/














































































































































Текст
Series. С.
Vol. IX. Fascicle 3.
PAL/EONTOLOGIA SINICA
Board of Editors
V. K, Ting (Chairman), T. C. Chow (Secretary), A. W. Grabau, J. S. Lee, Y. C. Sun, С. C. Young, T. H. Yin,
Fossil Proboscidea from China
BY
A. TINDELL HOPWOOD.
Department of Geology, British Museum (Natural History), London.
With Plates I—VIII.
Published by the Geological Survey of China
Peiping (Peking) Feb. 1935.
PUBLISHED FEBRUARY 1934.
A.-B. Hasse W. Tullbergs boktryckeri
Esscke ab. Stockholm 1935
429921
CONTENTS.
Page.
Preface......................................................................... 5
Introduction.................................................................... 9
Description of species....................................................... 13
Trilophodon connexus.................................................... 14
Trilophodon wimani....................................................... 19
Trilophodon spectabilis.................................................. 30
Serridentinus mongoliensis .............................................. 31
Serridentinus gobiensis.................................................. 32
Serridentinus florescens................................................. 32
Platybelodon granger!.................................................... 33
Tetralophodon exoletus................................................... 35
Tetralophodon (?) sinensis............................................... 40
Mastodon americanus ..................................................... 43
Mastodon borsoni......................................................... 46
Mastodon sp. indet....................................................... 48
Pentalophodon sinensis .................................................. 57
? Pentalophodon sinensis................................................ 61
Mastodontoidea incertae sedis............................................ 64
Stegodon officinalis .................................................... 73
Stegodon zdanskyi ....................................................... 75
Stegodon orientalis ..................................................... 77
Stegodon orientalis grangeri............................................. 82
Stegodon sinensis....................................................... 85
Stegodon aff, bombifrons................................................ 86
Archidiskodon cf planifrons.............................................. 88
Palaeoloxodon tokunagai ................................................. 92
Palaeoloxodon cf namadicus............................................... 95
Mammuthus primigenius.................................................... 98
The accompanying Fauna and the Age of the Deposits ........................... 101
List of works consulted...................................................... 105
Explanation of Plates......................................................... 109
PREFACE.
THE PALAEONTOLOGY OF CHINA WAS UNKNOWN UNTIL 1853 WHEN
Davidson wrote a short note on some fossils obtained in Shanghai by W.
Lockhart. The specimens were sent to Daniel Hanbury who gave them to the
British Museum. They were mostly invertebrate but among them were a few teeth.
One of these teeth (regd. 29007) was in later years described by Busk as a tooth of
E. armeniacus Falc. (Busk, 1868). Thus from the very first, fossil Proboscidea have
been known to occur in China.
Sir Richard Owen, who had referred to Lockhart's specimen in his address to the
British Association in 1858, was able in 1870 to describe a collection of fossil mam-
mals received from Mr. R. Swinhoe, at that time British Consul in Formosa. He described
two new species of Stegodon, S. orientalis and 5. sinensis, which later authors have
referred to the Indian species S. insignis and S. cliftii respectively.
In the following year, Gaudry, when dealing with the collection made by the
famous French Missionary Pere David, recorded a toothless jaw of an undetermined
species of elephant. Schlosser (1903, p. 42) showed that the specimen was probably
of the Mammoth, and remarked that although remains of Mammoth had often been
said to occur in China, no scientific study had yet been published. Even today, thirty
years later, this still seems to be the case, and the broken tooth described below is,
so far as I know, the first Mammoth molar to be recorded from China proper.
The first collection of any size was that taken back to Germany by Ferdinand,
Freiherr von Richthofen, and described by Professor Ernst Koken in 1885. This col-
lection contained specimens from several horizons, but they were all sufficiently close
together broadly to justify Koken's description of the whole as Upper Tertiary. The
Proboscidea were described under the names Mastodon perimensis var. sinensis nov.,
M. aff. pandionis, Stegodon cliftii, S. insignis, and S. aff. bombifrons (Koken, 1885,
pp. 6—16).
The next collection was that made by Dr. K. A. Haberer, who, during his journey
through China in 1899—1901, purchased great quantities of "dragons' bones and teeth”
from the native druggists. Professor Max Schlosser described the specimens in 1903.
255918
(Ш) 6 Palaontologia Sinica Ser. C.
There were very few remains of Proboscidea among the mammalian teeth, but, in so
far as they were represented, Professor Schlosser was able to identify Stegodon insig-
nis, Mastdon aff. latidens, 714. lydekkeri nov., and Mastodon sp. ex aff. pandionis
(Schlosser, 1903, pp. 42—49).
Twenty years after Haberer, Dr. J. G. Andersson in conjunction with the Geo-
logical Survey of China, and assisted by Dr. Otto Zdansky, made extensive collections
in various provinces of China. These collections, which are now preserved in the
Palaeontological Institute of Upsala University, were the first to be made under modern
conditions by which the localities were noted with scrupulous accuracy. They contain
a varied assortment of bones and teeth of Proboscidea which represent a dozen or
so species, most of them new.
Later collections were made by the Central Asia Expeditions of the American
Museum of Natural History. From this material five new forms have been described,
namely, Serridentinus gobiensis, S. mongoliensis, S. florescens, Platybelodon gran-
ger, and Stegodon orientalis grangeri (Osborn, 1924, 1929; Osborn & Granger
1931, 1932).
Additional collections have been made by the Geological Survey of China, as well
as by semi-private expeditions working in conjunction with the Survey, but the Pro-
boscidean remains have not been described in full, though certain authors (e. g. Boule
& Teilhard de Chardin, 1928) have referred to them incidentally when dealing with
a fauna.
In the present work species described by previous authors, and not represented
in the collections made by Dr. Andersson, are inserted. As a rule the original de-
scriptions are quoted and, where necessary, short comments added. Owing to the
uncertainty which attends efforts to deal with descriptions unaccompanied by speci-
mens, sections have been added on such previous records as "Stegodon insignis".
This has been done in preference to making definite redeterminations based on in-
sufficient knowledge of the specimens concerned. The object in making these addi-
tions has been to gather all the available information into one place. Advantage has
also been taken of the presence of a few specimens in the collections of the Geo-
logical Department of the British Museum (Natural History) to supplement the material
available for study. In this manner it has proved possible to add sections on Stegodon
orientalis, and on the Mammoth. Any specimen of which the registered number is
quoted is in the British Museum, all the others are in the Palaeontological Institute of
Upsala University.
In conclusion I wish to express my thanks to Professor Carl Wiman for his never-
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 7
failing courtesy and patience in face of the many delays to which this paper has been
Subjected, as well as for his kindness in entrusting me with the material for descrip-
tion; to Dr. Birger Bohlin, through whose mediation the specimens were sent tome;
and to Dr. Otto Zdansky for many publications on the Tertiary and Quaternary mam-
mals of China. To all three I am also indebted for their hospitable welcome to Up-
sala some years ago. Acknowledgements and thanks are also due to Dr. W. D. Lang,
F. R. S. Keeper of Geology in the British Museum (Natural History), for permission
to undertake this work, as well as to Dr. Guy E. Pilgrim, formerly of the Geological
Survey of India, for the opportunity to study the Indian species referred to in the
following pages. Finally I have to thank Father Teilhard de Chardin for discussing the
age of the various deposits with me when he was in London in 1932.
A. Tindell Hopwood.
Department of Geology, British Museum (Natural History), London.
INTRODUCTION.
I. The Morphology of the Mastodon Tooth.
Any tooth referable to the old collective genus Mastodon has certain characters
which are common to all other teeth classified under that head. Each tooth belonging
to the "intermediate molar” series consists of three or more transverse ridges composed
of two cusps apiece, an anterior cingulum, and a heel. It has long been the custom
to number the ridges from before backwards, and so we find references to the first,
second, or third ridge as the case may be. Another character is that the inner cusps
of the upper teeth, and the outer cusps of the lower, receive more wear than their
fellows. For this reason they are more strongly built, and in the course of evolution
are the first to be provided with additional strengthening structures. These stronger
cusps are known as the pretrite cusps, whereas the weaker ones are known as the
post-trite cusps.
A system of notation for the different cusps was arrived at by combining the idea
of strong and weak cusps, with that of strong and weak numbers. In this system the
strong, pretrite, cusps are distinguished by odd numbers, and the weaker, post-trite,
cusps by even numbers. The first cusp is the pretrite cusp of the first ridge, and
the second is the post-trite cusp of the same ridge; similarly the third and fourth cusps
are the pretrite and post-trite cusps of the second ridge, and so on in sequence.
This system has the great advantage of being entirely free from theory. It is
purely empirical, and capable of indefinite extension. For example, if a species were
to be discovered with twenty cross-ridges, the cusps of the seventeenth ridge would
be indicated by the numbers 33 and 34. Moreover, given the number (N) of the
cusp the ridge may be determined by employing the formula -y- for petrite cusps,
N
and 2 for post-trite cusps. Thus in the imaginary species just mentioned, cusp 39
39+1
belongs to the twentieth ridge since - 20 and cusp 14 belongs to the seventh
ridge since у = 7.
2—42992 A. Tindell Hop wood.
(Ill) 10 Palaontologia Sinlca Ser. C.
Anorner conveniioti wriicii is not without its uses is this. Ordinarily a simple
cusp consists of two cones, and is separated from its fellow by a fairly deep cleft.
This means that the ridge consists of four cones arranged in two pairs, thus, CC:CC.
In this formula 'C' stands for one of the ’’primary cones” and the colon for the cleft.
Later species have the cones variously divided. This, too, can be represented by means
of a formula in which 'c' stands for one secondary cone, and the representatives of
each primary cone are enclosed in brackets. For example, a tooth of the Indian species
Synconolophus corrugates (Pilgrim) has the formula
CC: (cc) (cc)
C (cc): C (cc)
(cc) С: C (cc)
which indicates that the first cusp has the two primary cones, the second has both
cones divided; the third cusp has the inner cone divided, whereas in the fourth cusp
the outer cone is divided and in the fifth and sixth cusps the outer cones alone have
undergone division.
An extension of this method is used when dealing with the teeth of Stegodon.
In these animals the ridges are made up of a succession of elements, each of which
consists of a varying number of large and small mammillae; the latter are represented
by M and m respectively, and suitable combinations placed within brackets indicate
the composition of each element. Examples of formulae of this type are given in the
account of Stegodon orientalis.
II. Systematic Arrangement.
The modern systematic treatment of the Proboscidea is due very largely to Pro-
fessor Henry Fairfield Osborn aided by a small band of assistants. Up to the present,
the results of their labour have been published only in part, mostly in the form of
summaries with very little evidence adduced in support. So far as possible, I have
made full use of the information already published, and also of much that is not yet
generally available. Professor Osborn has most generously kept me fully informed of
the progress of his studies, and has also given me the further advantage of being able
to read advance copies of his chapters as soon as page proofs were available.
It was inevitable under these conditions that the present paper should owe much
to Professor Osborn, and I wish to express to him my very real gratitude for all the
help given me in this way. At the same time, it must not be assumed that he is
responsible for any of the opinions given concerning the relationships of the different
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
species or groups unless such an opinion is definitely quoted as being his. Throughout,
I have tried to make it quite clear when I have adopted his views, and, in justice to
him, any statement made without his name being attached should be attributed to me.
Statements attributed to Osborn without reference to a published paper are from letters,
or from the proofs already mentioned. It is another instance of Professor Osborn's
generosity that he has allowed me to make this use of them.
Broadly speaking, the outcome of all this work has been the establishment of the
fact that not all elephants belong to the genus Elephas, any more than all mastodonts
belong to Mastodon. This, of course, was known to Dr. Hugh Falconer, and to those
who succeeded him, but Professor Osborn was the first to place it on a firm syste-
matic basis. Recognition of this fact has necessitated the making of many new genera,
and the resuscitation of many others long forgotten. In considering the problems in-
volved, the International Rules for Zoological Nomenclature have been adhered to only
so far as it seemed advisable. For example, Mastodon is used for the American
Mastodon despite the prior claims of Mammal; on the other hand, the generic name
Mammuthus is used for the Mammoth because it appears to be the first genus to
have Elephas primigenius specified as the genotype, whereas Professor Osborn uses
Mammonteus, a modification of Mamonteum which he claims to be Camper's generic
name for the same animal. By so doing I have attempted to make of the Rules a
useful servant, rather than to allow them to become a blind, unreasoning, master.
In the general systematic arrangement of the work, I have made use of other
authors' results but chiefly of Osborn's. The names for the families, however, have
been constructed on orthodox lines by adding the termination-/^ to the name of the
typical genus. Osborn employs descriptive names such as 'LongirostrinaT, 'Brevirostrinae',
and 'RhynchorostrinaT, but they do not seem to have any advantage over names formed
in the usual way.
The genera used in this work, and their approximate relationships to each other,
are shown in the following table,
PalaeomastodontidK
'Mastodonloidea
Proboscidea
Mastodontidae
.Dibunodontidae
Stegodontidae
.Elephantoidea
Elephantidae
(Trilophodon
"Serridentinus"
(Platybelodon
Tetralophodon
. Mastodon
. Pentalophodon
fitegolophodon
\Stegodon
(Archidiskodon
I Palceoloxodon
{Mammuthus
DESCRIPTION OF SPECIES.
Family PAL/FOMASTODONTID/E Andrews.
1906. Cat. Tert. Vert. Fayum, p. 130.
Diagnosis. - "In the skull the nasals shortened and the external nares somewhat
shifted back from the end of the snout. Mandibular ramus with elongated spout-like
symphysis, projecting beyond the skull. A single pair of tusks (i.2.) in both upper
and lower jaws. Upper incisors in form of downwardly directed tusks, with a band of
enamel on their outer side; lower incisors procumbent and continuing the upper sur-
face of the spout-like symphysis. Premolars replacing milk-molars in both upper and
lower jaws; molars with not less than three transverse ridges.” (Andrews, loc. tit).
Remarks. — This family, which is often spoken of as the Trilophodontidae, is
the equivalent of Osborn's sub-family, the Longirostrinae.
Genus TRILOPHODON Falconer & Cautley.
1817. Mastodon Cuvier, Regne Animal, I, p. 232.
1837. Gomphotherium Burmeister, Handb. Naturg., 11, p. 795. Genus ccelebs.
1841. Qamphotherium Gloger, Gemeinniitz. Naturg., I, p. 119.
1846. Trilophodon Falconer & Cautley, Fauna Antiqua Sivalensis, pt. i, p. 54.
1884. Tetrabelodon Cope, Proc. Amer. Phil. Soc., XXII, p. 5.
1895. Qamphotherium Gloger, Thomas, Ann. Mag. Nat. Hist., (6), XV, pp. 191, 192.
Diagnosis. — "Dentium molarium 3, utrinque intermediorum coronis colliculis 3."
(Falconer, 1857, p. 316).
This needs amendment as follows.
Longirostrine, bunolophodont, angusticoronate mastodonts with three transverse
ridges in the intermediate molars. Lower tusks flattened from above downwards, but
not expanded at their tips.
Genotype. — Mastodon angiistidens Cuvier.
Remarks. — Burmeister's genus Gomphotherium was originally diagnosed thus,
"Stosszahne in beiden Kiefern besass die gleichfalls untergegangene Gatt. Gompho-
therium". (Burmeister, loc. tit.), but he mentioned no species as belonging to this
genus. The late Dr. О. P. Hay (Hay, 1923, p. 109), regarded this as coming under
(Ill) 14
Palceontologia Sinica
Ser. C.
Opinion 46 of file International Commission on Zoological Nomenclature and adopted
it as a valid genus with genolectotype M. angustidens Cuvier (cf. Cope & Matthew,
1915, expl. to pl. cxx). Since the American Mastodon occasionally has mandibular
tusks, it is clear that Burmeister's diagnosis does not distinguish Gomphotherium from
Mastodon under which he expressly mentions “Ohiothier”, i. e., Mastodon americanus
On this we may regard Gomphotherium as invalid, because the diagnosis is inadequate.
Not only is it invalid, but it also antedates Gamphotherium Gloger. Oldfield Thomas
(1895, p. 189) regards this latter as a new generic name, and (op. tit., p. 191) italicises it
as a name which is not a simple synonym of an earlier name. The spelling, however,
is quite clearly either an error of transcription, or a lapsus calami, or even a misprint,
and the word should be written Gomphotherium, thus becoming a synonym of Gompho-
therium Burmeister. The next name, Trilophodon, was originally proposed for the
species M. ohioticus and M. angustidens. Of these the former is the genotype of
Mastodon, hence the latter only remains in Trilophodon and may be regarded as the geno-
type of that genus. For these reasons Trilophodon is here adopted as a valid.genus
to include all the bunolophodont proboscideans grouped round Mastodon angustidens.
Trilophodon first occurs in the Burdigalian of Europe as a migrant from an unknown
region. The species Tri. cooperi from the Gaj series of Baluchistan may be a little
earlier than Tri. angustidens from the Orleans Sands, but the difference, if any, is
not more than the time comprised in the interval between the transition from Aquitanian
to Burdigalian, Dr. G. E. Pilgrim places the Gaj in the Lower Burdigalian, whereas,
from a study of the Proboscidea, I am inclined to regard it as the transition period
between the Aquitanian and Burdigalian. Other primitive species occur* in Egypt,
and in Kenya, but in each locality the deposits are not older than Burdigalian, and
may be younger. Thus, despite the probability that Trilophodon derives from part
of the group of species which Andrews called Palceomastodon, there is at present no
evidence to indicate where the change took place. The Chinese species do not help,
for, so far as they are known, they are all too advanced to serve as connecting links
between Palceomastodon and Trilophodon.
TRILOPHODON CONNEXUS sp. nov.
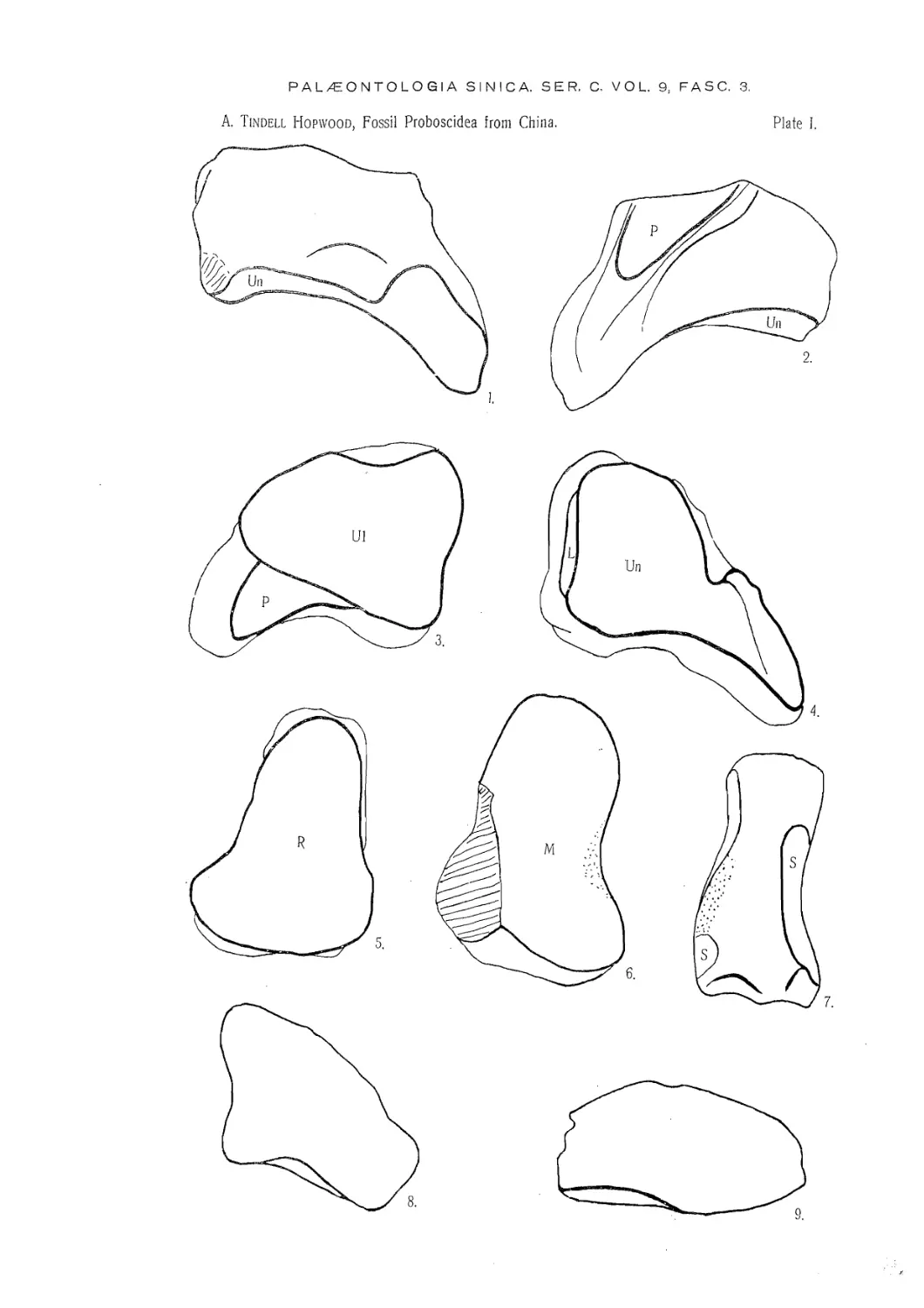
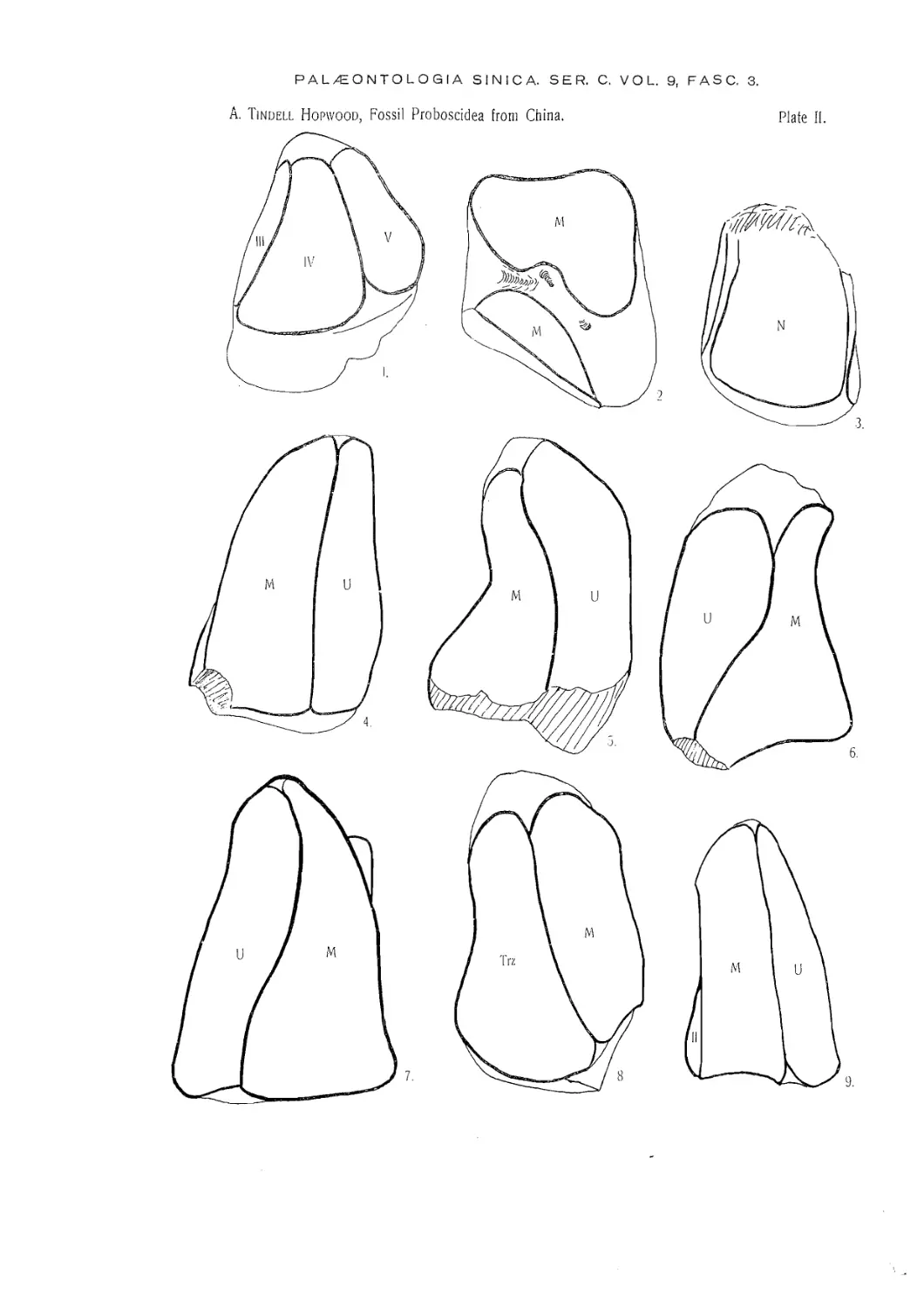
(Plate V).
Diagnosis. — A Trilophodon in which the dentition is angusticoronate, bunodont,
and moderately hypsodont; pretrite and post-trite cusps of upper molars, normal or
sub-normal to the long axis of the tooth; valleys wide, but obstructed by the pretrite
accessory structures; basal cingulum strong: lower molars with the pretrite cusps oblique,
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 15
and the post-trite cusps normal, to the long axis; pretrite cusps with anterior or posterior
buttresses, or both; valleys completely closed by vallecular conules.
Holotype. — A left mandibular ramus containing the part-worn second molar, and
the third molar just entering into wear.
Material. — A left fourth upper premolar; an unworn third upper molar; a left
mandibular ramus with the third milk-molar, third premolar, and first molar; a right
fourth lower premolar; a very worn left lower first molar; a left second lower molar,
unworn but very badly crushed; another specimen of the same tooth, uncrushed but
very worn.
Dimensions. —
Tooth Length Breadth Height Index
Upper P4 i 38.4 33.8 17.5 88
М3 ii 123 60.5 i 50.7 i 49.2
Lower Dm3 iii 71 26 i 24 iii ?
P3 iv 22 13.7 12 61.8
Ml v 85 44 iii 40.5 iii 51.8
P4 vi 41.3 30.9 17.2 74.8
М2 vii 102 51 iii 50
М2 viii 102 49.8 iii 31 iii 48.8
М3 ix 145.5 51.8 ii 45 i 35.6
Of these teeth, iii, iv, v are those in the smaller mandible, whereas viii, ix are
those in the holotype ramus. The remainder are isolated specimens.
Horizon. — Miocene.
Locality. — Kansu, Sining Fu, SW 20 li, Shui Ch'iian P'u, SE 5 li, Tiao Kou.
UPPER DENTITION.
Description. — There are only two teeth of the upper dentition in the collec-
tion, namely, the fourth premolar, and the third molar. The former is a small, bilophodont
tooth, almost quadrate in outline, but somewhat wider behind than it is in front. The
specimen is half worn, and hence details of the cusp structure are all but obliterated.
The posterior ridge consists of two cusps of which the pretrite is stouter than the post-
trite. The anterior ridge has a similar construction, but the pretrite cusp has both
anterior and posterior buttresses. There is a strong basal cingulum, which is weakest
on the buccal surface.
The third upper molar is trilophodont with a strong cingulum and talon. Cusps
1, 3, 5, 6 are slightly oblique to the long axis of the tooth, whereas the second and
fourth cusps are definitely normal to it. Each cusp consists of two cones, the inne
(Ill) 16
Palceontologia Sinlca
Ser. C.
being much smaller than the outer and occasionally divided at the summit, e. g., cusp.
4. Each of the pretrite cusps has one or two accessory conules anteriorly, and posteriorly;
sometimes, e. g., cusp. 5, the posterior conules are missing. On the post-trite side
the accessory structures are not definite cusps or conules, but rather buttresses formed
by outgrowths of the inner cones of each cusp. The talon consists of three stout
obtuse cones, two of which are on the pretrite side. The anterior cingulum is com-
plete, and is continued round on to the pretrite and post-trite sides as far as the second
ridge. It is broken away behind that point, but traces which remain prove that it
formed a cingule at the post-trite entrance to the second valley, and that on the pretrite
side it continued right back to the talon. It is coarsely nodular, especially in the front
of the tooth where it receives the anterior accessory buttress of the first cusp.
LOWER DENTITION.
The third lower milk molar is trilophodont, with an anterior cingulum, and a talonid.
The third and fifth cusps have been lost; those which remain are worn. Whereas all
the post-trite cusps are normal to the long axis of the tooth, the first cusp is oblique.
It has a strong anterior accessory structure which is seen in cross-section owing to
wear. In the centre of the first valley is the base of an independent column or valle-
cular cone. The talonid is a single stout cone standing on a cingular shelf. There
was a weak cingulum anteriorly, and at either end of the first ridge. It did not continue
past this point on the post-trite side, nor was there a cingule at the post-trite end of the
second valley. The conditions on the pretrite side are not known.
The third lower premolar consists of a large, compressed, main cusp with a single
conule in the front and two in the rear. It has the usual pear-shaped outline of these
teeth, and there is nothing to distinguish it from the corresponding tooth of other
Trilophodonts at the same stage of evolution.
The fourth lower premolar is a two-rooted, bilophodont tooth which is narrower
in front than behind. Each ridge has two cusps, the postero-pretrite being provided
with anterior and posterior accessory buttresses. The valley is closed by a strong valle-
cular conule situated on the middle line of the tooth. Other conules in the same rela-
tive position occur in front of the protolophid and behind the metalophid. In the latter
instance they form part of the talonid. There are strong anterior, pretrite, and posterior
cingula.
The first lower molar is trilophodont, with a slight anterior buttress and cingulum,
and a talonid of a single stout median cone. The cusps have a large outer and a
small inner cone. Those on the pretrite side of the tooth are oblique to the long axis;
Vol. IX. .4. Tindell Hopwood: Fossil Proboscidea from China 17
those on the post-trite side are normal to it. Each valley contains a large vallecular
connle, but in addition the pretrite cusps tend to develop accessory buttresses which
may be either anterior (cusp 5) or anterior and posterior (cusps 1, 3) in position. The
post-trite cusp have no accessory structures. There is an obscure, smooth pretrite
cingulum, and cingules occur at either entrance to each valley.
The second lower molars are practically identical with the first in structure. As
one would expect they are somewhat stouter, and have the features of the ridges and
accessory structures better defined. Apart from that, however, the only point of differ-
ence is in the cingulum, which, in these teeth, has a strong nodular lateral buttress
on the pretrite side. The second, fourth, and sixth cusps consist of a large external
cone and a very small inner one which arises as a bud from the outer, and which is
only indicated by a deep groove at the summit of the cusp. In other words, the inner
cone is not completely developed. The first, third, and fifth cusps have the inner
cones larger, and their development is practically complete. There are no post-trite
accessory structures. On the pretrite side the first and third cusps have each one
anterior and two posterior accessory conules. In each valley is a large vallecular
conule.
The third lower molar, whilst it retains the same general plan as the others, differs
from them in many points of detail. None of the post-trite cusps shows any trace of
an internal cone, and the internal cones of the pretrite cusps are not so well developed
as they are in the unworn second molar. The vallecular cone is only present in the
first valley; the second and third valleys are completely closed by buttresses accessory
to the cusps, and not by structures arising from the valley floor. Otherwise the struc-
ture of this tooth is the same as that of the first and second molars.
Discussion. — The stout columns which arise independently from the floors of
the transverse valleys of the lower molars make it quite certain that these teeth belong
to the genus Trilophodon in the restricted sense in which Osborn (1923, p. 2) uses
it. It is interesting to note that the upper molar has not these independent vallecular
cusps, and that the third lower molar has one in the first transverse valley only. Hence
we would seem to be dealing with a form which combines in itself the characters of
two genera in the upper and lower dentition. The upper molar, if I interpret Osborn’s
diagnosis rightly, should be referred to the genus Serridentinus, and the lower to
Trilophodon. This is evident from the following quotation of part of Osborn’s dia-
gnosis of Serridentinus: “Clearly distinguished from Trilophodon by the serrate crests
ascending on the outer cones of the lower molars and on the inner cones of the upper
molars; true trefoils, i. e. intermediate conules in the center of the valleys, observed
3—429921 A. Tindell Hopwood.
(Ill) 18
Palceontologia Sinica
Ser. C.
in ail species of Trilophodon, are wanting/ (Osborn, 1926, p. 2.) Comparison with
the third upper molar proves that this diagnosis applies exactly, whereas the lower
molars each have the ’’intermediate conules in the center of the valleys”.
It is, of course, open to argument that the upper and lower teeth here united as
a single species are really representative of two. Such a view does not commend it-
self to me for two reasons. First, all the teeth were obtained from a single locality,
and, so far as is known, from the same bed. Secondly, a similar instance occurs among
the proboscideans from the Bugti beds of Baluchistan. The specimens from those beds,
which were collected, described, and figured by Forster Cooper (1922) have "serri-
dentine” upper molars (cf. Cooper, 1922, pl. I, figs. 1, 2) and ”trilophodontine” lower
molars (op. cit., pl. II, fig. 1). Hence the doubts expressed by the late W. D. Matthew
(Osborn, 1924 a, p. 5) appear to have considerable justification.
Of all the primitive Trilophodonts hitherto described, that which approaches most
nearly to Tri. connexus is the form from Baluchistan mentioned above, a third upper
molar of which (M 12180 Brit. Mus. Geol. Dept.) is nearly indistinguishable from the
third upper molar of Tri. connexus. The Indian tooth is wider than the Chinese, the
breadth indices being 64 and 50.7 respectively. Moreover, the Indian tooth is more
advanced since the accessory structures are stronger, and more distinct, and the third
ridge is better developed. On the other hand, the Chinese tooth is more hypsodont,
for, although the two teeth are of approximately the same height, the Indian tooth is
the broader of the two.
This connexion between the Indian and Chinese species is of interest when
the resemblance between the Indian and European forms is considered. It is true that
the lower teeth of Tri. connexus differ from those of the Indian species in that they
have pretrite accessory buttresses in addition to the vallecular conules, but they agree
with lower teeth from Sansan in this respect. Thus the Chinese species has features
in common with both the European and Indian species, but differs from them
both in varying degree. On the balance, there is every indication that they were all
derived from a common stock, and are members of a widely distributed group of
animals radiating from a single centre. This is considered in more detail in a memoir
on certain Indian species which is in preparation for "Palceontologia Indica."
Trilophodon sendaicus Matsumoto, from the Tatsunokuchi formation of Japan is
a more advanced species without the vallecular columns of Tri. connexus. Another
Japanese species, "Hemimastodon" annectans Mats, is from the Hiramaki Formation,
and so probably of Burdigalian age. It is a smaller species with broader upper molars
(breadth index of M’ 66 67 %), but the unworn M:i figured by Matsumoto (1926,
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 19
pl. i, fig. 2; pl. ii, fig. 2) shows other points of difference as well. The cusps are
not so high, the accessory buttresses are not so complicated, and the vaiieys are iiot
so narrowed, nor so obstructed, as they are in T. connexus.
When comparison is made with the trilophodont species of America, it is found
that they are all more advanced than Tri. connexus. They all have four ridges and
a talonid, or rudimentary pentalophid, in the third lower molar, and the narrowest of
them, Tri. hicksi, Tri. lulli, Tri. osborni, have a breadth index of 40. Moreover,
they are all larger species (Cook, 1922, p. 8) and more complicated in their structure.
So far, the only species of Trilophodon known to occur in America have been
obtained from Pliocene deposits, and their relationship among themselves is not very
clear. Apparently the most simple form is Tri. willistoni Barbour, but, since it was
found in deposits of Upper Pliocene age, it does not throw much light on the question
of the early proboscidean immigrants into the New World.
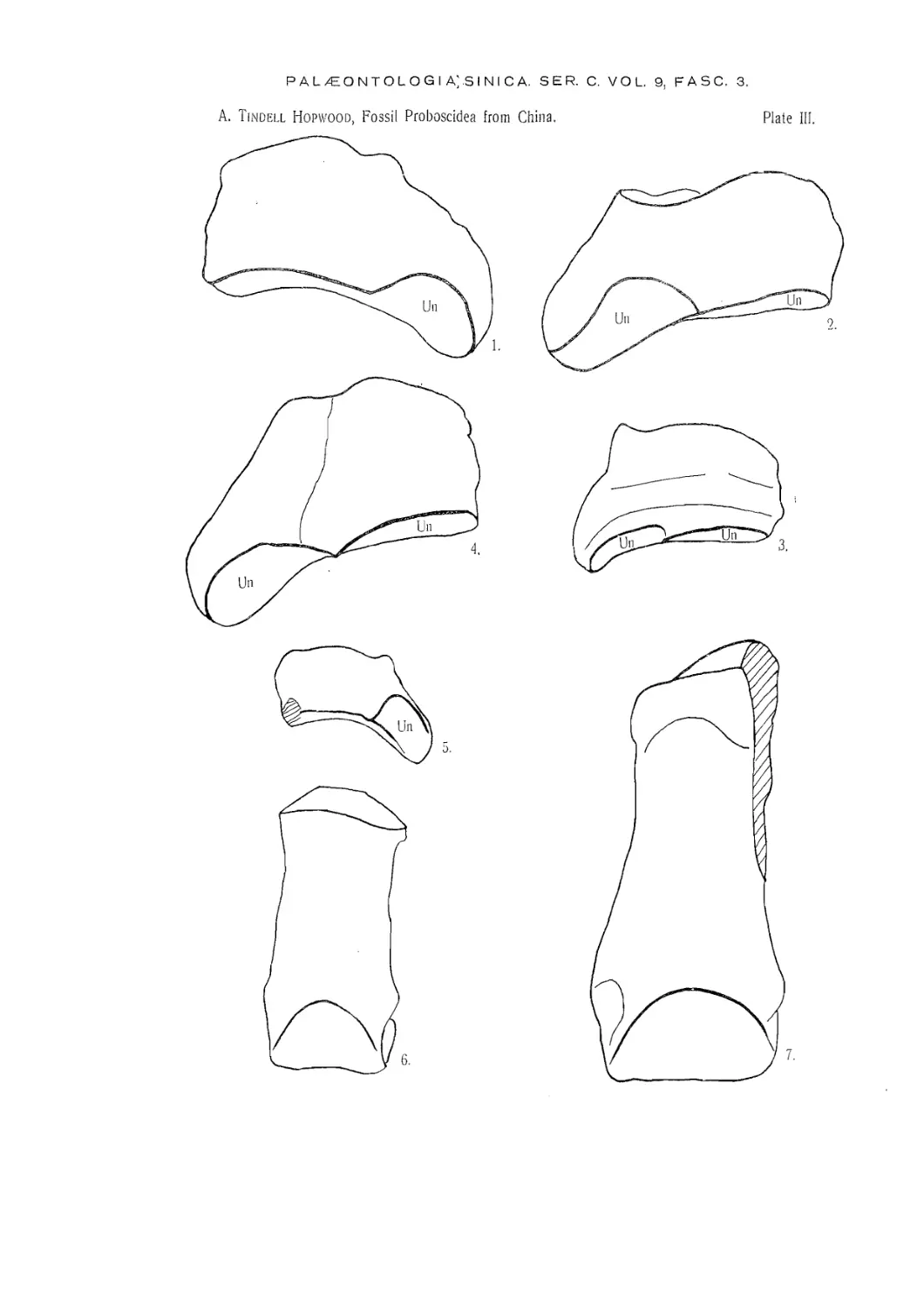
TRILOPHODON WIMANI sp. nov.
(Plates I, III, V, VI).
Diagnosis. — A Trilophodon of medium size with sub-tectiform ridges in
the upper molars; pretrite cusps sub-normal in the upper, oblique in the lower,
molars; post-trite cusps normal in both upper and lower molars; upper molars with,
strong pretrite accessory buttresses, and smaller post-trite ones; lower molars without
the vallecular cones of Tri. connexus, but with strong pretrite, and incipient post-trite
buttresses; cement fairly plentiful.
Holotype. — К palate with the right first molar very worn, second molars half
worn, third molars just entering into wear; the first molar on the left hand side is
missing, and the right third molar has been broken off just behind the second ridge.
Material. — In addition to the holotype; a left maxilla with the teeth from the
second premolar to the first molar inclusive; a right second upper molar almost unworn;
right mandible with the second molar worn, the third badly crushed and partly worn;
a worn, broken, and crushed right first lower molar; a left second lower molar in the
same condition as the preceding, and probably derived from the same individual; an
isolated, worn, left second lower molar, probably derived from the same individual as
the mandible; an unworn left third lower molar; three fourth premolars, two from the
upper, and one from the lower jaw; one milk incisor; and fragments of two others;
the tip of an adult incisor; fragments of the first, second, and third lower molars in
varying states of wear; the anterior ridge of an upper molar.
(Ill) 20
Palceontologia Sinica
Ser. C.
Upper Dentition. Length Breadth Height Index
Holotype. Left М2 107 67 iii 51 iii 62.6
М3 151 72 ii 64 ii 47.7
Right М2 103 65 iii 49 iii 63.1
М3 ? 69 i 63 ii —
Maxilla. Left P2 18? 19? ? —
P3 33 33 21 100
P4 41 39.5 28 96.3
Ml 81 54 iii 42 66.7
Isolated P4 46.5 41 29 88.2
P4 42.5 38 ? 89.4
М2 120 67 ii 55 iii 55.8
Lower Dentition.
Mandible. Right М2 103 71 iii ? —
М3 180 ? ? —
Isolated P4 50 35 28.5 70
М3 185 75 ii 67 ii 45.4
Although the remaining specimens are so broken that systematic measurements
are not feasible, yet they afford certain dimensions which are inserted in their appro-
priate places in the text.
Horizon. — No stratigraphical details as to the age of the beds whence these
fossils were derived is available, but, from their structure, one would expect them to
be of Sarmatian age. In this connexion, it is important to note that Dr. H. S. Pear-
son, when she described Listriodon gigas, the only other mammal known from Chuan
Tou Kou, says that the fragments: "show affinity with the small bunodont Listriodon
species of the European Lower and Middle Miocene . . .”, so that we should prob-
ably not differ very much concerning the age of the beds (Pearson, 1928, p. 1).
Another factor which helps in determining the probable age of these teeth is their
general agreement in evolutionary characters with certain undescribed teeth from the
Chinji beds of the Salt Range, Punjab, India. These beds are regarded by Dr. G. E.
Pilgrim as being of Upper Vindobonian age.
Localities. — Kansu, Ping Fan Hsien, SE 100 li, Hsien Shui Ho E 3 li, Chuan
Tou Kou; Sining Hsien from city SE 35 li, Hsin T’ien P'o N 3 li, Pa P'an Shan.
The latter is the locality of the holotype.
Vol. IX.
A. Tindell Hop wood: Fossil Proboscidea from China
21
Description. — The upper dentition of this species is represented by the tip of
л,-. Z.Z-1..U —z4 tu £____ il.------z4 1л ли
an auuiL uiLidui, dim uic uiccn-iccui 110111 Lilt: icouiiu picinuidi Lu tile Hinn iiiuidi. nu
these are in good condition, except the second premolar and first molar. The lower
dentition, of which the cheek-teeth from the fourth premolar to the third molar are present,
is not so well preserved. Most of the teeth are very worn, and others are crushed
laterally.
UPPER DENTITION.
The second premolar is badly broken. It is apparently circular, or sub-circular
in outline. All other details are lost.
The third premolar has a protoloph of two unequal cusps of which the post-trite
is the larger. The pretrite cusp has both anterior and posterior nodular buttresses. The
metaloph is represented by three or four conelets which stand on the strong, warty
cingulum. Both cusps of the protoloph are divided into two unequal cones.
The fourth premolar is bilophodont, and rounded in plan. The protolophs of all three
specimens are composed of two cusps, each of which is sub-divided into two or three
conules, the second cusp is stronger and more stoutly conical in the unworn tooth
than the first, but as wear proceeds the relative size changes owing to the anterior
and posterior buttresses of the first cusp coming into play. Both cusps of the metaloph
are of equal stoutness, but the pretrite cusp is strengthened by anterior and posterior
trefoil buttresses. In one specimen the fourth cusp has a very strong posterior buttress.
The cingulum is strong and warty; there is a well defined posterior talon.
The first molar is trilophodont. The second and fourth cusps have been broken off,
and the first and third are badly damaged. Each of the remaining cusps is made up of
two cones, a larger, outer, and a smaller, inner one. All the pretrite cusps have anterior
and posterior buttresses which are accessory to the outer cones of the cusps. The
post-trite cusps have much smaller buttresses, each of one or two conules, accessory
to the inner cones of the cusps. The fourth cusp has anterior and posterior buttresses,
whereas the sixth cusp has only an anterior buttress. The cingulum is strong and
nodular. It forms a posterior shelf remarkable for the number of stout, conical ex-
crescences with which it is covered. This nodular tendency is very noticeable in other
places on the tooth. The crevices at the bases of the cusps, both pretrite and post-
trite, are filled with small, but distinct, conical nodules. The valleys were originally
filled with cement. Unfortunately this has all fallen away.
The most complete specimen of the second molar is a single tooth which has
(Ill) 22
Palceontologia Sinica
Ser. C.
fallen out of a palate, and which retains the roots intact. It is large and massive,
trilophodont, with a talon at either end.
Cusps 1 and 3 are each made up of two cones, a large one externally and a smaller
one internally. The summit of the fifth cusp has been broken away so that the struc-
ture is not determinable. All the pretrite cusps have anterior and posterior accessory
buttresses which differ among themselves in structural details. That on the anterior
surface of the first cusp is a stout conule bearing on its inner surface a heavy ridge
which passes downwards and inwards to join the anterior talon. Doubtless this ridge,
when unworn, resembled that on the posterior surface of the fifth cusp in showing some
four or five cushion-like nodules on its occlusal surface. The anterior buttress of the
third cusp and the posterior one of the first resemble each other in being simple stout
conules, but the former is more massive than the latter. The anterior conule of the
fifth cusp is also very massive. The posterior conules of the third and fifth cusps are
unusually small. The small size of the latter may be due to the fact that the mamillated
talon would, to some extent, take over its functions as wear proceeded, and, by the
time that the tooth was half worn, would have replaced it altogether. It is interesting
to note that the outer face of the third cusp is produced backwards, as though in
compensation for the small size of the accessory structure.
The post-trite cusps are of simple construction. Each consists of a large outer,
and small inner cone. The fourth cusp is more complicated than the others; its inner
cone is composed of two unequal conules. Another feature of the same cusp is the
presence of anterior and posterior folds which, though slightly developed, would serve
as accessory structures. There is a similar structure on the anterior face of the sixth cusp.
The posterior talon begins two-thirds of the way up the fifth cusp. It forms a
sloping buttress which passes downwards and inwards to terminate rather less than half
way up the sixth cusp. On the occlusal surface are five sub-equal mammillae. The
anterior talon was probably of similar construction, except that it sloped away more
rapidly to the base of the second cusp. There it joined a fairly broad, beaded, shelf,
which passes across the front of the first cusp to the buccal surface of the tooth. This
cingulum merges into the buccal surface of the first cusp, and does not extend as far
as the mouth of the first valley. There are no lateral cingula, but each valley has a
definite cingule at either end.
The presence of cement has already been mentioned in the description of the first
upper molar. It is present in quantity in the second, in which tooth it is wholly
retained on the pretrite side, and, although most of it has been chipped away on the
other side, there is no doubt that the valleys were originally half full of this tissue.
Vol. IX. A. Tindell Hop-wood: Fossil Proboscidea from China 23
The differences between the tooth just described, and those of the holotype palate
are so slight as to be negligible. The only one of any importance is the stronger
internal cingulum of the holotype. Ail the cement has disappeared from the palate,
and no traces remain.
The third molar has four ridges and a talon. The first, third, and fifth cusps
have the usual structure of two unequal cones, but the seventh differs considerably
in that there is one large outer cone with three conules on its inner side. Each of
the first three post-trite cusps has both cones divided at the summit by a groove, but
this does not extend very far down on the sides of the cones; the eighth cusp is not
divided in this manner. The first and third cusps have anterior and posterior but-
tresses; that which is anterior is truly accessory, whereas that which is posterior is a
fold which forms part of the outer cone. The fifth and seventh cusps have
anterior conules closely pressed against the outer cone. In addition, a buttress from
the outer cone of the fifth cusp passes between this accessory conule and the inner
cone of the cusp forming a strong nodular ridge in the post-trite half of the valley.
The fourth cusp has small anterior and posterior accessory buttresses applied to the
inner cone. The second and sixth cusps have similar buttresses but only on the posterior
surface of the inner cones. The eighth cusp has an anterior buttress, but none posterior.
The lack of an anterior buttress to the sixth cusp is probably to be accounted for
by the manner in which the buttress from the fifth cusp passes over to the pretrite
side.
The anterior and internal cingula are similar to those of the second molars. Both the first
and second valleys have their post-trite ends occupied by small cingules. The talon
is a stout nodular ridge.
The two ridges of the right molar agree with this tooth, but it is interesting to note
that the anterior buttress of the third cusp tends to pass over to the post-trite side in
the manner already described for the fifth cusp of the left tooth.
The upper incisor is represented only by a fragment of the tip. It is about
195 mm long, has a maximum width of 51 mm, and a maximum depth of 70 mm.
The upper surface is plane and covered by a band of enamel which is 35 mm wide
at a point 63 mm from the tip. In cross-section the tooth is U-shaped; the inferior
face passes forwards and upwards in a graceful curve to meet the upper surface. On
the extreme tip is a small flattened area due to wear.
The larger of the two fragmentary milk incisors measures 40.5 X 26.5 mm near
the tip, which has a large irregular plane of wear parallel to the greatest diameter of
the tusk.
(Ш) 24
Palxontologia Sinica
Ser. C.
Г Н1ППП ГМ7МТТТТГЛ\Т
bV7VVbl\ L7bl\ 11UV71N,
The fourth premolar is an elongate, bilophodont tooth in which the metalophid is
oblique to the long axis. Each cusp is made up of two cones. The first and third cusps have
anterior and posterior accessory buttresses applied to their outer cones. The fourth
cusp has similar, but smaller, buttresses applied to its inner cone. There are strong
anterior and posterior talonids, as a well as a definite cingule at the pretrite mouth of the
transverse valley. As is usual in these teeth, the first ridge is taller than the second.
The first molar is trilophodont, with a high, detatched talonid. One specimen is worn,
crushed, and broken, so that no details of its construction that are of any value remain.
The other specimen, the anterior half of a tooth, retains cusps 1, 2 and 4, with the
anterior surface of the third cusp; its maximum width is 47.5 mm. Each cusp retains signs
of the original two cones, and the pretrite cusps show their accessory buttresses. The
posterior buttress of each pretrite cusp sends small spurs across to the post-trite side
of the valley, and resembles in this respect the anterior buttress of the fifth cusp of
the third upper molar. A large pressure mark obliterates the anterior buttress and cingulum
except for the extreme pretrite end. There is a large smooth cingule at the pretrite entrance
to the first valley. The enamel of this tooth is noticeably smooth and highly polished.
There is no satisfactory specimen of the second molar, but the two examples
of that tooth in the collection agree in showing a pronounced trefoil swelling' on the
anterior face of the sixth cusp.
The third molar is a trilophodont tooth with a large, bluntly triangular, talonid. The
three ridges are each composed of two cusps, of which the first four have a large outer,
and small inner cone. The second, fourth, and sixth cusps each have a posterior
accessory conule, but none anteriorly. The first and third cusps have anterior and
posterior accessory buttresses, whereas the fifth cusp has an anterior one only. The
talonid is massively built of stout, blunt, cones arranged in the form of a narrow U,
which is set obliquely to the long axis of the tooth. That is to say, the valley con-
tained by the cusps opens obliquely towards the buccal surface. As one would expect
the pretrite limb is the stouter. It is composed of three cones, the largest being in
front and the smallest behind. The post-trite side has three slender cusps where it
joins the pretrite side, and terminates on the buccal margin in a large, somewhat clumsy,
mass composed of three or four bosses. The anterior portion of the cingulum has
been lost, except for a small fragment which would seem to show that it resembled
that of the second molar. Any cingules which there may have been at the pretrite
ends of the valleys have been weathered away, but they persist on the post-trite side:
that between the protolophid and the metalophid is very strong.
Vol. IX.
Л. Tindell Hopwood: Fossil Proboscidea from China
25
Only slight traces of cement remain, but they are in positions which prove tha
originally a considerable quantity of this substance must have been present.
OSTEOLOGY.
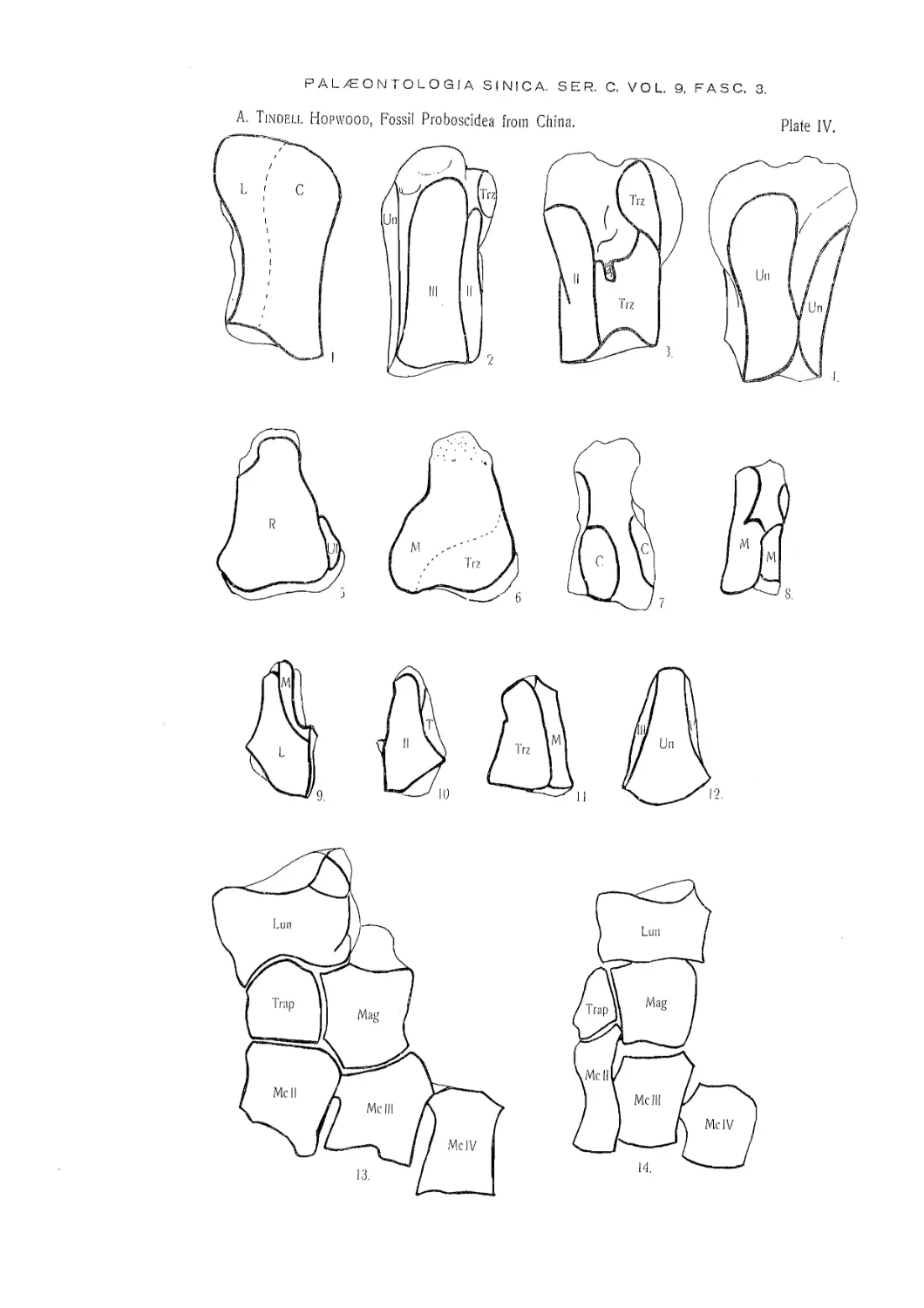
Eight bones found at Chuan Ton Kou and Pa P'an Shan are referred to this species
because none other is yet known from those localities. The five bones from Chuan
Ton Kou are, the proximal end of a small tibia, a crushed and broken lumbar vertebra,
a left cuboid wanting the posterior portion, a left meso-cuneiform, and a left unciform.
The three from Pa P’an Shan are, a left cuneiform, a left lunar damaged on the an-
tero-external angle, and a worn, broken almost square bone which is probably a small
right cuneiform.
Os lunatum. — The whole of the upper surface is occupied by the concavo-
convex radial facet; similarly the entire distal surface is occupied by the facet for the
magnum. The ulnar facet has disappeared from the outer face through fracture. On
the inner face of the bone are two facets. One forms a narrow band in the middle
of the superior margin, and the other, which is on the inferior margin and has been
much abraded, probably extended from the anterior-inferior corner backwards for three
quarters of the length of the margin. Both facets articulated with the scaphoid.
Os cuneiforms. — The cuneiform is a flattened, square, bone with a very large
decurved external process. On the upper, or proximal, surface there is a large facet
for the ulna. This facet is convex from side to side, and gently concave from behind
forwards; this concavity increases as it passes outwards so that the outer half of the
facet is concave, whereas the inner half is convex. On the under, or distal, surface
is a large facet for articulation with the unciform. This facet is slightly concave from
side to side, but from back to front it is gently concave behind and equally convex
in front. The postero-external corner is produced outwards into a long tongue which
is twisted. The inner portion is concave from side to side; it faces downwards, and
slightly inwards. The outer portion, which is convex from side to side as well as
from back to front, faces downwards and outwards.
Os unciforme. — The unciform is broken at its outer extremity, as well as on
the under side. It is roughly wedge-shaped, convex on the superior surface, and con-
cave on the inferior surface. The proximal facet, which articulates with the cuneiform,
occupies the whole of the upper surface. It is convex from side to side, as well as
from back to front, in its posterior portion. The anterior portion is gently concave.
All the hinder part of the inferior facet has been broken off, but the anterior part is
strongly convex on either side, and markedly concave in the centre. The median con-
4—429921 A: Tindell Hopwood.
(Ill) 26
Palceontologia Sinica
Ser. C.
cavity articulates with the fourth metacarpal, and the inner and outer surfaces with
the third and fifth metacarpals respectively. On the inner surface, which forms the
base of the wedge, there is, at its superior margin, a large abraded facet for articula-
tion with the magnum. It is not known whether there was a corresponding facet on
the inferior margin because of the extensive damage in that region.
Tibia. — The proximal end of the right tibia has undergone a considerable amount
of distortion through crushing. In its present state, the medial articular surface
occupies the whole of the internal condyle; it is nearly circular, and is concave
in all directions. The lateral facet looks upwards and outwards; it is transversely
oval, and concave in all directions. Between the two facets is the tibial spine, a stout
ridge which traverses. the whole of the proximal surface of the tibia from back to
front. Apparently, this bone derives from an animal not yet fully grown, for, in the
process of crushing, the epiphysis has been partly separated from the shaft. The same
crushing has so distorted what remains of the shaft as to make it unworthy of de-
scription.
Os cuboideum. — This was originally a flattened triangular bone, but the posterior
third is missing. On its convex superior surface there is a double facet, of which the
larger, internal, portion articulates with the calcaneum, and the smaller, external, portion
articulates with the navicular. On the under .surface is a large facet divided into two
parts for the fourth and fifth metatarsals. That for the fourth is slightly concave and
faces downwards, whereas that for the fifth is almost plane and faces downwards, and
slightly outwards. On the inner surface at the antero-inferior corner, there is a small
semi-circular facet for articulation with the adjacent cuneiform.
Os mesocuneiforme. — The meso-cuneiform is a triangular bone in which the
width from side to side is greater than the depth from back to front. All the edges,
and most of the surfaces, are deeply abraded, and the characters of the articulations
are consequently obscure. From the remains, it is clear that the superior and inferior
facets are convex in front, and concave behind. The facets on either side are inde-
cipherable.
Dimensions. — The principal dimensions of the various bones described are as
follow,
Os lunatum.
mm.
Greatest depth from back to front ................................................... 86
Greatest width from side to side....................................................... ?
Greatest height........................................................................ 57
Maximum diameter of ulnar facet........................................................ ?
Vol. IX, A. Tindell Hopwood: Fossil Proboscidea from China 27 .
Os cuneiforme.
Greatest depth from back to front ...................................................... 73
Greatest width from side to side....................................................... 136
Greatest height (anterior surface)..................................................... 44
Width of ulnar facet ................................................................... 86
Depth of ulnar facet ................................................................... 59
Os unciforme.
Greatest depth from back to front ............................................................ 77
Greatest width from side to side........................................................ 85
Greatest height (anterior surface)...................................................... 50
Tibia.
Maximum width across condyles.......................................................... 155
Greatest depth from back to front ...................................................... 97
Os cuboideum.
Greatest width from side to side............................................................. 104
Greatest height (anterior surface)..................................................... 43
Os mesocuneiforme.
Greatest depth from back to front ...................................................... 86
Greatest width from side to side........................................................ 88
Greatest height (anterior surface)...................................................... 49
Discussion. — So far as it is possible to compare the indices of this species with
those of T. connexus, the cheek teeth are broader in T. wimani. Thev are also
larger. The fundamental difference between the two species, however, is the presence
of vallecular conules in the lower dentition of T. connexus, and the absence of those
elements from the lower teeth of T. wimani. Not only does this confirm the fact
that the teeth are referable to separate species, but, so far as is at present known, it
proves that the two species are not members of the same genetic lineage. Nor has
this species any close connexion with T, (Serridentinus) florescens Osborn from the
Pliocene Khunuk formation of Mongolia, which is regarded by its discoverer as a de-
velopment of T. (S.) mongoliensis (Osborn, 1924 a, p. 1; 1929 a, p. 6). Neither of
these species has any accessory buttresses on the post-trite cusps.
In the Chinji beds of India, there are one or two species in approximately the
same evolutionary stage as this one from China. They are not yet described, but a
second lower molar in the Geological Department of the British Museum (Natural
History) which has been worn to approximately the same amount as the corresponding
tooth in the mandibular ramus of T. wimani, is almost indistinguishable. The tooth
in question bears the registered number M14014.
(Ill) 28
Palcsontologia Sinica
Ser. C.
The presence of vallecular conules in the European species T. angustidens, and
their absence from T. wimani is sufficient to separate the two species. Another animal
in the European Miocene has been described by Schlesinger (1917) as "Mastodon
(Bunolophodon) angustidens forma subtapiroidea nova forma". Between this and
T. wimani there are several differences, the chief of them being the presence of post-
trite accessory buttresses in the latter. Pending more complete stratigraphical cor-
relation, it is difficult to say whether there is any connexion between the two. The
type locality of T. a. subtapiroidea is Eibiswald in Steiermark, the lignites of which
are usually regarded as of Lower Helvetian age. The exact horizon of the specimens
from China is not given, but from their evolutionary stage I should judge it to be
Sarmatian.
The Japanese species T. sendaicus Matsumoto (1926, p. 9) is said by its author
to come from Pontian deposits, and thus would be younger geologically than T. wi-
mani. It is more simple, has lower crowned teeth, and no post-trite accessory but-
tresses. Matsumoto mentions the possibility that his species may be a geographical
and geological mutant of T. angustidens, using that name in its collective sense, but
it is clear that T. sendaicus is not a member of that lineage which includes T. wimani.
Comparison with the American species is not of much value until Professor Osborn
has published the results of his researches, for the various forms are often highly
collective.
Comparative studies on the bones referred to this species have been hampered
by lack of material for direct examination, as well as by the scarcity of good figures
and adequate descriptions in the literature. This is largely owing to the uncertainty
attending efforts to identify bones, an uncertainty arising from their plasticity and their
reaction to various influences, individual, as well as racial and environmental.
The lunar has been compared with that from Locality 77 described on page 65,
with a lunar from Sansan referred to T. angustidens (regd. 33592 a), and with one
from the Siwaliks figured by Falconer and Cautley (1847, pl. 1, fig. 7. regd. M3200).
The antero-external convexity of the radial surface is rather less than in the bone from
Sansan, and compares rather with that from the Siwaliks, although the bone itself is
very much smaller. One feature in which the lunar of T. angustidens differs from
that of T. wimani is in its breadth, which is greater than its depth, whereas in the
latter, as well as in the other two bones the depth is in excess of the breadth. The
inner surface of the lunar of T. wimani is more excavated than in the bones from
Loc. 77, and the Siwaliks, but less so than that from Sansan, and it has the superior
marginal facet better developed than either the bone from Loc. 77, or from Sansan.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 29
No isolated cuneiform bone of undoubted reference to one or other of the various
species of "Mastodon" was available for comparison. Al! those in the British Museum
collections are either from some species of elephant, or else, as is the case with those
figured by Falconer and Cautley (op. cit.), they are so large as to make it almost
certain that they were derived either from a stegodont, or else from an elephant. The
only exception is a left cuneiform from Missouri (regd. 17211) supposed to belong to
the American Mastodon. In addition to this last a right cuneiform of the Mammoth
(regd. 45114) from Ilford Essex has been used.
Compared with these two bones, that from China is noticeably shallower in an
antero-posterior direction, and intermediate in anterior thickness. The inferior facet
resembles that of the Mammoth, rather than that of the American Mastodon, whereas
that for the pisiform is closer to the type found in the latter. The lateral process in
T. wimani is longer, and more slender than in either of the other species.
The unciform was compared with one of T. angustidens from Sansan, and one
of the American Mastodon. The specimens are registered 17258, and 33592 b, re-
spectively. All three bones are damaged. The superior facet of the bone from China
is proportionately wider than it is in either of the other specimens, and it is also less
concave antero-posteriorly. Its facet for the fourth metacarpal is more deeply concave
anteriorly, and apparently that for the fifth metacarpal is more nearly in the same plane.
Both specimens in the British Museum have a distinct area of rough bone separating
the superior facet from that for the fifth metacarpal, but it would seem to be very
much reduced in T. wimani. The facets for the magnum in T. wimani are too
abraded for comparison; they are separated by a fairly deep concavity, which is inter-
mediate between the very deep trough seen in T. angustidens, and the almost plane
surface of M. americanus.
The cuboid is too broken to allow of any comparisons, but the mesocuneiform
is interesting. Apparently this bone is rarely found, probably because of its relatively
small size. Among the elephants, it, and its fellows, are long, narrow, wedge-shaped
bones, the depth being approximately twice the width, but in T. wimani, the width
is actually greater than the depth. No specimens other than those of Palceoloxodon
antiquus, and Mammuthus primigenius were available for comparison, and beyond
the statement that it is the smallest of the three cuneiforms in the pes of the American
Mastodon (Warren, 1852, p. 50). I have been unable to trace any information con-
cerning this bone in the Mastodontoidea.
The trivial name is in honour of Professor Carl Wiman to whose- energy and
enthusiasm much of our knowledge of the fossil faunas of China is due.
(Ill) 30
Palceontologia Sinica
Ser. C.
TRILOPHODON SPtCi ABLIS sp. nov.
(Plate VI.)
Diagnosis. — A Trilophodon of medium size with hypsodont, angusticoronate
dentition; pretrite cusps markedly oblique, post-trite cusps normal, to the long axis;
accessory vallecular cusps median, completely obstructing the valleys; cingulum absent,
except in front; cement plentiful.
Holotype. — К left lower third molar, partly worn and lacking the talonid.
Dimensions. — Length 176 mm, breadth 65 mm, height of second ridge in excess
of 65 mm. These measurements are taken from the holotype without any allowances
for breakage or wear.
Horizon. — Unknown.
Locality. — Unknown. The label on the specimen reads, “Sent febr. 1923 by
Rev. Beinhoff. Said to have come from Sian.”
Description. — The summits of the first six cusps have all been removed by
wear; those of the seventh and eighth cusps were broken off before burial.- Everything
behind the fourth ridge has been lost.
The pretrite cusps are distinguished by their extreme obliquity to the longitudinal
axis, which is so great that the outer cones alternate with those of the post-trite cusps.
Each cusp is composed of two cones, though, from the fifth cusp, it appears that the
inner, smaller, cone was divided at the summit. The post-trite cusps are normal to
the long axis, and composed of two cones. The only exception is the eighth cusp,
which is slightly oblique. This condition is very common in the ridge next to the
talonid in all species of mastodonts.
The accessory structures are restricted to the pretrite side almost without excep-
tion. The first cusp has a tall cone anteriorly, which forms a buttress merging with
the strongly nodular antero-external cingulum. Behind the same cusp is a large valle-
cular cone, or group of cones, which wears with a trefoil section. The third and
fifth cusps have each a posterior vallecular cone, but no anterior cone. There is a
small cone closely adpressed to the anterior surface of the inner cone of the sixth
cusp. Apart from this, the post-trite cusps have no accessory structures. There is no
cingulum other than that just mentioned. All the valleys are filled with cement.
Discussion. - In certain respects, this is the most advanced of the Trilophodon
group. None other has such hypsodont teeth, and so copious a supply of cement,
and in none are the pretrite cusps so oblique to the long axis of the tooth. The
species which, although much more primitive, shows the greatest superficial resemb-
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
31
lance to this one is that described by Lydekker (1884; pl. iv, figs. 3, 8; pl. v, figs.
1, la) as Mastodon angustidens var palceindica. That species is hypsodont, angusti-
coronate, and, as seen in specimens collected since Lydekker wrote, well provided with
cement, but an anlysis of its cusps shows that it cannot be regarded as an ancestor
of Trilophodon spectabilis. Another form with high slender cones is found in the
Perim Island beds, but this again appears to be a convergent resemblance and so far
as is known the two species are not related.
Genus SERRIDENTINUS Osborn.
1923. Serrldentituis Osborn, Amer. Mils. Novitates, No. 99, p. 2.
Diagnosis. — "Based upon the genotypic species Mastodon productus Cope
M. floridanus Leidy, M. obscuras Leidy, and Serridentinus simplicidens Osborn be-
low, characteristic of the southeast coast of the United States from Maryland to Flo-
rida; clearly distinguished from Trilophodon by the serrate crests ascending on the
outer cones of the lower molars and on the inner cones of the upper molars; true
trefoils, i. e. intermediate conules in the center of the valleys, observed in all species
of Trilophodon, are wanting.” (Osborn, loc. cit.)
Remarks. — The three species which follow are all referred to this "genus” by
their author. In view of the fact that none of them is represented in the present
collection, his determination has been retained; it is not certain, however, that the
"genus” is well founded, or that it is more than a section of Trilophodon.
SERRIDENTINUS MONGOLIENSIS Osborn.
1924. Serridentinus mongoliensis Osborn, Amer. Mus. Hovitates, No. 148, pp. 1—3, text-fig. 1.
Diagnosis. - "Ridgecrest formula of 5. mongoliensis: Dp472 Ml3 М2; М3;.
In each typical ridgecrest (e. g., the metalophid) three conelets and two trefoil conules
attached to the external cones. Molar proportions as in S. productus.”
Syntypes. — "Amer. Mus. 19152. A series of right inferior grinding teeth, p4
(dp4), m2, m3; also one left grinder, ml------; in juvenile condition, dp4 greatly
worn, ml partly worn, m2—3 embedded in the jaw.”
Horizon. — Loh Formation, ? Lower Miocene.
Locality. — Loh, Mongolia.
Remarks. — This species is compared with 5. productus (Cope). It is said to
be smaller and less progressive.
(Ш) 32
Pakeontologia Sinica
Ser. C.
crnninrKiTiMi те лггчптттктете _______________
JE.tWL'ClNUlNUO UVJDUJNOIO V75UU111.
1932. Serridentinus gobiensis Osborn, Amer. Mus. Novitates, No. 537, pp. 11—13, text-fig. 8.
Diagnosis. — ’’Extreme length of ramus from tip of incisor tooth to posterior
border 109 cm. Extension of lower incisor beyond alveolar border 12.5 cm. Greatest
diameter of tusk at alveolar border 5 cm. M2 a-p. 12.5 cm; tr. 8.5 cm. M3 a-p.
19.5 cm; tr. 8.7 cm. The lower border of the ramus, from the alveolar edge to the
angle of the jaw, is a nearly straight line which is set off at an angle of about 15е
from the plane of the molar crowns. Lower tusks rounded on the lower and outer
surface and somewhat flattened on the lingual face. M2 with three ridges, M3 with
four ridges and a heel. Serrated spur-crests or molar borders. Molar pattern extre-
mely simple and lophodont.”
Horizon. — Tung Gur Beds, Lower Pliocene.
Locality. — About 40 miles southeast of Iren Dabasu, Inner Mongolia.
Remarks. — This species is said to have its molars more simply constructed than
either 5. mongoliensis or 5. florescens.
SERRIDENTINUS FLORESCENS Osborn.
1929. Serridentinus florescens Osborn, Amer. Mas. Novitates, No. 393, p. 6, text-fig. 7.
Type. — ’’Amer. Mus. 21615. Second inferior molar of the right side, r.M2,
crushed laterally. Length of type crown 131 e mm, breadth of tritolophid 45-mm.”
Horizon. — "Khunuk formation, Pliocene, possibly equivalent to the Hung Kureh
beds of the Tsagan Nor region.”
Locality. — Kholobolchi Nor region, Mongolia.
"Specific Characters. — The specific name florescens refers to the remarkable
florescence or blossoming out of the external trefoil spurs into broad anteroposterior
plates crowned with four to five conelets which greatly exceed in prominence those
of any other species of Serridentinus. The length of the type crown (13 le mm)
exceeds that of r.M2 in 5. mongoliensis (108 mm); the breadth of the tritolophid is
45-mm and is greatly contracted by lateral crushing. 5. florescens appears to represent
a progressive stage in the blunt-coned series of Serridentinus, beyond 5. mongoliensis
of the Lower to Middle Miocene of Loh, out of which stage it has evolved.”
Remarks. — The only other species comparable with this in the development of
the accessory crests is one from the Upper Chinji horizon of India, which is usually
regarded as of Upper Vindobonian age. rfhat species, which has not yet been de-
scribed, is smaller, wider, and lower in the crown than 5. florescens.
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
33
Genus PLATYBELODON Borissiak.
1928. Platybelvdott Borissiak, Ann. Soc. paleont. Russte, VII, p. 119.
Diagnosis. — "Upper jaw with flat and wide fore part, incisors reduced and not
reaching the front border of the jaw; the mandible with long and wide spoon-shaped
symphysis, the incisors being wide and flat; the bunolophodont cheek teeth show a
ridge-formula similar to that of Trilophodon but somewhat more complicated” (о/ш?.,р. 120).
Genotype. — Platybelodon danovi Borissiak, 1928.
PLATYBELODON GRANGER! Osborn.
1929. Amebelodon grangeri Osborn, Natural History, XXIX, p. 16.
1931. Platybelodon grangeri Osborn, Amer. Mus. Novitates, No. 470, pp. 6—10, text-figs. 1—3.
Diagnosis. — ’’Type exceeding Platybelodon danovi in size, mandibular dimen-
sions in ratio of 5 to 4; in pre-mandibular dimensions in ratio of 8 to 5. Anterior
portion of mandible greatly broadened; second inferior incisors correspondingly broa-
dened, flattened, and relatively shortened; post-mandibular proportions as in P. danovi.”
(1931, p. 6.)
Holotype. — "Amer. Mus. 26200 (Field No. 812). Lower jaw, back part weathered
out; grinding teeth missing. A large fully adult individual” (op. cit.).
Horizon. — Tung Gur Pliocene.
Locality. — Tairum Nor Basin, Mongolia.
Genus TETRALOPHODON Falconer & Cautley.
1835. Mastodon, Каир, Descr. Oss. Foss. (4), p. 65.
1847. Tetralophodon Falconer & Cautley, Fauna Antiqua Sivalensis, pl. xlii.
1877. Bunolophodon Vacek, Abhand. k.-k. geol. Reichs., VII, Heft 4, p. 45.
1884. Tetrabelodon Cope, Proc. Amer. Phil. Soc., XXII, p. 5.
1886. Mastodon, Lydekker, Cat. Foss. Mam. Brit Mus., IV, p. 61.
Diagnosis. — ’’Dent, molar. 3, utrinque intermediorum coronis colliculis 4 (raro5).”
(Falconer, 1857, p. 316.)
This definition included ”Mastodon sivalensis", now referred to Pentalophodon
as the genotype. The diagnosis of Tetralophodon may be rewritten thus, —
Longirostrine, bunolophodont, angusticoronate mastodonts with four transverse
ridges in the intermediate molars. Cusps not alternating.
Genotype. — Mastodon longirostris Каир (Hay, 1923, p. 111).
Remarks. — Hay, in the* paper previously quoted, does not recognise Tetra-
lophodon as being anything more than a synonym of Gomphotherium. It has already
5—429921 A. Tindell Hopwood.
(1П) 34
Palceontologia Sinica
Ser. C.
been shown when discussing the validity of Trilophodon that Gomphotherium is
inadequately diagnosed, and that it cannot be adopted for that reason. Another objec-
tion to the use of that genus is that to do so would be to ignore the principle whereby
a genus ccelebs has no status. If Opinion 46 is generally adopted, and genotypes are
selected in order to validate genera to which the original author failed to allocate spe-
cies, all the systematic work of specialists for the last thirty years, and more, will have
to be revised so as to allow of the resuscitation of genera which they had discarded
on the grounds that no species were quoted. This would make confusion worse con-
founded, and would postpone the desired finality in Zoological Nomenclature to the
Greek Kalends. It is probably wiser, therefore, to ignore Opinion 46, and to continue
to sink such genera as hitherto. Tetralophodon is not a synonym of Gomphotherium,
but the first-named sub-genus which contains the four-ridged, bunolophodont mastodons,
and therefore the proper name to be employed. In this memoir it is raised to full
generic rank.
The early history of this genus is still obscure. Osborn (1924 c, p. 2) suggests
that his sub-genus Lydekkeria may be an ancestral form, but this is definitely based
on a misinterpretation of the type species M. falconeri Lydekker. Osborn gives the
horizon as Chinji, which he correlates with the Miocene of Saint Gaudens, France.
There is no evidence for this statement. Lydekker (1877, 1880, 1885) is never more
precise concerning the locality than, ’’the Potwar district, Punjab”. Pilgrim (1910)
gives the locality as Hasnot. The specimen was collected by Theobald in the years
just before 1877. From these three facts it is quite certain that the holotype of M.
falconeri did not come from the Chinji beds, that it probably was found in the Dhok
Pathan, and that it is certainly not earlier than the Pontian of Europe.
Apart from the uncertainty about M. falconeri, there is another reason why it
cannot be regarded as ancestral to Tetralophodon, namely that a mastodont with four
complete ridges in the lower molars is found in the Chinji beds. This proves that
four-ridged forms were already in existence in the Upper Vindobonian, and that even
if Osborn were correct in his dating of the genotype of Lyddekeria it would be con-
temporary with species in which the lower "intermediate molars” had four transverse ridges.
Actually none of the Iridian species was concerned in the evolution of the Tetra-
lophodon of Europe and China, for the area to the South of the Himalayas was isolated
from the beginning of the Miocene to the very top of the Pliocene, and formed a
local centre of evolution unaffected by outside influences. The reason that the same,
or very closely related, genera are found to the North and South of the Himalayas is
that in pre-Miocene times migration was easier, and the precursors of those genera
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
35
were able to enter India. When they were cut off from their fellows they continued
to evolve along much the same lines, though with local differences, and our know-
ledge at present is not enough to allow us to separate them from their cousins to the
North. Hence species which probably belong to different genera are referred to the
same genus, and a source of difficulty arises which can only be removed with the
advance of knowledge.
TETRALOPHODON EXOLETUS sp. nov.
(Plate VI.)
Diagnosis. — A Tetralophodon of large size with hypsodont, angusticoronate
dentition. Third lower molars with 61/, ridges and talonid, intermediate lower molars
with 4 to 47'2 ridges. Milk dentition with wrinkled enamel, and pronounced cingulum;
permanent teeth with smooth enamel and no cingulum.
Holotype. — A partly erupted and worn right lower third molar with part of
the mandibular ramus still attached.
Paratype. — A broken right mandibular ramus with the worn fourth deciduous
molar, the partly erupted third premolar, the fourth premolar still in its crypt, and the
first molar about to erupt.
Material. — A third upper left deciduous molar, incomplete behind; one lower
second deciduous molar; two lower third deciduous molars; one partly worn left lower
fourth deciduous molar; one left lower third premolar.
Dimensions. —
Upper Dentition. — Length Breadth Height Index
Left Dm3 ? 43 ii 27.5 ii —
Lower Dentition.
Isolated Dm2 27.5 18 19 65.4
Dm3 61 36 ii est. ? 59
Dm3 59 est. 38 ii 27 ii 64.5
Dm4 98 54 iv 38 55
Paratype Dm4 87 52 iv — 57.7
P3 38 30 22.5 79
Isolated P3 39 31.5 22.5 80.8
Paratype P4 — — — ,—
Ml 120? 55? 45? 45.8?
Holotype М3 272(295?) 102 iii 73.5 iv 34.5?
(Ill) 36
Palceontologia Slnica
Ser. C.
The holotype is broken both in front and behind; the first molar of the paratype
is in such a position that the dimensions can only be estimated; the fourth premolar
of the paratype is badly distorted through deposition of crystalline calcite in and about
the partly calcified germ.
Horizon. — Pontian.
Locality. — Shansi, Рао Te Hsien, Chi Chia Kou, Sang Chia Liang Kai, Loc.
43; holotype and paratype. Loc. 110(2) in the Province of Shansi; deciduous den-
tition. Shansi, PaoTe Chou, NE 20 li, P'u Fang Yen, N P/Ji, Ch’i Tsu Kou; isolated Dm4.
Description. — The third left upper milk molar is partly worm; the dentine of
the first two cusps is exposed, and the enamel of the third and fourth cusps is abraded.
The third ridge, consisting of the fifth and sixth cusps, has been broken away. Both
the post-trite cusps are normal to the long axis. The third cusp is slightly oblique,
and, from the position of the dentine lake of the first cusp, it appears that that cusp
is also oblique. It is not rotated to the same extent as the third cusp because of the
greater pressure exercised on the first ridge during the development of the tooth. Each
cusp consists of two cones, the outer is larger than the inner; any indications of finer
divisions of the cusps have been destroyed by wear. There are suggestions of accessory
folds in the enamel of the pretrite cusps, but they are hardly distinguishable from coarse
wrinkles. A strong nodular cingulum surrounds the first ridge on three sides, and
terminates in a cingule at either entrance to the first valley; the post-trite cingule is
stronger than the pretrite.
The second lower milk molar has a high, laterally compressed, crown composed
of two cones side by side, and a talonid of three blunt cones arranged in a semi-
circle. The crown is approximately twice the height of the talonid. In front of the
tooth is a small conule, closely pressed against the pretrite cone, and forming a buttress
to it. Another conule is wedged into the crevice between the postero-internal surfaces
of the two main cones. The median cone of the talonid is smaller than the other
two, which are of about equal size.
The third lower milk molar is represented by two complete teeth and three frag-
ments. It has three ridges, the anterior is the narrowest, and the median the widest.
Each cusp has two cones, and tends to form accessory folds which may be derived
from either cone. The first, third, fifth, and sixth cusps may all be more or less oblique
to the long axis of the tooth. There is a median vallecular conule in the first valley,
and a similar, more compressed element in the narrower second valley. The post-trite
half of the first valley is very wide with a convex floor; all the other valleys are narrow,
and their sides meet at a sharp angle at the bottom.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 37
The anterior and posterior talonids are compressed triangular masses, each made
up of two cones side by side, situated in the centre line. Apart from these there is
neither cingulum, nor cingules. As in the upper tooth, the enamel is everywhere
coarsely sculptured and wrinkled.
Two of these teeth have parts of the anterior ridge of the successional tooth firmly
cemented into the crypts beneath them. To detach these is impossible owing to their
fragile nature, and the hardness of the matrix. In their present condition they do not
yield much information concerning their structure.
The two specimens of the fourth lower milk molar differ in various details; the
isolated specimen is more advanced than that in the paratype jaw. Both have four
ridges with the pretrite cusps oblique to the long axis of the tooth. The cusps are
built up of two cones each. On the pretrite side the accessory structures arise in two
different ways, but always in conjunction with the external pretrite cone. In the first
two valleys are vallecular conules, connected to the external cones of the first and third
cusps by a buttress. In the third and fourth valleys the accessory structures are but-
tresses posterior to the external cones of the fifth and seventh cusps. There is a ten-
dency for the post-trite cusps to form trefoils, but this is achieved by the lateral com-
pression, and production antero-posteriorly, of the inner cones of the cusps; not by
the addition of accessory structures. The talonid forms a half ridge with a posterior
pretrite buttress; it is made up of four cones arranged in two cusps, both oblique to
the long axis of the tooth. There is a strong anterior and labial cingulum, with large
cingules at the pretrite entrances to the first and second valleys.
This description of the fourth milk molar is taken from the isolated, better pre-
served specimen. The tooth in the paratype is somewhat more simple in its construc-
tion, notably in the talonid, which appears to have been a buttress posterior to the
seventh cusp. Similarly, the accessory structures, now worn flat, do not seem to have
been so large, nor were the cingulum and cingules so well developed; moreover, the
tendency for the post-trite cusps to wear with a trefoil pattern is not so marked. Another
difference is in the enamel, which is smooth in the isolated tooth, except on the cin-
gulum where it is rugose, and which appears to have been wrinkled in the paratype.
The two specimens of the lower third premolar are identical in structure. Each
is bilophodont, narrower in front than behind, and well provided with accessory struc-
tures. The post-trite cusps are stouter than the pretrite, which alone have anterior and
posterior buttresses. Cusps 1 and 2 are irregularly triangular, and laterally compressed,
whereas the third and fourth cusps are composed of two cones placed side by side.
The first cusp has a strong anterior fold; its posterior buttress is made of two stout
(Ill) 38
Palceontologia Sinica
Ser. C.
COnulcS. The third CUSp haS anterior and posterior folds, the latter being nOdillar and
continuous with the posterior cingulum. The anterior cingulum is a high compressed
cone situated between the two cusps of the first ridge.
The last lower premolar is unknown. It is in position beneath the fourth milk
molar of the paratype, but calcification of the germ does not seem to have been very
far advanced at the time of death, and subsequent changes due to the deposition of
calcite in the crypt have destroyed the tooth.
The first lower molar is still in the crypt, and, although it is beginning to emerge,
it cannot have pierced the gum at the time the animal died. The tooth consists of
four ridges with a talonid. Each of the post-trite cusps shows a tendency for the outer
cone to divide into two, a tendency which is also shared by the inner cone of the
second cusp. None of these cusps has any accessory structures. All the pretrite cusps
show the same tendency to divide either, or both, cones at the summit. In addition,
the outer cone forms a posterior fold which passes inwards and downwards to the
middle line of the tooth, and which may be divided to form one or more conules
In the first valley there is also a low, stout, vallecular cusp. There is no post-trite
cingulum, nor is there a pretrite cingulum at the hinder end of the tooth; the antero-
pretrite part is concealed under the bone.
The third lower molar, which is incomplete, consists of six ridges and a talonid.
Each cusp is composed of two cones which are nearly equal in size. The most anterior
complete post-trite cusp, the fourth, shows signs of wearing with a trefoil section; this
is not due to accessory structures, but to an antero-posterior extension of the inner
cone. Similar extension of the inner cone is seen in all the post-trite cusps. The
pretrite cusps also wear with a trefoil pattern, but this is achieved differently. The
external lobe is formed by the outer cone of the cusp; the anterior lobe by the inner
cone, which is somewhat in advance of the outer, and, hence, the cusp is oblique
whereas the posterior lobe is formed by a large posterior accessory cone. The talonid
consists of three cones, two pretrite, and one post-trite; its extreme posterior portion
is lost, but there seems to have been a cone on the median lise of the tooth.
The only accessory structures are the posterior cones already mentioned, and they
are formed from a posterior fold on the outer cones of the pretrite side by division.
There are vestiges of cingules on the pretrite side, but none on the other. The an-
terior cingulum, if present, has been lost. There was little or no cement.
Discussion. — Although this species appears to be very close to T. spectabilis,
the resemblances are due to convergent adaptation rather than to actual relationship.
Both species are adapted for dry plains vegetation and hence both have high-crowned
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 39
teeth, with fairly sharp ridges. These resemblances are not of the same importance
as the differences. Thus, the pretrite accessory structures of T. exoletus are formed
from the outer cones of the cusps by division of the posterior fold, but in T. spec-
tabilis they are median and of vallecular origin. Again, the extreme obliquity of the pretrite
cusps, and copious cement, of \he latter, contrast with the slight obliquity, and practical
absence of cement in the former. These factors, especially the differences in the accessory
structures, and the disposition of the cusps, render any close relationship very unlikely.
Of the Indian mastodonts, the most like T. exoletus is a species represented by
a broken tooth (regd. 14746) in the British Museum. The tooth was figured by Fal-
coner & Cautley (F. A. S., pl. iv, figs 4, 4 a) as an upper tooth of Mastodon latidens,
but Lydekker (1886, p. 59) refers to it as a lower tooth of M. perimensis. This
specimen is not so hypsodont as the first molar of the Chinese species, and the ridges
are not so sharp and compressed. On the other hand, the pretrite accessory structures
are formed in exactly the same way in both species. There is no material with which
the holotype may usefully be compared.
T. longirostris is definitely more simple and primitive than T. exoletus. The ridges
of the third molars are lower and fewer, and the valleys wider. In addition, the
accessory structures differ in the two species, and the post-trite trefoils are more
strongly developed in T. exoletus than they are in the majority of the teeth
referred to T. longirostris. Despite these differences, however, there is evidence to
suggest that certain animals which lived in Europe, and whose remains are currently
referred to T. longirostris, were related to, though more primitive than, T. exoletus.
к left third lower molar from Eppelsheim (regd. 19435 b) is especially instruc-
tive. The tooth has four and a half ridges; a cone posterior to the half ridge has
been broken off. The post-trite cusps, notably the sixth and eighth, are of exactly
the same nature as those in the holotype of T. exoletus, the only difference being in
the height. The third and fourth ridges show the same tendency to curve, and the
posterior accessory buttresses on the pretrite side have strong resemblances to those of
the Chinese species. There is even closer agreement between the first molars of the
two species. T. exoletus is the more hypsodont, but the general structure of the tooth
is the same as in T. longirostris. Of all the teeth referred to T. longirostris, however,
those which are nearest to the Chinese specimens are from Maragha, Persia (cf. Schle-
singer, 1917, pl. xiv, fig. 4).
The American species which bear most resemblance to T. exoletus are T. cam-
pester (Cope), T. elegans (Hay), and T. (Morillia) barbouri Osborn. From all these
it is to be separated by its larger size, by its ridge formula, and by the small breadth
(1И) 40 Paloeontologia Sinica Ser. C.
index of the third molar. These differences are common to all three species; others
such as presence or absence of cement, development of the trefoils, and height of the
ridges, are individual.
Thus, T. exoletus has its closest affinities with the species found in the Pontian
of Europe, but little or none with those in the Pliocene of America. It is more advanced
than any of the forms with which it has been compared.
TETRALOPHODON ? SINENSIS (Koken).
1885. Mastodon perimensis var. sinensis Koken, Palceont. Abhand., Ill, 34 (7), pl. xii (vii), fig. 1.
1924. Tetr. (Lydekkeria) perimensis var. sinensis, Osborn, Amer. Mus. Novitates, No. 154, p. 2.
Diagnosis. — After pointing out the characters common to Mastodon
perimensis and his new specimen, Koken (op. cit., pp. 36—37) continues,
"Ausser einigen, vielleicht individuellen Eigenheiten im Detail der Kauflache. die
sich schwer in Worte fassen lassen und die man am besten aus einem Vergleich der
Abbildungen erkennt, unterscheidet sich aber der chinesische Molar durch den starker
entwickelten basalen Theil der Krone, wodurch der Eingang in die Thaler hbher zu
.liegen kommt, durch die noch schiefere und von aussen nach innen concave Abkauungs-
flache und besonders durch die im Verhaltniss zur Breite bedeutendere Lange. Dazu
kommt eine grossere Einfachkeit der Querjoche und geringere Verminderuhg der Breite
nach vorn zu”.
Holotype. — A second right lower molar lacking the last ridge (figd. Koken, loc.
city The legend opposite the plate says in error that the tooth is from the left; its
true position is correctly stated in the text.
Dimensions. — Lengthestimated at 180 mm, actually 151mm; greatest breadth, 68 mm.
Horizon. — Unknown.
Locality. — Yunnan.
Remarks. —Lydekker (1886, p. 57) dissented from Koken's views, and expressed the
opinion that the tooth from China was "probably specifically distinct” from M. perimensis.
Schlosser (1903, p. 47) added nothing to our knowledge, but made the sugges-
tion that the fragmentary upper teeth which he called Mastodon lydekkeri were per-
haps the same species. *
Osborn (loc. cit.) placed the species in a new sub-genus Lydekkeria, of which
Mastodon jalconeri Lydekker is the type. On general morphological grounds he suggested
that it is of Miocene age.
Koken discusses the question of the position of the tooth in the dental series very
carefully, and from his arguments, as well as from the figure, it is clear that the spe-
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 41
cimen came from the lower jaw. The increase in width from before backward is evi-
dence that it is an intermediate molar, and from its size it can only be the second.
The missing posterior portion was probably a large, ridge-like, talonid resembling that
seen in certain teeth of Tetralophodon, longirostris from Eppelsheim.
Without examining the original specimen, or a good cast, it is not feasible to make
any definite statement concerning the relationships of this species, but it seems certain
that it is an advanced member of the T. longirostris group, and that its age is Pliocene.
Family MAST0D0NT1M.
Diagnosis. — Brevirostrine, zygolophodont Proboscidea with or without a premolar
succession; accessory structures absent or rudimentary.
Remarks. — This family includes the two sub-families employed by Osborn which he
terms Mastodontinae and Zygolophodontinae typified byM americanusem^Zygolophodon
borsoni respectively. It is known as far back as the Upper Sarmatian, but before then its
history is blank. Professor Osborn (1925, p. 24) says, " It now appears probable that
the diminutive Palceomastodon.......gave rise to the giant mastodon (Mastodon
americanus) so abundant in the forests of eastern North America..... If this
line of descent is confirmed, it will furnish one of the romances of palaeontology and
of animal migration”.
The discovery in China of teeth indistinguishable from those of the American
Mastodon carries that species back to Pontian times, and specimens of Mastodon from
the Pontian of Pikermi prove the occurrence of the line in Southern Europe at that
date. Even then the premolar succession had been suppressed.
Species such as M. pyrenaicus Lartet from the European Miocene have some
connexion with the later zygolophodont forms, but in what manner has not yet been
determined.
Genus MASTODON Cuvier.
1799. Mammut Blutnenbach, Handb. Naturg., Ed. 6, p. 698.
1817. Mastodon Cuvier, Regne Animal, I, p. 232.
Diagnosis. — Zygodont Proboscidea of large size. Intermediate molars with
three ridges; third molars with four ridges and a heel. Premolar succession normally
suppressed.
Genotype. — Mastodon giganteam Cuvier (op. cit., p. 233), which is the same
6—429921 A. Tindell Hopwood.
(Ill) 42
Pakeontologia Sinica
Ser. C.
as blephas americanus Kerr (1792, p. 11G) and Mammal ohioticmn Blumenbach
(loc. cit.).
Remarks. — To retain Cuvier's generic name instead of Blumenbach's, is to follow
convenience rather than rule. The generic term Mammut and the vernacular Mam-
moth are so much alike that one or other should be suppressed in the interests of
clarity. It is very much easier to discourage the use of an unfamiliar name than a
well-known popular one, so that, whilst acknowledging Blumenbach’s prior claim, I
have given the preference to Cuvier.
The systematic Zoology of the zygodont Proboscidea is very confused, nor has
the creation of several genera without adequate material to indicate their inter-relation-
ships helped to clarify the position. Vacek (1877) used the genus Mastodon strictly
in the Cuvierian sense, and sub-divided it into two groups, Zygolophodon and Bano-
lophodon, including in the former, M. tapiroides Cuv., M. pyrenaicus Lartet, "Form
von Baltavar", ”M. Turicensis von Pikermi”, M. BorsoniHays, and M. ohioticus. Of
these species Matthew (1918, p. 200) selected M. tapiroides as the genotype. Osborn
(1926) has taken M. borsoni as genotype, but under the rules of priority this
selection is inadmissable.
The holotype of M. tapiroides is the hinder part of a tooth figured by Cuvier
(1821, pl. iii, fig. 6). It is said to come from Montabuzard, but this has always been
disputed, and the specimen is now lost. Moreover, there has always been considerable
doubt whether the specimen in question was not a milk-molar of M. angastidens.
Finally, there is no such species as M. tapiroides Cuvier. The phrase in that author
reads, ”je donnerai ........ au second ......... (le nom) de mastodonte. tapiroide”,
that is to say, Cuvier employed a vernacular name. The first mention of M. tapiroi-
des is in Desmarest (1822, p. 386) who latinised Cuvier’s vernacular, and took as his
type the specimen figured by Cuvier. Hence, the genotype of Zygolophodon is M.
tapiroides Desmarest, spec, indet, and thus Zygolophodon remains an indeterminate
genus.
Nevertheless, it is evident,, that this group does contain two genera of which one
typified by M. turicensis has a premolar succession. This is Turicius Osborn (1926).
The other one, which has no premolar succession, includes the zygodont form from
Pikermi, a similar species from Baltavar, and M. borsoni Hays. Until Osborn has
published his revision of the Proboscidea it is as well to follow Vacek, to place them
in one genus, but to term it Mastodon s. s. instead of Zygolophodon.
When teeth only are available for study, the history of this genus seems
to afford a close parallel to that of Dinotherium. So far as can be seen, the only
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea front China
43
changes during the passage of time are an increase of size, with slight modification of
minor details. In each case only one species is well known; al! the others are re-
presented by a few teeth. This makes comparison very difficult, and renders conclu-
sions drawn from a study of the less known species somewhat unreliable.
MASTODON AMERICANS (Kerr).
(Plate VI).
1792. Elephas americanus Kerr, Animal Kingdom, p. 116.
1799. Mammut ohioticwni Blumenbach, Handb. Naturg., Ed. 6, p. 698.
Diagnosis. — A Mastodon of large size which differs from M. borsoni in the
smaller size of the internal cones of the cusps, and in the lesser width of the summits
of unworn ridges.
Material. — A left mandibular ramus with the deciduous incisor, and three deck
duous molars; the crown of Dml is broken off, Dm2 is complete, Dm3 is a part-formed
germ, none of the teeth is fully erupted: five specimens of the lower fourth deciduous
molar: a broken second lower molar': an entire, but partly worn, third lower molar: a
fourth upper deciduous molar just entering wear: an unworn first upper molar: the two
anterior ridges of a third upper molar: numerous fragments and broken teeth, as well
as a malformed, pathological, germ of a deciduous molar?
Dimensions. —
Upper Dentition, Length Breadth Height Index
Left Dm4 77.5 56 iii 35.5 iii 72.3
Ml 97 64 iii 45 ii 66
Lower Dentition.
Left Dm2 26 16 ? 61.5
Dm3 56.5 ? ? ?
Dm4 80 ? ? ?
Dm4 (3?) 69 50 iii 31 iii 72.5
Right 81 54 iii 37 iii 66.6
78 54 iii 36 iii 69.2
70 49 iii ? 70
78 ? ? ?
М2 ? 86 iii 59 iii —
Horizon. — Lower Pliocene, Pontian, specimens from Shansi (Loc. 49) and Shensi
(Loc. 51); Pontian, fragments from Shansi, Hsi Kou fide Teilhard; not known, prob-
ably Pontian, specimen from Chihli (Loc. 47), fide Teilhard.
(Ill) 44
Palceontologia Sinica
Ser. C.
Localities. — Shansi, Рао Те Hsien, Chi Chia Kou, Yang Mu Kou, (Loc. 49);
Shansi, Wu Hsiang Hsien, NW 18 li, Chu Chia Wa, S y, li, Hsi Kou; Shensi, Fit
Ku Hsien, Wu Lan Kou, Pei Hou Kou, (Loc. 51); Chihli, Hsuan Hu Hsien, from
city 40 li, Chao Chia Wan, from village SE, Huang Tui Kou, (Loc. 47).
Description. — The fourth upper deciduous molar has three ridges with a well-
marked basal cingulum on all sides of the tooth. All the cusps are tectiform, com-
pressed, and with sharp summits. The pretrite cusps are separated from the post-trite
by comparatively deep clefts. On the pretrite side, the outer cones are very much
larger than the inner, and are furnished with slight accessory folds before and behind
which pass downwards and inwards from their summits, across the inner cones, to the
centre of the tooth. On the post-trite side there are no folds, and the inner cones
of the cusps are more nearly equal in size to the outer. The enamel is rough and
coarse, and the cingulum is strongly beaded.
The cusps of the first upper molar are less tectiform, and more serrate than those
of the preceding tooth. Each cusp is composed of two cones which may be more or
less divided at the summits. The pretrite cusps have each a very large outer cone
which is never divided at the summit, but their inner cones tend to be produced
transversely, and to have the summit serrate. The post-trite cusps resemble the pre-
trite, though the fourth cusp is composed of four cones, alternately large and small,
with a large one on the labial side.
Each pretrite cusp has anterior and posterior accessory folds. That which is an-
terior to the first cusp arises at the summit, and passes downwards and inwards to-
wards the base of the second cusp; there it merges with the cingulum. The posterior
fold is similar and meets the anterior fold of the third cusp at the bottom of the
valley. The anterior fold of the third cusp starts someway below the summit, forming
a buttress with a rounded mammillate top. The posterior fold is similar to the corre-
sponding fold of the first cusp. Similarly, the anterior and posterior folds of the fifth
cusps agree with the same folds of the third cusp. The post-trite cusps are devoid
of folds, but local thickenings in the enamel replace them to a certain extent.
The cingulum is strongly developed, especially before and behind; its width
may be as much as 10 mm. It is warty and pustulose, but does not show any ten-
dency to form cusps.
The enamel as a whole is coarse and rough, with horizontal striae at the base of
the cusps. It is covered with granular, vermiculate, markings.
The only other specimen from the upper dentition of this species consists of the
anterior two ridges of a large tooth, which, from its size, is regarded as the third
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
45
molar. This piece is broken and weathered. The whole of the enamel from the front
of the first and second cusps has been lost, as well as large pieces from the labial
and lingual sides. The cleft between the cusps is deep and pronounced; it is displaced
from the centre of the tooth towards the labial side. The pretrite cusps are wider than
the post-trite in consequence of this displacement. Each cusp has slight enamel folds;
small cingules occur at the entrances of the valleys; the enamel is covered with coarse
vermiculate markings.
The lower milk incisor is laterally compressed; in cross-section it measures 7x12
mm at the tip. The distal surface of the tip is concave, and the mesial convex. It is
enclosed in a sheath of enamel, which passes under a covering layer of cement at a
distance of 40 mm from the tip. The total length of the tusk is 93 mm.
The anterior milk-molar, the second, has had the crown broken, and all details
of its construction are lost. That part which remains is remarkable for the great size
of the pulp cavity seen in section.
The third milk-molar is bilophodont, with strong anterior and posterior talons.
The ridges are sharp; the cusps are separated by deep clefts; the cones are beaded
at the summit and the ridges are serrated. The inner cones have accessory emanel
folds on the first and third cusps, that posterior to the; first cusp is the strongest. The
folds do not obstruct the valleys. Both the anterior and posterior heels are broad and
shelf-like, with numerous conical beads on their margins. The enamel has a granular
surface, and there is no sign of cement.
Apart from the fact that it is trilophodont, the fourth milk molar differs very little
from the third. Each ridge becomes progressively wider from front to back. The
inner cones of the pretrite cusps are much reduced, and wrapped round by the ena-
mel folds of the outer.
The remainder of the lower molars are all broken, but each has the same cha-
racters'. The pretrite cusps have very weak inner cones which are tending to disappear
and to be replaced in their functions by the accessory folds of the outer cones; the
valleys are open, with occasional cingules at the pretrite entrances, and the posterior
cingula are more or less shelf-like and strongly beaded. The post-trite cusps have
stronger inner cones, each with two or three conules at the summit, and are devoid
of enamel folds. Where the ridges are still unworn, they are narrow at the summit.
Discussion. — The discovery of this species in a deposit which appears to be of Pon-
tian, or at least lower Pliocene, age was wholly unexpected. Its presence raises questions of
evolution and distribution which are not easy to solve, and which require consideration
of much more material than is at present available before they can finally be disposed of.
(Ill) 46
Palceontologla Sinica
Ser. C.
Schlesinger (1922) has given a careful account of ail the reported occurrences of
M. americanus in Europe, and, so far as possible, elucidated the horizons in which
they were found. He has also described a new subspecies M. tapiroides americanus
(op. cit., pp. 87—93) from the Lower Pontian of Hungary which he regards as leading
on to M. americanus prcetypica, a new form from the Hungarian Levantine deposits.
The latter (op. cit., pp. 115—116) is distinguished from the North American animals
by the greater width of the unworn ridges at the summit, and the less specialised
nature of the teeth. Specimens in the British Museum (regd. Geol. Dept. 33590,
33591) prove that M. tapiroides americanus is found in the Lipper Sarmatian deposits1
of St. Gaudens. These teeth, with three full ridges and a well developed, but somewhat
smaller, fourth ridge in the third upper molar, take the line back still further in time.
The specimens from China differ from those found in Hungary. They are not
to be distinguished from teeth found in the Pleistocene of Kentucky, and hence are
referred to the typical M. americanus. On this account I am inclined to regard Eastern
Asia as the centre of dispersal of this species.
The more primitive form appears in Europe in beds which are younger than those
Asiatic beds in which the typical form is found, and thus is to be regarded as a migrant
from the East. In North America M. americanus is not known earlier than the Plei-
stocene, which means that it is an invader from elsewhere. Thus, to East and West
of China there are signs of invasions of a species which is found there at an earlier
date than elsewhere. In other words, the species first arose in, or near, China.
From this it follows that M. tapiroides americanus and M. americanus prcetypica
are not truly ancestral to M. americanus, but represent ancestral forms which had
wandered from their place of origin, and which had, so far as we know at present,
reached the Western limit of their dispersal. Until earlier faunas in Asia are disco-
vered, they are most valuable as morphological stages in the evolution of a species
which, in its final stage, has always attracted attention by its apparent isolation.
MASTODON BORSONI Hays.
(Plate VII.)
1834. Mastodon borsoni Hays, Trans. Amer. Phil. Soc., n. s., IV (3), p. 334.
Diagnosis. — A zygodont mastodont of large size; pretrite cusps of the molar
teeth with unusually strong inner cones; ridge-crests very wide; lateral slopes of the
pretrite cusps steep. ,
Vol. IX. A, Tindell Hopwood: Fossil Proboscidea from China 47
Material. — A left third lower molar.
Лг'шлир/лг) n I Dnn+ln 171 Rrnnrlik Q7 ii I-lnirrhl- ЙО iii It-irlnv О
l^ci i oci ooiui co. buigui л i л , uiuiulu <j i n} iitigiu цц ишк,л uu.u.
Horizon. — Unknown.
Locality. — Shansi, Ho Shun Hsien, SW 120 li, Niu T’sao Kou.
Description. — The tooth is from an adult animal; both the first and second
ridges have the dentine exposed, and the enamel of the third ridge is abraded. It
consists of four ridges and a talonid. Each cusp has the inner cone distinctly marked
off from the outer, and although this is most noticeable on the post-trite side it is
almost equally clear on the pretrite side. The accessory folds in the enamel of the
outer cone of the pretrite cusps are relatively weak: the slope of the labial surface of
the same cones is steep. Anteriorly there is a cingulum which is reduced, and there
is a strong cingule at the pretrite entrance to the first valley. The talonid is stout
and shelf-like: it is composed of four conules with obtuse, flattened tops.
Remarks. — The separation of this species from its congener M. americanus
has always been a matter of some difficulty. So long as M. borsoni was only known
from the Pliocene of Europe, and M. americanus from the Pleistocene of America all
was well; lack of appreciation of the real distinctions was overlooked when it was
possible to separate the two on geographical and stratigraphical grounds. The first
to attempt their separation on purely morphological evidence was Mme. Marie Pavlow
(1894, 1901), who, as a result of her studies on the fossil proboscideans of Russia,
was led to the conclusion that both M. borsoni and M. ohioticus (americanus) were
found in the Pliocene deposits of that country.
Mme Pavlow based her identifications on such features as the height of the crown,
the presence or absence of a talon or of a cingulum, and the depth of the fissure
between the pretrite and post-trite cusps. Unfortunately, all these characters are most
variable in both species and insufficient for use in diagnosis. Other characters were
adopted by Schlesinger (1922, pp. 93—132, etc.) and these seem to enable the distinc-
tion between the unworn teeth of the two species to be made without any great
difficulty. In the case of worn teeth, the task is harder, and increases with the degree
of wear, until in the end they are indistinguishable. The following is quoted from
Schlesinger (op. cit., p. 167).
»B) Jochzahl an M3 4—4x, an M3 4x; Bauverhaltnisse wie bei M. tapiroides nur iiber-
trieben tapiroid, Sperrleisten schwacher, nie hockerShnlich. An den Jochen fallt pratriterseits die
ganz ausserordentlich schwache Entfaltung des Nebenhiigels auf, der meist gar nicht isoliert ist
........ M. americanus Pen.
a) Nebenhiigel noch als sblche kenntlich, Jochgratbreite massig .... f. prcetypica fkhles.
(Ill) 48 Palanotologia Sinica Ser. C.
b) Nebenhugel meist fehlend, zu einer ’Nebenleiste’ verkiimmert, Jochgrate iibermassig
eng f. typica autornm.
C) Charactere wie bei /14. americanus, die Nebenhfigel der pratriten Halften aber ausser-
gewohnlich stark, breit und wohlisoliert, Jochgrate demgemass sehr breit, Neigung der pratriten
Aussenhange der Joche steil ..... M. borsoni Hays.»
The intermediate molars are separable by the same means, but they have only three
ridges each. The main possibility of confusion lies jn the separation of /И. borsoni
and M. a. prcetypica, though the slope of the outer surfaces of the pretrite cusps
should be of importance.
The identification of the tooth under consideration rests on the large size of the
inner cone of the fifth cusp, on its differences from the third lower molar of M. ame-
ricanus figured by Pavlow (1894, pl. i, fig. 3), and its general agreement with the
tooth from Piedmont figured by Gastaldi (1861, pl. vii, figs. 9, 10). Except that it
appears to be more hypsodont, this tooth also displays the same general characters as
those from the Pliocene of France described and figured by Lortetand Chantre (1878,
pl. xi, figs. 1, 3, 4; pl. xii).
Until more is known of the fauna of which the Chinese remains of M borsoni
form part, speculation as to the route by which it migrated is unjustifiable. In Europe
the species is restricted to deposits which are conveniently grouped as being of Levantine
age; that is to say, it is not found before the Middle Pliocene and dies out in the
Upper Pliocene. Even its most easterly localities in Russia appear to be within these
limits although their exact stratigraphical position needs confirmation.
Under this head I am inclined to include the fragments which Schlosser (1903,
p. 45, pl. xiv, figs. 5, 7) described as Mastodon aff. latidens. They are characterised
by a tapiroid structure, lack of cement and accessory conules. In all the essential points
such as breadth, height, separation of the ridges, enamel characters, and structure of
the cusps, there is close agreement between the two specimens. In fact, apart from
individual differences, Schlosser's description could be applied equally well to either
specimen.
MASTODON sp. indet.
(Plate II.)
Material & Localities. — Three groups of bones of the manus and pes as follow,
Locality 44. The distal epiphysis of the right ulna, together with the somewhat
abraded right magnum and unciform.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 49
Locality 49. The anterior half of a much damaged left magnum, also a crushed,
and otherwise damaged, left third metacarpal lacking its distal epiphysis.
Locality 52. A left ectocuneiform.
Horizon. — All these localities are of Pontian age.
Dimensions. —
Ulna. Mm
Antero-posterior diameter of articulation .............................................. 60
Lateral diameter......................................................................... 92
Os magnum.
Greatest depth from back to front..................................................... 82
Anterior width of superior facet........................................................ 45
Posterior width of superior facet ....................................................... 61
Greatest height (posterior) ........................................................... 70
Anterior height ...................................................................... 46
Os unciforme.
Greatest depth from back to front....................................................... 80
Greatest width from side to side ....................................................... 90
Greatest height (posterior) ............................................................ 75
Anterior height (internal surface) ..................................................... 48
Os ectocuneiforme.
Greatest depth from back to front....................................................... 70
Greatest width from side to side ....................................................... 66
Greatest height (anterior).............................................................. 38
Metacarpal III.
Greatest depth of proximal epiphysis ................................................... 94
Greatest width of proximal epiphysis ................................................... 69
Width of facet for magnum (anterior) ................................................... 44
Width of facet for unciform (centre).................................................... 32
Description. — All the bones are more or less crushed, and their margins have
been abraded to a greater or less extent, with the consequence that many of the more
interesting features have been obliterated, arid the measurements can only be regarded
as approximate.
Os magnum. — The fragment from Locality 49 is almost twice the size of the
complete bone from Locality 44. Their anterior widths are in the ratio 7:4, and the
latter figure takes no account of the fractured inner surface of the larger bone. This
piece has a concave upper surface. The superior facet for the unciform is very large,
plane, and practically at right angles to the proximal articulating surface. The inferior
unciform facet is narrow and band-like. Of the metacarpal facets, that for the third
7-429921 A. Tindell Hopwood.
(Ill) 50
Palceontologia Sinica
Ser. C.
metacarpal is concave from side to side, whereas the other, for the second metacarpal
is convex in the same direction.
The entire bone from Locality 44 is remarkable among bones of this type because
of its great posterior height compared with its depth from back to front. The lunar
facet is convex posteriorly, and very slightly concave anteriorly; its inner margin is
gently sinuous, and there is a tendency towards concavity of the outer margin. The
facet for the trapezoid on the upper half of the inner surface is much narrower in the
centre than it is at either end; it is somewhat convex behind, and concave in front.
The inferior facet for the same bone has disappeared through abrasion. On the outer
surface, the cuneiform facet is much higher behind than in front, and is confined to
the upper portion of the bone. The facets on the distal surface for the second and
third metacarpals are separated by an angle which crosses the surface obliquely. This
has the effect of excluding the facet for the second metacarpal from the anterior margin,
and of rendering it triangular with its base at the back of the bone. This facet is
very much on the side of the bone; it faces inwards, and but slightly downwards,
and is wholly visible when the bone is looked at from the side.
Os unciforme. — The unciform from Locality 44 is probably part of the same
carpus as the magnum just described. It has the general transverse wedge shape
so commonly found among the Proboscidea, which is due to the convex upper,
surface curving downwards to meet the more grossly convex under surface. The
cuneiform surface is gently concave from back to front in its anterior portion, and con-
vex in the same direction behind. The under, or distal, surface is composed of three
facets inclined to each other at obtuse angles. Of these, the inner, which articulates
with the third metacarpal is the smallest. The next, for the fourth metacarpal is tri-
angular, with the apex behind, and almost the entire anterior margin for base; it is
gently concave. The facet for the fifth metacarpal is irregular, triangular, rounded, and
somewhat less in area than that for the fourth. It is gently concave antero-posteriorly,
and faces downwards, outwards, and slightly backwards. The facet on the inner sur-
face, which articulates with the magnum, is exactly the same shape as the correspond-
ing facet in the magnum from the same locality, with the exception that it is the
mirror image of that facet. It is restricted to the upper part of the inner surface of
the unciform.
Ulna. — The distal epiphysis of the ulna is crushed and distorted. Its articu-
lating surface is convex antero-posteriorly, and transversely it is convex at either side,
with a concave depression in the centre. In its total characters it agrees with the distal .
articulating surfaces of the same bone obtained from the Siwaliks and elsewhere.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 51
Metacarpale III. — The proximal end of the third metacarpal bears fourfacets, two
superior, and two lateral. The former are for the magnum and unciform, whereas the
latter are for the second and fourth metacarpals. The facet for the second metacarpal
has parallel sides, its length (56 mm) is about two and a half times its height (23 mm);
in shape it is a parallelogram with the upper side displaced to the rear. The facet for
the magnum is the largest of the four; it is convex from back to front, gently con-
cave from side to side, and tapers slightly as it passes from before backwards. A
similar description applies to the unciform facet, but its breadth (31 mm) is less than
that of the facet for the magnum (46 mm). In its outline, the facet for the fourth
metacarpal is an irregular ellipse with its long axis (65 mm) parallel to the long axis
of the head of the metacarpal; it is wide (30 mm) and faces outwards and slightly
downwards.
Os ectocuneiforme. — The ectocuneiform is a low, square, bone, in which the
area of the inferior surface is greater than that of the superior surface, and hence the
bone has the form of a truncated pyramid. The proximal facet, which articulates with
the navicular, is flat and narrower behind than7 in front. The distal facet is also flat
transversely in both parts, but has a slight antero-posterior concavity. The facet for the
third metatarsal is approximately four times as wide as that for the fourth; it is parallel
to the proximal surface, whereas the facet for the fourth metatarsal, which faces down-
wards and slightly outwards, is inclined to the proximal and distal surfaces at a small angle.
On the inner surface there are two facets for articulation with the mesocuneiform.
They are long and narrow, one superior and one inferior, and are separated by a deep
rugose groove. The outer surface bears on its inferior margin a long facet for the
cuboid. This facet occupies about half the height of the face; the other half is taken
up by a deep groove with rugose sides and floor.
Remarks. — All these specimens were found in localities, which from their fauna
are certainly of Pontain age. The localities are in the neighbourhood of Pao-Te-Hsien,
and that district has yielded Tetralophodon exoletus as well as Mastodon americanus.
Most of the bones are smaller than those of M. americanus preserved in the Geological
Department of the British Museum; on that account they might be referred to T.
exoletus, but the presence of the two species in the district enjoins caution, and it
is preferable to determine the bones as Mastodon sens. lat. without being more precise.
The only bone of M. americanus available for comparison is the unciform, of which
two isolated specimens, one much larger than the other, are in the collections of the
British Museum. The other bones of this species are either in a mounted skeleton, or
else, as in the case of the ulna, they have lost the epiphysis necessary for comparison.
(Ill) 52 Palazontologia Sinica Ser. C.
(~''1 nrvinnpjnz.n l-inr- л1пл клля A J n „-,’41-. J". 41л ГЧ- k/M-iAr. L * u л rt л 1 л л л 1 1 J-* л ГЛ АП .vmll лп
uuiupaiDUii нас aidu uccu HldUC Willi ULiici uuuco UUU1 VJllllCdt lucdlUitd, ad well dd
with remains of T. angustidens, and various other bones from different localities.
The large fragment of a magnum from Locality 49 differs from that from Locality
44, not only in its size, which is greater, but also in the fact that both facets on the
under surface share in the formation of the antero-inferior margin of the bone, whereas
in the latter specimen the facet for the second metacarpal is excluded from that margin.
In this respect the former specimen agrees with that from Locality 77, described below
but in the characters of the facet for the third metacarpal it differs from the bones
from Localities 44 and 77. In those two bones the facet is gently convex from side
to side, but in the bone from Locality 49 the same facet is concave from side to side.
Compared with the magnum from Locality 77, the bone from Locality 49 is
slightly wider in proportion to its length from back to front. The difference is not
very great, approximately three per cent, but it is very distinct when the two bones
are placed side by side. It is caused by the different proportion of the proximal
facet which is occupied by the posterior, wider, convex part. In this respect, as in
many others, the bone from Locality 77 seems to be the more advanced; it owes its
greater antero-posterior depth to the way in which the antero-externai angle is produced
forwards. The trapezoidal, and unciform facets of the smaller bone are more simple
than they are in the bone from Locality 77. For example, the trapezoidal facet is
gently sinuous, but its anterior portion is not deeply excavate, neither does it come
into contact with the metacarpal facet, nor is the pit for the interosseous ligament so
restricted in area and so deep as in the latter bone. Similarly there is practically no
inferior unciform facet, and the heights of the anterior and posterior moieties of the
superior unciform facet are much more unequal than in that bone.
When this small magnum is compared with a very large bone from Pikermi (regd.
M10122), it is seen to be narrower, to have a more sinuous trapezoidal margin, and
to be lower in proportion to its depth from back to front. It also has less complicated
facets on the outer and surfaces, but the metacarpal facets in both specimens are alike
in that that for the third nearly (M10122), or entirely (Loc. 49), excludes that for the
second metacarpal from the antero-inferior margin:
The unciform from Locality 44 has been compared with the left unciform of
Trilophodon wimani described above (p. 25), a left unciform of Trilophodon angus-
tidens from Sansan (regd. 33592 b), and two unciforms of Mastodon americanus from
Missouri (regd. 17211, 17193) both of them from the left carpus.
This specimen is relatively deeper from back to front than that of T. wimani, and
its posterior height is greater; correlated with these differences is the shape of the facet
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
53
for the magnum, which is higher and shorter than in T. wimani. Another difference
tvllirh nricno frnm Итпсо итппсг nmnnrl-innr io in oncrln knlnrnnn Ilin ntTwimal Und
,»nuvu U11UVU 11U111 U1VJV IIVIIO 10 ill Uli'' UVLVVWll 111V vauu
distal surfaces of the bones; this angle is greater in the specimen from Locality 44,
and since the pit for the interosseous ligament is elongate, and approximately parallel
to the distal margin of the inner surface, it follows that its direction is more oblique
in that bone than it is in the corresponding bone of T. wimani; it is also deeper.
The central part of the distal surface, the facet for the fourth metacarpal, is the more
concave in T. wimani. Damage to one or other of the bones prevents further
comparisons.
The posterior margin of the distal surface of the unciform of T. angustidens is
broken, otherwise it is perfect. It is relatively deeper from back to front than the
specimen from Locality 44, from which it also differs in that the proximal facet is not
so extended laterally, and hence there is a roughened area interposed between the
proximal and distal surfaces. On the internal surface, th,e bone from China has the
deeper pit for the interosseous ligament, and the limits of the pit are more sharply
defined; furthermore, the long axis of the pit is straight, and not concavo-convex as
it is in T. angustidens. Another difference observeable on the,inner surface is the
greater height of the superior magnum facet in proportion to its antero-posterier dia-
meter, on the bone from Locality 44, when compared with the corresponding facet in
T. angustidens. The inferior facet in the unciform of the latter is the more concave.
The facets for the fourth and fifth metacarpals correspond fairly well in both specimens
Viewed from the front, the two bones are seen to differ considerably. Whereas that
from Locality 44 is plane, or gently concave, from above downwards and has the
antero-internal margin almost perpendicular to the superior and inferior margins, the
unciform of T. angustidens has the anterior surface divided into two by a ridge parallel
to the inferior margin; each portion of the face above, and below the ridge is con-
cave. In addition, the antero-internal margin is very oblique; the angle between it and
the superior margin is approximately 60°.
The two specimens of the left unciform of Mastodon americanus form part of
the series of remains purchased from Dr. Albert Koch by the Trustees of the British
Museum in 1844. They differ considerably in size, and the larger one (regd. 17193)
is better preserved than the smaller (regd. 17258) in which the angles between the
various surfaces are badly abraded. The specimen from China is much smaller than
either: its depth back to front is 89% of its'maximum width, against 85 % in 17193
and 95 % in 17258, but whereas its depth is intermediate between the two American
specimens its height is less than either. Expressed in terms of the depth, the height
(Ill) 54
Palceontologia Sinlca
Ser. C.
of the Chinese specimen is 94 %, that of 17193 is 100 %, and that of 17258 is 107 %
None of these variations is big enough to affect the characters of the individual faces
of the three bones at all seriously. The proximal facet of the bone from Locality 44
agrees with the corresponding facet of 17193 better than it does with that of 17258
in its lateral sweep. This is because the proximal surface of 17258 is separated from
the distal surface by a roughened area similar to that in the unciform of Trilophodon
angustidens, whereas in the other two bones these surfaces are in contact. On the
anterior surface, the agreement is with 17258 rather than with 17193 in which the
maximum height is proportionally greater, the angle marking the junction of the anterior
margins of the facets for the third and fourth metacarpals, is more pronounced, and
the centre of the inferior margin more concave. When the interior, distal, surface is
examined, there are points of agreement now with one, now with the other, of the
American specimens, and sometimes differences from both. The facet for the second
metacarpal looks inward more than it does in 17193, and resembles 17258 in this
respect. The facet for the third metacarpal, which is gently concave, also resembles
that of 17258 rather than the deeply concave facet of 17193. On the other hand, the
sub-triangular facet for the fifth metacarpal is more like that of 17193 in shape, but
it differs from both 17193 and 17258 in its position, which is more downwards, and
not so much outwards. The greatest differences are between the three inner surfaces,
although in outline they all are of much the same shape. In the sum of characters the
inner face of the bone from Locality 44 Is intermediate between 17258 and 17193.
In the former, the superior facet for the magnum is very narrow in front when
compared with the bone from China, and still more so when compared with 17193.
This bone is also peculiar in the very shallow depression which takes the place of the
pit for the interosseous ligament, a pit which is elongate and oblique in the- Chinese
specimen, and deep and semicircular in 17193. The pit in the last specimen has its
superior and anterior walls .formed by an articulation which probably accommodated a
ateral peg from the magnum similar to that described below on an unidentified trape-
zoid from Locality 77 (p. 66).
The third metacarpal has been compared with the specimen from Locality 77 des-
cribed below (p. 67) and an imperfect specimen of the same bone of Mastodon ameri-
canus from Big Bone Lick, Kentucky (regd. M165). All three are from the left manus.
In comparison with the bone from Locality 77, that from Locality 49 is shorter
from back to front and wider from side to side. The facet for the second metacarpal
is wider from above downwards in proportion to its depth from back to front, and
has its superior and inferior margins parallel instead of convergent posteriorly. Both
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 55
bones have the facet for the magnum narrowing as it passes from front to back, but
tU _______x x--1—„ xt,„ 1___________ _c г___i:x.. ЛЛ ti__ ___ _1--x -----11__
LUC dlilUUllL U1 Ldpct 15 1C55 Ill lilt UU11C IfUlll bULdHLy 1 liey ШС dUUUL cquttliy
convex from back to front, but the concavity from side to side is greater in the
bone from Locality 77. Similar remarks apply to the facets for the unciform. The
facets for the fourth metacarpal bear the same relationships to each other as those for
the second metacarpal, that is to say, the facet on the bone from Locality 49 is higher
in proportion to its depth than that on the bone from Locality 77, and has its superior
and inferior margins more nearly parallel.
So far as the damaged state of the metacarpal of /14. americanus allows com-
parisons to be made, it appears to be a large edition of the bone from Locality 49.
The most noteworthy differences are in the lateral convexity of the facet for the magnum,
and the lateral concavity of that for the unciform. The facet for the fourth metacarpal
does not face downwards to the same extent as that in the bone from Locality 49,
but since that bone is crushed it is not altogether certain that the facet in question
is in its true position.
The best figure of an ectocuneiform is that given by Dietrich in his account of
Elephas antiquus recki (1916, pl. vii, fig. 7}. No actual bones were available for
comparison; the only fossil specimens in the British Museum are those of the Upnor
Elephant, and they are mounted in the skeleton. Andrews figures (1928, pl. viii, fig.
3 A) the left ectocuneiform from below. Both figures show a bone which is longer
from back to front than the Chinese specimen, and which have their facets differently
arranged. Thus Dietrich figures two facets for the cuboid compared with one in the
present specimen, and Andrews indicates that the facet for the fourth metacarpal has
about half the antero-posterior depth of that for the third, whereas in the specimen
from Locality 52 it is about three quarters the depth. The general tendency of the
cuneiform bones in the proboscidean tarsus seems to be one of reduction, but the
evidence is scattered and not very definite.
Family D1BUN0D0NT1D/E.
Diagnosis. — Brevirostrine, bunolophodont, Proboscidea in which the petrite and
post-trite cusps are arranged in alternating series. Intermediate molars with four or
five transverse ridges in the later forms, and three ridges in the earlier ones.
Remarks. — The two best-known genera in this family are Pentalophodon and
Dibunodon typified by Mastodon sivalensis Falconer & Cautley, and /14. arvernensis
Croizet & Jobert respectively. Another genus is Synconolophus Osborn. This last is
(HI) 56
Palceontologia Sinica
Ser. C.
unique, for, although it has only been recognised of recent years, its evolutionary
history has been traced from the Kamlial (Helvetian) of India to the Dhok Pathan
(Pontian). The earliest stage is the species described and figured by Lydekker (1884,
pp. 19—29, pl. iv, figs. 3, 8; pl. v, figs 1, la) as Mastodon angustidens var. palce-
indica and the genus culminates in Synconolophns corrugatus (Pilgrim) from the Attock
District of the Punjab. Professor Osborn does not agree as to the earliest term of the
series, but careful analysis of numerous teeth proves it to be correctly identified.
The importance of this series lies in the fact that it indicates the type of tooth
to be looked for when trying to find an ancestor of the outher genera. It also sug-
gests that even so far back as the Helvetian the main features of the later genera
were being laid down.
Genus PENTALOPHODON Falconer.
1857. Pentalophodon Falconer, Quart. Jour. Geol. Soc. XIII, p. 314. Genus caelebs.
1865. Pentalophodon Falconer, Quart. Jour. Geol. Soc., XXI, p. 262.
Diagnosis. — Bunolophodont Proboscidea in which there are four to five ridges
in the intermediate molars; petrite and post-trite cusps arranged in alternating series.
Genotype. — Mastodon sivalensis Cautley 1836, p. 294.
Remarks. — This name was originally used by Falconer as indicating a theore-
tical stage in the evolution of the Proboscidea from Mastodon to Stegodon. He later
decided that M. sivalensis came very close to this theoretical stage, and placed it
under the sub-generic heading Pentalophodon. After that all specimens from India
with any tendency to five ridges in the intermediate molars, and with alternating cusps,
were usually grouped with M. (Pentalophodon) sivalensis. This led to confusion
when dealing with a long series of teeth, for it is now evident that there was more
than one species involved, and that not all the intermediate molars have five ridges.
If, however, Falconer’s original series of specimens, now in the Geological Departe-
ment of the British Museum, be examined, it will be found that, with the notable
exception of one skull (regd. M2877, figd. Falc. & Cant, 1847, pls. xxxii, xxxiii) they
all show the same characters, and that the specimens which Falconer & Cautley figured
in the ’’Fauna Antiqua Sivalensis” as Mastodon sivalensis all belong to one species.
The confusion found in the works of later authors is largely due to the misinterpreta-
tion of Falconer’s work by Lydekker (cf. Lydekker, 1880), who included under this
species other forms which Professor Osborn has separated as a distinct genus, Syn-
conolophus (Osborn, 1929, p. 9).
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China oT
Cautley's best specimen (Cautley, 1836, pl. xi, figs 2, 3), appears to have been
lost many years ago. It is not mentioned by Falconer or Lydekker, nor is it figured
in the ’Fauna Antiqua Sivalensis", so that we are deprived of the valuable informa-
tion which would be afforded by a re-examination of the fossil itself, and are forced
back upon the very crude figure which accompanies the paper. There does not seem
to be any doubt, so far as the figure is reliable, that specimens such as that registered
M. 2874 Brit. Mus. Geol. Dept. (Falc. & Caul., 1847, pl. xxxvii, figs, 8, 8 a) are
correctly referred, and that from them we may learn all the characters of the species.
This is important, for the following specimen from Shansi has only four ridges id
the intermediate molars, yet the third upper molar differs but little from that of P.
sivalensis, and it is because of the agreement between the third upper molars that the
specimen is referred to Pentalophodon.
PENTALOPHODON SINENSIS sp. nov.
(Plate VII).
Diagnosis. — A Pentalophodon which is more primitive than the genotype. Inter-
mediate molars with four ridges and a strong talon. Anterior buttress of third upper
molar stronger, accessory columns not so highly developed.
Holotype. — The left second and third upper molars of one individual. The
second molar is extensively worn, and wear has just exposed the dentine of the first
ridge of the third molar.
Material. — In addition to the holotype, a left lower intermediate molar; either
the fourth deciduous molar, or else the first permanent tooth, very worn.
Dimensions. —
Upper Dentition. Length. Breadth. Height. Index.
Holotype М2 146 83 iii ? 57
М3 216 92 iii 66 iii 42,7
Lower Dentition.
Ml 114 63 iv 55
Horizon. — Unknown: Pliocene?
Localities. — Shansi, Yu She Hsien, NW 20 li, San Chuang, NE 1 li.........locality of
holotype; Shansi, Yu She Hsien, NW 20 li, Ni Ho Chang, E 1 li, Kuan Tao Kou.
Description. — The best preserved tooth is the third molar of the holotype. This
has six ridges which gradually decrease in height and complexity from before back-
wards. All the pretrite cusps, except the eleventh, are made up of two sub-equal cones;
8—429921 A. Tindell Hopwood.
(Ill) 58
Palaontologia Sinica
Ser. C.
the eleventh alone has three. Each of them, except the first, is slightly oblique to the
long axis of the tooth, and has its inner cone set in advance of the inner cone of
the post-trite cusp. In the first cusp, the. inner cone is decidedly smaller than the outer,
and is placed slightly to the rear of it; hence, the first cusp is oblique to the long
axis, but in the inverse sense to all the other pretrite cusps. There is an alternative
interpretation, namely, that , the first cusp is oblique in the same manner as the other
pretrite cusps, and that the cone which is here regarded as the inner cone of the cusp
is really an accessory column situated postero-internal to the outer cone of the cusp.
I do not think this can be correct, because the cone in question is so closely applied to
the outer cone of the cusp, and their general relations are exactly the same as those between
all the other inner and outer cones of the pretrite cusps, apart from their relative positions.
The post-trite cusps are not so regular in their construction as the pretrite. The
second cusp has both cones of about the same size; the fourth, sixth, and eighth
have the outer cone rather larger than the inner, and showing a groove at the summit;
he tenth cusp is one large, undivided cone; and the twelfth, two cones of about the
same size. All the post-trite cusps, except the second, are normal to the long axis of the
tooth. The second cusp is oblique, and has its outer cone in advance of the inner.
In this instance there is no alternative explanation.
The anterior cingulum is relatively stout and nodular on the pretrite side of the
tooth. It receives a very large, stout, columnar buttress, which is anterior to the first
cusp and accessory to its outer cone. That part of the buttress which is next the cone
has the appearance of being a large pillar, which was divided at the summit. As
pointed out above, it would be possible to interpret this element as the inner cone of
the first cusp. The other accessory structures are few in number. There are two
cones postero-accessory to the second cusp. They give it a concave posterior surface,
such as is frequently seen in the second cusp of many other primitive proboscideans.
The inner of these accessory cones has every appearance of being an independent
structure of vallecular origin, but the outer has developed through the backward ex-
pansion of the outer cone of the cusp, with which it is fused in its lower part, and
from which it is separated by a slight groove at the summit. A cone antero-internal
to the third cusp is in contact with the postero-internal accessory column of the second
cusp. There is also a column situated betwen the inner cones of the fourth and fifth
cusps; a backward extension, and tendency to divide at the summit of the inner cone
of the sixth cusp, seems to show that this column is accessory to the fourth cusp.
There are no other accessory cones or columns. Between the eleventh and twelfth
cusps is a small, stout, cone which forms the talon.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 59
Most of the details of the. construction of the second molar have been destroyed;
all that remains is the ground plan of the last two ridges. Those in front have dis-
appeared almost completely. The section of the fifth cusp suggests that it was made
of two cones without any accessory structures whereas the trefoil section of the seventh
cusp with the relative sizes of the lobes, indicates the presence of two cones, with a
third postero-internal accessoiy to them. Both the sixth and eighth cusps had the
usual two cones without any accessory structure. The talon was strong and nodular,
but it did not form an incipient ridge, and cannot have been separated from the fourth
ridge by a valley.
The skull of P. sinensis was shortened from back to front. This is shown by
the manner in which the occlusal surfaces of the upper teeth are inclined to one another.
The only specimen of a lower tooth in the collection is either a fourth deciduous
molar, or else the first of the true molars, probably the latter. It is well preserved,
but so worn that only the cross-sections of the cusps at the extreme base are
visible. Each of the petrite cusps is oblique, and each has the outer cone slightly
behind the inner, so that they all slope the same way. The post-trite cusps are all
normal to the long axis.
The accessory structures are represented by lobes in the enamel figures. There
is one lobe, almost circular in outline, behind the inner cone of the first cusp; another,
which is compressed obliquely, is behind the corresponding cone of the third cusp.
In a similar manner there is another bay behind the inner cone of the second cusp.
The seventh cusp is prolonged across the tooth behind the eighth cusp, showing that there
was a strong posterior buttress, or talonid. From the shape of these bays there can
be little doubt that the column behind the first cusp was of vallecular origin. Those
behind the second and third cusps, on the contrary, were probably part of the inner
cones of the cusps and derived from them by budding. The enamel was smooth on
the lingual and labial surfaces of the tooth, but where it borders the transverse valleys
it is more or less festooned. This is best seen on the post-trite side of the tooth.
Discussion. — The specimens of P. sivalensis used for comparison with this
species were a third left upper molar (M 2857) and a second right upper molar (40776).
These were chosen as being the best preserved specimens available; the third molar
is figured in the ’’Frjuna Antiqua Sivalensis”, pl. xxxvi, figs 6, 6 a.
When the two species are placed side by side, it is evident that P. sivalensis
has the shorter tooth (194 mm), and since it has a ridge formula of 61/, that the
ridges are closer together than they are in P. sinensis. It has the accessory structures
behind the inner cones of the post-trite better developed too. There is a distinct
(1П) 60
Palceontologia Sinica
Ser. C.
column behind the sixth cusp, and one beginning to bud off from the eighth. That
is, the budding off of accessory structures has proceeded back one ridge farther in
P. sivalensis than in P. sinensis. Another sign of progress is seen in the talon.
This consists of three cones, instead of the single cone flanked by slight swellings of
P. sinensis. On the other hand, the anterior buttress, even allowing for wear and
damage due to the development of a large pressure mark, was definitely smaller.. Other
differences are to be seen in the greater obliquity of the pretrite cusps, and the deve-
lopment of small cingules at the entrance of the valleys of P. sivalensis. The presence
of a small amount of cement in P. sinensis, and its apparent absence from P. sivalensis
is not of any great significance, for experience shows how easily mastodon teeth lose
this tissue either by weathering or through some other agency.
The second upper molar of P. sinensis is so worn that comparisons between it
and the corresponding tooth of P. sivalensis have very little value. It is evident
however, that the talon was only a posterior buttress and not. a fifth ridge similar to
that seen in the specimen of P. sivalensis regd. 40776. Another specimen from India
(16363) which is slightly more worn than the type of P. sinensis, and which formed
part of Cautley’s original material (Cautley, 1836, pl. xi, fig. 11) shows quite definitely
that it had five ridges, and that the pretrite cusp of the fifth ridge was oblique to the
long axis of the tooth. Behind this fifth ridge are traces of a small talon.
The lower tooth of P. sinensis is hardly to be distinguished from that of P.
sivalensis except by means of its lower ridge formula. That there are no other spe-
cimens in the collection is a pity, for comparison of the unworn teeth would yield
interesting differences in various points detail. Such would be a comparison of the
structure of the pretrite cones, or buttresses, accessory to the main cusp.
Since all previous authors have emphasised the close connexion between P. siva-
lensis and Dibanodon arvernensis, it is important to make a similar comparison
between that species and P. sinensis. Provided that they are examined with due care,
here is no possibilty of confusing the two. Apart from the fact that P. sinensis is
larger than D. arvernensis, and has more ridges in the third upper molar, they may
be separated by the differences in the pretrite accessory columns. Whether the spe-
cimens of D. arvernensis are from England (teeth regd. 28997, M 1983, M 7596,
M 9138, M 13713), France (Lortet & Chantre, 1878, pls. i-vi), Italy (Weithofer, 1890),
Austria-Hungary (Schlesinger. 1917, 1922), or elsewhere in Europe, the upper molars
alb have accessory conules of vallecular origin between each of the pretrite cusps, and
it is largely due to the manner in which these columns join up with cusps during
wear that the teeth appear to have the pretrite cusps alternating with the post-trite.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 61
Actually the dislocation of the two cusps in any one ridge is smaller than it is in
Pentalophodon.
In P. sinensis, on the other hand, the accessory columns are practically limited
to the first four cusps of the third upper molar, and the alternation of the cusps is
due to a definite dislocation of the pretrite and post-trite halves of the ridge, whereby
the pretrite half has its inner cone in front of the inner cone of the post-trite half.
There are certain teeth of D. arvernensis which show this feature as, for example, a
left first upper molar from the Red Crag of England (regd. M9139), but such teeth
are not common.
Lower teeth of the two species in the same state of wear are practically indistin-
guishable; such differences as are observable might quite well be regarded as individual
only, and until better specimens of P. sinensis are available nothing is gained by lis-
ting such differences.
The actual relationship between Dibunodon and Pentalophodon is very close indeed.
The lower teeth of the various species from Europe and India differ only in their ridge
formulae; their accessory structures, to which I attach great weight and importance, have
the same characters, and arise in the same way. Teeth from the upper jaw, however,
differ in just the same characters, and further work is needed to solve the problem of
whether the vallecular origin ascribed to the accessory columns of Dibunodon is correct.
This is further discussed in the memoir on certain Indian mastodonts previously men-
tioned.
? PENTALOPHODON SINENSIS.
(Plates II, III.)
Material. — A left cuneiform and right second metacarpal.
Locality. — Shansi, Yu She Hsien, NW 20 li, Ni Ho Chang, E 1 li, Kuan
Tao Kou.
Dimensions. —
Os cunei forme. mm
Greatest depth from back to front ................................................... 150
Greatest width from side to side..................................................... 231
Greatest height (antero-external) ...................................................... 93
Width of ulnar facet ................................................................ 159
Depth of ulnar facet ............................................................... 121
(Ill) 62
Palceontologia Sinica
Ser. C.
ЛЛ nf’n Л ZI 1л 11
JUUHLIW jt/U-tC J J.
Length ................................................................................... 225
Greatest depth of proximal end ........................................................... 129
Greatest width of proximal end exceeds..................................................... 90
Depth of distal end..................................................................... 112
Greatest width of distal end........................................................... 117
Width of distal articular surface ........................................................ 92
Description. — The cuneiform is an oblong bone, deeper than wide, with a
massive, decurved, rounded process attached to its outer surface. The proximal, su-
perior, facet for the ulna occupies the whole of the upper surface. It is convex from
side to side, and concave from before backwards; before the concavity reaches the
anterior margin it passes into a gentle convexity, hence the anterior third of the facet
is convex. Owing to a similar passage from a convex to a concave surface, that part
of the facet which is situated above the external process is gently concave in all di-
rections. The outer half of the posterior margin of this facet is in contact with the
superior margin of the facet for the pisiform. The latter faces directly backwards; it
is widest at its external end. On the under, or distal, surface is a large facet for
articulation with the unciform. This facet is slightly concave from side to side; the
posterior concavity is greater than the anterior. From back to front the facet is gently
concave behind and equally convex in front. The outer side is produced into a long
tongue which is twisted so as to face forwards and slightly downwards. On the inner sur-
face of the bone are two long, narrow facets for articulation with the lunar. They
are placed one above the other, with a deep trough for the interosseous ligaments
between them, and the superior facet is much the narrower.
The second metacarpal is a very massive bone with a sub-triangular head, and
a semi-cylindrical distal extremity. Apart from a large splinter broken away from the
internal corner of the proximal ehd, it is complete.
There are four facets on the head. That for the trapezium is on the internal
lateral surface: it is very small (33x22 mm), situated on the anterior third of the
superior margin of the bone, and has an indistinct continuation, only a few millimetres
wide, along that margin. The facet for the trapezoid faces upwards and inwards: it is
only half as wide posteriorly as it is in front, is almost plane from back to front, but
fairly deeply concave from side to side. The facet for the magnum is separated from
that for the trapezoid by a curved, somewhat oblique ridge; its surface is irregular.
From back to front it is first of all concave, and then convex, whereas from side to
side it is convex in varying degrees. Allowing for a certain amount of twist, this facet
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 63
looks directly upwards. The facet for the third metacarpal is on the external lateral
surface: it is rectangular, parallel sided, and well-developed. The shaft of the bone is
only slightly triangular. The distal articulation is very large. It has an almost obsolete
median keel on its posterior portion, and two well-marked areas indicating the position
of the sesamoid bones.
Remarks. — These two bones are from the same locality as the referred speci-
men of P. sinensis, and, presumably, from the same horizon. Although there is every
reason to believe that they belong to that species, they seem to be rather too large,
and because I felt some doubt in the matter I have kept them apart from the teeth.
Their size may be judged from the fact that the metacarpal is as big as, and in certain
measurements, bigger than the corresponding bone in the right manus of the Upnor
Elephant (Andrews, 1928, p. 12), and that the cuneiform is as large as the largest
specimens in the British Museum. For purposes of comparison, use has been made
of the right, second metacarpal of the Upnor Elephant (this bone is not in the mount;
it is registered Ml 1156). The same bone from an imperfect manus from Grays, Essex
(regd. 18259), the cuneiform from the same manus (regd. 18246), and a cuneiform
from Clacton referred to Archidiskodon meridionalis (regd. 28007). The cuneiform
has also been compared with the corresponding bone in Trilophodon wimani, with
a cuneiform of the Mammoth from Ilford, Essex (regd. 45114), and with one of the
American Mastodon (regd. 17211).
Apart from its very much greater size, the cuneiform differs from that of T. wi-
mani in many details. The antero-posterior convexity of the ulnar facet is much flatter,
and the posterior concavity of the same facet is more marked; also the width of the
facet is less in proportion to the depth from before backwards. The curves of the
inferior surface are stronger, and the length of the lateral process is less than half the
width of the unciform facet. The process itself is less decurved, and the change in
direction of its articular surface is not so abrupt; moreover, the process is relatively
stouter.
The cuneiform agrees fairly well with the corresponding bone of M. americanus.
It is somewhat thicker, with a larger lateral process, and bigger facets for articulation
with the lunar. The pisiform facet does not extend so far along the lateral process.
On the inferior surface, the articulating area of the lateral process faces forward, whereas
in the American Mastodon it is directed downwards.
The only undoubted cuneiform of Palceoloxodon antiquus (regd. 18246) has the
lateral process restored in plaster. It differs from the specimen from China in many
particulars. Thus, it is relatively thicker; the antero-posterior concavity of the proximal
(Ill) 64
Pcdceontologia Sinica
Ser. C.
siirfarp ic сггряЬг and fhp kfpral rnnvpvifv iq Ipqq- flip liinHpr hsH nf flip nisifnrm farpf
------- G------,------- *“'—*“* ---, LMV XKUVAVX - ----J-----------
is higher, and more rounded; the inferior facet is more triangular, and not so square,
in outline; and the inferior facet for the lunar is restricted to the antero-inferior corner
of the inner surface, where it forms a semi-circular area at right angles to the distal facet.
The cuneiform of Archidiskodon meridionalis has a much smaller ulnar facet,
which is more deeply concave antero-posteriorly, and less convex transversely. In both
these details it agrees with that of Pal. antiquus. The pisiform facet agrees with that
of T. wimanl and M. americanus in the extent to which it is produced downwards
and sideways along the lateral process, instead of being restricted to the body of the
bone as in the specimen from China. Viewed from the front, the cuneiform bones
of ? P. sinensis and A. meridionalis agree fairly well, the main differences consist in
the smaller amount of the inferior facet of the lateral process which is visible in the
latter. The facets for the lunar are much stronger in the cuneiform from China than
they are in A. meridionalis, which agrees with Pal. antiquus in this respect, and also
to a less extent with the Mammoth.
On the whole, the cuneiform of the Mammoth agrees with that of ?P. sinensis
far more closely than any other with which the latter was compared. Apart from the
difference in size, the main distinctions are in the restricted area of the facets for the
lunar, the manner in which the inferior facet on the lateral process faces downwards
as much as it does outwards, and the smaller lateral process. In the other characters
the cuneiform of the Mammoth resembles that from China very closely.
The main differences between the metacarpal from China and those of Palceoloxodon
antiques with which it has been compared lie in the relationships of the facets for the
carpal bones. These will be appreciated most readily by reference to the drawings on
plate II. So far as the lateral facets for articulation with the adjacent metacarpals are
concerned, those on the bones registered 18259, and Ml 1156, are narrower and more
restricted in area than they are on the Chinese specimen.
The great size of these bones does not imply that the shoulder height of the
animal was equivalent to that of the Upnor Elephant. It must be remembered that
the mastodonts, so far as their bodily proportions are known to us, were longer, and
lower than the elephants. ,
MASTODONTO1DEA Invert» Sedis.
(Plates II, III, IV.)
Indeterminate remains in this collection are noted here either for the sake of placing
the localities on record, or else because of their intrinsic interest. The front two ridges
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from. China 65
of a large, very worn, lower molar come from Honan, Meng-Chin-Hsien, East 40 li,
Yang-Chia-Kou, Erh-Liang-Kou, Wa Kou; from the same locality are numerous frag-
ments of tusks. Another fragment bears the label, Shansi, Fen-Yang, Western side of
Fen-Но. Two pieces of tusks are from Yunnan, Lan-P'ing Hsien, from city NW 65 li,
La-Chi-Ching. Lastly, two pieces are labelled, "Tarntj in Tall, Ostra Sunnit, 60 li NE
om Hallong OSSO, Mongoliet”.
Under this head are also included the specimens described by Koken (1885, p. 37,
pl. vii, fig. 2) and Schlosser (1903, p. 48, pl. xiv, fig. 4) as Mastodon sp. ex aff.
Pandionis Falc. The first is a very worn anterior ridge of a permanent molar; the
second is a premolar.
An incomplete hand from Locality 77 appears to be proboscidean in nature, al-
though the carpal bones interlock to a much greater extent than is usual in this group.
The specimen consists of the lunar, trapezoid, and magnum, the proximal ends of the
second and third metacarpals, and the entire fourth metacarpal, all from the left limb.
A detailed description follows.
Associated Carpal and Metacarpal Bones. Locality 77.
Os lunatum. — The lunar bone is nearly triangular in plan. The whole of the
upper surface is occupied by the radial facet, which has for section a sigmoidal curve
since it is concave from before backwards in its posterior moiety, and convex in the
same sense in front. The ulnar facet, which has much the same anterior convexity
and posterior concavity, is oval in contour; it truncates the antero-external corner of
the bone, and makes an angle of just over 90° with the radial facet. The borders of
the two facets are in close contact, and form the outer side of the triangular top of the bone;
The lower, or distal, surface has the same section as the upper, since it is wholly
occupied by the facet for the magnum, and is convex from before backwards in front
and concave in the same direction behind. Not only is the lunar wider than the
magnum on which it rests, but it has also been displaced inwards so that it almost
entirely covers the trapezoid, except for a small area at the posterior end of the bone
which is all that remains of the scaphoid facet. In addition the outer half of the upper
facet of the magnum, normally covered by the lunar, is uncovered. Hence, when the
three bones are articulated, and seen from the front, the whole of the trapezoid is
covered by the lunar, the outer third of which overlaps on. to the magnum. There
are narrow facets for the scaphoid and cuneiform on the antero-internal, and antero-
external margins respectively; each is approximately at right angles to the facet just
described.
9—429921 A. Tindell Hopwood.
([[[) 66
Palxontologia Sinica
Ser. C.
Os trapezoideum. — The upper, proximal, facet of the trapezoid is in the shape
of an oblique triangle of which the base is formed by the anterior margin, and the
longest side by the complex facet for the magnum. The facet is concave in a direction
parallel to its longest side, and gently convex parallel to its anterior border in front.
Consequently it is twisted. The antero-external corner is deflected sharply downwards
so as to allow it to follow the under surface of the lunar. The articulation with the
scaphoid is restricted to the narrow, tongue-like, posterior portion of the facet, but
there is no line of demarcation between it and the portion which articulates with the
lunar. The facet for the trapezium looks inwards and downwards. It is rhomboidal
in outline but has the upper third extended both anteriorly, and posteriorly. Of these
extensions the posterior is the larger.
The facet for the magnum is both large and complicated. It is divided into an-
terior and posterior portions by a deep notch. The anterior part, which is nearly as
deep as the bone itself, is plane with a slight elevation posteriorly where a peg of
bone projects to fit into a corresponding depression in the magnum. The hinder portion
is concave from behind forwards, and passes obliquely inwards from the front portion.
It faces outwards and upwards.
The distal facet, which articulates with the second metacarpal, is triangular in
outline, gently convex from side to side, and gently concave from back to front. The
bone articulates with the second metacarpal, but with none other.
Os magnum. — As in many proboscideans the upper surface of the magnum is
boldly convex behind, and flat in front. As pointed out above, the lunar nearly co-
vers the trapezoid, and in consequence of this lateral movement it leaves little less
than half the upper surface of the magnum to be covered by the cuboid. The limits
of the two areas thus covered are not indicated on the facet. The union with the
trapezoid is very close and the facet is the negative of the one on the trapezoid al-
ready described. Owing to this, the inner margin of the upper surface of the bone
is sinuous, and the bone itself is much wider at the back than it is in front. The facet
for the unciform is double. The superior portion is plane and vertical; it is roughly
oblong, but somewhat constricted in the middle. The inferior portion is a narrow
strip along the lower margin of the outher face of the bone.
The facets for the second and third metacarpals are on the inferior, distal, sur-
face. They form a fairly sharp angle with each other, and the width of that for the
third is twice the width of that for the second. They are concave from front to back,
but, whereas that for the second is plane, or even slightly concave, transversely, that
for the third is gently convex in the same direction.
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
67
Metacarpalia. — The proximal ends of the second and third metacarpals, and
the whole of the fourth, are preserved. They are unusually stout, and their general
proportions are very massive.
There are four facets at the upper end of the second metacarpal. One, which is
relatively small and semi-circular, is on its inner face; this is for the trapezium. The
second facet, which is for the trapezoid, is on the upper surface of the bone; it extends
from the front to the back, occupies rather more than two-thirds of the available area,
is gently concave from side to side, and is equally convex from back to front. A third
facet, which accommodates the magnum, occupies the remainder of the upper surface;
it is gently convex in both directions. On the outer surface of the bone, and forming
an angle of about 120° with the previous facet, is a smooth elongate, more or less
oval, articulation which is for the third metacarpal.
The third metacarpal is of the same type as the second, and has four facets at
its upper end. On the inner surface is a long, narrow, gently concave facet which
articulates with the second metacarpal. The proximal surface is formed by the facets
for the magnum and unciform. Both are narrower behind than in front, convex from
before backwards, slightly concave from side to side, and more concave in front than
behind. That for the magnum is half as wide again as that for the unciform, and the
two facets meet at an angle of about 120°.
The fourth metacarpal is complete. It is short and stumpy, expanded laterally,
and compressed antero-posteriorly, with the upper, proximal, end produced backwards
to form a stout triangular beak. Its upper end is occupied by a gently convex facet
for the unciform. The anterior border of the facet is convex, the inner border is
nearly straight, and the outer border is gently concave. Beneath this facet on either
side of the bone are facets for contact with the adjoining metacarpals. That for the
third is the widest and makes an angle of about 110° with the unciform facet, whereas
the narrower surface for the fifth is almost at right angles to it. The shaft is roughly
oblong in section; its anterior surface is deeply concave from above downwards, and
gently convex from side to side. The distal end consists of a semi-cylindrical facet
for articulation with the phalangeal bone. There are no signs of facets for the sesa-
moid bones on its posterior surface.
The Dimensions of the various bones are as follow.
Os lanatiim. inm
Greatest depth from back to front........................................................... 115
Greatest width from side to side ........................................................... 102
(Ill) 68
Palceontologia Sinica
Ser. C.
mm
Greatest height........................................................................ 65
Maximum diameter of ulnar facet........................................................ 50
Os Trapezoidenm.
Greatest depth from back to front .................................................... 95
Length of superior outer border ....................................................... 90
Greatest diameter of scapho-lunar facet ............................................. 93
Greatest width from side to side .................................................... 65
Greatest height........................................................................ 49
Greatest diameter of metacarpal facet................................................ 75
Os Magnum.
Greatest depth from back to front .................................................. 112
Anterior width of superior facet ...................................................... 60
Posterior width of superior facet .................................................... 77
Greatest height (posterior)............................................................ 94
Width of facet for metacarpal Ill (anterior).......................................... 43
Width of facet for metacarpal IV (anterior) ........................................... 22
Metacarpal II.
Greatest depth from back to front ..................................................... 84
Greatest width from side to side....................................................... 70
Greatest width of trapezoidal facet.................................................... 52
Greatest width of facet for magnum .................................................... 21
Metacarpal III.
Greatest depth from back to front ................................................. 109
Greatest width from side to side .................................................... 70
Width of facet for magnum (anterior)................................................. 44
Width of facet for unciform (anterior).............................................. 26
Metacarpal IV.
Length .................................;............................................. 145
Greatest depth of proximal end ....................................................... 103
Greatest width of proximal end ........................................................ 78
Width of distal end.................................................................... 70
Depth of distal end................................................................. 65
Width of middle of shaft .............................................................. 74
Greatest depth of middle of shaft ..................................................... 45
Discussion. — Despite the richness of the collections in the British Museum (Na-
tural History), there are almost no bones of "Mastodon" with which these specimens
can be compared. The series from the Siwaliks was made in the beginning of the
Nineteenth Century, and nothing is known of the localities where the individual speci-
mens were found. Two bones from Europe have definite localities. They are a
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
69
left lunar of Trilophodon angustidens from Sansan, and a left magnum of "Mastodon
pentelicT from Pikermi. In addition, three specimens from the Siwaliks have been
chosen for comparison with the Chinese specimens. These are a left lunar and tra-
pezoid, and a right magnum; they are figured in the Fauna Antiqua Sivalensis on
pl. 1, figs. 7, 26, and pl. li, fig. 5 respectively. In case the specimens described above
should belong to the Elephantidae, they have also been compared with Andrew's de-
scriptions of the Upnor Elephant (Andrews, 1928, pp. 8—12, pls. iii—v), as well as
with Dietrich's descriptions of the hand of Palceoloxodon antiquus recki (Dietrich,
1916, pp. 43—59, pls. iii—vi).
In the sum of its characters the lunar agrees best with the specimen from Sansan
(regd. 33592a), but has the various features such as the antero-external convexity of
the radial surface, and the posterior concavity of the under surface more strongly
marked, it differs in being longer from back to front than it is wide. The speci-
men 33592a confirms Dietrich’s observation (1916, p. 44) that in Tri. angustidens
the lunar is broader than long. Another difference in the bone from Sansan is the
greater concavity of the outer surface from back to front, as well as of the inner sur-
face from above downwards. The ratio of height to breadth is approximately 58 %
in both cases.
There are several well marked differences between the lunars from the Siwaliks
(regd. M3200) and China. In the former, the anterior convexity of both proximal and
distal surfaces is less pronounced, so that, when seen from in front, the radial facet
is barely visible. In the Indian specimen, too, the depth is very little greater than the
breadth (117:115), and the ratio of height to breadth is 69 %. The acutely triangular
form of the ulnar facet contrasts with the broad oval on the Chinese specimen.
Compared with the corresponding bone in P. antiquus this bone agrees with the
Upnor specimen in having the depth greater than the width, whereas Dietrich records
the two dimensions as practically equal in P. antiquus recki, as well as in the Eu-
ropean form. It also agrees with the Upnor Elephant in the relationships of the radial
and ulnar facets, and differs but little in the ratio height: breadth, which in the Upnor
specimen is 59.5 %. In the first of these characters it differs from the East African
form, which has the radial and ulnar facets at a relatively obtuse angle so that the
latter looks outwards and upwards. In P. a. recki the ratio height: breadth is also
59.5%.
The trapezoid is peculiar because of the extent to which it is covered by the lunar,
and because of the manner in which the proximal and external facets are twisted. The
specimen which makes the nearest approach to it in this respect is one from the Siwa-
(Ill) 70
Palceontologia Sinica
Ser. C.
liks (regd. M3215). The latter is the larger bone, and it has a flatter proximal facet.
The anterior half of the facet for the magnum is an upright oblong, instead of the
almost square portion on the Chinese bone. It is separated from the posterior section
by a veiy deep notch which prevents the two parts from coming into contact in the
manner shown in the other specimen. There is an approach to the condition whereby
the posterior portion of the facet in the bone from locality 77 faces outwards and up-
wards, but it is not so pronounced, nor is the facet so high in proportion to its length
from back to front. The facet for the trapezium is much larger than in the specimen
described above. This is due to the great development of the postero-superior exten-
sion until it is nearly the same size as the remainder of the facet. In consequence
the posterior half of the facet for the trapezium is separated from the metacarpal facet
by a deep narrow cleft, and the large roughened area seen on the postero-inferior
section of this face in the Chinese specimen has almost disappeared. The metacarpal
facet on the Indian specimen has a long, tongue-like hinder portion, and its anterior
expansion is more rounded than in the Chinese form, in which this facet is approxi-
mately triangular. The peg of bone on the outer surface is not developed in spe-
cimen M3215.
The magnum has peculiarities corresponding to those seen in the trapezoid. Its
sinuous inner margin corresponds exactly to the sinuous outer margin of that bone.
This margin is responsible for the unusually narrow apperance of the bone seen from
in front. The ratio width to depth is just under 54 % in front whereas at the back
it is 69 %. This compares with 67,5 % and 74 % respectively in the Upnor Elephant.
The specimen from Pikermi (regd. M 10122) is a much larger bone in which the breadth
and depth are almost equal (122 and 126 mm respectively), and which has the trape-
zoidal margin almost straight. Its greatest height (118 mm posteriorly) is relatively
greater, the percentage ratios height to depth are 84 % and 94 % in the Chinese and
Greek specimens respectively. The internal facet for the trapezoid is very much the
same shape in the two specimens, but the manner in which it is disposed differs
considerably. Each facet consists of an anterior and a posterior portion. In both the
anterior portion is concave from back to front, and the posterior portion, which is
practically plane, faces inwards. The Chinese specimen is the more complicated, for
in it the anterior portion is not only more deeply concave, but it also extends the
whole height of the inner side of the bone, and comes into contact with the facet for
the second metacarpal. The development of the facet has been carried so far that it
has arms both above and below the pit for the interosseous ligament. That on the
upper margin of the pit has been carried down into the pit wall so that, in that region,
Vol. IX. A. Tindell Hoopwood: Fossil Proboscidea from China 71
the facet is convex from above downwards and provision is afforded for the articula-
tion with the projecting peg of bone on the trapezoid.' There is none of this elabora-
tion in the specimen from Pikermi. The anterior portion of the facet for the trape-
zoid does not establish contact with the metacarpal facet, and there is no provision
for any peg on the trapezoid. The posterior portion of the facet is wider in the Chinese,
and higher in the Greek specimen, but whereas in specimen M 10122 it faces inwards,
in the Chinese bone it faces inwards and downwards. Another interesting difference
between the two bones is the development of an additional facet for the trapezoid
on the postero-inferior angle of the inner face of M 10122. This is not present
in the Chinese specimen. The metacarpal facets also differ to some extent, for whereas
those in the bone from Locality 77 are parallel sided, that for the third in specimen
M 10122 widens anteriorly and nearly excludes the facet for the second metacarpal
from the anterior margin of the bone (cf. Andrews, 1928, p. 10.).
The bone from the Siwaliks (regd. 17852) is much wider despite its sinuous trape-
zoidal margin. Unfortunately it is not in very good condition, and in consequence
the arrangement of the facets for the metacarpals and unciform is not easy to decipher.
The chief interest however lies in the manner in which the anterior portion of the
trapezoidal facet is modified so as to articulate with a peg from the side of that bone.
This is achieved in exactly the same way as in the bone from China, but the height
of this portion is less, and so it does not make contact with the metacarpal facet.
This specimen agrees with the majority of proboscidean magnums, and not only
with that of Palceoloxodon antiquus, in being broader than high in front, whereas
it is higher than broad behind. On the other hand it differs from both the Upnor
Elephant, and P. a. recki, as well as from several other species, in that the facet for
the second metacarpal has parallel sides, and that there is no tendency for it to be
excluded from the anterior margin of the bone by a greater development of that for
the third metacarpal.
The proximal ends of the metacarpal bones vaiy considerably among different
individuals as well as among the species of proboscideans, so that it not possible to
speak of a normal type. This is well shown by the figures published by Dietrich,
(1916, pl. vi.) and is further illustrated by those which accompany this paper.
Family STEGODONTID/E.
The animals included in this family have skulls which resemble those of the true elephants
but which are more primitive. They have very long sockets for the tusks, and the grinding
(П1) 72 Palceontologia Sinica Ser. C.
teeth and palate are well below the plane of the occipital condyles. The grinding teeth remain
brachyodont throughout the whole of their evolutionary history, but they parallel the teeth
of the true elephants in showing a progressive increase in the number of ridges. This
is especially true of the third molars. With this increase in the number of ridges, ajjd
its accompanying increase of length, there is an ever increasing curvature of the occlusal
surface which reaches its maximum in certain specimens referred to 5. airawana and.
5. insignis. Owing to the >short palate, it was impossible for the whole of the very
long teeth to be accommodated in the upper jaw at one time. The curvature of the
crown allowed the tooth to follow a more or less circular path which brought it from
a position practically parallel to the plane of the occiput to the correct position for
mastication. There is an ever increasing amount of cement, and the ridges show an
increasing number of mammillae on their crests.
This sub-family comprises two groups of animals. One, with compressed, tech-
form, ridges is given the generic name Stegodon Falconer & Cautley. The other, in
which the ridges are blunter, and composed of rounded conules, is known as Stego-
lophodon Schlesinger. Both genera occur in India, but, so far as is known, Stegodon
is the only genus found in China.
Genus STEGODON Falconer & Cautley.
1847. Fauna Antiqua Sivalensis, pls. xlii, xliv.
Diagnosis. — "Dentium molarium 3 utrinque intermediorum coronis complicata
colliculis hypisomeris (e. g.778),mammillatis, tectiformibus. Praemolares nondum obser-
vati”. (Falconer, 1857, p. 318).
Genotype — Mastodon elephantoides Clift.
Remarks. — The name Stegodon first appears engraved at the foot of plate xlii
of the "Fauna Antiqua Sivalensis”. On that plate and the next there is a series of
figures of proboscidean skulls which are grouped under various heads in the legend.
Figures 12—24 are referred to Elephas, with three sub-heads, namely, 12—15 Siego-
don, 16—18 Loxodon, and 19—24 Elasmodon. Figures 12—15 all occur on plate
xliii; taken in order, the following names are engraved against each. — fig. 12, E.
cliftii; fig. 13, E. bombifrons; fig. 14, E. ganesa; fig. 15, E. insignis. According to
the modern school of priority-purists this necessitates the selection of E cliftii. as
genotype as a matter of course. Fortunately, we can do so and adhere to the author's
original intention, for, in his later writings Falconer expressly says that Stegodon corre-
sponds "with the forms collectively designated Mastodon elephantoides by Clift”,
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
73
(1857, p. 314). The exact determination of Mastodon elephantoides Clift is dealt
ТТН-Ил Kxr Dmf/lf'r-nr ЛЛоКпГИ f11 Til (S' Л И /4 П£Ш/4 Ил1
vv 1111 uj i WlLOdUl vouvi 11 ill mo 1U1 LHLUUIHig 1 1 UUUdLlUCU ШСШМЦ , Uuvi hui
be entered into here, except to point out that the better known name Elephas (Stego-
don) cliftii Falconer & Cautley is a synonym.
There are three species of Stegodon represented in this collection, but there is
not a single complete tooth. In order to amplify the material, 1 have added to one
species such specimens as are in the British Museum. These include the type spe-
cimens of Owen’s Stegodon orientalis. The two new species are represented by three
scraps, one for one species, and two for the other.
STEGODON OFFICINALIS sp. nov.
(Plate VII).
Diagnosis. — A Stegodon with the ridge-crests widely spaced, with four to five
mamillae on each ridge; first and second ridges of lower molar divided by a median
cleft; conules of upper molars not united into ridges.
Holotype. — The first two ridges of an unworn lower molar.
Material. — The unworn last ridge and talon of a third molar. Probably from
the upper jaw.
Dimensions.—The holotype has the following dimensions: — Maximum length,
measured on the pretrite side, 95 mm. Maximum width, measured at the second ridge,
99 mm. Maximum height, measured on the post-trite side of the second ridge,
*58 mm.
The second fragment is 81 mm long, and has a maximum width and height of
101 mm and 51 mm respectively.
Horizon. — Unknown.
Locality. — "Bought in a medicine shop, Hanchow. Said to have come from
Szechuan.”
Description. — The anterior ridge is divided into two cusps by a deep median
cleft. The post-trite cusp has two large cones with a third, smaller one, between them.
The tips of the cones of the pretrite cusps have, with one exception, been broken off.
The second ridge is also divided by a median cleft, but this is not so prominent as
in thk first ridge. The pretrite cusp has three cones, and the post-trite has two. The
ridges are much narrower at the top than at the base, this is especially true of the
second hence the labial and lingual surfaces of the tooth slope inwards to a very
marked extent. The anterior surface of the second ridge is nearly perpendicular,
10-429921 A. Tindell Hopwood.
(Ill) 74
Palceontologia Sinica
Ser. C.
whereas the posterior surface slopes away at a fairly steep angle. This difference in
the slope of the two surfaces gives the ridge the appearance of being tilted forwards,
a sign of derivation from a lower tooth. There is a prominent anterior buttress of
three cones which passes across the front of the tooth towards the post-trite margin.
The space between the largest cone of the buttress and the antero-pretrite corner is
occupied by two cones, of which the outer is the smaller. There are no lateral cin-
gula, nor are there cingules at the entrances to the valleys. No cement is present.
The second fragment consists of the last ridge and talon of a large tooth, which
is doubtfully regarded as a third upper molar. The ridge consists of five cones which
are not fused to one mass, but are separated by definite clefts. One cleft is deeper
than the others; hence the cones are divided into two groups, one with three cones
and the other with two. A large part of the talon is covered by cement, above which
there rise the tips of four cones. Three of these are of about the same size but the
fourth, which is smaller, gives the impression of being formed by budding off the
inner surface of one of the outer cones. Behind everything is a coarse basal swelling.
It is not possible to state with certainty which side is pretrite and which post-trite. From
general principles, the pretrite cusp, or portion, should be slightly more advanced than
the post-trite. Applying this to the present specimen the cone formula would be
ccccc
pretrite £ qc q post-trite
The gap between the two post-triteXones of the talon is fyll of cement, which
hides the smaller cone one would except to find there.
The ridges are upright, with their anterior and posterior surfaces sloping at about
the same angle; the valley between them is narrow, and partly filled with cement.
Discussion. — There is no proof that these two specimens are correctly associated.
At the same time it is evident from their preservation that they are both derived from
the same deposit, which appears to have been of a lignitic, or peaty nature. They
indicate the existence of a species which has hitherto been unknown and it is prefe-
rable to assume an unproven connexion, instead of making two "species”, one of which
might have to be relegated to the synonymy at a later period.
In placing the species with Stegodon rather than in Stegolophodon, 1 have had
regard to the characters of the ridges of the holotype. One of the chief differences
between the two genera is that whereas in Stegodon the cones are united into ridges,
in Stegolophodon they remain discrete. Admittedly the cones in the fragment of an
upper tooth are still divided by deep clefts, but they show the first stages of union,
Vol, IX, A. Tindell Hopwood: Fossil Proboscidea from China 75
and the cones in each half of the ridge are more closely joined to each other than
thp twn halvps nf fhp rid ops
This species is the most primitive yet recorded from China. In the division of the
lower ridges into two cusps, as well as in the division of the upper ridges and the
slight displacement of the two halves, it retains characters which derive from a masto-
don! ancestry. Traces of this inheritance are never entirely lost among the Stegodon-
tidae and they persist with more or less regularity among the elephants. There are
several species in a similar stage of evolution among the fauna of the Siwalik deposits
of India, but they have not yet been described. So far as is at present known, the
Indian species have blunter cones than the Chinese form, and their ridges are not
so high.
The trivial name refers to the chance that these specimens were purchased in the
shop of a Chinese druggist.
STEGODON ZDANSKYI sp. nov.
* (Plate VII).
Diagnosis. — A Stegodon of very large size, with eight to ten mammillae on
each ridge-crest; anterior ridge still preserving the two cusps of the earlier mastodont
ancestors, posterior ridges foreshadowing the structure of the plates found in the teeth
of the Elephantidae.
Holotype. — The first four ridges of a right third lower molar.
Dimensions. — Length 191, Breadth 128 iv, Height 71 iv.
Horizon. — Unknown.
Locality, — Bought in a Medicine Shop, Shanghai.
Description. — The first ridge of this specimen is divided into two cusps by a
prominent mesial cleft. Each cusp, especially the pretrite one, is divided into two
cones by a cleft which is not so prominent as that which separates the cusps. The
cones themselves are again divided at the summit into two or three mammillae by
grooves, which do not proceed very far down the anterior and posterior surfaces. None
of the other ridges show this sub-division; they are all divided, but in a very different
manner.
Each of the posterior ridges has a large cone at either end, with one or two
mammillae at the summit. Between these cones is a long, roof-like portion with a
mammillated crest. The number of mammillae varies with the manner of counting. If
(lit) 76
Palceontologia Slnlca
Ser. C.
the main eminences alone are reckoned, there are from four to six; if each tiny sub-
division, or indication of one, is counted, this number is increased.
The crown of the tooth is curved to the right, and, seen from the side,, it is very
slightly concave. The ridges have a forward pitch; that is to say, their anterior faces
are steeper than the posterior. All these are marks of a right lower molar.
Anteriorly there is a relatively slender cingulum, which receives a nodular buttress
passing downwards and inwards from the inner cone of the pretrite cusp of the first
ridge. There are no lateral cingules at the entrances of the valleys. A small quantity
of cement is present.
Discussion. — This species is the largest Stegodon yet recorded. Apparently it
has nothing to do with the Indian species hitherto described; they are all smaller, have
the ridges closer together, and more copious cement. An undescribed Indian species,
in which the third upper molar is 243 mm long and 181 mm wide, is of about the
same size, but its ridges are more of the type of Stegolophodon stegodontoides (Pil-
grim), and it is probably referable to the same genus as that species.
Apart from its size, the most interesting feature of 5. zdanskyi is the curious
mixture of elephantine and mastodontine features in its ridges. In its composition of
two cusps, each made up of two cones, and in the relations between the pretrite cusp,
anterior buttress, and the cingulum, the first ridge is, fundamentally, that of any masto-
dont of the bunolophodont, or primitive zygodont, type. All the other ridges are es-
sentially elephantine in their structure. They each have a large cone at either end,
and a long, mammillated, roof-like portion in the centre. If the two fissures which
divide the tooth in this manner are deep enough, the partly worn to*oth would show
a tripartite enamel figure. This tripartite division is characteristic of the elephants,
though it is also shown by some, at least of the Stegodon group (cf. Soergel, 1912,
p. 8, fig. 2). In most cases it consists of two outer rings with an ellipse between
them; exactly the type of figure which could arise during the wearing down of a lower
tooth of 5. zdanskyi.
All the other remains of Stegodon from China hitherto described (Owen, 1870;
Koken, 1885; Schlosser, 1903; et alii) belong to more advanced species, and, by a
general consensus of opinion, the various writers, other than Owen, have referred them
to such species as S. bombifrons, S. ganesa, or 5. insignis.
The trivial name is given in honour of Dr 0. Zdansky of the Egyptian Univer-
sity, Cairo, who is well known for his valuable researches on the fossil mammals of
China.
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
77
STEGODON ORIENTALIS Owen.
un\
(ridic v и.)
1870. Stegodon orientalis Owen, Quart. Jour. Geol. Soc., XXVI, p. 421, pl. xxviii, figs. 1—4.
1885. Stegodon insignis Falconer et Cautley, Koken, Palceont. Abh., Ill, p. 14, pl. xi, fig. 8.
1903. Stegodon insignis Falconer, Schlosser, Abh. buyer. Akad. Wiss., XXII, p. 44, pl. xiv, fig. 10.
Diagnosis. — A Stegodon which resembles S. insignis Falconer & Cautley, but
differs from it in having the ridges more widely spaced, in the lesser development of
the cement, and in the greater lateral curvature of the lower molars, as well as the
greater convexity of the occlusal surface of the upper molars.
Syntypes. — Two fragments regd. 41926, 41927, Brit. Mus. Geol. Dept
Material. — Three specimen sin the Palaeontological Institute of Upsala University
may belong to this species. They are a broken Dm2, the hinder part of a first upper
molar, and a battered fragment of a true molar.
In addition to the syntypes, there are eight specimens in the Geological Depart-
ment of the British Museum, as well as a cast of another which is probably in the
Museum of the Itoyal Hungarian Geological Survey. These specimens are a right
fourth lower milk molar regd. M13986; a fragment of a lower tooth showing
two ridges regd. M12209; five pieces of lower teeth regd. M10222, M10223 —
two pieces, M10224 — two pieces; a first upper molar regd. M10270; a broken
third upper molar, with the first six ridges remaining, regd. M10249; the cast is of
the last six ridges of a right second lower molar, it is regd. M2015.
Dimensions. — Only the dimensions of the complete teeth are given in this table.
Upper Dentition.
Right Ml (M10270) ...................................
Length Breadth Height Index
146 68.6 iv 39 iv 47
Lower Dentition.
Right Dm4 (Ml3986)
154 65 v 43.5 vi 42.2
Horizon. — Probably all the material was obtained from Pleistocene deposits
though Lydekker (1886, p. 95) gives the horizon of M2015 as Pliocene (?), and the
late Dr C. W. Andrews assigned a similar age to M10270. The latter, however, stil
has a quantity of yellow matrix resembling loess adherent; for this reason among others
I have placed it in the Pleistocene.
Localities. — The specimens in the Upsala Museum have the following localities —
Fukien, Ning Hua Hsien, Lung Ti Tsun, Pei Shan Tung; Honan, Hien Chih Hsien,
NW 15 li, Shi Lang Tsun, Hsi Kou; Yunnan, Lan P'ing Hsien, from city NW 65 li,
La Chi Ching, Communicated by Mr C. Gimbel.
(IH) 78 Palceontologia Sinica Ser. C.
TLlO 1 11 j-i ЛО f-lnri Г-ГХ Л Г* I *-*-! Л Г- 1 *-i 4-ll Л D ♦*< J-inl-l АЛ..ЛЛ..МХ л«*Л нл 4- ЛЛ /4 Л-С< И < 4- л Т'Ч-» Л
mt lUVClULlvD U1 LUV О pCLUUCI Id III LUC UilUdil lUUdCUUl die IIUL du UCIlIlllC. 1111
syntypes are said to be ’’from a cave, near the city of Chung King Foo, in the pro-
vince of Sze Chuen”; the others are from ”Se Chuen”, M10222, M10223, M10224;
"Yen Ching K’ou near Wanhsien, Szechuen", M12209; "near Wan Hsun, Szechuen”,
Ml0270; "Szechuen”, Ml0249; "Chengtu", Ml3986; the cast is of a specimen obtained
"during the journey of Count Bela Szechenyi from the Pliocene (?) of Kansu, North-
west China” (Lydekker, 1886, p. 95.)
Description. — The only specimen of an anterior milk molar consists of the first
three ridges, together with the anterior talon and a part of the fourth ridge. The tips
of the ridges are only just touched by wear, but, since the tooth was taken out of
the ground, it has been artificially abraded in such a manner as to smooth off all the
fractured surfaces, and also to destroy all details of the anterior talon. As preserved,
the specimen is 31.5 mm long and 26 mm wide at the third ridge. Apparently the
second and third ridges are each composed of four cones; a large one at either end,
and two smaller ones betweeen them. There is no evidence to show whether the
outer cones had more than one mammilla at the summit. The enamel has coarse ver-
tical wrinkles. The former presence of cement is assumed, but cannot be proved.
The fourth lower milk-molar consists of seven ridges with a talonid at either end.
It is from the right hand side of the mandible, and has the right, or labial, surface
strongly concave. Each ridge is relatively high and compressed, and has eight or
more mammillae on its crest The exact number of mammillae varies considerably. If
everything which in any way resembles one be counted, the total may be as much
as twelve, as in the second ridge. If, however, the smaller ones are ignored, and
only those elements which are separated by furrows which extend some way down
the anterior and posterior surfaces of the ridge are counted, then the number varies
between seven and nine. The various components of the ridge have the enamel strongly
wrinkled, especially towards the base, and a strong tendency to sub-division at the sum-
mit. This leads to complications. The cement was stripped from the posterior surface
of the fourth ridge, and it was then seen that the ridge is made up of six cones,
which are divided at the top to yield a maximum of ten mammillae. Expressing the
relative sizes of the mammillae by large and small letters, and placing those belonging
to each cone within round brackets the structure of the ridge may be expressed as a
formula, thus,
(M) (mm) (mMM) (M) (M) (MM).
In the same way, the sixth ridge yields the formula
(Mm) (M) (Mm) (mMm) (MM) (M).
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 79
These two examples seem to indicate that the number of mammillae bears no fixed
relationship to the number of cones, and makes it doubtful whether the mammillae are
of much use for systematic purposes.
At its maximum development, the cement practically filled the valleys, leaving the
tips of the mammillae exposed. Its method of formation was such that the full amount
was not developed until just before the ridge pierced the gum. For this reason there
is very little on the last ridge and talonid, but it increases in quantity until it is thickest
on the third ridge, beyond which it again diminishes owing to wear. The third ridge
seems to have been exposed when the animal died so that some of the cement had
been worn away, and the top of the ridge uncovered. Hence, to obtain an accurate
picture of the maximum extent of the cement on any one ridge, it is necessary to
combine the thickness seen on the third ridge with the height seen on the fifth.
Two pieces are referred to the first lower molar; they are registered M10223 a, b.
Of these the first consists of three and a half ridges from about the centre of the tooth.
Most of the cement had been cut away with a knife before the specimen reached the
British Museum. Two of the ridges are practically undamaged; since they are con-
tiguous, their mammillary formulae are of interest for comparison with each other. The
anterior ridge shows various degrees of subdivision of the elements, thus,
(M) (M) (mMM) (MM) (mM).
All the elements of the posterior ridge are discrete and, with one exception, undivided.
The formula is
(Mm) (M) (M) (M) (M) (M) (M) (M).
The second specimen shows a noteworthy irregularity of construction in the an-
terior talonid, and in the first and second ridges. The third and fourth ridges are
practically normal, except for the fact that the outermost cone on the labial, or pre-
trite, side is separated from the rest of the ridge by a deep cleft. The mammillaiy
formulae are,
First ridge. - (M) (M) (M):(M) (m) (M).
Second do. — (M) (mmmmm): (m) (M),
Third do. - (M): (MmMMMMM).
Fourth do. - (M): (MMMmMMMM).
The anterior talonid is closely applied to the first ridge. It is a typical, nodular,
mastodont anterior cingulum and buttress in which the various cones and nodules have
become more slender and pointed, and fused together for the greater part of their
(Ill) 80 Palceontologia Sinica Ser. C.
length. There is a pointed, flattened, cone fastened against the centre of the anterior
face of the second ridge.
Two more fragments (regd. M10224) represent the third lower molar. One, which
consists of two ridges from the centre of the tooth, has the ridges close together,
with a deep, narrow valley between them. The ridges are sinuous: they have the
tips of the mammillae developed as backwardly directed points. The other is made
up of the last two ridges and talonid. These ridges, also, are sinuous, but they are
farther apart, and more ruggedly irregular in their construction. In the curvature of
the ridges, both these specimens agree with the tooth from Kansu identified by
Lydekker as 5. insignis (Lydekker, 1886, p. 95; cast. regd. M2015), but they differ from
it in having the ridges closer together.
Upper Dentition.
The upper dentition is represented by one complete first upper molar (regd.
M10270) and two pieces of similar teeth. One piece, in the British Museum, is re-
gistered M10222, the other is in the Palaeontological Museum of Upsala University.
In addition to these three specimens, there is a partial third molar, with the first six
ridges preserved. This specimen is also in the British Institute (regd. M10249).
The complete specimen of the first molar is half worn; most of the anterior talon
has been destroyed by the combined effects of wear and pressure against the preceding
tooth; the lingual cone of the posterior talon has been lost by fracture. There are six
ridges in the crown, and a talon at either end. The enamel figures expand from
either end as they approach the middle, and are defined by somewhat wavy enamel
from three to four millimetres in thickness. A curious feature of the second and
third ridges is the difference in shape of the two ends. One end of the enamel
figure is square, and the other rounded. Although the first ridge is influenced by
the anterior talon, it, too, shows a difference between the two ends. Both are round,
but that which corresponds to the square ends of the second and third ridges is not
so pointed as the other. There is no sign of a median cleft to any of the ridges, nor
do the enamel figures indicate that the cones at the ends of the ridges are distinct
from the other parts of the ridge, for the contraction of the figures of ‘the fourth and
fifth ridges in the appropriate positions is no greater than the contractions indicating
the limits between the other elements.
The posterior talon consists of a well developed ridge with a strong posterior
nodular, ’’cingulum”. It still retains a partial covering of cement after that substance
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 81
has disappeared from the rest of the crown. The last ridge and talon are slightly
sinuous.
The specimen from Yunnan (Pal. Inst. Upsala) consists of the last three ridges
and talon. All the ridges are more or less sinuous, and, fairly well covered by ce-
ment, though that substance is being worn away. The talon is still covered by ce-
ment; it is single, ridge-like, straight, and the valley which separates it from the last
ridge is not so wide as the valleys between the true ridges.
The remaining specimen (regd. M10222) which is referred to a first upper molar
consists of the last five ridges. This tooth was not worn, and has no cement pre-
served,. though there is no certain p^of that any had ever been deposited. The ridges
are straight, or curved, and the last one is very much distorted. Each ridge has the
two outer cones well separated from the remainder of the ridge. The formulas are as
follows, the anterior ridge being placed first,
(M) (МММ) (МММ) (M).
(M) (МММ) (mMMM) (М).
(М) (М) (mm) (тММ) (М) (М).
(М) (ММт) (ММ) (М).
(М) (МтМ) (ММ) (М).
Since there is no cement, this specimen illustrates the wall-like (Mauerartig) character
of the upper ridges when contrasted with the roof-like (Dachartig) character of the
lower ridges (Janensch, 1911).
The third upper molar, of which only the first six ridges are preserved in the
single specimen (regd. M10249) in the collection, has the occlusal surface strongly
convex. The anterior talon is almost destroyed by pressure against the second molar,
and by the effects of wear. The first three ridges all have some of the dentine ex-
posed; the first right across the tooth, the second one long lake and four small ones,
and the third six small lakes which correspond to the individual cones. Each of the
first two ridges shows, by a constriction of the enamel, that it is divided into two
unequal parts, but the constrictions are on different sides of the long axis. The length
of the specimen is 148 mm, and its maximum width 88 mm.
Discussion. — The syntypes of the species have not been redescribed, since
nothing additional to Professor Owen's original description has come to light. They
are very fragmentary, but confirm the identification of the new material as S. orientalis.
Authors writing subsequently to Owen have, with the exception of Osborn, united
this species with Falconer and Cautley's species 5. insignis. So long as the syn-
11—429921 A. Tindell Hopwood.
(Ш) 82
Paloeontologia Sinica
Ser. C.
types were the only specimens known, there was much to he. said for adopting this
course, but once better material became available it was evident that Owen saw more
clearly than his successors, and that 5. orientalis and S. insignis are indeed separate
species. Moreover, they are easily separated when complete teeth are available from
the mandible, and the separation of the upper molars is only a little less difficult.
The former species is a late survival of a more primitive type than the latter: it has
less cement developed, and the ridges are wider apart. On the other hand, it has
progressed in two directions which were not taken by 5. insignis: it has the greater
degree of hypsodonty, and the curvature of the crowns of both the upper and lower
teeth is more pronounced. From this latter circumstance it may safely be assumed
that the skull and mandible had continued to shorten. On the balance, therefore, it
is probably correct to regard 5. orientalis as more advanced than 5. insignis. In
other words, a species which was originally the more primitive, has passed the other
which was originally the more advanced in the struggle for existence. The more
advanced form became extinct, whereas the more primitive form, whilst retaining some
of its early characters, such as the wide-spaced ridges, progressed in other ways such
as the hypsodonty of the ridgecrests until in those respects it exceeded the other.
In certain of his preliminary publications, Prof. Osborn quoted the number of cones
in the cross ridges of the teeth of certain species of Stegodon (cf. Osborn, 1926, p. 16
& c). This number appeared to vary so considerably as to be of very little systematic
value. In studying the teeth of 5. orientalis, an attempt was made to test this with
the result that the formula, given above in the description of the specimens, was found
to vary in an apparently illogical manner, not only as between tooth and tooth, but
also as between the adjacent ridges of the same tooth. When any one factor can be
shown to fluctuate in this manner, it is evident that it cannot be used for systematic
purposes, however interesting it may be from the adaptational standpoint. On the other
hand, it is fair to quote the amount of sub-division of the original cones in general terms,
such as, "cones four to five in number”, "cones fairly numerous”, or "very numerous”,
when it is desired to give some idea of the evolutionary stage of the animal. There is no
doubt that the more primitive forms have fewer mammillae than the more advanced.
STEGODON ORIENTALIS GRANGERI Osborn.
(Plate VII.)
1929. S. orientalis grangeri Osborn, Amer. Mas. Novitates, No. 393, p. 16, fig. 16.
Diagnosis — "The sub-species Stegodon orientalis grangeri is more primitive
than the type of 5. orientalis--------the ridge-crests are less elevated and wider
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea front China
83
apart at the base and seem to be even more primitive than those of the S. insignis
type; the cranium is much smaller and simpler than that of S. insigrus-ganesa anti
resembles in its contour rather that of S. bombifrons.” (Osborn, 1929, p. 17.)
Syntypes. — The left third upper and lower molars of the same individual. Regd.
18714 Amer. Mus. Nat. Hist.
Material. — The only material available for the purposes of study is two speci-
mens which are, so far as is known, from the same locality as the type material in
the American Museum of Natural History. They are a first right lower molar registered
M12209, and two ridges of another tooth which also is registered M12209. Both
specimens were presented to the British Museum by J. L. Smith, Esq., in 1922.
Horizon. — Professor Osborn gives the horizon of the syntypes as Pliocene. The
horizon of the specimens in the British Museum is given as Pliocene?
Locality. — The syntypes are from "Yen-ching-kao, near Wanhsien, Province of
Sze-chuan”. The two specimens in the British Museum are from ”Yen-ching-k’ou, near
Wanhsien, Szechuen”. Allowing for variations in the transcription of Chinese place-
names, it seems probable that these localities are the same.
Description. — The first lower molar has lost the anterior talonid, and the labial
surfaces of the first, third, and fourth ridges; in addition, the first and second ridges
are just touched by wear. The tooth itself is bean-shaped, wider behind than in front,
and concave on the labial surface. It consists of six ridges with independent, ridge-
like talonids in front and behind. Each ridge has both the anterior and posterior sur-
faces concealed by plentiful deposits of cement so that the detailed structure is not
visible. The formula of the fifth ridge as revealed on removal of the cement is
(Mm) (mMniM) (mM) (M) (mM).
Neither this ridge, nor any of the others except the first has any trace of a median
division into two cusps similar to those found in the mastodonts. The first ridge,
however, is divided by a pronounced median cleft; removal of the cement showed its
formula to be
(MMm) (M): (MMMM).
Both talonids are detached from the adjacent ridges and might be regarded as inci-
pient ridges, but, since they have no roots of their own, and are supported by the root
of the nearest ridge, it is, perhaps, better to term them heels, or talonids. Apart from
this, the posterior talonid, which alone is preserved entire, has all the characters of a
small ridge.
(Ill) 84
Palieontologia Sinica
Ser. C.
the tips of the mammillae. It also fills the valleys to a depth of about 10 mm. As
in the fourth deciduous molar of 5. orientalis described above, the maximum develop-
ment is not reached until the tooth is cutting the gum, and it keeps pace with the
forward motion of the tooth. This is the explanation of the slight amount on the
posterior ridges, and the correspondingly open valleys, as compared with the heavy
deposits on the third and fourth ridges as well as in the second and third valleys.
The first and second ridges, with the intervening valley, show a decreased amount
owing to the wear to which the cement has been subjected. The specimen is 186
mm long, 87 mm wide, and 49 mm high. It has a breadth-length index of 46.9 %.
The fragment agrees very well with this tooth in all its main characters. The
formulae which express the construction of the ridges differ, but this has no great
significance.
Discussion. — The diagnosis, quoted from Osborn, contains all that has been
published concerning this variety. There was a mention in a previous paper (Matthew
& Granger, 1923, p. 567, figs. 3—6), but, apart from the photographs, no details
were given, and the material was referred to the typical 5. orientalis.
There can be no doubt that this form is more primitive than the type, since the
number of ridges is less, and the valleys are wider, but this does not necessarily mean
that the horizon is Pliocene. Matthew and Granger say that, ”The age of the fauna
is provisionally placed as Upper Pliocene on account of the abundance of Stegodon
remains and absence of any higher type of Proboscidean but its final correlation is
left open for the present” (op. cit., p. 563). On a later page (p. 597) they say that
the abundance of this form, the absence of elephants, and the presence of Chalicothe-
rium are ’’the only observed indications of Pliocene age; for the most part the fauna
appears to be quite closely related to modern species and might well be considered
Pleistocene”.
When estimating the age of any fauna, it is essential that the new arrivals be
given due weight, and that their presence should not be overshadowed by the survi-
vals of an earlier period. If, as Matthew and Granger say, the remainder of the fauna
from Yen-ching-kao is of a modern type which, but for the presence of two 'Pliocene'
genera, and the absence of a ’Pleistocene' genus, would be regarded as Pleistocene,
then, in my opinion, it would be preferable to regard the horizon as Lower Pleistocene.
Matsumoto (1915, p. 23) regards the specimens from Sze-chuan as belonging to
the younger Pliocene. He compares the fauna with that of the 'Siwaliks', the Narbada
alluvium, and with Java. In the result he concludes that, ’’the Stegodon-Vivm of Sze-
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 85
chuan is probably nearer to that of Siwalik than to that of Narbada and of Java. This
J., „ ____ Ci_____________________„о Tkin
idLL yiClllb d ItdbUll 1U1 icicuiug inc OLtgUMJfciauiia kjl utA^Aiuaii ao i uvixiit . mid
comparison and result is based on false assumptions as to the age of the Indian fauna.
The species quoted by Matsumoto are of all ages from the Dhok Pathan, or Pontian,
to the Pinjor and Boulder Conglomerate, which are certainly of Upper Pliocene and
Lower Pleistocene age, and may even be a little later. Again the statement that the
Szechuan fossils belong to the Pliocene is open to question.
Previous Records of Stegodon from China.
The species of Stegodon included under this head are extracted from the litera-
ture. Under each is given a suggestion as to its probable identification, but the old
names have been allowed to stand owing to doubts arising either from inadequate
descriptions or figures. Stegodon sinensis Owen is included here because of the lack
of suitable material for comparison.
STEGODON SINENSIS Owen.
1870. Stegodon sinensis Owen, Quart. Jour. Geol. Soc., XXVI, p. 417, pl. xxvii.
Original Description. — ’’The tooth in question is the second upper molar idS
of the type series) from the right side. Its crown, in a length of three inches, is di-
vided into five transverse ridges.....with ... ridged and wrinkled ... enamel. ...” /
"In the present tooth the first or foremost ridge is defined by a cleft on the outer
side of the tooth, but not on the inner side; here the abraded surfaces of ridges 1
and 2 are blended by wear into a common hollow field of smooth dentine. There is
a slight constriction near the part where the worn surface of the first ridge blends
with that of the second; and this constriction, which may be detected in the succeeding
ridges, I take to be a trace of that stronger one which more completely divides the
transverse coronal ridge in the molars of better Mastodons into an inner and an outer
part. A well-marked tubercle projects at the outer side of the base of the first ridge,
near the interspace between that and the second ridge. The second field of abrasion
although it broadens inwards to the common hollow, shows, before losing its indi-
viduality, a similar indication of constriction, or reciprocal inbending of the enamel
boundary. The same indication, though feeble, is obvious in the succeeding ridges,
which, by the unequal working of the lower grinder, show a broader field of dentine
as they pass inward. The third and fourth ridges, which are entire, show their slightly
undulated course from the outer to the inner side, which is lower and more worn.
(Ill) 86 Pakeontologia Sinica Ser. C.
The ridge at first inclines a little backward, then, at the indication of constriction,
bends forward, and finally resumes the transverse course to the inner, lower, and more
worn side of the tooth."
Remarks. — Owen says that, "the conformity of the Chinese molar with the
grinders of Mastodon elephantoides is close”. Koken (1885, p. 11), discusses this
resemblance at some length and comes to the conclusion that 5. sinensis is synony-
mous with S. cliftii (= M. elephantoides). The corresponding tooth of S. elephan-
toides is unknown, and so few of the teeth hitherto referred to that species are cor-
rectly identified, that it is preferable to leave the matter as Owen left it. Two milk
teeth bought in Shanghai probably belong to the same species, but they are so worn
as to be all but unidentifiable.
"STEGODON aff. BOMB1FRONS Falconer”.
1885. Stegodon aff. botnbifrons Falconer, Koken, Palceont. Abhand. Ill, p. 40, pl. xii, fig. 3.
Remarks. — This specimen consists of the last three ridges and talonid of a third
lower molar. It differs from 5. officinalis in that it is narrower, has the ridges closer
together and more compressed, and has about twice as many mammillae on each ridge.
According to Koken it occupies a position intermediate between 5. bombifrons and
5. insignis.
Family ELEPHANTID/E,
As used in this memoir, the family Elephantidse includes all those elephants which
are not included among the Stegodontidae. In this respect I differ from Professor
Osborn (Osborn, 1925, 1929, &c), who divides the family Elephantidse into at least
four sub-families, the Stegodontinae, Loxodontinas, Mammontinse, and Elephantinae.
The reasons which have lead him to this conclusion have not yet been published, but
he has informed me that much of the evidence is derived from a study of the cranial
proportions, as well as from the dental characters?1 Professor Osborn's results are so
revolutionary that one hesitates to adopt them without careful study of the facts on
which they are based. In consequence, I have kept to the more general division into
Elephants and Stegodonts, giving each group the rank of a family.
In one respect this was the only course open, for nearly all the specimens are
isolated teeth, or jaws. There is only one skull, which is so damaged as to make
accurate measurements and calculations impossible. The material studied, therefore is
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 87
not of the type needed in making any attempt to estimate the validity, or otherwise,
of Osborn's classification.
All the teeth of Chinese elephants hitherto described have been placed in the single
genus Elephas. To continue this practice seems to be unnecessarily conservative, for
there is a large, and ever-growing, body of literature which proves that Elephas is,
in its wider sense, a collective genus. This literature goes back to the ’’Fauna Antiqua
Sivalensis” in which Falconer and Cautley first sub-divided Elephas into three sections,
Stegodon, Loxodon, and Elasmodon. In view of the work of authors such as Pohlig
it is, perhaps, surprising that the prefence of several genera has not been more widely-
recognised. Not that finality could have been reached, for the difficulties attendant on
attempts to identify isolated teeth, which are all one usually has to study, are very
great; this is probably the reason which has prompted authors to err on the side of
caution, rather than to commit themselves to concepts which were not clearly defined,
and which, in the majority of instances, they had no opportunity of testing for themselves.
Genus ARCHIDISKODON Pohlig.
1825. Elephas, Nesli, N. Giorn. Litt. (Pisa), XI, p. 211.
1857. Loxodon, Falconer, Quart. Jour. Geol. Soc., LVII, p. 318.
1881. Elephas, Leith Adams, Monogr. Brit. Foss. Elephants, p. 181.
1886. Elephas, Lydekker, Cat. Foss. Mamm. Brit. Mus., IV, p. 107.
1888. Archidiskodon, Pohlig, Nova Acta Leop.-Carol. Deutsch. Akad., LIII, pp. 138, 252.
1912. Archidiscodon, Schlesinger, Jahrb. k.-k. geol. Relchs., LXII, p. 149.
1921. Elephas, Soergel, Geol. Palceont. Abh., XVIII, p. 17.
Diagnosis. — Elephants with brachyodont, laticoronate, parsilamellate, pachyganal
dentition; cement plentiful.
Genotype. — Elephas meridionalis Nesti.
Remarks. — This genus is very distinct and widely distributed. The genotype
is often found in Europe, especially in France and Italy; A. planifrons, a characteristic
fossil in the Lower Pleistocene of India, is also recorded from Europe; and several species
have been described in North America. Indeed, Osborn (1932) has recently described
a new variety of the genotype as A. meridionalis nebrascensis subsp. nov. It is clear
that there is a wide gap in the known distribution of the genus, and that the forms
from the Old and New Worlds are separated by most of Asia. Hitherto no species
of Archidiskodon has been described from the Far East, and the specimen described
below is the first evidence that this gap may be expected to close with the march of
knowledge.
(Ill) 88
Palceontologia Sinica
Ser. C.
ARCHIDISKODON cf. PLANIFRONS (Falconer & Cautley).
(Plate VII.)
1846. Elephas planifrons Falc. & Caut„ «Fauna Antiqua Sivalensis», Text, pt. i, pp. 11, 38
pl. ii. fig. 5 a.
Diagnosis. — An Archidiskodon with nine to ten plates in the third upper molars;
lamellar frequency 3—4.
Material. — A third upper molar.
Dimensions. — See table below.
Horizon. — Lower Pleistocene.
Locality. — Shansi, Ch’in Hsien, S 5 li, Sung Chia Kou, S 4 li, Hung Ch'ii Kou.
Description. — The seventh plate of this well-preserved tooth is in wear. The
anterior talon, and most of the first two plates have been broken off.
Before it was broken, this specimen had three complete enamel figures, and four
others in varying stages of development; now only one complete figure remains. This
follows a slightly sinuous course: at its labial extremity its hinder surface is concave,
but as it passes across the tooth it curves forward and then becomes straight. The
next two plates show three rings, produced transversely; the rings are of about equal
size in the first plate, whereas in the second the labial ring is smaller than either of the
other two. Behind these plates is another in which the tips of the digitations are worn,
and also damaged post mortem. There appear to be seven or eight of these struc-
Dimensions of
Species Locality j । Tooth Plates Length • Breadth Height
A. cf. planifrons. Hung Ch’ii Kou. M3 xi 8 x 232 94 iii 103 vii
P. foktinagai. Shanghai, Drug 3M x7 oo 149 80 i 210 iv
Store. M3 xl2x 223 83 i 210 vii
Locality 50. x lOx 256 75 iv ?
x9 x 242 | 78.5 iii ?
P. cf. namadicus. Ching Kou. M2 XI 11 X 223 96 v ?
"China”; (regd. 29007). M3 oo9x 166 101 180
Ching Kou. ,M XI12 X 243 72 vi ?
M2 Xi 12 x 244 72 vii ?
Locality 6. | M, x 13x 292 70 iii ?
Locality 8. M3 x 14 x • 333 79 vi 140 vii .
M. primigenius. Upper Yenesei. 3M x 25 x 242 84 viii 1 135 XTX
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
89
tures. The enamel is relatively thin for so primitive a tooth. Its average thickness is only
3.5—4 mm. It is thrown into fine folds, or wrinkles, which have a tendency to form festoons
here and there, more especially in the centre of the tooth. There is no true loxodont sinus.
The cement is very thick. Owing to some abnormality its wear is irregular, and
behind the third plate it has become deeply excavated. This is entirely due to wear,
since the surface of the cement at the bottom of the pit is highly polished by friction.
Discussion. — The material from India which Falconer identified as belonging to
this species, shows a considerable amount of variation in certain characters, but until
quite recently this variation was regarded as being within the limits of the species,
and Falconer's opinions were not questioned. In 1928, however, Professor Matsumoto
said that there were not only two species, but also two genera involved, and separated
Leith-Adamsia sivalikiensis gen. et sp. nov. (Matsumoto, 1928). The type specimens
of the new species are in the British. Museum (regd. 36695, M3070) and are figured
in the ’’Fauna Antiqua Sivalensis” (pl. xi, fig. 4; pl. xiv, fig. 8). Whether Matsumoto
is correct or not is open to question, but his paper makes it important to decide what
A. planifrons really is. In other words, we must determine which is the type speci-
men, a matter of some dispute.
According to Lydekker (1886, p. 100), it is the skull figured by Falconer and
Cautley on plates ix, x, of the ’’Fauna Antiqua Sivalensis”. Osborn does not agree
with this, but takes the specimen figured on plate ii, fig. 5 a, and described on pages
11 and 38 of part 1 of the text of the same work as his lectotype. It is more im-
Elephants’ Teeth.
Lamellar Frequency (Calculated) L-F. Mea- sured Length 4-La- mellae LengthH-Breadth Ena- mel thick- ness Digita- tions Entire Figu- res Development of Figures
232:100:: 8 = 3.4 3.25 232 4- 8 = 29.0 — 3.5-4 7-8? 3 Three equal rings.
149:100:: 7 = 4.6 4.5 149 7 = 21.3 — 2 ? — Lat. an. med. lam.
223:100:: 12 = 5.4 4.5—5 223 4-12 = 18.6 — 2 ? 3 Lat, an. med. lam.
256:100:: 11 = 4.3 4.5 2564-11=23,? 256-?75 = 3.4' 2 4? 1 8 ?*>open ovals.
242:100:: 10 = 4.1 4.5—5 242 4- 10 = 24.2 242-? 79 = 3.oi 2 ? 7 ?->almost true loxodonty.
223:100:: 11 = 4.9 6 223 4-11=20.3 — 2.5-3 ? 10 —
166:100:: 9 = 5.4 5-5.5 166 4- 9 = 18.4 — 3 5? . 6 Lat. an. med. lam.
243:100:: 14 = 5.8 5.5-6 243 4-14=17.4 2434-72 = 3.4 2 4-5 6 Lat. an. med. lam.
244:100:: 14 = 5.7 5.5-6 244 4-14 = 17.4 244-? 72 = 3.4 2-2.5 4 6 Lat. an. med. lam,
292:100:: 14 = 4.8 5-5.5 292 4- 14 = 20.9 292-? 70 = 4.2 1.5-2.5 4-6 3 Irregular.
333:100::15 = 4.5 3.5-6 3334-15 = 22.2 3334-79 = 4.2 ? 5-7 ? ?
1 242:100:: 26 = 10.7 10.5 242 4-26 = 9.3 242-? 84 = 2.9 1-1.5 1-2-1 14? f Lat an. med. lam.
12—429921 A. Tindell Hopwood,
(Ш) 90 Palceontologia Slnica Set. C.
portant that these two specimens should belong to the same species, than it is to find
out which of them is the technical lectotype, for, if one were broad and the other
narrow, then the latter would raise question of the validity of Leith-Adamsia in addition.
As it is, both have broad crowns so that they would be referred to A. planifrons by
Matsumoto, as well as by other authors, and they can be used to supplement each
other. Moreover, there can be no doubt that the specimen from Shansi, which also
has a broad crown, belongs to A. planifrons.
There are various differences between this tooth and Osborn's lectotype. The
exact breadth of the latter is unknown, for the specimen does not appear to have been
figured in the uncut state, and if the breadth was ever taken no record is now avail-
able. The enamel is well displayed, not only at the grinding surface, but also in the
section. It is thickest near the top of the plates, and thinnest at the bottom. The
thickness varies between four and six millimetres, and so is greater than in the Chinese
specimen. Moreover, the enamel on the lectotype is only slightly wavy, and never
crimped, or wrinkled, as in the other tooth. On the unworn portions, each plate is
clearly defined by the formation of the cement. Each plate stands up as a low,
rounded, ridge separated from its fellows by a very distinct groove. Owing to the
somewhat greater amount of cement in the Chinese tooth, this separation of the plates
is not so sharp. In both these characters, of the enamel and the cement, the tooth
from Shansi is the more advanced.
If the lamellar indices are compared, both specimens are in the same evolutionary
stage, since both have a lamellar index of 3, and troth are more primitive than Ly-
dekker’s type which has a lamellar index of 4. The agreement in the lamellar indices
of the Chinese tooth and Osborn’s lectotype is more significant than their differences,
for reference to the complete series in the British Museum shows that A. planifrons
varies considerably in such features as the crimping of the enamel, and that, in our
present state of knowledge, the variation is best regarded as individual.
This specimen, which might have been of interest when considering the place of
origin and dispersal of the genus, does not afford the help required for it is not yet
possible to compare its horizon with that of the specimens known from India and Eu-
rope. Until such information is available one cannot say whether A. planifrons ap-
peared earlier in the Siwaliks or in China.
Genus PAL/EOLOXODON Matsumoto.
1847. Elastnodon. Falconer & Cautley, »Fauna Antiqua Sivalensis», pl. xlii.
1857. Euelephas Falconer, Quart. Jour. Geol. Soc., XIII, p. 318. /
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 91
1888. Elephas, Polilig, Nova Acta Leop.-Carol. Deutsch. Akad., LIII.
1924. Palceoloxodon Matsumoto, Journ. Geol. Soc. Tokyo, XXXI, p. 257. (issued in September.
Japanese only.)
1924. Sivalikia Osborn, Amer. Mus. Novitates, No. 152, p. 2. (Issued December.)
1926. Palceoloxodon Matsumoto, Sci. Repts. Tohoku Imp. Univ., (2), X, pt. i, p. 2.
Diagnosis. — Elephants with hypsodont, angusticoronate to laticoronate, endioganal
dentition; enamel usually strongly crimppd; enamel figures with a slight mesial expan-
sion, but a true loxodont sinus rudimeriiary or absent. Cranium high, domed, with
a powerful frontal crest; premaxillae broad and diverging at the distal end, into which
the relatively straight, slightly upcurved tusks are inserted.
Genotype. — Elephas namadicus naumanni Makiyama, 1924.
Remarks. — The name of this genus antedates that proposed by Professor Os-
born by three months. It is satisfactory to find that two authorities, working inde-
pendently are agreed that those species grouped round Elephas namadicus Falconer
& Cautley belong to a distinct genus, but it would have been better if Professor Matsu-
moto had taken the Indian species as his genotype, instead of a variety about which
little is known.
The most striking feature of Palceoloxodon is the great frontal crest, which has
attracted attention ever since P. namadicus was first discovered. Thus Falconer writes,
"The species is especially interesting from the form of the cranium, which is so grotes-
quely constructed that it looks like the caricature of an elephants head in a periwig”.
(Falconer, 1868, p. 435). Apart from anything else, this crest differentiates Palceol-
oxodon from all other known members of the Elephantidse.
According to most students of the Proboscidea, any genus which is founded on
E. namadicus would also include the European species E. antiquus. Professor Os-
born, however, has recently (Osborn, 1931)- separated them by taking the latter as the
type of his ’’provisional genus" Hesperoloxodon. It appears from his paper that the
main distinction is the presence of the crest in the species P. namadicus, and P.
melitensis, and its apparent absence in P. antiquus, or at least in the specimen
which is the holotype of P. antiquus italicus Osborn. The evidence is not satisfactory,
for the vertex of the skull of P. a. italicus was destroyed before it reached the
American Museum of Natural History, and all one can rely on are photographs (de Lo-
renzo & d'Erasmo, 1927; Osborn, 1931). Moreover, in making his comparisons with
other skulls, Professor Osborn seems to have been misled by the gross measure
ments For example, he says that, ”As to Bathycephatf .... P. namadicus (592:728
mm) about equals L. africana (710:800 mm), while Pal. ant. italicus is much more
(Ill) 92
Palceontologia Sinica
Ser. C.
hathvrpnhalir " inn ri/ n 9Л\ If flip ratin? within thp hrarlrpf? hp pvnrpsspd as
I f-' * ДД V1*V TY1VA1111 IAAV k/iuv^^vvv Ж-/
percentages, it is clear that in this feature P. namadicus (81.2 %) is closer to the
Italian form of P. antiquus (81,8 %) than it is to the African elephant (88.7 %).
Although he does not give any figures, Professor Osborn says that, "The cranial
proportions [of P. a. italicus] .... are much nearer those of the Indian elephant
than of the African elephant. This bathycephaly is, however, a parallelism rather than
a point of affinity, because the very broad rostrum of Pal. ant. italicus presents an
extreme difference from the very narrow rostrum of Elephas indicus” (op. cit., pp. 20—21)
This argument can hardly be applied when comparing E. antiquus and E. namadicus,
for, not only do the cranial proportions agree so far as they have been published, but
also both species have the same type of wide rostrum, as well as the same type of
molar. Indeed the molars resemble each other so closely as to be distinguishable only
with difficulty.
Such far-reaching agreement in the characters of the two species counts for more
than the presence or absence of the frontal crest, and I prefer to keep both species in
the same genus until there are more certain grounds for separating them.
Palceoloxodon has long been known from China, but it has never been studied
in any detail. It also occurs in Japan and has formed the subject of several commu-
nications by Matsumoto and others. In addition to the new species P. tokunagai
Mats. (Matsumoto, 1929 b, p. 8), three sub-species of P. namadicus have been distin-
guished; these are P. n. naumanni Makiyama P. n. namadi Makiyama (Makiyama
1924, p. 264 & c.), andP, n. yabei Mats. (Matsumoto, 1929 a, p. 4).
PAI..T.OI.OXODON TOKUNAGAI Matsumoto.
(Plate VIII).
1929. Loxodonta (Palceoloxodon.) tokunagai Matsumoto, Sci. Repts. Tohoku Imp. Univ., (2), XIII,
pt. i, p. 7.
Diagnosis. — A Palceoloxodon with a lamellar index of 472—5; enamel figures
showing a loxodont sinus when well worn,
Holotype. — The third left lower molar described, but not figured, by Matsu-
moto in the above paper, and now in the Museum or Education, Ochanomizu, Tokyo, •
Japan. \
Material. — The incomplete second and third upper molars from either side.
The second molars very worn, and the third molars with the anterior lamellae showing
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
93
complete enamel figures. These four teeth have every appearance of belonging to the
same individual, though, in view of their history, this cannot be certain.
An incomplete lower jaw with both second molars in full wear. The jaw is
somewhat crushed, lacks the ascending rami, and is broken at the symphysis.
Dimensions. — See table on pp. 88—89.
. Horizon. — Lower Pleistocene? According to Matsumoto (op. cit.), this species
may belong to the upper Pliocene in Japan, but he does not give any stratigraphical
evidence in support of this opinion.
Locality. — The upper teeth were bought in Shanghai. The mandible is from
Locality 50.
Description. — The left second upper molar retains five and a half plates, together
with most of the posterior talon,; whereas the corresponding tooth from the right side
only has three and a half plates and the talon. In each tooth the less worn plates
curve backwards, but as the amount of wear increases the plates straighten until those in
front are almost straight. Each plate has a well marked median expansion, which
causes each enamel figure to take on an irregularly lozenge-shaped outline. The
enamel is relatively thick and crimped. Each loxodont sinus is so arranged that the
posterior angle of one touches the anterior angle of its neighbour. According to the posi-
tion chosen for the measurement, the lamellar frequency varies between four and a
half and five plates in a standard length of ten centimeters.
Of the third upper molars, the left has the anterior talon in full wear, and the
four succeeding plates showing various stages in the development of the enamel figures.
The right tooth has the anterior talon well worn, and the six following plates in various
stages from the abrasion of the tops of the digitations to the fully developed enamel
figures. None of the plates in the latter tooth shows a really definite loxodont sinus,
the most distinct being that in the third plate, and the plates are separated from their
neighbours by at least 4 mm of cement. The regular backward curve of the plates
seen in the preceding tooth is absent, and the curvature is irregular. For example,
the lingual end of the first plate curves backwards, whereas the labial end curves for-
wards. In the second plate there is a tendency to reverse this, and in the third plate
the labial end definitely curves backwards. The fifth plate is not so worn, but it has
the lingual end of the enamel figure curving forwards, and the labial end curving back-
wards. The other, left hand, tooth shows an even greater amount of irregularity.
The anterior talon has the centre bent backwards, as well as either end. The first
plate has an enamel figure of two ovals, of which the lingual is concave behind, but
is situated in advanced of the labial one. The second, third, and fourth plates are
(Ill) 94 Palceontologia Sinica Ser. C.
an aunormal in a soi'iiewhat similar iiiaiiner; that is to say, the second, which shows
a median lamella between two lateral annuli, has the median lamella branched so that,
taking the plate as a whole, the lingual half is in advancfTof the labial. The others
are in the same condition. Both the right and left teeth show the same enamel
characters as the two second upper molars.
The lower molars show certain individual differences which are recorded in the
table of measurements on pp. 88—89. The left has been damaged posteriorly, in addition
to losing the anterior talonid, and having a large amount of plaster inserted in the
neighbourhood of the sixth plate. Since the damage suffered by the right tooth is
restricted to a bruising of the hinder end, and the breaking away of the anterior talonid,
that tooth is here described.
The plates wear with a crescentic enamel figure which has the concavity facing
forwards. The enamel figures are narrow, and parallel-sided at first, but, as wear
goes on, they widen in the centre until they show a fairly distinct lozenge outline.
In the beginning there is a thick layer of cement between the plates, then the loxo-
dont sinuses come into contact, but later on the cement again separates the plates
throughout the whole width of the tooth. For example, the sixth and seventh plates
are separated by cement; the third, fourth and fifth, which are more worn, are in
contact, and the first and second which are still more worn, are again separated by
cement. This tooth is somewhat irregularly worn, especially posteriorly, and it might
be thought that this irregular wear was concerned in the varying positions of the
plates relative to each other, but since the left tooth also shows this variation, and
is normally worn, it is clear that the irregularity has nothing to do with it. The enamel
is moderately thick, probably a little less so than in the upper molars, and crimped
The lamellar frequency varies between 41/, and 5 */, according to where it is measured.
The mandible is more or less deformed by crushing; more especially is this true
of the right branch in which the molar has been displaced downwards and inwards.
The symphysis is not crushed or distorted. It is very short with a slight beak, most,
of which has been broken off.
Discussion. — These specimens have been referred to this species on account of
ther very distinct loxodont sinuses, and low lamellar frequencies. Neither P. nama-
dicus, P. antiquus, nor P. ausonius has the lozenge-shaped enamel figures found in
P. tokunagai, nor has any one of them so low a lamellar index as 4,5. It is true
that Professor Osborn quotes an index of 4 for the third lower molar of P. antiquus
(Osborn, 1931, p. 13, expl. to fig. 10) but he measures this index in an unusual
manner and so arrives at the figure of 6 for the third upper molar, and 4 for the
Vol. IX.
A. Tindell Hopwood: Fossil Proboscidea from China
95
corresponding lower molar. Thanks to his excellent figures, it is possible to meausure
the grinding surface in the usual way; this gives a lamellar index of 5,5 in both jaws.
In so far as the lamellar frequency is an indication of the evolutionary stage of
on elephant, it would seem that P. Tokunagai is the most primitive species of Palce-
oloxodon yet discovered, and that the teeth from China are more primitive than those
from Japan, which have a frequency of 5. To check this conclusion stratigraphically
is not yet possible, and no attempt is made here to distinguish the Chinese teeth as
a separate mutation. Moreover, nothing is yet known about the individual variation
of this species.
PAL/EOLOXODON cf. NAMADICUS (Falconer & Cautley).
(Plate VIII.)
1868. Elephas armeniacus Falconer, Busk, Quart. Jour. Geol. Soc. XXIV, p. 498.
1886. Elephas namadicus Falconer & Cautley, Lydekker, Cat. Foss. Mamm. Brit. Mus., IV
p. 169.
Material. — A left maxilla with the first and second milk molars; a badly broken
skull, lacking most of the brain case and the vertex, with the second upper molars in
full wear; two fragments of upper molars, each consisting of four or five plates; the
last nine plates and talon of a left third upper molar (Busk, loc. cit., regd. 29007),
a mandible, nearly complete, with the second molars in full use; a right mandibular
ramus with the second molar not yet fully erupted and part of the germ of the third
molar in the crypt; a damaged, isolated, right third lower molar.
Dimensions. — See table on pp. 88—89.
Horizon. — Middle Pleistocene?
Localities. — For convenience of reference, the localities are arranged in the same
order as the list of specimens. Locality 54; Honan, Kung-Hsien, Ching-Kou, from
village S 5 li; "said to have come from Sian"; Honan, Hsinan Hsien, from city N 12
li, Hsia-Ying-Kou; "China”; Honan, Kung-Hsien, Ching-Kou, from village S 5 li; Loca-
lity 6; Locality 8.
Description. — The anterior milk molar is worn flat. It had two plates in addi-
tion to anterior and posterior talods. The enamel was moderately crimped, thin, and
well covered with cement.
The succeeding tooth had five plates and two talons. Even after allowance has
been made for loss of height in front owing to wear, the anterior portion was lower
than the posterior. Externally, the surface of the enamel is more strongly wrinkled at
([[[) 96
Palaiontologia Sinica
Ser. C.
the base of the plates than it is at the top. Although each plate was originally covered
by cement it is clear that the cement was not thick enough to form a'smooth
outer coating which completely hid the plates, but that the position of each plate was
plainly shown by sharp folds in the cement.
All three specimens of the permanent upper teeth may be dealt with together,
since each is incomplete, and they supplement each other in many respects. The first
thing one notices about the teeth is the sharp angle which the grinding surface makes
with the transverse plane of the plates, and the extent to which the anterior enamel
surfaces of the plates are exposed. In the fragment from Sian the enamel is exposed
for as much as 17 mm. Because of this, the grinding surfaces appear to be built up
of a series of steps. In the earlier stages of wear, the enamel figures are separated
by 7 mm of cement, but, as wear proceeds their median. expansions, or loxodont
sinuses, establish contact with each other. It then seems (specimen from Hsia-Ying-
Kou) as though they separate again in the last stages, though this is not the case in
the specimen from Sian. The figures themselves are relatively narrow, with an indi.
stinct median expansion, and with the arms on either side directed backwards. In
this character, too, tfie tooth from Sian differs from the others, for it has the fourth
ridge turned forwards. All three of the isolated specimens differ from the teeth in
the broken skull in their lower lamellar index, though they agree in the shape of the
enamel figures, as well as in the thickness of the enamel.
Of the two teeth in the skull, only that in the right maxilla is at all complete.
It is in full wear, and has had the first plate and talon broken off. The enamel figures
have the same general characters as the teeth previously described, but the plates are
not so 'stepped', nor is the enamel so strongly crimped. The lamellar frequency is 6,
compared with 5 in the other specimens. \
The lower second molars show a fairly narrow range of variatiotK in the lamellar
frequency, and in the length-breadth ratio. Those in the mandible from Ching-Kou
have the plates closer together than they are in the specimen from Locality 6, and the
teeth are relatively broader. All the teeth have band-like enamel figures, with square
ends and slight median expansions, which are most pronounced on the posterior sur-
faces of the plates. In its thickness and amount of crimping the enamel is compa-
rable with that of the upper molars.
A somewhat intractable matrix, and not very much wear, make it difficult to
compare the third lower molar with the other specimens of lower teeth, but the avail-
able details seem to indicate that it is comparable with the tooth from Locality 6,
rather than with those from Ching-Kou, although it is slightly the more primitive.
Vol. IX. A. Tindell Hopwood: Fossil Proboscidea from China 97
Thus, the calculated lamellar frequency is slightly less, and the measured frequency,
which can only be taken on the side of the tooth varies between as little as 3,5 in
the centre, near the root, and 6 in the same part of the tooth, but taken at the tops
of the plates. This variation in the measured frequency suggests that a comparison
of the calculated frequencies is preferable. It is due to the tapering section of the
plates when seen from the side, and the manner in which their summits are placed
close together, so that the plates radiate like the sticks of a fan.
Discussion. - - The species Palceoloxodon namadicus, as at present interpreted,
appears to be very variable, even when the "typical” form the Narbada allu-
vium is considered without reference to specimens from extra-Indian localities which
are alleged to belong/to the same species. To take one character as an example;
the lamellar frequency varies from about 4 in a mandible registered M 3100 a (figd.
F. A. S., XII c, figs 4, 4 a) to just over 5 in the mandibular rami registered M3099
(figd. F. A. S., XII d, figs. 3, 3 a). This suggests that the sub-division practised by
Japanese authors is not desirable until a thorough revision of the Indian material has
been undertaken, and pending that revision I have not attempted to apply this sub-
division to the specimens from China.
In his latest monograph, Dr. Pilgrim places the Narbada deposits in the Middle
Pleistocene without any qualification other than the statement that his correlations with
Europe are approximate (Pilgrim, 1932, p. 8), and he has maintained this date over a
long period. If, then, the Middle Pleistocene age be accepted, and the species varies
to the extent quoted above, it is unwise to attempt to date deposits of which little
or nothing is known by means of a supposed mutational series. The Middle Pleist-
ocene age tentatively assigned to the specimens from China is entirely dependent on
the fact that the Indian teeth are said to be of the same age.
V
Genus MAMMUTHUS, Burnett.
1798. Elephas, Cuvier, Mem. Inst. Paris, II,- p. 21.
1830. Mammuthus Burnett, Q. Jour. Sci. Lit. Art., XXVIII (1829), p. 352.
1837. Dicyclotherium Geoffrey, C. R. Acad. Sci. Paris, IV, p. 119.
1841. Cymatotherium Каир, Akten der Urwelt, p. 11.
1888. Polydiskodon Pohlig, Nova Acta Leop.--Carol. Deutsch. Akad., till, pp. 138, 252.
1924. Mammonteus Osborn, Amer. Mas. Novitates, No. 152.
Diagnosis. — Elephants with medicoronate, hypsodont, densilamellate, endioganal
dentition; third molars with 24 or more plates.
Genotype. — Mammuthus borealis Burnett loc. cit.
13-429921 A. Tindell Hopwood.
(Ill) 98
Palaontologia Sinica
Ser. C.
Remarks. With the progress of research on the Proboscidea, if has become
ever clearer that the Mammoth is distinct from the Indian Elephant, which typifies the
genus Elephas. Professor Osborn has sought to give expression to this result by
reviving an alleged genus of Peter Camper’s and emending the name. If I do not
adopt the same name, it is because Camper was not referring to the Mammoth, and
because he was using the word 'Mamonteum’ as a vernacular. In the section ”De
ossibus Mamonteis” Camper (1788, p. 259) uses 'Mamonteum’ in an adjectival sense
throughout. Not only so, but he also makes it quite clear that he is referring to an
animal from America, and Pallas contributes a foot-note on p. 261 in which he explains
that whereas in Russia the term 'Mammontean bones' is commonly applied to the bones
of Elephants found in superficial deposits, it has suited Camper to apply the name to
bones found in America. With so much evidence as to Camper's meaning, it is no
possible to argue that the phrases on p. 251 of the same work refer to the Mammoth
as distinct from the Elephant.
Several names have been applied to the Mammoth, but the first that is valid, in
so far as it possesses a genotype, appears to be Mammuthus Burnett, 1830. Under
"Elephantidse, Elephant-kind” he groups the following, —
Genera. Elephas. Species. Indicus. Indian Elephant. Africanus. African.
Mammuthus. Borealis. Fossil Mammoth. Meridionalis.
Mastodon. Giganteum. Gigantic. Angustidens. Lesser.
From this table it is clear that he used the word Mammoth in the sense generally
accepted today, and did not apply it to the American Mastodon after the manner of
the majority of English and American authors at the beginning of the nineteenth cen-
tury, [cf. Cuvier, Ossemens foss., Ed. 1, Vol. II, Art. Sur le grand mastodonte), and
that his Mammuthus borealis is the equivalent of Elephas primigenius Blum.
Not only do the teeth of the Mammoth differ from those of all other elephants in the
number and structure of the plates, but the proportions of the skull are also different. The
skull is distinguished by its extreme foreshortening, as well as by a high, pointed vertex.
MAMMUTHUS PRIMIGENIUS (Blumenbach).
1799. Elephas primigenius Blumenbach, Handb. Naturg. Ed. 6, p, 697.
1830. Mammuthus borealis Burnett, Q. Jour. Sci. Lit. Art., XXVIII, p. 352.
Vol. IX.
A, Tindell Hopwood: Fossil Proboscidea from China
99
1837. Dicyclotherium primigenius Geoffroy, C. R. Acad. Sci. Paris, IV, pp. 119—120; p. 121, fig. 1.
1841. Cymatotherium antiquum Каир, Akten der Urwelt, pp. 11—15, pl. iv.
1845. Elephas primigenius, Falconer & Cautley, Fauna Antiqua Sivalensis, pl. i.
1888. Polydiskodon primigenius Pohlig, Nova Acta Leop.-Carol. Deutsch. Akad., LIU, pp. 138, 252.
1903. Elephas primigenius, Schlosser, Abh. buyer. Akad. Wiss., XXII, p. 43.
Diagnosis. — A Mammuthus in which the third molars have 24—27 plates;
enamel figures very narrow, with parallel sides; lamellar frequency progressing from
7 to 12.
Material. — A complete, partly worn, left third upper molar regd. M10941; part
of an upper molar just entering into wear, regd. M14102.
Dimensions. — See table on p.
Horizon. - Pleistocene. j
Localities. — Upper Yenesei Basin,VN. W. Mongolia (M10491); Chang-te-ho, on
the Yellow River, near Chang-te-fu, Hon|n (M14102).
Description. — The tooth from Mongolia is very fresh, but weathered. In its
general condition, it is hardly to be distinguished from teeth obtained from Alaska
and Siberia. The labial and lingual surfaces are slightly convex and concave respec-
tively. The occlusal surface, which has the shape of a long pear, is somewhat twisted
in addition to being convex antero-posteriorly and concave from side to side. The
plates are slightly sinuous, most of them have the ends directed backwards and are
gently concave in the centre in front. The enamel is not very much crimped, but
such as it is the crimping is very fine.
Seen from the side, one of the most noticeable features is the sharp angle, about
45°, between the occlusal surface and the base of the crown. There are seven roots,
or groups of roots, on the lingual side, and five on the labial. The root supporting
the first plate and talon is represented by a fractured stump.
The fragment from Honan is only sufficiently worn to show the cross-sections of
the digitations of the first plate and talon. It is noticeably narrower than the tooth
from Mongolia, and has the plates more widely spaced.
Discussion. — In its general characters, such as the thickness and degree of
crimping of the enamel and the shape of the enamel figures, the tooth from Mongolia
agrees with specimens from Alaska (regd. M13240) and Siberia (regd. M12357), but
there are certain differences which may be more than individual. Briefly, the Mongo-
lian tooth is shorter and broader than that from Siberia and has a higher lamellar
frequency. This suggests that there may have been a race in Mongolia which was
more highly specialised than others which lived farther North.
(1И) 100
Palceontologia Sinica
Ser. C.
From the details published hitherto (Osborn, 1924, 1930), it is not possible to
make any detailed comparisons between the sub-species ”Mammontens" primigenius
compressus and the Mongolian specimen. Osborn says that his form from Indiana
has 27 plates in the third upper molar, but it is not certain that this total does not
include the anterior and posterior talons. No dimensions of the teeth are given, so
that there is no opportunity of making comparisons on a statistical basis.
Numerous ribs and broken limb-bones from Ollan, Mongolia, are referred to this
species.
THE ACCOMPANYING FAUNAS AND THE AGE OF THE
DEPOSITS. x
Although so much descriptive work has beert^one on the fossil mammalian faunas
of China, very little has been written about the age of the beds whence the fossils
were derived. In only one or two cases have stratigraphical papers been accompanied
by faunal lists. It is no part of the function of a specialised memoir to attempt to
remedy this by giving extensive series of names of species found in this or that lo-
cality, but it is essential to provide some background against which the results may
be considered. For a long time I have made lists of the species described in Palceon-
tologia Sinica under their localities, and in this way endeavoured to form a picture
of the successive faunas. From these lists it has been possible to estimate the age of
any one deposit, and these estimates have been utilised under the heading ’’Horizon”
when describing the proboscidean remains. Many of the localities have nothing re-
corded against them except one mastodon or elephant, although others have quite an
extensive fauna. The latter have been extracted in the lists which follow, but, in order
to save space, genera only are quoted. In one or two instances the faunal list is quoted
from another author; then it is accompanied by a reference.
i. Trilophodon wimani occurs with Llstriodon gigas.
ii. Tetralophodon exoletus occurs with Ictltherium, Hycena, Indarctos?, Dlcerorhinus, Chi-
lotherium, Hipparion, Chleuastochoerus, Palceotragus.
iii. Pentalophodon sinensis is recorded from Kuan Tao Kou near Ni Ho Chang in the
district of Yu She Hsien, province of Shansi, as well as from San Chuang in the same district.
No other mammals are recorded from these localities, but Hipparion occurs at several other
places in the same district.
iv. Mastodon americanus was found associated with the following genera, — Proputorius,
Plesiogulo, Ultra, Melodon, Ictitherium, Lycycena?, Hycena, Machairodus, Fells, Dlcerorhinus,
Chilotheriwm, Diceratherium, Hipparion, Chleuastochcerus, Propotamochcerus, Cervocerus, Pro-
capreolus, Honanotherlum, Palceotragus.
v. Mastodontoidea incertae sedis from Locality 77. The foot bones so described were
accompanied by Hipparion, Eostyloceros, and Honanotherlum.
vi. Stegodon orientalis has been found associated with Cervus and Siphneus.
vii. S. orientalis grangeri was associated with Aeluropus, Ursus, Arctonyx, Cyon, Fells,
(Ill) 102
Palaiontologia Sinica
Ser. C.
Rhizomys, Tapirus, Sus, Proboselaphus, Bibos, two genera of monkeys, a chalicothere, and se-
veral other forms (Matthew & Granger, 1923).
viii. Elephas cf. primigenius in western Mongolia was accompanied by Rhinoceros, Bos,
Equus, Myospalax, Lagomys, Cervas, Oazella, Lupus, Meles, Cameliis (Teilhard de Chardin, 1923).
From these lists it is clear that there is little speculation in assuming the Pontian
age of the fossils identified as Tetralophodon exoletus, and Mastodon americanus.
The general agreement between the accompanying faunas and that of Pikermi is so
close that it is impossible for the beds to be regarded as other than homotaxial. Lists
vi, vii, viii, on the other hand, are not so distinctive. The last is undoubtedly Plei-
stocene, but whether the other two are Upper Pliocene, or Lower Pleistocene, does
not appear. Somewhat similar considerations apply to i, iii and v. In i both the
mastodont and Listriodon are of a type which elsewhere are found in beds of Miocene
(Sarmatian) age, and one assumes that age for the localities in which these remains
were discovered, but further information is necessary to comfirm this. The same is
true of iii. There are beds of Pliocene age in the Yu She Hsien district of Shansi,
for which reason a Pliocene age is assigned to Pentalophodon sinensis, and this is
confirmed to some extent by the morphology of the teeth, but other mammals from
the same locality would be welcome for the light they should throw on the question.
Locality 77, on the other hand, seems definitely to be of Pontian age, and the greatest
need is for teeth whereby the Proboscidea might be determined.
Certain species of the Tertiary Proboscidea of China have more definite resem-
blances to those found in Europe and America than others, and some are comparable
to the species found in India. Sufficient is known about the various forms to justify
a tentative synthesis which should endeavour to elucidate some of the migrations of
this group during the later Tertiary, from the Miocene up to the early Pleistocene.
Although this review is incomplete, there is nothing in the undescribed Indian series
which will run counter to it.
The most primitive form discovered in China is Trilophodon connexus which has
affinities with Tri. cooperi Osborn from Baluchistan, as well as with the European
Tri. angustidens. Of the three species the last is the more advanced and farthest
away from the ancestral line. Tri. connexus is the most primitive and, apart from its
greater hypsodonty, would do fairly well as the representative of a stage a little earlier
than Tri. cooperi. Other primitive/species are known from Moghara in the Libyan
Desert, but they do not appear t® belong to the same lineage as the Chinese species.
A third lower molar in the British Museum (regd. M14075) is shorter, with narrower
valleys, and a more complex talonid. It is in a fragment of mandible which indicates
Vol. IX.
/1. Tindell Hopwood: Fossil Proboscidea from China
103
a much stouter ramus. The general agreement in the disposition of the cusps and
their accessory structures is probably a sign of the primitive nature of both species.
Another species showing some affinity with a form occurring in India is Tetra-
lophodon exoletus from the Pontian. It seems to be related to an unnamed species
which occurs in the Upper Pliocene of India, and-may, or may not, be ancestral
to that species.
The third form which is interesting in this connexion is Mastodon americanus.
The genus Mastodon is found in the Pontian of Europe and China, at the top of the
Pliocene in India, and in the Pleistocene of America. This indicates that its centre of
dispersal was certainly not in either of the two last areas, but that it is to be sought
in Eurasia.
Insufficient is known of the Stegodon tidae of China to make a comparison with
those of India very profitable. At present it seems as though they occur earlier in
India than in China, where they are a late invasion from the South. Palceoloxodon,
on the other hand, may be more recent in India than in China.
The conclusion at which one arrives after a long study of Asiatic proboscidean
remains is that India has been isolated from China from early Miocene times, and
that until nearly the close of the Pliocene it was to all intents and purposes cut
off from the main migratory movements which were taking place to the North. Not
until the Pliocene was drawing to an end was there any interchange of fauna between
China and India. This interchange, which even then was restricted in its range, took
place along a route at present unknown, but which may have been by way of Indo-
China and Burma.
LIST OF WORKS CONSULTED.
Adams, A. L, 1881, Monograph of the British Fossil Elephants. Palaontogr. Soc. London,
volume for 1881.
Andrews, C. W. 1906, Catalogue of the Tertiary Vertebrata of the Fayum, Egypt. 4to, London,
Brit. Mus. (Nat. Hist.).
—, & Cooper, C. F., 1928, The. Upnor Elephant, 4to, London, Brit. Mus. (Nat. Hist).
Blumenbach, J. F., 1799, Handbuch der Naturgeschichte, Ed. 6, 8vo, Goettingen.
Borissiak, A., 1928, On a new Mastodon from the Chorak beds (Middle Miocene) of the Kuban
region. Platybelodon Danovi n. gen. n. sp., Ann. Soc. Paleont. Russ., VII, pp. 105—
120, pl. viii.
Boule, M., & Teilhard de chardin, 1928, ’’Paleontologie” in Le Paleolithique de la Chine, Arch.
Inst. Paleont. Humaine. Mem. 4.
Burmeister, С. H. C., 1837, Handbuch der Naturgeschichte, 8vo, Berlin.
Busk, G., 1868, Additional Remarks (to Has the Asiatic Elephant been found in a Fossil State?
by A. Leith Adams.), Quart. Jour. Geol. Soc., XXIV/ pp. 498—499.
Camper, P., 1788, Complementa Varia Acad. Imp. Scient. Petropolitanae communicanda, ad clar.
celeb, Pallas; Auctore Petro Camper, Nova Acta Acad. Scient. Imp. Petropol., II, pp.
240—265, pls. viii, ix.
Cautley, P. T., 1836, Note on teeth of the Mastodon a dents etroites of the Sewalik Hills, Jour
Asiatic Soc. Bengal, V, pp. 294—295.
Cook, H. J., 1922, Two new Bunomastodonts from Colorado. Proc. Colorado Mus. Nat. Hist.,
IV, No. 1.
Cooper, C. F., 1922, Miocene Proboscidea from Baluchistan. Proc. Zool. Soc. London, pp.
609—626, pls. i—iv.
Cope, E. D., 1884, The Extinct Mammalia of the Valley of Mexico, Proc. Amer. Phil. Soc.,
XXII, pp. 1-47, pl. i.
----, & Matthew, W. D., 1915, Tertiary Mammalia and Permian Vertebrata. Monogr. Amer.
Mus. Nat. Hist., Monogr. 2.
Cuvier, G., 1816—1817, Le Regne Animal, 4 vols, 4to, Paris.
----, 1821, Recherches sur les Ossements Fossiles, Ed. 2, 5 vols, 4to, Paris.
----, 1799, Memoire sur les especes d’elephans vivants et fossiles. Mem. Inst. Paris, II, pp.
1—22, pls. ii—vi.
Desmarest, A. G., 1822, Mammals in ’’Encyclopedie Methodique”, 4to, Paris.
Dietrich, W. O., 1916, Elephas antiquus Recki n. f. aus dem Diluvium Deutsch-Ostafrikas. Arch
Biontol. Berlin, IV. Heft. 1, pp. 1—80, pls. i—viii.
Falconer, H., 1857, On the Species of Mastodon and Elephant occurring in the Fossil State in
Great Britain. Part I, Mastodon, Quart. Jour. Geol. Soc., XIII, p. 307—360, pls. xi, xii,
----, 1865, The same. Part II, Elephant (Imperfect, Idem)., XXI, pp. 253—332.
—-, 1868, Palaeontological Memoirs and Notes of the late Hugh Falconer, A M., M. D..........
14—429921 A. Tindell Hopwood.
106
List of works consulted
Compiled and edited by Charles Murchison, M. D.. F. R. S. Vol. I. Fauna Antiqua
Sivalensis. 8vo. London.
Falconer, H, & Cautley, P. T., 1846, Fauna Antiqua Sivalensis, 64 pp. Text, Atlas fo. pls. i-xcii,
London.
Gastaldi, B., 1861, Cenni sui vertebrati fossili del Piemonte. Mem. Accad. Sci. Torino, (2).
XIX, pp. 19—84, pls. i—x.
Geoffroy saint Hilaire, 1837, Encore cet ecrit sur le Sivatherium considere sous le point de
vue de ses revelations contestees relativement a la Philosophic Zoologique. C. R.
Acad. Sci. Paris, IV, pp. 113—122.
Glooer, C. L., 1842, Gemeinniitziges Hand-und Hilfs-buch der Naturgeschichte. I, 8vo, Breslau.
Hay, 0. P., 1923, Characteristics of Sundry Fossil Vertebrates, Pan-Amer. Geol., XXXIX, pp.
101—120, pls. viii—ix.
Hays, L, 1834, Descriptions of the Inferior Maxillary Bones of Mastodons. Trans. Amer. Phil.
Soc., (2), IV, pp. 317—340, pls xx-xxix.
Janensch, W., 1911, Die Proboscidier-Schadel der Trinil-Expeditions-Sammlung, in Die Pithe-
canthropus-Schichten auf Java, 4 to, Leipzig, pp. 151 — 195, pls. xxi-xxv.
Kaup, J. J., 1835, Description des Ossements Fossiles. 4to, Darmstadt.
----, 1841, Akten der Urwelt, 8vo, Darmstedt.
Kerr, R., 1792, Animal Kingdom. 4to, London.
Koken, E., 1885, Ober fossile Saugethiere aus China. Palceont. Abhand., Ill, pp. 31—113 (1—
85), pls. vi—xii (i—vii).
Lortet, L., & Chantre, E., 1878, Recherches sur les Mastodontes. Arch. Mas. Lyon, II, pp.
285—311, pls. i—xvi bis.
de Lorenzo, G., & d’ERASMO, G., 1927, L’Elephas antiquus nell’ Italia Meridionale. Atti R.
Accad. Sci. Napoli, (2), XV]], No. 11.
Lydekker, R., 1877, Notices of New or Rare Mammals from the Siwaliks. Rec. Geol. Surv. India.,.
X, pp. 76-83.
----, 1880, Additional Siwalik and Narbada Proboscidea. Palceont. Indica, (10), I, pt. 5.
----, 1884, Additional Siwalik Perissodactyla and Proboscidea. Palceont. Indica, (10), III, pt. 1.
----, 1885, Catalogue of the Siwalik Vertebrates in the Indian Museum, 8vo, Calcutta.
----, 1886, Catalogue of the Fossil Mammals in the British Museum, IV, 8vo, London, Brit.
Mus. (Nat. Hist.)
Makiyama,-------, 1924. Notes on a Fossil Elephant from Totomi. Mem. Coll. Sci Kyoto Imp.
Univ., Ser. В, I, pp. 255—264. (Not seen. Reference from Osborn).
Matsumoto, H., 1915, On some fossil Mammals from Sze-Chuan, China. Sci. Rept. Tohoku
Imp. Univ., (2), III, pp. 1—28.
----, 1924, Preliminary Note on Fossil Elephants in Japan. Jour. Geol. Soc. Tokyo, XXXI, pp.
255. (Japanese only).
----, 1926, On two new Mastodonts and an Archetypal Stegodont of Japan. Sci. Rept. Tohoku
Imp. Univ., (2), X, pp. 1—11, pls. i—v.
----, 1928, On Leith-Adamsia siwalikiensis, a new generic and specific name of Archetypal Ele-
phants. Jap. Jour. Geol. Geogr., V, p. 213.
----, 1929 a, On Loxodonta (Palceoloxodon) namadica (Falconer and Cautley) in Japan. Sci.
Rept. Tohoku Imp. Univ., (2), XIII, pp. 1—5, pls. i—vi.
----, 1929 b, On Loxodonta (Palceoloxodon) Tokunagal Matsumoto, with Remarks on the Descent
of Loxodontine Elephants, Idem, pp. 7—11, pl. vii.
List of works consulted.
107
Matthew, W. D., 1918, Contributions to the Snake Creek Fauna. Bull. Amer. Mus. Nat. Hist.,
XXXV11I, Art. vii, pp. 183-229.
Matthew, W. D., & Granger, W., 1923, New Fossil Mammals from The Pliocene of Sze-Chuan,
China. Bull. Amer. Mus. Nat. Hist. XLVIII, Art. xvii, pp. 563—598.
Naumann, E., 1881, Ober japanische Elephanten der Vorzeit. Palaontographica, XXVIII. pp.
1—39, pls. i—vii.
Nesti, F., 1825, N. Giorn. Litt. (Pisa), XI, p. 211. (Not seen. Reference from ’’Index Animalium”).
Osborn, H. F., 1923, New Subfamily, Generic, and Specific Stages in the Evolution of the Pro-
boscidea. Amer. Mus. Novitates. No. 99.
----, 1924 a, Serridentinus and Baluchitherium, Loh Formation, Mongolia. Idem. No. 148.
—, 1924 b, Parelephas in relation to Phyla and Genera of the Family Elephantidas. Idem.
No. 152.
----, 1924 c, Additional Generic and Specific Stages in the Evolution of the Proboscidea. Idem.
No. 154.
----, 1925, Final Conclusions on the Evolution, Phylogeny, and Classification of the Probos-
cidea, Proc. Amer. Phil. Soc. LXIV, pp. 17—35.
----, 1926, Additional New Genera and Species of the Mastodontoid Proboscidea. Amer. Mus.
Novitates, No. 238.
----, 1929 a, New Eurasiatic and American Proboscidea, Idem, No. 393.
----, 1929 b, The Revival of Central Asiatic Life. Natural History, XXIX, pp. 3—16.
----, 1931, Palceoloxodon antiquus italicus sp. nov., Final Stage in the ’’Elephas antiquus”
Phylum. Amer. Mus. Novitates, No 460.
----, 1932, The Elephas Meridionalis Stage Arrives in America. Proc. Colorado Mus. Nat. Hist.
XI, No. 1.
----, & Granger, W., 1931, The Shovel-tuskers, Amebelodontins, of Central Asia. Amer. Mus.
Novitates, No. 470.
----, 1932, Platybelodon grangeri, Three Growth Stages and a new Serridentine from Mongolia
Idem. No 537.
Owen, R., 1870, On Fossil Remains of Mammals found in China. Quart. Jour, Geol. Soc., XXVI,
pp. 417—434, pls. xxvii—xxix.
Pavlow, M, 1894, Les Mastodontes de la Russie, Mem, Acad. Sci, St. Petersburg, [viii], I, fasc.
3, pp. 1—43, pls. i—iii.
----, 1901, Nouvelles Trouvailles de Mastodon Borsoni Lart, au Sud de la Russie. Anna. geol.
min. Passie, V, livr. 2—3, pp. 1—17, pl. i.
Pearson, H. S., 1928, Chinese Fossil Suidse, Palceont. Sinica, Ser. С, V. fasc. 5.
Pilgrim, G. E., 1910, Preliminary Note on a Revised Classification of the Tertiary Freshwater
Deposits of India. Pec. Geol. Surv. India, XL, pp. 185—204.
Pohlig, H., 1888, Dentition und Kranologie des Elephas antiquus Falc. Nova Acta Leop.-Carol.
Deutsch. Akad., LIU, pp. 1—279, pls. i—x.
Schlesinger, G., 1912, Studien fiber die Stammesgeschichte der Proboszidier. Jahrb. k.-k. geol.
Peichs., LXII, pp. 87—182, pls. vi—vii.
----, 1917, Die Mastodonten des k.-k. Hofmuseums, Denkschr. Naturh. Hofmus. Wien, I.
----, 1922, Die Mastodonten der Budapester Sammlungen. Geologica Hungarica, II, fasc. 1.
Schlosser, M., 1903, Die Fossilen Saugethiere Chinas. Abhand. bayer. Akad. Wiss., II CL, XXII,
pp. 1—221, pls, i—xiv.
Soergel, W., 1912, Elephas trogontherii Pohl., und Elephas antiquus Falc., Palceontographica,
LX, pp. 1—114, 8 tables, pls. i—iii.
----, 1921, Elephas Columbi Falconer. Geol. Palceont. Abhand., N. F., XIV, Heft 1—2.
108 List of works consulted.
Teilhard de Chardin, 1923, Cenozoic Vertebrate Fossils of E. Kansu and Inner Mongolia. Bull.
Geol. Soc. China, II, nos. 3—4, pp. 1—3.
Thomas 0., 1895, An Analysis ol the Mammalian Generic Names given in Dr. C. W. L. Gloger’s
’’Naturgeschichte” (1841). Ann. Mag. Nat. Hist., (6), XV, pp. 189—193.
Warren, J. C., 1852, The Mastodon Giganteus of North America. 4to Boston.
Weithofer, K. A., 1890. Die Fossilen Proboscidier des Arnothales in Toskana. Beitr. Palaont.
Osterr.-Ung., VIII, pp. 107—240, pls. i—xv.
Vacek, M., 1877, Ober Osterreichische Mastodonten. Abhand. k.-k. geol. Reichs, VII, Heft. 4.
EXPLANATION OF PLATES.
PLATE I.
PLATE I.
Trilophodon wimani sp. nov.
Fig. 1. Left Cuneiform. Anterior aspect.
2. The same. Posterior aspect.
3. The same. Superior aspect.
4. The same. Inferior aspect.
5. Left Lunar. Superior aspect.
6. The same. Inferior aspect.
7. The same. Internal aspect.
8. Left Unciform. Anterior aspect.
9. Left Cuboid. Anterior aspect.
L., facet for Lunar; facet for Magnum; P., facet for Pisiform; S., facet for Scaphoid
Ul., facet for Ulna; Un., facet for Unciform.
Note. — The drawings on plates I—IV are al] semi-diagrammatic; dotted areas represent abrasions; shaded
areas are fractures. AH the drawings on one plate are to the same scale, but the scale varies from plate
to plate.
PAL/EONTOLOGIA SINICA. SER. C. VOL. 9, FASC. 3.
A. Tindell Hopwood, Fossil Proboscidea from China.
Plate I.
8.
о
PLATE II.
PLATE К.
Mastodon sp. indet.
Fig. 1. Right Unciform. Inferior aspect.
2. The same. Internal aspect.
3. Left Ectocuneiform. Superior aspect.
4. Left Third Metacarpal. Superior aspect.
? Pentalophodon sinensis sp. nov.
8. Left Second Metacarpal. Superior aspect.
Mastodontoidea incertae sedis.
9. Left Third Metacarpal. Superior aspect.
Palaeoloxodon antiquus.
6. Right Third Metacarpal. Superior aspect.
(Pleistocene of Upnor, Kent. Regd. Ml 1156. Brit. Mus. Geol. Dept.)
7. The same.
(Pleistocene of Grays, Essex. Regd. 18248. Brit. Mus. Geol. Dept.)
Mammuthus primigenius.
5. Left Third Metacarpal. Superior aspect.
(Pleistocene of Slade Green, Kent. Regd. 23720e. Brit. Mus. Geol. Dept.)
M, facet for Magnum; M, facet for Navicular; Trz., facet for Trapezoid; //, ///, IV,
V, facets for Metacarpals II—V.
PALZEONTOLOGIA SINICA. SER. C. VOL. 9, FASC. 3.
15—42992]. A, Tindell Hopwood.
PLATE 1П.
PLATE III.
Trilophodon wimani. sp. nov.
Fig. 5. Left Cuneiform. Anterior aspect.
? Pentalophodon sinensis sp. nov.
2. Left Cuneiform. Anterior aspect.
7. Right Second Metacarpal. Anterior aspect.
Mastodontoidea incertae sedis.
6. Left Fourth Metacarpal. Anterior aspect.
Archidiskodon meridionalis.
1. Left Cuneiform. Anterior aspect.
(Lower Pleistocene of Clacton, Essex. Regd. 28007. Brit. Mus. Geol. Dept.)
Palaeoloxodon antiquus.
3. Right Cuneiform. Anterior aspect.
(Middle Pleistocene, Grays, Essex. Regd. 18246. Brit. Mus. Geol. Dept.)
Mammuthus primigenius.
4. Right Cuneiform. Anterior aspect.
(Upper Pleistocene, Ilford, Essex. Regd. 45114. Brit. Mus. Geol. Dept.)
Un., facet for Unciform.
palzeontologialsinica. SER. C. VOL. 9, FASC. 3.
PLATE IV.
PLATE IV.
Mastodontoidea incertae sedis.
Fig. 1. Left Magnum. Superior aspect.
2. The same. Inferior aspect.
3. The same. Internal aspect.
4. The same. External aspect.
5. Left Lunar. Superior aspect.
6. The same. Inferior aspect.
7. The same. External aspect.
8. Left Trapezoid. Internal aspect.
9. The same. Superior aspect.
10. The same. Inferior aspect.
11. Left Second Metacarpal. Superior aspect.
12. Left Fourth Metacarpal. Superior aspect.
13. Diagram of the above bones when articulated. Anterior aspect. Not to scale.
Palaeoloxodon antiquus recki.
14. Diagram to illustrate the usual relations of the same bones in the proboscidean
carpus. Adapted from Dietrich, 1916, pl. VIII, fig. 1. Not to scale.
C., facet for Cuneiform; L., facet for Lunar; Lun.., Lunar; M., facet for Magnum; Mag.,
Magnum; Me. //, Me. Ill, Me. V, facets for second, third, and fifth Metacarpals; T.,
facet for Trapezium; Trap., Trapezoid; Trz., facet for Trapezoid; Ul., facet for Ulna;
Un., facet for Unciform; //, ///, V, facets for second, third, and fifth Metacarpals.
PALZEONTOLOGIA SINICA. SER. C. VOL. 9, FASC. 3.
A. TfNHFH RnDU/nnn Rnooif г___ Г' L • - _
PLATE V.
PLATE V,
Trilophodon connexus sp. nov.
Fig. 1. Left mandibular ramus with second and third molars. Holotype.
2. Left upper third molar.
Trilophodon wimani sp. nov.
3. Palate with second and third molars. Holotype.
Figs. 1, 3, are y2 natural size. Fig. 2 is natural size.
PA L/EONTOLOG IA SINICA. SER. C. VOL. 9, FASC. 3.
Plate V.
A. Tindell Hopwood, Fossil Proboscidea from China.
PLATE VI.
PLATE VI.
Trilophodon wimani sp. nov.
Fig. 1. Left lower third molar.
Trilophodon spectabilis sp. nov.
2. Left lower third molar, lacking talonid. Holotype.
Tetralophodon exoletus sp. nov.
3. Right lower third molar. Holotype.
4. Right mandibular ramus with fourth deciduous molar, third premolar, and
first molar. Paratype.
Mastodon americanus.
5. Left ramus of juvenile mandible with the deciduous dentition.
All figures are natural size.
PALZEONTOL.O Gl A S I N I C A. SER. C. VOL. 9, FASC. 3.
Plate VI.
A. Tindell Hopwood, Fossil Proboscidea from China.
1C-429921, Л, Tindell Hopwood.
PLATE VII.
PLATE VII.
Mastodon borsoni.
Fig. 1. Left lower third molar.
Pentalophodon sinensis sp. nov.
2. Second and third left upper molars. Holotype.
Stegodon officinalis sp. nov.
3. Anterior two ridges of an unworn lower molar. Holotype.
4. Last ridge and talon of an unworn upper (?) molar.
Stegodon zdanskyi sp. nov.
5. First four ridges of a third lower molar. Holotype.
Stegodon orientalis.
6. First upper molar, Regd. M10270. Brit. Mus. Geol. Dept.
Stegodon orientalis grangeri.
7. Right lower first molar. Regd. M12209. Brit. Mus. Geol. Dept.
All figures are '/, natural size.
PALZEONTOLOGIA
SER. C. VOL. 9, FASC. 3.
A. Tindell Hopwood, Fossil Proboscidea from China.
Plate VII.
2.
PLATE VIII.
PLATE VIII.
Archidiskodon cf. planifrons.
Fig. 1. Upper third molar.
Palaeoloxodon tokunagai.
2. Portion of right mandibular ramus with the second molar.
Palaeoloxodon cf. namadicus.
3, Juvenile left maxilla with the first and second deciduous molars.
4. Part of a left upper third molar. Regd. 29007. Brit. Mus. Nat. Hist. The
first proboscidean recorded from China.
Mammuthus primigenius.
5. Left upper third molar. Regd. M10491. Brit. Mus. Nat. Hist.
All figures, except fig. 3, are natural size. Fig. 3 is natural size.
P A L /Е О N Т О L О G I A St Гм iCA. SER. С. VOL. 9, FASC. 3.
A. Tindell Hopwood, Fossil Proboscidea from China.
Plate VIII.
Palceoloxodon cf. namadicus °
* Mammuthus primigenius °
Elephantidaz
- Фек-Ж - °
~ ' 7^r^^S^<a? - r=^4W^N£5!fv
«) ° - е°1М^^К7^ ~ °
IE . • i 1 • э е^Бёйё!-Н 1нгж^^<
айМ^^г - - IH’ZS^W °
Е |j?
Ш
-ft- 4a 11
* Mastodon borsoni )ST *“ “H ®
* Mastodon SP- о
^S-HcSsS Mastodoniidte
* Pentalophodon sinensis •"
? Pentalophodon sinensis °
unodoniidce
* Stegodon officinalis ~ °
* Stegodon zdanskyi ~ S'S* й| К" К °
* Stegodon orientalis £{4<°
* Stegodon orientalis grangeri ~ °
Stegodon sinensis
Stegodon aff. bombifrons
Stegodontidae
* Archidiskodon cf. planifrons ~ U_ Jg^c
Palceoloxodon tok.anagai 4“^ -\l
* Trilophodon connexus С Щ ~ £
Trilophodon wimani ^;§g - С Кг^ ) * ШГ^ГТ^ *
Trilophodon spectabilis *- К- =§> °
Serridentinus mongoliensis ©
Serridentinus gobiensis x^nw - -н-s^ о
Serridentinus /lorescens о
Platybelodon grangeri * 'Э’ЙёЗё °
*’ Tetralophodon exoletus ~ “ fg[~ ! 1 ] Ц? Й$т*Ф
Tetralophodon (?) sinensis °
Pal<EomastodontidcE
* Mastodon americanus -3 fM ~ "H "
ж w
*tb Й
д
©Чй
> й
Я? ф
йч л
Ж W
ед й
«Г й
$1 й
ah й
)5f 4Ь
"и"
Л
М ffr
₽П
и
W. Л
Л
ж
ж
ж
ль й
W д
М
№ '!
Й9 £
Й W
ЛЁ Ж
W Й
® i
&r 4t
& Ж
№ -А
и t
й
йй
в
ffi
f