




















































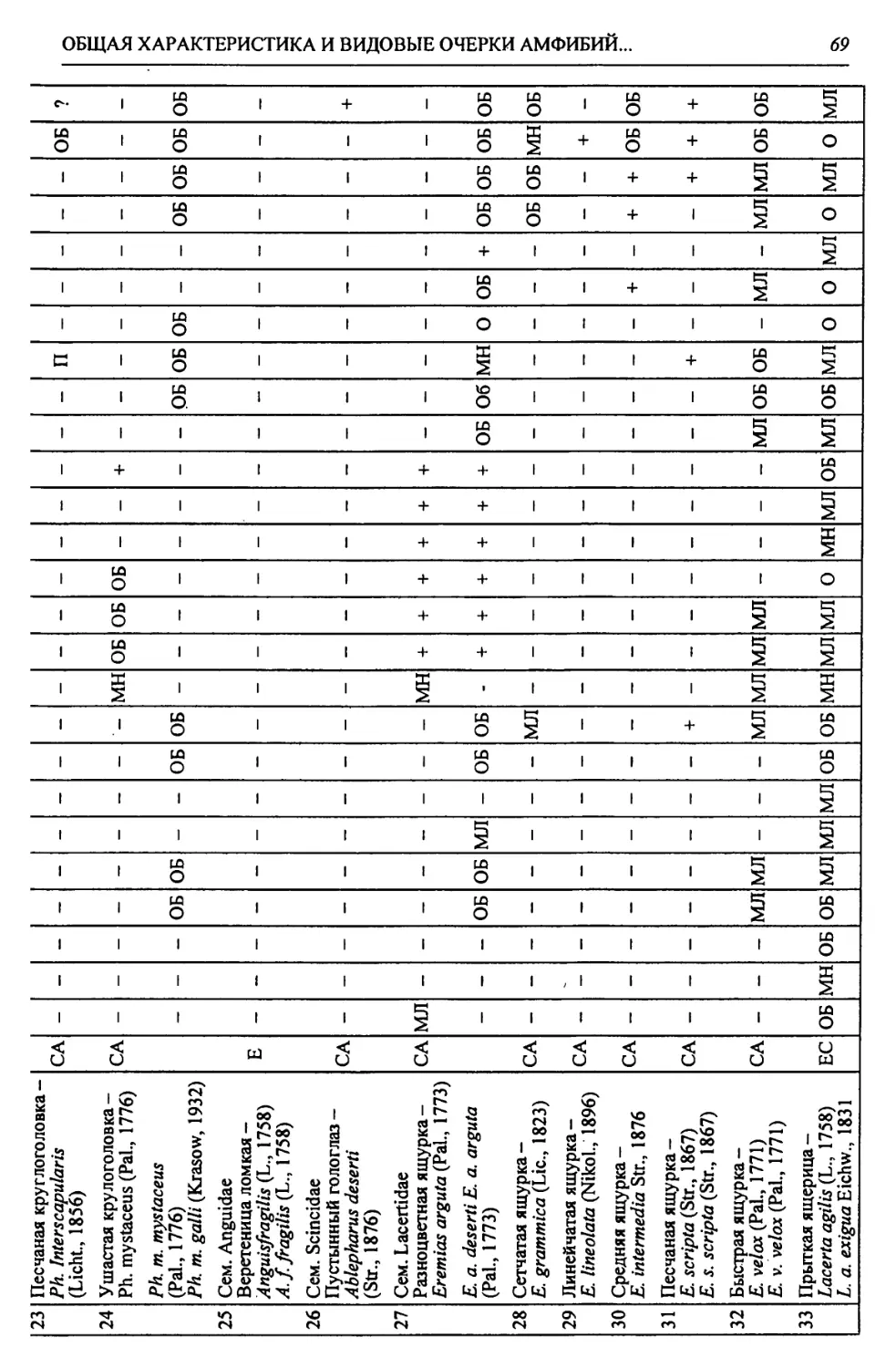
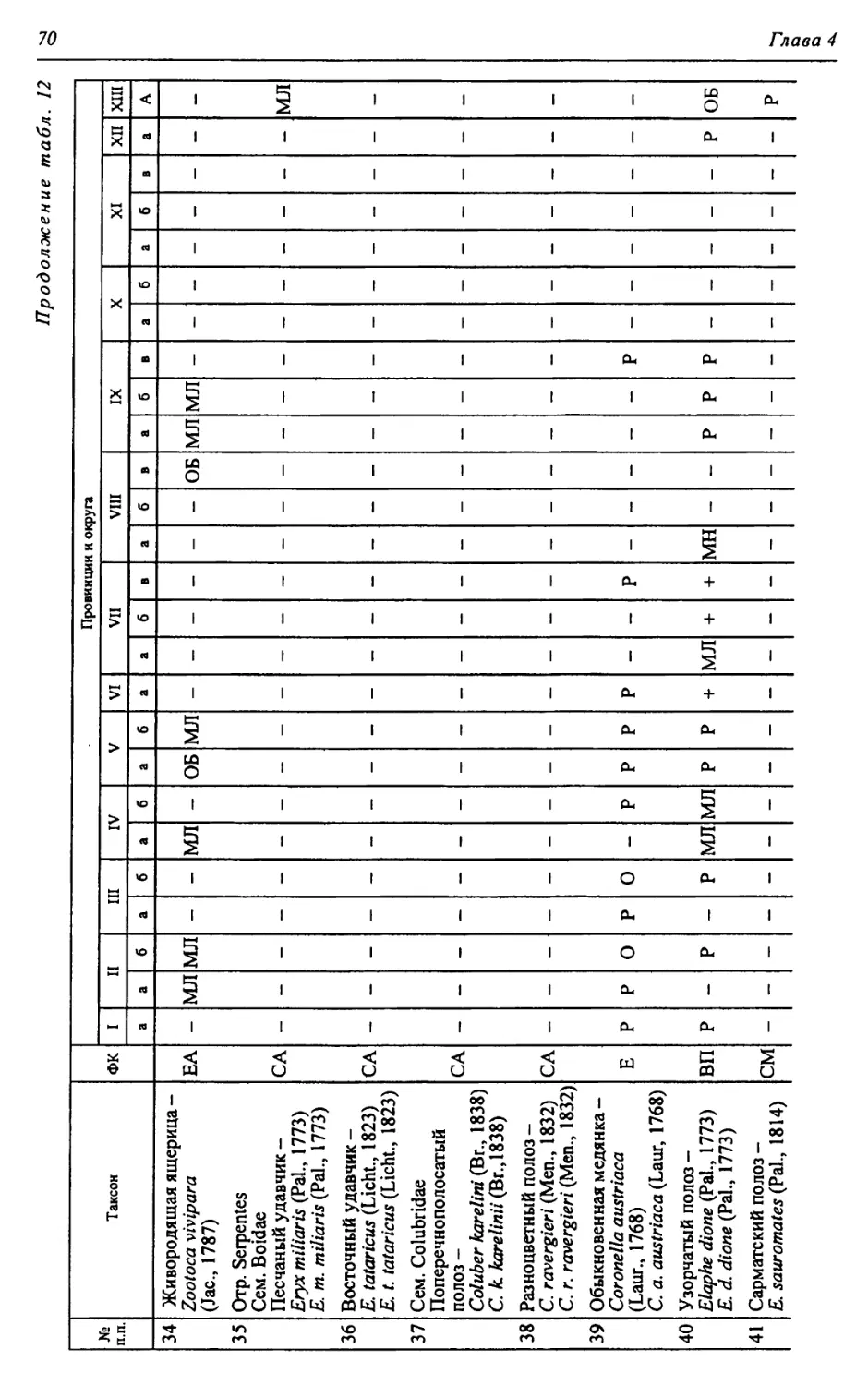
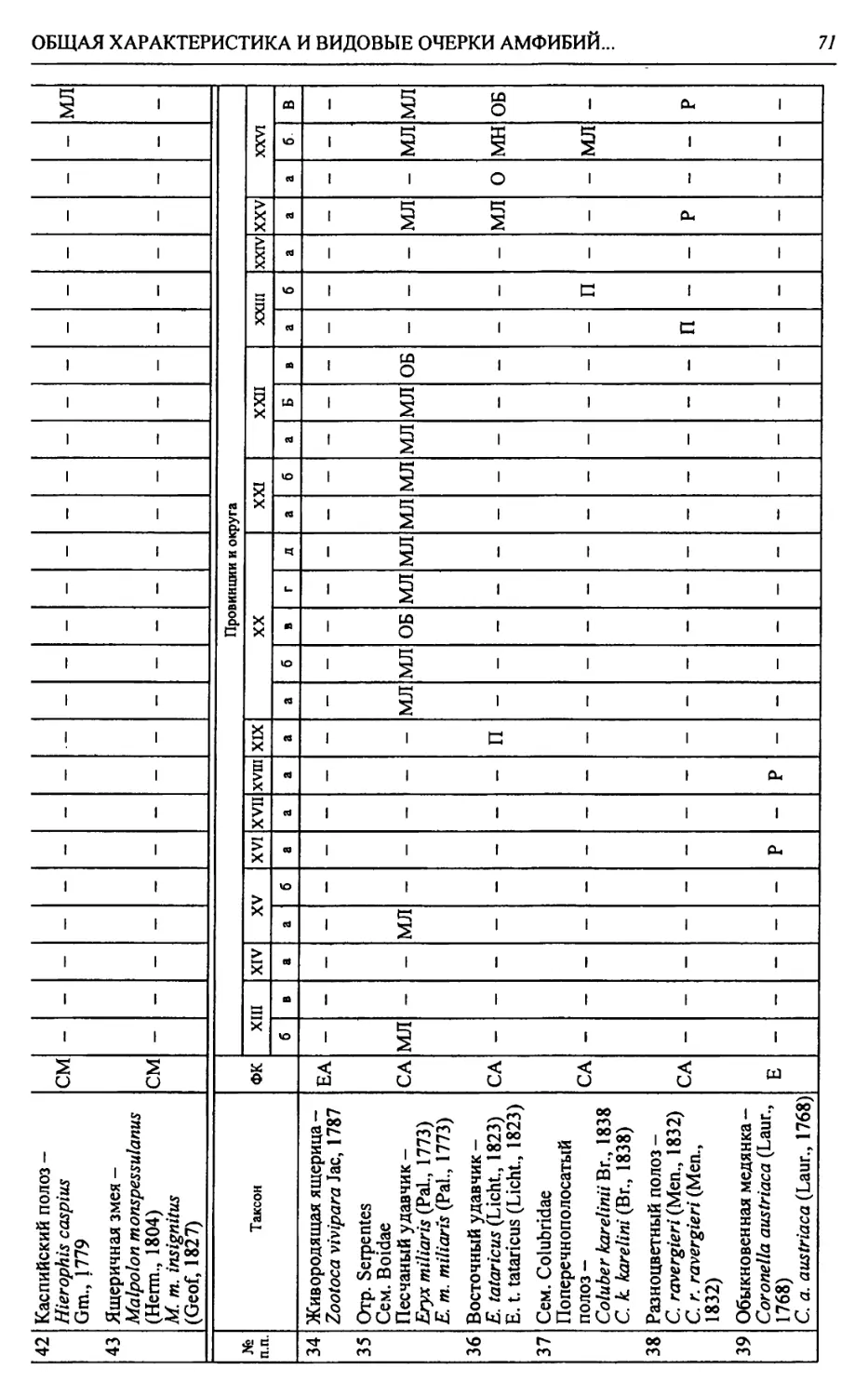
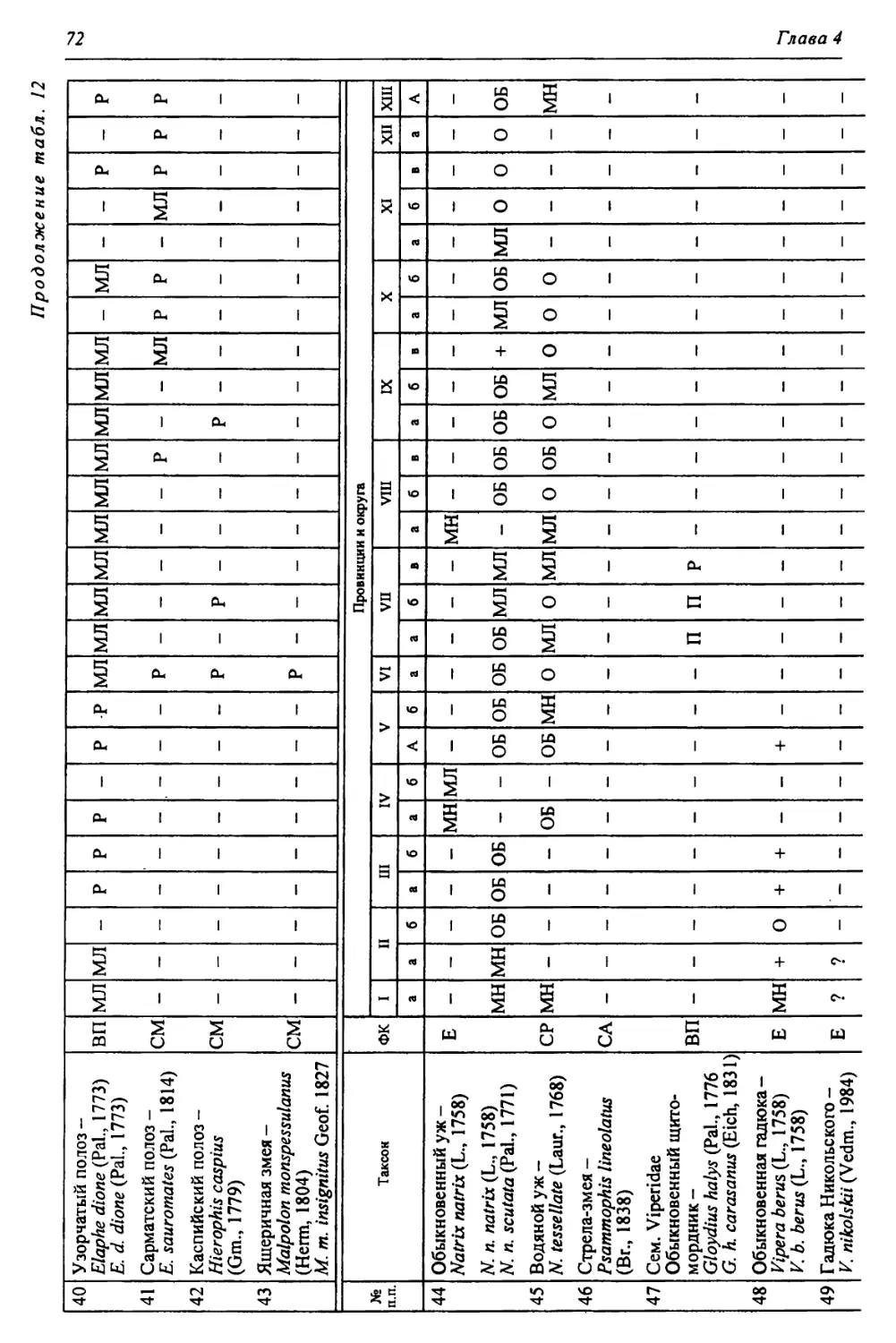





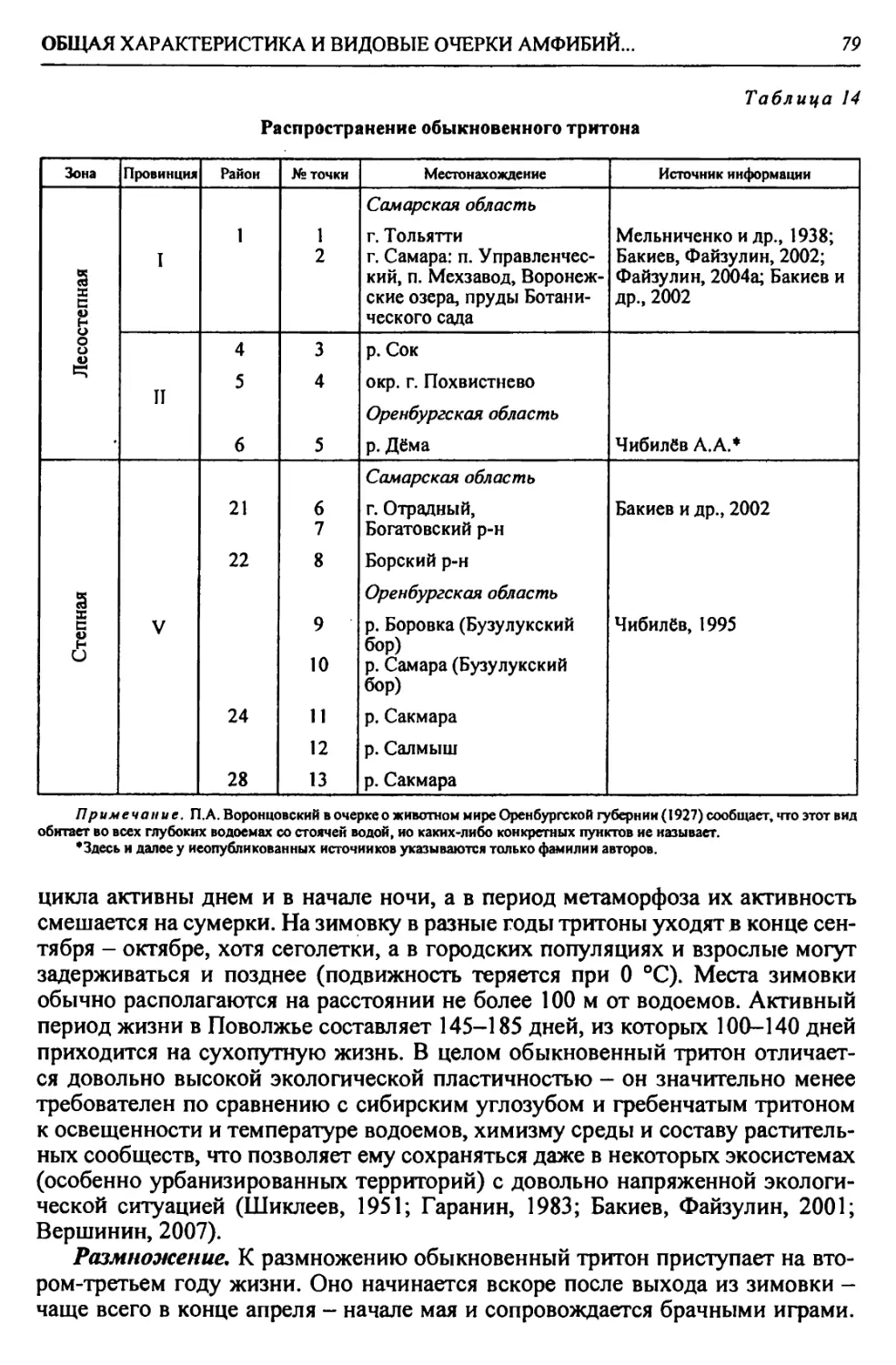





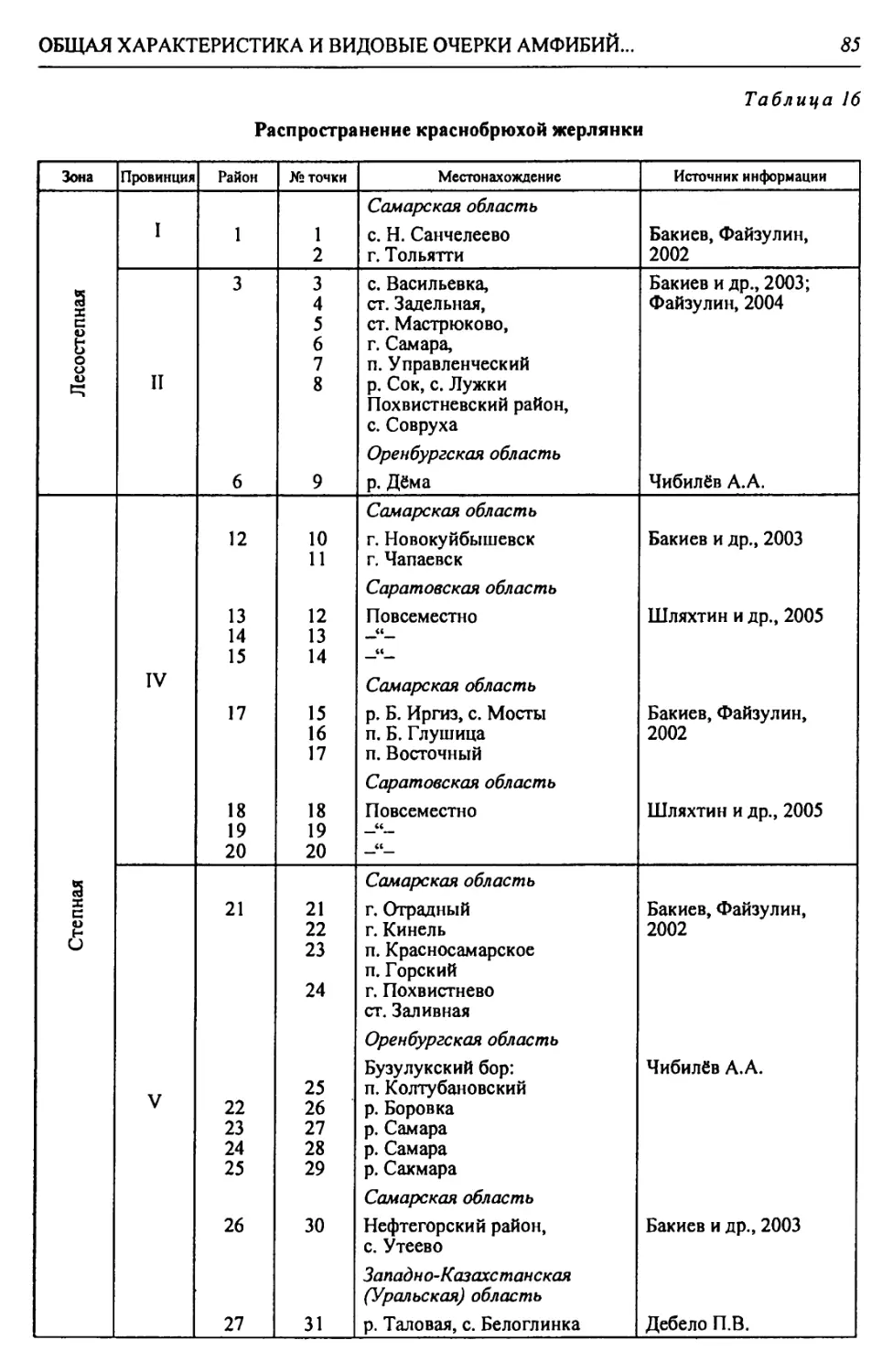
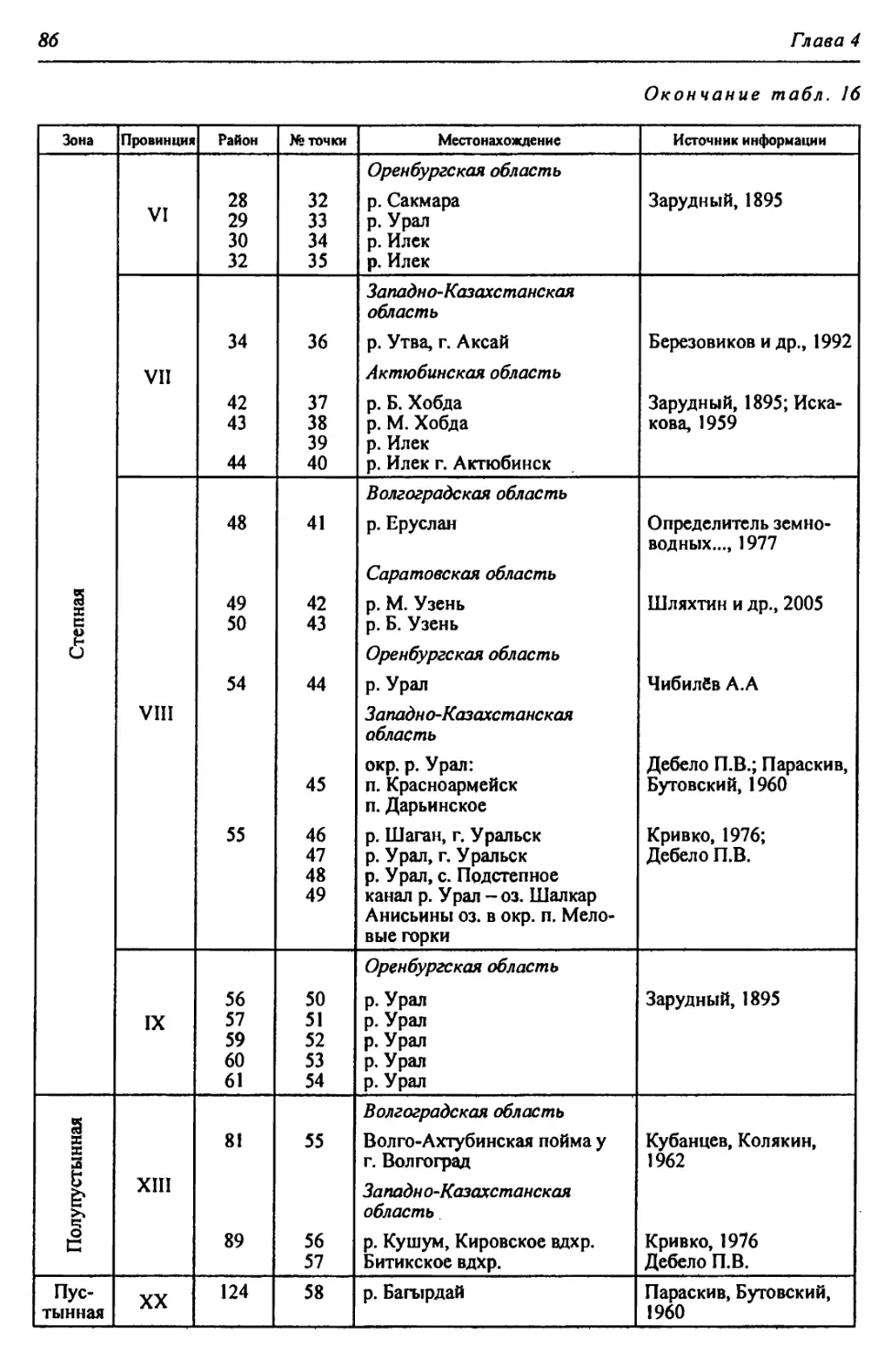






























































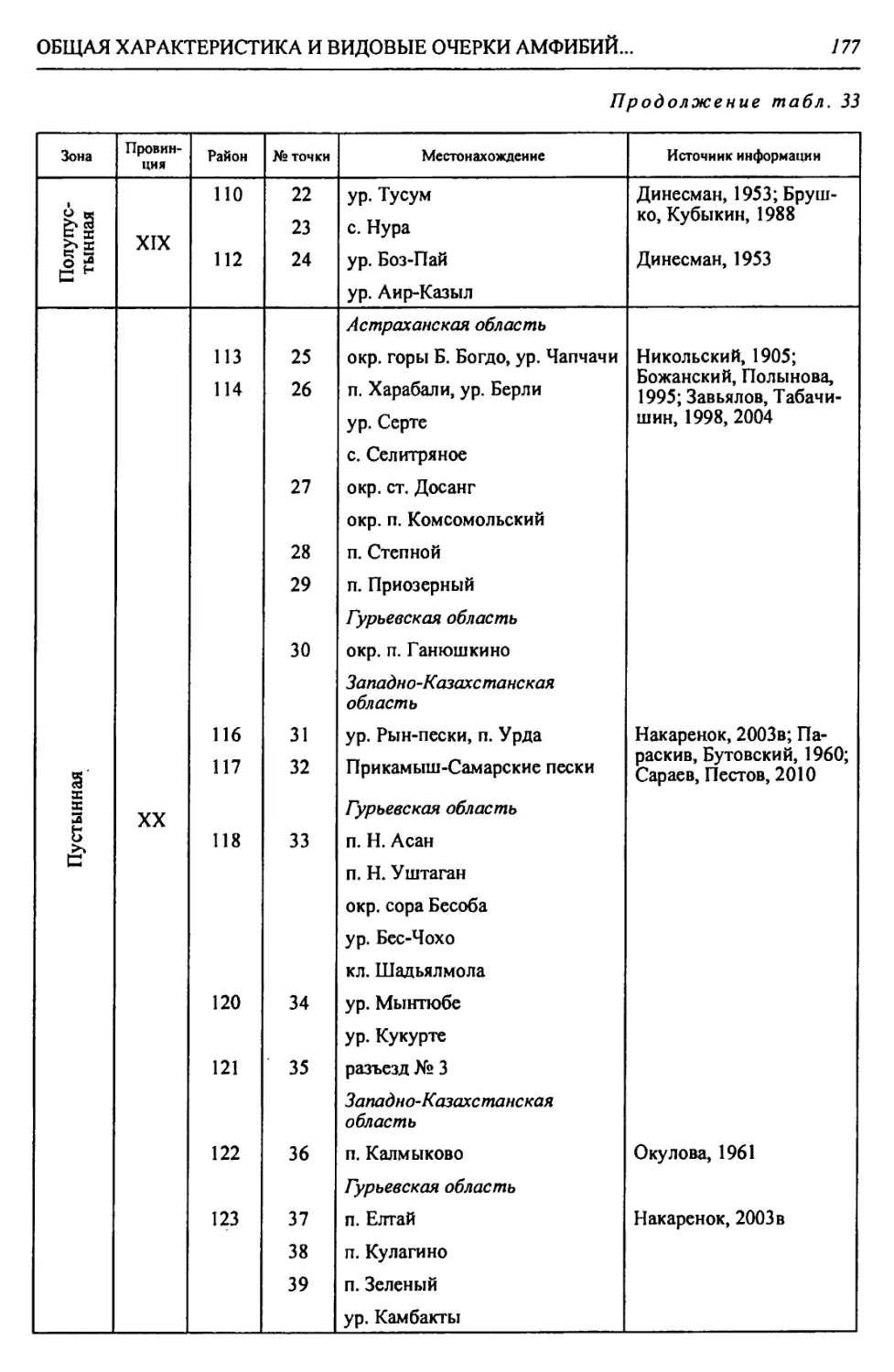




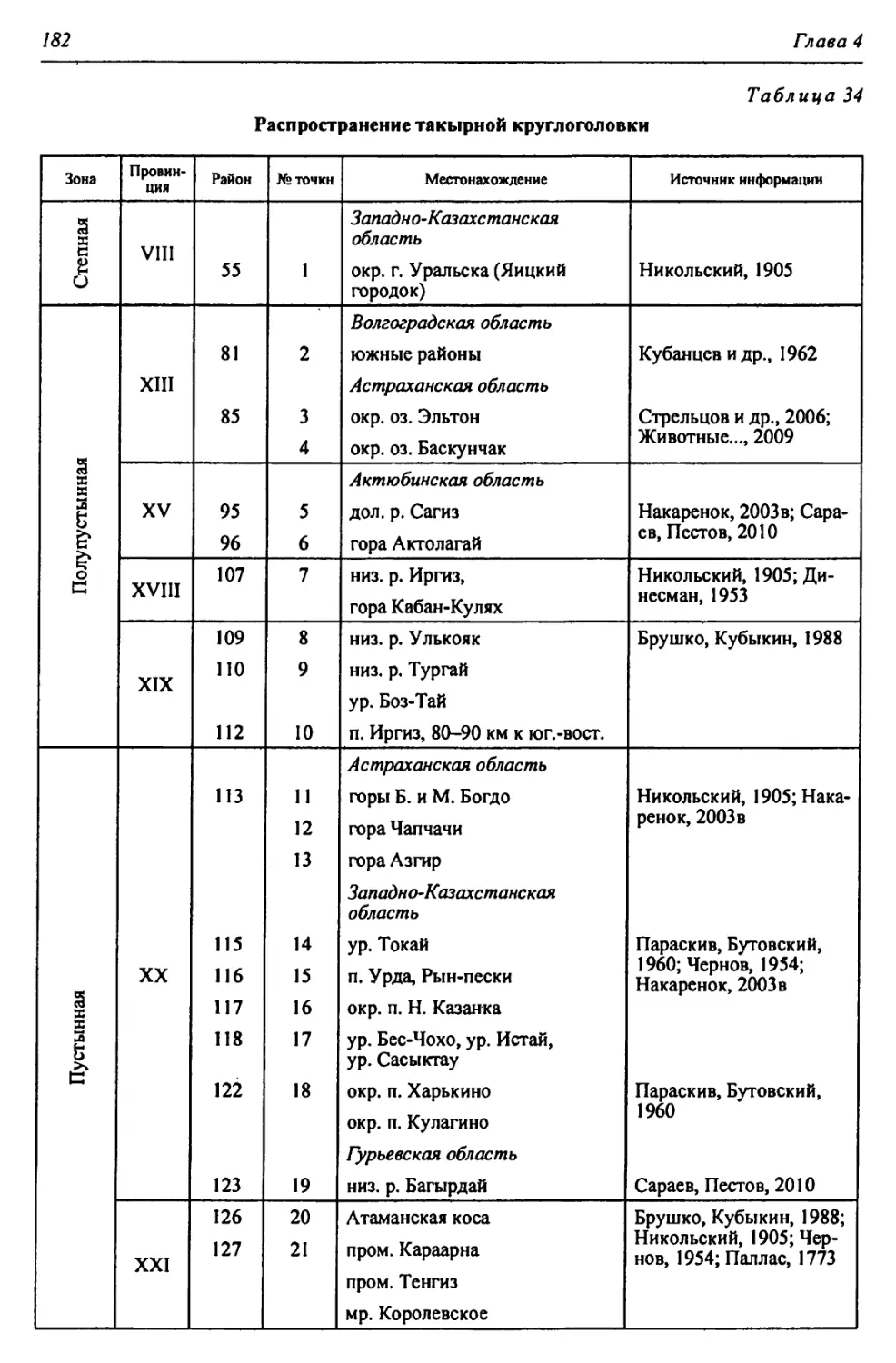
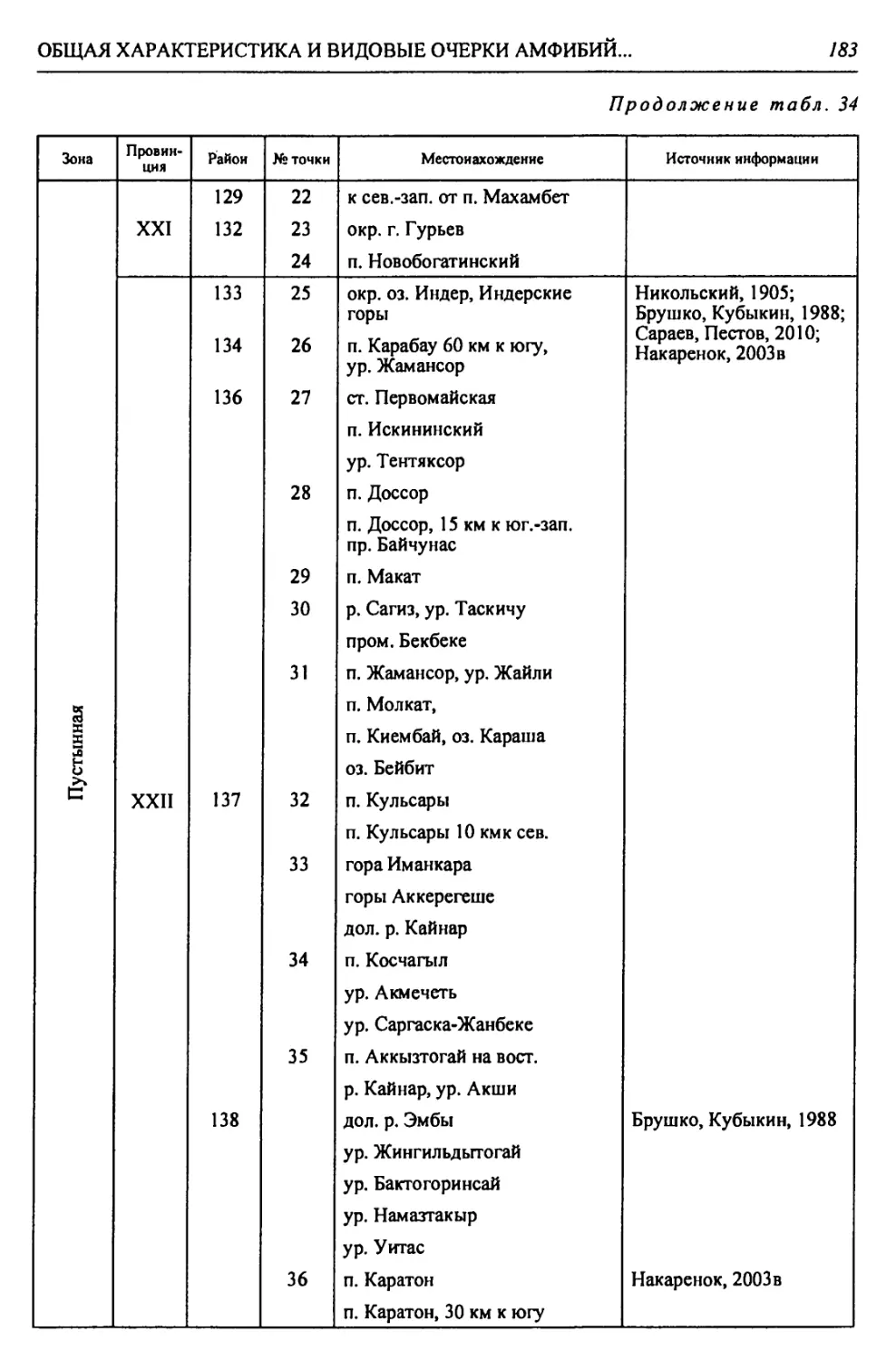
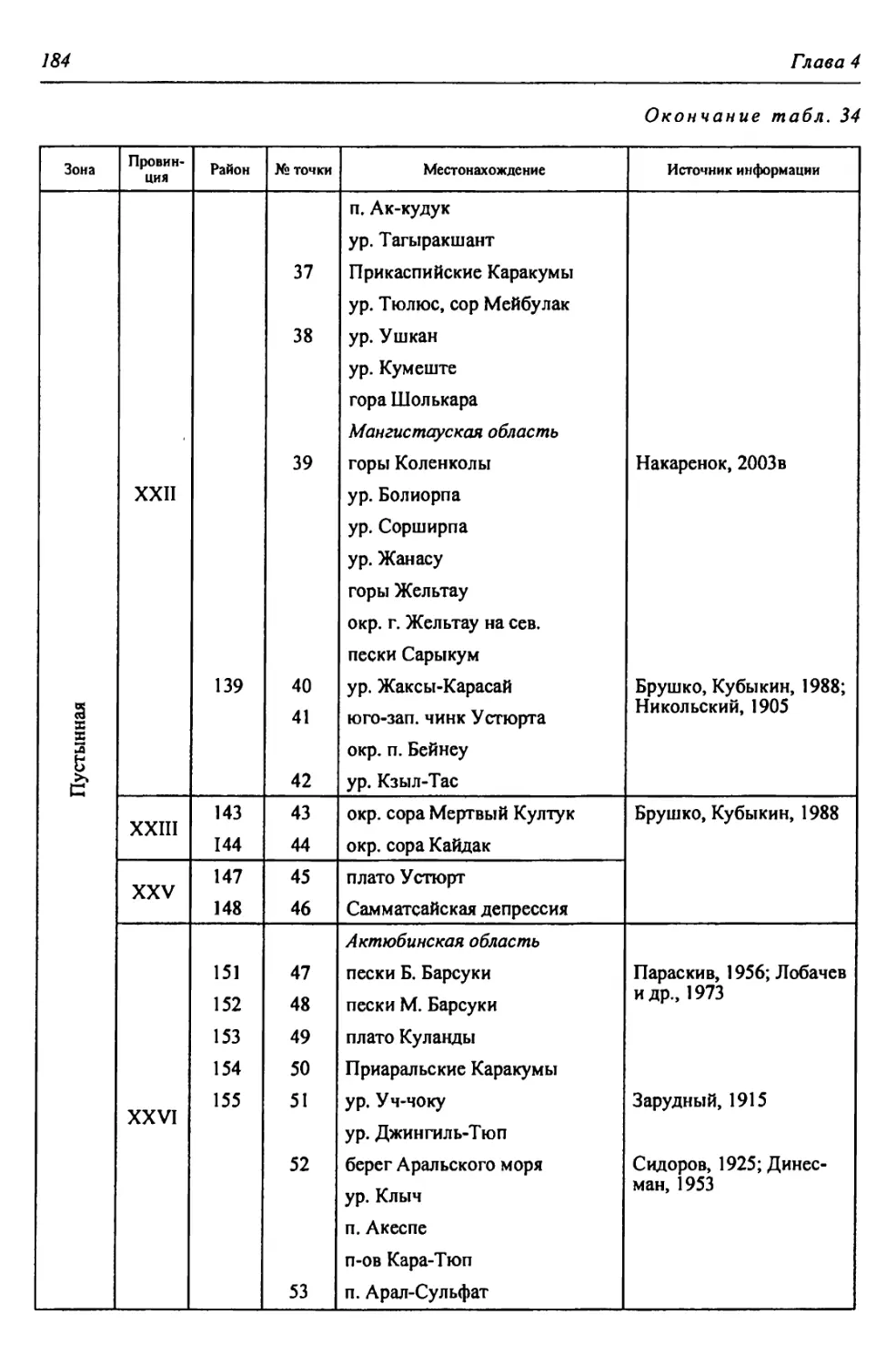









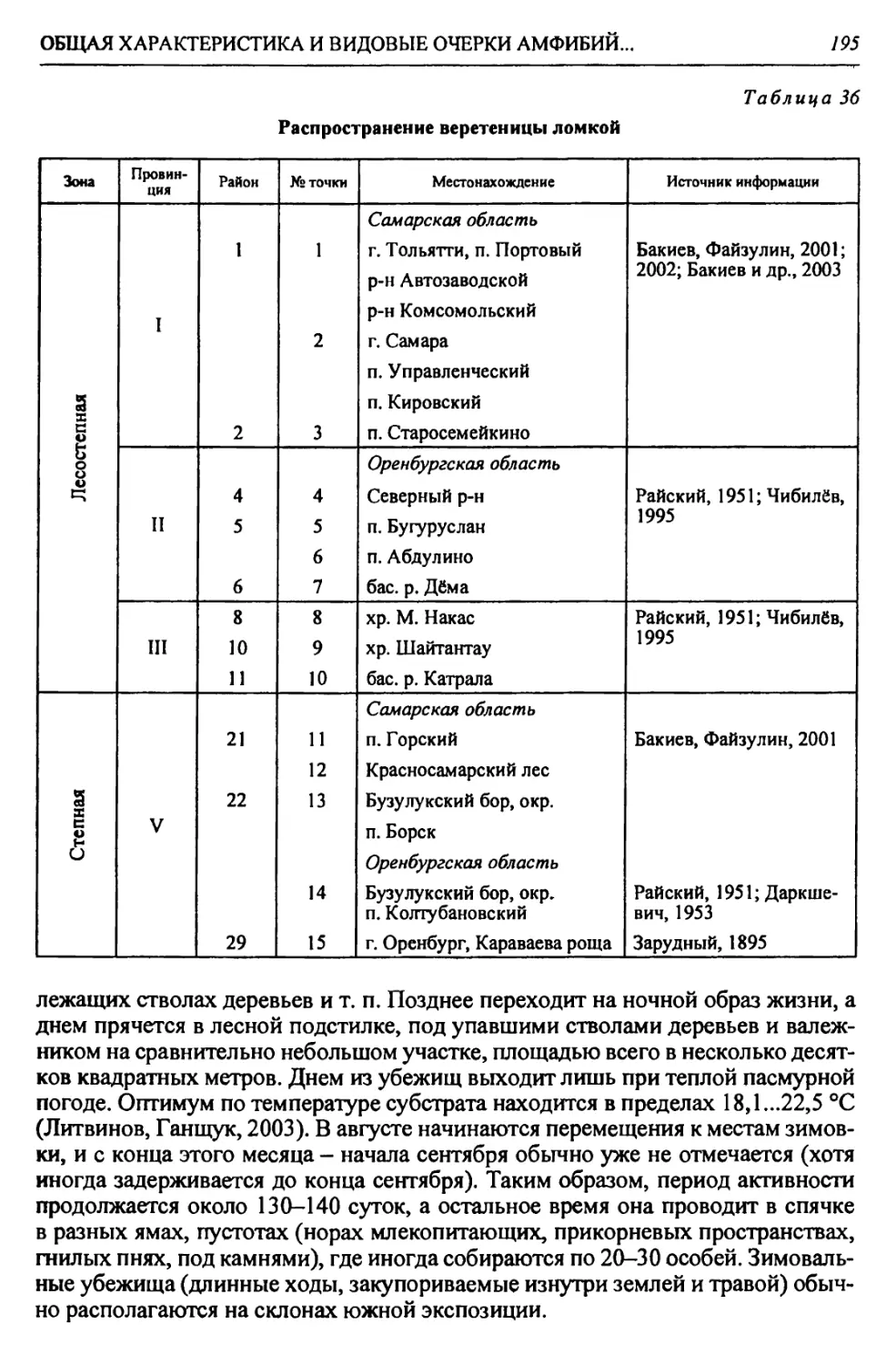




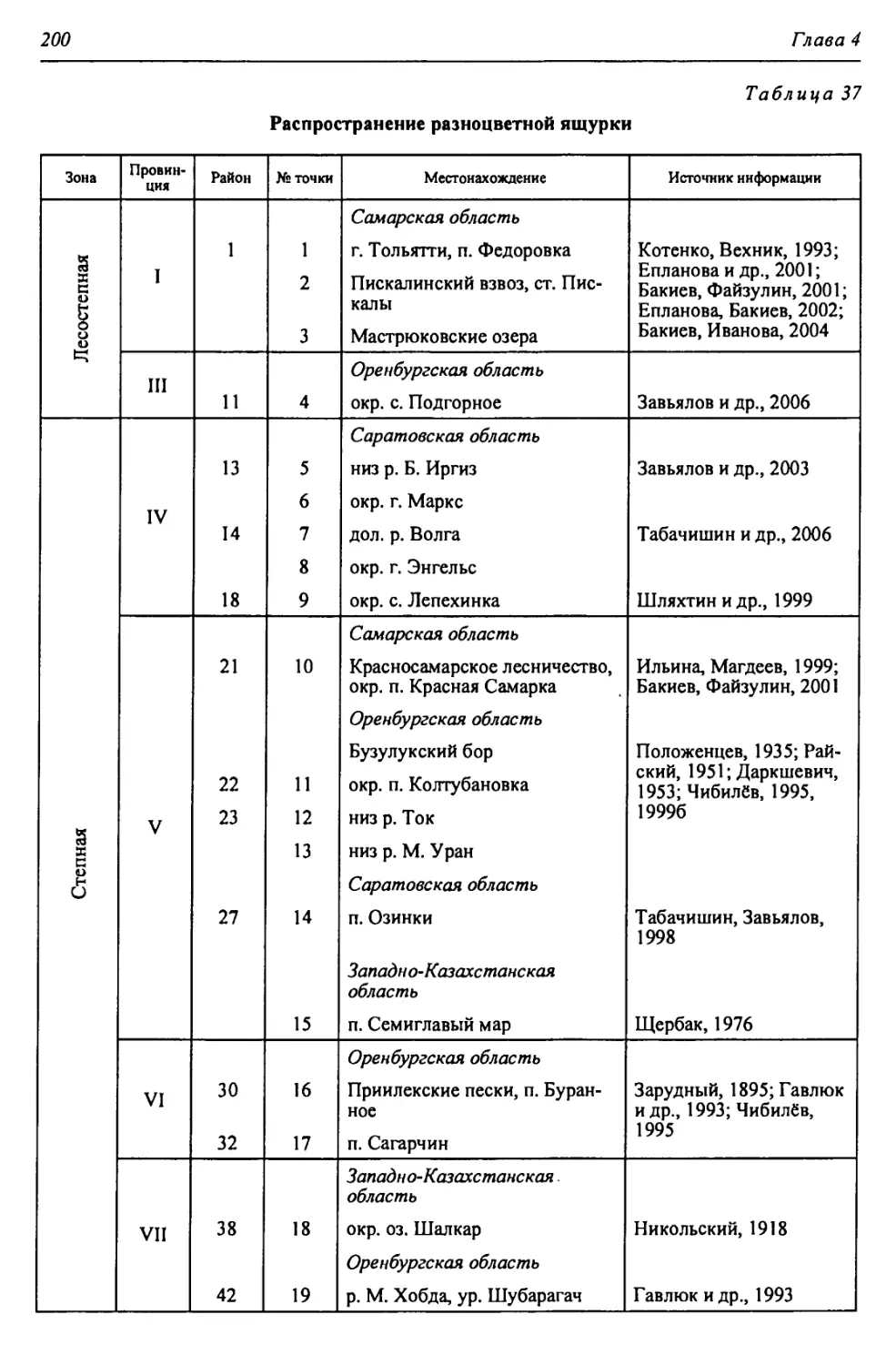
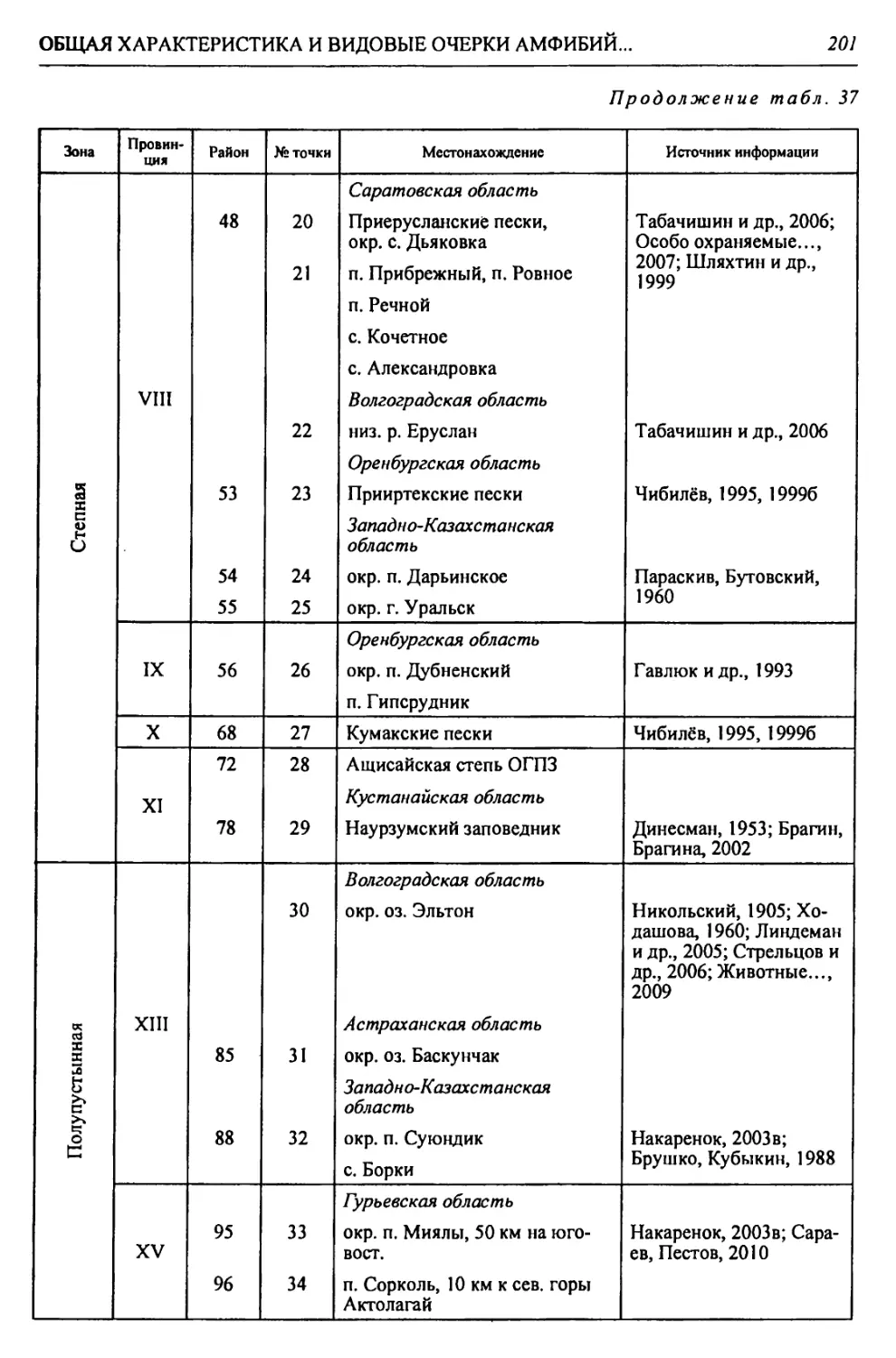







































































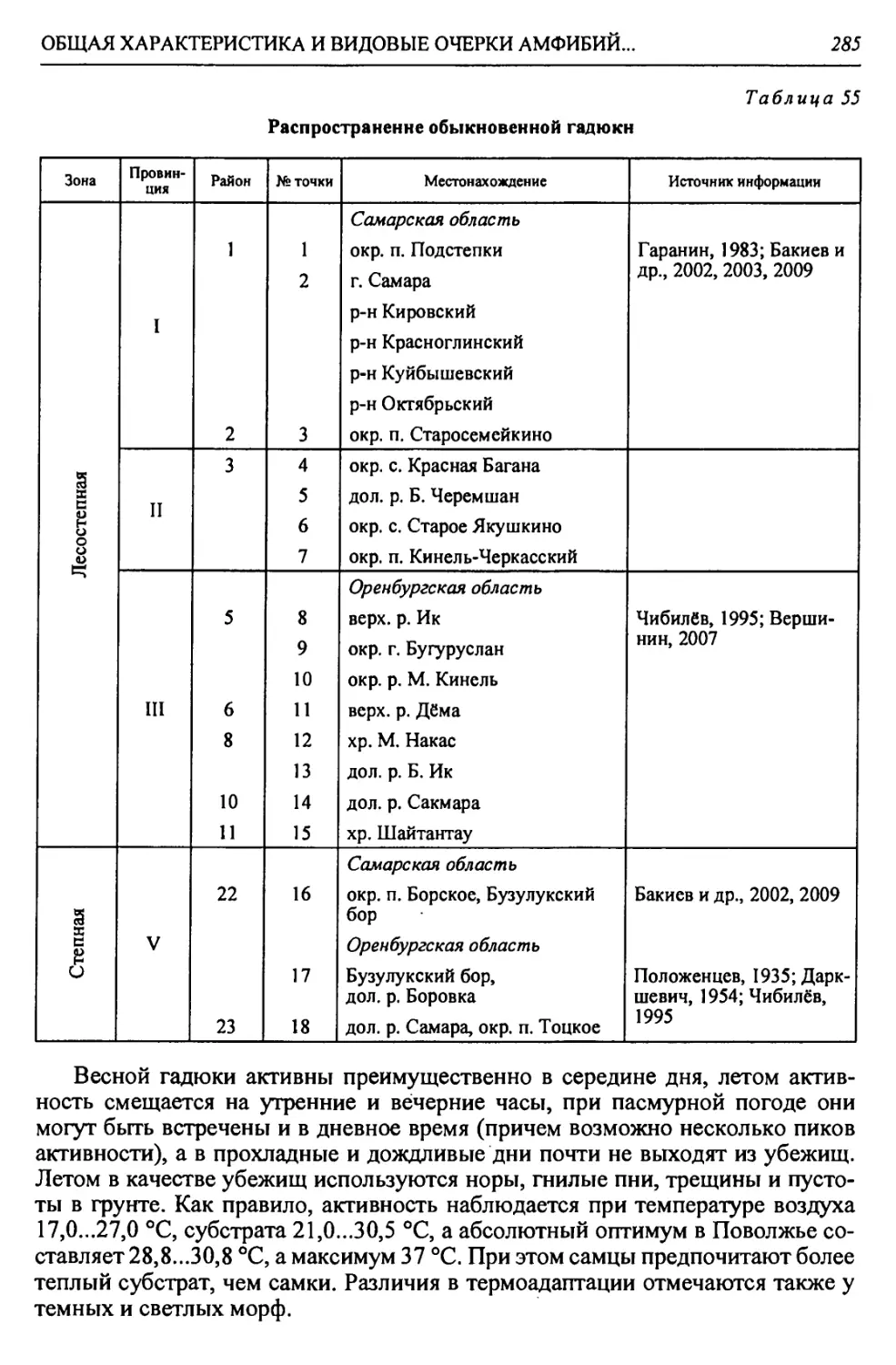



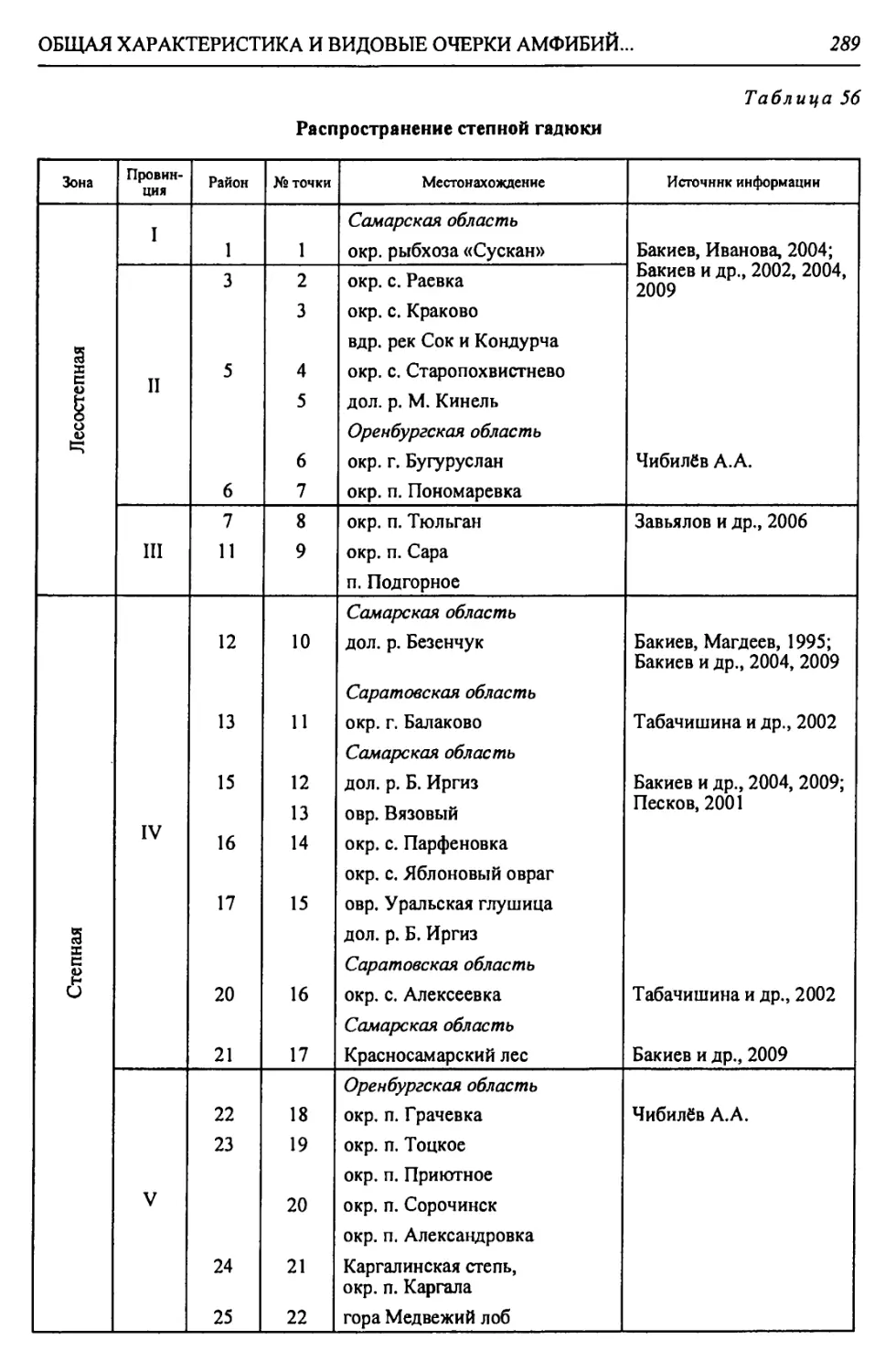
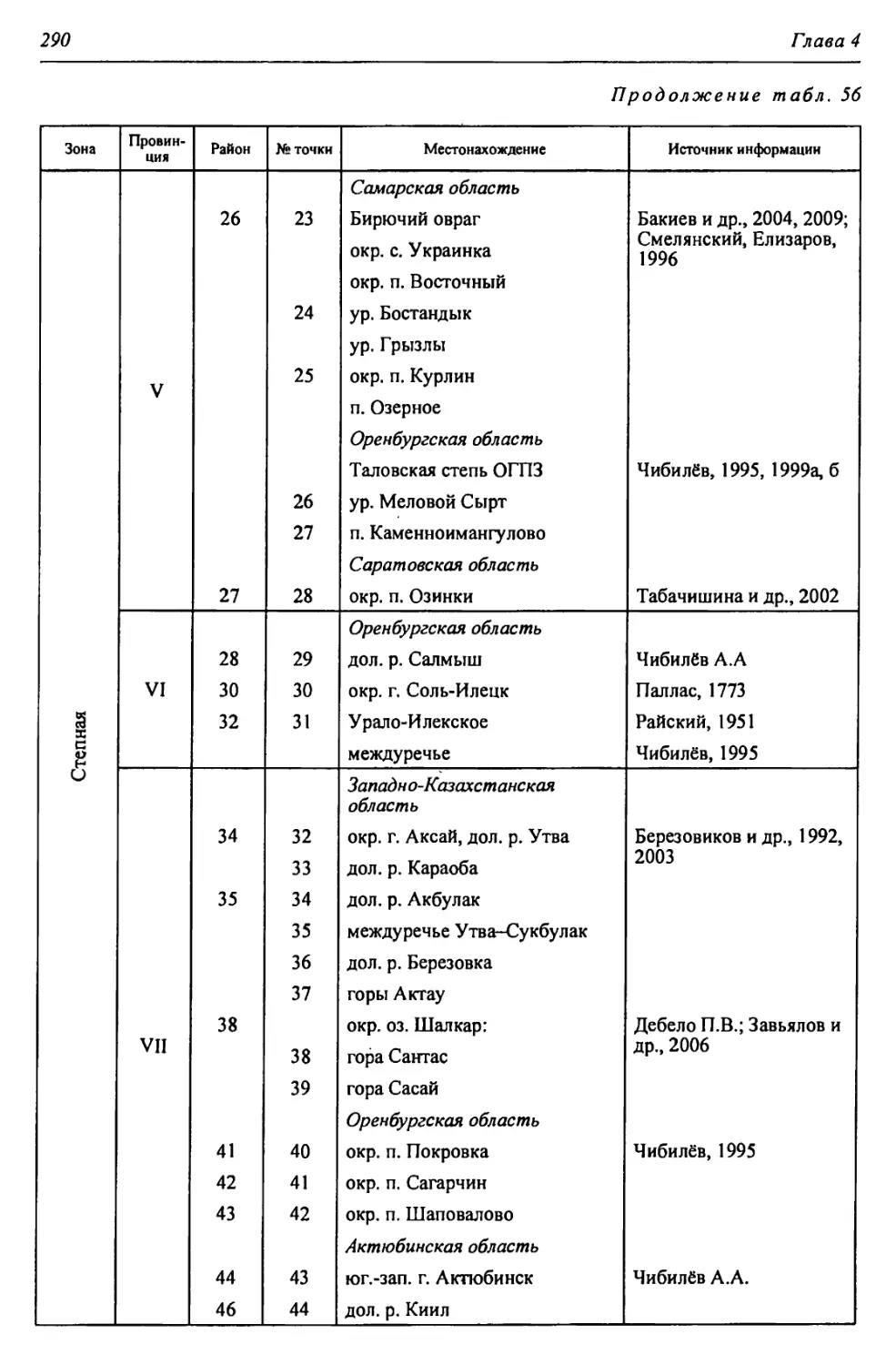























































Автор: Чибилёв А.А. Дебело П.В.
Теги: общая экология биоценология гидробиология биогеография экология и биогеография охрана живой природы биология зоология рептилии амфибии
ISBN: 978-5-7691-2363-4
Год: 2013




П.В. ДЕБЕЛО
А.А. ЧИБИЛЁВ
АМФИБИИ
И РЕПТИЛИИ
УРАЛО-
КАСПИЙСКОГО
РЕГИОНА
•НИИ
|«|ЙЙЫШГ|М ми
о
Дими*пн>| | » ।
' II Н. | |Ц I
fr-HWW ' I' •
•>ГУ»*М
Туйма>ы
ркт^Орьскнн
• 1а1ьятги
©Бугуруслан
( 14 'р ><Ь
I И|>
О \
И<И1 Ф
Ь • । аново
нН*1
htfCIU
©к?
Белебрй
*1 tang
I IРАМАНЬ
ЧЫГ ’•>
о
ку«нк|и ,ч>
>
О - САМАРА
Нокй^Дбы шевс к
УРАЛЬСК
оз. Шалкар
УФА
а
С терлйтймак V.
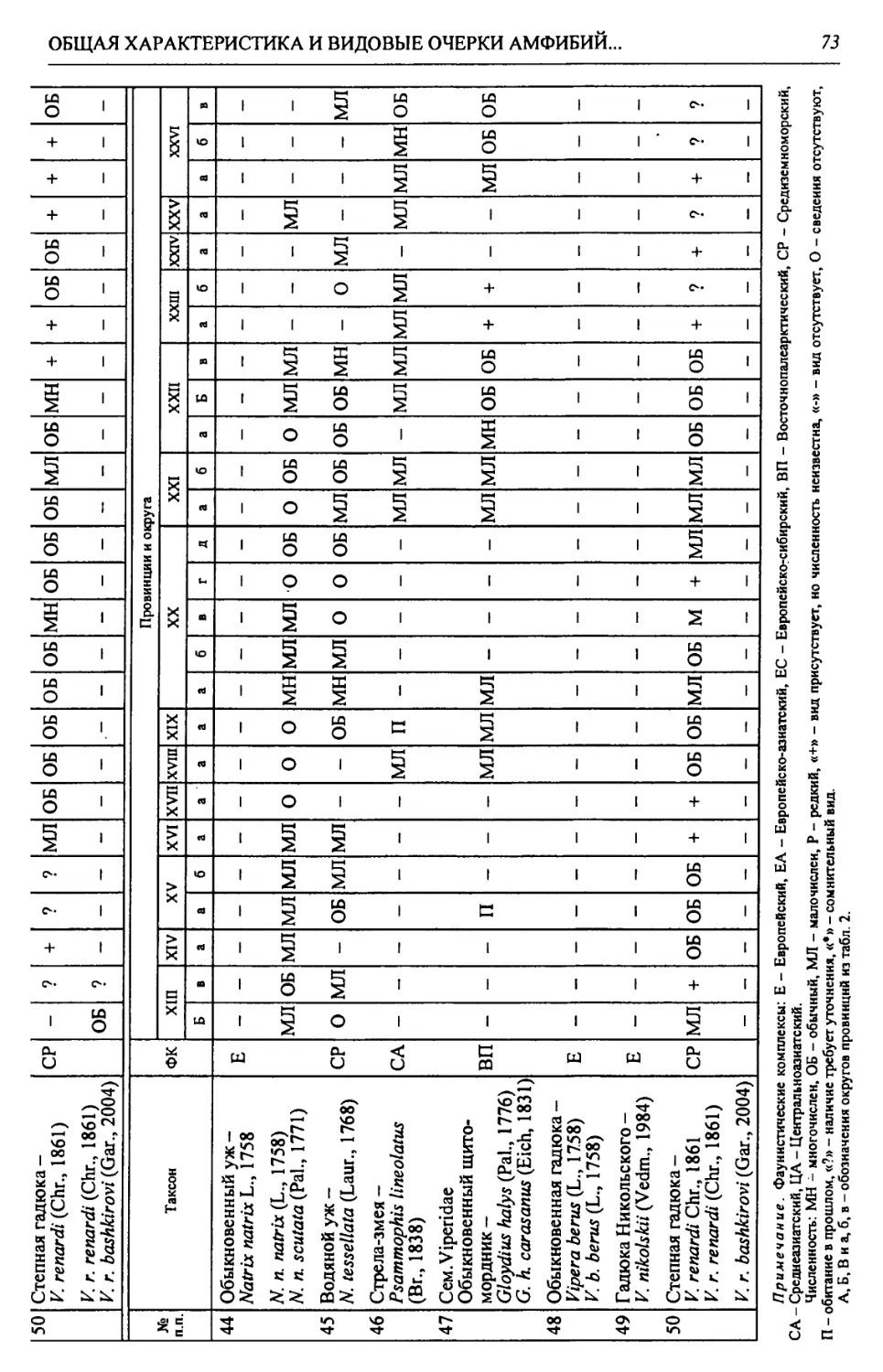
^.’ЭИшимбай
Салйват
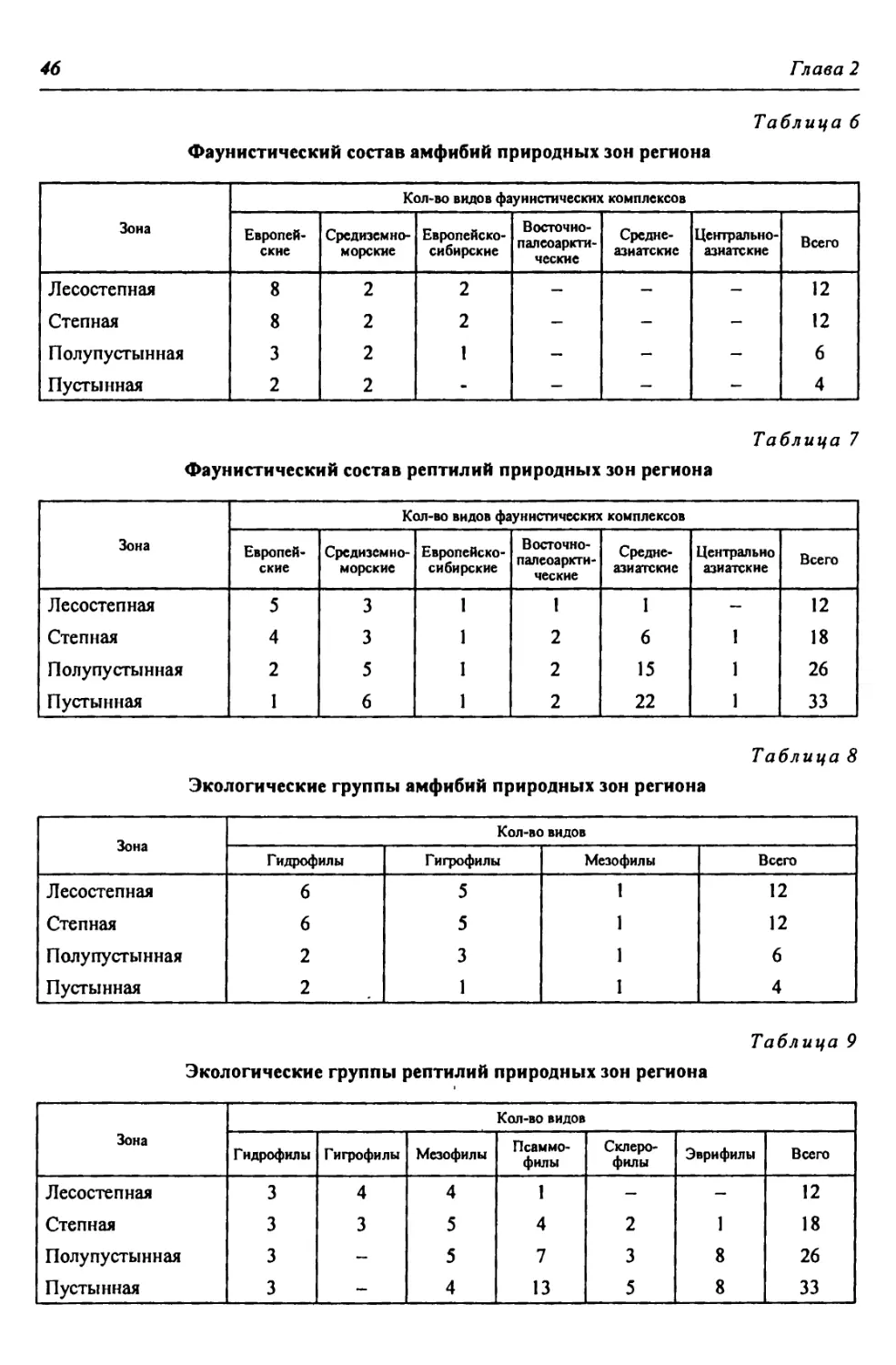
iy
1
1
Нойртройик
Лк ПОЩ11КК
J / ' ' е
’ хСатха
г
। арга jhhckoF* «у.
Златоуст
V Q
Миасс
}
Л ИрнклинсКоА
7 вдхр.
СГ
МаНнгтогррс*
oi 1 -in ' р
й
П Л А I (’
етроацм
•у
<в«Жетыколь г
то®2^
Вер^^юбол ьс кое
вдхрк
Т У Р Г А
о чздмугш
•V 7 ЧЕЛЯБИНСК
Верхнеуральское
вдхр»
КОС
О -
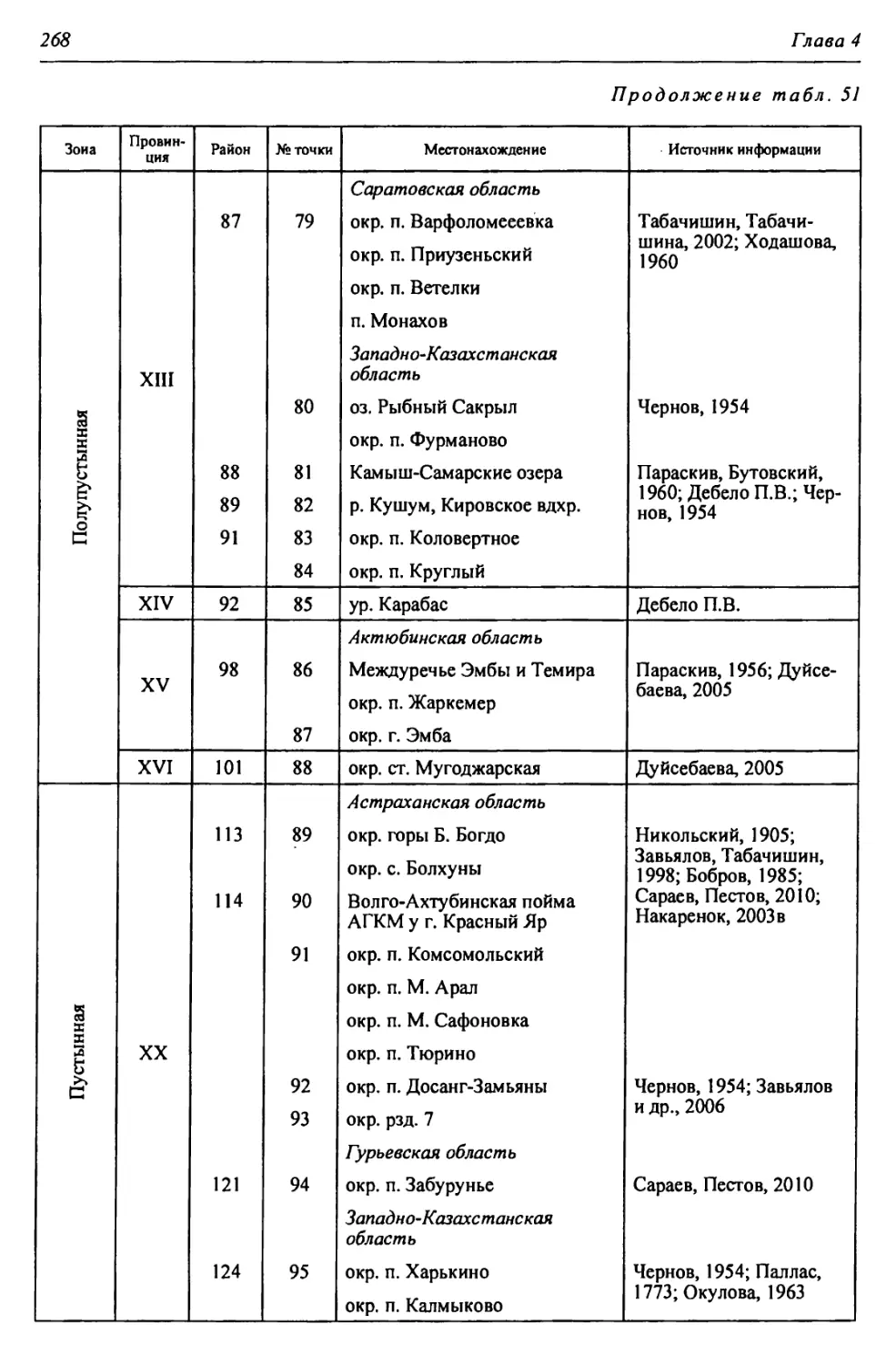
Рудный
Каратомарское
вдхр.
5
КУРГАН
о < ( apt II- он •
РОССИЙСКАЯ АКАДЕМИЯ НАУК • УРАЛЬСКОЕ ОТДЕЛЕНИЕ
ИНСТИТУТ СТЕПИ
П.В. Дебело, А.А. Чибилёв
АМФИБИИ И РЕПТИЛИИ
УРАЛО-КАСПИЙСКОГО РЕГИОНА
Серия: Природное разнообразие Урало-Каспийского региона
Том III
ЕКАТЕРИНБУРГ, 2013
RUSSIAN ACADEMY OF SCIENCES • URAL BRANCH
INSTITUTE OF STEPPE
P. V Debelo, A.A. Chibilyov
THE AMPHIBIANS AND REPTILIANS
OF THE URAL-CASPIAN REGION
Natural Diversity in the Ural-Caspian Region Series
Volume III
YEKATERINBURG, 2013
УДК 574/597
ББК 28.08.28.693.32
Д25
Ответственный редактор
кандидат биологических наук Е.В. Барбазюк
Рецензенты
доктор биологических наук В.Л. Вершинин
кандидат биологических наук А.Г. Бакиев
Дебело П.В., Чибилёв А.А.
Д25 Амфибии и рептилии Урало-Каспийского региона / П.В. Дебело, А.А. Чибилёв. Серия:
Природное разнообразие Урало-Каспийского региона. Т. III. - Екатеринбург: РИО УрО
РАН, 2013.-400 с.
ISBN 978-5-7691-2363-4
Дан обзор амфибий и рептилий Урало-Каспийского региона, который в прошлом со-
ставлял основную часть Оренбургского края. Эта территория представляет собой единое
информационное пространство, определенное классическими трудами естествоиспытате-
лей XVIII—XIX вв. П.И. Рычкова, П.С. Палласа, Э.А. Эверсманна и многих других иссле-
дователей, изучавших территорию современного Заволжья, Южного Урала и Западного
Казахстана как единое целое.
Проанализирована история формирования фауны амфибий и рептилий. Для всех ви-
дов даны краткие очерки с рисунками и фотографиями.
Монография предназначена для работников природно-ресурсных органов, студентов
биологических и географических специальностей, учителей школ, натуралистов и краеве-
дов.
УДК 574/597
ББК 28.08.28.693.32
Dcbclo P.V., Chibilyov А.А.
The Amphibians and reptilians of the Ural-Caspian Region / P.V. Dcbclo, A.A. Chibilyov.
Natural Diversity in the Ural-Caspian Region Series. Vol. 3. - Yekaterinburg: Ural Branch of
Russian Academy of Sciences, 2013. - 400 p.
This book provides a review of amphibians and reptiles of the Ural-Caspian region which
was the main part of the Orenburg region in the past. This area represents an integrated information
space having being defined in the classical works of naturalists Rychkov, Pallas, Eversmann in
18-19th centuries and many other researchers who studied the territory of modern Trans-Volga,
Southern Urals and Western Kazakhstan as a whole.
We analyzed the history of fauna formation for amphibians and reptilians. For all species
brief species accounts with figures and photographs are given.
Monograph is intended for agencies of natural resources, biology students, high school
teachers, naturalists and local historians.
Executive Editor
Candidate of Biological Sciences E.V. Barbazyuk
Reviewers
Doctor of Biological Sciences V.L. Vershinin
Candidate of Biological Sciences A.G. Bakiev
ISBN 978-5-7691-2363-4
© Дебело П.В.,Чибилёв A.A., 2013
© Институт степи УрО РАН, 2013
©РИО УрО РАН, 2013
© Debelo P.V., Chibilev А.А., 2013
© Institute of Steppe, UB RAS, 2013
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ....................................................... 7
Глава 1. КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА
РЕГИОНА........................................................ 8
1.1. Основные зональные и региональные ландшафтные различия. 8
1.2. Лесостепная зона...................................... 14
1.3. Степная зона.......................................... 15
1.4. Полупустынная зона..................................... 16
1.5. Пустынная зона........................................ 17
Глава 2. ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ РАСПРО-
СТРАНЕНИЯ И ЭКОЛОГИЧЕСКИЕ ГРУППЫ АМФИБИЙ И РЕПТИЛИЙ
УРАЛО-КАСПИЙСКОГО РЕГИОНА..................................... 18
Глава 3. ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ В
УРАЛО-КАСПИЙСКОМ РЕГИОНЕ................................ 48
Глава 4. ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ И
РЕПТИЛИЙ УРАЛО-КАСПИЙСКОГО РЕГИОНА...................... 58
Глава 5. ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ АМФИБИЙ И
РЕПТИЛИЙ РЕГИОНА....................................... 299
СПИСОК ЛИТЕРАТУРЫ ............................................. 308
CONTENTS
FOREWORD.............................................................. 7
Chapter 1. A BRIEF PHYSICAL AND GEOGRAPHIC CHARACTERISTICS OF THE
REGION................................................................. 8
1.1. The main zonal and regional landscape differences........ 8
1.2. Forest and Steppe Zone.................................. 14
1.3. Steppe Zone............................................. 15
1.4. Semi-Desert Zone ..................................... 16
1.5. Desert Zone........................................ 17
Chapter 2. THE HISTORY OF FAUNA FORMATION, CURRENT DISTRIBUTION
AND ECOLOGICAL GROUPS OF AMPHIBIANS AND REPTILIANS IN
THE URAL-CASPIAN REGION ............................................ 18
Chapter 3. AMPHIBIAN AND REPTILIAN STUDIES IN THE URAL-CASPIAN
REGION................................................;............... 48
Chapter 4. GENERAL CHARACTERISTICS AND SPECIES ACCOUNTS OF AMPHI-
BIANS AND REPTILIANS IN THE URAL-CASPIAN REGION.................. 58
Chapter 5. IMPORTANT ISSUES FOR CONSERVATION OF AMPHIBIANS AND
REPTILIANS IN THE URAL-CASPIAN REGION ............................... 299
REFERENCES ......................................................... 308
ПРЕДИСЛОВИЕ
Читателю предлагается очередной том из серии монографий Института
степи Уральского отделения РАН «Природное разнообразие Урало-
Каспийского региона». Он посвящен эколого-географическому обзору
батрахо- и герпетофауны обширного региона, расположенного между доли-
ной средней и нижней Волги на западе и Тургайской ложбиной на востоке.
Южная граница рассматриваемого региона доходит до северного побережья
Каспийского и Аральского морей. На севере он ограничен лесостепной зоной
Заволжья, Приуралья, Южного Урала и Зауралья. Выбор данной территории
в обозначенных пределах обусловлен не столько естественными рубежами,
сколько границами бывшей Оренбургской губернии, территория которой в
XVIII-XIX вв. именовалась «Оренбургским краем» или «Урало-Каспийским
краем».
На протяжении трех веков в данном регионе работали выдающиеся оте-
чественные естествоиспытатели: П.И. Рычков, П.С. Паллас, И.И. Лепехин,
А.И. Левшин, Э.А. Эверсманн, Н.А. Северцов, Г.С. Карелин, Н.А. Зарудный,
С.С. Неуструев и др. Это способствовало формированию единого информа-
ционного пространства, которое в настоящее время объединяет ряд областей
Западного Казахстана, Нижнего и Среднего Поволжья, Южного Урала Рос-
сийской Федерации.
В геоморфологическом отношении регион охватывает большую часть При-
каспийской низменности, Сыртовое Заволжье и Приуралье, оконечность Юж-
ного Урала, Зауралье, западную часть Тургайского плато, Мугоджары, Эмбен-
ское плато. В ландшафтном отношении на территории региона представлены
южная лесостепь, равнинные, холмисто-увалистые и низкогорные ландшафты
степной зоны, полупустыни и пустыни.
Обобщая и анализируя имеющуюся информационную базу по батрахо- и
герпетофауне обширной территории, авторы учитывали, что к западу и севе-
ру от Урало-Каспийского региона расположены два крупных центра изучен-
ния амфибий и рептилий Евразии (Тольятти, Институт экологии Волжского
бассейна РАН (А.Г. Бакиев) и Екатеринбург, Институт экологии растений и
животных УрО РАН (В.Л. Вершинин)), что в некоторой степени определи-
ло направление и характер нашей работы. Авторы надеются, что настоящая
монография будет способствовать новым зоогеографическим исследованиям
в Урало-Каспийском регионе, экосистемы которого сейчас испытывают зна-
чительную природную и антропогенную трансформацию.
Глава 1
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ
ХАРАКТЕРИСТИКА РЕГИОНА
1.1. ОСНОВНЫЕ ЗОНАЛЬНЫЕ И РЕГИОНАЛЬНЫЕ
ЛАНДШАФТНЫЕ РАЗЛИЧИЯ
Значительная протяженность региона с севера на юг и с запада на вос-
ток определяет исключительное разнообразие его ландшафтов, отража-
ющих природные условия в зональном, провинциальном и локальном
аспектах.
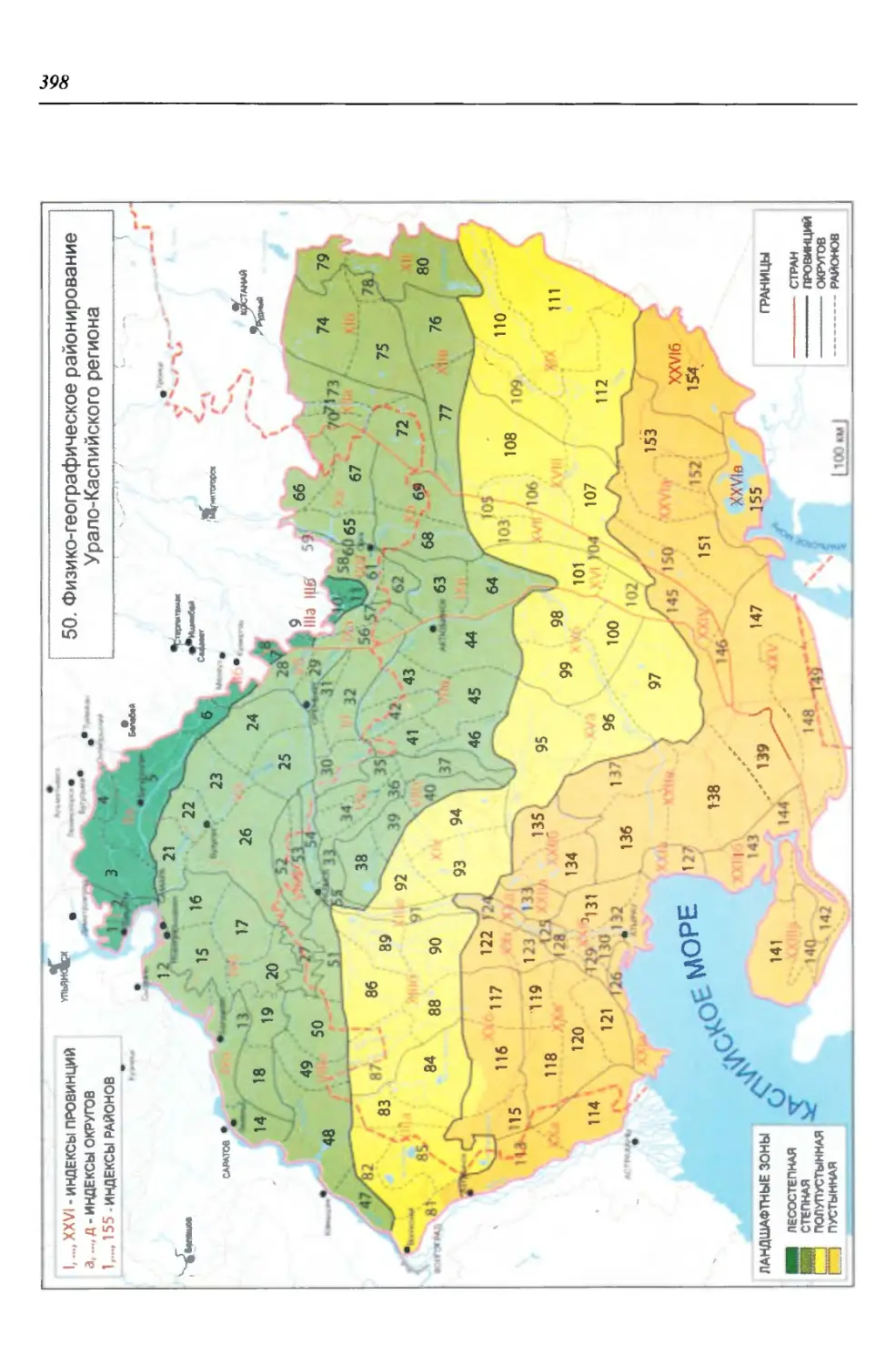
Природных зон (основных и промежуточных) в пределах региона выде-
ляют четыре: лесостепную, степную, полупустынную и пустынную. Грани-
цы между ними обусловлены в основном климатическими особенностями
(табл. 1), но физиономически они наиболее четко проявляются в сменах зо-
нальных типов растительности и почвенного покрова на плакорах (Мильков,
1986; Чибилёв, 1998).
Урало-Каспийский регион в авторском представлении охватывает терри-
торию от среднего и нижнего течения р. Волги на западе до Тургайской лож-
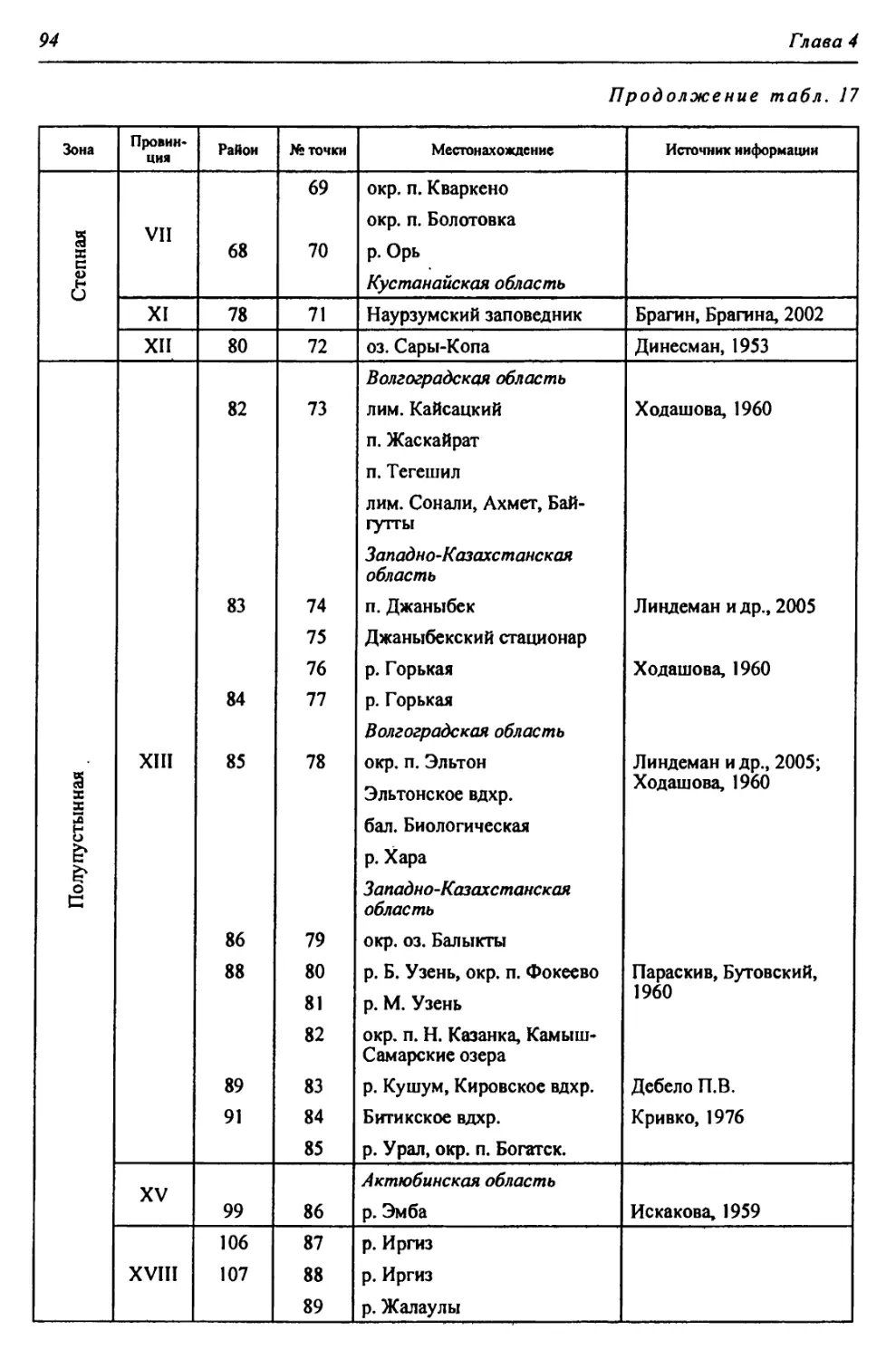
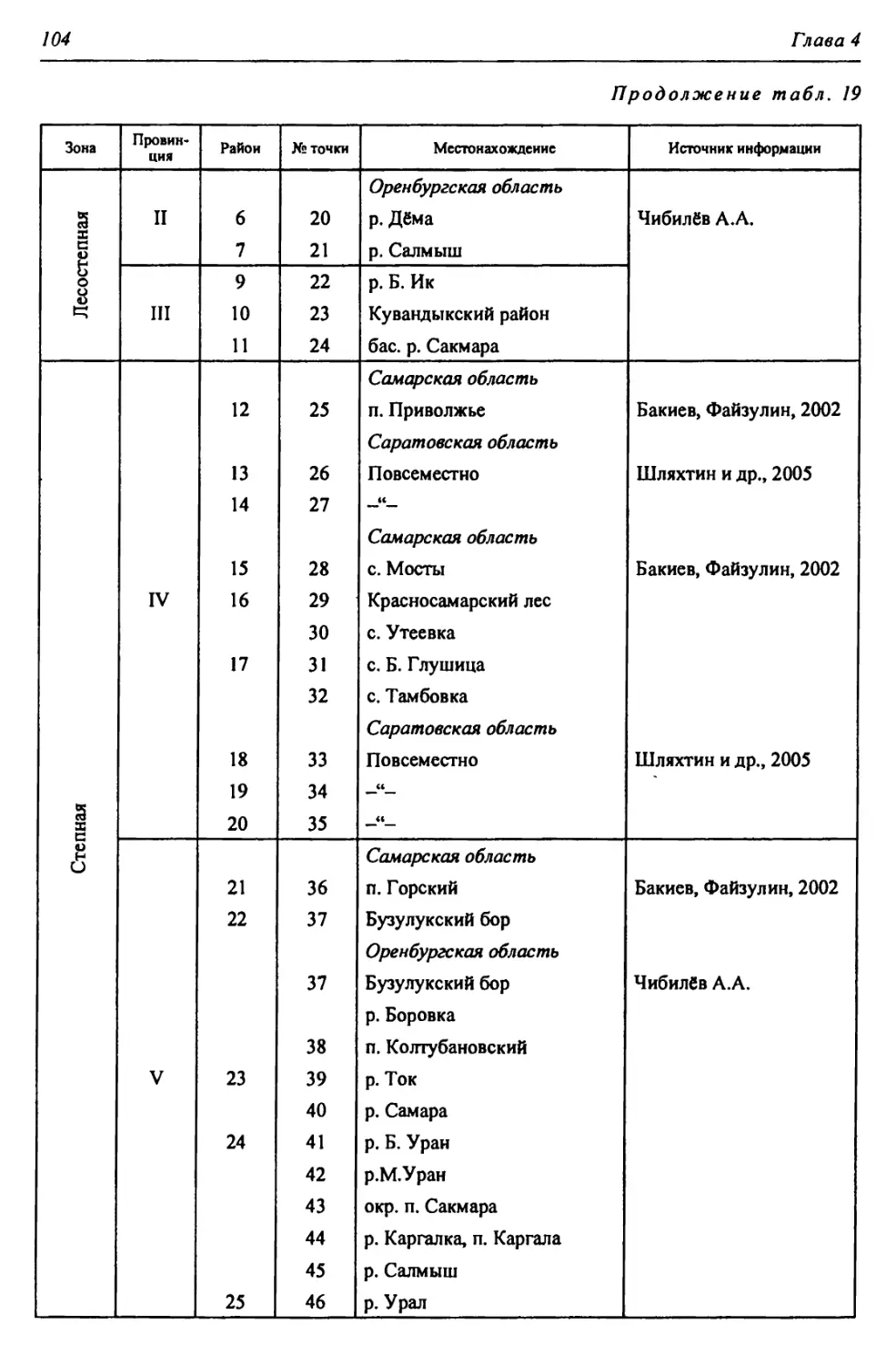
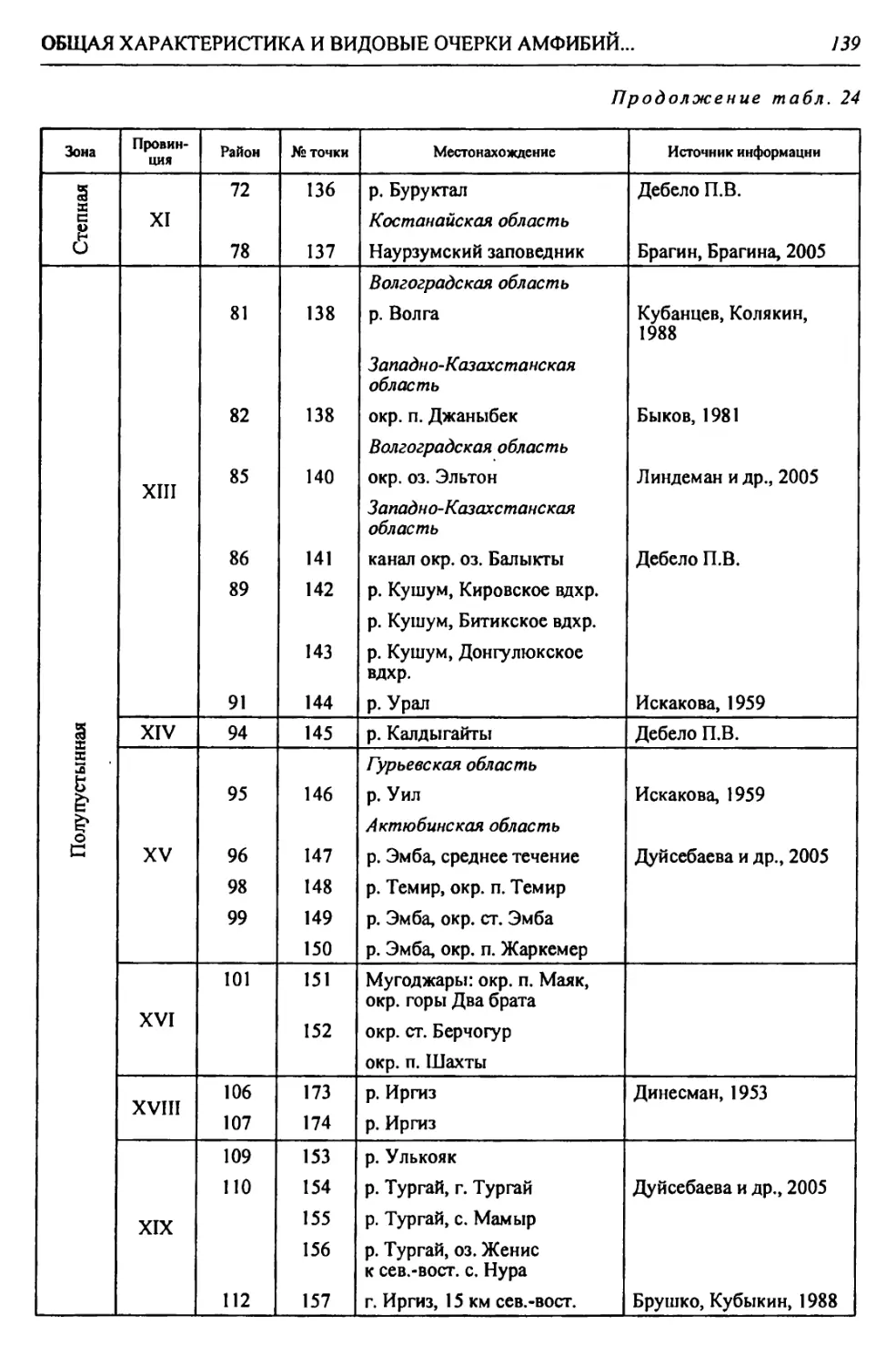
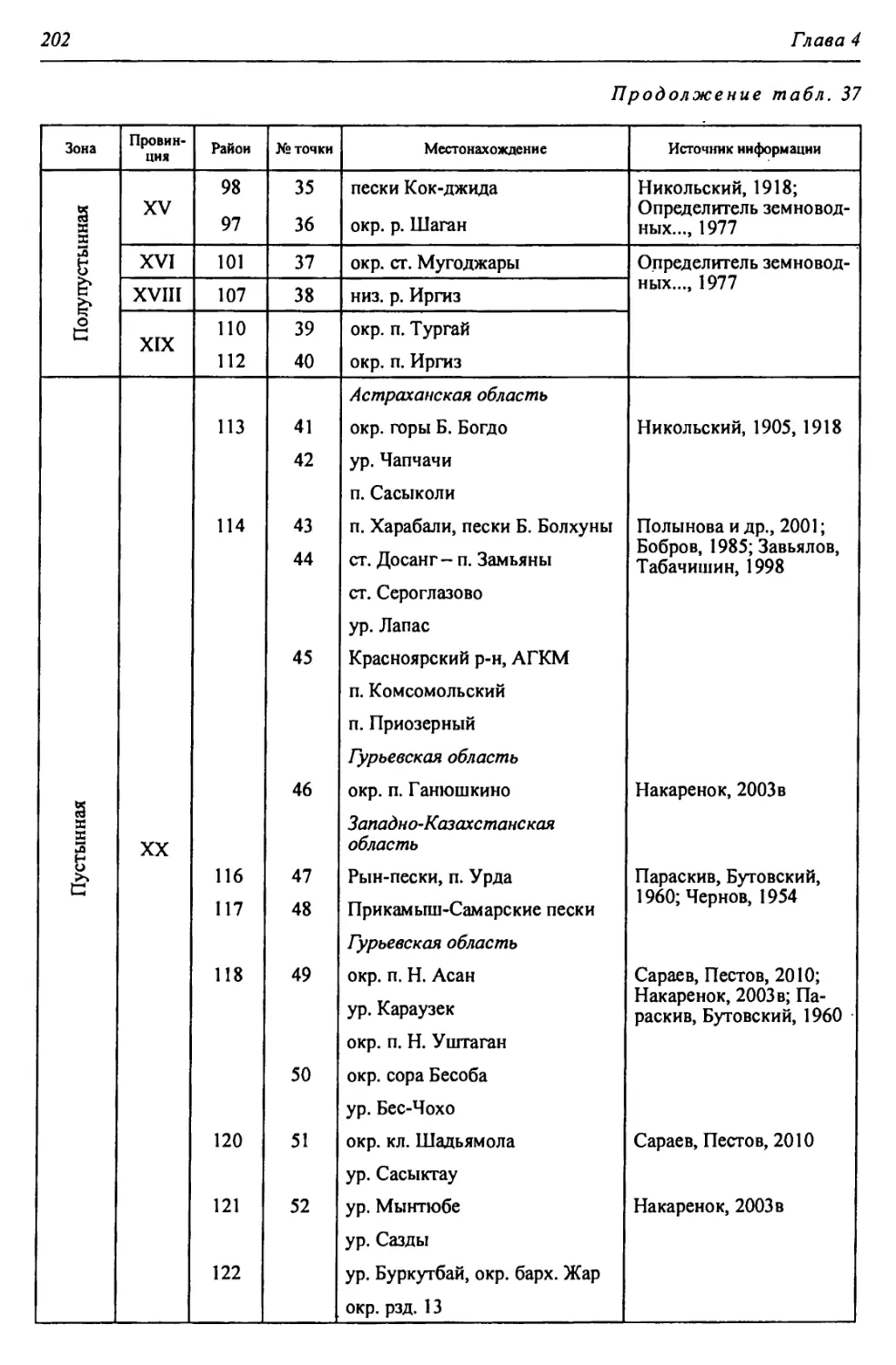
Таблица 1
Биоклиматическая характеристика природных зон
Зона Годовой радиацион- ный баланс, ккал/см 2т Термичес- кие условия: температура теплого и холодного месяцев, среднегодо- вая, °C Условия увлажнения Кол-во осадков, мм Почвы Зональные растительные сообщества
Коэфф, увлажнения Высоцкого - Иванова Индекс сухости
Лесо- степь* 30-36 +2,5; +20; —17 0,99- 0,60 1,0-1,5 400- 450 Типичные и выщелоченные черноземы с темно-серыми лесными Богато-разно- травные степные с массивами на- горных лесов
Степь 36-40 +4,5; +22; -14 0,59- 0,30 1,5-2,0 350- 400 Обыкновенные и южные чер- ноземы, темно- каштановые и каштановые Разнотравно- типчаково-ко- выльные, типич- ные злаковые, южные
Полу- пустыня 40-45 +24; -13; 5,2-5,9 0,29- 0,13 2,0-3,0 300- 350 Светло-кашта- новые с солон- цами и солонча- ками Злаковые. Слож- ная мозаика из степных злаков и прутняка
Пустыня 45-50 +25; -7; 6,4-7,2 0,12- 0,00 >3,0 200- 300 Бурые и серо-бу- рые пустынные Полынные и со- лянковые
В пределах региона.
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕГИОНА
9
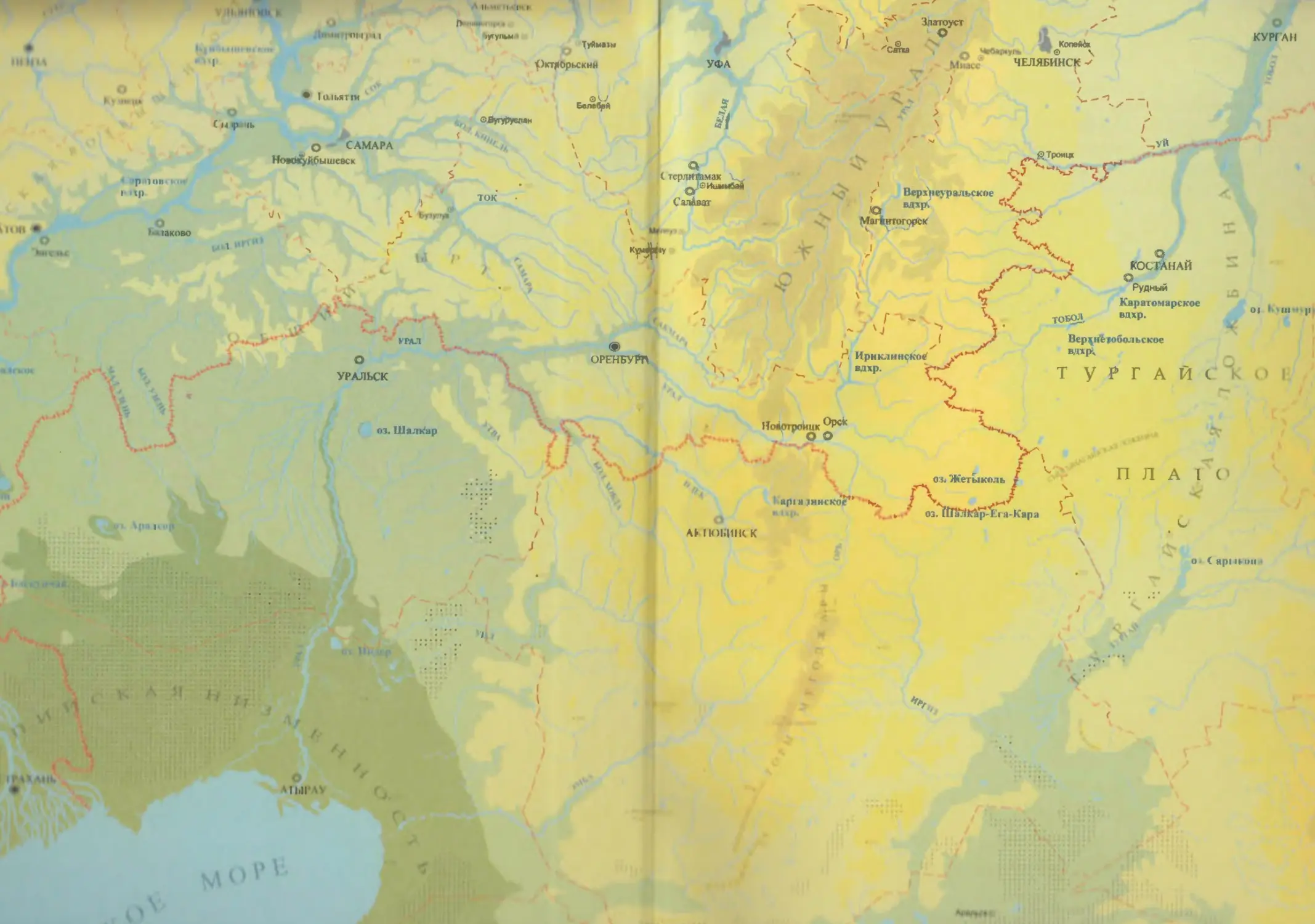
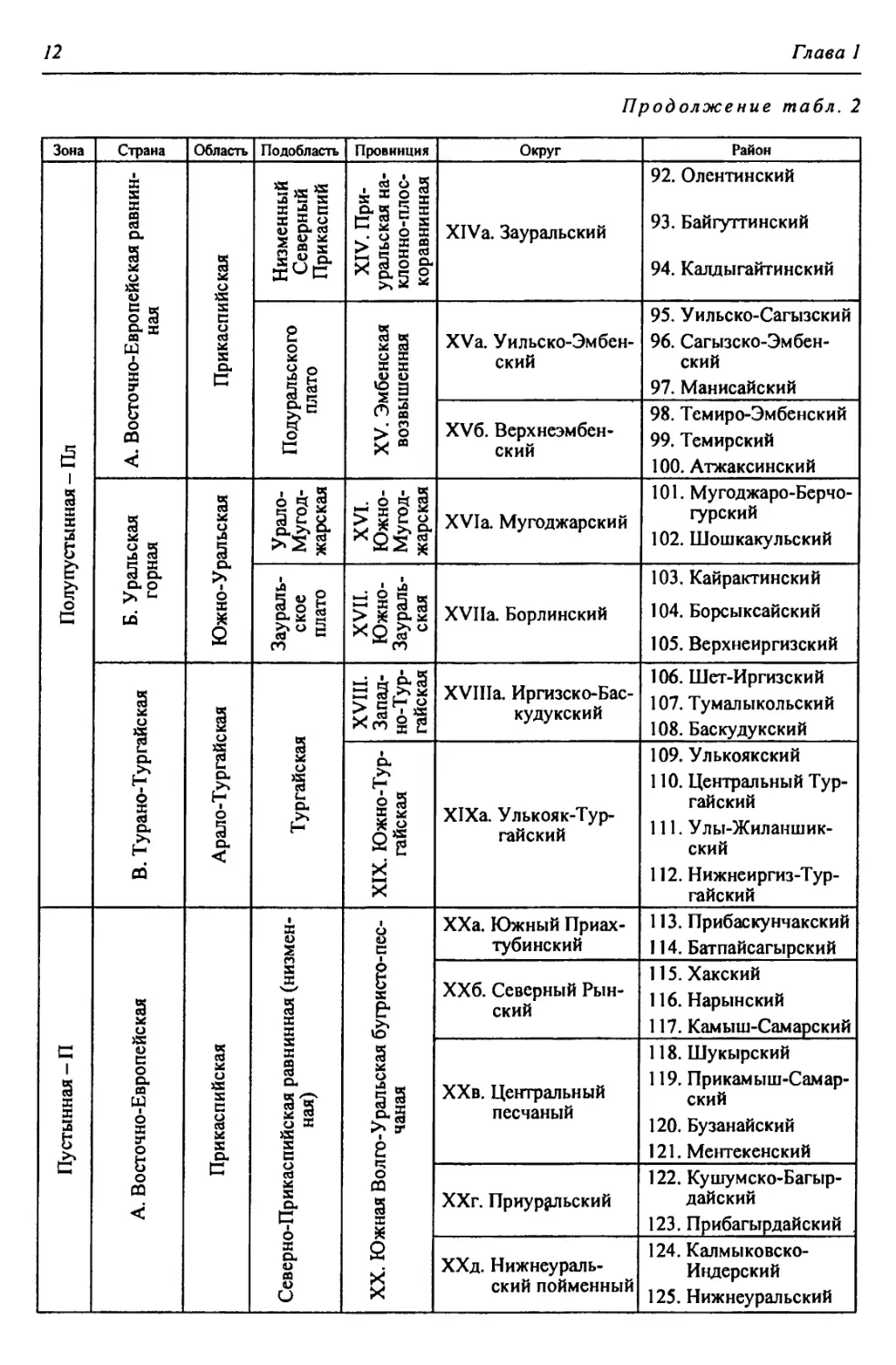
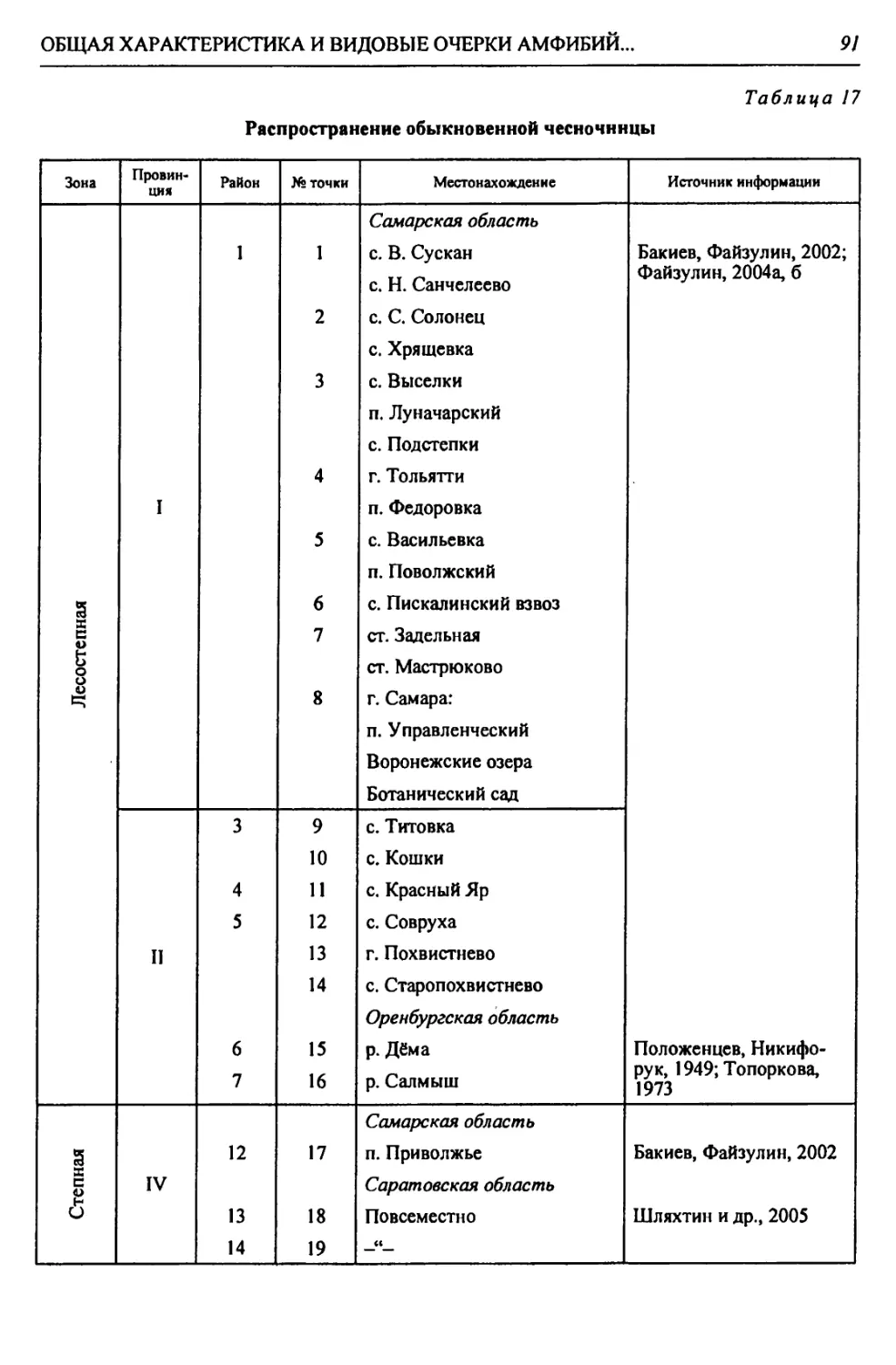
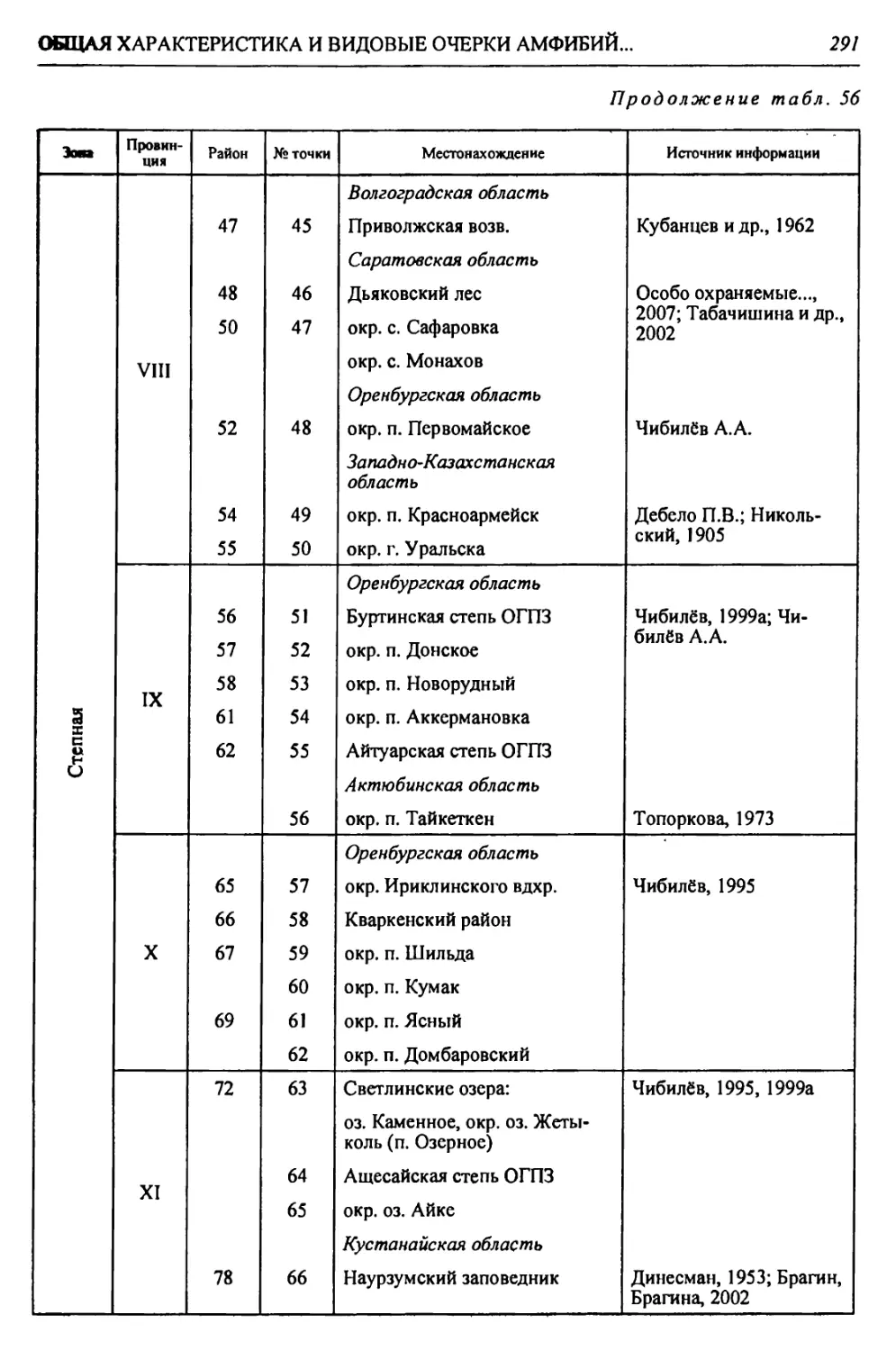
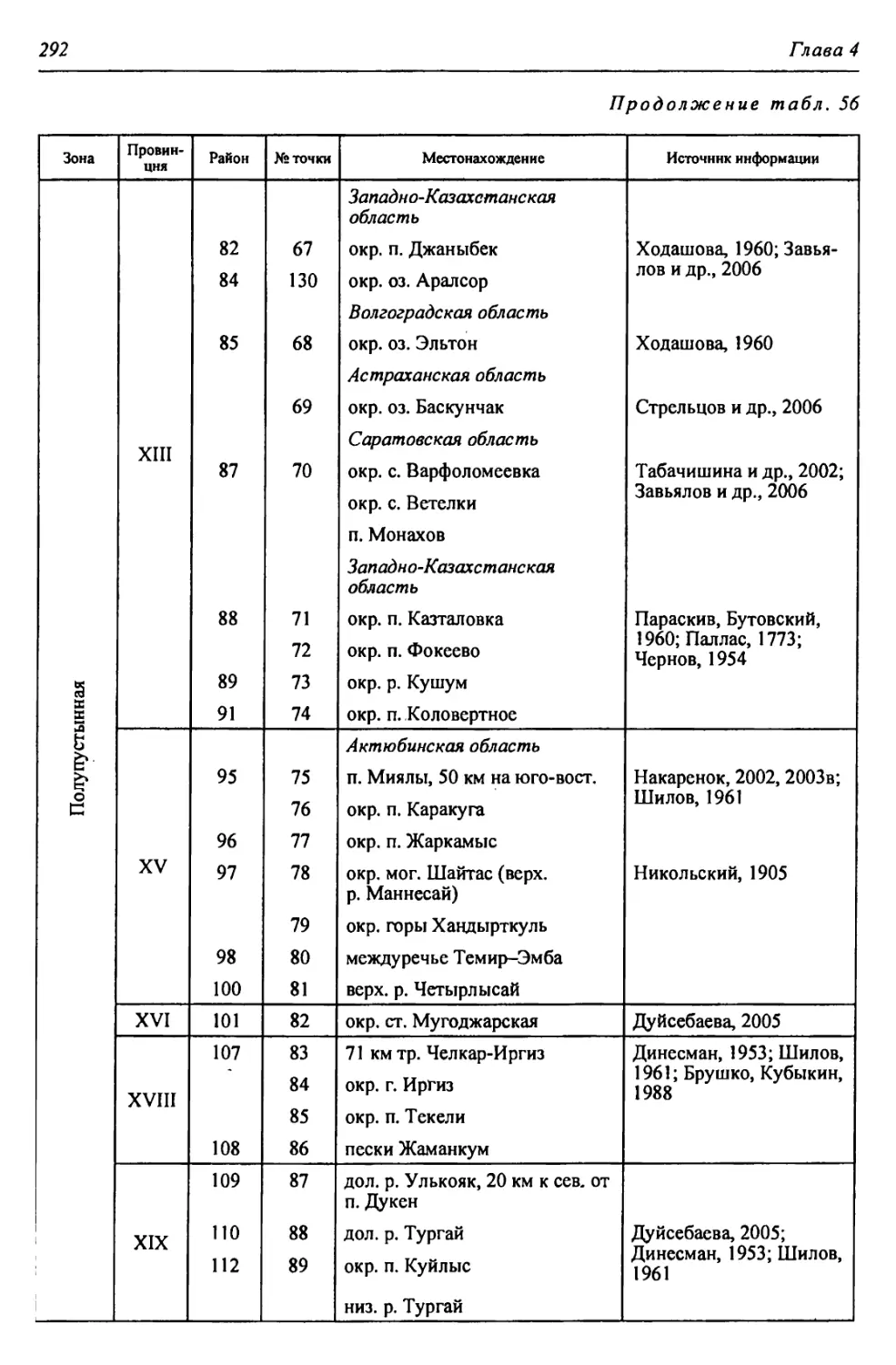
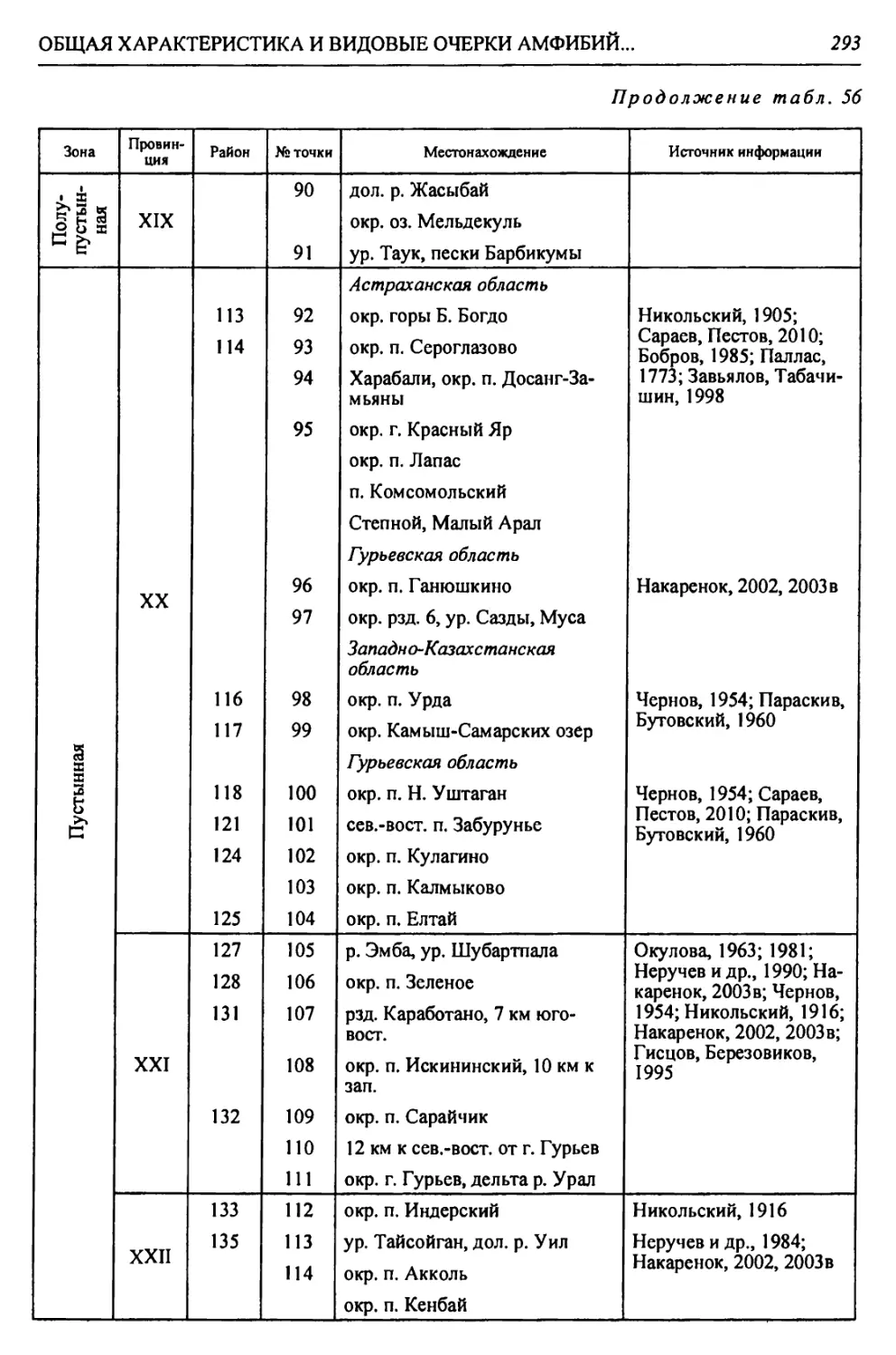
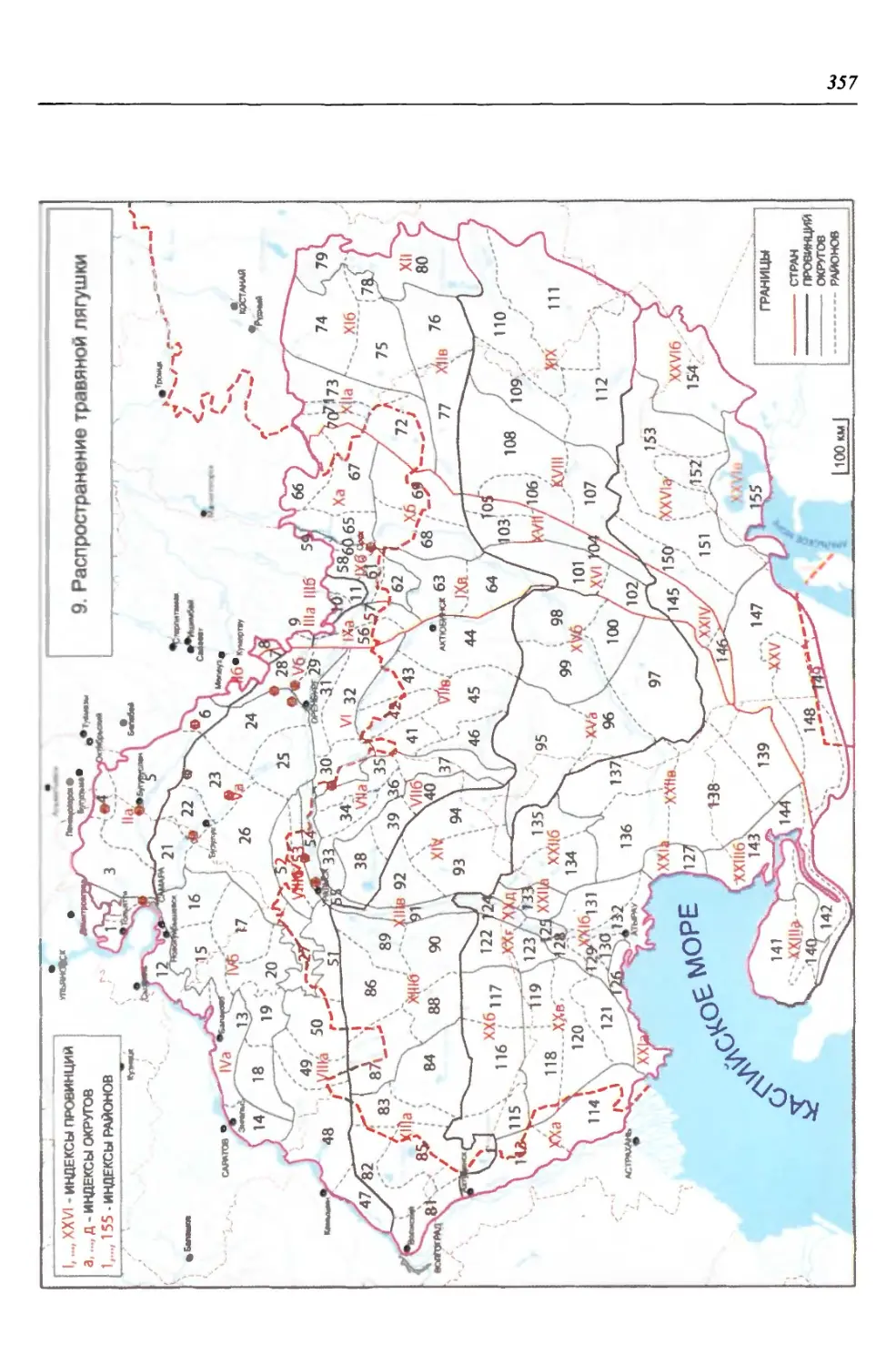
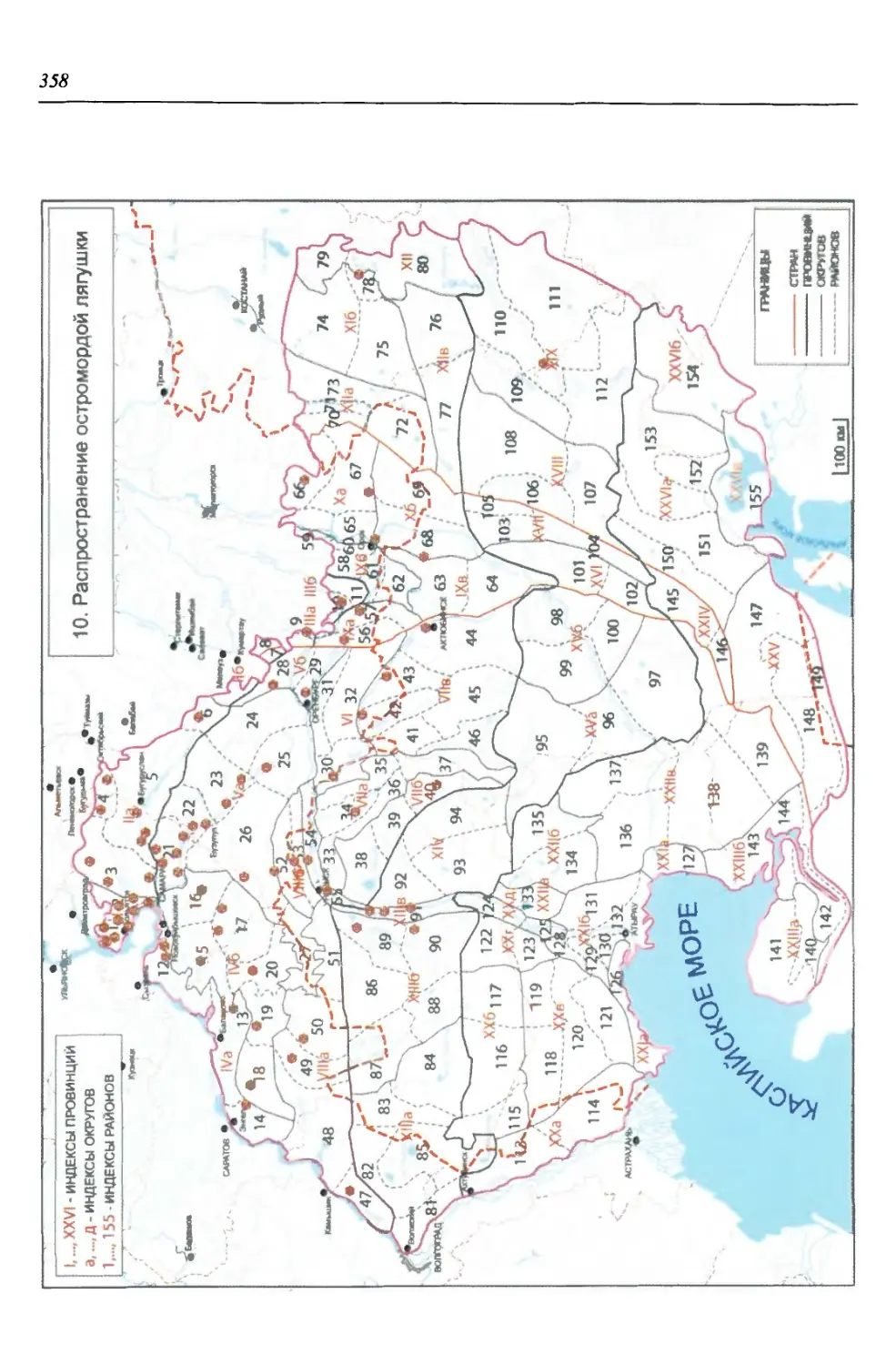
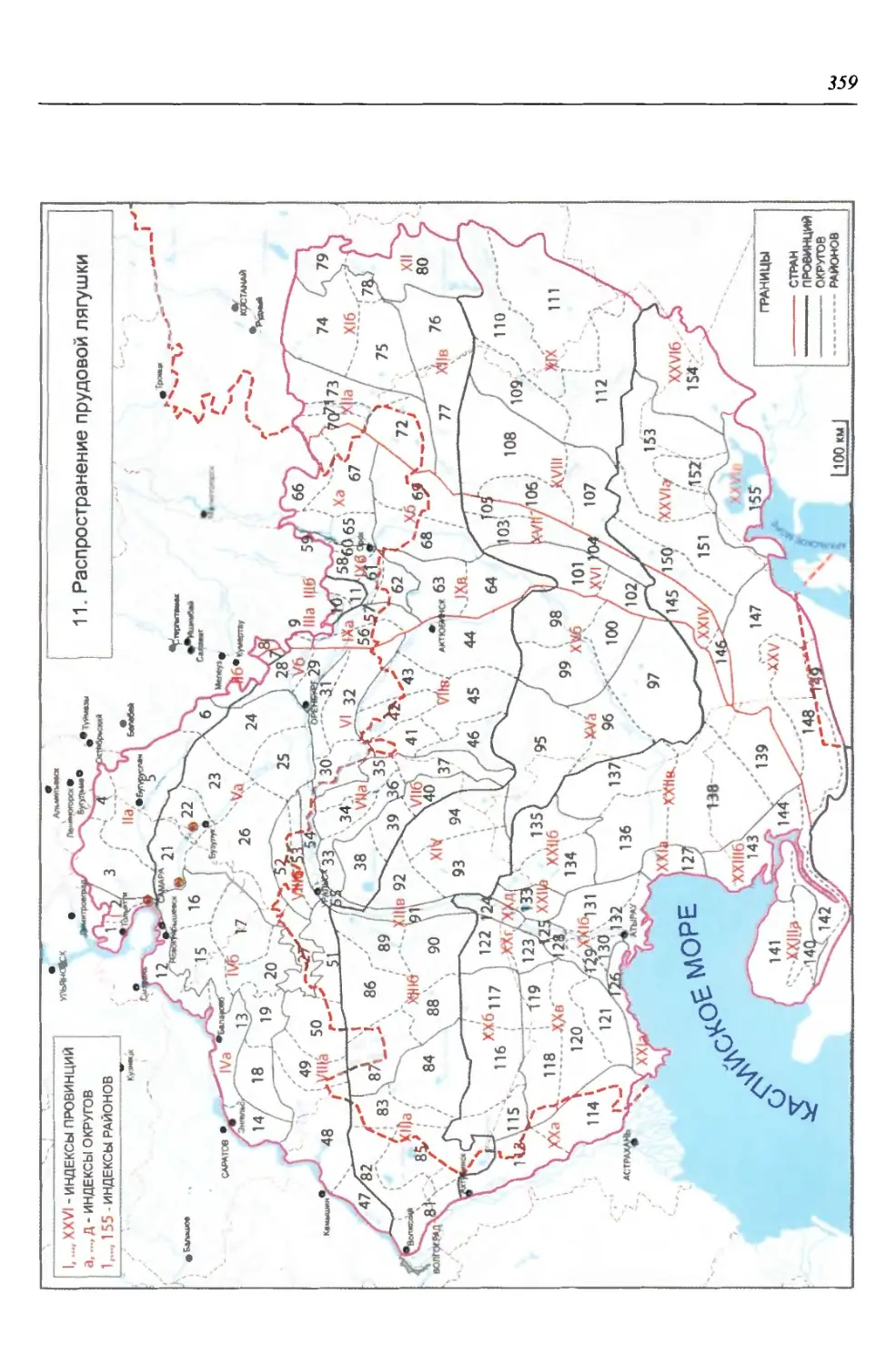
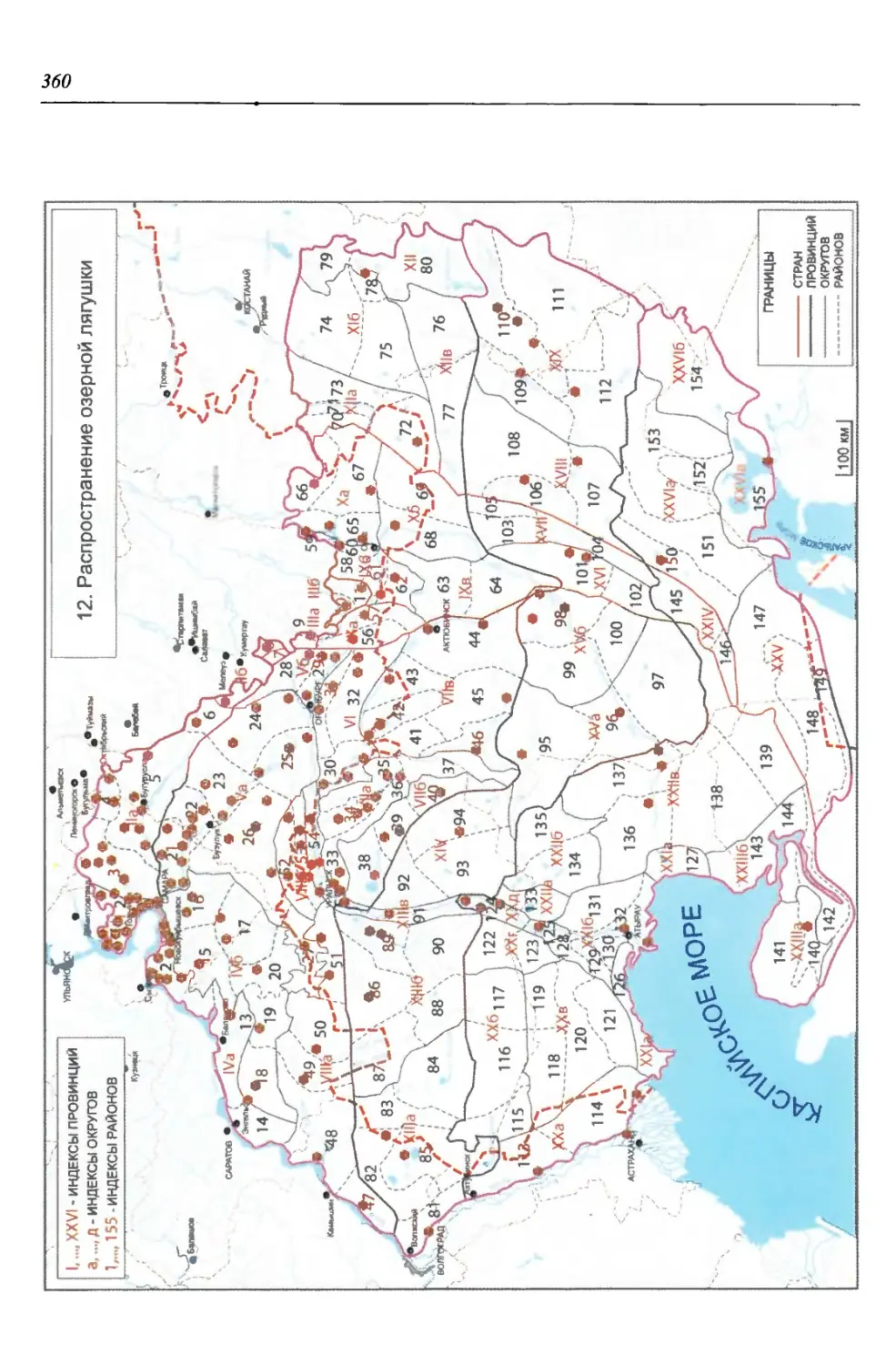
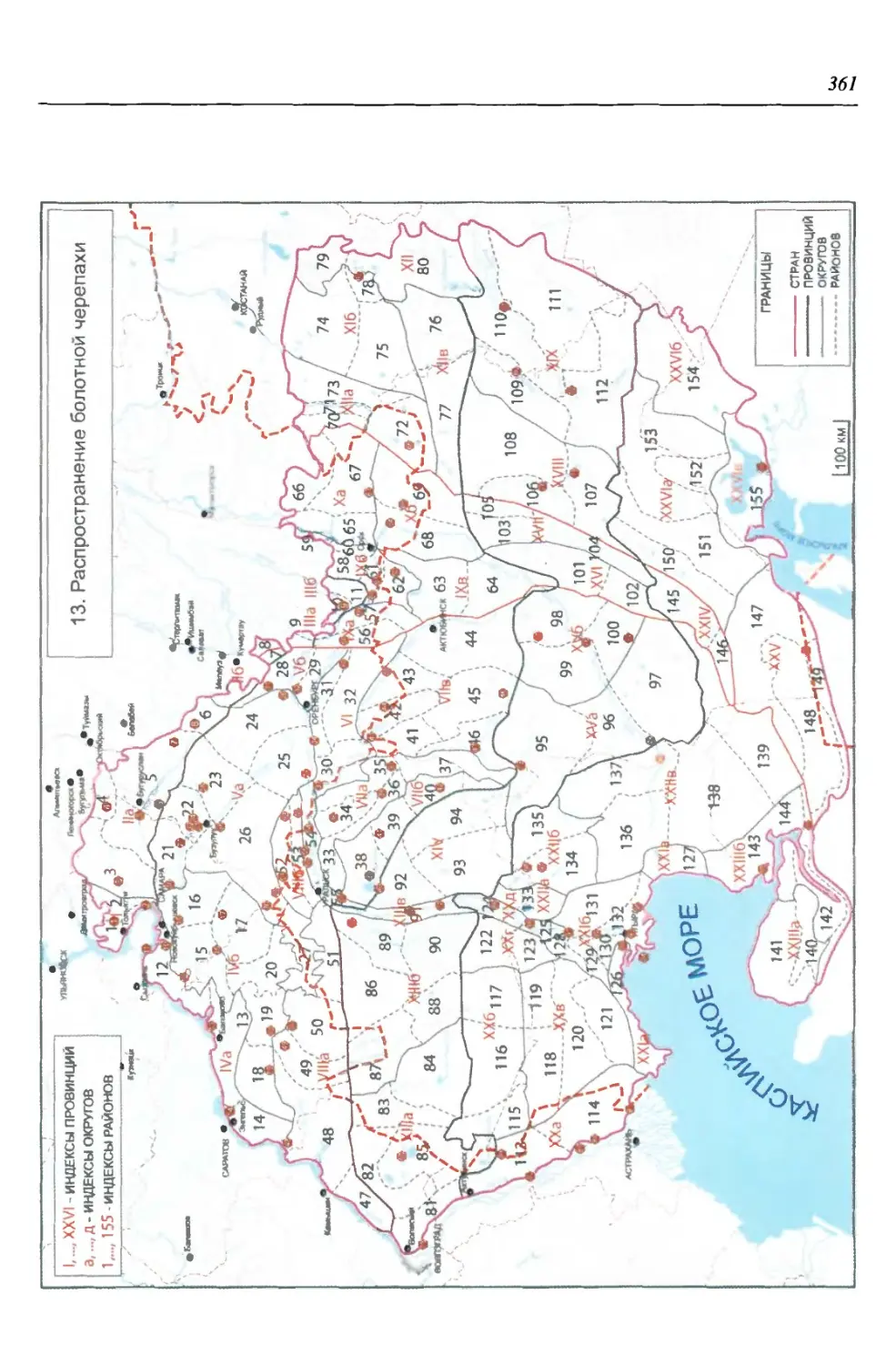
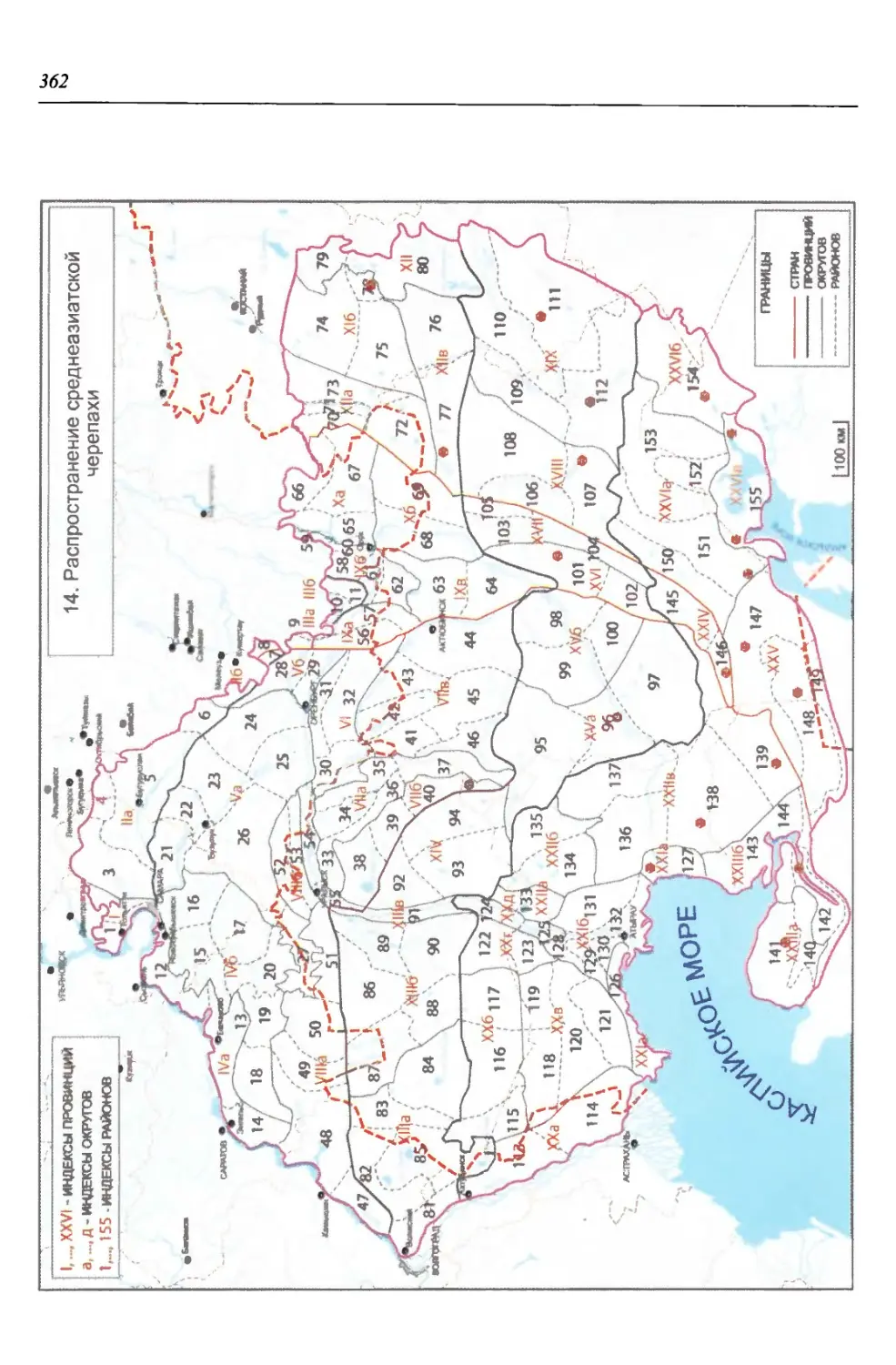
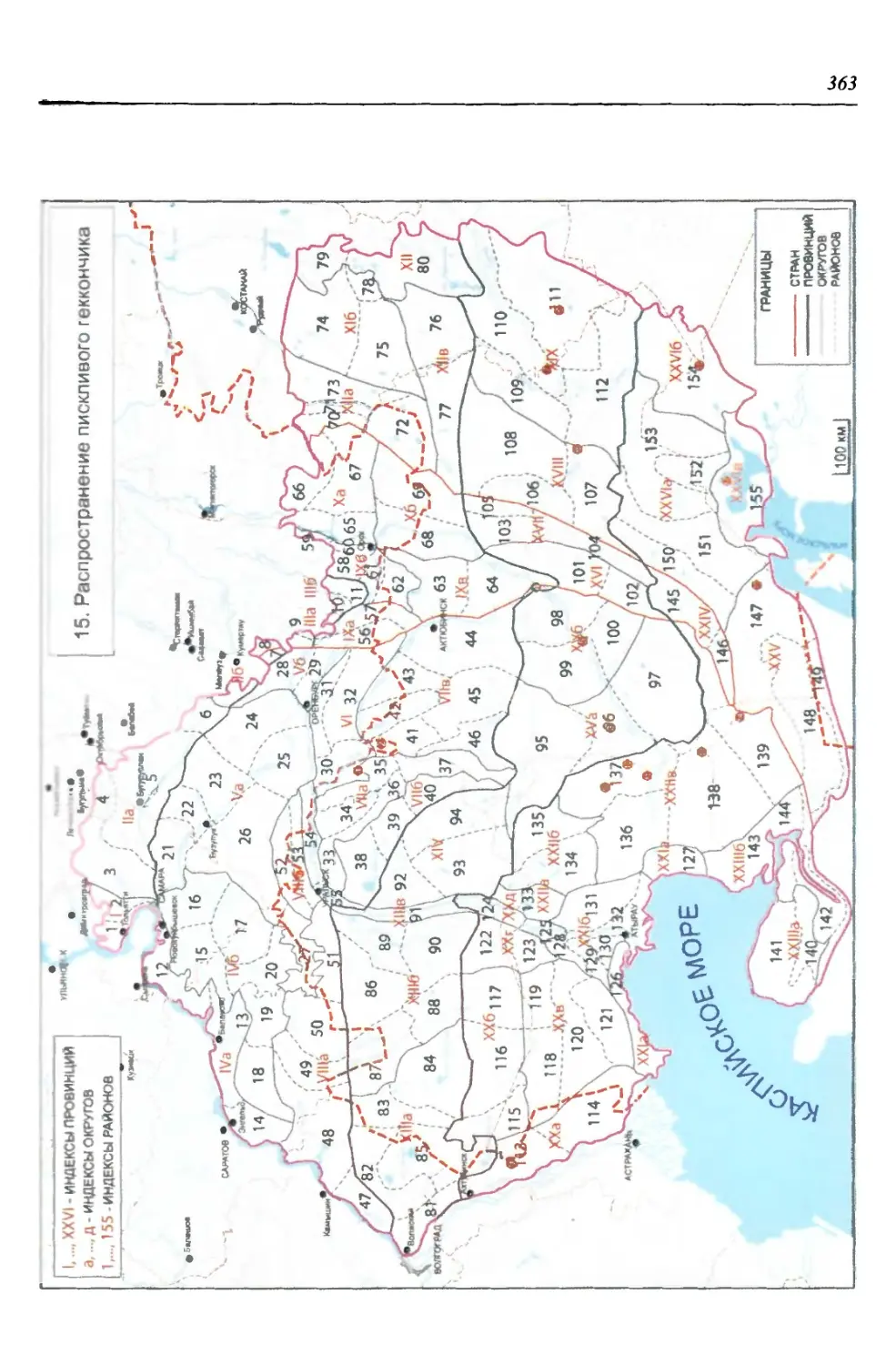
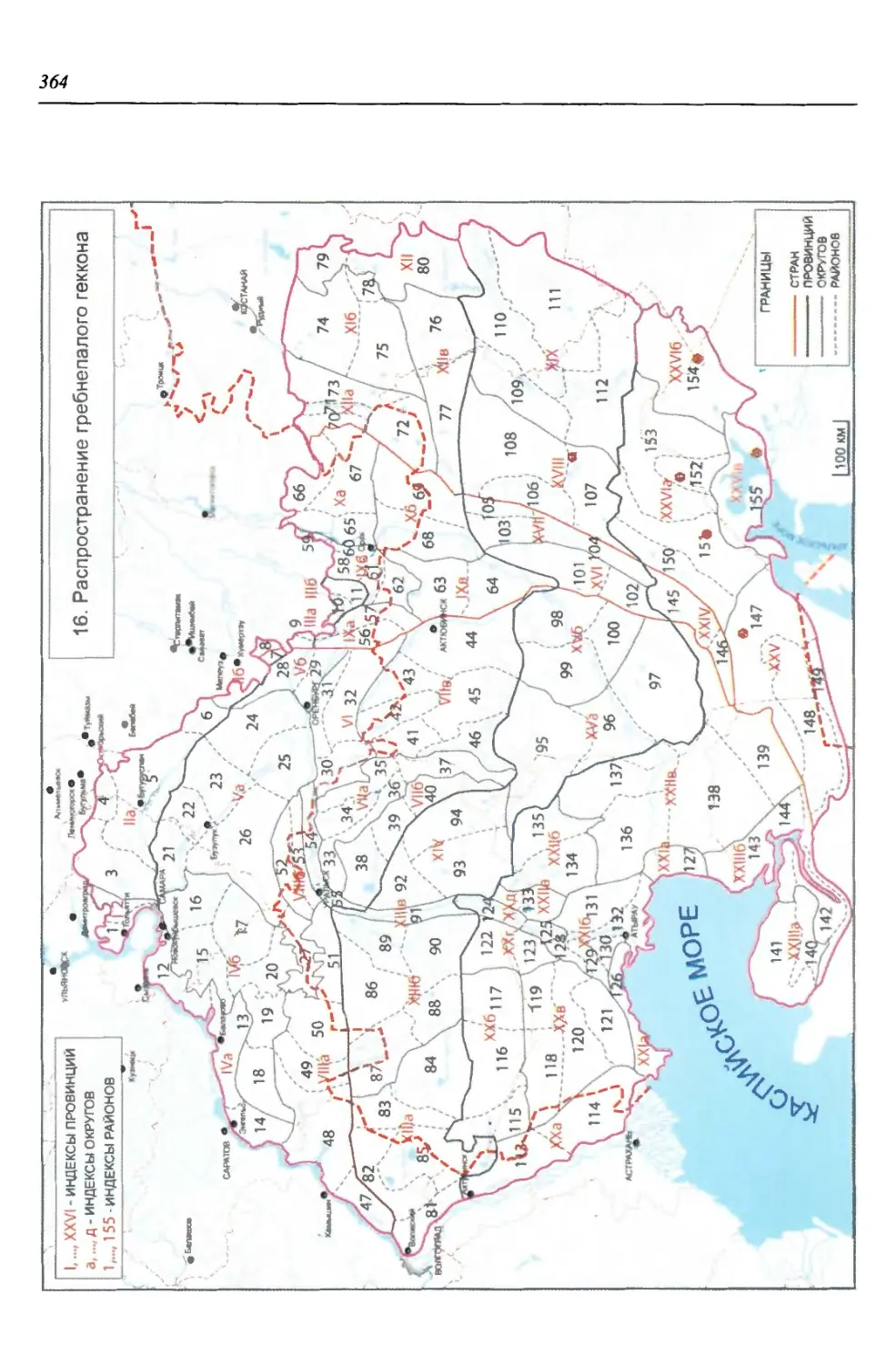
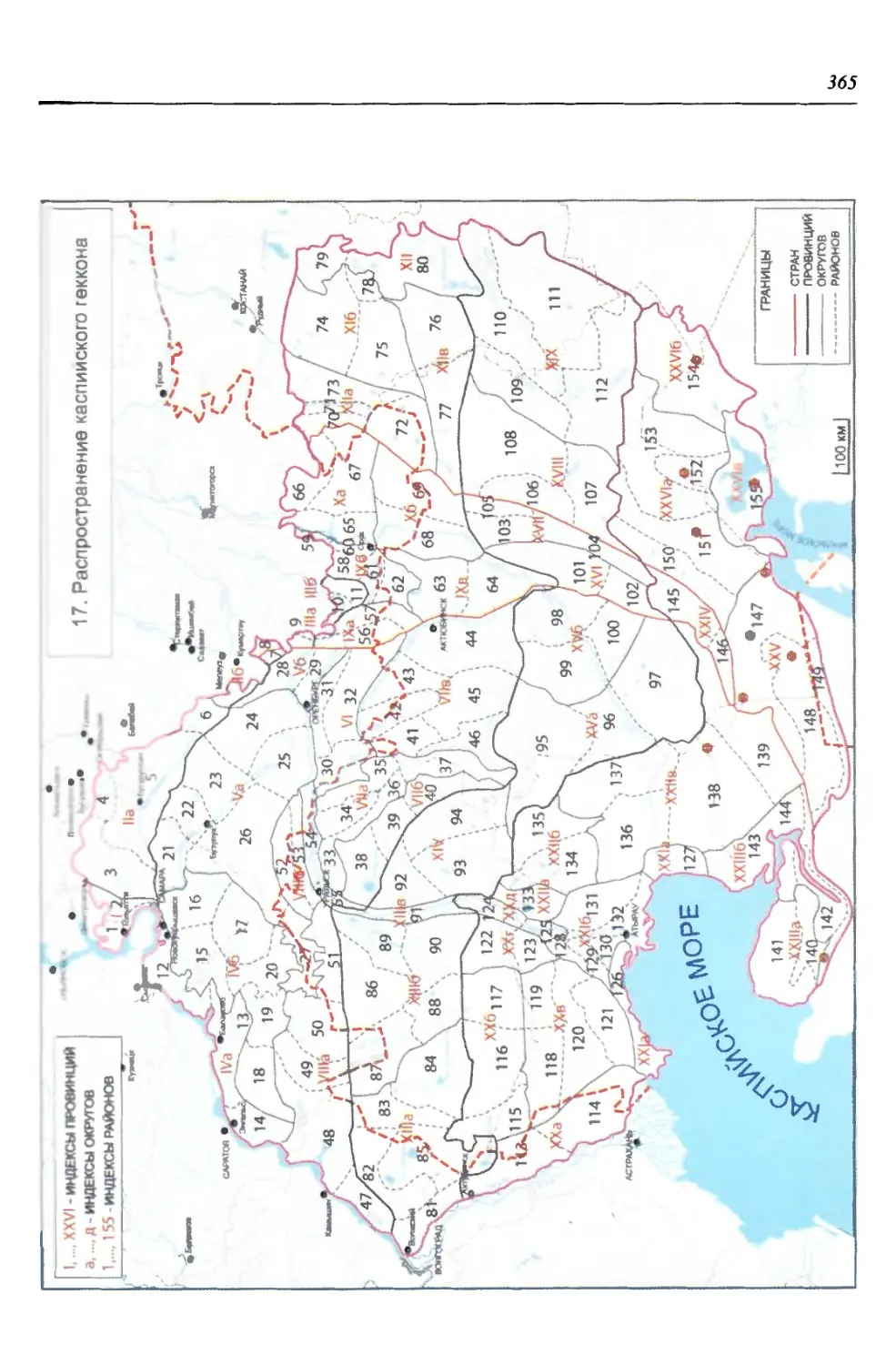
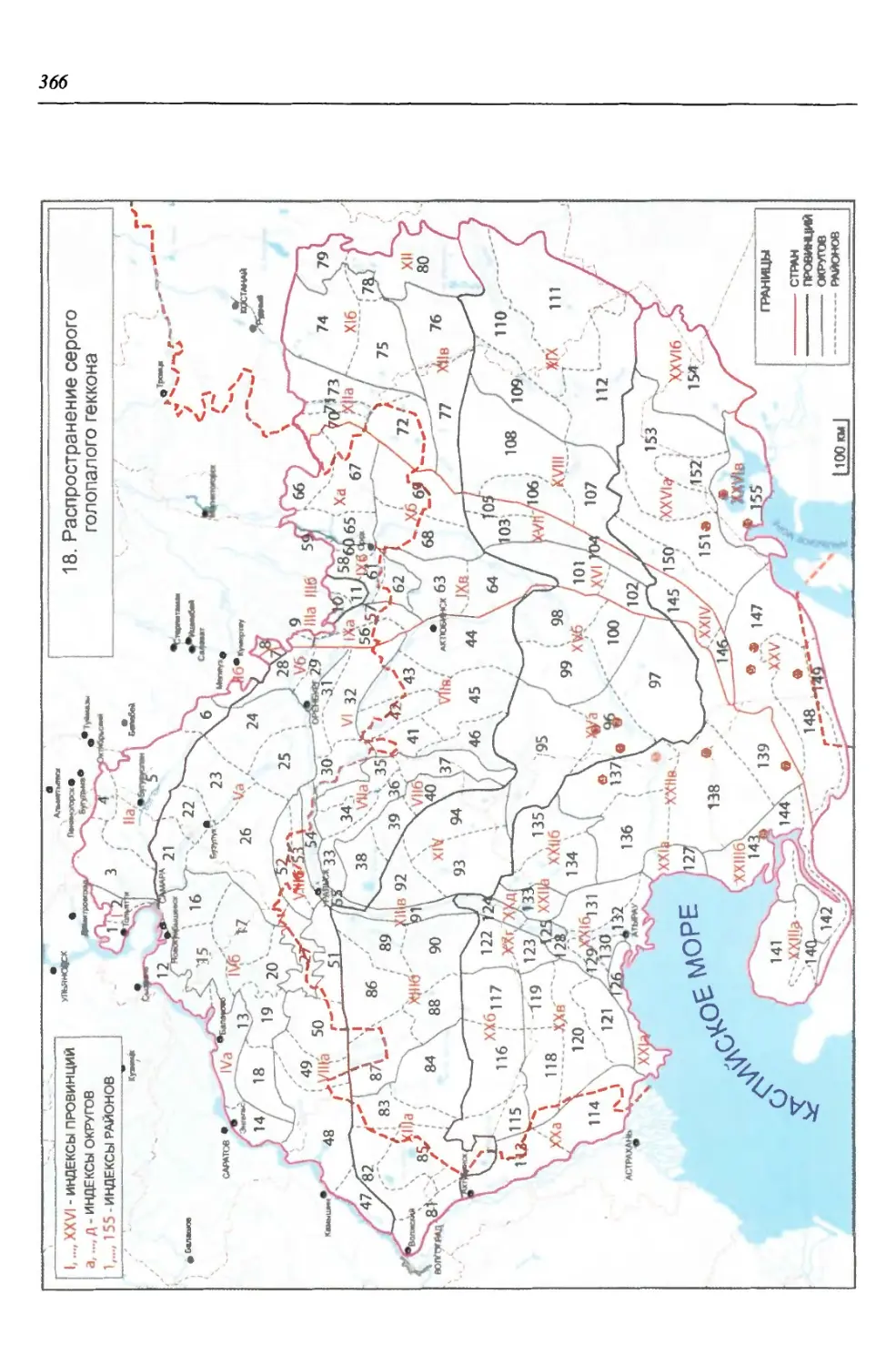
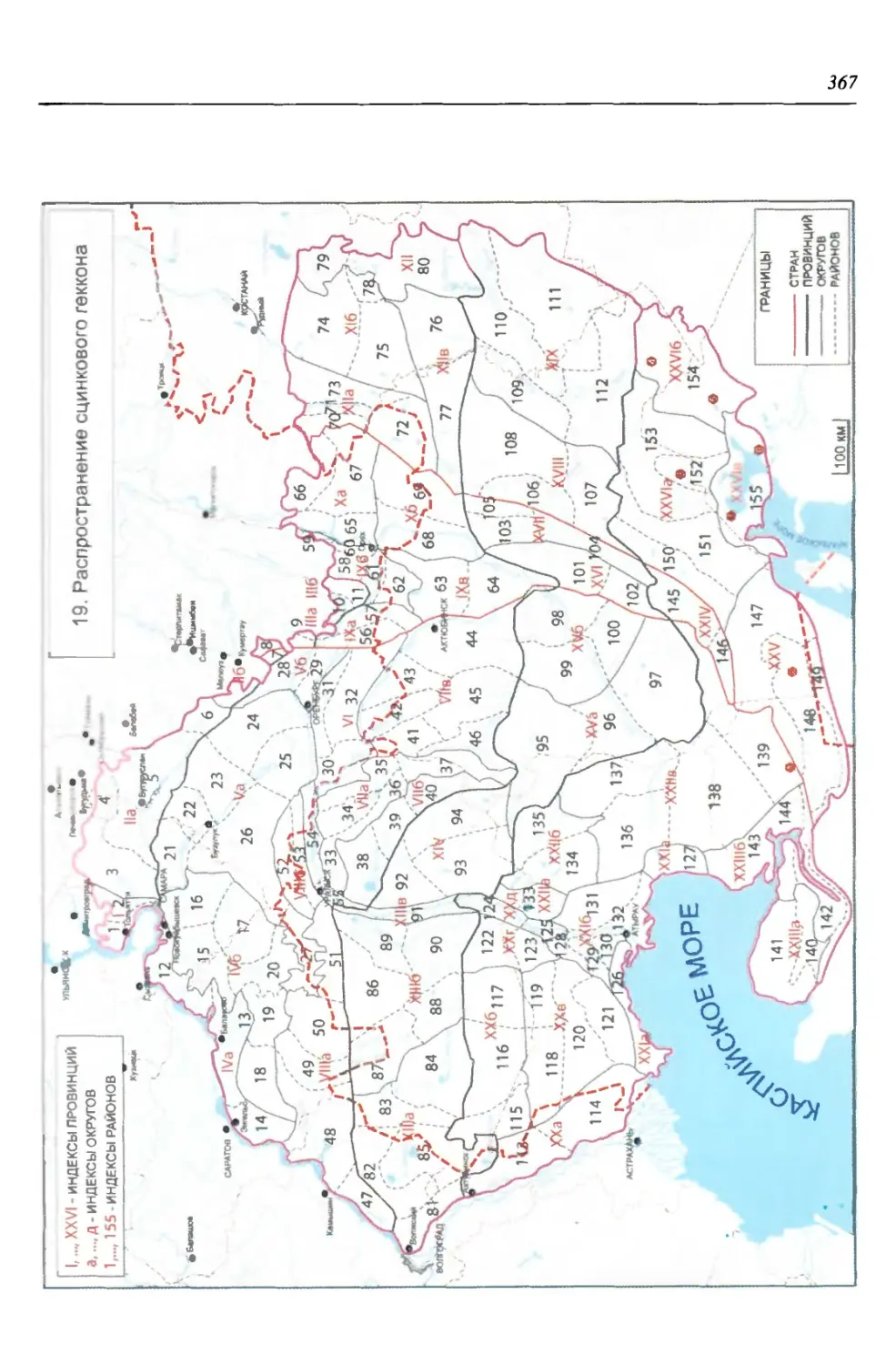
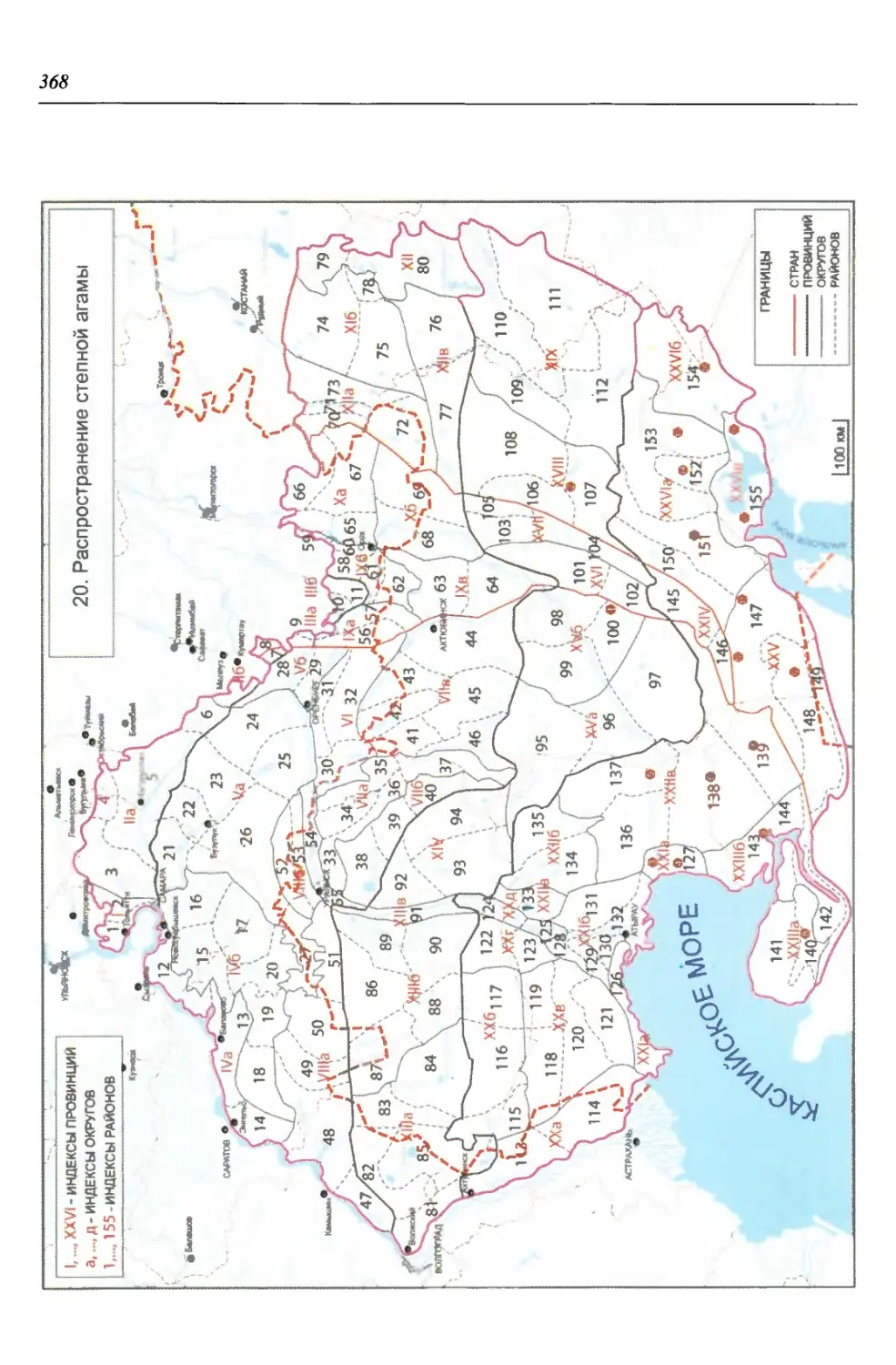
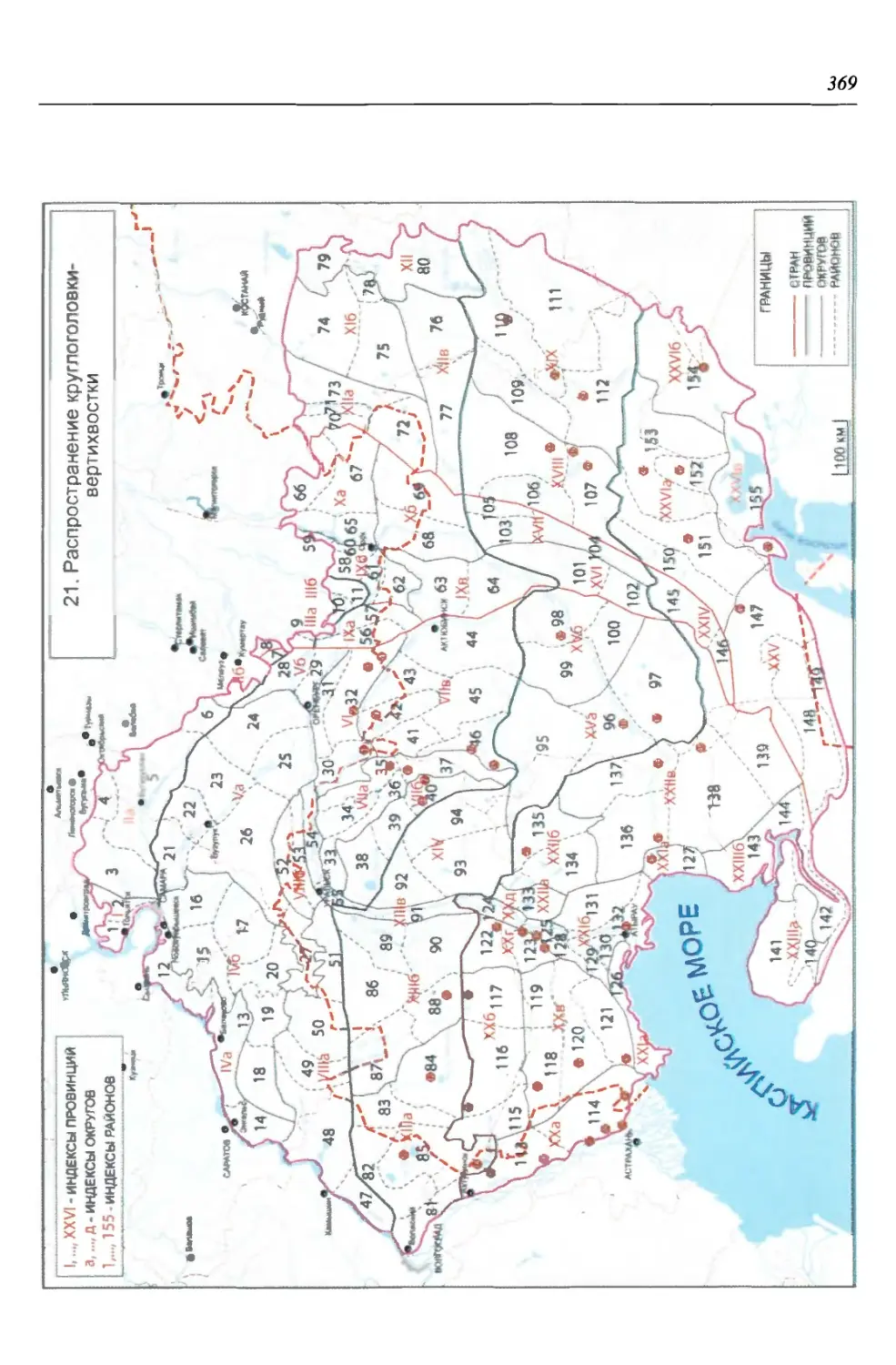
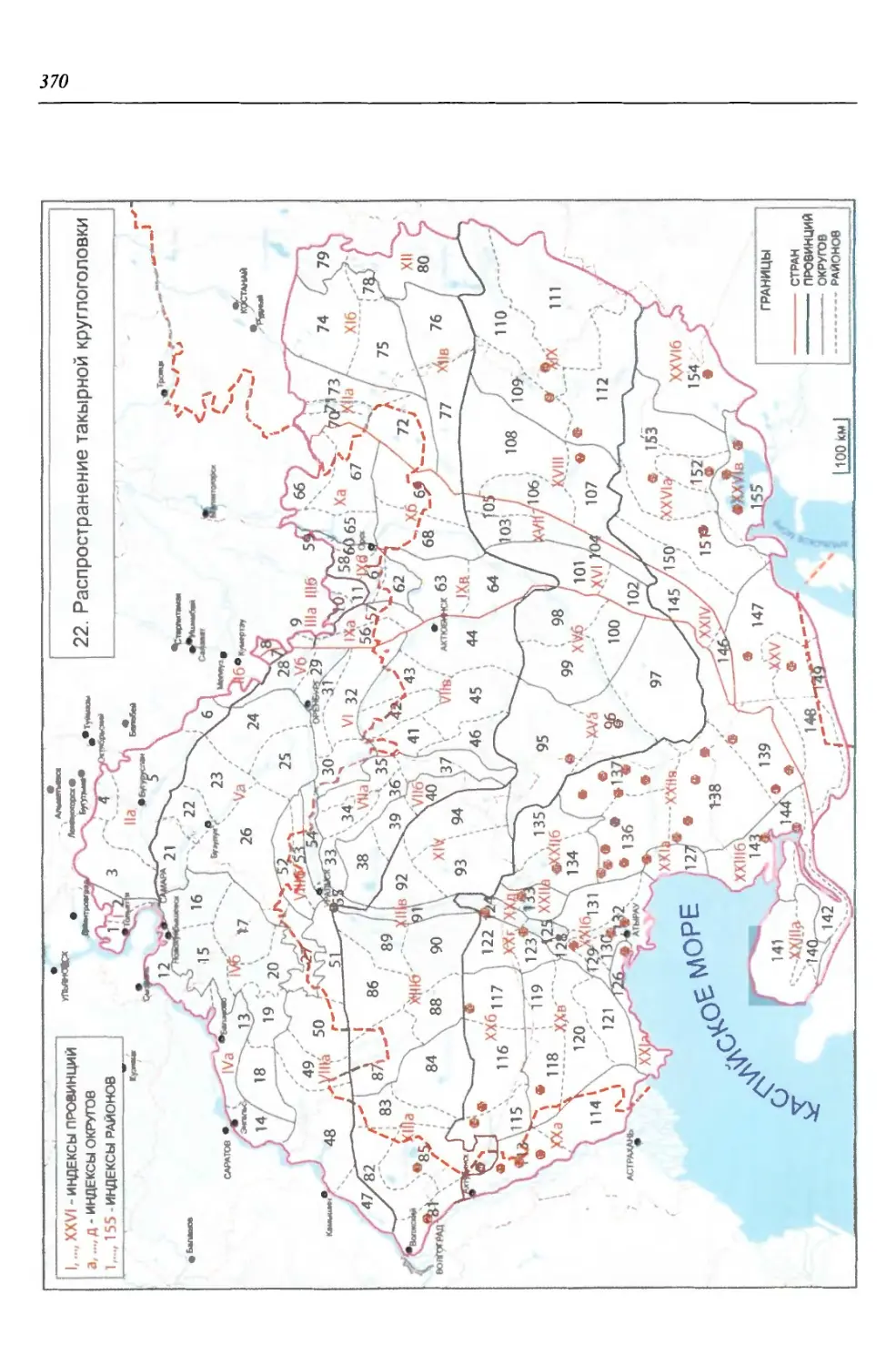
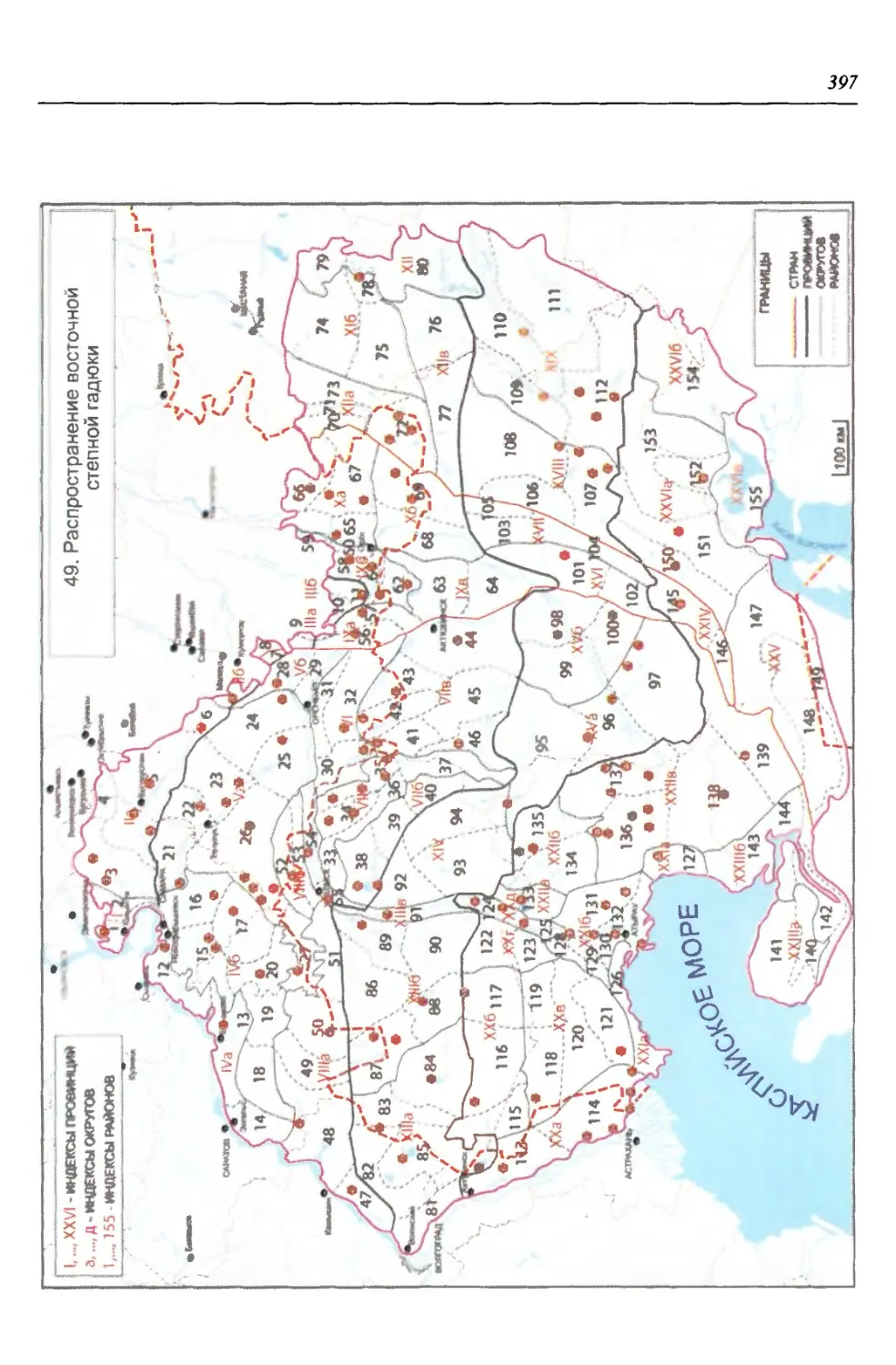
бины на востоке. Его во многом уникальная в физико-географическом отноше-
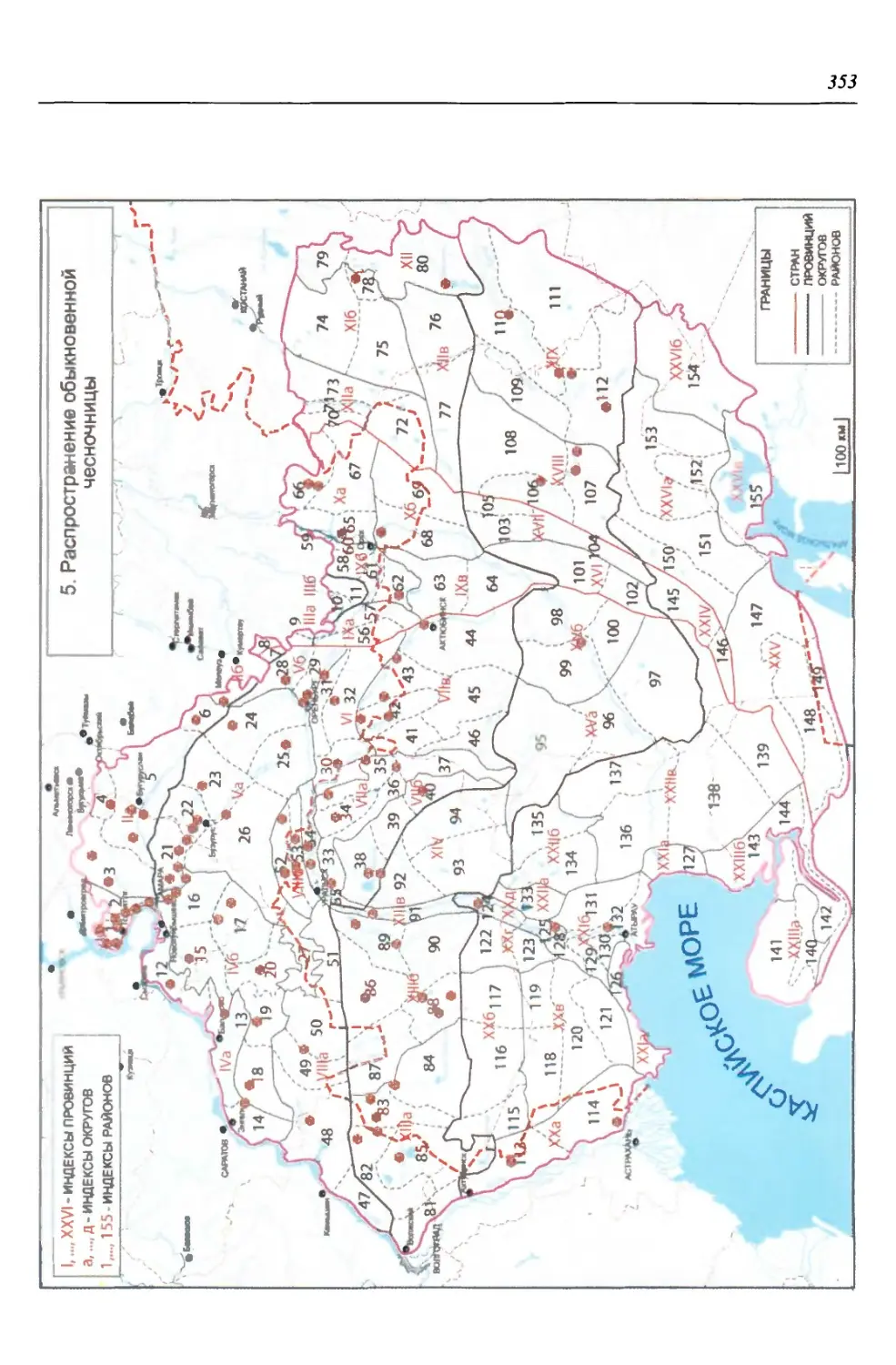
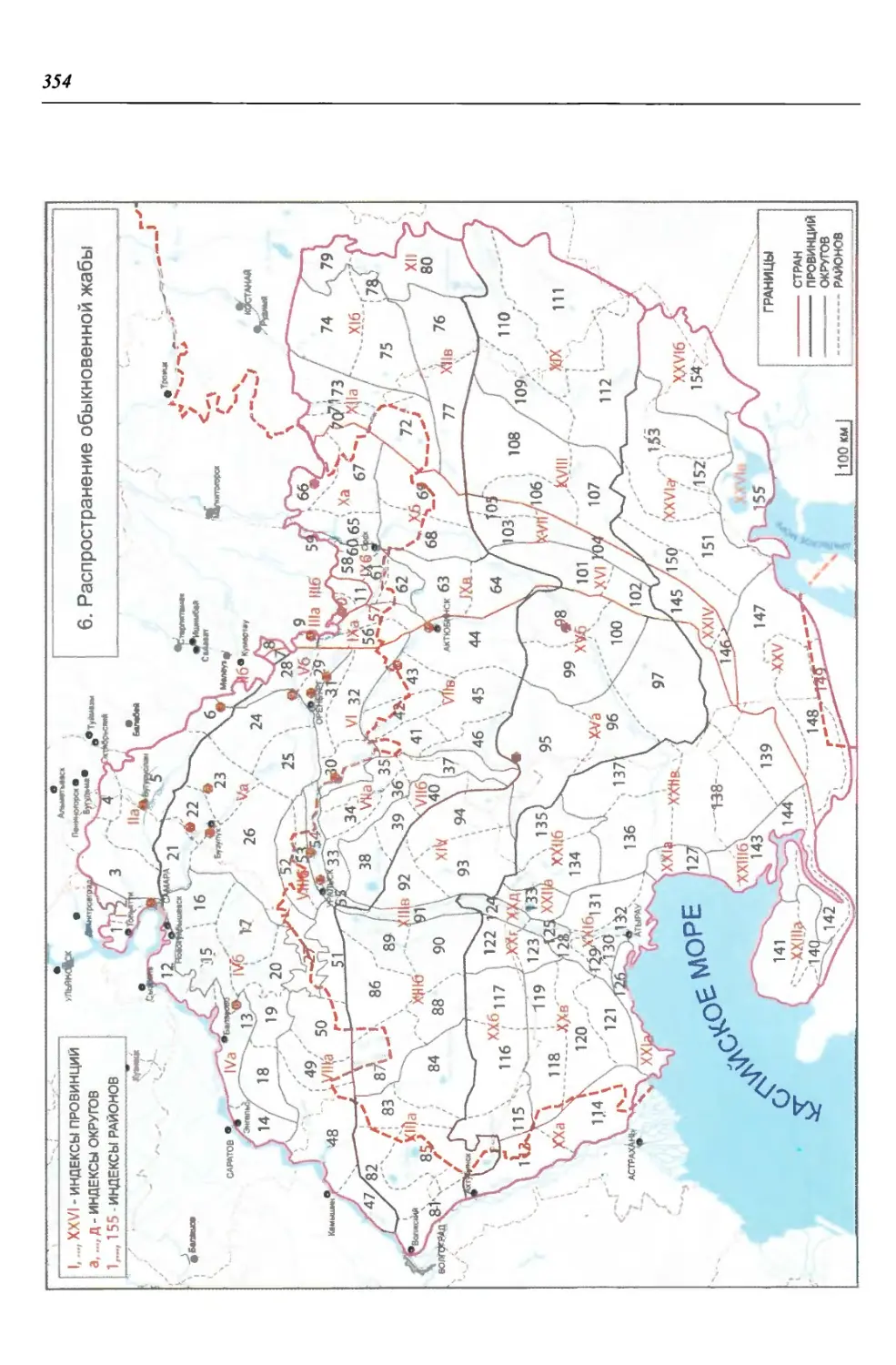
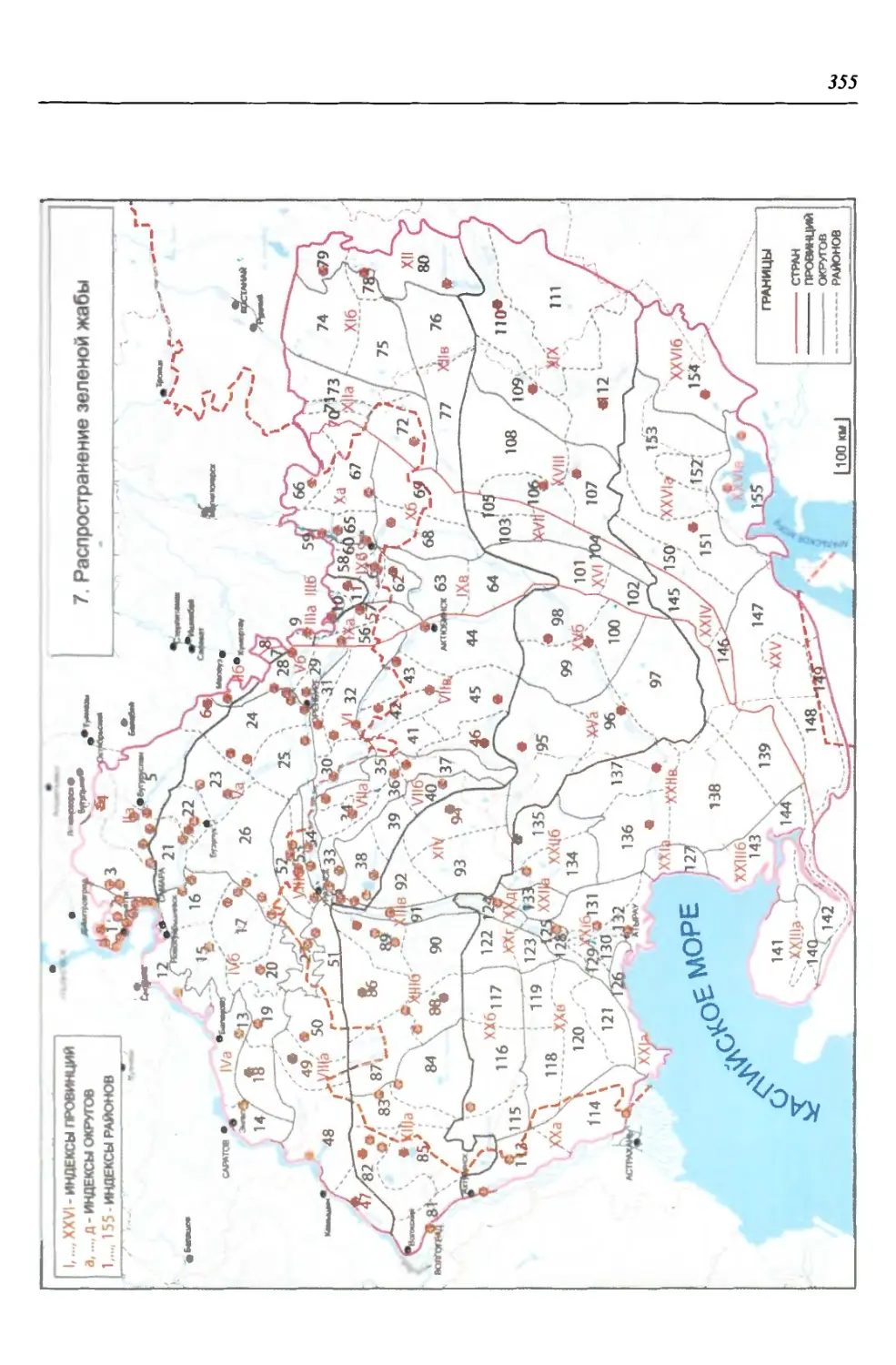
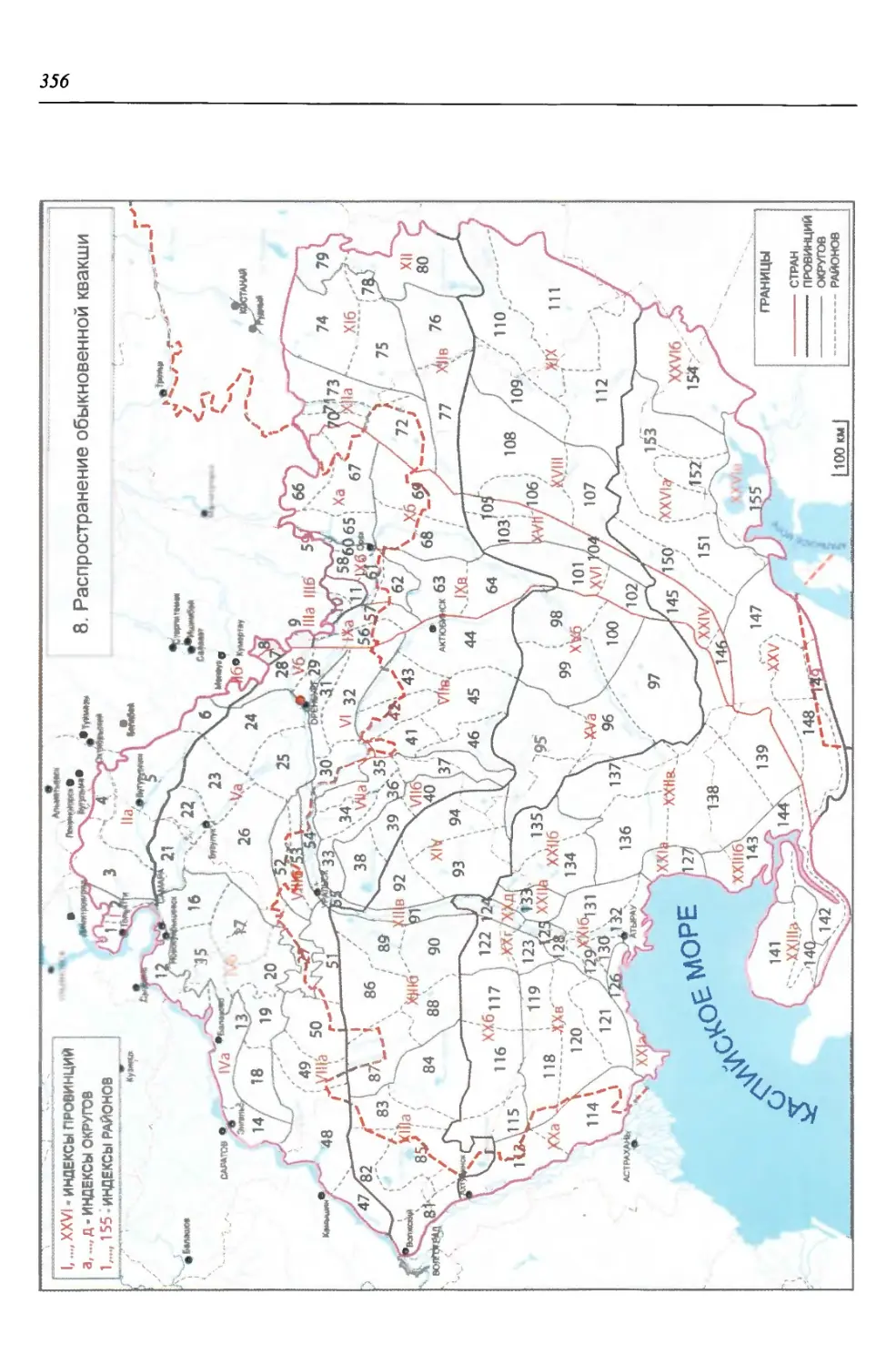
нии территория нами (Чибилёв, Дебело, 2006) разделена на 155 ландшафтных
районов, объединенных в 52 округа и 26 провинций (табл. 2).
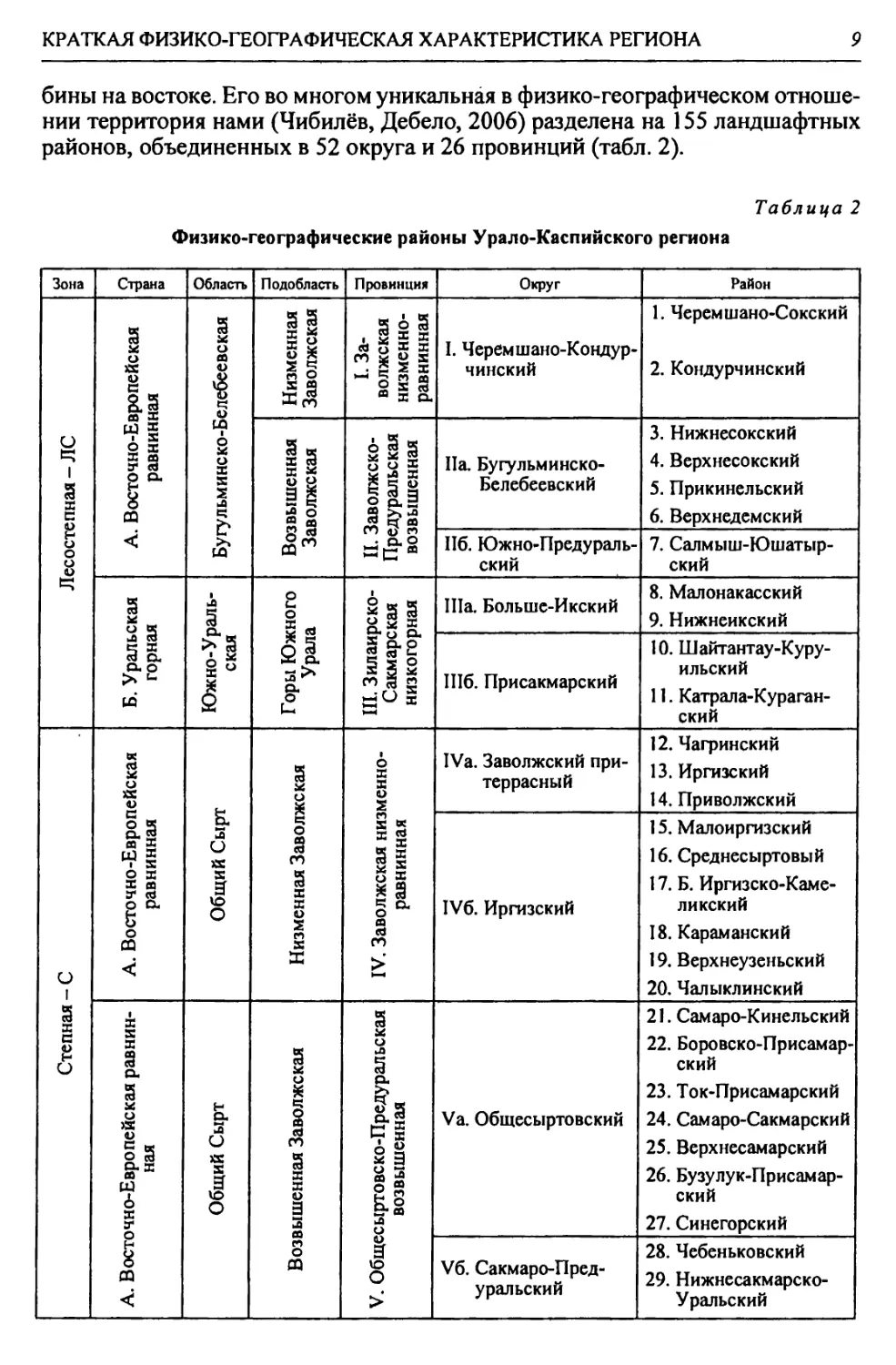
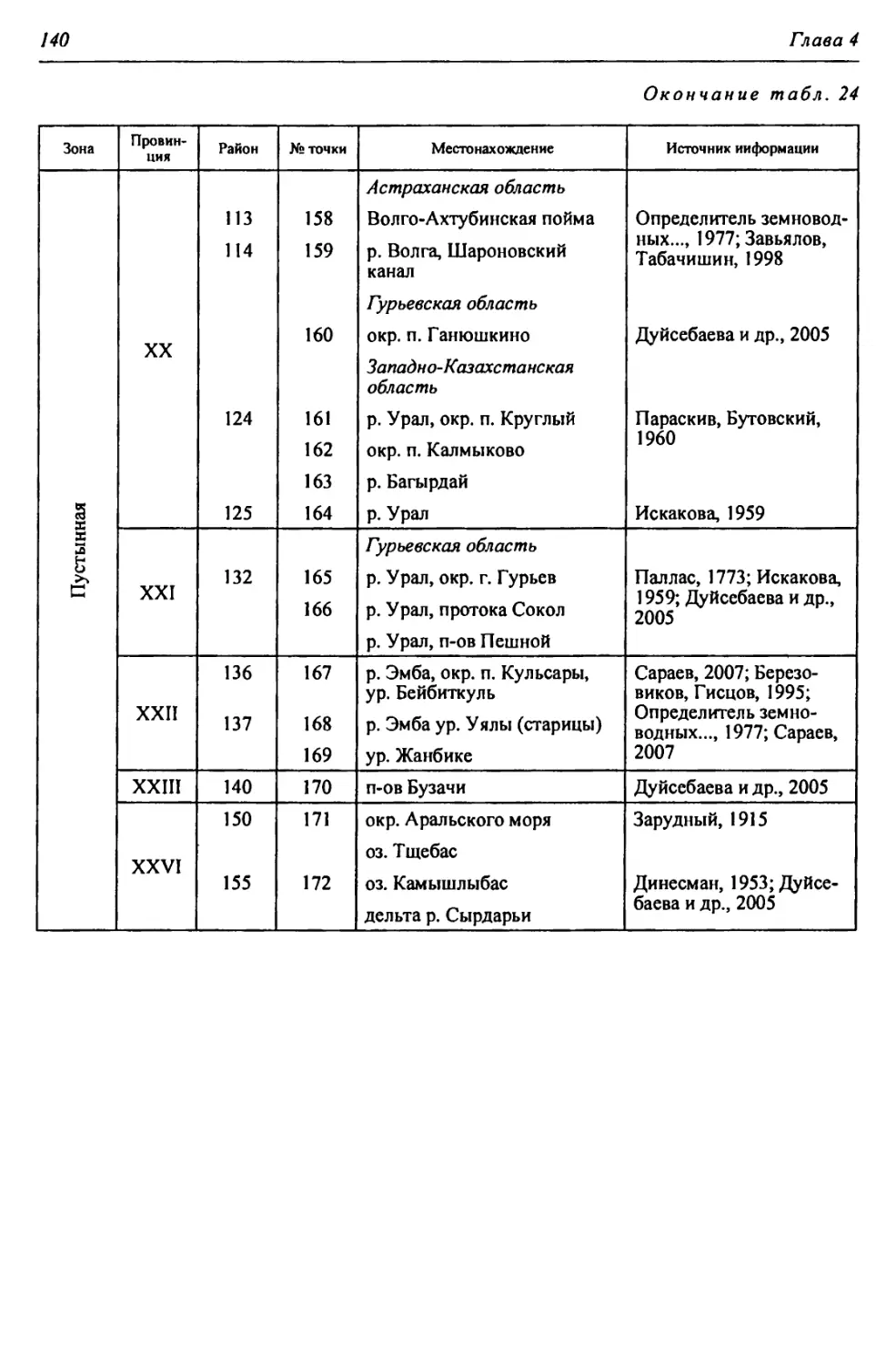
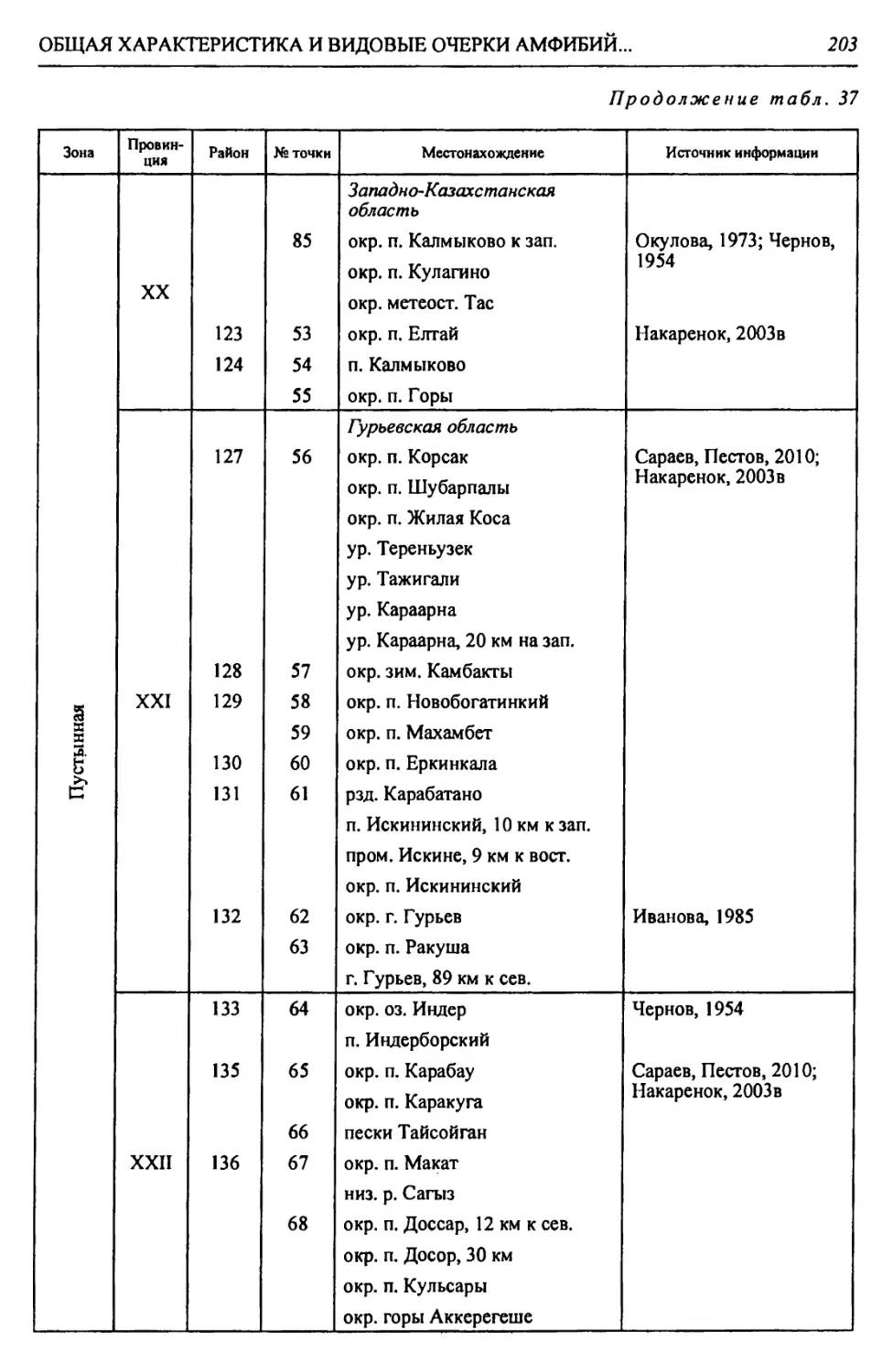
Таблица 2
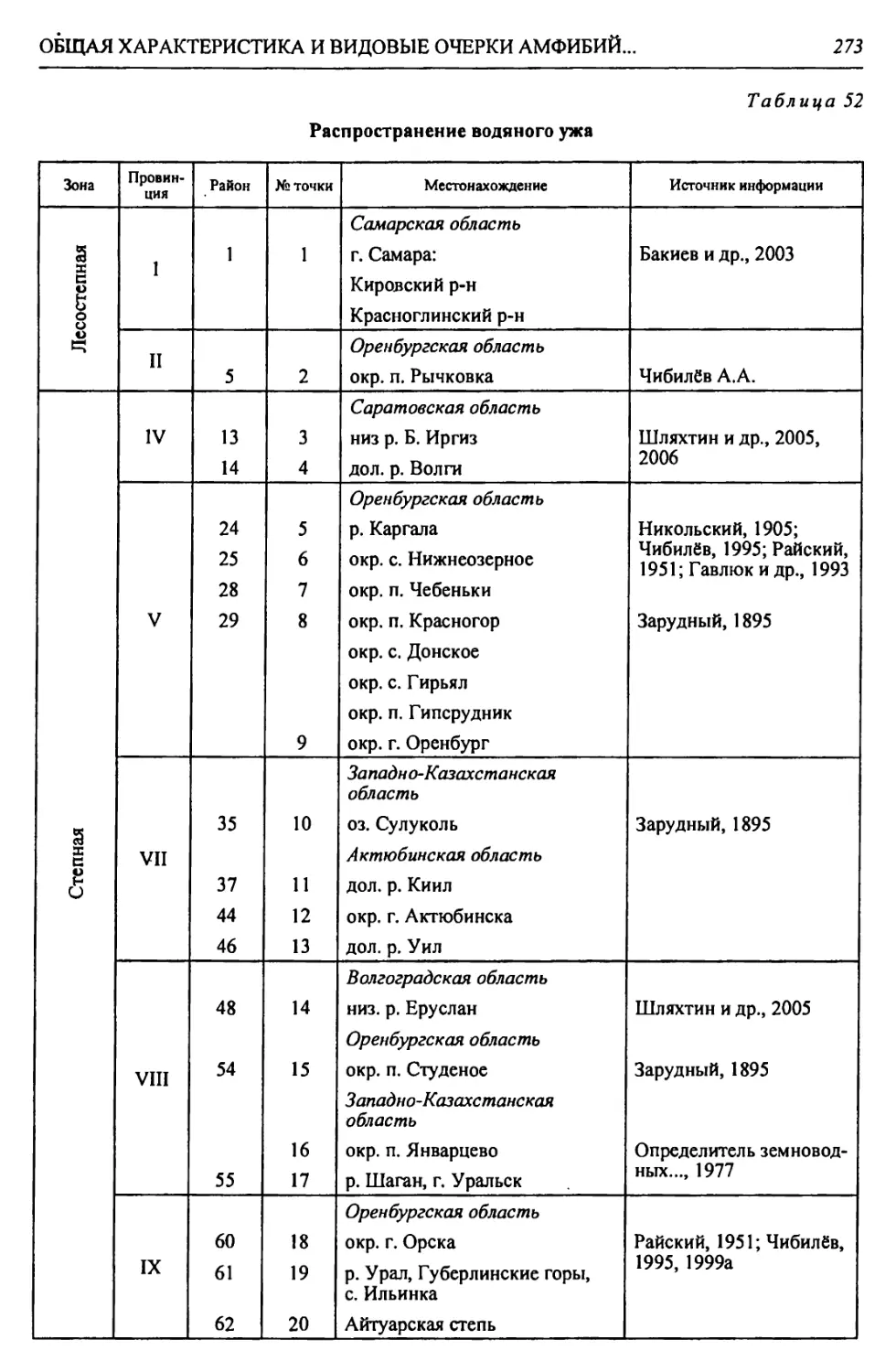
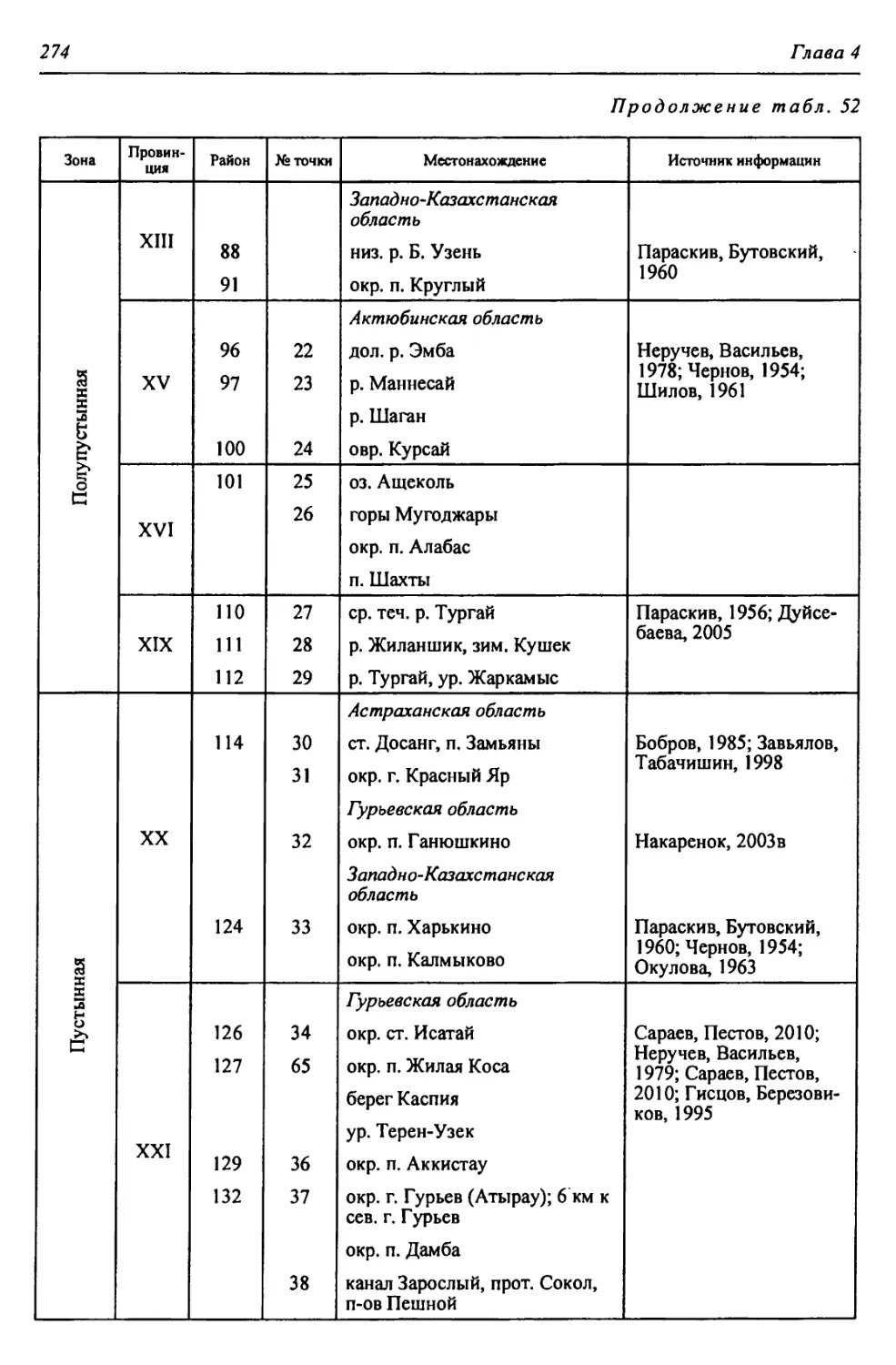
Физико-географические районы Урало-Каспийского региона
Зона Страна Область Подобласть Провинция Округ Район
Лесостепная - Л С А. Восточно-Европейская равнинная Бугульминско-Белебеевская Низменная Заволжская I. За- волжская низменно- равнинная I. Черемшано-Кондур- чинский 1. Черемшано-Сокский 2. КондурчинскиЙ
Возвышенная Заволжская II. Заволжско- Предуральская возвышенная Па. Бугульминско- Белебеевский 3. НижнесокскиЙ 4. Верхнесокский 5. Прикинельский 6. Верхнедемский
Пб. Южно-Предураль- ский 7. Салмыш-Юшатыр- ский
Б. Уральская горная Южно-Ураль- ская Горы Южного Урала III. Зилаирско- Сакмарская низкогорная П1а. Больше-Икский 8. Малонакасский 9. Нижнеикский
Шб. ПрисакмарскиЙ 10. Шайтантау-Куру- ильский 11. Катрала-Кураган- ский
Степная - С А. Восточно-Европейская равнинная Общий Сырт Низменная Заволжская IV. Заволжская низменно- равнинная IVa. Заволжский при- террасный 12. Чагринский 13. Иргизский 14. Приволжский
IV6. ИргизскиЙ 15. МалоиргизскиЙ 16. Среднесыртовый 17. Б. Иргизско-Каме- ликский 18. Караманский 19. Верхнеузеньский 20. ЧалыклинскиЙ
А. Восточно-Европейская равнин- ная Общий Сырт Возвышенная Заволжская V. Общесыртовско-Предуральская возвышенная Va. Общесыртовский 21. Самаро-Кинельский 22. Боровско-Присамар- ский 23. Ток-Присамарский 24. Самаро-Сакмарский 25. Верхнесамарский 26. Бузулук-Присамар- ский 27. Синегорский
V6. Сакмаро-Пред- уральский 28. Чебеньковский 29. Нижнесакмарско- Уральский
10
Глава 1
Продолжение табл. 2
Зона Страна Область Подобласть Провинция Округ Район
Степная - С А. Восточно-Европейская равнинная Общий Сырт ! Возвышен- ная Заволж- ская 1 VI. У рало- Ил екс кая возвышен- ная Via. Подуральский 30. Нижнеилекский 31. Донгуз-Приураль- ский 32. Донгуз-Буртинский
Подуральского плато VII. Подуральско-Илекская возвышенная Vila. Утвинский 33. Федоровский 34. Утвинский 35. Актауский 36. Приречный 37. Верхнеутвинско- Баянасский
VII6. Шалкаро-Жак- сыбаевский 38. Шал карский 39. Шидертинский 40. Калдыгайты- Булдуртинский
VIIb. Илекско-Хоб- динский 41. Илекско-Утвинский 42. Большехобдинский 43. Илекско-Хобдин- ский 44. Алгинский 45. Киил-Уильский 46. Киильский
Северный Прикас- пий 1 VIII. Южно-Сыр- товая Villa. Заволжско- Уральский 47. Приволжский 48. Еру с л ано-Торгу н- ский 49. Еруслано-Болыпе- узеньский 50. Болыиеузеньский 51. Предсыртовый
Степная - С А. Восточно-Ев- ропейская равнин- ная Общий Сырт Северный При- каспий VIII. Южно-Сыр- товая VIII6. Сыртово- Приуральский 52. Чаганско-Кинделин- ский 53. Рубежинско-Иртек- ский
VIIIb. Уральский долинный 54. Илекско-Уральский 55. Уральско-Бударин- ский
Б. Уральская горная Южно-У рал ьская Южно-У ральская горная IX. Урало-Мугоджар- ская низкогоорная 1Ха. Буртинский 56. Буртинско-Конду- ровский 57. Зиянчуринско- Киялыбуртинский
1X6. Саринско-Губер- линский 58. Саринский 59. Таналыкско-Урата- зымский 60. Ирендыкско-Орский
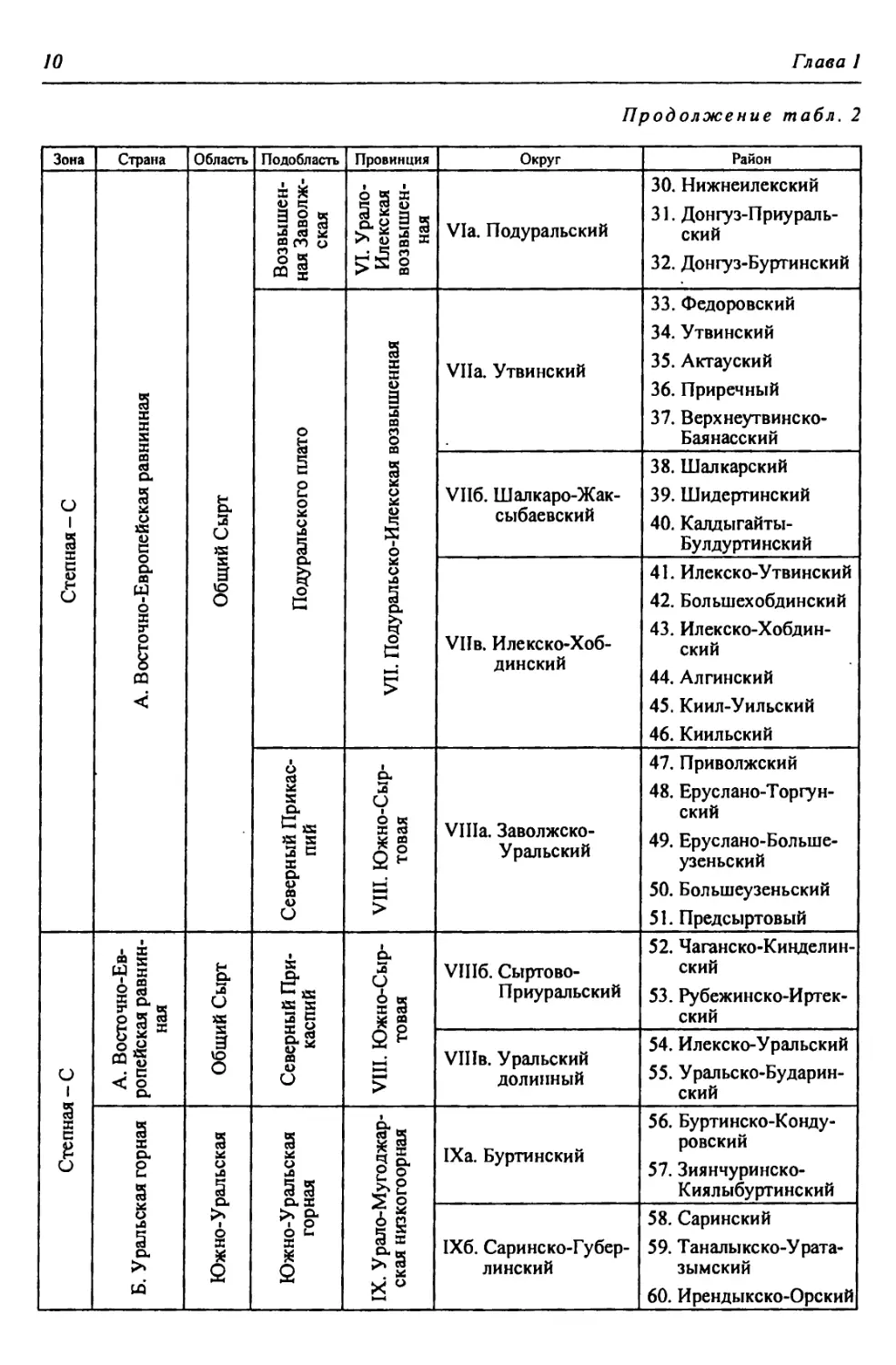
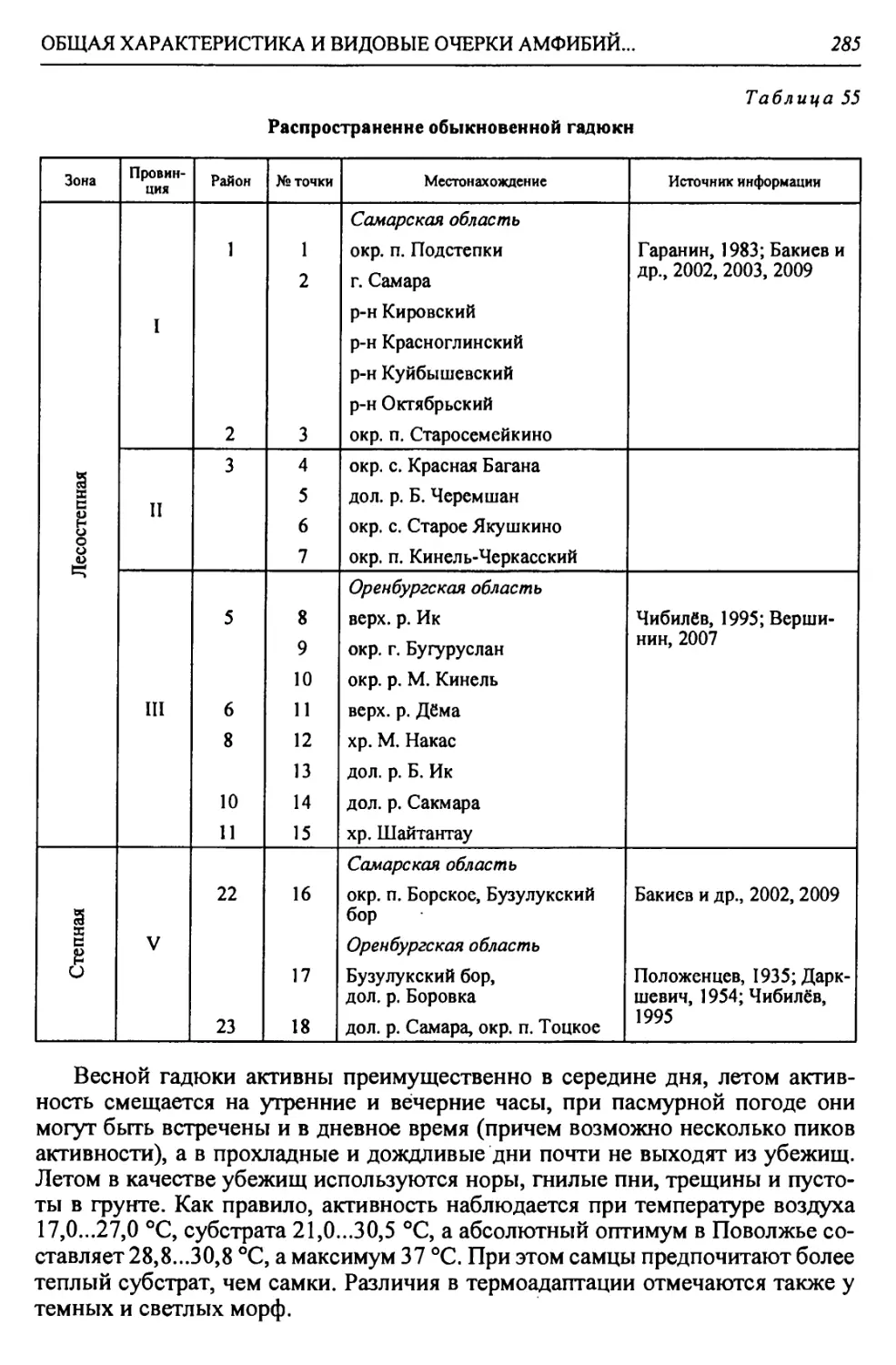
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕГИОНА
//
Продолжение табл. 2
Зона Страна Область Подобласть Провинция Округ Район
Степная - С Б. Уральская горная 1 Южно-У ральская Южно- Уральская горная IX. Урало- Мугоджар- ская низко- горная 61. Губерлинский 62. Алимбет-Каргалин- ский
1Хв. Северо-Мугод- жарский 63. Каргалинский 64. Илекско-Эмбенский
Заураль- ское Плато X. Урало- Тоболь- ская Ха. Суундук-Жарлин- ский 65. Суундук-Ириклин- ский 66. Верхнесуундукский 67. Жарлинский
Хб. Орь-Кумакский 68. Нижнеорьский 69. Верхнекумакско- Ушкотинский
В. Турано-Тургайская Арал о-Тур гайская Тургайская XI. Западно-Тургайская Х1а. Притобольский 70. Сынтасты-Верхне- тобольский 71. Шортанды-Тоболь- ский 72. Шалкар-Жетыколь- ский 73. Притобольский
XI6. Верхне-Иргиз- ско-Улькоякский 74. Верхнеулькоякский 75. Улькоякско-Кабыр- гинский
Х1в. Верхне-Тургай- ский 76. Северо-Тургайский 77. Сапсынагашский 78. Наурзумский
XII. Цен- трально- Тургайс- кая XII.а. Тургайский ложбинный 79. Кушмурунский 80. Сарьюзекский
Полупустынная - Пл А. Восточно-Европейская равнинная Прикаспийская Низменный Северный Прикаспий XIII. Северная Волго-Уральская плоскорав- нинная ХШа. Западный Вол- го-Уральский 81. Приахтубинский 82. Джаныбекский 83. Торгунско-Урдин- ский 84. Аралсорский 85. Эльтонский
XIII6. Восточный Волго-Ураль- ский 86. Чижинско-Балык- тинский 87. Приузеньский 88. Нижнеузеньский 89. Северный Урало- Ку шумский 90. Южный Урало- Кушумский
XIII6. Лбищенский 91. Бударинско-Калмы- ковский
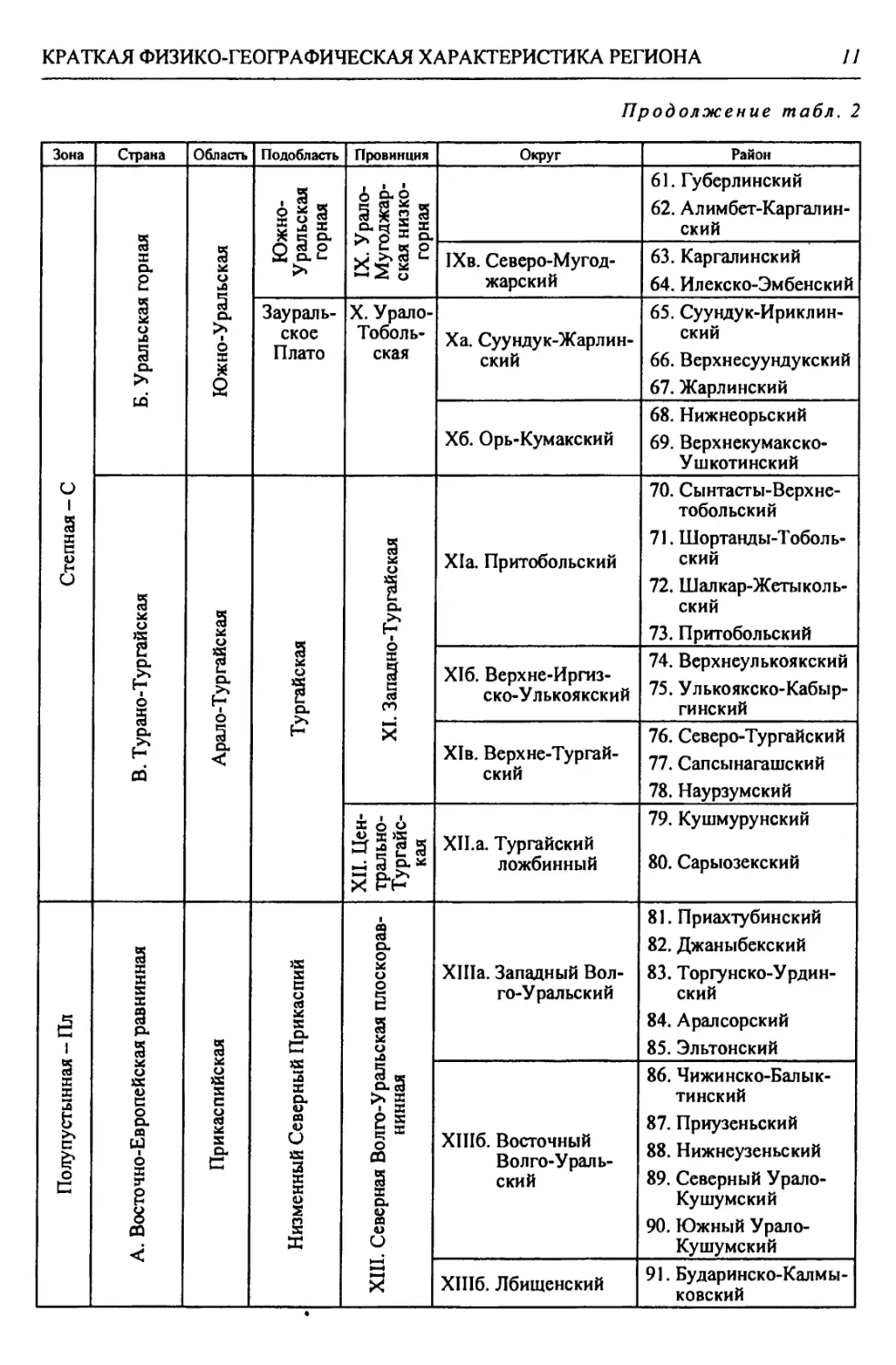
Пустынная - П Полупустынная - Пл 2 X в>
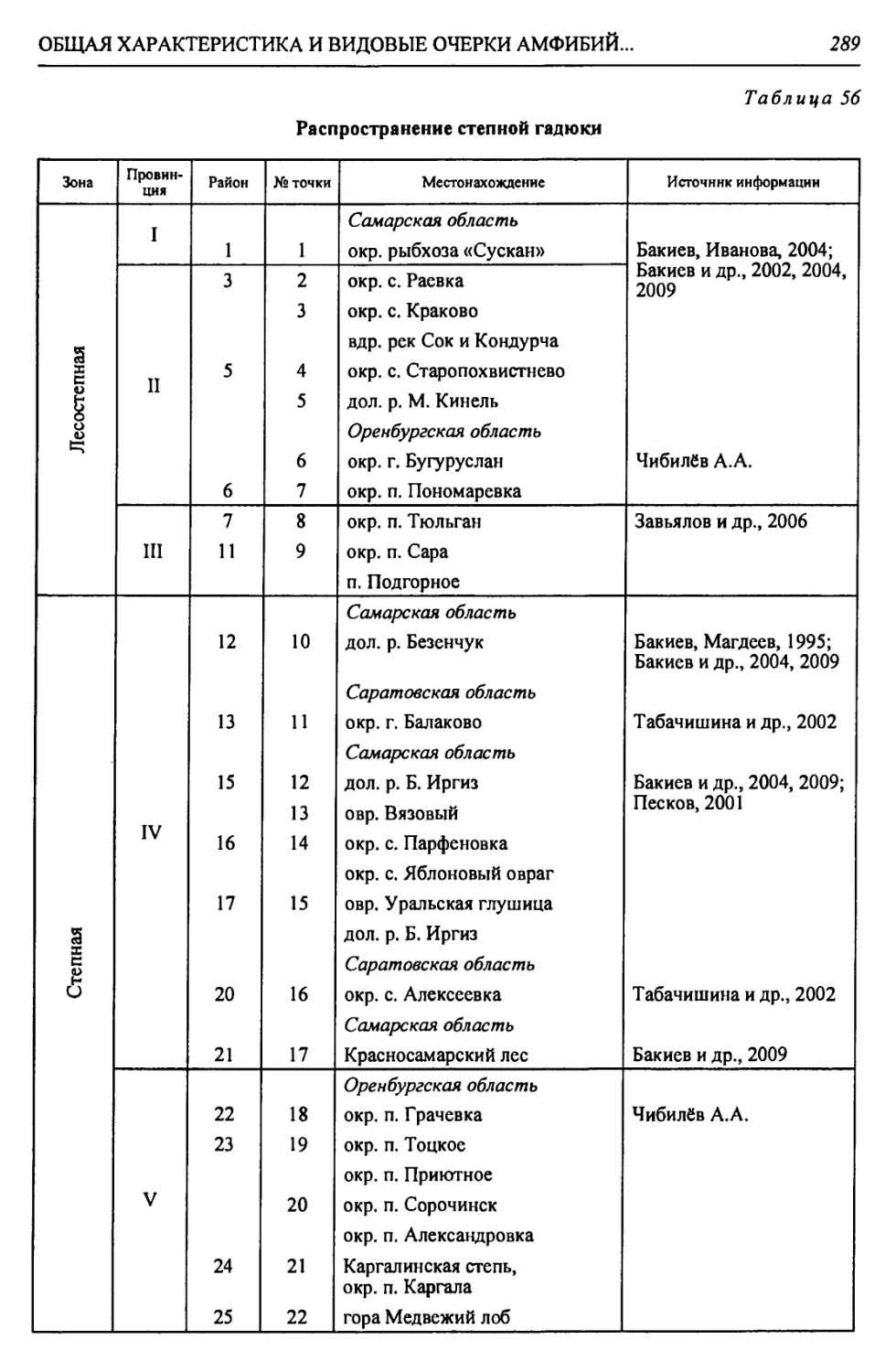
А. Восточно-Европейская В. Турано-Тургайская Б. Уральская горная А. Восточно-Европейская равнин- ная 1 Страна
Прикаспийская Арало-Тургайская Южно-У ральская Прикаспийская | Область
Северно-Прикаспийская равнинная (низмен- ная) Тургайская Заураль- ское плато Урало- Мугод- жарская Подуральского плато Низменный Северный Прикаспий | Подобласть
XX. Южная Волго-Уральская бугристо-пес- чаная XIX. Южно-Тур- гайская XVIII. Запад- но-Тур- гайская XVII. Южно- Заураль- ская XVI. Южно- Мугод- жарская XV. Эмбенская возвышенная XIV. При- уральская на- клонно-плос- коравнинная | Провинция
ХХд. Нижнеураль- ский пойменный ХХг. Приуральский ХХв. Центральный песчаный ХХб. Северный Рын- ский ХХа. Южный Приах- тубинский XIXa. Улькояк-Тур- гайский XVIIIa. Иргизско-Бас- кудукский XVIIa. Борлинский XVIa. Мугоджарский XV6. Верхнеэмбен- ский XVa. Уильско-Эмбен- ский XlVa. Зауральский jXd^Q |
124. Калмыковско- Индерский 125. Нижнеуральский 122. Ку шумско-Багыр- дайский 123. Прибагырдайский 118. Шукырский 119. Прикамыш-Самар- ский 120. Бузанайский 121. Ментекенский 115. Хакский 116. Нарынский 117. Камыш-Самарский 113. Прибаскунчакский 114. Батпайсагырский 109. Улькоякский 110. Центральный Тур- гайский 111. Улы-Жиланшик- ский 112. Нижнеиргиз-Тур- гайский 106. Шет-Иргизский 107. Тумалыкольский 108. Баскудукский 103. Кайрактинский 104. Борсыксайский 105. Верхнеиргизский 101. Мугоджаро-Берчо- гурский 102. Шошкакульский 98. Темиро-Эмбенский 99. Темирский 100. Атжаксинский 95. Уильско-Сагызский 96. Сагызско-Эмбен- ский 97. Манисайский 92. Олентинский 93. Байгуттинский 94. Калдыгайтинский | Район |
Продолжение табл.
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕГИОНА
13
Окончание табл. 2
Зона Страна Область Подобласть Провинция Округ Район
Пустынная - П А. Восточно-Европейская Прикаспийская Северно-Прикаспийская равнинная (низменная) XX.I Северо-Кас- пийская приморс- ко-равнинная ХХ1а. Волго-Эмбен- ский 126. Волго-Уральский 127. Урало-Эмбенский
XXI6. Устьуральский 128. Аксайский 129. Жалтырский 130. Баксайский 131. Курайлинский 132. Устьуральский
XXII. Урало-Эмбенская 1 плоскоравнинная । ХХПа. Индерский 133. Индерский
ХХПб. Нижнеуиль- ский 134. Устьуильский 135. Тайсойганский 136. Сагыз-Эмбенский
ХХПв. Приэмбенский 137. Кайнаро-Сагыз- ский 138. Прикаспийско- Каракумский 139. Подчинковый
XXIII. Бузачинская плоскоравнинная XXIIla. Бузачинский 140. Центральный Бузачинский 141. Бузачинский приморский 142. Примангышлак- ский
ХХП16. Култукский 143. Мертвокултукский 144. Кайдакский
Б. Ураль- ская горная Южно- Уральская Заураль- ское Плато XXIV. Ша- ги райская XXIVa. Шагырай- Донгузтауский 145. Шагырайский 146. Донгузтауский
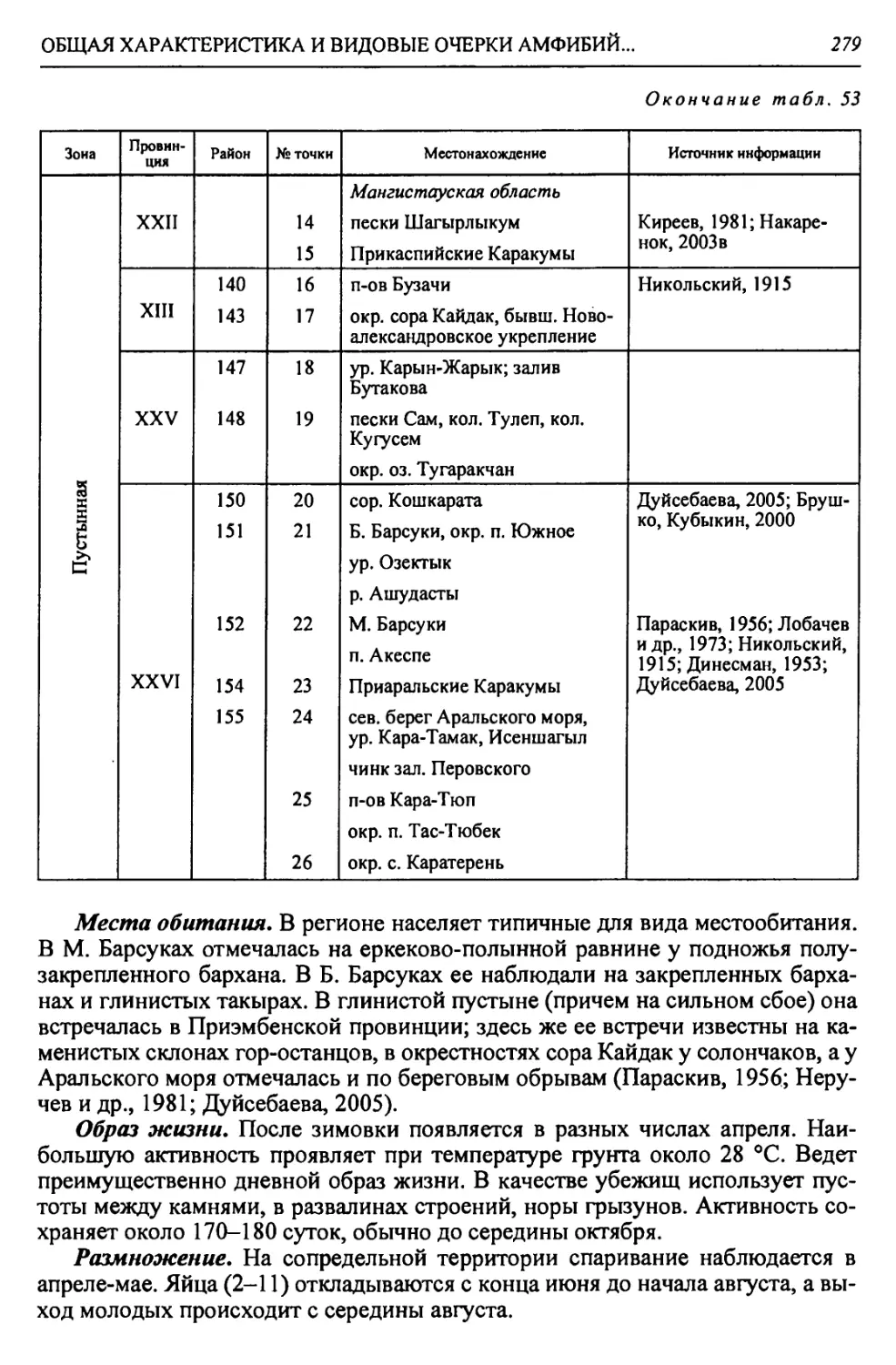
В. Турано-Тургайская Арало-Тургайская Приаральская XXV. Предус- тюртская XXVa. Северный Устюртский 147. Северный Устюртский 148. Сам-Матсайский 149. Подчинковый
XXVI. Северно-При- аральская XXVIa. Северо-запад- ный Приараль- ский 150. Шалкарский 151. Большебарсукский 152. Малобарсукский 153. Куландинский
XXVI6. Северо-вос- точный При- аральский 154. Приаральско- Каракумский
XXVIb. Приморский 155. Приморский
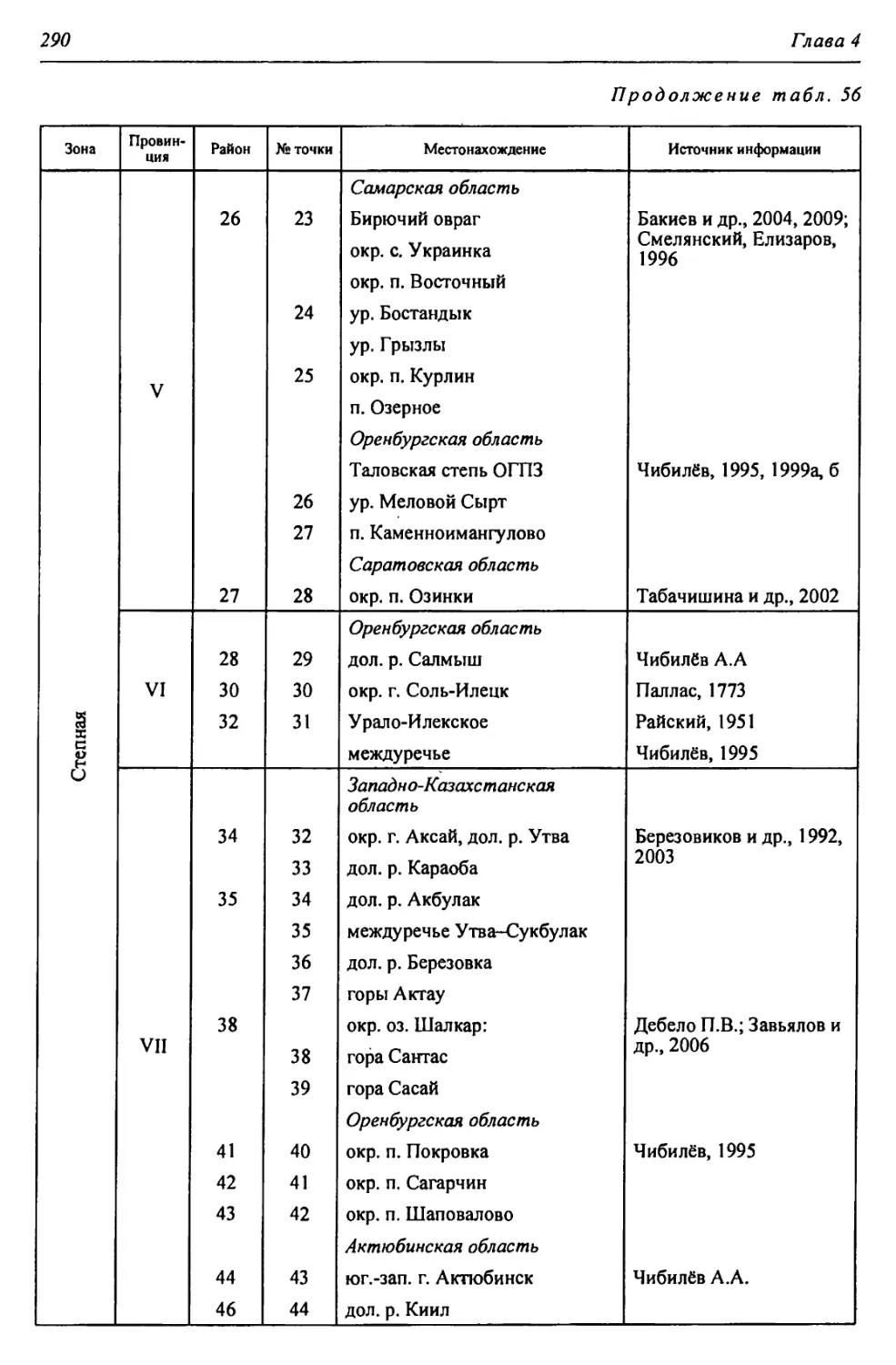
14
Глава 1
1.2. ЛЕСОСТЕПНАЯ ЗОНА
Наиболее характерной особенностью европейского лесостепного ланд-
шафта является чередование массивов преимущественно широколиственных
водораздельных лесов с безлесными междуречными пространствами луговых
степей.
На Восточно-Европейской равнине в пределах региона южная граница
зоны простирается немногим южнее долины р. Самары (р. Чапаевки), низо-
вий р. Б. Кинель, затем, на основе анализа современной и прошлой лесис-
тости водораздельных равнин, ее граница нами устанавливается по долине
р. М. Кинель, верховьям Б. Кинеля, истоку Салмыша(р. Неть), устью Б. Юша-
тыри, низовьям Б. Ика, устью Куруила, по Сакмаро-Уральскому водоразделу
южнее Кувандыка, верхней части бассейнов Киндерли и Чебаклы и верховьям
Губерли, включая северо-западную окраину Саринского плато.
В то же время, по мнению Л.С. Берга (1947), к лесостепи следует отнести
территорию Бузулукского бора, откуда граница зоны образует выступ к северу
за р. Б. Кинель и далее следует к г. Стерлитамаку. Ф.Н. Мильков (1986) южную
границу зоны проводит по южному склону Общего Сырта, устью р. Илека, ок-
рестностям п. Переволоцка, верховьям р. Салмыша, окрестностям п. Шарлык,
низовьям р. Юшатыри и далее в юго-восточном направлении.
Западная часть нашей территории в основе имеет сложную морфострук-
туру - Мелекесскую мульду с примыкающей частью Ульяновско-Саратовс-
кого прогиба, заполненную мощной толщей континентальных и морских па-
леозойских, мезозойских и более молодых отложений. В четвертичное время
эта территория испытала относительное опускание, что вызвало накопление
значительных по мощности преимущественно акчагыльских песчано-гли-
нистых отложений и их последующее неоднократное переотложение реч-
ными потоками. Медленное врезание русла р. Волги началось уже во время
акчагыльской регрессии, а заметно увеличилось (до 80-100 м) в послеап-
шеронскую эпоху. В конечном счете большая часть территории оказалась в
пределах погребенной (третичной) и современной долин Волги, к которым
примыкают связанные с ними абразионные и денудационные выровненные
поверхности, постепенно переходящие в Высокое Заволжье. В результа-
те последующих движений акчагыльская поверхность оказалась на высоте
120-165 м, ее абразионно-аккумулятивные террасы - 120-180 м, а одновре-
менно формировавшиеся денудационные поверхности на высоте 180-290 м.
В образовании современной морфоскульптуры основную роль сыграли эро-
зия, денудация и аккумулятивное выравнивание депрессии плейстоценовым
аллювием.
Большая часть территории в основе своей имеет Волго-Уральскую ан-
теклизу, кристаллический архейско-протерозойский фундамент которой об-
разует крупный Татарский свод. Восточнее к нему примыкают небольшие
куполовидные поднятия и осложненные разрывами и сбросами синклиналь-
ные структуры Предуральского прогиба, а затем в Уральской горной стране -
система узких асимметричных субмеридиональных складок Западно-Ураль-
ского поднятия и более крупных осложненных разрывами синклинориев и ан-
тиклинориев Центрально-Уральского поднятия. Кристаллический фундамент
здесь перекрыт морскими и континентальными отложениями, среди которых
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕГИОНА
15
наибольшее развитие получили отложения перми, ставшие важнейшими лан-
дшафтообразующими породами района. На западе Предуральского прогиба
пермские отложения образуют пологие складки, которые в его восточной час-
ти и далее в пределах Урала сменяются крупными осложненными разрывами
антиклиналями и синклиналями, сложенными палеозойскими и мезозойскими
породами. Длительная денудация в палеогене привела к формированию здесь
обширного пенеплена, на границе с неогеном испытавшего валообразные бло-
ковые поднятия с амплитудой от 200-250 м в пределах Татарского свода и до
500 м в области складчатого Урала. На западе это привело к возникновению
крупной прямой морфоструктуры (участка приподнятого третичного пенеп-
лена) холмисто-увалистой Бугульминско-Белебеевской возвышенности, а на
востоке - мелкосопочных и низкогорных массивов, граница между которыми -
важнейший ландшафтный рубеж края - проходит по осевой части Предураль-
ского прогиба.
Отмеченные особенности геологического строения и формирования круп-
ных орографических структур привели к формированию в пределах лесостеп-
ной зоны региона трех физико-географических (ландшафтных) провинций, две
из которых располагаются в пределах Восточно-Европейской страны и одна в
Южноуральской области Уральской горной страны.
1.3. СТЕПНАЯ ЗОНА
Понятие «степь» - широкое, емкое, его содержание со временем неод-
нократно менялось, что сказывалось на выделении границ степной зоны.
В физико-географическом плане степь представляет собой особый тип лан-
дшафта умеренного пояса, который отличается безлесием водоразделов, где
на черноземных, темно-каштановых и каштановых почвах господствуют сооб-
щества засухо- и морозоустойчивой (ксерофитной) многолетней травянистой,
преимущественно разнотравно-злаковой и злаковой растительности (Чибилёв,
1998).
При такой трактовке степь в регионе представляет собой широкую полосу,
простирающуюся от р. Волги через Общий Сырт, Подуральское плато, Юж-
ный Урал и Зауралье до Тургайского прогиба. Ее южная граница в общем пла-
не определяется Предсыртовым и Предуральскими уступами, на Урале - се-
верной окраиной Мугоджар, а в Зауралье - верховьями рек Иргиз, Улькояк
и Тургай. Точнее она прослеживается по южной и восточной окраине При-
волжской возвышенности, затем по широте п. Караоба (в междуречье Горь-
кой и М. Узеня), севернее п. Казталовки, Ал-Гая, через Богатыреве, Шильную
Балку, южнее Подтяжек, обходит Чижинско-Дюринские разливы севернее
оз. Бирказан, далее пересекает водораздел рек Кушума и Урала между п. Сквор-
киным и Янайкиным, обходит с юга (в 15-20 км) оз. Шалкар, направляется к
п. Джамбейты, Кара-Тюбе и, огибая с севера и востока пески Караганды-Кум,
выходит к верховьям рек Уила и Темира. Восточнее граница степи определена
недостаточно четко и в ряде случаев остается спорной. Казахстанскими авто-
рами далее она проводится к ст. Эмба, верховьям одноименной реки, северной
окраине Мугоджар, верховьям рек Иргиз, Улькояк, низовьям р. Сарыозек, об-
ходя с юга оз. Сарыкопа к рекам Тургай и Жалдома.
16
Глава 1
Таким образом, на территории региона степная зона простирается в преде-
лах трех физико-географических стран - Восточно-Европейской равнинной,
Уральской горной и Турано-Тургайской столовой, в пределах которых выде-
лено девять провинций.
1.4. ПОЛУПУСТЫННАЯ ЗОНА
Прошло более 100 лет с тех пор, как Н.А. Димо и Б.А. Келлер (1907) ввели
в научный обиход этот термин для характеристики природных условий об-
ширной территории, расположенной между степью и пустыней. Однако до
настоящего времени нет общепринятой трактовки отличительных признаков
полупустыни, а соответственно ее структуры и границ.
Вместе с тем ряд исследователей здесь отмечает тесное переплетение эле-
ментов, свойственных как степям, так и пустыням, что привело к появлению
таких терминов, как «опустыненная степь» или «остепненная пустыня», в
принципе отрицающих самобытность этой зоны. Своеобразие переходного,
экотонного характера данной зоны применительно к интересующей нас тер-
ритории хорошо отмечено В.С. Залетаевым (1989), писавшим, что в условиях
Северного Прикаспия, в Тургайской ложбине и Северном Приаралье сформи-
ровался особый «тип географически детерминированной геоэкотонной систе-
мы», включающей несколько парцелл, которые при смене фазы увлажнения
изменяют направление развития биоценоза то в сторону опустынивания, то
остепнения. Этот тип структурной организации геоэкотонной экосистемы на-
зван им «флуктуирующим» с обратимыми, периодически латеральными сдви-
гами парцеллярных границ.
Эта особенность в определенной мере затрудняет установление границ био-
ма, причем преимущественно у его северных пределов — на границе со степью.
Наиболее характерный признак полупустыни - мозаичность, которая проявля-
ется как в малых формах рельефа и геохимическом режиме, так и в почвенном
и растительном покровах. В связи с этим северная граница зоны на западе ре-
гиона принята нами по рубежу, установленному В.В. Ивановым (1958) между
типчаковой и пустынной степью, а южная - в основном по рубежу послед-
ней на светло-каштановых почвах. У западных пределов региона ее проводят
от Волги на широте Ахтубинска, затем несколько восточнее оз. Баскунчак к
ж.-д. станции Сайхин, пескам Джас-кус-Кум (севернее Урды), отсюда извилис-
той линией по кромке песков Нарын до района Камыш-Самарских озер и далее
на п. Саралжин и зим. Бескудук. Восточнее, огибая чернополынную пустыню
немногим севернее п. Калмыкове, она поднимается до п. Каленого на Урале.
В Зауралье граница спускается к югу до п. Базар-Тюбе, от которого направля-
ется к оз. Итмурун-Куль и низовьям р. Жаксыбай, проходит по северной кром-
ке песков Бийрюк, пересекает р. Уил вблизи песков Баркин и выходит в до-
лину р. Эмба у песков Кок-Джиде и Джага-Кулак. Далее она проходит между
хр. Шошкакуль и плато Шагырай, затем по восточной окраине Зауральского
(Примугоджарского) плато и, огибая с севера пески Б. и М. Барсуки, выходит
к низовьям рек Тургая и Жингылды-озека.
Таким образом, на территории региона полупустынная зона простирается в
пределах трех физико-географических стран - Восточно-Европейской равнин-
КРАТКАЯ ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕГИОНА
17
ной, Уральской горной и Турано-Тургайской столовой, в пределах которых
выделяем семь провинций.
1.5. ПУСТЫННАЯ ЗОНА
Неопределенность трактовки термина «полупустыня» или даже отрицание
целесообразности выделения ее в самостоятельную зону затрудняют установ-
ление границ следующей - пустынной - зоны, которые в пределах региона
разными авторами проводятся по-разному. Нами северная граница пустыни
принимается по линии г. Ахтубинск, восточнее оз. Баскунчак, Сайхин, север-
нее п. Урды, по кромке песков Нарын, южнее Аралсора к низовьям рек Б. и
М. Узеней, немногим севернее п. Калмыкове на р. Урал, к низовьям р. Уил,
п. Сагыз на реке с одноименным названием, п. Кулакши на р. Эмба и далее по
западной кромке плато Донгузтау, Шагырай, Шалкарской и Иргизской депрес-
сий и широте низовий рек Иргиз, Тургай и Жингылдыозек.
Таким образом, на территории региона пустыня расположена в преде-
лах трех физико-географических стран - Восточно-Европейской равнинной,
Уральской горной и Турано-Тургайской столовой, в пределах которых по осо-
бенностям ландшафтов выделено шесть провинций.
Глава 2
ИСТОРИЯ ФОРМИРОВАНИЯ,
СОВРЕМЕННАЯ ГЕОГРАФИЯ РАСПРОСТРАНЕНИЯ
И ЭКОЛОГИЧЕСКИЕ ГРУППЫ АМФИБИЙ И РЕПТИЛИЙ
УРАЛО-КАСПИЙСКОГО РЕГИОНА
В истории биосферы нашей планеты появление тетрапол стало одним из
важнейших узловых моментов эволюции ее биоты. Анализ палеонтоло-
гических рядов форм, морфологической преемственности комплексов
признаков и морфогенетической преемственности свидетельствует, что «кор-
ни» современных амфибий ведут к общему архетипу, сложившемуся на уров-
не древних кистеперых рипидистий (Воробьева, 1992). Последующая адаптив-
ная радиация разных филетических групп в амфибиотические, а затем в более
сухопутные экологические условия обеспечила им выход на сушу, по-види-
мому, еще в девоне, поскольку в карбоне уже различаются несколько линий
морфологических и экологических типов амфибий (Терентьев, 1961). К одной
из групп - карбоновым микрозаврам - восходит палеонтологическая история
хвостатых амфибий, а биологический тип бесхвостых амфибий сформировал-
ся несколько позднее.
Считалось, что с перми до середины мезозоя потомки мелких палеозойских
амфибий были малочисленны и, по-видимому, поддерживали свое существо-
вание преимущественно в горных резервациях, где были до известной степени
защищены (Шмальгаузен, 1964). Вместе с тем остатки тетрапод, найденные
в меловых отложениях Кызылкумов, свидетельствуют, что амфибии вместе с
рептилиями были довольно многочисленными и в специфических экологичес-
ких нишах окраинных сильно опресненных приморских лиманов, прилегаю-
щих болот и проток этого периода в условиях климата, относительно близкого
к тропическому (Несов, 1981).
В основном с современным по строению скелетом хвостатые и бесхвостые
амфибии встречаются уже в отложениях юрского периода, хотя их адаптивная
радиация началась, по-видимому, в поздней перми - триасе. С этого времени
или начала мелового периода прослеживается эволюция круглоязычных, а с
конца мела становятся известными и представители Pelobatidae. Представите-
ли семейств Buffbnidae и Hylidae, строение которых в значительной степени
соответствует современным формам, известны с палеоцена, хотя, возможно,
они существовали уже в меловом, а обособились еще в юрском периоде. Ис-
копаемые Ranidae известны с эоцена, а современные виды появились в начале
миоцена (Кэррол, 1992).
Происхождение пресмыкающихся (Reptilia) обычно связывают с примитив-
ными стегоцефалами (близкими к палеозойским антракозаврам), адаптивная
радиация которых в разные наземные условия привела также к формированию
многочисленных морфологических и экологических типов этих животных.
Наиболее древней специализированной группой, несомненно, являются че-
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
19
репахи (Testudines), геологическая история которых берет начало в перми
от каких-то Anapsidae, но уже в мелу они были представлены не менее чем
10 семействами, в том числе Emydidae (Зоогеография палеогена Азии, 1974).
От палеозойских диапсидных рептилий берут начало ящерицы (Sauria). По-
явившись в конце перми, они уже в начале триаса достигли значительного рас-
цвета. Однако к концу этого периода их разнообразие существенно снизилось
и вновь эта группа начала расцветать лишь в конце юры - мелу, когда впервые
возникли представители современных семейств.
Наиболее древними из семейств нашего региона, по-видимому, являют-
ся гекконовые (Gecconidae), формирование чьих исходных форм относится к
верхней юре и нижнему мелу Юго-Восточной Азии. С мелового периода, ве-
роятно, начинается геологическая история веретениц (Anguidae), варановых
(Waranidae) и агамовых (Agamidae). Считается, что большинство мезозойских
групп рептилий без существенных изменений «пересекло» границу кайнозоя
(Татаринов, 1981), на ранних этапах которого к ним добавились представители
сцинковых (Scincidae) и настоящих ящериц (Lacertidae), найденные к настоя-
щему времени остатки которых известны с эоцена.
Змеи (Serpentes), вероятно, также сформировались в верхней юре, хотя па-
леонтологические остатки их представителей обнаруживаются только с мела
и палеогена.
Таким образом, представители палеогена принадлежали в основном к тем
систематическим группам, которые существуют и в настоящее время, хотя ран-
нетретичные виды, несомненно, являются наиболее архаичными формами.
Ограниченность, а по ряду групп и районов полное отсутствие палеогер-
петологических сведений существенно затрудняют воссоздание как дальней-
шей истории формирования герпетофауны Северной Евразии в целом, так и
истории освоения территории нашего региона населяющими ее в настоящее
время амфибиями и рептилиями. Вместе с тем некоторые пробелы в ходе это-
го процесса может помочь восполнить палеогеографическая история региона,
основные этапы которой представляются (Чибилёв, Дебело, 2006) следующим
образом.
Наиболее древней частью региона является участок Восточно-Европей-
ской платформы, Урал и Зауральский пенеплен, которые на путь континен-
тального развития вступили в конце палеозоя - мезозое, преимущественно во
второй половине мелового периода, когда на смену существовавшей здесь с
триаса мезофитной флоре пришли покрытосемянные растения и на пространс-
твах Северной Еврази начались формирование и обособление трех климати-
ческих зон. Учитывая древность происхождения амфибий и рептилий, вполне
логично предположить, что с этого времени и началось заселение территории
региона древнейшими тетраподами.
Во многом сходной с позднемезозойской была обстановка и в палеоге-
не, к началу которого на территории региона господствовали вечнозеленые
тропические леса. Позднее в регионе и на смежных пространствах Восточно-
Европейской равнины установился умеренно влажный, а на юге Урала и Ка-
захстанской равнине (отделенной от Зауральского пенеплена трансгрессией в
районе Тургайского залива) - субтропический климат, в условиях которых в
засушливых районах получили распространение жестколиственные формации:
«полтавской флоры», а в более влажных - широколиственные леса с участи-
20
Глава 2
ем вечнозеленых форм «тургайской флоры». Это же время характеризуется
значительным расширением площади региона, поскольку в результате текто-
нических движений континентальные условия установились на значительной
части современного Общего Сырта, а также на востоке и юго-востоке регио-
на, что, несомненно, способствовало расширению области обитания древних
форм амфибий и рептилий. Во всяком случае уже к эпохе олигоцена относятся
наиболее древние из известных в регионе остатки сухопутных черепах, обна-
руженные в Тургайской впадине и Приаралье (Яковлева, 1964). Расселению
амфибий и рептилий, должно быть, способствовали также теплые и влажные
климатические условия, сложившиеся после регрессии среднеэоценного моря
и формирование озерно-аллювиальных ландшафтов с густой речной сетью на
освободившейся от моря территории.
Благоприятные (субтропические) условия в Заволжье и Предуралье (как и
на большей части Русской равнины) были и в начале миоцена, когда ту ргайская
флора достигла расцвета. Однако уже в комплексе средне-верхнего миоцена в
Предуралье и на Общем Сырте роль теплолюбивых флористических элемен-
тов снижается, выпадают и влаголюбивые виды, более широкое распростра-
нение получают холодоустойчивые широколиственные и мелколиственные, а
на Южном Урале смешанные (из широколиственных и хвойных пород) леса
и травянистые сообщества. На равнинах Прикаспия это время характеризует-
ся расширением площади травянистых сообществ, перемежающихся с мезо-
фильными лесами (лесостепные условия) и группировками саванного типа. На
территории Туранской равнины в условиях умеренно влажного климата по-
явились степные ценозы, хотя доминирующими по-прежнему оставались ши-
роколиственные (каштаново-дубовые) леса. На смежной территории Средней
Азии началась смена лесных и влажно-саванновых биоценозов сухо-саванно-
выми, степными (с вкраплениями небольших лесных массивов), полупустын-
ными и пустынными, которые затем стали постепенно завоевывать Прикаспий
(Федорович, 1946). Очевидно, к данному времени и относится начало форми-
рования современной герпетофауны Средней Азии и Казахстана.
В конце миоцена - начале плиоцена на значительной части региона леса
распались на отдельные массивы и господствующими постепенно станови-
лись степные формации, которые в кинельское время сменились холодной ле-
состепью. Эпоха среднего плиоцена способствовала дальнейшему развитию
на юге региона степных и полупустынных формаций, которые к концу плио-
цена (судя по спорово-пыльцевому комплексу) в окрестностях современного
г. Уральска и в бассейне р. Еруслан были доминирующими. У северных гра-
ниц региона в широколиственных лесах появились первые элементы таежной
флоры (пихта, ель, сосна), что позволяет флору этого времени отнести уже к
европейской флористической области. В Центральном Урале это время харак-
теризуется вымиранием теплолюбивых миоценовых элементов, хотя в целом
данный район стал убежищем (рефугиумом) для ряда форм, сейчас тяготею-
щих к широколиственным лесам. Для Орского Зауралья это время примеча-
тельно чередованием лесных, лесостепных и степных сообществ.
Акчагыльская эпоха (3,0-1,5 млн лет назад) характеризовалась неод-
нократными изменениями очертания, площади, климата региона и соответс-
твенно флористических и фаунистических комплексов. В максимальную фазу
трансгрессии, когда море занимало все современное низкое Заволжье, на юге
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
21
Общего Сырта доминировали степные сообщества, севернее сменяющиеся
смешанными и хвойными (таежными) лесами. В центральной части Южного
Урала господствовали лесные (таежные, смешанные, лиственные) сообщест-
ва с небольшими участками степных. В Зауралье чередовались лесные, лесо-
степные и степные ценозы. Во время наступившей затем глубокой регрессии
(Жуков, 1945) на значительной части Северного Прикаспия сформировались
открытые ландшафты. Изменение климата в апшероне (бассейн которого был
меньше акчагыльского и представлял собой в значительной степени опрес-
ненный лиман, благоприятствовавший распространению амфибиотических
форм) привело сначала к появлению обедненных смешанных лесов, затем к
преобладанию мелколиственных сообществ, а позднее к увеличению площади
безлесных пространств. В конечном счете в результате дальнейшей диффе-
ренциации растительного покрова всего неогенового периода, по образному
выражению К.К. Маркова (1960), произошло «великое остепнение» большей
части региона, лишь в горах которого сохранились лиственные леса с очагами
теплолюбивых форм. Южнее (в аридных и экстрааридных условиях) образова-
лись ландшафты полупустынь и пустынь с разреженной ксерофитной травяно-
кустарничковой растительностью с редкими оазисами древесных сообществ
(Дашкевич, 1969).
В целом окончание эпохи плиоцена считается временем формирования в
южной части пояса широколиственных лесов основного ядра современного
европейского лесного фаунистического комплекса (Завьялов и др., 2002), а для
юга региона предположительно и началом возникновения современного гер-
петологического комплекса (Зерова, Чхиквадзе, 1984). В европейских отложе-
ниях этой эпохи (или даже более ранних) к настоящему времени обнаружены
остатки 21 вида хвостатых и бесхвостых амфибий, из которых почти половину
(40%) фаунистического комплекса составляют предковые или близкие к ним
формы (Боркин, 1984). К плиоценовому времени относятся и более 20 мес-
тонахождений древних герпетофаунистических комплексов Восточно-Евро-
пейской платформы, в том числе расположенное на сопредельной территории
Нижегородской области Апастово. Здесь обнаружены остатки Pliobatrachus
cf. langhae, Bufo verrucosissimus (вымершие), В. bufo, Rana sp., а также Natrix
natrix и N. sp. В других лесных и реконструируемых как лесные местонахож-
дениях Поволжья найдены остатки еще ряда видов, в том числе Triturus cf.
alpestris, Ranomorphus similis, Bufo belogoricus (вымершие), Pelobates fuscus,
Rana ridibunda, R. lessonae, а также Anguis fragilis, Lacerta agilis, Eryx sp., Colu-
ber cf. viridiflavus и неопределенные зеленые жабы, бурые лягушки, гадюки
(табл. 3,4).
В состав лесостепных комплексов, которые, как отмечено ранее, также
были распространены в пределах региона, помимо значительной части ука-
занных видов входили вымершие здесь формы (Mioproteus sp., Latonia sp.,
Pelobates syriakus, Eupelobates sp., Bufo planus, B. albua, Rana yalpugiensis, Li-
ventsovkia jucunda, Varanus sp., Pseudopus apodus, Sauria incerta sedis, Coluber
aff. ravergieri, Elaphe cf. rufodorsata, Psammophius sp.) и современные виды,
представленные Bufo viridis, В, raddei, Lacerta viridis, Coronella austriaca, Ela-
phe aff. dione, E. longissima, Natrix tesselata, Vipera berus, К ursini. Часть этих
видов характерна и для хапровского фаунистического комплекса Восточно-Ев-
ропейской равнины, для которого наиболее вероятен степной палеоландшафт.
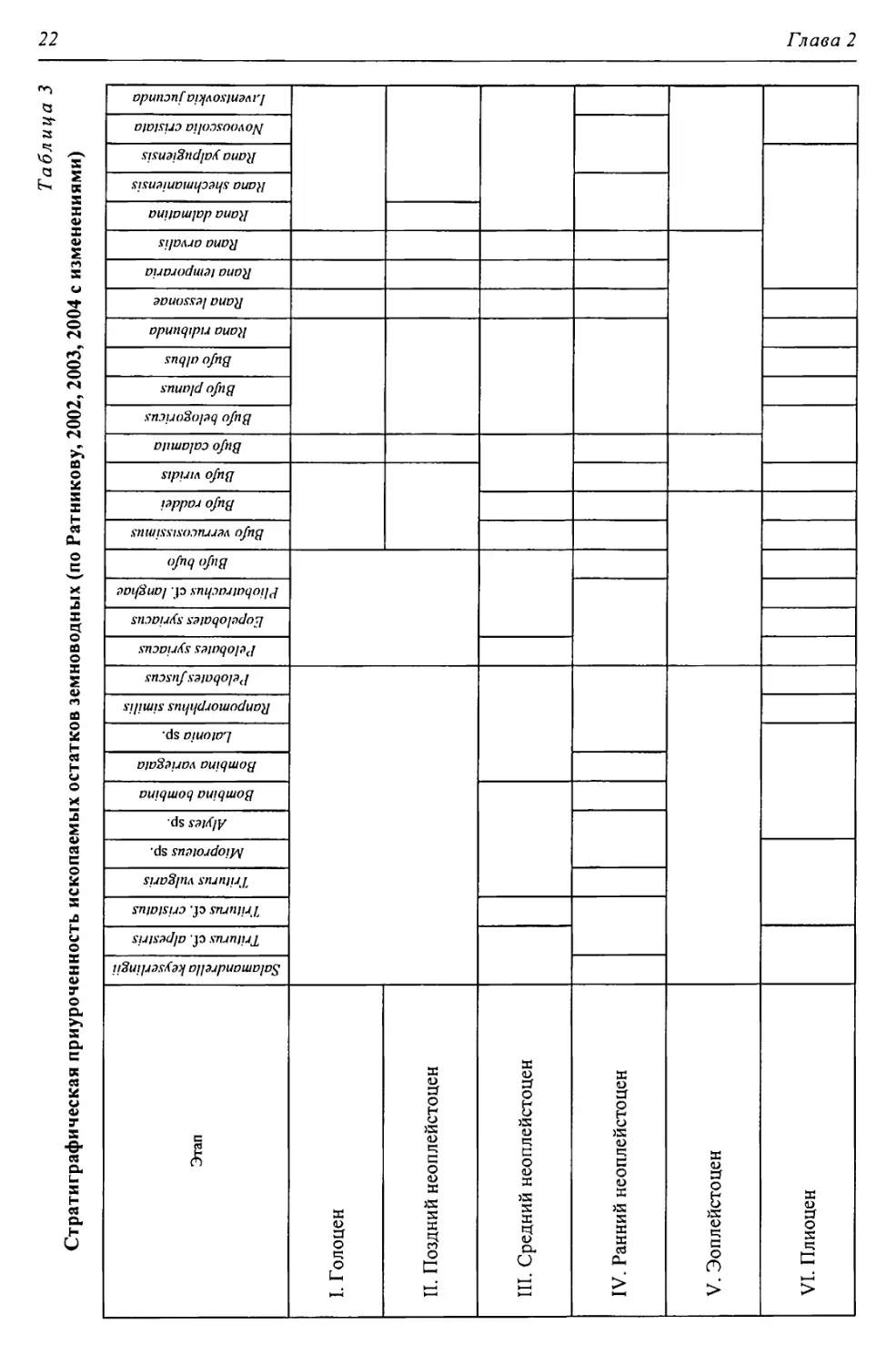
Таблица 3
Стратиграфическая приуроченность ископаемых остатков земноводных (по Ратникову, 2002, 2003, 2004 с изменениями)
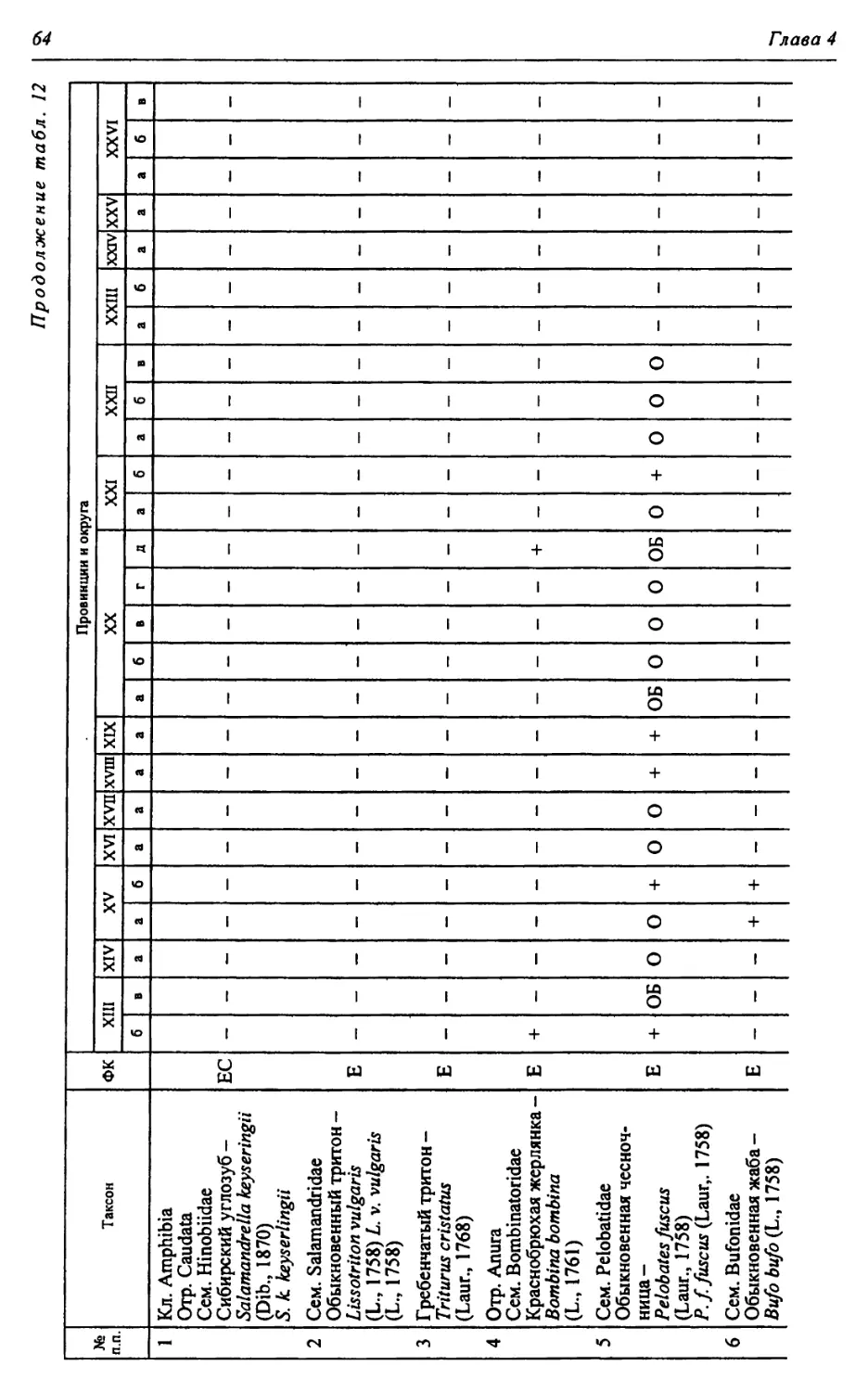
Этап | Salamandrella keyseriingii | | Triturus cf. alpestris | | Triturus cf. cristatus | | Triturus vulgaris | | Mioproteus sp. | | Alyles sp. | | Bombina bombina | | Bombina variegata | | Latonia sp. | | Ranpomorphhus si mil is | | Pelobates fuscus | | Pelobates syriacus | | Eopelobates syriacus | | Pliobatrachus cf. langhae | | Bufo bufo | | Bufo verrucosissimus | Bufo raddei 1 Bufo viridis | Bufo calamita 1 Bufo belogoricus | Bufo planus | Bufo albus | Rana ndibunda 1 Rana lessonae | Rana temporaria | Rana arvalis 1 Rana dalmatina | Rana shechmaniensis 1 Rana yalpugiensis | No vooscolia crislata | Liventsovkia jucunda
I. Голоцен
II. Поздний неоплейстоцен
III. Средний неоплейстоцен
IV. Ранний неоплейстоцен
V. Эоплейстоцен
VI. Плиоцен
Глава 2
Таблица 4
Стратиграфическая приуроченность ископаемых остатков пресмыкающихся (по: Ратников, 2002, 2003, 2004 с изменениями)
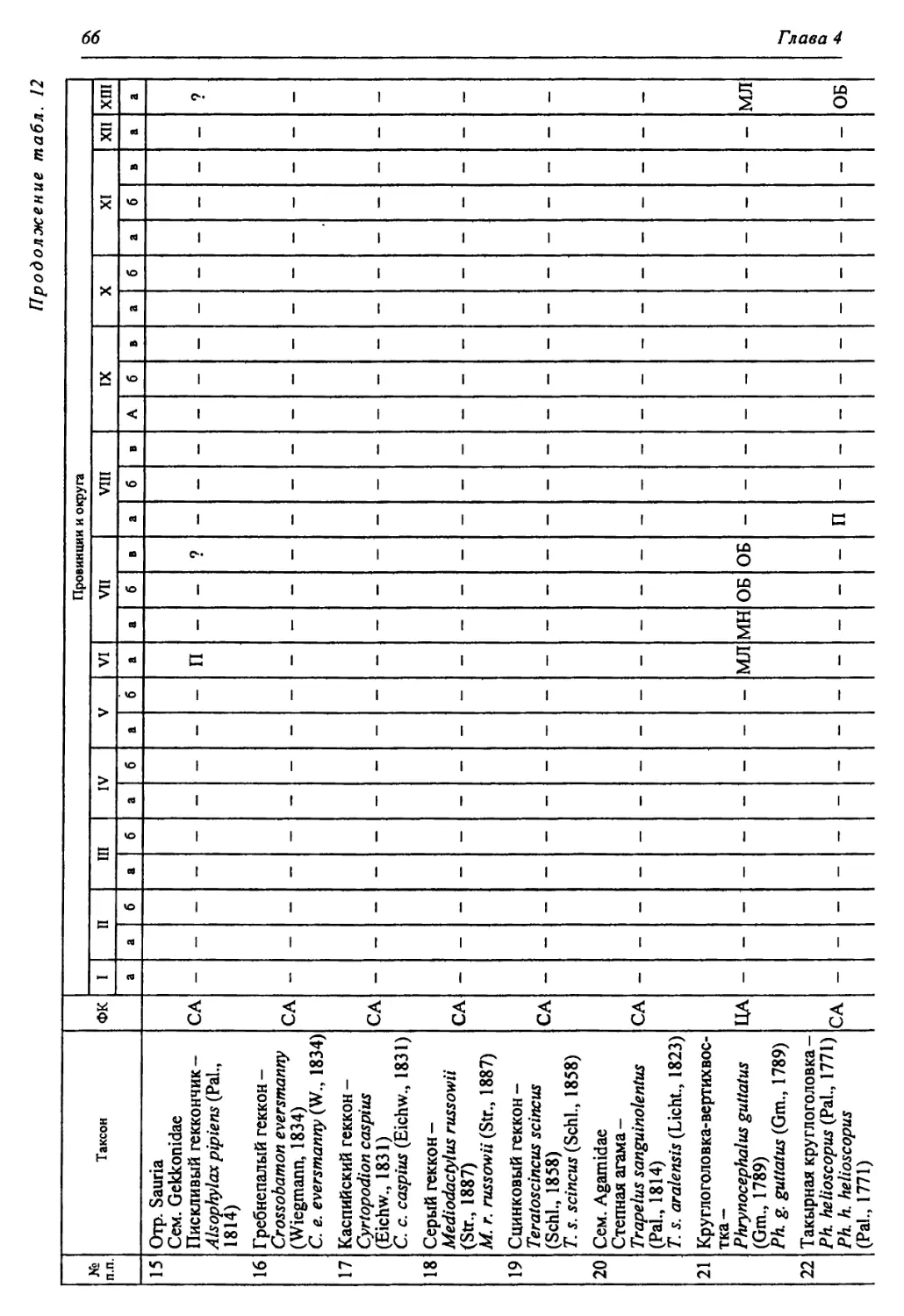
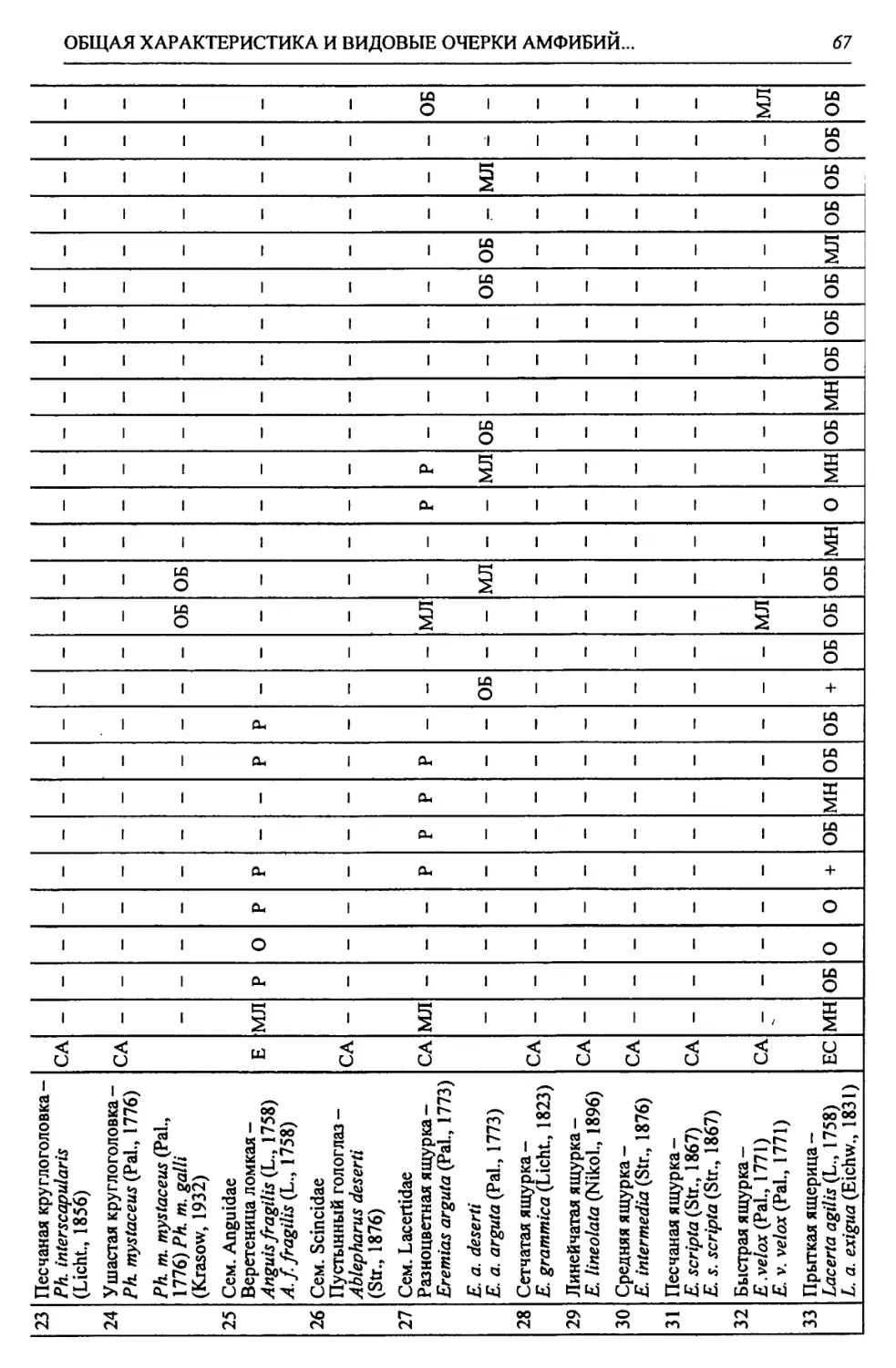
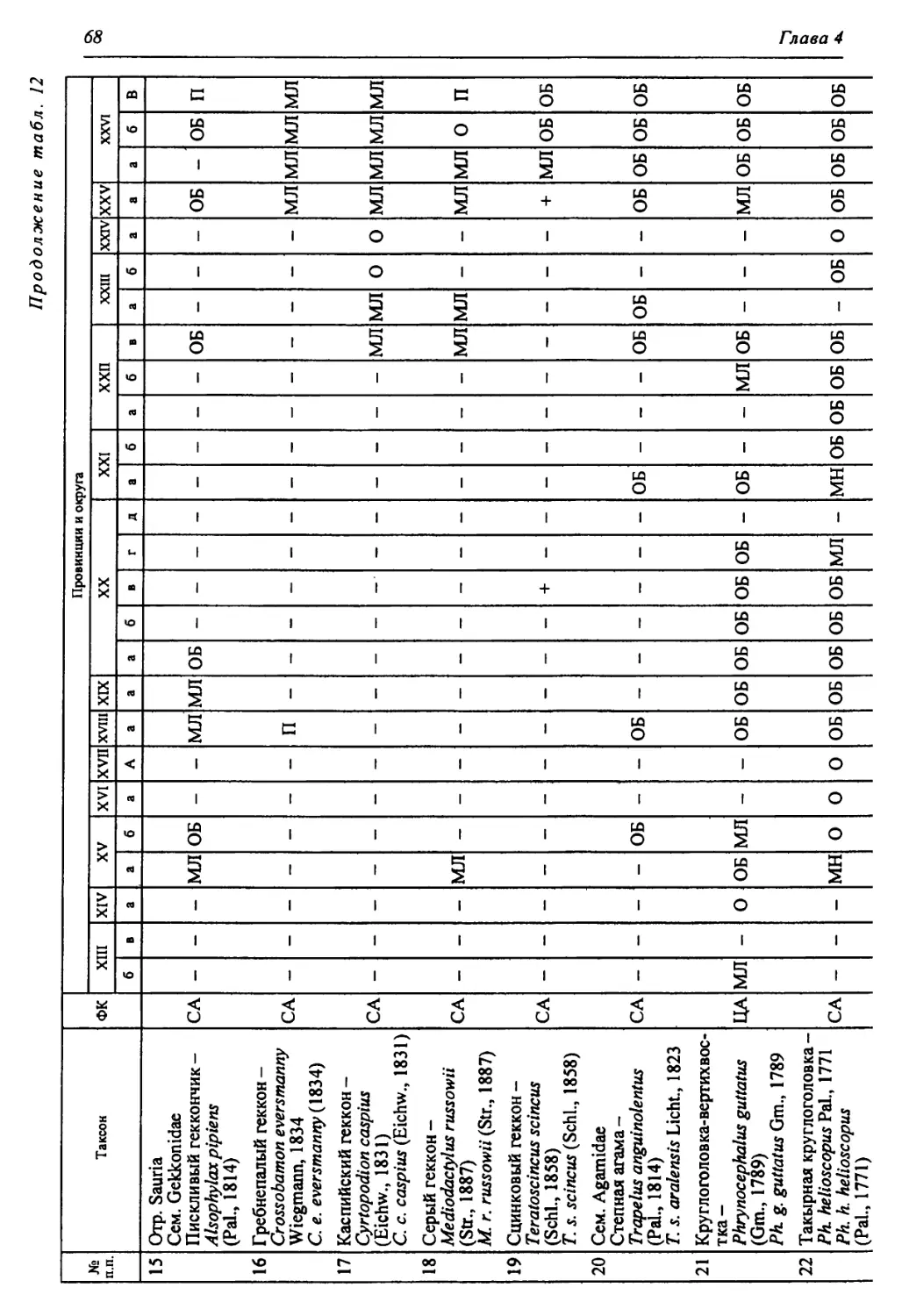
Этап ds ЯПиГМОД Eseodopus apodus Anguis fragUis Lacerta agilis Lacerta vivipara Lacerta vindis Eremias aff. argula Sauna insertasedis •ds xOj Coluber aff. ravergieri Coluber cf. viridijlavus Coronella austriaca Elaphe dione Elaphe longissima Psammorphis sp. Telescopus sp. Natrix natrix Natrix tesselata Vipera berus Vi per a ursini
I. Голоцен
II. Поздний неоплейстоцен
III. Средний неоплейстоцен
IV. Ранний неоплейстоцен
V. Эоплейстоцен
VI. Плиоцен
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
24
Глава 2
Таким образом, получившая к этому времени широкое распространение на
Восточно-Европейской платформе плиоценовая эпоха - время окончания тре-
тичного периода - характеризуется большим количеством позднее вымерших
форм, а значительная часть современных видов сейчас не встречается на тер-
ритории региона (Ратников, 2002, 2004). Считается также, что в этот период
в основном завершилось становление всего степного фаунистического комп-
лекса и активизировались адаптивные механизмы, способствующие расшире-
нию видового спектра не только семиаридных, но и аридных местообитаний
(Завьялов и др., 2002).
Наступление четвертичного периода ознаменовалось усилением динамич-
ности всех природных процессов, сопровождающихся чередованием ледни-
ковых и межледниковых эпох, плювиальных и ксеротермических условий,
трансгрессий и регрессий Каспия, что сказывалось на изменении всех элемен-
тов гидроклиматического цикла и облика ландшафтов, их границ и границ
более крупных региональных подразделений. В связи с этим Г.У. Линдберг
(1955) четвертичный период образно назвал временем «великого переселения
фаун и флор».
Всего в плейстоцене получили развитие 4-5 крупных оледенений, разде-
ленных межледниковыми эпохами, с общей продолжительностью ледниково-
межледниковых циклов в 100-200 тысяч лет (Дергачев, 1998).
Территория региона ледниками не покрывалась, хотя во время максималь-
ного днепровского оледенения его «языки» достигали широты Волгограда, а
во время окского - устья р. Кама. Вместе с тем близость ледников привела к
формированию своеобразных перигляциальных условий с холодным и сухим
климатом. В северной части региона (к югу до 53° с.ш.) складывались крио-
семигумидные условия с ландшафтами, сопоставимыми с современной тунд-
рой, южнее в Прикаспии (до 48^49° с.ш.) - криосемиаридные с ландшафтами
холодной степи и постоянной мерзлотой до широты ст. Азгир в Казахстане.
Южный Урал и Зауралье представляли собой в основном холодную сосно-
во-лиственично-березовую лесостепь. Однако, согласно палеоботаническим
данным, даже в период максимального днепровского оледенения по западным
предгорьям Южного Урала и среднему течению реки Урал сохранились широ-
колиственные леса, хотя и в угнетенном состоянии (Гричук, 1950).
Последним было валдайское оледенение, эпоха которого характеризова-
лась экстремально суровыми континентальными условиями, причем за весь
кайнозой (Величко, 1973). В центральной возвышенной части Урала похолода-
ние вызвало значительное расширение площади высокогорных сообществ. На
склонах Урала, в Предуралье и Зауралье господствующими стали лиственнич-
ные, сосновые и березовые леса с сопутствующими бореальными степными
элементами, а остатки широколиственных лесов сохранились, по-видимому,
лишь в возвышенной части Общего Сырта. Эти и отмеченные выше участки,
возможно, стали теми «микрорефугиями», где могли сохраниться некоторые
представители плиоценовой герпетофауны, «вымерзающие» на перигляциаль-
ных равнинах, представляющих собой открытую холодную степь с неболь-
шими разреженными перелесками по долинам рек. Южнее Общего Сырта
располагалась лесостепь, плакорные пространства которой были заняты пре-
имущественно степными ксерофитами и галофитами, а депрессии - лесами
таежного типа с примесью мелколиственных пород. Далее к югу находились
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
25
перигляциальные степи (с галерейными придолинными лесами), а затем полу-
пустыни и пустыни, «вступившие в свои права» еще с конца плиоцена (Федо-
рович, 1983).
В конечном счете в позднеледниковое время на обширной территории Рус-
ской равнины сформировался «гиперзональный» (в понимании А.А. Величко,
1973) комплекс тундровых, лесных и степных элементов и группировок, соот-
ношение между которыми менялось в связи с колебаниями климата в сторону
как потепления, когда распространение получали лесные сообщества, так и
похолодания, когда господствующими становились перигляциальные тундро-
степи. Последнее, наиболее значительное, распространение перигляциальные
сообщества получили в позднем дриасе (Dr-З) - времени перехода от поздне-
ледниковой растительности к послеледниковой (Хотинский и др., 1991). Пере-
гляциальные условия в целом приводили к обеднению биоразнообразия, хотя
в рефугиумах состав растительного и животного мира, по-видимому, обога-
щался. Биоразнообразие увеличивалось и в аридных районах, где при улучше-
нии обеспечения влагой менялся и тип ландшафта.
Существенные изменения ландшафтов происходили и в межледниковые
эпохи (фазы климатических оптимумов), когда условия становились более
благоприятными. Все эти изменения сказывались на динамике природных
комплексов и обусловливали периодические сдвиги их границ то к северу, то
в южном направлении, что довольно четко прослеживается при анализе лите-
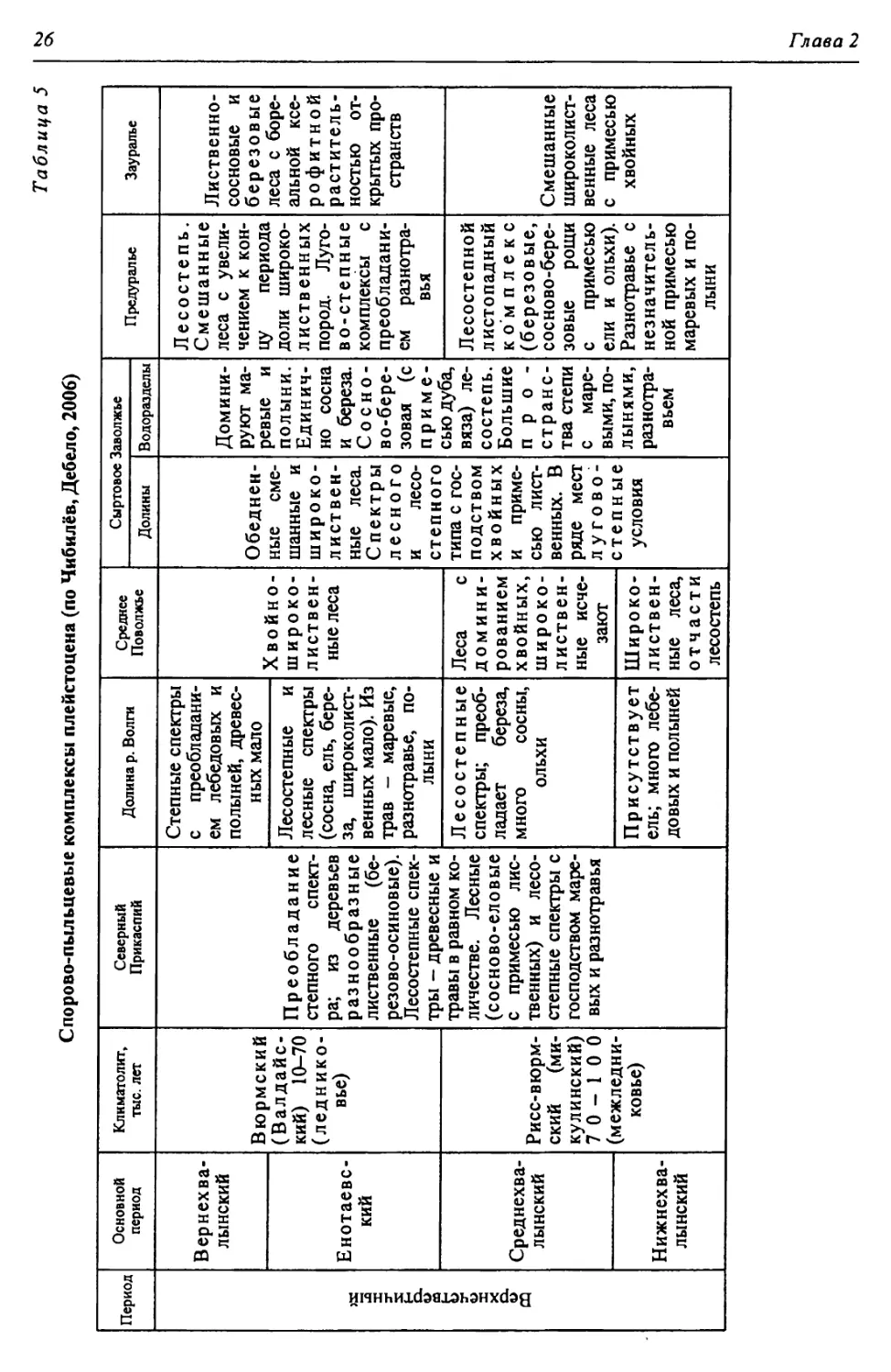
ратурных данных по изменению состава спорово-пыльцевых комплексов из
разных частей региона (табл. 5). В этой обстановке попеременного увеличе-
ния увлажнённости и аридизации, наступления криоаридных и термических
условий генеральная направленность развития биоты региона проявлялась в
постепенном усложнении ее зональной структуры и в периоды климатических
оптимумов она постепенно приобретала черты современной.
В конечном счете в днепровско-валдайскую (микулинскую) межледнико-
вую эпоху в связи с завершением формирования крупных региональных форм
рельефа, усилением западного переноса циклонов и изменением обводнённос-
ти территории (хвалынские трансгрессии с остановками на высотах +50, 20 и
0 м над уровнем моря) зональная структура и пространственное положение
ландшафтных зон постепенно приближались к современным. В это же время
начали складываться и современные зональные типы ландшафтов (Величко,
1977; Марков, 1977), причем основная последовательность событий в позд-
неледниковье в Заволжье (по материалам Бузулукского бора) была примерно
такой же, как в более западных районах Восточно-Европейской платформы, в
Западной Сибири и на сопредельной территории Казахстана (Климанов, 1994;
Кременецкий и др., 1998).
Приведенные материалы показывают, что наиболее существенные измене-
ния условий происходили в пределах современных лесной и лесостепной зон.
В степях Восточно-Европейской платформы в начале четвертичного периода
условия были более стабильны, что сказалось на сохранности местонахож-
дений и составе палеогерпетологических комплексов. В эпоху эоплейстоце-
на степные сообщества характеризуются остатками Pelobates sp., Bufo raddei,
В. cf. calamita, Rana cf. arvalis, неопределенными остатками других лягушек,
а также Lacerta cf. agilis, Eryx sp., Elaphe cf. longissima, Natrix sp. и неопреде-
лимыми остатками каких-то ужовых змей, что свидетельствует о некоторой
Таблица 5
Спорово-пыльцевые комплексы плейстоцена (по Чибилёв, Дебело, 2006)
Период Основной период Климатолит, тыс. лет Северный Прикаспий Долина р. Волги Среднее Поволжье Сыртовое Заволжье Предуралье Зауралье
Долины Водоразделы
Верхнечетвертичный Вернехва- ЛЫНСКИЙ ВюрмскиЙ (Валдайс- кий) 10-70 (леднико- вье) Преобладание степного спект- ра; из деревьев разнообразные лиственные (бе- резово-осиновые). Лесостепные спек- тры - древесные и травы в равном ко- личестве. Лесные (сосново-еловые с примесью лис- твенных) и лесо- степные спектры с господством маре- вых и разнотравья Степные спектры с преобладани- ем лебедовых и полыней, древес- ных мало Хвойно- широко- листвен- ные леса Обеднен- ные сме- шанные и широ ко- листвен- ные леса. Спектры лесного и лесо- степного типа с гос- подством хвойных и приме- сью лист- венных. В ряде мест л у г о в о - степные условия Домини- руют ма- ревые и полыни. Единич- но сосна и береза. Сосно- во-бере- зовая (с приме- сью дуба, вяза) ле- состепь. Большие п р о - странс- тва степи с маре- выми, по- лынями, разнотра- вьем Лесостепь. Смешанные леса с увели- чением к кон- цу периода доли широко- лиственных пород. Луго- во-степные комплексы с преобладани- ем разнотра- вья Лиственно- сосновые и березовые леса с боре- альной ксе- рофитной раститель- ностью от- крытых про- странств
Енотаевс- кий Лесостепные и лесные спектры (сосна, ель, бере- за, широколист- венных мало). Из трав - маревые, разнотравье, по- лыни
Среднехва- лынский Рисс-вюрм- ский (ми- кулинский) 70-100 (межледни- ковье) Лесостепные спектры; преоб- ладает береза, много сосны, ольхи Леса с домини- рованием хвойных, ш иро ко- листвен- ные исче- зают Лесостепной листопадный к омп л е к с (березовые, сосново-бере- зовые рощи с примесью ели и ольхи). Разнотравье с незначитель- ной примесью маревых и по- лыни Смешанные широколист- венные леса с примесью хвойных
Нижнехва- лынский Присутствует ель; много лебе- довых и полыней Широко- листвен- ные леса, отчасти лесостепь
Глава 2
Среднечетвертичный Ательский Р и с с к и й 1 00-2 2 5 Моско вс- кий 100-110 Одинцовс- кий 110-175 Днепровс- кий 175-220 Лихвинский 225-370 Лесостепные и степные спектры с господством маре- вых. Ксерофитные степные спектры (маревые, реже по- лыни, злаки), дре- весных мало (со- сна, береза, ольха) Степные спект- ры с господством маревых; древес- ных мало Смена тундро- вой расти- тельности хвойно-бе- р езо в ы - ми, затем хвойными (преиму- щественно сосновы- ми и тем- нохвой- н ы м и ) лесами. Тундро- вый Сухостеп- ной спектр с маре- выми, по- лынями и разнотра- вьем. Не- большие массивы сосны, ели, березы с примесью широко- листвен- ных пород; папорот- ники, пла- уны Лесостепь. Преимущес- твенное развитие степ- ных ксеротермических комплексов - маревые с примесью полыней и раз- нотравья. Из древесных - сосна, ель, липа Сложные комплексы лиственич- но-сосново- березовых лесов и боре- ально-ксеро- фитной рас- тительности открытых пространств. Светлохвой- ные леса. Ле- состепь. Со- сновые леса с примесью лиственни- цы, пихты и широко- лиственных пород
Верхнеха- зарский
Черноярс- кий Холодная лесостепь-лесотундра: дре- весные - ель, сосна, береза, при потеп- лении - широколиственные, смешан- ные; травянистая - маревые, полыни; степь
Нижнеха- зарский Холодная лесотундра с доминировани- ем травянистой растительности, дре- весная развита слабо (сосна, береза, ель), при потеплении распространя- лись темнохвойные открытые массивы с маревыми и полынями
Нижнечетвертичный Сингаль- ский М и н - дельский (окский) 370-605 Гюнцский 605-965 Доледни- ковый 965- 2070 Преимущественно лесостепные, реже лесные Лесостепные и лесные спектры (с преобладанием сосны, ели, бе- резы, маревых и разнотравье) ре- ликты - единицы Лесной комплекс с преоб- ладанием хвойных (преиму- щественно сосны). Мезофиль- ный лес- ной комп- лекс Преоб- ладают хвойные, хвойно- широко- лиственные, березовые леса. Без- лесные участки с марево-по- лынными и лугово- степными комплек- сами Домини- рует ком- плекс ма- ревых и полыней. Из дре- весных - сосна, ель, бе- реза Лесные и лесо- степные комп- лексы с учас- тием реликтов. В лесах до- минируют хвойные (сосна, ель), лиственные занимают подчиненное положение. Среди трав - маревые, полыни, раз- нотравье Темнохвой- ные леса с примесью сосны. Сме- ш а н н ы е широко- лиственные леса. Бедные бореальные леса. Бога- тые смешан- ные хвойно- лиственные леса
Астраханс- кий Лесостепные спек- тры
Верхнеба- к и н с к и й Нижнеба- кинский Преимущественно травянистые спек- тры. Лесные спек- тры (сосна, ель, ольха, береза), по- лыни, маревые
Тюрканс- кий Преобладают тра- вы; сосна, ель, мхи, папоротники
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
28
Глава 2
перестройке степного герпетокомплекса. К этому времени относятся остатки
ящерицы, предположительно определенные как L. agilis, и из местонахожде-
ния Аккулаево в Башкирии (Хабибуллин, 2002).
Эпоха раннего неоплейстоцена (тираспольский фаунистический комп-
лекс) представлена сообществами, которые формировались преимущественно
в условиях степных и лесостепных, в меньшей степени лесных ландшафтов.
В состав степных герпетокомплексов этого времени входили Pelobates fuscus,
Р. syriacus, Bufo raddei, Rana cf. ridibunda, R. arvalis, Eremias aff. arguta, Na-
trix natrix, Vipera ursine, а также некоторые другие ящерицы и гадюки (по не-
определенным остаткам). В состав лесных сообществ входили Salamandrella
keyseriingii, Triturus cf. cristatus, Pelobates sp., Bombina variegata, Bufo bufo,
B. verrucosissimus, Rana arvalis, R. temporaria, R. schechmaniensis, R. lessone,
Anguis fragilis, Lacerta agilis, Elaphe dione, Natrix natrix, Vipera cf. berus, а так-
же позднее вымершие Pliobatrachus cf. langnae. Лесостепные герпетокомплек-
сы были богаче и кроме большинства отмеченных лесных и степных видов
включали Triturus vulgaris, Bombina bombina, Natrix cf. tesselata.
Эпоха среднего неоплейстоцена, представленная сингильским и хазарским
фаунистическими комплексами (известна в значительной степени по серии
местонахождений на реке Волга в районе п. Черный Яр - Нижнее Займище
Астраханской области), выражена в основном современными видами, в том
числе Bufo raddei и В. verrucosissimus, ныне не обитающими в пределах реги-
она, и L. agilis.
Jipn позднего неоплейстоцена (микулинское межледниковье, валдайское
время), представленного верхнепалеолитическим фаунистическим комплекса-
ми, сформировавшимися в лесостепных и лесных ландшафтах, помимо отме-
ченных уже современных видов характерны также Coronella austriaca, Emys
orbikularis, Rana dalmatica (ныне обитающая за пределами региона), а также
Eremias arguta, ареал которой сейчас расположен южнее местонахождения ис-
копаемых остатков (Еласы).
Приведенные материалы свидетельствуют, что наибольшим видовым раз-
нообразием отличается герпетологический комплекс плиоцена, со времени
которого количество видов лишь неуклонно сокращается. Так, если в эпоху
плиоцена в целом на Восточно-Европейской платформе обитало 45 видов ам-
фибий и рептилий, то в нижнем неоплейстоцене - 23, а рубеж верхнего нео-
плейстоцена преодолели лишь 16 (Ратников, 2002). Из этих форм обыкновен-
ная и сирийская чесночницы, зеленая жаба, озерная и прудовая лягушки, лом-
кая веретеница, обыкновенный и водяной ужи населяли Восточно-Европейс-
кую равнину с позднего плиоцена, а серая жаба, по-видимому, является еще
более древней формой. Столь длительное существование ряда видов, очевид-
но, свидетельствует, что темпы эволюции амфибий и рептилий существенно
уступают скорости эволюции высших тетрапод. Вместе с тем за это время на
Восточно-Европейской равнине появился ряд новых видов - как эволюционно
более молодых, так и осваивавших ее пространства в голоцене.
Завершение плейстоцена и переход к современной эпохе (голоцену) чаще
всего проводят на уровне 11,5-10,0 тыс. лет назад (окончание Dr-З и начало
РЬ). Это по времени совпало с половецким потеплением и позднехвалынской
(мангышлакской) регрессией Каспия (уровень которого опускался до -51 м
ниже уровня моря (Величко, 1973). По-видимому, именно в это время про-
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
29
изошло мощное расселение среднеазиатских псаммофильных и других пус-
тынных видов рептилий (ушастой круглоголовки, вертихвостки, песчаного
удавчика и др.) в Северный Прикаспий и Предкавказье (Неручев, Васильев,
1978; Накаренок, 20036). На севере региона это сопровождалось усилением
роли березовых и хвойных (с примесью широколиственных пород) лесов, ко-
торые, оттеснив к северу таежные сообщества, достигли предгорий западного
склона Урала. Наступившее вскоре похолодание замедлило расширение лес-
ной зоны, но более существенно проявилось в аридизации лесостепи, степи и
формировании после отступления моря ниже нулевой отметки маршевых, а
затем криоаридных ландшафтов, сменившихся со временем условиями сухой
степи и полупустыни.
Во время термического оптимума голоцена (атлантического периода) су-
щественное улучшение гидротермических условий (Рычагов, 1993) привело к
формированию на севере региона (как и на значительной части Русской равни-
ны) лесостепных условий, при которых восточно-европейские леса (где более
существенную роль стали играть широколиственные породы) вышли на водо-
разделы, хотя в основном эти ценозы по-прежнему оставались связанными с
интерзональными элементами рельефа (Динесман, 1977; Климанов, Немкова,
1988). В предгорной части западных склонов Урала получили распространение
смешанные и широколиственные (преимущественно липовые) леса, его цент-
ральная возвышенная часть оказалась занятой в основном хвойными и мелко-
лиственными породами, а на восточном склоне и по Зауральному плато более
континентальные условия способствовали сохранению лиственично-сосново-
березовой лесостепи (Чигуряева, Яхимович, 1965). На Орь-Илекском между-
речье и примыкающей территории Западно-Сибирской низменности влияние
оптимума сказалось слабее, хотя степные формации и здесь стали более высо-
котравными, приобрели лугово-степной облик (Динесман, 1977; Кременецкий
и др., 1994). Небольшие лесные массивы встречались также в степях южных
отрогов Общего Сырта и примыкающих пространствах Северного Прикаспия,
ландшафты которого постепенно эволюционировали от полупустынно-пус-
тынных до степных (Киселева, 1982). Вместе с тем по наиболее благоприятным
местообитаниям небольшие участки древесной растительности (преимущест-
венно из сосны и березы) доходили до полупустынь Северного Прикаспия, юг
которого вскоре оказался во власти очередной трансгрессии (Рычагов, 1993).
Считается, что в это время произошло существенное обогащение сложившего-
ся к этому моменту ядра фаунистического комплекса позвоночных животных
региона причерноморско-понтийскими степными, европейскими лесостепны-
ми, крымско-кавказскими горно-степными, казахстанско-южносибирскими
степными, а также каспийско-туранскими и другими пустынными азиатскими
элементами (Завьялов и др., 2003).
Переход от теплого и влажного атлантического периода к теплому сухо-
му суббореалу сопровождался резкой аридизацией климата, произошедшей
практически синхронно на громадных пространствах евразиатских степей.
В результате полупустыни и сухие степи Прикаспия сменились пустынями
(Иванов, 1989), а на освободившихся при махачкалинской регрессии про-
странствах началось формирование эоловых форм рельефа. Севернее ариди-
зация проявилась лишь в упрощении структуры степных сообществ (умень-
шилось обилие сложноцветных и злаков), снижении доли липы и некоторых
30
Глава 2
других гидрофильных лесных видов и возрастании роли дуба. Более сущест-
венные изменения произошли в Зауралье, где уменьшились площадь березо-
вых колков и ареалы широколиственных пород (Кременецкий и др., 1994).
Наступившая затем эпоха среднего суббореала (так называемый «полтав-
кинский микроплювиал» бронзы) ознаменовалась восстановлением на значи-
тельных пространствах Северного Прикаспия лесостепи, разнотравно-злако-
вых степей, а также возрастанием доли степных и уменьшением пустынных
видов животных и их численности (Иванов, Васильев, 1995). Севернее этому
времени соответствует наступление новой лесной фазы - более широкого рас-
пространения дубрав (с участием липы, вяза, бука), с которой, по-видимому,
началось формирование современной лесостепи Русской равнины (Фёдоров,
1978). Поздний суббореал, судя по полинологическим данным (Болиховская,
1990), характеризовался своеобразным импульсным иссушением юга региона,
а севернее аридизация проявилась в меньшей степени и лесостепь, например,
сохранила многие особенности экосистем раннего голоцена.
Примерно 2,7-2,5 тысяч лет назад начался последний, заключительный,
период голоцена - субатлантический, в котором также происходили значи-
тельные изменения климата, сопровождающиеся подвижками растительного
покрова и перестройкой структуры экосистем, особенно на юге региона, испы-
тавшего на себе ряд новокаспийских трансгрессий и регрессий, приводивших
к разрыву ареалов ряда видов, а затем их вторичной интерградации. Раннесуб-
атлантическое время, начавшееся некоторым повышением влажности (пре-
имущественно на юге региона), закончилось усилением континентальности
климата, сопровождающегося уменьшением роли широколиственных пород
деревьев и установлением господства сосны и лиственницы с елью, а также
ксерофитизацией травянистых сообществ. В то же время в пределах Южного
Урала динамика природной обстановки не выходила за рамки степных - су-
хостепных условий (Рысков, Демкин, 1997).
Среднесубатлантическое время, наступление которого совпадает с нача-
лом современной эры, характеризуется как «малый климатический оптимум
голоцена» (Климанов, 1994). Его начальная (межплювиальная) фаза опреде-
лялась мезофилизацией степной растительности Прикаспия и расширением
площади лесных сообществ, которые по долинам рек проникали далеко на юг.
Сходные процессы протекали и в Зауралье, где начались возрождение степных
боров, закрепление песков, расширение площади лимнофильных экосистем
(Николаев, 1999). Начавшееся в IV в. н.э. потепление, которое достигло макси-
мума в X-XI вв. (ксеротермическая фаза по Николаеву, 1977), способствова-
ло ксерофитизации степей, сокращению площади лесов и замене в них части
широколиственных элементов ивняками, а также усилению регрессии Каспия
(уровень которого упал до -40 м - дербентская регрессия) и Аральского моря
(которое перешло в стадию так называемого «оксийского болота»).
Почти все последнее тысячелетие относится к позднесубатлантическому
времени, значительная часть которого приходится на «малый ледниковый пе-
риод», характеризующийся возрастанием суровости климата, определившего
уменьшение значимости широколиственных пород и березы, и расширением
площади сосновых лесов. В степной зоне начало этого времени ознаменовано
значительным развитием лугово-разнотравных сообществ и некоторым уве-
личением облесенности территории, что придало этой части региона облик
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
31
кустарниковых и редколесных савваноидов, южнее сменяющихся полупус-
тынными сообществами. В XV в. в аридной зоне своеобразный средневековый
плювиал по сути дела завершился, и с XVI в. здесь начинается новая волна
ксерофитизации растительного покрова, получившая прогрессирующее разви-
тие в XVIII и XIX в. К концу этого периода лесные сообщества здесь почти
вытесняются, широкое распространение получает травянистая растительность
с доминированием ксерофитов, а побережья Арала и Каспия опустыниваются
(Абрамова, 1994).
В Рын-песках эпоха XVI-XIX вв. отличалась большей увлажненностью,
что обусловило закрепление песков, увеличение облесенности и улучшение
обводнения Камыш-Самарских озер (Иванов, Васильев, 1995). Севернее в это
время начались формирование комплекса кустарниковой степи по многим па-
динам, развитие древесно-кустарниковых зарослей по берегам рек и в круп-
ных лиманах. В конечном счете, пойменные леса по рекам Иргиз, Еруслан,
Б.и М. Узени и их менее значительные массивы по рекам Горькая, Хара, Лан-
цуг и Паника сомкнулись с вековыми дубравами южных отрогов Общего Сыр-
та, описанных Э.А. Эверсманном и Г.С. Карелиным. На легких почвах здесь
сформировались березово-осиновые и частично сосновые леса, которые вос-
точнее, по-видимому, продолжались по Подуральскому и Урало-Эмбенскому
плато (о чем свидетельствуют реликтовые лески, сохранившиеся до настояще-
го времени в некоторых речных долинах). Широкое распространение лесных
сообществ и их значительная площадь позволяют некоторым авторам утверж-
дать, что Северный Прикаспий на том этапе приобрел в определенной степени
лесостепной облик (Абрамова, Турманина, 1982). Эти же процессы обусло-
вили подъем уровня Каспия (до -23-24 м), Аральского моря (ново-аральская
трансгрессия), а также значительное изменение видового состава и соотноше-
ния эколого-генетических типов животных Северного Прикаспия, которые
постепенно приобрели черты, характерные для современного этапа (Иванов,
Васильев, 1995).
Считается, что к середине XIX в. «малый ледниковый период» окончился и
началась новая, продолжающаяся по настоящее время, климатическая эпоха -
потепление, сопровождающееся уменьшением атмосферной увлажненности и
ксерофитизацией растительного покрова, хотя на общем фоне аридизации на-
блюдались как более сухие (1915-1935; 1970-1990), так и относительно влаж-
ные (1875-1915; 1940-1960) периоды с более короткими, преимущественно
кратными брикнеровскому, циклами.
Со времени начала голоцена, но особенно в последнее тысячелетие, фор-
мирование внешнего облика ландшафтов региона и структуры их биоты шло
под все возрастающим воздействием антропогенных факторов (пасторальной
дигрессии, вырубки лесов, распашки земель и общего освоения территории),
которые снивелировали их многие первобытные черты, особенно на юге регио-
на. У его северных пределов меньшая засушливость позднего голоцена наряду
с особенностями хозяйственного освоения территории определили большую
стабильность структуры природных биоценозов и их биоты, которые сохрани-
ли целый ряд особенностей экосистем раннего голоцена. В целом же голоцен,
очевидно, следует рассматривать как время преимущественной стабилизации
ареалов всех позвоночных животных, медленного их расселения и освоения
новых территорий, своеобразного заполнения ценотических вакуумов, об-
32
Глава 2
разевавшихся в «малом ледниковом периоде» (Кривенко, 1991 2002; Белик,
2000). Это как будто подтверждается составом герпетологического комплекса
Поволжья, дополненного лишь Lacerta vivipara, остатки которой отмечены в
двух пещерных местонахождениях Башкирии (Ратников, 20046; Хабибуллин,
2002).
Имеющиеся к настоящему времени палеонтологические и палеогеографи-
ческие материалы позволяют наметить важнейшие этапы в «естественной ис-
тории» разных таксонов амфибий и рептилий, предположительно воссоздать
историю освоения региона населяющими его современными видами и воспол-
нить некоторые пробелы в динамике их ареалов.
Хвостатые амфибии (Caudata, Urodela) в регионе известны по двум семейс-
твам, из которых наиболее примитивными являются углозубы (Hynobiidae),
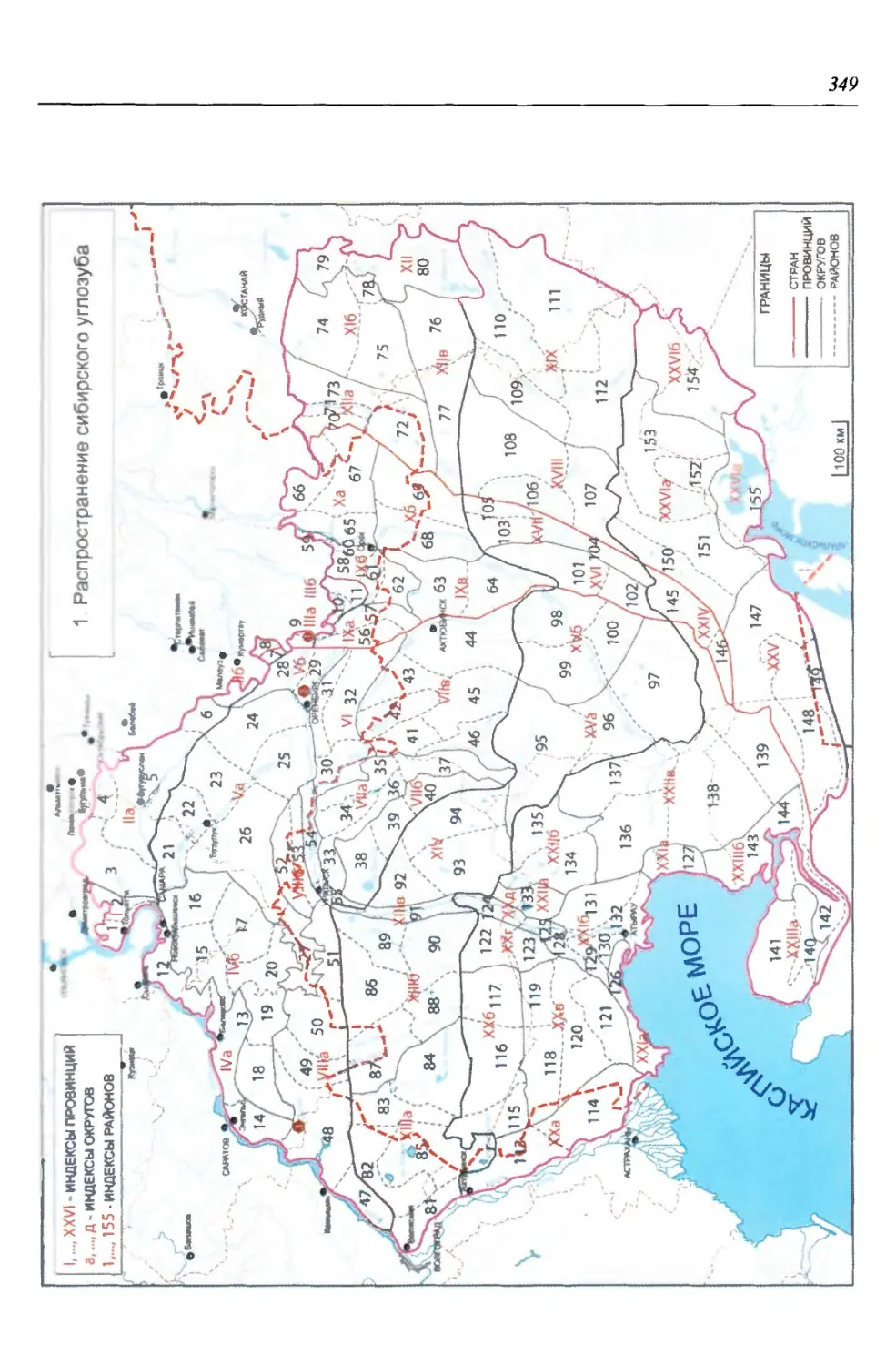
представленные сибирским углозубом {Salamandrella keyseriingii). Предпола-
гается, что центром дифференциации этого семейства являются горы Цент-
рального Китая, откуда еще в доледниковое время шло расселение его видов,
в том числе и предка сибирского углозуба. Считается, что современный си-
бирский углозуб - это геологически молодой вид, который, вероятно, сфор-
мировался в первой половине плейстоцена в Центральной Азии и Восточной
Сибири, откуда после отступления ледника шло заселение его современного
ареала (Сибирский углозуб, 1994). Однако находка ископаемых остатков вида
в ранненеоплейстоценовых отложениях (мучкапский горизонт тираспольско-
го фаунистического комплекса) Восточно-Европейской платформы (Ратников,
2002) позволяет по-иному взглянуть на историю формирования ареала вида.
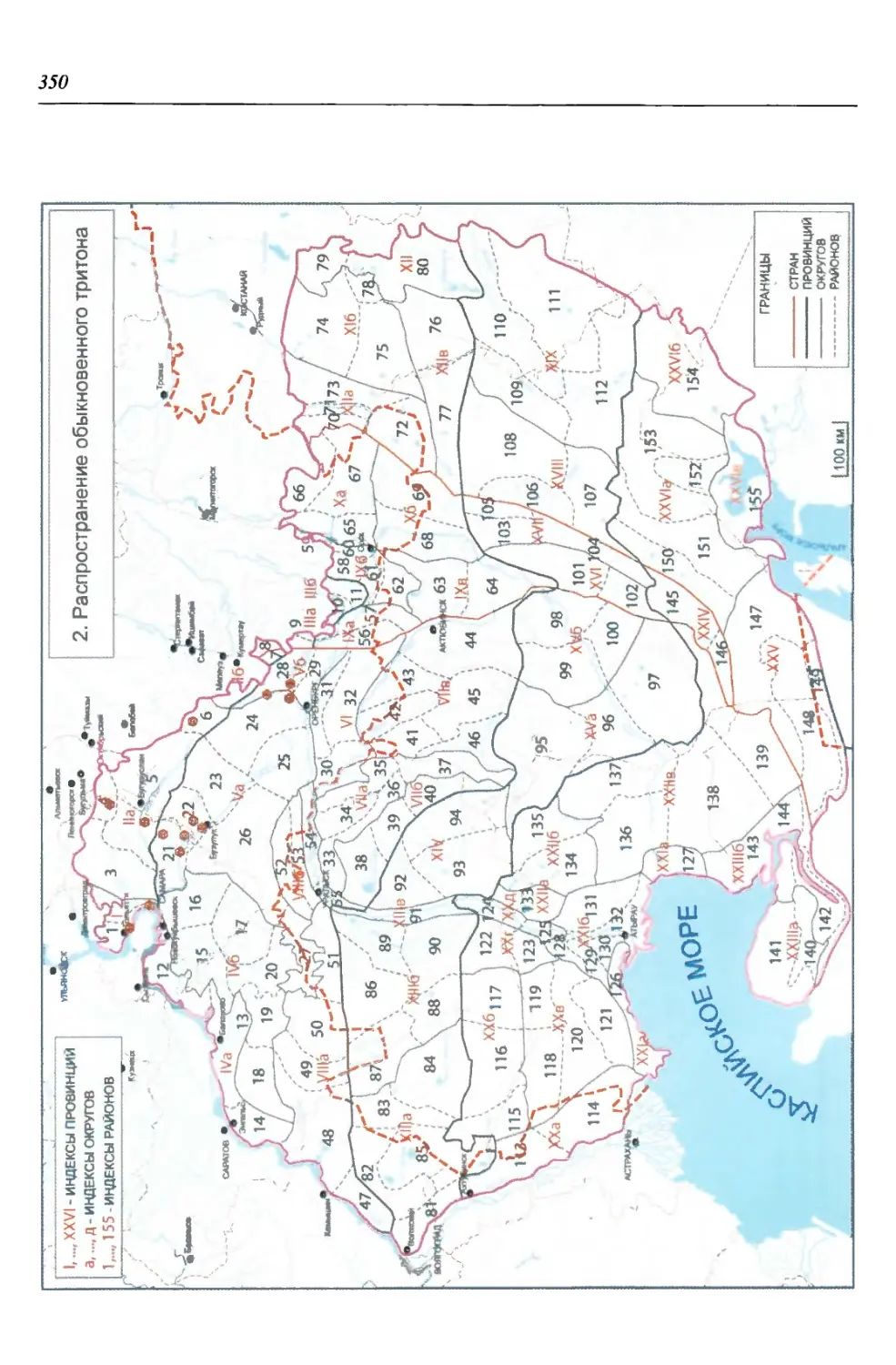
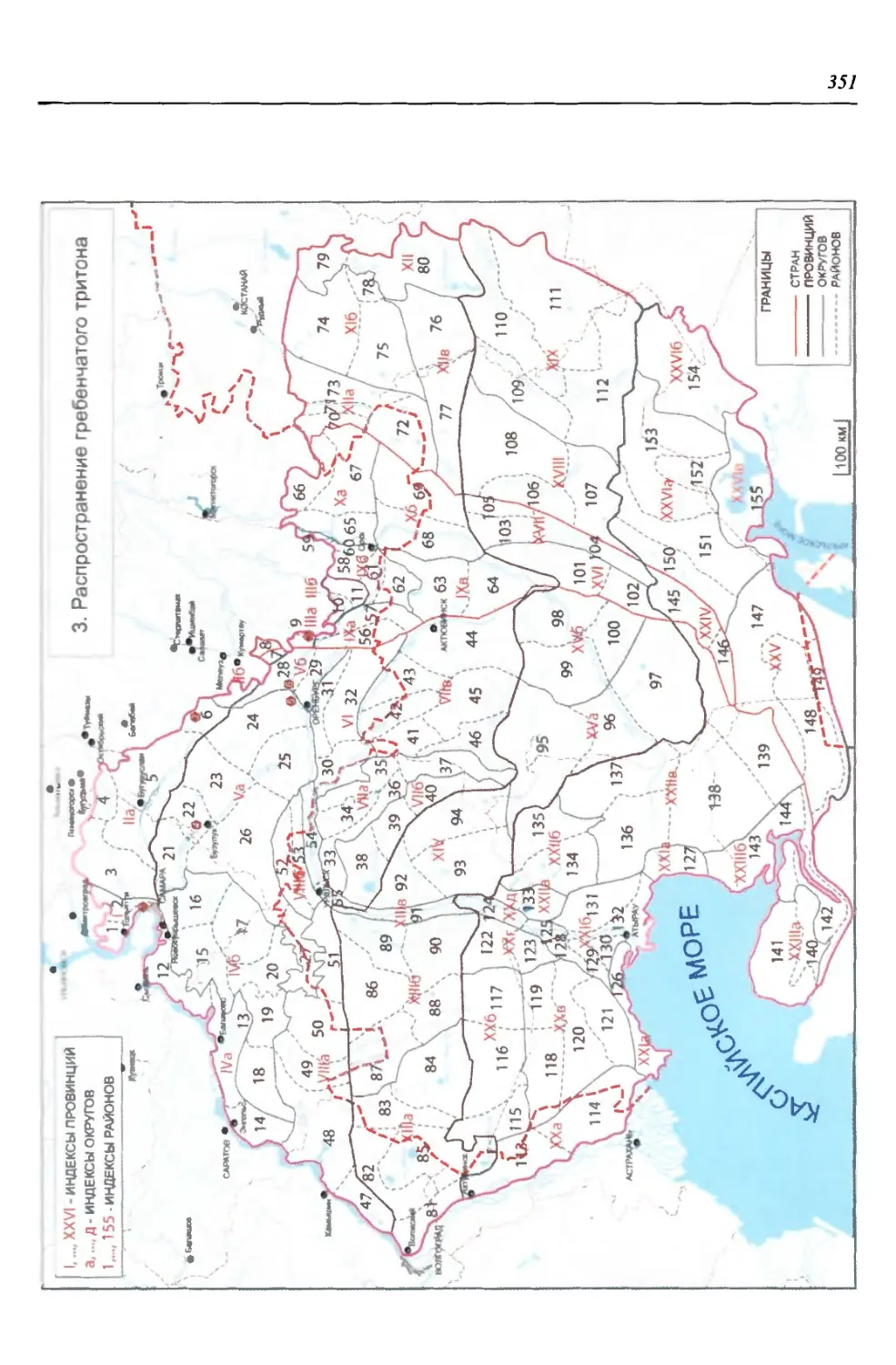
Саламандровые (Salamandridae) - одно из эволюционно наиболее продви-
нутых семейств хвостатых амфибий. В регионе оно представлено двумя совре-
менными видами тритонов: гребенчатым {Triturus cristatus) и обыкновенным
(Г. vulgaris). Древнейшие представители этого рода известны с миоцена, а тер-
риторию региона начали осваивать как минимум с плиоцена, поскольку имен-
но к этому времени отосятся ископаемые остатки Т. cf. alpestris, обнаружен-
ные на сопредельной территории Нижегородской области в местонахождении
Апастово. Из современных видов ископаемые остатки Т. cf. cristatus известны
из лесных ассоциаций Ильинского горизонта, а обыкновенного тритона - из
лесостепного герпетокомплекса мучкапского горизонта нижнего неоплейс-
тоцена. Остатки этих трех видов прослеживаются также в лесостепном лих-
винском горизонте сингильского фаунистического комплекса среднего нео-
плейстоцена Восточно-Европейской платформы, а остатки Т. cf. cristatus - из
среднего неоплейстоцена Башкирии (Ратников, 2002). Отмеченные изменения
обстановки, несомненно, сказывались на размерах и очертаниях ареалов этих
видов, современную конфигурацию которые приобрели с наступлением по-
следнего сухого периода.
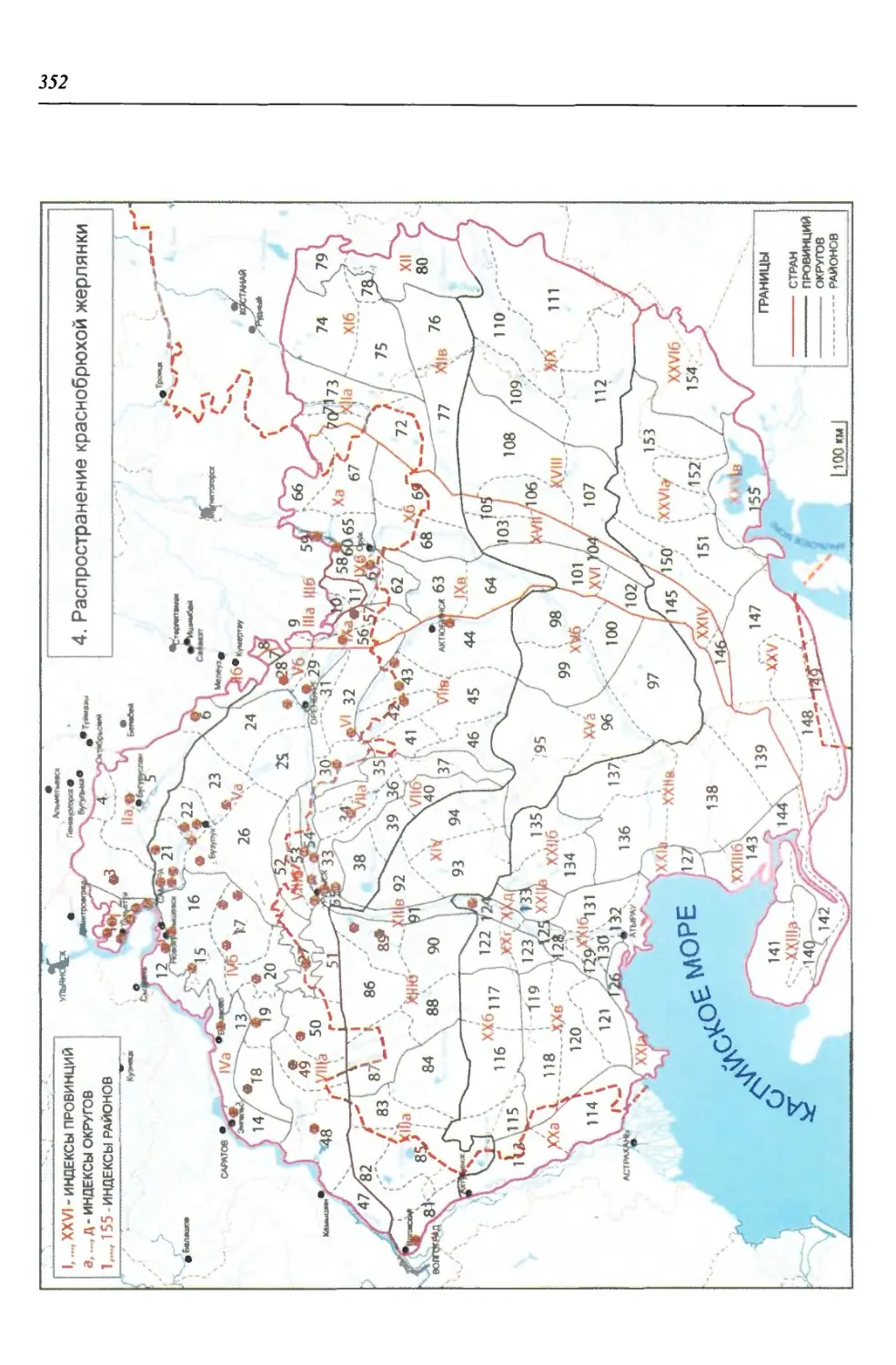
Среди бесхвостых амфибий одной из древнейших групп являются жерлян-
ки (Bombinatoridae), сформировавшиеся предположительно в Юго-Восточной
Азии, откуда впоследствии дивергировали на восточную и западную ветви.
Считается, что краснобрюхая жерлянка или близкий к ней вид {В. cf. bombi-
na), известные из нижнего плиоцена Казахстана, уже в доледниковое время
заселяли обширную территорию Сибири и Европы, хотя на сопредельной тер-
ритории Восточно-Европейской равнины ископаемые остатки жерлянок ста-
новятся известны из лесостепного фаунистического комплекса ильинского, а
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
33
затем мучкапского горизонтов нижнего неоплейстоцена. В ледниковое время
жерлянки, по-видимому, сохранились в Придунайском рефугиуме, откуда
впоследствии пошло их расселение. Продвижение краснобрюхой жерлянки на
территорию региона, в Предуралье и к юго-западным склонам Урала, вероят-
но, началось с наступлением голоцена, а максимума достигло в атлантическое
время. Формирование современных очертаний ареала началось с наступлени-
ем субатлантического похолодания и (если учесть, что в этом периоде выделя-
ется до 12 крупных климатических и соответствующих им фитоценотических
фаз) происходило в очень динамичных условиях (Боркин, 1984; Юшков, 1997;
Ратников, 2002; Вершинин, 2007).
Квакши (Hylidae) Палеарктики, по-видимому, уже на ранних этапах эво-
люции дивергировали на восточную и западную ветви. Европейские квакши в
ископаемом состоянии прослеживаются из позднего миоцена, но на Восточно-
Европейской равнине они становятся известны лишь для позднего плестоце-
на-голоцена Украины (Боркин, 1984). Таким образом, литературные сведения
о распространении квакши (Hyla arborea) в Поволжье и Предуралье пока не
подтверждены палеонтологическими материалами.
Представители семейства чесночницы (Pelobatidae) в ископаемом состоят
нии прослеживаются с конца мелового периода, рода Pelobates - с олигоцена
Монголии. Некоторые современные представители рода, возможно, сформи-
ровались на юго-востоке Европы или сопредельной территории Азии, а в пре-
делах Восточно-Европейской платформы они становятся известны начиная с
молдавского (сирийская чесночница) и урывского (обыкновенная чесночница)
фаунистических комплексов плиоцена. Их остатки далее обнаружены в отло-
жениях эоплейстоцена, ильинского и мучкапского горизонтов тираспольского
фаунистического комплексов, затем в лихвинском и среднерусском горизон-
тах сингильского и хазарского фаунистических комплексов среднеплейсто-
ценового местонахождения Татарстана (обыкновенная чесночница), а также
микулинском и валдайском горизонтах верхнего неоплейстоцена и ряде место-
нахождений со степными и лесостепными ассоциациями голоцена, в том числе
с низовий Волги (Средняя Ахтуба, Черный Яр - Нижнее Займище). Считается,
что после окончания ледникового периода на равнинах Восточно-Европейс-
кой платформы и Урала обыкновенная чесночница появилась в атлантическую
эпоху с распространением широколиственных лесов, а современные очерта-
ния ее ареал приобрел во время прохладной субатлантической стадии (Юш-
ков, 1997; Ратников, 2002; Вершинин, 2007), чему, несомненно, способствова-
ло сведение лесов в Северном Прикаспии, на Общем Сырте и Подуральском
Плато.
1?скопаемые остатки представителей семейства жабы (Bufonidae) известны
с начала третичного периода (палеоцена), хотя возможно существовали уже в
мелу. На Восточно-Европейской равнине они прослеживаются с молдавско-
го фаунистического комплекса плиоцена, причем в одесских катакомбах уже
принадлежат современным видам. Остатки обыкновенной (Bufo bufo) и зеле-
ной (Bufo viridis) жаб известны с лесостепных ассоциаций урывского фаунис-
тического комплекса плиоцена и прослеживаются вплоть до голоцена, в том
числе по местонахождениям на сопредельной с регионом территории - Апас-
тово, Еласы, Б. Тиганы, Красный Бор, Средняя Ахтуба и Черный Яр - Нижнее
Займище. Здесь и в ряде других местонахождений обнаружены остатки и иных
34
Глава 2
жаб, впоследствии вымерших или отступивших за пределы прежних мест на-
ходок. Считается, что В. bufo была широко распространена в Северной Евра-
зии уже в доледниковое время. С наступлением ледника она, расселившись по
разным рефугиумам, образовала несколько форм, в том числе наиболее моло-
дую, возникшую в Придунайской котловине В. b. bufo. Современные очерта-
ния ареал этого вида начал приобретать после отступления ледника в процес-
се иррадиации его из рефугиума на запад, север и восток, включая и Сибирь.
Зеленая жаба, по-видимому, появилась в аридных ландшафтах европейского
Средиземноморья или Средней Азии. Северных пределов региона после оле-
денения она, скорее всего, достигла в теплую и сухую суббореальную стадию
голоцена, а после некоторого сокращения в прохладном суббореале ее ареал
вновь стал расширяться на север и восток (Гаранин, 1983; Ратников, 2002; Вер-
шинин, 2007).
Древнее происхождение имеют и лягушки (Ranidae), ископаемые остатки
которых известны с эоцена, а современных видов - с начала миоцена. Центром
формирования зеленых лягушек, по-видимому, была Европа, где они известны
с олигоцена. Бурые лягушки, вероятно, являются восточно-азиатскими вида-
ми, которые в Европу проникли в период образования олигоценового сухо-
путного моста на месте Тургайского моря. Из зеленых лягушек самый древ-
ний вид - прудовая (Rana lessonae), известная из олигоценовых отложений
Западной Европы, а озерная лягушка (R. ridibunda), возможно, - миоценовый
вид. На Восточно-Европейской равнине оба вида достоверно известны из лес-
ных ассоциаций урывского фаунистического комплекса плиоцена и просле-
живаются почти во всех последующих фаунистических комплексах (включая
и голоценовые) из сопредельных поволжских местонахождений - Апастово,
Березовка, Б. Тиганы, Еласы, Коминтерн, Красная Лука, Средняя Ахтуба,
Черный Яр - Нижнее Займище. Третий вид - гибридная съедобная лягушка -
R. (esculenta) sp. - в ископаемом состоянии известна из лесных ассоциаций ми-
кулинского горизонта верхненеоплейстоценового местонахождения Б. Тиганы
(Татарстан).
Из представителей западной группы бурых лягушек - остромордая (R. аг-
valis), вероятно, ранее (эоцен) отделившаяся от общего филетического ствола,
и травяная (R. temporarid), являющаяся геологически более молодым видом,
современную область обитания в Европе, скорее всего, начали осваивать с
олигоцена. В окрестностях региона их наиболее древние остатки обнаруже-
ны в отложениях раннего плиоцена северо-восточного Казахстана, в восточ-
но-европейских местонахождениях неопределенные- остатки каких-то бурых
лягушек становятся известны из лесных ассоциаций урывскоге-фауццстичес-
кого комплекса плиоцена, а остатки Rana cf. arvalis — из эоплейстоценовых
отложений. Позднее остатки этих видов встречались в лесных ассоциациях
почти всех фаунистических комплексов нижнего, среднего и верхнего нео-
плейстоцена, а также голоцена из местонахождений сопредельных территорий
Поволжья - Березовка, Б. Тиганы, Еласы, Коминтерн, Красная Лука, Черный
Яр - Нижнее Займище. Формирование их современных ареалов, несомненно,
происходило в послеледниковое время расселением предположительно из вос-
точно-европейского рефугиума.
История рептилий, как известно, начинается в перми палеозоя, хотя совре-
менные таксоны появились намного позднее. Черепахи, достигшие значитель-
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ... 35
ного расцвета уже в мелу, в эоценовую эпоху палеогена были представлены
как обитателями теплых (тропических и субтропических) водоемов, так и су-
хопутными (хотя и архаичными) формами. С конца палеогена - верхнеоли-
гоценового времени на сопредельной азиатской территории известны остатки
морских (Chelonia aralis), пресноводных (Clemys kasachstanica) и сухопутных
(Testudo aralensis, Protestudo alba и др.) видов (Зоогеография палеогена Азии,
1974). Наиболее древней в регионе формой является пресноводная черепаха,
имеющая европейско-средиземноморские «корни». Представители рода Emys
в Европе известны уже с отложений эоцена, а в составе восточно-европейских
палеогерпетологических комплексов остатки каких-то черепах отмечаются с
начала плиоцена. Считается, что болотная черепаха {Emys orbicularis) - вид,
эндемичный для Средней Азии и Восточного Ирана, - сформировалась во вто-
рой половине плиоцена в Причерноморье, расселяясь откуда в плейстоцене
она освоила значительную часть Восточно-Европейской равнины (Хозацкий,
1949, 1958, 1982; Богданов, 1963; Ратников, 2002). Предполагается, что на
Южном Урале она впервые могла появиться уже в раннем плейстоцене (Ха-
бибуллин, 2002), хотя палеонтологическими материалами это не подтвержде-
но. Современные очертания ее ареал приобрел после окончания оледенения.
К эпохе плиоцена относят и возникновение в Передней Азии каспийской че-
репахи (Mauremys caspica), которая, возможно, еще в историческое время оби-
тала по северному побережью Каспия. Вместе с тем род Mauremys, по-види-
мому, образовался на Дальнем Востоке, откуда позднее расселился на запад.
Остатки сухопутной среднеазиатской черепахи (Agrionemys horsfieldii) - вида,
эндемичного для Средней Азии и Восточного Ирана, - в регионе известны
из олигоценовых палеогерпетологических комплексов Тургайской впадины и
Приаралья, а современные очертания ее ареал приобрел к началу четвертично-
го времени -т. е. имеет голоценовый возраст (Яковлева, 1964).
Формирование современных семейств ящериц (Sauria) началось в конце
юры - мелу, и без существенных изменений многие их группы пересекли гра-
ницу кайнозоя (Татаринов, 1981).
Наиболее древним, по-видимому, является семейство гекконовые (Gecco-
nidae), сформировавшееся в Юго-Восточной Азии, откуда его представители
впоследствии расселялись на север и запад, освоив пустынные и горные место-
обитания на юге региона и сопредельных пространствах Казахстана и Средней
Азии. В регионе это семейство представлено пятью современными видами,
из которых три - гребнепалый, серый, каспийский {Crossobamon eversmanni,
Gymnodactylus russowi, Cyrtopodion caspius) эндемичные (автохтонные), а два-
пискливый геккончик, сцинковый геккон (Alsophylax pipieus, Teratoscincus
scincus) - субэндемичные виды Средней Азии и Восточного Ирана (причем
гребнепалый и сцинковый гекконы входят в состав ядра территории). Считает-
ся, что образование современного герпетокомплекса этого региона могло на-
чаться не ранее миоцена, возможно, с его второй половины или раннего плио-
цена (Чернов, 1949; Богданов, 1963). Территорию исследуемого нами региона
пискливый геккончик населяет, по-видимому, со времени мангышлакской рег-
рессии, а современные очертания ареалы других видов приобрели во время
новокаспийских трансгрессий и регрессий.
Семейство агамовые (Agamidae) известно из отложений мелового време-
ни, а формирование населяющих регион современных видов - степной агамы
36
Глава 2
{Trapelus agilis) и круглоголовок, вероятно, началось в конце палеогена. Счи-
тается, что круглоголовка-вертихвостка {Phrynocephallus guttatus) имеет цен-
тральноазиатское происхождение и в Среднюю Азию проникла, скорее всего,
уже в четвертичное время. Автохтонным среднеазиатским видом признается
песчаная круглоголовка {Ph. interscapularis), а степная агама, такырная и ушас-
тая круглоголовки {Ph. helioscopus, Ph. mystaceus) - являются субэндемиками
Средней Азии и Восточного Ирана. Территорию нашего региона они начали
осваивать по мере формирования здесь континентальных условий после от-
ступления морских вод.
Семейство варановых (Varanidae) известно с мелового периода. В про-
шлом его ареал был значительно шире и остатки Varanus sp. известны, напри-
мер, из лесостепных ассоциаций урывского фаунистического комплекса плио-
цена Восточно-Европейской равнины (Ратников, 2002). В настоящее время к
южным рубежам региона почти вплотную примыкает ареал единственного
представителя этого рода - серого варана {Varanus griseus), имеющего сахаро-
синдские корни.
Семейство веретеницевых (Anguidae) известно с мелового периода, а ряд
современных видов - из эоценовых и более поздних отложений запада Евра-
зии. На Восточно-Европейской равнине остатки веретеницы ломкой {Anguis
fragilis) прослеживаются в лесных ассоциациях плиоцена, а также Ильинско-
го, лихвинского, микулинского и валдайского горизонтов нижнего, среднего и
верхнего неоплейстоцена и голоцена смежных районов Поволжья (Березовка,
Еласы и Б. Тиганы). К плиоценовому времени относятся и остатки обитаю-
щего сейчас в Калмыкии и Дагестане желтопузика (Сухов, 1972; Чхиквадзе,
Сухов, 1977; Гаранин, 1983, 1999; Ратников, 2002).
Сцинковые (Scincidae) на сопредельной территории известны с эоценовых
отложений (Терентьев, 1961). Единственный современный представитель в
регионе - пустынный гологлаз {Ablepharus deserti) - считается автохтонным
среднеазиатским видом, современные очертания ареала которого здесь сфор-
мировались, очевидно, в голоцене.
Настоящие ящерицы (Lacertidae) известны с эоцена Западной Европы, а род
Lacerta на сопредельной территории Башкирии и Восточно-Европейской рав-
нины - с миоцен-плиоценовых отложений. В пределах Восточно-Европейской
равнины остатки прыткой ящерицы {Lacerta agilis, L. aff. agilis, L. cf. agilis) -
европейскосредиземноморского вида - прослеживаются с урывского и хап-
ровского фаунистических комплексов плиоцена, одесского фаунистического
комплекса эоплейстоцена, петропавловского, Ильинского и мучкапского гори-
зонтов тираспольского фаунистического комплекса нижнего неоплейстоцена,
среднерусского горизонта хазарского фаунистического комплекса среднего
неоплейстоцена, микулинского и валдайского горизонтов палеолитического
фаунистического комплекса верхнего неоплейстоцена и голоценовых отложе-
ний, в том числе из сопредельных местонахождений Б. Тиганы (Татарстан),
Башкирии, а также Средней Ахтубы и Черный Яр - Нижнее Займище Аст-
раханской области (Ратников, 2002; 20046; Хабибуллин, 2002). Современные
очертания ареала этого вида в регионе и его ближайших окрестностях сформи-
ровались в процессе послеледникового расселения восточной группы популя-
ций из Кавказского рефугиума (Прыткая ящерица, 1996). Из местонахождений
Б. Тиганы и Средняя Ахтуба известны и остатки зеленой ящерицы {L. viridis),
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
37
современный ареал которой простирается к западу от Полтавы. Живородящая
ящерица (Zootoca vivipara = Lacerta vivipara) известна из двух голоценовых пе-
щерных местонахождений Башкирии (Хабибуллин, 2002). Вероятно, это наи-
более молодая форма, возникшая при образовании тайги, а затем освоившая
более южные части лесного пояса (Гаранин, 1983).
Наиболее древние ископаемые остатки ящурок (Eremias) на территории
бывшего СССР обнаружены в отложениях позднего плиоцена (Зерова, Чхик-
вадзе, 1984). В регионе они представлены шестью видами, в том числе три -
линейчатая, средняя и сетчатая (Е. lineolata, Е. intermedia, Е. grammica) - эн-
демичными и три - разноцветная, песчаная и быстрая (£. arguta, Е. scripta,
Е. velox) - субэндемичными для Средней Азии и Восточного Ирана (Богда-
нов, 1963). Из них разноцветная ящурка (Е. arguta) известна с микулинского
горизонта верхнего неоплейстоцена местонахождения Еласы, расположенно-
го несколько севернее современного ареала вида, другие из более отдаленных
пунктов за пределами региона.
Змеи (Serpentes), вероятно, в самостоятельную группу обособились в верх-
ней юре, хотя палеонтологические остатки их представителей обнаруживают-
ся только в отложениях мелового периода.
Современные представители семейства ложноногих, или удавов (Boidae)
встречаются в отложениях с миоценового времени, а остатки какого-то пред-
ставителя рода удавчики (Eryx sp.) известны с миоцен-голоцена Европы, в том
числе с урывского фаунистического комплекса плиоцена Восточно-Европейс-
кой равнины. Современные виды - восточный и песчаный удавчики (£. (atari-
cus, Е. miliaris), несомненно, имеющие азиатское происхождение, территорию
региона освоили после ее освобождения от морских вод.
Представители семейства ужовых (Colubridae) известны с позднего эоцена,
а виды рода стройные полозы (Coluber cf. viridiflavus, С. aff. ravergieri), медян-
ка (Coronella aff. austriaca), лазающие полозы (Elaphe aff. dione u sp.) и настоя-
щие ужи (Natrix natrix, N. sp.) из лесных и лесостепных ассоциаций урывского
фаунистического комплекса плиоцена Восточно-Европейской равнины. Здесь
же из современных представителей разноцветный полоз, обыкновенная ме-
дянка или близкие к ней виды прослеживаются с урывского фаунистического
комплекса плиоцена, а также в лесных ассоциациях микулинского и валдайс-
кого горизонтов с верхнего неоплейстоцена, хотя в Центральной Европе извес-
тны с позднего миоцена (Ратников, 2002). В послеледниковое время из евро-
пейских рефугиумов медянка расселилась, очевидно, в суббореальную стадию
голоцена (Юшков, 1997; Хабибуллин, 2001). Наиболее древние палеонтоло-
гические находки лазающих полозов (Elaphe) встречаются с раннемиоцено-
вых отложений Европы. На Восточно-Европейской равнине узорчатый полоз
или близкий к нему вид (Elaphe aff. dione, Е. dione), имеющий, по-видимому,
восточно-палеарктические корни прослеживается из лесостепных ассоциаций
плиоцена, затем в лесных герпетокомплексах ильинского горизонта нижнего
неоплейстоцена, в том числе на смежной территории Поволжья (местонахож-
дение Березовка). В ряде местонахождений среднерусского горизонта хазарс-
кого фаунистического комплекса среднего неоплейстоцена Поволжья отмечен
близкий вид - эскулапов полоз (Е. longissima), ныне обитающий за пределами
этого региона. Европейско-средиземноморский сарматский (четырехполосый)
полоз (£. quatorlineata) также, вероятно, имеет плейстоценовый возраст.
38
Глава 2
Ящеричная змея [Malpolon monpessulanus), по-видимому, плейстоценовый
вид, имеющий средиземноморские корни. Представители рода настоящие ужи
[Natrix natrix, N. tesselata) известны с плиоцена, а на Восточно-Европейской
равнине практически на всем протяжении от урывского фаунистического ком-
плекса плиоцена до голоценовых отложений включительно на сопредельной
территории поволжских местонахождений - Березовка, Б. Тиганы, Еласы,
Средняя Ахтуба, Черный Яр - Нижнее Займище, Красный Бор и в пещерах Ле-
меза. Современные очертания их ареалы приобрели в постгляциальное время
(Сухов, 1972; Гаранин, 1983; Ратников, 2002, 20046; Вершинин, 2007). Стре-
ла-змея [Psamophis lineolatus), вероятно, третичный вид, сформировавшийся в
Средней Азии.
Древнейшие остатки современных гадюковых змей (Viperidae) встречают-
ся с миоцен-плиоценовых отложений Палеарктики. Наиболее древней среди
евразиатских гадюк признается степная [Vipera ursini), имеющая, очевидно,
европейско-среднеазиатские корни, в Поволжье известна с плиоценовых от-
ложений, а обыкновенная гадюка [V. berus), по-видимому, является самым мо-
лодым видом, сформировавшимся в конце ледникового периода в рефугиумах
Центральной Европы из северных популяций неогенового правида, который
с потеплением расселился в лесной и лесостепной зонах. К миоценовому вре-
мени относится и образование современного рода щитомордников (Серебров-
ский, 1937).
Приведенные материалы дают основание предполагать, что наиболее мно-
гочисленные и разнообразные составы батрахологических и герпетологичес-
ких фаунистических комплексов региона были в третичном периоде - неогене
(миоцене) - времени расцвета тургайской флоры. Однако последующие изме-
нения очертаний региона и облика его ландшафтов, вызванные похолодани-
ем на севере и ксерофитизацией на юге, несомненно, должны были сказаться
на составе этих фаунистических комплексов уже в конце третичного периода
(плиоцене), что и подтверждается вымиранием в это время ряда архаичных
тепло- и влаголюбивых видов (см. табл. 3, 4). Этот процесс продолжался и в
плейстоцене, чему способствовало усиление динамичности всех природных
процессов в регионе и на сопредельной территории. Среди вымерших форм
практически все входили в состав лесных или лесостепных ассоциаций: Муо-
proteus sp., Pliobatrachus cf. langhae, Eopelobatus sp., Latonia sp., Liventsovkia
jacunda, Bufo planus [Bufo albus}, Ranomorphus similis, Elaphe cf. rufodorsata,
Coluber aff. ravergiere и некоторые другие.
У ряда видов отмечено отступление границ ареала, преимущественно в
южном направлении. Наименьший сдвиг ареала произошел, по-видимому, у
разноцветной ящурки, которая отступила только в пределы степной зоны По-
волжья (хотя изолированный очаг остался в районе Самарской луки), ящерич-
ной змеи - в Предкавказье и у камышовой жабы, сместившейся западнее. Юж-
нее, в пояс широколиственных, смешанных и отчасти хвойных лесов Европы
и Кавказа, перешли ареалы альпийского тритона, желтобрюхой жерлянки, кав-
казской жабы, прыткой лягушки и эскулапова полоза; в районы с засушливым
климатом южнее дельты р. Дунай - сирийской чесночницы; в пояс песчаных
пустынь южного Казахстана и Средней Азии - среднеазиатской черепахи и ва-
рана; в предгорья и горный пояс этого района - разноцветного полоза [Coluber
aff. ravergieri). Наибольший интерес, несомненно, вызывает изменение ареала
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
39
монгольской жабы, которая в настоящее время обитает на юге Восточной Си-
бири и Дальнем Востоке.
Считается, что у ряда обитателей лесного пояса Северной Евразии - си-
бирского углозуба, серой и зеленой жабы, озерной лягушки и обыкновенной
гадюки - в последние тысячелетия ареал значительно расширился и, по-ви-
димому, продолжает увеличиваться. Наряду с этим у гребенчатого тритона
и обыкновенной чесночницы произошло некоторое сокращение ареала в его
южной части - у первого вида в связи с наступлением сухого и теплого кли-
мата, а у чесночницы - с наступлением прохладной субатлантической стадии.
Наряду с более или менее четко выраженными изменениями границ области
распространения у многих видов продолжались заполнение ценотического ва-
куума и усложнение «кружева» ареала.
В последнее тысячелетие формирование облика ландшафтов и соответс-
твенно состава и структуры их биоты шло под все возрастающим воздействи-
ем антропогенных факторов: вырубки лесов, пасторальной дигрессии, затем
распашки земель и общего освоения территории, которые сейчас оказывают
решающее влияние как на биоту в целом, так и на состав и состояние батра-
хологических и герпетологических комплексов отдельных регионов. Вместе
с тем у северных пределов региона меньшая засушливость позднего голоце-
на наряду с особенностями хозяйственного освоения территории определили
большую стабильность природных биоценозов, которые на слабо трансфор-
мированных участках сохранили многие особенности экосистем раннего голо-
цена. Степная, полупустынная, а в последние десятилетия и пустынная зоны
испытали значительное воздействие антропогенного пресса вследствие выруб-
ки лесов на Общем Сырте и в Северном Прикаспии, увеличения масштабов
пасторальной дигрессии в Волго-Уральских песках, Предуралье и примыка-
ющих пространствах глинистой полупустыни, а затем и земледельческого ос-
воения территории. Во время первого этапа земледельческого освоения были
выборочно распаханы крупные массивы черноземов, темно-каштановых почв
и некоторые крупные падины в южной части степного пояса. На втором этапе
завершилась распашка почти всех удобных в технологическом плане земель, а
затем были распаханы и условно пахотнопригодные земли (с пониженным аг-
роэкологическим потенциалом и большой комплексностью) вплоть до южной
границы полупустыни, что обусловило широкое распространение антропоген-
ного опустынивания.
В последние годы в связи со сменой формы организации сельскохозяйс-
гвенного производства и как следствие переводом части пахотных земель в
залежь, а также с уменьшением пастбищной нагрузки наметился новый этап
в землепользовании, в процессе которого замедлилось опустынивание регио-
на и, возможно, положено начало процессу восстановления его ландшафтов и
эиоразнообразия. Это настоятельно требует объективной оценки изменений в
состоянии экосистем региона, составе и структуре их фаунистических комп-
пексрв^выявления закономерностей этих процессов, что позволит разработать
эффективныёПчетоды оценки состояния окружающей среды, обеспечить ее мо-
ниторинг и оптимизировать использование биоресурсов региона при сохране-
нии его биологического разнообразия.
Отмеченное позволяет принять для региона несколько измененную схему
периодизации истории развития батрахо- и герпетофауны Восточно-Европей-
40
Глава 2
ской равнины, предложенную В.Ю. Ратниковым (2002), в которой целесооб-
разнее выделить шесть последовательных этапов со специфическими комп-
лексами амфибий и рептилий.
- Неогеновый - охватывает время формирования исходного типа батра-
хо- и герпетологического фаунистических комплексов региона, для которого
характерны множество впоследствии вымерших форм (см. табл. 3,4) и появле-
ние ряда современных видов, часть которых в настоящее время не обитает на
исследуемой территории (в том числе Bufo raddei и Bufo verrucosissimus).
- Эоплейстоценовый - характеризующийся резким обеднением видово-
го состава фаунистических комплексов, в которых в небольшом количестве
представлены молодые современные {Bufo raddei) и некоторые новые {Егух
sp.) виды.
- Раннеплейстоценовый - отмечающийся присутствием ряда довольно
многочисленных вымирающих форм {Pliobatrachus cf. langhae, Natrix sp.forma
А и В), постоянным присутствием и доминированием значительного количест-
ва современных видов при сохранении ряда ныне здесь не обитающих.
- Среднеплейстоценовый - характеризующийся небольшим количеством
вымирающих форм, доминированием в основном современных видов, в соста-
ве которых еще сохраняются ныне не обитающие на исследуемой территории
представители.
- Позднеплейстоценовый - с фаунистическими комплексами, которые
почти во всех местонахождениях представлены современными видами, оби-
тающими на исследуемой территории. Характерно резкое увеличение находок
Bufo viridis, которые ранее были редкими.
- Голоценовый - с фаунистическими комплексами только из современных
видов, ныне обитающих на изучаемой территории. Границы их ареалов очень
динамичны.
В последнее тысячелетие фауна испытывает все возрастающее воздействие
антропогенных факторов, что сказывается на численности видов и их распре-
делении внутри ареала.
В настоящее время батрахо- и герпетофауна региона представлена 50 ви-
дами, из которых 12 относят к амфибиям и 38 к рептилиям (Дебело, Чибилев,
2012). Их распространение здесь выглядит следующим образом.
В лесостепной зоне региона, занимающей низменное и возвышенное Са-
марское Заволжье, а также юго-западные отроги Южного Урала, установлено
обитание 12 видов амфибий и такого же количества рептилий. В формировании
батрахологического комплекса региона основная роль (66,6%) принадлежит
европейским видам (обыкновенному и гребенчатому тритонам, краснобрю-
хой жерлянке, обыкновенной чесночнице, серой жабе, обыкновенной квакше,
травяной и прудовой лягушкам) и по 2 вида (по 16,7%) считаются предста-
вителями средиземноморского (зеленая жаба и озерная лягушка)^ европей-
ско-сибирского (сибирский углозуб и остромордая лягушка) фаунистических
комплексов. Герпетологический комплекс^обраЗуют 5 (41,7%)европейских
(веретеница ломкая, обыкновенная м^дяйка, обыкновенныйужтобыкновенная
гадюка и гадюка Никольского)>>^5%) средиземноморских (болотная чере-
паха, степная гадюка, водяной уж) и по одному (по 8,3%) широко распростра-
ненному европейско-сибирскому (прыткая ящерица), европейско-азиатскому
(живородящая ящерица), восточно-палеарктическому (узорчатый полоз) и
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
41
среднеазиатскому (разноцветная ящурка) виду. В целом в состав фаунистичес-
кого комплекса лесостепной зоны региона входят: европейских - 13 (54,1%)
видов, средиземноморских - 5 (20,8), европейско-сибирских - 3 (12,5), евро-
пейско-азиатских - 1 (4,2), восточно-палеарктических - 1 (4,2), среднеазиатс-
ких-! (4,2%).
Вместе с тем обитание двух видов здесь известно лишь по одному пункту:
- разноцветной ящурки на небольшом (около 40 га) участке надпойменной
террасы Волги в районе Самарской Луки;
- сибирского углозуба, известного в прошлом из окрестностей Богославска
в бассейне реки Сакмары.
Кроме того, здесь восточных рубежей ареала достигает гадюка Николь-
ского (если будет подтвержден ее видовой статус), а в прошлом достигала
обыкновенная квакша. Таким образом, основное ядро герпетофауны региона
составляют виды европейского происхождения.
По характеру использования территории (жизненного пространства) зем-
новодных и пресмыкающихся обычно делят на лесных, обитателей открытых
пространств и водоемов (Гаранин, 1983; Гаранин, Щербак, 1989; Бакиев и др.,
2004). Все амфибии, учитывая их привязанность к водоемам в период размно-
жения, являются гидрофильными видами. Однако после завершения размно-
жения наиболее тесно к водоемам оказывается привязанной лишь озерная ля-
гушка. Это фактически полуводная форма (которая от воды обычно удаляется
не дальше нескольких метров), что позволило ей освоить все пресные водоемы
региона, за исключением горных ручьев и речек (где она местами встречается
лишь в тихих заводях). Только в водоемах и по их берегам встречается так-
же прудовая лягушка, хотя севернее в лесной части ареала эта привязанность
не столь четко выражена. К этим видам экологически близка и краснобрюхая
жерлянка, которая благодаря тесной привязанности к пресноводным водоемам
освоила территорию региона вплоть до пустынной зоны. Эти экологически
близкие виды составляют особую группу гидрофильных амфибий.
В меньшей (хотя и в разной) степени привязаны к водоемам гребенчатый
тритон, травяная лягушка и сибирский углозуб. Первый из них населяет лес-
ные массивы по берегам крупных и постоянных водоемов. Основным место-
обитанием травяной лягушки являются сырые луга и лесные массивы по бе-
регам рек. Подобными локальными местообитаниями она проникает и южнее.
В сходных местах - заболоченных участках речных пойм, перемежающихся
с сырыми лугами и древесно-кустарниковыми зарослями у южных пределов
ареала, встречался и сибирский углозуб.
Однако большинство амфибий региона являются гигрофильными видами.
Из них обыкновенный тритон, обыкновенная жаба, чесночница, а в прошлом
и квакша в лесостепи явно тяготеют к сырым и тенистым пойменным лист-
венным лесам с зарослями кустарников. Здесь, а также в островных колковых
лесах и лесополосах обитает и остромордая лягушка. Наиболее засухоустой-
чивый, по сути мезофильный вид - зеленая жаба, которая в регионе населя-
ет разреженные лесные массивы, а также поймы рек, где придерживается в
основном открытых биотопов, причем иногда на значительном удалении от
воды.
Из рептилий гидрофильными видами в регионе являются болотная черепа-
ха, водяной уж и в меньшей степени обыкновенный уж, населяющий не толь-
42
Глава 2
ко побережья водоемов, но и другие, преимущественно лесные, увлажненные
биотопы в их ближайших окрестностях. Возможно, обыкновенного ужа следу-
ет считать промежуточной формой между гидрофилами и типичными гигро-
филами, к числу которых относятся живородящая ящерица, веретеница, обык-
новенная гадюка и гадюка Никольского. Среди рептилий значительно больше
мезофильных форм, к числу которых относятся прыткая ящерица, медянка,
узорчатый полоз и степная гадюка. Достигающую здесь северных пределов
распространения разноцветную ящурку можно считать ксерофильным видом.
В целом комплекс амфибий и рептилий лесостепной зоны составляют: гидро-
филы - 9 (37,5%) видов; гигрофилы - 9 (37,5), населяющие в основном увлаж-
ненные лесные сообщества; мезофиллы - 5 (20,8), населяющие преимущест-
венно экотоны - пограничные местообитания окраин лесов, опушки и поляны;
ксерофилы - 1 (4,2%) вид, населяющий небольшой участок надпойменной
террасы Волги.
В степной зоне региона установлено обитание 30 видов, в том числе
12 амфибий и 18 рептилий (без гадюки Никольского, распространение которой
здесь еще точно не выяснено). Из них южных пределов распространения здесь
достигают сибирский углозуб, обыкновенный и гребенчатый тритоны, обык-
новенная квакша, травяная и прудовая лягушки, веретеница ломкая, живоро-
дящая ящерица и приближаются к южной границе ареала обыкновенная жаба
и медянка. Кроме того, здесь появляется ряд новых видов - среднеазиатская
черепаха, круглоголовки вертихвостка, такырная и ушастая, быстрая ящурка и
обыкновенный щитомордник.
Среди амфибий, достигающих южных пределов распространения и «вы-
бывающих» из состава батрахологического комплекса региона, 5 видов имеет
европейское происхождение и один относится к европейско-сибирскому фау-
нистическому комплексу (углозуб). Среди «выбывающих» рептилий один вид
имеет европейские «корни» и один - евразиатские. В целом из состава фау-
нистического комплекса степной зоны региона южных пределов распростра-
нения достигают 6 европейских видов и по одному европейско-сибирскому и
евразиатскому.
Из достигающих в степной зоне северных пределов распространения реп-
тилий 4 вида имеют среднеазиатское происхождение (среднеазиатская чере-
паха, такырная и ушастая круглоголовки, быстрая ящурка), один - централь-
ноазиатское (круглоголовка-вертихвостка) и один - восточнопалеарктическое
(обыкновенный щитомордник). Таким образом, в целом комплекс амфибий и
рептилий степной зоны региона составляют: европейские виды - 12 (40,0%),
среднеазиатские - 6 (20,0), средиземноморские - 5 (16,7), европейско-сибир-
ские - 3 (10,0), восточнопалеарктические - 2 (6,7), европейско-азиатские -
1 (3,3), центральноазиатские - 1 (3,3%).
Среди видов, достигающих в степной зоне южных пределов ареала, 4 яв-
ляются гидрофилами и 4 гигрофилами, населяющими преимущественно ув-
лажненные лесные биотопы. Из видов, достигающих северных пределов рас-
пространения, к эвритопным мезофиллам можно отнести (с известной долей
условности) практически лишь щитомордника, а остальные 6 (наряду с раз-
ноцветной ящуркой) - типичные ксерофилы, среди которых 4 вида (средне-
азиатская черепаха, круглоголовки вертихвостка и ушастая и быстрая ящурка)
связаны с локальными песчаными массивами (псаммофилы) и 2 вида (писк-
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
43
ливый геккончик, такырная круглоголовка) являются эвритопными, населя-
ющими преимущественно участки с каменистыми или плотными грунтами.
В целом комплекс амфибий и рептилий степной зоны составляют: гидро-
филы - 9 (30,0%) видов, гигрофилы - 8 (26,7), населяющие преимущественно
сообщества пойменных и островных лесов, мезофилы - 6 (20,0), эвритопные
виды - 1 (3,3), ксерофилы - 6 (20,0%), преимущественно псаммофилы.
Таким образом, если исключить азональные (в широком смысле этого тер-
мина) водные и околоводные (сибирского углозуба, озерную, прудовую, травя-
ную и остромордую лягушек, гребенчатого тритона, краснобрюхую жерлянку,
обыкновенную чесночницу, болотную черепаху, обыкновенного и водяного
ужей), лесные (обыкновенную квакшу, обыкновенного тритона, живородящую
ящерицу, медянку, обыкновенную гадюку), населяющие островные песчаные
массивы псаммофильные виды (среднеазиатскую черепаху, круглоголовок
вертихвостку и ушастую, быструю ящурку), то окажется, что собственно степ-
ные плакорные ландшафты населяют лишь 6 видов (20,0% общего видового
состава), в том числе зеленая жаба, такырная круглоголовка, прыткая ящерица,
узорчатый полоз, обыкновенный щитомордник и степная гадюка.
В полупустынной зоне региона установлено обитание 32 видов, в том чис-
ле 6 (18,8%) амфибий и 26 (81,2%) рептилий. Среди амфибий половина со-
става (50,0%) имеет европейское происхождение (краснобрюхая жерлянка,
обыкновенная чесночница, обыкновенная жаба), 2 вида (33,3) относятся к сре-
диземноморскому (зеленая жаба, озерная лягушка) и 1 (16,7%) - к европейско-
сибирскому (остромордая лягушка) фаунистическим комплексам. Основу гер-
петологического комплекса образуют 15 (57,7%) среднеазиатских и 5 (19,2%)
средиземноморских видов. В составе среднеазиатских 9 видов достигают в
регионе северных пределов распространения. Это каспийский, грепнепалый
и серый голопалый гекконы, степная агама, сетчатая и песчаная ящурки, пес-
чаный и восточный удавчики и стрела-змея. Среди них 5 видов являются эв-
ритопными и 4 населяют преимущественно псаммофитные местообитания.
В целом в составе среднеазиатских видов доминируют псаммофилы (8 видов),
остальные (7 видов) населяют преимущественно участки с каменистыми или
плотными глинистыми грунтами. В составе вторых, помимо широко распро-
страненных болотной черепахи, водяного ужа и степной гадюки, появляются
эвритопные сарматский и каспийский полозы, достигающие здесь северно-
восточных пределов ареала. Из европейских видов остаются лишь 2 (7,7%) -
широко распространенный обыкновенный уж и достигающая южных преде-
лов распространения медянка. Восточнопалеарктический комплекс (7,7%)
представляют широко распространенные узорчатый полоз и обыкновенный
щитомордник и по одному виду относятся к европейско-сибирскому (прыткая
ящерица) и центральноазиатскому (вертихвостка) комплексу.
В целом в состав фаунистического комплекса полупустынной зоны регио-
на входят: среднеазиатских - 15 (46,9%) видов, средиземноморских - 7 (21,9),
европейских - 5 (15,6), европейско-сибирских - 2 (6Д), восточнопалеарктичес-
ких - 2 (6,2), центральноазиатских - 1 (3,2%).
По характеру использования жизненного пространства среди амфибий
2 вида - озерная лягушка и краснобрюхая жерлянка - являются типичными
гидрофилами (последняя здесь приближается к южной границе ареала). Тес-
нее к водоемам становится привязана обыкновенная чесночница, которая
44
Глава 2
встречается лишь на участках с мягким грунтом по кромке водоемов. Гигро-
фильные обыкновенная жаба и остромордая лягушка достигают здесь южной
границы ареала, и только зеленая жаба достигает в полупустыне оптимума,
освоив практически все пригодные местообитания, чему в известной мере спо-
собствовало хозяйственное освоение территории человеком.
Из рептилий гидрофильные виды в регионе - болотная черепаха, водяной
уж и в меньшей степени обыкновенный уж, который, однако, в полупустыне
уже практически не выходит за пределы речных пойм. С мезофильными ста-
циями (площадь которых в полупустыне существенно уменьшилась) по-пре-
жнему связаны прыткая ящерица, медянка, узорчатый полоз, степная гадюка и
обыкновенный щитомордник. Все остальные представители практически ксе-
рофилы, среди которых 7 видов населяют локальные псаммофитные сообщес-
тва и 11 видов в регионе являются эвритопными, населяющими разные, пре-
имущественно с глинистыми, реже каменистыми грунтами, местообитания.
В целом комплекс амфибий и рептилий полупустынной зоны составляют: гид-
рофилы -5(15,6%) видов, гигрофилы (околоводные) - 3 (9,4), мезофиллы -
6 (18,7), псаммофилы - 7 (21,9), склерофилы - 3 (9,4), эвритопные - 8 (25,0%).
Если учесть, что в этом комплексе 9 видов встречаются в водоемах и на их
побережье, 1 вид - в лесных сообществах, а 7 - в основном в островных песча-
ных массивах, то количество видов, населяющих преимущественно зональные
полупустынные сообщества, по сравнению с таковым степной зоны возрастает
более чем в 2 раза и достигает 15 (46,9% общего видового состава).
В пустынной зоне региона установлено обитание 37 видов, в том числе
4 (10,8%) амфибий и 33 (89,2%) рептилий. Среди амфибий 2 вида имеют ев-
ропейское происхождение (жерлянка, чесночница) и 2 - средиземноморское
(зеленая жаба и озерная лягушка). Основу герпетологического комплекса об-
разуют 22 среднеазиатских и 6 средиземноморских видов. В составе первых
6 видов здесь достигают северных пределов ареала (сцинковый геккон, песча-
ная круглоголовка, средняя и линейчатая ящурки, поперечнополосатый и раз-
ноцветный полозы). В составе средиземноморского комплекса новым видом
становится ящеричная змея, отмеченная недавно на левобережье Волги. Вмес-
те с тем южных пределов распространения в регионе достигает широкого рас-
пространенная прежде степная гадюка. Из европейских видов остается лишь
широко распространенный обыкновенный уж, из европейско-сибирских -
прыткая ящерица, доходящая здесь до южных пределов распространения. Вос-
точные палеаркты по-прежнему представлены широко распространенными
узорчатым полозом и обыкновенным щитомордником, а центральноазиатский
комплекс - круглоголовкой вертихвосткой. В целом в состав фаунистическо-
го комплекса пустынной зоны региона входят: среднеазиатские - 22 (59,5%)
вида, средиземноморские - 8 (21,6), европейские - 3 (8,1), восточнопалеар-
ктические - 2 (5,4), европейско-сибирские - 1 (2,7), центральноазиатские -
1 (2,7%).
По характеру использования жизненного пространства среди амфибий ти-
пичными гидрофилами являются краснобрюхая жерлянка (достигающая здесь
южных пределов распространения) и озерная лягушка, освоившая в регионе
и водоемы с повышенной минерализацией (как, например, опресненные лагу-
ны Аральского моря, примыкающие к устью р. Сыр-Дарья). Южной границы
ареала достигает здесь и обыкновенная чесночница, населяющая преимущес-
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
45
твенно прибрежные участки пресных водоемов с рыхлым грунтом. Наиболее
адаптированной к условиям пустыни оказалась зеленая жаба, ночной образ
жизни которой и способность использовать для размножения временные водо-
емы позволили ей широко расселиться в пределах региона.
Среди рептилий гидрофильный комплекс представляют болотная черепа-
ха, водяной и обыкновенный ужи, населяющие почти исключительно долины
Волги, Урала и Эмбы. Преимущественно мезофильных стаций речных долин
и их ближайших окрестностей придерживаются прыткая ящерица и степная
гадюка, а эвритопные мезофилы узорчатый полоз и обыкновенный щитомор-
дник проникают и в типичные пустынные стации с колониями грызунов. Ос-
тальные представители являются типичными ксерофилами, в составе которых
как отражение господства в регионе песчаных ландшафтов доминируют псам-
мофилы, в свою очередь дифференцирующиеся на обитателей развеваемых,
полузакрепленных и закрепленных песков (среднеазиатская черепаха, гребне-
палый геккон, сцинковый геккон, круглоголовка-вертихвостка, песчаная круг-
логоловка, ушастая круглоголовка, сетчатая ящурка, линейчатая ящурка, пес-
чаная ящурка, быстрая ящурка, песчаный удавчик, поперечнополосатый полоз,
ящеричная змея). Преимущественно с каменистыми и щебнистыми склонами
гор-останцов, скалами и береговыми обрывами связана специализированная
группа склерофилов (пискливый геккончик, серый геккон, разноцветный по-
лоз, палласов полоз, каспийский полоз). Глинистую пустыню населяют эври-
топные ксерофилы (каспийский геккон, степная агама, такырная круглоголов-
ка, пустынный гологлаз, разноцветная, средняя ящурки, восточный удавчик,
стрела-змея). В целом, комплекс амфибий и рептилий пустынной зоны состав-
ляют: гидрофилы - 5 (13,5%), гигрофилы - 1 (2,7), мезофиллы - 5 (13,5), псам-
мофилы - 13 (35,2), склерофилы - 5 (13,5), эвритопные - 8 (21,6%).
Эти материалы показывают, что в типичных пустынных ландшафтах реги-
она обитает 26 видов, или 70,3% его фаунистического комплекса амфибий и
рептилий.
Наряду с отмеченными ряд изменений в фаунистических комплексах этих
групп наблюдается и с запада на восток (табл. 6-9).
Среди амфибий 6 видов (50% видового состава) являются широко распро-
страненными и встречаются на всем протяжении или значительной части со-
ответствующих природных зон региона. Из них обыкновенный тритон, серая
жаба и озерная лягушка считаются монотипическими видами, сибирский уг-
лозуб и остромордая лягушка представлены одним (номинативным), а зеленая
жаба 2 подвидами.
Остальные 6 видов представляют европейский фаунистический комплекс,
из состава которого прудовая лягушка в регионе достигает центральной час-
ти Волжско-Уральского междуречья (возможно, этих же пределов достигает и
съедобная лягушка). До юго-западных склонов Урала в прошлом простирался
ареал обыкновенной квакши. До Уральских гор простирается основная часть
ареала еще 4 видов, хотя локально они проникают и в Зауралье, где достигают:
краснобрюхая жерлянка - 60° в.д., гребенчатый тритон - Курганской области,
обыкновенная чесночница - Тургайской депрессии, а травяная лягушка - ни-
зовий р. Иртыш. Из них гребенчатый тритон, жерлянка и прудовая лягушка
считаются монотипическими видами, а чесночница, квакша и травяная лягуш-
ка представлены номинативными подвидами.
46
Глава 2
Таблица б
Фаунистический состав амфибий природных зон региона
Зона Кол-во видов фаунистических комплексов
Европей- ские Средиземно- морские Европейско- сибирские Восточно- пал соаркти- ческие Средне- азиатские Центрально- азиатские Всего
Лесостепная 8 2 2 — — - 12
Степная 8 2 2 - - — 12
Полупустынная 3 2 1 — — - 6
Пустынная 2 2 - - — — 4
Таблица 7
Фаунистический состав рептилий природных зон региона
Зона Кол-во видов фаунистических комплексов
Европей- ские Средиземно- морские Европейско- сибирские Восточно- палеоаркти- ческие Средне- азиатские Центрально азиатские Всего
Лесостепная 5 3 1 1 1 - 12
Степная 4 3 1 2 6 1 18
Полупустынная 2 5 1 2 15 1 26
Пустынная 1 6 1 2 22 1 33
Таблица 8
Экологические группы амфибий природных зон региона
Зона Кол-во видов
Гидрофилы Гигрофилы Мезофилы Всего
Лесостепная 6 5 1 12
Степная 6 5 1 12
Полупустынная 2 3 1 6
Пустынная 2 1 1 4
Таблица 9
Экологические группы рептилий природных зон региона
Зона Кол-во видов
Г ндрофилы Г игрофилы Мезофилы Псаммо- филы Склеро- филы Э ври фи л ы Всего
Лесостепная 3 4 4 1 - - 12
Степная 3 3 5 4 2 1 18
Полупустынная 3 - 5 7 3 8 26
Пустынная 3 — 4 13 5 8 33
ИСТОРИЯ ФОРМИРОВАНИЯ, СОВРЕМЕННАЯ ГЕОГРАФИЯ...
47
У рептилий, отличающихся повышенной гетерогенностью состава, распро-
странение имеет более сложный характер. Среди них лишь несколько видов
являются широко распространенными, ареалы которых выходят за пределы
региона (прыткая ящерица, круглоголовки вертихвостка и такырная, узорча-
тый и сарматский полозы). У остальных видов в регионе имеется несколько
довольно четко выделяющихся естественных рубежей.
Долина р. Волги с востока ограничивает основную часть ареала ящеричной
змеи (которая лишь недавно проникла на левобережье), с запада - ареалы пис-
кливого геккончика, щитомордника, а в недалеком прошлом и степной агамы.
Кроме того, здесь проводятся границы между номинативными и восточными
подвидами обыкновенного ужа, степной гадюки и, возможно, номинативно-
го и кавказского подвидов быстрой ящурки. По мнению некоторых авторов,
волжская долина ограничивает также распространение гадюки Никольского
(Ефимов и др., 2008) и приморской солончаковой формы разноцветной ящур-
ки (Иванова, 1985).
Долина р. Урал ограничивает распространение стрелы-змеи, каспийско-
го (желтобрюхого) полоза, подвидов ушастой круглоголовки и разноцветной
ящурки (у которой в Волжско-Уральском междуречье наблюдается интергра-
дация номинативного и западного подвидов). Южнее, ее продолжением, явля-
ется восточное побережье Каспия, ограничивающее распространение на запад
ряда пустынных среднеазиатских видов.
Уральские горы - Мугоджары на севере региона ограничивают основную
часть ареала болотной черепахи (локально в нескольких пунктах известна и в
Зауралье), а на юге смещают ареал сарматского полоза.
Турано-Тургайская депрессия ограничивает проникновение на восток ве-
ретеницы, болотной черепахи и на запад - гребнепалого геккона, восточного
удавчйка, разноцветного полоза и, возможно, сетчатой круглоголовки.
Таким образом, фаунистический комплекс амфибий и рептилий, а также
распространение их видов определяются историческими и комплексом совре-
менных факторов.
В герпетогеографическом плане территория региона относится к Пале-
арктической области. Его северная лесостепная часть включается в состав
переходной лесостепной провинции Бореальной Европейско-Сибирской под-
области, а остальная - степной и пустынной Туранской провинциям аридной
Средиземно-Центральноазиатской подобласти (Щербак, 1981).
Глава 3
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ
И РЕПТИЛИЙ В УРАЛО-КАСПИЙСКОМ РЕГИОНЕ
Земноводные и пресмыкающиеся известны людям с древнейших времен.
Сведения о них нашли отражения в разных легендах, поверьях, магичес-
ких и религиозных символах, аллегориях, имитационных действиях, по-
нятиях, представлениях, средствах народной медицины и т.п. (Ананьева и др.,
1998; Атлас пресмыкающихся..., 2004; Кузьмин, 1999).
Первые сведения об этой группе животных Оренбуржья приводятся в из-
вестной «Топографии Оренбургской...» 1762 г., в которой за 10 лет до зна-
менитых академических экспедиций дано обстоятельное (по тем временам)
описание природы Прияицких степей. В этой работе П.И. Рычков (цит. по:
1999 г. С. 150, 151) со слов «служилых людей» сообщает, что «черепахи в
Каспийском море в довольном числе находятся,... тако же и около реки Янка
по обеим сторонам в озерах и в степных речках... их много,... токмо мелкие,
и весьма изредка такие попадают, которых скорлупа больше ординарной та-
релки». По мнению В.И. Гаранина и А.Т. Бакиева (2004), здесь, по всей ви-
димости, говорится о трех видах черепах: среднеазиатской (Agrionemys hors-
fieldi}, каспийской (Mauremys caspia) и болотной (Emys orbicularis}, которых
автор не различал. Определенно к болотной черепахе относятся следующие
слова П.И. Рычкова: «...сказывают, ежели она человека уязвит, то многие от
того умирают. От них же молодым уткам и гусям случается вред, а особливо,
когда они заплывают к их гнездам; ...а в змеях и ящерицах ...чтоб какие особ-
ливости были - поныне неизвестно». Есть еще стрела-змея, которая в броске
якобы пронзает человека. Кроме того, в разделе «Птицы знатные» сообщается,
что аист или стерх, который при Оренбурге изредка встречается «залетом из
Хивы», питается змеями и лягушками. Учитывая, что в данной работе не при-
меняется бинарная номенклатура, то, согласно схеме периодизации развития
герпетологии в России, предложенной Л.Я. Боркиным (2003), ее следует от-
нести к произведениям долинеевского периода.
Первые научные (в современном смысле) батрахо- и герпетологические
данные по региону появились в результате экспедиционных исследований
оренбургских и астраханского отрядов, организованных Академией наук.
Оренбургскими академическими отрядами руководили известные естество-
испытатели того времени П.С. Паллас (1741-1810 гг.), И.И. Лепехин (1740-
1802 гг.), И.П. Фальк (1727-1774 гг.), а одним из астраханских отрядов -
С.Г. Гмелин (1745-1774 гг.).
П.С. Паллас детальное изучение природы бывшего Оренбургского края на-
чал в июне 1769 г. на пути вдоль Самарской укрепленной линии к Оренбургу.
Из окрестностей Самары он впервые описал Coluber melanis - встречающуюся
в регионе черную форму гадюки (систематическое положение которой уточ-
няется до настоящего времени), С. berus (по-видимому, молодые особи чер-
ной формы обыкновенной гадюки) и С. natrix (обыкновенного ужа), а также
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ...
49
простых зеленых и серых ящериц (Lacerta agilis). Далее, вверх по р. Самаре
в окрестностях современных с. Алексеевка (Кинельский р-н) и с. Борское им
была отмечена болотная черепаха (Паллас, 1773), сохранившаяся здесь и до
настоящего времени (Бакиев, 2001а).
Из Оренбурга в июле этого года он совершил поездки к Илецкой соли,
Орской крепости, а затем через Илек (23.07) и г. Уральск (27.07-12.08) напра-
вился к Каспийскому морю. Для г. Уральска и его ближайших окрестностей
(п. Б. Чаган) из интересующей нас группы животных отмечены лишь «пест-
рые лягушки» (зеленые жабы), которые встречаются как в жилых постройках,
так и в норах «сей сухой страны», а для южнее расположенного п. Бударино
(13.08) - и болотные черепахи. В окрестностях п. Калмыкове (16-17.08) он
наблюдал «великое множество змей», которые находились в тростниковых за-
рослях по руслу р. Багырдая, а по Индерским горам (20.08) - «много пригожих
ящериц особливого рода» (Lacerta helioscopa, L. velox, L. cruenta). Для окрест-
ностей г. Гурьева (25-31.08) отмечается, что «Яик и все его рукава наполнены
лягушками и есть между ними превеликие, особливого рода, кои так же водят-
ся в р. Волге». Здесь много водилось и водяных змей (Coluber hudris et scuta-
tus). Возвратившись в г. Уральск, он обследовал его окрестности (06-17.09),
а затем через Чернореченскую и Разсыпную станицы отправился в г. Уфу.
В 1770 г. нижнее течение р. Урал, побережье Каспия и прилегающие террито-
рии были обследованы частью отряда П. Палласа под руководством студента
Н. Соколова.
По материалам этих экспедиций на территории региона отмены также ак-
жилан - узорчатый полоз (Coluber dione), который «по соленым местам в сте-
пях около Каспийского моря» водился; пятенный уж (Anguis miliarisT), кото-
рый «находится около Каспийского моря», и лукавая ящерица (Lacerta arguta),
«обитающая около Каспийского моря и по соседственным оному пескам».
Второй раз территорию региона П.С. Паллас посетил в 1770 г., когда была
обследована его северная часть. Основной целью экспедиции 1773 г. стало
изучение природы Волго-Уральского междуречья (хотя часть его отряда со
студентом В. Зуевым направлялась и в Индерские горы). Проследовав через
города Бугуруслан, Бузулук, Сорочинск и Уральск 24.05.1773 г., исследова-
тели направились от р. Урал в степь - на юго-запад к р. Кушум, по этой реке
впоследствии были отмечены проворные ящерицы зеленого цвета и песчаные
ящерицы, которые были «тут весьма обычны». Далее экспедиция пересекла
Камыш-Самарскую депрессию, Рын-пески и 08.06.1773 г. достигла берегов
р. Волги у Ахтубы.
Полностью все батрахо- и герпетологические материалы, собранные в
разных путешествиях П.С. Паллас обобщил в «Zoographia Rosso - Asiatica»
(1814 г.), где были описаны обитающие в регионе чесночница, оба вида жаб,
озерная, прудовая и травяная лягушки, гребенчатый тритон, краснобрюхая
жерлянка, прыткая ящерица, веретеница, а также Coluber scutatus (в настоя-
щее время признается восточным подвидом обыкновенного ужа - Natrix natrix
scutata), С. hydris - водяной уж (N. tesselata), С. cupreus — обыкновенная медян-
ка (Coronella austriaca), С. dione - узорчатый полоз (Elaphe dione), С. prestes,
С. berus — вероятно, разные формы обыкновенной гадюки - и С. melanis.
Руководителем второго оренбургского отряда академических экспедиций
был И.И. Лепехин. Его отряд обследовал Заволжье, затем по широте Крас-
50
Глава 3
ный Яр (выше Астрахани) - Яманхалинка пересек Волго-Уральские пески и
23.08.1769 г. достиг г. Гурьева. Во время этого перехода им были отмечены
ящерицы, змеи и ряд других животных, но подробнее природу «прияицких
степей» И.И. Лепехин не описывает, считая, что «их как естественные, так и
гражданские редкости» достаточно полно охарактеризовал П.С. Паллас. Пе-
резимовав в Табынске, И.И. Лепехин в 1770 г. обследовал верховья р. Урал
и сопредельные пространства. Для этой территории в «Дневных записках...»
(1772) упоминаются лягушки, зеленая жаба, веретеница, обыкновенная гадю-
ка, обыкновенный и водяной ужи, однако без указания конкретных мест их
встречи. На Урале им впервые была обнаружена медянка, которую он описал
и изобразил на рисунке, не указав, к сожалению, места добычи (Юшков, Во-
ронов, 1994).
Третий академический отряд И.П. Фалька летом 1770 г. пересек Волго-
Уральское междуречье по маршруту Сарепта (близ современного г. Волго-
града) - Камыш-Самарские разливы - низовья Б. и М. Узеней - Уральск, а в
1771 г. обследовал Южный Урал. Собранные на этих маршрутах сведения о
прыткой и живородящей ящерицах, обыкновенном уже и гадюках (С. berus и
С. prestes) приводятся в «Полном собрании сочинений ученых путешествий по
России... И.П. Фалька», подготовленных к печати И.Г. Георги (Фальк, 1824).
В его собственной работе для региона указываются гадюки С. melanis (Сама-
ра), С. berus, С. prestes (р. Волга), а также болотная черепаха и квакша (Georgi,
1800; цит. по: Бакиев и др., 2004).
Западные и южные районы обследовал астраханский отряд академичес-
кой экспедиции С.Г. Гмелина, который в 1769 г. изучал часть Рын-Песков, а в
1772 и 1773 г. совершил плавание вдоль восточных берегов Каспийского моря.
Во время этих путешествий была собрана богатая зоологическая коллекция,
включающая и представителей герпетофауны.
Долгое время после окончания работ академических экспедиций в регионе
не предпринималось сколько-нибудь значительных исследований. Лишь спус-
тя полвека - в начале 20-х гг. XIX столетия, начался новый этап, связанный
главным образом с деятельностью Э.А. Эверсмана (1794-1860 гг.), Г.С. Каре-
лина (1801-1872 гг.) и в меньшей мере ряда других естествоиспытателей.
Изучению природы региона Э.А. Эверсманн посвятил около 30 лет. Первой
его работой, содержащей сведения по герпетофауне региона, была «Путешес-
твие из Оренбурга в Бухару», изданная в 1823 г. в Берлине и Лондоне. Более
подробно материалы, собранные им совместно с К. Пандером во время этого
путешествия в составе «миссии А. Негри» в 1820-1821 гг. (переданные Бер-
линскому университету), были обработаны Лихтенштейном и опубликованы в
его специальной работе (Lichtenstein, 1823; цит. по: Богданов, 1965) в качестве
приложения к берлинскому изданию «Путешествия...» Э. Эверсманна. Отде-
льные наблюдения о герпетофауне, собранные во время этого путешествия,
содержатся и у Мейендорфа (1826; цит. по: Терентьев, 1957).
В последующие годы наиболее значительные исследования им проведе-
ны в 1825-1826 гг. на побережье Каспийского и Аральского морей и плато
Устюрт, в 1827 г. - по Общему Сырту и Рын-пескам (совместно с Г.С. Каре-
линым). В 1829 г. Э.А. Эверсманн посещал Индерские горы, низовья рек Б. и
М. Узени, горы Бестау и Чапчачи, низовья Волги, северное побережье Каспия и
уральскую дельту. Этому путешествию посвящена специальная работа «Пуге-
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ...
51
шествие от Казани по разным местам Оренбургской и Астраханской губерний
и по берегам Каспийского моря в 1829 г.», опубликованная в Вестнике Казан-
ского университета (1830-1832 гг.) (цит. по: Чибилёв, 1993). В 1830-1840-е гг.
по заданию Э.А. Эверсманна в разных частях региона сборы коллекций прово-
дил препаратор П. Романов (Чибилёв, 1993). Собранные в этих путешествиях
материалы по распространению нескольких видов ящериц, змей и болотной
черепахи были опубликованы в двух специальных работах на латинском язы-
ке (Eversmann, 1834, 1844; цит. по: Чибилёв, 1995; Бакиев, Файзулин, 2001).
Возможно, поэтому в «Естественной истории Оренбургского края...» 1840 г.
(Эверсманн, 1949) рептилиям посвящено лишь несколько строк. В ней, в час-
тности, отмечается, что змей «в южных степях водятся много, но их видно
только весною».
Более 30 лет в регионе провел Г.С. Карелин, обстоятельно изучивший
Приуралье, Волжско-Уральское междуречье и восточное побережье Каспия.
Результаты его исследований обобщены в многотомной «Урало-Казачьей фа-
уне», которая, однако, как и большая часть коллекций, погибла при пожаре в
г. Гурьеве, где автор жил последние 16 лет. Часть герпетологической коллек-
ции Г.С. Карелина находится в Зоологическом институте РАН и музее Мос-
ковского государственного университета. Ряд сведений о распространении
17 видов пресмыкающихся, собранных на п-ове Мангышлак и в Закаспийском
крае, приводится в дневниках «Экспедиции для осмотра северо-восточных
берегов Каспийского моря в 1832 г.» (Карелин, 1883), а о змеях, собранных
Г.С. Карелиным во время этих путешествий, - и в заметке, опубликованной
в 1838 г. В. Брандтом (цит. по: Терентьев, 1957). В 1830-1840-е гг. природа
северного и восточного побережий Арала и сопредельных территорий Север-
ного Прикаспия и Средней Азии была объектом исследований Э. Эйхваль-
да, А. Лемана, А. Кайзерлинга, в 1850-1870-е гг. - сотрудников экспедиции
К.М. Бэра и Н.А. Северцова, А. Беккера, И. Ауэрбаха. Их богатые герпетоло-
гические коллекционные материалы, хранящиеся в основном в Зоологическом
институте РАН, использовались при подготовке практически всех региональ-
ных и общих сводок по животному миру (Эйхвальд, 1841; Брандт, 1852; Беккер,
1866; Кесслер, 1871; Северцов, 1873; Аленицин, 1987; цит. по: Никольский,
1905; Параскив, 1956). Некоторые сведения о местах находок пресмыкаю-
щихся по западной части региона - «степям Киргизким, Киргиз-Кайсацким,
на реках Урале...» - содержатся в книге И. Двигубского «Опыт естественной
истории всех животных Российской империи» (Гады, или животные пресмы-
кающиеся, 1832; цит. по: Терентьев, 1957).
Важный вклад в изучении батрахо- и герпетофауны северной части реги-
она внес известный зоолог Л.П. Сабанеев. Он в работе, посвященной позво-
ночным животным Пермской и Оренбургской губерний (1874), характеризует
места обитания ставших известных к тому времени амфибий и рептилий, их
распространение, образ жизни, а также показывает морфологические отличия
остромордой и травяной лягушек. Кроме того, со слов местных жителей он
сообщает о существовании на Урале каких-то еще неизвестных крупных ужей
или полозов.
За последние десятилетия XIX в. известны зоологические экспедиции на
п-ова Мангышлак и Бузачи А. Остроумова, во время которых он собирал и
пресмыкающихся. Их список и некоторые особенности распространения при-
52
Глава 3
водятся в работе, опубликованной в 1889 г. (цит. по: Сидоров, 1925). Тургай-
ская область, а также верхнее и среднее течение р. Эмба были обследованы
П.П. Сушкиным, который попутно с орнитологическими наблюдениями соб-
рал значительную коллекцию пресмыкающихся, переданную впоследствии
Зоологическому институту АН и обработанную А.М. Никольским (1899).
Своеобразный итог изучения амфибий и рептилий региона к концу XIX —
началу XX столетий подведен в работах Н.А. Зарудного (1895, 1915). Во время
многочисленных экспедиций на громадной территории, простирающейся от
Южного Урала до Индийского океана, эти животные собирались им попут-
но, но достаточно тщательно и были во многих случаях новыми для науки -
если не для систематики, то для зоогеографии. Для региона он указывает
11 видов амфибий и 15 видов рептилий. Всего же с учетом сопредельных
территорий (и изменений в систематике) Н.А. Зарудным описано не менее
16 форм 13 видов амфибий и 93 формы 88 видов рептилий. Из описанных им
форм 5 А.М. Никольский назвал в его честь, причем 2 из них сохраняют свое
название и в настоящее время; не исключено, что может быть восстановлен в
правах и ряд других форм (Гаранин, 2000).
Батрахо- и герпетологические материалы по северной окраине региона
(Каргалинской степи в окрестностях г. Оренбурга) содержатся также в работе
В.А. Линдгольма, причем наиболее подробные сведения приводятся по фено-
логии, биотопическому размещению, питанию и врагам степной гадюки (Lind-
holm, 1902; цит. по: Никольский, 1918).
По «Гадам Арала» на начало XX в. имеется обстоятельная работа В.С. Ел-
патьевского (1903), при написании которой помимо его собственных матери-
алов использованы и герпетологические сборы Л.С. Берга (Параскив, 1956).
В 1883,1885и 1886г. на островах и в окрестностях Аральского моря, в Закаспий-
ской области (совместно с Н.А. Зарудным) и по маршруту Аральское море-Те-
мир-Илецк-Оренбург большая коллекция рептилий и амфибий была собрана
А.М. Никольским. Результатом обработки многочисленных собственных сбо-
ров, материалов зоологического кабинета Петербургского университета, Зоо-
логического музея АН и отдельных лиц (Казнакова, Липского, Борщевского
и др.) стала обстоятельная монография «Пресмыкающиеся и земноводные
Российской империи» (Никольский, 1905), которая стала первой капитальной
сводкой по амфибиям и рептилиям нашей страны, где нашли отражение все
известные на тот момент материалы. Издание этой монографии, а затем «Оп-
ределителя...» (Никольский, 1907) сыграло важную роль в развитии дальней-
ших исследований в этой области на территории всей страны. Позже данные
работы А.М. Никольским были расширены до трехтомной монографии серии
«Фауна России и сопредельных стран» (1915, 1916, 1918). В ней подытожены
результаты изучения амфибий и рептилий России почти за полтора столетия:
дано описание морфологии всех известных на то время видов и подвидов,
характеризуется их распространение как в пределах страны, так и на приле-
гающей территории, описывается их образ жизни, приведены определитель-
ные таблицы и краткие диагнозы всех видов, здесь приводится также общий
зоогеографический обзор фауны земноводных и пресмыкающихся страны и
рассматриваются некоторые палеогеографические гипотезы формирования ее
батрахо- и герпетофауны. В итоге эти работы, с одной стороны, ознаменова-
ли окончание периода оформления батрахологии и герпетологии как самосто-
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ...
53
ягельных научных дисциплин в нашей стране, а с другой - дали своеобраз-
ный толчок дальнейшему развитию исследованиям этих классов животных
в России.
К этому времени относится и выход работы И.В. Васильева, в которой на
основе собственных материалов и данных Эйхвальда (1841), А. Лемана (1852),
Н.А. Северцова (1873) и А. Остроумова (1889, 1890) приводятся сведения о
распространении и некоторых особенностях биологии 20 видов рептилий
Мангышлака и сопредельных территорий (цит. по: Параскив, 1956). Краткая
информация об амфибиях и рептилиях региона дана также в соответствующих
разделах томов издания «Россия», в том числе «Среднее и Нижнее Поволжье
и Заволжье» (Гаврилов, Ососков, 1901); «Урал и Приуралье» (Гриневецкий,
Клюге, 1914). Этими работами по сути заканчивается период инвентаризации
батрахофауны и герпетофауны региона и составления ее первого зоогеографи-
ческого обзора.
В течение ряда последующих десятилетий, практически включая и сере-
дину XX столетия, уточнялось распространение отдельных видов в пределах
уже в основном очерченных ареалов, выяснялись особенности их биотопичес-
кого распределения, экологии, численность и биоценотическая роль в разных
экосистемах и регионах. При этом значительная часть материалов собиралась
попутно при проведении других исследований, а некоторые сведения сообща-
лись в краеведческих очерках и заметках.
На начальном этапе этого периода сведения о встречах амфибий и репти-
лий в реках Б. Иргиз, Самара и оз. Лебяжье (Боровое) приводится в «Работах
Волжской биологической станции» (Бенинг, 1913, 1926; Редько, 1915), о фе-
нологии лягушек и ящериц и встрече гадюки в черте г. Самара в «Дневниках
Самарской природы» за 1916 г. (Щербиновский, 1919), а о распространении
отдельных видов - в краеведческом очерке «Животный мир Самарского края»
(Хрущова, 1924). С 1906 г. периодически, а в 1920-х гг. более регулярно бе-
рега р. Урал, озера и болота в форштадтских лугах Оренбурга обследовались
П.А. Воронцовским. Результаты его зоологических экскурсий были опуб-
ликованы в «Трудах общества по изучению Киргизского края» (Воронцовс-
кий, 1922), а позднее некоторые сведения о видовом составе амфибий и реп-
тилий Оренбуржья сообщаются в очерке «Животный мир» (Воронцовский,
1927).
В 1920-1921 гг. коллекцию пресмыкающихся попутно собрали сотрудни-
ки Аральской научно-промысловой экспедиции, которая впоследствии была
обработана С.А. Сидоровым (1925), что позволило уточнить распространение
некоторых видов в регионе. В 1928 г. на территории Бузулукского и Пугачев-
ского уездов бывшей Самарской губернии В.С. Бажанов (1930) выявил пре-
бывание 5 видов земноводных и 7 видов пресмыкающихся (ошибочно указана
зеленая ящерица). Состав этого комплекса и характер распространения ряда
видов впоследствии были уточнены П.А. Положенцевым (1935) и Я.Н. Дарк-
шевичем (Даркшевич и др., 1940). Для г. Самары и его окрестностей сведения
о земноводных и ящерицах приводятся также в работе А.И. Мельниченко с
соавт. (1938), о некоторых особенностях распространения в регионе 10 видов
амфибий и 12 видов рептилий - в книгах «Животный мир Среднего Повол-
жья» (Положенцев, 1937, 1941), а о фауне Приерусланской степи - в работе
И.Б. Волчанецкого и Н.П. Яльцева (1934).
54
Глава 3
Итоги проведенных в 1920-1930-х гг. исследований подведены в соответ-
ствующих главах подготовленного Зоологическим институтом АН коллектив-
ного труда «Животный мир СССР», в I томе которого опубликован общий очерк
герпетофауны страны (Терентьев, Чернов, 1949). Все отмеченные в регионе
виды амфибий и рептилий были описаны, а их выявленные к этому времени в
общих чертах ареалы проиллюстрированы картами распространения в «Опреде-
лителе...» П.В. Терентьева и С.А. Чернова (1949). Эта книга наряду с первыми
изданиями (1936,1940) некоторое время была наиболее полной для территории
СССР сводкой по указанным группам животных, заменив отчасти устаревшие
работы А.М. Никольского (Терентьев, 1957; Гаранин, Бакиев, 2004).
Своеобразным рубежом в эколого-фаунистических исследованиях репти-
лий и амфибий региона стала вторая половина 40-х и 50-е гг. XX столетия.
Результаты исследований этого времени на Урале обобщены в сводке «Жи-
вотный мир Урала» (Шварц и др., 1951), ряде статей и соответствующих раз-
делах физико-географических очерков Чкаловской (Оренбургской) области
(Райский, 1951, 1956), Бузулукского бора (Даркшевич, 1953, 1956), Среднего
Поволжья (Мильков, 1953), Куйбышевской области (Шиклеев, 1951) и Ниж-
него Поволжья (Нижнее Поволжье..., 1948). Материалы, собранные участни-
ками комплексных экспедиций АН СССР в Волго-Уральском междуречье, по
среднему и нижнему течению р. Урал, а также в Тургайской области и При-
аралье, отражены в обзорных работах Л.Г. Динесмана (1953) и С.А. Чернова
(1954). В это же время значительную коллекцию пресмыкающихся на Ман-
гышлаке и сопредельной территории собрали И.А. Долгушин, К.П. Параскив,
Д.И. Чекменев; в бассейнах рек Тургая и Джиланчика, песках Б. Барсуки, на се-
веро-западном берегу Аральского моря и вблизи г. Уральска - В.С. Бажанов и
М.Д. Бирюков; в междуречье Волга-Урал - участники комплексной экс-
педиции Института зоологии АН КазССР (П.М. Бутовский, А.И. Агапова,
М.С. Шакирзянова, Ф.Л. Евдаков, Б.В. Муханов), а в окрестностях оз. Шал-
кар - Серов Н.П. Все эти материалы обобщены в монографиях «Пресмыкаю-
щиеся Казахстана» (Параскив, 1956), «Земноводные Казахстана» (Искакова,
1959), региональном очерке по Западному Казахстану (Параскив, Бутовский,
I960), а по Заволжью - приводятся в работах Б.С. Кубанцева и др. (1962) и
К.С. Ходашовой (1960).
Очерки фауны амфибий и рептилий степной и лесной зон страны приводят-
ся также в III и IV томах «Животного мира СССР» (Чернов, 1950; 1953; Терен-
тьев, 1953), а некоторые сведения о распространении и особенностях экологии
затрагиваются в очерке о жизни прыткой ящерицы (Даревский, 1946), «Очер-
ках по биологии земноводных» (Банников, Денисова, 1956), соответствующих
разделах монографии «Лягушка» (Терентьев, 1950, 1957) и учебного пособия
«Герпетология» (Терентьев, 1961). Важную роль в унификации последующих
исследований сыграла работа Л.Г. Динесмана и М.П. Калецкой (1952) с обзо-
ром и методическими указаниями по количественному учету земноводных и
пресмыкающихся.
Значительным расширением масштабов батрахологических и герпетоло-
гических исследований в стране характеризуются 60-70-е гг. XX в. При этом
в ряде районов наряду с уточнением состава фауны и границ распространения
отдельных видов (особенно на периферии ареалов) существенно расширились
экологические исследования. Большинство экологических работ этого периода
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ...
55
посвящено определению численности фоновых видов, рассмотрению их цено-
тических связей, биотопического распределения, а также изучению структуры
и динамики их популяций, особенностей использования ими разных биотопов,
их развития и питания.
Все более популярными становятся исследования регуляции жизненного
цикла, межвидовых отношений, питания, роста и развития. При этом начинают
широко применяться математические методы, амфибии и рептилии все чаще
используются как модельные объекты при физиологических, биохимических,
морфологических и других экспериментах. Определенную роль в разверты-
вании этих работ сыграл организованный в 1960-х гг. Герпетологический ко-
митет СССР и созываемые им Всесоюзные герпетологические конференции.
К этому времени сформировались батрахологические и герпетологичес-
кие центры в ЗИНе АН СССР, ИПЭЭ АН СССР, ИЭ УрО АН СССР, ИЗ АН
КазССР, группы в Казанском, Куйбышевском, Нижегородском, Саратовском
университетах и системе противочумных учреждений.
Одними из первых работ этого периода стали статьи В.И. Гаранина (1964 а, б)
об изменении численности земноводных востока европейской части СССР,
их распространении, численности и динамике структуры их ареала в регионе.
В дальнейшем по результатам более чем 20-летних наблюдений этим автором
прослежена динамика численности ряда видов (в том числе на северной окраине
нашего региона), предпринята попытка выявить основные факторы, влияющие
на численность ряда видов амфибий и рептилий, а также объяснить механизмы
их действия (Гаранин, 1971,1977). В материалах I Всесоюзной герпетологичес-
кой конференции (Ленинград, 1964 г.) публикуются материалы С.А. Шиловой
(1964) о питании обыкновенной чесночницы в Бузулукском бору.
Материалы по фауне, распространению и экологии ряда амфибий и репти-
лий приводятся также в IV томе «Жизни животных» (Банников, 1969), а затем
в справочнике-определителе «Земноводные и пресмыкающиеся СССР» (Бан-
ников и др., 1971), снабженном картами с уточненными границами ареалов
видов, в том числе обитающих на территории Урало-Каспийского региона.
Территория нашего региона затрагивается в монографии «Ящурки Палеар-
ктики» (Щербак, 1974), поскольку здесь проходят границы распространения
ряда видов и подвидов этой группы рептилий (в частности отмечается отсутс-
твие или скудность данных по Самарской и Оренбургской областям). Матери-
алы по региону имеются также в одной из первых видовых монографий (Прыт-
кая ящерица, 1964), подготовленной в рамках Международной биологической
программы (IBP) и программы «Человек и биосфера» (МАВ): кадастр встреч
вида, в частности, включает около 40 точек на территории Урало-Каспия.
Большинство ставших известными к этому времени мест встреч амфибий и
рептилий нашло отражение на картах, иллюстрирующих распространение ви-
дов в «Определителе земноводных и пресмыкающихся фауны СССР» (1977).
Сведения об некоторых особенностях экологии амфибий и рептилий, обитаю-
щих на территории Саратовской области, приводятся Р.А. Девишевым (1976),
а о 10 видах амфибий и рептилий, нуждающихся в особой охране на террито-
рии Куйбышевской (Самарской) области, сообщается в статье В.М. Шапош-
никова (1978). В это же время начато систематическое изучение герпетофауны
Северо-Восточного Прикаспия сотрудниками Нижегородского университета
(В.В. Неручевым). Попутно герпетологические материалы в разных частях
56
Глава 3
региона собирали также сотрудники Института зоологии АН КазССР и про-
тивочумной службы Э.М. Ауэзов, А.П. Гисцов, Б.М. Губин, Е.С. Ташибаев,
Р.Т. Шаймарданов, А.С. Климов, О.В. Панкратов, С.И. Смирнов.
Временем наиболее интенсивного развития в истории батрахо- и герпето-
логических исследований страны в XX в. стали 80-е гг., хотя часть резуль-
татов публиковались уже в 1990-е гг. - в «постсоветский» (по Л.Я. Боркину,
2003) период, когда условия для деятельности научных учреждений и вузов
заметно ухудшились. В работах этого этапа большое внимание уделяется изу-
чению состояния популяций разных видов амфибий и рептилий, динамике их
численности, обсуждаются роль некоторых видов в экосистемах, возможности
их использования и проблемы сохранения в антропогенных ландшафтах (Ки-
реев, 1981, 1984; Неручев и др., 1984, 1985, 1986, 1989, 1990; Горелов, 1990;
Березовиков и др., 1992; и др.). Тогда же выходит ряд обобщающих сводок по
фауне отдельных частей региона и сопредельных территорий (Гаранин, 1983;
Щербак, Голубев, 1986), совершенствуется также «Методика полевых иссле-
дований экологии амфибий и рептилий» (Шляхтин, Голикова, 1986).
В духе этих идей и традиций продолжились батрахо- и герпетологические
исследования в регионе и в 1990-е гг., хотя в начале десятилетия их масштабы
заметно сократились, поскольку нарушились связи по линии научных учреж-
дений и между вузами. На этом фоне особую значимость приобрели инициати-
ва Института экологии Волжского бассейна РАН по проведению конференции
герпетологов Поволжья в 1995 г, открывшая цикл герпетологических конфе-
ренций на базе ИЭВБ РАН с публикацией сборников их материалов (1995,
1999 г.), а также организация издания сборников научных трудов «Актуальные
проблемы герпетологии и токсинологии», в которых приняли участие специ-
алисты как центральных научных учреждений, так и региональных групп из
Тольятти, Саратова, Астрахани и Оренбурга (всего в этих изданиях опублико-
вано более 30 работ по фауне, распространению, экологии и некоторым про-
блемам охраны амфибий и рептилий региона).
Подобные материалы публикуются в бюллетене «Самарская лука», а с
1997 г. - в сборниках молодых ученых «Проблемы общей и прикладной эколо-
гии», издаваемых Саратовским университетом. Батрахо- и герпетологические
сведения по региону содержатся также в материалах ряда научных симпозиу-
мов, в том числе «Степи Северной Евразии», «Проблемы охраны и рациональ-
ного использования природных экосистем и биологических ресурсов», а также
в «Вестнике Зоологии» и других периодических изданиях.
По ряду областей региона обобщаются материалы по амфибиям и рептили-
ям в связи с подготовкой и последующим изданием Красных книг: Саратовс-
кой в 1996 г.; Волгоградской - 1996; Оренбургской - 1998; Самарской - 1999,
а также Казахстана - 1991, 1996 г. Сведения о распространении и некоторых
особенностях экологии ряда видов амфибий и рептилий региона приводятся
также в обзорных сводках «Земноводные и пресмыкающиеся» (Ананьева и
др., 1998), «Земноводные бывшего СССР» (Кузьмин, 1999), «Ящерицы пус-
тынь Казахстана» (Брушко, 1995).
Важным событием в истории изучения амфибий и рептилий региона
(и всей страны) стал созыв Первого съезда Герпетологического общества
им. А.М. Никольского, открывшего цикл симпозиумов, выполняющих
роль своеобразных координаторов батрахо- и герпетологических исследо-
ИСТОРИЯ ИССЛЕДОВАНИЙ ФАУНЫ АМФИБИЙ И РЕПТИЛИЙ...
57
ваний страны. В издании материалов этого, затем второго (совмещенного с
XII съездом Европейского герпетологического общества) и последующего
съездов приводится ряд сведений по фауне и экологии низших тетрапод ре-
гиона. Практически в это же время в Саратовском университете организовано
издание сборников научных трудов «Современная герпетология», основу ко-
торых составляют региональные материалы.
Наряду с этим продолжалось издание сборников научных трудов «Акту-
альные проблемы герпетологии и токсинологии», сборников трех конферен-
ций герпетологов Поволжья (из которых третья прошла как Международная
конференция «Змеи Восточной Европы»), а также бюллетеня «Самарская
лука», содержащих современную информацию о систематике, распростране-
нии и образе жизни видов, обитающих в нашем регионе.
Сведения по разным аспектам затрагиваемых проблем публиковались также
в материалах ряда международных симпозиумов: «Степи Северной Евразии»
(2000, 2003, 2006, 2009 г.), «Проблемы изучения и охраны биоразнообразия»
(2001 г.), Международной молодежной конференции (2003 г.), межвузовских и
ведомственных сборниках «Фауна и экология животных» (2002 г.), «Природное
наследие России» (2004 г.), сборниках региональных и вузовских конференций
«Самарский край в истории России» (2001 г.), «Животный мир Южного Урала
и Северного Прикаспия» (2000 г.), «Экологические проблемы бассейнов круп-
ных рек», «Биоразнообразие и биоресурсы Среднего Поволжья и сопредельных
территорий» (2002 г.), Волжского и Оренбургского университетов, периоди-
ческих изданиях («Герпетологический вестник», «Поволжский экологический
журнал», «Selevinia»), а также в некоторых краеведческих работах.
Батрахо- и герпетологические материалы обобщались в ряде публикаций,
посвященных подготовке к изданию Красных книг: Красной книги РФ (2001),
Самарской (2009), Саратовской (2006), Волгоградской (2004), Астраханской об-
ластей (2004) РФ и Западно-Казахстанской области (Дебело, Булатова, 1999).
Развитию исследований рептилий и амфибий способствовало также из-
дание учебных пособий: «Герпетология» (2001, 2002 гг.), «Земноводные и
пресмыкающиеся Самарской области» (2001 г.), «Земноводные и пресмыка-
ющиеся Самарской и Ульяновской областей» (2002 г.), «Животный мир Сара-
товской области» (2005 г.), «Животный мир Казахстана» (2005 г.), а также по
ряду сопредельных территорий.
Из обобщающих работ этого периода наиболее значительными являются
«Земноводные и пресмыкающиеся» (Ананьева и др., 1998), «Атлас пресмы-
кающихся Северной Евразии» (2004), «Конспект фауны земноводных и пре-
смыкающихся России» (Кузьмин, Семенов, 2006), «Змеи Волжско-Камского
края» (Бакиев и др., 2004), «Змеи Самарской области» (Бакиев и др., 2009),
региональные сводки «Материалы к кадастру амфибий и рептилий бассейна
Средней Волги» (2002) и «Амфибии и рептилии Урала» (Вершинин, 2007).
В настоящее время группы специалистов или отдельные зоологи, изучаю-
щие амфибий и рептилий (в том числе попутно), имеются почти во всех рос-
сийских областях региона и Казахстана. Их основные публикации приведены
в библиографическом списке, хотя, к сожалению, часть работ оказалась недо-
ступной авторам и не все они упомянуты в настоящем обзоре.
Глава 4
ОБЩАЯ ХАРАКТЕРИСТИКА
И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ И РЕПТИЛИЙ
УРАЛО-КАСПИЙСКОГО РЕГИОНА
В соответствии с принятой в настоящее время системой земноводных и
пресмыкающихся России их современные представители включаются в
следующую классификацию:
Царство Annimalia - Животные
Тип Chordata - Хордовые
Подтип Craniota - Черепные или Vertebrata - Позвоночные
Класс Amphibia - Земноводные
Класс Reptilia - Пресмыкающиеся.
С учетом представлений о номенклатуре и таксономическом статусе той
или иной систематической группы современных земноводных региона отно-
сят к 12 видам, пресмыкающихся - к 38. Среди амфибий 5 видов являются
монотипическими и 7 - политипическими, представленными 6 номинативны-
ми и 1-2 (номинативным и локальным) подвидами, которые объединяются в
8 родов и 7 семейств, относящимися к 2 отрядам. Среди них подавляющее
большинство (9) видов относится к отряду бесхвостых, на долю которых при-
ходится 66,6% общего видового состава этой группы низших тетрапод реги-
она. В числе современных пресмыкающихся 11 видов - монотипические и
27 - политипические, представленные 18 номинативными, 5 локальными и
4 (номинативными и локальными) подвидами, которые объединяются в
24 рода и 10 семейств, отнесенных к 3 отрядам (табл. 10, 11).
Порядок расположения таксонов в классах, их объем и названия приня-
ты по сводке D.R. Frost (2002) с учетом изменений и дополнений для амфи-
Табяица 10
Таксономический и количественный состав батрахо- и герпетофауны
Урало-Каспийского региона
Класс Отряд Кол-во видов в Кол-во в регионе
мировой фауне Российской Федерации видов родов семейств
Земновод- Хвостатые 400 6 3 3 2
ные Бесхвостые 4000 22 9 5 5
Итого 2 4500 28 12 8 7
Черепахи 250 6 2 2 2
Пресмыка- ющиеся Ящерицы 4300 99 20 12 5
Змеи 3000 71 16 10 3
Итого 3 7000 176 38 24 10
Всего 5 11500 204 50 32 17
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
59
Таблица 11
Перечень видов земноводных и пресмыкающихся Урало-Каспийского региона
Класс I. AMPHIBIA LINNAEUS, 1758 - ЗЕМНОВОДНЫЕ
Отряд I. CAUDATA OPPEL, 1811— ХВОСТАТЫЕ
Сем. 1. Hynobiidae Соре, 1860-Углозубовые
Род 1. Salamandrella Dybowski, 1870 - сибирские углозубы
1. Salamandrella keyseriingii (Dybowski, 1870) - сибирский углозуб
Сем. 2. Salamandridae Goldfuss, 1820-Саламандровые
Род 2. Lissotriton Bell, 1839 - лиссотритоны, тритоны
2. Lissotriton vulgaris (Linnaeus, 1758) - обыкновенный тритон
Номинативный подвид L. vulgaris vulgaris (Linnaeus, 1758)
Род 3. Triturus Rafinesque, 1815 - тритоны
3. Triturus cristatus (Laurenti, 1768) - гребенчатый тритон
Отряд IL ANURA RAFINESQUE, 1815 - БЕСХВОСТЫЕ
Сем. 3. Bombinatoridae Gray, 1825-Жерлянки
Род 4. Bombina Oken, 1816 - жерлянки
4. Bombina bombina (Linnaeus, 1761) - краснобрюхая жерлянка
Сем. 4. Pelobatidae Bonaparte, 1850-Чесночницы
Род 5. Pelobates Wagler, 1830 - чесночницы
5. Pelobates fuscus (Laurenti, 1768) - обыкновенная чесночница
Сем. 5. Bufonidae Gray, 1825 - Жабы
Род 6. Bufo Laurenti, 1768 - жабы
6. Bufo bufo (Linnaeus, 1758) - обыкновенная жаба
7. Bufo viridis (Laurenti, 1768) - зеленая жаба
Сем. 6. Hylidae Gray, 1825 - Квакши
Род 7. Hyla Laurenti, 1768 - квакши
8. Hyla arborea (Linnaeus, 1758) - обыкновенная квакша
Сем. 7. Ranidae Rafinesque, 1814-Лягушки
Род 8. Rana Linnaeus, 1758 - лягушки
9. Rana temporaria (Linnaeus, 1758) - травяная лягушка
10. Rana arvalis (Nilsson, 1842) - остромордая лягушка
* Rana esculenta (Linnaeus, 1758) - съедобная лягушка
11. Rana lessonae (Camerano, 1882) - прудовая лягушка
12. Rana ridibunda (Pallas, 1771) - озерная лягушка
Класс II. REPTILIA LINNAEUS, 1758-ПРЕСМЫКАЮЩИЕСЯ
Отряд III. TESTUDINES BATSCH, 1788 - ЧЕРЕПАХИ
Сем. 8. Emydidae Rafinesque, 1815-Американские пресноводные черепахи
Род 9. Emys Dumeril, 1806 - болотные черепахи
13. Emys orbicularis (Linnaeus, 1758) - болотная черепаха
Номинативный подвид Е. о. orbicularis Linnaeus, 1758
Сем. 9. Testudinidae Batsch, 1788 - Сухопутные черепахи
Род 10. Agrionemys Khozatsky et Mlynarski, 1966 - среднеазиатские черепахи
14. Agrionemys horsfieldii (Gray, 1844) - среднеазиатская черепаха
Подвид A. h. kasakhstanica Chkhikvadze, 1988 -казахстанская черепаха
Надотряд Squamata, Oppel, 1811 - Чешуйчатые
Отряд IV. S AURIA MCCARTHNEY, 1822 - ЯЩЕРИЦЫ
Сем. 10. Gekkonidae Gray, 1825-Гекконы, или цепкопалые
Род 11. Alsophylax Fitzinger, 1843 - североазиатские геккончики
15. Alsophylaxpipiens (Pallas, [1814]) - пискливый геккончик
60
Глава 4
Продолжение табл. 11
Род 12. Crossobamon Boettger, 1888 - гребнепалые гекконы
16. Гребнепалый геккон - Crossobamon eversmanny (Wiegmann, 1834)
Род 13. Cyrtopodion Fitzinger, 1843 -тонкопалые гекконы
17. Cyrtopodion caspius (Eichwald, 1831)- каспийский геккон
Номинативный подвид С. с. caspius (Eichwald, 1831)
Род 14. Mediodactylus Szczerbak et Golubev, 1977 - средиземноморские тонкопалые гекконы
18. Mediodactylus russowii (Strauch, 1887) - серый геккон
Род 15. Teratoscincus Strauch, 1863 - сцинковые гекконы
19. Teratoscincus scincus (Schlegel, 1858) - сцинковый геккон
Номинативный подвид Т. s. scincus (Schlegel, 1858)
Сем. 11. Agamidae Spix, 1825 - Агамовые
Род 16. Trapelus Cuvier, 1817 - равнинные агамы
20. Trapelus sanguinolentus (Pallas, [1814]) - степная агама
Род 17. Phrynocephalus Каир, 1825 - круглоголовки
21. Phrynocephalus guttatus (Gmelin, 1789) - круглоголовка-вертихвостка
Номинативный подвид Ph. g. guttatus (Gmelin, 1789)
22. Phrynocephalus helioscopus (Pallas, 1771)-такырная круглоголовка
Номинативный подвид Ph. h. helioscopus (Pallas, 1771)
23. Phrynocephalus interscapularis Lichtenstein, 1856 - песчаная круглоголовка
24. Phrynocephalus mystaceus (Pallas, 1776) - ушастая кругологоловка
Номинативный подвид Ph. т. mystaceus (Pallas, 1776)
Подвид Ph. m. galli Krasowsky, 1932
Сем. 12. Anguidae Gray, 1825-Веретеннциевые
Род 18. Angius Linnaeus, 1758 - веретеницы
25. Anguis fragilis Linnaeus, 1758 - веретеница ломкая
Номинативный подвид A.f. fragilis Linnaeus, 1758
♦Род 19. Pseudopus Merrem, 1820 - панцирные веретеницы
♦26. Pseudopus apodus (Pallas, 1775) - желтопузик, или глухарь
Номинативный подвид Р. a. apodus (Pallas, 1775)
Сем. 13. Scincidae Oppel, 1811 - Сцинковые
Рбд 20. Ablepharus Fitzinger in Eversmann, 1823 - гологлазы
27. Ablepharus deserti Strauch, 1876 - пустынный гологлаз
Сем. 14. Lacertidae Bonaparte, 1831 - Настоящие ящерицы
Род 21. Eremias Fitzinger in Wiegmann, 1837 - ящурки
28. Eremias arguta (Pallas, 1773) - разноцветная ящурка
Номинативный подвид E.a.arguta (Pallas, 1773)
29. Eremias lineolata Nikolsky, 1896 - линейчатая ящурка
30. Eremias intermedia (Strauch, 1876) - средняя ящурка
31. Eremias grammica (Lichtenstein, 1823) - сетчатая ящурка
32. Eremias scripta (Strauch, 1867) - песчаная, или полосатая ящурка
Номинативный подвид Е. s. scripta (Strauch, 1867)
33. Eremias velox (Pallas, 1771) - быстрая ящурка
Номинативный подвид Е. v. velox (Pallas, 1771)
Род 22. Lacerta Linnaeus, 1758 - настоящие ящерицы
34. Lacerta agilis Linnaeus, 1758 - прыткая ящерица
Подвид L. a. exigua Eichwald, 1831 - восточная прыткая ящерица
Род 23. Zootoca Wagler, 1830 - лесные ящерицы
35. Zootoca vivipara Jacquin, 1787 - живородящая ящерица
Номинативный подвид Z. v. vivipara Jacquin, 1787
Отряд V. SERPENTES LINNAEUS, 1758-ЗМЕИ
Сем. 15. Boidae Gray, 1825-Ложноногие, или Удавы
Род 24. Eryx Daudin, 1803 - удавчики
36. Eryx miliaris (Pallas, 1773) - песчаный удавчик
Номинативный подвидЕ т. miliaris (Pallas, 1773)
37. Eryx tataricus (Lichtenstein, 1823) - восточный удавчик
Номинативный подвид Е. t. tataricus (Lichtenstein, 1823)
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
61
Окончание табл. 11
Сем. 16. Colubridae Oppel, 1811 -Ужеобразные
Род 25. Coluber Linnaeus, 1758 - стройные полозы
38. Coluber karelinii Brandt, 1838 - поперечнополосатый полоз
Номинативный подвид С. k. karelinii Brandt, 1838
39. Coluber ravergieri Menetries, 1832 - разноцветный полоз
Род 26. Coronella Laurenti, 1768 - медянки
40. Coronella austriaca (Laurenti, 1768) - обыкновенная медянка
Номинативный подвид С. a. austriaca (Laurenti, 1768)
Род 27. Elaphe Fitzinger In Wagler, 1833 - лазающие полозы
41. Elaphe dione (Pallas, 1773) - узорчатый полоз
42. Elaphe sauromates (Pallas, [1814]) - сарматский или паласов полоз
Род 28. Hierophis Fitzinger In Bonaparte, 1832 - гиерофисы
43. Каспийский, или желтобрюхий полоз - Hierophis caspius (Gmelin, 1779)
Род 29. Malpolon Fitzinger, 1826 - ящеричные змеи
44. Malpolon monspessulanus (Hermann, 1804) - ящеричная змея
Подвид М. т. insignitus (Geoffroy, 1809)
Род 30. Natrix Laurenti, 1768 - настоящие ужи
45. Natrix natrix (Linnaeus, 1758) - обыкновенный уж
ПодвидУ п. scutata (Pallas, 1771)
46. Natrix tessellata (Laurenti, 1768) - водяной уж
Род 31. Psammophis Boie In Fitzinger, 1826 - песчаные змеи
47. Psammophis lineolatus (Brandt, 1838) - стрела-змея
Сем. 17. Viperidae Laurenti, 1768 - Гадюковые
Род 32. Gloydius Hoge Romano-Hoge, 1981 - щитомордники
48. Gloydius halys (Pallas, 1776) - обыкновенный щитомордник, или щитомордник
Палласа
Подвид G. h. carasanus (Eichwald, 1831)
Род 33. Vipera Laurenti, 1768 - гадюки
49. Vipera (Pelias) berus (Linnaeus, 1758)-обыкновенная гадюка
Номинативный подвид V. (Pelius) b. berus (Linnaeus, 1758)
50. Vipera nikolskii Vedmederja, Grubant et Rudaeva, 1986 - гадюка Никольского, или
лесостепная гадюка
51. Vipera (Pelias) renardi (Christoph, 1861) - восточная степная гадюка
* Сомнительный таксон.
бий, предложенных С.Л. Кузьминым, Д.В. Семеновым (2006), и рептилий -
Н.Б. Ананьевой с соавт. (Атлас пресмыкающихся..., 2004).
В повидовых очерках принята следующая рубрикация:
- латинское (научное) название с указанием автора, года описания и обще-
принятое русское название;
- значение таксона для сохранения биоразнообразия, его структура;
- распространение;
- места обитания;
- образ жизни;
- питание, биоценотическая роль;
- размножение;
- численность, тенденции ее изменения;
- основные лимитирующие факторы;
- охрана;
- региональный статус.
Обобщенные сведения о распостранении и численности населяющих тер-
риторию региона видов и подвидов по разным ландшафтным провинциям и
округам приведены в табл. 12.
Таблица 12
Видовой состав, распространение и численность амфибий и рептилий Урало-Каспийского региона
№ п.п. Таксон ФК Провинции и округа
I II III IV V VI VII VIII IX X XI XII ХШ
a а б а б а б а б а а б в а б в а б в а б а б в а а
1 Кл. Amphibia Отр. Caudata Сем. Hinobiidae Сибирский углозуб - Salamandrella keyseriingii (Dub., 1870) S. к. keyser- iingii EC п п ?
2 Сем. Salamandridae Обыкновенный тритон - Lissotriton vulgaris (L., 1758) L. v. vulgaris (L., 1758) E МЛ МЛ - — - - — мл п? -
3 Гребенчатый тритон - Triturus cristatus (Laur., 1768) E P Р — - р — - р р -
4 Отр. Anura Сем. Bombinatoridae Краснобрюхая жерлянка - Bombina bombina (L., 1761) E МЛ МН О о О мл мл мл мл МЛ МЛ О мл МЛ О мл МЛ мл о О О О О О - МЛ
5 Сем. Pelobatidae Обыкновенная чесноч- ница- Pelobates fuscus (Laur., \15%) P.f fuscus (Laur., 1768) E ОБ ОБ мл О О ОБ мл ОБ ОБ ОБ ОБ мл ОБ О ОБ ОБ О 4- О ОБ 4- о О 4- 4- ОБ
6 Сем. Bufonidae Обыкновенная жаба - Bufo bufo (L., 1758) E МЛ МЛ мл мл мл 4- О МЛ МЛ МЛ О о МЛ 6 О МЛ -
Глава 4
7 Зеленая жаба - Bufo viridis (Laur., 1768) CP ОБ ОБ ОБ ОБ 4- ОБ ОБ ОБ ОБ МН ОБ МЛ МЛ МН ОБ мн ОБ ОБ мл ОБ ОБ мл мл - - ОБ
В. v. viridis (Laur., 1768) В. v. turanensis (Hem., 1978) - ОБ мл -
8 Сем. Hylidae Обыкновенная квакша - Hyla arborea (L., 1758) H. a. arborea (L., 1758) E П
9 Сем. Ranidae Травяная лягушка - Rana temporaria (L., 1758) Л t. temporaria (L., 1758) E ? ? 0 О О - - Р Р р — - - - - р? - П — - — - - - — -
10 Остромордая лягушка - Rana arvalis (Nil., 1842) R. a. arvalis (Nil., 1842) EC MH MH 0 МН МН МН ОБ ОБ МН ОБ ОБ мл ОБ ОБ МН мн мн ОБ О МН ОБ О О ОБ - мл
11 Прудовая лягушка - Rana lessonae (Cam., 1882) E p ? Р -
12 Озерная лягушка - Rana ridibunda (Pal., 1771) CP MH MH ОБ ОБ ОБ мн ОБ МН МН МН МН ОБ ОБ МН мн мн ОБ ОБ ОБ мн Об ОБ О ОБ О мн
13 Кл. Reptilia Отр. Testudines Сем. Emididae Болотная черепаха - CP МЛ МЛ МЛ МЛ МЛ ОБ МЛ МН МН мн ОБ МН ОБ ОБ мн ОБ ОБ МЛ О мл мн МЛ О МЛ О мл
Emys orbicularis (L., 1758) E. o. orbicularis (L., 1758)
14 Сем. Testudinidae Среднеазиатская черепаха - Agrionemys horsfieldii (Gray, 1844) A. h. kasakhstanica (Ckhikv., 1988) CA - ? ? ? - - — — - - - п — - мл - -
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИИ...
Продолжение табл. 12
№ п.п. Таксон ФК Провинции и округа
XIII XIV XV XVI xvn xvin XIX XX XXI ххп XXIII XXIV XXV XXVI
б в а а б а а а а а б в г д а б а б в а б а а а б в
1 Кл. Amphibia Отр. Caudata Сем. Hinobiidae Сибирский углозуб - Salamandrella keyseringii (Dib., 1870) S. к. keyserlingii EC -
2 Сем. Salamandridae Обыкновенный тритон - Lissotriton vulgaris (L., 1758) L. v. vulgaris (L„ 1758) E -
3 Гребенчатый тритон - Triturus cristatus (Laur., 1768) E -
4 Отр. Anura Сем. Bombinatoridae Краснобрюхая жерлянка - Bombina bombina (L., 1761) E + 4-
5 Сем. Pelobatidae Обыкновенная чесноч- ница- Pelobates fuscus (Laur., 1758) P.f fuscus (Laur,. 1758) E + ОБ О О 4- О О 4- 4- ОБ О О О ОБ О 4- О О о -
6 Сем. Bufonidae Обыкновенная жаба - Bufo bufo (L., 1758) E - — — 4- 4- -
Глава
7 Зеленая жаба - Bufo viridis Laur., 1768 В. v. viridis (Laur., 1768) CP MH MH МЛ мл мл 00 00 - - ОБ МЛ 0 0 ОБ - ОБ 0 мл мл -
В. v. turanensis (Hem. 1978) - - - - — ОБ ОБ ОБ 0 ОБ
8 Сем. Hylidae Обыкновенная квакша - Hyla arborea (L., 1758) H a. arborea (L., 1758) E
9 Сем. Ranidae Травяная лягушка- Rana temporaria (L., 1758) R. t. temporaria (L., 1758) E
10 Остромордая лягушка- R. arvalis (Nils., 1842) R. a. arvalis (Nils., 1842) EC ОБ ОБ мл
11 Прудовая лягушка - R. lessonae (Cam., 1882) E -
12 Озерная лягушка- R. ridibunda (Pal., 1771) CP MH MH ОБ ОБ ОБ мл 0 мл мл МН 0 — ОБ МН - МН — мл ОБ мл - - - МЛ — 7
13 Кл. Reptilia Отр. Testudines Сем. Emydidae Болотная черепаха- Emys orbicularis (L., 1758) E. o. orbicularis (L., 1758) CP ОБ MH 0 МЛ МЛ мл 0 мл ОБ ОБ 0 0 7 мн мл ОБ 0 мл мл 0 мл 0 мл - МЛ
14 Сем. Testudinidae Среднеазиатская чере- паха- Agrionemys horsfieldii (Gray, 1844) A. h. kasakhstanica (Ckhikv., 1988) CA - — МЛ мл О п 0 мл МЛ - — - - - мл - — 0 мл мл + мл ОБ ОБ ОБ МЛ
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
Продолжение табл. 12
№ п.п. Таксон ФК Провинции и округа
I П III IV V VI VII VIII IX X XI XII ХШ
a a 6 a 6 a 6 a 6 a a 6 в a б в А б в а б а б в а а
15 Огр. Sauria Сем. Gekkonidae Пискливый геккончик - Alsophylax pipiens (Pal., 1814) CA П 9 ?
16 Гребнепалый геккон - Crossobamon eversmanny (Wiegmann, 1834) C. e. eversmanny (W., 1834) CA
17 Каспийский геккон - Cyrtopodion caspius (Eichw., 1831) C. c. caspius (Eichw., 1831) CA
18 Серый геккон - Mediodactylus russowii (Str., 1887) M. r. russowii (Str., 1887) CA
19 Сцинковый геккон - Teratoscincus scincus (Schl., 1858) T. s. scincus (Schl., 1858) CA
20 Сем. Agamidae Степная агама - Trapelus sanguinolentus (Pal., 1814) T. s. aralensis (Licht., 1823) CA -
21 Круглоголовка-вертихвос- тка- Phrynocephalus guttatus (Gm., 1789) Ph. g. guttatus (Gm., 1789) ЦА - МЛ MH ОБ ОБ - МЛ
22 Такырная круглоголовка- Ph. helioscopus (PaL, 1771) Ph. h. helioscopus (Pal., 1771) CA - п ОБ
Глава 4
23 Песчаная круглоголовка - Ph. interscapularis (Licht., 1856) CA -
24 Ушастая круглоголовка- Ph. mystaceus (Pal., 1776) CA -
Ph. m. mystaceus (Pal., 1776) Ph. m. galli (Krasow, 1932) -
25 Сем. Anguidae Веретеница ломкая - Anguis fragilis (L., 1758) A.f.fragilis (L„ 1758) E МЛ p 0 p p - - p P —
26 Сем. Scincidae Пустынный гологлаз - Ablepharus deserti (Str., 1876) CA -
27 Сем. Lacertidae Разноцветная ящурка- Eremias arguta (Pal., 1773) CA МЛ - - - p p P p - —
E. a. deserti E. a. arguta (Pal., 1773) - ОБ
28 Сетчатая ящурка - E. grammica (Licht, 1823) CA -
29 Линейчатая ящурка - E. lineolata (Nikol., 1896) CA -
30 Средняя ящурка - E. intermedia (Str., 1876) CA -
31 Песчаная ящурка- E. scripta (Str., 1867) E. s. scripta (Str., 1867) CA -
32 Быстрая ящурка - £ .ve/ох (Pal., 1771) E. v. ve/ox(Pal., 1771) CA -
33 Прыткая ящерица - Lacerta agilis (L., 1758) L. a. exigua (Eichw., 1831) EC MH ОБ 0 0 + ОБ MH ОБ ОБ +
ОБ ОБ -
— МЛ — — р р — — — — — — — — ОБ
— — МЛ — — мл ОБ — — — ОБ ОБ МЛ — —
— мл - МЛ
ОБ ОБ ОБ МН О мн ОБ МН ОБ ОБ ОБ МЛ ОБ ОБ ОБ ОБ
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
Продолжение табл. 12
№ п.п. Таксон ФК Провинции и округа
XIII XIV XV XVI XVII XVIII XIX XX XXI ххп XXIII XXIV XXV XXVI
б в а а б а А а а а б в г д а б а б в а б а а а б в
15 Отр. Sauria Сем. Gekkonidae Пискливый геккончик - Alsophylax pipiens (Pal., 1814) CA - - — МЛ ОБ - - МЛ МЛ ОБ - - - — — - - - ОБ — - - ОБ — ОБ П
16 Гребнепалый геккон - Crossobamon eversmanny Wiegmann, 1834 C. e. eversmanny (1834) CA п МЛ МЛ МЛ МЛ
17 Каспийский геккон - Cyrtopodion caspius (Eichw., 1831) C. c. caspius (Eichw., 1831) CA - МЛ МЛ О О мл мл мл МЛ
18 Серый геккон - Mediodactylus russowii (Str., 1887) M. r. russowii (Str., 1887) CA мл - мл мл — - мл мл О п
19 Сцинковый геккон - Teratoscincus scincus (Schl., 1858) Г. s. scincus (Schl., 1858) CA 4- + мл ОБ ОБ
20 Сем. Agamidae Степная агама - Trapelus anguinolentus (Pal., 1814) T. s. aralensis Licht., 1823 CA - - — - ОБ - — ОБ — — - — — - ОБ - — - ОБ ОБ - - ОБ ОБ ОБ ОБ
21 Круглоголовка-вертихвос- тка— Phrynocephalus guttatus (Gm., 1789) Ph. g. guttatus Gm., 1789 ЦА МЛ - О ОБ МЛ — — ОБ ОБ ОБ ОБ ОБ ОБ — ОБ — - МЛ ОБ - — - МЛ ОБ ОБ ОБ
22 Такырная круглоголовка- Ph. helioscopus Pal., 1771 Ph. h. helioscopus (Pal., 1771) CA - - — МН О О О ОБ ОБ ОБ ОБ ОБ МЛ - МН ОБ ОБ ОБ ОБ — ОБ О ОБ ОБ ОБ ОБ
Гпава 4
23 Песчаная круглоголовка - Ph. Interscapularis (Licht., 1856) CA П ОБ ?
24 Ушастая крулоголовка - Ph. mystaceus (Pal., 1776) CA - МН ОБ ОБ ОБ + -
Ph. m. mystaceus (Pal., 1776) Ph. m. galli (Krasow, 1932) — - — ОБ ОБ - - ОБ ОБ - - — - - - - - ОБ ОБ ОБ - - ОБ ОБ ОБ ОБ
25 Сем. Anguidae Веретеница ломкая - Anguisfragilis (L., 1758) A//ragi7w(L., 1758) E -
26 Сем. Scincidae Пустынный гологлаз - Ablepharus deserti (Str., 1876) CA - 4-
27 Сем. Lacertidae Разноцветная ящурка - Eremias arguta (Pal., 1773) CA МЛ МН + + + + + + -
E. a. deserti E. a. arguta (Pal., 1773) - — - ОБ ОБ МЛ - ОБ ОБ - + + + + + + ОБ Об МН О ОБ + ОБ ОБ ОБ ОБ
28 Сетчатая ящурка - E. grammica (Lie., 1823) CA МЛ - ОБ ОБ МН ОБ
29 Линейчатая ящурка - E. lineolata (Nikol., 1896) CA + —
30 Средняя ящурка - E. intermedia Str., 1876 CA - + - + + ОБ ОБ
31 Песчаная ящурка- E. scripta (Str., 1867) E. s. scripta (Str., 1867) CA + - + + + +
32 Быстрая ящурка - E. velox (Pal., 1771) E. v. velox (Pal., 1771) CA — - — МЛ МЛ - - — МЛ мл МЛ МЛ — — - — МЛ ОБ ОБ - МЛ - мл МЛ ОБ ОБ
33 Прыткая ящерица - Lacerta agilis (L., 1758) L. a. exigua Eichw., 1831 EC ОБ MH ОБ ОБ МЛ мл мл ОБ ОБ мн МЛ мл О мн мл ОБ МЛ ОБ МЛ О О МЛ О мл О МЛ
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
Продолжение табл. 12
№ п.п. Таксон ФК • Провинции и округа
I II III IV V VI VII VIII IX X XI ХП XIII
a a 6 a 6 a 6 a б a a 6 в a 6 в a 6 в a 6 a 6 в a A
34 Живородящая ящерица - Zootoca vivipara (Jac., 1787) EA - МЛ МЛ - - МЛ - ОБ МЛ - - - — - - ОБ МЛ мл -
35 Отр. Serpentes Сем. Boidae Песчаный удавчик - Eryx miliar is (Pal., 1773) Е. т. miliaris (Pal., 1773) CA - МЛ
36 Восточный удавчик - Е. tataricus (Licht., 1823) Е. t. tataricus (Licht., 1823) CA -
37 Сем. Colubridae Поперечнополосатый полоз - Coluber karelini (Вг., 1838) С. к. karelinii (Вг., 1838) CA -
38 Разноцветный полоз - С. ravergieri (Men., 1832) С. г. ravergieri (Men., 1832) CA -
39 Обыкновенная медянка - Coronella austriaca (Laur., 1768) С. a. austriaca (Laur, 1768) E P P 0 P 0 — p P p P P p
40 Узорчатый полоз - Elaphe dione (Pal., 1773) E. d. dione (Pal., 1773) ВП P - p - p МЛ МЛ P p + МЛ 4- 4- MH — — p p p — - - - - P ОБ
41 Сарматский полоз - E. sauromates (Pal., 1814) CM - P
Глава
42 43 Каспийский полоз - Hierophis caspius Gm., 1779 Ящеричная змея - Malpolon monspessulanus (Herm., 1804) M m. insignitus (Geof, 1827) CM CM мл
№ п.п. Провинции и округа
Таксон ФК XIII XIV XV XVI XVII xvni XIX XX XXI XXII ххш XXIV XXV XXVI
б в а а б а а а а а б в г д а б а Б в а б а а а б. в
34 Живородящая ящерица - Zootoca vivipara Jac, 1787 EA
35 Отр. Serpentes Сем. Boidae Песчаный удавчик - Eryx miliaris (Pal, 1773) E. m. miliaris (Pal., 1773) CA МЛ — - МЛ — - — - - МЛ мл ОБ мл МЛ МЛ МЛ МЛ МЛ ОБ - - - МЛ - мл мл
36 Восточный удавчик - E. tataricus (Licht., 1823) E. t. tataricus (Licht., 1823) CA п - МЛ О мн ОБ
37 Сем. Colubridae Поперечнополосатый полоз - Coluber karelinii Br., 1838 С. E karelini (Br., 1838) CA - п мл -
38 Разноцветный полоз - C. ravergieri (Men., 1832) C. r. ravergieri (Men., 1832) CA - п Р - Р
39 Обыкновенная медянка - Coronella austriaca (Laur., 1768) C. a, austriaca (Laur., 1768) E Р Р
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
П родолжение табл. 12
40 Узорчатый полоз - Elaphe dione (Pal., 1773) Е. d. dione (Pal., 1773) ВП MJI МЛ - p P P - p P МЛ МЛ МЛ мл мл мл мл МЛ МЛ мл - мл - — р — р
41 Сарматский полоз - E. sauromates (Pal., 1814) CM - p р мл р р — мл р р р
42 Каспийский полоз - Hierophis caspius (Gm., 1779) CM p р - р
43 Ящеричная змея - Malpolon monspessulanus (Herm, 1804) M. m. insignitus Geof. 1827 CM p -
№ п.п. Провинции и округа
Таксон ФК I II III IV V VI VII VIII IX X XI XII XIII
a a 6 a 6 a 6 A 6 a a б в а б в а б В а б а б в а А
44 Обыкновенный уж - Natrix natrix (L., 1758) E — - — — - MH МЛ мн -
N. n. natrix (L., 1758) N. n. scutata (Pal., 1771) MH MH ОБ ОБ ОБ — - ОБ ОБ ОБ ОБ мл мл - ОБ ОБ ОБ ОБ 4- мл ОБ МЛ О О О ОБ
45 Водяной уж - N. tessellate (Laur., 1768) CP MH — — — — ОБ - ОБ MH 0 МЛ О мл мл О ОБ О МЛ 0 О О — — — - МН
46 Стрела-змея - Psammophis lineolatus (Br., 1838) CA -
47 Сем. Viperidae Обыкновенный щито- мордник - Gloydius halys (Pal., 1776 G. h. carasanus (Eich, 1831) ВП - П п р -
48 Обыкновенная гадюка- Vipera berus (L., 1758) V b. berus (L., 1758) E MH 4- 0 4- 4- 4- -
49 Гадюка Никольского - V. nikolskii (Vedm., 1984) E ? ?
Глава 4
50 Степная гадюка - V. renardi (Chr., 1861) CP - ? + ? ? МЛ ОБ ОБ ОБ ОБ ОБ МН ОБ ОБ ОБ мл ОБ МН + + ОБ ОБ + + + ОБ
V. г. renardi (Chr., 1861) И г. bashkirovi (Gar., 2004) ОБ ? -
№ п.п. Провинции и округа
Таксон ФК ХШ XIV XV XVI XVII XVIII XIX XX XXI XXII ххш XXIV XXV XXVI
Б в а а б а а а а а б в г д а б а Б в а б а а а б в
44 Обыкновенный уж - Natrix natrix L., 1758 E -
N n. natrix (L., 1758) N n. scutata (PaL, 1771) МЛ ОБ МЛ МЛ МЛ МЛ О О О МН МЛ мл О ОБ О ОБ О МЛ мл - — - МЛ - - -
45 Водяной уж - N. tessellata (Laur., 1768) CP О МЛ - ОБ МЛ мл - - ОБ мн мл О О ОБ мл ОБ ОБ ОБ мн - О МЛ - - - мл
46 Стрела-змея - Psammophis lineolatus (Br., 1838) CA - МЛ П — - - - - мл МЛ - МЛ мл МЛ мл - МЛ МЛ мн ОБ
47 CeM.Viperidae Обыкновенный щито- мордник - Gloydius halys (PaL, 1776) G. h. carasanus (Eich, 1831) ВП — - — П - - - мл мл мл — - — - мл мл МН ОБ ОБ + + - - мл ОБ ОБ
48 Обыкновенная гадюка - Vipera berus (L., 1758) V. b. berus (L., 1758) E -
49 Гадюка Никольского - V. nikolskii (Vedm., 1984) E -
50 Степная гадюка - V. renardi Chr., 1861 V. r. renardi (Chr., 1861) CP мл + ОБ ОБ ОБ + + ОБ ОБ мл ОБ м + МЛ мл мл ОБ ОБ ОБ + ? + ? + ? ?
V. r. bashkirovi (Gar., 2004) -
Примечание. Фаунистические комплексы: Е - Европейский, ЕА - Европейско-азиатский, ЕС - Европейско-сибирский, ВП - Восточнопалеарктический, СР - Средиземноморский,
СА - Среднеазиатский, ЦА - Центральноазиатский.
Численность: МН — многочислен, ОБ — обычный, МЛ — малочислен, Р — редкий, «+» — вид присутствует, но численность неизвестна, «-» — вид отсутствует, О - сведения отсутствуют,
П - обитание в прошлом,«?» - наличие требует уточнения, «•» - сомнительный вид.
А, Б, В и а, б, в - обозначения округов провинций из табл. 2.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
74
Глава 4
Класс 1. AMPHIBIA LINNAEUS, 1758 - ЗЕМНОВОДНЫЕ
Отряд 1. CAUDATA OPPEL, 1787 - ХВОСТАТЫЕ
Семейство 1. Hynobiidae Соре, 1860 - Углозубые
Род 1. Salamandrella Dybowski, 1870 - сибирские углозубы
1. Сибирский углозуб - Salamandrella keyseriingii (Dybowski, 1870)
Значение таксона для сохранения биоразнообразия. Единственный
представитель уникального среди земноводных рода. За время его изучения
описывалось несколько подвидов, валидность которых, однако, признается не
всеми авторами. В последних сводках вид считается политипическим, в соста-
ве которого на территории России и смежных районов Казахстана (на осно-
вании различий в морфологии, генетике и экологии) выделяют два подвида,
в том числе номинативный S. К. keyseriingii (Dybowski, 1870), известный на
территории региона. Наряду с этим в его составе довольно четко проявляются
межпопуляционные различия по ряду морфологических признаков (окраске,
длине тела, конечностей и хвоста, длине и ширине головы, пропорциям тела,
количестве бороздок по бокам тела и др.), особенностям размножения, типам
населяемых биотопов и некоторым другим деталям экологии (Кузьмин, Мас-
лова, 2005; Кузьмин, Семенов, 2006).
Распространение. Вид отличается самым широким распространением
среди современных земноводных. Его нынешний ареал охватывает Северо-
Восточную Европу, Урал, Западную, Среднюю и Восточную Сибирь, Камчат-
ку, Курилы, Сахалин. На севере граница ареала местами выходит за пределы
полярного круга (в зону тундры), а на юге по долинам рек (обычно занятых
интразональными ландшафтами) достигает Средней Волги, Южного Урала,
Алтае-Саянской горной страны, Забайкалья, Маньчжурии и Северной Кореи.
Вместе с тем палеонтологические данные (Ратников, 1989, 2002) свиде-
тельствуют о его былом обитании значительно южнее современной границы
в европейской части ареала. Это существенно подкрепляет концепцию релик-
тового характера европейских популяций, а не их формирования в результате
расселения вида из Сибири. Последняя точка зрения не согласуется и с мигра-
ционными способностями вида, которых явно недостаточно для преодоления
свыше 1000 км за 30 лет (Григорьев, 1984).
В свете этих данных не такими уж невероятными оказываются сведения
о находке вида в Дьяковском лесном массиве Краснокутского района Сара-
товской области (коллекция ЗИН РАН, экз. № 5592, 1990 г.). Следует иметь
в виду, что в эпоху «малого ледникового периода» (не говоря уже о более
ранних временах) Общий Сырт был занят вековыми дубравами (они описаны
Э.А. Эверсманом и Г.С. Карелиным), которые смыкались с обширными пой-
менными дубово-вязовыми лесами по рекам Иргиз, Еруслан (Дьяковский лес),
Б. и М. Узени и менее значительными массивами по рекам Горькая, Хара,
Ланцуг, Паника и другим, заходящим в пределы Прикаспийской низменнос-
ти. Широкое распространение лесных сообществ и их значительная площадь
дают основание считать, что Северный Прикаспий в это время приобретал в
определенной степени лесостепной облик (Абрамова, Турманина, 1982). На-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 75
чавшееся с середины XIX столетия потепление климата и последующая общая
аридизация региона неоднократно прерывались относительно влажными пе-
риодами. Последнее, наиболее значительное, увлажнение территории региона
приходится на 1875-1915 гг. - период, когда вид, верятно, ставший в это время
более или менее обычным, был отмечен в бассейне р. Сакмара «у Богославска»
(Сабанеев, 1874) и в пойме р. Урал в окрестностях г. Оренбурга Н.П. Навозо-
вым 1-28.05.1911 (экз. № 2439 из коллекции отдела герпетологии ЗИН РАН)
(Сибирский углозуб, 1994). Следует отметить также, что в коллекции ЗИН есть
экземпляр, доставленный Могульским «откуда-то из Киргизской степи». Ве-
роятнее всего этот экземпляр также происходит «из окрестностей Оренбурга,
где сибирский углозуб был достаточно обычен» (Искакова, 1959. С. 13). Эти
животные доставлялись также Н.А. Зарудному (1895), но откуда-то из горного
Урала, скорее всего из «Челябинского уезда».
В свете приведенных материалов вероятным кажется и предположение,
что сибирский углозуб мог сохраниться и в другом подходящем месте реги-
она-Бузулукском бору, где основная последовательность событий и смен рас-
тительного покрова в позднеледнековье и голоцене была примерно такой же,
как и на смежной территории Восточно-Европейской равнины (Кременецкий
и др., 1998). Возможно, и здесь, как и на остальной территории европейского
ареала, углозуб «просто не попадался» зоологам. Последующая естественная
аридизация территории, негативные последствия которой усугубились все
возрастающим воздействием антропогенных факторов, по всей вероятности, и
стали основной причиной отступления границы его ареала к северу. Известную
современную южную границу ареала вида сейчас проводят на широте верхо-
вий р. Урал у г. Магнитогорска, что по меридиану примерно на 250 км севернее
г. Оренбурга (Вершинин, 2007), а также на территории Санарского и Карагай-
ского боров Челябинской области (Чибилёв, 2005). Если учесть, что областью
дифференциации семейства углозубов и, по-видимому, местом формирования
сибирского углозуба являются Центральная Азия и Восточная Сибирь, то это
дает основание относить вид к сибирскому фаунистическому комплексу, хотя
до общепринятого решения вопроса о центре происхождения, вероятно, более
правильным будет считать его европейско-сибирским видом. С учетом отме-
ченного возможное (в прошлом) обитание вида в регионе в соответствии с лан-
дшафтным районированием территории (здесь и далее по: Чибилёв, Дебело,
2006) представляется следующим образом (табл. 13, карта 1).
Места обитания. Сибирский углозуб населяет в основном влажные хвой-
ные, смешанные и лиственные леса, где предпочитает притеррасные низинные
болота и (в меньшей мере) поймы крупных и средних рек с перемежающими-
ся лесами, болотами и лугами. В лесостепи Южного Урала, непосредственно
примыкающей к нашему региону, он встречается на заболоченных участках
лугов, в мелколиственных лесах, а также по остаточным водоемам с обильной
прибрежной кустарниковой растительностью среди сосновых боров. Подоб-
ные местообитания имеются и в Бузулукском бору, что позволяет поставить
вопрос о возможности интродукции вида в этот массив, значительной части
которого в 2007 г. придан статус национального парка. Этому в определенной
степени может способствовать высокая экологическая пластичность вида, в
частности его способность размножаться даже в небольших водоемах, особен-
ности питания (как на личиночной стадии так и взрослого животного), раз-
76
Глава 4
Таблица 13
Места обитания сибирского углозуба
Зона Провинция Район № точки Местонахождение Источник информации
Лесостепная ш 9 1 окр. Богославска в бас. р. Сакмары Сабанеев, 1874
V 29 2 окр. г. Оренбурга Сибирский угло- зуб, 1994
Степная VIII 48 3 Дьяковский лес по р. Еруслан Краснокутс- кого р-на Саратовской обл.
множения, значительная толерантность к трансформации мест обитания под
воздействием рекреационной нагрузки, а также некоторые другие особеннос-
ти экологии. Эти животные местами встречаются по придорожным канавам, в
парках, окрестностях поселков и городов с затененными лесистыми или кус-
тарниковыми зарослями, хотя как типично лесной вид углозуб плохо перено-
сит уплотнение поверхностного слоя опада под действием рекреации и неко-
торых других антропогенных факторов.
Образ жизни. Взрослые животные в начале водной фазы жизненного цик-
ла активны днем, по мере прогревания водоема активность сдвигается на су-
меречное время, а затем и ночное. В сухопутную фазу животные активны в
сумерки и ночью, а днем прячутся в разных укрытиях. Личинки активны круг-
лосуточно. На Урале в сентябре-октябре углозуб уходит на зимовку. Зимует в
трухлявых пнях, деревьях, валежнике, под опавшей листвой, в норах и прикор-
невых пустотах деревьев, обычно небольшими группами или по одиночке на
расстоянии до 500 м от водоема. Уникальной особенностью вида является его
способность переносить низкие температуры (известны находки в вечной мер-
злоте и «оживание» после оттаивания). Выход из зимовки на Урале отмечается
в первой-третьей декадах апреля в зависимости от хода развития весенних яв-
лений (Ананьева и др., 1998; Кузьмин, 1999; Вершинин, 2007).
Размножение начинается вскоре после выхода с зимовки (еще во время
таяния льда или сразу после его разрушения). На Урале период икрометания
иногда растягивается до 19.06, что, по-видимому, объясняется микроклима-
тическими особенностями мест зимовки и размножения (даже в пределах
одного местообитания), возрастными и физиологическими особенностями
животных. Обычно откладывается 2 икряных мешка, содержащие от 30 до
120 яиц каждый. В условиях технического загрязнения плодовитость снижает-
ся и возрастает число аномалий развития. Эмбриональное развитие в условиях
Свердловской области продолжается 19-65 дней, личиночное - 26-83, общая
продолжительность развития составляет 55-120 суток и с 19.07 личинки начи-
нают выходить на сушу, где завершается метаморфоз. Половозрелым стано-
вится в возрасте 3 лет (Вершинин, 2007).
Питание. В воде личики питаются мелкими ракообразными, насекомыми
и их личинками. Основу пищи взрослых составляют дождевые черви, немато-
ды, многоножки, паукообразные, насекомые, их личинки и другие беспозво-
ночные, состав которых может изменяться в зависимости от биотопа.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
77
Численность. В естественных экосистемах на большей части ареала это
обычный вид. В лесных экосистемах пригородов Урала численность зависит
от степени трансформации среды, в связи с чем ее показатель может исполь-
зоваться для диагностики ранних стадий нарушения внешне благополучных
сообществ.
Лимитирующие факторы. Загрязнение окружающей среды различными
полютантами. В пригородных лесопарковых зонах крупных центров интен-
сивная рекреация.
Охрана. На большой части ареала специальных мер охраны не требуется,
но в ряде областей заслуживают охраны краеареальные популяции.
Региональный статус. Редкий краеареальный вид у южных пределов рас-
пространения. Занесен в Красную книгу сопредельной Челябинской области
(III категория. Редкий, малочисленный вид на периферии ареала).
Семейство 2. Salamandridae Goldfuss, 1820 - Саламандровые
Род 2. Lissotriton Bell, 1839 - лиссотритоны
2. Обыкновенный тритон - Lissotriton vulgaris (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода на территории России. Политипический вид, в составе которо-
го выделяют до 7 подвидов; из них на территории России известны 2. Евро-
пейско-сибирскую часть ареала населяет номинативный подвид L. v. vulgaris
(Linnaeus, 1758), обитающий и в пределах региона.
Отличается высокой экологической пластичностью, образуя многочислен-
ные популяции, морфологические (длина тела, хвоста, конечностей и их соот-
ношение, пятнистость, наличие полос, окраска тела и др.), фенологические и
генетические отличия которых определяются конкретными условиями сущес-
твования (Ананьева и др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает большую часть Европы, Запад-
ной Сибири и Кавказа (европейский фаунистический комплекс). Северная гра-
ница распространения вида достигает Скандинавии, Карелии, Архангельской,
Тюменской, Пермской, Кемеровской областей и юго-запада Красноярского
края. Южная граница номинативного подвида на Восточно-Европейской рав-
нине простирается от низовьев Дуная и Днепра к среднему течению р. Дон,
затем проходит через Волгоградскую, Саратовскую, Оренбургскую и Челя-
бинскую области, окрестности Новосибирска и Барнаула на юго-запад Крас-
ноярского края.
В прошлом на Восточно-Европейской равнине граница проходила несколь-
ко южнее (Ратников, 2002), а восточнее изолированные очаги якобы были из-
вестны на северном побережье Каспийского моря (в ЗИН РАН имеется экзем-
пляр, доставленный В. Хлебниковым из окрестностей г. Астрахани), нижнем
течении р. Эмба, области ее верховий, северном побережье Аральского моря,
Мангышлаке (в Зоологическом музее Казанского университета имеется экзем-
пляр с этикеткой, указывающей добычу вида в этом пункте) и в нижнем тече-
нии р. Или, недалеко от места впадения в оз. Балхаш; предполагалось также
обитание вида в области низовий р. Илек (Искакова, 1959; Определитель зем-
78
Глава 4
новодных..., 1977; Гаранин, 1983). Впрочем, все герпетологи, позднее рабо-
тавшие в этих районах, сомневались в достоверности этих сведений, объясняя
приведенные указания в основном путаницей в этикетах.
Точнее в пределах региона и его ближайших окрестностях южная граница
вида представляется следующим образом: у западного рубежа региона он из-
вестен из окрестностей Волгограда, но после зарегулирования р. Волги здесь
уже не встречается (Кубанцев, Колякин, 1988). В Саратовской области грани-
ца следует по долине Волги, откуда он предположительно заходит в низовья
Еруслана, Б. и М. Иргиза (Шляхтин и др., 2005). На левобережье Самарской
области граница ареала проходит через г. Самару, затем, очевидно, по пой-
ме р. Самары (находки известны в Кинельском, Богатовском и Борском райо-
нах) (Бакиев, Файзулин, 2001, 2002) к Бузулукскому бору и далее к низовьям
р. Сакмара в окрестностях Оренбурга (Райский, 1951; Чибилёв, 1995). Восточ-
нее южный рубеж проходит приблизительно по среднему течению р. Урал до
лесостепных районов Челябинской области, в пределах которой этот тритон
встречается во многих водоемах, расположенных в лиственных и смешанных
лесах (Чибилёв, 2005).
В отмеченных пределах встречи обыкновенного тритона известны в г. То-
льятти, в каждом третьем административном районе г. Самары, Кинельском,
Богатовском и Борском районах Самарской области, затем в Бузулукском и
ряде смежных районов Оренбургской области (табл. 14, карта 2). Прежние ука-
зания на обитание вида в бассейне р. Сакмары современными материалами не
подтверждаются, что скорее всего свидетельствует о его исчезновении. В гео-
графическом отношении его конкретные местонахождения известны в 4 (36,4%
из 11) ландшафтных районах лесостепной и 4 (5,8% из 69) степной зоны.
Места обитания. Обыкновенный тритон в пределах региона явно тяготе-
ет к пойменным лиственным (осиновым, березовым, дубово-липовым) лесам и
зарослям кустарников, иногда садам и паркам в пригородах даже крупных го-
родов (Самары), если поблизости есть пригодные для размножения водоемы.
Весной он обычно встречается в мелких стоячих или слабо проточных вре-
менных и постоянных водоемах - старицах, прудах, ручьях и даже канавах с
pH = 5,6-9,0 и значительным химическим загрязнением. На суше придержива-
ется тенистых влажных мест, скрываясь в лесной подстилке, кучах валежника,
трухлявых пнях, под отставшей корой упавших деревьев, карягами; открытых
пространств (даже влажных лугов) избегает. Подобная узкая специализация
в выборе мест обитания является одной из причин сомнений в достоверности
данных о встречах вида в изолированных очагах южнее сплошной границы его
ареала, хотя в прошлом облесенность территории региона, как это отмечалось
ранее, была более значительной.
Образ жизни. Весной на сопредельной территории у северной границы
региона обыкновенные тритоны обычно появляются во второй половине апре-
ля, реже в начале мая, когда воздух прогревается до 8... 10 °C, а вода до 4... 12
°C и более. В это время они держаться на водоемах и активны круглосуточ-
но. С окончанием периода размножения переходят к наземному образу жизни
(в июле, а в некоторых городских популяциях в августе), во время которого ак-
тивность смещается на сумеречное и ночное время. Днем на поверхности три-
тоны отмечаются лишь в период миграции или при дождливой погоде; обыч-
но же укрываются в разных убежищах. Личинки в водной фазе жизненного
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
79
Таблица 14
Распространение обыкновенного тритона
Зона Провинция Район № точки Местонахождение Источник информации
Самарская область
3 X X § I 1 1 2 г. Тольятти г. Самара: п. Управленчес- кий, п. Мехзавод, Воронеж- ские озера, пруды Ботани- ческого сада Мельниченко и др., 1938; Бакиев, Файзулин, 2002; Файзулин, 2004а; Бакиев и др., 2002
о о о 4 3 р. Сок
t=; II 5 4 окр. г. Похвистнево Оренбургская область
6 5 р. Дёма Чибилёв А.А?
Самарская область
21 6 7 г. Отрадный, Богатове кий р-н Бакиев и др., 2002
22 8 Борский р-н
3 Оренбургская область
X с 5 V 9 10 р. Боровка (Бузулукский бор) р. Самара (Бузулукский бор) Чибилёв, 1995
24 11 р. Сакмара
12 р. Салмыш
28 13 р. Сакмара
Примечан ие. П.А. Воронцовский в очерке о животном мире Оренбургской губернии (1927) сообщает, что этот вид
обитает во всех глубоких водоемах со столчен водой, ио каких-либо конкретных пунктов ие называет.
’Здесь и далее у неопубликованных источников указываются только фамилии авторов.
цикла активны днем и в начале ночи, а в период метаморфоза их активность
смешается на сумерки. На зимовку в разные годы тритоны уходят в конце сен-
тября - октябре, хотя сеголетки, а в городских популяциях и взрослые могут
задерживаться и позднее (подвижность теряется при О °C). Места зимовки
обычно располагаются на расстоянии не более 100 м от водоемов. Активный
период жизни в Поволжье составляет 145—185 дней, из которых 100-140 дней
приходится на сухопутную жизнь. В целом обыкновенный тритон отличает-
ся довольно высокой экологической пластичностью - он значительно менее
требователен по сравнению с сибирским углозубом и гребенчатым тритоном
к освещенности и температуре водоемов, химизму среды и составу раститель-
ных сообществ, что позволяет ему сохраняться даже в некоторых экосистемах
(особенно урбанизированных территорий) с довольно напряженной экологи-
ческой ситуацией (Шиклеев, 1951; Гаранин, 1983; Бакиев, Файзулин, 2001;
Вершинин, 2007).
Размножение. К размножению обыкновенный тритон приступает на вто-
ром-третьем году жизни. Оно начинается вскоре после выхода из зимовки -
чаще всего в конце апреля - начале мая и сопровождается брачными играми.
80
Глава 4
К икрометанию самки приступают, когда вода прогревается до 10 °C. В свя-
зи с этим в городских популяциях откладка икры начинается несколько рань-
ше, чем в загородных. Обычно на хорошо прогреваемых участках водоемов с
обильной придонной растительностью в условиях Волго-Камского края самки
откладывают 70-140 икринок. Эмбриогенез продолжается 14-20 сут, а еще
спустя 60-70 сут - в августе-сентябре - сеголетки, завершившие метаморфоз,
появляются на суше. В городских популяциях Урала отмечено удлинение сро-
ков личиночного и общего развития, что, вероятно, связано с ингибирующим
действием полютантов. Известны случаи и перезимовки личинок (Гаранин,
1983; Вершинин, 2007).
Питание. Личинки сначала питаются мелкими ракообразными, а в даль-
нейшем спектр их кормов расширяется за счет моллюсков, насекомых, их ли-
чинок и других водных беспозвоночных. Взрослые потребляют в пищу червей,
почвенных личинок и имаго разных членистоногих, состав которых в разных
биотопах может несколько отличаться. В свою очередь личинок и взрослых
обыкновенных тритонов потребляют хищные водные насекомые, зеленые
лягушки, ужи, ряд водоплавающих, околоводных и некоторых других видов
птиц, водяная полевка.
Численность. В окрестностях Волгограда обыкновенный тритон еще в се-
редине прошлого столетия был малочислен (Кубанцев, Колякин, 1988), а сей-
час на этой широте известны лишь единичные случаи встреч (Кубанцев, 2003).
Севернее в области Среднего Поволжья он был сравнительно обычен, а мес-
тами и многочислен (Гаранин, 1983). Сейчас в г. Самаре он наиболее обычен
в Красноглинском районе, где ежегодно отмечается по 5-10 особей, в других
районах встречается единично, а в г. Тольяти отдельные особи встречаются
не каждый год (Бакиев и др., 2003). В лесопарковых зонах городов Среднего
Урала он местами и сейчас по количеству уступает лишь лягушкам, хотя в
большинстве случаев численность городских популяций невысока, а в Челя-
бинской области он уже определенно малочислен (Чибилёв, 2005; Вершинин,
2007). В Бузулукском бору и долине р. Дёмы в связи с ограниченностью мест
обитания количество его невелико. В области нижнего течения р. Сакмары
даже в прошлом тритон был немногочисленным видом (Райский, 1951), а сов-
ременное состояние этой популяции не известно.
Таким образом, современную численность популяций в регионе в целом
следует рассматривать как незначительную, что соответствует их статусу
краеареальных.
Лимитирующие факторы. Загрязнение водоемов разными полютантами,
усиление рекреационной нагрузки, способствующей значительной трансфор-
мации (особенно иссушению) местообитаний, обезлесение ландшафтов; в ряде
мест, по-видимому, отлов животных.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На большей части остального ареала номинативного подвида
специальных мер охраны не требуется. Региональные популяции заслуживают
охраны как краеареальные.
Региональный статус. Малочисленный краеареальный вид у южных пре-
делов распространения. Включен в Красную книгу Самарской области (2009),
Приложение III Бернской конвенции (виды, эксплуатация которых регулиру-
ется в соответствии с требованиями конвенции).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
81
Род 3. Triturus Rafinesque, 1815 - тритоны
3. Гребенчатый тритон - Triturus cristatus (Laurenty, 1768)
Значение таксона для сохранения биоразнообразия. Один из двух видов
рода, обитающих на территории России. Раньше считалось, что Т. cristatus
включает четыре подвида, которые сейчас рассматриваются как отдельные
виды надвида Triturus cristatus подрода Triturus', видовое название Triturus
cristatus в настоящее время относится к прежнему номинативному подвиду,
населяющему и территорию региона. Отличается значительной экологической
пластичностью, образуя ряд популяций, морфологические, фенологические и
генетические отличия которых обусловлены конкретными условиями сущест-
вования (Ананьева и др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает значительную часть Централь-
ной и Восточной Европы, Урала и сопредельную территорию Западной Сиби-
ри (европейский фаунистический комплекс). Его северная граница на Восточ-
но-Европейской равнине прослеживается от Калининградской области до юга
Карелии, затем идет по Вологодской, Костромской, Пермской (г. Соликамск),
Свердловской и Курганской областям. Южная граница прослеживается от
горных районов Карпат через Молдавию (Кишинев), Одесскую, Херсонскую,
Кировоградскую, Харьковскую и Луганскую области Украины, затем через
Белгородскую, Пензенскую, Саратовскую, Самарскую, Оренбургскую, Челя-
бинскую и Курганскую области России. В прошлом на Восточно-Европейской
равнине граница проходила несколько южнее (Ратников, 2002). Существует
ряд указаний и на более южное современное распространение вида, что требу-
ет уточнения южной границы ареала в целом.
Точнее в пределах региона южная граница ареала прослеживается сле-
дующим образом: у западной границы региона находки вида известны из
окрестностей Саратова (данные 1878 г., позднее не подтвержденные) (Кузь-
мин, 1999), в п. Управленческий Красноглинского района г. Самары (Бакиев,
Файзулин, 2002), Бузулукском бору, выше по долинам рек Самара и Сакмара,
низовьям р. Большой Ик (Зарудный, 1895; Райский, 1951; Чибилёв, 1995) и
затем в Челябинской области у Аргазинского водохранилища (Чибилёв, 2005).
В отмеченных пределах встречи гребенчатого тритона известны у п. Управ-
ленческий Красноглинского района г. Самары, в верховьях р. Дёма (Понома-
ревский район), Бузулукском бору, по долинам рек Сакмара и Б. Ик (в преде-
лах Оренбургский области) (табл. 15, карта 3). В географическом отношении
его конкретные местонахождения известны в 3 (27,3% из 11) ландшафтных
районах лесостепной и 3 (4,3% из 69) степной зоны.
Места обитания. В пределах региона населяет преимущественно круп-
ные массивы пойменных лесов (по рекам Самаре, Боровке, Сакмаре и др.),
где придерживается мест у опушек, полян и вырубок поблизости от крупных
и глубоких стариц и других стоячих водоемов (чем и отличается от обыкно-
венного тритона); в окрестностях г. Самары встречается в пригородной лесо-
парковой зоне, где, однако, избегает загрязненных мест. Весну и начало лета
проводит преимущественно в воде, а на сушу выходит во второй половине
лета, чаще всего с середины июля. С этого времени обычно придерживается
увлажненных мест, где скрывается в трухлявых пнях, под гнилыми стволами
82
Глава 4
Таблица 15
Распространение гребенчатого тритона
Зона Провинция Район № точки Местонахождение Источник информации
I 1 1 г. Самара: п. Управ- Бакиев, Файзулин, 2002;
ленческий Бакиев и др., 2003;
Лесостепная II 6 2 р, Дёма Чибилёв, 1995
III 9 3 р. Б. Ик
22 4 Бузулукский бор Райский, 1951; Чибилёв, 1995
Степная V 24 5 р. Сакмара Зарудный, 1895; Рай- ский, 1951; Чибилёв,
1995
28 6 р. Сакмара
упавших деревьев, кучами валежника, в норах грызунов, ямах с опавшей лис-
твой, корневых ходах и т. п. В этих же убежищах тритоны могут и зимовать.
Сеголетки после выхода из водоемов обычно далеко не уходят и держатся в
подходящих укрытиях всю осень; здесь же они могут оставаться и зимовать.
Образ жизни. Весной гребенчатый тритон появляется при температуре
воздуха 9... 10 °C и воды 6...8 °C (т. е. после вскрытия водоемов), что обычно
наблюдается во второй половине апреля. Всю весну и начало лета проводит
в воде, где, как правило, наибольшую активность проявляет в дневные часы.
После окончания периода размножения (чаще всего с середины июля) начина-
ется выход на сушу. С этого времени активность смещается на темное время
суток, а днем он укрывается в убежищах. Личинки в водной фазе жизненного
цикла активны в светлое время суток, а в период метаморфоза их активность
постепенно смещается на ночные часы.
На суше гребенчатые тритоны сохраняют активность до конца октября,
когда температура воздуха снижается до 6...4 °C и начинаются регулярные
ночные заморозки (хотя подвижность сохраняется и при температуре около
0 °C). С этого времени обычно и начинается их зимовка, на которую трито-
ны скапливаются в подходящих местах зачастую с жабами и жерлянками (ли-
чинки могут зимовать и в замерзающих водоемах). Активный период жизни
в Поволжье в разные годы составляет 123-215, в среднем около 153 дней, из
которых в воде этот тритон проводит около 120 и до 90 суток приходится на
сухопутную жизнь (Определитель земноводных..., 1977; Гаранин, 1983; Чи-
билёв, 1995; Бакиев, Файзулин, 2001; Вершинин, 2007).
Размножение. Половой зрелости гребенчатый тритон достигает на третий
год жизни. Как правило, через 3-6 дней после выхода из зимовки - чаще всего
в третьей декаде апреля - самцы идут в водоем, где вскоре приобретают брач-
ный наряд. Для нереста гребенчатые тритоны (в отличие от обыкновенных)
обычно концентрируются в боле,е глубоких местах водоемов (по сравнению с
другими тритонами гребенчатый отличается более высокой требовательнос-
тью к качеству воды). Здесь, при температуре воды выше 10 °C, после ритуаль-
ного ухаживания на листочках, ветках и других плавающих в воде предметах
самка откладывает 150-200 икринок. Эмбриогенез продолжается 13-18 суток,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
83
а еще спустя 3 месяца - в конце лета (в августе) завершается метаморфоз личи-
нок. Часть из них (в отличие от личинок обыкновенного тритона) может долго
задерживаться в воде (Гаранин, 1983).
Питание. Личинки в основном питаются мелкими ракообразными.
В дальнейшем спектр их пищи расширяется за счет крупных планктонных
форм ракообразных, мелких ползающих беспозвоночных, личинок насекомых
и других водных обитателей. Взрослые тритоны во время обитания в водоемах
питаются в основном мелкими моллюсками, водными насекомыми и их ли-
чинками, могут поедать также икру и личинок разных видов амфибий, иногда
и молодых тритонов. В наземную фазу жизненного цикла потребляют глав-
ным образом дождевых червей, слизней, других моллюсков, насекомых и их
личинок. В свою очередь личинок и взрослых гребенчатых тритонов поедают
болотная черепаха, ужи, серая жаба, ряд водоплавающих, околоводных и не-
которых других видов птиц.
Численность. Спорадическое распространение на окраине ареала опреде-
ляет его повсеместную незначительную численность, которая в 4-6 раз ниже,
чем у обыкновенного тритона.
Лимитирующие факторы. Комплекс природных ландшафтно-клима-
тических условий на границе ареала. В последние десятилетия значительно
возросла роль антропогенных факторов - промышленного и пестицидного за-
грязнения, обмеления и эвтрофикации водоемов, что приобрело особую значи-
мость в связи с высокой чувствительностью вида к качеству воды.
Охрана. Обитает и охраняется на территории национального парка «Бу-
зулукский бор». На значительной части остального ареала состояние популя-
ций ухудшается в связи с усилением воздействия антропогенных факторов.
Особого внимания заслуживают все популяции региона, которые относятся к
категории краеареальных.
Региональный статус. Занесен в Красную книгу МСОП 1996 г. (в кате-
гории LRcd - таксон низкого риска, зависимый от сохранения), Самарской
(2009), Оренбургской (1998), Саратовской (2006) областей, а также сопредель-
ных Челябинской области, Башкортостана и Татарстана. В Западной Европе
охраняется Бернской конвенцией (Приложение II - виды животных, для кото-
рых требуются специальные меры охраны).
Отряд 2. ANURA RAFINESQUE, 1815 - БЕСХВОСТЫЕ
Семейство 3. Bombinatoridae Gray, 1825 - Жерлянки
Род 4. Bombina Oken, 1816 - жерлянки
4. Краснобрюхая жерлянка -Bombina bombina (Linnaeus, 1761)
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей рода, обитающих на территории России. Монотипический вид, ко-
торый вместе с тем проявляет значительную географическую, биотопическую
и индивидуальную изменчивость в размерах тела (возрастают в направлении
юго-запад), окраске и рисунке спины, брюшка и конечностей. Географичес-
кая изменчивость усиливается интрогрессией генов желтобрюхой жерлянки,
84
Глава 4
в связи с чем южные популяции оказываются более полиморфны (Ананьева и
др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает значительную часть Централь-
ной и Восточной Европы (европейский фаунистический комплекс). Северная
граница ареала на Восточно-Европейской равнине проходит от Калининграда
через Белоруссию (Оршу), Псковскую, Московскую, Нижегородскую, Перм-
скую области и Башкирию к северной горно-лесной части Челябинской области.
Южная граница следует от Карпат через север Крыма, Тамань, Краснодарский
и Ставропольский края, Ростовскую, Волгоградскую, Западно-Казахстанскую
(Уральскую), Актюбинскую, Оренбургскую и Челябинскую области. Точнее
в пределах региона и его ближайших окрестностях южная граница прослежи-
вается следующим образом: у западной границы региона жерлянка известна в
Волго-Ахтубинской пойме южнее г. Волгограда (Кубанцев, Колякин, 1988).
По р. Волге граница, очевидно, отступает к северу и в Заволжье жерлянка ста-
новится известной лишь в прудах и водохранилищах Палласовского водовод-
ного канала и лужах-разливах вдоль него (Животные..., 2009). Далее ее обита-
ние известно во всех районах Саратовской области (Шляхтин и др., 2005). На
сопредельной территории Казахстана нами отмечена в верховьях р. Таловая
(окр. п. Белоглинка). Вероятно, по р. Деркул (или прудам в верховьях балок
Предсыртового уступа) граница ее ареала выходит к долине р. Урал немногим
южнее г. Уральска (п. Круглозерное), отсюда по каналу она попала в хвалын-
ский рукав Урала - Кушум, по которому в 1970-1980 гг. нами прослежива-
лась в Кировском и Битикском водохранилищах. К настоящему времени, не-
сомненно, проникла южнее, как минимум до Донгелекского водохранилища.
В многоводные годы по каналам выносится на орошаемые лиманы вплоть до
Чижинских и Балыктинских разливов, где формируются временные популя-
ции. По пойме р. Урал в 1950-е гг. П.М. Бутовским прослежена до п. Б. Ча-
ган. Южнее по пойме ее обитание неизвестно, но не исключено, что местами
жерлянка может встречаться почти до широты п. Калмыкове, поскольку этим
же автором обнаружена в древнем рукаве Урала - р. Багырдай (Параскив, Бу-
товский, 1960). Здесь же по опросным сведениям вблизи п. Антоново она была
известна и в начале 1970-х гг. Несомненно, что из р. Урал жерлянка попадала
в Урало-Шалкарский канал и р. Барбастау. Восточнее известна на Утва-Илек-
ском междуречье (Березовиков и др., 1992), по р. Илек (вверх до ур. Биш-Та-
мак) и его притокам Б. и М. Хобда (Зарудный, 1895), откуда граница следует к
верховьям р. Орь, затем по долине этой реки и р. Урал в Челябинскую область
(Кузьмин, 1999; Вершинин, 2007; Чибилёв, 2005).
В очерченных пределах краснобрюхая жерлянка обитает в ряде районов
Самарского левобережья, большинстве северных и центральных районов Са-
ратовского Заволжья, большинстве западных и центральных районов Орен-
буржья, северных и примыкающих к р. Урал районах Западно-Казахстанской
(Уральской) и северо-западных районах Актюбинской областей. В географи-
ческом отношении ее конкретные местонахождения известны в 4 (36,4% из 11)
ландшафтных районах лесостепной, 32 (46,4% из 69) степной, 2 (6,3% из 32)
полупустынной и 1 (2,3% из 43) пустынной зон (табл. 16, карта 4).
Места обитания. В регионе жерлянка находится у южных пределов рас-
пространения. Здесь она встречается не только в лесостепной и степной зо-
нах, но проникает в полупустыню и даже пустыню. В связи с этим она теснее
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
85
Таблица 16
Распространение краснобрюхой жерлянки
Зона Провинция Район № точки Местонахождение Источник информации
Самарская область
I 1 1 2 с. Н. Санчелеево г. Тольятти Бакиев, Файзулин, 2002
Лесостепная II 3 3 4 5 6 7 8 с. Васильевка, ст. Задельная, ст. Мастрюково, г. Самара, п. Управленческий р. Сок, с. Лужки Похвистневский район, с. Совруха Бакиев и др., 2003; Файзулин, 2004
Оренбургская область
6 9 р. Дёма Чибилёв А.А.
Самарская область
12 10 И г. Новокуйбышевск г. Чапаевск Бакиев и др., 2003
Саратовская область
13 14 15 12 13 14 Повсеместно Шляхтин и др., 2005
IV Самарская область
17 15 16 17 р. Б. Иргиз, с. Мосты п. Б. Глушица п. Восточный Бакиев, Файзулин, 2002
Саратовская область
18 19 20 18 19 20 Повсеместно а и Шляхтин и др., 2005
3 Самарская область
X Е О и 21 21 22 23 24 г. Отрадный г. Кинель п. Красносамарское п. Горский г. Похвистнево ст. Заливная Бакиев, Файзулин, 2002
Оренбургская область
V 22 23 24 25 25 26 27 28 29 Бузулукский бор: п. Колтубановский р. Боровка р. Самара р. Самара р. Сакмара Чибилёв А.А.
Самарская область
26 30 Нефтегорский район, с. Утеево Бакиев и др., 2003
Западно-Казахстанская (Уральская) область
27 31 р. Таловая, с. Белоглинка Дебело П.В.
86
Глава 4
Окончание табл. 16
Зона Провинция Район № точки Местонахождение Источник информации
VI 28 29 30 32 32 33 34 35 Оренбургская область р. Сакмара р. Урал р. Илек р. Илек Зарудный, 1895
VII 34 42 43 44 36 37 38 39 40 Западно-Казахстанская область р. Утва, г. Аксай Актюбинская область р. Б. Хобда р. М. Хобда р. Илек р. Илек г. Актюбинск Березовиков и др., 1992 Зарудный, 1895; Иска- кова, 1959
Степная VIII 48 49 50 54 55 41 42 43 44 45 46 47 48 49 Волгоградская область р. Еруслан Саратовская область р. М. Узень р. Б. Узень Оренбургская область р. Урал Западно-Казахстанская область окр. р. Урал: п. Красноармейск п. Дарвинское р. Шаган, г. Уральск р. Урал, г. Уральск р. Урал, с. Подстепное канал р. Урал - оз. Шал кар Анисьины оз. в окр. п. Мело- вые горки Определитель земно- водных..., 1977 Шляхтин и др., 2005 Чибилёв А.А Дебело П.В.; Параскив, Бутовский, 1960 Кривко, 1976; Дебело П.В.
IX 56 57 59 60 61 50 51 52 53 54 Оренбургская область р. Урал р. Урал р. Урал р. Урал р. Урал Зарудный, 1895
Полупустынная XIII 81 89 55 56 57 Волгоградская область Волго-Ахтубинская пойма у г. Волгоград Запади о-Казахстанская область р. Кушум, Кировское вдхр. Битикское вдхр. Кубанцев, Колякин, 1962 Кривко, 1976 Дебело П.В.
Пус- тынная XX 124 58 р. Багырдай Параскив, Бутовский, I960
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
87
(особенно на юге) привязана к пресным водам речных долин. В поймах рек
Сакмары, Самары и Урала она населяет преимущественно старицы и поймен-
ные озера, где придерживается хорошо прогреваемых мелководий с зарослями
мягкой водной растительности. По долине р. Урал после высоких паводков
часто остается в «отмирающих» (лишь периодически наполняемых) старицах,
межгривных понижениях, которые вскоре (как это отмечено еще Н.А. Заруд-
ным (1895)) приобретают облик заболоченных лугов, в колодцах-копанках, ка-
навах, ямах и подобных местах. По небольшим степным речкам и некоторым
балкам обитает в прудах, небольших запрудах, иногда в копанках или ямах
с кустарниковыми зарослями. На участке, напоминающем типичную степ-
ную речку с прибрежными зарослями рогоза, отмечена по каналу из р. Урал к
оз. Шалкар. По р. Кушум наблюдалась в водохранилищах, где придерживалась
мелководных заливов с мягкой водной растительностью. По ирригационным
каналам выносится на орошаемые лиманы, где после сброса воды некоторое
время держится в заболоченных понижениях с луговой растительностью (от-
мечена по Кировско-Чижинскому каналу). В окрестностях г. Уральска насе-
ляет пруды рыбхоза, а до 1990-х гг. обитала в наиболее чистой части город-
ского накопителя сточных вод - Анисьиных озерах (возле Меловых горок).
Встречается в некоторых определенно загрязненных водоемах рекреационных
зон городов Самары (п. Управленческий), Уральска (р. Чаган), Оренбурга. Со-
лоноватой воды не переносит, в связи с чем в устьевых Кушумских озерах и
степных речках Зауралья не встречалась.
Образ жизни. Сроки весеннего пробуждения жерлянок зависят от хода
развития весенних явлений. В обычные годы на широте городов Саратов -
Уральск они, как правило, появляются в третьей декаде апреля, в ранние - на-
чале второй декады этого месяца, а в поздние - в начале мая, когда воздух
прогревается до 10 °C, а вода до 7. У северных пределов региона эти сроки
смещаются соответственно на 7-10 дней. Весенне-летний период проводит в
воде с температурой от 10 до 30 °C (предпочтительнее 21), хотя иногда пере-
мещается из одного водоема в другой. Жерлянки активны днем, но в брачный
период пик вокальной активности смещается на вечерние часы. На зимовку
жерлянки обычно уходят во второй половине сентября - первой половине ок-
тября, проводя в активном состоянии около 200 суток. Зимуют они чаще всего
на суше в норах грызунов, пустотах почвы, ямах (образуя иногда значительные
скопления, в том числе с другими видами амфибий), закапываются в мягкий
грут по берегам водоемов, часть особей зимует на дне водоемов, зарываясь в
отложения ила.
Размножение. Половой зрелости жерлянки достигают на второй-третий
год жизни. К размножению они приступают спустя 10-15 дней после выхода
из зимовки, что чаще всего наблюдается в конце апреля - мае, а разгар «тока» -
когда температура воды достигает 14... 16 °C. По этой причине в более глубоких
и менее прогреваемых водоемах размножение начинается несколько позднее
и в целом брачный период (даже в одном районе) оказывается сильно растя-
нут (в Волжско-Камском крае спаривающиеся жерлянки отмечались со второй
декады мая до начала июля). Плодовитость сравнительно невелика - обычно
300-450 икринок откладывается несколько раз небольшими порциями среди
водных растений. Эмбриогенез продолжается 4-10 дней. Первая свежеотло-
женная икра вблизи г. Уральска (с. Дарьинское) отмечена 3 мая, а 7 мая здесь
88
Глава 4
уже наблюдали головастиков длиной 3,5-5,0 мм. В р. Багырдай 30 июля они
достигали 15 мм в длину. Примерно при таких или немногим более крупных
размерах - спустя 51-74 дня начинается метаморфоз, пик которого обычно
приходится на июль-август. После завершения метаморфоза сеголетки около
2 месяцев остаются в воде.
Питание. Головастики питаются в основном водорослями, простейши-
ми и мелкими ракообразными. С завершением метаморфоза основу питания
составляют личинки насекомых (преимущественно двукрылых), моллюски,
другие водные беспозвоночные; иногда поедают головастиков других амфи-
бий. Довольно разнообразна и сухопутная добыча, в составе которой преоб-
ладают насекомые (жуки, полужесткокрылые, двукрылые) и многоножки,
иногда поедаются дождевые черви и некоторые другие беспозвоночные, хотя
в целом возможности добывания наземных кормов у жерлянок более ограни-
ченны, чем у лягушек. В свою очередь икру и головастиков жерлянки в боль-
шом количестве поедают болотные черепахи, а взрослых жерлянок - зеленые
лягушки, ужи, многие водоплавающие и околоводные птицы и млекопи-
тающие.
Численность. На большей части регионального ареала в прошлом жер-
лянка была сравнительно обычна, а местами даже многочисленна и в целом
уступала по численности лишь зеленым лягушкам. В настоящем времени на
Нижнем Поволжье она немногочисленна (Кубанцев, 2003). Не бывает она
особо многочисленной и в Волжско-Камском крае, где по данным учетов со-
ставляет не более 33% общего количества отлавливаемых амфибий (Шикле-
ев, 1951; Гаранин, 1983). В условиях городов Самары и Тольятти встречается
ежегодно, но лишь в виде единичных особей (Бакиев и др., 2003). В Оренбург-
ской части ареала в середине прошлого столетия она была довольно обычна,
а местами даже многочисленна по долинам всех рек области (Райский, 1951).
В ее северной части существенного изменения численности, по-видимому, не
произошло, и сейчас в долине р. Дёма на сопредельной территории Башкор-
тостана она характеризуется прежними показателями (Баянов, 1995). В то же
время по долинам Урала, Сакмары и Илека в связи со значительным снижени-
ем уровня паводка, при котором талые воды практически не выходят в пойму,
численность жерлянки резко сократилась, поскольку исчезли основные места
ее размножения - мелководные и заболоченные межгривные луговые пони-
жения. Подобное прослеживается и в казахстанской части бассейна р. Урала.
Таким образом, в целом по региону краснобрюхую жерлянку следует считать
малочисленным видом.
Лимитирующие факторы. Разрушение и пересыхание водоемов, суро-
вые морозы, негативные последствия которых усиливаются индустриальным
загрязнением водоемов и рекреацией.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». Состояние популяций на остальной части региона особых мер
по охране не требует, достаточно поддержания естественного состояния лан-
дшафтов.
Региональный статус. Малочисленный вид, населяющий специфические
местообитания. Занесена в Красную книгу МСОП (категория LRcd - таксон
низкого риска, зависимый от сохранения). Охраняется Бернской конвенцией
(Приложение II).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 89
Семейство 4. Pelobatidae Bonaparte, 1850 - Чесночницы
Род 5. Pelobates Wagler, 1830 - чесночницы
5. Обыкновенная чесночница — Pelobatesfuscus (Laurenti, 1768)
Значение таксона для сохранения биоразнообразия. Один из двух видов
рода, населяющих территорию России. Политипический вид, представлен-
ный двумя подвидами. В России и в пределах региона обитает номинативный
подвид Р. f. fuscus (Laurenti, 1768), в составе которого по генетическим дан-
ным (размерам генома) выделяют две географические популяции, из которых
территорию региона населяет восточная (Ананьева и др., 1998; Боркин и др.,
20036; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает значительную часть Западной,
Центральной и Восточной Европы, Урала, Западной Сибири и Западного Ка-
захстана (европейский фаунистический комплекс). Северная граница ареала
на Восточно-Европейской равнине проходит от Балтийского побережья Эс-
тонии через Ленинградскую, Вологодскую, Костромскую, Нижегородскую,
Пермскую, Свердловскую, Тюменскую и Курганскую области. Южная гра-
ница следует от Карпат через Крым, Краснодарский и Ставропольский края,
окрестности г. Грозного, междуречье Кумы и Терека, Черные земли Калмы-
кии, по долине Нижней Волги, в Волго-Уральском междуречье по северной
кромке Камыш-Самарской озерной депрессии, низовьям р. Кушум, долинам
рек Урала и Эмбы, к низовьям р. Тургай, затем по Тургайской депрессии, Кус-
танайской, Курганской и Тюменской областям. Точнее в пределах региона и
его ближайших окрестностях южная граница ареала прослеживается следую-
щим образом: в дельте Волги известна в Астраханском заповеднике (Кузьмин,
1999), выше по течению - в Волго-Ахтубинской пойме и примыкающих буг-
ристых песках Волго-Уральского междуречья (Завьялов, Табачишин, 1998),
территории Богдинско-Баскунчакского заповедника (Стрельцов и др., 2006),
в Волгоградской области (Кубанцев, Колякин, 1988), низовьях р. Ерусла-
на, откуда поворачивает на восток и примерно по 50-й параллели выходит к
р. М. Узень, далее по долине этой реки следует до Камыш-Самарских озер
(окр. поселков Н. Казанка, Фокеево), затем к низовьям р. Кушум и долине
р. Урал, по которой спускается до взморья. В Урало-Эмбенском междуречье
найдена у озер Шалкар, Сорколь, в долине р. Калдыгайты, где-то в области
среднего или верхнего течения р. Эмбы добывалась Н.А. Северцовым (экз. в
коллекции ЗИН РАН). Находки вида в этих пунктах позволяют допустить его
обитание и в верховьях других речек, стекающих с Подуральского плато и
образующих несколько связанных массивов раннехвалынских аллювиально-
дельтовых песчаных отложений. Восточнее Уральского хребта известна по
нижнему течению р. Иргиз, в ур. Жалаулы (100 км южнее р. Иргиз), в пес-
ках Иргиз-Кум, на протоке Сарыузек (между реками Иргиз и Тургай), затем
по р. Тургай, на оз. Сары-Копа (Динесман, 1953), в Наурзумском заповеднике
(Брагин, Брагина, 2002) и далее на север в Курганской и Тюменской областях.
Старые сведения о находках личинок чесночницы в окрестностях Аральского
моря, низовьях р. Сырдарьи (и даже в окрестностях г. Ташкента и по Чуйской
долине) оказались ошибочными - головастики при переисследовании оказа-
лись гигантскими головастиками озерной лягушки (Ананьева и др., 1998).
90
Глава 4
В очерченных границах чесночница известна на всей территории Самар-
ского (Бакиев, Файзулин, 2001, 2002), Саратовского (Шляхтин и др., 2005) и
северной части Волгоградского Заволжья (Кубанцев, Колякин, 1988), в запад-
ных и центральных районах Оренбуржья, почти по всей Западно-Казахстан-
ской, Актюбинской и западной части Кустанайской области (Чибилёв, 1995;
Искакова, 1959; Динесман, 1953; Параскив, Бутовский, 1960; Брагин, Брагина,
2002). В географическом отношении ее конкретные местонахождения извес-
тны в 6 (54,5% из 11) ландшафтных районах лесостепной, 43 (62,3% из 69)
степной, 13 (40,6% из 32) полупустынной и 5 (11,6% из 43) пустынной зон
(табл. 17, карта 5).
Места обитания. В Бузулукском бору и более северной части ареала
встречается как в смешанных, так и широколиственных лесных массивах. На
большей части остального ареала привязана к долинам крупных и небольших
рек с хорошо выраженными поймами, по которым повсеместно проникает
в степную зону, полупустыню (реки Б. и М. Узени, Кушум) и пустыню (по
р. Урал до взморья). По этой причине распространение вида на большей части
региона имеет ленточный характер. Несколько реже встречается чесночница в
котловинах крупных постоянных озер.
Во всех этих случаях возможность ее существования определяется наличи-
ем пресных вод (с низкой минерализацией и отсутствием нитратов) и рыхлого
грунта, в котором она может относительно легко закапываться (плотных каме-
нистых и глинистых грунтов определённо избегает). Эта особенность опреде-
ляет наиболее частые встречи чесночниц на участках с рыхлой почвой вбли-
зи уреза воды стариц, разных канав и копаней, причем как с травянистыми,
так и кустарниковыми зарослями. Вместе с тем на обрабатываемых огородах
и дачных участках с регулярным поливом в окрестностях г. Оренбурга она
встречалась от воды на расстоянии более 200 м. Из Волго-Ахтубинской поймы
проникает в примыкающие закрепленные бугристые пески Волго-Уральско-
го междуречья. Не переносит чесночница и засоления грунта, в связи с чем в
Тургайской депрессии, например, встречается по берегам только пресных озер
или их крупных пресных плесов (Динесман, 1953). Наличие легкого хорошо
промытого грунта в ряде случаев позволяет чесночнице существовать почти
у уреза соленых вод. В таких условиях чесночница обнаружена нами вдоль
стока пресной воды из системы водоснабжения п. Сары-Умыр в оз. Шалкар, а
также на конусе выноса в устье одной из балок, открывающейся в соленое оз.
Сорколь (30 км восточнее оз. Шалкар).
Образ жизни. Из зимовки в окрестностях г. Уральска (с. Дарьинское)
чесночницы выходили в начале третьей декады апреля (Искакова, 1959), а у
северной границы региона в Волжско-Камском заповеднике преимуществен-
но в конце апреля - начале мая, причем несколькими днями раньше в быст-
рее прогреваемых биотопах (Гаранин, 1983). Обычно это наблюдается, ког-
да температура воздуха превышает 12-14 °C, а при 16... 18 °C в окрестностях
с. Дарьинское они уже встречались в значительном количестве. Ко времени
размножения чесночницы собираются на мелководных, быстрее прогрева-
емых участках нерестовых водоемов, где проводят весь этот период. В это
время они активны почти круглые сутки. По окончании периода размножения
взрослые чесночницы выходят на сушу и расселяются по типичным место-
обитаниям. На суше они бывают активны в вечерние сумерки и начале ночи,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
91
Таблица 17
Распространение обыкновенной чесночницы
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
1 1 с. В. Сускан с. Н. Санчелеево Бакиев, Файзулин, 2002; Файзулин, 2004а, б
2 с. С. Солонец с. Хрящевка
3 с. Выселки п. Луначарский с. Подстепки
4 г. Тольятти
I 5 п. Федоровка с. Васильевка п. Поволжский
3 6 с. Пискал и некий взвоз
X с А) 7 ст. Задел ьная
ь о Q ст. Мастрюково
8 г. Самара: п. Управленческий Воронежские озера Ботанический сад
3 9 с. Титовка
10 с. Кошки
4 11 с. Красный Яр
5 12 с. Совруха
II 13 г. Похвистнево
14 с. Старопохвистнево Оренбургская область
6 7 15 16 р. Дёма р. Салмыш Положенцев, Никифо- рук, 1949; Топоркова, 1973
Самарская область
3 12 17 п. Приволжье Бакиев, Файзулин, 2002
X X IV Саратовская область
5 13 18 Повсеместно Шляхтин и др., 2005
14 19
92
Глава 4
Продолжение табл. 17
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
15 16 20 21 с. Владимировка п. Рубежное, Яицкие озера Бакиев, Файзулин, 2002; Файзулин, 2004а, б
22 с. Белозерки
IV 17 23 окр. с. Б. Глушица
24 р. Б. Иргиз, окр. с. Тепловка Саратовская область
18 25 Повсеместно Шляхтин и др., 2005
19 26
20 27
Самарская область г. Кинель:
21 28 29 п. Горский Красносамарский лес Файзулин, 2004а, б; Бакиев, Иванова, 2004
30 с. Богатое
31 ст. Заливная
Борский район:
S X 32 оз. Лебяжье
с о п. Лесное
0 22 33 Бузулукский бор Оренбургская область
V 34 Бузулукский бор: д. Елшанка п. Колтубановский р. Боровка Даркшевич, 1953
23 35 р. Ток Чибилбв А.А.
24 36 р. Ток
25 37 р. Самара
28 38 р. Б. Ик
39 р. Сакмара г. Оренбург:
29 40 пойма р. Урал, Форштадт- ские луга, Воронцовский, 1922; Дебело П.В.
41 о. Мира
30 42 р. Илек Зарудный, 1895
VI 31 43 р. Урал
32 44 р. Киялы-Буртя
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
93
Продолжение табл. 17
Зона Провин- ция Район № точки Местонахождение Источник информации
Западно-Казахстанская область
34 45 р. Утва, окр. г. Аксай Березовиков и др., 1992
35 46 р. Березовка ГП-16, пруд Коншубай,
47 р. Илек
36 48 верх. р. Утва
38 49 оз. Шалкар, п. Сары-Умыр, Дебело П.В.
50 оз. Сорколь
40 51 р. Калдыгайты Актюбинская область Кривко, 1976
42 52 р. Илек Зарудный, 1895
53 р. Б. Хобда
54 р. М. Хобда
43 55 р. Илек
44 56 р. Илек, г. Актюбинск Волгоградская область
1 VII 48 57 бас. р. Еруслан Определитель земновод- ных..., 1977
с л Саратовская область
о 49 58 Повсеместно Шляхтин и др., 2005
50 58
Оренбургская область
52 59 р. Чаган Чибилвв А. А.
60 р. Иртек
53 61 р. Чаган
62 р. К ин дел я
Западно-Казахстанская область
54 63 64 пойма р. Урал, п. Красноар- мейск с. Дарьинское Дебело П.В.; Параскив, Бутовский, 1960; Иска- кова, 1959
55 65 р. Урал, окр. п. Подстепное Актюбинская область Дебело П.В.
62 66 р. Куагач, п. Кимперсай Оренбургская область Топоркова, 1973
65 67 р. Урал Чибилёв А.А.
66 68 р. Суундук
94
Глава 4
Продолжение табл. 17
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная VII 68 69 70 окр. п. К варке но окр. п. Болотовка р. Орь Кустанайская область
XI 78 71 Наурзумский заповедник Брагин, Брагина, 2002
XII 80 72 оз. Сары-Копа Динесман, 1953
Полупустынная XIII 82 83 84 85 86 88 89 91 73 74 75 76 77 78 79 80 81 82 83 84 85 Волгоградская область лим. Кайсацкий п. Жаскайрат п. Тегешил лим. Сонали, Ахмет, Бай- гутты Западно-Казахстанская область п. Джаныбек Джаныбекский стационар р. Горькая р. Горькая Волгоградская область окр. п. Эльтон Эльтонское вдхр. бал. Биологическая р. Хара Западно-Казахстанская область окр. оз. Балыкты р. Б. Узень, окр. п. Фокеево р. М. Узень окр. п. Н. Казанка, Камыш- Самарские озера р. Кушум, Кировское вдхр. Битикское вдхр. р. Урал, окр. п. Богатск. Ходашова, 1960 Линдеман и др., 2005 Ходашова, 1960 Линдеман и др., 2005; Ходашова, 1960 Параскив, Бутовский, 1960 Дебело П.В. Кривко, 1976
XV 99 86 Актюбинская область р. Эм ба Искакова, 1959
XVIII 106 107 87 88 89 р. Иргиз р. Иргиз р. Жалаулы
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
95
Окончание табл. 17
Зона Провин- ция Район № точки Местонахождение Источник информации
а 3 ПО 90 91 окр. п. Тургай р. Тургай, 105 км сев.-вост., Динесман, 1953; Бруш- ко, Кубыкин, 1988;
XIX г. Иргиз
1 112 92 пески Иргизкум Динесман, 1953
Е 93 р. Сары-Озек
Астраханская область:
113 94 Богдинско-Баскунчакский заповедник Стрельцов и др., 2006
§ а а ё XX 114 95 Красноярский р-н, АГКМ Западн о-Казахстанская область Завьялов, Табачишин, 1998
с 124 96 р. Урал, окр. п. Круглый Параскив, Бутовский, 1960
Гурьевская область
XXI 125 132 97 98 р. Урал р. Урал, окр. г. Гурьев Искакова, 1959
затем в предутренние часы, а к 6 часам большинство особей уходит в укрытия
(норы грызунов, другие пустоты почвы, подстилку), или закапываются в грунт.
В Наурзуме А.Н. Формозов находил много чесночниц в старых гнездах во-
дяных крыс, расположенных в воде в 10-40 м от берега (Динесман, 1953).
В пасмурную погоду на поверхности иногда встречаются и днем. В целом на
активность чесночницы основное влияние оказывают влажность воздуха у
поверхности почвы и температура. На зимовку чесночницы уходят в сентяб-
ре-октябре, когда температура воздуха понижается до 9 °C. Взрослые особи
зимуют в норах грызунов, других пустотах земли, колодцах или зарываются в
грунт до 1,5-2,0 м; сеголетки чаще всего зарываются вблизи уреза воды. Ак-
тивный период жизни продолжается до 170, чаще всего 130-150 дней, а зимнее
оцепенение длится около 200-220 дней.
Питание. Головастики первое время питаются детритом, а затем переклю-
чаются в основном на водоросли, реже потребляют листья ряски и некоторых
других высших растений, простейших, коловраток, рачков и других мелких
водных беспозвоночных. Взрослые кормятся в основном разными наземными
насекомыми: жуками (особенно жужелицами), в окрестностях с. Дарьинское -
чернотелками и листоедами, муравьями, двукрылыми, гусеницами бабочек, а
также пауками, дождевыми червями и некоторыми другими беспозвоночными
(Искакова, 1959; Параскив, Бутовский, 1960; Топоркова, Меныциков, 1974; Га-
ранин, 1983). В свою очередь обыкновенную чесночницу иногда поедают щука,
окунь, взрослые зеленые лягушки, но гораздо чаще ужи, степная гадюка, а так-
же околоводные, хищные и некоторые другие виды птиц и хищные млекопита-
ющие. Известен и каннибализм в форме личинка-личинка (Кузьмин, 1999).
96
Глава 4
Численность. В прошлом на большей части региона обыкновенная чес-
ночница была сравнительно обычной, местами многочисленной, а в пойме
средней Волги одним из массовых видов земноводных. Однако с затоплением
поймы и исчезновением типичных местообитаний численность ее заметно сни-
зилась, хотя и остается на уровне «обычна» (Гаранин, 1983). Подобное, очевид-
но, произошло и в зоне сооружения Саратовского и Волгоградского водохра-
нилищ. На внепойменной территории этой части региона она никогда не была
многочисленной, а с ее освоением стала редкой и встречается спорадически
(Кубанцев, Колякин, 1988; Кубанцев, 2003). На незарегулируемом участке в
Волго-Ахтубинской пойме она и сейчас остается сравнительно обычной - на
водоемах ее численность оценивается в 17,5±0,69 ос/га, а на примыкающих, за-
крепленных бугристых песках, - 22,6±0,43 ос/га (Завьялов, Табачишин, 1998).
По-видимому, близкий уровень плотности характерен и для расположенной се-
вернее песчаной Приахтубинской части Прибаскунчакского района, а восточ-
нее на плотных грунтах Богдинско-Баскунчакского заповедника она встречает-
ся в незначительном количестве и лишь спорадически (Стрельцов и др., 2006).
В Волго-Уральском междуречье по поймам рек Самары, Б. и М. Узеней и в
Камыш-Самарской озерной системе в 1950-е гг. была сравнительно обычной, а
в конце 1960-х гг. в связи с сильным обмелением рек и почти полным пересы-
ханием озер численность ее, несомненно, значительно уменьшилась. На Волж-
ско-Узеньской равнине сравнительно обычной она бывает лишь в многовод-
ные годы (Ходашова, 1960; Линдеман и др., 2005). По р. Уралу в окрестностях
г. Оренбурга в конце XIX и начале XX в. чесночница встречалась довольно час-
то, а в 1921 г. (после засушливого года) даже при настойчивых поисках с тру-
дом удалось найти лишь несколько маленьких экземпляров (Зарудный, 1895;
Воронцовский, 1922). Подобное заметное снижение численности в этом райо-
не, вызванное продолжающейся в течение ряда последних лет засухой, наблю-
дается и в настоящее время. В целом же по Оренбургской области в последние
десятилетия обыкновенная чесночница чаще всего встречается по поймам рек
в Первомайском, Ташлинском, Илекском, Оренбургском, Соль-Илецком, Ак-
булакском и Домбаровском районах (Чибилёв, 1995). Ниже по течению р. Ура-
ла в 1950-е гг. чесночница была довольно многочисленной как в окрестностях
г. Уральска (с. Дарьинское), так и значительно южнее - у с. Круглого (Иска-
кова, 1959; Параскив, Бутовский, 1960). В 1970-е гг. на этом отрезке нижнего
течения р. Урала у п. Богатск в пойме реки в ловчие канавки с цилиндрами в
июне за ночь попадало по 5-20 особей (Кривко, 1976). Сейчас здесь с усыхани-
ем поймы количество чесночниц, видимо, также уменьшается.
Восточнее по долине р. Утвы чесночница обычна, но на примыкающей тер-
ритории Карачаганакского нефтегазового месторождения (роща возле УПК-16
в верховье р. Березовка) в 1990 г. за все лето отмечено лишь 8 особей, хотя в
отдельных местах на пруду Коншубай наблюдалось до 4-6 головастиков на
1 м2 поверхности водоема. Низкая численность вида здесь, очевидно, связана с
нефтяным загрязнением водоема, а возможно, и действием газовых грифонов
(Березовиков и др., 1992, 2003). По рекам Илек, Б. и М. Хобда в конце XIX в.
чесночница была сравнительно обычна (Зарудный, 1895); сейчас в этой части
региона она чаще всего встречается лишь в Соль-Илецком районе, что видимо,
вызвано загрязнением р. Илек промышленными сбросами с высокой концент-
рацией тяжелых металлов.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
97
В Зауралье чаще всего этот вид встречается в Кваркенском и Домбаровском
районах Оренбуржья (Топоркова, 1973; Чибилёв, 1995), а южнее - в Наурзуме,
на оз. Сары-Копа, бассейнах рек Иргиз и Тургай она, несомненно, испытыва-
ет значительные колебания в связи с изменением обводненности территории
и степени минерализации водоемов. Таким образом, на значительной части
региона за прошедшее столетие с начала более или менее регулярных наблю-
дений численность обыкновенной чесночницы заметно снизилась в связи с
трансформацией ее типичных местообитаний и изменением обводненности
территории.
Лимитирующие факторы. Обмеление и пересыхание водоемов, суровые
морозы (вызывающие массовую гибель головастиков и взрослых особей), не-
гативные последствия которых усиливаются трансформацией местообитаний,
индустриальным загрязнением водоемов и рекреацией; местами автотранс-
порт (Кузьмин, 1999; Файзулин, 20046).
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор», Оренбургского, Наурзумского, Богдинско-Баскунчакского запо-
ведников и Иргиз-Тургайского резервата. Состояние популяций на остальной
части региона особых мер по охране не требует, достаточно поддержания ес-
тественного состояния ландшафтов и предупреждения загрязнения водоемов
промышленными стоками, пестицидами, отходами.
Региональный статус. Типичный представитель батрахофауны региона с
сокращающейся численностью. Охраняется Бернской конвенцией (Приложе-
ние II).
Семейство 5. Bufonidae Gray, 1825 - Жабы
Род 6. Bufo Laurenti, 1768 - жабы
6. Обыкновенная жаба — Bufo bufo (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Один из примерно
10 видов комплекса Bufo bufo, систематические отношения между которыми
во многом остаются неясными. Обитающих на территории СССР серых жаб
раньше относили к трем подвидам, которые сейчас рассматриваются как са-
мостоятельные, географически изолированные виды отмеченного комплекса.
Видовое название Bufo bufo сейчас применяется лишь к выделяемому ранее
номинативному подвиду В. b. bufo (Linnaeus, 1758) с хорошо выраженными
межпопуляционными отличиями (которые, однако, по мнению ряда исследо-
вателей, не достигают подвидового уровня) в размерах тела, паротид, барабан-
ной перепонки, встречаемости крупных бугорков на коже, рисунку, окраске
тела и некоторым другим признакам (Ананьева и др., 1998; Кузьмин, 1999).
Распространение. Ареал вида охватывает северо-запад Африки, почти
всю Европу, юг Западной и Центральной Сибири и север Казахстана (европей-
ский фаунистический комплекс). Северная граница ареала на Восточно-Евро-
пейской равнине проходит от южной части Кольского полуострова через Ар-
хангельскую и Пермскую области, затем через Средний Урал, Красноярский
край и Иркутскую область Сибири (немного не доходя до оз. Байкал). Южная
граница следует от северной части Молдавии через Украину, Воронежскую,
98
Глава 4
Саратовскую, Оренбургскую, Челябинскую области и по северу Казахстана
достигает Алтая и Саян.
Точнее в пределах региона и его ближайших окрестностях южная граница
ареала прослеживается следующим образом: в Саратовской области известна
из окрестностей г. Вольска на правобережье и в Заволжье в долине р. Б. Ир-
гиз (Определитель земноводных..., 1977 ), затем в Бузулукском бору (Чибилёв,
1995), по долине р. Урала в окрестностях г. Уральска, выше по течению этой
реки до устья р. Илека, по его долине до ур. Биштамак, в верховьях рек Уила
и Темира (Зарудный, 1895). В Зауралье находки известны в Брединском со-
сновом бору Челябинской области (Чибилёв, 2005), затем в сходных условиях
Болотовского, Аландского и других реликтовых сосновых боров Кваркенского
района Оренбуржья. Восточнее на этой широте обыкновенная жаба появляет-
ся уже только в бассейне р. Иртыша (Искакова, 1959).
В очерченных пределах находки обыкновенной жабы известны в черте
г. Самары, саратовском Заволжье по долине р. Б. Иргиз, в Бузулукском бору,
бассейне р. Б. Кинель в окрестностях г. Бугуруслана, по долинам р. Сакмары
и ее притоков Салмыша и Б. Ика,.в долинах Урала и Илека на территории
Оренбургской области, а также в Западно-Казахстанской и Актюбинской об-
ластях. В географическом отношении ее конкретные местообитания известны
в 5 (45,5% из 11) ландшафтных районах лесостепной, 13(18,8% из 69) степной
и 2 (6,3% из 32) полупустынной зон (табл. 18, карта 6). Вместе с тем новей-
шие находки обитание вида подтверждают лишь в северной части региона, а
по пункатам вдоль южной границы ареала (рекам Б. Иргиз, Урал, Илек, Уил,
Темир) такие сведения отсутствуют. Возможно, в этой части она уже исчезла,
поскольку в окрестностях г. Уральска не найдена даже при специальных ис-
следованиях (Искакова, 1959; Параскив, Бутовский, 1960).
Места обитания. На сопредельной территории Среднего Урала и Волж-
ско-Камского края основным местообитанием вида являются глухие боры-зе-
леномошники, влажные еловые, смешанные и мелколиственные (березняки,
осинники) леса, а также хорошо облесенные поймы рек, в сходных условиях
встречается эта жаба в лесопарковых зонах городов и разных населенных пун-
ктов. Именно по поймам Сакмары, Б. Кинеля, Салмыша, Б. Ика она распро-
странена в лесостепи, а по Уралу, Самаре и Волге (до Вольска) проникла и
в степную зону. Местообитания по р. Иргизу, в окрестностях г. Уральска, по
рекам Илеку, Уилу и Темиру, вероятно, - реликты малого ледникового пе-
риода, когда облесенность территории была более значительной. Начавшаяся
позднее ксерофитизация региона, развитию которой в значительной степени
способствовали вырубка лесов и зарегулирование стока большинства рек, ве-
роятно, и привели к исчезновению краеареальных популяций на юге степной
зоны. В итоге к настоящему времени местообитания вида сохранились лишь в
г. Самаре, Бузулукском бору, окрестностях г. Бугуруслана, по р. Сакмаре и ее
притокам, а также в Кваркенских борах; обитание вида по широтному отрезку
р. Урала требует подтверждения.
Образ жизни. Обыкновенная жаба является преимущественно наземным
видом и в водоемах встречается лишь в период размножения. Из зимовки она
выходит во второй половине апреля-мае, когда воздух прогревается до 10 °C и
более. Ко времени размножения собирается в водоемы, где проводит весь этот
период, будучи активной практически круглосуточно. После окончания икро-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
99
Таблица 18
Распространение обыкновенной жабы
Зона Провин- ция Район № точки Местонахождение Источник информации
1 1 Самарская область
3 1 г. Самара, ст. Пятилетка Мельниченко и др., 1938; Бакиев, Файзулин, 2002
X с Оренбургская область
о и 5 2 р. Б. Кинель, окр. г. Бугу- руслан Чибилёв, 1995
6 3 р. Салмыш
111 9 10 4 5 р. Б. Ик хр. Шайтантау
Саратовская область
IV 13 6 р. Б. Иргиз. Определитель земновод- ных..., 1977
Оренбургская область
22 7 Бузулукский бор, р. Боровка Зарудный, 1895; Чи- билёв, 1995
V 23 8 Грачевский район, р. Ток
24 9 р. Сакмара
26 10 р. Самара, г. Бузулук
29 11 р. Урал, г. Оренбург
3 X с а VI 30 31 12 13 р. Илек р. Урал
и Актюбинская область
VII 43 14 р. Илек Зарудный, 1895
44 15 р. Илек
Западно-Казахстанская область
VIII 54 16 р. Урал
55 17 р. Урал, окр. г. Уральск Дебело П.В.
X 66 18 Оренбургская область окр. п. Кваркено. Чибилёв, 1995
• х Актюбинская область
Полу пусты ная XV 95 98 19 20 р. Уил р. Темир Зарудный, 1895
100
Глава 4
метания взрослые особи выходят на сушу и рассредоточиваются по прилегаю-
щей к нерестовым водоемам территории. На суше активность жаб смещается
на вечерние и ночные часы с максимумом около полуночи, хотя в пасмурную
и дождливую погоду часть особей активна и в дневные часы. Обычно днем они
прячутся в разных убежищах - под корнями и упавшими деревьями, в норах,
разных ямах или густой траве. Сеголетки активны днем и вечером.
На зимовку в Волжско-Камском крае обыкновенная жаба уходит в основ-
ном в сентябре, хотя последние иногда задерживаются почти до конца октября.
Зимует она преимущественно небольшими группами на суше в разных норах
и других пустотах, под упавшими деревьями, в трухлявых пнях; часть осо-
бей иногда задерживается в водоемах. Активный период жизни в большинстве
случаев продолжается 140-180, а зимнее оцепенение около 200-220 суток.
Размножение, Половой зрелости обыкновенная жаба достигает на треть-
ем-четвертом году жизни. По достижении этого возраста вскоре после выхо-
да из зимовки у северных пределов региона в Волжско-Камском крае обычно
в конце апреля - начале мая (когда вода прогревается до 8... 10 °C) взрослые
особи собираются в мелководных стоячих или слабо проточных водоемах с
зарослями погруженной растительности. Здесь в прибрежной части водоема
(на глубине до 1 м) чаще всего на подводные растения несколькими шнурами
откладывается 1200-7200 икринок. Период эмбрионного развития при темпе-
ратуре воды 10...20 °C продолжается до 20 суток. Личинки в разных водоемах
развиваются 45-95 суток. После завершения метаморфоза - с конца июня до
середины сентября (иногда только на следующий год) - сеголетки выходят на
сушу и вскоре начинается их расселение от водоемов.
Питание. Головастики питаются разными водными беспозвоночными.
Взрослые особи потребляют в основном ползающих наземных беспозвоноч-
ных - муравьев, жуков, других взрослых насекомых и их личинок, пауков,
слизней и других моллюсков, причем количество и размеры их с возрастом
увеличиваются. В свою очередь личинок и сеголеток обыкновенной жабы пот-
ребляют зеленые лягушки, а взрослых также ужи, гадюки, околоводные и не-
которые другие птицы и млекопитающие, хотя в целом благодаря ядовитым
железам обыкновенные жабы имеют не очень много врагов.
Численность. Несмотря на обширность ареала, обыкновенная жаба как
сравнительно стенотопный вид в целом отличается невысокой численностью,
особенно в южной лесостепи, где в среднем насчитывается лишь 0,57 экз. на
10 км маршрута (Шошева, 1985). Поскольку эта подзона является основным
местообитанием вида на территории региона, то в целом она здесь (даже в
лучшем случае) малочисленна, а на остальной части - редка. Невысокая чис-
ленность обыкновенной жабе была свойственна и ранее (Зарудный, 1895; Рай-
ский, 1951), в последние десятилетия она заметно снизилась, а ряд южных по-
пуляций, по-видимому, исчез.
Лимитирующие факторы. Наиболее существенное значения для вида
имеют обезлесение региона и уменьшение его обводненности, что способству-
ет ксерофитизации территории. Оставшиеся водоемы испытывают усиленное
техногенное загрязнение, их побережья находятся под достаточно высоким
прессом рекреационной нагрузки, что снижает успешность размножения вида.
В садах и на огородах численность снижается из-за интенсивной агротехники
и применения ядохимикатов. Местами на численность вида заметное влияние
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
101
оказывает автотранспорт. Все это при невысокой способности вида к синант-
ропотизации и определяет снижение его численности и сокращение площади
ареала.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной территории региона необходимы сохранение мес-
тообитаний, предупреждение загрязнения водоемов и водоохранных зон.
Региональный статус. Малочисленный краеареальный вид. Занесена в
Красные книги Самарской области (2009) и сопредельных территорий Баш-
кортостана и Татарстана. Охраняется Бернской конвенцией (Приложение III -
виды, эксплуатация которых регулируется в соответствии с требованиями
Конвенции).
7. Зеленая жаба - Bufo viridis (Laurenti, 1768)
Значение таксона для сохранения биоразнообразия. Это центральный
член комплекса зеленых жаб, включающий ряд диплоидных и полиплоидных
форм, систематические отношения между которыми во многом остаются не-
ясными (Писанец, Щербак, 1979). Считается, что популяции европейской час-
ти ареала этого комплекса представлены диплоидными формами, а восточные
преимущественно полиплоидны. Выделяется несколько (по старым данным
семь) подвидов, однако статус большинства из них требует уточнения. Приня-
то считать, что восточноевропейскую часть региона населяет номинативный
подвид В. v. viridis Laurenti, 1768, а равнинную часть Средней Азии -туранская
жаба В. v. turanensis Hemmer, Schmidtler et Bohme, 1978. Однако граница меж-
ду ними проводится довольно условно по «южным степям и полупустыням
Западного Казахстана». Возможно, центральная часть региона представляет
собой своеобразную зону интерградации подвидов, атипичные В. v. viridis и
В. V. turanensis населяют соответственно лишь северо-запад и юго-восток реги-
она. Возможность подобной ситуации подтверждается тем, что морфометри-
ческие показатели особей уральской популяции не укладываются в пределы,
свойственные номинативному подвиду (Топоркова, 1981).
Различные популяции различаются высокой географической, биотопичес-
кой и индивидуальной изменчивостью в размерах тела, конечностей, паротид,
сочленовных бугорков, величине и форме зеленых пятен, общей окраской,
величиной и расположением бугорков и шипиков на коже, причем значения
многих из диагностических признаков перекрываются (Ананьева и др, 1998;
Кузьмин, 1999; Пескова, 2003; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает северную Африку, большую
часть Европы, южную Сибирь, Переднюю, Среднюю и часть Центральной
Азии до Китая и Монголии (средиземноморский фаунистический комплекс).
Северная граница ареала на Восточно-Европейской равнине проходит от Бал-
тики через Чудское озеро, Псковскую, Вологодскую, Пермскую области, за-
тем через Средний Урал, Челябинскую, Курганскую, Павлодарскую, Семипа-
латинскую, Восточно-Казахстанскую области и Алтай в Монголию. Южная
граница проходит через Переднюю Азию, хребет Копетдаг, Памиро-Алай и
Тянь-Шань. Территория региона, таким образом, полностью входит в ареал
вида. Вместе с тем распространен он здесь далеко не равномерно, а в ряде
мест отсутствует. Конкретные местонахождения зеленой жабы известны в
102
Глава 4
10 (90,9% из 11) ландшафтных районах лесостепной зоны, 55 (79,7% из 69)
степной, 20 (62,5% из 32) полупустыни и 14 (32,6% из 43) пустыни (табл. 19,
карта 7).
Места обитания. Зеленая жаба является одним из самых эвритопных и
пластичных видов земноводных Палеарктики, что обусловлено ее высокой за-
сухоустойчивостью, способностью использовать для икрометания временные
водоемы, меньшей подверженностью негативному влиянию внешней среды, а
также ночным образом жизни, который в большинстве случаев ведут взрослые
особи (Ананьева и др., 1998; Вершинин, 2010). Она населяет смешанные и ши-
роколиственные леса, лесостепную, степную, полупустынную и пустынную
зоны материка.
В северной лесостепной части региона зеленая жаба обитает в разреженных
лесах, по дорогам и опушкам проникает в глубь плотных лесных массивов,
встречается по песчаным руслам лесных речек, постоянно обитает в поймах
средних и крупных рек, где преимущественно придерживается незаливаемых
грив, остепненных склонов надпойменных террас, других открытых биотопов,
причем порой на значительном удалении от воды (Гаранин, 1983).
В степной зоне она является постоянным обитателем долин рек Волги,
Самары, Урала, их крупных (Сакмары, Илека, Ори) и менее значимых (Б. и
М. Иргиза, Еруслана, Чапаевки, Тока, Кумака, Чагана, Утвы и др.) притоков и
Уила, где придерживается окрестностей стариц, участков с медленным течени-
ем, защищенных от постоянного волнобоя участков водохранилищ. На более
мелких степных речках и по балкам основное местообитание (и своеобразные
стации переживания) - небольшие водохранилища (Черновское, Кумакское) и
степные пруды (их только в Оренбургской области насчитывается более трёх
тысяч). На пересыхающих речках она придерживается в основном участков с
постоянными плесами. В небольших балках и других временных водотоках
наблюдалась у пересыхающих луж, где на некоторое время задерживаются та-
лые или скапливаются дождевые воды. Населяет зеленая жаба также котлови-
ны пресных озер (Наурзум, Сары-Копа, Светлинские), отмечена на опреснен-
ном участке солоноватого озера Шалкар (в месте впадения канала из р. Урал)
и на песчаном участке его берега у п. Сары-Умыр. Известна также в бывшем
накопителе сточных вод г. Уральска (Анисьины озера) и прудах биологичес-
кой очистки (балка Вишневая).
У южных пределов степной зоны существенную роль в пространственном
распределении зеленой жабы начинает играть такой важный биоценотический
компонент сухостепных ландшафтов, как малый суслик, в поселениях кото-
рого, по мнению ряда исследователей, обитает не менее 75-80% численности
природных популяций вида (Варшавский и др., 1985). Значительно чаще, чем
в лесостепной зоне, встречается жаба и в поселениях человека, где держится
в погребах и хозяйственных постройках. Кстати, г. Уральск (бывший Яицкий
городок), пос. Чаганский и его окрестности признаются типовой территорией
подвида (Паллас, 1773; цит. по: Паллас, 2006), а окрестности расположенного
в 40 км от этих пунктов п. Переметное признаются типовой территорией этого
вида, описанного как Bufo cursor И. Лепехиным (1772).
В полупустыне основными постоянными местообитаниями зеленой жабы
являются травянистые поймы рек Волги, Урала и Эмбы. В Волго-Уральском
междуречье важную роль играют долины Б. и М. Узеней, Ку шума, Горькой и
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
ЮЗ
Таблица 19
Распространение зеленой жабы
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная I 1 2 1 2 3 4 5 6 7 8 *9 10 11 Самарская область с. В. Сускан с. Н. Санчелеево с. Хрящевка с. Выселки п. Луначарский п. Подстепки п. Приморский г. Тольятти, Центральный район, ул. Баныкина п. Портовый Автозаводской район, пром- зона, жилая зона п. Федоровка с. Васильевка п. Поволжский с. П искал и некий взвоз ст. Задельная ст. Мастрюкова г. Самара: п. Управленческий Воронежские озера пруды Ботсада п. Старосемейкино с. Красный Яр Бакиев, Файзулин, 2002; Файзулин, 2004а
II 3 4 5 12 13 14 15 16 17 18 19 с. Кошки с. Елховка р. Шешма с. Савруха г. Похвистнево п. Октябрьский п. Тимашово г. Отрадный р. Сарбай, г. Кинель Файзулин, 2004а; Баки- ев, Файзулин, 2002
104
Глава 4
Продолжение табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная II 6 7 20 21 Оренбургская область р. Дема р. Салмыш Чибилбв А.А.
III 9 10 11 22 23 24 р. Б. Ик Кувандыкский район бас. р. Сакмара
S X с IV 12 13 14 15 16 17 18 19 20 25 26 27 28 29 30 31 32 33 34 35 Самарская область п. Приволжье Саратовская область Повсеместно Самарская область с. Мосты Красносамарский лес с. Утеевка с. Б. Глушица с. Тамбовка Саратовская область Повсеместно <с Бакиев, Файзулин, 2002 Шляхтин и др., 2005 Бакиев, Файзулин, 2002 Шляхтин и др., 2005
и О V 21 22 23 24 25 36 37 37 38 39 40 41 42 43 44 45 46 Самарская область п. Горский Бузулукский бор Оренбургская область Бузулукский бор р. Боровка п. Колтубановский р. Ток р. Самара р. Б. Уран р.М.Уран окр. п. Сакмара р. Каргалка, п. Каргала р. Салмыш р. Урал Бакиев, Файзулин, 2002 Чибилев А.А.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
105
Продолжение табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
26 47 С. Б. Черниговка п. Восточный с. Украинка Бакиев, Файзулин, 2002
48 с. В. Ростоши Смелянский, Елизаров, 1996
п. Кошкин
49 ур. Синий Сырт
V ур. Грызл ы
Западно-Казахстанская область
27 50 р. Деркул Оренбургская область Дебело П.В.
28 51 р. Сакмара Чибилбв А.А.
п. Саракташ Воронцовский, 1922
29 52 г. Оренбург, Форштадские луга Дебело П.В.
be 53 о. Мира
Я Я С 30 54 р. Илек Зарудный, 1895
р О VI 31 55 р. Донгуз Березовиков и др., 1992
32 56 р. Черная Чибилбв А.А.
Западно-Казахстанская область
33 57 р. Барбастау Дебело П.В.
34 58 р. Утва р. Акбулак г. Аксай Березовиков и др., 1992
VII 35 59 р. Березовка пруд Коншубай прудуУКПГ-16
36 60 р. Утва р. Караоба
37 61 верх. р. Утва
38 62 оз. Шалкар у п. Сарыумыр Дебело П.В.
63 оз. Шалкар, канал из р. Урал
40 64 р. Калдыгайты п. Егендыколь
106
Глава 4
Продолжение табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
Актюбинская область
42 65 р. Илек Зарудный, 1895
66 р. М. Хобда
VII 43 67 68 р. Илек р. Б. Хобда
44 69 р. Илек, г. Актюбинск
45 70 р. Уил
46 71 р. Уил
Волгоградская область
47 72 р. Волга Кубанцев, Колякин, 1988
48 73 р. Еруслан Саратовская область Кубанцев, 2003
49 74 Повсеместно Шляхтин и др., 2005
50 75
Западно-Казахстанская область
51 76 п. Переметное, Лепехин, 1772
5 р. Деркул
53 77 р. Ембулатовка, п. Рожков, п. Петров Дебело П.В.
р. Быковка, п. Чеботарев
VIII 78 р. Рубежка
79 п. Рассвет
54 80 п. Красный Свет р. Урал п. Красноармейск
81 с. Дарвинское Искакова, 1959
55 82 г. Уральск Паллас, 1773; Дебе- ло П.В.
83 Анисьины озера
84 п. Подстепное оз. Едучее п. Круглоозерное Биостанция ЗКУ
85 п. Б. Чаган
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
107
Продолжение табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
IX 56 57 59 60 61 62 86 87 88 89 90 91 Оренбургская область пойма р. Урал р. Урал р. Урал г. Орск р. Урал Актюбинская область р. Куагач п. Тайкеткен п. Кимпсрсай Чибилёв А.А. Топоркова, 1973
Степная X 65 66 67 68 92 93 94 95 Оренбургская область Ириклинское вдхр. р. Суундук п. Болотовка р. Кумак р. Орь
XI 72 78 96 97 р. Буруктал, Светлинские озера Кустанайская область Наурзумский заповедник Дебело П.В. Динесман, 1953; Брагин, Брагина, 2002
XII 79 80 98 99 р. Сары-Озек оз. Сары-Копа Искакова, 1959 Никольский, 1899; Ди- несман, 1953
Полупустынная XIII 81 82 83 84 100 101 102 103 104 105 Волгоградская область Волго-Ахтубинская пойма лим. Кайсацкий Западно-Казахстанская область п. Джаныбек, ДжаныбекскиЙ стационар Волгоградская область п. Жаскайрат п. Тегешил лим. Сонали, Ахмет, Бай- гутты р. Горькая р. Горькая Кубанцев, Колякин, 1988; Кубанцев, 2003 Ходашова, 1960 Параскив, Бутовский, 1960; Быков, 1981 Линдеман и др., 2005; Ходашова, 1.960
108
Глава 4
Продолжение табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
85 106 п. Эльтон окр. оз. Эльтон бал. Биологическая р. Хара
Западно-Казахстанская область
86 107 окр. оз. Балыкты
XIII 87 108 р. Б. Узень, п. Фокеево Параскив, Бутовский, 1960
109 р. М. Узень, п. Н. Казанка
88 ПО Камыш-Самарские озера р. Ку шум: Дебело П.В.
89 111 Кировское вдхр.
3 X 112 Битикское вдхр.
я н 113 Донгу люкс кое вдхр
Q 91 114 р. Урал, окр. п. Кожехарово Параскив, Бутовский,
t? 1960
О С XIV 94 115 р. Калдыгайты, окр. п. Еген- дыколь Дебело П.В.
95 116 р. Уил Гурьевская область Искакова, 1959
XV 96 117 р. Эмба (ср. течение)
98 118 р. Темир
99 119 р. Эмба
Актюбинская область
XVIII 106 120 р. Иргиз Динесман, 1953
107 121 р. Иргиз
109 122 р. Улькояк
XIX НО 123 р. Тургай
112 124 р. Иргиз, ур. Жал аулы
Астраханская область
лынная 113 125 Вол го-Ахту би некая пойма Определитель земновод- ных..., 1977
XX 126 Богдинско-Баскунчакский Стрельцов и др., 2006
заповедник
114 127 Красноярский р-н АГКМ Завьялов, Табачишин, 1998
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
109
Окончание табл. 19
Зона Провин- ция Район № точки Местонахождение Источник информации
XX Западно-Казахстанская область
116 128 п. Урда, ур. Копа Параскив, Бутовский, 1960; Искакова, 1959
124 129 р. Урал, п. Круглый
125 130 р. Урал Гурьевская область
3 X X XXI 131 132 131 132 р. Урал, проток Курайли р. Урал, окр. г. Гурьев, г. Гурьев, 35-40 км вост, дельты р. Урал Определитель земновод- ных..., 1977; Искакова, 1959; Варшавский и др., 1985; Гисцов, Березови- ков, 1995
о К 134 133 р. Уил Искакова, 1959
XXII 135 136 134 135 р. Уил р. Эмба
137 136 р. Эмба
151 137 пески Б. Барсуки
154 138 Приаральские Каракумы Лобачев и др., 1973
XXVI 155 139 140 Аральское море, сев. побе- режье окр. г. Аральск Зарудный, 1915; Сидо- ров, 1925; Динесман, 1953
других менее значимых рек. После спада паводковых вод русла их обычно пе-
рекрываются плотинами, обеспечивающими поддержание уровня по наиболее
глубоким плесам (своеобразным стациям переживания вида), а на р. Кушу-
ме сооружены четыре водохранилища с системой каналов, поддерживающих
существование популяций Чижинских, Дюринских, Балыктинских разливов и
лиманов Урало-Кушумского междуречья.
В Уральско-Эмбенском междуречье основными местообитаниями являют-
ся долины рек Калдыгайты, Уил, Темир, а в Зауралье - Иргиз и Тургай; в мень-
шей степени она оказывается привязана к другим водотокам, большинство ко-
торых летом пересыхает или сильно осолоняется (Улькояк и др.). Повышенная
минерализация водоемов в ряде случаев - один из важнейших лимитирующих
факторов, определяющих здесь характер распространения вида.
В полупустыне более четко проявляется привязанность зеленых жаб к по-
селениям малого суслика, в норах которого постоянно сохраняется до 85-90%
численности природных популяций вида (Варшавский и др., 1985), который
вне их поселений встречается несравненно реже или даже единично. Здесь
важную роль в сохранении популяций по-прежнему играют населенные пун-
кты, в постройках, погребах которых, на приусадебных участках, колодцах и
на орошаемых плантациях овощных и бахчевых культур обитает значительное
количество жаб. Существенно усложняли «кружево ареала» в этой зоне мно-
гочисленные животноводческие «точки» с сопутствующими им колодцами и
но
Глава 4
копанями. В ряде мест благоприятную роль играют придорожные кюветы, где
вода иногда может сохраняться до середины лета.
Однако наиболее ярко адаптационные способности зеленых жаб прояв-
ляются в зоне пустыни. Здесь они обитают не только у пресных, но и слабо
минерализованных (до 11%) водоемов - весенних разливов на солончаках и
такырах, в солоноватых заливчиках и у мест выхода пресных источников в
Аральском море, что для региона было отмечено еще Н.А. Зарудным (1895,
1915). Л.Г. Динесман (1953) эту способность вида существовать в условиях
повышенной солености Аральского моря объяснил выработкой адаптации в
процессе длительного постепенного осолонения некогда пресного озера, а в
р. Улькояк и других соленых водоемах изменение минерализации происходит
резко и быстро, что оказывается недостаточным по времени для выработки
приспособления, или даже если бы приспособление возникло, оно вряд ли со-
хранилось. У пресных водоемов наиболее стабильные условия обитания харак-
терны для долин рек Волги и Урала. В Волго-Уральском междуречье вблизи
Ахтубинской поймы зеленая жаба населяет закрепленные бугристые пески и
межбугровые депрессии (Завьялов, Табачишин, 1998), а по уральским рукавам
она проникает на восточную окраину Волго-Уральских песков и в зауральское
левобережье.
В пустыне наиболее ярко проявляется и способность вида использовать
для икрометания и временные пресные водоемы, даже такие, которые обра-
зуются в колее автотранспорта после дождей. Здесь же возрастает и значение
биоценотических партнеров, в роли которых в пустыне выступают песчанки -
в их норах находят убежище до 90-95% обитающих здесь природных популя-
ций (Варшавский и др., 1985). Определенную роль в сохранении природных
популяций в пустыне играют постоянные населенные пункты и временные
поселения (с сопутствующими скважинами, колодцами и копанями), их по-
явление благоприятно сказалось на расширении спектра осваиваемых видом
биотопов как в Приаральских Каракумах (Лобачев и др., 1973), так и в пустын-
ной зоне в целом.
Образ жизни. Зеленая жаба вне периода размножения ведет исключитель-
но наземный образ жизни. Из зимовки она выходит несколько раньше других
видов земноводных, когда воздух прогревается до 8-10, а вода до 3,5 °C. На
юге региона это обычно происходит в середине или даже начале марта, а у его
северных пределов - в начале, середине или даже конце апреля. В зависимости
от хода развития весенних явлений жабы сразу же или через одну-две недели
начинают концентрироваться в местах размножения, где и проводят весь этот
период на мелководных, хорошо освещенных и прогреваемых участках водо-
емов. В период размножения они активны почти круглосуточно, а после его
завершения активность сохраняется в основном в сумерки и ночью (хотя на
юге региона активность иногда наблюдается и в дневные часы). После окон-
чания размножения жабы выходят на сушу, но на юге региона в жаркие дни
они отмечаются и в водоемах. Днем они скрываются в норах, разных пустотах
почвы, под корнями и другими лежащими предметами (причем иногда неболь-
шими группами) или зарываются в мягкий грунт. В населенных пунктах но-
чью собираются на освещенных электричеством местах, где охотятся на при-
влеченных светом насекомых - в г. Астрахани на дворе 50x40 м 16.06.1972 г.
в таких условиях насчитали 110 охотящихся жаб (Варшавский и др., 1985).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 111
В зависимости от хода развития осенних явлений в зимнюю спячку зеленая
жаба уходит в разное время - обычно при снижении температуры воздуха до
6...8 °C. На севере региона это чаще всего происходит в конце сентября - пер-
вой половине октября, на юге - в конце октября - начале ноября. Таким обра-
зом, активный период жизни продолжается до 220, чаще всего 180-200 дней,
а зимнее оцепенение - около 150 дней. Зимуют жабы в норах, пустотах земли
(на глубине до 3 м), ямах, колодцах, погребах, подвалах, иногда зарываются в
рыхлую почву. На юге ареала известны случаи зимовки в водоемах.
Размножение. Половой зрелости зеленая жаба достигает на третьем-чет-
вертом году жизни. В этом возрасте после выхода из зимовки жабы концент-
рируются в пресных или солоноватых водоемах со стоячей или слабопроточ-
ной водой - старицах рек, прудах, мелководных заливах озер и водохранилищ,
заболоченных лугах, различных временных водоемах (придорожных кюветах,
канавах, дождевых лужах, лужах у скважин, колодцев, в местах водопоя скота,
весенних разливах талых вод и т. п.). О начале размножения свидетельствуют
брачные «трели», которые на юге региона становятся слышны с начала апреля,
а на севере с начала мая. В постоянных водоемах брачный период продолжа-
ется около месяца, во временных, особенно в пустынной зоне, его продолжи-
тельность сильно растянута и зависит от времени выпадения обильных дождей
и образования луж. Это иногда наблюдается лишь в середине августа, и пери-
од размножения таким образом увеличивается до 150-170 суток. Нерестятся
жабы обычно на заросших водной растительностью мелководьях (чаще всего
глубиной до 0,5 м), когда температура воды достигает 9... 18 °C. Во временных
водоемах икра часто откладывается на грунт. Кладка обычно состоит из 2 тя-
жей длиной 0,7-7,0 м, в которых насчитывается от 2 тыс. до 30 тыс. икринок.
Эмбриональное развитие при температуре воды более 10 °C продолжается
3-11 суток и уже с середины апреля - начала мая появляются головастики. Их
развитие обычно продолжается 45-70 суток, а во временных водоемах пустын-
ной зоны сокращается до 21-25 суток. Конкретные сроки отдельных фаз про-
цесса размножения в разных пунктах региона и его ближайших окрестностях
приведены в табл. 20.
После завершения метаморфоза молодые особи некоторое время держатся
в водоемах (в глубоких значительно дольше), а затем выходят на сушу, что в
условиях Казахстана чаще всего наблюдается во время дождей.
Питание. Головастики питаются преимущественно детритом и водорос-
лями, а животные корма (простейшие, коловратки, мелкие ракообразные) пот-
ребляют в меньшем количестве. Во время интенсивного метаморфоза питание
прекращается и вновь возобновляется, когда молодые (с небольшим рудимен-
том хвоста) выходят на сушу. В это время они питаются двукрылыми, паука-
ми, жуками, а взрослые переходят на ползающих беспозвоночных, в основном
на муравьев и другие фоновые виды. В желудках особей, выловленных в под-
валах, было много многоножек, червей, мокриц; у обитателей огородов — слиз-
ней, медведок, чернотелок, совок; в других местах кроме муравьев отмечались
саранчевые, клопы, мухи, медведки, жужелицы, чернотелки, щелкуны и дру-
гие насекомые; водные беспозвоночные отмечались лишь в весенний период
жизни. В свою очередь взрослых и сеголеток зеленых жаб поедают лягушки,
ужи, гадюки, околоводные и некоторые другие птицы и млекопитающие; из-
вестен и каннибализм.
112
Глава 4
Таблица 20
Сроки размножения и развития зеленых жаб
Сроки
Пункт икрометания регистрации голо- вастиков метаморфоза Примечание
г. Казалинск 07-10 ИЮЛЯ Массовый выход
Аральское море 28 мая - 29 июня 16 августа
г. Гурьев (Атырау) Конец мая - 18 июня Июнь - массовый выход
р. Урал п. Круглый 13-15 июня
п. Урда 19 июня
р. Иргиз 8-12 июля
р. Тургай Август
Наурзум Июнь Июль-август
оз. Шалкар Июль-август
р. Кушум Кировское вдхр. Июль-август Конец июня
Окр. г. Уральска Анисьины озера Конец июня
р. Урал с. Дарьинск Конец июня
Саратовская обл. Июль*август
Башкирия 03 мая Июнь Конец июня - июль Массовый выход
Численность. Зеленая жаба в регионе наиболее распространенный вид
земноводных и на большей части его территории довольно обычна, а местами
даже многочисленна.
Наиболее ранние сведения о ее численности имеются для г. Оренбуга, в ок-
рестностях которого в конце XIX - начале XX в. по пойме р. Урала встречалась
«очень часто, а местами в изобилии», обычной была по р. Илек, его притокам
и р. Утва (Зарудный, 1895; Воронцовский, 1922). В это же время «довольно
обыкновенною» она была на северном побережье Аральского моря и «часто
попадалась» на небольших его островах (Елпатьевский, 1903; Зарудный, 1915;
Сидоров, 1925). Существенных изменений численности здесь и, по-видимому,
на остальной территории региона не происходило до середины прошлого сто-
летия. Во всяком случае для Оренбуржья это подтверждается наблюдениями
А.П. Райского (1951), а для Приаралья Л.Г. Динесмана (1953). По материалам
последнего автора зеленая жаба на берегу моря была немногочисленна, но у
колодцев и луж по сухим долинам их было очень много; севернее по долинам
рек Иргиз, Тургай, в ур. Жалаулы они были сравнительно обычны (хотя в по-
пуляциях численно преобладали молодые и сеголетки), а у соленых водоемов
(р. Улькояк, оз. Сарыкопа) встречались единично. В целом численность зеле-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
113
ных жаб в этом районе «подвержена сильным годовым изменениям», связан-
ным с усыханием и осолонением водоемов. Западнее в окрестностях г. Гурьева
в июне 1966 г. на одной из луж плотность достигала 8-20 ос/м2, а всего в ко-
лее протяженностью 2,5 км насчитывалось около 70 тысяч молодых (Варшав-
ский и др., 1985). Немногим севернее в пойме по нижнему течению р. Урал
(п. Круглый) во второй декаде июня плотность достигала 8-12 ос/м2, а в Волж-
ско-Уральском междуречье близ п. Урда (ур. Копа) месяцем позже насчиты-
валось 2-7 ос/м2 (Параскив, Бутовский, 1960). Вполне обычной зеленая жаба
была в зеленых зонах населенных пунктов, на огородах и плантациях северной
части нижнего Поволжья, нередко здесь встречалась в степи, а в Волгограде
на 1 км маршрута вечером насчитывалось 15-20 особей. Однако уже к нача-
лу 1960-х гг. численность ее повсеместно заметно упала, в оврагах и балках
ее удавалось обнаружить очень редко и лишь местами, а в городах она стала
встречаться лишь изредка (Кубанцев, Колякин, 1988).
В последующие десятилетия усиление освоения территории, несомненно,
сказалось на динамике численности вида, хотя в разных частях региона оно
было неоднозначным. В пустыне и полупустыне в 1970-1980-е гг. появились
постоянные и временные населенные пункты и многочисленные животновод-
ческие точки с сопутствующими скважинами, колодцами, колонками, водопой-
ными пунктами, что способствовало не только усложнению «кружева ареала»
вида, но и увеличению его численности (Быков, 1981). Однако уже в 1990-е гг.
многие временные поселения были заброшены, а запруды разрушены. На
состоянии полусинантропных популяций негативно сказалась неумеренная
химизация сельскохозяйственных угодий. Зарегулирование поверхностного
стока на временных водотоках, сооружение здесь многочисленных небольших
запруд благотворно повлияли на численность вида в степной и полупустын-
ной зонах, но вместе с тем поставило под угрозу существование популяций
Камыш-Самарской, Кушумской, Уильской и других устьевых озерных систем.
Негативные последствия этого на фоне усиливающейся засушливости климата
и уменьшения уровня паводков (что особенно заметно в последние годы) при-
вели к пересыханию и осолонению многих прежних мест массового размно-
жения, что повлияло на численность вида; в связи с обмелением Аральского
моря и обсыханием его северной части многие популяции здесь, по-видимо-
му, исчезли. На состояние ряда популяций негативно воздействует разработ-
ка нефтяных и газовых месторождений. Так, по наблюдениям на территории
Карачаганакского нефтегазоконденсатного месторождения (на границе Орен-
бургской и Западно-Казахстанской областей) в местах выхода грифонов и на
действующих буровых площадках зеленые жабы не отмечались, сюда они воз-
вращались лишь спустя несколько лет после рекультивационных работ. В зоне
влияния промысла на пруду Коншубай (бассейн р. Березовка) численность
вида была в 1,5 раза меньше, а смертность в 3 раза выше, чем на контрольной
площадке по относительно чистой р. Утве (Березовиков и др., 1992). На состо-
янии ряда популяций негативно сказывается урбанизация.
Современное состояние численности вида в регионе и на некоторых сопре-
дельных участках по далеко неполным сведениям представляется следующим
образом: в лесостепной зоне региона зеленая жаба в целом остается сравни-
тельно обычным видом; на сопредельной территории Альметьевского района
Татарстана на 1 км береговой линии р. Елга было обнаружено 15 «поющих
114
Гпава 4
самцов» (Гаранин, 1983), в Зиянчуринском районе Башкортостана средняя
плотность летом 2004 и 2005 г. составляла 37,6 ос/га (Габитова, Юмагулова,
2006), весьма обычна она была и в населенных пунктах. Вместе с тем в горо-
дах она немногочисленна - только в г. Тольятти ежегодно регистрируется по
5-10 особей, а в г. Самаре она встречается единично (Бакиев и др., 2003). Юж-
нее в степной зоне Самарской (Куйбышевской) и Оренбургской областей зеле-
ная жаба является довольно обычным видом вдоль р. Самары, в Бузулукском
бору, окрестностях Оренбурга, хотя здесь в пойме рек Сакмары и Урала (как и
ниже по течению этой реки) в связи с низким паводком ряд известных прежде
мест ее размножения, вероятно, исчез. По-видимому, на таком же уровне нахо-
дится численность вида у северо-восточных пределов Оренбуржья в Кваркен-
ском районе, поскольку в естественных биоценозах сопредельной территории
музея-заповедника «Аркаим» в Челябинской области насчитывалось в сред-
нем 3,7 ос/100 м2, а в черте научного городка до 39 ос/ 100 м2 (Чибилёв, 2005).
В Утва-Илекском междуречье в 1989 г. по долине р. Березовка плотность
достигала 12,4 ос/га, а по р. Утве - 6,7. В 1990 г. в этом районе по р. Утва в
окрестностях г. Аксай насчитывали 14,5 ос/км, а в пруду Коншубай (зона про-
мысла КТКМ) - 10,7 (Березовиков и др., 1992).
В окрестностях г. Уральска очень многочисленной зеленая жаба была в
наиболее осветленной части городского накопителя сточных вод - Анисьиных
озерах. Здесь в окрестностях аграрного института в 1970-1980 гг. при выходе
сеголеток на 1 м2 берега мы насчитывали до 14-35 особей (сейчас с прекраще-
нием сброса они пересыхают). В пойме р. Урала этого района (у озер Едучего,
Песчаного) численность была значительно ниже - до 3 ос/м2. На оз. Шалкар в
эти годы А.М. Кривко (1976) и нами вблизи устья канала из р. Урал и в окрес-
тностях п. Сары-Умыр насчитывалось до 20 ос/м2.
В полупустыне по северной части Нижнего Поволжья зеленая жаба сей-
час малочисленна (Кубанцев, 2003; Линдеман и др., 2005), в небольшом ко-
личестве встречается и на территории Богдинско-Баскунчакского заповедника
(Стрельцов и др., 2006). В восточной части Волго-Уральского междуречья на
Кировском водохранилище по р. Кушум во время выхода молодых числен-
ность достигала 2-10 ос/м2 (Кривко, 1976).
Южнее - в Волго-Ахтубинской пойме на территории Астраханского
НГКМ - плотность в среднем составляла 27,4±0,4 ос/га, в примыкающих за-
крепленных бугристых песках Волго-Уральского междуречья 10,00±0,44, а в
межбугровых понижениях 4,30±0,65 ос/га (Завьялов, Табачишин, 1998). На
остальной территории пустыни в связи с усилением аридности численность в
целом, по-видимому, существенно уменьшилась, а некоторые популяции по-
бережья Аральского моря исчезли или сильно деградировали.
Лимитирующие факторы. Разрушение, обмеление, пересыхание и осоло-
нение водоемов, трансформация типичных местообитаний в процессе урбани-
зации и рекреационного освоения территории. Негативное влияние оказывают
неумелое и нерациональное использование удобрений и пестицидов на полях,
плантациях овощных и бахчевых культур, огородах и приусадебных участках,
а также загрязнение водоемов техногенными сбросами и отходами. В ряде
мест много жаб гибнет под колесами автотранспорта (Файзулин, 20046). Тем
не менее зеленая жаба отличается высоким потенциалом синантропизации,
что позволяет ей осваивать новые местообитания.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 115
Охрана. Обитает и охраняется на территории национального парка «Бу-
зулукский бор», Оренбургского, Наурзумского, Богдинско-Баскунчакского за-
поведников и Иргиз-Тургайского резервата. На остальной территории региона
угрозы существованию вида нет. Здесь достаточно поддержания естественно-
го состояния ландшафтов и предупреждения загрязнения водоемов техноген-
ными сбросами, пестицидами и отходами.
Региональный статус. Типичный представитель батрахофауны региона.
Охраняется Бернской конвенцией (Приложение II).
Семейство 6. Hylidae Gray, 1825 - Квакши
Род 7. Hyla Laurenti, 1768 - квакши
8. Обыкновенная квакша - Hyla arborea (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Это центральный
член комплекса видов Hyla arborea, отношения между которыми в ряде случа-
ев остаются спорными. Обитавших на территории СССР квакш раньше отно-
сили к трем подвидам, которые сейчас на основании биохимических, поведен-
ческих и экологических различий рассматриваются и признаются (не всеми)
как самостоятельные виды отмеченного комплекса. Видовое название Hyla
arborea сейчас применяется к выделяемому ранее номинативному подвиду,
который считается политипическим видом, состоящим, по-видимому, из че-
тырех подвидов. Европейскую часть России населяет номинативный подвид
Н. a. arborea (Linnaeus, 1758), отдельные популяции которого отличаются раз-
мерами, рисунком, окраской тела и некоторыми другими признаками (Анань-
ева и др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2002).
Распространение. Ареал вида охватывает Западную, Центральную, юг
Восточной Европы и Кавказ (европейский фаунистический комплекс). На Вос-
точно-Европейской равнине современная северная граница ареала проходит от
Балтийского моря через южную Литву, Белоруссию (Гомель), Брянскую, Кур-
скую, Белгородскую, Харьковскую, Днепропетровскую и Донецкую области.
Вместе с тем, по сведениям И.П. Фалька (1786; цит. по: Гаранин, 1983),
квакша в прошлом встречалась на р. Урале, И.Г. Георги (цит. по: Николь-
ский, 1918) сообщал о ее встречах по Волге и Уралу, П.С. Паллас (1814) и
Э. Эйхвальд (цит. по: Гаранин, 1983) указывали ее для Волги, а по сведени-
ям Н.А. Зарудного (1895), квакша в очень небольшом числе попадалась в
Караваевой роще около г. Оренбурга. Кроме того, имеются сообщения о ее
встречах и на сопредельных территориях, в том числе юго-западных склонах
Урала (Сабанеев, 1874). Все это дает основание предполагать, что в недале-
ком прошлом квакша обитала на Волге и Южном Урале, но затем исчезла
(карта 8).
Предполагается, что основной причиной исчезновения вида у восточных
пределов его распространения, в том числе на территории нашего региона,
стала общая аридизация климата, негативные последствия которой усугубля-
ются вырубкой лесов, исчезновением водоемов и некоторыми другими видами
антропогенной деятельности. Учитывая, что населявший прежде восточную
часть ареала номинативный подвид Н. a. arborea (Eichwald, 1830) находится
116
Глава 4
в состоянии, близком к угрозе исчезновения, а возможно, что в 1980-е гг. он в
России полностью исчез, предпринимаются попытки его реинтродукции (Ана-
ньева и др., 1998; Пестов и др., 2003).
Семейство 7. Ranidae Rafinesque, 1814 - Лягушки
Род 8. Rana Linnaeus, 1758 - лягушки
9. Травяная лягушка -Rana temporaria (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Это центральный
член обширной группы бурых лягушек, в чей состав на разных этапах раз-
вития систематики включались формы, часть из которых сейчас рассматри-
вается как самостоятельные виды. Главным образом это обусловлено широ-
ким развитием в группе параллельного диморфизма по разным направлениям
(В.Г. Ищенко, 1978):
- редукции спинного рисунка;
- наличию (или отсутствию) светлой спинной полосы;
- степени развития крапчатости, пигментации и ряду других, что и сейчас
затрудняет определение границ некоторых таксонов.
В настоящее время травяная лягушка признается самостоятельным полити-
пическим видом, в составе которого распознается четыре подвида, в том числе
номинативный R. t. temporaria (Linnaeus, 1758), населяющий территорию ре-
гиона. Вместе с тем и он отличается сильной межпопуляционной изменчивос-
тью, которая проявляется в размерах и окраске тела в целом и отдельных его
частей, биологическом предпочтении и других особенностях экологии, диа-
пазон которых (фенотипический спектр) значительно больше, чем у других
видов группы (Ананьева и др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2006;
Вершинин, 2007).
Распространение. Ареал вида охватывает большую часть Европы и Урала
до Западной Сибири на востоке - это типичный представитель европейско-
го фаунистического комплекса на территории Восточной Европы. Северная
граница ареала проходит от Баренцева моря к северному побережью Канда-
лакшского залива, Белому морю, затем через Республику Коми и окрестности
г. Воркуты к Полярному Уралу, далее вдоль р. Обь к низовьям р. Иртыш и
через Курганскую область к окрестностям г. Петропавловска (Северо-Казах-
станская область).
Южная граница проходит от среднего течения р. Днестр через Одесскую,
Николаевскую, Запорожскую и Днепропетровскую области к г. Харькову, за-
тем через Белгородскую, Курскую, Воронежскую, Пензенскую, Самарскую,
Оренбургскую, Уральскую, Костанайскую и Кокчетавскую области к окрест-
ностям г. Петропавловска. Точнее в пределах региона и его ближайших окрес-
тностях она прослеживается по находкам у с. Клиновка Шигонского района
(правобережье Волги) Самарской области (Бакиев, Файзулин, 2002), окрес-
тностях г. Самары (Гаранин, 1983), в Бузулукском бору (Даркшевич, 1953),
верховьях р. Самара у с. Мамалаевка (Зарудный, 1895), по среднему течению
р. Урал в окрестностях г. Уральска (экземпляр от Кайзерлинга в коллекции
ЗИНа), низовьях р. Илека, окрестностях г. Оренбурга (Зарудный, 1895), вблизи
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
117
Таблица 21
Распространение травяной лягушки
Зона Провин- ция Район № точки Местонахождение Источник информации
т Самарская область
5 1 1 г. Самара Гаранин, 1983
е о 4 2 р. Сок Вершинин, 2007
и о о о II Оренбургская область
5 3 р. Б. Кинель Чибилёв А.А.
6 4 р. Дёма, окр. г. Бугуруслан
22 5 Бузулукский бор Даркшсвич, 1953
23 6 р. М. Кинель Вершинин, 2007
7 окр. п. Сорочинск Бенинг, 1926
V 24 8 с. Мамалаевка Линдгольм, 1902; цит. по: Гаранин, 1983
9 р. Салмыш Зарудный, 1895
со X X 29 10 р. Сакмара Эверсман, 1949
5 VI Западно-Казахстанская область
30 11 р. Илек Искакова, 1959
VIII 54 12 р. Урал, окр. с. Дарьинское
55 13 окр. г. Уральск
IX 60 14 р. Урал, окр. г. Орск Определитель земновод- ных..., 1977
г. Орска (Определитель земноводных..., 1977) и далее уже за пределами реги-
она в Челябинской области (Вершинин, 2007).
В очерченных пределах травяная лягушка известна из окрестностей г. Са-
мары, двух пунктов Уральской области, а остальные встречи были приуро-
чены к территории Оренбургской области. В географическом отношении ее
конкретные местонахождения известны в 4 (36,4% из 11) ландшафтных райо-
нах лесостепной и 8 (11,6 % из 69 ) степной зоны (табл. 21, карта 9). Вместе с
тем последние сводки не подтверждают обитание вида в окрестностях городов
Самары (Файзулин, Вехник, 2001; Бакиев, Файзулин, 2002), Уральска, а возле
с. Дарьинского она еще в 1950-е гг. отмечалась лишь однажды (Искакова,
1959); нет сведений о текущем распространении вида и в Оренбургской об-
ласти, хотя с учетом прошлого состояния популяций и современного состо-
яния среды обитания с известной долей вероятности можно допустить, что
она сохранилась в Бузулукском бору, по долинам рек Сакмара, Салмыш и в
верховьях р. Дёма - т.е. преимущественно в лесостепной зоне. Таким образом,
южная граница ареала вида на большей части региона отступила к северу на
100-150 км. Основные изменения, по-видимому, произошли во второй поло-
вине XX столетия, хотя в окрестностях Оренбурга эта лягушка не отмечалась
уже в его первой половине (Воронцовский, 1922; Райский, 1951).
118
Глава 4
Места обитания. На сопредельной территории Волжско-Камского края
и Урала травяная лягушка населяет хвойные, смешанные и лиственные леса,
причем встречается как в глубине лесных массивов, так и в зарослях кустарни-
ков по берегам озер и болот. В смешанных массивах она отмечалась и в Бузу-
лукском бору. Вместе с тем у южных пределов ареала основными местообита-
ниями вида являются речные долины Самары, Сакмары, Урала и некоторых их
притоков, где она встречалась по сырым лугам с хорошо развитой травянистой
растительностью, древесно-кустарниковыми зарослями по берегам рек, ста-
риц или в местах выхода грунтовых вод и других местах с повышенной (до
90%) влажностью. В подходящих условиях встречается и в разных антропо-
генных модификациях ландшафтов - в парках, садах, огородах. За прошедшее
столетие в связи с вырубкой лесов и хозяйственным освоением пойм условия
обитания вида здесь существенно изменились, что обусловлено иссушением
типичных местообитаний, нарушением благоприятных для него микроклима-
тических условий, негативно сказавшихся на состоянии большинства краеаре-
альных популяций в регионе.
Образ жизни. Травяная лягушка более влаголюбивый вид (по сравнению
с симтопической остромордой), в связи с чем теснее связана с водоемами, что
ярче всего проявляется на юге ареала. Зимует она преимущественно в неболь-
ших проточных водоемах (непромерзающих речках, ручьях, прудах, канавах)
с достаточной концентрацией кислорода, но избегает крупных рек; некоторые
особи зимуют в норах, ямах с растительным опадом и подобных местах.
Из зимовки выходит довольно рано, при среднесуточной температуре воз-
духа около 2 °C, воды 3...4 °C и разрушении ледяного покрова, что в регионе
обычно происходит в начале-середине апреля. Вскоре она приступает к раз-
множению, по завершению которого рассредоточивается по типичным место-
обитаниям. В это время лягушки наиболее активны в сумерки (прячась днем
под валежником, кустами, в густой траве и т. п.), хотя в затененных и сырых
местах, а также во время дождя и после него, могут быть встречены и днем
(в целом доля дневной активности выше весной и осенью). В водной фазе
взрослые особи активны в теплое время суток. При возвратах холодов актив-
ность снижается и они прячутся под водой. Предпочитаемые температуры
взрослых особей на суше 17...20 °C. Чаще в светлое время суток попадаются
сеголетки и неполовозрелые особи, особенно при миграциях.
На зимовку травяная лягушка уходит позднее других видов лягушек -
обычно в середине октября, когда начинаются регулярные заморозки, а сред-
несуточная температура воздуха приближается к 0 °C; мелкие особи иногда
задерживаются и позднее. Некоторые особи зимуют поодиночке, чаще они об-
разуют небольшие скопления, но в местах с хорошей аэрацией периодически
собирается по несколько тысяч особей. Во время зимовки способны перено-
сить снижение концентрации кислорода до 0,5 мг/л. Зимовка продолжается
160-180 дней, а активный период - около 200 дней (больше, чем у других ви-
дов лягушек).
Размножение. Половой зрелости травяная лягушка достигает в возрасте
2-3 лет. После зимовки первые активные лягушки появляются при температу-
ре воды 3...4 °С,т. е. когда водоемы еще полностью не освобождаются ото льда.
Через несколько суток, когда температура воды превышает 5 °C, начинается
спаривание, пик которого достигается при 14 °C. Икрометание в отдельных
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
119
микропопуляциях продолжается до 10-14 суток, хотя в разных частях водоема
в связи с разной скоростью его прогревания растягивается почти на месяц. Еще
больше могут различаться сроки размножения в одном и том же водоеме в раз-
ные годы. Конкретные сведения о сроках размножения в регионе отсутствуют,
но на смежной территории Жигулевского заповедника, очевидно в запоздалую
весну, первые кладки икры отмечались с конца апреля, хотя в севернее распо-
ложенной Нижегородской (Горьковской) области они известны с конца марта
(Гаранин, 1983). Кроме того, на Урале в Свердловской области начало размно-
жения датируется 13 апреля - 10 мая, а окончание 21 апреля - 19 мая (Верши-
нин, 2007). Икрометание обычно происходит на заросших мелководных (до
80 см) участках озер, стариц и заливчиках рек, прудов, в разных ямах, канавах,
лужах талой или дождевой воды. Икра (до 4000) откладывается одной порцией
в виде плотного комка, иногда сотнями самок на одном участке.
Эмбриональное развитие при температуре воды 9... 18 °C продолжается
5-15, в других условиях до 30 дней, личинки развиваются до 60 дней, а общая
продолжительность периода развития составляет 70-75 дней. С учетом этого
появления молодых в регионе можно ожидать уже в первой декаде июня.
Питание. Головастики питаются в основном детритом, водорослями и вы-
сшими растениями, в меньшем количестве потребляется животная пища. Сего-
летки чаще всего потребляют мелких сухопутных членистоногих (коллембол,
личинок двукрылых и др.), но по мере развития кормовой спектр постепенно
расширяется. Основу питания взрослых на сопредельной территории Урала
составляют насекомые (жужелицы, двукрылые, слоники, чешуекрылые, чер-
нотелки, листоеды, муравьи и др.), в меньшей степени другие членистоногие
(Gastropoda, Arachnidae) (Топоркова, 1973); в лиственных лесах потребляются
также моллюски, дождевые черви. Состав кормов существенно меняется по
годам, сезонам и местам обитания в связи с изменением климатических усло-
вий. В свою очередь икру и личинок травяной лягушки потребляют хищные
водные насекомые и взрослые особи других земноводных. Сеголеток и взрос-
лых лягушек едят хищные рыбы, ужи, гадюки, а также многие околоводные и
некоторые другие виды птиц и млекопитающих. Известно и явление канниба-
лизма.
Численность. В пределах основной части ареала травяная лягушка очень
обычный, во многих местах наиболее массовый вид амфибий. В регионе в про-
шлом обычной она признавалась только для Бузулукского бора (Даркшевич,
1953), а по рекам Урал, Илек, Сакмара и Самара еще в конце XIX - начале
XX столетия встречалась в небольшом («ничтожном... сравнительно с ос-
тромордой лягушкой») количестве (Зарудный, 1895; Линдгольм, 1902; цит.
по: Гаранин, 1983; Бенинг, 1926; Райский, 1951). В пойме р. Урал в пределах
Уральской области у с. Дарьинское даже при специальных исследованиях за
2,5 месяца был найден только один экземпляр (Искакова, 1959). К настоящему
времени популяции по рекам Урал, Илек, нижнему течению Сакмары, вер-
ховьям Самары и в г. Самаре, по-видимому, исчезли. По остальным, извест-
ным в прошлом, местообитаниям сведений, подтверждающих существование
вида, нет, хотя, возможно, в небольшом количестве эта лягушка сохранилась
в Бузулукском бору, по среднему течению р. Сакмара, а также в долинах рек
Салмыш и Дёма. На сопредельной территории Зиянчуринского района Баш-
кортостана в лесах междуречья Саплаяк и Асель на площадке 400 м2 учтено
120
Глава 4
7 особей (Габитова, Юмагулова, 2006), а в Башкирском заповеднике на 10 км
маршрута отмечалось 5,66 экз. (Шошева, 1985).
Лимитирующие факторы. Основной причиной сокращения ареала вида
в регионе стала вырубка зрелых пойменных лесов по рекам Урал, Сакмара
и Самара, способствующая, наряду с хозяйственным освоением территории
(сенокошением, выпасом скота, особенно опасными в засушливые годы) и
рекреацией, иссушению типичных местообитаний и нарушению благоприят-
ных микроклиматических условий; на антропогенно модифицированной тер-
ритории неблагоприятно сказывается и общее техногенное загрязнение среды
обитания.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной территории региона необходимы комплекс мер,
предупреждающих деградацию пойменных ландшафтов по Уралу, Сакмаре,
Самаре и другим рекам, выявление и контроль состояния существующих по-
пуляций.
Региональный статус. Редкий краеареальный вид. Занесена в Красную
Книгу Оренбургской (1998), Самарской (2009) областей и сопредельного Баш-
кортостана (2004). Охраняется Бернской конвенцией (Приложение Ш).
10. Остромордая лягушка—Rana arvalis (Nilsson, 1842)
Значение таксона для сохранения биоразнообразия. Представитель об-
ширной группы бурых лягушек, из состава которой на территории России оби-
тает четыре вида. Таксономические отношения между некоторыми формами
не совсем ясны, что обусловлено широким развитием в группе параллельного
полиморфизма и естественной гибридизацией. На большей части ареала это
хорошо обособленный вид, в котором выделяются два-три подвида (реаль-
ность некоторых рядом авторов оспаривается), в том числе номинативный
Rana a. arvalis (Nilsson, 1842), населяющий территорию региона. Вместе с тем
и в его пределах наблюдается довольно значительная географическая и топи-
ческая изменчивость, которая проявляется в пропорциях задних конечностей
(длинноногие, коротконогие), рисунке и окраске спинной и брюшной сторон
тела, наличии (отсутствии), форме и размерах дорсомедиальной и дорсолате-
ральной полос и затылочного пятна, структуре кожи спины, биотопическом
предпочтении и некоторых других особенностях экологии; в целом это один
из самых экологически пластичных видов бесхвостых амфибий России (Ищен-
ко, 1978; Ананьева и др., 1998; Кузьмин, 1999; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает большую часть Западной, Цен-
тральной и Восточной Европы, Западной Сибири, юг Средней Сибири (до
Якутии) и Северный Казахстан (это типичный европейско-сибирский вид).
В пределах восточной части ареала северная граница распространения прохо-
дит от Карелии через север Архангельской области, окрестности г. Воркуты,
Полярный Урал и Таймыр к северному побережью оз. Байкал и юго-западной
Якутии. Южная граница прослеживается от Дунайских плавней через Никола-
евскую область, окрестности г. Херсона, Запорожское водохранилище, Донец-
кую, Ростовскую, юг Волгоградской области, окрестности городов Уральска,
Актюбинска, пески Тусум (на р. Тургай), окрестности Караганды, Семипала-
тинска, оз. Алаколь, Алтай, Туву и Забайкалье к юго-западу Якутии.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
121
В пределах региона и его ближайших окрестностях точнее южная граница
прослеживается от нижнего течения р. Медведица (притока Дона) в направ-
лении г. Быково на левобережье Волги (Кубанцев, Колякин, 1988) или к юж-
ным рубежам Волгоградской области (Кузьмин, Семенов, 2006). В Заволжье
обитание этой лягушки известно в Самарской, Саратовской и Оренбургской
областях. По долине р. Урал в 1950-е гг. отмечалась до г. Уральска (Параскив,
Бутовский, 1960). Однако здесь она, по-видимому, обитала и южнее, посколь-
ку вскоре была найдена в окрестностях поселков Владимировка, Бударино и
Чапаеве (125 км южнее г. Уральска), а так же по древнему рукаву Урала -
р. Кушум на Кировском водохранилище (Кривко, 1976); здесь самцы в брач-
ном наряде неоднократно отмечались и нами. По р. Кушум она, несомненно,
проникла и южнее (во всяком случае до Битикского водохранилища), а по ка-
налам «вышла» на орошаемые лиманы (по Тайпакскому каналу прослежена до
широты п. Батурине).
На левобережье р. Урал отмечена по р. Утва (Чингирлау) в окрестностях
г. Аксая (Березовиков и др., 1992; 2003). Возможно, обитает в верховьях рек,
стекающих с Подуральского плато, поскольку найдена в р. Калдыгайты, на
которой прослежена почти до п. Каратюбе (Кривко, 1976). Восточнее отме-
чалась по р. Илек у г. Актюбинска и несколько выше него в ур. Биш-Тамак
(Зарудный, 1895), а также у п. Кимперсай, Тайкеткен и Степной, в долине
р. Орь у г. Орска и п. Кумак на одноименной реке (Топоркова, 1973). Отсюда
граница, резко поворачивая на юго-восток, следует к Наурзуму и пескам Ту-
сум на р. Тургай (Динесман, 1953; Брагин, Брагина, 2002), а затем к озерам
Шоиндыкуль и Тениз (Искакова, 1959).
В очерченных пределах ареал вида охватывает всю Самарскую и Орен-
бургскую области, северную часть Саратовской, Уральской, Актюбинской и
значительную часть Кустанайской областей. В географическом плане конк-
ретные местонахождения остромордой лягушки известны в 8 (72,7% из 11)
ландшафтных районах лесостепной, 37 (53,6% из 69) степной и 4 (12,5% из
32) полупустынной зон (табл. 22, карта 9). Вместе с тем существование ряда
краеареальных популяций, особенно на юго-востоке региона, где существенно
увеличилась аридность климата, новыми материалами не подтверждено.
Места обитания. Остромордая лягушка, обитающая на юге тундры, в
лесной, лесостепной и степной зонах, а местами проникающая в полупусты-
ню, - довольно эвритопный вид. Она населяет леса разного типа (в Предуралье
и на Урале преимущественно лиственные), в которых чаще всего держится
по опушкам, полянам и просекам. В лесостепной зоне основными местооби-
таниями являются островные, колковые и пойменные леса, полезащитные и
придорожные лесные полосы, древесно-кустарниковые заросли по берегам
водоемов и заливные высокотравные пойменные луга. В степную зону она
проникает уже почти исключительно по сырым участкам пойменных лесов,
прибрежным древесно-кустарниковым зарослям и высокотравным лугам пре-
имущественно первой и второй пойменных террас. По р. Урал, придерживаясь
подобных биотопов, она проникает и в полупустыню. В открытой степи она
отмечается лишь по сырым луговинам у пресных (Наурзумские и др.) озер,
речек, карасу и копаней. В полупустынном Волго-Уральском междуречье по
р. Кушум, водохранилищам и отходящим каналам ее можно встретить на сы-
рых прибрежных луговинах и заболоченных участках орошаемых лиманов.
122
Глава 4
Таблица 22
Распространение остромордой лягушки
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
1 1 с. В. Сускан с. Н. Санчелеево Бакиев, Файзулин, 2002
2 с. Хрящевка с. Выселки
3 п. Приморский
4 г. Тольятти
п. Портовый
т п. Федоровка
5 с. Васильевка
6 с. Пискалинский взвоз ст. Мастрюково ст. Задельная
7 г. Самара
5 п. Управленческий
X 5 2 8 с. Старый Буян
б о 9 с. Старосемейкино
1 п. Водино
3 10 р. Коцдурча, с. Кошки
11 п. Шентала
12 с. Алакаевка
4 13 д. Р. Байтуган Оренбургская область Файзулин, 20046
II 14 с. Андреевка Самарская область Чибилёв А.А.
5 15 г. Похвистнево Бакиев, Файзулин, 2002
16 р. Сарбай
17 п. Тимашево
6 18 р. Дёма Положенцев, Ханисла- мов, 1942
Оренбургская область
III 9 19 р. Б. Ик Вершинин, 2007
10 20 р. Сакмара
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
123
Продолжение табл. 22
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
12 21 г. Новокуйбышевск Бакиев и др., 2003
22 г. Чапаевск
Саратовская область
13 23 Повсеместно Шляхтин и др., 2005
14 24
15 25 а
Самарская область
IV 16 26 р. Чапаевка, с. Каменный Брод Бакиев, Файзулин, 2002
с. Тепловка
17 27 р. Б. Иргиз, с. Мосты
28 с. Тамбовка с. Августовка Саратовская область
18 29 Повсеместно Шляхтин и др., 2005
3 X 19 30
с S 20 31
о 21 32 33 Самарская область г. Отрадный Красносамарский лес Гаранин, 1983; Бакиев и др., 2003; Бакиев, Фай- зулин, 2002
34 п. Горский
35 Богатовский р-н
36 р. Самара, оз. Лебяжье
V 22 37 Оренбургская область Бузулукский бор р. Боровка
38 р. Самара, с. Колтубановка Чибилёв А.А.
23 39 р. Самара Вершинин, 2007
40 р. М. Уран
25 41 р. Самара
Самарская область
VI 26 42 п. Восточный Бакиев, Файзулин, 2002
с. Украинка
124
Глава 4
Продолжение табл. 22
Зона Провин- ция Район № точки Местонахождение Источник информации
Оренбургская область
28 43 р. Сакмара Вершинин, 2007
VI 29 44 г. Оренбург, Форштадтские луга Воронцовский, 1922
30 45 р. Илек Зарудный, 1895
Западно-Казахстанская область
34 46 р. Утва, г. Аксай Березовиков и др., 1992
VII 40 47 р. Калдыгайты Актюбинская область Кривко, 1976
42 48 р. Илек Зарудный, 1895
43 49 р. Илек
44 50 р. Илек, г. Актюбинск
Волгоградская область
47 51 р. Волга, г. Быково Кубанцев, Колякин, 1988
Саратовская область
3 X с 49 50 52 53 Повсеместно _и Шляхтин и др., 2005
к О VIII 52 54 Оренбургская область Повсеместно Чибилёв А.А.
53 55 _«
Западно-Казахстанская область
54 55 56 57 р. Урал, с. Дарвинское р. Урал, г. Уральск Параскив, Бутовский, 1960; Кривко, 1976
Оренбургская область
IX 56 57 58 59 р. Урал р. Урал Чибилёв А.А.
61 60 р. Орь, окр. г. Орска
66 61 р. Суундук
67 62 п. Болотовка
р. Кумак, п. Кумак
X 68 63 Актюбинская область р. Куагач, п. Тайкеткен п. Кимперсай п. Степное Топоркова, 1973
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
125
Окончание табл. 22
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная XI 78 64 Кустанайская область Наурзумский заповедник: оз. М. Дксуат, Б. Аксуат, Кенсуат, Умурбек, Жар- Куль, Туркобай, Шегенбай, Акужан, Жаркуль, Карасу, Наурзум-Карасу, Катантал Динесман, 1953; Брагин, Брагина, 2002
Полупустыня XIII 89 90 91 65 66 67 68 69 Западн о-Казахстане кая область р. Кушум, Кировское вдхр. р. Кушум, Тайпакский канал р. Урал, п. Владимировка п. Бударино п. Чапаево Кривко, 1976 Дебело П.В.
XIX 110 70 Актюбинская область р. Тургай, пески Тусум Динесман, 1953
В Тургайской области она отмечалась на сыром берегу у пресноводной стари-
цы р. Тургай с густой мезофильной растительностью (осокой и т. п.). Как общая
закономерность во всех местообитаниях степной и полупустынной зон с про-
движением к югу наблюдается усиление привязанности вида к гидроморфным
биотопам.
Повсеместно она встречается (но значительно реже чем в естественных
биотопах) по обрабатываемым сельхозугодиям, садам, паркам и в населенных
пунктах. Вместе с тем по сравнению с другими видами лягушек остромор-
дая отличается более быстрой и лучшей адаптацией к антропогенизации сре-
ды, ее техногенному загрязнению. Это в значительной степени обусловлено
высокой устойчивостью эмбрионов к кислородному стрессу, низкой кожной
проницаемостью для многих веществ и соответственно низкой способностью
к биоаккумуляции, наиболее ярко проявляющимися у морфы «Striata», что оп-
ределяет увеличение частоты ее встречаемости на территориях естественных
и искусственных геохимических аномалий, в том числе на сильно техногеннно
модифицированных ландшафтах (Вершинин, 2007).
Образ жизни. На большей части ареала вне периода размножения ведет
преимущественно наземный образ жизни, причем по сравнению с травяной
лягушкой населяет более сухие места. Однако на юге региона, как и в других
местах у южных пределов распространения, наблюдается более тесная при-
вязанность к водоемам. Большая мезофильность вида проявляется и в выборе
мест зимовки, для которых обычно используются норы грызунов, другие пус-
тоты в земле, ямы с большим количеством опавшей листвы, трухлявые пни и
т. п., где она переносит более низкие температуры, чем зимующая в воде тра-
вяная лягушка. Вместе с тем в долине р. Урал у с. Дарьинского известна зимов-
ка и в непромерзающей старице (Параскив, Бутовский, 1960).
126
Глава 4
Сроки выхода из зимовки определяются ходом развития весенних явле-
ний, и в одном и том же районе они в разные годы могут различаться почти
на месяц. Первые особи обычно пробуждаются, когда воздух прогревается до
10... 13 °C, а местами еще сохраняется снег. Так в долине р. Урал у с. Дарвин-
ского в некоторые годы она отмечается в начале апреля, а в 1956 г. недавно
вышедшие на поверхность лягушки замечены в первых числах мая (Искако-
ва, 1959; Параскив, Бутовский, 1960), когда температура воздуха превысила
13 °C. В окрестностях г. Уральска она обычно появляется во второй половине
апреля (Кривко, 1976). Севернее - в Волго-Камском крае - первые встречи
чаще всего приходятся на третью декаду апреля (Гаранин, 1983).
После выхода из зимних убежищ лягушки довольно быстро (в течение не-
скольких дней) концентрируются в местах размножения. В это время они ак-
тивны практически круглые сутки и лишь при опасности скрываются на дне
водоема. По завершению размножения взрослые особи покидают водоемы и
рассредотачиваются по типичным местообитаниям, где придерживаются срав-
нительно небольших (0,2-0,3 га) участков. Здесь они активны преимуществен-
но в утренние и вечерние часы, но по сравнению с другими бурыми лягушками
чаще встречаются и днем. В дневное время взрослые особи в окрестностях
г. Уральска обычно располагались в 3-5 м вдоль береговой кромки водоема.
У молодых особей повышенная активность чаще всего наблюдалась в середи-
не дня.
Активные особи в окрестностях г. Уральска и с. Дарьинского отмечались
до конца сентября (обычно исчезают с наступлением первых заморозков), но в
отдельные годы зимовка начинается позднее. В Волжско-Камском крае в раз-
ные годы последние встречи регистрировались 8.09-25.10 (в 1951 г. в середине
ноября) и период активности, таким образом, продолжается 142-214, в сред-
нем около 170 дней (Гаранин, 1983).
Размножение, Половой зрелости остромордая лягушка достигает на треть-
ем-пятом году. В этом возрасте через несколько дней после выхода из зимов-
ки лягушки начинают концентрироваться на мелководных (до 30 см) откры-
тых участках водоемов (на которых вода быстрее прогревается до 5... 10 °C),
где через несколько дней начинается спаривание. Места размножения подоб-
ны таковым у травяной лягушки - это старицы, разливы рек, другие поймен-
ные водоемы, разные пруды, канавы, некоторые лесные (преимущественно
временные) водоемы, крупные лужи и т. п. На каждом отдельном водоеме
размножение продолжается до 25-28 суток, во время которого зачастую не-
сколько самок одновременно откладывают по 200-3000 икринок. Обычно это
происходит в конце апреля или начале мая - на несколько дней позже, чем у
травяной лягушки, что наряду с отличиями в местах нереста способствует реп-
родуктивной изоляции этих видов (хотя нередки случаи образования смешан-
ных пар). У ряда популяций кладки охраняются самцами. Продолжительность
эмбрионального развития находится в обратной пропорциональной зависи-
мости от температуры и других факторов среды: в норме оно завершается за
4-10 дней, а при похолодании затягивается до 34 суток. Личиночное развитие
на Южном Урале в норме занимает около 48 дней, а при менее благоприятных
условиях иногда растягивается до 120 суток. Общий срок развития остромор-
дой лягушки чаще всего продолжается 60-100 дней, а при благоприятных ус-
ловиях сокращается до 55. В ряде природных популяций скорость развития
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
127
и роста молодых зависит и от их плотности (Ананьева и др., 1998, Кузьмин,
1999; Вершинин, 2007). В связи с разной скоростью развития выход сеголеток
растягивается до 2 месяцев. У южных границ ареала на Кировском водохрани-
лище по р. Кушум они отмечались в первой половине июня (Кривко, 1976), а у
северных пределов региона в Волжско-Камском крае - с конца июня до конца
июля (Гаранин, 1983). После завершения метаморфоза сеголетки рассредото-
чиваются по типичным местообитаниям.
Питание. Головастики остромордой лягушки питаются в основном во-
дорослями, листьями высших растений и детритом, в небольшом количест-
ве потребляют водных беспозвоночных. Недавно закончившие метаморфоз
сеголетки кормятся преимущественно членистоногими, а в составе кормов
взрослых особей преобладают наземные беспозвоночные более 130 видов: на-
секомые (жуки, гусеницы бабочек, перепончатокрылые, прямокрылые и др.),
пауки, моллюски, дождевые черви и другие, состав и соотношение которых
меняется в зависимости от места обитания и сезона. В свою очередь яйца и
головастиков остромордой лягушки поедают головастики травяной лягушки,
которые вылупляются ранее. Сеголеток и взрослых особей потребляют не-
которые хищные рыбы, зеленые лягушки, ужи, гадюки, ряд водоплавающих,
околоводных и других птиц и млекопитающих; известны также разные формы
каннибализма.
Численность. Остромордая лягушка в центральной части ареала доволь-
но обычна, а в отдельных районах, как, например, в Волжско-Камском крае,
является наиболее массовым видом. В тоже время состояние ряда популяций,
особенно краеареальных, менее благополучно. Так, в полупустынных волго-
градских районах Нижнего Поволжья даже в прошлом она была немногочис-
ленна, а с середины XX столетия встречается лишь эпизодически (Кубанцев,
Колякин, 1988; Кубанцев, 2003). Более обычна она на побережье Кировского
водохранилища по р. Кушум, где во время выхода сеголеток на 1 м2 насчиты-
валось по 5-10 особей (Кривко, 1976). Восточнее - в Тургайской области -
она определенно малочисленна, поскольку отмечена лишь на одной старице
р. Тургай у песков Тусум (Динесман, 1953).
По долине р. Урал в окрестностях г. Уральска в начале 1950-х гг. была
сравнительно немногочисленна, но уже в 40 км выше по течению этой реки
у с. Дарьинского становилось многочисленной - ловчие банки на грызунов,
установленные на влажном лугу у оз. Чаганак, за два дня почти на половину
заполнялись этими лягушками (Параскив, Бутовский, 1960). В «достаточном
количестве» в 1989 г. она отмечалась по р. Утва немногим севернее предыду-
щих пунктов, но на сопредельной территории Карачаганакского НГКМ была
редка, а в зоне его промышленного загрязнения (р. Березовка, пруд Коншубай)
отсутствовала (Березовиков и др., 1992, 2003). Сравнительно обычен этот вид
был в р. Илек, по которому прослежен до окрестностей г. Актюбинска (Заруд-
ный, 1895; Искакова, 1959), затем в верховьях р. Куагача (Топоркова, 1973).
В Зауралье остромордая лягушка наиболее многочисленной бывает на озерах
Наурзумского заповедника (Динесман, 1953; Брагин, Брагина, 2002).
В лесных и пойменных биотопах Самарской области и сопредельной тер-
ритории Татарстана эта лягушка является одним из самых многочисленных
видов земноводных (Гаранин, 1983). Довольно обычна, а на прибрежных и
пойменных лугах многочисленна она в западной части степного Оренбуржья
128
Глава 4
(Зарудный, 1895; Воронцовский, 1922; Райский, 1951; Чибилёв, 1995), места-
ми (например у с. Болотовки в верховье р. Суундук) часто встречается у севе-
ро-восточных рубежей региона (Ищенко, 1978), а также в бассейне р. Убаган
и его окрестностях (Искакова, 1959). В лесостепной зоне на пойменных лугах
р. Саплаяк (Зиянчуринский район Башкортостана) вблизи границ Оренбург-
ской области численность ее достигала 9,4 экз. на 100 м2 (Габитова, Юмагуло-
ва, 2006), что в пересчете дает 940 особей на 1 га. Для сравнения отметим, что
численность в 250-300 ос/га, отмечаемая летом на Южном и Среднем Урале,
считалась очень высокой (Определитель земноводных..., 1977).
Остромордая лягушка довольно толерантна к антропогенным преобразова-
ниям территории, а на Южном и Среднем Урале она даже численно преобла-
дает в антропогенных ландшафтах (Вершинин, 2007). Высокая толерантность
позволяет ей существовать и на урбанизированной территории. Так, в Самар-
ской области она встречается в парковой зоне практически всех городов, а в
г. Самаре в половине административных районов, где местами (в Красноглин-
ском районе) ежегодно встречается до 5-10 особей (Бакиев и др., 2003). Вмес-
те с тем, как отмечалось ранее, на территории Карачаганакского НГКМ она
встречается редко, а в местах с повышенным техногенным загрязнением (пруд
Коншубай) отсутствует (Березовиков и др., 1992, 2003).
Лимитирующие факторы. Уничтожение и разрушение типичных мес-
тообитаний в результате урбанизации, хозяйственного освоения территории
(сенокошения, перевыпаса скота, особенно опасных в засушливые годы) и
рекреации. На антропогенно модифицированой территории неблагоприятно
сказывается общее техногенное и пестицидное загрязнение, неумелое приме-
нение удобрений, а в ряде мест движение автотранспорта и бессмысленное
уничтожение людьми.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор», Оренбургского и Наурзумского заповедников. На большей час-
ти остального ареала в регионе угрозы существованию вида нет. В качестве
общих мер охраны необходимы комплекс мероприятий по предупреждению
деградации типичных местообитаний и контроль за состоянием краеареаль-
ных популяций.
Региональный статус. На большей части региона весьма обычный вид.
Охраняется Бернской конвенцией (Приложение III).
*Съедобная лягушка - Rana esculenta (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Один из восьми
представителей обширного рода, населяющих территорию России. Является
центральным видом комплекса зеленых лягушек (Rana esculenta). Согласно
современным взглядам, она представляет собой «гибридогенный комплекс»,
сформировавшийся путем гибридизации R. lessonae и R. ridibunda (клептон -
категория, характер которой не совпадает с биологической концепцией вида).
Эта форма характеризуется сложными генетическими механизмами, включа-
ющими так называемое полуклональное наследование, полиплоидию, сущест-
вование одно- и двуполых популяций, образующих вместе с родительскими
видами разные варианты популяционных систем. На территории России сей-
час выявлены семь вариантов популяций этих форм, в которых гибриды могут
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
129
сосуществовать и в смешанных группировках с одним или двумя родительски-
ми видами, чаще с R. lessonae. Это обусловлено слабой воспроизводительной
способностью гибридов, что не позволяет им полностью поглотить или вытес-
нить родительские виды (Александровская, Быков, 1979; Боркин и др., 2003а;
Кузьмин, Семенов, 2006).
Еще сравнительно недавно на смежной территории Волжско-Камского
края указывалось на обитание только двух видов зеленых лягушек и отсутс-
твие гибридов в местах совместного обитания родительских видов (Гаранин,
1983). Таким образом, складывался своеобразный «волжский парадокс», ког-
да (в отличие от Центральной Европы) в зоне симпатрии зеленых лягушек
гибридные формы отсутствовали. Однако вскорее стало известно о находке
гибридной формы в окрестностях г. Казани, хотя это было установлено лишь
по морфологическим параметрам, применение которых при определении
вида представляет некоторые трудности (Ананьева и др., 1998; Галеева и др.,
2002). Немногим позднее в рамках программы по изучению амфибий Волж-
ского бассейна с помощью метода проточной ДНК-цитометрии достоверно
доказано, что на территории этого региона, в том числе Татарстана (Раифский
участок Волжско-Камского заповедника), Ульяновской и правобережной час-
ти Самарской области, обитают все три вида зеленых лягушек (Боркин и др.,
2003).
Учитывая симпатрическое обитание родительских форм в пределах наше-
го региона, вполне реально допустить существование гибридной формы и в
его пределах (доказательство этого, по-видимому, лишь вопрос времени).
11. Прудовая лягушка - Rana lessonae (Camerano, 1882)
Значение таксона для сохранения биоразнообразия. Один из восьми
представителей обширного рода, обитающих на территории России. Относит-
ся к комплексу зеленых лягушек (Rana esculenta). Морфометрические индексы
вида характеризуются относительно низкой географической изменчивостью, в
связи с чем подвиды не выделяются. Вместе с тем имеются достоверные меж-
популяционные различия в размерах задних конечностей, рисунке и окрас-
ке (особенно спинной поверхности) тела (Кузьмин 1999; Кузьмин, Семенов,
2006). По другой точке зрения, она может быть представлена номинативным
подвидом R. I. lessonae (Camerano, 1882), распространенным у нас и на боль-
шей части остального ареала (Ананьева и др., 1998).
Распространение. Ареал вида охватывает почти всю Западную, Цент-
ральную и Восточную Европу до Среднего Поволжья, что позволяет отнести
вид к европейскому фауанистическому комплексу. На Восточно-Европейской
равнине северная граница ее ареала от Эстонии проходит через окрестности
г. Санкт-Петербурга, юг Вологодской, Кировскую области, Татарстан и
Башкортостан к г. Белорецку на Южном Урале. Южная граница от низовий
р. Дунай проходит через Николаевскую, Днепропетровскую, Харьковскую,
Белгородскую, Воронежскую, Тамбовскую, Пензенскую, Самарскую области
и Башкортостан к г. Белорецку в верховьях р. Белой (Кузьмин, 1999; Галеева и
др., 2002; Вершинин, 2007).
В пределах региона распространение вида требует уточнения. Достоверно
его обитание установлено в окрестностях п. Управленческий Красноглинс-
130
Глава 4
Таблица 23
Распространение прудовой лягушки
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
Лесостепная I 1 1 г. Самара, п. Управлен- ческий Бакиев, Файзулин, 2002; Файзулин, 2004
Степная V 21 2 Красносамарское лесни- чество, окр. с. Красная Самарка, п. Горский Оренбургская область Бакиев, Файзулин, 2002; Файзулин, 2004
22 3 Бузулукский бор Чибилёв, 1995
кого района г. Самары, а также в окрестностях с. Красная Самарка и п. Гор-
ного (Красносамарское лесничество) Кинельского района Самарской облас-
ти (Бакиев, Файзулин, 2002; Файзулин, 2004а). Приводимые ранее сведения
этих авторов (Бакиев, Файзулин, 2001) о встречах прудовой лягушки в Став-
ропольском, Красноярском, Нефтегорском, Богатовском и Борском районах
Самарской области в их последних работах не подтверждаются. Восточнее она
отмечалась в водоемах Оренбургской части Бузулукского бора, а предполо-
жительно указывалась и для сопредельных лесостепных районов Оренбуржья
(Чибилёв, 1995). Вместе с тем в сводке «Животный мир Оренбургской облас-
ти» она наряду с озерной лягушкой указывается как широко распространен-
ный в области вид, обитающий повсеместно, где имеются реки, пойменные
и степные озера, мелкие речушки (Гавлюк и др., 1993), что в свете последних
данных о распространении вида не соответствует действительности (табл. 23,
карта 11).
Места обитания. В Волжско-Камском крае населяет преимущественно
смешанные и лиственные леса, по приречным массивам которых проникает
в лесостепную и степную зоны. Обитает в основном в небольших, но посто-
янных озерах, старицах, болотах, прудах и других искусственных водоемах с
густой травянистой растительностью; реже встречается в тихих заводях не-
больших речек и ручьев, а иногда и в частично пересыхающих водоемах, от-
куда в ряде случаев взрослые лягушки мотуг мигрировать в сохранившиеся
соседние водоемы. С водой связана не так тесно, как озерная лягушка, что
позволяет ей удаляться от водоемов на значительные расстояния. В более за-
сушливых местах у южной границы ареала она встречается лишь в водоемах
и по их берегам.
Образ жизни. Прудовая лягушка сравнительно теплолюбивая форма. Пос-
ле окончания зимовки взрослые особи обычно появляются во второй половине
апреля при температуре воды 8 °C и более, летом встречаются в водоемах, про-
гревающихся до 27 °C. На протяжении большей части теплого времени года
имеет полифазовый ритм активности, четкая цикличность наблюдается лишь
в период размножения, а осенью она активна только днем. Первую половину
дня активна обычно на суше, а затем лягушки уходят в воду. Осенью неактив-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
131
ными становятся при понижении температуры до 9 °C, на зимовку прудовые
лягушки обычно уходят в сентябре - начале октября, сначала взрослые, затем
сеголетки. В целом период активности продолжается до 164, а зимовка около
200 дней. Зимуют они в основном в воде, реже на суше в норах или закапыва-
ясь в рыхлую почву под корнями деревьев.
Размножение. Половой зрелости прудовая лягушка достигает на втором-
третьем году жизни. В этом возрасте через 10-15 дней после выхода из зимов-
ки (когда вода прогревается выше 8 °C) начинают проявляться первые при-
знаки брачной активности. Интенсивные брачные концерты свидетельствуют
о начале спаривания, которое обычно приходится на вторую декаду мая. На
отдельных водоемах икрометание, как правило, продолжается около месяца,
но в целом по Южному Уралу и Волжско-Камскому краю оно завершается
лишь к концу июня. Откладка икры (порционно, до 2-3 тыс.) происходит на
заросших подводными растениями мелководных участках водоемов, где днем
температура воды достигает 15... 19 °C и более. Эмбриональное развитие про-
должается 4-12 суток, а личиночное 47-77 (иногда затягиваясь до 135) и обыч-
но завершается в июле-августе, иногда продолжается до октября; часть голо-
вастиков зимует. Сеголетки обычно остаются в родном водоеме, хотя часть из
них расселяется и в другие.
Питание. Головастики питаются в основном водорослями. Сеголетки в
большом количестве потребляют двукрылых и их личинок. В составе кормов
взрослых доминируют сухопутные беспозвоночные, как правило, животные
с дневной активностью: преимущественно жуки, муравьи, другие перепонча-
токрылые, двукрылые, в меньшей степени потребляются моллюски, дождевые
черви, ракообразные, мальки рыб, головастики и сеголетки других видов ам-
фибий. В свою очередь головастиков и взрослых прудовых лягушек поедают
хищные рыбы, озерные лягушки, ужи, черепахи, водоплавающие, околовод-
ные, хищные и ряд других птиц и млекопитающих. Известны и разные формы
каннибализма.
Численность. Все популяции региона являются локально краеареальны-
ми, в связи с чем общая численность вида, вероятно, незначительна; в услови-
ях окрестностей п. Управленческий Красноглинского района г. Самары еже-
годно отмечается лишь около 10 особей (Бакиев и др., 2003).
Лимитирующие факторы. На большей части ареала на состояние вида
негативно влияют осушение и разрушение водоемов в процессе хозяйствен-
ного освоения, урбанизации и инфраструктурного обустройства территории.
Смертность в остающихся водоемах повышает их техногенное и биоцидное
загрязнение. В регионе, по-видимому, действуют весь комплекс факторов, оп-
ределяющих существование вида на границе ареала, а также вытеснение дру-
гим, более приспособленным видом.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной части региона в качестве общих мер необходимы
комплекс мероприятий по предупреждению деградации типичных местооби-
таний и контроль за состоянием популяций.
Региональный статус. В регионе малочисленный и, по-видимому, сокра-
щающийся вид. Включена в «Приложение...» к Красной книге Самарской об-
ласти (2009). На сопредельной территории занесен в Красную книгу Башкор-
тостана. В Европе охраняется Бернской конвенцией (Приложение III).
132
Глава 4
12. Озерная лягушка - Rana ridibunda (Pallas, 1771)
Значение таксона для сохранения биоразнообразия. Один из восьми
представителей обширного рода, населяющих территорию России. Относится
к комплексу зеленых лягушек (Rana esculent а). Это, вероятно, политипический
вид со сложной структурой, что затрудняет разработку его таксономии. По
старым данным (Определитель земноводных..., 1977) популяции большей час-
ти ареала (в том числе нашего региона) относятся к номинативному подвиду
R г. ridibunda Pallas, 1771. По современным представлениям подвиды у озер-
ной лягушки не выделяются, хотя в популяциях проявляется тенденция уве-
личения размеров генома к северу, размеров тела - к юго-западу и от равнин
к высокогорьям; есть также различия в рисунке, степени развития фоновой
окраски спины и некоторых других признаках. Ряду популяций южной части
ареала иногда придается статус самостоятельных видов, хотя по другой точке
зрения их диагностические признаки не соответствуют современным требова-
ниям не только к видам, но даже к подвидам (Ананьева и др., 1998; Кузьмин,
Семенов, 2006).
Распространение. Ареал вида охватывает обширную территорию от Фран-
ции до Восточного Казахстана (средиземноморский фаунистический комп-
лекс). Северную границу в пределах Восточно-Европейской равнины прово-
дят от г. Риги примерно через окрестности городов Пскова, Санкт-Петербурга,
Твери, Ярославскую, Костромскую и Пермскую области, затем вблизи горо-
дов Уфы и Белорецка на Урале, восточнее через Курганскую область и Северо-
Восточный Казахстан. Восточнее (до г. Якутска) ряд популяций образовался в
результате интродукции. Южная граница проводится от Балкан через Турцию,
Закавказье, Иран, Афганистан и южный Таджикистан в северо-западный Ки-
тай. Территория региона, таким образом, полностью входит в ареал вида, хотя
распространен он здесь далеко не равномерно, а на значительных пространс-
твах полупустыни и пустыни отсутствует.
Конкретные местонахождения озерной лягушки известны в 11 (100% из 11)
ландшафтных районах лесостепной зоны, 56 (81,2% из 69) степной, 17 (53,1%
из 32) полупустыни и 10 (23,3% из 43) пустыни (табл. 24, карта 12).
Места обитания. Озерная лягушка является одним из наиболее широко
распространенных видов земноводных Западной Евразии, где населяет доволь-
но широкий комплекс биотопов смешанных и лиственных лесов, лесостепной,
степной, полупустынной и пустынной зон.
В лесостепной и степной зонах обитает в стоячих и проточных водоемах,
постоянно или временно связанных с речными долинами (по которым про-
никает в глубь лесных и горных массивов). В горных речках избегает участ-
ков с быстрым течением и обрывистыми или плоскими, но голыми берегами.
По рекам Волга, Самара, Урал, их крупным (Сакмара, Илек, Орь, Бузулук) и
менее значительным (Б. и М. Иргизы, Еруслан, Чапаевка, Ток, Кумак, Чаган,
Утва и др.) притокам, а также в р. Уил озерная лягушка придерживается пре-
имущественно тихих заводей, затонов, мелководных заливов и других мест,
укрытых от волнобоя, с негустой прибрежной и водной травянистой расти-
тельностью, редкими кустарниками или рыхлыми наносами, в подходящих
местах встречается и на постоянных плесах небольших пресноводных степ-
ных речек и речушек (Ембулатовка, Быковка, Рубежка, Утва и др.). Во всех
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
J33
Таблица 24
Распространение озерной лягушки
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
1 1 2 с. В. Сускан с. Н. Санчелеево с. Хрящевка с. Выселки Бакиев, Файзулин, 2002; Бакиев и др., 2003; Фай- зулин, 2004
3 п. Луначарский п. Подстепки
4 п. Приморский
5 г. Тольятти, Центральный р-н, ул. Баныкина
п. Портовый
6 п. Федоровка
7 с. Васильевка
п. Поволжский
8 с. Пискалинский взвоз ст. Задельная
3 1 ст. Мастрюково, Мастрюков- ские озёра
X X о 9 п. Волжский
1 г. Самара:
10 п. Управленческий ст. Пятилетка, Воронежские озера Ботанический сад п. Рубежное, Яицкие озера
2 11 с. Елховка
12 с. Сгаросемейкино п. Водино
13 с. Старый Буян п. Висловка с. Красный Яр
14 п. Алексеевка п. Советы
3 15 с. Кошки
16 с. Чекалино
II 17 г. Сергиевск п. Суходол с. Старое Вечканово
134
Глава 4
Продолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
18 с. Боровка
19 с. Лужки
4 20 с. Иваново-Подбельское
21 с. Красный Яр д. Русский Байтуган Оренбургская область
22 п. Северное Самарская область Чибилёв А.А.
5 23 с. Совруха Бакиев, Файзулин, 2002
II 24 с. Абдул-Завод
« с. Старопохвистнево
cd X с 25 г. Похвистнево
1 26 р. Сарбай, п. Тимашево
о о е Оренбургская область
27 окр. г. Бугуруслан п. Михайловка Чибилёв А.А.
28 Абдулинский р-н, р. Ик
6 29 р. Дема
7 30 р. Сал мы in
8 31 п. Троицк п. Тюльган
III 9 10 32 33 р. Б. Ик р. Сакмара
11 34 р.Губерля р. Чебакла, с. Хмелевка
Самарская область
12 35 36 р. Кривуша п. Маяк Бакиев, Файзулин, 2002; Файзулин, 2004
37 г. Чапаевск
5 38 п. Бсзенчук, Саратовское
5 IV вдхр.
б 39 окр. с. Иовокиевка
40 окр. с. Кандеевка
41 с. Александровка с. Натальино
42 п. Приволжье
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
135
Продолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
43 р. Чагра, с. Новотулка с. Владимировка р. Свинуха Саратовская область
13 44 Повсеместно Шляхтин и др., 2005
14 45 «с
15 46 а
Самарская область
IV 16 47 р. Чапаевка, с. Каменный Брод Файзулин, 2004
48 с. Тепловка
17 49 р. Б. Иргиз, с. Мосты
50 с. Б. Глушица Саратовская область
18 51 Повсеместно Шляхтин и др., 2005
19 52
20 53 и
5 X Самарская область
С £ . о 21 54 55 г. Отрадный г. Кинель с. Бобровка Бакиев, Файзулин, 2002; Файзулин, 2004
56 п. Красносамарское
57 п. Октябрьск
58 п. Горский
59 с. Богатое
с. Заливное
V 60 Борский р-н, оз. Лебяжье
61 с. Петровка п. Лесное Оренбургская область
22 62 Бузулукский бор: п. Колтубановский р. Боровка, Даркшевич, 1953; Чи- билёв, 1995; Чиби- лёв А.А.
63 Грачевский р-н, р. Ток
23 64 р. Ток
65 р. Самара
136
Гпава 4
Продолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
66 р. Самара, п. Тоцкое-2
24 67 р. Ток
68 Октябрьский р-н, р. Ток
69 р. Каргала
25 70 окр. п. Новосергиевка Воронцовский, 1922
71 окр. п. Переволоцк
72 р. Урал
26 73 р. Таналык
74 р. Бузулук
75 Ташлинский р-н, р. Бузулук
V 76 Первомайский р-н, р. Чаган
Западно-Казахстанская область
27 77 р. Таловая, окр. с. Бело- глинка Дебело П.В.
Оренбургская область
28 78 р. Сакмара Чибилёв А.А.
3 29 79 г. Оренбург, Форштадтские луга Воронцовский, 1922; Дебело П.В.
X с о 80 о. Мира
О 81 п. Чебеньки Чибилёв А.А.
п. Черный Отрог
30 82 р. Илек
83 окр. п. Илек
Западно-Казахстанская область
84 р. Илек, окр. п. Карачаганак Дуйсебаева и др., 2005
VI 31 85 окр. п. Успенка Оренбургская область Черновское вдхр. Вершинин, 2007; Чи- билёв А.А.
86 р. Донгуз, п. Ивановка
32 87 Беляевский р-н, р. Буртя
88 р. У рта-Бу ртя
Западно-Казахстанская область
VII 33 89 р. Барбастау, п. Чаганский Дебело П.В.
34 90 91 низ. р. Утва р. Утва, окр. г. Аксай Зарудный, 1895; Бе- резовиков и др., 1992; Дуйсебаева и др., 2005
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
137
П родолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
окр. п. Пугачеве окр. п. Пролетарка
92 р. Карабау, п. Григорьевка р. Ащесай, п. Аксу
93 р. Акбулак, р. Сукбулак, р. Шыбынды (Чибенды)
35 94 оз. Сулуколь Зарудный, 1895
36 95 оз. Шошкала (Чушкалы)
38 96 оз. Шалкар, п. Сары-Умыр Дебело П.В.
VII 39 97 р. Олеиты
98 р. Булдурты
40 99 р. Калдыгайты Актюбинская область
42 100 р. Илек Зарудный, 1895; Иска-
101 р. Б. Хобда, р. М. Хобда кова, 1959; Дуйсебаева и др., 2005
43 102 р. Илек
44 103 р. Илек, г. Актюбинск
5 104 окр. ст. Кандагач
X с 45 105 р. Уил
н и 46 106 р. Киил
Волгоградская область
47 107 р. Волга Кубанцев и др., 1962
48 108 р. Волга, р. Еруслан Саратовская область
49 109 р. М. Узень Шляхтин и др., 2005
50 ПО р. Б. Узень
Западно-Казахстанская область:
VIII 51 111 р. Чижа 2 Дебело П.В.
112 р. Деркул, п. Переметное Оренбургская область
52 113 р. Чаган Чибилёв А.А.
53 114 р. Иртек р. Кинделя
Западно-Казахстанская область
115 р. Ембулатовка, п. Чесноко- во, п. Петрово, п. Рожково Дебело П.В.
138
Глава 4
Продолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
116 р. Быковка, п. Чеботарево р. Рубежка, п. Рассвет
117 п. Чиров р. Шатан, п. Фурманов
54 118 119 п. Облавка р. Урал, окр. п. Красноар- мейск оз. Зимник, оз. Кивалино с. Чулпан Исакова, 1959; Парас- кив, Бутовский, 1960
VIII 120 с. Дарьинск
55 121 окр. г. Уральск: р. Шатан, пруды рыбхоза Никольский, 1905; Иска- кова, 1959; Дебело П.В.
122 окр. п. Подстепное оз. Песчаное оз. Едучее
123 окр. п. Меловые горки Анисьины озера
124 Биостанция ЗКУ
13 X С р. р. Урал, окр. п. Круглоозер- ное
О 56 125 Оренбургская область Буртинская степь ОГПЗ Чибилёв, 1999а
57 126 р. Урал
59 127 р. Урал
60 128 р. Урал
IX 62 129 р. Урал Айтуарская степь ОГПЗ Актюбинская область
130 п. Тайкеткен п. Кимперсай р. Куагач Топоркова, 1973
Оренбургская область
65 131 Ириклинское вдхр. Чибилёв А.А., Топорко-
132 р. Урал, окр. г. Орск ва, 1973
X 66 133 р. Суундук, окр. п. Боло- товка
67 134 р. Кумак Чибилёв А.А.
68 135 р. Орь
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
139
Продолжение табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
5 72 136 р. Буруктал Дебело П.В.
X 5 XI Костанайская область
о 78 137 Наурзумский заповедник Брагин, Брагина, 2005
Волгоградская область
81 138 р. Волга Кубанцев, Колякин, 1988
Западно-Казахстанская область
82 138 окр. п. Джаныбек Волгоградская область Быков, 1981
XIII 85 140 окр. оз. Эльтон Линдеман и др., 2005
Западно-Казахстанская область
86 141 канал окр. оз. Балыкты Дебело П.В.
89 142 р. Кушум, Кировское вдхр. р. Кушум, Битикское вдхр.
143 р. Кушум, Донгулюкское вдхр.
91 144 р. Урал Искакова, 1959
i XIV 94 145 р. Калдыгайты Дебело П.В.
ас 3 Гурьевская область
1 95 146 р. Уил Искакова, 1959
Актюбинская область
с XV 96 147 р. Эмба, среднее течение Дуйсебаева и др., 2005
98 148 р. Темир, окр. п. Темир
99 149 р. Эмба, окр. ст. Эмба
150 р. Эмба, окр. п. Жаркемер
XVI 101 151 Мугоджары: окр. п. Маяк, окр. горы Два брата
152 окр. ст. Берчогур окр. п. Шахты
XVIII 106 173 р. Иргиз Динесман, 1953
107 174 р. Иргиз
109 153 р. Улькояк
ПО 154 р. Тургай, г. Тургай Дуйсебаева и др., 2005
XIX 155 р. Тургай, с. Мамыр
156 р. Тургай, оз. Женис к сев.-вост. с. Нура
112 157 г. Иргиз, 15 км сев.-вост. Брушко, Кубыкин, 1988
140
Глава 4
Окончание табл. 24
Зона Провин- ция Район № точки Местонахождение Источник информации
Астраханская область
113 114 158 159 Волго-Ахтубинская пойма р. Волга, Шароновский канал Гурьевская область Определитель земновод- ных..., 1977; Завьялов, Табачишин, 1998
XX 160 окр. п. Танюшкино Дуйсебаева и др., 2005
Западно-Казахстанская область
124 161 р. Урал, окр. п. Круглый Параскив, Бутовский,
162 окр. п. Калмыкове 1960
163 р. Багырдай
5 X 125 164 р. Урал Искакова, 1959
X н Гурьевская область
ь С XXI 132 165 166 р. Урал, окр. г. Гурьев р. Урал, протока Сокол р. Урал, п-ов Пешной Паллас, 1773; Искакова, 1959; Дуйсебаева и др., 2005
ххп 136 137 167 168 169 р. Эмба, окр. п. Кульсары, ур. Бейбиткуль р. Эмба ур. У ялы (старицы) ур. Жанбике Сараев, 2007; Березо- виков, Гисцов, 1995; Определитель земно- водных..., 1977; Сараев, 2007
XXIII 140 170 п-ов Бузачи Дуйсебаева и др., 2005
150 171 окр. Аральского моря Зарудный, 1915
XXVI 155 172 оз. Тщебас оз. Камышлыбас дельта р. Сырдарьи Динесман, 1953; Дуйсе- баева и др., 2005
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
141
случаях предпочтение отдается водоемам (или их участкам) с пологими гли-
нистыми, илистыми и тинистыми берегами. В поймах рек предпочитает зре-
лые и отмирающие старицы, притеррасные озера и заболоченные участки. На
степных речках и балках довольно быстро осваиваются небольшие водохра-
нилища, запруды и соединяющие их каналы. Населяет озерная лягушка так-
же котловины пресных озер (Сулуколь, Чушкалы), отмечена на опресненном
участке солоноватого озера Шалкар (в месте впадения канала из р. Урала) и
на временных лужах песчаного участка его берега у п. Сары-Умыр. Известна
также в наиболее чистой части бывшего накопителя сточных вод г. Уральска
(Анисьины озера у п. Меловые горки).
Озерная лягушка отличается высокой степенью синантропизации, что поз-
воляет ей заселять подходящие места во многих городских территориях - она
встречается во всех административных районах городов Самары, Тольятти,
ряде мест Оренбурга, Орска, Уральска и некоторых других.
В полупустыне основными постоянными местообитаниями озерной ля-
гушки являются транзитные поймы рек Волги, Урала и Эмбы. В Волжско-
Уральском междуречье важную роль играют долины Б. и М. Узеней, Кушума,
наиболее глубокие плесы и водохранилища которых выполняют роль своеоб-
разных стаций переживания вида. В Урало-Эмбенском междуречье обитание
вида известно по пресным старицам и плесам в долинах рек Калдыгайты, Уила
и Темира, а в Зауралье - Иргиза и Тургая. Высокая степень минерализации во-
доемов здесь выступает основным лимитирующим фактором, определяющим
распространение вида.
В зоне пустыни основными местообитаниями озерной лягушки являют-
ся водоемы Волго-Ахтубинской поймы и низовий Урала, где она встречает-
ся не только по основному руслу, но и по протокам и старицам. Окрестности
г. Гурьева, кстати, признаются типовой территорией вида в Восточной Европе
(Паллас, 1773; цит. по: Паллас, 2006).
По р. Эмбе она проникает вниз до окрестностей п. Кульсары, где отмечает-
ся по некоторым старицам. Встречалась озерная лягушка также в опресненных
лагунах Аральского моря, примыкающих к устью р. Сырдарья.
Практически все отмеченные местообитания связаны с речными долина-
ми. Вместе с тем на сопредельной территории Мангыстауской области озерная
лягушка встречалась также в небольшом изолированном водоеме, питаемом
родниками в каньоне Саура, что свидетельствует об исключительно высокой
способности вида к расселению.
Образ жизни. Озерная лягушка обычно ведет полуводный образ жизни и
от воды почти никогда не удаляется дальше нескольких метров (чаще всего в
пределах 3-4 прыжков). Однако в полупустыне Заволжья расселяющиеся осо-
би встречались и в нескольких километров от каналов (Линдеман и др., 2005).
После зимовки активные особи появляются при температуре воздуха не ниже
10 °C и воды 5 °C. На юге региона это наблюдается в начале, а у его северных
пределов - в конце апреля или начале мая.
Весной и осенью взрослые лягушки активны в основном днем, летом ак-
тивность проявляется почти круглосуточно (с пиками в утренние и вечерние
часы). По сравнению с взрослыми лягушата охотятся в течение всего светлого
времени суток с почти одинаковой активностью. Осенью активность прекра-
щается при снижении температуры воды до 6...9 °C. Зимовка на севере региона
142
Глава 4
обычно начинается в конце сентября - начале октября, иногда задерживаясь
до начала-середины ноября. Продолжительность активного периода жизни
на севере региона составляет в среднем 140, на юге около 170 дней. Зимуют
озерные лягушки чаще всего зарываясь в ил на дне водоемов, причем обычно
небольшими группами. Известны случаи зимовки и на суше в норах и дру-
гих пустотах грунта. Продолжительность зимовки на юге региона составляет
180-190, на севере около 220 дней.
Размножение. Половозрелой озерная лягушка становится на третьем году
жизни. В этом возрасте обычно через несколько дней (иногда до месяца) пос-
ле выхода из зимовки, когда вода прогревается выше 10... 12 °C, начинается
концентрация лягушек на открытых, хорошо прогреваемых мелководных
(30-100 см) участках водоемов с богатой травянистой растительностью. О на-
ступлении времени размножения свидетельствует начало «брачных концер-
тов», сроки которых зависят от хода развития весенних явлений. Брачный пе-
риод сильно растянут и даже на одном и том же водоеме может продолжаться
до 1,5-2,5 месяцев (на некоторых водоемах брачные концерты оканчиваются
лишь с наступлением заморозков).
Сроки начала откладки икры также зависят от метеоусловий, но обычно
приходятся на время, когда вода прогревается до 15... 18 °C. Икра выметыва-
ется одновременно или порциями (до 7 раз). Общее количество икринок, от-
кладываемых одной самкой, колеблется от 1,1 до 16,5 тысяч. Эмбриональное
развитие продолжается 3-10 (при понижении температуры затягивается до 18)
дней, и уже с середины мая появляются головастики. Их развитие в разных
условиях продолжается от 50 до 100 суток, и в связи с растянутостью икроме-
тания они встречаются до конца августа. Иногда их в процессе метаморфоза
встречают и поздно осенью, а в ряде случаев личинки зимуют. Первые сего-
летки появляются в июле, но основная масса личинок заканчивает метаморфоз
в августе-сентябре.
Питание. Головастики питаются детритом, диатомовыми, зелеными и
другими водорослями-обрастателями, высшими растениями; в меньшей степе-
ни потребляются животные: простейшие, коловратки, мелкие ракообразные,
черви и другие виды. По ходу онтогенеза разнообразие пищи возрастает, но ос-
нову питания постоянно составляют бентосные организмы. В составе кормов
сеголеток преобладают моллюски, ракообразные, клещи, пауки, равнокрылые,
перепончатокрылые, двукрылые, а у взрослых особей - наземные насекомые:
жуки, перепончатокрылые, двукрылые, бабочки; в меньшем количестве пот-
ребляются пауко-, ракообразные, моллюски, дождевые черви. В ряде случаев,
особенно в искусственных водоемах, потребляются разные виды амфибий (на
всех стадиях развития), рыба, мышевидные грызуны, насекомоядные; извест-
ны и разные формы каннибализма, который у озерной лягушки развит силь-
нее, чем у других наших земноводных. В свою очередь икру, головастиков,
сеголеток и взрослых озерных лягушек поедают многие виды рыб, черепахи,
ужи, гадюки, водоплавающие, околоводные и некоторые другие виды птиц и
млекопитающих.
Численность. Озерная лягушка практически во всех характерных для
вида местообитаниях была и остается одним из наиболее многочисленных ви-
дов земноводных. Первые представления о ее численности в регионе мы на-
ходим у П.С. Палласа (1773; цит. по: Паллас, 2006), который, экскурсируя в
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
143
окрестностях г. Гурьева в августе 1769 г., отмечал, что «Яик и все его рукава
наполнены лягушками и есть между ними особливого рода, кои также водят-
ся в Волге». Подобное определение количества, по-видимому, можно было
употребить и для многих других водоемов региона в XVIII-XIX столетиях.
Во всяком случаев в конце XIX - начале XX в. она была «очень обыкновенна»
по всем подходящим местам на всем пространстве исследуемого Н.А. Заруд-
ным (1895) Оренбургского края - т. е. по рекам Урал, Илек, Утва, Уил, Киил,
Темир, а на озерах Сулуколь, Чушкалы, реках Ащесай, Чимбеды (притоках
Утвы, Илека), по утверждению этого автора, она жила «массами». На водо-
емах форштадтских казаков в окрестностях г. Оренбурга очень обыкновенной
она была и в начале XX в. (Воронцовский, 1922). У южных пределов реги-
она достаточно обычной она была и по всей дельте р. Сырдарья (Сидоров,
1925).
Вероятно, на достаточно высоком уровне на большей части региона чис-
ленность сохранялась и в середине XX в. Во всяком случае как обычный вид
она отмечена для Бузулукского бора (Даркшевич, 1953), как самый распро-
страненный и всем хорошо известный вид указывается для остальной террито-
рии Оренбургской области (Райский, 1951). Очень высокой была численность
по р. Урал, вблизи с. Дарьинское, но южнее у п. Круглый лягушек встречалось
уже значительно меньше (Параскив, Бутовский, 1960). Регулярно наблюдалась
озерная лягушка на пресных старицах рек Иргиз и Тургай в окрестностях од-
ноименных городов, а единично и по берегам этих степных рек с повышенной
минерализацией (Динесман, 1953).
Однако, по-видимому, наиболее многочисленной она была по долине Вол-
ги. Так, в 1960-х гг. на территории Волгоградской области в окрестностях
с. Политотдельское на 1 км маршрута учитывалось до 490 особей (Косарева,
1970), а по озерам и притокам Волго-Ахтубинской поймы в 1960-х и начале
1970-х гг. численность ее достигала 200-1600 особей на 1 км маршрута. Вместе
с тем за последующие 10 лет численность вида здесь сократилась почти в 5 раз
и на тех же маршрутах насчитывалось уже по 96-435 особей на 1 км (Кубан-
цев, Колякин, 1988). Выше по течению реки в Волжско-Камском крае, а также
в долинах рек Урал, Сакмара, Салмыш, Ик, по старицам и пойменным озерам,
поросшим водной растительностью, численность лягушек достигала 7,2 экз. на
100 м береговой линии, а на озерах и прудах 50,6-97,5 экз. на 100 м2 (Гара-
нин, 1983). В окрестностях г. Уральска в это время на берегах водоемов поймы
р. Урала с прибрежными зарослями травянистой растительности насчитыва-
лось 2-7 особей на 1 м2 (Кривко, 1976).
В последние десятилетия численность озерных лягушек заметно снизи-
лась, но на общем фоне уменьшения количества других видов земноводных
они почти повсеместно остаются самыми многочисленными, особенно в райо-
нах с повышенным техногенным процессом. В значительной степени этому
способствует ее более высокая толерантность к техногенной трансформации
окружающей среды, в частности в условиях городских агломераций. В итоге
она оказалась наиболее обычным видом в условиях городов Самары и Толь-
ятти, где ежегодно отмечается более 10 особей (Бакиев и др., 2003) и Урала
(Вершинин, 2007). В естественных биотопах лесостепной зоны по р. Б. Ик ее
численность достигает 9,4 экз. на 100 м береговой линии (Габитова, Юмагу-
лова, 2006). Южнее - в дачной зоне г. Оренбурга (о. Мира) на участке с при-
144
Глава 4
брежной травянистой растительностью по р. Урал на таком же маршруте пос-
тоянно отмечается 12,5-15,6 особей. Довольно обычна озерная лягушка в зоне
Карачаганакского ГКМ. Здесь в условно чистой р. Утва (Чингирлау) в 1989-
1990 гг. ее численность достигала 865,5 особей на гектар, в отдельных местах
(роще у ГП-16) была еще выше - 1170 особей на гектар, хотя в черте промыш-
ленной зоны (р. Березовка) снижалась до 250 особей на гектар, а в условиях
наиболее загрязненного пруда Коншубай (L = 6 км) она не отмечена вовсе
(Березовиков и др., 1992). В Волго-Ахтубинской пойме в пределах АГКМ по
водоемам и их окрестностям численность достигает 260,0+0,09 особей на гек-
тар (Завьялов, Табачишин, 1998). В массе она обитает на мелководьях дельты
р. Урал (Гисцов, Березовиков, 1995).
Лимитирующие факторы. Разрушение, обмеление, пересыхание и осо-
лонение водоемов, трансформация местообитания в процессе урбанизации и
рекреационного освоения территории. Негативное влияние оказывает сильное
загрязнение водоемов техногенными сбросами и отходами, резко снижающи-
ми выживаемость личинок и молодых. В ряде мест существенное воздействие
оказывает отлов лягушек с разными (в том числе с коммерческими) целями.
Тем не менее сооружение искусственных водоемов (в том числе технического
назначения) способствует расселению вида.
Охрана. Обитает и охраняется на территории Бузулукского национального
парка, Оренбургского заповедника, Иргиз-Тургайского резервата. На осталь-
ной территории региона особых мер по охране вида не требуется. Достаточно
поддержания естественного состояния ландшафтов и предупреждения загряз-
нения водоемов техногенными сбросами, пестицидами и отходами.
Региональный статус. Фоновый вид. Охраняется Бернской конвенцией
(Приложение III).
Класс 2. REPTILIA LINNAEUS, 1758 - ПРЕСМЫКАЮЩИЕСЯ
Отряд 3. TESTUDINES BATSCH, 1788 - ЧЕРЕПАХИ
Семейство 8. Emydidae Rafinesque, 1815 -
Американские пресноводные черепахи
Род 9. Emys Dumeril, 1806 - болотные черепахи
13. Болотная черепаха - Emys orbicularis (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, обитающий на территории России. Политипический вид;
в настоящее время выделяют 13 подвидов, из которых 3 известны на террито-
рии России и смежных районов Казахстана, в том числе номинативный Е. о.
orbicularis (Linnaeus, 1758), населяющий территорию региона. Наряду с этим
в его составе наблюдаются некоторые межпопуляционные отличия по ряду
морфологических признаков (форме, окраске, пятнистости панциря, голове и
конечностям), особенностям размножения и некоторым другим деталям эко-
логии (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семе-
нов, 2006).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
145
Распространение. Ареал вида охватывает большую часть Европы, Юж-
ного Урала, Северо-Западную Африку, Переднюю Азию, Кавказ, Прикаспий,
Приаралье, Тургай (восточно-средиземноморский фаунистический комплекс).
Северная граница ареала на Восточно-Европейской равнине проходит через
Литву, северную Белоруссию, Смоленскую область, среднюю Волгу, Татарс-
тан (р. Б. Черемшан), юг Башкортостана и Челябинской области (Брединский
район) откуда, поворачивая на юго-восток, следует к Аральскому морю и ни-
зовьям р. Сырдарья. Южная граница проходит через Переднюю Азию, Ирак,
Прикаспийские районы Ирана, западную Туркмению, Мангышлак и по Пре-
дустюртью выходит к Аральскому морю.
Территория региона, таким образом, почти полностью включается в ареал
вида, за исключением северо-западной части Тургайского плато. В географи-
ческом отношении его конкретные местонахождения известны в 6 (54,5% из
11) ландшафтных районах лесостепной, 46 (66,7% из 69) степной, 14 (43,7% из
32) полупустынной и 12 (27,9% из 43) пустынной зон (табл. 25, карта 13).
Места обитания. Болотная черепаха обитает преимущественно в лесо-
степной и степной зонах региона, а в полупустыне и пустыне встречается в ос-
новном в поймах рек. Как правило, населяет слабопроточные или бессточные
водоемы с заросшими пологими берегами.
В лесостепном Высоком Заволжье, низкогорном Предуралье и Урале,
а также по долинам рек Самара, Сакмара и Урал чаще всего населяет зарас-
тающие старицы и пойменные озера, а иногда и тихие заводи рек на участ-
ках со слабыми течениями. На большей части степной зоны и примыкающих
пространствах полупустыни она встречается в постоянных плесах и прудах
небольших рек (верховья рек Б. Иргиз, Б. и М. Узени), мелких притоков Ура-
ла (Бурте, Утве, Рубежке, Ембулатовке и др.), Илека (Б. и М. Хобдах), Уила
(Киила), речках, стекающих с Подуральского плато (Шидерты, Калдыгайты) и
Мугоджар. По р. Кушум освоила Кировское водохранилище. Местами встре-
чается в изолированных степных озерах (Сулуколь в верховьях р. Утва) и даже
небольших болотцах (Черепашье болото в Буртинской степи Оренбургского
заповедника). В полупустыне и пустыне она связана почти исключительно
со слабопроточными или замкнутыми пойменными водоемами в долинах рек
Волга, Урал, Эмба, а также Иргиз, Тургай и Улькояк.
Образ жизни. Активный образ жизни болотная черепаха обычно ведет
лишь в теплое время года, а зимой, зарываясь в ил водоема, впадает в оцепене-
ние. Зимовка в северной части региона продолжается с конца октября - начала
ноября до конца апреля - середины мая (около 180-190, на юге, по-видимому,
160-170 дней). Весной черепахи обычно появляются, когда воздух прогревает-
ся до 6... 14 °C, а вода до 5... 10 °C. Большую часть жизни они проводят в воде,
но довольно часто (особенно в жаркие дни) отдыхают на кочках, наводных
предметах (корягах и т.п.) или даже гнездах водоплавающих птиц, выходят и
на сушу. Активны днем и в сумерки. Период активности продолжается около
180 суток, хотя на юге региона в теплые годы отдельные особи активность
могут сохранять и зимой.
Размножение. К размножению болотные черепахи приступают в возрасте
5-8 лет. Спаривание наблюдается в конце апреля - мае. В регионе, по-види-
мому, бывает одна кладка. В пойме р. Урал у п. Круглый выход самок для
откладки яиц наблюдался с 15 июня, а в неволе здесь две самки 19 и 21 июня
146
Гпава 4
Таблица 25
Распространение болотной черепахи
Зона Провин- ция Район № точки Местонахождение Источник информации
I I 1 2 Самарская область с. Васильевка г. Самара Бакиев, Файзулин, 2002; Никольский, 1915; Баки- ев, 2001а
Лесостепная II 3 4 5 6 3 4 5 6 7 8 р. Кондурча р. Сок, р. Сургут, оз. Голу- бое р. Б. Кинель, окр. с. Старо- похвистнево Оренбургская область р. М. Кинель р. Садак р. Дйма Зарудный, 1895; Бакиев, Файзулин, 2001 Чибилбв, 1995
III 10 9 р. Сакмара (хр. Шайтантау)
Степная IV 12 13 14 15 16 17 18 19 10 11 12 13 14 15 16 17 18 19 Самарская область окр. п. Безенчук р. Волга, Васильевские о-ва р. Чагра, окр. с. Новотулка Саратовская область р. Волга р. Волга, окр. г. Энгельс р. Б. Иргиз Самарская область р. Чапаевка, лиманы р. Б. Иргиз, с. Мосты Саратовская область верх. р. Еруслан верх. р. М. Узень Бакиев, 2001а; Бакиев, Файзулин, 2002 Шляхтин и др., 2005 Шапошников, 1978 Завьялов и др., 2003
V 21 22 20 21 22 23 24 Самарская область р. Самара, с. Алексеевка ст. Заливная с. Борское Бузулукский бор: р. Самара, р. Шелехметь Оренбургская область Бузулукский бор, с. Колтуба- новка, Опытное лесничество Бакиев, 2001а; Бакиев, Файзулин, 2002 Райский, 1951
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
147
П родолжение табл. 25
Зона Провин- ция Район № точки Местонахожден ие Источник информации
V 23 24 25 26 28 29 25 26 27 28 29 30 31 32 р. Ток р. В. Каргалка р. С. Каргалка р. Урал р. Самара р. Бузулук руч. Ялга Самарская область р. Таловая, р. Балабанка Оренбургская область р. Салмыш р. Сакмара Чибилбв, 1995 Линдгольм, 1902 (цит. по: Гаранин, 1983) Чибилев, 1995 Смелянский, Елизаров, 1996 Чибилев, 1995
Степная VI 30 31 32 33 34 35 р. Илек р. Урал р. Бурля, р. Киялыбуртя руч. Джамылчигай руч. Тузлыколь Черепашье болото (Буртинс- кая степь) руч. Караагач
VII 34 35 36 38 39 40 41 42 36 37 38 39 40 41 42 43 44 Западно-Казахстанская область р. Утва оз. Сулуколь р. Утва р. М. Анкаты р. Б. Анкаты (оз. Шалкар) р. Солянка (ур. Карабас) р. Шидерты (п. Джамбейты) р. Калдыгайты Актюбинская область р. Б. Хобда р. М. Хобда р. Илек р. Уил р. Киил Березовиков и др., 1992; Зарудный, 1895 Параскив, Бутовский, 1960; Дебело П.В. Зарудный, 1895
148
Глава 4
Продолжение табл. 25
Зона Провин- ция Район № точки Местонахождение Источник информации
§ VIII 48 49 50 52 53 54 55 48 49 50 51 52 53 54 55 56 57 Саратовская область р. Волга (п. Ровное) р. М. Узень р. Б. Узень Оренбургская область р. Чаган р. Иртек, р. Кинделя р.Иртек, р. Кинделя Западно-Казахстанская область р. Урал окр. п. Январцево р. Ембулатовка в окр. п. Красноармейск р. Рубежка, п. Раздольное р. Урал Завьялов и др., 2003 Чибилёв, 1995 Параскив, Бутовский, 1960; Дебело П.В. Параскив, 1956
Е <5 IX 56 57 61 62 58 59 60 61 62 Оренбургская область р. Урал Буртинская степь ОГПЗ р. Урал Айтуарская степь ОГПЗ Актюбинская область р. Куагач р. Каргала Райский, 1951; Чибилёв, 1995, 1999а Топоркова, 1973
X 67 68 69 63 64 65 Оренбургская область р. Кумак, р. Карабутак р. Орь, р. Камсак р. Кумак Чибилёв, 1995
XI 72 78 66 67 р. Буруктал Костанайская область Наурзумский заповедник Брагин, Брагина, 2002
Полупустынная XIII 81 85 89 91 68 69 70 71 72 Волгоградская область р. Волга р. Соленая Куба р. Б. Сморогда Западно-Казахстанская область р. Кушум, Кировское вдхр. р. Урал, п. Бударино р. Урал, п. Круглый Кубанцев и др., 1962; Ходашова, 1960; Живот- ные..., 2009 Дебело П.В. Паллас, 1773 Параскив, 1956
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
149
Продолжение табл. 25
Зона Провин- ция Район № точки Местонахождение Источник информации
Актюбинская область
а XV 95 96 98 73 74 75 р. Уил р. Эмба, ур. Жанбике р. Темир Неручев и др., 1990; Не- ручев, Васильев, 1978
X X 99 76 р. Эмба
ё 100 77 р. Маннисай
С е; о С XVIII 106 107 78 79 р. Иргиз р. Иргиз Зарудный, 1895; Нака- ренок, Неручев, 2001; Динесман, 1953; Шилов,
109 80 р. Улькояк 1961
XIX по 81 р. Тургай, ур. Тусум
112 82 р. Тургай
Астраханская область
113 83 Волго-Ахтубинская пойма Чернобай, 2006
84 Босдинско-Баскунчакский заповедник Стрельцов и др., 2006
114 85 86 Волго-Ахтубинская пойма окр. ст. Досанг Завьялов, Табачишин, 1998; Определитель земноводных..., 1977; Бобров, 1985
XX Гурьевская область
87 рукав р. Волги, окр. п. Та- нюшкино, оз. у разъезда 7 Сараев, Пестов, 2010
к Западно-Казахстанская область
Я X X 124 88 р. Урал, п. Калмыкове Окулова, 1963
1 Гурьевская область
с 125 89 р. Урал, окр. п. Индербор- ский
126 132 90 91 92 п. Забурунье, ер. Белужий р. Урал, окр. п. Махамбет окр. п. Енбекши Рычков, 1999; Николь- ский, 1915; Сараев, Пестов, 2010; Гисцов, Березовиков, 1995
XXI 93 окр. г. Гурьев, ур. Казган, канал Зарослый
94. п-ов Пешной
95 зал. Каменный Култук
XXII 134 96 97 р. Уил, окр. п. Райгородок п. Карабау, 25 км сев.-вост. Накаренок, 2003в; Неручев, Васильев, 1978
137 98 р. Эмба, ур. Жанбике
150
Глава 4
Окончание табл. 25
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXIII 143 144 99 100 Мангистауская область зал. Мертвый Култук Предустюртьс Определитель земновод- ных..., 1977
XXV 149 101 Актюбинская область Предустюртье Зарудный, 1915; Оп- рсделитель земновод- ных..., 1977
XXVI 155 102 дельта р. Сырдарья, побе- режье Аральского моря
отложили 16 и 11 яиц. В окрестностях оз. Шалкар массовый выход самок
для откладки яиц наблюдался 24 июня 1949 и 1951 г., причем из долин рек
Б. и М. Анкаты в одном и том же месте на равнину они начинали подниматься
по довольно крутым лессовым берегам не ранее 11-12 часов дня (Параскив,
1956; Параскив, Бутовский, 1960). Выход самок для откладки яиц на незатоп-
ляемые участки речных долин в Оренбуржье наблюдал Н.А. Зарудный (1895),
по сведениям которого здесь «яйца кладутся в числе 9-20 во второй половине
мая, в июне и первой половине июля». У северных пределов региона в Волж-
ско-Камском крае откладка яиц продолжается с начала июня почти до конца
июля (Гаранин, 1983).
Яйца (партиями по 3-4 штуки, всего до 20) откладываются в ямку, обычно
вырытую в сухом хорошо прогреваемом грунте. Инкубационный период здесь
продолжается 60-110 суток и в августе-сентябре появляются первые молодые
(Топоркова, 1973), хотя большинство их остается под землей до следующей
весны.
Питание. Болотная черепаха потребляет корма преимущественно живот-
ного происхождения. На суше она кормится саранчевыми, разными жуками,
клопами, другими насекомыми, мокрицами; в воде ловит насекомых, рако-
образных, моллюсков, головастиков и лягушек, мальков рыб. На оз. Шалкар
по наблюдениям Н.П. Серова черепахи чаще всего встречались в местах кон-
центрации колюшки (Параскив, 1956). В р. Ембулатовка (1,5 км от п. Крас-
ноармейск) нами черепахи дважды ловились на удочку с наживкой из дож-
девого червя. Потребляют также водоросли, высшие водные и околоводные
растения.
Численность. Первые публикации по региону показывают, что в
XVIII столетии на значительной части территории болотная черепаха была
если не многочисленным, то во всяком случае довольно обычным видом. Так,
П.И. Рычков в 1762 г. сообщал (1999), что «черепахи в Каспийском море в
довольном числе находятся, также около реки Яика (Урала) по обеим сторо-
нам в озерах и степных речках их много...». Для противоположной - северо-
западной части региона - П.С. Паллас (1773 г.), направляясь вдоль р. Самары
(Самарской укрепленной линии) к г. Оренбургу, отмечал, что в окрестностях
«Алексеевского пригорода» на территории современного Кинельского района
в подходящих местах есть «много не только рыбы, но и выхухолей и чере-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
151
пах», а в окрестностях Борского находится «много наполненных черепахами
непроточных болотин» (цит. по: Бакиев, 2001а). Исключение, по-видимому,
составляли популяции северной окраины региона, которые, являясь краеаре-
альными, изначально (в связи с локальностью местообитаний) не могли быть
многочисленными. Подобная ситуация сохранялась и более века спустя, когда
Н.А. Зарудный (1895) «в небольшом числе» находил болотных черепах «в бо-
лоте около Голубого озера, по рекам Сургут и Сок; редки они были также по
р. Салмыш и нижнему течению р. Сакмара, не часто встречались и по долине
р. Урал между городами Орском и Оренбургом, но ниже по течению между
городами Илецком и Уральском встречались уже несколько чаще». Вместе с
тем «чрезвычайно обыкновенна болотная черепаха была в долинах р. Илек и
его притоков Сары-Хобда, Кара-Хобда и Улу-Хобда, а также по рекам Утва
(Чингирлау), Термир, Уил, Киил и многим притокам этих рек, «массами» жила
она на многих степных озерах, лежащих в стороне от речных долин (например
оз. Сулуколь)».
Довольно высокой на многих этих водоемах численность была и в первой
половине XX столетия. Во всяком случае П.А. Положенцев (1935), описывая
герпетофауну Бузулукского бора, сообщает, что она «довольно обыкновенна в
непересыхающих, глубоких и больших озерах Борового, Опытного, Колтубан-
ского и, по-видимому, других лесничеств». Немногим позднее А.П. Райский
(1951) отмечал, что «много ее было по левым притокам р. Урал (Буртя, Бурля
и т.д.)», «не редко» она встречалась и в долине р. Илека; обычной оставалась и
в Бузулуском бору (Даркшевич, 1953).
Довольно «значительной» была численность болотной черепахи и по ниж-
нему течению р. Урал. Так, у п. Круглый при тонении береговым неводом
только на одной из стариц было отловлено около 20 особей. Определенно мно-
гочисленными были черепахи и по руслам впадающих в оз. Шапкар, реках
Б. и М. Анкаты, где Н.П. Серов наблюдал их «массовый выход» на места от-
кладки яиц. Несомненно, сохранилась она и в реках, стекающих с Подураль-
ского плато, поскольку отмечалась на р. Шидерта (Параскив, 1956; Параскив,
Бутовский, 1960), а также в Мугоджарах и по «пресноводным старицам» рек
Тургай, Улькояк и Иргиз (Динесман, 1953; Шилов, 1961).
Несколько неопределенным к этому времени было состояние аралморс-
кой популяции. Старые сведения о обитании болотной черепахи в р. Сырда-
рья базировались в основном на факте добычи ее Н.А. Северцовым в середине
XIX столетия. Н.А. Зарудный (1915) во время исследования низовий р. Сырда-
рья в 1911 г. отмечал черепаху всего один раз, и, по его мнению, она уже тогда
находилась здесь в «периоде окончательного вымирания» (С. 115). Других до-
стоверных сведений о добыче черепах в этой реке, а также у южных пределов
региона на п-ове Мангышлак (где она в прошлом отмечалась А. Остроумовым)
к 1950-1960-м гг., насколько нам известно, нет.
Последняя треть XX и начало XXI в. характеризуется повсеместным сниже-
нием численности вида, что особенно ярко проявилось в наиболее освоенной
северо-западной части региона. Так, в Самарской области в последние годы
ее пребывание достоверно установлено не более чем в 20 пунктах, а общая
численность оценивается лишь в несколько сот особей (Бакиев, 2001а; Бакиев,
Файзулин, 2001). В связи с этим уже с 1970-х гг. этот вид стали относить к
категории редких и указывать на необходимость занесения его на страницы
152
Глава 4
областной Красной книги (Шапошников, 1978; 1996; Бакиев, Маленев, 1999;
Шапошников и др., 2001; Бакиев, Файзулин, 2001), что и было сделано при ее
издании.
Большая часть Саратовского Заволжья характеризуется как территория
предполагаемого или вероятного обитания вида и лишь по р. Волге и в юго-
восточной части (верховьях рек Еруслана, Б.и М. Узеней) обитание указывает-
ся как достоверное; здесь эти животные являются обычными, а местами мно-
гочисленными (Завьялов и др., 2003).
Более благоприятная ситуация сложилась на территории Оренбуржья.
Здесь малочисленным этот вид является лишь в северных районах по Высоко-
му Заволжью, Предуралью (по рекам Садак, Б. и М. Кинели, Салмыш, Сакма-
ра) и, по-видимому, в бассейне р. Суундук в Зауралье, откуда встречи черепах
известны также из сопредельной территории Челябинской области (Чибилёв,
2005) и в бассейне р. Кумак. На большей части остальной территории Орен-
бургской области (за исключением окрестностей городов и крупных населен-
ных пунктов) болотные черепахи повсеметно обычны, а ряд районов отличает-
ся довольно высокой плотностью населения. К их числу относятся:
- Верхнебузулукский, особенно по крупным озеровидным плесам ручья
Ялга (Тоцкий р-н);
- Чаганский, особенно в низовьях рек Таловой, Балабанки и верховьях
Чагана;
- Илекско-Иртекский - по р. Урал, в районе низовий рек Илека, Кинделя,
Иртека;
- Буртинский, особенно по речкам Киялыбуртя, Бурля, Джамылчигай и
в верховьях ручья Тузлукколь. В этом районе особый интерес представляет
карстово-родниковое Черепашье болото в Буртинской степи у госзаповедника
«Оренбургский», где на площади около 150 м2 насчитывалось до 18 взрослых
и более 40 молодых особей;
- Орь-Камсакский, включающий низовье р. Орь и ее притока Камсак;
- Кумакский, включающий среднее течение р. Кумак и бассейн его прито-
ка р. Карабутак (Чибилёв, 1995, 1999а, б).
Сравнительно обычной до конца XX столетия болотная черепаха была и
южнее в пределах казахстанской части ареала, хотя засухи и суровые морозы
последних лет негативно сказались на ее численности, особенно за пределами
долин рек Урал, Эмба и Тургай. На территории Карачаганакского нефтегазо-
конденсатного месторождения причиной исчезновения вида, очевидно, стало
техногенное загрязнение р. Березовки и пруда Коншубай, хотя в соседней бо-
лее чистой р. Утве черепахи наблюдаются «в достаточном количестве» (Бе-
резовиков и др., 1992, 2003). Обычна она и в дельте р. Урал - на п-ове Пеш-
ном численность составляет 1-3 ос/км береговой линии (Гисцов, Березовиков,
1995).
В Волго-Ахтубинской пойме черепахи довольно обычны, а местами много-
численны (Стрельцов и др., 2006), а на водоемах и их окрестностях в пределах
Астраханского ГК месторождения их численность достигает 10,10±0,47 экз/га
(Завьялов, Табачишин, 1998). Важно также и то, что численность ее здесь в
течение многих лет остается стабильной (Кубанцев, 2003).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
153
Таким образом, в целом по региону болотная черепаха является сравни-
тельно обычным (нередким) видом, хотя численность ее, особенно в послед-
ние годы, заметно сокращается.
Лимитирующие факторы. Разрушение естественных местообитаний
при зарегулировании стока рек, застройке, хозяйственном и рекреационном
освоении береговой линии, техногенное загрязнение, беспокойство в местах
откладки яиц, гибель в ставных рыболовных орудиях, отлов местным населе-
нием и туристами. Однако устройство гидротехнических сооружений на ряде
степных рек способствует сохранению вида.
Охрана. Обитает и охраняется на территории национального парка «Бу-
зулукский бор», государственных заповедников «Оренбургский», «Наурзумс-
кий», «Богдинско-Баскунчакский» и Иргиз-Тургайского резервата. Поскольку
на большей части остальной территории региона реальной угрозы существова-
нию вида нет, достаточно поддержания естественного состояния ландшафтов
и предупреждения загрязнения водоемов техногенными сбросами, пестицида-
ми и отходами; в ряде мест целесообразно создание заказников местного зна-
чения, запрещение отлова.
Региональный статус. Характерный для герпетофауны региона вид с
тенденцией сокращения численности или полного исчезновения вблизи круп-
ных населенных пунктов; включен в список МСОП (категория низкого риска),
приложение Бернской конвенции, занесен в Красную книгу Самарской облас-
ти (2009), а также сопредельных Татарстана, Башкортостана и Челябинской
области.
*Род Mauremys Gray, 1869 - водные черепахи
*Каспийская черепаха - Mauremys caspica (Gmelin, 1774)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода в России. Политипический вид, на территории России пред-
ставленный номинативным подвидом М. с. caspica (Gmelin, 1774).
Распространение. Ареал вида охватывает Южную Европу, Переднюю
Азию, юго-запад Туркмении, Кавказ и Дагестан. В российском Прикаспии на-
селяет разные водоемы, в том числе прибрежные участки Каспийского моря,
где сейчас достигает низовий р. Терек (Определитель земноводных..., 1977).
Вместе с тем по старым данным (Georgi, 1800) эта черепаха в прошлом оби-
тала и на Нижней Волге, хотя еще А.М. Никольский (1905) считал эти сведе-
ния ошибочными. Кроме того, в приведенном ранее сообщении П.И. Рычкова
(1762; цит. по: 1999), по мнению В.И. Гаранина (1983), речь идет о трех видах
черепах, в том числе каспийской (обитавшей ранее в приморской полосе вбли-
зи устья р. Урал), которых автор не различал.
Окончательное решение этого вопроса, по-видимому, окажется возмож-
ным лишь с привлечением палеогерпетологических материалов, которые к на-
стоящему времени не известны.
154
Глава 4
Семейство 9. Testudinidae Batsch, 1788 - Сухопутные черепахи
Род 10. Agrionemys Khozatsky et Mlynarski,
1966 - среднеазиатские черепахи
14. Среднеазиатская черепаха -Agrionemys horsfieldii (Gray, 1844)
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, известный на территории Евразии. Политипический вид,
представленный тремя подвидами, в том числе казахстанской черепахой A. h.
Kazakhstanica Ckhikvadze, 1988, населяющей территорию региона (Ананьева и
др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает северо-западный Китай, значи-
тельную часть Пакистана, Афганистана, Ирана, Среднюю Азию и Казахстан
к северу до низовий р. Тургай (среднеазиатский фаунистический комплекс).
Впрочем, детали его распространения в ряде мест, в том числе в регионе, оста-
ются слабо изученными, а отчасти не признанными.
Обычно (вслед за А.М. Никольским (1905; 1916)) в регионе и его ближай-
ших окрестностях граница ареала проводится (Параскив, 1956; Определитель
земноводных..., 1977) по восточному побережью Каспийского моря, низовьям
р. Эмбы, средним Мугоджарам, низовьям рек Иргиз, Тургай, северному При-
балхашью и Алакульской котловине к хр. Тарбагатай. Между тем Н.А. Заруд-
ный (1895) сообщает о находке этой черепахи на полпути между г. Орском и
форпостом Карабутак (в бассейне р. Кумак) примерно на меридиане большой
излучины р. Орь. Восточнее этих мест нами (А.А. Чибилёвым) в 1977 г. она
отмечена в 120 км южнее п. Светлый, т. е. вблизи поворота р. Иргиза на юг.
В 2005 г. две одиночные особи этих черепах нами же наблюдались в песках
Кугузюк-Кум (бассейне р. Калдыгайты), а в 1980-1989 гг. они найдены в На-
урзуме (Брагин, Брагина, 2002). Все эти пункты находятся в степной зоне, что
в некоторой степени реабилитирует старое название вида степная черепаха.
С учетом этих находок северная граница постоянного распространения
казахстанской черепахи прослеживается от низовий р. Эмба, по кромке мас-
сивов аллювиально-дельтовых отложений у подножья Предуральского усту-
па до р. Калдыгайты, затем восточнее (обходя с юга Северные Мугоджары) к
р. Кумак, верховьям рек Иргиз, Наурзум, низовьям р. Тургай и далее к север-
ному Прибалхашью.
В очерченных пределах распространение этой черепахи выглядит сле-
дующим образом: в степной зоне Приуралья сейчас она обитает в песчаном
массиве Кугузюк-Кумы (А.А. Чибилёв), в Зауралье известна в низовьях р. Ку-
мак (Н.А. Зарудный), а также в Сапсынагашской депрессии (А.А. Чибилёв).
В 1980-е гг. была найдена в п. Наурзум, близ оз. М. Аксуат и на бархане На-
урзумского бора (Брагин, Брагина, 2002). В полупустынной зоне отмечалась в
Средних Мугоджарах (Н.А. Зарудный), низовьях рек Иргиз и Тургай. В пус-
тыне рапространена более широко - от Бузачи, Мертвого Култука (Определи-
тель земноводных..., 1977), Прикаспийских Каракумов (Сараев, Пестов, 2010)
до юга Предуральского плато, северо-западных и северных чинков Донгузтау
и Сам-Матсайской депрессии, юго-западной кромки Б. Барсуков, северо-за-
падного и северного побережья Аральского моря и Приаральских Каракумов
(Шилов, 1961). В географическом отношении ее конкретные местонахождения
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
155
известны в 4 (5,8% из 69) ландшафтных районах степной, 5 (15,6% из 32) полу-
пустынной и 11 (25,6% из 43) пустынной зон (табл. 26, карта 14).
Таблица 26
Распространение среднеазиатской черепахи
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная VII 40 1 Западно-Казахстанская область пески Кугузюк-Кумы Чибилёв А.А.
X 68 2 Оренбургская область окр. городов Орск, Карабу- так Зарудный, 1895
XI 77 78 3 4 Актюбинская область Сапсынагашская ложб. Кустанайская область Наурзумский заповедник Чибилёв А.А. Брагин, Брагина, 2002
Полупустынная XV 96 5 Актюбинская область ср. течение р. Эмба Определитель земновод- ных..., 1977; Зарудный, 1895; Динесман, 1953; Шилов, 1961
XVI 101 6 Средние Мугоджары
XVIII 107 4 низ. р. Иргиз
XIX 111 112 8 9 междуречье Тургай-Жилан- шык низ. р. Тургай ур. Тауп ур. Жаманкум оз. Аткайколь Чибилёв А.А. Определитель земно- водных..., 1977; Шилов, 1961
Пустынная XXI 127 10 Гурьевская область низ. р. Эмба Неручев, Васильев, 1978; Накаренок, Неру- чев, 2001; Шилов, 1961; Определитель земновод- ных..., 1977 Накаренок, 2003; Ши- лов, 1961; Определитель земноводных..., 1977
XXII 138 139 11 12 Прикаспийские Каракумы ур. Карашунгыл окр. мог. Ушкан окр. п. Аккудук ур. Комек ур. Устибукай Мангистауская область Преду тюртье гора Токсанбай сев.-зап. и сев. Чинки ур. Мол куду к
156
Глава 4
Окончание табл. 26
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXIII 141 143 13 14 п-ов Бузачи окр. Мертвого Култука Сараев, Пестов, 2010
XXIV 146 15 Актюбинская область юж. чинк Донгузтау, Север- ный Устюрт Параскив, 1956
XXV 147 148 16 17 гора Таксанбай дол. рек Аксай, Тасай Сам-Матсайская депрессия; ур. Асматай-Матай, Косбу- лаксор Шилов, 1961
XXVI 151 154 155 18 19 20 21 юг.-зап. кромка Б. Барсуков Приаральские Каракумы сев.-зап. побережье Араль- ского моря, п-ов Каратюп окр. г. Аральска Определитель земновод- ных..., 1977
Места обитания. В пределах региона населяет преимущественно пес-
чаные, реже глинистые пустыни и полупустыни с разреженной травянистой
растительностью и зарослями тамарикса и саксаула. На Мангышлаке и Бузачи
наблюдалась на приморских песках у подножья чинков. Преимущественно в
песках отмечались эти черепахи в низовьях рек Иргиз и Тургай, в Приараль-
ских й Прикаспийских Каракумах, на побережье Аральского моря, местами в
Предустюртье.
Вместе с тем в окрестностях Мертвого Култука по соровым понижениям
и чинкам Северного Устюрта они обитают и на участках с суглинистыми поч-
вами. У северных пределов ареала в Приуралье находки приурочены к масси-
вам древних дельтово-аллювиальных отложений в низовьях рек, стекающих
с Подуральского плато. В Сапсынагашской депрессии Тургайского плато они
также встречались на участках с легкими почвами и разреженной псаммофит-
ной растительностью.
Образ жизни. Среднеазиатские черепахи зимоспящие животные. Из зим-
ней спячки у южных пределов региона они выходят в конце марта - апреле.
Весной активны почти в течение всего светлого времени суток, иногда пере-
мещаясь за день на расстояние до 2 км. Летом пики активности смещаются
на утренние и вечерние часы, а середину дня (с 13 до 17 ч) они проводят в
убежищах - неглубоких собственных норах или расширенных норах песчанок,
тушканчиков, ушастых ежей, располагающихся обычно под кустами полыней,
саксаула, жузгуна и других растений. В летнее время черепахи, накопив доста-
точное количество жира, залегают в убежищах. Здесь часть из них в состоянии
оцепенения проводят 2-3 месяца и в сентябре вновь выходят на поверхность.
У подавляющего большинства особей летнее состояние покоя обычно пере-
ходит в зимнюю спячку и общая продолжительность состояния оцепенения
достигает 8-9 месяцев.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
157
Размножение. Половозрелыми черепахами становятся в 10-13 лет. До-
стигнувшие этого возраста особи вскоре после выхода из спячки приступают к
спариванию, которое продолжается почти 2 месяца - с середины апреля почти
до середины июня. Формирование яиц занимает около 10 дней и уже с первых
чисел мая начинается их откладка. На сопредельной территории Казахстана
за весь период размножения двумя-тремя порциями в специально вырытые в
сухом грунте ямки самки откладывают по 5-12 яиц. Эмбриональное развитие
продолжается 80-110 суток, и уже в первой декаде августа появляются первые
молодые. Их выход продолжается до второй половины октября, но на повер-
хности они чаще всего появляются лишь весной следующего года (Параскив,
1956; Ананьева и др., 1998).
Питание. Кормятся черепахи в основном листьями, побегами, семенами
и плодами эфемеров, многолетних трав и кустарников, изредка потребляются
членистоногие, почвенные олигохеты, а в отдельных случаях и мелкие позво-
ночные; при случае черепахи пьют воду. В свою очередь молодых и взрослых
черепах поедают лисицы, волки, вороны и крупные хищные птицы.
Численность. В пределах региона эта черепаха наиболее обыкновенна на
северном побережье Аральского моря (ур. Кульмес, Туранглы, Байкун, п-ов
Каратюб), в Приаральских Каракумах и низовьях р. Тургай (ур. Тауп, Жаман-
кум, Аткайколь). Довольно высокая численность отмечалась на побережье
Каспийского моря у Форт-Шевченко, в долинах некоторых речек, ручьев и
балок с редкими зарослями тамарикса, а также на склонах песчаных гряд с
разреженной злаково-полынной растительностью, где насчитывалось по 5-12,
в среднем до 10 особей на 1 га. По кромке Б. Барсуков она отмечалась еди-
нично, редкой была и на Северном Устюрте (Параскив, 1956; Шилов, 1961). В
этом районе лишь у чинка Донгузтау наблюдалась высокая встречаемость - до
10 особей на 1 км маршрута (Сараев, Пестов, 2010). Севернее в пределах степ-
ной зоны она обнаружено по 1-2 экземпляра.
Лимитирующие факторы. Коренные изменения местообитаний в про-
цессе хозяйственного освоения территории, местами пресс хищных млекопи-
тающих и птиц, а также сбор с коммерческими целями (Кубыкин, Брушко,
1994).
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резервата
и Устюртского заповедника. Особых специальных мер по охране вида на ос-
тальной части региона не требуется, достаточно поддержания естественного
состояния ландшафтов и ограничение промысла.
Региональный статус. Характерный для герпетофауны региона краеаре-
альный вид у северных пределов распространения; включен в Список Между-
народной конвенции о торговле видами дикой фауны и флоры (СИТЕС, При-
ложение II).
158
Глава 4
Надотряд SQUAMATA, OPPEL, 1811-ЧЕШУЙЧАТЫЕ
Отряд 4. SAURIA. MCCARTHNEY, 1822 - ЯЩЕРИЦЫ
Семейство 10. Gekkonidae Gray, 1825-Гекконы, или цепкопалые
Род И. Alsophylax Fitzinger, 1843 - североазиатские геккончики
15. Пискливый геккончик - Alsophylaxpipiens (Pallas, [1814])
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, населяющий территорию региона. Монотипический вид,
хотя в его пределах наблюдаются довольно существенные межпопуляционные
различия в рисунке на голове, туловище, хвосте и конечностях. Почти всю
территорию заселяют представители западной группы популяций, отличаю-
щиеся более мелкими размерами, слабее выраженными спинными бугорками
и их упорядоченностью в ряды, некоторыми средними значениями количес-
твенных признаков и особенностями экологии, в частности предпочитанием
местообитаний и некоторыми другими (Щербак, Голубев, 1986; Ананьева и
др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает прикаспийские районы Ирана,
северный Афганистан, северо-запад Китая и Монголии, Среднюю Азию, зна-
чительную часть Казахстана и примыкающие районы Нижнего Поволжья и
Приуралья (субэндемик Средней Азии и Восточного Ирана). Большая часть
территории района, таким образом, входит в ареал вида, где проходит его се-
верная граница (во многом еще четко не определенная). На крайнем северо-за-
паде ареала она прослеживается по находкам на горе Б. Богдо (в Астраханской
области), откуда вид был впервые описан П.С. Палласом. Позднее здесь его
находили Беккер и Ауэрбах (Кесслер, 1871; цит. по: Чернов, 1954), затем в
1887 г. в окрестностях этой горы его отмечали А. Харузин и К. Сатунин, а в
1888 г. где-то «в степях Букеевской орды» он был найден Н. Калугиным (Чер-
нов, 1954; Параскив, Бутовский, 1960). Последний пункт на карте ареала вида
указывается в центре Волжско-Уральского междуречья (Щербак, Голубев,
1986), где нет подходящих местообитаний. Учитывая привязанность вида к
останцовым горам, а также то, что ханская ставка Букеевской орды находилась
в Урде, логично предположить, что и эта находка была сделана в Прибаскун-
чакском или Приэльтонском районах (в пределах которых помимо «крупных»
островных гор Б. и М. Богдо, Улаган, Чапчачи имеется и множество останцов
меньших размеров), но никак не в центре Волжско-Уральского междуречья.
Таким образом, единственным достоверным местом обитания вида на западе
ареала, где он отмечается и в настоящее время (Смирнов и др., 1985; Стрель-
цов и др., 2006), является гора Б. Богдо, поскольку многолетние поиски этого
вида в подобных окрестных биотопах результатов не дали.
Другим пунктом, где достоверность обитания вида подтверждена несколь-
кими исследователями, были меловые горы Алмас-Тау в верховьях р. Чибенды
(Шыбынды) в Утва-Илекском междуречье. Здесь пискливый геккончик был
найден в 1882 г. Н.А. Зарудным (1895), а позднее его обитание в этих местах
подтверждено А.М. Никольским (1915). Сравнительно недалеко от этого пун-
кта в 1976 г. он был отмечен и в меловых горах в 4 км восточнее с. Троицкое
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
159
Соль-Илекского района Оренбургской области (Чибилёв, 1995). Однако во вре-
мя специального обследования потенциально пригодных местообитаний этого
вида в верховьях р. Шыбынды, по оврагу Акбулак (у с. Троицкое), а также в
прилегающих обнажениях песчаников и меловых отложений по г. Ишкаргану,
проведенных во второй половине 1990-х гг., ни сам пискливый геккончик, ни
следы его жизнедеятельности обнаружены не были, в связи с чем вид внесен в
список исчезнувших животных Оренбуржья (Давыгора, 2001). С учетом этих
сведений ближайшие (около 600 км) к горе Б. Богдо местообитания вида на-
ходятся в Урало-Эмбенском междуречье на горах Иман-Кара, Аккерегешин,
Каражар, Жельтау, Коленкала, ур. Бесбай, чинке Донгузтау и плато Актолгай
(Неручев, Васильев, 1978; Смирнов и др., 1985; Щербак, Голубев, 1986; Нака-
ренок, Неручев, 2001; Накаренок, 2003в; Сараев, Пестов, 2010).
Восточнее пискливый геккончик отмечался в 4 км западнее г. Эмба (Актю-
бинская область), низовьях р. Иргиз (Lehman, 1852; цит. по: Динесман, 1953),
на правобережье и в верховьях р. Тургай, затем в Центральном Казахстане,
Северном Прибалхашье, Алакольской котловине (Параскив, 1956) и далее по
северо-западному Китаю до Монголии. Южная граница полностью находится
в пределах пустынной зоны.
В очерченных пределах пискливый геккончик не образует сплошных по-
селений, а встречается лишь в ограниченных по площади изолированных оча-
гах (рефугиумах). В Северном Прикаспии его существование, по-видимому,
является следствием нижнехвалынской и последующих трансгрессий и рег-
рессий Каспийского моря, в результате которых они оказались разделенными
непригодными для обитания вида глинистыми, песчаными и солончаковыми
пустынями (Неручев, Шатилович, 1985; Накаренок, 2003в). В итоге в настоя-
щее время ареал вида только на горе Б. Богдо (возможно, и в Приэльтонском
районе); в Приэмбенской провинции, окрестностях городов Эмбы и Иргиза
и по р. Тургай встречается в полупустыне, а остальная часть его ареала на-
ходится в пустынной зоне. В целом в географическом отношении его конк-
ретные местообитания известны в 2 (2,9% из 11) ландшафтных районах степ-
ной зоны, 6 (18,8% из 32) полупустынной и 9 (20,9% из 43) пустынной зонах
(табл. 27, карта 15).
Места обитания. Пискливый геккончик - типичный обитатель полупус-
тынных и пустынных ландшафтов и даже в местах прежних встреч в степной
зоне придерживался полупустынных местообитаний. В северной части регио-
на встречается в основном на эродированных склонах останцовых холмов или
плато с выходами песчаников, мела или покрытых грубообломочным матери-
алом. Для этих стаций, как правило, характерна скудная растительность из по-
лыней, злаков или низкорослых кустарников. Южнее долины р. Эмбы селится
также на глинисто-щебнистых и песчаных равнинах с обрывами известняков,
пухлых гипсоносных глин или лессовых отложений с обилием трещин, щелей,
других пустот, в том числе нор роющих животных.
Образ жизни. Пискливый геккончик из зимней спячки на сопредельной
территории Казахстана выходит в конце марта - начале апреля. Активен пре-
имущественно в ночные часы, хотя в пасмурную погоду встречается и днем,
чаще всего среди лучше прогреваемых каменистых осыпей. Днем, как прави-
ло, укрывается в различных убежищах. Активный образ жизни продолжается
около 200 суток и заканчивается в разных числах октября.
160
Глава 4
Таблица 27
Распространение пискливого геккончика
Зона Провин- ция Район № точки Местонахождение Источник информации
3 X VI Западно-Казахстанская область
с о 30 1 гора Алмас-Тау (исчез ?) Зарудный, 1895; Чи- билёв, 1995; Давыгора, 2001
О VII 41 2 с. Троицкое (исчез ?)
Актюбинская область
густынная XV 96 98 99 3 4 5 горы Актолагай окр. г. Эмба, 4 км к зап. р. Эмба, побережье Сараев, Пестов, 2010; Щербак, Голубев, 1981; Накаренок, Неручев, 2001
£ XVIII 107 6 окр. г. Иргиз Динесман, 1953; Па-
к XIX ПО 111 7 8 низ. р. Тургай низ. р. Жиланчик раскив, 1956; Щербак, Голубев, 1986
Астраханская область
XX 113 9 окр. г. Б. Богдо Паллас, 1814 (цит. по: Чернов, 1954); Божанс- кий, Полынова, 1998
Гурьевская область
137 10 11 12 дол. р. Кайнар дол. р. Эмба гора Иман-Кара гора Кой-Кара Неручев, Васильев, 1978; Смирнов и др., 1985; Накаренок, Неру- чев, 2001; Накаренок, 2003е; Сараев, Пестов, 2010
3 ур. Боздык
X X XXII гора Бесбай гора Аккерегешин
е* 138 13 гора Каражар ур. Бесбай Мангистауская область горы Жельтау
139 14 чинк Донызтау горы Коленкела
XXV 147 15 ур. Каска-жол у сев.-зап. берега Аральского моря Параскив, 1956
XXVI 154 16 Приаральские Каракумы Лобачев и др., 1973
155 17 ур. Уш-Чоку Зарудный, 1915
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
161
Размножение. Половой зрелости геккончики достигают на вторую весну
после появления на свет. В этом возрасте спустя 3-4 недели после выхода из
зимней спячки начинается спаривание, а с конца мая - откладка яиц, которая
продолжается до июля. Яйца находили в рыхлой меловой пыли под обрывами
и в растрескавшемся пухляке на глубине 10-15 см (Смирнов и др., 1985). Мо-
лодые геккончики появляются в июле-августе, а спустя 2-3 месяца залегают
в спячку.
Питание. Питается преимущественно насекомыми. В желудках 10 особей,
добытых в регионе, по встречаемости и массе преобладали чешуе- и жестко-
крылые (Смирнов и др., 1985). В других районах Казахстана отмечались также
прямокрылые, двукрылые и их личинки, причем большинство (70%) оказалось
дневными формами (Параскив, 1956).
Численность. В связи с ночным образом жизни сведения о численности
вида довольно ограничены. Вместе с тем известно, что в 1860-1870-х гг. на
горе Б. Богдо он не являлся редкостью и встречался в довольно значительном
количестве - зоолог Беккер за 3 дня поймал более 20 особей. Позднее числен-
ность его, по-видимому, существенно сократилась, поскольку зоологи, посе-
щавшие Б. Богдо в первой половине XX в. его не упоминали (Чернов, 1954).
Сокращение численности здесь продолжалось и позднее, причем главным об-
разом в связи с разрушением типичных местообитаний (Ананьева, 2001). Сей-
час на Б. Богдо пискливый геккончик обычный, хотя и немногочисленный вид
(Стрельцов и др., 2006). Локальная плотность на разных участках достигает
18-40 (в среднем 29 ос/га), а общая численность оценивается в 1,0-1,5 тыс.
особей (Божанский, Полынова, 1998). На остальной территории региона он,
по-видимому, наиболее многочисленен в Приэмбенской провинции, где по
меловым обрывам и крутым пухляковым склонам останцов за час находили
3-10 экземпляров (Смирнов и др., 1985), в то время как в других местах обыч-
но отмечались единичные экземпляры (Зарудный, 1915; Динесман, 1953; Па-
раскив, 1956; Щербак, Голубев, 1981; 1986; Лобачев и др., 1973; Накаренок,
2003в). Вместе с тем в некоторых локальных местообитаниях плотность его
населения достигает максимальных величин, характерных для рептилий Ка-
захстана (Брушко, 1993).
Лимитирующие факторы. Хозяйственное освоение территории, часто
сопровождающееся разрушением типичных местообитаний (разработка карь-
ера на горе Б. Богдо).
Охрана. Обитает и охраняется на территории Богдинско-Баскунчакского
государственного заповедника и Иргиз-Тургайского резервата; на остальной
территории специальных мер охраны не предпринималось.
Региональный статус. Занесен в Красную книгу Российской Федерации
(Ананьева, 2001) как редкий узкоареальный вид (категория 3) и Красную кни-
гу Астраханской области (2004). Для казахстанской части ареала это харак-
терный немногочисленный представитель герпетофауны у северных пределов
распространения.
162
Глава 4
Род 12. Crossobamon Boettger, 1888 - гребнепалые гекконы
16. Гребнепалый геккон - Crossobamon eversmanny (Wiegmann, 1834)
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, населяющий территорию региона. Политипический вид,
в регионе представлен номинативным подвидом С. е. eversmanny (Wiegmann,
1834). Популяции различаются деталями рисунка на голове, боках, конечнос-
тях, некоторыми другими признаками и особенностями экологии. У северных
популяций довольно четко прослеживается клинальная изменчивость в чис-
ле чешуй вдоль нижней стороны тела, вокруг середины туловища и на горле
(Щербак, Голубев, 1986).
Распространение. Ареал вида охватывает северные районы Ирана, Аф-
ганистана, значительную часть Средней Азии и южного Казахстана (эндемик
Средней Азии и Восточного Ирана). Северная граница ареала прослеживается
от восточного побережья Каспийского моря через песчаный массив Ак-тюбе
на п-ове Мангышлак, низовья р. Иргиз, Приаральские Каракумы, окрестности
г. Аральска и далее на восток через Муюнкумы до границы Киргизии, откуда
поворачивает к южным рубежам.
В очерченных пределах этот геккон отмечался в песках у родника Кин-Тыч-
ке на Устюрте (Параскив, 1956), в Б. и М. Барсуках (Брушко, 1993), низовьях
р. Иргиз (Lehman, 1852; цит. по: Никольский, 1905), Приаральских Каракумах
(Лобачев и др., 1973; Брушко, 1993), ряде пунктов на побережье Аральского
моря и в окрестностях г. Аральска (Зарудный, 1915; Динесман, 1953). В гео-
графическом отношении его конкретные местонахождения известны в одном
(3,1 % из 32) ландшафтном районе полупустынной и 5 (11,6% из 43) районах
пустынной зон (табл. 28, карта 16).
Места обитания. Вид является типичным псаммофилом с высокой сте-
пенью специализации. Как правило, населяет слабо закрепленные жузгуном и
верблюжей колючкой холмы и их незаросшие вершины, реже встречается на
их склонах и у основания. Участков с уплотненными глинистыми, песчаными
и лессовыми почвами, особенно закрепленных растительностью, определенно
Таблица 28
Распространение гребнепалого геккона
Зона Провин- ция Район № точки Местонахождение Источник информации
Полупустын- ная XVIII 107 1 низовья р. Иргиз Lehman, 1852 (цит. по: Никольский, 1905)
XXV 147 2 плато Устюрт Параскив, 1956
Пустынная XXVI 151 152 154 3 4 5 пески Б. Барсуки . пески М. Барсуки Приаральские Каракумы Лобачев и др., 1973; Брушко, 1993; Зарудный, 1915; Динесман, 1953
155 6 побережье Аральского моря, окр. г. Аральска
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
163
избегает (Параскив, 1956; Брушко, 1993). Обычно встречается с другим псам-
мофилом - сцинковым гекконом.
Образ жизни. Гребнепалый геккон из зимней спячки чаще всего выходит
в апреле (Мангышлак, Муюнкумы) и сохраняет активность почти до конца
сентября (около 170-180 суток). Это типичное сумеречно-ночное животное -
из укрытий выходит около 21 часа и возвращается в них до восхода солнца.
За это время он проходит значительное расстояние (обследуя норки, другие
углубления, взбираясь на кустики), удаляясь от убежищ на 100-150 м. Днем
укрывается в норках (прямой наклоненный ход длиной 70-110 см с расшире-
нием на глубине 30-40 см), которые чаще всего роет сам или использует нор-
ки крупных жуков (навозников), в которых устраивает свои отнорки. На зиму
выход из норок забивается песчаной пробкой.
Размножение. Половозрелым этот геккон становится на следующий год
после выхода из яиц. К спариванию приступает с середины апреля, но у неко-
торых особей этот процесс продолжается до начала июня. Яйца обычно откла-
дываются несколькими порциями по 1-2 штуки в июне-июле. Период инкуба-
ции продолжается 45-53 дня и уже с конца июля появляются молодые.
Питание. Основу пищевого рациона гребнепалого геккона составляют на-
секомые, главным образом жуки, перепончатокрылые, ночные бабочки и их
гусеницы; реже потребляются другие членистоногие.
Численность. На сопредельной региону территории Казахстана в ти-
пичных местообитаниях средняя плотность составляет 5,5, максимальная
15,0 ос/га (Брушко, 1993).
Лимитирующие факторы. Комплекс абиотических и биотических факто-
ров на границе ареала, а также хозяйственное освоение территории (в первую
очередь выпас скота и инфраструктурное обустройство, которые в наиболь-
шей степени сказываются на состоянии песчаных пустынь).
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резерва-
та и Устюртского заповедника. На остальной территории региона необходимо
поддержание естественного состояния типичных местообитаний вида (регули-
рование выпаса и т. п.).
Региональный статус. Типичный представитель герпетофауны пустынь у
северных пределов распространения.
Род 13. Cyrtopodion Fitziger, 1843 - тонкопалые гекконы
17. Каспийский геккон - Cyrtopodion caspius (Eichwald, 1831)
Значение таксона для сохранения биоразнообразия. Один из двух
представителей обширного рода гекконов, населяющий территорию регио-
на. Политипический вид, в регионе представленный номинативным подви-
дом С. с. caspius (Eichwald, 1831). В его пределах наблюдаются некоторые
межпопуляционные отличия по числу чешуй вдоль тела, поперек брюшка,
на голове, а также в размерах спинных бугорков, некоторых других морфо-
логических признаках и особенностях экологии (Щербак, Голубев, 1986;
Ананьева и др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов,
2006).
164
Глава 4
Таблица 29
Распространение каспийского геккона
Зона Провин- ция Район № точки Местонахождение Источник информации
Полупустын- ная XXII 138 1 Мангистауская область горы Жельтау горы Коленкелы Киреев, 1981; На- каренок, 2003в
XXIII 142 2 окр. Форт-Шевченко Смирнов и др., 1985; Параскив, 1956
Пустынная XXV 147 148 3 4 5 6 сев. чинк Устюрта Устюрт: окр. сора Карын- Жарык ур. Кугусем сев.-зап. окр. Аральского моря окр. сора Асматай-Матай Параскив, 1956; Брушко, Кубыкин, 1988
XXVI 151 152 154 155 7 8 9 10 пески Б. Барсуки пески М. Барсуки Приаральские Каракумы ур. Кок-Арал Параскив, 1956 Брушко, 1993; Щербак, Голубев, 1986
Распространение. Ареал вида охватывает прикаспийские районы Азер-
байджана и Ирана, северо-запад Афганистана, значительную часть Средней
Азии и юго-запад Казахстана (эндемик Средней Азии и Восточного Ирана).
Северная граница его ареала проходит от северо-востока Каспия к среднему
течению р. Эмбы, северному побережью Аральского моря, Приаральским Ка-
ракумам, затем через центральную часть Кызылкумов к предгорьям Памиро-
Алая и далее к южным рубежам. Таким образом, в состав ареала вида входит
только южная часть исследуемого нами региона. Точнее в ближайших .окрес-
тностях и на территории региона граница ареала прослеживается по находкам
этих гекконов у Форт-Шевченко на п-ове Мангышлак, северном чинке плато
Устюрт, останцовых горах по р. Эмба, в Б. и М. Барсуках и Приаральских Ка-
ракумах, у сора Асматай-Матай, северо-западного побережья Аральского моря
и на его бывшем острове Кок-Арал. Эти пункты по сути являются и единствен-
ными известными местами обитания в регионе (табл. 29, карта 17).
Места обитания. Это один из наиболее эвритопных видов гекконов, на-
селяющий самые различные местообитания. В Казахстане встречается по рав-
нинам и предгорьям, обычно у нагромождений камней, развалин сооружений,
в постройках, пустотах обрывов и норах грызунов (Параскив, 1956; Брушко,
1993). Южнее набор местообитаний еще более разнообразен.
Образ жизни. На зиму впадает в спячку, во время которой проводит около
150 суток - как правило, с конца октября по апрель. Убежищем на этот период,
а затем и в летнее время им служат разные естественные пустоты, а также норы
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
165
грызунов. После выхода из спячки активны преимущественно в сумеречные и
ночные часы, хотя весной и в условиях рассеянного света нередко появляются
и днем. Летом при рассеянном свете они чаще всего только начинают охотится
за добычей, быстро и ловко передвигаясь по наклонным, вертикальным и по-
толочным поверхностям.
Размножение. Половой зрелости эти гекконы достигают к середине сле-
дующего после выхода из яиц года. Вскоре после выхода из зимней спячки у
них начинаются брачные турниры самцов, а через несколько дней и спарива-
ние. Яйца (по 1-2 штуке) откладываются в конце мая - июне - июле, причем
в одно гнездо (в песке, гнездах муравьев, лессе) яйца могут откладывать не-
сколько самок. Выход молодых (на Мангышлаке) начинается с конца июня и
продолжается до августа-сентября, а спустя примерно 2-3 месяца они залега-
ют в спячку.
Питание. Основу пищевого рациона составляют насекомые (прямокры-
лые, жуки, перепончатокрылые, клопы, некоторые бабочки и их гусеницы) и
другие членистоногие, иногда поедает и мелких ящериц. В свою очередь он
является добычей для змей, ряда хищных птиц и млекопитающих.
Численность. В целом по Казахстану этот вид отличается довольно высо-
ким уровнем численности. Его плотность в типичных местообитаниях состав-
ляет в среднем 77,3 экз/га, а в оптимальных условиях достигает 185,0 экз/га
(Брушко, 1993). У северных пределов ареала это очень редкий вид - в Приэм-
бенской пустыне на крутых склонах с выходами рухляка насчитывали лишь
1,0 ос/10 км (Накаренок, 2003в).
Лимитирующие факторы. У северных пределов, по-видимому, комплекс
факторов, действующий на краеареальные популяции; на остальной террито-
рии - естественные враги.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На большей части ареала высокая и стабильная численность особых мер охра-
ны не требует; для краеареальных популяций достаточно сохранения естест-
венного состояния ландшафтов.
Региональный статус. Характерный представитель герпетофауны регио-
на, населяющий специфические местообитания у северной границы ареала.
Род 14. Mediodactylus Szczerbak et Golubev,
1977 - средиземноморские тонкопалые гекконы
18. Серый геккон - Mediodactylus russowii (Strauch, 1887)
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей обширного рода, населяющий территорию региона. Политипичес-
кий вид, в регионе представленный номинативным подвидом С. г. russowii
(Strauch, 1887), в составе которого проявляются существенные межпопуляци-
онные различия в размерах, окраске, щитковании тела и некоторых особеннос-
тях экологии. Популяции нашего региона (как самые северные) отличаются
максимальными размерами тела, наибольшим количеством преанальных пор,
у них возрастает частота встречаемости дополнительных спинных бугорков; у
популяций, населяющих заросли саксаула, рисунок на спинной стороне тела
166
Глава 4
становится очень сложным, а у обитателей построек поперечные полосы зачас-
тую сохраняются только на туловище и хвосте (Щербак, Голубев, 1986; Ана-
ньева и др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает восточное Предкавказье, север-
ные районы Ирана, Афганистана, северо-запад Китая, Среднюю Азию и зна-
чительную часть территории Казахстана (эндемик Средней Азии и Восточного
Ирана). Его северная граница от залива Мертвый Култук в северо-восточной
части Каспийского моря проходит к низовьям рек Эмба и Кайнар, по западной
кромке плато Донузтау к северному Приаралью, затем через пески Арыскум к
западному побережью оз. Балхаш и далее по р. Каратал, предгорьям Джунгар-
ского Алатау и долине р. Или уходит в пределы Китая. Таким образом, ареал
вида охватывает только южную часть исследуемого нами региона.
Более точно пределы распространения этого геккона здесь прослеживают-
ся по его находкам на побережье залива Мертвый Култук, останцовых горах
Кой-Кара, Иман-Кара, горам Аккерегешын, Сары-Нияз, плато Ак-Толгай, в
долинах рек Кайнар, Эмба, Шаган (у кромки плато Донгузтау), ур. Жарбулак
на Устюрте, ур. Кос-Казах, Тюп-Каран и Турангли на северо-западном и север-
ном побережье Аральского моря. Южнее в очерченных пределах серый геккон
отмечался также в горах Жельтау, Коленкелы, балке Жаман-Карасай, а также
у сора Асматай-Матай в Предустюртье. Таким образом, по останцовым горам
вид заходит в полупустыню, где встречается в одном (3,1% из 32) ландшафт-
ном районе, а основная часть его ареала находится в 8 (18,6% из 43) районах
пустынной зоны (табл. 30, карта 18).
Места обитания. Серый геккон - широко распространенный и эвритоп-
ный пустынный вид. Однако у северных пределов ареала, где он частично за-
ходит в зону полупустыни (в Приэмбенской провинции), и на сопредельной
территории Устюрта, Прикаспия и Приаралья становится более стенотопным
и селится в основном на выходах песчаников, мела, известняков, сланцев, реже
по растрескавшимся пухляковым гипсоносным глинам на крутых склонах хол-
мов, по обрывам сухих русел рек и временных водотоков, где придерживается
в основном нагромождений обломочного материала. Довольно часто поселя-
ется в пустотах и трещинах стен разных строений, несколько реже встречается
в норах грызунов, пустующих муравейника^.
Образ жизни. Активный образ жизни начинается после выхода из зимней
спячки, что чаще всего наблюдается в начале апреля. С этого времени с наступ-
лением сумерек гекконы выходят из убежищ, охотятся ночью, а с рассветом
деятельность прекращается и животные скрываются в убежищах (естествен-
ных пустотах и норах животных). Ранней весной и в прохладные осенние дни
чаще встречается днем. Обычно живут поодиночке, но неполовозрелые особи
встречаются и небольшими группами (по 3-4 экз.). Активность продолжается
около 160 суток, и в сентябре гекконы залегают в спячку (выбирая более глу-
бокие убежища), которая продолжается около 200 суток.
Размножение. Половой зрелости серый геккон (как и другие виды), по-
видимому, достигает на втором году после появления на свет. Спаривание (по
наблюдениям на сопредельной территории) отмечалось с середины мая, а от-
кладка яиц - с начала июня. Период откладки яиц продолжается около 45 дней
(до середины июля), а позднее самки со зрелыми яйцами уже не встречались.
Яйца (по 1-2 шт. в 2 приема) обычно откладываются в хорошо прогреваемых
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
167
Распространение серого голопалого геккона
Таблица 30
Зона Пронин* ция Район № точки Местонахождение Источник информации
Полупустынная XV 96 1 2 3 Гурьевская область долина р. Кайнар горы Актолагай междуречье Сагиз-Кайнар ур. Жантеректау Накаренок, Неручев, 2001; Смирнов и др., 1985; Накаренок, 2003в
Пустынная XXII 137 138 139 4 5 6 7 гора Койкара гора Иманкара гора Аккерегешын гора Сарынияз ур. Жанбике по р. Эмба ур. Уитас Мангистауская область горы Жельтау гора Коленкелы балка Жаман-Карасай окр. п. Бейнсу Неручев, Василь- ев, 1978; Смирнов и др., 1985 Накаренок, 2003 в; Сара- ев, Пестов, 2010 Накаренок, 2003в; Па- раскив, 1956
XXIII 143 8 окр. сора Мертвый Култук Щербак, Голубев, 1986
XXV 147 148 9 10 11 Устюрт: ур. Жарбулак окр. сора Карын-Жарык окр. сора Асматай-Матай Брушко, Кубыкин, 1988
XXVI 151 155 12 13 14 пески. Б. Барсуки ур. Тюп-Караган ур. Турангли ур. Кос-Казах Параскив, 1956; Брушко, Кубыкин, 1988; Заруд- ный, 1915
естественных пустотах, а также в ходах муравейников, норах грызунов. Инку-
бация продолжается около 50 суток, и уже в начале августа появляются моло-
дые, которые примерно месяц спустя залегают в спячку.
Питание. Основу кормового рациона вида составляют насекомые, состав
которых меняется в зависимости от местообитания и сезона года. Весной в
добыче преобладают жуки и перепончатокрылые, летом - прямокрылые и рав-
нокрылые, а к осени увеличивается количество перепончатокрылых, клопов
и мух (спектр значительно шире, чем у других видов - за счет более крупных
размеров и повышенной дневной активности). В свою очередь серых гекконов
добывают некоторые змеи и хищные птицы.
Численность. В основной части ареала серый геккон является обычным
или многочисленным видом. Так, на сопредельной территории в Казахстане
168
Глава 4
средняя плотность составляет 104,0, а максимальная 130,0 экз/га (Брушко,
1993). В регионе он встречается гораздо реже - на меловых обрывах Приэм-
бенских гор-останцов численность определена в 1,0, их защебненных склонах
7,0, крутых склонах с выходами рухляка 13,0 ос/10 км (Накаренок, 2003в), а на
обширных пространствах он отсутствует вовсе, в связи с чем общая числен-
ность его в регине, очевидно, невелика.
Лимитирующие факторы. Комплекс абиотических и биотических факто-
ров у северных пределов ареала; хозяйственное освоение территории, сопро-
вождающееся разрушением типичных местообитаний, ремонт глинобитных
сооружений.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На остальной территории региона достаточно поддержания естественного со-
стояния ландшафтов.
Региональный статус. В европейской части ареала, вероятно, исчез, в свя-
зи с чем занесен в Красную книгу Российской Федерации (2001) в категории О.
В регионе характерный немногочисленный вид у северной границы ареала.
Род 15. Teratoscincus Strauch, 1863 - сцинковые гекконы
19. Сцинковый геккон - Teratoscincus scincus (Schlegel, 1858)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода, обитающий на территории региона. Политипический вид, в
пределах региона представлен номинативным подвидом Т. s. scinus (Schlegel,
1858). В его составе имеются межпопуляционные отличия по рисунку взрос-
лых особей; количество чешуй вокруг середины туловища и поперек головы
перекрывается (Щербак, Голубев, 1986; Ананьева и др., 1998; Атлас пресмы-
кающихся.., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает прикаспийские районы Ирана,
север Афганистана, Пакистана, Белуджистана, северо-запад Китая, Среднюю
Азию и Южный Казахстан (субэндемик Средней Азии и Восточного Ирана).
Его северная граница прослеживается от Мангышлака через плато Устюрт
(в окрестностях п. Бейнеу и сора Асматай-Матай), северное побережье Араль-
ского моря (п-ов Каратюп), Приаральские Каракумы, Арыскумы, пески по ре-
кам Каратал, Или (его типовая территория) и предгорьям Тянь-Шаня в Китай;
южная соответственно - по песчаным массивам Црана и Афганистана. Таким
образом, сцинковый геккон населяет только самую южную часть нашего реги-
она в пределах пустынной зоны. Его конкретные местонахождения здесь из-
вестны в песках у п. Бейнеу, Сам на Устюрте, п-ове Каратюп, М. Барсуках, в
окрестностях г. Аральска и Приаральских Каракумах (табл. 31, карта 19).
Места обитания. Это типичный псаммофильный вид. В Казахстане сцин-
ковый геккон чаще всего поселяется на склонах песчаных гряд и бугров с ред-
кой травянистой растительностью, а также на слабо закрепленных или голых
песках (даже оказавшихся внутри населенных пунктов) и лишь изредка отме-
чается на прилегающих к пескам участках плотных грунтов.
Образ жизни. Зимний период (около 200 суток) сцинковые гекконы про-
водят в спячке в норах, которые роют сами, или селятся с другими живот-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
169
Распространение сцинкового геккона
Таблица 31
Зона Провин- ция Район № точки Местонахождение Источник информации
XXIII 139 I ур. Копа в окр. п. Бейнсу Брушко, Кубыкин, 1988
XXV 148 2 пески Сам у сора Асматай- Матай Параскив, 1956
S 152 3 пески М. Барсуки Неручев, Васильев, 1978
X 154 4 Приаральские Каракумы, Щербак, Голубев, 1986
в сев. пески Коимбай
tz? XXVI 5 кол. Ильля-Кудук, 100 км к сев-вост. г. Аральска Динесман, 1953
155 6 окр. г. Аральска, ПЧС Щербак, Голубев, 1986
7 п-ов Каратюп Динесман, 1953
ними (чаще всего крупными насекомыми или тушканчиками). Здесь же они
впоследствии проводят и дневные часы. Из зимних убежищ в северной части
ареала выходят в первых числах апреля. Взрослые гекконы обычно живут по-
одиночке. Активны ночью; на поверхности появляются с наступлением суме-
рек и охотятся примерно с 21 до 5-6 часов, не уходя далеко от норы. Активный
период продолжается 150-160 суток и прекращается с наступлением осени,
поскольку уже в первых числах сентября животные залегают в спячку.
Размножение. Половозрелым сцинковый геккон становится через 18-
20 месяцев после выхода из яиц. Спаривание начинается в конце апреля и про-
должается до середины мая. Начало откладки яиц на сопредельной территории
отмечено в первой половине июня, и продолжается она приблизительно до
конца июля. Яйца откладываются порционно (по 1-3 шт.) и, по-видимому, не
менее 3 раз. Инкубационный период продолжается около 2 месяцев, и с сере-
дины августа начинают появляться молодые, выход которых иногда растяги-
вается до середины сентября.
Питание. Основу пищевого рациона сцинкового геккона составляют
крупные насекомые и их личинки: жуки (чернотелки, жужелицы), прямо-, пе-
репончатокрылые (муравьи), а также другие членистоногие. В свою очередь
он служит добычей змей, хищных птиц и млекопитающих.
Численность. В краеареальных популяциях, очевидно, невелика. В целом
же для Казахстана он сейчас сравнительно обычен - средняя плотность до-
стигает 6,5, а максимальная 16,5 ос/га (Лобачев и др., 1973; Брушко, 1993); в
прошлом во многих местах Казахстана считался многочисленным (Параскив,
1956).
Лимитирующие факторы. В основном лишь сильный перевыпас скота
и (локально) внедорожная техника. В последние годы вид стал объектом тор-
говли и наряду с некоторыми другими видами предметом вывоза за границу
(Брушко, 1993; Брушко, Кубыкин, 1994).
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На остальной части региона достаточно под держания естественного состояния
ландшафтов и регулирования возможных заготовок.
170
Глава 4
Региональный статус. Характерный представитель герпетофауны пусты-
ни у северных пределов распространения.
Семейство 11. Agamidae Spix, 1825 - Агамовые
Род. 16. Trapelus Cuvier, 1817 - равнинные агамы
20. Степная агама - Trapelus sanguinolentus (Pallas, [1814])
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель обширного рода, обитающий на территории региона. Политипичес-
кий вид, в регионе представлен восточно-прикаспийским подвидом Т. s. ага-
lensis Lichenstein, 1823 (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004).
В то же время рядом аторов (Кузьмин, Семенов, 2006) этот вид рассматрива-
ется лишь как северный подвид (Г. agilis sanguinolentus Oliver, 1804) западно-
азиатской прыткой агамы, в составе которого таксономическая особенность
аральских популяций вообще не находит подтверждения.
Распространение. Ареал вида охватывает восточное Предкавказье, север-
ные районы Ирана, Афганистана, Пакистана, северо-запад Китая (Джунгарию),
Среднюю Азию и Южный Казахстан (субэндемик Средней Азии и Восточного
Ирана). Его северная граница в прошлом достигала низовий р. Волги (Кир-
гизские степи в окрестностях г. Астрахани, откуда имеется экземпляр в кол-
лекции МГУ; Никольский, 1905). Сейчас она прослеживается от восточного
побережья Каспия к Прикаспийским Каракумам, оконечности Подуральского
плато южнее среднего течения р. Эмба, обходит плато Шагырай, затем следует
к низовьям р. Иргиз, Приаральским Каракумам, далее к р. Сары-су, Северному
Прибалхашью, окрестностям г. Аягуз и в Китай. Его южные рубежи от прикас-
пийбких районов Ирана простираются к северо-западу Китая.
Таким образом, ареал вида охватывает лишь южную пустынную часть ис-
следуемого нами региона, где он местами (по останцовым массивам) проника-
ет в зону полупустынь.
Точнее в настоящее время в регионе и его ближайших окрестностях се-
верная граница прослеживается по находкам на Мангышлаке, Бузачах, п. Жи-
лая Коса в низовьях р. Эмба, севере Прикаспийских Каракумов, южной око-
нечности Подуральского плато у рек Майнисай, Атжаксы, низовьях р. Иргиз
и Приаральских Каракумах (Параскив, 1956; Определитель земноводных..,
1977; Накаренок, Неручев, 2001). В очерченных пределах степную агаму на-
ходили также по западному и восточному чинкам северного Устюрта, в Сам-
Матсайской котловине, ур. Каратюп, Б. и М. Барсуках, плато Куланды, в ряде
мест Приаральских Каракумов, ур. Ильля-Кудук и в окрестностях г. Аральска
(табл. 32, карта 20).
Места обитания. В пределах региона степная агама является эвритопным
видом, населяющим пустынные и полупустынные сообщества. Вместе с тем
она явно предпочитает закрепленные песчаные и петрофитные группировки
с кустарниковыми и полукустарниковыми зарослями (жузгуном, терескеном,
саксаулом и др.); несколько реже встречается по окраинам слабо закреплен-
ных травянистой растительностью песков (в том числе у населенных пунктов
и по обочинам дорог). Засоленных глинистых и сырых солончаковых пустынь
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
171
Таблица 32
Распространение степной агамы
Зона Провин- ция Район № точки Местонахождение Источник информации
[©пу- стын- ная XV 100 1 Актюбинская область юг Подуральского плато Определитель земновод-
'-g' XVIII 107 2 бас. р. Иргиз ных..., 1977
Гурьевская область
XXI 127 3 ур. Кара-арна пром. Тенгиз
4 п. Жилая Коса
137 5 Приэмбенская пустыня Накаренок, 2003в
138 6 Прикаспийские Каракумы, сев-зап. кромка п. Косчагыл окр. п. Кульсары, 37 км к югу окр. п. Тургызба окр. некр. Карашунгыл окр. Толтурсора окр. некр. Токсанкелы окр. некр. Ушкан Определитель земновод- ных..., 1977; Неручев, Васильев, 1978; Нака- ренок, 2003в; Сараев, Пестов, 2010
5 окр. некр. Айнамыз
я я п. Аккудук, ур. Утибукай
S XXII некр. Темирорта
с 139 7 горы Колекелы ур. Ыргызбай ур. Буранкуль ур. Кумтюбе метеост. Джугара бол. Жаман-Карасай ур. Молорпа ур. Шагырлыкум гора Таксанбай ур. Мынсуалмас чинк Донызта кол. Тасастау зап. чинк Устюрта
140 8 п-ов Бузачи Параскив, 1956
XXIII 143 9 окр. Мертвого Култука Определитель земновод- ных..., 1977
172
Глава 4
Окончание табл. 32
Зона Провин- ция Район № точки Местонах ожден не Источник информации
§ XXV 147 148 10 11 12 плато Сев. Устюрт ур. Молкудук вост, чинк Устюрта Сам-Матсайская депрессия Неручев, Васильев, 1978 Параскив, 1956; Брушко,
я я 151 13 пески Б. Барсуки 1993, 1995
1 152 14 пески М. Барсуки Определитель земновод-
с 153 15 плато Куланды ных..., 1977
XXVI 154 155 16 17 18 Приаральские Каракумы п-ов Карапоп окр. г. Аральска Лобачев и др., 1973 Динесман, 1953
явно избегает, в связи с чем отсутствует на приморских равнинах и внутрен-
них солончаковых депрессиях.
Образ жизни. Из зимней спячки (в которой проводит около 150-160 су-
ток) у северных рубежей ареала степные агамы выходят в конце марта, реже в
начале апреля. Весной активны весь день, летом пики активности смещаются
на утренние и вечерние часы; самцы днем (между 13 и 16 часами) часто наблю-
даются на кустарниках, деревьях или других возвышениях, а самки и молодые
укрываются в убежищах - естественных пустотах или норах животных (пес-
чанок, тушканчиков, ежей и др.). Агамы отличаются территориальной привя-
занностью и свои участки охраняют от конкурентов. Активный образ жизни
продолжается около 200 суток, до начала или середины октября, а затем они
скрываются в зимовочных убежищах (глубоких расселинах, пустотах под кам-
нями, норах большой песчанки или других животных).
Размножение. Половой зрелости степные агамы достигают в возрасте
около 2 лет; они моногамы. Спаривание чаще всего начинается вскоре после
выхода из спячки со второй половины апреля и продолжается до конца мая.
Яйца (4-18) за сезон откладываются дважды - со второй декады мая до по-
следних чисел июля (или первых чисел августа) обычно в нору или специально
выкапываемую самкой ямку. Инкубация, вероятно, продолжается 50-60 дней,
и уже в июле появляются молодые первой генерации; из вторых кладок выход
продолжается до поздней осени.
Питание. Основу пищевого рациона в Казахстане составляют насекомые:
жуки (чернотелки, усачи, бронзовки), прямо-, перепончатокрылые, клопы, ба-
бочки, их личинки и другие членистоногие; в небольшом количестве поедают-
ся также листья и цветки растений. Весной местами наблюдается концентрация
агам вдоль хорошо прогреваемого асфальта дорог со скоплениями насекомых.
В свою очередь агамы (преимущественно молодые особи) являются жертвами
разных хищников.
Численность. На большей части ареала степная агама является сравни-
тельно обычным, а местами даже многочисленным видом. Так, по материалам
З.К. Брушко (1993) в Казахстане плотность населения вида в среднем составля-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
173
ст 8,5 экз/га, а в наиболее оптимальных местообитаниях достигает 12,0 экз/га.
Для сравнения отметим, что в Средней Азии численность в 10 экз/га считается
очень высокой (Ананьева и др., 1998).
Это в отдельные годы позволяет ей считаться доминантным видом в верх-
нехвалынской полосе ,ландшафтов Северного Прикаспия (Накаренок, 2003а,
б), хотя чаще всего она здесь не входит даже в число содоминантов (Неручев
и др., 1989). В других случаях важную роль играет конкуренция экологичес-
ки близких видов. Так, в Приаральских Каракумах, на значительной части ко-
торых агама считается массовым видом, на закрепленных бугристых песках
при совместном обитании ее с сетчатой ящуркой численность агам составляет
лишь 0,4 экз/га (Лобачев и др., 1973). На побережье Аральского моря довольно
многочисленна, что установлено еще Н.А. Зарудным (1915). Л.Г. Динесман
(1953) здесь отмечал в среднем одну особь на километр маршрута.
Лимитирующие факторы. Комплекс факторов у границы распростране-
ния вида, хозяйственная деятельность, в основном перевыпас скота, местами
транспорт.
Охрана. Обитает и охраняется на территории Устрюртского заповедника.
На остальной территории специальных мер охраны не требуется; достаточно
поддержания естественного состояния ландшафтов.
Региональный статус. Характерный представитель герпетофауны у се-
верных пределов распространения.
Род 17. Phrynocephalus Каир, 1825 - круглоголовки
21. Круглоголовка-вертихвостка - Phrynocephalus guttatus
(Gmelin, 1789)
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода круглоголовок, населяющий территорию района. По-
литипический вид, но структура его недостаточно разработана, в связи с чем
статус и систематическое положение отдельных форм требуют уточнения. По
современным представлениям всю территорию региона населяет номинатив-
ный подвид Ph. g. guttatus (Gmelin, 1789), в пределах которого, однако, наблю-
даются довольно существенные межпопуляционные различия. Так, у вертих-
восток из Волгоградской области отмечаются отличия от основных параметров
фолидоза подвида по количеству чешуи поперек шляпки, от теменного до
межчелюстного щитков, числу подпальцевых пластинок и черных полос снизу
хвоста, что, по-видимому, определяется их окраинным положением в пределах
западной части ареала (Шенброт, Семенов, 1987; Завьялов, Табачишин, 1999;
2000а; 2004). Несомненно, определенные различия (особенно в окраске и ри-
сунке тела) свойственны и популяции самой северной «оренбургской» части
ареала, площадь которой несколько увеличилась в последние десятилетия.
Распространение. Ареал вида охватывает обширную территорию, про-
стирающуюся от Волго-Донского междуречья и западного Предкавказья через
Казахстан и север Узбекистана до северо-западных районов Китая (централь-
но-азиатский фаунистический комплекс). Его северная граница от правобе-
режья р. Волга на широте г. Волгограда простирается в Волжско-Уральском
174
Гпава 4
междуречье по окрестностям оз. Баскунчак, Эльтон, п. Урды и северной кром-
ке Волжско-Уральских песков до п. Калмыкове на р. Урал, Урало-Эмбенском
междуречье по р. Уил, затем примерно по 49-й параллели в низовьях р. Иргиз,
оз. Сары-Копа, по рекам Тургай, Сары-Су и далее через южное Прибалхашье
и Зайсанскую котловину в северо-западный Китай.
В западном Предуралье в конце XIX - начале XX в. севернее 49-й парал-
лели вертихвостка зоологами не отмечалась, в том числе таким известным
фаунистом, как Н.А. Зарудный, в течение ряда лет изучавшим эту часть ре-
гиона. Вместе с тем в 1972-1974 гг. нами (Чибилёв, 1995) вертихвостка была
отмечена в песках по долине р. М. Хобда (бас. р. Илек), затем в разные годы
на правом берегу р. Илек у с. Покровка, в ур. Крутые Горки Соль-Илецкого
района, у п. Кайрат Акбулакского района и других песчаных массивах Илек-
ского района Оренбургской области. В 1955 г. она была найдена в верховь-
ях р. Уртабуртя у с. Рыбаковка Мортукского района Актюбинской области
Ю.А. Дубровским (1967).
Позднее вертихвостка отмечалась также в песчаном массиве Аккумы, пес-
ках у оз. Сорколь (верховья р. Калдыгайты), песках Шагыл у оз. Сулуколь (вер-
ховья р. Утвы), ур. Шубар-Агач в бассейне р. М. Хобда, правобережной части
поймы р. Илек и по среднему течению р. Уртабуртя (левобережный приток
р. Урал), что лишь в 25 км севернее известной находки Ю.А. Дубровского
(1967). Считается, что вид расселяется вниз по долине р. Илек, хотя в 1991 г.
пески у поселков Буранный и Линевка еще не достигались (Давыгора, 1992;
1995).
Если допустить, как это делает А.В. Давыгора, что в отмеченные пункты
вертихвостка «пришла» после 1880-х гг. (время исследования Н.А. Зарудного),
то придется признать, что примерно за 70 лет она не только преодолела более
200 км по прямой, но и пересекла при этом такую крупную (для этого вида) пре-
граду, как р. Илек, в то время как вниз по долине этой реки (где также имеются
крупные песчаные массивы) она почему-то продвинулась на гораздо меньшее
расстояние. В связи с этим, по-видимому, более правдоподобным будет допу-
щение, что и в прошлом вертихвостка обитала севернее долины р. Уил, но в
силу меньших размеров песчаных массивов и их большей закрепленности она
была гораздо малочисленнее и оказалась «не замеченной» зоологами. Ариди-
зация климата и мощная пастбищная дигрессия 1970-1990-х гг., несомненно,
способствовали увеличению ее численности и некоторому расширению ареала,
но не в таких масштабах, как это может показаться на первый взгляд. В других
пунктах у северных пределов ареала сколь-нибудь существенных изменений
границ не выявлено, если не считать находки Ю.А. Дубровским (1967) этой
круглоголовки в 70 км севернее п. Иргиз в низовьях одноименной реки и у
с. Мамыр Актюбинской области (Брушко, Кубыкин, 1988).
Южная граница ареала от низовий р. Волга прослеживается вдоль побе-
режья Каспийского моря до области низовий р. Эмба. Далее она несколько
неопределенна, поскольку, по свидетельству Эйхвальда, на восточном побе-
режье Каспия вертихвостка встречалась до Балканского залива (Никольский,
1905). Кроме того, в коллекции ЗИН АН имеется экземпляр от Н.А. Северцо-
ва с этикеткой, на которой значится, что он добыт в 1876 г. в горах Каратау
на Мангышлаке. Эти сведения до настоящего времени никем не подтверж-
дены и, как отмечает известный исследователь герпетофауны Казахстана
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
175
К.П. Параскив (1956), «находка даже на Мангышлаке по общим соображениям
маловероятна».
Достоверными на настоящий момент являются лишь находки в песках у
оз. Аккуль в дельте р. Эмба (Неручев, Васильев, 1978), Кызыл-Кумах, непо-
далеку от Аральского моря (Определитель земноводных..., 1977), низовьях
р. Сырдарья в окрестностях г. Кызыл-Орда, затем по бассейну р. Или и далее
в Китае (Параскив, 1956).
В очерченных пределах региона круглоголовка-вертихвостка встречается
на всем пространстве Волжско-Уральских песков (где впервые была отмечена
И.И. Лепехиным, считаются типовой территорией вида), большинстве песча-
ных массивов Урало-Эмбенского междуречья, Зауралья и вблизи Аральского
моря. В географическом плане ее встречи известны в 7 (10,1 % из 69) ландшаф-
тных районах степной зоны, 10 (31,3 % из 32) полупустыни и 21 (48,8% из 43)
пустынной зоны (табл. 33, карта 21).
Места обитания. Круглоголовка-вертихвостка - типичный псаммофил.
Вместе с тем в разных частях региона в его распространении проявляются
некоторые особенности, а в ряде случаев и явно выраженные биотопические
предпочтения. Так, в древнеаллювиальных песках Приаралья она населяет
котловины выдувания с полынно-злаковыми и вершины бугров с полынно-
эфемеровыми сообществами. Севернее в песках по низовьям рек Иргиз и Тур-
гай ее типичными местообитаниями являются полынно-злаковые ассоциации
на буграх (Динесман, 1953).
В Приэмбенской провинции вертихвостка встречается в основном в долин-
ных (пойменных) песчаных массивах дюн с редкими кустарниками, окраинах
барханов, по рекам Уил, Сагиз, Эмба, местами здесь даже заходит на кромки
пухлых солончаков. В песках междуречий отмечена на мозаично сочетающих-
ся участках со сбитой вторичной сорной растительностью и полуразвеваемы-
ми песками (часто возникающими в результате хозяйственной деятельности);
на голых барханах отсутствует, исчезает и при закреплении песков (Накаре-
нок, 2003в).
Севернее, в степном Предуралье, населяет экстразональные массивы буг-
ристых и выровненных слабозакрепленных песков, где они перемежаются с
развеваемыми участками и куртинами полукустарниковой и кустарниковой
растительности, а в котловинах придерживаются днищ и склонов южной экс-
позиции (Давыгора, 1992,1995; Чибилёв, 1995).
В центральной части Волжско-Уральских песков она придерживается в
основном окраин бугристых, грядово-бугристых и барханных массивов (где
травянистая, полукустарниковая и кустарниковая растительность не образу-
ет сколь-нибудь сомкнутого покрова) и участков разбитых (особенно мелко-
очаговых) песков, переходящих в слабозакрепленные и закрепленные масси-
вы -т. е. здесь довольно четко проявляется экотонный эффект (Чернов, 1954;
Параскив, 1956; Окулова, 1981; Накаренок, 2003а). Вместе с тем по западной
окраине междуречья при переходе от Волго-Ахтубинской поймы к солонцам,
межбарханным понижениям с редкой полынно-злаковой растительностью и
участками закрепленных бугристых песков подобного явления не наблюдает-
ся (Завьялов, Табачишин, 1998, 2004).
В деталях пространственного распределения вида, возможно, определен-
ную роль играет и конкуренция со стороны ушастой круглоголовки. Во всяком
176
Глава 4
Таблица 33
Распространение круглоголовки-вертихвостки
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная V 30 32 35 37 40 1 2 3 4 5 6 7 8 Оренбургская область р. Илек, с. Покровка ур. Крутые горки р. Илек, ст. Сагарчин р. Уртабуртя с. Междуречье Актюбинская область с. Рыбаковка, Мартукский р-н Западно-Казахстанская область ур. Шагыл у оз. Сулу коль пески у оз. Сорколь ур. Аккумы ур. Карагаш Чибилбв, 1995 Давыгора, 1992, 1995 Дубровский, 1967 Давыгора, 1992, 1995 Дебело и др., 2000
VII 42 46 9 10 11 Оренбургская область р. М. Хобда, ур. Шубарагаш Актюбинская область пойма р. Киил р. Уил, ур. Баркин Чибилёв А.А.
Полупустынная XIII 84 85 88 12 13 14 15 Западно-Казахстанская область окр. сора Хаки Астраханская область окр. оз. Эльтон окр. оз. Баскунчак, п. Н. Бас- кунчак Западно-Казахстанская область окр. п. Эдельсор Животные..., 2009; Ходашова, 1960 Завьялов, Табачишин, 2004; Стрельцов и др. 2006 Накаренок, 2003 в
XV 96 97 98 16 17 18 Актюбинская область дол. р. Эмбы, ур. Толагай пески Аккум-Сагиз ур. Кок-джиде Неручев и др., 1994; Накаренок, Неручев, 2001; Параскив, 1956; Завьялов и др., 2005
XVIII 107 108 19 20 21 ур. Иргиз-Кум п. Иргиз п. Текели 70 км. к сев. от п. Иргиз Никольский, 1905; Ди- несман, 1953 Дубровский, 1976
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
177
Продолжение табл. 33
Зона Провин- ция Район № точки Местонахождение Источник информации
Полупус- тынная XIX по 112 22 23 24 ур. Тусум с. Нура ур. Боз-Пай ур. Аир-Казыл Динесман, 1953; Бруш- ко, Кубыкин, 1988 Динесман, 1953
Пустынная XX 113 114 116 117 118 120 121 122 123 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 Астраханская область окр. горы Б. Богдо, ур. Чапчачи п. Харабали, ур. Берли ур. Серте с. Селитряное окр. ст. Досанг окр. п. Комсомольский п. Степной п. Приозерный Гурьевская область окр. п. Танюшкино Западно-Казахстанская область ур. Рын-пески, п. Урда Прикамыш-Самарские пески Гурьевская область п. Н. Асан п. Н. Уштаган окр. сора Бесоба ур. Бес-Чохо кл. Шадьялмола ур. Мынтюбе ур. Кукурте разъезд № 3 Западно-Казахстанская область п. Калмыкове Гурьевская область п. Елтай п. Кулагине п. Зеленый ур. Камбакты Никольский, 1905; Божанский, Полы нова, 1995; Завьялов, Табачи- шин, 1998, 2004 Накаренок, 2003в; Па- раскив, Бутовский, 1960; Сараев, Пестов, 2010 Окулова, 1961 Накаренок, 2003 в
178
Глава 4
Окончание табл. 33
Зона Провин- ция Район № точки Местонахождение Источник информации
XXI 127 132 135 40 41 42 дельта р. Эмба, оз. Аккуль окр. г. Гурьева (Атырау) пески Тайсоган Неручев, Васильев, 1978; Накаренок, 2003в; Брушко, Кубыкин, 1988; Сараев, Пестов, 2010; Неручев и др., 1984; Брушко, 1995; Киреев, 1981
136 43 п. Сарматшагыл п. Карабау п. Тасшагыл ур. Бек-Беке
XXII п. Кульсары п. Аккызтогай
5 X 137 44 п. Аралтюбе
3 g ур. Тогалай
к 138 45 дол. р. Эмба горы Жельтау
Актюбинская область
XXV 147 46 Сев. Устюрт ст. Челкар Определитель земно- водных..., 1977; Брушко, 1995; Динесман, 1953
151 47 ур. Б. Барсуки
152 48 ур. М. Барсуки
XXVI 153 49 плато Куланды
154 50 Приаральские Каракумы
155 51 п-ов Куланды
случае у северной окраины песков, где конкурент отсутствует, вертихвостка
забегает и на ближайшие барханы. В окрестностях п. Урда она довольно обык-
новенна и в сосновом бору (Параскив, Бутовский, 1960).
Приведенные примеры показывают, что несмотря на кажущуюся стенотоп-
ность у этого вида в естественных ландшафтах, особенно у северных пределов
ареала, проявляется довольно высокая экологическая пластичность.
Повсеместно распространена вертихвостка и в антропогенных модифика-
циях ландшафтов региона - от сильно выбитых скотом окрестностей населен-
ных пунктов и колодцев до колеи проселочных дорог, причем иногда на участ-
ках, измененных выпасом, вид находит более оптимальные условия обитания.
Внося определенные изменения в структуру псаммофитных ландшафтов, мы
тем самым способствуем расселению вертихвостки внутри песчаных масси-
вов. Однако чрезмерность антропогенного воздействия со временем может
негативно сказаться как на распространении, так и на численности не только
этого вида, но и всего герпетокомплекса региона.
Образ жизни. Из зимней спячки (в состоянии которой проводит примерно
200 суток) вертихвостка начинает выходить во второй половине апреля, но
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
179
массовый выход иногда затягивается на всю первую декаду мая. После выхода
активна в течении всего светлого времени суток, а затем преимущественно
утром (в 6-11 часов) и вечером (17-21). Дневные часы проводит либо в норках
(наклонный ход, длинной 10-20 см, с небольшим расширением), обычно вы-
рытых у основания кустика, или просто закапываясь в песок. Территориальна,
причем большей оседлостью отличаются самки. Осенью деятельна до второй
половине сентября - первой половине октября. Общая продолжительность пе-
риода активности, таким образом, достигает 150-160 суток.
Размножение. Половой зрелости вертихвостки достигает на втором году
жизни (через 12-13 месяцев). В этом возрасте вскоре после выхода из спячки,
а чаще всего с первой половины мая у них начинается брачный период. От-
кладка яиц начинается в конце мая и заканчивается в июле. Предполагается,
что в Волжско-УральскОм междуречье в течении одного сезона бывает 2 или
даже 3 кладки (Чернов, 1954), хотя сильная растянутость репродуктивного пе-
риода, возможно, связана с разновременным созреванием фоликул у разных
возрастных групп. В одной кладке бывает 1-3 (чаще всего 2-3) яйца, которые
помещаются в норку или специально вырытую ямку на глубине 5-8 см. Сего-
летки появляются во второй половине июля - августе, на юге ареала, видимо,
и позднее.
Питание. Вертихвостка отличается широким спектром пищевых объек-
тов, хотя повсеместно преобладают перепончатокрылые (муравьи), жуки, а
местами и прямокрылые; в меньшей степени потребляются гусеницы бабочек,
клопы, пауки и двукрылые; довольно часто (до 33%) в желудках отмечаются
растительные остатки (листья, семена, кусочки стеблей злаков).
Численность. Какой-либо географической закономерности (в зональном
плане) в распространении вертихвостки в регионе проследить не удается,
поскольку разные исследователи применяли различные методы учета числен-
ности.
У западной кромки Волжско-Уральских песков в 1980-е гг. на надпоймен-
ной террасе р. Волги численность достигала 2 особей на 10 км маршрута (Не-
ручев и др., 1990). В этих же местах в 1991 и 1994 г. на закрепленных мелко-
бугристых и бугристых песках, примыкающих к Волго-Ахтубинской пойме,
ее плотность была определена в 0,75 ос/га, а в песчаном массиве Берли (в ок-
рестностях п. Харабали) - 3,75 ос/га (Божанский, Полынова, 1995). Немногим
южнее - на территории АГКМ в Красноярском районе в мае 1998 г. на солон-
цовых участках плотность составляла 1,0 ос/га, на выровненных участках в
межгрядовых понижениях - 13,6, в бугристых песках барханного типа - 24 и в
закрепленных бугристых песках - 38,5 ос/га (Завьялов, Табачишин, 1998).
У северных пределов - в окрестностях озера Баскунчак в небольшом изо-
лированном песчаном массиве Шигерты численность вертихвостки определя-
ется в количестве 4 особи за 1 час поисков; несколько чаще она встречалась
в песках Шкили (Стрельцов и др., 2006). В Урдинском бору в 1958 г. на 1 км
маршрута в среднем встречалась одна особь (Параскив, Бутовский, 1960).
В центральной части Волжско-Уральских песков в 1963-1980 гг. на учас-
тках пастбищ по плотным с очагами сбоев пескам численность достигала
1,0 особей на 10 км маршрута, соответственно на закрепленных перевеянных
песках - менее 1,0, в песках с очагами развевания - 3,0-4,0 и полузакреплен-
ных песках - 27,0 особей на 10 км маршрута (Неручев и др., 1981).
180
Глава 4
У восточной кромки Волжско-Уральского междуречья в злаково-полын-
ной пустыне в сочетании с бугристыми песками на широте п.Антоново-Кал-
мыково-Харькино в 1959-1960 гг. численность в среднем составляла 1,16 осо-
бей на 10 км маршрута (Окулова, 1963). На песчаных участках поймы р. Урал
вертихвостка не отмечалась.
В Урало-Эмбенском междуречье в низовьях р. М. Хобда (бас. р. Илека)
в 1972-1974 гг. на 1 км маршрута насчитывалось не более 2, а в низовьях
р. Илека - около 3 особей (А.А. Чибилёв). В целом в этих и других пунктах
Оренбургской области вертихвостка считается малочисленным видом - все-
го насчитывается до тысячи особей (Давыгора, 1998). Южнее в песках Ак-
кумы (верховьях р. Калдагайты) в 1990-е гг. она была сравнительно обычна
(А.А. Чибилёв), а в районе верховий р. Уртабуртая в 1955 г. считалась много-
численной (Дубровский, 1967).
В бассейне р. Уила состояние численности в прошлом не известно, но в
1980-е гг. в песках собственно поймы этой реки численность достигала 6 осо-
бей, а по надпойменной террасе - 9 ос/10 км (Неручев и др., 1990). Южнее
на зарастающих с очагами развевания участках песков Аккум-Сагиз в пос-
ледние годы ее численность достигала 20, а на закрепленных с кустарниками
60 ос/10 км. В то же время в полузакрепленных стациях песков Толагай и При-
каспийских Каракумов она не превышала 1-2 ос/10 км (Накаренок, 2003в).
В Зауралье по полынно-эфемеровым сообществам окрестностей Аральско-
го моря в 1947 г. ее плотность достигала 4,0 особи в полынно-злаковых - 6,6,
в песках низовий р. Иргиз - 3,0, а в массиве Тусум (в бассейне р. Тургай) -
23,0 особи на 1 км маршрута (Динесман, 1953). Довольно обычной считается
она и в Приаральских Каракумах (Лобачев и др., 1973).
В целом же для Казахстана средняя плотность составляет 3,3, а максималь-
ная - 8,9 ос/га и оценивается как низкая (Брушко, 1993).
Лимитирующие факторы. Для краеареальных популяций комплекс фак-
торов, действующих на границе ареала, и перевыпас.
Охрана. Обитает и охраняется на территории Устюртского и Богдинско-
Баскунчакского заповедников. На остальной части региона состояние попу-
ляций применения особых мер охраны не требует; достаточно поддержания
естественного состояния ландшафтов.
Региональный статус. Характерный для региона, спорадически (у север-
ных рубежей ареала лишь на изолированных островных участках) встречаю-
щийся вид. Занесена в Красную книгу Волгоградской области (2004).
22. Такырная круглоголовка -
Phrynocephalus helioscopus (Pallas, 1771)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Политипический
вид, но структура его недостаточно разработанна, в связи с чем систематичес-
кое положение некоторых форм на уровне подвидов требует уточнения. По
современным представлениям территорию региона населяет номинативный
подвид Ph. h. helioscopus (Pallas, 1771), впервые списанный из «Индерской
степи». Вместе с тем в его составе проявляются довольно существенные меж-
популяционные отличия в окраске и рисунке верха тела, которые изменяются
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
181
в зависимости от цвета субстрата (Параскив, 1956; Ананьева и др., 1998; Атлас
пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида простирается от низовий р. Волги, вос-
точных (прикаспийских) районов Кавказа, Турции через северный Иран,
Среднюю Азию и Казахстан до северо-западных районов Монголии и Ки-
тая (субэндемик Средней Азии и Восточного Ирана). Его северная грани-
ца проходит от юга Волгоградской области через верховья р. Б. Сморогда к
г. Уральску (Яицкий городок), области среднего течения р. Эмба, низовьям
рек Иргиза, Тургая, далее вблизи Карсакпая, по северу Бетпак-далы, окрест-
ностям городов Аягуз и Семипалатинск уходит в Китай и Монголию. Южная
граница от северо-восточных районов Турции проходит по северному Ирану
и далее на восток к Китаю и Монголии. Таким образом, значительная часть
ареала вида охватывает территорию исследуемого региона. Здесь северные ру-
бежи ареала прослеживаются по находкам в южных районах Волгоградской
области (Кубанцев и др., 1962), окрестностях г. Уральска (экз. коллекции ЗИН
от Кайзерлинга), в Приэмбенской пустыне между р. Кайнар и Прикаспийски-
ми Каракумами (Неручев, Капустина, 1981), низовьях рек Иргиз (г. Иргиз),
Улькояк и Тургай (Динесман, 1953; Брушко, Кубыкин, 1988; Животные...,
2009).
В очерченных пределах региона локальное распространение такырной
круглоголовки выявлено в 1 (1,4% из 69) ландшафтном районе степной зоны,
8 (25,0% из 32) районах полупустыни и 26 (60,5% из 43) пустынной зоны
(табл. 34, карта 22).
Места обитания. Типовой территорией вида является «Индерская степь»,
откуда эта круглоголовка впервые была описана П.С. Палласом. Здесь она
населяет довольно широкий спектр биотопов, проявляя себя типичным эври-
топным видом. Вместе с тем наиболее типична она здесь по глинистым по-
лынно-солянковым стациям, такырам, такыровидным участкам и окраинам со-
ров. Восточнее в полосе верхне- и нижнехвалынских ландшафтов основными
местообитаниями вида становятся глинистые полынно-биюргуновые стации,
откуда она проникает в долины рек Сагиза, Кайнара, Эмбы, а местами и на
супесчаные сбои (Неручев, Капустина, 1981; Неручев и др., 1981, 1990; Нака-
ренок, 2003 в). В глинистой полупустыне только среди биюргуна и биюргуна
с итсигеком она встречается также в низовьях рек Иргиз, Улькояк и Тургай.
Южнее в этой части региона (вплоть до побережья Аральского моря) количес-
тво биотопов, занимаемых такырной круглоголовкой, постепенно увеличива-
ется: в глинистой пустыне она населяет как минимум четыре биотопа и три
среди массивов древнеаллювиальных песков, хотя наиболее обычной является
натакырах и окраинах соров (Динесман, 1953).
По участкам с плотными суглинистыми и песчаными грунтами (аши-
кам) она проникает в Волжско-Уральские пески. Детали ее распространения
здесь (особенно в центральной части) во многом остаются неясными, хотя по
межгрядовым понижением она достигает окрестности п. Урды. По-видимому,
в сходных местообитаниях и по соровым депрессиям в прошлом отмечалась в
окрестностях гор Б. и М. Богдо, Чапчачи, Азгыр; недавно подтверждено оби-
тание у оз. Баскунчак (Никольский, 1905; Чернов, 1954; Параскив, Бутовский,
1960; Стрельцов и др., 2006). В последние десятилетия важным фактором, спо-
собствующим расселению вида, здесь стали дороги, существенно изменяющие
182
Глава 4
Таблица 34
Распространение такырной круглоголовки
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная VIII 55 1 Западно-Казахстанская область окр. г. Уральска (Яицкий городок) Никольский, 1905
Волгоградская область
81 2 южные районы Кубанцев и др., 1962
XIII 85 3 4 Астраханская область окр. оз. Эльтон окр. оз. Баскунчак Стрельцов и др., 2006; Животные..., 2009
Полупустынна] Актюбинская область
XV 95 96 5 6 дол. р. Сагиз гора Актолагай Накаренок, 2003в; Сара- ев, Пестов, 2010
XVIII 107 7 низ. р. Иргиз, гора Кабан-Кулях Никольский, 1905; Ди- несман, 1953
109 8 низ. р. Улькояк Брушко, Кубыкин, 1988
XIX ПО 9 низ. р. Тургай ур. Боз-Тай
112 10 п. Иргиз, 80-90 км к юг.-вост.
Астраханская область
113 11 12 горы Б. и М. Богдо гора Чап чачи Никольский, 1905; Нака- ренок, 2003в
13 гора Азгир
Западно-Казахстанская область
3 X XX 115 116 117 14 15 16 ур. Токай п. Урда, Рын-пески окр. п. Н. Казанка Параскив, Бутовский, 1960; Чернов, 1954; Накаренок, 2003в
3 В 118 17 ур. Бес-Чохо, ур. Истай, ур. Сасыктау
с 122 18 окр. п. Харькино окр. п. Кулагине Гурьевская область Параскив, Бутовский, 1960
123 19 низ. р. Багырдай Сараев, Пестов, 2010
XXI 126 127 20 21 Атаманская коса пром. Караарна пром. Тенгиз мр. Королевское Брушко, Кубыкин, 1988; Никольский, 1905; Чер- нов, 1954; Паллас, 1773
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
183
Продолжение табл. 34
Зона Провин- ция Район № точки Местонахождение Источник информации
129 22 к сев.-зап. от п. Махамбет
XXI 132 23 окр. г. Гурьев
24 п. Новобогатинский
133 25 окр. оз. Индер, Индерские Никольский, 1905;
горы Брушко, Кубыкин, 1988;
134 26 п. Карабау 60 км к югу, ур. Жамансор Сараев, Пестов, 2010; Накаренок, 2003в
136 27 ст. Первомайская п. Искининский ур. Тентяксор
28 п. Доссор
п. Доссор, 15 км к юг.-зап. пр. Байчунас
29 п. Макат
30 р. Сагиз, ур. Таскичу пром. Бекбеке
31 ; п. Жамансор, ур. Жайли
5 п. Мол кат,
к к п. Киембай, оз. Караша
в оз. Бейбит
К XXII 137 32 п. Кульсары п. Кульсары 10 кмк сев.
33 гора Иманкара горы Аккерегеше дол. р. Кайнар
34 п. Косчагыл ур. Акмечеть ур. Саргаска-Жанбеке
35 п. Аккызтогай на вост.
р. Кайнар, ур. Акши
138 дол. р. Эмбы ур. Жингильдьггогай ур. Бактогоринсай ур. Намазтакыр ур. Уитас Брушко, Кубыкин, 1988
36 п. Каратон п. Каратон, 30 км к югу Накаренок, 2003 в
184
Глава 4
Окончание табл. 34
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXII 139 37 38 39 40 41 42 п. Ак-кудук ур. Тагыракшант Прикаспийские Каракумы ур. Тюлюс, сор Мейбулак ур. Ушкан ур. Кумеште гора Шолькара Мангистауская область горы Коленколы ур. Болиорпа ур. Сорширпа ур. Жанасу горы Жельтау окр. г. Жельтау на сев. пески Сарыкум ур. Жаксы-Карасай юго-зап. чинк Устюрта окр. п. Бейнеу ур. Кзыл-Тас Накаренок, 2003в Брушко, Кубыкин, 1988; Никольский, 1905
XXIII 143 144 43 44 окр. сора Мертвый Култук окр. сора Кайдак Брушко, Кубыкин, 1988
XXV 147 148 45 46 плато Устюрт Самматсайская депрессия
XXVI 151 152 153 154 155 47 48 49 50 51 52 53 Актюбинская область пески Б. Барсуки пески М. Барсуки плато Куланды Приаральские Каракумы ур. Уч-чоку ур. Джингиль-Тюп берег Аральского моря ур. Клыч п. Акеспе п-ов Кара-Тюп п. Арал-Сульфат Параскив, 1956; Лобачев и др., 1973 Зарудный, 1915 Сидоров, 1925; Динес- ман, 1953
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
185
некоторые физические параметры рельефа и повышающие экологическую
значимость территории (Неручев, Накаренок, 1999).
В ряде мест она осваивает участки полузакрепленных и закрепленных пес-
ков, в том числе в приморской части побережья Каспия, где отмечалась на
буграх среди солончаков, как, например, у Атаманской косы (Параскив, Бу-
товский, 1960). В прошлом ее встречи были известны по всему восточному
побережью, за исключением недавно освободившейся приморской полосы.
У Аральского моря она встречается на берегу как среди крупнозернистого пес-
ка, мелкой гальки, так и на участках плотных солонцеватых песков (Зарудный,
1915; Динесман, 1953). В окрестностях Индерского озера такырная круглого-
ловка освоила также долины оврагов, соленых ручьев и речушек, по которым
спускается на щебнистое побережье (Чернов, 1954).
Таким образом, в пределах региональной части ареала такырная кругло-
головка может быть встречена почти везде, за исключением сыпучих песков,
скальных участков Устюрта, мокрых соров приморской полосы и некоторых
других непригодных для вида местообитаний.
Высокой биотопической пластичностью и способностью занимать новые
места отличается эта круглоголовка и на сопредельной территории ареала (Па-
раскив, 1956; Шенброт, Антонова, 1981; Брушко, 1993, 1995).
Образ жизни. После зимней спячки у южных пределов региона иногда
появляется в первой, чаще всего во второй половине марта, на севере в кон-
це этого месяца или в разных числах апреля. В это время активна в течение
всего светлого времени суток, а летом преимущественно в утренние (с 7-8) и
вечерние (с 17) часы. От других видов отличается способностью переносить
большие перепады суточных температур, хотя в ранние утренние часы срав-
нительно малоподвижна; понижение активности наблюдается также при пас-
мурной и дождливой погоде. Летом, в наиболее жаркое время дня, укрывается
в тени под кустиками, в разных пустотах грунта, собственных норках (устраи-
ваемых у основания кустиков, под камнями, коркой такыра) или норах других
животных (чаще всего песчанок, тушканчиков, крупных насекомых). В подоб-
ных убежищах (собственные отнорки углубляются до 20-25 см) она и зимует.
На зимовку у северных пределов региона уходит в разных числах октября, на
юге преимущественно в конце октября, иногда задерживаясь до середины но-
ября. Общая продолжительность периода активности, таким образом, достига-
ет 180-200, а зимней спячки - 160-180 суток.
Размножение. Половозрелыми такырные круглоголовки становятся на
втором году жизни. К спариванию в Приэмбенской провинции они приступа-
ют в апреле, а в конце этого месяца - начале мая в яйцеводах самок отмеча-
лись крупные яйца (Неручев и др., 1989а). Вместе с тем в Волжско-Уральских
песках в это время отмечались самки только с увеличенными фолликулами, а
сформированные яйца здесь наблюдались лишь в конце мая (Чернов, 1954).
Судя по этим данным, массовая кладка яиц в регионе проходит в конце мая -
июне, а в июле размножение прекращается. За это время самки дважды откла-
дывают по 2-5 яиц. Инкубационный период продолжается около 40 суток, и в
июле повсеместно появляются первые молодые, выход которых растягивается
более чем на месяц (на сопредельной территории до второй декады августа).
Питание. Основу пищевого рациона такырных круглоголовок составляют
насекомые. Среди них на большей части ареала доминирующими являются
186
Глава 4
муравьи (до 70-90% по числу экземпляров), жуки и прямокрылые, в меньшем
количестве потребляются гусеницы бабочек, личинки жуков, паукообразные,
изредка и растительная пища (листья, семена). В свою очередь эти круглого-
ловки потребляются крупными пресмыкающимися, а также хищными птица-
ми и млекопитающими.
Численность. В соответствующих местообитаниях на большей части аре-
ала такырная круглоголовка является сравнительно обычным, а в Приэмбенс-
ких ландшафтах и некоторых других местообитаниях доминирующим видом.
В Приэмбенской провинции по слаботрансформированным супесчаным ста-
циям ее численность достигает 25-30, в глинистой пустыне - 21, на надпой-
менных террасах рек Эмбы и Кайнара, соответственно 16 и 10, а по краю сора
в Прикаспийских Каракумах - 5 особей на 10 км маршрута. На этой широте в
Волжско-Уральском междуречье по ашикам среди полузакрепленных песков
ее численность достигала И ос/10 км (Неручев и др., 1981; 1990; Накаренок,
2003в). В Астраханской части массива встречается на отдельных солончаках,
где плотность достигает 1,1-2,5 ос/га (Божанский, Полынова, 1998).
В разных сообществах Приаральских пустынь численность еще более вы-
сокая - по древнеаллювиальным пескам достигает 8-80, а в глинистой пусты-
не - 6-80 особей на 10 км маршрута, снижаясь к северным пределам распро-
странения до 5 особей на 10 км маршрута (Динесман, 1953). По-видимому, на
столь же высоко^ уровне ее численность в Приаральских Каракумах сохраня-
ется и в последние десятилетия, где она считается массовым видом (Лобачев
и др., 1973).
На побережье Аральского моря «очень обыкновенной» она была как в на-
чале XX столетия (Зарудный, 1915; Сидоров, 1925), так и в середине, когда
численность достигала 7 экз/км (Динесман, 1953). Следует отметить, что на
близко расположенном от побережья о-ве Барсакельмес ее плотность достига-
ла 82-116, а после выхода молодых - 156-170 экз/га. Вместе с тем в среднем
по Казахстанской части ареала ее плотность составляет лишь 5,1, а максималь-
но 14,2 экз/га (Брушко, 1993, 1995).
Лимитирующие факторы. У северной границы ареала весь комплекс
факторов, ограничивающий обитание вида за пределами области распростра-
нения; в его основной части, по-видимому, неумеренный выпас.
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резер-
вата, Устюртского и Богдинско-Баскунчакского заповедников. На остальной
территории состояние популяций особых мер охраны не требует.
Региональный статус. Номинативный подвид внесен в аннотированный
перечень таксонов и популяций, нуждающихся в особом внимании к их состо-
янию в природной среде (Приложение к Красной книге РФ, 2001) в связи со
снижением численности в низовьях Волги. Занесена в Красную книгу Астра-
ханской области (2004). В казахстанской части ареала характерный для регио-
на вид у северных пределов распространения.
23. Песчаная круглоголовка -
Phrynocephalus interscapularis Lichtenstein, 1856
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода круглоголовок, населяющий территорию региона. Мо-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
187
нотипический вид. Несмотря на значительную площадь ареала, существенных
межпопуляционных отличий в морфологии не выявлено (Ананьева и др., 1998;
Атлас пресмыкающихся..., 2004).
Распространение. Ареал вида охватывает северные районы Ирана, Афга-
нистана, Среднюю Азию и юг Казахстана (эндемик Средней Азии и Восточ-
ного Ирана). Северная граница определяется Прикаспийскими и Приаральски-
ми Каракумами и долиной р. Сырдарьи. Южная граница проходит по северу
Ирана и Афганистана, затем по южным рубежам пустыни Каракум к среднему
течению реки Амударьи и далее по окраине песков Кызылкум к р. Сырдарья.
В очерченных пределах обитание песчаной круглоголовки в изолирован-
ном очаге Прикаспийских Каракумов (Определитель земноводных..., 1977) но-
вейшими наблюдениями не подтверждается (Неручев, Васильев, 1978; Сараев,
Пестов, 2010), в связи с чем, по-видимому, единственным местом обитания
вида в регионе сейчас являются Приаральские Каракумы (карта 23). С обмеле-
нием Аральского моря и соединением ряда его островов (где она отмечалась
Н.А. Зарудным, 1915) с сушей, возможно, появилась и в Приморском районе.
Места обитания. Песчаная круглоголовка, как свидетельствует название
вида, - типичный псаммофил. Она заселяет сыпучие барханы, развеваемые и
бугристые, почти лишенные растительности пески; местами встречается и на
песчаных наносах такыров. В Приаральских Каракумах чаще всего ее можно
встретить в виде изолированных поселений на островках барханов, возле ко-
лодцев, иногда отделенных один от другого десятками километров (Лобачев и
др., 1973). Обычно придерживается вершин барханов, хотя иногда заходит на
участки более закрепленных песков.
Образ жизни. Песчаная круглоголовка в Казахстане - слабо изученный
вид, в связи с чем представление о ее жизненном цикле можно составить в
основном по материалам из сопредельной территории. Из зимней спячки вы-
ходит в конце марта. Активна в утренние и вечерние часы, днем укрывается в
норках (наклонный ход длинной до 20 см) или закапывается в песок. Терри-
ториальна, занимает участки площадью до 200 м2. На зимовку уходит в конце
октября - ноябре. Активный период, таким образом, продолжается около 210,
а зимняя спячка - 150 суток.
Размножение. Половозрелыми песчаные круглоголовки становятся к кон-
цу первого года жизни. Откладка яиц происходит со второй половины мая до
начала августа. За это время самки откладывают не менее 4 раз по одному
яйцу. Молодые появляются в конце июня - августе.
Питание. Основу пищевого рациона составляют муравьи, жуки и терми-
ты, в меньшем количестве потребляются саранчевые, клопы, мухи, а также
паукообразные.
Численность. В соответствующих местообитаниях Приаральских Кара-
кумов песчаная круглоголовка является сравнительно обычным видом - на
островках барханов и возле колодцев их численность достигает до 20,5 экз/га
(Лобачев и др., 1973). В более крупных песчаных массивах Казахстана ее плот-
ность достигает в среднем 37,2, максимум 100,0 экз/га (Брушко, 1993).
Лимитирующие факторы. Отсутствие соответствующих местооби-
таний.
Охрана. Состояние популяций на основной части ареала особых мер охра-
ны не требует.
188
Глава 4
Региональный статус. Вид на северной окраине ареала в пределах огра-
ниченных изолированных местообитаний.
24. Ушастая круглоголовка-Phrynocephalusmystaceus (Pallas, 1776)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Политипический
вид, представленный несколькими подвидами, количество и границы ареалов
которых остаются до конца не выясненными. По одной точке зрения, популя-
ции европейской части ареала до Волжско-Уральских песков включительно
относятся к номитативному подвиду Ph. т. mystaceys (Pallas, 1776), восточнее
сменяющемуся Ph. т. galli Krasowsky, 1932, отличающемуся более длинным
хвостом, конечностями и размером ушной складки кожи. По иной, менее рас-
пространенной, точке зрения, в пределах европейско-среднеазиатской час-
ти ареала дискретные формы не выделяются и все ее популяции относятся к
номинативному подвиду. Вместе с тем при любой трактовке структуры вида
признается, что он представлен рядом географически изолированных популя-
ций, распространение которых определяется наличием подвижных песчаных
массивов (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Се-
менов, 2006).
Крупнейшей в регионе является волжско-уральская популяция, населя-
ющая единый в генетическом отношении песчаный массив, расположенный
между низовьями рек Волги и Урала. В Урало-Эмбенском междуречье рас-
пространение вида также связано с единой в прошлом (хвалынской) песча-
ной аллювиально-дельтовой равниной (расположенной у подножья Под-
уральского плато), сейчас разделенной на ряд изолированных очагов. Едина
по генезису и популяция зауральской части региона, населяющая песчаные
массивы в низовьях рек Иргиза и Тургая, а также Приаральские Каракумы.
Несколько обособлены популяции Северного Устюрта и сопредельной тер-
ритории Мангышлака, что, несомненно, сказывается на особенностях их
экологии.
Распространение. Ареал вида охватывает пески Восточного Прикавка-
зья, севера Ирана, Афганистана, северо-запада Китая, Средней Азии, Прикас-
пийской низменности (это типовая территория вида, откуда он впервые был
описан П.С. Палласом) и Южного Казахстана (субэндемик Средней Азии и
Восточного Ирана). Его северная граница следует по кромке Астраханских, а
затем Волжско-Уральских песков до широты п. Кулагине на р. Урал. Восточ-
нее еще недавно она определялась находками в низовьях р. Уил, при слиянии
рек Эмбы и Темира, а затем (обходя Мугоджары с юга) в низовьях р. Иргиз,
окрестностях оз. Сары-Копа в Тургайской депрессии, на среднем течении
р. Сарысу, южном Прибалхашье и окрестностях оз. Алаколь (Определитель
земноводных..., 1977). Вместе с тем нами в 1990-х гг. она найдена в песчаном
массиве Аккумы в верховье р. Калдыгайты, песках ур. Караагаш в верховье
р. Булдурты и в бугристых песках на правобережье р. Б. Хобда на территории
Актюбинской области Казахстана (Чибилёв, 1995), что примерно на 200 км
севернее известных ранее рубежей. В ур. Аккумы и Караагаш она отмечалась
и позднее (Дебело и др., 2000). В других частях региона изменения очертания
северной границы ареала нам не известны. Южная граница ареала проходит
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
189
по северным районам Ирана, Афганистана и северо-западу Китая. Таким обра-
зом, ушастая круглоголовка населяет только южную часть региона.
В очерченных пределах она обитает в Волжско-Уральском междуречье (за
исключением сырой солончаковой приморской полосы), песчаных массивах
Предуралья (где заходит в степную зону), п-ове Бузачи, Северном Устюрте,
Б. и М. Барсуках, низовьях рек Иргиз и Тургай, Приаральских Каракумах и на
побережье Аральского моря. В целом ее обитание известно в 2 (2,9% из 69)
ландшафтных районах степной зоны, 6 (18,7% из 32) полупустыни и 21 (48,8%
из 43) пустынной зоны (табл. 35, карта 24).
Таблица 35
Распространение ушастой круглоголовки
Зона Провин- ция Район № точки Местонахождение Источник информации
Западно-Казахстанская область
3 X VII 40 1 ур. Караагаш Чибилёв, 1995
с 2 ур. Аккумы Дебело и др., 2000
42 3 Актюбинская область окр. р. Б. Хобда Чибилёв А.А.
Западно-Казахстанская
XIV область
94 4 окр. р. Калдыгайты Чибилёв А.А.
Актюбинская область
3 X XV 95 5 р. Сагиз, п. Каракуга Накаренок, 2003в
X 96 6 ур. Тогалай Неручев, Васильев,
ё 98 7 междуречье Эмбы-Темира 1978; Никольский, 1905; Динесман, 1953
& ч* XVIII 107 8 ур. Бооз-Тай
о С ПО 9 пески Тусум
10 окр. оз. Сары-Копа
XIX 112 11 окр. г. Иргиз
12 ур. Иргиз-Кум низ. р. Тургай
Астраханская область
113 13 окр. горы Богдо Никольский, 1905; Боб-
окр. п. Лапас, Харабали ров, 1985; Божанский, Полынова, 1998; Завья-
3 114 14 окр. п. Селитряное лов, Табачишин, 1998
X X XX 15 ст. Досанг, п. Замьяны
Й 16 г. Красный Яр, АГКМ
Е 17 окр. ст. Бузан окр. п. Степной окр. п. Комсомольский окр. п. Приозерный
190
Глава 4
Продолжение табл. 35
Зона Провин- ция Район № точки Местонахождение Источник информации
Гурьевская область
18 окр. п. Танюшкино
Западно-Казахстанская область
116 19 Рын-пески, п. Урда Параскив, Бутовский,
ур. Караузек Гурьевская область 1960
118 20 п. Н. Асан, 11 км на юго-вост, окр. сора Бесоба ур. Бес-Чохо кл. Шадьялмола ур. Жаскайрат Сараев, Пестов, 2010
21 ур. Тауп окр. п. Н. Уштаган
120 22 ур. Ортакыр ур. Кокмай
23 ур. Сазды
3 X М 24 ур. Алтай
X ё XX ур. Бегайдар
с 25 ур. Бикшек ур. Жалпакшагыл ур. Балт ур. Мынтюбе
121 26 окр. ст. Исатай
27 ур. Сахаба ур. Барсы ур. Мерген ур. Аманколь
122 28 окр. п. Калмыкове ур. Салыкмула ур. Акмула ур. Катя-чага ур. Колбай Гурьевская область Окулова, 1963
123 29 30 окр. п. Кулагине метеостанция Тас, окр. п. Зеленое Чернов, 1954; Сараев, Пестов, 2010
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
191
Окончание табл. 35
Зона Провин- ция Район № точки Местонахождение Источник информации
128 31 зим. Комбакты
XXI 130 32 окр. п. Новобогатинский
132 33 окр. г. Гурьев (Атырау)
135 34 35 окр. п. Карабау окр. п. Тайсойган мог. Акмола Параскив, 1956; Брушко, Кубыкин, 1988
ххи 137 36 37 пески Аккум-Сагиз пески Толагай Неручев, Шатилович, 1985
3 Мангистауская область
X X 3 138 38 Прикаспийские Каракумы Неручев и др., 1990
в Е 139 39 Сев. Устюрт Параскив, 1956; Брушко, 1993
XXIII 140 40 п-в Бузачи Параскив, 1956
XXV Актюбинская область
148 41 ур. Сам
151 42 пески Б. Барсуки
152 43 пески М. Барсуки
XXVI 154 44 Приаральские Каракумы
155 45 46 ур. Агыспе окр. г. Аральск Лобачев и др., 1973; Сидоров, 1925; Динес- ман, 1953
Места обитания. Ушастая круглоголовка - типичный псаммофил, насе-
ляющий преимущественно сухие песчаные массивы междуречий. Вместе с тем
в Приэмбенской провинции она обитает также на надпойменных террасах рек
Уила и Сагиза, где условия обитания существенно не отличаются от окружа-
ющей пустыни, а местами (когда существование на окружающей территории
уже невозможно) проникает в пойму рек (пески Толагай по р. Эмбе).
В целом по Северному Прикаспию наблюдается довольно четкая привязан-
ность вида к характерным и для южных песчаных пустынь участкам рыхлых
развеваемых песков, в связи с чем распространение даже в крупных песчаных
массивах (как, например, в Волжско-Уральском междуречье) носит очаговый
характер.
На некоторых территориях подобные стации (иногда с разреженной вто-
ричной растительностью) формируются на местах сильного сбоя, вызванно-
го перевыпасом, вблизи колодцев, при прокладке дорог или другой деятель-
ности, поддерживающей существование очагов развевания и в связи с этим
являющейся важным элементом в формировании «кружева ареала». В не-
которых местах ушастая круглоголовка встречается и на участках слабо за-
крепленных песков с редкой кустарниковой и травянистой растительностью.
192
Глава 4
В этом плане исключительное зрелище представляют собой круглоголовки
на фоне стелющегося можжевельника казацкого в песках Аккумы; здесь же
их можно наблюдать и на фоне верхушек берез, засыпаемых движущимися
барханами.
На побережье Аральского моря ушастая круглоголовка занимает участки
древнеаллювиальных песков с очень бедной растительностью - преимущест-
венно на вершине песчаных бугров; котловин выдувания или их закрепленных
склонов определенно избегает, но к концу лета (с отмиранием растительности)
расселяется несколько шире, особенно молодые особи. Севернее в песках Ир-
гизкум, Аир-Кызыл и Тусум живет в нижней пологой части больших котловин
выдувания, причем в последнем случае среди редких растений, у основания
погибших стеблей которых они обычно устраивают свои норки. Таким обра-
зом, даже на небольшой территории региона этот типичный псалмофил прояв-
ляет определенную эвритопность.
Образ жизни. Из зимней спячки в ранние вёсны у южных пределов реги-
она она появляется в конце марта, обычно же в начале апреля, а у северной
границы ареала во второй половине этого месяца. Территориальна и очень
агрессивна при защите индивидуальных участков. Половозрелые особи нор
обычно не роют, а на ночь и при необходимости зарываются в песок. Моло-
дые особи, а иногда и беременные самки роют норки, вблизи которых посто-
янно и находятся. Норки представляют собой прямой наклонный ход длиной
50-90 см, с расширением в зоне увлажненного песка (на глубине 25-50 см).
Весной активна с 8-9 часов утра в течение всего светлого времени суток, ле-
том активность проявляется несколько раньше, но в жаркую часть дня зары-
вается в песок или укрывается в норке. Осенью становится активной лишь
при температуре почвы выше 20 °C. В целом период активной жизни продол-
жается около 180-190 суток, а в начале октября она уже залегает в спячку (на
170-180 суток).
Размножение. Половой зрелости ушастые круглоголовки достигают к
концу второго года жизни. На сопредельной территории они спариваются
со второй половины апреля до начала июля. Яйца (по 2-6) откладываются
дважды - с середины мая до конца июня, а затем во второй половине июля.
Первые молодые выходят из яиц в конце июля, а в массе они появляются с
середины августа. Молодые чаще всего селятся вблизи мест расположения
кладок.
Питание. Основу пищевого рациона составляют насекомые и их личин-
ки: разные жуки (преимущественно навозники) и прямокрылые, в меньшем
количестве потребляются муравьи, клопы, двукрылые и другие беспозвоноч-
ные, изредка мелкие ящерицы, цветки и зеленые части растений. В свою оче-
редь круглоголовки добываются различными хищными птицами и млекопи-
тающими.
Численность. В различных частях региона она неодинакова и к тому же
существенно изменяется по годам.
В Волжско-Уральских песках в окрестностях п. Урда (Нарынский мас-
сив - типовая территория вида) в июне 1952 г. на 1 км маршрута встреча-
лось 2-3 особи (Параскив, Бутовский, 1960). По восточной кромке междуре-
чья, вблизи п. Калмыкове в злаково-полынной пустыне, перемежающейся с
участками бугристых песков, по результатам весеннее-летних учетов 1958-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
193
1960 гг. в среднем отмечалось лишь 0,48 экз/км, хотя на отдельных участках
численность их была значительно выше - в ур. Салыкмула 21.04.1959 г. на
1 км маршрута насчитывалось 9 особей, а 30.05.1960 г. здесь же за 1,5 часа из
норок было выкопано И круглоголовок (Окулова, 1963). На западной кром-
ке междуречья в районе ст. Досанг - п. Замъяны в июне 1979 г. численность
достигала 10 ос/га, в начале 1990-х гг. - 23,44, в других популяциях 1,9-
11,1 ос/га (Бобров, 1985; Божанский, Полынова, 1998). Весной 1998 г. пример-
но в этих же местах на территории Астраханского газоконденсатного место-
рождения (Красноярский район) в бугристых песках барханного типа плот-
ность достигала 17,3±0,55, в закрепленных бугристых песках - 11,2±0,65 ос/га
(Завьялов, Табачишин, 1998). В то же время в некоторых местообитаниях она
практически исчезла, что связано с зарастанием песков (Дунаев, Жбанова,
1998).
В Урало-Эмбенском междуречье в разные годы в песках Аккумы числен-
ность достигала 6,8-9,5 ос/км, в ур. Караагаш 5,0 ос/км (Чибилёв А.А.). Срав-
нительно обычной является она в песках по рекам Уилу, Сагизу, Эмбе и юж-
нее в массиве Сам на Устюрте (Неручев, Васильев, 1978; Неручев и др., 1981,
1990). Обычным видом она считается и в Приаральских Каракумах (Лобачев
и др., 1973).
В целом по песчаным массивам Казахстана ее средняя плотность составля-
ет 25,4, максимальная 47,5 экз/га, а после выхода молодых она увеличивается
до 75,5 экз/га (Брушко, 1993).
Лимитирующие факторы. У северных пределов комплекс факторов,
действующих на окраине ареала. На приморской полосе и по Устюрту, оче-
видно, высокая минерализация грунта. На остальной территории - отсутствие
подходящих местообитаний.
Охрана. Обитает и охраняется на территории Устюртского заповедника и
в низовьях Волги. На большей части остальной территории региона специаль-
ных мер охраны, по-видимому, не требуется.
Региональный статус. Занесена в Красную книгу Астраханской области
(2004). На остальной территории характерный для региона вид, населяющий
специфические местообитания у северной границы ареала.
*Сетчатая круглоголовка - Phrynocephalys reticulates Eichwald, 1831
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода круглоголовок. Политипический вид, представленный
тремя подвидами. Западную часть его ареала населяет Ph. г. bannikoviTlaxexsVy,
Rustamov et Shammakov, 1976.
Распространение. Населяет территорию Туркмении, Узбекистана и не-
которые южные районы Казахстана. На территории региона известна для се-
верного побережья Аральского моря, «кажется, только по одному экземпляру
в музее АН», найденному В.С. Елпатьевским (1903, цит. по: Сидоров, 1925).
Другие зоологи, позднее работавшие в данной районе, эту круглоголовку не
отмечали, хотя ее типичные местообитания - покрытые мелкой щебенкой и
разреженной растительностью - по всему Устюрту распространены довольно
широко.
В связи с этим вид нами рассматривается как сомнительный.
194
Глава 4
Семейство 12. Anguidae Gray, 1825 - Веретеннциевые
Род 18. Angius Linnaeus, 1758 - веретеницы
25. Веретеница ломкая -Anguis fragilis Linnaeus, 1758
Значение таксона для сохранения биоразнообразия. Один из двух видов
рода, обитающий на территории региона. Политипический вид, представлен-
ный тремя подвидами, отношения между которыми и границы ареалов выяс-
нены недостаточно. По мнению одних исследователей, территорию региона
населяет номинативный подвид A. f fragilis Linnaeus, 1758, по другой точке
зрения - A. f colchicus Nordmann, 1840, реальность существования которого
рядом авторов вообще подвергается сомнению (Ананьева и др., 1998; Атлас
пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает Центральную и Южную Евро-
пу, Переднюю Азию, Северный Иран, Кавказ, большую часть Восточной Ев-
ропы и Западную Сибирь к востоку до р. Тобол (европейский фаунистический
комплекс). Его северная граница на Восточно-Европейской равнине примерно
от широты г. Архангельска постепенно смещается к 60-й параллели на Сред-
нем Урале, а затем к р. Тобол. Южная граница от Молдавии прслеживается к
излучине р. Дон, затем к р. Волга, по долине которой поднимается до устья
р. Самара, далее по долине этой реки выходит к окрестностям г. Оренбурга, по
р. Сакмара следует к островным борам на границе Оренбургской и Челябинс-
кой областей и долине р. Тобол. Таким образом, ареал вида охватывает только
северную часть региона (Определитель земноводных..., 1977; Чибилёв, 1995,
1998; Вершинин, 2007), хотя, предположительно, веретеница может обитать и
в низовьях волжских притоков в пределах Саратовского левобережья (Завья-
лов и др., 2003, 2006).
В очерченных пределах встречи веретеницы известны в черте городов Са-
мара и Тольятти, Красноярском, Кинельском и Борском районах Самарской
области (Бакиев, Файзулин, 2001,2002; Бакиев и др., 2003), оренбургской части
Бузулукского бора, окрестностях г. Оренбурга и лесостепных Северном, Тюль-
ганском, Саракташском, Шарлыкском, Кувандыкском районах Оренбургской
области (Зарудный, 1895; Райский, 1951; Даркшевич, 1953; Чибилёв, 1995).
В целом по региону веретеница отмечена в 8 (72,7% из 11) ландшафтных райо-
нах лесостепной и 5 (7,2% из 69) степной зоны. Ее конкретные месторождения
отражены в табл. 36 и на карте 25.
Места обитания. В регионе населяет нагорные дубово-липовые и пой-
менные мелколиственные (преимущественно березово-осиновые) леса, где
придерживается вырубок, просек, слабо заросших гарей, окраин опушек, по-
лян и границы леса вдоль лугов и полей. В Бузулукском бору встречалась на
участках со смешанным древостоем. В окрестностях г. Оренбурга Караваева
роща представляет собой участок высокой поймы р. Сакмара, где с луговыми
участками перемежаются небольшие тополевые рощи с сопутствующим ком-
плексом кустарников. На сопредельной территории Челябинской область она
встречается по островным реликтовым борам.
Образ жизни. После зимней спячки появляется в конце апреля - начале
мая и (по наблюдениям на сопредельной территории Волжско-Камского края)
первое время мало активна - чаще всего греется на солнце на тропинках, пнях,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
195
Таблица 36
Распространение веретеницы ломкой
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
1 1 г. Тольятти, п. Портовый Бакиев, Файзулин, 2001;
р-н Автозаводской 2002; Бакиев и др., 2003
т р-н Комсомольский
1 2 г. Самара
п. Управленческий
3 X п. Кировский
Е О 2 3 п. Старосемейкино
& 8 Оренбургская область
О 4 4 Северный р-н Райский, 1951; Чибилев,
II 5 5 п. Бугуруслан 1995
6 п. Абдулино
6 7 бас. р. Дёма
8 8 хр. М. Накас Райский, 1951; Чибилёв,
III 10 9 хр. Шайтантау 1995
И 10 бас. р. Катрала
Самарская область
21 11 п. Горский Бакиев, Файзулин, 2001
12 Красносамарский лес
3 X 22 13 Бузулукский бор, окр.
Е О V п. Борек
О Оренбургская область
14 Бузулукский бор, окр. Райский, 1951;Даркше-
п. Колтубановский вич, 1953
29 15 г. Оренбург, Караваева роща Зарудный, 1895
лежащих стволах деревьев и т. п. Позднее переходит на ночной образ жизни, а
днем прячется в лесной подстилке, под упавшими стволами деревьев и валеж-
ником на сравнительно небольшом участке, площадью всего в несколько десят-
ков квадратных метров. Днем из убежищ выходит лишь при теплой пасмурной
погоде. Оптимум по температуре субстрата находится в пределах 18,1...22,5 °C
(Литвинов, Ганщук, 2003). В августе начинаются перемещения к местам зимов-
ки, и с конца этого месяца - начала сентября обычно уже не отмечается (хотя
иногда задерживается до конца сентября). Таким образом, период активности
продолжается около 130-140 суток, а остальное время она проводит в спячке
в разных ямах, пустотах (норах млекопитающих, прикорневых пространствах,
гнилых пнях, под камнями), где иногда собираются по 20-30 особей. Зимоваль-
ные убежища (длинные ходы, закупориваемые изнутри землей и травой) обыч-
но располагаются на склонах южной экспозиции.
196
Глава 4
Размножение. Половозрелыми веретеницы становятся на третьем году
жизни. В этом возрасте вскоре (на Южном Урале приблизительно через полме-
сяца) после выхода из спячки начинается спаривание, что приходится пример-
но на середину мая. Для веретеницы характерно яйцеживорождение и спус-
тя примерно 90 суток - в конце июля - начале августа появляются молодые.
Плодовитость в Волжско-Камском крае составляет 7-17, на западном склоне
Южного Урала 5-26 молодых. Они более подвижны и отличаются меньшей
привязанностью к территориям, хотя зачастую зимуют вместе со взрослыми.
Питание. Питаются беспозвоночными: преимущественно дождевыми
червями, голыми слизнями (которые для большей части птиц и рептилий не-
доступны или несъедобны), многоножками, кивсяками, мокрицами, а также
насекомыми и их личинками (чаще всего гусеницами бабочек); иногда поеда-
ются и мелкие рептилии. В свою очередь веретеницы входят в состав кормово-
го рациона некоторых хищных птиц и млекопитающих.
Численность. В пределах региона веретеница, как и любой другой вид,
обитающий на периферии ареала, малочисленна, особенно в антропогенно
модифицированных местообитаниях. В окрестностях г. Оренбурга (Каравае-
ва роща) она редко встречалась уже в конце XIX столетия (Зарудный, 1895).
В Оренбургской области сравнительно обычна она была, по-видимому, лишь в
Бузулукском бору (Даркшевич, 1953), где относительно стабильной ее числен-
ность сохраняется и в настоящее время. В целом по области к концу XX сто-
летия общее количество веретениц ориентировочно не превышало 200 особей
(Чибилёв, 1998) и, к сожалению, снижается.
В Самарской области за последние десятилетия относительно стабильной
численность остается лишь в Красносамарском лесничестве, а в лесопарковой
зоне, городов Самары и Тольятти она снизилась более чем в 5 раз и сейчас
единичные особи здесь встречаются не каждый год (Бакиев, Файзулин, 2001;
Бакиев и др., 2003). Малочисленна веретеница также в приграничных районах
Башкирии (Шошева, 1985) и Челябинской области (Чибилёв, 2005).
Лимитирующие факторы. Комплекс факторов, действующих на перифе-
рии ареала; хозяйственное и рекреационное освоение территории (особенно в
пригородной лесопарковой зоне), бессмысленное уничтожение людьми.
Охрана. Обитает и охраняется на территории национального парка «Бу-
зулукский бор». Занесена в Красную книгу Оренбургской (1998), Самарской
(2009), Саратовской (2006), сопредельной Челябинской области и Башкор-
тостана.
Региональный статус. Редкий краеральный вид, находящийся у южных
пределов распространения.
*Род 19. Pseudopus Меггеш, 1820 - панцирные веретеницы
*26. Желтопузик, или глухарь - Pseudopus apodus (Pallas, 1775)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода, встречающийся на территории России. Политипический вид,
для которого П.С. Палласом типовой теорией указана «нарынская степь» меж-
ду Волгой и Уралом.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
197
Этим названием обозначается северо-западная часть Волжско-Ураль-
ских песков в окрестностях п. Урда (бывшая Ханская ставка), представляю-
щая собой сложный комплекс перемежающихся широких (5-20 км) песчаных
гряд-нарынов и долиноообразных понижений (ашиков), издавна называемая
«нарынская степь». Позднее обитание в этих местах желтопузика не подтвер-
дилось, в связи с чем типовой территорией вида была принята долина р. Терек.
В «Определителе земноводных...» (1977) эти отстоящие друг от друга поч-
ти на 500 км пункты ошибочно оказались объединены как «нарынская степь»
(Северный Кавказ). В связи с этим вид включаем в так называемый «теневой
список региона».
Семейство 13. Scincidac Oppel, 1811 - Сцинковые
Род 2Ъ. Ablepharus Fitzinger In Eversmann, 1823 - гологлазы
27. Пустынный гологлаз -Ablepharus deserti Strauch, 1876
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, известный на территории региона. Монотипический вид,
представленный рядом изолированных популяций (Ананьева и др., 1998; Ат-
лас пресмыкающихся...,2004).
Распространение. Ареал вида охватывает часть Средней Азии и Южно-
го Казахстана. Его северная граница достигает дельты р. Сырдарья. Здесь, а
также на одном из островов Аральского моря близ Ак-бике этот вид в 1914 г.
был найден Н.А. Зарудным (1915). По низовьям р. Сырдарьи позднее он отме-
чался и другими исследованиями, в связи с чем эту территорию, по-видимому,
можно считать районом постоянного обитания вида (карта 26). Кроме того, в
коллекции ЗИН АН есть экземпляр пустынного гологлаза с Устюрта (без точ-
ного указания места находки), доставленный в 1843 г. Манизером. Возможно,
что этот экземпляр был обнаружен в казахстанской части Устюрта (Параскив,
1956).
Семейство 14. Lacertidae Bonaparte, 1831 - Настоящие ящерицы
Род 21. Eremais Fitzinger In Weigmann, 1837 - ящурки
28. Разноцветная ящурка - Eremias arguta (Pallas, 1773)
Значение таксона для сохранения биоразнообразия. Один из представите-
лей обширного рода, обитающий на территории региона. Политипический вид,
представленный шестью подвидами, в том числе двумя в регионе, различающи-
мися размерными показателями, параметрами фолидоза, рисунком и окраской
верхней стороны тела (которые во многом определяются характером населяе-
мого субстрата), числу бедренных пор, числу чешуй вокруг хвоста, их форме
и некоторым другим признакам, причем по длине тела и количеству щитков
наблюдается клинальная изменчивость с запада на восток и с севера на юг.
Считается, что всю европейскую часть ареала на восток до р. Урала насе-
ляет Е. d. deserti Gmelin, 1789, который восточнее сменяется номинативным
198
Глава 4
подвидом Е. a. arguta, описанным П.С. Палласом (1773) из песков Кок-джи-
да Урало-Эмбенского междуречья (Щербак, 1993; Атлас пресмыкающихся...,
2004; Кузьмин, Семенов, 2006). Вместе с тем детальное изучение фенетичес-
ких различий в зоне вторичной интерградации этих подвидов, возникшей в го-
лоцене в Северном Прикаспии при сокращении акваторий моря (и восстанов-
лении целостности разорванного ранее ареала вида) показало существование
здесь более сложной структуры ареала.
Наиболее близки по фенетическому спектру популяции Урало-Эмбен-
ского междуречья (кроме песков Тайсойган в низовьях р. Уил) и дельты
р. Эмба, связь между которыми, по-видимому, не прерывалась во время неод-
нократных хвалынских трансгрессий Каспийского моря. К этим группиров-
кам довольно близкой оказывается популяция молодой приморской равнины,
простирающейся на запад практически до волжской дельты. В свою очередь
эта группировка по некоторым фенам оказалась ближе и к популяции Волжс-
ко-Уральских песков. Несколько изолированно от всех них стоит выборка из
песчаного массива Тайсойган. Данные группировки различаются также био-
топической приуроченностью, деталями экологии, отношением к антропоген-
ным факторам и как результат плотностью населения (Кудакина, 1981; Ива-
нова, 1985). Не исключено, что и выше по р. Уралу структура границы между
подвидами также имеет гораздо более сложный характер. Это подтверждается
особенностями полиморфизма ящурок из северо-восточной окраины Волжс-
ко-Уральских песков вблизи п. Калмыкове. Здесь фенетический облик «пес-
чаных микропопуляций» Е. a. deserti сближается с обликом микропопуляций
Е. a. arguta, повсеместно приуроченным к песчаным почвам, в то время как
обитатели глинистых почв имеют черты типичного подвида. В связи с этим
формирование Е. a, deserti предполагается на основе «песчаной формы», близ-
кой к номинативному подвиду, что впоследствии позволило виду заселить но-
вые территории (Окулова, 1973). Севернее в Волжско-Уральском междуречье
западную границу зоны интерградации двух подвидов проводят по меридиа-
ну Камыш-Самарских озер до государственной границы Казахстана и России
(Щербак, 1974), а затем скорее всего она следует к Бузулукскому бору, пос-
кольку особи из Поволжья являются типичными представителями Е. a. deserti
(Епланова, Бакиев, 2002; Епланова и др., 2003). Восточнее зона интерградации
подвидов, вероятно, охватывает всю территорию ареала в пределах Оренбург-
ской области вплоть до Предуралья.
Распространение. Ареал вида охватывает южную часть Восточно-Евро-
пейской равнины, Кавказ, Среднюю Азию, большую часть Казахстана и за-
падные районы Китая и Монголии (субэндемик Средней Азии и восточного
Ирана). Его северная граница от восточных рубежей Румынии проводилась че-
рез Украину, Курскую, Воронежскую и север Волгоградской области к долине
р. Волги. В начале XX столетия северным пределом распространения ящур-
ки здесь считалось устье р. Саратовки, а затем район Жигулей (Положенцев,
1937, 1941).
В Заволжье ее распространение В.С. Бажановым (1930) несколько неоп-
ределенно указывалось для Пугачевского уезда бывшей Самарской губернии,
восточнее она отмечалась в Бузулукском бору (Положенцев, 1935; Райский,
1951, 1956; Даркшевич, 1953), затем по пескам и песчаным почвам в низовьях
р. Илека и по среднему течению р. Урала (Зарудный, 1895; Шварц и др., 1951).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
199
В этой части ареала, по-видимому, самое северное местообитание отмечено в
окрестностях с. Подгорное Кувандыкского района (Завьялов и др., 2006). Вос-
точнее в середине XX столетия граница проводилась через Наурзум (Динес-
ман, 1953), далее примерно по 51-й параллели до окрестностей оз. Алаколь и
Зайсан в Китай (Параскив, 1956).
В исследованиях последующих лет установлено, что в Заволжье разно-
цветная ящурка обитает по поймам рек Волги и Еруслана, а также в их меж-
дуречье, затем в низовьях р. Б. Иргиз (Завьялов и др., 2003, 2006; Табачишин
и др., 2006). Однако самая северная из известных в настоящее время не только
в регионе, но и во всем ареале популяция обитает в окрестностях г. Тольятти
между селами Федоровка и Пискалинский взвоз. Еще одна новая популяция
этой ящурки недавно обнаружена в Красносамарском лесничестве Кинель-
ского района Самарской области и на окраине этого леса у с. Красная Самар-
ка (Котенко, Вехник, 1993; Ильина, Магдеев, 1999; Бакиев, Файзулин, 2001;
Епланова, Бакиев, 2001, 2002; Епланова и др., 2001; Бакиев, Иванова, 2004).
Несколько неопределенной до настоящего времени остается ситуация с
(условно ее назовем) «пугачевской» популяцией. Территория, которую ранее
занимал бывший Пугачевский уезд, в настоящее время относится не только к
Пугачевскому, но и Балаковскому, Духовницкому, Ивантеевскому, Краснопар-
тизанскому, Марксовскому и Прелюбскому административным районам Са-
ратовской области, а также четырем - Большеглушицкому, Большечернигов-
скому, Пестравскому и Хворостянскому районам Самарской области. Из всех
этих районов двух областей находки вида по данным публикаций последних
лет (Табачишин, Завьялов, 1998) известны только в Озинском и Перелюбском
районах Саратовской области. По мнению Г.В. Еплановой с коллегами (2001),
возможно, именно здесь, а не на территории современного Пугачевского райо-
на В.С. Бажановым (1930) и была отмечена разноцветная ящурка. Однако
В.Г. Табачишин с коллегами (2006) считают, что В.С. Бажанов при составле-
нии кадастра встреч вида, очевидно, использовал данные о его регистрации в
пойме р. Волги в пределах прежнего Пугачевского уезда, где пойменные пес-
чаные участки более типичны.
Восточнее разноцветная ящурка постоянно отмечается на территории
Бузулукского бора, затем по изолированным песчаным массивам в низовьях
рек Ток и М. Уран (впадающих в р. Самару), а также в Прииртекских пес-
ках (Чибилёв, 1995, 1999а), низовьях р. Илек, найдена в районе излучины
р. М. Хобда, ур. Шурбагач, по р. Урал у п. Дубенский и Гипсрудник Беляевс-
кого района, Кумакских песках и Ащисайском участке Оренбуржского степ-
ного заповедника (Гавлюк и др., 1993; Чибилёв, 1995, 1999а; Давыгора, 1998).
В Казахстанской части региона находки новых местообитаний в последние
годы за пределами ранее установленной границы нам не известны. Южнее
ареал простирается до северо-восточных районов Турции, Северного Ира-
на, Афганистана и Китая. Таким образом, ареал вида охватывает большую
часть нашего региона. В очерченных пределах разноцветная ящурка обитает в
2 (18,2% из И) ландшафтных районах лесостепной зоны, 18 (26,1% из 69)
степной, 10 (31,3% из 32) полупустынной и 29 (67,4% из 43) пустынной зоны.
Ее конкретные места обитания приведены в табл. 37 и на карте 27.
Места обитания. Разноцветная ящурка генетически пустынно-степной
(полупустынный в современной трактовке) вид, сформировавшийся в услови-
200
Глава 4
Таблица 37
Распространение разноцветной ящурки
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
[есостепная I 1 1 2 3 г. Тольятти, п. Федоровка Пискалинский взвоз, ст. Пис- калы Мастрюковские озера Котенко, Вехник, 1993; Епланова и др., 2001; Бакиев, Файзулин, 2001; Епланова, Бакиев, 2002; Бакиев, Иванова, 2004
III И 4 Оренбургская область окр. с. Подгорное Завьялов и др., 2006
Саратовская область
13 5 низ р. Б. Иргиз Завьялов и др., 2003
IV 6 окр. г. Маркс
14 7 дол. р. Волга Табачишин и др., 2006
8 окр. г. Энгельс
18 9 окр. с. Лепехинка Шляхтин и др., 1999
Самарская область
21 10 Красносамарское лесничество, окр. п. Красная Самарка Ильина, Магдеев, 1999; Бакиев, Файзулин, 2001
Оренбургская область
3 к к V 22 23 11 12 13 Бузулукский бор окр. п. Колтубановка низ р. Ток низ р. М. Уран Положенцев, 1935; Рай- ский, 1951; Даркшевич, 1953; Чибилёв, 1995, 19996
о О 27 14 Саратовская область п. Озинки Табачишин, Завьялов, 1998
Западно-Казахстанская область
15 п. Семиглавый мар Щербак, 1976
Оренбургская область
VI 30 16 Приилекские пески, п. Буран- ное Зарудный, 1895;Гавлюк и др., 1993; Чибилёв,
32 17 п. Сагарчин 1995
Западно-Казахстанская область
VII 38 18 окр. оз. Шалкар Оренбургская область Никольский, 1918
42 19 р. М. Хобда, ур. Шубарагач Гавлюк и др., 1993
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
201
Продолжение табл. 37
Зона Провин- ция Район № точки Местонахождение Источник информации
Саратовская область
48 20 21 Приерусланские пески, окр. с. Дьяковка п. Прибрежный, п. Ровное п. Речной с. Кочетное с. Александровка Табачишин и др., 2006; Особо охраняемые..., 2007; Шляхтин и др., 1999
VIII 22 Волгоградская область низ. р. Еруслан Оренбургская область Табачишин и др., 2006
5 X 53 23 Прииртекские пески Чибилёв, 1995, 19996
с V б Западно-Казахстанская область
54 55 24 25 окр. п. Дарвинское окр. г. Уральск Параскив, Бутовский, 1960
Оренбургская область
IX 56 26 окр. п. Дубненский п. Гипсрудник Гавлюк и др., 1993
X 68 27 Кумакские пески Чибилёв, 1995, 19996
72 28 Ащисайская степь ОГПЗ
XI 78 29 Кустанайская область Наурзумский заповедник Динесман, 1953; Брагин, Брагина, 2002
Волгоградская область
30 окр. оз. Эльтон Никольский, 1905; Хо- дашова, 1960; Линдеман и др., 2005; Стрельцов и др., 2006; Животные..., 2009
к cd XIII Астраханская область
X X ьй 85 31 окр. оз. Баскунчак
в с Западно-Казахстанская область
5 с 88 32 окр. п. Суюндик с. Борки Накаренок, 2003в; Брушко, Кубыкин, 1988
Гурьевская область
XV 95 33 окр. п. Миялы, 50 км на юго- вост. Накаренок, 2003в; Сара- ев, Пестов, 2010
96 34 п. Сорколь, 10 км к сев. горы Актолагай
202
Глава 4
П родолжение табл. 37
Зона Провин- ция Район № точки Местонахождение Источник информации
5 а а XV 98 97 35 36 пески Кок-джида окр. р. Шаган Никольский, 1918; Определитель земновод- ных..., 1977
XVI 101 37 окр. ст. Мугоджары Определитель земновод-
ь XVIII 107 38 низ. р. Иргиз них..., 1977
о С XIX ПО 112 39 40 окр. п. Тургай окр. п. Иргиз
Астраханская область
113 41 окр. горы Б. Богдо Никольский, 1905, 1918
42 ур. Чапчачи п. Сасыколи
114 43 44 п. Харабали, пески Б. Болхуны ст. Досанг- п. Замьяны ст. Сероглазово ур. Лапас Полынова и др., 2001; Бобров, 1985; Завьялов, Табачишин, 1998
45 Красноярский р-н, АГКМ п. Комсомольский п. Приозерный Гурьевская область
к 46 окр. п. Танюшкино Накаренок, 2003в
cd X X 3 и XX Западно-Казахстанская область
Q С 116 117 47 48 Рын-пески, п. Урда Прикамыш-Самарские пески Гурьевская область Параскив, Бутовский, 1960; Чернов, 1954
118 49 окр. п. Н. Асан ур. Караузек окр. п. Н. Уштаган Сараев, Пестов, 2010; Накаренок, 2003 в; Па- раскив, Бутовский, 1960
50 окр. сора Бесоба ур. Бес-Чохо
120 51 окр. кл. Шадьямола ур. Сасыктау Сараев, Пестов, 2010
121 52 ур. Мынтюбе ур. Сазды Накаренок, 2003в
122 ур. Буркутбай, окр. барх. Жар окр. рзд. 13
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИИ...
203
Продолжение табл. 37
Зона Провин- ция Район № точки Местонахождение Источник информации
Западно-Казахстанская область
85 окр. п. Калмыкове к зап. окр. п. Кулагине Окулова, 1973; Чернов, 1954
XX 123 53 окр. метеост. Тас окр. п. Елтай Накаренок, 2003в
124 54 п. Калмыкове
55 окр. п. Горы
Гурьевская область
127 56 окр. п. Корсак окр. п. Шубарпалы окр. п. Жилая Коса ур. Тереньузек ур. Тажигали ур. Караарна ур. Караарна, 20 км на зап. Сараев, Пестов, 2010; Накаренок, 2003в
128 57 окр. зим. Камбакты
g XXI 129 58 окр. п. Новобогатинкий
х х 59 окр. п. Махамбет
3. S 130 60 окр. п. Еркинкала
Е 131 61 рзд. Карабатано п. Искининский, 10 км к зап. пром. Искине, 9 км к вост. окр. п. Искининский
132 62 окр. г. Гурьев Иванова, 1985
63 окр. п. Ракуша г. Гурьев, 89 км к сев.
133 64 окр. оз. Индер п. Индерборский Чернов, 1954
135 65 окр. п. Карабау окр. п. Каракуга Сараев, Пестов, 2010; Накаренок, 2003в
66 пески Тайсойган
XXII 136 67 окр. п. Макат низ. р. Сагыз
68 окр. п. Доссар, 12 км к сев. окр. п. Досор, 30 км окр. п. Кульсары окр. горы Аккерегеше
204
Глава 4
Окончание табл. 37
Зона Провин- ция Район № точки Местонахождение Источник информации
69 окр. п. Косчагыл дельта р. Эмбы
137 70 дол. р. Сагиз, ур. Камбай
71 дол. р. Кайнар, ур. Акша г. Иманкара, 30 км к сев. ур. Мадьяр ур. Брлик гора Кой-Кара
72 дол. р. Эмбы, ур. Жанбике ур. Бактогоринсай ур. Намазтакыр ур. Уитас
XXII ур. Акмечеть ур. Тугыракшан
Кромка: ур. Тугарышкан, ур. Буранкуль
окр. сора Толтуртуз
138 73 ур. Кумшете Брушко, Кубыкин, 1998
cd К Я ур. Айнамас
В ур. Ушкан
с 74 Мангистауская область окр. г. Жельтау окр. ур. Сорширпа ур. Деримжанмола Накаренок, 2003в
139 75 окр. п. Бейнеу ур. Жанасу
XXIII 143 76 86 окр. р. Сары-Чеганак, пром. Тенгиз ур. Прорва
144 77 окр. сора Кайдак Никольский, 1905
XXIV 146 78 ур. Донгузтау Богданов, 1965
XXV 148 79 окр. кол. Кюше пески Сам Брушко, Кубыкин, 1988
153 80 между городами Аральском и Иргизом Динесман, 1953
XXVI 154 81 Приаральские Каракумы Лобачев и др., 1973
155 82 сев. берег Аральского моря Зарудный, 1915
83 окр. г. Аральска Сидоров, 1925
84 низ. р. Сырдарья Динесман, 1953
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
205
ях Центрального Казахстана (Щербак, 1974). Вероятно, именно эта особен-
ность исходных ландшафтов и определила высокий уровень полиморфизма
вида, а также его эвритопность.
В регионе наиболее близка к исходной форме популяция, по-видимому, из-
начально населяющая территорию возвышенной полупустынной Эмбенской
провинции с типичными для местности солонцеватыми глинистыми, сугли-
нистыми, мергелистыми, а местами и древнедельтовыми песчаными и супес-
чаными отложениями (Чибилёв, Дебело, 2006), занятыми полынно-солянково-
эфемеровыми пустынными группировками (Бабаев и др., 1986). Отступление
Хвалынского моря и формирование обширной дельтово-морской глинистой,
суглинистой и супесчаной равнины несомненно способствовали расширению
ареала вида в Урало-Эмбенском, а затем и Волжско-Уральском междуречье.
Последующая трансгрессия привела к изоляции этих популяций, а специфич-
ность условий западной части ареала, по-видимому, и определила появление
здесь Е. a. deserti. Объединение этих двух участков ареала в результате после-
дующей регрессии способствовало возникновению обширной зоны интергра-
дации двух подвидов, охватывающей значительную часть Волжско-Уральско-
го междуречья, заселенного особым фенотипом (Иванова, 1985).
По-видимому, уже в пределах ареала исходной формы начались ее диффе-
ренциация и формирование двух условных экоморф.
- «Солончаковая», населяющая дельтово-лиманные ландшафты, изобилу-
ющие сорами и солончаками. Формирование обширной полосы приморских
солончаковых пустынь во время неоднократных регрессий моря способствова-
ло расширению ареала этой формы, и сейчас она является «пионерной» на наи-
более молодой новокаспийской солончаковой приморской равнине (Неручев и
др., 1990; Накаренок, 2003в).
- «Песчаная». Ее существенные фенетические отличия от популяции
глинистых пустынь скорее всего свидетельствуют о раннем обособлении от
исходной формы, а ограниченность этого типа местообитаний оставила ее в
ранге «несостоявшегося подвида» (типа Е. a. deserti). Так в наиболее общих
чертах представляются гипотетическая история освоения территории региона
разноцветной ящуркой и причины ее эврито'пности.
В настоящее время в исходных (приэмбенских) ландшафтах разноцветная
ящурка явно предпочитает глинистые пустыни со слабо сбитыми полынно-би-
юргуновыми, полынно-солянковыми и эфемеровыми сообществами, а на учас-
тках с сильным сбоем почти исчезает. Значительно реже она начинает встре-
чаться и у нижней кромки раннехвалынской полосы, где типичные глинистые
и суглинистые пустыни постепенно замещаются песчано-полынными на су-
песях. Здесь же она проникает на отдельные участки речных долин, природ-
ные условия которых при углублении и иссушении, вызванном отступлением
моря, перестали существенно отличаться от окружающих пространств.
В полосе верхнехвалынских ландшафтов она по-прежнему наиболее
обычна на глинистых участках, но поскольку они здесь имеют ограниченное
распространение, то основным (фоновым) местообитанием здесь становится
песчано-полынная пустыня. Здесь ящурка несколько чаще встречается и на
пухлых солончаках по разным депрессиям. По этим своебразным экологи-
ческим руслам она выходит и на просторы пухлых приморских солончаков
с сарсазаном (Неручев и др., 1990; Накаренок, 2003в). В зоне интерградации
206
Глава 4
двух подвидов в северо-восточной части Волжско-Уральского междуречья
(п. Калымово) по северной кромке песчаного массива она населяет преиму-
щественно злаково-полынные и полынные сообщества на глинистых участках,
а южнее встречается уже исключительно в песчаной злаково-полынной пусты-
не, перемежающейся с бугристыми песками. Явное предпочтение полынных
сообществ, здесь, по-видимому, связано с небольшой сомкнутостью их покро-
ва и обилием саранчевых, составляющих основу ее питания. Населяются здесь
и переходные участки от глинистой к типичной песчаной пустыне (Окулова,
1963).
У северной кромки песков на Прикамыш-Самарской равнине эта ящурка
встречается исключительно на закрепленных участках. В центральную часть
массива она проникает в основном по ашикам, вдоль которых заселяет как
закрепленные участки, так и полузакрепленные пески с очагами развевания и
окраины барханов (Чернов, 1954; Параскив, Бутовский, 1960; Неручев и др.,
1981).
У западной кромки Волжско-Уральского междуречья на территории Ас-
траханского ГКМ она населяет преимущественно закрепленные бугристые
пески и выровненные участки в межбугровых понижениях, значительно реже
встречается на солонцах и бугристых песках барханного типа (Завьялов, Таба-
чишин, 1998). В бугристых песках эта ящурка отмечалась и несколько север-
нее - в окрестностях горы Богдо, оз. Баскунчак, п. Урда (Рын-пески); пример-
но в этой же части региона наблюдалась также среди лесных посадок и на
межполосных пространствах (Чернов, 1954; Параскив, Бутовский, 1960).
В Волгоградской области разноцветная ящурка мозаично встречается в
разных, но преимущественнно полупустынных местообитаниях (Кубанцев и
др., 1962). В окрестностях оз. Эльтон предпочитает микроповышения с пус-
тынным типом растительности (Ходашова, 1960; Быков, 1981). У северных
границ этой области и в Саратовском Заволжье ее обитание связано главным
образом с участками закрепленных и полузакрепленных песков в долине рек
Волги, Еруслана, а также их междуречья (Табачишин и др., 2006).
В районе Самарской луки популяция занимает лишь небольшой (до 40 га)
участок надпойменной террасы, где ящурки держаться на сравнительно крутом
(до 50°) склоне южной экспозиции, сложенном сухим, хорошо прогреваемым
желто-серым песком, занятым разряженными (покрытие 20-40%) житняково-
разнотравными и полынно-лебедовыми сообществами с участием галофитов.
Здесь они явно предпочитают укрепленные корнями растений стации, на неук-
репленных местах встречались значительно реже, а на участках с древостоем
не отмечались вовсе (Ильина, Магдеев, 1999; Епланова и др., 2001). Восточ-
нее - в долине р. Самара, наоборот, она отмечена лишь в Красносамарском и
Бузулукском лесных массивах. Вместе с тем, по наблюдениям в Бузулукском
бору, она и здесь встречается лишь на участках с террасированными или кру-
тосклонными бугристыми песчаными дюнами с разреженным сосновым дре-
востоем, что обеспечивает хорошую прогреваемость грунта. Определенную
роль здесь играют и детали микрорельефа, в часности глубоко врезанные в
песчаном грунте колеи дорог, увеличивающие инсоляцию и способствующие
его лучшей прогреваемости. В низовьях рек Ток, М. Уран и на Иртекско-Рубе-
жинской равнине эта ящурка связана с открытыми степными песчаными мас-
сивами. В последнем случае она населяет полузакрепленные среднебугристые
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
207
пески, сформировавшиеся в низовьях рек Иртек, Ембулатовка, Выковка, Ру-
бежка и простирающиеся почти до г. Уральска.
В низовьях р. Иртек, Б. и М. Хобда разноцветная ящурка населяет песча-
ные и супесчаные террасы с бугристо-песчаным типом местности, в пределах
которых выделяется несколько довольно крупных (у п. Привольное, ст. Са-
гарчин) массивов кучевых и барханных песков с разреженной псаммофитной
растительностью. Южнее она обитает в массивах полузакрепленных бугристо-
гривистых и грядово-барханных песков Караагаш и Аккумы. Популяции этих
и приилекских песков генетически скорее всего ближе к уильской, в прошлом
эти пески составляли единый аллювиально-дельтовый массив, простирающий-
ся вдоль Предсыртового Уступа.
Довольно широко распространена разноцветная ящурка и в Зауралье. Вмес-
те с тем наиболее разнообразны ее местообитания в южной приаральской части
региона. Здесь она населяет как типичные для номитативного подвида участ-
ки глинистой пустыни (занятые биюргуном с полынью и одним биюргуном),
так и ячеистые пески (с тремя полынными и адраспановым сообществами).
Севернее в низовьях рек Иргиз и Тургай она встречается уже только в двух
(полынном и биюргуновым), а в Наурзуме в одном (полынно-разнотравном)
степных сообществах (Динесман, 1953). Общим для всех этих местообитаний
является разряженность растительного покрова (не затрудняющая передви-
жения) и примерно одинаковый температурный режим поверхности грунта.
В долине р. Тургая она, кроме того, встречается в соответствующих местах по
берегам озер и среди прибрежных зарослей тамарикса (Лобачев и др., 1973).
Образ жизни. Зимние убежища на сопредельной территории у южных
границ региона разноцветная ящурка покидает в первой декаде марта или не-
многим позднее. Севернее в центральной части Волжско-Уральских песков
(окрестности п. Сасыктау) и на территории Волгоградской области первые
особи появляются в конце этого месяца или начале апреля, а в самой северной
популяции в Самарской области во второй-третьей декаде апреля. Массовый
выход обычно наблюдается через 1-2 недели, что в значительной степени оп-
ределяется ходом развития весенних явлений.
Весной максимум активности наблюдается с 10 до 16 часов при темпера-
туре почвы выше 15 °C, а летом активность смещается на утренние и вечерние
часы; наиболее жаркое время дня она проводит в собственных норках (наклон-
ный ход длиной до 70 см и глубиной до 35 см) или в других убежищах (тре-
щинах в почве, пустотах под камнями, норах разных животных). Предпочита-
емые ящурками температуры воздуха на пятисантиметровой высоте находятся
в пределах от 31,3 до 35,1 °C (Литвинов, Ганщук, 2003). Территориальна, зани-
мает участки площадью до 180 м2.
Осенью активность снижается, и со второй половины сентября она зале-
гает в спячку, хотя в теплые годы задерживается до первой половины октяб-
ря. Таким образом, активный период на севере региона продолжается около
150-170, а на юге до 210-220 суток, зимняя спячка соответственно около 200
и 150 суток (Параскив, Бутовский, 1960; Кубанцев и др., 1962; Епланова, 2001,
2004; Табачишин и др., 2006).
Размножение. Брачный период начинается вскоре после массового выхо-
да из зимних убежищ. У южных пределов региона спаривание начинается при-
мерно в середине апреля, у северных границ в Самарской области - в третьей
208
Глава 4
декаде этого месяца (Епланова, 2004) и на большей части ареала продолжатся
до конца июня (т. е. около 2 месяцев). Кладка яиц в Волжско-Уральских песках
начинается в первой половине мая, а в конце июня - начале июля происходит
вторая. В кладках здесь отмечали до 7, в Волгоградской и Саратовской облас-
ти до 10, а в Самарской области 3-5 (но более крупных) яиц; меньше яиц и во
второй кладке. Инкубационный период продолжается 25-30 суток. Массовый
выход сеголеток наблюдается с начале июня и продолжается почти до конца
сентября (ур. Сасыктау Волжско-Уральских песков, 1938 г.).
Питание. Основу пищевого рациона вида составляют насекомые, пре-
имущественно жуки, бабочки и перепончатокрылые, в меньшем количестве
потребляются клопы, двукрылые и прямокрылые, хотя доля последних при
их массовом размножении увеличивается (Волжско-Уральские пески). Кроме
того, потребляются пауки, мокрицы и моллюски, в единичных случаях поеда-
ются мелкие ящерицы, изредка вегетативные части растений и семена. В свою
очередь ящурок поедают змеи, хищные птицы и млекопитающие.
Численность. В связи со значительными размерами ареала численность в
разных частях региона существенно различается. Вместе с тем даже в перифе-
рических популяциях местами она может достигать значительной величины и
в целом может быть сопоставима с таковой в центральной части ареала. Так,
по кромке закрепленных и полузакрепленных Приерусланских песков мак-
симальная численность достигает 367,5 ос/га, а на антропогенно-преобразо-
ванных местообитаниях и неудобях - 77,0-96,7 ос/га (Шляхтин и др., 1999;
Табачишин и др., 2006; Табачишина и др., 2006а). Довольно многочисленна
она была в полупустынных и пустынных сообществах Заволжья (Ходашова,
1960; Кубанцев и др., 1962). Однако с 1970-х гг. численность ее здесь резко
сократилась и сейчас она становится обычной лишь в Эльтонской котловине
(Линдеман и др., 2005; Стрельцов и др., 2006). В окрестностях горы Богдо в
прошлом она встречалась в «поразительном количестве» (Никольский, 1905).
Немногим южнее, на территории Астраханского ГКМ, сравнительно много-
численна она и сейчас. Здесь по наиболее типичным для района местообита-
ниям в закрепленных бугристых песках и ашиках между ними в 1998 г. на-
считывалось соответственно 78,6 и 40,8 ос/га, хотя на солонцах и бугристых
барханах она встречалась значительно реже - соответственно 23,8 и 8,4 ос/га
(Завьялов, Табачишин, 1998). Впрочем численность ящурки в этом районе ис-
пытывает довольно значительные колебания. Так, если в 1979 г. в окрестнос-
тях ст. Досанг (ур. Батпайсагыр) по разным местообитаниям насчитывалось
всего 1,1-3,3 ос/га (Бобров, 1985), то в 1997 г. их количество увеличилось до
20,4 ос/км (Дунаев, Жбанова, 1998).
Сравнительно многочисленной в середине прошлого столетия она была
и в Прикамыш-Самарских бугристых песках (Параскив, Бутовский, 1960).
В центральной части Волжско-Уральского песчаного массива роль ее снижа-
ется, но и здесь она остается довольно обычной, а местами и многочисленной:
на плотных песках численность ее составляла 0,4-0,5 ос/км, а на участках за-
крепленных и полузакрепленных песков с очагами развевания - 1,0-2,4 ос/км
(Неручев и др., 1981). На восточной кромке песков Волжско-Уральского
междуречья в окрестностях п. Калмыкове в глинистой злаково-полынной и
полынной пустыне ее плотность составляла 1,24 и 3,26 ос/км, а в песчаной
злаково-полынной в сочетании с бугристыми песками - 2,4 ос/км (Окулова,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
209
1963). Сейчас здесь она встречается значительно реже - 0,4 ос/км (Накаренок,
2003 в).
Неодинакова численность вида и в краеральных популяциях северной час-
ти Волжско-Уральского междуречья. Наиболее обычна она в районе Самар-
ской луки (Епланова и др., 2001), хотя в связи с ограниченностью площади
места обитания общая численность вида здесь незначительна. По-видимому,
всегда немногочисленной была ящурка в сосновых лесах по р. Самара, во вся-
ком случае так охарактеризовывал состояние популяции Бузулукского бора в
середине прошлого столетия Я.Н. Даркшевич (1953). В изолированных Ниж-
нетокском, Малоурановском и Иритекском песчаных массивах в разные сезо-
ны 1990-х гг. ее насчитывали по 4,5-6,0 ос/км (А.А. Чибилёв), а в Приураль-
ных бугристых песках в окрестностях п. Дарьинское она отмечалась единично
(Параскив, Бутовский, 1960).
В Урало-Эмбенском междуречье наиболее обычна она в Приилекских пес-
ках и, что не менее важно, количество ее здесь относительно стабильно. Общая
численность популяций Оренбургской области ориентировочно оценивается в
несколько тысяч особей (Давыгора, 1998). Южнее в массивах Караагаш и Ак-
кумы она отмечается несколько чаще (А.А. Чибилёв). По р. Уил условия оби-
тания ящурки, по-видимому, благоприятны, поскольку здесь она встречается
по 2,3-2,6 ос/км. Значительно реже она наблюдается по рекам Сагиз (1,6 ос/км)
и Кайнар, причем в последнем случае по надпойменной террасе численность
достигает 1,0, а в пойме лишь 0,2-0,4 ос/км (Неручев и др., 1990). Вместе с тем
локально численность здесь достигает 11,0 ос/км. На приморской солончако-
вой равнине в среднем насчитывали 0,6, по краям соров - 0,2, а локально (на
местах буровых площадок) численность достигала 5,0 ос/км. В Прикаспийских
Каракумах она определенно малочисленна - 0,1 ос/км (Накаренок, 2003в).
В Зауралье наиболее высокая численность разноцветной ящурки наблюда-
ется в Приаральских пустынях, особенно в их песчаных вариантах, где достига-
ет 12-23 ос/км, в то время как в глинистых насчитывали лишь по 0,5-0,6 ос/км.
Мало ее встречается в низовьях рек Иргиз, Тургай и в Наурзумском заповед-
нике, причем лишь локально - на полынно-песчаных и песчано-ковыльных
участках и опушках Наурзумского бора и Сапсына (Динесман, 1953; Брагин,
Брагина, 2002). Впрочем, местами по берегам озер и в прибрежных зарослях
тамарикса по р. Тургай она отмечалась несколько чаще - соответственно 3,0
и 3,7 ос/га. В Приаральских Каракумах эта ящурка считается обычным видом
(Лобачев и др., 1973). В целом по Казахстану она относится к видам со средней
категорией плотности - 7,9 экз/га, хотя в оптимальных биотопах плотность
достигает 50,0 ос/га (Брушко, 1993).
Лимитирующие факторы. В основном перевыпас, нерегулируемое дви-
жение транспорта и другие антропогенные факторы, способствующие раз-
рушению местообитаний, формированию на их месте сбоев и развеваемых
песков.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор», Оренбургского (Ащисайский участок), Наурзумского, Устюрт-
ского, Богдинско-Баскунчакского заповедников и Иргиз-Тургайского резерва-
та. На большей части ареала в регионе специальных мер охраны не требуется,
достаточно поддержания естественного состояния ландшафтов и контроля со-
стояния численности.
210
Глава 4
Региональный статус. По северу региона редкий, ограниченно распро-
страненный вид. Занесен в Красные книги Оренбургской (1998), Самарской
(2009) и Саратовской (2006) областей, а также в Приложение III Бернской кон-
венции.
29. Линейчатая ящурка - Eremias lineolata Nikolsky, 1896
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода, населяющий территорию региона. Монотипический
вид; межпопуляционные различия проявляются в числе чешуек вокруг сере-
дины туловища, количестве бедренных пор, окраске верхней стороны тела
(Ананьева и др., 1998).
Распространение. Ареал вида охватывает восточные районы Ирана, север
Афганистана, Среднюю Азию и юг Казахстана (эндемик Средней Азии и Вос-
точного Ирана). В пределах региона найдена лишь в Приаральских Каракумах
(карта 28), в связи с чем детали распространения и экологии практически не
изучены.
30. Средняя ящурка - Eremias intermedia (Strauch, 1876)
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода, населяющий территорию региона. Монотипический
вид; межпопуляционные различия проявляются в количестве брюшных щит-
ков, чешуй вокруг хвоста, незначительном увеличении числа чешуй вокруг
середины туловища с севера на юг (клинальная изменчивость), уменьшении
числа горловых чешуй и бедренных пор у периферических популяций (Щер-
бак, 1974; Ананьева и др., 1998; Атлас пресмыкающихся..., 2004).
Распространение. Ареал вида охватывает северо-восток Ирана, большую
часть равнинной Средней Азии и юг Казахстана (эндемик Средней Азии и
Восточного Ирана). Его северная граница еще недавно проводилась от сора
Мертвый Култук на восточном побережье Каспийского моря по Северному
Устюрту, северному побережью Аральского моря, югу Приаральских Караку-
мов и северному Прибалхашью к бассейну оз. Алаколь (Определитель земно-
водных..., 1977). В последние десятилетия в Приаральских Каракумах она пе-
ренесена на 100-150 км севернее (Лобачев и др., 1973; Брушко, 1995). Южная
граница проводится от северо-востока Ирана по предгорьям Средней Азии к
оз. Алаколь. Таким образом, ареал вида захватывает только южную пустын-
ную часть нашего региона. Здесь ящурка отмечалась у сора Мертвый Култук
(территория лектотипа вида), на северном Устюрте, у северо-западных бере-
гов Аральского моря, песках Б. Барсуки, Приаральских Каракумах, ур. Ильля-
Кудук и между г. Аральском и дельтой р. Сырдарья (табл. 38, карта 29).
Места обитания. На большей части ареала это типичный псаммофил,
населяющий закрепленные участки песков и межбарханные понижения с та-
мариксом, саксаулом и другими кустарниками, местами встречается и на со-
лончаковых грунтах с разреженной растительностью. В ур. Ильля-Кудук на-
блюдалась в песках, поросших жузгуном, полынью и кияком.
Образ жизни. В регионе не изучен. На сопредельной территории Казахста-
на из зимних убежищ выходит в первой декаде апреля и активна до середины
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
211
Распространение средней я щурки
Таблица 38
Зона Провин- ция Район № точки Местонахождение Источник информации
XXIII 143 1 окр. сора Мертвый Култук Параскив, 1956
XXV 147 2 Северный Устюрт
й X X 151 3 пески Б. Барсуки Никольский, 1915
3 в 154 4 Приаральские Каракумы Лобачев и др., 1973
£ XXVI 5 ур. Ильля-Кудук Динесман, 1953
155 6 сев.-зап. побережье Араль- ского моря Параскив, 1956
октября (около 190 суток). Территориально, причем отдельные особи живут на
значительном расстоянии друг от друга, придерживаясь определенного убе-
жища, в основном нор песчанок, сусликов, других животных и естественных
пустот в грунте. Летом активны утром и вечером, а наиболее жаркое время дня
проводят в тени кустарников или норках. Зимует в норах грызунов.
Размножение. Половозрелыми становятся в возрасте 12-14 месяцев.
К спариванию приступают в конце апреля. Кладка яиц (дважды за сезон по
2-6 шт.) начинается в середине июня и продолжается почти до конца июля.
Выход молодых длится с начала августа до середины сентября.
Питание. В ранневесенний период потребляет в основном муравьев; позд-
нее чаще всего добываются прямокрылые, жуки (особенно чернотелки), бабоч-
ки и их личинки. Реже потребляются другие членистоногие, мелкие ящерицы
и растительная пища. В свою очередь наряду с другими ящурками она отлав-
ливается крупными рептилиями, хищными птицами и млекопитающими.
Численность. В пустынях Казахстана эта ящурка относится к видам со
средней категорией плотности - 8,2-1,6 ос/га (Брушко, 1993). В Приаральских
Каракумах - к категории обычных видов (Лобачев и др., 1973).
Лимитирующие факторы. Комплекс абиотических и биотических фак-
торов, действующих у северной границы ареала; разрушение местообитаний в
результате деятельности человека.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На остальной территории региона специальных мер по охране вида не требу-
ется, достаточно поддержания естественного состояния ландшафтов.
Региональный статус. Стенотопный краеральный вид с ограниченной
численностью.
31. Сетчатая ящурка - Eremias grammica (Lichtenstein, 1823)
Значение таксона для сохранения биоразнообразия. Один из представи-
телей обширного рода ящурок, населяющий территорию региона. Монотипи-
ческий вид. Все выявленные различия популяций ниже различий подвидового
ранга. Наиболее вариабельно количество бедренных пор, а также спинных и
горловых чешуй (Щербак, 1974; Ананьева и др., 1998; Атлас пресмыкающих-
ся..., 2004).
212
Глава 4
Распространение. Ареал вида охватывает территорию Южного Казах-
стана, большую часть Средней Азии, северные районы Ирана, Афганистана
и северо-запад Китая (эндемик Средней Азии и Восточного Ирана). Его се-
верная граница прослеживается от побережья Каспийского моря через Се-
верный Устюрт, Б. Барсуки, низовья р. Иргиз (где почти достигает 49° с.ш.),
Приаральские Каракумы и далее по среднему течению р. Сары-су, северному
Прибалхашью и окрестностям оз. Алаколь в Китай. Южные рубежи ареала оп-
ределяются находками в северных районах Ирана, Афганистана и Китае. Таким
образом, ареал вида захватывает только южную, полупустынную и пустынную
части региона. В его пределах находки вида известны на Северном Устюрте у
сора Асматай-Матай, в песках Б. и М. Барсуки, Иргизкум в низовьях р. Ир-
гиз, Приаральских Каракумах, севернее г. Аральска и в низовьях р. Сырдарья
(табл. 39, карта 30).
Места обитания. Это типичный псаммофил, населяющий главным обра-
зом вершины и склоны песчаных бугров и барханов с разреженной травянистой
и кустарниковой растительностью. На побережье Аральского моря это древне-
аллювиальные кучевые пески и прибрежные дюны, обычно занятые разрежен-
ными сообществами полыни с примесью ксерофитных злаков и разнотравья.
В песках Иргизкум отмечена только в наиболее развеваемой части массива,
опять же лишь с отдельными куртинами полыни, злаков и разнотравья (Динес-
ман, 1953). В Приаральских Каракумах она предпочитает слабо закрепленные,
проросшие белым саксаулом и жузгуном бугристые пески, хотя встречается
также на закрепленных и развеваемых песках (Лобачев и др., 1973).
Образ жизни. Весной из зимовки на сопредельной территории Казахстана
выходит в разных числах первой половины апреля, когда температура грун-
та превышает 16 °C. В это время, а затем и осенью, активна в течение всего
светлого времени суток, летом активность смещается на утренние и вечерние
часы. В наиболее жаркое время и на ночь обычно скрывается в норках (каж-
дый раз новой) или зарывается в сыпучий песок. Норки, вырытые у основания
растений, как правило, представляют собой слегка наклонный ход длиной до
100 см; зимние более глубокие. Территориальна, агрессивна и составляет кон-
куренцию не только совместно обитающей быстрой ящурке, но и ушастой
круглоголовке и степной агаме. Осенью активность снижается, и в конце сен-
тября - первой половине октября она уходит в спячку, в состоянии которой
проводит около половины года.
Размножение. Молодые из ранних кладок половой зрелости достигают
к лету следующего года, а из поздних, видимо, лишь после второй зимовки.
Спаривание начинается вскоре после выхода из спячки и сопровождается дра-
ками между самцами и ухаживаниями за самками. С апреля по август бывает
2 (вторая в середине июля), а, возможно, и 3 кладки по 4-6 яиц. Выход моло-
дых из первых кладок начинается с середины июля, из поздних продолжается
до сентября.
Питание. Основу пищевого рациона составляют разнообразные мелкие
беспозвоночные, преимущественно жуки (личинки, куколки, имаго), пере-
пончатокрылые (в основном муравьи), бабочки (и их гусеницы), прямокры-
пые, двукрылые и, кроме того, пауки и скорпионы. Поедает также молодых
ящериц других и своего вида, листья и плоды растений. В поисках пищи за
день обследует территорию площадью около 0,5 га. В свою очередь сама
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
213
Распространение сетчатой ящурки
Таблица 39
Зона Провин- ция Район № точки Местонахождение Источник информации
Полу- пустын- ная XIX 112 1 2 Актюбинская область пески Иргизкум низ. р. Тургай Никольский, 1915 Динесман, 1953
Пустынная XXV 148 3 пески Сам у сора Асматай- Матай Параскив, 1956
XXVI 151 152 154 155 4 5 6 7 8 9 пески Б. Барсуки пески М. Барсуки Приаральские Каракумы ур. Ильля-Кудук п. Аралсульфат низ. р. Сырдарья Лобачев и др., 1973; Динесман, 1953
является объектом охоты крупных рептилий и ряда хищных птиц и млеко-
питающих.
Численность. В пустынях Казахстана сетчатая ящурка в целом является
видом со средней категорией плотности. В типичных местообитаниях ее плот-
ность в среднем составляет 12,6, а максимальная - 22,1 экз/га (Брушко, 1993).
В бугристых песках Приаральских Каракумов, где она считается массовым ви-
дом, плотность составляет 7,6, а на развеваемых и закрепленных песках - со-
ответственно 2,6 и 2,0 экз/га (Лобачев и др., 1973). В окрестностях г. Аральска
наиболее высокая численность отмечалась в прибрежных дюнах, где на 1 км
маршрута насчитывалось 29 особей, несколько реже - 27 ос/км - она встреча-
лась в полынниках на кучевых песках, а в других сообществах была значитель-
но ниже - 4-11 ос/км, что сопоставимо с численностью в Иргизкумах - 6 ос/км
(Динесман, 1953).
Лимитирующие факторы. Комплекс абиотических и биотических факто-
ров, действующих у северных пределов ареала; локально сильный перевыпас.
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резерва-
та. На большей части ареала состояние популяции особых мер охраны не тре-
бует, достаточно поддержания естественного состояния ландшафтов.
Региональный статус. Стенотопный вид у северных пределов ареала.
32. Песчаная, или полосатая ящурка - Eremias scripta (Strauch, 1876)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Политипический
вид, в регионе представленный номинативным подвидом Е. s. scripta (Strauch,
1876) (описан по сборам Г.С. Карелина и А. Лемана; лектотип добыт Г.С. Ка-
релиным в 1837 г.). В его пределах наблюдаются межпопуляционные отличия
по характеру рисунка верхней поверхности туловища (в популяциях региона
214
Глава 4
Распространение песчаной ящурки
Таблица 40
Зона Провин- ция Район № точки Местонахождение Источник информации
Полу- пусты- ня XVIII 112 I Актюбинская область пески Иргизкум Динесман, 1953
XXII 137 2 окр. р. Эмба Параскив, 1956
151 3 пески Б. Барсуки Динесман, 1953
Пус- тынная XXVI 154 155 4 5 ур. Ильля-Кудук п. Аралсульфат
6 сев. побережье Аральского моря
отмечается наибольшее количество темных особей), количеству чешуи вокруг
9-10-го кольца хвоста и некоторыми особенностями экологии (Щербак, 1974;
Ананьева и др., 1998; Атлас пресмыкающихся..., 2004).
Распространение. Ареал вида включает северные района Ирана, Афга-
нистана, Пакистана, Среднюю Азию и юг Казахстана (субэндемик Средней
Азии и Восточного Ирана). Его северная граница прослеживается от р. Эмба
через пески Б. Барсуки, Иргизкум, окрестности г. Аральска и далее по северу
Муюнкумов, южному Прибалхашью и бассейну оз. Алаколь. Южная граница
проводится по песчаным массивам северных районов Ирана, Афганистана и
Пакистана. Таким образом, ареал вида захватывает только южную часть ре-
гиона (табл. 40, карта 31). Подробнее в его пределах граница распростране-
ния вида прослеживается по находкам А.Н. Северцова где-то в районе сред-
него течения р. Эмба, затем в песках Б. Барсуки (экз. ЗИН РАН, Гожев), в
песках Иргизкум по низовьям р. Иргиз, ур. Ильля-Кудук в 20 км к юго-востоку
г. Аральска и окрестностях п. Аралсульфат (Параскив, 1956; Динесман, 1953).
Считалось, что южнее этих пунктов полосатая ящурка населяет почти все
пески (причем всюду является довольно обычной), хотя конкретные местона-
хождения не назывались (Параскив, 1956). Однако в ряде последующих работ
по ящерицам Казахстана (Неручев, Васильев, 1978; Неручев и др., 1981, 1990;
Брушко, 1993,1995) ее обитание в Северном Прикаспии и на Мангышлаке уже
не указывается.
Места обитания. На большей части ареала населяет преимущественно
полузакрепленные пески с кустарниковой, полукустарниковой и эфемерной
травянистой растительностью. В Приаралье она встречалась в сообществах
тамарикса, жуз1уна, верблюжей колючки и полыни с участием ксерофитного
разнотравья, злаков и небольшими пятнами (несколько квадратных метров)
чистого, незаросшего песка. В Иргизкумах отмечена на участках, поросших
полынью, жузгуном и молочаем с примесью злаков (Динесман, 1953).
Образ жизни. В регионе не изучен. На сопредельной территории активна
с начала апреля до конца сентября, иногда задерживаясь до середины октября.
Это наиболее термофильный вид ящурок с одновершинным пиком активнос-
ти. При опасности «ныряет» в песок или скрывается в убежищах - норках раз-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
215
ных животных или собственных, которые роет у основания растений. Хорошо
передвигается по кустам. Активна около 170-180 суток, в зимней спячке про-
водит 180-190 суток.
Размножение. Позднее, чем у других пустынных ящурок. Спаривание на-
чинется в апреле, кладка яиц (дважды по 1-2 шт.) продолжается со второй
половины мая до начала августа. Молодые встречаются с конца июля по сен-
тябрь. Половой зрелости достигает после первой зимовки, часть особей после
второй.
Питание. Основу пищевого рациона составляют мелкие беспозвоночные,
преимущественно насекомые (жуки, равно-, перепончато-, прямокрылые, ба-
бочки и др.); изредка потребляется растительная пища. В свою очередь ящурка
становится добычей более крупных рептилий, а также хищных птиц и млеко-
питающих.
Численность. В пустынях Казахстана эта ящурка относится к видам со
средней категорией плотности. В типичных местообитаниях ее плотность в
среднем составляет 13,0 экз/га, максимум - 26,6 (Брушко, 1993). По наблю-
дениям Л.Г. Динесмана (1953), в Каракумах средняя численность составляла
6,8 ос/км, а в Иргизкумах - 12,0.
Лимитирующие факторы. Комплекс абиотических и биотических факто-
ров, действующих у северной границы ареала.
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резерва-
та. На остальной территории региона, по-видимому, специальных мер охраны
не требуется, достаточно поддержания естественного состояния ландшафтов.
Региональный статус. Стенотопный краеареальный вид с незначитель-
ной численностью.
33. Быстрая ящурка -Eremias velox (Pallas, 1771)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода ящурок, населяющий территорию региона. Полити-
пический вид, в регионе, по-видимому, представленный лишь номинативным
подвидом Е. v. velox (Pallas, 1771), который отличается исключительно высо-
кой вариабельностью признаков, особенно рисунка спинной стороны тела и
количеством чешуй вокруг середины туловища (Щербак, 1974; Ананьева и
др., 1998; Атлас пресмыкающихся..., 2004; Чирикова, 2004; Кузьмин, Семе-
нов, 2006). Детальное изучение размеров и рисунка тела показало, что только
в Приаралье по этому признаку выделяется около 10 локальных популяций
(Чирикова, 2003). Возможно, западная часть региона, примыкающая к нижней
Волге, представляет собой зону интерградации номинативного и кавказского
подвидов.
Распространение. Ареал вида охватывает нижнее Поволжье, восточную
часть Кавказа, север Ирана, Афганистана, Среднюю Азию, большую часть Ка-
захстана и северо-запад Китая (субэндемик Средней Азии и Восточного Ира-
на). Его северная граница в пределах региона проводится от южных районов
Волгоградской области (Кубанцев и др., 1962; Щербак, 1974) по окрестностям
оз. Баскунчак (Стрельцов и др., 2006), Прихакской низменности (Животные...,
2009), северной кромке Волжско-Уральских песков вблизи п. Урда и Новая
Казанка к р. Урал в районе оз. Индер (Параскив, Бутовский, 1960; Окулова,
216
Глава 4
1963). Указание на распространение вида на север до Приерусланских песков
в дальнейшем не подтвердилось.
В Урало-Эмбенском междуречье в конце XIX столетия она проводилась в
области низовий р. Илек (Зарудный, 1895), хотя уже к середине XX столетия
обитание вида здесь было поставлено под сомнение Н.П. Райским (1951), а в
его более поздних работах (Райский, 1954, 1956) быстрая ящурка для этого
района не упоминается вовсе. Достоверно не найдена здесь и позднее. Восточ-
нее рубежи распространения вида определяются его находками в окрестностях
с. Подгорное (Завьялов и др., 2006), низовьях р. Иргиз, ур. Тусум по р. Тургай
(Динесман, 1953), в северном Прибалхашье, у оз. Алаколь, а затем в Китае
(Параскив, 1956). Южная граница от Закавказья прослеживается по северным
районам Ирана, Афганистана и далее в Китае. Таким образом, ареал вида ох-
ватывает большую часть региона.
В очерченных пределах быстрая ящурка населяет примыкающие к Волго-
Ахтубинской пойме пески в окрестностях ст. Досанг и г. Красный Яр (Щер-
бак, 1974), центральную (п. Н. Уштан) и северную (п. Урда, Новая Казанка)
части Волжско-Уральского массива. В Урало-Эмбенском междуречье обита-
ет в окрестностях оз. Индер, принятых за типовую территорию вида, откуда
он впервые был описан П.С. Палласом; отсюда же описан и неотип вида, до-
бытый С.А. Черновым (Щербак, 1974). Обитание вида по нижнему Илеку в
известной монографии «Ящурки Палеарктики» (Щербак, 1974) вообще отри-
цается, поскольку оно, якобы, основано на ошибочном определении Н.А. За-
рудным разноцветной ящурки (Eremias arquta). Вместе с тем примерно в этом
районе быстрая ящурка была отмечена немного южнее в песчаных массивах у
оз. Сорколь и Аккумы (Давыгора, 1995). Далее она обитает по р. Уилу (пески
Тайсойган), в междуречье Темира и Эмбы, ниже по течению этой реки, в При-
каспийских Каракумах, на Северном Устюрте (Неручев, Васильев, 1978; Неру-
чев и др., 1981, 1990), в Приаралье, Б. Барсуках, по рекам Иргиз (Иргизкумы),
Тургаю (ур. Тусум) и в Приаральских Каракумах (Динесман, 1953; Брушко,
1993, 1995). Из этого списка несколько неожиданно выделяется находка в об-
ласти среднего течения р. Урал у с. Подгорное (Завьялов и др., 2006). Кроме
этого пункта быстрая ящурка обнаружена в одном (1,5% из 69) ландшафтном
районе степной зоны, 7 (21,9% из 32) полупустынных и 7 (16,3% из 43) райо-
нах пустынной зоны (табл. 41, карта 32).
Места обитания. В предполагаемом центре формирования эта ящурка
является типичным мезофильным склеробионтом. На большей части наше-
го региона она населяет преимущественно закрепленные и полузакреплен-
ные пески, хотя местами не избегает солончаковых, плотных глинистых и
щебнистых участков. Это проявляется уже в расположенном ближе к центру
формирования вида Приаралье. Здесь на побережье Аральского моря и в При-
аральских Каракумах она обитает по прибрежным дюнам, глинистым обры-
вам, каменистым россыпям, у основания останцов (где глинистый грунт сме-
няется песчаным), по сухим долинам весенних водотоков, на песках с кус-
тарниками (в сообществах тамарикса, жузгуна и полыней с участием злаков),
несколько реже встречается на задернованных травянистой растительностью
песках и участках глинистой пустыни (причем почти исключительно на коло-
ниях большой песчанки). Севернее, в полупустынных песках Иргизкум и Ту-
сум, благоприятных местообитаний значительно меньше, и здесь она населяет
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
217
Распространение быстрой ящурки
Таблица 41
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесо- степ- ная III 11 1 Оренбургская область с. Подгорное Завьялов и др., 2006
Степная VII 40 1а 2 Западно-Казахстанская область окр. оз. Сорколь ур. Аккумы Давыгора, 1995, 2001
Полупустынная XIII 84 85 3 4 5 Прихакская низменность Астраханская область окр. оз. Баскунчак пески Шигреты карьер у п. Н. Баскунчак Стрельцов и др., 2006 Животные..., 2009
XV 95 96 98 6 7 8 9 Актюбинская область окр. п. Каракуга гора Кой-Кара гряда Актолагай междуречье Эмбы и Темира Неручев, Васильев, 1978
XIX ПО 112 10 11 ур. Тусум ур. Иргизкум Динесман, 1953 Чирикова, 2004
Пустынная XX 113 114 115 116 117 118 121 12 13 14 15 16 17 18 19 20 21 Астраханская область гора Б. Богдо пески Б. Болхуны окр. п. Харабали, ур. Серте окр. ст. Досанг окр. ст. Сероглазово ур. Батпайсагыр окр. п. Красный Яр п. Приозерный п. Комсомольский Западно-Казахстанская область сев.-зап. окр. Волжско-Ураль- ских песков Рын-пески, п. Урда окр. п. Новая Казанка Гурьевская область окр. п. Новый Уштаган окр. сора Бесоба окр. п. Забурунье Чернов, 1954; Стрельцов и др., 2006; Щербак, 1974; Накаренок, 2003в; Бобров, 1985; Дунаев, Жбанова, 1998 Определитель земновод- ных..., 1977; Параскив, Бутовский, 1960 Чернов, 1954; Сараев, Пестов, 2010
218
Гпава 4
Окончание табл. 41
Зона Провин- ция Район № точки Местонахождение Источник информации
133 22 окр. оз. Индер Чернов, 1954
135 23 ур. Тайсойган окр. п. Тайсойган Неручев и др., 1984
24 окр. п. Карабау окр. м. Рахиммола ур. Акмола окр. п. Тасшагыл Чирикова, 2004; Сараев, Пестов, 2010
136 25 окр. п. Комсомольский пески Бек-Беке Неручев и др., 1984
окр. п. Доссор
137 26 гора Койкара окр. п. Аккызтогай окр. п. Аралтюбе Неручев, Васильев, 1978; Накаренок, 2003в; Сараев, Пестов, 2010
XXII окр. п. Акмечеть ур. Уитас р. Эмба, ур. Жанбике пески Тогалай окр. п. Казбек, ур. Брлик
5 X 138 28 ур. Тюлюс
X 3 окр. п. Косчагыл
В & окр. кл. Карашунгыл
1-и 29 окр. п. Аккудук Мангистауская область ур. Комек ур. Устибукай Щербак, 1974; Чирико- ва, 2004
139 30 ур. Буранкуль пески Шагырлыкум Устюрт
XXIII 144 31 окр. кол. Кугусем окр. сора Кайдак
Актюбинская область
XXV 148 151 32 33 пески Сам пески Б. Барсуки окр. ст. Челкар Неручев, Васильев, 1978; Чирикова, 2004; Лобачев и др., 1973; Динесман, 1953
154 34 Приаральские Каракумы
155 35 окр. г. Аральск
XXVI 36 сев. побережье Аральского моря
37 п-ов Куланды
38 низ. р. Сырдарья
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
219
лишь участки древнеаллювиальных песков с жузгуном и полынью с участием
ксерофильного разнотравья и злаков (Динесман, 1953). В долине р. Тургай со
своеобразным сочетанием биотипов быстрая ящурка встречается также по бе-
регам озер и в прибрежных зарослях тамарикса (Лобачев и др., 1973).
Западнее она отмечается в песках Сам (Устюрт), Прикаспийских Караку-
мах, на останце Кой-Кара и по долине р. Эмба. В прошлом была известна так-
же в окрестностях сора Кайдак; в настоящее время здесь и в сходных условиях
Приморской солончаковой равнины встречи не известны.
В северо-восточном Прикаспии она появляется лишь на верхнехвалынской
равнине, причем исключительно на умеренно сбитых участках, где формиру-
ется вторичный комплекс псаммофильных видов рептилий. Выше по р. Эмбе
встречается не только на антропогенно модифицированных участках, но и в ее
долине и междуречье с Темиром (Неручев, Васильев, 1978; Неручев и др., 1986,
1990). С пойменными и надпойменными песками связана быстрая ящурка и в
бассейне р. Уила (массив Тайсойган), где держится преимущественно на учас-
тках с полукустарниковыми и кустарниковыми зарослями. В песчаном массиве
Аккумы (верховья р. Калдыгайты) она встречается на поросших казацким мож-
жевельником окраинных участках, а у оз. Сорколь отмечена на южных склонах
песчаных бугров, заросших полынком (Давыгора, 1995). В окрестностях оз. Ин-
дер (типовая территория вида) она наблюдалась только на песчано-глинистых,
почти лишенных растительности обрывистых берегах спускающихся к озеру
оврагов и даже в непосредственной близости от воды, не избегая сильно засо-
ленных и покрытых солевыми выпотами грунтов (Чернов, 1954).
В Волжско-Уральском междуречье для быстрой ящурки также характерен
широкий набор местообитаний. У северной кромки песков в районе Камыш-
Самарских озер (окрестности п. Н. Казанка) она населяет периферийные учас-
тки, хорошо закрепленные жузгуном, тамариксом и полынью. В Нарынском
массиве по окрестностям п. Урды придерживается более или менее заросших
песчаных бугров, задернованных межбугровых котловин, встречается также
среди подлеска в сосновом лесу. Вблизи п. Н. Уштаган наблюдалась на аши-
ках. У оз. Баскунчак она отмечена в песках Шигреты, Шкили и в песчаном ка-
рьере у п. Н. Баскунчак. Довольно широкий набор местообитаний, причем на
ограниченной территории, она осваивает в окрестностях горы Б. Богдо - учас-
тки бугристых песков, лесополосы тамарикса, выращенные на супесях, песча-
ный карьер и скалы на юго-восточном склоне горы (Чернов, 1954; Параскив,
Бутовский, 1960; Стрельцов и др., 2006). Встречается также в незакрепленных
и слабо закрепленных песках у ст. Досанг и окрестностях г. Красный Яр (Щер-
бак, 1974; Бобров, 1985).
Образ жизни. Весной из зимних убежищ в регионе выходит в разных чис-
лах апреля. В это время, а затем и осенью активна в наиболее теплое время
дня. Летом активность смещается на утренние и вечерние часы, а днем она
укрывается в тени кустарников, трещинах почвы, пустотах под камнями, но-
рах животных (в которых обычно устраивает собственные отнорки) или собс-
твенных, которые представляют собой слегка наклонный ход длиной около
25 и глубиной до 10 см в наиболее сухом слое песка, устроенный у основания
жузгуна, тамарикса или других кустарников. Территориальна, но иногда две
самки или самка с молодой особью уживаются в одной норке. При совместном
обитании наблюдается конкуренция с сетчатой ящуркой. В зимнюю спячку
220
Глава 4
обычно залегает в последних числах октября, хотя иногда задерживается до
начала ноября. Период активности, таким образом, продолжается около 200, а
зимней спячки 160-170 суток.
Размножение. Половой зрелости молодые первой генерации достигают
весной следующего года, из поздних кладок, очевидно, после второй зимовки.
Начало размножения сопровождается драками между самцами и своеобраз-
ным ухаживанием за самками. Спаривание начинается в конце апреля-нача-
ле мая, кладка яиц (2, возможно, 3 раза за сезон по 2-6 шт., в краеареальных
популяциях по 2-4 шт.) происходит с конца мая до середины июля, а выход
молодых в конце июля - августе.
Питание. Основу пищевого рациона составляют мелкие беспозвоночные,
преимущественно насекомые, среди которых преобладают жуки, перепонча-
токрылые (муравьи), прямокрылые; в меньшей степени потребляются двукры-
лые, чешуекрылые, их личинки, а также другие членистоногие и мелкие яще-
рицы; изредка плоды и семена. Добычу обычно ловят на поверхности почвы,
а насекомых и на ветках растений. В свою очередь сами ящурки становятся
добычей более крупных рептилий, хищных птиц и зверей.
Численность. Среди пустынных ящериц Казахстана быстрая ящурка от-
носится к видам с низкой категорией плотности - ее средняя плотность со-
ставляет 4,6, максимальная 10,0 экз/га (Брушко, 1993). В пределах региона
в наиболее типичных местообитаниях в окрестностях Аральского моря на
1 км маршрута насчитывалось: в прибрежных дюнах 14 особей, а в песках с
кустарниками и полукустарниками - 3-8, хотя в небольших колониях песча-
нок в пересчете на 1 км маршрута этот показатель достигал 40-80. В песках
ур. Иргизкум в среднем на 1 км маршрута насчитывалось 4-11 (максимально в
сообществах жузгуна), а в ур. Тусум 12-16 ос/км (Динесман, 1953). В долине
р. Тургай ее плотность по берегам озер достигала 4,0, а в прибрежных зарослях
тамарикса - 0,5 ос/га (Лобачев и др., 1973).
В Урало-Эмбенском междуречье наиболее многочисленна быстрая ящурка
в песках Аккум-Сагиз, где плотность достигает 10,0-12,5 ос/км, и приуильских
песках Тайсойган, где по надпойменной террасе на 1 км в среднем насчитыва-
лось 2,7, в пойме - 0,3, а по долине р. Эмба лишь 0,1 ос/км. В Прикаспийских
Каракумах она определенно малочисленна - до 0,1 ос/км (Неручев и др., 1990;
Накаренок, 2003в).
В Волжско-Уральских песках по окрестностям п. Урды на 1 км маршрута
встречалось 3-4 особи (Параскив, Бутовский, 1960), а вблизи ст. Досанг - 5.
(Бобров, 1985). В других отмеченных ранее пунктах междуречья она, по на-
блюдениям С.А. Чернова (1954), в середине XX столетия не составляла ред-
кости, хотя встречалась в меньшем количестве, чем разноцветная ящурка; в
окрестностях оз. Баскунчак за последние годы наблюдается реже примерно
в 5-10 раз (Стрельцов и др., 2006), а в окрестностях ст. Досанг к 1997 г. она
полностью вытеснилась (Дунаев, Жбанова, 1998).
Лимитирующие факторы. Преимущественно разрушение местообита-
ний в процессе хозяйственной деятельности (перевыпас, бесконтрольный рост
сети дорог в пустыне, внедорожное движение транспорта).
Охрана. Обитает и охраняется на территории Устюртского и Богдинско-
Баскунчакского заповедников. На остальной территории ареала достаточно
поддержания естественного состояния ландшафтов.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 221
Региональный статус. Характерный, спорадически встречающийся в ре-
гионе вид.
Род 22. Lacerta Linnaeus, 1758 - настоящие ящерицы
34. Прыткая ящерица -Lacerta agilis Linnaeus, 1758
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода настоящих ящериц, населяющих территорию реги-
она. Политипический вид, в регионе представленный восточным подвидом
L. a. exigua Eichwald, 1831, в пределах которого довольно отчетливо прояв-
ляются межпопуляционные отличия в длине туловища и хвоста, количестве
чешуй вокруг середины тела, числе и характере расположения задненосовых и
преанальных щитков, числе бедренных пор, рисунке и окраске тела, причем по
ряду признаков изменения показателей проявляются в направлениях в общих
чертах, совпадающих с направлениями расселения вида из предполагаемого
«кавказского» центра его формирования (Прыткая ящерица, 1976; Ананьева и
др., 1998; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает большую часть Европы, Запад-
ной Сибири, Казахстана, горы восточной части Средней Азии и Прибайкалье
с примыкающими районами Монголии и Китая (средиземноморский фаунис-
тический комплекс). Его северная граница на Восточно-Европейской равнине,
Уралу и Западной Сибири проходит немногим севернее 60-й параллели, затем
постепенно снижаясь достигает примерно 52°с.ш. в Прибайкалье. Южная грани-
ца проводится от Крыма, по Северному Кавказу, северному побережью Каспия
до низовий р. Эмба, долине этой реки, Северному Устюрту, Б. Барсукам, низо-
вьям рек Иргиза и Тургая, среднему течению р. Сары-Су, северу Муюнкумов,
Прибалхашью и предгорьям среднеазиатских хребтов. Точнее в регионе и его
ближайших окрестностях она прослеживается по находкам в окрестностях горо-
дов Астрахань и Красные Яры (Прыткая ящерица, 1976; Завьялов, Табачишин,
1998), ст. Досанг (Бобров, 1985), оз. Баскунчак (Стрельцов и др., 2006), кромке
Волжско-Уральских песков у п. Урда, районе Камыш-Самарских озер, п. Кал-
мыкове, рукава Баксай, низовьях р. Урал у г. Гурьева (Чернов, 1954; Параскив,
Бутовский, 1960; Окулова, 1963; Брушко, Кубыкин, 1988), низовьях и долине
р. Эмба (Неручев, Васильев, 1978), долине р. Шатан, севернее Б. Барсуков, низо-
вьях рек Иргиза и Тургая (Динесман, 1953; Параскив, 1956; Лобачев и др., 1973).
Таким образом, прыткая ящерица населяет Заволжье, Волжско-Уральское
междуречье в основном севернее песков, Общий Сырт, Предуралье, Урало-
Эмбенское междуречье, Южный Урал, Мугоджары и зауральскую (тургай-
скую) часть региона, где отмечена в 8 (77,7% из 11) ландшафтных районах
лесостепной зоны, 40 (58,0% из 69) степных районах, 20 (62,5% из 32) районах
полупустыни и 23 (53,5% из 43) районах пустынной зоны. Ее конкретные мес-
тонахождения приводятся в табл. 42 и на карте 33.
Места обитания. Прыткая ящерица - широко распространенный эври-
топный вид, обитающий в лесной, лесостепной, степной, полупустынной и
пустынной зонах. Вместе с тем наибольшее число местообитаний в регионе
она освоила, по-видимому, в лесостепной зоне, где встречается как в лесных,
так и открытых луговых и степных сообществах.
222
Глава 4
Таблица 42
Распространение прыткой ящерицы
Зона Провин- ция Район № точки Местонахождение Источник информации
1 1 Самарская область с. В. Сускан Н. Санчелеево Бакиев, Файзулин, 2002
2 с. Хрящевка с. Выселки п. Луначарский п. Подстенки
3 п. Приморский
4 г. Тольятти, ул. Баныкина Автозаводской район:
п. Портовый п. Федоровка
5 п. Волжск
Мастрюковские озера
6 с. Царевщина
I с. Васильевка
7 п. Поволжский
cd X С 8 с. Пискалинский взвоз
ё Q ст. Задельная
О и (=: 9 ст. Мастрюково г. Самара:
Красноглинский район Ботанический сад Воронежские озера Яицкие озера
2 10 с. Елховка с. Ст. Буян, п. Висловка с. Царевщина
И п. Старосемейкино п. Водино с. Алакаевка
3 12 с. Советы
13 с. Алексеевка
п 14 с. Титовка
11 15 п. Шептала
16 с. Кошки
17 р. Сок, с. Чекалино
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
223
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
18 р. Сок, с. Лужки
19 р. Сарбай, п. Тимашево
20 г. Сергиевск, п. Суходол
21 р. Сок, с. Старое Вечканово
22 р. Шешма, с. Боровка
4 23 с. Красный Яр
5 24 д. Русский Байтуган
с. Абдул-Завод
Я X Е ё II 25 с. Совруха с. Старопохвистнево
о <и 26 г. Похвистнев
27 Оренбургская область г. Бугуруслан Вершинин, 2007; Чи-
28 окр. п. Рычковка билёв А.А.
29 окр. п. Матвеевка
30 окр. п. Абдулино
6 31 окр. п. Пономаревка
7 32 окр. п. Шарлык
III 11 33 окр. с. Подгорное Завьялов и др., 2006
Самарская область
12 34 г. Новокуйбышевск, п. Маяк Бакиев, Файзулин, 2002
35 г. Чапаевск
п. Б. Тамылово
36 п. Безенчук с. Новокиевка
37 с. Александровка с. Наталино
X Е IV с. Конуевка
£ 38 п. Приволжье с. Владимировка с. Новотулка Саратовская область
39 окр. р. М. Иргиз окр. г. Балаково Определитель земновод- ных..., 1977; Шляхтин и
13 40 др., 2005
14 41 окр. г. Энгельс
42 окр. п. Александровка
15 43 дол. р. Б. Иргиз
224
Глава 4
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
Гб 44 с. Каменный брод Бакиев, Файзулин, 2002
45 р. Чапаевка
46 с. Белозерки р. Самара, с. Съезжее с. Утеевка
17 47 с. Мосты
IV 48 п. Б. Гдушица с. Августовка с. Тамбовка Саратовская область
18 49 Повсеместно Шляхтин и др., 2005;
19 50 сс Завьялов и др., 2006
20 51 <с
Самарская область
21 52 г. Отрадный Бакиев, Файзулин, 2002
3 53 г. Кинель
X Е О' с. Бобровка
О 54 п. Красносамарское Красносамарский лес
55 п. Горский
56 с. Богатое
с. Заливное
оз. Лебяжье
V 57 с. Петровка п. Лесное Оренбургская область
22 58 Бузулукский бор п. Колтубановский р. Ток Зарудный, 1895; Чи- билбв А.А.
23 59 р. Самара
25 61 окр. р. В. Каргалка р. Янгиз окр. истока р. Самара
26 62 п. Курманаевка
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
225
Продолжение табл. 42
Зона Провин- ция Район № точки Местонах ождение Источник информации
Самарская область
63 64 с. Верхнесъезжее с. Б. Черниговка с. В. Росташи с. Украинка п. Восточный п. Кошкин Бакиев, Файзулин, 2002; Смелянский, Елизаров, 1996
V 27 65 ур. Грызлы Саратовская область окр. п. Озинки (с. Модин) Шляхтин и др., 2005; Завьялов и др., 2006
Оренбургская область
28 66 р. Сакмара, п. Октябрьский Зарудный, 1895; Чи- билёв А.А.
29 67 окр. г. Оренбург о. Мира Дебело П.В.
VI 30 68 р. Илек, п. Буранное Зарудный, 1895
§ Западно-Казахстанская область
£ 33 69 п. Барбастау Дебело П.В.
34 70 р. Утва, окр. п. Григорьевка Березовиков и др., 1992
71 п. Пугачеве
г. Аксай
35 72 р. Березовка, пр. Коншубай
VII 36 73 п. Алгабас
37 74 обочина шоссе Уральск-Ак- тюбинск Прыткая ящерица, 1976
38 75 г. Сантас, окр. оз. Шалкар Дебело П.В.
39 76 п. Джамбейты Актюбинская область Прыткая ящерица, 1976
42 77 п. Акраб
44 78 окр. г. Актюбинск
Саратовская область
48 79 окр. п. Дьяковка Шляхтин и др., 2002
VIII 80 Волгоградская область п. Палласовка с. Тараново Завьялов и др., 2006
226
Глава 4
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахожден не Источник информации
Степная VIII 52 54 55 81 82 83 84 85 86 87 88 89 90 Оренбургская область окр. п. Первомайский окр. п. Ташла п. Новоуральское Западно-Казахстанская область п. Январцево п. Красноармейск п. Рубежка п. Дарвинское г. Уральск пойма р. Урал окр. оз. Едучее дол. р. Деркул п. Донецкое п. Владимировка Чибилёв А.А. Дебело П.В.; Прыткая ящерица, 1976
IX 57 60 62 91 92 93 Оренбургская область п. Новоуралец г. Орск Актюбинская область п. Батамшинский Прыткая ящерица, 1976; Вершинин, 2007 Топоркова, 1973
X 65 66 68 72 94 95 96 97 98 Оренбургская область окр. Ириклинс кого вдхр. п. Кваркено низ. р. Орь гора Сарыоба оз. Б. Обалыколь оз. Каменное Чибилёв А.А. Давыгора и др., 2006; Дебело П.В.
XI 78 99 Кустанайская область Наурзумский заповедник Динесман, 1953; Брагин, Брагина, 2002
XII 80 100 ур. Казан-Босы оз. Сары-Копа
Полупустыня XIII 82 85 101 102 Западно-Казахстанская область п. Джаныбек Астраханская область окр. оз. Баскунчак Параскив, Бутовский, 1960
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
227
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
Запади о-Казахстанская область
87 103 окр. п. Фурманове
88 104 окр. п. Новая Казанка
105 окр. п. Фокеево
89 106 р. Кушум, окр. Кировского вдхр. Дебело П.В.
107 окр. Битикского вдхр.
91 108 окр. п. Янайкино Окулова, 1963
XIII 109 окр. п. Кожехарово
1 ПО окр. п. Чапаеве окр. п. Мергенево окр. с. Сугандык с. Ерментай с. Даутпай окр. с. Бесагаш окр. п. ЕсенсаЙ окр. п. Верблюжий завод Накаренок, 2003в
ё & с: 111 окр. п. Кулагино
XIV 92 112 р. Солянка, ур. Карабас Дебело П.В.
о С 95 113 Гурьевская область п. Миялы, 50 км к юго-вост. Актюбинская область Шилов, 1961
96 114 ур. Тубускен окр. п. Жаркам ыс ур. Жолуткуль Прыткая ящерица, 1976
97 115 окр. р. Маннисай
XV 116 окр. с. Шантас окр. с. Байсуат окр. ст. Кзылжар окр. п. Таскичу окр. ст. Тогузкен
98 117 междуречье Темира и Эмбы
99 118 ст. Караулкельды
100 119 окр. п. Жаркамыс, 10 км вост.
kvi 101 102 120 121 горы Мугоджары горы Шошкакуль Динесман, 1953; Бруш- ко, Кубыкин, 1988
228
Глава 4
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
104 122 окр. п. Карабутак
XVII 123 п. Карабутак, юж. ур. Талдыкум
XVIII 107 124 окр. г. Текели окр. оз. Джансангир
X в >> 109 ПО 125 126 низ. р. Улькояк, окр. с. Дукен низ. р. Тургай Прыткая ящерица, 1976; Накаренок, 2003в
ь & 112 127 пески Иргизкум
о С пески Бос-Тай
XIX 128 устье р. Иргиз
129 низ. р. Тургай
130 овр. оз. Мельдекуль ур. Жабысай ур. Жалаулы
Астраханская область
113 131 132 Волго-Ахтубинская пойма окр. п, Б. Богдо Чернов, 1954; Сараев, Пестов, 2010
114 133 окр. п. Сасыколи окр. п. Селитряное
134 окр. г. Красный Яр
135 окр. п. Комсомольский окр. п. Приозерный
Западно-Казахстанская область
лынная 115 136 ур. Теректы Параскив, Бутовский, 1960
XX 116 137 Рын-пески, окр. п. Урда
С 117 138 окр. Камыш-Самарских озер Гурьевская область
121 139 окр. п. Забурунье
140 окр. п. Аккистау
Западно-Казахстанская область
124 141 окр. п. Круглый
142 окр. п. Калмыкове окр. п. Харькино окр. с. Шимкудук окр. с. Косчеген
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
229
Продолжение табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
Гурьевская область
XX 125 143 144 окр. п. Горы окр. с. Копкудук Прыткая ящерица..., 1976
145 окр. с. Айгыр
127 146 окр. п. Жилая Коса окр. пром. Ботахан окр. оз. Бейбит
128 147 ур. Шубарпала
129 148 окр. с. Киралы
XXI 130 149 150 окр. с. Миндский низ. р. Баксай Брушко, Кубыкин, 1988; Сараев, Пестов, 2010
151 ур. Тайлы
132 152 ’ 153 окр. с. Яманхалинка окр. п. Сарайчик Неручев и др., 1984, 1990; Сараев, Пестов, 2010
154 окр. г. Гурьев (Атырау) окр. п. Искине, 20 км к юту
'стынная 133 134 135 155 156 157 окр. п. Индерский окр. п. Карабау, 27 км к зап. дол. р. Сагиз, ур. Кимбай Прыткая ящерица, 1976; Неручев, Васильев, 1978; Сараев, Пестов, 2010
с 158 окр. п. Карабау мог. Рахиммола
136 159 окр. п. Караколь ур. Тайсойган
160 дол. р. Эмба, ур. Дюсеке
161 ур. Кзылжар
XXII 162 окр. оз. Бартылдакты
137 163 ерик Доссор гора Аккерегешин окр. п. Кульсары Накаренок, 2003 в; Ши- лов, 1961
164 окр. Тентяксора, ур. Мол кат р. Эмба, ур. Жанбике
165 дол. р. Кайнар
166 горы Актолагай дол. р. Сагиз Мангистауская область
139 167 чинк С. Устюрта Сараев, Пестов, 2010; Неручев и др., 1984, 1990
230
Глава 4
Окончание табл. 42
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXIV 146 168 Актюбинская область горы Донгузтау Шилов, 1961
XXVI 150 151 153 155 169 170 171 172 173 окр. оз. Копа-Сор окр. г. Чел кар оз. Сасык-Коль окр. р. Жабысай окр. г. Аральск Никольский, 1905; Прыткая ящерица, 1976
В лесных сообществах северной части региона придерживается опушек,
полян, вырубок, гарей, участков редколесья, окраин галерейных, байрачных
и островных лесов, в Бузулукском бору - полян, в долинах Волги, Урала и
некоторых других рек предпочитает высокие участки прирусловых валов, не-
затапливаемые в половодье участки грив с зарослями кустарников, склоны
надпойменных террас, а также разные лесопосадки. В открытых биотопах она
отмечается практически повсеместно (за исключением заболоченных участ-
ков): в пойменных и суходольных лугах, вариантах степных сообществ и их
антропогенных модификациях - сельскохозяйственных угодьях, пустырях,
парках, кладбищах, карьерах, склонах балок и оврагов. В западной (заволж-
ско-предуральской) части степной зоны региона она населяет все лесные со-
общества (которые здесь в основном приурочены к речным долинам), заросли
степных кустарников, лесопосадки, речные долины, побережья озер и прудов,
целинную степь (кроме участков солончаков), окраины сельхозугодий, участ-
ки с пересеченной местностью, особенно получающие дополнительное увлаж-
нение (и как следствие с хорошо развитой травянистой и кустарниковой расти-
тельностью), карьеры, обочины дорог, населенные пункты и сопутствующие
им элементы инфраструктуры. Восточнее - в Наурзумском бору она встреча-
лась по лесным опушкам, березовым и осиновым колкам, балкам, берегам озер
и карасу — всюду, где есть мезофитное разнотравье. В окрестной типчаковой
степи она также населяет лишь наиболее влажные, пониженные участки рель-
ефа, прорезанные ложбинами стока с мезофитной растительностью, изредка
пырейные западинки среди черной полыни. У оз. Сары-Копа прыткая ящери-
ца была обнаружена на берегу среди луговин морского камыша, зарослей та-
марикса и на поросших пыреем бидаяках (Динесман, 1953; Брагин, Брагина,
2002). По окраинам прибрежного разнотравно-злакового пояса отмечена на
Светлинских озерах. На возвышенном Урало-Тобольском плато отмечалась
по пятнам злаково-разнотравных группировок на склоне р. Сарыоба и кромке
балки у ее подножья, занятой разнотравно-злаковыми сообществами с приме-
сью полукустарничков (Давыгора и др., 2006). В широко распространенных
здесь степных сообществах с ковылем Лессинга и на солонцах отсутствует.
Южнее в полупустыне, с ее своеобразным экотонным обликом (опреде-
ляемым мозаичностью малых форм рельефа и комплексностью почвенного и
растительного покрова), пространственное распределение ящериц существен-
но изменяется. Наиболее четко это проявляется на плакорах, в то время как в
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
231
долинах Волги, Урала, Эмбы, Б. и М. Узеней, Уила и некоторых других рек эти
изменения выражены слабее. Основными местообитаниями здесь становятся
гигромезофильные и мезофильные сообщества пойменных лугов и побережий
водоемов. На плакорах Волго-Уральского междуречья она населяет разные,
порой причудливо извивающиеся руслоподобные понижения, небольшие за-
падины, а также озеровидные депрессии (падины, лиманы), занятые лугово-
степными сообществами с разнотравьем и кустарниками. С продвижением к
югу она встречается также в злаково-полынных комплексах с кустарниками и
сопутствующим разнотравьем, а местами и на возвышениях с сообществами
черной полыни. Вместе с тем после многоснежных зим, когда большинство
депрессий длительное время залито талыми водами, часть их популяций, по-
видимому, перемещается на возвышения в сообщества черной полыни, где на-
блюдается значительно чаще (Ходашова, 1960; Животные..., 2009).
У южных пределов зоны на границе с Волжско-Уральскими песками прыт-
кая ящерица встречается лишь в речных долинах и по глинистым участкам со
злаково-полынными сообществами, причем иногда занимает очень небольшие
(0,25 га) понижения и рвы среди полыней, выделяющиеся более богатой рас-
тительностью (Окулова, 1963). В окрестностях оз. Баскунчак она отмечалась у
тростниковых зарослей на берегу пруда в Кардонной балке (Стрельцов и др.,
2006).
В Урало-Эмбенском междуречье основными местообитаниями являются
долины рек Уил, Сагиз, Кайнар, Эмба, Майнисая и ряда временных водотоков,
где она обитает по злаково-разнотравным и кустарниковым зарослям, обсох-
шим тальвегам русел и саям, на берегах прудов, по орошаемым угодьям, среди
полукустарничковой и кустарничковой растительности на кладбищах (Шилов,
1961; Неручев, Васильев, 1978). Вне речных долин она местами встречается
также по северным (с остепненным разнотравьем) склонам некоторых бугров
и останцов (Неручев и др., 1986, 1990).
В Зауралье в долине р. Иргиза прыткая ящерица отмечалась в пойме и по
склонам русла среди зарослей софоры, полыней, осок и морского камыша, в
песках Иргизкум - по берегам озер, поросших полынью, пыреем, осокой с
примесью разнотравья, в ур. Жалаулы - по склонам русла с зарослями тавол-
ги, тамарикса, пырея, а отдельные особи и среди тростника морского камыша
и ситника. В долине р. Улькояка она встречалась по склонам русла и в пойме
с зарослями тамарикса, солодки и полыни. По р. Тургай населяет пойму, а по
ложбинам стока выходит за ее пределы, где наблюдается также в небольших
западинках с более богатой мезофильной растительностью (Динесман, 1953).
В пустыне распространение прыткой ящерицы еще более ограничено.
Здесь ее местообитания почти исключительно приурочены к долинам рек.
В западной части Волжско-Уральского междуречья она отмечается в основ-
ном по берегам водоемов и их окрестностям, откуда местами проникает в ос-
тепненные межбугровые депрессии (Завьялов, Табачишин, 1998). По задерно-
ванным участкам песков они встречалась и в окрестностях п. Урды. Детали
распространения в пойме р. Урал хорошо иллюстрирует отрывок из повидо-
вого очерка этой ящерицы известной работы К.П. Параскива и П.М. Бутовско-
го (1960. С. 154): «близ Каспия населяет места с более влажными условиями
существования; лиманы, берега разливов, дельтовые луга, огороды, пониже-
ния со злаковой растительностью, пространства, заросшие тростником и часто
232
Глава 4
затопляемые водой. В последнем случае поселяется почти вплотную к урезу
воды и здесь, в сыром грунте, роет норки. Несмотря на то что вода часто затоп-
ляет тростники, ящерицы все же живут здесь».
На левобережье р. Урал она населяет в основном прирусловые участки до-
лин рек Уил, Сагиз, Кайнар, Эмба, их древние дельтовые протоки; по доли-
нам временных водотоков проникает в горный массив Донгузтау, подходит к
оз. Индер и чинкам Северного Устюрта. В сходных условиях встречается она
на западе Приаралья в Шалкарском районе, пересекаемом многочисленными
временными водотоками, заканчивающимися в бессточных озерах (Копа-Сор
и др.), а также по ложбинам северной части Б. Барсуков и плато Куланды.
Ранее в Приаралье прыткая ящерица, по-видимому, имела более широкое рас-
пространение, свидетельством чего является существование изолированного
очага вблизи г. Аральска (Прыткая ящерица, 1976).
Образ жизни. Из зимней спячки на юге региона прыткая ящерица выходит
в конце марта - начале апреля, на севере (в Волжско-Камском крае) обычно в
последних числах апреля, на Южном Урале в конце апреля - начале мая (как
правило, когда воздух прогревается до 7...9 °C) - т. е. сроки пробуждения к
северу запаздывают почти на месяц. Примерно на такое же время сроки про-
буждения могут различаться в разные годы в одном и том же районе (в Волж-
ско-Камском крае, например, от 11.04 до 13.05), что зависит от хода развития
весенних явлений и состояния погоды (Кубанцев и др., 1962; Гаранин, 1983;
Шляхтин и др., 2005; Вершинин, 2007).
Первое время после спячки ящерицы малоактивны, но по мере потепления
активность повышается и оптимума в Поволжье достигает при температуре
грунта 28,4...45,3 °C, и температуре тела 29,6...33,90 °C (Литвинов, Ганщук,
2003). Весной, а затем и осенью ящерицы активны весь день и в убежищах
скрываются только с наступлением сумерек. Летом активность смещается на
утренние и вечерние часы, а при пасмурной погоде - к середине дня. Наиболее
жаркое время года часть особей проводит в летней спячке.
Убежищем служат временные и постоянные собственные норы (наклон-
ные ходы длиной до 70 см, которые устраиваются преимущественно в легких
и средней плотности грунтах), норы других животных, пустоты и расселины
в грунте, кучах камней, строительных материалов и мусора, нагромождения
хвороста и т. п.; в этих же наиболее глубоких убежищах и зимует, забивая вход
листьями и травой.
Прыткая ящерица - территориальный вид, занимает участки площадью до
250-300 м2(в зависимости от обеспеченности кормом) с радиусом индивиду-
альной активности до 15 м, хотя иногда самцы за короткий срок перемещаются
до 200 м. Более значительные перемещения происходят при резких изменени-
ях условий обитания и расселении молодых.
В зимнюю спячку на севере региона уходит во второй половине августа -
начале сентября, иногда задерживаясь до конца месяца. Южнее это явление
сдвигается на более поздние сроки - в Саратовской области на вторую по-
ловину сентября - октябрь, в Волгоградской - на октябрь, а на сопредельной
региону территории южного Казахстана задерживается почти до середины
ноября. Таким образом, на севере региона период активности продолжается
около 110-140, а зимней спячки 220-250 суток, на юге, соответственно около
180-190 и 160-180 суток.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
233
Размножение. Половой зрелости прыткая ящерица достигает на третьем
году жизни. В этом возрасте спустя несколько дней после выхода из спячки
начинается спаривание, а с конца мая - начала июня - откладка яиц, кото-
рая продолжается и в июле. В одной кладке насчитывается до 15 яиц, причем
максимальное их количество откладывают наиболее крупные особи. Кладки
обычно располагаются в рыхлом грунте или пустотах под камнями, пнями, как
правило, на южных или восточных хорошо инсолируемых участках склонов,
с южной стороны кустов, пней, стволов деревьев и т. п. Инкубационный пе-
риод продолжается 50-55 дней и в июле-августе появляются молодые первой
генерации, в сентябре-октябре - второй. Спустя некоторое время после вылуп-
ления сеголетки расселяются на свободные участки, иногда преодолевая при
этом до 1200 м (Прыткая ящерица, 1976).
Питание. Основу пищевого рациона прыткой ящерицы составляют мас-
совые насекомые, среди которых преобладают жестко-, перепончато- и че-
шуекрылые, в меньшем количестве потребляются прямо-, дву-, полужестко-
и равнокрылые. Кроме того, в небольшом количестве потребляются черви,
брюхоногие моллюски, паукообразные и многоножки, в редких случаях по-
едаются небольшие позвоночные, в основном пресмыкающиеся (известен и
каннибализм), листья, цветки и ягоды. Наблюдаются довольно существенные
сезонные и суточные изменения рациона (в соответствии с биологическими
циклами ящериц и температурой окружающей среды), а также предпочтение
в выборе пищи молодых и взрослых особей, самцов и самок (т. е. половозрас-
тная структура популяции влияет на спектр потребляемых кормов). В урба-
низированных условиях состав кормов существенно не различается (Прыткая
ящерица, 1976; Шляхтин и др., 2006; Вершинин, 2007). В свою очередь прыт-
ких ящериц добывают более крупные рептилии, ряд хищных птиц и млекопи-
тающих, случайно амфибии.
Численность. На большей части ареала в регионе прыткая ящерица яв-
ляется довольно обычным, а местами многочисленным видом. По лесостеп-
ной зоне, как известно, ее плотность в среднем (при пересчете на 1 га) состав-
ляет 173,0, максимально 386,0 ос/га, в степной зоне соответственно 404,0 и
925,0 ос/га, а в полупустыне в среднем 256,0 ос/га (Прыткая ящерица, 1976).
Эти материалы, дополненные сведениями из разных частей региона, сущест-
венно уточняют наши представления о численности вида в ареале. Так, в до-
лине р. Утва в конце лета - начале осени 1989 г. ее плотность составляла 19,3,
а по р. Березовка (на территории Карачаганакского месторождения) 7,6 ос/га.
Летом и осенью 1990 г. здесь же в среднем насчитывалось 3,5 (3,1-3,8) и 6,2
(9,0-5,3) ос/га (Березовиков и др., 1992). На участке со слаборазвитой щеб-
нистой почвой и разреженным растительным покровом Тургайского плато
(г. Сарыоба) в июле численность ее достигала 25 ос/га (Давыгора и др., 2006).
В северной части Волжско-Уральской полупустыни в окрестностях п. Ко-
жехарово (на р. Урал) на засоленных почвах со злаково-полынными комп-
лексами на 1 км маршрута летом 1953 г. насчитывалось 2-4 особи, а на ме-
жах полезащитных полос до 20 особей. Южнее, по окраине поймы р. Урал у
п. Круглый, в этом же году ее численность достигала 5-8 особей на 100 м2 (Па-
раскив, Бутовский, 1960). В то же время вблизи п. Калмыково по глинистым
участкам со злаково-полынными сообществами на 1 км маршрута учитыва-
лось лишь 0,70, а в полынниках - 0,52 особи (Окулова, 1963).
234
Глава 4
В Урало-Эмбенском междуречье по долинам рек Уил, Сагиз и Кайнар чис-
ленность была еще ниже - соответственно 0,4, 0,6 и 0,2-0,3 ос/км (Неручев и
др., 1990). По р. Тургай на берегу озер с тростниковыми зарослями плотность
составляла 0,7 ос/га, а в прибрежных зарослях тамарикса - 1,0 (Лобачев и др.,
1973).
В зоне пустыни по северной кромке Волго-Уральских песков на задерно-
ванных участках в окрестностях п. Урды в июне 1953 г. на 1 км маршрута
отмечалось до 20 особей (Параскив, Бутовский, 1960). Южнее у г. Красный
Яр в пойме р. Волга на берегах водоемов плотность достигала 27,10±0,17, а в
межбугровых депрессиях лишь 0,70±0,31 ос/га (Завьялов, Табачишин, 1998).
На противоположной окраине междуречья вблизи г. Гурьева у п. Сарайчик в
мае 1950 г. в неглубокой канаве на 1 км отмечалось до 25-30 ящериц (Чернов,
1954).
Приведенные материалы свидетельствуют, что на большей части регио-
нального ареала состояние популяций вполне благополучное, но в некоторых
локальных группировках Урало-Эмбенского междуречья численность близка
к нижнему пределу устойчивости (что при ухудшении условий может привес-
ти к гибели).
Лимитирующие факторы. Аномальные климатические условия: ранние
заморозки (вызывающие гибель молоди), суровые зимы (гибель при промер-
зании почвы), половодье, засуха, а также полное разрушение или сильная
трансформация условий обитания в процессе хозяйственной деятельности и
рекреации; в ряде мест хозяйственное освоение территории способствует си-
нантропизации вида.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор», Оренбургского, Наурзумского и Богдинско-Баскунчакского за-
поведников, Иргиз-Тургайского резервата. На остальной территории региона
специальных мер охраны не требуется, достаточно поддержания естественно-
го состояния ландшафтов.
Региональный статус. На значительной части региона краеареальный
вид, находящийся у южных пределов распространения; включен в Приложе-
ние II Бернской конвенции.
Род 23. Zootoca Wagler, 1830 - лесные ящерицы
35. Живородящая ящерица —Zootoca vivipara Jacquin, 1787
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, обитающий на территории региона. Политипический вид,
внутривидовая таксономия которого разработана еще недостаточно. В регио-
не он представлен «восточной» хромосомной формой номинативного подвида
Z. V. vivipara Jacquin, 1787, в пределах которого существует как региональ-
ная, так и постепенная клинальная изменчивость ряда морфологических при-
знаков. Наиболее четко клинальная изменчивость проявляется в уменьшении
количества чешуй вокруг середины туловища, причем в направлении и с юга
на север, и с запада на восток. Межпопуляционные отличия заметны также
по фолидозу головы (особенно типам соединения предлобных щитков), числу
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
235
бедренных пор, рисунку и окраске тела (Орлова, 1973; Ананьева и др., 1998;
Атлас пресмыкающихся..., 2004; Куприянова и др., 2003; Кузьмин, Семенов,
2006; Вершинин, 2007).
Распространение. Ареал вида охватывает практически всю лесную полосу
Северной Евразии и сопредельные пространства тундры, лесостепной и степ-
ной зон (широко распространенный евразиатский вид). Его северная граница
на Восточно-Европейской равнине прослеживается от Кольского полуострова
к окрестностям г. Воркуты, Полярному Уралу, затем восточнее к устью р. Обь,
низовьям р. Енисей, месту слияния рек Лена и Алдан, Шантарским островам и
о. Сахалин. Южная граница от Закарпатья на восток проводится в основном по
рубежу между лесостепью и степью. Таким образом, ареал вида захватывает
только северную часть региона. Более точно в его пределах и окрестностях
южная границы ареала прослеживается по находкам у п. Пристанное Сара-
товского района одноименной области (Шляхтин и др., 1999; Завьялов и др.,
2003), полустанка Зелененький южнее г. Чапаевска Самарской области (Ша-
пошников, 1999а), затем по долине р. Сакмара в Красносамарском лесничестве
(Кирилов, 1999), у п. Заливное Богатского района и в Бузулукском бору (Рай-
ский, 1951; Бакиев, Файзулин, 2002). По долинам р. Самара она приближается
к р. Урал, в пойме которого находки известны в окрестностях г. Уральска (еще
от Кайзерлинга; цит. по: Никольский, 1905), 60 км к северу от г. Уральска
вблизи п. Красноармейское-Спартак и Облавка (Брушко, Кубыкин, 1988), у
п. Нижнеозерный, в Караваевой роще возле г. Оренбурга (Зарудный, 1895).
Восточнее в Оренбургской области граница прослеживается по долине р. Ура-
ла (захватывая и облесенные поймы его левых притоков, а также территорию
Буртинского и Айтуарского участков Оренбургского степного заповедника),
затем уже в пределах Челябинской области (Чибилёв, 1999а, б; Вершинин,
2007).
К северу до границ региона в Самарской области живородящую ящери-
цу отмечали в долине р. Сок, бассейне р. Б. Кинель у п. Похвистнево (Баки-
ев, Файзулин, 2002). В Оренбургской области помимо Бузулукского бора и
окрестностей г. Оренбурга ее встречи известны в долине р. В. Каргалка, по
уремам в долине р. Сакмара и ее притока Салмыш (Райский, 1951; Чибилёв,
1995; Вершинин, 2007). В целом по региону обитание живородящей ящерицы
выявлено в 3 (27,3% из 11) ландшафтных районах лесостепной и 11 (15,9%
из 69) районах степной зоны. Ее конкретные местонахождения приведены в
табл. 43 и на карте 34.
Места обитания. Живородящая ящерица - широко распространенный
лесной вид, придерживающийся преимущественно влажных местообита-
ний. В южной части Волжско-Камского края населяет преимущественно
широколиственные леса с полянами, вырубками и редколесьем вблизи водо-
емов. По рекам Урал, Сакмара и Салмыш селится преимущественно в тенис-
тых пойменных (уремных) местах, вблизи г. Оренбурга в Караваевой роще
наблюдалась на участках нижнего уровня луговой поймы, перемежающейся
с тополевыми и ветловыми рощами. Из поймы р. Урал проникает в облесен-
ные устьевые участки его притоков, галерейные ольшаники и сырые овраж-
но-балочные березово-осиновые заросли. В долине р. Самара отмечалась в
смешанных лесных массивах Красносамарского лесничества и Бузулукско-
го бора. В прошлом, по-видимому, имела более широкое распространение,
236
Глава 4
Таблица 43
Распространение живородящей ящерицы
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная II 3 5 7 1 2 3 Самарская область р. Сок, п. Лужки окр. п. Абдул-Завод Оренбургская область бас. р. Салмыш Бакиев, Файзулин, 2002 Чибилёв, 1995; Верши- нин, 2007
IV 12 4 Самарская область окр. п. Зелененький Шапошников, 1999а
Степная V 21 22 24 25 28 29 5 6 7 8 9 10 11 Красносамарское лесничество окр. с. Заливное Оренбургская область Бузулукский бор, окр. п. Колту- бановский р. В. Каргалка р. Сакмара р. Урал п. Нижнеозерный р. Сакмара г. Оренбург, Караваева роща Кирилов, 1999; Бакиев, Файзулин, 2002 Райский, 1951; Чиби- лёв А.А. Никольский, 1905 Зарудный, 1895; Чи- билёв, 1999а
VIII 54 55 12 13 14 Западно-Казахстанская область окр. п. Облавка дол р. Урал, окр. п. Красноар- мейск, п. Спартак дол. р. Урал у г. Уральск Березовиков и др., 1992; Брушко, Кубыкин, 1988 Никольский, 1905
IX 57 62 15 16 Оренбургская область Буртинская степь ОГПЗ Айтуарская степь ОГПЗ Чибилёв, 1999а
о чем свидетельствуют реликтовые местообитания в островных лесах вблизи
г. Чапаевска.
Образ жизни. Из зимней спячки ящерица выходит в конце апреля - нача-
ле мая. Первоначально, а затем в пасмурную прохладную погоду наибольшая
активность проявляется при прояснении. Максимальная активность наблю-
дается при температуре субстрата 20,6...28,7 °C, а с повышением несколько
снижается (Литвинов, Ганщук, 2003). Территориальна, обычно занимает учас-
ток площадью 8-10 м2. В его пределах, как правило, держится у поваленных
стволов и старых пней, где находится основное убежище или несколько вре-
менных укрытий. В их качестве используются норы грызунов, межкорневые
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
237
пустоты, пространства за отставшей корой на пнях и сухих деревьях, дупла,
трещины в грунте и т. п. Поселяясь вблизи человеческого жилья, укрывается
в кучах бревен, древесного мусора, заброшенных строениях. При опасности
может прятаться и в водоемах. В спячку уходит в конце августа - сентябре,
сначала самки, а затем сеголетки с самцами. Таким образом, период активнос-
ти продолжается около 150—160, а зимней спячки - около 200 суток.
Размножение. Половой зрелости достигает через 2 года. В этом возрасте
вскоре после выхода из зимней спячки начинается брачный период, сопровож-
дающийся драками между самцами. В регионе ящерица яйцеживородяща и оп-
лодотворенные яйца (6-12) задерживаются в организме самки на 70-90 дней.
Молодые обычно появляются во второй половине июля - начале августа и до
ухода на зимовку держатся группами (выводками).
Питание. Основу пищевого рациона вида составляют «мягкие» корма:
паукообразные, дождевые черви, двукрылые, жуки, чешуекрылые и их личин-
ки; несколько меньше потребляется прямокрылых, перепончатокрылых, мно-
гоножек, моллюсков, клещей и растительной пищи. Пища добывается как на
земле, так и на стволах и ветках деревьев (на высоте до 3 м). В свою очередь
живородка служит объектом добычи более крупных рептилий, хищных птиц
и млекопитающих.
Численность. В целом по региону живородящая ящерица чаще всего ха-
рактеризуется как малочисленный, «изредка» встречающийся вид. Однако на
облесенных территориях Богатовского, Борского, Камышлинского, Кошкин-
ского, Сергиевского и Шенталинского районов Самарской области на гектар
без учета сеголеток численность местами достигает 5-10 особей (Бакиев, Фай-
зулин, 2001). Конкретные данные о численности на единицу площади или мар-
шрута на остальной территории региона отсутствуют, хотя, судя по масшта-
бам отлова для исследований и коллекционирования, в ряде мест (особенно по
пойме р. Урала) она еще сравнительно обычна. На сопредельной территории
Башкирского заповедника ее численность в среднем составляет 0,36 ос/10 км,
в том числе по пойменным уремам 0,5, а в смешанных лесах 0,08 ос/10 км
(Шошева, 1985).
Лимитирующие факторы. Комплекс абиотических (аридизация) и био-
тических факторов, действующих на периферии ареала, разрушение и транс-
формация местообитаний при хозяйственном и рекреационном освоении тер-
ритории.
Охрана. Обитает и охраняется на территории национального парка «Бу-
зулукский бор» и Оренбургского заповедника. На остальной территории спе-
циальных мер охраны не требуется, достаточно поддержания естественного
состояния ландшафтов.
Региональный статус. Вид у южной границы ареала в специфических
местообитаниях. Занесена в Красные книги Саратовской (2006) и Самарской
(2009) областей.
238
Глава 4
Отряд 5. SERPENTES LINNAEUS, 1758 - ЗМЕИ
Семейство 15. Boidae, Gray, 1825 - Удавы, или Ложноногие
Род 24. Eryx Daudin, 1803 - удавчики
36. Песчаный удавчик - Eryx miliaris (Pallas, 1773)
Значение таксона для сохранения биоразнообразия. В соответствии
с принятой нами системой классификации рептилий (Ананьева и др., 1998;
Атлас пресмыкающихся..., 2004) на территории региона обитает 2 вида удав-
чиков - песчаный и восточный. Песчаный удавчик политипический вид, в ре-
гионе представленный номинативным подвидом Е. т. miliaris (Pallas, 1773),
отличающимся в среднем большим числом брюшных (165-192) и подхвосто-
вых (19-40 пар) щитков. Четких межпопуляционных различий внутри подвида
не выявлено - установлено лишь наличие 2 типов окраски тела: типичной и
меланистической (Определитель земноводных..., 1977; Сараев, Пестов, 2010).
Распространение. Ареал вида охватывает восточное Предкавказье, низо-
вья р. Волги, юго-западные части Казахстана и Средней Азии, а также северные
районы Ирана и Афганистана (среднеазиатский фаунистический комплекс).
В регионе северных пределов распространения песчаный удавчик, по-ви-
димому, достигает на юго-востоке Волгоградской области (Кубанцев и др.,
1962), поскольку сведения о его якобы единичных встречах на юге Саратовс-
кого левобережья (Шляхтин, Голикова, 1986; Шляхтин, 1996) позднее досто-
верными находками не подтверждены (Табачишина и др., 2004; Завьялов и
др., 2005). Восточнее его распространение определяется встречами по север-
ной кромке Волжско-Уральских песков вблизи п. Урда, островных песках в
районе Камыш-Самарских озер (Параскив, Бутовский, 1960), п. Джантемир на
широте п. Калмыкове (Окулова, 1963), п. Елтая, Индерского озера (Чернов,
1954), низовьях р. Сагиз на широте п. Макат (Неручев и др., 1984), у гряды
Актолагай (Накаренок, 2002), горы Токсанбай (Сараев, Пестов, 2010), на Се-
верном Устюрте в песках Сам (Дуйсебаева, 2005), у северо-западного побе-
режья Аральского моря, южной части Приаральских Каракумов, окрестностях
г. Аральска и в песках Муюнкумы; Южная граница ареала проводится через
северные районы Ирана и Афганистана и далее по южной кромке Каракумов и
Кызылкумов к Муюнкумам (Определитель земноводных..., 1977).
Таким образом, ареал вида охватывает только южную часть региона.
В очерченных пределах песчаный удавчик населяет Волжско-Уральское меж-
дуречье, область низовий рек Сагиза и Эмбы, Прикаспийские Каракумы, пес-
ки Северного Устюрта, Мангышлака, северного побережья Аральского моря
и Приаральские Каракумы. Его конкретные местонахождения известны в
4 (12,5% из 32) ландшафтных районах полупустынной и 17 (39,5% из 43) пус-
тынной зоны (табл. 44, карта 35)
Места обитания. Песчаный удавчик в центральной части ареала типич-
ный псаммофил, а в регионе населяет не только разные типы песков, но и учас-
тки с рыхлым супесчаным грунтами, а местами и глинистым.
По западной кромке Волжско-Уральских песков в пределах АГКМ населя-
ет как бугристые барханные пески, так и закрепленные бугристые. На задер-
нованных песках он отмечался также в окрестностях п. Урда, а в центральной
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
239
Таблица 44
Распространение песчаного удавчика
Зона Провин- ция Район № точки Местонахождение Источник информации
Волгоградская область
5 85 1 Юго-восточные р-ны Кубанцев и др., 1962
X X 3 в XIII Западно-Казахстанская область
ё? § 88 2 Прикамыш-Самарские ост- ровные пески Параскив, Бутовский, 1960; Окулова, 1963
С 90 3 окр. п. Джантемир
XV 96 4 горы Актолагай Накаренок, 2002
Астраханская область
114 5 окр. п. Лапас ст. Досанг - с. Замьяны Бобров, 1985; Завьялов, Табачишин, 1998
6 окр. г. Красный Яр, АГКМ окр. п. Комсомольский
Западно-Казахстанская область
116 7 Рын-пески: окр. п. Урда ур. Бармак Гурьевская область Параскив, Бутовский, 1960
гынная XX 118 120 8 9 ур. Бес-Чохо ур. Майтюбе окр. п. Кукурте Сараев, Пестов, 2010; Кубыкин, Брушко, 1988; Чернов, 1954; Окулова, 1963
Б С 122 10 окр. п. Басчикан ур. Сары-Иван ур. Салыкмула
11 окр. п. Елтай
123 12 окр. п. Зеленое, 25-30 км зап.
Западно-Казахстанская область
124 13 окр. п. Калмыкове, ур. Камбакты Чернов, 1954; Накаре- нок, 2002, 2003 в
Гурьевская область
XXI 127 14 15 ур. Аккуль ур. Каратон Неручев, Васильев, 1978; Никольский, 1905
132 16 окр. п. Новобогатинский окр. г. Гурьев (Атырау)
240
Глава 4
Окончание табл. 44
Зона Провин- ция Район № точки Местонахождение Источник информации
133 136 17 18 окр. п. Индерборский ур. Караша ур. Кзыл-Жар Чернов, 1954; Накаре- нок, 2002
дол. р. Эмба, 10 км к сев. п. Кульсары
окр. п. Кульсары
19 окр. п. Комсомольский ур. Мадиар Щербак, 1974; Неручев и др., 1984
20 ур. Брлик ур. Сагиз ур. Испулай ур. Бек-Беке
XXII 137 21 гора Кой-Кара гора Иман-Кара гора Аккерегешин ур. Жусалы ур. Аралтюбе ур. Уялы (Акмечеть) ур. Уитас Накаренок, 2003 в; Неру- чев и др., 1984; Неручев, Васильев, 1978; Нака- ренок, 2003в; Сараев, Пестов, 2010
22 ур. Намазтакыр ур. Жанбике
138 23 окр. п. Мунайлы ур. Каратон ур. Ушкан Неручев, Васильев, 1978; Киреев, 1981
24 горы Жельтау ур. Жанасу Мангистауская область
139 25 гора Токсанбай Сараев, Пестов, 2010;
XXV 148 26 пески Сам, Устюрт Дуйсебаева, 2005; Лоба- чев и др., 1973
154 27 Приаральские Каракумы
155 28 сев. берег Аральского моря
XXVI 29 ур. Каскаджол сев.-зап. берег, ур. Тас-Тюбе
30 окр. г. Аральск Параскив, 1956
31 низ. р. Сырдарьи Динесман, 1953
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
241
части песков встречался на участках с очагами сбоев. На северо-восточной ок-
раине массива придерживается преимущественно песчаных островов (часто с
колониями песчанок), а вблизи п. Джантемир заходит и в глинистую полынную
пустыню (с поселениями малого суслика). На юго-востоке Волжско-Уральско-
го междуречья в окрестностях п. Новобогатинского он чаще всего придержи-
вается барханных песков на границе с полынниками (где обычно охотится).
В этом же районе, в окрестностях ст. Первомайская, песчаные удавчики жи-
вут на рыхлых с поверхности песчанистых почвах в полынно-солянковой пус-
тыне между сорами. Следует напомнить, что именно эти места на побережье
Каспийского моря вблизи г. Гурьева (откуда песчаный удавчик впервые был
описан П.С. Палласом и где он позднее отмечался Г.С. Карелиным), признаны
типовой территорией вида.
В окрестностях Индерского озера он, по-видимому, обитает в пустынно-
солянковых стациях, а не в песках, расположенных на правом берегу р. Урал
в 12 км от его поймы. По долине р. Эмба и ее надпойменным террасам встре-
чается как в песках, так и среди супесчаной и глинистой полынно-солянковой
пустыни. На Устюрте единственным местом его обитания является песчаный
массив Сам. На участках с древними аллювиальными песками он отмечается
также и на побережье Аральского моря.
Образ жизни. У южной границы региона из зимних убежищ выходит в
марте, на севере - со второй декады апреля. Весной и осенью активен в днев-
ные часы, летом - в сумерки и ночью, проползая за это время свыше 0,5 км.
По наблюдениям в Волжско-Уральском междуречье за 1,5-2,0 часа до наступ-
ления темноты из песков он направляется к ближайшим полынникам (где пре-
имущественно и ловит добычу), а утром возвращается в барханы или укрыва-
ется в норах песчанок или малых сусликов. Норы грызунов использует и как
зимние убежища, в которых на севере ареала укрывается уже в сентябре, на
юге - в октябре. Период активности, таким образом, продолжается 200-180, а
зимней спячки 180-160 суток.
Размножение. Половой зрелости песчаные удавчики достигают на треть-
ем-четвертом году жизни. Судя по состоянию гонад, самцы, добытые в Волж-
ско-Уральском междуречье в конце мая - начале июня, по-видимому, уже при-
нимали участие в размножении. Молодые (8-11) на сопредельной территории
Мангышлака отмечались со второй пятидневки июля, в июле они появляются
и у северных пределов ареала в Волгоградской области; в то же время у самки,
добытой 21 августа на западном побережье Аральского моря, в яйцеводах еще
находились яйца (Чернов, 1954; Параскив, 1956; Кубанцев и др., 1962).
Питание. Основу пищевого рациона молодых составляют насекомые и
мелкие ящерицы. Взрослые добывают грызунов (в том числе песчанок, сусли-
ков), ящериц (разноцветных ящурок и др.) и мелких птиц.
Численность. Судя по количеству мест регистрации, песчаный удавчик
наиболее обычен в Приэмбенской провинции (Сараев, Пестов, 2010). У за-
падной окраины Волжско-Уральского междуречья в закрепленных песках на
территории АГКМ его плотность достигает 2,40±0,21 ос/га, а среди бугрис-
тых песков барханного типа - 1,10±0,06 (Завьялов, Табачишин, 1998). Немно-
гим севернее в окрестностях ст. Досанг его численность в песках составляла
1,0 экз/км (Бобров, 1985), а в начале 1990-х гг. - 0,3 (Божанский, Полыно-
ва, 1998). У северной окраины Волжско-Уральских песков в окрестностях
242
Глава 4
п. Урды и Новой Казанки он определенно редок (Параскив, Бутовский, 1960).
По северо-восточной кромке этого массива на широте п. Калмыкове - Харь-
кино в злаково-полынной глинистой и злаково-полынной песчаной пустыне
в сочетании с бугристыми песками его численность достигала 0,2 ос/10 км
(Окулова, 1963). Южнее он становится более обычным (Чернов, 1954), а на
пустынных пастбищах с очагами сбоев его численность достигает 2,0 ос/10 км
(Неручев и др., 1981). По наблюдениям последних лет на основной части аре-
ала вида в регионе в Гурьевской области песчаный удавчик может быть встре-
чен практически повсеместно, однако численность его везде невелика (Сараев,
Пестов, 2010). В Прикаспийских Каракумах плотность достигает 1,0 ос/10 км
(Накаренок, 2003 в).
Лимитирующие факторы. По-видимому, в основном комплекс абиоти-
ческих и биотических факторов, действующий на границе ареала. Хозяйствен-
ное освоение территории.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
Занесен в Красную книгу Астраханской области (2004). На остальной террито-
рии состояние популяций проведения специальных мер охраны не требует.
Региональный статус. Характерный малочисленный представитель гер-
петофауны региона у северных пределов распространения.
37. Восточный удавчик - Eryx tataricus (Lichtenstein, 1823)
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей рода, обитающий на территории региона. Политипический вид,
представленный в регионе номинативным подвидом Е. I. tataricus (Lichten-
stein, 1823) (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004). Вместе с
тем, по мнению А.А. Токаря (1990, цит. по: Дуйсебаева, 2005), он представляет
собой лишь одну из вариаций песчаного удавчика.
Распространение. Ареал вида охватывает юг Казахстана, большую часть
равнинной Средней Азии, север Ирана, Афганистана, Пакистана, Индии, запад
Китая и юг Монголии (среднеазиатский фаунистический комплекс). Его север-
ная граница еще недавно проводилась от дельты р. Урал примерно через район
излучины р. Эмба в окрестностях п. Кульсары (Неручев, Васильев, 1978; Неру-
чев и др., 1990), Северный Устюрт, Иргиз-Тургайское междуречье, окрестнос-
ти г. Моинты, оз. Алаколь и далее в Китае (Параскив, 1956). Переопределение
экземпляров, найденных в Северном Прикаспии, и тщательное изучение новых
находок показали, что самой западной точкой обитания восточного удавчика
в регионе является урочище Озектык, расположенное между сорами Донге-
лек и Косбулак на Северном Устюрте (Дуйсебаева, 2005). Поскольку система-
тическая принадлежность удавчиков из Приаральских Каракумов и низовий
р. Тургай не подвергалась ревизии, то восточнее северная граница ареала в
регионе проводится к низовьям р. Тургай, где «вероятно, именно этот вид был
обнаружен П.П. Сушкиным» (Динесман, 1953). Таким образом, ареал вида за-
хватывает только самую южную часть исследуемого нами региона. В очерчен-
ных пределах обитание восточного удавчика в окрестностях Аральского моря
(принятых за типовую территорию вида), вблизи г. Аральска и в ур. Ильля-
Кудук (Динесман, 1953) косвенно подтверждается новейшей находкой вблизи
этих пунктов в дельте р. Сырдарьи (Дуйсебаева, 2005). Систематическая при-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
243
Распространние восточного удавчика
Таблица 45
Зона Провин- ция Район № точки Местонахождение Источник информации
Полу- пус- тынная XIX 112 1 Актюбинская область низовья р. Тургай Динесман, 1953
XXV 147 2 ур. Озектык Дуйсебаева, 2005
154 3 Приаральские Каракумы Лобачев и др., 1973
Пус- тынная XXVI 155 4 5 сев. берег Аральского моря окр. г. Аральска Определитель земновод- ных..., 1977
6 ур. Ильля-Кудук Динесман, 1953
7 низ. р. Сырдарья Дуйсебаева, 2005
надлежность удавчиков из Приаральских Каракумов (Лобачев и др., 1973) и
низовий р. Тургай к восточному виду пока не подвергается сомнению.
Таким образом, восточный удавчик в пределах региона известен в одном
(3,1% из 32) ландшафтном районе полупустынной и, по крайней мере, 3 (7,0%
из 43) районах пустынной зоны. Его конкретные местонахождения отражены
в табл. 45 и на карте 36.
Места обитания. В Казахстане населяет «полынные степи», глинистые,
лессовые и песчаные пустыни. В окрестностях г. Аральска отмечен на песках,
поросших полынью, песчаной осочкой и молочком, а в ур. Ильля-Кудук - на
песках с жузгуном, тамариксом, полынью и пыреем (Динесман, 1953).
Образ жизни. На сопредельной территории активен с первой декады мая
до середины сентября. Взрослые обычно охотятся ночью (проползая до 1 км),
но при пасмурной погоде встречаются и днем, хотя чаще всего это время про-
водят укрывшись в норах или прикорневых пустотах; в подобных, но более
глубоких укрытиях они и зимуют. Молодые активны преимущественно утром
и вечером.
Размножение. Восточный удавчик - яйцеживородящий вид. Молодые
(5-34) появляются в июне-июле или в августе - начале сентября.
Питание. Взрослые животные питаются преимущественно ящерицами и
грызунами, иногда добывают мелких птиц. Молодые особи в условиях Казах-
стана питаются в основном чернотелками и саранчевыми.
Численность. В центральной части ареала является сравнительно обыч-
ным (Ананьева и др., 1998), а в Приаральских Каракумах - «массовым» видом
(Лобачев и др., 1973).
Лимитирующие факторы. Комплекс факторов, действующих на границе
ареала; хозяйственное освоение территории.
Охрана. Обитает и охраняется на территории Устюртского заповедника (?).
Региональный статус. Характерный представитель герпетофауны с огра-
ниченным распространением у северных пределов ареала.
244
Гпава 4
Семейство 16. Colubridae Oppel, 1811 -Ужеобразные
Род 25. Coluber Linnaeus, 1758 - стройные полозы
38. Поперечнополосатый полоз Coluber karelinii Brandt, 1838
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей обширного рода, населяющий территорию региона. Политипичес-
кий вид, в регионе представленный номинативным подвидом Е. к. karelinii
Brandt, 1838.
Распространение. Ареал вида охватывает юг Казахстана, большую часть
равнинной Средней Азии, а также северные районы Ирана, Афганистана, Па-
кистана и Индии (среднеазиатский фаунистический комплекс). Его северная
граница проводится от побережья Каспийского моря у залива Мертвый Кул-
тук (в окрестностях бывшего укрепления Ново-Александровска, где этот по-
лоз добывался А. Леманом) к южной оконечности Приаральских Каракумов и
северу Муюнкумов. В пределах региона она определяется отмеченными выше
находками у Мертвого Култука (Параскив, 1956) и на юге Приаральских Кара-
кумов (Лобачев и др., 1973), которые являются единственными известными к
настоящему времени пунктами обитания вида (карта 37).
Места обитания. В Казахстане обитает на задернованных песках, меж-
бугровых впадинах, у границы региона на Мангышлаке отмечался в долине
пересохшей речки Агашты на мелком галечнике среди редких кустиков чия,
полыни и терескена (Параскив, 1956).
Образ жизни. На севере ареала активен со второй половины апреля по
ноябрь (около 200 суток). Весной и осенью максимальная активность проявля-
ется в дневные часы, летом ведет преимущественно сумеречно-ночной образ
жизни, днем укрываясь в трещинах, промоинах почвы и норах разных живот-
ных. При пасмурной погоде может быть встречен в любое время суток.
Размножение. Откладка яиц (3-9) в ценральной части ареала наблюдается
во второй половине июня, молодые появляются в августе.
Питание. Основу пищевого рациона составляют пресмыкающиеся, пре-
имущественно ящерицы, реже потребляются грызуны и мелкие птицы.
Численность. В Казахстане встречается редко.
Лимитирующие факторы. Комплекс факторов у северных пределов рас-
простанения.
Охрана. Специальные меры не предпринимались.
Рациональный статус. Малочисленный краеареальный вид с ограничен-
ным распространением.
39. Разноцветный полоз - Coluber revergieri Menetries, 1832
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей обширного рода, населяющий территорию региона. Политипичес-
кий вид, представленный в регионе номинативным подвидом С. г. revergieri
Mentries, 1832.
Распространение. Ареал вида охватывает восточное Закавказье, Перед-
нюю и Среднюю Азию на восток до северо-западного Китая, юг Казахстана
(среднеазиатский фаунистический комплекс). Его северная граница от Каспия
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
245
проводится по северо-западному побережью Аральского моря и далее через
Прибалхашье и окрестности оз. Алаколь в Китай. Точнее в пределах регио-
на она определяется находками на п-ове Бузачи, откуда этот полоз доставлен
Г.С. Карелиным (коллекция АН), в северо-западной части Аральского моря он
добывался В.Д. Аленициным (Сидоров, 1925), а затем Г. Никольским, Сати-
ным и В.С. Бажановым (Параскив, 1956); С.А. Сидоров (1925) указывал также,
что этот полоз иногда попадался «на правом берегу Сырдарьи, в ее низовьях
и на оз. Камышлыбаш», но позднее его здесь не находили. Эти пункты и по
настоящее время являются единственными известными местонахождениями
вида в регионе (карта 38).
Места обитания. В Казахстане населяет участки глинистой и щебнистой
пустыни, задернованные и закрепленные пески с редкими кустами терескена
и тамарикса.
Образ жизни. Активен с начала апреля до октября-ноября (около 190—
200 суток), причем утром и вечером. В качестве убежищ использует трещины
и пустоты в грунте, под камнями, а также норы разных животных.
Размножение. В июне-июле откладывает 5-10 яиц, из которых в конце
августа - начале сентября появляются молодые.
Питание. Основу пищевого рациона составляют мелкие позвоночные:
ящерицы, грызуны, птицы.
Численность. В Казахстане повсюду сравнительно редкая змея.
Лимитирующие факторы. Комплекс факторов на границе ареала.
Охрана. Обитает и охраняется на территории Устюртского заповедника (?).
На остальной территории специальные меры охраны не предпринимались.
Региональный статус. Малочисленный краеареальный вид у северных
пределов ареала с изолированными очагами распространения.
Род 26. Coronella Laurenti, 1768 - медянки
40. Обыкновенная медянка - Coronella austriaca (Laurenti, 1768)
Значение таксона для сохранения биоразнообразия. Единственный
представитель рода, населяющий территорию региона. Политипический вид,
в регионе представленный номинативным подвидом С. a. austriaca (Laurenti,
1768). Подвидовая структура изучена недостаточно, но во всех описаниях от-
мечается изменчивость окраски и рисунка тела (Ананьева и др., 1998), а также
количества брюшных щитков и отношения длины туловища к длине хвоста
(Табачишин и др., 1996).
Распространение. Ареал вида охватывает большую часть Европы (на се-
вер до 62° с.ш.), Кавказа, Южный и Средний Урал, юг западной Сибири и со-
предельную территорию Казахстана (европейский фаунистический комплекс).
Это один из наиболее северных видов пресмыкающихся. На Восточно-Евро-
пейской равнине его северная граница от низовий р. Западная Двина просле-
живается в окрестностях оз. Ильмень, г. Кострома, по северу Нижегородской
области, 58-й параллели на Урале и далее к р. Тобол. В Сибири долиной этой
реки достигает окраин Восточно-Уральского поднятия, а затем Мугоджар в
районе правых притоков р. Иргиз и верховий р. Эмба. Южная граница от вое-
246
Глава 4
точного Предкавказья, обходя Черные Земли, достигает р. Волга, по долине
которой поднимается до Самарской Луки (возможно обитание в устьевых
участках левобережных притоков), затем прослеживается по нижнему тече-
нию р. Самары до Красносамарского леса, верховьям р. Б. Иргиз и долиной
р. Илек подходит к Мугоджарам.
В Волжско-Уральском междуречье и Предуралье по вековым дубравам
Общего Сырта (описанным Э.А. Эверсманом и Г.С. Карелиным) и пойменным
дубово-вязовым лесам по рекам Б. Иргиз, Еруслан, Б. и М. Узени, Подураль-
скому плато медянка, по-видимому, еще недавно распространялась почти до
Прикаспийской низменности, что косвенно подтверждается наличием изоли-
рованных (реликтовых) очагов в верховьях рек Б. Иргиза и Эмбы. В настоящее
время ареал медянки охватывает только северную часть исследуемого нами
региона. Здесь ее обитание известно в 6 (54,5% из 11) ландшафтных районах
лесостепной, 8 (11,6% из 69) районах степной и 2 (6,2% из 32) полупустынной
зон (табл. 46, карта 39).
Места обитания. Населяет преимущественно лесные сообщества. В Са-
марской области обитает в лиственных и смешанных лесах, где предпочитает
разреженные, хорошо прогреваемые участки, поляны, опушки, вырубки, про-
секи, обочины дорог, притеррасные участки поймы (Бакиев и др., 2004, 2009).
Подобных мест придерживается и в лесных массивах на севере Оренбургской
области, а в Бузулукском бору отмечалась на старых, захламленных порубоч-
ными остатками вырубках и в зарослях степных кустарников, где больше все-
го мелких грызунов (Даркшевич, 1953; Чибилёв, 1995, 1998). В окрестностях
г. Оренбурга ее встречи известны в поймах рек Сакмара, В. и Н. Каргалки,
а в Большечерниговском районе - на заросших кустарниками склонах балок.
В казахстанской части ареала становится более гигрофильной и помимо лес-
ных и кустарниковых зарослей встречается на окраинах влажных лугов и забо-
лоченных участков (Параскив, 1956).
Образ жизни. После зимовки на юге Волжско-Камского края появляется
во второй декаде апреля - мае. Это вид преимущественно дневной (10-19 ча-
сов) активностью, но при жаркой погоде встречается и в ночные часы. Обычно
это время проводит в укрытиях: норах, пустотах в грунте, под камнями или
упавшими деревьями. Территориальна, чаще всего занимает участок радиу-
сом 60-80 м, хотя изредка при поисках корма мигрирует и на более значи-
тельные расстояния. Довольно мирная змея, но защищаясь становится агрес-
сивной. На зимовку уходит во второй половине сентября - начале октября.
Период активности, таким образом, продолжается около 150 суток, а зимней
спячки 200.
Размножение. Половозрелой медянка становится на третьем-четвертом
году жизни. В этом возрасте вскоре после выхода из зимовки начинается спа-
ривание, а спустя 70-80 суток - в июле-августе появляются 2-15 детенышей; в
условиях неволи возможны 2 приплода в год.
Питание. Основу пищевого рациона составляют разные ящерицы (ме-
дянка преимущественно заурофаг), мышевидные и насекомоядные. В Бузу-
лукском бору потребляются крупные жуки - майский жук, июньский хрущ
и другие вредители сосны. В редких случаях отлавливаются иные пресмыка-
ющиеся, амфибии, грызуны, известны случаи каннибализма. В свою очередь
медянка является добычей ряда хищных птиц и млекопитающих.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
247
Таблица 46
Распространение обыкновенной медянки
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная I 1 2 1 2 3 4 5 Самарская область окр. рыбхоза «Сускан» г. Тольятти: лесопарковая зона Центральный р-н окр. с. Федоровка, Писка- линский взвоз г. Самара, п. Управленчес- кий с. Старая Бинарадка Бакиев и др., 2001 Бакиев, Иванова, 2004 Бакиев и др., 2009
II 4 5 6 6 7 8 9 Оренбургская область окр. с. Рычковка Самарская область окр. с. Старопохвистнево Оренбургская область с. Покровка верх. р. Дёма Чибилёв, 1995 Бакиев и др., 2001 Чибилёв, 1995, 1998
III 8 10 хр. М. Накас
Степная IV 17 11 Самарская область с. В. Росташи Бакиев и др., 2002,2009; Бакиев, Маленев, 1999 Даркшевич, 1953 Никольский, 1905 Зарудный, 1985; Райс- кий, 1951
V 2! 22 24 29 12 13 14 15 16 Красносамарский лес Бузулукский бор, Борский Р-н Оренбургская область Бузулукский бор дол. р. В. Каргалка дол. р. Н. Каргалка окр. г. Оренбург, пойма р. Сакмара
VI 32 17 дол. р. Илек
VII 44 18 Актюбинская область окр. г. Актюбинск, с. Родни- ковое Параскив, 1956
IX 64 19 хр. Мугоджары
Полупус- тынная XVI 101 20 хр. Мугоджары дол. р. Кундуз Дубровский, 1967
XVIII 106 21 ур. Кайракты ур. Бала-Талдык
248
Глава 4
Численность. В целом по региону медянка немногочисленный вид. На
территории Оренбургской области по ориентировочной оценке ее численность
не превышает 100—150 особей (Чибилёв, 1998). В Самарской области наиболее
высокая численность отмечена в Красносамарском лесничестве, где достигает
2 экз/га (Бакиев и др., 1996,2004, 2009). По мнению ряда исследователей, бла-
годаря скрытому образу жизни показатели ее численности, как правило, везде
занижены.
Лимитирующие факторы. Комплекс факторов на границе ареала, сведе-
ние лесов, преследование и уничтожение из-за ошибочного представления о
ядовитости.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной территории региона специальных мер не предпри-
нималось.
Региональный статус. Малочисленный краеареальный вид с ограничен-
ным распространенном у южных пределов ареала. Занесена в Красную книгу
Оренбургской (1998), Самарской (2009), смежной Челябинской области, Баш-
кортостана и Татарстана; охраняется Бернской конвенцией.
Род 27. Elaphe Fitzinger, 1833 - лазающие полозы
41. Узорчатый полоз -Elaphe dione (Pallas, 1773)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Внутривидовая
структура изучена недостаточно, но в свете последних данных (Смирнова и
др., 2003) его, по-видимому, следует признать политипическим видом, особи
западной (до Алтая) части ареала которого относятся к номинативному подви-
ду Е. d, dione (Pallas, 1773). В его пределах отмечается высокая изменчивость
окраски и рисунка тела, кроме того, в направлении с севера на юго-восток на-
блюдаются некоторое увеличение числа брюшных щитков и уменьшение под-
хвостовых; других, значимых в подвидовой систематике отличий, не выявлено
(Табачишин и др., 1996; Завьялов, Табачишин, 2000).
Распространение. Ареал вида от левобережья Украины через Поволжье,
Кавказ, Иран, Среднюю Азию, Казахстан, юг Сибири и Монголию простира-
ется до Приморья, Кореи и Северного Китая (восточно-палеарктический фау-
нистический комплекс). Его северная граница в ближайших окрестностях ре-
гиона прослеживается по территории Ульяновской (г. Дмитровград) области,
Башкортостана (г. Мелеуз, хр. Ирендык), югу Челябинской области (Бакиев
и др., 2004, 2009; Хабибулин, 2004; Чибилёв, 2005), далее по Казахстану к
окрестностям г. Семипалатинска (Параскив, 1956) и югу Сибири к низовьям
р. Амур (Определитель земноводных..., 1977). Южная граница от Восточной
Украины следует через Закавказье, север Ирана и Среднюю Азию в Северный
Китай. Таким образом, тарритория региона полностью оказывается в пределах
ареала вида.
В пределах региона узорчатый полоз встречается на всем протяжении По-
волжья, большей части Волжско-Уральского междуречья, в Предуралье, При-
эмбенской провинции, Северо-Восточном Прикаспии, Северном Устюрте,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
249
Мугоджарах, по Тургаю и в Приаралье. В целом его пребывание установлено
в 3 (27,3% из 11) ландшафтных районах лесостепной, 18 (26,1 % из 69) степной,
13 (40,6% из 32) полупустынной и 19 (44,2% из 43) пустынной зон. Конкрет-
ные местообитания приведены в табл. 47 и на карте 40.
Таблица 47
Распространение узорчатого полоза
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная I 1 1 2 Самарская область г. Тольятти, лесопарковая зона г. Самара, Студеный овраг, Лысая гора Бакиев и др., 2004, 2009; Бакиев, Магдеев, 1995
II 7 3 Оренбургская область Тюльганский р-н Чибилёв, 1995
III 10 4 хр. Шайтантау
IV 13 15 5 7 Саратовская область окр. г. Балаково Пугачевский р-н Табачишина и др., 2005
Степная V 24 25 26 27 29 8 9 10 11 6 12 Оренбургская область Общий Сырт: верх. р. Самара верх р. Каргалка ур. Котлы низ. р. Погромка верх. р. Бузулук Саратовская область окр. п. Первомайский Оренбургская область окр. г. Оренбург гора Гребени гора Маячная с. Гирьял Зарудный, 1895; Чи- билёв, 1995 Табачишина и др., 2005 Чибилёв А.А.
VI 31 13 р. Донгуз
VII 34 38 40 44 14 15 16 17 18 Западно-Казахстанская область ср. течение р. Утва низ. р. Утва, ур. Ак-Тепе окр. оз. Шалкар р. Калдыгайты, ур. Куагаш Актюбинская область верх р. Илек Березовиков и др., 1992, 2003; Зарудный, 1895 Дебело П.В. Зарудный, 1895
250
Глава 4
П родолжение табл. 47
Зона Провин- ция Район № точки Местонахождение Источник информации
VIII 48 19 Саратовская область окр. п. Ровное (о. Круглый) Шляхтин, 1996;Табачи- шина и др., 2005
Степная IX 56 61 62 64 20 21 22 23 Оренбургская область Буртинская степь, ОГПЗ низ. р. Губерля окр. с. Белошапка дол. р. Чебакла дол. р. Кинделя Айтуарская степь, ОГПЗ Актюбинская область горы Мугоджары, к сев.-вост. от горы Эмба Чибилёв, 1999а Давыгора, 1998 Чибилёв, 1995, 1999а, б Дусейбаева, 2005
XII 80 24 верх. р. Сары-Тургай Никольский, 1905
Полупустынная XIII 84 85 87 88 91 25 26 27 28 29 30 31 32 Западно-Казахст анская область окр. Арал-Сора Волгоградская область окр. оз. Эльтон Астраханская область окр. оз. Баскунчак Саратовская область р. Б. Узень окр. с. Ворфоломеевка окр. п. Приузеньский Западн о-Казахстанская область низ. р. Б. Узень низ. р. М. Узень окр. п. Коловертное Ходашова, 1960 Параскив, Бутовский, 1960 Стрельцов и др., 2006; Косарева, 1950 Табачишина и др., 2005 Параскив, Бутовский, 1960; Чернов, 1954
XV 97 98 33 34 Актюбинская область ур. Жанасу ур. Кумжарган п. Жагабулак междуречье Эмбы и Темира Дуйсебаева, 2005; Ни- кольский, 1905
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
251
Продолжение табл. 47
Зона Провин- ция Район № точки Местонахождение Источник информации
ынная XVI 101 102 35 36 Мугоджары, окр. п. Алабас хр. Шошкакуль Брушко, Кубыкин, 1988; Дуйсебаева, 2005; Ди- несман, 1953
8 XVIII 107 37 Иргизский р-н
С ё4 О С XIX 111 112 38 39 ур. Жыланшыктурме низ. р. Тургай
гтынная XX 113 114 117 118 122 123 125 127 40 41 42 43 44 45 46 47 48 Астраханская область гора Б. Богдо гора Чапчачи окр. п. Селитряное окр. г. Красный Яр, АГКМ Западно-Казахстанская область окр. Камыш-Самарских озер Гурьевская область окр. п. Н. Уштаган Западно-Казахстанская область окр. п. Калмыкове, п. Харькино Гурьевская область окр. п. Орлик окр. п. Будене Никольский, 1905 Завьялов, Табачишин, 1998 Чернов, 1954 Ралль, 1935 Окулова, 1963, 1985
С XXI 129 129 132 49 50 50 51 52 53 54 окр. п. Ракуша ур. Тажигали п. Прорва, ур. Кзылжар окр. протоки Баксай окр. п. Махамбет окр. п. Зеленый окр. п. Сарайчик г. Гурьев; Аэропорт, карьер окр. г. Гурьев дельта р. Урал Сараев, Пестов, 2010; Накаренок, 2003в Чернов, 1954
XXII 133 135 136 55 56 57 окр. оз. Индер ср. теч. р. Сагиз окр. п. Жамансор р. Эмба, ур. Айренкуль окр. п. Кульсары Брушко, Кубыкин, 1988; Гисцов, Березовиков, 1995; Чернов, 1954 Сараев, Пестов, 2010
252
Глава 4
Продолжение табл. 47
Зона Провин- ция Район Ns точки Местонахождение Источник информации
Пустынная XXII 137 138 58 59 60 61 62 63 окр. п. Мукур ур. Мадьяр низ. р. Сагиз, ур. Тентяксор оз. Бартылдакты гора Аккерегешин ур. Брлик гора Иманкара дол. р. Эмба, ур. Намазтакыр ур. Казбек, гора Койкара ур. Жанбике окр. сора Есенжол мог. Аламола, ур. Акмечеть ур. Шубарпала горы Шолькара: ур. Ушкан ур. Бекжал горы Жельтау, окр. хр. Жель- тау ур. Белиорна Брушко, Кубыкин, 1988 Сараев, Пестов, 2010 Накаренок, 2003 в
XXIII 143 64 Мангистауская область окр. бывш. форта Новоалек- сандровск Никольский, 1905
XXVI 153 154 155 65 66 67 68 Актюбинская область окр. ст. Саксаульская Приаральские Каракумы берег Аральского моря окр. оз. Камышлыбаш Динесман, 1953 Лобачев и др., 1973 Динесман, 1953 Дуйсебаева, 2005
Места обитания. Узорчатый полоз - широко распространенный вид, в ре-
гионе населяющий разные ландшафты лесостепной, степной, полупустынной
и пустынной природных зон. В условиях лесостепной зоны региона он населя-
ет сильно расчлененные речными долинами оврагами и лощинами предгорье
хр. Накас и мелкосопочные участки хр. Шайтантау с небольшими массивами
редкостойных березово-осинных лесков и густых зарослей кустарников, пере-
межающихся с расположенными преимущественно на южных склонах учас-
тками каменистой степи. В сходных условиях он встречается и южнее, уже в
степной зоне на территории Губерлинского мелкосопочника, сильно расчле-
ненного реками Губерля, Чебакла, Киндерля и их притоками (этот участок, по-
видимому, является основным местообитанием .вида в Оренбуржье). На терри-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
253
тории Предуралья (Буртинская степь ОГПЗ) и Общего Сырта придерживается
преимущественно расчлененных холмисто-увалистых и останцовых массивов
с каменистой и кустарниковой степью.
В южных степях Предуральского плато, его отрогов и Предсыртового усту-
па все известные поселения узорчатого полоза приурочены к речным долинам
(Илека, Карахобды, Калдыгайты, Утвы, Б. Узеня), а также Волжской пойме,
где береговые обрывы представляют собой элементы расчлененного рельефа;
в сходных условиях отмечен он нами среди разреженных злаково-полынных
зарослей на берегу оз. Шалкар возле устьевой части впадающего в него канала
из р. Урал (обрывистые берега которого заняты колонией береговых ласточек).
На берегу р. Б. Узень полозы наблюдались на участке разнотравной степи с
зарослями тростника. По долинам рек Б. и М. Узеней полозы отмечались на
всем протяжении полупустынной зоны вплоть до Волжско-Уральских песков.
В этом районе их встречи известны также в окрестностях оз. Баскунчак (у Кар-
донной балки с прудом), Аралсора, а также по равнинной глинистой полупус-
тыне вблизи п. Коловертное. Восточнее он отмечен в условиях возвышенного
Темир-Эмбенского междуречья, Мугоджар, хр. Шошкакуль, а также по рекам
Иргиз, Тургай и Жыланшик.
В условиях пустынной зоны на западе Волжско-Уральского междуре-
чья узорчатый полоз отмечался по эродированным склонам горы Б. Богдо, у
г. Чапчачи, а также в закрепленных бугристых песках, межбугровых пониже-
ниях заволжской волнисто-грядовой равнины (АГКМ) и у окрестных водо-
емов Волго-Ахтубинской поймы. Местами на западе междуречья (окрестности
п. Н. Уштаган), но чаще всего в районе Камыш-Самарской озерной депрессии,
проникает вглубь песков, где держится по берегам озер и у колодцев.
По северо-восточной кромке Волжско-Уральских песков полозы обита-
ют в глинистой пустыне, где придерживаются полынных, в меньшей степени
злаково-полынных сообществ, а в песках и здесь они отмечались лишь у ко-
лодцев. По долине р. Урал и его древним рукавам (Баксай) полозы населяют
почти все биотопы: полынные, полынно-солянковые и солянковые сообщест-
ва, встречаются в карьерах, а также поблизости от лесных полос. По мнению
С.А. Чернова, здесь, «вероятно, у них нет сколько-нибудь постоянных убежищ
и района охоты. Поохотившись в каком-либо ограниченном районе несколько
дней, узорчатый полоз, по всей вероятности, перемещается в другой». И еще -
«по-видимому, он может совершать значительные передвижения и оставаться
более или менее продолжительное время в том биотопе, где в данный момент
имеется некоторая концентрация животных, которыми он питается, и где есть
подходящие для него убежища в виде старых нор грызунов, нор береговой
ласточки, различных искусственных сооружений и т. д.» (1954. С. 150).
Следует отметить также, что именно эти «солончаковые степи около Кас-
пийского моря», откуда в 1770 г. «красивая сия и безвредная змейка» была
доставлена П.С. Палласу участником его экспедиции студентом Соколовым,
по-видимому, и нужно считать типовой территорией вида, поскольку по
р. Иртыш, как показали исследования А.Г. Бакиева с соавторами (2009), он
был найден почти год спустя.
На левобережье р. Урал узорчатый полоз отмечался в Индерских горах,
низовьях р. Сагиз, в горах Аккерегешин, Иманкара, Шолькара, Жельтау, а
в северо-восточном Прикаспии на солончаковой равнине у бывшего форта
254
Глава 4
Ново-Александровска. В Приаралье его встречи известны на плато у ст. Сак-
саульская, в Приаральских Каракумах, на берегу Аральского моря вблизи
оз. Камышлыбаш в низовьях р. Сырдарья.
Наряду с отмеченными пунктами естественных местообитаний узорчатый
полоз населяет и их антропогенные модификации. Его встречи известны в чер-
те г. Сакмара, а на сопредельной территории и в г. Саратове. Вместе с тем все
отмеченные популяции как естественных, так и антропогенных местообита-
ний являются изолированными, локальными, и, вероятно, нигде за исключени-
ем Губерлинского мелкосопочника и Приэмбенской провинции не занимают
сколь-нибудь значительной площади, на которой вид образовывал бы более
или менее сплошные поселения.
Образ жизни. После зимней спячки у южных пределов региона первые
полозы появляются в первой половине марта, в Саратовской области - 18-
24 марта, а у северной границы региона - с 5 апреля по первые числа мая
(в случае поздней и холодной весны). Однако массовый выход происходит
несколько позднее - на юге Поволжья по второй декаде апреля, в Саратов-
сой области - во второй половине апреля - первой декаде мая, т. е. примерно
3 недели спустя. Это вид с дневным образом жизни, причем весной активность
имеет типичный одновершинный характер, а в летнее время она смещается
на утренние и вечерние часы. Вместе с тем даже в самое жаркое время года,
по наблюдениям на юге Астраханской области, активные полозы в ночные
часы не отмечены. Днем они быстро передвигаются по земле, хорошо лазают
по ветвям деревьев и кустарников, хорошо плавают. В качестве убежищ ис-
пользует норы разных животных (в том числе береговых ласточек), пустоты
в земле, искусственных сооружениях, под камнями, а по берегам водоемов и
густые тростниковые и рогозовые заросли. В зимнюю спячку на севере реги-
она уходит во второй половине сентября - первой половине октября, на юге
активность сохраняется до конца октября - начала ноября. В целом на севере
региона период активности продолжается около 200 суток, на юге 240, зимняя
спячка соответственно 160 и 120 суток.
Размножение. Вскоре после выхода из зимовки, чаще всего в первой по-
ловине мая у полозов начинается спаривание. Яйца формируются довольно
быстро и уже в конце мая - начале июля поступают в яйцеводы (Чернов, 1954).
Откладка яиц (в регионе по 3-16 шт.) происходит в конце июня - первой по-
ловине июля, инкубационное развитие при температуре 31,9 °C продолжается
23-28 (в среднем 27) суток, и уже в конце июля - августе появляются молодые
(Завьялов, Табачишин, 20006; Шляхтин и др., 20056).
Питание. Узорчатый полоз - хищник, охотящийся на всех животных,
которых способен проглотить. Из пресмыкающихся он потребляет мелких
грызунов: мышей, полевок и других, вплоть до молодых малых сусликов и
водяных крыс. Из птиц в регионе объектом добычи являются жаворонки, бере-
говые ласточки, золотистые щурки и каменки. По-видимому, обилие и легкая
доступность пищи (в виде яиц и птенцов) привлекают полозов к колониям бе-
реговых ласточек, что наблюдалось на волжском острове Круглом (Завьялов и
др., 2000) и нами у оз. Шалкар. В низовьях р. Урал в окрестностях п. Сарайчик
узорчатый полоз встречался только в тех местах, где в большом числе живут
прыткие ящерицы, «составляющую здесь если не единственную, то во всяком
случае основную пищу». Кроме них в Волжско-Уральском междуречье полоз
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
255
потребляет разноцветных ящурок (по встречаемости стоят на первом месте) и
такырных круглоголовок (Чернов, 1954; Параскив, Бутовский, 1960). Помимо
отмеченных видов полозы добывают амфибий (зеленую жабу, озерную лягуш-
ку), рыбу и насекомых. В свою очередь они являются объектами охоты ряда
видов хищных птиц и млекопитающих; известны и явления каннибализма (Ба-
киев и др., 2009).
Численность. Учитывая локальность распространения общая численность
узорчатого полоза в регионе, очевидно, невелика, хотя местами известны оча-
ги с повышенной плотностью.
В лесостепной зоне на Лысой горе и в Студеном овраге (в черте г. Самара)
по разным источникам плотность населения вида составляет до 0,1 экз/км мар-
шрута (Бакиев и др., 2009). На территории Оренбуржья, занимающей юг лесо-
степной (Тюльганский р-н, хр. Шайтантау) и северную часть степной зоны, в
большинстве пунктов (по-видимому, за исключением Губерлинского мелко-
сопочника) узорчатый полоз встречается единично (Чибилёв, 1995), а общая
численность здесь ориентировочно определяется в несколько сот особей (Да-
выгора, 1998). На обитаемых участках разнотравно-типчаковой степи Балаков-
ского и Пугачевского районов Саратовской области плотность достигает 0,8-
1,2 экз/га, в соответствующих стациях Александрово-Гайского района - 1,6,
местами в долине р. Б. Узень - 1,6 и на о. Круглый в средней зоне Волгоград-
ского водохранилища - 28,4 экз/га (Табачишина и др., 2005; Завьялов, Табачи-
шин, 20006; Шляхтин и др., 2005). Конкретные данные о численности вида в
долинах рек Илек, Карахобда, Утва и у оз. Шалкар отсутствуют, но складыва-
ется впечатление, что он здесь определенно немногочислен.
В полупустынной зоне более или менее постоянные поселения, вероятно,
имеются лишь в долинах рек Б. и М. Узеней и Эмба; в окрестностях оз. Бас-
кунчак «за три года сделана только одна находка» (Стрельцов и др., 2006), в
Мугоджарах этот вид вообще известен по единичным встречам (Дуйсебаева,
2005).
В зоне пустыни узорчатый полоз более многочислен. Во всяком случае по
западной кромке Волжско-Уральских песков у п. Красный Яр (АГКМ) в за-
крепленных бугристых песках его плотность достигала 18,20±0,35 экз/га, по ок-
рестным водоемам Волго-Ахтубинской поймы - 11,70±0,29 и лишь в межбуг-
ровых понижениях снижалась до 1,70±0,14 экз/га (Завьялов, Табачишин, 1998).
Не представлял ранее особой редкости узорчатый полоз у восточной кромки
песков в Приуралье (Чернов, 1954), а в 1958-1960 гг. здесь его численность
составляла 0,5-0,7 ос/10 км (Окулова, 1985). В низовьях р. Эмба, где известно
наибольшее количество мест его встречи в пустынной зоне, он считается обыч-
ным видом (Неручев, Васильев, 1978), но сейчас в Гурьевской области числен-
ность его «относительно невелика» (Сараев, Пестов, 2010); а в Приаралье он
становится определенно редким (Лобачев и др., 1973). В целом по региону об-
щая численность вида, очевидно, не превышает несколько тысяч особей.
Лимитирующие факторы. По-видимому, в основном преследование и
уничтожение из-за ошибочного представления о ядовитости при хозяйствен-
ном освоении территории.
Охрана. Обитает и охраняется на территории Оренбургского, Баскунчак-
ско-Богдинского заповедников и Иргиз-Тургайского резервата. На остальной
территории региона специальных мер охраны не предпринималось.
256
Глава 4
Региональный статус. Малочисленный спорадически втречающийся вид.
Занесен в Красные книги Оренбургской (1998), Самарской (2009), Саратовс-
кой (2006) областей и сопредельного Башкортостана.
42. Палласов (сарматский) полоз - Elaphe sauromates (Pallas, [1814])
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Монотипичес-
кий вид, который ранее рассматривался в качестве подвида четырехполосого
полоза. Его видовой статус восстановлен на основе аллозимного анализа, что
подтверждено филогенетическими исследованиями на молекулярном уровне
(Атлас пресмыкающихся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает большую часть южной Европы,
Кавказа, северные районы Малой Азии, Ирака, Ирана, а также прикаспийс-
кие и приаральские районы Средней Азии и Казахстана (средиземноморский
фаунистический комплекс). Его северная граница на Восточно-Европейской
равнине от Молдовы проходит через южную Украину, Ростовскую и Астра-
ханскую области, Северный и Северо-Восточный Прикаспий, Предустюртье,
Северный Устюрт, северное побережье Аральского моря и Приаральские Ка-
ракумы, откуда, поворачивая на юго-запад, выходит к восточному побережью
Каспийского моря в окрестностях Кара-Богаз-Гола. Таким образом, ареал вида
охватывает только южную часть региона.
В его пределах до недавнего времени самым северным местообитанием
вида были окрестности оз. Баскунчак, где он был найден еще в конце XIX в.
(Никольский, 1905). С тех пор этот полоз никем из работавших здесь зооло-
гов не отмечался около 50 лет (Ходашова, 1960). Затем в 2002, 2003 и 2005 г.
вблизй этого пункта 2 особи были отловлены на юго-восточном склоне горы
Б. Богдо (причем одна ближе к оз. Баскунчак) и еще один экземпляр отловлен
в 0,5 км восточнее Богдо (Стрельцов и др., 2006). Южнее этого полоза в конце
1940-х гг. И.С. Даревский с помощником добывали в окрестностях ст. Досанг
(Чернов, 1954).
Далее этот вид отмечался примерно в 600 км восточнее в Приэмбенской
провинции (Неручев, Васильев, 1978). Существующий разрыв ареала не-
давно заполнила находка этого полоза в окрестностях г. Гурьева, о которой,
основываясь на опросных сведениях, сообщал Р.А. Кубыкин (1994). Одна-
ко, по мнению Ф.А. Сараева и М.В. Пестова (2010), современное обитание
вида в этом районе маловероятно и, следовательно, нуждается в подтверж-
дении.
В восточной части региона палласов полоз населяет Предустюртье, Север-
ный Устюрт, окрестности сора Кайдак, п-ов Бузачи, Приаралье и Приараль-
ские Каракумы (Кубыкин, 1994; Дуйсебаева, 2005; Сараев, Пестов, 2010). Его
крайние северные находки известны из окрестностей ст. Опорная, гор Шоль-
кара и Жельтау, на северном чинке Устюрта, в устье оврага Аксай и его ок-
рестностях, затем на южной оконечности Б. Барсуков, северном побережье
Аральского моря и в Приаральских Каракумах. В очерченных пределах ре-
гиона его конкретные местонахождения известны в одном (3,1% из 11) ланд-
шафтном районе полупустыни и 13 (30,2% из 43) районах пустынной зоны
(табл. 48, карта 41).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
257
Распространение палласова полоза
Таблица 48
Зона Провин- ция Район № точки Местонахождение Источник информации
Полупус- тынная XIII 85 1 Астраханская область окр. оз. Баскунчак Чернов, 1954; Ходашо- ва, 1960; Стрельцов и др., 2006
XX ИЗ 2 гора Б. Богдо юго-вост. окр. г. Б. Богдо Чернов, 1954; Божанс- кий, Полынова, 1998
114 окр. ст. Досанг
XXI 132 Гурьевская область окр. г. Гурьев (Атырау) Кубыкин, 1994а
138 5 6 сев-зап. Прикаспийских Каракумов окр. п. С ары камы с окр. ст. Опорная ур. Тугаракшан Панкратов, 1989; Ку- быкин, 1994; Неручев, Васильев, 1978; Сараев, Пестов, 2010
7 гора Шолькара Мангистауская область
XXII 8 горы Жельтау
иная 139 9 гора Тамды ур. Молорпа ур. Орта-Шагырлы Киреев, 1981; Нака- ренок, Неручев, 2001; Сараев, Пестов, 2010; Панкратов, 1989
10 ур. Мынсуалмаз
с 11 оз. Туракчан гора Токсанбай овр. Аксай окр. овр. Аксай Шилов, 1961
142 12 окр. п. Тиген кл. Аузорка Кубыкин, 1994
XXIII 143 13 окр. бывш. Новоалександ- ровска Никольский, 1905
144 14 окр. сора Кайдак Кубыкин, 1994
147 15 Сев. Устюрт: 18 км сев. Лобачев и др., 1973
метеост.
окр. п. Туруш
XXV 16 ур. Косбулаксор окр. ур. Косбулаксор окр. кол. Куштайкудук окр. ур. Тогураксор
258
Глава 4
Окончание табл. 48
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXV 148 17 18 пески Сам, окр. п. Туруш пески Сам, п. Туруш 30 км к зап. метеост. Дукен ур. Ак-Жигит ур. Матайкум кол. Ушкудук Неручев, Васильев, 1978 Кубыкин, 1994 Шилов, 1961
XXVI 151 154 155 19 20 21 22 юг Б. Барсуков Приаральские Каракумы п-ов Куланды окр. зал. Тущебас окр. п-ова Каратюб Ерофеев, 1986; Лобачев и др., 1973; Кубыкин, 1994; Дуйсебаева, 2005; Шилов, 1961
Места обитания. Сарматский полоз в целом не только широко распро-
страненный, но и эвритопный вид. Однако в регионе он занимает довольно
ограниченный круг местообитаний. Гора Б. Богдо и ряд других, упомянутых
ранее, где встречается этот вид, представляют собой отдельные соляные ку-
пола или небольшие останцовые «хребтики» с эродированными склонами,
«поющими скалами», карстовыми воронками, каменистыми россыпями. В по-
добных местообитаниях встречается полоз и по чинкам Северного Устюрта.
Здесь же, как и на сопредельной территории, он отмечен по песчаным руслам
временных водотоков Аксай, Туранглы и в Босагинских оврагах.
В Б. Барсуках населяет бугристые полузакрепленные пески с жузгуном,
полынями и злаками, встречается также на супесчаных участках с эфемеро-
выми сообществами. В Сам-Матсайской депрессии на Устюрте обитает как
на плотных закрепленных песках, так и в полынно-биюргуновой глинистой
пустыне с сильным засолением. На побережье Аральского моря отмечался по
берегам высохшего залива среди терескена и тамарикса, здесь же и у Кайда-
ка не избегает и соровых участков. Характерной особенностью вида является
привязанность к колониям песчанок, которыми он питается и в норах которых
находит укрытие. На западе ареала не избегает близости с человеком и селится
в садах и виноградниках, хотя в целом по региону его места обитания еще мало
затронуты деятельностью человека (Кубыкин, 1994; Ерофеев, 1986; Ананьева
и др., 1998).
Образ жизни. После зимовки на поверхности появляется в начале или се-
редине апреля. Весной активен в течение всего светлого времени суток, а ле-
том активность смещается на утренние и вечерние часы. Агрессивен, хорошо
лазает по кустам и обрывам. Днем укрывается в норах грызунов, прикорневых
пустотах, под камнями; здесь же и зимует. Активность продолжается до конца
сентября - начала октября (около 170-180 суток).
Размножение. В регионе практически не изучено. Известно лишь, что
яйца (4—16) откладываются в июне-июле, а молодые появляются в августе-
сентябре.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
259
Питание. Основу пищевого рациона составляют крупные и средней вели-
чины грызуны (песчанки и др.), реже потребляются птицы и их яйца, изредка
ящерицы (Панкратов, 1989; Ананьева и др., 1998).
Численность. В Нижнем Поволжье и на большей части казахстанского
участка ареала встречается единично (Кубыкин, 1994; Стрельцов и др., 2006)
н лишь в некоторых пунктах несколько чаще: в хр. Жельтау у Северного Ус-
тюрта в конце 1970-х гг. плотность составляла 0,2-0,7 ос/га (Киреев, 1981), на
равнинах, примыкающих к Прикаспийским Каракумам, и по останцам севе-
ро-западного Устюрта (в зависимости от характера биотопов, численности и
видового состава грызунов) она достигала 0,3-1,8-2,0 ос/га (Панкратов, 1989),
чаще по 2-3 взрослые особи отмечались в колониях большой и краснохвостой
песчанок по Большим Барсукам (Ерофеев, 1986).
Лимитирующие факторы. Хозяйственное освоение территории и сопутс-
твующее уничтожение человеком из-за ошибочного представления о ядови-
тости, местами отлов для содержания в неволе.
Охрана. Обитает и охраняется на территории Устюртского и Богдинско-
Баскунчакского заповедников.
Региональный статус. Редкий вид с сокращающейся численностью. Зане-
сен в Красную книгу Казахстана (1996), Красную книгу Астраханской области
(2004) и Аннотированный перечень таксонов и популяций, нуждающихся в
особом внимании к их состоянию в природной среде (Приложение к Красной
книге Российской Федерации - 2001). Находится под охраной Бернской кон-
венции.
Род 28. Hierophis Fitzinger In Bonaparte, 1832 - гиерофисы
43. Каспийский, или желтобрюхий полоз -
Hierophis caspius (Gmelin, 1779)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода в герпетофауне региона. Монотипический вид, до недавнего
времени рассматривающийся в качестве подвида Coluber jugularis (Атлас пре-
смыкающихся..., 2004).
Распространение. Ареал вида охватывает европейский юго-восток, Пред-
кавказье и Нижнее Поволжье, откуда заходит в пределы Казахстана (среди-
земноморский фаунистический комплекс). Его северная граница на Восточ-
но-Европейской равнине от Молдавии проводится через Южную Украину к
Поволжью. До недавнего времени считалось, что здесь она достигает Крас-
нокутского и Новоузеньского районов Саратовской области (Девишев, 1976;
Шляхтин, Голикова, 1986; Шляхтин, 1996). Однако в последующие годы,
несмотря на специальные поиски в этих районах и на сопредельной террито-
рии, каспийский полоз не обнаружен (Шляхтин и др., 2005, 2006). В связи с
этим считается, что этот вид в Заволжье не проникает севернее устья р. Хара у
оз. Эльтон. Кроме того, в этих местах он отмечался в песках Шкили Ахтубин-
ского района (Табачишина и др., 2004,20066), окрестностях оз. Баскунчак и на
юго-восточном склоне горы Б. Богдо (Линдеман и др., 2005; Стрельцов и др.,
2006).
260
Глава 4
Таблица 49
Распространение каспийского полоза
Зона Провин- ция Район № точки Местонах ождение Источник информации
А Волгоградская область
Полупу< тынная XIII 85 1 2 устье р. Хара в окр. оз. Эльтон окр. оз. Баскунчак Табачишина и др., 2006а; Завьялов и др., 2006; Ходашова, 1960; Стрельцов и др., 2006
Астраханская область
i X 3 XX 113 114 3 4 гора Б. Богдо, Богдинские полезащитные полосы пески Шкили окр. г. Красный Яр (Волго- Ахтубинская пойма) Сараев, Пестов, 2010; Стрельцов и др., 2006; Чернов, 1954; Табачи- шина и др., 20066
В Гурьевская область
к 118 5 окр. п. Н. Асан окр. ур. Бес-Чохо окр. кл. Шадьялмола Ралль, 1935; Островских и др., 2010
XXII 133 6 окр. п. Индерборский Сараев, Пестов, 2010
Наиболее восточным пунктом, куда проникал этот полоз, считался неболь-
шой массив холмистой полынно-эбелековой степи Бес-Чохо, вдающийся в
Волжско-Уральские пески (Ралль, 1935; Чернов, 1954). Однако в последние
годы получены данные о более широком распространении вида как в районе
Бес-Чохо, так и за ее пределами - на горе М. Богдо и даже в окрестностях
п. Индерборский на левом берегу р. Урала, примерно в 300 км восточнее (Ос-
тровских и др., 2010; Сараев, Пестов, 2010). Эта находка заставляет по-иному
взглянуть на сообщение Г.С. Карелина (1883) о встрече «большой змеи у Гу-
рьева» под названием Coluber caspius «желтопузик», которое считали отно-
сящимся к описанию нередко встречающегося в низовьях р. Урал узорчато-
го полоза (Чернов, 1954). Южнее по р. Волга этот полоз отмечался в районе
Богдинских полезащитных полос, а также в долине между Волгой и Ахтубой
(Определитель земноводных..., 1977).
В очерченных пределах конкретные местообитания известны в одном
(3,1% из 32) ландшафтном районе полупустынной и 4 (9,3% из 43) пустынной
зоны (табл. 49, карта 42).
Места обитания. Основными местообитаниями вида в регионе являются
склоны возвышающихся над грядово-холмистой равниной соляных куполов
с обнажениями песчаников, известняков, солей и широко развитым карстом,
склоны долин рек (Хара), оврагов и другие стации со сложным микрорелье-
фом и зарослями кустарников, полезащитные полосы.
В кустарниковых зарослях, перемежающихся с луговыми участками, оби-
тает этот вид и в Волго-Ахтубинской пойме. В «древней степи Бес-Чохо» он
отмечался среди разреженных полынно-кокпековых зарослей в карстовой зоне
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
261
и поблизости от кладбищных сооружений (Ралль, 1935; Чернов, 1954; Табачи-
шина и др., 20066; Островских и др., 2010).
Образ жизни. После зимней спячки появляется в апреле и сохраняет ак-
тивность до октября (около 180 суток). Весной активен днем, летом преиму-
щественно утром и вечером. В качестве убежищ использует норы грызунов,
трещины в грунте, пустоты среди камней, развалины различных сооружений.
Размножение. Спаривание происходит вскоре после выхода из спячки. На
сопредельной территории яйца (6-18) откладываются в июне-июле, молодые
появляются в сентябре.
Питание. Основу пищевого рациона в регионе составляют ящерицы, за-
тем грызуны, птицы, реже потребляются насекомые и другие животные.
Численность. У северных пределов распространения встречается спора-
дично и единичными особями: на кромке песков Шкили насчитывали до 2 экз.
на 1 км маршрута, а в устье р. Хара - 0,7 экз/км (Табачишина и др., 20066). Бо-
лее обычен на юго-восточном склоне горы Б. Богдо, где за 3 сезона отмечено
более 30 особей (Стрельцов и др., 2006). За пределами этих пунктов встреча-
ется еще реже-до 1,0 ос/10 км (Накаренок, 2003в). По мнению В.Ф. Чернобая
(2006), в Волго-Ахтубинской пойме он находится на грани исчезновения.
Лимитирующие факторы. Комплекс факторов на границе ареала, но,
скорее всего, хозяйственное освоение территории, сопровождающееся унич-
тожением этих змей из-за их агрессивности.
Охрана. Обитает и охраняется на территории Богдинско-Баскунчакского
заповедника. На остальной территории специальные меры охраны не предпри-
нимались.
Региональный статус. Редкий, спорадически встречающийся вид у се-
верной границы ареала. Занесен в Красную книгу Казахстана (1996), Астра-
ханской (2004), Волгоградской (2007) областей и Аннотированный перечень
таксонов и популяций, нуждающихся в особом внимании к их состоянию в
природной среде (Приложение к Красной книге Российской Федерации, 2001).
Находится под охраной Бернской конвенции.
Род 29. Malpolon Fitzinger, 1826 - ящеричные змеи
44. Ящеричная змея -Malpolon monspessulanus (Hermann, 1804)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода на территории региона. Политипический вид, в регионе пред-
ставленный подвидом М. т. insignitus (Geoffroy, 1809) (Атлас пресмыкающих-
ся..., 2004; Кузьмин, Семенов, 2006).
Распространение. Ареал вида охватывает Северную Африку, Переднюю
и Южную Азию, Кавказ и значительную часть Южной Европы (средиземно-
морский фаунистический комплекс). Изолированный участок ареала занимает
восточную часть Калмыкии и сопредельные районы Ставропольского края и
Астраханской области, откуда, по-видимому, сравнительно недавно ящерич-
ная змея проникла на территорию региона. В его пределах впервые была обна-
ружена И.С. Деревским в конце 1940-х гг. в окрестностях ст. Досанг (Чернов,
1954), а затем нами (А.А. Чибилёвым) наблюдалась на горе и в окрестностях
262
Глава 4
Таблица 50
Распространение ящеричной змеи
Зона Провин- ция Район № точки Местонахождения Источник информации
Полу- пус- тынная XIII 85 1 Астраханская область окр. горы Б. Богдо Чибилёв А.А.
а X я н XX 113 114 2 3 Карстовые ландшафты в окр. оз. Баскунчак Волго-Ахтубинская пойма окр. ст. Досанг Определитель земно- водных..., 1977; Чернов, 1954; Божанский, Полы- нова, 1998
ь К Западно-Казахстанская область
115 4 окр. горы М. Богдо (Жамантау) Чибилёв А.А.
горы Б. Богдо в Астраханской области (2006, 2008 г.) и на горе М. Богдо (Жа-
мантау) в Бокейординском районе Западно-Казахстанской области (2000,
2011 г.) (табл. 50, карта 43).
Места обитания. В пределах региона встречалась в области приволжских
бэровских бугров, где невысокие гряды бугристо-барханных и полузакреплен-
ных бугристых песков перемежаются с плоскодонными ложбинами, занятыми
белополынными, белополынно-прутняковыми и чернополынными (на солон-
цах) сообществами.
Образ жизни. В связи с незначительным числом встреч практически не
изучен.
Численность. Повсеместно невысока. На территории Астраханского ГКМ
даже при специальных поисках не отмечена (Завьялов, Табачишин, 1998).
Лимитирующие факторы. Повсеместно хозяйственное освоение терри-
тории, как правило, сопровождающееся разрушением типичных местообита-
ний (Ананьева и др., 1998).
Охрана. Специальные меры не предпринимались.
Региональный статус. Малочисленный краеареальный вид у северных
пределов распространения. Занесена в Красную книгу Астраханской области
(2004) и Аннотированный перечень таксонов и популяций, нуждающихся в
особом внимании к их состоянию в природной среде (Приложение к Красной
книге Российской Федерации, 2001).
Род 30. Natrix Laurenti, 1768 - настоящие ужи
45. Обыкновенный уж - Natrix natrix (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей обширного рода, населяющий территорию региона. Политипичес-
кий вид; описано до 13 подвидов, в том числе номинативный N. п. natrix (Lin-
naeus, 1758) и восточный N. п. scutata (Pallas, 1771), обитающие на территории
региона. Вместе с тем, пределы их распространения разными авторами указы-
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
263
ваются неодинаково. По принятой нами системе (Атлас пресмыкающихся...,
2004) большую часть европейского ареала (на восток до р. Волги) населяет
номинативный подвид, который в Заволжье сменяется восточным подвидом.
Тем не менее В.Л. Вершинин (2007) считает, что номинативный подвид насе-
ляет и всю территорию Урала. Однако исследования последних лет показали,
что обыкновенные ужи с признаками, характерными для номинативного под-
вида, преобладающие главным образом в правобережной части Поволжья, по
территории Саратовской области проникают и в Заволжье. На большей же его
части доминирует форма с признаками N. п. scutata, в свою очередь места-
ми проникающая и на правобережье р. Волги (Бакиев и др., 2004; Трохимен-
ко, Бакиев, 2001; Табачишин и др., 1996). Вместе с тем в последней работе
А.Г. Бакиев с соавторами (2009) восточную форму считает внутриподвидовой
формой номинативного подвида.
Таким образом, территория исследуемого нами региона в пределах Волж-
ско-Уральского междуречья является своеобразной зоной интерградации под-
видов, границы которой еще не определены и требуют специальных иссле-
дований (Кузьмин, Семенов, 2006). При этом не следует выпускать из вида,
что р. Урал признается типовой территорией У. п. scutata. Наряду с этим у
обыкновенного ужа проявляются довольно существенные межпопуляционные
отличия по цвету, рисунку, выраженности формы затылочных пятен и некото-
рых других признаков (Морозенко и др., 2003).
Распространение. Ареал вида охватывает большую часть Европы, севе-
ро-запад Африки, Западную Азию, Кавказ, значительную часть Казахстана и
Южной Сибири до северо-запада Монголии й Китая (европейский фаунисти-
ческий комплекс). Его северная граница от юга Феноскандии прослеживается
в области верховий р. Печора, нижнего течения р. Иртыш, по среднему те-
чению р. Обь, окрестностям г. Красноярска и верховью р. Лена к оз. Байкал,
затем по Забайкалью уходит в Монголию. Южная граница от Западной Азии
проходит по Ирану и Северному Китаю в Монголию. Таким образом, террито-
рия региона полностью вписывается в пределы видового ареала. Сейчас здесь
он известен в 4 (36,4% из 11) ландшафтных районах лесостепной, 35 (50,7% из
69) степной, 11 (34,4% из 32) полупустынной и 9 (20,9% из 43) пустынной зон
(табл. 51, карта 44).
Места обитания. Обыкновенный уж населяет разные типы местообита-
ний, как правило, расположенные по берегам водоемов или поблизости от них,
но в значительной степени увлажненные (гидрофильные, гигрофильные).
В лесостепной зоне Волжско-Камского края, Оренбуржья и Урала это бе-
рега рек, ручьев, озер, прудов, водохранилищ, болот, а также опушки, поляны
и лесосеки смешанных, широко-, узколиственных (березняки, осинники) ле-
сов, заросли кустарников, луга низкой части поймы и гривы, причем часть из
них используется круглогодично, другие периодически как места кормежки,
размножения или зимовки.
В степной зоне западной части региона более или менее сплошные (не-
прерывные) поселения он образует лишь в поймах рек Волга, Самара, Урал,
Сакмара и Илек, откуда по элементам овражно-балочных систем с луго-
во-болотными урочищами и галерейными лесками выходит на сопредель-
ные плакоры (как, например, в Буртинском и Айтуарском участках Орен-
бургского заповедника). Заселяет эта змея и долины небольших степных
264
Глава 4
Таблица 51
Распространение обыкновенного ужа
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная I 1 2 1 2 3 4 5 6 7 8 9 10 Самарская область с. В. Сускан с. Н. Санчелеево с. Хрящевка с. Выселки п. Луначарский с. Подстепки п. Приморский г. Тольятти: р-н ул. Баныкина п. Портовый п. Федоровка с. Пискалинский взвоз ст. Задельная ст. Мастрюково п. Волжский г. Самара: п. Управленческий Воронежские озера Ботанический сад п. Рубежное Яицкие озера с. Васильевка п. Старосемейкино п. Водино п. Висловка с. Старый Буян Бакиев, Файзулин, 2002; Бакиев и др., 2003, 2004, 2009
II 3 11 12 13 14 15 16 с. Кошки окр. г. Сергиевска р. Сок, с. Чекалин с. Лужки п. Красный Яр с. Алакаевка с. Советы п. Алексеевка окр. п. Грачевка
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
265
Продолжение табл. 51
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная II 5 17 18 19 20 21 с. Боровка окр. д. Русский Байтуган с. Абдул-Завод с. Старопохвистнево г. Похвистнев р. Сарбай
Степная IV 12 13 14 15 16 17 18 19 20 22 23 24 25 26 27 28 29 30 31 32 33 34 35 с. Александровка с. Натальино с. Кануевка окр. п. Безенчук окр. с. Новокиевка г. Новокуйбышевск г. Чапаевск Саратовская область окр. г. Балаково окр. р. Б. Караман дол. р. Волги Самарская область р. Чагра, с. Новотулка окр. п. Маяк южнее г. Новокуйбышевска Саратовская область Повсеместно сс сс Табачишин, Табачиши- на, 2002 Бакиев, Файзулин, 2002; Бакиев и др., 2003, 2004, 2009 Табачишин, Табачиши- на, 2002
V 21 36 37 38 39 40 41 Самарская область окр. г. Кинель с. Бобровка п. Красносамарское, Красноса- марский лес окр. п. Горский пойма р. Самара окр. с. Богатое с. Заливное, оз. Лебяжье г. Отрадный окр. с. Петровка Бакиев, Файзулин, 2002; Бакиев и др., 2004, 2009; Трохименко, 2003
266
Глава 4
Продолжение табл. 51
Зона Провин- ция Район № точки Местонахождение Источник информации
22 42 окр. п. Борское п. Лесное Оренбургская область
43 Бузулукский бор: окр. п. Колтубановский Линдгольм, 1902; цит. по: Никольский, 1905; Чибилёв А.А.
44 низ. р. Ток
23 45 дол. р. Ток
24 46 дол. р. В. Каргалка
47 дол. р. Салмыш Самарская область
V 26 48 г. Нефтегорск, окр. с. Съезжее (дол. р. Самара) Бакиев и др., 2002, 2004, 2009
окр. с. Утеевка
49 окр. с. Украинка окр. с. В. Ростоши Оренбургская область
50 Таловская степь ОГПЗ Чибилёв, 1999а
5 х Саратовская область
с е 27 51 окр. с. Модин Завьялов и др., 2006
о 29 52 Оренбургская область окр. г. Оренбург о. Мира Дебело П.В.
VI 30 53 окр. Илецкого укр. дол. р. Илек Паллас, 1773; Николь- ский, 1905, 1918
Западн о-Казахст анская область
33 54 окр. п. Барбастау Дебело П.В.
34 55 дол. р. Утва дол. р. Караоба Зарудный, 1895; Березо- виков и др., 1992
35 56 ур. Актау
VII 57 окр. оз. Сулуколь Зарудный, 1895
37 58 оз. Чушкалы
38 59 дол. р. М. Анката, окр. п. Рыбцех Дебело П.В.
60 окр. оз. Шалкар
40 61 р. Калдыгайты юж. п. Егендыколь
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
267
Продолжение табл. 51
Зона Провин- ция Район № точки Местонахождение Источник информации
Степная VII 44 45 46 62 63 64 Актюбинская область верх. р. Уил р. Уил дол. р. Киил Никольский, 1905; Па- раскив, 1956
VIII 48 50 51 54 55 65 65а 66 67 68 69 70 Саратовская область окр. п. Ровное окр. п. Дергачи Западно-Казахстанская область р. Деркул п. Красноармейск, ст. Зимник, ст. Кивалино, дол. р. Ембула- товка дол. р. Выковка, окр. п. Спар- так окр. п. Подстепное, оз. Едучее окр. г. Уральска, пруды рыб- хоза Табачишин, Табачиши- на, 2002; Табачишин и др., 1996 Дебело П.В.
IX 56 60 62 71 72 73 Оренбургская область Буртинская степь ОГПЗ окр. г. Орска Айтуарская степь, ОГПЗ, р. Куагач Чибилёв, 1999а; Опреде- литель земноводных..., 1977; Топоркова, 1973
Полупустынная XIII 81 82 83 85 86 74 75 75а 76 77 78 Волгоградская область дол. р. Волга Западно-Казахстанская область Джаныбекский стаи. АН р. Горькая Волгоградская область окр. оз. Эльтон Астраханская область окр. оз. Баскунчак Западно-Казахстанская область окр. оз. Балыкты Определитель земновод- ных..., 1977 Ходашова, 1960; Живот- ные..., 2009 Стрельцов и др., 2006 Ходашова, 1960
268
Глава 4
Продолжение табл. 51
Зона Провин- ция Район № точки Местонахождение Источник информации
Полупустынная XIII 87 88 89 91 79 80 81 82 83 84 Саратовская область окр. п. Варфоломееевка окр. п. Приузеньский окр. п. Ветелки п. Монахов Западно-Казахстанская область оз. Рыбный Сакрыл окр. п. Фурманово Камыш-Самарские озера р. Кушум, Кировское вдхр. окр. п. Коловертное окр. п. Круглый Табачишин, Табачи- шина, 2002; Ходашова, 1960 Чернов, 1954 Параскив, Бутовский, 1960; Дебело П.В.; Чер- нов, 1954
XIV 92 85 ур. Карабас Дебело П.В.
XV 98 86 87 Актюбинская область Междуречье Эмбы и Темира окр. п. Жаркемер окр. г. Эмба Параскив, 1956; Дуйсе- баева, 2005
XVI 101 88 окр. ст. Мугоджарская Дуйсебаева, 2005
Пустынная XX 113 114 121 124 89 90 91 92 93 94 95 Астраханская область окр. горы Б. Богдо окр. с. Болхуны Волго-Ахтубинская пойма АГКМ у г. Красный Яр окр. п. Комсомольский окр. п. М. Арал окр. п. М. Сафоновка окр. п. Тюрино окр. п. Досанг-Замьяны окр. рзд. 7 Гурьевская область окр. п. Забурунье Западно-Казахстанская область окр. п. Харькино окр. п. Калмыкове Никольский, 1905; Завьялов, Табачишин, 1998; Бобров, 1985; Сараев, Пестов, 2010; Накаренок, 2003 в Чернов, 1954;Завьялов и др., 2006 Сараев, Пестов, 2010 Чернов, 1954; Паллас, 1773; Окулова, 1963
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
269
Окончание табл. 51
Зона Провин- ция Район № точки Местонахождение Источник информации
Пустынная XXI 129 132 96 97 98 99 100 Гурьевская область окр. Баксанской станции окр. п. Сарайчик окр. п. Редут окр. г. Гурьев пр. Сокол Чернов, 1954 Чернов, 1954; Сараев, Пестов, 2010 Киреев, 1981
XXII 134 138 101 102 окр. п. Крабау Мангистауская область хр. Жельтау
XXV 147 103 Актюбинская область окр. кол. Казалы Параскив, 1956
речек, где придерживается постоянных плесов, водохранилищ (Варфоломе-
евское, Кировское и др.) многочисленных запруд и за пределы их узких до-
лин почти не выходит. В связи с этим «кружево ареала» здесь становится
гораздо реже.
В степном Зауралье по сведениям А.П. Райского (1951) обыкновенный уж
также встречался (хотя и редко), но конкретные данные по этой части региона,
как и по Тургайскому плато, отсутствуют.
В полупустынной и пустынной зонах (в связи с их большей ксерофитнос-
тью) основным местом обитания являются долины рек Волги и Урала, где этот
уж теснее привязан к слабопроточным и стоячим водоемам. Здесь он встре-
чается преимущественно в прирусловых (галерейных) лесах, влажных лугах
(особенно граничащих с лесами или имеющими вкрапления кустарников), за-
болоченных участках вблизи пойменных озер и стариц (особенно с прибреж-
ными тростниковыми или рогозовыми зарослями и богатыми лягушками и
рыбой).
В полупустынном Волжско-Уральском междуречье он встречается в ок-
рестностях оз. Баскунчак (Кардонная балка), по рекам Горькая, Б. и М. Узени,
Кушум (с русловыми водохранилищами), на оз. Рыбный, Сакрыл, Балыкты и
устьевых Камыш-Самарских озерах. При разливах р. Урал отмечается и по его
протокам (Баксай и др.).
В Урало-Эмбенском междуречье его обитание известно по долинам рек
Уил, Киил, в верховьях Эмбы, Темира и их междуречье. В прошлом по севе-
ро-восточному Прикаспию он, вероятно, имел более широкое распростране-
ние, о чем свидетельствуют изолированные местообитания в горах Жельтау и
на Северном Устюрте. В Северном Приаралье обыкновенный уж не известен,
хотя Т.Н. Дуйсебаева (2005) не исключает возможности его встречи в районе
низовий рек Иргиз и Тургай.
Обыкновенный уж хорошо уживается на антропогенно-модифицирован-
ных и урбанизированных территориях, причем часто ведет себя как хорошо
выраженный синантроп, встречаясь не только в населенных пунктах сельского
270
Глава 4
типа, где поселяется на огородах, в строениях, кучах бытового и строительно-
го мусора, но и в черте городов, особенно на дачных участках. Вместе с тем
в типичных местообитаниях зоны промысла и промышленного загрязнения
Карчаганакского нефтегазоконденсатного месторождения отсутствует, хотя
по долине недалеко протекающей р. Утва встречается в достаточном количес-
тве (Березовиков и др., 2003).
Образ жизни. Выход из зимних убежищ на юге региона начинается в пер-
вых числах марта, в Саратовской области со второй его половины (наиболее
ранняя встреча у п. Ровное зарегистрирована на 09.03.1995 г.), а в Волжско-
Камском крае - с конца первой декады апреля - т. е. примерно на месяц позже.
Однако массовый выход происходит позднее и зависит от хода развития весен-
них явлений. На юге Саратовской области это наблюдается в третьей декаде
марта - начале апреля, на ее севере - в разных числах второй половины апреля,
а в Волжско-Камском крае - в конце апреля-начале мая, что совпадает со вре-
менем прогревания почвы до 12 °C.
Весной, а затем осенью ужи активны преимущественно в середине дня,
летом пик активности смещается на утреннее (7-12 часов) и вечернее (17-19)
время. На ночь и при неблагоприятной погоде ужи укрываются в кучах листвы,
мусора, нагромождениях камней, норах животных, причем, избегая перегрева,
они из одних убежищ могут переходить в другие, с более благоприятными
условиями (оптимальной считается температура в 25...31 °C).
В Волжско-Камском крае с конца августа начинаются перемещения ужей
к местам зимовки, а в спячку здесь они залегают в конце сентября - октябре,
задерживаясь в отдельные годы до первых чисел ноября (04.11.1996 г.). На юге
региона уход в зимовку запаздывает примерно на месяц.
Таким образом, период активности в Волжско-Камском крае в среднем
продолжается 124 (максимум 169) дня, что примерно совпадает с продолжи-
тельностью безморозного периода и активности его объектов питания. В усло-
виях Саратовской области период активности обычно продолжается с середи-
ны марта до середины октября - т. е. около 200 дней. Зимуют ужи под старыми
пнями, в прикорневых пустотах, норах животных, кучах мусора, стогах сена,
разных строениях, в том числе жилых (Гаранин, 1983; Табачишин, Табачиши-
на, 2002; Бакиев и др., 2004, 2009; Шляхтин и др., 2005).
Размножение. Брачный период начинается вскоре после выхода из зи-
мовки, но чаще всего в конце апреля - первой половине мая. Сроки откладки
яиц в разные годы несколько изменяются. Так, в Саратовской области кладка
проходила с 13 июня по 24 июля, а несколько южнее, в Волжско-Уральском
междуречье, в начале 1950-х гг. у обследованных самок в конце июня - начале
июля были только готовые или почти готовые к откладке яйца. Одна самка в
пределах региона и его ближайших окрестностях откладывает до 30 яиц, но
часто в одно место яйца откладывают несколько самок. Продолжительность
инкубации зависит от конкретных погодных и микроклиматических условий,
но обычно молодые появляются со второй половины августа - самая ранняя
дата 18 июля 1966 г. (Параскив, Бутовский, 1960; Гаранин, 1983; Шляхтин и
др., 2005; Бакиев и др., 2009).
Питание. Основу пищевого рациона обыкновенного ужа в регионе со-
ставляют земноводные: озерная и остромордая лягушки, обыкновенная чес-
ночница и молодняк зеленой жабы. Помимо них в составе пищи отмечалась
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
271
рыбья и лягушачья икра, прыткая и живородящая ящерицы, мелкие птицы и
их яйца, мыши и полевки, рыба (интересный случай нападения ужа на кара-
ся на мелководье вблизи впадения Кардонной балки в оз. Баскунчак описан
А.Б. Стрельцовым с соавторами, 2006). В небольшом количестве потребляют-
ся беспозвоночные: жуки (чернотелки, жужелицы, водолюбы, плавунцы, дол-
гоносики), клопы, саранчовые, личинки, слизни и некоторые другие. В свою
очередь и сам обыкновенный уж служит добычей ряда рыб, амфибий, репти-
лий, птиц и млекопитающих.
Численность. В типичных местообитаниях западной части региона он
является сравнительно обычным, а местами (локально) и многочисленным
видом. Так на трансекте 5x200 м, идущей по берегу р. Волги около грани-
цы Волжского и Ставропольского районов Самарской области отмечено
35 особей, что в пересчете составляет 350,0 экз/га (Бакиев и др., 2009). Южнее
в пойме Волгоградского водохранилища вблизи п. Ровное в мае 1996 г. его
численность составляла 12,0 экз. на 1 км береговой линии, а на пологом склоне
речной террасы - 3,0 экз/км (Табачишина, Табачишин, 2002). В Волго-Ахту-
бинской пойме на водоемах и их окрестностях вблизи г. Красный Яр весной
1998 г. плотность достигала 29,70±0,16 экз/га (Завьялов, Табачишин, 1998).
Здесь наблюдается явная тенденция к повышению численности, что связано
с сооружением искусственных водоемов, уменьшением химизации овощного
производства и разрушением животноводческой структуры (Кубанцев, 2003;
Чернобай, 2006). Как «значительная» оценивалась его численность и по доли-
не р. Урала (Параскив, Бутовский, 1960).
В северной части Волжско-Уральского междуречья на территории Орен-
бургской области он постоянно считается обычным видом (Райский, 1951;
Чибилёв, 1995). Южнее в Саратовском Заволжье у Ворфоломеевского водо-
хранилища на р. Б. Узень его плотность в 1996 г. составляла 0,52 экз/га (Та-
бачишина, Табачишин, 2002). Невысока его численнось и ниже по течению
этой реки, хотя иногда, как, например, в 1953 г. на оз. Рыбный Сакрыл, его
бывает довольно много. Несколько реже попадался он на берегу оз. Балыкты,
а по р. Горькая за 1950-1954 гг. отмечался лишь дважды (Ходашова, 1960).
Определенно редок он и в окрестностях оз. Баскунчак, где периодически от-
мечались лишь несколько особей (Стрельцов и др., 2006). Восточнее в Ка-
захстане, за исключением примыкающей к р. Урал территории, он повсемес-
тно считается малочисленным или редким видом (Параскив, 1956, Окулова,
1963).
Лимитирующие факторы. Разрушение и коренное преобразование ти-
пичных местообитаний в процессе хозяйственного и рекреационного осво-
ения территории, местами автотранспорт. На территории Карачаганакско-
го нефтегазоконденсатного месторождения загрязнение пруда Коншубай и
р. Березовка, где ужи исчезли, хотя на протекающей поблизости р. Утва встре-
чаются регулярно (Березовиков и др., 1992, 2003). Возможно, это связано и с
особенностями их трофики. По мнению Д.В. Магдеева (1999), на Самарской
Луке, «питаясь водными амфибиями, ужи аккумулируют в своем организме
значительные количества канцерогенных веществ, что ведет к изменениям фи-
зиологии внутренних органов и сказывается на репродуктивных способностях
особей из “загрязненных” популяций, которые со временем элиминируются»
(С. 196).
272
Глава 4
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор», Оренбургского и Богдинско-Баскунчакского заповедников. На
остальной территории специальных мер по охране не предпринималось.
Региональный статус. Типичный представитель герпетофауны региона,
населяющий специфические местообитания.
46. Водяной уж - Natrix tessellata (Laurenti, 1768)
Значение таксона для сохранения биоразнообразия. Один из двух пред-
ставителей обширного рода, населяющих территорию региона. Монотипичес-
кий вид. Выделяемым некоторыми исследователями по особенностям окраски
подвидам N. t. hydrus (Pallas, 1771), N. t. elaphoides Brandt, 1793, N. t. heinroth
Hecht, 1930 сейчас таксономического значения не придается; изменчивость
этого признака и различия в количестве брюшных, подхвостовых, предглаз-
ничных и заглазничных щитков рассматриваются как межпопуляционные
(Табачишин и др., 1996; Атлас пресмыкающихся..., 2004; Кузьмин, Семенов,
2006).
Распространение. Ареал вида охватывает большую часть Центральной и
Южной Европы, север Африки, Кавказ, Малую, Переднюю, Среднюю Азию
и Южный Казахстан до северо-западных регионов Китая на востоке и Пер-
сидского залива на юге (европейский фаунистический комплекс). Его север-
ная граница на Восточно-Европейской равнине прослеживается от Молдавии,
по югу Украины, северу Самарской Луки, югу Башкирии, среднему течению
р. Урал, а затем в Казахстане по среднему течению рек Тургай и Сарысу, Се-
верному Прибалхашью, окрестностям оз. Алаколь и далее в Китае, откуда по-
ворачивает на юг к побережью Персидского залива. Таким образом, большая
часть региона (за исключением его северо-восточной части) входит в пределы
ареала вида.
В регионе и его ближайших окрестностях более точно северная граница
ареала прослеживается по следующим пунктам: в окрестностях п. Старая Май-
на и по долине р. Б. Черемшан в Ульяновской области, городах Бирск и Ишим-
бай в Башкирии, окрестностях п. Чебеньки на р. Сакмара, поселках Донское,
Гирьял и Гипсрудник Беляевского района (Гавлюк и др., 1993), п. Ильинка
Кувандыкского района, подножья Губерлинских гор и у г. Орск Оренбургской
области (Райский, 1951; Чибилёв, 1995,1999а).
Южнее этих пунктов водяной уж обитает по долинам рек Волга, Самара,
Урал, Илек, Эмба, в Волжско-Уральском и Урало-Эмбенском междуречьях,
Мугоджарах, Северном Устюрте, побережье Каспийского моря, Северном
Приаралье и на побережье Аральского моря. Его конкретные местонахож-
дения известны в 2 (18,2% из 11) ландшафтных районах лесостепной зоны,
16 (23,2% из 69) степной, 8 (25,0% из 32) полупустынной и 15 (34,9% из 43)
пустынной зоны (табл. 52, карта 45).
Места обитания. Водяной уж - типичный обитатель побережий водо-
емов. В Самарской области чаще всего он населяет обрывистые, скалистые
берега рек, причем от воды уходит не далее 300 м (Бакиев и др., 2009). В Са-
ратовском и Волгоградском Заволжье он, по-видимому, встречается лишь по
устьевым участкам крупных притоков Волги - Б. Иргиз, Еруслан (Шляхтин
и др., 2005, 2006), а ниже по течению, особенно в Волго-Ахтубинской пойме,
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
273
Таблица 52
Распространение водяного ужа
Зона Провин- ция Район № точки Местонахождение Источник информации
5 X с ё о о 1 1 1 Самарская область г. Самара: Кировский р-н Красноглинский р-н Бакиев и др., 2003
о II 5 2 Оренбургская область окр. п. Рычковка Чибилбв А.А.
IV 13 14 3 4 Саратовская область низ р. Б. Иргиз дол. р. Волги Шляхтин и др., 2005, 2006
V 24 25 28 29 5 6 7 8 9 Оренбургская область р. Каргала окр. с. Нижнеозерное окр. п. Чебеньки окр. п. Красногор окр. с. Донское окр. с. Гирьял окр. п. Гипсрудник окр. г. Оренбург Никольский, 1905; Чибилёв, 1995; Райский, 1951; Гавлюк и др., 1993 Зарудный, 1895
Степная VII 35 37 44 46 10 11 12 13 Западно-Казахстанская область оз. Сулу коль Актюбинская область дол. р. Киил окр. г. Актюбинска дол. р. Уил Зарудный, 1895
VIII 48 54 55 14 15 16 17 Волгоградская область низ. р. Еруслан Оренбургская область окр. п. Студеное Западно-Казахстанская область окр. п. Январцево р. Шаган, г. Уральск Шляхтин и др., 2005 Зарудный, 1895 Определитель земновод- ных..., 1977
IX 60 61 62 18 19 20 Оренбургская область окр. г. Орска р. Урал, Гу берлинские горы, с. Ильинка Айтуарская степь Райский, 1951; Чибилёв, 1995, 1999а
274
Глава 4
Продолжение табл. 52
Зона Провин- ция Район № точки Местонахождение Источник информации
XIII Западно-Казахстанская область
88 91 низ. р. Б. Узень окр. п. Круглый Параскив, Бутовский, 1960
Актюбинская область
5 X XV 96 97 22 23 дол. р. Эмба р. Маннесай Неручев, Васильев, 1978; Чернов, 1954; Шилов, 1961
£ р. Шаган
& 100 24 овр. Курсай
Ь-< 101 25 оз. Ащеколь
кн XVI 26 горы Мугоджары окр. п. Алабас п. Шахты
XIX по 111 27 28 ср. теч. р. Тургай р. Жиланшик, зим. Кушек Параскив, 1956; Дуйсе- баева, 2005
112 29 р. Тургай, ур. Жаркам ыс
Астраханская область
114 30 31 ст. Досанг, п. Замьяны окр. г. Красный Яр Гурьевская область Бобров, 1985; Завьялов, Табачишин, 1998
XX 32 окр. п. Танюшкино Накаренок, 2003в
Западно-Казахстанская область
3 X 124 33 окр. п. Харькино окр. п. Калмыкове Параскив, Бутовский, 1960; Чернов, 1954; Окулова, 1963
X Гурьевская область
о £ XXI 126 127 34 65 окр. ст. Исатай окр. п. Жилая Коса берег Каспия ур. Терен-Узек Сараев, Пестов, 2010; Неручев, Васильев, 1979; Сараев, Пестов, 2010; Гисцов, Березови- ков, 1995
129 36 окр. п. Аккистау
132 37 окр. г. Гурьев (Атырау); 6 км к сев. г. Гурьев
окр. п. Дамба
38 канал Зарослый, прот. Сокол, п-ов Пешной
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
275
Окончание табл. 52
Зона Провин- ция Район № точки Местонахождение Источник информации
133 39 окр. п. Индерборский оз. Индер Чернов, 1954
135 40 р. Уил, п. Коныстану Сараев, Пестов, 2010
136 41 дол. р. Эмба ур. Кзыл-Жар ур. Кой-Сарай ур. Дюсеке
5 XXII 137 138 42 43 р. Эмба, ур. Акмечеть р. Эмба, ур. Жанбике ур. Торенан Неручев, Васильев, 1978; Сараев, Пестов, 2010; Накаренок, 2003в; Шилов, 1961
а а 44 ур. Бали
В ур. Шолькара
с 139 45 горы Жельтау сев. чинк, ур. Жимбулак мог. Шайтас
XXIV 146 46 ур. Донгузтау
154 47 Приаральские Каракумы Лобачев и др., 1973
155 48 окр. п. Куланды Динесман, 1953
XXVI 49 50 п-ов Кара-Тюп оз. Камышлыбаш окр. п. Раим р. Сырдарья Дуйсебаева, 2005
встречается лишь непосредственно около воды (Завьялов, Табачишин, 1998).
Его более тесная по сравнению с таковой обыкновенного ужа привязанность к
водоемам в дельте Волги отмечалась еще А.Н. Никольским (1905).
Отчетливо выраженное предпочтение высоким обрывистым берегам, сло-
женным песчаниками и меловыми отложениями с сетью трещин и корневых
ходов, наблюдалось и по среднему течению р. Урал (Чибилёв, 1995). Ниже по
течению этой реки водяной уж встречается преимущественно по пойменным
озерам и старицам, а в Волжско-Уральском междуречье он отмечался в низо-
вьях р. Б. Узень и по Камыш-Самарским озерам (Параскив, Бутовский, 1960).
Привязанность к речным долинам замечена и в Урало-Эмбенском междуречье,
где он обнаружен по долинам рек Уил, Эмба и Маннесай. В низовьях послед-
ней он поселяется у запруд с обильной прибрежно-водной растительностью и
небольших оставшихся плесиков по пересыхающему руслу, откуда, по-види-
мому, попадает в колонии песчанок (Шилов, 1961; Неручев, Васильев, 1978;
Сараев, Пестов, 2010). Интересной представляется и находка погибшего ужа в
соленом оз. Индер (Чернов, 1954).
276
Глава 4
В Зауралье водяной уж населяет долины рек Тургай, Жыланшик и зарос-
шее рогозом озеро Камышлыбаш в низовьях р. Сырдарья (Дусебаева, 2005).
Встречается этот уж и на побережье Каспия, где его можно наблюдать как
на песчано-галечниковой прибрежной полосе с зарослями солянок, так и на ка-
менистых участках. Сходных мест придерживается он и на побережье Араль-
ского моря (Параскив, 1956).
Образ жизни. Из зимней спячки в регионе выходит в разных числах второй
половины апреля - начале мая, когда температура воздуха превышает 10 °C.
Обычно активен в течение всего светлого времени суток (7-21 час), что опре-
деляется его более высокой термофильностью по сравнению с таковой обыкно-
венного ужа (оптимальной считается температура субстрата 20...27 °C) и тем,
что при температуре воздуха выше 28 °C большую часть времени он проводит
в воде (в том числе в реках с быстрым течением и в море, удаляясь от берега на
несколько километров). В качестве укрытий использует расщелины в грунте,
пустоты под камнями, пнями, деревьями, норы грызунов, корневые ходы.
Со второй половины августа активность водяных ужей снижается, а в кон-
це сентября - начале октября они (сначала самцы) уходят на зимовку. Период
активности, таким образом, продолжается около 160-180 суток, что несколько
меньше, чем у обыкновенного ужа. Зимует на территории Самарской области,
по-видимому, главным образом в карстовых пустотах, других подобных мес-
тах, также в разных подземных убежищах, нередко скоплениями в несколько
сот особей (иногда совместно с обыкновенным ужом).
Размножение. Половозрелым водяной уж становится на третьем-четвер-
гом году жизни. В этом возрасте, вскоре после выхода из зимовки, обычно в
конце апреля - начале мая при температуре воздуха около 18 °C начинается
спаривание, ко времени которого ужи собираются в пары или образуют «брач-
ные хороводы», в которых участвуют до 15-20 особей. Яйца (4-23) откладыва-
ются в июне - начале июля; инкубация в естественных условиях продолжается
40-52 суток; и с конца июня начинается выход сеголеток, который продолжа-
ется и в сентябре (Кубанцев и др., 1962; Бакиев и др., 2004, 2009; Шляхтин и
др., 2005).
Питание. Основу пищевого рациона водяных ужей в большинстве водо-
емов региона составляет рыба (до 60-93% по массе), видовой состав и разме-
ры которой существенно меняются по сезонам, в зависимости от численности,
доступности пищевых объектов и активности ужей. Кроме того, потребляются
земноводные (преимущественно озерные лягушки, чесночница, реже зеленые
жабы, жерлянки и их личинки), в ряде мест являющиеся основной добычей, а
также водные беспозвоночные и их личинки. В Предустюртье, где свойствен-
ная водяным ужам пища отсутствует, он перешел на питание мышевидными
грызунами и большими песчанками, в колониях которых (причем иногда вда-
ли от свойственных этим змеям местообитаний) иногда встречается (Шилов,
1961). В свою очередь водяные ужи имеют большое количество естественных
врагов - главным образом хищных птиц и млекопитающих, реже он служит
объектом добычи крупных рыб, некоторых земноводных и пресмыкающихся.
Численность. В типичных местообитаниях водяной уж является обычным,
а местами и многочисленным видом. Так, по Волге в районе Самарской Луки у
мест зимовки, в конце апреля - начале мая 1991-2000 гг. его плотность достиг-
ла 20-30 экз/га (Бакиев и др., 2009). Южнее, по средней зоне Волгоградского
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
277
водохранилища, она составляла 3-17 ос/га (Шляхтин и др., 2005), а в Волго-
Ахтубинской пойме весной 1998 г. - 4,90±0,19 экз/га (Завьялов, Табачишин,
1998). В последние годы численность его здесь постоянно увеличивается, чему
способствует создание искусственных водоемов (Кубанцев, 2003).
По среднему течению р. Урал в Переволоцком районе Оренбургской об-
ласти на участке обрывистого берега протяженностью 300 м в мае 1962 г. было
насчитано более 90 только убитых ужей (Чибилёв, 1995). Усеяные трупами во-
дяных ужей берега р. Урал отмечались и в других местах по среднему течению
этой реки. Вместе с тем за его пределами даже в начале XX в. водяной уж был
редок (Райский, 1951).
Численность водяных ужей увеличивается во влажные, особенно следую-
щие один за другим, годы. Так в 1950-1951 гг. по долине р. Урал у п. Круглый
ужей было сравнительно немного, во влажном 1952 г. их численность заметно
увеличилась, а в следующем влажном 1953 они были очень многочисленны
(Параскив, Бутовский, 1960). Обычным в начале 1950-х гг. водяной уж был
и в низовьях р. Урал (Чернов, 1954), по руслам пересыхающих рек Шаган и
Маннесай, а местами на небольших прудах по их течению очень многочис-
ленным (Шилов, 1961); весьма обычен этот вид и по долине р. Эмба (Неручев,
Васильев, 1978).
В Зауралье численность, по-видимому, подвержена довольно значитель-
ным колебаниям. Во всяком случае по р. Тургай на протяжении почти всего
XX столетия он не наблюдался, а в мае 2002 г. здесь отмечена его довольно
высокая плотность; частые встречи регистрировались в это время и в дельте
р. Сырдарьи (Дуйсебаева, 2005). На побережье Каспийского и Аральского
морей водяной уж, по-видимому, немногочислен, а на остальной-территории
представлен лишь небольшими изолированными популяциями.
Лимитирующие факторы. Действие комплекса факторов у северных пре-
делов ареала. Застройка, хозяйственное и рекреационное освоение типичных
местообитаний, необоснованное преследование людьми.
Охрана. Обитает и охраняется на территории Оренбургского заповедника.
На остальной территории региона специальные меры охраны не применялись.
Строительство новых гидросооружений в ряде мест способствует расширению
местообитаний.
Региональный статус. Характерный для региона представитель герпето-
фауны, населяющий ограниченные местообитания. Занесен в Красные книги
Саратовской (2006), Самарской (2009) областей и сопредельной Башкирии.
Род 31. Psammophis Boie In Fitzinger, 1826 - песчаные змеи
47. Стрела-змея - Psammophis lineolatus (Brandt, 1838)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель обширного рода, населяющий территорию региона. Монотипичес-
кий вид.
Распространение. Ареал вида охватывает южные районы Казахстана,
Среднюю Азию, юго-запад Монголии и Китая, север Пакистана, Афганис-
тана, Ирана и юго-восточное Закавказье (среднеазиатский фаунистический
278
Глава 4
комплекс). Его северная граница от окрестностей г. Гурьева прослеживается
вблизи п. Макат, через низовья р. Сагиз, гряду Актолагай, среднее течение
р. Эмба, горы Шолькара, Жельтау, пески Шогырлыкум (Сараев, Пестов, 2010),
Б. Барсуки, низовья р. Тургай, пески Арыскум, Муюнкумы, северное Прибал-
хашье, Зайсанскую котловину (Параскив, 1956; Брушко, Кубыкин, 2000) и да-
лее в Китай и Монголию. Южная граница проходит по пустыням Пакистана,
Афганистана и Ирана. Таким образом, ареал вида охватывает только южную
часть региона и все отмеченные пункты (вплоть до низовий р. Тургай) опреде-
ляют его северные пределы распространения в исследуемом регионе. Южнее
стрела-змея известна на п-ове Бузачи, в окрестностях сора Кайдак, по нижней
Эмбе, Северному Устюрту, побережью Аральского моря и в Приаральских Ка-
ракумах. Конкретные местонахождения приведены в табл. 53 и на карте 46.
Таблица 53
Распространение стрелы-змеи
Зона Провин- ция Район № точки Местонахождение Источник информации
Полу- пустын- ная XVIII 107 1 Актюбинская область окр. с. Жайсанбай Брушко, Кубыкин, 1989
XIX 112 2 низ. р. Тургай Никольский, 1915
Гурьевская область
XXI 127 3 ур. Теренозек Брушко, Кубыкин, 2000
131 4 окр. п. Искининский
132 5 22 км к сев.-вост. от г. Гурьева
136 6 низ. р. Сагиз, окр. п. Макат окр. п. Кенбай гора Кой-Кара Сараев, Пестов, 2010; Неручев и др., 1981, 1984
7 окр. оз. Бартылдакты ур. Брлик
3 X им гора Аккерегешин
►С 8 окр. п. Косчагыл
£ XXII 137 9 10 окр. п. Кульсары горы Актолагай п. Аралтюбе ур. Жанбике Неручев, Шатилович, 1985; Сараев, Пестов, 2010; Брушко, Кубыкин, 1988, 2000
11 ур. Кыргынтюбе окр. сора Есенжал
138 12 горы Шолькара горы Жельтау Неручев и др., 1981, 1984; Сараев, Пестов, 2010
13 горы Коленкелы Прикаспийские Каракумы
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
279
Окончание табл. 53
Зона Провин- ция Район № точки Местонахождение Источник информации
Мангистауская область
XXII 14 пески Шагырлыкум Киреев, 1981; Накаре-
15 Прикаспийские Каракумы нок, 2003в
140 16 п-ов Бузачи Никольский, 1915
XIII 143 17 окр. сора Кайдак, бывш. Ново- александровское укрепление
147 18 ур. Карын-Жарык; залив Бутакова
XXV 148 19 пески Сам, кол. Тулеп, кол. Кугусем
окр. оз. Тугаракчан
3 я 3 В 150 151 20 21 сор. Кошкарата Б. Барсуки, окр. п. Южное Дуйсебаева, 2005; Бруш- ко, Кубыкин, 2000
£ XXVI 152 154 22 23 ур. Озектык р. Ашудасты М. Барсуки п. Акеспе Приаральские Каракумы Параскив, 1956; Лобачев и др., 1973; Никольский, 1915; Динесман, 1953; Дуйсебаева, 2005
155 24 сев. берег Аральского моря, ур. Кара-Тамак, Исеншагыл
чинк зал. Перовского
25 п-ов Кара-Тюп окр. п. Тас-Тюбек
26 окр. с. Каратерень
Места обитания. В регионе населяет типичные для вида местообитания.
В М. Барсуках отмечалась на еркеково-полынной равнине у подножья полу-
закрепленного бархана. В Б. Барсуках ее наблюдали на закрепленных барха-
нах и глинистых такырах. В глинистой пустыне (причем на сильном сбое) она
встречалась в Приэмбенской провинции; здесь же ее встречи известны на ка-
менистых склонах гор-останцов, в окрестностях сора Кайдак у солончаков, а у
Аральского моря отмечалась и по береговым обрывам (Параскив, 1956; Неру-
чев и др., 1981; Дуйсебаева, 2005).
Образ жизни. После зимовки появляется в разных числах апреля. Наи-
большую активность проявляет при температуре грунта около 28 °C. Ведет
преимущественно дневной образ жизни. В качестве убежищ использует пус-
тоты между камнями, в развалинах строений, норы грызунов. Активность со-
храняет около 170-180 суток, обычно до середины октября.
Размножение. На сопредельной территории спаривание наблюдается в
апреле-мае. Яйца (2-11) откладываются с конца июня до начала августа, а вы-
ход молодых происходит с середины августа.
280
Глава 4
Питание. Стрела-змея - узкоспециализированный герпетофаг - питает-
ся главным образом ящурками, агамами, круглоголовками; в меньшей мере
другими видами. Основным способом охоты является подкарауливание. Мо-
лодые питаются насекомыми. В свою очередь стрела-змея добывается бо-
лее крупными рептилиями, хищными птицами и некоторыми млекопитаю-
щими.
Численность. В казахстанской части ареала считалась сравнительно обыч-
ным видом (Параскив, 1956), а в Приаральских Каракумах - даже массовым
(Лобачев и др., 1973). В последние годы численность, по-видимому, сократи-
лась, поскольку в Северном Приаралье, например, за 2 полевых сезона (май-
июнь 2002 и 2003 г.) встречено лишь 4 особи (Дуйсебаева, 2005). В Приэмбен-
ской пустыне ее численность определена в 0,5-1,0 ос/10 км, а в Прикаспийских
Каракумах 1,0 (Накаренок, 2003 в).
Лимитирующие факторы. Хозяйственное освоение территории (сельхоз-
работы, автотранспорт), уничтожение людьми.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На остальной территории состояние популяций принятия особых специальных
мер охраны не требует.
Региональный статус. Узкоспециализированный представитель герпето-
фауны пустынь; северные популяции краеареальные.
Семейство 17. Viperidae Laurenti, 1768 - Гадюковые змеи, или гадюки
Род 32. Gloydius Hoge et Romano-Hoge, 1981 - щитомордники
48. Обыкновенный щитомордник, или щитомордник Палласа -
Gloydius halys (Pallas, 1776)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода, населяющий территорию региона. Политипический вид; в ре-
гионе представлен подвидом G. h. caraganus (Eichwald, 1831), отличающимся
светлой окраской и рисунком темных полос на туловище (Атлас пресмыкаю-
щихся..., 2004).
Распространение. Ареал вида простирается от низовий р. Волги и юго-
восточного Закавказья через Казахстан, Среднюю Азию и юг Сибири до побе-
режья Тихого океана (восточно-палеарктический фаунистический комплекс).
Его южная граница проводится от юго-восточных районов Азербайджана че-
рез север Ирана, Афганистана и Китай. Северная до настоящего времени четко
не определена. По старым данным она проводилась от Саратовского Заволжья
(Шляхтин, Голикова, 1986) немногим южнее верховий р. Утва (Чингирлау),
ур. Куагаш в верховьях р. Калдыгайты, окрестностях г. Актюбинска (Николь-
ский, 1905), вблизи г. Иргиза (Динесман, 1953), ур. Акмин на р. Тургай и далее
через окрестности г. Семипалатинска по югу Сибири к тихоокеанскому по-
бережью. Считалось, что к югу от этих пунктов щитомордник в регионе рас-
пространен повсеместно (Параскив, 1956). Однако последующими исследова-
ниями его обитание в Саратовском Заволжье не подтверждено (Табачишина
и др., 2004), нет также новых сведений о его обитании в верховьях рек Утва,
Калдыгайты и у г. Актюбинска.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 281
Достоверно распространение щитомордника в регионе в последние годы
определяется находками в окрестностях ст. Досанг, Индерских горах, на ос-
танцовых горах по р. Эмба — Аккерегешин, Иманкара, Актолагай, Шолька-
ра, Жельтау, чинке Донызтау и в его окрестностях (Накаренок, 2003 в; Сараев,
Пестов, 2010), 71 км к северу ст. Чел кар и долине р. Жыланшик (Дуйсебаева,
2005). Южнее его встречи в регионе известны в окрестностях г. Гурьева, по
р. Эмбе, в Прикаспийских Каракумах, окрестностях сора Кайдак (бывшее ук-
репление Новоалександровск), п-ове Бузачи, Северном Устюрте, Б. Барсуках,
северо-западном и северном побережьях Аральского моря, Приаральских Ка-
ракумах и окрестностях г. Аральска. В целом его конкретные местообитания
известны в 3 (4,4% из 69) ландшафтных районах степной зоны, 5 (15,6% из 32)
полупустыни и 13 (30,2% из 43) районах пустынной зоны (табл. 54, карта 47).
Таблица 54
Распространение щитомордника
Зона Провин- ция Район № точки Местонахождение Источник информации
Западно-Казахстанская область
5 X VII 37 1 верх. р. Утва (исчез?) Никольский, 1905
к н 40 2 ур. Куагаш (исчез?)
О Актюбинская область
44 3 окр. г. Актюбинск Параскив, 1956
3 XV 97 4 окр. р. Шатан Никольский, 1905
X X XVIII 107 5 71 км к сев. от ст. Челкар Дуйсебаева, 2005
ё & 110 6 р. Тургай, ур. Акмин Параскив, 1956
п* Q XIX 111 7 окр. р. Жыланшик Смирновский, 1963
С 112 8 окр. г. Иргиз Динесман, 1953
Астраханская область
XX 114 9 окр. ст. Досанг Накаренок, 2003в
10 окр. г. Красный Яр Чернов, 1954
Гурьевская область
XXI 127 11 80 км к юго-зап. от п. Куль- Брушко, Кубыкин, 1998;
в? сары Сараев, Пестов, 2010;
cd X X 132 12 окр. г. Гурьев Чернов, 1954; Неручев и др., 1984
в 133 13 Индерские горы
£ 136 14 гора Аккерегешин гора Иман-Кара
XXII 15 окр. п. Косчагыл окр. п. Кульсары
137 16 горы Актолагай Сараев, Пестов, 2010
17 гора Каражар
282
Глава 4
Окончание табл. 54
Зона Провин- ция Район № точки Местонахождение Источник информации
138 18 19 гора Шолькара горы Жельтау Накаренок, 2003в; Па- раскив, 1956
139 20 Прикаспийские Каракумы
XXII 21 Мангистауская область ур. Кызылкуз чинк Донызтау Зап. Устюрт Параскив, 1956; Завья- лов и др., 2006
3 X X i XXIII 140 144 22 23 п-ов Бузачи окр. сора Кайдак
С 151 24 пески Б. Барсуки Лобачев и др., 1973
154 25 Приаральские Каракумы
155 26 п-ов Кара-Тюп Сидоров, 1925
XXVI 27 п-ов Куланды Дуйсебаева, 2005
28 окр. родн. Каска-Джо л ур. Достан окр. п. Екеспе
29 окр. г. Аральск Динесман, 1953
Места обитания. Щитомордник не только широко распространенный, но
и эвритопный вид. Его типовой (и типичной) территорией являются Волжс-
ко-Уральские пески в окрестностях г. Красный Яр; в песчаных массивах он
отмечался также у северных пределов своего распространения в верховьях
рек Калдыгайты и Утва, затем в Прикаспийских и Приаральских Каракумах,
Б. Барсуках, причем везде в этих биотопах он придерживается закрепленных
злаково-полынными сообществами участков или колоний песчанок.
По-видимому, вторым по значимости местообитанием в регионе являются
поросшие полынями и биюргуном каменистые склоны останцовых гор, а так-
же карстовые воронки и разные ниши в Индерских горах, чинки Донызтау и
подобные им глинисто-песчаные обрывы коренного берега Аральского моря.
Не избегает он засоленных биотопов в окрестностях сора Кайдак и по обсох-
шему дну Аральского моря, занятых полынно-кермековыми группировками.
Все эти местообитания он явно предпочитает глинистым пустыням, хотя из-
редка встречается и в них, как по р. Эмба, так и на побережье р. Жиланшик
(Чернов, 1954; Неручев, Васильев, 1978; Дуйсебаева, 2005).
Образ жизни. После зимовки появляется в разных числах апреля. Весной
активен в течение всего светлого времени суток, летом преимущественно в су-
мерки и ночью, хотя в пасмурную погоду выходит и днем. В качестве убежищ
используются норы, пустоты в грунте, прячется среди камней или развалин;
временными укрытиями могут служить и кусты. Период активности обычно
продолжается до середины октября (хотя иногда задерживается до первой де-
кады ноября)-т. е. около 180-200 суток.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
283
Размножение. Щитомордник - яйцеживородящий вид. Спаривается он в
апреле-мае, молодые (3-14) появляются с конца июля.
Питание. Основу пищевого рациона взрослых особей составляют мелкие
позвоночные животные, преимущественно грызуны (полевки, мыши, хомячки,
тушканчики), птицы, ящерицы (ящурки, круглоголовки); молодые (первого и
второго годов жизни), вероятно, питаются в основном насекомыми и другими
беспозвоночными. В свою очередь щитомордников потребляют более круп-
ные рептилии, некоторые хищные птицы и звери.
Численность. По-видимому, существенно изменяется по годам. Так, в ок-
рестностях Аральского моря в 1920-1921 гг. он попадался «довольно редко»
(Сидоров, 1925), в 1945 и 1946 гг. вблизи г. Аральска, а в 1946 г. и вблизи
г. Иргиз (у северной границы ареала) щитомордников было очень много, а уже
год-два спустя (в 1947-1948 гг.) они опять попадались довольно редко (Динес-
ман, 1953). В последние годы, судя по контексту сообщения Т.Н. Дуйсебаевой
(2005), они здесь не многочисленны, хотя и не представляют собой редкости.
Сравнительно обычным видом считается щитомордник и в Приаральских Ка-
ракумах (Лобачев и др., 1973).
В западной части ареала в 1950 г. много щитомордников было в Индерских
горах, где за часовую экскурсию перед заходом солнца на песчано-глинистом
северном берегу озера было отмечено девять особей (Чернов, 1954). Судя по
количеству мест обитания, не представляют они особой редкости в песках и
по останцовым горам в Приэмбенской провинции, хотя в глинистой пустыне
встречаются чрезвычайно редко (Неручев, Васильев, 1978; Накаренок, 2003в;
Сараев, Пестов, 2010). В Волжско-Уральском междуречье он, возможно, уже
исчез, хотя в 1990-е гг. имелись достаточно достоверные (но документально не
подтвержденные) сведения о его встречах в районе Богдинско-Баскунчакского
заповедника (Божанский, Полынова, 1998).
Лимитирующие факторы. В основном хозяйственное освоение террито-
рии, сопровождающееся уничтожением змей.
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резерва-
та. На остальной территории региона специальные меры охраны не применя-
лись.
Региональный статус. Характерный представитель пустынной герпето-
фауны, населяющий изолированные специфические местообитания у север-
ных пределов ареала.
Род 33. Vipera (Pelias) Laurenti, 1768 - гадюки
49. Обыкновенная гадюка - Vipera (Pelias) berus (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Политипический
вид, в регионе представленный номинативным подвидом V. b. berus (Linnaeus,
1758), в пределах которого наблюдаются некоторые межпопуляционные раз-
личия в окраске тела (цветовые морфы), количестве брюшных и подхвостовых
щитков (Табачишин и др., 1996; Морозенко и др., 2003; Атлас пресмыкающих-
ся..., 2004; Кузьмин, Семенов, 2006).
284
Глава 4
Распространение. Ареал вида охватывает значительную часть Евразии от
Атлантики до долины р. Лены и северо-восточного Китая (европейский фау-
нистический комплекс). Его северная граница от побережья Баренцева моря
на Кольском полуострове постепенно смещается к 60° с.ш. в долине р. Енисей,
затем несколько южнее, но по долине р. Лена вновь продвигается до 60-й па-
раллели. К югу распространена практически до границ степной зоны. Таким
образом, обыкновенная гадюка населяет только северную часть исследуемого
нами региона.
Южная граница ареала вида в регионе и его ближайших окрестностях в
1970-е гг. проводилась по лесным территориям в окрестностях г. Волгограда
(Кубанцев, 2003), сейчас она прослеживается по находкам вблизи г. Хвалынс-
ка Саратовской области, затем в черте г. Самары, долинах р. Самара у п. Бор-
ский Самарской области (Бакиев и др., 2009), р. Боровки в Бузулукском бору,
р. Самары у п. Тоцкое, долинах рек М. Кинель и Б. Кинель вблизи г. Бугурус-
лан, затем в верховьях р. Ик в Абдулинском и р. Демы в Шарлыкском районах
Оренбургской области, хр. М. Накас (Даркшевич, 1953; Райский, 1951; Чи-
билёв, 1995; Вершинин, 2007) и далее уже за пределами региона в Башкирии.
Кроме отмеченных, по сути пограничных, пунктов встречи обыкновен-
ной гадюки известны в окрестностях поселков Подстепки, Кинель-Черкас-
ский (Гаранин, 1983), сел Старое Якушкино и Красная Богана в верховьях
р. Б. Черемшан Самарской области (Бакиев и др., 2009). К настоящему вре-
мени в пределах региона обыкновенная гадюка известна в 8 (77,7% из 11)
ландшафтных районах лесостепной и 2 (2,9% из 69) степной зоны (табл. 55,
карта 48).
Места обитания. Обыкновенная гадюка - типичный обитатель лесных
и лесостепных ландшафтов, где придерживается преимущественно окраин
опушек и полян с кустарниками, зарастающих вырубок и гарей, просек, до-
рог, хорошо прогреваемых стаций по берегам рек, озер и подобных экотонов.
В пределах региона практически все известные пункты встреч приурочены
к долинам рек Волга, Самара, Б. и М. Кинели, Сок, Б. Черемшан, Ик, Дёма,
Боровка и речек, стекающих с хребтов Накас и Шайтантау. Здесь она насе-
ляет поросшие кустарниками побережья пойменных озер, влажные поймен-
ные участки с травянисто-кустарниковой растительностью, пойменные луга и
выпасы. В целом выбор местообитаний помимо степени влажности биотопов
(лимитирующей распространение вида на юг), их инсоляции и наличия ук-
рытий определяется также обеспеченностью кормом и степенью беспокойс-
тва. Разрушение местообитаний и постоянное беспокойство приводят, с одной
стороны, к инсуляризации ареала, распадающегося на изолированные участки,
а с другой - к вытеснению гадюк в непригодные для деятельности человека
угодья (овраги, свалки, пустоши, развалины строений и т. п.), что уже четко
прослеживается в лесопарковой зоне г. Самары и на Урале, которые становят-
ся своеобразными рефугиями вида в антропогенно трансформированной среде
(Чибилёв, 1995; Бакиев и др., 2004, 2009; Вершинин, 2007).
Образ жизни. Весной в регионе обыкновенная гадюка появляется в ап-
реле-начале мая, когда местами еще сохраняются значительные пятна снега.
Обычно это совпадает с переходом среднесуточных температур через диапа-
зон от 0 до 5 °C. Первыми из зимних убежищ выходят самцы, спустя несколько
дней самки, а затем молодые особи.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
285
Таблица 55
Распространение обыкновенной гадюки
Зона Провин- ция Район № точки Местонахождение Источник информации
Лесостепная I 1 2 1 2 3 Самарская область окр. п. Подстепки г. Самара р-н Кировский р-н Красноглинский р-н Куйбышевский р-н Октябрьский окр. п. Старосемейкино Гаранин, 1983; Бакиев и др., 2002, 2003, 2009
II 3 4 5 6 7 окр. с. Красная Багана дол. р. Б. Черемшан окр. с. Старое Якушкино окр. п. Кинель-Черкасский
III 5 6 8 10 11 8 9 10 11 12 13 14 15 Оренбургская область верх. р. Ик окр. г. Бугуруслан окр. р. М. Кинель верх. р. Дема хр. М. Накас дол. р. Б. Ик дол. р. Сакмара хр. Шайтантау Чибилёв, 1995; Верши- нин, 2007
Степная V 22 23 16 17 18 Самарская область окр. п. Борское, Бузулукский бор Оренбургская область Бузулукский бор, дол. р. Боровка дол. р. Самара, окр. п. Тонкое Бакиев и др., 2002, 2009 Положенцев, 1935; Дарк- шевич, 1954; Чибилёв, 1995
Весной гадюки активны преимущественно в середине дня, летом актив-
ность смещается на утренние и вечерние часы, при пасмурной погоде они
могут быть встречены и в дневное время (причем возможно несколько пиков
активности), а в прохладные и дождливые дни почти не выходят из убежищ.
Летом в качестве убежищ используются норы, гнилые пни, трещины и пусто-
ты в грунте. Как правило, активность наблюдается при температуре воздуха
17,0...27,0 °C, субстрата 21,0...30,5 °C, а абсолютный оптимум в Поволжье со-
ставляет 28,8...30,8 °C, а максимум 37 °C. При этом самцы предпочитают более
теплый субстрат, чем самки. Различия в термоадаптации отмечаются также у
темных и светлых морф.
286
Глава 4
Обыкновенная гадюка - территориальная форма и чаще всего держится на
участках от 500-800 м2 до 1,5-4,0 га, совершая незначительные перемещения,
хотя во время сезонных миграций отдельные особи иногда могут перемещать-
ся до 2-5 км.
К осени активность гадюк снижается, и уже во второй половине сентября -
первой половине октября они залегают в спячку, которую проводят в грудах
хвороста, прикорневых и других пустотах грунта, норах животных, карстовых
воронках и подобных местах ниже уровня промерзания грунта, иногда образуя
скопления в несколько сот особей. Период активности, таким образом, про-
должается около 160 дней, в среднем около 140 (Гаранин, 1983; Бакиев и др.,
2004, 2009).
Размножение. Половой зрелости обыкновенная гадюка достигает в возрас-
те 4-5 лет. Достигшие этого возраста особи спустя 2-4 недели после выхода из
зимовки приступают к спариванию. Сроки его наступления и продолжитель-
ность (чаще всего около 2 недель) зависят главным образом от особенностей
биотопа и его микроклиматических условий (обычно после перехода средних
температур через 10 °C). Эмбриональное развитие продолжается 2,5-3,0 неде-
ли, и во второй половине июля - начале августа в Волжско-Камском крае появ-
ляется от 4 до 23 молодых. Размножается обыкновенная гадюка не ежегодно, в
разных частях ареала выявлены 2-, 3- и 4-летние циклы.
Питание. Основу пищевого рациона обыкновенной гадюки в регионе
(около 80%) составляют мелкие млекопитающие (преимущественно полевки,
мыши, замлеройковые), в меньшей мере потребляются амфибии (практически
все местные виды), ящерицы, ужи, изредка в составе пищи отмечаются насе-
комые, слизни и растительные остатки. В свою очередь обыкновенные гадюки
добываются рядом птиц, млекопитающих и других более крупных рептилий
(известно и явление каннибализма).
Численность. В связи с некоторыми трудностями определения в полевых
условиях обыкновенной гадюки, гадюки Никольского (видовая самостоятель-
ность которой некоторыми авторами отрицается) и восточной степной гадюки
старые данные о численности вида использовать не представляется целесо-
образным. В связи с этим отметим лишь, что в черте г. Самары (лесопарко-
вая зона у Горелого хутора) в 1993-2001 гг. плотность этого вида достигала
40-50 экз/га (локально до 125), что в переводе на данные маршрутного учета
составляет 20-25 экз/км (Бакиев и др., 2009).
Лимитирующие факторы. Комплекс факторов, действующих у южной
границы ареала. Разрушение местообитаний, хозяйственное и рекреационное
освоение территории, уничтожение человеком.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной территории специальных мер охраны не предпри-
нималось.
Региональный статус. Типичный представитель герпетофауны реги-
она у южных пределов ареала. Занесена в Красную книгу Самарской облас-
ти с категорией 5 Б как условно редкий вид со снижающейся численностью
(2009).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
287
50. Гадюка Никольского, или лесостепная гадюка -
Vipera nikolskii Vedmederja, Grubant and Rudaeva, 1986
Значение таксона для сохранения биоразнообразия. Один из представи-
телей комплекса Vipera berus, населяющий территорию региона. В качестве
самостоятельного вида выделена сравнительно недавно (Грубант и др., 1973;
Ведмедеря и др., 1986), хотя ряд герпетологов считает ее лишь подвидом обык-
новенной гадюки V. berus nikolskii или даже внутриподвидовой формой номи-
нативного подвида V. b. berus. Из региональных герпетологов гадюку Николь-
ского самостоятельным видом считают саратовские специалисты (Табачишин
и др., 1995, 1996; Шляхин и др., 1995, 2001; Завьялов, Табачишин, 1997; Та-
бачишина и др., 2004). Однако при дальнейших исследованиях оказалось, что
указанные в первоначальном диапазоне признаки ненадежны, а особенности
биологии и экологии не вполне соответствуют биологической и экологической
концепции вида. В итоге накопившиеся к настоящему времени данные пока-
зывают, что лесостепная гадюка имеет, скорее всего, недостаточные отличия
от обыкновенной гадюки для присвоения ей таксономической категории ви-
дового ранга и является лишь подвидом V. b. Nikolskii или иной внутривидо-
вой формой V. berus (Бакиев, Кренделев, 1999; Бакиев и др., 2000, 2004, 2009;
Песков и др., 2003). Это подтверждают и результаты анализа последователь-
ностей гена цитохрома в митохондральном ДНК, согласно которому «гадю-
ка Никольского, по-видимому, не может считаться отдельным видом, а имеет
более низкий уровень отличий (вероятно, подвидовой) от обыкновенной га-
дюки». Кроме того, состав пептидов ядовитого секрета гадюк волжского бас-
сейна типичен для V. b. berus и имеет различия лишь в количестве некоторых
пептидных компонентов (Старков, Уткин, 2003). Все это позволяет считать,
что гадюки, по крайней мере, Самарской области, представлены популяциями,
совмещающими в себе признаки V. b. berus и V. b. Nikolskii (Колябина и др.,
2003; Павлов и др., 2004; Бакиев и др., 2009; Маленев и др., 2009). Возможно
также, что это свидетельствует о наличии широкой интрогрессивной гибриди-
зации между этими двумя формами, в связи с чем они, видимо, широко обме-
ниваются генами и представлены по большей части гибридными и смешанны-
ми популяциями (Зиненко, 2003).
Распространение. По литературным данным (Ананьева и др., 1998), ареал
вида охватывает «лесные и лесостепные районы Украины и европейской части
России к югу от линии Канев-Курск-Тамбов-Бузулук, а по влажным долинам
рек проникает в степные районы Саратовской и Самарской областей (евро-
пейский фаунистический комплекс). Позднее, при уточнении распространения
вида, граница на востоке ареала от оренбургского г. Бузулука была перенесена
к г. Бугульма (Татарстан) и далее продвинута «до предгорий Южного и Сред-
него Урала» (Мильто, 2003; Павлов и др., 2004). По другой версии (Ефимов
и др., 2008), восточным пределом распространения вида V. nikolsKii является
долина р. Волга, причем район ее Самарской Луки является зоной вероятной
гибридизации V. berus и V. nikolskii.
Таким образом, таксономический статус и распространение меланистичес-
кой формы гадюк региона до настоящего времени остаются неопределенными
и требуют дальнейшего изучения, поскольку описанный ареал обыкновенной
гадюки в регионе может оказаться зоной интерградации двух видов (подви-
288
Глава 4
дов?). В связи с этим неясными, а во многом неопределенными становятся и
сведения об образе жизни, экологии и численности гадюки Никольского.
Региональный статус. Гадюка Никольского в качестве самостоятельно-
го вида V. nikolsKii включена в Красную книгу Российской Федерации с ка-
тегорией 4 как неопределенный по статусу малоизученный вид (Божанский,
2001). Это обстоятельство стало достаточным основанием для включения вида
в Красную книгу Саратовской области (2006).
51. Восточная степная гадюка, или гадюка Ренарда -
Vipera (Pelias) renardi (Christoph, 1861)
Значение вида для сохранения биоразнообразия. Один из представителей
обширного рода, населяющий территорию региона. До недавнего времени счи-
талась восточным подвидом полиморфного комплекса V. ursini-V. и. renardi.
Сейчас признается самостоятельным политипическим видом, в регионе пред-
ставленным двумя подвидми - номинативным V. г. renardi (Christoph, 1861)
и гадюкой Башкирова V. г. bashkirovi (Garanin et al., 2004), отличающимися
от других размерами, степенью проявления меланизма, количеством брюш-
ных, подхвостовых, верхнее- и нижегубных щитков, а также некоторыми осо-
бенностями экологии (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004;
Павлов, 1997; Кузьмин, Семенов, 2006; Бакиев и др., 2009). Вместе с тем спе-
цифичен ряд метрических и меристических признаков у локальных популяций
Нижнего Поволжья (Табачишин и др., 1996; Кармышев, Табачишин, 2003), что
требует дополнительных исследований.
Распространение. Ареал вида охватывает лесостепную, степную, полу-
пустынную зоны Европы, Кавказа, Казахстана и Средней Азии, а по речным
долинам проникает и в пустынную зону (средиземноморский фаунистический
комплекс). Его северная граница на Восточно-Европейской равнине прово-
дится от Молдавии по северной Украине и Поволжью к месту слияния рек
Волги и Камы (острова Спасского архипелага), окрестностям г. Кумертау
(Башкирия), г. Верхнеуральска на Урале, Кустаная, Петропавловска, Семипа-
латинска в Казахстане и долиной р. Иртыш уходит в северо-западный Китай.
Южная граница от Молдавии проводится через Крым, северные районы Кав-
каза, северное побережье Каспийского моря, примерно по долине реки Эмбы
к Мугоджарам, затем по северному побережью Аральского моря, предгорь-
ям Улытау, северному Прибалхашью к оз. Алаколь и по предгорьям Средней
Азии в Китай (Параскив, 1956; Ананьева и др., 1998; Бакиев и др., 2004, 2009;
Вершинин, 2007; Табачишина и др., 2002). Таким образом, большая часть
региона включается в ареал вида, за исключением Волжско-Уральских пес-
ков, Предустюртья, Северного Устюрта, северного Приаралья и Приараль-
ских Каракумов. В целом местообитания вида установлены в 6 (54,5% из 11)
ландшафтных районах лесостепной, 41 (59,4% из 69) степной, 32 (53,1% из
32), полупустынной и 22 (51,2% из 43) пустынной природных зон (табл. 56,
карта 49).
Места обитания. Как свидетельствует название вида, его основными мес-
тообитаниями являются открытые равнинные и горные степные ландшафты,
а также сопредельные территории лесостепной, полупустынной и пустынной
зон.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
289
Распространение степной гадюки
Таблица 56
Зона Провин- ция Район № точки Местонахождение Источник информации
I 1 1 Самарская область окр. рыбхоза «Сускан» Бакиев, Иванова, 2004;
Лесостепная II 3 5 6 2 3 4 5 6 7 окр. с. Раевка окр. с. Кракове вдр. рек Сок и Кондурча окр. с. Старопохвистнево дол. р. М. Кинель Оренбургская область окр. г. Бугуруслан окр. п. Пономаревка Бакиев и др., 2002, 2004, 2009 Чибилёв А.А.
III 7 11 8 9 окр. п. Тюльган окр. п. Сара п. Подгорное Завьялов и др., 2006
Степная IV 12 13 15 16 17 20 21 10 11 12 13 14 15 16 17 Самарская область дол. р. Безенчук Саратовская область окр. г. Балаково Самарская область дол. р. Б. Иргиз овр. Вязовый окр. с. Парфеновка окр. с. Яблоновый овраг овр. Уральская глушица дол. р. Б. Иргиз Саратовская область окр. с. Алексеевка Самарская область Красносамарский лес Бакиев, Магдеев, 1995; Бакиев и др., 2004, 2009 Табачишина и др., 2002 Бакиев и др., 2004, 2009; Песков, 2001 Табачишина и др., 2002 Бакиев и др., 2009
V 22 23 24 25 18 19 20 21 22 Оренбургская область окр. п. Грачевка окр. п. Тонкое окр. п. Приютное окр. п. Сорочинск окр. п. Александровка Каргалинская степь, окр. п. Каргала гора Медвежий лоб Чибилёв А.А.
290
Глава 4
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
26 23 Бирючий овраг окр. с. Украинка окр. п. Восточный Бакиев и др., 2004, 2009; Смелянский, Елизаров, 1996
24 ур. Бостандык ур. Грызлы
V 25 окр. п. Курлин п. Озерное Оренбургская область Таловская степь ОГПЗ Чибилёв, 1995, 1999а, б
26 ур. Меловой Сырт
27 п. Каменноимангулово Саратовская область
27 28 окр. п. Озинки Табачишина и др., 2002
Оренбургская область
28 29 дол. р. Салмыш Чибилёв А.А
VI 30 30 окр. г. Соль-Илецк Паллас, 1773
i 32 31 Урало-Илекское Райский, 1951
е а междуречье Чибилёв, 1995
О Западно-Казахстанская область
34 32 33 окр. г. Аксай, дол. р. Утва дол. р. Караоба Березовиков и др., 1992, 2003
35 34 дол. р. Акбулак
35 междуречье Утва-Сукбулак
36 дол. р. Березовка
37 горы Актау
VII 38 38 окр. оз. Шалкар: гора Сантас Дебело П.В.; Завьялов и др., 2006
39 гора Сасай Оренбургская область
41 40 окр. п. Покровка Чибилёв, 1995
42 41 окр. п. Сагарчин
43 42 окр. п. Шаповалово Актюбинская область
44 43 юг.-зап. г. Актюбинск Чибилёв А.А.
46 44 дол. р. Киил
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
291
П родолжение табл. 56
Зоя» Провин- ция Район № точки Местонахождение Источник информации
VIII 47 48 50 52 54 55 45 46 47 48 49 50 Волгоградская область Приволжская возв. Саратовская область Дьяковский лес окр. с. Сафаровка окр. с. Монахов Оренбургская область окр. п. Первомайское Западно-Казахстанская область окр. п. Красноармейск окр. г. Уральска Кубанцев и др., 1962 Особо охраняемые..., 2007; Табачишина и др., 2002 Чибилев А.А. Дебело П.В.; Николь- ский, 1905
Степная IX 56 57 58 61 62 51 52 53 54 55 56 Оренбургская область Буртинская степь ОГПЗ окр. п. Донское окр. п. Новорудный окр. п. Аккермановка Айтуарская степь ОГПЗ Актюбинская область окр. п. Тайкеткен Чибилёв, 1999а; Чи- билёв А.А. Топоркова, 1973
X 65 66 67 69 57 58 59 60 61 62 Оренбургская область окр. Ириклинского вдхр. Кваркенский район окр. п. Шильда окр. п. Кумак окр. п. Ясный окр. п. Домбаровский Чибилёв, 1995
XI 72 78 63 64 65 66 Светлинские озера: оз. Каменное, окр. оз. Жеты- коль (п. Озерное) Ащесайская степь ОГПЗ окр. оз. Айке Кустанайская область Наурзумский заповедник Чибилёв, 1995, 1999а Динесман, 1953; Брагин, Брагина, 2002
292
Глава 4
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
>шная XIII 82 84 85 87 88 89 91 67 130 68 69 70 71 72 73 74 Западно-Казахстанская область окр. п. Джаныбек окр. оз. Аралсор Волгоградская область окр. оз. Эльтон Астраханская область окр. оз. Баскунчак Саратовская область окр. с. Варфоломеевка окр. с. Ветелки п. Монахов Западно-Казахстанская область окр. п. Казталовка окр. п. Фокеево окр. р. Кушум окр. п. Коловертное Ходашова, 1960; Завья- лов и др., 2006 Ходашова, 1960 Стрельцов и др., 2006 Табачишина и др., 2002; Завьялов и др., 2006 Параскив, Бутовский, 1960; Паллас, 1773; Чернов, 1954
ь Ё? £ о К XV 95 96 97 98 100 75 76 77 78 79 80 81 Актюбинская область п. Миялы, 50 км на юго-вост, окр. п. Каракуга окр. п. Жаркамыс окр. мог. Шайтас (верх, р. Маннесай) окр. горы Хандырткуль междуречье Темир-Эмба верх. р. Четырлысай Накаренок, 2002, 2003в; Шилов, 1961 Никольский, 1905
XVI 101 82 окр. ст. Мугоджарская Дуйсебаева, 2005
XVIII 107 108 83 84 85 86 71 кмтр. Челкар-Иргиз окр. г. Иргиз окр. п. Текели пески Жаманкум Динесман, 1953; Шилов, 1961; Брушко, Кубыкин, 1988
XIX 109 ПО 112 87 88 89 дол. р. Улькояк, 20 км к сев. от п. Дукен дол. р. Тургай окр. п. Куйлыс низ. р. Тургай Дуйсебаева, 2005; Динесман, 1953; Шилов, 1961
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
293
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
• я 90 дол. р. Жасыбай
R Й Я О S S XIX окр. оз. Мельдекуль
с £ 91 ур. Таук, пески Барбикумы
Астраханская область
113 114 92 93 94 окр. горы Б. Богдо окр. п. Сероглазово Харабали, окр. п. Досанг-За- мьяны Никольский, 1905; Сараев, Пестов, 2010; Бобров, 1985; Паллас, 1773; Завьялов, Табачи- шин, 1998
95 окр. г. Красный Яр окр. п. Лапас п. Комсомольский Степной, Малый Арал Гурьевская область
XX 96 окр. п. Танюшкино Накаренок, 2002, 2003 в
97 окр. рзд. 6, ур. Сазды, Муса
Западно-Казахстанская область
116 117 98 99 окр. п. Урда окр. Камыш-Самарских озер Чернов, 1954; Параскив, Бутовский, 1960
3 я я Гурьевская область
3 в к* 118 121 124 100 101 102 окр. п. Н. Уштаган сев.-вост. п. Забурунье окр. п. Кулагино Чернов, 1954; Сараев, Пестов, 2010; Параскив, Бутовский, 1960
103 окр. п. Калмыково
125 104 окр. п. Елтай
XXI 127 128 131 105 106 107 108 р. Эмба, ур. Шубартпала окр. п. Зеленое рзд. Каработано, 7 км юго- вост. окр. п. Искининский, 10 км к Окулова, 1963; 1981; Неручев и др., 1990; На- каренок, 2003 в; Чернов, 1954; Никольский, 1916; Накаренок, 2002, 2003в; Гисцов, Березовиков, 1995
зап.
132 109 окр. п. Сарайчик
ПО 12 км к сев.-вост. от г. Гурьев
111 окр. г. Гурьев, дельта р. Урал
133 112 окр. п. Индерский Никольский, 1916
XXII 135 ИЗ 114 ур. Тайсойган, дол. р. Уил окр. п. Акколь окр. п. Кенбай Неручев и др., 1984; Накаренок, 2002, 2003в
294
Глава 4
Окончание табл. 56
Зона Провин- ция Район № ТОЧКИ Местонахождение Источник информации
136 115 окр. п. Макат
116 низ. р. Сагиз
117 п. Жамансор
118 дол. р. Эмбы, ур. Кзылжар окр. оз. Бартылдакты
119 ур. Тургузба ур. Дюсеке ур. Брлик
ххп ур. Мадьяр
5 окр. п. Камбай
I I 3 120 плато Аккерегеше
Й с“ 137 138 121 122 гора Иманкара окр. р. Кайнар Шилов, 1961; Накаре- нок, 2003 в
139 123 ур. Намазтакыр пески Актам аймагут
124 окр.п. Косчагыл
125 сев. чинк Устюрта
XXIV 145 126 п. Карабулак, 10 км к сев. Шилов, 1961; Дуйсебае-
150 127 окр. п. Каир ва, 2005
XXVI 151 128 окр. ст. Тогыз Б. Барсуки, окр. п. Бегимбет
152 129 окр. ст. Саксаульская
В условиях лесостепи Волжско-Камского края степная гадюка населяет це-
линные степные участки, залежи, опушки лиственных лесов и окраины колков,
примыкающие к ним луговые и остепненные пространства, кустарниковые
луга, а также облесенные склоны речных и овражно-балочных систем. Вместе
с тем в ряде мест здесь (окр. рыбхоза «Сускан») и в некоторых южнее распо-
ложенных островных лесах (Красносамарском лесу) она помимо свойствен-
ных виду открытых ксерофильных биотопов проникает в более гигрофильные
сообщества полян, опушек, просек, зарастающих вырубок, а также молодых
посадок (Гаранин, 1983; Бакиев и др., 2004, 2009).
В степях Самарской и Саратовской областей обитание связано с участка-
ми целинных степей и старых залежей, где. придерживается биотопов разных
типов, но преимущественно стаций со сложным микрорельефом: расчленен-
ных каменистых склонов сыртов, сухих склонов балок, оврагов, речных до-
лин, прибрежных участков степных речек, чаще всего с зарослями кустарни-
ков, разреженных островных и байрачных лесков с прогалинами, полянами и
зарастающими вырубками. В наиболее освоенных земледельческих районах
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
295
вытеснена на обочины дорог, в овраги и другие малопригодные для хозяйс-
твенной деятельности человека места (Смелянский, Елизаров, 1996; Бакиев и
др., 2001, 2004, 2009; Песков, 2001; Шляхтин и др., 2006).
На территории Оренбургской области наиболее благоприятны условия в
пределах типичной и южной степи, особенно в карстовых районах Предуралья,
изобилующих трещинами и пустотами. В Каргалинской степи и на террито-
рии сохранившихся целинных массивов Таловского, Буртинского, Айтуарско-
го и Ащисайского участков Оренбургского степного заповедника также чаще
всего населяет пересеченные сырты и горные балки со скальными выходами
по бортам (Чибилёв, 1999а, б). В Зауралье обычно встречается по останцам
и в местах выхода на поверхность кварцитов и песчаников, местами прида-
ющих территории холмисто-останцовый облик. В Наурзумской степи встре-
чи, как правило, приурочены к сообществам пырея и черной полыни (Шилов,
1961).
На Утва-Илекском междуречье степная гадюка встречается по долинам
степных речек Березовка, Караоба, Акбулак, Утва, а также по меловым ос-
танцам гор Актау (Березовиков и др., 1992). В окрестностях оз. Шалкар ос-
новные местообитания приурочены к склонам соляных куполов Сантас (самое
змеиное в округе место) и Сасай, реже отмечается среди чернополынников
на сопредельной территории и в верховьях балок, впадающих в реки Б. и
М. Анкаты и озеро.
На полупустынных равнинах Волжско-Уральского междуречья степная
гадюка населяет в основном водораздельные комплексы с господством чер-
нополынных сообществ, несколько реже встречается в злаково-разнотравных
понижениях (западины, падины), а местами (у п. Круглый) и в биюргунниках.
В окрестностях оз. Эльтон и Баскунчак ее встречи приурочены к склонам реч-
ных долин и балок с преобладанием остепненных участков или с зарослями
кустарников (Параскив, Бутовский, 1960; Ходашова, 1960; Стрельцов и др.,
2006). У восточных пределов междуречья (окрестности п. Калмыкове) основ-
ными местообитаниями считаются полынные и злаково-полынные сообщества
глинистой полупустыни; характерным обитателем она является и в песчаной
полупустыне, хотя встречается значительно реже (Окулова, 1963).
В Урало-Эмбенском междуречье, как правило, населяет долины и поймен-
ные урочища по рекам Уил, Сагиз и Эмба: берега, сухие староречья, солонча-
ковые луга, прибрежные заросли тростника и тамарикса (Неручев, Васильев,
1987; Накаренок, Неручев, 2001). В верховьях р. Маннесай она отмечалась в
колониях большой песчанки и на курганчике малого суслика; в сходных усло-
виях на солонцах у колоний малого суслика, степной пеструшки, других по-
левок ее встречи известны в Зауралье в низовьях рек Иргиз, Улькояк и Тургай
(Шилов, 1961).
Пустынную зону региона степная гадюка населяет лишь по переферии. По
западной кромке Волжско-Уральских песков почти повсеместно она отмеча-
ется в основном в пределах надпойменной террасы. В окрестностях г. Крас-
ный Яр основными местообитаниями являются межбугровые депрессии, хотя
встречи известны также по закрепленным бугристым пескам и на солонцах
(Завьялов, Табачишин, 1998).
По межбугровым депрессиям (ашикам) в Рын-песках (окрестностях
п. Урда) она проникает в глубь массива, где встречается также в понижениях
296
Глава 4
с ивняками и осокорем. По северной и южной кромкам Волжско-Уральского
массива изредка встречается на задернованных песках и очень редко между со-
рами, где придерживается солянковых сообществ. Здесь она отсутствует лишь
в барханных и разбитых песках (Чернов, 1954; Параскив, Бутовский, 1960),
хотя в местах с очагами сбоев и развевания она все же встречается (Неручев
и др., 1983). По шлейфу песков Актамаймогут и кромке песчаных массивов
Жаманкумы и Барбикумы встречи известны также в Предустюртье и Зауралье
(Шилов, 1961).
Образ жизни. Из зимних убежищ степная гадюка выходит, когда темпе-
ратура воздуха превышает 6...9 °C, что на юге региона приходится на конец
марта - начало апреля, а на севере на разные числа апреля, т. е. запаздывает
до 15-20 дней; примерно на такое же время сроки выхода могут различаться
и в одном районе в разные годы в зависимости от хода развития весенних яв-
лений. Массовый выход происходит, когда температура воздуха превышает
12 °C, - т. е. во второй половине апреля (Кубанцев и др., 1962; Бакиев и др.,
2004, 2009; Песков, 2001; Шляхтин и др., 2006; Табачишина и др., 2002).
В условиях эксперимента добровольный минимум для приземного воздуха
оказался равен 8,7 °C; для субстрата - 10,8 °C, а максимум соответственно
32,6 и 43,5 °C, диапазон оптимальных температур определен в 15,0...29,0 °C
(Литвинов, 2004). Это практически совпадает с материалами полевых на-
блюдений. В Поволжье активные гадюки встречались на субстрате с тем-
пературой 4,5...43,5 °C (Песков и др., 2003), а у северной кромки Волжско-
Уральских песков (окрестности п. Калмыкове) при температуре воздуха, °C:
в апреле-мае-18,0...31,5, июне-июле-21,0...28,0, сентябре-октябре-14,0...27,0.
Ранней весной активность имеет однопиковый характер, который примерно
с середины мая сменяется двухпиковым. В июне-июле максимальная актив-
ность наблюдается в 9-12 и 15—18 часов, хотя в пасмурную погоду она актив-
на и днем. Днем же обычно укрывается в нежилых норах разных животных,
других полостях в грунте, стогах и подобных местах. На ритмику активности
помимо температуры влияют также сила ветра, характер растительности, обес-
печенность кормом и ряд других факторов.
С началом осени активные степные гадюки наблюдаются в течение все-
го светлого времени суток; с этого же времени начинается их концентрация
около мест зимовки. В спячку на севере региона залегают во второй половине
сентября, на юге - в первой половине октября, хотя отдельные особи иногда
задерживаются и позднее (почти на месяц) (Бакиев и др., 2004,2009; Шляхтин
и др., 2006).
Размножение. Половозрелыми степные гадюки, по-видимому, становят-
ся на втором-третьем году жизни. В этом возрасте спустя 2-3 недели после
выхода из зимовки начинается спаривание, в отдельных популяциях продол-
жающееся 7-10 дней, но в целом этот период оказывается растянут. В Волж-
ско-Уральском меадуречье он приходится на вторую половину мая - первую
половину июня. В это время образует скопления численностью более десятка
особей (гора Сантас). Количество эмбрионов достигает 20. Период эмбрио-
нального развития в условиях Саратовской области продолжается около 100 су-
ток. Выход сеголеток начинается с середины июля (в Волжско-Уральском
междуречье наиболее ранняя находка отмечена 10.07.1953 г.) и продолжается
до середины сентября (Параскив, Бутовский, 1960; Шляхтин и др., 2006).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
297
Питание. Основу пищевого рациона степной гадюки составляют мелкие
грызуны (преимущественно полевки, мыши) и ящерицы (прыткая, ящурки);
в ряде специфических местообитаний яйца и птенцы мелких наземногнездя-
щихся видов птиц, а также амфибии и рептилии. Повсеместно широко потреб-
ляются насекомые (особенно прямокрылые, временами чешуекрылые, жуки),
реже паукообразные. Их соотношение несколько изменяется по сезонам и ста-
циям, что свидетельствует о значительной трофической пластичности вида.
Молодые особи кормятся в основном прямокрылыми насекомыми и мелки-
ми ящерицами. В свою очередь для степной гадюки характерно и «обилие ес-
тественных врагов» - это преимущественно хищные птицы, млекопитающие,
крупные рептилии.
Численность. Сведения о ней довольно противоречивы даже по одним и
тем же районам. В связи с этим по некоторым пунктам приводим данные лишь
последних лет. В Самарской области даже на локальных участках с макси-
мальной плотностью (в Большечерниговском, Кинельском и Ставропольском
районах) встречаемость весной и осенью не превышает 3-4 экз/га, а в летние
месяцы 1-2 экз/га (Бакиев, Гафарова, 1999; Бакиев и др., 2009). На значитель-
ной части Саратовской области численность степных гадюк не превышает
2-5 экз/га, хотя в разных стациях межузеньской равнины в 1998 г. она состав-
ляла 1,2-9,4, на участках с травянисто-кустарниковой растительностью по ов-
ражно-балочным системам отрогов Общего Сырта (Дергачевский, Перелюб-
ский, Озинский районы) - 5-11, а в 2001 г. местами она достигала 17 экз/га
(Табачишина и др., 2002; Шляхтин и др., 2006). В Оренбургской области на
участках заповедника степная гадюка является обычным видом, а в карсто-
вой зоне Предуралья местами численность достигает 7-10 экз/га (Чибилёв,
1999а; данные последних лет). В Зауралье ее встречаемость значительно ниже,
хотя и здесь она может достигать значительной величины - на сопредельной
территории музея-заповедника «Аркаим» в Челябинской области, например,
12,7 экз/га (Чибилёв, 2005).
В полупустыне Волжско-Уральского междуречья в середине прошлого сто-
летия степная гадюка считалась обыкновенной, а местами, как, например, в ни-
зовьях р. Б. Узень (окрестности п. Фокеево), и очень многочисленной (Параскив,
Бутовский, 1960). В комплексах Волжско-Узеньского водораздела, по наблюде-
ниям К.С. Ходашевой (1960), в это время численность испытывала значительные
колебания в зависимости от влажности года. Так, в сухом 1950 г. встречаемость
здесь достигала 5,0 ос/10 км маршрута, во влажном 1952 г. - 1,0, а в средние по
влажности - 3,0-4,0 ос/10 км. В окрестностях оз. Эльтон эти показатели соот-
ветственно равнялись 20,0; 5,0 и 7,0-11,0 ос/10 км. На специфических участках
поймы р. Б. Узень в окрестностях Александрова Гая плотность и сейчас дости-
гает значительных величин - 4,0-9,0 ос/га (Кармышев, Табачишин, 2003), а на
межузеньской равнине - 5,6 ос/км (Табачишина и др., 2002). На восточной кром-
ке междуречья в окрестностях п. Калмыкове в глинистой полынной полупусты-
не в весенне-летние сезоны начала 1950-х гг. численность в среднем достигала
3,2 ос/10 км, в злаково-полынных сообществах - 2,8, а в песчаной полупусты-
не 0,6 ос/10 км, хотя весной (апрель-май) змеи встречались несколько чаще: в
глинистой полупустыне - 0,86 экз/га, а в песках - 0,24 (Окулова, 1963, 1981).
В Урало-Эмбенском междуречье степная гадюка наиболее обычна по речным
долинам Уила, Сагиза и Эмбы, а в Зауралье в низовьях рек Иргиз и Тургай.
298
Глава 4
В пустынной зоне степная гадюка встречается в основном по кромке пес-
ков, хотя по ашикам местами проникает и в глубь массивов. По Волго-Ахту-
бинской надпойменной террасе в окрестностях г. Красный Яр в межбугровых
депрессиях ее плотность составляет 8,70±0,09 экз/га, на закрепленных буг-
ристых песках 1,20±0,18, на солонцах 0,97±0,16 (Завьялов, Табачишин, 1998),
вблизи ст. Досанг в песчаной степи 0,2 экз/га (Бобров, 1985). На территории
Богдинско-Баскунчакского заказника известны ее лишь единичные встречи
(Стрельцов и др., 2006), хотя на сопредельной территории Рын-песков в ок-
рестностях п. Урда в межбугровых депрессиях с зарослями ивняка и осокоря,
а также у Камыш-Самарских озер она не представляла особой редкости (Па-
раскив, Бутовский, 1960). В Волжско-Уральских песках немногочисленна - до
1-2 экз/10 км (Неручев и др., 1981). В Северо-Восточном Прикаспии, судя по
количеству пунктов встреч, наиболее обыкновенна степная гадюка по рекам
Сагиз (до 6,0 экз/10 км) и Эмба (Накаренок, 2003 в; Сараев, Пестов, 2010).
Лимитирующие факторы. Хозяйственное (распашка земли, сенокоше-
ние, выжигание травы, выпас скота, использование ядохимикатов) и рекреаци-
онное освоение территории, урбанизация, истребление людьми, коммерческие
заготовки. После многоснежных зим затопление мест зимовки.
Охрана. Обитает и охраняется на территории Оренбургского, Наурзумско-
го и Богдинско-Баскунчакского заповедников, Иргиз-Тургайского резервата.
На остальной территории специальных мер охраны не предпринималось.
Региональный статус. Включена в Красную книгу МСОП (2000), Сара-
товской (2006) и Самарской (2009) областей, Аннотированный перечень таксо-
нов и популяций животных, нуждающихся в особом внимании к их состоянию
в природной среде России, Приложение III и Приложение IСИТЕС. Находится
под охраной Бернской конвенции (Приложение 2).
Глава 5
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ
АМФИБИЙ И РЕПТИЛИЙ РЕГИОНА
Современный состав фауны амфибий и рептилий региона является
результатом длительного исторического развития. Как уже отмеча-
лось, ядро ее исходного типа сформировалось в плиоцене, к концу
которого активизировались адаптивные механизмы, способствующие расши-
рению видового спектра фауны региона. Эти процессы продолжались и в плей-
стоцене, чему способствовало усиление динамики всех природных явлений в
регионе, приведшее к вымиранию ряда архаичных и существенному измене-
нию ареала «молодых» форм.
Современный облик фауна региона постепенно приобретала в голоцене,
который рассматривается как время преимущественной стабилизации аре-
алов, медленного расселения и освоения новых территорий, своеобразного
заполнения ценотического вакуума, образовавшегося в результате действия
природных факторов. Последним «штрихом» преимущественного их дейс-
твия, по-видимому, является исчезновение на Восточно-Европейской равнине
обыкновенной квакши.
Со времени начала голоцена, но особенно в последние тысячелетия, фор-
мирование ландшафтов региона и структуры их биоты идет под все возраста-
ющим воздействием антропогенных факторов: вырубки лесов, пасторальной
дигрессии, затем распашки земель и общего освоения территорий, которые
снивелировали их многие первобытные черты. Вместе с тем у северных пре-
делов региона меньшая засушливость позднего голоцена наряду с особеннос-
тями хозяйственного освоения территории определили большую стабильность
природных биоценозов, которые на целинных участках сохранили многие осо-
бенности экосистем раннего голоцена, исчезнувшие в степной и полупустын-
ной зонах.
Во время первого этапа земледельческого освоения территории в лесо-
степной и степной зонах были выборочно распаханы крупные массивы чер-
ноземных и темно-каштановых почв, а также некоторые крупные падины в
полупустыне. На втором этапе оказались распаханы практически все удобные
в технологическом плане, а затем и условно пахотнопригодные земли, что
обусловило широкое распространение антропогенного опустынивания.
Наряду с этим в регионе сооружен ряд крупных водохранилищ на Волге,
Урале, Кушуме, зарегулирован сток практически всех степных речек, созданы
Урало-Кушумская, Иргизская и около десятка менее значительных водоканаль-
ных систем. Регион стал и ареной крупного лесохозяйственного эксперимента,
результат которого - создание лесополос вдоль Волги и Урала, многочисленных
полезащитных и придорожных насаждений, проложение тысяч километров ав-
томагистралей, линий электропередач, связи, трасс трубопроводов, многочис-
ленные инженерные сооружения; в регионе создан мощный промышленный
(преимущественно нефтегазовый и горно-металлургический) комплекс.
300
Глава 5
Совокупное действие этих факторов привело к сокращению, а в ряде слу-
чаев и уничтожению на значительной территории типичных местообитаний
животных, их загрязнению и чрезмерной эксплуатации ресурсов живой приро-
ды, что настоятельно требует разработки комплекса природоохранных меро-
приятий, снижающих воздействие человека на мир животных.
Воздействие это, как известно, бывает прямым и косвенным. Прямое воз-
действие на амфибий и рептилий региона проявляется в их добыче для разных
целей и уничтожении, чаще всего бессмысленном. Истребляются в основном
гадюки, щитомордник и стрела-змея как ядовитые животные, в меньшей степе-
ни преследуются из-за ошибочного представления об их ядовитости медянка и
узорчатый полоз, а из-за агрессивности некоторые другие полозы. В ряде сель-
ских районов вредным считается обыкновенный уж, хотя большинству населе-
ния известны его неядовитость и хорошо выраженные отличительные призна-
ки. Отмеченное массовое уничтожение водяных ужей, вероятнее всего, связано
с их некоторым сходством с обыкновенной гадюкой, отличие от которой не
известно местным жителям (хотя, возможно, здесь имеет место бессмысленное
уничтожение безобидных животных). Общие масштабы уничтожения змей в
регионе с учетом значительных размеров его территории по ориентировочным
расчетам составляют не менее тысячи особей в год. В ряде случаев беспричин-
но уничтожаются также лягушки, чесночницы и ящерицы. Наиболее действен-
ным мероприятием по снижению масштабов такого воздействия, по-видимому,
является усиление экологического просвещения населения.
Более существенны масштабы специальной добычи амфибий и рептилий.
Их постоянными потребителями являются медицинские и другие учебные за-
ведения естественного профиля и научно-исследовательские учреждения, ис-
пользующие разные виды в учебном процессе и как коллекционный материал.
В значительном количестве змей (в основном массовых видов) добывают для
нужд медицинской промышленности. Перспективно в этой сфере использова-
ние секрета кожных желез ряда видов жаб и лягушек. По ориентировочным
расчетам для всех этих целей ежегодно в регионе добывается до 100-150 ты-
сяч экземпляров разных видов.
В последнее время амфибии и рептилии стали играть важную роль как
объекты коммерческой торговли. По-видимому, несколько тысяч особей ряда
видов, особенно экзотических, продается для содержания в частных коллекци-
ях. Однако при недостаточном уровне специальной подготовки заготовителей,
коллекционеров и отсутствии надлежащего ухода значительная часть содер-
жащихся в неволе животных гибнет. В еще большем количестве (очевидно,
сотни тысяч) некоторые виды заготавливаются для нужд пищевой промыш-
ленности (деликатесы).
Для предупреждения негативных последствий нерегулируемого промыс-
ла необходимы ограничение частного коллекционирования; разумное лицен-
зирование добычи с периодической сменой мест отлова при учете состояния
численности популяций; в районах с достаточно высокой численностью змей
создание специализированных питомников, стационаров или мобильных ла-
бораторий по эксплуатации животных непосредственно в природе. В особом
внимании нуждаются краеареальные популяции малочисленных видов, что
позволит стабилизировать их численность и обеспечит возможность их хо-
зяйственного использования.
276
Глава 4
В Зауралье водяной уж населяет долины рек Тургай, Жыланшик и зарос-
шее рогозом озеро Камышлыбаш в низовьях р. Сырдарья (Дусебаева, 2005).
Встречается этот уж и на побережье Каспия, где его можно наблюдать как
на песчано-галечниковой прибрежной полосе с зарослями солянок, так и на ка-
менистых участках. Сходных мест придерживается он и на побережье Араль-
ского моря (Параскив, 1956).
Образ жизни. Из зимней спячки в регионе выходит в разных числах второй
половины апреля - начале мая, когда температура воздуха превышает 10 °C.
Обычно активен в течение всего светлого времени суток (7-21 час), что опре-
деляется его более высокой термофильностью по сравнению с таковой обыкно-
венного ужа (оптимальной считается температура субстрата 20...27 °C) и тем,
что при температуре воздуха выше 28 °C большую часть времени он проводит
в воде (в том числе в реках с быстрым течением и в море, удаляясь от берега на
несколько километров). В качестве укрытий использует расщелины в грунте,
пустоты под камнями, пнями, деревьями, норы грызунов, корневые ходы.
Со второй половины августа активность водяных ужей снижается, а в кон-
це сентября - начале октября они (сначала самцы) уходят на зимовку. Период
активности, таким образом, продолжается около 160-180 суток, что несколько
меньше, чем у обыкновенного ужа. Зимует на территории Самарской области,
по-видимому, главным образом в карстовых пустотах, других подобных мес-
тах, также в разных подземных убежищах, нередко скоплениями в несколько
сот особей (иногда совместно с обыкновенным ужом).
Размножение. Половозрелым водяной уж становится на третьем-четвер-
гом году жизни. В этом возрасте, вскоре после выхода из зимовки, обычно в
конце апреля - начале мая при температуре воздуха около 18 °C начинается
спаривание, ко времени которого ужи собираются в пары или образуют «брач-
ные хороводы», в которых участвуют до 15-20 особей. Яйца (4-23) откладыва-
ются в июне - начале июля; инкубация в естественных условиях продолжается
40-52 суток; и с конца июня начинается выход сеголеток, который продолжа-
ется и в сентябре (Кубанцев и др., 1962; Бакиев и др., 2004, 2009; Шляхтин и
др., 2005).
Питание. Основу пищевого рациона водяных ужей в большинстве водо-
емов региона составляет рыба (до 60-93% по массе), видовой состав и разме-
ры которой существенно меняются по сезонам, в зависимости от численности,
доступности пищевых объектов и активности ужей. Кроме того, потребляются
земноводные (преимущественно озерные лягушки, чесночница, реже зеленые
жабы, жерлянки и их личинки), в ряде мест являющиеся основной добычей, а
также водные беспозвоночные и их личинки. В Предустюртье, где свойствен-
ная водяным ужам пища отсутствует, он перешел на питание мышевидными
грызунами и большими песчанками, в колониях которых (причем иногда вда-
ли от свойственных этим змеям местообитаний) иногда встречается (Шилов,
1961). В свою очередь водяные ужи имеют большое количество естественных
врагов - главным образом хищных птиц и млекопитающих, реже он служит
объектом добычи крупных рыб, некоторых земноводных и пресмыкающихся.
Численность. В типичных местообитаниях водяной уж является обычным,
а местами и многочисленным видом. Так, по Волге в районе Самарской Луки у
мест зимовки, в конце апреля - начале мая 1991-2000 гг. его плотность достиг-
ла 20-30 экз/га (Бакиев и др., 2009). Южнее, по средней зоне Волгоградского
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
277
водохранилища, она составляла 3-17 ос/га (Шляхтин и др., 2005), а в Волго-
Ахтубинской пойме весной 1998 г. - 4,90±0,19 экз/га (Завьялов, Табачишин,
1998). В последние годы численность его здесь постоянно увеличивается, чему
способствует создание искусственных водоемов (Кубанцев, 2003).
По среднему течению р. Урал в Переволоцком районе Оренбургской об-
ласти на участке обрывистого берега протяженностью 300 м в мае 1962 г. было
насчитано более 90 только убитых ужей (Чибилёв, 1995). Усеяные трупами во-
дяных ужей берега р. Урал отмечались и в других местах по среднему течению
этой реки. Вместе с тем за его пределами даже в начале XX в. водяной уж был
редок (Райский, 1951).
Численность водяных ужей увеличивается во влажные, особенно следую-
щие один за другим, годы. Так в 1950-1951 гг. по долине р. Урал у п. Круглый
ужей было сравнительно немного, во влажном 1952 г. их численность заметно
увеличилась, а в следующем влажном 1953 они были очень многочисленны
(Параскив, Бутовский, 1960). Обычным в начале 1950-х гг. водяной уж был
и в низовьях р. Урал (Чернов, 1954), по руслам пересыхающих рек Шаган и
Маннесай, а местами на небольших прудах по их течению очень многочис-
ленным (Шилов, 1961); весьма обычен этот вид и по долине р. Эмба (Неручев,
Васильев, 1978).
В Зауралье численность, по-видимому, подвержена довольно значитель-
ным колебаниям. Во всяком случае по р. Тургай на протяжении почти всего
XX столетия он не наблюдался, а в мае 2002 г. здесь отмечена его довольно
высокая плотность; частые встречи регистрировались в это время и в дельте
р. Сырдарьи (Дуйсебаева, 2005). На побережье Каспийского и Аральского
морей водяной уж, по-видимому, немногочислен, а на остальной-территории
представлен лишь небольшими изолированными популяциями.
Лимитирующие факторы. Действие комплекса факторов у северных пре-
делов ареала. Застройка, хозяйственное и рекреационное освоение типичных
местообитаний, необоснованное преследование людьми.
Охрана. Обитает и охраняется на территории Оренбургского заповедника.
На остальной территории региона специальные меры охраны не применялись.
Строительство новых гидросооружений в ряде мест способствует расширению
местообитаний.
Региональный статус. Характерный для региона представитель герпето-
фауны, населяющий ограниченные местообитания. Занесен в Красные книги
Саратовской (2006), Самарской (2009) областей и сопредельной Башкирии.
Род 31. Psammophis Boie In Fitzinger, 1826 - песчаные змеи
47. Стрела-змея - Psammophis lineolatus (Brandt, 1838)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель обширного рода, населяющий территорию региона. Монотипичес-
кий вид.
Распространение. Ареал вида охватывает южные районы Казахстана,
Среднюю Азию, юго-запад Монголии и Китая, север Пакистана, Афганис-
тана, Ирана и юго-восточное Закавказье (среднеазиатский фаунистический
278
Глава 4
комплекс). Его северная граница от окрестностей г. Гурьева прослеживается
вблизи п. Макат, через низовья р. Сагиз, гряду Актолагай, среднее течение
р. Эмба, горы Шолькара, Жельтау, пески Шогырлыкум (Сараев, Пестов, 2010),
Б. Барсуки, низовья р. Тургай, пески Арыскум, Муюнкумы, северное Прибал-
хашье, Зайсанскую котловину (Параскив, 1956; Брушко, Кубыкин, 2000) и да-
лее в Китай и Монголию. Южная граница проходит по пустыням Пакистана,
Афганистана и Ирана. Таким образом, ареал вида охватывает только южную
часть региона и все отмеченные пункты (вплоть до низовий р. Тургай) опреде-
ляют его северные пределы распространения в исследуемом регионе. Южнее
стрела-змея известна на п-ове Бузачи, в окрестностях сора Кайдак, по нижней
Эмбе, Северному Устюрту, побережью Аральского моря и в Приаральских Ка-
ракумах. Конкретные местонахождения приведены в табл. 53 и на карте 46.
Распространение стрелы-змеи
Таблица 53
Зона Провин- ция Район № точки Местонахождение Источник информации
Полу- пустын- ная XVIII 107 1 Актюбинская область окр. с. Жайсанбай Брушко, Кубыкин, 1989
XIX 112 2 низ. р. Тургай Никольский, 1915
Гурьевская область
XXI 127 3 ур. Теренозек Брушко, Кубыкин, 2000
131 4 окр. п. Искининский
132 5 22 км к сев.-вост. от г. Гурьева
136 6 низ. р. Сагиз, окр. п. Макат окр. п. Кенбай гора Кой-Кара Сараев, Пестов, 2010; Неручев и др., 1981, 1984
7 окр. оз. Бартылдакты ур. Брлик
5 X им гора Аккерегешин
►С 8 окр. п. Косчагыл
с XXII 137 9 10 окр. п. Кульсары горы Актолагай п. Аралтюбе ур. Жанбике Неручев, Шатилович, 1985; Сараев, Пестов, 2010; Брушко, Кубыкин, 1988, 2000
11 ур. Кыргынтюбе окр. сора Есенжал
138 12 горы Шолькара горы Жельтау Неручев и др., 1981, 1984; Сараев, Пестов, 2010
13 горы Коленкелы Прикаспийские Каракумы
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
279
Окончание табл. 53
Зона Провин- ция Район № точки Местонахождение Источник информации
Мангистауская область
XXII 14 пески Шагырлыкум Киреев, 1981; Накаре-
15 Прикаспийские Каракумы нок, 2003в
140 16 п-ов Бузачи Никольский, 1915
XIII 143 17 окр. сора Кайдак, бывш. Ново- александровское укрепление
147 18 ур. Карын-Жарык; залив Бутакова
XXV 148 19 пески Сам, кол. Тулеп, кол. Кугусем
окр. оз. Тугаракчан
в 150 151 20 21 сор. Кошкарата Б. Барсуки, окр. п. Южное Дуйсебаева, 2005; Бруш- ко, Кубыкин, 2000
£ XXVI 152 154 22 23 ур. Озектык р. Ашудасты М. Барсуки п. Акеспе Приаральские Каракумы Параскив, 1956; Лобачев и др., 1973; Никольский, 1915; Динесман, 1953; Дуйсебаева, 2005
155 24 сев. берег Аральского моря, ур. Кара-Тамак, Исеншагыл
чинк зал. Перовского
25 п-ов Кара-Тюп окр. п. Тас-Тюбек
26 окр. с. Каратерень
Места обитания. В регионе населяет типичные для вида местообитания.
В М. Барсуках отмечалась на еркеково-полынной равнине у подножья полу-
закрепленного бархана. В Б. Барсуках ее наблюдали на закрепленных барха-
нах и глинистых такырах. В глинистой пустыне (причем на сильном сбое) она
встречалась в Приэмбенской провинции; здесь же ее встречи известны на ка-
менистых склонах гор-останцов, в окрестностях сора Кайдак у солончаков, а у
Аральского моря отмечалась и по береговым обрывам (Параскив, 1956; Неру-
чев и др., 1981; Дуйсебаева, 2005).
Образ жизни. После зимовки появляется в разных числах апреля. Наи-
большую активность проявляет при температуре грунта около 28 °C. Ведет
преимущественно дневной образ жизни. В качестве убежищ использует пус-
тоты между камнями, в развалинах строений, норы грызунов. Активность со-
храняет около 170-180 суток, обычно до середины октября.
Размножение. На сопредельной территории спаривание наблюдается в
апреле-мае. Яйца (2-11) откладываются с конца июня до начала августа, а вы-
ход молодых происходит с середины августа.
280
Глава 4
Питание. Стрела-змея - узкоспециализированный герпетофаг - питает-
ся главным образом ящурками, агамами, круглоголовками; в меньшей мере
другими видами. Основным способом охоты является подкарауливание. Мо-
лодые питаются насекомыми. В свою очередь стрела-змея добывается бо-
лее крупными рептилиями, хищными птицами и некоторыми млекопитаю-
щими.
Численность. В казахстанской части ареала считалась сравнительно обыч-
ным видом (Параскив, 1956), а в Приаральских Каракумах - даже массовым
(Лобачев и др., 1973). В последние годы численность, по-видимому, сократи-
лась, поскольку в Северном Приаралье, например, за 2 полевых сезона (май-
июнь 2002 и 2003 г.) встречено лишь 4 особи (Дуйсебаева, 2005). В Приэмбен-
ской пустыне ее численность определена в 0,5-1,0 ос/10 км, а в Прикаспийских
Каракумах 1,0 (Накаренок, 2003 в).
Лимитирующие факторы. Хозяйственное освоение территории (сельхоз-
работы, автотранспорт), уничтожение людьми.
Охрана. Обитает и охраняется на территории Устюртского заповедника.
На остальной территории состояние популяций принятия особых специальных
мер охраны не требует.
Региональный статус. Узкоспециализированный представитель герпето-
фауны пустынь; северные популяции краеареальные.
Семейство 17. Viperidae Laurenti, 1768 -Гадюковые змеи, или гадюки
Род 32. Gloydius Hoge et Romano-Hoge, 1981 - щитомордники
48. Обыкновенный щитомордник, или щитомордник Палласа -
Gloydius halys (Pallas, 1776)
Значение таксона для сохранения биоразнообразия. Единственный пред-
ставитель рода, населяющий территорию региона. Политипический вид; в ре-
гионе представлен подвидом G. h. caraganus (Eichwald, 1831), отличающимся
светлой окраской и рисунком темных полос на туловище (Атлас пресмыкаю-
щихся..., 2004).
Распространение. Ареал вида простирается от низовий р. Волги и юго-
восточного Закавказья через Казахстан, Среднюю Азию и юг Сибири до побе-
режья Тихого океана (восточно-палеарктический фаунистический комплекс).
Его южная граница проводится от юго-восточных районов Азербайджана че-
рез север Ирана, Афганистана и Китай. Северная до настоящего времени четко
не определена. По старым данным она проводилась от Саратовского Заволжья
(Шляхтин, Голикова, 1986) немногим южнее верховий р. Утва (Чингирлау),
ур. Куагаш в верховьях р. Калдыгайты, окрестностях г. Актюбинска (Николь-
ский, 1905), вблизи г. Иргиза (Динесман, 1953), ур. Акмин на р. Тургай и далее
через окрестности г. Семипалатинска по югу Сибири к тихоокеанскому по-
бережью. Считалось, что к югу от этих пунктов щитомордник в регионе рас-
пространен повсеместно (Параскив, 1956). Однако последующими исследова-
ниями его обитание в Саратовском Заволжье не подтверждено (Табачишина
и др., 2004), нет также новых сведений о его обитании в верховьях рек Утва,
Калдыгайты и у г. Актюбинска.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 281
Достоверно распространение щитомордника в регионе в последние годы
определяется находками в окрестностях ст. Досанг, Индерских горах, на ос-
танцовых горах по р. Эмба - Аккерегешин, Иманкара, Актолагай, Шолька-
ра, Жельтау, чинке Донызтау и в его окрестностях (Накаренок, 2003 в; Сараев,
Пестов, 2010), 71 км к северу ст. Челкар и долине р. Жыланшик (Дуйсебаева,
2005). Южнее его встречи в регионе известны в окрестностях г. Гурьева, по
р. Эмбе, в Прикаспийских Каракумах, окрестностях сора Кайдак (бывшее ук-
репление Новоалександровск), п-ове Бузачи, Северном Устюрте, Б. Барсуках,
северо-западном и северном побережьях Аральского моря, Приаральских Ка-
ракумах и окрестностях г. Аральска. В целом его конкретные местообитания
известны в 3 (4,4% из 69) ландшафтных районах степной зоны, 5 (15,6% из 32)
полупустыни и 13 (30,2% из 43) районах пустынной зоны (табл. 54, карта 47).
Таблица 54
Распространение щитомордника
Зона Провин- ция Район № точки Местонахождение Источник информации
Западно-Казахстанская область
3 X с н VII 37 40 1 2 верх. р. Утва (исчез?) ур. Куагаш (исчез?) Никольский, 1905
О Актюбинская область
44 3 окр. г. Актюбинск Параскив, 1956
3 XV 97 4 окр. р. Шатан Никольский, 1905
X X ё & XVIII 107 5 71 км к сев. от ст. Челкар Дуйсебаева, 2005
ПО 6 р. Тургай, ур. Акмин Параскив, 1956
п* Q XIX 111 7 окр. р. Жыланшик Смирновский, 1963
С 112 8 окр. г. Иргиз Динесман, 1953
Астраханская область
XX 114 9 окр. ст. Досанг Накаренок, 2003в
10 окр. г. Красный Яр Чернов, 1954
Гурьевская область
[иная XXI 127 132 11 12 80 км к юго-зап. от п. Куль- сары окр. г. Гурьев Брушко, Кубыкин, 1998; Сараев, Пестов, 2010; Чернов, 1954; Неручев и др., 1984
ё 133 13 Индерские горы
£ 136 14 гора Аккерегешин гора Иман-Кара
XXII 15 окр. п. Косчагыл окр. п. Кульсары
137 16 горы Актолагай Сараев, Пестов, 2010
17 гора Каражар
282
Глава 4
Окончание табл. 54
Зона Провин- ция Район № точки Местонахождение Источник информации
138 18 19 гора Шолькара горы Жельтау Накаренок, 2003 в; Па- раскив, 1956
139 20 Прикаспийские Каракумы
XXII 21 Мангистауская область ур. Кызылкуз чинк Донызтау Зап. Устюрт Параскив, 1956; Завья- лов и др., 2006
§ X i XXIII 140 144 22 23 п-ов Бузачи окр. сора Кайдак
С 151 24 пески Б. Барсуки Лобачев и др., 1973
154 25 Приаральские Каракумы
155 26 п-ов Кара-Тюп Сидоров, 1925
XXVI 27 п-ов Куланды Дуйсебаева, 2005
28 окр. родн. Каска-Джол ур. Достан окр. п. Екеспе
29 окр. г. Аральск Динесман, 1953
Места обитания. Щитомордник не только широко распространенный, но
и эвритопный вид. Его типовой (и типичной) территорией являются Волжс-
ко-Уральские пески в окрестностях г. Красный Яр; в песчаных массивах он
отмечался также у северных пределов своего распространения в верховьях
рек Калдыгайты и Утва, затем в Прикаспийских и Приаральских Каракумах,
Б. Барсуках, причем везде в этих биотопах он придерживается закрепленных
злаково-полынными сообществами участков или колоний песчанок.
По-видимому, вторым по значимости местообитанием в регионе являются
поросшие полынями и биюргуном каменистые склоны останцовых гор, а так-
же карстовые воронки и разные ниши в Индерских горах, чинки Донызтау и
подобные им глинисто-песчаные обрывы коренного берега Аральского моря.
Не избегает он засоленных биотопов в окрестностях сора Кайдак и по обсох-
шему дну Аральского моря, занятых полынно-кермековыми группировками.
Все эти местообитания он явно предпочитает глинистым пустыням, хотя из-
редка встречается и в них, как по р. Эмба, так и на побережье р. Жиланшик
(Чернов, 1954; Неручев, Васильев, 1978; Дуйсебаева, 2005).
Образ жизни. После зимовки появляется в разных числах апреля. Весной
активен в течение всего светлого времени суток, летом преимущественно в су-
мерки и ночью, хотя в пасмурную погоду выходит и днем. В качестве убежищ
используются норы, пустоты в грунте, прячется среди камней или развалин;
временными укрытиями могут служить и кусты. Период активности обычно
продолжается до середины октября (хотя иногда задерживается до первой де-
кады ноября)-т. е. около 180-200 суток.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
283
Размножение. Щитомордник - яйцеживородящий вид. Спаривается он в
апреле-мае, молодые (3—14) появляются с конца июля.
Питание. Основу пищевого рациона взрослых особей составляют мелкие
позвоночные животные, преимущественно грызуны (полевки, мыши, хомячки,
тушканчики), птицы, ящерицы (ящурки, круглоголовки); молодые (первого и
второго годов жизни), вероятно, питаются в основном насекомыми и другими
беспозвоночными. В свою очередь щитомордников потребляют более круп-
ные рептилии, некоторые хищные птицы и звери.
Численность. По-видимому, существенно изменяется по годам. Так, в ок-
рестностях Аральского моря в 1920-1921 гг. он попадался «довольно редко»
(Сидоров, 1925), в 1945 и 1946 гг. вблизи г. Аральска, а в 1946 г. и вблизи
г. Иргиз (у северной границы ареала) щитомордников было очень много, а уже
год-два спустя (в 1947-1948 гг.) они опять попадались довольно редко (Динес-
ман, 1953). В последние годы, судя по контексту сообщения Т.Н. Дуйсебаевой
(2005), они здесь не многочисленны, хотя и не представляют собой редкости.
Сравнительно обычным видом считается щитомордник и в Приаральских Ка-
ракумах (Лобачев и др., 1973).
В западной части ареала в 1950 г. много щитомордников было в Индерских
горах, где за часовую экскурсию перед заходом солнца на песчано-глинистом
северном берегу озера было отмечено девять особей (Чернов, 1954). Судя по
количеству мест обитания, не представляют они особой редкости в песках и
по останцовым горам в Приэмбенской провинции, хотя в глинистой пустыне
встречаются чрезвычайно редко (Неручев, Васильев, 1978; Накаренок, 2003в;
Сараев, Пестов, 2010). В Волжско-Уральском междуречье он, возможно, уже
исчез, хотя в 1990-е гг. имелись достаточно достоверные (но документально не
подтвержденные) сведения о его встречах в районе Богдинско-Баскунчакского
заповедника (Божанский, Полынова, 1998).
Лимитирующие факторы. В основном хозяйственное освоение террито-
рии, сопровождающееся уничтожением змей.
Охрана. Обитает и охраняется на территории Иргиз-Тургайского резерва-
та. На остальной территории региона специальные меры охраны не применя-
лись.
Региональный статус. Характерный представитель пустынной герпето-
фауны, населяющий изолированные специфические местообитания у север-
ных пределов ареала.
Род 33. Vipera (Pelias) Laurenti, 1768 - гадюки
49. Обыкновенная гадюка - Vipera (Pelias) berus (Linnaeus, 1758)
Значение таксона для сохранения биоразнообразия. Один из предста-
вителей обширного рода, населяющий территорию региона. Политипический
вид, в регионе представленный номинативным подвидом V. b. berus (Linnaeus,
1758), в пределах которого наблюдаются некоторые межпопуляционные раз-
личия в окраске тела (цветовые морфы), количестве брюшных и подхвостовых
щитков (Табачишин и др., 1996; Морозенко и др., 2003; Атлас пресмыкающих-
ся..., 2004; Кузьмин, Семенов, 2006).
284
Глава 4
Распространение. Ареал вида охватывает значительную часть Евразии от
Атлантики до долины р. Лены и северо-восточного Китая (европейский фау-
нистический комплекс). Его северная граница от побережья Баренцева моря
на Кольском полуострове постепенно смещается к 60° с.ш. в долине р. Енисей,
затем несколько южнее, но по долине р. Лена вновь продвигается до 60-й па-
раллели. К югу распространена практически до границ степной зоны. Таким
образом, обыкновенная гадюка населяет только северную часть исследуемого
нами региона.
Южная граница ареала вида в регионе и его ближайших окрестностях в
1970-е гг. проводилась по лесным территориям в окрестностях г. Волгограда
(Кубанцев, 2003), сейчас она прослеживается по находкам вблизи г. Хвалынс-
ка Саратовской области, затем в черте г. Самары, долинах р. Самара у п. Бор-
ский Самарской области (Бакиев и др., 2009), р. Боровки в Бузулукском бору,
р. Самары у п. Тоцкое, долинах рек М. Кинель и Б. Кинель вблизи г. Бугурус-
лан, затем в верховьях р. Ик в Абдулинском и р. Демы в Шарлыкском районах
Оренбургской области, хр. М. Накас (Даркшевич, 1953; Райский, 1951; Чи-
билёв, 1995; Вершинин, 2007) и далее уже за пределами региона в Башкирии.
Кроме отмеченных, по сути пограничных, пунктов встречи обыкновен-
ной гадюки известны в окрестностях поселков Подстепки, Кинель-Черкас-
ский (Гаранин, 1983), сел Старое Якушкино и Красная Богана в верховьях
р. Б. Черемшан Самарской области (Бакиев и др., 2009). К настоящему вре-
мени в пределах региона обыкновенная гадюка известна в 8 (77,7% из 11)
ландшафтных районах лесостепной и 2 (2,9% из 69) степной зоны (табл. 55,
карта 48).
Места обитания. Обыкновенная гадюка - типичный обитатель лесных
и лесостепных ландшафтов, где придерживается преимущественно окраин
опушек и полян с кустарниками, зарастающих вырубок и гарей, просек, до-
рог, хорошо прогреваемых стаций по берегам рек, озер и подобных экотонов.
В пределах региона практически все известные пункты встреч приурочены
к долинам рек Волга, Самара, Б. и М. Кинели, Сок, Б. Черемшан, Ик, Дёма,
Боровка и речек, стекающих с хребтов Накас и Шайтантау. Здесь она насе-
ляет поросшие кустарниками побережья пойменных озер, влажные поймен-
ные участки с травянисто-кустарниковой растительностью, пойменные луга и
выпасы. В целом выбор местообитаний помимо степени влажности биотопов
(лимитирующей распространение вида на юг), их инсоляции и наличия ук-
рытий определяется также обеспеченностью кормом и степенью беспокойс-
тва. Разрушение местообитаний и постоянное беспокойство приводят, с одной
стороны, к инсуляризации ареала, распадающегося на изолированные участки,
а с другой - к вытеснению гадюк в непригодные для деятельности человека
угодья (овраги, свалки, пустоши, развалины строений и т. п.), что уже четко
прослеживается в лесопарковой зоне г. Самары и на Урале, которые становят-
ся своеобразными рефугиями вида в антропогенно трансформированной среде
(Чибилёв, 1995; Бакиев и др., 2004, 2009; Вершинин, 2007).
Образ жизни. Весной в регионе обыкновенная гадюка появляется в ап-
реле-начале мая, когда местами еще сохраняются значительные пятна снега.
Обычно это совпадает с переходом среднесуточных температур через диапа-
зон от 0 до 5 °C. Первыми из зимних убежищ выходят самцы, спустя несколько
дней самки, а затем молодые особи.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
285
Таблица 55
Распространение обыкновенной гадюки
Зона Провин- ция Район № точки Местонахождение Источник информации
i в О о о I 1 2 1 2 3 Самарская область окр. п. Подстепки г. Самара р-н Кировский р-н Красноглинский р-н Куйбышевский р-н Октябрьский окр. п. Старосемейкино Гаранин, 1983; Бакиев и др., 2002, 2003, 2009
II 3 4 5 6 7 окр. с. Красная Багана дол. р. Б. Черемшан окр. с. Старое Якушкино окр. п. Кинель-Черкасский
III 5 6 8 10 11 8 9 10 11 12 13 14 15 Оренбургская область верх. р. Ик окр. г. Бугуруслан окр. р. М. Кинель верх. р. Дема хр. М. Накас дол. р. Б. Ик дол. р. Сакмара хр. Шайтантау Чибилёв, 1995; Верши- нин, 2007
Степная V 22 23 16 17 18 Самарская область окр. п. Борское, Бузулукский бор Оренбургская область Бузулукский бор, дол. р. Боровка дол. р. Самара, окр. п. Тонкое Бакиев и др., 2002, 2009 Положенцев, 1935; Дарк- шевич, 1954; Чибилёв, 1995
Весной гадюки активны преимущественно в середине дня, летом актив-
ность смещается на утренние и вечерние часы, при пасмурной погоде они
могут быть встречены и в дневное время (причем возможно несколько пиков
активности), а в прохладные и дождливые дни почти не выходят из убежищ.
Летом в качестве убежищ используются норы, гнилые пни, трещины и пусто-
ты в грунте. Как правило, активность наблюдается при температуре воздуха
17,0...27,0 °C, субстрата 21,0...30,5 °C, а абсолютный оптимум в Поволжье со-
ставляет 28,8...30,8 °C, а максимум 37 °C. При этом самцы предпочитают более
теплый субстрат, чем самки. Различия в термоадаптации отмечаются также у
темных и светлых морф.
286
Глава 4
Обыкновенная гадюка - территориальная форма и чаще всего держится на
участках от 500-800 м2 до 1,5-4,0 га, совершая незначительные перемещения,
хотя во время сезонных миграций отдельные особи иногда могут перемещать-
ся до 2-5 км.
К осени активность гадюк снижается, и уже во второй половине сентября -
первой половине октября они залегают в спячку, которую проводят в грудах
хвороста, прикорневых и других пустотах грунта, норах животных, карстовых
воронках и подобных местах ниже уровня промерзания грунта, иногда образуя
скопления в несколько сот особей. Период активности, таким образом, про-
должается около 160 дней, в среднем около 140 (Гаранин, 1983; Бакиев и др.,
2004, 2009).
Размножение. Половой зрелости обыкновенная гадюка достигаете возрас-
те 4-5 лет. Достигшие этого возраста особи спустя 2-4 недели после выхода из
зимовки приступают к спариванию. Сроки его наступления и продолжитель-
ность (чаще всего около 2 недель) зависят главным образом от особенностей
биотопа и его микроклиматических условий (обычно после перехода средних
температур через 10 °C). Эмбриональное развитие продолжается 2,5-3,0 неде-
ли, и во второй половине июля - начале августа в Волжско-Камском крае появ-
ляется от 4 до 23 молодых. Размножается обыкновенная гадюка не ежегодно, в
разных частях ареала выявлены 2-, 3- и 4-летние циклы.
Питание. Основу пищевого рациона обыкновенной гадюки в регионе
(около 80%) составляют мелкие млекопитающие (преимущественно полевки,
мыши, замлеройковые), в меньшей мере потребляются амфибии (практически
все местные виды), ящерицы, ужи, изредка в составе пищи отмечаются насе-
комые, слизни и растительные остатки. В свою очередь обыкновенные гадюки
добываются рядом птиц, млекопитающих и других более крупных рептилий
(известно и явление каннибализма).
Численность. В связи с некоторыми трудностями определения в полевых
условиях обыкновенной гадюки, гадюки Никольского (видовая самостоятель-
ность которой некоторыми авторами отрицается) и восточной степной гадюки
старые данные о численности вида использовать не представляется целесо-
образным. В связи с этим отметим лишь, что в черте г. Самары (лесопарко-
вая зона у Горелого хутора) в 1993-2001 гг. плотность этого вида достигала
40-50 экз/га (локально до 125), что в переводе на данные маршрутного учета
составляет 20-25 экз/км (Бакиев и др., 2009).
Лимитирующие факторы. Комплекс факторов, действующих у южной
границы ареала. Разрушение местообитаний, хозяйственное и рекреационное
освоение территории, уничтожение человеком.
Охрана. Обитает и охраняется на территории национального парка «Бузу-
лукский бор». На остальной территории специальных мер охраны не предпри-
нималось.
Региональный статус. Типичный представитель герпетофауны реги-
она у южных пределов ареала. Занесена в Красную книгу Самарской облас-
ти с категорией 5 Б как условно редкий вид со снижающейся численностью
(2009).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ... 287
50. Гадюка Никольского, или лесостепная гадюка -
Vipera nikolskii Vedmederja, Grubant and Rudaeva, 1986
Значение таксона для сохранения биоразнообразия. Один из представи-
телей комплекса Vipera berus, населяющий территорию региона. В качестве
самостоятельного вида выделена сравнительно недавно (Грубант и др., 1973;
Ведмедеря и др., 1986), хотя ряд герпетологов считает ее лишь подвидом обык-
новенной гадюки V. berus nikolskii или даже внутриподвидовой формой номи-
нативного подвида V. b. berus. Из региональных герпетологов гадюку Николь-
ского самостоятельным видом считают саратовские специалисты (Табачишин
и др., 1995, 1996; Шляхин и др., 1995, 2001; Завьялов, Табачишин, 1997; Та-
бачишина и др., 2004). Однако при дальнейших исследованиях оказалось, что
указанные в первоначальном диапазоне признаки ненадежны, а особенности
биологии и экологии не вполне соответствуют биологической и экологической
концепции вида. В итоге накопившиеся к настоящему времени данные пока-
зывают, что лесостепная гадюка имеет, скорее всего, недостаточные отличия
от обыкновенной гадюки для присвоения ей таксономической категории ви-
дового ранга и является лишь подвидом V. b. Nikolskii или иной внутривидо-
вой формой V. berus (Бакиев, Кренделев, 1999; Бакиев и др., 2000, 2004, 2009;
Песков и др., 2003). Это подтверждают и результаты анализа последователь-
ностей гена цитохрома в митохондральном ДНК, согласно которому «гадю-
ка Никольского, по-видимому, не может считаться отдельным видом, а имеет
более низкий уровень отличий (вероятно, подвидовой) от обыкновенной га-
дюки». Кроме того, состав пептидов ядовитого секрета гадюк волжского бас-
сейна типичен для V. b. berus и имеет различия лишь в количестве некоторых
пептидных компонентов (Старков, Уткин, 2003). Все это позволяет считать,
что гадюки, по крайней мере, Самарской области, представлены популяциями,
совмещающими в себе признаки V. b. berus и V. b. Nikolskii (Колябина и др.,
2003; Павлов и др., 2004; Бакиев и др., 2009; Маленев и др., 2009). Возможно
также, что это свидетельствует о наличии широкой интрогрессивной гибриди-
зации между этими двумя формами, в связи с чем они, видимо, широко обме-
ниваются генами и представлены по большей части гибридными и смешанны-
ми популяциями (Зиненко, 2003).
Распространение. По литературным данным (Ананьева и др., 1998), ареал
вида охватывает «лесные и лесостепные районы Украины и европейской части
России к югу от линии Канев-Курск-Тамбов-Бузулук, а по влажным долинам
рек проникает в степные районы Саратовской и Самарской областей (евро-
пейский фаунистический комплекс). Позднее, при уточнении распространения
вида, граница на востоке ареала от оренбургского г. Бузулука была перенесена
к г. Бугульма (Татарстан) и далее продвинута «до предгорий Южного и Сред-
него Урала» (Мильто, 2003; Павлов и др., 2004). По другой версии (Ефимов
и др., 2008), восточным пределом распространения вида V. nikolsKii является
долина р. Волга, причем район ее Самарской Луки является зоной вероятной
гибридизации V. berus и V. nikolskii.
Таким образом, таксономический статус и распространение меланистичес-
кой формы гадюк региона до настоящего времени остаются неопределенными
и требуют дальнейшего изучения, поскольку описанный ареал обыкновенной
гадюки в регионе может оказаться зоной интерградации двух видов (подви-
288
Глава 4
дов?). В связи с этим неясными, а во многом неопределенными становятся и
сведения об образе жизни, экологии и численности гадюки Никольского.
Региональный статус. Гадюка Никольского в качестве самостоятельно-
го вида V. nikolsKii включена в Красную книгу Российской Федерации с ка-
тегорией 4 как неопределенный по статусу малоизученный вид (Божанский,
2001). Это обстоятельство стало достаточным основанием для включения вида
в Красную книгу Саратовской области (2006).
51. Восточная степная гадюка, или гадюка Ренарда -
Vipera (Pelias) renardi (Christoph, 1861)
Значение вида для сохранения биоразнообразия. Один из представителей
обширного рода, населяющий территорию региона. До недавнего времени счи-
талась восточным подвидом полиморфного комплекса V. ursini-V. и. renardi.
Сейчас признается самостоятельным политипическим видом, в регионе пред-
ставленным двумя подвидми - номинативным V. г. renardi (Christoph, 1861)
и гадюкой Башкирова V. г. bashkirovi (Garanin et al., 2004), отличающимися
от других размерами, степенью проявления меланизма, количеством брюш-
ных, подхвостовых, верхнее- и нижегубных щитков, а также некоторыми осо-
бенностями экологии (Ананьева и др., 1998; Атлас пресмыкающихся..., 2004;
Павлов, 1997; Кузьмин, Семенов, 2006; Бакиев и др., 2009). Вместе с тем спе-
цифичен ряд метрических и меристических признаков у локальных популяций
Нижнего Поволжья (Табачишин и др., 1996; Кармышев, Табачишин, 2003), что
требует дополнительных исследований.
Распространение. Ареал вида охватывает лесостепную, степную, полу-
пустынную зоны Европы, Кавказа, Казахстана и Средней Азии, а по речным
долинам проникает и в пустынную зону (средиземноморский фаунистический
комплекс). Его северная граница на Восточно-Европейской равнине прово-
дится от Молдавии по северной Украине и Поволжью к месту слияния рек
Волги и Камы (острова Спасского архипелага), окрестностям г. Кумертау
(Башкирия), г. Верхнеуральска на Урале, Кустаная, Петропавловска, Семипа-
латинска в Казахстане и долиной р. Иртыш уходит в северо-западный Китай.
Южная граница от Молдавии проводится через Крым, северные районы Кав-
каза, северное побережье Каспийского моря, примерно по долине реки Эмбы
к Мугоджарам, затем по северному побережью Аральского моря, предгорь-
ям Улытау, северному Прибалхашью к оз. Алаколь и по предгорьям Средней
Азии в Китай (Параскив, 1956; Ананьева и др., 1998; Бакиев и др., 2004, 2009;
Вершинин, 2007; Табачишина и др., 2002). Таким образом, большая часть
региона включается в ареал вида, за исключением Волжско-Уральских пес-
ков, Предустюртья, Северного Устюрта, северного Приаралья и Приараль-
ских Каракумов. В целом местообитания вида установлены в 6 (54,5% из 11)
ландшафтных районах лесостепной, 41 (59,4% из 69) степной, 32 (53,1% из
32), полупустынной и 22 (51,2% из 43) пустынной природных зон (табл. 56,
карта 49).
Места обитания. Как свидетельствует название вида, его основными мес-
тообитаниями являются открытые равнинные и горные степные ландшафты,
а также сопредельные территории лесостепной, полупустынной и пустынной
зон.
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
289
Распространение степной гадюки
Таблица 56
Зона Провин- ция Район № точки Местонахождение Источник информации
I 1 1 Самарская область окр. рыбхоза «Сускан» Бакиев, Иванова, 2004;
Лесостепная II 3 5 6 2 3 4 5 6 7 окр. с. Раевка окр. с. Кракове вдр. рек Сок и Кондурча окр. с. Старопохвистнево дол. р. М. Кинель Оренбургская область окр. г. Бугуруслан окр. п. Пономаревка Бакиев и др., 2002, 2004, 2009 Чибилёв А.А.
III 7 11 8 9 окр. п. Тюльган окр. п. Сара п. Подгорное Завьялов и др., 2006
Степная IV 12 13 15 16 17 20 21 10 11 12 13 14 15 16 17 Самарская область дол. р. Безенчук Саратовская область окр. г. Балаково Самарская область дол. р. Б. Иргиз овр. Вязовый окр. с. Парфеновка окр. с. Яблоновый овраг овр. Уральская глушица дол. р. Б. Иргиз Саратовская область окр. с. Алексеевка Самарская область Красносамарский лес Бакиев, Магдеев, 1995; Бакиев и др., 2004, 2009 Табачишина и др., 2002 Бакиев и др., 2004, 2009; Песков, 2001 Табачишина и др., 2002 Бакиев и др., 2009
V 22 23 24 25 18 19 20 21 22 Оренбургская область окр. п. Грачевка окр. п. Тонкое окр. п. Приютное окр. п. Сорочинск окр. п. Александровка Каргалинская степь, окр. п. Каргала гора Медвежий лоб Чибилёв А.А.
290
Глава 4
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
Самарская область
26 23 Бирючий овраг окр. с. Украинка окр. п. Восточный Бакиев и др., 2004, 2009; Смелянский, Елизаров, 1996
24 ур. Бостандык ур. Грызлы
V 25 окр. п. Курлин п. Озерное Оренбургская область Таловская степь ОГПЗ Чибилёв, 1995, 1999а, б
26 ур. Меловой Сырт
27 п. Каменноимангулово Саратовская область
27 28 окр. п. Озинки Табачишина и др., 2002
Оренбургская область
28 29 дол. р. Салмыш Чибилёв А.А
VI 30 30 окр. г. Соль-Илецк Паллас, 1773
i 32 31 Урало-Илекское Райский, 1951
е а междуречье Чибилёв, 1995
О Западно-Казахстанская область
34 32 33 окр. г. Аксай, дол. р. Утва дол. р. Караоба Березовиков и др., 1992, 2003
35 34 дол. р. Акбулак
35 междуречье Утва-Сукбулак
36 дол. р. Березовка
37 горы Актау
VII 38 38 окр. оз. Шал кар: гора Сантас Дебело П.В.; Завьялов и др., 2006
39 гора Сасай Оренбургская область
41 40 окр. п. Покровка Чибилёв, 1995
42 41 окр. п. Сагарчин
43 42 окр. п. Шаповалове Актюбинская область
44 43 юг.-зап. г. Актюбинск Чибилёв А.А.
46 44 дол. р. Киил
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
291
Продолжение табл. 56
Зои» Провин- ция Район № точки Местонахождение Источник информации
VIII 47 48 50 52 54 55 45 46 47 48 49 50 Волгоградская область Приволжская возв. Саратовская область Дьяковский лес окр. с. Сафаровка окр. с. Монахов Оренбургская область окр. п. Первомайское Западно-Казахстанская область окр. п. Красноармейск окр. г. Уральска Кубанцев и др., 1962 Особо охраняемые..., 2007; Табачишина и др., 2002 Чибилёв А.А. Дебело П.В.; Николь- ский, 1905
Степная IX 56 57 58 61 62 51 52 53 54 55 56 Оренбургская область Буртинская степь ОГПЗ окр. п. Донское окр. п. Новорудный окр. п. Аккермановка Айтуарская степь ОГПЗ Актюбинская область окр. п. Тайкеткен Чибилёв, 1999а; Чи- билёв А.А. Топоркова, 1973
X 65 66 67 69 57 58 59 60 61 62 Оренбургская область окр. Ириклинского вдхр. Кваркенский район окр. п. Шильда окр. п. Кумак окр. п. Ясный окр. п. Домбаровский Чибилёв, 1995
XI 72 78 63 64 65 66 Светлинские озера: оз. Каменное, окр. оз. Жеты- коль (п. Озерное) Ащесайская степь ОГПЗ окр. оз. Айке Кустанайская область Наурзумский заповедник Чибилёв, 1995, 1999а Динесман, 1953; Брагин, Брагина, 2002
292
Глава 4
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
>шная XIII 82 84 85 87 88 89 91 67 130 68 69 70 71 72 73 74 Западно-Казахстанская область окр. п. Джаныбек окр. оз. Аралсор Волгоградская область окр. оз. Эльтон Астраханская область окр. оз. Баскунчак Саратовская область окр. с. Варфоломеевка окр. с. Ветелки п. Монахов Западно-Казахстанская область окр. п. Казталовка окр. п. Фокеево окр. р. Кушум окр. п. Коловертное Ходашова, 1960; Завья- лов и др., 2006 Ходашова, 1960 Стрельцов и др., 2006 Табачишина и др., 2002; Завьялов и др., 2006 Параскив, Бутовский, 1960; Паллас, 1773; Чернов, 1954
ь Ё? £ о К XV 95 96 97 98 100 75 76 77 78 79 80 81 Актюбинская область п. Миялы, 50 км на юго-вост, окр. п. Каракуга окр. п. Жаркамыс окр. мог. Шайтас (верх, р. Маннесай) окр. горы Хандырткуль междуречье Темир-Эмба верх. р. Четырлысай Накаренок, 2002, 2003в; Шилов, 1961 Никольский, 1905
XVI 101 82 окр. ст. Мугоджарская Дуйсебаева, 2005
XVIII 107 108 83 84 85 86 71 кмтр. Челкар-Иргиз окр. г. Иргиз окр. п. Текели пески Жаманкум Динесман, 1953; Шилов, 1961; Брушко, Кубыкин, 1988
XIX 109 ПО 112 87 88 89 дол. р. Улькояк, 20 км к сев. от п. Дукен дол. р. Тургай окр. п. Куйлыс низ. р. Тургай Дуйсебаева, 2005; Динесман, 1953; Шилов, 1961
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
293
Продолжение табл. 56
Зона Провин- ция Район № точки Местонахождение Источник информации
л i 90 дол. р. Жасыбай
О и я XIX окр. оз. Мельдекуль
с £ 91 ур. Таук, пески Барбикумы
Астраханская область
113 114 92 93 94 окр. горы Б. Богдо окр. п. Сероглазово Харабали, окр. п. Досанг-За- мьяны Никольский, 1905; Сараев, Пестов, 2010; Бобров, 1985; Паллас, 1773; Завьялов, Табачи- шин, 1998
95 окр. г. Красный Яр окр. п. Лапас п. Комсомольский Степной, Малый Арал Гурьевская область
XX 96 окр. п. Танюшкино Накаренок, 2002, 2003в
97 окр. рзд. 6, ур. Сазды, Муса
Западно-Казахстанская область
116 117 98 99 окр. п. Урда окр. Камыш-Самарских озер Чернов, 1954; Параскив, Бутовский, 1960
с5 я я Гурьевская область
3 в с 118 121 124 100 101 102 окр. п. Н. Уштаган сев.-вост. п. Забурунье окр. п. Кулагине Чернов, 1954; Сараев, Пестов, 2010; Параскив, Бутовский, 1960
103 окр. п. Калмыкове
125 104 окр. п. Елтай
XXI 127 128 131 105 106 107 108 р. Эмба, ур. Шубартпала окр. п. Зеленое рзд. Каработано, 7 км юго- вост. окр. п. Искининский, 10 км к Окулова, 1963; 1981; Неручев и др., 1990; На- каренок, 2003 в; Чернов, 1954; Никольский, 1916; Накаренок, 2002, 2003в; Гисцов, Березовиков, 1995
зап.
132 109 окр. п. Сарайчик
ПО 12 км к сев.-вост. от г. Гурьев
111 окр. г. Гурьев, дельта р. Урал
133 112 окр. п. Индерский Никольский, 1916
XXII 135 ИЗ 114 ур. Тайсойган, дол. р. Уил окр. п. Акколь окр. п. Кенбай Неручев и др., 1984; Накаренок, 2002, 2003в
294
Глава 4
Окончание табл. 56
Зона Провин- ция Район № ТОЧКИ Местонахождение Источник информации
136 115 окр. п. Макат
116 низ. р. Сагиз
117 п. Жамансор
118 дол. р. Эмбы, ур. Кзылжар окр. оз. Бартылдакты
119 ур. Тургузба ур. Дюсеке ур. Брлик
ххп ур. Мадьяр
5 окр. п. Камбай
I I 3 120 плато Аккерегеше
Й С* 137 138 121 122 гора Иманкара окр. р. Кайнар Шилов, 1961; Накаре- нок, 2003 в
139 123 ур. Намазтакыр пески Актамаймагут
124 окр.п. Косчагыл
125 сев. чинк Устюрта
XXIV 145 126 п. Карабулак, 10 км к сев. Шилов, 1961; Дуйсебае-
150 127 окр. п. Каир ва, 2005
XXVI 151 128 окр. ст. Тогыз Б. Барсуки, окр. п. Бегимбет
152 129 окр. ст. Саксаульская
В условиях лесостепи Волжско-Камского края степная гадюка населяет це-
линные степные участки, залежи, опушки лиственных лесов и окраины колков,
примыкающие к ним луговые и остепненные пространства, кустарниковые
луга, а также облесенные склоны речных и овражно-балочных систем. Вместе
с тем в ряде мест здесь (окр. рыбхоза «Сускан») и в некоторых южнее распо-
ложенных островных лесах (Красносамарском лесу) она помимо свойствен-
ных виду открытых ксерофильных биотопов проникает в более гигрофильные
сообщества полян, опушек, просек, зарастающих вырубок, а также молодых
посадок (Гаранин, 1983; Бакиев и др., 2004, 2009).
В степях Самарской и Саратовской областей обитание связано с участка-
ми целинных степей и старых залежей, где. придерживается биотопов разных
типов, но преимущественно стаций со сложным микрорельефом: расчленен-
ных каменистых склонов сыртов, сухих склонов балок, оврагов, речных до-
лин, прибрежных участков степных речек, чаще всего с зарослями кустарни-
ков, разреженных островных и байрачных лесков с прогалинами, полянами и
зарастающими вырубками. В наиболее освоенных земледельческих районах
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
295
вытеснена на обочины дорог, в овраги и другие малопригодные для хозяйс-
твенной деятельности человека места (Смелянский, Елизаров, 1996; Бакиев и
др., 2001, 2004, 2009; Песков, 2001; Шляхтин и др., 2006).
На территории Оренбургской области наиболее благоприятны условия в
пределах типичной и южной степи, особенно в карстовых районах Предуралья,
изобилующих трещинами и пустотами. В Каргалинской степи и на террито-
рии сохранившихся целинных массивов Таловского, Буртинского, Айтуарско-
го и Ащисайского участков Оренбургского степного заповедника также чаще
всего населяет пересеченные сырты и горные балки со скальными выходами
по бортам (Чибилёв, 1999а, б). В Зауралье обычно встречается по останцам
и в местах выхода на поверхность кварцитов и песчаников, местами прида-
ющих территории холмисто-останцовый облик. В Наурзумской степи встре-
чи, как правило, приурочены к сообществам пырея и черной полыни (Шилов,
1961).
На Утва-Илекском междуречье степная гадюка встречается по долинам
степных речек Березовка, Караоба, Акбулак, Утва, а также по меловым ос-
танцам гор Актау (Березовиков и др., 1992). В окрестностях оз. Шалкар ос-
новные местообитания приурочены к склонам соляных куполов Сантас (самое
змеиное в округе место) и Сасай, реже отмечается среди чернополынников
на сопредельной территории и в верховьях балок, впадающих в реки Б. и
М. Анкаты и озеро.
На полупустынных равнинах Волжско-Уральского междуречья степная
гадюка населяет в основном водораздельные комплексы с господством чер-
нополынных сообществ, несколько реже встречается в злаково-разнотравных
понижениях (западины, падины), а местами (у п. Круглый) и в биюргунниках.
В окрестностях оз. Эльтон и Баскунчак ее встречи приурочены к склонам реч-
ных долин и балок с преобладанием остепненных участков или с зарослями
кустарников (Параскив, Бутовский, 1960; Ходашова, 1960; Стрельцов и др.,
2006). У восточных пределов междуречья (окрестности п. Калмыкове) основ-
ными местообитаниями считаются полынные и злаково-полынные сообщества
глинистой полупустыни; характерным обитателем она является и в песчаной
полупустыне, хотя встречается значительно реже (Окулова, 1963).
В Урало-Эмбенском междуречье, как правило, населяет долины и поймен-
ные урочища по рекам Уил, Сагиз и Эмба: берега, сухие староречья, солонча-
ковые луга, прибрежные заросли тростника и тамарикса (Неручев, Васильев,
1987; Накаренок, Неручев, 2001). В верховьях р. Маннесай она отмечалась в
колониях большой песчанки и на курганчике малого суслика; в сходных усло-
виях на солонцах у колоний малого суслика, степной пеструшки, других по-
левок ее встречи известны в Зауралье в низовьях рек Иргиз, Улькояк и Тургай
(Шилов, 1961).
Пустынную зону региона степная гадюка населяет лишь по переферии. По
западной кромке Волжско-Уральских песков почти повсеместно она отмеча-
ется в основном в пределах надпойменной террасы. В окрестностях г. Крас-
ный Яр основными местообитаниями являются межбугровые депрессии, хотя
встречи известны также по закрепленным бугристым пескам и на солонцах
(Завьялов, Табачишин, 1998).
По межбугровым депрессиям (ашикам) в Рын-песках (окрестностях
п. Урда) она проникает в глубь массива, где встречается также в понижениях
296
Глава 4
с ивняками и осокорем. По северной и южной кромкам Волжско-Уральского
массива изредка встречается на задернованных песках и очень редко между со-
рами, где придерживается солянковых сообществ. Здесь она отсутствует лишь
в барханных и разбитых песках (Чернов, 1954; Параскив, Бутовский, 1960),
хотя в местах с очагами сбоев и развевания она все же встречается (Неручев
и др., 1983). По шлейфу песков Актамаймогут и кромке песчаных массивов
Жаманкумы и Барбикумы встречи известны также в Предустюртье и Зауралье
(Шилов, 1961).
Образ жизни. Из зимних убежищ степная гадюка выходит, когда темпе-
ратура воздуха превышает 6...9 °C, что на юге региона приходится на конец
марта - начало апреля, а на севере на разные числа апреля, т. е. запаздывает
до 15-20 дней; примерно на такое же время сроки выхода могут различаться
и в одном районе в разные годы в зависимости от хода развития весенних яв-
лений. Массовый выход происходит, когда температура воздуха превышает
12 °C, - т. е. во второй половине апреля (Кубанцев и др., 1962; Бакиев и др.,
2004, 2009; Песков, 2001; Шляхтин и др., 2006; Табачишина и др., 2002).
В условиях эксперимента добровольный минимум для приземного воздуха
оказался равен 8,7 °C; для субстрата - 10,8 °C, а максимум соответственно
32,6 и 43,5 °C, диапазон оптимальных температур определен в 15,0...29,0 °C
(Литвинов, 2004). Это практически совпадает с материалами полевых на-
блюдений. В Поволжье активные гадюки встречались на субстрате с тем-
пературой 4,5...43,5 °C (Песков и др., 2003), а у северной кромки Волжско-
Уральских песков (окрестности п. Калмыкове) при температуре воздуха, °C:
вапреле-мае-18,0...31,5,июне-июле-21,0...28,0, сентябре-октябре-14,0...27,0.
Ранней весной активность имеет однопиковый характер, который примерно
с середины мая сменяется двухпиковым. В июне-июле максимальная актив-
ность наблюдается в 9-12 и 15—18 часов, хотя в пасмурную погоду она актив-
на и днем. Днем же обычно укрывается в нежилых норах разных животных,
других полостях в грунте, стогах и подобных местах. На ритмику активности
помимо температуры влияют также сила ветра, характер растительности, обес-
печенность кормом и ряд других факторов.
С началом осени активные степные гадюки наблюдаются в течение все-
го светлого времени суток; с этого же времени начинается их концентрация
около мест зимовки. В спячку на севере региона залегают во второй половине
сентября, на юге - в первой половине октября, хотя отдельные особи иногда
задерживаются и позднее (почти на месяц) (Бакиев и др., 2004,2009; Шляхтин
и др., 2006).
Размножение. Половозрелыми степные гадюки, по-видимому, становят-
ся на втором-третьем году жизни. В этом возрасте спустя 2-3 недели после
выхода из зимовки начинается спаривание, в отдельных популяциях продол-
жающееся 7-10 дней, но в целом этот период оказывается растянут. В Волж-
ско-Уральском междуречье он приходится на вторую половину мая - первую
половину июня. В это время образует скопления численностью более десятка
особей (гора Сантас). Количество эмбрионов достигает 20. Период эмбрио-
нального развития в условиях Саратовской области продолжается около 100 су-
ток. Выход сеголеток начинается с середины июля (в Волжско-Уральском
междуречье наиболее ранняя находка отмечена 10.07.1953 г.) и продолжается
до середины сентября (Параскив, Бутовский, 1960; Шляхтин и др., 2006).
ОБЩАЯ ХАРАКТЕРИСТИКА И ВИДОВЫЕ ОЧЕРКИ АМФИБИЙ...
297
Питание. Основу пищевого рациона степной гадюки составляют мелкие
грызуны (преимущественно полевки, мыши) и ящерицы (прыткая, ящурки);
в ряде специфических местообитаний яйца и птенцы мелких наземногнездя-
щихся видов птиц, а также амфибии и рептилии. Повсеместно широко потреб-
ляются насекомые (особенно прямокрылые, временами чешуекрылые, жуки),
реже паукообразные. Их соотношение несколько изменяется по сезонам и ста-
циям, что свидетельствует о значительной трофической пластичности вида.
Молодые особи кормятся в основном прямокрылыми насекомыми и мелки-
ми ящерицами. В свою очередь для степной гадюки характерно и «обилие ес-
тественных врагов» - это преимущественно хищные птицы, млекопитающие,
крупные рептилии.
Численность. Сведения о ней довольно противоречивы даже по одним и
тем же районам. В связи с этим по некоторым пунктам приводим данные лишь
последних лет. В Самарской области даже на локальных участках с макси-
мальной плотностью (в Большечерниговском, Кинельском и Ставропольском
районах) встречаемость весной и осенью не превышает 3-4 экз/га, а в летние
месяцы 1-2 экз/га (Бакиев, Гафарова, 1999; Бакиев и др., 2009). На значитель-
ной части Саратовской области численность степных гадюк не превышает
2-5 экз/га, хотя в разных стациях межузеньской равнины в 1998 г. она состав-
ляла 1,2-9,4, на участках с травянисто-кустарниковой растительностью по ов-
ражно-балочным системам отрогов Общего Сырта (Дергачевский, Перелюб-
ский, Озинский районы) - 5-11, а в 2001 г. местами она достигала 17 экз/га
(Табачишина и др., 2002; Шляхтин и др., 2006). В Оренбургской области на
участках заповедника степная гадюка является обычным видом, а в карсто-
вой зоне Предуралья местами численность достигает 7-10 экз/га (Чибилёв,
1999а; данные последних лет). В Зауралье ее встречаемость значительно ниже,
хотя и здесь она может достигать значительной величины - на сопредельной
территории музея-заповедника «Аркаим» в Челябинской области, например,
12,7 экз/га (Чибилёв, 2005).
В полупустыне Волжско-Уральского междуречья в середине прошлого сто-
летия степная гадюка считалась обыкновенной, а местами, как, например, в ни-
зовьях р. Б. Узень (окрестности п. Фокеево), и очень многочисленной (Параскив,
Бутовский, 1960). В комплексах Волжско-Узеньского водораздела, по наблюде-
ниям К.С. Ходашевой (1960), в это время численность испытывала значительные
колебания в зависимости от влажности года. Так, в сухом 1950 г. встречаемость
здесь достигала 5,0 ос/10 км маршрута, во влажном 1952 г. - 1,0, а в средние по
влажности - 3,0-4,0 ос/10 км. В окрестностях оз. Эльтон эти показатели соот-
ветственно равнялись 20,0; 5,0 и 7,0-11,0 ос/10 км. На специфических участках
поймы р. Б. Узень в окрестностях Александрова Гая плотность и сейчас дости-
гает значительных величин - 4,0-9,0 ос/га (Кармышев, Табачишин, 2003), а на
межузеньской равнине - 5,6 ос/км (Табачишина и др., 2002). На восточной кром-
ке междуречья в окрестностях п. Калмыков© в глинистой полынной полупусты-
не в весенне-летние сезоны начала 1950-х гг. численность в среднем достигала
3,2 ос/10 км, в злаково-полынных сообществах - 2,8, а в песчаной полупусты-
не 0,6 ос/10 км, хотя весной (апрель-май) змеи встречались несколько чаще: в
глинистой полупустыне - 0,86 экз/га, а в песках - 0,24 (Окулова, 1963, 1981).
В Урало-Эмбенском междуречье степная гадюка наиболее обычна по речным
долинам Уила, Сагиза и Эмбы, а в Зауралье в низовьях рек Иргиз и Тургай.
298
Глава 4
В пустынной зоне степная гадюка встречается в основном по кромке пес-
ков, хотя по ашикам местами проникает и в глубь массивов. По Волго-Ахту-
бинской надпойменной террасе в окрестностях г. Красный Яр в межбугровых
депрессиях ее плотность составляет 8,70±0,09 экз/га, на закрепленных буг-
ристых песках 1,20±0,18, на солонцах 0,97±0,16 (Завьялов, Табачишин, 1998),
вблизи ст. Досанг в песчаной степи 0,2 экз/га (Бобров, 1985). На территории
Богдинско-Баскунчакского заказника известны ее лишь единичные встречи
(Стрельцов и др., 2006), хотя на сопредельной территории Рын-песков в ок-
рестностях п. Урда в межбугровых депрессиях с зарослями ивняка и осокоря,
а также у Камыш-Самарских озер она не представляла особой редкости (Па-
раскив, Бутовский, 1960). В Волжско-Уральских песках немногочисленна - до
1-2 экз/10 км (Неручев и др., 1981). В Северо-Восточном Прикаспии, судя по
количеству пунктов встреч, наиболее обыкновенна степная гадюка по рекам
Сагиз (до 6,0 экз/10 км) и Эмба (Накаренок, 2003в; Сараев, Пестов, 2010).
Лимитирующие факторы. Хозяйственное (распашка земли, сенокоше-
ние, выжигание травы, выпас скота, использование ядохимикатов) и рекреаци-
онное освоение территории, урбанизация, истребление людьми, коммерческие
заготовки. После многоснежных зим затопление мест зимовки.
Охрана. Обитает и охраняется на территории Оренбургского, Наурзумско-
го и Богдинско-Баскунчакского заповедников, Иргиз-Тургайского резервата.
На остальной территории специальных мер охраны не предпринималось.
Региональный статус. Включена в Красную книгу МСОП (2000), Сара-
товской (2006) и Самарской (2009) областей, Аннотированный перечень таксо-
нов и популяций животных, нуждающихся в особом внимании к их состоянию
в природной среде России, Приложение III и Приложение IСИТЕС. Находится
под охраной Бернской конвенции (Приложение 2).
Глава 5
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ
АМФИБИЙ И РЕПТИЛИЙ РЕГИОНА
Современный состав фауны амфибий и рептилий региона является
результатом длительного исторического развития. Как уже отмеча-
лось, ядро ее исходного типа сформировалось в плиоцене, к концу
которого активизировались адаптивные механизмы, способствующие расши-
рению видового спектра фауны региона. Эти процессы продолжались и в плей-
стоцене, чему способствовало усиление динамики всех природных явлений в
регионе, приведшее к вымиранию ряда архаичных и существенному измене-
нию ареала «молодых» форм.
Современный облик фауна региона постепенно приобретала в голоцене,
который рассматривается как время преимущественной стабилизации аре-
алов, медленного расселения и освоения новых территорий, своеобразного
заполнения ценотического вакуума, образовавшегося в результате действия
природных факторов. Последним «штрихом» преимущественного их дейс-
твия, по-видимому, является исчезновение на Восточно-Европейской равнине
обыкновенной квакши.
Со времени начала голоцена, но особенно в последние тысячелетия, фор-
мирование ландшафтов региона и структуры их биоты идет под все возраста-
ющим воздействием антропогенных факторов: вырубки лесов, пасторальной
дигрессии, затем распашки земель и общего освоения территорий, которые
снивелировали их многие первобытные черты. Вместе с тем у северных пре-
делов региона меньшая засушливость позднего голоцена наряду с особеннос-
тями хозяйственного освоения территории определили большую стабильность
природных биоценозов, которые на целинных участках сохранили многие осо-
бенности экосистем раннего голоцена, исчезнувшие в степной и полупустын-
ной зонах.
Во время первого этапа земледельческого освоения территории в лесо-
степной и степной зонах были выборочно распаханы крупные массивы чер-
ноземных и темно-каштановых почв, а также некоторые крупные падины в
полупустыне. На втором этапе оказались распаханы практически все удобные
в технологическом плане, а затем и условно пахотнопригодные земли, что
обусловило широкое распространение антропогенного опустынивания.
Наряду с этим в регионе сооружен ряд крупных водохранилищ на Волге,
Урале, Кушуме, зарегулирован сток практически всех степных речек, созданы
Урало-Кушумская, Иргизская и около десятка менее значительных водоканаль-
ных систем. Регион стал и ареной крупного лесохозяйственного эксперимента,
результат которого - создание лесополос вдоль Волги и Урала, многочисленных
полезащитных и придорожных насаждений, проложение тысяч километров ав-
томагистралей, линий электропередач, связи, трасс трубопроводов, многочис-
ленные инженерные сооружения; в регионе создан мощный промышленный
(преимущественно нефтегазовый и горно-металлургический) комплекс.
300
Глава 5
Совокупное действие этих факторов привело к сокращению, а в ряде слу-
чаев и уничтожению на значительной территории типичных местообитаний
животных, их загрязнению и чрезмерной эксплуатации ресурсов живой приро-
ды, что настоятельно требует разработки комплекса природоохранных меро-
приятий, снижающих воздействие человека на мир животных.
Воздействие это, как известно, бывает прямым и косвенным. Прямое воз-
действие на амфибий и рептилий региона проявляется в их добыче для разных
целей и уничтожении, чаще всего бессмысленном. Истребляются в основном
гадюки, щитомордник и стрела-змея как ядовитые животные, в меньшей степе-
ни преследуются из-за ошибочного представления об их ядовитости медянка и
узорчатый полоз, а из-за агрессивности некоторые другие полозы. В ряде сель-
ских районов вредным считается обыкновенный уж, хотя большинству населе-
ния известны его неядовитость и хорошо выраженные отличительные призна-
ки. Отмеченное массовое уничтожение водяных ужей, вероятнее всего, связано
с их некоторым сходством с обыкновенной гадюкой, отличие от которой не
известно местным жителям (хотя, возможно, здесь имеет место бессмысленное
уничтожение безобидных животных). Общие масштабы уничтожения змей в
регионе с учетом значительных размеров его территории по ориентировочным
расчетам составляют не менее тысячи особей в год. В ряде случаев беспричин-
но уничтожаются также лягушки, чесночницы и ящерицы. Наиболее действен-
ным мероприятием по снижению масштабов такого воздействия, по-видимому,
является усиление экологического просвещения населения.
Более существенны масштабы специальной добычи амфибий и рептилий.
Их постоянными потребителями являются медицинские и другие учебные за-
ведения естественного профиля и научно-исследовательские учреждения, ис-
пользующие разные виды в учебном процессе и как коллекционный материал.
В значительном количестве змей (в основном массовых видов) добывают для
нужд медицинской промышленности. Перспективно в этой сфере использова-
ние секрета кожных желез ряда видов жаб и лягушек. По ориентировочным
расчетам для всех этих целей ежегодно в регионе добывается до 100-150 ты-
сяч экземпляров разных видов.
В последнее время амфибии и рептилии стали играть важную роль как
объекты коммерческой торговли. По-видимому, несколько тысяч особей ряда
видов, особенно экзотических, продается для содержания в частных коллекци-
ях. Однако при недостаточном уровне специальной подготовки заготовителей,
коллекционеров и отсутствии надлежащего ухода значительная часть содер-
жащихся в неволе животных гибнет. В еще большем количестве (очевидно,
сотни тысяч) некоторые виды заготавливаются для нужд пищевой промыш-
ленности (деликатесы).
Для предупреждения негативных последствий нерегулируемого промыс-
ла необходимы ограничение частного коллекционирования; разумное лицен-
зирование добычи с периодической сменой мест отлова при учете состояния
численности популяций; в районах с достаточно высокой численностью змей
создание специализированных питомников, стационаров или мобильных ла-
бораторий по эксплуатации животных непосредственно в природе. В особом
внимании нуждаются краеареальные популяции малочисленных видов, что
позволит стабилизировать их численность и обеспечит возможность их хо-
зяйственного использования.
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ АМФИБИЙ...
301
Косвенное воздействие антропогенных факторов проявилось в процессе
освоения территории. Наиболее значительные изменения ландшафтов и со-
става фауны произошли в северной части региона, где уничтожены обшир-
ные дубравы по Общему Сырту, еще в историческое время смыкавшиеся с
пойменными лесами по рекам Волго-Уральского междуречья, Подуральского,
Урало-Эмбенского плато и Мугоджар (что придавало всей этой территории
лесостепной облик).
Сейчас южная граница лесостепи отступила на рубежи г. Самары, долины
р. М. Кинель, истокам р. Салмыш, низовым р. Б. Ик и верховьям р. Губерля -
т. е. более чем на 200-300 км, а остальная территория приобрела степной об-
лик с реликтовыми Бузулукским и Красносамарским борами и рядом неболь-
ших лесков, ставших своеобразными рефугиями для обитавших здесь лесных
форм. Существенные изменения произошли и в структуре пойменных лесов
по рекам Самаре, Сакмаре и Уралу, которые потеряли «уремный» облик, стали
более осветленными и мезофильными.
Это сопровождалось изменением границ и «кружева ареалов» всех лес-
ных видов, свидетельством более широкого распространения которых явля-
ются находки сибирского углозуба в Дьяковском лесу, обыкновенной жабы
по долинам рек Б. Иргиз, Илек, Уил и Темир; травяной лягушки по р. Илек,
живородящей ящерицы в окрестностях г. Чапаевска, обыкновенной медянки в
верховьях р. Б. Иргиз, по р. Илек и в Мугоджарах.
Наиболее значительные изменения структуры фауны амфибий и рептилий
произошли в междуречье Волги и Урала, особенно в низменном лесостепном
Заволжье. С зарегулированием Волги практически полностью исчезли поймен-
ные ландшафты - места обитания более десятка гидрофильных и мезофиль-
ных видов. Почти полностью освоена первая надпойменная терраса, сейчас
занятая пригородными плантациями овощных культур, огородами и дачными
участками с регулярным поливом. Это способствовало значительному увели-
чению численности чесночницы и зеленой жабы, которые стали здесь массо-
выми видами; временные водоемы заселяют также озерная лягушка и красно-
брюхая жерлянка. На дачных участках массовым видам стала также прыткая
ящерица; кучи мусора, компостные ямы и разные строения привлекают сюда
обыкновенных ужей (места кормежки, размножения, зимовки), на периферий-
ные участки дачных массивов иногда проникают гадюки и узорчатые полозы,
вытесненные в неудобные для хозяйственного использования участки пересе-
ченной местности и заросли кустарников.
На второй и третьей террасах помимо дачных участков значительные пло-
щади (распаханность территории достигает 50-60%) заняты посевами зерно-
вых и технических культур. На посевах собственных видов практически нет,
но в них на 10-20 м местами проникает прыткая ящерица. Естественная рас-
тительность территории представлена небольшими массивами дубрав, остеп-
ненных сосновых боров и примыкающих к ним вторичных березово-осиновых
и кустарниковых зарослей (облесенность достигает 12%); обитатели открытых
местообитаний ютятся на опушках, полянах, пойменных лугах, небольших
участках луговой степи по склонам и окраинам полей, обочинам дорог и дру-
гих инфраструктурных элементов. В этих сильно фрагментированных, локаль-
ных (по сути островных) стациях сейчас и сохранились прежде значительно
шире распространенные лесные виды и обитатели открытых пространств.
302
Глава 5
Сходные изменения произошли и в Высоком Заволжье. Распаханность тер-
ритории здесь достигает 50% общей площади, а лесистость - 14% в Самарской
области и 5-18% в разных административных районах Оренбургской. Вместе
с тем здесь гораздо слабее развито поливное земледелие и более крупные мас-
сивы занимают посевы зерновых и технических культур, что сказывается на
распространении связанных с этими местообитаниями видов. Менее освоена
территория Зилаирско-Сакмарской низкогорной провинции Урала, где сейчас
сохранились основные местообитания всех лесных видов.
Значительные изменения в распространении и численности амфибий и
рептилий произошли в староосвоенных степях между Волгой и Уралом. Как
и севернее, наиболее существенно изменилась структура фауны в низком За-
волжье и на Приволжской граде, где распаханы практически все удобные в
технологическом плане площади. Здесь большие площади также занимают
поливные земли, хотя преобладают массивы зерновых культур с соответству-
ющими фаунистическими комплексами. Лесная растительность (около 1%
площади) представлена байрачными колками и небольшими рощами по эле-
ментам овражно-балочной сети и долинам рек. Крупнейшими из них являются
Дьяковский лес по р. Еруслан и массив по р. Чапаевка, где помимо обычных
видов известны встречи сибирского углозуба и серой жабы. Распространению
зеленой жабы и прыткой ящерицы способствуют придорожные и полезащит-
ные лесополосы.
Почти полностью распаханы выровненные водоразделы по Общему Сыр-
ту, Урало-Илекскому и Подуральскому плато, сейчас занятые в основном
крупными массивами посевов зерновых культур. Естественные сообщества
здесь сохранились по наиболее возвышенным, сильно расчлененным (местами
с обликом мелкосопочника) грядам сыртов с холмами-останцами и выходами
кварцитов и конгломератов, военным полигонам, долинам рек, местами вдоль
границ полей севооборотов, байрачных лесков, элементов овражно-балочной
и инфраструктурной сети, ставших своеобразными рефугиями и для зональ-
ных сообществ. По песчаным массивам здесь отмечается ряд южных форм
амфибий и рептилий. Эта территория несколько больше (до 2-4%) облесена.
Наиболее значительные массивы представлены Бузулукским и Красносамар-
ским борами, а также лесами в поймах рек Самара, Сакмара, Урал, Илек и
некоторых степных речках. По ним в этой части региона проходят южные ру-
бежи ареалов обыкновенного и гребенчатого тритонов, серой жабы, веретени-
цы, медянки, водяного ужа; они являются важными местообитаниями зеленой
жабы, жерлянки, обыкновенного ужа, прыткой ящерицы.
Пойменные леса и луга - основные местообитания большинства этих ви-
дов и в Урало-Мугоджарской низкогорной провинции. На участках ее горных
петрофитных степей сохранились крупные популяции поперечнополостного
полоза, здесь появляется разноцветная ящурка. В Зауралье по сохранившимся
степным сообществам наибольшее распространение получили прыткая яще-
рица и степная гадюка. На распространение амфибий и рептилий всей степной
зоны региона большое влияние оказало зарегулирование стока рек. Формиро-
вание прибрежных зарослей по волжским водохранилищам и в устьях впада-
ющих в них рек расширило зону обитания озерной лягушки, ужей, болотной
черепахи, а на побережье - зеленой жабы, чесночницы, жерлянки и прыткой
ящерицы; в зоне горного Ириклинского водохранилища по р. Урал это воз-
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ АМФИБИЙ...
303
действие оказалось значительно меньше. Существенно расширилось распро-
странение и усложнилось «кружево ареала» всех этих видов по руслам кана-
лов и запрудам на степных речках и балках.
Наряду с этим в пределах степной зоны начало проявляться негативное
воздействие разработки нефтегазовых месторождений и предприятий горнодо-
бывающего комплекса. Так, в водоемах зоны Карачаганакского нефтегазового
месторождения в месте сброса нефтепродуктов резко снизилась численность
большинства водных обитателей, а озерная лягушка вообще исчезла на этом
участке (Березовиков и др., 2003). Негативное воздействие предприятий гор-
но-металлургического комплекса проявляется в основном в пределах Орского
и Актюбинского промышленных узлов и выражается в разрушении типичных
местообитаний при разработке карьеров и загрязнении водоемов стоками с по-
вышенным содержанием тяжелых металлов.
В полупустынной и пустынной зонах важную роль в распространении ам-
фибий и рептилий сыграли создание животноводческих «точек» с колодца-
ми, копанями, артезианскими скважинами, а также появление разведочных и
буровых скважин, ряду которых сопутствуют временные водоемы. В Волж-
ско-Уральском и Урало-Эмбенском междуречьях это привело к усложне-
нию «кружева ареала» зеленой жабы и прыткой ящерицы, а в Приаральских
Каракумах - зеленой жабы, водяного ужа и некоторых гекконов (Лобачев и
др., 1973). Вместе с тем животноводческое освоение территории способство-
вало развитию пастбищой дигрессии, что негативно сказалось на численности
почти всех зональных видов.
В долинах рек Волги и Урала широкое развитие получило орошаемое зем-
леделие, массивы которого стали очагами высокой и (что также немаловажно)
стабильной численности чесночниц, зеленых жаб, озерных лягушек, ужей, а
местами некоторых других видов.
Создание лесных посадок в Заволжье способствовало увеличению числен-
ности быстрой ящурки в окрестностях горы Б. Богдо, а в посадках Баксайской
опытной станции - прыткой ящерицы и узорчатого полоза. С разработкой ка-
рьера на горе Б. Богдо оказались разрушенными местообитания пискливого
геккончика.
Важным элементом инфраструктурного обустройства территории яв-
ляются дороги. В лесу и на пойменных лугах они выполняют роль своеоб-
разных экологических русел, способствующих проникновению в глубь
сплошных массивов серой жабы, прыткой ящерицы; полевые степные до-
роги способствуют расселению степной гадюки, временные водоемы в при-
дорожных кюветах - расселению амфибий и приводных рептилий, а также
служат местом их кормежки и размножения, а пруды в ряде случаев со вре-
менем становятся местами их постоянного обитания. Образующиеся после
дождя в колеях автотранспорта лужи в пустыне иногда становятся местом
массового размножения зеленых жаб. Дороги с твердым покрытием (при-
влекающие насекомых) служат местами кормежки ряда ящериц, а в пусты-
не они наряду с грунтовыми являются важным фактором, определяющим
пространственное рассредоточение многих видов рептилий (Неручев и др.,
1999).
Вместе с тем на дорогах в местах с высокой численностью амфибий и реп-
тилий обычно, а при массовой миграции особенно, гибнет значительное ко-
304
Глава 5
личество животных (преимущественно малоподвижных видов), что требует
создания специальных направляющих линий и переходов (проводов).
На всей территории региона проявляется и своеобразный эффект синан-
тронизации ряда видов амфибий и рептилий. В его северной части известны
случаи поселения в хозяйственных и жилых постройках ужей. Зеленые жабы
являются постоянными обитателями погребов, хозяйственных и жилых по-
строек на всей территории региона, что для Яицкого городка (Уральска) и его
окрестностей отмечено еще П.С. Палласом. Этот вид довольно многочислен
и в условиях крупного города, где успешно охотится на насекомых, привле-
каемых ночью электрическими фонарями. На юге региона в глинобитных
строениях и на их развалинах довольно обычны разные гекконы. Повсеместно
в садах, на дачных участках и некоторых других антропогенно-модифициро-
ванных местообитаниях массовым видом стала прыткая ящерица, численность
которой местами значительно превосходит таковую в естественных биотопах.
В парковых зонах городов определенное влияние на видовой состав и чис-
ленность представителей герпетофауны оказывают рекреационная нагрузка,
особенно уплотнение почвы, вытаптывание растительности, благоустройство
территории. Впрочем все эти факторы действует комплексно и последствия
зависят от общей степени урбанизации территории.
Отмеченные негативные последствия воздействия антропогенных факто-
ров на фауну амфибий и рептилий региона настоятельно требуют усиления
их охраны. Наиболее действенной формой охраны является создание сети
ООПТ, в которой по принципу ландшафтной репрезентативности и прави-
лу провинциального ряда должны быть представлены все ландшафтные эле-
менты, образующие основу единого ландшафтно-экологического каркаса
территории.
В регионе сейчас имеются следующие категории ООПТ: заповедники, ре-
зерваты, национальный парк, заказники и памятники природы. Высший уро-
вень охраны (категория 1 по классификации МСОП) обеспечивают заповед-
ники. В настоящее время на территории региона имеются четыре заповедника
(Богдинско-Баскунчакский, Оренбургский, Наурзумский, Устюртский), близ-
кий по режиму Иргиз-Тургайский резерват, национальный парк «Бузулукский
бор», а также несколько десятков заказников и сотни памятников природы,
охраняемых на местном уровне. Представить информацию о всех ООПТ не-
возможно, в связи с этим ограничимся данными о заповедниках, резервате и
национальном парке (табл. 57).
Материалы показывают, что в региональных ООПТ высшего ранга пред-
ставлены 10 (83,3% общего состава) видов амфибий (все за исключением си-
бирского углозуба и квакши) и 32 (84,2%) рептилий (все за исключением ло-
кально распространенных).
Вместе с тем среди амфибий обыкновенная чесночница, зеленая жаба, ост-
ромордая и озерная лягушки, тритоны, обыкновенная жаба, травяная и прудо-
вая лягушки (видьг, связанные преимущественно с лесными местообитаниями)
наиболее широко представлены лишь в одном - Бузулукском бору. Из репти-
лий преобладают болотная черепаха, разноцветная ящурка, прыткая ящерица
и степная гадюка; в 3 ООПТ отмечены такырная круглоголовка, узорчатый по-
лоз и обыкновенный уж, в двух - 7 видов, в том числе среднеазиатская черепа-
ха, пискливый геккончик, гребнепалый геккон, вертихвостка, быстрая ящурка,
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ АМФИБИЙ...
305
Таблица 57
Распространение амфибий и рептилий по ООПТ Урало-Каспийского региона
№ п.п. Вид Богднн- ско-Бас- кунчак- ский Оренбург- ский Наурзум- ский Иргиз- Тур ган- ский Устюр- ский Нацио- нальный парк «Бу- зулукский бор»
1 Обыкновенный тритон Амф ибии — — — 4-
2 Гребенчатый тритон - - — - - 4-
3 Краснобрюхая жерлянка - - — - - 4-
4 Обыкновенная чесночница 4- 4- 4- 4- — 4-
5 Обыкновенная жаба - - - - - 4-
6 Зеленая жаба 4- 4- 4- 4- — 4-
7 Травяная лягушка - — - — - 4-
8 Остромордая лягушка — 4- 4- - 4-
9 Прудовая лягушка — - — - — 4-
10 Озерная лягушка - 4- - 4- — 4-
11 Болотная черепаха Репп 4- гилии 4- 4- 4- — 4-
12 Среднеазиатская черепаха - - - 4- 4- -
13 Пискливый геккончик 4- - - 4- - -
14 Гребнепалый геккон — — — 4- 4- —
15 Каспийский геккон - - - — 4- -
16 Серый геккон — - - — 4- -
17 Сцинковый геккон - - - - 4- —
18 Степная агама — - — — 4- —
19 Круглоголовка-вертихвостка 4- — - — 4- -
20 Такырная круглоголовка 4- - — 4- 4- —
21 Ушастая круглоголовка - - - - 4- -
22 Веретеница ломкая - - - — - 4-
23 Разноцветная ящурка 4- 4- 4- 4- 4- 4-
24 Сетчатая ящурка - - — 4- - -
25 Средняя ящурка - - - - 4- -
26 Песчаная ящурка - — - 4- — -
27 Быстрая ящурка 4- — - - 4- -
28 Прыткая ящерица 4- 4- 4- 4- — 4-
29 Живородящая ящерица - 4- - — — 4-
30 Песчаный удавчик - - - — 4- —
31 Восточный удавчик - - - — 4- -
32 Разноцветный полоз — - — - 4- -
306
Глава 5
Окончание табл. 57
№ п.п. Вид Богдин- ско-Бас- кунчак- ский Оренбург- ский Наурзум- ский Иргиз- Турган- ский Устюр- ский , в >?х О 3 U5 V - s х - у а. X S м 5 Х С г?
33 Обыкновенная медянка — — - - - 4-
34 Полоз узорчатый 4- 4- — 4- - —
35 Палласов полоз 4- — - - 4- —
36 Каспийский полоз + - - - - —
37 Обыкновенный уж 4- 4- - — - 4-
38 Водяной уж - 4- - - - —
39 Стрела-змея - - - - 9 -
40 Обыкновенный щитомордник - - - 4- - -
41 Обыкновенная гадюка - - - - — 4-
42 Степная гадюка 4- 4- 4- 4- - 4-
Примечание. «+» - вид присутствует, «-» - вид отсутствует, «?» - требуется подтверждение.
живородящая ящерица и палласов полоз, а остальные (преимущественно узко-
или краеареальные) - только в одном. В целом незначительное (в 1-2 ООПТ)
представительство имеет 31 вид (62,8% общего состава), что явно недостаточ-
но для обеспечения устойчивого состояния их популяций.
Важную роль в сохранении ряда видов животных играет занесение их на
страницы Красной книги. В настоящее время в Красные книги разного уровня
занесены 23 вида, обитающие на территории региона, в том числе 4 амфибии
(33,3% общего состава) и 19 (50,0%) рептилий (табл. 58).
Из них в Красную книгу высшего ранга - МСОП - занесены 2 вида (4,0%
общего состава) - краснобрюхая жердянка и болотная черепаха (категория
низкого риска, зависят от охраны). Вместе с тем на территории как региона,
так и Российской Федерации и Республики Казахстан в целом эти виды до-
вольно обычны или во всяком случае нередки и существенной угрозы их су-
ществованию нет.
В Красную книгу РФ занесены и 8 видов рептилий (21,1% их общего со-
става), в том числе серый голопалый геккон (как, вероятно, исчезнувший в ев-
ропейской части ареала), пискливый геккончик (категория 3), гадюка Николь-
ского (категория 4), а также занесенные в Аннотированный перечень таксонов
и популяций животных, нуждающихся в особом внимании к их состоянию
в природной среде на территории РФ (Ананьева, Даревский, 2001) такырная
круглоголовка, ящеричная змея, степная гадюка, сарматский (четырехполо-
сый) и каспийский (желтобрюхий) полозы. Два последних вида занесены так-
же в Красную книгу Казахстана, причем каспийский полоз с категорией 3, а
сарматский с категорией 4.
Среди областей Российской Федерации наибольшее число видов занесено
в Красные книги Самарской, Астраханской и Оренбургской областей. Редки-
ми в 3 областях считаются широко распространенные разноцветная ящурка,
узорчатый полоз и степная гадюка. Остальные представители, встречающи-
ВАЖНЕЙШИЕ ПРОБЛЕМЫ СОХРАНЕНИЯ ФАУНЫ АМФИБИЙ...
307
Таблица 58
Амфибии и рептилии Урало-Каспийского региона на страницах Красных книг
№ п.п. Вид Красная книга*
МСОП 1 g Астра- ханской обл. Волго- градской обл. Сара- товской обл. Самар- ской обл. Орен- бургской обл. bd
1 Гребенчатый тритон / \мфиби и — — 4- 4- —
2 Краснобрюхая жерлянка 4- - — — - - — —
3 Серая жаба - — - — — 4- — —
4 Травяная лягушка - — - — - 4- 4- -
5 Болотная черепаха Р 4- ептили и — — 4- — —
6 Пискливый геккончик — ?+ - - - — - -
7 Серый геккон - 4- - — - - - —
8 Круглоголовка-вертихвостка - - - 4- — - 4- -
9 Такырная круглоголовка - 4- 4- - — - - -
10 Ушастая круглоголовка — — 4- — — — - —
11 Веретеница ломкая - — - — 4- - 4- —
12 Разноцветная ящурка - - — — 4- 4- 4- —
13 Живородящая ящерица - - - - — 4- - -
14 Песчаный удавчик - ?+ 4- — - - - -
15 Обыкновенная медянка - — - - - 4- 4- -
16 Узорчатый полоз - - - - 4- 4- 4- —
17 Сарматский (четырехполо- — 4- 4- 4- — — — 4-
18 сый) полоз Каспийский (желтобрюхий) — 4- 4- 4- — — — 4-
19 полоз Ящеричная змея — 4- 4- — — - — -
20 Водяной уж - - - - 4- 4- - -
21 Обыкновенная гадюка - - - - - 4- - -
22 Гадюка Никольского — 4- - - - 4- - -
23 Степная гадюка - 4- 4- - 4- 4- - —
Итого: 2 8 8 3 5 12 7 2
*В пределах региона.
еся в равном соотношении (по 8 видов), в 1 или 2 областях являются в ос-
новном краеареальными, что заставляет обратить внимание на состояние их
популяций.
СПИСОК ЛИТЕРАТУРЫ
Абрамова Т.А. Изменения природной среды некоторых регионов Евразии за два последних
тысячелетия // Вести. МГУ. Сер. геогр., 1994. № 5. С. 59-66.
Абрамова Т.А., Турманина В.И. Реконструкция климатических изменений последнего тыся-
челетия в Прикаспии // Изв. ВГО, 1982. Вып. 3. С. 208-214.
Агарков С.Г., Кононова Н.К, Савинова С.С., Хмелевская Л.В. Циклические колебания ат-
мосферной циркуляции северного полушария в XX столетии // Изв. АН СССР. Сер. геогр., 1980.
№ 5. С. 24-34.
Адрианов В.В., Лапаева И.В. Растительный и животный мир Волго-Уральских песков в ус-
ловиях динамики техногенных ландшафтов // Метеорол. и гидрол., 2005. Вып. 3. С. II1-115.
Александровская Т.О., Быков А.В. Особенности распределения зеленых лягушек в европей-
ской части СССР И VII Всесоюз. зоогеогр. конф.: тез. док л. М.: Наука, 1979. С. 279-281.
Алексеев С.К, Стрельцов А.Б., Константинов Е.Л., Устюжанина О.А. Земноводные и реп-
тилии окрестностей озер Баскунчак // Богдинско-Баскунчакский заповедник и его роль в сохра-
нении биоразнообразия севера Астраханской области. Астрахань, 2004. С. 75-76.
Ананьева Н.Б. Пискливый геккончик Alsophylax pipiens (Pallas, 1814) // Красная книга Рос-
сийской Федерации. Животные. М.: ACT; Астрель, 2001. С. 328-329.
Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся.
Энциклопедия природы России. М.: ABF, 1998. 576 с.
Ананьева Н.Б., Даревский И.С. Класс Пресмыкающиеся - Reptilia // Аннотированный пе-
речень таксонов и популяций животных, нуждающихся в особом внимании к их состоянию в
природной среде (Приложение 3 к Красной книге РФ) // Красная книга Российской Федерации.
Животные. М.: ACT; Астрель, 2001. С. 853-855.
Атлас пресмыкающихся Северной Евразии (таксономическое разнообразие, географичес-
кое распространение и природоохранный статус). Н.Б. Ананьева, Н.Л. Орлов, Р.К. Халиков.
СПб.: Зоол. ин-т, 2004. 232 с.
Бабаев А.Г., Зонн И.С., Дроздов Н.Н., Фрейкин З.Г. Пустыни. М.: Мысль, 1986. 398 с.
Бажанов В.С. Список гадов Бузулукского и Пугачевского уезда бывшей Самарской гу-
бернии, собранных в 1928 году // Средне-Волжская краевая станция защиты растений. Бюл. за
1926-1928 гг. Самара: Средне-Волжское краевое изд-во, 1930. 68 с.
Бакиев А.Г. Болотная черепаха Emys orbikularis в Самарской области (региональная сводка)
// Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр. Тольятти, 2001а. Вып. 5.
С. 3-18.
Бакиев А.Г История изучения фауны рептилий Самарской области: XVIII век // Самарский
край в истории России: Матер, юбил. науч. конф. Самара, 20016. С. 271-272.
Бакиев А.Г, Гафарова ЕВ. О состоянии охраны гадюк в Среднем Поволжье // Самарская
Лука, 1999. № 9, 10. С. 187-190.
Бакиев А.Г., Иванова М.А. Земноводные и пресмыкающиеся Самарской области в коллек-
ции Тольяттинского краеведческого музея И Актуальные проблемы герпетологии и токсиколо-
гии: Сб. науч. тр. Тольятти, 2004. Вып. 7. С. 23-24.
Бакиев А.Г., Кренделев В.В. Сравнение топографии внутренних органов гадюк обыкновен-
ной и Никольского И Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр. Толь-
ятти, 1999. Вып. 3. С. 60-62.
Бакиев А.Г., Магдеев Д.В. К вопросу о фауне змей Самарской Луки // Самарская Лука, 1995.
№ 6-95. С. 225-228.
Бакиев А.Г., Маленев А.А. К обоснованию перечня пресмыкающихся для занесения в Крас-
ную книгу Самарской области // Актуальные проблемы герпетологии и токсикологии: Сб. науч,
тр. Тольятти, 1999. С. 62-71.
Бакиев А.Г., Маленев А.Л. Пресмыкающиеся Среднего Поволжья. Тольятти, 1996. 25 с.
Бакиев А.Г., Песков А.Н. Медянка и ящеричная змея в Астраханской области И Актуальные
проблемы герпетологии и токсикологии: Сб. науч, тр., 2006. Вып. 9. С. 13-14.
Бакиев А.Г, Ратников В.Ю. История формирования ареала узорчатого полоза Elaphe dione
и современное распространение вида в Волжском бассейне // Изв. СНЦ РАН. Спец. вып. Акту-
альные проблемы экологии, 2003. Вып. 2. С. 313-316.
СПИСОК ЛИТЕРАТУРЫ
309
Бакиев А.Г., Файзулин А.И. Земноводные и пресмыкающиеся Самарской области: Методич.
пособ. Самара: ОРФ «Самарская Лука», 2001. 68 с.
Бакиев А.Г., Файзулин А.И. Материалы к кадастру земноводных и пресмыкающихся Самар-
ской области // Матер, к кадастру амфибий и рептилий бассейна Средней Волги. Н. Новгород:
Экоцентр «Дронт», 2002. С. 97-132.
Бакиев А.Г., Магдеев Д.В., Песков А.Н. Данные о распространении и экологии медянки в Са-
марской области // Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр. Тольятти,
1996. Вып. 2. С. 72-73.
Бакиев А.Г., Гранин В.И., Литвинов Н.А., Ратников В.Ю. Змеи Волжско-Камского края.
Самара: Изд-во СНЦ РАН, 2004. 182 с.
Бакиев А.Г, Маленев А.А., Зайцева О.В., Шуршина И.В. Змеи Самарской области. Тольятти:
ООО «Кассандра», 2009. 170 с.
Бакиев А.Г„ Маленев А.Л., Песков А.И, Гриднев Д.В. Морфологическая характеристика га-
дюк из лесопарковой зоны г. Самара// Актуальные проблемы герпетологии и токсикологии: Сб.
науч. тр. Тольятти, 2000. Вып 4. С. 3-8.
Бакиев А.Г., Баринов В.Г., Песков А.Н. и др. Составление видового списка пресмыкающихся
Бузулукского бора и Красносамарского леса // Актуальные проблемы герпетологии и токсико-
логии: Сб. науч. тр. Тольятти, 1996. Вып. 2. С. 73-74.
Бакиев А.Г., Быков Е.В., Вехник В.П. и др. Животные // Растения, животные и грибы Крас-
ной книги Самарской области. Тольятти: ИЭВБ РАН, 2000. С. 11-23.
Бакиев А.Г., Кривошеев В.А., Епланова Г.В. и др. К вопросу о состоянии охраны пресмыка-
ющихся в Самарской и Ульяновской областях // Актуальные проблемы герпетологии и токсико-
логии: Сб. науч. тр. Тольятти, 2001. Вып. 5. С. 18-22.
Бакиев А.Г., Кривошеев В.А., Файзулин А.И. и др. Земноводные и пресмыкающиеся крупных
городов Самарской и Ульяновских областей // Биоразнообразие и биоресурсы Среднего Повол-
жья и сопредельной территории. Казань, 2002. С. 105-106.
Бакиев А.Г, Маленев А.Л., Песков А.Н. и др. Змеи Среднего Поволжья и их распространение
в регионе // Вопросы герпетологии: Матер. I Съезда герпетологического о-ва им. А.М. Николь-
ского. Пущино; Москва, 2001. С. 22-24.
Бакиев А.Г, Маленев А.Л., Песков А.Н. и др. К фауне пресмыкающихся Богдино-Баскунчак-
ского заповедника // Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр. Тольят-
ти, 2005. Вып. 8. С. 3-5.
Бакиев А.Г, Файзулин А.И., Кривошеев В.А. и др. Земноводные и пресмыкающиеся, обита-
ющие на городских территориях в Самарской и Ульяновской областях // Актуальные проблемы
герпетологии и токсикологии: Сб. науч. тр. Тольятти, 2003. Вып. 6. С. 3-9.
Банников А.Г. Земноводные. Пресмыкающиеся //Жизнь животных. М.: Просвещение, 1969.
Т. 4, ч. 2. С. 35-92.
Банников А.Г, Денисова М.Н. Очерки по биологии земноводных. М.: Учпедгиз., 1956. 168 с.
Банников А.Г, Даревский И.С., Рустамов А.К Земноводные и пресмыкающиеся СССР. М.:
Мысль, 1971. 308 с.
Баянов М.Г. Земноводные. Пресмыкающиеся. Животный мир Башкортостана. Уфа: Китап,
1995. С. 211-222.
Белик В.П. Птицы степного Придонья. Ростов-на-Дону, 2000. 376 с.
Бенинг А.Л. Материалы по гидрофауне придаточных систем реки Волги. Материалы по гид-
рофауне р. Б. Иргиз // Работы Волж. биол. станции. Саратов, 1913. Т. IV, № 5. С. 1-50.
Бенинг А.Л. Материалы по гидрофауне придаточных систем реки Волги. Материалы по гид-
рофауне реки Еруслан И Работы Волж. биол. станции. Саратов, 1921. Т. V, № 4-5. С. 255-331.
Бенинг А.Л. Материалы по гидрофауне придаточных систем реки Самара // Работы Волж.
биол. станции. Саратов, 1926. Т. IX, № 1-2. С. 71-110.
БергЛ.С. Географические зоны Советского Союза. Т. 1. М.: Геогрфгиз, 1947. 398 с.
Березовиков Н.Н., Хроков В.В., Брушко З.К. Влияние разработки Карчаганакского нефте-
газоконденсатного месторождения на животный мир Западного Казахстана // Selevinia, 2003.
С. 123-137.
Березовиков Н.Н., Хроков В.В., Брушко З.К. и др. Современное состояние фауны позвоноч-
ных животных на территории Карчаганакского нефтегазоконденсатного месторождения // Эко-
логия Карачаганака: Матер, науч.-практ. конф. Алма-Ата, 1992. С. 78-81.
Бобров В.В. Материалы по фауне и населению пресмыкающихся Северо-западного Прикас-
пия // Вопросы герпетологии: VI Всесоюз. герпетол. конф. Л.: Наука, 1985. С. 29-30.
310
СПИСОК ЛИТЕРАТУРЫ
Богданов О.П. Экология пресмыкающихся Средней Азии. Ташкент: ФАН, 1965. 258 с.
Божанский А.Т. Гадюка Никольского Viperia nikolskii Vedmederja, Grubant et Rudaeva, 1986
//Красная книга Российской Федерации. Животные. М.: ACT; Астрель, 2001. С. 348-349.
Божанский А.Т., Полынова Г.В. Предварительные результаты герпетологического райони-
рования песчаных пустынь Астраханской области //Тез. докл. I Конф, герпетологов Поволжья.
Тольятти, 1995. С. 5-7.
Божанский А.Т., Полынова Г.В. Проект регионального списка рептилий Красной книги Ас-
траханской области // Проблемы сохранения биоразнообразия аридных регионов России: Ма-
тер. между нар. науч.-практ. конф. Волгоград: Изд-во ВолГУ, 1998. С. 57-59.
Болиховская Н.С. Палиноиндикация изменений ландшафтов Нижнего Поволжья в пос-
ледние 10 000 лет // Каспийское море. Вопросы геологии и геоморфологии. М.: Наука, 1990.
С. 52-68.
Большаков В.Н., Вершинин В.Л. Амфибии и рептилии Среднего Урала. Екатеринбург: УрО
РАН, 2005. 128 с.
Боркин Л.Я. Европейско-дальневосточные разрывы ареалов у амфибий и рептилий СССР и
сопредельных стран //Тр. Зоол. ин-та АН СССР. Л., 1984. Т. 124. С. 55-88.
Боркин Л.Я. Краткий очерк развития герпетологии в России // Московские герпетологи. М.:
КМК, 2003. С. 7-33.
Боркин Л.Я., Литвинчук С.Н, Розанов О.М. и др. Гибридогенный комплекс Rana esculenta’.
существует ли «волжский парадокс»? И Тез. докл. III Конф, герпетологов Поволжья. Тольятти,
2003а. С. 7-12.
Боркин Л.Я., Литвинчук С.Н., Розанов О.М. и др. Распространение двух криптических форм
обыкновенной чесночницы (Pelobates fuscus) на территории Волжского бассейна//Тез. докл. III
Конф, герпетологов Поволжья. Тольятти, 20036. С. 3-6.
Брагин Е.А., Брагина Т.М. Фауна Наурзумского заповедника. Рыбы, земноводные, пресмы-
кающиеся, птицы, млекопитающие (аннотированные списки видов). Костанай: Костанайский
дом печати, 2002. 60 с.
Брушко З.К. Эколого-фаунистический обзор ящериц, населяющих пустыни Казахстана
И Selevinia, 1993. № 1. С. 19-36.
Брушко З.К. Ящерицы пустынь Казахстана. Алматы: Конжык, 1995. 231 с.
Брушко З.К, Кубыкин Р.А. Каталог герпетологической коллекции Института зоологии Ака-
демии наук КазССР. Алма-Ата: Наука, 1988. 42 с.
Брушко З.К, Кубыкин Р.А. Земноводные и пресмыкающиеся // Красная книга Казахской
ССР. Животные. Алматы: Гылым, 1991. С. 263-295.
Брушко З.К, Кубыкин Р.А. Распространение и экология стрелы-змеи (Psammophis lineola-
tum Brandt, 1838) в Казахстане И Selevinia, 2000. С. 130-137.
Быков А.В. Фауна и распределение амфибий и рептилий в окрестностях Джаныбекского
стационара// Вопросы герпетологии (Сб. статей). Л., 1981. С. 26.
Варшавский С.Н., Крылова КТ., ШиловаМ.Н., КамневП.И. Об особенностях распростране-
ния и численности зеленой жабы в субаридных и аридных ландшафтах И Вопросы герпетологии
(Матер. VI Всесоюз. герпетол. конф.). Л., 1985. С. 41-42.
Васильев Н.Ф., Неручев В.В., Кудакина Е.Ю. Население рептилий в условиях пустынных
пастбищ Северного Прикаспия И Новые проблемы зоологич. науки и их отражение в вузовском
преподавании: Тез. докл. Ставрополь: ГПИ, 1979. Ч. 11. С. 223-224.
Ведмедеря В.И., Грубант В.Н., РудаеваА.В. К вопросу о названии черной гадюки лесостепи
европейской части СССР И Вести. ХГУ, 1986. № 288. С. 83-85.
Величко А.А. Природный процесс в плейстоцене. М.: Наука, 1973. 256 с.
Величко А.А. Опыт палеогеографической реконструкции природы верхнего плейстоцена
для территории Восточной Европы и СССР // Изв. АН СССР. Сер. геогр., 1977. № 4. С. 28-44.
Вершинин В.Л. Амфибии и рептилии Урала. Екатеринбург: УрО РАН, 2007. 170 с.
Вершинин В.Л. Топография морфологических аномалий и формирование брачных пар у
жаб Bufo bufo L., 1758 и Bufo viridis Laur., 1768 // Экология, 2010. № 4. С. 305-309.
Вилкина Е.А., Табачишин В.Г., Шляхтин В.Г. Особенности питания островной популяции
узорчатого полоза (Elaphe dione) острова Круглый средней зоны Волгоградского водохранили-
ща//Совр. герпетология, 2000. Вып. 1. С. 66-68.
Волчанецкий И. Б., Ялъцев Н.П. К орнитофауне Приерусланской степи // Уч. зап. Сар ГУ У,
1934. Т. 11, вып. 1.С. 63-93.
Воробьева Э.И. Проблемы происхождения наземных позвоночных. М.: Наука, 1992. 344 с.
СПИСОК ЛИТЕРАТУРЫ
311
Воронцовский П.А. Амфибии окрестностей г. Оренбурга//Тр. О-ва по изучению Киргизско-
го края. Оренбург, 1922. Вып. 3. С. 26-30.
Воронцовский П.А. Животный мир. Сельскохозяйственные районы и земельные нормы
Оренбургской губернии. Оренбург: Оренполиграфпром, 1927. 108 с.
Габитова ГМ., Юмагулова Г.Р, Видовой состав и численность бесхвостых амфибий на тер-
ритории Зиянчуринского района республики Башкортостан И Матер. Междунар. науч.-практ.
конф. «Природное наследие России в XXI веке». Уфа, 2008. С. 101-102.
Гавлюк Э.В., Давыгора А.В,, Руди В.Н. Животный мир Оренбургской области (позвоноч-
ные). Оренбург: Изд-во ОГПИ, 1993. 48 с.
Гаврилов Н.Г., Ососков П.А. Растительный и животный мир. Россия: Полное географи-
ческое описание нашего отечества. Т. 6. Среднее и Нижнее Поволжье и Заволжье. СПб., 1901.
С. 69-110.
Галеева Д.Н., Гаранин В.И., Замалетдинов Р.И., Павлов А.В. Материалы к кадастру зем-
новодных и пресмыкающихся Республики Татарстан И Материалы к кадастру амфибий и
рептилий бас. Средней Волги. Н. Новгород: Межд. соц.-экол. союз, Экоцентр «Дронт». 2002.
С. 186-221.
Гаранин В.И. Изменения численности земноводных востока европейской части СССР
//Совр. проблемы изучения динамики численности популяций животных: Матер, сов. М., 1964а.
С. 26-27.
Гаранин В.И. Материалы по распространению и численности амфибий Волжско-Кам-
ского края // Природные ресурсы Волжско-Камского края: животный мир. М.: Наука, 19646.
С.127-133.
Гаранин В.И. К вопросу о динамике численности земноводных и пресмыкающихся // При-
родные ресурсы Волжско-Камского края. Животный мир. Казань, 1971. Вып. 3. С. 79-83.
Гаранин В.И. Класс Земноводные. Класс Пресмыкающиеся, или рептилии И Животный мир
Башкирии. Уфа, 1977. С. 252-259.
Гаранин В.И. Земноводные и пресмыкающиеся Волжско-Камского края. М.: Наука, 1983.
176 с.
Гаранин В. И. О границах ареалов амфибий и рептилий в Волжском бассейне И Тез. докл. II
Конф, герпетологов Поволжья. Тольятти, 1999. С. 13-15.
Гаранин В.И. Вклад Н.А. Зарудного в герпетологию И Животный мир Южного Урала и Се-
верного Прикаспия: Тез. докл. IV per. конф. Оренбург: Изд-во ОГПУ, 2000. С. 19-20.
Гаранин В.И., Бакиев А.Г. К истории изучения низших наземных позвоночных Волжско-
Камского края И Самарская Лука, 2004. № 14/04. С. 222-234.
Гаранин В.И., Щербак Н.Н Изучение биотопов И Руководство по изучению земноводных и
пресмыкающихся. Киев, 1989. С. 111-117.
Гисцов А.П., Березовиков Н.Н. Современное состояние животного мира дельты р. Урал
//Животный мир Южного Урала и Северного Прикаспия. Оренбург, 1995. С. 7-9.
Горелов М.С. Земноводные и пресмыкающиеся И Природа Куйбышевской области. Куйбы-
шев: Кн. изд-во, 1990. С. 365-379.
Горелов М.С. О находке ящурки разноцветной (Eremias arguta) в Самарской области И Са-
марская Лука, 1991. № 1. С. 132.
Горелов М.С. Животный мир И Самарская область. Самара: ЗАО «Самарский информаци-
онный концерн», 1999. С. 39-57.
Горелов М.С., Магдеев Д.В., Павлов С.И. Состояние популяции гадюки обыкновенной на
территории Самарской области // Самарская Лука, 1992. № 3. С. 171-181.
Григорьев О.В. О скорости расселения сибирского углозуба в связи с формированием запад-
ной части его ареала//Тез. докл. VIII Всесоюз. зоогеогр. конф. М.: Наука, 1984. С. 36-37.
Гриневицкий Б.Б., Клюге Г.А. Растительный и животный мир И Россия: полное географи-
ческое описание нашего отечества. Т. 5. Урал и Приуралье. СПб.: Изд-во А.Ф. Девриена, 1914.
С. 97-125.
Гричук В.П. Растительность Русской равнины в нижне- и среднечетвертичное время И Тр.
Ин-та географии АН СССР, 1950. Вып. 46. С. 5-202.
Грубант В.Н., Рудаева А.В., Ведмедеря В.И. О систематической принадлежности черной
формы обыкновенной гадюки // Вопросы герпетологии (Сб. статей). Л., 1973. С. 68-71.
Давыгора А.В. Современное состояние и долговременные изменения герпетофауны Степ-
ного Предуралья И Животный мир Южного Урала и Северного Прикаспия: тез. докл. регион,
конф. Оренбург: Изд-во ОГПИ, 1995. С. 88-94.
312
СПИСОК ЛИТЕРАТУРЫ
Давыгора А.В. Узорчатый полоз // Красная книга Оренбургской области. Оренбург: ОКИ,
1998. С. 82-83.
Давыгора А.В. Реликтовое нахождение живородящей ящерицы (Lacerta vivipara) в Орен-
бургском степном Предуралье //Животный мир Южного Урала и Северного Прикаспия. Орен-
бург: Изд-во ОГПУ, 2000. С. 25-29.
Давыгора А.В. Новые данные о герпетофауне Оренбуржья и сопредельных территорий
// Биоразнообразие и биоресурсы Урала и сопредельных территорий. Оренбург: ИПК «Газпром-
печать», 2001. С. 258-260.
Давыгора А.В., Толин С.Л. Болотно-луговое урочище на ручье Тузлуколь - уникальное лан-
дшафтно-фаунистическое дополнение к системе участков заповедника «Оренбургский» // Про-
блемы сохранения и восстановления степных экосистем. Оренбург, 1999. С. 13-14.
Давыгора А.В., Русаков А.В., Дебело П.В. Материалы к фауне горы Сарыоба И Проблемы
геологии, охраны окружающей среды и управление качеством экосистем. Оренбург: ИПК ГОУ
ОГУ, 2006. С. 98-100.
Даревский И.С. К биологии прыткой ящерицы // Природа, 1946. № 9.
Даревский И.С. Герпетология И Развитие биологии в СССР. М.: Наука, 1967. С. 245-249.
Даркшевич Я.Н. Бузулукский бор (научно-популярная монография). Чкалов: Чкаловское
кн. изд-во, 1953. 88 с.
Даркшевич Я.Н. В Бузулукском бору // По родному краю: Сб. краеведческих очерков. Чка-
лов: Чкаловское кн. изд-во, 1956. С. 173-186.
Даркшевич Я.Н, Кнорре Е.П., Лаченков С.Т. Бузулукский бор. Чкалов, 1940. 17 с.
Дашкевич З.В. Палеогеография. Л.: Изд-во ЛГУ, 1969. 152 с.
Дебело П.В., Булатова КБ. Животные Западно-Казахстанской области. Уральск: РИО
ЗКГУ, 1999.212 с.
Дебело П.В., Чибилёв А.А. Некоторые эколого-географические особенности распростра-
нения амфибий и рептилий аридных и семиаридных ландшафтов Урало-Каспийского региона
И Степи Северной Евразии: Матер. VI междунар. симп. Оренбург: ИПК «Газпромпечать», 2012.
С. 203-208.
Дебело П.В., Фомин В.П., Мазяркина Т.Н. Заповедное Приуралье (особо охраняемые объек-
ты). Уральск: Полиграфсервис, 2000. 56 с.
Девишев Р.А. Позвоночные животные родного края И Природа и люди. Саратов: Изд-во
СарГУ, 1976. С. 177-200.
Дергачев В.А. О крупномасштабных природных процессах И Изв. РГО, 1998. Вып. 6.
С. 58-76.
Димо НА., Келлер Б.А. В области полупустыни: почвенные и ботанические исследования на
юге Царицинского уезда Саратовской губернии. Саратов, 1907. 587 с.
Динесман Л.Г. Амфибии и рептилии юго-востока Тургайской столовой страны и Северного
Приаралья // Тр. Ин-та географии АН СССР. Зоогеография и экология наземной фауны Казах-
стана. М.: Наука, 1953. 186 с.
Динесман Л.Г. Биогеоценозы степей в голоцене. М.: Наука, 1977. 160 с.
Динесман Л.Г., Калецкая М.П. Методы количественного учета амфибий и рептилий И Ме-
тоды учета численности и географического распределения наземных позвоночных. М.: Изд-во
АН СССР, 1952. С. 329-341.
Дубровский Ю.А. Новые находки рептилий в степях Казахстана И Бюл. МОИП. 1967.
Вып. 1.С. 36-37.
Дуйсебаева Т.Н. Новые находки амфибий и рептилий в Приаралье и сопредельных районах
Казахстана. Змеи (Reptilia: Squmata: Serpentes) И Selevinia, 2005. С. 49-56.
Дуйсебаева Т.Н., Березовиков Н.Н., Брушко З.К, и др. Озерная лягушка (Rana ridibunda Pal-
las, 1771) в Казахстане: изменение ареала в XX столетии и современное распространение вида
И Современная герпетология, 2005. Т. 3/4. С. 28-59.
Дунаев Е.А., Жданова Ю.М. О динамике населения ящериц Астраханских пустынь
И Проблемы сохранения биоразнообразия аридных регионов России. Волгоград, 1998. С. 125-
127.
Елпатъевский В.С. Гады Арала//Изв. Туркестанского отдела РГО. Науч, результаты Араль-
ской экспедиции. Вып. IV. 1903.
Епланова Г.В. Особенности репродуктивной биологии разноцветной ящурки Eremias arguta
на северном пределе ареала // Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр.
Тольятти, 2004. Вып. 7. С. 71-74.
СПИСОК ЛИТЕРАТУРЫ
313
Епланова ГВ., Бакиев А.Г. К истории исследования ящериц Самарской области И Самарс-
кий край в истории России: Матер, науч, конф, Самара, 2001. С. 273-274.
Епланова Г.В., Бакиев А.Г. Таксономический статус разноцветной ящурки Eremias arguta на
северном пределе распространения И Selevinia, 2002. С. 298-299.
Епланова Г.В., Бакиев А.Г, Бэр С.В. К эколого-морфологической характеристике разно-
цветной ящурки на северном пределе распространения И Тез. докл. III Конф, герпетологов По-
волжья. Тольятти, 2003. С. 16-18.
Епланова Г.В., Бакиев А.Г., Лысенко Т.М. О распространении и распределении разноцвет-
ной ящурки Eremias arguta (Pallas, 1773) в Среднем Поволжье, особенно в Самарской области
И Актуальные проблемы герпетологии и токсикологии: сб. науч. тр. Тольятти, 2001. С. 50-56.
Ерофеев А.В. Краткое сообщение о четырехполосом полозе// Редкие животные Казахстана.
Алма-Ата, 1986. С. 180.
Ефимов Р.В., Завьялов Е.В., Табачишин В.Г. Аспекты экологической сегрегации и техноло-
гия видовой идентификации гадюковых змей (Reptilia, Viperidae, Vipera) в Поволжье на основе
генотипирования // Поволжский эколог, журн., 2008. № 2. С. 147-153.
Животные глинистой полупустыни Заволжья (конспект фаун и экологические характерис-
тики / Под ред. А.А. Тишкова. М.: КМК, 2009. 164 с.
Жизнь животных. Т. 5. Земноводные и пресмыкающиеся / Под ред. А.Г. Банникова. М.:
Просвещение, 1985. 399 с.
Жуков М.М. Плиоценовая и четвертичная история севера Прикаспийской впадины // Про-
блемы Западного Казахстана. Т. 2. М.: Изд-во АН СССР, 1945. 236 с.
Завьялов Е.В. Герпетофауна севера Нижнего Поволжья // Матер, межрегион, науч.-практ.
конф. Чебоксары, 2002. С. 37-39.
Завьялов Е.В., Табачишин В.Г. Распространение популяции круглоголовки-вертихвостки
(Sauria, Agamidae, Phrynocephalus guttatus'} на севере Нижнего Поволжья // Современная герпе-
тология: Сб. науч. тр. Саратов: Изд-во СарГУ, 2000а. Вып. 1. С. 40^17.
Завьялов Е.В., Вилкина Е.В., Табачишин В.Г. Хищничество узорчатого полоза Elaphe dione в
отношении островных популяций береговой ласточки Riparia riparia в средней зоне Волгоград-
ского водохранилища // Рус. орнитол. журн. Экспресс вып., 2000. № 116, С. 18-19.
Завьялов Е.В., Табачишин В.Г. Распространение и особенности биологии Vipera nikolskii в
северной части Нижнего Поволжья // Проблемы общей биологии и прикладной экологии, 1997а.
Вып. 1.С. 168-170.
Завьялов Е.В., Табачишин В.Г. Особенности биологии и морфологическая характеристика
болотной черепахи (Emys orbicularis) в северной части Нижнего Поволжья // Проблемы общей
биологии и прикладной экологи, 19976. С. 49-54.
Завьялов Е.В., Табачишин В.Г Современное состояние популяций амфибий и рептилий на
территории Астраханского газоконденсатного месторождения // Проблемы экологической безо-
пасности Нижнего Поволжья в связи с разработкой и эксплуатацией нефтегазовых месторожде-
ний с высоким содержанием сероводорода. Саратов, 1998. С. 142-146.
Завьялов Е.В., Табачишин В.Г. Современное распространение и таксономический статус
популяций круглоголовки-вертихвостки (Phrinocephalus guttatus, Reptilia, Agamidae) в Нижнем
Поволжье // Тез. докл. II Конф, герпетологов Поволжья. Тольятти, 1999. С. 22-26.
Завьялов Е.В., Табачишин В.Г. Узорчатый полоз (Elaphe dione Pallas, 1773) в Прикаспийс-
кой низменности // Животный мир Южного Урала и Северного Прикаспия: тез. докл. и матер.
IV регион, конф. Оренбург, 20006. С. 31-34.
Завьялов Е.В., Табачишин В.Г. Распространение, некоторые аспекты морфологии и эколо-
гии круглоголовки-вертихвостки - Phrinocephalus guttatus (Agamidae, Sauria) на юго-востоке
европейской части России // Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр.
Тольятти, 2004. Вып. 7. С. 75-84.
Завьялов Е.В., Табачишин В.Г, Шляхтин Г.В. Современное распространение и морфологи-
ческая характеристика степной гадюки (Vipera ursinii) в Поволжье // Вопросы герпетологии (Сб.
статей). Пущино; Москва, 2001. С. 101-104.
Завьялов Е.В., Табачишин В.Г., Шляхтин ГВ. Современное распространение рептилий на
севере Нижнего Поволжья И Современ. герпетология: Сб. науч. тр. Саратов: Изд-во СарГУ,
2003. Т. 2. С. 52-67.
Завьялов Е.В., Табачишин В.Г, Шляхтин Г.В. и др. Генезис природных условий и основные
Направления современной динамики ареалов животных на севере Нижнего Поволжья // Поволж-
ский эколог, журн., 2004. № 3. С. 252-276.
314
СПИСОК ЛИТЕРАТУРЫ
Завьялов Е.В., Табачишин В.Г., Шляхтин Г.В. и др. Фондовые коллекции в системе монито-
ринга герпетофауны И Каталогизация зоологических коллекций. Вып. 2. Саратов: Изд-во Сара-
товского ун-та, 2006. 96 с.
Завьялов Е.В., Шляхтин Г.В., Табачишин В.Г. и др. Генезис природных условий и основные
направления современной динамики ареалов животных на севере Нижнего Поволжья. Сообщ.
111. Генезис фауны и флоры в четвертичное время. Плейстоцен // Поволжский эколог, журн.,
2002. №3. С. 217-235.
Завьялов Е.В., Шляхтин Г.В., Табачишин В.Г. и др. Генезис природных условий и основные
направления современной динамики ареалов животных на севере Нижнего Поволжья И Поволж-
ский эколог, журн., 2003. Ns 2. С. 119-146.
Залетаев В.С. Экологически дестабилизированная среда (экосистемы аридных зон в изме-
няющемся гидрологическом режиме). М.: Наука, 1989. 150 с.
Зарудный Н.А. Экскурсия на озеро Сулюк-Куль // Природа и охота, 1883. № 5. С. 1-16.
Зарудный Н. Материалы для фауны амфибий и рептилий Оренбургского края И Bui. Soc.
impcr. des Naturalistes de Moscou, 1895. (Nov. Scric, 1896. T. 5, Ns 3. C. 361-370.)
Зарудный НА. Поездка на Аральское морс летом 1914 г. Гады Арала// Изв. Туркестанского
отдела РГО, 1915. Т.П, вып. ГС. 113-125.
Зерова Г. А., Чхиквадзе В.М. Обзор кайнозойских ящериц и змей СССР И Изв. Ан ГССР. Сер.
биол., 1984. Т. 10, Ns 5. С. 319-325.
Зиненко А.И. Особенности морфологии Vipera berus (Linnaeus, 1758) и Vipera nikolskii (Vc-
dmedrja, Grubant et Rudaeva, 1986) - следствие интрогрессивной гибридизации? // Змеи Восточ-
ной Европы. Тольятти: ИЭВБ РАН, 2003. С. 20-22.
Змеи Восточной Европы: Матер, междунар. конф. Тольятти: ИЭВБ РАН, 2003. 96 с.
Иванов В.В. Степи Западного Казахстана в связи с динамикой их покрова. М.: Изд-во АН
СССР, 1958. 228 с.
Иванов И.В. Эволюция почв степной зоны как индикатор изменения климатических усло-
вий в голоцене // Палеоклиматы позднеледниковья и голоцена. М.: Наука, 1989. С. 67-75.
Иванов И.В., Васильев И.Б. Человек, природа и почвы Рын-песков Волго-Уральского меж-
дуречья в голоцене. М.: Интеллект, 1995. 264 с.
Иванова Н.Н. К изучению популяций разноцветной ящурки (Eremias arguta) в Северном
Прикаспии И Вопросы герпетологии. (Матер. VI Всесоюз. герпетол. конф.). Л., 1985. С. 83-84.
Ильин В.IO., Смирнов Д.Г., Титов СВ. Стациальная приуроченность и относительная чис-
ленность пресмыкающихся в полупустыне Западного Казахстана И Тез. докл. I Конф, герпето-
логов Поволжья. Тольятти, 1995. С. 20-22.
Ильина Н.С., Магдеев Д.В. Биотопичсское распределение ящурки разноцветной {Eremias
arguta) в Самарской области // Тез. докл. II Конф, герпетологов Поволжья Тольятти, 1999.
С. 25-26.
Искакова К.М. Земноводные Казахстана. Алма-Ата: Изд-во АН Каз. ССР, 1959. 92 с.
Ищенко В.Г. Динамический полиморфизм бурых лягушек фауны СССР. М.: Наука, 1978.
148 с.
Калябина С.А., Йогер У., Орлов Н.Л., Винк М. Филогения и систематика гадюковых змей
комплекса «.Vipera berusv> И Змеи Восточной Европы. Тольятти: ИЭВБ РАН, 2003. С. 22-24.
Карелин Г.С. Экспедиция для осмотра северо-восточных берегов Каспийского моря в 1832 г.
//Зап. РГО по общей географии. Т. 10. М., 1883. 198 с.
Кармышев Ю.В., Табачишин В.Г. Эколого-морфологическая характеристика крымских и
нижневолжских популяций степной гадюки (Vipera (Renardi) ursini) И Змеи Восточной Европы.
Тольятти: ИЭВБ РАН, 2003. С. 26-29.
Каталогизация зоологических коллекций. Вып. 2. Фондовые коллекции в системе монито-
ринга герпетофауны / Е.В. Завьялов, В.Г. Табачишин, Г.В. Шляхтин и др.; Под рсд. докт. биол.
наук Е.В. Завьялова. Саратов: Изд-во Саратовского ун-та, 2006. 96 с.
Киреев В.А. Земноводные и пресмыкающиеся хребта Жельтау // Вопросы герпетологии
(Тез. докл. V Всесоюз. герпетол. конф.). Л., 1981. С. 64-65.
Киреев В.А. О северной границе серого голопалого геккона Gymnodactylus russowi Strauch //
Экология и фаунистика амфибий и рептилий сопредельных стран. Л.: Наука, 1984. С. 141-142.
Кириллов Л.А. Гельминтофауна настоящих ящериц (Lacertidae, Lacerta) Бузулукского бора
и Краспосамарского лесничества И Тез. докл. II Конф, герпетологов Поволжья. Тольятти, 1999.
С. 26-27.
Киселева Н.К. Эволюция биогеоценозов Прикаспия в голоцене. М.: Наука, 1982. 112 с.
СПИСОК ЛИТЕРАТУРЫ
315
Климанов В.А. Особенности изменения климата Северной Европы в позднеледниковье и
голоцене И Бюл. МОИП. Отделение геологии, 1994. Т. 69. С. 58-62.
Климанов В.А., Немкова В.К. Изменение климата Башкирии в голоцене И Палеоклиматы
голоцена европейской территории СССР. М.: 1988. С. 45-51.
Книга генетического фонда фауны Казахской ССР. Позвоночные животные / Под ред.
Е.В. Гвоздева. Алма-Ата: Наука, 1989. 216 с.
Колякин НН Распределение и численность пресмыкающихся в северных районах Нижнего
Поволжья И Тез. докл. Всесоюз. совещ. по проблемам кадастра и учета животного мира. Уфа,
1989. Т. 2. С. 57-58.
Кононова НК. Циркуляционные характеристики климатических экстремумов И Палеокли-
маты позднеледниковья и голоцена. М.: Наука, 1989. 146 с.
Копылов П.Е., Бакиев А.Г. Об укусах людей гадюками в Самарской области И Актуальные
проблемы герпетологии и токсинологии: Сб. науч. тр. Тольятти, 2001. Вып. 5. С. 57-62.
Косарева Н.А. Рептилии юга Сталинградской области И Уч. зап. Сталинградского гос. пед.
ин-та. Вып. 2. Зоология, ботаника, химия. Сталинград, 1950. С. 227-240.
Косарева Н.А. Материалы по экологии озерной лягушки в Волгоградской области И Вопро-
сы морфологии, экологии и паразитологии животных. Волгоград, 1970. С. 50-56.
Котенко Т., Вехник В. О находке разноцветной ящурки (Eremias arguta) в районе Жигулей
Вести, зоологии, 1993. № 1. С. 42.
Красная книга Астраханской области / Под ред. Ю.С. Чуйкова. Астрахань: Изд-во Нижне-
волжского центра экологического образования, 2004. 356 с.
Красная книга Волгоградской области. Волгоград, 2004. 172 с.
Красная книга Казахстана. Т. 1. Животные. Ч. 1. Позвоночные. Алматы: Конжык, 1996. 324 с.
Красная книга Оренбургской области. Животные и растения. Оренбург: ОКИ, 1998. 176 с.
Красная книга Российской Федерации. Животные. М.: ACT; Астрель, 2001. 860 с.
Красная книга Самарской области. Т. 2. Редкие виды животных / Под ред. чл.-корр. РАН
Г.С. Розенберга и проф. С.В. Саксонова. Тольятти: ИЭВБ РАН; «Кассандра», 2009. 332 с.
Красная книга Саратовской области: Грибы. Лишайники. Растения. Животные. Саратов:
Изд-во Торгово-промышленной палаты Саратовской обл., 2006. 528 с.
Крашенников И.М. Из истории развития ландшафтов Южного Урала И Географические ра-
боты. М.: ОГИЗ, 1954. С. 103-128.
Кременецкий К.В., Беттер Т., Климанов В.А. и др. История растительности и климата Бу-
зулукского бора в позднеледниковье и голоцене и ее палеогеографическое значение И Изв. АН.
Сер. геогр., 1998. № 4. С. 60-74.
Кременецкий К.В., Тарасов П.Е., Черкинский А.С. История островных боров Казахстана в
голоцене И Бот. журн., 1994. Т. 79, № 3. С. 13-26.
Кривенко В.Г. Водоплавающие птицы и их охрана. М.: Агропром, 1991. 272 с.
Кривенко В.Г Современный статус водоплавающих птиц России с позиций природных и
антропогенных воздействий И Многолетняя динамика численности птиц и млекопитающих:
Матер, междунар. симп. Казань: Новое знание, 2002. С. 51-77.
Кривко А.М. К распространению и экологии амфибий Северного Прикаспия И Биологичес-
кие науки. Алма-Ата: Изд-во КазПИ, 1976. Вып. 3. С. 46-48.
Кубанцев Б.С. Земноводные и пресмыкающиеся северных районов Нижнего Поволжья И
Тез. докл. III Конф, герпетологов Поволжья. Тольятти, 2003. С. 33-36.
Кубанцев В.С., Колякин Н.Н. Состав и распределение земноводных в северных районах
Нижнего Поволжья И Животный мир Предкавказья и сопредельных территорий. Ставрополь,
1988. С. 54-59.
Кубанцев Б.С., Колякин Н.Н. Распределение и численность пресмыкающихся в северных
районах Нижнего Поволжья И Матер. Всесоюз. сов. по проблемам кадастра и учета животного
мира. Уфа: БКИ, 1989. Ч. III. С. 280-282.
Кубанцев Б.С., Уварова В.Я., Косарева Н.А. Животный мир Волгоградской области. Назем-
ные позвоночные животные. Волгоград: Кн. изд-во, 1962. 192 с.
Кубыкин Р.А. Современное распространение и численность четырехполосого полоза Elaphe
qutorlineata (Reptilia, Colubridae) в Казахстане И Selevinia, 1994. № 1. С. 61-64.
Кубыкин Р.А., Брушко З.К. Новые сведения по распространению пресмыкающихся в Казах-
стане И Бюл. МОИП. Отд. биол., 1989. Т. 94, вып. 3. С. 33-36.
Кубыкин Р.А., Брушко З.К. О промысле амфибий и рептилий в Казахстане И Selevinia, 1994.
№2. С. 78-81.
316
СПИСОК ЛИТЕРАТУРЫ
Кубыкин Р.А., Брушко З.К К современному распространению восточного удавчика - Erix
tataricus (Lichtenstain, 1823) в Казахстане И Зоологические исследования в Казахстане. Алматы,
2002. С. 173-174.
Кудакина Е.Ю. О географической изменчивости разноцветной ящурки (Eremias arguta) в
Северном Прикаспии И Вопросы герпетологии (Сб. статей). Л., 1981. С. 75.
Кузьмин С.Л. Земноводные бывшего СССР. М.: КМК, 1999. 298 с.
Кузьмин С.Л., Маслова И.В. Земноводные российского Дальнего Востока. М.: КМК, 2005.
434 с.
Кузьмин С.Л., Семенов Д.В. Конспект фауны земноводных и пресмыкающихся России. М.:
КМК, 2006. 139 с.
Куприянова Л.А., Завьялов Е.В., Табачишин В.Г. Живородящая ящерица Zootoca vivipara
(Lacertidae) на севере Нижнего Поволжья: цитогенетический анализ и экология размножения
//Тез. докл. III Конф, герпетологов Поволжья. Тольятти, 2003. С. 36-38.
Кэррол Р. Палеонтология и эволюция позвоночных. Т. 1. М.: Мир, 1992. 280 с.
Лепехин И. И. Дневные записки путешествия по разным провинциям Российского государс-
тва в 1768-1769 годах. Т. I. СПб., 1771. 538 с.
Линдберг ГУ. Четвертичный период в свете биогеографических данных. М.; Л.: Изд-во АН
СССР, 1955. 336 с.
Линдеман Г.В., Абатуров Б.Д., Быков А.В., Лопушков В.А. Динамика населения позвоноч-
ных животных заволжских полупустынь. М.: Наука, 2005. 254 с.
Литвинов Н.А. Термобиологические исследования//Змеи Волжско-Камского края. Самара:
Изд-во СНЦ РАН, 2004. С. 109-146.
Литвинов Н.А., Ганщук С.В. Температурные условия обитания ящериц Волжско-Камского
края // Тез. докл. III Конф, герпетологов Поволжья. Тольятти, 2003. С. 42-44.
Лобачев В.С, Чугунов Ю.Д., Чуканина И.Н. Особенности герпетофаупы Северного Приара-
лья И Вопросы герпетологии (Тез. докл. III Всесоюз. герпетол. конф.). М., 1973. С. 116-118.
Магдеев Д.В. Анализ состояния популяций амфибий и рептилий Самарской Луки // Самар-
ская Лука на пороге третьего тысячелетия: Матер, докл. «Состояние природного и культурного
наследия Самарской Луки». Тольятти: ИЭВБ РАН; ОСИП «Парквей», 1999. С. 191-200.
МагдеевД.В., Бакиев А.Г. Изучение фауны гадюк Самарской области // Актуальные пробле-
мы герпетологии и токсинологии: Сб. науч. тр. Тольятти, 1995. Вып. 1. С. 38-40.
Маленев А.Л., Бакиев А.Г. Пресмыкающиеся Самарской области И Науч. инф. сб. ИЭВБ.
1991-1996. Тольятти, 1997. 84 с.
Маленев А.Л., Бакиев А.Г, Песков А.Н., Вехник В.Л. Гадюка обыкновенная Vipera berus
(Linnaeus, 1758) И Красная книга Самарской области. Т. 2. Редкие виды животных. Тольятти:
ИЭВБ РАН; ООО «Кассандра», 2009. С. 247.
Марков КК. Палеогеография (историческое землеведение). М.: Изд-во МГУ, 1960. 268 с.
Марков КК. Плювиальные условия. Рельеф и ландшафты. М.: Изд-во МГУ, 1977.
С. 14-28.
Материалы к кадастру амфибий и рептилий бассейна Средней Волги / Под ред. М.В. Песто-
ва. Н. Новгород: Междунар. социально-экол. союз; экоцентр «Дронт», 2002. 222 с.
Матросов А. Н., Кузнецов А.А. Размещение и численность наземных позвоночных живот-
ных в Волго-Уральских песках //Животные в природных экосистемах. Н. Новгород: НПУ, 1994.
С. 107-117.
Мельниченко А., Положенцев П., Куликова М., Королева К. Куйбышев и его окрестности
как места для школьных экскурсий по зоологии // Уч. зап. Гос. пед. и учит, ин-та. Куйбышев:
Куйбышев, изд-во, 1938. Вып. 1.С. 158-167.
Мильков Ф.Н. Среднее Поволжье: физико-географическое описание. М.: Изд-во АН СССР,
1953.262 с.
Мильков Ф.Н. Физическая география: учение о ландшафте и географическая зональность.
Воронеж: Изд-во ВГУ, 1986. 328 с.
Мильто КД. Распространение и морфологические особенности черной лесостепной гадю-
ки // Змеи Восточной Европы. Тольятти: ИЭВБ РАН, 2003. С. 56-57.
Морозенко Н.В. Эколого-морфологическая структура и фенетический анализ популяций
обыкновенного ужа (Reptilia; Colubridae, Natrix natrix)'. Автореф. дис. ... канд. биол. паук. Сара-
тов: ФГОУ ВПО «Саратовский ГАУ», 2003. 20 с.
Морозенко Н.В., Шляхтин Г.В., Завьялов Е.В. Изменчивость окраски обыкновенного ужа
(Natrix natrix) в Нижнем Поволжье //Змеи Восточной Европы. Тольятти, 2003. С. 56-57.
СПИСОК ЛИТЕРАТУРЫ
317
Накаренок Е.Г. Экологические аспекты формирования герпетофауны Северного Прикаспия
и тенденции ее современного развития: Дис. канд. биол. наук. Н. Новгород: НГПУ, 2002. 200 с.
Накаренок Е.Г. Влияние четвертичных трансгрессий Каспия на современный состав и раз-
мещение герпетофауны Северного Прикаспия И Тез. III конф, герпетологов Поволжья. Тольят-
ти, 2003а. С. 53-55.
Накаренок Е.Г. Распространение и особенности экологии круглоголовки-вертихвостки
(Phrynocephalus guttatus) песчаных пустынь Северного Прикаспия И Поволжский экол. журн.,
20036. № 1.С. 77-81.
Накаренок Е.Г., Неручев В.В. О редких видах рептилий Северного Прикаспия // Биоразно-
образие и биоресурсы Урала и сопредельных территорий. Оренбург: ИПК «Газпромпечать»,
2001. С. 295-296.
Неручев В.В., Васильев НФ. Фауна рептилий Северо-восточного Прикаспия // Вестн. зоол.,
1978. №6. С. 36^11.
Неручев В.В., Капустина С.Ф. Такырная круглоголовка (Phrynocephalus helioscopus) в при-
эмбинских пустынях // Вопросы герпетологии (Сб. статей). Л., 1981. С. 90-91.
Неручев В.В., Накаренок Е.Г. О современном состоянии герпетофауны низовий Волги и
юга Волго-Уральского междуречья И Тез. докл. I Конф, герпетологов Поволжья Тольятти, 1995.
С. 41—43.
Неручев В.В., Накаренок Е.Г. О влиянии дорожной сети па состав и численность населения
рептилий Приволжской пустыни И Тез. докл. II Конф, герпетологов Поволжья Тольятти, 1999.
С.4М2.
Неручев В.В., Шатилович О.А. О некоторых закономерностях формирования герпетофау-
ны Северо-Восточного Прикаспия И Вопросы герпетологии. (Матер. VI Всесоюз. герпетолог,
конф.). Л., 1985. С. 147-148.
Неручев В.В., Арженкова Н.Г, Панфилова Т.А. О современном составе и размещении гер-
петофауны Гурьевской области И Тез. докл. сов. по проблеме кадастра и учета животного мира.
Уфа: 1989а. Ч. 3. С. 289-291.
Неручев В.В., Арженкова Н.Г, Шатилович О.А. Герпетофауна речных долин Северного
Прикаспия И Наземные и водные экосистемы: Межвуз. сб. Горький: Изд-во ГГУ, 1990. С. 26-33.
Неручев В.В., Козырева О.А., Накаренок Е.Г. К фепогеографической характеристике попу-
ляций разноцветной ящурки (Eremias arguta) в Северном Прикаспии И Вопросы герпетологии
(Сб. статей). Киев, 19896. С. 171-172.
Неручев В.В., Кудакина Е.Ю., Васильев НФ. Влияние выпаса на фауну и население реп-
тилий в пустынях Северного Прикаспия И Наземные и водные экосистемы. Горький, 1983.
С. 67-72.
Неручев В.В., Накаренок Е.Г, Лебединский А.А. Пресмыкающиеся пустынь Северного При-
каспия И Животный мир Южного Урала и Северного Прикаспия: Тез. докл. Оренбург: Изд-во
ОГПИ, 1995. С. 94-96.
Неручев В.В., Шатилович О.А., Иванова Н.Н. Современная динамика герпетофаупы юга
Урало-Эмбипского междуречья // Наземные и водные экосистемы: Межвуз. сб. Горький: Изд-во
Горьковского ГУ, 1986. С. 75-81.
Неручев В.В., Ширяев А.Ф., Накаренок Е.Г Герпетофауна песков Северного Прикаспия
И Животные в природных экосистемах: Межвуз. сб. науч. тр. Н. Новгород: Изд-во ННПУ, 1994.
С. 127-136.
Неручев В.В., Васильев Н.Ф., Кудакина Е.Ю., Смирнов А.Н. Исторические и антропогенные
факторы в современном распространении рептилий в Северном Прикаспии // Тез. докл. VII Все-
союз. зоогеогр. конф.: М.: Наука, 1979. С. 212-214.
Неручев В.В., Ширяев А.Ф., Капустина С.Ф., Кудакина ЕЮ. Фауна и ландшафтные комп-
лексы рептилий юга Урало-Эмбенского междуречья // Наземные и водные экосистемы: Межвуз.
сб. Горький, 1984. С. 86-91.
Несов Л.Р. Амфибии и рептилии в экосистемах мела Средней Азии И Вопросы герпетоло-
гии (Сб. статей). Л., 1981. С. 91-92.
Несов Л.А., Хозацкий Л.И. Черепахи мезозоя СССР // Вопросы герпетологии (Сб. статей).
Л„ 1977. С. 157-159.
Нижнее Поволжье: физико-географическое описание / Отв. ред. А.А. Григорьев. М.; Л.:
Изд-во АН СССР, 1948. 136 с.
Николаев В.А. Наследие ксеротермических эпох плейстоцена в природе степей Казахстана.
Рельеф и ландшафты. М.: Изд-во МГУ, 1977. С. 181-189.
318
СПИСОК ЛИТЕРАТУРЫ
Николаев В.А. Ландшафты азиатских степей. М.: Изд-во МГУ, 1999. 288 с.
Никольский А.М. Пресмыкающиеся и амфибии, собранные П.П. Сушкиным в Тургайской
области И Bull. Soc. Nat. Moscow. Nouv. ser., 1899.
Никольский A. M. Летние поездки натуралиста. СПб., 1900. 240 с.
Никольский А.М. Пресмыкающиеся и земноводные Российской Империи (Herpctologia Ros-
sica) И Записки АН. Т. XVII, № 1. СПб., 1905. 442 с.
Никольский А.М. Определитель пресмыкающихся и земноводных Российской Империи.
Харьков: Русская типография и литография, 1907. 182 с.
Никольский А.М. Пресмыкающиеся (Reptilia). Фауна России и сопредельных стран: Chelo-
nia и Sauria. Т. 1. СПб., 1915. 534 с.; Т. 2. 1916. 610 с.
Никольский А.М. Земноводные (Amphibia). Фауна России и сопредельных стран. Пг.: Ти-
погр. АН, 1918.310 с.
Окулова Н.М. Ландшафтные особенности фауны позвоночных животных па северной окра-
ине Волго-Уральских песков//Зоол. журн., 1963. Т. XLII, вып. 6. С. 882-892.
Окулова Н.М. К биологии круглоголовок Западного Казахстана // Вопросы герпетологии
(Сб. статей). Л., 1964а. С. 50-51.
Окулова Н.М. Некоторые черты биологии разноцветной ящурки в Западном Казахстане //
Вопросы герпетологии (Сб. статей). Л., 19646. С. 56-58.
Окулова Н.М. Популяционный полиморфизм разноцветной ящурки на северо-восточной
окраине Волго-Уральских песков // Вопросы герпетологии (Сб. статей). М., 1973. С. 135-137.
Окулова Н.М. К биологии степной гадюки (Vipera ursini) в Западном Казахстане // Вопросы
герпетологии (Сб. статей). Л., 1981. С. 93-94.
Окулова Н.М. Узорчатый полоз (Elaphe dione) и песчаный удавчик (Eryx miliaris) в Западном
Казахстане//Вопросы герпетологии (Матер. VI Всесоюз. герпетол. конф.). Л., 1985. С. 153.
Определитель земноводных и пресмыкающихся фауны СССР. А.Г. Банников, И.С. Даревс-
кий, В.Г. Ищенко и др. М.: Просвещение, 1977. 416 с.
Орлова В.Ф. Географическая изменчивость живородящей ящерицы И Вопросы герпетоло-
гии (Сб. статей). Л., 1973. С. 137-139.
Особо охраняемые природные территории Саратовской области И Комитет охраны окру-
жающей среды и природопользования Саратовской области / Под ред. В.З. Макарова. Саратов:
Изд-во СарГУ, 2007. 300 с.
Островских С.В., Пестов М.В., Шапошников А.В. К вопросу о распространении каспийско-
го полоза - Hierophis caspius (Gmelin, 1789) в Волго-Уральском междуречье // Герпетологичес-
кие исследования в Казахстане и сопредельных странах. Алматы, 2010.
Павлов А.В. К биологии степной гадюки на севере ареала И Чтения памяти В.А. Попова.
Казань, 1997. С. 24-26.
Павлов А.В., Гаранин В.И, Бакиев А.Т. Обыкновенная гадюка Vipera berus (Linnaeus, 1758)
И Змеи Волжско-Камского края. Самара: Изд-во СамНЦ РАН, 2004. С. 49-61.
Паллас П.С. Путешествие по разным провинциям Российской Империи. 4.1. СПб.: 1773.
658 с. (Цит. по Паллас П.С. Путешествия по разным провинциям Российской империи. Избран-
ные главы. Уральск: Изд-во ТОО «Оптима», 2006. 220 с.)
Панкратов О.В. Численность и биотопическое распределение четырехполосого полоза в
Западном Казахстане И Вопросы герпетологии (Сб. статей). Киев, 1989. С. 183-184.
Параскив К.П. Пресмыкающиеся Казахстана. Алма-Ата: Изд-во АН Каз. ССР, 1956. 228 с.
Параскив К.П., Бутовский П.М. О фауне земноводных и пресмыкающихся Западного Ка-
захстана//Тр. Ин-та зоол. АН Каз. ССР. Алма-Ата, 1960. Т. 13. С. 148-159.
Песков А.Н. Степная гадюка в Самарской области: история изучения распространения и
распределения // Самарский край в истории России: Матер, юбилейн. науч. конф. Самара, 2001.
С. 276-278.
Песков А.Н, Бакиев А.Г., Маленев А.Н. Vipera melanis (Pallas, 1771): что за гадюки обитают
в городской черте Самары? // Тез. докл. III Конф, герпетологов Поволжья. Тольятти, 2003. С. 62-64.
Пескова Т.Ю. Внутрипопуляционный полиморфизм окраски зеленой жабы // Актуальные
проблемы герпетологии и токсикологии: Сб. науч. тр. Тольятти, 2003. Вып. 6. С. 90-91.
Пестов М.В., Лебединский А.А., Ануфриев В.М. и др. Проект по реинтродукции номинатив-
ного подвида обыкновенной квакши (Hyla arborea arborea) на территории России И Тез. докл.
III Конф, герпетологов Поволжья. Тольятти, 2003. С. 64-66.
Писанец Е.М., Шербак Н.Н. Систематика зеленых жаб (Amphibia, Anura) фауны СССР Н
Вести, зоол., 1979. № 4. С. 11-16.
СПИСОК ЛИТЕРАТУРЫ
319
Положенцев П.А. К фауне млекопитающих и гадов Бузулукского бора И Материалы по
изучению природы Среднего Поволжья. Москва; Куйбышев: Куйбышев, краевое изд-во, 1935.
Вып. 1.С. 77-96.
Положенцев П.А, Классы Пресмыкающиеся и Земноводные И Животный мир Среднего По-
волжья (полезные и вредные животные). Куйбышев: Кн. изд-во, 1937. С. 91-99.
ПоложенцевП.А. Классы Пресмыкающиеся и Земноводные //Животный мир Среднего По-
волжья (полезные и вредные животные). Изд-во 2-е. Куйбышев: ОГИЗ, 1941. С. 103-114.
Положенцев П.А., Никифорук К.С. Животный мир Башкирии. Кл. Пресмыкающиеся. Зем-
новодные. Уфа: БГИ, 1949. С. 126-131.
Положенцев П.А., Ханисламов М.Г. К вопросу о фауне амфибий и рептилий Башкирской
АССР//Тр. Башкирского с/х ин-та. Уфа, 1942. Вып. 3. С. 143-147.
Полынова ГВ., Кудрявцева К.А., Соломатина Т.А. Некоторые аспекты экологии богдин-
ской популяции пискливого геккончика И Богдинско-Баскунчакский заповедник и его роль в
сохранении биоразнообразия севера Астраханской обл. Астрахань: АГТУ, 2004. С. 76-87.
Полынова Г.В., Полынова О.Е., Вулич Т.А. Особенности биотопического распределения
разноцветной ящурки {Eremias arguta deserti) на территории Богдинско-Баскунчакского го-
сударственного заповедника // Вопросы герпетологии (Матер. I Съезда герпетол. о-ва им.
А.М. Никольского). Пущино; Москва, 2001. С. 239-241.
Прыткая ящерица/Отв. ред. А.В. Яблоков. М.: Наука, 1976. 374 с.
Пуляев А.И. Роль заповедника в сохранении биологического разнообразия Оренбуржья И
Проблемы сохранения и восстановления степных экосистем. Оренбург, 1999. С. 17-20.
Разноцветная ящурка / Под ред. Н.Н. Щербака. Киев: Наук, думка, 1993. 238 с.
Райский А.П. Животный мир Чкаловской области И Очерки физической географии Чкалов-
ской области. Чкалов: Чкаловское кн. изд-во, 1951. С. 157-209.
Райский А.П. О животных Чкаловской области И По родному краю (краеведческие очерки).
Чкалов: Чкаловское кн. изд-во, 1954. С. 71-89.
Райский А.П. О животных Чкаловской области И По родному краю: Сб. краеведческих очер-
ков. Изд-во 2-е. Чкалов: Чкаловское кн. изд-во, 1956. С. 187-215.
Раллъ Ю.М. Древняя степь «Бес-Чохо» в Волго-Уральских песках // Природа, 1935. № 4.
С. 55-60.
Ратников В.Ю. Некоторые замечания о находках герпетофауны в четвертичных отло-
жениях Восточной Европы И Вопросы герпетологии (Сб. статей). Киев: Наук, думка, 1989.
С. 208-209.
Ратников В.Ю. Герпетофауна верхнеплейстоценового местонахождения Еласы в бассей-
не Волги И Актуальные проблемы герпетологии и токсикологии: Сб. науч. тр. Тольятти, 2001.
Вып. 5. С. 81-88.
Ратников В.Ю. Позднекайнозойские земноводные и чешуйчатые пресмыкающиеся Вос-
точно-Европейской равнины И Тр. НИИ геологии Воронеж, гос. ун-та. Вып. 10. Воронеж:
Изд-во ВГУ, 2002. 138 с.
Ратников В.Ю. Некоторые особенности ископаемых остатков змей из позднекайнозойских
отложений Восточно-Европейской равнины И Змеи Восточной Европы. Тольятти: ИЭВБ РАН,
2003. С. 67-70.
Ратников В.Ю. К истории фауны ящериц Волжского бассейна // Актуальные проблемы
герпетологии и токсинологии: Сб. науч. тр. Тольятти, 2004а. Вып. 7. С. 103-111.
Ратников В.Ю. К истории формирования офидиофауны Волжско-Камского края И Змеи
Волжско-Камского края. Самара: Изд-во СамНЦ РАН, 20046. С. 21-27.
Ратников В.Ю. Ископаемые остатки современных видов земноводных и чешуйчатых пре-
смыкающихся как материал для изучения истории их ареалов И Тр. НИИ геологии Воронеж. ГУ.
Вып. 59. Воронеж: Изд-во ВГУ, 2009. 92 с.
Редько Б.А. Поездка на озеро Лебяжье Самарского уезда с целью выяснения причин гибели
в нем рыбы // Работы Волжской биол. станции, 1915. Т. 5, № 2. С. 89-101.
Россия. Полное географическое описание нашего отечества Т. 5. Урал и Приуралье. СПб.,
1914. С. 120-121.
Рысков Я.Г., Демкин В.А. Развитие почв и природной среды степей Южного Урала в голо-
цене. Пущино: ОНТИ ПНЦ РАН, 1997. 166 с. <
Рычагов Г.И. Уровенный режим Каспийского моря за последние 10 000 лет И Всстн. МГУ.
Сер. геогр., 1993. № 2. С. 38-49.
Рычков П.И. Топография Оренбургской губернии (1762). Уфа: Китап, 1999. 360 с.
320
СПИСОК ЛИТЕРАТУРЫ
Сабанеев Л. Позвоночные Среднего Урала и географическое распространение их в Перм-
ской и Оренбургской губерниях. М., 1874. 204 с.
Сараев Ф.А. Новые данные о находках озерной лягушки в Западном Казахстане И Selevinia,
2007. С. 172.
Сараев Ф.А., Пестов М.В. К кадастру рептилий Северного и Северо-восточного Прикас-
пия // Герпетологические исследования в Казахстане и сопредельных странах. Алматы, 2010.
С. 172-191.
Серебровский П.В. Очерк третичной истории наземной фауны СССР // Животный мир
СССР. М., Л.: Изд-во АН СССР, 1937. Т. 1. С. 11-78.
Сибирский углозуб {Salamandrella Keyserlingii Dybowski, 1870): зоогеография, системати-
ка, морфология / Под ред. С.Н. Литвинчука, Л.Я. Боркина. М.: Наука, 1994. 368 с.
Сидоров С. Амфибии и рептилии Арала// Бюл. МОИП. Отд. биолог., 1925. Т. ХХХШ, вып.
1-2. С. 188-200.
Смелянский И.Э., Елизаров А.В. О проектируемом степном заказнике «Синий Сырт» в Са-
марской области И Самарская Лука., 1996. № 7-96. С. 104-123.
Смирнов С.И., Шкунов В.Ф., Кудакина Е.И. Гекконы Северного Прикаспия // Вопросы гер-
петологии (Матер. VI Всесоюз. герпетол. конф.). Л., 1985. С. 195-196.
Смирнова Ю.А., Рябов С.А., Ананьева Н.Б. Изучение внутривидовой структуры в комплексе
Е. dione - Е. bimaculata с использованием молекулярных маркеров РАПД // Змеи Восточной
Европы: Матер, междунар. конф. Тольятти: ИЭВБ РАН, 2003. С. 81-82.
Смирновский Б.Н. Ядовитые змеи Казахстана. Алма-Ата: Казсельхозгиз, 1963. 64 с.
Старков В.Г., Уткин 1О.Н. Новые данные о видовой принадлежности гадюк Самарской
области//Тез. докл. III Конф, герпетологов Поволжья. Тольятти, 2003. С. 81-82.
Стрельцов А.Б., Константинов Е.Л., Алексеев С.К, Устюжина О.А. О герпетофауне Бог-
динско-Баскупчакского заповедника и одноименного заказника // Вести. Калужского ун-та,
2006. № 2. С. 30-32.
Сухов В.П. Позвоночные - Vertebrata (мелкие) // Фауна и флора Аккуласва (опорный разрез
среднего акчагыла - среднего апшерона Башкирии). Уфа, 1972. С. 119-139.
Табачишин В.Г, Завьялов Е.В. Распространение и таксономический статус разноцветной
ящурки {Eremias arguta) в северной части Нижнего Поволжья И Вести, зоол., 1998. № 32 (4).
С. 51-59.
Табачишин В.Г, Завьялов Е.В. Распространение и особенности биологии узорчатого полоза
(Colubridae, Reptilia) в Поволжье//Герпетол. вести., 2000. № 3-4. С. 14-23.
Табачишин В.Г., Табачишина И.Е. Распространение и особенности экологии обыкновенно-
го ужа {Nartix natrix) на севере Нижнего Поволжья // Поволж. экол. журн., 2002. № 2. С. 179—
183.
Табачишин В.Г, Завьялов Е.В., Табачишина И.Е. Пространственное размещение разноцвет-
ной ящурки -Eremias arguta (Pallas, 1773) на севере ареала в Поволжье // Соврем, герпетология,
2006. Вып. 5-6. С. 117-124.
Табачишина И.Е., Табачишин В.Г, Завьялов Е.В. Морфоэкологическая характеристика
нижневолжских популяций степной гадюки {Vipera ursini) И Поволж. экол. журн., 2002. № 1.
С. 76-81.
Табачишин В.Г, Завьялов Е.В., Табачишина И.Е., Шляхтин Г.В. Ужеобразные и гадюко-
вые змеи юго-востока европейской части России // Зоологическое исследование регионов Рос-
сии и сопредельных территорий. Н. Новгород: Изд-во Нижегородского гос. пед. ун-та, 2002.
С. 157-158.
Табачишина И.Е., Табачишин В.Г, Завьялов Е.В. Эколого-фаунистическая характеристика
пресмыкающихся севера Нижнего Поволжья // Актуальные проблемы герпетологии и токсико-
логии: Сб. науч. тр. Тольятти, 2004. Вып. 7. С. 129-132.
Табачишина И.Е., Табачишин В.Г, Завьялов Е.В. Пространственное размещение и тенден-
ции изменения численности узорчатого полоза {Elaphe dione) на севере ареала в Поволжье И
Поволж. экол. журн., 2005. № 3. С. 277-280.
Табачишина И.Е., Табачишин В.Г., Завьялов Е.В. Разноцветная ящурка - Eremias arguta
(Pallas., 1773) И Красная книга Саратовской области. Саратов: Изд-во Торгово-промышленной
палаты, 2006а. С. 365-366.
Табачишина И.Е., Табачишин В.Г, Завьялов Е.В. Современное распространение каспийс-
кого полоза {Hierophis caspius (Gmelin, 1779)) на севере Нижнего Поволжья и сопредельных
территорий // Поволж. экол. журн., 20066. № 1. С. 91-94.
СПИСОК ЛИТЕРАТУРЫ
321
Табачишин В.Г., Шляхтин Г.В., Завьялов Е.В, Распространение и морфометрическая харак-
теристика гадюки Никольского (Vipera nikolskii Vedmederja, Grubant et Rudaeva) в Саратовской
области //Тез. докл. I Конф, герпетологов Поволжья. Тольятти, 1995. С. 54—55.
Табачишин В.Г., Шляхтин Г.В., Завьялов Е.В. и др. Морфометрическая дифференциация
к таксономический статус пресмыкающихся сем. Colubridae и Viperidae И Фауна Саратовской
области: Сб. науч. тр. Саратов: СарГУ, 1996. Т. I, вып. 2. С. 39-70.
Татаринов Л.П. Современные данные о происхождении ящериц И Вопросы герпетологии:
Автореф. докл. Л.: Наука, 1981. С. 131-133.
Терентьев П.В. Лягушка. М.: Сов. наука, 1950. 342 с.
Терентьев П.В. Земноводные - Amphibia И Животный мир СССР. М.; Л.: Изд-во АН СССР,
1953. Т. 4. С. 220-227.
Терентьев П.В. Материалы к истории отечественной герпетологии И Тр. Ин-та истории ес-
тествозн. и техники. М.; Л.: Изд-во АН СССР, 1957. Т. 16, вып. 3. С. 97-122.
Терентьев П.В. Герпетология. М.: Высш, шк., 1961. 336 с.
Терентьев П.В., Чернов С.А. Краткий определитель пресмыкающихся и земноводных
СССР. Л.: Учпедгиз, 1940. 184 с.
Терентьев П.В., Чернов С.А. Определитель земноводных и пресмыкающихся. М.: Сов. на-
ука, 1949. 340 с.
Тертышников М.Ф. Стации, численность, биомасса И Разноцветная ящурка. Киев: Наук,
думка, 1993. С. 114-119.
Тихонов В.Е. Ритмичность метеорологических факторов и биоклиматический потенциал в
степной зоне Урала // Вопросы степеведения. Оренбург: ИС УрО РАН, 2000. С. 93-101.
Топоркова Л.Я. Амфибии и рептилии Урала // Фауна европейского Севера, Урала и Запад-
ной Сибири. Свердловск, 1973. С. 84-117.
Топоркова Л.Я., Меныциков А.П. К экологии чесночницы обыкновенной на северо-
восточном пределе ее ареала // Фауна Урала и Европейского Севера. Свердловск, 1974.
С. 46-50.
ТопорковаЛ.Я. Морфологические особенности зеленой жабы (Bufo virdis) Южного Урала//
Вопросы герпетологии (Сб. статей). Л., 1981. С. 133-134.
Трохименко Н.М. К морфологии обыкновенного ужа в Самарской области И Тез. докл.
III Конф, герпетологов Поволжья. Тольятти, 2003. С. 82-84.
Трохименко Н.М., Бакиев А.Г. История изучения фауны ужеобразных змей (Colubridae) Са-
марской области // Самарский край в истории России: Матер, юбилейн. науч. конф. Самара,
2001. С. 274-276.
Файзулин А.И. Земноводные в фондовой коллекции Института экологии Волжского бассей-
на РАН (материалы к каталогу) // Актуальные проблемы герпетологии и токсинологии. Вып. 7.
Тольятти: ИЭВБ РАН, 2004. С. 141-151.
Файзулин А.И. Антропогенный фактор как причина гибели земноводных (Amphibia) на тер-
ритории Среднего Поволжья //Актуальные проблемы герпетологии и токсинологии: Сб. науч,
тр. Тольятти, 20046. Вып. 7. С. 152-154.
Файзулин А.И. Земноводные г. Тольятти И Актуальные проблемы герпетологии и токсино-
логии: Сб. науч. тр. Тольятти, 2005. № 8. С. 183-187.
Файзулин А.И., Вехник В.П. Травяная лягушка (Rana temporaria) - вид, включенный в Крас-
ную книгу Самарской области И Актуальные проблемы герпетологии и токсинологии: Сб. науч,
тр. Тольятти, 2001. Вып. 5. С. 101-104.
Фальк И.П. Записки путешествия академика Фалька. Полное собрание ученых путешествий
по России. Т. 6. СПб., 1824. 546 с.
Федоров П.В. Плейстоцен Понто-Каспия. М.: Наука, 1978. 166 с.
Федорович Б.А. Вопросы палеогеографии равнин Средней Азии. Динамика и закономернос-
ти рельефообразования пустынь. М.: Наука, 1983. С. 31^5.
Флеров К.К., Беляева Е.И., Яновская Н.М., Гуреев А.А. Зоогеография палеогена Азии. М.:
Наука, 1974. 304 с.
Хабибуллин В.Ф. Фауна пресмыкающихся Республики Башкортостан. Уфа: Изд-во БашГУ,
2001. 128 с.
Хабибуллин В.Ф. К истории формирования современной фауны пресмыкающихся Башки-
рии //Зоол. журн., 2002. Т. 81., № 3. С. 342-349.
Хабибуллин В.Ф. К офидиофауне Республики Башкортостан И Актуальные проблемы герпе-
тологии и токсинологии: Сб. науч. тр. Тольятти, 2004. Вып. 7. С. 155-156.
322
СПИСОК ЛИТЕРАТУРЫ
Хабибуллин В.Ф., Онучина Л.С. О питании прыткой ящерицы (Lacera agilis L.) на Южном
Урале // Животный мир Южного Урала и Северного Прикаспия. Тез. докл. Оренбург, 2000.
С. 83-84.
Ходашова К.С. Природная среда и животный мир глинистых полупустынь Заволжья. М.:
Изд-во АН СССР, 1960.32 с.
Хозацкий Л.И. История фауны черепах СССР в свете палеографии И Тр. II Всесоюз. гсогр.
съезда. М„ 1949. Т. 3. С. 220-230.
Хозацкий Л.И. Современное и прошлое распространение черепах в СССР//Проблемы гео-
графии суши. Львов: Изд-во Львовского ун-та, 1958. С. 319-324.
Хозацкий Л.И. Земноводные и пресмыкающиеся // Четвертичная система. Стратиграфия
СССР. М.: Недра, 1982. С. 248-262.
Хотинский И.А., Алешинская З.В., Гуман М.А. Новая схема периодизации ландшафтно-кли-
матических изменений в голоцене//Изв. АН СССР. Сер. геогр., 1991. № 3. С. 30-42.
Хрущева К.А. Животный мир Самарского края // Краеведение. Природа и население Самар-
ского края. Самара: Губиздат, 1924. С. 135-149.
Чернобай В.Ф. Фаунистический обзор наземных позвоночных природного парка «Волго-
Ахтубинская пойма» // Волго-Ахтубинская пойма - природно-ресурсный потенциал: Науч. ст.
Волгоград, 2004. С. 130-141.
Чернов С.А. Пресмыкающиеся (Reptilia) и земноводные (Amphibia). Зоографический и эко-
логический обзор фауны СССР И Животный мир СССР. М.; Л.: Изд-во АН СССР, 1937. Т. 1.
С. 326-346.
Чернов С.А. Основные черты истории фауны пресмыкающихся Средней Азии И Тр. II Все-
союз. геогр. съезда. М., 1949. Т. 3. С. 230-237.
Чернов С.А. Пресмыкающиеся. Земноводные // Животный мир СССР. Зона степей. М.; Л.:
Изд-во АН СССР, 1950. Т. 3. С. 140-161.
Чернов С.А. Эколого-фаунистический обзор пресмыкающихся юга междуречья Волга-Урал
//Тр. Зоол. ин-та АН СССР. Л., 1954. Т. 16. С. 137-159.
Чернов Ю.И. Природная зональность и животный мир суши. М.: Мысль, 1975. 222 с.
Чернов С.А. Четырехполосый полоз. Краткое сообщ. // Редкие животные Казахстана. Алма-
Ата: Наука, 1986. С. 180.
Чибилёв А.А. В глубь степей. Очерки об естествоиспытателях Оренбургского края. Екате-
ринбург: УИФ «Наука», 1993. 120 с.
Чибилёв А.А. Земноводные и пресмыкающиеся Оренбургской области и их охрана. Матери-
алы для Красной книги Оренбургской области. Екатеринбург: УрО РАН, 1995. 45 с.
Чибилёв А.А. Обыкновенная медянка // Красная книга Оренбургской области. Оренбург:
ОКИ, 1998. С. 82.
Чибилёв А.А. Герпетофауна госзаповедника «Оренбургский» // Тез. докл. II Конф, герпето-
логов Поволжья. Тольятти, 1999а. С. 55-56.
Чибилёв А.А. Герпетофауна Оренбургской области и прилегающих территорий // Тез. докл.
II Конф, герпетологов Поволжья. Тольятти, 19996. С. 56-57.
Чибилёв А.А., Дебело П.В. Ландшафты Урало-Каспийского региона. Оренбург: ИС УрО
РАН: Печатный дом «Димур», 2006. 264 с.
Чибилёв Е.А. Амфибии и рептилии Челябинской области. Миасс, 2005. 17 с.
Чигуряева А.Н., Яхимович В.Л. О флоре и растительности Южного Предуралья в среднеак-
чагыльско-голоценовое время // Антропоген Южного Урала. М.: Наука, 1965. С. 164-187.
Чигуряева А.А., Жидовинов Н.Я., Мичурин В.Г. Изменения растительности и климата на
юго-востоке европейской части СССР в четвертичное время И Вопр. ботаники Юго-Востока,
1988. Вып. 6. С. 53-79.
Чирикова М.А. Особенности рисунка ящурки быстрой (Eremias velox Pallas, 1771) из При-
аралья И Актуальные проблемы герпетологии и токсинологии: Сб. науч. тр. Тольятти, 2003.
Вып. 6. С. 109.
Чирикова М.А. Материалы по изменчивости быстрой ящурки Eremias velox Pallas, 1771
(Reptilia, Sauria) в Казахстане П Selevinia, 2004. С. 24-34.
Чхиквадзе В.М., Сухов В.П. Земноводные и пресмыкающиеся из четвертичных отложений
Красного Бора (р. Кама) // Вопросы герпетологии (Сб. статей). Л., 1977. С. 227-228.
Шапошников В.М. Животные Куйбышевской области, нуждающиеся в особой охране И
Вопросы лесной биогеоценологии, экологии и охраны природы в степной зоне. Куйбышев,
1978. Вып. 3. С. 120-131.
СПИСОК ЛИТЕРАТУРЫ
323
Шапошников В.М. О пересмотре границы распространения живородящей ящерицы в Са-
марской области // Тез. докл. II Конф, герпетологов Поволжья. Тольятти, 1999а. С. 60-61.
Шапошников В.М. О разноцветной ящурке, обитающей в Самарской области И Тез. докл.
II Конф, герпетологов Поволжья. Тольятти, 19996. С. 62-64.
Шапошников В.М., Магдеев Д.В., Бакиев А.Г. и др. Красная книга Самарской области:
земноводные и пресмыкающиеся И Вопросы герпетологии (Матер. I Съезда герпетол. о-ва им.
А.М. Никольского). Пущино; Москва, 2001. С. 341-342.
Шварц С.С., Павлинин В. И., Данилов Н.Н. Амфибии, или земноводные, и рептилии, или
пресмыкающиеся // Животный мир Урала (наземные позвоночные). Свердловск, 1951. С. 9-14.
Шенброт Г.И., АнтоноваГ.С. Сравнительная экология такырной и сетчатой круглоголовок
в условиях их совместного обитания на юге Бухарской области (Узбекская ССР) И Вопросы
герпетологии (Сб. статей). Л., 1981. С. 156-157.
Шенброт Г.И., Семенов Д. В. Современное распространение и систематика круглоголов-
ки-вертихвостки - Phrynocephalus guttatus (Reptilia, Agamidae) // Зоол. журн., 1987. Т. 66, № 2.
С. 259-272.
Шиклеев С.М. Земноводные (амфибии). Природа Куйбышевской области. Куйбышев: ОГИ,
1951. С. 288-289.
Шилов М.Н. Заметки о некоторых рептилиях Северного Приаралья П Тр. Ин-та зоол. АН
КазССР, 1961. Т. 15. С. 170-177.
Шилова С.А. О питании чесночницы в южных лесных массивах // Вопр. герпетологии (Сб.
статей). Л., 1964. С. 77-78.
Шляхтин Г.В. Класс Рептилии. Reptilia // Красная книга Саратовской области. Саратов:
Изд-во «Детская книга», 1996. С. 211-214.
Шляхтин Г.В., Голикова В.Л. Методика полевых исследований экологии амфибий и репти-
лий. Саратов: Изд-во СарГУ, 1986. 78 с.
Шляхтин Г.В., Завьялов Е.В., Табачишин В.Г. Обыкновенный уж (эколого-морфологичес-
кая характеристика)//Актуальные проблемы: Ст. науч. тр. герпетологии и токсинологии. Толь-
ятти, 1996. Вып. 2. С. 54-66.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Распространение пресмыкающихся сем. Vi-
peridae и Colubridae на территории Саратовской области И Тез. докл. I Конф, герпетологов По-
волжья. Тольятти, 1995. С. 61-63.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Распространение ящериц и современное со-
стояние их популяций в северной части Нижнего Поволжья П Тез. докл. II Конф, герпетологов
Поволжья. Тольятти, 1999. С. 66.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Экология питания обыкновенного ужа (Natrix
natrix) на севере Нижнего Поволжья И Современная герпетология: Сб. науч. тр. Саратов, 2005а.
Т. 3/4. С. 111-116.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Экология питания прыткой ящерицы (Lacerta
agilis) на севере Нижнего Поволжья И Современная герпетология: Сб. науч. тр. Саратов, 2006а.
Т. 5/6. С. 93-99.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Водяной уж - Natrix tessellata (Laurenti, 1768)
И Красная книга Саратовской области. Саратов: Изд-во торгово-промышленной палаты, 20066.
С. 369-370.
Шляхтин ГВ., Табачишин В.Г, Завьялов Е.В. Восточная степная гадюка Vipera (Pelias) re-
nardi (Christoph., 1861) И Красная книга Саратовской области. Саратов: Изд-во торгово-про-
мышленной палаты, 2006в. С. 371-372.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В. Характеристика рациона краснобрюхой жер-
лянки Bombina bombina (Linnaeus, 1758), его сезонная динамика на севере нижнего Поволжья //
Современная герпетология: Сб. науч. тр. Саратов, 2009. Т. 9, вып. 3/4. С. 27-32.
Шляхтин Г.В., Рузанова И.Е., Любущенко С.Ю., Завьялов Е.В. К уточнению южной гра-
ницы распространения гадюки Никольского (Vipera nikolskii) на юго-западе России // Вопросы
герпетологии (Сб. статей). Пущино; Москва: МГУ, 2001. С. 347-349.
Шляхтин ГВ., Табачишин В.Г, Завьялов Е.В., Табачишина И.Е. Животный мир Саратовс-
кой области. Амфибии. Рептилии. Саратов: Изд-во СарГУ, 20056. 116 с.
Шляхтин Г.В., Табачишин В.Г, Завьялов Е.В., Табачишина И.Е. Редкие и исчезающие виды
амфибий и рептилий, рекомендуемые к внесению во второе издание Красной книги Саратовс-
кой области // Поволжский экологический журнал. Спец, вып., 2006г. С. 78-83.
Шмальгаузен И.И. Происхождение наземных позвоночных. М.: Наука, 1964. 272 с.
324
СПИСОК ЛИТЕРАТУРЫ
Шошева И.В. Биотопическое размещение и численность амфибий и рептилий Башкир-
ского заповедника И Вопросы герпетологии (Матер. VI Всесоюз. герпетол. конф.). Л., 1985.
С. 237-238.
Щербак Н.Н. Ящурки Палеарктики. Киев: Наук, думка, 1974. 246 с.
Щербак Н.Н. Основы герпетогеографического районирования территории СССР И Вопро-
сы герпетологии (Сб. статей). Л., 1981. С. 157-158.
Щербак Н.Н. Ареал // Разноцветная ящурка. Киев: Наук, думка, 1993. С. 9-21.
Щербак Н.Н., Голубев М.Л. Новые находки земноводных и пресмыкающихся в Средней
Азии и Казахстане // Вести, зоологии, 1981. Вып. I. С. 70-72.
Щербак Н.Н., Голубев М.Л. Гекконы фауны СССР и сопредельных стран. Определитель.
Киев: Наук, думка, 1986. 232 с.
Щербак Н.Н., Котенко Т.Н, Тертышников М.Ф. Разноцветная ящурка. Киев: Наук, думка,
1993.238 с.
Щербиновский Н. Дневники Самарской природы 1916 года. Самара, 1919. 146 с.
Эверсман Э. Естественная история Оренбургского края. Вступление в подробную естест-
венную историю Оренбургской губернии или общий взгляд на край Оренбургский в отношении
к произведениям природы (1840) И Оренбургские степи в трудах П.И. Рычкова, Э.А. Эверсмана,
С.С. Неуструева. М., 1949. С. 205-261.
Юшков Р.А. География и экология амфибий и рептилий Камского Предуралья (Пермской
области): Автореф. дис. ... канд. биол. наук. М.: МГУ, 1997. 26 с.
Юшков Р.А., Воронов ГА. Амфибии и рептилии Пермской области: Предварительный ка-
дастр. Пермь: Изд-во Перм. ун-та, 1994. 158 с.
Яковлева И.Д. Пресмыкающиеся Киргизии. Фрунзе: Гылым, 1964. 274 с.
Яхимович В.Л. Корреляция антропогенных отложений Предуралья, Западного и Восточного
склонов Южного Урала // Антропоген Южного Урала. М.: Наука, 1986. С. 140-143.
325
Сибирский углозуб - Hynobius keyserlingii
Обыкновенный тритон - Lissotriton vulgaris
326
Гребенчатый тритон Trituris cristatus
Краснобрюхая жерлянка - Bombina bombina
327
Обыкновенная чесночница - Pelobates fuscus
Обыкновенная жаба Bufo bufo
328
Зеленая жаба - Bufo viridis
Обыкновенная квакша - Hyla arborea
329
Травяная лягушка - Rana temporaria
Остромордая лягушка - Rana arvalis
330
Озерная лягушка Rana ridibunda
Болотная черепаха - Emys orbicularis
331
Среднеазиатская черепаха - Agrionemys horsfieldi
Пискливый геккончик -Alsophylaxpipiens
332
Каспийский геккон - Cyrtopodion caspius
Серый геккон - Mediodactylus russowi
333
Сцинковый геккон - Teratoscincus scincus
Степная агама - Trapelus agilis sanguinolentas
334
Круглоголовка-вертихвостка Phrynocephalus guttatus
Такырная круглоголовка - Phrynocephalus helioscopus
335
Песчаная круглоголовка - Phrynocephalus interscapularis
Ушастая круглоголовка - Phrynocephalus mystaceus
336
Веретеница ломкая - Angius fragilis
Желтопузик, или глухарь - Pseudopus apodus
337
Разноцветная ящурка - Eremias arguta
Линейчатая ящурка - Eremias lineolata
338
Средняя ящурка - Eremias intermedia
Сетчатая ящурка - Eremias grammica
339
Быстрая ящурка Eremias velox
Прыткая ящерица - Lacerta agilis
340
Живородящая ящерица Zootoca vivipara
Песчаный удавчик - Eryx miliaris
341
Восточный удавчик Eryx tataricus
Желтобрюхий (каспийский) полоз - Coluber caspius
342
Поперечнополосатый полоз - Coluber karelini
Разноцветный полоз - Coluber ravergieri
343
Обыкновенная медянка — Coronella austriaca
Узорчатый полоз Elaphe dione
344
Сарматский (Палласов) полоз - Elaphe sauromates
Ящеричная змея Malpolon monspessulanus
345
Обыкновенный уж - Natrix natrix
Водяной уж - Natrix tesselata
346
Стрела-змея Psammophis lineolatus
Обыкновенный щитомордник, или щитомордник Палласа - Gloydius halys
347
Обыкновенная гадюка - Vipera berus
Гадюка Никольского, или лесостепная гадюка Vipera nikolskii
348
Восточная степная гадюка Vipera renardi
349
350
352
358
360
362
364
Научное издание
Петр Васильевич Дебело
Александр Александрович Чибилёв
АМФИБИИ И РЕПТИЛИИ УРАЛО-КАСПИЙСКОГО РЕГИОНА
Серия: Природное разнообразие Урало-Каспийского региона
Том III
Рекомендовано к изданию
ученым советом Института степи
и президиумом УрО РАН
Редактор М.О. Тюлюкова
Технический редактор Е.М. Бородулина
Корректоры Г.Н. Старкова, Н.В. Каткова
Компьютерная верстка Н.С. Глушковой
НИСО УрО РАН № 14(13)—38. Подписано в печать 25.11.13.
Формат 70x100 1/16. Бумага типографская. Печать офсетная.
Усл. печ. л. 32,25. Уч.-изд. л. 32. Тираж 200 экз. Заказ № 548.
Оригинал-макет подготовлен в РИО УрО РАН.
620990, Екатеринбург, ул. Первомайская, 91.
Отпечатано в типографии
«Уральский центр академического обслуживания».
620028, Екатеринбург, ул. Крылова, 27.