/




































































































































































































































































Текст
АКАДЕМІЯ НАУК УКРАЇНСЬКОЇ РСР
ІНСТИТУТ БОТАНІКИ ім М.Г. ХОЛОДНОГО
Ю.Р. Шеляг-Сосонко
ЛІСИ ФОРМАЦІЇ ДУБА ЗВИЧАЙНОГО НА ТЕРИТОРІЇ УКРАЇНИ ТА ЇХ ЕВОЛЮЦІЯ
ВИДАВНИЦТВО «НАУКОВА ДУМКА»
КИЇВ — 1974
634.9 Ш 45
УДК.58І.526.42:582.632. 2(477):58І.55
У монографії вперше для території всієї України подано геоботанічну характеристику 43 асоціацій чистих дубових лісів 8 дуба звичайного. Описується їх поширення, екологічні умови, фітоценотична будова, флористичний склад та продуктивність. Велика увага приділяється висвітленню та розробці теоретичних питань геоботаніки: геоботанічному районуванню, фітоценотичній ролі видів, фітоценотипній будові ареалів, класифікації та еволюції досліджуваних лісів.
Розрахована на геоботаніків, ботаніко-географів, ландшафтознавців, викладачів кафедр ботаніки університетів і педвузів та працівників лісового господарства.
Відповідальний редактор д-р біологічних наук Є.М.Брадіс
Рецензенти
д-р біологічних наук С.С.Харкевич,
д-р біологічних наук С.М.Стойко
Редакція загальної біології
Ш
40502-301
166-74
М221 (04)-74
© Видавництво "Наукова думка", 1974 р.
ВСТУП
Розробка наукових основ охорони та перетворення природи в метою поліпшення природного середовища і кращого використання природних ресурсів розглядається в Директивах XXIV з’зду КПРС по IX п’ятирічному плану (1971 - 1975 рр.) розвитку народного господарства СРСР як одне з основних завдань дальшого наукового прогресу.покликаного забезпечити в новій п'ятирічці значне піднесення матеріального і культурного рівня радянського народу. Необхідною умовою успішного виконання цього завдання є проведення найширших геоботанічних досліджень рослинного покриву країни, особливо гуотонаоеле- них територій,, до яких належить Україна.
Ліси з перевагою дуба звичайного (Quercus robur L.) на території України становлять близько 26% усієї площі лісів республіки, тобто 8432 тис. га. Із заходу на схід масиви .їх про- стлглись на 1300 км, а з півночі на південь на 500 км. Поширені в південній частині широколистяноліоової, лісостепової та в північній частині степової зон дубові ліси займають найбільш круті еро- зійнонебезпечні схили долин, ярів і погорбованих межиріч. Тому вони мають винятково важливе грунтозахисне та водоохоронне значення. Разом з тим, оскільки дубові ліси ростуть на родючих землях і на більшій частині території в головним джерелом деревини, вони, як жодна інша лісова формація, відчули на собі вплив господарської діяльності людини. Забезпечення раціональної експлуатації цих лісів і виявлення найбільш цінних і унікальних в науковому відношенні масивів, які ще збереглися, стає важливим народногосподарським і науковим завданням, що вимагає детального фітоценотичного дослідження.
За ІЗО-річний період вивчення дубових лісів України було опубліковано близько 150 праць. З них майже половина праць належить представникам екологічного напрямку, які не висвітлюють асоціаційного та флористичного складу формації. Не торкаються вони й голов-
3
них теоретичних проблем, поставлених ще в працях А.М.Краснова, Г.І.Танфільева, Й.К.Пачоського, - походження неморальної рослинності, розвитку !!, взаємовідношення зі степовою рослинністю та 1н.
У працях представників фітоценологічного напрямку дубові ліси описано повніше, проте в більшості випадків не наведено повних списків флористичного складу описаних асоціацій. Крім того, вони переважно присвячені вивченню невеликих регіонів. Ці роботи виконані на різному методичному рівні, багато в чому застаріли і не дають достатньо ясного, цілісного уявлення про дубові ліси республіки.
Головним завданням автора було дослідити сучасний стан дібровного ценотичного комплексу України та подати його детальну фітоцено- тичну характеристику. Об'єктом дослідження були дубові ліси з дуба звичайного та ліси, в які він входить як складова частина: грабово- дубові, липово-дубові, сосново-дубові та ясеново-дубові на території всіх областей України.Всього описано 1718 пробних ділянок, з них 1329 були закладені в корінних і тривалопохідних дубових лісах 8 добре виявленим ярусом підліска. Вони й лягли в основу написання монографії. Решту 387 пробних ділянок закладено в змішаних лісах і використано для додаткової характеристики перших. Зібраний фактичний матеріал дозволив виявити і охарактеризувати 43 корінні та ква- зікорінні асоціації дубових лісів. Дослідження провадились деталь- номарщрутним методом з попереднім аналізом наявних в лісгоспзагах матеріалів. Розмір пробної ділянки дорівнював 0,5 га і тільки в деяких випадках, головним чином для реліктових дібров - 0,25 га. Ми згодні 8 думкою ряду дослідників (Грейг-Смит, 1967; Василевич, 1969), що методика геоботанічних досліджень, яка тепер широко застосовується, значною мірою субпєктивна. Ми частково позбавились суб'єктивізму за рахунок використання планових матеріалів лісовпорядження і максимально можливого, часто серійного закладання пробних ділянок. Виділити та охарактеризувати асоціацію на підставі кількох описів ми не вважали за можливе. Проведені нами серійні описи (120- 150 пробних ділянок однієї асоціації) показали, що в ряді випадків у виділених таксонів досить значна синекологічна амплітуда і відносно невелике (21% загальної кількості видів асоціації) флористичне ядро постійних видів.
Значна увага в монографії приділяється також вирішенню теоретичних питань фітоценології, таких, як геоботанічне районування, класифікація, фітоцеяотипи та будова ареалу.
4
ІСТОРІЯ ВИВЧЕННЯ ДУБОВИХ ЛІСІВ УКРАЇНИ
У вивченні дубових лісів України чітко простежуються, на наш погляд, чотири періоди: І) від початку ХУШ ст. і до 1890 р.; 2) з 1890 р. до 1917 р.; 3) з 1917 р. до 1930 р.; 4) з 1930 р. до наших днів. Розглядаючи роботи, що належать до того чи іншого періоду, зупинимось тільки на огляді найбільш важливих праць, пов”язаних з вивченням дубових лісів.
Перший період
Дослідження в цей час мали епізодичний характер і охоплювали невеликі території. Одержані дані стосувались найбільш загальних питань флори та господарського стану лісів. Необхідно відзначити праці Фогеля (1836), Г.Білецького (1840), В.М.Черняєва (1858) та В.М.Туцевича (1880).
В праці Фогеля висвітлена загальна характеристика поширення лісів в повітах Волинської губернії та розглянуто їх господарський стан. Автор відзначає, що дубові ліси є переважно в південній частині губернії, і обмежується лише загальними зауваженнями, наприклад: ”В Волинській губернії ростуть переважно дубові Й соснові ліси, між якими знаходяться клен, ясен, в"яз і зрідка ялина” (с.223).
Г.Білецький описує місцерозташування, межі, рельєф, грунти і переважаючі породи окремих дач Кролевецького повіту Чернігівсько! губернії. Ліси, як і в попередній праці, характеризуються дуже стисло.
Значним кроком вперед є праця В.М.Черняєва (1858), в якій розрізняються дубові ліси, створені ранньою та пізньою формами дуба, звертається увага на поширення їх в зв“язку з рельєфом місцевості та характером грунтів. Автор вже виділяє в дубових лісах високостовбурні дерева, дерева середньої висоти, низькі дерева та чагарникові породи. Вперше зупиняється на супутниках дуба.
В цілому праці першого періоду мають значення переважно для історії науки.
Другий період
Дослідження цього періоду планомірніші, зростає й кількість їх. Вони становлять для нас інтерес насамперед з точки зору загальної характеристики місцевості, оскільки разом з рослинністю подано не менш розгорнутий огляд геологічної будови, рельєфу та грунтів. в деяких з них вже є перші спроби не тільки загального опису
5
широколистяних або дубових лісів в цілому, але й виділення окремих асоціацій або типів.
Цей період починається відомими монографіями А.М. Краснова (1893, 1894а), в яких автор поряд з характеристикою степової рослинності дає загальне уявлення про широколистяні ліси Лівобережного лісостепу України. А.М.Краснов вперше чітко висловив думку про тісний взаємозв'язок широколистяних лісів з рельєфом місцевості. Він писав: "Високі праві береги річок, добре дреновані системами ярів і балок, заселені лісовою рослинністю. Ця остання найбільш багата в піднятих пунктах краю. Навпаки, чим рівніша місцевість, чим менш порізана ярами та балками, чим ближча до рівня Дніпра, тим різкіший і характерніший її степовий характер" (1894а, с.48).
Велику увагу цим питанням приділяє також Г.І.Танфільєв (1894, 1896, 1902). В першій праці він наводить списки видового складу Чорного лісу (тепер Чорноліоький лісгоспзаг Кіровоградської області) та заплавних лісів р.Деркулу. Так само як А.М.Краснов, Г.І.Тан- фільєв приходить до висновку про зв"язок лісів з піднесеними вододілами та високими, порізаними ярами берегами річок. На його думку, відсутність лісів в степу і на солонцях пояснюється одним - засоленістю грунтів. В наступних працях Г.І.Танфільєв продовжує розвивати погляд про приналежність лісів до верхівців річок і, крім того, вперше відзначає для лісів Леонтьєвого байрака (Торезь- кий район Донецької області) граб звичайний (Oarpinus betuius L.). Надзвичайно важливим для розуміння історичного розвитку лісів є висунуте ним положення, що сучасні ліси є збіднілими залишками пліоценових лісів, які пережили льодовиковий період на території сучасного лісостепу. Висловлювання А.М.Краснова і Г.І.Танфільєва і досі не втратили свого значення.
У 1901 і 1913 рр. з"являються праці В.І.Талієва, в яких подається загальний флористичний склад окремих масивів дубових лісів в межах сучасних Харківської та Ворошиловградської областей, а також їх весняний аспект. На відміну від А.М.Краснова і Г.І.Танфіль- єва, В.І.Талієв вважав, що в цих районах степ сформувався на місці лісу, знищеного людиною. Топографічні умови, на його думку, прискорюють або сповільнюють цей процес.
Цікавою в геоботанічному відношенні є стаття П.Прокорова (1905), яка, крім флористичного списку, містить ще й типи дубових насаджень Чорного лісу - рівнинного лісостепу, пологих схилів, великих хвилястих схилів і тальвегів. У 1907 р. характеристику б
цього ж лісу дав В.Доктуровський. Його праця є зразком найбільш повного геоботанічного опису даного періоду, тому наведемо невеликий уривок з неї: “Тут старе насадження складається з дуба звичайного (Quercus robur),береста (Ulmus campestris), клена польового (Acer campestre) і граба звичайного (Carpinus betulus), В надзвичайно густому й високому підліску трапляються ясен звичайний (Fraxinus excelsior), КЛЄН звичайний (Acer platanoides), клен польовий (A. campestre^ клен татарський (a. tataricum), берест (Ulmus campestris) , лідина звичайна (Corylus avellana), бруслина європейська (Kuonymus europaea). Трав’яниста рослинність має буйний характер. Взагалі помічається перевага переліски багаторічної (Mercurialis perennis), купини лікарської (Polygonatum officinale), зірочника лісового (Stellaria holostea), копитняка європейського (Asarum europaeum), маренки запашної (Asperula odo- rata)” (с.235). .
В 1913 р. з’являється важлива в теоретичному відношенні праця Г.М.Висоцького "Про діброви Європейської Росії та їх області", де вперше обгрунтовано поділ дубових лісів на області Й райони та подано їх класифікацію. Г.М.Висоцький виділив чотири області дубових лісів (Заволзьку, Центральну, або Середню, Задніпровську та Південну передгірну), кожну з яких поділив на три райони. На такому ботаніко-географічному фоні він розрізняє десять формацій дубняків: І) пристеповий дубняк; 2) підборовий пристеповий дубняк; 3) дубняк з чагарниковим підліском; 4) липбвий дубняк; 4а) липовий дубняк з ясенем; 5) підборовий липо-дубняк;, 6) польовокленовий дубняк; ?) грабовий дубняк; 8) гірський дубняк; 9) куртинний дубняк або гирнець; 10) плавневий або в’язовий дубняк. Класифікація Г.М.Висоцького географічних областей дубових лісів не втратила свого значення і на сьогодні.
В 1914 р. виходить праця Г. Степуніна з переліком видового складу деревостою і травостою лісів у верхів’ях р. Міуоу (Торезь- кий лісгоопзаг Донецької області), які належать до трьох типів: тальвегів, плато та ярів.
Завершують другий період праці Й.К.Пачоського (1910, 1914, 1915). В двох перших автор не виділяє асоціацій дубових лісів, обмежується лише загальною короткою характеристикою окремих їх ділянок. У роботі 1915 р. Й.К. Пачоський всі ліси Херсонської губернії залежно .від їх місцезнаходження поділяє на два тили: ліси плато і ліси знижених місць. Серед першого типу він розрізняє чор7
нолісся та діброви, а серед другого - ліси високих схилів, ліси надзаплавної другої тераси та ліси заплав. На підставі наведених в книзі описів Чорного лісу та деяких ділянок лісів Поділля мохно встановити, що йдеться про діброви ліщиново-яглицеву, ліщиново-зі- рочникову та польовокленово-зірочникову. З низки основних положень, висунутих Й.К.Пачоським , для вирішення ряду проблем, повиваних з формуванням дубових лісів, трансгенезом їх та взаємовідношеннями із степової) рослинністю, особливо важливими, на наш погляд, є такі: І) флора Поділля, Бесарабії і власне лісостепу є автохтонною і вихідною для- формування рослинності Полісся і степової зони; 2) вона має третинний (пліоценовий) вік; 3) під час льодовикового періоду ліси не були цілком знищені, а збереглись у захищених місцях Поділля, Бесарабії та Придніпровської височини; 4) найбільш універсальною причиною безлісся степів є боротьба між трав'янистою і лісовою рослинністю; 5) появу всієї рослинності в даній місцевості можна зрозуміти лише виходячи з розвитку та зміни попередньої флори.
Таким чином, дослідження другого періоду мають значення в зв"язку з вивченням питань загальнотеоретичного характеру. Описів конкретних ділянок дубових лісів було дуже мало і виконані вони на методичному рівні, який не відповідав сучасним вимогам.
Третій період
Після перемоги Жовтневої революції і встановлення на Україні Радянської влади починається розгортання планомірних досліджень дубових лісів, головним чином науковими силами товариств природознавців і кафедр ботаніки. В цей час на Україні поряд з фітоцено- тичним напрямком сформувався екологічний. Ці напрямки базуються на різних принципах та методиці, тому розглянемо їх окремо .
Фітоценотичний напрямок. Саме представники цього напрямку продовжили розробку широких теоретичних проблем, поставлених у працях А.М.Краснова, Г.І.Танфільєва, Г.М.Висоцького і Й.К.Пачоського.
Однією з перших з'явилась стаття Є.М.Лавренка (1925), в якій вказувалось на зростання граба в Міуоькому лісництві (Торезький лісгоспзаг) та подано характеристику дубових лісів, представлених
1 Для зручності екологічний напрямок розглянуто в четвертому періоді. 8
такими асоціаціями, дібровами: польовокленово-перлівковою (Q. aceroso (campestris) - meIleosum (pictae) , татарськокленово-грясти- цевою ( Q. aceroso(tararici)-dactilosun) і татарськоклеяово- горобейниковою (Q. aceroso (tararlci)-lithospermosuM). В Пізніших працях цього періоду головна увага приділяється закономірностям розподілу, віку і географічному аналізу флори лісів, серед них і дубових (Лавренко, 1926, 1927). По відношенню до рельєфу Є.М.Лав- ренко (1926, 1927) виділяє три типи лісів: байрачні, вододільні та заплавні. На думку автора, вони являють собою збіднілі залишки пліоценових лісів, які перехили льодовиковий період на території сучасного лісостепу, зокрема на Донецькому кряжі.
В 1927 р. виходить праця І.Матківського, в якій подаються короткі відомості загального характеру про байрачні ліси північно- східної частини Одеської області. В цьому а році дві статті опублікував М.І.Котов (1927а, б). Перша з них присвячена описові аспектів дубових лісів околиць Харкова та характеристиці асоціацій дібров ліщиново-зірочникової ( Queraetua coryloso-stellarlosun) і ліщиново-яглицевої ( Q.coryloso-aegopodlosun). в другій праці наводиться опис асоціації діброви орлякової (Q.pteridiosum).
В 1929 р. цей хе автор опублікував статтю, присвячену флористичним особливостям дубових, грабових та інших лісів центральної частини Хмельницької області.
Дві статті у 1930 р. опублікував Є.Юавренко (1930а,б). В першій з них подано загальний огляд рослинності України та геоботанічне районування, в другій розглядаються реліктові центри широколистяних лісів Європи та Азії.
В 1931 р. Г.Ф.Морозов у відомій книзі "Вчення про типи насаджень" наводить опис Чорного лісу за П. Прохоровим (1905).
Таким чином, в третій період, на відміну від другого, збільшується роль вузькорегіональних досліджень. Описи стають повнішими і більш диференційованими, проте це переважають праці, які містять лише флористичні списки окремих масивів дубових лісів. В цей період з'являються узагальнюючі праці О.В.Фоміна (1925), Є.М.Лав- ренка (1926, 1927, 1930а) та Ю.Д. Клеопова (1930). Можна без перебільшення сказати, що вони заклали основи сучасного геоботанічного районування України і розглянули головні проблеми походження, віку та формування її рослинного покриву.
9
Четвертий період
В роботах цього періоду наводяться описи не тільки загального характеру, але вже встановлюються асоціації на підставі сучасної або близької до сучасної методики. Так, у 1933 р. виходить велика стаття Ю.Д.Клеопова з характеристикою таких асоціацій дібров: та- тарськокленово-бугилової ( Q.acerO8o(tatarici)-anthri8cosuni),татар- оькокленово-перлівково! (Q. ас, (tatarici)-meliooeum (pictae), грястицевої ( Q.dactylosum), кінсько-часникової ( Q.al- Ііагісаии), крушиново-лісовокуцоніжкової ( Q.rhamnoso-brachypodio- eum(silvatici), бородавчастобруслиново-тонконогової ( Q.euonymo- 8о(7елгисоаае)-р0О8ит(патоі:а1іа),п0ЛЬ0В0КЛЄН0В0-ЯГЛИЦЄВ0Ї (Q.ac. (campes tris)-.aegopodiosum), польовокленово-бородавчастобруслино- во-маренкової (Q.ас,(campestrisj-euonyrnoao-asperuloaum) та польовокленово-бородавчастобруслиново-грястицевої ( Q.ac.(сашре- strisj-euonymoso-dactylosum), а також ряду асоціацій змішаних дубових лісів. У 1934 р. цей же автор опублікував характеристику асоціацій рослинності Лівобережжя Середньої Наддніпрянщини, в тому ЧИСЛІ Дібров костяницево-розхідникової ( Q.ruboso-glechomosum), татарськокленово-конвалієвої ( Q.ac.(tatarici)-convallariosuin), свидиново-конвалієвої (Q.thelycrardoso-convallariosuja), кропивної ( Q.urticosum), татарськокленово-фіалкової (Q.ac.(tatarici)-violosum), татарськокленово-грястицевої (Q.ac,(tatarici)-dactylosum), ліщи- ново-грястицевої (Q,COrylOSO-dactylosum),BiniHHOBO-KyUOHiMKOBOI (Q,c,-brachypodiosum(silvatici), ліщиново-орлякової (Q.c.-pte- ridiosuia), ліщиново-гірськоосокової (Q.c.-caricosum(wontanae), ліщиново-яглицевої (Q.c.-aegopodiosum), ліщиново-волосистоосокової . (Q,ce~tfaricosuin(pilosae), глідово-грястицевої (Q,crataegoso~da- ctylosum). Кожна з перелічених асоціацій характеризується лише щодо видового складу та грунтів. Відомостей про вік деревостану, висоту його, зімкненість крон, діаметр дерев та проективне покриття травостою немає. Крім геоботанічного опису рослинності, в обох статтях Ю.Д.Клеопов приділяє велику увагу історії її розвитку.
Є.М.Лавренко та Ф.Я.Левіна (1,934) дають коротку характеристику асоціацій дібров орлякової (Qa^terldlosum) та веснівкової (Q.madantbemosum) з ОКОЛИЦЬ ВІННИЦІ.
В 1935 р. виходить велика стаття В.Шафера (Szafer, 1935), в якій автор,користуючись методикою Й.Браун-Бланке, наводить опис асоціації діброви подільської (Q.podolicum) з території Тернопільської області та розглядає історію лісів Західного Поділля.
10
Дві праці присвячено окремим лісовим масивам Вінницької області. Перша з них- Ю.Х.Молотківського (1936) з описом асоціацій діброви ліщиново-волосистоосокової (Q.coryloso-сагісояшп (рііовае), друга- Ф.О.Гриня (1936)з характеристикою асоціацій дібров чагар- никово-гірськоосокової (Q.fruticoso-caiicosuo(Dontanae), ліциново- яглицевої (q.coryloao-aegopodioauin), ліщиново-гірськоосоково-пле- уроцієвої ( Q.c.-caricoswa(tnontanae)-pleurozlo3uni та смовдевої (Q.peucedanosum). Ю.Х.Молотківський вперше на Україні, крім видового складу асоціацій дубових лісів, подав зімкненість дерево- стану, висоту його та бонітет.
В статті Г.І.Потапенка (1937) для Савраноького лісу (Балтський лісгоспзаг Одеської обл.) наводиться коротка характеристика асоціацій дібров польовокленово-волосистоосокової (Q.aceroso(campeatria)- caricosum(pilosae), польовокленово-типчакової (^.ас.(сащревЬгів)- featucosum(sulcatae) та татароькокленово-тонконогово! (Q.ac.(ta- tarici)-poosun(nemojcalia). В цьому Ж році М.Коотинюком та К.Вечореком (Koatyniuk, wieozorek,1937) була описана для околиць Моршина Стрийського району Львівської області асоціація діброви трясучковидноосокової (Q.oaricosuM(brizoidia).
У 1938 р. Ю.Д.Клеопов та Є.М.Лавренкр на'підставі зібраного на той час на Україні матеріалу публікують перелік головних груп асоціацій рослинності України.
У 1940 р. М.І.Котов подав опис дубових (без виділення асоціацій) і дубово-грабових лісів південної частини Вінницької області. У цьому ж році М.І.Котов разом з Є.Д.Карнаух опублікував статтю про рослинність заповідників Донецької області, яка містить коротку характеристику асоціацій діброви чагарниково-трав"янисто! (Q. £ruticoso-herboBum), а також ясеново-дубових та грабово-дубових лісів.
Велика праця проблемного характеру Ю.Д.Клеопова "Основане чертн развития флорн широколиственннх лесов европейской части, СССР* виходить у 1941 р. Геоботанічних описів дубових лісів в ній немає, проте значення її для розуміння еволюції та флористичних особливостей дібров важко переоцінити. Б.Д.Клеопов вперше показав, що флора широколистяних лісів Європейської рівнини СРСР в історико- генетичному відношенні являв собою надзвичайно складний комплекс, в утворенні якого брали участь різні генетичні, історичні, міграційні та ценотичні елементи,
У 1948 р. Г.І.Білик опублікував статтю, яка містить коротку характеристику байрачних лісів між Дністром та Бугом. Він виділив
асоціації чистих дубових лісів: скумлієву ( Q.cotinosum),кизилово- глухокропивну (Q.cornoso-lamiosum(laevigatae), татарськокленово- кропивну (q.ac.(tatarici)-urticosum), татарськокленово-гравілато- ву (q.ac. (tatarici)-geosum), татарськокленово-горобейникову (q.ao,(tatarici)-.lithosperniosua)).
В 1950 р. з’являється докладна стаття Ф.О.Гриня, присвячена лісам Тернопільської області, яка містить характеристику двох груп асоціацій дубових лісів - діброви горобейникової (Querceta lithospermosa) та гірськоосокової (Q. caricos a (montanae). В цьому ж році характеристику асоціації діброви заплавної (Q.inun- datum)3 території Закарпатської області опублікував В.0.Повари Іцин.
Простий перелік асоціацій дубових лісів Львівської області наводить М.Г.Косець (1953). Це діброви:волосистоосокова (Q.caricosum (pilosae), куцоніжкова (Q.brachypodiosum), копитнякова (Q.asaro- sum ), підлісникова (Q.saniculosum), маренкова (Q.asper uloaum), трясучковидноосокова (qecaricosum(brizoidia) та квасеницева ( Q.oxalidosum).
В колективній монографії "Рослинність Закарпатської області УРСР" Ф.О.Гринь (1954) дає детальну характеристику дубових лісів Закарпаття, представлених конвалієво-фіалковою (Q.convallarioso- violosum), трясучковидноосоковою (Q.caricosum (brizoidis) та го- логадючниковою (Q. filipenduloaum (denudati) дібровами. Групи асоціацій автор виділив за екологічними ознаками.
У 1955 р. вийшла стаття В.О.Поварніцина з детальною характеристикою діброви ліщиново-волосистоосокової ( Q.corylosо-сагісо- •um(pilosae) на території Ровенської області. Того ж року була надрукована велика праця А.ЬБарбарича, в якій подано опис трьох асоціацій дубово-грабових лісів на території Полісся.
Стаття Ф.О.Гриня 1957 р. присвячена проблемі лісових реліктів. В ній міститься і коротка характеристика двох асоціацій кленово- липово-дубових лісів Сумської області.
У 1958 р. Д.Я.Афанасьєв подав коротку характеристику дубових лісів заплави Дніпра, серед яких він розрізняє діброви трав’янисту та трав’Яниото-злакову. В тому ж році з’явилась велика стаття К.Ю.Голгофської, присвячена байрачним лісам Деркульоького району Ворошиловградської області. В статті описано такі типи дубняка: кленово-липовий, ясеновий, ясеновий змішано-чагарниковий, липово- бруслиновий, липовий яглицевий, в’язовий кропивний, липовий на терасах, чагарниковий та дубняк на піщаних схилах.
У монографії В.О.Поварніцина "Ліси Українського Полісся" (І959) описано групи асоціацій грабових дубняків - злакового (Q.graminosum), грабового ( Q.oarpinosum^ крушинового ( Q.frangulosum),
12
ліщиново-осокового (Q.coryloso-caxicosum), ліщиново-маренкового (Q.o.-asperulosum), ліщиново-яглицевого (Q.c.-aegopodiosum), зеленчукового (Q.galeobdolosum), орлякового (Q.pteridiosum), яглицевого (Q.aegopodiosum), жіночопапоротевого (Q.dryopterlosum), трав'янистого (Q.herbosum) та групи асоціацій заплавних дібров - рокитникової (Q.cytisosum), злакової ( Q.graminosum), калинової ( q.viburnosum) та кущової ( Q.frutloosum).
Невелика стаття З.Н.Горохової (1960) містить перелік кількох асоціацій дубових лісів Чернівецького лісгоСпзагу - дібров мертво- покривної, волосистоосокової, маренкової, яглицевої та діброви апо- зерисової.
М.А.Бухало (1961) в невеликій статті дав коротку характеристику лісів Золочівського району Львівської області (діброви копит- някова, яглицева, квасеяицева, лісовоосокова). В цьому ж році М.І.Алексієнко описала кленово-дубову з липою, ясеново-дубову та дубову асоціації на території Харківської області.
У 1962 р. О.Н.Моляка подала опис діброви татарськокленово- зірочникової (Q.ac.(tatjarici)-stellariosum) для заплави Росі. Стаття Л.В.Климентова (1962) містить тільки загальновідомі факти і за методичним рівнем відповідав працям другого періоду.
В статті Л.С.Балашова (1963) наводяться описи трьох асоціацій дубових лісів річки Снов, дібров - ліщиново-трав'янисто! (q.coryloso- herbosum), свидиново-ліщиново-трав"янистої (Q.thelycranioso-coryloso- herbosum) та крушинОво-трав"янистої (Q.franguloso-herbosum).
С.О.Мулярчук (1965) подає характеристику дубових лісів Наддес- нянської вододільної рівнини (Чернігівська обл.), представлених дібровою ліщиново-трав"янистою (Q.coryloso-herbosiim). Невелика стаття М.І.Котова (1965) містить опис асоціацій дібров конвалієвої (Q.convallariosum) та ліщиново-яглицевої (Q.coryloso-aegqpodio- sum) на території Ізюмського л1спгоспзагу Харківсько! області.
У 1967 р. опубліковано статтю В.С.Ткаченка, присвячену заплавним лісам Сіверського Дінця. В ній в перелік асоціацій дубових лісів, дібров - конвалієвої, розхідникової, хвилівникової, вербозіл- лєвої, ранньоосокової, костяницевої, підмаренникової, бородачевої та грястицевої. В тому ж році А.І.Кузьмичов навів загальну характеристику дубових лісів Волинської височишцпредставлених дібровами барвінковою та маренковою.
С.М.Стойко (1968) опублікував коротку характеристику дібров волосистоосоково-зірочникової (Q.caricoso(pilosae)-etellarios4m), злакової (Q.graminoBui»), орлякової (Q.pteridioeum), трясучковид-
ноосокової (Q.caricc8uw(br|zoidls), копитняково-маренкової (Q.asaio- so-asper uloaum), конвалієво-фіалкової (Q.convallarioso-violoeuin),
оизоожинової (Q.caesii) та калюжницево-гадючникової (Q.calthoso- tilipendulosum). Стаття С.О.Мулярчука (1968) містить досить детальну характеристику таких асоціацій, дібров - коотяницевої (Q.ruboeum), ліщиново-яглицевої ( Q.coryloao-aegopodiosum); ліщиново-копитнякової (Q.c.-asarosum), ліщиново-медункової (Q.c.-pulmonariosum) та бруо- линово-трав'Янистої (Q.euonymoso-herbosum),
1969 р. з'являється стаття О.П.Мринського, де є загальна характеристика дубових лісів північно-західної частини Полтавської області, представлених Дібровами ліщиново-яглицевою (Q.ooryloso-aego- podioeum), ліщиново-Зірочниковою (Q.c.-stellariosum) та ліщиново- трав'Яниотою (Q.o.herbosum). С.О.Мулярчук та Л.С.Балашов (1969) подали характеристику таких асоціацій заплавних лісів Чернігівської області,дібров - ліщиново-конааліевоІСЧ.о.-соотаІіагіовиш), ліщино- во-трав"яниотої (Q.o.herbosum) та овидиново-трав'Янистої (Q.thelycra- nioso-herboautn). Праця С.М.Стойка (1969) містить загальні відомості про дубові ліси Карпат а переліком їх асоціацій, а також ряд теоретичних положень щодо їх класифікації, поширення та районування. Виділені цим автором асоціації М.А.Голубець та К.А.Малиновоький (1969) наводять в статті, присвяченій класифікації рослинності Українських Карпат.
С.О.Мулярчук (1970) в монографії про рослинність Чернігівської області вміщує опис асоціацій дубових лісів, наведених у згаданих вище роботах цього автора.
У 1971 р. вийшла колективна монографія по вивченню лісів Ук- раїни^В ній Ф.О.Гринь на підставі своїх попередніх праць подав детальний, з переліком флористичного окладу, опис груп асоціацій дубових лісів: горобейникових (Querceta libhospermosa), фіалкових (Q.ylolosa), яглицевих (Q.aegopodiosa), гірськоосокових (Q.cari- еosa(montanae), орлякових (Q.pteridiosa) та Тряоучковидно-
осокових ( Q.oarioosa(brisoidis),
Неважко помітити, що розглянуті нами праці можна поділити на три групи. В працях першої групи,крім подання фактичного матеріалу, розв'язуються важливі теоретичні питання. Таких праць небагато (Клеопов, 1933, 1934, 1941; Згаїег, 1935; Клеопов, Лавренко, 1938; Гринь, 1957, 1971; Голубець, Малиновоький, 1968, 1969; Стойко, 1969). До другої групи належать праці, що містять загальні відомості про дубові ліси або короткі описи окремих асоціацій (таких більшість), і до третьої - де подано більш детальну характеристику асоціацій дубових лісів.
14
Характеристики асоціацій дубових лісів, наведені в перелічених працях, хибують на відсутність повних списків видового складу асоціацій. Перелічуються, як правило, лише види, що часто зустрічаються, а видів характерних, але менш рясних, немає. Такі матеріали дають деяке уявлення про асоціацію, але не дозволяють аналізувати II. Повні списки рослин наведено тільки в кількох працях (Клео- пов, 1933; Szafer, [935; Гринь, 1936, 1971; Koatyniuk.Wieczorek, 1937; Котов, 1940; Поварніцин, 1959). Слід зазначити, що відсутність в більшості праць даних про оклад деревостану 1 особливо підліска виключав можливість порівняння виділених асоціацій.
З методичного боку необхідно відзначити відсутність загальноприйнятого критерію для виділення асоціацій si змішаним дерево- станом. В.0.Поварніцин (1959), наприклад, до грабових дібров відносить насадження, в складі деревостану яких граб трапляється поодиноко або становить 30-50%, тоді як С.О.Мулярчук (1968) насадження, в яких участь граба незначна, до грабових дібров не відносить. Те саме спостерігається і при виділенні в"язових, ясенових, липових та інших дібров. Крім того, слід відзначити, відсутність єдиного критерію для виділення асоціацій та єдиного обсягу таксонів, які виділяють. Так, К.О.Голгофська (1958) виділяє асоціації за характером деревостану, характером підліску або травостою і навіть - рельєфу..
Як правило, фітоценотичну роль видів деревного ярусу автор не бере до уваги і вважається, що для виділення нової асоціації досить присутності або відсутності видів. За такою ж ознакою виділяються й групи асоціацій. О.Н.Моляка (1962) виділяв асоціації в залежності від домінантних видів усіх ярусів і співдомінантних видів головного. С.О.Мулярчук та Л.С.Балашов (1969) враховують, крім, того, співдомінанти і другорядних ярусів. В.О.Поварніцин (1955, 1959) виділяє асоціації а врахуванням домінантних видів усіх ярусів або двох-трьох. М.І.Кооець (1953) бере до уваги домінантні види лише деревного та травпянистого ярусів. Не дотримується одного принципу й Ф.О.Гринь. Все це призводить до того, що скласти уявлення про ранг описуваних таксонів немає змоги. Даних, які’ свідчили б про кількість пробних ділянок, на підставі яких подано описи асоціацій, автори, як правило, не наводять. Для ряду праць характерний низький методичний рівень. Наприклад, М.І.Алексєєнко, характеризуючи байрачні ліси, пише: "Досліджені нами ліси... представлені асоціаціями, в складі яких характерні для південної експозиціїїбиег- cu0 robur, Ulmus foliacea, Pyrus communia; в підліску: Acer tataricum, види бруслини... В трав"янистоМу покриві: Роа nemoralia,
15
Viola odorata, Ballota nigra, Urtioa dioica та ін. На менш сухих схилах (північна експозиція і по тальвегах балок) в складі асоціацій: Quercus robur, fraxinus excelsior, Tilia cordata..."(1961, 0.378). Які асоціації описано і що вони собою являють, лишається незрозумілим. Подібним схематизмом позначені описи лісової рослинності К.Ю.Голгофської (1958). Особливо показова в цьому відношенні стаття 1.В.Климентова (1962), в якій характеристику лісів обмежено переліком кількох звичайних видів деревостану та чагарників, а травостій не згадується. В більшості праць групи асоціацій не виділено або вони виділяються на підставі різних показників, тому доводиться мати оправу з таксонами різного обсягу навіть у одного й того ж автора. Наприклад, В.О.Поварніцин (1959) виділяє групи асоціацій на підставі типу грунту, геоморфології території або рослинності.
Екологічний напрямок. Виник в зв'язку з практичними потребами лісового господарства. Головними таксономічними одиницями лісівницької класифікації, за Д.В.Воробйовим (1953, 1967), є типи - лісової ділянки, лісу і деревостану. Асоціація розглядається як допоміжна одиниця і практично не виділяється. Найголовнішою ознакою для визначення типу лісу і лісової ділянки та їх виділення є рослини-індикатори (Воробйов, 1967). Рослинність виділених таксонів представники цього напрямку описують скорочено. Все це робить неможливим (за незначним винятком) встановлення асоціацій на підставі таких матеріалів.
Основоположник екологічного напрямку Є.Б.Алексєєв розробив систему типів лісу, опубліковану 1925 р. За цим автором, усі суходольні типи лісу об'єднуються в чотири групи: бори, субори, груди і діброви. До- грудів він відносить змішані широколистяні насадження з двоярусним деревоотаном на сірих грунтах, а до дібров - широколистяні з одноярусним деревостаном на деградованих чорноземах.
З'являються праці про ліси Полісся Д.В.Воробйова (1927, 1928) та П.С.Погребняка (1927, 1928), в яких вони поряд з іншими лісами описують свіжий, вологий та сирий груди.
У 1931 р. виходять друком праці про ліси Поділля П.С.Погріб- няка і П.П.Кожевникова. Крім грудів, для цієї території вони описують сухі, свіжі та вологі в'Язові діброви, що відрізняються від грудів трав'янистим покривом та відсутністю граба.
Великий інтерес викликають праці П.П.Кожевникова (1936, 1937, 1939), в яких висвітлюється характеристика берестових та кленово- дипово-дубових лісів.
Робота 0.1.Бельгарда (1939) присвячена описові байрачних лі-
16
сів, розташованих на південний схід від Дніпропетровська. В наступні роки автор неодноразово повертається до вивчення лісів південно-східної частини України (Бельгард, 1940, 1950, 1960), він вважається кращим їх знавцем.
З наведених у перелічених роботах описів видно, що О.Л.Бельгард описав діброви: ліщиново-зірочникову, ліщиново-яглицеву, лі- щиново-грястицеву, ліщиново-волосистоооокову, ліщиново-конвалієву, кленово-розхідникову, кленово-конвалієву, кленово-грястицеву, кле- ново-куничникову, кленово-волосистоосокову, кленово-фіалкову, кле- ново-дібровнотонконогову, терново-фіалкову та крушиново-молінієву. Встановлені цим автором таксони мають детальну екологічну та найбільш детальну фітоценотичну характеристику, в зв"язку з чим праці його можна віднести і до фітоценотичного напрямку.
У 1941 р. вийшла узагальнююча монографія П.С.Погребняка "Типологія і екологія лісу", перевидана 1955 р.в ширшому обсязі під назвою "Основи лесной типологии". В цих працях автор об"єднує груди і діброви, за Є.Б.Алексєєвим (1928), в групу дібров - дуже сухих, сухих, свіжих, вологих і сирих.
Велика праця О.Б.Іукова (1949) присвячена переважно лісогосподарській характеристиці дубових лісів України.
У 1953 р. опубліковано монографію Д.В.Воробйова, де він дає загальну схему типів лісів європейської частини СРСР та їх характеристику. Ця праця є значним досягненням екологічного напрямку, так само як і праці Є.Б.Алексєєва (1928) та П.С.Погребняка (1955), вона значною мірою сприяла його розвитку.
У 1954 р. Д.Д.Лавриненко повторює схему, запропоновану П.С.По- гребняком.
Серед робіт, присвячених типології лісів окремих регіонів України, слід відзначити праці О.М.Гаврусевича (1959), В.В.Осич- нюка (1960), Б.Ф.Остапенка (1961), В.М.Данька (1961, 1964, 1965), В.В.Воробйова, І.П.Федця (1968), І.П.Федця, М.С.Улановського (1968) та ін.
Монографія "Ліси Української РСР" С.С.П"ятницького та П.П.Ізюм- ського вийшла друком 1965 р. Один з головних розділів її - "Типи лісу" - фактично повторює відомі вже матеріали і не позбавлений ряду помилок, про іцо ми вже писали (Брадіс, Шеляг-Сосонко, 1968).
Загалом праці екологічного напрямку у відношенні вивчення рослинності характерні тим, що: І) описи рослинності обмежуються коротким схематизованим переліком звичайних, видів без урахування І?
їх фітоценотичної ролі; 2) наведені в списках види в індикаторами родючості та ступеня зволоження грунту і служать основою для виділення типів лісу. Звичайно перелічують характерні (Погребняк, 1955), найбільш звичайні або переважаючі (Воробйов, 1953) види. Проте, на наш погляд, ступінь індикаторної цінності видів доцільно було б визначати залежно від трапляемості, рясності та регіональних особливостей.
Праці цього напрямку мають важливе практичне значення для лісового господарства республіки.
X X
X
Отже, якщо розглядати асоціацію як таксономічну одиницю, яка об"єднуе однорідні за своїми істотними рисами фітоценози, виділяючи при цьому короткочасні сукцесійні стадії в ранзі неоасоціацій, то всі 85 асоціацій чистих дубових лісів, відбитих у літературі, на підставі наведеного вище аналізу можна поділити на п"ять груп.
До першої групи ввійдуть таксони, встановлені на підставі короткочасних сукцесійних стадій рослинності. Як правило, в перелічених працях вказівок на походження їх немає. Це такі умовні асоціації (неоасоціації), діброви: травпяниста, мертвопокривна, апозериоова, розхідникова, бородачева, вербозіллвва, барвінкова, смовдева, кропивна, койвалієво-фіалкова, ранньоосокова, трав'янисто-злакова, грястицева, кінськочасникова, ліщиново-трав"яниота, ліщиново-орлякова, бруслиново-трав"яниста, бруслиново-дібровно- тонконогова, крушиново-куцоніжкова, татарськокленово-дібровнотон- коногова ,татароькокленово-бугилова, татарськокленово-грястицева, татароькокленово-фіалкова, татарськокленово-куничникова, татар- ськокленово-кропивна, костяницево-розхідникова, свидиново-трав"я- ниота, кизилово-глухокропивна, глідово-грястицева, орлякова, тра- в"яниото-гілокомівва, терново-степо-кущова, кущово-трав"яниста, кущово-гравілатова, кущово-кропивна, кущово-костяницева, кущово- злакова, різнотравно-кущово-злакова, різнотравно-кущово-типчако- ва і терново-фіалкова. До цієї ж групи належать майже всі типи, описані К.Ю.Голгофською (1958). Як показали наші дослідження, всі фітоценози без ярусу підліска, без виразної переваги одного-двох видів у травостої, з перевагою в підліску бруслини бородавчастої (Kuonyaus verrucosa flcop.), терену звичайного (Prunus spinosa L.), Глоду одноматочкового (Crataegus aonogyna Jacq.) або в тра-
18
востої - грястиці збірної (Dactylis glomerata L.)t тонконога дібровного (Роа nemoralis L.), вербозілля лучного (Lysimachia nummularia L.), КІНСЬКОГО часнику звичайного ( АІІІагІа petiolata (М. В.) С a v a r a et Grande), бугили ЛІСОВОЇ (Anth- riscus silvestris (Le) H o f f me), розхідника звичайного ( Glechorna hederacea L.), кропиви ДВОДОМНОЇ (Urtica dioica L.), глухої кропиви гладенької (Lamium laevigatum L.)t орляка звичайного (Pteridium aquilinum K u h n.), фіалки шершавої (Viola hirta L.), осоки ранньої (Carex praecox S c h r e b.) та КОСТРИЦІ борознистої (Festuca sulcata Hack*) мають антропогенне походження.
До другої групи ввійдуть таксони неясного таксономічного рангу, встановити його неможливо через неповноту описів. До цієї групи відносяться такі умовні асоціації, діброви: трясучковидно-осо- кова, гологадючникова, крушинова, зеленчукова, яглицева, жіночопа- поротева, кущова, волосистоосокова, маренкова, конвалієва, хвилівни- кова, костяницева, веснівкова, горобейникова, подільська, кущово- гірськоосокова, кущово-кореневищноосокова, скумпієва, заплавна, ква- сеницева і копитнякова. Ця група, безумовно, неоднорідна. В ній можна передбачити такі варіанти: а) під виглядом асоціацій описано таксони вищого рангу; б) описані таксони відповідають рангові асоціації; в) в ранзі асоціації описано сукцесійні стадії. Найможливі- шим прикладом першого варіанту є діброва подільська, другого - діброва трясучковидноосокова і третього - діброва кущова. Баз детального опису асоціації або хоч переліку переважаючих видів ярусів не можна встановити, що являє собою виділений таксон. Наприклад, діброву конвалієву можна розглядати як сукцесійну стадію (якщо немає підліска) або як різні асоціації (діброви крушиново-конвалієва, татарськокленово-конвалієва та ін.).
До третьої групи ввійдуть таксони, встановлені на підставі фрагментів асоціацій. Це такі умовні асоціації, як діброви ліщино- во-медункова, лісовокуцоніжкова, підлісникова, калинова.
До четвертої групи ввійдуть таксони в ранзі субасоціацій, що розглядаються в літературі як асоціації. Наприклад, діброви ліщино- во-яглицево-волосистоосокова, ліщиново-гірськоосоково-плеуроцієва, копитняково-маренкова і калюжницево-гадючникова. Останні дві оубасо- ціації мають до того ж неясні таксони.
В п"яту групу ввійдуть таксони, що відповідають своєму рангові і встановлені на достатньому методичному рівні. Це такі асоці19
ації, діброви: крушиново-молінієва, ліщиново-маренкова, ліщиново- копитнякова, ліщиново-зірочникова, ліщиново-яглицева, ліщиново-воло- систоосокова, ліщиново-конвалієва, ліщиново-гірськоосокова, та- тарськокленово-зірочникова, татарськокленово-горобейникова, татар- ськокленово-конвалієва, татарськокленово-волосистоосокова, татар- ськокленово-перлівкова і свидиново-гірськоосокова. Відомостей про поширення їх по території всієї України та про регіональні особливості їх в літературі немає.
Таким чином, з 85 асоціацій чистих дубових лісів, описаних за останні 65 років, 42 являють собою короткочасні сукцесійні стадії, 21 має неясний таксономічний ранг, 4 - фрагменти t ще 4 мають фактично ранг субасоціацій; лише 14 асоціацій можна вважати корінними асоціаціями дубових лісів, що відповідають своєму рангові.
Ми виділили та охарактеризували 43 корінні асоціації дубових лісів з дуба звичайного, діброви: крушиново-трясучковидноосокова ( Quercetum franguloso-caricosum(brizoidis), крушиново-молінієва (Q.franguloso-moliniosum), крушиново-конвалієва (Q.fr angui os о- convallariosum), ліщинбво-трясучковидноосокова (Q.coryloso-carico- sum(brizoidis), ліщиново-квасенішвва ( Q.coryloso-oxalidosum), ліщиново-чоловічопапоротева ( Qtcoryloso-dryopteriosum(filicis masis) ліщиново-жіночопапоротева (Q.coryloso-athyriosum(filicis
feminae), ліщиново-веснівкова (Q.coryloso-madanthemosum), Ліщиново-Зірочникова (Q.coryloso-stellariosum), ліщиново-копитня- кова (Q.coryloso-asarosum), ліщиново-маренкова (Q.coxyloso- asperulosum), ліщиново-яглицева ( Q.coryloso-aegopodiosum), ліщи- ново-зеленчукова (Q.coryloso-galeobdolosum), ліщиново-волосисто- осокова (Q*ooryloso-caricosum(pilosae), ліщиново-гірськоосокова (Q,coryloso-caricosum(montanae)9 ліщиново-конвалієва ( Q.coryloso- convallariosum), ліщиново-парвськоосокова (Q.coryloso-caricosum (brevicollis ),ліщиново-горобейникова (Q. coryloso-lithospermosum), свидиново-зірочникова (Q^thelycranioso-stellariosum), свидиново- яглицева (Q.tbelycranioso-aegopodiosum), свидиново-гірськоосоко- ва ( Q.thelycranioso-caricosum(montanae), СВИДИНОВО-КОНвалієва (Q<thelycranio8o-oonvallarioeum),CBHflHH0B0-iiapBCbK00C0K0Ba ( Q.the- lyoranioeo-oaricoswn (brevicollie), татарськокленово-зіроч- никова (Q.aceroso(tatarici)“Stellariosum)9 татарськокленово-ягли- цева ( Q.aceroso(tatarici)Taegopodiosum), татарськокленово-волосисто- осокова (Q.aceroso(tatarici)-carioosuiD (pilosae), татарськокле- ново-кореневищноосокова ( Q.aceroso(tatarici)-caricosum(rhiziiiae), 20
татарськокленово-гірськоосокова (4.aceruso(tatarici)-cariaoeum (montanae), татарськокленово-конвалієва ( q,aoeroso(tatariai)- convallariosum), татарськокленово-рябоперлівкова ( Q.aceroso(tata- rici)-melicosum (pictae), татарськокленово-горобейникова (Q.aceroso (tatar ici) -li t hosper mosum), татарськокленово-м І кал І євоооокова (Q. aceroso (tatarici)~carico8um(michelii), кизилово-парвськоосокова (Q.cornoso-caricosum(brevicolLis), кизилово-зірочникова (Q.cornoso- stellariosum), кизилово-конвалієва (Q.cornoso-convallariosum), кизилово-волосистоосокова (Q.cornoso-caricosum(pilosae), кизи- лово-кореневищноосокова ( Q.cornoso-curicosum(rhizinae), кизилово- горобейникова ( Q.cornoso-lithospexmosuiD), кизилово-мікелієвоосо- кова ( Q.cornoso-caricosum(michelii), скумпієво-конвалієва (Q.oo- tinoso-convallariosum), скумпієво-рябоперлівкова (Q.cotinoso-
melicosum(pictae), скумпієво-пірчастокуцоніжкова (Q.cotinoso- brachypodiosum(pinnati) t скумпієво-мікелієвоосокова (Q.cotinoso- caricosum(miohelii).
Підводячи підсумки вивчення дубових лісів України за 130 років, необхідно відзначити таке.
Праць, спеціально присвячених вивченню дубових лісів всієї України, в літературі немає. Описуються ліси окремих територій, що не охоплюють навіть головних природно-історичних районів України. Наприклад, немає зовсім даних про чисті дубові ліси Західного і Правобережного Полісся України, майже не вивчені дубові ліси Придніпровсько! височини, практично відсутні дані про дубові ліси других терас лісостепу України. Саме тому в узагальнюючих працях дубові ліси не знайшли належного висвітлення.
Згадані роботи написані з різною метою і на різному теоретичному та методичному рівні. Звичайно автори не мали єдиного погляду на найбільш загальні методичні і теоретичні питання. Все це ускладнює або робить неможливим інтерпретацію зібраних матеріалів.
Спостерігається поступовий відхід від вирішення проблемних питань, накреслених в працях А.М.Краснова, Г.І.Танфільєва, Й.К.Па- чоського, Є.М.Лавренка та Ю.Д.Клеопова.
ПРИРОДНО-ІСТОРИЧНІ УМОВИ території дослідження
Українська РСР займав південно-західну частину Російської рівнини, Українські Карпати та Кримські гори. На півночі вона межує з Білоруською РСР, на сході - з Російською РСР, на південному заході - з Молдавською РСР, а на заході - з Польщею, Чехооловаччиною.
21
Румунією та Угорщиною. Загальна площа її дорівнює 603 тис.км2.
Географічне положення України зумовило надзвичайну різноманітність її природних умов, зокрема й рослинності. Характер та поширення дубових лісів залежить насамперед від клімату, геологічної будови, рельєфу та грунтів.
Клімат
Клімат, як відомо, зумовлює зональність рослинного покриву. На Україні з півночі на південь дубові ліси послідовно беруть участь у створенні рослинного покриву широколиотянолісової, лісостепової та північної частини степової зон, таким чином, через клімат поширення дубових лісів обмежується тільки в південному напрямку. В межах кожної зони формується певний набір асоціацій дубових лісів. Найчіткіше це видно в плакорних лісах. Наприклад, на Правобережній частині України ллакорна асоціація широколистянолісової зони - діброва ліщиново-волосистоооокова - в плакорних умовах лісостепової зони змінюється на асоціацію діброву свидиново-гірсько- осокову. В межах окремих регіонів формування та топографія асоціацій визначаються на загальному кліматичному фоні мікрокліматом, в свою чергу зумовленим рельєфом місцевості.
Клімат впливає на формування певних асоціацій дубових лісів та їх продуктивність, що виявляється не тільки в широтному, але й у меридіанальному напрямку. Так, з посиленням континентальності клімату асоціація діброва свидиново-гірськоосокова, поширена в плакорних умовах західного лісостепу, змінюється на тій самій широті у східній частині лісостепу на асоціацію діброву ліщиново-волосисто- ооокову. Дуже чітко вплив клімату простежується і на флористичному складі дубових лісів. При просуванні з півночі на південь дубові ліси поступово бідніють на види бореального клімату, до яких належать рамішія однобока (Hamisohia secunda (L.) G a r c k e}, грушанка круглолиста (Pyrola rotundifolis L.), веснівка дволиста (Majanthemum trifolium (L.)F.w.s c h в i d t). При просуванні з заходу на схід вникають види атлантичного та субатлантичного Клімату: бук лісовий (Fagus silvatica Ь.), скополія карніолійська (Scopolia carniolica J а о k ,), черешня (Cerasus avium Moeno h.), граб звичайний (Carpinus betulus L.), клен несправжньо- платановий (Acer pseudoplatanus L.), апозерис смердючий (Aposeris foetida (L.) Cass) та ін. Відповідно в першому випадку спостерігається проникнення видів аридного клімату - осоки Мікелі (Carex
22
mlchelii Host.), залізняка бульбистого (РЬІошіа tuberosa L.) та ін., а в другому - видів континентального клімату - скереди сибірської (Crepis sibirica L.), серпію звичайного (Serratula coronata L.) та ін. Зміна флористичного складу охоплює не тільки кліматично заміщуючі асоціації, але й різні частини ареалу однієї асоціації.
Продуктивність дубових лісів в залежності від клімату зменшується від Iа бонітету в широколистянолісовій зоні до ІУ-Уа бонітету в степовій.
У кліматичному відношенні територія поширення дубових лісів на Україні відноситься Б.П.Алісовим (1956) до південно-західної під- області Атлантико-континентальної лісової кліматичної області та західної підобласті Атлантико-континентальної степової області. Границя між ними збігається приблизно з південною межею лісостепової зони. Головні риси клімату території зумовлені впливом Атлантичного океану, Середземного моря та материка. Клімат України І.Є.Бучинський (1963) характеризує як перехідний між м”яким кліматом Західної Європи та континентальним кліматом східних областей європейської частини СРСР. Для західних районів України характерні більш м"яка зима та прохолодне літо, ніж у східних районах. Так, за даними М.І.Гук, І.К.Половка, Г.Ф.Прихотька (1958), в західних районах широколистянолісової зони середня температура січня становить від -2 до -4°С, а середня температура липня +І8°С. Середня річна кількість опадів 600-750 мм. У східних районах зони ці показники відповідно дорівнюють -7°, +І9°С та 520-560 мм. В лісостеповій зоні оередньосічнева температура в західній частині -5°, а в східній -8°С, відповідно середньолипнева температура становить +19° та +20°. Кількість опадів із заходу на схід зменшується від 680 до 500 мм. В степовій зоні (байрачний степ) в західних районах серед- ньосічнева температура дорівнює -4°, а в східних -7°. Середи олип- нева температура становить відповідно +21 та +22°С. Річна кількість опадів із заходу на схід майже не змінюється і становить 410- 450 мм (в окремі роки різниця між західними та східними районами України може досягати 500 мм). Південна межа поширення дубових лісів збігається з ізогієтою 300 мм.
Геологічна будова
На території поширення дубових лісів В.Г.Бондарчук (1959) виділяє такі геоструктурні елементи: Український кристалічний щит, Воронезький кристалічний масив, Волино-гПодільська плита,
23
Поліський масив, Галицько-Волинська западина, Дніпровсько-Донецька западина, Донецький кряж, Прикарпатський передовий прогин, Карпати і Закарпатська западина. В орографічному відношенні Українському кристалічному щиту відповідають Житомирське Полісся і Придніпровська височина; Воронезькому кристалічному масиву - Середньоруська височина; Волино-Подільській плиті - Волино-Подільська височина; Поліському масиву - Західнополіська низовина; Галицько-Волинській западині - Сансько-Бузька рівнина; Дніпровсько-Донецькій западині - Придніпровська низовина і Полтавська рівнина; Прикарпатському передовому прогину - Прикарпатська височина і Закарпатській западині - Закарпатська низовина.
На формування дубових лісів безпосередньо впливають відклади, що залягають близько до поверхні і підстелюють грунт або служать для нього материнською породою.
Архейська і протерозойська групи відкладів. За даними В.Г.Бондарчука (1946, 1949, 1959), ці групи представлені кристалічними породами, різко відмінними за складом і поширенням від порід пізнішого походження. Породи цієї групи, утворюючи докембрійську товщу, беруть участь у створенні кристалічного фундаменту платформної частини УРСР. На більшій частині території вони перекриті товщею осадових порід і на денну поверхню виходять лише в окремих районах, - переважно на території Житомирської області, а також в долинах Південного Бугу, Росі та їх приток. Представлені гранітами, а на Овруцькому кряжі - кварцитами. На продуктах руйнування їх формуються дерново-підзолисті грунти, що зумовлюють проникнення в дубові ліси бореальних видів і просування їх у зону лісостепу.
Палеозойська група. Відклади цієї групи порід, за даними В.Г.Бондарчука (1946, 1959), на території України дуже поширені, особливо на захід і схід від Українського кристалічного щита. Палеозойська група представлена товщами окремих стратиграфічних систем, з яких дубові ліси пов’язані переважно з силурійською, девонською та кам’яновугільною системами.
Силурійські відклади поширені на захід від Українського кристалічного щита, від Полісся до Чорного моря. Відслонюються вони в долині подільської частини Дністра та його лівих приток. Представлені силурійські відклади глинистими сланцями або мергелями.
Девонські відклади утворюють великі поля на захід та схід від Українського кристалічного щита. Відслонення їх зустрічаються в південній частині Донбасу і в долині подільської частини Дністра
24
та його приток. Девонські відклади представлені сланцями та пісковиками .
На силурійських і девонських породах формуються малопотужні дерново-карбонатні грунти, на яких ростуть реліктові дубові ліси з значною участю субсередземноморських видів.
Кам"яновугільні відклади трапляються в Карпатах та Дніпровсько- Донецькій западині, Галицько-Волинській западині і на Донецькому кряжі. На території останнього вони виходять на денну поверхню і представлені шарами сірих глинистих і піщаних сланців, пісковиків, вапняків та вугільних пластів. На продуктах руйнування їх формуються малопотужні чорноземи з дубовими лісами базифільного характеру.
Мезозойська група. Відклади мезозойського віку на Україні, за даними згаданих вище авторів, поширені нерівномірно. Представлені вони Тріасовою, Юрською та Крейдовою системами, з яких дубові ліси пов"язані лише з останньою.
Серед осадових порід крейдяні відклади особливо поширені і виходять на денну поверхню на Поділлі, Західному Поліссі, а також в долинах Сейму, Пола, Ворскли та Сіверського Дінця. Представлені крейдяні відклади вапняками, крейдою і мергелями, на яких формуються дерново-карбонатні та чорноземні грунти. Крейдяні породи зумовлюють просування реліктових дубових лісів субсередземноморсько- го характеру далеко на північ від межі їх суцільного ареалу. Особливо добре цей взаємозв"язок простежується на території Поділля й Донбасу. Так, на Поділлі на північних схилах річкових долин та межиріч, крейдяні відклади яких перекриті лесом, ростуть дубово- грабові ліси. Останні на таких самих схилах, але позбавлених покриву кайнозойських порід, заміщаються дубовими лісами з низкою субсередземноморських видів.
Кайнозойська група. Відклади цієї групи характерні для всієї території України. Відслонення їх трапляються в долинах усіх річок, в ярах та балках; представлені третинною та четвертинною системами.
Третинні відклади поділяються на палеогенові та неогенові. Палеогенові відклади, за даними В.Г.Бондарчука (1959) та П.К.За- морія (1961), в межах Українського кристалічного щита трапляються лише у вигляді окремих островів. Найчастіше вони відслояюються в долинах річок Дніпровсько-Донецької западини і Донецького кряжа. Складають їх глауконітові піски, пісковики, мергелі та глини. На породах цього віку дубові ліси ростуть рідко. Участь субсередвем-
25
номорських видів в них більша, порівняно з дубовими лісами на ле- оах.
Неогенові відклади більш поширені, ніж попередні, і трапляються на території усіх геоструктурних елементів. Найцікавішими для нас є відклади полтавської серії, осадонакопичення яких відбувалось в умовах регресії і висихання мілководь олігоценового та міоценового морів. Полтавська серія відкладів складається з кварцових пісків, на яких формуються дерново-підзолисті грунти, різко відмінні від суцільних масивів чорноземів, які їх оточують.
Ділянки з виходом на поверхню полтавських пісків - класичний приклад залежності сучасної природної рослинності від геологічних порід. Так, тут можна згадати масив дубового лісу, описаний нами в підзоні байрачного степу на території Біловодського лісництва Дер- кульського дослідного лісгопзагу. Масив займав верхню частину крутого південно-західного схилу глибокої балки. Строкаті й червоно- бурі глини з плащем з лесів, які звичайно вкривають полтавські піски, на цій ділянці розмиті і грунти сформувались безпосередньо на полтавських пісках. На таких ділянках формується асоціація татар- ськокленово-конвалієва діброва з низкою борових видів. На сусідніх схилах, в аналогічних умовах, але з покривом строкатих або червоно-бурих глин, що збереглися на чорноземних грунтах, формується асоціація татарськокленово-рябоперлівкова діброва з низкою субсе- редземноморських видів.
На території Галицько-Волинської западини і на Волино-Поділь- ській плиті неогенові відклади представлені тортонськими літотамні- євими вапняками і сарматськими вапняками, осадонакопичення яких відбувалось в умовах Сарматського моря. На них формуються дерново- карбонатні грунти. Вапняки зумовлюють острівне просування асоціації діброви свидиново-гірськоосокової у північно-західному напрямку за межі її суцільного поширення.
Четвертинні відклади поширені скрізь. Лише на території Донецького кряжа вони були змиті і зустрічаються зрідка. Звичайно їх прийнято поділяти на постпліоценові, плейстоценові та голоценові.
Постпліоценові відклади представлені червоно-бурими глинами (на балтських пісках, строкатих глинах), які утворились в кінці пліоцену - на початку четвертинного періоду в континентальних умовах (Заморій, 1954, 1961; Бондарчук, 1959; Ворона, 1969). Поширені вони на території Лівобережного Полісся, Поділля, Прикарпаття і Закарпаття лише островами. На решті території, за винятком По26
лісся та Донецького кряжа, трапляються майже всюди. Гливи дуже щільні, в"язкі і складаються з 60-78% кремнезему і 8-12% глинозему з включеннями кристалів гіпсу. Потужність червоно-бурих глин (0,5- 40 м) залежить від умов залягання. На них формуються чорноземи. Для поширення дубових лісів ці глини мають винятковий інтерес. З ними пов"язане крайнє східне знаходження реліктових дубових лісів з підліском з дарену справжнього (Cornus mas L.), які ми відносимо до субсередземноморських дібров.
Плейстоценові відклади представлені різними генетичними типами континентальних осадів, значно поширених. Складаються вони з льодовикових, флювіогляціальних, лесових та елювіально-делювіальних відкладів.
Льодовикові відклади на ератичній частині території УРСР представлені мореною, потужність якої становить в середньому 2-4 м. Морена складається з валунних суглинків, пісків і глин (Веклич, 1958; Бондарчук, 1959; Маринич, 1963). Дубові ліси ростуть тільки на карбонатній морені, яка залягає на глибині 1,5-2 м. Якщо карбонатна морена лежить глибше, дубові ліси заміщуються дубово-сосновими та сосновими.
Флювіогляціальні відклади, за даними згаданих авторів, дуже поширені. Для дубових лісів мають значення надморенні флювіогляціальні відклади дніпровського і валдайськогогляціалів. Складаються вони з пісків, рідше супісків та суглинків з галькою. Потужність відкладів звичайно не перевищує 2-5 м. Вони сприяють формуванню ацидофільних дубових лісів з участю бореальних видів.
Лесові відклади вкривають майже суцільним шаром мало не 70% території зростання дубових лісів УРСР. Тільки в Поліссі і на Донецькому кряжі вони є окремими невеликими островами. Лес в північних та західних районах УРСР містить близько 14 піщаної, 70 пилуватої і 18% глинистої фракцій. В більш південних районах вміст глинистої фракції збільшується до 25-30%. Характерним для лесової формації в її карбонатність (Соколовський,І96І; Мельник, Чугунний, 1969). Дубові ліси, які ростуть на лесовій формації в плакорних умовах, зональні. їх можна розглядати як клімаксові угруповання, якщо прийняти теорію клімаксу з поправками Р.УІттекера («hittabet, 1953), тобто розглядати клімакс як частково стале угруповання, що найбільн стійко пристосувалось до максимального перетворення оточуючих ресурсів в біологічну продукцію. Як функціонуюча система, що розвивалась в тісному зв"язку з середовищем, угруповання є структуряо- 27
функціональною моделлю, пристосованою до всіх факторів оточення, в якому вона існує.
Елювіально-делювіальні відклади поширені переважно на Донецькому кряжі і являють продукт руйнування дочетвертинних порід. На них формуються чорноземні та темно-сірі опідзолені грунти, на яких росте більша частина дубових лісів кряжа.
Голоценові відклади представлені алювієм заплавних терас. Вони складаються з різнозернистих пісків з прошарками гальки та глини в нижній частині. Потужність їх становить 2-25 м (Бондарчук, 1959; Заморій, 1961; Цись, 1962; Маринич, 1963). Заплавні алювіальні відклади наймолодші з усіх розглянутих. У дубових лісах, що ростуть на них, реліктових видів немає. Флористичний склад цих лісів найбільш нівельований і складається переважно з неморальних видів.
Рельєф
По відношенню до рослинності рельєф виступає насамперед як фактор, що перерозподіляє тепло і вологу на поверхні землі.
На початку четвертинного періоду, за даними В.Г.Бондарчука (1959) та І.Л .Соколовського (1965), платформна частина УРСР мала рівнинну, мало розчленовану поверхню і лише долини Дністра та Прута врізувались на глибину до 200 м. Нерівності колишнього рельєфу були згладжені товщею червоно-бурої глини. Підняття, що почалося на початку четвертинного періоду, збільшило базис ерозії і тим самим призвело до утворення більшістю річок постійних долин і до дальшого розчленування поверхні. В плейстоцені діяльність льодовика викликала утворення моренних рівнин, а діяльність флювіогляціальних вод - зандрових рівнин та флювіогляціальних долинних рівнин. З льодовиковою епохою пов”язане й утворення лесових рівнин. Підняття, яке почалося після дніпровського зледеніння, викликало підйом південно-західної частини Поділля та Прикарпаття на 150 м, Середнього Придніпров"я - на 100 м, Полісся і Дніпровсько-Донецької западини - на 25 м (Соколовоький, 1965). В цей період, як і в голоцені, головними рельєфоутворюючими факторами були ерозійні процеси, які переважно й створили сучасний рельєф. Характерною рисою його є чергування долин і вододільних просторів. Природно, що ділянки, які зазнали більш інтенсивного епейрогенічного підняття, висохли в неогені і плейстоцені першими. Ерозійні процеси були на них інтенсивніш!, що призвело до більш значного вилуговування поверхневих горизонтів. Такі території заселялись лісами в першу 28
чергу. Зпейрогенічні підняття в історичній перспективі сприяють поширенню лісів, а різниця клімату і геоморфології веде до їх диференціації.
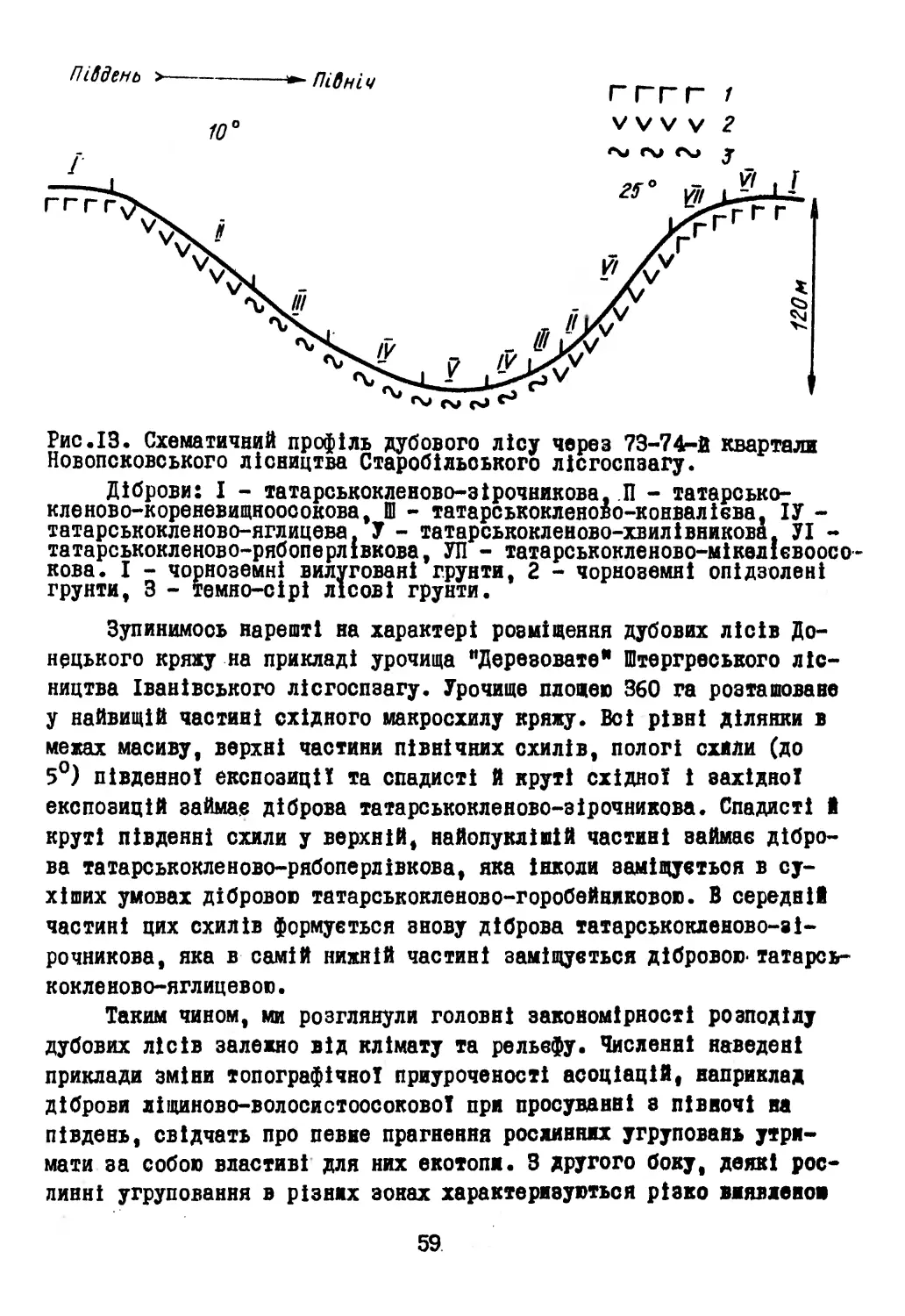
На території поширення дубових лісів (рис.І) розрізняють такі головні орографічні одиниці: Поліська низовина, Волино-Подільська височина, Карпати, Закарпатська низовина, Придніпровська височина, Лівобережна низовина, Середньоруська височина і Донецький кряж (Бондарчук, 1949; Бондарчук та ін., 1959; Геренчук, 1960; Цись, 1962).
Рис.1. Схематична карта орографічних районів території дослідження: 1 — Волинське Полісся, 2 — Мале Полісся, 3 — Житомирське Полісся, 4 — Київське Полісся, 5 — Чернігівське Полісся, 6 — Новгород-Сіверське Полісся, 7 — Волинська височина, 8 — Розточчя, 9 — Опілля, 10 — Гологоро-Кременецький кряж, 11 —
Західне Поділля, 12 — Східне Поділля, 13 — Товтровий кряж, 14 —
Покуття, 15 — Хотинська височина, 16 — Карпати, 17 — Прикарпатська височина, 18 — Закарпатська низовина, 19 — Придніпровська височина, 20 — Придніпровська терасова рівнина, 21 — Полтавська рівнина, 22 — Середньоруська височина, 23 — Донецький кряж.
Абсолютні висоти Поліської низовини на території УРСР нижче 200 м. Піднесеність вище 200 м спостерігається лише на більшій частині Житомирського Полісся, Малого Полісся, південній і захід29
ній частинах Новгород-Сіверського Полісся. Загалом Українське Полісся має плоску вирівняну поверхню, розчленовану неглибокими долинами річок. Виняток становлять різко піднесена Словечансько-Ов- руцька височина, абсолютна висота якої дорівнює 320 м (Бондарчук, 1949; Цись, 1962). З цією височиною і піднесеними ділянками повмя- зані дубові ліси.
Волино-Подільська височина з півночі обмежена Поліссям, а з півдня - Прикарпаттям. Східну межу цієї височини К.І.Геренчук (1950, 1960), В.Г.Бондарчук (1959) та П.М.Цись (1962) проводять по західному краю відкритої частини Українського кристалічного масиву, лінії: Шепетівка - Старокостянтинів - Могилів-Подільський. Волино-Подільську височину Мале Полісся поділяє на Волинську та Подільську височину. Перша з них має абсолютні максимальні висоти в межах 255-360 м і характеризується плоскими, трохи хвилястими межиріччями, друга піднесена в середньому на 300-400 м над р.м. Характерною рисою рельєфу Подільської височини є чергування глибоких долин (60-180 м) з плоскими межиріччями. На її території орографічно виділяються: Розточчя, Опілля, Гологоро-Кременецький кряж, Західне Поділля, Східне Поділля, Товтровий кряж, Покуття і Хотинська височина.
Карпати в орографічному відношенні звичайно поділяють на власне Карпати, Прикарпатську височину та Закарпатську низовину. В Карпатах дубових лісів немає. Прикарпатська височина має абсолютні висоти 300-500 м і характеризується чергуванням витягнутих в бік Дністра вододілів а широкими долинами (Цись, 1962, 1968). Закарпатська низовина рівна, з коливаннями абсолютних висот 105-120 м (Цись^ 1968).
Придніпровська височина на півночі межує з Українським Поліссям, а на заході * а Волино-Подільською височиною. На сході її омивав Дніпро. Абсолютна висота - близько 200 м, у західній частині - до 300-380 м. Для більшої частини височини характерними є плоскі, дещо хвилясті межиріччя. Річки і балки мають неглибокі долини. Ближче до Дністра рельєф стає розчленованішим, долини річок - каньяоноподіоними. Глибина розчленування поверхні перевищує 80- 100, а у Дністра 180-200 м. Середні нахили в цьому районі досягають 18-22°. Подібний дуже розчленований, тільки з більш згладженими формами, рельєф має придніпровська південно-східна частина Придніпровської височини (Бондарчук, 1949; Геренчук, 1960; Цись, 1962).
Лівобережна низовина на півночі межує з Українським Поліссям, на півдні - з Донецьким кряжем, на сході - з Середньоруською височиною, а на заході - з Придніпровською височиною. Абсолютні висоти її 140-180 м, ближче до Дніпра вони знижуються до 100 м, а біля Середньоруської височини становлять близько 210 м. Межиріччя ближчі до Дніпра плоскі, слаборозчленовані. Глибина розчленування становить 30-60 м (Бондарчук, 1949; Геренчук, 1960; Цись, 1962). З розчленованими ділянками рельєфу і пов"язані дубові ліси.
Середньоруська височина займає східну частину України.Поверхня височини являє собою хвилясту рівнину з коливаннями абсолютних висот від 65 до 258 м. Середні висоти дорівнюють 180-210 м. Характерною рисою території є густа сітка ярів та балок, на окремих межиріччях рельєф горбастий. Середні нахили на височині дорівнюють 9-11°.
Донецький кряж являє собою типовий пенеплен. Середні абсолютні висоти його дорівнюють 180-220 м, максимальна висота - 367 м. Рельєф сильно розчленований, з амплітудою висот 100-150 м. Межиріччя двох типів - пологосхилові та грядово-улоговинні. Середні нахили на Донецькому кряжі дорівнюють 15-20° (Бондарчук, 1949; Цись, 1962).
На території описаних орографічних одиниць дубові ліси пов"я- зані переважно з Долинами річок, вододілами та балками.
Долини річок утворились під впливом ерозійних процесів, які розвивались в зв"язку з епейрогенічними підняттями. Головним елементом долин є тераси. Формування дубових лісів залежить не стільки від висотного рівня терас, скільки від їх екологічного режиму і характеру породи, що вкриває тераси. Коли друга борова і третя (однолесова) тераси морфологічно не виявлені й об'єднуються в одиц терасовий рівень, дубові ліси на них різні. Значно більше значення має наявність на терасах морени, гальки або нижньоплейстоцено- вих пісків, якщо вони залягають близько від поверхні. Якщо лесо- вий покрив досить потужний, то дубові ліси практично однакові на всіх лесових терасах і вододілах, вкритих лесом. Варіювання лісів в цьому випадку залежить від особливостей рельєфу. Отже, значна різниця простежується завжди міх заплавними дубовими лісами (перша тераса), дубовими лісами другої тераси (борова) і дубовими лісами лесових терас (третя і вище).
В заплавах річок дубові ліси пов”язані з середніми екологічними умовами 1 зустрічаються звичайно в центральній частині нал-
31
лава. У притерасній та прирусловій частинах вони ростуть лише в заплавах річок високого рівня південної смуги лісостепової та пів~ нічної половини степової зон.
На території України дубові ліси найбільш поширені в долинах Дністра, Дніпра, Росі, Сейму, Сули, Ворскли, Псла та Сіверського Дінця.
Високі корінні береги річок досить різко переходять у вододільні простори, на яких дубові ліси пов"язані з хвилястим та горбастим рельєфом. Плоскорівнинні простори позбавлені дубових лісів. Таким чином, зростання дубових лісів в зоні лісостепу зумовлене не стільки абсолютною висотою місцевості, скільки характером рельєфу. Причиною такого розподілу лісів, на думку Ю.Д.Клеопова (1935), є гідрологічний режим плоскорівнинних вододілів лісостепу. Ми вважаємо, що підтвердити це може такий приклад. На Подільській височині плоскорівнинне межиріччя Стрипа - Серет (висота 330-350 м) було зайняте степовою рослинністю, представленою формаціями ковили найкрасивішої (stipeta pulcherrimae) та довголистої (stipeta longifollae). В той же час вододіли з горбастим рельєфом, розташовані значно нижче, вкриті дубовими або дубово-грабовими лісами.
Балки є характерним елементом рельєфу лісостепової і степової зон України. Сучасні балки, за М.І.Дмитрієвим (1958), невеликі. Довжина їх не перевищує 2-3 км, а ширина - кількох сотень метрів. Давні балки мають довжину до 10 км і ширину до І км. Глибина їх сягає 60-80 м. Частіше зустрічаються балки глибиною 30-40 м, з шириною тальвега 40-50 м та крутизною схилів 10 - 40°. Вище балки переходять у пологі прибалочні схили. Більшість балок галузиться.
Балки можна розглядати як класичний приклад залежності розподілу асоціацій дубових лісів від характеру рельєфу. Найбільш теплими і сухими в балках є верхні частини схилів південної експозиції. З ними цов"язана асоціація діброва татарськокленово-горобей- никова. Найбільш вологими є тальвеги балок, в умовах яких формується асоціація діброва татароькокленово-яглицева. Чим стримкіші схили, тим більша різниця в грунті, характері зволоження та мікрокліматі їх верхніх і нижніх частин і тим різкіша диференціація дубових ліоів. В балках зміна асоціацій часто відбувається вже на протязі 20-30 м. Аналогічна картина спостерігається і на високих крутих правих схилах долин. Велика розчленованість рельєфу, інтенсивні процеси змиву створюють на таких схилах різноманітні умови, 32
мікроклімат тощо. Амплітуда коливань цих умов в окремих випадках може бути дуже значною, що видно і з розподілу рослинності. Наприклад, на вододілі і схилах дністровського каньйону, на ділянці, розташованій на території, коефіцієнт зволоження якої відповідає широколистянолісовій зоні (Тернопільська обл.), на північних схилах каньйону ростуть дубово-грабові ліси, а верхня частина стрімких схилів південної експозиції вкрита дібровою кизилово-горобей- никовою. Закінчуючи розгляд залежності поширення дубових лісів від рельєфу, необхідно відзначити, що найбільші масиви їх в межиріччях знаходяться на Середньоруській височині і піднятій частині Українського Полісся. Південна межа Дубових лісів збігається з вододілами, абсолютні висоти яких не нижчі 200 м. По долинах річок вони трапляються й далі на південь.
Грунти
Дубові ліси - формація відносно вимоглива до грунтових умов - на підзолистих або бідних відмінах дерново-підзолистих грунтів не ростуть. В першому випадку вони витісняються ялиновими, а в другому - сосновими лісами. В кліматичних умовах України, за нашими даними, дубові ліси пов"язані з п"ятьма генетичними типами грунтів, а саме: дерново-підзолистими, дерново-опідзоленими алювіальними, сірими лісовими (сірими опідзоленими), дерново-карбонатними (перегнійно-карбонатними) і чорноземними. Така досить значна екологічна амплітуда дубових лісів (щодо зволоження, то вони займають грунти від дуже сухих до сирих) зумовлює велику різноманітність їх асоціаційного складу і продуктивності. Практично на території України зростають корінні дубові ліси від Iа до У® класу бонітету. Зменшення їх продуктивності нижче Ш класу бонітету повезене не з бідністю грунтів на елементи живлення, а з нестачею вологи. Так, на Поліссі (широколистяноліоова зона), де коефіцієнт зволоження вище 1,0, корінних дубових лісів нижче Ш класу бонітету немає. Це пояснюється тим, що з менш багатих грунтів вони витісняються сосновими лісами і, таким чином, не утворюють низькобоні- тетних деревостанів. У степовій зоні, де дуб не має конкурентів, на багатих, але сухих чорноземних грунтах утворюються деревоста- ни ІУ-Уа класу бонітету. Отже, на Україні виявлена значною мірою лише кліматична межа ареалу дібров. Едафічні фактори не створюють меж, а визначають тільки їх топографічне розташування.
Як відомо, грунтоутворюючий процес не є самостійним і неза-
33
лежним. Його найважливішою рушійною силою є життєдіяльність рослинності. Дубові ліси по-різному впливають на цей процес. Різниця впливу визначається різницею кліматичних умов, грунтсутворюючих і підстилаючих порід та рельєфу, тобто умов, в яких відбувається одночасний розвиток лісу і грунту. Здійснюється вплив лісу на грунт безпосередньо через опад і кореневі системи. В середньому в ягли- цевій діброві віком 50-100 років кількість опаду становить 4-5 т/га на рік, а коренів у грунті - близько 38 т/га. Всього за 200 років у ліс! залишається і впливає на грунт 650 т/га органічної речовини (Зони, 1954; Ремезов, 1962).
Винос продуктів розкладу органічної маси залежить від кількості атмосферних опадів і наявності доступного для рослин кальцію. В однакових кліматичних умовах на багатих кальцієм грунтоут- ворюючих породах під дубовими лісами формуються менш вилуговані та багатші грунти, ніж на кислих або бідних на кальцій породах під хвойними лісами. Ця різниця виступає тим яскравіше, чим більш виявлені риси гумідного клімату і, за інших однакових умов, вона більша у глинистих відмінах грунтів, ніж у піщаних. Саме такий тип залежності між дубовими лісами і грунтом характерний для території Українського Полісся. В степовій і деякою мірою лісостеповій зонах дубові ліси, навпаки, віддають перевагу менш карбонатним породам з більш вилугованими грунтами на них, чим і пояснюється зв"язок їх з підвищеними та розсіченими ділянками рельєфу.
Дерново-підзолисті грунти формуються під дубовими лісами на плоскорівнинних межиріччях Полісся, Прикарпаття та других терасах річок лісостепової і степової зон. Характерною рисою цих грунтів є значна зруйнованість силікатної частини верхніх елювіальних горизонтів (НЕ и Е) грунту з виносом більш рухомих магнію, заліза, кальцію та алюмінію і накопиченням кремнієвої кислоти. Ці процеси виявлені у дерново-підзолистих грунтів під дубовими лісами значно сильніше, ніж у всіх Інших типів грунтів під Ними, але слабіше, ніж у аналогічних грунтів під сосновими лісами, в комплексі в якими вони зустрічаються. Порівняно з усіма Іншими грунтами під дубовими лісами дерново-підзолисті грунти менш структурні (практично безструктурні), менш гумусовая! і кисліші. За даними ряду дослідників (Вернандер и др., 1951; За- сульська, Захарченко, 1969), ступінь насиченості їх основами гумусово-елювіального горизонту найменший і дорівнює 60-63%. За нашими даними*, в цьому горизонті міститься 1,74-5,48% гумусу. В
1 Аналізи грунтів під дубовими лісами виконано в хіміко-ана- літичній лабораторії Інституту ботаніки ім.М.Г.Холодного АН УРСР.
34
розподілі його за профілем для цих грунтів характерне різке падіння вмісту гумусу при переході до елювіального горизонту - 0,70%. У солевій витяжці гумусово-елювіального горизонту рН дорівнює 3,25- 5,46, із зменшенням в ілювіальному горизонті до 4,65 - 5,63 і в материнській породі до 5,31 - 7,42. Ілювіальний горизонт, на відміну від сірих лісових грунтів, слабо ущільнений і складається із звивистих червоно-бурих або сірувато-іржавих шарів псевдофібр та білястих або жовтуватих прошарків. Потужність гумусово-елювіального горизонту у дерново-підзолистих грунтів становить 12 - 36 см.
Дерново-підзолисті грунти загалом порівняно з сірими мають нижчу потенціальну родючість. Горизонт їх виносу лежить біля поверхні, а винос складає 80% обмінних основ. Проте, як показано для дерново-підзолистих грунтів Тульських засік (Пряхин,1960),знизані кальцієм гумінові речовини при недостатності поглинутих основ і більшій кислотності дерново-підзолистих грунтів більш рухомі, ніж у сірих лісових грунтів, що й забезпечує досить високу активну родючість цих грунтів під дубовими лісами. Особливо різко збільшується під дубовими лісами потенціальна й активна родючість цих грунтів у випадках, коли вони на глибині 90-150 см підстелені крейдою або мергелем, як на територіях Волинського і Малого Полісся. Завдяки наявності кальцію різко зростає роль процесів акумуляці^результатом чого є збільшення потужності гумусовоелювіального горизонту до 40-50 см і збільшення вмісту гумусу в ньому до 4,85 - 6,91%. Морфологічна різниця між генетичними горизонтами профілю цього варіанта дерново-підзолистих грунтів майже не виявлена. При близькому до поверхні заляганні крейди профіль горизонтів за будовою нагадує профіль карбонатних чорноземів•
Одержані нами дані показали, що дерново-підзолисті грунти під дубовими лісами належать до олабо- та середньопідволистих типів. За механічним складом перші бувають від глинисто-піщаних до пилувато-супіщаних, а другі - від супіщаних до легкосуглинистих. За ступенем оглеєності грунти бувають глеюватими, глеєвими і лове рхнево-оглеєними. На дерново-слабопідзолистих піщаних грунтах ростуть дубові ліси, у флористичному складі яких зустрічаються олігомезотрофні види, такі, як веснівка дволиста, ожика волосиста (Luzuia pilosa (L.) w і 1 1 d .), береза бородавчаста (Betula verrucosa E ь г ь.) та ін. Топографічно й екологічно ці ліои безпосередньо повиязані з дубово-сосновими лісами. Дубові ліси на дерново-середньопідзолистих глеєвих грунтах топографічно та 35
едафічно пов"язані з варіантом вільхових лісів на бідних грунтах.
Дернові опідзолені алювіальні грунти сформувались під заплавними дубовими лісами. За зайнятою площею вони є найменш поширеними і трапляються головним чином в заплавах Дністра, Сіверського Дінця, Псла та Ворскли. Від дерново-підзолистих грунтів вони відрізняються відсутністю елювіального горизонту, а від лучних - наявністю присипки в нижній частині гумусового горизонту. За профілем заплави гумусовий горизонт цих грунтів дуже варіює. В прирусловій частині він сірий або світло-сірий, супіщаний, потужністю від 15 до 25 см. Вміст гумусу в ньому дорівнює 2,46 - 3,65%, рН 5,3 - 5,8. Нижче йде досить потужний піщаний, з слідами шаруватості перехідний горизонт, який поступово світлішає. У високих грив він має білуваті елювіальні плями з початковими стадіями ілювіальних процесів у нижній частині. Ближче до прируслової частини потужність гумусового горизонту збільшується до 50-70 см, вміст гумусу - до 8,65 - 10,42%, рН 5,9-6,3. Колір горизонту стає тЬмно-сірим і навіть чорним. Перехідний горизонт в нижній частині оглеєний. Дубові ліси на таких грунтах екологічно і топографічно пов"язаяі з варіантом вільхових лісів на багатих грунтах.
Сірі лісові грунти найбільш поширені під дубовими лісами на території Опілля та Розточчя, в лісостеповій зоні і на Волинській височині. Окремими островами вони зустрічаються також і на Поліссі, Прикарпатській височині і знижених елементах рельєфу степової зони. Залежно від ступеня виявленості елювіального процесу сірі лісові грунти поділяються на темно-сірі, сірі та світло-сірі. За І.В.Тюріним (1966), темно-сірі лісові грунти є першою, а світло-сірі - останньою ланкою в генетичному ряді переходу від вилугованих чорноземів до дерново-підзолистих грунтів.
На думну О.С.Скородумова (1954), для елювіального ряду грунтів важливим показником є ступінь виносу та накопичення кальцію. За його даними, горизонт акумуляції поглиненого кальцію світло-сірих лісових грунтів дорівнює 7-8 см. У сірих опідзолених грунтів він отановить 12 см, а у темно-сірих - 19 см. У опідзолених чорноземів цей горизонт у 2-3 рази більший, ніж у темно-сірих грунтів. Не менш показовим є і ступінь насиченості цих грунтів основами. Так, за даними Н.Б.Вернандер та ін. (1951), а також Т.М.Засуль- ської, І.Г.Захарченко (1969) , для світло-сірих опідзолених грун-
36
тів вона дорівнює 67-70%, а для сірих опідзолених грунтів 70-74%, для темно-сірих опідзолених 76-84% і для опідзолених чорноземів 86-92%.
Світло-сірі лісові грунти мають добре виявлений білуватий елювіальний горизонт, пластинчастої структури і дуже щільний, досить багатий на колоїди, з горіхувато- призматичною структурою - ілювіальний. Морфологічні ознаки світло- сірих лісових грунтів найбільш чітко виявлені у відмінах важливого механічного складу. У відмінах легкого механічного складу вони майже не виявлені, і практично ці грунти важко відрізнити від дерново- підзолистих, з якими на Поліссі вони пов"язані цілим рядом проміжних форм.
Вміст гумусу і рН гумусово-елювіального горизонту світло-сірих лісових грунтів, за нашими даними, дорівнюють відповідно 1,76-3,65% і 4,5-5,65. За механічним складом ці грунти представлені відмінами від піщано-суглинистих до важкосуглинистих. Як і для дерново-підзолистих грунтів, для них характерне різке зменшення вмісту гумусу за профілем вже з незначної глибини.
Більшість асоціацій дубових лісів, пов"язаних з дерново-підзолистими грунтами, росте і на світло-сірих лісових грунтах, проте у флористичному складі їх майже немає індикаторів суборевих еда- фотопів. Відповідно дубові ліси на цих грунтах екологічно і топографічно пов"язані не з дубово-сосновими, а з буковими і дубово-грабовими лісами в західній частині України і з дубово-липовими лісами - в східній.
Сірі лісові грунт и,на відміну від світло-сірих, мають темніше забарвлення, більш потужний, нерідко до 40 см, структурний гумусово-елювіальний горизонт і морфологічно слабо виявлений елювіальний. Ілювіальний горизонт дуже щільний, червонувато-бурого кольору, в верхній частині горіхуватої структури, дуже багатий на колоїди. Вміст гумусу і рН гумусово-елювіального горизонту цих грунтів, за нашими даними, відповідно дорівнюють 2,08 - 5,41% і 4,60 - 5,95.3а механічним складом грунти представлені відмінами від середньосуглинистих до важкосуглинистих. Дубові ліси на оірих опідзолених грунтах топографічно та екологічно ще можуть бути пов"язані з дубово-грабовими і дубово-липовими лісами, однак цілком позбавлені індикаторів суборевих едафотопів. Характерне для них переважання мезотрофяих, нейтрофільних, неморальних європейських видів.
37
Темно-сірі лісові грунти мають структурний гумусово-елювіальний горизонт потужністю 35-48 см. Елювіального горизонту тут немає. Морфологічно процес елювіальності виявлений лише в борошнистій присипці кремнезему. Ілювіальний горизонт досить потужний - 65-85 см, горіхувато-призматичної структури, темно-коричневого або червонувато-буруватого кольору. Вміст гумусу у гумусово-елювіальному горизонті цих грунтів, за нашими даними, дорівнює 3,65-6,81%. Зменшення вмісту гумусу з глибиною виступає не так різко, як у попередніх грунтів. Ці грунти, як правило, також менш кислі. У верхньому горизонті рН їх дорівнює в західних областях 4,60-5,18, у східних 5,34 - 6,47. За механічним складом вони представлені відмінами від середньосуглинистих до глинистих. Дубові ліси на темно-сірих лісових грунтах, як правило, топографічно й екологічно з іншими формаціями вже не пов"язані. В них переважають мезо- і мегатрофні, нейтрофільні неморальні види, з домішкою базифільних неморальних (субсередземноморських).
Чорноземні опідзолені грунти під дубовими лісами пбширені на хвилястих і горбастих межиріччях південної частини Правобережного лісостепу, в центральній і південній частинах Лівобережного лісостепу і верхній частині схилів балок степової зони. Як вже відзначалось, в них ще більш послаблені елювіальні та посилені акумулятивні процеси. Морфологічно це виявляється в розвитку потужнішого, ніж у темно-сірих лісових грунтів, гумусово-елювіального горизонту, який Досягає 50-85 см. У сильно опідзолених відмін його розрізняються два підгоризонти, за наявністю слабкої щільності і борошнистої присипки у другому з них. Ілювіальний горизонт формується, але виявлений він значно слабше, ніж у темно-сірих лісових грунтів, менш щільний і має світло-бурий колір. Вміст гумусу в першому підгоризонті гумусово- елювіального горизонту цих грунтів, за нашими даними, становить 5,21-11,34%, в другому - 3,74-5,98%. Реакція гумусово-елювіального горизонту слабокисла, рН 5,26-6,85. За механічним складом вони представлені відмінами від середньосуглинистих до важкогли- нистих. Опідзолені чорноземи пов"язані з темно-сірими лісовими грунтами низкою поступових переходів, що ускладнює віднесення їх до того чи іншого типу.
Чорноземи вилуговані - остання ланка елювіальних грунтів, на яких зростають діброви. Від звичайних (типових) чорноземів вони відрізняються тим, що під гумусовим горизон38
том є ще горизонт, позбавлений карбонатів кальцію. Елювіальний горизонт у цих грунтів не розвивається. Звичайно можна відзначити лише сліди вмивання колоїдів у некапілярних шпаринах. Сліди елювіального процесу виявляються наявністю в кореневих ходах слабовиявле- ної присипки, яка,можливо, має фітолітарне походження. Вміст гумусу верхнього шару вилугованих чорноземів, за нашими даними,дорівнює 6,36-10,66%. З глибиною кількість його поступово зменшується. Цим вилуговані чорноземи відрізняються від усіх попередніх грунтів. У гумусовому горизонті рН дорівнює 6,38-7,25 і практично не змінюється з глибиною. За механічним складом вилуговані чорноземи представлені відмінами від важкосуглинистих до важкоглинистих грунтів. Дубові ліси на опідзолених і вилугованих чорноземах на території України топографічно й.екологічно повпязані з чагарниковими степовими формаціями і аж ніяк не повпязані з іншими лісовими формаціями. У їх флористичному складі переважають базифільні неморальні субсередземноморські види.
Дерново-карбонатні грунти під дубовими лісами трапляються зрідка на території Волино-Подільської височини і південно-східної частини Середньоруської височини. Приурочені вони до верхніх і середніх частин схилів річкових долин та балок і формуються, як правило, на крейді або мергелях, що залягають на глибині 20-50 см. Дерново-карбонатні грунти мають гумусовий і перехідний горизонти. Гумусовий горизонт структурний, з високим вмістом (12,46-34,18%) гумусу, колір його має відтінки від темно-сірого до чорного. В нижній частині цього горизонту вже трапляється щебінка материнської породи. Реакція гумусового горизонту слаболужна, рН його 7,05-7,36. Із збільшенням потужності верхнього горизонту до 60-110 см в умовах більшої кількості опадів (500-600 мм за рік) і при водному режимі промивного типу дерново-карбонатні грунти поступово опідзолюються, вміст гумусу в них знижується і вони поступово перетворюються на дерново-підзолисті.
Питання про походження сірих лісових грунтів має принципове значення для вирішення проблем, повлязаних з формуванням рослинного покриву, взаємовідношенням лісу і степу та геоботанічним районуванням.
Зараз більшість грунтознавців, як відомо, додержуються поглядів С.І.Короноького (І887),який вважав, то сірі лісові грунти виникли з чорноземних, які опідзолились. В.В.Докучаєв (1889) стояв на протилежних позиціях, він розвивав думку про самостійне ут- 39
вороння сірих лісових грунтів лід дубовими лісами. Після критичного перегляду поглядів прихильників С.І.Коржинського (Танфильев, 1894, 1896; Полннов, 1916; Глинка, 1927; Прасолов, 1939,та ін.) такого ж висновку дійшов і І.В.Тюрін (1966). Докази самостійного походження сірих лісових грунтів, які наводить останній автор, полягають в тому, що процес грунтоутворення відбувається значно швидше, ніж вважали раніше, що між ступенем опідзолеяості і вмістом гумусу немає певної кореляції, що сірі грунти мають меншу потужність елювіального горизонту, ніж гумусовий горизонт поширених тут чорноземів, що в умовах лісостепу під дубовими лісами можуть акумулюватись в значних масштабах гумати кальцію. Останнім часом на підставі аналізу літературних даних Г.І.Дохман (1968) визнала, що за незмінних кліматичних умов сірі лісові грунти під дубовими лісами можуть довго зберігатись без дальшого опідзолювання, проте автор заперечує можливість самостійного їх формування. З першим можна погодитись повністю. Те, що дубові ліси в певних умовах не опід- золюють чорнозему, відзначає багато дослідників (Соловьев, 1951; Зони, 1954, 1961; Надеждин, 1954; Зозулин, 1955; Мигунова, 1960; Афанасьева ,1966, та ін.). Про це свідчать і описані нами під перестійними дубовими насадженнями вилуговані чорноземи. Друге твердження,на наш погляд, не можна вважати остаточним. Є.С.Мигунова (1960) з цього приводу пише: "Наявність великих запасів перегною, в окладі якого багато вільних гумінових кислот, а також чудово виявленої водотривкої зернистої структури в залісених чорноземах, з одного боку, і тривалого перебування багатьох лісових масивів на чорноземах, з другого, свідчить про те, що і перегній, і структура залісених чорноземів являють сучасне утворення, що виникло в умовах зростання лісу на чорноземі, а не є реліктом, що був тут колись*(с.182). До подібного висновку на підставі численних даних порівняльного вивчення чорноземів під дубовими лісами і степовою рослинністю дійшла і Є.А.Афанасьева (1966). На її думку, лісові чорноземи значно поширеніші, ніж здається, оскільки подекуди їх відносять до темно-сірих лісових грунтів, а розорані лісові чорноземи не відрізняють від чорноземів, що утворились під трав"я нистою рослинністю. Можливо, ці висновки надто категоричні, але, у всякому разі, якщо ми визнаємо циклічні зміни рослинності, спричинені змінами клімату або епейрогенічними рухами, то маємо визнати і можливість утворення сірих лісових грунтів та чорноземів під дубовими лісами. Крім того, слід враховувати при вирішенні цього питання 1 проградацію грунтів.
40
ОСНОВНІ ЗАКОНОМІРНОСТІ ПОШИРЕННЯ ДУБОВИХ ЛІСІВ НА УКРАЇНІ
В кінці валдайського зледеніння почалось врізання уступів другої тераси. До того рельєф був рівнішим, ніж тепер. Вирівнювання його відбулося за рахунок відкладів лесової та флювіогляціальної товщі потужністю 3-6 м. Крім того, інтенсивні епейрогенічні рухи, які почалися після дніпровського зледеніння, підняли до сучасного рівня,за даними І.Л.Соколовського (1965), південно-західну частину Подільської височини та Прикарпаття на 150 м, Придніпровську височину - на 50-100 м, Подільську височину - на 25 м і Середньо- руську височину - на 50-100 м.
Природно, що підняття супроводжувалось зниженням базису ерозії та посиленням розчленування рельєфу. Наприкінці валдайського льодовикового періоду - на початку голоцену річки в долинах протікали на 5-60 м вище, ніж тепер,і води в них, завдяки таненню льодовика, було значно більше, а в умовах вирівняного і, отже, сповільненого стоку відбувалось підтоплення і підвищення рівня грунтових вод значних територій. Тому формувались грунти болотяного ряду або чорноземи, насичені карбонатами. Природно, що грунти гідрогенного та переходового (елювіально-гідрогенвого) рядів сприяли формуванню трав"яної рослинності і перешкоджали поширенню дубових лісів, повезених з грунтами елювіального ряду. Останні формувались на ділянках найбільш підвищеного і розчленованого рельєфу. Такі ділянки розташовані, як вже зазначалось, вздовж річкових долин, на Подільській, Придніпровській та Середньоруській височині. Саме з ними пов"язане поширення сірих опідзолених грунтів лісостепу України. Такий зв"я- зок між широколистяними лісами, грунтами і висотою місцевості простежується, за даними М.В.Водовозової (1964), 1 в східних районах’ Руської рівнини. Один з перших гіов"язав широколистяні ліси з подільським островом цих грунтів А.І.Иабоких (1912), який вважав, що з цього центра в післяльодовиковий період і розселились вони. Слід додати, що на Україні таких центрів було кілька (Лавренко, 1930а; Гринь, 1949, 1957; Артюшенко, 1970).
На нашу думку, дубові ліси під час валдайського льодовикового періоду займали сучасні верхні частини схилів річкових долин 1 балок. Крутизна схилів мала бути такою, що виключала інтенсивне накопичення лесу, але й не надто великою, оскільки на малопотужному лесовому покриві формується степова рослинність. Цим умовам 41
відповідають схили крутизною 10-30°. Одержані нами дані свідчать, що на території Подільської височини схили південних експозицій подібної крутизни зайняті дубовими лісами тільки при наявності лесо- вого покриву або іншої грунтоутворюючої породи потужністю не менш як 30-40 см. Грунти на схилах східної та західної експозицій вилуговуються більше, тому для них досить шару грунтоутворюючої породи потужністю 15-20 см. На пологіших схилах, де, природно, потужність лесового покриву значна і залягання карбонатів глибоке, сформувались сірі лісові грунти. Як правило, це схили західних або північних експозицій. На більш крутих схилах з лесовим покривом меншим як І м і карбонатними породами, що підстелюють лес, сформувались чорноземні грунти. Це переважно південні схили. В зв’язку з такою диференціацією грунтового покриву залежно від рельєфу спостерігається і диференціація неморальних реліктів. Види, пов’язані головним чином з тінистими кззофільними буковими або липовими лісами, пережили валдайський льодовиковий період в дубових лісах, що росли на сірих лісових грунтах. Це граб звичайний, лунарія оживаюча (Lunaria rediviva Lt), гусимець пужниковий (Arabis turrita L.), страусове перо звичайне (Matteuccia struthiopteris (L.) T o d a r о), ХВОЩ великий (Equisetum majus G a r s#), вероніка Тіньова (Veronica umbrosa 11, Вф), шавлія клейка (Salvia glutinosa L.), СКОПОЛІЯ карнІОЛІЙСЬка (Scopolia carniolica J a c q.), Зірочник гайовий (Stellaria nemorum L*), костриця висока (Festuca sitissima A 1 1.) та ІН.
Види, повпязані переважно із світлими ксеромезофільними дубовими лісами, пережили валдайське зледеніння в дубових лісах на чорноземних грунтах. Це дерен справжній (Cornus mas L.), скумпія звичайна ( Cotinus ooggygria S с о р.),лазурник трилопатевий (Laser trilobum (L.) В o r k h*)* ластовень ВИТКИЙ ( Vincetoxicum scandens S o m a.et L e v.), жабриця гарна (Seseli elegans S c h i s 0 h ke), фізосперм Дана (Pbysospermum danaa (M.B.) Schischk.), живокіст кримський ( Symphytum tauricum f i 1 l d.)ia ін. З цих місцевих рефугіумів відбулось радіальне розселення широколистяних лісів (Кац, 1955). Крім цих місцевих рефугіумів, до збереглись дуже збіднені широколистяні ліси, були й значно віддалені (Балкани, Талиш, Колхіда) . В них широколистяні ліси збереглись майже повністю. Такі рефугіуми мали, мабуть, значення флорогенетичних центрів.
42
Розселення широколистяних лісів в місцевих рефугіумів звичайно пов"язують з потеплінням клімату. Проте, на наш погляд, не лише суворий клімат перешкоджав розселенню дубових лісів. Ще однією причиною були флювіогляціальні та лесові відклади, що вкривали межиріччя. Дубові ліси не могли поселитись на них, доки менш вибагливі види, такі, як сосна звичайна (Pinus silvestris L.) і береза бородавчаста, не підготували для них більш сприятливі грунтові умови. Панування на початку голоцену березових 1 соснових лісів на Україні і відбивають результати спорово-пилкових аналізів, наведені в роботах М.Кочвари (Koczwara,I928),С.Кульчинського (Kulczyn- ski, 1930?, Д.К.Зерова (1933,1934,1938,1950.1952). С.Толпи (Тоїра, 1935), В.Тимракевича (Tymrakiewicz,I935), К.Костинюка (Kostyniuk, 1938), О.Т.Артюшенко (1957, 1960, 1970), Г.О.Пашкевич (1963) і Д.К.Зерова, О.Т.Артюшенко, (1961, 1966).
П.Д.Клеопов (1941) на підставі аналізу флори широколистяних лісів дійшов висновку, що в ранньому голоцені на території України був поширений лісостеп типу сучасного Західносибірського. Рослинність в цей час була представлена березово-сосновими лісами і степами. Цей погляд С.Д.Клеопова поділяли Д.К.Зеров (1950,1952) на О.Т.Артюшенко (1957, 1960, 1970). Справді,пилкові спектри в нижніх горизонтах містять головним чином пилок сосни, берези і травнистих видів родин, в яких переважають ксерофіти. Однак у нас немає підстав припускати панування змішаних березово-соснових лісів. На території Полісся переважали в той час соснові ліси, а березові зустрічались окремими острівцями на важчих за механічним складом грунтах. На території лісостепу на вододілах переважали березові ліси, а на других терасах - соснові. Переважання у спектрах пилку сосни пояснюється тим, що досліджувались болота в долинах річок. Виняток становлять тільки Західне Поділля та Передкарпаття, де другі тераси не мають піщаних відкладів і, отже, соснові ліси тут мали дуже обмежене поширення. Виходячи з того, що в ранньому голоцені рельєф був більш вирівняний і враховуючи характер відкладів, ми вважаємо, що на Поліссі на межиріччях були поширені не ліси і степи, а соснові ліси з оотепненим травостоєм. Частина цих видів, наприклад гадючник шестипелюстковий (Filipendula bexapetala G і 1 і b.), шавлія лучна (Salvia pratensis Ь.), тимофіївка степова (Phleua phleoides К a r s t e п.), конюшина гірська (Trifolium montana L.), волошка сумська (Centaurea suaensis K а 1 e n.), вероніка сива (Veronica incana L.)( келеріЯ сиза (Koeleria glau-
43
са ПО.) , вероніка колосиста (Veronica siccata L.), лещиця висока (Gypsophila altissima L.), нечуйвітер синяковидний (Hieracium echioides L u о n. ex N. P.), тонконіг вузьколистий (Poa anguati- folia L.),збереглася в соснових лісах Полісся й досі (Шеляг-Сосон- ко, 1966а; Шеляг-Сосонко, Давиденко, 1969). Цілком імовірно, що клімат у ранньому голоцені був сухий, континентальний і нагадував сучасний клімат північного Казахстану та півдня Західного Сибіру. В цих районах на піщаних грунтах тепер ростуть соснові ліси, травостій і підлісок яких утворюють степові види: карагана кущова (Oaragana frutex (L.)G. К о о h.), шипшина найколючіша (Нова spinosissima L.), КИЗИЛЬНИК чорноплодий (Cotoneaster melanooarpa L о d d.), ковила пірчаста (Stipa pennata L.), лещиця висока, вероніка сива, підмаренник справжній (Galium verum L.), смілка поникла (Silene nutans L.), гадючник шестипелюстковий, вероніка колосиста, костриця борозниста, тимофіївка степова, тонконіг вузьколистий (Сукачев, 1948; Карамншева, 1961; Крнлов, Речан, 1965). Як видно з наведених: переліків, значна частина видів соснових лісів Полісся, Казахстану і Західного Сибіру є спільною.
На території лісостепу в ранньому голоцені на межиріччях були поширені остепнені березові ліси і степи. Аналогічні березові ліси, за даними Г.В.Крилова та Н.Г.Салатової (1950), Г.В.Крилова (1953, 1961), Г.А.Глумова (1960), Є.І.Лапшина (1963), поширені тепер в лісостеповій зоні Західного Сибіру. В їх травостої трапляються залізняк бульбистий, горицвіт весняний (Adonis vernalis L.), оман шершавий (Inula hirta L.), люцерна серповидна (Medicago falcata Ь.>, ДЗВОНИКИ сибірські (Campanula sibirica L.), рутвиця мала*(Thalictrum minus L.), куцоніхка Пірчаста (Brachypodium pinnatum (І.) p.b.) та ін. Всі названі види ростуть і в описа¬
них нами дубових ліоах.
Березові ліси на початку голоцену займали, очевидно, плоско- рівнинні, олабодреновані вододіли. В міру диференціації рельєфу,зумовленої епейрогенічними рухами, відбувалась диференціація рослинного покриву в напрямку вичленування степів і мезофільних березових лісів з одночасним зменшенням остепнених березняків. Цей процес пов"язаний з посиленням загальної дренованості території і, отже, вичленуванням, з одного боку, сухіших ділянок і, а другого, - більш вилугованих ділянок.Таким чином.дані спорово-пилкових аналізів (Firbas,1949; Нейштадт,І957; Пьявченко, 1958) та аналіз флори свідчать про те, що в ранньому голоцені існував на території Єв-
44
ропи й Азії суцільний пояс остепнених соснових і березових лісів.
В кінці раннього голоцену дубові ліси із рефугіумів починають виходити на вододіли. Вони витіснили в першу чергу березові ліси в сприятливих для цих едафотопах, тому корінних березняків на Україні не лишилось. Значно пізніше дубові ліси почали витісняти ліси соснові,та лише з найбагатших едафотопів. Існує загальноприйнятий погляд, що цей процес був викликаний потеплінням клімату, яке почалося на початку середнього голоцену. На нашу думку, клімат обумовлює лише потенціальну можливість розселення. Для реалізації її потрібні були ще дві умови: наявність грунтового покриву та диференційований рельєф. Ю.Д.Клеопов прямо зазначає, що Придніпровська терасова низовина позбавлена лісів через галогідрогенний генезис чорноземів цієї території, зумовлений її рослинністю. За нашим глибоким переконанням, яке засноване на викладених в попередніх розділах даних, і зараз на загальному фоні кліматичних умов лісостепу, практично однакових для певного району, розподіл степів і дубових лісів визначає характер рельєфу, отже - епейрогенічні рухи. При цьому слід враховувати вплив конкуренції. Відсутність лісу на багатьох ділянках лісостепу пояснюється не тим, що він не може рости за даних екологічних умов, а конкуренцією між лісом та степом.
За даними обліку лісового фонду на І.І 1966 р., площа лісів України з переважанням дуба звичайного становила 1420 тис.га,або 26% всієї вкритої лісом площі республіки. З неї близько 45% припадає на чисті дубові ліси, близько 30% - на грабово-дубові ліси, поширені переважно на Подільській і Придніпровській височині, близько 10% - на сосново-дубові ліси на Поліссі та других терасах річок лісостепової зони, і близько 10% - на липово-дубові 1 ясеново-дубові, ліси, поширені переважно на Середиьоруській височині, та близько 5% - на кленово-дубові, вільхово-дубові, смереково- дубові й буково-дубові ліси.
Твердження П.П.Кожевникова (1937), що на Середиьоруській височині кленово-липово-дубові ліси складають майже 80% площі лісів, безпідставне П.П.Кожевников до цих лісів відносив фітоценози з поодинокою домішкою липи серцелистої (Tilia cordata м і 1 1,) чи клена звичайного (Acer platanoidee L.), з чим не можна погодитись.
За зонами площі чистих дубових лісів розподіляються в такий
45
спосіб: у широколистянолісовій зоні їх знаходиться близько 25% t в лісостеповій - близько 70%, в степовій лише близько 5%. Найбільші масиви дубових лісів площею від І до 8 тис.га трапляються в південній частині Житомирського Полісся, на Овруцькому кряжі, Серед- ньоруській височині та зрідка на Придніпровській височині. На території Закарпатської низовини та Прикарпатської височини переважають масиви дубових лісів площею від 100 до 700 га. Дещо менша площа їх на Волино-Подільській височині. На Поліссі, за винятком Житомирського та південної частини Волинського Полісся, переважають ділянки площею декілька гектарів. На більшій частині Придніпровської височини звичайно трапляються масиви площею 300-1000 га. На Полтавській рівнині площа окремих масивів значно менша, не більше як 400-500 га. На Донецькому кряжі переважають ділянки розміром 200- 400 га. В степовій зоні масиви дубових лісів мають площу 50-300 га.
В розподілі масивів дубових лісів чітко простежується приуроченість їх до найбільш високих і розсічених територій. Геоботаніки розглядають цю закономірність як прояв зародків вертикальної зональності (Лавренко, Прозоровский, 1939; Корчагин, 1954; Водовозо- ва, 1964). Проти цього, однак, заперечує В.Б.Сочава (1948). На його думку, в середніх широтах Руської рівнини вертикальної зональності немає. Ми вважаємо, що з першою думкою можна погодитись, якщо взяти до уваги те,що зазначена залежність виявляється лише в межах однієї зони і, крім висоти,вона зумовлюється і мірою розчленованості рельєфу. Тому зв"язок абсолютних висот з розміщенням рослинних поясів на рівнині чітко не виявляється і, як відмічає М.В.Водовозова (1964), досі не з"ясований. Окрім загальних вказівок, відсутні дані такого характеру і для рівнинної частини України.
На території Полісся найбільші основні масиви дубових лісів ростуть на межиріччях, розташованих вище 190 м над рівнем моря; на Прикарпатській височині оптимум їх поширення лежить в межах абсолютних висот 300-450 м; на Подільській височині - вище від 90 м, в долині Дністра до 350 м на межиріччях; на Придніпровській височині - 220 - 310 м; на Донецькому кряжі - 250-370 м; на Полтавській рівнині - 160 - 200 м; на Середиьоруській височині (в межах України) 170 - 220 м в північно-східній частині і 190 - 230 м в східній.
За межами України дубові ліси, за даними М.В.Водовозової (1964), на Середньоруській височині найбільш поширені на ділянках з абсолютною висотою 240-250 м. Внутрішньозональні зміни рослинності най46
більш виявлені на Подільській височині, де значна амплітуда абсолютних висот (близько 300 м) при загальному сприятливому для лісів кліматі призводить до закономірної зміни одних формацій іншими. Досить чітко така зміна простежується і на Прикарпатській височині. На решті території спостерігаються лише зміни, що не виходять за межі формації і зумовлені коливаннями не стільки абсолютної, скільки відносної висоти.
На Подільській височині ми виділяємо кілька висотних смуг лісової рослинності, які закономірно змінюють одна одну. Розміщуються вони згори донизу так: букові ліси-дубово-грабові—дубові (Шеляг- Сосонко, 1970а). Цей висотний ряд відбиває і ступінь опідзоленості грунтів від світло-сірих лісових під буковими лісами до темно-сірих лісових під дубовими. Посилення процесів опідзолювання пояснюється збільшенням кількості опадів з висотою і посиленням дренування, а отже, й процесів виносу.
На ділянках з глибоко розчленованим рельєфом (Опілля, Розточчя, Західне Поділля) спостерігається висотна диференціація і між асоціаціями дубових лісів. Згори донизу вони утворюють такий ряд дібров: ліщиново-маренкова — ліщиново-зірочникова—-ліщиново-ягли- цева. Перші дві асоціації займають денудаційні частини рельєфу, а остання - акумулятивні. Наведений ряд значною мірою зумовлений також зміною відносної висоти. Однак наявність деяких видів букових лісів, таких, як апозерис смердючий, купина кільчаста (Polyganatue verticiiiatun а 1 1.) і бук лісовий у першій асоціації свідчить про диференціацію, пов'язану si зміною абсолютної висоти.
Таким чином, найвищі та найстарші за віком ділянки рельєфу з малопотужними четвертинними відкладами займають букові ліси, а наймолодші з більш потужними четвертинними відкладами - дубові. Такий характер розміщення лісів пояснюється кліматичними та гідрологічними факторами; з кліматичних найбільший вплив має збільшення з висотою кількості опадів. На висоті 450 м за рік в середньому випадає на 90-120 мм опадів більше, ніж на висоті 300 м. Збільшення кількості опадів посилює інтенсивність опідзолювання грунтів і призводить до витіснення дубових лісів буковими і грабовими. Крім того, з висотою збільшується і відносна вологість повітря, що також сприяє посиленню позиції букових лісів. Однак на ділянках в плоским слабодренованим рельєфом, де елювіальний процес послаблений висхідними або боковими стоками слаболужних грунтових вод, дубові ліси збереглись. Залежно від міри виявлення дії гідрологічних факторів вони можуть змінюватись навіть степовою рослинністю.
47
За крутизною та експозицією схилів, на яких вони ростуть, дубові ліси розміщуються таким чином. В північній частині Волино-По- дільської височини, де рельєф більш вирівняний, ніж в південній, і кількість опадів більша, дубові ліси займають невисокі опуклі з плоскими вершинами ділянки рельєфу зі схилами, що мають крутизну 5 - 20° та відносну висоту 20 - 50 м. В цих умовах неширокі (100- 500 м) верхівки цих ділянок з досить потужним лесовим покривом займає діброва ліщиново-маренкова. Вона ж, як правило, займає верхню частину схилів північної експозиції, а в середній частині змінюється ліщиново-зірочниковою дібровою і в нижній - ліщиново-яг- лицевою. Верхню частину крутіших схилів південної експозиції займає діброва ліщиново-волосиотоосокова, а більш пологих - ліщиново- зірочникова. Вона ж займає і середню частину схилів. В нижній частині схилів розміщується ліщиново-яглицева діброва, яка біля підніжжя змінюється ліщиново-копитняковою. Якщо рельєф менш розсічений і ширина плоских ділянок межиріч збільшується, то діброва ліщиново-маренкова спочатку заміщується ліщиново-яглицевою, а при ще більшій ширині межиріч - дібровою свидиново-гірськоосоковою. Ця зміна асоціацій від ліщиново-маренкової діброви до свидиново- гіроькоосокової пояснюється послабленням елювіальних процесів (рис. 2).
В південній частині Подільської височини(Придністров"!) переважає горбаотий рельєф з вузькими гребенястими вододілами і схилами
Рис. 2. Схематичний профіль дубового лісу в 5-му кварталі Сковородського лісництва Старокостянтинівського лісгоспзагу. Діброви: I — ліщиново-мареякова, II — ліщиново-зірочникова, III — ліщиново-яглицева, IV — ліщиново-копитнякова. 1 — світло- сірі лісові грунти, 2 — сірі лісові грунти.
крутизною від 20 до 40° і відносною висотою 60-200 м. В цих різко відмінних від решти території екологічних умовах плоскі верхівки вузьких гребенів займав діброва кизилово-зірочникова, верхівки широких гребенів (більш як 200-300 м) - дубово-грабові ліси,
48
що пояснюється кращими умовами зволоження широких гребенів» Верхню частину північних схилів займає діброва ліщиново-конвалієва, нижче якої розташовується діброва ліщиново-зірочникова і ще нижче - ліщиново-яглицева. Менш круті північні схили або круті, але в вузьких балках, зайняті звичайно дубово-грабовими, а іноді й буковими лісами. Верхню, найбільш пологу частину південного схилу з крутизною до 5° займає діброва ліщиново-волосистоосокова, опуклу, найбільш суху верхню частину схилу - діброва кизилово-горобейникова, що в середній частині схилу заміщується дібровою кизилово-мікелієвоосоковою. Нижче від останньої розташовується діброва кизилово-зірочникова, яка біля підніжжя переходить у діброву ліщиново-копитнякову (рис.З).
Рис. 3. Схема синтетичного профілю дубового лісу в 23-му кварталі, Нирківського лісництва Бучацького лісгоспзагу (південний схил) та в 55-му кварталі Пановецького лісництва Кам’янець-Подільського лісгоспзагу (північний схил). Діброви: I — кизилово-зірочникова, II — ліщиново-конвалієва, III — ліщиново-зірочникова, IV — ліщиново-яглицева, V — ліщиново- волосистоосокова. VI — кизилово-волосистоосокова. VII — кизилово- горобейникова, VIII — кизилово-мікелієвоосокова, IX — ліщиново-копитнякова. 1 — сірі лісові грунти, 2 — дерново-карбонатні грунти, 3 — темно-сірі лісові грунти.
На території Полісся формування дубових лівів обмежується наявністю в грунтоутворюючих породах кальцію. На більшій частині території вони трапляються в комплексі з сосновими та вільховими лісами. Перші приурочені до верхніх частин рельєфу з найбільш легкими та бідними дерново-підзолистими грунтами, а останні - до найбільш низьких ділянок рельєфу з потенціально найбагатшими, але заболоченими грунтами. Дубові ліси займають проміжне положення і по- в"язані з достатньо дренованими найбагатшими і важкими відмінами
49
дерново-підзолистих грунтів. Перехід від дубових лісів до соснових або вільхових у північній і середній смузі Полісся досить часто відбувається вже на протязі кількох десятків метрів при змінах висоти рельєфу на кілька метрів. Звичайно це ділянки якоїсь однієї асоціації. В південній смузі Полісся та на Овруцькому кряжі в зв"язку з більшою диференціацією рельєфу цей перехід виявлений більш різко. Дубові ліси ростуть тут великими масивами, і зміна їх асоціацій спостерігається вже залежно від характеру рельєфу в межах того самого масиву. Рельєф цієї частини звичайно слабохвилястий, з відносною висотою гребенів 5-10 м і крутизною схилів 3-8°. У прирічковій частині він більш розчленований. Відносні висоти тут збільшуються до 30-40 м, а крутизна схилів до 15-25°.
У зв"язку з тим, що при просуванні на північ екологічні умови для дубових лісів погіршуються, спостерігається зміна їх асоціацій. У північній частині Полісся найпоширеніші діброви ліщиново- веснівкова та ліщиново-квасеницева, а в південній - ліщиново-во- лосистоосокова та ліщиново-трясучковидноооокова. Перша з них займає дещо підвищені плоскі ділянки з найбіднішими для дубових лісів грунтами і безпосередньо пов"язана з дубово-сосновими лісами. Друга асоціація займає незначні зниження, що добре дренуються, і пов”яза- на з вільховими лісами. В південних районах або на найбільш багатих для дубових лівів Полісся грунтах її заміщує діброва ліщиново-копит- някова. Третя асоціація займає верхні частини пологих схилів і вершини невисоких плоских горбів, тобто пов"язана з найбільш сухими в умовах Полісся грунтами, на яких ростуть дубові ліси. Четверта асоціація в південних районах займає дещо знижені ділянки, а при просуванні на північ переходить на більш підвищені ділянки та пологі схили. Порівняно з попередніми трьома асоціаціями вона займає найменш дреновані оглеєні і глеєві дерново-підзолисті грунти. Дальший розвиток процесу оглеєння і заторфованості викликає заміну її дібровою крушиново-трясучковидноосоковою, а в крайньому вираженні в ряді дубових лісів - дібровою крушиново—молінієвою. Діброви ліщиново—з ірочникова та ліщиново-яглицева, що також трапляються на Поліссі, розташовуються нижче асоціації діброви ліщиново-волосисто- осокової, але вище від усіх останніх, за винятком діброви ліщиново- веснівкової (рис.4,5,6).
На Поліссі, в зв"язку з значною його протяжністю з заходу на схід, на розподілі асоціацій дубових лісів відбивається також вплив збільшення континентальності клімату. Так, діброва ліщиново-трясуч- 50
ковидноосокова, яка панує на території Волинського і Житомирського Полісся, виклинюється в західній частині Чернігівського Полісся (Городнянський лісгоспзаг), тут трапляються лише 1! фрагменти. Східна межа ареалу цієї діброви збігається з ізотермою січня, що дорівнює 7°С. При просуванні з заходу на схід зі складу дубових лісів зникає низка середньоєвропейських видів, таких, як граб звичайний (Придеснянське плато), клен несправжньоплатановий (Волинське Полісся), осока трясучковидна ( Carex brizoides Juslen) та ін.
Рис. 4. Схема синтетичного профілю лісів Гущинського лісництва Любомльського лісгоспзагу: I — сосновий ліс, II — діброва ліщиново-веснівкова, III — діброва ліщиново-квасеницева, IV — вільховий ліс, V — діброва ліщиново- яглицева. 1 — дерново-слабопідзолисті грунти, 2 — дерново-середньопідзолисті грунти.
Рис.5. Схематичний профіль лісів 5-6-го кварталів Григурського лісництва Житомирського лїсгоспзагу: 1 — сосновий ліс, II — діброва ліщиново-копитнякова, III — діброва ліщиново-волосистоосокова, IV — діброва ліщиново-трясучковидноосокова. 1 — дерново-слабопідзолисті грунти, 2 — дерново-середньопідзолисті грунти.
На території лісостепової зони в умовах кліматичної рівноваги як для дубових лісів, так і для степової рослинності зміни в той чи інший бік визначаються ступенем розчленованості рельєфу. Посилення континентальності клімату при просуванні з заходу на
51
Рис. 6. Схематичний профіль лісів 60-65-го кварталів Дзержинського лісництва Бердичівського лісгоспзагу:
1 — дубово-сосновий ліс орляковий, II — діброва ліщиново-трясучковидноосокова, III — діброва крушиново-трясучковидноосокова, IV —
діброва крушиново-молінієва. 1 — дерново-слабопідзолисті грунти, 2 — дерново-середньопідзолисті грунти.
схід зумовлює зміни взаємовідносин дубових лісів з іншими лісовими формаціями. Якщо в західній та центральній частинах лісостепу вони екологічно і топографічно повпязані з дубово-грабовими лісами, то в східній частині лісостепу - з липово-дубовими. Паралельно йде процес збіднення дубових лісів на середньоєвропейські види. Зникають, наприклад, черешня, граб евичайний, клен несправжньопла- тановий, проліска дволиста (Scilla bifolia L.), рівноплідник рут- вицелистий (Іаорулии thalictroides L.), клопогін смердючий (Cimicifuga foetida L.), осока парвська (Carex brevicollis DC.) та 1н.
Являє інтерес, безумовно, поширення асоціацій дубових лісів в зв^язку зі змінами клімату з півночі на південь 1 з заходу на схід та рельєфними особливостями території лісостепу. Розглянемо, наприклад, для порівняння, дубові ліси Придніпровської і Середньо- руської височини. В північній частині Придніпровської височини переважають вододіли з слабохвилястим рельєфом. Звичайно крутизна схилів становить 3-8°, а відносні висоти - кілька метрів. Грунто- утворююча порода - лес, підстелений гранітною щебінкою. В цих умовах плоскі, дещо опуклі вершини широких вододілів займає діброва татарськокленово-гірськоосокова, а вужчі - діброва ліщиново- гірськоосокова. В останньому випадку це пояснюється інтенсивнішим розвитком процесів елювіальності. Верхні частини найкрутіших північних схилів (8-10°) займав діброва ліщиново-маренкова (грунти під цими лісами найбільш опідзолюються), а більш пологі - діброва ліщиново-зірочникова. На верхній частині більш крутих південних схилів росте діброва ліщиново-волосистоосокова, а на пологіших -
52
ліщиново-зірочникова. Нижче від останньої розміщується діброва ліщиново-яглицева, а найбільш низькі ділянки займав діброва ліщиново- копитнякова (рис.7).
Рис. 7. Схематичний профіль дубового лісу 34-40-го кварталів Козятинського лісництва Хмільницкого лісгоспзагу.
Діброви: I — ліщиново-яглицева, II — ліщиново-маренкова, III —
ліщиново-гірськоосокова, IV — ліщиново-зірочникова, V — ліщиново- копитнякова, VI — татарськокленово-гірськоосокова. 1 — сірі лісові грунти, 2 — світло-сірі лісові грунти, 3 — темно-сірі лісові грунти.
Таким чином, в умовах слабохвилястого рельєфу головне значення має не експозиція схилів, як на півдні Подільської височини, а ступінь протікання еловіальних процесів, зумовлених характером рельєфу.
На південній частині височини, на вододілах з слабохвилястим рельєфом, схили якого не перевищують 3-5°, плоскі широкі підвищені ділянки займає діброва татарськокленово-горобейникова, пологі схили - діброва ліщиново-яглицева, а найбільш знижені - ліщиново- конвалієва.
Рис. 8. Схематичний профіль дубового лісу через 59-й квартал Котовського лісництва Котовського лісгоспзагу.
Діброви: I — татарськокленово-горобейникова, II — татарськокленово-конвалієва, III — татарськокленово-зірочникова, IV — татарськокленово-яглицева, V — татарськокленово-мікелієвоосокова. 1 —
чорноземні вилуговані грунти, 2 — темно-сірі лісові грунти, 3 —
чорноземні опідзолені грунти.
53
Зміна діброви татарськокленово-гірськоосокової в аналогічних умовах рельєфу дібровою татарськокленово-горобейниковою пояснюється аридизацією клімату. Розподіл двох останніх асоціацій залежить від ступеня вологості грунту.
Інший характер має розподіл асоціацій на ділянках з горбастим рельєфом, де схили більш як 15-20°,а відносна висота 150-250 м.Тут вершини плоских вододілів займає діброва татарськокленово-горобейни- кова, верхню частину північного схилу - діброва татарськокленово- конвалієва, середню - татарськокленово-зірочникова, а нижню і дно балки - татарськокленово-яглицева. Верхню прибалочну частину південного схилу займає татарськокленово-зірочникова діброва, трохи нижче розташовується діброва татарськокленово-горобейникова. На найбільш опуклих частинах схилів панує діброва татарськокленово- мікелієвоосокова. Нижче, в середній частині схилу розташовується знову діброва татарськокленово-горобейникова, яка заміщується та- тарськокленово-зірочниковою, а ще нижче - татарськокленово-яглице- вою (рис.8). Залежно від крутизни схилів та ширини балки асоціації можуть зміщуватися. Загалом відзначимо, що збільшення крутизни південних схилів і ширини балон веде до того, що асоціації а пануванням у травостої яглиці звичайної (Aegopodiua podagraria L.) та зірочника лісового (stellaria holoetea L»), зміщуються донизу, а поширюються асоціації з пануванням у травостої горобейника пурпурово- голубого ( LithospernosuB purpureo-coexuleum L») і осоки Мікелі. Зменшення крутизни схилів викликає зворотний процес і навіть веде до зникнення діброви татарськокленово-мікелієвоосокової. Наведений розподіл асоціацій пояснюється особливостями мікроклімату і характером зволоження різних частин схилів.
Західні й східні схили із збільшенням аридності мікроклімату на східних схилах займають проміжне положення між північними та південними схилами. За характером рослинності дуже круті східні схили відповідають приблизно спадистим південним схилам.
На Полтавській рівнині та Середньоруській височині в зв"яз- ку із збільшенням континентальнеєті клімату низка розглянутих асоціацій заміщується Іншими, при збереженні закономірності розміщення їх залежно від рельєфу. В північній та центральній частинах території плоскі широкі вододіли (ширше 300 м) займає діброва ліщи- ново-яглицева, вужчі - татарськокленово-зірочникова і зовсім вузькі (20-60 м) - ліщиново-волосистоосокова. Пояснюється це тим, що грунти широких плоских вододілів менше вилуґовуються і, таким чи-
54
ном , є багатшими порівняно з вужчими вододілами. Зовсім вузькі вододіли дренуються такою мірою, що на перший план виступає вже фактор висушування. Отже, розподіл першої та другої асоціацій зумовлений інтенсивністю елювіальних процесів, а останньої - характером зволоження. Цей фактор діє і в першому випадку, але менше. Верхню частину північних пологих схилів займає діброва ліщиново-маренкова, середню - ліщиново-зірочникова і нижню - ліщиново-яглицева. Верхню частину північних спадистих або крутих схилів до найбільш опуклої їх частини займає діброва ліщиново-зірочникова, нижче розміщується ліщиново-волосистоосокова, ще нижче - знову ліщиново-зірочникова, якої може й не бути, і в нижній частині схилу - ліщиново-яглицева. Верхню частину південних пологих схилів займає діброва ліщиново- волосистоосокова, середню - ліщиново-зірочникова і нижню - ліщиново- яглицева. На крутих південних схилах діброва ліщиново-волосистоосокова займає верхню і середню їх частини, а в широких балках з дуже крутими схилами частково й нижню (рис.9). В південній частині височини плоскі, широкі, трохи підвищені ділянки вододілів займає діброва ліщиново-волосистоосокова, яка в зв'язку з загальною ари- дизацією клімату заміщає діброву ліщиново-яглицеву. До верхньої частини північних пологих схилів приурочена діброва ліщиново-зірочникова, а до середньої та нижньої - ліщиново-яглицева. На спадистих і крутих північних схилах у верхній та середній частинах поширена діброва ліщиново-волосистоосокова і лише нижче - ліщиново- зірочникова і ліщиново-яглицева. На південних схилах найкрутіші верхню і середню частини їх займає діброва татарськокленово-зі- рочникова, нижче якої розміщується діброва ліщиново-волосистоосокова і вже в притальвежній частині - ліщиново-яглицева (рис.10). На дуже крутих південних схилах в їх верхній опуклій, найбільш сухій частині з'являється діброва татарськокленово-рябоперлівкова.
Неабиякий інтерес являє формування і характер розміщення асоціацій дубових лісів в однакових кліматичних умовах лісостепової зони, які, проте, різко відмінні від розглянутих едафічних умов, - на другій терасі і в заплаві. В призаплавній частині другої тераси, де найбільш виявлений гривиотий характер рельєфу і дерново- підзолисті піщані грунти, панують соснові ліси, серед яких окремими плямами на найбільш родючих ділянках трапляється діброва ліщиново- волосистоосокова. В напрямку до третьої тераси рельєф згладжується, а грунти стають багатшими, а за механічним складом більш важкими. Збільшується вміот гумусу і потужність гумусово-елювіального гори-
55
зонту, поступово починають формуватись світло-сірі, а потім і сірі лісові грунти, характерні для третьої тераси. Вершини грив стають пологіними і ширшими. В таких умовах формується діброва ліщиново- гірськоосокова, а діброва ліщиново-волосистоосокова зміщується на схили грив. Виє біля третьої тераси рівні, дещо підвищені ділянки займає діброва ліщиново-зірочникова, а знижені - ліщияово-копитня- кова. На міжгривних зниженнях росте діброва ліщиново-конвалієва (рис. 11).
Рис.9. Схематичний профіль дубового лісу через 5-й і 9-й квартали Юнаківського лісництва Сумського лісгоспзагу. Діброви: I — ліщиново-яглицева, II — ліщиново-зірочникова, III —
ліщиново-волосистоосокова, І — темно-сірі лісові грунти, 2 — сірі лісові грунти.
Рис. 10. Схематичний профіль дубового лісу через 20-й квартал Коломацького лісництва Октябрьського лісгоспзагу. Діброви: I — ліщиново-волосистоосокова, II — ліщиново-зірочникова, III — ліщиново-яглицева, IV — татарськокленово-зірочникова. 1 — чорноземні опідзолені грунти, 2 — темно-сірі лісові грунти.
В заплаві формування і розподіл дубових лісів залежить від міри виявленості процесів заплавності й седиментації. В прирусловій частині заплави, де рельєф дуже гривистий, а групти представлені найбільш легкими і бідними відмінами дернових опідзолених,
56
Рис. 11. Схематичний профіль дубового лісу через 20-й і 24-й квартали Низовського лісництва Сумського лісгоспзагу:
I — сосновий ліс, II — діброва ліщиново-волосистоосокова, III —
діброва ліщиново-гірськоосокова, IV — діброва ліщиново-конвалієва, V — діброва ліщиново-зірочникова, VI — діброва ліщивово-копитнякова. 1 — дерново-слабопідзолисті грунти, 2 — світло-сірі лісові грунти.
вершини грив займав діброва ліщиново-волосистоосокова по річках північної та середньої частин лісостепової зони і татарськокленово-зірочникова - по річках південної частини лісостепової зони. Схили грив і невисокі гриви в першому випадку займав діброва ліщиново- зірочникова, а в другому - татарськокленово-конвалівва. Відповідно в неглибоких міхгривних зниженнях формуються діброви ліщиново-ко- питнякова та ліщиново-кірказонова, а в глибших - крушиново-конвалієва. В центральній частині заплави більш вузькі і високі гриви займає діброва ліщиново-конвалієва, а нижчі й ширші (50-100 м) - ліщиново-яглицева. Схили грив і неглибокі міжгривиі зниження в відносним перевищенням до І м займає діброва ліщиново-конвалієва, а глибші міжгривні зниження - крушиново-конвалієва. В притерасній частині заплави переважає діброва ліщиново-яглицева (рис.І2).
Рис.12. Схематичний профіль дубового лісу через 59-60-й квартали Совєтського лісництва Кременчуцького лісгоспзагу.
Діброви: І - татарськокленово-зірочникова, П - татарськокленово-конвалівва, Ш - крушиново-конвалієва, ІУ - ліщиновр-хвилівнн- кова, У - ліщиново-яглицева, УІ - ліщиново-конвалієва. І - дернові опідзолені грунти.
57
В степовій зоні існування і розподіл дубових лісів визначаються характером рельєфу. Ерозійні форми рельєфу сприяють додатковому вволохенню грунтів за рахунок стікаючих з прилеглих вододілів поверхневих і грунтових вод, що в свою чергу веде до більшого вилуговування грунтів. Крім того, балки захищають ліси від безпосереднього впливу сухих вітрів і високих температур. Ліси займають схили балок, для яких характерна надзвичайна різноманітність екологічних умов, чим і пояснюється велика різноманітність асоціацій дубових лісів на одиниці площі. Асоціації розташовуються у вигляді вузьких смуг, що закономірно змінюються в напрямку від тальвега до прибалочного схилу. Ширина їх звичайно становить 10-80 м.
Як правило, верхні положисті привододільні схили балок займає діброва тарарськокленово-зірочникова, яка на вершинах північних відрогів балок заміщується дібровою татарськокленово-конвалієвою. Ця х асоціація займає середню частину схилів північної експозиції. Нижче від неї, на північному схилі розміщується діброва татарськокленово- яглицева, а вище - у верхній, найбільш опуклій частині схилу - діброва татарськокленово-кореневищноосокова. Південні схили крутизною до 20° у верхній і середній частинах займає діброва татарськокленово - рябоперлівкова. На більш крутих схилах у верхній опуклій частині формується діброва татарськокленово-мікелієвоосокова або татарськокле- яово-горобейникова. В таких випадках вище і нижче розташовується попередня асоціація. В нижній частині схилу формується діброва та- тарськокленово-кореневищноосокова або татароькокленово-зірочникова, а це нижче - татароькокленово-яглицева. На східних t західних спадистих схилах широких балок переважають діброви татарськокленово- а і рочникова, татароькокленово-рябоперл і вкова, татарськокленово-кон- валієва та татарськокленово-яглицева, розташовані у зазначеній послідовності (рис.ІЗ). Західні й східні схили вузьких балок та вершини їх за характером рослинності відповідають північним схилам. Крім перелічених асоціацій, в глибоких балках у середній і нижній частинах північних схилів трапляється ще діброва ліщиново-волооисто- осокова, а інколи і дубово-липові ліси.
При порівнянні лісів наведеного профілю з більш північними і західними (задніпровськими) байрачними лісами можна відзначити їх особливості. В північних байраках з'являються вже діброви ліщиново- зірочникова і ліщиново-яглицева. В західних байраках діброва татар- оьжокленово-кореневищноосокова зникає, проте підвищується роль діброви татарськокленово-горобейникової.
58
Рис. 13. Схематичний профіль дубового лісу через 73-74-й квартали Новопсковського лісництва Старобільського лісгоспзагу.
Діброви: I — татарськокленово-зірочникова, II — татарськокленово-кореневищноооокова, III — татарськокленово-конвалієва, IV —
татарськокленово-яглицева, V — татарськокленово-хвилівникова, VI —
татарськокленово-рябоперлівкова, VII — татарськокленово-мікелієвоосокова. 1 — чорноземні вилуговані грунти, 2 — чорноземні опідзолені грунти, 3 — темно-сірі лісові грунти.
Зупинимось нарешті на характері розміщення дубових лісів Донецького кряжу на прикладі урочища "Дерезовате" Штергреського лісництва Іванівського лісгоспзагу. Урочище площею 360 га розташоване у найвищій частині східного макросхилу кряжу. Всі рівні ділянки в межах масиву, верхні частини північних схилів, пологі схили (до 5°) південно! експозиції та спадисті й круті східної і західної експозицій займає діброва татарськокленово-зірочникова. Спадисті І круті південні схили у верхній, найопуклішій частині займає діброва татарськокленово-рябоперлівкова, яка інколи заміщується в сухіших умовах дібровою татарськокленово-горобейняковою. В середній частині цих схилів формується знову діброва татарськокленово-зірочникова, яка в самій нижній частині заміщується дібровою- татарсь- кокленово-яглицевою.
Таким чином, ми розглянули головні закономірності розподілу дубових лісів залежно від клімату та рельєфу. Численні наведені приклади зміни топографічної приуроченості асоціацій, наприклад діброви ліщиново-волосистоосокової при просуванні з півночі на південь, свідчать про певне прагнення рослинних угруповань утримати за собою властиві для них екотопи. З другого боку, деякі рослинні угруповання в різних зонах характеризуються різко виявленою
59
тенденцією до зміни екотопів. Це, наприклад, діброва ліщиново-яг- лицева, поширена від супіщаних дерново-підзолистих грунтів лісової зони до глинистих опідзолених чорноземів лісостепової зони. Виходячи з цього всі асоціації дубових лісів України ми поділяємо на стеноекотопні (діброви крушиново-молінієва, кизилово-горобейникова, татарськокленово-горобейникова та ін.) та евріекотопні (діброви лі- циново-яглицева, ліщиново-конвалієва та ін.).
ГЕОБОТАНІЧНЕ РАЙОНУВАННЯ ТЕРИТОРІЇ ПОШИРЕННЯ ЛІСІВ ФОРМАЦІЇ ДУБА ЗВИЧАЙНОГО (Querceta roboris) НА УКРАЇНІ
Диференціація рослинного покриву і всієї географічної оболонки Землі є наслідком складної взаємодії різних процесів. В основі їх, на думку М.І.Будики (1948), О.О.Григор"єва і М.І.Будики (1956),лежить насамперед зміна кількості тепла та вологи, а точніше - співвідношення кількості тепла і вологи. Засновник вчення про зони В.В.Докучаев (1948),так само як І Л.С.Берг (1947,1952), при встановленні зон дотримувався комплексного підходу. Тепер під комплексним районуванням звичайно розуміють фізико-географічне (ландшафтне, або природне) районування (Гвоздецкий, 1959; Исаченко, 1965). За С.В.Ко- лесниковим (1959), таке районування являє собою класифікацію ландшафтів на регіональній основі, що має відбивати різні угруповання реально існуючих територіальних комплексів.
Крім фізико-географічного,звичайно розрізняють ще галузеві районування за одним якимось ведучим фактором. Таким районуванням в геоботанічне. Галузеві районування - частина загального фізико- географічного районування. В зв'язку 8 цим виникає питання про спів відношення одиниць фізико-географічного та геоботанічного районування. Ю.Д.Цинзерлінг вважав, що "... геоботанічне районування повинно вбігатись з районуванням природно-географічним, тобто за природними ландшафтами* (1934, о.295). Погляди Ю.Д.Цинзерлінга поділяють С.Я .Соколов (1940), В.Б.Сочава (1952), О.В.Кумінова (1960), П.Д.Ярошенко (1961) та ін. Порівнянними ці поняття й одиниці вважав П.Г.Іоаченко (1956), але він додає, що це не означає їх повного територіального суміщення. Відсутність повного збігу меж фізико- географічного і геоботанічного районування відзначав також 1 Ф.М.Мільков (1956). Правильну позицію в цьому питанні, на наш погляд, займав ІЛ.Тумаджанов, який робить такий висновок: "Хоча геоботанічне районування і в невід"ємною частиною загального ландшафт-
60
ного районування, яке1побудоване на спільному обліку різних ознак природних ландшафтів, проте далеко не завжди і не на всіх градаціях слід чекати повного збігу відповідних щаблів супідрядних таксономічних одиниць" (1963, с.9). Наш досвід зіставлення таксономічних одиниць геоботанічного і фізико-географічного районування України і границь виділених таксонів підтверджує, що повного збігу між ними немає. Це в загальних рисах пояснюється тим, що єдність За одним фактором і відсутність за низкою інших обумовлюється різним ходом і характером притаманних їм процесів, тому при комплексному підході часто доводиться орієнтуватись на опосередковані показники або віддавати перевагу одному чи двом з них. Як правило, це - грунти, рослинність або рельєф.
З питанням, яке ми розглянули, пов"язане питання про ознаки, які мають бути покладені в основу при виділенні геоботанічних регіонів: зовнішні умови чи сама рослинність. Ясно й чітко на нього відповів О.П.Шенников: "... при районуванні слід виходити з ознак самої рослинності і тільки рослинності" (1940, с.23). Про це писали також Є.М.Лавренко (1947а, 1950, 1968), М.О.Прозоровський і
С.М.Тюремнов (1967), М.О.Гвоздецький (1967) та ін. Головним доказом на користь такого підходу є те, що в протилежному випадку геоботанічне районування не буде відрізнятись істотно від фізико-географічного, а ботанічні об'єкти опиняться на другому плані.
З першого положення безпосередньо випливає і друге, сформульоване Є.М.Лавренком (1947а): необхідно грунтуватись на облікові складу едифікаторів, в першу чергу плакорних формацій. Звідси ми робимо також принципово важливий висновок: межі таксонів великих таксономічних одиниць слід проводити по границі суцільного ареалу зональних едифікаторів, виділяючи території а фрагментарним поширенням їх як самостійні округи, але вже інших таксонів. За третім положенням, висунутим В.Б.Сочавою (1952), в основу геоботанічного районування треба покласти класифікацію рослинності, а за четвертим, висунутим П.Д.Ярошенком (1961), - і карту рослинності.
Звичайно при геоботанічному районуванні користуються такою системою таксономічних одиниць: область або зона, підобласть, провінція, підпровінція, округа, підокруга 1 район. За цією системою, за винятком останньої одиниці, було проведено природне геоботанічне районування всієї території СРСР. В.Б.Сочава (1968) запропонував іншу систему, а саме: планетарний пояс, геоботанічний 61
пояс, система типів рослинності, світи типів рослинності, геоботанічна область, зона рослинності, провінція, макрофітоценохора, фіто- ценохора, мезофітоценохора, мікрофітоценохора і елементарна фіто- ценохора. В цій системі геоботанічний пояс відповідає об'єднанню домініонів Є.М.Лавренка (1968), а геоботанічна область наближається аа своїм змістом до "країни", прийнятої географами. Вищі таксономічні одиниці геоботанічного районування і класифікації рослинності є загальними. Системи близькі до найбільш поширеної, розробленої в Ботанічному інституті ім. В.Л.Комарова AH СРСР, опубліковані М.О.Про- еоровським і С.М.Тюремновим (1967) та С.М.Тюремновим і П.П.Жудовою (1967).
Нами прийнято такі таксономічні одиниці; район, округа, під- провінція, провінція, зона та домініон.
Район розглядається як основна одиниця геоботанічного районування. Характерним для нього є однорідний рослинний покрив, на підставі якого і виділяють його (Прозоровский і Тюремнов, 1967). Варіювання рослинності всередині району, як цілком вірно відзначає М.А.Голубець (1967), має бути меншим, ніж між районами.
Округа відзначається, за Є.М.Лавренком (1947а, 1968), певними сполученнями типологічних одиниць рослинного покриву, які залежать від грунтово-геоморфологічних умов. М.О.Прозоровський і С.М.Тюремнов (1967) пропонують виділяти її за перевагою в плакорних умовах будь-якої однієї рослинної формації. З цією пропозицією не можна погодитись, бо вона викличе виділення дуже великих територіальних одиниць. Наприклад, через перевагу формації дуба звичайного весь Лівобережний лісостеп України треба було б віднести до однієї округи. На нашу думку, при виділенні округи необхідно грунтуватись на групах асоціацій і співвідношенні формацій плакорних та непла- корних.
Підпровінція характеризується відмінами в складі співдоміную- чих видів плакорних формацій (Лавренко, 1947а, 1968). Межі підпро- вінцій, як t провінцій, на нашу думку, доцільно встановлювати по границі суцільного поширення едифікатора або субедифікатора планерної формації, виділяючи територію острівного поширення їх в ранзі округ другої підпровінції або провінції.
Провінція, на думку Є.МЛавренка (1947а, 1968), виділяється 8а складом плакорних формацій.
Область Є.М.Лавренко (1947а, 1968) встановлює за перевагою на плакорах зонального типу рослинності з супровідними типами 62
позаплакорної рослинності. В такому розумінні "область" часто збігається з "зоною", принаймні на території України, але вона вужча, ніж "зона" в трактовці М.О.Прозоровського і С.М.Тюремнова (196?). Області Є.М.Лавренко (1968) об"єднує в домініони. Вся територія СРСР належить до Голарктичного домініону.
Схема геоботанічного районування території дослідження
Перше геоботанічне районування України провів О.В.Фомін (1925-1933), який виділив лісову та степову зони і лісостепову під- зону. Кожна зона поділялась на райони, що являли собою географічні або іноді зональні категорії і в наш час не відповідають ніяким таксонам геоботанічного районування. Більш детальне й обгрунтоване геоботанічне районування України, яке послужило основою для всіх наступних районувань, розробили Є.М.Лавренко (1927, 1930а, 1938а) та Є.М.Лавренко t П.С.Погребняк (1930). Взявши за основу зональний поділ О.В.Фоміна, Є.М.Лавренко в перших двох працях поділив усю територію на 15 геоботанічних районів, які наближаються до округ в сучасному розумінні. В пізнішій праці Є.М.Лавренко (1938а) поділяє територію України на три провінції. Найдетальніше районування України дано в праці Є.М.Лавренка та П.С.Погребняка (1930). В ній автори на території України виділяють 27 районів. В багатьох випадках межі районів майже збігаються з межами округ, виділених пізніше Г.І.Біликом та Є.М.Брадіо (1962). Геоботанічне районування інших авторів (Кожевников, 1936; Котов, Гринь, 1953) по суті аналогічне районуванню Є.М.Лавренка (1938а).
Прийняте в наш час геоботанічне районування України в своїх головних рисах розроблене Є.М.Лавренком (19476). Лісостепову під- зону він виділив як самостійну зону (область), в межах кожної зони виділив провінції, підпровінції та округи. Описувана в цій праці територія, за районуванням Є.М.Лавренка, поділяється на 3 області, 4 провінції, б підпровінцій та 17 округ.
Пізніші доповнення були внесені Г.І.Біликом і Є.М.Брадіс (1962), які прийняли в основному ту саму систему таксономічних одиниць і виділили в більшості випадків аналогічні таксони. Загалом вони уточнили або переглянули границі ряду округ t виділили чотири нові. За найбільшу таксономічну одиницю автори прийняли "країну", прийняту географами. Тим самим вони відійшли від двох головних принципів геоботанічного районування - зонально-провін- ціального та районування за самою рослинністю, замінивши їх геоморфологічним принципом, з чим ми не можемо погодитись.
63
Детальне геоботанічне районування окремих невеликих територій за останні двадцять років подавали М.І.Косець (1953), Є.М.Брадіс (1957), В.В.Осичнюк (1958), Г.В.Козій (1963), Ю.Р.Шеляг-Сосонко (19666) та І.В.Артемчук (1967). Районування відносно великих територій опублікували В.О.Поварніцин (1959), М.А.Голубець, К.А.Ма- линовський, С.М.Стойко (1965), Г.І.Білик (1970) і Ю.Р.Шеляг-Сосон- ко, Г.С.Куковиця (1971).
На підставі критичного перегляду опублікованих даних геоботанічного районування України і зібраних матеріалів з рослинності республіки ми пропонуємо наведену нижче схему геоботанічного районування (рис.14). Вона розроблена на основі викладених на початку розділу теоретичних положень та схем геоботанічного районування Є.М.Лавренка (19476,в,г), Г.І.Білика і Є.М.Брадіс (1962).
І. Європейська широколистянолісо- в а зона займає Полісся,північно-західну частину Середньорусь- кої височини і Карпати. На південь від неї лежить Євразійська лісостепова зона, відносно якої між геоботаніками, географами і грунтознавцями розходжень немає, тому встановлення південної і західної меж широколистянолісової зони має принципове значення для будь-якого районування. Проте єдиного погляду на межі між зазначеними зонами ще немає. Географи (Борг, 1947, Мильков, 1950, 1964; Ланько та ін., 1959; Геренчук, 1968; Ланько, 1968) і грунтознавці (Прасолов, 1939; Вернандер та ін., 1951; Андрющенко та ін., 1967) північну межу лісостепу проводять по південній межі Полісся. Отже, 2, 6, 7, 8 і П-ту округи (див.рис.І4) вони відносять до зони лісостепу. Обгрунтуванням північної межі лісостепової зони, на думку географів (Берг, 1947; Мильков, 1952, 1956), є головний ландшафтний рубіж, з яким збігаються південна межа ялини і лінія суцільного поширення сірих лісових грунтів на лесах. Цей рубіж характеризується зміною позитивного балансу вологи на негативний і звичайно встановлюється на підставі низки інтегральних кліматичних індексів, що виявляють зв'язок між кліматичними факторами і зональним поширенням ландшафту. Найбільш вдалим з них вважають коефіцієнт зволоження і радіаційний індекс сухості (Исаченко, 1965).
Згідно з даними М.М.Іванова (1948), лісостепова зона лежить в межах території, річний коефіцієнт зволоження якої дорівнює 0,99- 0,60. З північною границею лісостепу цей коефіцієнт збігається лише на території Лівобережної України. На Правобережжі коефіцієнт зво-
64
Рис. 14. Схематична карта геоботанічного районування досліджуваної території. 1 — межі зон, 2 — межі провінцій, 3 — межі підпровінцій, 4 —
межі округ. I. Європейська широколистянолісова зона. A. Східноєвропейська широколистянолісова провінція: а) Поліська підпровінція — 1. Західнополіська, 2. Центральнополіська, 3. Придніпровсько-Поліська, 4. Східнополіська і 5. Малополіська округи; б) Середньоруська підпровінція — 6. Кролевецько-Глухівська округа; в) Подільсько-Бесарабська підпровінція —
7. Волинська лесова, 8. Західноподільсько-Покутська, 9. Вінницько- Могилів-Подільська і 10. Прутсько-Дністровська округи. B. Середньоєвропейська широколистянолісова провінція: г) Західноукраїнська підпровінція — 11. Кременецько-Хотинська округа; д) Карпатська підпровінція — 12. Самбірсько-Стрийська, 13. Калусько-Вашківцівська і 14. Закарпатська притисенська округи. II. Європейсько- Сибірська лісостепова зона. C. Східноєвропейська лісостепова провінція: е) Подільсько-Придніпровська підпровінція — 15. Західноподільська, 16. Старокостянтинівсько-Білоцерківська, 17. Козятинсько-Канівська, 18. Бершадьсько-Котовська, 19. Голованівсько-Новомиргородська і 20. Чигиринсько-Знаменська округи; ж) Середньодніпровська підпровінція — 21. Лівобережнопридніпровська терасова, 22. Сумсько-Гадяцька і 23. Харківсько-Зміївська округи. III. Євразійська степова зона. D. Східноєвропейська (понтична) провінція: з) Чорноморсько-Азовська підпровінція — 24. Придністровська, 25. Бузько-Дніпровська, 26. Присамарська, 27. Куп’янсько-Старобільська та 28. Донецька байрачні округи.
ложення, наприклад, для Тернополя, розташованого в лісостеповій зоні, дорівнює 1,2 (Попов, 1958). Не збігаються з західною частиною межі лісостепової зони і гідротермічний та плювіотермічний коефіцієнти. Таким чином, зазначена вище межа між зоною змішаних 65
лісів 1 лісостеповою зоною на Правобережній Україні не відповідав головному ландшафтному рубежеві. Не збігається на території Україні південна межа зони змішаних лісів і з межею острівного поширення ялики європейської (Picea abies (L.)Kajisten). Остання проходить на 50-150 км далі на північ від південно! межі зони змішаних лісів. Не можна прийняти південну межу ялини європейсько! за межу між вказаними зонами і для сусідньої з Україною території Польщі,де вона на більшій частині зони змішаних лісів не росте.
Північна межа суцільного поширення сірих лісових грунтів збігається з межею, яка розділяв зону широколистяних лісів і зону лісостепу по лінії Володимир-Волинський - Кролевець. Але цей показник для розмежування зазначених зон не може бути прийнятий, тому що північну межу лісостепу визначав чергування сірих лісових та чорноземних грунтів.Саме такий характер мав границя, яку розглядаємо на ділянці Шепетівка - Конотоп. На захід від Шепетівка сірі лісові грунти чергуються з опідзоленими чорноземами, на яких ростуть ліси. Ділянки степової рослинності, що збереглися на цій території, пов"язані виключно зі схилами різної крутизни, на вододілах їх немає. Як правило, це схили південних румбів-з малопотужними дерново-карбонатними грунтами. Не можна прийняти за межу між широколистянолісовою і лісостеповою зонами й північну межу суцільного поширення лесу. Ці зони збігаються на правобережній частині України і не збігаються на лівобережній. Таким чином, ні з позицій ландшафтного районування, ні з позицій геоботанічного районування не можна погодитись з пропозицією географів і грунтознавців проводити північну межу лісостепу по північній окраїні Полісся. Різниця між позицією вищезгаданих дослідників та нашою пояснюється двома причинами. По-перше, Полісся вони розглядають як зональну одиницю, отже, все, що лежить далі на південь, повинно відноситись до лісостепу. З цим не можна погодитись, тому ЩО Полісся утворилось на місці тектонічного зниження Руської рівнини і своєрідність його ландшафту зумовлена геоморфологічними умовами. По-друге, зазначені дослідники не виділяють зони широколистяних лісів, яка на території України була розірвана льодовиком і збереглась на рівнині лише у вигляді двох язиківгохідного (&-та округа) та західного (7, 8, 9, 10 і П-та округи). Східна межа західного язика широколистяних лісів є водночас і межею лісостепової зони. Ми проводимо її по лінії більш або менш суцільного поширення степової та лісової рослинності. Тому межиріччя Стрипа-Серет, де в плакорних умовах є
6S
лише одна ділянка ковилового степу (формації ковили найкрасивішої - Stipeta pulcherrimae і ковили довголистої - Stipeta longifollae), рослинність якої зумовлена особливостями гідрологічного режиму в умо вах близького до поверхні залягання карбонатних порід, відносимо також до широколистянолісової зони.
А. Східноєвропейська широколистянолісова провінція характеризується на Україні пануванням лісів з едифікатором першого рангу - дуба звичайного і едифікатором другого рангу - сосни звичайної (Pinus silvestris L.).Значну роль відіграє в лісах семіедифікатор - граб звичайний (Carpinus betulus Lt), який далі на схід зникає. На підставі різної ролі в лісах перелічених видів Є.МЛавренко (19476) поділяє провінцію на три підпровінції: Поліську, Середньоруську і Подільсько-Бесарабську.
а) В Поліській підпровінції на межиріччях за зайнятою площею переважають соснові та дубово-соснові ліси. Менш поширені дубові ліси. Залісненість у західній частині території складає 35, в східній - 18—20%. Крім лісів, зустрічаються болота, які займають близько 5%,і луки, на які припадає майже 8% всієї території. Серед боліт переважають евтрофні, але значне поширення в західній частині мають мезотрофні та оліготрофні болота. Луки переважно заплавні (болотисті, торфові та справжні), менш поширені суходольні (пус- тищні й торф’янисті).
І. Західнополіська округа. Східну межу її ми проводимо на 100-120 км. західніше, ніж за схемою, В.О.Поварніцина (1959) і Г.І.Білика та Є.М.Брадіс (1962). Пояснюється це тим, що на території, яку розглядаємо, переважають дубово-соснові, дубові та дубово- грабові ліси, характерні для Центральнополіської округи, до якої мр цю територію відносимо. Західнополіська округа характеризується перевагою соснових лісів, представлених переважно сосняками дов- гомошними, зеленомошними та сфагновими. Значно меншу роль відіграють дубово-соснові та дубові ліси, які трапляються в комплексі з вільховими та зрідка вільхово-ялиновими лісами. Для дубових лісів найбільш характерними є діброва ліщиново-квасеницева і менше - ліщиново-веснівкова. В південній частині округи трапляються діброви ліщиново-копитнякова, ліщиново-зірочникова та ліщиново-маренко- ва. Характерною для округи є найбільша на Україні заболоченість, що дорівнює 11% (Брадіс, Лихобабіна, 1969). За площею серед боліт переважають евтрофні болота (Брадіс, 1969). Крім них, значні площі займають мезотрофні й оліготрофні лісові та рідколісні сфагнові 67
болота. Цікаво відзначити, що своєрідність рослинного покриву округи виявляється 1 в заплавних луках, представлених переважно болотистими луками формації осоки стрункої (Cariceta gracilia ) та лепешняку великого ( Glycerleta maximae). Менш поширені торфянисті луки, представлені формацією щучника дернистого ( Deschampsleta caes- pitosae). Таких великих площ, як у Західнополіській окрузі, перелічені формації не утворюють ніде. З флористичного боку для заплавних луків округи характерними є середньоєвропейські види, такі, як трясучка середня (Bliza media 1>«)та гребінник звичайний (Cynosurus cristatus L.), а також надзвичайна бідність на бобові.
2. Центральнополіська округа характеризується перевагою дубово-соснових та дубових лісів. Значну роль відіграють і їх похідні березові ліси. Серед дубових лісів найбільш характерною та поширеною є діброва ліщиново-трясучковидяоосокова. Менш поширеною, але також характерною є діброва крушиново-трясучковидноосокова. Крім них, трапляються діброва ліщиново-зірочникова, ліщиново-конвалієва і в південній частині округи - ліщиново-волосистоосокова. Болота займають близько 3,5% території і належать майже виключно до евтроф- них. Заплавні луки двох найбільших річок округи представлені переважно справжніми луками (формації мітлиці білої - Agrostideta albae , лисохвосте лучного - Alopecureta pratensis і костриці лучної Festucata pratensis ) і в меншій мірі болотистими і торф'янистими (Шеляг-Сосонко, Кузьмичов, 1967).
3. Придніпровсько-Поліська округа. В.О.Поварніцин (1959) її не вибіляє, а Г.І.Білик і Є.М.Брадіс (1962) виділяють лише на підставі рослинності заплави Дніпра та його другої тераси і,отже, встановлюють округу не за плакорною рослинністю. Практично в даному випадку округа відповідав геоботанічному району, з чим не можна погодитись.
До Придніпровсько-Поліської округи ми відносимо територію, на якій переважають дубово-соснові та соснові ліси. Дубові ліси трапляються зрідка. Найбільш поширеними серед них є діброви ліщиново- веснівкова та ліщиново-зірочникова. Болота переважно евтрофні, займають 2,5% площі.
4. Східнополіська округа окреслена нами в загальноприйнятих межах. Найбільш поширені ліси її - соснові та дубово-соснові. В минулому значну роль відігравали й дубові ліси, які тепер значно вирубані. Представлені вони головним чином дібровами ліщиново-яглице- вою, ліщиново-копитияковою та ліщиново-зірочниковою. Характерними 68
для округи є липово-дубові ліси, а також те, що в західній її частині проходить східна межа ареалу осоки трясучковидяої, а майже посередині - східна межа граба звичайного. Болота займають близько 4% площі округи, вони майже виключно евтрофні (Брадіс, Лихобабіна, 1969).
5. Малополіська округа також прийнята нами без змін загальноприйнятих меж. В рослинному покриві цієї округи переважають соснові та дубово-соснові ліси. Дубові ліси займають незначну площу, найпоширенішими серед них є діброви ліщиново-трясучковидноосокова та ліщиново-яглицева. З флористичного боку ці ліси характеризуються наявністю ряду середньоєвропейських видів, наприклад астранції великої (Astrantla major ь.). Заболоченість Малополіської округи становить близько 5%.
б) Середньоруська підпровінція. В рослинному покриві її переважають дубові ліси. Значно рідше трапляються дубово-соснові та липово-дубові ліси. Болота й луки мало поширені. На території України до цієї підпровінції належить лише одна округа.
6. Кролевецько-Глухівська округа окреслюється нами в загальноприйнятих межах. Дубові ліси округи представлені переважно дібровами ліщиново-яглицевою та ліщиново-зірочниковою. Серед липово-дубових лісів переважають липово-дубові ліси волосистоосокові. Середньоєвропейських видів у лісах округи немає. Характерною рисою рослинності є дві невеликі (близько 2 га) екстразонально-азональні ділянки степової рослинності, представленої формацією костриці борознистої ( Festuceta sulcatae), Дуки переважно справжні (формації сто- колосу безостого - Brometa inermis, МІТЛИЦІ білої - Agrostideta albae, костриці лучної -Festucata pratensis та ін.) і болотисті (формації осоки стрункої - Oariceta gracilis та бекманії звичайної - Beckmannieta cruciformis ).
в) Подільсько-Бесарабська підпровінція. Рослинний покрив її утворюють дубові та дубово-грабові ліси. В середньому залісненість становить 11-12%. Болота і луки займають близько 2% площі. Характерною рисою підпровінції є наявність екстразонально-азональних ділянок степів на стрімких схилах.
7. Волинська лесова округа належить до найбільш спірних щодо зональної та провінціальної належності. Є.М.Лавренко (1947а) відніс її до Поліської підпровінції широколистянолісової зони. Г.І.Білик
і Є.М.Брадіс (1962) виділили її як лісостепову, а в номенклатурному списку залишили в Поліській підпровінції. До лісостепу відно69
сять цю округу грунтознавці й географи. І.С.Амелін (1962) та Г.В.Ко- зій (1963) вважають її лісовою. Таким чином, нез"ясованими лишаються два питання: І) що являв собою ця територія за характером рослинності; 2) чим треба керуватись при віднесенні округ до підпровінції - принципом територіальної цілісності чи принципом близькості рослинного покриву.
В наш час на Волинській височині переважають дубово-грабові ліси та їх похідні грабові ліси. Значно менш поширені дубові та дубово-соснові ліси. Останні пов"язані з другими терасами річок і їх можна не брати до уваги. Найхарактернішими серед дубових лісів в діброви ліщиново-яглицева та ліщиново-маренкова. Степова рослинність майже не збереглась. За даними С.Мацко (Маско, 1938),на стрімких схилах трапляються лише окремі екземпляри степових видів, зростання яких обумовлене едафічними факторами. Клімат Волинської височини мав явно виявлений "лісовий1* характер. Річна кількість опадів становить 600-650 мм, середня температура липня дорівнює І8°С. Коефіцієнт зволоження І,1-І,2 (Бучинский, 1958). Головним доказом на користь думки про лісостеповий характер території Волинської височини є поширені подекуди глибокі малогумусові чорноземи. Генезис їх не зовсім ясний. На нашу думку, вони могли сформуватись і під дубовими лісами, оскільки на Волинській височині є головна для цього умова - близьке до поверхні залягання крейди. Дослідження дубових лісів височини, які збереглися, показали, що в їх складі повністю відсутні асоціації союзу Quercion pubescenti-petraea, характерні для лісостепу. Це дає нам підстави розглядати Волинську лесову округу як лісову, а не лісостепову.
Плакорна рослинність Волинської лесової округи подібна до рослинності Західноподільсько-Покутської округи і різко відрізняється від рослинності Поліської підпровінції. Територіально Волинська лесова округа відділена від Західноподільсько-Покутської округи вузькою смугою Малополіської округи. Геоморфологічно територія останньої - знижена рівнина водно-льодовикового походження, яка розсікав генетично єдину височину на дві частини - Волинську і Подільську височини. За характером рельєфу, грунтами та рослинністю вони єдині, тому відносити Волинський лесовий острів до Поліської підпровінції вважаємо недоцільним.
8. Західноподільсько-Покутська округа виділяється нами. Характерними для неї в дубово-грабові, дубові та острівні букові ліси. Дубові ліси представлені переважно дібровами ліщиново-яглице- 70
вою, ліщиново-зеленчуковою," ліщиново-зірочниковою, ліщиново-воло- систоосоковою та ліщиново-конвалієвою. З’являється на території округи і діброва свидиново-гіроькоосокова, характерна для лісостепової частини Подільської височини. В південній частині округи на схилах збереглися невеликі ділянки степової рослинності, представленої формаціями костриці борознистої - Festucata sulcatae, осоки низької - Oariceta humilis t ковили волосистої -Stipeta capillatae (Шеляг-Сосонко, Байрова, 1965).
9. Вінницько-Могилів-Подільська округа окреслюється нами в загальноприйнятих межах. В рослинному її покриві переважають дубово-грабові ліси та їх похідні - грабові. Значно рідше трапляються дубові ліси, найхарактернішою асоціацією яких є діброва свидино- во-гірськоосокова. В придністровській частині округи, так само як
і в попередній, на стрімких схилах ростуть дубові ліси класу асоціацій Querceta comosa. На території округи проходить східна межа середньоєвропейських видів, таких, як рівноплідник рутвицелио- тий (laopyruia thalictroides L.), кадила мелісолисте ( Melittis inelissophyllum L.) j скополія карніолійська.
10. Прутсько-Дністровська округа виділяється нами. Рослинність її представлена дубовими лісами з дуба звичайного і дуба скельного (Quercus petraea L і e b 1.) та дубово-грабовими. Дубових овидиново-гіроькоосокових лісів, характерних для попередньо! округи, немає. Найбільш поширені діброви ліщиново-волооиотоооокова та ліщиново-яглицева.
В. Середньоєвропейську широколистянолісову провінцію на рівнинній частині території України Є.М.Лавренко (19476), а також Г.І.Білик і Є.М.Брадіс (1962) не виділяють, хоча в літературі (Qajewski, 1937) є вказівки на доцільність віднесення деяких рівнинних районів західної частини України до цієї провінції. Як відомо, ця провінція характеризується пануванням на рівнині І в гірському лісовому поясі лісів союзу Fagion (Domin, 1952» Tuxen, 1937» 1955; Moor, 1938, 1960; KnapP» 1948; Lebrun et alM 1949; Klika 1955» Oberdorfer, 1957; Medwecka-Korпав, Kornad, Pawlowakl, 1959; Borza, 1963; Braun-Blanquet, 1964; Sod, 1964; Holub et al,, 1967; Passarge, 1968X Тому східну границю Середньоєвропейської провінції доцільно проводити по межі суцільного поширення букових лісів. На Україні, крім Карпат, суцільні масиви їх трапляються на Розточчі, Гологоро-Кременецькому кряжі, в Опіллі та на Хотинській височині (Косец, 1947; Горохова, 1960; Заверуха, 1963). Цю тери-
71
торію, на нашу думку, слід віднести до Середньоєвропейської провінції, виділивши її в окрему підпровінцію, яку пропонуємо назвати Західноукраїнською.
г) Західноукраїнська підпровінція характеризується поширенням дубово-грабових, букових та в невеликій кількості дубових і дубово- соснових лісів. До неї належить тільки одна Кременецько-Хотинська округа. На схід від цієї округи букові ліси трапляються лише у вигляді окремих островів на підвищених ділянках. До границі суцільного поширення їх 40-60 км. У флористичному відношенні букові ліси Кременецько-Хотинської округи стоять ближче до карпатських букових лісів, ніж до острівних букових лісів Західноподільсько-Покутської округи.
Саме на території Кременецько-Хотинської округи проходить межа більшості видів, характерних для карпатських дубових лісів: ялиці білої(АЬіев alba М і 1 1.),пренанта пурпурового ( Prenantbea purpurea L.), ожики лісової ( Luzula silvatica (Н u d s.) G a u d.), купини кільчастої ( Polygonatum verticillatum (L.) A 1 1.),кремпя- ника гарного ( Telekia speciosa (S о b r e b.) B a u m g,), білоцвіту весняного ( Leucojun vernum ь.) та ін. Трапляються в межах цієї округи також невеликі ділянки ацидофільних дубових лісів з дуба скельного та дуба звичайного союзу оцегсіоп robori-pe- traea, характерного для Північної рівнини Середньої Європи (Гихеп 1937, 1955; Braun-Blanquet, Tiixen, 1943: Braun-Blanquet, 1964). Загалом у дубових , дубово-грабових і букових лісах цієї округи досить звичайними є середньоєвропейські види: апозерис смердючий, аконіт молдавський (Aconitum moldavicum Нас k.), астран- ція велика, осока трясучковидна, карпезій пониклий (Oarpesium dernuum L.), осот клейкий (Oirsium erisithales (J а с k.) S c o p.), фітеума куляста (Phyteuma orbiculare L.), багаторядник Брауна (Polystichum braunii F e e), живокіст серцевидний (Symphytum cordatum w . K.) та інші, які далі на схід не зустрічаються або зустрічаються дуже рідко. Степова рослинність на території Кременецько- Хотинської округи пов"язана виключно 8 місцями, де близько від поверхні (до 30-40 см) залягають карбонатні породи, тобто поширення її зумовлене едафічними факторами і аналогічне поширенню степових угруповань на сусідній території Польщі (Kozlowska, 1931; Slawinski, 1952; Izdebski, 1958, 1959; Medwecke-Kornas', Kor- nas,I959), ЩО Відноситься, за Й.Браун-Бланке (Braun-Blanquet, 1964), до Середньоєвропейської (Центральноєвропейської) провінці ї.
72
В межах Кременецько-Хотинської округи трапляються описані нами заплавні та суходольні луки, представлені формаціями райграса високого - Arrhenathereta elatior та трищетинника жовтуватого - Trlseteta flavescentis, які належать ДО порядку Arrhenatheretalia, характерного для Середньоєвропейської провінції (тііхеп, 1937і Кпарр, 1948; Oberdorfer, 1957S Hundt, 1953* Medwecka-Kornas, Kornas', Paw- lowski, "1959» Passaxge, 1959, 1964* Matuszkiewioz, 1962* Borza,196J, 1966; soo,1964). Лише в цій окрузі на степових схилах трапляються в"язіль звичайний ( Coronilla coronata L.), очиток іспанський ( Sedum hispanicum L.)і скорзонера іспанська ( Scorzonera hispanloa L.), які мають середземноморсько-середньоєвропейський тип ареалу (Шеляг-Сосонко, Куковиця, 1970а).
11. Кременецько-Хотинська округа виділена нами. Рослинність її представлена буковими, дубово-грабовими, хвойно-широколистяними і дубовими лісами. Останні утворені дібровами ліщиново-маренковою, ліщиново-яглицевою, ліщиново-зірочниковою та ліщиново-трясучковид- ноосоковою. Степова рослинність складена формаціями осоки низької - Cariceta humilis, ковили довголистої -Stipeta longifoliae і ковили найкрасивішої - Stipeta pulcherrimae.
д) Карпатська підпровінція приймається нами без змін її за- гальноприйнятих-меж. В рослинному покриві її переважають букові та ялинові ліси, рідше трапляються дубові, грабово-дубові та ялицеві. Крім лісів, широко представлені справжні і пустищні суходільні луки. Г.І.БІлик і Є.М.Брадіс (1962) в межах підпровінції виділяють три округи; Передкарпатську, Східнокарпатську гірську та Закарпатську притисенську.М.А.Голубець,К.А.Малиновоький.С.М.Стой- ко (1965), залишивши першу і останню округи, поділили Східнокарпатську гірську округу на 5 округ. Лісів, досліджуваних нами, в них немає, тому на них не зупиняємось. Передкарпатську округу ми поділяємо на дві округи - Самбірсько-Стрийську та Калусько-Вашків- цівську. Залісненість їх становить 25-30%.
12. Самбірсько-Стрийська округа характеризується перевагою в рослинному покриві дубових лісів. Значно менше поширені тут дубово-грабові ліси. Букові, ялинові та яглицеві трапляються зрідка
в підгір“ї. Найпоширенішою серед дубових лісів є діброва ліщиново- трясучковидноосокова, рідше трапляються діброви ліщиново-волосистоосокова, ліщиново-зірочникова та ін. З флористичного боку для них характерна наявність значної кількості середньоєвропейських гірських видів, таких, як тирлич ваточниковидний (Gentiana моїарі- 73
adea L.), купина кільчаста, пренант пурпуровий, сугайник австрійський (DoxonXcum austriacum J a c q.), аконіт волотистий ( Aconitum paniculatuia l а в.)та ін. Порівняно з сусідньою ця округа значно більша заболочена (близько 2% території). Значна кількість асоціацій болотної рослинності її (Calamagrostis lanceolata - Carex appropinquata - Sphagnum teres; Oalamagrostis LanceoLata-Oxycocaus quadcipetalua - Sphagnum teres та ІН.) в сусідній окрузі не трапляється. Значно більші площі, порівняно з наступною округою, займають і луки, представлені головним чином класом формацій справжніх і болотистих луків (Шеляг-Сосонко, 1963а,б).
13. Калусько-Вашківцівська округа від попередньої відрізняється перевагою в рослинному покриві букових лісів. Значно менше поширені ялицеві, ялинові та дубово-грабові ліси. Дубові ліси трапляються лише в західній частині округи.
14. Закарпатська притисенська округа приймається нами без змін її загальноприйнятих меж. Переважають в окрузі дубові ліси з дуба звичайного, найбільш характерною серед них є діброва ліщино— во-тряоучковидноооокова. Флористично від цієї ж асоціації, поширеної на території Самбірсько-Стрийської округи, діброва ліщиново- трясучковидноосокова відрізняється наявністю клена татарського (Acer tataricum L.), глоду колючого (Crataegus oxyacantha L.), гордовини (Viburnum Lantana L») та інших більш термофільних видів.
П. Європейсько-Сибірська лісостепова зона займає східну частину Подільської та більшу частину Придніпровської і Середньоруської височини і Лівобережної низовина. Південну межу зони проводимо за районуванням Г.І.Білика і Є.М.Брадіс (1962), з відхиленням у західній та найбільш східній частинах її. В останній межу слід перенести на 40-60 км на північ, оскільки тут поширені байрачні дубові ліси з типовими для них асоціаціями. В звнязку з цим ніяк не можна погодитись з пропозицією Ф.М.Мількова (1950, 1964) проводити південну межу лісостепу в смузі звичайних чорноземів, тобто на 60-250 км півден- півдеиь, ніж за розглянутими схемами геоботанічного та фізико-географічного районування України. Твердження Ф.М.Мількова (1950, 1964), що на цій території є ділянки вододільних лісів, наші дослідження не підтвердили. У всіх випадках зростання дубових лісів на вододілах маємо справу з середбалочними вододілами, а не міжбалочними, що не може бути підставою для перенесення межі зони в південному напрямку.
74
Рослинність лісостепової зони представлена головним чином лісами (11%). Домінують серед них дубові, дубово-грабові та дубово- липові ліси. На других терасах трапляються дубово-соснові й соснові ліси. Луки і болота займають близько 4% території і збереглися лише в заплавах річок. Болота майже виключно евтрофні, переважають трав"яні, менш поширені трав"яно-мохові. Луки крупнозлакові справжні, рідше болотисті та остепнені. Степова рослинність збереглась лише на непридатних для розорювання схилах та на заповідній ділянці "Михайлівська цілина". Представлена вона формаціями костриці борознистої - Featuoeta auloatae, стоколосу прибережного - Broaeta riparii (у східній частині), ковили волосистої - Stipeta capillatae, тонконога вузьколистого -Poeta anguatifoliae та осоки низько! - Cariceta Ьшаі1іа(в західній частині). Є.М.Лавренко (І947в), а також Г.І.Білик t Є.М.Брадіо (1962) весь лісостеп України відносять до Середньодяіпровської підпровінції Східноєвропейської лісостепової провінції. Ми вважаємо за доцільне в Середньодніпровській підпровінції залишити лише лісостеп Лівобережної рівнини, а лісостеп Подільської і Придніпровської височин виділити в окрему Подільсько- Придніпровську підпровінцію, на тій підставі, що на плакорах цієї території поширені дубово-грабові ліси, які далі на схід трапляються вже окремими островами на найвищих прирічкових правобережних ділянках. Із східною межею виділеної підпровінції збігається і східна границя суцільного поширення черешні. В межах цієї підпровінції майже не трапляються липово-дубові ліси, значно поширені на Лівобережній Україні. Таким чином, на схід від Подільсько- Придніпровської підпровінції відбувається зміна головних співдомі- нуючих видів плакорних формацій, на підставі чого, за Є.М.Лаврен- ком (1947а, 1968), і виділяються підпровінції.
е) Подільсько-Придніпровська підпровінція характеризується перевагою дубово-грабових та дубових лісів.
15. Західноподільська округа. До неї відносимо територію, обмежену на сході лінією острівного поширення букових лісів, що відповідає Товтровому кряжеві. На схід від кряжу в межах підпровінції немає формації осоки низької. На території округи переважають дубово-грабові ліси. Для дубових лісів найбільш характерною є діброва свидиново-гірськоосокова, крім неї трапляються діброви ліщи- ново-яглицева, ліщиново-волосистоосокова, ліщиново-копитнякова та деякі інші.
16. Старокостянтинівсько-Білоцерківська округа приймається
75
нами баз змін її загальноприйнятих меж*. В ній переважають дубові ліси, представлені дібровами свидиново-гірськоосоковою, ліщиново- гірськоосоковою, ліщиново-яглицевою та ліщиново-зірочниковою. Дубово-грабові ліси трапляються значно рідше, ніж у попередній окрузі.
17. Козятинсько-Канівська округа прийнята нами в межах, запропонованих Г.І.Біликом (1970). В окрузі переважають дубово-грабові та дубові ліси. Найбільш характерні серед останніх діброви ліщиново-гірськоосокова і татарськокленово-гірськоосокова.
18. Бершадьоько-Котовоька округа в загальних рисах відповідає Дністровсько-Бузькій окрузі на схемах Г.І.Білика і Є.М.Брадіс (1962) та Г.І.Білика (1970). Різниця полягає тільки в тому, що північну межу округи ми проводимо на 30-40 км далі на північ та дещо змінюємо південно-східну межу. В окрузі переважають дубові ліси з дуба звичайного і дуба скельного. Грабово-дубові ліси поширені мало. Характерними асоціаціями дубових лісів в діброви свиди- ново-парвськоосокова та кизилово-парвоькоооокова, які далі на охід вже не трапляються.
19. Голованівоько-Новомиргородська округа частково відповідає Правобережній південно-західній окрузі на схемах Г.І.Білика і Є.М.Брадіс (1962) та Г.І.Білика (1970). Східну межу її ми проводимо порівняно із зазначеними схемами на 70-120 км далі на захід. Переважають в окрузі дубові ліси, представлені дібровою татар- оькокленово-горобейниковою.
20. Чигиринеько-8Йєменська округа виділяється нами. Переважають в окрузі дубово-грабові та дубові ліси. Характерною рисою останніх є поширення асоціацій класу дубових лісів кизилових (Querceta
о ох поза).
ж) Середньодніпровська підпровінція. На її території переважають дубові та липово-дубові ліси. Г.І.Білик (1970) запропонував поділити цю територію на дві підпровінції: Лівобережно-Придніпровську та.Середньоруську лісостепову, відповідно до поділу, запропонованого географами (Виленкин, Никитин, 1968; Демченко, 1968). Останні виділили Середньоруську лісостепову ландшафтну провінцію переважно на підставі геоморфологічних особливостей території, що цілком правомірно. Проте з позицій геоботанічного районування виділення цієї підпровінції потребує дальшого вивчення. На території
^Г.І.Білик (1970) перейменував цю округу на Північноподіль- ську, що не можна визнати вдалим, тому що більша частина її лежить за межами Подільської височини.
76
Середньодніпровської підпровінції серед лісової рослинності не спостерігається змін головних сгіівдомінуючих видів плакорних формацій, тому цю підпровінцію ми приймаємо в раніше встановлених межах.
21. Лівобережнопридніпровська терасова округа окреслюється нами в загальноприйнятих межах. За площею в природній рослинності округи переважають луки і болота. Характерною рисою є значне поширення засолених луків та трав"яних заплавних боліт. Дубові ліси трапляються переважно на других терасах і представлені дібровами ліщиново-гірськоосоковою, ліщиново-жіночопапоротевою та ліщиново- чоловічопапоротевою. Відсутність дубових лісів на більш високих терасах пояснюється надзвичайно рівнинним характером рельєфу, який уповільнює знесолення грунтів.
22. Сумсько-Гадяцька округа виділяється нами. В ній переважають липово-дубові та дубові ліси. Найбільш поширена асоціація останніх - діброва ліщиново-яглицева. Значно поширені луки та болота, а на других терасах річок - соснові та дубово-соонові ліси*
23. Харківсько-Зміївська округа виділяється нами. Границі її лише частково відповідають Харківській окрузі схеми Г.І.Білика (1970). Переважають в окрузі дубові ліси, найбільш поширеною і характерною асоціацією яких є діброва ліщиново-волосистоосокова. З асоціацій, які не трапляються в попередній окрузі, можна відзначити діброви татарськокленово-мікелієвоосокову та татарсько- кленово-горобейникову. Липово-дубові ліси порівняно 8 попередньою округою поширені значно менше і займають не вододіли, а схили балок. Болота і луки мало поширені.
Ш. Євразійська степова зона в смузі поширення лісів відповідає південній частині Придніпровської височини, Лівобережній низовині, Середньоруській височині і Донецькому кряжеві. Залісненість її 3-4%, ліси майже виключно дубові. Є.М.Лавренко (І947в) відносить степову зону України до Азовської підпровінції Східноєвропейської провінції.
24. Придністровська байрачна округа виділяється нами. Вона частково відповідає Дністровсько-Бузькій окрузі схеми Г.І.Білика і Є.М.Брадіс (1962). В рослинному покриві переважають дубові ліси, представлені класами асоціацій дібров татарськокленових
( Querceta aceroso(tatarici) та скумпієвих ( Querceta ootinoea). Крім лісів, на схилах балок поширені різнотравяо-типчаково-ковкло- ві степи.
77
25. Бузько-Дніпровська округа приймається нами, за винятком західної границі, без змін. Останню ми переносимо на 70-90 км на захід. Серед найбільш поширених і характерних асоціацій дубових лісів округи можна відзначити діброви татарськокленово-зірочникову, татарськокленово-яглицеву та татарськокленово-рябоперлівкову.
26. Присамарська байрачна округа виділяється нами. Межі її частково відповідають Дніпровсько-Донецькій окрузі схеми Г.І.Бі- лика і Є.М.Брадіс (1962), які відносять до неї територію аж до Азовського моря. На нашу думку, цю територію доцільно виділити в окрему Приазовську округу. Найбільш поширеною і характерною асоціацією дубових лісів Присамарської байрачної округи є діброва та- тарськокленово-зірочникова.
27. Кулмянсько-Старобільська байрачна округа, межі якої, за винятком північно-західної, відповідають Лівобережнопридонецькій окрузі схеми Г.І.Білика і Є.М.Брадіс (1962). Запропонована цими авторами назва округи, на нашу думку, не зовсім вдала, тому що в ній
не відбито історично і географічно обумовлену назву території "Ста- робільщина". За площею в окрузі переважає діброва татарськоклено- во-зірочникова. З характерних, хоча менш поширених асоціацій, слід відзначити діброви татарськокленово-горобейникову, татарськоклено- во-мікелієвоосокову, татарськокленово-кореневищноосокову та татар- ськокленово-рябоперлівкову. Флористичною особливістю дубових лісів округи Є наявність В НИХ ПОИТИЧНИХ (вербозілля Кільчасте - Lysimachia verticiiiaris s р г e n g,# ластовень виткий) та сибірських (скереда сибірська та серпій увінчаний) видів. Крім Куп"ян- сько-Старобільської байрачної округи, в досліджуваних лісах ці види трапляються лише на території Донецької байрачної округи.
28. Донецька байрачна округа в своїх межах відповідає Донецькій лісостеповій окрузі схеми Г.І.Білика та Є.М.Брадіс (1962),з тією різницею, що західну границю її ми проводимо на 70-80 км далі на схід.Так само як і Волинська лесова округа, це одна з найбільш спірних щодо зональної приналежності. Є.М.Лавренко (1926) розглядав її як лісостепову, оскільки, на його думку, тут є значна кількість дубових лісів та вододільні масиви. Проте автор відзначає, що вододільні масиви виявлені в більшості випадків недостатньо різко, це скоріше великі байрачні масиви, що перетворюються на вододільні.У праці 1947 р. Є.М.Давренко вже не підкреслює лісостеповий характер округи 1 в номенклатурному списку відносить !! до степової зони. Лісостеповий характер рослинності округи підкреслювали
78
Є.М.Лавреяко та П.С.Погребняк (1930), Ю.Д.Клеопов (1933) та ія. Протилежної думкй дотримувався П.П.Кожевников (1937). Таке розходження поглядів пояснюється переходовим характером рослинності Донецького кряжу, яка має більш мезофільні риси, ніж оточуюча рослинність рівнини, але більш ксерофільні, ніж рослинність типового лісостепу. Залісненість округи становить 5-6%, вона більша, ніж в інших округах степової зони, але менша,, ніж у лісостеповій зоні.
Поширення лісів на кряжі зумовлене значною абсолютною висотою місцевості, що перевищує 300 м. Кращому розвиткові лісів сприяють і безкарбонатні породи (сланці та пісковики), які на кряжі переважають. На території Донецької байрачної округи дійсно поширені більші масиви лісів, ніж у прилеглій степовій зоні. Проте, якщо порівняти ліси округи з лісами не тільки степової, а й лісостепової зони, то виявиться, що звичайно найбільші масиви дубових лісів Донецького кряжа перевищують найбільші масиви дубових лісів степової зони у 4-6 разів, але вони у 8-Ю разів менші, ніж найбільші масиви дубових лісів лісостепової зони. Чітко виявлені вододільні масиви трапляються лише в Теплинському та Маяцькому лісництвах Слов'янського лісгоспзагу Донецької області. Розташовані вони на високому правому березі Сіверського Дінця, зростання лісів на цьому вододілі пояснюється, на нашу думку, безпосереднім впливом водної артерії.
Аналогічна залежність виявлена і розглянута нами і для букових лісів Поділля. На самому Донецькому кряжі вододіли, які в двох- трьох випадках в Сніжнянському і Торезькому лісництвах Торезького лісгоспзагу були зайняті дубовими лісами, належать скоріше до вододілів великих середбалочних систем. Нарешті, важливим показником зональної приналежності рослинного покриву може бути, на наш погляд, поширення певних асоціацій. Для дубових лісів Донецької байрачної округи найбільш характерними в діброви татарськокленово-зі- рочникова, татарськокленово-рябоперлівкова та татарськокленово-го- робейникова. Всі вони - типові асоціації байрачних лісів степової зони. Навпаки, діброва ліщиново-волосистоосокова, широко представлена в південній частині Лівобережного лісостепу, на Донецькому кряжі не росте. З флористичного боку в лісах округи трапляються види, характерні як для лісів широколистянолісової та лісостепової зон (граб звичайний, гніздівка звичайна - Neottia nidus-avls в і с h), так і для байрачних дібров, наприклад карагана кущова, солонечник естрагоновидний (Galatella dracunculoides (Ь а ш.) Я e e в), грудниця волохата (Linoayrls villosa D, О.) та ін. Тому ми вва-
79
маємо, що хоч дубовим лісам Донецької байрачної округи й притаманні риси переходового характеру, проте асоціаційний склад їх свідчить про близкість цих лісів до дубових лісів степової зони. Виходячи з того, що дубові ліои в зональними, ми схильні розглядати рослинність округи в зональному відношенні як степову. Аналіз степової рослинності округи за тим самим принципом, що й лісової, тепер ускладнений. Вона дуже порушена і майже не збереглася.
ФІТОЦЕНОТИПИ
Основним завданням геоботаніки як науки є вивчення властивості рослин асоціюватись, тобто утворювати фітоценози. Становлення фітоценозу являє собою боротьбу рослин за існування, з одного боку, з абіотичними факторами, а з другого - поміж собою. Відбувається природний відбір рослин з біологічними властивостями, що дають їм можливість за даних екологічних умов відігравати певну роль у створенні фітоценозу. В деяких випадках для вотановлення ролі індивідууму серед тих або інших видів досить прямого спостереження. Природно, що популяціям одного й того ж виду в різних екологічних умовах, а також різним видам належить різна роль у створенні фітоценозів і вони мають різний характер зв'язків між собою. Вивчення останніх дав можливість виявити фактори, які формують фітоценоз і виявити значення кожної популяції в створенні фітоценозу.
В основу фітоценотичних класифікацій радянської геоботанічної школи покладено фітоценотипи. Аналіз їх дає можливість зрозуміти взаємозв"язки між видами в ценозі, що зумовили його організацію. Отже, проблема фітоценотипів є однією а найважливіших теоретичних проблем сучасної геоботаніки. Цілком природно, що з появою відомих праць Г.М.Висоцького (1915), Й.К.Пачооького (1917), Й.Браун-Бланке (Bxaun-Blanquet, 1921), О.П.Ільїнського (1921),А.Я.Гордяг1на(1922), Г.І.Поплавоької (1924), В.М.Сукачова (1926), Й.Браун-Бланке, Ю.Павіля- ра ( Bsaun-Blanquet, Pavlllard, 1928), в яких сформульовано перші головні положення цієї проблеми, їй а кожним роком приділяється все більша увага. Лише за останні ЗО років цієї проблеми в тій чи іншій мірі торкались Є.М.Лавренко (1940, І947д, 1959), С.Я.Соко- Ібв (1947), Б.О.Биков (1949, 1957, 1960, 1966а), І.Х.Блюменталь, Т.А.ПопОва (1950), Ю.ПавіЛяр (Pavillaxd,I954), М.В.Марков (1958, 1968), І.Х.Блюменталь, Л.О.Кіршсова (1960), Т.І.Іоаченко, К.І.Рач- ківоька (1961), В.С.Порфир’вв (1961, 1968), В.Д.Лопатін (1963, 1966), Х.Х.Трас (1963), А.О.Ніценко (1965), Т.О.Работнов (1966),
80
Ю.Р.Шеляг-Сосоико (1968а, 19686, 1969а) та ін. Проте, незважаючи на значні успіхи, в проблемі фітоценотипів є багато нерозв"язаних питань, спірних, часом протилежних і неузгоджених положень. Такий отан склався, на нашу думку, через: І) відсутність визначення єдиного розуміння самого поняття “фітоценотип"; 2) виділення фітоце- нотипів на підставі різних ознак і неоднакового розуміння термінів і виділених одиниць; 3) відсутність таксономічних одиниць класифікації фітоценотипів; 4) недостатню розробку класифікацій. Розглянемо перелічені причини в зазначеній послідовності.
Поняття "фітоценотип". Теоретично геоботаніки в це поняття вкладають певний якісний зміст і розуміють під ним міру і характер впливу рослин на склад та будову фітоценозів. Але фактично цього положення додержується тільки частина авторів. Інші (Блюменталь, Петровичева, 1951; Блюменталь, Попова, 1960; Номоко- нов, 1959; Блюменталь, Кирикова, 1960; Буторина, 1963; Афанасьєв, 1964) ототожнюють його з рясністю і трапляємістю. Проте зведення ролі рослин у створенні фітоценозів до суто кількісного значення не можна прийняти* Правомірним є лише виділення фітоценотипів за їх роллю в житті ценозу. При такому підході в розумінні фітоценотипів за А.О.Ніценком склалося два напрямки. Більшість геоботаніків застосовує поняття Ифітоценотипн до видових популяцій, тобто один і той самий вид відносять до різних фітоценотипів. Фіто- ценотипи в такому розумінні А.О.Ніценко (1965) пропонує назвати популяційними. Прихильники другого напрямку (Виооцький, 1915; Па- чоський, 1917; Ільїнський, 1921; Раменський, 1938; Лопатін, 1963, 1966 ; Ніценко, 1965; Работнов, 1966) застосовують це поняття до видів, отже, кожен вид може належати лише до одного фітоценотипу. За А.О.Ніценком (1965) це видові фітоценотипи.
З уявленням про популяційні фітоценотипи можна погодитись повністю, але поняття видових фітоценотипів потребує уточнення. А.О.Ніценко (1965) підкреслює, що в такому разі види поділяють не за фактичною роллю їх у фітоценозі, а за тим, яку роль вони можуть відігравати в сприятливих умовах в межах однієї кліматичної області. Якщо так, то поняття "фітоценотип11 застосовується вже не до видів, а до їх найбільш типових у фітоценотичному відношенні популяцій, в межах певного кліматичного регіону. Отже, фітоценотипи в такому розумінні краще називати не видовими, а регіональними. Ми вважаємо, що під видовими фітоценотипами слід розуміти види, популяції яких займають однакові фітоценотичні позиції в межах їх
81
географічних ареалів. Таким чином, в тлумаченні поняття "фітоцено- типп намітились три напрямки: популяційний, регіональний та видовий.
Поняття "фітоценотип" в популяційному значенні фактично не визначене. Формулювання, яке дає С.Я.Соколов: "Фітоценотип є сукупність індивідів, які мають однакові фітоценотичні властивості" (1947, с.152), на думку А.О.Ніценка (1965), розходиться з його трактуванням, бо йдеться не про властивості тієї чи іншої популяції, а про позицію, яку вона займає в даний момент.
Визначення Б.0.Викова: "...під "фітоценотипом" треба розуміти сукупність видів, що займають однакове і при тому найбільш стійке й типове для них положення в фітоценозах" (1966а, с.1234), - лише частково відповідає змістові, який вкладає в нього автор. Воно припускає віднесення до фітоценотипу видів, тоді як в статті мова переважно йде про віднесення до фітоценотипу популяцій видів.
Ми пропонуємо таке формулювання цього поняття: "Популяційним фітоценотипом є група видових популяцій, яка, залежно від їх біологічних особливостей і відношення до умов середовища, з одного боку, і відносин з іншими групами видових популяцій, з другого, характеризується однаковою роллю та позиціями в створенні фітоценозів". Популяційний фітоценотип є показником цих відношень. Для поняття "видовий фітоценотип" ми пропонуємо таке визначення: "Видовим фітоценотипом є група видів, яка характеризується більш або менш однаковою кількістю популяційних фітоценотипів, що займають однакові позиції".
Принципи виділення фітоценотипів, термінологія та зміст. На думку В.М.Сукачова (1926), виділення фітоценотипів має грунтуватись на рясності, обсязі та просторі, зайнятому видом, характері використання видом середовища та його конкурентної здатності. Б.О.Би- ков (1949, 1957) головними показниками вважає щільність зростання рослин, характер розміщення їх в угрупованнях та життєву форму. За І.Х.Блюменталем із співавторами (1950, 1951, 1960), Л.І.Номоконовим (1959) та Д.Я.Афанасьєвим (1964), головними показниками є градації рясності або проективного покриття І трапляемооті рослинГформально- статистичний підхід). Внаслідок чого до одного й того ж фітоценотипу відносять види, які характеризуються різною роллю в складанні фітоценозу. Наприклад, І.Х.Блюменталь та О.Л.Петровичева (1951) до одного фітоценотипу в одній формації разом з трав"яними видами віднесли чагарники й лишайники, у яких різні життєва форма, біологічні та екологічні особливості, а також фітоценотична роль.
82
На нашу думку, виділення фітоценотипів може грунтуватись лише на принципі групування рослин за їхнім значенням в створенні середовища, або,інакше кажучи, на підставі єдності зайнятих у фітоценозі позицій. Зовнішнім проявом вказаного принципу є конкурентна здатність виду як вияв його еколого-біологічних властивостей, біологічна форма, життєвість, синузіальна належність, рясність та ступінь взаємопбв"язаності. Як показали одержані нами дані (Шеляг-Сосонко, Куковиця, 1969, 19706) вивчення міжвидових зв’язків в асоціації діброви ліщиново-волосистоосокової у фітоценозах з виробленою ценотичною структурою, ступінь взаємолов"я- заності видів травостою різний. Пояснюється він не тільки і не стільки міжвидовими відносинами видів, які його складають, скільки відносинами з едифікатором і конфектором першого рангу.
Поряд з різним підходом до виділення фітоценотипів немає єдності і в тлумаченні готовних понять та одиниць. Найбільш істотною є розбіжність поглядів на поняття "домінанта11 й "едифікатор”, що переконливо показано В.С.Порфир”євим (1968). Так, І.Х.Блюмен- таль і О.Л.Петровичева (1951), А.Скамоні (Scamoni, 1955) та М.В.Мар- ков (1958, 1962, 1968) вважають ці терміни синонімами. Цю думку активно відстоює М.В.Марков, посилаючись на роботу А.Я.Гордягіна (1922), де вперше у вітчизняній літературі вжито термін "домінанта” в якісному значенні. Ще далі йде Б.О.Биков (1957, 1966а), який термін "едифікатор" приймає в загальному значенні і відносить до едифікаторних не тільки популяції головних синузій фітоценозу, але й другорядних.Ми не можемо погодитись з такими поглядами на розглянуті терміни. М.В.Марков має рацію в тому, що А.Я.Гордягін надавав домінантам не тільки кількісне, але і. якісне значення. Проте термін "домінанта" А.Я.Гордягін розумів ширше, ніж Г.І.Поплав- ська (1924) термін "едифікатор" (вона ввела його у вітчизняну літературу).Ми вважаємо, що необхідно погодитись з думкою Є.М.Лаврен- ка (1959), П.Д.Ярошенка (1961) та Х.Х.Траса (1963), які розуміють термін "домінанта" ширше, ніж термін "едифікатор". Отже, домінанти, на нашу думку, повинні поділятися на едифікатори і конфекто- ри, тобто рослини, що переважають у другорядних ярусах.
Таксономічні одиниці класифікації фітоценотипів. Практично цей бік проблеми фітоценотипів не розроблений. Більшість авторів ( Вхаид-Blanquet, Pavillard, 1928;Раменський, 1938; Соколов, 1947; Бьпсов, 1949, 1957, 1966а; Блюменталь, Попова, 1951; Pavil- lard, 1954; Номоконов, 1958; Лавренко, 1959; Исаченко f Рачковская,
83
1961; Ильинская, 1963; Афанасьев, 1964; Работнов, 1966) наводять простий перелік фітоценотипів з коротким їх визначенням. Наскільки можна зрозуміти з тексту, вони виділяють головні фітоценотипи і групи фітоценотипів. Обсяг фітоценотипів у різних авторів або навіть у одного і того ж автора неоднаковий.
Цікаву спробу внести деякий порядок в це питання зробив А.0.НІценко(1965).Хоч він і не розглядав таксономічні одиниці, проте в своїй класифікації видових фітоценотипів розрізняв групи типів,підгрупи типів І типи. Групи типів автор виділяв на підставі ролі видів у створенні фітоценозів, а дальший поділ провадить на підставі особливостей виділених груп типів, внаслідок чого вони можуть бути постійними, переходовими або тимчасовими. Інакше кажучи, він виділяв власне не підтипи, а варіанти групи типів. Ще раніше це було зроблено Г.І.Поплавською (1924), коли вона запропонувала розрізняти едифікатори автохтонні й дигресивні. Х.Х.Трас (1963) запропонував розрізняти домінанти за розміром вкритої індивідуумом площі, розміром ділянки, зайнятої популяцією виду, силою впливу на інші компоненти фітоценозу і екотоп, здатністю утворювати чисті або змішані фітоценози і т.п. Ми погоджуємось з такою глибокою і різнобічною диференціацією фітоценотипів, але вважаємо, що в даному випадку мова йде про варіанти одного і того ж фітоцено- типу залежно від екологічних, морфологічних, сезонних та інших особливостей виду і сукцесійних стадій. Таким чином, сучасний стан вивчення фітоценотипів потребує розробки їх класифікації.
Цілком зрозуміло, що класифікації популяційних, регіональних і видових фітоценотипів можуть мати неоднакове число таксономічних одиниць різного рангу. Виділення таксонів у них може грунтуватись на різних принципах, але в будь-якому випадку необхідно строго дотримуватись супідрядності прийнятих одиниць, а одиниці одного рангу не повинні перекриватися за вибраною ознакою.
Розуміючи всю складність цього питання, ми пропонуємо поки що для класифікації популяційних фітоценотипів такі таксономічні одиниці.
Тип. Головна таксономічна одиниця. Виділяється на підставі позиції, яку видова популяція займає у фітоценозі, впливу її на середовище й інші видові популяції. Всі популяції одного виду, з однаковою роллю у складанні фітоценозів ми називаємо фітоценотип- ною популяцією. Фітоценотипні популяції різних видів, які займають у фітоценозах однакові позиції, належать до одного типу. Кожен вид може мати одну або кілька фітоценотипних популяцій.
84
Група типів. Наступна за рангом одиниця, що об’єднує типи на підставі взаємозв'язку і взаємообумовленості фітоцено- типних популяцій.
Порядок типів. Об"єднує групи типів за ступенем їх значення.
Клас типів. Об’єднує порядки типів по відношенню їх ролі до фітоценозу в цілому.
Об “єднання типів. Найвища за рангом одиниця, до якої входять всі типи. Залежно від розуміння фітоценотипу вона може мати три варіанти: І) об’єднання популяційних фітоценотипів, 2) об’єднання регіональних і 3) об’єднання видових фітоценотипів. Перелічені таксономічні одиниці виявлені лише в першому варіанті. У двох інших деякі з них випадають. Пояснюється це їх недостатньою розробленістю і різним тлумаченням фітоценотипів у цих варіантах. Наприклад, у другому і третьому варіантах випадають фітоценотипи- деструктатори і адвентивні, тому що жоден вид не може лише руйнувати фітоценози або траплятись тільки випадково і ніде не роста цілком закономірно. Можливо, доцільно виділити варіанти в. самостійні об’єднання і розробити таксономічні одиниці для кожного з них окремо.
Варіанти типів. Ми вважаємо їх не таксономічною одиницею. До них пропонуємо відносити популяції, які займають у фітоценозах однакові позиції, але відрізняються життєвою формою, розмірами, продуктивністю, характером !х зумовленості, рясністю, розвитком у часі і т.п. Виділення фітоценотипів на підставі цих факторів, особливо останнім часом (Трасо, 1963; Ниценко, 1965; Бнков, 1966а) , є скоріше поділом фітоценотипів за особливостями фітоценозів, які вони утворюють, а не за характером їх ролі у створенні фітоценозів. Наприклад, за Б.А.Виковим (1966а), домінантні популяції фісташки справжньої (Pistacia vera L.) та ялини сибірської ( Picea obovata L e df)відносяться до різних фітоценотипів. На наш погляд, це невірно, тому що кожна з цих популяцій у відповідних екологічних умовах займає однакові фітоце- нотичні позиції. В боротьбі за існування вони набули властивостей, які дозволили їм зайняти екологічну нішу, в якій популяція саме цього, а не іншого виду найбільш інтенсивно використовує зовнішнє середовище і зумовлює оклад фітоценозу. Фітоценотип є показником ролі виду або його популяції у створенні фітоценозу, а не показником особливостей її серед аналогічних фітоценотипів, але у фітоце85
нозів різних екологічних умов. Тому, якщо названі види вважати різними фітоценотипами, то це буде суперечити самій ідеї фітоценотипів. Крім того, це призвело б до виділення величезної КІЛЬКОСТІ фітоценотипів. Лише різних типів едифікаторів треба було б розрізняти стільки, скільки існує різних екологічних умов. На підставі викладеного ми вважаємо, що розглянуті види належать до одного і того ж фітоценотипу, але є різними його варіантами. Варіанти типів - найбільш поширена група. Більш або менш повний огляд її потребує розгляду багатьох показників і детального знання біології видів, тому зупинимось лише на деяких варіантах типів.
Насамперед доцільно поділити варіанти типів на дві групи: І) варіанти, які можуть мати всі фітоценотипи і 2) варіанти, які мають тільки певні фітоценотипи.
Перша група. До неї ввійдуть автохтонні та дигресивні, кліматичні й едафічні, постійні та тимчасові, нано-, мікро-, мезо-, макро-, і мегафітні, темпоральні і консервативні, еврітопні й стенотопні варіанти фітоценотипів. Доцільно розрізняти варіанти 1 за ступенем виробленості ценозів: агломераційні та семіасоційні і взагалі варіант сукцесійних стадій в широкому розумінні.
Друга група. Домінанти можуть бути патулекторними, дензектор- ними, конекторними, нано-, мікро-, мезо-, макро- і мегагрегідни- ми, гіпо-, суб-, і гіпердомінантними, еврітопнодомінантними і стенотопнодомінантними. Асектатори можуть бути різних рангів, залежно від рясності і трапляємості. Слід розрізняти конфектори й асектатори також за ступенем пов"язаності з едифікатором, залежно від чого вони матимуть різні класи зв"язку.
Крім перелічених варіантів, популяційні фітоценотипи конфек- торів та асектаторів можуть бути справжніми і похідними. Справжні - коли популяція займає відповідну до біології її індивидів фі- •гоценотипну позицію. Похідні - коли фітоценотична позиція популяції не відповідає біології індивидів. Наприклад, асектаторна фі- тоценотипна популяція дуба звичайного є похідною, а груші звичайної (Pyrua communis L.) - справжньою. Мабуть, похідні популяційні фітоценотипи можна розрізняти починаючи з субедифікаторів. Це положення має практичне значення. Воно вказує на те, що на території поширення асектаторної популяції, яка є похідною від едифі- каторної популяції, екологічні умови дозволяють створити чисті або змішані культури цього виду.
Сучасний стан проблеми фітоценотипів дає нам можливість за-
86
Класифікація популяційних фітоценотипів
Об’єднання типів
Клас типів
Порядок типів
Група типів
Тип
Популяційні фітоценотипи
Фітоценобіонти
Домінанти
Едифікатори
Едифікатор I рангу
Едифікатор II рангу
Конедифікатор
Співедифікатор
Субедифікатор
Семіедифікатор
Конфектори
Конфектор I рангу
Конфектор II рангу
Субконфектор
Співконфектор
Конфектор дегратогенний
Асектатори
Асектатори едифікаторні
Асектатор едифікаторний ординарний
Асектатор едифікаторний субординарний
Асектатор едифікаторний дегратогенний
Асектатори конфекторні
Асектатор конфекторний ординарний
Асектатор конфекторний субординарний
Асектатор конфекторний дегратогенний
Фітоценоабіонти
Адвентні
Деструктатори
Деструктатор ординарний
деструктатор остаточний
Солітарії
Солітарій I рангу
Солітарій II рангу
Індиферентні
Індиферентні I рангу
Індиферентні II рангу
Класифікація видових фітоценотипів
Об’еднання типів
Група типів
тип
Видові фітоценотипи
Монотипні
Валіденти
Медієнти
Дебіленти
Політипні
Абсолютенти
Партикуленти
Інтермісенти
87
пропонувати класифікації популяційних та видових фітоценотипів.
З розглянутих позицій дубові ліси з дуба звичайного мають едифікаторну популяцію І рангу і едифікаторну популяцію П рангу. До перших належать популяції дуба звичайного, які утворюють головну синузію на вододілах, а до других - популяції з дуба звичайного, що утворюють головну синузію в заплавах річок та на других терасах. Звичайно до однієї асоціації доводиться відносити фітоценози, утворені обома едифікаторними популяціями. Структурно та фізіономічно вони подібні, але мають більш значні відміни у флористичному складі асектаторів та в екології, ніж фітоценози однієї едифікаторної популяції. Цілком ймовірно, що в процесі дальшого вивчення доцільно буде виділити їх у самостійні асоціації, поєднавши в популяційні формації.
Конедифікаторів у досліджуваній формації немає. Співедифіка- тори на території Західного Поділля і Прикарпаття представлені популяціями бука лісового і лише на території Прикарпаття - ялиці білої. На Поліссі, у лісостеповій і степовій зонах трапляються співедифікаторні популяції ясена звичайного (Fraxinus excelsior L.), а переважно на Поліссі - популяції сосни звичайної та вільхи клейкої ( Alnus glutinosa (Ь.) G a e r t п.).
Як субедифікатори в дубових лісах переважно Лівобережної України трапляються популяції клена звичайного і липи серцелистої, а в дубових лісах Полісся - популяція сосни звичайної.
Семіедифікатори в дубових лісах лісової зони і більшої частини лісостепової зони представлені популяціями граба звичайного, а в лісостеповій і степовій зонах - клена польового (Acer campestre Ь.), в"яза гладенького (Ulmus laevis Раї 1,) і в"яза шорсткого (Ulmus scabra M і 1 1,7.
Конфекторів І рангу в дубових лісах України немає.
Конфектори П рангу представлені популяціями клена татарського, яглиці звичайної, копитняка європейського ( Asarum europaeum L.), осоки парвської, осоки гірської ( Carex montana І».), ліщини звичайної (Corylus avellana L.) Та ІН.
Співконфекторні популяції можуть мати всі види, які утворюють популяції конфекторів П рангу. В цій ролі вони трапляються в перехідних екологічних умовах.
В ролі субконфекторів зустрічаються популяції яглиці звичайної, осоки гірської, осоки волосистої та барвінку малого (vinea minor L.).
88
Конфектори дегратогенні в дубових лісах України представлені лише популяціями горобини звичайної (Sorbus aucupar ia L.),
Асектатори едифікатори! ординарні в дубових лісах представлені популяціями клена польового, клена звичайного, берези бородавчастої, граба звичайного, черешні, ясена звичайного та ін.
В ролі асектаторів едифікаторних субординарних бувають популяції черешні, сосни звичайної та береста (Ulmus foliacea G і - і і ь.).
До асектаторів конфекторних ординарних і асектаторів конфек- торних субординарних належать популяції більшості видів, що створюють ярус травостою і підліску.
В ролі, деструктаторів ординарних трапляються популяції граба звичайного.
Деструктатори, остаточні за мірою ролі, яку вони відіграють у побудові фітоценозу, можуть бути конфекторами або асектатора- ми. В дубових лісах до них належать популяції карагани кущової, скумпії звичайної, квасениці звичайної (Oxalis acetosella L.) та ін.
До індиферентних фітоценотипів належать популяції бур"янів. Таким чином, в дубових лісах України кожен з фітоценотипів представлений такою кількістю видових фітоценотичних популяцій: едифікатори І рангу - І, едифікатори П рангу - І, співедифікатори - 4, субедифікатори - 3, семіедифікатори - 4, конфектори П рангу - 44, співконфектори - 45, субконфектори - 4, конфектори дегратогенні - І, асектатори едифікатори! ординарні - 24, асектатори едифікатори! субординарні - 3, асектатори конфекторні ординарні і асектатори конфекторні субординарні - 302, деструктатори ординарні - І, деструктатори остаточні - 58 та індиферентні - 87.
Значно важче проаналізувати дубові ліси з позицій видового розуміння фітоценотипів, тому що при цьому треба знати, яку роль відіграє вид в межах усього ареалу. Попередньо, без врахування групи видів, невластивих дубовим лісам, ми одержали такі дані: валідентів - немає; інтермісентів - немає; медіанти - цибуля ведмежа (Allium ursinum L.) і барвінок малий; дебіленти - арум плямистий (Arum maculatum L,), жовтушник лісовий (Er ув imum silvaticum М. В.), медунка вузьколиста (Pulmonaria angustifolia L.), груша звичайна та ін.; абсолютенти - ялиця біла, береза бородавчаста, граб звичайний, бук лісовий, сосна звичайна, осика (Populus tremula L.), дуб звичайний, дуб скельний і липа серцелиста; парти-
89
куленти - дерен справжній, свидина кров"яка, крушина ламка, ліщина звичайна, осоки волосиста, кореневищна, парвська і трясучковидна, зірочник лісовий, перлівка ряба, куцоніжка пірчаста, горобейник пурпурово-голубий, копитняк європейський, клен татарський, яглиця звичайна, маренка запашна (Asperula odorata L,), конвалія звичайна (Oonvallaria majalia L.), веснівка дволиста та ін. Отже, в дубових лісах переважають партикуленти і дебіленти. В групі звичайних видів до перших належить 151, а до других - 64 види. Слід вважати, що в групі рідкісних видів це співвідношення змінюється в бік зростання дебілентів.
Таким чином, розглянута нами в теоретичному плані проблема фітоценотипів характеризується багатогранністю своїх аспектів, і твердження, висловлені нами, далеко не вичерпують всієї різноманітності, важливості і глибини її. Особливо плідним для дальшого розвитку проблеми є, на наш погляд, вивчення хімічної взаємодії рослин. Дослідження останніх років показали, що алелопатичний вплив властивий більшості видів рослин (Гродзинский, 1965). Виділення рослин значно впливають на екологічні умови місцезростання і, отже, сприяють усталенню зайнятих позицій чи захвату нових. В зв"язку з цим великий інтерес являє запропонований А.М.Гродзин- ським (1964) поділ рослин за токсичністю виділень їх на три групи: І) рослини з дуже токсичними виділеннями; 2) рослини з менш токсичними виділеннями; 3) рослини з малотоксичними виділеннями. Виявилось, що едифікатори мають, як правило, слаботоксичні виділення. Вони створюють загальний фон алелопатичних виділень фітоценозу, для деяких рослин навіть сприятливий, через що кожен фітоценоз має свій певний флористичний склад. Навпаки, виділення рослин, які ростуть окремими екземплярами і є асектаторами, дуже токсичні. Цікаві дані одержали О.Ланге і Г.Канцов (tange, Kanzow, 1965). Вивчаючи популяції цибулі ведмежої, що в широколистяних лісах Європи створюють трав”янистий ярус, вони дійшли висновку, що причиною пригнічення рослин інших видів є, можливо, дія виділених цибулею КОЛІНІВ.
АРЕАЛ ДУБА ЗВИЧАЙНОГО ТА ЙОГО БУДОВА
Взаємовідносини рослин з умовами середовища складаться не довільно, - певні вади потребують лише певних умов. Цілком зрозуміло, що зводити поширення виду лише до екологічних умов, хоч вони й важливі, не можна. Сучасний ареал рослин обумовлений дією
90
історичних, топографічних t антропогенних факторів. При цьому кожен з цих факторів може мати більше значення для певного виду, ніж усі інші, і тим самим зумовлювати межі його поширення. Отже, доцільно говорити про історичну, екологічну (кліматичну, едафічну), фіто- ценотичну і антропогенну границі ареалу. Всі фактори, крім екологічних, не дають можливості виду зайняти територію з придатними для нього екологічними умовами. В природі трапляються в більшості випадків саме такі види, ареали яких ми відносимо до типу неповних. Якщо перелічені фактори не впливають, то вид, при наявності достатнього для цього часу, займає територію з сприятливими для свого існування екологічними умовами. Такий тип ареалу ми називаємо повним. Межі зумовлені дією лише найнесприятливіших для дальшого поширення виду екологічних факторів. Змінюються вони в напрямку з півночі на південь швидше, ніж з заходу на схід, тому ареал звичайно має вигляд видовженого еліпса.
При аналізі сучасного географічного ареалу виду ми виходимо в таких положень: І) якщо вид досяг своєї екологічної межі, то він має пригнічений вигляд, дерева, наприклад, набувають габітусу кущів; 2) якщо вид на межі свого ареалу має властивий для нього габітус 1 високу життєвість, то дальшому його поширенню перешкоджають не екологічні, а якісь Інші фактори; 3) значну роль у поширенні виду відіграє конкуренція; 4) ареал виду-едифікатора має звичайно більш складну будову, ніж ареал виду, що є асектатором; 5) в різних частинах свого ареалу вид має границі різного характеру; 6) ареали різних видів мають неоднакову кількість фітоценотишшх частин.
Здебільшого вид представлений кількома популяційними фітоце- нотипами. Кожна фітоценотипна популяція в межах географічного ареалу виду займає певну територію, на якій вона, через умови, що склалися, є більш характерною, ніж інші фітоценотипні популяції цього виду. Таким чином, доцільно виділяти частини географічного ареалу виду, в яких він представлений популяціями, що займають у фітоценозах найбільш типове для цієї території положення. Такі частини ми пропонуємо називати хоріонами. Географічний ареал кожного виду буде поділятись на стільки хоріонів, скільки вид має популяційних фітоценотипів.
За даними Г.Мейзеля, Є.Єгера, біВаЙнерта (Meusel, Jagex, Weinert, 1965), крайня південно-західна відмітка границі ареалу Дуба звичайного лежить в Португалії, недалеко від Лісабона, близь91
ко 39° північної широти. На північ ареал дуба займає Португалію, Кантебрійські гори, Іспанію, всю Францію, Ірландію і Англію, за винятком їх крайньої північно-західної приморської смуги. Далі північна межа його ареалу проходить в Норвегії біля 62° північної широти, вздовж південного узбережжя Швеції та Фінляндії і доходить до Ленінграда. Південна частина ареалу на території Середньої Європи охоплює узбережжя Середземного моря, Апеннінський півострів, усю Югославію, північну частину Греції (де межа ареалу доходить майже до 40° північної широти), всю Болгарію і узбережжя Чорного моря. Далі на схід межа ареалу проходить біля Кривого Рога, на північ від Запоріжжя і Донецька, доходить до Волгограда, де, ва даними О.Ф.Кірєєва (1964), в заплаві Волги росте найпівденніша діброва. Від Волгограда вона йде на Уральск і по р. Уралу досягає біля м. Кувандика східної відмітки. На Уралі межа, за даними П.Л.Гор- чаківського (1968), проходить на захід від Білорецька на Красно- уфімськ, звідки повертає на захід. На півночі по заплавах річок дуб звичайно доходить до Кірова, Костроми і Череповця (Норин, 1954; Чеведаев, 1963). Найбільш південні ексклави дуба знаходяться на Кавказі та в Криму.
В горах дуб звичайний підіймається до висоти: в Піренеях - 1200, в Альпах - 1400, в Карпатах - 700 і на Кавказі - 500 м.
Ареал дуба звичайного, який описується нами, ми поділяємо на 9 хоріонів: І) едифікаторогенний першого порядку - Ер 2) едифіка- торогенний другого порядку - Eg, 3) співедифікаторогенний - Сд, 4) субедифікаторогенний - Сб, 5) конфекторно-дегратогенний - Кд, 6) асектаторогенний - А, 7) оолітаріогенний - С, 8) дегратогенно- природний - Дп, 9) дегратогенно-синантропний - Де (рис.15),
Хоріон Ej займає більшу частину Ірландії, Англії, Північної рівнини, лісостепової зони України, Середньоруської височини і західну частину Заволжя. На цій території популяції дуба звичайного в плакорних умовах відіграють роль едифікатора і утворюють головні масиви лісів (Кожевников, I939;Taneley, 1939; Напалков, 1951; Bubnet, Relnhold, 1953; Passacge, 19571 Pasearge, Hofmann, 1968, ia ін.).
Хоріон Е2 займає всю південну частину ареалу від Португалії до Уралу, вузьку смугу в Приураллі і від Уралу до Дніпра на північ від хоріона Ej. В південній смузі хоріону на території Португалії, Іспанії та Італії ліси з дуба звичайного в звнязку з наявністю в умовах макротермного клімату сильних конкурентів трапля-
92
Рис.15. Ареал дуба звичайного та його будова. 1 — межа ареалу дуба звичайного, 2 — межа хоріонів: Е1 — едифікаторогенний I порядку. Е2 — едифікаторогенний II порядку, Сд —
співедифікаторогенний, Сб — субедифікаторогенний, А — асектаторогенний та Дс — дегратогенно-синантропний.
ються лише в горах серед каштанових 1 дубових лісів, створених іншими видами цього ж роду на буроземних і буроземно-підзолистих грунтах, які добре дренуються і підстелюютьоя кислими породами 8 вмістом гумусу в верхньому горизонті 6% і рН 5,2 - 5,7 (Вгаїш- Blanquet,1952, 1967; Dantaa Barreto, 1958). На схід, на території Югославії, Угорщини, Болгарії, Румунії та Молдавії дуб звичайний в північніших районах створює ліси лише в депресіях рельєфу, в заплавах річок і в нижній частині гірського поясу на темяо-сірих лісових, дерново-карбонатних і чорноземних грунтах серед зональних лісів з дуба пухнатого, бургундського (б.селгіл Ь.^скельного та 1н.., а в південніших районах - лише в заплавах річок (Sorf, 1943, 1958; Giavac, 1962; Радков, Минков, 1963; Гейдемая, Остапенко, 1964; Pudear u-S ох ос ©anu, Poppva-»Oucuf 1966j Pa^oovschi, Donifa, 1967). На території України та РРФСР дубові ліси в межах розглянутого хоріону повиязані виключно з балками та долинами річок І на плакорах не трапляються. В Приураллі, в зв"язку із зростаючою
93
континентальні сію клімату, ці ліси поступаються місцем липовим та дубово-липовим і займають лише схили хребтів, долин річок і балок, які краще прогріваються, з темно-сірими лісовими і чорноземними опідзоленими грунтами (Крашенинников, 1941; Марков, 1948; Бельгард, 1950; Напалков, 1951; Александрова, 1954; Игошина, 1961; Киреев, 1964). В північній смузі цього хоріону на території СРСР більш холодний клімат сприяє поширенню ялинових лісів, і дубові ліси трапляються окремими островами лише на багатих грунтах. На території Середньої Європи клімат сприяє формуванню букових лісів, а дубові зустрічаються на сірих лісових і дерново-карбонатних грунтах, ЩО краще прогріваються і підстелені лесом (Bubner, Belahold, 1953; Buhl, 1956).
Хоріон Сд займає північну частину Ірландії, Англії, більшу частину Франції, ФРН, Данії, південну частину Норвегії, Швеції, Прибалтику і підзону південної тайги. На цій території популяції дуба звичайного виступають переважно в ролі співедифікатора, в межах Радянського Союзу і Скандінавії вони беруть участь у формуванні дубово-ялинових лісів, а на решті території хоріона - дубово- букових (Цинзерлинг , 1934; Ильинский, 1939; Тапвіеу, 1939» Алехии, І947; Bubnex, Beinhold, 1953; Llndquist, 1959» Sjors, 1965).
Хоріон Сб займає поліську частину України, Білорусію, середню і північну частину Польщі. Популяції дуба звичайного в його межах виступають в ролі співедифікатора, утворюючи другий ярус в СОСНОВИХ лісах ( Matuszkiewlcz, 1947; Matuszkiewicz, Polakowska, 1955; Поварніцин, 1959; Szafer, 1959; Юркевич, Гельтман, 1965). Формування цього хоріону пояснюється не кліматичними, як в попередніх випадках, а едафічними факторами.
Хоріон А займає найбільш північну частину ареалу від Англії до Уралу. В його межах популяції дуба звичайного в нлакорних умовах трапляються у вигляді окремих деров або кущів і роль едифікатора можуть відігравати лише в заплавах річок (Смирнова, 1928; Цинзерлинг, 1934; Игошина, 1943; Зубарев, 1958; Llndquist, 1959; Таскаева, 1963). В південній частині ареалу на території Радянського Союзу цього регіону немає, на території Середньої Європи він виявлений дуже слабо і не має чітких границь.
Хоріони Дп та Кд займають дуже вузьку смугу, шириною 5- 15 км на Південному Уралі і на схемі не виділені. За даними К.І.Ха- мітова (1956), в цьому районі на висоті 590-820 м дуб звичайний має вигляд чагарника.
94
Хоріон С виявлений лише на південній межі границі ареалу від Уралу до Молдавії. Ширина його вимірюється кількома сотнями метрів.
Хоріон Де займав досить значну територію, кліматичні умови якої не дають можливість даному видові зростати. Не зустрічається він лише через конкуренцію з боку інших едифікаторів.
В межах хоріону фітоценотипна популяція займає місця з різними екологічними умовами, наче "переборюючи" їх в процесі перетворення в умови місцезростання. Роль її у створенні фітоценозів визначається загальними екологічними та історичними особливостями цієї території. За межами даного хоріону популяції виду займають у фітоценозах вже інші позиції, на підставі чого виділяється наступний хоріон. Проте це не означав, що в межах нового хоріону не можуть траплятися популяції виду з такою ж позицією у фітоценозах, як і в попередньому хоріоні (йдеться про територію, на якій фітоценотипна популяція найбільш поширена). Таким чином, крім хоріонів, необхідно розрізняти популяційні фітоценотипи ареали виду. Під таким ареалом ми розуміємо територію, зайняту даною фіто-г ценотипною популяцією, на якій вона найпоширеніша та трапляється хоч би зрідка. Як правило, популяційний фітоценотипний ареал більший за відповідний хоріон, проте для невеликої групи видових монотипних фітоценотипів їх географічний ареал, хоріон і популяційний фітоценотипний ареал збігаються.
Топографія популяційного фітоценотипного ареалу складна. Для більшості видів (щодо інших фітоценотипних популяцій виду) можна виділити чотири частини такого ареалу: І) однотипну, в якій поширена лише певна фітоценотипна популяція; 2) переважаючу, в якій, крім неї, трапляються також інші популяції, але вона переважав; 3) змішану, де певна фітоценотипна популяція поширена досить рівномірно разом з іншою; 4) випадкову, в якій ця фітоценотипна популяція трапляється зрідка, а переважає інша.
ФІТОЦЕНОТИЧНА КЛАСИФІКАЦІЯ ДУБОВИХ ЛІСІВ УКРАЇНИ
У фітоценології класифікація рослинності є однією з центральних проблем, яка за рівнем свого значення відповідає проблемі асоційованості рослин. Вона є синтезом наших знань в галузі фітоценології і разом з тим - знаряддям дальшого пізнання фітоценозів. Тому цілком закономірна увага, яку приділяють розробці цієї проб-
95
леми радянські та зарубіжні геоботаніки. Ми дозволимо собі не робити огляду їх робіт з цього питання, тому що повне зведення з класифікації рослинності було нещодавно опубліковано В.Д.Алексаядровою (1969).
На наш погляд, в проблемі класифікації рослинності можна виділити три кардинальних кола питань, пов"язаних: І) з ознаками, за якими виділяють об'єкти класифікації; 2) обсягом таксономічних одиниць; 3) принципами, які кладуть в основу класифікації.
Головним об"єктом класифікації ми вважаємо фітоценоз, точніше пробні ділянки. За визначенням В.М.Сукачова (1961), фітоценоз, або рослинне угруповання, - будь-яка конкретна ділянка рослинності, на певному протязі однорідна за складом, оинузіальною структурою, будовою та характером взаємодії між рослинами та між ними і середовищем. З цього визначення випливають, як неодноразово підкреслював автор (Сукачев, 1926, 1931, 1938, 1961), такі головні положення: І) якщо між рослинами немає зазначеної взаємозалежності, то немає і фітоценозу; 2) до фітоценозів належать посіви та інші штучно створені сполучення рослин, якщо між ними існує зазначена взаємодія; 3) фітоценоз - поняття конкретно-територіальне, він є головною одиницею рослинності і підлягає дальшій типізації.
Всю різноманітність фітоценозів у природі можна звести до певних типів. Тили, що об'єднують фітоценози, однорідні за своїми суттєвими рисами,В.М.Сукачов (1931) вважає асоціацією. Найбільш повне визначення асоціації знаходимо в працях автора 1957 та 1961 рр.: "Рослинна асоціація (тип фітоценозу) є головною таксономічною одиницею у фітоценології, об"єднує всі фітоценози, що беруть однорідну участь в акумуляції і трансформації речовини та енергії на поверхні землі, або точніше її фітогеосфери. Відповідно до цього вони характеризуються головним чином однорідним видовим складом, однорідною оинузіальною структурою, що відбиває відповідний оклад екологічних типів рослин, t однорідним складом факторів середовища, які впливають на фітоценотипи процес” (1961, с.38). Таким чином, "рослинна асоціація" є одночасно і флористичне, і екологічне, і фітоценотичне поняття. За критерій однорідності фітоценозів, що належать до однієї асоціації, В.М.Сукачов (1929, 1931, 195?, 1961) пропонує обрати ознаки, в яких безпосередньо відображаються процеси трансформації речовини та енергії: І)видовий склад та екологічні властивості видів, наскільки вони нам відомі; 2) синузіальну структуру угруповання і будову синузій; 3) фактори
96
зовнішнього середовища, в тій мірі, в якій вони можуть бути враховані при маршрутному дослідженні.
Добре помітними екологічними ознаками і відмінами структури та будови має відрізнятися асоціація і на думку В.В.Мазинга та Х.Х.Траса (1963). Останнім часом в широкому теоретичному аспекті питання однорідності фітоценозів розглянув В.І.Василевич (1969, 1971). Ми цілком погоджуємось з його пропозицією застосовувати при класифікації рослинності пробні ділянки, розмір яких відповідає ценоквантові, тобто найменшій ділянці рослинного покриву, до якої можна вжити кількісну характеристику.
З наведених трьох груп ознак при виділенні асоціацій найбільші труднощі виникають при оцінці однорідності фітоценозів за екологічними показниками. Пояснюється це головним чином складністю оцінки екологічних умов, яку звичайно роблять не прямими методами, а за наявною рослинністю та екологічною амплітудою обнєктів, що підлягають класифікації, через що вимога однорідності факторів середовища в фітоценозах однієї асоціації є відносною. В природі трапляються численні випадки, коли екологічні амплітуди різних асоціацій перекриваються і фітоценози крайніх умов однієї й тієї ж асоціації в екологічному відношенні стоять ближче до фітоценозів іншої асоціації, ніж до своєї.
. До однієї асоціації ми відносимо фітоценози, які мають однакові едифікатори, субедифікатори і конфектори, однакову синузіаль- ну будову, майже однаковий флористичний склад з ядром детермінант- них видів та більш або менш однорідними умовами місцезростання, принаймні в межах певної зони. Однорідність екотопу встановлювалась переважно на підставі грунтів та характеру рельєфу.
З наведеного визначення асоціації і неодноразових роз’яснень її обсягу видно, що В.М.Сукачов (1926, 1931, 1938, 1957, 1961), по- перше, розглядав асоціацію як нижчу фітоценотичну одиницю і, по- друге, в дуже вузькому значенні. Слід зупинитись на поясненні цього. Фактичний матеріал, одержаний на підставі описів фітоценозів однієї асоціації в різних природно-історичних регіонах, свідчить про значну неоднорідність асоціацій в різних частинах її ареалу. Ця неоднорідність має чітко виявлений закономірний характер і виявляється в певних змінах загального флористичного складу, проективного покриття і появи співконфекторів і субконфекторів при загальній незмінній структурі асоціації та однаковому складі видів, що визначають її головні риси. Такі фітоценози з несуттєвими відмї-
97
нами ми вважаємо за доцільне виділити як дрібніші одиниці: суб- асоціації, варіанти асоціацій та фації. Менші за асоціацію одиниці виділяють представники цюріхської школи фітоценологів(КІІка, І955;3сашопі, 1955; Pawlowska, 1959; Bjcaun-Blanquet, 1964; So8, 1964; Fukarek, 1967). Простежується тенденція виділення менших, ніж асоціація, таксономічних одиниць і серед радянських геоботаніків (Шенников, 1964; Камншев, 1966; Голубец, Малиновский, 1967). Різниця полягає лише в обсязі цих таксонів.
В.М.Сукачов (1931) розрізняв корінні та похідні асоціації. На нашу думку, виділення їх має базуватися на поглядах про інваріантну структуру асоціацій і сполучені з нею змінні структури. Останні, за В.Б.Сочавою (1968), відповідають різним серійним і антропогенним модифікаціям. Короткочасовопохідні суксцесійні стадії підносити до рангу асоціацій ми вважаємо недоцільним. Краще виділяти їх в ранзі неоасоціацій і відносити до корінної асоціації, на місці якої вони сформувались. Асоціація та відповідні неоасоціації створять епіасоціацію в розумінні В.Б.Сочави.
Щодо обсягу формації, так само як і асоціації, єдиної думки не існує. Багато дослідників, застосовуючи цю таксономічну одиницю, трактують її по-своєму. Пояснюється це трьома головними причинами. По-перше, недостатньою розробкою низки теоретичних положень геоботаніки, наприклад оцінки видів залежно від їхньої ролі у фітоценозі, на чому засноване встановлення деяких таксономічних одиниць, серед них і формації. Оскільки немає єдиного погляду на таке поняття, як "едифікатор", неминуча й розбіжність у трактуванні поняття "формація". По-друге, використанням поняття "формація" в розумінні, далекому від вжитого О.П.Шенніковим (1964). Наприклад, Л.Р.Лаасимер (1958) виділяє формацію на підставі фізіономічних ознак 1 відносить до цієї таксономічної одиниці такі таксони, як сухі та помірновологі суходольні ліси. По-третє, протиріччям, що намічається між принципом супідрядності таксономічних одиниць та змістом таксонів, які відносяться до них.
Останнє протиріччя В.Д.Александрова (1968) пояснює такими обставинами, у полідомінантних угруповань виділення формації за домінантним принципом носить формальний характер, як щодо обсягу, так і природності одиниць. Окрім того, деякі види утворюють однорідні одиниці й відзначаються екологічною і фітоценотичною єдністю, тобто створюють фітоценози, близькі між собою в екологічному та морфологічному відношеннях, і навпаки, є види з ширшою екологічною 98
амплітудою, ніж подібна амплітуда вищої таксономічної одиниці. На луках деякі види не є панівними з року в рік. Тому Б.М.Міркін (1968, 1970) вважає, що від формації як таксономічної одиниці слід відмовитись. Звичайно з розглянутого протиріччя виходять завдяки встановленню надвидових або середвидових формацій. Першим скористались Є.М.Брадіс (1956, 1963), С.Я.Соколов (1962), а другим - П.Д.Ярошенко (1961). Ми вважаємо, що для лісового типу рослинності відмовлятися від формації як таксономічної одиниці поки що немає можливості, хоча в цілому запропонований Б.М.Міркіним домїнанто- детермінантний принцип класифікації рослинності безумовно приваблює і дає можливість уникнути згаданого протиріччя.
На питання, пов"язані з принципами, покладеними в основу класифікаційних побудов, також немає єдиного погляду. Основний вододіл, на нашу думку, тут проходить між екологічними (фітотипологіч- ними) і фітоценотичними (в широкому розумінні) класифікаціями. Більшість геоботаніків заперечують проти виключно екологічних побудов як таких, що не відповідають самому об"єктові класифікації (Ильинский, 1937; Шенников, 1938; Сукачев, 1944; Лавренко, 1963). В екологічних класифікаціях одна і та сама асоціація потрапляє в різні підрозділи одного таксономічного рангу, що несумісне з правилом “поділу обсягу понять".
З головними принципами класифікацій тісно пов’язане питання про їх природність чи штучність. За В.М.Сукачовим (1931), штучні класифікації базуються на одній будь-якій ознаці, хоча б випадковій, але в даний момент для нас важливій. Природні класифікації засновані на сукупності багатьох ознак. Цю ж думку розвиває і
A. Г.Долуханов (1959), за яким природна класифікація лісових фітоценозів повинна будуватись на даних всебічного вивчення їх складу, синузіальної структури, динаміки та ритміки життєвих процесів.
B. Д.Александрова (1969) природними вважає всі класифікації і пропонує розрізняти лише ступінь їх природності.
Найбільш природними класифікаціями Б.А.Биков (19666) вважає генетичні. До них відносить свою класифікацію і Б.П.Колесников (1956, 1958а, 19586, 1961, 1966), хоча вже в першій з наведених праць відзначає, що в поняття "генезис" вкладено не флорогенез і ступінь історичної спорідненості об"єктів, що класифікуються, а зміст сукцесій під час формування певних фітоценозів. Підкреслюючи цю різницю, Б.П.Колесников (19586) називає свою класифікацію онтогенетичною, тобто застосовує термін "онтогенез" до лісоутворю— 99
ючих процесів, що відбуваються яа конкретних ділянках території протягом півтора-двох сторіч. Тим самим тип лісу він обмежує не лише в просторі, але і в часові. Ця класифікація, на нашу думку, не викликає заперечень, проте термінологія, якою оперує автор, не зовсім вдала; Насправді йдеться про динаміку фітоценозу, тобто об'єднання в один тип лісу всіх його серійних і антропогенних сук- цесійних стадій, пов"язаних між собою часто лише єдністю території та едафотопу, тому говорити про онтогенез можна лише умовно. За думкою О.П.Шенникова (1958), в цій класифікації відображена лише еволюція рослинного покриву в масштабі дигресія і демутація корінних форм рослинності. В.М.Сукачов ще в 1944 р. застерігав від застосування терміну "онтогенез" до рослинності як неправомірного. Отже, класифікація Б.П.Колосникова не є генетичною. На наш погляд, її слід відносити до еколого-фітоценологічних класифікацій, найбільш поширених в Радянському Союзі. До них належить, наприклад,переважна більшість регіональних класифікацій лісів європейської частини СРСР. Ці класифікації найбільш розроблені й зручні. На їх основі Можливий перехід до генетичної класифікації.
Якісно новим етапом є побудова біоценотичної класифікації, що випливає з уявлень В.М.Сукачова (1942, 1945, 1964) про біогеоценоз як якісно відмінне від фітоценозу явище природи з притаманною лише йому взаємодією складаючих його компонентів. В напрямку створення цієї класифікації особливо плідно працюють М.В.Диліс (1964) та ОЛ.Уткін (1968). На їх думку, бхогеоценотична класифікація лісів може дати більш глибоку і різнобічну характеристику природи лісу, ніж фітоценотична, і може стати основою для узгодження поглядів рівних шкіл, з чим погоджується, відзначають вони, більшість геоботаніків. Щодо цього існує цікава думка (Krajin, - цит. за Шен- яиковим, 1962), згідно з якою пропонується розрізняти три рівні в розробці класифікаційної проблеми. Перший становлять топологічні класифікації, що відповідають нижчому рівню знань про природу рослинних угруповань. Другий - фітоценотичні класифікації за ознаками рослинності. Вони потребують більш різнобічних відомостей про рослинність. Третій - біогеоценотична класифікація, заснована на розумінні кожного тилу фітоценозу як члена біогеоценозу. Зараз ми перебуваємо на другому рівні. Найближчим завданням є розробка філоценогенетичної класифікації.
100
Головні класифікації рослинного покриву України
На Україні ботаніки користуються трьома класифікаціями; екологічною, еколого-фітоценогенетичною і еколого-фітоценотичною.
Екологічна лісівницька класифікація, розроблена Є.В.Алексє- євим (1928), згодом творчо була доповнена П.С.Погребняком (1941, 1955), О.Л.Бельгардом (1950)* та Д.В.Воробйовим (1953). В основу своєї класифікації Є.В.Алексввв (1928) поклав два провідних екологічних фактори - зволоження і трофність. Зміни цих факторів в напрямку збільшення вологості та багатства грунту утворюють ряди едафічної сітки; гігрогенний ряд - сухі, свіжі, вологі, сирі і мокрі; трофогенний ряд - бори, субори, складні субори та груди (діброви). Кожна з клітин сітки являє собою єдність гігротолу 1 трофо- топу та відповідає типові лісової ділянки або едатопу. Останній, за Д.В.Воробйовим (1953), об”єднує типи лісу, які схожі за родючістю грунту і заміщуються кліматично, географічно та ареогенетич- но. Всі гігротопи одного едафотопу об'єднуються в чотири групи типів лісової ділянки. Вони об'єднують усі вододільні ліси України. Найменшою класифікаційною одиницею є тип деревостану. Принципова відмінність цієї класифікації від фітоценотичної полягає в тому, що в ній класифікація рослинності підміняється класифікацією екотопів. Якщо фітоценологи припускають існування певної амплітуди екологічних умов під однорідними фітоценозами, то Є.В.Алексеєв кладе в основу встановлення таксономічних одиниць однорідність екотопу і припускав певні коливання однорідності фітоценозів, які часто перевищують міру обсягу асоціації. Тому представники цього напрямку вивчають рослинність лише для її господарської оцінки і виділення однорідних екотопів, що потребують застосування одних і тих самих лісогосподарських заходів. На практиці представники екологічного напрямку встановлюють типи за індикаторними видами, які є провідною ознакою. Допоміжними ознаками вважаються рельєф і грунти. Таким чином, класифікація Є.В.Алексєєва-П.С.Погребняка має на меті виділення і згрупування не фітоценозів і навіть не однорідних екологічних умов, а скоріше екологічних умов, однорідних за лісо- рослинним ефектом.
І Класифікацію О.Л.Бельгарда можна так само віднести й до еколого-фітоценотичних, оскільки вона базується не тільки на екологічних показниках, але й на синузіальній будові, біоморфах та флористичному складі фітоценозів.
101
Еколого-фітоценогенетична класифікація розроблена М.А.Голубцем і К.А.Малиновським (1967, 1968, 1969) для рослинності Українських Карпат. Вона є найбільш деталізованою і являв значне досягнення фітоценотичної школи на Україні. В основу класифікації авторами покладено найбільш цінні, стосовно до своєрідної рослинності Карпат, принципи деяких вітчизняних і зарубіжних класифікацій. В ранзі головної таксономічної одиниці прийнята асоціація. Дальше об'єднання у таксономічні одиниці вищих рангів провадиться за ознаками корінної рослинності. Похідні асоціації об'єднуються з корінною асоціацією, на місці якої вони виникли. Тим самим, на думку авторів, розкриваються генетичні зв"язки між корінними та похідними асоціаціями, отже,вони розуміють генезис за Б.П.Колосниковим,а не як філоценогенез. Ми вважаємо, що ця класифікація належить до групи еколого-фітоценотичних. Таксономічні одиниці найвищого рангу виділяються авторами за життєвими формами. В цілому ми погоджуємося з принципами, покладеними в основу розглянутої класифікації. Зупинимось лише на трьох, дискусійних, на наш погляд, моментах.
По-перше, тип рослинності автори приймають в дуже великому обсязі. Наприклад, до виділеного ними типу трав"яної рослинності доводиться віднести разом з альпійськими і субальпійськими луками’ степову та частину болотяної рослинності t т.п., тобто в межах тільки України один тип об'єднує різну в екологічному і генетичному відношеннях рослинність, проти чого обгрунтовані заперечення висунув Є.М.Лавренко (1940, 1956, 1959). Це стосується і підтипу рослинності, який виділяється за життєвими формами, але не завжди. Наприклад, підтипи - "високотрав"я", "різнотравні луки" і "прионігові лучки", як видно, встановлені за різними показниками.
По-друге, потребує уточнення і запропоноване М.А.Голубцем та К.А.Малиновським виділення формацій. "Формація, - пишуть вони,- являє собою об"єднання монодомінантних і кондомінантних оубформа- цій однієї домінанти едифікаторної синузії (наприклад, формація ялини європейської об"єднує субформації чистих ялинових лісів, кедрово-ялинових, буково-ялинових лісів та ін.)" (1969, с.240). На нашу думку, до субформації слід відносити фітоценози, складені едифікатором і субедифікатором, наприклад липово-дубові ліси.Фітоценози, створені двома і більше едифікаторами (кондомінантні), ми вважаємо за доцільне виділити в ранзі формації, як, наприклад, буково-дубові ліси.
102
По-третє, досвід показує, що класифікація довготривалопохід- ної рослинності дуже зручна для наукової і практичної мети. Тому відмова від систематизації асоціацій довготривалопохідної рослинності внесе нові ускладнення і кінець кінцем виникне потреба створення паралельних класифікацій. Крім того, при такому підході губиться оуть асоціації як таксономічної одиниці, яка об'єднує всі однорідні фітоценози, тому що припускають віднесення однієї і тієї самої асоціації до кількох корінних асоціацій.
Еколого-фітоценотична класифікація на Україні дуже поширена. Для її побудови багато зробили Є.М.Лавренко (1928, 1940), Ю.Д.Кле- опов і Є.М.Лавренко (1938), Д.К.Зеров (1938), Г.І.Білик, (1954, 1963), Д.Я.Афанасьєв та ін. (1956), Є.М.Брадіс (1956, 1961, 1963), В.О.Поварніцин (1957, 1959), Д.Я.Афанасьєв (1959, 1968), В.І.Ко- мендар (1966), Ф.О.Гринь (1971) та ін.
Детальної еколого-фітоценотичної класифікації для всіх типів рослинності України поки що немає. Є тільки узагальнююча класифікаційна схема, запропонована Д.Я.Афанаоьєвим та ін. (1956), і класифікації окремих типів рослинності (Афанаоьев, 1968; Брадіо, 1969; Гринь, 1971; Поварніцин, 1971), більш чи менш деталізовані. На відміну від попередньої класифікації названі автори приймають вищу таксономічну одиницю (тип) в значно меншому обсязі. Як самостійні типи рослинності вони виділяють ліси, чагарники, отепи, луки, фриганоподібні півчагарнички1, болота, солончаки і пустища. На наш погляд, типи рослинності пустища та фриганоподібні півчагарнички обгрунтовані недостатньо, бо не відповідають прийнятому авторами принципу, за яким встановлюється тип рослинності.
До лісового типу рослинності автори згаданої класифікаційної схеми відносять три класи формацій - хвойні ліси, широколистяно- хвойні та широколистяні. Класи формацій автори встановлюють на підставі подібності екологічних особливостей рослинного покриву, що визначається близькими умовами місцезростання. Це не зовсім так, бо за екологічними ознаками формація, сосни кримської (Pineta pallae іапае) ближче стоїть до формації дуба пухнатого ( Querceta pubescentis), яка належить до іншого класу формацій.ніж до формації ялини європейської (Piceeta abies) - одного в нею класу формацій.
Очевидно, авторами враховувались і біоморфи, з чим цілком
Термін "фриганоподібні півчагарнички" невдалий, оскільки вія означає життєву форму, а не тип рослинності.
103
можна погодитися. Групи формацій автори виділяли на підставі подібності біоморф. Проте, цього принципу вони не скрізь дотримуються. Так, у класі формацій широколистяних лісів розрізняють групи формацій; дубових лісів, букових, широколистяно-змішаних, дрібно- листяних, заплавних, заболочених і т.п. Через такий підхід в одній групі формацій заплавних лісів опинились різні в біоморфоло- гічному відношенні формації - в"язово-дубова (Queioeto - Uliieta) та верби білої (Sallceta albae) і, з другого боку, відсутні інші формації заплавних лісів, наприклад вільхи клейкої (Alneta glutinosae), які автори віднесли до групи формацій заболочених лісів. Крім того, в"язово-дубову формацію, віднесену до групи формацій заплавних лісів, одночасно слід було б віднести до групи формацій широколистяно-змішаних лісів. Застосування таких різнорідних ознак, що перекриваються при виділенні таксономічних одиниць одного рангу, безумовно, не виправдовує себе. Не відповідав обраному авторами принципові і виділення формацій. Вони пишуть: "Формації об'єднують групи асоціацій із спільними едифікаторами панівних ярусів" (Афаяаоьев та ін., 1956, с.65) і в той же час виділяють їх на підставі видів, що належать до другого, не панівного ярусу (наприклад, в"язово-дубова формація).
На підставі зазначеного вище та одержаних нами матеріалів ми пропонуємо класифікацію досліджуваних лісів.
Фітоценотична класифікація дубових лісів з дуба звичайного
В основу класифікації нами покладені: І) життєві форми рослин; 2) фітоценотична роль видів, яка виявляється в певному сину- зіальному складі фітоценозу, життєвості, рясності та є наслідком певних взаємовідношень з іншими видами, що вирішуються в умовах певного екологічного середовища; 3) постійні види.
При створенні класифікації ми дотримувалися правила поділу поняття, можливості виділення таксономічних одиниць рівних рангів на підставі різних показників та принципу оубординаційної класифікації.
Таксономічні одиниці класифікації такі: клас типів, група типів, тип, клас формацій, група формацій, формація, субформація, клас асоціацій, група асоціацій, асоціація, субасоціація, варіант асоціації та фація. З них асоціація, формація 1 тип - головні. Фація, варіант асоціації та субасоціація належать до системи внут- 104
рішньоасоціаційних таксономічних одиниць. Фація розглядається нами як найменша таксономічна одиниця, що об"єднує однорідні за всіма основними ознаками фітоценози. Від інших фітоценозів тієї самої асоціації вони відрізняються наявністю групи регіональних постійних видів, тобто тих, що мають регіональний 8-ІО-й клас постійності. Далі ми будемо розрізняти спільні абсолютно постійні види, тобто такі, що мають 8-ІО-й клас постійності, і спільні відносно постійні види, які мають 5-7-й клас постійності. З числа останніх ми виділяємо регіональні постійні види, що обумовлюється неоднорідністю групи постійних видів. Одні з них трапляються епізодично на всій території поширення асоціацій, наприклад коручка широколиста (Epipaotia latifolia (L.).A 11.), ооока отягнута (Carex oontgua Н о р р e)t ранник вузлуватий (Sorophularia nodooa L.), а інші - лише в її окремих частинах, де вони мають 10—й клас постійності. Можливо, що перші менш повпязані з регіональними змінами екологічних умов асоціації, ніж другі. Таким чином, індикаторна цінність перших видів обумовлена амплітудою екологічних умов, яка виходить за межі міри подібності асоціації, а других - амплітудою екологічних умов в межах подібності асоціації.
Варіант асоціації об"єднує фітоценози, пов"язані з різними екологічними умовами, наприклад, ділянки дубового лісу ліщиново- яглицевого на плакорах та в заплавах річок.
Субасоціація об"єднує фітоценози зі спільними співконфекто- рами або субконфекторами.
Асоціацію ми приймаємо в розумінні В.М.Сукачова (1961), з тим лише уточненням, що тимчасові антропогенні і вікові сукцесійні стадії виділяємо в ранзі неоасоціації. Асоціація (корінна або тривало- похідна) разом з неоасоціацією (тимчасові сукцесійні стадії) утворює епіасоціацію. Неоасоціації відносимо до асоціацій, на місці яких вони виникли, тим самим досягаємо відбиття динаміки і наступності. Таке об’єднання важливе t з господарського погляду, тому що практично ми виділяємо однотипні в межах екологічної амплітуди асоціацій ділянки. Як правило, иеоасоціація відрізняється від асоціації, на місці якої вона виникла, лише складом конфекторів, тому, якщо не надавати цьому показнику чисто формального значення, різниця між ними не виходить за межі міри подібності асоціації. Кожний вірно встановлений такоон асоціації, крім спільної синузіаль- ної будови, буде мати ще кілька спільних абсолютно постійних видів. Разом з конфекторами вони утворюють флористичне ядро асоціа-
105
ці!, яке в індикатором її екологічної амплітуди. Відсутність спільних абсолютно характерних видів свідчить про методичну помилку або про те, що ми маємо справу з порушеними фітоценозами, ще більше порушення яких веде до появи спільних абсолютно характерних антропогенних видів t зміни конфектора. В останньому випадку будемо мати неоасоціацію.
Група асоціацій виділяється нами на підставі кількох ознак. В.М.Сукачов (1934) вважав, що кожна з груп асоціацій повинна характеризуватися певним типом рельєфу, певним зв'язком з умовами середовища та продуктивністю. Такої ж думки М.А.Голубець та К.А.Ма- линовський (1969), але В.М.Сукачов будує свої ряди залежно від прямо діючих екологічних факторів і не обмежує їх обсягу, а М.А.Голубець та К.А.Малиновський обмежили групу асоціацій трьома ступенями трофності і п"ятьма ступенями вологості. Крім того, в групу асоціацій вони включили і всі похідні асоціації, що утворились на її місці. Нам здається, що підхід цих авторів принципово вірний, але не слід було б обмежувати кількість груп асоціацій вузькими рамками трофності та вологості, оскільки у випадку появи нових асоціацій, що відрізняються порівняно в описаними більшою оліготроф- ністю або евтрофнїстю, цілком можливий перехід раніш розподілених асоціацій з однієї групи в іншу.
До однієї групи асоціацій ми пропонуємо відносити асоціації, ярус підліску яких утворений конфектором одного і того ж виду та близькими екологічно і генетично конфекторами трав“яниотого ярусу аі спільними абсолютно і відносно постійними видами. Ступінь спільності флористичного складу асоціацій встановлювався за С.Кульчин- ським (Kulczynski, 1940). Одержані величини спільності (подібності) порівнюваних асоціацій зводились в матрицю, за якою будували дендрит. На підставі останнього виділяли однорідні групи, які розглядаються нами як групи асоціацій. Методика побудови й аналізу дендриту докладно описана в роботах Я.Фалінського (Failtiski, 1960) та В.І.Василевича (1969).
Клас асоціацій об"єднує асоціації, деревний яруо яких складений одним t тим самим едифікатором та конфектором чагарникового ярусу.
Субформація об"єднує асоціації, деревний ярус яких складений одним і тим самим едифікатором та субедифікатором.
Формація об'єднує асоціації, едифікаторний ярус яких складений одним і Тим самим едифікаторним видом або підвидом чи близькою в екологічно-генетичному відношенні групою видів одного роду.
106
Група формацій об'єднує формації, едифікаторний ярус яких складений едифікаторами, що належать до одного роду, до однієї біоморфи і близькі аа ареалом.
Клас формацій об'єднує формації з едифікаторами, що належать до однієї біоморфи типу "світлолюбна широколистяна порода".
Тип рослинності об'єднує формації, утворені едифікаторами однієї біоморфи типу '‘широколистяна літньо-зелена порода”, тобто цю таксономічну одиницю ми встановлюємо в звуженому обсязі, як це пропонує Є.М.Лавренко (1959).
Група типів об"єднує формації, едифікатори яких належать до однієї життєвої форми типу "літньо-зелена листяна порода".
Клас типів об"єднує всі ліси, тобто формації з едифікаторами.
Виходячи з розглянутих положень та одержаних матеріалів ми пропонуємо класифікаційну схему дубових лісів України.
Клас типів рослинності - ліси (silvae).
Група типів рослинності - літньо-зелені лиотяні ліси(Silvae aeatif oliolignosa).
Тип рослинності - широколистяні літньо-зелені ЛІСИ (silvae folioaeetilignosa).
Клас формацій - широколистяні літньо-зелені світлолюбні ліси (Silvae lucidiphilus latifolioaestilignosa).
Група формацій - дубові літньо-зелені ліси (Silvae quercetae aestіlignosa).
Формація - дубові ліси з дуба звичайного (Querceta roboris).
Субформації: грабово-дубова (Oarpiaeto-Queroeeta), липово- дубова (Tilieto-Querceeta), кленово-дубова ( Acereto-Queroeeta) та чисто дубове (Queroe- eta roboris)^ .
Остання оубформація поділяється на такі одиниці:
А. Клас асоціацій - дубові ліси крушинові (Querceta frangulosa).
І. Група асоціацій - дубові ліси крушиново-тря- сучковидноосокові (Querceta franguloeo-oaricosa (brizoidis).
АсоціацІ.Ї.: діброва крушиново-трясучковидноосокова (Quercetun franguloeo-caricoeua(brizoidis) та діброва круши- НОВО-молІНІвва (Quercetum franguloeo-mollnioeum).
!усі субформації, за винятком останньої, досить повно описані Ф.О.Гринем (1971).
107
П. Група асоціацій- дубові ліси крушиново- конвалієві ( Querceta fran- guloso-convallarloea).
Асоціація - діброва крушиново-конвалієва (Quercetum
~ frauguloao-convallariosuia) е
Б» Клас асоціацій - дубові ліси ліщинові (Querceta corylosa).
1. Група асоціацій - дубові ліси ліщиново-
трясучковидноосокові (Querceta corylosо_сагіао- sa (brizoidis).
Асоціацій діброва ліщиново-трясучковидноосокова (Quercetum coryloso-caricoaum(brizoidifl) та діброва ліщи- ново-квасеницева (Quercetum coryloso-oxalidosum).
IV. Група асоціацій - дубові ліси ліщиново-чо-
ловічопапоротеві (Querceta coryloso-dryopteriosa).
Acoi^iauiJj діброва лїщиново-чоловічопапоротева (Quercetum coryloao-dryopteriosum(filicis таз)діброва ліщи*- ново-жіночопапоротава (Quercetum coryloso-athyrlosuio (filiole feminae) та діброва ліщиново-веснівкова (Quercetum coryloso-majanthemoeum).
V, Група асоціацій - дубові ліси ліщиново-зі-
рочникові (Querceta corylos o-atellariosa).
дібРова ліщиново-зірочникова ( Querceta coryloso- atellariosum), діброва ліщиново-копитнякова (Quercetum coxyloso-asaroaum), діброва ліщино¬
во-маренкова (Quercetum coryloso-asperuloeum), Діброва ліщиново-яглицева (Quercetum coryloso- aegopodioaum), діброва ліщиново-зеленчукова (Quercetum coryloso-galeobdolosum), діброва ліщиново-волосистоосокова (Quercetum coryloso- caricoa um(pilosae).
УІ< Трупа асоціацій - дубові ліси ліщиново-гір- CbKOOCOKOBi (Querceta corylos с-с аг іс os а( montanae ) ф Асоціації^ Діброва ліщиново-гірськоосокова (Quercetum corylo- ао-сагісоаum(montanae), діброва ліщиново- конвалієва (Quercetum coryloso-convallariosum)«
108
УП. Група асоціацій - дубові ліси ліщиново- парвськоосокові (Querceta coryloso-caricosa(brevi- collis).
Асоціація^- діброва ліщиново-парвськоосокова (Quercetum corylos o-caricosum(brevicollis).
УШ. Група асоціацій - дубові ліси ліциново- горобейникові (Querceta coryloso-lithospermosa).
Асоціація - діброва ліщиново-горобейникова (Quercetum corylos о- lit hosp er mos шп) .
В. Клас асоціацій - дубові ліси свидинові (Querceta thelyora- niosa).
IX. Група асоціацій - дубові ліси свидиново- аірочникові (Querceta tbe- lycranioso-stellariosa).
AcogiaiUX? Діброва свидиново-зірочникова (Quercetum thelycra- nioso-stellariosum), діброва свидиново-яг- ЛИцева (Quercetum thelycranioso-aegopodiosum).
X» Група асоціацій - дубові ліси свидиново-гір- ськоосокові (Querceta the- lycranioso-caricosum(montana).
Асодіації^ діброва свидиново-гірськоосокова (Quercetum thely- cranioao-caricosum(montanae) та діброва сви- диново-конвалівва (Quercetum thelycranioso-conval- lariosun),
XI. Група асоціацій - дубові ліси свидиново- парвськоооокові (Querceta thelycranioso- caricosa(brevicollis).
Дсоціація^- діброва свидиново-парвськоосокова (Quercetum the- lycranioso-cariooBum(brevicollis).
Г. Клас асоціацій - дубові ліси татарськокленові (Querceta acerosa(tatarioi)e
ХП. Група асоціацій - дубові ліси татарськокле- ново-зірочникові (Querceta aceroso(tatarici)- atellariosa).
109
АооціацП^ діброва татарськокленово-зірочникова (Quercetum acero8o(tataEici)-stellariosum), діброва татар- ськоклвново-яглицева (Quercetum aceroso(tatarici)- aegopodioaum), діброва татарськокленово-волосис- TOOCOKOBa ( Quercetum aceroso (tatarici)-caricosum (pilosae), діброва татарськоклбново-коранбвищ- ноооокова ( Quercetum aoeroso(tatarici)-caricosum (rhizinae).
XHL Група асоціацій - дубові ліси татарсько- кленово-гірськоосокові (Querceta aceroso(tatari- сісагісоаа(montanae).
Асоціації; діброва татарськокленово-гірськоосокова ( Quercetum acer oso(tatarioi)~caricosum(montanae), діброва татарзькокленово-конвалюва (Quercetum aceros o(tatarlol)-convallariosum).
XIV. Група асоціацій - дубові діси татарсько-
кленово-рябоперлівкові (Querceta acerosо(tatа- rici)-'melicosa(pictae) е
Асоціація^- діброва татарськокланово-рябоперлівкова (Quercetum ~ aceroso (tat arici)-meliooaum(pictae).
XV. Група асоціацій - дубові ліси татарсько-
кленово-горобейникові (Querceta aceroso(tatari- ci)-lithospermosa).
ДсоціацЦ^ Дарова татарськокленово-горобейникова (Quercetum aceroso(tatarioi)-lithospermosum, діброва татарськоклаково-мікеліевоосокова (Quercetum aceroso(tatarlci)-caricosum(michelii).
Д. Клас асоціацій - дубові ліси кизилові (Querceta comosa).
XVI. Група асоціацій - дубові ліси кизилово-
парвськоосокові (Querceta cornoao-caricoaa(bre- vicollis).
AgogiagH^ діброва кизилово-парвськоосокова ( Quercetum oorno- so-caricosum (brevicollis), діброва кизи- лово-зірочникова (Quercetum cornoso-stellario- sum), діброва кизилово-конвалієва (Quercetum ЇІ0
согао0О-оагісоашп(р11о8ае), діброва кизилово1- волосистоосокова ( Quercetum cornos о—o&rloosum fcilosae), діброва кизилово-кореневищноосокова (Quercetum aceroeo(tatarici)-oaricosum(rhizlnae).
ХУП. Група асоціацій- дубові ліси кизилово- горобейникові (Querceta cornoso-litbospernosa).
Асоціації: діброва кизилово-горобойникова (Quercetum coxnoso- Litbospermosum), діброва кизилово-мікелієвоосо- кова (Quercetum aornoso-caxicosum(mlchelii).
Е. Клас асоціацій - дубові ліси скумпієві (Querceta ootlnosa).
ХУ1П. Група асоціацій - дубові ліси скумпієво-коя- валієві (Querceta cotinos o-convallariosa).
Асоціації^ діброва скумпієво-конвалієва (Quercetum cotinoeo- convallariosua), діброва скумпівво-рябоперлівкова (Quercetum cotinoso-welicosum(piotae).
ХІХ.Г рупа асоціацій - дубові ліси скумпієво-пір- частокуцоніжкові (Querceta cotinoso-brachypodioea (pinnati).
Асоціації; діброва скумпієво-пірчастокуцоніжкова ( Quercetum ootinoso-braohypodiosuB>(pinnati), діброва скум- піево-мікеліевоосокова ( Quercetum ootlnoso-oari- cosum(michelii).
В запропонованій нами схемі таксони класів асоціацій розташовуються в порядку зростання ксеротермності умов І родючості грунту, так само як і групи асоціацій. Кожний з виділених класів асоціацій має свій ареал, трапляється в певних екологічних умовах і характеризується певним флористичним складом. Все це дає можливість подану класифікаційну схему легко трансформувати в ординаційву систему еколого-фітоценотичних рядів В.М.Сукачова.
ФІТОЦЕНОТИЧНА ХАРАКТЕРИСТИКА ДУБОВИХ ЛІСІВ З ДУБА ЗВИЧАЙНОГО НА ТЕРИТОРІЇ УКРАЇНИ
Дубові ліси з дуба звичайного являють собою світлі монодомі- нантні з одноярусним деревостаном фітоценози,ступінь участі едифікатора в яких становить не менте ніж 80$ загальної кількості
стовбурів.За висотою едифікатор перевищує більшість своїх супутників 1 завжди формує перший ярус,в якому разом з ним звичайно трапляються ясен звичайний, сосна звичайна (Полісся) та бук лісовий (Поділля). Нижче розташовуються поодинокі дерева граба звичайного, липи серцелистої, клена звичайного, клена польового, черешні та інших видів. Метрично вони відповідають другому ярусові, який як структурна одиниця не формується. Крім ярусу деревостану, у дубових лісів добре розвинутий ярус підліску і травостою. Моховий ярус формується лише в деяких асоціаціях на вологих і сирих дерново-підзолистих грунтах.
Дубові ліси мають два екологічні оптимуми поширення. Перший з них, в межах ареалу граба звичайного і бука лісового, розташований в північній частині Західної і Середньої Європи (Міндленд- ська, Ріонська і Північна низовини) з її великими,погано дренованими рівнинами, а другий - в межах ареалу липи серцелистої - в лісостеповій і степовій зонах Радянського Союзу. Перший з них зумовлений едафічними, а другий - кліматичними факторами. Па території України виявлено обидва оптимуми. Перехідний від атлантичного до континентального клімат республіки, в якому конкурентна здатність ліооутворюючих широколистяних порід Середньо! Європи різко послаблена (бук лісовий, дуб скельний, дуб пухнатий і граб звичайний), а порід Східної Європи (липа серцелиста) ще не виявлена, сприяє поряд з наявністю погано дренованих територій Полісся, Прикарпаття і Закарпаття формуванню дубових лісів з дуба звичайного. Саме тому вони на досліджуваній території займають великі площі й представлені такою великою кількістю асоціацій. За- лежно^від екологічних особливостей і флористичного складу асоціації можна поділити на три еколого-генетичні ланки: І) ацидофільну - дубові ліси на бідних і дуже кислих грунтах (дерново-підзолистих); 2) нейтрофільну - дубові ліси на середньобагатих кислих і слабокислих грунтах (дерново-підзолистих та сірих лісових);
3) базифільяу - дубові ліси на багатих нейтральних і лужних грунтах (чорноземних). До першої ланки належить 5, до другої - 18 і до третьої - 15 асоціацій. Асоціації першої ланки поширені в ши- роколистянолісовій зоні. В їх флорі важливу роль відіграють боре- альні види. Асоціації другої ланки поширені в лісостеповій зоні. У флорі їх панують неморальні балтійські види, під якими маємо на увазі види мезотермної групи, і асоціації третьої ланки, поширені в степовій зоні, де переважають неморальні субсередземноморські види коеромезотермно! еколого-генетичної групи.
112
Дубові ліси України представлені асоціаціями, спільними з більшістю асоціацій районів їх ареалу. Найбільше вони відрізняються від дубових лісів південно-західної частини ареалу (південь Франції, північ Португалії та Іспанії), з якими у них близько 15$ спільних видів і немає жодної спільної асоціації. Менше відрізняються вони від дубових лісів крайньої південно-східної частини ареалу,з якими не мають жодної спільної асоціації, проте більшість видів у них спільні. З дубовими лісами Кавказу вони мають одну спільну асоціацію і близько 70% спільних видів. Три асоціації і майже 75% видів спільні у них з дубовими лісами північної частини Західної та Середньої Європи. Більшість асоціацій і близько 90% видів в спільними у цих лісів з аналогічними лісами центральної та південно-східної частини Середньої Європи. З територією Молдавії та Середньоруської височини вони мають спільні майже всі асоціації та види поширені там.
Фітоценологи франко-швейцарської геоботанічної школи відносять дубові ліси до двох класів: Quercetea robori-petraeae і Querco-Kagetea (Braun-Blanquet, Tiixen, 1943; bebrun, Noirfalise, Helnemann, Vanden Bergben, 1949; Braun-Blanquet, 1952, 1964; Kli- ka, 1955; Tiixen, 1955; Oberdorfer,1957; Holub, Hejny, Mo.raveo, Neuhausl, 1967).
Перший клас об"вднув ацидофільні ліси (дубові, букові, каштанові, дубово-березові та дубово-ооонові) на дуже кислих (рН 3,2- 5,2) і бідних, слабо дренованих піщаних дерново-підзолистих або буроземних опідволених грунтах атлантичного і оубатлантичного клімату. Представлений він одним порядком. Характерними видами класу і порядку є ацидофільні бореальні і атлантичні неморальні види: осока кульконоона (Carex pilulifera L.), щучник звивистий (Desohampsia flexuosa 1:1 п.), перестріч лучний (Меіащрулит pratense L.),нечуйвітер савойський (Ніегасіии sabaudun L.), ОрЛЯК звичайний (Pteridiua aquilinum Kuhn.), Teuoriua scorodonia В., Lonicera periclymenun L, та ін. В межах класу дубові ліси входять до порядку Quercetalla robori-petraeae та двох союзів Queroion occidentale (Dantas Barreto, 1958; Braun-Blanquet, 1967) та Чмегоіоп robori-petraeae (Tuxen, 1957» 1955; Klika, 1955; Oberdorfer, 1957; Pukarek, 1961, та ін.). Дубових лісів, що належать до першого союзу, характерними видами якого з стокротки лісові (Bellis eyivestris ь.), анемона трилиста (Anemone trifolia L.), горлянка західна (A;Juga occidentalis L.)
та ін., на Україні зовсім немає. Дубові ліси другого союзу трапляються й на Україні.
Другий клас об"єднує євросибірські ліси і чагарники на багатих глинистих кислих, нейтральних та лужних (рН 5,1 - 7,2) добре дренованих буроземних, сірих лісових та дерново-карбонатних грунтах атлантичного і континентального клімату. Представлений клас кількома порядками. Характерними видами його є нейтрофільні неморальні балтійські види, такі, як копитняк звичайний, цибуля ведмежа, яглиця звичайна, фіалка лісова (viola sylvestris ь а ш.), гравілат міський (Geum urbanum L.) та ін. Дубові ліси в межах класу належать переважно до союзу Quercion pubescenti-petraeae порядку Quercetalia pubescenti-petraeae,проте деяка частина їх потрапляє і в союз Сагріпіоп порядку Fagetalia. На Україні трапляються ліси обох союзів.
Останнім часом П.Якуч (Jakuce,1960) виділив з класу Queroc- Fagetea новий клас Quercetea pubescenti-petraeae, до якого ВІДНІС найбільш ксеротермофільні літньо-зелені ліси Субоередземномор"я, півдня Середньої та Східної Європи на дерново-карбонатних і чорноземних грунтах. Клас включає два порядки, один з яких (Orno-Coti- netalia) об'єднує субсередземноморські ліси,а другий (Quercetalia petraeae-pubescentis)-більш континентальні і частково менш ксеро- фільні середньоєвропейські та східноєвропейські дубові ліси. Дубові ліси з дуба звичайного належать головним чином до союзу Аоегі (tatarico)-Querclon останнього порядку ( Jakuce, 1961} Зоо, 1964, Fekete, 1965). Трапляються вони й на Україні. До характерних видів класу належать базифільні неморальні субсередземноморські види: куцоніжка пірчаста, дерен справжній, ясенець білий(Dictamnus albus L.), оман верболистий (Inula salicina L.), смовдь оленяча (Peucedanum cervaria (L.) O u s s.), маруна ЩИТК0В8 (Pyrethrum corymbosum (L.) ii 1 1 d.), горобейник пурпурово-голубий, лазурник трилопатевий, дуб пухнатий, гордовина та ін.
На нашу думку, П.Якуч (Jakucs, 1960, 1961) виділив новий клас цілком обгрунтовано. Проте автор виявив непослідовність, звівши клас Querco-Fagetea в ранг групи класів, через що змушений був виділити ще один клас для мезофільних неморальних лісів. Проте новий клас, який він назвав Carpino-Pagetea, об"єднує лише західноєвропейські та середньоєвропейські мезофільні ліси на родючих грунтах. Аналогічні східноєвропейські ліси вія не включає, отже, виникає необхідність виділити новий, третій клас, а це досить пробле114
матичне питання, тому то ці ліси порівняно а першими не мають практично характерних видів і відрізняються лише біднішим флористичним складом. Отже,доцільніше було б не зводити клас Quexco-Eagetea до рангу групи класів, а залишити його в попередньому ранзі,лише виділити з його складу ксеротермофільніші ліси. Далі ми ці класи будемо розглядати саме так.
Дубові ліси України представлені шістьма класами асоціації, дубовими лісами: крушиновими (Querceta fxangulosa), ліщиновими ( y.coxyloea), свидиновими ( Q.thelycraniosa), татарськокленовими (Q.acerosa (tatarici) 1 скумпієвими (Q.ootlnoaa). З загальної площі 620 тис.га, яку вони займають,приблизно 60% належить дубовим лісам ліщиновим, майже 20%-татарськокленовим, близько 10%-свиди- новим і близько 10%-крушиновим. Дубові ліси кизилові займають приблизно 2-3 тис.га, а скумпієві - близько 1 тис.га. Центральне місце серед перелічених класів асоціацій в системі еколого-фітоцено- тичних рядів займає клас асоціацій дубових лісів ліщинових, який представлений найбільшим числом асоціацій і відповідав оптимальним умовам зростання дубових лісів. Від нього в бік заболочування і збідніння грунтів відходить клас асоціацій дубові ліси крушинові, а в бік зростання родючості грунтів - клас асоціацій дубові ліси свидинові. Три інші класи асоціацій створюють ряд збільшення багатства і сухості грунтів.
Дубові ліси крушинові
Центр поширення цих лісів на слабодренованих атлантичних t субатлантичних низинах західної та північно-західної Європи (південно-східна частина Великобританії, Франція, Бельгія, Нідерланди, ФРН, НДР), він досягає північно-західної частини Чехооловаччи- ни та півдня Польщі. Відзначено окремі місцезнаходження їх в горах (Вогези, північний макросхил Альп), де вони ростуть на плоских слабо дренованих ділянках нижнього гірського поясу (Koztowska, 1936; Steffen, 1936; Walter, 1950; Matuskiewicz, 1952; Onno,1954; Scamoni, 1954, 1964; Noixfalise, 1955; Bxeton,1956; Eassarge, 1956, 1957, 1962, 1960; Knoxzex, 1957; Mraz, 1958; Duxin, Gehu, 1959; Krausoh, 1960; Schubext, 1960; Wiedenxotb, 1964; Sohindlex, 1965; Neubausl, Neuhauslova-Novotn^, 1966, 1967; Mikyaka, 1968; Passarge, Hofmann,l968). На території Радянського Союзу такі ліси описані лише в західній та північно-західній частинах України, південній частині Білорусії і на МологоЧИекснинському межиріччі
115
(Kostyniuk, Wieczorek, 1937; Гринь, 1954; 1971; Корчагин, Селя- нинова-Корчагина, 1957; Юркевич, 1960; Шеляг-Сосонко, 1961, 19706; Юркевич, Гельтман, 1965; Шеляг-Сосонко, Горохова, 1969). Пов"язані вони з кислими (рН 3,2 - 5,0) дерново-підзолистими глеевими і буроземними прихованопідзолистими грунтами, тобто формуються в екстремальних для дуба звичайного грунтових та кліматичних умовах 1 містять ацидофільні, бореальні, атлантичні та гігрофільні види.
Дубові ліси крушинові на території України представлені двома групами асоціацій - крушиново-трясучковидноосоковими та крушиново-конвалієвими. Перші пов"язані з найбільш вологими, бідними й кислими глинисто-піщаними та супіщаними дерново-підзолистими грунтами плакорів. Другі в екологічному і флористичному відношеннях досить різко відрізняються від них. Вони займають багатші і сухіші дерново-підзолисті грунти в позаплакорних умовах. Усі асоціації зазначених груп, через вузьку синекологічну конфекторну амплітуду крушини ламкої ( Frangula alnus м і 1 1.), належать до сте- ноекотопних. Досить близькі вони за трофніотю екотопу, але істотно відрізняються за вологістю, створюючи ряд від сирих до свіжих дубових лісів.
І. Група асоціацій дубові ліси крушиново-трясучковидноосокові представлена дібровою крушиново-молінієвою та крушиново-трясучко- видноосоковою, що належать до ацидофільної ланки. Перші займають найбільш бідні і найменш дреновані для дубових лісів грунти. Характерною рисою лісів цієї групи асоціацій є значна участь в їх складі бореальних видів, таких, як щучник дернистий ( Deschampsia caespitosa (L.) Р. В.), молінія голуба (Molinia coerulea (L.) Moene h), перестріч звичайний (Melampyrum vulgatum Реї s.), Pleurosium schreberi (1111 d.) Mi t t„ Dicranum rugosum H e d w.,Polytrichum commune H e d w., одинаряик європейський (Trientalis europaea L.), чорниця ( Vaccinium myrtillus L.), ожика волосиста, веснівка дволиста, грушанка круглолиста, орляк звичайний, перстач прямостоячий (Potentilla erecta (L.) H a m p $ та ін.; ацидофільних, оліготрофних та олігомезотрофних (більшість перелічених бореальних видів), гігрофільних і мезогігрофільних, таких, як ситник розлогий (Juncus effusus L.), гадючник голий (Filipendula denudata (P r e s 1.) F r i t s c h вербозілля звичайне (Lysimachia vulgaris L.), жовтець повзучий (Ranunculus repens L.), верба попеляста (Salix cinerea L.), вільха клейка,
116
вербозілля лучне, молінія голуба та ін. Базифільних та ксерофіль- них видів немає. З числа середньоєвропейських видів до їх складу на Прикарпатті входять; тирлич ваточниковидний (Gentiana asclepi- ad e a L.), купина кільчаста (Polygonatun verticillatua A 1 1.),
осока трясучковидна(Oarex brizoides J u s 1 e n.) та ін.
Діброва крушиново-моліиієва
( Quercetum franguloso-noiiniosuD) поширена в лісовій зоні: на Поліссі та в Прикарпатті (Стрийський район Львівської області, Бердичівський та Городнянський лісгоспзаги). По других терасах лівобережних приток Дніпра вона заходить і в північну частину лісостепу (Прилуцький та Переяолав-Хмельницький лісгоспзаги). Ці ліси не утворюють великих масивів, а трапляються зрідка невеликими ділянками розміром до 0,5-2 га.Займають вони плоскі, блюдцеподібні, слабо дреновані зниження з дерново-середньопідволистими глеєвими з ознаками заболочування грунтами і в рельєфі розташовуються вище вільхових, але нижче решти асоціацій дубових або сосново-дубових лісів. Деревостани їх невисокої зімкненості, суховершинні, зі скривленими стовбурами, вкритими лишайниками. Все це створює загальний сіруватий, ніби позбавлений життя вигляд.
Деревостан у віці 70-100 років має зімкненість крон 0,7-0,8, середню висоту 16-20 м, діаметр дерев 22-30 см і Ш-ІУ клас бонітету. Створюють його дуб звичайний 8 участю вільхи клейкої, берези бородавчастої, осики та ялини європейської. Останній вид має 2-й клас постійності і трапляється на Прикарпатті. На Поліссі 8 4-м класом постійності трапляється сосна звичайна (Pinus silvestris L.\ Підлісок добре виявлений. Він має зімкненість 0,3-0,5 1 середню висоту І,6-2,5 м. Створює його крушина ламка (0,2-0,5) з участю горобини звичайної (+ - 0,І)\ бруслини бородавчастої (+ - 0,1)верби попелястої, свидини кров"яної (ШЬеІусгапіа sanguinea (L.) У о - u г хД вовчих ягід звичайних (Dapbne mezereun L.) та ожини звичайної (Bubus caesius L.). Останні п"ять видів мають 2-3-й клас постійності.
Трав"яно-чагарничковий ярус виявлений добре і має проективне покриття 30-50%. Створюють його молінія голуба (25-45%), чорниця (+ - 15%),орляк звичаиний(+ - 10%).веснівка дволиста(+ - 5%).костяниця (Bubus saxatilis L.) (+ - 1%).вербозІдля звичайне(+ - 1%), куничник тростиновий ( Calamagrostia arundinacea (L.) Botb), безщитник жіночий (Athyriun flllx-femina (L.)B o t h), перстач
:Нами прийнята десятибальна шкала постійності видів; перший клас - 0-10%, другий клас - П-20%, третій клас - 21-30% , четвертий - 31-40%. п"ятий - 41-50% і т.п.
117
прямостоячий ( Potentilla erecta (L.) н а ш p e),золотушник звичайний (Bolidago virgaurea L.), одинарник європейський, конвалія звичайна, ситник розлогий, ожика волосиста та перестріч лучний. З видів, що мають 2-4-й клас постійності, трапляються горлянка повзуча (Ajuga reptans Ь.), пахуча трава звичайна (Anthoxan- thum odoratum L,), КОСТРИЦЯ велетенська ( Festuoa gigantea (L.) V i 1 І.), гадючник в"язолистий ( Filipendula ulmaria (L.) Maxim.), дріоптерис ЧОЛОВІЧИЙ (Dryopteris filix-maa(L.) S o h o t t ) та остистий (Dryopteris spinulosa (Mi 1 l,)K u- n t 2 e ), ХВОЩ ЛІСОВИЙ ( Equisetum silvaticum L.), ЗНІТ дрібно- квітковий ( Epilobium parviflorum (s c h r e b.) DC.), купина лікарська (polygonatum offioinale (L.) AU.), зірочник лісовий, гравілат річковий (Geum rivale L.), ранник вузлуватий, грушанва круглолиста, осоки трясучковидна, волосиста (Загех pilosa Scop.) та бліда (G.pallescens L,), гравілат міський та ін. Моховий яруо виявлений погано.Він має проективне вкриття 10—20% і створений переважно Polytrlobum commune H e d w., Polytricbum formosum H e d w., Dicrdnum rugosum H e e w. та іншими видами.
Видова насиченість асоціації дорівнює 72 видам і є найнижчою серед дубових лісів.
Діброва крушиново-тряоучковид- ноооокова( Quercetum franguloso-carloosum (brisoidis) помирена лише в західній частині лісової зони: на Прикарпатті, в Західному і Правобережному Поліссі (Косівський, Коломийський та Надвірнянський райони Івано-Франківської області; Золочівський, Овруцький, Соснівський та Бердичівський лісгоспзаги). Трапляється вона зрідка також невеликими (до 1-3 га) ділянками. Ці ліси займають рівнинні,плоскі зниження з сирими або вологими дерново-підзолистими глеєвими задернованими грунтами. В рельєфі вони розташовуються трохи вище попередньої асоціації, але нижче діброви лі- щиново-трясучковидноосокової (рис.16)1.
Деревоотан у віці 40-90 років має зімкненість крон 0,6-0,8, висоту 15-22 м, діаметр дерев 20-38 см і П-Ш клас бонітету. Створює його дуб звичайний з участю осики (+ - 0,2), берези бородавчастої (+ - 0,1), Вільхи клейкої, Інколи - сосни звичайної.
1 Рис.16-35 див. на вклейці між стор.128 - 129.
118
Ярус підліску виявлений добре і мав зімкненість 0,2-0,7. Його створюють крушина ламка (0,2-0,7) з незначною домішкою ліщини звичайної(* - 0,1).горобини звичайної, верби попелястої, черемхи звичайної (Padua racemosa (Lam.)Gilib,), ожини шорсткої (Bubus hirtus W. К.) та дроку красильного (Geniata tinctoria L,). Останні чотири види мають 2-3-й клас постійності.
Травостій звичайно густий з проективним покриттям 40-80%. Переважає в ньому осока трясучковидна (26-65%), яка в більшості випадків створює суцільний фон. Разом з нею ростуть куничаик тростиновий (+ - 5%), молінія голуба (+ - 5%), орляк звичайний!* - б%), копитняк звичайний (+ - 1%), щучник дернистий (+ - 1%), вербозілля звичайне!* - 1%). веснівка дволиста (+ - 1%), перстач прямостоячий (+ - 1%), вероніка дібровна (Veronica chamaedrys L.), підмаренник весняний (Gallum vernum s с о р.), ожика волосиста, перестріч звичайний, квасениця звичайна, зірочник лісовий, одинарник звро- пейський, горлянка повзуча, безщитник жіночий, конвалія звичайна та види з 2-4-м класом постійності: астрагал солодколистий (Astragalus glycyphyllus L.), осока волосиста (Carex рііова Sco р»), осока бліда (Carex pallescens Ь.), підмаренник проміжний (Gallum intermedium 8 о h u 1 c,), звіробій чотиригранний ( Hypericum quadrangulum L.), просянка розлога ( Milium effusum L.), суховершки звичайні (Pr irnella vulgaris L.), яглиця звичайна, буквиця лікарська, дріоптерис чоловічий, сугайник австрійський, герань лісова (Geranium silvaticum L.), гадючник ГОЛИЙ, жовтець повзучий, костриця велетенська, мітлиця тонка (Agrostis tenuis S і b t h.), медунки темна (Pulmonaria obscura Dumos t.) та м"якенька (P.mollissima K e r n.), зозулинець Фукоа (Orchis fuschii D r u d e), осока стягнута, золотушник звичайний, фіалка лісова та ін.
Неважко помітити, що в розглянутій асоціації порівняно з попередньою підвищується роль спільних для обох асоціацій неморальних видів, з'являється низка нових видів (копитняк звичайний, осока трясучковидна, зірочник лісовий, фіалка лісова та ін.) та помічається зниження роді або зникання бореальних та гігрофільних видів (ситник розлогий, молінія голуба та ін.). В групі постійних з'являється один вид (підмаренник весняний) з групи неморальних субсередземноморських видів з широкою синекологічною амплітудою. Екологічна ємкість асоціації збільшується, завдяки чому П видова насиченість зростає до 10І виду.
119
П. Група асоціацій дубові ліси крушиново-конвалієві представлена лише однією асоціацією, яка закінчує ряд збільшення сухості і багатства грунтів в межах класу. Порівняно а попередніми асоціаціями вона є і найбільш термофільною.
Діброва крушиново-конвалієва ( Quercetum franguloso-convallariosum) поширена в лісостеповій зоні Лівобережжя УРСР (Прилуцький, Мринський, Переяслав-Хмельниць- кий, Кременчуцький та Красноградський лісгоспзаги). Трапляються ці ліси невеликими ділянками в заплавах і на других терасах Дніпра та його приток, де займають міжгривні зниження або невисокі плоскі гриви прируслової та центральної частини заплави або других терас. В заплавах ці ліси займають вологі супіщані дернові опідзолені з різким перемінним зволоженням грунти, а їй других терасах - глинисто-піщані та супіщані слабопідзолисті.
Деревостан у віці 60-90 років має зімкненість крон 0,7-0,9, висоту 18-23 м, діаметр дерев 24-40 см і П-Ш клас бонітету. Створює його дуб звичайний з незначною домішкою осики (+ - 0,1) та видів з 2-4-м класом Постійності - берези бородавчастої, липи серцелистої, груді звичайної (Pyrus оопшншіе L.), в"яз1в гладенького (Ulmus laevis Раї 1.) та шорсткого (U.soabra М і 1 1.), сосни звичайної та ін.
Підлісок розвинений добре і має зімкненість 0,3-0,7. Крім крушини ламкої (0,2 - 0,5), його створюють свидина кров"яна (+ - 0,2), клен татарський (+ - 0,1), ожина звичайна (+ - 0,1), бруслини бородавчаста та європейська (Euonymus europaea L.), глід український ( Orataegus ucrainica Р о j а в k.),жостір проносний (Bhanmus cathaxtica L.), черемха звичайна та горобина звичайна. Чотири останні види мають 2-4-й клас постійності.
Травостій з проективним покриттям 25-60% створений конвалією звичайною (20-55%) з участю гравілату міського (+ - 1%), кропиви дводомної (Urtica dioica L.) (+ - 1%), дзвоників персиколистих ( Oarnpanula persicifolia L.) та ріпчаотовидних (O.rapunouloides L.), пахучіш звичайної (Clinopodium vulgare Ь,), дутня ягідного (Cucu- balus baccifer L.), торилЮу ЯПОНСЬКОГО ( Torilis Japonica (Я o ut t.) DC.), фіалки шершавої, буквиці лікарської, осоки стягнутої, грястиці збірної, коручки широколистої, костриці велетенської, золотушника звичайного, вероніки дібровної та видів з 2-4-м класом постійності, до яких належать холодок лікарський (Asparagus officinalis L.), куцоніжка лісова (Brachypodium 120
silvaticum (H u d s.) P. В.), ВПЯЗІЛЬ барвистий (Coronilla varia Ь.), 8НІТ Гірський (Epilobium montanum L.), чини чорна (Lathyrus niger (L.) В e r n h.) та весняна ( L.vernus (L.) в e r n h.), міцеліс стінний (Mycells muralis (L.) H c h b.), материнка звичайна (Origanum vulgare L.), медунка темна (Pulmonaria obscura D u - m o r t.), регнерія ообача (Roegnerla canina (L.) N e v s k i ), ломиніс прямий (Clematis recta ь.), гірча клинописта (seiinum oarvifolia L,), серпій різнолистий ( Serratula heterophylla (L.) Des f.), фіалка дивна (Viola mirabilis L.), бугила лісова, ку- ничник тростиновий, осоки бліда та пальчаста (Carex digitata L.), герань криваво-червона, просянка роалога, вербозілля звичайне, яглиця звичайна, орляк звичайний, маруна щиткова, ранник вузлуватий, СМОВДЬ Гірська (Peucedanum oreoselinum (L.) Moench), аконіт дібровний (Aconitum nemorosum M.B.), хвощ зимовий (Equisetum hiemale L.)( стародуб пруський (Laserpitlum pruthenicum L.), конюшина альпійська ( Trifolium alpestre Ь.),перестріч гребінчастий (Melampy- rum cristatum L.) та ін.
Видова насиченість асоціації становить 130 видів.
Від попередніх ця асоціація відрізняється досить різко. Вона мав свій відокремлений ареал і формується в найбільш сухих і теплих для всього класу асоціацій кліматичних умовах. Далі на південь асоціації цього класу вже не трапляються. Для асоціації характерна перевага сціофільних неморальних балтійських видів (яглиці звичайної, астрагалу солодколистого, конвалії звичайної, коручки широколистої, чини весняної, медунки темної та ін.) та значна участь світлолюбних неморальних субоередземномороьких мезокоерофільних видів (ломиніс прямий, в"я8Іль різнобарвний, купина лікарська, маруна щиткова, регнерія собача,*фіалка шершава та ін.). Роль 1 кількість бореальних та гігрофільних видів у ній різко падає.
В еколого-генетичяому відношенні асоціація має цікаву особливість. За типом грунтів і деякими бореальними видами вона ще по- в"язана з попередніми,проте переважають у ній неморальні балтійські та неморальні субсередземноморські види, 1 в цьому відношенні вона повпязана з дубовими лісами нейтрофільної 1 базифільної ланок. Пояснюється це тим, що ліси, які належать до цієї асоціації, формуються виключно в позаплакорних умовах других терас, де вони займають найбідніші та найлегші для всього ряду дубових лісів других терас грунти. Останні можна розглядати як аналоги поліських грунтів, але в зовсім інших кліматичних умовах,- лісостепової зони,- 121
які зумовлюють формування нейтрофільної та базифільної ланок дубових лісів. Отже, флористичний склад асоціації зазнав впливу екстразональних грунтів і зонального клімату, по й зумовило його різноманітність.
Дубові ліси ліщинові
Найбільш широкий ареал цих лісів пояснюється широкою оинеко- логічною конфекторною амплітудою ліцияи звичайної, яка майже збігається з оинекологічною едифікаторною амплітудою дуба звичайного. В звпязку в цим багато асоціацій класу є евріекотопними. Сте- ноекотопні лише асоціації, які займають крайні положення, або асоціації з вузькою синекологічною конфекторною амплітудою ярусу травостою, незалежно від їх місця в екологічному ряді.
На території Західної (Атлантичної) і Середньої (Центральної) Європи дубові ліси ліщинові трапляються в південній і середній частинах Великобританії, північній, північно-західній та середній частинах Франції, в Бельгії, Нідерландах, ФРН, НДР, Чехословач- чині, Польщі та Данії. Острівні місцезнаходження цих лісів спостерігаються в північно-західній частині Іспанії та Португалії (південно-західні Піренеї), в північній частині Великобританії, найбільи південних частинах Норвегії та Швеції, в Австрії, Угорщині й Румунії. В Піренеях і Альпах вони піднімаються до висоти 800 м ( tfowinski, 19295 Меиаєі, 19355 Kosloweka, 19365 Steffan, 19365 Ujvaxoei, 19411 Balaze, 1943s 800', 19*35 Matuszkiewlcz, 1W, 19525 Vanden Baxghen, 1953, 1955, 19565 Pas sarge, 1953, 1956, 1957, 19625 Onno, 19545 Michalko, 1957» Juxko,1958; Iteaz, 19585 Hofaann, 19595 Izdebaka, 19595 Eljalkowskl, 1961» Fukaxek, 19615 Isdebaki, 19625 MikySka, 1963, 19685 Meuhausl, Keubauslo- vA-Hovotaa, 1964, 1968, 19695 flcamoni, 1964> Tanghe, 1964j Wie- dearoth, 1964j Sjoxs, 1965s Lamorisea, 1966, Pupcaru-Soxcceauu, Fopova-Oucu, 1966j Bxaun-Blanquet, 1967).
На території Радянського Союзу ці ліси поширені в широко- листянолісовій та на більшій частині лісостепової зони і доходять до Уралу. Острівні місцезнаходження їх є на Кавказі, у хвойнолі- оовій 1 степовій зонах і практично збігаються з ареалом дуба звичайного (Коновалов, 1929, 1949; Морозов, 1931а; Цинзерлинг, 1934; Сукачев, 1934; Марков, 1935; Szafex, 1935; Гринь, 1936, 1950,1954; Koetyniuk і Vleozoxek, 1937; Кожевников, 1937, 1939; Крашенинников
122
1941; Алехин, 1947; Бельгард, 1950; Напалков, 1951, 1966; Елагян, 1953; Прилипло, 1954; Юркевич, 1960; Шеляг-Сосонко, 1961, 1966а, 19696, 19706, 1971а, І97І6; Соколова, 1963; Юркевич, Гельтмая, 1965; Кайрюкштис, 1966; Карписонова, 1967; Нешатаев, 1967; Бара- банщиков, 1968; Дохман, 1968; Шеляг-Сосонко, Горохова, 1969; Попов, Вакуров, 1970; Сенкевич та ін., 1970).
В західній частині ареалу ліщинові дубові ліси повлязані переважно з кислими і олабокислими (рН 3,5-6,2) дерново-підзолистими, буроземними прихованопідзолистими та світло-сірими лісовими грунтами, рідше трапляються на нейтральних і слаболужнйх дерново- карбонатних грунтах з рН 6,5-7,2. У східній частині ареалу вони займають світло-сірі, сірі й темно-сірі лісові грунти, рідше трапляються на дерново-підзолистих, дерново-карбонатних і чорноземних опідзолених грунтах. У флорі лісів переважають неморальні балтійські види, проте значна участь, особливо в західних і північних районах ареалу, також бореальних видів.
На Україні дубові ліси ліщинові поширені переважно в широко- листянолісовій і лісостеповій зонах. В лісовій зоні вони займають вододільні простори з найбільш багатими грунтами і трапляються в комплексі з дубовими лісами крушиновими, дубово-сосновими (Полісся) та дубово-грабовими (Прикарпаття, Розточчя, Опілля). В лісостеповій зоні вони займають в західній частині схили межирічних пагорбів, а в східній - схили балок і плакори широких межиріч. В першому районі вони поширені в комплексі з дубово-грабовими, а в другому - а липово-дубовими лісами, заміщуючись на південь і в ксеротермічніших місцезнаходженнях дубовими лісами татарськоклено- вими.
Дубові ліси ліщинові представлені 6 групами асоціацій, які створюють ряд дібров від сирих до сухих 1 від найбільш оліготроф- них (для дубових лісів) до мегатрофних. Центральне місце серед них займав група асоціацій дубові ліси ліщиново-зірочников'і, пов’язана з середніми для всього класу асоціацій екологічними умовами та більшістю інших груп асоціацій. Від неї, в системі едафо-фіто- цетичних рядів В.М.Сукачова, в бік більшого зволоження і збіднення грунтів розташовується група асоціацій дубові ліси ліщиново- кіночопапоротеві. Завершує ряд збільшення зволоження група асоціацій дубові ліси ліщиново-трясучковидяоосокові. Порівняно з центральною групою асоціацій сухіші грунти займав група асоціацій дубові ліси ліщиново-гіроькоосокові. Продовжує ряд збільшення су-
123
іості, але з обмеженішою амплітудою багатства грунтів,група асоціацій дубові ліси ліщиново-парвськоосокові. Найбільш сухі та багаті грунти в класі асоціацій займає група асоціацій дубові ліси ліщиново-горобейникові. Найбільш поширеною в центральна група асоціацій, на яку припадав близько 60% площі лісів цього класу.Найменш поширені групи асоціацій дубові ліси ліщиново-жіночопапоро- теві, ліщиново-парвськоосокові та ліщиново-горобейникові, які становлять лише близько 10% площі лісів цього класу.
Ш. Група асоціацій дубові ліси ліщиново-трясучковидноосокові складається з дібров ліщиново-трясучковидноосокової та ліщиново- квасеницевої. Перша з них займав більш вологі та менш дреновані грунти. В екологічному та флористичному відношенні вона дуже близька до дубових лісів крушиново-тряоучковидноосокових і поєднує дубові ліси ліщинові з дубовими лісами крушиновими. Характерною особливістю лісів цієї групи асоціацій в значна участь бореальних, ацидофільних та гігрофільних видів, хоча в порівнянні а попередніми лісами у них відзначається посилення позицій неморальних балтійських видів. Ці ліси належать до ацидофільної еколого-генетич- ної ланки. Асоціації їх стеноекотолні.
Діброва ліщиново-трясучковидно- 0 0 0 к 0 в а ( Queicetum coiyloao-caricoaum(brizoidi8) поширена лише в широколиотянолісовій зоні (Закарпатська низовина, Прикарпаття, Розточчя, Мале Полісся, Західне та Правобережне Полісся), де вона характерна для більшості ліогоопзагів. Найбільш східні ділянки її трапляються в Городнянському лісгоспзагу Чернігівської області. На Прикарпатті і в південній частині Полісся діброва лі- щиново-тряоучковидноооокова пов"язана а великими плоскими слабо- дренованими ділянками, а в північній частині Полісся переходить на пологі схили. Основні площі цих лісів зосереджені в Прикарпатті та Житомирській області, де окремі масиви їх займають кілька десятків і навіть сотень гектарів і де вони територіально переважають над усіма іншими асоціаціями дубових лісів. Зростають ліси цієї асоціації на дерново-слабо- або оередньопідзолистих глинисто-піщаних та супіщаних глеєвих грунтах.
Деревостан у віці 50-160 років мав зімкненість крон 0,6-0,9, висоту 18-28 м, діаметр дерев 26-54 см і П, рідше І або Ш клас бонітету. Створює його дуб звичайний з незначною домішкою постійних видів,таких, як береза бородавчаста, граб звичайний і осика, та випадкових (клен звичайний, вільха клейка, граб звичайний та
124
ін.). Неважко помітити, що порівняно з попередніми асоціаціями деревостан характеризується посиленням ролі більш вибагливих до грунтів широколистяних мезотрофних видів. В асоціаціях наступних груп їх значення буде все більше зростати,а значення менш вимогливих дрібнолистяних та хвойних порід все зменшуватиметься доти, доки вони не випадуть зі складу деревостану асоціацій нейтрофільної ланки.
Підлісок в цілому виявлений добре. Він мав висоту 2,5-3,5 м 1 зімкненість 0,3 - 0,7.Створюють його ліщина звичайна(0,2 - 0,6), крушина ламка (+ - 0,2), горобина звичайна (постійні види); вовчі ягоди звичайні, бруслини європейська і бородавчаста, черемха звичайна, свидина кров"яна, ожина звичайна та інші випадкові види, які трапляються на найбільш дренованих і багатих едафотопах.
Травостій густий і має проективне покриття 35-75%. Основний фон його створює осока трясучковидна (20-70%), яка на найбільш вологих з оглеєними грунтами ділянках створює суцільне покриття. В числі постійних видів травостою зустрічаються веснівка дволиста (+ - 5%), квасениця звичайна (+ - 5%), зірочник лісовий (+ - 5%), ожика волосиста (+ - 1%), вербозілля звичайне (+ - 1%), щучник дернистий, підмаренник весняний, просянка розлога, одинарник європейський (абсолютно постійні), яглиця звичайна (+ - 10%), маренка запашна (+ - 5%), безщитник жіночий (+ - 5%), осока волосиста (+ - 5%), куничник тростиновий (+ - 1%), дріоптерис гребенястий (Dryopteris oristata (L.) G в а у ),медунка темна (+ - 1%), Жов- тець кашубський ( Ranunculus cassubicus L,), горлянка повзуча, дріоптерис чоловічий, костриця велетенська, зеленчук жовтий ( Galeobdolon luteum н u d в,), підмаренник проміжний, чина весняна, вербозілля лучне, молінія голуба, перстач прямостоячий та фіалка лісова (відносно постійні).. Регіонально постійними видами травостою є тирлич ваточниковидний та купина кільчаста (середньоєвропейські монтанні види, що зустрічаються лише в Прикарпатській частині ареалу асоціації). Із видів з 2-4-м класом постійності трапляються костяниця (Rubus saxatillis L.), копитняк європейський буквиця лікарська, конвалія звичайна, чемериця Лобелієва (Varatru» lobeilanum в e r n h.), зніт гірський, гравілат міський, ситник розлогий, перестріч звичайний, вороняче око звичайне (Paris qua- drifolia L.), купина багатоквіткова (Polygonatua multiflonua (L.) A 1 1.),купина лікарська,орляк звичайний, підлісник європейський (Sanicula europaea ь.), ранник вузлуватий, золотушник звичайний, вероніка дібровна та ін.
125
Видова насиченість асоціації становить 149 видів.
Таким чином,з числа постійних видів 45% становлять борвальні; гігрофілів, за винятком вербозілля звичайного, немає. В складі неморальної балтійської групи з'явились види з вужчою, ніж у аналогічних видів першої асоціації попереднього класу, синекологіч- ною амплітудою (яглиця звичайна, осока волосиста, костриця велетенська, копитняк європейський, жовтець кашубський, зірочник лісовий, підмаренник проміжний, купина багатоквіткова, медунка темна, маренка запашна, чина весняна, фіалка лісова, анемона дібровна, підлісник європейський та ін.).
Діброва ліщиново-квасеницева
( Quercetum coryloso-oxalidoaua)поширена на Поліссі, Малому Поліссі та Розточчі (Ніццький, Любомльський, Киверцівський, Золочів- ський, Пастерівський,. Львівський, Малинський, Овруцький, Старо- костянтинівський ліогоспзагь). Найхарактернішою вона є для півні чно^захі дно ї частини Західного Полісся, де займає рівнинні або трохи підвищені плоскі ділянки. На решті території ці ліси звичайно трапляються на середніх та нижніх частинах пологих схилів північної, західної та східної експозицій або в незначних зниженнях. Ця асоціація не створює великих масивів і трапляється досить рідко. Грунти під нею вологі, дерново-оередньопідзолиоті, глеюваті.
Деревоотан у віці 60-150 років має зімкненість крон 0,7-0,9, висоту 16-30 м, діаметр дерев 26-52 см і П-Ш клас бонітету. Крім дуба звичайного, його створюють граб звичайний, сосна звичайна, осика, вільха клейка, береза бородавчаста та ясен звичайний. Останні три види мають 2-й клас постійності.
Підлісок розвинений добре і мав зімкненість 0,3-0,6. Постійними видами його є ліщина звичайна (0,2 - 0,5), крушина ламка (+ - 0,1), бруслина бородавчаста, вовчі ягоди звичайні та черемха звичайна, а випадковими - бруслина європейська, свидина кро- в"яна, горобина звичайна та ожина неоійська (Rubus nessensis W, Н а 1 1.).
Травостій має проективне покриття 25-45%. Ценозоутворююча роль конфектора ярусу виявлена досить слабко, що відбивається в його проективному покритті (15-25%) та значній ролі інших видів. Пояснюється це тим, що квасениця звичайна належить до типових сціо- фітів. Оптимум синекологічної конфекторної амплітуди її відповідає ялиновим, ялицевим та гірським буковим лісам. Тому в світлих
126
дубових, вибагливих до грунтів,лісах конфекторна роль цього виду проявляється слабо 1 забезпечується лише в північній частині їх ареалу на бідних, але досить дренованих грунтах. Постійно супроводжують конфектор копитняк європейський (+ - 10%), маренка запашна (+ - 10%), веснівка дволиста (+ - 5%), зірочник лісовий (+ - 5%), яглиця звичайна (+ - 1%), дріоптерис гребенястий (+ - 1%), горлянка повзуча, куцоніжка лісова, осока лісова (Carex silvatica в u а в.), осока стягнута, конвалія звичайна,, зеленчук жовтий, підмаренник ресняний, чина весняна, вербозілля звичайне, ожика волосиста, просянка розлога, міцеліо стінний, підлісник європейський, купина лікарська, медунка темна, костяниця та фіалка лісова. Рідше (2-4-й клас постійності) трапляються актея колосиста (Actaea spicata L.), цирцея звичайна (Circaea lutetiana L.), бутень запашний (Cbaexophyllua агornatіочи L.), жовтець кашубський ( Ranunculus cassubicus L.) пахучка звичайна (Ollnopodiun vulgare Ь.), горлянка повзуча, дріоптерис чоловічий, щучник дернистий, бугила лісова, гравілат річковий, ооока трясучковидна, дзвоники ріпчаотовидні.орляк звичайний, ооока волосиста, зніт гірський, перестріч звичайний, хвощ лісовий, костриця велетенська, кадило меліоолисте, вороняче око звичайне, одинарник європейський, ранник вузлуватий, фіалка дивна та ін.
Видова насиченість асоціації становить ПО видів.
Незважаючи на те, що конфектором ярусу травостою описаної асоціації є бореадьний вид, значення неморальних балтійських видів у ній порівняно з попередньою зростає, що пов"язане з більшою роздільною здатністю екотопу (краща дренованість) та послабленою конкурентною здатністю квасениці звичайної.
ІУ. Група асоціацій дубові ліси папоротеві представлена трьома стеноекотопними асоціаціями: дібровами ліщиново-веснівковою, ліщииово-жіночопапоротевою та ліщиново-чоловічопапоротевою. Об'єднує їх те, що вони займають найбільш бідні для дубових лісів дерново-підзолисті грунти, але не вологі, а свіжі. В цих вузьких межах вони і створюють екологічний ряд. За ступенем зволоження остання асоціація наближається до ліщиново-жіночопапоротевої діброви, але формується в більш південних умовах. За ступенем трофяості грунтів перша асоціація близька до свіжих дубово-соснових лісів. Формується вона в північній частині ареалу формації.
Роздільною здатністю для виявлення конфекторної ролі веснівки дволистої, дріоптериса чоловічого та безщитника жіночого є не-
127
достатня, щоб ці види були витіснені конфекторами наступних асоціацій, родючість грунтів, якої, проте, досить щоб вони не були витіснені менш вибагливими до родючості грунту конфекторами дубово-соснових лісів, наприклад орляком звичайним. Конфектори попередньої групи асоціацій (квасениця звичайна та осока трясучковидна) переходять в ранг асектаторів, що зумовлюється кращою дрено- ваяістю грунту.
Діброва ліщиново-веснівкова відноситься нами до ацидофільної ланки дубових лісів. Дві останні асоціації цієї групи,незважаючи на те, що вони ростуть на бідних дерново-підзолистих грунтах, розглядаємо як початкові асоціації нейтрофільної ланки. Вони характеризуються ще значною роллю бореальних і ацидофільних видів, але мають вже, хоча і незначну, перевагу неморальних балтійських і низку неморальних оубсередземноморських видів з широкою синекологіч- ною амплітудою. Ареал їх, порівняно з першою асоціацією, зміщений на південь. По ярах та других терасах річок вони заходять в північну частину лісостепу. В екологічному та флористичному відношеннях вони дуже близькі між собою.
Діброва ліщиново-веснівкова (Quercetum coryloeo-majenthemoeum) поширена лише на Поліссі та Малому Поліссі (Золочівський, Кременецький, Новоград-Волинський, Любомльський, Йепетівський, Святошинський, Городнянський та Ям- пільоький лісгоспзаги), де займає рівні, трохи підвищені ділянки або пологі схили. Великих масивів ці ліси не утворюють, хоча трапляються досить часто. Звичайно окремі ділянки їх не перевищують 1-2 га і зустрічаються в комплексі з дубово-сосновими лісами, з якими вони пов"яаані екологічно і флористично. Останнє виявляється в низці спільних видів, до яких належать: квасениця звичайна, охика волосиста, купина лікарська, вороняче око звичайне, горлянка повзуча, костяниця, куцоніжка лісова, чина весняна, сосна звичайна, чорниця, одинарних європейський, грушанка круглолиста, ра- мішія однобока, орляк звичайний, веснівка дволиста та ін.
Деревоотан у віці 50-160 років має зімкненість крон 0,7-0,9, висоту 14-25 м, діаметр дерев 18-40 ом і Ш та рідше П клас бонітету. Крім едифікатора, абсолютно постійними видами його є береза бородавчаста (+ - 0,1) та ооона звичайна (+ - 0,1). У багатших еда- фотопах зрідка трапляється граб звичайний, а в найбільш вологих - вільха клейка.
Підлісок добре сформований і має зімкненість 0,2-0,6. До по-
128
стійних видів його належать ліщина звичайна (0,2 - 0,6), крушина ламка (+ - 0,1), бруслина бородавчаста та горобина.
Травостій досить розріджений, з проективним покриттям 20-40%« Ценозоутворююча роль конфектора виявлена слабо, тому що визначається не кліматичними,а едафічними факторами» Травостій утворюють веснівка дволиста (15-30%), квасениця звичайна (+ - 5%), костяниця (+ - 5%), конвалія звичайна (+ - 1%), суниці лісові (Eragaria vesca L.) (+ - i%)t підмаренник весняний (♦ - 1%), ожина волосиста (+ - 1%), перлівка поникла (Melica nutans L.) (+ - I%)t орляк звичайний (4- - 1%),молочай ребристий (Euphorbia angulata J а с q.), перстач білий (Potentila alba L.), горлянка повзуча, буквиця лікарська, осока стягнута, пахучка звичайна, перестріч звичайний, міцеліс стінний, купина лікарська, зірочник лісовий, одинарник європейський, вероніка дібровна та фіалка лісова. Менш поширені підмаренник північний ( Calium boreale L.), печіночниця звичайна ( Hepatica nobilis G a r s a u 1 t.), ЛІЛІЯ лісова ( Lilium marta- gon і,.), вероніка лікарська ( Veronica officinalis L.), конюшина альпійська ( Trifolium alpestre L.), очиток звичайний (Sedum telep- hium L.), осока пальчаста ( Carex digitata L.), яглиця звичайна, копитняк європейський, безщитник жіночий, куцоніжка лісова, куничник тростиновий, осока трясучковидна, осока бліда, дріоптерис чоловічий, ломиніс прямий, зніт гірський, зеленчук жовтий, гравілат міський, нечуйвітер зонтичний ( Hieracium umbellatum L.), чина весняна, підмаренник проміжний, герань криваво-червона, вербозілля звичайне, звіробій чотиригранний, перстач прямостоячий, медунка темна, тонконіг дібровний, підлісник європейський та ін. Крім перелічених трав"янистих видів, в ярусі травостою постійно присутній і типовий конфектор ялинових лісів чорниця (+ - 1%).
Видова насиченість асоціації дорівнює 123 видам. Порівняно з наступними асоціаціями групи діброва ліщиново-веснівкова характеризується відсутністю в числі постійних видів травостою мезогіг- рофітів. Помітно посилюється і роль неморальних субсередземномор- ських видів, що становлять майже 25% усього видового складу постійних видів, проте всі вони мають невелику рясність. Провідне значення наложить бореальним видам, які складають близько 40% видів асоціації (решта - неморальні види).
Діброва ліщиново-жіночопапоро- Т е В а ( Quercetum coryloso-athyriosum(filicis feminae) поширена в південних, на межі з лісостеповою зоною, районах По129
лісся (Цуманський, Новоград-Волинський, Мринський та Прилуцький лісгоспзаги), де займав нижні частини схилів прирічкових ярів та балок або міжгривні зниження невисоких грив других терас. Зустрічається вона зрідка і невеликими (до 0,5 - 2 га) вузькими смугами шириною 20-40 м і довжиною 0,1-2 км. Грунти під цією асоціацією вологі дерново-середньопідзолисті супіщані, часом світло- сірі лісові.
Деревостан у віці 60-110 років має зімкненість крон 0,7-0,9, висоту 22-26 м, діаметр дерей 26-40 см і П клас бонітету. У створенні його беруть участь дуб звичайний (0,7-0,9), береза бородавчаста (+ - 0,1), осика (+ - 0,1) та липа серцелиста. На відміну від попередньої асоціації відсутня сосна звичайна.
Підлісок виявлений добре і має зімкненість 0,3-0,5. З групи постійних видів його створюють: ліщина звичайна (0,2-0,5), крушина ламка (+ - 0,1), ожина звичайна (+ - 0,1), бруслина бородавчаста, горобина звичайна, глід український ( Crataegos ucrainica Р о j ai k) іа свидина кров"яна.Крім них зустрічаються ще бруслина європейська ожина несійська та черемха звичайна.
Травостій має проективне покриття 35-55% і створений безщит- ником жіночим (20-30%), конвалією звичайною (+ - 10%), орляком звичайним (+ - 10%), веснівкою дволистою (+ - 5%), костяницею (+ - 5%), підмаренником весняним (+ - 1%), горлянкою повзучою, астрагалом солодколистим, буквицею лікарською, куничником тростиновим, пахучкою звичайною,коручкою широколистою,кострицею велетенською, чиною чорною, вербозіллям звичайним, перлівкою пониклою, медункою вузьколистою, гравілатом міським, знітом гірським, ранником вузлуватим, перстачем білим, веронікою дібровною, торилісом японським та дзвониками персиколистими. Серед випадкових видів трапляються борщівник сибірський (Heracleum sibiricum L.), ві- халка гілляста (Anthericum ramosum L,), наперстянка великоцвіта (Digitalis grandiflora М і 1 1.), герань Робертова (Geranium го- bertianum !>.), ластовень лікарський (Vincetoxicum officinale L.), кропива дводомна, осока гірська, фіалка дивна, одинарник європейський, чина весняна, ожика волосиста, парило звичайне (Agrimonia eupatoria L.), перестріч звичайний, просянка розлога, міцеліс стінний, вороняче око звичайне, вероніка лікарська, тонконіг дібровний, купина лікарська, перстач прямостоячий, мітлиця тонка, медунка темна, жовтець кашубський, актея колосиста, підлісник європейський, зірочник лісовий, копитняк європейський, дріоптериси чоловічий та остистий, очиток звичайний та 1н.
130
Видова насиченість асоціації дорівнює 104 видам.
Порівняно з лісами попередньої асоціації серед постійних видів діброви ліщиново-жіночопапоротевої вже з’являються мезогігро- філи, різко зростав роль і кількість балтійських неморальних видів. Бореальні види складають лише близько 20% всіх видів або майже 26% групи постійних видів.
Діброва ліщиново-чоловічопапо- р О т е в a (Quercetum coryloso-dryopterioaum (filiois masis) поширена на Розточчі та в південній частині Полісся, по других терасах річок заходить і в північну смугу лісостепової зони (Ра- дехівський, Івано-Франківський, Володимир-Волинський, Клеванський, Житомирський, Фастівський, Богуславський та Мринський лісгоспза- ги). Пов’язана вона з середніми або нижніми частинами схилів вузьких прирічкових ярів та балок або з плоскими невисокими гривами других терас і розташовується безпосередньо над дібровою ліщиново- жіночопапоротевою. Грунти під нею свіжі, дерново-підзолисті, супіщані (рис.17). Деревостан у віці 80-140 років має зімкненість крон 0,6-0,9, висоту 22-26 м, діаметр дерев 26-48 см і П та рідше Ш клас бонітету. Порівняно з деревостаном попередньої асоціації серед постійних видів його з’являється черешня, а серед випадкових - вільха клейка та сосна звичайна.
Підлісок з зімкненістю крон 0,2-0,6 утворює ліщина звичайна (0,2-0,5), крушина ламка (+ - 0,1), бруслина бородавчаста, черемха звичайна, горобина звичайна, ожина звичайна, глід український, свидина кров’Яна та інші види з 2-4-м класом постійності.
Травостій густіший, ніж у попередньої асоціації і має проективне покриття 30-55%. Його складають дріоптерис чоловічий (15-35%), копитняк європейський (+ - 10%), безщитник жіночий (+ - 10%), яглиця звичайна (+ - 5%), конвалія звичайна (+ - 5%), гравілат міський (■*• - 1%), веснівка дволиста (+ - 1%), медунка темна (+ - i%)t костяниця (+ - 1%), актея колосиста, астрагал солодколистий, ку- цоніжка лісова, ломиніс прямий, пахучка звичайна, костриця велетенська, підмаренник весняний, перлівка поникла, вороняче око звичайне, тонконіг дібровний, орляк звичайний, чистець лісовий (stachya sylvatica L.), фіалка дивна, горлянка повзуча, зніт гірський, дзвоники персиколисті, звіробій гірський (HypericUJB montanum ь.), перстач прямостоячий, медунка вузьколиста, грушанка круглолиста, ториліс японський. З випадкових видів трапляються:
ІЗІ
бугила лісова, підлісник європейський, дзвоники ріпчастовидні, ластовень лікарський, осока трясучковидна, цирцея звичайна, хвощ лісовий, зніт гірський, зірочник лісовий, вербозілля звичайне, чина весняна, кадило мелісолисте, зеленчук жовтий, печіночниця звичайна, купина багатоквіткова та ін.
Видова насиченість асоціацій становить 124 види.
У. Група асоціацій дубові ліси ліщиново-зірочникові складається з шести асоціацій, центральне місце серед яких займає діброва ліщиново-зірочникова. На більш сухих грунтах росте діброва ліщиново-волосистоосокова, а на вологіших - діброва ліщиново-яг- лицева. Інші три асоціації заміщують центральну переважно в північній та західній частинах ареалу цієї групи асоціацій. В більш вологих умовах формується діброва ліщиново-копитнякова, дуже близька в цьому відношенні до діброви ліщиново-яглицевої, а в сухіших - діброва ліщиново-зеленчукова. Проміжне положення займає діброва ліщиново-маренкова. Найбільш поширеними є перші три асоціації, які займають близько 60% площі дубових лісів лісостепової зони. Вони виявляють значно меншу варіабельність по відношенню до вологи грунтів,ніж до їх родючості.Особливістю поширення цих асоціацій є майже повний збіг їх південної межі, яка проходить по лінії: Новопсковськ - Куп"янськ - Слов"янськ - Красноград - Олександрія - Умань - Могилів-Подільоький.
Діброва ліщиново-ягдицева( Quexcetue coxyloao-aegopodiosum) поширена переважно в лісостеповій зоні, де трапляється майже в усіх лісництвах. В степову зону вона заходить лише по тальвегах балок в північно-східній частині зони (Харківська та Донецька обл.). В широколистянолісовій зоні на території Прикарпаття, Розточчя, Опілля, Західного Поділля, Малого Полісся і особливо північної частини Полісся ці ліси трапляються зрідка і невеликими ділянками (Косівський район Івано-Франківської області; Золочівський, Бродівський, Дубровицький та Овруцький лісгоспзаги). Більш характерні вони для південної частини Полісся, особливо Волинської височини. В межах лісової зони діброва ліщиново-яглицева займає рівні, дещо підвищені, добре дреновані ділянки або верхні частини пологих 1 нижні частини крутих схилів з свіжими й вологими дерново-слабопідзолистими глинисто-піщаними та супіщаними, рідше світло-сірими лісовими грунтами. В лісостеповій зоні ці ліси трапляються часто і є одними з найхарактерніших. Зростають вони на різних елементах рельєфу - в балках, на плакорах та в долинах річок.
132
В північній частині лісостепової вони діброва ліщиново-ягли- цева займає звичайно верхню частину слабопологих схилів горбистих вододілів. Зі збільшенням їх крутизни та просуванням на південь вона зміщується все нижче і в південній смузі зони формується лише в тальвегах балок. Так само впливає на її розміщення і експозиція. На південних схилах ці ліси спускаються нижче, ніж на північних. Звичайно окремі ділянки цієї асоціації мають ширину кілька десятків метрів і довжину від кількох сотень метрів до 5-Ю і більше кілометрів. Грунти під ними - свіжі або вологі сірі лісові суглинисті, рідше - темно-сірі лісові.
В середній та південній смузі лісостепової зони на широких і слаборозчленованих межиріччях Лівобережжя ці ліси займають плоскі, найбільш знижені ділянки плакорів з чорноземними сильноопідзоле- ними грунтами.
В заплавах річок дубові ліси ліщиново-яглицеві трапляються на території всієї зони. Займають вони невисокі плоскі гриви прируслової або міжгривні зниження центральної та притерасної частин з дренованими опідзоленими грунтами; значно рідше трапляються в міжгривних зниженнях других терас з дерново-слабопідзолистими супіщаними грунтами (рис.18).
Деревостан у віці 60-160 років має зімкненість крон 0,6-1,0, висоту 18-30 м, діаметр дерев 20-52 см і І-П клас бонітету. Створюють його дуб звичайний з участю ясена звичайного (+ - 0,2), липи серцелистої (+ - 0,2), клена звичайного (+ - 0,1) (постійні види); берези бородавчастої, осики, в"яза шорсткого, клена польового, черешні, груші звичайної, клена неоправжньоплатанового (Acer pseudoplatanys L.),граба звичайного, яблуні лісової, в"яза гладенького та інших випадкових видів.
Підлісок має зімкненість 0,2-0,7. Крім ліщини звичайної (0,2- 0,7), до нього входять бруслини європейська та бородавчаста, свидина кров"яна, черемха звичайна, роза щитконосна (Вона corymbifera В о r k h.j, крушина ламка, клен татарський, гордовина (Viburnum lantana L.) та глід несправжньо-кривостовпчиковий (Crataegus рае- udokyrboetyla К 1 о k.). Останні п"ять видів мають 2-4-й клас постійності.
Травостій з проективним покриттям 25-60% створює яглиця звичайна (14-45%) з участю таких постійних видів: копитняк звичайний (+ - 10%), зірочник лісовий (+ - 10%), маренка запашна (+ - 10%), осока волосиста (+ - 10%), гравілат міський (+ - 5%), медунка тем133
на (+ - 5%), конвалія звичайна (+ - 5%), кропива дводомна (+ - 5%), переліска багаторічна (Mercurialis perennis L.) (+ - 5%), купина багатоквіткова (+ - 1%), фіалка дивна (+ - 1%), куцоніжка лісова, дзвоники ріпчастовидні, ториліс японський, кадило мелісолисте, розхідник шорсткий ( Glechoma hirsuta W. К.), чистець лісовий, перлівка поникла, грястиця збірна. Регіонально постійні види представлені гордовиною (Придніпровська височина), осокою трясучковид- ною (Правобережне Полісся) та бутнем запашним (західні райони УРСР). До випадкових видів належать: жовтець золотистий (Banuncu- lus auricomus L.), шоломниця висока (Scutellaria aitізsima L.), празелень звичайна (Lapsana communis L.), актея колосиста, горлянка повзуча, бугила лісова, астрагал солодколистий, безщитник ЖІНОЧИЙ, КІНСЬКИЙ часник черешковий (Alliaria petiolata Cavata et Grand e).осока стягнута, осока лісова, пахучка звичайна, дріоптерис чоловічий, зніт гірський, коручка широколиста, костриця велетенська, зеленчук жовтий, підмаренник весняний, борщівник сибірський, чина чорна, вербозілля лучне, веснівка дволиста, вороняче око звичайне, тонконіг дібровний, квасениця звичайна, просянка розлога, міцеліс стінний, жовтець кашубський, маруна щиткова, фіалка лісова, підлісник європейський, печіночниця звичайна, парило звичайне, хвощ лісовий, дзвоники персиколисті, звіробій чотиригранний, герань лісова, бутень запашний, в"язіль барвистий, підмаренник проміжний, зубниця бульбиста, лілія лісова, стародуб пруський та ін.
Видова насиченість асоціацій - 241 вид, проте з них тільки 88 видів мають 2-ІО-й клас постійності.
В діброві ліщиново-яглицевій порівняно з дібровами ліщиново- папоротевими серед групи постійних немає бореальних видів, вони трапляються лише в невеликій кількості в групі непостійних видів. Помітно вменшується роль папоротей, характерних для асоціацій лісової зони, і навпаки, зростають роль і кількість неморальних балтійських мезо- і мегатрофів, серед яких, крім спільних з попередньою асоціацією, з"являються клен звичайний, ясен звичайний, переліска багаторічна, ранник вузлуватий та ін. Роль неморальних субсередземноморських видів практично не змінюється.
Діброва ліщиново-зірочникова (Quercetum ooryloso-stellariosum), як і попередня, трапляється в усіх лісових масивах лісостепової зони, де звичайно розміщується над дубовими лісами ліщиново-яглицевими. Вона пов"язана з 134
більш сухими, а отже, більш піднесеними та краще дренованими ділянками рельєфу. В широколистянолісовій зоні ліщиново-зірочни- кова діброва поширена дещо більше, ніж діброва ліщиново-яглицева (Симбірський та Стрийський райони Львівської області, Косівський та Снятинський райони Івано-Франківської області, Золочівський, Любомльський, Володимир-Волинський, Маневицький, Камінь-Кашир- ський, Степанський, Овруцький, Житомирський, Малинський, Коростен- ський, Новоград-Волинський, Іванківський, Городнянський та Глу- хівський лісгоспзаги). В зазначеній зоні ця діброва займає невисокі плоскі вершини вододілів або верхні частини похилих схилів південної експозиції, спадистих та крутих схилів північної та західної експозицій. На крутих південних схилах вона займає їх нижні частини. В північній частині лісостепу ці ліси пов’язані з верхніми та середніми частинами спадистих схилів північних напрямків та нижніми частинами схилів південних напрямків. У південній смузі лісостепу на схилах південної експозиції вони вже не ростуть, а на спадистих та крутих схилах північної експозиції переходять в їх нижні частини. Найбільш південні ділянки таких лісів описані нами в північно-східній частині степової зони (Старобільський та Куп"ян- ський лісгоспзаги), де вони займають нижні частини північних схилів глибоких ярів. На плакорах та в заплавах річок ці ліси трапляються рідше, ніж дубові ліси ліщиново-яглицеві, але на других терасах річок вони представлені значно краще. На Поліссі та на других терасах ліщиново-зірочникова діброва займає свіжі дерново-, слабо- та середньопідзолисті глинисто-піщані та супіщані грунти і в північній частині лісостепу та на Волинській височині - сірі лісові суглинисті грунти, а в південній частині лісостепу - темно-сірі лісові і чорноземні сильноопідзолені глинисті грунти (рис.19).
Деревостан у віці 50-130 років має зімкненість крон 0,7-1,0, висоту 17—27 м, діаметр дерев 18-40 см та П, рідше І або Ш клас бонітету. Створюють його ті самі види, що й у попередньої асоціації, але з меншою участю ясена звичайного.
Підлісок має зімкненість 0,2-0,7. Складають його ліщина звичайна (0,2-0,6) свидина кров"яна (+ - 0,2), бруслина бородавчаста (+ - 0,1) та інші відмічені для попередньої асоціації види, але з більшою участю клена татарського (+ - 0,1).
Травостій має проективне покриття 30-55%. Абсолютно постійними видами його є зірочник лісовий (15-35%), яглиця звичайна (+ - 10%), копитняк європейський (+ - 10%), медунка темна (+ - 5%), чина
135
весняна (+ - 1%), фіалка дивна (+ - 1%), гравілат міський та купина багатоквіткова. Відносно постійні види, так само як і випадкові, переважно спільні а попередньою асоціацією. Серед них збільшується тільки роль і кількість неморальних субсередземноморських видів, наприклад, стають постійними ториліс японський, розхідник шорсткий, фіалка шершава та ін.Ступінь конфекторної ролі зірочника лісового дуже змінюється, що пояснюється збігом синекологічних амплітуд деяких видів (яглиця звичайна, осока шорстка) з більшою конкурентною здатністю (краще розвиненою кореневою системою, більшою висотою тощо).
Видова насиченість асоціації становить 218 видів.
Діброва ліщиново-волосистоосокова (Queroetun coryloso-caricoeum(piloaae) поширена на тій самій території, що Й дві попередні асоціації, але в рельєфі займає ще сухіші та вищі місця і розміщується беапосередньо над дібровою ліщиново-зірочниковою. П.С.Погребняк (1955) відносить цю асоціацію до сухого типу дібров, що, на наш погляд, не зовсім вірно. Доцільніше відносити її до свіжих дібров.
На Поліссі (Камінь-Каширський, Дубровицький, Іванівський та Словечанський лісгоспзаги) ліси зазначеної асоціації займають найбільш підвищені та дреновані серед дубових лісів опуклі ділянки межиріч або верхні частини спадистих та крутих схилів південної та східної експозицій. На пологих схилах північної експозиції вони, на відміну від попередніх, не ростуть. Окремі ділянки цих лісів у північній частині Полісся мають площу менше І га і трапляються дуже рідко. В південній частині Полісся дубові ліси ліщи- ново-волосистоосокові стають досить звичайними і окремі ділянки їх мають розміри до кількох гектарів. Особливо великі ділянки (5- 20 га) трапляються в Житомирському лісгоспзагу.
Для території Прикарпаття та Західного Поділля ці ліси не характерні, тому що на багатих грунтах зазначених регіонів в умовах вологого клімату вони заміщуються дубово-грабовими волосистоосоко- вими лісами. Грунти під ними на території широколистянолісової зони свіжі дерново-середньопідзолисті, глинисто-піщані та супіщані, рідше - світло-сірі лісові.
На території лісостепової зони на Правобережжі найбільш великі ділянки (близько 1-2 га) дубових лісів ліщиново-волосистоосо- кових трапляються на других терасах річок дніпровського басейну (Фастівський та Білоцерківський лісгоспзаги), де вони займають вер136
шини та схили грив. На межиріччях ці ліси трапляються рідко (Тернопільський, Ізяславський, Старокостянтинівський, Вінницький, Звенигородський та Уманський лісгоопзаги). Головні масиви їх зосереджені на Лівобережжі, де в північній смузі лісостепової зони вони пов"язані з найбільш крутими та сухими схилами різних експозицій, а в південній смузі - з середніми і нижніми частинами спадистих і крутих схилів північної та західної експозицій і тільки як виняток - з нижніми частинами схилів південно! експозиції. Інакше кажучи, при просуванні на південь відбувається заміна топографічних позицій асоціації, що зумовлює збереження однакових екологічних умов. Грунти, як правило, в лісостеповій зоні сірі або темно-сірі лісові суглинисті. На плоскі рівні межиріччя діброва ліщиново-волосис- тоосокова виходить у південно-східній частині Черкаської та Полтавської областей, проте найбільші масиви її розміщені на вододілах лісостепової частини Харківської області, де вона за площею переважає усі інші асоціації формації і створює останні плакорні ліси. Грунти під ними в цій частині ареалу сильноопідзолені чорноземи (рис.20).
Деревостан у віці 50-140 років має зімкненість крон 0,6-1,0, висоту 18-30 м, діаметр дерев 18-42 см і П-1 клас бонітету. Постійними видами його, крім дуба звичайного, є тільки липа серцелиста (+ - 0,1) та клен звичайний. Ясен звичайний в цю групу вже не входить, тому що в північній частині ареалу асоціації грунти для нього надто бідні, а в південній - надто сухі. В західній частині він поступається місцем грабові звичайному. Таким чином, постійно цей вид трапляється лише в центральній та південно-східній частинах ареалу асоціації. Інші види деревостану виявляють таку закономірність: береза бородавчаста і осика характерні для ділянок асоціації, поширених на Поліссі, тоді як клен польовий - для лісостепу. Крім перелічених видів трапляються зрідка черешня, груша звичайна, вільха клейка та ін.
Підлісок за зімкненістю та видовим складом подібний до попередньої асоціації. В більш південних районах на багатих та сухих грунтах постійними супутниками ліщини звичайної виявляється свидина кровпяна (+ - 0,1) і клен татарський, в північніших районах на бідних супіщаних грунтах - горобина звичайна та крушина ламка, а в межах всього ареалу асоціації - бруслина бородавчаста. Зрідка трапляються бруслина європейська, глід український, вовчі ягоди звичайні, шипшина щитконосна, ожина звичайна, черемха звичайна та інші види з І-2-м класом постійності.
137
Травостій мав проективне покриття 25-50%. Порівняно з попередньою асоціацією ценозоутворююча роль його конфектора виявлена краще (20-45%). Група постійних видів майже та сама, що й у діброви ліщиново-зірочникової (яглиця звичайна, маренка запашна, копитняк європейський, астрагал солодколистий, куцоніжка лісова, дзвоники ріпчастовидні, осока стягнута, конвалія звичайна, гравілат міський, розхідник шорсткий, чина весняна, перлівка поникла, переліска багаторічна, просянка розлога, медунка темна, зірочник лісовий, ранник вузлуватий, фіалка шершава та фіалка дивна), але з меншим проективним покриттям.
Закономірність розподілу групи випадкових видів така сама, як у видів верхніх ярусів. Наприклад, печіночниця звичайна та підлісник європейський лов"язані тільки з ділянками асоціації, розташованими в західних районах. Веснівка дволиста, квасениця звичайна, ожика волосиста, орляк звичайний та костяниця - з ділянками асоціації північних районів на найбільш бідних грунтах, а горошок горохоподібний (Viola pisiformis L,), підмаренник пухнатоногий ( Gallum dazypodum К 1 о k.), борщівник сибірський, звіробій шорсткий (Hyperioum hirsutum Ь.), шоломниця висока та ін. - з ділянками асоціації південних районів на багатих грунтах. Костриця велетенська, вербозілля лучне та безщитник жіночий трапляються в найбільш вологих екотопах асоціації і, навпаки, - осоки Мікелі та кореневищна (Oarex rhizina в 1 у t t.),фіалка шершава - в найсухіших.Крім перелічених видів,до цієї групи входять ще осока пальчаста,купини багатоквіткова І широколиста ( Polygonatum latifolium D e a f.)( фіалка лісова, маруна щиткова, жовтець золотистий, отоколос Бене- кена (Zarna benekeni Ь і n d ш.), підмаренники весняний і проміжний, чина чорна, актея колосиста, зеленчук жовтий, горлянка повзуча, коручка широколиста, буквиця лікарська, медунка м'якенька, ввіробої звичайний та чотириграний, мїцеліс стінний, смрвдь гірська, золотушник звичайний, хвощ лісовий, молочай мигдалевид- ний, бутень ароматний, ломиніо прямий, куничник тростиновий,дзвоники персиколисті, лілія лісова, заяча капуста велика та ін.
Видова насиченість асоціації - 213 видів.
Діброва ліщиново-копитнякова (Quercstum oaryloso-asarosuii) поширена в Прикарпатті, на Поділлі, в середній та південній частинах Полісся і в північній частині Середньоруської височини, тобто на території широколиотяноліоо- вої та в північній частин! лісостепової зон (Галицький район івано-
138
Франківської області, Золочівський, Любомльський, Малайський, Но- воград-Волинський, Житомирський, Іванківський, Бучацький, Тернопільський, Забаровський, Хмільницький, Кам"янець-Подільський, Старокостянтинівський, Ізяславський, Іллінецький, Фастівський, Пере- яслав-Хмельницький, Богуславський, Прилуцький та Роменський лісгоспзаги). В південну частину лісостепу вона заходить тільки по тальве- гах глибоких балок (Балтський, Вовчанський та Слов"янський лісгоспзаги). На других терасах ліси цієї асоціації займають міжгривні зниження з найбільш багатими свіжими дерново-слабопідзолистими суглинковими грунтами. В балках та на ділянках з хвилястим рельєфом вони звичайно ростуть в нижній частині схилів з сірими лісовими або зрідка дерново-карбонатними грунтами. Ця асоціація, як і дві наступні, має більш вузький, ніж попередні асоціації дубових ліщино- во-зірочникових лісів, ареал, який в загальних рисах відповідає перехідній смузї між широколистянолісовою та лісостеповою зонами.
Деревостан у віці 50-130 років має зімкненість крон 0,7-1,0, висоту 19-28 м, діаметр дерев 20-50 см і П, рідше І клас бонітету. До постійних видів його належать тільки дуб звичайний, осика (+ - 0,2) та липа серцелиста (+ - 0,1). В екотопах домішується: в найбільш вологих - вільха клейка, в найбідніших - береза бородавчаста, в багатих - ясен звичайний, черешня, граб звичайний, клен звичайний та ін.
Підлісок загалом має більш високу зімкненість, ніж у попередньої асоціації. Постійними видами його є ліщина звичайна (0,2-0,6), свидина кров"яна (+ - 0,2), клен татарський (+ - 0,1) та бруслина бородавчаста. Крім них, трапляються бруслина європейська, жостір проносний, крушина ламка, шипшина собача, черемха звичайна, ожина сиза, гордовина, глід Липоького (Crataegos Іірвкуі Klo k.) та ін.
Травостій з проективним покриттям 30-50% утворює копитняк європейський (15-30%), а з числа постійних видів - яглиця звичайна (+ - 20%), зірочник лісовий (+ - 10%), конвалія звичайна (+ - 5%), гравілат міський (+ - 5%), медунка темна (+ - 5%), перлівка поникла (+ - 1%), куцоніжка лісова, чини весняна і чорна, вороняче око звичайне, купина широколиста, ранник вузлуватий, веснівка дволиста, фіалка дивна, актея колосиста, костриця велетенська, осока стягнута, маренка запашна, дзвоники ріпчатовидні, ториліс японський, пахучка звичайна, фіалка лісова та чистець лісовий. Група випадкових видів складається з тонконога дібровного, осок гірської та пальчастої, фіалки шершавої, підлісника європейського, орляка 139
звичайного, горлянки повзучої, купини лікарської, грястиці збірної, цирцеї звичайної, печіночниці звичайної, зніту гірського, дріоптериса чоловічого, наперстянки великоцвітої, кадила мелісолисто- го, медунки вузьколистої, чистецю прямого (Stachys recta Ь.)та ін.
Видова насиченість асоціації - 170 видів.
Діброва ліщиново-маренкова (Quercetum ooryloeo-asperulosua) трапляється переважно в західній частині широколистянолісової та лісостепової зон на території Прикарпаття, Розточчя, Опілля, Волині, Поділля та південної частини Західного Полісся (Самбірський район Львівської області, Рогатинський, Калу ський райони Івано-Франківської області, Золочівський, Радехів- оький, Пастерівський, Львівський, Чортківський, Тернопільський, Кременецький, Старокостянтинівський, Маневицький, Володимир-Волин- ський та Любомльський лісгоспзаги). На Лівобережній Україні вона поширена в північно-східній частині Середньоруської височини (Сумський, Романський та Галицький лісгоспзаги). Характерною особливістю її є зв"язок з найбільш високим для дубових лісів місцепо- ложеннями, вище якого на заході вона заміщується дубово-грабовими та буковими лісами. Великих масивів ці ліси ніде не утворюють, а трапляються невеликими ділянками 1-2 га. Займають вони вершини ме- жирічних горбів або верхні частини західних схилів із свіжими світло-сірими та сірими лісовими суглинистими або дерново-середяьопід- зодиотими супіщаними (на Поліссі) грунтами.
Деревоотан у віці 60-180 років має зімкненість крон 0,8-1,0, висоту 21-35 м, діаметр дерев 22-60 см і І-П клас бонітету. Відмінною рисою його,порівняно з попередньою асоціацією,є постійна присутність осики (+ - 0,1), берези бородавчастої (+ - 0,1) та граба звичайного.
Підлісок більш іустий (0,3 - 0,7) і з краще виявленою, ніж у попередніх асоціацій, роллю ліщини звичайної (0,3 - 0,7). Крім неї, постійно трапляються також свидина кров"яна та бруслина бородавчаста. Значно рідше (2-4-й клас постійності) зустрічаються бруслина європейська, вовчі ягоди звичайні, глід несправжньо-криво- стовпчиковий, жимолость пухната (Lonicera xyloeteum L.), крушина ламка, черемха звичайна, горобина звичайна, ожина сиза, жостір проносний, шипшина собача та ін.
Травостій має проективне покриття 30-55%. Міра ценозоутво- рюючої ролі його конфектора невелика (15-30%). Пояснюється це тим, що оптимум синекологічної конфекторної амплітуди цього виду відпо-
140
відає буковим лісам і в світлих дубових лісах його конкурентна здатність послаблена. Абсолютно постійними видами травостою .є яглиця (+ - 10%), копитняк європейський (+ - 10%), медунка темна (+ - 5%), веснівка дволиста (+ - 1%), куцоніжка лісова, актея колосиста, чина весняна, просянка розлога, вороняче око звичайне, дріоптерис чоловічий та купина багатоквіткова. Більшість відносно постійних та випадкових видів спільні з попередньою асоціацією. Різниця полягає лише в значно більшій участі в цих лісах середньоєвропейських видів: апозериса смердючого, осоки трясучковидної, бука лісового, купини кільчастої, дуба скельного та астранції великої, що наближає їх до діброви ліщиново-трясучковидноосокової.
Видова насиченість асоціації - 185 видів.
Діброва л 1 щ и н о в о - з є л є н ч у к о в а ( Quercetum ooryloao-galeobdoloaum) поширена в західній частині широколистянолісової та лісостепової зон на території Опілля, Розточчя, Поділля, Західного та Правобережного Полісся (Золочівський, Львівський, Любомльський, Дубровицький, Клеванський, Новоград-Волинський, Ізяславський, Бучацький та Вінницький лісгоспзаги). Трапляється вона досить рідко і невеликими (до І га) ділянками, що займають незначні плоскі піднесення межиріч або похилі та спадисті північні схили в дерново-слабопідзолистими супіщаними та світло- сірими суглинистими грунтами.
Деревостан у віці 100-190 років має зімкненість крон 0,6-0,8, висоту 26-31 м, діаметр дерев 34-86 см 1 П клас бонітету. Утворює його дуб звичайний з участю липи серцелистої (+ - 0,1), граба звичайного (+ - 0,1), осики (+ - 0,1), клена звичайного, явора, ясена звичайного та берези бородавчастої.
Підлісок із зімкненістю 0,2-0,6 складається 8 ліщини звичайної (0,2 - 0,6), свидини кровияної, бруслини бородавчастої, черемхи звичайної та інших видів з І-З-м класом постійності.
Травостій має проективне покриття 30-40%, з нього близько половини припадає на долю розхідника жовтого, тобто його конфек- торна здатність в цій асоціації виявлена слабо. Постійні супутники конфектора - яглиця звичайна (+ - 10%), копитняк європейський (+ - 10%), маренка запашна (+ - 5%), веснівка дволиста (+ - 5%), медунка темна (+ - 1%), зірочник лісовий (+ - 1%), підлісник європейський (+ - 1%), актея колосиста, горлянка повзуча, безщит- ник жіночий, дріоптерис чоловічий, гравілат міський, розхідник шорсткий, чина весняна, перлівка поникла, просянка розлога, во-
141
роняче око звичайне, жовтець кашубський, купина багатоквіткова, міцел іс стінний, фіалка лісова, фіалка дивна, тонконіг дібровний та підмаренник проміжний. Випадково трапляються цибуля ведмежа (Allium ursinum L.), апозерио смердючий, костриця велетенська, конвалія звичайна, грястиця збірна, осоки лісова та бліда, печіночниця звичайна, костяниця, чина чорна, підмаренник весняний, герань темна (Geranium phaeum L.) та ІН.
Видова насиченість асоціації - 106 видів.
УІ. Група асоціацій дубові ліси ліщиново-гірськоосокові представлена дібровами ліщиново-конвалієвою та ліщиново-гірськоосоко- вою. Перша з них займає дещо вологіші екотопи, ніж друга, і трапляється інколи на більш багатих грунтах, тобто має широку екологічну амплітуду. Порівняно з попередньою групою асоціацій в цій асоціації світлолюбні неморальні субсередземноморські види представлені значно краще (куцоніжка пірчаста, перстач білий, дзвоники персиколисті, ластовень лікарський, клен татарський, медунка вузьколиста та ін.). З другого боку, в цих лісах в ще й бореальні види (орляк звичайний, веснівка дволиста, куничник тростиновий, костяниця та ін.). Така різноякісність цих лісів пояснюється тим, що вони ростуть в умовах лісостепового клімату, який сприяє розвиткові базифільних дібров з субсередземноморськими видами, але на бідних едафотопах (другі тераси річок), що сприяють розвиткові ацидофільних дібров з бореальними видами.
В процентному відношенні всі види, що створюють, наприклад, діброву ліщинбво-гірськоосокову, розподіляються таким чином. Якщо з загальної кількості видів асоціації вилучити види з першим класом постійності, то залишиться І18 видів, з них 44% - неморальні субсередземноморські,40% - неморальні балтійські і 16% - бореальні. Якщо взяти до уваги, що в другій групі є 9 видів з дуже широкою синекологічною амплітудою, що відповідає водночас амплітуді як неморальних балтійських, так і неморальних субсередземноморсь- ких видів, то кількість видів першої групи збільшиться на 4%. Таким чином, за флористичним аналізом дубові ліщиново-гірськоосокові ліси слід віднести до базифільної ланки, а на підставі особливостей едафотопу - до ацидофільної. Подвійна позиція цих лісів і щодо вологості грунту. Наявність в них значної кількості мезо- ксерофітів, таких, як ломиніс прямий, куцоніжка пірчаста, самосил гайовий ( Teucrium chamaedrys L.), свідчить про сухі грунти.
142
Проте в цих же лісах разом з мезокоерофітами трапляються мезогі- грофіти (безщитник жіночий, костриця велетенська, чистець лісовий) і навіть гігрофіти (вербозілля звичайне). Причини цього криються в грунтово-геоморфологічних особливостях других терас. Через відносно неглибоке залягання грунтової води та легкі, ивидко пересихаючі з поверхні грунти створюються вже при незначній зміні мікрорельєфу дуже різноманітні умови зволоження.
Діброва ліщиново-конвалієва (■Quercetum coryloso-convallarioaum) поширена переважно в північних та середніх районах лісостепової зони, але трапляється й північніше та південніше. Крайні північні місцезнаходження асоціації розташовані в південній частині Полісся (більшість лісгоспзагів) та на Словечанському кряжі, а південні - в північній частині Кіровоградської (Голованівський лісгоспзаг) та Донецької (Словенський лісгоспзаг) областей. Трапляється вона досить часто, але невеликими ділянками. В межах основного ареалу дубові ліси ліщиново-конвалієві ростуть на других терасах річок, де займають середньови- сокі плоскі гриви та схили високих грив з дерново-олабо- і се- редньопідзолистими супіщаними грунтами. На таких самих грунтах ці ліси трапляються і на території Полісся, де пов"язані з рівними вершинами вузьких горбів прирічкової смуги. В північно-західній частині лісостепу вони займають західні та круті північні схили з дерново-карбонатними грунтами (рис.21).
Деревостан у віці 50-160 років має зімкненість крон 0,7-0,9, висоту 17-26 м, діаметр дерев 20-64 см та П-Ш клас бонітету. Постійними видами його є дуб звичайний, осика (+ - 0,1), липа серцелиста (+ - 0,1) та береза бородавчаста. В багатих трофотопах ростуть ясен звичайний, а в бідних - сосна звичайна. В західних районах постійно трапляється граб звичайний.
Підлісок частіше густий (0,4 - 0,7). Утворює його ліщина звичайна (0,3-0,6) з участю клена татарського (+ - 0,2), бруслини бородавчастої (+ - 0,1), свидини кров"яної (+ - 0,1), крушини ламкої, горобини звичайної, ожини звичайної, черемхи звичайної, глоду українського та інших видів з І-З-м класом постійності. Травостій має проективне покриття 30-50%. Конфекторна роль конвалії звичайної виявлена добре. Звичайно їй належить більше половини проективного покриття травостою. З числа постійних видів до травостою входять копитняк європейський (+ - 5%), осока гірська (+ - 5%), зірочник лісовий (+ - 5%), куцоніжка лісова (+ - 1%),
143
гравілат міський (+ - 1%), перлівка поникла (+ - 1%), медунка темна (+ - 1%), орляк звичайний (+ - 1%), чини весняна і чорна, па- хучка звичайна, розхідник шорсткий, тонконіг дібровний, веснівка дволиста, костяниця, грястиця збірна, ранник вузлуватий, буквиця лікарська, осоки пальчаста і стягнута, яглиця звичайна, ториліс японський, фіалка дивна, зірочник лісовий та підмаренник весняний. Характерною рисою травостою е висока насиченість регіонально постійними видами, починаючи від бореальних (веснівка дволиста, охи- ка волосиста, одинарник європейський) і закінчуючи неморальними оубсередземноморськими (гордовина, осока парвська, кадило мелісо- лиоте, перлівка ряба, медунка вузьколиста, шоломниця висока та ін.).
Видова насиченість асоціації - 197 видів.
Діброва ліщиново-гірськоосоко- В а (Quercetum с or yl os o-oario os um (montanae) поширена виключно в лісостеповій зоні на других терасах річок. Трапляється вона значно частіше, ніж попередня, і більшими, до 2-5 га, ділянками. На другій терасі Росі, Ірпеня, Дніпра ця асоціація за площею перевищує інші асоціації дубових лісів. Значно рідше вона трапляється на другій терасі Південного Бугу, Псла та Сули (Тульчинський, Вінницький, Хмільницький, Сумський та Роменський лісгоспзаги). Діброва ліщиново-гіроькоооокова займав верхні частини грив і розташовується вище діброви ліщиново-волооистоосокової. Грунти під нею свіжі дерново-слабо- і середньопідзолисті глинисто-піщані та супіщані. Звичайно на найвищих гривах над лісами цієї асоціації розташовуються соснові насадження. Екологічно асоціація виявлена добре і, на відміну від попередньої, належить до стеноекотопних.
Деревостан у віці 50-120 років має зімкненість крон 0,7-1,0, виооту 17-25 м, діаметр дерев 20-38 ом і П-Ш клас бонітету. Утворюють Його ті види, що й у попередній асоціації, але без участі оосни звичайної та ясена звичайного (більш вирівняний трофотоп).
Підлісок має зімкненість 0,2-0,5. За видовим складом він подібний до підліску попередньої асоціації (ліщина звичайна, клен татарський, бруслина бородавчаста, крушина ламка, жостір проносний та ін.).
Травостій має проективне покриття 25-55% і складається з осоки гірської (15-40%), конвалії звичайної (+ - 10%), орляка звичайного (+ - 5%), пахучки звичайної (+ - 1%), астрагалу солодколистого, куничника тростинового, перлівки пониклої, тонконога дібровного, купини лікарської, парила звичайного, ториліоу японсько-
144
го, вероніки дібровної, ластовня лікарського, серпію різнолистого, ломиноса прямого, маруни щиткової, грястиці збірної, в"язелю різнобарвного, герані криваво-червоної, гравілату міського, підмаренника, весняного, медунок вузьколистої та м”якенької, чин чорної та весняної, фіалок дивної та шершавої, конюшини альпійської, смовді гірської (Peucedanum oreoselinum М о e nc h,)ia інших видів з I-4-м класом постійності. Від травостою попередньої асоціації він відрізняється наявністю численнішої групи постійних виді> та краще виявленою роллю неморальних субсередземноморських видів.
Видова насиченість асоціації - 199 видів.
УП. Група асоціацій дубові ліси ліщиново-парвськоосокові складаються тільки з однієї асоціації, яка характеризується відсутністю бореальних та ацидофільних видів серед групи постійних. Роль неморальних субсередземноморських видів в межах нейтрофільної ланки асоціацій виявлена найкраще. Це перша асоціація; у якої роль конфекто- ра травостою належить неморальному субсередземноморському видові з субсередземноморським типом ареалу і з диз'юнкціями на Паннонській низовині та на Кавказі. Вона належить до групи стеноекотопних і ряду свіжих дібров нейтрофільної ланки (рис.22).
Діброва ліщиново-парвськоосоко- В а ( Quercetum coryloso-cariooflum(brevicollis) зустрічається тільки в Придністровських районах південно-західної частини лісостепової зони (Бершадський та Могилів-ПоДільський лісгоспзаги). Займає вона верхні частини пологих схилів північної експозиції з сірими лісовими грунтами. Окремі ділянки її мають площу близько 1-2 га.
Деревостан у віці 60-100 років має зімкненість крон 0,8-1,0, висоту 18-23 м, діаметр дерев 22-30 см і П клас бонітету. За видовим складом він багатший, ніж деревостани всіх попередніх асоціацій. До його окладу, крім дуба звичайного, входять граб звичайний (+ - 0,2)., ясен звичайний (+ - 0,1), липа серцелиста(+ - 0,1%), клен польовий, черешня, груша звичайна, береза бородавчаста, клен звичайний, дуб скельний та інші види з І-З-м класом постійності.
Підлісок (0,3-0t5) створюють ліщина звичайна (0,2-0,3), бруслина бородавчаста (+ - 0,1), бруслина європейська, глід український, дерен справжній, городовика, клен татарський, свидина кров"яна та інші види з І-З-м класом постійності.
Травостій звичайно густий (40-60%). Створює його осока парв- ська (25-40%) з участю осоки волосистої (+ - 15%), копитняка єв145
ропейського (+ - 5%), зірочника лісового (+ - 5%), конвалії звичайної (+ - 5%), зеленчука жовтого (+ - 1%), медунки темної (+ - 1%), горошку лісового (vicia silvatica L.), астрагалу солодколистого, дзвоників ріпчастовидних, грястиці збірної, гравілату міського, чин чорної та весняної, жовтецю кашубського, шоломниці високої, тонконога дібровного, пахучки звичайної, маренки запашної, горлянки женевської, фіалки дивної, купини багатоквіткової, ранника вузлуватого, горошку горохоподібного, підмаренників проміжного та весняного, звіробою гірського, купини широколистої, стоко— лосу Бенекена, печіночниці звичайної, фіалки шершавої, перлівки пониклої, куцоніжки лісової, горлянки повзучої, веснівки дволистої та інших видів з І-4-м класом постійності.
Видова насиченість асоціації - 75 видів, що пояснюється як незначним поширенням її, так і вузькою екологічною амплітудою.
Характерною рисою флористичного складу асоціації є порівняно велика кількість середньоєвропейських видів, наприклад граба звичайного, черешні, дуба скельного, барвінку малого, перлівки одноквіткової (Melica uuiflora R e t z.). Переважають у складі асоціації типові мезофільні мезо- і мегатрофні неморальні балтійські види (56%), значну участь беруть також неморальні субсеред- земноморські види (28%).
УШ. Група асоціацій дубові ліси горобейникові.як і попередня, представлена тільки однією асоціацією. Ліси, що належать до неї, створюють південну межу ареалу класу асоціацій дібров ліщинових і є серед них найбільш термофільними.
Діброва лїщиново-горобейникова (Quetcetum coxyloao-llthospermosum) поширена тільки в південно- західній частині лісостепової зони (Балтський лісгоспзаг) і на Донецькому крижі (Словиянський лісгоспзаг). У першому районі вона займає знижені ділянки рівних та широких вододілів з темно-сірими лісовими грунтами, а в другому - верхні частини середбалочних спадистих схилів південних напрямків з дерново-карбонатними грунтами. Топографічно, екологічно і флористично вона безпосередньо пов"я- зана з дібровою татарськокленово-горобейниковою, в комплексі з якою і трапляється невеликими ділянками. Асоціація належить до групи стеноекотопних і ряду сухих дібров (рис.23).
Деревостан у віці 80-120 років має зімкненість крон 0,7-0,9, висоту 16-24 м і Ш, рідше ІУ бонітет. Постійними видами його є дуб звичайний, липа серцелиста (+ - 0,1), черешня, клен звичайний та 145
ясен звичайний. Рідне трапляються клен польовий, берест, груша звичайна , в"яз шорсткий та інші види з I-2-м класом постійності.
Підлісок загалом виявлений краще, ніж у попередньої асоціації. Його створюють ліщина звичайна (0,3-0,б), клен татарський (0,1-0,2) бруслина бородавчаста, свидина кров"яна, глід кривочашечковий ( Orataegus curvisepala Li nd m.), городовина (постійні види), бирючина звичайна, карагана кущова, терен звичайний, глід український, шипшина щитконосна, бузина чорна (Sambucus nigra L.) та інші випадкові види.
Травостій досить густий (35-45%). Утворюють його горобейник пурпурово-голубий (25-35%), конвалія звичайна (+ - 5%), кінський часник звичайний (-і- - 5%), гравілат міський (+ - 5%), купина широколиста (+ - 1%), осока Мікелі (+ - 1%), лопух дібровний (Агобіит nemorosum L e j e u n .), очиток великий (Sedum maximum Sute r.), холодок тонколистий ( Asparagus tenuifollus L a m.), астрагал солодколистий, куцоніжка пірчаста, дзвоники персиколисті, дзвоники ріпчастовидні, осока стягнута, костриця велетенська, чина чорна, празелень звичайна, міцеліо стінний, маруна щиткова, шоломниця висока, осока гірська, ториліс японський, регнерія собача, жовтець золотистий, фіалка шершава, в''язіль барвистий, отоколос Зенекена, горошок горохоподібний, ластовень лікарський, дутень ягідний та інші види з І-4-м класом постійності.
Видова насиченість асоціації - 108 видів.
В еколого-флористичному відношенні розглянута асоціація об'єднує нейтрофільні та базифільні дубові ліси. Вона займав найбагатші для всього класу грунти, подіо'ні до грунтів дубових лісів базифіль- ної ланки. Роль конфектора її травостою належить типовому субсеред- земноморському виду. У флористичному окладі асоціації, якщо не брати до уваги 8 видів, наявність яких зумовлена антропогенним впливом (кінський часник звичайний, кропива дводомна, мняточник чорний, чистотіл звичайний та ін.), переважають неморальні субсередземно- морські (55%) та неморальні балтійські (45%) види. Бореальні види, на відміну від попередньої асоціації, в якій вони входили до групи випадкових видів, повністю відсутні. Натомість вперше стають постійними з більш термофільної групи неморальних субсередземномор- ських видів півники злаколисті (Xrls graminea L.), горобейник пурпурово-голубий, осока Мікелі, перлівка ряба ( Melica piota к о с ь.) та ін. Крім того, різко збільшується,порівняно з попередніми асоціаціями, кількість і роль ксеро- і мезоксерофітів (ка147
рагана кущова, куцоніжка пірчаста, горобейник пурпурово-голубий, терен звичайний та ін.). На підставі зазначеного ми відносимо розглянуту асоціацію до зональних базифільних дубових лісів.
Дубові ліси свидинові
Дубові ліси свидинові мають досить обмежений ареал. Пояснюється це тим, що фітоценотичний оптимум свидини кров"яної лежить в умовах субатлантичного клімату на багатих свіжих та вологих грунтах в перехідній смузі між термофільними і мезофільними широколистяними лісами. На території Середньої Європи вони поширені в південній частині Словаччини, ФРН, північній частині Угорщини і Румунії (Зоо', 1943; Simon, 1957; Jurko, 1958; Papp» Szodfiidt, 1967). На території Радянського Союзу вони відомі з Подільської і Придніпровської височин (Szafer, 1935; Гринь, 1950; Шеляг-Сосонко, 1961).
На Україні дубові ліси свидинові безпосередньо пов"язані з дубовими лісами ліщиновими, цілий ряд асоціацій яких вони заміщують на більш родючих грунтах і в порівнянні з якими вони характеризуються більшою однорідністю екологічних умов і флористичного складу.Серед них вже немає асоціацій на бідних і кислих грунтах з участю ацидофільних бореальних видів 5-ІО-го класу постійності, але немає і ксеро-мезотермофільних асоціацій, аналогічних до дубових лісів ліщиново-горобейникових. Поширені дубові ліси свидинові в межах ареалу дубово-грабових лісів, там вони трапляються в долинах річок або на широких слаборозсічених межиріччях, несприятливих для зростання останніх. Грунти під ними сірі та темно-сірі лісові суглинки, з рН 5,2-6,8. У флористичному складі цих лісів переважають неморальні балтійські види.
Представлені дубові ліси свидинові трьома групами асоціацій. Центральне місце серед них займає група асоціацій дубові ліси сви- диново-зірочникові. Від неї в бік незначного збільшення сухості та родючості грунтів розташовується група асоціацій дубові ліси свидиново-гірськоосокові, по відношенню до якої група асоціацій дубові ліси свидиново-парвськоосокові займає багатші грунти. За зайнятою площею друга група асоціацій складає близько 80% лісів класу.
IX. Група асоціацій дубові ліси свидиново-зірочникові представлена дібровами свидиново-зірочниковою та свидиново-яглицевою. Остання порівняно з першою займав більш вологі грунти.
148
Порівняно з іншими групами асоціацій даного класу вони характеризуються меншою роллю неморальних субсередземноморських видів, представлених головним чином більш мезофільними видами з північнішим ареалом (розхідник шорсткий, шоломниця висока, ластовень лікарський, фіалки шершава і дивна та ін.). З другого боку, присутність неморальних субсередземноморських видів і високий клас постійності відрізняють дану групу від дубових лівів ліщиново-зіроч- никових.
Діброва свидин, о во-яглицева( Qoercetua
thelycranioso-aegopodioaum) поширена в північній і менше в південній частині лісостепової зони (Чортківський, Жовтневий, Старо- костянтинівський та Голованівський лісгоспзаги). По заплавах річок вона заходить і в західну частину широколистянолісової зони (заплава Дністра). Трапляються ці ліси зрідка і невеликими ділянками. Вони займають незначні плакорні зниження із свіжими темно-сірими лісовими суглинистими грунтами, в заплавах річок - знижені ділянки центральної частини із дерновими опідзоленими грунтами.
Деревоотан у віці 50-140 років має зімкненість крон 0,8-1,0, висоту 20-27 м, діаметр дерев 24-56 см, І-П клас бонітету (і складається з дуба звичайного, липи серцелистої, явора, ясена звичайного, граба звичайного, клена польового, черешні, груші звичайної та яблуні лісової ( Malus silvestris м і і 1.).Останні чотири види мають 3-4-й клас постійності.
Підлісок добре виявлений. Він має висоту 2-2,7 м, зімкненість 0,3-0,7 і велику видову насиченість, його утворюють свидина кров"я- на (0,2-0,6), крушина ламка (+ - 0,2), глід Липського(+ - 0,1), клен татарський (+ - 0,1), ліщина звичайна, бруслина бородавчаста, гордовина, ожина сиза, бруслина європейська, черемха звичайна, вовчі ягоди звичайні, калина звичайна та інші види. Останні п"ять видів мають 2-4-й клас постійності.
Травостій з проективним покриттям 30-55% утворюють яглиця звичайна (20-45%), розхідник шорсткий (+ - 10%), зірочник лісовий (+ - 10%), конвалія звичайна (+ - 5%), кропива дводомна (+ - 5%), фіалка лісова (+ - 5%), фіалка дивна (+ - 5%), копитняк європейський, астрагал солодколистий, куцоніжка лісова, осока стягнута, цирцея звичайна, коручка широколиста, костриця велетенська, лілія лісова, гравілат міський, вороняче око звичайне, просянка розлога, ранник вузлуватий, ториліо японський та жовтець кашубський. З видів 2-4-го класу постійності трапляються любка дволиста (riatanthe- 149
xa bifolia R i c h ), медунка м"якенька (Pulmonaria mollissima K e r n.), фіалка шершава, вероніка дібровна, чистець лісовий, шоломниця висока, підлісник європейський, тонконіг дібровний, купина багатоквіткова, купина широколиста, купина лікарська, кунич- ник тростиновий, перстач білий, стоколос Бенекена, безщитник жіно- чий, лазурник трилопатевий, маруна щиткова, чина лісова, пахучка звичайна, грястиця збірна, наперстянка великоцвіта, дріоптерис чоловічий, зніт гірський, хвощ лісовий, зеленчук жовтий,костриця велетенська, горобейник пурпурово-голубий, переліска багаторічна, перлівка поникла та ін.
Видова насиченість асоціації становить ІІЗ видів.
Таким чином, у створенні асоціації, особливо в групі видів з 5-І0-м класом постійності, беруть участь насамперед мезофільні неморальні балтійські види. Значно поступаються перед ними неморальні субсередземноморські види (гордовина, клен татарський, холодок тонколистий, півники злаколисті, лазурник трилопатевий, горобейник пурпурово-голубий, ластовень лікарський та ін.), більшість з яких має 2-4-й клас постійності. Участь бореальних видів незначна. В екологічному відношенні переважають мезофіти. Гігрофіти та гігро- мезофіти- жеруха недоторкана (Cardamine impatiens L.), осока лісова , хвощ лісовий, гадючник голий, гравілат річковий (Geum rivale 1.), вербозілля звичайне-представлені погано і зустрічаються зрідка. Ще рідше трапляються мезоксерофіти і ксерофіти.
Діброва свидиново-зірочникова (Quercetum tbelycranioso-stellariosun) поширена В північній смузі лісостепу t лише як виняток - в південній (Прилуцький, Сумський, Богуславський, Звенигородський, Корсунь-Шевченківський та Красно- градський лісгоспзаги). Ці ліси трапляються зрідка 1 невеликими ділянками на межиріччях і в заплавах річок. В першому випадку їх яайтиповішим місцезнаходженням є верхні частини пологих південних схилів або рівні ділянки широких межиріч із свіжими сірими лісовими суглинистими грунтами, у другому - низькі широкі гриви центральної частини заплави з дерновими опідзоленими грунтами.
Деревостан у віці 60-110 років має зімкненість крон 0,8-0,9, висоту 20-25 м, діаметр дерев 24-38 см і П клас бонітету. Крім едифікатора, його створюють липа серцелиста, граб звичайний, берест, в"яз шорсткий, клен звичайний, яблуня лісова, груша звичайна, ясен звичайний та клен польовий (+ - 0,2). П"ять останніх видів мають 2-3-й клас постійності.
150
Підлісок мав зімкненість 0,2-0,4 і виявлений гірше, ніж у попередніх асоціацій. Створює його свидина кров"яна (0,2-0,3) з участю клена татарського (+ - 0,1), ліщини звичайної, бруслини бородавчастої, гордовини та інших видів, характерних для попередньої асоці- аці ї.
Травостій має проективне покриття 30-50%. Крім зірочника лісового (15-45%), до нього входять яглиця звичайна (+ - 10%), конвалія звичайна (+ - 10%), розхідник шорсткий (+ - 5%), фіалка дивна (+ - 5%), купина багатоквіткова (+ - 1%), медунка темна (+ - 1%), астрагал солодколистий, тонконіг дібровний, шоломниця висока, ториліс японський, фіалка шершава, хвилівник звичайний, грястиця збірна, лілія лісова, гірча клинолиста, зеленчук жовтий, костриця велетенська, коручка широколиста, ранник вузлуватий та інші види з 2-4-м класом постійності, спільні з попередньою асоціацією.
Видова насиченість асоціації - 83 види.
Порівняно з дібровою свидиново-зірочниковою ця асоціація характеризується зменшенням ролі гігрофітів і незначним посиленням ролі мезоксерофітів.
X. Група асоціацій дубові ліси свидиново-гірськоосокові представлена двома асоціаціями: дібровами свидиново-конвалієвою та сви- диново-гірськоосоковою. Перша займає більш вологі й бідні грунти і безпосередньо пов"язана з дібровою свидиново-зірочниковою, майже так само, як і з дібровою свидиново-гірськоосоковою. Ліси цієї групи є зональними для центральної частини західного лісостепу.
Діброва овидиново-коявалієва ( Dercetum bhelycraoioso-oonvallaciosum) поширена В ПІВНІЧНІЙ і середній смузі лісостепу (Іллінецький, Богуславський, Переяслав- Хмельницький, Голованівоький, Мринський та Прилуцький лісгоспзаги). Зустрічається вона звичайно на других терасах річок у неглибоких міжгривних зниженнях з дерновими слабопідзолистими супіщаними грунтами. В плакорних умовах вона займав рівні ділянки, рідше схили з темно-сірими лісовими суглинистими грунтами.
Деревостан у віці 50-90 років має зімкненість крон 0,8-1,0, висоту 15-25 м, діаметр дерев 20-32 см і П-Ш клас бонітету. Крім дуба звичайного, його створюють береза бородавчаста, осика, липа серцелиста, черешня, в"яз шорсткий, груша звичайна, граб звичайний, клен польовий та інші види з I-2-м класом постійності.
Підлісок виявлений добре 1 має зімкненість 0,3-0,5.Створюють його свидина кров"яна (0,2-0,4), крушина ламка (+ - 0,1), бруслина 151
бородавчаста, ожина сиза (+ - 0,1), жостір проносний, клен татарський, глід український, бруслина європейська, жимолость пухнаста, гордовина, шипшина щитконосна та ін. Останні шість видів мають 2-3-й клас постійності.
Травостій з проективним покриттям 30-55% складається з конвалії звичайної (20-45%), осоки гірської (+ - 10%), куцоніжки пірчастої (+ - 1%), астрагалу солодколистого, пахучки звичайної, грястиці збірної, перстачу білого, ранника вузлуватого, торилісу японського, вероніки дібровної, фіалки шорсткої, медунки вузьколистої, осоки стягнутої, тонконога дібровного, куничника тростинового костриці велетенської, купин багатоквіткової і лікарської, дужня ягідного, кропиви дводомної та інших видів з I-4-м класом постійності.
Видова насиченість асоціації - 132 види.
Діброва свидиново-гірськоосоко- в а (Quercetum thelycranioso-caricoeujB (montanae) поширена в північній і середній смузі Правобережного лісостепу. Її можна вважати типовою асоціацією дубових лісів для території Поділля та західної частини Придніпровської височини, де вона займає найбільш опуклі ділянки плоских межиріч із свіжими темно-сірими лісовими суглинистими грунтами і де окремі її масиви мають площу до кількох десятків гектарів. Фрагменти асоціації трапляються t на других терасах, тут вона.приурочена до найбільш багатих грунтів. Найбільш східні ділянки її зустрічаються в Київській (Богуславській лісгоспзаг) та Черкаській (Корсунь-Шевченківський лісгоспзаг) областях. Таким чином, діброва свидиново-гірськоосокова завершує екологічний ряд розподілу асоціацій дубових лісів плакорів Поділля 1 других терас залежно від міри зростання родючості грунтів (рис. 24).
Деревостан у віці 50-180 років має зімкненість крон 0,8-1,0, висоту 19-29 м, діаметр дерев 20-60 см і І-Ш клас бонітету. Крім дуба звичайного, його створюють береза бородавчаста, граб звичайний, черешня, липа серцелиста, в"яз шорсткий, осика, ясен звичайний, груша звичайна та клен звичайний. Останні шість видів мають 3-4-й клас постійності. Порівняно з деревостаном попередньої асоціації в його складі помітно більшу участь беруть граб звичайний, черешня і значно меншу - липа серцелиста.
Підлісок виявлений добре і має зімкненість 0,3-0,7. Створений він свидиною кров’яною (0,2-0,6) з участю бруслини європейської (+ - 0,2), бруслини бородавчастої (+ - 0,1), глоду несправжньо- кривостовпчикового (+ - 0,1), клена татарського, ліщини звичай152
ної, гордовини, крушини ламкої, ожини звичайної, глоду Липського, шипшини собачої, жимолості пухнасто! та інших видів з I-2-м класом постійності.
Травостій з проективним покриттям 30-60% складається а осоки гірської (20-40%), конвалії звичайної (+ - 15%), копитняка європейського (+ - 5%), куцоніжки пірчастої (+ -5%), гравілату міського (+ - 5%), яглиці звичайної (+ - 5%), буквиці лікарської (+ - 1%), пахучки звичайної (+ - 1%), чини чорної (+ - 1%), астрагалу солодколистого, грястиці збірної, наперстянки великоцвітої, суниць лісових, підмаренника весняного, чини весняної, тонконога дібровного, молочаю ребристого, перстачу білого, медунки м”якенької, маруни щиткової, торилісу японського, вероніки дібровної, ластов- ня лікарського, фіалки шершавої, фіалок лісової та дивної, стоко- лосу Бенекена, актеї колосистої, горлянки повзучої, коручкн широколистої, жовтецю кашубського, кропиви дводомної, очитка великого (постійні види); стародуба пруського (Lasarpitiuii prutheaicum Ь.), смовді Любименка ( Peucedanum lubimeukoanum К о t о ▼ .)» медунки темної, костяниці, оману верболистого, ранника вузлуватого, конюшини альпійської, розхідника шорсткого, борщівника сибірського, празелені звичайної, вербозілля лучного, веснівки дволистої, зніту гірського, просянки розлогої, купини багатоквіткової, бугили лісової, цирцеї звичайної, ломиноса прямого, дріоптериса чоловічого, костриці велетенської, звіробою шорстокого, гадючника шестипелюсткового, підмаренника проміжного, медунки вузьколистої, герані криваво-червоної, куцоніжки лісової, костриці різнолистої, дзвоників ріпчастовидних, дутня ягідного та інших випадкових видів.
Видова насиченість асоціації - 162 види.
Конфектор ярусу травостою цих лісів є світлолюбним видом і має вузьку синекологічну амплітуду, його легко витісняють інші види при незначному збільшенні затіненості, вологості або сухості грунту. Тому в травостої переважають світлолюбні види, серед них і види плейстоценового комплексу, такі, як куничник тростиновий, суниці лісові, клопогін смердючий та медунка м"якенька, або види з широкою синекологічною амплітудою.
XI. Група асоціацій дубові ліси свидиново-парвськоосокові представлена лише однією асоціацією з вузькою синекологічною амплітудою.
Діброва свидиново-парвськоосо- к о в а (Quercetum thelycraoioso-caricosun(brevicollls) поши153
рена в південній частині Вінницької області (Бершадський лісгосп- заг). Вона займав широкі, рівні або дещо хвилясті межиріччя зі свіжими чорноземними сильноопідзоленими або темно-сірими лісовими суглинистими грунтами. Окремі масиви її мають кілька десятків гектарів, але трапляються в двох-трьох місцях (41-56-й квартали Бершадського лісництва).
Деревоотан у віці 60-100 років має зімкненість крон 0,8-0,9, висоту 22-25 м, діаметр дерев 28-32 см і П клас бонітету. Створюють його ті самі види, що й попередню асоціацію, але із значно більшою участю ясена звичайного (0,1-0,2).
Підлісок має зімкненість 0,3-0,5 і, крім конфектора, складається з ліщини звичайної (+ - 0,1), бруслини бородавчастої (+ - 0,1) глоду Липського, гордовини, бруслини європейської та інших видів з І-3-м класом постійності.
Травостій з проективним покриттям 35-60% створюють осока парв- ська (20-45%), копитняк європейський (+ - 10%), осока волосиста (+ - 10%), зірочник лісовий (+ - 10%), перлівка ряба (+ - 5%), розхідник шорсткий (+ - 1%), медунка темна (+ - 1%), яглиця звичайна, астрагал солодколистий, конвалія звичайна, грястиця збірна, зеленчук жовтий, гравілат міський, півники злаколисті, чина чорна, переліска багаторічна, купина широколиста, шоломниця висока, фіалки дивна і шершава, стоколос Бенекена, звіробій шорсткий, ранник вузлуватий, купина багатоквіткова, куцоніжка лісова, дзвоники ріп- частовидні, коручка темно-червона (Epipactia atrirubens (Н о f f и.) 8 e hui і,), бутень пмянкий ( Chaerephyllum teaulum L.), холодок тонколистий, чина весняна, лілія лісова, вероніка широколиста, маренка запашна та інші види з I-4-м класом постійності.
Видова насиченість асоціації становить лише 53 види, що пояснюється незначною площею досліджуваних лісів і однотипністю умов.
Дана асоціація в межах класу завершує ряд зростання термофіль- ності при незмінній вологості грунту. Цим і пояснюється велика кількість балтійських неморальних видів. Бореальних видів, які в незначній кількості І в невисоким класом постійності траплялись в попередніх асоціаціях, тут майже немає. З неморальних субсередзем- номорських видів, крім уже названих, абсолютно постійними стаюті, осока парвська, півники злаколиоті і перлівка ряба. Таким чином, цю асоціацію ми розглядаємо як таку, що завершує ряд нейтрофіль- них дубових лісів.
154
Дубові ліси татарськокленові
Конфектор ярусу підліску цих лісів мав відносно вузьку сине- кологічну амплітуду, яка відповідає континентальному і субконти- нентальному ксеротермічному кліматові південної частини ареалу едифікатора. В цій смузі він створює ярус підліску до самої межі лісів на більшій частині території Східної Європи і витісняється лише в південно-східній частині ареалу дуба звичайного більш стійкими до континентального клімату видами, такими, як вишня кущова (Cerasus fruticosa (Pali ,)G.W о r о n. ), шипшина корична ( Rosa cinna- fflomea !>.), акація жовта (Caragana arborescsns L а в.) та ІН. В Середній Євроці, вже на території Середньо-Дунайської низовини клен татарський витісняється більш термофільними дереном справжнім, бирючиною звичайною і скумпією звичайною. Отже, дубові ліси татарсько- кленові поширені в південно-східній частині Середньої Європи на території Угорщини та Румунії (Balazs, 1943} 806, 1943» 1958, 1960, 1963} Jakucs, Fekete, 1957} Zolyoal, 1957} z61yoni, Jakucs, 1957« Jakucs, 1960, 1961} Maths, Kovacs, 1962} Feketa, 1965} Jar6,1965« Michalko, D&atko, 1965} Pu^oaru-Soroceanu, Popova-Cucu, 1966« Pa^oovschi Doni^S, 1967} Purcelean, 1968).
На території Радянського Союзу вони зустрічаються в південній частині лісостепової зони і в смузі різнотравно-типчаково-ковилових степів степової зони (Бельгард, 1950; Напалков, 1951; Фурсаев,І952; Александрова, 1954; Киреев, 1964; Барабанщиков, 1968; Попов, Ваку- ров, 1970; Сенкевич та ін., 1970).
На Україні дубові ліси татарськокленові поширені в лісостеповій зоні. Північна межа їх проходить по лінії: Суми - Прилуки - Козятин - Чортків. На Поділлі вони трапляються лиже в південно- східній частині його, з решти території витісняються дубовими лі-- сами свидиновими. Представлені ці ліси чотирма групами асоціацій, дубовими лісами: татарськокленово-зірочниковими, татарськокленово- гірськоосоковими, татарськокленово-рябоперлівковими та татароько- кленово-горобейниковими. Причому перша група займає центральне місце. Від неї в бік незначного збільшення сухості і бідності грунтів розташовується друга група, а в бік більшого багатства, що супроводжується зростанням сухості грунту, - третя. Четверта група завершує ряд збільшення сухості і багатства грунтів, а разом з тим і ряд зростання ролі неморальних субсередземноморських видів. За площею перше місце займають дубові ліси татарськоклеяово-зіроч- никові та дубові ліси татарськокленово-горобейникові. їм належить
155
близько 70% усіх лісів класу. Дубові ліси татарськокленові займають темно-сірі лісові, дерново-карбонатні або чорноземні опідзоле- ні грунти з рН від 5,5 у перших до 7,4 у останніх. У флорі цих лісів переважають неморальні субсередземноморські та неморальні балтійські види, тобто, порівняно з лісами попереднього класу вони характеризуються дальшим збільшенням кількості перших та зростанням їх ролі.
ХП. Група асоціацій дубові ліси татарськокленово-зірочникові представлена чотирма асоціаціями. Центральне місце серед них займав діброва татарськокленово-зірочникова з широкою екологічною амплітудою. Від неї в бік більшого зволоження і багатства грунтів розташовується діброва.татарськокленово-яглицева, а в бік більшої сухості грунтів - діброва татарськокленово-волосистоосокова. Подальше зростання сухості й багатства грунтів призводить до формування діброви татарськокленово-кореневищноосокової.
Діброва татарськокленово-яглицева (Quercetum aceroso(tatarici)~aegopodio8uin) поширена в південній частині лісостепової зони і особливо в різнотравно-типчако- во-ковиловій підзоні степової зони, де вона трапляється в усіх ліс- госпзагах. Північна межа II поширення проходить по лінії: Могилів- Подільський - Умань - Полтава - Богодухів. Ці ліси займають майже виключно нижні Частини схилів глибоких балок та їх тальвеги зі свіжими й вологими чорноземними сильноопідзоленими або темно-сірими лісовими суглинистими грунтами. В кожній з балок вони тягнуться невеликими вузькими і довгими ділянками площею кілька гектарів (рис.25).
Деревостан у віці 60-90 років мав зімкненість крон 0,8-0,9, висоту -17-26 м, діаметр дерев 20-38 см і продуктивність П-Ш класу бонітету. Створений він дубом звичайним з участю ясена звичайного (+ - 0,2), клена польового (+ - 0,2), липи серцелистої (+ - 0,2), клена звичайного (+ - 0,2), черешні, яблуні лісової, вияза шорсткого та інших видів. Чотири останніх мають 2-3-й клас постійності.
Підлісок має висоту 3-5 м і зімкненість 0,2-0,6. Створює його клен татарський (0,2 - 0,5) з участю ліщини звичайної (+ - 0,1), свидини кровияної (♦ - 0,1), бруслини бородавчастої, бруслини європейської, гордовини, глоду несправжньо-кривостовпчикового, черемхи звичайної, ГЛОДУ обманливого ( Grataegus fallacina к 1 о kj, шипшини щитконосної, дерену справжнього та ін. Чотири останні види мають 2-3-й клас постійності.
156
Травостій з загальним проективним покриттям 35-65% утворює яглиця звичайна (25 - 45%). З числа абсолютно постійних видів до нього належать зірочник лісовий (+ - 10%), копитняк європейський (+ - 1%), гравілат міський (+ - 1%), медунка темна (+ - 1%), чистець лісовий (+ - 1%), фіалки дивна (+ - 1%) та шершава (+ - і%), просянка розлога, маруна щиткова, шоломниця висока, ранник вузлуватий, тори- ліс японський. Відносно постійні види представлені вербозіллям лучним (+ - 5%), конвалією звичайною (+ - 5%), розхідником шорстким (+ - 1%), купиною багатоквітковою, латуком стріловидним (bactuca sagittata W а 1 d , et Ki t , ), регнерією собачою, чи- ною весняною, тонконогом дібровним, празеленню звичайною, переліо- кою звичайною, дзвониками ріпчастовидними, грястицею збірною, астрагалом солодколистим, осокою стягнутою та бугилою лісовою.З регіонально постійних видів можна відзначити вербозілля кільчасте (Донецький кряж) і купину широколисту (Придніпровська височина). З випадкових видів трапляються хвилівник звичайний (Aristolochia clematitis ь.), чемериця чорна ( Veratrua aigrua L. ), перлівки висока (Melica altissima І».) і ряба, холодок тонколистий, горошок горохоподібний, очиток великий, костриця велетенська, буквиця лікарська, лілія лісова, дутень ягідний, звіробій шорсткий, борщівник сибірський, зеленчук жовтий, підмаренник пухнатоногий, фіалка запашна, бутень п"янкий, ломиніс прямий, аконіт Роговича, куцоніжка лісова та ін.
Видова насиченість асоціації - 136 видів.
Діброва татарськокленово-зіроч- н и к о в а ( Quercetum aoeroso(tatarici)-etellarloBum) поширена від середньої частини лісостепової зони і до південної межі лісової. В південній частині свого ареалу ці ліси трапляються в кожному лісовому масиві. Окремі ділянки їх тягнуться вузькими смугами вздовж схилів балок, розташовуючись безпосередньо над дібровою татарськокленово-яглицевою. Найбільші масиви цих лісів характерні для Донецького кряжа, де вони займають по кілька десятків гектарів. У північній частині ареалу ці ліси трапляються значно рідше (Тульчинський, Уманський, Звенигородський та Прилуцький ліс- госпзаги). На цій території вони пов"язані з верхніми та середніми частинами сильноспадистих і крутих схилів, східної та західної, рідше - південної експозицій. В міру просування на південь згадані ліси, зберігаючи подібні до попередніх умови зволоження, займають вже прибалочні схили та схили північної експозиції. При цьо-
157
му на схилах, що знаходяться всередині великих балок, вони ростуть у верхній частині їх, а на інших - в середній та нижній. На схилах річкових долин, де умови вологіші, ніж в балках, вони займають такі самі позиції, як у північній частині ареалу. Грунти під дібровою татарськокленово-зірочниковою - від сухих і свіжих темно-сірих лісових до чорноземних дуже опідзолених суглинистих.
Деревостан у віці 50-100 років має зімкненість крон 0,6-0,9, висоту 14-26 м, діаметр дерев 16-40 см і Ш, рідше П і ІУ клас бонітету. Крім дуба звичайного, до його складу входять: клен польовий (+ - 0,2), ясен звичайний (4 - 0,2), липа серцелиста (+ - 0,1), груша звичайна, граб звичайний (Подільська та Придніпровська височини), черешня, клен звичайний, впяз шершавий, яблуня лісова, берест та інші види з I-2-м класом постійності.
Підлісок виявлений добре і має зімкненість 0,3-0,6. Утворюють його клен татарський (0,2-0,5), бруслина бородавчаста (+ - 0,1), свидина кров”яна (4- -0,1), жостір проносний, гордовина (Подільська та Придніпровська височини), ліщина звичайна, глід несправжньо- кривостовпчиковий, бруслина європейська, черемха звичайна та інші види з І-З-м класом постійності.
Травостій менш однорідний, ніж в попередній асоціації. Він має проективне покриття 30-50%. На конфектор припадає 20-40%. Абсолютно постійними видами травостою є тонконіг дібровний (+ - 15%), фіалки шершава (+ - 1%) та дивна (+ - 1%), грястиця збірна (+ - 1%), гравілат міський (+ - 1%), купина багатоквіткова (4 - 1%), медунка темна (4 - 1%), осока стягнута, маруна щиткова, ранник вузлуватий та шоломниця висока. До відносно постійних видів належать: яглиця звичайна (+ - 5%), осока волосиста (+ - 5%), конвалія звичайна (4 - 5%), горобейник пурпурово-голубий (+ - 5%), перлівка ряба (4 - 5%), астрагал солодколистий, куцоніжка лісова, дзвоники ріпчастовидні, персиколисті та кропиволисті ( Сатрапиla trachelium L.), пахучка звичайна, підмаренник пухнатоногий, буквиця лікарська, латук отріловидний, чина весняна, звіробій шорсткий, регнерія собача, перлівка поникла, ранник вузлуватий, ториліс японський, горошок горохоподібний, хвилівник звичайний та стоколос Бенекена. Якщо проаналізувати цю групу видів, то неважко помітити, що яглиця звичайна, астрагал солодколистий, хвилівник звичайний, конвалія звичайна та ранник вузлуватий пов”язані з максимальними для асоціації умовами зволоження. В найменш вологих едафотопах вони відсутні. Тут з'являються горобейник пурпурово-голубий і перлівка ряба. З
158
північною частиною ареалу пов’язане поширення осоки волосистої і перлівки пониклої, а з південною - латука стріловидного, регнерії собачої, горошку горохоподібного та ін. Серед регіонально постійних видів можна відзначити граб звичайний, ясенець голостовпчико- вий ( Dictamnus gymnostylis Stev.), вербозілля кільчасте, фізо- сперм Дана (Донецький кряж) та деякі інші. Випадково трапляються горлянка женевська, кінський часник черешковий, аконіт дібровний, бутень п’янкий, осока Мікелі, бугила лісова, віхалка гілляста, лопух дібровний, холодок тонколистий, копитняк європейський, маренка запашна, в"язіль барвистий, осока пальчаста, бутень ягідний, коручка широколиста, борщівник сибірський, празелень звичайна, чина чорна, очиток великий, жовтеці золотистий і кашубський, медунка м’якенька, зеленчук жовтий, переліска багаторічна, просянка розлога, купина лікарська, ластовень лікарський, осока гірська, чемериця чорна та ін.
Видова насиченість асоціації - 196 видів.
Отже, при порівнянні з попередньою асоціацією видно, що зі складу флористичного ядра описуваної асоціації зникають найбільш гігрофільні види (бугила лісова, вербозілля лучне, чистець лісовий), зате з’являються більш ксерофільні-горобейник пурпурово-голубий та перлівка ряба. Одночасно послаблюється роль неморальних балтійських видів та посилюється - неморальних субсередземно- морських.
Діброва татарськокленово-воло- систоосокова (Queioetua aceroso(tat&£lci)-oarlcoeua (pilosae) поширена в південній смузі лісостепової зони (уманський, Звенигородський, Смілянський, Кремлівський, Полтавський, Вовчанський, Жовтневий та Гутянський лісгоспзаги) в зв’язку з тим, що екологічна амплітуда осоки волосистої відносно термічності клімату менша, ніж у клена татарського. Ці ліси займають верхні та середні частини пологих і спадистих схилів південних, рідше - західних або східних напрямків з сухими або свіжими темно- сірими лісовими або чорноземними оильноопідзоленими суглинистими грунтами. Трапляються ці ліси зрідка і за площею значно поступаються перед попередніми, по відношенню до яких займають більш сухі й теплі схили.
Деревостан у віці 50-140 років має зімкненість крон 0,7-0,9, висоту 16-25 м, діаметр дерев 20-50 см t П-Ш клас бонітету. Крім дуба звичайного, його створюють клен польовий (+ - 0,2), липа сер-
159
целиста, ясен звичайний та інші види, спільні з попередньою асоціацією.
Підлісок виявлений добре і має зімкненість 0,3-0,6. Він складається з клена татарського (0,2-0,4), ліщини звичайної (+ - 0,2), бруслини бородавчастої (+ - 0,1), гордовини (+ - 0,1), бирючини звичайної (+ - 0,2), свидини кров"яної, глоду українського та інших видів.
Травостій з проективним покриттям 30-60% створює осока волосиста (25-50%), а з числа абсолютно постійних видів - зірочник лісовий (1-10%), медунка темна (+ - 5%), копитняк європейський (+ - 1%), гравілат міський (+ - 1%), розхідник шорсткий (+ - 1%), дзвоники ріпчастовидні, чина весняна, перлівка поникла, тонконіг дібровний, купина багатоквіткова, маруна щиткова, фіалки шершава та лісова. Відносно постійні та випадкові види травостою представлені підмаренником пухнатоногим, астрагалом солодколистим, дзвониками кропиволистими, зеленчуком жовтим, конвалією звичайною, осокою кореневищною, купиною широколистою, серпієм різнолистим, очитком великим, ранником шишкуватим та іншими спільними с попередньою асоціацією видами.
Видова насиченість асоціації становить 86 видів.
Діброва татарськокленово-коре- невищноосокова ( Quercetum aceroao(tatarici)-caricoaum (rhialnae) поширена лише в східній частині степової і південній смузі лісостепової зон (Вовчанський, Жовтневий, Гутянський, Куп"ян- ський та Старобільський ліогоспзаги). Пояснюється це тим, що основний ареал конфектора ярусу травостою лежить на території Західного Сибіру і Східної Європи. В південній смузі лісостепової зони ці ліси займають верхні частини сильноспадистих і крутих схилів південної експозиції. В міру просування на південь вони зміщуються спочатку на середні частини схилів південної експозиції, а біля своєї межі - на верхні частини північно-західних та північно-східних схилів. Ростуть ці ліси на сухих чорноземних сильноопідзоле- них суглинистих або дерново-карбонатних грунтах. Окремі масиви їх, як правило, менші І га. Більші зустрічаються лише на території Вов- чанського лісгоспзагу (рис.26).
Деревостан у віці 50-90 років має зімкненість крон 0,7-0,9, висоту 12-20 м, діаметр дерев 16-32 см 1 Ш-ІУ клас бонітету. Створює його дуб звичайний 8 участю ясена звичайного (+ - 0,1), груші звичайної, липи серцелистої та клена польового. Всі вони ма-
160
ють лише 2-4-й клас постійності, чим ця асоціація відрізняється від описаних раніше.
Підлісок виявлений добре і має зімкненість 0,3-0,5. Крім кон- фектора (0,3-0,5), його створюють бруслина бородавчаста (+ - 0,1), свидина кровмяна, жостір проносний, ліщина звичайна, глід криво- чашечковий ( Crataegos curvisepala L і n d mj, бирючина звичайна та інші види з І-3-м класом постійності. З перелічених видів ліщина звичайна трапляється в найвологіших, а бирючина звичайна - в най- сухіших екотопах, які посідає діброва татарськокленово-кореневищно- осокова.
Травостій має проективне покриття 30-55%, з них на осоку кореневищну припадає 20-50%. Абсолютно постійними видами його є зірочник лісовий (+ - 10%), копитняк європейський (+ - 1%), осока Мі- келі (+ - 1%), розхідник шорсткий (+ - 1%), перлівка ряба (+ - 1%), тонконіг дібровний (•*• - 1%), медунка темна (+ - 1%), астрагал солодколистий, грястиця збірна, підмаренник пухнатоногий, гравілат міський, чина весняна, маруна щиткова, горошок горохоподібний, в’язіль барвистий і фіалка шершава; відносно постійними - осока волосиста (+ - 5%), буквиця лікарська, дзвоники персиколисті та ріп— частовидні, пахучка звичайна, звіробій шорсткий, купина багатоквіткова, регнерія собача, ториліс японський, вероніка дібровна, шоломниця висока, ранник вузлуватий, очиток звичайний, фіалка дивна. З регіонально постійних видів можна відзначити ясенець голостовпчи- ковий та ластовень виткий (Донецький кряж). Випадкові види представлені фіалкою запашною (Viola odorata L.)f яглицею звичайною, хвилівником звичайним, куцоніжкою лісовою, конвалією звичайною, дзвониками кропиволисуими, борщівником сибірським, празеленню звичайною, переліскою багаторічною, стоколосом Бенекена, перлівкою пониклою, горошком горохоподібним, серпієм різнолистим та ін.
Видова насиченість асоціації - 94 види.
Роль світлолюбних неморальних субсередземноморських видів у даній асоціації найбільш виявлена. Хоча в число постійних її видів все ще входять типові неморальні балтійські види (копитняк європейський, чина весняна, зірочник лісовий та ін.), кількісно переважають перші. Більше того, серед абсолютно постійних видів вже є перлівка ряба, осока Мікелі та в’язіль барвистий, які не входили в групу постійних видів попередніх асоціацій. Загалом асоціація добре виявлена екологічно і флористично. Як найбільш гігрофільні (вербозілля лучне), так і найбільш ксерофільні (залізняк бульбис-
161
гий, горобейник пурпурово-голубий) види цієї групи асоціації є для неї випадковими.
ХШ. Група асоціацій дубові ліси татарськокленово-гірськоосоко- ві представлена двома асоціаціями. З них діброва татарськокленово- гірськоосокова займав більш сухі місцеположення, ніж діброва та- тарськокленово-конвалієва. Проте ця різниця незначна, тим більше, що синекологічна амплітуда конвалії звичайної більша за синеколо- гічну амплітуду осоки гірської.
Діброва татарськокленово-конва- л і є в a (Quercetum aoeroao(tatarici)-convallaxioaum) поширена в межах всього ареалу класу асоціацій. Найбільш північні її місцезнаходження трапляються в Сумському, Прилуцькому та Чортків- ському лісгоспзагах, а найбільш південні майже збігаються з межею поширення лісів. В північній частині свого ареалу ці ліси безпосередньо пов"язані з дубовими лісами ліщиновими, серед яких в екологічному відношенні вони найбільш близькі до діброви ліщиново-воло- систоосокової, останню вони заміщують у верхніх частинах схилів південної експозиції з багатшими темно-сірими лісовими грунтами. В південній частині лісостепу та в степовій зоні ці ліси пов"язані з верхньою частиною спадистих і крутих схилів південних напрямків з сухими темно-сірими лісовими або чорноземними дуже опідзоленими суглинистими грунтами. Цікаво відзначити, що в степовій зоні вони можуть займати верхні та середні частини крутих схилів південної експозиції, але тоді грунти під ними дерново-слабопідзолисті глинисто-піщані, тобто більш холодні. Крім межиріч, діброва татарсь- кокленово-конвалієва трапляється в лівобережних заплавах річок південної частини лісостепової зони на найбільш високих гривах прируслової частини з дерновими опідзоленими глинисто-піщаними грунтами та на гривах других терас з дерново-підзолистими супіщаними грунтами. В заплавних умовах вони завершують еколого-фітоценотич- ний ряд дубових лісів. Усі наступні асоціації їх в заплавах уже не трапляються. Великих масивів дубові татарськокленово-конваліеві ліси не створюють ніде.Звичайно площа їх ділянок не перевищує 1-2 га (рис. 27).
Деревоотан у віці 50-110 років має зімкненість крон 0,7-1,0, висоту 15-24 м, діаметр дерев 20-38 см та Ш, рідше ІУ клас бонітету. Більшість видів його, за винятком дуба звичайного та липи серцелистої (+ - 0,1), мають 2-3-й клас постійності. Пояснюється такий низький клас постійності досить великою екологічною амплі-
162
тудою асоціації, завдяки чому кожен з супутніх видів трапляється лите в певному діапазоні, наприклад, черешня - лише в західній частині ареалу асоціації, ооика - на других терасах річок, клен польовий - в південній частині ареалу.
Підлісок виявлений добре й мав зімкненість 0,2-0,7. Створюють його клен татарський (0,2-0,6) з участю ліщини звичайної (+ - 0,1), глоду несправжньо-кривостовпчикового (+ - 0,1), бруслини бородавчастої (+ - 0,1), свидини кров'яної (+ - 0,1), бруслини європейської, гордовини, терену звичайного та інших видів з I-2-м класом постійності.
Травостій мав проективне покриття 30-55%. До абсолютно постійних видів його, крім конфектора (20-50%), належать: зірочник лісовий (+ - 10%), грястиця збірна (+ - 1%), гравілат міський (+ - 1%), розхідник шорсткий (+ - 1%), купина багатоквіткова (+ - 1%), фіалка дивна (+ - 1%), маруна щиткова, ториліс японський, фіалка шершава. Відносно постійні види представлені купиною широколистою (+ - 5%), тонконогом дібровним (+ - 1%), осокою Мікелі (+ - 1%), осокою стягнутою, холодком тонколистим, астрагалом солодколистим, буквицею лікарською, куцоніжкою пірчастою, дзвониками персиколисти- ми, дзвониками ріпчастовидними, ломиносом прямим, пахучкою звичайною, борщівником сибірським, чиною чорною, ранником вузлуватим, шоломницею високою, чемерицею чорною та ластовнем лікарським, а випадкові - яглицею звичайною, бугилою лісовою, віхалкою гіллястою, осокою гірською, дзвониками кропиволистими, фіалкою запашною, копитняком європейським, просянкою розлогою, переліскою багаторічною, перлівкою рябою, чиною весняною, півниками злаколисти- ми, латуком стріловидним, звіробоєм шорстким, фіалкою високою, маренкою запашною, осокою волосистою, коручкою широколистою, оманом верболистим, золотушником звичайним, дзвониками широколистими, купиною лікарською, медункою темною, регнерією собачою, дутнем ягідним, бутнем п'янким та ін. З числа регіонально постійних видів зустрічаються холодок тонколистий, купина широколиста, гордовина (Придніпровська височина) та ластовень виткий (Донецький кряж).
Видова насиченість асоціації - 183 види.
Порівняно з асоціаціями попередньої групи в цій асоціації в числі постійних з'являється ряд світлолюбних мегатрофних мезоморфних видів високого різнотрав'я (борщівник сибірський, чемериця чорна), які, можливо, входили в плейотоценовий комплекс.
163
Діброва татарськокленово-гірсь- КОО сокова ( Quercetum aceroso(tatarici) - сагіcosum(montanae} поширена в південній і середній смузі лісостепової зони в західній частині ареалу дубових лісів татарськокленових (Вінницька та північна частина Одеської обл.), де на багатших та більш сухих грунтах вона заміщує дубові ліси свидиново-гірськоосокові. Дубові ліси татарськокленово-гірськоосокові займають верхні частини пологих t спадистих схилів південної експозиції в сухими та свіжими темно-сірими лісовими грунтами в північній частині ареалу і середні частини менш крутих схилів тієї х або східної експозиції в південній частині ареалу з чорноземними опідзоленими грунтами. Крім того, вони трапляються і на других терасах правобережних річок (Хмільницький та Богуславський лісгоспзаги), де ростуть на вершинах грив з найбільш сухими і багатими в цих умовах дерново-слабопідзолистими супіщаними грунтами. Отже, ці ліси завершують еколого-фітоценотичний ряд дубових лісів на других .терасах річок. Всі наступні асоціації на них вже не трапляються (рис. 28).
Деревостан у віці 50-100 років має зімкненість крон 0,7-0,9, висоту 18-23 м, діаметр дерев 24-36 см і П-Ш клас бонітету. Створює його дуб звичайний з участю липи серцелистої (+ - 0,1), черешні, граба звичайного, клена польового, груші звичайної та яблуні лісової (постійні види). Крім того, трапляються береза бородавчаста, клен звичайний, в"яз шорсткий та ясен звичайний, що мають І-З-й клас постійності.
Підлісок виявлений добре і має зімкненість 0,3-0,6. Крім клена татарського (0,2-0,5), до нього належать: свидина кров"яна (+ - 0,1), ліщина звичайна, бруслина бородавчаста, глід Липського, гордовина та інші види 8 І-4-м класом постійності.
Травостій з проективним покриттям 25-45% створює осока гірська (20-35%) з участю медунки м”якенької (+ - 5%), пахучки звичайної (+ - 1%), суниць лісових (+ - 1%), гравілату міського (+ - 1%), чини чорної (+ - 1%), перстачу білого (+ - 1%), буквиці лікарської, куцоніжки пірчастої, ломиноса прямого, маруни щиткової, торилісу японського, вероніки дібровної, фіалки шершавої (абсолютно постійні види), конвалії звичайної (+ - 5%), осоки Мікелі (+ - 1%), ластовня лікарського (+ - 1%), горлянки женевської (ajuga genevensis L.), парила звичайного (Agrimonia eupatoria L.), само- сила гайового, холодна тонколистого, астрагалу солодколистого, смовді оленячої, дзвоників персиколистих, молочаю ребристого (eu - 164
phorbia angulata Jaoq.),розхідника шорсткого, празелені звичайної, чемериці чорної, купини широколистої, очитку великого, підмаренника весняного, фіалки давно! та інших відносно постійних та випадкових видів.
Видова насиченість асоціації - 155 видів.
ХІУ. Група асоціацій дубові ліси татарськокленово-рябопер- лівкові представлена лише однією асоціацією, яка займає проміжне положення між попередніми, загалом більш мезофільними асоціаціями класу, і наступними - більш ксерофільними. Цю групу можна розглядати як таку, що завершує ряд сухих дубових лісів. Наступна група асоціацій починає ряд дуже сухих дібров,
Діброва татароькокленово-рябо- п. ерлівкова (Quecoebuu aceroso (tat аг io i) - melioosum (pictae) поширена в степовій зоні (в усіх ліогоспаагах) і лише в східній та західній частинах України заходить у лісостепову зону (Одеська та Харківська обл.). Північна межа її поширення проходить по лінії Котовоьк - Олександрія - Краоноград - Харків. В більш північних районах ці ліси займають верхні частини спадистих і крутих схилів балок південної експозиції, а в південних - середні частини крутих схилів південної експозиції або верхні частини крутих і сильноспадиотих схилів східної та західної експозицій. Грунти під ними дуже сухі чорноземні опідзолені суглинисті або дерново- карбонатні. Ліси трапляються часто, але невеликими довгими вузькими смугами розміром до 1-3 га. В вологіших умовах вони звичайно заміщуються дубовими лісами татарськокленово-зірочниковими, а в сухіших - дубовими лісами татарськокленово-горобейниковими (рис.29).
Деревостан у віці 50-90 років має зімкненість крон 0,6-0,9, висоту 10-18 м, діаметр дерев 18-32 см і ІУ-У клас бонітету. Постійними видами його, крім дуба звичайного, є ясен звичайний (+ - 0,2), клен польовий (+ - 0,1), липа серцелиста (* - 0,1) та груша звичайна.
Підлісок виявлений добре і має зімкненість 0,3-0,6. Створюють його клен татарський (0,2-0,5), карагана кущова (+ - 0,2), бруслина бородавчаста (+ - 0,1), жостір проносний, свидина кров"яна, бирючина звичайна, терен звичайний, глід неоправжньо-кривостовп- чиковий та інші види з І-З-м класом постійності.
Травостій має проективне покриття 25-50% і складається з перлівки рябої (20-40%), зірочника лісового (+ - 10%), осоки Мікелі (+ - 5%), тонконога дібровного (+ - 5%), в"язелю барвистого (+ - 165
1%), грястиці збірної (+ - 1%), молочаю несправжньо-волохатого, гравілату міського, залізняка бульбистого, маруни щиткової, горошку горохоподібного та фіалки шершавої (абсолютно постійні види). Відносно постійні види представлені горобейником пурпурово-голубим (+ - 10%), грястицею збірною (+ - 1%), розхідником шорстким (+ - 1%), купиною багатоквітковою (+ - 1%), астрагалом солодколистим, дзвониками персиколистими та кропиволистими, осокою стягнутою пахучкою звичайною, підмаренником пухнатоногим, звіробоєм шорстким чиною весняною, медункою темною, латуком стріловидним, регнерією собачою, шоломницею високою, торилісом японським та барвінком тра- в"янистим (Vinea herbacea w. К.), Із регіонально постійних видів травостою слід відзначити солонечник естрагоновидний, який зустрічається лише в східній частині ареалу асоціації на ділянках з солонцюватими грунтами., Групу випадкових видів складають фіалка висока (Viola elatior F т.), СТОКОЛОС 6Є30СТИЙ (Zerna inermis (L е у s s.)L i n d m.),дзвоники болонські (Campanula bononiensis Ь.), м"яточник чорний (Ballota nigra L,), ломиніс цілолистий ( Clematis integrifolia Ь.), вероніка дібровна, гадючник шестипелюстковий, суниці лісові, осока кореневищна, коііитняк європейський, хвилівник звичайний, буквиця лікарська, борщівник сибірський, ранник вузлуватий, фізосперм Дана, стоколос Бенекена, горлянка женевська, фіалка дивна, рутвиця мала та ін.
Видова насиченість асоціації - 137 видів.
Екологічно 1 флористично розглянута асоціація виявлена добре. Характерною рисою її є те, що роль конфектора ярусу травостою (вперше в межах класу) відіграв типовий субсередземноморський світлолюбний вид з вузьким ареалом, північна межа суцільного поширення якого не виходить за границю південної смуги лісостепової зони. Разом з ним серед постійних видів вперше з'являється низка інших оубсередвемяомороьких світлолюбних ксеро- та мезоксерофільних видів - карагана кущова, залізняк бульбистий 1 барвінок трав"яни- стий. В цілому, якщо не брати до уваги грястиці збірної і тонконога дібровного, присутність яких в числі постійних видів зумовлена, на нашу думку, впливом людини, флористичне ядро асоціації на 75% створюють неморальні субсередземноморські види,і лише 25% припадає на неморальні балтійські види 8 широкою синекологічною амплітудою (астрагал солодколистий, медунка темна, зірочник лісовий, липа серцелиста та ін.).
ХУ. Група асоціацій дубові ліси татарськокленово-горобейни- 166
кові об’єднує дві найбільш коеро- і термофільні асоціації класу, дуже близькі між собою. З них діброва татарськокленово-мікелієво- осокова займає дещо сухіші місцеположення, ніж діброва татарсько- кленово-горобейникова. Конфектори ярусу травостою обох асоціацій є ксерофітами. За вимогами до термічності клімату вони дещо перевищують клен татарський, поотупаючись перед останнім щодо кріо- тичності та континентальності клімату. Внаслідок цього названі асоціації трапляються лише в західній та центральній частинах південної смуги ареалу дубових лісів татарбькокленових.
Діброва татарськокленово - горобейник ова ( Quercetum aceroso(tatarici)-lithospermosum (purpureo-coerulei) поширена в степовій та південній частині лісостепової зони (в усіх лісгоспзагах на Донецькому кряжі, Придніпровській та Середньоруській височинах). Північна межа її проходить по лінії Кам”янець-Подільський - Брацлав - Шпола - Богодухів. Най- північніша острівна ділянка цієї асоціації описана нами біля села Михайлівна Лебединського району Сумської області. Біля своєї північної межі ця асоціація займає виключно верхні найопукліші частини схилів південної експозиції з крутизною 15-30°, що мають дуже сухі чорноземні опідзолені грунти, або рівнозначні за умовами місцеположення, наприклад, нижні частини схилів на ділянках з близьким заляганням гальки, що веде до значного підсушення грунтів. Біля південної границі лісостепової зони ліси асоціації, крім схилів, займають на Правобережжі ще й широкі рівні межиріччя з чорноземними сильпоопідзоленими грунтами. На таких ділянках вони ростуть великими масивами і за зайнятою площею переважають уоі інші асоціації. Наприклад, у Полтавському та Голованів- ському лісгоспзагах на них припадає майже 70% площі найбільших лісових урочищ. Саме дубові ліси татарськокленово-горобейникові є останніми вододільними (плакорними) лісами на Правобережжі України. В південній частині свого ареалу вони в межах ареалу дерну справжнього (Придніпровська височина) займають верхні частини тільки слабкоспадистих схилів південної експозиції. На крутіших схилах ці ліси зміщуються на східну та західну експозиції і займають верхні та середні частини, а на дуже крутих - і нижні. Пояснюється це тим, що з південних експозицій їх витісняють дубові ліси кизилові. На Лівобережжі після перерви на Дніпровській низовині зазначені ліси в південній частині їх ареалу, як і в північній, займають верхні частини схилів південної експозиції, знижую167
чись по дуже крутих схилах до їх нижніх частин. Трапляються вони в цьому районі в кожному лісовому масиві й утворюють на південних схилах верхню вузьку смугу байрачних дібров. Крім того, вони займають і вузькі сильнодреновані середбайрачні гребені південних експозицій з чорноземними опідзоленими грунтами (рис. ЗО).
Деревостан у віці 50-100 років мав зімкненість крон 0,6-0,9, висоту 10-22 м, діаметр дерев 16-38 см і ІУ, рідше Ш або У клас бонітету. Характерною особливістю його є відсутність постійних супутників едифікатора, в зв"язку з тим, що асоціація формується в екстремальних умовах, несприятливих для неморальних мезофільних видів Руської рівнини. Регіонально постійними видами є черешня (Придніпровська височина), ясен звичайний (північна частина ареалу асоціації) та липа серцелиста (Харківська обл.). Випадково трапляються граб звичайний, берест, в"яз корковий (Ulmua suberoaa Moench.), груша звичайна, клен польовий та інші види з I-2-м класом постійності .
Підлісок має зімкненість 0,3-0,7 і складається з клена татарського (0,2-0,6), бруслини бородавчастої (+ - 0,1), глоду обманливого (+ - 0,1), терену звичайного (+ - 0,1), глоду несправжньо- кривостовпчикового, свидини кров"яної (+ - 0,1), жостіру проносного, бирючини звичайної, гордовини, шипшини повстистої та інших видів з І-З-м класом постійності. Регіонально постійними видами підліску в дерен справжній (Придніпровська височина) та ліщина звичайна (північні райони ареалу асоціації).
Травостій має проективне покриття 25-60%, з них на горобей- ник пурпурово-голубий припадав 20-55%. Група абсолютно постійних видів представлена гравілатом міським (+ - 1%), буквицею лікарською, медункою м"якенькою, маруною щитковою, торилісом японським та фіалкою шершавою. Більш численна група відносно постійних видів, до якої входять осока Мікелї (+ - 10%), конвалія звичайна (+ - 10%) бугила лісова (+ - 5%), куцоніжка пірчаста (+ - 5%), перлівка ряба (+ - 5%), купина широколиста (+ - 5%), грястиця збірна (+ - 0,1%), залізняк бульбистий (+ - 1%), собача кропива п’ятилопатева (їлопи- rus gulnquelobatua G і і і ь.), холодок тонколистий, астрагал солодколистий, дзвоники персиколисті, осока стягнута, пахучка звичайна, півники злаколисті, латук стріловидний, чина чорна, чемериця чорна, купина лікарська, фіалка дивна, стоколос Бенекена та оман верболистий. Така досить значна кількісна різниця між двома групами видів пояснюється більшою порушеністю і ксерофітизацією 168
цих лісів яа південних схилах, внаслідок чого зі складу їх зникав ряд видів (перш за все холодок тонколистий, півники злаколисті, латук стрїловидний, чемериця чорна, дутень ягідний та лопух дібровний), абсолютно постійних для менш порушених ділянок. Мабуть, постійним видом цих лісів був і ломиніс цілолистий, який зараз в них майже не трапляється. З регіонально постійних видів, крім тих, що зазначали вже для попередніх асоціацій, необхідно вказати перлівку одноквіткову, яка проникла на Україну з дубових та букових лісів Середньої Європи, молочай несправжньо-волохатий (ендем західної частини Придніпровської височини) та ясенець голостовпчи- ковий (Донецький кряж). З випадкових видів трапляються вероніка волотиста (Vexonica paniculata L.), котяча м"ята паннонська (Nepeta pannonica L.), перлівка висока, латук Ше (Laotuoa ohai- xli у 1 1 1. ), віхалка гілляста, дзвоники скупчені, осока кореневищна, герань криваво-червона, золотушник звичайний, просянка розлога, переліски яйцевидна та багаторічна, фіалка висока, горошок горохоподібний та ін.
Видова насиченість асоціації - 192 види.
Діброва татарськокленово-міке- ліввоосокова (Quercetum acero8o(tatarici)-oariooBum (michelii) поширена в степовій зоні (в більшості лісгоспзагів) і в південній частині лісостепової зони (в Тульському, Звенигородському, Полтавському та Гутянському лісгоспзагах). Північна межа її майже збігається з межею попередньої асоціації. В лісостеповій зоні ці ліси займають верхні частини найбільш крутих схилів південної експозиції з дуже сухими чорноземними слабопідзолистими суглинистими грунтами. В степовій зоні вони пов"язані з верхньою частиною спадистих і сильноспадистих схилів південної експозиції та верхніми і середніми досить опуклими і добре дренованими частинами крутіших схилів тієї ж експозиції з чорноземними вилугованими суглинистими грунтами. Таким чином, у цієї асоціації при просуванні на південь не спостерігається змін щодо експовиції й крутизни схилів. Трапляється вона зрідка і невеликими (розміром до І га) ділянками у вигляді дуже вузьких (10-30 м) смуг, розташованих вище попередньої асоціації (рис .31).
Деревостан у віці 60-90 років має зімкненість крон 0,7-0,9, висоту 10-20 м, діаметр дерев 18-30 см і ІУ-У клас бонітету. Характерною рисою його є те, що всі супутники едифікатора (клен польовий, липа серцелиста, в"яз корковий, ясен звичайний та груша зви169
чайна) мають клас постійності нижче 5-го, що пояснюється формуванням асоціації в екстремальних умовах.
Підлісок має зімкненість 0,3-0,6. Створює його клен татарський (0,2-0,5) з участю бруслини бородавчастої (+ - 0,1), жості- ра проносного, гордовини, терену звичайного, глоду Дилського,карагани кущової, вишні кущової ( Cerasus fruticosa W о r о пф), свидини кровияної та інших видів з І-З-м класом постійності.
Травостій з проективним покриттям 30-50% створює осока Мікелі (20-35%) та ряд абсолютно постійних і випадкових видів. Перші представлені куцоніжкою пірчастою (+ - 5%), грястицею збірною, гравілатом міським (+ - 5%), буквицею лікарською, залізняком бульбистим, маруною щитковою та фіалкою шершавою. Другі - перлівкою рябою (+ - 10%), зірочником лісовим (+ - 5%), холодком тонколистим, дзвониками персиколистими та ріпчастовидними, осокою стягнутою, па- хучкою звичайною, в"язелем барвистим, молочаєм несправжньо-волохатим, гадючником шестипелюстковим, суницями лісовими, розхідником шорстким, латуком стріловидним, купиною широколистою, репне- рією собачою, очитком звичайним, шоломницею високою, торилїоом японським, горошком горохоподібним, бутнем бульбистим, барвінком тр.ав"янистим, ластовнями лікарським та витким, підмаренником пух- натоногим, стоколосом Бенекена, рутвицею малою та дивиною волотистою (Verbascum lyohnitis L.). Регіонально постійними є гордовина, купина широколиста та ластовень виткий. Більшість випадкових видів спільна з попередньою асоціацією.
Видова насиченість асоціації - 145 видів.
Таким чином, клас асоціацій дубових лісів татарськокленових представлений низкою асоціацій від типово мезофільних до тилово ксерофільних. Екологічно і флористично серед них найкраще виявлені асоціації, конфектори ярусу травостою яких перебувають в даному класі в екстремальних умовах. Значно гірше виявлені асоціації, наприклад, діброва татарськокленово-зірочникова. У них синеколо- гічна амплітуда конфекторів ярусу травостою не обмежується, хоча б за одним з провідних екологічних факторів, синекологічною амплітудою конфектора ярусу підліска.
Незважаючи на досить значну різницю між крайніми асоціаціями, всі асоціації класу належать до ланки базифільних дубових лісів, повпязапих безпосередньо, як вже відзначалось, з аналогічними лісами Угорщини і Румунії. На схід від України дубові ліси татар- ськокленові стають флористично все біднішими та одноманітнішими.
170
З їх складу випадають всі середньоєвропейські види, а в міру просування на схід, спочатку в басейні Дону, а потім Волги, зникають і деякі європейські види (клен польовий, бруслина європейська, ясен звичайний, печіночниця звичайна та ін.). Натомість з'являється невелика кількість сибірських видів, з яких в лісах даного класу на території східної частини України трапляються скереда сибірська та серпій увінчаний - види, характерні для західносибірських остепнених березових лісів.
Дубові ліси кизилові
Кизил або дерен справжній характеризується вузькою синеколо- гічною амплітудою, яка відповідає теплому клімату південної частини Середньої Європи. Оптимум синекологічної конфекторної амплітуди його виходить за межі синекологічної едифікаторної амплітуди дуба звичайного і відповідає синекологічній едифікаторній амплітуді дуба пухнатого. Тому дерен справжній створює підлісок в лісах з дуба пухнатого і в ліси з дуба звичайного заходить тільки в південній середньоєвропейській частині їх ареалу - на території східної частини Франції, південної частини ФРН, в Чехословаччині, Угорщині та Румунії. В цих районах дубові ліси кизилові займають південні схили передгірського і низькогірського поясів на висоті 250-700 м (Gauckler, 1938; гбіуоші» Jakuca, I957;Jucko, 1958; Jakuca, 1960, I96I;Fekete, 1965). Найбільш північні острівні місцезнаходження цих лісів відомі на території Бельгії, де дерен справжній разом з бирючиною звичайною і ліщиною звичайною утворює підлісок в дубових лісах на вапняках крутих схилів річкових долин (Vanden Barghen, 1955). На території Радянського Союзу дубові ліси кизилові трапляються в Молдавії, на Поділлі і на Придніпровській височині. Острівні місцезнаходження їх відзначено в Криму і на Кавказі на висоті 350-500 м (Цнрина, 1925; Грудзинская, 1953; Елагин, 1953; Гейдеман, Остапенко, 1964; Львов, 1964). Ростуть дубові ліси кизилові на нейтральних і слабколухних дерново-карбонатних або чорноземних вилугованих і опідзолених грунтах з рН 6,4. - 7,4. У їх флорі переважають неморальні субоередземноморські види з участю неморальних балтійських видів.
На Україні ці ліси трапляються тільки у вузькій придністровській смузі від західної межі Івано-Франківської області до північної частини Одеської (Бучацький, Чортківський, Кам"янець-По- дільський, Могилів-Подільоький, Бершадьський та Балтський лісгосп- ваги) та у східній частині Придніпровської височини в трикутнику
І7Т
між містами Чигирин - Сміла - Знам"янка. В Придністров"ї вони займають дуже круті і навіть урвисті схили (35-60°) долини Дністра та його притоків і балок, де на південних схилах утворюють смуги шириною 60-300 м (рис.32). На плакорах та схилах інших експозицій ці ліси заміщуються дубовими лісами ліщиновими або татарськокленови- ми. На Придніпровській височині вони займають найбільш круті (15- 25°) південні схили горбів межиріч, де пов"язані безпосередньо з дубовими лісами татарськокленовими.
До початку господарської діяльності людини дубові ліси кизилові в Придністров"ї були досить поширеними. Після суцільних рубок головного користування вони не відновлюються і тепер знищені майже повністю. У порівнянні з лісами попереднього класу асоціацій для дубових лісів кизилових характерне зростання ролі неморальних суб- середземноморських видів. Серед останніх в група видів, північно- східна суцільна межа ареалу яких проходить на Україні саме в цих лісах. Далі на схід, на території Радянського Союзу ці види трапляються лише в Криму і на Кавказі. До них належать дерен справжній, берека ( Sorbus torminalia Crantz.), чемерник червонуватий (Heileborus purpurascens W.K.) та клокичка периста ( Staphylea pinnata L.). Всі вони в фітоценотично вірними ксеротермофільним лісам оубатлантичного клімату.
Щодо клімату і родючості грунту асоціації цього класу характеризуються практично однаковими вимогами, а за умовами зволоження грунту створюють ряд від сухих до дуже сухих дібров. У найбільш гіг- ротопних асоціацій класу до числа постійних видів входять ксерофіти, яких в числі постійних видів не було у найбільш гігротопних асоціацій попереднього класу. В дубових лісах кизилових найбільш гігрофільні види представлені типовими мезофітами (копитняк європейський, ліщина звичайна, яглиця звичайна, зірочник лісовий та ін.), а в дубових лісах татарськокленових - гігромезофітами (костриця велетенська, чистець лісовий та ін.). Неморальні субсередзем- номорські види в асоціації з найменш виявленою їх роллю становлять майже 55%, тобто на 10% більше, ніж у відповідній асоціації попереднього класу.
Дубові ліси кизилові представлені двома групами асоціацій. З них дубові ліси кизилово-парвоькоосокові займають більш вологі грунти, ніж дубові ліси кизилово-горобейникові. Всі їх асоціації в ствноекотопними.
ХУІ. Група асоціацій дубові ліси кизилово-парвськоосокові 172
об'єднує п"ять екологічно дуже близьких асоціацій з найменш виявленою роллю неморальних субсередземноморських видів. Центральне місце серед них займав діброва кизилово-парвськоосокова. Від неї в бік незначного збільшення сухості грунту розташовується діброва кизилово-зірочникова, а в бік більш вилугованих грунтів - діброва кизилово-волосистоосокова. Вологіші, ніж під лісами центральної асоціації, грунти займав діброва кизилово-конвалієва, а багатші - кизилово-кореневищноосокова.
Специфічною рисою лісів зазначених асоціацій є наявність в їхньому складі видів, які виявляють ценотичні зв"язки з буковими лісами Середньої Європи на багатих грунтах. Це такі види, як явір, шавлія клейка, зубниця залозиста ( Dentaria glandulosa w. к.) та молочай мигдалевидний ( Euphorbia amygdaloidea L.), В.Шафер (£za- fer, 1930), враховуючи значне поширення цих видів у букових та ялицевих лісах Карпат, відніс їх до гірського елементу флори, що вказує на більш тісні флористичні зв"язки з буковими лісами дубових лісів кизилових, ніж дубових лісів татароькокленових. Мабуть, зміна континентальності клімату послаблює фітоценотичні зв"язки в більшій мірі, ніж зміни температури.
Діброва к и з и л о-в о - к о н в а л і є в a (quercetum cornoso-oonvallariosum) поширена у вузькій придністров¬
ській смузі на всьому протязі ареалу класу. В північних районах вона займав нижні частини крутих схилів південної експозиції або верхні частини західних схилів з сухими дерново-карбонатними грунтами. В південних районах ця асоціація займає плоскі неширокі верхівки горбів та верхні частини їх схилів північної та західної експозиції. Розміри окремих ділянок не перевищують І га.
Деревоотан у віці 50-90 років має зімкненість крон 0,8-1,0, висоту 14-20 м, діаметр дерев 18-24 см і Ш клас бонітету. Створює його дуб звичайний з участю ясена звичайного (+ - 0,2), липи серцелистої (+ - 0,2), граба звичайного, клена польового, береки, черешні, клена звичайного, дуба скельного та явора. Останні чотири види мають 2-3-й клас постійності.
Підлісок виявлений добре, має зімкненість 0,3-0,5 і висоту 3-4 м. Створюють його дерен справжній (0,2-0,4),клен татарський (+ - 0,1), бруслина оородавчаота, свидина кров"яна, гордовина, глід український,жимолость пухнаста та інші види з I-4-м класом постійності.
Травостій має проективне покриття 25-40%. До його складу вхо- 173
дать конвалія звичайна (20-30%), осока волосиста (+ - 10%), зірочник лісовий (+ - 5%), ломиаіс прямий (+ - 1%), копитняк європейський (+ - 1%), грястиця збірна (+ - 1%), горобейник пурпурово- голубий (+ - 1%), холодок тонколистий, дзвоники ріпчастовидні, чина чорна, медунка темна, купина широколиста, купина багатоквіткова, шоломниця висока, фіалки шершава та дивна, горлянка повзуча, астрагал солодколистий, осока гірська, зеленчук жовтий, перлівка поникла, маруна щиткова, а з числа випадкових видів: чемериця чорна (Veratrum nigrum L.), ласкавець серповидний ( Bupleurum falcatum ь.), дзвоники кропиволисті, маренка запашна, кадило мелі- солисте, смовдь оленяча, тонконіг дібровний, медунка м”якенька, дзвоники персиколисті, купина лікарська, пахучка звичайна, кадило мелісолисте, підмаренник весняний, перлівка ряба, впязіль барвистий, наперстянка великоцвіта, осока парвська, осока стягнута, осока Мікелі, маренка запашна та інші види з І-З-м класом постійності .
Видова насиченість асоціації - 96 видів.
Діброва кизилово-парвськоосоко- В а ( Quercetum cornoso-oaricoBum(brevicollis) поширена тільки В центральній частині Придністров"я (Кам"янець-Подільоький та Моги- лів-Подільоький лісгоспааги). Займав вона середні частини дуже крутих охилів південної експозиції або верхні частини схилів західної експозиції в дерново-карбонатними та чорноземними вилугованими грунтами.
Деревоотан у віці 50-80 років має зімкненість крон 0,8-0,9, висоту 18-22 м, діаметр дерев 18-32 ом і Ш клас бонітету.За видовим окладом він подібний до деревостану попередньої асоціації,відрізняється тільки меншою участи липи серцелистої та ясена звичайного і дещо більшою - клена звичайного.
Підлісок (0,3-0,6) утворюють дерен справжній (0,2-0,4), ліщина звичайна (+ - 0,1), бруслина бородавчаста (+ - 0,1), свидина кро- в"яна (+ - 0,1), гордовина, глід Липського, жостір проносний та інші види з І-З-м класом постійності.
Травостій (35-45%) складається з осоки парвської (20-30%), конвалії звичайної (+ - 15%), осоки волосистої (+ - 10%), копитняка європейського (+ - 5%), зеленчука жовтого (+ - 1%), дзвоників ріпчастовидних (+ - 1%), чини весняної (+ - 1%), печіночниці звичайної (+ - 1%), купини широколистої (+ - 1%), грястиці збірної, перлівки пониклої, медунки темної, куцоніжки лісової, пере-
174
ліски багаторічної, маруни щиткової, горлянки женевської, розхідника шорсткого, очитка великого та інших видів, спільних з попередньою асоціацією.
Видова насиченість асоціації дорівнює 81 виду.
Діброва кизилово-кореневищноосо- к о в a (Quercetum cornoBo-caricosum(rhiziaae) зустрічається зрідка переважно на Придніпровській височині в лісах Чорноліського лісгоспзагу. Займає вона нижні частини південних схилів з чорноземними опідзоленими грунтами.
Деревостан у віці 70-80 років має зімкненість крон 0,8-0,9, висоту 15-22 м, діаметр дерев 28-32 см і Ш-ІУ клас бонітету. До нього, крім дуба звичайного, входять ясен звичайний (+ - 0,1), граб звичайний, клен польовий, липа оерцелиста, гордовина та інші види з І-З-м класом постійності.
Підлісок (0,2-0,б) складають дерен справжній (0,2-0,5), клен татарський (+ - 0,2), гордовина, глід несправжньо-кривостовпчико- вий, свидина кров"яна, бруслина бородавчаста та ін.
Травостій має проективне вкриття 25-60% і складається 8 осоки кореневищної (20-40%), осоки волосистої (+ - 5%), зірочника лісового (+ - 5%), тонконога дібровного (+ - 5%), дзвоників кро- пиволистих (Campanula txaohelium І>.) (+ - J%) t чини чорної (+ - 1%), грястиці збірної (+ - 1%), перлівки пониклої, пахучки звичайної, самосилу гайового, шоломниці високої, гравілату міського, тори лі су японського, маруни щиткової, аконіту кущистого, буквиці лікарської, підмареннику весняного, медунки м”якенької, фіалок дивної та шершавої, оману верболистого, горошку горохоподібного, купини широколистої, чини весняної, переліски багаторічної, дзвоників персиколистих, осоки пальчастої, медунки темної, холодку тонколистого, астрагалу солодколистого, звіробою жорсткого, куцоніжки лісової, осоки стягнутої, конюшини альпійської, купини багатоквіткової, горобейника пурпурово-голубого, празеленя звичайного та інших видів з I-5-м класом постійності.
Видова насиченість асоціації - 79 видів.
Діброва кизилово-зірочнякова ( Quercetum ooxnoso-stellarioeum) трапляється зрідка в північній частині придністровського ареалу класу та на Придніпровській височині. Займає вона верхівки вузьких вододільних гребенів та нижні частини стрімких схилів південної експозиції з дерново-карбонатними грунтами (Придністров"я) або верхні частини крутих схилів
175
західної та східної експозицій (Придніпровська височина) з чорноземними опідзоленими грунтами.
Деревостан у віці 60-120 років мав зімкненість крон 0,7-0,9, висоту 16-25 м, діаметр дерев 22-96 см і Ш класу бонітету. Складають йогОіКрім дуба звичайного, клен польовий (+ - 0,2), ясен звичайний (+ - 0,1), липа серцелиста, граб звичайний, груша звичайна, клен звичайний черешня та інші види з I-4-м класом постійності.
Підлісок із зімкненістю 0,3-0,6 складається з дерену справжнього (0,2-0,4), клена татарського (+ - 0,1), бруслини бородавчастої (+ - 0,1), гордовини,бирючини звичайної, свидини кровняної, глоду українського та інших видів з I-4-м класом постійності.
Травостій має проективне покриття 30-45%, з якого 15-35% складає зірочник лісовий. Крім нього, постійно трапляються копитняк європейський (+ - 5%), осока волосиста (+ - 5%), астрагал солодколистий (+ - 1%), грястиця збірна (+ - 1%), тонконіг дібровний (+ - 1%), шоломниця висока (+ - 1%), фіалка дивна (+ - 1%), дзвоники ріпчастовидні, осока стягнута, гравілат міський, розхідник шорсткий, чина чорна, перлівка поникла, купина багатоквіткова, медунка темна, фіалка шершава, стоколос Бенекена, холодок тонколистий, горлянка женевська, осока пальчаста та горобейник пурпурово- голубий. З випадкових видів ростуть осока Мікелі, зеленчук жовтий, переліска багаторічна, Купина широколиста, печіночниця звичайна, жовтець золотистий, ранник вузлуватий, дзвоники кропиволисті, па- хучка звичайна, зубниця бульбиста, осока кореневищна, підмаренник весняний, фіалка запашна, печіночниця звичайна, ломиніс прямий та ін.
Видова насиченість асоціації - 91 вид.
Діброва кизилово-волосистоосо- к о в a (Quercetum cornoso-carioosum(pilosao) характерна для всієї території, яку займають дубові ліси кизилові. Трапляється вона, як і всі попередні асоціації класу, досить рідко і невеликими ділянками розміром до І га. Останні звичайно розташовуються у верхній частині опадистих або середній частині крутих схилів південної експозиції з чорноземними опідзоленими суглинками.
Деревостан у віці 60-110 років має зімкненість крон 0,6-0,9, висоту 17-24 м, діаметр дерев 28-36 см і Ш клас бонітету. До складу його входять ті самі види, що й у попередній асоціації, але з дещо більшою участю ясена звичайного (+ - 0,2) та липи серцелистої (+ - 0,1).
176
Підлісок виявлений добре і має зімкненість 0,2-0,6. Складають його дерен справжній (0,2-0,5), клен татарський (+ - 0,1), бруслина бородавчаста (+ - 0,1), глід український, ліщина звичайна, свидина кровпяна, жостір проносний, гордовина та інші види з І-3-м класом постійності.
Травостій більш густий (35-50%), ніж у попередньої асоціації. Крім осоки волосистої (15-40%), абсолютно постійними видами його є зірочник лісовий (+ - 10%), горобейник пурпурово-голубий (+ - 5%), тонконіг дібровний (+ - 5%), конвалія звичайна (+ - 1%), розхідник шорсткий (+ - 1%), копитняк європейський, дзвоники пер- сиколисті, грястиця збірна, чина чорна, чина весняна, перлівка поникла, медунка темна, шоломниця висока, фіалки шершава та дивна. Відносно постійно трапляються астрагал солодколистий, зубниця залозиста, ластовень лікарський, пахучка звичайна, дзвоники ріпчао- товидні, зеленчук жовтий, звіробій жорсткий, маруна щиткова, купина багатоквіткова, горошок горохоподібний, переліска багаторічна та стоколос Бенекена. Більшість випадкових видів є спільна з попередньою асоціацією.
Видова насиченість асоціації - 9? видів.
Як це видно з наведених описів, усі ценотаксони розглянутої групи асоціацій формуються в дужо близьких екологічних умовах і па реважна більшість їх видів спільні. Тому різниця між ними полягає не стільки у флористичному окладі, який відбиває істотні зміни око логічних умов, скільки в зміні фітоценотичної ролі видів та їх постійності.
ХУП. Група асоціацій дубові ліси кизилово-горобейникові складається з двох екологічно близьких асоціацій, пов"язаних з дуже сухими та теплими екстремальними для дуба звичайного екологічними умовами, в яких він утворює невисокі, розріджені, з кривими стовбурами деревоотани.
Діброва кизилово-горобейникова (Quercetum oornoao-lithospeiMosun) в придністровських північних районах займає середні й верхні найбільш опуклі частини дуже крутих та урвистих південних схилів з дерново-карбонатними грунтами. В придністровських південних районах та на Придніпровській височині вона повпязана з менш крутими схилами, що мають чорноземні вилуговані грунти. Порівняно з іншими асоціаціями класу діброва кизилова-горобейникова є найбільш поширеною, окремі ділянки її на дністровських схилах досягають кількох гектарів.
177
Деревостан у віці 60-90 років має зімкненість крон 0,5-0,8, висоту 8-16 м, діаметр дерев 18-28 ом і ІУ-У, зрідка Уа клас бонітету. Абсолютно постійним видом тут е тільки дуб звичайний. Відносно постійно трапляються клен польовий (+ - 0,1), ясен звичайний (+ - 0,1) та в"яз норковий (Ulmus suberosa M о e n с h .). З інших видів (2-4-й клас постійності) наявні граб звичайний, груша звичайна, черешня та берека (Sorbus torminalis (L.) с r а п і г).
Підлісок густий (0,4-0,?). Створюють його дерен справжній (0,3-0,5), клен татарський (+ - и,2), бруслина бородавчаста (+ - 0,1), гордовина (+ - 0,1), терен звичайний, жостір проносний, свидина кров"яна та іншівидди з I-4-м класом постійності.
Травостій мав проективне покриття 30-50%, з якого на горо- бейник пурпурово-голубий припадав 20-35%. До числа його постійних видів належать осока Мікелі (+ - 1%), купина широколиста (+ - 1%), ластовень лікарський (+ - 1%), фіалка шершава (+ - 1%), грястиця збірна (+ - 1%), ласкавець серповидний, смілка широколиста (Silene latifolia Bendle et В r i t t.), холодок тонколистий, дзвоники ріпчьстовидні та болонські, ломиніс прямий, в"язіль барвистий, гравілат міський, оман верболистий, чина чорна, маруна щиткова, шоломниця висока, чистець прямий, очиток великий, самосил гайовий, фіалка дивна та стоколос Бенекена. Регіонально постійним видом в ясенець білий (придністровська північна частина ареалу). Випадкові види представлені суницями зеленими (Fragaria viridis Duo h.), жовтушником сіруватим (Brysimua canescens В o t h.), айстрою степовою (Aster anellus !>.), рутвицею малою, підмаренником справжнім^ буквицею лікарською, осокою волосистою, дзвониками персиколистами, осокою гірською, зеленчуком жовтим, півниками злаколистими та угорськими, горлянкою женевською, перлівкою одноквітковою, конюшиною альпійською та гірською, купиною лікарською, залізняком бульбистим та ін.
Видова насиченість асоціації - 126 видів.
Діброва кизил о в о - м і к е л і є в о о с о - к о в а ( Quercetun cornoso-caricoswn(michelii) поширена на тій самій території, що й попередня асоціація, порівняно з якою займає більш опуклі й сухі частини схилів з малопотужними дерново- карбонатними або чорноземними вилугованими грунтами.
Деревостан у віці 50-100 років має зімкненість крон 0,5-0,8, висоту 8-18 м, діаметр дерев 18-34 см і ІУ-Уа клас бонітету. Як 1 в попередньої асоціації, абсолютно постійних супутників едифі-
178
катор не має, є лише види з 3-5-м класом постійності: клен польовий, граб звичайний, ясен звичайний, вняз корковий, берека та черешня»
Підлісок густий (0,4-0,7), утворюють Мого дерен справжній (0,3- 0,5), свидина кровияна (+ - 0,2), гордовина (t - 0,1), клен татарський (+ - 0,1), бруслина бородавчаста, глід несправжньо-кривостовп- чиковий, жостір проносний, жимолость пухнаста та інші види з І-З-м класом постійності.
Травостій має проективне покриття 30-45%. Ценозоутворююча здатність його конфентора виявлена гірше, ніж у попередній асоціації, що виявляється у невисокому проективному покритті осоки Мікелі (15-26%),досить строкатому видовому складі та більшій ролі постійних видів: виязелю барвистого (+ - 10%)8 горобейника пурпурово-голубого (+ - 10%), куцоніжки пірчастої (+ - 5%), тонконога дібровного (+ - 5%), віхалки гіллястої (4- - 1%), грястиці збірної (+ - 1%), розхідника шорсткого (+ - 1%), чистецю прямого (+ - 1%), сто- колоса Бенекена (+ - 1%) та ластівня лікарського (+ - 1%). Меншу роль у складі травостою відіграють холодок тонколистий, астрагал солодколистий, ласкавець серповидний, дзвоники болонські та перси- колисті, горошок горохоподібний, пахучка звичайна, чина чорна, смовдь оленяча, фіалка запашна, купина широколиста, купина лікарська, маруна щиткова, шоломниця висока, очиток великий, герань криваво-червона, фіалка шершава, самосил гайовий, рутвиця мала, осока кореневищна., підмаренник справжній, оман верболистий, зірочник лісовий та інші види з I-4-м класом постійності. Регіонально постійними видами є.чина паннонська (Latbyrus panno пісна G a r a k е ) та ясенець білий (Придністровпя).
Видова насиченість асоціації - 120 видів.
Отже, порівняно з асоціаціями попередньої групи в розглянутій з числа постійних зникають типові неморальні балтійські види. Разом з цим посилюється роль і збільшується кількість неморальних субсередземноморських видів, таких, як ласкавець серповидний, чистець прямий, самосил гайовий та ін.). У групі постійних видів на них припадає близько 80% видового складу, решту складають неморальні балтійські види з широкою екологічною амплітудою. Група випадкових видів представлена переважно типовими мезофільними неморальними балтійськими видами (яглиця звичайна, копитняк європейський, підмаренник проміжний, медунка темна, жовтець кашубський та ін.), найбільш мезофільними неморальними оубоередземноморськими
179
(горошок горохоподібний, горлянка женевська, латук Ше, празелень звичайна та ін.) видами, які не в лісовими (молочай кипарисовид- ний, буркун лікарський, звіробій звичайний та ін.)і неморальними субсередземноморськими видами, характерними для лучних степів (герань криваво-червона, півники угорські, люцерна серповидна, рутвиця мала, конюшина альпійська, конюшина гірська, залізняк бульбистий та ін.).
Дубові ліси скумпієві
Синекологічна конфекторна амплітуда скумпії звичайної майже не збігається з синекологічною едифікаторною амплітудою дуба зви-* чайного. Скумпія звичайна, як і дерен справжній, не переносить континентального клімату, але більш вимоглива до тепла. В досліджуваних дубових лісах України вона перебуває в екстремальних умовах і легко витісняється з менш сухих і теплих екотопів кленом татарським. За даними зарубіжних фітоценологів (Rikli, I943;Braun-Blan- quet, 1952} Jakucs, 1955» 1959, 1960, 1961j Zolyoad., Jakucs, 1957} Soo', 1958, 1960, 1963, 1964, 1968; Jekete, . 1959} Радков, Минков, 1963; Borza, Bo^caiu, 1965; Pa^covschi.Donifi, 1967; Велчев, Ганчев, Бондев.Кочев, 1969; Пенев, Маринов, Горелков, На- умов, 1969), скумпія звичайна є найтиповішим конфектором ксеротер- мобазифільних (доломіти, вапняки) субсередземноморських лісів з дуба пухнатого, бургундського ( Quercus cenis L.) і фрайнетто (О, frainetto Т е а.), ясена білоцвітого (Fraxinus ornus L.), граба східного ( Carpinus orientalis М і 1 1.) та сосни чорної (Pinus nigra L.). Характерними видами цих лісів, крім названих едифікаторів, є види з субсередземноморським, середземно-субсеред- земноморським та балканським типом ареалу. Всі вони, за винятком кількох видів, наприклад, холодна тонколистого, чистецю прямого, переліски яйцевидної (Mercurialis ovata s t e r n b.et H o p p e) в досліджуваних лісах не трапляються. З другого боку, в західних субсередземноморських лісах немає низки видів, що ростуть в лісах класу з пептичним, понтично-південносибірським і понтично-півден- носибірсько-середньоазіатським типом ареалу, таких, як ломиніс ці- лолистий, ясенець голостовпчиковий, карагана кущова, молочай несправжньо-волохатий, жовтушник лісовий, вероніка волотиста ( Vero- nloa panlculata L,),
Дубові ліси скумпієві в Середній Європі поширені на території Угорщини і Румунії роб» 1964; Borza, 1963 ) на більш теплих, ніж 180
попередній клас асоціацій, схилах. В Радянському Союзі вони трапляються в Молдавії та в Придністровській частині України. Острівні місцезнаходження відомі в Криму, Донецькій області, на Кавказі, (Царина, 1925; Елагин, 1953; Гейдеман, Остапенко, 1964; Шеляг-Сосонко, Собко, 1970). Приурочені вини до слабколужних дерново-карбонатних і чорноземних опідзолених або вилугованих грунтів з рН 6,7-7,6. У флорі їх переважають неморальні субсередземноморські види з участю степових.
На Україні дубові ліси скумпієві трапляються лише в степовій зоні на межі з Молдавією (Котовський лісгослзаг) та на "Горах Ар- тема" (Слов"янський лісгослзаг). У першому випадку на південних схилах глибоких і -широких балок вони створюють верхню кромку лісу при переході до безлісних плакорних ділянок, заміщуючись нижче по схилу дубовими лісами татароькокленовими. На "Горах Артема” ці ліси займають круті й високі (100-150 м) крейдяні схили корінного берега Сіверського Дінця на ділянці довжиною 7-9 км. На бідніших едафотопах (крейдяні відслонення) вони заміщуються сосновими лісами, а на більш вологих (північні схили, менш круті східні та західні, нижня частина південних схилів) - дубовими лісами татарсько- кленовими .
Схили, зайняті дубовими лісами скумпієвими,дуже прогріваються. Дуб звичайний перебуває на них в екстремальних умовах. Він дуже суховершить, має скривлені стовбури і утворює розріджені невисокі деревостани. Синекологічна амплітуда класу надто вузька. Всі асоціації його належать до дуже сухих мегатрофних базифільних дібров і виявляють варіабельність лише в цих межах, займаючи порівняно з попереднім класом ще сухіші та менш вилуговані грунти. На більш сухих та лужних грунтах дубові ліси з дуба звичайного вже не рос-' туть. Навіть у найбільш мезофільної асоціації класу в числі постійних видів травостою типових неморальних балтійських мезофітів майже немає. Так само немає в них і групи видів букових лісів, яка трапляється в лісах попереднього класу.
Максимально можливого для формації значення в дубових лісах скупієвих досягають степові види, наприклад горицвіт весняний (Adonis vernalis L.),пирій середній (Elytrigia intermedia(H o s t.) N e v s k і), різак звичайний (Faloaria vulgaria В e r n h.), грудниця волохата (Linosyris villosa DO.), перлівка трансільванська (Melica transsilvanioa S o h u л.), костриця борозниста та ін., в тому числі чагарники (мигдаль степовий Amygdalus nana L., 181
вишня кущова та карагана кущова). В цьому відношенні ліси даного класу е аналогом континентальних дубових лісів південно-східної частини ареалу формації, які вони заміщують в умовах субконтинен- тального клімату. Другою характерною рисою їх е ще значніше збіднення видового складу деревостану і спрощення його структури (повна відсутність ознак другого ярусу), збільшення видового складу, ускладнення структури ярусу підліску (самостійну синузію формують степові чагарники).
До класу дубових лісів скумпієвих належать дві групи асоціацій. З них дубові ліси скумпіево-рябоперлівкові формуються в менш сухих умовах, ніж дубові ліси скумпієво-мікеліевоосокові.
ХУШ. Група асоціацій дубові ліси скумпіево-рябоперлівкові представлена дібровами скумпієво-конвалієвою та скумпієво-рябопер- лівковою. Перша по відношенню до другої займає більш вологі едафо- топи з вищим вмістом гумусу в верхньому горизонті (до 24%). Другу асоціацію, як? формується в більш вологих умовах порівняно з наступними, можна розглядати як центральну.
Діброва скумпієво-конвалієва (Quercetum ootinoao-convallariosum) поширена в обох районах, проте більше на "Горах Артема". Пов"язана вона з верхніми та середніми частинами дуже крутих схилів східних і західних напрямків а дерново-карбонатними або чорноземними опідзоленими грунтами. Площа окремих ділянок цих лісів не перевищує 1-2 га.
Деревостан у віці 60-200 років має зімкненість крон 0,4-0,8, висоту 10-19 м, діаметр дерев 24-60 см і ІУ-У клас бонітету. Крім едифікатора, до складу його входять ясен звичайний (+ - 0,1), липа серцелиста, клен польовий, берест, клен звичайний та ін. З них лише два перші види трапляються постійно.
Підлісок висотою 2,5-3,5 м досить густий (0,5-0,7).Створюють його скумпія звичайна (0,4-0,5), клен татарський (+ - 0,2), свидина кров"яна (+ - 0,2), карагана кущова (+ - 0,1), бруслина бородавчаста (+ - 0,1), жостір проносний, глід обманливий, бирючина звичайна, терен звичайний, ліщина звичайна, шипшина повстиста (Нова tomentosa Smlt.) та інші види з I-2-м класом постійності.
Травостій має проективне покриття 35-45%. Характерною рисою його є добре виявлена ценозоутворююча роль конвалії звичайної (30- 40%) і значна кількість постійних видів з незначним проективним покриттям. З них відзначимо лише найхарактерніші, такі, як перлівка ряба (♦ - 5%), віхалка гілляста, грястиця збірна, осока ко-
182
реневищна, рагнерія собача, дзвоники персиколисті, холодок тонколистий, осока Мікелі, осока пальчаста, в"язіль барвистий, підмаренник пухнатоногий, гравілат міський, латук стріловидний, астрагал солодколистий,купина лікарська, шоломниця висока, зірочник лісовий, золотушник звичайний, стоколос Бенекена, очиток звичайний, вероніка волотиста, дутень ягідний, фіалка шершава, порізник проміжний (Libanotis inteimedia H u р г.) та смілка широколиста (Silene latifolia (Mill.)Bendle et Bi i t t.). Привертав увагу той факт, що в групі постійних видів ще присутні неморальні види з широкою синекологічною амплітудою (осока пальчаста, золотушник звичайний), які заходять навіть в ялинові ліси. Це лише підкреслює те, що скумпія звичайна в ролі конфектора в даній асоціації перебуває в екстремальних умовах. Посилення гігро- і кріотичнооті факторів середовища призводить вже до заміщення його конфекторами інших видів.
Видова насиченість асоціації - 87 видів.
Діброва окумпівво-ряболерлівко- в а ( Quercetua cotinoBo-aeiiooeum(piotae) поширена на "Горах Артема", де займав верхні та середні частини крутих і дуже крутих схилів південної, рідше східної та західної експозицій з дерново- карбонатними грунтами.
Деревостан у віці 70-160 років має зімкненість крон 0,4-0,6, висоту 9-18 м, діаметр дерев 28-50 см і У-Уа клас бонітету. Постійними видами в лише дуб звичайний, ясен звичайний та берест.
Підлісок густий (0,4-0,7). Створюють його скумпія звичайна (0,3-0,5), бруслина бородавчаста (+ - 0,2), карагана кущова (+ - 0,1), свидина кров"яна (+ - 0,2), вишня кущова, бирючина звичайна, терен звичайний, клен татарський та інші види з I-4-м класом постійності.
Травостій має проективне покриття 20-40%. До його окладу входить перлівка ряба (15-30%), а з групи постійних видів осока Міке- лі (+ - 5%), конвалія звичайна (+ - 5%), в"язіль барвистий (+-1%), купина лікарська (+ - 1%), купина багатоквіткова, підмаренник пухнатоногий, шоломниця виоока, очиток звичайний, смілка широколиста, барвінок трав'янистий, ластовень лікарський (абсолютно постійні види), віхалка гілляста, материнка звичайна, ласкавець серповидний, молочай несправжньо-волохатий, дзвоники персиколисті та ріп- частовидні, грястиця збірна, гравілат міський, латук стріловидний, тонконіг дібровний, маруна щиткова, чиотець прямий та фіалка шер183
шава (відносно постійні види). Регіонально постійними є рутвиця смердюча (Thaliotrum foetidum L.) та ясенець голоотовпчиковий. Випадково трапляються деревій паннонський (Achillea раппопіса Scheele ), ушанка несправжньоушанкова (Otites pseudootites К 1 о К.), жовтушник прямий ( Srysimum strictum G.-M.-s с h.), підмаренник напіводягнений ( Gallum semiamictum К 1 о k.),порізник Проміжний ( Libanotis intermedia Вир г.), нечуйвітер отруйний, люцерна серповидна, залізняк бульбистий, пужник голий (Turritis glabra L.) стоколос Бенекена, зірочник лісовий, рутвиця мала, очиток великий, вероніка волотиста (Veronica paniculata L.), горлянка женевська,кінський часник черешковий, астрагал солодколистий, осока кореневищна, куцоніжка лісова та ін.
Видова насиченість асоціації - 82 види.
XIX. Група асоціацій дубові ліси скумпієво-мікелієвоосокові обвєднує дві найбільш ксеротермофільні асоціації, з яких діброва окумпієво-мікелієвоосокова займав більш вологі едатопи, ніж діброва скумпієво-пірчастокуцоніжкова. Поширені вони лише в північно- західній частині степової зони на межі а Молдавією (Павлівське ліо- ництво Котовського ліогоопзагу). В цих лісах неморальні балтійські види до числа постійних не входять і належать до групи випадкових^. Головна роль належить неморальним субсередземноморським та степовим мегатрофним базифійьним ксеро- І ксеромезофільним видам а північною межею ареалу, яка проходить у лісостеповій зоні України. Лише в лісах цієї групи асоціацій із степових видів постійно присутні горицвіт весняний, мигдаль степовий, дзвоники скупчені ( Campanula glomerata L.),пирій середній, костриця борозниста, перлівка трансільванська, тимофіївка степова, стоколос бев- остий та ряд видів, які ще не втратили лісових зв"яаків, а саме: ферульник ЛІСОВИЙ fPerulago silvatica (Bees.) В e io h b,), горобейник лікарський (Lithospermum offioinale L.), котяча м"ята паннонська та ін. Найхарактернішою рисою цих лісів в наявність в їх складі окремих екземплярів дуба пухнатого. З цими лісами повезена північно-східна межа його ареалу.
Діброва скумпієво-мікелієвоосо- К о В a (Quercetum cotinoso-carlcosum (miohelil) займає верхні.
1 Абсолютно постійні види гравілат міський та бруслина бородавчаста мають таку широку синекологічну амплітуду, що в рівній мірі можуть розглядатись як характерні види неморальних ксеротер- мофільних і неморальних мезофільних лісів.
184
зрідка середні частини найбільш крутих (20-30°) в даній місцевості схилів південних напрямків та привододільні схили вужчих балок з чорноземними вилугованими глинистими грунтами. Як правило, окремі ділянки її мають ширину 15-30 м, довжину 50-200 м і чергуються з степовими узліссями та куртинами дуже порушених дубових рідколісь (рис.33).
Деревостан у віці 60-140 років має зімкненість крон 0,4-0,8, висоту 7-Ю м, діаметр дерев 16-30 см і У® клас бонітету. Створюють його, крім дуба звичайного, лише три види, з яких постійно трапляється лише дуб скельний. Два інші види (липа серцелиста та ясен звичайний) мають 2-3-Й клас постійності.
Підлісок (зімкненість 0,4-0,6) складається зі скумпії звичайної (0,3-0,5), клена татарського (+ - 0,1), мигдалю степового, бруслини бородавчастої, терену звичайного, жостеру проносного, глоду несправжньо-зігнутостбвпчикового, гордовини, шипшини повстистої та видів з І-З-м класом постійності.
Травостій з проективним покриттям 25-40% створює конфектор (15-35%), а а числа постійних видів - куцоніжка пірчаста (+ - 5%), гравілат міський (+ - 1%), холодок тонколистий, дзвоники персико- листі, грястиця збірна, гадючник шестипелюстковий, суниці зелені, півники угорські, латук стріловидний, маруна щиткова, рутвиця мала, барвінок трав"янистий, горицвіт весняний, віхалка гілляста, буквиця лікарська, дзвоники скупчені, молочай несправжньо-волохатий, пирій середній, ферульник лісовий, підмаренник справжній, горобейник лікарський, купина лікарська, очиток звичайний, регнерія собача, чистець прямий, конюшина альпійська, ластовень лікарський, стоколос безостий, фіалка шершава, самосил гайовий та маточник чорний. З групи випадкових видів ростуть горошок тонколистий (Vicia tenuifolia В о t h.), валеріана пагононосна (Valeriane stolonlfera с e e г n.), азинеума сірувата (Azyneuma canescens fw.K.) a r і в e b. et з o n e n k), різак звичайний, вероніка волотиста, собача кропива сизувата, герань криваво-червона, наперстян* ка великоцвіта, чина паннонська, смовдь гірська, материнка звичайна, ломиніс цілолистий, кінський часник черешковий, дуТеиь ягідний та ін.
Видова насиченість асоціації - 105 видів.
Діброва скумпієво-пірчастокуцо- н і ж к о в а ( Quercetum cotinoeo-brachypodiosum(piimati) поширена лише на границі з Молдавією. Вона займає привододільні 185
ділянки та верхні частини схилів південної експозиції з чорноземними вилугованими грунтами. Порівняно з лісами попередньої асоціації окремі ділянки її мають не меншу площу і більш порушені. Після рубок головного користування вони, як і ліси попередніх асоціацій цього класу, не поновлюються (рис.34).
Деревостан у віці 50-60 років має зімкненість крон 0,4-0,8, висоту 6-8 м, діаметр дерев 18-20 см і продуктивність уа класу бонітету. Створює його дуб звичайний, інколи з незначною участю дуба скельного.
Підлісок (зімкненістю 0,4-0,6) складається зі скумпії звичайної (0,3-0,5), мигдалю степового (+ - 0,1), глоду несправжньо-кри- востовпчикового, жостіра проносного, терену звичайного, клена татарського, бруслини бородавчастої, гордовини, вишні кущової, дереву справжнього та ін. Пмять останніх видів мають 2-4-й клас постійності .
Травостій має проективне покриття 40-55%. Постійними видами його є куцоніжка пірчаста (30-45%), пирій середній (1-5%), гравілат міський (+ - 1%), горицвіт весняний, парило звичайне, віхалка гілляста, холодок тонколистий, буквиця лікарська, дзвоники болонські, скупчені, персиколисті та ріпчастовидні, осока Мікелі, в"язіль барвистий, грястиця збірна, бутень бульбистий, ферульник лісовий, костриця борозниста, гадючник шестипелюстковий, суниці зелені, підмаренник справжній, латук стріловидний, чина паннонська, горобей- ник лікарський, перлівка висока, перлівка трансільванська, котяча мпята паннонська, тимофіївка степова, купина лікарська, маруна щиткова, очиток звичайний, чистець прямий, самосил гайовий, ториліс японський, конюшина альпійська, рутвиця мала, барвінок трав"янис- тий, ластовень лікарський та стоколос Бенекена. Рідше (2-4-й клас постійності) трапляються дивина східна (Verbascum orientale М.В.), нечуйвітер отруйний ( Ніегасішп virosum раї 1.), оман шершавий, грудниця волохата, валеріана пагононосна, вероніка волотиста,перстач темний ( Potentilla obscura wi 11 d.), фіалка шершава, го- робейник пурпурово-голубий , материнка звичайна, ломиніс прямий, ломиніс цілолиотий, люцерна серповидна, тонконіг дібровний, маточник чорний, наперстянка великоквіткова, смовдь гірська, чина чорна, різак звичайний та ін.
Видова насиченість асоціації - 77 видів.
Таким чином, усі специфічні риси лісів класу досягають в лісах асоціації свого максимального виявлення. Деревостан складений
186
практично одним едифікатором. Неморальних балтійських мезофільних видів немає. Кількість видів, однаково характерних для мезофільних і ксеротермофільних лісів (з урахуванням кінського часнику черешкового, м"яточника чорного і грястиці збірної), складав всього близько 8%. На степові види припадає майже 24% і на неморальні суб- середземноморські види - 68%. Серед останніх 46% складають види, що трапляються і на лучних степах, останні 22% - суто лісові. На екологічні групи види поділяються так: ксерофіти і ксеромезофіти становлять 82%, мезофіти - 18%, облігатні мегатрофи - 86% і факультативні мегатрофи - 14%. Фітоценотипна синекологічна амплітуда само! куцоніжки пірчастої дуже широка. В ролі конфектора вона виступає в західносибірських березових лісах (Крнлов, 1953; Глумов, 1960; Лапшина, 1963), в дубових і соснових лісах Криму (Троицкий, 1936; Посохов, 1961) і Субсередземномор"я (Gauoklex, 1938; Jakucs,1955, 1961; Зоб, 1963; Fekete, 1965; Пенев та ін., 1969). Крім того, вона може бути також едифікатором степових фітоценозів ( Koczwara, 1931; Gauoklex, 1938; Зоб, 1947; 1949, 1959; StawiAski, 1952;
Шифферс, 1953; Klika, 1954; Привалова, 1956; Boxza,1963), чим відрізняється від усіх попередніх конфекторів. Отже, цю асоціацію ми розглядаємо як таку, що завершує природний ряд формування ксеротермофільних дубових лісів з дуба звичайного. Ліси з більш ксеро- фільним травостоєм, наприклад з кострицею борознистою, ми відносимо до похідних.
ЕВОЛЮЦІЯ ДУБОВИХ ЛІСІВ
Як ми вже зазначали, головне флористичне ядро досліджуваних лісів складають неморальні види. Існують дві протилежні гіпотези щодо їх походження. Більшість дослідників ( Snglax, 1879; Кучие- цов, 1920; Криштофович, 1936, 1946, 1958; Гросогейм, 1948; Попов, 1949; Pawiowska, 1959; Szafex, 1964 та ін.) обстоюють приполярне (Гренландія, Аляска, Новосибірські острови) походження неморальної флори, звідти пізніше, на їх думку, вона мігрувала на південь. А.М.Криштофович (1958) вважає, що ця флора виникла на базі гренландської флори, яка існувала у крейдяному періоді на сучасних островах Північного льодовикового океану і на початку третинного періоду поширилась на південь, створивши суцільний трансконтинентальний пояс неморальної рослинності, який проходив через усю Канаду, Скандинавію, Урал та Сибір. У складі гренландської флори панували Metasequoia, Pterospexmites, Txochodendxoides, але траплялись види 187
t широколистяних лісів, представлені родами клен (Acer), платан (Platanus), в**ЯЗ (Ulnus), вільха (Alnus), горіх (Juglans) і, можливо, дуб (Quercus). Перед кінцем існування гренландської флори на півночі та північному сході на її основі сформувалась неморальна флора, яку А.М.Криштофович (1936) назвав тургайською. В цей час (кінець еоцену - середина олігоцену) на території Російської рівнини була поширена полтавська лісова вічнозелена тропічна флора. В середині олігоцену тургайська флора почала проникати й на Україну, де вона стала в міоцені переважаючою (Криштофович, 1938; Гричук, 1961; Щекина, 1962). В її складі панували представники таких родів, як вільха (Alnus), платан (Platanus), секвойя (Sequoia), бук (Pagus), болотний кипарис (Taxodium), каштан (Castana), береза (Betula), горіх (Juglans), дзельква (Zelkova), клен (Acer), фікус (Picus) та ін. Рід дуб (Quercus) в цей час на Україні, за даними А.М.Криштофовича та Т.М.Байковоької (1951, 1965), був представлений вічнозеленими цільнолиотими видами, а також листопадними із туполопатевим листям з менш диференційованою листовою пластинкою ( Quercus deuterogona) і більш диференційованою (Quercus pseu- dorobur). Вони були досить поширеними, оскільки пилок їх у відкладах нижнього міоцену складає 5-12% (Ротман, 1956).
Послідовно й аргументовано обстоюючи гіпотезу приполярного походження неморальної флори, М.Г.Попов (1949) висуває на доказ її такі положення: І) не всі арктотретинні види Західної Європи або Передньої Азії мають родинні зв"язки у Південно-Східній Азії; 2) Західна Європа та Передня Азія мають свої ендемічні роди, які за мезофільністю не поступаються перед найближчими до них східноазі- атськимн; 3) існуюча в поширенні видів секційна смугастість свідчить проти міграції арктотретинних видів зі сходу Азії. "Таким чином, - твердить М.Г.Попов (1949, с.125), - ми можемо сформулювати загальний висновок: з третинної Арктики на захід Євразії поширилась не менш багата арктотретинна флора, ніж у Східну Азію або Північну Америку... ".Якщо прийняти ці погляди як робочу гіпотезу, то слід визнати що всі найбільш близькі до тургайських сучасні євразійські широколистяні ліси належать до вторинних центрів походження.
За іншою гіпотезою,неморальна флора виникла автохтонно в Пів- дейно-Східній Азії з елементів тропічної та субтропічної флори (Краснов, 18946; Комаров, 1908; Diels, 1929; Вульф, 1944; Федоров, 1957; Тахтаджяи, 1957, 1970; Васильєв, 1961,та ін.). Цей процес 188
відбувався у горах, де Й зараз спостерігаються поступові переходи від помірної до тропічної флори. На захід неморальна флора мігрувала на початку третинного періоду по гірських хребтах. На користь цієї гіпотези, за даними згаданих дослідників, свідчать такі факти:
1) флористична бідність арктичної викопної неморальної флори;
2) більш рання у часові поява елементів неморальної флори в Південно-Східній Азії; 3) наявність на цій території цілих філогенетичних ланок неморальної флори; 4) відсутність сприятливих умов для міграції неморальної флори з півночі на південь; 5) відсутність у складі сучасної флори змішаних лісів Далекого Сходу представників третинної флори північних районів; б) походження листопадних видів дуба від вічнозелених предків, родинні звиязки яких тяжіють до півдня, а не до півночі.
Розселення неморальної флори з цього центру походження, згідно з даними А.Л.Тахтаджяна (1970), в загальних рисах закінчилося у крейдяному періоді. В цей час, вважав автор, вже існувала верхньокрейдяна Бореальна область, яка трансформувалась далі в третинну Бореальну область, до складу якої входили такі роди,як клен (Acer), граб (Carpinus), каштан (Castanea), чорниця ( Vaccinium), карія (Сагуа), калопанакс (Acanthopanax), в"яз (Ulmus), бук (Fagus), липа (Tilia),дерен ( Cornus) та ін. Збіднені залишки цих лісів і дали початок сучасним широколистяним лісам.
На нашу думку, в доказах високоширотного походження неморальної флори більше гіпотетичного, ніж в доказах низькоширотного походження. У всякому разі, не викликав сумніву, що з сучасних неморальних лісів найбільш близькими до третинних неморальних лісів за флористичним окладом та фітоценотичною будовою в ліси південно- східної частини Азії. В.Б.Сочава (1946) відносить ці ліси до манч- журської фратрії і вважав, що найбільш характерними рисами їх є плюрідомінантність, багатоярусність, велика кількість ліан, багатий видовий склад чагарників і бідний - трав"яного ярусу. Саме ці риси були притаманні третинним лісам, дальший розвиток яких, за Є.М.Синською (1933), йшов в напрямку інтенсивного збіднення видами верхніх ярусів і зведення багатьох домінантів до одного. Збіднення складних Полі домінантних угруповань відбувалось у зв"язку з погіршенням клімату та міграціями. Однією з форм цього процесу в виділення консоціацій, які з часом диференціюються на пануючі та підпорядковані. Далі йде процес звільнення підпорядкованих кон- соцій, обмін або інкумбація їх, зміна просторового положення і т.д
189
Флористичні зв"язки сучасної флори дубових лісів України з манчжурськими лісами простежуються в таких спільних родах, як граб (Carpinus), дуб ( Quercus), бруслина (Buonyaus), липа (Silia), клен (Acer), ліщина (Corylus), в"яз ( ЇЛаив), свидина (Thelyirania) та багато інших. Вони входили до складу тургайської флори, розрив суцільного євразійського поясу якої відбувся в кінці третинного періоду. Внаслідок цього утворився ряд регіонів, в яких неморальна флора далі розвивалась самостійно. За Є.М.Лавренком (19386), це такі регіони: Середньоєвропейський, Карпатсько-Північнобалканський, Колхідський, Талишський та Манчжурський. З них два перші дали переважну більшість сучасних неморальних видів дубових лісів України. Вихідними видами були дуже поширені в нижньому міоцені викопні форми тилу Quercus pseudorobur, Carpinus grandis, Corylus cf. avellana та ін. Відбитки листя їх нагадують існуючі тепер види. Ця близька .до сучасної широколистяна флора відступила в пліоцені на південь під прикриття Альпійсько-Балканської гірської системи, де й закінчилось формування багатьох її видів. Цілком можливо, що в середньому пліоцені на цій території існувала більшість сучасних видів. В усякому разі В.Шафер (Szafer, 1938) для пліоцену передгір"їв Карпат наводить тис ягідний (laxus baccata), ялину білу (Picea abies), граб звичайний ( Carpinus botulus), фізаліс звичайний fPnysalle alkekengi), ліщину звичайну {Corylus cf.avellana) та ІН. Ю.Єнтіо-Шаферова pentys-Szaferowa, 1958) на підставі вивчення анатомічної будови викопних горішків граба з пліоценових відкладів підніжжя Карпат прийшла до висновку, що за анатомо-морфологіч- ними ознаками вони тотожні з горішками сучасного граба звичайного. В пліоцені виник і дуб звичайний. А.М.Криштофович (1941) для відкладів цього періоду з України наводить листя дуба типу дуба звичайного. Б.Стефанов (stefanoff, 1928) за вихідну форму його вважає Quercus roburoides, дуже поширений у Болгарії в кінці пліоцену. Такої самої думки дотримуються В.П.Малєєв (1935) та Ю.Клео- пов (1941). Останнім часом Ю.Меніцький (1969) переглянув це положення. Базуючись на тому, що Quercus pseudorobur знайдено в Європі та Західному Сибіру в міоцені, тобто значно раніше, ніж Quercus roburoides, вія вважає, що дуб звичайний виник з Quercus pseudorobur в районах,північніших за Болгарію. Численні знахідки викопних форм дуба звичайного в кінці пліоцену - на початку плейстоцену яа території від Каспію до Середньої Європи підтверджують цей висновок.
190
Серед сучасної неморальної флори дубових лісів можна виділити дві великі групи, одну з них складають мезотермні, а другу - ксеромезотермні види. Останні являють собою кверцетальний ценоеле- мент Ю.Д.Клеопова (1941) в широкому розумінні. Перша група видів повпязана з неморальними лісами, що ростуть на північ від Альпійської гірської системи, а друга - з неморальними лісами субсередземноморських районів.
Види мезотермної групи є дериватами тінявого тургайського комплексу. Більшість їх виникла ще до розриву його суцільного поясу і тепер вони мають екоклави в Західному Сибіру (наприклад,копитняк європейський, костриця лісова, дзвоники кропиволисті, яглиця звичайна), Східному Сибіру (костриця велетенська, чина весняна та ін.), на Далекому Сході і навіть в Японії (маренка запашна, просянка розлога, осока отягнута, конвалія звичайна, актея колосиста, купина багатоквіткова та ін.).
Види ксеромезотермної групи виникли з тургайського комплексу в області Древнього Субсередземномор"я дещо пізніше. Дубові ліси з дуба звичайного поглинули їх в кінці пліоцену, коли ці ліси під впливом, похолодання відступили в субсередземноморські райони. Мабуть, процес цей відбувався у передгірському поясі Балкан та Південних Карпат, де дубові ліси створювали розріджені деревостани, під які проникли ксеромезотермні види, такі, як осока парвська, горобейник пурпурово-голубий, смовдь оленяча, скумпія звичайна, дерен справжній, півники злаколисті, холодок тонколистий, купина широколиста, гордовина, берека та ін. Вище формувалися мезотермні фітоценози дубових лісів з участю мезофільних, але більш світлолюбних, ніж типові мезотермні види першої групи, видів типу чини чорної, перстачу білого, підмаренника весняного, осоки гірської, фіалки дивної та ін.
Бореальві види дубових лісів теж представлені двома групами: мікротермною тіньолюбною та иікротермною світлолюбною. Перша група видів, генетичні зв"язки якої ще не достатньо 8"ясован1, походить, мабуть, від бореального темнохвойного комплексу, який виник не пізніше третинного періоду. За О.І.Толмачовим (1954), видо- утворюючим центром бореальної флори були гори Східного Сибіру, на схилах їх в міоцені сформувалась темнохвойна тайга. З похолоданням вона поширилася на рівнину й утворила сучасний пояс темнохвойних лісів. На початку плейстоцену цілий ряд видів цих лісів був поглинутий дубовими лісами: квасениця звичайна, веснівка дволиста, 191
грушанка круглолиста, одинарник європейський, ожика волосиста, рамішія однобока, чорниця та ін. Крім темнохвойних лісів, перелічені види трапляються ще в гірських букових лісах Європи та хвойно-широколистяних лісах Далекого Сходу, що дало підставу В.Б.Соча* ві (1946) віднести перші три види до неморальних, точніше до ква- зібореальних форм залишків пліоценових змішаних тінявих лісів.
З третинними широколистяними лісами пов"язув походження вказаних видів В.М.Васильєв (1958).
На наш погляд, процес видоутворення мікротермної тіньолюбної групи видів відбувався в холодостійкішій частині тургайського комп лексу, яка в неогені трансформувалася у вихідний бореальний комплекс. Досі від останнього вбереглися тільки види, що входили в трав"яно-чагарничкові синузії. Первинні види едифікаторних сину- зій не збереглися. В кінці третинного періоду вони були замінені типовими едифікаторами темнохвойних лісів тайги. Саме тому едифікатори цих лісів мають обмеженіший ареал, ніж поглинуті ними види вихідного бореального комплексу. Завдяки цьому останні також є досить звичайними видами гірських букових і змішаних далекосхідних лісів. Першого максимального розвитку види мікротермної тіньолюбно! групи на території Російської рівнини досягли, за даними П.І.Дорофєєва (1963), в мінделі. З того часу роль їх на території України в холодні кліматичні періоди посилюється, а в теплі - послаблюється.
Друга група видів виникла в близьких до першої групи кліматичних умовах, але в світлих бореальних лісах гірських пасм північно-східної Азії в кінці третинного періоду, і на територію України проникла в плейстоцені. До неї належать лілія лісова, борщівник сибірський, грястиця збірна, береза бородавчаста, осика та ін. В загальних рисах ця група видів являє собою плейотоценовий флориотичний комплекс І.П.Крашеніннікова (1951), найбільш характерною рисою якого була колись слабка диференційованість в загальних екологічних умовах, а також в складі лісових і напіввідкритих (типу степових і лучних) ценозїв, що поєднувалось із загальними суворими кліматичними умовами. Ю.Д.Клеопов (1941) більшу частину видів плейотоценового флористичного комплексу відносить до бету- лярного ценотичного елементу. Види, що належать до нього, в сучасний період пов"язані із західносибірськими корінними березовими лісами, які, на думку Г.В.Крилова (1957), сформувалися на початку плейстоцену і проникли в Європу в лихвінсько-дніпровський міжльодовиковий період.
192
Початкові етапи утворення синузій сучасних дубових лісів відносяться до кінця пліоцену - початку плейстоцену. В цей час на території України і суміжних територій неморальна рослинність, за даними В.П;Гричука (1950) і В.Шафера (szafer, 1953), поступається місцем хвойним лісам. В.Шафер ( Szafer, 1953, с.40) описує межу між пліоценом і плейстоценом так: "Виявляється вона в раптовому зникненні в пилковому профілі всіх теплолюбних пліоценових рослин і в рівній мірі раптовій зміні в пилковому дощі дерев, серед яких тепер починає панувати пилок хвойних дерев, головним чином родів pinus типу Silvestris, Picea...". З цим періодом і пов"язані початкові етапи проникнення на територію України видів мікротермної групи.
В наступних лихвінсько-дніпровському та дніпровсько-валдайсько- му інтергляціалах на спорово-пилкових діаграмах виразно виступає чергування трьох фаз, що відповідають пануванню двох рослинних комплексів. На початку і в кінці інтергляціалу спостерігається максимум пилку сосни та берези, а в середній частині інтергляціалу - широколистяних порід. При цьому поступово біднішає видовий склад пилкового спектра, через відмирання або зникнення з певної території термофільних видів. Так, за В.Шафером (Szafer, 1953), під час дніпровського гляціалу відмерли Tsuga europaea, Oarpinus ші- піша, vitis ludwigi та ін. З території Європи зникла Abies fra- seri, Liriodeniron tulipifera, а з території Польщі - Picea ошохіса, Pinus subgen. Haploxylon та ін. Це характерне в загальних рисах і для рослинності інтергляціалів України (Артюшенко, Хриотофорова, Карева, 196?; Паришкура, 1967; Артюшенко, 1970).
Незважаючи на наявність у пилковому спектрі інтергляціалів третинних рестанців, які свідчать про клімат м"якіший, ніж клімат голоцену, зональний розподіл рослинності під час максимуму пилку широколистяних порід був близьким до сучасного. Відповідно зональний розподіл рослинності та характер її на початку і в кінці кожного інтергляціалу наближались до раннього голоцену, для якого характерне панування пилку сосни та берези. Таким чином, починаючи з кінця пліоцену відбувається збіднення неморальної флори за рахунок зникнення її термофільного ядра і посилення ролі мікротерм- них світлолюбних, мікротермних тіньолюбних та ксерсмезотермних видів. Синузії двох перших груп флори сформувались у плейстоцені, а останньої - в кінці пліоцену.
В ранньому голоцені в бореальний час на території Російської 193
рівнини були поширені соснові та березові ліси (Нейштадт, 1957; Пьявченко, 1958; Хотинский, 1969). На Україні спорово-пилкові спектри, які вказують на поширення в той час подібних лісів, відомі з багатьох районів (Koczwara, 1928; Kulczyiiski, 1930,• Tymraklewicz, 1935; Тоіра, 1935; Зеров, 1938, 1950; Koatyniuk, 1938; Артю- шенко, 1957, 1960, 1970; Зеров, Артюшенко, 1961, 1966, Пашкевич, 1963). Наявність в них пилку полинів, лободових та ефедри свідчить про те, що клімат у ранньому голоцені був сухий і відносно теплий. Від сучасного він. відрізнявся значно меншою кількістю опадів і відносно меншою різницею температур. В таких кліматичних умовах розподіл мікротермного світлолюбного та мікротермного тіньолюбного комплексів визначався в першу чергу рельєфними особливостями території. На початку середнього голоцену першими на межиріччях почали формуватися, на наш погляд, дубові ліси з синузіями, утвореними видами мікротермної світлолюбної та ксеромезотермної груп. Як вже зазначалось, вони поширились на місці остепнених березових та частково соснових лісів. На півдні спочатку це були діброви татарськокленово-кореневищноосокова, татарськбкленово-рябо- перлівкова, татарськокленово-горобейникова, а згодом - діброви та- тарськокленово-яглицева, татарськокленово-зірочникова та ін. На півночі в цей час поширювались діброви крушиново-молінієва та крушиново-конвалієва. Пізніше, в міру зволоження клімату, яке супроводжувалось вилуговуванням грунтів, поширилися дубові ліси з синузіями, утвореними видами мезотермної групи. Представлені вони були переважно дібровами ліщиново-яглицевою, ліщиново-зірочниковою, лі- щиново-волосистоосоковою та ін. Мабуть, на той же час припадав і поширення в північній частині України дубових лісів з мікротермною тіньолюбною групою видів - дібров ліщиново-веснівкової та ліщиново- квасеницевої. Останніми, вже на початку пізнього голоцену, поширилися дубові ліси з участю неморальних мезотермних видів атлантичного та субатлантичного клімату. Це такі види, як бук лісовий, граб звичайний, купина кільчаста, явір, осока трясучковидна, пренант пурпуровий та ін.
Отже, на Україні в голоцені в фазу змішаних лісів з елементами дубового лісу (середній голоцен) і в наступну фазу змішаних лісів з вологолюбними породами (пізній голоцен), встановлених Д.К.Зеровим (1950, 1952), можна виділити кілька стадій розвитку дубових лісів. Першою за часом є стадія розвитку та поширення дубових лісів з березою бородавчастою, другою - дубових лісів з ли194
пою серцелистою, третьою - дубових лісів з грабом звичайним і четвертою - дубових лісів з буком лісовим. Перші дві стадії розвитку лісів проходили на всій території України, третя - тільки на території поширення граба звичайного і четверта - бука лісового. Такому розвиткові в часі дубових лісів відповідає сучасне поширення перелічених деревних порід. Нами (19696) було показано, що пилок будь-якого виду анемофільних деревних порід з’являється в голоцені тим раніше, чим далі на схід цей вид поширений. Дійсно, східна межа берези бородавчастої по відношенню до східної межі липи серцелистої віддалена на 4500 км, відповідно липи серцелистої відносно граба звичайного - на 2900 км і граба звичайного відносно бука лісового - на 700 -км.
Крім зазначених чотирьох стадій, можна виділити ще п”яту стадію розвитку чистих дубових лісів. Максимум розвитку її, мабуть, відповідає максимумові пилку дуба звичайного на спорово-пилкових діаграмах. У широколистянолісовій та більшій частині лісостепової зон України максимум кількості пилку дуба звичайного наставав пізніше від максимуму кількості пилку липи серцелистої. В південніших районах має спостерігатись протилежна закономірність.
Таким чийом, широколистяні ліси в західній частині України (Поділля) проходили в часі такі стадії розвитку: березово-дубові, липово-дубові, дубові, грабово-дубові, буково-дубові та букові ліси. При проходженні цих стадій відбувалась трансформація грунтів - від вилугованих чорноземів до світло-сірих лісових. Грунти і ліси межиріч з дуже розсіченим рельєфом пройшли ці стадії швидше і тепер перебувають в останній стадії. Стадій березово-дубових та липово- дубових лісів на Поділлі не збереглося. Вони зараз повністю витіснені стадіями чисто дубових та грабово-дубових лісів. У східній частині України (Середньоруська височина) широколистяні ліси пройшли тільки три стадії - від березово-дубових до липово-дубових лісів. Під час дніпровсько-валдайського інтергляціалу п"ять стадій розвитку проходили всі широколистяні ліси України. Цілком зрозуміло, що кожній з перелічених стадій відповідав розвиток в дубових лісах певної групи видів. Наприклад, під час першої стадії спостерігалось посилення позицій мікротермної світлолюбної групи, видів, під час стадії чистих дубових лісів - неморальної ксеротер- мофільної, а під час стадії липово-дубових лісів - неморальної ме- зотермної групи видів.
Стадія березово-дубова відповідає початкові фази змішаних
195
лісів з елементами дубового лісу, встановленої Д.К.Зеровим (1950, 1952), яка синхронізується з підфазою ранньої частини теплого часу, за Ф.Фірбасом (Firbas, 1949), або з періодом поширення теплолюбних порід,за О.Сьродонем (&eodori, 1959), або з шостою зоною,за М.І.Ней- штадтом (1957). Стадію чисто дубову та липово-дубову можна синхронізувати відповідно з серединою та кінцем фази змішаних лісів, за Д.К.Зеровим, підфазою середньої частини теплого часу - за Ф.Фірбасом, з початком періоду панування теплолюбних порід, - за О.Сьродонем та з п"ятою зоною, за М.І.Нейштадтом. Стадії грабово-дубова та буково-дубова синхронізуються відповідно з фазою змішаних лісів з вологолюбивими породами за Д.К.Зеровим, з підфазою пізнього теплого часу - за Ф.Фірбасом, з періодом панування теплолюбних порід- за О.Сьродонем і з четвертою зоною - за М.І.Нейштадтом. Під час розвитку грабово-дубової та буково-дубової стадій, як видно на спорово-пилкових діаграмах, спостерігається зменшення кількості пилку дуба звичайного та ліщини звичайної, що є наслідком витіснення дуба звичайного грабом звичайним та буком лісовим. Крім того, в період, що відповідає другій половині буково-дубової стадії, Ф.Фір- бао (Firbas, 1949), О.Т.Артюшенко (1957, 1960, 1970), М.І.Нейштадт (1957), С.Сьродонь (inodori, 1959) відзначають зменшення кількості пилку широколистяних порід, пов"язане не тільки з діяльністю людини, а й з деяким похолоданням. О.Сьродонь (1959, с.527) з цього приводу пише: "Суворі зими, що повторюються, завдають місцями значної шкоди цілим ялицево-буковим деревоотанам, змінюючи їх територіальне розміщення. Весняні заморозки на межі ареалу бука лісового є, цілком вірогідно, головною причиною його відступу з висунутих на схід ділянок". Зникнення в другій половині пізнього голоцену з території Західного Полісся пилку бука лісового простежується на багатьох спорово-пилкових діаграмах (lymrakiewioz, 1935; Артюшенко, 1957, 1970). Г.О.Пашкевич (1963) знайшла пилок цього виду і в більш східних районах Полісся. Мабуть, сучасні кліматичні зміни сприяють зникненню на території Полісся вологолюбних неморальних видів і посиленню позицій мікротермних тіньолюбних видів.
З вивченням розвитку дубових лісів тісно пов"язане питання про вік їх неморальної флори в області міграційних флор, до якої належить територія України. Згідно з поглядами Й.К.Пачоського (1910, 1915), під час льодовикового, періоду клімат на території сучасної лісостепової зони України не був настільки суворий, щоб перешкодити зростанню широколистяних лісів. На його думку, вони 196
зникли в цей період лише з межирічних плато. Отже, за Й.К.Пачось- ким, принаймні частина неморальної флори України має третинний вік. З сучасних українських ботаніків з цим висновком погоджуються М.В.Клоков (1963), Б.В.Заверуха (1964), О.М.Дубовик (1964) та Г.К.Смик (1965). Ю.Д.Клеопов (1930, 1941) на підставі аналізу палеоботанічних, педологічних та ареалогічних даних прийшов до висновку про дніпровсько-валдайський інтергляціальний вік реліктової флори широколистяних лісів України. Цей висновок обстоювали Д.К.Зе- ров (1950, 1952, 1963), О.Т.Артюшенко,(1957, 1960, 1970), Д.К.Зе- ров, О.Т.Артюшенко (1961) та ін. За даними цих дослідників, широколистяні ліси під час валдайського гляціалу могли зберегтися на більшій частині території сучасної лісостепової зони і навіть в південній частині Полісся на ділянках з сильно погорбованим рельєфом, під захистом високих схилів гідрографічної мережі. Одним з перших на такий характер залежності між рельєфом та реліктовими видами неморальних лісів вказав Й.К.Пачоський.
Ми не можемо погодитись з висновком М.Я.Каца (1955), що заплавні дубові ліси лісостепу старіші, ніж дубові ліси на схилах річкових долин. По-перше, цей висновок не узгоджується з відсутністю у флорі заплавних дубових лісів видів з розірваним ареалом. По-друге, як ми вже відзначали, заплави річок є найбільш молодими геологічними утвореннями, на яких зростають дубові ліси. На перших стадіях свого розвитку заплави інтенсивно затоплювались і виповнювались піщаними осадами, а в цих умовах дубові ліси не могли рости.
Дальший розвиток поглядів,згідно з якими під час льодовикових періодів були суворі кліматичні умови, належить Г.б.Гросоету (1935, 1958, 1961, 1962, 1966, 1967). На його думку, клімат навіть валдайського льодовикового періоду виключав можливість вростання широколистяних лісів на території Російської рівнини, Південного Уралу і Західного Сибіру. Ці ліси льодовикову епоху пережили на Далекому Сході, Кавказі, в Криму і за Карпатсько-Альпійською дугою. Отже, на рівнинній частині України ці види є реліктами вологого і теплого часу голоцену. Свої висновки цей автор будує на численних палеогеографічних та палеоботанічних даних літературних джерел. Найбільш важливими з них, на наш погляд, є дані про існування на території України під час дніпровського та валдайського гляціалів мерзлоти, що доходила майже до Чорного та Азовського морів і наявність яко! відкидає можливість існування широколистяних лісів на цій території з третинного та льодовикового часів. Це дуже важливе заперечен-
197
ия зростання лісів не враховує проте, по-перше, значного коливання кліматичних умов залежно від рельєфу місцевості, а, по-друге, кліматичного мінімуму, для можливого збереження решток широколистяних лісів, який значно нижчий від кліматичного мінімуму північної межі сучасного їх ареалу. Не узгоджується цей висновок t з швидким поширенням широколистяних лісів під час алередського інтерстадіалу на території не тільки сучасного лісостепу, але й Полісся. Враховуючи віддаленість цих територій від Карпатського рефугіуму, незначну швидкість розселення широколистяних порід і коротку тривалість цього інтерстадіалу, можна дійти висновку про наявність на території лісостепу 1 південної частини Полісся місцевих рефугіумів неморальної флори. Останнім часом Г.О.Пашкевич (1970) одержала спорово-пилкові спектри, які свідчать про те, що в плейстоцені широколистяні ліси з рівнинної частини України не зникали повністю і росли принаймні на території Приазов"я. На користь можливого існування під час льодовикового періоду в лісостеповій зоні України неморальної флори свідчить такий факт. Ми вже зазначали, що другі тераси річок мають неовюрмський вік. Отже, дубові ліси, які досить Поширені на них в лісостеповій зоні, згідно з поглядами Г.Є.Грос- сета, повинні були б мати у своєму складі неморальні види з розірваним ареалом. Проте в цих лісах, так само як 1 в заплавних, реліктові неморальні види не трапляються.
Таким чином, розглянуте питання не може вважатись остаточно розв’язаним. Воно потребує дальшого вивчення Із застосуванням нових підходів і методів.
ЛІСОГОСПОДАРСЬКА ХАРАКТЕРИСТИКА ДУБОВИХ ЛІСІВ ТА ПЕРСПЕКТИВИ ЇХ ОХОРОНИ
Ліси України займають близько 13% території республіки, тоді як залісненість по Радянському Союзу в цілому становить 34%. За даними обліку лісового фонду, на І.І 1966 р. на Україні вкрита лісом площа становила 7190 тис.га. Це на 1240 тис-га більше, ніж у 1914 р. За 60 років експлуатації лісів до Великої Іовтневої революції (1854 - 1914 рр.) площа їх, за даними А.Г.Солдатова (1960), зменшилась на 30,5%.
Лісогосподарська характеристика
За значенням українські ліси віднесено до І і П груп. Ліси І групи становлять 34,1% усієї площі лісів держлісфонду. В них
198
забороняються суцільні рубки головного користування. Ліон П групи становлять 65,9% і належать до експлуатаційних. Площа лісів з перевагою дуба звичайного дорівнює 1420 тис.га або 26% всієї вкритої лісом площі республіки. За лісоросдинними зонами, прийнятими на Україні (Тюков, 1960; Пятницкий, Изюмский, 1966), ці ліси розподіляються нерівномірно - в Карпатах знаходяться лише близько 7% всієї площі дубових лісів,на Поліссі - 18, в Лісостепу - 64 і в Степу - 11%.Із згаданої вище площі дубові ліси, віднесені до І групи .становлять 680 тис.га,або 28% всіх лісів цієї групи, що свідчить про винятково важливу протиерозійну та водоохоронну їх роль. Пояснюється це тим, що переважна частина дубових лісів рооте на високих сильно розчленованих правих берегах річок лісостепової та байраках степової зон України. За класифікацією І.В.Тюріна (1949), за водоохоронним і захисним значенням дубові ліси належать до І і П класів.
За групами віку дубові ліси розподіляються таким чином. Молод- няки займають 764 тис.га,або 53,8%; середньовікові - 418 тис.га, або 29,4%, достигаючі - 124 тис.га, або 8,7% і спілі та перестійні - ІІЗ тис.га, або 7,9%. Загальний запас дуба високостовбурного дорівнює І07 240 тио.м3, а дуба низькостовбурного - 2923 тис.м3.
В умовах високої густоти населення, розвиненої промисловості і відносно невеликої залісненості проблема підвищення продуктивності дубових лісів набуває особливого значення. Лісові господарства України досягли в цьому значних успіхів.£ усіх лісах кожні 10 років провадять ліоовпорядкування.Основні заходи пов"язані з доглядом, експлуатацією та лісовідновленням, здійснюють відповідно з планом розвитку лісового господарства УРСР, внаслідок чого з року в рік збільшується продуктивність лісів і зменшується обсяг рубок головного користування. Всі зрубувані лісосіки своєчасно заліснюються, а кількість зрубаної за всіма видами рубок деревини дорівнює середньому приростові деревини в лісах республіки. З кожним роком за рахунок лісових культур, створюваних на невкритих лісом схилах ярів, балок, на пісках та інших незручних для сільськогосподарського користування землях, збільшується площа лісів, у тому числі й дубових.
В середньому по республіці дуб високостовбурний має П клас бонітету, а дуб низькостовбурний - Ш. Серед асоціацій найбільшою продуктивністю відзначаються ті, що формуються в свіжих та вологих екотопах (діброви ліщиново-трясучковидноосокова, ліщиново-яглицева.
199
ліщиново-маренкова, ліщиново-зірочникова, свидиново-гірськоосокова, татарськокленово-яглицева та ін.), а найменшо» - які формуються в дуже сухих екотопах (більшість асоціацій дубових лісів татарсько- кленових та всі асоціації дубових лісів кизилових і скумпієвих). Середній запас спілих насаджень асоціацій першої групи, поширених переважно на території широколистянолісової та більшій частині лісостепової зон, дорівнює 210-230 м3/га, а насаджень асоціацій другої групи, поширених на території степової та південної частини лісостепової зон - 120—ІВО м3/га. За цим показником вони поступаються перед сосновими, буковими та ялиновими лісами. Значно вищі запаси кращих дубових насаджень по окремих зонах. Так, за даними лісовпорядних партій, кращі насадження в степовій зоні на території То- резького лісгоспзагу віком 80 років при повноті 0,8 мають запас 210 м3/га;в лісостеповій зоні на території Романського лісгоспзагу віком ПО років при повноті 1,0 - 508 м3/га, на території Іллінець- кого лісгоспзагу віком 70 років при повноті 1,0 - 440 м3/га; в широколистянолісовій зоні на території Бердичівського лісгоспзагу при повноті 0,8 - 290 м3/га. За даними С.М.Стойка (1960), кращі дубові насадження Закарпатської області мають запас 500-600 м3/га. Наведені дані свідчать про високі потенціальні можливості дубових насаджень. Повністю використати їх перешкоджають, на наш погляд, - переважно невисока повнота насаджень, випас худоби в лісах, заміна дуба звичайного малоцінними породами, невідповідність створених культур типові умов місцезростання, ураження шкідниками лісу, перевищення розміру розрахункової лісосіки, досить висока питома вага порослевих насаджень, недостатнє використання природного поновлення та ін.
Повнота насаджень впливає не тільки на продуктивність, але й на якість лісопродукції, тому що умови зростання в зімкнутих насадженнях сприяють кращому формуванню ділової деревини. Зниження повноти деревостану спілих насаджень дуба звичайного лише на 0,1 призводить до втрати 40-60 м3/га цінної деревини. Тому заходи по створенню високоповнотнях деревостанів є одним з найбільш важливих завдань лісового господарства. Середня повнота дуба високостовбурного дорівнює 0,72, низькостовбурного - 0,66. Загалом найнижча вона у дуже сухих дубових лісів степової зони і найвища - у свіжих і вологих дубових лісів лісостепової зони, Прикарпаття та Волино- Подільської височини. В окремих лісгоспзагах Й досі площа низько- повнотннх насаджень дещо перевищує площу високоповнотних. Наприк200
лад, у Дубровицькому лісгоспзагу дубові насадження з повнотою 0,3- 0,5 становлять 31,6%, а з повнотою 0,9 - 1,0 - лише 0,8%; в Аітир- ському лісгоспзагу відповідно 7 і 3%,Полтавському - 15 і 7%, Котов- ському 16 і 3% і т.п. Отже, тільки через зниження повноти деревостану втрати деревини становлять близько 30%.
Випасання худоби в дубових лісах в господарському відношенні не в доцільним. Більшість конфекторів ярусу травостою і супутніх видів належать до малоцінних або неїстівних і не можуть мати істотного значення для годівлі худоби. Випасання худоби завдає лісовому господарству значної шкоди, тому що худоба знищує підріст та підлісок, а такок ущільнює грунт, порушуючи тим самим його екологічний режим. Під час дощу верхні горизонти грунту швидко перезволожуються, а в сухий період швидко пересихають, що сприяв висушуван- нюгрунту і ксерофітизації трав'янистого покриву. Ущільнення погіршує також азотний баланс грунту, при цьому різко знижуються процеси азотфіксації та нітрифікації. Через погіршання еколого-бі©логічного грунтового режиму і суто механічне пошкодження рослинності формуються сукцесійні стадії фітоценозів,
В класі асоціацій дубових лісів ліщинових інтенсивний випас викликає спочатку розрідження ліщинового підліску та заміну його на бруслиновий з бруслини бородавчастої, а пізніше - повне знищення підліску. Цей процес супроводжується задернінням грунту. Панівними в травостої північної частини ареалу класу є костриця червона ( Festuca .rubra L.), а південної - тонконіг вузьколистий. Ліси набувають паркового характеру (рис.35). Площа таких лісів тепер різко зросла і продовжує збільшуватись. В цілому ряді лісництв, наприклад Дорогинському Фастівського лісгоспзагу, Іржавському Мрин- ського лісгоспзагу, Старосалтівському Вовчанського лісгоспзагу, Білозерському Переяслав-Хмельницького лісгоспзагу паркові діброви займають вже десятки кварталів. Є навіть випадки майже повного знищення природного травостою в лісах всього лісництва (Часовеняр- ське лісництво Слов'янського лісгоспзагу).
В класі асоціацій дубових лісів татарськокленових підлісок із клена татарського заміщується підліском з глоду українського та обманливого, шипшини повстистої та інших видів. В травостої починають панувати тонконіг дібровний, грястиця збірна, кінський часник черешковий, а в найбільш сухих умовах - костриця борозниста. Приріст дуба різко знижується , він починав суховершити і гине. Деревостани розріджуються, з'являється поверхневий стік і розви201
вається ерозія. В умовах степової зони такі ділянки надалі ставів господарчо непридатними. Особливо шкідливі наслідки випасання худоби в верхніх частинах південних схилів балок, де навіть незначне порушення біологічної рівноваги призводить до швидкого розрідження деревостанів та змиву грунтового покриву.
Через недостатні в минулому заходи в догляді за лісом, застосування невідповідних до біологічних особливостей дуба як світлолюбної породи суцільних лісосічних рубок дуб заміщують менш цінні породи. Особливо різко цей процес виявлено на Правобережжі України, де внаслідок експансії граба звичайного на місці дубових лісів дуже поширились грабняки, що призводить до зниження продуктивності лісу ІУ класу віку на 60-110 м3/га. Зараз досить частими є випадки, коли граб віком 15-30 років вже повністю витіснив підлісок і створив густий полог зімкненістю 1,0. В багатьох лісництвах Прикарпаття, Поділля, Західного та Правобережного Полісся такі ділянки займають більше половини площі дібров. Природним шляхом дуб звичайний на них вже не поновиться. На нашу думку, в зв"язку з експансією граба звичайного, повне вирубування підліску, широко застосовуване лісогосподарствами на ділянках, відведених під рубки головного користування, недоцільне. Підлісок в дубових лісах виконує дуже важливу біологічну функцію "страва" цілісності фітоценозу. Тому, якщо орієнтуватись на природне поновлення дубових лісів, яке на жаль, не проводять, підлісок необхідно зберігати. Значно рідше, переважно у вологих екотопах, відбувається заміна дуба звичайного на осику.
В більшості ліогоопзагів степової і лісостепової зон площа, зайнята дубовими лісами, за останні роки збільшується через створення нових культур дуба. На жаль, в лісгоспзагах широколистяно- лісової зони вона скорочується, тому що на місці дубових лісів створюють культури сосни звичайної і навіть ялини європейської. Так, в Кам"янець-Подільоькому лісгоспзагу за останні 10 років близько 10% площі вирубаних дубових лісів було зайнято культурами ялини європейської 1 близько 5% - культурами сосни звичайної. Ще гірший стан у поліських ліогоопзагах (Дубровицькому, Цумансько- му, Словечанському, Малиноькому та ін.) , де під культури сосни звичайної відводять 15 - 30% площі вирубаних дубових лісів. Причина цього полягає в недоліках методичних оцінок типів умов місцезростання, коли за наявністю поодиноких екземплярів видів, які є Індикаторами суборевих умов, наприклад орляка звичайного, ожи- 202
ки волосистої, підмаренника, весняного, ці умови оцінюють як відносно біллі, придатні лише для культур сосни звичайної.
Викликане господарською діяльністю, зокрема випасанням худоби, порушення біологічної рівноваги і послаблення життєвості насаджень призводить до посилення інвазій шкідників лісу. Особливо великі площі дібров пошкоджуються в лісостеповій і степовій зонах дубовою зеленою листовійкою та зимовою л"яденицею. За останні 10 років в окремі роки в Полтавському лісгоспзагу було пошкоджено від
2 до 9 тис. га або від 19 до 80% площі всіх дубових лісів; в Чигиринському - від 3 до 7 тис.га, в Кременчуцькому - від 1,5 до
3 тис.га, в Звенигородському - від 1,5 до 11,5 тис.га, в Іванівоь- кому - від 1,5 до 4 тис.га і т.п. За даними В.Ф.Самарина (1969), при ураженні листя зазначеними шкідниками на 80% середньорічний приріст знижується на 31%, що становить 1,76 м3 абсолютної втрати деревини на І га насаджень.
Незважаючи на низку заходів, спрямованих на раціональніше використання лісів і скорочення рубок головного користування, в деяких лісгоспзагах широколистянолісової І лісостепової зон останні все ж перевищують розрахункову лісосіку. Так, по Кам"янець- Подільському лісгоспзагу таке перевищення становить 30%, по Іранському лісгоспзагу - 20%, по Звенигородському ліогоопаагу - 25%. А це призводить до зниження віку рубок головного користування, встановленого генеральним планом.
Хоча площа порослевих низькопродуктивних дубових лісів поступово зменшується, проте в ряді лісгоспзагів питома вага їх все ще досить висока. Практично всі ліси сухих і дуже сухих екотопів, зосереджені переважно в південній смузі лісостепової та північній смузі степової зон, - малопродуктивні порослеві насадження. Заміна їх насіннєвими пов"язана з значними труднощами і витратами. В окремих лісгоспозагах більш північних районів низькопродуктивні порослеві насадження становлять 2-4%.
Поновлення дубових лісів на Україні майже повністю засноване на штучному лісовідновленні. Пояснюється це не відсутністю в достатній кількості насіннєвого підросту дуба звичайного. Як показали одержані нами дані, більшість асоціацій дубових лісів, на які припадає близько 70% їх площі, має потенціальну можливість самопоновлення. Добре проходить відновлення в малопорушених насадженнях дібров лішиново-яглицевої, ліщиново-зірочникової, лі- щиново—зеленчукової, ліщиново-волосистоооокової, свидиново-гір- 203
ськоооокової, татарськокленово-яглицевої та татарськокленово- зірочникової. В середньому на одному гектарі в перелічених асоці. аціях нараховується близько 15-30 тисяч екземплярів підросту дуба звичайного віком 2-5 років 1 до 1-5 тисяч віком 6-Ю років. Максимальна кількість підросту досягає 200 тисяч екземплярів на І га. Задовільно проходить відновлення в насадженнях дібров ліщи- ново-трясучковидноосокової, ліщиново-маренкової, татарськокле- ново-волосиотоосокової, ліщиново-гірськоооокової та ліщиново-конвалієвої. В середньому на одному гектарі в них нараховується 7-9 тисяч екземплярів підросту дуба звичайного віком 2-5 років. Проте під впливом випасання худоби більша частина підросту гине, а на ділянках, де він лишається, заходів для збереження його звичайно не вживають,і згодом ці ділянки відводять під культури. Погано протікає відновлення тільки в насадженнях дібров татарсько- кленово-кореневищноосокової, татарськокленово-рябоперлівкової татарськокленово-горобейникової та у всіх асоціацій дубово-кизилових і дубово-скумпієвих лісів. Проте і в цих лісах при застосуванні лише вибіркових рубок головного користування і проведенні всіх заходів догляду за лісом можна одержати достатню кількість надійного насіннєвого підросту для самовідновлення. Мабуть, частину площ дубових лісів треба обов'язково поновлювати за рахунок природного насіннєвого підросту дуба звичайного. Це необхідно робити тому, що штучні насадження дубових лісів біологічно менш стійкі, ніж природні, і повна заміна останніх культурами може призвести до небажаних наслідків. Раціональне використання дубових лісів означає, в першу чергу, підвищення їх продуктивності і не стільки збільшення площ, зайнятих дубовими лісами (це не завжди можливо і доцільно), скільки зростання запасів деревини на одиницю площі. Останнє має стати принциповою основою розширеного поновлення лісів. Для цього необхідно в першу чергу усунути відзначені вище недоліки. Низькопродуктивні порослеві низько- повнотні та малоцінні насадження необхідно повністю реконструювати, що поки провадиться в недостатньому обсязі. Випасання худоби в лісах необхідно заборонити.
На практиці слід точно розрівняти категорії дубових лісів з задовільним та незадовільним поновленням. В перших з них треба орієнтуватись на природне ліоопоновлення і лише в другій - переважно на штучне. Реалізація тільки цих заходів підвищить продуктивність дубових лісів удвічі. Не менш перспективне 204
також введення змішаних культур дуба з високопродуктивними і цінними екзотами. За даними І.Н.Гегельського (1960), Б.Й.Логінова, М.І.Гордієнко (1963), І.В.Агаркова, Б.М.Махмета, М.І.Шендрикова (1963), найбільш придатними можуть бути дуб червоний, модрина європейська, горіх чорний та ін. Такі насадження продуктивніші від чисто дубових на 30-50%.
Перспективи охорони дубових лісів
Необхідною умовою раціонального використання дубових лісів є науково обгрунтоване виділення об'єктів охорони їх як частини єдиного біологічного комплексу. По республіці в цьому напрямку за останні роки досягнуто великих успіхів. У 1960 р. прийнято Закон про охорону природи; у 1967 р. при Раді Міністрів УРСР створено Державний Комітет по охороні природи, який провів велику роботу по виявленню та охороні ряду об'єктів; створено два нові заповідники і т.д. На жаль,в жодному заповіднику на Україні немає дубових лісів. Лише дуже незначна частина таких лісів охороняється в заказниках місцевого значення.
Науковий підхід до розв"язання проблеми охорони дубових лісів повинен, на нашу думку, здійснюватись на підставі комплексної оцінки їх як з точки зору господарського значення і можливості поновлення, так і з точки зору наукової цінності. Оскільки всі дубові ліси в тій чи іншій мірі тепер змінені і надалі в міру інтенсифікації природокористування вони підлягатимуть це більшим змінам, виділення особливо цікавих в науковому відношенні об"єктів набував першочергового значення. Якщо виходити з цих позицій, то охороні підлягають добре збережені дубові ліси найбільш типові для різних зон, окремі ділянки асоціацій на границі кліматичних, едафічних та фітоценотичних меж, реліктові, ендемічні та рідкісні асоціації. Охорона таких об'єктів має першочергове значення для з’ясування широких теоретичних питань; пов'язаних з реконструкцією клімату минулих часів, пізнанням центрів формування флори і рослинності, встановленням їх віку,- вивченням міграційних шляхів і генетичних зв'язків, закономірностей формування структури та взаємозв'язків фітоценозів і нарешті - пізнанням процесів видоутворення та еволюції рослинного світу, бо процес видоутворення у рослин відбувається в популяціях, які складають рослинні угруповання. Виходячи з цього ми пропонуємо зробити заповідними такі ценотаксони дубових лісів.
205
1. Найбільи типові та високопродуктивні дубові ліси, що підлягають охороні.
Широколистянолісова зона: І) квартали 35-37 і 58-65 Дзер- жинського лісництва Бердичівського лісгоспзагу; 2) квартали 37- 63 Піщевського лісництва Новоград-Волинського лісгоспзагу; 3) квартали 3-7 Григуроького лісництва та квартали 82-96 Корабельного лісництва Житомирського лісгоспзагу; 4) квартали 84-91 Гущин- оького лісництва Любомльського ліогоспааіу. В цих кварталах добре збереглися найбільш типові для Полісся дубові ліси П класу бонітету віком 9U-I40 років, які належать до дібров ліщиново-тря- сучковидноосокової, ліщиново-волосистоосокової та ліщиново-веснів- кової.
Лісостепова зона: І) квартали 55-70 Улашківського лісництва Чортківського лісгоспзагу; 2) квартали 18-55 Савранського і квартали 19-37 Слюоарівоького лісництва Балтоького лісгоспзагу; 3) квартали 20-45 Голочаноького лісництва Голованівського лісгоспзагу; 4) квартали 22-48 Ольшаницького лісництва Богуславсько- го лісгоспзагу; 5) квартали 72-95 Іскрівського лісництва Полтавського лісгоспзагу; б) квартали 71-85 Вовчиноького лісництва і квартали 20-43 Коломацького лісництва Жовтневого лісгоспзагу. В цих кварталах збереглися найбільш типові для Подільської,Придніпровської та Середньоруської височин дубові ліси І-П класу бонітету віком 80-120 років, вони належать до дібров свидиново- гірськоосокової, ліщиново-волосистоосокової, ліщиново-гірсько- осокової, діброви ліщиново-яглицевої, ліщиново-зірочникової, татарськокленово-яглицевої та татарськокленово-горобейникової.
Степова зона: І) квартали 12, ІЗ, 30-33, 50, 71 Новопсковсь- кого лісництва Старобільського лісгоспзагу; 2) квартали 5-8, 39 Біловодського лісництва Деркульського лісгоспзагу; 3) квартали 3-9 Сніхиянського лісництва Торезького лісгоспзагу; 4) квартали 56-63 Штергреського лісництва Іванівського лісгоспзагу. В цих кварталах збереглись найбільш типові й продуктивні ліси Ш-ІУ класу бонітету віком 70-90 років, що належать до дібров татарськокленово-аірочникової, татарськокленово-рябоперлівкової, татарськокленово-мікелієвоосокової та татарськокленово-горобей- никової.
2. Найбільш рідкісні дубові ліри на межі свого ареалу, що підлягають охороні.
Широколистянолісова зона: І) квартал 28 Грицівського ліс206
ництва Старокостянтинівського лісгоспзагу (південна межа діброви ліщиново-кваоеницевої); 2) околиці о.Шепаровців Коломийського району Івано-Франківської області (південно-східна межа діброви ліщиново-трясучковидноосокової з участю ряду гірських видів); 3) квартали 4-18 Іржавського лісництва Мринського лісгоспзагу (південна межа діброви ліщиново-жіночобезщитникової); 4) квартал 19 Моложавського лісництва Городнянського лісгоспзагу (східна межа діброви ліщиново-трясучковидноосокової).
Лісостепова зона: І) квартали 41-52 Бершадьського лісництва Бершадьського лісгоспзагу (східна межа дібров свидиновб-парвсько- осокової та ліщиново-парвськоосокової); 2) квартали 39 - 46 , 67, 68 Старосалтівського лісництва Вовчанського лісгоспзагу (добре збереглись ділянки діброви татарськокленово-кореневищноосокової)|
3) квартал 82 Шполянського лісництва Звенигородського лісгоспзагу (північна межа діброви татарськокленово-мікелієвоосокової);
4) квартал 14 Будейського лісництва Балтського лісгоспзагу (південна межа діброви ліщиново-гірськоосокової).
Степова зона: І) квартали 45-53 Красноградського лісництва Красноградського лісгоспзагу (південна межа діброви крушиново- конвалієвої); 2) квартали 43-45 Великобурлуцького лісництва Ку- п"янського лісгоспзагу (південна межа дібров ліщиново-зірочни- кової та ліщиново-яглицевої); 3) квартал 57 Новопсковоького лісництва Старобільського лісгоспзагу (південна межа діброви ліщино- во-волосистоосокової).
3. Реліктові та ендемічні дубові ліси, що підлягають охороні. До реліктових належать усі асоціації дубово-кизилових та ду- бово-скумпієвих лісів, які, можливо, мають третинний вік. Саме вони найбільш насичені рідкісними субсередземноморськими видами з розірваним ареалом. У багатьох з них східна, північна та південна межа ареалу пов"язані з цими лісами. Після рубок головного користування вони не поновлюються, бо ростуть на дуже крутих і сухих схилах південної експозиції. Щоб зберегти їх, необхідно зробити заповідними майже всі ділянки, насамперед на крутих схилах:в Нирківському лісництві (квартали 6-2Ї) Бучацького лісгоспзагу; Мурованокуриловецькому лісництві (квартали 45-52) Могилів-Подільського лісгоспзагу; Дмитріевському лісництві (квартали 3-29) Чорноліського лісгоспзагу; Пановецькому лісництві (квартали 55-56) Кам"янець-Подільського лісгоспзагу; Смілянсько- му лісництві (квартали І0І-І02) Смілянського лісгоспзагу та в Павлівському лісництві (квартали 34-40) Котовського лісгоспзагу.
207
XXX
На території України дубові ліси поширені в широколистяно- лісовій, лісостеповій та степовій зонах. В геоботанічній літературі дубові ліси звичайно розуміють широко і відносять до них буково-дубові,яглицево-дубові, вільхово-дубов і, ясеново-дубові, кленово-дуо'ові, липово-дубові, сосново-дубові, грабово-дубові, кленово-липово-дубові і чисто дубові ліси. Найбільш поширені сосново-дубові, грабово-дубові, липово-дубові та чисто дубові ліси, які складають близько 90% всієї площі дубових лісів республіки, що становить 1420 тисяч га.
Сосново-дубові ліси трапляються на Поліссі та на других терасах річок лісостепової і почасти степової зон. Ростуть вони на найбідніших для дубових лісів дерново-підзолистих супіщаних грунтах і є проміжною ланкою між дубово-сосновими та чистими дубовими лісами. Представлені сосново-дубові ліси класами асоціацій ліщинових та крушинових. Сосново-дубові ліси ліщинові трапляються в південній частині Полісся і на других терасах річок лісостепової та отепової зон Лівобережжя України, на Правобережжі їх майже немає. Найпоширенішими є: сосново-дубові ліси ліщиново-орляко- ві ( Pineto-Quercetua оолу1ово-рЬелі<11овилі),ліщиново-ввонівкові (P.-Q.coryloso-majantheBoaum), ліщиново-костяницеві (P.-Q.corylos o-rubosum (saxatilis)), лІЩИНОВО-ЗіроЧНИКОВІ (P.-Q.coryloso- stelleriosum), ЛІЩИНОВО-ЯГЛИЦЄВІ (P. Q. ooryloso-aegopodiosum), ліщиново-копитнякові (P.-Q.ooryloso-asarosum). Сосново-дубові ліси крушинові трапляються в південній та середній частинах Українського Полісся та на других терасах річок північної частини лісостепу. Представлені вони сосново-дубовими лісами: крушиново- чорницевими (Pineto-Qusrcetim fxaaguloso-vaociniosum),крушиново- костяницевими (P,-Q.franguloso-rubosw»(saxatilis), крушиново- орляковими (P.-Q.fraj3galoso-pteridioaua) та крушиново-конвалієвими ( P.-Q.fxaaguloso-ooovallariosuK). Характерною рисою видового складу перелічених асоціацій дубово-соснових лісів є незначне переважання бореальних видів,таких, як ожика волосиста, грушанка круглолиста, орляк звичайний, сосна звичайна, веснівка дволиста, чорниця, одинарник європейський, квасениця звичайна, рамішія однобока, костяниця, куничник тростиновий, вербозілля звичайне, перстач прямостоячий та ін.
208
Грабово-дубові ліси поширені в Карпатах, на Поділлі, Придніпровській височині та Поліссі. Південна межа їх ареалу проходить через Котовський, Чорноліський та Полтавський лісгоспзаги, а східна - через Прилуцький та Корюківський лісгоспзаги. Ростуть грабово-дубові ліси на широко- та вузькогорбистих межиріччях з світло-сірими та сірими лісовими грунтами. Найпоширенішими з них е: грабово-дубові ліси волосистоосокові (Cacpineto-Quercetum са- ricosum(piloaae), яглицеві (C.-Q.aegopodiosuni) та зірочникові ( c.-Q.stellarioauB). В цих лісах, на відміну від сосново-дубових, переважають неморальні види, серед яких найбільш постійними, крім едифікатора, семіедифікатора та конфектора, є бруслина бородавчаста, свидина кров"яна, ліщина звичайна, клен звичайний, липа серцелиста, медунка темна, куцоніжка лісова, фіалка лісова, просянка розлога, чина весняна, маренка запашна, конвалія звичайна, гравілат міський, зеленчук жовтий, купина багатоквіткова, ранник вузлуватий, копитняк європейський, дріоптерис чоловічий, переліска багаторічна, зубниця бульбиста, підлісник європейський, костриця велетенська, коручка широколиста, астрагал солодколистий та ін. Бореальні види трапляються в незначній кількості в північній частині ареалу грабово-дубових лісів.
Липово-дубові ліси поширені переважно в лісостеповій зоні Лівобережжя України. Острівні ділянки їх трапляються на Прикарпатті, Поліссі та Придніпровській височині. Ростуть вони на пла- корах із світло-сірими, сірими та- темно-сірими лісовими грунтами. Біля південної межі свого ареалу (північна частина степової зони) липово-дубові ліси ростуть тільки в нижній частині північних схилів глибоких ярів. Представлені вони липово-дубовими лісами ВОЛОСИСТООСОКОВИМИ ( Tilieto-Quercetuxn caricosumCpilosae), зірочниковими (C.-Q.stellarіоашв), яглицевими ( T.-Q.aegop odiosum), липово-дубовими лісами лішиново-маренковими (T.-Q.coryioso- asperiliosum), ліщиново-волосистоосоковими (T.-Q.coryloeo-oairicoeuB pilosae), ліщиново—яглицевими (T.-Q.ooiyloso-aegopodioaum) та
ліщиново-зірочниковими ( T.-Q.coryloeo~stellariosiue).jepeBOCTaH липово-дубових лісів, як і грабово-дубових, двохярусний із зімкненістю крон 0,1-1,0. Перший ярус створює дуб звичайний, інколи з участю ясена звичайного, рідко - берези бородавчастої. В другому ярусі переважає липа серцелиста з домішкою клена звичайного. Ярус підліску зімкненістю 0,2-0,3 формується тільки у чотирьох останніх асоціацій. Крім ліщини звичайної, до нього входять 209
бруслина бородавчаста, свидина кров"яна, бруслина європейська, черемха звичайна, клен татарський та ін. Травостій має проективне покриття 20-45%. Крім вищенаведених конфекторів, його створюють майже виключно неморальні види, такі, як куцоніжка лісова, перлівка поникла, тонконіг дібровний, астрагал солодколистий, чина весняна, копитняк європейський, конвалія звичайна, зубниця бульбиста, гравілат міський, переліска багаторічна, купина багатоквіткова, фіалка дивовижна, ранник вузлуватий, просянка розлога, дзвоники кропиволисті, медунка темна та ін.
Чисто дубові ліси поширені на Поліссі, Поділлі, в Карпатах, на Придніпровській і Середньоруській височинах та на Донецькому кряжі. Ростуть вони на плакорах, в ярах та долинах річок. За флористичним складом, генезом та екологічними умовами ці ліси поділяються на три еколого-генетичні ланки: ацидофільну, нейтрофільну та базифільну.
Ацидофільні дубові ліси формуються на дерново-підзолистих грунтах, які підстелюються архейськими та протерозойськими гранітами, мореною або флювіо-гляціальними відкладами. Поширені вони в північній та північно-західній частинах ареалу формації і характеризуються переважанням бореальних (дріоптерис гребенястий, ожика волосиста, веснівка дволиста, квасениця звичайна, грушанка круглолиста, підлісник європейський, орляк звичайний, рамішія однобока, чорниця, фегоптерис дубовий та ін.) та участю неморальних балтійських видів (дуб звичайний, костриця велетенська, ясен звичайний, зеленчук жовтий, переліска багаторічна, жимолость пухната, фіалка лісова, печіночниця звичайна, яглиця звичайна, медунка темна, зірочник лісовий та ін.). До цієї ланки належить б асоціацій.
Нейтрофільні дубові ліси формуються на сірих лісових грунтах, підстелених лесом, або на дернових опідзолених грунтах, підстелених голоценовим алювієм. Вони займають середню частину ареалу формації і створені переважно балтійськими неморальними мезо- трофними видами (клен звичайний, яглиця звичайна, цибуля ведмежа, копитняк європейський, астрагал солодколистий, осоки пальчаста і волосиста, лінійна звичайна, зубниця бульбиста, медунка темна, бруслини європейська та бородавчаста, зірочник лісовий, липа серцелиста,підлісник європейський та ін.). До цієї ланки належить І? асоціацій.
Базифільні дубові ліси формуються на чорноземних і дерново- 210
карбонатних грунтах, підстелених силурійськими та девонськими сланцями, крейдою, червоно-бурими глинами або лесом. Поширені вони в південній частині ареалу формації і характеризуються переважанням неморальних субсередземноморських базифільних мега- трофних видів (клен татарський, віхалка гілляста, півники зла- колисті, перлівка ряба, чистець прямий, берест, горошок горохоподібний, барвінок трав'янистий, клен польовий, ооока Мікелі, дерен справжній, скумпія звичайна, ясенець білий, горобейник пурпурово-голубий, гордовина, ластовень виткий, купина широколиста, лазурник трилопатевий, смовдь оленяча та ін.). До цієї ланки належить 20 асоціацій.
На Україні проходить південна межа всіх 43 асоціацій субформації дубових лісів з дуба звичайного, східна межа 15 асоціацій та північна межа 28 асоціацій. За класифікацією франко-швей- царської школи всі асоціації дубових лісів належать до трьох класів рослинності - Quercetea toboximpoCiaeaa, QuorcoaFageCea та Querceta a pubesceati-petraeae.
В досліджуваних лісах трапляється близько 530 видів вищих рослин - близько 25% усього видового окладу рослинності на території їх поширення. З числа названих 230 видів є звичайними, в їх складі 140 видів - основне флористичне ядро дубових лісів. 210 видів поширені мало і 90 - невластиві дубовим лісам.
Всі види, що ростуть в дубових лісах, належать до 69 родин та 295 родів. При цьому до 7 родин (Складноцвіті, Розові, Злакові, Жовтецеві, Зонтичні, Губоцвіті, Бобові) входять близько 47% усіх видів; до 14 родин (Ранникові, Осокові, Маренові, Лілійні, Гвоздикові, Хрестоцвітні, Дзвоникові, Шорстколисті, Багатоніж- кові, Молочайні, Первоцвіти!, Орхідні, Фіалкові, Жимолостеві) - 33% видів і до решти 48 родин - 20% видів. За кількості видів на першому місці стоять Складноцвіті (50), на другому - Розові (38) і на третьому - Злакові (33). Серед групи звичайних видів на першому місці стоять Розові (24), на другому - Злакові та Губоцвіті (по 14) і на третьому - Складноцвіті (ІЗ), а серед групи малопоширених видів відповідно-Складноцвіті (21), Жовтецеві (14) та Розові і Зонтичні (по II).,Серед родів найбільшу кількість видів мають Осока (16), Підмаренник (II) та Дзвоники (9).
Серед групи конфекторів, яка охоплює 26 видів, переважають Осокові (6), Злакові (3), Багатоніжкові, Деренові та Лілійні
211
(по 2). Таким чином, роль родин так само як і родів в різних групах видів неоднакова.
Екологічний аналіз видів показав, що в дубових лісах України найгірше представлені гігрофіти,які становлять близько 2% постійних видів (вербозілля звичайне, гадючники в"язолистий та голий, вільха клейка та ін.). Дещо більше в цих лісах ксерофітів. Вони становлять близько 4% видового складу постійних видів (горо- бейник пурпурово-голубий, залізняк бульбистий, осока Мікелі, карагана кущова, шавлія лучна та ін.). Значно краще представлені в дубових лісах мезогігрофіти, які становлять 11% (куничник тростиновий, вербозілля лучне, розрив-трава звичайна, безщитник жіночий та ін.) та мезоксерофіти - 21% (буквиця лікарська, дзвоники персиколисті, ломиніс прямий, фіалка шершава, барвінок трав'янистий та ін.). Переважають мезофіти - 62% (яглиця звичайна, зірочник лісовий, маренка запашна, копитняк європейський, підлісник європейський та ін.).
За поділом щодо родючості грунту в дубових лісах найгірше представлені оліготрофні види, які, за винятком сосни звичайної, в дубових лісах не ростуть. Олігомезотрофнпх видів налічується близько 10% (веснівка дволиста, орляк звичайний, вероніка лікарська, ожика волосиста та ін.). Ще більша участь (39%) мегатроф- них (смілка широколиста, фіалки шершава та запашна, кропива дводомна, ломиніс цілолистий, гордовина,шоломниця висока, переліска багаторічна та ін.) видів. Переважають мезотрофні види, які складають 51% (яглиця звичайна, зірочник лісовий, маренка запашна, куцоніжка лісова, орляк звичайний, фіалка лісова та ін.).
Щодо кислотності грунту, то найгірше представлені ацидофіт- ні види,вони становлять близько 6% (ожика волосиста, куничник тростиновий, вероніка лікарська, орляк звичайний, віснівка дволиста, перстач прямий та ін.). Ацидонейтрофітні (горлянка повзуча, дріоптерис чоловічий, крушина ламка, золотушник звичайний, горобина звичайна та ін.) та базифітні (гордовина, горобейник пурпурово-голубий, перлівка ряба, осока Мікелі, ломиніс цілолистий, куцоніжка пірчаста, купина широколиста, шоломниця висока та ін.) види становлять по 25%. Найкраще представлені нейтрофітні види, - 44% (медунка темна, клен звичайний, копитняк європейський, осоки стягнута та волосиста, дзвоники ріпчастовидні, яглиця звичайна, бруслина європейська, ясен звичайний, жовтець золотистий та ін.).
212
Якщо розглядати екологічні групи видів дубових лісів в межах зон, то співвідношення їх буде іншим. В широколистянолісовій зоні на території Полісся базифітних, ксерофітних і мегатрофних видів в дубових лісах практично немає. Мезоксерофітні складають 7%, гігрофітні та мезогігрофітні-тільки 3%, ацидофітні та ацидо- нейтрофітні - 4%, ксерофітні і мезоксерофітні - 37%, базифітні - 44%, нейтрофітні - 52% і мезофітні - 55%.
У формуванні дубових лісів України брали участь види двох філоценогенетичних та одного селектоценогенетичного комплексів. Неморальний філоценогенетичний комплекс в дубових лісах представлений мезо- та ксеромезотермною еколого-ценогенетичними групами, бореальний-мікротермною тінелюбною та мікротермною світлолюбною зколого-ценогенетичними групами. А плейстоценовий селек- тоценогеяетичний комплекс представлений мезофільними мікротерм- ними альпійською та лучною, а також ксеромезотермною степовою світлолюбними еколого-ценогенетичними групами. Кожна з еколого- ценотичних груп складається з певної кількості історичних свит рослинності.
Дубові ліси мають винятково важливе водоохоронне, протиерозійне та кліматичне значення. З одного боку, вони сприяють накопиченню снігу та збільшенню опадів, а з другого - регулюють сток з басейнів річок і збільшують водні ресурси території, вкритої лісом. Характерно, що дубові ліси збільшують водні ресурси в усіх зонах України, проте в південній частині їх ареалу, яка лежить в посушливих районах, позитивний вплив лісів на річковий сток помітніший. Крім того, дубові ліси - могутній фактор боротьби з ерозією грунтів. На схилах долин, балок та горбів, де вони ростуть, різко зменшується або й зовсім припиняється поверхневий сток дощових та снігових вод і збільшується підземний. Отже, дубові ліси запобігають змиванню грунту, замулюванню річок, сприяють збільшенню запасів грунтових вод та покращанню гідрологічного режиму території, що має надзвичайно важливе значення для сільського господарства республіки.
Не можна забувати і про естетичне, рекреаційне та оздоровче значення дубових лісів, особливо для південних районів України, де інші ліси не ростуть природно.
Неабияку цінність мають дубові ліси і як джерело високоцін- ної міцної деревини, а також дубильної (дуб звичайний, вільха клейка, граб звичайний, скумпія звичайна та ін.), лікарської 213
(конвалія звичайна, крушина ламка, любка дволиста, дріоптерис чоловічий, липа серцелиста, осока парвська, цибуля ведмежа, різні види шипшини, суниць та ін.) та харчової (груша звичайна, дерен справжній, черешня, яблуня лісова, ліщина звичайна, гриби та ін.) сировини. Все це свідчить про те, що питання охорони дубових лісів, їх раціонального використання та відновлення в умовах все більш інтенсивного природокористування стають надзвичайно актуальними і важливими. В майбутньому, в міру інтенсифікації природокористування, значення їх буде все зростати, тому вже зараз необхідно розпочати роботи по комплексному вивченню дубових лісів та прогнозуванню раціонального їх використання, розміщення і природоохоронного значення на найближчі 100 років.
Рис. 16. Діброва крушиново- трясучковидноосокова. Дзержинське лісництво Бердичівського лісгоспзагу Житомирської області. (Всі фото Г.С. Куковиці).
Рис. 17. Діброва ліщиново-чоловічопапоротева. Левківське лісництво Житомирського лісгоспзагу Житомирської області.
Рис. 18. Діброва ліщиново-яглицева. Диканське лісництво Полтавського лісгоспзагу Полтавської області.
Рис. 19. Діброва ліщиново-зірочнико-
ва. Іскрівське лісництво Полтавського
лісгоспзагу Полтавської області.
Рис. 20. Діброва ліщиново-волосистоосокова. Корабельне лісництво Житомирського лісгоспзагу Житомирської області.
Рис. 21. Діброва ліщиново-конвалієва. Ічнянське лісництво Прилуцького лісгоспзагу Чернігівської області.
Рис. 22. Діброва ліщиново-парвськоосокова. Мурованокуриловецьке лісництво Могилів-Подільського лісгоспзагу Вінницької області.
Рис. 23. Діброва ліщиново-горобейникова. Савранське лісництво Балтського лісгоспзагу Одеської області.
Рис. 24. Діброва свидиново-гірськоосокова. Немирівське лісництво Ілінецького лісгоспзагу Вінницької області.
Рис. 25. Діброва татарськокленово-яглицева. Котовське лісництво Котовського лісгоспзагу Одеської області.
Рис. 26. Діброва татарськокленово-кореневищноосокова. Коломакське лісництво Жовтневого лісгоспзагу Харківської області.
Рис. 27. Діброва татарськокленово-конвалієва. Шполянське лісництво Звенигородського лісгоспзагу Черкаської області.
Рис. 28. Діброва татарськокленово-гірськоосокова. Брацлавське лісництво Тульчинського лісгоспзагу Вінницької області.
Рис. 29. Діброва татарськокленово-рябоперлівкова. Новопсковське лісництво Старобільського лісгоспзагу Ворошиловградської області.
Рис. 30. Діброва татарськокленово-горобейникова. Голованівське лісництво Голованівського лісгоспзагу Кіровоградської області.
Рис. 31. Діброва татарськокленово-мікелієвоосокова. Павлівське лісництво Котовського лісгоспзагу Одеської області.
Рис. 32. Дубовий ліс кизиловий на схилі Дністра. Нирківське лісництво Бучацького лісгоспзагу Тернопільської області.
Рис. 33. Діброва скумпієво-мікелієвоосокова. Павлівське лісництво Котовського лісгоспзагу Одеської області.
Рис. 34. Діброва скумпієво-пірчастокуцоніжкова. Павлівське лісництво Котовського лісгоспзагу Одеської області
Рис. 35. Дубовий ліс паркового характеру. Фастівське лісництво Фастівського лісгоспзагу Київської області.
ЛІТЕРАТУРА
А г а р к о з И.В., М а х м е т Б.М.. Шандри- к о в Н.И. Орех грецкий в лесных культурах Винницкой области. - В кн.: Разведение и возобновление леса. Госоельхозиздат УССР, К., 1963.
Александрова В.Д. Краткий очерк растительности вдоль Дона от Воронежа до Ростова-на-Дону. - Тр.Бот.ин-та, 1954, оер.З, геоботаника, 9.
Александрова В.Д. Об очередных аадачах в области классификации растительности. - В кн.: Основные проблемы современной геоботаники. "Наука", Л., 1968.
Александрова В.Д. Классификация растительности. "Наука", Л., 1969.
Алексеев Е.В. Типы украинского леса. Правобережье. Изд-во УАН, К., 1928.
Алексеенко М.И. К характеристике типов лесной растительности Харьковской области. - В кн.: Природные ресурсы Левобережной Украины и их использование, 2. Изд-во Харьковского унта, Харьков, 1961.
Алехин В.В. Растительность и геоботанические районы Московской и сопредельных областей. МОИП, М.. 1947.
Алисов Б.П. Климат СССР. Изд-во МГУ, М., 1956.
Аме л і н І.С. Про зональну приналежність рівнинних територій Львівської і суміжних з нею областей. - Укр.бот.журн.,1952,
Андрущенко Г.О.. Вернаядер Н.В., Гринь Г.С., Канаш О.П., Кис 1 л ь В.Д., Скори- н а С.С., Р е о е з о В.М. Карта грунтів Української PCP. К., 1967.
Артемчук И.В. Опыт геоботанического районирования Советской Буковины. - В кн.: Материалы межвузовской конференции по геоботаническому районированию СССР. Изд-во МГУ, М., 1967.
Артюшенко О.Т. Історія розвитку рослинності Західноукраїнського Полісся в пізньольодовиковий та післяльодовиковий час на основі спорово-пилкових досліджень. - Укр.бот.журн., 1957, H, І.
Артюшенко О.Т. До історії розвитку рослинності Лівобережного Лісостепу УРСР В ПІЗНЬО- І ПІСЛЯЛЬОДОВИКОВИЙ часйс - Укр.бот.журн». I960, 17, 3.
Артюшенко А.Т. Растительность лесостепи и степи Украины в четвертичном периоде. "Наукова думка”. K.r 1970.
Артюшенко U.Т., Христофорова Т.Ф., Карева О.В. Міндель-рісське міжльодовикове торфовище в околицях с.Крукеничі Львівської області. - Укр.бот.журн., 1967, 24t4» Афанасьев Д.Я. Деревно-чагарникова рослинність заплави Поліського Дніпра. - Укр.бот.журн., 1958, 15, І.
215
Афанасьев Д.Я. Заплавні луки Поліського Дніпра. їх поліпшення і раціональне використання. Вид-во АН УРСР, К., 1959.
Афанасьев Д.Я. Фітоценотипи дніпровських лук. - Укр.бот.журн., 1964, 21, 4. *
Афанасьев Д.Я. Природні луки УРСР. - В кн.: Рослинність УРСР. "Наукова думка", К., 1968.
Афанасьев Д.Я., Б і л и к Г.І.. Б p а д і c Є.М., Гринь Ф.О. Класифікація рослинності Української PCP. - Укр. бот.жури., 1956, ІЗ, 4.
Афанасьева Е.А. Черноземы Среднерусской возвышенности. "Наука", М.-Л., 1966.
Балашов Л.С.Загальна характеристика рослинності долини р.Снов. - У кн.: Питання фізіології, цитоембріології і флори України. Вид-во АН УРСР, К., 1963.
Барабанщиков А.С. Дубовые леса Саратовской области. - Тр.Саратов. с.-х. ин-та, 1958, IS.
Барбарич A.I. Флора і рослинність Полісся Української PCP. - В кн.: Нариси про природу і сільське господарство Українського Полісся. Вид-во КДУ, К., 1955.
Белецкий Г. Сведения о лесах Кролевецкого уезда Чер-~ ниговской губернии. - Лесной журн., 1840. 4.4, І.
Бельга рд О.Л. Надсамарські байрачні ліси. - 36.робіт біол.фак-ту Дніпропетровського ун-ту,1939, 9, 2.
Бель гард А.Л. Байрачные леса бывшей порожистой части Днепра. - Сб.работ биол.фак-та Днепропетровского ун-та, 1940, 18, 3.
Бельгард А.Л. Лесная растительность юго-востока УССР. Изд-во КГУ, К., 1950.
Бельгард А.Л. О дубовых лесах степной зоны Украины. - Науч.зап. Воронежского лесного ин-та, I960, 20.
Берг Л.С. Географические зоны Советского Союза, 1. Госиздат географической литературы, М., 1947.
Берг Л.С.Географические зоны Советского Союза, 2_. Госиздат географической литературы, М.. 1952.
Б і лик Г.І. Рослинність Дністро-Бузького байрачного степу. - Бот.журн. АН УРСР, 1948, 5, 2.
Б і л и к Г.І. До класифікації галофільної рослинності УРСР. - Бот.журн. АН УРСР, 1954, II, 4.
Б і л и к Г.І. Рослинність засолених грунтів України. "Наукова думка", К., 1963.
Б і л и к Г.І. Детальне геоботанічне районування Лісостепу Української PCP. - Укр.бот.журн., 1970, 22, 3.
Б і л и к Г.І., Б p а д і c Є.М. геоботанічне районування Української PCP. - Укр.бот.журн.. 1962, 19, 4.
Блюменталь И.X., Попова Т.А. Геоботанический очеок Формации овсяницы джимильской Festuoa djimiiensie Boise, et Bal. на Северо-Западном Кавказе. - Тр .ленинградского о-ва естествоисп., 1950. 20, 3.
Блюменталь И.Х., Петровичева О.Л. Геоботанический очерк высокогорных мелкоосоковых пастбищ северо-западного Кавказа. - Уч.зап. ЛГУ,сер.биол. наук, 1951, 143. 30.
Блюменталь И.Х., Кирикова лТГ. Геоботанический очерк формации остреца ложнопырейного ( Agrорутum pseudo- agToggum^D i(i а ) F і a n с h ,). - Уч.зап. ЛГУ, сер.биол.на-
Б о‘н д’а р’ч у к В.Г. Геологічна структура УРСР. Вид-во АН УРСР та КДУ, К., 1946.
216
Бондарчук В.Г. Геоморфологія УРСР. "Радянська школа", К., 1949.
Бондарчук В.Г. Генетичні типи та стратиграфія четвертинних відкладів Української PCP. - Геологічний журн., 1958,
Бондарчук В.Г. Геологія України. Вид-во АН УРСР, К • ț 1959 •
Бондарчук В.Г., В е к л и ч М.Ф., Р о м о д а- нова А.П., Соколовський І.Л. Геоморфологічне районування Української PCP і Молдавської PCP. - ДАН УРСР, 1959,
Б р а д і с Є.М. Про класифікацію рослинності боліт Української PCP. - Укр.бот.журн., 1956, ІЗ, 3.
Б р а д і с Є.М. Рослинність східної частини Малого Полісся та питання ботаніко-географічного районування західних областей УРСР. - Укр.бот.жури., 1957, И, 4.
Б р а д и с Е.М. Растительный покров болот Башкирской АССР,- Тр.Ин-та биологии АН СССР, 1961, £7.
Брадис Е.М. Принципы и основные единицы классификации болотной растительности. - Уч. зап.Тартусского ун-та. Тр. по ботанике, 1963, 145, 7.
Б pva д ł с Є.М. Рослинний покрив боліт УРСР. - В кн.: Рослинність УРСР. "Наукова думка", К., 1969.
Б р а д і с Є.М., Лихобабіна Є.П. Торфово-бп- лотний фонд Української PCP та його районування. - В кн.: Мате- ріали^іУ |д||ЯУ Українського ботанічного товариства. "Наукова дум-
’ б’р а д и с Е.М.. Шеляг-Сосонко D.P. П я т - ницкий С.С., Изюмский П.П. Леса Украинской СССР. (Рецензія.) - Укр.бот.журн., 1968, £5, 1«
Б у д ы к'о м.И. і) закономерностях поверхностного Физико- географического процесса. - Метеорология и гидрология, 1948, 4.
Буторина Т.Н. Эколого-ценотическии анализ кустарничково-травяного яруса лесных ассоциаций. - В кн.: Типы лесов Сибири. Изд-во АН СССР, М., 1963.
Бухало М.А. Флора і рослинність Гологір. - Доп. та повідомлення Львівського ун-ту, 1961, 9.
Бучинский И.Е. Районирование Украины по степени увлажнения. - Тр.Укр. н.-и. гидрометеорол. ин-та, 1958, 13.
Бучинский И.Е. Климат Украины в прошлом, настоящем и будущем. Сельхозиздат УССР, К., 1963.
Быков Б.А. 3 диФика торы растительных формаций Советского Союза. - Вести. АН КазССР, 1949, 3.
Быков Б.А. О составлении флоры эдификаторов (Picea eohxenkiana и Arteniaia incana как эдификаторы). - Бот.журн., 1954, 39» 4.
Бы ков Б.А. Геоботаника. Изд-во АН КазССР, Алма-Ата, 1957.
Быков Б.А. Доминанты растительного покрова Советского Союза. I. Изд-во АН КазССР, Алма-Ата, I960.
Быков Б.А. Проблема эдификаторов растительного покрова. - Бот.журн., 1966а, 51, 9. „ .
Быков Б.А. На пути к генетической классификации растительности. - Изв.АН КазССР, сер.биол., 19666,4.
Василевич В.И. Принципы выделения растительных ассоциаций. - Бот. журн., 1968. Sâ , 6.
- Василевич В.И. Статистические методы в геоботанике. "Наука", Л., 1969.
217
Василевич В.И. Некоторые вопросы теории классификации фитоценотических объектов. - Тез.докл. Совещания по классификации растительности. "Наука" Л., 1971.
Васильев В.Н. Происхождение флоры и растительности Дальнего Востока и Восточной Сибиои. - В кн.: Материалы по истории флоры и растительности СССР,8. Изд-во АН СССР» М.-Л.,1958.
Васильев В.Н. О миграционных гипотезах и видообразовании. - Бот. журн.. 1961, 46, II.
Веклич М.Ф. Четвертинні відклади правобережжя Середнього Дніпра. - Тр.Ін-ту геол.наук, сер.геоморфол. та четвер- іинної геології, 1958, 3.
Веклич М.Ф. Стратиграфия лессовой формации Украины и соседних стран. "Наукова думка", К.. 1968.
В е л ч е в В., Г а н ч е в С., Бо н де в И., К о- ч е в X. Проект за легенда на геоботаническата карта на HP България. - Изв. на ботан. ин-т. 1965. 19.
Вериандер Н.Б., Г о д л и н МЛ., С а м- б у р Г.И.. Скорина С.А. Почвы УССР. Госиздат сельскохозяйственной литературы УССР, К. -Харьков, 1951.
Викторов С.В., Востокова Е.А., В ы- ш и в к и н Д.Д. Введение в индикационную геоботанику. Изд-во МГУ, М., 1962.
Викторов С.В., Востокова Е.А., Вышив- к и н Д.Д. Некоторые вопросы теории геоботанических индикационных исследований. - Тр.Московского о-ва испыт. природы, 1964, 8,
Виленкин В.Л., Никитин В.Н. Сумская лесостепная область западных отрогов Среднерусской возвышенности. - В кн.: Физико-географическое районирование Украинской ССР. Изд-во КГУ, К., 1968•
Водовозова М.В. Высотная дифференциация растительности центральной полосы Русской равнины. - Вот.жури., 1964, 49,
Воробйов Д.В. Типи лісу та лісові асоціації Українського Правобережного Полісся. - Труди з лісової досвідної справи на кра і н і, 1927, 7.
Воробйов Д.В. Типи лісу та лісові асоціації* Українського Лівобережного Полісся. - Труди з лісової досвідної справи на Україні. 1928. 10.
Воробьев Д.В. Типы лесов Европейской части СССР. Изд-во АН УССР. К., 1953.
Воробьев Д.В. Методика лесотипологических исследований. "Урожай", К., 1967.
Воробйов Д.В., Фе д е цьІ.Ф. Класифікація типів лісу Українських Карпат. - В кн.: Природні умови та природні ресурси Українських Карпат, "наукова думка , К.» 1968.
Ворона В.О. Червоноколірна формація Лівобережжя Середнього Придніпров"я. - В кн.: Стратиграфія УРСР» 2. Антропоген. "Наукова думка". К., 1969.
Вульф Е.В. Историческая география растений. Изд-во АН СССР. М•—Л •, 1944.
Высоцкий Г.Н. 0 дубравах в Европейской России и их областях. - Лесной журн., 1913, ДЗ. 1-2.
Высоцкий Г.Н. Ергеня. Культурно-фитологический очерк.- Тр.Бюро по прикл. бот.. 1915, І0-ІІ.
Гаврусевич А.Н. К вопросу о типах леса Львовской области. - Науч.тр. Львовского лесотехн, ин-та, 1959, 4.
Гвоздецкий Н.А. Физико-географическое районирование СССР для целей сельского хозяйства. - В кн.: Материалы к Ш съезду Географического общества СССР. Изд-во ГО СССР, Л., 1959.
218
Гвоздецкий Н. А. Геоботаническое и комплексное природное районирование СССР. - В кн.: Материалы межвузовской конференции по геоботаническому районированию. Изд-во МГУ, М.,
Гегельський І.Н.Досвід розведення дубя бореального на Україні. - Наук.пр.лісогоспод. фак-ту УАСГН,1960,1^,7.
Гейдеман Т.С., Остапенко Б.Ф. лесовод- ственно-экологическая и геоботаническая характеристика зональных типов леса. - В кн.: Типы леса и лесные ассоциации Молдавской ССР."Картя Молдовеняска". Кишинев. 1964.
Геренчук К.Й. Геоморфология Подолии. - Уч.зап.Черновицкого ун-та, сер.геол.-геогр.наук, 1950, 8, 2.
Геренчук К.И. Тектонические закономерности в орографии и речной сети Русской равнины. Изд-во МГУ, М., I960.
Геренчук К.И. Область Волынской возвышенности. - В кн.: Физико-географическое районирование Украинской ССР. Изд-во КГУ і К • у 1968 •
Глинка К.Д. Почвоведение. "Новая деревня", М.-Л., 1927 Глумов Г.А. Естественные леса южной части Лесостепи Зауралья. - Тр.Ин-та биол. Уральского филиала АН СССР, I960. 19.
Голгофская К.Ю. Типы байрачних лесов района Дер- кульской научно-исследовательской станции по полезащитному лесоразведению. - Тр. Ин-та леса, 1958, 39, I.
Голубец М.А. О принципах геоботанического районирования горных стран (на примере Украинских Карпат). - В кн.: Материалы межвузовской конференции по геоботаническому районированию СССР. Изд-во МГУ. М., 1967.
Голубец М.А..М алиновский К.А. Принципы классификации и классификация растительности Украинских Карпат. - Бот.журн., 1967, 52, 2.
Голубець М.А.,М алиновоький К.А. Рослинність. - В кн.: Природа Українських Карпат. Вид-во Львівського ун-ту, Львів, 1968.
Голубец М.А., Малиновский К.А. Класси- ^икация^растительности Украинских Карпат. - Проблемы ботаники.
* Г о’л~у*б е ц М.А., Малиновский К.А., С т о 1- к о С.М. Геоботаническое районирование Украинских Карпат.-Докл. и сообщ. Львовского отд. географ, о-ва УССР, 1965.
Гордягин А.Я. Растительность Татарской республики. - В кн.: Географическое описание Татарской республики, 1.ГИЗ Казань, 1922.
Горохова З.Н. Леса Черновицкого лесхоза и пути повышения их продуктивности. - Науч.ежегод. Черновицкого ун-та за 1959 г., Черновцы, I960.
Горчаковский П.Л. Растения Европейских широколиственных лесов на восточном пределе их ареала. - Тр.Ин-та экологии растений и животных. Свердловск, 1968, 59.
Грейг-Смит П. Количественная экология растений. "Мир", Й., 1967.
Григорьев А.А., Будыко М.И.О периодическом законе географической зональности. - ДАН СССР, 1956, ПО, I.
Гринь Ф.О. Лісова рослинність Немирівського району. - Журн. Ін-ту ботаніки АН УРСР, 1936, 8.
Гринь Ф.О. Реліктові елементи у флорі Диканських лісів. - Бот. журн. АН УРСР, 1949, £, 2. _
Гринь Ф.О. Нарис лісової рослинності Тернопільської області. - Бот.журн. АН УРСР, 1950, 2, І.
219
Г р и и ь Ф.О. Дубові ліси. - В кн.: Рослинність Закарпатської області УРСР. Вид-во АН УРСР, К., 1954.
Гринь Ф.О. Рефугіум міжльодовикових лісових реліктів на Сумщині. - Укр.бот.журн., 1957, L4, І.
Гринь Ф.О. Дубові та широколистяно-дубові ліси. - В кн.: Рослинність УРСР. Ліси. "Наукова думка", К., І9?І.
Гричук В.П. Растительность Русской равнины в нижнє- и среднечетвертичное время. - Тр.Ин-та географии АН СССР, 1950, 46.
Гричук в.П. Ископаемые флоры четвертичных отложений и стратиграфическое расчленение плейстоцена на основании флористических материалов. - В кн.: Материалы Всесоюзного совещания по изучению четвертичного периода.1.Изд-во АН СССР, М., 1961.
Гродзінський А.М. Рослинність Білоцеркївщини. - В^кн.:^Білоцерківщина. Білоцерківське краєзнавче товариство, ’Гродзі нський А.М. Питання взаємовпливу між спів- компонентами фітоценозу. - Укр.бот.журн., 1964, 21, 3.
Гродзинский А.М. Аллелопатия в жизни растений и их сообществ. "Наукова думка" К., 1965.
Гроссгейм А.А. Растительный покров Кавказа. Изд-во МОИП, М., 1948.
Г р о с с е т Г.Э. О возрасте реликтовой флоры равнинной европейской части СССР. - Землеведение, 1935, 37, 3.
Г Р О С С Є Т Г.Э. Распространение рас Cornua sanguines DC. и история широколиственных лесов европейской части СССР. - Бюл. Московского о-ва испыт. природы, отд. биол., 1958, 63 4.
Г р о с с е т Г.Э. Колебания границ между лесом и степью в голоцене в свете ууения о смещении зон. - Бюл. Московского о-ва иопыт. природы, отд.биол., 1961, 66. 2.
Г р о о с е т Г.Э. Возраст термофильной реликтовой флоры широколиственных лесов Русской равнины, Южного Урала и Сибири в связи с палеогеографией плейстоцена и голоцена. - Бюл. Московского о-ва испыт. природы, отд. биол.. 1962. 67, 3.
Г р о с о е т Г.Э. Антигляциализм в ботанической географии.^- Бюл. Московского о-ва испыт.природы, отд.биол., 1966, ’ Гроосет Г.Э. Пути и время миграции лесных Крымско- Кавказских видов на территории Одесской равнины и последующие изменения их ареалов в связи о эволюцией ландшафтов. - Бюл.Московского о-ва испыт.природы, отд. биол., 1967, 72, 5.
Гр у д з и н с к а я И.А. Широколиственные леса предгорий северо-западного Кавказа. - В кн.: широколиственные леса предгорий северо-западного Кавказа. Изд-во АН СССР, М., 1953.
Гук МЛ., П о л о в к о I.K., Прихотько Г.Ф. Клімат Української PCP. "Радянська школа", К., 1958.
Данько В.М. Лісорослинні .умови в центральному Донбасі. - Наук. пр. Уко.н.-д. ін-ту лісов.господ., 1961. 21.
Данько В.М. Типы поименных лесов Донецкой и Луганской областей. - Лесоведение и лесоводство, 1964, I.
Данько В.М. Классификация типов лесорастительных условий байрачних лесов Старобельщины. - Лесовед. и агролесомелиорация, 1965, 3.
Демченко М.А. Харьковская лесостепная область юго- западных отрогов Среднерусской возвышенности. - В кн.: Физико- географическое районирование Украинской ССР. Изд-во КГУ, К.,
Дмитриев Н.И. Рельеф Харьковской области. - Уч.зап. Харьковского ун-та, 97. Тр.геогр. ф-та, 1958, 4.
220
Доктуровский В. О растительности Черного леса ^Херсонской губернии). - Тр.Бот.сада Юрьевского ун-та, 1907, 8, Докучаев В.В. Реферат и подробный критический разбор геоботанических исследовании профессора Коржинского в бассейне Камы и части Волги. - Тр.Вольного эконом.о-ва, 1889, 3.
Докучаев В.В. Учение о зонах природы. Йзд-во АН СССР, М., 1948.
Долуханов А.Г. Вопросы естественной классификации лесных ценозов. - Тр.Тбилисского бот.ин-та АН ГССР, 1959, 20.
Дорофеев П.И. Новые данные о плейстоценовых флорах Белоруссии и Смоленской области. - В кн.: Материалы по истории флоры и растительности СССР, 1963.
Д о X м а н Г.И. Лесостепь европейской части СССР. - Московское о-во испыт. природы, отд.бот., 1968, 14.
Дубовик О.Н. Основные черты развития флоры Донецкой Лесостепи. Автореф. канд. дис. К., 1964.
Д ы л и с Н.В. Принципы построения классификации лесных биогеноценозов. - В кн.: Основы лесной биогеноценологии. "Наука“, М., 1964.
Елагин И.Н. Дубовый лес крайней западной части северного склона Кавказского хребта. - В кн.: Широколиственные леса северо-западного Кавказа. Изд-во АН СССР, М., 1953.
Елагин И.Н. Типы леса нагорной части Теллермановско- го опытного лесничества и их хозяйственное значение. - В кн.: Биогеоценотические исследования в дубравах лесостепной зоны. Изд-во АН СССР, N.. 1963.
Жуков А.Б. Дубравы УССР и способы их восстановления. - В кн.: Дубравы СССР, J. Гослесбумиздат, М.-Л.. 1949.
Заверуха Б.В. Нарис рослинності Кременецьких гір,- В кн.: Питання фізіології, цитоембріології і флори України. Вид- во АН УРСР, К., 1963.
Заверуха Б.В. Реліктові і ендемічні рослини Кременецьких гір та необхідність їх охорони. - В кн.: Охороняйте рідну природу. "Урожай", К., 1964.
2 Заморій П.К. Червонобурі глини УРСР. - Пр.КДУ, 1954, Заморій П.К. Четвертинні відклади Української PCP.
Вид-во КДУ, K., 1961.
Засульська Т.М., Захарченко І.Г. Грунти Київської області. "Урожай", К., 1969.
З е р о в Д.К. Стратиграфія торфовищ України як одно з джерел до четвертинної історії її рослинності та клімату. - Четвертинний період, 1933, 5.
З е р о в Д.К. Час та умови розвитку сфагнових боліт північно-західної України. - Журн. Ін-ту бот. АН УРСР, 1934, 2(10).
З е р о в Д.К. Болота УРСР. - Рослинність і стратиграфія. Вид-во АН УРСР, К., 1938.
З е р о в Д.К. Основные черты послеледниковой истории растительности Украинской ССР. - Тр.конф. по споро-пыльцевому анализу 1948 Г. Изд-во МГУ, М., 1950.
3 е р о в Д.К. Нарис розвитку рослинності на території Української PCP у четвертинному періоді на основі палеоботанічних досліджень. - Бот.жури. АН УРСР, 1952, 2, 4.
З е р о в Д.К. Хвощ великий ( Kquisetua œadue Gar а.) і хвощ строкатий (Egulsetum variegatum S о h 1 е і о b.) в околицях Києва. - Укр.бот. журн., 1963, 20, 6.
З е р о в Д.К., Артюшенко А.Т. История растительности Украины со времени максимального оледенения по данным спорово-пыльцевого анализа. - Четвертичный период, 1961. 13.
221
3 е р о в Д.К., Ар. тюшенко О.Т. Спорово-пилкові дослідження озерно-болотних відкладів Овруцького кряжа. - В кн.: палеогеографічні умови території України в пліоцені та антропогені. ’’Наукова думка", К.. І9ь6.
Зозулин Г.М. Взаимоотношения лесной и травянистой Растительности в Центрально-Черноземном госзаповеднике. - Тр. ентрально-Черноземного госзап., 1955, 3.
Зони С.В. Влияние леса на почвы. Изд-во АН СССР. М.. 1954. ’
3 о н н С.В. Краткие методические указания к изучению почв при лесотипологических исследованиях. - В кн.: Методические указания к изучению типов леса. Изд-во АН СССР. М., 1961.
Зубарев В.М. Дуб черешчатый на северных пределах его распространения. - Сообт. ин-та леса, 1958, 9.
Иванов Н.Н. Ландшафтно-климатические зоны земного шара. - Зап.геогр. о-ва, 1948, £.
Игошина К.Н. Остатки широколиственных ценозов среди пихтово-еловой тайги Среднего Урала. - Бот.журн., 1943, 28. 4.
Игошина К.Н. Опыт ботанико-географического районирования Урала на основе зональных флористических групп. - Бот. журн., 1961, 4g, 2.
Ильинская С.А. Изучение синузиальной структуры лесных сообществ. - В кн.: Типы лесов Сибири. Изд-во АН СССР, М., 1963.
Ильинский А.П. Опыт формирования подвижного равновесия в сообществах растений. - Изв.Главк .бот.сада, 1921, go, 2.
Ильинский А.П. Растительность земного шара. Изд-во АН СССР, М.-Л., 1937.
ИльинскиИ А.П. К вопросу о северной границе хвойношироколиственных лесов в европейской части РСФСР. - В кн.; Президенту АН СССР акад. В.Л.Кбмарову к 70-летию со дня рождения и 45- летию научной деятельности. Изд.-во АН СССР, Л., 1939.
Исаченко А.Г. Учение о ландшафте и современная геоботаника. - В кн.: Академику В.Н.Сукачеву к 75-летию со дня рождения. М.-Л., 1956.
Исаченко А.Г. Основы ландшафтоведения и физико-географическое районирование. "Высшая школа", М., 1965.
Исаченко Т.И., Рачковская Е.И. Основные зональные типы степей Северного Казахстана. - Тр.Бот.ин-та, 1961, сер.З, геоботаника, 13.
Йайрюкщтис А.А. Леса ЛитССР. - В кн.: Леса СССР, g. "Наука", М., 1966. ~
Камышев Н.С. Принципы установления, наименования и выделения в природе растительных ассоциаций. - Бот.журн., 1966, 51, 10.
Карамышева З.В. Растительность северо-восточной части Центрально-Казахстанского мелкоеопочника (в пределах Павлоградской области). - Тр.Бот.ин-та, 1961, сер.З, геоботаника,13.
Карписонова Р.А. Дуоравы лесопарковой зоны Москвы. "Наука", М., 1967.
Кац Н.Я. О центрах консервации, времени и условиях расселения некоторых широколиственных пород в европейской части СССР в валдайскую и послевалдайскую эпохи. - Тр.Комиссии по изучению четвертич. периода. 1955, lg..
Киреев А.Ф.К типологии нагорных и байрачных лесов севера Волгоградской области. - Тр.Воронежского с.-х. ин-та, 1964, 16 »
К л е о п о в Ю.Д. До Історії рослинного вкриття України. Четвертинний період, 1930, 1-2.
222
К л е о п о в Ю.Д. Рослинне вкриття півдепно-західної частини Донецького кряжа. - Вісн. Київського бот.саду, 15, 1933.
К л е о п о в Ю.Д. Геоботанічний ескіз Лівобережжя Середньої Наддніпрянщини. - Журн. Ін-ту бот. АН УРСР, 1934, 2 (10).
Клеопов Ю.Д. Про геоморфогенетичні мотиви розвитку рослинного вкриття УРСР. - Журн. Ін-ту бот.АН УРСР, 1935, 5 (ІЗ).
Клеопов Ю.Д. Основные черты развития флоры широколиственных лесов европейской части СССР. - В кн.: Материалы по истории флоры и растительности СССР, I. Изд-во АН СССР, М.-Л., 1941.
Клеопов Ю.Д., Лавренко Є.М. Легенда до карти рослинності УРСР. - Геооот. зб. 1938, 2.
Климентов Л.В. Растительность Одесской области. - Тр. Одесского ун-та, 1962, сер.геол, и геогр. наук, 152, 10.
Клоков М.В. Основные этапы развития равнинной флоры европейской части СССР. - В кн.: Материалы по истории флоры и растительности СССР, 4. Изд-во АН СССР, М.-Л., 1963.
Кожевников П.П, Типи лісу та лісові асоціації Поділля. - Всесоюэ. н.-д. ін-т лісов.госп. та агромеліор. Сер.наук, видань, 1931, 10. „
Кожевников П.П. Природно-історичні райони, екотопи і типи насаджень лісокультурної зони України. - В кн.: Лісові культури. Лісостепова частина УРСР. Вид-во колгоспної і радгоспної літератури УРСР, К.-Полтава. 1936.
Кожевников П.П. Екологічний нарис дубових лісів Лівобережного Лісостепу УРСР. - Тр.Н-д. ін-ту бот.Харківського
о ж ê в н и к о в П.П. Дубовые леса лесостепи европейской части СССР. - Тр.Всесоюз.н.-и. ин-та леей.хоз., 1939, I.
К о з і й Г.В. Флора і рослинність західних областей України. - Пр.Бот.саду Львівського ун-ту, 1963.
Колесник С.В. Современное состояние учения о ландшафтах. - В кн.: Материалы к ш съезду Геогр. о-ва СССР. Изд-во ГО СССР, Л«, 1959.
Колесников Б.П. Кедровые леса Дальнего Востока. - Тр.Дальневосточного филиала АН СССР, 1956, Z, 4.
Колесников Б.П. Состояние советской лесной типологии и проблема генетической классификации типов леса. - Изв. СО АН СССР, 1958а, 2.
Колесников Б.П. О генетической классификации типов леса и задачах лесной типологии в восточных районах СССР. - Изв. СО АН СССР, 19586, 4.
Колесников Б.П. Генетическая классификация леса и ее задачи на Урале. - Тр.Ин-та биол. АН СССР, 1961, 27.
Колесников Б.П. Генетическая классификация типов леса и некоторые ближайшие задачи ее совершенствования. - Тр.Казахского н.-и. ин-та лесн.хоз., 1966, 5» 5.
Комаров В.Л. Введение к флорам Китая и Монголии. - Тр.СПб.»бот. сада, 1908, gȘ, 1,2.
Комендар В.и. Форпосты горных лесов. "Карпаты", Ужгород. 1966.
Коновалов Н.А. Типы леса подмосковных опытных лесничеств ЦЛОС. - Тр. по лесному опытному делу Центр, лесной опытной станции, 1929, 5.
Коновалов Н.А. Очерк широколиственных лесов центральной лесостепи. - Уч.зап. Уральского ун-та, 1949, 10.
Коркинский С.И. Предварительный отчет о почвенных и геоботанических исследованиях.- Тр.о-ва естествоисп. при Казанском ун-те, 1887, 16, 6.
223
К о р ч а г ин А.А. Вертикальная зональность на Русской равнине и на Среднем Тимане в частности. - Вот. журн.,1954, 22,6.
Корчагин А.А. Некоторые важнейшие современные задачи изучения растительных сообществ. - В кн.: Основные проблемы современной геоботаники. "Наука", Л., 1968.
Корчагин А.А., Селянинов а-К о р ч а- г и н а И.В. Леса Молого-Шекснинского междуречья. - Тр.Дарви- новского гос. заповедника, 195?, 4.
К о с е ц ь МЛ. Букові ліси Західного Поділля. - Бот.журн. АН УРСР, 1947, А, 3-4.
К о с е ц ь МЛ. Нарис лісової рослинності Львівської області УРСР. - Бот.журн. АН УРСР, 1953, 10,4.
Котов М.И. Опыт стационарного изучения растительности харьковских лесов. - Наук. зап. Харківської науково-дослідної кафедри ботаніки, 1927а, JL.
Котов М.І. Ботанічно-географічний нарис Тростянецького комбінату Сумської округи. - Тр.сільськогоспод.бот., 19276, 1,3.
Котов М.І. Основні риси рослинності Проскурівської округи в зв"язку з грунтами. - В.кн.: Матеріали дослідних грунтів України, І9г9, 3.
Котов М.І. Геоботанічний нарис південної частини Вінниць кої області. - Бот.журн. АН УРСР, 1940, І, 2.
Котов М.І. Рослинність Ізюмського лісгоспу на Лівобережжі Сіверського Дінця у Харківській області. - Укр.бот.журн.,
Котов МЛ., Гринь Ф.О. Добір деревних та чагарникових порід для полезахисних смуг Української PCP. - Бот.журн. АН УРСР, І §53, 10, І.
Котов М.І., Карнаух 6.Д. Рослинність заповідників Донецької області. - Бот.журн. АН УРСР, 1940,1, 3-4.
Краснов А.Н. Рельєф, растительность и почвы Харьковской гуо. - Докл .Харьковского о-ва сельск.хозяйств, 1893.
Краснов А.Н. Ботанико-географический очерк Полтавской губернии. СПб., 1894а.
К р а о и о в А.Н. Из поездки на Дальний Восток Азии. Заметки о растительности Явы, Японии и Сахалина. - Землеведение, 18946,
2, 3 •
Крашенинников И.М. Основные зональные типы растительности и главнейшие ассоциации БашкАССР. - В кн.: Природные jeoggoH БашкирокоОССР, I. Растительность Башкирской АССР. Изд-во
Краше’н ин ников И.М. Основные пути развития растительности Южного Урала в связи с палеогеографией северной Евразии в плейстоцене и голоцене. - В кн.: Географические работы. Геог- рафгив, М., 1951.
Криштофович А.Н. Основные пути развития флоры Авии. - Уч.зап. Ленинградского ун-та, оер.геол.-почв.-геогр., 1936, , 2 •
Kp и и т о ф о в и ч А.Н. Миоценовая флора Украины и ее связи через Урал с третичной флорой Азии. - В кн.: Памяти акад. А.В.Фомина. Изд-во АН УССР, К.. Î938.
.Криштофович А.Н. Палеоботаника. Изд-во АН СССР, М.-Л.. 1941.
Криштофович А.Н. Эволюция растительного покрова в геологическом прошлом и ее основные факторы. - В кн.: Материалы по истории флоры и растительности СССР. Изд-во АН СССР, М.-Л., 1946.
Криштофович А.Н. Происхождение флоры ангарской суши. - В кн.: Материалы по истории флоры и растительности СССР,
3. Изд-во АН СССР, М.-Л.. 1958.
224
Криштофович A.H.t Байковская Т.Н. Сар- иатские растения из Амвросиевки в Донецком бассейне. - В кн.: Паняти акад. А.Д.Архангельского. Изд-во АН СССР, М.-Л., 1951«.
Криштофович А.Н., Байковская Т.Н. Сарматская флора Крынки. "Наука”, М.-Л.д 1965.
Крылов А.Г., Речам С.П. Леоорастительное районирование и типы леса. - В кн.: Леса горного Алтая. "Наука”, М., Крылов Г .В. Березовые леса Томской области и их типы. Язд-во АН СССР, Новосибирск, 1953.
Крылов Г .В. Природа лесов Западной Сибири и направление аспользования и улучшения лесных богатств. - Тр.по леснрму хоз-ву Зап.-Сиб.филиала АН СССР, 1957, 3.
Крылов Г.В. Леса Западной Сибири. Изд-во АН СССР, М., [961.
Крылов Г.В., Крылов А.Г. Леса Западной Сибири. - В кн.: Леса СССР, 4. "Йаука**, М., 1969.
Крылов Г.В., Салатова Н.Г. Леса Западной Сибири. Новосибирское областное госиздательство, Новосибирск, 1950.
Кузнецов Н.И. Курс географии растений. Изд-во Таври- іеского губернского земства, Симферополь, 1920.
Кузьмичов А.І. Ліси волинського лесового плато. - Укр.бот.журн. 1967, 24, 2.
Куминова А.В. Растительный покров Алтая. Изд-во АН СССР, Новосибирск, I960.
Лаасимер Л.Р.Геоботаническое районирование Эстонской ССР. - Бот.журн., І958. 43, 3.
Лавренко Е.М. Ботанико-географические исследования между р.Миуоом и р.Кальмиусом. - Тр.с.-х. опыта.учрежд. Дона и Сев.Кавказа. Ростов-на-Дону, 1925.
Лавренко Е.М. Леса Донецкого кряжа. - Почвоведение, 1925, 3-4.
Лавренко Ё.М. Рослинність України. - Вісн.природознавства, 1927, 1-2.
Ла в р^е^н ко Є.М. Болота України. - Вісн. природознав-
*Л а в’р е н к о Є.М. Нарис рослинності України. - В кн.: Грунти України. "Радянський селянин," Харків, І930а.
Лавренко Е.М. Лесные реликтовые (третичные) центры между Карпатами и Алтаем. - Іурн. Русск. бот.о-ва, 19306, 15, 4.
Лавренко Є.М. Характеристика ботаніко-географічних районів УРСР. - В кн.; Флора УРСР, I. Вид-во АН УРСР, К., 1938а.
Лавренко Е.М. История флоры и растительности СССР по данным современного распространения растений. - В кн.: Растительность СССР, I. Изд-во АН СССР, М.-Л Л 1938 б.
Лавренко Е.М. Степи СССР. - В кн.: Растительность СССР, 2. Изд-во АН СССР, М.-Л., 1940.
Лавренко Е.М. Принципы и единицы геоботанического районирования. - В кн.: Геоботаническое районирование СССР. Изд-во АН СССР, М.-Л., 1947а.
Лавренко Е.М. Европейская широколиственная область. - В^кнл Геоботаническое районирование СССР. Изд-во АН СССР, М.-Л., Лавренко Е.М. Евразиатокая степная область. - В кн.: Геоботаническое районирование СССР. Изд-во АН СССР, М.-Л., 1947в.
Лавренко Е.М. Основные черты ботанико-географического разделения СССР и сопредельных стран. - Проблемы ботаники. М.-Л., ’Лавренко Е.М. Степи и сельскохозяйственные земли на
225
месте степей. - В кн.: Растительный покров СССР, 2. Изд-во АН СССР. М.-Л., 1956.
Лавренко Е.М. Основные закономерности растительных сообществ и пути их изучения. - В кн.: Полевая геоботаника. I. Изд-во АН СССР, М.-Л., 1959. ’
Лавренко Е.М. О работе А.А.Ниценко "О некоторых спорных вопросах теории геоботаники". - Бот. журн., 1963, 48, 8.
Лавренко Е.М. Об очередных задачах изучения географии растительного покрова в связи с ботанико-географическим районированием СССР. - В кн.: Основные проблемы современной геоботаники. "Наука", Л., 1968.
Лавренко Є.М. , Л е в і н а Ф.Я. Острівне находите бореальної рослинності в околицях Вінниці. - Вісн. Київського бот. саду, 1934, 17.
Лавренко Є.М.. Погребняк П.С. Матеріали до детальної геоботанічної районізації України. - Вісн. прикладної ботаніки, 1930, 5-6.
Лавренко Е.М., Прозоровский А.В. Растительность Европейской части СССР. - В кн.: Почвы СССР, 1. Изд-во АН СССР, М.-Л., 1939.
Лавриненко Д.Д. Типы лесов Украинской ССР. Гослео- бумиздат, М.-Л., 1954.
Ланько А.И. Новгород-Северское Полесье. - В кн.: Физико-географическое районирование Украинской ССР. Изд-во КРУ, К.,
Ланько А.И., М а р и н и ч А.М., Попов В.П., Порывкина А .В.. Ц ы с ь П.Н. Физико-географическое районирование Украинской ССР для целей сельского хозяйства. - В кн.: Материалы к Ш съезду Геогр. о-ва СССР. Изд-во ГО СССР, Л.,
Лапшина Е.И. Березовые леса лесостепи юго-востока Западной Сибири. - Тр.Центр. Сиб. бот.сада, 1963, 6.
Л о г г и н о в Б.И., Гордиенко М.И. Взаимоотношения амурского бархатного дерева и других пород в культурах УССР.- В кн.: Разведение и возобновление леса. Госсельхозиздат УССР, К.,
1963.
Лопатин В.Д.К вопросу о взаимосвязях между ценотипами растений-здификаторов и их ареалами. - ДАН СССР, 1963, 148, 4.
Лопатин В.Д., Ниценко А.А. О фитоценотипах.- Бот.журн., 1966, 51, 4.
Львов ПТл. Леса Дагестана. Дагкнигоиздат, Махачкала,
1964.
М а з и н г В.В., Трасс Х.Х. Развитие некоторых теоретических проблем в работах эстонских геоботаников. - Бот.журя.„ Í963, 4S, 3.
Малеев В.П. Обзор дубов Кавказа в их систематических и географических отношениях и в связи с эволюцией группы Robur. - Бот.журн., 1935, 20, 2-3.
м а р и н и ч А.М. Геоморфология Южного Полесья. - Изд-во КГУ, К., Í963.
Марков М.В. Лес и степь в условиях Закамья. - Уч.зап. Казанского ун-та, 1935, 25, 2. ,
“і94§ к 0 в М,В* ?астительяооть Татарии. Татгосиздат, Ка-
*М а р*к о в М.В. 0 доминантах фитоценоза по работам советских геоботаников. - Бот.журн., 1958, 43, 4.
Марков М.В. Общая геоботаника. "Высшая школа", М., 1962.
Марков М.В. Некоторые соображения о путях изучения эди- фикаторной роли доминантов и кондоминантов фитоценозов. - Уч.зап. Пермского пед.ин-та, 1968, Ät.
226
Матвеева Е.П. Луга Советской Прибалтики. ’’Наука“, Л., 1967.
Матковський І. Дністро-Ягорлицький байраковий степ. - Зап.Одеського наук.т-ва, 1927, I.
Мельник ВЛ., Чугунний Ю.Г. Речовинний склад відкладів лесової формації. - В кн.: Стратиграфія УРСР,П.Антропоген. “Наукова думка“, К., 1969.
Меницкий Ю.л. Предки и эволюция кавказских и переднеазиатских дубов подсекции Quercus. -Бот.журн., 1969, 54, II.
Мигунова Е.С. О характере почвообразования под пологом широколиственных древесных насаждений в черноземной зоне. - Науч.докл. высшей школы. Биол.науки. I960 I.
Мильков Ф.Н. Лесостепь Русской равнины. Изд-во АН СССР, М., 1950.
Мильков Ф.Н. К анализу ландшафтных (физико-географических) рубежей на Русской равнине. - Изв.Всесоюз.геогр. о-ва, 1959, £4,1.
Мильков Ф.Н. Физико-географические провинции и типы местности. - В кн.: Лесостепь и степь Русской равнины. Изд-во АН СССР, М., 1956.
Мильков Ф.Н. Природные зоны СССР. “Мысль”, М., 1964.
Миркин Б.М. Критерии доминантов и детерминантов при классификации фитоценозов. - Бот.журн., 1968, £3, 6.
Миркин Б.М. Узловые вопросы курса общей геоботаники. Изд-во Башкирского госуниверситета, Уфа, 1970.
Молотківський Ю.Х. Геоботанічне дослідження дубового насадження в Цибулівському лісі. - Журн. 1н-ту бот .АН УРСР, 1936, 8 (16).
М о л я к а О.Н. Деревно-чагарникова рослинність заплави р.Рось та її приток. - Укр.бот.журн., 1962. 19, 2.
Морозов Г.Ф. Учение о типах насаждений. Сельхозиздат, М.-Л.. 1931•
мринський О.П. До характеристики лісової рослинності пониззя р.Удай. - Укр.бот.журн., 1969, 2£, 4.
Мулярчук С.0. Рослинність наддеснянської вододільної рівнини. - Укр.бот.журн.. 1965, 22, 2.
Мулярчук C.Ö. Ліси Чернігівщини. II. Листяні ліси. - Укр.бот.журн., 1968, 25, 2.
K 197$ л я Р 4 У к V.O. Рослинність Чернігівщини. “Вища школа”, ** М у*л ярчук С.О., Балашов Л.С. Ліси Чернігівщини. Деревно-чагарникова рослинність заплав. - Укр.бот.журн., 1969, 25 і 5 *
Набоких А.И. Состав и происхождение различных горизонтов некоторых южно-русских почв и грунтов, - В кн.; Сельское хозяйство и лесоводство за 1906 - 1911 г. СПб., I9II.
Надеждин Б.В. Материалы к изучению влияния лесных насаждений на черноземные почвы. - Сб.работ Центр.музея почвоведения. 1954, I.
Напалков Н.В. Дубравы центральной лесостепи и Нижнего Поволжья. - В кн.: Дубравы СССР, £. Гослесбумиздат, М.-Л., 19 51 •
Напалков Н.В. Леса Чувашской, Татарской АССР и Ульяновской области. - В кн.: Леса СССР, 2. "Наука", М., 1966.
Нейштадт М.И. История лесов и палеогеография СССР в голоцене. Изд-во АН СССР, М., 1957.
Нешатаев Ю.н. Опыт крупномасштабного геоботанического картирования лесостепной дубравы "Лес на Ворскле". - Уч.зап. Ленинградского ун-та, 1967, сер .биол.наук, 50. 331.
227
Ниц е нк о А.Л. О фитоценотипах. - Бот.журн., 1965, 50, б •
Ниценко А.А.О критериях выделения растительных ассоциаций. - Бот.журн., 1966, 54. 8.
Номоконов JI .ИТЛуговые фитоценотипы Енисейской поймы и способы их выделения. - Тр.Вост.-Сиб.филиала АН СССР, 1958,
Номоконов Л.И. Пойменные луга Енисея. Изд-во АН СССР, М., 1959.
Норин Б.Н. Некоторые данные о произрастании дуба в Западном Предуралье. - Бот.журн., 1954, 3£. 3.
О с и ч н ю к В.В. Геоботанічне районування Середнього Побужжя. - Наук.щорічник Київського ун-ту за 1957 р. Біол.фак.,1958.
О с и ч я ю к В.В. Лісова рослинність Середнього Побужжя. - ВІсн. Київського уя-ту, I960, 2, сер.біол., 2.
Остапенко Б.Ф. Классификация типов леса Северной Буковины. - Тр.Харьковского с.-х. ин-та, 1961, 3£).
Пари шкура С.І. Спорово-пилкова характеристика антропогенових відкладів опорного розрізу в м.Прилуки. - Укр.бот.журн., 1967, 24, 6.
Пачоский И.К. Основные черты развития флоры юго- западной России. - Зап.Новороссийского о-ва< естествоиоп. Прил. к 24-му тому. Херсон, 1910.
Пачоский И.К. Очерк растительности Бессарабии. Кишинев, 1914.
Пачоский И.К. Описание растительности Херсонской губернии, 1. Леса. Издание естественно-исторического музея, Херсон, Пачоский И.К. Описание растительности Херсонской губернии. 2. Степи. Издание естественно-исторического музея, Херсон, 1917.
Пашкевич Г.0. Історія рослинності Житомирського Полісся в голоцені за даними спорово-пилкових досліджень. - Укр. бот.журн., 1963, 2Q, 6.
Пашкевич Г.О. Розвиток рослинності північно-східного Приазов"я в пізньому пліоцені-антропогені. - Укр.бот.журн., 1970, “** П е н е в К., Маринов М., Горелков Д., Наумов 3. Типове гора в България. София, 1969.
Поварніцин В.О. Ліси Закарпаття. - Бот.журн. АН УРСР, 1950, 2, 3.
Поварніцин В.О. Лісова рослинність поліської частини Ровенської області. - Бот.журн. АЙ УРСР, 1955, 12, І.
Поварніцин В.О. Принципи класифікації лісово! рослинності Української PCP. - Укр.бот.журн., 1957, 14, 4.
Поварніцин В.О. Ліси українського Полісся. Вид-во АН УРСР, K«, 1959.
Поварніцин В.О. Світлохвойні ліси. - В кн.: Рослинність УРСР. Ліси. "Наукова ^іумка", K.t 1971.
Погребняк П.С. Лісорослинні умови Правобережного Полісся. - Тр.з лїсов.досв. справи на Україні. Харків, 1927 7.
Погребняк П.С. Лісорослинні умови Лівобережної Полісся України. - Тр. з лїсов.досв. справи на Україні. Харків, 1928, 8.
Погребняк П.С. Лісорослинні умови Поділля. - Все- союз. н.-д. ін-т лісов.госп.та агролісомеліор., 1931, сер.наук, вид., 10.
Погребняк П.С. Типология и экология леса. - В кн.; Погребняк П.С., Ш м и д т В.Э., Калужский Н.И., Готшалк Ю.Ф.,
228
Вербицкий Л.Н. Основы лесной типологии. Изд-во АН УССР, К., 1941.
Погребняк П.С. Основы лесной типологии. Изд-во АН УССР, К., 1955.
П о л ы н о в Б.Б. К вопросу о лесной растительности как факторе почвообразования. - Русский почвовед, 1916, 16 - 18.
Поплавская Г.И. Опыт фитосоциологического анализа растительности целинной заповедной степи. Аскания-Нова. - Журн. Русек. бот. о-ва. 9, 1924.
Попов В.П. Агроклиматическое районирование УССР. - В кн.: Вопросы агроклиматического районирования СССР. Гидрометео- издат. М., 1958.
Попов В.В., Вакуров А.Д. Леса Мордовской АССР. - В кн.: Леса СССР. 5. "Наука", М., 1970.
Попов М.Г. Очерк растительности и флоры Карпат. - Изд. Московского о-ва испыт. природы отд. бот., М., 1949, 5.
Порфирьев В.С. Темнохвойно-широколиственные леса северо-востока Татарии. - Уч.зап. Казанского пед.ин-та, 1950, 2..
Порфирьев В.С. Елово-широколиственные леса Райфы. - Тр.о-ва естествоиспыт.природы при Казанском ун-те, 1961, JL2JL, 9«
Порфирьев В.С. Опыт классификации хвойно-широколиственных лесов Волхско-Камского края. - Бот.журн., 1964, 42, 2.
Порфирьев В.С. К проблеме изучения доминантов и эди- Їикаторов растительного покрова. - Уч.зап. Пермского пед. ин-та, 968, 64.
Посохов П.П. Экологический очерк лесов горного Крыма.- Бот.журн., 1961, 46, 4.
Потапенко Г.І. Нарис рослинності Савранського лісу.- Тр.Одеського ун-ту. Біологія, 1937, 2.
Прасолов Л.И. Черноземы как тип почвообразования. - В кн.: Почвы СССР, 1. Иэд-во АН СССР, М.-Л., 1939.
Привалова Л.А. Растительный покров восточного нагорья Крыма и его хозяйственное использование. - Тр.Никитского бот.сада, 1956, 26.
Прилипко Л.И. Лесная растительность Азербайджана. Изд-во АН АзССР, Баку, 1954.
Прозоровский Н.А., Тюремнов С.Н. Вопросы геоботанического районирования. - В кн.: Материалы межвузовской конференции по геоботаническому районированию СССР. Иэд-во ’Прохоров П. Ботанико-географические письма из русской лесостепи. - Почвоведение, 1905, 4.
I960 Пряхин и,в< ТУ^кие засеки. Гослесбумиздат, М.-Л.,
Пьявченко Н.И. Торфяники русской лесостепи. Изд-во АН СССР, М., 1958.
Пятницкий С .С., И аі в м с к и й П.П. Леса Украинской ССР. - В кн.: Леса СССР’_а_. "Наука", М., 1966.
Работнов Т.А. Об эколого-биологическом и ценотичес- ком своеобразии видов на примере луговых растений. - Тр.Москов- ского о-ва испыт. природы, отд. биол., секция бот., 1956, 22-
Р а д к о в И., Минков И. Дъбовите гори в България. Варна. 1963.
Раменский Л.Г. Введение в комплексное почвенно-геоботаническое исследование земель. Сельхозгиз, M.t 1938.
Ремезов Н.П. Динамика взаимодействия широколиственного леса с почвой. - В кн.: Проблемы почвоведения. Изд-во АН СССР. М., 1962.
Ротман Р.Н. Спорово-пилковий склад бурого вугілля Шесте- ринецького родовища Дніпровського басейну УРСР. - ДАН УРСР 1956 6.
229
С а ма р і н В.Ф. Зниження приросту деревини в заплавних дібровах, пошкоджених листовійками. - Вісн. с.-г. науки, 1959, 2.
Сенкевич А.А., Бондаренко Н.Я., Щербакова Л.Б.. Каргов В.А. леса юго-востока европейской части СССР (куйбышевская, Оренбургская, Саратовская. Ростовская, Волгоградская. Астраханская области и Калмыцкая АССР). - В кн.: Леса СССР, .2.. "Наука", М., 1970.
С и в с к а я Е.Н. Основные черты эволюции лесной растительности Кавказа в связи с историей видов, I и П. - Бот.журн., 1933,
’ Ô ко р о д у м о в А.С. Почвы Черного леса. Изд-во АН УССР, К., 1954.
СмирноваЗ.Н. Лесные ассоциации северо-западной части Ленинградской области. - Тр.Петергофского ест.-науч.ин-та, 1928,5.
Смык Г.К. О реликтовых растениях Овручско-Словечанского кряжа. - В кн.: Природная обстановка и фауна прошлого, 2. "Наукова думка". К., 1965.
Соколов С.Я. Принципы геоботанического районирования.- Тр.Бот.ин-та, сер.З, геоботаника, 1940, 4.
Соколов С.Я. Фитоценотические типы. - ДАН СССР, 1947, 55, 2.
Соколов С.Я. Таксономия лесных ассоциаций. - Проблемы ботаники. М.-Л., 1962, 6.
Соколова В.А. Лесостепная растительность юго-западной части Рязанской области. - Науч.докл.высшей школы. Биол.науки, 1963, 4.
Соколовский И.Л. О различных условиях залегания, составе и свойствах лессовых пород западных областей УССР. - Тр. I Украинского гидрология.совещания. Изд-во АН УССР, К., 1961.
Соколовский И.Л. Четвертичные движения земной коры на территории Украины. - Материалы по четвертичному периоду. • К УП конгрессу IKQOA. "Наукова думка", К., 1965.
Солдатов А.Г. Ліси України до Великої Ховтневої соціалістичної революції. - В кн.: Ліси України. Вид-во УАСГН, К., I960.
Соловьев П.Е. Влияние полезащитных лесных полос на Їизико-химические свойства предкавказских черноземов. - Вести. ГУ, 1951, II.
С о ч а в а В.Б. Вопросы флорогенеза и филоценогенеза маньчжурского смешанного леса. - Материалы по истории флоры и растительности СССР. Изд-во АН СССР, М.-Л., 1946.
Сочава В.Б. Некоторые данные об инверсии растительных ассоциаций в связи с вопросом об инверсиях растительности вообще.- Уч.зап. ЛГУ. Геогр.фак.,1948, 6, 2.
Сочава В.Б. Основные положения геоботанического районирования. - Бот.журн.. 1952, 37, 3.
Сочава В.Б. Растительные сообщества и динамика природных систем.-Докл. Ин-та географии Сибири и Дальнего Востока, 1968, 20.
Степунин Г. Леса в верховьях Миуса. - Лесной жури.,
'Стойко С.М. Гірські діброви Карпат та їх відновлення. - В зб.: Лісове господарство Карпат. Вид-во УАСГН. К., I960.
Стойко С.М. Дубові ліси Українських Карпат та їх типологічна класифікація. - В кн.: Природні умови та природні ресурси Українських Карпат. "Наукова думка", К., 1968.
Стойко С.М. Дубовые леса Карпатской горной системы. - Автореф. докт. дис. К., 1969.
Сукачев В.Н. Растительные сообщества. "Книга", М.-Л., 1926.
230
Сукачев В.Н. Сущность типов леса как растительной ассоциации. - Тр. по лесному опытному делу, 1929, 1/75/.
Сукачев В.Н. Руководство к исследованию типов лесов. Сельхозгиа, Л., 1931.
Су к а ч е в В.Н. Дендрология с основами леоной геоботаники. Гослестехиздат. Л., 1934.
Сукачев В.Н. Главнейшие понятия из учения о растительном покрове. - В кн.: Растительность СССР, £. Изд-во АН СССР. М.-Л.. 1938.
Сукачев В.Н. Идеи развития в фитоценологии. - Советская ботаника, 1942, 1-2.
Сукачев В.Н. О принципах генетической классификации в биоценологии. - Журн. общей биол., 1944, 5 4.
Сукачев В.Н. Биогеоценология и фитоценология. - ДАН СССР, 1945, 4?, 6.
Сукачев В.Н. Кокчетавские горные леса. - Землеведение, 1948, 2 /42/.
Сукачев В.Н. О некоторых основных вопросах фитоценологии. - Проблемы ботаники, М.-Л., 1950, I.
Сукачев В.Н. Некоторые общие теоретические вопросы фитоценологии. - Вопросы ботаники,1. М.-Л., Í954.
Сукачев В.Н. Общие принципы и программа изучения типов леса. - В кн.: Сукачев В.Н., Зонн С.В., Мотовилов Г.П. Методические указания к изучению типов леса. Изд-во АН СССР, М., 1957.
Сукачев В.Н. Общие принципы и программы изучения типов леса. - В кн.: Методические указания к изучению типов леса. Изд-во АН СССР, М., 1961.
Сукачев В.Н. Основные понятия лесной биогеоценологии. - В кн.: основы лесной биогеоценологии. "Наука". М., 1964.
Т а л и е в В.I. Контакт леса и степи в Валковском уезде харьковской губернии. - Тр. о-ва испыт. природы при Харьковском ун-те, 1901, 36, 2•
Пі лиев ВЛ. Введение в ботаническое исследование Харьковской губ*, Харьков, 1913.
Танфильев Г.И. Пределы лесов на юге России .-Тр.экспед., снаряженной Леся, департаментом под рук. проф.Докучаева, ^’Т^а н’ф и л ь е в Г.И. Доисторические степи Европейской России. - Землеведение, 1896, 2, 2.
Танфильев Г.И. Главнейшие черты растительности России. - В кн.: Варминг Е. Распределение растении. СПб.. 1902.
Таскаева Н.Я. Дубняки юго-запада Пермской области. - Бюл. Московского о-ва испыт. природы, отд. биол., 1963, 6§, I.
Тах ta дія н А.Л. К вопросу о происхождении умеренной флоры Евразии. - Бот. журн., 1957, 42, И.
Тахтаджян А.Л. Происхождение и расселение цветковых растений. "Наука", Л., 1970.
Ткаченко В.С. Лісова рослинність заплави Сів.Дінця. - Укр.бот. журн., 1967, 24 2.
Толмачев А.Й. К истории возникновения и развития темнохвойной тайги. Изд-во АН СССР, М.-Л., 1954.
Трасс Х.Х.О типологии доминантов растительных сообществ. - Бюл.Московского о-ва испыт. природы, отд. биол., 1963, 5.
Тумаджанов И.И. Опыт дробного геоботанического районирования северного склона Большого Кавказа. Изд-во АН ГССР. Тбилиси, 1963. ’
Туцевич В.Н. Гайсинское лесничество Подольской губернии. - Лесной жури., 1880, 6-7.
231
Тюков С.Ю. Розміщення лісів Української PCP по природах районах. - В кн.: Ліси України. Вид-во УАСГН, К., I960.
Тюремнов С.Н., Ж у д о в а П.П. Принципы геоботанического районирования, проводимого вузами в помощь народному хозяйству. - В кн.: Материалы межвузовской конференции по геоботаническому районированию СССР. Изд-во МГУ, М., 1967.
Тюрин И.В. Опыт классификации лесных площадей водоохранной зоны по их водоохранной защитной роли. - Тр.Всесоюз. н.-и. ин-та лесн.хоз.,1949, 26.
Тюрин И.В. К вопросу о генезисе и классификации лесостепных и "лесных" почв. - В кн.: Тюрин И.В. Вопросы генезиса и плодородия почв. "Наука", М., 1966.
Уткин А.И., Д ы л и с Н.В. Принципы построения биогео- ценотической классификации лесов. - Лесоведение, 1968, 3.
Ф е д є ц И.Ф., Улановский М.С. Типологическая классификация дубовых лесов Лесостепи и Полесья УССР. - В кн.: Дубравы Советского Союза и повышение их производительности. "Урожай*, К., 1968.
Федоров Ан.А. Флора юго-западного Китая и ее значение для познания растительного мира Евразии. - Комаровские чтения, —'* Ф о*г е л ь . Сведения о лесах в Волынской губернии. - Лесной журн., 1836, I. 2.
Фомин А.В. Краткий очерк естественных ботанико-географических районов. - Технико-экономическое право, 1925, 4-5.
Фомин А.В. Схзматическая карта ботанико-географических районов Украины. - Материалы по картографии Украины, J. Харьков, Фу рс а ев А.Д. Естественные леса в пределах трассы государственной лесной полосы Саратов - Камышин. - Уч.зап. Саратовского ун-та. 1952, 29.
Хамитов К.И. О вертикальных границах распространения широколиственных пород на Южном Урале. - Сб.тр. Поволжского лесотехнического ин-та, 1956. 51.
Хотинский Я.А. Корреляция голоценовых отложений и абсолютная хронология схемы Блитта - Сернандера. - В кн.: Голоден. К УШ конгрессу INQUA. "Наука", М., 1969.
Циязерлинг Ю.Д. География растительного покрова северо-запада европейской части СССР, 4. Изд-во АН СССР, Л., 1934.
Ц и с ь П.М. Геоморфологія УРСР. - Вид-во Львівського ун-ту, Львів, 1962.
ц и с ь П.М. Геоморфологія і неотектоніка. - В кн.: Природа Українських Карпат. Вид-во Львівського ун-ту, Львів, 1968.
Цырина Т.С. Ливенские дубки. - Зап.Крымского о-ва ес- тествоисп. и любит, природы, 1925, £.
Чеведаев А.А. дуб, его свойства и значение. Гослесбум- издат, М., 1963.
Черняев В.М. О лесах Украины. М., 1858.
Шеляг-Сосонко Ю.Р. Ліси долини верхньої течії р.Дністра. - Укр. бот.журн., 1961, 12. 6.
Шеляг-Сосонко Ю.Р. Рослинність боліт долини верхнього Дністра. - Укр.бот.журн., 1963а, 20, І.
Шеляг-Сосонко Ю.Р. Лучна рослинність долини верхнього Дністра. - В кн.: Питання фізіології, цитоембріології і флори України. Вид-во АН УРСР, K., 19636.
Шеляг-Сосонко Ю.Р. Ліси межиріччя Десна - Сейм.- Укр.бот.журн., І9бба, 22, 3.
Шеляг-Сосонко Ю.Р. До зональної та провінціаль- ної приналежності Придеснянських відрогів Середньоросійської височини. - Укр.бот. журн., 19666, 22.. 4.
232
Шеляг-Сосонко ю;р. Ареал виду та його розчленування залежно від факторів середовища. - Укр.бот.журн., 1967,24,7. 4.
Шеляг-Сосонко Ю.Р. Класифікація фітоценотипів. - Укр.бот.журн., 1968а, 5, 4.
Шеляг-Сосонко Ю.Р., Б.А.Быков. Проблема здифика- торов растительного покрова. Рецензия. - Бот.куря., 19686, 53, I.
Шеляг-Сосонко Ю.Р. Популяційні фітоценотипи! частини ареалів дуба звичайного (Quexcus robur L.) та липи серцелистої (Tilia coxdafca Mil 1.;, — Укр.бот.журн., І9б8в, 25. 2.
Шеляг-Сосонко Ю.Р. Проблема фитоценотипов. - Бот.журн., 1969а, 54, 7.
.Шеляг-Сосонко Ю.Р. Липово-дубові ліси Західного та Правобережного Полісся УРСР. - Укр.бот.журн., 19696, 26.6.
' Шеляг-Сосонко Ю.Р. Висотна диференціація рослинного покриву Поділля. - Укр.бот.журн.. 1970а, 27, 4.
Шеляг-Сосонко Ю.Р. Дубові ліси Полісся України.- Укр.бот.журн., 19706, 27, І.
Шеляг-Сосонко Ю.Р. Дубовые леса Подолии. - Бот. журн., 1971а, 55, 4.
Шеляг-Сосонко Ю.Р. Дубові ліси других терас лісостепу України. - Укр.бот.журн., І97І6, 2&, 2.
Шеляг-Сосонко Ю.Р., Б а Гр о в a P.O. Нарис рослинності Подільського Покуття. - Укр.бот.журн., 1965. 22,5.
Шеляг-Сосонко Ю.Р., Горохова 8.Н. Дубові ліси Передкарпаття. - Укр.бот.журн., 1969, 26, 2.
Шеляг-Сосонко Ю.Р., Давиденко 1.0. Ліси смуги підтоплення та дренування захисної зони Київського водосховища та прогнозування їх зміни. - Укр.бот.журн., 1969< 26, 4.
Шеляг-Сосонко Ю.Р., Кузьмичов А.І. Заплавні луки р.Горині. - Укр.бот.журн., 1967 24, 3.
Шеляг-Сосонко Ю.Р., Куковица Г.С. 0 характере межвидовой сопряженности травянистого покрова лещино- во-волосистоосоковой дубравы. - В кн.: Количественные методы анализа растительности. Материалы П всесоюз. совещания. "Применение количественных методов при изучении структуры растительности". Тарту 1969»
Іеляг-Сосонко Ю.Р., Куковиця Г.С. Нові відомості до флори Розточчя та Опілля.-Укр.бот.жур., 1970а. 22,6.
Шеляг-Сосонко Ю.Р., Куковиця Г.С. До методики вивчення міжвидової асоці Иованоеті трав"яного покриву дібров. - Укр.бот.журн., 19706, 22. 6.
Шеляг-Сосонко Ю.Р., Куковица Г.С. Геоботаническое районирование запада Украины. - Бот.журн., 19.71, 5§, 10.
Шеляг-Сосонко Ю.Р., С о б к о В.Г. Нові місцезростання рутвиці смердючої (Thallctrum foetidua Ł.) на Україні. - Укр.бот.журн., 1970, 22, 6.
Шенников А.П. луговая растительность СССР. - В кн.: Растительность СССР, І. Изд-во АН СССР, М.-Л., 1938.
Шенников А.П. Принципы геоботанического районирования. - Тр. Ин-та ботаники АН СССР, сер.З, геоботаника, 1940, 4.
Шенников А.П. К созданию единой естественной классификации растительности. - Проблемы ботаники. Изд-во АН СССР, М.-Л., 9б2‘Л н н и к о в А.П. Введение в геоботанику. Изд-во ЛГУ, Л., 1964.
233
Шиффере Е.В. Растительность Северного Кавказа и его природные угодья. Изд-во АН СССР, М.-Л., 1953.
Щекина Н.А. Спорово-пыльпевые комплексы третичных отложений Украинской ССР. - В кн.: К I междунар.палинологической конференции.(Таксон США.) Докл. советских палинологов. М., 1962. ' ■ 6 р к е в и я ІІ.Д.'Дубравы БССР. Изд-во АН БССР, Мииск, I960.
Юрк є ви ч И.Д., Гельтман В.С. География, типология и районирование лесной растительности Белоруссии."Наука и техника11, Минск. 1965. , „ 1М ,
Ярошенко П.Д. Геоботаника. Изд-во АН СССР, М.-Л., 1961. , , ,
Balázs F. Nagykároly es Erdőd kornyékének erdői» - Acta Ge ob o tanie a Hugn», 1943» J?» 2»
Borza A, Pflanzengesellschaften der rumänischen Karpaten» - Biologie9 Boán. 1963, 18, 11»
Borza A» Cercetări asupra florei și vegetației din Clm- pia Română I« - Contribuții botanice, 1966, 1»
Borza A.» Boșoaiu N. Introducere in studiul covorului vegetal» Edit» A RPR, București, 1963.
Braun-Blanquet J» Prinzipien einer Systematik der Pflanzengesellschaften auf floristischer Grundlage» - Bas Jahrb. der St» Gall» Naturwissenschaft» Ges» 1921, 52, 2, Braun-Blanquet J. Les groupements végétaux de la Erance Méditerranéenne. Macabet, 1952» Braun-Blanquet J. Pflanzensoziologie. Wien - N»Y» 1964.
Braun-Blanquet J» Vegetationsskizzen aus dem Baskenland mit Ausblicken auf das Heitere Ibero-Atlantikum. II Teil. - Vegetatio, 1967, 14, 1-4.
Braun-Blanq uzezt J., Pavillard J. Vocabulaire de Sociologie Végétale. Montpellier, 1928,
Braun - Blanque t J», T ii x e n R.Übersicht der höheren Vegetationseinheiten Mitteleuropas. - Communication, 1943, 83.
Breton R. Recherches phytosooiologiques dans la région de Dijon (2* partie). - Ann. de L’In -tut Nat. de la Reche,ehe Agronomique, Serie, A. Annales Agronomiques, 1956» 7, 4.
Dantas Barreto R.R. Os carvalhais da Serra da Penda. Estudo fitosociologico. - Agronomia lusitana, 1958, ЗД» 2.
Diels L. Pflanzengeographie. Berlin - Leipzig, 1929. Domin K, The Beech Forest of Czechoslovakia. - In: Veröffentl. des Geobot. In-tes RÜbel in Zürich. 1932, 8.
D u r і n L., G і h u J. - M. Contribution а 1*étude de la végétation des sources de l’Oise. - Bull, de la Soc.Bot. de France, 1959» 106, 5-6.
E n g le r A. Versuch einer Entwicklungsgeschichte der Pflanzenwelt, insbesondere der Florengebiete, seit der Tertiarpe- riode, 1, 2. Leipzig, 1879.
Palins ki J.B. Zastosowanie taksonomii wrocławskiej do fitosocjologii. - Acta Soc. Bot. Pol,. I960, 29. 3» Fekete G, Angaben zur ZÖnologie der meosischen Schwarzföhrenwälder. - Acta Bot. Acad. Sc. Hung., 1959» 5, 3-4.
Fekete G. Die Waldvegetation im GödöllŐer Hügelland. Budapest, 1965. z
Fijałkowski D. Rezerwat; leśny "Bachus” koło Chełma» - Ann. Univ. M.O. - Skł., 1961, 14.
F i r b a s F. Spät-und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen. Jena, 1949. 1.
234
F u к а г е к F. Die Vegetations des Dard- und ihre Geschichte, - Pflanzensoz. 1961, 12.
Fukarok F. Fitos ocJologia. Warszawa, 1967.
Gajewski W. Klementy flory Polskiego Podola. - Planta Polonica, 1957, 5.
Gauckler K. Steppenheide und Steppenheidewald der Fränkischen Alb in pflanzensoziologischer, ökologischer und geographischer Betrachtung. - Berichte der Bayer. Bot. Ges, zur Krf. der heim. Flora, 1958, 23.
G 1 a V а с V. Osnovno f itocenoloâko r eső lan jen Je nizin- • skih suma u Posavini. - Sumarski list, 1962, g§, 1.
Hofmann G, Die Wälder des Meininger Muschelkalkgebietes. - Feddes Repert., Beihf 1959» 138, 3,
Holub J., H e J n y S., Moravec J.. Neuhäusl R. Übersicht der höheren Vegętątionseinheiteę der Tschechoslowakei. - Rozp. CAV? Rada mat. prirod^ved,1967. 77.
Hundt R. Die Äuenwlesen an Kibe, Saale und Mulde. - Nova Acta Leopoldina, 1958, gg. 135.
Izdebska M. Badania fitosосJologiczne w lasach leśnictwa Zemborzyce, - Ann. Univ. M.C. - Ski., Sect. 0., 1959, 1J,
I z d e b s к i* K. Zbiorowiska z roślinnością kserotermięsna w Rudniku K.Lublina i.Dobożku k. Łaszczowa. - Acta Soc. Bot. Pol,, 1958, 22, 4.
Izdebski K. Badania fitosocJologiczne i florystycz- ne w rezerwacie Stawska Góra pod Chełmem. - Ann. Univ. M.C. - Skł., Seo. С..1959» 15. ,
Izdebski K. Grady na Roztoczu Środkowym. - Якої. Polska, Ser.,A., 1962, 10, 18.
Jakucs P. GeoBotanische Untersuchungen und die Karstaufforstung in Nordungarn. - Acta Bot, Acad. Sc. Hung., 1955» 2, 1-2*
J а к u с э Р. Uber die ostbalkanischen Flieder - Busohwäl- der. - Acta Bot. Acad. Sc. Hung., 1959, W*
Jakucs P. Nouveau classement oenologique des bois de chênes xśrоthermes (Quercetea pubescenti-petraeae cl.nova) de L’Kurope. - Acta Bot. Acad. So. Hung.. I960, 6, 3*4.
Jakucs P. Die phytozbnologischen Verhältnisse der Flaumeichen Buschwälder Südostmitteleuropas. Budapest. 1961, Jakucs P., Fekete G, Karstbuschwald des Nordöstlichen Ungarischen Mittelgebirges (Quer eus pubescens. - Prunus mahaleb nova ass.). - Acta Bot, Acad. Sc. Hung., 1957, 3» 3*4.
Járó Z, Beiträge zu den St and örtlichen und b обе nk ländlichen Verhältnissen der Waldgesellschaften, - In: G,Fekete. Die Waldvegetation im GödöllÓer Hügelland. Budapest, 1965.
JentyâsSzațerowa J. Znaczenie materiałów czwartorzędowych dla badan nad ewolucja historyczna roślin; - . Kosmos, Ser. A, 1958. 7, 1,
J u г к oz A. Podne ekologicke pomory a lesné spoloöenstva PodunaJskeJ nifiny, Bratislava, 1958.
Klika J, 25 Years of phytocenological investigation of our xerothermic vegetation. - Vegetatio, 1954, $-6,
Klika J. Nauka о rostlinnych spoleiensEvech. Praha, 1956.
Knapp R. Die Pflanzengesellschaften Mitteleuropas. Stuttgart, 194В.
Knörzer К.-Н. Die Pflanzengesellschaften der Walder im nördlichen Reinland zwischen Niers und Nieder-Rbein und experimentelle Untersuchungen über den Einfluß einiger Ваше- Arten auf ihre Krautschicht. Düsseldorf, 1957,
235
Koczwara M, 2 badań pyłkowych nad torfowiskami Podola, - Kosmos, 1928, 1,
Koczwara M. Zespoły stepowe Podola Pokuckiego. - Prace Geograf., 1931» là»
KostyniuK M. Analiza pyłkowa dwóch torfowisk w okolicy Rudek i Jambora. - Kosmosț 193», ,
Kostyniuk M., Wieczorek K, Zespoły lesne okolicy Morszyna. - Kosmos, 1957, 62, 5*
Kozłowska A. Elementy genetyczne i pochodzenie flory stepowej_ Polski• - Bull, de 1’Acad. Pol. des Sc. et des Lettres. Ser. в, 1951» i t t
Kozłowska A. Charakterystyka zespołów lesnycn Pogórza Cieszyńskiego. - Prace Biol,, 1936, 1.
K r a u s c h H.D. Die Pflanzenwelt des Spreewaldes. Wittenberg, I960. z
Kulczyński S. Stratygrafia torfowisk Polesia, - Prace Biura Meljor. Polesia, Brześć, 1930, 1» 2.
Kulczyński S.Torfowiska Çolesia. £, Krakow, 1940* Lamorisse R. Recherches géographiques sur la foret des Palanges (Aveyron), - Bull, Soc. Lanqued. de Geograf.,1966, 27> 3e
Lange O.L., K a n z o w H. Wachs turns he mm ung an höheren Pflanzen durch abgetötete Blätter und Zwiebeln von Allium ursi- num, - Flora oder allgem. bot. Zeit, Abt. B, 1965, 156» 1*
L e b r un Ja, Noirfalise A., H e in e m a n n P., VandanBerghen C* Lee assooiationa végétales de Belgique, - Bull, de la Soc, Royale de Botan, de Belg,, 1949» 82, 1,
Lindquist<B, Forest Vegetation Belts in Southern Scandinavia, - Acta Hprti Gotoburgensis, 1959, 22. z
Macko S. Roslinnośó projektowanych rezerwatów na Wołyniu, -Ochrona1 Przyrody, 193B» 17. ,
M h t h 6 I,,K o y a c s M, A gyöngyösi Sárhege vegetációja. - Botanikai Köziemen,, 1962, 49, 3-4.
Matuszkiewicz W. Zespoły lesne południowego Polesia. - Ann, Univ. M.C. - Ski., Sect. E, 1^47, 2, 5.
Matuszkiewicz W. Zespoły lesne Białowieźsklego Parku Narodowego. - Ann. Univ, M.K. - Skł, Sect. C.,1952, Suppl, 6,
Matuszkiewicz W. Dypkusja nad systemem zbiorowisk roślinnych Europy Zachodniej i Środkowej, - Wiad, Bot«, 1962 , 6, 5.
Matuszkiewicz WMP olakowa M, Materiały do fitosocjologicznej systematyki borow mieszanych w Polsce, - Acta Soc, Bot. Pol., 1955» 2£» 2.,
MedweckarKornas А., К o r n a ś J. Zespoły stepów i suchych muraw, - Szata roślinna Polski. Warszawa, 1959» 1. .
MedweckarKornas А., К o r n a s J,, Pawłowski xB. Przegląd ważniejszych zespołów roślinnych Polski. - Szata roślinna Polski. Warszawa, 1959, 1.
Meusel H, Die Waldtypen des Grabfels und ihre Stellung Innerhalb der Wälder zwischen Main und Werra, - Beih. zum Bot,, Centr., Abt. B, 1935, 55. 1.
Meusel H., Ja ger E., Weinert E, Vergleichende Chorologie der Zentraleuropäischen Flora. Jena, 1965,
Michalko J, Geobotaniokó pomery pohoria Vihorlat, Bratislava, 1957,
Michalko J. Dźatko M, Fitocenologioká a ekologická Charakteristika rostlinnÿch spoloÖenstiev lesa Dubaik pri Seredi. - Biolog, práce SAV, 1965, II, 5.
236
M і к y é к a R. Lesy v Zálabí Vychodoőeská niziny, - Fito- cen, studie. Rozp. CAV, Hada mat.z prirod. ved.,1965, 73» 15»z
M і к у ă к a R. Bezkolenkove brèzove' doubravy a гладіlnne bfeziny. - In: Vegețace USSR, A 2, Geobotanická mapa USSR, 1. Geské zemé Nakiad. CAV, Praha, 1968.
Moor M, Zur Systematik der Fagetalia. - Ber. de Schweiz. Bot. Ges., 1938» 48.
Moor M, Zur Systematik der Qherceto-Fagetea. - Mitteil, der Flor.-;soz. Arbeitsgemeinschaft N.F., I960, 8.
M r a z K. Beitrag zur Kenntnis der Stellung des Potentil- lo-Quercetum. - Archiv für Forstv., 1958, 7, 9.
z Neuhäusl R., Ne uhäuslova-Novot - n á Z. VegetationsVerhältnisse am südrand des Schemnitzer Gebirges. - Biolog, pra'ce, SAV, 1964, 10, 4.
Neuhäusl R. NeuhSuslova-NoTOtna 8. Geobotanická Charakteristika lesa "Vidrholec” u Prahy. - Preslla. Roőn., 1966, 2Ê, 3.
Neuhäusl R., Neuhauslova-Novotná Z Syntax on omis ehe Revision der azidophilen Eichen * und Eichenmisch- Wälder im westlichen Teile der Tschechoslowakei. - Folia geo- bot. et phytotax,,1967, 2, 2.
Neuhäusl R., Neuhâuslovâ-Nov.ot- n á Z. Mesophile und subxerophile Waldgesellschaften Mittelböh* mens. - Folia geobot. et phytotax., 1968, 3, 3t
Neuhäusl R.. Neuhäus lova-Novöt- n á Z. Die Laubwaldgesellschaften des östlichen Teiles der Elbeebene, Tschechoslowakei. - Folia geobot. et phytotax., 1969» 5, J.
Noirfalise A. La végétation forestière. - In: Lebrun J.,Noirfalise A. et Saugnez N. Sur l’extension du district fluviatile dans la basse vallée de la Meuse belge. - Bull, de la Soc. Roy. de Botan. de Belg., 1955»,§7, 1•
Nowiński M. Zespoły roślinne Puszczy Sandomierskiej Kosmos, S.A., 1929, 54» 1*2.
Oberdörfer E. Süddeutsche PflanzengeselIsöhaften, - Pflanzensoziologie, 1957» Ifi*
0 n n p M. Vergleichende Studien über die natürliche Waldvegetation Osterreiches und der Schweiz. - In: Festschrift für E.Aichinger., I. Wien, 1954.
Papp L., Szodfridt I. Die Vegetation und die ökologischen Verhältnisse des Naturschutzareals bei Kunbaracs.— Acta. Bot. Acad. Sc. Hung., 1967, 1?» 3~4.
Passarge H. Waldgeaellschaften des mitteldeutschen Trockengebietes. - Archiv für Forstw., 1953» 2, 2-J, Passarge H. Die Wälder von Magdeburgerforth (NW * Fläming). - Wiessenschaft. Abhandl., 1956, 18.
Passarge H. Waldgesellschaften des nördlichen Havellandes. - Wiessenschaft. Abhandl., 1957, 27.
Passarge H. Pflanzengesselscharten zwischen Trebel, Grenz-Bach und Peene (O-Mecklenburg). - Feddes Report., Beib., 1959» 138, 3.
P~s sarge H. Waldgesellschaften des Eichenwaldgebietes von Sw-Mecklenburg und der Altmark. - Archiv für Forstw., 1962, 2, 2.
Passarge H. Pflanzengesellschaften des nordostdeutschen Flachlandes. I. - Pflanzensoz., 1964, 13.
Passarge 11. Neue Vorschläge zur Systematik nordmitteleuropäischer Waldgesellschaften. - Feddes Repert., 1968. 27j 1»
Passarge H., Hofmann G. Ff Lanzengesselischaf- ten des nordostdeutschen Flachlandes. П. -Pflanzensoz., 1968, *]£,
237
Pațcovschi S.j-Doniț ă N. Vegetația lemnoasa din ailvostepa României. Edit, AR3R, 1967.
Pavillard J, aur la valeur sociologique des espèces en phytosociologie. - Bull, de la Soc. Botan, de France. 1954. 101, 3-4.
Pawłowska S. Charakterystyka statystyczna i elenen- ty flory Polskiej. - Szata roślinna Polski, Warszawa, 1959, 1* Puroelean Șt. Grupele de tipuri de pădure fundamentale din România. - Rev. pădurilor., 1968, ÔJ9 10.
Pus car u - S о r oc eanu E.,Popova-Cu- o u A. Metode de cercetare vegetației cu applicații la teritoriul Republicii Socialiste România. Edit, Științ., București,
Rikli M. Das Pflanzenkleid der Mittelmeer1ander , X» Bern. 1943.
R u b n e r K., Reinhold F. Das natürliche Waldbild Europas. Berlin, 1955*
Rühl A. Über die linksrheinischen Wärmeliebenden Trockenwälder, - Allgem. Forst- und Jagdzeit,, 1956, 127, 11-12, Scąmoni A. Die Waldvegetation des ünterspreewaldes. - Archiv, für Forstw,, 1954, 3, 3-4*
Scamoni A. Einführung in die praktische Vegetationkunde. Berlin, 1955.
Scamoni A. VegetationsStudien im Waldschutzgebiet "Fauler Ort" und in den angrenzenden Waldungen, - Feddes Repert., Beih., 1957» 157, 2.
Scamoni A. Grundwasserbedingte Birken - Steileichen- und Steileichen-Buchenwälder, - Inj Vegetationskarte der Deutschen Demokratischen Republik. Berlin, 1964.
Schindler W. Natürliche Waldgesellschaften am Sonnenhübel (Königsholz) bei Zittan, - Archiv für Forstv., 1965, 11» 8.
Schubert R. Die zwergstrauchreichen azidophilen Pflanzengesellschaften Mitteldeutschlands, - Pflanzesoziologie, I960, 2.
Simon T. Die Wälder des Nördlichen Alföld. Budapest,1957* S j ö r s H, Forest Regions. - Inj The Plant Cover of Sweden. Acta Phytogeogr. Suec., 1965, §0. >
Sławiński W. Zespoły kserotermiczne okolic Kazimierza nad Wisła, - Ann. Univ. M.C.-Skł., 1952. 6.
S o 6 R.A nyírségi erdők a növényszövetkezetek rendszereben.- Aota Geobçt. Hung., 1943, 5, 2.
S о о R. Revue systématique des associations végétales des environs de Kolozsvár. - Acta Geobot. Hung., 1947» 6, 1,
S o é R. Les associations végétales de la Moenne-Transylvanie. - Acta Geobot. Hung., 1949» ê, 2.
Sod R. Die Wälder des Alföld. - Acta Bot, Acad.Sc. Hung о 1958, 4, 3-4..
S 0 0 R. Systematische Übersicht der pannonischen Pflanzengesellschaften. II. - Acta Bot, Acad.Sc. Hung.,1959, 5, 3—ł.
S o 6 R. Magyarország erddtarsulasainak és erdotipusalnak áttekintése. - Az Erdó', I960, Eve 9*
S o 0 fi. Systematische Übersicht der pannonischen Pflanzengesellschaften. VI. Die Gebirgswälder. II, - Acta Bot. Acad. Sc. Hung., 1965, 2, 1-2.
800 R. A Magyar flora és vegetáció rendszertani - Növeny- f Öldralzi ^kézikönyve, Budapest, ’1964.
8 0 ó R. Neu Übersicht der höheren zönologischen Einheiten der ungarischen Vegetation. - Acta Bot, Acad. Sc. Hung., 1968, lib 5t4. z
0 r o d o n A, Zarys historycznego rozwoju szaty roślinnej
238
Polski w późnym glacjale i postglacjale. - Szata roślinna Polski. Warszawa, 1959» 1•
Stefanoff B. Uber einige rezente und fossile Eichen- arten in Bulgarien. - Mitteil. Bulg. Bot. Ges., 1928, 2.
Steffen H. Ostpreußen Eichenwälder. - Beih. zum Bot. Centr., Abt. B., 1936, 55, 1.
Szafer W. Element górski we florze niżu polskiego. - Rozpr. Wydz, mat.-przyr. Pols. Akad. Umiejętn., Dz, B, 1930,
S z a f e 1? W. Las i step na Zachodniem Podolu. - Rozp. Wydz. mat. - przyr. Pola. Acad. Umiejetn, Dz. Bt 1935, 71, 2.
Szafer W. Eine plipzäne Rióra in Krościenko am Dunajec. - Bull. Intern, de IіAcad, Pol, des Scienc, et des Lettr., Ser. B,, 1938, 1.
Szafer W. Stratygrafia plejstocenu w Polsce na podstawie florystycznej. - Rocznik Pols. Tow, Geolog,, 1953, 22, 2. z Szafer W. Szata roślinna Polski Niżowej, - Szata roślinna Polski. Warszawa, 1959, 2,
Szafer W. Ogólna geografia roślin. Warszawa, 1964.
T a n g h e M, Contribution a 1’etude de la végétation forestière de la Haute Belgique. Note 1 : La Chênaie-érabliére eutrophe a caractère montagnard des versants froides de l’Arden- ne occidentale. - Bull, de la Soc, Roy. de Botan. de Belg.. 1964, 2Z*
T a n s 1 e y A.G. The British islands and their vegetation« University press, Cambridge, 1939.
T o ł p a S, Zatorfowiania jeziorne na południowej krawędzi Polesia. - Acta Soc. Bot. Pol., 1935» 12, 1. T
T ü X e n R, Die PflanzengesellscHaften Nordwestdeutschlands. - Mitteil, der Flor.-soziologischen Arbeitsgemeinschaft in Niedersen, 1937» З»
Taxen R, Das System der nordwestdeutschen Pflanzengesell schäften. - Mitteil, der Flor. - soz. Arbeitsg., F, 1955, 5.
Tymrakiewioz W. Stratygraf ja torfowisk krasowych poludn, Polesia i połn. Wołynia. - Kosmos .1935» 60, 3.
Újvárosi M, A Sojólódi-erdtf vegetaoISja. - Aota Geobot. Hung., 1941,.4, 1.
V ą n ćL e n Bêrghen C. Aperçu aur la vegetation de la region de Lebbeke. - Bull, de la Soc. Roy« de Botan. de Belg., 1953» ß6» 1. z
Van dze n Berghen 0. Etude sur les irraidationa de plantes méridionales dans la vallee de la Meuse Wallonne. - Bull, de la Soc, Roy, de Botan, de Belg., 1955» 87» 1.
Vanden Berghen C, Etude sur la forets située« au nord de Virton (Belgique méridionale). - Bull, de la Soc. Roy. de Botan. de Belg,, 1956» 8§, 1.
Walter K, Die Pflanzengeaellschaften im Grundwassera- chaclengebiet Lathen - Dörpen am Dortmund - Ems - Seitenkanal« - Mitteil, der Turing. Böt. Ges., 1950, 2, 3.
Whit taker R,H. A consideration of climax theory! the climax as a population and pattern. - Ecol. Monogr., 1953» 22.« Wiedenroth E.4Í. Vegetations Untersuchungen im Par- thegebiet, ein Beitrag zur Kenntnis des Landșchaftschaushaltea Nordwestsachsens. - In: Meusel H., Schubert R. Vegetationskundli- che Untersuchungen als Beiträge zur Lösung von Aufgaben der Landeskultur und Wasserwirtschaft« Halle-Wittenberg, 1964.
Zoloyml B, Der Tatarenahorn-Eichen - Lösswald der zona len Waldsteppe (Acereto tatarici-Quercetum). - Acta Bot. Acad, Sei Hung., 1,957» 3» 3-4.
Zólyomi B., J а к u c s P. Neue Einteilung der Assoziationen der Quercetalia pubescentis-petraeae Ordnung im panno- nischen Eichenwaldgebiet, - Ann. Hist.-Nat. Musei Nation. Hung., 1957, 8,
ЗМІСТ
Вступ 3
Історія вивчення дубових лісів України 5
Природно-історичні умови території дослідження 21
Основні закономірності поширення дубових лісів на Україні 41
Геоботанічне районування території поширення лісів формації дуба звичайного (Querceta roboris) на Україні 60
Фітоценотипи 80
Ареал дуба звичайного та його будова 90
Фітоценотична класифікація дубових лісів України 95
Фітоценотична характеристика дубових лісів з дуба звичайного на території України 111
Дубові ліси крушинові 115
Дубові ліси ліщинові 122
Дубові ліси свидинові 148
Дубові ліси татарськокленові 155
Дубові ліси кизилові 171
Дубові ліси скумпієві 180
Еволюція дубових лісів 187
Лісогосподарська характеристика дубових лісів та перспективи їх охорони 198
Література 215
Юрий Романович Шеляг-Сосонко
ЛЕСА ФОРМАЦИИ ДУБА ОБЫКНОВЕННОГО НА ТЕРРИТОРИИ УКРАИНЫ И ИХ ЭВОЛЮЦИЯ (на украинском языке)
Друкується за постановою вченої ради Інституту ботаніки ім. М.Г. Холодного АН УРСР
Редактор В.І. Середа
Художній редактор M.І. Возний
Оформлення художника І.Й. Анікін
Технічний редактор Р.Е. Хазірова
Коректор С.В. Лісіцина
Підписано до друку 15.V74 р. БФ 30237. Папір офс. №2, 60×901/16. Ум.-друк. арк. 16.0. Облік.-вид. арк. 17,0. Тираж 700. Зам. №4-377 Вид. №144. Ціна 1 крб. 81 коп.
Видавництво "Наукова думка". Київ, Репіна, 3.
Київська друкарня наукової книги Республіканського виробничого об’єднання "Поліграфкнига" Держкомвидаву УРСР. Київ, Репіна, 4.