано, что 940 дней потребовалось бы для диффузии 1 мг сахарозы на расстояние 1 м через заполненную водой трубку с поперечным сечением 1 см2 из 10%-ного раствора в чистую воду. Следовательно, для ежедневного перемещения 150-200 л воды от корней к побегам или нескольких сотен граммов сахаров из листьев необходимо участие механизмов более быстрых, чем диффузия. Рассмотрим пути и механизм транспорта наиболее важных из передвигающихся веществ, а также некоторые факторы, от которых зависит транспорт на большие расстояния.
ТРАНСПОРТ ВОДЫ
Транспорт воды обсуждается в главе 13, а в этой главе ток воды будет рассмотрен в качестве переносчика для передвижения вверх растворенных веществ. В жаркий солнечный день 100—150 л воды
302
Глава 11
11.1. КОНЦЕНТРАЦИИ РАСТВОРЕННЫХ"ВЕЩЕСТВ В КСИЛЕМНОМ И ФЛОЭМНОМ СОКЕ ЯСЕНЯ ПЕНСИЛЬВАНСКОГО
10 ОКТЯБРЯ, МГ/МЛ (Von Die, Willemse, 1975)
Наименование показателей
Ксилемный сок Флоэмный сок
Сухое вещество 1,4 220,0
Сахароза 0,128 140,0
Калий 0,177 2,1
Магний 0,0115 0,077
Натрий 0,004 0,037
Кальций 0,018 0,049
Фосфор 0,005 0,052
рн 4,9-5,0 7,5
может перемещаться по корням и стволу в листья большого дерева. Это движение сока происходит путем массового перетекания по ксилеме (строение ее обсуждалось в главе 2). При передвижении воды в радиальном направлении из почвы к ксилеме корней она пересекает массу живых клеток, вероятно, передвигаясь в основном по клеточным стенкам (апопласту). В листьях вода выходит из ксилемы в стенки клеток мезофилла. Все продольное передвижение происходит через просветы мертвых сосудов и трахеид. В ксилеме вода почти всегда движется вверх. Но в ксилеме нет клапанов или других устройств для предотвращения тока вниз, и в определенных условиях направление движения воды может меняться. Движущей силой обычно является градиент водного потенциала от почвы до листьев, возникающий в результате испарения воды. В связи с этим скорость тока воды регулируется в основном интенсивностью транспирации, хотя в этом участвуют и другие факторы.
СОСТАВ КСИЛЕМНОГО СОКА. Ксилемный сок (пасока) деревьев представляет собой разбавленный раствор минеральных солей и органических веществ, в том числе азотистых соединений, углеводов, ферментов и регуляторов роста, особенно цитокининов и гиббереллинов (табл. 11.1). Концентрация и состав ксилемного сока подвержены суточным и сезонным колебаниям. Дневное увеличение скорости тока воды, вызванное сильной транспирацией в полдень, снижает концентрацию растворенных в соке веществ.
Углеводы. В пасоке содержание углеводов зимой и ранней весной высокое, в конце весны и начале лета оно снижается до минимума. У груши оно начинает увеличиваться в конце лета, достигая максимума зимой. Основные сахара ксилемного сока — сахароза, глюкоза и фруктоза. Особенно высоко содержание сахара в пасоке клена сахарного, содержащего так много сахарозы (1—7%), что из него получают кленовый сироп. Значительные количества фруктозы и глюкозы содержит березовый сок. (Истечение сока у березы и клена обсуждается в главе 13).
Передвижение веществ
303
Соединения азота. Большая часть используемого деревьями азота восстанавливается в корнях до органических соединений и передвигается в побеги с ксилемным соком. Для пасоки растений разных семейств характерны различные соединения азота, но большая его часть присутствует в виде аминокислот и амидов, сообщалось лишь о следах нитратного и аммонийного азота. Концентрация и состав соединений азота изменяются в течение года. В соке вяза идентифицирована 21 аминокислота. Содержание аминокислот постепенно уменьшалось с начала весны до лета. В соке яблони летом основными были аспарагиновая кислота и глутамин. В сентябре содержание аспарагиновой кислоты уменьшалось, а содержание аргинина увеличивалось, и в течение всей зимы он был доминирующим компонентом. Глутамин был самой важной аминокислотой у семи видов сосны, изученных Барнесом (1963), но у других изученных видов преобладали другие аминокислоты. Несмотря на то, что этот автор (1963) обнаружил лишь 20 мг/л азота в ксилемном соке сосен, он рассчитал, что в сосновом древостое благодаря высокой интенсивности транспирации около 50 кг/га год азота может передвигаться в кроны с ксилемным соком. (Более подробно азотное питание рассмотрено в главе 9.)
Другие компоненты. Кроме углеводов и органических азотсодержащих соединений пасока обычно содержит значительные количества различных минеральных веществ и небольшие количества регуляторов роста (особенно цитокининов и гиббереллинов), ферментов и разнообразных соединений. Минеральные вещества составляют относительно небольшую фракцию сухого вещества ксилемного сока деревьев в конце зимы — начале весны. В ксилемном соке травянистых растений они часто являются единственным значительным растворенным компонентом. (Транспорт солей обсуждается в следующем разделе, а транспорт регуляторов роста - в конце этой главы.)
ТРАНСПОРТ МИНЕРАЛЬНЫХ ВЕЩЕСТВ
Минеральные вещества передвигаются из корней в побеги в ксилемном соке, перемещающемся путем массового перетекания с транспирационным током. Существует также значительное латеральное передвижение минеральных веществ между ксилемой и флоэмой (рис. 11.1). Минеральные вещества, поднимаясь вверх, диффундируют из ксилемы, предположительно, вследствие того, что они используются в зонах роста вдоль пути. Клетки верхушек стеблей, камбиальные зоны и другие метаболические активные участки накапливают минеральные веш етва, в результате чего возникают градиенты, вызывающие передвижени к этим участкам.
ВОСХОДЯЩИЙ ТОК. Скорость транспорта минеральных веще, i в регулируется в основном интенсивностью транспирации и потому в деревьях, сбросивших листья, она намного ниже, чем в облиственных. Удивительно, что не все минеральные вещества передвигаются вверх по ксилеме с одной и той же скоростью: передвижение некоторых ионов, по-видимому, замедляется из-за адсорбции на стенках элементов ксилемы. Примером медленно передвигающихся ионов служат каль-
304
Глава 11
Рис. 11.1. Пути восходящего транспорта радиоактивного калия у ивы. Кора у левой ветви отделена от древесины слоем парафинированной бумаги (прирост К в листьях-8,7 ч/млн.), правая ветвь оставалась нетронутой (прирост К в листьях - 9,9 ч/млн.). В коре, отделенной от корней, обнаружено очень мало калия, но в условиях нормального контакта его было много. Этот эксперимент показывает, что передвижение вверх происходит в основном по ксилеме, но имеется также и латеральное передвижение из древесины в кору ( Stout, Hoagland, 1939). Цифры на рисунке - концентрация К, ч/млн. (числитель - в коре; знаменатель - в древесине): а — кора отделена; б — контроль
ций, железо и цинк. Некоторые группы растений, вероятно, удерживают в корнях больше определенных ионов, чем другие группы. Например, некоторые разновид. ности винограда при выращивании на концентрированных растворах хлорида натрия перемещают в побеги меньше солей, чем
другие. Как отмечалось в главе 10, клетки обладают высокой избирательностью в отношении накопления ионов. Корневые системы также достаточно избирательны, но хотя бы следы почти каждого иона просачиваются из окружающей корни среды через ионные барьеры корней и обнаруживаются в свободном пространстве или апопласте листьев.
КРУГОВОРОТ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. Ксилемный сок - основной источник минеральных веществ и азота для зон роста побегов, но важную роль в минеральном питании играет также циркуляция по флоэме. Так, растущие листья могут получать минеральные вещества как через ксилему, так и через флоэму. Постоянное передвижение минеральных веществ в листья с транспирационным током в течение вегетационного периода вызывает накопление солей, которое иногда причиняет вред листьям растений, растущих на солонцах или слишком обильно удобренных. Такие элементы, как кальций и магний, а на засоленных почвах и натрий, обычно накапливаются в течение вегетационного периода. Концентрации таких мобильных элементов, как азот, фосфор и калий, часто уменьшаются при пересчете на сухой вес (см. рис. 10.3). Для мобильных элементов общим является их передвижение из старых, метаболически менее активных тканей, в молодые листья, развивающиеся плоды и другие метаболически активные зоны. Кроме того, осенью в больших масштабах происходит переход минеральных веществ из стареющих листьев в ветви (рис. 11.2).
Заметное количество солей теряется из свободного пространства или апопласта листьев вследствие вымывающего действия дождя (см. главу 10). Некоторые виды деревьев теряют соли, выделяя их через особые железы. Часть солей теряется при гуттации с водой, выдавливаемой из гидатод под действием корневого давления. Эксперименты с мечеными соединениями, например 32р, показывают, что при введе-
Передвижение веществ
305
Возвращение 6 почву вымыванием и при листопаде
Минеральные и ореанические вещества почвы
л -
Почвенный раствор
0
К листьям по ксилеме
Диффузия и массовое перетекание
Через хору -
К листьям и из них по флозме
Обмен между ксилемой и флозмой
Передвижение по флоэме,в основном вниз
Ксилемный сок Поверх- корней корней
ность —
корней
Накопление в клетках корней
Передвижение в органической и неорганической форме, вверх к побегам преимущественно массовым перетеканием в ксилеме
Ианы поглощенные^ почвенными коллоидами
Потери вымыванием
Рис. 112. Круговорот минеральных веществ в дереве и их возвращение в почву путем вымывания и при листопаде (Kramer, 1969, с разрешения Мс Graw-НШ Rook Company)
нии в листья они часто передвигаются по флоэме вниз к корням, где часть просачивается наружу, а часть переносится в ксилему и движется по ней'вверх.
Мобильность различных ионов широко варьирует. Буковак и Витвер (1957) разделили их на высокоподвижные (К, Rb, Na, Mg, Р, S и Cl), ионы со средней подвижностью (Fe, Mg, Zn, Си и Мо) и относительно подвижные (Са, Li, Sr, Ва и В). Однако подвижность ионов промежуточной группы различается по видам, фазам развития и в зависимости от количества элемента в растении. Например, Си, Zn и S подвижны только при высоких концентрациях, их мобильность невысока в растениях, страдающих от дефицита этих элементов. Концентрация неподвижных элементов может быть достаточной в старых листьях в то время, как у молодых листьев того же растения наблюдаются симптомы дефицита. Наоборот, высокоподвижные элементы, такие, как N, Р и К, часто отводятся из более старых листьев, вызывая у них симптомы дефицита, тогда как у молодых листьев никаких признаков недостаточности не наблюдается. Это следует учитывать при отборе проб листьев для анализа с целью выявления недостаточности минеральных веществ.
РЕГУЛЯЦИЯ ПОГЛОЩЕНИЯ МИНЕРАЛЬНЫХ ВЕЩЕСТВ. На уровне растения потребление минеральных веществ регулируется преимущественно скоростью, с которой ионы перемещаются из почвы через корни
306
Глава 11
и поступают в корневую ксилему. На нее влияет и скорость передвижения воды в ксилему корня. Быстрый ток воды перемещает ионы в почве к корням и облегчает их передвижение по свободному пространству и симпласту по направлению к ксилеме корня, Интенсивный транспирационный ток снижает концентрацию солей в ксилеме корня и способствует их переходу в пасоку.
На тканевом уровне накопление ионов, по-видимому, связано с биосинтезом белка, и сила впитывания поэтому выше в растущих тканях. Так, молодые листья часто получают ионы из более старых тканей, а по мере старения отдают ионы молодым листьям, плодам и другим метаболически активным тканям. Часто утверждают, что растительные гормоны участвуют в регуляции передвижения солей. Однако трудно отличить влияние быстрого использования солей растущими тканями от действия большого количества гормонов, образующихся в таких тканях. Вероятно, примером действия гормонов на поглощение ионов могут служить такие случаи, как вызванное ауксином выделение Н + в листовой подушечке или потеря К+ замыкающими клетками устьиц, вызванная абсцизовой кислотой. Утверждается также, что в присутствии абсцизовой кислоты уменьшается транспорт ионов в корневую ксилему без снижения скорости поглощения ионов корнями.
ПЕРЕДВИЖЕНИЕ ОРГАНИЧЕСКИХ ВЕЩЕСТВ
Изучение передвижения органических веществ в растениях началось с работ Теодора Гартига. Он описал в 1837 г. ситовидные трубки, а в 1859 г. открыл, что при их прокалывании можно получить сок. В результате этих исследований он обеспечил методологическую основу для многих дальнейших исследований по передвижению веществ. В растениях перемещается множество органических соединений, в том числе углеводы, азотсодержащие вещества, регуляторы роста, витамины, вирусы и различные биоциды - гербициды, фунгициды и инсектициды. Большая часть органических соединений перемещается по флоэме, как показывают опыты с кольцеванием, анализы флоэмного сока и эксперименты с радиоактивньпми метками. Растущее растение можно рассматривать как интегральную систему потребителей питательных веществ, конкурирующих между собой в отношении направления передвижения и количества углеводов и азотсодержащих веществ, двигающихся к различным тканям в зависимости от нужд потребителей и значения разных источников снабжения. Таким образом, любое обсуждение вопроса о передвижении веществ требует количественной характеристики происходящего роста и места, где он наблюдается.
УГЛЕВОДЫ. Количество передвигающихся углеводов превышает общее количество всех других растворенных веществ. Углеводы передвигаются преимущественно вниз, но отмечается и значительное движение вверх. Их источниками служат фонды запасных углеводов и продукты текущего фотосинтеза. Количество передвигающихся веществ зависит от потребности в уг л ев одах и близости растущей ткани к источникам снабжения. У большинства древесных растений углеводы передвигаются в виде сахарозы. У немногих семейств важными коМпонен
Передвижение веществ
307
тами флоэмного сока являются рафиноза, стахиоза, вербаскоза. Саха-роспирт сорбит обнаружен у яблони и вишни, маннит — у ясеня. Редуцирующие сахара во флоэмном соке не встречаются. Библейская "манна небесная” может происходить из разных источников, но имеющаяся в продаже манна (медвяная роса) добывается из содержащего маннит флоэмного сока ясеня манного при его подсочке.
У молодых голосеменных растений умеренной зоны до распускания почек главным потребителем углеводов являются обычно корни (рис. 11.3). После распусканйя почек происходит обратное передвижение, когда большее количество углеводов движется к развивающимся побегам. В это время рост камбия является второстепенным потребителем притекающих углеводов. После окончания сезонного роста побегов основными потребителями углеводов являются растущий камбий и корни, возобновляющие рост. Этот обобщенный сезонный ход событий может сильно изменяться в год обильного плодоношения, когда репродуктивные ткани монополизируют основную массу имеющихся углеводов. При одновременном росте корней и побегов соотношение остающихся в побегах и передвигающихся в корни ассимилятов зависит от относительного роста каждого из этих потребителей и их близости к источникам углеводов.
Например, после экспозиции с 14СОз сеянцев ели белой у быстрорастущих растений побеги сохраняли больше продуктов фотосинтеза (70%), чем у медленнорастущих (55%). У медленнорастущих растений больше 14С передвигалось в корни. Количество ассимилятов, передвигавшихся в корни, изменялось также по мере развития корней. У сеянцев сосны смолистой и веймутовой с плохо развитыми корневыми системами передвижение продуктов фотосинтеза было намного слабее, чем у сеянцев с обширными, быстро разрастающимися корневыми системами.
Значение отношения между отдачей и потреблением углеводов показано путем чередования в условиях эксперимента относительной силы поглощения у источников и у потребителей. Например, перед цветением и во время цветения побеги винограда были сравнительно независимы один от другого в отношении снабжения продуктами фотосинтеза. При увеличении потребности в углеводах у одного из побегов, вызванном стимуляцией роста под действием гиббереллина, произошло компенсирующее передвижение ассимилятов из соседнего побега. Аналогично понижение количества доступных ассимилятов в побеге или в его части, вызван-ноезатемнением или дефолиацией, приводило к передвижению ассимилятов из близлежащих листьев. Такое компенсирующее передвижение показано как у одного побега, так и между побегами. Напротив, удаление потребителя увеличивает доступность углеводов для прилегающих к нему потребителей. Эти опыты хотя и проводились в искусственно регулируемых условиях, но они показывают, что временное затенение или повреждение некоторых листьев насекомыми или болезнями воздействуют на характер передвижения продуктов фотосинтеза.
Углеводы, перемещенные из листьев в корни проростков в начале вегетационного периода, могут затем передвигаться вверх к мощным потребителям, таким, как быстро развивающиеся побеги. Например, у 3-летних сеянцев сосны
Рис. П.З. Сезонные изменения передвижения 14С в корни 3-летних деревьев сосны веймутовой (Nelson, 1964)
1961г, Меся
jpg Глава 11
веймутовой, побеги которой выдерживались в атмосфере с 1^СС>2 в июне, концентрация в корнях ассимилированного при фотосинтезе увеличивалась от 1% через 8 ч до 10% после 1 месяца. В течение следующего месяца, однако, концентрация ; 14с в корнях уменьшалась до 4%, но к октябрю она снова увеличивалась до 14%. Уменьшение концентрации в корнях между первым и вторым месяцами после фотоассимиляции указывало на наличие передвижения 1*с из корней обратно в побеги. По сравнению с этим, у 8-летних деревьев сосны смолистой запасы углеводов в корнях имели сравнительно небольшое значение для роста побегов- Когда ; ток по флоэме блокировали кольцеванием главного ствола на уровне земли перед ?. началом вегетационного периода, предотвращая передвижение по флоэме запасных ? углеводов из корней в надземные ткани, последующий рост побегов подавлялся лишь незначительно.
Имеются доказательства того, что по мере увеличения размера де- >. ревьев и удлинения пути, по которому происходит передвижение ве- f ществ от побегов к корням, растущие ткани верхушки кроны все мень- < ше зависят от запасов углеводов, находящихся в нижней части ствола и в корнях. Таким образом, значение нижцих ветвей для верхней части > стебля неуклонно уменьшается по мере роста дерева.
Рост побегов. На рост побегов используются как запасные углеводы, так и продукты текущего фотосинтеза. При этом их соотношение изменяется в зависимости от вида, генетической природы, типа побегов и расположения побегов на стволе. Растущие побеги покрытосеменных растений используют продукты текущего фотосинтеза в дополнение к утилизации запасных углеводов в начале роста. У голосеменных источники вновь образующихся углеводов, используемых на рост побегов, изменяются по временам года.
Так, в начале мая, когда у побегов сосны смолистой в центральных районах Висконсина (США) раскрываются верхушечные почки, они начинают мобилизовывать углеводы, в основном продукты фотосинтеза из однолетней хвои. К 1 июня, когда побеги находятся на стадии ’’свечки” (междоузлия хорошо вытянуты, а хвоя вдоль междоузлий относительно недоразвита) , однолетняя хвоя поставляет большую часть продуктов текущего фотосинтеза разрастающимся побегам, тогда как 2-3-летняя хвоя дает меньше. Снабжение побегов продуктами текущего фотосинтеза из старой хвои снижается в середине августа. К этому времени полностью развившаяся хвоя текущего года замещает старую хвою в качестве основной фотоассимилирующей ткани и поставщика углеводов для роста различных других тканей. Обнаружено также, что передвижение меченых ассимилятов из старой хвои к растущим побегам сосны смолистой бывает наибольшим в период быстрого удлинения новой хвои.
Старая хвоя - самый важный источник метаболитов для роста побегов сосны смолистой. Козловский и Винжет (1964) изучали роль запасных веществ ветвей, стволов и корней, а также роль резервов вместе с продуктами текущего фотосинтеза хвои.для роста побегов 8-летних деревьев сосны смолистой. С этой целью исследователи удаляли листву, блокировали флоэму кольцеванием, применяли различные комбинации этих методов до начала вегетационного периода. Дефолиация и кольцевание, отдельно или вместе, уменьшали рост побегов- Рост побегов в результате этих воздействий снижался в таком порядке: кольцевание ветвей + удаление хвои>кольцевание у основания главного ствола + удаление хвои > удаление хвои > кольцевание ветвей > кольцевание у основания ствола. Участие в росте побегов запасных веществ (из тканей кроме старой листвы) изменялось так: ветви > главный ствол > корни (табл. 11.2).
Передвижение веществ
309
11.2. РАСЧЕТ ДОЛИ РАЗНЫХ ЧАСТЕЙ РАСТЕНИЙ В ОБЩЕМ СУХОМ ВЕЩЕСТВЕ И ПРИРОСТЕ ГЛАВНЫХ ВЕРХУШЕЧНЫХ ПОБЕГОВ (Kozlowski, Winget, 1964)
Участие в приросте отдельных частей растения
Сухой вес побегов, % контроля
Хвоя + главный ствол + ветви + корни 100,0
)&оя:
на целом дереве 87,6
на целом дереве минус корни 75,8
на ветвях 66,7
Ветви 5,7
Главный ствол:
со старой хвоей
после удаления старой хвои
Главный ствол + корни после удаления старой хвои
Корни:
со старой хвоей на дереве
после удаления старой хвои
13,9
4,9
6,7
13,6
1,8
Направление передвижения углеводов от отдельных листьев разнолистных побегов покрытосеменных растений изменяется по мере того, как побег разрастается и на нем продолжает увеличиваться количество новых листьев. Очень молодые листья получают углеводы от зрелых листьев, расположенных ниже. Развившиеся до некоторой степени листья и получают и отдают углеводы. Полностью развившиеся листья только отдают углеводы. С разрастанием листа гетерофильного побега характер передвижения углеводов изменяется от движения вверх до движения в обоих направлениях и, наконец, когда выше его оказываются несколько хорошо развитых листьев, к преимущественному движению вниз. В итоге, когда листья стареют фотосинтез и отток ассимилятов уменьшаются. Такое передвижение углеводов показано для тополя и яблони Ларсоном, Гордоном (1969), Хансеном (1967). Большая часть передвигающихся вверх углеводов используется на развитие сосудистых и механических тканей стебля.
Растущие плоды часто видоизменяют характер оттока углеводов из листьев разнолистных побегов. Плоды — мощные потребители углеводов, и они часто монополизируют имеющиеся углеводы до такой степени, что вместо нисходящего передвижения наблюдается двустороннее. При этом часть продуктов фотосинтеза направляется вверх, в плоды из листьев, находящихся ниже, а часть передвигается вниз от листьев, расположенных на ветви выше.
Согласно Кридеману (1969) быстрорастущие побеги апельсина получают большие количества углеводов. Но как только листья текущего цикла роста (самой близкой по времени вспышки роста) или
310
Глава 11
предыдущих циклов полностью развиваются, они становятся поставщиками углеводов в основном для расположенных рядом растущих плодов. Зрелые листья, находящиеся на некотором удалении от растущего побега, поставляют углеводы преимущественно корням.
Камбиальный рост. Зона камбия — относительно диффузная меристема и слабый потребитель углеводов. Поэтому она не очень успешно конкурирует с мощными потребителями — быстро удлиняющимися побегами и корнями или разрастающимися репродуктивными структурами. Тем не менее активный камбий, покрывающий древесину главного ствола, всю систему ветвей и главные корни, потребляет за вегетационный период большие количества углеводов. Они используются на образование новых инициальных клеток камбия и на прирост ксилемы и флоэмы. Так как большая часть новых инициальных клеток отделяется путем псевдопоперечндго деления, происходит избыточное образование и потеря многих новых клеток. Выживают инициальные клетки, имеющие лучший контакт с лучами, что свидетельствует о сильной конкуренции за углеводы и воду.
У многих лиственных видов активность камбия на участках ниже почек начинается до их раскрытия, поэтому камбиальный рост в начале вегетационного периода должен зависеть от запасов углеводов. В частности, значительные количества запасных углеводов используются на образование флоэмы, начинающееся очень рано и часто предшествующее образованию ксилемы, иногда на несколько недель (см. главу 3) . Такой ранний камбиальный рост сопровождается истощением резервных углеводов в запасающих тканях, таких, как клетки осевой и дучевой паренхимы. У голосеменных растений на камбиальный рост в начале вегетационного периода используются, помимо запасных углеводов, продукты текущего фотосинтеза. Например, у 3, 5 и 15-летних деревьев сосны смолистой заметные количества 14С концентрировались в дифференцирующихся трахеидах вскоре после экспозиции целых деревьев или отдельных веток с 14СО2. Когда Дикман-и Козловский (1970) выдерживали в 14СС>2 разновозрастную хвою 20-летних деревьев сосны смолистой в середине июня, значительная доля 14С быстро включалась в структурные соединения ксилемы, что еще раз подчеркивает раннюю утилизацию продуктов текущего фотосинтеза на камбиальный рост.
Фонд углеводов для роста камбия используется неравномерно во времени и пространстве. У многих видов умеренной зоны углеводы используются на камбиальный рост в течение лишь немногих месяцев года. И даже в это время углеводы используются периодически вследствие конкуренции камбиального роста с более мощными потребителями углеводов и вследствие влияния на снабжение углеводами колебаний климатических факторов, нападения насекомых, болезней и т.д. У многих тропических видов углеводы используются на рост камбия в течение всего года, но скорость мобилизации углеводов подвержена сезонным колебаниям. Иногда тропические деревья, у которых прирост камбия происходит ежемесячно, образуют в течение года многочисленные кольца роста, что свидетельствует о неравномерной утилизации углеводов.
Использование углеводов также сильно изменяется в зависимости от высоты ствола и в разных ветвях. Например, у сеянцев и у мощных,
Передвижение веществ
311
растущих на открытом месте взрослых деревьев больше углеводов используется на камбиальный рост в нижней части ствола по сравнению с Верхней. У угнетенных и очень старых деревьев утилизация углеводов при росте в основном ограничена верхней частью ствола, что подтверждается частым выпадением колец древесины в нижней части ствола.
У голосеменных деревьев мутовки ветвей верхней части кроны являются основным источником продуктов текущего фотосинтеза для камбиального роста. Например, у типичных деревьев сосны смолистой большую часть углеводов, используемых на камбиальный рост, дают ветви верхней трети кроны. Такие относительно незатененные ветви имеют высокую фотосинтезирующую способность и короткий путь передвижения углеводов в главный ствол. Относительное значение углеводов^ образуемых другими ветвями, изменяется в зависимости от условий роста, возраста деревьев, конкурентных отношений и других факторов. У растущих на открытом месте деревьев сохраняющиеся на нижней части ствола ветви дают камбиальному росту этой части важный источник снабжения углеводами. В сомкнутых насаждениях в условиях жесткой конкуренции естественное опадение нижних ветвей или потеря ими мощности ограничивают источники углеводов для камбиального роста ветвей верхней части ствола. Приток углеводов для камбиального роста из отдельных ветвей уменьшается по мере их угнетения. У голосеменных, например, стареющие нижние ветви покрываются сверху все новыми и новыми мутовками ветвей, в результате они все больше затеняются и уменьшается их фотосинтетическая активность. В этих нижних, угнетенных, ветвях меньше углеводов, пригодных для оттока и использования на камбиальный рост, по сравнению с ветвями, расположенными выше на стволе.
Как отмечалось ранее, у многих взрослых деревьев нижние, угнетенные, ветви не участвуют в снабжении углеводами роста главного ствола. Такие ’’отрицательные” ветви могут даже не давать достаточно углеводов для образования ксилемы по всей длине самой ветви. Например, у взрослых деревьев дугласии в основании нижних ветвей часто было на 9—10 ксилемных колец меньше, чем в главном стволе у места отхождения этой ветви. Ветви сосны ладанной в нижней половине кроны, имеющие менее трех веточек, или ветви нижней четверти кроны менее чем с пятью веточками совсем не дают углеводов для роста главного ствола.
Значение разновозрастной хвои как источника углеводов для камбиального роста изменяется в зависимости от времени года. По данным Гордона и Ларсона (1968, 1969), у молодых деревьев сосны смолистой большая часть весенней древесины в старых междоузлиях стволов образовывалась во время удлинения побегов и на ранних этапах развития хвои, а новая хвоя в это время не давала заметных количеств ассимилятов на камбиальный рост. На образование весенней древесины скорее всего использовались углеводы, оттекающие из старой хвои, в основном однолетней. Новая, почти полностью развившаяся хвоя начинает давать большие количества углеводов на образование ксилемы. В это же время наблюдалось изменение направления, в котором передвигались углеводы из старой хвои: теперь транспорт происходил главным образом в корни и очень мало углеводов двигалось вверх, к новым ветвям. Усиленное снабжение углеводами после
312 Глава 11
созревания новой хвои коррелировало с утолщением вторичных стенок трахеид, характерным для развития поздней древесины. Ближе к концу вегетационного периода новая хвоя снабжала углеводами новые почки и камбиальный рост верхних междоузлий ствола, а старая хвоя поставляла углеводы для роста камбия нижних междоузлий. В конце сезона, когда камбий уже не был заметным потребителем, хвоя всех возрастов снабжала углеводами корни, и в паренхимных тканях по всему дереву накапливались запасные углеводы.
Репродуктивный рост. Растущие плоды являются мощными потребителями, использующими большие количества запасных и (или) вновь образованных углеводов. Например, у бука много запасных углеводов передвигалось в развивающиеся цветки и плоды главным образом из древесины ветвей. Камерон и Борст (1938) обнаружили, что у плодоносящих деревьев авокадо содержание крахмала в ветвях в течение лета и осени было намного ниже, чем у неплодоносящих деревьев. Запасы крахмала осенью и в начале зимы явно уменьшались из-за потребностей растущих плодов. Предпочтительное использование углеводов на репродуктивный рост было показано Риго и Дависом (1959) для пяти сортов персиковых деревьев, плодоносящих в разное время. У каждого сорта содержание крахмала в плодоносящих ветвях уменьшалось перед сбором урожая плодов и увеличивалось после него. На запасы углеводов сильно влияет периодичность плодоношения. Например, содержание крахмала в листьях мандариновых деревьев, образующих немного плодов, было примерно вдвое выше, чем в листьях деревьев с обильным урожаем. Молодые и старые плоды на одном и том же дереве также конкурируют за углеводы. В случаях, когда на деревьях оставляли зрелые апельсины, в течение нескольких месяцев до сбора урожая отмечались четкая периодичность плодоношения и уменьшение размеров очередного урожая. Происходило это, вероятно, потому, что поздний сбор уменьшал количество углеводов, которыми располагает дерево для последующего урожая.
На пятую или шестую неделю после полного цветения содержание крахмала в плодоносящих плодовых веточках яблони быстро падало приблизительно до 1/3 содержания в неплодоносящих веточках
11.3. СОДЕРЖАНИЕ КРАХМАЛА В ПЛОДОНОСЯЩИХ
И НЕПЛОДОНОСЯЩИХ ПЛОДОВЫХ ВЕТОЧКАХ ЯБЛОНЬ СОРТА ЛАНДСБЕРГЕР-РЕНЕТ В 1969 Г., МГ/Г СЫРОГО ВЕСА (Grochowska, 1973)
Плодовые веточки Время после полного цветения, недели
-2 0 3 6 9 12 15 18
Неплодоносящие 3,56 1,96 2,81 4,50 4,48 4,46 4,51 4,19
Плодоносящие 3,59 2,08 3,05 1,19 3,10 2,29 2,65 3,78
Разность 0,03 0,12 0,24 3,31 1,38 2,17 1,86 0,41
Примечание. Наименьшая достоверная разность при 1%-ном уровне значимости равна 0,29.
Передвижение веществ
313
Даты
Рис. 11.4. Кривые роста пяти листьев винограда. Стрелками указаны даты начала оттока углеводов из листьев (Hale, Weaver, 1962)
Рис. 11.5. Три этапа развития побега винограда. Стрелками показано основное направление передвижения ассимилятов из различных листьев (Hale, Weaver, 1962)
(табл- 11.3). Это быстрое истощение запасов совпадает с сильным увеличением содержания гиббереллина в развивающихся семенах. Продолжительный приток гормонов из растущих семян, по-видимому, стимулирует передвижение углеводов в быстро растущие плоды.
Наряду с мобилизацией резервов, репродуктивные структуры используют также большие количества продуктов текущего фотосинтеза. Мокицуки (1962) подвергал действию 14СС>2 плодоносящие яблони через различные промежутки времени после цветения. В начале вегетационного периода основная масса ассимилятов поступала в вегетативные ткани, но в конце лета большая часть их передвигалась в плоды. Хансен (1967) показал, что яблоки получали до 90% 14С от соседних листьев. Косточковые плоды особенно интенсивно потребляют углеводы в процессе развития. Например, персики и абрикосы являются такими мощными потребителями, что они успешно конкурируют за продукты текущего фотосинтеза с расположенными рядом развивающимися листьями. Это контрастирует с ситуацией у цитрусовых, где более мощными потребителями являлись молодые, быстро растущие вегетативные ткани.
По данным Халя и Вавера (1962), у быстро растущих побегов винограда передвижение углеводов протекало по типу, описанному для свободно растущих побегов, за исключением того, что оно было видоизменено развитием кистей плодов (рис. 11.4 и 11.5). Вначале молодой лист получал углеводы от листьев, расположенных ниже. Когда он
314
Глава II
Рис. 11.6. Направление передвижения ассимилятов из листьев карии пекан в различные моменты вегетационного периода. Тип 1 - ассимиляция без оттока; тип П - передвижение в обоих направлениях; тип Щ - базипетальное передвижение (Davis, Sparks, 1974). Этапы развития:
1 — развитие побегов; 2 — опыление; 3 — завязывание плодов; 4 — увеличение плодов; 5 — затвердевание скорлупы; 6 - развитие ядра; 7 - созревание плодов;
8 — листопад
разрастался наполовину, начинался отток углеводов из него в новые и недоразвитые вышерасположенные листья. Этот транспорт, направленный строго вверх, продолжался лишь 1—2 дня, после чего часть продуктов фотосинтеза передвигалась также вниз, к плодам. Такой характер передвижения сохранялся только 2—3 дня, а затем транспорт происходил целиком' вниз. Главную функцию листа - снабжение растущих кончиков побегов углеводами — теперь принимали на себя молодые листья, расположенные ближе к верхушке побега.
На ранних этапах развития побегов винограда, когда ассимиляты передвигались в кончик побега из первого листа, кисть цветков была слабым потребителем, и такое положение сохранялось до конца цветения. Однако вслед за завязыванием плодов (около 10 дней после полного цветения) кисть ягод становилась доминирующим потребителем. В это время кончик побега делался несколько более слабым потребителем углеводов, так как скорость его роста уменьшалась. Когда начиналось развитие плодов, большие количества углеводов передвигались в кисть из листьев, расположенных как выше, так и ниже.
По исследованиям Дэвиса и Спаркса (1974), в удлиняющихся плодоносящих побегах карии пекан передвижение углеводов из различных листьев изменялось в зависимости от характера развития вдоль оси побега и от разной мощности плодов как потребителей углеводов (рис. 11.6). Начиная с раскрытия почек и в течение разрастания листвы углеводы из листьев не передвигались. Незрелые листья частично снабжали сами себя, но также зависели и от углеводов, накопленных в предыдущем сезоне. Полностью развившийся лист начинал поставлять углеводы вверх и вниз, пока не созревал вышерасположенный лист (рис. 11.6, тип II), после чего углеводы из нижнего листа передвигались только вниз (рис. 11.6, тип Ш). Этот характер передвижения сохранялся по мере последовательного созревания листьев, все выше расположенных на стебле, и в конце концов все листья отдавали углеводы. В начале развития побега карии из многих листьев происходил отток углеводов вверх и вниз, свидетельствуя о том, что как незрелые листья, так и женские цветки были мощными потребителями продуктов текущего фотосинтеза. Доля
Передвижение веществ
315
листьев, поставлявших углеводы в обоих направлениях, во время разрастания плода намного уменьшалась. Однако во время быстрого развития ореха углеводы из нижних листьев вновь передвигались как вверх, так и вниз^ После созревания плода происходил еще один сдвиг и все листья отдавали'углеводы только вниз. В тот год, когда плодоношения не было или оно было слабым, и после полного развития всех листьев на ветке преобладало передвижение углеводов вниз.
На некоторых этапах развития репродуктивные структуры голосеменных растений являются очень мощными потребителями углеводов. По данным Дикмана и Козловского (1970, 1973), шишечки сосны смолистой в первый год развития растут мало и, следовательно, являются относительно слабыми потребителями углеводов. На второй год развития быстро растущие шишки потребляют углеводов больше, чем растущие побеги, с которыми они конкурируют. Мобилизация углеводов из старой, хвои репродуктивными структурами (однолетними шишечками и двухлетними шишками) почти в 3 раза превышала мобилизацию углеводов побегами. Характер источников углеводов для роста двухлетних шишек подвержен сезонным изменениям. В начале вегетационного периода важную роль играют запасные углеводы, потому что в штате Висконсин сухой вес шишек начинает увеличиваться в середине апреля, когда еще холодно и интенсивность фотосинтеза невелика. К маю шишки получают продукты текущего фотосинтеза. В начале и середине периода роста шишек наиболее важным источником ассимилятов для них является однолетняя хвоя (та, которая развилась в ходе предыдущего вегетационного периода), а 2 и 3-летняя хвоя играет все менее важную роль. Передвижение углеводов в шишки из хвои, развивающейся в текущем году, приобретает значение лишь в конце вегетационного периода после ее созревания. До того времени новая хвоя удерживает большую часть фиксированного ею 14С. Значение фотосинтеза самих зеленых шишек сравнительно невелико.
РЕГУЛЯТОРЫ РОСТА. Регуляторы роста всех классов, за исключением этилена, обнаружены как в ксилеме, так и во флоэме. Передвигаются они, по-видимому, на большие расстояния. Меченые регуляторы роста передвигались из листьев с такой же скоростью, что и другие органические соединения, а вещества, стимулирующие цветение, быстро переходят из листьев в кончики стеблей. Ауксин, гиббереллин, абсцизовая кислота и цитокинины (см. главу 15) передвигаются из корней по ксилеме. Таким образом, нет сомнений в наличии транспорта регуляторов роста на большие расстояния. Однако физиологическое значение такого дальнего транспорта неясно, так как регуляторы роста могут образовываться во всех органах растений. Поэтому представляется возможным, что синтезом in situ объясняется большинство результатов, приписываемых транспорту из корней и других отдаленных тканей. Для оценки значения транспорта на большие расстояния необ-димо дальнейшее изучение источников и потребителей.
ГЕРБИЦИДЫ. Мобильность отдельных гербицидов в растениях различная. Передвижение их может происходить через симпласт или апо-пласт. В то время как 2, 4-дихлорфеноксиуксусная кислота и 2, 4, 5-трихлорфеноксиуксусная кислота передвигаются в основном по флоэме, триазины (например, симазин, атразин, пропазин) и производные мочевины свободно передвигаются только по ксилеме. Еще одна группа гербицидов, в том числе 2, 3,6-трихлорбензойная кислота, пикло-
316
Глава 11
рам и гидразид малеиновой кислоты, свободно перемещается как по ксилеме, так и по флоэме. Гербициды последней группы могут мигрировать из флоэмы в ксилему и циркулировать по растению наподобие фосфора. На передвижение гербицидов влияют многие факторы, включающие их химический состав и количество, вид растения, ткани, в которые их ввели, мощность различных потребителей и факторы, окружающей среды.
По флоэме гербициды перемещаются вместе с углеводами. Нанесенный на листья 2,4-Д проникает через кутикулу, поступает в сим-пласт, достигает в конце концов ситовидных трубок и далее движется с током ассимилятов к мощным потребителям, часто обходя зрелые ткани, являющиеся источниками ассимилятов. Крафт и Ямажук (1964) показали, что ^^С-меченый 2, 4-Д легко передвигался из зеленых (но не желтых) листьев. Передвижение зависит от мощности потребителей, т.е. от скорости роста различных тканей, поэтому иногда преобладает движение вниз к корням, а иногда в основном к растущим побегам, цветкам или плодам.
ВИРУСЫ. Вирусные инфекции возможны только при их прямом введении в определенные ткани, обычно при участии насекомого-переносчика. Все вирусы могут проникать в царенхимные клетки. Некоторые вирусы, по-видимому, адаптированы к определенным тканям, а другие (например, вирусы мозаики) могут инфицировать большинство живых клеток растения. У основной части более специализированных вирусов инфицирование зависит от флоэмной ткани, хотя некоторые, видимо, передвигаются по ксилеме. Передвижение инфицирующих агентов через клетки паренхимы или между ними и ситовидными элементами происходит, вероятно, через плазмодесмы, движение из одного ситовидного элемента в другой — через поры в ситовидных пластинках. Систематическое распространение возбудителей вирусных болезней по флоэме возможно двумя путями:. 1) вирионами, пассивно накапливающимися в молодых ситовидных элементах; 2) связанными с мембранами их промежуточными формами, которые реплицируются в ситовидных элементах или в клетках паренхимы, находящихся в контакте с ситовидными элементами, и в других тканях, поддерживающих репродукцию вирусов.
Возбудитель вирусной инфекции перемещается по флоэме с током ассимилятов. Поскольку флоэма пронизывает все растение, а транспорт происходит в соответствии с характером изменений потребителей, вирусная инфекция может легко распространиться по всему растению. Инфекция, вызванная вирусами, передвигающимися только по флоэме, не может распространяться за окольцованную часть ствола. В противоположность этому инфекция, вызванная вирусами мозаики, может обойти кольцо через клетки паренхимы в ксилеме окольцованной части ствола. Поскольку инфекционный возбудитель вирусной болезни, передвигаясь по растению, может оставить некоторые его части неинфици-рованными, вполне вероятно, что возбудитель переносится пассивно. Некоторые вирусные инфекции причиняют серьезный вред флоэме, другие нет.
Скорость распространения вирусных инфекций по различным тканям сильно варьирует. По клеткам паренхимы возбудитель передвигается
317
Передвижение веществ
очень медленно, но когда он достигает сосудистых тканей, движется намного быстрее. Транспорт по флоэме в листьях и стеблях происходит быстро; например, вирус курчавости в черешках свеклы движется со скоростью до 150 см/ч. Однако из корней перемещаемые по флоэме вирусы передвигаются очень медленно. Корни являются постоянными потребителями углеводов, поэтому передвижение из них по флоэме весной наиболее интенсивно протекает в тех корнях, которые содержат запасы углеводов. Вирусные инфекции, распространяющиеся с транспирационным током, могут передвигаться по ксилеме со скоростью приблизительно до 30 см/мин.
ПЕРЕДВИЖЕНИЕ ВЕЩЕСТВ ОТ РАСТЕНИЯ К РАСТЕНИЮ
Многие древесные растения образуют с другими растениями ассоциации, характер которых может быть от паразитического до симбиотического. Способность болезнетворных грибов получать углеводы от растений-хозяев подверждается в книгах по фитопатологии и выходит за пределы нашего труда. Ниже будут кратко рассмотрены несколько примеров растительных ассоциаций, в которых происходит перенос углеводов, воды, минеральных веществ и различных других соединений от одного растения к другому.
СИМБИОТИЧЕСКИЕ И ПАРАЗИТИЧЕСКИЕ АССОЦИАЦИИ. Как подчеркивалось в главах 2 и 10, между многими древесными растениями и микоризными грибами существуют симбиотические отношения. При этом грибы получают углеводы и воду от хозяина и играют важную роль в усилении поглощения растением-хозяином минеральных веществ. Выдерживая сеянцы в атмосфере с 14qq2, Мелин и Нильсон (1957) показали, что меченые продукты фотосинтеза быстро передвигались в ассоциированные с растением микоризные грибы. Хотя имеются некоторые свидетельства в пользу передвижения углеводов также и из микоризных грибов к хозяину, вероятно, превалирует передвижение от хозяина к грибам.
В ряде работ показано, что микориза усиливает поглощение и перенос минеральных веществ к хозяину. Например, работа Мелина и Нильсона (1950) продемонстрировала, что гифы грибов поглощали радиоактивный фосфор и переносили его к сеянцам сосны. Позже (1955) эти авторы показали, что грибы могли переносить и другие вещества из среды к сеянцам. Режим минерального питания у микоризных ассоциаций древесных растений проанализировал Бовен (1973). Корни с микоризой описаны в главе 2, их роль в поглощении минеральных веществ - в главе 10.
Некоторые содержащие хлорофилл растения, такие, как омела, паразитируют на древесных растениях, получая от них углеводы и воду. Карликовые виды омелы ( Arceuthob ium spp.) обычно считают способными к ограниченному фотосинтезу, но Миллер и Токер (1975) показали, что у" Arceuthobium tsugense фотосинтез давал лишь часть углеводов, необходимых для дыхания и роста. Гуль и Леонард (1964) показали, что карликовые омелы получают большую часть углеводов от хозяев, тогда как зеленые омелы (Phoradendron spp.) сами синтезируют
318
Глава 11
большую часть потребляемых ими углеводов и лишь очень небольшие количества получают от хозяина. Эти исследователи показали также, что фиксированный углерод как у карликовых, так и у зеленых омел не передвигается к хозяину.
Омелы получают от растения-хозяина также воду, на что указывает поглощение ими красителя, введенного хозяину. Марк и Рид (1971) показали, что для побегов карликовой омелы градиент водного потенциала благоприятен, даже если растение-хозяин испытывает сильный водный стресс. Карликовые виды омелы считают одним из наиболее опасных болезнетворных агентов для лесных пород на западе США. Они не только замедляют рост деревьев, но часто и убивают их. Механизм замедления роста хозяина омелами не совсем выяснен, но в него входит, вероятно, лишение хозяина углеводов, воды и минеральных веществ. Такое лишение дерева материалов, необходимых для роста, приводит в конечном счете к повреждению кроны и уменьшению фотосинтезирующей поверхности до размеров, недостаточных для поддержания роста дерева-хозяина.
СРАСТАНИЕ КОРНЕЙ. В, главе 2 отмечалось, что значительное передвижение воды, углеводов, минеральных веществ, гормонов, биоцидов, микроорганизмов и грибных спор происходит от одного дерева к другому через сросшиеся корни. Поэтому многие экземпляры деревьев не являются полностью независимыми.
Живые пни. Большинство пней погибает через 1—2 года после рубки дерева. Однако сообщество деревьев может быть так эффективно связано сросшимися корнями, что если срубить все деревья, кроме одного, то оставшиеся корневые системы могут выжить благодаря поступлению углеводов и регуляторов роста из кроны этого единственного дерева. У пней, сросшихся с живыми деревьями, часто длительное время продолжается камбиальный рост. По данным Айса (1972), в выборочно вырубленных насаждениях дугласии 45% пней росли после рубки, а многие оставались живыми даже через 32 года после рубки. От деревьев-доноров 65% живых пней находились менее чем в 2 м, но некоторые доноры поддерживали существование живых пней на расстояниях почти 10 м. Распределение радиального прироста у живых пней и у сросшихся деревьев со срезанной верхушкой было сходным. До высоты пня прирост ксилемы был обычно нерегулярным, наиболее сильным — выше основных мест срастаний. Выше по стволу отмечался равномерный прирост по всей окружности, но с высотой постепенно уменьшался.
Значение передвижения питательных веществ между деревьями варьирует в зависимости от класса крон сросшихся партнеров, Айс (1972) обнаружил 4 типа связи камбиального роста пней и деревьев-доноров:
1. Когда срубали кодоминантное дерево в соединении доминантного дерева с кодоминантным, камбиальный рост пня медленно снижался, и за 3—4 года доходил до 20-30% первоначального уровня. Рост камбия доминантного донора немного усиливался, что отражало уменьшение конкуренции.
2. Если сросшиеся деревья были сходного размера и одного класса (например, оба были доминантными, или когда срубали доминантное дерево в соединении доминантного с кодоминантным), камбиальный рост пня быстро снижался. После первого года следовало несколько лет слабого роста камбия. В дальнейшем камбиальный рост пня усиливался и достигал приблизительно уровня 20—30% до
?>О1Э
Передвижение веществ 319
рубки. Камбиальный рост дерева*донора впоследствии усиливался, возможно, благодаря пониженной конкуренции.
3. При рубке доминантного дерева в соединении доминантного с угнетенным или доминантного с промежуточной формой, пень жил лишь несколько лет. Камбиальный рост угнетенного дерева-донора снижался, пока пень был еще жив, и медленно увеличивался после гибели пна.
4. Если пень от срубленного доминантного дерева из соединения доминантного .дерева с угнетенным оставался живым, камбиальный рост угнетенного дерева-донора был ограниченным. Снижение конкуренции сопровождалось усилением роста большинства остальных деревьев, независимо от того, были ли они сросшимися или нет.
Эти эксперименты показали, что сросшиеся корнями угнетенные деревья получают часть или все необходимые для корней нижней части ствола углеводы от доминантных деревьев, с которыми они срослись. Такие отношения могут отсрочить гибель сросшихся корнями угнетенных деревьев, растущих в густых древостоях.
Передвижение гербицидов. Многие гербициды легко передвигаются от одного дерева к другому через сросшиеся корни. Например, на плантации 30-летних деревьев сосны веймутовой 43% деревьев, не подвергавшихся обработке гербицидом, погибли из-за передвижения сульфамата аммония от обработанных этим химикатом деревьев через срастания корней. В сходной ситуации погибала половина остававшихся деревьев сосны смолистой. С учетом этого явления в лесоводстве Айс (1972) подчеркивает, ,что для видов, у которых легко срастаются корни, использовать арборициды при прореживании и прочистке можно только в молодых насаждениях, у которых еще не произошло обширное срастание корней. Наоборот, передвижение гербицидов через сросшиеся корни может способствовать устранению нежелательных видов.
Передача болезнетворных и гнилостных организмов. Возбудители некоторых наиболее опасных болезней деревьев перемещаются через сросшиеся корни. К ним относятся; Fomes pini у дугласии, Armillaria mellea у ели, Endothia gyrosa у дуба черного, Poria weirii у дугласии., Ceratostomella ulmi у вяза, Ceratocystis fogacearum у дубов. Сросшиеся пни были источником заражения Polyporus schwemitziiдерева, служившего донором углеводов. Была обнаружена также связь между числом деревьев вяза, пораженных голландской болезнью, и расстоянием между ними. Ввиду большого распространения инфекции через сросшиеся корни деревьев, расположенных ближе 8 м друг от друга, многие авторы рекомендуют размещать вязы на расстоянии не менее 9 м.
НЕСРОСШИЕСЯ РАСТЕНИЯ. Перенос различных веществ между близко расположенными деревьями возможен, даже если их корни не срослись. Например, гербициды иногда переносятся к деревьям, корни которых не срослись. Введенные в пни одних видов радиоактивные изотопы впоследствии обнаруживались в других видах, с которыми они не были связаны через сросшиеся корни. Смит (1976) предполагает, что корневые выделения играют роль в круговороте минеральных веществ. Такие наблюдения служат доказательством важности выделения корнями минеральных веществ и последующего поглощения их другими растениями. Аналогично эти вещества переносятся грибами, образующими микоризу, и ризосферными организмами.
Многие аллелопатические соединения, освобождающиеся из деревьев и других растений вследствие вымывания, улетучивания, выделения
Глава 11
320
и разложения непосредственно или в результате действия микроорганизмов, часто подавляют прорастание семян и рост прилегающих растений. Например, в калифорнийских пустынях проростки креозотового куста погибают от соседства со взрослыми растениями того же вида, вероятно, из-за токсического действия веществ, выделяемых корнями взрослых растений.
МЕХАНИЗМ ФЛОЭМНОГО ТРАНСПОРТА
Одна из наиболее спорных и нерешенных проблем в физиологии растений — вопрос о механизме передвижения растворенных веществ по флоэме. Вардлав (1974) озаглавил свой обзор по передвижению веществ так: ’’Флоэмный транспорт: физический, химический или невозможный”. Данная проблема трудна ввиду большого объема транспорта растворенных веществ. Она еще более усложняется тем, что вещества могут двигаться по флоэме вниз или вверх и вместе с сахарами передвигается множество веществ, таких, как минеральные элементы, регуляторы роста, витамины и вирусы. В этом разделе будут кратко рассмотрены те аспекты анатомии флоэмных тканей, которые важны для понимания механизмов передвижения. Будут также обсуждены различные гипотезы, выдвинутые для объяснения дальнего транспорта растворенных веществ по флоэме.
СИТОВИДНЫЕ ТРУБКИ ПОКРЫТОСЕМЕННЫХ. У покрытосеменных растений ряд проводящих элементов флоэмы - членики ситовидных трубок соединены концами с образованием длинных ситовидных трубок. Термин ’’сито” относится к торцевым стейкам, пронизанным группой пор, через которые соединяются протопласты смежных члеников ситовидных трубок. Ситовидные пластинки — части стенки, несущие ситовидные участки с большими порами, — обычно находятся на концах члеников ситовидных трубок.
Протопласты молодых члеников ситовидных трубок содержат все клеточные компоненты, встречающиеся в норме у живых клеток растений. На заключительных этапах дифференциации членики ситовидных трубок теряют многие из этих компонентов, в том числе тонопласт, микротрубочки, рибосомы и диктиосомы. У одних видов ядро в ходе дифференциации полностью исчезает, у других остатки дегенерировавшего ядра остаются долгое время. Членики ситовидных трубок у многих видов содержат белковое вещество, называемое Р-белком или слизью, которое может встречаться в виде тяжей, распространяющихся по всему просвету клетки, или занимать строго пристенное положение.
Каллоза в ситовидных участках зрелых проводящих ситовидных трубок имеется в небольших количествах или отсутствует. По мере старения ситовидных трубок количество каллозы увеличивается и после их отмирания ситовидные пластинки могут содержать очень большие количества каллозы. Постепенно она исчезает из старых, неактивных клеток. Оболочки члеников ситовидных трубок обычно рассматриваются как первичные. Членики ситовидных трубок почти неизменно сопровождаются паренхимными клетками (клетками-спутниками), с которыми ситовидные трубки связаны структурно и функционально.
Передвижение веществ
321
СИТОВИДНЫЕ КЛЕТКИ ГОЛОСЕМЕННЫХ. Проводящие элементы флоэмы голосеменных растений, названные ситовидными клетками, по форме и размерам похожи на трахеиды. В процессе дифференциации ситовидные клетки дезорганизуются и теряют многие из своих компонентов. В них нет Р-белка. Оболочки у ситовидных клеток только первичные, за исключением растений семейства?inасеае, у которых имеется также вторичная оболочка. Аналогом клеток-спутников покрытосеменных растений у голосеменных служат белковые клетки, называемые также клетками Страсбургера.
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ ПРОВОДЯЩИХ ЭЛЕМЕНТОВ. У большинства видов покрытосеменных растений ситовидные элементы живут недолго и участвуют в передвижении веществ только в течение того сезона, когда они образовались. У нескольких родов они остаются живыми и функционируют более 1 года. Например, ситовидные трубки винограда и тюльпанного дерева функционируют не менее двух сезонов. При этом в противоположность активности в течение первого лета отсутствует активность зимой. Ситовидные трубки липы могут оставаться функционирующими 5-10 лет. У покоящихся, неактивных ситовидных трубок на ситовидных пластинках и ситовидных участках находятся обильные отложения каллозы. Весной, когда ситовидные трубки становятся активными, каллоза постепенно растворяется. Реактивация флоэмы начинается прежде всего рядом с камбием. Флоэму, становящуюся активной, можно отличить от покоящейся по тонким отложениям каллозы и заметному образованию Р-белка у первой. После отмирания ситовидных элементов связанные с ними клетки-спутники или белковые клетки также погибают, что указывает на тесную взаимосвязь между ними.
У ряда видов покрытосеменных и голосеменных передвижение углеводов может наблюдаться ранней весной, так как первыми функционирующими ситовидными элементами являются те, которые последними возникли в предыдущем вегетационном периоде и перезимовали в виде недифференцированных образований. Они достигают зрелости в начале весны и могут транспортировать углеводы до реактивации камбия. Такие перезимовавшие ситовидные элементы не теряют способности к передвижению веществ до тех пор, пока не образуются новые, функционирующие ситовидные трубки. У разных вечнозеленых двудольных и у голосеменных часть флоэмы остается активной, транспортируя углеводы в течение двух вегетационных периодов. У некоторых покрытосеменных, например у клена ясенелистного и белой акации, неактивные ситовидные элементы и сопровождающие их клетки спадаются, но у других, таких как тюльпанное дерево, орех и липа, их структура остается без изменений. Ситовидные трубки древесных однодольных должны оставаться функционирующими в течение всей жизни этих растений.
ДВИЖУЩИЕ СИЛЫ ТРАНСПОРТА. Наиболее важные соединения, такие, как сахароза и, возможно, некоторые другие сахара, накапливаются во флоэме в концентрациях, превышающих концентрации в окружающих тканях. Это объясняется ’’нагрузкой” флоэмы, т.е. выделением растворенных веществ во флоэму путем активного транспорта с затратой метаболической энергии. Для объяснения движущей силы, вызывающей передвижение растворенных веществ после их попадания
322
Глава 11
в ситовидные трубки, выдвинуто несколько гипотез. Они основаны в общем на одном из двух противоположных представлений: 1) растворенные молекулы передвигаются за счет энергии метаболизма независимо от воды; 2) растворенные молекулы движутся с водой как перетекающий раствор.
Диффузия. Передвижение газов и даже растворенных веществ на расстояния в несколько миллиметров можно объяснить диффузией, но за счет этого процесса нельзя отнести передвижение веществ на многие метры. В ряде опытов показано, что во флоэме существуют градиенты концентрации сахарозы, что ближе к источникам флоэма содержит сахарозы больше, чем ближе к потребителям, и что передвижение сахарозы происходит по градиенту концентрации. Однако эти градиенты , слишком малы, чтобы объяснять известную скорость передвижения , сахарозы диффузией.
Движение протоплазмы. Имеется предположение, что движение j протоплазмы может участвовать в транспорте растворенных веществ. * Куртис (1935) придерживается этой теории на том основании, что она предусматривает одновременное передвижение различных веществ в разных направлениях, а также учитывает потребности живых клеток. Главное возражение против механизма передвижения веществ на основе движения протоплазмы заключается в том, что такое движение не отвечает количественным требованиям для объяснения известной скорости транспорта на большие расстояния. К тому же существование того типа движения протоплазмы, который встречается в некоторых клетках, не подтверждено для зрелых ситовидных элементов.
Электроосмос. Электроосмотический ток часто принимали в качестве движущей силы передвижения растворов. Спаннер (1958), например, выдвинул теорию о передвижении ионов калия, приводимом в действие; энергией метаболизма. Для этого необходимо, чтобы у значительного числа ситовидных пластинок ряда проводящих элементов поры были закупорены Р-белком и чтобы ситовидные пластинки представляли собой мембраны, хорошо приспособленные для развития электроосмотических сил. Вероятно, разница электрических потенциалов поперек ситовидной пластинки поддерживается активным поглощением ионов калия через мембрану ситовидной трубки выше пластинки и потерей их ниже ситовидной пластинки. Энергию для передвижения, по-видимому, поставляют макроэнергетические соединения, например АТФ, которые поступают из клеток-спутников.
Против теории электроосмотического тока выдвинут ряд возражений. К’наиболее важным из них относятся: 1 ) этот процесс не может обеспечить транспорт как катионов, так и анионов, которые, как известно, также передвигаются по флоэме; 2) количество находящегося в обращении К+ слишком велико; 3) система требует для рециркуляции калия слишком много энергии. Например, у деревьев для рециркуляции необходимого количества калия потребовалось бы больше энергии, чем имеется вообще в распоряжении всей флоэмы.
Трансцеллюлярные тяжи и цитоплазматический насос. Тин (1962) обнаружил тяжи протоплазмы, протянутые по всей длине ситовидных* элементов. Он наблюдал также частицы, передвигающиеся в обоик направлениях по ситовидным элементам и через поры в ситовидных
Передвижение веществ
323
пластинках в прилегающие ситовидные элементы. В 1969 г. этот исследователь выдвинул предположение, что тяж с диаметром приблизительно 5 мкм состоит из нескольких трубочек диаметром около 1 мкм, причем остальное его пространство заполнено жидкостью, содержащей подвижные частицы. По его представлениям, нити белка в стенке тяжа испытывают ритмические сокращения и растяжения, перемещая содержимое трубочек путем перистальтического нагнетания.
О существовании трансцеллюлярных тяжей велись многочисленные дискуссии. Согласно Эверту (1977) в норме Р-белок не существует в виде трансцеллюлярных тяжей. 'Эверт с соавторами (1969) сделали вывод, что тяжи Р-белка фибриллярной сети представляют собой артефакты, вызванные повреждением клеток. Отсутствие доказательств о наличии трансцеллюлярных тяжей в члениках ситовидных трубок, основанных на изучении их развития, являются серьезным аргументом против теории Тина.
Ток под давлением. Мюнх (1930) высказал предположение о существовании тока растворенных веществ через флоэму от подающего конца (источника) к получающему концу (потребителю). По этой теории требуется: 1) чтобы ситовидные трубки имели избирательно проницаемые мембраны и функционировали как осмотические системы; 2) они должны допускать продольный ток; 3) чтобы на подающем конце имелся ’’нагружающий” механизм для растворенных веществ, а на потребляющем конце — механизм ’’разгрузки”; 4) должен существовать градиент уменьшающегося тургорного давления от источника к потребителю для того, чтобы действовать в качестве движущей силы.
В современной форме эта теория предполагает, что в листьях паренхимные клетки флоэмы накапливают сахара, получаемые из клеток мезофилла и транспортируют их в ситовидные трубки. Такая ’’нагрузка” флоэмы требует затрат метаболической энергии. Высокая концентрация сахаров снижает осмотический потенциал в ситовидных трубках, вызывая приток воды из смежных тканей и развитие высокого тургорного давления. Удаление сахаров (’’разгрузка” флоэмы) различными потребителями, такими, как меристемы и запасающие ткани, снижает осмотический потенциал в ситовидных трубках. В результате происходят вытекание воды и массовое перетекание растворенных веществ по ситовидным трубкам от источника к потребителю. Таким образом, от энергии метаболизма зависит не действительное передвижение от источника к потребителю, а процессы ’’нагрузки” и ’’разгрузки”, делающие передвижение чувствительным к изменениям температуры и к ингибиторам дыхания. Милбурн (1975) проанализировал несоответствия в теории тока под давлением и высказал, мысль о том, что она реально применима только к отдельным ситовидным трубкам. Однако при существовании мощного потребителя ситовидные трубки работают в унисон как единая система.
Среди возражений, выдвинутых против этой теории, укажем на следующие: быструю закупорку слизью или каллозой ситовидных пластинок в разрезанных тканях; трудность объяснения передвижения в обоих направлениях; затруднения при работе такого механизма у растений в условиях водного стресса, а также сомнения в существовании градиента тургорного давления во флоэме. В настоящее время считается,
124 Глава 11
•гго закупорка ситовидных пластинок является артефактом, вызванным рн’цюзанием флоэмы. Очевидно, эта теория не допускает одновременного передвижения в обоих направлениях по ситовидным трубкам, но допускает возможность тока в двух направлениях по различным пучкам и, может быть, по различным ситовидным трубкам. Проблему дну стороннего тока обсуждали Циммерман и Милбурн (1975). Эта юория предусматривает также возможность изменения направления пе-|кднижения. Например, очень молодой лист функционирует как по1рсбитель, поскольку он использует сахара, но позже он начинает служить источником, производящим сахара. Запасающие ткани могут функционировать как потребители к концу вегетационного периода, но становятся источниками углеводов для роста корней почек и камбия ранней весной, до того, как появляются продукты фотосинтеза.
Водный стресс уменьшает скорость передвижения, но, как утверждаю! исследователи, некоторое передвижение наблюдается даже в рас- I гениях с увядшими листьями до тех пор, пока существуют потребители ] органических веществ, т.е. пока происходит рост. Кажется вероятным, что массовое перетекание может продолжаться, пока осмотический гкпенциал флоэмного сока ниже водного потенциала ксилемы. Имелись также сомнения относительно существования градиента тургорного давления в ситовидных трубках. Несомненно, однако, что сок выступает через надрезы, сделанные во флоэме различных травянистых и древесных видов. Кроме того, много литров сока в день часто вытекает hi стеблей пальм и агав при удалении их соцветий. По исследованиям Циммермана и Милбурна (1975), поведение тлей также указывает на i vшествование давления во флоэме. Некоторые виды тлей вводят свои xiНютки в ситовидные трубки и питаются флоэмным соком, который поступает в них под давлением и через тела насекомых выделяется в виде медвяной росы. Когда тлю отделяют от хоботка, последний служит трубкой, через которую часто в течение нескольких дней продолжает вытекать флоэмный сок, в отличие от надрезов, которые могут быстро закупориваться. Такие разносторонние доказательства оставляют мало сомнений в том, что массовое перетекание растворенных веществ происходит через флоэму, хотя относительно некоторых важных деталей in лается много неясного.
И ГУЛЯЦИЯ ФЛОЭМНОГО ТРАНСПОРТА
Понимание механизма транспорта по флоэме имеет важное значение, но еще важнее понять, как контролируется распределение метаболитов. Другими словами, чем регулируется количество питательных веществ, доставляемых в различные конкурирующие органы и ткани, и как осуществляются изменения, необходимые на разных стадиях развития? В прорастающих семенах питательные вещества передвигаются И’1 запасающих тканей в различные зоны роста, в том числе в развивающиеся листья до начала в них фотосинтеза. Затем направление транспор гп меняется и продукты фотосинтеза передвигаются из листьев к кончикам корней и стеблей. У зрелых листопадных древесных расте
Передвижение веществ
325
ний весной питательные вещества сначала передвигаются из запасных тканей ствола и ветвей в кончики стеблей. Затем, по мере разраста-ния новых листьев, последние поставляют питательные вещества еще более новым листьям, камбию и корням. Развивающиеся плоды и семена становятся мощными потребителями, к которым питательные вещества передвигаются за счет вегетативного роста. После прекращения роста и созревания плодов и семян питательные вещества начинают накапливаться в запасающих органах. В разгар вегетационного периода относительная мощность различных потребителей часто распределяется следующим образом: плоды и семена молодые листья и кончики стеблей зрелые листья камбий корни запасающие ткани.
Решающим является вопрос: что определяет относительную мощность различных потребителей и тем самым контролирует распределение питательных веществ между конкурирующими органами и тканями? Представляется что флоэма обладает значительной избыточной способностью к транспорту веществ, так как частичное удаление почти не влияет на рост растения. Кроме того, процесс передвижения веществ происходит в широком диапазоне температур и водного стресса.
Распределение питательных веществ, по-видимому, контролируется в первую очередь масштабами их поступления и относительной величиной различных потребителей. При этом большие потребители при получении питательных веществ более активны по сравнению с меньшими. Важное значение имеет также расстояние от источника до потребителя: обычно потребители снабжаются от ближайшего источника. В результате молодые листья имеют преимущество перед камбием ствола, а в нижнюю часть ствола питательных веществ доставляется меньше, чем в верхнюю, несущую крону. Корни оказываются в наиболее невыгодном положении по мере перемещения фотосинтезирующей поверхности вверх с ростом дерева. Следует помнить, что корни могут быть и наиболее мощными потребителями в период, когда не растут другие части дерева. Например, корни начинают расти весной, до начала роста побегав и стволов. Следовательно, в начале сезона корни являются самыми мощными потребителями, и к ним направляются большие количества текущих продуктов фотосинтеза (см. рис. 11.3). К тому же корни деревьев умеренной зоны продолжают расти позже, чем побеги и стволы (см. главу 3). Таким образом, в конце лета — начале осени и вновь ранней весной корни являются более мощными потребителями, чем побеги и стволы.
Считают, что в определении мощности потребителей играют роль регуляторы роста, или гормоны. Например, передвижение метаболитов обычно усиливается при введении цитокинина или гиббереллина в определенный участок. Неясно, действуют ли гормоны прямо, например, ускоряя ’’нагрузку” или ’’разгрузку” ситовидных трубок, или косвенно, воздействуя на относительную мощность источников и потребителей. В некоторых случаях развитие шюда, по-видимому, увеличивает интенсивность фотосинтеза, но неизвестно, вызвано ли это увеличение использованием питательных веществ развивающимся плодом или гормональным стимулом из плода. Возможно, имеются оба пути.
ЗНАЧЕНИЕ ВОДЫ И ПРОЦЕСС ТРАНСПИРАЦИИ
Распространение деревьев на земной поверхности, где температурные условия делают возможным рост растений, зависит главным образом от водоснабжения. На большинстве лугов и в пустынях могли бы расти леса при условии достаточного количества и благоприятного сезонного распределения осадков. На других больших пространствах произрастают только редкие древостои вследствие ограниченного количества воды. Экологическое значение воды обусловлено ее физиологической ролью. Достаточное водоснабжение так же необходимо для успешного произрастания растений, как фотосинтез и другие биохимические процессы, участвующие в синтезе питательных веществ и их преобразовании в новые ткани. Важным моментом в водном режиме растений является поддержание достаточно высокого содержания воды и тургора, чтобы сделать возможным нормальное функционирование физиологических и биохимических процессов, связанных с процессами роста. Это находится под контролем относительной скорости поглощения и потери воды.
ЗНАЧЕНИЕ ВОДЫ. Значение достаточного водоснабжения для роста древесных растений показал Цанер(1968), работа которого содержит обширный обзор литературы. По сообщению этого автора, до 80 % изменений роста в толщину деревьев во влажных районах (и до 90 % в засушливых) можно отнести за счет разного количества осадков и водного стресса. Действительно, возможность описания прошлых климатических условий по ширине годичных колец деревьев (’’дендрохронология”) полностью доказана для определенных засушливых районов, а иногда и для влажных областей. (Более подробно этот вопрос будет рассмотрен в главах 13 и 16.)
Значение воды в жизни древесных растений можно показать, перечислив наиболее важные ее функции, которые можно разделить на четыре группы: 1. Вода важный компонент протоплазмы, составляющий 80—90 % сырой массы активно растущих тканей. 2. Вода — растворитель, в котором передвигаются газы, соли и другие растворимые вещества в клетки и из клеток и из одного органа в другой. 3. Вода участвует в качестве реагента в фотосинтезе и процессах гидролиза. 4. Вода необходима для поддержания тургора. Определенная степень тургесцентности имеет существенное значение для растяжения клеток, для открывания устьиц' и для поддержания формы молодых листьев и других слабо одревесневших структур.
Для полного понимания водного режима растений нужно принимать во внимание состояние почвенной и атмосферной влаги. Проанализируем вначале два взаимосвязанных аспекта водного режима растений. Один из них рассматривает водообмен клеток и тканей внутри растения, а другой — водный режим растения в целом. Водный режим растения включает в себя потерю воды в процессе транспирации, поглощение воды, подъем ксилемного сока и внутренний водный баланс дерева. В теплые солнечные дни транспирация часто превышает поглощение, вызывая водный дефицит, потерю тургора и конкуренцию за воду между различными частями дерева. Происходящее в результате этого перераспределение воды обусловливается водным потенциалом клеток и тканей.
Значение воды и процесс транспирации
327
ВОДНЫЙ РЕЖИМ КЛЕТКИ. Водный режим растений контролируется прежде всего водным режимом клеток, поэтому необходимо рассмотреть структуру и функции клетки в связи с передвижением воды.
Строение клетки. Живые растительные клетки состоят из протопластов, окруженных более или менее жесткими стенками. Эти стенки ограничивают изменение объема клеток, особенно в более старых тканях, где клеточные стенки лигнифицированы. В зрелой клетке протопласт состоит из большой центральной вакуоли, окруженной тонким слоем прижатой к стенке цитоплазмы, в которой расположены ядро и ряд других органелл, в том числе пластиды и митохондрии (рис. 12.1). Электронная микроскопия обнаруживает также многие другие структуры в цитоплазме: рибосомы, пероксисомы, диктиосомы, микротрубочки и сложную систему мембран, известную под названием эндо-
Рис. 12.1. Электронная микрофотография паренхимной клетки ксилемного луча сосны смолистой, показывающая ее основные структуры:
1 — клеточная стенка; 2 — вакуоль; 3 — хлоропласт; 4 — митохондрия; 5 — липидная капелька; 6 — микротельце; 7 — трахеида; 8 — ядро; 9 — эндоплазматическая сеть; 10 — плазмодесмы (Фото D, Neuberger)
328
Глава 12
плазматической сети. В активных клетках большие количества воды связаны с белковыми структурами. Поверхностные мембраны протопластов (плазмалемма и тонопласт) проницаемы для воды, но относительно непроницаемы для растворенных веществ. В результате ионы и растворимые органические вещества накапливаются в вакуолях, создавая осмотический потенциал в пределах от —5 до —50 бар. Часто протопласты соседних клеток соединяются друг с другом тяжами цитоплазмы (плазмодесмами), образуя непрерывную систему, так называемый симпласт. Размеры вакуолей колеблются от очень маленьких палочковидных или сферических структур в меристематической ткани до больших центральных вакуолей, занимающих более 50 % объема зрелых паренхимных клеток. Вакуоли заполнены водой, в которой растворены самые разнообразные ионы и органические молекулы.
Передвижение воды. Происходит оно по градиенту уменьшающейся свободной энергии или молекулярной активности, часто выражаемому в виде разности водных потенциалов. Если разность свободной энергии или потенциалов создается каким-либо внешним агентом, например давлением или силой тяжести, то передвижение называют массовым потоком. Примерами подобных процессов являются ток воды в трубах под давлением, вытекание водных струй под действием силы тяжести, подъем ксилемного сока в растениях, вызываемый испарением воды побегами. Если передвижение происходит в результате беспорядочного движения молекул за счет их собственной кинетической энергии, например при испарении, то этот процесс называется диффузией. Осмос — пример диффузии, вызываемой разностью водных потенциалов на двух сторонах мембраны, обычно вследствие разности концентраций поперек мембраны.
Терминология. Свободная энергия системы или компонента системы характеризует ее способность совершать работу. Абсолютную величину свободной энергии воды измерить трудно, но легко определить разность между свободной энергией чистой воды и воды в растворе, например в клеточном соке. Эту разность, которую раньше называли сосущей силой или дефицитом диффузионного давления, теперь обычно называют водным потенциалом. Вывод формулы этого потенциала обсуждается в работах Крамера (1969), Крамера с соавторами (1966) и Слуге- ц? ра (1967). Связь потенциала с терминологией, применявшейся раньше, можно показать следующим образом: 7
сосущая сила = осмотическое давление тургорное давление;
дефицит диффузионного давления = осмотическое давление - тургорное дав- , ление;
водный потенциал ф — осмотический потенциал + потенциал давления.
+ % ? (12-D
где _ потенциал воды в клетке; ф$ (иногда пишут Фц) ~ осмотический потенциал; фр — потенциал давления; иногда к этому добавляют матричный потенциал (обозначаемый также )
для учета силы, с которой вода связывается твердыми телами. Вели--
Значение воды и процесс транспирации
329
чины 0S и Фт отрицательны, а положительна,
за исключением редких случаев отрицательного тургорного давления или в ксилемном соке при натяжении сосудов. Сумма чисел в правой части уравнения отрицательна, за исключением полной тургесцентно-сти клеток, когда она становится равной нулю. Ситуации, складывающиеся в клетках при различном тургоре, можно показать на следующих примерах (величины выражены в барах):
ъ >
тургесцентная 0 — -18+(+18);
частично тургесцентная -10 = -19 + (+9); (12.2)
вялая -20 = -20 + (0).
Повышение с возрастанием тургора осмотического потенциала 0 s от —20 до —18 бар вызывается разбавлением клеточного сока вследствие того, что при этом увеличивается объем клетки. Передвижение водаг обусловливается водным потенциалом, но осмотический потенциал и потенциал давления также имеют большое значение. Потенциал давления, или тургорное давление, является особенно важным при растяжении клеток, а понижение осмотического потенциала может частично компенсировать уменьшение водного потенциала (см. главу 13).
ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ВОДНОМ РЕЖИМЕ РАСТЕНИЙ. Одним из важных представлений, способствующих пониманию водного режима растений, является рассмотрение передвижения воды через почву, в корни, по растению и выделение ее в атмосферу как ряда тесно взаимосвязанных процессов. Это представление иногда называют концепцией непрерывной системы почва—растение—атмосфера. Следует подчеркнуть необходимость рассмотрения всех аспектов водного режима при изучении водного баланса растений. Эта концепция приводит к представлению об аналогии передвижения воды в системе почва — растение — атмосфера с протеканием электрического тока в проводящей системе. Поэтому передвижение воды можно записать уравнением, аналогичным закону Ома:
разность водных потенциалов
поток =------------------------------------ . (12.3)
сопротивление
Эту концепцию можно применить к непрерывному стационарному потоку воды через растение следующим образом:
0 - 0
w почвы г поверхности корня поток = ----------------------------------- =
Г почвы
0 0 0 ксилемы- 0 клеток листа
в поверхности2<орня_ _ксилемьд _ _________________у_____________
, , г корня г ксилемы
0 ... 0
-Листа_______воздуха_____ (12.4)
Г + г
листа*
воздуха
330
Глава 12
Устьичные отверстия межклетники листа
мезофилл
Атмосфера
Кутикула
Переход из жидкой фазы в пароадриз-ную
Рис. 12.2. Упрощенная схема, показывающая ток воды через систему почва - растение - атмосфера по аналогии с электрическим током через ряд сопротивлений и емкостей
Ксилема
Накопление воды S тканях
, Запас почвенной влаги
Почва Корень
концепция полезна также пои
Концепция непрерывности способствует разработке полезной унифицированной теории, в которой передвижение воды через почву, корни, стебли и листья и ее испарение в атмосферу можно изучать с точки зрения движущих сил и сопротивлений, действующих на каждом этапе. Эта анализе способов, при помощи кото
рых разные растения и факторы внешней среды действуют на передвижение воды, влияя на движущие силы или на сопротивления, а иногда и на то и другое. Например, высыхающая почва вызывает как повышение сопротивления току воды по направлению к корням, так и уменьшение движущей силы или водного потенциала. Недостаточная аэрация увеличивает сопротивление току воды через корни, повышение температуры листьев и воздуха усиливает транспирацию, так как увеличивает градиент давления водяного пара (движущую силу) от листа к атмосфере. Закрывание устьиц повышает сопротивление диффузии водяного пара из листьев.
Концепция непрерывности облегчает также моделирование передвижения воды, как это показано, например, на рис. 12.2. Модели могут быть различными: от модели для отдельного устьица до модели для целого древостоя. Авторы моделей Микель, Шарп (1973),Вагонер, Рифшнайдер (1968) надеются, что они смогут предсказывать поведение растений в широком диапазоне условий внешней среды, однако до того, как это станет возможным, потребуется гораздо больше информации.
Следует предупредить читателей, что концепция непрерывности является очень упрощенной, так как она предполагает наличие условий устойчивого состояния, которые редко встречаются у растений. К тому же сложность заключается в том, что передвижение воды в жидкой фазе пропорционально разности водных потенциалов, тогда как движение в парообразной фазе пропорционально градиенту давления водяного пара. Более важно то, что при точном применении аналогии закона Ома к водному току в растениях сопротивление току должно изменяться в зависимости от его скорости, а это маловероятно.
ТРАНСПИРАЦИЯ
Транспирация — это потеря растениями воды в виде пара. В своей основе она является процессом испарения. Однако она до некоторой
Значение воды и процесс транспирации
331
степени контролируется структурой растения и поведением устьиц наряду с физическими факторами, которые влияют на испарение со свободной водной поверхности. Транспирация — наиболее важный фактор водного режима растений, так как испарение водь! создает энергетический градиент, который является причиной передвижения воды по растению. В связи с этим она определяет скорость поглощения воды и подъема ксилемного сока и вызывает почти ежедневно водный дефицит листьев. Одно отдельно стоящее дерево может испарять 200-400 л в день, а лиственный лес во влажных Южных Аппалачах теряет 42—55 см воды в год. Несколько сотен килограммов воды расходуют растения на каждый килограмм выработанного сухого вещества, причем около 95 % этой воды просто прокодит через растение и теряется при транспирации.
Иногда утверждают, что транспирация полезна^ потому что она охлаждает листья, вызывает подъем ксилемного сока и усиливает поглощение и передвижение минеральных веществ. Например, Гейтс (1965) придает большое значение охлаждающему эффекту транспирации. Все это отчасти правильно, но транспирация не является жизненно необходимым процессом. Листья, развивающиеся при полном солнечном освещении, редко повреждаются из-за повышения температуры, которое отмечается при ослаблении транспирации вследствие завядания. Использование воды растущими тканями могло бы вызывать медленный подъем ксилемного сока даже если бы не было транспирации. Вода передвигается к верхушкам высоких деревьев, а транспирация только увеличивает скорость и количество передвигающейся воды. Поглощение и передвижение солей, по-видимому, усиливаются благодаря транспирации, но у многих растений наблюдается пышный рост в затененных влажных местообитаниях, где транспирация очень невелика.
С другой стороны, быстро транспирирующие растения теряют так много воды в солнечные дни, что клетки молодых побегов и листьев теряют тургор и завядают, устьица закрываются, понижая фотосинтез, а рост ослабевает или прекращается. (Вредное влияние водного стресса будет подробнее рассмотрено в главах 13 и 16.) Мы рассматриваем транспирацию как неизбежное зло: неизбежное вследствие строения листьев, и зло, так как она часто создает водный дефицит и вызывает повреждение и гибель от обезвоживания.
ПРОЦЕСС ТРАНСПИРАЦИИ. Вследствие большого значения транспирации для всего водного режима растений сущность этого процесса и влияющие на него факторы заслуживают тщательного рассмотрения. Повторим, что транспирация в своей основе является физическим процессом испарения, который контролируется физическими факторами. Однако транспирация — это и физиологический процесс, который находится под влиянием таких внутренних факторов, как строение и расположение листьев, поведение устьиц. Транспирация обычно происходит в две стадии: испарение воды из клеточных стенок в межклетники и диффузия водяного пара в наружную атмосферу.
Транспирация как физический процесс. Скорость испарения воды с какой-либо поверхности зависит от притока энергии, которая может использоваться на испарение, от градиента давления пара, или его концентрации, являющегося движущей силой процесса, и от сопротивле
332
Глава 12
ний на пути диффузии водяного пара. Наибольшее количество водяного пара выделяется через устьица, некоторая часть проходит через эпидерму листьев и его кутикулярный покров, а часть выделяется через чечевички в коре ветвей и побегов древесных растений.
Испарение г/см 2 . с, можно выразить простым уравнением:
с — с е - е
воды возд воды возд
Е =---------------------„„----------------------, (12.5)
возд возд
где своды и свозд — концентрации водяного пара соответственно на поверхности воды и в основной массе воздуха, г/мл; еводы и евозд — соответствующие давления водяного пара в миллиметрах ртутного столба или в миллибарах; г возд сопротивление пограничного
слоя, преодолеваемое диффундирующими молекулами, с/см.
Транспирация Т в значительной степени контролируется сопротивлением листьев, поэтому для ее описания нужно ввести дополнительный член:
£ __ £ . р _ р
листа возд листа возд
- —_ или -
листа возд листа возд
(12.6)
где с и ет„та - соответственно концентрация и давление водяно-JlrlGTd JlMCld
го пара на испаряющих поверхностях внутри листа; г листа —дополнительное сопротивление диффузии в листе.
Это уравнение показывает, что интенсивность транспирации в граммах воды на квадратный сантиметр транспирирующей поверхности пропорциональна с, разности концентраций водяного пара, или е, разности
евозд.
( га
) (Сопротивление
J пограничного ) слоя воздуха )
} 0,1-3,0
(Устьичное < сопротивление) 1,0-10
(Мен.клет') HUKU) ’ листа
(Кутикулярное сопротивление) 10-> 100
(Сопротивление листа)
Общее сопротивление
! (Сопротивление мезофилла) очень низкое
Рис. 12.3. Схема, показывающая сопротивление диффузии водяного пара из листа. Интенсивность транспирации прямо пропорциональна крутизне градиента давления водяного пара (от «’^иста до евозд) и обратно пропорциональна сопротивлениям, с/см. Устьичное и кутикулярное сопротивления широко варьируют у разных видов и в зависимости от степени гидратации и могут быть значительно выше, чем указано здесь
Чнвчение воды и процесс транспирации 333
давлений пара между испаряющими поверхностями в листе и в основной массе наружного воздуха, разделенной на сумму сопротивлений диффузии ( г листа + гвозд ) в с/см. Схема
сопротивлений показана на рис. 12.3. Компоненты уравнения (12.5) будут рассмотрены после краткого обзора энергетического баланса.
Расходование энергии в процессе транспирации- Энергия, подводимая к фитоценозу или к отдельным листьям, поступает в виде прямой солнечной радиации, радиации, отражаемой и излучаемой почвой и окружа-щсй растительностью, и потока теплоты (адвекции) от окружающей среды. Поступившая энергия рассеивается тремя путями: излучением, конвекцией содержащего тепла и расходованием скрытой теплоты при испарении воды (транспирации). Энергетический баланс листа складывается следующим образом:
S + (7 — г S — А ± Н ± Е + А = 0, (12 7)
нетто-радиация
где нетто-радиация - фактическая радиация, имеющаяся в распоряжении листьев (она состоит из S-общей солнечной радиации, плюс длинноволновой радиации из внешней среды, минус rS ~ отражаемой листьями радиации, минус R - радиации, излучаемой листьями); И — теплообмен с окружающей средой путем конвекции и адвекции; Е - скрытая теплота, теряемая при транспирации; А — энергия, используемая на метаболические процессы, в основном на фотосинтез. Последняя составляет только 2—3 % общей энергии, и обычно ею можно пренебречь. Все перечисленные величины могут сильно изменяться, поэтому энергообмен отдельных листьев довольно сложен. При одинаковых температурах листа и воздуха вся тепловая энергия затрачивается на излучение и транспирацию. Если закрывание устьиц останавливает транспирацию, то тепловая энергия может рассеиваться излучением и переносом тепла, но обычно существует динамическое равновесие, в котором действуют все три механизма. По мнению Идсо и Вакера (1967), отдельные листья и растения теряют больше тепла излучением, чем посредством конвекции или транспирации, но для фитоценозов наибольшее значение имеет потеря тепла при транспирации. Иногда в результате переноса энергии путем адвекции от окружающей среды к небольшой изолированной массе растительности интенсивность транспирации превышает величину, объяснимую с точки зрения падающей радиации. Это так называемый оазисный эффект. Ночью листья часто охлаждаются путем излучения в атмосферу, вследствие чего происходит приток к ним содержащейся и скрытой теплоты и выпадение росы.
Градиент давления пара между листом и атмосферой. Как указывалось выше, движущей силой диффузии водяного пара из растений является разность давления или концентрации пара между тканями растения И воздухом: Элиста — Своэд ИЛИ ^листа — ^возд-
Эта разность зависит от двух переменных: давления, или концентрации, пара на испаряющих поверхностях и от соответствующих величин в окружающей массе воздуха.
334
Глава 12
Рис. 12.4. Влияние повышения температуры на градиент дав-ления водяного пара (ГДП) между листом и атмосферой при условии насыщения воздуха внутри листа водяным паром и 60 %-ной относительной влажности наружной атмосферы при любой температуре: 1 — давление водяного пара в листе; 2 — давление водяного пара в атмосфере; 3 — давление водяного пара при постоянной относительной влажности
12.1. ВЛИЯНИЕ ПОВЫШЕНИЯ ТЕМПЕРАТУРЫ НА ДАВЛЕНИЕ ВОДЯНОГО ПАРА В ЛИСТЬЯХ И НА ГРАДИЕНТ ДАВЛЕНИЯ ПАРА МЕЖДУ ЛИСТЬЯМИ
И АТМОСФЕРОЙ ПРИ ПОСТОЯННОЙ ОТНОСИТЕЛЬНОЙ ВЛАЖНОСТИ 60 %*
Температура, °C Давление пара в ткани, мбар Давление пара в атмосфере при 60 %-ной относительной влажности, мбар Градиент давления пара, мбар
10 12,27 7,36 4,91
20 23,37 14,02 9,35
30 42,43 25,45 16,98
* Предполагается, что давление пара в тканях листа равно давлению пара над водой в точке насыщения, так как его понижение под влиянием содержащихся в клетке растворенных веществ составляет всего лишь около 3 % при осмотическом потенциале-40 бар. См. также рис. 12.4.
На давление пара на испаряюйщх поверхностях клеток влияют в основном температура и водный потенциал этих поверхностей. Если водный потенциал на поверхности клеток принять за нуль (тургесцентные клетки), то можно считать, что давление пара соответствует точке насыщения при данной температуре. Ниже приведены данные о влиянии повышения температуры листа и воздуха при постоянной абсолютной влажности на градиент давления пара между листом и атмосферой (см. также рис. 12.4 и табл. 12.1) :
Температура листа и воздуха, °C. . . . 10 20 30
Относительная влажность воздуха при постоянной абсолютной влажности, % ............... . 80 44 25
Давление пара, мбар: на испаряющей поверхности листа . 12,27 23,37 42,43
в атмосфере при указанной температуре . 9,81 10,10 10,50
Градиент давления пара между листом и атмосферой . 2,46 13,27 31,93
1начекие воды и процесс транспирации
335
Из приведенных данных видно, что повышение температуры от 10 до 30°С приводит к увеличению давления пара более чем в 3 раза. Таким образом, даже небольшие изменения температуры листа могут вызвать значительные изменения интенсивности транспирации, если ЛВОЗД И г листа остаются постоянными.
Влияние уменьшения *Атиста на давление водяного пара незначительно: понижение листа до —40 бар уменьшает гра-
диент давления пара всего лишь приблизительно на 5 % при 50 %-ной относительной влажности и 30°С. Поэтому умеренные изменения водного потенциала или осмотического потенциала листьев являются несущественными по сравнению с другими факторами, вызывающими изменение Де, и слабо влияют на транспирацию. Обычно предполагаем, что водный потенциал на поверхности клеток мезофилла сходен с водным потенциалом всей ткани, но это, по-видимому, не всегда правильно. Некоторые исследователи сообщают, что влажность в межклетниках быстро транспирирующих растений эквивалентна —100 бар или ниже.
Давление пара в массе окружающего растения воздуха зависит от температуры и влажности (см. табл. 12.1 и 12.2). Фактическое количество воды, содержащейся в атмосфере, называют абсолютной влажностью и обычно выражают в миллиграммах воды на литр воздуха. Более часто содержание влаги в воздухе выражают в виде относительной влажности, показывающей процент насыщения при данной температуре. Относительная влажность не является хорошей мерой условий атмосферной влаги в отношении испарения или транспирации, потому что при повышении температуры массы воздуха относительная влажность уменьшается, а давление пара увеличивается.
Как видно из табл. 12.1, интенсивность испарения с влажной поверхности при 3(К>С и 60 %-ной относительной влажности более чем в 3 раза выше, чем при ЮОС и такой же относительной влажности. Причиной увеличения испарения и транспирации с повышением температуры является то, что давление водяного пара в листьях увеличивается быстрее, чем в ненасыщенном воздухе, а не понижение относительной влажности, как это часто ошибочно считают. Несомненно, более точной будет оценка условий атмосферной влажности в окружающей среде, например в Камере для выращивания растений, по величине дефицита давления пара, т. е. разности между существующим давлением атмосферного пара и давлением пара, насыщающего воздух при данной температуре.
СОПРОТИВЛЕНИЯ НА ПУТИ ВОДЯНОГО ПАРА. Небольшие количества водяного пара выделяются через кору, преимущественно через чечевички. Некоторое количество выделяется через эпидерму, но около 90 % водяного пара выходит через устьица, так как сопротивление диффузии через устьица (если они открыты) гораздо ниже, чем сопротивление диффузии через эпидерму. Существуют два типа сопротивлений диффузии водяного пара: сопротивления, связанные со свойствами листьев, и внешнее сопротивление пограничного слоя воздуха, примыкающего к листовой поверхности.
336
Глава 12
12.2. СОПРОТИВЛЕНИЕ ПЕРЕДВИЖЕНИЮ ВОДЯНОГО ПАРА И ДВУОКИСИ УГЛЕРОДА ЧЕРЕЗ ПОГРАНИЧНЫЙ СЛОЙ Г «КУТИКУЛУ Г ,
а с
ОТКРЫТЫЕ УСТЬИЦА Г * И СОПРОТИВЛЕНИЕ МЕЗОФИЛЛА ДЛЯ СО, Г
s I т
(HOLMGREN И ДР., 1965)
Сопротивление диффузии, с/см
Вид водяного лара COj
r a r s r rs r m
Betula verrucosa 0,80 ^92 83 1,56
Quercus robur 0,69 6,70 380 11,30 9,6
Acer platanoides 0,69 4,70 85 8,00 7,3
Helianthus annuus 0,55 0,38 — 0,65 2,4
12.3. ОБЩАЯ И КУТИКУЛЯРНАЯ ТРАНСПИРАЦИЯ ЛИСТЬЕВ (ПОВЕРХНОСТЬ ВКЛЮЧАЕТ ОБЕ СТОРОНЫ ЛИСТЬЕВ) РАЗНЫХ ГРУПП РАСТЕНИЙ ПРИ СТАНДАРТНЫХ УСЛОВИЯХ ИСПАРЕНИЯ, МГ Н,О / ДМ2/ч i f
(LARCHER, 1975)
Транспирация при от-
Кутикулярная транспирация
Вид крытых устьицах при закрытых устьицах % от общей l В
Древесные растения i
Betula pendula 780 95 12,0 '
Fagus silvatica 420 90 21,0
Picea abies 480 15 3,0
Pinus silvestris 540 13 2,5 .'
Rhododendron ferrugineum 600 60 10,0
Травянистые растения
Coronilla veria 2000 190 9,5 Л.
Stachysrecta 1800 180 ю,о
Oxvtropis pilosa . 1700 100 6,0
Сопротивления листьев. Наружная поверхность эпидермы обычно покрыта слоем кутина, который относительно непроницаем для воды. Толщина и проницаемость этого слоя сильно различаются у листьев разных видов и у листьев, развивающихся при различных условиях внешней среды. Кутикула часто бывает покрыта налетом воска, который, по-видимому, выделяется через клеточную стенку и кутикулу и накапливается на наружной поверхности (см. главу 8, роль воска в кутикуле обсуждалась в той же главе). Кутикулярное сопротивление широко варьирует и обычно значительно повышается при дегидратации листьев (табл. 12.2, 12.3).
При открытых устьицах устьичное сопротивление г s настолько меньше кутикулярного, что большая часть воды испаряется через устьица.
и процесс транспирации ЗУ1
Н ионной фактор, влияющий на устьичное сопротивление, - степень hiнрыгости устьичных отверстий, которая контролируется в значительном степени интенсивностью света и водным потенциалом листьев и в ММ1ЫНСЙ мере - влажностью воздуха и концентрацией СО2- Виншер и Котовский (1971) считают, что повышение температуры в интер-Hrtiir or 20 до 40°С тоже увеличивало устьичное сопротивление у деревьям некоторых видов. Можно также различать сопротивление диффу-1ии водяного пара в межклетниках и в клетках мезофилла. Сопро-i«мление в межклетниках, измеренное диффузионным порометром, «мимется, по-видимому, существенным при широко открытых устьицах. Значение сопротивления мезофилла дебатировалось в течение нескольких десятилетий. Еще в начале этого столетия Ливингстон и Браун (1912) сообщили, что интенсивность транспирации часто уменьша-пйгь без заметного изменения степени открытости устьиц. Ливингстон приписывал это явление втягиванию менисков в клеточных стенках ио время периодов усиленной транспирации и назвал это начинающим-i и подсыханием. Существование повышения сопротивления мезофилла шрицалось некоторыми исследователями, однако о нем недавно сообщили Симс (1963); Витеман и Коллер (1964). Джервис и Слейтер ( !‘П0) относят :это повышение за счет пониженной водопроводимости, мы «ываемой слоем кутикулы, который покрывает наружную поверхность клеточных стенок мезофилла. Другие исследователи не обнаружили сопротивления клеточных стенок мезофилла у некоторых травянистых видов и сделали вывод, что давление водяного пара на испаряющих поверхностях равно его давлению в точке насыщения. Устьичное и кутикулярное сопротивления действуют параллельно, поэтому их можно ншисать так:
L = 2_+_L; rs= rm + П + Гр , (128)
г г г
I с S.
где г J — общее сопротивление листа; г с — кутикулярное сопротивление; г s — устьичное сопротивление; r m сопротивление клеточных стенок мезофилла; г*. - сопротивление в межклетниках; г & — сопротивление устьичных щелей. Схема различных сопротивлений в листе показана на рис. 12.3.
Внешние сопротивления. Внешнее сопротивление зависит в значи-юльной степени от размера, формы листа и от скорости ветра. Свойства листьев обсуждаются в разделе о внутренних факторах, влияющих на транспирацию. Усиление движения воздуха оказывает прямое действие, увеличивая транспирацию путем удаления пограничного слоя водяного пара, окружающего листья в неподвижном воздухе, и понижая г В03д в уравнении (12.6). Ветер действует также косвенно, уменьшая транспирацию вследствие охлаждения листьев и понижения с в уравнении (12.6). Наибольший эффект наблюдается при низ-ких скоростях ветра (рис. 12.5). Кнор (1967) указывает, что хотя слабый ветер увеличивает транспирацию листьев при низкой освещенности, но при более высоком уровне радиации, когда они становятся
338
Глава 12
Скорость 6empat м/ч
Скорость ветра, см/с
Рис. 12.5. Влияние повышения скорости движения воздуха на интенсивность транспирации горшечных экземпляров подсолнечника, выращиваемых в освещенной солнцем теплице:
Гр растений, обдуваемых ветром; контрольных растений в неподвижном воздухе; наибольший эффект наблюдается при очень низкой скорости ветра; скорость I миля/ч равна 44,7 см/с (Martin, Cements, 1935)
Рис. 12.6. Кривые, показывающие теоретический ’ ход обмена скрытой теплоты j отдельного листа путем транспирации при разных скоростях ветра и различной j«-нетто-радиации ^(Rд) • При высоком уровне радиации ветер может понижать тран- г спирацию, охлаждая листья, при низком Уровне он может усиливать транспира- J цию, поставляя энергию (Knoerr, 1967) : £
Рис. 12.7. Влияние повышения скорости ветра на транспирацию молодых горшечных экземпляров субальпийских растений в продуваемом тоннеле при температуре воздуха 20°С, температуре почвы 15°С и интенсивности света 30000 лк. Эвапорация измерялась при тех же условиях при помощи зеленого атмометра Пайча (Tranqufflini, 1969). С увеличением скорости ветра эвапорация возрастала, транспирация у большинства видов понижалась:
1 - ольха (Alnus viridis); 2 - лиственница (Larix decidua); 3 - ель (Picea aides); 4 - сосна кедровая (Pinus cemhra); 5 - рябина (Sorbus aucuparia); 6 — рододендрон (Rhododendron femigineutn); 7 — эвапорация
1начение воды и процесс транспирации
339
гоплее воздуха, ветер может уменьшать транспирацию, охлаждая листья (рис. 12.6). Однако фактическое проведение растений может быть довольно различным.
Девис с соавторами (1974) сообщили о значительных различиях в реакции сеянцев трех видов в продуваемом тоннеле при искусственном освещении от 2Н7ОО до 35500 лк и скорости движения воздуха ,5,8—26 м/с. Интенсивность транспирации как при ветре, так и в спокойном воздухе была: ясень американский клен сахарный сосна смолистая. Ветер усиливал транспирацию
сеянцев ясеня при всех скоростях, понижал транспирацию клена и существенно не влиял на сосну. Устьица листьев клена быстро закрывались под действием негра, а устьица ясеня оставались открытыми до тех пор, пока не было значительного обезвоживания.
Транквилини (1969) также сообТцил о различиях между видами в австрийской зоне верхней границы распространения леса. Транспирация лиственницы европейской и ольхи зеленой была выше, чем в спокойном воздухе, при всех скоростях ветра вплоть до 20 м/с, а у ели европейской, сосны кедровой европейской и Rhododendron fernigineum при 10 м/с наблюдалась понижение интенсивности транспирации — от умеренного до сильного (рис.,12.7). Исследователь приписал это уменьшение транспирации закрыванию устьиц, вызванному скорее местным подсыханием поверхности листьев, чем общим обезвоживанием растений, которые во время экспериментов подвергались действию лишь небольшого дефицита насыщения водой. Видовые различия, отмеченные в данной работе, были, по-видимому, связаны с разным поведением устьиц и величиной кутикулярной транспирации. По другим сообщениям, хотя ветер и вызывает немедленное закрывание устьиц у R. ferrugineum,OH слабо влияет на степень открытости уегьиц сосны кедровой, и любое понижение фотосинтеза у последней должно вызываться изменением освещенности хвои.
ВНУТРЕННИЕ ФАКТОРЫ, ВЛИЯЮЩИЕ НА ТРАНСПИРАЦИЮ
Интенсивность транспирации в основном находится под контролем физических факторов, но на нее влияет ряд внутренних факторов, действующих как на движущие силы, так и на сопротивления.
ПЛОЩАДЬ ЛИСТЬЕВ. Общая площадь листьев значительно влияет на потерю воды отдельнымй~растёниями. Растения с большой листовой поверхностью обычно транспирируют больше, чем растения с малой поверхностью. Однако при удалении с дерева половины листьев не обязательно пропорционально сократится интенсивность транспирации. Миллер (1938) приводит данные ряда экспериментов, в которых удаление части листьев уменьшало транспирацию в расчете на одно дерево, по повышало интенсивность транспирации на единицу листовой поверхности. Такое повышение произошло в результате усиления движения воздуха и лучшего освещения оставшихся листьев, а также более высокого отношения поверхности корней к поверхности листьев. Некоторые виды древесных растений, например креозотовый куст, сбрасывают большую часть листьев при водном стрессе, значительно сокращая транспирирующую поверхность. Это происходит также и у некоторых мезофитных видов, например у конского каштана и даже у тюльпанного дерева. Скручивание и свертывание завядающих листьев также уменьшает открытую поверхность и повышает сопротивление диффузии водяного пара, особенно если большинство устьиц находится на внутренней поверхности скрученного листа.
Некоторые виды сбрасывают листья раньше, чем другие, и, вероятно, они извлекают из почвы меньше воды в начале осени, чем виды, кото
340
Глава 12
рые сохраняют листья значительно дольше, например дубы. Если действительно в этом существуют значительные различия, то имело бы смысл преобразовать растительный покров на водоразделах в пользу видов с минимальной интенсивностью транспирации осенью и таким образом увеличить сток в это время года. Некоторые соображения были также высказаны относительно возможности дефолиации деревьев на водоразделах в конце лета для уменьшения транспирации и увеличения запаса воды. Дефолиация может сократить рост, но это было бы несущественно там, где накопление воды более важно, чем производство древесины. Удаление подлеска из лавра и рододендрона на водоразделе в Кауита (Северная Каролина) значительно усилило сток, но в последующие годы запас воды уменьшался вследствие восстановления растительности.
12,4. ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ НА ЕДИНИЦУ ЛИСТОВОЙ ПОВЕРХНОСТИ И НА ОДИН СЕЯНЕЦ СОСНЫ ЛАДАННОЙ И ЛИСТВЕННЫХ ПОРОД С 22 АВГУСТА ПО 2 СЕНТЯБРЯ (СРЕДНИЕ ВЕЛИЧИНЫ ДЛЯ ШЕСТИ СЕЯНЦЕВ КАЖДОГО ВИДА)
Наименование показателей Сосна Тюльпан- Дуб
ладанная ное дерево красный
Транспирация, г/с у т/дм 2 5,08 9,76 12,45
Транспирация, г/сут на один сеянец ,106,70 59,10 77,00
Средняя площадь листьев одного дерева, дм^ 21,00 6,06 6,18
Средняя высота деревьев, см 34,00 34,00 20,00
Имеются большие различия в транспирации на единицу листовой поверхности среди разных видов (табл. 12.4), Однако эти различия могут быть обманчивыми, потому что неодинаковая общая площадь листьев может компенсировать разницу в интенсивности на единицу листовой поверхности. Например, как видно из табл. 12,4, интенсивность транспирации на единицу листовой поверхности сосны ладанной ниже, чем у некоторых лиственных пород, но общая поверхность листвы одного сеянца сосны настолько больше, что потеря воды на один сеянец оказывается также больше. Таким образом, одно дерево сосны ладанной может терять за день столько же воды, сколько и дерево лиственной породы с кроной сходного размера.
ОТНОШЕНИЕ КОРНИ / ПОБЕГИ. Отношение корней к побегам или, точнее, поглощающей поверхности к транспирирующей имеет большее значение, чем одна только площадь листовой поверхности, так как если поглощение отстает от транспирации, то возникает водный дефицит, устьица закрываются и транспирация уменьшается. Паркер (1949) обнаружил, что интенсивность транспирации на единицу листовой поверхности у сеянцев дуба красного и сосны ладанной, растущих на влажной почве, повышалась с увеличением отношения сухого веса корней к поверхности листьев (рис. 12.8). Перейра и Козловский (1977) установили, что у сеянцев клена сахарного с большой поверхностью листьев развивался более сильный водный стресс, чем у сеянцев после частичной дефолиации. Деревья с обширной, сильно разветвлен-
iNMtKue воды и процесс транспирации
341
Рис, 12.8. Влияние увеличения отношения корни / нпПеги иа интенсивность транспирации сеянцев цриитных растений. На оси ординат отложены । риммы воды, теряемой на 1 дм* листовой поверх-шнт и, на оси абсцисс — вес корней, разделенный на шишцщь листьев, дмА С увеличением отношения корни / побеги транспирация у бирючины не изме^ HHiuK'i», так как количество корней, по-видимому, ни разу не сокращалось настолько, чтобы влиять ин |рннспирацию (Parker, 1949):
I бирючина; 2 — сосна ладанная; 3 — сосна смо-шстон; 4— можжевельник виргинский
Сухой Вес корней/ площадь листьев
Ной корневой системой гораздо лучше переносили засуху, чем деревья с поверхностной или слабо разветвленной корневой системой.
Потеря корней во время выкопки сеянцев - серьезная проблема, потому что наиболее обычной причиной гибели пересаживаемых сеянцев является засыхание из-за отсутствия эффективной поглощающей поверхности. Подрезка и ’’скручивание” сеянцев в грядках проводится для создания компактной, сильно разветвленной корневой системы, которую можно выкопать с минимальным повреждением. По сообщениям, выживаемость сеянцев дугласии и сосны желтой, пересаженных в районе с засушливым летом, повышалась при хорошо развитой корневой системе и большой величине отношения корни J побеги. Аллен (1955) установил, что подрезка саженцев сосны болотной до длины 12,5 см уменьшала отпад после пересадки. На практике обычно применяется обрезка надземной массы пересаживаемых деревьев и кустарников для того, чтобы скомпенсировать потерю корней в процессе пересадки. Однако сокращение транспирирующей поверхности уменьшает и фотосинтезирующую поверхность, что нежелательно. Исключение составляют случаи, когда сохранение содержания воды важнее потери фотосинтетической способности.
РАЗМЕР И ФОРМА ЛИСТЬЕВ. Размер и форма листьев влияют на интенсивность транспирации на единицу поверхности. По сообщению Слаутера (1967), сопротивление воздуха rQ для листьев шириной 10 см почти в 3 раза больше, чем для листьев шириной 1 см.[кроме того, небольшие, глубокорассеченные листья и сложные листья с маленькими листочками обычно бывают холоднее, чем крупные листья, так как их более тонкие пограничные слои способствуют более быстрому переносу тепла.ДГибалс с соавторами (1964) установили, что в спокойном воздухе широкие листья значительно теплее хвои сосны, подвергающейся действию такой же падающей радиации. Нобель (1976) обнаружил, что крупные теневые листья Hyptis emoryi нагревались гораздо сильнее, чем небольшие световые листья, а это должно усиливать транспирацию. Однако тонкий пограничный слой и более низкая вели- / чина гвозд мелких листьев также являются более благоприятными^ для потери водяного пара, так что эти два эффекта имеют тенденцию,/ взаимно уравновешивать свое влияние на перенос скрытой теплоты
342
Глава 12
и на транспирацию в том случае, если устьичное сопротивление имеет сходную величину.
ОРИЕНТАЦИЯ ЛИСТЬЕВ. Большинство листьев растет таким образом, чтобы располагаться более или менее перпендикулярно наиболее яркому свету, проникающему к ним. Это хорошо заметно у покрывающих стены лиан и у одиночных деревьев, на наружной поверхности которых имеется сплошная мозаика листьев, четко расположенных так, чтобы поглощать как можно больше света. С другой стороны, листья некоторых видов, например, Silphium и дуба гладкого, а также хвоя сеянцев сосны болотной в травянистой фазе располагаются вертикально. Такая ориентация, очевидно, уменьшает поглощение энергии и понижает температуру листьев в полдень, что в свою очередь может сократить потерю воды, хотя еще не было доказано, что такое необычное расположение лцстьев действительно имеет жизненно важное значение. Иглы большинства сосен располагаются пучками и затеняют друг друга, что понижает фотосинтез, а возможно, и транспирацию. Опускание и скручивание, характерные для вялых листьев, также уменьшают количество получаемой радиации. Завядание служит признаком водного стресса, но оно способствует сокращению дальнейшей потери воды, достаточному, вероятно, для продления жизни. По сообщениям, вызываемые ветром изменения ориентации листьев сосны кедровой сильно понижают фотосинтез.
ПОВЕРХНОСТЬ ЛИСТЬЕВ. Мы уже упоминали о значении кутикулы и воска для увеличения сопротивления потере воды через эпидерму. Суммарное действие плотного покрова из волосков, имеющегося у некоторых типов листьев, является менее определенным. Следовало бы ожидать, что опушение будет увеличивать сопротивление пограничного слоя г а , понижая при этом как потерю тепла, так и выделение водяного пара в подвижный воздух. Однако недавно появилось сообщение Элерингера с соавторами (1976) о том, что диффузия СО2 в опушенные листья Encelia farinosa не является замедленной по сравнению с диффузией в неопушенные листья Е. californica. Если г fl не повышается для СО2, то оно, по-видимому, не будет повышаться и для водяного пара. Живые волоски могут усиливать транспирацию вследствие увеличения испаряющей поверхности.
Альбедо, или отражательная способность, поверхности листьев существенно влияет на их температуру. Растительность обычно отражает 15-25 % падающей радиации. По сообщению Биллингса и Мориса (1951), листья пустынных растений обычно отражают света больше, чем листья растений менее освещаемых местообитаний. Белые, густо опушенные листья пустынного кустарника Encelia farinosa отражают приблизительно вдвое больше радиации, чем зеленые неопушенные листья Е. californica^ происходящей из влажного прибрежного района. Отражая так много света, Е. farinosa отличается пониженным нетто-фотосинтезом. По сообщению Миллера (1938), после обработки бордоской жидкостью транспирация уменьшается, вероятно, вследствие понижения температуры листьев, вызванного белым налетом, который при этом образуется на листьях.
30/3
'Значение воды и процесс транспирации
343
АНАТОМИЧЕСКОЕ СТРОЕНИЕ ЛИСТЬЕВ, Структура листьев может сильно изменяться под влиянием внешней среды, в которой они развиваются, У световых листьев по сравнению с теневыми мельче клетки, меньше поверхность и промежутки между жилками, толще слой кутина, Они имеют большую толщину, благодаря наличию дополнительных слоев палисадных клеток (рис. 12.9). Верхние листья обычно имеют более ксероморфную структуру, чем нижние. Вообще листья растении сухих местообитаний сходны со световыми листьями (ксероморфный тип)., а растений влажных мест — с теневыми листьями (мезоморфные । ины, рис. 12.10).
Часто высказывалось предположение { что потеря воды листьями мезоморфного типа с тонким слоем кутина и крупными межклетниками (например, листьями тюльпанного дерева, дуба красного или клена), больше, чем у растений с более толстыми и более кутинизиро-нанными ксероморфными листьями (такими, как у магнолии или падуба). Максимов (1929) показал, что у хорошо увлажняемых растений это совсем не обязательно. Более поздние данные по этому вопросу приведены в табл. 12.5. Как Ilex glabra, так и Gordonia lasianthus имеют толстые, сильно к утилизированные листья. Однако интенсивность транспирации на единицу листовой поверхности при хорошем увлажнении у них выше, чем у дубов, кленов или тюльпанного дерева.
Рис. 12.9. Поперечные разрезы листьев одиночного дерева клена сахарного (Weaver и Cements, 1938; по разрешению издательства Me Graw-Hill): а — с южной стороны; б — из центральной части кроны
344
Глава 12
Рис. 12.10. Поперечные разрезы листьев:
а — дуба малого; б — бука; листья дуба малого ксероморфмого типа, с толстой кутикулой, двойным слоем палисадных клеток, сильно развитой обкладкой пучков и высоким отношением внутренней поверхности к наружной; листья бука — мезоморфного типа, они тоньше, с тонкой кутикулой и одним слоем палисадных клеток, с меньшей внутренней поверхностью, обращенной к межклетникам (Kramer, Kozlowski, i960)
2)
По сообщению Свенсона (1943), падуб тусклый в солнечный день транспири-ровал интенсивнее, чем сирень, колеус или табак, а в облачный день интенсивность транспирации падуба была ниже, чем у сирени. Эти различия связаны, по крайней мере частично, с большей величиной отношения внутренней поверхности к наружной у ксероморфных и световых листьев вследствие более сильного развития палисадной ткани по сравнению с теневыми и мезоморфными листьями. Свенсон (1943) определил отношение внутренней поверхности листьев к наружной у падуба тусклого, табака и колеуса, которое оказалось равным соответственно 12,9; 7,1 и 4,6. Бб^ьшая величина отношения внутренней поверхности к наружной обеспечивает большую испаряющую поверхность на единицу внешней листовой поверхности и при хорошем водоснабжении поддерживает, вероятно, межклеточное пространства в состоянии почти полного насыщения. Однако если'Подсыхание почвы вызывает водный дефицит и закрывание устьиц, то транспирация ксероморфных листьев бывает, как правило, ниже, чем у мезоморфных, вследствие того, что кутикулярная транспирация первых очень мала. По сообщениям, внутреннее заполненное воздухом пространство занимает от 20 до 70 % общего объема листа. При этом самая низкая величина наблюдается у световых листьев, а наиболее высокая у теневых.
Приведенное выше высказывание основано на предположении о том, что испарение внутри листа происходит главным образом из стенок клеток мезофилла. Майднер (1975, 1976) предположил, что зна-
Значение воды и процесс транспирации
345
12.5- ИНТЕНСИВНОСТЬ ЛЕТНЕЙ ТРАНСПИРАЦИИ РАЗЛИЧНЫХ ВИДОВ ДРЕВЕСНЫХ РАСТЕНИЙ (ВСЕ СЕЯНЦЫ ВЫРАЩИВАЛИСЬ НА ПОЧВЕ ПРИ ВЛАЖНОСТИ, БЛИЗКОЙ К ПОЛЕВОЙ ВЛАГОЕМКОСТИ)
Место \ Месяц Про-ДОЛ- Число Сред- Автор няя
Вид жи- рас- интен-
тель- тений сив-
ность, ность
сут транспирации,
г/дм2
сут
Liriodendron tulip if era Колумбус, Огайо Август 1 7 10,11 Meyer (19 32)
Liriodendron tulipifera Дарем, Северная Каролина Август * 3 4 11,78 Caughey (1945)
Quercus alba То же 3 4 14,21
Quercus rubra « 3 4 12,02 »J
Quercus rubra Файет, Миссури Июль 14 6 8,1 Biswell (1935)
Acer saccharum То же 14 6 12,2
Acer negundo »> V , 14 6 6,4
Platanus ocddentalis ‘ 14 6 8,8
Pinus taeda Дарем, Север-Август нал Каролина 3 4 4,65 Caughey (1945)
Clethra alnifolia То же 3 4 9,73 »*
Ilex glabra •» »♦ 3 4 16,10
Myrica cerifera >> 3 4 10,80 tL
Gordonia lasianthus Июль 23 4 17,77 »»
Liriodendron tulipifera Август-сентябрь 12 6 9,76 Kramer (неопубл.)
Quercus rubra *> То же 12 6 12,45 JI
Pinus taeda ♦» 12 6 5,08
чительная часть воды испаряется с внутренних поверхностей клеток эпидермы, особенно в непосредственной близости от устьиц. При этом упускается из внимания возможность того, что внутренние поверхности эпидермальных клеток бывают кутинизированы, например нижняя эпидерма листьев груши. Появляется все больше данных о том, что тургор замыкающих клеток может изменяться независимо от тургора мезофилла. Однако представляется невероятным, чтобы внутренняя поверхность эпидермы, обращенная к воздушным полостям внутри листа, была достаточно велика, чтобы представлять собой глав-
346 t Глава 12
нуй испаряющую поверхность. Вопрос об относительном значении разных испаряющих поверхностей в листьях нуждается в дальнейших исследованиях.
УСТЬИЦА
Большая часть воды, теряемой растениями, выделяется через расположенные на листьях устьица (см. рис. 2.3), и наибольшее количество двуокиси углерода, используемой при фотосинтезе, проникает тем же путем. Несмотря на то, что устьичные отверстия обычно занимают не более 1 % поверхности листа, диффузия водяного пара через устьица часто достигает 50 % интенсивности испарения со свободной водной поверхности. Это происходит благодаря тому, что размеры устьиц и промежутков между ними обеспечивают их функционирование в качестве эффективных путей для диффузии газов. (Более подробная информация об устьицах приведена в главе 2.)
Размер устьичных отверстий регулируется тургором замыкающих клеток, увеличиваясь с повышением тургора и уменьшаясь с его понижением. Устьица открываются на свету или в ответ на низкую концентрацию двуокиси углерода в межклетниках, закрываются в темноте. Они закрываются также и при потере тургора вследствие обезвоживания. Существует некоторая неясность относительно механизма, при помощи которого изменения освещенности или концентрации двуокиси углерода вызывают изменения тургора замыкающих клеток и степени открытости устьиц. Однако представление о том, что тургор замыкающих клеток изменяется в результате изменений в соотношении крахмала и сахара, было вытеснено гипотезой о накоплении или потере ионов, главным образом калия, как причине изменения тургора.
В последние годы показано, что соприкосновение эпидермы с сухим воздухом вызывает закрывание устьиц тургесцентных листьев некоторых видов. Таким образом, закрывание устьиц может вызываться обезвоживанием листьев, или дегидратацией замыкающих клеток, и, по-видимому, соседних клеток эпидермы. Кроме того, как сообщили Лавлор и Милфорд (1975), устьица листьев сахарной свеклы могут оставаться открытыми при выдерживании во влажном воздухе даже в том случае, когда в мезофилле существует водный дефицит. Высказано предположение, что у некоторых древесных растений, подвергающихся действию ветра, устьица закрываются скорее из-за подсыхания поверхности листьев, чем из-за обезвоживания всей ткани. Данные Халла, Кауфмана (1975, 1976) также показывают, что низкая влажность вызывает закрывание устьиц, но этот эффект уменьшают низкая концентрация СО2 и высокая температура.
Циклическое открывание и закрывание устьиц с периодами от 15 ? до 120 мин наблюдалось у травянистых растений, страдающих от водно- 5, го стресса. Сообщалось о циклических устьичных движениях у цитрусо-вых деревьев, растущих в садах. Физиологическое значение этой циклич- | ности точно не известно, однако Кован (1972) предположил, что она £ оптимизирует противоречивые потребности поглощения двуокиси угле- * рода и ограничения потери воды. Дальнейшее усложнение внесли наблю-
iNnwoNur воды и процесс транспирации
347
дгния, показавшие, что на поведение устьиц влияют регуляторы роста, особенно абсцизовая кислота. Было высказано предположение, что у рис гений, страдающих от водного стресса, устьица закрываются вслед-сгние уменьшения содержания цитокининов и увеличения количества цбсцизовой кислоты. Дэвис и Козловский (1975) сообщили, что обработка листьев ясеня американского, клена сахарного и каламондина абсцизовой кислотой понижала транспирацию. Представляется вероятным, что водный стресс является причиной закрывания устьиц вслед-ствис потери тургора замыкающими клетками, прежде чем он может им звать изменение концентрации регуляторов роста, достаточное для loro, чтобы оказать соответствующее действие.
Устьичные реакции изменяются в зависимости от возраста растений и условий внешней среды. Известно, что устьица чутко реагируют на интенсивность света, концентрацию СО2 и водный стресс, но на их ответную реакцию влияет предшествующая ситуация. При старении листьев их устьица становятся менее чувствительными и полуденное устьичное сопротивление может увеличиться в несколько раз, так как они не открываются полностью.
Такое поведение хорошо заметно у некоторых злаков. Имеются также сообщения о том, что устьица растений, выращенных в теплицах и в специальных камерах, закрываются при более низком уровне водного стресса (более высоком подлом потенциале листьев), чем устьица растений, выросших в поле. Все эти эксперименты были проведены на травянистых растениях, но не на древесных. Но утверждению Миллера (1938), устьица большинства травянистых растений, растущих при благоприятных условиях, можно разделить на три группы: 1) типа люцерны, которые открываются утром и закрываются вечером; 2) типа картофеля, которые всегда открыты, за исключением нескольких часов после захода солнца; 3) типа ячменя (куда входят многие злаки), которые начинают закрываться вскоре после открывания. Нам неизвестно, делались ли попытки классифицировать таким же образом поведение устьиц древесных растений, но подобные исследования могли бы представлять интерес.
Данные, обобщенные Лопушинским (1969), указывают на значительные различия в реакции устьиц на водный стресс среди разных видов хвойных, при этом устьица сосны обладают наибольшей чувствительностью. В экспериментах с 2-летними сеянцами исследователь обнаружил, что у сосен желтой и скрученной, а также у ели Зигельмана устьица закрывались при более слабом водном стрессе (более высоком Фу), чем у сеянцев дугласии и пихты великой (табл. 12.6). Рун-нинг (1976) также сообщил о различиях среди хвойных в отношении связи между водным стрессом листьев и открыванием устьиц. По данным Федерера (1977), устьица закрывались у березы при —15 бар, у тополя при —17 бар, у дуба при —21 бар и у вишни при —23 бар. Он считает различия между видами более важными, чем различия, связанные с местообитанием или размером дерева. Хинкли (1978) сообщил, что в засушливое лето устьица клена сахарного были закрыты 50 % времени, дуба белого и бархатистого около 20 % и дуба красного приблизительно 30 %. Часто трудно оценить значение быстрого закрывания устьиц у растений, страдающих от водного стресса, потому что польза от сохранения воды может быть сведена на нет уменьшением фотосинтеза.
348
Глава 12
Рис. 12.11. Зависимость интенсивности транс* пирации от степени открытости устьиц при различной потенциальной транспирации* Числа на кривых показывают интенсивность испарения с фильтровальной бумаги атмометров, выраженную в мг воды/ч на 25 см2 испаряющей поверхности. Транспирация также выражена в мг воды/25 см2 листовой поверхности. При низкой интенсивности эвапорации и потенциальной транспирации степень открытости устьиц намного слабее влияет на интенсивность транспирации, чем при потенциально высокой интенсивности (St&lfelt, 1932)
12.6. СОДЕРЖАНИЕ ВОДЫ, ОТНОСИТЕЛЬНОЕ СОДЕРЖАНИЕ ВОДЫ (ОТНОСИТЕЛЬНАЯ ТУРГЕСЦЕНТНОСТЬ) И ВОДНЫЙ ПОТЕНЦИАЛ ПРИ ЗАКРЫВАНИИ УСТЬИЦ У НЕКОТОРЫХ ХВОЙНЫХ ДЕРЕВЬЕВ (Lopushinsky, 1969)
Вид Содержание, % Относитель- Водный по-
---------------------ное содержа- тенциал, бар
сухого вещества ВОДЫ ние воды, %
Сосна желтая 31,3 68,7 84,4 -16,5
Сосна скрученная 29,4 70,6 81,6 -14,6
Пихта великая 17,9 82,1 82,2 -25,1
Дугласия 21,4 78,6 85,0 -19,0
Ель Энгельмана 32,8 67,2 86,1 -16,0
УСТЬИЧНОЕ РЕГУЛИРОВАНИЕ ТРАНСПИРАЦИИ. В начале этого столетия было сделано немало попыток определить влияние частичного закрывания устьиц на интенсивность транспирации. На исследователей, работавших в этой области, оказали влияние эксперименты Брауна и Экомба (1900), которые проводились в спокойном воздухе, где сопротивление пограничного слоя г а было равно устьичному сопротивлению г Их результаты показали, что большие изменения степени открытости устьиц слабо влияют на интенсивность транспирации. Однако неоднократно было показано, что в подвижном воздухе, где г а невелико, имеется четкая связь между степенью открытости устьиц и транспирацией. В настоящее время пришли к соглашению о том, что хотя частичное закрывание устьиц слабо влияет на транспирацию в спокойном воздухе, оно сильно понижает ее интенсивность в подвижном воздухе, рис. 12.11,
ИЗМЕРЕНИЕ СТЕПЕНИ ОТКРЫТОСТИ УСТЬИЦ. Понимание того, что степень открытости устьиц значительно влияет как на потерю водя-
Значение воды и процесс транспирации
349
кого пара, так и на поглощение СО2, стимулировало интерес к измерению устьичных отверстий. Был использован ряд методов, в том числе прямое наблюдение под микроскопом, сдирание кусочков эпидермы и фиксация gx в чистом спирте, приготовление отпечатков эпидермы при помощи коллодия или пленок силиконового каучука. Другой метод включает определение времени, требуемого для инфильтрации жидкости с разной вязкостью, например бензола, парафинового масла и керосина. Фри и Валкер (1967) описали метод инфильтрации под давлением, используемый для игл хвойных. Этот метод показывает, открыты ли устьица или закрыты, но не дает возможности вычислить сопротивление листьев.
В 1911 г. Дарвин и Пертц использовали порометр, измеряющий протекание газа через листья под небольшим давлением. Были описаны многочисленные модификации, в том числе модели, пригодные для использования в полевых условиях. Однако эти порометры измеряют поток газа, являющийся в своей основе процессом диффузии под давлением, и если давление слишком велико, то это может вызвать закрывание устьиц. Это привело к разработке диффузионных порометров, которые измеряют диффузию газа через листья. Слатир и Джервис (1966) описали диффузионный порометр с использованием закиси азота, который позволяет непрерывно регистрировать устьичное сопротивление.
В настоящее время сделано много измерений при помощи различных модификаций описанного Канемази с соавторами (1969) прибора, в котором используется датчик влажности, соединенный с электронной цепью и счетчиком для определения изменений влажности в манжете, помещаемой над листом на несколько секунд. Эти приборы могут быть оснащены электронным хронометром, что повышает точность измерения. Порометры, измеряющие поток газа, невозможно использовать для исследования листьев с устьицами только на одной стороне и жилками, которые препятствуют латеральной диффузии газа (гетеробарический тип).
ВЗАИМОДЕЙСТВИЕ ФАКТОРОВ, ВЛИЯЮЩИХ НА ТРАНСПИРАЦИЮ
Основными факторами внешней среды, влияющими на транспирацию, являются: интенсивность света, дефицит давления водяного пара в воздухе, температура, ветер и водоснабжение корней. К внутренним факторам относятся: площадь листьев, расположение и структура листьев, поведение устьиц и эффективность действия поглощающей поверхности корней. Существуют сложные взаимодействия между различными факторами. Эти взаимодействия можно суммировать по их влиянию на различные члены уравнения (12.6).
Например, изменения интенсивности света влияют на гписта через степень открытости устьиц и на елиста, изменяя его температуру. Температура воздуха влияет как на елиста, так и на евоэд или на Де. Напомним, что хотя повышение температуры определенной массы воздуха от 20 до 30°С уменьшает относительную влажность, оно значительно
350
Глава 12
увеличивает давление водяного пара в воздухе. Поэтому повышение температуры усиливает транспирацию, так как увеличивает градиент давления пара be между листом и атмосферой, но не потому, что оно сопровождается уменьшением относительной влажности воздуха. При постоянной температуре изменение влажности атмосферы аналогично влияет на транспирацию, изменяя £возд и Де между листом и атмосферой. Мартин (1943) обнаружил тесную связь между транспирацией растений в темноте при постоянной температуре и давлением пара в атмосфере. Коль и Декер (1973) также установили, что транспирация является линейной функцией Д е.
Ранее уже упоминалось о том, что ветер усиливает транспирацию, унося водяной пар, содержащийся в пограничном слое, и понижая Гвозд’ и чт0 он Действует косвенно, уменьшая транспирацию посредством охлаждения листьев и снижения елиста- Другие стороны действия ветра на транспирацию — усиление вентиляции межклетников вслед- -ствие сгибания листьев и ускорение движения воздуха через амфисто-матические листья (имеющие устьица на обеих сторонах) имеют, по-видимому, второстепенное значение. Однако некоторые исследователи обнаружили, что сгибание листьев тополя вызывает значительный массовый поток газа через них и понижает общее сопротивление приблизительно на 25 %.
Расположение листьев влияет на их освещение солнечными лучами и температуру, что в свою очередь влияет на елисТЬев- Вертикально расположенные листья й самое жаркое время дня получают энергии меньше, чем горизонтальные листья, а листья, собранные группами, например пучки сосновых игл, получают меньше света, чем расположенные по отдельности. Различия внутренней геометрии и объема межклетников могут влиять на сопротивление в межклетных пространствах г • , толщина кутина влияет на г^а степень открытости устьиц — на г s (см. рис. 12.3).
Следует подчеркнуть, что изменение одного из влияющих на тралспи-^ рацию факторов не обязательно вызывает пропорциональное измене- I. ние транспирации, интенсивность которой контролируется не одним г; , каким-либо фактором. Примером может служить тот факт, что легкий $ ветер имеет тенденцию усиливать транспирацию, понижая г воздтно I может и уменьшать ее, если при этом лист охлаждается достаточно ( сильно, чтобы понизилось Подобным образом (см. рис. 12.11) *
ЛИСТа £
частичное закрывание устьиц оказывает меньшее влияние в спокойном .. воздухе, где гВОздуха велико (2) по сравнению с чем в подвиж- I ном воздухе, где гВ03д низкое (0,1). Действие различных факторов s на интенсивность транспирации можно объяснить с точки зрения их { влияния на разность давления пара между листом и воздухом (Д е = ~ £„-,„) и на величину сопротивлений на пути диффузии К
4 ЛИСТД ВОЗД
пара в листе и в воздухе (г + г возд). р
Приток воды к корням также влияет на транспирацию, потому что:;
351
Значение воды и процесс транспирации
Рис. 12.12. Влияние понижающейся влажности почвы на интенсивность транспирации сеянцев дуба (Bourdeau, 1954) :
1 — белого; 2 — малого; 3 — мерилендского; 4 - красного; 5 — серповидного; 6 — влажность завядайия
О -2 -Ь -6 -8 -Ю -12 -14 -16 -18 М
Водный потенциал почвы, бар
Рис. 12.13. Влияние понижения водного потенциала почвы на интенсивность транспирации сеянцев:
1 — сосны желтой; 2 — сосны скрученной; 3 — ели Зигельмана; 4 — дугласии; 5 — пихты великой; более быстрое снижение транспирации у сосен происходит вследствие того, что их устьица начинают закрываться при более высоком водном потенциале листьев, чем устьица пихты великой и дугласии (см. табл. 1217) (Lopushinski, Klock, 1974)
недостаточное водоснабжение вызывает обезвоживание растения и закрывание устьиц. В почве при влажности близкой к полевой влаго-смкости вода передвигается в корни быстрее и интенсивность транспирации контролируется в значительной степени атмосферными факторами, за исключением наступающего время от времени полуденного завя-дания интенсивно транспирирующих растений, вызываемого большим сопротивлением корней. При снижении содержания воды в почве приток воды к корням становится ограничивающим фактором и интенсивность транспирации уменьшается. Было показано разными исследователями от Гартинга и Хенеля в XIX в. до Козловского (1949), Боур-да (1954), Леме (1956), Слатира (1956), Яксена с соавторами (1973), что понижение влажности почвы вызывает уменьшение транспирации деревьев (рис. 12.12). Лопушинский и Клок (1974) установили, что интенсивность транспирации некоторых хвойных начинала уменьшаться при потенциале почвенной влаги — 1 или - 2 бар, но понижение при 10 бар было выражено гораздо сильнее у сосен желтой и скрученной, чем у дугласии или пихты великой (рис. 12.13). По сообщению Рингита (1952), низкая влажность почвы так сильно понижала транспирацию масличных пальм, что деревья больше испаряли в дождливый сезон, чем в засушливый, несмотря на то, что атмосферные факторы были более благоприятны для транспирации во время засухи. Паркером (1949) показано, что изменение отношения поверхности корней к поверхности листьев влияет на транспирацию слабо укорененных
352
Глава 12
сеянцев древесных растений, но это влияние было незначительным у видов с густыми корневыми системами. Холодная почва и высокое содержание солей также понижают поглощение воды.
ИЗМЕРЕНИЕ ТРАНСПИРАЦИИ
Первые количественные измерения потери воды растениями были, по-видимому, сделаны Стивеном Гейлсом до 1727 г. Он измерял потерю воды, взвешивая выращенные в горшках виноград, яблони, лимоны и различные травянистые растения. Ряд исследований был проведен во второй половине XIX в., из которых наиболее известны работы Хенеля, опубликованные в 1881 и 1884 гг. Ранние работы по транспирации обобщил Рабер (1937), а Миллер (1938) проанализировал методы измерения транспирации.
В последние годы произошли существенные изменения взглядов на то, какие измерения транспирации могут быть полезными. Первоначально наибольшее внимание уделялось проведению измерений потери воды отдельными растениями, отдельными листьями или побегами. В настоящее время привлекает все больший интерес определение испарения воды растительным покровом или фитоценрзами. Измерения транспирации отдельных листьев или веток имеют большое значение в физиологических исследованиях, но они не могут быть достоверно экстраполированы для вычисления потери воды целыми растениями или фитоценозами. Тем не менее эти методы будут рассмотрены, так как часто только они имеются в распоряжении исследователя.
ГРАВИМЕТРИЧЕСКИЕ МЕТОДЫ. Со времен Гейлса (1727) и до настоящего времени исследователи выращивают растения в сосудах и измеряют потерю воды, взвешивая сосуды через определенные промежутки времени. Массу почвы, в которой растет растение, необходимо помещать в водонепроницаемый сосуд и пополнять запас почвенной влаги так часто, чтобы водоснабжение не становилось ограничивающим фактором. Размеры исследуемых растений зависят от величины сосуда, который можно взвесить. Ваймаер (1927) измерял транспирацию небольших деревьев, выращенных в баках с почвой весом около 450 кг. Нутман (1941) использовал весы грузоподъемностью более 225 кг и чувствительностью 25 г для измерения потери воды кофейными деревьями. Однако во многих ранних исследованиях сосуды были, по-видимому, слишком малы для того, чтобы получать достоверные результаты.
Полученные этим методом данные полезны для таких физиологических исследований, как сравнение потери воды отдельными деревьями. Однако является спорным, насколько приложимы результаты этих исследований к деревьям, растущим в нормальных условиях внешней среды. Обычно растения выращивают на почве, поддерживая влажность около полевой влагоемкости, вместо того, чтобы подвергать их действию циклов подсыхания и увлажнения, характерных для полевых условий. К тому же растения обычно выдерживаются в ненормальных атмосферных условиях. Один из способов приблизиться к естественным условиям — на участках, где эти растения нормально произрао
Значение воды и процесс транспирации
353
тают, устанавливать сосуды в ямы так, чтобы их верхний край был на одном уровне с поверхностью почвы. Так делали Бисвел (1935) и Хольх (1931) при исследовании транспирации сеянцев древесных растений на солнце и в тени в штате Небраска. Усовершенствованный метод — использование лизиметров — приборов, вмещающих большие массы почвы, на которой можно выращивать растения, и при помощи которых можно точно измерять изменения содержания воды. Некоторые типы лизиметров описал Таннер (1967). Вероятно, самый большой из лизиметров для древесных растений был сконструирован Фритшеном с соавторами (1973): он вмещал монолит почвы 3,7 х х 3,7 х 1,2 м, в котором была укоренена дугласия высотой 28 м. Лизиметр установили в лесу, где росло это дерево. Контейнер, почва и дерево имели массу 28900 кг, а чувствительность измерительного аппарата составляла 630 г.
ИЗМЕРЕНИЕ ТРАНСПИРАЦИИ СРЕЗАННЫХ ПОБЕГОВ. Вследствие ограниченных размеров растений, выращиваемых в сосудах, и сроков измерений, а также из-за требуемых для этого расходов многие ученые определяли потерю воды отделенными листьями или побегами. Обычно измерения проводятся в течение нескольких минут после срезания, так как интенсивность транспирации имеет тенденцию к понижению, хотя иногда в первые минуты после срезания возможно временное увеличение (эффект Л.А. Иванова). Этот метод использовали многие исследователи. Славик (1974) подробно обсуждает этот метод, а Франк и Магалис (1965) — его недостатки. Рингот (1952) сравнил транспирацию отдельных побегов и горшечных сеянцев и установил, что интенсивность, полученная при измерении потери воды листьями за 2-минутные интервалы, была выше, чем у горшечных экземпляров. Различия были выражены сильнее у светолюбивых растений, например у масличной пальмы, чем у растущих в тени. Исследователь сделал вывод, что взвешивание отдельных частей растений позволяет оценить влияние физических факторов на испарение, тогда как взвешивание интактных горшечных растений дает результаты, характеризующие взаимодействие растения с внешними факторами, влияющими на транспирацию. Халеви (1956) обнаружил, что в осенние дни с умеренной погодой интенсивность транспирации срезанных листьев и горшечных растений апельсина была сходной, а в жаркие сухие дни последние испаряли на 25—50 % слабее, чем отделенные листья. Погрешность изме-
Рис. 12.14. Интенсивность транспирации отделенных ветвей:
1 — сосны желтой; 2 — пихты великой; числа на кривых показывают относительное содержание воды (относительную тургесцент-ность) в момент полного закрывания устьиц; устьица сосны желтой при водном стрессе закрываются быстрее устьиц пихты великой, а кутикулярная транспирация у хвои сосны происходит менее интенсивно, чем у пихты великой (Lopushinski, 1969)
Время, ч
354
Глава 12
Рис. 12.15. Ход изменений в течение дня:
1 - транспирации; 2 — скорости протекания пасоки Данные получены для сосны алепской н Тэмпе, Аризона (Decker и Skau, 1964)
рений транспирации срезанных листьев увеличивается из-за больших отклонений отдельных листьев одного и того же растения и листьев разных растений одного вида. Этот метод, несмотря на его недостатки, использовался для определения различий в интенсивности
транспирации листьев, расположенных на разной высоте и на северной и южной сторонах деревьев, а также для сравнения разных видов (см. табл. 12.8). Хиген и другие исследователи использовали разновидности f этого метода для измерения потери воды срезанными листьями и для определения момента, когда устьичная транспирация прекращается и поте- $ ря воды происходит в основном через кутикулу (рис. 12.14). ?
ОБЪЕМНЫЕ МЕТОДЫ. Другой метод изучения потери воды — из- ! мерение поглощения воды срезанным листом или веткой. К сожале- f нию, интенсивность транспирации срезанной ветки может быть совсем Г иной, чем у неотделенной, так как ветка на растении конкурирует за воду со всеми остальными ветвями, а ее водоснабжение зависит также от скорости поступления воды через корни. Кроме того, поглощение воды отделенными листьями и ветками часто уменьшается вследствие закупорки проводящей системы воздухом и обрывками тканей. Более достоверных результатов можно достичь при помощи порометров, в которые погружают всю корневую систему, если растения выращивались в водной культуре. Погружение в воду корневых систем, выросших в почве, может сопровождаться ослаблением поглощения воды, водным дефицитом и закрыванием устьиц^ Робертс (1977) успешно > измерил транспирацию деревьев сосны обыкновенной высотой до ; 16 м, помещая срезанные основания стволов в резервуары с водой и поддерживая верхушки в их первоначальном положении в пологе.
УЧЕТ ВЫДЕЛЯЕМОГО ВОДЯНОГО ПАРА. Многие измерения тран- ... спирации выполнены путем определения изменения содержания воды в воздухе, просасываемом через приемник, в который помещают ис- ; следуемый материал. Приемники, обычно изготавливаемые из пластмассы, имеют разные размеры — от кювет, вмещающих один лист, до пластмассовых палаток, достаточно больших, чтобы окружить не- г большие деревья. В открытых системах определяют разность между ( содержанием воды в воздухе, входящем в приемник и выходящем из него, а в закрытых системах измеряют увеличение содержания вдды. Использовались многие методы для измерения изменений содержания
Значение воды и процесс транспирации .
355
воды в воздухе, проходящем над частями растений, в том числе: поглощение пара некоторыми гигроскопическими веществами, применение термометров с влажным и сухим шариками, или термопар, коронного гигрометра, микроволнового гигрометра, резисторных гигрометров и инфракрасных газоанализаторов, регистрирующих количество водяного пара.
Использование приемников исключает погрешности, вызываемые отделением листьев или ветвей, но этот метод создает до некоторой степени искусственную внешнюю среду вокруг листа или растения, помещенного в приемник. Изменения температуры листьев и воздуха, скорости ветра и влажности могут вызвать существенные различия между интенсивностью транспирации, измеренной этими методами и у полностью открытых растений. Один из способов регуляции внешних условий в камерах - обеспечить непрерывную компенсацию изменений влажности и концентрации СО£, как это делали Келлер, Сатит (1964) и Мосс (1963).,
Короткий период измерений потери воды устраняет некоторые погрешности, вызываемые длительным пребыванием листьев в приемнике. Гриф и Вент (1965) описали способ применения стандартного датчика влажности с хлоридом лития для измерения изменения влажности в приемнике для одного листа. Некоторые исследователи установили соответствие между интенсивностью транспирации, измеренной методом взвешивания, и при использовании разновидности этого прибора. Измерения транспирации, требующие менее 1 мин, можно также сделать при помощи дифференциального психрометра. Хлоркобальтовым методом измеряют выделение водяного пара за время, необходимое для стандартного изменения окраски куска бумаги, пропитанной хлоридом кобальта и прижатой к листу. Однако интенсивность транспирации, измеренная этим методом, не соответствует величинам, полученным гравиметрическими методами, так как листья в лечение значительного времени находятся в ненормальной атмосфере.
СКОРОСТЬ ПРОТЕКАНИЯ ПАСОКИ. Некоторые исследователи пытались определять интенсивность транспирации измерением скорости протекания пасоки по стволам деревьев, используя некоторые разновидности метода тепловых импульсов, описанного Хибером и Шмидтом (1937). Ладефогед (1963) применил диатермическую машину в качестве источника тепла и сообщил о быстрых реакциях на прохождение облаков и порывы ветра и о надежных указаниях на направления сезонных изменений. Он измерял поглощение воды стволами деревьев, срезанных и помещенных в резервуары с водой, и на основании корреляции между поглощением и скоростью протекания пасоки у этих деревьев подсчитывал методом экстраполяции интенсивность транспирации интактных деревьев. Декер и Скау (1964) установили хорошую корреляцию между одновременно измеренными значениями скорости передвижения пасоки и интенсивностью транспирации (рис. 12.15). Свенсон и Лей (1966) рассмотрели преимущества и недостатки этого и других методов учета транспирации и сделали вывод, что хотя по скорости протекания пасоки трудно определить транспирацию, но эти измерения полезны тем, что показывают, когда транспирация начинается утром, как быстро она увеличивается и как влияют на нее из
Глава 12
менения атмосферных условий. Некоторые данные этих авторов указывают на измеримое запаздывание понижения скорости протекания пасоки около основания древесного ствола после того, как дождь вызывал уменьшение транспирации. Подобное запаздывание наблюдается утром, когда транспирация на верхушке дерева быстро возрастает, по-видимому, благодаря способности стволов деревьев запасать воду. Лейтон (1970) обобщил информацию относительно метода тепловых импульсов.
ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ \
Выполнены многочисленные измерения интенсивности транспирации деревьев и кустарников различных видов и разного возраста при всевозможных условиях. Некоторые результаты более ранних исследований обобщил Рабер (1937), а многие другие содержатся в серии статей Пизека и его сотрудников, на которые ссылается Лархер (1975). Приведем несколько примеров. В целом пределы изменений интенсивности транспирации древесных и травянистых растений сходны. Существуют большие различия этого показателя у отдельных особей и у листьев разного возраста и расположения на одном и том же растении. Интенсивность транспирации на единицу листовой поверхности у сосны ладанной ниже, чем у широколистных видов, а у падуба и гардении, имеющих кожистые листья, интенсивность транспирации такая же, как у деревьев с мезоморфными листьями, например у тюльпанного дерева и дуба красного. Результаты определения транспирации сеянцев двух листопадных твердолиственных пород и сосны ладанной (см. табл. 12.4) показывают, что хотя на единицу листовой поверхности лиственные транспирировали приблизительно в 2 раза интенсивнее, чем сосна, транспирация в расчете на один срянец такого же размера была больше у сосны, так как она имела большую листовую поверхность. Уже давно Грум (1910) обнаружил, что интенсивность транспирации на единицу листовой поверхности у хвойных ниже, чем у лиственных, а в расчете на одно дерево сходного размера — одинакова.
Рис. 12.16. Сезонный ход транспирации горшечных сеянцев вечнозеленых и листопадных деревьев в Дареме (Северная Каролина) :
1 — тюльпанное дерево; 2 — сосна ладанная
Значение воды и процесс транспирации
357
Сезонная цикличность транспирации листопадных и вечнозеленых видов показана на рис. 12.16. В этом эксперименте у сосны ладанной зимняя транспирация составляла в среднем около 10 % ее интенсивности в середине лета. В другом эксперименте Козловский (1943) измерял транспирацию вечнозеленых и листопадных видов в конце декабря и в начале января (табл. 12,7) и установил, что интенсивность транспирации сосны ладанной ниже, чем у обнаженных ветвей листопадных растений. Обе серии измерений проводились в Дареме (Северная Каролина), но погода во время исследований Козловского была, вероятно, необычно холодной. Интенсивность зимней транспирации хвойных в Небраске (Вавер, Могенсен, 1919) и Ленинграде (Иванов, 1924) составляла менее 1 % интенсивности летней транспирации. Однако в этих исследованиях зимние температуры были, по-видимому, гораздо ниже обычно наблюдаемых в Северной Каролине, где часто бывают теплые солнечные дни. Известно, что низкие температуры сильно понижают транспирацию у хвойных. Даже в некоторых тропических районах наблюдаются сезонные циклы транспирации вследствие изменения количества осадков, влажности воздуха и почвы. Например, Ринг от (1952) обнаружил большие сезонные колебания транспирации у масличных пальм, растущих в Заире.
При измерении транспирации неизбежны погрешности, поэтому у читателей могут возникнуть сомнения относительно значения каких-либо приведенных конкретных величин интенсивности этого процесса. Однако некоторые данные являются более достоверными, чем можно было ожидать. Например, интенсивность летней транспирации горшечных сеянцев тюльпанного дерева составляла в среднем 10,1 г/дм^-сутв Колумбусе (Огайо) и 11,7 г/дм2 - сут в Дареме (Северная Каролина). Сравнение транспирации, измеренной двумя методами и четырьмя разными исследователями (табл. 12.8), показывает, что из шести видов древесных растений у березы была самая высокая интенсивность транспирации, у ели — самая низкая. Губер (1953) также отметил соответствие относительной интенсивности транспирации многих видов, из-
12.7. ИНТЕНСИВНОСТЬ ЗИМНЕЙ ТРАНСПИРАЦИИ (С 22 ДЕКАБРЯ ПО 5 ЯНВАРЯ) НА ЕДИНИЦУ ПОВЕРХНОСТИ СЕЯНЦЕВ ХВОЙНЫХ И ШИРОКОЛИСТВЕННЫХ ДРЕВЕСНЫХ РАСТЕНИЙ (Kozlowski 1943)
Вид Интенсивность транспирации, г/дм2’ , сут Вид Интенсивность транспирации, г/дм2 ’ сут
Prunus laurocerasus* Piniis taeda* 1,65 0,40 Liriodendron tulipifera** 0,76
Acer saccharum**
0,26 Quercus alba** 0,84
*Ha единицу поверхности листьев.
** На единицу поверхности побегов и ствола.
Глава 12
НОСИТЕЛЬНАЯ ИНТЕНСИВНОСТЬ ТРАНСПИРАЦИИ РАЗЛИЧНЫХ ’НЫХ ПОРОД (Hubei, 1953)
Ис след ов атель
Эйд мал* Губер** Пизек и Картель-ер и** Польстер*
618 541 740
282 \ 468 (377-559) 512 460
268 379 (218-541) 137 372
[Я 130 — — 94
100 100 (64-136) 100 100
181 134 (118-150) 133 . 139
щица 310 409 (341-476) 212 212
е получены с горшечными растениями.
..те получены методом быстрого взвешивания срезанных ветвей.
юй в разные дни. Принципиальные трудности возникают при \ках экстраполировать результаты измерений, выполненных на Чих или отдельных листьях, на целые древостои.
Чя ВОДЫ ФИТОЦЕНОЗАМИ
‘доводов, садоводов и агрономов больше интересует количество Л теряемой фитоценозами, чем отдельными растениями. Потеря Ч лесом, фруктовым садом, полем или лугом включает как транс-рию растений, так и испарение с поверхности почвы. Совокуп-этих процессов обычно называют эвапотранспирацией.
ЛТОДЫ ИЗМЕРЕНИЯ. Раньше делались попытки вычислить поте-Лды лесом по среднесуточной интенсивности транспирации на еди-7 листовой поверхности и площади листьев на единицу поверхности J4i. Используя этот метод, Польстер (1950) вычислил интенсивность Чпирации германских лесов в пределах от 5,3 мм/сут для дугласии мм для березы до 2Д мм для сосны обыкновенной. В середине ’Л столетия лесоводы и экологи обратились к другим методам. С определения потери воды земной поверхностью наиболее часто уЧзуются четыре основных метода: 1) применение уравнения вод-? баланса; 2) применение уравнения энергобаланса; 3) аэродинами-метод (перетекания водяного пара); 4) подсчет по эмпири-'jW формулам на основании метеорологических данных или интен-:^сти эвапорации. Полезные данные были также получены при по-лизиметров, например в Голландии и в Сан-Димасе (Калифор-или на регулируемых водоразделах, как, например, в Африке,
Лцарии, Колорадо, Северной Каролине и Западной Виргинии.
акторы, контролирующие, эвапотранспирацию. испаре-^сды почвой и транспирацию растений можно рассматривать как
Значение воды и процесс транспирации 359
альтернативные пути передвижения воды в массу воздуха. Как упоминалось выше, испарение зависит от притока энергии, требуемой для превращения воды в пар, от разности давления пара между испаряющей поверхностью и массой воздуха и от сопротивлений на пути диффузии пара. Выделение водяного пара растительностью (транспирация) усложняется регуляцией этого процесса растением наряду с внутренним сопротивлением и различными свойствами испаряющих поверхностей. Несмотря на это, эвапотранспирацию ,7?-можно описать, несколько модифицируя уравнение (12.6):
С — С е — е
внутр. возд. внутр. возд.
Е =-----—-------------- =---------------------
внутр, возд, внутр. возд. (12,9)
Индекс ’’внутр” относится к внутренней части пути между испаряющей поверхностью и поверхностью почвы или растений. Во влажных почвах г внутр можно пренебречь, а СвнуТр - это поверхность почвы, но при ее высыкании испаряющая поверхность отходит в глубь почвы, г внутр становится значительным, а интенсивность эвапорации снижается.
Если эпидерма сухая, то испаряющие поверхности находятся внутри листьев, но в фитоценозах их поверхность иногда принимают за испаряющую поверхность. Однако это является сильным упрощением, потому что как атмосферные условия, так и состояние листьев изменяются от нижней части растительного покрова до его верхушки. Вагонер и Райфснидер (1968) разработали модель для оценки взаимодействия факторов, влияющих на испарение внутри полога, если известны падающая радиация, сопротивления и площадь листьев на разных уровнях.
Количество воды, испаряемой с единицы земной поверхности, зависит прежде всего от имеющейся в наличии энергии. Поскольку в ясный летний день на испарение может быть использовано 320 380 Гкал/см^, а для испарения 1 г воды требуется 570 кал, максимально возможная испаряемость составляет приблизительно 6 мм в день. Однако иногда падающая энергия дополняется адвекцией (горизонтальным потоком) энергии от окружающей среды, как это происходит у свободно стоящих деревьев или на возделываемых полях, окруженных пустыней (так называемый ’’оазисный эффект”). Несмотря на то, что площадь листьев фитоценозов обычно в 4—6 раз больше поверхности почвы, на которой они растут, потеря воды не может превышать испарения с влажной почвы или со свободной водной поверхности, получающей такое же количество энергии.
ВЛИЯНИЕ ИЗМЕНЕНИЙ РАСТИТЕЛЬНОГО ПОКРОВА. Лесоводы, гидрологи и агрономы интересуются влиянием изменений состава, высоты и густоты растительности в фитоценозах на интенсивность потери воды. Часто утверждают, что скорость испарения воды сходна у всех типов сомкнутого растительного покрова, в том числе у лугов, сельскохозяйственных культур и лесов, до тех пор, пока почва остается влажной. Однако, как предупреждает Ридер (1957), это утверждение следует принимать с осторожностью, так как альбедо и внутреннее
360
Главе 12
12.9. ИСПАРЕНИЕ ВОДЫ ЛЕСОМ И ПРИМЫКАЮЩИМ ЛУГОМ, РАСТУЩИМИ НА ВЛАЖНОЙ ПОЧВЕ, MM/ГОД (Rutter, 1968)
Тип леса Лес Примыкающий луг Луг: лес
Picea sitchensis 800 416 0,52
Picea abies 579 521 0,90
Сметанный хвойно-лиственный 861 696 0,81
Смешанный лиственный — 0,8-1,0
Eucalyptus niphophila — — 1,0
сопротивление у разных типов растительности различны. Светлоокрашенный покров, по-видимому, теряет меньше воды. По мнению Руте-ра (1968), устьичное сопротивление в лесных фитоценозах, вероятно, несколько выше, чем в сообществах травянистых растений. Однако поверхность лесного полога часто бывает более неровной, чем у низкорослых сельскохозяйственных растений, а это должно приводить к большей турбулентности, уменьшая гвн^,по сравнению с этим показателем в культурах с однородной поверхностью. Несмотря на это, большинство исследований показывает небольшие различия между видами древесных растений и между лесами и лугами до тех пор, пока водоснабжение не является ограничивающим фактором. Потеря воды лугами составляет около 80 % или более от потери воды лесами. При недостатке воды в почве эвапотранспирация снижается сильнее у лугов, чей у лесов, отчасти, возможно, из-за более поверхностной корневой системы. Результаты некоторых исследований приведены в табл. 12.9.
ПРОРЕЖИВАНИЕ. Прореживание лесных древостоев может понижать эвапотранспирацию или не изменять ее, Кнорр (1965) изучал истощение запасов почвенной воды под насаждениями пихты великолепной с покрытием от 50 до 100 % и обнаружил максимальное поглощение воды древостоями с 70—80 %-ным покрытием, Гевлет и Гиберт (1961) установили, что усиление притока воды с водораздела в Кауита (Северная Каролина) было почти пропорционально сокращению площади оснований стволов. Частично это увеличение могло быть вызвано меньшим перехватом осадков (интерцепцией) прореженными древостоями. Удаление растительности подлеска также, по-видимому, уменьшает потерю воды. Влияние прореживания лесных насаждений бывает, очевидно, различным, хотя по сообщению Гиберта (1967), результаты 30 исследований показывают, что обычно уменьшение лесного покрова увеличивало запас воды, а возобновление леса совровождалось его уменьшением.
ИЗМЕНЕНИЯ ВИДОВОГО СОСТАВА. Влияние изменения видового состава лесного насаждения на запас воды усложняется вследствие интерцепции осадков и различиями в глубине корневых систем и продолжительности периода транспирации. Большинство экспериментов, на которые ссыпается Рутер (1-968), показывают небольшие различия между видами, по крайней мере летом. Измерения, проведенные в Дании и Голландии, указывают на то, что, как и следовало ожи-
WHM воды и процесс транспирации
361
Нйн>, потеря воды за год больше у хвойных, чем у листопадных пород. И одном из экспериментов в Каунта (Северная Каролина) через 15 лет Нт не преобразования спелого,лиственного леса на двух эксперимен-нньных водоразделах в насаждений сосны веймутовой ежегодный сток нииштился на 20 %. Уменьшение стока наблюдалось ежемесячно, но пяипольшее — во время покоя и в начале вегетационного периода, когда индекс листовой поверхности составлял 9,9 для сосны и менее 1 для' лиственных пород. Вследствие этого происходили большая интерцеп-цин и последующее испарение с поверхности сосны по сравнению с этими показателями голых лиственных деревьев. Результаты измерения <»Н»ка показывают, что общие потери воды путем интерцепции и транспирации в течение вегетационного периода были также больше у сосны.
ОТНОСИТЕЛЬНЫЕ ПОТЕРИ ВОДЫ ПУТЕМ ЭВАПОРАЦИИ И ТРАНСПИРАЦИИ. В тех местах, где запас воды имеет большое значение, представляют большой интерес относительные количества воды, теряемой путем эвапорации и транспирации. На всей территории Соединенных Штатов около 1/4 всех осадков уходит в виде стока, а 3/4 воз-н pin дается в атмосферу посредством эвапотранспирации. В предыдущих разделах было отмечено, что, по-видимому, нет больших различий и потере воды разными типами растительности, растущей на влажной почве. Однако при высыхании почвы могут приобретать значение различия в глубине расположения корней и в регулировании растениями грпнспирации. Утверждают, что как в Центральной долине Калифорнии, так и в южной части Калифорнии превращение Чапарраля в луг ’ишчительно увеличивает сток воды. Сплошная рубка сосны скрученной в Колорадо увеличивала сток приблизительно на 30 %, а удаление всей древесной растительности с водораздела в Кауита (Северная Каролина) — более чем на 70 % в первый год. Это увеличение было выражено сильнее на склонах, обращенных на север. Влияние удаления лесно-i о покрова на сток зависит от суммы и сезонного распределения осадков и от количества осадков, выпадающих за 1 раз во время грозы. Следовательно, сток не был точным показателем интенсивности транспирации. Фактическая потеря воды посредством транспирации в лесах Кауита была, вероятно, значительно больше количества, измеренного по увеличению стока после сплошной рубки. Патрик с соавторами (1965) измерил истощение запасов почвенной влаги под древостоями, росшими на почве, покрытой пластмассовой пленкой для предотвращения потери воды путем испарения. Было установлено, что транспирация с апреля по октябрь включительно соответствовала 41 см у 21 -летнего насаждения сосны ладанной и 37,2 см у древостоя дуба и карии. По сообщению Стольфальтэ (1963), испарение с поверхности почвы под открытым насаждением ели европейской составляло более 20 % общей потери воды, тогда как Рутер (1968) установил, что только 8 % терялось путем эвапорации под древостоем сосны обыкновенной. Баумгартнер (1967), применив метод энергетического баланса, вычислил количество воды, испаряемой почвой под лесами, лугами и сельскохозяйственными культурами, которое оказалось равным 10, 25 и*45 % общей потери воды.
Деревья могут иногда приживаться в районах с ограниченным количеством осадков, но они отмирают при истощении запасов влаги
362
Глава 12
в почве. Многие насаждения лесных деревьев были созданы в прерии и на равнинах в 1880-х и 1890-х годах. Большая часть этих деревьев росла хорошо несколько лет, но затем они начали отмирать, так как исчерпали все запасы почвенной влаги и не могли выжить за счет осадков, выпадающих в течение засушливых периодов.
МЕТОДЫ УМЕНЬШЕНИЯ ТРАНСПИРАЦИИ. Ущерб, наносимый чрезмерной транспирацией, вынуждает искать пути сокращения потери воды. Усилия исследователей сосредоточены на решении трех проблем. Уменьшение транспирации после пересадки давало бы возможность растениям сохранять тургор до восстановления их корневых систем. Сокращение транспирационных потерь во время засух помогло бы выживанию растений с наименьшим ущербом, а уменьшение транспирации растительного покрова на водоразделах увеличило бы запас воды, которая пригодилась бы для других целей. Используемые методы заключаются в основном в применении водонепроницаемых покрытий или материалов, вызывающих закрывание устьиц. Вещества, предназначенные для уменьшения транспирации, часто называют антитранспирантами.
Ряд первоначальных исследований был основан на погружении или опрыскивании надземных частей сеянцев различными веществами: эмульсией латекса, соединениями поливинила, полиэтилена и винил акрилата. Наблюдалось некоторое уменьшение транспирации и повышение выживаемости, однако Ален (1955) предложил провести дальнейшее исследование этих покрытий, перед тем как рекомендовать их применение. Лей и Козловский (1974), Дэвис и Козловский (1975) проанализировали недостатки, встречающиеся при использовании анти транспирационных покрытий сеянцев древесных растений. По-видимому, их эффективность зависит от вида и фазы развития растения и от атмосферных условий в течение периода исследований. По мнению некоторых исследователей, антитранспиранты не уменьшают повреждения вечнозеленых деревьев при температуре воздуха ниже точки замерзания, но могут быть эффективными веской, когда температура воздуха повышается, а почва остается холодной или замерзшей.
Польза от нанесения пленок на растущие растения ограничивается тем, что по мере увеличения листовой поверхности необходимы повторные обработки, а также из-за того, что вещества, непроницаемые для водяного пара* непроницаемы и для двуокиси углерода и понижают интенсивность фотосинтеза. В настоящее время пленки полезны только там, где сокращение транспирации более важно, чем высокая интенсивность фотосинтеза. Примером может служить их использование на плодовых деревьях для увеличения размеров созревающих плодов и улучшения их лежкости после уборки.
Представляется более перспективным применение веществ, вызывающих закрывание устьиц, так как их частичное закрывание в большей мере уменьшает транспирацию, чем фотосинтез. Это утверждение основано на наблюдениях Гастра (1959), согласно которым сопротивление мезофилла поступлению двуокиси углерода значительно ^выше сопротивлений, влияющих на выход водяного пара. Таким Образом, большое повышение г $ должно слабее влиять на поглощение двуо
Значение воды и процесс транспирации
363
киси углерода, чем на потерю воды. В опытах Слатира и Бируцена (1964) закрывание устьиц фенилмеркур ацетатом понижало транспирацию больше, чем фотосинтез. С другой стороны, имеются сообщения о том, что фотосинтез и транспирация уменьшались в равной степени при закрывании устьиц у хлопчатника, томатов и сосны ладанной.
Одним из. наиболее перспективных соединений, вызывающих закрывание устьиц, оказался фенилмеркурацетат, который, по сообщению Вегенера и Браудо (1967), значительно понижал транспирацию целого соснового древостоя. Однако нежелательно добавлять еще большие количества ртутных соединений- в нашу и так уже загрязненную внешнюю среду. К тому же, как сообщили Вайзель с соавторами (1969), фенилмеркурацетат повреждает листья березы. Келлер (1966) обратил внимание на то, что фенилмеркурацетат и образующее пленку соединение поливинила задерживали фотосинтез и рост корней сеянцев ели, причем первый повреждал и хвою. Имеются также сообщения о том, что фенилмеркурацетат замедляет вегетативный рост чая. После того как было обнаружено, что абсцизовая кислота тормозит открывание устьиц, ее испытали в качестве антитранспир анта. Дэвис и Козловский (1975) установили, что эта кислота понижает транспирацию сеянцев клена сахарного, ясеня американского и каламондина приблизительно на 60 %, причем некоторое уменьшение сохранялось до 21 дня после обработки. Токсических эффектов не наблюдалось. В другой серии экспериментов* силиконовое покрытие оказалось более эффективным средством сокращения транспирации сеянцев ясеня американского и сосны смолистой, чем абсцизовая кислота. Однако был сделан вывод о том, что пленкообразующие антитранспиранты, по-видимому, непригодны для обработки некоторых голосеменных, так как они понижали фотосинтез на 95 % по меньшей мере в течение 12 дней. Следовательно, необходимы дальнейшие исследования для разработки эффективных методов регулирования транспирации. (Влияние антитранспирантов на фотосинтез было рассмотрено в главе 5.)
ТРАНСПИРАЦИОННЫЙ КОЭФФИЦИЕНТ. Представляет большой интерес количество воды, требуемое для выработки единицы сухого вещества и иногда называемое транспирационным коэффициентом. Миллер (1938) обобщил многие данные относительно транспирационного коэффициента, который колеблется от 200 до 1100 г воды на 1 г сухой массы у разных сортов сельскохозяйственных растений. Самый низкий транспирационный коэффициент (около 50) был отмечен для ананаса. Слатир (1967) указывает, что транспирационный коэффициент может доходить до 5000 в полупустынях, где растения расположены на большом расстоянии друг от друга и относительно мало энергии улавливается листьями. Для деревьев данные немногочисленны. Польс-тер (1950) подсчитал, что в ГДР количество воды, требуемой для выработки 1 г сухого вещества, колеблется от 170 у бука и дугласии до 300 г у сосны обыкновенной, 317 у березы и 344 у дуба черешчатого. Кох (1957) утверждает, что отношение фотосинтеза к транспирации складывается более благоприятно при облачной погоде, чем при ясной солнечной, потому что транспирация понижается в большей степени, чем фотосинтез. Транквилини (1969) проделал интересные вычисле
364
Глава 12
ния так называемой продуктивности транспирации (отношения фотосинтеза к транспирации) при различной скорости ветра. Это отношение было сходным при всех скоростях ветра у лиственницы и ели. Потеря' воды уменьшалась сильнее, чем фотосинтез, у сосны кедровой европейской, рябины и рододендрона. При более высоких скоростях ветра интенсивность фотосинтеза рододендрона понижалась быстрее, чем транспирация. Фотосинтез ольхи твердой уменьшался быстрее, чем потеря воды, при самой низкой скорости ветра.
ЭФФЕКТИВНОСТЬ ИСПОЛЬЗОВАНИЯ ВОДЫ. Термин эффектив-. ность использования воды (сокращенно ЭИВ) все шире применяется для описания соотношения между продуктивностью растений и расходованием воды. По определению Бегга и Турнера (1976), ЭИВ — это отношение выработанного сухого вещества к количеству воды, израсходованной путем эвапотранспирации. Этот показатель включает в себя потерю воды как при транспирации, так и при эвапорации, поэто-„му он дает характеристику условий, существующих в поле и в лесу, более реалистичную, чем транспирационный коэффициент.
Как указывалось выше, трудно что-дибо сделать для изменения интенсивности эвапотранспирации, так как она контролируется в основном приходящей энергией, которая может расходоваться на испарение воды. Однако в земледелии* эффективность использования воды может быть повышена увеличением густоты растительного покрова, чтобы лишь небольшая часть радиации доходила до почвы, а эвапорация оставалась минимальной. При таких условиях максимально возможная часть падающей энергии может использоваться на фотосинтез и образуется наибольшее количество сухого вещества на единицу испаренной воды. Иногда максимальное образование сухого вещества менее важно в лесном хозяйстве, чем другие аспекты, например, приток воды с облесенных водоразделов. Как указывалось в одном из предыдущих разделов, сток часто усиливается при прореживании или полном удалении всех деревьев. Это происходит в значительной степени потому, что глубоко укоренившиеся деревья извлекают воду с гораздо большей глубины, чем травянистая растительность с более поверхностной корневой системой. Несмотря на широкие исследования, еще не решен вопрос относительно того, какой тип растительного покрова лучше всего использовать на определенных водоразделах, чтобы свести до минимума эрозию и максимально увеличить долгосрочный запас воды.
ПОГЛОЩЕНИЕ ВОДЫ, ПОДЪЕМ КСИЛЕМНОГО СОКА И ВОДНЫЙ БАЛАНС
Транспирация - доминирующий процесс в водообмене растений, так как она и значительной степени определяет скорость поглощения и передвижения воды по растению и вызывает водный дефицит. Непрерывное поглощение достаточного количества воды для возмещения ее потери при транспирации имеет существенное значение для роста и даже для выживания растений.
ПОГЛОЩЕНИЕ ВОДЫ
Большая часть поглощаемой растениями воды поступает в жидком миде через их корни, но небольшое количество поглощается через листья и даже через побеги.
ПОГЛОЩЕНИЕ ЧЕРЕЗ ЛИСТЬЯ И СТЕБЛИ. Количество воды, поглощаемой через листья и стебли, невелико, но этому процессу уделяется большое внимание. Ранние литературные данные, обзор которых сделал Миллер (1938), показывают, что в листья многих растений могут проникать значительные количества воды. Кутикула при увлажнении становится довольно проницаемой, что делает возможным внесение удобрений через листья (см. главу 10). Некоторое количество минеральных питательных веществ и, по-видимому, воды поглощается через чечевички и другие щели в коре и даже через листовые рубцы. Поглощение росы и ее значение для водного режима растений было рассмотрено Стоуном (1957). Стоун и Фовеллс (1955) показали, что в тепличных экспериментах роса повышала выживаемость сеянцев сосны желтой. Однако существуют большие разногласия относительно ее значения в полевых условиях. Некоторые исследователи считают, что настоящая роса, т. е. атмосферная влага, конденсирующаяся на листьях, не имеет большого практического значения вследствие малого количества воды, поставляемого этим процессом. Другие подсчитали, что максимально возможное количество росы, осаждающейся за ночь, составляет менее 0,5 мм, а сообщения о более высоких величинах были вызваны погрешностями измерений, в частности включением воды, испаряемой почвой. Считают также, что роса имеет значение не только для выживания, но даже для роста трав и древесных растений. Гиндел (1966, 1973) утверждает даже, что сосна алеппская поглощает влагу из воздуха и передает ее в корни, где она выделяется в почву, увеличивая содержание воды в прикорневой зоне. Однако это повышение влажности происходит, возможно, потому, что более холодная затененная почва под деревьями действует как ’’ловушка” водяного пара.
Поглощение воды сеянцами сосны из насыщенной атмосферы было исследовано Стоуном с соавторами (1950). Слатир (1956) наблюдал поглощение воды из насыщенного воздуха корнями или побегами сеянцев сосны и ее передвижение через сеянцы, если создавался большой градиент водного потенциала. По сообщениям, вода поглощалась из воздуха, транспортировалась по растению и выделялась в почву.
Процесс противоположный поглощению через листья — вымывание минеральных веществ из листьев дождем или при орошении посред-
366 Глава 13
13.1. ОТНОСИТЕЛЬНАЯ ПРОНИЦАЕМОСТЬ ДЛЯ ВОДЫ И 32Р КОРНЕЙ ВИНОГРАДА РАЗНОГО ВОЗРАСТА (ИЗМЕРЕНИЯ ПРОВОДИЛИСЬ ПРИ ГРАДИЕНТЕ ДАВЛЕНИЯ 660 МИЛЛИБАР) (ПО Queen, 1967)
Зона и состояние корней Относительная проницаемость
для ВОДЫ для 32Р
Корни текущего сезона (растущие) Терминальные 8 см, удлиняющиеся, неразветвленные, 1,0 1,0
неопро бковев шие Неопробковевшие с удлиняющимися ответвлениями 155,0 75,0
(покоящиеся) Главная ось и ответвления, покоящиеся и частично опробковевшие до окончанйя удлинения Главная ось и ответвления покоящиеся и частично 545,0 320,0
65,0 35,0
опробковевшие Корни предшествующих сезонов (сегменты с ответе-
пениями) Сильно опробковевшая главная ось с многими корот- 0,2 0,04
кими опробковевшими ответвлениями (неразвет- вленные сегменты) Сильно опробковевшие, с толстой корой и относительно небольшим цилиндром ксилемы: интактные 0,2 0,02
после снятия коры 290,0 140,0
ством дождевания. Выпадение росы и тумана также вызывает выщелачивание. По некоторым исследованиям лиственные растения теряют летом больше питательных веществ из-за их вымывания, чем хвойные, однако у последних этот процесс продолжается и зимой. Если растворимые вещества могут вымываться, то, очевидно, может происходить поглощение воды и растворенных веществ, когда жидкость находится на поверхности листьев и градиент водного потенциала является благоприятным.
ПОГЛОЩЕНИЕ ЧЕРЕЗ КОРНИ. Корневые системы древесных многолетних растений состоят из корней, находящихся на всех стадиях развития, — от тонких, недавно образовавшихся, неопробковевших кончиков диаметром менее 1 мм до старых, одревесневших корней, покрытых толстых слоем коры. Кроме того, корни часто бывают видоизменены из-за наличия микоризных грибов. В результате у разных корней наблюдаются большие различия в проницаемости для воды и солей (табл, 13.1). На рис. 13.1 показаны ткани, через которые должна проходить вода, чтобы проникнуть в молодые корни тюльпанного дерева. Стенки эпидермальных и коровых клеток на этой стадии состоят преимущественно из целлюлозы, но стенки клеток эндодермы уже начинают утолщаться. Полоски опробковевшей ткани (пояски Каспа-ри) образуются на радиальных стенках и, как предполагают, понижают их проницаемость для воды и растворенных веществ. Однако недавние исследования свидетельствуют о том, что эндодерма не всегда является непроницаемым барьером для воды и растворенных
плющение воды, подъем ксилемного сока и водный баланс
367
Рис. 13.1. Поперечный разрез молодого корня тюльпанного дерева на расстоянии около 0,6 мм от верхушки (выделяется толстой слой коровой паренхимы, окружающей стелу) (Popham, 1952, с разрешения автора)
веществ. Например, когда молодые корешки образуются в перицикле и пробиваются через эндодерму, возникают щели, по которым могут легко проникать вода и растворенные вещества, пока эндодермы бо-I ковых и родительских корней не будут соединены друг с другом. 1 Предполагают также, что вода и растворенные вещества могут пере-| двигаться через плазмодесмы в клеточных стенках эндодермы. Ка-। ковы бы ни были детали, вероятно, значительные количества воды 1 и ионов пересекают эндодерму на расстоянии многих сантиметров от
верхушки корней травянистых, а возможно, и древесных растений.
При старении корней эпидерма, корневые волоски и часть коры отмирают вследствие образования пробкового камбия в наружной
368
Глава 13
части коры (рис. 13.2). В дальнейшем, в результате деятельности камбия, утрачивается даже эндодерма и корень составляют ксилема, камбий, флоэма и опробковевшие слои на наружной поверхности флоэмы. Раньше предполагали, что вода и растворенные вещества передвигаются от клетки к клетке через вакуоли клеток, расположенных между поверхностью корня и ксилемой. Эксперименты Штругера (1949) показали, что движение воды может в значительной мере происходить по клеточным стенкам. Эта точка зрения находит все большую поддержку в последние годы. Она подтверждается данными, свидетельствующими о том, что сопротивление току воды через неопробковевшие клеточные
Рис. 13.2. Поперечный разрез более старого корня тюльпанного дерева, у которого' произошло слущивание большей части наружной паренхимы. На поверхности корня развивается слой опробковевшей ткани (Popham, 1952, с разрешения автора)
Поглощение воды, подъем ксилемного сока и водный баланс
стенки значительно ниже сопротивления току через протопласты. Это мнение подвергает сомнению Ньюмэн (1976), который утверждает, что вода передвигается в основном по симпласту. Пояски Каспари, по-видимому, препятствуют передвижению воды через стенки клеток эндодермы, однако, как указывалось выше, существует, вероятно, значительный массовый поток через разрывы в эндодерме, которая не всегда представляет собой сплошной барьер.
Появляется все больше доказательств того, что значительное количество воды поглощается через старые опробковевшие корни древесных растений. У недавно посаженных сеянцев деревьев мало или отсутствуют неопробковевшие корни, при этом некоторые остаются живыми в течение многих месяцев без образования новых корней. Цитрусовые в Калифорнии и сосны на юго-востоке США почти не имеют зимой растущих корней, но они должны поглощать большие количества воды в солнечные дни, чтобы возместить транспирационные потери. Крамер и Буллок (1966) установили, что летом менее 1 % поверхности корней под древостоями тюльпанного дерева и сосны ладанной не было покрыто пробкой, а Хад (1967) обнаружил заметное уменьшение новообразования корней у яблони и сливы в течение лета. Все три автора сделали вывод, что значительные количества воды и солей должны поглощаться через опробковевшие корни (см. табл. 13.1). Более подробная информация о корневых системах приведена в главе 2.
Сопротивление корней. Наряду с изменениями сопротивления потоку воды, вызываемыми созреванием корней, существуют труднообъяснимые изменения наблюдаемого сопротивления корней, связанные с интенсивностью транспирации и потока воды через корни. Некоторые исследователи сообщают об уменьшении наблюдаемого сопротивления корней при усилении транспирации и потока воды, а другие этого не наблюдали. Физус и Крамер (1975) пытались объяснить противоречивые данные тем, что вклад осмотического и массового (гидравлического) потока в общий поток воды по растению изменяется при увеличении его скорости. Имеются также сообщения о суточных изменениях наблюдаемого сопротивления корней с минимумом в полдень и максимумом около полуночи. Эти циклы связаны, по-видимому, с сигналами, поступающими из побегов, так как они устанавливаются в соответствии с условиями чередования света и темноты, в которых находятся побеги. Исследование Бунка (1978) травянистых растений также показывает, что наблюдаемое сопротивление корней зависит от тех или иных воздействий на побеги. Таким образом, условия эксперимента могут существенно влиять на результаты измерений наблюдаемого сопротивления корней.
МЕХАНИЗМЫ ПОГЛОЩЕНИЯ ВОДЫ
В соответствии с терминологией Реннера (1915) существуют два механизма поглощения воды: активное поглощение, обычно наблюдаемое у слабо транспирирующих растений, и пассивное, которое преобладает у интенсивно транспирирующих растений и обеспечивает ббль-
Глава 13
370
шую часть поглощения воды древесными растениями. Необходимо подчеркнуть, что всякое поглощение воды происходит по градиентам водного потенциала — от среды обитания корней до ксилемы. Различие между активным и пассивным поглощением заключается в способе создания этих градиентов.
АКТИВНОЕ ПОГЛОЩЕНИЕ. Корни растений, произрастающих на теплой, хорошо аэрируемой, влажной почве, действуют как осмометры, когда растения транспирируют медленно, потому что накопление солей в ксилемном соке понижает его осмотический потенциал, а следовательно, и водный потенциал, который становится меньше водного потенциала почвы. Возникающая в результате этого диффузия воды, направленная внутрь, создает корневое давление, благодаря которому происходят гуттация и ’’плач”, наблюдаемый при поранении некоторых растений (например, березы и винограда). Делались попытки объяснить корневое давление активной секрецией воды или процессом электроосмоса, однако простая осмотическая теория дает, по-видимому, достаточное объяснение. Нет даже доказательств того, что в растениях происходит активный транспорт воды, а электроосмос, вероятно, не может вызвать передвижение такого объема воды, который вытекает из корневых систем после срезания стеблей.
Основные проблемы заключаются в объяснении того, как концентрируются ионы в ксилеме и что служит барьером для ионов, внутри от которого они накапливаются. Возможно, что паренхимные клетки, граничащие с сосудами ксилемы, действительно выделяют ионы в мертвые сосуды. Однако, по предположению некоторых авторов, часть растворенных веществ поступает из протопластов клеток, дифференцирующихся в элементы ксилемы. Эндодерму обычно характеризуют как барьер для ионов или избирательно проницаемую мембрану в корнях. Аткинс (1916) предполагал, что вся кора может функционировать в качестве многоклеточной, избирательно проницаемой мембраны. До сих пор не объяснены суточные колебания корневого давления, о которых сообщалось уже более 100 лет назад (рис. 13.3). Нет также вполне адекватного объяснения того, что у некоторых видов, в том числе у большинства хвойных, редко или никогда не обнаруживается корневое давление, хотя наблюдения Лери (1965) наводят на мысль о возможной причине этого. Этот исследователь не наблюдал ’’плача” у пней здоровых сеянцев сосны ладанной, ели сизой или клена сахарного, тогда как пни клена красного, березы, тюльпанного дерева и винограда обильно выделяли пасоку. Однако он обнаружил, что апикальные отрезки корней тех трех видов, у которых не наблюдалось ’’плача” из пней, выделяли измеримое количество сока, содержащего
много солей. Таким образом, если не удается выявить корневое давление, то это не является результатом отсутствия накопления солей в ксилемном соке. Возможно, что высокое внутреннее сопротивление продольному передвижению воды в корнях в сочетании с тенденцией сока просачиваться под давлением из более старых участков корней препятствуют передаче в побеги какого бы то ни было давления, развиваемого в апикальных зонах корней. Вит с соавторами (1958) и Димблеб (1952) также сообщали о ’’плаче” отрубленных корней
Поглощение воды, подъем ксилемного сока и водный баланс
371
Март Zf Время су тон
Рис. 13.3. Суточные колебания ’’плача” и корневого давления корневых систем подсолнечника, выдержанных 6 дней при постоянной температуре (/7н - полночь; Пд — полдень). Во время эксперимента растения находились на разбавленном в 2 раза питательном растворе Хоаглоща (Vaadia, 1960)
хвойных. Обнаружено, что корневое давление развивается у покоящегося винограда даже при отсутствии растущих корней.
ПАССИВНОЕ ПОГЛОЩЕНИЕ. Когда интенсивность транспирации увеличивается и возникает натяжение в ксилемном соке, начинается массовый приток воды. При этом уносятся соли, накопившиеся в пасоке корней, и уменьшается значение осмотического передвижения воды. Корни становятся пассивной поглощающей поверхностью, через которую вода всасывается благодаря массовому потоку, создаваемому транспирирующимй побегами. Вероятно, что практически все поглощение воды транспирирующими растениями, как древесными, так и гравянистыми, происходит пассивно. Некоторые авторы, например, Руфельт (1956) и Броувер (1965), утверждают, что активное и пассивное поглощение действуют параллельно, даже у быстро транспири-рующих растений. Однако они не учитывают того, что быстрый ток воды через корни разбавляет раствор солей в ксилеме корня и нарушает осмотический градиент, от которого зависит активное поглощение.
КОРНЕВОЕ И СТЕБЛЕВОЕ ДАВЛЕНИЕ
С незапамятных времен люди наблюдали выделение сока из поврежденных растений. На Дальнем Востоке добывали сок из пальм для приготовления сахара и вина. По мнению Эвелина (1670), в Англии и в контитентальной Европе давно проводилась подсочка березы, а сок использовался для различных целей, в том числе в пивоварении. Интерес с использованию березового сока вновь повысился на Украине. Первые европейцы, побывавшие в Канаде и Новой Англии, заметили, что индейцы делали надрезы на стволах клена и выпаривали сок для получения сахара. В Мексике испанские конкистадоры обнаружили, что туземцы собирали сладкий сок из агавы и сбраживали его для получения спиртного напитка ’’пульке”. К сожалению, авторы ранних работ объединяли без разбора все случаи подсочки и ’’плача” независимо от
372
Глава 13
Март
Л прель Даты
5
Рис. 13.4. Давление, одновременна измеренное у березы черной (а) и клена красного (в) : 1 — в стеблях; 2 — корневое; корневое давление у березы превышает давление в стебле, причем оба изменяются почти одновременно; у клена корневое давление обычно отсутствует, даже если в стеблях существует положительное давление
их происхождения. Вилер (1893) перечислил почти 200 видов, относящихся ко многим родам, но его список включал примеры настоящего корневого давления, истечения сока из ран, гуттации и даже выделений железистых волосков. Необходимо различать истечение сока, вызываемое корневым давлением, как например, у винограда и березы, и
вызываемое давлением в стеблях (у клена), или вследствие поранения (у пальм).
КОРНЕВОЕ ДАВЛЕНИЕ. Такое давление не распространено среди деревьев умеренной зоны, встречается оно главным образом весной, пока листья не распустились и транспирадия невелика. Однако Паркер (1964) наблюдал обильный ’’плач” у березы вишневой в Новой Англии в октябре-ноябре, после листопада. После засушливого лета ’’плача” не было.
Кларк (1874) исследовал более 60 видов древесных растений в Массачусетсе и обнаружил ’’плач” только у немногих из них, в том числе у клена, березы, ореха, хмелеграба и винограда. Гальс (1727) впервые опубликовал данные об ' измерении корневого давления у винограда, равного 1 бар, а Кларк (1874) и Мервин и Лион (1909) сообщили, что корневое давление у березы составляет 2-3 бара. Давление в стебле и корневое давление у клена и березы показаны на рис. 13.4. По данным Кларка (1874) береза бумажная диаметром 37,5 см вырабатывала 28 л сока в день и 675 л в течение весны, а Енсон (1944) сообщил о выходе 20-100 л сока от деревьев березы бумажной диаметром 20-38,5 см. Выход сока не был пропорционален диаметру: некоторые небольшие деревья v вырабатывали сока больше, чем более крупные. Истечение сока прекращалось, С когда распускались листья и усиливающаяся транспирация создавала отрицательное давление (натяжение) в ксилемном соке. Содержание сахара в березовом соке (преимущественно редуцирующего) составляет около 1,5 %, т. е. ниже, % чем в кленовом соке. <!
Боллард (1960) составил обширный обзор литературы о составе j, пасоки и отметил, что состав играет важную роль в передвижении минеральных веществ и органических азотистых соединений, синтезируе-
Поглощение воды, подъем ксилемного сока и водный баланс
373
Гис. 13.5. Сезонные изменения концентрации в ксилемном соке лещины, растущей в Бельгии (Denaucr - De Smet, 1967):
I калия; 2 — кальция; 3 — распускание листьев; 4 — старение листьев; 5 — листопад
мых в корнях. Пасока некоторых древесных растений содержит также цитокинины с гиббереллины. Дальнейшие исследования покажут, вероятно, что эти вещества широко распространены. Содержание минеральных веществ в пасоке варьирует в зависимости от снабжения
корней доступными для них солями и от времени года (см. рис. 10.9 и 13.5). Некоторые исследователи пытались использовать состав пасоки для определения потребности растений в минеральных питательных веществах. Однако более полезную информацию можно получить при
помощи анализа листьев.
ГУТТАЦИЯ. Наиболее распространенное доказательство корневого давления у травянистых растений — выделение капелек жидкость на краях или кончиках листьев. Количество выделяемой жидкости колеблется от нескольких капель до многих миллилитров, а состав — от почти чистой воды до разбавленного раствора органических и неорганических веществ .Гуттация обычно происходит через похожие на устьица отверстия в эпидерме, называемые гидатодами, которые располагаются около окончаний жилок. В тропических влажных лесах гуттация обычно происходит ночью. У древесных растений умеренной зоны она встречается редко, так как необходимое для этого процесса сочетание тепла, влажной почвы и сырого воздуха наблюдается не так часто, как в тропиках. Бюсин и Мюнх (1931) приводят несколько примеров гуттации побегов деревьев. Рабер (1937) наблюдал вытекание сока из листовых рубцов лиственных деревьев в Луизиане после листопада, а Фризнер (1940) сообщил о выделении жидкости пневой порослью клена красного в феврале в Индиане. Имеется также сообщение о выделении жидкости корнями и корневыми волосками древесных растений. Это явление вызывается, вероятно, корневым давлением, поэтому его можно назвать корневой гуттацией. О гуттации у хвойных не сообщалось, как и следовало ожидать, вследствие отсутствия у них корневого давления. Однако можно вызвать искусственную гуттацию, подвергая их корневую систему внешнему давлению.
Гуттация не имеет существенного значения для растений. Иногда осадок солей, остающихся при испарении гуттационной воды, повреждает край листа. Утверждают также, что гуттационная жидкость служит путем для проникновения патогенных организмов. Гуттацию мож
Глава 13
374
но считать просто побочным результатом повышения гидростатического давления у медленно транспирирующих растений.
ИСТЕЧЕНИЕ КЛЕНОВОГО СОКА. Это явление заслуживает особого внимания, так как на получении кленового сока основана важная отрасль промышленности в северо-восточной части США, интересно оно и в физиологическом отношении. По ряду причин можно предположить, что истечение происходит независимо от корневого давления. Это подтверждается и экспериментально. Так, измерительные приборы, прикрепленные к корням клена, часто показывают отрицательное давление, когда давление у стеблей оказывается положительным (см. рис. 13.4). Более убедительным является тот факт, что у отрезков стволов и ветвей, отделенных от деревьев клена, наблюдается истечение сока, если их снабжать водой и подвергать воздействию температур выше или ниже точки замерзания. Обширные исследования истечения кленового сока, проведенные Кларком (1874, 1875) более 100 лет назад, все еще сохраняют свое значение. В Массачусетсе истечение кленового сока может происходить в любое время от октября до апреля при смене морозных точен теплыми днями. Вытекание сока прекращается при сохраняющейся температуре выше или ниже нуля. Оно прекращается весной, когда ночные температуры не опускаются ниже точки замерзания. При этом обычно оно приостанавливается в полдень и не возобновляется до следующего утра, пока температура не поднимется выше нуля. Отсутствие знаний о суточных колебаниях давления сока в стеблях деревьев от положительного до отрицательного привело к ошибкам в интерпретации экспериментальных данных (Kramer, 1940). В противоположность истечению сока у клена, ’’плач” березы и винограда, вызываемый корневым давлением, ускоряется при нагревании почвы до тех пор, пока усиливающаяся вследствие распускания листьев транспирация не приведет к прекращению корневого давления. Истечение сока обычно начинается прежде всего на южной стороне деревьев, но впоследствии происходит и из отверстий, высверленных на северной стороне стволов. Сок выделяется из заболони.
В Вермонте более 60 % истечения сока происходит между 9 ч утра и полуднем. Вследствие его зависимости от погоды истечение сока обычно происходит с перерывами, и в течение одной весны может наблюдаться от 2-3 до 10-12 "периодов работы". Некоторые производители кленового сиропа применяют в настоящее время вакуумные насосы для усиления истечения сока. Иногда урожай за сезон может быть увеличен более чем втрое, так как при помощи насоса можно добывать сок в те дни, когда нормально он обычно не выделяется или выделяется слабо. По сообщению Брауна с соавторами (1937), средний выход сока без применения вакуума составляет 38—75 л/дерево за сезон, изредка деревья дают 150 л. Содержание сахара колеблется от 0,5 до 7 или даже до 10, обычно составляет 2—3 %. В пальмовом соке сахара содержится намного меньше.
Тейлор (1956) в результате обширных исследований показал существование больших различий в содержании сахара в соке, полученном от разных деревьев и в разных древостоях. Это указывает на возможность селекции деревьев на высокий выход сахара. Морсели и другие исследователи (1978) сообщили, что у высокопродуктивных деревьев клена лучи более многочисленны и крупнее, чем у низкопродуктивных. Концентрация сахара, как правило, в начале сезона невелика, затем быстро возрастает до максимума, к концу сезона постепенно умень
Поглощение воды, подъем ксилемного сока и водный баланс
375
шается. Наряду с сахарозой и небольшим количеством глюкозы кленовый сок содержит немного неорганических солей, азотистые соединения (пептиды и аминокислоты), амилазы и неидентифицированные органические компоненты. Характерный вкус, усиливающийся при нагревании, приписывают некоторым аминокислотам. Сахар образуется из накопленного в течение предшествующего лета крахмала, который превращается в сахарозу в конце осени — начале зимы. Активность ферментов, участвующих в этом превращении, по-видимому, повышается под влиянием низких температур. По мнению Саутера с соавторами (1973), сахароза выделяется в ксилему, создавая высокую концентрацию в пасоке. Потеря сахара при подсочке, очевидно, безвредна, так как многие деревья подсачивались десятилетиями без видимых повреждений. Подсчитано, что подсачиваемые клены выделяют менее 10 % общего сахара. Эта потеря слишком мала, чтобы иметь значение, если только условия не будут очень неблагоприятными для фотосинтеза в следующее лето. Деревья на бедной или сухой почве обычно дают меньше сока, чем растущие на плодородной, влажной почве. Выход сахара зависит от фотосинтеза, и этому благоприятствуют большие хорошо освещенные кроны. Известно, что удобрения также повышают урожай. Деревья, предназначенные для добывания сока, должны быть расположены дальше друг от друга, чем выращиваемые для получения древесины, и, как утверждают, придорожные деревья вырабатывают больше сока. По сообщениям, дефолиация деревьев летом значительно уменьшает выход сиропа следующей весной.
Причиной истечения кленового Сока является, по-видимому, давление в стеблях, вызываемое повышением температуры после низких, но необязательно морозных температур. Одно из более современных объяснений было предложено Саутером и его сотрудниками (1971), работавшими в Гарвардском лесу. Согласно его толкованию двуокись углерода накапливается в межклетниках в течение дня и возникающее в результате этого давление выдавливает сок из ран. Ночью СО2 поглощается, понижая давление и вызывая восходящее передвижение из корней воды, которая вновь заполняет сосуды ксилемы.
ДРУГИЕ ПРИМЕРЫ ДАВЛЕНИЯ В СТЕБЛЯХ. Двумя другими растениями, которые дают большое количество сока, имеющего практическое значение, являются пальмы и агавы. В тропических районах Индии и других азиатских стран пальмовый сок использовался в качестве источника сахара, вероятно, до того как начали выращивать сахарный тростник. Его также сбраживают для изготовления пальмового вина. По последним сообщениям, ежегодная выработка составляет более 150 тыс. т сахара, получаемого главным образом из кокосовой, финиковой и пальмировой пальм. По сообщению Молиша (1902), изучавшего этот процесс на Яве, истечение сока, обычно вызываемое срезанием соцветия, может поддерживаться в течение недель или даже месяцев посредством неоднократного срезания и поколачивания по стеблю. Сок получают также из пальмировых пальм, делая надрезы на коре, причем этот прием можно повторять из года в год. Если срезать центральную почку финиковой пальмы, то истечение сока прекращается через несколько недель, и пальма отмирает. Дэвис (1961) предполагал, что корневое давление имеет большое значение у пальм, но
Глава 13
376 впоследствии ои сообщил, что оно наблюдается редко и, вероятно, не играет роли в истечении пальмового сока. По-видимому, сок вытекает из флоэмы. Содержащийся в нем сахар мобилизуется для использования развивающимися соцветиями или верхушками стеблей.
Высокое давление, обнаруженное в стеблях многими авторами, является, вероятно, результатом поранения. Приводились данные о давлении экссудации в отверстиях, высверленных в стеблях крупных кактусов, сосны замечательной, грецкого ореха и дуба, но это явление приписывается скорее действию поранения, чем корневому давлению. Иногда из трещин и других ран на деревьях вытекающий сок сбраживается дрожжами и бактериями, вызывая ’’слизетечение”. По сообщению Картера (1945), у вязов слизетечение происходит вследствие пропитывания водой ядровой древесины (так называемой мокрой гнилью, вызываемой деятельностью бактерий). У деревьев с гниющим ядром развивается такое высокое давление в стволе, что оно может выбросить бур из пробуравливаемых отверстий. В некоторых случаях газ, выделяющийся из отверстий в стволе, загорается при поджигании, так как он содержит метан, вырабатываемый гнилостными организмами.
СМОЛОТЕЧЕНИЕ. Один из наиболее важных примеров выделений, вызываемых поранением, — истечение из сосен живицы, которая служит основой подсочки. Однако живица вытекает из особых смоляных ходов и ее истечение не связано с истечением сока, обсуждавшимся в этой главе. Истечение латекса из пораненных стеблей каучуконосных деревьев — другой важный пример экссудации (см. главу 8).
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОГЛОЩЕНИЕ ВОДЫ
Во влажной почве скорость поглощения воды контролируется в основном двумя факторами: интенсивностью транспирации, поскольку она в значительной степени определяет величину водного потенциала в ксилеме корня, и эффективностью деятельности корневых систем как поглощающих поверхностей. При высыхании почвы доступность воды начинает ограничиваться вследствие понижения водного потенциала и гидравлической проводимости. Аэрация и температура почвы, концентрация и состав почвенного раствора также иногда ограничивают поглощеие воды.
ПОЧВЕННАЯ ВЛАГА. Скорость поглощения воды зависит от крутизны градиента водного потенциала от почвы до корней. При высыхании почвы вода становится все менее доступной, так как ее потенциал уменьшается, а сопротивление ее передвижению по направлению к корням повышается (рис. 13.6). Легкодоступной водой часто считают ту, которая находится между полевой влагоемкостью и влажностью устойчивого завядания. Полевая влагоемкость — это содержание воды через несколько дней после того, как почва была увлажнена и нисходящее передвижение воды стало очень медленным. Влажность устойчивого завядания — это содержание воды, при котором растения остаются завядшими, пока почва не будет вновь увлажнена. Из рис. 13.6 видно, что количество легкодоступной воды гораздо больше в почвах с мел
Поглощение воды, подъем ксилемного сока и водный баланс
377
ким механическим составом, чем в грубозернистых почвах. Как полевая влагоемкостъ, так и влажность устойчивого завядания представляют собой не физические константы, а только общепринятые участки на кривой зависимости водного потенциала от содержания воды. Обычно считают, что влажность устойчивого завядания наблюдается приблизительно при -15 бар, но это объясняется тем, что для определения данного показателя использовались подсолнечник или сходные с ним мезофиты. Слатир (1957) обнаружил, что водный потенциал при сильном завядании бирючины достигал -70 бар, значения меньше —15 бар были установлены и для других растений.
Одно время считали, что почвенная вода может быть доступной или недоступной для растений. От этого представления отказались вследствие того, что оно неприемлемо как теоретически, так и практически.
Многие исследования с сеянцами древесных растений, выращенными в
сосудах, показали, что вода по мере понижения водного потенциала почвы постепенно становится менее доступной (см., например, рис. 12.12).
Остается неясным вопрос об относительном сопротивлении току воды через почву и корни (Г1ЮЧВ в сравнении с лКОрН) в высыхающей почве. Известно, что сопротивление току воды через почву быстро
возрастает при высыхании почвы вследствие сокращения поперечного
сечения, доступного для перетекания воды, так как уменьшается толщина пленок воды и нарушается ее непрерывность. Иногда при высыхании почвы происходит, по-видимому, усадка почвы и корней, вызывающая уменьшение передвижения воды от почвы к корням. Гарднер (1960) и Кован (1965) подсчитали, что сопротивление передвижению воды через почву будет превышать сопротивление корней при водном потенциале почвы -1 или —2 бар, а сопротивление почвы будет оказывать наибольшее влияние на передвижение воды по направлению к корням. Однако Ньюмэн (1969) установил, что фактическая плотность корней обычно значительно больше той, которую принимали в расчет Гарднер и Кован. При более высокой плотности корней воде не нужно передвигаться через почву так далеко или так быстро, как считалось раньше. Поэтому сопротивление почвы не станет, вероятно, ограничивающим фактором для поглощения воды до тех пор, пока водный потенциал почвы не приблизится к влажности устойчивого завя-
дания. Вероятно, доступность почвенной влаги обычно больше зависит от крутизны градиента водного потенциала от почвы к корням
Рис. 13.6. Кривые зависимости матричного потенциала суглинков от содержания воды в почве:
1 — влажность устойчивого завядания;
2 — суглинок тяжелый пылеватый (по Richards и Weaver, 1944); 3 —
легкий суглинок (по Wadleigh и др., 1946); 4 — полевая влагоемкостъ
178
Глава 13
(^орн~ Аючв)’ чемо1 ; сопротивления почвы (г nQqB).
КОНЦЕНТРАЦИЯ И СОСТАВ ПОЧВЕННОГО РАСТВОРА. Водный потенциал почвы обусловливают поверхностные силы, которые удерживают воду в капиллярах и на поверхностях (матричный потенциал (ф^,и осмотические силы, создаваемые растворенными веществами (осмотический потенциал ф$). Если осмотический потенциал ниже —2 или —3 бар, то рост растений обычно замедляется даже на почвах с содержанием воды, близким к полевой в л аго емкости. Однако чрезмерная засоленность обычно наблюдается только в засушливых районах, где эвапотранспирация значительно превышает количество осадков, а на покрытых лесом территориях встречается редко. Иногда эта проблема возникает в отношении плодовых деревьев в засушливых областях, где поливные воды часто содержат избыточные количества солей. Представляет также определенный интерес выявление древесных растений, пригодных для выращивания в прибрежных зонах, где соленые брызги вызывают повреждения.
АЭРАЦИЯ ПОЧВЫ. Рост и физиологическая активность корней часто понижаются из-за недостатка кислорода. Наиболее резко это проявляется в затопленных почвах, однако хронический, но умеренный недостаток кислорода часто существует в тяжелых глинистых почвах, что ограничивает проникновение корней вглубь и, возможно, поглощение минеральных питательных веществ. Было даже высказано предположение, что плохая аэрация ограничивает заселение прерий древесными растениями. Затопление почвы водой обычно резко понижает ее поглощение (рис. 13.7), так как увеличивает сопротивление току воды в корни. Однако существуют большие различия между видами по их устойчивости к затоплению. Кипарис болотный и нисса водная растут неограниченно долго на затопленной почве, ивы буйно разрастаются на почве, насыщенной водой, многие виды выдерживают ежегодные периоды затопления, но некоторые виды, например дерен и тюльпанное дерево, быстро отмирают на насыщенной водой поЛве.
Недостаточная аэрация не только уменьшает поглощение воды и солей, но и понижает синтетическую активность корней. Имеются данные, свидетельствующие о том, что корни, по крайней мере некоторых видов, синтезируют цитокинины и гиббереллины. Установлено также, что такие азотистые органические вещества, как амиды и аминокислоты, синтезируются в корнях многих древесных растений, в том числе яблони и других видов розоцветных. Поэтому вероятно, что недостаточная аэрация и холодная почва могут вызывать ослабление роста побегов, уменьшая их снабжение регуляторами роста и органическими азотистыми соединениями, а также замедляя поглощение воды и солей.
ТЕМПЕРАТУРА ПОЧВЫ. Многие авторы считают холодную почву важным экологическим фактором. Пониженная доступность воды в холодной почве на больших высотах над уровнем моря может влиять на растительность и на расположение верхней границы распространения леса. Плохо дренированные почвы медленно нагреваются весной. Имеются данные, что холодные почвы европейских верховых болот ограничивают рост растений. По сообщению Камерона (1941),апельсиновые деревья часто завядают зимой в Калифорнии вследствие мед-
Поглощение воды, подъем ксилемного сока и водный баланс
379
Рис. 13.7. Изменение интенсивности транспирации, свидетельствующее о влиянии затопления почвы на поглощение воды. В результате водного стресса, вызванного пониженным поглощением воды, сеянцы теряли большую часть листьев, но дуб лировидный после осушения почвы вновь покрывался листвой и быстро оправлялся (Parker, 1950):
1 — дуб красный; 2 — дуб лировидный; 3 — дуб каштановый; 4 — затопление; 5 — осушение
ленного поглощения воды из холодной почвы. Существуют значительные различия между видами в отношении влияния низкой температуры на поглощение воды. Козловский (1943) установил, что при понижении температуры почвы от 15 до 5°С поглощение воды у сосны ладанной сокращалось сильнее, чем у сосны веймутовой. Вообще у видов из теплого климата обнаруживается большее Снижение поглощения, чем у видов из холодного климата (рис. 13.8). Цитрусовые деревья больше страдают от холодной почвы, чем древесные породы из более холодного климата, но между разиовидностями и видами существуют, вероятно, различия.
Холодная почва уменьшает поглощение воды двумя путями: прямо — вследствие понижения проницаемости корней для воды, и косвенно, увеличивая вязкость воды, что замедляет ее передвижение как через почву, так и через корни. В результате совместного действия этих факторов сопротивление току воды через корни приблизительно удваивается при понижении температуры от 25 до 5°С. Пониженную проницаемость при низких температурах некоторые авторы относят за счет перехода мембранных липидов из жидкого состояния в твердое. Относительно влияния высоких температур почвы на поглощение воды данные немногочисленны.
Существуют и вторичные температурные эффекты — уменьшение растяжения корней и изменение их метаболизма, но это, по-видимому, имеет гораздо меньшее значение, чем прямое влияние температуры на сопротивление водному току.
380
Глава 13
Рис. 13.8. Влияние охлаждения почвы на поглощение воды двумя южными и двумя северными видами сосен, измеренное по интенсивности транспирации- Поглощение из холодной почвы снижалось сильнее у южных видов, чем у северных (Kramer, 1942) :
1 — веймутова; 2 — смолистая; 3 — Эллиота; 4 — ладанная
ПРОТЯЖЕННОСТЬ И ЭФФЕКТИВНОСТЬ РАБОТЫ КОРНЕВЫХ СИСТЕМ. Успешное поглощение любым растением воды и минеральных веществ зависит от протяженности и проницаемости корнав. Как отмечалось в главе 2, большинства деревьев
корневые системы про-
стираются за пределы проекции ветвей и проникают в почву так глубоко, как это допускают аэрация и физическая структура почвы. В сомкнутых древостоях почва полностью занята корнями на глубину от 25—30 см до нескольких метров. По данным Виганса (1936), 18-лет-ние яблони на хорошо аэрируемой лессовидной почве в Небраске поглощали воду с глубины более 10 м. В противоположность этому у груш на плохо дренированной наносно- глинистой почве в Орегоне около 90 % корней находились в верхнем 1-метровым слое, а Коль (1937) обнаружил в сосновых и дубовых лесах Северной Каролины 90 % корней в верхнем горизонте 12,5 см. Несмотря на относительно небольшое количество корней, расположенных глубоко в почве под лесными насаждениями, почва часто высыхает до влажности устойчивого завядания. Руссель (1973) приводит интересные данные о развитии корней в связи
с поглощением минеральных веществ и воды.
Эффективность работы корней зависит от размера поверхности, находящейся в контакте с почвой, и от проницаемости их поверхности. Очевидно, корневые системы с многочисленными мелкими- ответвлениями должны быть более эффективными, чем состоящие из небольшого количества крупных, слабо разветвленных корней. Микоризы, по-видимому, повышают эффективность поглощения .минеральных веществ благодаря тому, что гифы далеко распространяются в почве, увеличивая таким образом поглощающую поверхность. Они также долго сохраняют активную поглощающую систему более старых корней после их опробковения, Влияние микоризных корней на поглощение воды определить труднее. По сообщению некоторых исследователей, наличие микоризы повышает засухоустойчивость сеянцев древесных растений. Кроме того, сопротивление передвижению воды через корни сои уменьшается благодаря наличию микоризы. Как отмечалось выше, значительное поглощение воды и минеральных веществ происходит через опробковевшие корни, несмотря на их относительно низкую проницаемость.
Поглощение воды, подъем ксилемного сока и водный баланс
381
ПОДЪЕМ КСИЛЕМНОГО СОКА
Существование высоких наземных растений стало возможным только после того, как развилась сосудистая система, обеспечивающая возможность быстрого передвижения воды к транспирирующим побегам, Наземным растениям высотой более 20—30 см трудно существовать без сосудистой системы, за исключением растений, произрастающих в наиболее влажных местообитаниях. Это объясняется тем, что передвижение воды от клетки к клетке путем диффузии происходит слишком медленно для того, чтобы предотвратить обезвоживание надземной массы транспирирующих растений. На важность этой проблемы для деревьев указывает тот факт, что в жаркий летний день 200 л воды или более может пройти от корней к испаряющей поверхности листьев на высоту 20, 30 или даже 100 м. Гейле (1727), определивший поглощение и потери воды, писал: ’’Все три последних эксперимента показывают, что заполненные соком капиллярные сосуды обильно всасывают влагу; однако у них не хватает силы, чтобы продвинуть ее дальше без помощи потеющих листьев, которые способствуют ее продвижению”. Объяснение Гейлса предвещало наше современное толкование, несмотря на то, что он не понимал, как может транспирация ’’способствовать ее продвижению”. К концу XIX столетия Бем, Сакс и Страс-бургер пришли к заключению, что потеря воды создает натяжение, вызывающее подъем сока, хотя и у них недоставало одного существенного факта для полного объяснения. Заключительный шаг сделали Аскенази (1895), Диксон и Жоли (1895), которые показали, что вода, заключенная в узких трубках, какими являются элементы ксилемы, обладает очень большой силой сцепления и может подвергаться натяжению.
Теория сцепления для объяснения подъема ксилемного сока хотя и существует с конца XIX в., была воспринята неохотно, и авторы учебников до сих пор ощущают необходимость отстаивать ее. Нежелание - согласиться с этой теорией отчасти происходит, вероятно, от почти интуитивного мнения, что трудно поверить в возможность натяжения воды, а также от сомнений относительно того, можно ли сохранять хрупкую, напряженную систему в качающемся древесном стволе. Теория сцепления основана на следующих предпосылках: 1) вода имеет большие внутренние силы сцепления, и если ее заключить в тонкие трубки со смачивающимися стенками, например в элементы ксилемы, то она может выдержать растяжение от 30 до, вероятно, 300 бар; 2) вода в растении образует непрерывную систему в насыщенных водой клеточных стенках от испаряющих поверхностей листьев до поглощающей поверхности корней; 3) когда вода испаряется из какой-либо части этой системы, главным образом из листьев, уменьшение водного потенциала на испаряющих поверхностях вызывает передвижение воды из ксилемы к испаряющим поверхностям; 4) благодаря взаимному притяжению между молекулами воды потеря воды создает натяжение в ксилемном соке, которое передается через непрерывные столбики воды до корней, где понижает водный потенциал и вызывает приток воды из почвы. Таким образом (как указывалось в разделе
382
Глава 13
’’Пассивное поглощение”), у транспирирующих растений поглощение воды непосредственно регулируется интенсивностью транспирации.
Недостаточно доказано, что вода может выдержать натяжение, необходимое для того, чтобы притягивать воду к верхушкам высоких деревьев. Теоретически межмолекулярные силы притяжения в воде чрезвычайно велики. Уршпрунг (1915) измерил натяжение в клетках аннулюса спорангия папоротника, оказавшееся равным 315 бар, Бриггс (1949) показал, что натяжение в воде, подвергнутой действию центробежной силы, составляет 223 бар. По мнению Грениджа (1954), наибольшее натяжение в деревьях должно составлять в среднем только около 30 бар. Однако измерения потенциала ксилемного сока, выполненные при помощи барокамеры, показали существование давления до 80 бар, а для преодоления как силы тяжести, так и сопротивления току при передвижении воды к верхушке дерева высотой 100 м достаточно натяжения всего лишь около 20 бар. Установлено, что многие водные столбики разрываются. Мильбурн и Янсон (1966) сообщили, что при использовании усилительной аппаратуры они обнаруживали ’’щелчки” при разрыве водных столбиков в стеблях, находящихся под давлением. Однако пузырьки, образующиеся вследствие кавитации, состоят из водяного пара к обычно легко рассасываются яри уменьшении натяжения. Данные Кларка и Гибса (1957) показывают, что летом приблизительно до 50 % воды в стволах некоторых деревьев может быть замещено газом (рис. 13.9). Однако проникающий воздух занимает только крупные сосуды и не блокирует всю проводящую систему. Неоднократно было показано, что нанесенные на стебель горизонтальные зарубки с противоположных сторон на расстоянии 15 см друг от друга не препятствуют подъему пасоки, потому что непрерывность водных столбиков сохраняется в более коротких элементах ксилемы и происходит радиальный ток вокруг зарубок. Однако сопротивление току при этом повышается.
Другая сложность в теории сцепления заключается в том, что замерзание может вытеснять растворенные газы из раствора, а образующиеся
13.9. Сезонные изменения содержания в стволах березы желтой (Clark, Gibbs, 1957):
а — воды (определенной в дисках, вырезанных из основания, середины и верхушки стволов) ; б — воды и газа, вычисленных в виде процентов от объема свежеср убиенных стволов: 1 — вода; 2 — древесина; 3 — газ; заметно уменьшение содержания воды и увеличение содержания газа в середине лета, в течение периода быстрой транспирации, а также осеннее повышение после листопада
Поглощение воды, подъем ксилемного сока и водный баланс
383
пузырьки могут блокировать большинство или все элементы ксилемы. Однако, по сообщению Гамеля (1967), после оттаивания замороженных отрезков стволов и ветвей тсуги сопротивление току воды не увеличивалось, хотя у покрытосеменных замораживание вызывало повышение этого сопротивления. Автор предположил, что у голосеменных пасока изолирована в трахеидах, находящихся под давлением, и после оттаивания мельчайшие газовые пузырьки вновь растворяются, но этого не происходит в крупных сосудах кольцесосудистых покрытосеменных. Циммерман (1964) указывает, что у кольцесосудистых деревьев (например, у дуба и ясеня) старые сосуды остаются заполненными воздухом, но каждую весну, до полного распускания листьев, образуются новые сосуды. Рассеяннососудистые виды ведут себя, вероятно, подобно голосеменным: у них ксилема никогда не блокируется газовыми пузырьками. У некоторых пород, например у березы и винограда, корневое давление может способствовать растворению газа и заполнению сосудов соком вновь. Вероятно, у многих кольцесосудистых деревьев хотя крупные сосуды никогда вновь и не заполняются, но более мелкие сосуды поздней древесины никогда не бывают заблокированы и продолжают выполнять функцию проведения воды. Необходимо иметь также в виду, что в большинстве стволы имеют большой излишек проводящей способности и могут утрачивать значительную часть без значительного ущерба. Чтобы Передвинуть воду вверх, против силы тяжести, требуется напряжение —0,1 бар на каждый метр плюс некоторое напряжение для преодолевания сопротивления восходящему току в ксилеме вследствие трения.
Коннор с соавторами (1977) ссылаются на измерения, в которых градиент водного потенциала в стволе был, по-видимому, меньше необходимого минимума. Однако их обширные исследования, проведенные на эвкалиптах высотой 45 и 75 м, показали значения градиентов водного потенциала гораздо ниже теоретически ожидаемой величины - 0,1 бар/м, за исключением тех случаев, когда листва была увлажнена. Был обнаружен также вертикальный профиль осмотического потенциала около 0,1 бар/м в ранние послеполуденные часы, но значительного вертикального градиента устьичного сопротивления не было. Вследствие различий в гидростатическом давлении между деревьями разной высоты листья 75-метровых деревьев подвергались действию напряжения по крайней мере -7,5 бар, а 45-метровых -4,5 бар. С увеличением высоты деревьев эвкалипта размеры листьев уменьшаются, а толщина кутикулы становится больше.
Сложностям, присущим теории подъема пасоки при помощи сил сцепления, придается, по-видимому, слишком большое значение. Во всяком случае, как давно указывал Реннер (1912), теория сцепления представляет собой только одно из возможных объяснений эффективной взаимосвязи поглощения воды и транспирации. Непрерывные столбики воды, простирающиеся от листьев до корней, обеспечивают механизм обратной связи, при помощи которого взаимно контролируются изменения интенсивности потери воды и ее поглощения. Этот механизм важен для выживания транспирирующих растений, несмотря на то, что его значением пренебрегали некоторые критики теории сцепления.
384
Глава 13
ВОДОПРОВОДЯЩАЯ СИСТЕМА
Вода из корней в листья передвигается по специализированной проводящей ткани, называемой ксилемой или древесиной, обеспечивающей путь наименьшего сопротивления. Однако, как упоминалось выше, вода должна пересечь ряд слоев живых клеток, чтобы поступить в ксилему корня, а в листьях она опять должна пройти через несколько клеток, прежде чем достигнет испаряющих поверхностей. В ксилеме она передвигается по мертвым элементам. Ксилема состоит из клеток древесной паренхимы и лучей, волокон, трахеид и (у покрытосеменных) сосудов, но вода передвигается в основном по трахеидам у хвойных и по сосудам у покрытосеменных, так как они оказывают наименьшее сопротивление току воды. Трахеиды — это одиночные клетки длиной до 5 мм и 30 мкм в диаметре, вода должна проходить через тысячи клеточных стенок, передвигаясь вверх по стеблям. Несмотря на то, что это передвижение облегчается благодаря многочисленным порам ( в стенках трахеид, сопротивление току воды через трахеиды хвойных и короткие сосуды рассеяннососудистой древесины значительно боль- ? ше, чем через длинные сосуды кольцесосудистых видов. Сосуды покрытосеменных -- это сложные трубчатые структуры, образуемые ’ путем растворения поперечных стенок и протопластов большого ко- J личества клеток, расположенных конец к концу. В результате возникают трубки диаметром от 20 до 700—800 мкм и длиной от нескольких сантиметров до многих метров. i
СЛОЖНОСТЬ ПРОВОДЯЩЕЙ СИСТЕМЫ. Было бы большим упро-щением рассматривать ксилему как простое собрание трубок. На самом деле, как указывают Козловский и Винчет (1963), она нередко образует удивительно сложную систему, в которой пасока проходит неожиданными путями. Например, ксилема растет обычно так, что вода чаще передвигается по спирали, чем вертикально. У некоторых голосеменных путь восходящего тока оказывается более спираль- > ным, что можно объяснить структурным косослоем ксилемы. Это, вероятно, вызывается характером расположения окаймленных пор I в трахеидах. По сообщению Томаса (1967), в кизиле краска передвигается по спирали приблизительно на 90° окружности на каждый метр подъема. Рудинский и Вите (1959) утверждают, что спиральный характер подъема сока обеспечивает более эффективное распределение воды по всем частям кроны, чем строго вертикальный. Это, по-видимому, обусловлено результатами наблюдений, показавших, что дуб белый, в котором сок передвигается вверх почти вертикально, меньше повреждается от болезни увядания, чем дуб болотный, в котором транспирационный ток проходит по спирали и расходится в верхней части. Путь подъема сока представляет значительный интерес в связи с инъекцией химических препаратов для защиты от насекомых и грибов.
Длина сосудов и строение древесины. Многочисленные исследования длины сосудов выполнены посредством определения расстояния, на которое можно было продавить через стебли ртуть, суспензии крахмала или туши. Длина сосудов рассеяннососудистых видов составляет несколько сантиметров, а у кольцепоровых она может достигать не
Помещение воды, подъем ксилемного сока и водный баланс 385
скольких метров. У некоторых кольцесосудистых деревьев весной дифференцируются сосуды, которые тянутся без перерыва от верхушки до основания (табл. 13.2). У большинства вьющихся растений сосуды большого диаметра и очень длинные, образующие чрезвычайно эффективные проводящие системы. Длинные сосуды кольцесосудистых пород обычно закупориваются пузырьками газа, тиллами и др., поэтому их эффективная длина к концу вегетационного периода уменьшается, а через 1—2 года они в большинстве случаев утрачивают функцию проведения сока.
Казалось бы, более низкое сопротивление передвижению воды по крупным сосудам кольцесосудистых видов должно давать им большое преимущество, однако некоторые наиболее высокие деревья на Земле относятся к хвойным, у которых передвижение воды происходит по трахеидам длиной не более нескольких миллиметров. Каждый тип строения древесины имеет свои достоинства и недостатки. Рассеяннососудистая структура обеспечивает лучшую защиту от обширного блокирования транспорта, так как газовые пузырьки не могут легко распространяться, но она создает и более высокое сопротивление току воды. Низкое сопротивление -току воды кольцесосудистой структуры особенно важно в длинной, тонкой виноградной лозе, но большой диаметр сосудов делает эти стебли более уязвимыми в отношении блокирования из-за кавитации, если они подвергаются натяжению. Губер (1935), подробно рассмотрев значение двух типов проводящих систем, пришел к заключению, что кольцесосудистый тип древесины также биологически целесообразен, как и рассеяннососудистая структура листопадных пород, у которых новые сосуды образуются каждой весной до распускания листьев. Однако он высказал сомнение, сможет
13.2- ДЛИНА СОСУДОВ В СТВОЛАХ РАЗЛИЧНЫХ ЛИСТВЕННЫХ ДЕРЕВЬЕВ (Greenidge, 1952)
Вид Длина сосудов, см мини- макси- средняя мальная мальная
Рассеянно сосудистые
Acer saccharum 81 94 88,0
Betula lutea 86 142 119,0
F agus grandifolia 488 556 302,0
Populus tremuloides 101 132 122,0
Alnus rugosa 86 122 105,0
Кольцесосудистые
Quercus rubra 853 1524 94,8*
ITaxinus americana 772 1829 97,1*
Ulmus americana 518 853 94,6*
♦Среднее расстояние, пройденное воздухом, % от высоты дерена.
386
Глава 13
ли успешно функционировать кольцесосудистая древесина у вечнозеленых растений, так как в конце каждого вегетационного периода она в значительной степени утрачивает водопроводимость. В подтверждение этой точки зрения он указывает на то, что вечнозеленые дубы Средиземноморского района являются рассеяннососудистыми, а листопадные дубы имеют кольцесосудистую структуру.
ЭФФЕКТИВНОСТЬ ПРОВЕДЕНИЯ ВОДЫ. По мнению Губера, существуют два способа оценки эффективности водопроводящей системы. Первый заключается в определении удельной проводимости, которая показывает объем воды, передвигающейся за единицу времени при данном давлении через отрезок стебля определенной длины и поперечного сечения (литров в час, на 1 см2 поперечного сечения, на 1 бар, на расстояние 1 м). Другой показатель — относительная проводящая поверхность, т. е. отношение проводящей поверхности (поперечного сечения элементов ксилемы) к транспирирующей поверхности.
По сообщению Фармера (1918), удельная проводимость стволов хвойных гораздо ниже, чем у покрытосеменных, а Губер (1956) приводит следующие величины удельной проводимости: 20 для хвойных, 65—128 для листопадных широколиственных деревьев, 236—1273 для виноградных лоз, а для корней еще больше. Имеются данные, что удельная проводимость ветвей и побегов ниже, чем у ствола, и что удельная проводимость ствола уменьшается от основания к верхушке. Это уменьшение компенсируется значительным повышением относительной проводящей поверхности от основания к верхушке ствола. Относительная проводящая поверхность, выраженная в квадратных миллиметрах ксилемы на 1 г сырой массы листьев, возрастает от 0,02
13.3. СКОРОСТЬ ПЕРЕДВИЖЕНИЯ ВОДЫ В КСИЛЕМЕ, ОПРЕДЕЛЕННАЯ РАЗЛИЧНЫМИ МЕТОДАМИ
Исследователь Метод Материал Скорость, м/ч
Greenidge (1958) Кислый фуксин Acer saccharum 1,5 — 4,5
** Fagus grandifolia 3,6 - 4,2
Ulmus americana 4,3 - 15,5
Huber, Schmidt Термоэлектри- Quercus pedun- 43,6 (макс.)
(1937) ческий culata
То же Fraxinus 25,7 (макс.)
excelsior
Ulmus effusa 6,0 (макс.)
Juglans cinerea 3,79 (макс.)
Liriodendron 2,62 (макс.)
tulipifera
in” Хвойные Менее 0,5
Kutz, Riker Quercus 27,5 — 60 (макс.)
(1955) 86 . ellipsoidalis
Rb Quercus macrocarpa 27,5 — 60 (макс.)
Decker, Shau Термоэлектри- Juniperus 0,25 (макс.)
(1964) ческий ostej sperma
Owston и др. 32 рj Pinus contorta 0,1 - 0,8
Moreland (1950) 32 р Pinus taeda 1,2
Поглощение воды, подъем ксилемного сока и водный баланс
387
Рис. 13.10. Изменение относительной водопроводящей поверхности по сравнению с листовой поверхностью вдоль главного ствола и в ветвях 6-летней пихты одноцветной (в сотых долях квадратных миллиметров поперечного сечения ксилемы на 1 г сырого веса хвои). Относительная проводимость увеличивается от основания к верхушке, но она ниже в местах прикрепления мутовок ветвей (числа светлым шрифтом), чем между узлами (жирный шрифт) (Huber, 1928)
у водных растений примерно до 0,5 у деревьев и 3,4 у несуккулентных пустынных растений. Пример изменения относительной проводящей поверхности с высотой дерева приведен на рис. 13.10. Губер сделал вывод, что передвижение воды по здоровым растениям ограничивается не сопротивлением ксилемы, а сопротивлением, с которым встречается вода, проходя через живые клетки и листьев, — вывод, с которым согласны авторы этой книги. Робертс (1977) подсчитал, что половина сопротивления приходится на компонент почв — корень.
Сопротивление радиальному передвижению воды в корнях от почвы до ксилемы велико, но сопротивление продольному току в ксилеме корней древесных растений ниже, чем в стеблях. Стоун (1975) установил, что проводимость корней сосны смолистой была в 50 раз больше, чем стеблей, причем с увеличением расстояния от основания ствола проводимость повышалась. В корнях не наблюдалось спирального передвижения, несмотря на то, что оно обычно происходит в стволах. Низкое сопротивление продольному току воды через корни деревьев контрастирует с высоким сопротивлением, наблюдаемым в корнях некоторых злаков. Сообщают, что у некоторых деревьев в ксилеме, снабжающей листья, имеется суженная зона вследствие уменьшения количества и диаметра сосудов, что повышает сопротивление притоку воды в листья.
Ж
СКОРОСТЬ ПЕРЕДВИЖЕНИЯ СОКА. Различия в удельной проводимости между кольце- и рассеяннососудистыми видами выявляется при измерении скорости протекания сока (табл. 13.3). Некоторые измерения выполнены при помощи инъекции красок, например кислого фуксина или радиоактивных метчиков. При инъекции красок приходится губить стебли для определения расстояния, на которое передвинулся краситель. Измерения при помощи красок и радиоактивными метчиками имеют тот недостаток, что скорость передвижения во вскрытой и неповрежденной ксилеме может быть неодинаковой. Наиболее
достоверные результаты измерения скорости протекания пасоки получаются, по-видимому, при использовании термоэлектрического метода (тепловых импульсов), разработанного Губером и его сотрудниками. Несмотря на то, что этот метод может давать заниженную оценку ско-
388
Глава 13
Рис. 13.11. Термоэлектрическая установка. Нагревающий элемент помещают под кору рядом с камбием или в отверстие, высверленное в наружной части ксилемы. Точки 1 и П - термопары или термисторы, используемые для измерения температуры протекающего сока. Точки 17 и Ш использованы Daum (1967) для определения глубины, до которой происходит проведение воды. Детали методики описаны Swanson (1962):
1 — нагревающий элемент; 2 — кора; 3 — активная ксилема; 4 — неактивная ксилема
Рис. 13.12. Скорость протекания воды (м/ч) в различных частях дуба в полдень, измеренная методом тепловых импульсов. Скорость тока уменьшается по направлению к верхушке, так как относительная проводимость (отношение поперечного сечения ксилемы к листовой поверхности) повышается (см. рис. 13.10). У березы скорость увеличивается по направлению к верхушке вследствие понижения относительной проводимости (Huber, 1956, по Huber and Schmidt, 1937)
Поглощение воды, подъем ксилемного сока и водный баланс
389
рости протекания сока, его использовали многие исследователи для измерения суточных колебаний скорости восходящего тока у деревьев. В этом методе поднимающуюся пасоку нагревают при помощи маленького электрического элемента, вставленного под кору или в заболонь. Измеряют время, необходимое для того, чтобы нагретая пасока достигла термопары или термистора, установленного в древесине выше нагревающего элемента. Даум (1967) повысил чувствительность метода, помещая нагреватель под кору (рис. 13.11). Несколько примеров полученной скорости тока приведено в табл. 13.3. Распределение скоростей в разных частях дерева показано на рис. 13.12. У хвойных и рассеяннососудистых видов скорость невелика, потому что вода передвигается по нескольким годичным кольцам, тогда как у кольцесосудистых она быстро поднимается всего лишь по одному или двум годичным кольцам (рис. 13.13). Данный метод безвреден, поэтому его можно использовать в длительной серии измерений.
НАПРАВЛЕНИЕ ПРОТЕКАНИЯ ПАСОКИ. Мы предполагали до сих пор, что вода всегда передвигается вверх. Обычно это правильно, потому что передвижение воды происходит по направлению к участкам с более низким водным потенциалом, который наблюдается в транспирирующих листьях. Однако вода может передвигаться вниз, если возникает обратный градиент, как было показано Джоном Реем в 1669 г., Стивеном Гейлсом в 1727 г. и другими ранними исследователями. Обратный ток, который вызывал Слатир (1956) в сеянцах сосны и другие исследователи у разных растений, указывает на то, что нет или почти нет дополнительного сопротивления передвижению воды Ери перемене нормального восходящего тока на нисходящий. Даум (1967) показал наличие нисходящего тока в одной ветви с одновременно восходящим током в другой, более освещенной ветви двухвершинного дерева ясеня.
Рис. 13.13. Суточные колебания скорости подъема сока, измеренной методом тепловых импульсов, (Schubert, 1939) :
1 - лиственница; 2 - бук; 3 - ель
390
Глава 13
ПОПЕРЕЧНОЕ СЕЧЕНИЕ СТЕБЛЕЙ, УЧАСТВУЮЩЕЕ В ПОДЪЕМЕ ПАСОКИ. Размеры общего поперечного сечения стволов деревьев, участвующего в проведении воды, различны. У большинства видов центральная часть ствола превращается в ядро и становится физиологически неактивной. Даже не вся заболонь участвует в проведении сока, хотя существуют весьма различные мнения по этому вопросу. По крайней мере, у некоторых видов кольцесосудистых деревьев, в том числе у каштана, ясеня и дуба красного, вода передвигается, по-видимому, по самому наружному годичному кольцу, хотя некоторая часть может проходить по поздней древесине предыдущего года (Румбольд, 1920). Каней и Козловский (1975) сообщили, что в сеянцах ясеня американского вода передвигается по крупным сосудам ранней древесины кольца текущего года, а в сеянцах клена сахарного — по крупным сосудам данного года и в двух третях годичного кольца предшествующего года. Исследования Козловского и Вингета (1963) также показывают, что передвижение воды ограничено одним или двумя самыми наружными годичными кольцами у сосны смолистой, дуба белого и дуба крупноплодного. Имеются сообщения о восходящем передвижении краски по концентрическим цилиндрам поздней древесины у некоторых видов, но это подвергалось сомнению. Характер передвижения сока у хвойных и рассеяннососудистых древесных растений более сложен вследствие участия в этом процессе большого числа годичных колец. У покоящихся сеянцев хвойных вплоть до 4-летнего возраста вода передвигается по всем годичным кольцам, причем по самому наружному кольцу менее быстро, чем по второму. Через крупные трахеиды ранней древесины проходит больше воды, чем через более мелкие поздней древесины. У более старых хвойных деревьев существуют, по-видимому, значительные различия в ширине проводящей воду заболони (некоторые данные о числе колец, входящих в состав заболони, приведены в табл. 2.3). По сообщению Свенсона (1967), у сосны скрученной и ели Энгельмана вода наиболее быстро передвигается в древесине на расстоянии 15—20 мм от периферии, а в наружном 5-миллиметровом слое древесины восходящий ток идет слабо. Это удивительно, так как следовало ожидать, что наиболее быстрое проведение воды происходит в одном или двух самых наружных годичных кольцах, которые, по-видимому, непосредственно связаны с транспирирующими листьями. Анатомическое строение проводящей системы нуждается, очевидно, в дальнейшем изучении.
УСТОЙЧИВОСТЬ К ПОВРЕЖДЕНИЯМ. Деревья имеют, по-видимому, большой запас прочности во до проводящей системы и могут выдерживать разрушение значительной части ее поперечного сечения. Наблюдалось, что деревья дуба, у которых 50 % окружности было уничтожено пожаром, росли в течение последующих 10 лет так же, как и соседние неповрежденные деревья. При этом поврежденные деревья быстро образовывали новую ксилему, ориентированную так, чтобы получился эффективный путь вокруг ран (рис. 13.14). Такая тенденция к переориентации ксилемы и флоэмы вокруг поранений часто наблюдается при спиральном кольцевании деревьев. По сообщениям, горизонтальная зарубка на глубину более половины древесного ствола не вызыва-
Поглощение воды, подъем ксилемного сока и водный баланс
391
Рис. 13.14. Изменение ориентации ксилемы и флоэмы, окружающих рану, вызванную пожаром (Jemison, 1944):
1 _ сгоревшая зона; 2 — сохранившаяся зона; 3, 4 — корни, соединенные новыми проводящими тканями с оставшимися в живых
Рис.' 13.15. Схемы, показывающие влияние горизонтальных зарубок на стволе сосны на восходящее передвижение сока, меченного 3Zp (Postlethwait, Rogers, 1958):
а, б, в, г — зарубки, доходившие до половины ствола. "Р вводился в точке р, участки, обозначенные пунктиром, показывают его путь
ла заметного водного дефицита в листьях, но две параллельные зарубки на стволе создавали водный дефицит в надземных частях (рис. 13.15).
Некоторые так называемые сосудистые заболевания (вилт), например болезни увядания дуба и мимозы, вертициллез вяза и клена, голландская болезнь вяза, происходят вследствие того, что патогенные организмы блокируют прохождение воды через ксилему. Вряд ли мицелий сам по себе действительно блокирует ксилему. Более вероятно, что патогенные организмы вырабатывают токсины, которые повреждают живые клетки, примыкающие к элементам ксилемы, и вызывают образование камеди и каллусов, закупоривающих ксилему и вызывающих повреждение из-за обезвоживания. Тальбус (1968) указывает, что повреждения от так называемых сосудистых заболеваний являются сложными и не все они вызываются единственной причиной. Сверлящие насекомые также вызывают частичное повреждение ксилемы, вероятно, вследствие того, что их ходы являются причиной обезвоживания. В некоторых случаях насекомые способствуют также внедрению грибов, что вызывает дополнительное повреждение проводящей системы.
ВОДНЫЙ БАЛАНС
Рост древесных и травянистых растений чаще ослабляется вследствие водного дефицита, чем из-за какого-либо другого отдельного
392
Глава 13
фактора. Обширные литературные данные, обобщенные Цанером (1968), показывают наличие корреляции между ростом растений в высоту и толщину и количеством доступной воды: 70—80 % изменений ширины годичных колец во влажных районах и 90 % в сухих местообитаниях можно отнести за счет различий в напряженности водного режима. Степень водного стресса в растениях определяется относительной скоростью поглощения и потери воды, поэтому водный дефицит могут вызывать или слишком медленное поглощение, или слишком быстрая потеря воды, или чаще всего — сочетание того и другого. Таким образом, изучение факторов, влияющих на поглощение воды и на транспирацию, имеет большое значение, так как способствует пониманию внутреннего водного баланса, который в свою очередь влияет на физиологические процессы и условия, контролирующие количество и качество роста.
Под водным стрессом понимают такие условия, при которых клетки не вполне тургесцентны, а водный потенциал значительно ниже нуля. Первые видимые признаки водного стресса — закрывание устьиц, завядание листьев и молодых стеблей и прекращение роста. Существует много и других незаметных, но важных последствий, которые будут обсуждены позже. Степень водного стресса часто оценивают количественно при помощи таких показателей, как дефицит насыщения, относительное содержание воды или лучше всего — водный потенциал.
ПРИЧИНЫ ВОДНОГО СТРЕССА. Использование таких понятий, как водный баланс и водное хозяйство, подчеркивает возможность рассматривать внутренний водный режим растений наподобие бюджета, в котором содержание воды (баланс) определяется относительной скоростью поглощения (приходом) и потери воды (расходом).
Отставание поглощения от потери воды. Точно установлено, что содержание воды в листьях и стеблях растений часто заметно понижается около полудня при солнечной погоде (рис. 13.16). По сообщениям многих исследователей, это проявляется и в уменьшении диаметра ствола (рис. 13.17). Гиббс (1935) тщательно исследовал суточные изменения содержания воды в древесине стволов березы (табл. 13.4).
Он установил, что содержание воды достигает максимума около восхода солнца, понижается утром и в полдень, повышается после полудня и вечером. Такой режим, характерный, по-видимому, для многих растений яри теплой солнечной погоде, указывает на то, что стволы
13.4. СУТОЧНЫЕ КОЛЕБАНИЯ СОДЕРЖАНИЯ ВОДЫ В ДРЕВЕСИНЕ СТВОЛОВ БЕРЕЗЫ (Gibbs, 193S)
Показатель Дата
24 августа 2S августа
Время, часы суток 5 13 19 5 13 19
Погода Ясно Ясно, жарко Ясно Ясно Небольшая облачность Ясно
Содержание годы, % к сухому весу 65 54 58 59 50 53
Поглощение воды, подъем ксилемного сока и водный баланс
393
Рис. 13.16, Суточные колебания (Ackley, 1954) :
1 — содержание воды; 2 — дефицита насыщения в листьях груши
Vqcw суток
деревьев служат местом запасания воды, которая извлекается, когда транспирация превосходит поглощение, и вновь пополняется при противоположной ситуации.
Причина колебаний содержания воды заключается в более низком сопротивлении оттоку воды из тургесцентных растительных тканей по сравнению с сопротивлением поглощению воды корнями (рис. 13.18). Поэтому утром, когда транспирация усиливается, поглощение не ускоряется до тех пор, пока понижающийся водный потенциал не вызовет достаточно большого натяжения в ксилемном соке, чтобы преодолеть сопротивление току воды по ксилеме и еще большее сопротивление радиальному передвижению воды из почвы в ксилему корня. Тем временем вода удаляется из тканей, оказывающих слабое сопротивление ее оттоку, какими являются паренхимные клетки листьев и стеблей, что создает водный дефицит, иногда достаточно сильный, чтобы вызвать временное завядание листьев. По сообщению Хелквис-та с соавторами (1974), у ели ситхинской, растущей на влажной почве в Шотландии, водный потенциал листьев составляет от —12 до — 15 бар, а тургор уменьшается на 40 % от максимального.
То, что передвижение воды из древесного ствола отстает от ее потери листьями, показывают наблюдения Вагнера и Турнера (1971), согласно которым сжатие стволов сосны на высоте груди запаздывает приблизительно на 2 ч по сравнению с уменьшением водного потенциала листьев. Цаер (1971) наблюдал аналогичное отставание у дуг-
Рис. 13.17. Записи дендрографа, показывающие сжатие стволов сосны смолистой днем после их набухания ночью (Пд - полдень; Пн — полночь). Верхняя запись показывает отсутствие ежедневного сжатия и набухания ствола в течение последних дней недели, когда преобладала пасмурная и дождливая погода . (Kozlowski, 1968)
Время суток
Рис. 13.18. Соотношение между поглощением воды (2) и транспирацией (1):
а — ясеня американского; б — сосны ладанной; сеянцы выращивались в сосудах с автоматическим поливом, что позволяло измерять поглощение воды; измерения проводились через каждые 2 ч (Kramer, 1937)
ласии. Это отставание указывает на наличие значительного сопротивления току воды через ствол и ветви и по направлению к испаряющим поверхностям листьев. К концу дня, когда температура понижается и устьица закрываются, транспирация быстро уменьшается, но вода продолжает поглощаться до тех пор, пока водный потенциал в расте-нии не повысится приблизительно до величины водного потенциала ‘ г почвы. На это может потребоваться вся ночь. После высыхания почвы в последующие ночи полного оправления не происходит, затем наступает устойчивое завядание. Длительный сильный водный дефицит вы- ( зывает отмирание вследствие обезвоживания.
Полуденный водный дефицит и временное завядание вызываются тем, что транспирация временно превышает поглощение даже у растений, растущих на почве при влажности равной полевой влагоемко-сти. Длительное завядание происходит вследствие слабого поглощения воды, вызванного высыханием почвы, а иногда низкой температурой или недостаточной аэрацией, препятствующими возмещению воды, теряемой при транспирации. При усилении водного стресса устьица закрываются, уменьшая потерю воды, и деревья с сильно ку-тинизированными листьями, например маслина и сосна ладанная, могут выдерживать длительные периоды водного стресса с минимальным ущербом.
Конкуренция за воду внутри растения. В течение вегетационного периода разные части деревьев и крупных трасянистых растений часто конкурируют за воду. Вследствие различий в степени затенения и концентрации растворенных веществ разные части побегов теряют
Поглощение воды, подъем ксилемного сока и водный баланс
395
воду с различной скоростью, и в них создается выраженный в разной степени водный дефицит и неодинаковый водный потенциал. Это бывает особенно важно при высыхании почвы, когда те части растений, в которых создается самый низкий водный потенциал, получают воду за счет более старых тканей. Несмотря на то, что на растениях, страдающих от водного стресса, молодые листья могут завядать первыми, они обычно отмирают последними. Водный стресс ускоряет старение, вероятно, отчасти потому, что он уменьшает поступление цитокининов и изменяет баланс регуляторов роста в листьях. Нижние, затененные листья тоже страдают из-за недостатка воды, так как они вырабатывают углеводов меньше, чем верхние, лучше освещенные листья, и поэтому менее способны к осмотической-конкуренции за воду. Таким образом, обезвоживание может быть одной из причин отмирания нижних, затененных ветвей деревьев.
По мнению Халмерса и Ьильсона (1978), потребность развивающихся плодов персика в углеводах И возрастающий водный стресс понижают рост ветвей и плодоношение персиковых деревьев. Твердо установлено существование конкурент дии между листьями и плодами: у плодов наблюдается ослабленный рост или даже сморщивайие в полдень, когда во всем дереве развивается водный дефицит (рис. 13.19). Это сокращение размеров было установлено для плодов лимона, апельсина, груши, вишен и других плодов, а также для шишек голосеменных. Однако обнаружено, что молодые апельсины диаметром меньше 35 мм не теряют воду в течение периодов стресса. Это согласуется с результатами наблюдений о неспособности листьев отнимать воду от молодых коробочек хлопчатника, хотя у более старых коробочек происходит ежедневное полуденное сокращение. Эти данные соответствуют общей концепции, согласно которой более молодые части растений получают или сохраняют воду за счет более старых частей.
Колебания содержания воды и водного потенциала, наблюдаемые в различных частях транспирирующих растений и даже в разных частях крупных листьев, создают серьезную проблему отбора проб для измерения этих величин.
ДЛИТЕЛЬНЫЕ ИЗМЕНЕНИЯ СОДЕРЖАНИЯ ВОДЫ. Более 50 % общей сырой массы дерева состоит из воды. Содержание ее широко варьирует в разных частях дерева и зависит от вида растений, возрас-
Время суток
Рис. 13.19. Полуденное (Яд) сокращение диаметра плодов вишни после их расширения ночью (Пн), когда исчезал водный стресс. Стрелками показано время полива (Kozlowski, 1968)
396
Глава 13
Рис. 1320. Сезонные изменения содержания воды в древесине хвойных и лиственных пород в Восточной Канаде. Содержание воды в древесине хвойных изменяется меньше, чем в древесине лиственных пород (см. рис. 13.9). Изменение содержания воды в наружной части заболони:
1 ~~ пихты бальзамической; 2 - сосны Банкса; 3 — тополя осинообразного; изменение содержания воды в наружной части ядра;
4 — тополя осинообразного; 5 — сосны Банкса
та, места обитания и времени года. Содержание воды в хорошо развитой ядровой древесине обычно гораздо ниже, чем в заболони (рис. 13.20). Кария составляет исключение, так как ее ядровая древесина постоянно содержит больше воды, чем заболонь во все времена года. Пытались уменьшить содержание воды, не отделяя в течение нескольких дней после рубки облиственную верхушку. Эти попытки оказались безуспешными для карии, хотя у березы этот прием сильно понижал влажность стволов. Вследствие необычного распределения воды в древесине карии ее трудно высушить без раскалывания. По сообщению Гиббса (1939), у сосны Банкса и ели белой ядровая древесина очень сухая. В ядре пихты* бальзамической и тсуги часто имеются влажные зоны, в которых наблюдаются суточные или сезонные изменения содержания воды. Паркер (1954) сообщил, что у дугласии и можжевельника виргинского ядро содержит меньше воды, чем у сосны желтой и пихты великой.
По сообщению Овингтона (195$), содержание воды обычно увеличивается от основания к вершине деревьев, до Ито (1955) обнаружил у Castanea crenata уменьшение содержания воды от основания вверх. Люксфорд (1930) установил, что у секвойи наибольшее количество воды содержится у основания ствола и оно понижается по направлению к верхушке. В заболони, наоборот, самая низкая влажность наблюдается у основания, максимальная — у вершины.
Сезонные изменения содержания воды. У некоторых видов древесных растений наблюдаются большие сезонные колебания содержания воды в стволах. Эти изменения интересны не только в физиологическом отношении, но имеют и большое практическое значение, так как они влияют на скорость высыхания, на плавучесть бревен и на транспортные расходы.
В Восточной Канаде бревна березы и тополя в мае - начале июня имеют такую большую плотность, что их невозможно сплавлять. В сентябре плотность уменьшается на 60 — 75 %, так как часть воды замещается воздухом. Содержание воды и плотность заболони и сырого ядра пихты бальзамической остаются высокими в течение всего лета (табл. 13.5). Обычно наибольшие сезонные колебания содержания воды наблюдаются у лиственных пород, но Ито (1955) установил, что в Японии большие сезонные изменения влажности древесины происходят у сосны густоцветной, чем у каштана городчатого, причем ми-
Поглощение воды, подъем ксилемного сока и водный баланс
397
13.5. СОДЕРЖАНИЕ ВОДЫ И ГАЗА В СТ ВОДАХ НЕКОТОРЫХ ВИДОВ ДРЕВЕСНЫХ РАСТЕНИЙ В РАЗНОЕ ВРЕМЯ ГОДА (Gibbs, 1935 г.)
Й
I I I I > I I I i ) I I I I J I 1 I ) ) I I I I I I 1 I I J I I i i ) I I I t ! f I I I ) I I I I I I t 1 I I I I I t I I I I I I I I i I I ( f I I I I I I I I i 1
398
Глава 13
нимум у обоих видов приходится на август. Гиббс приводит данные Р. Гартига и Э. Мюнха, согласно которым, в Европе у хвойных наблюдаются значительные сезонные колебания содержания воды, тогда как сам Гиббс обнаружил у хвойных в Восточной Канаде довольно слабые изменения (см. рис. 13.21).
Типичный характер сезонных изменений содержания воды у рассеяннососудистых пород (у березы) показан на рис. 13.9. В Восточной Канаде наибольшее содержание воды в стволах берез, тополей, некоторых осин и ив достигается весной, перед самым распусканием листвы. В течение лета содержание воды понижается до минимальной величины перед листопадом, затем увеличивается осенью, после того как опадение листьев уменьшит транспирацию, но лишь до тех пор, пока достаточно сильное охлаждение почвы не будет препятствовать поглощению воды. У некоторых видов наблюдается второе понижение в конце зимы, по-видимому, вследствие затрудненного поглощения воды из холодной почвы, а после оттаивания почвы содержание воды увеличивается до максимального, но до распускания листьев. Среди отклонений от описанного хода изменений влажности стволов можно указать следующие: у ясеня американского и ильма белого не наблюдается осеннего повышения, у клена серебристого и бука максимум содержания воды достигается осенью, причем бук резко отличается от всех изученных видов отсутствием весеннего повышения содержания воды. Данные для более мягкого климата немногочисленны, но зимнее понижение менее вероятно там, где почва не замерзает.
Методы пересчета содержания воды. Уместно обсудить здесь методы пересчета содержания воды. Обычно его выражают в процентах к сырому или сухому весу, однако оба метода имеют некоторые недостатки. Пересчет на сырой вес — неточен, так как сырой вес сочных тканей, например листьев и молодых стеблей, может сильно колебаться по дням и даже по часам. Кроме того, большие изменения содержания воды на единицу ткани приводят к необычно маленьким изменениям процентного содержания на сырой вес. Как указывают Куртис и Кларк (1950), при уменьшении содержания воды в листьях пустынного кустарника Nicotiana glauca с 85 до 80 % в пересчете на сырой вес они теряли 25 г воды на первоначальные 100 г сырого веса, или 29,4 % исходного содержания воды. При пересчете на сухой вес листьев содержание воды понижалось с 566 до 400 % (при этом сухой вес не изменялся) . К сожалении), сухой вес также не является постоянной величиной, потому что фотосинтез, дыхание и передвижение веществ могут быстро его изменять, а в течение длительных периодов значительно увеличивается сухая масса вследствие утолщения клеточных стенок. Например, установлено, что сухой вес листьев груши увеличивался почти на 150 % с июня по август, тогда как количество воды оставалось без изменений (рис. 13.21), в результате сильно уменьшилось содержание воды на единицу сухого веса.
Сходные результаты получили Козловский и Клаузен (1965) для листьев дуба эллипсоидального. В начале вегетационного периода процентное содержание воды увеличивалось в шишках ели белой и лиственницы американской в результате усиленного поглощения воды. В дальнейшем оно уменьшалось вследствие того, что сухая масса шишек быстро увеличивалась, тогда как количество воды в них изменялось
Поглощение воды, подъем ксилемного сока и водный баланс
399
Рис. 1321. Сезонные изменения сухого веса, содержания воды и количества воды, поглощаемой для достижения насыщения листьев груши (Acklev, 1954):
1 — общий вес при насыщении; 2 — вес воды в полевых условиях; 3 — сухое вещество; 4 — вода, поглощенная при насыщении
незначительно. Таким образом, изменение процентного содержания воды отражает изменения в содержании как сухого вещества, так и воды.
Масон и Маскель (1928), а также Денни (1932) попытались получить более подходящую основу для пересчета изменений в содержании воды и других веществ путем экстрагирования листьев в разбавленной НС1 для удаления
из них легко гидролизуемых соеди-
нений, в результате чего остаются только клеточные стенки и другие стойкие компоненты. Это уменьшает ежечасные колебания, вызываемые фотосинтезом, дыханием и оттоком веществ, но не устраняет длительных изменений, связанных с утолщением клеточных стенок. Какой бы метод ни использовали исследователи, они должны помнить о не
достатках, присущих этим методам.
ВЛИЯНИЕ ВОДНОГО СТРЕССА. Водный дефицит изменяет анатомию, морфологию, физиологию и биохимию выращиваемых растений. В сухих местообитаниях произрастают деревья меньших размеров, их листья обычно мельче, толще и сильнее кутинизированы, диаметр сосудов ранней древесины часто бывает меньше, а клеточные стенки толще и более лигнифицированы. Как уже упоминалось, прирост деревьев тесно коррелирует с наличием воды. Водный стресс задерживает деление клеток в меньшей степени, чем их растяжение. Вследствие этого размеры растений уменьшаются при недостатке воды в основном из-за того, что для растяжения клеток необходим некоторый минимальный уровень тургора. Тургор имеет также большое значение для регулирования степени открытости устьиц, для распускания листьев и цветков и для движений частей растений, например для никти-настического складывания листочков мимозы. Другое важное следствие пониженного тургора — ослабление фотосинтеза, вызываемое закрыванием устьиц, влиянием на механизм фотосинтетического процесса и уменьшением листовой поверхности (см. главу 5).
Исследования, проведенные преимущественно на травянистых растениях, показали, что водный стресс действует на многие процессы, идущие с участием ферментов, в том числе на дыхание, темновые реакции фотосинтеза, образование хлорофилла, углеводный и азотный обмен (см. главы 7 и 9), влияет он и на органеллы (например, на митохонд
400
Глава 13
рии и рибосомы), на структуру мембран. Водный стресс влияет также на банане регуляторов роста: содержание цитокинина в побегах уменьшается, а содержание абсцизовой кислоты и ауксина увеличивается. Синтез гиббереллина в корнях тоже может сокращаться. Большая часть этих исследований была выполнена на травянистых растениях, но результаты, вероятно, в равной степени приложимы и к древесным растениям.
Связь водного стресса с устойчивостью к насекомым и болезням. Существуют доказательства того, что поражение деревьев сверлящими насекомыми, живущими во внутренней коре или в наружной части древесины, бывает более сильным в засушливые годы, чем в годы с незначительным водным стрессом. Как утверждает Вите (1961), гораздо более сильное заражение сосны желтой жуками бывает у деревьев, страдающих от водного стресса, чем у хорошо увлажненных. Имеются также сообщения, что степень и продолжительность водного стресса существенно влияют на восприимчивость сосны ладанной к поражению южным сосновым лубоедом. Очевидно, высокое давление живицы в деревьях, не испытывающих стресса, препятствует вторжению сверлильщиков. Сходная ситуация встречается у пих^ы одноцветной, в которой поселяются короеды при водном стрессе — 20 бар, но не при —15 бар вследствие более сильного истечения смолы у деревьев, меньше страдающих от стресса.
Имеются также данные, свидетельствующие о том, что водный стресс благоприятствует проникновению в растения некоторых грибов. По сообщению Бира (1959), гриб, вызывающий рак ивы, вторгается в кору только когда относительное содержание воды в ней ниже 80 %. Сходную зависимость автор обнаружил у возбудителя рака коры тополя. Паркер (1961) сделал обзор литературы по связи между содержанием воды и болезнями растений и пришел к выводу, что развитие возбудителей рака коры обычно коррелирует с низким содержанием воды. Утверждают также, что повреждение растений Fomes annosus усиливается при водном стрессе. Некоторые грибы, заражающие листья, например возбудители мучнистой росы, причиняют большой вред при сухой погоде, однако распространение многих грибов усиливается в сырую погоду, когда поверхность листьев остается влажной в течение многих часов, что создает условия для прорастания спор грибов, например возбудителя парши яблони. (Влияние водного стресса на вегетативный и репродуктивный рост рассматривается более подробно в главе 16.)
Благоприятное действие водного стресса. При определенных условиях умеренный водный стресс может улучшать качество растительных продуктов, даже если он уменьшает вегетативный рост. Это положение справедливо для гваюлы, у которой наблюдается большое повышение содержания каучука при умеренном водном стрессе, несмотря на то, что сырой вес одного растения уменьшается. Установлено, что качество яблок, груш, персиков и слив улучшается под влиянием водного стресса. По сообщениям, содержание масла в маслинах повышается при водном стрессе, хотя общий урожай, вероятно, уменьшается. Утверждают, что водный стресс вызывает увеличение содержания алкалоидов в некоторых лекарственных растениях, но, по некоторым данным, содержание алкалоида в Cinchona ledgeriana понижается. Можно
Поглощение воды, подъем ксилемного сока и водный баланс
401
также согласиться с тем, что повышенное количество толстостенных элементов ксилемы, которые образуются в деревьях, подвергающихся действию водного стресса, в некоторых случаях является благоприятным, так как в результате этого увеличивается плотность древесины. Интересны дальнейшие исследования влияния водного стресса на качество древесины.
ЗАСУХОВЫНОСЛИВОСГЬ
В течение многих лет термин засухоустойчивость использовался для обозначения способности растений переносить засуху. Однако этот термин имеет недостаток: при обсуждении так называемой засухоустойчивости часто смешивают фактор внешней среды, засуху, с реакцией растений на недостаток воды, который лучше описывать как водный дефицит или водный стресс. Мы предпочитаем пользоваться термином засуховыносливость, потому что он более точно характеризует реакцию растений на засуху. Необходимо подчеркнуть, что засуха — это метеорологическое явление, обычно описываемое как период без осадков, достаточно продолжительный для того, чтобы вызвать истощение запасов почвенной влаги и ослабление роста растений. Засуха может быть долговременной, как в засушливых районах, сезонной, как в областях со строго определенными влажными и сухими временами года, или случайной -- в наиболее влажных зонах. Продолжительность периода без дождей, необходимая для создания засушливых условий, зависит от влагоемкости почвы и скорости эва-потранспирации и в меньшей степени — от типа растительности. Даже в таких влажных районах, как Западная Европа и юго-восток США, засухи обычно вредны для растений.
Засуха — это фактор внешней среды, вызывающий водный дефицит или водный стресс в растениях. В технике и в физических науках стресс обозначается как ’’напряжение”, и наблюдаемый в растениях .эффект следовало бы скорее называть не водным стрессом, а ’’напряжением воды”. Однако в физиологии растений термин водный стресс используют как для характеристики водного дефицита во внешней среде, так и в. отношении водного дефицита растений. Водный стресс, или водный дефицит, растений начинает развиваться при водном потенциале листьев —2 или —3 бар или когда тургор клеток становится значительно ниже его максимальной величины.
Водный дефицит растений и стресс всегда сопровождают засуху, но встречаются они и в другое время - - вследствие слишком быстрой транспирации или когда поглощению воды препятствуют холодная почва, избыток солей в почвенном растворе или повреждение корневых систем. Большинство растений подвергается действию временного водного дефицита и стресса в полдень при жаркой солнечной погоде даже в тех случаях, когда они растут на почве с влажностью близкой к полевой влагоемкости или на разбавленном питательном растворе. Например, Хелквист с соавторами (1974) обнаружили заметный водный дефицит в верхних ветвях ели ситхинской, растущей на влажной почве в мягком климате Шотландии. Верхние, более осве
Глава 13
щенные части древесных крон обычно подвергаются действию более сильного стресса, чем нижние.
Последствия водного дефицита, вызываемого засухой или другими причинами, так же важны для роста лесных, фруктовых и декоративных древесных растений, как и для однолетних травянистых сельскохозяйственных культур. Способность переносить засуху зависит от ряда фенологических, морфологических и физиологических факторов. Фермеры и лесоводы, так же как и экологи и физиологи, знают, что деревья одних видов переносят засуху с меньшим ущербом, чем деревья других видов. Например, сосна желтая переносит засуху лучше, чем пихта великая; дубы малый и мерилендский растут в местообитаниях слишком сухих для дубов красного и белого, а сосна ежовая растет в более сухих местах, чем сосна ладанная. Некоторые виды из полуза-сушливых районов, например Larrea, Laurus, маслина и некоторые акации, переносят сильное обезвоживание во время длительных засух, а Cornus florida сильно повреждается засухой. Некоторые расы сосны ладанной лучше переносят водный стресс, чем другие, а дугласии из удаленных от моря территорий легче переносят обезвоживание, чем прибрежные расы. Имеются сообщения и о других различиях между популяциями из разных местообитаний. Например, Ладигес (1974) обнаружил, что сеянцы Eucalyptus viminalis из районов с малым количеством осадков более устойчивы к повреждению от высыхания, чем из районов с большим количеством осадков. Сосны замечательная, ладанная и ежовая, по-видимому, лучше всего переносят обезвоживание при атмосферных условиях, сходных с их естественной средой обитания. Лар хер (1975) собрал данные о засуховыносливости самых разнообразных древесных и травянистых растений.
ПРИЧИНЫ ЗАСУХОВЫНОСЛИВОСТИ. Растения выживают в районах с недостаточным количеством осадков благодаря тому, что они избегают засухи, так как обладают морфологическими или физиологическими особенностями, дающими им возможность не подвергаться обезвоживанию, или удлинять этот срок, или потому, что они могут выносить высыхание. )
Избегание засухи. Растения, избегающие засуху встречаются в районах со строго определенными засушливыми сезонами. К ним относятся пустынные эфемеры с таким коротким циклом развития, что они f завершают его в течение нескольких недель после зимних дождей, и растения, созревающие в начале лета до высыхания почвы. К эфемерам относятся главным образом однолетние растения, хотя избегание засухи может иметь значение и для некоторых многолетних растений •’ в средиземноморском климате.
Избегание или отсрочка высыхания. Повреждающая степень обез- воживания при пониженном поступлении воды в корни может быть отсрочена несколькими путями. Наиболее понятный способ заключается в запасании большого количества воды в мясистых корнях или стеблях. Однако такая способность свойственна немногим видам, к числу которых относятся кактусы, способные накапливать много воды и хорошо регулирующие интенсивность транспирации. Несмотря на то, что в стволах деревьев запасается много воды, ее масса незначи
Поглощение воды, подъем ксилемного сока и водный баланс
403
тельна по сравнению с транспирационными потерями древесных растений.
У многих растений из засушливой зоны или из районов с продолжительной летней засухой имеются сильно кутинизированные листья и очень низкая интенсивность транспирации после закрывания устьиц. По сообщению Опенхаймера (1951), в Израиле такие растения, как це-ратония стручковая, лавр, маслина, сосна алеппская и Arbutus andrache, имеют очень низкую интенсивность транспирации при исчерпании запасов почвенной влаги, тогда как миндаль и инжир слабо регулируют свою транспирацию. Кауль и Крамер (1965) установили, что устьица падуба (Ilex cornuta, var Burfordii) закрывались при гораздо более низком водном Дефиците листьев, чем устьица азалии (Rhododendron poukhanensis). Транспирация срезанных ветвей вначале была более быстрой у азалии, что говорит о лучшей регуляции как кутикулярной, так и устьичной транспирации у падуба. У растений, страдающих от стресса, транспирация часто сокращается посредством сбрасывания листьев, однако это уменьшает и фотосинтез.
Корневые системы. Большинство исследователей признают, что глубокие, широко простирающиеся корневые системы имеют большое значение для отсрочки повреждения от высыхания. Опенхаймер (1951) считает это важным свойством растений, произрастающих в Израиле. Утверждают, что быстрое разрастание корней - главная причина того, что некоторые расы сосны ладанной в Техасе лучше противостоят обезвоживанию, тогда как регулирование транспирации играет второстепенную роль. Установлено, что большие корневые системы имеют большое значение для приживаемости пересаживаемых сеянцев дугласии и сосны желтой на тихоокеанском северо-западе США. По наблюдению Сато (1956), степень развития корневых систем — важный фактор выживания сеянцев хвойных в Японии. Фовелс и Кирк (1945) сделали вывод, что уменьшение поглощающей системы вследствие ее повреждения при выкопке и посадке является важной причиной отпада саженцев сосны желтой. Следует отметить особое значение выращивания саженцев с мощными корневыми системами, способных быстро возобновлять рост после пересадки.
Важность глубокой корневой системы признается всеми лесоводами и садоводами, причем первые уделяют особое внимание различиям в начальной стадии развития корней сеянцев древесных растений как причине разной выживаемости. Например, кипарис болотный и береза желтая приживаются только на влажной почве, так как из-за поверхностных корней они не способны переносить летнюю засуху, тогда как большинство суходольных видов имеют довольно глубокие корневые системы. Неспособность сеянцев сосны выживать под пологом леса, где сеянцы лиственных пород растут хорошо, можно объяснить, хотя бы частично, развитием в условиях затенения более поверхностных корневых систем, что делает их более чувствительными к вредному действию летней засухи.
Способность переносить обезвоживание. Другой крайностью является устойчивость к высыханию, объясняемая, способностью протоплазмы подвергаться чрезвычайно сильному обезвоживанию без необратимых повреждений. Выдающиеся примеры такой выносливости встречаются
Глава 13
404
среди мхов и лишайников, но лишь немногие цветковые растения также могут обезвоживаться до воздушно-сухого состояния. Некоторые древесные растения засушливых районов, например креозотовый куст, полынь, акации, кустарники средиземноморского маквиса и калифорнийского чаларраля, обладают значительной выносливостью протоплазмы к обезвоживанию. Считают, что у дубов малого и мерилендского протоплазма более вынослива, чем у дубов белого или красного, так как они переносят степень высыхания смертельную для других дубов, хотя и не обладают более обширными корневыми системами или лучшей способностью регулировать транспирацию. Следует отметить, что корни этих дубов, по-видимому, более устойчивы к обезвоживанию, чем побеги, в результате чего после гибели побегов от высыхания возникает корневая поросль. В противоположность этому наблюдения Брикса (1960) и Лезели (1965) показали, что корни сосны отмирают раньше побегов. Парис и Феррель (1966) установили, что сеянцы дугласии из внутренних районов более устойчивы к обезвоживанию, чем сеянцы из прибрежных. Паркер (1968) считает способность переносить высыхание важным признаком древесных пород.
Основы устойчивости к обезвоживанию. При понижении содержания воды до критического уровня выживание растений зависит от степени обезвоживания, которую способна выдержать протоплазма, не претерпевая необратимых повреждений. В этом отношении между разными видами имеются, по-видимому, большие различия. По сообщениям, листья миндаля могут высыхать без повреждения до 70 %-ного дефицита насыщения, маслина — до 60 %, а инжир — только до 25 %. Существуют также и сезонные различия. Листья креозотового куста, образовавшиеся при влажной погоде, имеют большие размеры и легко
Месяцы
195И
5
Рис. 13.22. Сезонные изменения выносливости к обезвоживанию двух видов хвойных. Выносливость к дегидратации повышается зимой и понижается в течение вегетационного периода. Ель может подвергаться дегидратации без повреждения до более низкого содержания воды, чем сосна (Pisek, Laichei, 1954): а — сосна кедровая европейская; б --ель обыкновенная; 1 — без повреждения; 2 — незначительное повреждение; 3 - 10 % повреждений; 4 — 1/3 погибшие
Поглощение воды, подъем ксилемного сока и водный баланс
405
повреждаются под влиянием водного дефицита, а мелкие листья, которые сформировались при сухой погоде, могут быть подсушены до 50 %-ного дефицита насыщения. Установлено, что у некоторых видов устойчивость к обезвоживанию повышается зимой одновременно с холодостойкостью, а затем понижается весной (рис. 13.22).
Одно время связанная вода считалась важным фактором засухо-и холодоустойчивости, однако наибольшее количество воды связывается в клеточных стенках и не играет большой физиологической роли. Высокое осмотическое давление также считалось важным приспособлением к водному стрессу. Благоприятное влияние осморегуляции на водный стресс было вновь показано на сельскохозяйственных культурах, но оно может иметь значение и для некоторых древесных растений. Например, по сообщению Осонуби и Дэвиса (1978), накопление солей является важным фактором поддержания тургора у испытывающего водный стресс дуба черешчатого, но не у березы.
Западноевропейские и советские исследователи придают особое значение протоплазматическим и биохимическим изменениям, повышающим устойчивость к обезвоживанию. Левит (1972) придает значение образованию — SH-групп, а Штокер и Росс (1956) приписывает особую роль изменениям структуры протоплазмы. Стадельман (1971) рассмотрел методы измерения проницаемости и вязкости протоплазмы в связи с устойчивостью к обезвоживанию. В настоящее время особое внимание уделяется устойчивости тонких структур к повреждению вследствие обезвоживания.
Как утверждают Паркер (1956, 1968), Бег, Турнер (1976) и Козловский (1976), существует несколько причин засуховыносливости. К ним относятся факторы, которые удлиняют срок обезвоживания (глубокие корневые системы, толстая кутикула и хорошее регулиро-! вание транспирации), а также способность протоплазмы переносить ( высушивание. Левит (1972) подробно рассмотрел эти факторы и пришел к выводу, что способность избегать обезвоживание имеет большее значение, чем способность выносить его. Несмотря на то, что способность выдерживать обезвоживание часто имеет большое значение для приживаемости сеянцев и для выживания дикой растительности, ни современное лесное хозяйство, ни садоводство не могут быть рентабельными в районах, где бывают продолжительные и сильные засухи, без искусственного орошения.
ЗАКАЛИВАНИЕ ПРОТИВ ЗАСУХИ. Известно, что растения, подвергавшиеся действию водного стресса, меньше повреждаются засухой, чем растения, не испытавшие до этого недостатка воды. Например, если внезапно перенести горшечные растения из затененных, влажных условий внешней среды на полное солнечное освещение, то листья часто повреждаются, даже если растения хорошо поливать. Это обычно относят за счет меньшей кутинизации и больших промежутков между жилками листьев, выросших в тени, а также более низкого отношения корней к побегам. Рок (1973) установил, что у ежедневно поливаемых сеянцев сосны замечательной интенсивность устьичной и кутикулярной, транспирации была более высокой, чем у поливаемых реже, хотя у них не различалось отношение корни / побеги. Сеянцы, поливаемые ежедневно, испытывали более сильный водный стресс и хуже укоренялись
Глава 13
406
после пересадки, чем поливаемые менее часто, вероятно, вследствие того, что последние лучше регулировали транспирацию.
При выращивании травянистых и древесных растений для пересадки их в поле сеянцы для улучшения приживаемости обычно закаливают. Для этого сеянцы выставляют на солнце и уменьшают частоту полива. На лесных питомниках часто подрезают или выдергивают корни сеянцев, чтобы создать компактные, сильно разветвленные корневые системы. Эти приемы также вызывают умеренный водный стресс.
Имеются также доказательства, что протоплазматические изменения, возникающие при водном стрессе, благоприятствуют выживанию. Например, по некоторым исследованиям, фотосинтетический аппарат креозотового куста, выращенного в сухом местообитании, более устойчив к водному стрессу, чем у выросшего на влажном месте. Это явление, вероятно, аналогично ’’привыканию” мембран хлоропластов креозотового куста к изменению температуры.
ИЗМЕРЕНИЕ ВОДНОГО СТРЕССА
Еще в начале столетия экологи и физиологи понимали необходимость количественного измерения водного стресса растений, но не было еще соответствующих методов. Были проведены многочисленные измерения осмотического давления, но и после того, как стали понятны преимущества измерения водного потенциала (который тогда называли сосущей силой или дефицитом диффузионного давления), не было удовлетворительных методов для его определения. Вследствие этого многие ранние исследования водного режима были неубедительными, поскольку не проводились количественные измерения степени стресса, которому подвергались растения. В последние два десятилетия разработаны надежные методы, поэтому не оправдано отсутствие оценки водного стресса растений в экспериментах, для которых имеет значение состояние воды в растениях.
ПРОБЛЕМА ОТБОРА ПРОБ. При любом методе важен сбор типичных и сравнимых проб. Листья — это чувствительный индикатор водного стресса, но состояние воды в них сильно изменяется в зависимости от возраста, времени суток, освещенности и расположения на растении. Возможны большие ошибки при сравнении проб р азног о возраста или собранных в разное время суток. Необходимо также позаботиться о том; чтобы предотвратить изменения состояния воды во время между сбором проб и измерением.
ПРЯМОЕ ИЗМЕРЕНИЕ СОСТОЯНИЯ ВОДЫ. Уже упоминалось о погрешностях, присущих измерению и пересчету содержания воды на сырой или сухой вес. Вследствие этого некоторые исследователи выражают содержание воды в тканях в процентах от ее содержания при максимальном тургоре. Штоккер, который ввел этот метод, назвал величину, вычисленную по нижеприведенному уравнению, водным дефицитом (часто его называют дефицитом насыщения водой ДНВ):
вес при полном тургоре — сырой вес
Водный дефицит =--------------------------------- ,
вес при полном тургоре - абс. сухой вес
Поглощение воды, подъем ксилемного сока и водный баланс
407
Рис. 13.23. Связь между водным дефицитом и водным потенциалом разновозрастных листьев дерена из двух местообитаний (Knipling, 1967) :
а — у ручья; б — на водоразделе; 1 — май; 2 -- июнь; 3 — июль; 4 — август; более сильные возрастные изменения отмечены у листьев деревьев, произрастающих на водоразделе, чем у листьев хорошо увлажняемых деревьев около ручья
Ватерлей, видоизменир уравнение Шток кера, вычислил показатель, который он назвал относительной тургесцентностью:
Относительная сырой вес — абс, сухой вес
тургесцентность, =----------------------------------------х 100.
или относитель- вес при полном тургоре — абс. сухой ное содержа- вес
ние воды
Относительную тургесцентность часто предпочитают называть относительным содержанием воды (ОСВ). Два названных показателя взаимно дополняют друг друга: ОСВ = 100 — ДНВ. Точная методика, подходящая для получения наиболее достоверных результатов, изменяется в зависимости от видов растений. Ватерлей предложил использовать вырезанные из листьев диски, а Клаузен и Козловский (1965), Гармс и Грегор (1962) считают целесообразным использовать для некоторых видов хвойных целые иглы. Гевлет и Крамер (1963) установили, что для некоторых древесных пород целые листья дают лучшие результаты, чем диски.
Использование дефицита насыщения водой, или относительного содержания воды, дает возможность следить за изменениями содержания воды при помощи простейшей аппаратуры. К сожалению, определенный водный дефицит не означает один и тот же водный потенциал в листьях разного возраста, разных видов и местообитаний (рис. 13.23). Водный стресс растений, измеренный в процентном отношении, нельзя также сравнивать с напряжением почвенной воды.
ИЗМЕРЕНИЕ ВОДНОГО ПОТЕНЦИАЛА. Вероятно, единственный наиболее удовлетворительный метод определения состояния воды в растении и почве — измерение водного потенциала, поскольку передвижение воды из почвы в растение и внутри растения контролируется ее потенциалом. Это измерение можно сделать, используя мето
Глава 13
ды равновесных жидкостей, термопарные психрометры или барокамеру Сколандера.
Метод окрашивания не требует дорогостоящего оборудования и может использоваться в поле, но для некоторых видов он дает большие погрешности. Метод барокамеры, рекомендуемый Сколандером и его сотрудниками, также пригоден для полевых исследований. Этим методом измеряют водный потенциал ксилемы, но обычно предполагают, что он почти равен водному потенциалу клеток листьев. Бауер (1967) установил, что результаты измерения водных потенциалов тиса и подсолнечника в барокамере сходны с величинами, полученными психрометрическим методом. В опытах с рододендроном этого не отмечалось. Кауфман (1968) также обнаружил большее соответствие между двумя методами для одних видов, чем для других.
Для решения многих задач наиболее пригодны измерения при помощи термопарных психрометров, так как этот прибор измеряет состояние свободной энергии воды в системе. Кроме того, после гибели пробы можно определить осмотический потенциал и вычислить тургор-ное давление. К сожалению, наличие солей на поверхности листьев приводит к ошибкам, поэтому для листьев, выделяющих соли, по-види-мому, более точен метод барокамеры. Оборудование для термопарных психрометров дорогостоящее, к тому же его трудно использовать в поле. В настоящее время психрометры доступны для измерений водного потенциала несрезанных листьев и корней, почвы in situ, а также отделенных частей растения и образцов почвы.
Водный потенциал — это мера состояния свободной энергии воды в растениях и почве. Он контролирует передвижение воды и, вероят- ) но, является лучшим простым показателем состояния воды в растении. Однако растяжение клеток обусловливается их тургором (по- 1 тенциалом давления); поэтому осмотический потенциал также может иметь значение. В связи с этим при изучении роста и растяжения клеток желательно или даже необходимо измерение как водного потенциала, так и осмотического потенциала для оценки тургора (потенциала давления). Объемный метод определения давления, разработанный Тире и Гаммелем (1972), использовали Роберт и Кнор ; (1977) для определения величины различных компонентов водного потенциала в некоторых видах деревьев. У пяти изученных видов наблюдалось постоянное увеличение отношения тургорного давления к водному потенциалу с лета до осени. х
КОСВЕННЫЕ МЕТОДЫ ОЦЕНКИ СОСТОЯНИЯ ВОДЫ. Делались попытки охарактеризовать состояние воды в растениях косвенными методами. Самым старым методом является, вероятно, наблюдение за степенью завядания растений (слабое, умеренное или сильное). Однако при этом не учитывается тот факт, что листья одних растений вянут после очень небольшой потери воды, тогда другие не завядают даже при сильном обезвоживании. Преждевременное закрывание устьиц — надежный показатель усиливающегося водного стресса у многих, но не у всех видов. Диффузионные порометры позволяют легко проводить измерения, но не обеспечивают количественной оценки водного стресса. Уменьшение толщины листьев, диаметра стеблей и пло
Поглощение воды, подъем ксилемного сока и водный баланс
дов также указывает на развитие водного стресса. Создание бета-из-мерительных приборов для определения изменений массы листьев позволяет контролировать быстрые изменения содержания воды, а использование измерителей деформации (тензометров) дает возможность точно и быстро определять изменения размеров стеблей и плодов. Некоторые исследователи установили корреляцию между давлением выделения живицы и водным стрессом у сосен. По-видимому, давление латекса также может быть показателем водного стресса. Однако ни один из этих методов не дает количественной оценки состояния воды в растениях, что абсолютно необходимо во всех исследованиях, связанных с изучением влияния воды на процессы жизнедеятельности и рост растений.
Следует подчеркнуть, что состояние воды в растении, или водный баланс, регулируется сложными взаимодействиями, в которых участвуют почва, растение и атмосфера. К тому же, поскольку водный стресс существенно влияет на все физиологические процессы, засуха является фактором внешней среды,* который наиболее часто ограничивает рост деревьев. Поэтому следует подчеркнуть особое значение изучения водного режима растений для лучшего понимания того, как водный стресс замедляет рост и каким образом можно свести к минимуму это неблагоприятное действие.
ФИЗИОЛОГИЯ СЕМЯН
Рост древесного растения обычно начинается с прорастания самого важного органа размножения, семени. Существенная часть семени - живой зародыш, поэтому основное внимание при обработке и хранении семян уделяется созданию условий, обеспечивающих жизнеспособность зародыша, хорошую всхожесть семени.
Возобновление роста зародыша и его развитие в новый, независимый сеянец включают наиболее важные процессы, которые являются предметом изучения физиологии растений. К ним относятся: поглощение воды, усвоение питательных веществ, синтез ферментов и гормонов, азотный и фосфорный метаболизм, передвижение веществ и ассимиляция.
СТРОЕНИЕ И СОСТАВ СЕМЯН
Семена древесных растений различаются размерами, формой, цветом и строением. Размеры бывают от едва видимых, например у оксидендрума древовидного и рододендрона, до таких, как у кокоса, вес которых достигает иногда 9 кг. Поверхность оболочки семян также очень разнообразна: она может быть совершенно гладкой и очень грубой, шероховатой. Семена иногда снабжены дополнительными образованиями, такими, как крылатки, присемянники, выступы, шины, । бугорки, волоски. у
Нормальное семя — это оплодотворенная зрелая семяпочка, кото- £ рая содержит зародыш, запасные питательные вещества (лишь очень * редко они отсутствуют) и защитную оболочку или оболочки. Термин семя не всегда ограничивается этим определением. Охотнее его используют в функциональном смысле как единицу рассеивания, распрост-ранения. В этом смысле термин семя применим к сухим, односемян- ? ? ным или редко дву- многосемянным плодам, также и к настоящим се-менам. Например, сухие односемянные плоды (крылатки ильма, оре- шек и плюска бука) обычно, хотя и неточно, относят к семенам. :
Зародыш — это миниатюрное растение, имеющее одну или более семядолей (первых листьев), зародышевые почки, гипокотиль (часть стебля) г и корешки (зачаточные корни) (рис. 14.1). Размер зародыша значительно варьирует у семян разных видов. У некоторых видов — это зачаточная структура, у других он почти полностью занимает семя (рис. 14.2).
Зародыши семян бамбука и пальмы имеют одну семядолю, поэтому классифицируются как однодольные. Зародыши большинства древесных покрытосеменных имеют две семядоли, у голосеменных,4^ зависимости от вида, возможны от 2 до 18 семядолей. '
Питательные вещества в семенах могут быть локализованы в семядолях или в тканях, окружающих зародыш, у покрытосеменных — в эндосперме. Нормальный эндосперм по числу хромосом триплоиден, так как образуется после соединения диплоидного ядра и спермия. Эндосперм — основная запасающая питательные вещества ткань семян многих двудольных видов. У семян некоторых растений, таких, как айлант, некоторые питательные вещества запасаются в эндосперме,
411
Физиология семян
Рис. 14.1. Зрелое семя сосны Ламберта (Anonymous, 1948) :
а—б — внешний вид в двух планах; в — продольный разрез; г — зародыш: 1 — кожура семени; 2 — нуцеллус; 3 — почечка зародыша; 4 — семядоли; 5 — эндосперм; 6 — полость зародыша; 7 — первичный корешок; 8 — подвесок; 9 — микропиле
другие — в семядолях. В семенах голосеменных питательные вещества запасаются прежде всего в мегагаметофите (женском гаметофите), который окружает зародыш. Мегагаметофит по числу хромосом гаплоиден й отличается по происхождению от эндосперма, хотя и выполняет те же функции.
Семенная кожура, защищающая зародыш от высыхания или от поражения вредителями, состоит из наружной твердой оболочки, тесты и тонкой внутренней пленчатой оболочки. Имеются значительные различия в свойствах кожуры семян. Например, у тополя и ивы теста очень мягкая, а у боярышника, падуба и большинства бобовых она очень
Рис. 142. Строение семени (Anonymous, 1948) :
а — аралии, большой эндосперм и маленький зародыш; б — тсуги, большой з'аро-дыш, окруженный эндоспермом; в — ирги канадской, эндосперма нет, зародыш почти целиком заполняет полость семени; 1 — наружная кожура семени; 2 — внутренняя кожура семени; 3 — эндосперм; 4 — семядоля; S — первичный корешок; 6 — микропиле
412
Глава 14
14.1. ОТНОСИТЕЛЬНОЕ СОДЕРЖАНИЕ В СЕМЕНАХ ДЕРЕВЬЕВ УГЛЕВОДОВ, ЖИРОВ И БЕЛКОВ (Mayer, Poljakoff — Mayber, 1953; Anonyrruis, 1948)
Вид Содержание в воздушно-сухих семе-
нах, %
углеводов жиров белков
Клен серебристый 62,0 4,0 27,5
Каштан конский 68,0 5,0 7,0
Каштан 42,0 3,0 4,0
Дуб черешчатый 47,0 3,0 3,0
Дуб белый 58,4 6,8 7,4
Дуб красный 34,5 22,8 —
Тунговое дерево 5,0 21,0 62,0
Сосна веймутова 4,8 35,4 30,2
Сосна австралийская 4,5 31,7 35,2*
твердая. У ильма и внутренняя и наружная оболочки семян пленчатые. Простая кожура семян голосеменных может быть твердой, как у сосны, или мягкой, как у пихты.
Семена содержат разные количества питательных веществ в форме углеводов, жиров и белков. Крахмал в семенах обычно откладывается в виде гранул. Липиды находятся в жировых каплях. Около 80 % белков семян содержится в запасающих органеллах, остальные 20 % распределены в ядрах, митохондриях, пропластидах, микросомах и в цитоплазме. Соотношение углеводов, жиров и белков различно у разных видов, но обычно преобладают углеводы или липиды. Семена белого дуба и клена сахарного заслуживают внимания из-за высокого содержания в них углеводов, в семенах сосен и тунга содержится много жиров (табл. 14.1). Иногда бывают большие различия в составе семян различных видов одного рода. Например, семена клена сахарного богаты углеводами, а семена клена ясенелистного содержат много липидов и белков. Некоторые белки семян, такие, как ферменты, и нуклеопротеиды, обладают метаболической активностью, но большая их часть не активна. Кроме белков, в семенах содержатся и другие азотистые соединения — свободные аминокислоты и амиды. Из амидов чаще всего встречаются глутамин и аспарагин. В семенах содержатся в разных количествах минеральные вещества, фосфорсодержащие соединения (фосфаты, нуклеотиды, фосфолипиды, нуклеопротеины), нуклеиновые кислоты, алкалоиды, органические кислоты, фитостеролы, пигменты, фенольные соединения, витамины и гормональные регуляторы роста.
Химический состав семян генетически детерминирован, но относительное содержание тех или иных веществ заметно изменяется под влиянием окружающих условий и в зависимости от питания родительского дерева. Были обнаружены значительные различия в химическом составе зародышей и мегагаметофитов семян сосны Банкса, собранных из разных географических районов (табл. 14.2). Климат влияет на интенсивность обмена углеродных и азотистых соединений до прорастания и во время начальных его этапов.
Физиология семян
413
14,2. ВАРИАЦИИ В ХИМИЧЕСКОМ СОСТАВЕ МЕГАГАМЕТОФИТА И ЗАРОДЫША СЕМЯН СОСНЫ БАНКСА, СОБРАННЫХ В РАЗЛИЧНЫХ ГЕОГРАФИЧЕСКИХ РАЙОНАХ (Durzan, Chalupa, 1968)
Нейле Харбор (Новая Шотландия) Смоки Хиллс (Онтарио) Висконсин Дейле (Висконсин) Марл Лейк (М читан) Рейндеер и- Лейк (Саскачеван)
Гаметофит Сухой вес, мг/100 гамето- 200,80 152,30 255,40 224,40 178,40
фитов Влажность, % на сырой вес 5,51 6,25 6,41 4,92 5,88
Растворимые сахара, % на 3,36 3,93 '3,21 3,13 4,14
сухой вес: В том числе: стахиоза 1,88 2,08 1,83 1,84. 2,39
раффиноза 0,53 0,96 0,51 0,50 0,90
сахароза 0,95 0,89 0,87 0,79 1,12
Растворимый белок, 17,80 19,60 16,60 17,20 19,20
мкм/кг порошка Зародыш Сухой вес, мг/100 зароды- 33,40 22,30 41,70 39,90 22,60
шей Влажность, % на сырой вес 4,39 4,09 4,37 4,33 4,18
Растворимые сахара, % на 8,95 9,58 7,87 8,86 12,56
сухой вес: В том числе: стахиоза 4,08 4,57 3,75 4,58 5,61
раффиноза 2,13 2,61 1,84 1,88 3,14
сахароза 2,74 2,40 2,28 2,40 3,81
Растворимый белок, 36,00 39,10 35,60 36,60 38,90
мкм/мг порошка Длина зародыша, мм 2,83 2,48 2,98 3,04 2,37
ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ СЕМЯН
Одна из самых характерных особенностей семян — широкая вариабельность продолжительности жизни, которая может быть от нескольких дней до нескольких десятилетий и даже столетий. Известно, например, что семена люпина из арктической тундры, которые были погребены в замороженном иле на протяжении 10 тыс. лет,хорошо проросли в лаборатории. Семена таких древесных пород, как тис, тополь, ильм, ива, дуб, гикори, береза и конский каштан, содержат много воды и недолговечны. Быстро теряют всхожесть семена многих тропических растений. К ним относятся представители таких родов: Theobroma, Coffea, Cinchona, Erythroxylon, Lit chia, Monterzuma, Macadamia, Hevea, Thea и Cocos. Однако подбором подходящих температуры и влажности во время хранения семян можно продлить срок их жизни от нескольких недель или месяцев до года.
Семена постепенно стареют и переходят от одной стадии нарушения жизнедеятельности к другой. Начальные симптомы старения семян
414
Глава 14
заключаются в неспособности прорастать и увеличении чувствительности к микроорганизмам. По мере ухудшения состояния семян с возрастом у них при прорастании появляются короткие корешки, а семядоли не могут пробиться через оболочку семян. В конце концов семена погибают. Скорость старения семян контролируется в основном влажностью среды, от которой зависит содержание воды в семени, и температурой, которая влияет на биохимические процессы. Гарингтон (1972) ссылается на два полезных правила, оба убедительны и применяются независимо друг от друга. Одно из них касается влияния на старение семян влажности в сухих семенах, другое — температуры: 1. При увеличении содержания воды в семенах на 1 % (в диапазоне от 5 до 14 % исходной их влажности) срок жизни семян сокращается в 2 раза. При содержании воды в семенах ниже 5 % старение часто ускоряется из-за самоокисления липидов. При содержании воды в семенах выше 14 % прорастание семян часто снижается из-за поражения их грибами. 2. При увеличении температуры на каждые 5°С срок жизни семян уменьшается в 2 раза. Это правило обычно применимо при повышении температуры от 0 до 50°С.
ТЕСТИРОВАНИЕ СЕМЯН
В связи с продолжительностью жизни семян возникает необходимость проверки их жизнеспособности и качества. Знание процента всхожести семян необходимо не только в исследованиях условий их хранения, но и для того, чтобы правильно определять нормы посева. Результаты, полученные при тестировании семян, зависят от окружающих условий и использованного оборудования, поэтому проверку качества семян необходимо проводить в стандартных условиях разработанными методами. Если работы осуществляются хорошо подготовленными работниками, эти методы обеспечивают высокую степень точности и воспроизводимости, где бы они ни применялись. В настоящее время правилами Американской ассоциации семенно-контрольных станций используются следующие тесты: 1) пробный тест для разделения семян на чистые семена, семена других культур, семена сорняков и инертный материал; 2) проверка на вредность семян сорняков; 3) проверка на всхожесть. Правила Международной ассоциации испытания семян содержат три вышеупомянутых теста и еще несколько тестов на качество семян, включающие такие показатели состояния семян, как качество материнских растений, чистопородность вида и данной культуры, содержание воды, местопроисхождение или место сбора семян, единица веса семян и гомогенность пробы семян.
Прямая проверка всхожести семян часто занимает много времени. Поэтому были сделаны попытки определить жизнеспособность семян методом прижизненного окрашивания индиго-кармином и при помощи солей селена и теллура. Кроме того, пробовали использовать рентгенографию и методы, основанные на большем выделении сахара из мертвых семян по сравнению с живыми. Эти методы не получили широкого распространения, и в настоящее время руководящими лабораториями по контролю семян официально приняты лишь два теста.
Физиология семян
415
ТЕТРАЗОЛИЕВЫЙ ТЕСТ. Наиболее широко применяемый тест был описан Лаконом (1949, 1954). Живые клетки выявляются в результате восстановления бесцветной соли тетразолия. Используются несколько разных солей тетразолия, но наиболее часто 2, 3, 4-трифенил-тетразолий хлористый. В присутствии дегидрогеназ бесцветный тегра-золий восстанавливается в живых клетках до нерастворимого розового или красного формазана.
На протяжении многих лет семеноводы сталкивались с некоторыми сложностями при использовании тетразолиевого теста: 1) трудность окрашивания некоторых семян; 2) необходимость разрезания семян, чтобы наблюдать изменения их окраски; 3) слабая согласованность в некоторых случаях со всхожестью семян, особенно тех, которые обладают низкой способностью прорастать; 4) отсутствие единства в интерпретации окрашивания; 5) трудоемкость, превышающая нормы, которые установлены для стандартных тестов на всхожесть. Некоторые из этих проблем были преодолены. Тем не менее тетразолиевый тест применяется лишь для покоящихся семян, которые нельзя прорастить обычными методами. Американская ассоциация семенно-контрольных станций утвердила этот тест для покоящихся семян Fraxinus sp., Mains sp., Prnnns, Pyrus sp. и Finns cembra. Международная ассоциация испытания семян санкционировала применение тетразолиевого теста для семян кипарисовика туеобразного, кипариса болотного, девяти видов сосны и всех видов 17 других родов.
ТЕСТ ВЫДЕЛЕНИЯ ЗАРОДЫША. Изолированный из семени зародыш помещают на влажную фильтровальную бумагу и проверяют на прорастание при обычной интенсивности света и при комнатной температуре (не выше 24°С). Живые зародыши обычно прорастают в течение 2 недель. Этот тест утвержден Американской ассоциацией для восьми видов сосны и ряда других видов.
ТИПЫ ПРОРАСТАНИЯ СЕМЯН
Прорастание семян можно рассматривать как начало роста зародыша, в результате чего происходят разрыв оболочки семени и появление молодого растения. Рост зародыша требует и деления клеток и растяжения, причем у одних видов сначала происходит деление клеток, у других вначале наступает растяжение клеток. Например, деление клеток предшествует их растяжению во время роста зародыша семян Pinus thunbergii. У лавровишни, однако, в эмбриональных органах деление и растяжение клеток начинаются почти одновременно. Запасные питательные вещества в семени поддерживают растущий зародыш до тех пор, пока развернутся листья — фотосинтезирующая система — и разовьются корни, способные поглощать воду и минеральные вещества. Таким образом, молодое растение становится физиологически самостоятельным.
В процессе прорастания семени корешок удлиняется и проникает в почву. У некоторых древесных растений, таких, как большая часть видов голосеменных, бук, кизил, белая акация, ясень и многие виды клена, семядоли пробиваются наверх при помощи растущего гипокотиля
416
Глава 14
Рис. 14.3. Стадии прорастания семян (переснято из разных источников) :
а — желудя дуба белого, у которого семядоли остаются под землей (подземное) ;
б — клена красного, у которого семядоля пробиваются на поверхность почвы (надземное)
(надземное прорастание). У других видов, таких, как дуб, орех, конский каштан, семядоли остаются под землей, в то время как эпико- ? тиль растет вверх и развивает листья (подземное прорастание) (рис. 14.3 и 14.4). •
В самом начале прорастания делятся все клетки зародыша, когда = сформируется проросток, деление клеток локализуется в кончиках ; побега и корня. После прорастания семени начинают последовательно ! формироваться листья, узлы и междоузлия из апикальных меристем. Побеги могут возникать из апикальных меристем в пазухах листьев, обеспечивая молодое растение системой ветвей. Корневая апикальная меристема формирует стержневой, или первичный, корень. Часто боко- • вые, или вторичные, корни возникают в новой апикальной меристеме в перицикле стержневого корня.
Инс. 14.4. Надземное прорастание Семян сосны (фото t-Regis Paper Со.)
Физиология семян
417
ВЛИЯНИЕ ОКРУЖАЮЩИХ УСЛОВИЙ НА ПРОРАСТАНИЕ СЕМЯН
Быстрое прорастание семян очень желательно, так как при этом уменьшается угроза повреждения семян насекомыми, грибами или неблагоприятными условиями, а также поедания их птицами илитры-зунами. Из окружающих факторов, контролирующих прорастание семян, очень важными являются вода, температура, свет, кислород и различные химические вещества.
ВОДА. Семена, выходя из состояния покоя, должны поглотить некоторое количество воды для того, чтобы у них возобновились физиологические процессы, связанные с прорастанием. Например, дыхание семян значительно возрастает при небольшом возрастании гидратации выше критического уровня. Абсолютное количество воды, необходимое для начала прорастания, относительно невелико, как правило, не превышает 2—3-кратного веса семян. Растущий проросток нуждается в большом количестве воды, и эта потребность возрастает с усилением транспирации. Все жизнеспособные семена, за исключением тех, которые обладают непроницаемой оболочкой, при прорастании могут поглощать воду из почвы при полевой влагоемкости. На подсыхающих почвах скорость прорастания и конечный процент проросших семян снижается. Например, Кауфман (1969) показал,'что почвенный водный дефицит —2,3 и —4,7 бара значительно ингибировал прорастание семян цитрусовых. В отсутствии водного дефицита (0 бар) через 31 день всходило 88 % семян, в то время как при —2,3 бар и —4,7 бар прорастало соответственно лишь 23 и 3 % семян.
Влияние влажности почвы на прорастание семян зависит от их принадлежности к определенным видам. Когда влажность почвы становилась ниже полевой влагоемкости, прорастание семян сосны густоцветной и кипарисовика туполистного снижалось. При этом семена кипари-совика были более чувствительны к подсушиванию. Например, всхожесть семян кипарисовика туполистного понижалась примерно на 6 % на каждый бар снижения водного потенциала почвы. В этих условиях у сосны густоцветной всхожесть семян падала лишь на 2,5 %. Влияние водного стресса на прорастание семян часто зависит от температуры. Например, при температуре около 38°С водный стресс был решающим фактором в самом начале прорастания семян мескитового дерева. Однако при 29°С на рост зародыша сильнее влияла температура, а водный стресс становился лимитирующим фактором только в конце процесса прорастания.
ТЕМПЕРАТУРА. Из состояния покоя семена выводятся низкими температурами, а для индуцирования быстрого прорастания необходимы значительно более высокие температуры. Семена с нарушенным покоем могут расти и при низкой температуре, но для начала ростовых процессов требуется больше времени. Прорастание любой партии семян может происходить в широком температурном диапазоне, но в его пределах имеется оптимум, при котором в кратчайший срок достигается самый высокий процент всхожести. Минимальная, оптимальная и максимальная температуры прорастания семян значительно различаются у, разных видов. У видов умеренной зоны они обычно ниже, чем у тропических. Температуры прорастания также значительно зави
418 Глава 14
сят от места сбора семян. Оптимальные температуры бывают различными и для семян, собранных с разных деревьев одного и того же вида.
Семена многих видов часто прорастают одинаково хорошо в широких пределах температурной шкалы. Семена сосны скрученной широкохвойной прорастали примерно с одинаковой скоростью при 20 и 30°С, у семян сосны Банкса не отмечалось заметных различий в скорости прорастания при 15, 21 и 27©С при непрерывном освещении. Есть данные о том, что массовое появление проростков ели Зигельмана и сосны скрученной широкохвойной было и при 16°, и при 25°С, но меньше всего при 35°С. Однако при 16°С проростки ели появились на 2 дня раньше, чем проростки сосны. У некоторых видов семена прорастают в узком температурном диапазоне, который можно изменить только применением предпосевной обработки семян. Например, семена клена платановидного прорастали лучше всего при 5 и 10°С. Всхожесть семян клена ясенелистного составляла 67 % при температурах от 10 до 25°С, и лишь 12 % при температуре между 20 и 25°С.
Семена многих видов хорошо прорастают при постоянной температуре, но обычно все-таки необходимы суточные температурные колебания. Так, семена ясеня маньчжурского увлажняли и выдерживали при низкой температуре, а затем при 25°С. При этом некоторое количество семян прорастало, а при постоянной температуре 8°С прорастание существенно тормозилось. При содержании семян в течение 20 ч при 8°С и затем при температуре до 25° на протяжении 4 ч прорастание их значительно ускорялось. Прорастание семян сосны густоцветной также ускорялось под влиянием суточных колебаний температур.
РАДИАЦИЯ. Большинство семян не чувствительно к свету и прорастает одинаково хорошо как в темноте, так и на свету. Семена некоторых видов требуют для прорастания свет. Так, для ели необходимо освещение всего в 0,08 лк, для березы 1 лк, для сосны 5 лк. Но есть и такие виды, для прорастания семян которых необходимо освещение до 100 лк. Интенсивность освещения оказывает относительно небольшое влияние на прорастание, но часто длина дня и длина волны усиливают этот эффект. Стимулирующее действие света на прорастание обусловлено увеличением потенциала ростовых процессов зародыша.
Длина дня. Для массового прорастания семян большинства светочувствительных видов древесных растений необходим 8— 12-часовой фотопериод. Прерывание темнового периода короткими световыми вспышками, а также увеличение температуры обычно оказывают то же действие, что и удлинение экспозиции на свету. Можно привести ; несколько примеров различных требований к фотопериоду при про- •* растании семян разных видов. У тсуги восточной 8 или 12-часовой день вызывал максимальное прорастание семян, с увеличением дня до 14 или 20 ч дополнительный эффект не наблюдался. Семена эвкалипта хорошо прорастали при 8-часовом дне, семена березы — при 20-часовом фотопериоде. Семена дугласии, однако, прорастали на длинном, или 16-часовом, дне, но совсем не прорастали при 8-часовом фотопериоде.
Длина волны. Прорастающие семена большого числа видов травянистых и древесных растении чувствительны к свету определенной длины волны. Это характерно для таких древесных покрытосеменных, как Alnus inocuma, Artemisia monosperma, Betula pubescens,
Физиология семян
419
14.3. ВЛИЯНИЕ ЧЕРЕДОВАНИЯ БЛИЖНЕЙ И ДАЛЬНЕЙ КРАСНОЙ РАДИАЦИИ НА ПРОРАСТАНИЕ СЕМЯН СОСНЫ ВИРГИНСКОЙ (Toole etal., 1961)
Характер радиации Прорастание, % Характер радиации Прорастание, % .
Темновой контроль 4 К + ДК + К 94
К 92 К + ДК + К + ДК 3
К + ДК 4 К + ДК + К + ДК + К 93
Примечание. К- ближняя; ДК - дальняя красная радиация.
Fraximus mandshurica и Ulmus americana. Среди голосеменных к свету определенной длины волны при прорастании семян чувствительны: Abies, Picea и Pinus (Р. thunbeigii, Р. strobus, Р. palustris, Р. silvestris, Р. taeda и Р. virginiana).
Реакция на длину волны при прорастании контролируется фитохромной пигментной системой. Ближний красный свет стимулирует прорастание, дальний ингибирует его. Если ближний (650 нм) и дальний красный (730 нм) свет дают последовательно, то способность семян прорастать зависит от последней длины волны. Стимулирующее влияние ближнего красного света на прорастание нивелируется, если после него дают освещение дальним красным светом. Если же после дальнего красного света давали ближний, активность прорастания усиливалась (табл. 14.3). Стимулирующее влияние красного света зависит часто от температуры или длительности снабжения водой. Например, установлено, что прорастание стимулированных красным светом семян сосны виргинской ускорялось больше после 20-дневного намачивания в воде при 25°C, чем после 1-дневного намачивания. Прорастание семян стимулировалось красным светом сильнее, если перед облучением они поглощали воду при 5 °C, а нелтри 25°С.
Радиация
ДК К к+дк к+дк+к к+дк+к+дк +ДК+К+ДК + К
Контроль (теннота)
Дыхание Митозы Прорастание
25 50 2S 50 75 100 25 50 75 100
Qo2 °/о %
Рис. 14.5. Обратимое действие красной К и дальней красной ДК радиации на скорость дыхания, появление митозов в зародышах и на процент прорастания семян сосны обыкновенной (Nyman, 1961)
420
Глава 14
У светочувствительных семян метаболическая активность и митоз в зародышах стимулировались ближним красным светом и ингибировались дальним. Это было продемонстрировано Найманом (1961) при переменном действии ближнего и дальнего красного света на дыхание, митоз и прорастание семян сосны обыкновенной (рис. 14.5). Время ответной реакции каждого из исследованных процессов было разным. Значительное усиление дыхания индуцировалось красным светом после намачивания семян в течение 24 ч, митотическая активность стимулировалась после 36-часового набухания. Первичные корни появлялись только после 48-часового намачивания.
Фитохром — это рецептор света, белково-пигмеитный комплекс, который распределяется в зеленых растениях в низких концентрациях. Он обычно содержится больше всего в меристематических тканях. Свет действует на фитохром Ф, превращает его из неактивной формы с максимумом поглощения в красной части спектра (660 нм) в активную форму с максимумом поглощения в дальней красной части (730 нм). В зрелык семенах часто присутствует Ф660. Семёна, подготовленные к прорастанию обработкой красным светом, могут утратить способность к прорастанию под влиянием обработки дальней красной радиацией, которая вновь приводит к превращению Ф730 в Ф660.
Трансформацию фитохрома можно выразить так;
Разрушение
Темновое превращение
биологический
Фдк о/пвет
ик
Фитохром, кроме участия в прорастании семян, влияет также на цветение, растяжение стебля, фототропизмы, ’’движение сна” листьев (никтинастии) и на ориентацию хлоропластов в клетках.
КИСЛОРОД. Дыхание необходимо на ранней фазе прорастания семян, поэтому снабжение кислородом влияет на прорастание. Семена обычно нуждаются в более высоких концентрациях кислорода для прорастания, чем это требуется для последующего роста проростков. Относительно высокая потребность в кислороде у семян некоторых видов растений объясняется наличием кожуры, являющейся барьером для диффузии кислорода в семена. Удаление кожуры семян сосны смолистой или выдерживание интактных семян при высокой концентрации кислорода значительно увеличивало скорость поглощения кислорода. Выдержи
вание семян, освобожденных от кожуры, при высокой концентрации кислорода еще больше ускоряло дыхание.
Кислород играет роль первичного акцептора электронов в дыхании. У некоторых видов он может участвовать в инактивации одного и более ингибиторов. Прорастание изолированных зародышей березы повислой и бородавчатой тормозилось при обработке семян водными экстрактами, такое ингибирование частично снималось выдерживанием зародышей на свету. Оказалось, что интактная кожура семени тормозила про-
Физиология семян
421
растание в темноте, снижая доступ кислорода ниже критического уровня, однако зародыш без семенной кожуры прорастал при низкой концентрации кислорода. Следовательно, высокая потребность зародыша в кислороде обусловлена семенной кожурой.
Замачивание семян в течение нескольких часов ускоряет прорастание, но продолжительное замачивание индуцирует повреждение и потерю жизнеспособности многих семян, главным образом вследствие сниженной концентрации и доступности растворенного кислорода. Степень влияния замачивания семян на прорастание значительно отличается у разных видов растений. Замачивание семян некоторых видов в течение 3—5 дней не снижало всхожести, но замачивание в течение 10 дней значительно ухудшало прорастание, удлинение срока до 30 дней убивало семена. Семена растений, произрастающих в долинах (например ниссы и болотного кипариса), обладали малой потребностью в кислороде и могли выдерживать продолжительное погружение в воду, не теряя всхожести. Не обнаружено существенного влияния на прорастание у 6 видов растений долин после замачивания семян в течение 32 дней.
СРЕДЫ ДЛЯ ПОСЕВА СЕМЯН. Прорастание семян и укрепление проростков в естественных условиях в значительной мере зависят от их физических свойств, температуры, доступности воды и минеральных веществ. Почва является хорошей средой для посева семян благодаря высокой способности к фильтрации воды, хорошей аэрации и тесному контакту между частицами почвы и семенами. Остатки растений и лесная подстилка менее пригодны для посева, так как они медленно прогреваются, препятствуют проникновению корней и контакту семян с минеральными веществами почвы, быстро высыхают и затеняют маленькие проростки. Сфагнум, обладающий высокой водоудерживающей способностью, часто пригоден в качестве среды для посева, но после их прорастания он может заглушить молодые растения. Разлагающаяся древесина также прекрасный естественный субстрат для прорастания семян лесных деревьев из-за ее способности поглощать и удерживать влагу.
ХИМИЧЕСКИЕ ВЕЩЕСТВА. Некоторые химические вещества (например, инсектициды, фунгициды, гербициды и удобрения) иногда препятствуют восстановлению леса, подавляя прорастание семян, или из-за их токсичности для молодых сеянцев, или вследствие того или другого. Установлено, что такие широко используемые химикаты, как гексахлорциклогексан и фунгицид N-(трихлорометил) тио-4-циклогек-сан-1,2—дикарбоксимид (каптан), повреждают корни и подавляют рост молодых проростков. Триазиновые гербициды, такие, как 2-хлор-4 (этиламино) -6 (изопропиламино) -с-триазин (атразин) и 2-хлор-4,6-бис (этиламин) -с-триазин (симазин), не влияют на прорастание семян, но токсичны для только что появившихся проростков. Другие гербициды в сравниваемых дозах, такие, как N-1-нафтилфталевая кислота (нафталам), 2-хлораллилдиэтил-дитиокарбамат (СДЕС), S-этил дипропил тиокарбамат (ЕРТС), N, М-диаллил-2-хлорацетамид (СДАА) и 2,4-дихлорфеноксиуксусная кислота (2,4-Д), по-разному ингибируют прорастание семян и проростки в ранней фазе роста. Сильно подавляли прорастание семян также 2,4-Д и СДАА.
422
Глава 14
В то время как некоторые гербициды убивают проростки, другие вызывают ненормальное их развитие, например скручивание, сморщивание или срастание семядолей, хлороз, искривление и подавление роста различных листовых придатков. Обнаружено, что у сосны смолистой особенно чувствительны к повреждению семядоли первичная и вторичная хвоя. Токсически действующие гербициды влияют на жизненно важные процессы и непосредственно повреждают клетки и ткани. Действие гербицидов в значительной мере зависит от методов их применения, так как проростки при этом могут их поглощать по-разному. Токсичность гербицидов, остающихся на поверхности почвы, мала, но она увеличивается, если гербицид проникает в почву и концентрация его становится очень высокой, если он в растворе или в виде суспензии контактирует с растительными тканями. Высокая токсичность многих гербицидов при попадании их в почву может резко снижаться вследствие частичного испарения, разрушения их микроорганизмами и лишайниками, необратимого абсорбирования почвой.
Прорастание семян й рост молодых проростков ингибируются не только применяемыми химикатами, но также и многими естественно образующимися в растениях метаболитами, которые выделяются в почву. Такие аллелопатические соединения выделяются корнями и надземными органами. К группе таких естественно образующихся веществ относятся фенолы, кумарины, хиноны, терпены, эфирные масла, алкалоиды и органические цианиды. Аллелопатические соединения имеют важное экологическое значение, так как они влияют на смену сообществ, на доминирование видов и на динамику растительности, на разнообразие видов, структуру растительных сообществ и их продуктивность. (Действие аллелопатических веществ обсуждается в главе 17.)
ФИЗИОЛОГИЯ ПРОРАСТАНИЯ СЕМЯН
Основной момент в прорастающем семени — начало роста зародыша и его превращение в самостоятельное растение. Когда начинается прорастание, включаются многие процессы, при этом происходит: 1) гидратация семени; 2) увеличение скорости дыхания; 3) активация ферментов; 4) увеличение количества аденозинфосфатов; 5) увеличение количества нуклеиновых кислот; 6) усвоение запасных питательных веществ и транспорт растворимых продуктов к зародышу, где синтезируются клеточные компоненты; 7) активация деления клеток и их растяжение; 8) дифференциация клеток в ткани и органы. Точный порядок ранних изменений не ясен, наблюдаются наложения одного этапа на другой, но первым обязательным этапом всегда является поглощение воды. Увеличение гидратации связано с растяжением и делением клеток в точках роста, так же как и с освобождением гормонов, которые стимулируют образование ферментов и их активность. При набухании увеличивается сырой вес семени и одновременно вследствие окисления субстратов и некоторой утечке их из семян снижается сухой вес. Когда появляются корни и начинают поглощать минеральные вещества, а семядоли или листья становятся фотосинтетически
Физиология семян
423
Рис. 14.6. Поглощение воды разными частями семени сосны Ламберта во время стратификации (Stanley, 1958) :
1 — зародыш; 2 — все семя; 3 — гаметофит; 4 — кожура семени (теста и внутренняя мембрана)
активными, сухой вес проростков вновь увеличивается и достигает исходного уровня, а затем даже превосходит первоначальный вес семян.
ГИДРАТАЦИЯ. Вода должна всасываться семенем, чтобы увеличивалась оводненность протоплазмы и приходили в движение звенья метаболических процессов, связанных с прорастанием. При поглощении воды размягчаются твердые покровы семени, а набухание зародыша приводит к разрыву кожуры, что позволяет первичному корешку пробиться наружу. Крахмальные зерна, запасные белки и жиры водонерастворимы, поэтому вода, вероятно, поглощается очень рано начинающими функционировать органеллами и наружными мембранами запасающих органоидов. В конце начального периода набухания абсорбция воды семенами часто прекращается на несколько часов или дней, затем этот процесс возобновляется и во время появления корешков происходит очень активно.
Количество абсорбированной воды различными частями семени во время прорастания значительно варьирует. Зародыш семени сосны Ламберта, который был самым маленьким его компонентом, поглощал воду (в % на сухой вес) больше всего. Мегагаметофит поглощал воды меньше и совсем мало абсорбировала воду семенная кожура (рис. 14.6). На скорость и характер поглощения воды зародышем воздействуют факторы, нарушающие покой семян. Например, поглощение воды зародышами семян сосны Ламберта возрастало при увеличении периода стратификации. Скорость и количество всасываемой воды зависят от природы кожуры семени, размеров и химического состава семян и от температуры всасываемой воды.
ДЫХАНИЕ. Скорость дыхания у сухих семян незначительна, но при намачивании их она усиливается и, как правило, коррелирует со скоростью поглощения воды. В ходе активации дыхания поглощение кислорода семенем в целом, отдельно его зародышем или гаметофитом быстро становится равным и может продержаться на этом уровне в течение нескольких дней. Затем скорость дыхания вновь резко возрастает.
Рис. 14.7. Изменения количества аденозинмонофосфата (АМФ), аденоэиндифос-фата (АДФ), аденозинтрифосфата (АТФ) и общего содержания аденозинфосфа-тов^в процессе стратификации и прорастания сосны желтой (Ching, Ching, а — гаметофит; б — зародыш или проросток; 1 — появление корешка; 2 — появление семядолей; 3 — всего; 4 — АТФ; S — АДФ; 6 — АМФ; 7 — стандартные отклонения
В процессе дыхания происходит распад органических составных веществ семени, прежде всего крахмала, сахаров, жирных кислот и триглицеридов, что обеспечивает большое количество энергии в форме АТФ, необходимой для синтеза ферментов, участвующих в превращениях резервных веществ в запасающих тканях семян, и для формирования клеточных компонентов проростков (см. также главу 6).
В живых клетках семян имеются митохондрии, которые продуцируют АТФ, и ферментные системы, синтезирующие АДФ. В период стратификации при низкой температуре семян сосны желтой общее содержание аденозинфосфатов (АФ) увеличивалось в зародыше в 7, в гаметофите в 6 раз. Во время прорастания общее количество аденозинфосфатов возрастало в 20 раз (рис. 14.7). В семенах клена сахарного уровень содержания АТФ в зародыше, изолированном из сухих семян,
Физиология семян
425
14.4. ДЕЙСТВИЕ СТРАТИФИКАЦИИ ПРИ 5°C НА СОДЕРЖАНИЕ АТФ, АДФ, АМФ И ЭНЕРГЕТИЧЕСКИЙ ЗАРЯД В СЕМЕНАХ КЛЕНА САХАРНОГО (Simmonds, Dumbroff, 1974)
Период стратификации, дней АТФ Аденилаты, нмоли/г сухого веса энергети- ряд
АДФ АМФ
0 46,9 ±12,6 180,2 ±34,6 690,8 ±31,1 0,15
1 137,9± 13,4 163,3 ±24,7 194,9 ±15,0 0,44
5 238,9 ±27,9 119,4 ±14,1 57,3 ±14,1 0,72
10 327,2± 31,7 98,1 ±17,0 60,2 ± 14,0 0,78
20 354,3 + 63,2 99,8±21,0 63,6 ±18,3 0,78
40 416,3 ±48,8 92,1± 11,5 63,2+12,2 0,80
75 456,3 ±70,7 91,4 ±29.3 56,8 ±11,8 0,83
был относительно низким, но он быстро возрастал в течение первых 10 дней стратификации при 5°С, а затем продолжал увеличиваться, но с меньшей скоростью (табл. 14.4).
Высокий энергетический заряд (ЭЗ) очень важен при завершении периода покоя семян вследствие той роли, которую выполняет АТФ в дыхании (см. главу 6). Энергетический заряд — это сумма молей фракции АТФ плюс половина молей фракции АДФ, поделенные на сумму молей фракции АТФ, АДФ и АМФ:
[АТФ]+1/2 [АДФ] энергетический =____________________________
заряд [АТФ] + [АДФ] + [АМФ]
Было показано (см. табл. 14.4), что во время стратификации семян клена сахарного резко возрастал ЭЗ и что даже при высоком уровне АТФ зародышевая ось удлинялась в ответ на действие гиббереллиновой кислоты и кинетина лишь когда энергетический заряд был не ниже 0,8.
ЦИКЛ ФЕРМЕНТОВ. В процессе прорастания семян заметно возрастает синтез и активность ферментов, хотя некоторые из них и сохраняли активность в период покоя. В основном активность ферментов увеличивается в результате их перехода из неактивной формы в активную. При набухании семян многие ферменты синтезируются вновь. Одновременно с этим существующие ферменты разрушаются, поэтому происходит интенсивная смена ферментных систем. Для разных семян характерны определенные ферментные системы, соответствующие составу содержащихся субстратов, но они непременно включают такие ферменты, которые усваивают запасные питательные вещества, и ферменты, участвующие в гидролизе компонентов клеточных стенок.
Как уже отмечалось, важную роль в регулировании синтеза ферментов в семенах выполняют гормоны. Например, гиббереллин стимулировал синтез амилазы, присутствующей в крахмалистых семенах, липазы, содержащейся в семенах хлопка, изоцитратлиазы и РНК-полимера-
426
Глава 14
зы в семенах лещины. Такие наблюдения подчеркивают важность активации гормонов как первого момента после поглощения семенами воды и завершения стратификации.
ФОСФОРНЫЙ МЕТАБОЛИЗМ. К фосфорсодержащим соединениям в составе семян относятся нуклеотиды, нуклеиновые кислоты, фосфолипиды, фосфорные эфиры сахаров и фитин. Значительный интерес к фосфорному метаболизму в прорастающих семенах объясняется связью между фосфором и переносом энергии в ходе метаболизма. Когда зародыши вишни охлаждали для нарушения периода покоя, фосфаты транслоцировались из семядолей и появлялись в зародышевой оси в виде сахарофосфатов, богатых энергией нуклеотидов и нуклеиновых кислот. В клетках неохлажденных зародышей, однако, накапливались неорганические фосфаты. Эти эксперименты показали, что прекращение покоя семян сопровождается фосфатным метаболизмом и увеличением запасов энергии в зародыше (см. главу 6).
НУКЛЕИНОВЫЕ КИСЛОТЫ. Для роста сеянцев необходим синтез белков. Нуклеиновые кислоты играют важную роль в сохранении и считывании генетической информации в клеточных ядрах, так же как и в использовании такой информации в синтезе белка. В общем в процессе прорастания содержание нуклеиновых кислот в запасающих тканях снижается, а в зародыше увеличивается. Характер метаболизма нуклеиновых кислот и синтеза белков несколько различен у разных семян и даже в группе семян одного вида растений.
По некоторым исследованиям, в процессе прорастания семян сосны смолистой количество нуклеиновых кислот резко возрастало в зародыше, а в мегагаметофите не отмечалось почти никаких изменений. Прекращение покоя семян охлаждением или обработкой гиббереллином сопровождалось увеличением синтеза нуклеиновых кислот и ферментных систем. Добавление абсцизовой кислоты, гормона, который индуцирует покой, ингибировало образование нуклеиновых кислот ( Villiers, 1975). В зародышах семян дугласии содержание ДНК несколько увеличивалось в процессе стратификации, но удваивалось в стадии появления первичного корня и возрастало на 450 % к концу прорастания. Синтез РНК в проростках также усиливался, а в мегагаметофите количество РНК снижалось. Увеличение содержания растворимых нуклеотидов сопровождалось возрастанием количества РНК. Изменения в содержании ДНК, РНК и растворимых нуклеотидов указывают на то, что после прекращения покоя происходит синтез нуклеиновых кислот (см. рис. 14.10).
УСВОЕНИЕ, ПЕРЕДВИЖЕНИЕ И ИСПОЛЬЗОВАНИЕ ЗАПАСНЫХ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ. Процесс прорастания семян вызывает значительные изменения в метаболизме запасных веществ в тканях. Клетки, которые исходно синтезировали нерастворимые крахмал, белки и липиды, во время развития семени внезапно приступают к гидролизу этих веществ. Во время формирования семян питательные вещества транспортируются в зпасающие ткани, а при переходе к прорастанию растворимые продукты гидролиза транслоцируются к меристематическим тканям проростка. При этом необходима существенная активация и деактивация ферментов и превращение ’’вклада” в ’’источник”. Это снижает сухой вес запасающих тканей. Например, су-
Физиология семян
427
200
100
О
30
20
10 О
0 2 4 6 8 10 12 0 2 4 6 8 10 12 14 16 18 | Стратификация при 5 °C | Прорастание при 1S-20°C*\
Дни
Рис. 14.8. Изменения сырого и сухого веса зародыша проростков в процессе стратификации и прорастания сосны желтой (Ching, Ching, 1972) :
1 — мегагаметофит; 2 — зародыш проростков; 3 — всего; 4 — появление первичного корня; 5 — появление семядолей; 6 — стандартные отклонения
хой вес мегагаметофита семени сосны желтой резко снижался за счет распада запасных веществ при прорастании (рис. 14.8).
Углеводы. Во время начальных стадий прорастания нерастворимые крахмал и запасные сахара превращаются в растворимые сахара, при
Рис. 14.9. Распределение крахмала в семядолях желудей дуба бархатного (Vozzo, Young, 1975):
а — клетки покоящихся желудей; б — клетки стратифицированных желудей; в — клетки прорастающих желудей
Дни прорастания
Рис. 14.10. Изменение веса и структуры зародыша, мегагаметофита и всего семени дугласии в течение прорастания (ВС - воздушно-сухие семена ; С - стратифицированные семена) (Ching, 1966):
1 — семя; 2 — проросток; 3 — гаметофит; 4 — кожура
этом возрастает активность амилазы и фосфорилазы. Растворимые сахара транспортируются из эндосперма в ткани семядолей и к растущим участкам зародыша. Клетки тканей семядолей покоящихся желудей дуба бархатного содержат много гранул крахмала, в то время как семядоли прорастающих желудей крахмала содержат их мало (рис. 14.9).
Липиды. Запасные липиды в семенах сначала гидролизуются до глицерина и жирных кислот за счет активации липазы. Некоторые из жирных кислот вновь используются в синтезе фосфолипидов и гликоли-
Физиология семян
429
Продолжение рис. 14.10.
пидов, которые необходимы как компоненты органелл и мембран. Однако большинство жирных кислот превращается в ацетил-кофер-мент А, а затем в сахара путем превращений в гликолитическом цикле (см. рис. 6.20). В семенах Simmondsia chinensis воскоподобные вещества играют роль запасных питательных веществ во время прорастания, Структурные изменения в семени дугласии во время прорастания показаны на рис. 14.10. Липиды, основные запасные питательные вещества, первоначально составляли 48 и 55 % на сухой вес гаметофита и зародыша соответственно. В течение ранних стадий прорастания со-
Продолжение рис. 14.10.
держание липидов в гаметофите сильно снижалось и использовалось при развитии зародыша. Сухой вес зародыша увеличивался на 600 %, а сухой вес мегагаметофита падал на 70 %.
Белки. Запасные белки в период прорастания разрушаются протеолитическими ферментами до растворимых азотистых соединений, которые затем используются различными частями проростка. Запасные белки уменьшаются с возрастанием количества аминокислот и амидов, за счет которых происходит синтез новых белков в растущих частях зародыша. Лишь небольшое количество азотистых веществ накапливается в запасающих тканях, так как в процессе быстрого синтеза новых
Физиология семян
431
белков в развивающемся зародыше расходуются имеющиеся азотистые соединения.
Сухие семена содержат лишь некоторые белки и пептидазы, значительная часть их появляется в процессе прорастания. Ферменты с протеолитической активностью, локализованные в семядолях или эндосперме, не отличаются от тех, что обнаруживаются в других растительных тканях.
ПОКОЙ СЕМЯН
Зрелые семена многих древесных растений, попав в соответствующие условия, сразу же прорастают, но семена большинства видов проходят через стадию покоя. Это означает, что они не прорастают сразу после созревания даже если их посеять в самые благоприятные условия. Покой семян может быть для растений благоприятен и неблагоприятен. Для прохождения стадии покоя семенам многих растений умеренной зоны требуется продолжительное охлаждение, поэтому они не прорастают до наступления весны. Это обеспечивает молодым растениям безопасность, так как раннее прорастание привело бы проростки к гибели от морозов. Семена некоторых дикорастущих растений могут оставаться в состоянии покоя в почве на протяжении многих лет, и период прорастания таких семян растягивается на годы. Такой длительный покой способствует закреплению и выживанию видов, даже если случайно очень рано появившиеся некоторые проростки данного года будут убиты засухой или морозом. Другое преимущество покоя семян в жарких и сухих областях заключается в том, что они прорастают в очень короткий влажный период года. У семян некоторых пустынных растений ингибиторы, содержащиеся в оболочке, препятствуют прорастанию. Однако ингибиторы выщелачиваются обильными дождями во влажную почву, вследствие чего семена прорастают лишь при достаточном для укоренения растений содержании воды в почве. С другой стороны, работники питомников рассматривают покой семян как отрицательное явление, так как они стремятся быстро получать семена хорошей всхожести, чтобы иметь большой и однородный выход проростков. Причины покоя семян и методы нарушения его имеют физиологическое и практическое значение.
ПРИЧИНЫ ПОКОЯ. Знание причин покоя семян часто способствует целесообразному применению соответствующих приемов для преодоления состояния покоя отдельных партий семян. Покой семян обусловлен рядом причин: 1) незрелостью зародыша; 2) непроницаемостью оболочек семени; 3) механическим сопротивлением оболочек семени растущему зародышу; 4) блокированием метаболических процессов в зародыше; 5) комбинацией первого и второго условий; 6) вторичным покоем.
Семенная кожура и покой семян. Распространенная причина покоя -непроницаемость оболочек для воды и кислорода. Покой семян, связанный со свойствами оболочек, особенно характерен для семян растений семейства Leguminosae, таких, как белая акация, гледичия и церцис. Проблема оболочек семян также характерна для можжевельника вир
432
Гллвв 14
гинского, липы, сосны веймутовой и яблони. Природа непроницаемости семенных оболочек у разных видов различна. У гледичии и белой акации для всасывания воды и поглощения кислорода семенная кожура является барьером. У сосны веймутовой и ясеня зеленого кожура проницаема для воды, но препятствуют газообмену. Кожура семян яблони задерживает поглощение кислорода, создавая его недостаточность, что препятствует высокой скорости дыхания, необходимой при прорастании зародыша.
Считается, что семенная кожура некоторых видов механически препятствует прорастанию зародыша, и он полностью развивается внутри семени. Однако Виллере (1972) утверждает, что причины этого явления во многих опубликованных случаях иные, возможно, это обусловлено физиологическим покоем зародыша.
Покой зародыша. Иногда зародыш не созревает до конца вегетацион- : ного периода и требует времени для дозревания (хранения при благо- приятных условиях), чтобы достичь определенной стадии развития и ; быть готовым к прорастанию (например, семена калины, падуба и | гинкго). Однако наиболее распространен такой тип покоя семян, при котором морфологически зрелый зародыш не может возобновить рост и прорастать. Этот *ип физиологического покоя зародыша распростра- ; нен среди покрытосеменных у видов яблони, сирени, дуба, каштана, ; кизила, гикори, груши и явора. У голосеменных физиологический • покой зародыша встречается у некоторых видов сосны, кипариса болотного, дугласии, тсуги, можжевельника, лиственницы, ели и пихты.
Состояние физиологического покоя зародыша, так же как и спящих почек, проходит, по-видимому, в две стадии, одна из которых — слабый и обратимый покой — постепенно переходит в глубокий покой, который необратим в тех же условиях, которые его вызвали. Семена неко- . торых видов Fraxinus, нормально переходящие в состояние покоя, начинают прорастать сразу же, если их собрать и посеять до того, как
Сезон • 1 11 Лето । Зима i Лето 1 Зима | Весна L 1 ।
Процесс Одразова- ‘ Рост зародыша, 1 | ние ! изменение -запасных веществ, 1 дение ] 1 ' й 1 семян । разрушение кожуры семени - I эаробыша j । < ! | . |
Тип ПОКОЯ 1 1 Вступление |О покои i 1 1 ! * I । Незрелый зародыш 1 ।
4 1 к i J ! § Ограничение газообмена кожурой семени । । j=
*- 1 1 ,1 { 1 а Необходимость 6 охлаждении зародыша L о i i ! ! | 1 i ! ! * ।
Рис. 14.11. Многообразие механизмов покоя семян ясеня обыкновенного (Villiers, 1975)
433
Физиология семян
они успевают пройти через фазу высушивания. Иногда неспособность семян прорастать обусловлена воздействием нескольких типов покоя. У некоторых видов Rosa прорастанию препятствуют как механическое сопротивление мощного перикарпия увеличивающемуся в размерах зародышу, так и покой семян, который возникает под влиянием ингибиторов роста. Такой ’’удвоенный покой” известен также для семян боярышника, можжевельника виргинского, тиса, липы, дерена, маклюры яблоконосной, скумпии американской, гамамелиса виргинского, сосен Сабина, кедровой европейской и белоствольной. Покой семян падуба обусловлен как незрелостью зародыша, так и твердой кожурой семян. Некоторые представления о сложности механизма покоя семян у определенных видов можно получить из анализа причин покоя семян ясеня обыкновенного - (рис. 14.11). Этот вид образует сухие односемянные, покоящиеся плоды, которые нормально прорастают не раньше как на второй год после опадения плодов. Зародыш в это вемя морфологически полностью сформирован, но для того, чтобы прорасти, он должен увеличиться в. размерах в 2 раза относительно исходной величины за счет насыщения водой. Росту незрелого зародыша препятствуют оболочки плода, которые затрудняют поглощение воды и кислорода. Поэтому наружные слои оболочек должны быть разрушены. Но даже полностью выросший зародыш находится в состоянии покоя и не может прорасти из семени до тех пор, пока оно не подвергнется охлаждению в течение нескольких месяцев. В связи с недостаточным развитием зародыша и ограниченным поступлением кислорода семена не могут прорасти в первую весну. Необходимое воздействие низкими температурами осуществляется лишь в течение второй зимы, и семена прорастают после своего формирования только на вторую весну. Прорастание может быть задержано и на более длительный срок, если охлаждение во время второй зимы будет недостаточным, чтобы началось разрушение оболочек.
Гормоны и покой семян. Весь физиологический покой семян, вероятно, находится под контролем ростовых гормонов. Начало покоя часто связывают с накоплением ингибиторов, а нарушение покоя — со сдвигом в балансе гормонов в пользу стимуляторов роста, которые начинают перекрывать действие ингибиторов. Сходство состояния покоя зародышей семян и покоя почек подчеркивается тем, что физиологические карликовые проростки, образующиеся из неохлажденных зародышей, очень похожи на ветви плодовых деревьев с розеточной формой роста после мягкой зимы. Кроме того, и покой зародышей и покой почек могут быть нарушены охлаждением, экзогенными стимуляторами роста и длинным фотопериодом.
Ингибиторы. Различные ингибиторы прорастания семян у многих видов древесных растений содержатся во всех частях семян и плодов, включая зародыш, нуцеллус, тесту и перикарпий. Абсцизовая кислота (АБК), вероятно, наиболее хорошо известный самый сильный ингибитор в комплексе подобного действия веществ. Она содержится в семенах персиков, ясеня и яблони. У некоторых видов, таких, как ясень, содержание АБК во время охлаждения семян снижается. У других видов АБК не исчезает при охлаждении, но ее ингибирующее действие может быть преодолено такими стимуляторами роста, как гиббереллин
434 Глава 14
или цитокинин. Например, охлаждение, которое ускоряло прорастание ясеня обыкновенного, не снижало активности АБК в зародыше или эндосперме, но оно стимулировало активаторы роста до начала прорастания. К другим естественно образующимся ингибиторам прорастания относятся этилен, аммиак, кумарин и цианистый водород.
Стимуляторы роста. Важность стимуляторов роста в преодолении покоя семян может быть иллюстрирована несколькими примерами. Как уже ранее упоминалось, в процессе нормального дозревания или после искусственного охлаждения некоторых семян уровень стимуляторов роста возрастает, хотя количество ингибиторов не всегда снижается. Выщелачивание семян, которое, как предполагается, стимулирует прорастание вследствие вымывания ингибиторов, часто вызывает формирование карликовых проростков, вероятно, за счет удаления и стимуляторов роста. Например, примененный гиббереллин индуцировал ферментативную активность и рост зародыша у лещины. У клена белого цитокинины, а не гиббереллины, нарушали покой семян, У других видов, таких, как персик и клен сахарный, гиббереллины и цитокинины могли преодолеть покой семян. Эти эксперименты подчеркивают сложность и изменчивость специфического гормонального регулирования покоя зародышей у разных видов. Кан (1975) подчеркивает возможность гормональных взаимоотношений в покоящихся семенах и предлагает приемлемую модель, в которой гиббереллины имеют первостепенное значение в переходе семян к прорастанию, а роль ингибиторов и цитокининов рассматривается как вторичная, так как их действие вполне предотвратимо.
НАРУШЕНИЕ ПОКОЯ. У многих видов покой семян может быть нарушен приемами, направленными на изменение соотношения между ингибиторами и стимуляторами роста и увеличивающими их механическое сопротивление по отношению к растущему зародышу. Эффективность применяемого воздействия зависит от глубины и характера покоя семян. У одних видов покой семян легко нарушить любой обработкой, семена других видов отзываются лишь на какой-нибудь один, специальный прием. Но есть и такие виды, семена которых не удается вывести из состояния покоя ни одним из известных и широко применяемых методов.
Приемы, используемые для нарушения покоя зародыша, обычно также направлены на то, чтобы сдвинуть соотношение между гормонами в пользу стимуляторов роста. Это может сопровождаться либо снижением уровня эндогенных ингибиторов, либо увеличением уровня стимуляторов роста. Так, по исследованиям ряда авторов, охлаждение семян персика и ясеня резко снижало уровень ингибитора (АБК). Однако снйжение уровня ингибиторов не всегда объясняет прекращение покоя семян, которые были обработаны охлаждением. Брадбером (1958) показано, что воздействие низкими температурами нарушает покой семян лещины вследствие синтеза гиббереллина. Гамильтон и Карпентер (1975) обнаружили, что активность ингибитора роста при охлаждении семян Elaeagnus не снижалась, но ростовые стимуляторы, которые появлялись, чтобы нейтрализовать действие ингибитора, синтезировались в зародыше во время дозревания при низкой температуре.
Физиология семян
435
Дозревание. У семян, обладающих лишь средней глубиной покоя зародыша, период дозревания проходит в условиях сухого хранения. Во время дозревания устойчивость семян к окружающим условиям увеличивается. Температурный диапазон для, прорастания становится значительно шире по сравнению с температурами для свежесобранных семян, которые могут прорастать либо при очень низких, либо при очень высоких температурах. Специфические требования к температурному и световому режиму в процессе дозревания постепенно исчезают.
Скорость дозревания зависит от времени года и вида растений.. Даже у генетически однородной популяции семян, которые дозревают в одних и тех же окружающих условиях, отдельные семена бывают разной степени зрелости. У некоторых видов семян при сухом хранении происходит множество различных физических и химических изменений. По крайней мере часть из них является результатом изменений в кожуре семени, которая изменяет свою силу натяжения или увеличивает проницаемость для воды и газов. В других случаях изменения происходят в зародыше или в окружающих тканях. У семян с незрелыми зародышами происходят дальнейшие морфологические преобразования. У некоторых видов ясеня морфологически сформировавшийся зародыш, прежде чем начать прорастать, должен увеличиться в размерах. Биохимические изменения в процессе дозревания включают: уменьшение количества запасных липидов, углеводов и белков; возрастание метаболической активности; изменение гормонального баланса таким образом, чтобы стимуляторы роста преобладали над ингибиторами.
Стратификация. Покой семян обычно нарушается после хранения их в течение 30—120 дней при температурах 1—5°С, при обильной аэрации и влажности. Этот прием воспроизводит до некоторой степени естественные зимние условия. Термин стратификация употребляют в случаях, когда семена помещают между слоями материалов, хорошо удерживающих влагу (сфагнум, опилки, песок). Затем создают низкую температуру. Низкая температура и влажность — необходимые условия для хранения семян слоями. Таким образом, при стратификации семена увлажняют, помещают в пластиковые мешки и хранят при низкой температуре. После такой обработки их можно высевать в землю, но если после этого семена подсушить, то они могут перейти в состояние очень глубокого вторичного покоя.
Химические вещества. Покой зародышей семян часто нарушается такими химическими веществами, как гибберелловая кислота й цитокинины. У видов с относительно слабым покоем зародыша (сосен ладанной, ежовой и густоцветной, лиственниц субальпийской и западной) окислительные вещества, например перекись водорода, стимулировали дыхание и ускоряли прорастание. Однако перекись водорода имеет практические ограничения при стимуляции прорастания некоторых семян с покоящимся зародышем. Например, было показано, что при обработке семян веймутовой сосны концентрированной перекисью водорода у одних партий семян удавалось стимулировать прорастание, а другие семена погибали, вследствие этого конечный процент всхожести этих партий семян снижался.
Скарификация. Покой семян с непроницаемой оболочкой может быть нарушен намачиванием в концентрированной серной кислоте
436
Глава 14
в течение 15—60 мин. Проницаемость кожуры семян можно увеличить прокалыванием или обработкой ее наждаком.
ФИЗИОЛОГИЯ МОЛОДЫХ ПРОРОСТКОВ
Как у голосеменных, так и у покрытосеменных растений семядоли играют первостепенную физиологическую роль в ростовых процессах проростков. Семядоли, которые остаются в земле (у ореха, дуба), служат прежде всего запасающими органами. Наземные семядоли некоторых древесных растений также запасают значительные количества углеводов. У других видов, например у сосны и дерена, в семядолях накапливаются лишь небольшие количества углеводов, но они становятся фотосинтетически активными вскоре после появления на поверхности почвы. У видов, семена которых лишены эндосперма (безбел-ковые семена), семядоли приспособлены и для запасания и для фотосинтеза (например, у березы и белой акации). Если в семенах имеется эндосперм, то семядоли служат не только как запасающие органы, они поглощают запасные питательные вещества из эндосперма и переносят их к растущим осевым органам. Так, эндосперм не играл существенной роли в ограниченном раннем росте проростков ясеня зеленого. Однако у семядолей, которые были в контакте с эндоспермом, оказались большая скорость растяжения и увеличение сухого веса, так же как и более замедленный распад белков и липидов, по сравнению с семядолями, отделенными от эндосперма. Скорость роста интактных проростков была значительно выше, чем у проростков, выросших из изолированных зародышей (табл. 14.5).
В процессе развития наземные семядоли семян некоторых видов последовательно выполняют функции запасания и переноса питательных веществ и фотосинтеза. Это требует радикальных изменений в ферментативной активности, как уже упоминалось ранее. В фазу запасания семядоли наполнены запасными питательными веществами (углеводами, белками, липидами), которые распадаются во время прорастания и используются в течение раннего роста проростков. Типы питательных веществ, запасаемых в семядолях, у разных видов растений различны. Например, семядоли клена перистолистного* содержат больше белков, а в семядолях айланта высочайшего откладываются большие запасы липидов. Зародышевые семядоли клена перистолистного и белой акации содержат значительные количества углеводов, а в семядолях айланта высочайшего и ясеня зеленого углеводы откладываются в небольших количествах (рис. 14.12).
В семядолях откладываются в разных количествах в запас также и минеральные вещества. Зародышевые семядоли безбелковых семян клена красного и белой акации содержат огромные количества питательных элементов, которые перемещаются к быстро развивающимся осевым органам в течение ранних стадий роста проростков. Напротив, в небольших, тонких семядолях ясеня зеленого откладываются в запас незначительные количества питательных веществ.
Переходная стадия развития семядолей характеризуется интенсивным ростом, распадом и утилизацией запасных питательных веществ
Физиология семян
437
14.5. ИЗМЕНЕНИЕ ВЕЛИЧИНЫ 10-ДНЕВНЫХ СЕМЯДОЛЕЙ ЯСЕНЯ ПЕНСИЛЬВАНСКОГО И СОДЕРЖАНИЯ В НИХ СОСТАВНЫХ ВЕЩЕСТВ У ИНТАКТНЫХ ПРОРОСТКОВ И У ПРОРОСТКОВ С УДАЛЕННЫМ ЭНДОСПЕРМОМ ИЗМЕРЕНИЯ ПРОВОДИЛИСЬ ЧЕРЕЗ КАЖДЫЕ 2 ДНЯ (Marshall, Kozlowski, 1976)
I I I I
I I I I I I t I I I I I I I I I I I I I I I I I I I I I i i ( I I I I I I I I I I I i f I I I I I I I I I I I I I I I i !
* t 1 О t О
o, П, м °° еГ <4 U' o'
I i I I
I I
. «lit i/lNnC^Q’tO’tenO 'J 04 to чЧ tn
0'0'000000000
I i t I t
O^Os'f^O’Tat^N’t о" о о о о о о о о' о о
. t 1 |„|
^7^ о © o'
СП ГП 40 СП 00 ГН ОС Tf —< ич
..*11
О О tn, tn tn О in О О О О
SO OS 00 СЧ чч Os 'J- чч izi rJ »П
о’ О О чч* г-Г чч т-J гГ 4 г; -
+1+ I + It I + I + I
I I I i I I
Достоверных различий нет. ** Достоверность Р 0,95. *** Достоверность Р<0,01.
438
Глава 14
Дни дня; 4 - ясень
б
в меристематических тканях, дифференциацией замыкающих клеток устьиц, выходом семядолей из кожуры семени и синтезом хлорофилла. Семядоли безбелковых семян прорываются через оболочки раньше, чем у семян с семядолями, погруженными в эндосперм. Очень важная фотосинтетическая стадия начинается с появления видимого фотосинтеза и заканчивается старением семядолей. Результаты нескольких исследований продемонстрировали определенную связь между фотосинтезом надземных семядолей и ранним развитием проростков как у голосеменных, так и у покрытосеменных. После прорастания семян сосны проростки представляют собой систему конкурирующих потребителей углеводов.
Развивающийся проросток — это также сложная система с участками синтеза углеводов, которые в онтогенезе сеянца перемещаются от семядолей к первичной, а затем к вторичной хвое. Существует тесная зависимость роста позднее развивающихся листьев от способности к синтезу необходимых для роста веществ ранее появившихся листовых зачатков. Отсюда развитие первичной хвои зависит от вклада семядолей, а развитие вторичной хвои, в свою очередь, зависит от вклада, внесенного первичной хвоей. Молодой проросток на стадии семядолей функционирует с максимальной потребностью в питательных веществах для ппстй. поэтому он очень чувствителен к действию факторов среды. В
Физиология семян
в 1 1 1 I 1 1 I 1 f 1 1 1 1 f 1 1 1 s 1 1 1 i 1 1 1 1 1 I I I I 1 1 1 1 I 1 1 i 1 1 1 1
О 1 1 1
ь 1 1 *л 1
х 1 1 N |
R 1 1 1
0 s 1 c > 1 1 I I I
Q 1.1 1
3 i « 1 1 i p* । i
X । ^* । f
U
О 1 © j ।
Q
X о !3i8!
Л 1 1 1
X I I 1
1 I I
0 1 1 1
X 1 1 1
S' 1 1 1
X III
m 1 1*1
c. 1 1 ™ 1
M
R 1 I 1
X 1 1 1
« 1 1 1
Ы III"
t; 1 1 1
о I 1 1
ч 1 1 1
ч 1 1 1
s 1 . I X 1
M 1 1 1
и 1 1 1
H 1 1
>0 0 00
oocotj r—
xjp<7\t-H t—
E x'u-
440
Глава 14
14.7. ВЛИЯНИЕ ИНТЕНСИВНОСТИ СВЕТА НА ЗАЛОЖЕНИЕ И РОСТ ПЕРВИЧНОЙ ХВОИ СОСНЫ СМОЛИСТОЙ (Kozlowski, Borger, 1971)
1 I I I I I I I I I I 1 f I ) I I I I I I I I I I I l i I I 1 J I I I I I I I I I I I I I I
1 I I I I I I I I I I I I I I I i I I ) I ! ) I t I I I I I I I I I I I
<u !
S 1
о 00 X 1
СП СП
м X 1
й я ° 1 1
сП СП 5 !
40
© © о | ?
и 1
о (
X
и S а । 1
С4 g 1 i
СП g. 1 ’i
l| r
-н 00 X '
© О 40 X
и 1
§ 1
ex 1
° 1 i
1 1 -H I ' i
© о 5? Si , 1
S' ' «
ET 1
« t
X i
щ 1
о !
и 1 i 8 । I
о g | ।
ю сч 1 jq V) гч ё s ! : t ’
<ч •* &
'и Q 8
'и <ч § ем д & ГЧ В сх о Среда
tr> ’о d . 1 ' ' '-
г» 1—1 X ci 1
2 X S !. ? '
и о СП 40 -8
X X X меч еле п
5 ! X R О R О со О О о s § | °* S
н ьп S К I 1
Физиология семян
441
Рис. 14.13. Влияние повторного освещения проростков СОСНЫ СМОЛИСТОЙ на удлинение первичной хвои после различных сроков выдержки в темноте (Sasaki, Kozlowski, 1970)
начале онтогенеза окружающие условия сильно влияют на заложение всех органов, кроме ранее сформированных зачатков первичной хвои, и на рост всей первичной хвои, включая и сформированные ранее. От наличия продуктов фотосинтеза в семядолях зависит заложение лишь немногих первичных хвоинок. Например, низкая температура и малая интенсивность света на стадии семядолей препятствовали дополнительному заложению первичной хвои сосны смолистой (табл. 14.6 и 14.7). Когда же после длительной инкубации при низкой температуре или малой интенсивности света проростки помещали в благоприятные условия, начинали быстро формироваться зачатки первичной хвои, которые затем полностью разворачивались (рис. 14.13).
У покрытосеменных рост и развитие проростков на стадии семядолей также необычайно чувствительны к стрессовым условиям, так как они влияют на синтез веществ, необходимых для роста. Большая часть углеводов, используемых в процессе роста, у некоторых древесных покрытосеменных с выходящими на поверхность почвы семядолями поступает именно из семядолей до тех пор, пока не разовьются листья. Маршал и Козловский (1974) показали, что продукты фотосинтеза из семядолей экспортировались к растущим тканям листьев. Лишь полностью развернувшиеся листья лиственных пород приобретали большее значение, чем семядоли в синтезе питательных веществ, кото-
442
Глава 14
14.8. СУХОЙ ВЕС РАЗЛИЧНЫХ ЧАСТЕЙ СЕЯНЦЕВ БЕЛОЙ АКАЦИИ ПОСЛЕ УДАЛЕНИЯ СЕМЯДОЛЕЙ У ПРОРОСТКОВ РАЗНОГО ВОЗРАСТА
(Mar&all, Kozlowski, 1976)
Часть растения Возраст про- Сухой вес, м, проростков, росших без семядолей, дни
КОВ, дней 2 4 6 8 10 контроль
Корни 14 2,1 2,2 3,2 4,4 7,1 22,7
21 5,6 9,8 7,8 8,7 14,2 45,4
28 18,5 19,4 12,1 20,6 27,7 59,5
Гипокотили 14 1,0 1,4 1,7 1,8 1,9 3,0
21 1,8 1,9 2,1 2,1 2,9 4,9
28 2,6 3,2 23 2,8 3,2 7,6
Листва 14 2,6 2,9 4,2 5,0 4,4 11,4
21 8,0 11,0 9,7 8,6 14,0 22,6
28 16,0 17,0 14,6 24,8 23,7 71,2
рые транспортировались из них ко всем растущим органам. Важность здоровых, фотосинтетически активных семядолей для развития проростков доказывается опытами, в которых их рост подавляют затенением, применяют ингибиторы фотосинтеза или их совсем удаляют. В разное время после прорастания семян клена перистолистного, айланта высочайшего, ясеня зеленого, белой акации и ильма американского семядоли удаляли или обрабатывали раствором ДХММ [ 3-(3,4-дихлор-фенил)-1,1-диметилмочевины], который подавляет фотосинтез. Когда удаляли семядоли в течение первых 10 дней после появления зародышевого корешка, у всех видов прекращался рост корней, гипокотилей
Рис. 14.14. Фотосинтез семядолей проростков в разное время после прорастания (Marshall, Kozlowski, 1976а):
1 — айланта; 2 — ясеня зеленого; 3 — белой акации; 4 — ильма американского
443
Физиология семян
и листьев (табл. 14.8). Удаление или притенение семядолей в одинаковой мере ингибировало рост проростков, следовательно, в развитии проростка важную роль играет фотосинтез семядолей.
У разных видов широко варьирует скорость фотосинтеза семядолей и длительность периода, в течение которого они способны к фотосинтезу. Например, уровень фотосинтеза семядолей у ильма значительно ниже, чем у белой акации, айланта высочайшего или ясеня зеленого. У айланта высочайшего и ясеня зеленого фотосинтез поддерживается на высоком уровне дольше, чем' у ильма и белой акации (рис. 14.14). По сообщениям, у дерева какао надземные семядоли играют важную роль в запасании жиров, аминокислот и углеводов. Эти запасные питательные вещества поддерживают растущий проросток до тех пор, пока не развернутся листья и не начнется передвижение углеводов. У дерева какао семядоли представляют собой относительно не очень важный фотосинтезирующий орган.
Начало стадии старения характеризуется прогрессивным пожелтением семядолей, которое продолжается до их опадения. Функциональный жизненный цикл семядолей у разных видов различен. Например, по некоторый данным, надземные семядоли белой акации, шелковицы и черной ивы жили недолго (около 1 месяца). Более долговечными были семядоли тополя, хмелеграба и явора (около 2 месяцев). Еще дольше ’ жили семядоли аралии колючей, каркаса западного и граба каролинского (около 3 месяцев) и необычайно долго жили семядоли ясеня американского и магнолии (более 6 месяцев). Точно установить продолжительность жизненного цикла семядолей у различных видов трудно, так как он в значительной мере зависит от стрессовых условий. Например, засуха может индуцировать быстрое старение и опадение зрелых семядолей.
4»*
РЕГУЛЯТОРЫ РОСТА
Строение деревьев и других древесных растений обсуждалось в главе 2, а их рост — в главах 3 и 4. В этой главе излагаются вопросы, связанные с гормонами -специфическими веществами, регулирующими или контролирующими рост и развитие растений. Рост в упрощенном смысле — это увеличение размеров в результате деления и растяжения клеток, а развитие - это дифференциация клеток на различные ткани и органы, что в конечном итоге создает зрелое растение. Эти процессы относительно легко наблюдать у растущих растений начиная от прорастания семян. Первый шаг роста - это всасывание воды и набухание семян, за которыми следуют активация имеющихся и синтез новых ферментов, гидролиз запасных питательных веществ, их перемещение к точкам роста, синтез новых метаболитов и тканей (см. главу 14). Деление, растяжение и дифференциация клеток на различные структуры проростка создают растение, которое растет достигая зрелости, приобретает характерную для него форму, образует цветки, плоды и семена. Все это легко наблюдать, но трудно понять, каким образом бесчисленные физические и биохимические процессы, включая рост и развитие растений, контролируются и программируются так, что они проходят в определенной последовательности и необходимом количестве, чтобы возник пропорциональный, эффективно функционирующий организм. Происхождение и развитие формы живых организмов входит в понятие морфогенез.
ОСНОВНЫЕ ФАКТОРЫ, РЕГУЛИРУЮЩИЕ РОСТ И РАЗВИТИЕ
КРУГ ПРОБЛЕМ. Ранее упоминалось, что рост и развитие контролируются как внешними, так и внутренними факторами. К внешним относятся температура, свет, вода, кислород, углекислота и минеральное питание. Некоторые из этих факторов рассматривались в главах, посвященных фотосинтезу, минеральному питанию, водному режиму, и более подробно будут обсуждаться в главе 17, Как уже указывалось в главе 1, факторы внешней среды влияют на рост, изменяя физиологические процессы и условия их протекания. Например, ослабление тургора вследствие водного дефицита сокращает растяжение клеток, а отсюда и размеры растений. Но вследствие водного дефицита закрываются устьица и снижается поступление СО2, необходимого для фотосинтеза, что снижает снабжение углеводами и, как следствие этого, подавляет рост и развитие. Интенсивность, качество света и длительность освещения также очень важны для фотосинтеза, но они действуют на рост и цветение опосредованно, путем влияния на фитохромы, гормоны и светозависимые ферменты. Возникает вопрос, каким образом внешние условия, такие, как фотопериод, могут вызывать соответствующие ответные реакции клеток, например побудить меристемы к формированию зачатков цветков. Тот же вопрос возникает относительно действия внутренних регуляторов роста, таких, как ауксин.
В этой главе коротко обсуждаются внутренние факторы, которые контролируют рост и развитие. Они действуют на двух уровнях — на внутриклеточном и межклеточном. Внутриклеточный, контролирующий регулятор - это генетический механизм, или геном, заключенный в значительной степени в ДНК хромосом, контролирующий через матричную РНК типы синтезируемых белков и ферментов, которые, в свою очередь, контролируют структуру и поведение клеток. Пробле-
Регуляторы роста
445
мп контроля роста и развития усложняется тем фактом, что все клетки растения тотипотентные, т. е. каждая клетка содержит всю генетическую информацию, необходимую для создания цело-ю растения. В процессе созревания большинство клеток теряет спо-гобность использовать свои возможности, так как они подвергаются нииянию различного уровня распределения питательных веществ и юрмонов. Однако в культуре тканей клетки часто обретают способность создавать новые растения. Кусочек семядоли дугласии может образовать ткани каллуса, из которых затем формируется новое растение. Для растений, которые развиваются из оплодотворенного яйца или одной клетки в культуре тканей, необходим комплекс регулирующих систем, которые могут включать различные гены в нужное время, чтобы образовалась матричная РНК, необходимая для обеспечения ферментов, требуемых для синтеза различных материалов, используемых в построении разного типа клеток и тканей формирующихся рас-1сний, а затем выключать эти гены, когда отпадает необходимость в их работе. Регулирующее действие генов обычно объясняется соответственно схеме Жакобо и Моно, которая была создана на основании результатов, полученных в работах с микроорганизмами. Внутриклеточная регуляторная система многоклеточных организмов, таких, как деревья, значительно сложнее, чем у одноклеточных организмов, и не так понятна. Внеклеточная контролирующая система координирует активность отдельных клеток, так чтобы развивались различные ткани и органы растения. Она создает внутренние условия для развития клеток и включает снабжение органическими и неорганическими питательными веществами, гормонами, водой в определенном состоянии, в связи с локализацией клеток. Локализация клеток — важный фактор в определении их дальнейшей дифференциации. Например, клетки, отделившиеся от камбия внутрь, дифференцируются в ксилему, а клетки, отделившиеся наружу, дифференцируются во флоэму. Часть клеток листовых зачатков дифференцируется в эпидерму, часть — в мезофилл, другие клетки - в различные ткани проводящих пучков.
ДВА ТИПА КООРДИНИРУЮЩИХ СИСТЕМ. Известны две группы внеклеточных координирующих факторов в растениях - поступление метаболитов и . гормонов в необходимых соотношениях и количествах. К основным метаболитам относятся углеводы и азотистые соединения. Одна из интересных проблем, связанная с продуктивностью, заключается в том, чтобы понять, в какой мере образование углеводов в процессе фотосинтеза контролирует рост и в какой степени использование углеводов в процессе роста влияет на скорость фотосинтеза. Рост часто ограничивается недостатком азота, и до 1920 г. уделялось много внимания углеводно-азотному отношению как фактору, контролирующему развитие растений. Вследствие недостатка азота развиваются низкорослые древесные растения, которые часто образуют мало цветов. Избыток азота вызывает мощный вегетативный рост и у некоторых видов снижает плодоношение. Периодичность плодоношения у плодовых деревьев объясняют тем, что при обильном плодоношении основная часть питательных веществ идет на образование плодов, а в этот период как раз закладываются цветочные почки. Снижение оттока углеводов от верхушек ствола и ветвей путем кольцевания иногда
446
Глава 15
вызывает более раннее формирование цветочных почек и увеличение плодоношения у молодых деревьев. На протяжении многих лет роль питательных веществ затушевывалась усиленным вниманием к гормонам как регуляторам роста и развития. В последнее время вновь возник интерес к роли питательных веществ, но при этом учитывается, что на их распределение и утилизацию могут влиять гормоны. Таким образом, и снабжение питательными веществами и гормоны играют существенную роль в росте и развитии растений.
ПРИРОДНЫЕ ГОРМОНЫ
Гормонам приписывается значительная роль в координации ростовых процессов, поэтому рассмотрим основные группы растительных гормонов и некоторые призеры их деятельности. Термин гормон впервые был использован физиологами животных Бэйлисом и Старлингом (1904). Теперь он обычно применяется к определенным органи-
ческим соединениям, образуемым растениями и животными. В низких концентрациях (10'5 -<• 10*” М и ниже) они влияют на различные физиологические процессы, часто на расстоянии от места их синтеза. В XIX в. Сакс предположил существование веществ, формирующихся в корнях, стеблях и листьях, а Дарвин и Бойсен-Иенсен допускали, что стимул, распространяющийся вниз от верхушки проростка овса, вызывает фототропическое искривление колеоптилей. Вент в 1928 г. показал, что стимул — это специфическое вещество, теперь известное как ауксин, и разработал на этот гормон биопробу. После второй мировой войны исследования гормонов проводились очень интенсивно, что привело со временем к идентификации пяти типов регуляторов роста растений: ауксинов, цитокининов, гиббереллинов, этилена и ингибиторов роста, из которых наиболее широко известна абсцизовая кислота.
В этой главе рассматриваются природа и основные функции растительных гормонов. Их роль в процессах покоя, растяжения побегов, камбиальной активности и внутреннем координировании роста растений подробно обсуждается в главе 16.
АУКСИНЫ. Первыми идентифицированными растительными гормонами были ауксины, и на протяжении многих лет они были единственными, о существовании которых было известно. Открытие ауксина было сделано в ранних исследованиях, касающихся причин фототропи-ческих изгибов стеблей и черешков листьев. Эти работы впервые начал Дарвин (1880). В 1934 г. в результате дальнейших исследований была выделена из грибов индолил-3-уксусная кислота (ИУК). Из цветковых растений ИУК выделили в 1946 г. Ауксины обычно определяют как вещества, которые в низких концентрациях (10'5 М) стимулируют растяжение декапитированных колеоптилей овса или сегментов эпикотиля гороха. Другие природные индольные соединения проявляют небольшую стимулирующую рост активность. ИУК оказалась наиболее активным естественно синтезирующимся ауксином. Его формула выглядит так
Гг зудя торы роста
447
Н с сн2-соон
НС^ С---С
I II II
нс^ ,сн XNZ н н
Считают, что ауксин получается из аминокислоты триптофана, который в результате реакций декарбоксилирования и дезаминирования превращается в индолип-3 ацетальдегид, окисляющийся затем до ИУК. Это происходит главным образом в молодых, растущих тканях верхушек стебля, листьев, цветков, зародышей и кончиков корней. Транспорт ауксина строго полярен; он передвигается вниз от верхушек стебля и от молодых листьев. При этом создается градиент концентраций, снижающихся от верхушек стеблей к корням. Ауксин легко превращается в другие соединения, такие, как индолацетил, аспартат или глюкозидные соединения, быстро окисляется ИУК-оксидазой и пероксидазой. Таким образом, концентрация свободного ауксина в растительных тканях оказывается очень нцзкой. Ауксин также может быть связан с белками в цитоплазме, что затрудняет выявление четкой корреляции между ростом и содержанием ауксина.
Имеется большое количество синтетических соединений, способных оказать действие, подобное естественно синтезируемому ауксину. Среди них: индолилбутиловая кислота (ИБК), а-нафтилуксусная кислота (НУК), 2,4,6-трихлорбензойная кислота, дихлорбензойная кислота, (2,4-Д), и трихлорфеноксиуксусная кислота (2,4,5-Т). Последние два соединения в высоких концентрациях являются сильнодействующими гербицидами. В низких концентрациях их действие сходно с действием индолилуксусной кислоты.
Роль ауксина. Ауксин выполняет в растениях смешанные функции. В концентрации 10-5 М он стимулирует растяжение стебля. В той же самой концентрации он тормозит растяжение корня (рис, 15.1). Он вызывает апикальное доминирование за счет подавления развития пазушных почек стебля, но в относительно высоких концентрациях индуцирует заложение корней на стеблях. Благодаря этому синтетические ауксины ИБК и НУК широко используются в сельском хозяйстве и лесоводстве для стимуляции укоренения черенков. Ауксин участвует в фототропизме, направляя стебли и черешки к свету. На затененной стороне стебля концентрация ИУК становится более высокой, чем на освещенной, что приводит к большему растяжению клеток этой части стебля и изгибу его по направлению к свету. Он также участвует в геотропизме, т. е. росте стеблей вверх, а корней вниз в ответ на действие гравитации* Эта реакция точно так же является, по-видимому, результатом накопления большого количества ауксина на нижней стороне, вследствие чего подавляется растяжение клеток корня и он начинает расти вниз. Несимметричное распределение гиббереллиновой кислоты известно также в корнях, но там возможно взаимодействие между двумя гормонами. Пока не известно, почему существует дифференциальное распределение ауксина или других регуляторов роста в кор-
448
Глава 15
Мо/гярные концентрации андо/тлуксусной кислоты
Рис. 15.1. Реакция на разные концентрации ауксина:
1 - корней; 2 - побегов; эти различия могут объяснить положительный геотропизм стеблей и отрицательный геотропизм корней (Leopold, Thimann, 1949)
нях и стеблях в вертикальном направлении. Статолитная теория, которая приписывает это оседанию крахмальных зерен или других органелл на нижней стороне клеток, еще дискуссионна, но представляется наиболее вероятной.
У некоторых растений ауксины индуцируют цветение. НУК используется в коммерческих целях для стимуляции цветения ананаса. Эффект в этом случае, вероятно, косвенный. В настоящее время считается, что ауксин индуцирует образование этилена, который непосредственно влияет на цветение. У некоторых видов растений ауксин вызывает партенокарпию — образование плодов без семян — если его наносят на рыльце цветков. Он может быть также использован для образования после оплодотворения недоразвитых зародышей, приводящего к опадению цветков. Синтетический ауксин иногда используют для уменьшения количества плодов но есть, однако, примеры естественного опадения плодов, связанного с низким уровнем ауксина, и оно может быть предотвращено опрыскиванием растворами ауксина. Ауксин принимает участие, по-видимому, в отделении листьев и других частей растений, а также в возбуждении камбиальной активности. Так, по сообщениям, у деревьев ауксин, продуцируемый в распускающихся почках, стимулирует деление клеток камбия, но недавно стало известно, что при этом играют роль гиббереллины и некоторые цитокинины. С тех пор как научились идентифицировать гиббереллины, цитокинины и этилен в качестве регуляторов роста, стало ясно, что эффект, приписываемый первоначально исключительно ауксину, получается при взаимодействии нескольких ростовых гормонов. Это будет обсуждаться позднее.
Ауксины в роли гербицидов. Давно установлено, что ауксин, при-' мененный в концентрациях более высоких, чем он нормально обнару*’. живается в растительных тканях, угнетает растяжение, вызывает не-' нормальный рост (например, образование опухолевых тканей, скручи
Регуляторы роста
449
вание черешков, листьев, разрушение листовой пластинки), а еще более высокие концентрации губят растения. Наблюдения за этими явлениями позволили сделать вывод, что ауксины можно использовать в качестве гербицидов и арборицидов. Индолилуксусная кислота для этой цели не очень пригодна, так как она действует слишком быстро, но родственные ей синтетические соединения 2,4-Д и 2,4,5-Т очень эффективны в борьбе с нежелательной растительностью. Некоторые соединения действуют очень избирательно. Например, 2,4-Д в соответствующей концентрации убивает большинство лиственных пород деревьев, но не повреждает травы и зерновые культуры, 2,4,5-Т действует на некоторые сорняки и на большинство древесных растений.
Хвойные обычно более устойчивы к 2,4,5-Т, чем лиственные породы, поэтому его можно использовать для удаления конкурирующих кустарников с участков молодых насаждений хвойных. При этом хвойные повреждаются незначительно. Однако чувствительность к повреждению в значительной степени зависит от стадии роста побегов (рис. 15.2).
ГИББЕРЕЛЛИНЫ. Эти стимуляторы были открыты в Японии перед второй мировой войной, но на Западе о них узнали лишь после окончания войны. Они были обнаружены учеными при исследовании причин заболевания, называемого ’’дурные сеянцы”, у риса, которое характеризуется ненормальным ростом в высоту. Было установлено, что заболевание является результатом поражения грибком Gibberella fujikuroi (известном в несовершенной стадии развития как Fusarium monili-forme) и что экстракт из этого грибка вызывает ненормальное растяжение стебля риса. Активное вещество, выделенное из экстракта, было названо гиббереллином. Исследования, проведенные после второй мировой войны, показали, что гиббереллины широко распространены среди семенных растений, и более 50 из них было идентифицировано. Среди них гиббереллин К3, часто называемый ГК3, т. е. гибберелловая кислота 3, изучен лучше остальных. Его формула представлена ниже:
Н
НС Н хон н о Т I \
Хк Xй с=сн2
HOHC^VJc----с^ нсн
I ноос н сн;
Имеются данные, что гиббереллины значительно различаются по активности. Они представляют собой насыщенные кислородом дитерпены (см. главу 8) и синтезируются почти во всех органах растений, но особенно в листовых зачатках верхушек стеблей и в кончиках корней. Действие гиббереллинов на растения примерно так же многообразно, как и ауксинов. По Существу, если бы гиббереллины были открыты первыми, они заняли бы первое место в литературе, которое ныне занимает ауксин.
450
Гпава 15
Рис. 15.2. Чувствительность к 2, 4, 5, 5-Т сосны желтой и конкурирующих кустарников (земляничного дерева Меициза, додонеи, цеанотуса цельнолистного и ивы). Сосна желтая становится в конце лета значительно устойчивее по сравнению с конкурирующими кустарниками (Gratkowski, 1977):
1 — верхушки кустарников погибают; 2 — гибель кустарников;
3 — повреждение сосны желтой
ФИЗИОЛОГИЧЕСКАЯ РОЛЬ. Гиббереллины обычно индентифици-руют по их способности вызывать растяжение влагалища листа однодольных или междоузлий карликовых двудольных. Самое отличительное свойство гиббереллинов — их способность вызывать растяжение стебля, особенно у карликовых растений, включая и генетически обусловленные карликовые формы, а также у астры и зерновых, пораженных вирусом. Гиббереллины стимулируют как растяжение, так и деление клеток. Хотя ауксин тоже действует на растяжение стебля интактных растений, но гиббереллины в этом отношении значительно более эффективны. Наоборот, ауксин сильнее, чем гиббереллин, воздействует на растяжение отрезанных кусочков стебля, колеоптилей и гипокотилей. Комбинация этих двух стимуляторов роста оказывает синергическое действие на некоторые ткани, вызывая значительно большее растяжение, чем каждый из них отдельно. Ауксин обычно перемещается вниз, по флоэме, а гиббереллины, если они вносятся извне, перемещаются и вверх и вниз. Следовательно, они двигаются и по ксилеме, и по флоэме. Значительное количество ГК обнаружено в экссудате из корней винограда, в соке ксилемы яблони и груши.
Многие длиннодневные травянистые растения могут зацветать при коротком дне под влиянием обработки гиббереллином. Применение гиббереллина способно также вызвать выход в стрелку или преждевременное зацветание двулетних травянистых растений, которые нормально требуют периода низких температур, чтобы приступить к цветению. Использование гиббереллинов для стимуляции цветения хвойных рассматривается в ряде работ Фариса (1976), а также этого же автора с Куо (1977). Гиббереллин Кз часто используется, чтобы вызвать более раннее, обильное и длительное цветение у камелий. Он также увеличивает длину стебля у роз. Однако оказалось, что ГК ингибирует заложение цветков у древесных двудольных. У некоторых растений гиббереллин, так же как и ауксин, вызывает партенокарпию, но он может вызвать также партенокарпию у персиков, абрикосов, некоторых сортов яблонь, где ауксин оказывается не эффективным. По данным Вавера (1972), ГК оказала влияние на виноград сорта Delaware , но была не эффективна по отношению к другим сортам, хотя опять-таки уменьшала у них сморщивание ягод. Гиббереллин может заменить воз
Регуляторы роста 451
действие света, необходимого для нарушения покоя светочувствительных семян и при индуцирований распускания листьев,
Практический интерес представляет возможность использования гиббереллинов для стимулирования роста в высоту сеяйцев в питомниках. Обработка гиббереллином нарушает покой у большого числа нидов древесных растений, которые нуждаются в охлаждении для перехода к фазе цветения. Например, Лавендер с соавторами (1973) в полевых условиях обнаружили, что одновременно с повышением температуры почвы увеличивалась активность почек и содержание ГК н ксилемном соке растений дугласии. Когда ГК применялась к побегам сеянцев дугласии, растущих в холодной почве, они возобновляли рост побегов как только сеянцы пересаживали в теплую почву. Это привело авторов к предположению, что когда корневая система находится в холодной почве, возобновление роста задерживается вследствие замедленного транспорта из корней регуляторов роста, вероятно ГК. С другой стороны, Думброф и Браун (1976) упоминают о том, что в почках клена сахарного перед возобновлением роста не оказалось гиббереллинов, и они предположили, что появление гиббереллинов скорее сопровождает нарушение покоя, чем вызывает его. Необходимы дальнейшие исследования с использованием более совершенных технических средств.
Способы действия. Трудно объяснить разнообразие функций гиббереллинов и ауксинов. Проблема усложняется трудностью решения вопроса, одинаково ли действие ГК, даваемой извне и эндогенной ГК, а также все ли гиббереллины, экстрагируемые из растительных тканей, физиологически активны. У прорастающих семян ячменя и других зерновых культур ГК активирует имеющиеся ферменты, особенно а -амилазу, а также синтез а -амилазы и некоторых других гидролитических ферментов. Активация ферментов, по-видимому, происходит в результате изменения в проницаемости клеточных мембран. Синтез ферментов, вероятно, связан с синтезом матричной РНК. Джонс (1973) приводит результаты интересных исследований, проведенных на семенах лещины, у которых экзогенная ГК нарушала вторичный покой и стимулировала метаболизм ДНК и РНК.
Механизм действия гиббереллинов в целом растении анализировать значительно труднее. Некоторые из воздействий гиббереллинов на ростовые процессы приписываются увеличению количества ауксинов в результате стимуляции гиббереллином синтеза ауксинов или за счет торможения распада ауксинов. Однако это вызывает сомнения. Леопольд и Кри деман (1975) считают, что экспериментальные данные дают основания предположить, что ауксин и гиббереллины оказывают влияние на разные стороны ростовых процессов. Еще не совсем понятно, как влияет гиббереллин на растяжение клеток и каков его вклад в стимуляцию клеточного деления. Совершенно ясно, что необходимо поставить много экспериментов, чтобы до конца понять роль и механизм действия гиббереллинов.
ЦИТОКИНИНЫ. Эти соединения в основном стимулируют деление клеток, но они также участвуют и в процессах растяжения клеток, дифференциации тканей, цветения, развития плода, старения. Мерой активности цитокинина обычно является его влияние на увеличение
452
Глава 15
сырого веса тканей каллуса сои, или на торможение старения, о чем судят по количеству хлорофилла в целых тканях растений, особенно много их содержится в эндосперме семян молочной спелости в кончиках корней и в экссудатах ксилемы большого числа растений.
Химически цитокинины являются производными пурина — аденина. Первый природный цитокинин, который был идентифицирован в растениях, это зеатин, экстрагированный из семян кукурузы. Известно также большое число других соединений с цитокининовой активностью. Ниже даны структурные формулы зеатина и синтетического цитокинина — бензил амино пурина (бензиладенина):
НОНгС /Н нс-он
ос нс Ъ-снг-ын
H3CZ чснг-^н нЪ-сн (I N
Г Y 'он HCV%Z н
hr С Ч I н СИ
HCW
н
В соке ксилемы винограда обнаружено пять различных цитокининов и еще несколько в других древесных растениях. В лабораторных условиях цитокинины широко используются в комбинации с ауксином для стимуляции образования побегов с почками и листьями в культуре растительных тканей (Murashigej 1974). Они также могут прерывать покой светочувствительных семян, преодолевать апикальное доминирование пазушных почек у яблони и абрикоса и стимулировать образование клубней на столонах картофеля. Синтетический цитокинин индуцирует партенокарпию у инжира. Кроме того, применение цитокининов эффективно при стимуляции цветения некоторых растений при неблагоприятных условиях. По данным ряда авторов, значительное усиление активности цитокинина у клена сахарного, которое начиналось в конце февраля в период роста’ корней, не имело прямой связи с нарушением покоя (рис. 15.3). Концентрация цитокинина в ксилемном соке снижалась, когда корни подвергались действию неблагоприятных внешних условий, и это могло существенно влиять на развитие побегов. Быстрое старение листьев при водном стрессе или при затоплении может быть вызвано как торможением передвижения цитокинина из корней, так и прямым действием водного стресса.
Способность применяемых цитокининов задерживать старение листьев и вырезанных из них дисков хорошо известна и дает основание для некоторых выводов. Старение, очевидно, задерживается в результате синтеза гидролитических ферментов и усиления транспорта питательных веществ к местам их активного использования. Возможно, цитокинины играют основную роль в регулировании передвижения веществ. Это согласуется с их влиянием на апикальное доминирование пазушных почек и на ускорение образования клубней. Они, вероятно, играют также роль и в азотном метаболизме, на что указывает их участие в задерживании старения. Пока остается не ясным, насколько это обуслов-
Регуляторы роста
453
Рис. 15.3. Активность цитокинина и ингибитора (АБК) в почках клена сахарного в течение зимы. Содержание АБК в период, когда глубина покоя уменьшалась, значительно не снижалось. Увеличение содержания цитокининов в конце февраля и марта наблюдалось после уменьшения глубины покоя почек и, вероятно, происходило вследствие усиления роста корней, а не активности почек (Taylor, Dumbroff, 1975)
лено торможением гидролиза белков, а насколько усилением их синтеза. Место приложения действия цитокининов не известно. Хотя цитокинин и входит в состав транспортной РНК, все-таки сомнительно, чтобы эта фракция обладала какой-либо гормональной активностью.
ЭТИЛЕН. Несколько десятилетий назад стало известно, что этилен (£^2= СН?) оказывает существенное влияние на физиологические процессы, включая нарушение покоя у картофеля, ускорение созревания плодов, индуцирует эпинастию и опадение листьев. Оказалось, что индуцируемое ауксином цветение ананаса вызывается усилением образования этилена у обработанных ауксином растений. Первоначально этилен привлекал внимание физиологов главным образом в связи с исследованиями процесса созревания и поисками лучших условий хранения плодов. Он продуцируется созревающими плодами и уско
454
Глава 15
ряет дозревание бананов, цитрусовых, хурмы виргинской и некоторых других плодов. Концентрация этилена в хранилище должна поддерживаться на низком уровне, чтобы задержать созревание яблок и дефолиацию саженцев при хранении.
Возможность быстрых и точных измерений концентрации этилена при помощи газовой хроматографии вызвала большой интерес у исследователей. В результате этого прояснилась его роль в растении. Этилен стали рассматривать как унйкальный по действию регулятор роста в форме газа. Несмотря на многочисленные попытки исследовать биосинтез этилена, единого мнения о его происхождении нет. Вероятно, одним из предшественников этилена является метионин, но возможно также, что он получается из жирных кислот. Предполагаются и вне-ферментные пути его образования. Он возникает в различного рода тканях и на его появление влияют такие внешние факторы, как свет, температура, водный стресс, механическое повреждение и поражение грибами. Некоторые грибы образуют такое же большое количество этилена, как и пораженные ими ткани. Побитые или порезанные растительные ткани, ^особенно некоторых плодов, часто характеризуются большим содержанием этилена. Известно, что водный стресс усиливает образование этилена и стимулирует опадение листьев хлопка после возобновления полива. При затоплении почвы наблюдалась эпинастия (искривление) листьев травянистых и древесных растений вследствие образования этилена в корнях. При нормальных условиях этилен выделяется в почву. Однако при избыточном увлажнении почвы концентрация этилена возрастает в корнях вследствие его низкой растворимости в воде и он диффундирует в побеги, скапливаясь там в очень больших количествах. Этилен диффундирует так же легко, как и окись углерода, поэтому он быстро перемещается по межклетникам в растительных тканях. Он растворим в липидах, в связи с этим может концентрироваться в липидной фракции клеточных мембран, влияя на их проницаемость.
Как упоминалось ранее, этилен многофункционален. Он может ингибировать растяжение. Вместе с тем наблюдали усиление растяжения клеток и локального набухания стеблей сеянцев гороха под влиянием этилена. Наряду с этим он ингибировал рост боковых почек у гороха. Эти и другие наблюдения навели на мысль о том, что ингибиторная функция ауксина, по крайней мере частично, усиливается синтезом этилена. Действие ауксина и этилена на корни различно. Этилен нейтрализует нормальный геотропизм стебля некоторых видов растений так, что они занимают горизонтальное положение вместо вертикального. Если использовать этилен в качестве дефолианта для растительных подвоев, то их можно выкапывать раньше срока. Следует при этом учитывать, что вечнозеленые растения могут пострадать. Имеются данные о том, что этилен усиливает передвижение питательных веществ к растущим плодам персиков (Jarie, Chalmers, 1976). Созревание плодов иопадение листьев в основном ускоряются вследствие увеличения активности ферментов, но точный механизм действия этилена неизвестен. Предложены три гипотезы: 1) этилен присоединяется к металлсодержащим белкам, которые выполняют регуляторные функции; 2) он присоединяется к мембранам и изменяет их функционирование; 3) он влияет
Регуляторы роста 455
на РНК и синтез белка, контролируемый РНК. Возможно, что существуют все эти механизмы. Многие действия этилена нейтрализуются углекислотой. Пока в растениях не известна система, которая регулирует концентрацию этилена. Возможно, этилен, как газ, легко диффундирует через межклетники и быстро выделяется наружу, не накапливаясь в тканях до нежелательного уровня.
АБСЦИЗОВАЯ КИСЛОТА. В Англии во время исследований состояния покоя у деревьев было выделено вещество из листьев клена белого, которое вызывало развитие чешуек почек, если его наносили на кончики стебля. Это вещество было названо дормином. В США группа исследователей, изучая процесс опадения листьев, выделила два вещества, которые индуцировали этот процесс, и назвали их абсцизин 1 и абсцизин II. Когда абсцизин II был очищен, то оказалось, что он идентичен дормину и теперь известен как абсцизовая кислота, или АБК. АБК представляет собой сесквитерпен, который, по-видимому, является производным каротиноидов или, что более вероятно, получается из мевалоновой кислоты. Ее формула представлена ниже:
Н3С СН3^
НС
’ ! ОН I
н
сн3
соон
Абсцизовая кислота содержится, возможно, у всех семенных растений и, очевидно, синтезируется во всех частях растения. В противоположность другим гормонам, АБК главным образом ингибирует рост. Она накапливается (по крайней мере, у некоторых видов древесных растений) в условиях короткого фотопериода. Обработка некоторых многолетников растворами АБК индуцировала у них переход в состояние покоя. Однако это не является убедительным доказательством регуляторной роли эндогенной АБК в процессе покоя почек. Тейлор и Думброф (1975) показали, что изменения концентрации АБК в корнях и стеблях проростков клена сахарного не были связаны с нарушением покоя. Хотя сокращение фотопериода в конце лета совпадает с увеличением содержания АБК и с прекращением роста почек, все-таки не доказано, что накопление АБК индуцирует переход в состояние покоя.
Применение АБК может задержать прорастание семян и сохранить покой почек у клубней картофеля. Она обычно тормозит растяжение стебля и рост других органов, но в некоторых случаях АБК стимулирует образование корней. Экзогенная АБК также ускоряет старение отрезанных листьев и распад хлорофилла, несмотря на добавление цитокинина. Возможно, что некоторые признаки вырождения, которые возникают при старении, вызываются действием АБК. Однако Мильборо (1974) это действие АБК считает косвенным. Абсцизовая кислота ускоряет опадение листьев у некоторых травянистых растений. Это действие также до некоторой степени противоположно влиянию ГК и цитокинина на многие процессы, в том числе на опосредован
456
Глава 15
ный ГК синтез а-амилазы в семенах зерновых культур. Этот же исследователь в своем обстоятельном обзоре работ, касающихся вопросов химии АБК и влияния ее на физиологические процессы, не смог установить место приложения действий этого вещества.
Другое интересное наблюдение заключается в том, что АБК может влиять на движение устьиц. Обнаружено заметное увеличение АБК в листьях растений, страдающих от водного дефицита, вызванного почвенной засухой, затоплением и избытком солей, поэтому предполагается, что она стимулирует закрывание устьиц, которое наблюдается в этих условиях. Концентрация АБК обычно возвращается к нормальному уровню в течение 1-2 дней после устранения водного стресса. По данным Ловейса (1977), АБК синтезируется в хлоропластах (этот вопрос более подробно обсуждался в главе 13). Было высказано предположение, что снижение содержания цитокинина и, вероятно, ГК взаимодействует с увеличением АБК при регулировании устьичного движения у растений в условиях водного стресса. Однако это предположение подвергалось сомнению. Суждение о том, что накопление АБК во время водного стресса влияет на сбрасывание листьев у клена, также подвергается сомнению.
ДРУГИЕ ИНГИБИТОРЫ РОСТА. Наряду с абсцизовой кислотой в растениях обнаружены другие вещества, которые функционируют как ингибиторы. Наиболее важными из них являются фенольные соединения, такие, как коричная кислота, которая ингибирует активность ауксина. Монофенолы, применяемые в опытах, увеличивают активность ИУК-оксидазы и деструкцию ауксина, но ди- и полифенолы ингибируют деструкцию индолилуксусной кислоты. Имеются кроме того, некоторые довольно сбивчивые данные о влиянии фенолов на реакции проростков на свет. Флавоноиды также могут играть какую-то роль в реакциях листьев на свет. Некоторые лактоны, например кумарин, также считаются ингибиторами роста. Наиболее хорошо известны кумарин и прото-анемонин. Кумарин встречается в травах, обусловливая характерный запах свежескошенных трав. Он подавлял прорастание семян салата и растяжение корней. Предполагается, что танины являются основными антагонистами и регуляторами работы гиббереллинов. Возможно, что перечисленными соединениями список гормонов не исчерпывается. При дальнейших исследованиях могут быть обнаружены новые вещества.
ВЗАИМОДЕЙСТВИЕ ГОРМОНОВ
Когда-то предполагалось, что каждая группа гормонов выполняет одну основную функцию, но читатель знает, что один и тот же процесс может регулироваться более чем одним гормоном и что существуют важные взаимодействия между различными гормонами. Так, первоначально считалось, что ауксин вызывает цветение ананаса, однако оказалось, Что на цветение влияет этилен, образование которого стимулируется ауксином. Очень важное синергичное действие ауксина и цитокинина регулирует рост и дифференциацию клеток в культуре тканей и, возможно, при развитии почек. Сходное взаимодействие может возникать между ауксином и гиббереллином при растяжении стебля. Бывает также
457
регуляторы роста
и антагонистическое действие: например, влияние экзогенной АБК нейтрализует влияние экзогенной ИУК на отделение листьев и плодов.
Таким образом, процессы роста и развития часто, по-видимому, контролируются разными гормонами в соответствующих концентрациях, хотя эти гормоны действуют различными путями. В главе 16 обсуждается некоторая специфическая роль регуляторов роста в таких процессах, как покой, активность камбия, заложение цветков, а также влияние гормонов на апикальное доминирование и тем самым на отношение корней к побегам. Предпринималось много попыток сопоставить уровень содержащихся гормонов с этими явлениями. В результате была установлена слабая связь, а иногда она и вовсе не улавливалась. Эти выводы сделаны на основании сравнительных наблюдений за растениями, к которым применяли гормоны и которые не обрабатывались гормонами. При этом делалось сомнительное допущение, что экзогенно применяемый гормон оказывает такое же действие, как и эндогенный. Слишком мало информации о действии гормонов на молекулярном уровне. Предполагается, что они оказывают влияние на гены, которые регулируют синтез РНК, или действуют на один из предшественников РНК и таким образом влияют на уровень содержания некоторых ферментов, участвующих в различных процессах обмена веществ. Имеются некоторые примеры их непосредственного влияния на проницаемость мембраны. Необходимы дальнейшие биохимические исследования механизмов действия гормонов, но, по-видимому, трудности работы с древесными растениями расхолаживают большинство исследователей.
РЕГУЛЯТОРЫ РОСТА И БОЛЕЗНИ. Нередко допускается, что некоторые симптомы, которые появляются у растений, подвергаемых атаке патогенных организмов, могут быть вызваны чрезмерной гормональной активностью. Деформация листьев персиков при поражении Taphrina deformans, гипертрофия, вызванная корончатым галлом, корневая деформация у крестоцветных, называемая килой, и галлы, образуемые на деревьях ржавчиной Cronarium, часто приписываются действию высоких концентраций ауксина и, возможно, других регуляторов роста. Часто приводится пример ненормального роста риса, инфицированного Gibberella fujikuroi, обусловленного избытком гиббереллина. С другой стороны, допускается, что некоторые патогенные организмы редуцируют рост за счет снижения концентрации ауксина или гиббереллина. Другой симптом, известный как образование зеленых островков, и обусловленный сохранением хлорофилла в пораженных частях листа, напоминает зеленые участки у стареющих листьев, на которые попали капли цитокинина.
Многие виды* Патогенных и непатогенных бактерий и грибов вырабатывают ауксин и другие ростовые регуляторы. Концентрация этих веществ в больных тканях часто бывает выше, чем в здоровых. Поэтому невольно возникает искушение объяснить ненормальный рост избытком регуляторов роста, в том числе этилена, продуцируемых внедрившимся организмом. Однако не ясно, какая доля этого возросшего количества гормонов синтезируется патогенами, а какая часть продуцируется самой клеткой хозяина. Есть мнение, что избыток регуляторов роста, возникает скорее за счет изменений метаболизма клеток
458 , Глава /5
хозяина, чем в результате синтеза патогенами. Разнообразие признаков, связанных с поражением растений насекомыми, также допускает гормональную стимуляцию. Примером могут служить корневые галлы, образующиеся при внедрении нематод, и галлы насекомых, формирующиеся на листьях. Все они появляются в результате ненормального клеточного деления. Зеленые островки иногда формируются на листьях березы, пораженных лиственным пилильщиком. Содержание в них цитокинина, как правило, высокое, но происхождение большой концентрации цитокинина не ясно. Необходимы дальнейшие исследования, чтобы установить причинно-следственные отношения между концентрацией регуляторов роста и заболеваниями растений.
СОДЕРЖАНИЕ
От издательства................................................. 3
Предисловие..................................................... 4
Глава 1, Значение физиологии растений.......................... 6
Роль физиологии растений..................................... 7
Круг вопросов физиологии дерева.......................... . 9
Проблемы и перспективы...................................... 13
Глава 2. Строение.............................................. 17
Лист........................................................ 17
Ствол..................................................... 22
Строение древесины голосеменных............................. 29
Строение древесины покрытосеменных.......................... 33
Кора........................................................ 35
Корни....................................................... 41
Глава 3. Вегетативный рост................................... 54
Рост побегов............................................... 54
Типы побегов и способы их роста............................. 60
Рост побегов в тропиках.................................... 70
Камбиалытый рост............................................ 73
Рост корня.................................................. 87
Измерение и анализ роста.................................... 94
Глава 4. Репродуктивный рост................................... 99
Периодичность роста репродуктивных органов.................. 99
Половое размножение у покрытосеменных...................... 102
Половое размножение у голосеменных..........................115
Бесполое размножение........................................128
460
Содержание
Глава 5. Фотосинтез...............................................136
Пигменты хлоропластов......................................... 136
Факторы, влияющие на образование хлорофилла................... 141
Механизм фотосинтеза.......................................... 142
Изменения интенсивности фотосинтеза........................... 149
Факторы, влияющие на фотосинтез............................... 157
Глава 6. Ферменты, энергетика и дыхание................,..........181
Ферменты и энергетика......................................... 181
Дыхание....................................................... 186
Дыхание растений и их частей................................. 193
Факторы, влияющие на дыхание...................................203
Ассимиляция............................*.......................208
Глава 7. Углеводы................................. -............. 211
Классы углеводов....................................;......... 211
Превращения углеводов...................................... 215
Использование углеводов....................................... 218
Накопление углеводов.......................................... 220
Сезонная цикличность содержания углеводов. ................... 226
Осенняя окраска............................................... 227
Глава 8, Липиды, терпены и родственные им вещества............... 230
Липиды........................................................ 231
Воски, кутин и суберин....................:................... 233
Внутренние липидные слои...................................... 235
' Состав липидов.............................................. 235
Распространение липидов в древесных растениях................. 236
Изопреноиды, или терпеноиды................................... 237
Функции веществ вторичного происхождения в растениях.......... 243
Глава 9. Метаболизм азота и азотное питание...................... 245
Распределение и сезонные колебания содержания азота........... 245
Содержание
461
Основные соединения азота................................... 253
Потребность в азоте......................................... 259
Источники азота.......................... .................. 261
Круговорот азота............................................ 265
Глава 10. Минеральное питание и поглощение солей................. 269
Минеральное питание........................................... 269
Поглощение солей......................................'....... 291
Глава 11. Передвижение веществ................................... 301
Транспорт воды............................................... 301
Транспорт минеральных веществ.................................. 303
Передвижение органических веществ............................ 306
Передвижение веществ от растения к растению.................... 317
Механизм флоэмного транспорта.................................. 320
Регуляция флоэмного транспорта................................. 324
Глава 12. Значение воды и процесс транспирации................... 326
Транспирация................................................... 330
Внутренние факторы, влияющие на транспирацию.................. 339
Устьица........................................................ 346
Взаимодействие факторов, влияющих на транспирацию.............. 349
Измерение транспирации. t..................................... 352
Интенсивность транспирации.................................... 356
Потеря воды фитоценоэами....................................... 358
Глава 13. Поглощение воды, подъем ксилемного сока и водаый баланс . . 365
Поглощение воды................................................ 365
Механизм поглощения воды....................................... 369
Корневое и стеблевое давление.................................. 371
Факторы, влияющие на поглощение воды........................... 376
Подъем ксилемного сока......................................... 381
Водопроводящая система....................................... 384
452 Содержание
Водный баланс................................................ 391
Засуховыноспивость................................................ 401
Измерение водного стресса......................................... 406
Глава 14. Физиология семян........................................ 410
Строение и состав семян....................................... 410
Продолжительность жизни семян.................................. 413
Тестирование семян............................................. 414
Типы прорастания семян......................................... 415
Влияние окружающих условий на прорастание семян................ 417
Физиология прорастания семян............................... 422
Покой семян.................................................... 431
Физиология молодых проростков................................ 436
Глава 15. Регуляторы роста...................................... 444
Основные факторы, регулирующие рост и развитие................ 444
Природные гормоны.............................................. 446
Взаимодействие гормонов........................................ 456
Паул Крамер, Теодор Козловский
ФИЗИОЛОГИЯ ДРЕВЕСНЫХ ПОРОД
Редактор издательства З.С. Чугунова
Оформление художника В.Н. Тикунова
Художественный редактор К.П. Остроухое
Технический редактор Т.И. Ю р о в а
Корректоры Ж-А. Лобанова, И, Б- Шеманская
Вычитка Е.Н. Соколовой
ИБ № 1699