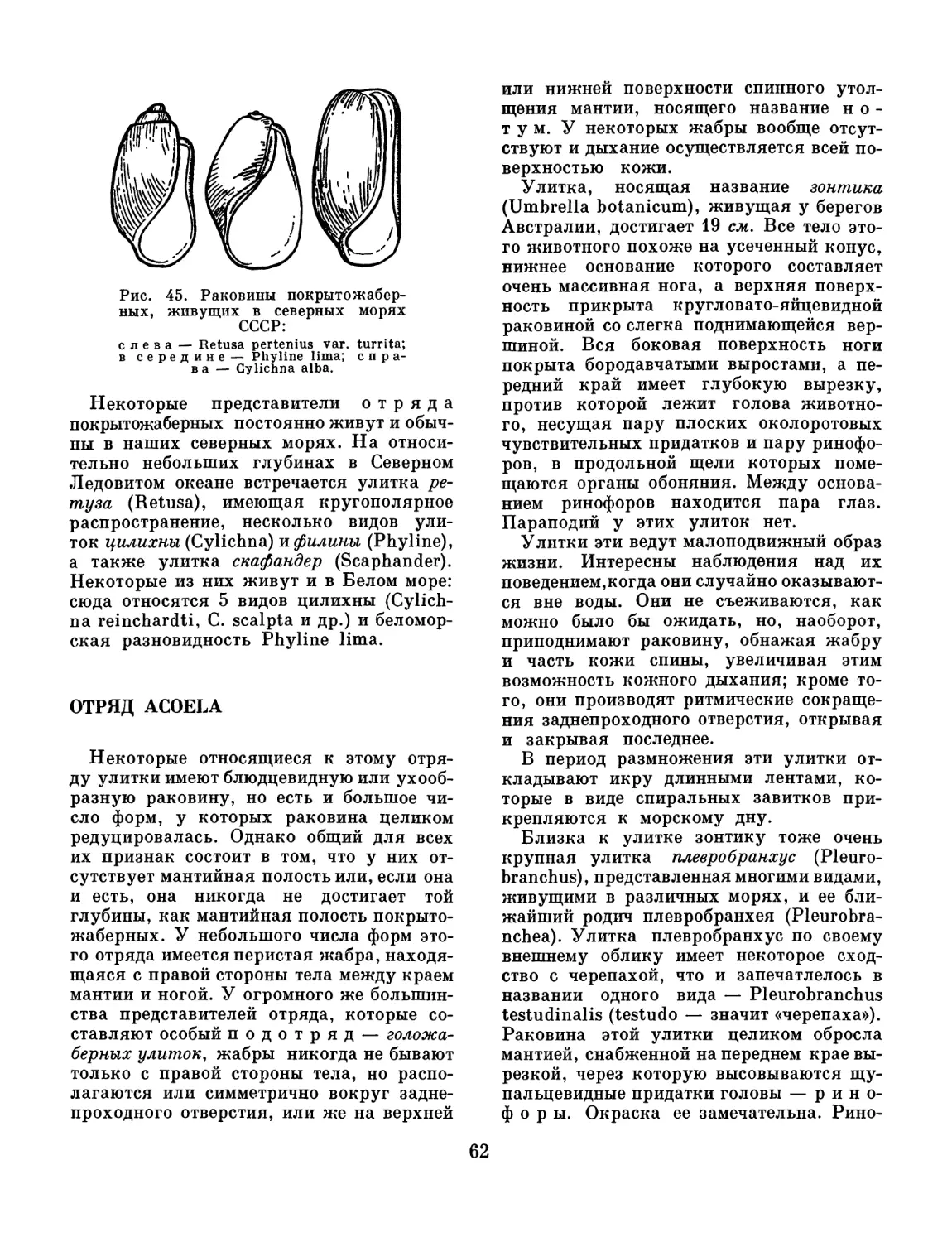
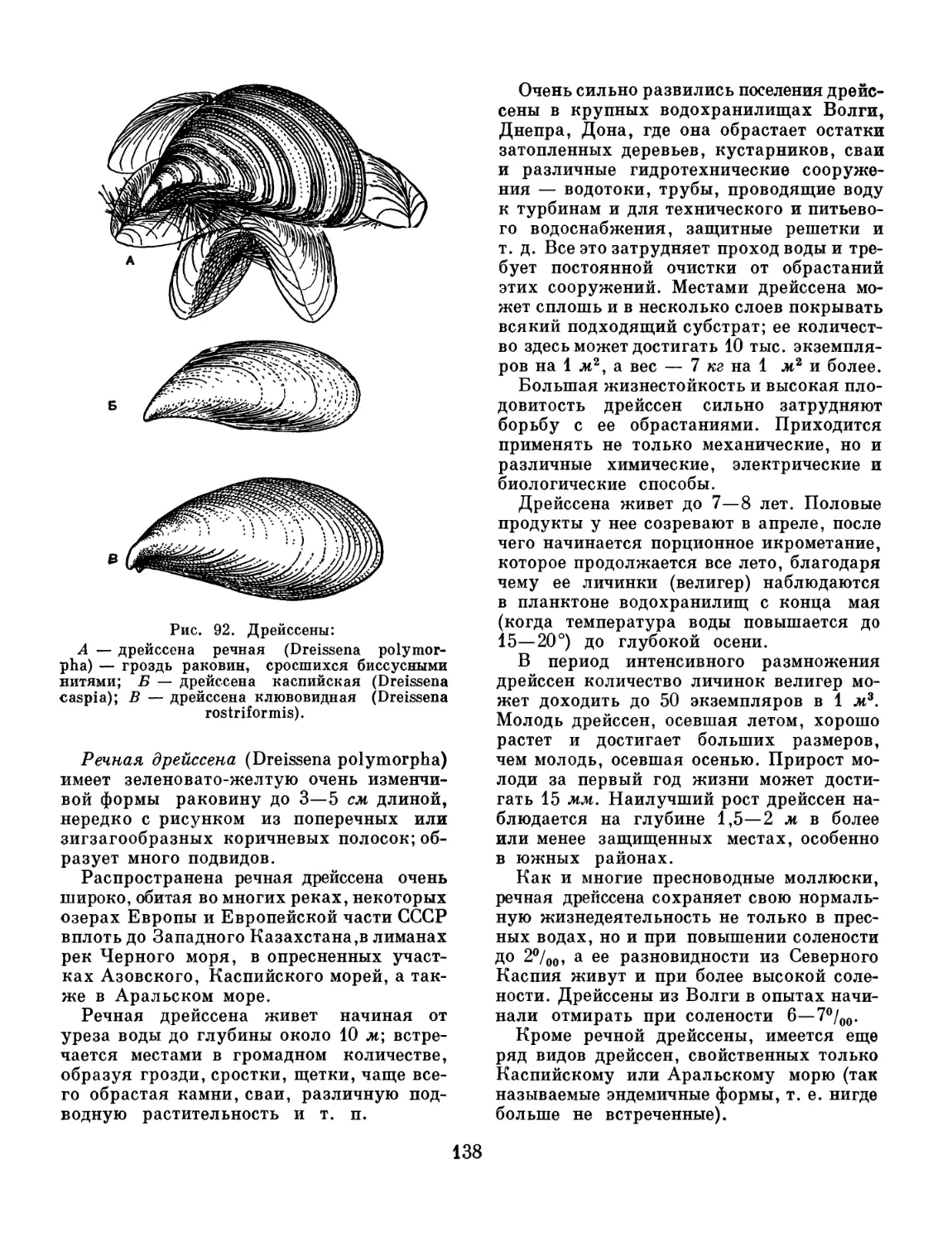
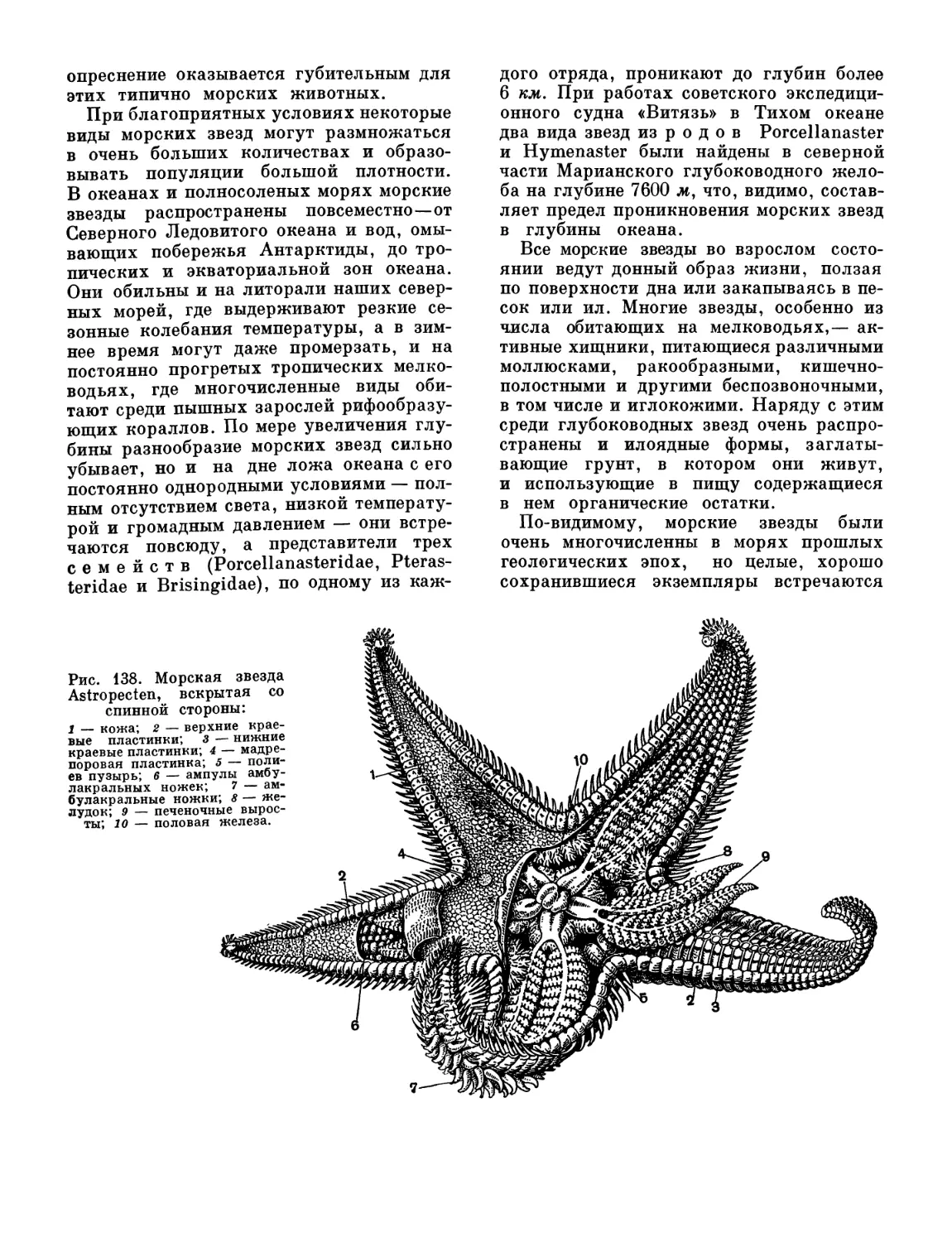
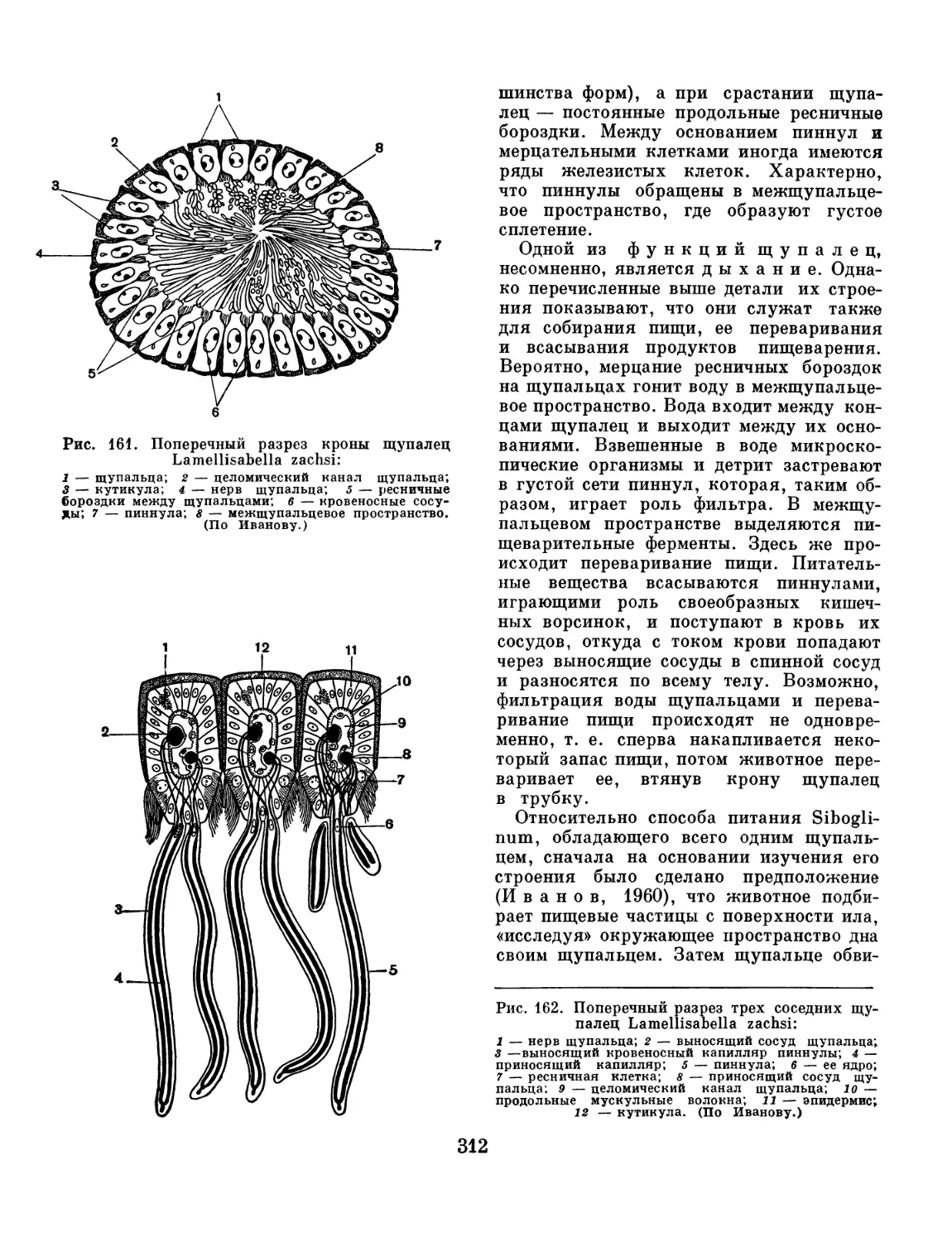
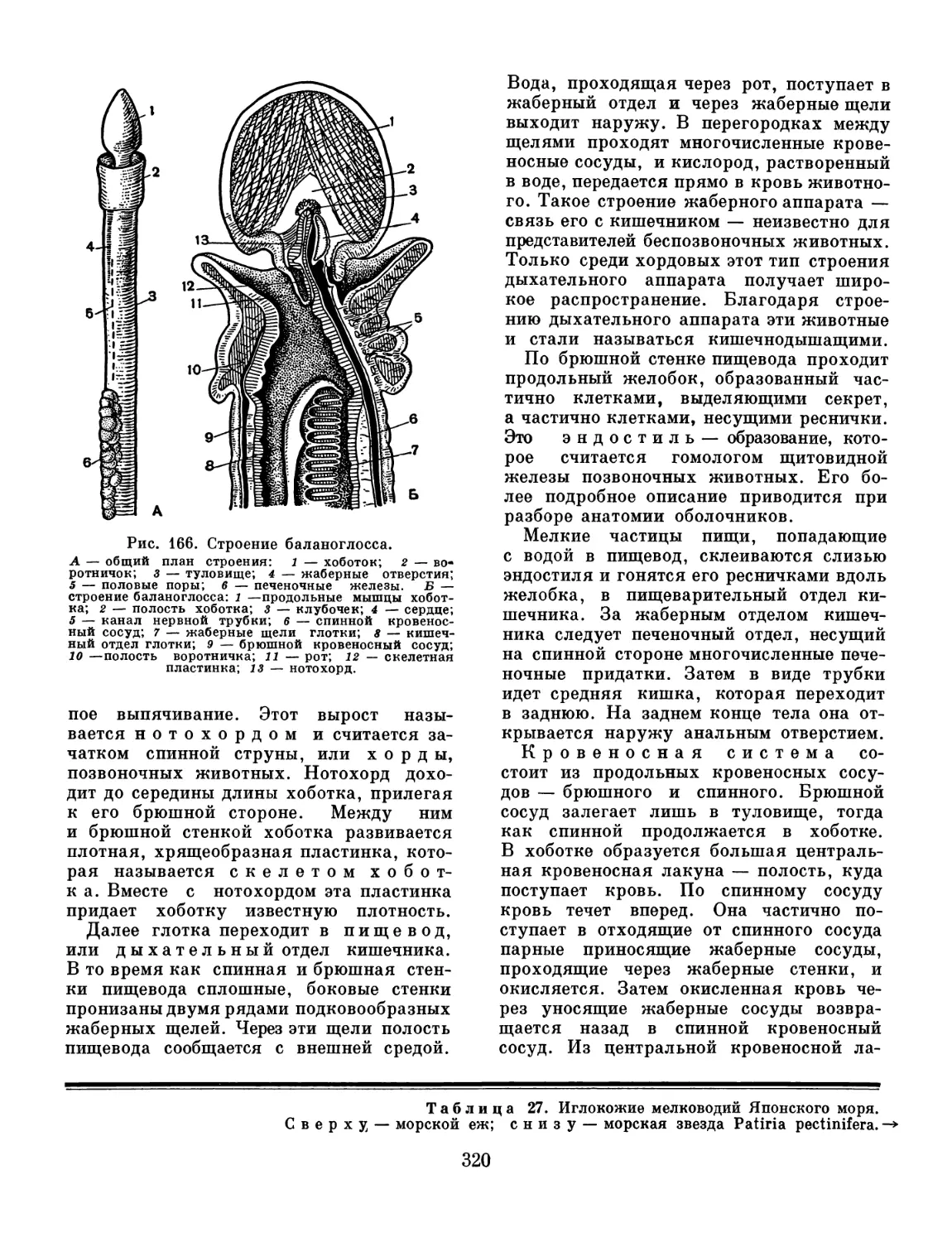
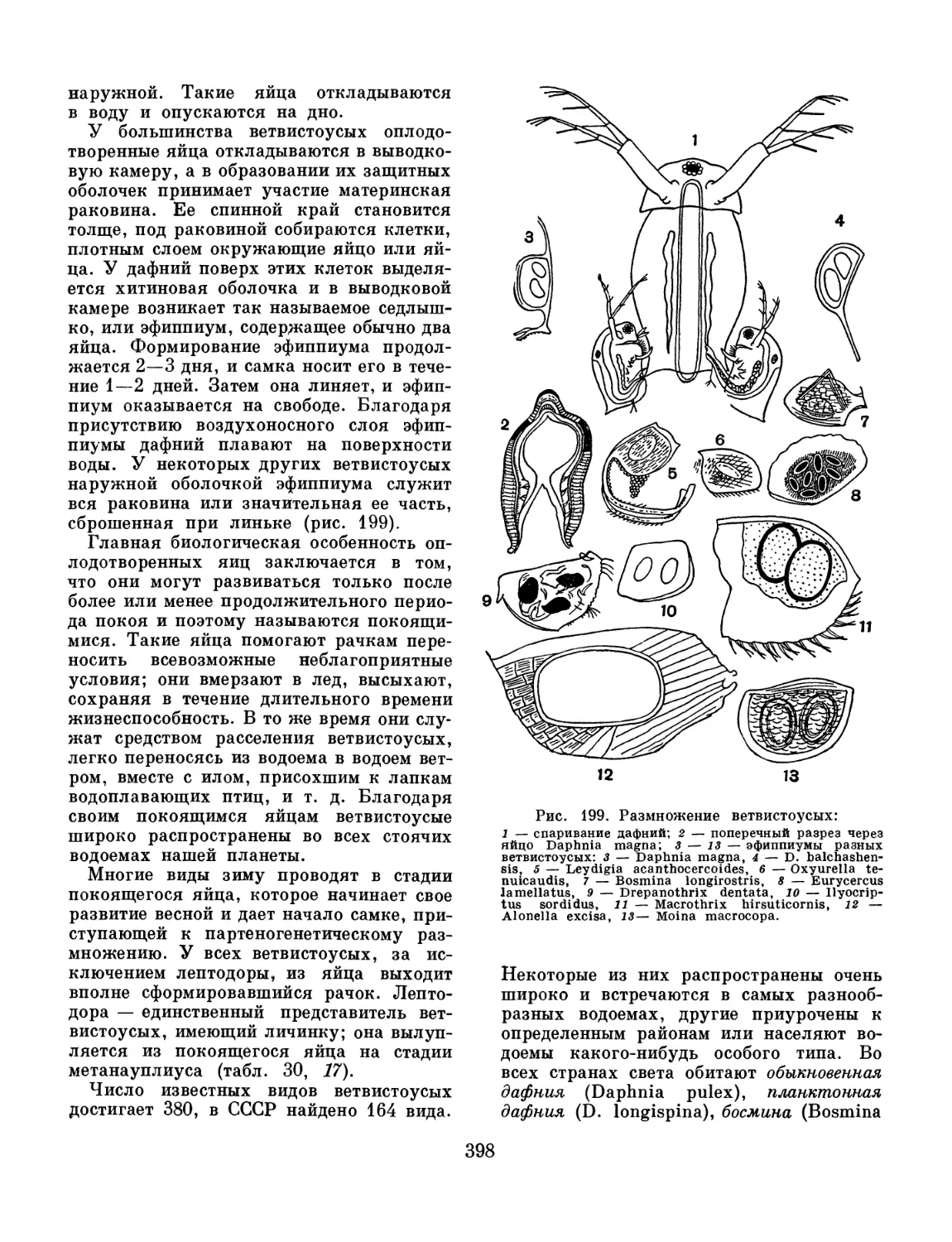
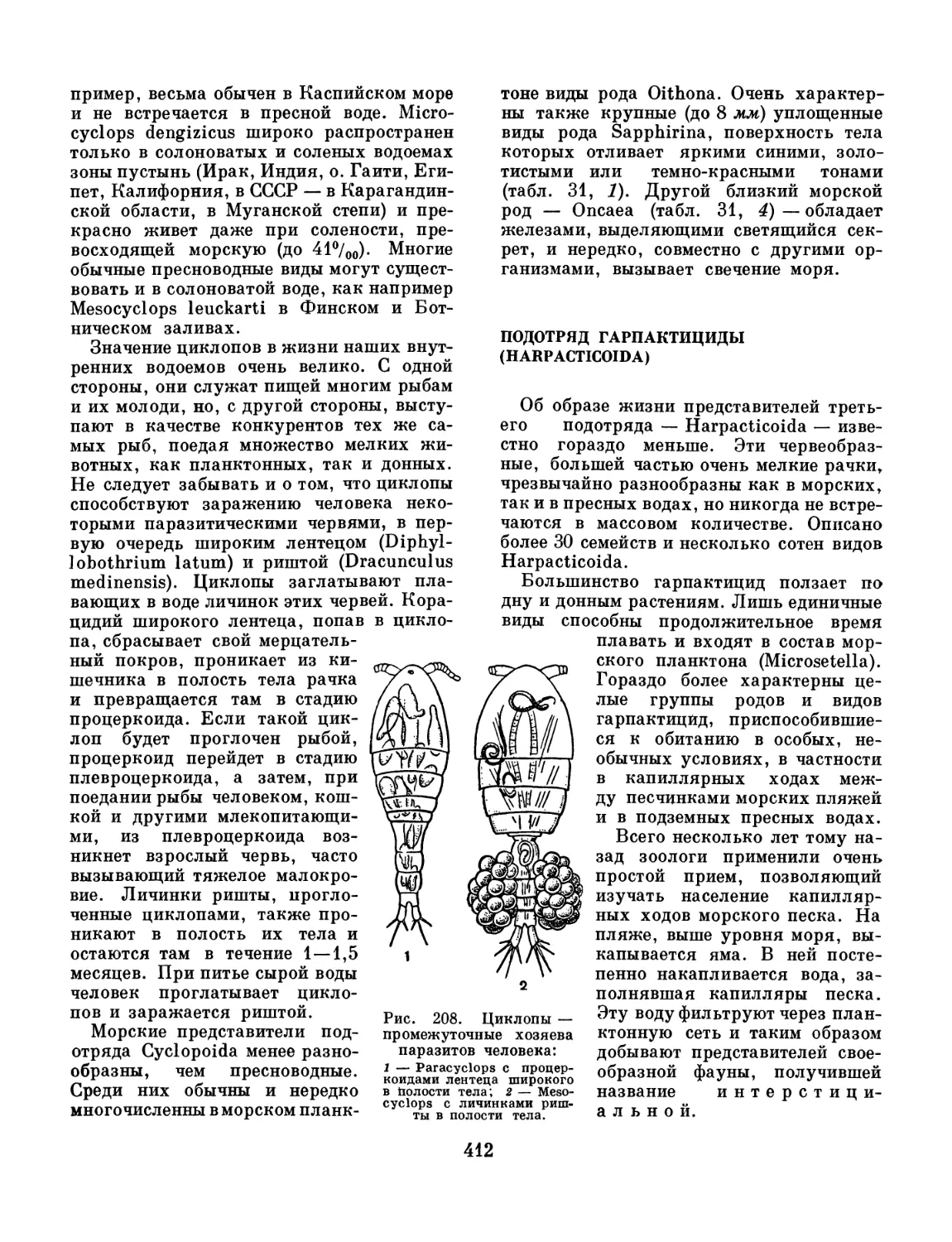
/










































































































































































































































































































































































































































































































































































































Автор: Зенкевича Л.А.
Теги: биология зоология животные ихтиология беспозвоночные академия наук ссср издательство просвещение серия жизнь животных
Год: 1968




Текст
жизнь
ЖИВОТНЫХ
2
ЖИЗНЬ ЖИВОТНЫХ
в
6
ТОМАХ
РЕДАКЦИОННАЯ КОЛЛЕГИЯ:
академик Л.А.ЗЕНКЕВИЧ (гл. редактор),
член-корреспондент АН СССР М.С. ГИЛЯРОВ,
профессора: А. Г. БАННИКОВ, Н. А. ГЛАДКОВ,
А. П. КУЗЯКИН,А. В. МИХЕЕВ,
С. П. НАУМОВ, Ф.Н.ПРАВДИН,
Т. С. РАСС * * * *
МОСКВА • 1968
БЕСПОЗВОНОЧНЫЕ
ПОД РЕДАКЦИЕЙ
действительного члена АН СССР
Л. А, ЗЕНКЕВИЧА * * * *
ИЗДATЕЛЬСТВО «ПРОСВЕЩЕНИЕ»
2
ТОМ
ЖИЗНЬ ЖИВОТНЫХ
59
Ж 71
В написании второго тома «Жизни животных»
участвовали:
доктора биологических наук
Я. А. Бирштейн. \Б. В. Властов\, М. С. Гиляров9
Л. А. Зенкевич, А. В. Иванов, П. В. Матекин,
3. А. Филатова;
кандидаты биологических наук
И. И. Акимушкин, Г. М. Беляев, 3. И. Баранова,
Н. Г. Виноградова, В. И. Зацепин
2-10-6
Моллюски
Mollusca
ТИП МОЛЛЮСКИ (MOLLUSCA)
ОБЩИЙ ОЧЕРК
Моллюски, или мягкотелые, составляют
ясно обособленную группу, и уже более
ста лет назад их стали рассматривать
как отдельный тип животных. В него
входят следующие классы: Брюхоногие
(Gastropoda) — около 85 тыс. видов,
Панцирные (Loricata) — около 1000 видов,
Беспанцирные (Aplacophora) — около 150
видов, Моноплакофоры (Monoplacopho-
га) — несколько видов, Двустворчатые
(Bivalvia) — около 15 тыс. видов, Лопа-
тоногие (Scaphopoda) — около 300
видов, Головоногие (Cephalopoda) — около
600 видов. Несмотря на огромное
разнообразие форм, все моллюски имеют ряд
характерных только для них признаков,
которые, во-первых, подчеркивают
общность и единство происхождения всей
этой группы, а во-вторых, указывают на
своеобразный путь ее эволюции. В
основном это билатерально симметричные нто-
ричнополостные животные. Их тело состоит
из головы, мешковидного несегментиро-
ванного туловища и ноги. Нога
представляет собой утолщенную и разросшуюся
брюшную стенку туловища. Для
моллюсков весьма характерна твердая
минеральная раковина, нередко покрывающая
все тело животного. С внутренней стороны
к раковине прилежит мантия — кожная
складка, которая свободно свисает со
спинной стороны туловища на его бока.
Пространство, образуемое между
стенками туловища и мантией, называется
мантийной полостью. Там лежат органы
дыхания — жабры, и туда открываются
наружные отверстия органов выделения
и анальное отверстие.
Лишь у немногих моллюсков может быть
отмечено метамерное расположение
некоторых наружных и внутренних
органов (жабр, органов выделения,
мышц). В большинстве же случаев они
не обладают ни наружной, ни внутренней
метамерией.
Форма и строение, а также функции
ноги, раковины, мантийной полости и
других органов моллюсков сильно
видоизменяются в пределах типа и могут быть
очень различны у представителей разных
классов; сильно видоизменяется и
общий облик животных в связи хотя бы
с тем, что многие моллюски, как
например брюхоногие — представители самого
обширного их класса, утрачивают
билатеральную симметрию. Раковина во
многих случаях, вместо того чтобы служить
прикрытием для мягких частей тела,
обрастает мантией и становится частично или
полностью внутренней, в связи с чем
обычно уменьшается в размерах, а иногда
может и полностью редуцироваться.
Наконец, нога у одних форм служит для
ползания, и тогда она имеет обычно более
или менее широкую подошву; у других
форм — это орган, при помощи которого
моллюск закапывается в грунт; у
третьих нога видоизменилась в орган
плавания; у четвертых такое значение
приобрела только задняя часть ноги, а
передняя видоизменилась в органы,
служащие главным образом для поимки
добычи и лишь отчасти для передвижения.
Наконец, имеются и такие моллюски,
которые во взрослом состоянии ведут
совершенно неподвижный образ жизни,
однажды на всю жизнь прикрепляясь
к тем или иным предметам, а это бывает
связано с более или менее сильной
редукцией ноги.
Однако, несмотря на все эти резкие
различия, можно установить единство
организаций всех моллюсков на
основании сравнительноанатомического их
изучения и особенно на основании данных их
развития. Так, например, те формы, у
которых резко нарушена билатеральная
симметрия благодаря спиральному
закручиванию раковины и смещению ряда органов,
проходят в своем развитии стадию дву-
стороннесимметричной личинки.
Раковина моллюсков обычно
состоит из кристаллов углекислой извести,
которые расположены в несколько слоев,
и из слоя органического вещества — кон-
хиолина. Из углекислой извести
образуются и другие скелетные элементы —
чешуйки, иглы и др., лежащие либо на
поверхности, либо в коже. Отметим, что
кальциевая раковина моллюсков образуется
на органическом, белковом, материале,
который в виде пластинок формируется
мантией. При наиболее полним развитии
раковины в ней можно различить
наружный, конхиолиновый, слой (периостра-
кум), средний, известковый, слой (ос-
тракум) и внутренний, тоже
известковый, слой (гипостракум). В то время
как при росте раковины остракум
нарастает только по своему краю,
внутренний слой (гипостракум) нарастает и в
толщину, обусловливая увеличение
толщины самой раковины по мере роста
моллюска. Гипостракум на раковинах
некоторых моллюсков состоит из тончайших
пластиночек извести, расположенных
параллельно поверхности самой раковины,
и имеет характерный перламутровый блеск,
получающийся благодаря неравномерному
отражению света от поверхностей этих
пластинок.
У всех моллюсков, за исключением
представителей одного класса, более или
менее явственно обособлена голова,
несущая ротовое отверстие. Кроме того, на
голове могут быть различные щупальце-
видные придатки и глаза; те и другие,
однако, имеются не у всех представителей
типа. Чувствительные щупальцевидные
придатки, как и глаза, могут развиваться
также у некоторых форм по краю мантии.
Глаза могут также находиться и на спине
или на концах щупалец. Из других
органов чувств обычно имеются различные
органы химического чувства и органы
равновесия — статоцисты (каждый статоцист
представляет собой пузырек, в стенке
которого находятся чувствительные
клетки, тогда как внутренняя полость
содержит одно крупное или много мелких
известковых телец). Давление в том или ином
участке на стенку пузырька дает
возможность моллюску ориентироваться в
пространстве.
Центральная нервная
система моллюсков обнаруживает
различную степень сложности у
представителей разных классов. У беспанцирных
и панцирных моллюсков, представителей
двух примитивных классов, она состоит
в основном из окологлоточного кольца
и отходящих от него двух пар нервных
стволов, из которых одна пара лежит
в ноге (педальные стволы), а другая
пара тянется вдоль по бокам туловища
(плевровисцеральные стволы);
педальные стволы соединены поперечными
перемычками между собой, а также с пле-
вровисцеральными стволами. Последние
также дугообразно переходят один в
другой на заднем конце тела. В некоторых
местах нервные стволы расширяются и
образуют как бы узелки — слабенькие
ганглии. Такой тип строения нервной
системы весьма примитивен.
Примитивность выражается здесь также и в том, что
нервные клетки рассеяны по всему ходу
нервных стволов, как это наблюдается,
например, у всех панцирных моллюсков.
У других моллюсков центральная нерв-
ная система уже ясно дифференцирована
на несколько пар связанных между собой
нервных узлов.
Ротовое отверстие у моллюсков ведет
обычно в объемистую ротовую полость с
мускулистыми стенками, называемую часто
глоткой. У входа в последнюю находится
часто парная или непарная челюсть. Кроме
того, чрезвычайно характерно для ротового
аппарата моллюсков наличие особого
органа — терки, или радулы, состоящей из
ленты с расположенными на ней рядами
зубчиков; число, форма и расположение
последних сильно видоизменяются в
различных систематических группах. Терка
лежит на особом выступе на дне ротовой
полости; внутри выступа находится хрящ,
дающий опору всему аппарату терки.
Благодаря особой мускулатуре терка может
перемещаться вперед или назад внутри
ротовой полости, а кроме того, может
несколько выдвигаться через ротовое
отверстие. При помощи зубчиков терки
моллюски могут соскабливать пищу с
поверхности различных предметов, например
водоросли с поверхности камней или со
стенок аквариумов. У некоторых хищных
моллюсков радула и глотка могут сильно
подаваться вперед, как бы выворачиваться
наизнанку; в этом случае зубцы радулы
служат для схватывания добычи. В
ротовую полость открываются обычно те или
иные железы, например слюнные, хотя
последние имеются не у всех моллюсков.
Секрет этих желез иногда содержит яд
и способствует скорейшему умерщвлению
добычи. Однако в тех случаях, когда
пищей служат микроскопические
водоросли или органический дотрит, который
приносится током воды, как это
происходит у ракушек, строение ротового
аппарата крайне упрощается и радула,
челюсти, а также и слюнные железы
подвергаются полной редукции. Ротовая полость
моллюсков обычно переходит в пищевод,
который иногда расширяется наподобие
зоба, а далее следует желудок, в который
открывается парная или непарная
пищеварительная железа, называемая часто
печенью. Желудок переходит в труб-
кообразную кишку, открывающуюся
заднепроходным отверстием наружу.
Заднепроходное отверстие у одних моллюсков
открывается на заднем конце тела, а у
других сдвинуто ближе к переднему концу
и лежит иногда сейчас же за головой,
открываясь в мантийную полость. В этом
случае кишечник образует резкий
петлеобразный изгиб. Моллюски обладают
развитой кровеносной системой, состоящей
из сердца, кровеносных сосудов и
специальных полостей — лакун, или с и-
н у с о в.
Сердце состоит у большинства форм из
одного желудочка и двух предсердий:
в последних собирается обогащенная
кислородом кровь, поступающая из органов
дыхания. При этом наблюдается
соответствие между числом настоящих, или
первичных, жабр и числом предсердий.
Первичными органами
дыхания моллюсков служат парные
внешние жабры, называемые ктенидиями,
которые лежат симметрично около
заднепроходного отверстия в мантийной
полости. Они представляют собой кожные
выросты, имеющие форму пера; внутри
такой жабры проходят приводящий и
отводящий кровь сосуды. Ктенидии связаны
обычно с особыми органами химического
чувства — осфрадиями, которые служат,
по-видимому, для опробования воды,
притекающей к жабрам. У многих моллюсков
из двух первичных ктенидиев остается
только один, тогда как другой
редуцируется, а иногда редуцируются и оба кте-
нидия. Дыхание может осуществляться и
через кожу, особенно через поверхность
мантии. Во многих случаях, кроме
ктенидиев, развиваются вторичные
жабры в виде различной формы придатков.
Вторичная полость тела,
или целом, который бывает ясно
выражен в зародышевом развитии моллюсков,
у взрослых форм сохраняется обычно
в виде двух участков: один представляет
собой окружающую сердце полость — так
называемую околосердечную сумку, или
перикардий, а другой составляет полость
половой железы. Остальные части
вторичной полости тела заполняются клетками
или сильно развитыми органами. Лишь
у представителей одного класса — моно-
плакофор — вторичная полость тела
развита хорошо и сохраняется у взрослого
животного в виде обширных отделов тела.
Органами выделения служат
парные почки, которые одним концом
сообщаются с полостью перикардия, а
другим концом открываются в
мантийную полость. Однако одна почка может
редуцироваться. Также и половые
железы — гонады — у одних, более
примитивных, форм сохраняются парные,
а у других — остается лишь одна
половая железа; последнее получается или
в результате слияния парных желез
в одну, или в результате редукции одной
из гонад. Более примитивной чертой для
моллюсков является также их
раздельнополость — черта, преобладающая у
моллюсков. Однако многие формы
утратили раздельнополость и стали
гермафродитами; соответственно чрезвычайно
усложнилось строение их полового
аппарата. Несмотря на гермафродитизм, как
правило, у моллюсков происходит
перекрестное оплодотворение.
В развитии моллюсков есть много
общего с развитием кольчатых червей.
Сходно с последними происходит у них
дробление яйца. Типичная для
моллюсков личинка, так называемый велигер,
или парусник, чрезвычайно сходна с
личинкой аннелид — трохофорой. Однако
вместо сегментации нижнего полушария
у велигера начинает расти на спинной
стороне горб — будущее туловище, на
вершине которого закладывается
личиночная раковинка. Название личинки
моллюсков связано с тем, что передний отдел
тела личинки здесь расширен и образует
парус, или велум, покрытый ресничками
и имеющий значение как орган плавания
личинки. Кроме паруса, характерны
довольно рано возникающие зачатки
некоторых органов, которых нет у аннелид:
зачаток раковины на спинной стороне и
зачаток ноги на брюшной стороне тела.
Представители типа моллюсков
известны уже в древнейших пластах земной
коры, содержащих остатки вымерших
организмов, а именно уже в
отложениях кембрийского и силурийского
периодов, причем в это время жили уже весьма
разнообразные представители разных
классов моллюсков. Этот факт ясно
указывает на то, что обособление типа
моллюсков в процессе эволюции должно было
произойти гораздо раньше, в те еще более
древние периоды существования Земли,
от которых не могли сохраниться
ископаемые остатки вымерших организмов. Это
обстоятельство лишает нас возможности
установить происхождение моллюсков
на основании преимущественно
палеонтологических данных и заставляет опираться
главным образом на материал
сравнительной анатомии и эмбриологии. Данные
эмбриологии особенно ясно указывают
на родство моллюсков с кольчатыми
червями. На это же родство указывает и то,
что моллюски относятся к вторичнополост-
ным животным, и то, что у них имеется
сообщение полости перикардия (т. е.
остатка целома) с наружной средой через
органы выделения, что весьма сходно
с чертами строения кольчатых червей.
Некоторые исследователи усматривают
сходство между кольчатыми червями и
моллюсками в том, что у примитивных
форм последних, как, например, у
беспанцирных, строение нервной системы
обнаруживает некоторые следы метамерии.
Эти исследователи склонны считать, что
моллюски происходят от членистых червей,
утративших затем эту членистость. Другие
исследователи считают, что и первые
древние моллюски были нечленистыми
животными и что они произошли от
нечленистых плоских червей.
Таким образом, в вопросе о
происхождении типа моллюсков остается еще
много невыясненного. Наиболее вероятно,
однако, их раннее обособление от общих
предков с кольчатыми червями, причем
эти формы были целомическими, т. е.
вторичнополостными животными. Были
ли они сегментированными, остается
неясным. Согласно одним взглядам
наиболее примитивны панцирные моллюски,
другие исследователи считают их,
наоборот, сильно специализированными.
Беспанцирных моллюсков
большинство исследователей рассматривает как
потомков древних панцирных моллюсков,
утративших в своей эволюции раковину
и соответственно изменившихся в ряде
других признаков. Большие разногласия
вызывает толкование организации моно-
плакофор—моллюсков с чертами наружной
и внутренней метамерии и, как уже было
10
отмечено, обладающих обширными
отделами вторичной полости тела. Одни
исследователи видят в них наиболее
примитивных моллюсков и считают их исходными
формами, другие все отмеченные нами
оригинальные черты их организации
считают возникшими вторично.
Подобно панцирным моллюскам, и
другие классы моллюсков рано обособились
в своем развитии от древних
родоначальников типа и стали развиваться как
отдельные ветви, отходящие от общего корня,
сохранив, однако, в своей организации
общие для всех моллюсков черты, которые
ясно указывают на единство
происхождения типа в целом. Представители
некоторых классов моллюсков весьма значимы
во многих отношениях. Так,
фильтрационный способ питания двустворчатых
приводит к тому, что многие из них,
пропуская через мантийную полость большое
количество воды со взвешенными в ней
минеральными и органическими частицами,
осаждают их, образуя на дне океанов
и пресноводных водоемов мощные осадки.
Двустворчатые же, некоторые
брюхоногие и головоногие являются важнейшими
компонентами пищи рыб, птиц и морских
млекопитающих, которые в свою очередь
представляют собой промысловых
животных. Представители этих классов
служили пищей для первобытных людей,
а некоторые из них это значение
сохранили и до наших дней (устрицы, мидии,
корбикулы, морские гребешки,
виноградная улитка, осьминоги).
Раковины двустворчатых и
брюхоногих являются сырьем для получения
перламутра, а некоторые из двустворчатых
производят жемчуг.
Многие виды брюхоногих, особенно
пресноводные и наземные,— промежуточные
хозяева трематод, нематод, цестод —
паразитических червей.
Велико значение моллюсков как
руководящих ископаемых. Особенно следует
отметить вымерших представителей
головоногих — аммонитов. Однако и
двустворчатые, так же как брюхоногие, тоже имеют
определенное значение: нахождение их
раковин может без ошибки указать,
представляют ли собой слои, содержащие эти
раковины, морские отложения, или они
сформировались в пресноводных
водоемах, или они, наконец, представляют
собой погребенные участки древней суши.
КЛАСС БЕСПАНЦИРНЫЕ (APLACOPHORA)
Едва лишь исполнилось полсотни лет,
как были открыты и более точно описаны
некоторые малозаметные морские
животные, которых мы теперь объединяем
в этот класс и которых прежде относили
то к иглокожим, то к червям. Только
в дальнейшем на основании изучения их
внутреннего строения и особенно нервной
системы было обнаружено их родство
с мягкотелыми, хотя они и лишены
раковины. Они особенно близки к группе
панцирных моллюсков, от которых они,
однако, во многом отличаются, почему их и
выделили в самостоятельный класс.
Тело беспанцирных, или, как их часто
называют, бороздчатобрюхих (Solenogaster),
моллюсков в большинстве случаев
цилиндрическое и сильно вытянуто в длину,
которая превышает ширину в несколько
(иногда во много) раз. Это и создает
некоторое внешнее сходство их с червями.
У некоторых форм оно слегка сплющено
с боков, а на спинной стороне несет
невысокий киль.
У большинства форм на брюшной
стороне проходит узкая бороздка, которая
начинается богатой железами ямкой,
выстланной мерцательным эпителием и
лежащей сейчас же за ртом. В бороздке обычно
находится тонкая, тоже покрытая
мерцательным эпителием кожная складка. Это
видоизмененная нога моллюска,
лишенная, однако, характерной для последней
сильно развитой мускулатуры. Наличие
мерцательных ресничек резко отличает
характер наружного покрова в этой части
11
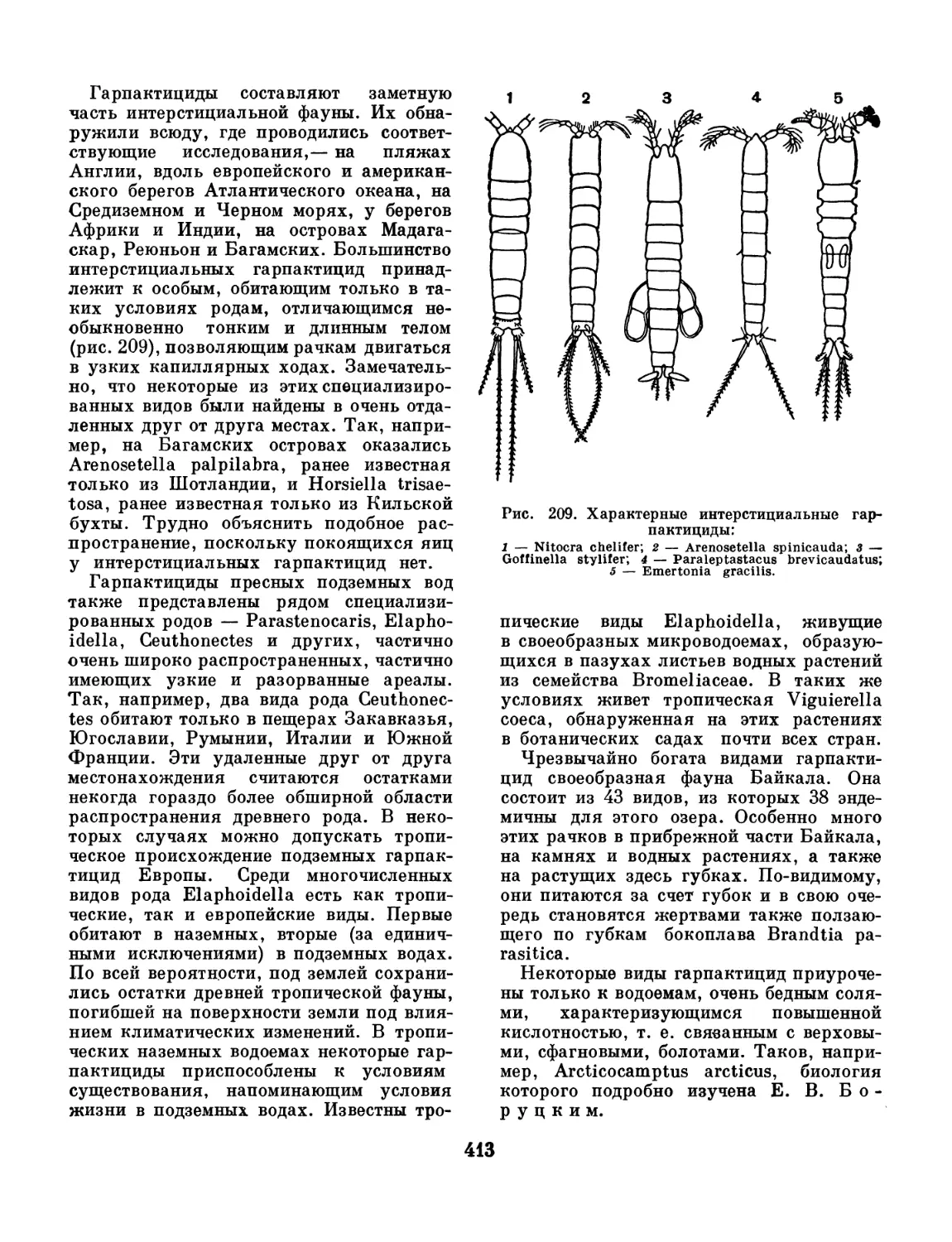
Рис. 1.
1 — хэтодерма (Chaetoderma nitidulum), la — головной
конец, вид со спинной стороны (увеличено); 2 — неоме-
нин (Neomenia), вид с брюшной стороны (слева) и
со спинной стороны (справа); 3,4 — беспанцирные
моллюски, живущие на колониях гидроидных полипов;
3 — ропаломения (Rhopalomenia aglaopheniae); 4 —
мизомения (Myzomenia).
тела, так как другие участки последнего
покрыты плотной кутикулой, достигающей
у некоторых форм значительной толщины,
благодаря чему тело в значительной
степени утрачивает свою гибкость.
Характерно и строение
покрова беспанцирных моллюсков:
свойственное им одеяние состоит из
известковых шипиков, чрезвычайно
разнообразных по форме. В одних случаях это
простые, заостренные на концах иглы, в
других — это чешуйки; то они остры,
как шило, то более или менее
притуплены, то заострены, как лезвие ножа,
то имеют вид лопатки или ложки или
загнуты крючком. Обычно они
направлены назад и придают шелковистый
глянец поверхности тела.
В связи с общей примитивностью
строения, в частности в связи с отсутствием
органов зрения и слуха, все
беспанцирные моллюски — чрезвычайно
малоподвижные животные, ведущие крайне
однообразный образ жизни. Их биология,
впрочем, изучена еще очень
недостаточно.
Беспанцирные моллюски встречаются на
различных глубинах, некоторые из них
обнаруживаются только на больших
глубинах — 3—4 тыс. м. Обитают либо в
илистых грунтах, питаясь детритом, либо на
стволах гидроидных и коралловых
полипов, объедая их верхушки.
У некоторых холодноводных видов
Арктики и Антарктики обнаружена забота
о потомстве: яйца откладываются в
камеру, которая представляет собой часть
клоаки; в этой камере и происходит
развитие яиц.
Беспанцирные моллюски встречаются
почти во всех морях земного шара от
Арктики до Антарктики. Число форм их
относительно невелико. Некоторые из них
встречаются и в наших северных морях.
Наиболее обычен здесь один вид,
относящийся к роду Хэтодерма
(Chaetoderma nitidulum). Этот небольшой
моллюск (длина тела до 81 мм) встречается
на глубинах 30—80 м и в Баренцевом,
и в Белом морях, и дальше на восток;
вид этот, по-видимому, имеет кругополяр-
ное распространение. Вдоль западного
побережья Европы встречается до
Средиземного моря, а вдоль берегов Азии
спускается к югу до Японского моря.
Для Баренцева моря известны также
и два других вида беспанцирных
моллюсков, относящихся к р о д у Пронеоме-
ния. Один из них (Proneomenia sluiteri)
достигает размера в 140 мм, тогда как
другой (Proneomenia thulensis) относится
к мелким формам (длина до 20 мм).
Пронеомении по сравнению с хэтодермой
более глубоководные формы и в
Кольском заливе живут на глубинах от 100 до
150 м.
Интересен способ передвижения в иле
представителей рода хэтодерма. Когда
животное оказывается на поверхности ила,
на котором его трудно различить благодаря
его маскирующей серо-белой окраске, оно
12
передвигается крайне медленно и
оставляет за собой характерный след в виде
елочки, получающийся благодаря маят-
никообразным колебаниям заднего конца
тела. Однако обычно эти моллюски
держатся, закопавшись в ил, так, что их
задний конец приходится в уровень
с поверхностью дна и над последним
выдаются лишь красные жаберные
придатки животного, совершающие
медленные ритмические колебания и тем
способствующие обмену обмывающей их
воды.
При малейшей тревоге животное с
молниеносной быстротой закрывает клоаку
и уходит на глубину нескольких
сантиметров.
При закапывании в грунт животное
сперва прочно закрепляет положение
заднего конца тела при помощи
вонзающихся в ил шипиков, а затем вытягивает
передний конец тела и вбуравливается
в ил. Достигнув определенной глубины,
передний конец вздувается, что
закрепляет тело в новом положении, после чего
закрывается клоака, иглы заднего конца
выпрямляются параллельно оси животного
и все оно подтягивается вперед.
Интересно, что расположение шипов,
которые направлены своими концами назад,
не дает животному возможности
обратного движения в его узком ходе, и этим
объясняются те характерные петли,
которые моллюск должен делать, чтобы снова
подняться на поверхность и выставить
наружу задний конец для дыхания.
Относительно рода пищи беспанцирных
моллюсков и способа ее захвата данных
очень мало; у многих форм при
исследовании содержимого кишечника он ока-
зался пустым; у обитателей ила, например
Chaetodermatidae, пищу составляет, по-
видимому, детрит, наряду с которым
в желудке находят обычно диатомовые
водоросли, корненожек, иногда
радиолярий, часто иглы губок и частицы
ракообразных, которые могут заглатываться
моллюском вместе с илом.
Период размножения у северных видов,
например у Chaetoderma nitidulum,
падает на холодное время года. У герма-
фродитных форм мужские половые
продукты созревают раньше, чем женские,
т. е. проявляется протандрический
гермафродитизм.
Из яйца выходит личинка, которая
свободно плавает в воде при помощи своего
ресничного покрова. При дальнейшем
развитии постепенно вырастает задний конец
тела личинки; внутри клеток наружного
покрова закладываются известковые ши-
пики и пластинки, покрывающие тело
с боков и со спины; большая часть
ресничного покрова атрофируется, и
личинка падает на дно.
В этой стадии у нее на спине можно
ясно различить 7 известковых щитков,
черепицеобразно налегающих друг на
друга и чрезвычайно сходных с раковинными
пластинками панцирных моллюсков.
В дальнейшем как эти щитки, так и
другие шипики, покрывающие тело личинки,
атрофируются и заменяются
окончательными иглами и шипиками, свойственными
взрослым формам. Появление в
личиночной жизни беспанцирных моллюсков
спинных пластинок, сходных с раковиной
панцирных моллюсков, дает основание
многим исследователям рассматривать этих
последних как предков беспанцирных.
Рис. 2. Известковые пгапики и
чешуйки кожного покрова
беспанцирных моллюсков.
Рис. 3. Стадии развития мизомении.
13
КЛАСС ПАНЦИРНЫЕ (LORICATA)
К панцирным моллюскам относятся
своеобразные морские животные,
населяющие главным образом приливо-отливную
зону различных морей, и все их
строение носит яркие черты приспособленности
к своеобразным условиям жизни в этой
среде, к жизни под ударами волн
морского прибоя. Их обтекаемая сверху
форма тела и их способность плотно
присасываться к поверхности камней дают
им возможность выдерживать натиск
самых сильных волн.
Большинство представителей этого
класса имеет удлиненноовальную форму
тела, выпуклая спинная сторона которого
прикрыта раковиной; раковина состоит
из восьми пластинок, черепицеобразно
налегающих одна на другую. При
отрывании моллюска от субстрата он обычно
свертывается кольцом на брюшную
сторону, выставляя наружу покрытую
раковиной более защищенную часть своего
тела. Что касается нижней, брюшной
стороны, то она, наоборот,
представляется как бы срезанной в одной плоскости,
и здесь прежде всего бросается в глаза
мускулистая нога с плоской подошвой:
нога двумя глубокими продольными
бороздками отчленяется от боковых частей
тела. Нога служит не только органом
передвижения, но также и для плотного
прикрепления животного к камням, скалам,
известковым водорослям и тому
подобным предметам. На переднем конце
выделяется голова в виде полукруглого диска,
отделенного от ноги поперечной
бороздкой. В центре головного диска лежит рот,
а сзади, над ногой, заднепроходное
отверстие. В продольных бороздках, которые
можно рассматривать как мантийную
полость, с каждой стороны расположен
более или менее длинный ряд жабр, и
в эти же бороздки открывается пара
половых и одна пара выделительных
отверстий. Что касается самой мантии, этого
типичного для моллюсков органа, то у
панцирных моллюсков она образуется
всей той частью их кожного покрова,
которая находится между жаберными
бороздками и краем раковины на спинной
стороне. Эта часть поверхности покрыта
известковыми иглами, что характерно как одна
из черт сходства этих моллюсков с
беспанцирными. Однако, в отличие от последних,
мантия здесь относительно хорошо развита
и разделена на верхнюю и нижнюю
половины, граница между которыми и
составляет внешнее очертание тела. У
большинства форм, нижняя поверхность мантии
обычно плотно прижимается к
поверхности того предмета, на котором находится
животное, и способствует более плотному
прикреплению его, что очень существенно
для донных обитателей зоны морского
прибоя.
Панцирные моллюски в общем много
крупнее беспанцирных, и, хотя их самые
мелкие формы меньше сантиметра,
некоторые наиболее крупные представители
этого класса достигают размеров в 20—
30 см и значительного веса — до
нескольких килограммов.
Во внутреннем строении обеих групп
также наблюдаются как черты сходства,
так и черты отличия. Нервная система
панцирных моллюсков состоит из четырех
продольных стволов, соединенных
поперечными перемычками; спереди эти
стволы замыкаются нервным кольцом,
окружающим глотку. Однако здесь нет
обособленного головного мозга, и
соответствующая ему по положению верхняя
часть глоточного кольца мало отличается
по характеру своего строения от других
участков нервных стволов. Рот ведет
в объемистую глотку, в которой лежат
язык и радула — орган, похожий на
рашпиль и состоящий из множества
расположенных рядами зубчиков. Посредством
этих зубчиков моллюск захватывает
различные мелкие пищевые частицы, как
например водоросли, корненожек и др.,
соскабливая их с поверхности скал или
камней, на которых он сидит. Кишка здесь
имеет иное строение, чем у беспанцирных.
Она сильно вытянута, образует много
извивов и снабжена парой крупных
пищеварительных желез. Кровеносная система
14
устроена весьма сложно, и, кроме сердца,
имеется сеть кровеносных сосудов и
полостей с весьма сложным циклом
кровообращения. Сердце лежит на средней
линии спинной стороны под последней
пластинкой раковины и имеет вид
вытянутого мешка, в который с обеих сторон
открывается пара предсердий.
Околосердечная сумка (перикардий) отделена от
половой железы, а последняя
отграничена от почек. От половой железы, также
лежащей на спинной стороне, отходят
два канала, через которые выводятся
наружу зрелые половые продукты — яйца
или сперматозоиды. Почки имеют довольно
сложное строение и одним своим концом
сообщаются с околосердечной сумкой,
а другим открываются наружу по бокам
тела.
Раковина панцирных моллюсков, как
и у других представителей типа,
выделяется мантией, и каждая из восьми
образующих ее пластинок состоит из
нескольких слоев. Внутренние слои построены
на 98% из углекислого кальция и имеют
лишь прослойки из органического
вещества; самый наружный тонкий слой, так
называемый периостракум, состоит,
наоборот, целиком из органического вещества
конхина, обладает эластичностью и
противостоит действию слабых кислот и
щелочей, предохраняя известковую часть
раковины от разрушения. Раковинные
пластинки пронизаны множеством каналов,
в которых расположены своеобразные
органы чувств, а края этих пластинок более
или менее погружены в кожный покров
спины. У многих форм нижний слой
раковинных пластинок выступает из-под
верхних слоев, образуя крыловидные или
другой формы выросты, служащие для
прикрепления мускулатуры. Все эти черты,
типичные для большинства
представителей класса, подверглись у ряда форм
изменениям, выражающимся прежде всего в-не-
которой редукции раковины, пластинки
которой уменьшаются в размерах и
частично или полностью обрастают сверху
мантией. Так, например, урода Крип-
топлакс отдельные пластинки раковины
хорошо видны снаружи, но они
уменьшены в размерах и отодвинуты одна от
другой, а все тело животного напоминает
по форме червя, так как сильно сужено
и вытянуто в длину; узкая нога теряет
форму широкого мясистого диска, как
у других панцирных моллюсков, и
приобретает сходство с ногой беспанцирных.
У других форм, как, например, у
изображенного на таблице одного из
представителей рода Криптохитон, мантия
целиком обрастает раковину, так что
последняя совершенно невидима снаружи.
При этом наблюдается и некоторое
упрощение всей формы тела, так как
исчезает ясная граница между его верхней
и нижней сторонами и оно приближается
по форме к яйцу, которое лишь слегка
сплющено на брюшной стороне. У всех
остальных представителей класса,
которых насчитывается несколько сот видов и
которые объединены во множество родов,
мантия как бы особым кантом по краю
тела ясно подразделяется на нижнюю
плоскую ее часть (между краевым кантом
и жаберной бороздкой) и верхнюю
выпуклую (между краевым кантом и раковиной).
Мантию часто называют пояском
благодаря ее форме и положению вокруг
тела. Кроме раковины, мантией
образуются также и все те разнообразные кожные
включения, которые для панцирных
моллюсков так же характерны, как и для
беспанцирных, и которые подобно раковине
состоят частью из органического вещества
конхина и частью из извести, причем
наблюдается чрезвычайное разнообразие в
соотношении того и другого. Интересно, что
на нижней стороне пояска развиваются
чаще всего ромбовидные, плоские, как бы
фарфоровые пластинки с острыми краями,
которые располагаются красивой
мозаикой, причем некоторые из них лежат
косо к поверхности тела, выступая наружу
своими краями. Зацепляясь своими краями
за поверхность субстрата, они служат для
более прочного закрепления моллюска на
поверхности камней или скал и как нельзя
лучше приспособлены к этой роли.
Другие кожные включения мантии, которые
развиваются преимущественно на ее
верхней поверхности, имеют форму
цилиндрических шипиков или тонких щетинок,
причем закругленное основание некоторых
из них часто бывает окружено кольцом
из конхина, через которое проходит волок-
15
нистый тяж, связанный с нервом.
Такие шипики имеют значение органов чувств.
Переходя к рассмотрению органов
чувств, прежде всего приходится
отметить отсутствие органов равновесия—ста-
тоцистов, глаз на голове и головных
щупалец. Для определения качества воды,
приносящей к жабрам необходимый для
дыхания кислород, животному служат
чувствительные клетки кожи, находящиеся
на стенках жаберных бороздок и на самих
жабрах; в глотке помещается хорошо
развитый орган вкуса; наконец, у тех форм,
у которых верхние слои раковинных
пластинок пронизаны каналами, имеются
своеобразные органы чувств, носящие
название эстетов—выросты мантийного покрова,
которые проникают в эти каналы до их
наружных отверстий и оканчиваются
округлой конхиновой крышечкой,
замыкающей канал. От более широких каналов
ответвляется несколько узких, каждый
из которых заполнен одной большой
клеткой; они также оканчиваются отверстиями,
закрытыми конхиновыми крышечками.
Как предполагают, эти органы служат для
восприятия давления воды, хотя точно
их значение еще не выяснено. Характерно
для панцирных моллюсков развитие
совершенно своеобразных раковинных глаз,
представляющих собой в основном как
бы видоизмененные эстеты, у которых
толстая конхиновая крышечка заменена
тонким, тоже конхиновым покровом;
Таблица 1. Панцирные A—12 и бороздчатобрюхие
A3—14) моллюски:
1 — Placiphorella stimpsoni:
2 — Ischnochiton hasodadensis;
3 — Lepidochiton aleuticus;
4 — Tonicella granulata;
5 — Mopalia middendorffii;
6 — Mopalia schrenkii;
7 — Acanthochiton rubrolineatus;
8 — Placiphorella uschacovi;
9 — Cryptochiton stelleri;
10 — Tonicella marmorea;
11 — Cryptoplax Japonica;
12 — Mopalia seta;
13 — Nematomenia flavens;
14 — Echinomenia corallophila.
под этим покровом лежит прозрачная
светопреломляющая линза, находятся
светочувствительные клетки и отлагается
красящее вещество — пигмент. По мере
роста раковинных пластинок у их края
постоянно образуются новые глаза, и это
продолжается почти в течение всей жизни
животного. Число глаз у одной особи
может превосходить 11 тыс.
Наибольшей интенсивности окраска
достигает на верхней поверхности тела,
которая часто окрашена в охряно-жел-
тые или коричневые тона, но наряду
с этим можно видеть разнообразный узор
из пятен всех цветов радуги: зеленых,
красных и др., что в сочетании со
скульптурой раковинных пластинок придает
животному очень красивый вид. Окраска
некоторых форм прекрасно скрывает их
в окружающей обстановке, как, например,
живущего в Средиземном море красного
хитона (Chiton rubicundus), который
совершенно неотличим от красных
водорослей (кораллин), на которых он живет
на глубине 50—60 м.
Будучи тесно связаны с дном, эти
животные населяют все моря нормальной
океанской солености, встречаясь на разных
глубинах — от береговой линии до
глубины в 4 тыс. м и более. При этом они всегда
живут лишь на твердом грунте и этим
существенно отличаются от беспанцирных,
некоторые формы которых, наоборот,
зарываются в ил. Большинство видов этого клас-
Рис. 4. Изящный хитон (Chiton elegans).
са населяет зону прибоя, и лишь немногие
обитают в глубинах моря, причем
большинство таких форм составляют мелкие виды с
довольно примитивным строением. В зоне
прилива, наоборот, встречаются наиболее
крупные формы, у которых можно видеть
самое мощное развитие раковины и
мантии, самые длинные цилиндрические иглы
и, наконец, самое сильное развитие
пластинок, служащих для прикрепления
кожной мускулатуры. Развитие
последней имеет особенное значение в связи с
необходимостью противостоять действию
волн и крепко присасываться к
поверхности камней или скал. Нога и нижняя
сторона мантии плотно прижимаются при
этом к поверхности камня и играют роль
мощной присоски. Чем сильнее бушует
прибой, тем крепче присасывается
моллюск; если все же волны срывают его,
он сворачивается кольцом на брюшную
сторону, подобно мокрице, и
развертывается после этого лишь через
продолжительное время. Исключение в группе
панцирных моллюсков представляют по
своему образу жизни лишь червеобразные
Cryptoplacidae, которые живут в щелях
на коралловых рифах, обнаруживая
в этом отношении некоторое сходство
с образом жизни многих беспанцирных.
За исключением двух видов, относящихся
к роду Cryptoplax, которые живут
у берегов Японии, представители этой
группы встречаются лишь в тропических
морях — в Вест-Индии и в западной части
Тихого океана, омывающего юго-восточное
побережье Азии и прилегающие к нему
острова.
Большинство панцирных моллюсков—
малоподвижные животные, но встреча-
ются среди них и такие формы, которые
передвигаются относительно быстро.
Фишер указывает, что он наблюдал однажды,
как хитон (Chiton fulvus) быстро полз
вверх по якорной цепи, но не отмечает,
однако, более точно, какова же была эта
скорость. Возможно, что наше
представление о подвижности панцирных
моллюсков вообще неправильно, так как они,
по-видимому, ночные животные и более
подвижны ночью, чем днем.
Многие из панцирных моллюсков
живут у верхней границы зоны прилива и без
вреда переносят пребывание вне воды.
Возможно, что при этом они сами
набирают воздух в жаберную полость; по
крайней мере, по свидетельству Блэнвиля,
можно видеть, как сильное внешнее
раздражение вызывает у этих форм
выбрасывание воды и воздуха из жаберных
бороздок. Хэдлей указывает, что один
австралийский хитон живет у самой верхней
границы зоны прилива, где вода может
покрывать его только случайно.
Пища панцирных моллюсков,
обитающих в зоне прилива и на малых
глубинах, обычно состоит из растений,
которыми обрастают скалы; по крайней мере
в их кишечнике часто находят диатомовые
водоросли, а у одного крупного
экземпляра криптохитона Стеллера (Crypto-
chiton stelleri), обитающего в
Беринговом море, Миддендорф находил в кишке
даже нити водорослей длиной до дюйма.
Наряду с растительноядными известны
некоторые виды, питающиеся морскими
корненожками, а один глубоководный вид
(Hanleya abyssorum) поедает губок.
Наличие органа чувств,
лежащего под радулой, указывает на
некоторую способность моллюсков к
вкусовому отбору.
Н. 3 и м р о т полагает, что у
некоторых форм в связи с этим возникает и
специальная приспособленность к
определенной пище, и связывает с этим тот
факт, что некоторые панцирные
моллюски встречаются, по-видимому, только на
определенных видах ракушек, как,
например, один вид хэтоплевры (Chaetop-
leura bullata), который живет на
раковинах Spondylus calcifer, или ишнохитон
малый (Ischnochiton exiquus), попадаю-
17
2 Жизнь животных, т. 2
щийся на морской жемчужнице. Однако
совершенно не доказано, что эта связь с
определенными видами ракушек
обусловлена тем, что лишь на их раковинах
указанные виды находят необходимую им пищу.
Панцирные моллюски — раздельнополые
животные, за исключением, может быть,
лишь одного вида. При простоте строения
органов размножения и отсутствии
специальных приспособлений к внутреннему
оплодотворению возможность последнего
здесь, как правило, исключена. Оба пола
выделяют свои продукты в воду, где и
происходит оплодотворение яиц. Однако
среди панцирных моллюсков имеется
один живородящий вид (Lorica (Callis-
tochiton) viviparus), у которого яйца
развиваются в половых органах самки.
Как происходит при этом оплодотворение
яиц, остается неизвестно. Яйца имеют
округлую форму и покрыты нежной
оболочкой, усаженной у некоторых видов
выпуклостями или шипами. Они
склеиваются слизью и откладываются в виде
длинных шнуров «икры», которые, однако,
быстро разбиваются волнами на
отдельные куски. У одного вида ишнохитона
(Ischnochiton magdalensis) такие шнуры
достигают 77 мм длины и содержат свыше
миллиона яиц. Период размножения
у разных видов падает на разные месяцы;
несколько изменяются, вероятно, сроки
размножения и в разных географических
районах. Некоторые формы, живущие у
берегов Калифорнии, откладывают «икру»
уже в феврале—марте, другие — в мае-
июне; у большинства европейских форм,
для которых известны сроки их
размножения, последнее захватывает или весну,
или также и лето. Интересно, что
некоторые виды выбрасывают половые продукты
лишь ночью, другие по утрам и притом
лишь в спокойной воде, в период между
отливным и приливным течением.
Наряду с этой типичной для панцирных
моллюсков формой
размножения среди них есть целый ряд видов,
которые держат отложенную икру в
жаберных бороздках, так что последние
превращаются таким образом в
выводковые камеры, внутри которых и
развивается молодь. Интересно, что в связи
с этим приспособлением, выработавшимся
путем естественного отбора, резко
уменьшается число яиц, откладываемых
самкой, так как при развитии в жаберных
бороздках резко сокращается и процент
гибели зародышей. Тогда как у видов,
откладывающих яйца прямо в воду, число
яиц в кладках чаще всего достигает в
среднем 1400—1600 штук, у видов,
выращивающих молодь под прикрытием тела матери,
наблюдалось не свыше 200, а у некоторых
лишь по 80 зародышей в каждой из двух
жаберных бороздок. Из яйца развивается
личинка, у которой еще до вылупления
по бокам тела образуются личиночные
глазки, на спинной стороне — зачатки
раковинных пластинок, а на брюшной
стороне — зачаток ноги. После некоторого
периода свободного существования в виде
плавающей личинки молодой моллюск
падает на дно и постепенно приобретает
характерную для взрослого форму,
причем параллельно исчезают характерные
для личинки органы, как, например,
личиночные глаза и др.
К сожалению, как и в отношении
большинства морских животных, нам еще очень
мало известны быстрота роста
панцирных моллюсков, продолжительность их
жизни, возраст, в котором они достигают
половой зрелости. Имеются лишь
немногие отрывочные данные для нескольких
видов. Криптохитон Стеллера растет
довольно быстро и уже на четвертом году
достигает размера в 270—330 мм, а
половозрелым становится уже на втором году
жизни. У одного из видов хитонов
(Chiton tuberculatus) установлена возможность
определения возраста по годичным следам
нарастания, сохраняющимся на
раковинных пластинках; таким методом на
большом количестве особей этого вида
определена средняя продолжительность жизни
этих хитонов в 8—9 лет, хотя некоторые
экземпляры доживают и до 12 лет.
Интересно, что самки этого вида отмирают
несколько раньше самцов, но вообще
смертность сильно возрастает уже на
седьмом году жизни. Старых хитонов
часто легко можно отличить от более
молодых также по степени разрушения их
раковины от механического действия
волн и песчинок, которые волны несут
с собой, а также от разрушающего дей-
18
ствия различных организмов, которые
поселяются на моллюсках. Такое
разрушающее действие оказывают,
например, некоторые водоросли, которые
проникают даже в пустые каналы эстетов,
и некоторые сверлящие животные.
Вообще раковины хитонов обрастают часто
разнообразными сидячими формами,
населяющими вместе с ними приливо-отливную
зону моря; здесь можно находить
различных мшанок, сидячие формы многоще-
тинковых кольчецов, усоногих
ракообразных, как например морских желудей,
и др. На одном экземпляре хитона
(Chiton tuberculatus) наблюдалось
однажды 125 экземпляров морского желудя,
которые густо покрывали спину
животного. Наблюдались также случаи
прятания в жаберных бороздках крупных
хитонов некоторых ракообразных из группы
равноногих раков. Некоторые
паразитические одноклеточные частью живут в
полости этих бороздок, частью проникают в
их стенки и разрушают кожный покров.
Однако в общем панцирные моллюски
имеют относительно мало врагов и,
вероятно, спасаются от нападения многих
хищников главным образом благодаря
очень плотному присасыванию к
поверхности камней или скал; у некоторых
видов способность присасывания
соединяется со скрытым образом жизни в
щелях между камнями (как например
виды Schizochiton), или в щелях
коралловых рифов (как виды Cryptoplax), или
среди разветвлений известковых
водорослей литотамниев (как виды Trachyde-
rmon и др.).
Жизнь у верхней границы приливо-
отливной зоны в самой полосе прибоя
также ограничивает число возможных
врагов. Известны лишь единичные случаи
находок хитонов в желудке некоторых
рыб, например камбал.
Некоторые панцирные моллюски
употребляются в пищу человеком, как
например один из хитонов (Chiton
tuberculatus) на Бермудских островах, крипто-
хитон Стеллера (Cryptochiton stelleri)
и иглобоки (Acanthopleura). В пищу идет
мясистая нога, а в некоторых случаях
и икра этих моллюсков. Блэнвиль
указывает, что в старину их употребляли также
как амулеты, якобы приносящие
исполнение всех желаний.
Из представителей класса
панцирных моллюсков, встречающихся в пределах
СССР, наиболее крупные формы живут
в дальневосточных морях. Так, в
Беринговом, Охотском и Японском морях на
скалах и на каменистых грунтах живет
крупный криптохитон Стеллера
(Cryptochiton stelleri), достигающий длины
150—180 мм. Его спина, целиком
обросшая мантией, под которой лежат
пластинки раковины, окрашена в
темно-коричневый цвет, и этот общий тон окраски как
нельзя лучше соответствует цвету скал,
на которых он сидит. В ряде пунктов
в определенное время года (весной или
осенью) этот вид в массовом количестве
поднимается в приливо-отливную зону.
С другой стороны, в Беринговом и
Японском морях встречаются также и мелкие
хитоны, как например формы, живущие
среди известковых красных водорослей
и тоже окрашенные в красный цвет.
Ряд видов хитонов встречается также
и в наших северных морях. В Кольском
заливе часто можно находить на зарослях
известковой водоросли литотамний
небольших (около 12 мм) хитонов красного
цвета (Trachydermon ruber), с беловатыми
пятнами, которых очень трудно отличить
среди ветвлений водоросли, дающей им
приют. Сходной окраской обладает и
другой, тоже часто встречающийся здесь на
литотамний вид — Tonicella marmorea,
который населяет также и прибрежную
зону Белого моря. На каменистом и
ракушечном грунтах, а отчасти и на
литотамний как в Белом море, так и в Кольском
заливе встречается также белый хитон
(Trachydermon albus) с более удлиненным
телом; наконец, к числу обычных для
Кольского залива форм должен быть
отнесен и один представитель рода Lepido-
pleurus (L. arcticus) с темно-желтой
раковиной, живущий на различных грунтах.
Фауна Черного моря весьма бедна
панцирными моллюсками. Более обычен
здесь один вид хитона (Chiton polii),
живущий под прибрежными камнями и
среди ракушника. Кроме того,
встречается еще несколько видов (Acanthochites
fascularis и ДО.)-
2*
19
КЛАСС БРЮХОНОГИЕ (GASTROPODA)
Улитки, или брюхоногие, составляют
наиболее богатый формами класс
мягкотелых, охватывающий около 85 тыс. видов.
Развитие этого огромного разнообразия
форм стоит в связи с чрезвычайно
большим разнообразием условий
существования, к которым приспособились в своей
эволюции представители брюхоногих: они
заселили не только прибрежную зону
океанов и морей, но завоевали также и
значительные глубины и область
открытого моря; часть их расселилась по
пресным водам, а некоторые группы
приспособились к жизни на суше и выработали
типичные черты приспособления к
наземному существованию, заселив даже пустыни
и горные вершины до области вечных
снегов. Наконец, часть наземных форм
вернулась в воду и заселила пресноводные
бассейны, смешавшись там с исконными
морскими выходцами, но сохранив как
одну из ярких черт своего прошлого
наземного существования легочный тип
дыхания.
Одним из характерных признаков
брюхоногих служит наличие у них
раковины, состоящей из цельного куска и
прикрывающей спину животного;
правильнее было бы сказать, что раковина
прикрывает здесь так называемый
внутренностный мешок, т. е. выдающееся на
спине мешковидное выпячивание, внутри
которого находится целый ряд органов.
Другим типичным признаком
брюхоногих служит то, что почти все из них,за
ничтожным исключением, утратили
билатеральную симметрию
многих внутренних органов. Характерно
и то, что кишечник всех брюхоногих
образует петлеобразный изгиб, в связи
с чем анальное отверстие лежит над
головой или сбоку от нее, на правой стороне
тела.
У большинства брюхоногих раковина
закручена в спираль, при этом обороты
спирали чаще всего лежат в разных
плоскостях. Такая спираль носит название
турбоспирали. Соответственно со
спиральной закрученностью раковины спирально
свит и внутренностный мешок. В
огромном большинстве случаев эта закручен-
ность бывает по движению часовой стрелки,
т. е. вправо, если смотреть на раковину
с заостренного конца; в более редких
случаях закручивание раковины и
внутренностного мешка бывает против движения
часовой стрелки, т. е. влево. По
направлению закручивания раковины и
различают правозакрученные ( д е к с и о-
тройные) и левозакрученные (л е о-
т р о п н ы е) раковины, причем те и
другие могут встречаться иногда у
представителей одних и тех же систематических
групп.
В зависимости от того, насколько пологи
или круты обороты спирали, в завцсимо-
сти от числа и формы этих оборотов
чрезвычайно разнообразны по своему виду
и раковины различных улиток. Иногда
завороты раковины, плотно прилегая друг
к другу, срастаются своими внутренними
частями, образуя цельный столбик,
называемый колонкой (columella), иногда
они отстают один от другого, благодаря
чему по оси раковины вместо сплошного
столбика образуется щель, так
называемый пупок. Наконец, в ряде случаев мы
видим у улиток как будто более простую
раковину в форме колпачка или блюдца,
но, как показывает история развития,
такие раковины у современных улиток
представляют результат упрощения
первоначально спирально закрученной
раковины.
Раковина брюхоногих покрыта тонким
слоем органического вещества,
составляющего наружный слой раковины — пери-
о с т р а к у м. Последний образует иногда
щетинковидные отростки, благодаря чему
раковина снаружи кажется мохнатой.
Прикрытая периостракумом известковая
часть раковины слагается из тонких
известковых пластинок, составляющих в
своей совокупности так называемый
фарфоровый слой, в котором в свою очередь
можно различить до трех слоев
известковых пластинок. У некоторых
(сравнительно небольшого числа) улиток внутрен-
20
Рис. 5. Вертикальный
распил раковины улитки
Buccinum undatum.
Виден осевой столбик A),
обороты завитка раковины
B) и ее наружное
отверстие — устье C).
няя поверхность раковины
выстлана еще блестящим
перламутровым слоем. Раковина
соединена с телом при помощи
мощного мускула, сокращение
которого и втягивает улитку
внутрь раковины.
Переходя к рассмотрению
мягких частей тела улиток,
прежде следует отметить
наличие у них более или менее
обособленной головы, несущей рот,
глаза и щупальца, а на
брюшной стороне — наличие
массивной мускулистой ноги с
широкой нижней поверхностью,
называемой подошвой.
Характерный для большинства улиток
способ
передвижения — это медленное
скольжение по субстрату на широкой
подошве ноги, причем само
передвижение осуществляется
благодаря волнам сокращения, пробегающим
по подошве ноги сзади наперед. Обильная
слизь, выделяемая при этом кожей,
смягчает трение и облегчает скольжение по
твердому субстрату. У некоторых улиток
в связи с переходом их к иному типу
движения изменяются и функция и
строение ноги, что указано в описании
отдельных подклассов и отрядов брюхоногих.
У многих улиток задняя часть ноги несет
на спинной поверхности особую роговую
или обызвествленную крышечку и, когда
улитка прячется в раковину, крышечка
плотно закрывает устье.
Непосредственно под раковиной,
одевая внутренностный мешок, находится
мантия, передний утолщенный край
которой свободно свисает над туловищем
животного и прикрывает образующуюся
под ним мантийную полость, в которую
открываются заднепроходное,
выделительное и половое отверстия.
В мантийной же полости находятся
и органы дыхания — чаще всего
одна перистая жабра, или ктенидий.
Сравнительно небольшое число дышащих
жабрами улиток имеет две такие жабры;
у улиток, относящихся к подклассу
легочных, жабры утрачиваются, а свод
мантийной полости функционирует как
легкое. Свободный край мантии
у некоторых улиток может
вытягиваться в более или менее
длинную трубку — сифон; в
этих случаях обычно и по краю
устья раковины образуется же-
лобообразный выступ, внутри
которого сифон и
помещается. В других случаях
свободный край мантии может
заворачиваться над краем
раковины, так что мантия как бы
выступает из-под раковины и
прикрывает ее сверху частично
или даже полностью.
В последнем случае раковина
становится внутренней и
обычно в той или иной мере
подвергается редукции.
Рот у улиток ведет в
объемистую ротовую полость, в
которой находится парная или
непарная челюсть и типичный для
большинства моллюсков орган — терка,
или радула. В ротовую полость
открываются протоки парных слюнных желез,
а у некоторых улиток — протоки и
других железок, например ядовитых или
желез, выделяющих кислоту. От ротовой
полости отходит тонкий пищевод, у
некоторых улиток расширяющийся в
объемистый зоб, а последний переходит в
желудок, в который открывается
пищеварительная железа («печень»). От желудка
начинается кишка, более короткая у
хищных улиток и более длинная у
растительноядных; кишка открывается наружу
заднепроходным отверстием внутри
мантийной полости. Кровеносная система
улиток не замкнута: сердце состоит из одного
желудочка и одного предсердия (у
немногих форм два предсердия) . В
предсердии собирается окисленная кровь из
жабры или легкого, откуда она
перегоняется в желудочек, а затем по
разветвляющимся головной и внутренностной аортам
разносится по телу. Сердце улиток лежит
внутри полости перикардия; с полостью
этой сообщаются и органы выделения—
почки, в редких случаях парные. Обычно
сохраняется лишь одна почка.
Нервная система улиток состоит
из пяти пар нервных узлов, или ганглиев:
21
пары церебральных ганглиев, пары
ножных, или педальных, ганглиев, пары
плевральных ганглиев, пары висцеральных
и пары париетальных ганглиев.
Церебральные, педальные и висцеральные
ганглии соединены между собой
поперечными тяжами — комиссурами, а, кроме
того, церебральные, педальные и
плевральные узлы взаимосвязаны друг
с другом нервными тяжами — коннекти-
вами. Такими же продольными нервными
тяжами, или коннективами, связаны
между собой плевральные и висцеральные
ганглии, а париетальные ганглии
представляют собой скопления нервных клеток
в определенных местах этих коннектив.
В связи с закручиванием
внутренностного мешка у улиток, относящихся к
подклассу переднежаберных, а также у
некоторых самых низших представителей двух
других подклассов (заднежаберных и
легочных улиток) образуется характерное
перекрещивание коннектив, идущих
между плевральными и висцеральными
ганглиями. У высших заднежаберных и
легочных этого перекреста нет. У многих
улиток — очень резкое сближение
различных ганглиев и соответственное
укорочение связывающих их коннектив.
Из органов чувств, кроме пары
глаз на голове и пары головных
щупалец, имеющих значение органов осязания,
у улиток развиты органы равновесия —
статоцисты, одна пара которых иннерви-
руется от церебральных ганглиев, хотя
и лежит в непосредственной близости
от педальных ганглиев. Статоцисты
представляют собой пару замкнутых
пузырьков, стенки которых выстланы
ресничными и чувствительными клетками, а
полость пузырьков содержит жидкость,
в которой плавает одно крупное или
много мелких зернышек углекислой
извести. Давление, которое эти зернышки
извести оказывают на ту или иную стенку
пузырька при различных положениях
улитки, позволяет ей ориентироваться
в пространстве. Улиткам присущи также
и органы химического чувства—о с ф р а -
д и й, лежащий у основания жабры и
служащий для опробования воды,
попадающей в мантийную полость, а у наземных
улиток —вторая пара головных щупалец,
которые имеют значение органов
обоняния. Кроме того, кожа улиток богата
чувствительными клетками.
Половые органы имеют
различное строение у представителей разных
подклассов, причем среди улиток есть
и раздельнополые и гермафродитные
формы. У последних строение полового
аппарата наиболее сложно. Оплодотворение
у большинства брюхоногих внутреннее.
Развитие яйца и образование из него
личинки (в е л и г е р) у огромного
большинства форм имеет большое сходство
с развитием многощетинковых червей.
Интересной и важной особенностью
развития примитивных брюхоногих следует
считать изменение положения раковины
и внутренностного мешка. До
определенных стадий развития у личинки вершина
внутренностного мешка и вершина
раковины нависают над головой; перед
оседанием личинки на дно внутренностный
мешок и раковина поворачиваются на 180°,
в результате чего их вершины нависают
над задней частью ноги. Это так
называемое личиночное закручивание и приводит
к тому, что оказываются перекрещенными
нервные стволы, идущие от головы в
туловище, а мантийная полость с жабрами
и анальным отверстием лежит теперь
впереди, над головой. У других
брюхоногих, не имеющих свободноплавающей
личинки, все это перемещение
внутренностного мешка и раковины совершается
внутри яйца или даже смещается на столь
ранней стадии эмбрионального развития,
что проявляется лишь в неравномерном
росте правой и левой сторон тела. Но
и в этих случаях мантийная полость
взрослого животного лежит впереди, а
кишка образует петлю.
Характерная для большинства
брюхоногих асимметрия возникает благодаря
тому, что башневидная турбоспиральная
раковина располагается так, что одним
своим краем прилежит к голове плотнее,
нежели другим; это вызывает сужение левой
части мантийной полости, редукцию жабры
и связанных с ней других органов.
Брюхоногие делятся на три подкласса:
1) переднежаберных (Prosobranchia);
2) заднежаберных (Opisthobranchia) и
3) легочных (Pulmonata).
22
ПОДКЛАСС ПЕРЕДНЕЖАБЕРНЫЕ (PROSOBRANCHIA)
Переднежаберные составляют наиболее
обширную и разнообразную группу
брюхоногих моллюсков. Почти у всех у них
хорошо развита раковина; у огромного
большинства представителей подкласса
она спирально закручена, а у некоторых
имеет форму колпачка или блюдца.
Типично для всех переднежаберных то, что
жаберная полость направлена вперед
и лежит на спинной стороне животного,
причем соответственно вперед направлена
и вершина настоящей жабры, или ктени-
дия.
У более примитивных представителей
подкласса, которые по своей организации
ближе стоят к древним брюхоногим, кте-
нидии, а равным образом и предсердия-
парные, тогда как у других
переднежаберных правый ктенидий и правое предсердие,
как правило, редуцируются. У некоторых
форм подвергается редукции также и
левый ктенидий, а как органы дыхания
функционируют вторичные жабры.
Правая почка, хорошо развитая у некоторых
более примитивных переднежаберных,
у других форм или подвергается полной
редукции, или преобразуется в часть
выводящих путей полового аппарата.
Животные, как правило,
раздельнополы, хотя к этому подклассу и
относится значительный ряд гермафродитных
форм. К очень характерным для всех
переднежаберных признакам относится так
называемый перекрест нервных стволов,
соединяющих пару плевральных ганглиев
с парой висцеральных (перекрест плевро-
висцеральных коннектив). Соответственно
висцеральный ганглий, лежащий,
например, на правой стороне тела, в
действительности представляет собой левый
висцеральный ганглий, переместившийся
на правую сторону.
Из органов чувств обычно
бывают развиты глаза, в большинстве
случаев довольно примитивного строения,
органы равновесия (статоцисты), пара
головных щупалец, а иногда и другие щу-
пальцевидные придатки и типичный для
моллюсков орган чувств, связанный с
настоящей жаброй,— осфрадий.
Рис. 6. Расположение важнейших органов в теле переднежаберных моллюсков.
А — двупредсердная пунктурелла (Puncturella noachina): l — голова; 2 — ктенидий; з — анальное
отверстие. Б — однопредсердная лужанка (Viviparus viviparus): 1 — церебральные ганглии; 2 — нервный тяж;
3 — раковинный мускул; 4 — ктенидий; 5 — мочеточник; 6 — задняя кишка; 7 — выводковая сумка
(матка) с эмбрионами; 8 — сердце; 9 — передняя кишка; 10 — печень.
23
Нога, как правило, хорошо развита
и у более примитивных форм имеет вид
массивного нерасчлененного органа с
широкой ползательной подошвой. Нога
обычно несет на себе роговую или обыз-
вествленную крышечку, при помощи
которой может плотно запираться отверстие
раковины, когда улитка в нее прячется.
У некоторых переднежаберных форма ноги
усложняется. Может обособляться ее
передняя часть (проподиум) или
задняя часть (м е т а п о д и у м); у
некоторых вокруг основания ноги
обособляется окружающая основание ноги
выступающая складка (эпиподиум).
В основном нога служит для ползания
и опоры на субстрате; у некоторых форм—
также для закапывания в грунт. Наконец,
у немногих она превратилась в орган
прикрепления, а у некоторых
переднежаберных, приспособившихся к
планктонному существованию, — в орган
плавания. Большинство переднежаберных
имеет хорошо развитую радулу. У
переднежаберных, питающихся детритом,
ротовой аппарат очень слаб — радулярная
пленка тонкая, зубы слабо хитинизирова-
ны, челюстей нет. Некоторые переднежа-
берные приспособились к питанию
фильтрационным способом или к хищничеству.
Некоторые приспособились к
определенному составу пищи, как например к
питанию губками или кишечнополостными.
Для некоторых хищных форм главную
добычу составляют двустворчатые или
иглокожие. Наконец, среди переднежаберных
мы встречаем также и ряд паразитов,
причем можно наблюдать все переходы
между хищническим образом питания,
с одной стороны, и внешним или
внутренним паразитизмом, с другой стороны.
Некоторые внутренние паразиты
настолько резко изменились в своей организации
под влиянием образа жизни, что
утратили заднепроходное отверстие, сердце
и ряд других органов, типичных для
брюхоногих.
Переднежаберные в огромном
большинстве своем обитают в морях, но часть их
перешла к жизни в пресных водах, а ряд
форм в условиях очень влажного климата
приспособился к жизни на суше. У нас
в стране наземные переднежаберные
встречаются на Кавказе и в Приморье Дальнего
Востока. Наконец, есть ряд видов,
которые ведут земноводный образ жизни.
В заключение мы хотим очень кратко
сказать о разнообразном использовании
улиток человеком, причем по сравнению
с другими улитками особенно широко
используются именно переднежаберные.
Улитки уже издавна имели большое
значение в хозяйственной деятельности
человека, главным образом, естественно, в
приморских странах; но уже очень давно
пример этих стран проник далеко в глубь
материков. Моллюски издавна служили
не только в качестве пищи, их раковины
не только использовались как украшения
или знаки отличия, но во многих
областях они сделались важнейшим мерилом
ценности и употреблялись в качестве
денег, а на некоторых островах Океании
это свое значение они сохраняли вплоть
до самого последнего времени.
Большие кучи раковин, «кухонные
остатки» первобытного населения многих
приморских стран (в Дании, Бразилии,
Франции, Англии, Португалии,
Северной Америке, Японии, а также у нас по
берегам Черного моря), так же как и
Рис. 7. Различные формы радулы у
переднежаберных улиток:
1 — Neritina — один поперечный ряд зубцов радулы;
2 — Patella — левая половина одного ряда зубцов;
3 — Bithynia — средний зубец и левая половина
ряда; 4 — Scalaria — средний зубец и правая половина
ряда; 5 — Mitra — средний зуб и левый боковой
зубец; 6 — Conus — один зубец радулы.
24
украшения, находимые в погребениях,
указывают на широкое потребление
переднежаберных наряду с ракушками еще
в глубокой древности. Судя по мифам,
улитки играли большую роль в жизни
народов древней Индии. В Передней Азии
пурпурная улитка, из которой
финикияне добывали красящее вещество,
уже с древних времен приобрела большое
хозяйственное значение; у греков
морские улитки находили применение и как
лечебное средство.
В качестве денег улитки служили
преимущественно в трех областях: на
островах Океании, в Африке и в Америке. На
островах Тихого океана особенной
известностью пользуются так называемые «пела»,
изготовленные обитателями
Соломоновых островов из обработанных и
нанизанных на шнуры раковин улитки
нассы; особенно искусно обработаны
бывают некоторые виды денег у папуасов
восточного берега Новой Гвинеи; они делают
из них цепочки многометровой длины.
В Африке раньше служили в качестве
денег маленькие улитки оливы (Oliva
nana), которых собирали на одном из
островов южнее устья Конго, но теперь
они уже давно вышли из употребления.
Напротив, очень распространены и теперь
так называемые деньги «каури» —
раковины многих видов ципрей (Сургеа
moneta, С. annulus, а возможно, что также
и др.), которые все обитают у
юго-восточных берегов Азии и служили сначала
в Китае и в Японии средством оплаты
и размена, а оттуда они были
перенесены в Индостан и Индокитай, где
они с начала нашей эры сделались
единственной валютой и даже сейчас не
совсем еще вышли из употребления. Отсюда
они в доисторический период
распространились даже вплоть до Северной Европы.
В Африку до озера Чад, в большую часть
Судана они проникли с Запада через
посредство венецианцев, голландцев и
англичан; на западном берегу Африки их
уже нет.
В Северной Америке, где раковины
.морского ушка, а также денталиумы широко
применялись в качестве монет и
украшений, заслуживают упоминания, как
особая достопримечательность, вампумы —
Рис. 8. Деньги «каури» и украшения
из раковин переднежаберных.
кожаные пояса, вышитые кусочками
раковин улитки буссикон (Bussycon) и
служившие символами и документами. Так,
например, на одном таком вампуме был
увековечен договор, относящийся к 1682 г.,
которым некий Лени-Ленапе передает
Вильяму Пенну нынешнюю
Пенсильванию.
Едва ли возможно перечислить всех
переднежаберных, которые употреблялись
и даже сейчас еще употребляются людьми
в качестве пищи; многие виды, например
литорины, букцинум, пателлы и др.,
находят и сейчас большой спрос и у
культурных народов. Пестрые раковины улиток
находят чрезвычайно разнообразное
применение. И сейчас раковины улиток
употребляются в виде нательных
украшений у народов Африки, Азии,
Америки, Австралии; европейцы из них
вырезают камеи, причем цветной гипо-
стракум, темно-коричневый у Cassis cameo,
желтый у С. rufa, розово-красный у Strom-
bus gigas, очень эффектно выступает на
белом фоне остракума. Наконец, раковины
трохус идут в качестве сырья для
пуговичного производства.
ОТРЯД ДРЕВНИЕ БРЮХОНОГИЕ
(ARCHAEOGASTROPODA)
Наиболее примитивны в подклассе
переднежаберных те представители о т-
р я д а древних брюхоногих, которые
сохранили некоторые черты строения,
свойственные первичным брюхоногим,
например две перистые жабры, лежащие
25
в мантийной полости, парные почки с
парой выделительных отверстий и парные
предсердия. Желудочек сердца здесь
обычно пронзен задней кишкой, а
расположение нервов в ноге имеет более или
менее явственный лестничный характер,
что также может рассматриваться как
признак примитивного строения. В
раковине этих улиток обычно имеется
щель, которая у одних форм тянется
от устья до середины последнего завитка,
у других расположена у верхушки блюд-
цеобразной раковины, а у третьих как бы
разбита на ряд отдельных отверстий
округлой формы. Ряд этих отверстий
тянется вдоль оборотов раковины, которые
сильно расширены и уплощены, так что
вся раковина приобретает форму уха,
определившую характерное для этих
моллюсков название морского ушка.
Эта группа наиболее примитивных
переднежаберных составляет несколько
семейств, причем некоторые из них
обитают в морских глубинах, например
представители рода плеуротомарий (Pleu-
rotomaria), живущие у берегов Японии,
Молуккских островов, а также в Вест-
Индии. Раковины у плеуротомарий имеют
прекрасный перламутровый блеск и у
некоторых видов достигают значительных
размеров (свыше 20 см).
Современные плеуротомарий
представляют собой остатки прежде богатого
формами рода; число ископаемых видов
его, живших со времен силура до
третичного времени, определяется в несколько
сотен, в настоящее время виды
плеуротомарий исчисляются единицами.
Гораздо более многочисленны в
настоящее время представители другого
семейства, так называемые улитки морские
ушки (Haliotidae), которые все
относятся к одному роду Haliotis, но
представлены сейчас многими десятками видов.
Они живут в Тихом океане, у его
азиатского и американского берегов и частично
у берегов Австралии, а также
распространены в Индийском океане, у восточных
берегов Африки и в Атлантическом—
у западных берегов этого континента,
откуда они проникли и в Средиземное
море. Своеобразная ухоподобная раковина
этих улиток, пронизанная рядом
округлых отверстий, о которых говорилось выше,
ярко окрашена снаружи, а с внутренней
стороны покрыта перламутровым слоем
с превосходным блеском и отливающим
всеми цветами радуги. Нога здесь
достигает чрезвычайно мощного развития,
с широкой подошвой, при помощи
которой улитка так крепко присасывается
к поверхности камней, что выдерживает
удары самых сильных волн; отделить ее
от субстрата можно только при помощи
долота, которое просовывают между
подошвой ноги и поверхностью камня или
скалы, на которой укрепился моллюск.
Основание ноги у этих улиток окаймлено
мощно развитой кожной складкой,
носящей название эпиподиума, край которой
усажен многочисленными щупальцевид-
ными придатками. На переднем конце,
несколько выше, чем щупальца головы,
находятся более короткие выросты,
несущие глаза примитивного строения. Сверху
над основанием ноги спускается край
мантии, прикрывающий сдвинутую влево
глубокую мантийную полость, где лежат две
жабры и куда открывается
заднепроходное отверстие и выводные отверстия почек.
От наружной поверхности мантии
отходят щупальцевидные отростки, которые
высовываются наружу через отверстия
в раковине.
Интересные наблюдения над морскими
ушками (Haliotis tuberculata), живущими
в аквариуме в Неаполе, приводит в
своих работах Н. 3 и м р о т.
«Если мы с большим усилием оторвем
улитку от стеклянной стенки аквариума,—
пишет он, —причем иногда отрываются и
куски скрепляющей стекла замазки,
которые остаются на подошве ноги, то
поверхность последней опрокинутого на спину
животного прежде всего изгибается внутрь,
а края подошвы несколько утолщаются.
Затем моллюск делает попытки
перевернуться и принять нормальное положение
подобно тому, как это наблюдается у
перевернутых на спину черепах. Концы ноги
вытягиваются то в одну, то в другую
сторону, ища точку опоры; как только
задний или передний конец ноги
наткнется на стеклянную стенку аквариума,
соответствующий участок подошвы
сейчас же прилипает к ней, а затем постепен-
26
но получает опору и вся подошва,
прилипающая к стеклу, и сам моллюск занимает
нормальное положение. Но совсем другой
результат получается в тех случаях,
когда в поисках точки опоры у
перевернутой на спину улитки и передний и
задний концы ноги одновременно
прикоснутся к стенке аквариума. Оба они плотно
присасываются к последней, но при
попытке выпрямиться, имея эти две точки
опоры, улитка падает обратно на спину,
хотя, казалось бы, возврат к нормальному
положению теперь мог бы осуществиться
легче всего. Не совсем ясны причины
этого явления, но, по-видимому, тогда как
механическое раздражение подошвы,
вызванное прикосновением ее к поверхности
того или иного предмета, вызывает
ответную реакцию присасывания, мощный
раковинный мускул, или аддуктор, может
сокращаться с полной силой лишь тогда,
когда раздражение действует на него
лишь с одной стороны, постепенно
распространяясь на соседние участки, тогда
как при раздражении, идущем от обоих
концов ноги, его действие ослабляется,
и трудно сказать, обусловлено ли это
характером самой мускулатуры или
возбуждающих ее нервов. Как только наша
улитка снова укрепится на стенке
аквариума и подошва ее плотно прилипнет
к стеклу, она принимается ползти, но
чрезвычайно медленно, передвигаясь едва
на 6 см в течение 5 минут».
Органами дыхания
морского ушка, как уже отмечалось, служат
парные жабры, лежащие в глубине
мантийной полости. Токи свежей воды входят
в мантийную полость через широкую щель,
сообщающую ее с наружной средой,
в передней части тела, и, омывая жабры,
выходят наружу через ряд описанных
выше округлых отверстий на спинной
стороне раковины.
По наблюдениям Зимрота над морскими
ушками, жившими в аквариуме, эти улитки
иногда выползают по стенке аквариума
выше уровня воды и принимают
своеобразное положение, широко раскрывают
отверстие жаберной полости и
приподнимают передний край раковины, так что
последняя становится под углом, иногда
почти перпендикулярно к плоскости ноги.
Зимрот ставит это явление в связь с часто
наблюдаемой и у других переднежаберных
склонностью подниматься над уровнем
воды, причем в это время дыхание
осуществляется у них, по-видимому, не только
через жабры, но и всей поверхностью кожи.
Однако в природных условиях виды
морского ушка населяют обычно более глубокие
слои сублиторальной зоны и только
изредка попадаются в мелкой воде во время
отливов, как, например, живущие у
тихоокеанского побережья виды, служащие
предметом промысла и встречающиеся
преимущественно на глубине нескольких
метров. Также и европейский вид, живущий
в Атлантическом океане у западных
берегов Франции и Пиренейского полуострова,
ловится только во время самых сильных
отливов.
Улитки сидят обычно, плотно
присосавшись, на камнях и скалах, и, для того
чтобы отделить их от субстрата,
пользуются обычно ножом или долотом. В США
обычно такое долото прикрепляют к
длинному шесту; при этом через деревянный
ящик с вставленным в дно стеклом
высматривают с лодки местоположение
моллюска, так как без этого приспособления
трудно обнаружить его под водой
благодаря ряби на поверхности. В Японии
морское ушко добывается
преимущественно водолазами, работающими в
скафандрах. Водолаз берет с собой корзину
и складывает в нее собранных моллюсков;
по данному им сигналу корзину поднимают
в лодку и, опорожнив ее, снова опускают
на дно, причем один и тот же ловец
работает под водой около трех часов.
Иногда лов производится и более
примитивно. Водолаз ныряет под воду без
скафандра, подчас на глубину до 6 м, только
защитив глаза очками, а уши заложив
тампоном, и собирает улиток вручную
в течение одной-двух минут. Если место
для лова выбрано удачно, он каждый раз
возвращается с добычей.
Морское ушко добывают и ради мяса,
которое считается ценным продуктом
питания, и ради раковин, которые
представляют высокосортное сырье для
различных изделий из перламутра. Иногда
в раковинах попадаются и ценные
жемчужины. Использование мяса морского
27
ушка как продукта питания издавна
практиковалось в Китае, а с конца XIX в.
получило широкое распространение и в
США, где, например, за один 1921 г.
добыча этого моллюска возросла до
1 481 170 фунтов. Промыслы эти развились
главным образом в Калифорнии, где
промысловое значение имеют два вида: Halio-
tis fulgens и Н. rufescens. Параллельно
с развитием промыслов морского ушка
в США они стали развиваться и в
Японии, где за один 1923 г. японскими
промышленниками было добыто у берегов
Японии, Кореи и Шань-Дуня 7 544 344 кг
живущего здесь промыслового вида Halio-
tis gigantea.
Этот же вид морского ушка благодаря
высокому качеству перламутра и
крупным размерам A0—20 см) издавна
добывался и ради его раковин по всему восточ-
ноазиатскому побережью Тихого океана,
а также в Австралии. Ценное значение
как перламутровое сырье, идущее для
инкрустационных работ и на пуговичные
изделия, имеют также раковины других
видов морского ушка. В Европе
промысел этих моллюсков не имеет, однако, столь
широкого размаха, как в США и в
Японии: кроме упомянутого выше вида Halio-
tis tuberculata, промысловое значение здесь
имеет и другой вид — Н. lamellosa, который
ловится на средиземноморском побережье
Европы.
Представители рода галиотис по
преимуществу относятся к океаническим
формам и не встречаются ни в Черном,
ни в Балтийском морях, где для них нет,
по-видимому, подходящих условий
солености. Отсутствуют они и в северных
морях, омывающих европейское
побережье СССР, но встречаются на Дальнем
Востоке у берегов Камчатки.
Из других представителей этой группы,
более примитивных по своему строению
переднежаберных, имеющих парные
жабры, пару предсердий и раковину со
щелью, через которую выходит ток воды
из жаберной полости, следует упомянуть
живущую в водах Советского Союза
улитку пунктуреллу (Puncturella noachina),
принадлежащую к довольно
многочисленному по числу форм семейству
Fissurellidae. Раковина этой небольшой
но размерам улитки (длина до 1,5 см)
имеет форму овального в своем основании
колпачка с радиально ребристой
наружной поверхностью и лишена
перламутрового слоя. Верхушка раковины имеет
зид загнутой назад зачаточной спирали,
а спереди от нее лежит узкая короткая
щель. Пунктурелла — довольно обычный
представитель фауны моллюсков наших
северных морей, так как имеет кру го-
полярное распространение. Она часто
попадается при драгировках с глубин от
35 до 350 м и живет на камнях или на
известковых водорослях литотамниях,
а иногда встречается и на песчано-илистых
грунтах.
Интересно, что очень близкая к нашей
пунктурелле форма, составляющая,
однако, особый подрод, живет в южной части
Тихого океана. Другие представители
семейства широко распространены в
морях северного и южного полушарий. Один
вид (Fissurella graeca) встречается и в
Черном море в районе Босфора.
Весьма сходны по внешности с
представителями предыдущего семейства, но
глубоко отличны от них анатомически
улитки, составляющие три близких с е-
м е й с т в a: Patellidae, Acmaeidae и Le-
petidae. Они все характеризуются
упрощенной симметричной раковиной,
имеющей форму колпачка или опрокинутого
вверх дном блюдца. Важным
анатомическим признаком, характерным для всей
этой группы, служит наличие у этих
улиток не двух, а лишь одного предсердия,
что стоит в связи со своеобразным
изменением у них и органов дыхания; у
представителей семейства Patellidae обе
жабры редуцируются, сохраняясь лишь
в виде рудиментов, но взамен них на
нижней поверхности мантии развивается
венчик листовидных придатков, которые
функционируют как вторичные жабры. В
семействе Acmaeidae настоящие
первичные жабры тоже редуцируются,
но взамен них образуется одна вторичная
жабра, а у представителей семейства
Lepetidae исчезновение жабр
компенсируется кожным дыханием всей
поверхности тела. Большинство улиток,
относящихся к этой группе, раздельнополы,
но некоторые виды гермафродитны, при-
28
чем образование мужских половых
продуктов предшествует образованию
женских (протерандрический гермафродитизм).
Яйца обычно откладываются прямо в
воду, где и происходит оплодотворение.
К семейству Patellidae относится
сравнительно небольшое число форм.
Типичным представителем группы может
служить улитка блюдечко (Patella),
красочное описание строения и образа жизни
которой дает Зимрот. «В
приливо-отливной зоне всех морей,— пишет он,— сидят
на скалах ленивые блюдцеобразные
улитки пателлы; здесь их исконная область,
где они живут, прочно присосавшись к
береговым скалам. Их раковина, имеющая
форму низкого конуса с эксцентрически
лежащей верхушкой, вплотную
пригоняется к поверхности субстрата даже
тогда, когда это шероховатая, размытая
шлаковая лава: в этом случае края
раковины становятся неровными и зубчатыми,
следуя за всеми неровностями
поверхности субстрата, что дает улитке
возможность плотно прижаться к ней. Когда
улитку тревожат, она прижимает свою
раковину к тому предмету, на котором она
сидит, с огромной силой, и уже Реомюр
сделал наблюдение, что для того, чтобы
преодолеть силу присасывания
обыкновенной пателлы (Patella vulgata),
необходимо приложить силу, равную 14—15 кг.
Если загнать клин между
раковиной и камнем и, действуя
им как рычагом, стараться
отделить животное от камня, то
нередко оно разрывается на две
части: нога и голова
остаются на камне, а раковина с
мантией и с большей частью
внутренностного мешка отрывается.
Но если улитка сидит,
приподняв раковину так, что голова
ее и боковые части тела
остаются открытыми, то
достаточно легкого бокового удара,
чтобы животное отделилось от
места своего прикрепления; этим
и пользуются обычно рыбы,
поедающие пателлу.
Долгое время считался
нерешенным вопрос о том, как
происходит прикрепление пател-
лы: приклеивается ли она выделением
ножных желез или удерживается
исключительно силой присасывания, благодаря
действию раковинного мускула.
Согласно последним исследованиям,
можно, по-видимому, считать
установленным, что сначала действительно
выделяется небольшое количество слизи из
множества одноклеточных кожных желез
подошвы ноги, но служит эта слизь не для
приклеивания улитки, а для заполнения
мелких щелей между подошвой и камнем.
После этого уже начинает действовать со
всей силой раковинный мускул,
кольцевая форма которого только спереди
нарушается небольшой выемкой, благодаря
чему он имеет сходство с подковой. Он
напрягается при каждой волне прибоя,
а также во все время отлива, пока
животное бывает открыто действию солнечного
света. Такое поведение пателлы повело
к своеобразному сожительству ее с
некоторыми животными, как например с
коловратками, которые на это время
укрываются под краями ее мантии, находя
здесь пристанище, где долго сохраняется
необходимая им влажная среда.
На том основании, что края раковины
бывают очень точно пригнаны как к
гладкой, так и к неровной поверхности
скалы, делали заключение, что пателла
никогда не меняет своего места и как бы
прикреплена к нему. Однако
такое заключение оказалось
ошибочным. Улитка часто
совершает ночные путешествия на
расстояние около метра.
Замечательно, что, двигаясь
определенным образом всегда в
левую сторону, она в конце
концов возвращается к исходной
точке своего пути и
укрепляется на старом месте точно
так же, как сидела там
раньше, — замечательный случай
ориентации в пространстве,
которому, правда, помогает при
движении равномерное
отклонение от прямой линии.
Животное возвращается к своему
прежнему месту подобно тому,
как заяц, преследуемый
собакой и обежавший большой круг,
Рис. 9.
Улитка-блюдечко (Patella algira):
l — подошва ноги; 2 —
мантия; з — вторичные
жабры; 4 — голова с парой
щупалец.
29
после многих сделанных им петель попадает
опять в свое логовище. Эта привязанность
к своему месту жительства, «чувство
дома», теперь подробно и разносторонне
исследована, и оказывается, что только
в том случае, если место, где улитка
живет, претерпело коренные изменения за
время ее отсутствия, она решается на
поиски нового. Ни в коем случае она не
селится где попало. При выборе места она
руководствуется потребностью в
достаточно насыщенном водяными парами воздухе;
поэтому она предпочитает расщелины
утесов, и именно их теневую сторону. Ночные
странствования служат преимущественно
для утоления голода. Улитка во время
своего передвижения объедает
поверхность скалы, и обглоданная полоска
выдает ее путь, потому что все время, пока
животное ползет, его радула находится
в действии.
Пищей улитке служат различные
организмы, обрастающие утесы, в первую
очередь микроорганизмы, которыми
питаются также и панцирные моллюски,
а попутно и небольшие растения, как уль-
вы и фукусы, но их она не отыскивает
нарочно, поедая главным образом все
то, что может сбрить по дороге с
поверхности камня своей радулой. Нам ничего
не известно о том, делает ли она при этом
какой-нибудь выбор пищи по вкусу. Для
такого способа питания радула с ее
толстыми крепкими лезвиями оказывается
прекрасно приспособленным
инструментом; ее прочные зубцы вполне
соответствуют своему назначению —
соскабливать обрастание скал, которое в
прибойной зоне держится на них так же крепко,
как и сама улитка. Эта работа, однако,
ведет к чрезвычайно быстрому
изнашивание) орудия, благодаря чему возникает
потребность в столь же быстром его
восстановлении. Поэтому влагалище радулы,
в котором образуются зубцы, необычайно
длинно, почти такой же длины, как все
тело животного, и закручено спиралью,
чтобы быть в состоянии уместиться в нем.
Питательность маленьких существ,
добываемых таким способом со скал,
по-видимому, довольно мала, что
обусловливает чрезвычайную длину кишечника па-
теллы, многократно изгибающегося и
открывающегося в дыхательную полость
на конце небольшого сосочка.
В большинстве случаев раковина пател-
лы малоприметна и покрыта таким же
обрастанием, как и камень, на котором улитка
сидит; раковина толста и прочна у видов,
обитающих в прибойной зоне, но
встречаются и другие формы раковины, как
например у маленькой Patella pellucida,
живущей у берегов Германии. Она
покинула скалы и переселилась на водоросли
фукусы. В связи с этим ее раковина
сделалась тонкой и прозрачной, с чудесными
переливчатыми зелено-голубыми
полосками, которые придают ее окраске
сходство с искрящейся на солнце водорослью».
Пателлиды широко распространены
в различных морях, причем
некоторые виды встречаются и в сильно
опресненных внутренних морях, как например
черноморская пателла (Patella pontica),
которая проникла и в Азовское море. Этот
вид живет на береговых скалах,
подверженных действию прибоя, в зоне которого
он обычно держится; но иногда эти
улитки всползают по скалам и выше уровня
воды, так что их лишь обдают брызги
прибойных волн. Наша черноморская
пателла, достигающая 35—40 мм длины,
употребляется в пищу в свежем виде под
названием «пателлиды». Съедобны также
и другие виды этого рода, как например
средиземноморская Patella coerulea и
живущая в Атлантическом океане Patella
vulgata.
* * *
Среди представителей семейства
Acmaeidae некоторые виды достигают
относительно больших размеров, до 7,5 см,
как, например, гигантская лоттия (Lot-
tia gigantea), живущая у западного
побережья Северной и Центральной Америки,
тогда как встречающиеся в наших
северных морях улитки рода Акмеа (Acmaea
testudinalis, A. virginea и A.
rubella) значительно меньше ее по
размерам; длина раковины у наиболее крупных
Acmaea testudinalis обычно не
превосходит 3 см. Этот вид весьма часто попадается
в Кольском заливе и Белом море на
глубинах от 0 до 8 м и по своему образу жизни
имеет много общего с пателлой, так как
тоже населяет зону морского прибоя. Дру-
30
гой вид, Acmaea virginea, живет в более
глубоких слоях — от 6 до 100 м.
Только что рассмотренные семейства
составляют как бы боковую ветвь
развития улиток, выделившуюся в отряде
древних брюхоногих, к которому относится
большое число форм со значительным
разнообразием строения и составляющих
много семейств. Однако у всех у них в той
или иной степени проявляются некоторые
черты, свойственные древним
брюхоногим, как например примитивная радула,
наличие у ряда форм двух предсердий и
парных почек, лестничное расположение
нервов в ноге и другие признаки.
* * *
Обширную группу в отряде древних
брюхоногих составляет семейство
улиток волчков, или трохид (Trochidae),
охватывающее около 50 различных родов
морских улиток и делящееся на ряд
подсемейств. У всех этих улиток правая
первичная жабра редуцирована, но
сохраняются два предсердия и парные почки.
Раковина здесь лишена щели и обычно
спирально закручена в форме
конусовидной башенки или башенки с уступами.
Очень своеобразна ее форма у улиток
рода ангария, живущих в
Индийском и смежных с ним областях Тихого
океана. У этих улиток невысокие обороты
раковинной спирали покрыты отростками
причудливой формы.
Обычно у всех этих улиток хорошо
развита кожная складка, окаймляющая
основание ноги (эпиподиум) и
несущая щупальцевидные придатки, которые
у рода трохус могут втягиваться в особые
влагалища. Эти придатки имеют также
значение органов осязания, как и пара
развитых головных щупалец трохид.
Зимрот дает очень красочное
описание действия этих
придатков у улитки каллиостомы
(Calliostoma): «В то время как
большинство водных улиток как
бы оберегает свои щупальца от
резких раздражений,
осторожно втягивая их или отводя в
сторону, трохиды ведут себя в
этом отношении гораздо
смелее, напоминая этим
головоногих. Примером может служить улитка
каллиостома. Щупальца у нее
длиннее, чем эпиподиальные осязательные
усики, окрашены в черный цвет и
постепенно заостряются к концам. Они
действуют гораздо свободнее, чем щупальца
других улиток, они могут сворачиваться
и даже изгибаться, как бич при ударе;
сходно, но лишь несколько слабее
выражено это и у эпиподиальных осязательных
придатков. Поэтому щупальца здесь более
походят на руки головоногих, чем на
щупальца других улиток. Сходство
усугубляется еще тем, что, когда они касаются
какого-нибудь твердого тела, они
пристают к его поверхности на короткое время,
как это типично для рук головоногих».
Из многочисленных представителей с е-
м е й с т в а трохид некоторые имеют
большое хозяйственное значение, как
например представители рода трохус
(Trochus), живущие в Индийском и
смежной с ним области Тихого океана.
Раковина этих улиток дает превосходного
качества перламутр, который используется
преимущественно для пуговичной
промышленности, причем наибольшее
промысловое значение имеет вид Trochus
niloticus, раковины которого добываются
на Сингапуре, в Красном море, на
островах Фиджи и в Новой Каледонии.
Раковины более мелкого
средиземноморского вида трохид — Trochus adriaticus —
употребляются лишь для ювелирных
изделий.
Наши северные моря богаты трохида-
ми: в Баренцевом море живет Trochus
occidentalis, несколько видов,
относящихся к роду соляриелла (Solariella) и к
роду Маргарита (Margarita), имеющих
кругополярное распространение.
В Черном море живет несколько видов
мелких улиток, относящихся
к роду гиббула (Gibbula),
которые держатся в области
сублиторали, а иногда и очень
близко к поверхности,
усеивая сплошь береговые скалы.
Один из этих видов, живущий
также и в восточной части
Средиземного моря (Gibbula albida),
постоянно встречается на
устричных банках.
Рис. 10. Каллиостома
(Calliostoma zizyphinus)
с щу пальцевидными
придатками на ноге.
31
Рис. И. Пресноводные улитки из семейства
неритид.
Слева направо: Theodoxus fluviatilis, T. tran-
sversalis (из р. Днестр), Т. pallasi (опресненные
участки Черного и Каспийского морей).
Близки к трохидам морские улитки,
относящиеся к семейству Turbinidae,
из которых наиболее интересен род
Turbo. Это по большей части весьма крупные
улитки с закрученной раковиной,
имеющей форму волчка и достигающей
размера 20 см и более. Внутреннюю
поверхность ее покрывает перламутровый слой
прекрасного качества и блеска, в связи
с чем эти улитки издавна служат объектом
промысла. Раковины их идут на
различные изделия из перламутра или
непосредственно употребляются как украшения;
в этом случае с них удаляются
наружные, не имеющие блеска, слои и
сохраняется один перламутровый слой. Из этого
рода на первом месте по своему
промышленному значению стоит вид Turbo olea-
rius, который вылавливается в
Юго-Восточной Азии, у берегов Австралии и
Новой Гвинеи и раковины которого достигают
веса в 2 кг.
Крупные виды турбо, живущие в
тропических морях, держатся обычно в
поверхностных слоях воды и часто
всползают выше ее уровня; они наиболее
деятельны ночью.
Такой полуземноводный образ жизни
в еще большей степени свойствен
некоторым представителям другого близкого
семейства неритид (Neritidae),
среди которых имеются обитатели и морей,
и пресных вод. Из числа перешедших
к полупресноводному или пресноводному
образу жизни улиток один вид (Theodoxus
pallasi) живет в опресненных участках
Каспийского и Черного морей, а
несколько видов заселяют реки, принадлежащие
к бассейнам этих морей, и реки бассейна
Аральского моря. Все это небольшие по
размерам улитки со своеобразной формой
раковины, которая может плотно
закрываться обызвествленной крышечкой.
Рисунок на верхней поверхности раковины
крайне изменчив и состоит либо из
желтовато-белых пятен на черном и
черно-зеленоватом фоне, либо из темных полос,
пробегающих по светлой серо-желтой
поверхности раковины.
Наиболее часто в Днестре, Южном
Буге, Днепре, Доне, а также в их
притоках и в некоторых озерах можно
встретить одного из представителей
пресноводных неритид — небольшую улитку Theo-
Таблица 2. Брюхоногие моллюски:
1 — Fissurella graeca,
2 — Acmaea cassis;
3 — блюдечко (Patella granulans);
4 — Acmaea testudinalis;
5 — Margarita;
6 — Perotrochus (Pleurotomaria)
hirasei:
7 — морское ушко (Haliotis tube-
rculata);
8 — Cypraea caurica;
9 — Tectus maximus,
10 — Cypraea tigris;
11 — Nerita spinosa;
12 — Delphinula rugosa;
13 — Purpura lapiilus;
14 — Vharonia tritonis;
15 — Natica vitellus;
16 — Strorabus gigas;
17 — Turitella terebra cerea;
18 — Cirsotrema perpexum;
19 — Murex ternispina;
20 — Murex scarpio,
21 — Murex saxatilis,
22 — Distarsio (Murex) anus;
23 — Galerita hungarica;
24 — Crepidula grandis;
25 — Terebra subulata.
doxus fluviatilis. Она держится обычно
близко к поверхности воды на
прибрежных камнях, спускающихся в воду корнях
деревьев или на стеблях тростника. Часто
эти улитки всползают и выше
поверхности воды и их можно находить в таком
положении даже в самое жаркое время
дня.
Как и у многих моллюсков, переход
к пресноводному образу жизни отразился
на характере развития этих улиток: они
утратили стадию свободноплавающей
личинки. Развитие зародышей здесь
происходит внутри яйцевых капсул, которые
откладываются самками иногда на
поверхность самой раковины.
Несмотря на то что неретиды уже
довольно далеко уклонились в своем
строении от наиболее примитивных
представителей отряда древних брюхоногих, они
сохранили еще некоторые характерные
для этих улиток признаки, как
например наличие двух предсердий.
Однако у некоторых семейств,
составляющих особую ветвь развития древних
брюхоногих и приспособившихся к
наземному существованию, мы встречаем уже
сердце с одним предсердием и наряду
с этим некоторые новые признаки,
носящие черты приспособления к наземному
образу жизни. Жабры здесь нацело
редуцируются, а свод мантийной полости
служит как легкое. Большинство видов этих
небольших по размерам улиток относится
к семейству гелицинид (Helicini-
dae), виды которого распространены в
двух разобщенных между собой областях:
в Центральной Америке и Вест-Индии,
а с другой стороны, в небольшой части
Восточной Азии, Австралии, на Зондских
островах и на островах Полинезии.
Улитки эти живут на влажной почве
тропических лесов, а частично и на деревьях,
сохраняя характерный признак
большинства переднежаберных — наличие
крышечки, запирающей отверстие
раковины,— признак, по которому их легко
можно отличить от наземных легочных
улиток.
ОТРЯД MESOGASTROPODA
У представителей отряда Mesogas-
tropoda в огромном большинстве случаев
имеется лишь одно предсердие и лишь
одна почка, а что касается органов
дыхания, то у водных обитателей они
представлены вторичными жабрами,
развивающимися на нижней поверхности мантии,
а у наземных видов жабры редуцированы
и дыхание осуществляется через
поверхность мантийной полости. Также в
строении нервной системы, радулы и половых
Таблица 3. Брюхоногие моллюски:
1 — Rapana thomasiana;
2 — Phalium strigatum;
3 — Bulla lignaria;
4 — Thatcheria mirabilis;
5 — Pyrula pernessa;
6 — Valva valva;
7 — Cassis cornuta;
8 — Pterocera rugosa;
9 — Strombus auris-dianae;
10 — Harpa ventricosa;
11 — Siliquaria cumingi;
12 — Конус (Conus marmoreus);
13 — Dorioconus aulicus;
14 — Conus ebraeus;
15 — Mitra episcopalis;
16 — Mitra cruentata;
17 — Oliva mustelina:
18 —Cymbium (Voluta) amporus;
19 — Tiara praestantissima;
20 — Mitra strictica.
3 Жизнь животных, т. 2
Рис. 12. Циклостома (Cyclostoma, или Pomatias
elegans) в двух фазах движения.
органов (причем в большинстве случаев
полы разделены) эти улитки представляют
более высокую ступень организации, чем
представители древних брюхоногих
(Archaeogastropoda). У них также обычно
сохраняется крышечка, иногда весьма
сложного строения.
Наиболее богаты видами наземных Meso-
gastropoda тропические и субтропические
страны.
Некоторые относящиеся к этой группе
улитки приспособились к жизни на
деревьях, как, например, живущие на
Филиппинских островах некоторые
представители рода Pterocyclus, обладающие
в связи с этим крайне тонкой и нежной
раковиной.
Несколько больше, чем о тропических
улитках, мы знаем о европейских формах
наземных переднежаберных, которые
в своем распространении на север в
Европе заходят гораздо дальше, чем в какой-
либо другой части света. Среди опавшей
листвы, в лесах под поваленными стволами
деревьев, а также во мху или на
известковых почвах среди камней можно
находить этих маленьких улиток, наличие
у которых крышечки показывает на их
принадлежность к переднежаберным.
Одни из них живут под опавшей листвой
и ползают, опираясь на всю подошву
ноги, как например широко
распространенная Acme polita или другой более
южный вид A. lineata; другие, как
например циклостомы (Cyclostoma elegans),
встречающиеся обычно на известковой
почве под камнями на хорошо
прогреваемых солнцем местах, перемещаются
своеобразным способом.
Вот как описывает способ передвижения
этих улиток Зимрот: «Заслуживает
внимания наличие у этой улитки плоского
округлого диска на конце рыльца,
давшего основание для ее названия
(циклостома — значит «круглорылая»). Этот
диск представляет собой как бы присоску
и помогает животному в его
поступательном движении. Если извлечь эту
зеленоватую улитку из-под опавшей листвы и,
положив в стакан, наполненный влажным
мхом, поманить морковью или кусочками
огурца, то можно увидеть в высшей
степени своеобразную форму передвижения.
Одна половина подошвы плотно
прилегает к стеклу и несколько расширяется.
Она остается неподвижной и удерживает
животное на месте, в то время как
другая половина отделяется от стекла и
вытягивается вперед, причем здесь не
образуются правильные поперечные
углубления подошвы, как у наземной легочной
улитки, но возникают неправильные
переливы волн сокращения, которые лучше
всего можно сравнить с волнующейся
под ветром поверхностью нивы; действие
этих переливов лежит в основе движения
у всех водных улиток. Когда же ей не
удается почему-либо продвинуться вперед
таким способом, она вытягивает рыльце,
присасывается его дисковым придатком
и, сокращая тело, подтягивается таким
образом вперед».
Кроме только что описанных наземных
форм, отряд Mesogastropoda включает
в себя еще большее разнообразие форм
морских улиток, приспособившихся
к чрезвычайно разнообразным условиям
обитания: среди них мы находим и
типичных обитателей прибрежной зоны, и
формы, населяющие более значительные
глубины, и улиток, приспособившихся
к планктонному существованию, наконец,
ряд форм, превратившихся в наружных
или внутренностных паразитов
различных морских беспозвоночных. К этому же
отряду относятся и некоторые семейства
пресноводных переднежаберных. Из всей
этой обширной группы мы рассмотрим
лишь некоторых наиболее интересных
представителей отряда.
Типичными обитателями прибрежной
зоны морей являются представители с е -
34
м е й с т в а литорин (Littorinidae), или
береговых улиток. Все они живут в
приливо-отливной зоне и часто остаются над
уровнем воды, оставаясь, однако,
в этом положении неподвижными, а
иногда и впадая как бы в спячку. При этом
тело улитки глубоко втягивается в
раковину, а между краем устья и
поверхностью скалы можно видеть образовавшуюся
кожистую пленку, причем в жаберной
полости не наблюдается того нормального
количества воды, которое обычно бывает
у этих улиток. Грей наблюдал, что улитка
пребывала в таком оцепенении в течение
целой недели, а затем, помещенная в
морскую воду, через несколько минут
вернулась к состоянию своей нормальной
активности.
Одна из самых обыкновенных и широко
распространенных береговых улиток —
Littorina littorea — живет в мелкой воде
или держится на сваях, водорослях,
камнях и часто сидит неподвижно в течение
долгого времени над уровнем воды.
Уползая снова в воду, она захватывает с собой
воздух, и, если ее потревожить вскоре
после погружения, можно видеть
пузырьки воздуха, поднимающиеся из воды.
Движения ее медленны; средней величины
экземпляр этой улитки при ползании по
стеклянной стенке аквариума двигался
со скоростью 0,5 мм в секунду. Пища
обыкновенных береговых улиток
состоит частью из растений, частью из
животных.
В Англии литорин сажают в устричные
ящики, чтобы они очищали их дно от
морских водорослей, наличие которых здесь
нежелательно, так как отмирающие
водоросли образуют отложения ила.
Обыкновенная береговая улитка
литорина относится к самым широко
распространенным моллюскам северного
полушария. В Балтийском море она доходит
до восточного берега Борнгольма и Рю-
гена, а дальше на восток для нее уже
оказывается недостаточной соленость воды,
хотя в одну из прошлых геологических
эпох она была распространена
повсеместно на территории так называемого Лито-
ринового моря, очертания которого лишь
немного отличались от современного
Балтийского. Ее много всюду на северном
и западном побережьях Европы вплоть
до Португалии. Встречается она также
в Адриатическом море, в Белом море
и в Атлантическом океане, у берегов
Гренландии и на северо-востоке Америки.
Интересно, что в Белом море, где, кроме
Littorina littorea, живут также два
других вида (Littorina rudis и L. palliata),
все эти виды, по наблюдениям К. М.
Дерюгина, спускаются на глубину до
10—12 му т. е. теряют свою
специфичность литоральных животных.
На французском побережье Ламанша
водятся три вида: Littorina obtusata,
населяющая зону фукусов; Littorina
littorea, обитающая несколько выше в зоне
ульвы, которая часто подсыхает,
оставаясь выше уровня воды; Littorina rudis,
селящаяся, как правило, выше приливо-
отливной черты. Эти виды, однако, часто
перемешиваются, и даже Littorina
obtusata встречается иногда вместе с Littorina
rudis.
Самцы и самки у Littorina rudis
заметно отличаются друг от друга:
раковина самки крупнее и с круглой формой
устья, тогда как у самцов устье более
угловато. Наблюдаются скрещивания
между разными видами. Икра Littorina
obtusata прикрепляется к грунту. Молодь
выходит из яиц уже в совершенно
развитом виде, похожая на взрослых
животных. Littorina rudis — живородящая
форма, и новорожденная молодь у нее
появляется вполне развитой. Littorina
littorea, напротив, при высоком стоянии воды
откладывает плавающую икру, которая
держится вблизи берегов и носит
название «неритической». Два или три яйца,
из которых каждое раз в 20 мельче яиц
живородящей Littorina rudis, вместе
заключены в светлую капсулу, окаймленную
дисковидным расширением наподобие
широких полей соломенной шляпы. У
личинок здесь хорошо развит парус, и они
представляют собой типичных
плавающих личинок велигеров. Выловленные
и помещенные в аквариум, через 6—11
дней они опускаются на дно и переходят
к донному существованию. Здесь мы
видим, таким образом, совершенно
различный характер развития личинок в
зависимости от зоны обитания взрослой улитки*
3*
35
Рис. 13.
J — кладка яиц Littorina obtusata; 2 —
свободноплавающая яйцевая капсула Littorina littorea; з — Ris-
soa costata (справа — в натуральную величину,
слева — увеличена); 4 —гидробия (Hydrobia steini); 5 —
байкальские улитки из семейства Hydrobidae
(слева — Kobeltocochlea maxima, высота раковины —
17 — 21 мм, вверху и внизу — Benedictia fragilis,
высота раковины — 40—50 мм)\ 6 — битиния (Bi-
thynia leachi).
Сходные наблюдения были сделаны над
литоринами, живущими на английском
побережье.
Литорина широко используется в
Англии как объект промысла и добывается для
питания, причем дешевизна и
доступность этой улитки обусловливает массовое
потребление ее преимущественно
малоимущими слоями населения.
Сходный с литоринами образ жизни
ведут маленькие улитки риссоиды (Ris-
soidae), имеющие удлиненную раковину
с клетчатой скульптурой на поверхности.
Некоторые из них встречаются и на
значительных глубинах, до 200 м. Виды,
населяющие прибрежную зону, обладают,
как и литорины, способностью набирать
воздух в мантийную полость, причем
захваченный пузырек воздуха у
зарывшихся в ил улиток время от времени
выталкивается наружу, но не отрывается
от животного, а остается как бы
связанным с ним слизистой нитью, к которой
пристают частицы ила. Имеются
указания, что относящаяся к этой группе
улитка Liliopa, живущая среди
водорослей Саргассова моря, случайно
оторвавшись от субстрата, сохраняет связь с ним
посредством аналогичной нити и
пользуется последней, чтобы снова добраться до
водоросли, наподобие того как пауки
пользуются выделенной паутиной.
Относительно близкое к литоринам
семейство составляют улитки гидробии
(Hydrobiidae), большинство которых
приспособилось к жизни в пресных водах или
в солоноватых водоемах, а некоторые
стали обитателями суши. Несколько
типичных представителей этого семейства из
рода гидробия (Hydrobia) живет на
территории СССР, причем один из них,
Hydrobia ventrosa, встречается в
солоноватых водоемах Средней Азии, Северного
Кавказа и Закавказья, а другой вид,
Hydrobia steini, живет в прудах и озерах,
относящихся к бассейнам Волги, Днепра
и Балтийского моря. Гидробии — это
маленькие улитки, башнеобразная
раковина которых достигает высоты лишь
4—5 мм и закрывается тонкой роговой
крышечкой.
Еще меньшими размерами отличается
другой представитель этого семейства —
ручьевая улитка (Bythinella adsharica),
живущая в Закавказье, высота раковинки
которой не превышает 1,6 мм, К этому
же семейству относятся и мелкие улитки
рода Paladilhia (Lartetia), которые живут
в ручьях внутри пещер в ракушечных
известняках Южной Германии и которые
образуют множество местных форм,
возникших в связи с изолированными
местообитаниями этих улиток в подземных
водах, куда они переселились, вероятно,
во время ледникового периода.
Значительно более крупные
представители семейства живут в Байкале (из р о-
д о в Kobeltocochlea и Benedictia) и
встречаются в реках Черноморского
бассейна на медленном течении (Lithoglyphus
naticoides); относительно крупны также
некоторые представители широко
распространенного у нас в пресных водах рода
битиния (Bithynia).
Bithynia tentaculata, пожалуй,наиболее
обычный и широко известный из обитате-
36
лей текучих и замкнутых водоемов
Европейской части СССР. Это небольшая, но
хорошо заметная улитка с
башнеобразной, заостряющейся к вершине раковиной
A0—12 мм высоты), которая может
закрываться обызвествленной крышечкой. Би-
тиний можно видеть часто в огромных
количествах на прибрежных камнях, в иле
или на водных растениях. Особенно
обычны они в пазухах листьев плавающего
у поверхности воды телореза, на одном
растении которого их иногда можно
собирать десятками. Как и у других
пресноводных улиток, в развитии битинии
выпадает стадия свободноплавающей
личинки. Оплодотворенные яйца улитка
откладывает кучкой, причем яйца так тесно
прилегают друг к другу, что соприкасающиеся
между собой яйцевые оболочки образуют
прямые грани, дающие ложное
представление о наличии между ними перегородок.
Вышедшая из яйца молодая битиния ведет
образ жизни, сходный с образом жизни
взрослых. Тогда как Bithynia tentaculata в
своем распространении на восток не
заходит далее бассейна Оби, другой, также
часто встречающийся в наших водоемах
более мелкий вид битинии, Bithinia leachi,
образующий ряд разновидностей,
проникает и дальше, вплоть до бассейна Тихого
океана. Наконец, в бассейне Амура
живет маленькая улитка Bithynia striata (до
10 мм), отличающаяся от своих
родственников тем, что ее раковинка украшена
продольными ребрышками.
К относительно крупным
представителям переднежаберных, заселившим
пресные водоемы всего земного шара, за
исключением Южной Америки, относятся
улитки живородки (Viviparidae). Несколь-
ко улиток из р о д а настоящих
живородок (Viviparus) обычны для наших рек
и замкнутых водоемов, озер, прудов
и даже болот. Наиболее крупными
размерами среди них отличается живородка
уссурийская (Viviparus ussuriensis),
живущая в бассейне Амура (высота раковины
до 60 мм), а из европейских видов —
широко распространенная у нас в
стоячих водоемах от озер до болот так
называемая лужанка (Viviparus contectus).
Относительно тонкостенная,
спирально завитая раковина этой улитки бледно-
коричневатой окраски с зеленоватым
отливом и тремя темно-коричневыми
полосами бывает сильно изменчива в
оттенках, то более светлой, то более темной
окраски. Устье раковины, когда улитка
втягивается в нее, очень плотно
закрывается роговой крышечкой с ясно
заметными концентрическими полосками.
Замечательно красива окраска и мягких
частей тела этой улитки, у которой на
общем темном фоне кожи рассеяны
золотисто-коричневые точки. На голове лу-
жанки довольно сильно выдается вперед
лопастевидное рыло, а по бокам его
расположены весьма длинные щупальца,
у основания которых находятся глаза.
Правое щупальце у самцов сильно
расширено и отличается закругленной
формой: оно играет роль копулятивного
органа. Оплодотворенные в теле самки яйца
развиваются в яйцеводе, в концевой части
которого при беременности можно
находить обычно более развитые стадии
зародышей, тогда как следующие за ними
зародыши находятся на более ранних
стадиях развития; это делает лужанку таким
удобным и излюбленным объектом для
эмбриологических исследований.
Размножение происходит,
по-видимому, почти в течение круглого
года, так как и весной, и летом,
и зимой можно находить самок
с вполне развитыми зародышами,
число которых колеблется от 12
до 25. Новорожденные лужанки,
которые сразу же начинают вести
такой же образ жизни, как и
взрослые, отличаются от последних не
только своими размерами, но
также и наличием на их раковине
Рис. 14. Пресноводная, или
живородка лужанка (Viviparus
contectus).
Рис. 15. Вальвата (Val
vata cristata), высунув
шаяся из раковины.
37
Рис. 16. Изменения формы
раковины у различных видов пресноводных
вальват.
Слева направо: Valvata piscinalis
v. fluviatilis (широко распространенный в
реках вид); Valvata brandti (Армения,
озеро Гокча); Valvata cristata (широко
распространенный вид в стоячих водоемах).
Рис. 17.
Байкалия
(Baicalia
stiedea).
конхиновых бахромчатых выростов,
расположенных в несколько рядов вдоль
завитков раковины, так что последняя
приобретает мохнатый вид. Впоследствии эти
бахромки отпадают, но некоторое время
еще после того у молодых лужанок
можно наблюдать на раковинке спиральные
ряды точек соответственно прежним
местам расположения этих конхиновых
выростов.
Замечательна выносливость лужанок
к низким температурам: плотно закрывая
раковину крышечкой и впадая в
оцепенение, они без вреда для себя могут
пережить замерзание воды и оставаться
вмерзшими в льдину.
Другой широко распространенный у нас
вид живородки, Viviparus viviparus,
населяет не только поемные водоемы в
долинах рек, но и сами реки, причем
речные формы отличаются более толстой
и прочной раковиной.
Наряду с живородками наши пресные
водоемы населены крошечными улитками
вальватами (Valvatidae), которые
живут на илистом грунте, а также на
водных растениях в реках, озерах и прудах.
Наиболее распространена Valvata
piscinalis, различные подвиды которой
населяют и реки, и замкнутые водоемы;
эти маленькие улитки, высота раковины
которых не превышает 8 мм, утратили
свойственные древним переднежаберным
первичные жабры, расположенные на
своде мантийной полости, но зато край
последней несет у них с каждой стороны по
одному щупальцеобразному отростку,
причем щупальцевидный придаток,
находящийся на левой стороне, усажен с обеих
сторон листочками и весь орган функцио-
нирует как жабра. В отличие от
раздельнополых живородок, вальваты —
гермафродиты. Яйца они откладывают в
коконах, которые приклеиваются к
субстрату: внутри кокона находится слизистая
масса, обволакивающая яйца, а
последние как бы подвешены на плотных тяжах.
У Valvata piscinalis, по наблюдениям А. Д.
Некрасова, таких яиц в коконе бывает
до 31, а у другого вида (Valvata cristata)
их бывает не больше 6, чаще же 1—3 яйца.
У этого последнего вида завитки
раковины располагаются почти в одной
плоскости, а на ряде других видов можно
наблюдать все переходы от
возвышающейся башнеобразной раковинки до плоско-
дисковидной формы последней.
Несколько видов вальват встречается
в Байкале, а некоторые виды
приспособились к жизни в пещерных водоемах.
Одна такая вальвата, выделенная в
самостоятельный род горация (Horatia),
живет в Рионской пещере у Кутаиси. Это
почти микроскопических размеров улитка
(высота раковины до 1,1 мм).
* * *
Из других представителей
пресноводных переднежаберных большой интерес
представляет семейство Micromelanii-
dae, охватывающее небольшое число
родов, из которых два встречаются на
Балканском полуострове, три в Каспийском
море, один в солоноватых водах Индии
и один род байкалия (Baicalia) в
Байкале, причем последний представляет
огромное разнообразие форм и делится на много
подродов, каждый с многочисленными
видами, размеры которых сильно
варьируют — от 3 до 20 мм\ остальные роды этого
семейства включают каждый весьма
ограниченное число видов.
Разнообразные представители другого,
тоже живущего в пресных водах
семейства улиток меланиид (Melaniidae)
широко распространены в Южной и Юго-
Восточной Азии, на Зондских островах,
на Мадагаскаре, в Африке, Центральной
и Северной Америке, тогда как у нас
встречается лишь небольшое число видов
этих улиток, относящихся к двум р о-
д а м — меланопсис (Melanopsis) и мела-
ния (Melania). Представители первого жи-
38
вут в реках, относящихся к
Черноморскому бассейну, и некоторые, как например
Melanopsis acicularis, живущие в
бассейне Днестра, Днепра и Южного Буга,
достигают значительных размеров (высота
раковины до 17—20 мм). Представителем
же второго рода у нас является
единственный вид Melania amurensis, живущий
в бассейне Амура и отличающийся
относительно крупными размерами (высота
раковины до 45 мм). Другие же
многочисленные виды этого рода широко
распространены от Африки до островов
Океании. Некоторые мел аниды живородящи,
как например живущая в африканском
озере Танганьика улитка тифобия
(Tiphobia), с причудливой формой раковины,
напоминающей раковины морских
брюхоногих.
* * *
Заканчивая обзор переднежаберных,
приспособившихся к жизни в пресных
водах, необходимо упомянуть о самых
крупных представителях этой
разнородной группы, входящих в семейство
ампулярий (Ampullariidae) и
распространенных по всему тропическому поясу
земного шара. Раковина их соответственно
толста и крепка. У некоторых видов она,
кроме того, сильно уплощена, иногда до
такой степени, что можно принять ее за
левозакрученную раковину, и только
правостороннее положение порошицы и
полового отверстия обнаруживает, что мы
имеем дело все же с правозакрученнои
улиткой. Очень своеобразно строение этих
улиток, приспособленное к земноводному
образу жизни и соответственно к дыханию
и атмосферным воздухом, и
растворенным в воде кислородом. Мантийная
полость разделена здесь перегородкой на
правую и левую части; в одной части
находится жабра, служащая для водного
дыхания, другая часть функционирует
как легкое. Край мантии,
ограничивающий отверстие, сильно вытянут и может
складываться в виде длинной трубки;
улитки выставляют ее открытым концом
над поверхностью воды. Таким образом,
улитки дышат некоторое время
атмосферным воздухом, находясь на неглубоком
месте под поверхностью воды.
Ампулярий способны, как и многие
другие брюхоногие, переносить долгую,
длящуюся годами спячку: при недостатке
влаги они плотно закрывают раковину
крышечкой. Эти улитки раздельнополы.
Яйца самка или откладывает вне воды,
или прикрепляет под водой к водным
растениям, и тогда их оболочка разбухает,
как у икры лягушек. На левой части
нашего рисунка можно видеть, как яйца
выходят из полового отверстия и, скользя
вдоль так называемой генитальной
складки, которая тянется до правого
переднего угла ноги, соединяются в
общую массу икры, как это показано на
правом рисунке.
* * *
Возвращаясь к тем представителям о т-
р я д a Mesogastropoda, которые типичны
как обитатели морского дна, прежде всего
отметим ряд форм, ведущих
малоподвижный или даже прикрепленный образ
жизни. В этом ряду форм мы встречаем
представителей различных семейств, довольно
далеко отстоящих друг от друга в системе.
Рис. 18. Слева — тифобия (Tiphobia horei);
справа— две последовательные фазы откладки икры
ампулярией (Ampullaria gigas).
Рис. 19. Крепидула
(Crepidula farnicata) —
вид раковины снизу.
39
Интересную по своей биологид группу
составляет семейство калиптреи
(Calyptraeidae), один из представителей
которого (Calyptraea sinensis) живет и
у нас в Черном море и весьма обычен
у берегов Крыма. Это маленькие улиточки
с плоскоконической раковинкой, которые
держатся на небольших относительно
глубинах. Половые железы их гермафродит-
ны, причем здесь наблюдается протеранд-
рический гермафродитизм, т. е. раньше
развиваются мужские половые продукты.
Сроки созревания и откладки яиц
различны в различных географических районах:
так, средиземноморские калиптреи у
берегов Неаполя размножаются с января по
апрель включительно; живущие в
Адриатическом море около Триеста
размножаются в марте и апреле, тогда как
размножение черноморских калиптреи
начинается лишь в мае и длится до сентября.
Интересно при этом проявление у
калиптреи так называемой заботы о потомстве,
выражающейся в том, что яйца,
откладываемые в особые капсулы, не бросаются
на произвол судьбы, а остаются под
прикрытием тела матери, которая закрывает
их своей раковиной и отчасти ногой.
К семейству калиптреи относится также
другая интересная улитка — крепидула
(Crepidula), или улитка-туфелька, как
можно ее назвать за некоторое сходство
ее раковины с туфлей, если смотреть на
раковину снизу.
У этих улиток раковина имеет
удлиненно-овальную форму с очень низкой
макушкой, которая обнаруживает едва
заметную спиральную закрученность,
а на нижней поверхности раковины
находится поперечная перегородка с прямым
или слегка вогнутым краем. Крепидулы
ведут неподвижный образ жизни,
прикрепляясь к различным предметам
посредством известковой пластинки, которая
выделяется ногой; при этом обычно
образуются длинные цепочки особей, сидящих
одна на другой. Часто они поселяются
на раковинах устриц, особенно у берегов
Северной Америки, откуда крепидула
была случайно завезена и к берегам
Европы.
Близка по своему положению в
системе к семейству калиптреи улитка тика
(Thyca), принадлежащая к
немногочисленному семейству капулид (Сари-
lidae). Эта улитка интересна тем, что
живет в качестве наружного паразита на
теле морских звезд. В связи с этим
раковина ее сильно уплощена, а на брюшной
стороне у нее для прикрепления к телу
хозяина развилась особая присоска,
окружающая ротовое отверстие. Для
высасывания соков из тела хозяина служит
длинный хобот, который прободает
стенку тела морской звезды.
* * *
Особо резкому изменению в связи с
неподвижным образом жизни подверглась
внешняя форма тела в семействе
верметиду или червеобразных улиток (Ver-
metidae). Раковина здесь имеет вид более
или менее вытянутой в длину, спирально
закрученцой трубки с круглым устьем,
которое может запираться роговой
крышечкой. На голове животного сидит
пара коротких щупалец, на наружной
стороне которых находятся глаза. Два щу-
пальцевидных придатка отходят также
от передней части ноги цад крышечкой,
тогда как ползательная подошва ноги
у этих форм редуцирована. Мантийная
полость хорошо развита и содержит
жабру, состоящую из множества узких
листочков. У верметид хорошо развита
радула, но при их неподвижном образе
жизни они питаются главным образом
органическим детритом. Очень
своеобразен способ, при помощи которого эти
улитки захватывают пищу. Ножная
железа этой улитки выделяет обильную
слизь, которая наподобие вуали нависает
над устьем. Через некоторое время эта
слизь втягивается и поедается вместе
с приставшим к ней детритом.
Принадлежащий к этому семейству род верметус
(Vermetus) представлен многими видами,
распространение которых приурочено к
теплым морям, а отчасти и к морям
умеренного пояса. Червеобразные улитки
раздельнополы. Самка откладывает яйца
в двояковыпуклых капсулах, которые
приклеиваются в ряд в концевой части
трубки. В каждой капсуле содержится
несколько яиц, из которых развивается
типичная для моллюсков личинка вели-
40
гер. Некоторые представители этого
семейства, как например виды другого р о-
д а тенагодус (Tenagodus), живут обычно
в теле морских губок.
Относительно близка к рассмотренному
семейству совсем крохотная улитка це-
кум (Caecum), величиной меньше
миллиметра. Правда, сходство здесь почти
ограничивается тем, что раковина ее,
образовав вначале короткий завиток, растет
затем в форме изогнутой дугой трубки
и открывается круглым устьем, которое
может закрываться роговой крышечкой.
Такая форма раковины сильно
затрудняет ползание улитки, однако
внутренностный мешок не заполняет здесь
внутренней полости раковины до самого завитка,
как у других брюхоногих, но даже
отделяется от завитка внутренней
перегородкой; завиток раковины в конце концов
отпадает, и раковина становится
похожей на миниатюрного денталиума, с той
только разницей, что конец ее трубки
замкнут. Характерно, что у этой улитки
передвижение осуществляется посредством
ресничного эпителия кожи.
* * *
К своеобразным обитателям дна моря,
но не в прибрежной зоне, а на
значительных глубинах относятся улитки,
составляющие особое семейство
Xenophoridae с единственным родом
ксенофора (Xenophora) и немногими
видами.
По форме раковины эти улитки
несколько похожи на калиптрею с ее несколько
уплощенной конусовидной раковиной, но
только они значительно крупнее, и
раковина их расширена по своему краю. При
этом наружная поверхность раковины
обычно усажена различными
чужеродными телами, сходными у каждой улитки,
но различными у разных индивидов. У
одних — это раковины ракушек, у других —
раковины улиток, у третьих — иглы
морских ежей, которые равномерно
выступают над краем раковины; с одного
определенного участка морского дна
обыкновенно вылавливают совершенно сходные
экземпляры, как это, например,
наблюдалось в работах глубоководной
экспедиции «Вальдивии».
Рис. 20. Верметус (Vermetus lumbricalis).
Полную противоположность
неподвижно прикрепленным донным формам Meso-
gastropoda представляют по своему
образу жизни пелагические киленогие улитки,
составляющие особую группу (Heteropo-
da). Сюда относятся три семейства:
Atlantidae, Carinariidae и Pterotracheidae.
Киленогие водятся только в южных
морях; Средиземное море еще богато ими,
так как оно вообще представляет собой
в некоторых отношениях как бы северный
форпост тропической зоны. Что касается
Северного моря, то в него заносятся
Гольфстримом только редкие единичные
представители этой группы. Как пелагические
обитатели водной толщи, почти все они
принадлежат к типичным планктонным
животным с телом
стекловидно-прозрачным, за исключением глаз и маленького
внутренностного мешка. Атланты,
которые, прячась в раковину, погружаются
при этом глубже в воду, имеют лиловую
и коричневую окраску; большая карина-
рия из Индийского океана имеет резкое
крупное коричневое пятно на теле. Как
у большинства пелагических животных
тропиков, области распространения
видов очень обширны, и близкие виды трудно
отличимы друг от друга, как, например,
близкие тихоокеанские формы и формы,
живущие в Атлантическом океане. Также
и роды имеют обширное распространение,
что затрудняет установление их
происхождения. Только у птерозомы, которая
похожа формой своего тела на скрипку, аре-
41
ал распространения более
ограничен Австралийским
морем, южной частью Тихого
и Индийским океаном.
Животное это представляет
большую редкость, и оно
попадается всегда только
единичными экземплярами, почему
и не удавалось долго
определить его место в системе, и
долгое время эту улитку
принимали за червя.
Размножение киле-
ногих отличается некоторыми
интересными особенностями.
Полы раздельны; яйца
выбрасываются в воду на
светлой цилиндрической нити,
кодец которой часто задерживается
внутри женского полового отверстия так,
что яйцо продолжает плыть за матерью.
Свободноплавающие личинки снабжены
парусом, который достигает большого
размера и с каждой стороны расщеплен
на две или три довольно крупные
лопасти. Естественно, что не всегда легко
установить принадлежность той или
другой личинки к определенному виду, хотя
по различию многих плоских атлантопо-
добных личиночных раковин,
встречающихся в разных частях тропиков, можно делать
вывод о различии и взрослых форм; это
служит подсобным средством для
исследования распространения видов.
О сроках размножения и о скорости
развития молоди мы знаем пока очень
мало. Но как и для других настоящих
пелагических животных, более или менее
длительная благоприятная погода, в
первую очередь слабые ветры или штили,
содействует наиболее обильному
размножению и ведет к колоссальному увеличе-
нию численности этих
животных. Такие скопления их
часто удавалось наблюдать.
Они удерживаются, пока
длится благоприятное
состояние ветра и течений.
Начавшийся шторм выбрасывает их
огромными массами на
берег, который тогда на целые
мили бывает засыпан
толстым слоем раковин одного
какого-нибудь вида. Жизнь
в скопищах улиток
протекает очень ровно; день и ночь
они находятся в
беспрестанном движении. Необычайна
выносливость и кажущаяся
безразличность к ранениям
у этих животных. Часто преследующий
хищник откусывает у них начисто
внутренностный мешок или переднюю часть
тела, а бывали и такие случаи, что
попадались особи с откушенной головой,
но с уже зажившей раной, бодро
продолжавшие плавать; иногда их даже
описывали как новую форму, думая, что они
лишь были попорчены консервирующими
средствами, что действительно часто
бывает.
Атланта (Atlanta), представитель с е-
мейства Atlantidae,— небольшая
улитка, которая вся умещается в
раковине величиной не более 1 см. Раковина ее
плоская, завитая в одной плоскости,
и только самое начало завитка нарушает
здесь асимметрию. В строении этого
животного все приспособлено к возможно
более равномерному распределению
тяжести при плавании. Гребневидное
расширение, находящееся на последнем
завитке, служит превосходным килем при
плавании в перевернутом положении, как
обычно и плавает эта улитка. Наибольшие
же изменения претерпела здесь нога. Ее
передняя часть сжата с боков и в виде
пластинки отвесно опущена книзу. Животное
плавает, двигая из стороны в сторону этим
плавником, а на заднем конце последнего
находится присоска — остаток ползатель-
ной подошвы. В ней сохранилась хорошо
развитая мускульная ткань, тогда как
в прозрачном плавнике лишь при
тщательном рассмотрении можно видеть диаго-
Рис. 21. Атланта (Atlanta
peronii).
Рис. 22. Птеротрахея (Pterotrachea sp.).
42
нально перекрещивающиеся мускульные
пучки. На задней части ноги, имеющей
вид обособленной лопасти, находится
крышечка — оперкулум. Таким образом,
плавник, присоска и задняя лопасть,
несущая крышечку, составляют каждый как
бы переднюю, среднюю и заднюю ноги.
Атланты, как и прочие киленогие,
живут в открытом океане; но часто они
с помощью своей присоски прочно
укрепляются на каком-нибудь плавающем
предмете и, если их потревожить,
втягиваются внутрь раковины, замыкаясь
крышечкой.
Совсем по-другому ведут себя карина-
рии (Carinariidae). Они значительно
крупнее: большинство форм достигает размера
человеческого пальца, а попадаются
и чрезвычайно крупные представители
семейства, как например некоторые
глубоководные формы, живущие в
Индийском океане и достигающие размера
почти 40 см. Однако большей частью карина-
рии живут близко к поверхности моря.
Раковина у этих улиток немногим больше,
чем у атланты, но ее короткий завиток
сразу расширяется, так что получается
форма, сходная с якобинским колпаком.
Раковина служит только для прикрытия
жаберных лепестков, которые и
помещаются в ней. Остальное же тело
вытянуто в длину, особенно вперед,
имеет вальковатую форму и
веретенообразно заострено с
концов. Плавник, в
большинстве случаев только у
самцов, имеет присоску,
служащую органом
прикрепления во время полового акта.
Тело уплотнено
расположенными в коже крепкими со-
единительноткаными
клетками, содержащими слизь.
Направленный вертикально
вверх плавник, сгибаясь под
косым углом то в одну, то
в другую сторону, сообщает
животному медленное, но
верное поступательное
движение. Хвост бьет из
стороны в сторону, и все
туловище, насколько позволяет ему
его малая плотность,
производит подобные же движения. Этими
толчками животное как бы бросается то туда,
то сюда, но все же плывет вперед, хотя
и без всякой грации. Как следует из этого
описания, животное может с одинаковым
удобством плыть также и назад, и
действительно у него наблюдали
передвижения в обоих направлениях.
Другой представитель семейства
каринарий— птерозома (Pterosoma) —
обладает особенной устойчивостью при
плавании благодаря форме тела. По бокам
тело ее позади головы крыловидно
расширено, и весь ее облик несколько
напоминает скрипку.
Однако наиболее изменилась в сторону
приспособления к плаванию птеротра-
хея (Pterotrachea) с ее наиболее
правильной веретенообразной формой тела. Эта
улитка, относящаяся к семейству
Pterotracheidae, совсем не имеет
раковины; ее серебристо-блестящий, украшенный
крапчатым узором внутренностный мешок
замкнут у заднего края, так что из него
выступают только жабры, снабженные
сверху и снизу кожистой оторочкой;
хвост часто бывает вытянут в виде
длинной нити с утолщениями в виде узелков
красного цвета. В коже лежат хорошо
развитые пучки продольных мускулов.
Поэтому животное плавает очень быстро,
двигаясь не только с
помощью плавника, но извиваясь
всем телом, как рыба. Пте-
ротрахеи — в высшей
степени хищные животные. Их
вытянутое хоботообразное
рыльце беспрестанно
поворачивается из стороны в
сторону в поисках добычи. В
связи с этой особенностью,
вероятно, находится высокая
степень развития их глаз и
органов равновесия,
чрезвычайно важных при
подвижном образе жизни этих
животных. Те и другие органы
достигают здесь наивысшей
степени развития среди всех
брюхоногих. Нервные
волокна идут от этого органа
равновесия к ножным
ганглиям и дальше к мозгу. Глаз
Рис. 23. Орган равновесия
птеротрахеи (схематизирован-
но):
1 — нерв; 2—бесструктурная
мембрана, покрывающая статоцист;
3,4 — ресничные клетки; 5 —
отолит; 6 и 8 — чувствительные
клетки; 7 — клетки поддерживающей
ткани.
43
очень велик, имеет форму шара с
усеченной частью и односторонне расширен
у основания, а сверху замкнут
полукруглой роговицей и снабжен линзой.
Удлиненная форма всего глаза обеспечивает
значительность расстояния между
линзой и сетчаткой. Последняя имеет на дне
своеобразную щель, помогающую,
вероятно, легче определять расстояние до
различных предметов. Пигмент,
выстилающий боковые стенки и дно глаза, имеет по
сторонам скважины — окна, которые
пропускают свет. Внутрь от них лежат
смежные зрительные клетки.
В заключение нужно упомянуть, что
птеротрахеи при встряхивании светятся,
особенно их внутренностный мешок,
который при раздражении животного
испускает красивый голубоватый свет.
* * *
Кроме киленогих, к пелагическому
образу жизни в открытом океане
приспособились улитки, относящиеся к родам
янтина (Janthina) и реклюзия (Recluzia).
Эти два рода, представленные
несколькими живущими на поверхности воды
видами, составляют особое семейство
Janthinidae.
По своему внешнему облику янтины
мало отличаются от других типичных
переднежаберных. Округлая или
конусообразная раковина янтин по своим
пропорциям несколько напоминает раковину
обыкновенной лужанки, однако
раковина их значительно тоньше и легче, что
связано с приспособлением янтин к
свободному плаванию. Для удержания на
поверхности воды янтины строят из
пузырьков воздуха вытянутый в длину
особый поплавок. При построении поплавка
пузырьки воздуха захватываются
подвижным выростом передней части ноги и
обволакиваются быстро затвердевающим на
воздухе секретом особых слизистых
желез. Сама улитка держится на нижней
поверхности такого поплавка, или
«плота», спиной книзу и в таком положении,
расправив у поверхности воды
раздвоенные щупальца, пассивно плавает по воле
течений, ожидая встречи с другими
животными, которые могут послужить ей
добычей. Пищей янтинам служат главным
образом хандрофоры Velella и Porpita, а
также и другие в изобилии плавающие на
поверхности воды животные. Отмечены
случаи каннибализма и поедания одним
видом янтин молоди других видов.
Глаза и органы
равновесия (статоцисты) у янтин отсутствуют.
Однако их щупальца, пронизанные сильно
развитой сетью нервных волокон,
обладают высокой чувствительностью к
изменению освещенности и служат им хорошо
развитым органом осязания. Раковина
большинства янтин, и особенно нижняя
сторона, обращенная кверху, окрашена
в яркий лилово-голубой цвет, который
делает янтин малозаметными на голубом
фоне поверхности воды.
Янтины — гермафродиты, но мужские
и женские половые продукты развиваются
у них в разное время (протерандрический
гермафродитизм). Один из видов янтин —
живородящий (J. janthina), другие виды
откладывают яйца в небольшие коконы,
которые прикрепляются моллюском
к нижней поверхности поплавка. Актив-
ноплавающие личинки янтин (в е л и г е-
р ы) некоторое время живут в планктоне.
Янтины — тепловодные животные. Они
широко распространены в тропических
и субтропических областях Мирового
океана. Дрейфуя вместе с течениями, они
образуют часто огромные стайные
скопления на поверхности воды. Подхваченные
течениями, стаи янтин периодически
выносятся в холодные районы океана, и тогда
их можно встретить у берегов Ирландии,
Японии или Новой Зеландии. Штормовые
ветры выбрасывают часто большое
количество янтин на берега, и тогда их
раковины окрашивают пляжи в яркие
голубые цвета.
* * *
Из числа донных улиток, к которым мы
опять возвращаемся, весьма интересен
род натика (Natica), или пупочная
улитка (из семейства Naticidae),
представленный многими видами. У нати-
ки толстая гладкая шаровидная раковина
с наростом на пупке. Нога имеет
отчлененный проподиум, который может
прикрывать голову. Снизу помещается
хорошо развитая крышечка — оперку-
44
лум. Животные имеют много особенностей.
Так, например, они выкапывают из ила
ракушек, которыми питаются. Это повело
к тому, что нога приспособилась
набирать воду. Она бесформенно разбухает,
причем проподиум выпячивается вперед;
за ним посредине лежит большой метапо-
диум, или собственно подошва, а сзади —
оперкулум. До сих пор не удалось
законсервировать улитку с ногой в разбухшем
состоянии, потому что при малейшем
раздражении она выпускает воду и тело
ее возвращается к обычной форме. Раньше
считали, что она набирает воду в ногу
через отверстие, находящееся на середине
или немного ближе к переднему краю
подошвы; у многих переднежаберных это
отверстие ведет к задней ножной железе.
Но у пупочной улитки на переднем крае,
где обыкновенно находится широкая щель,
куда открывается передняя ножная
железа, были обнаружены справа и слева
многочисленные маленькие отверстия. Они
ведут к каналам, которые тончайшим
образом разветвляются между
мускулатурой по всей толще их ноги и могут
замыкаться кольцевыми мускулами. Можно
предположить, что они образовались как
видоизменение желез переднего края
ноги. Вслед за набиранием воды отверстия
замыкаются, и общим сжатием кожно-му-
скульного слоя вода вгоняется в
расширившиеся разветвленные полости,
лежащие между мускулами ноги. Это
приспособление оказалось полезным при
откапывании ракушек, которые служат
пищей пупочным улиткам. Эти
улитки обволакивают свою добычу
разбухшими частями ноги. Дальнейшее
овладение ракушкой не менее своеобразно.
Над передней частью ноги натики
выступает дыхательный желобок —
сифон, наличие которого обеспечивает обмен
воды в жаберной полости даже тогда,
когда улитка закопалась; над передней же
частью ноги находятся щупальца и,
наконец, вытянутое хоботообразное рыльце.
Под ним, можно сказать на подбородке,
помещается имеющая форму плоского
блюдечка сверлильная железа: синяя
лакмусовая бумажка, когда мы ее прижимаем
к этой железе, краснеет, обнаруживая
кислую реакцию ее выделений. Кислота
Рис. 24. Натика (Natica josepliina),
схватившая ракушку:
1 — схваченная ракушка; 2 — задняя часть ноги; з —
передняя часть ноги; 4 — хобот; 5 — сифон; б —
начало разрушения раковины ракушки в результате
действия кислого выделения железы натики; 7 —
образовавшееся в стенке раковины ракушки отверстие,
через которое натика выедает мягкие части тела
ракушки.
эта растворяет известковое вещество
раковин ракушек (мы еще встретимся в
дальнейшем с подобными примерами); сначала
на раковине ракушки образуется
размягченный участок, на котором
кругообразными движениями принимается работать
радула, пока не возникнет отверстие
совершенно правильной круглой формы.
Тогда в него проникает хобот улитки и она
поедает мягкие части ракушки.
Способ откладки икры у натики
находится в полном соответствии со
способностью этой улитки к копанию в песке.
Икру она помещает в песочную ямку;
стенки этой ямки густо усажены яйцевыми
капсулами, помещающимися ряд над
рядом. Очевидно, этот кокон во время
откладки яиц улитка устраивает ногой, делая
ею вращательные движения и выделяя
слизь. Молодь появляется в виде личинок-
велигеров с парусом, разделенным на
четыре одинаковые лопасти, как мы это уже
видели у атлант; чем ближе к тропикам,
тем длиннее эти лопасти у личинок. При
этом раковина и оперкулум уже у
личинок имеют ряд признаков, по которым,
как например по строению крышечки,
можно определить систематическое
положение личинки.
45
Очень сильное развитие крышечки,
наблюдаемое здесь, служит
приспособлением для защиты от нападения
раков-отшельников, которые в связи со своей
потребностью в раковине улиток
обратились в злейших врагов брюхоногих
литоральной зоны. Бауер считает, что как
оборонительные приспособления против того
же врага у некоторых улиток, как
например у Conus, развились утолщения
последнего завитка раковины или
утолщенная наружная губа у многих
морских улиток (Cassidaria, Tritonium) и иглы,
которыми усажен край устья у Мигех
и которые часто превращаются в
настоящие пилки. Обрастание иглами не только
наружной губы, но и всей поверхности
раковины оказывает также хорошую
услугу при защите от другой группы
хищников, а именно от морских звезд, к
взаимоотношениям с которыми мы еще не раз
возвратимся. Эти нападения повели у Nas-
sa reticulata к выработке особого
защитного рефлекса: если дотронуться до
двух лоскутообразных щупалец на задней
части ноги, она начинает кувыркаться
с неимоверной быстротой.
Близки к пупочным улиткам
представители семейства ламеллярид (Lamel-
lariidae), которые, однако, развивались
в другом направлении. Мы можем считать
у них за исходные формы тех, которые
имеют шаровидную или более
приплюснутую уховидную раковину, как например
улитки Velutina и Sigaretus. У последней
тело еще сходно с телом натики, у велу-
тины же раковина более хрупка и
мантийный край загибается на нее, частью
покрывая ее собой. Это тот путь развития, по
которому пошли в своей эволюции голые
улитки с неуклюжим малоподвижным
телом, с внутренней раковиной, которая,
несмотря на свою хрупкость, все-таки
сохраняет полностью весь свой завиток;
таковы Lamellaria, Marsenia, Marseniop-
sis, Oncidiopsis и некоторые другие. Они
водятся во всех морях от Арктики до
Антарктики, но в своем существовании они
связаны не с ракушками, а с
оболочниками, именно с асцидиями. По
наблюдениям Берга, они прогрызают круглые
отверстия в толстой мантии асцидий,
откладывают туда свои яйца и закрывают
отверстие крышечкой из слизи.
Мы видели, что у предыдущей группы
сифон в связи с закапыванием в грунт
служил приспособлением,
поддерживающим связь между дыхательной полостью
и водной средой. У других улиток мы
уже видим сифон лежащим поверх
раковины, в которой для него образовалось
специальное отверстие; формы,
обладающие этой особенностью, объединяются
иногда общим названием сифоноротых.
* * *
Составляющие особое семейство
улитки церитии (Cerithiidae), или
игольные у литки у имеют крепкую длинную
башневидную раковину, украшенную
многочисленными шишечками. Более крупные
из них, обитательницы тропиков,
замечательны тем, что проникают в устья рек
и в болота. Во время засухи они,
замкнувшись крышечкой, подвешиваются при
помощи затвердевшей полоски слизи к
ветвям мангровых деревьев. Из
средиземноморских улиток более всех
вынослив к высыханию вид Cerithium vulga-
tum.
Из семейства церитии
относительно небольшое число видов
встречается и в Черном море, а некоторые из них,
как например Cerithiolum reticulatum
и Cerithium ponticum, заходят в Азовское
море. Наиболее обычен в Черном море
Cerithium vulgatum, постоянно встре-
Рис. 25. Свободноплавающая ли-
чинка-велигер натики с
разделенным на лопасти парусом.
46
чающийся на устричных банках.
Черноморские Cerithium откладывают яйца
шнурами в виде неправильной спирали,
причем размножение их происходит летом
и их личинки особенно многочисленны
в планктоне в июле и августе. Близкая
к этой группе маленькая с левозавитой
раковиной улитка Trifora обнаруживает
другую особенность: ее личинка с
помощью своего четырехлопастного паруса
плавает по морю долго после того, как
ее раковина уже сильно выросла.
* * *
Фарфоровая улитка, или ципрея,
относится к семейству Cypraeidae.
Внутренняя часть ее раковины тонка, как
бумага, в то время как наружная часть,
составляющая последний оборот завитка,
очень толста и как бы отполирована,
причем мантия прикрывает раковину сверху
целиком или в большей части. Хотя
раковина этой улитки хорошо известна и
раньше широко использовалась, образ жизни
этой улитки мало известен. По
наблюдениям Румфа, ципреи обыкновенно сидят,
зарывшись в песок, а во время новолуний
и полнолуний выползают оттуда и
держатся на скалах. Некоторые наблюдения
над ними производил в Неаполе Зимрот,
который пишет: «У относящейся к этому
семейству маленькой улитки Trivia все
мягкие части тела имеют
бледно-оранжевую или охряную окраску. Раковина
Сургаеа коричневатая, с двумя стертыми
беловатыми полосками и имеет ту же
окраску, что и у Trivia. Иначе обстоит
с мягкими частями тела. Подошва здесь
белесая с розоватым оттенком, а по
спинной части ноги проходят густо-черные
полосы, расположенные на светлом фоне,
что делает ее малозаметной. Мантийный
край, который завернут на раковину
и покрывает всю ее сверху, имеет
коричневую окраску, как и сама раковина,
которая образована мантией; только
окраска его несколько бледнее.
Интересно, что раздражение
прикосновением вызывает сильное сокращение
мантии, которая втягивается в отверстие
раковины; по окончании раздражения мантия
снова расширяется до прежнего объема,
но этот процесс происходит крайне мед-
Рис. 26. Стромбус (Strombus lentiginosis).
Рис. 27. Кассис (Cassis cornuta).
Рис. 28. Улитка бочонок (Dolium perdix).
47
ленно и заканчивается лишь в течение
1—2 дней. На поверхности мантии можно
видеть беловатые или серые бородавочки,
среди которых на некотором расстоянии
друг от друга возвышаются более светлые
выросты в форме острого шипа. Что
особенно с первого взгляда привлекало
внимание, так это яркая шеллаково-красная
окраска отдельных частей тела ципреи:
рыльца, наружных светочувствительных
щупалец, которые смыкались каждый раз,
когда их касалась тень, и сифона,
выступавших резко на общем, как было описано,
тусклом цвете туловища».
Улитки ципреи живут в теплых,
преимущественно тропических морях,
некоторые виды встречаются и в
Средиземном море, но уже не заходят в Черное
море.
Близкие к настоящим ципреям улитки,
составляющие особое подсемейство
Amphiperasinae, живут на коралловых
полипах и питаются ими.
Своеобразен характер передвижения
у видов Strombus и Pteroceras
представителей семейства Strombidae. У тех
и других раковина чрезвычайно тяжелая
и массивная, но, несмотря на тяжесть
раковины, животные довольно подвижны
благодаря мощному развитию и
расчленению их ноги, подобно тому как было
описано у атланты. Узкая, заостренная
рогатая крышечка посажена на
удлиненном метаподиуме; у подошвы особенно
активен передний край. Опираясь на эти
две отдаленные друг от друга точки,
улитка может сделать прыжок: за эти бойкие
скачки туда и сюда один из видов получил
название фехтовальщика. С такой
подвижностью связано и хорошее развитие
глаз. Интересно, что своей заостренной
крышечкой стромбиды могут пользоваться
как оружием, нанося ею значительные
раны, почему они и получили в
некоторых местах название чертова когтя.
* * *
Некоторые переднежаберные отличаются
чрезвычайно развитым хоботом,
равняющимся по длине всей раковине. Он может
полностью втягиваться, и тогда видно
обыкновенное ротовое отверстие. Сюда
относится большое число разнообразных
видов, ведущих хищный образ жизни. Мы
рассмотрим некоторые из них. Шлемовид-
ные улитки, или кассиды (Cassididae),
обитают преимущественно у тропиков; из
них один род кассидария (Cassidaria)
водится и в Средиземном море, которое
представляет северную границу их
распространения. Крупная улитка шишак
(Cassis cornuta) известна своим коротким
Таблица 4. Голожаберные моллюски:
1 — Coryphella rufibranchialis;
2 — Hermissenda crassicornis;
3 — Eubranchus (Galvina) flava;
4 — Tethys fimbria;
5 — Dendronotus arborescens;
6 — Hermaenia maculosa;
7 — Tritonia hombergi;
8 — Berghia coerulescens;
9 — Doris pilosa с кладкой;
10 — Doto coronata;
11 — Chromodoris coerulea;
12 — Triopha carpenteri.
завитком и длинным узким отверстием
между утолщенными гладкими устьевыми
губами. Шлемовидные улитки живут в
большинстве случаев на незначительной
глубине, вблизи берега, на песчаном
грунте, в который они целиком или почти
целиком закапываются, преследуя
ракушек, составляющих их постоянную и
излюбленную пищу. Отметим
оригинальную улитку пеликанья нога (Aporrhais
pes pelicani).
* * *
Улитки бочонки, составляющие с е-
м е й с т в о долиид (Doliidae), охотятся
главным образом на иглокожих, на
морских звезд и на морских огурцов
(голотурий), кожа которых содержит
известковые включения и образует часто прочный
известковый панцирь. В связи с этой
особенностью добычи слюна долиид
содержит кислоту. К эффектнейшим
открытиям нужно отнести случай, когда Т р о-
шель, взяв в руки Dolium, увидел, что
улитка выбросила струю слюны, причем
мрамор, на который она попала, вдруг
зашипел. Так же изумительно было
открытие, что слюнные железы долиид имеют
особое приспособление, вырабатывающее
серную кислоту, и что в их слюне
содержится от 3 до 4% свободной серной кисло-
ты; в слюне долиума были найдены также
и другие кислоты, например а с п а р а-
гиновая. В дальнейшем было
доказано, что кислота приводит иглокожих
в состояние оцепенения. Таким образом,
тут достигается сразу двойная цель:
ошеломление врага и разрушение его
защитного покрова. Хотя серная кислота и не
кажется тут наиболее подходящим
средством, поскольку она, соединяясь с
известью, дает трудно растворимый сульфат,
однако она делает покров, защищающий
звезду или морского ежа, хрупким и
рассыпающимся и уже не представляющим
препятствий для действия радулы.
К долиидам можно отнести, пожалуй,
и грушевидную улитку, или пирулу (Ру-
rula), хотя она и отличается от них.
Нога у нее маленькая, но зато ее
пятнистая мантия, сильно завернутая на
раковину, расширена почти в горизонтальном
направлении, т. е. в одной плоскости
с подошвой, и это расширение мантии
наряду с ногой служит как опора при
ползании. Грушевидная улитка живет в Ост-
и Вест-Индии. Род Dolium также
встречается в этих обеих областях, с ними
связаны через Средиземное море и
африканские формы. Сходное распространение
обнаруживают и близкие к ним улитки
тритониум (Tritonium); но они имеют
Таблица 5. Наземные моллюски:
1 — Jaminia potaniniana;
2 — Helicella candacharica;
3 — кустарниковая улитка (Bradybaena fruticum);
4 — янтарка (Succinea putris);
5 — Pup ilia muscorum;
6 — Perforatella bidens;
7 — полевой слизень (Deroceras agrestis);
8 — Arion subfuscus;
9 — Arion empiricorum;
10— Limax maximus;
11 — Arion maximus;
12 — Cochlicopa lubrica;
13 — Theba fruticum;
14 — Ena obscura;
15 — Iphigena ventricosa;
16 — Gepaea nemoralis;
17 — Zebrina cylindrica;
18 — Gigantomilax kollyi;
19 — Bradybaena maacki;
20 — Caucasotachea atrolabiata;
21 — Helix vulgaris;
22 — Vallonia costata;
23 — Philomycus bilineatus;
24 — Trigonochlamys imitatrix.
А Жизнь животных, т. 2
замечательную особенность: их различные
виды распространены одновременно и в
Ост- и в Вест-Индии. Связь между
обеими областями, очевидно, поддерживается
очень крупными плавающими личинками
с длинными лопастными в е л у м а м и.
Личинки тритониума достигают в длину
больше 1 см, а личинки долиума легко
распознаются по их раковине, особенно
по ее рогатой верхушке. Известно с
достоверностью, что эти личинки встречаются
в южной части Индийского и в
Атлантическом океане.
* * *
К о т р я д у Mesogastropoda относится
также своеобразная группа
переднежаберных, носящая название безъязычных
(Aglossa); у этих улиток мы не находим
ни челюстей, ни радулы, а у
принадлежащих к этой же группе некоторых
паразитных форм редуцируются и раковина,
и крышечка, причем и все их
анатомическое строение носит яркие черты
приспособления к паразитическому
существованию. У относящегося к этой группе с е-
м е й с т в а меланеллид (Melanellidae)
богатого видами и широко
распространенного рода меланелла (Melanella) мы
встречаем, с одной стороны, свободножи-
вущие формы и, с другой стороны, ряд
форм, в той или иной степени перешедших
к паразитическому существованию
преимущественно на иглокожих. Так,
антарктическая Melanella troglodytes живет в
полости мозолевидных утолщений на коже
морских ежей (рис. 31). В этих своеоб-
разных «пещерах» происходит и
откладка яиц, и выплод молоди. Еще неясно,
можно ли этот вид считать паразитом:
в его строении не обнаружено каких-либо
приспособлений для паразитарного образа
жизни, наблюдаемых у других моллюсков.
Например, Stilifer brychius (рис. 32)
и некоторые другие виды этого рода
обладают длинным хоботком, с помощью
которого они сосут соки из полости
тела иглокожих, сидя на поверхности их
кожи. Это настоящие эктопаразиты; их
раковина и внутренние органы не
подверглись редукции. У других представителей
семейства наблюдается дальнейшее
развитие облика паразитов. У этих форм
наблюдается редукция не только органов чувств,
но также и ноги с крышечкой, различных
частей пищеварительного аппарата и даже
таких органов, как сердце и почки. Зато
хобот достигает здесь огромных по
отношению ко всему телу размеров, а вокруг
его основания развивается подобие
воротника, как, например, у улитки мукрона-
лии (Mucronalia), живущей у берегов
Японии. У другого представителя того же
семейства — улитки стилифер (Stilifer),
живущей в Тихом и Индийском океанах,
у основания хобота образуется кожная
складка, которая охватывает раковину,
покрывая часть ее наподобие ложной
мантии. У третьего рода (Gasterosiphon),
живущего в Индийском океане, раковина
уже целиком покрыта такой ложной
мантией, которая срастается в трубку (сифон),
прободающую кожу хозяина. Однако
наибольшая степень изменений под влиянием
Рис. 29. Улитка «пеликанья
нога» (Aporrhais pes pelicani).
Рис. 30. Пирула (Pyrula).
паразитизма наблюдается у
представителей семейства Entoconchidae. Один
из относящихся к нему видов (Entocolax
ludwigi) встречается в наших северных
морях, где он живет в полости тела
голотурий Myriotrochus. Ни раковины, ни
сердца, ни почек не сохраняется у этих
паразитов. Характерно, однако, что они
раздельнополы и обнаруживают резкие
различия в строении самцов и самок. Тело
последних имеет вид удлиненного мешка,
на одном конце которого находится ротовое
отверстие, а на другом — короткая
трубка-сифон, посредством которого паразит,
находящийся в полости тела голотурии,
прикрепляется изнутри к стенке ее
кишки. Этот сифон ведет в объемистую
выводковую полость, куда откладываются яйца
в яйцевых капсулах, содержащих обычно
развивающихся личинок. В этой же
полости находятся обычно и карликовые
самцы, иногда до 46 штук в одной самке.
Строение самца еще более упрощено,
чем самок. Их тело представляет собой
продолговатый пузырек, внутри которого
помещаются семенник, рудиментарная
кишка и немногочисленные тяжи мышц
и соединительной ткани. Самцы медленно
ползают благодаря сокращению
слаборазвитой кожной мускулатуры.
Несмотря на столь резкие изменения
в организации взрослых форм, личинки
этих улиток сохраняют типичные черты
велигеров. Выход их из тела хозяина
происходит, вероятно, через сифон, когда
они еще находятся в яйцевых капсулах.
Через сифон они могут попасть в кишку
хозяина, а оттуда уже выходят наружу.
Карликовые самцы были обнаружены
также и у рода энтоконха (Entoconcha).
Интересно, что кишечник у обоих
последних родов представляет собой слепой
мешок и не имеет заднепроходного
отверстия.
ОТРЯД STENOGLOSSA
Нам остается рассмотреть довольно
обширную группу переднежаберных,
которая объединяет высших представителей
этого подкласса, относящихся к
обитателям морей.
Рис. 31. Глубоководный паразитический
моллюск меланелла пещерная (Melanella troglodytes)
в теле морского ежа.
Рядом с раковиной моллюска видны кучки яиц ме-
ланеллы.
Рис. 32. Паразитический моллюск стилифер
брихиус (Stilifer brychius) с длинным хоботом B)
на теле морского ежа A).
Огромное большинство из них
хищники, некоторые же питаются трупами
разных животных.
У этих улиток перламутровая структура
гипостракума давно исчезла. Рыльце
вытянуто в форме хобота, а основание его
глубоко впячивается в форме уха; радула,
строение которой можно видеть даже на
упрощенном рисунке нашего изображения,
носит типичные черты приспособления к
плотоядному питанию; в хоботе, чаще всего
4*
51
Рис. 33.
Паразитическая улитка сти-
лифер (Stilifer cele-
bensis) и ее
проникновение в тело
хозяина (морской
звезды Certonardoa semi-
regularis):
1 — общий вид
паразита; 2 — момент
погружения паразита под
кожу морской звезды;
3 — морская звезда с
вздутием на одном из
лучей, показывающим
местонахождение
паразита.
Рис. 34. Личинка
паразитической
улитки энтоконхи.
Рис. 35.
Паразитическая улитка Ento-
concha mirabilis в
полости тела голотурии
Lapidoplax digitata:
слева — общий вид
голотурии с паразитом;
справа — средняя
часть тела голотурии с
паразитом (при сильном
увеличении). 1 —
стенка тела голотурии; 2 —
5 кишка голотурии с
идущими вдоль нее
спинным (а) и брюшным
(б) кровеносными
каналами; 3 — кнопкооб-
разно вздутый передний
конец тела паразита;
4 — туловище паразита
с просвечивающими
половыми органами; 5 —
выводковая камера; 6 —
семенной пузырек.
в передней его части, находятся железы,
которые ошибочно принимались за
слюнные, но которые оказываются скорее
сверлильными железами, помогающими
продырявливать раковины двустворчатых
моллюсков, которыми преимущественно
питаются эти улитки. У всех у них хорошо
развит сифон, образованный
выпячиванием мантийного края. Крышечка в
большинстве случаев имеется.
Все улитки, относящиеся к этому
отряду, раздельнополы. Личинки
приспособлены к настоящему пелагическому образу
жизни и имеют хорошо развитый парус
с длинными лопастями.
К этому отряду относится очень большое
число форм, из которых мы рассмотрим
лишь некоторые. Несмотря на то что
раковины многих из этих улиток, особенно
тропических видов, имеют спрос у
любителей, образ жизни большинства из них
нам очень мало известен, хотя мы знаем,
что они ведут образ жизни хищников или
пожирателей трупов, а наличие длинного
сифона указывает на присущую им
способность закапываться в грунт.
* * *
Наиболее хорошо известны некоторые
улитки из семейства букцинид(Вис-
cinidae), относящиеся к р о д у букцинум
(Buccinum), который представлен и в
наших северных морях несколькими видами.
Один из них — Buccinum undatum —
достигает размера 80 мм и широко
распространен в арктических и субарктических
морях где живет на сравнительно
небольших пубинах — около 150 м и меньше.
Этот вчд весьма многочислен в Северном
море и в качестве съедобного моллюска
служит здесь объектом довольно
значительных промыслов. Букцинум часто
встречается здесь на устричных банках и вне
их на небольших глубинах, причем,
однако, не поднимается выше чем на глубину
нескольких метров ниже уровня воды во
время отливов. Питаются букцинумы как
живой добычей, так и падалью, но сами
в свою очередь часто становятся добычей
мелких акул.
Молодь этой улитки можно находить
в большом количестве летом и ранней
осенью; растут они довольно медленно и,
по-видимому, только на четвертый год
достигают размера 55 мм и больше, когда
они уже приобретают промысловую
ценность. При особо благоприятных
условиях букцинум достигает размера
в 120 мм.
Промыслы букцинума особенно развиты
в Англии, в Шотландии и Ирландии. Этих
52
улиток употребляют и как
пищу для людей (в вареном виде),
и в качестве наживки для
рыбной ловли. Там, где промыслы
букцинума поставлены на
широкую ногу, как, например, у
устья Темзы и в некоторых
других местах английского
побережья, лов идет круглый год.
С моторных ботов,
выезжающих в море, спускают
одновременно около 180—200 ловчих
корзин с приманкой, куда и
попадаются букцинумы. В
других местах этих улиток ловят
на крабов, которых просто
привязывают на веревке.
* * *
Очень близки к семейству
букцинид улитки нептуниды
(Neptunidae); к этому
семейству относятся самые крупные из
брюхоногих моллюсков наших
северных морей. Такова,
например, улитка Neptunea despecta,
раковина которой достигает
высоты 130 мм. Эта улитка живет
на глубинах от 25 до 300 м и
встречается у берегов
Гренландии, Шпицбергена, Новой
Земли и Кольского полуострова.
На раковинах как букцинума,
так и нептунии часто
поселяются актинии, а пустые раковины
этих улиток постоянно служат
приютом для
раков-отшельников.
В то время как
представители родов букцинид и неп-
шунид приурочены к северным
морям, многочисленные виды
рода Насса (Nassa)
распространены по всем морям, за
исключением Северного
Ледовитого океана. К широко
распространенным видам этого рода
относится маленькая улитка вер-
ша (Nassa reticulata), которая
широко известна рыбакам
нашего побережья Черного и
Азовского морей под разными
местными названиями: ее называ-
ют то ракушкой, то червем, то
пиявкой в связи с вредом,
который она приносит,
чрезвычайно затрудняя рыболовство в тех
местах, где ее особенно много.
Эта маленькая улитка,
достигающая размера 20—30 мм,
обладает чрезвычайно красивой
раковиной светло-желтой окраски
с проходящими по желтому
фону голубовато-черной и
коричневой полосками, причем вся
поверхность украшена слегка
извилистыми продольными
ребрами, которые пересекаются
глубокими спиральными
бороздками, иногда ярко
окрашенными.
Эта улитка отличается
большой подвижностью. Когда ей
случается упасть на спину, она
вытягивает вбок ногу,
щупальца и рыльце и быстро
переворачивается обратно. На
Атлантическом побережье и в
Средиземном море, где многочисленны
нассы, можно наблюдать, как
во время отлива многие из них
остаются лежать на берегу, и:
тогда видны только их длинные
сифоны, торчащие над песком и
как бы обнюхивающие
окружающий грунт; дело в том, что
на дне дыхательной трубки
находится осфрадиум—орган
обоняния. Как только улитка
почует добычу, она выбирается
наружу и направляется к ней.
Яркое описание улитки верши
дают Мейер и Мёбиус:
«Улитки верши — настоящие
мясоеды. Мы видели, как они
нападали на живую морскую
звезду, и она не могла их
стряхнуть, сколько ни извивалась.
Когда в аквариум бросают
мясо, они очень скоро чуют его
и, где бы они ни находились,
тотчас же начинают двигаться,
разыскивая его. Те, что
сидели у поверхности воды,
устремляются вниз, другие, которые
ползли вверх, поворачивают
Рис. 36. Радулы
хищных улиток:
сверху — Tritonium
undatum, с н и 8 у — Ми-
гех erinaceus.
Рис. 37. Нептуния
(Neptunea despecta) и бук-
цинум (Buccinum
undatum).
Рис. 38. Черноморская
улитка верша (Nassa
reticulata).
53
Рис. 39. Мурекс (Mu-
гех) спереди и тритони-
ум (Tritonium) сзади.
обратно. Многие при этом отрывают
ногу от стеклянной стенки и падают на
дно: таким образом они сразу
приближаются на значительную часть пути к
учуянной пище и дальше ползут к ней по дну.
Те, которые сидели на дне, закопавшись
в ил, выбираются из него и также ползут
к мясу.
Орган, которым улитка верша чует
мясо, помещается, по-видимому, в
дыхательной трубке. Улитки вытягивают ее и
водят ею из стороны в сторону. К мясу они
двигаются не прямым путем, но
отклоняясь то вправо, то влево, иногда даже
поворачивая обратно, но скоро замечают,
что удаляются от учуянной пищи, и
возвращаются к прежнему, более короткому
пути. Повеем их движениям можно
заключить, что они руководствуются не
световыми раздражениями, но раздражениями
другого рода, а именно запахами, которые
воздействуют на их орган обоняния.
Когда улитка, наконец, прикоснется к мясу,
по ее щупальцам и дыхательной трубке
проходит содрогание. Светло-красная
трубочка хобота высовывается изо рта и
начинает сверлить кусок мяса, погружаясь
в него.
Вскоре улитки верши со всего
аквариума собираются тесной толпой вокруг
куска мяса: каждая держится на своем
участке, и только поднятый кверху сифон
покачивается туда и сюда.
Иногда улитка верша пользуется своей
ногой, чтобы захватить и удержать пищу.
Одна насса только что нашла кусок мяса,
как появилась креветка Palaemon squilla
и ухватила этот же кусок своими
клешнями. Тогда улитка судорожно обхватила
кусок своей ногой и не выпускала его все
время, пока креветка оставалась тут же
и ела вместе с нею».
Улитка верша встречается повсюду
в Черном море на глубинах от 5 до 20 м,
а иногда и глубже. Многочисленна она
и в Азовском море. Питаясь, как хищник,
она может просверливать тонкие раковины
двустворчатых, на которых часто можно
находить следы ее работы в виде округлых
отверстий. Однако устричные раковины,
особенно взрослых устриц, для нее
слишком толсты, и потому она едва ли
причиняет вред устричным банкам. Надо
отметить, что улитки верши вредят
рыболовству, потому что нападают на живца
(т. е. наживленную на крючья приманную
рыбу) и быстро его съедают. Поэтому там,
где нассы особенно многочисленны, лов
крючками камбал и других рыб на
глубинах, населенных этой улиткой, очень
затруднителен. Интересны наблюдения,
что насса различно относится к соленому
и свежему живцу; первого она съедает
гораздо быстрее, нежели свежего, к
которому приступает только тогда, когда
он начинает раскисать.
* * *
Относительно близки к описанным
формам пурпурные улитки (Muricidae),
раковина которых по краю устья снабжена
более или менее длинным выступающим
вперед желобком и, кроме того, часто
бывает украшена длинными отростками.
Одним из характерных для всех этих улиток
признаком служит наличие особой
пурпурной железы в мантии. Это
хищные улитки, которые просверливают
раковины других брюхоногих, или
ракушек. Впрочем, один из относящихся сюда
видов (Murex fortispina), живущий у
берегов Каледонии, поступает иначе. Его
раковина снабжена зубцами, сидящими на
наружной губе устья, причем один из
зубцов достигает особенно сильного
развития; захватив небольшую ракушку,
54
улитка плотно зажимает ее между этим
зубцом и крышечкой, а затем с силой
втягивается в свою раковину, и ракушка
раздавливается, как орех. В Черном море
на зарослях филофоры обитает
представитель того же семейства Trophonop-
sis breviatus striata, с темно-красной
раковиной, имеющей защитное значение.
Семейство пурпурных улиток
широко распространено по всем морям
земного шара и имеет своих представителей
также и в наших водах.
Из дальневосточных представителей
этого семейства наибольшего внимания
заслуживает улитка рапана (Rapana),
страшный вредитель устричных банок,
в массе пожирающая устриц на
устричных банках Японского моря. В сороковых
годах нашего века рапана обнаружилась
в Черном море, где она также вредит.
Считают, что личинки рапаны проникли
в этот внутренний морской бассейн на
днищах кораблей, совершающих рейсы через
Средиземное море. Личинки могли
приютиться среди обрастателей. В настоящее
время рапаны в Черном море не редкость
и в ряде пунктов они, как уже
сообщалось, сильно вредят устричным банкам.
К вредителям устричных промыслов на
Атлантическом и Тихоокеанском
побережьях Северной Америки принадлежит
и другой представитель семейства — улитка
устричное сверло (Urosalpinx cinerea).
Несмотря на то что процесс просверливания
толстой устричной раковины у этой улитки
продолжается довольно долго, она
приносит значительный вред, уничтожая
главным образом устричную молодь.
Можно было наблюдать, что одна и та же
улитка нападала подряд на 5 молодых
устриц, а если принять во внимание
способность к быстрому размножению этого
вредителя (одна самка способна
произвести до 100 штук молоди за сезон
размножения), то понятна значительность
приносимого ею вреда. Жертвой ее
становятся, кроме устриц, также и мидии,
причем для того, чтобы просверлить
раковину последней, улитке необходимо от 18
до 36 часов.
Пурпурные улитки в древности служили
для получения краски пурпура. Теперь
искусство этой добычи сохранилось
только у некоторых рыбаков Средиземного
моря, которые этой краской красят свои
одежды. Своеобразие пурпурного вещества
состоит в том, что, будучи извлечено из
железы, оно имеет белый или
бледно-желтый цвет, причем некоторые виды
пурпура и мурекса дают разновидности
оттенков. Выставленное на солнечный свет,
пурпурное вещество становится сначала
лимонно-, потом зеленовато-желтым;
пройдя через стадию зеленого цвета, оно
становится наконец лиловым и, чем дальше
подвергается воздействию солнечных
лучей, тем более темнеет. Тот или иной
оттенок фиолетового зависит от способа
наложения краски и от толщины ее слоя;
таким образом, в руках умелого
красильщика находится большое разнообразие
тонов. Обычно красящее вещество с
помощью упругой кисточки берут с того
участка мантии, где оно вырабатывается, а
затем непосредственно наносят его на
материю, подлежащую окраске. Л а к а з-
Д ю т ь е, который был не только
зоологом, но и художником, нашел, что
пурпурное вещество могло бы найти
применение в фотографии. Он проделал в этом
направлении ряд опытов и пришел к
выводу, что пурпур может найти
применение при перенесении фотографических
изображений на батист или тонкие шелковые
ткани.
Пелагические личинки пурпурных
улиток совершенно особенно приспособлены
к своему образу жизни. Их конхиновая
раковина около устьевой оторочки имеет
вырез, предназначенный для длинного
лопастного паруса, вследствие чего этих
личинок принимали за взрослую форму
и ошибочно присваивали им название
рода Sinusigera.
У всех этих высших переднежаберных
моллюсков (Stenoglossa) пол,
по-видимому, строго разделен. Икра по большей
части состоит из изящных капсул очень
разнообразной формы: в виде урн,
бокалов, плоских коробочек, располагающихся
рядами или кучками. У букцинума они
плоские, двояковыпуклые, крепкие; их
большими массами прибивает к берегам,
и рыбаки называют их «морским мылом».
Пельзенеер доказал, что в образовании
одного из видов такой кладки принимают
55
участие выделения ножной железы. Яйца
скользят вверх по упоминавшемуся гени-
тальному желобку, ножная железа
выделяет вещество, из которого образуются
капсулы, а нога формирует и скрепляет
их. Вероятно, большинство видов
пользуется этим способом. Капсулы содержат
иногда много яиц — до 80, а иногда
больше. Развитие происходит очень по-разному.
Так, у букцинума только наиболее
сильные эмбрионы созревают до стадии велиге-
ра; обыкновенно созревает только один,
который питается своими более слабыми
и отстающими в росте собратьями. У нас-
сы, наоборот, обыкновенно созревают все
эмбрионы, что ведет к вредной для них
тесноте, вызывающей различные
деформации раковинки и крышечки.
От представителей семейства букцинид
значительно отличаются по внешности
улитки оливы (Oliva), которых выделяют
в особое семейство Olividae. Их
блестящая, как бы полированная раковина
имеет короткий завиток, но ее
последний оборот сильно увеличен и вытянут так,
что вся раковина приобретает валько-
ватую форму. В отличие от букцинид,
устье оливы не имеет вытянутого
желобком края. Улитки эти лишены крышечки;
голова их несет утолщенные у основания
щупальца, на внешней стороне которых
находятся глаза. Нога здесь хорошо
развита, с обособленной передней частью—
проподиум, а мантийный край несет
щупальцеобразные придатки. Оливы
живут исключительно в теплых морях,
преимущественно под тропиками, и в
некоторых местах встречаются в огромных
количествах. Они на долгое время
закапываются в мягкий грунт или песок,
находясь там в вертикальном положении:
наружу из субстрата слегка выступает
лишь притуплённая вершинка раковины.
Интересно, что у олив по шву между
оборотами раковины проходит глубокий
спиральный желобок, по которому, видимо,
к устью подходит чистая вода, свободная
от минеральной взвеси.
Так же как и оливы, исключительно
в тропических морях живет улитка арфа
(Награ) с ее сильно ребристой и красиво
окрашенной раковиной. Ребристость,
особенно сильно выраженная на последнем
обороте, представляет утолщения
наружной губы, указывающие здесь на очень
медленный рост раковины или на частые
приостановки его. Своеобразным
защитным приспособлением этой улитки
оказывается способность автоматически
отбрасывать задний конец ноги; это
производится посредством самоампутации,
и, когда при раздражении животное
очень быстро втягивается в свой
домик, острый край устья раковины
врезается в ногу именно там, где
заканчивается идущий поперек ноги синус
кровеносной системы.
Улитки арфы относятся к самым
красивым представителям брюхоногих.
* * *
В этом отношении с ними могут, однако,
соперничать и некоторые представители
семейства митрид (Mitridae),
распространение которого приурочено не
только к тропическим морям, но и к морям
умеренного пояса. Особенно красива
раковина улитки епископская шапка (Mit-
га episcopalis), достигающая размера
13 см. Однако представители этого
семейства, встречающиеся в морях умеренного
пояса, например средиземноморские виды,
не достигают столь крупных размеров и
яркой окраски, как обитатели тропиков.
Как ни сильно вытянута в высоту
раковина некоторых митрид, еще более
вытянута она у представителей семейства так
называемых винтовых улиток (Terebridae),
у которых она действительно напоминает
по форме узкий и длинный винт, особенно
когда отверстие раковины плотно закрыто
крышечкой. Интересно, что глаза у
некоторых из этих улиток сидят на щупаль-
цеобразных придатках.
Рис. 40. Олива (Oliva maura).
56
Наиболее богатый видами, относящийся
к этому семейству род теребра
(ТегеЬга) широко распространен в теплых
морях земного шара.
* * *
Своеобразной формой раковины
обладают также представители
многочисленного семейства улиток (Conidae) и
особенно богатый видами род конус
(Conus), представители которого населяют
теплые моря. Ее тяжеловесная раковина
начинается коротким уплощенным
завитком и оканчивается узкой длинной щелью
устья, причем вся раковина в целом имеет
почти правильную форму конуса.
Эти улитки особенно интересны тем,
что они обладают хорошо развитой
ядовитой железой, которая открывается с
правой стороны у основания хобота.
Жителям тех мест, где водится конус, хорошо
знакомо действие этой железы, и они
предостерегают европейцев, занимающихся
сборами улиток, о том, что укус конуса
вызывает и у человека резкое воспаление
на коже. По всей вероятности, этот яд
скорее должен служить целям поимки
добычи, чем самозащиты.
Об образе жизни этих улиток известно
относительно немного. Зимрот наблюдал,
как маленькая средиземноморская улитка
Conus mediterraneus закапывалась в грунт.
При этом работала главным образом
передняя часть ноги, тогда как осталь-
ная часть ноги оставалась неподвижной.
Лишь время от времени она как бы
подтягивалась назад вместе со всей раковиной.
Разные виды конусов благодаря
разнообразию и красоте узоров на их
раковинах очень ценятся коллекционерами: за
экземпляры некоторых видов германские
любители платили до 20 тыс. марок и
даже больше.
Большое разнообразие улиток рода
конус находится в связи со способностью
этих улиток к образованию местных форм.
Этот род, по-видимому, находится в
процессе интенсивного видообразования.
Один из видов, однако, прекратил свое
существование, так как он обитал лишь
на одном маленьком острове
Филиппинского архипелага, ставшем жертвой
вулканического извержения, — интересный
пример вымирания в результате действия
мощных геологических факторов.
Рис. 41. Конус (Conus textilis).
ПОДКЛАСС ЗАДНЕЖАБЕРНЫЕ (OPISTHOBRANCHIA)
Заднежаберные значительно уступают
переднежаберным улиткам в
многообразии форм, но все же составляют довольно
богатую видами группу брюхоногих.
Наиболее примитивные по своей организации
представители этого подкласса
сохранили в некоторых чертах сходство с
переднежаберными. Это сходство выражается
не только в чисто внешних признаках
формы тела или в наличии у них
спирально закрученной раковины с более или
менее возвышающимся завитком, но и в
некоторых анатомических чертах
строения их нервной системы, жаберного
аппарата и в других признаках. Однако
большая часть видов заднежаберных довольно
далеко уклонилась в своей эволюции от
исходных форм своих предков, которые,
как можно предполагать, имели типичные
черты переднежаберных.
У очень многих заднежаберных
раковина обрастает мантией и в той или иной
степени подвергается редукции. У
некоторых форм она сводится до небольшой
пластинки неправильной формы, которая
лежит под мантией, а у некоторых реду-
57
цируется полностью. Крышечка,
запирающая устье, есть только у очень немногих
более примитивных видов. Нога
видоизменяется очень сильно у многих
представителей подкласса. Известен ряд форм, у
которых нога развита чрезвычайно слабо,
а у некоторых она редуцируется вовсе.
У других, наоборот, боковые стороны
ноги разрастаются в широкие крыловидные
лопасти, так называемые параподии,
которые служат для плавания. Резким
изменениям подвергается также строение
органов дыхания.
Чаще всего в различных местах тела
заднежаберных расположены кожные
выросты — вторичные жабры, развившиеся
взамен утраченных настоящих ктенидиев.
Такие вторичные жабры чаще всего
образуются симметрично или вокруг
заднепроходного отверстия, или по бокам спины,
или на нижней стороне особого
утолщения мантии на спине животного. У
заднежаберных может быть отмечена общая
характерная черта во внешней форме их
тела: некоторая тенденция возврата к
билатеральной симметрии; эта черта
проявляется не только у плавающих форм,
ведущих планктонный образ жизни, но и у
форм, которые живут на морском дне и
передвигаются ползая, как другие
улитки. Заднепроходное отверстие у
некоторых заднежаберных находится на средней
линии спины и около него симметрично
располагаются вторичные жабры;
симметрично разрастаются параподии; все тело
у одних видов сильно вытянуто в длину и
сжато с боков, а у других, наоборот,
сплющено в направлении от спины к брюху и
приобретает общее внешнее сходство с
формой тела плоских червей турбеллярий.
Некоторый возврат к билатеральной
симметрии проявляется также и в
строении нервной системы: тогда как у
примитивных представителей подкласса, более
близких к переднежаберным, мы
встречаем еще типичный для последних
перекрест плевровисцеральных нервных
стволов, у других заднежаберных эта черта
сглаживается.
Из числа типичных для моллюсков
органов чувств, как правило, имеются
органы равновесия (статоцис-
ты); связанный с настоящей
жаброй (ктенидием) осфрадий
имеется у представителей отряда покрыто-
жаберных, к которому принадлежат более
примитивные формы подкласса.
Характерными для заднежаберных являются
участки кожи со скоплениями
чувствительных клеток, находящиеся на голове по
бокам рта и служащие, по-видимому, как
органы обоняния или вкуса. У ряда форм
участки чувствительных клеток
расположены на кожных выростах щупальцевид-
ной формы, которые сидят на голове в
числе одной или двух пар. Такие выросты
носят название ринофоров и
служат, по-видимому, органами
обоняния или химического
чувства. Как органы осязанияу
некоторых заднежаберных развиваются
щупальцевидные придатки
по бокам рта. Что касается глаз, то хотя
они имеются у большинства
заднежаберных, но имеют у этих улиток
второстепенное значение и обычно бывают прикрыты
кожей. Сердце у заднежаберных улиток
состоит из одного желудочка и одного
предсердия и лежит в перикардии.
Только у одного рода (Rodope) сердце
редуцировано. Непарная почка соединяется с
полостью перикардия, а ее наружное
выводное отверстие открывается на правой
стороне тела или у основания жабры.
Половые железы гермафродитны, и половой
аппарат более сложно устроен, чем у
переднежаберных .
Половая зрелость наступает обычно в
течение второго года жизни, и после
размножения заднежаберные улитки быстро
умирают.
Среди заднежаберных мы встречаем и
растительноядные формы, и хищников.
У большинства хорошо развита радула, а
у некоторых, кроме того, рот вооружен
кольцом шипиков или многочисленными
крючочками. Имеются слюнные железы
и пищеварительная железа, так
называемая печень, которая у некоторых
заднежаберных разветвляется на множество
отдельных веточек. Этот орган служит
для переваривания и усвоения пищи,
частицы которой захватываются клетками и
подвергаются внутриклеточному
перевариванию. Интересно, что мускулистый
желудок у некоторых заднежаберных несет
58
на внутренней поверхности твердые обыз-
вествленные пластинки, служащие для
лучшего размельчения пищи. Почти все
представители подкласса обитают в морях;
их распространение охватывает и
холодные моря земного шара, однако
большинство видов живет в теплых морях или
морях умеренного пояса. Один вид
встречается в Байкале, и несколько видов
приспособилось к жизни в устьях рек на
Малайских островах.
Часть заднежаберных обитает на
поверхности морского дна на песчаном или
илистом грунте, причем некоторые живут
на небольших глубинах, так что во время
отлива их можно находить у самой
границы воды среди зарослей водорослей.
Некоторые формы, которые, как правило,
держатся на дне, могут, однако, при
помощи развитых параподии подниматься
над грунтом и проплывать небольшое
расстояние. Другие заднежаберные, особенно
формы, составляющие особый отряд
крылоногих, относятся к типичным
планктонным животным; развиваясь иногда в
огромных количествах, они играют
значительную роль в круговороте
веществ моря, так как служат
пищей для ряда промысловых животных, как
например для некоторых китообразных.
ОТРЯД ПОКРЫТОЖАБЕРНЫЕ
(TECTIBRANCHIA)
У представителей отряда покрыто-
жаберных в большинстве случаев имеются
раковина и жаберная полость,
которая открывается на правой стороне
тела. Однако среди относящихся сюда
видов мы встречаем значительное
разнообразие форм, различающихся не только по
своему внешнему облику, но и по своему
образу жизни. Мы рассмотрим наиболее
характерных представителей отряда.
* * *
Наиболее примитивными из всех
заднежаберных и наиболее близкими по своему
облику к переднежаберным могут
считаться улитка актеон (Actaeon) и некоторые
другие представители того же
семейства Actaeonidae, которые сохранили
не только башневидную спирально
закрученную раковину, но и крышечку. Род
актеон включает небольшое число видов.
Наиболее известен из них Actaeon tor-
natilis, встречающийся на Атлантическом
побережье Европы и в Средиземном море.
Резко отличается от актеона по всему
своему облику улитка ацера (Acera bulla-
ta) из с е м е й с т в a Aceridae. Тело этого
животного удлиненно-цилиндрической
формы; голова приплюснута и как бы тупо
обрублена спереди. Мантия сзади несет
нитевидный придаток, который может
сильно вытягиваться и сокращаться.
Биологическое значение этого придатка не
выяснено, но по своему виду он
несколько напоминает хвостовой придаток,
имеющийся у некоторых киленогих улиток,
например у птеротрахеи. Тонкая
эластическая раковина ацеры яйцевидной
формы и относительно небольших размеров,
так что улитка уже не может целиком
спрятаться в нее. Крупные экземпляры
этого вида в вытянутом состоянии при
ползании достигают размера 4 см.
Хорошо развитая нога с широкими
кругловатыми лопастяхми по бокам служит не
только для ползания по субстрату, но и
для плавания. Если животное спокойно
лежит на дне или ползет, боковые лопасти
ноги загибаются вверх и прикрывают не
только бока тела, но и середину спины,
находя своими краями друг на друга и
закрывая также часть раковины. Но
достаточно вынуть улитку из воды или чем-
нибудь ее потревожить, как все туловище
настолько сильно сокращается, что
оказывается целиком скрытым под ножными
лопастями, и животное в таком состоянии
приобретает вид мягкого, покрытого
слизью шарика, на котором из-под
свернувшихся лопастей ноги можно видеть лишь
маленький участок раковины
треугольной формы. Об образе жизни этой
улитки М е й е р и М ё б и у с приводят
следующие данные. Более крупные
экземпляры ацер вылавливаются в течение зимы
и весны, тогда как летом, в июле, можно
находить лишь молодь 3—5 мм длиной,
а также пустые раковины среди отмершей
растительности, из чего можно заключить,
что ацеры живут лишь один год — от
59
Рис. 42. Актеон (Actaeon
tornatilis), вид со спинной
стороны.
Рис. 43. Ацера (Асега bullata) в спокойном положении
на водоросли: боковые лопасти ноги завернуты
на раковину A).
одной весны до другой. В Кильской
бухте, где изучалась жизнь этих животных,
они принадлежат к весьма обычным
формам, населяющим участки дна с илистым
грунтом, заросшим водорослями, и
держатся обычно среди отмершей
растительности, на которой они находят для себя
богатую пищу. При содержании в
аквариуме они охотно едят не только
растения, но также и мясо.
«Ацера, или улитка-шарик,— пишут
упомянутые исследователи,— почти
непрерывно находится в движении. То она
ползет по дну, то поднимается по стенкам
аквариума. Иногда она подвешивается
снизу к поверхности воды, несколько
криво изогнув при этом свое тело. При
ползании она то опускает, то
приподнимает голову и изгибает переднюю часть
туловища то направо, то налево, а
приподнятые вверх ножные лопасти то
смещаются вперед вместе с нижней частью
ноги, обнажая часть раковины, поверх
которой они лежат, то снова, отодвигаясь
назад, покрывают последнюю. Если эти
последние движения становятся более
частыми, чем обычно,— это признак того,
что наша улитка собирается плыть, а
такой способ ее передвижения можно было
бы назвать полетом в воде. Ее желтая
раковина скользит все быстрее и дальше
то вперед, то назад; движения переднего
конца тела становятся ритмическими, а
разгибания и обратные загибания ножных
лопастей становятся все сильнее и шире,
пока, наконец, их удары не отталкивают
всего тела от дна. Теперь уже животное
движется, все выше и выше
приподнимаясь в воде над дном и покачиваясь в
разные стороны, пока не повисает довольно
грациозно в прозрачной толще воды.
Когда эти движения достигают предела своей
интенсивности, нога делает по 2—3
сильных удара в секунду: при расхождении
лопастей ноги получается вогнутая с
нижней поверхности плоскость.
Одновременно передний конец тела сгибается вперед
или назад, и, пока это происходит,
животное несколько опускается в воде, но
новый удар по воде растянутых лопастей
ноги скачком вновь поднимает животное
вверх в косом направлении. После
нескольких минут такого быстрого
движения удары по воде становятся слабее и
улитка начинает падать ко дну. Часто,
прежде чем достигнуть последнего, она
делает несколько сильных ударов,
которые поднимают ее вверх, но уже не до
прежней высоты. Силы ее слабеют, она
падает на дно, бьет только краями
лопастей вверх, несколько раз помахивает
ими, а затем складывает их спокойно над
раковиной и переходит к ползанию».
В аквариумах ацеры откладывают яйца
иногда уже с января, тогда как в самой
Кильской бухте Мейер и Мёбиус лишь
в мае и июне находили их икру в таких
огромных количествах, что при помощи сачка
могли собирать на водорослях полными
горстями яйцевые шнуры этих улиток.
60
Характерны и наиболее эффектны из
отряда покрытожаберных улитки
морской заяц, или аплизия (Aplysia), и
близкая ей форма Dolabella; это самые
крупные из всех заднежаберных.
Некоторые из них достигают веса более 400 г.
Род аплизия широко распространен по
теплым морям земного шара; он включает
довольно большое число видов.
Чуть вогнутая раковина аплизии с
тонким внутренним известковым слоем
утрачивает у взрослых улиток ясную
спиральную закрученность, отчетливо
выраженную у молодых; ее с полным правом
можно рассматривать лишь как рудимент
настоящей раковины. На голове морского
зайца находятся две пары чувствительных
придатков, а по бокам тела
приподнимаются кверху параподии, закрывая с боков
середину спины. Часть ноги, которая
служит для ползания, сужена и сильно
вытянута в длину. Замечательно красива
окраска аплизии: то она бывает темно-
фиолетовой с разбросанными по этому
фону круглыми белыми пятнами, то эти
Рис. 44. Морской заяц, или аплизия (Aplysia
depilans):
А — вид сверху; 1 — раковина; Б — вид сбоку.
пятна рассеяны по охряно-желтому фону,
то преобладают грязно-серые и
желтоватые тона без резко очерченных пятен.
Аплизии могут хорошо плавать в течение
довольно долгого времени. При этом
параподии расправляются в стороны и по
ним пробегают спереди назад резкие
волнообразные сокращения, как на грудных
плавниках ската, и эти волны сокращения
охватывают весь свободный край обеих
параподии, затухая лишь там, где эти
последние срастаются над задним краем
мантии.
Аплизия может присасываться задней
частью своей ползательной подошвы к
стеклянной стенке аквариума, причем
тело ее свободно свешивается в воде под
любым углом наклона. Так она держится
довольно крепко, как если бы у нее на
заднем конце ноги была присоска; однако
как только улитка начинает ползать по
дну, видно, что никакой присоски у нее
нет. Возможность такого присасывания,
по-видимому, обусловлена сокращением
поперечных мускульных пучков,
расположенных в плотной хвостовой части ноги,
начинающейся сзади от места сращений
параподии. Будучи, как и все другие
заднежаберные, гермафродитами, аплизии
при спаривании могут поочередно играть
роль самца или самки; наблюдалось
образование цепочек из копулирующих особей;
в таких цепочках можно было насчитать
их по двенадцати штук. Аплизии
растительноядны и весьма прожорливы. Они
подобно змеям глотают пищу целиком,
а для размельчения заглоченной пищи у
них служат твердые пластинки,
выстилающие изнутри стенки мускулистого и
объемистого желудка. Зубцы
вывороченной вперед радулы захватывают пищу
снизу, а также справа и слева и при
обратном втягивании радулы тащат ее в
глотку косо по диагонали вверх и назад. Если
захваченный кусок пищи оказывается
слишком большим, улитка из рыгает его
посредством обратного движения зубцов,
причем незаглоченная пища выходит в
закрученном наподобие веревки виде. Как
на характерную особенность аплизии
можно указать на их способность выделять
при раздражении красящее вещество
пурпурного или белого цвета.
61
Рис. 45. Раковины покрыто
жаберных, живущих в северных морях
СССР:
слева — Retusa pertenius var. turrita;
в середине — Phyline lima;
справа— Cylichna alba.
Некоторые представители отряда
покрытожаберных постоянно живут и
обычны в наших северных морях. На
относительно небольших глубинах в Северном
Ледовитом океане встречается улитка
решу за (Retusa), имеющая кругополярное
распространение, несколько видов
улиток цилихны (Cylichna) и филины (Phyline),
а также улитка скафандер (Scaphander).
Некоторые из них живут и в Белом море:
сюда относятся 5 видов цилихны
(Cylichna reinchardti, С. scalpta и др.) и
беломорская разновидность Phyline lima.
ОТРЯД ACOELA
Некоторые относящиеся к этому
отряду улитки имеют блюдцевидную или ухооб-
разную раковину, но есть и большое
число форм, у которых раковина целиком
редуцировалась. Однако общий для всех
их признак состоит в том, что у них
отсутствует мантийная полость или, если она
и есть, она никогда не достигает той
глубины, как мантийная полость
покрытожаберных. У небольшого числа форм
этого отряда имеется перистая жабра,
находящаяся с правой стороны тела между краем
мантии и ногой. У огромного же
большинства представителей отряда, которые
составляют особый подотряд —
голожаберных улиток, жабры никогда не бывают
только с правой стороны тела, но
располагаются или симметрично вокруг
заднепроходного отверстия, или же на верхней
или нижней поверхности спинного
утолщения мантии, носящего название н о -
т у м. У некоторых жабры вообще
отсутствуют и дыхание осуществляется всей
поверхностью кожи.
Улитка, носящая название зонтика
(Umbrella botanicum), живущая у берегов
Австралии, достигает 19 см. Все тело
этого животного похоже на усеченный конус,
нижнее основание которого составляет
очень массивная нога, а верхняя
поверхность прикрыта кругловато-яйцевидной
раковиной со слегка поднимающейся
вершиной. Вся боковая поверхность ноги
покрыта бородавчатыми выростами, а
передний край имеет глубокую вырезку,
против которой лежит голова
животного, несущая пару плоских околоротовых
чувствительных придатков и пару ринофо-
ров, в продольной щели которых
помещаются органы обоняния. Между
основанием ринофоров находится пара глаз.
Параподий у этих улиток нет.
Улитки эти ведут малоподвижный образ
жизни. Интересны наблюдения над их
поведением,когда они случайно
оказываются вне воды. Они не съеживаются, как
можно было бы ожидать, но, наоборот,
приподнимают раковину, обнажая жабру
и часть кожи спины, увеличивая этим
возможность кожного дыхания; кроме
того, они производят ритмические
сокращения заднепроходного отверстия, открывая
и закрывая последнее.
В период размножения эти улитки
откладывают икру длинными лентами,
которые в виде спиральных завитков
прикрепляются к морскому дну.
Близка к улитке зонтику тоже очень
крупная улитка плевробранхус (Pleuro-
branchus), представленная многими видами,
живущими в различных морях, и ее
ближайший родич плевробранхея (Pleurobra-
nchea). Улитка плевробранхус по своему
внешнему облику имеет некоторое
сходство с черепахой, что и запечатлелось в
названии одного вида — Pleurobranchus
testudinalis (testudo — значит «черепаха»).
Раковина этой улитки целиком обросла
мантией, снабженной на переднем крае
вырезкой, через которую высовываются щу-
пальцевидные придатки головы — р и н о-
ф о р ы. Окраска ее замечательна. Рино-
62
форы оранжевого цвета, а грубые
щитообразные выпуклости спины окрашены в
коричневый цвет, но разных оттенков:
иногда они бывают светло-коричневыми,
у других животных, наоборот, они
совершенно темные, тогда как разделяющие их
бороздки выглядят как белоснежные
полоски, поверх которых проходит в
некоторых местах красивая карминно-красная
линия. Интересно, что эта окраска
связана с отложениями в коже различных
продуктов обмена веществ. Белые
полоски представляют собой отложения в
коже мочевой кислоты, а красные,
вероятно,— отложения мурексида,
продукта этой кислоты, тогда как
различные оттенки коричневого цвета отражают
различный ход процессов обмена веществ
при кожном дыхании.
Более бледно, но не менее красиво
окрашена улитка плевробранхея, которая
отличается от предыдущей формы также
и тем, что ее раковина целиком
редуцирована.
* * *
Подотряд голожаберные (Nudibran-
chia) представлен многими видами, со-
Рис. 46. А — улитка зонтик (Umbrella
mediterranean Б — плевробранхус (Pleurobranchus
testudinalis).
ставляющими свыше 30 различных
семейств. Однако в этом многообразии
могут быть отмечены две основные группы:
типичным представителем первой может
служить улитка дорис (Doris), а типичным
представителем второй — улитка эолис
(Aeolis). Каждый из этих родов
показывает один из основных путей эволюции
голожаберных; у других родов
обнаруживается тенденция развития в одном из
этих двух направлений.
У форм, относящихся к группе дорид,
на спине находится утолщение мантии
(нотум), на верхней поверхности которого
обычно и располагаются вторичные
жабры, которые, как правило, симметрично
группируются около заднепроходного
отверстия. Последнее расположено на
средней линии спины ближе к заднему концу,
и тело этих улиток в отношении внешних
черт строения как бы возвращается к
двусторонней симметрии, нарушенной лишь
тем, что половое и выделительное
отверстия лежат на правой стороне. В отличие
от дорид у улиток, более близких к роду
эолис, заднепроходное отверстие лежит на
правой стороне. Что касается жаберного
аппарата, то у некоторых форм жабры
полностью отсутствуют и дыхание
осуществляется через поверхность кожи; у других
вторичные жабры, как правило,
располагаются в два ряда по бокам спины, а
иногда и поперечными рядами. Кроме того,
имеются внутренние анатомические
отличия, в частности в строении радулы и
печени. В то время как у дорид печень имеет
строение цельного массивного органа, у
эолис она состоит, как правило, из трех
долек, делящихся на ветви, причем
разветвления печени заходят внутрь кожных
выростов на спине.
По своему образу жизни улитки дориды
относятся к чисто донным животным с
весьма слабой способностью к плаванию.
Род дорис (Doris), представленный
небольшим количеством видов, живущих в
Атлантическом океане и в Средиземном
море, характеризуется несколько
уплощенным телом, а мантийное утолщение на
его спине имеет неровную бородавчатую
поверхность и содержит многочисленные
известковые включения — элементы
кожного скелета. Сидящие венчиком вокруг
63
заднепроходного отверстия жабры, а
также прободающие спинную покрышку
щупальца при раздражении могут быстро
втягиваться, и животное в таком состоянии
приобретает особенную плотность,
компактность. Представители группы дорид
бывают крайне разнообразной окраски: от
матово-коричневой, сходной с окраской
грунта, на котором улитка живет, до
самой ярко-красной у тех форм, которые
населяют коралловые рифы в тропическом
поясе. Чудесную голубую окраску имеет
улитка глоссодорис (Glossodoris), живущая
в Средиземном море около Неаполя.
Из группы дорид многие виды обитают
и в наших северных морях: сюда
относится Acanthodoris pillosa, Onchidoris bila-
mellata, О. muricata и некоторые другие.
Acanthodoris pillosa — довольно обычный
обитатель каменистой литорали вдоль
берегов Кольского полуострова; встречается
и в Белом море. Эта небольшая улитка,
длиной до 30 мм, размножается в
середине лета, откладывая яйца в нижней
полосе литорали на водорослях фукусах.
Интересна отмеченная Ю. М.
Дерюгиным периодичность появления и
исчезновения этой улитки: в тех местах, где
она была в изобилии несколько лет назад,
нельзя было найти ни одного экземпляра,
а еще через ряд лет они опять появились
в массе. Виды Onchidoris встречаются
также вдоль побережья Кольского
полуострова и в Белом море. Doris bobretzkii
отмечен в Черном море.
Рис. 47.
1 — дорис (Doris
verrucosa); 2 — аканто-
дорис (Acanthodoris
pillosa), вид сбоку.
Рассматривая эолид и денд ропоту сов,
прежде всего следует остановиться на
характерных особенностях внешней формы
тела многих из этих улиток, придающих
им совершенно причудливый вид.
Последнее связано с развитием на их теле
кожных выростов, функционирующих как
вторичные жабры, но имеющих также и
другое значение, как мы увидим в
дальнейшем. Урода эолис (Aeolis) эти
кожные выросты расположены поперечными
рядами и так густо покрывают верхнюю
часть тела животного, что оно приобретает
некоторое внешнее сходство с метелкой.
Еще более своеобразную форму благодаря
кожным выростам приобретает
древовидная улитка (Dendronotus).
На переднем конце ее удлиненного тела,
а также двумя продольными рядами вдоль
спины расположены длинные и тонкие
ветвящиеся придатки кустовидной формы;
другие придатки в форме звездчатых
розеток сидят по краю углублений, из
которых высовываются кольцевидно
перетянутые щупальца. Интересно, что спинные
придатки в случае их отрыва могут быстро
регенерировать. Спина красивого красного
цвета, а придатки окрашены в светло-
коричневый цвет. Вообще окраска эолид,
так же как и дорид, поражает
наблюдателя своей красотой и изяществом. «До сих
пор с удовольствием вспоминается,—
пишет Зимрот,— как много лет назад,
будучи на Азорских островах, я обнаружил в
банке улитку идулию (Idulia, или Doto)
чудесного карминно-красного цвета:
особенно сверкали крупные спинные
придатки колбообразной формы, как бы
покрытые шишечками. И все же, несмотря на эту
окраску, мы долго не замечали этой
улитки, пока она сидела на красных
водорослях, которые были выловлены нами с
некоторой глубины. Окраска ее тела и
водорослей совершенно совпадала, а
спинные придатки очень походили
на спорангии, и таким образом
получалась весьма совершенная
покровительственная окраска.
Совершенно иное значение
имеет яркая окраска у эолис (Aeolis
papillosa). Основной цвет
спинных придатков здесь голубой,
а устье окружено широким жел-
Рис. 48.
I — эолис (Aeolis papillosa); 2 — дендронотус (Dendronotus frontosus); 3 — идулия (Idulia coronata).
тым кольцом. Животное с такой окраской
нельзя проглядеть. Это, наоборот,
угрожающая окраска, которая выявляет себя.
Но рыба, которая схватывает эту улитку,
сейчас же выплевывает ее, так как
получает ожог. Дело в том, что в концевых
частях придатков находятся такие же
защитные образования, как у
кишечнополостных, а именно стрекательные капсулы.
Первоначально думали, что эти
стрекательные капсулы образуются клетками
самой улитки, пока не было замечено, что
различные улитки держатся на колониях
определенных видов полипов и что во всех
случаях у улиток можно находить в
качестве средства защиты совершенно такие
же стрекательные капсулы, как те,
которыми обладают полипы, служащие пищей
для этих улиток. Загадка таким образом
разрешилась. Незрелые стрекательные
клетки кишечнополостных не
перевариваются в кишечнике улиток, но попадают
в разветвления печени, заходящие в
спинные придатки и открывающиеся на конце
наружу. Чужие стрекательные клетки как
бы вскармливаются в расширениях этих
печеночных ветвей; капсулы достигают
здесь своей зрелости и используются
улиткой как защитное оружие».
Некоторые представители группы эолид
встречаются и у нас в Баренцевом море:
улитка идулия (Idulia coronata), эолис
(Aeolis papillosa), корифелла (Coryphella
rufibranchialis, C. sarsi) и др. Idulia
coronata встречается и в Черном море.
Из тропических форм интересна своим
приспособлением к плаванию и своей
окраской улитка глаукус (Glaucus). Она
подвешивается к поверхности воды с
нижней стороны, вытягивая и распластывая
длинные спинные придатки,
расположенные группами на выростах тела, а газы,
скопляющиеся в ее кишечнике, придают
этому органу значение также и
гидростатического аппарата. В таком состоянии
улитка пассивно плавает у поверхности
моря, причем в ее окраске проявляется та
же закономерность, как и в окраске
многих морских рыб: сторона тела,обращенная
книзу (в данном случае — спина), имеет
у нее блестящую серебристую окраску,
тогда как обращенная кверху брюшная
сторона зелено-голубоватого цвета, как
морская вода, если смотреть на нее сверху.
5 Жизнь животных, т. 2
65
Таким образом, у этой улитки также
весьма совершенная покровительственная
окраска. Яйца, откладываемые этой
улиткой в виде спиральной ленты,
приклеиваются к колоколу парусника (Velella),
причем самые полипы предварительно
съедаются. Самые крупные дендронотусы
относятся к небольшому роду Tethys (
Fimbria). Т. fimbria (или leporina) живет в
Средиземном море и в Атлантическом
океане. Другой представитель — улитка
мелибе (Melibe rosea)— живет в Тихом и
Индийском океанах. Обе эти улитки
имеют и крайне причудливую форму тела:
их рыло окружено широкой
воронкообразной и бахромчатой по краю
лопастью — своеобразным ловчим аппаратом
для поимки добычи. Замечательна и
окраска тетис: вся она белого цвета с
черным узором, а спинные придатки имеют
красноватый оттенок. Образ жизни этой
улитки весьма разнообразен: она может и
плавать у поверхности, и ползать по дну,
и даже закапываться в грунт. По
наблюдениям Крумбаха, в Адриатическом
море бывает два периода, когда эта
улитка ведет пелагическое существование. «При
спокойной погоде,— пишет он,— можно
часто видеть тетис плавающей у
поверхности моря. Она подвешивается тогда
снизу к поверхности воды при помощи
подошвы ноги и ловит своим черпаком молодых
рыбок и всякую другую живность. Но это
подвешивание обусловлено не силой
поверхностного натяжения воды, а скорее
Рис. 49. Филлирое (Phyllirrhoe bucephalum).
эту улитку можно уподобить суденышку с
низкими бортами, образованными краями
ее ноги, причем спина здесь заменяет
киль. В таком положении она медленно
продвигается вперед, попеременно то
вытягивая, то сокращая свое тело... Когда
тетис хочет опуститься на дно или,
наоборот, подняться со дна к поверхности, она
плывет в толще воды посредством
волнообразных движений подобно
какой-нибудь планарии, а по дну она ползает
довольно живо, на широко
расправленной подошве своей ноги. В толще же
самого грунта она продвигается крайне
медленно. Способ и характер питания этой
улитки довольно хорошо изучен. У нее
совершенно нет радулы, а между тем она
свирепый хищник. На поверхности воды
она захватывает наряду с растительным
также и животный планктон, в том числе
и рыбью молодь. На дне она ловит
преимущественно ракообразных и офиур.
Ловчим аппаратом ей служит окружающий ее
рот воронковидный черпак, который
снаружи и особенно на внутренней
поверхности покрыт чувствительными
щупальцами; в стенке этого черпака, у основания
которого находится рот, проходят
радиальные и кольцевые мышцы. Когда тетис
ловит добычу у поверхности, она действует
своим ловчим аппаратом как вершей,
когда же она охотится на дне, она
прикрывает им добычу сверху. При этом
тетис, по-видимому, не отбирает добычу, а
залавливает все, что движется и что имеет
определенную консистенцию. Только уже
во рту делается отбор пищи, что можно
видеть из опытов кормления этой улитки.
Если ей дать в качестве корма мясо и
внутренности сепии, первоначально
захватывается и то и другое, но потом
внутренности выплевываются изо рта обратно...
Последнее имеет довольно забавный вид.
В мешке образуется складка, куда и
выплевывается неприятная пища со всеми
признаками отвращения... Подкармливая
тетис рыбьим мясом и мясом моллюсков,
можно было поддерживать ее
существование в течение месяцев».
Кладка икры имеет вид студенистой
спирально закрученной ленты. На конце
ее — комочек липкой слизи с пристающими
к ней песчинками, т. е. плавучий якорь,
66
который держит ленту с икрой в
определенном положении у дна. После полового
акта и откладки икры животные умирают.
Интересно отметить, что тетис
принадлежит к тем морским животным, которые
способны к свечению.
Совершенно своеобразная форма тела
и строение развилось у представителей
семейства Phyllirrhoidae в связи с их
настоящим планктонным существованием,
так как они никогда не опускаются на
дно. К этому семейству отряда
голожаберных принадлежит всего два рода улиток,
которые распространены исключительно в
теплых морях. Тело их сплющено с
боков и приобрело рыбообразную форму,
что особенно характерно выражено у
улитки филлирое (Phyllirrhoe). На голове
улиток этого семейства расположена пара
ринофоров, но жабры совершенно
редуцированы. Нога сведена до степени
рудимента (урода Cephalopyge) или
вовсе исчезла. Типичный представитель
этого семейства улитка филлирое, едва
достигающая длины 3 см, часто
попадается при планктонных ловах в
Средиземном море у поверхности воды, но ее легко
проглядеть из-за прозрачности ее тела.
Эта прозрачность настолько совершенна,
что через тело улитки можно читать без
всякого затруднения. В темноте ее тело
светится при раздражении; начинает оно
светиться и при опускании улитки в
пресную воду, а особенно сильное свечение
можно вызвать действием аммония. Тогда
все тело вместе с ринофорами испускает
яркий голубой свет, который затухает
вместе со смертью животного. Свечение
связано с раздражением периферических
нервных клеток кожи и с выделением
особого вещества, которое может
светиться и после смерти животного под действием
различных факторов, как например
пресной воды. Это вещество выделяется
особыми железистыми клетками.
Указание на то, каким образом
совершился переход некоторых эолид, ведущих
донный образ жизни, к планктонному
существованию, мы имеем в особенностях
образа жизни улитки Cephalopyge. Эта
улитка была найдена А. Куном
прикрепившейся к сифонофоре Halistemma, на
которой она держалась при помощи своей
рудиментарной ноги. Таким образом, здесь
также проявлялась характерная для
многих эолид связь с кишечнополостными, но
уже не с прикрепленными, а с
планктонными формами последних. Отсюда уже
мог быть сделан следующий шаг в
эволюции в сторону приспособления к
свободному планктонному существованию.
ОТРЯД КРЫЛОНОГИЕ (PTEROPODA)
В этом отряде значительно меньше
разнообразия форм, чем в предыдущих. Это
стоит в связи и с более однородным
характером среды, которую населяют эти
улитки, так как все они ведут планктонный
образ жизни. Характерным
анатомическим признаком улиток этого отряда
служит развитие пары более или менее
крупных плавников, которые
образуются из верхней части ноги. У всех у них
также хорошо развиты органы
равновесия — статоцисты, которые имеют
большое значение для этих животных,
свободно плавающих в водной толще. Все
крылоногие — гермафродиты, причем мужские
половые продукты созревают раньше, чем
женские (протерандрический
гермафродитизм).
Среди представителей этого отряда в
процессе его эволюции обособились две
ветви.
Одни формы, в большинстве
сохранившие свойственную и их предкам раковину,
приспособились исключительно к
питанию морским микропланктоном, что
отразилось на ряде черт их организации. В
месте соединения обоих плавников у них
лежит ротовое отверстие, окруженное гу-
бообразными лопастями ноги. Самая
голова с парой щупалец слабо обособлена от
туловища, челюстной аппарат и радула
развиты относительно слабо, но в желудке
имеются пластинки, служащие для
перетирания пищи. У некоторых форм,
относящихся к этой группе, вместо раковины
бывает стекловидно-прозрачный хрящевой
домик.
Другая группа крылоногих, наоборот,
совершенно утратила раковину и
приспособилась к хищному способу питания тоже
планктонными, но более крупными живот-
5*
67
Рис. 50. Спирателла, или лимацина (Spiratel-
1а, или Limacina) (Л) и Cavolinia tridentata (Б):
1 — раковина; 2 — общий вид спирателлы.
ными. Голова здесь довольно четко
обособилась от туловища и несет две пары
щупалец и часто весьма сложный ротовой
аппарат, приспособленный к поимке и
умерщвлению добычи. Радула здесь
хорошо развита. Глотка, которая у некоторых
форм может высовываться наподобие
хоботка, иногда вооружена
присасывательными придатками. Хорошо развитые
челюсти в виде пластинок с острыми зубцами
могут втягиваться в особые карманы или
высовываться вновь. Твердые пластинки
на внутренней стенке желудка здесь,
наоборот, отсутствуют. Характерно, что
личинки этих улиток первоначально имеют
раковину сперва блюдцевидной, а затем
трубковидной формы. Раковина, однако,
в дальнейшем развитии отбрасывается, как
и парус; на веретеновидном теле личинки
образуется три ресничных кольца, из
которых заднее сохраняется наиболее долго,
У представителей одного рода (Halopsiche)
молодь развивается внутри тела матери и
выходит наружу через разрыв ее тела,
вызывая этим ее гибель.
Крылоногие встречаются во всех морях,
но наибольшее разнообразие их форм
принадлежит тропическим морям. Они не
держатся только у поверхности моря, но
совершают обычно суточные миграции с
одних глубин на другие. Прозрачность их
тела не столь совершенна, как у Phylli-
rrhoe. Некоторым из них свойственна
также окраска или в блеклых, или в ярко-
красных тонах.
* * *
Группа крылоногих, сохранивших
раковину,— подотряд Thecosomata —
включает около 15 родов улиток. У всех
у них приспособление к питанию
микропланктоном выражено в образовании
весьма совершенного аппарата, служащего для
улавливания мельчайших планктонных
организмов. На заднем крае плавника
здесь находится «мерцательное поле», т. е.
участок, покрытый мерцательным
эпителием, суживающийся по направлению к
туловищу и доходящий до самого рта.
Движением ресничек мелкие планктонные
организмы и подгоняются ко рту.
У относящейся к этой группе улитки
спирателлы, или лимацины (Spiratella,
или Limacina), тонкая, почти прозрачная
раковина спирально закручена на левую
сторону. Раковина эта может закрываться
крышечкой, которая лежит на задней
лопасти ноги. Из относящегося к этому роду
небольшого числа видов в наших северных
водах встречаются два вида: один из них
(Limacina helicina) относится к числу хо-
лодноводных форм и встречается как в
Арктике, так и в Антарктике, тогда как
другой (Limacina retroversa) бывает у нас
в Баренцевом море гостем, приносимым
Гольфстримом из Атлантического океана.
У других крылоногих этой группы мы
встречаем уже более или менее отличную
от лимацины форму раковины: она может
быть конической, как например у
улитки крезеис (Creseis), иногда слегка
закрученной вперед или другой формы.
Окраска некоторых из этих улиток весьма
своеобразна.У улитки каволинии (Cavolinia
tridentata) раковина коричневатого цвета,
по краю крыловидных придатков ноги
проходит тускло-лиловая каемка,
обрамляющая коричневые поля — общую
основную окраску этих придатков. Каволинии
очень подвижные животные. 3 и м р о т
наблюдал, как некоторые из них, устав
плавать, опускались на дно, но затем опять
быстро поднимались, делая около НО —
115 ударов в минуту своими
плавательными лопастями.
Каволинии откладывают яйца в
простых стекловидно-прозрачных оболочках,
причем яйцевые шнуры не приклеиваются,
как у других морских брюхоногих, к
камням и другим предметам, а свободно
плавают на волнах, пока в них не
разовьются зародыши, которые затем выходят из
яйцевых оболочек и продолжают свободное
68
существование в планктоне. По
наблюдениям Гегенбауера, Cavolinia tri-
dentata откладывает в течение двух дней
около 200 яиц, тогда как другой вид —
Cavolinia gibbosa — 60—80 штук. Столько
же откладывает и крылоногая улитка.
На седьмой или восьмой день развития
зародыш с ресничным кольцом на
переднем и заднем концах тела уже покидает
свою яйцевую оболочку и, кружась
внутри яйцевого шнура, ищет из него выхода,
чтобы начать свободное существование.
Венчик ресничек на переднем конце
приобретает форму овала с двумя вырезами,
благодаря чему появляется уже знакомая
нам у других брюхоногих форма
двухлопастного паруса.
К группе раковинных крылоногих
обычно относят и ряд таких форм,
которые занимают как бы промежуточное
положение между этой группой и теми
крылоногими, которые утратили
раковину. Сюда можно отнести те виды, которые
лишены настоящей раковины, но у
которых развилась вторичная, «ложная»
внутренняя раковина (псевдокон-
х а), состоящая из «хрящевой» ткани. Сюда
относится, например, улитка цимбулия
(Cymbulia peronii), у которой плотная
студенистая раковина в форме деревянного
башмака покрыта мантией, улитка глеба
(Gleba cordata) и немногие другие.
Улитка глеба — одна из самых крупных
представителей крылоногих. Немногие виды
этого рода населяют тропическую зону
Атлантики, а также встречаются и в
Средиземном море. Характерно для этих
форм особенно мощное развитие
плавников, причем они срастаются сзади; у
улитки цимбулии в месте сращения их от
заднего края отходит длинный придаток
с тонким жгутообразным концом.
Характерно для этих форм также образование
хобота, на конце которого находится рот.
Хобот этот отличается большой
подвижностью, но совсем не служит целям
поимки добычи, так как эти улитки питаются
микропланктоном; последний
улавливается таким же ловчим ресничным
аппаратом, как и у других питающихся
микропланктоном крылоногих.
* * *
Группа так называемых голых, т. е.
лишенных раковины, крылоногих (Gym-
nosomata) включает в себя около 15
различных родов; некоторые представители
этой группы постоянно обитают и в
наших морях, как например улитка клио-
не (Clione limacina), часто встречающаяся
и в Баренцевом и в Белом морях.
Характерно, что, будучи типично
арктической формой, клионе приносится обычно
холодными течениями. Эта улитка —
прожорливый хищник, и главную пищу ее
составляет такая же, как и она, холодно-
водная форма — улитка лимацина
(Limacina helicina). В свою очередь сама клионе
составляет излюбленную пищу для
некоторых китов Как у хищника, у клионе
Рис. 51. Клионе
(Clione limacina).
Рис. 52. Глеба (Gleba coronata — Tiede-
mania neapolitana).
69
весьма совершенно развит ловчий аппарат
в виде расположенных вокруг рта шести
ротовых придатков (цефалоконий),
снабженных железами с клейким секретом,
удерживающим пойманную добычу. В
период спаривания две особи клионе
принимают вертикальное положение и,
соприкасаясь брюшными сторонами, взаимно
оплодотворяют друг друга. Через 20—
24 часа начинается откладка яиц. Кладка
состоит из сравнительно крупных клубков
или шнуров студенистой консистенции,
содержащих каждый большое число яиц.
У вылупившейся личинки имеется три
венчика ресничек; передний из них
соответствует морфологически парусу.
Распространение клионе так же, как и
служащей ей в качестве добычи лимацины, не
ограничивается холодными водами
северного полушария. Она встречается также
и в области Антарктики.
ОТРЯД МЕШКОЯЗЫЧНЫЕ
(SACOGLOSSA)
Заканчивая обзор заднежаберных,
укажем еще на один относительно бедный
формами отряд мешкоязычных улиток
(Sacoglossa), охватывающий всего около
15 родов с небольшим числом видов. Одни
из этих улиток имеют тонкую бесцветную
наружную раковину с маленьким
завитком и широким устьем, другие вовсе
лишены раковины. Все они, однако,
характеризуются значительным упрощением
радулы, передний конец которой находится
в слепом мешковидном углублении, чем и
обусловлено само название отряда
мешкоязычных. Большинство относящихся к этому
отряду форм обитает в теплых морях, где
они входят в состав донной фауны, но
некоторые виды захватывают в своем
распространении более северные широты, и в
частности встречаются на побережье Европы
в пределах СССР, как например
маленькая улитка лимапонция (Limapontia),
весьма обыкновенная на Мурманском побережье
Кольского полуострова, где она живет в
зоне литорали.
Особо следует отметить среди
моллюсков этого отряда бертелиний (Bertelinia).
Рис. 53. Двустворчатый брюхоногий
моллюск бертелиния (Bertelinia).
По внешнему облику их можно принять
за каких-нибудь представителей класса
двустворчатых, но не брюхоногих, так как
бертелиний имеют раковину, состоящую
из двух створок (рис. 53). Так же как и у
двустворчатых, створки раковины
бертелиний соединены лигаментом — тонкой
эластичной связкой, представляющей собой
видоизменение наружного органического
слоя. В противоположность
двустворчатым створки раковины не несут зубов,
однако, как у двустворчатых, с
внутренней стороны они стягиваются одной парой
мышц-замыкателей. Для двустворчатых,
правда, характерны две пары
мышц-замыкателей, но у некоторых из них может
быть лишь одна пара.
Несмотря на внешнее и отчасти
внутреннее сходство бертелиний с
двустворчатыми моллюсками, в основных чертах
строения тела они, конечно, настоящие
брюхоногие. У них хорошо развита голова со
щупальцами, типично для брюхоногих
устроена нога и, что, пожалуй, самое
главное, дыхание совершается с помощью
одной жабры, по своему строению иной,
нежели у двустворчатых. Радула и другие
органы пищеварения типичны для
мешкоязычных, что и заставляет относить
бертелиний к этому отряду брюхоногих
моллюсков.
Развитие бертелиний начинается с
формирования типичной для брюхоногих
личинки с шапочковидной раковиной. Од-
70
нако далее происходит как бы
расщепление краев раковины и независимый рост
каждой из ее сторон с соответствующими
изменениями и в форме мантии. Так и
формируется эта оригинальнейшая для
брюхоногих двустворчатая раковина, одна
из створок которой несет спирально закру-
ченную вершинку — трохоидную,
личиночную раковинку — след типичного для
брюхоногих начала развития.
Бертелинии стали известны науке
совсем недавно. Впервые они обнаружены у
берегов Японии, среди водорослей,
которыми, видимо, и питаются.
ПОДКЛАСС ЛЕГОЧНЫЕ (PULMONATA)
Легочные улитки представляют собой
группу, наиболее далеко уклонившуюся
в процессе эволюции от общего ствола
брюхоногих.
Все легочные улптки приспособились к
жизни или на суше, или в пресных водах,
а если некоторые их представители и
встречаются иногда в морях, то лишь в очень
сильно опресненных участках.
Раковина этих улиток чаще всего
спирально закручена и весьма разнообразна
по форме — от башневидной или валькова-
той до плоскодисковидной. У некоторого
относительно небольшого числа форм
раковина приобрела вид колпачка,
прикрывающего сверху все тело, как, например,
у некоторых улиток, приспособившихся
к жизни на быстром течении рек; у
других этот колпачок прикрывает лишь
небольшую часть тела и представляет
собой скорее рудимент настоящей раковины,
как мы это видим у многих наземных
улиток; наконец, у наземных же улиток мы
встречаемся со случаями полного
обрастания раковины мантией, иногда
сопровождающегося и полным исчезновением
раковины. Там, где раковина хорошо
развита и обнаруживает ясную спиральную за-
крученность, обычно она бывает закручена
направо; однако есть некоторые группы
легочных улиток, у которых раковины
закручены налево, а экземпляры с правоза-
крученной раковиной составляют
исключение. Устье раковины обычно остается
открытым, так как крышечка сохранилась
у представителей только одного семейства
(Amphibolidae). У небольшой, но древней
группы наземных легочных улиток клау-
зилиид (Clausiliidae) устье закрывается
особым клапаном раковины — к л а у з и-
л и е м, который опирается в устье на
сложную систему пластинок. Клаузилий
внешне напоминает крышечку
переднежаберных, однако он совсем иного
происхождения. Другой защитой от
неблагоприятных физических условий среды, как
например от засухи или холода, служит
затягивание отверстия раковины пленочкой
слизи, содержащей известь, которая
затвердевает на воздухе, — так называемой
эпифрагмой; между пленочкой и
глубоко втянутым внутрь телом улитки
остается обычно прослойка воздуха.
Насколько надежная защита создается таким
образом, показывают, с одной стороны,
опыты с действием низких температур
на садовых улиток. Под защитой эпифраг-
мы они оказались способными в течение
нескольких дней переносить температуру
в 110 и 120° ниже нуля, за исключением
тех экземпляров, у которых эпифрагма
была с трещинкой. С другой стороны,
известны примеры перенесения
наземными улитками довольно сильной жары
и засухи благодаря тому же
приспособлению. Обильному и быстрому выделению
слизи, необходимой для образования эпиф-
рагмы, способствуют так называемые «з у -
б ы» в устье, особенно характерные для
видов, обитающих в засушливых условиях.
У одних они представляют собой иногда
весьма многочисленные и сильные
выпуклости на внутренних стенках устья, у
других они выглядят как тонкие и острые
пластинки, уходящие по внутренней
стенке оборота далеко в глубь раковины. Все
эти образования при втягивании ноги
внутрь раковины давят на мягкие ткани и
выжимают слизистый секрет,
застывающий тотчас в пленку — эпифрагму.
71
К закупориванию устья при
наступлении неблагоприятных обстоятельств
прибегают иногда и водные легочные
улитки, которые тоже закрывают
отверстие раковины слоем слизи с воздушной
прослойкой между ним и своим телом; в
таком состоянии они даже иногда
вмерзают в лед и переживают зиму без вреда для
себя. Гораздо хуже защищены в этом
отношении безраковинные наземные улитки—
так называемые голые слизни. Сильная
засуха, яркое солнечное освещение в летний
зной, как равным образом и резкие холода,
заставляют их искать убежище под
различными прикрытиями,например под
слоями опавшей листвы, заползать в щели под
корой на гниющих пнях или прятаться
между комьями почвы,забираясь иногда
довольно глубоко под землю; там
сохраняется влага и температурные колебания менее
резки.
Для всех легочных улиток характерно
плавное скольжение на подошве ноги, в
передней части которой имеется сильно
развитая железа, выделяющая слизь.
Последняя смачивает подошву и предохраняет
ее кожный покров от повреждений,
уменьшая трение о твердую поверхность
субстрата. Само перемещение улитки вперед
происходит благодаря пробегающим по
подошве сзади наперед волнообразным
сокращениям, обусловленным
взаимодействием продольной и поперечной
мускулатуры. Передвигаясь вперед, улитка
вытягивает обычно свои щупальца, пользуясь
ими как органами осязания. У
пресноводных форм голова несет пару таких
щупалец, а у их основания расположена пара
глаз. У наземных улиток часто имеются
Рис. 54. Схема вдыхания и выдыхания воздуха
из легочной полости у виноградной улитки
(Helix pomatia):
1 — верхняя спинная стенка легочной полости; 2 —
положение нижней брюшной стенки легочной полости
при выдохе; 3 — положение нижней брюшной
стенки легочной полости при вдохе.
две пары щупалец, а у некоторых форм —
еще и третья пара щупальцевидных
придатков, расположенных по краям рта.
Глаза у наземных улиток расположены
иначе, чем у пресноводных,— на концах
щупалец. Из других органов чувств
обычно бывают развиты органы равновесия —
статоцисты. У водных форм имеется также
слаборазвитый осфрадий.
Характерная черта строения,
обусловившая само название подкласса, состоит
в устройстве органов дыхания,
в превращении мантийной полости в
легкое, причем более или менее узкое
отверстие, через которое полость последнего
сообщается с внешней средой, может
закрываться; на своде мантийной полости
развито густое сплетение сосудов. Жабра у
легочных улиток встречается лишь в виде
исключения. Таким образом, большинство
и пресноводных форм дышит атмосферным
воздухом, в связи с чем улитки должны
время от времени подниматься к
поверхности воды, набирая запас воздуха в свою
легочную полость. Сердце легочных улиток
состоит из одного желудочка и одного
предсердия. Нервные ганглии более или менее
явственно сконцентрированы в
окологлоточное кольцо.
Среди легочных улиток мы встречаем и
растительноядные, и всеядные, и хищные
виды. Хищные формы питаются другими
улитками, а некоторые — червями. У
легочных улиток хорошо развита радула, а
у растительноядных также и непарная
подковообразная челюсть. Хорошо развита
глотка, а зубцы на пластинках радулы
особенно длинны и заострены и напоминают
по форме клыки позвоночных. В глотку
открываются протоки обычно хорошо
развитых слюнных желез, а в мускулистый
желудок открывается пищеварительная
железа — печень. Кишка образует петлю
и открывается обычно около дыхательного
отверстия на правой стороне тела. Рядом
с заднепроходным отверстием находится
обычно и выходное отверстие единственной
почки, которая соединена с
околосердечной сумкой — перикардием. Особенной
сложности у легочных улиток достигает
половой аппарат. Половая железа здесь
гермафродитна — вырабатывает и
сперматозоиды и яйца. Отходящий от нее общий
72
проток разъединяется затем на
мужскую и женскую части
полового аппарата, причем та и
другая имеют ряд придаточных
образований. В женской части
образуются белковая и скорлуповая
железы, семеприемник, а иногда
и ряд других железистых
придатков. Семеприемник у наиболее
высокоорганизованных
представителей подкласса непосредственно
переходит в мужской копулятивный
орган. Для некоторых
характерно образование сперматофоров,
т. е. особых вместилищ для
семени. При спаривании оба
партнера взаимно оплодотворяют друг
друга, причем самому спариванию
обычно предшествует так
называемая любовная игра. У
некоторых форм во время спаривания в
тело партнера проникают особые
известковые тельца, так
называемые «любовные стрелы»,
служащие для полового возбуждения.
Яйца легочных улиток
откладываются или в общем студенистом
коконе той или иной формы (у
пресноводных форм), или
разъединенно, хотя и в общую кладку
(у наземных). Каждая яйцевая
клетка бывает окружена
значительным запасом питательного
материала, причем у некоторых форм
отношение между массой яйца и массой
окружающего его белка равно 1:8000
(Limax variegatus). Развитие, как
правило, проходит без образования
свободноплавающей личинки, и из яйца обычно
выходит почти сформированная улитка.
Легочные улитки делятся на два отряда.
ОТРЯД УЛИТОК С ГЛАЗАМИ
У ОСНОВАНИЯ ЩУПАЛЕЦ
(BASOMMATOPHORA)
Характерный признак представителей
этого отряда уже указывается в самом его
названии. Интересно отметить, что цаи-
более древние и во многих отношениях
наиболее примитивные легочные принадле-
Рис. 55. Герма-
фродитный
половой аппарат
слизня (Limax maxi-
mus):
l — гермафродитная
железа; 2 — ее
проток; з — белковая
железа, 4 —
яйцевод; 5 —
семеприемник; б —
семяпровод;
7—мускул,втягивающий мужской
копулятивный орган
жат к одному из семейств этого
отряда — семейству
сифонариид (Siphonariidae),
представители которого ведут амфибиоти-
ческий образ жизни на морском
побережье. Для сифонариид
характерна шапочковидная
раковина, очень похожая на раковину
морских блюдечек, дисковидная
присасывательная нога. Наряду
с легким — органом дыхания,
типичным для легочных, у них есть
и жабры. Некоторые
исследователи одну из жабр считают
рудиментом настоящих жабр, т. е.
ктенидиев переднежаберных;
другие же полагают, что все жабры
сифонариид вторичного
происхождения, развившиеся в связи с
переходом сифонариид к жизни в
постоянно влажной среде.
Своеобразными местообитаниями и
образом жизни отличаются
представители немногочисленного
семейства амфиболид (Amphibolidae),
распространенного в Восточной
Азии, Австралии и Новой
Зеландии. Эти улитки населяют сильно
опресненные участки моря в
различных бухтах, встречаются и в
устьях рек; это единственное
семейство легочных улиток,
сохранивших крышечку; другая их
характерная черта состоит в том, что
их легочная полость обычно наполнена
водой, хотя они и не имеют жабр.
Представителей отряда Basommatophora
можно встретить также среди обитателей
мангровых болот тропиков на островах
Зондского архипелага (Cassidula), среди
населения прибойной зоны, например
маленькую улитку Otina otis, живущую в Англии;
некоторые формы заселили карстовые
пещеры, как например улитка Zospeum spe-
laeum, утратившая в связи с пещерным
существованием глаза.
Лишь очень немногие представители
сидячеглазых обитают на суше. Это
единичные виды семейства эллобиид
(Ellobiidae) — карихиумы (Carichium
minimum), в то время как остальные эл-
лобииды солоноватоводные, эстуарные или
пресноводные виды. Карихиумы — очень
73
Рис. 56. Левозакрученные (леотроп-
ные) пресноводные улитки:
с л е в а—физа (Physa fontinalis);
справа — аплекса (Aplexa hypnorum).
Рис. 57. Прудовик обыкновенный
(Lymnaea stagnalis).
мелкие формы, они обитают в сырой лесной
подстилке и широко распространены в
северном полушарии.
Однако большинство представителей
отряда населяют пресные водоемы, и эти
формы представляют для нас наибольший
интерес как постоянные и обычные
обитатели наших рек, озер, прудов и ручьев.
В числе их мы встречаем прежде всего
представителей двух р о д о в—физа (Physa)
и аплекса (Aplexa), составляющих особое
семейство физид (Physidae),
которые особенно характерны тем, что их
раковина и все внутренние органы закручены
не на правую, а на левую сторону (лео-
тропные улитки). У физы тонкая
яйцевидная раковинка с коротким завитком,
прикрытая сверху длинными лопастевидными
отростками мантийного края. Улитки
этого рода распространены по всем частям
света, а у нас представлены лишь двумя
видами, один из которых (Physa
fontinalis) широко распространен в небольших
медленно текучих ручьях, озерах,
прудах почти по всей территории СССР, тогда
как другой вид (Physa acuta) населяет реки
и ручьи более южных районов
Европейской территории нашего Союза,
встречаясь в бассейне Дона, Днепра и в
Закавказье. Этот вид физы отличается
и более крупными размерами (высота
раковины 10—17 мм), чем первый (высота
раковины 4—6 мм). Род аплекса, имеющий
своих представителей в Европе, Азии и
Северной Америке, представлен у нас
одним видом (Aplexa hypnorum) —
небольшой улиткой (высота раковины 12—15 мм),
которая живет в болотах, небольших
пересыхающих лужах и очень сильно за-
росших ручьях, встречаясь и в
Европейской и в Азиатской частях СССР. Эти
улитки заканчивают свой жизненный цикл в
течение одного года. Их молодь можно
находить осенью, в сентябре, а к весне из
нее уже вырастают взрослые улитки,
которые в июне приступают к усиленному
размножению. Но уже к середине июля
нельзя найти ни одной аплексы в тех
местах, где еще в первую половину лета они
встречались массами.
Гораздо богаче видами представлено у
нас широко распространенное по всем
частям света семейство прудовиков—
самых обычных легочных улиток многих
прудов, озер, стариц. Из большого числа
видов, относящихся к этому семейству,
наиболее известен благодаря своим
крупным размерам обыкновенный прудовик
(Lymnaea stagnalis), некоторые более
крупные экземпляры которого достигают
размера 68—70 мм. С ранней весны до
поздней осени можно наблюдать этих
крупных улиток преимущественно в
прудах, небольших озерах или в затонах рек,
где особенно в тихий солнечный день они
оживленно ползают по дну или по
прибрежной растительности. Особенно много
их бывает в середине лета среди зарослей
кувшинок, или водяных лилий, на
подводных лугах роголистника, или урути,
на неглубоких местах. Одни из них
пасутся на зарослях этой растительности,
объедая листья урути или соскабливая
своей радулой с нижней поверхности
листьев кувшинок водоросли, а попутно и
различных мелких животных, которые
могут им попасться, так как улитки эти
всеядны. Другие, поднявшись к поверх-
74
ности водоема и подвесившись к ней снизу
широкой подошвой своей ноги, которая
удерживает животное благодаря
натяжению поверхностной пленки воды, медленно
и плавно скользят в таком положении,
оставляя за собой незаметную полоску
слизи. Дыхательное отверстие улитки,
ведущее в легочную полость, при этом чаще
всего бывает широко открыто. Прудовик
обыкновенный принадлежит к самым
прожорливым обитателям пресных вод,
поедая не только растения и животных,
которые могут ему попасться при его
медленном способе передвижения, но также и
трупы. При спаривании этих улиток оба
партнера взаимно оплодотворяют друг
друга. Икра откладывается в длинных
студенистых шнурах, которые приклеиваются
к самым различным предметам,
находящимся под водой и
имеющим гладкую твердую
поверхность. Иногда икра
приклеивается к раковине другой особи
того же вида. Заслуживает
внимания сложное строение
яйцевой кладки как пример
приспособления, характерного и
для других пресноводных
легочных улиток: это
приспособление выражается в снабжении
питательным материалом и
защитными оболочками
зародышей, которые все свое
развитие проходят в этих оболочках,
без стадии свободноплавающей
личинки. Само яйцо улитки
представляет собой сложное
образование, так как яйцевая
клетка погружена в массу
белка, находящуюся внутри
двойной оболочки. Такие яйца
включены в слизистую массу, которая
в свою очередь одета особой
капсулой, или коконом. От
внутренней стенки кокона отходит тяж,
прикрепленный другим концом
к наружной оболочке яйца,
которое оказывается таким
образом подвешенным к стенке
кокона. Число яиц в каждой
кладке варьирует в довольно
широких пределах, так же как
и сам размер яйцевого шнура.
Иногда можно насчитать до 275 яиц в одной
кладке. Болотный прудовик, как и другие
представители рода прудовиков,
отличается крайней изменчивостью, причем
Ьарьируют и размеры, и форма раковины,
и толщина ее, и окраска ноги и туловища
улитки. Наряду с крупными
представителями этого вида известны почти
карликовые формы, живущие в неблагоприятных
для них по комплексу условий водоемах,
как, например, одна разновидность в
горном озере Гокча (Армения),
достигающая всего 19,5 мм. Наряду с формами
обладающими крепкой твердостенной
раковиной, встречаются и формы с
чрезвычайно тонкой хрупкой раковиной, которая
ломается при малейшем давлении.
Варьирует также и форма устья и завитка;
окраска ноги и туловища
изменяется от сине-черной до пес-
чано-желтой. Эта широко
проявляющаяся изменчивость вида
привела в процессе его
эволюции к возникновению
большого числа местных
разновидностей, причем и до настоящего
времени далеко не во всех
случаях выяснено, когда мы имеем
дело с настоящими
географическими подвидами на обширном
ареале распространения этого
вида и когда — с узко
местными вариациями, связанными с
определенными условиями
существования в том или ином
водоеме.
Наряду с обыкновенным
прудовиком, постоянным
обитателем наших внутренних
водоемов, встречается и другой,
также крайне изменчивый вид—
ушастый прудовик (Radix au-
ricularia), затем прудовик
яйцевидный (Radix ovata), прудовик
болотный (Stagnicola palust-
ris) — виды, живущие в
стоячих и медленно текучих
водоемах, и некоторые другие.
Наибольший интерес представляет
небольшая улитка малый
прудовик (Galba truncatula), широко
распространенный по всей
территории нашего Союза. Этот вид
Рис. 58.
Внутривидовая изменчивость
раковины у обыкновенного
прудовика Lymnaea sta-
gnalis:
1 — типичная форма; 2 —
var. ampliata; 3 — var.
subula; 4 — var. arenaria;
5 — var. bodamica.
75
живет даже в небольших лужах, болотах и
родниках, встречаясь в соответствующих
условиях и по берегам рек и озер. Эта
маленькая улитка, безобидная сама по себе,
оказывается тем не менее одним из самых
вредных обитателей наших водоемов,
поскольку она служит промежуточным
хозяином для опасного паразита
нашего рогатого скота, а отчасти и человека —
червя из класса сосальщиков —
печеночного сосальщика, или фасциолы печеночной
(Fasciola hepatica). Соответственно в ряде
мест отмечалась прямая зависимость
заражения овец и крупного рогатого скота
фасциолезом от обилия в данной местности
малого прудовика, так как последний
бывает иногда заражен на 70%. Если принять
во внимание огромные количества этого
прудовика в местах водопоев скота, в
небольших лужах на окраинах болот и
других водоемов, становится понятным
его огромное значение в распространении
фасциолеза среди наших домашних
животных. Поэтому как одна из важных мер
борьбы с этим заболеванием
рекомендуется уничтожение этой улитки, для чего
весьма хорошим средством служит
известкование тех водоемов, где она встречается
в больших количествах. Это средство ведет
к быстрой гибели улиток, тогда как при
благоприятных условиях они
размножаются весьма интенсивно, достигая
половозрел ости уже к 6—7 месяцам при общей
продолжительности их жизни до 21
месяца. В яйцевой кладке малого прудовика
бывает от 4 до 25 яиц, а развитие
молоди улиток продолжается 10—20 дней.
Как интересную особенность некоторых
прудовиков (Lymnaea profunda, L. abys-
sicola, L. foreli) следует отметить
приспособление их к жизни на значительных
глубинах в больших озерах Швейцарии. В
этих условиях они уже лишены
возможности подниматься на поверхность для
захвата атмосферного воздуха, и их
легочная полость наполнена водой. В связи
с этим они дышат, используя
растворенный в воде кислород.
Своеобразным приспособлением к
жизни в горячих ключах на берегу Байкала
отличается вид Limnaea peregra,
образовавший в этих условиях особую
разновидность var. geyser icola.
К семейству прудовиков
принадлежит и другой, тоже широко
распространенный у нас род улитки — Myxas (Am-
phipeplea) с единственным у нас
европейским видом Myxas glutmosa. Другие
представители этого рода распространены в
Австралии, на Филиппинских,
Молуккских и Зондских островах. Это небольших
размеров улитка (высота раковины до
15 мм), отличающаяся от прудовиков
главным образом тем, что ее исключительно
тонкая и хрупкая раковина почти сплошь
покрыта мантией.
Эти улитки живут преимущественно в
поемных прудах и озерах, где иногда
их можно находить в огромных
количествах весной и в первую половину
лета.
Однако уже к середине лета прудовики
исчезают, так как их жизненный цикл
заканчивается в течение одного года.
Рис. 59. Различные виды прудовиков:
1 — Galba truncatula; 2 — Radix peregra; 3-Я.
ovata; 4 — Stagnicola palustris; 5 — S. stagnalis; 6 —
Radix auricularia.
Рис. 60. Левозакрученная легочная улитка
миратеста (Miratesta celebensis), у которой
развились вторичные жабры:
1 — глаз; 2 — ротовые лопасти; з — карман
щупальца, образованный двумя складками; 4 — жабры.
76
Помимо прудовиков, широко
распространено в наших прудах и старицах
богатое видами семейство катушек(Р1а-
norbidae), представленное у нас
несколькими родами, те или иные виды которых
встречаются и в Европейской и в Азиатской
частях Советского Союза. Другой, тоже
богатый видами род этого семейства —
Choanomphalus—имеет относительно
ограниченный ареал распространения —
Байкал и реку Ангару; наконец, на
Дальнем Востоке, в пойменных озерах
бассейна Амура, встречается еще один
представитель этого семейства — Glyptophysa
resvoi, тогда как остальные формы этого
семейства распространены в Австралии,
Африке, Южной Азии и в Новом Свете.
Некоторые из них обладают явной лево-
закрученной раковиной, как, например,
один африканский род (Isidora), один
индийский (Camptoceras) и один род с
острова Целебес (Miratesta). Последний интересен
также тем, что у этого представителя
легочных улиток имеется хорошо развитая жабра.
Наиболее крупна из встречающихся у нас
представителей семейства катушка
роговидная (Coretus corneus), часто обитающая в
прудах, небольших озерах, в затонах рек.
Закрученная в одной плоскости раковина
этой улитки достигает в диаметре 25—32 мм
и окрашена в оливково-коричневые или
коричневые тона разных оттенков, а нога и
туловище чаще всего сине-черного цвета.
Голова несет пару длинных нитевидных
щупалец, у основания которых с
внутренней стороны расположена пара глаз.
Одна из характерных особенностей
строения состоит в том, что дыхательное
Рис. 61. Катушка роговидная (Coretus
corneus).
Видны просвечивающие кровеносные сосуды (J).
Рис. 62. Улитки из семейства Ancylidae.
Слева направо: Acroloxus lacustris; Ancylus
fluviatilis; Pseudancylastrum dybowskii.
и половые отверстия лежат здесь, как
и у других представителей семейства, не
с правой, а с левой стороны. Эта
улитка обладает радулой и челюстями
значительно более слабыми, нежели у
прудовика; она держится большей частью
у дна, где легче найти мягкую пищу — ил,
отмершие растения, трупы животных. В
крови у катушки содержится
связывающий кислород гемоглобин, что позволяет
ей реже подниматься на поверхность для
дыхания атмосферным воздухом. Из
других более мелких видов катушек, которые
населяют наши водоемы, особенно
интересен вид Planorbis vortex, как
обладающий настолько тонкостенной, почти
прозрачной раковиной, что через нее
свободно можно наблюдать под лупой пульсацию
сердца и расположение некоторых
внутренних органов. Эта маленькая улитка
(диаметр раковины около 9—10 мм) часто
в массе попадается в стоячих водоемах,
преимущественно на нижней стороне
листьев кубышек и водяных лилий. Будучи
в большинстве своем крайне
изменчивыми, виды катушек приспособились к
существованию в весьма разнообразных
условиях: один из видов (Gyraulus gredleri
var. borealis) встречается даже в горячих
ключах на берегу Байкала при
температуре 31°. Некоторые виды хорошо переносят
вмерзание в лед (Planorbis planorbis) и в
таком состоянии переживают зиму.
Наконец, ряд видов приспособился к жизни
в небольших пересыхающих лужах:
улитки закапываются в ил или забираются под
дернины временно залитой водой луговой
растительности и, затянув устье раковины
тонкой пленкой, переживают
неблагоприятный для них период (таковы,
например, Planorbis planorbis, PI. rossmaes-
sleri, Segmentina hilida, PI. leucostoma).
77
Этой же способностью обладают и
некоторые мелкие прудовики, например Radix
peregra, G. truncatula и S. palustris, a
также битинии и валъваты из
переднежаберных.
Заканчивая обзор пресноводных
легочных улиток, остается упомянуть о весьма
распространенных в наших водоемах
маленьких улитках из семейства ан-
цилид (Ancylidae), раковина которых имеет
форму колпачка с невысокой вершиной,
несколько наклоненной назад; иногда она,
кроме того, и сдвинута вбок. Самая
крупная из этих улиток, Acroloxus lacustris,
достигает 7—8 мм длины, тогда как все
остальные представители семейства еще
меньших размеров. Acroloxus lacustris —
постоянный обитатель стоячих водоемов
почти на всей территории СССР и
встречается обычно на стеблях и листьях
водных растений, не поднимаясь, как
правило, выше уровня воды. Другой вид,
Ancylus fluviatilis, с яйцевидноовальной
формой устья и сильно сдвинутой назад
вершиной раковины, живет, наоборот,
только в текучих водоемах и охватывает
своим распространением реки
Европейской части СССР, распространяясь и
далее на запад. Наконец, несколько
видов, относящихся к р о д у Pseudancy-
lastrum, населяют Байкал и реку Ангару.
Для всех этих улиток характерен возврат
от легочного дыхания к дыханию
растворенным в воде кислородом: мантийная
полость у них более или менее редуцирована,
но на нижнем крае мантии находится ло-
пастевидный вырост, функционирующий
как вторичная жабра.
Водные легочные улитки
непосредственного значения в хозяйственной
деятельности человека не имеют, но косвенное
влияние некоторых их видов огромно. Они
служат промежуточными хозяевами для тех
или иных видов паразитических червей
сосальщиков (Trematoda), а некоторые из
последних наносят существенный вред
нашему животноводству. О жизненном цикле
печеночного сосальщика, который связан
с малым прудовиком (Galba truncatula),
было указано выше. Другие водные
легочные улитки служат промежуточными
хозяевами для паразитов и самого
человека, например некоторые виды
катушки (Planorbis pfeifferi), в которых
проходит некоторые стадии своего развития
такой опасный паразит, как кровяная
двуустка (Schistosomum bovis),
встречающийся не только у некоторых
домашних животных и обезьян, но и у
человека.
ОТРЯД УЛИТОК С ГЛАЗАМИ
НА КОНЦАХ ЩУПАЛЕЦ
(STYLOMMATOPHORA)
Принадлежащие к этому отряду улитки
почти все ведут наземный образ
жизни и составляют наиболее
высокоорганизованную группу среди всех
брюхоногих. Все они характеризуются
наличием пары втягивающихся щупалец, на
конце которых находятся глаза. Кроме
того, имеется еще одна пара более
коротких щупалец. У большинства стебельча-
тоглазых раковина развита очень хорошо.
В связи с существованием на суше этих
животных раковина тут служит не только
защитой мягкого тела от механических
повреждений, но и от врагов и от испарения.
Естественно, что упоминавшаяся нами
прежде эпифрагма одна не в состоянии
обеспечить надежную защиту от
избыточной потери влаги.
В связи с этой новой функцией
раковина наземных стебельчатоглазых может
весьма сильно видоизменяться в своей
структуре. У одних видов, живущих во
влажной среде, она слабо
кальцинирована, тонка и прозрачна. У других,
приспособившихся к жизни в очень сухом
климате, она, наоборот, имеет толстые стенки,
богата углекислым кальцием и нередко
ярко-белая, что способствует отражению
солнечных лучей. У скальных видов
раковина часто несет на поверхности то тонкие,
то толстые и высокие ребрышки. Ребра не
только придают раковине прочность, но и
представляют собой результат, так
сказать, «экономии материала»: чересчур
толстые стенки раковины — в толщину ребер—
сделали бы ее чрезмерно тяжелой,
сковывающей подвижность улитки.
Несомненно, что явным
приспособлением к сохранению нужного объема вла-
78
ги в теле сухоустойчивых видов служит
увеличение их размеров. Наиболее сухо-
устойчивые крымские зебрины или
среднеазиатские яминии, живущие на камнях и
кустарнике на солнцепеке, весьма крупны.
В этом случае большой объем мягких
частей тела и раковины обеспечивает
накопление большого количества воды, а
неизбежное испарение части ее относительно
меньше, чем испарение такого же
количества у мелких форм.
У многих стебельчатоглазых раковина
рудиментарна и частично или полностью
прикрыта мантией, а у некоторых форм
она совершенно редуцирована. К таким
совершенно безраковинным улиткам
относятся, например, представители
семейства онцидиид (Oncidiidae),
большинство которых живет в Индии, Австралии,
на Филиппинах, на Молуккских и на
Галапагосских островах, а один род (Oncidi-
ella) встречается в Африке, Европе и
Америке.
Большинство этих улиток ведет
земноводный образ жизни, населяя зону
морского прибоя, где они оказываются то
под водой, то вне ее, но некоторые виды
их завоевали и сушу, приспособившись к
жизни на значительных высотах над
уровнем моря и находя для себя убежище под
корой деревьев.
Полуземноводный образ
жизни ведут и некоторые другие
представители описываемого отряда, как,
например, улитка янтарка (Succinea putris),
названная так за янтарно-желтый цвет
своей раковины. Эта небольшая улитка
весьма обыкновенна у нас на сырых
местах в непосредственной близости от
водоемов, где она встречается на стеблях и
листьях прибрежной растительности, но ее
же можно наблюдать и на плавающих в
воде листьях кувшинок, или водяных
лилий. Вообще она не боится воды и
часто уходит под поверхность последней. С
другой стороны, ее можно встретить и
довольно далеко от водоемов на
кустарниках или на луговой растительности во
влажных местах.
Другие виды янтарок не так тесно
связаны с водоемами и встречаются
постоянно на растениях в садах, на опушках леса
и на лугах.
Настоящие наземные легочные улитки,
целиком приспособившиеся к жизни на
суше, составляют чрезвычайно обширную
группу, охватывающую свыше сорока
семейств, некоторые из которых
характеризуются чрезвычайно интенсивно идущим
у них в настоящее время процессом
видообразования. Ввиду невозможности охватить
в кратком очерке все это многообразие
форм мы остановимся на тех из них,
которые особенно интересны чертами своего
строения, особенностями образа жизни,
или тех, которые имеют хозяйственное
значение в жизни человека. Среди этих
улиток мы также встречаем ряд форм с более
или менее редуцированной раковиной,
причем различная степень редукции
раковины проявляется у представителей
семейств, часто довольно далеко отстоящих
друг от друга. Однако у большинства
наземных легочных улиток хорошо развита
раковина, относительно массивная у
некоторых форм, хотя если сравнивать
раковины наземных пульмонат с
раковинами морских переднежаберных
приблизительно того же размера, то первые, как
правило, уступают по своей массивности
последним. Это понятно, так как в воде
раковина теряет часть своего веса и
становится относительно более легким грузом,
чем на суше. Наибольшими размерами
отличаются раковины самых крупных из
наземных улиток, живущих в тропической
Африке и на юге этого континента,
принадлежащих к семейству ахатин
(Achatinidae). С ними могут отчасти
конкурировать лишь некоторые мадагаскар-
ские улитки (Clavator clavator), высота
раковины которых превосходит 10 см,
и южноамериканские, тоже по
преимуществу жители тропиков, улитки рода
Рис. 63. Улитка янтарка (Succinea putris).
79
строфохилус (Strophochilus popelairianus)
с очень толстой и массивной раковиной,
высотой до 14 см. О прочности раковины
этих улиток можно судить хотя бы уже по
тому, что их употребляли на табачных
плантациях в качестве утюгов для
разглаживания табачных листьев. Африканские
улитки ахатины (Achatina, Cochlitoma)
настолько тяжелы, что, когда они
собираются по нескольку штук на ветвях
деревьев или кустарников, ветки иногда
обламываются под их тяжестью. Один из
видов этих живородящих улиток
(Achatina fulica) был завезен с ее родины,
Восточной Африки, в Ост-Индию и на
Цейлон и прекрасно акклиматизировался в
этих местах; взрослые улитки выполняют
полезную роль ассенизаторов, поедая
главным образом разлагающиеся растительные
остатки, экскременты животных и
различные нечистоты. Однако молодь этого вида
нередко вредит различным культурным
растениям, поедая почки бананов,
различные плоды и клубнеплоды.
Полную противоположность этим
гигантам среди наземных легочных улиток
представляют, например, маленькие улитки из
семейства зонитид (Zonitidae), часть
которых живет в Южной и Центральной
Америке, на Сандвичевых и Багамских
островах, а некоторые роды
распространены в палеоарктической области. Таковы,
например, маленькая улитка витрея (Vit-
геа), некоторые виды которой имеют
совершенно прозрачную тонкую раковину,
не превышающую высотой одного
миллиметра, или некоторые виды широко
распространенного в различных частях света
рода зонитоидес (Zonitoides), тоже с
тонкой прозрачной раковиной. Один из
видов этой улитки (Zonitoides nitidus)
часто встречается и у нас
многочисленными сообществами на сырых лугах в
непосредственной близости от водоемов.
Почти настолько же, насколько
разнообразны размеры легочных улиток,
варьирует и форма их раковин — от почти
дисковидной, как, например, у живущей
в лесах Европы под камнями улитки Discus
(Patula) solarius и некоторых других, до
кубаревидной, как, например, у
виноградной улитки, или башневидной и даже уд-
линенно-веретеновидной, как, например,
Рис. 64.
Слева — клаватор
(Clavator clavator),
высота раковины
до 10,2 см.
Справа — строфохилус
(Strophochilus po-
pelairianus),
высота раковины до
14 см.
у европейских видов улитки клаузилии
(Clausilia laminata) или у вестиндского
вида аномы (Anoma tricolor) и многих
других. В пределах этих основных типов
и варьирует обычно форма раковины
наземных легочных улиток, и только у
относительно небольшого числа форм
раковины отличаются особенно причудливым
видом. Наконец, у ряда форм, у которых
раковина в той или иной мере
обнаруживает тенденцию к редукции, она
приобретает вид более или менее уплощенного
колпачка со слегка закрученным завитком, и
эта форма конвергентно проявляется у
представителей разных далеких друг от
друга семейств, как, например, у
живущей в странах Средиземноморья и на
Кавказе улитки даудебардии (Daudebardia),
у одного новозеландского вида (Schizoglos-
sa novoselandica), у тропических
представителей семейства янтарок (Suc-
cineidae) и некоторых других.
Если у многих морских
переднежаберных, как мы видели, на наружной
поверхности раковины развиваются различные
отростки и шипы, то у наземных пулъмонат
такие шипы образуются только по
внутреннему краю устья, тогда как наружная
поверхность раковины чаще всего бывает
гладкая или покрытая сравнительно мало
выступающей скульптурой в виде
ребрышек, струек и т. п. У небольшого числа
улиток поверхность раковины бывает
покрыта кожистыми выростами перио-
стракума, как, например, у одной
маленькой европейской улитки Acanthi-
nula aculeata, живущей в лесах среди
гниющих древесных остатков. Защита устья
выступающими по его краю зубцами у
некоторых улиток из семейства клаузилиид
усложняется образованием с внутренней
80
Рис. 65. Ахатина (Achatina mauritana).
стороны устья также нескольких
выступающих в полость раковины перегородок
и одной подвижной пластинки (так
называемого замочка, или клаузили-
у м а), сидящей на эластичном
стерженьке. Когда улитка прячется в раковину,
эта пластинка замыкает отверстие
последней. Наконец, в одном случае (у улитки
Thyrophorella, живущей на Принцевых
островах) известен своеобразный способ
замыкания устья: часть верхнего края
последнего выступает в виде лопасти,
которая подвижно сочленена с раковиной
и закрывает ее входное отверстие, когда
улитка прячется внутрь.
В связи с наземным существованием и
передвижением по суше у многих наземных
пульмонат видоизменилось строение их
подошвы: тогда как у одних волны
сокращения, перемещаясь сзади наперед,
охватывают всю подошву, у других улиток
они проходят лишь по средней ее части,
которая обособлена от боковых частей
двумя продольными бороздками, а иногда,
кроме того, и выделяется своей окраской.
Об обильном выделении слизи передней
ножной железой, а в некоторых случаях
и задней железой ноги, которую часто
неправильно называют хвостовой, уже
упоминалось. В связи с наземным
существованием изменилось отчасти и строение
кожного покрова, особенно у крупных
улиток, у которых кожа приобрела
характерный морщинистый вид. Эти морщинистые
утолщения кожи медленно пульсируют,
как это можно наблюдать, например, у
крупных слизней; эта пульсация имеет
значение в процессе кожного дыхания,
которое у наземных улиток дополняет
дыхание через легкое. С одним крупным
слизнем был проделан интересный опыт: он
был погружен в воду, пока у него не
проявились явные признаки удушения. После
того как он был снова положен на воздух,
постепенно восстановилась пульсация
кожных морщин на его спине, а затем уже
через продолжительное время открылось
его легочное отверстие. В другом опыте
легкое слизня Agriolimax заполнялось
расплавленным парафином, но это не
вело к полному прекращению дыхательных
процессов, которые осуществлялись при
этом через кожу. В связи с этим важно
отметить, что дыхательные движения,
выражающиеся в выдыхании и вдыхании
воздуха через легочное отверстие, так
называемый пневмостом, происходят не
ритмично, а через разные промежутки
времени в зависимости от потребности
животного. Открывание пневмостома и
выдыхание воздуха происходит,
по-видимому, тогда, когда в легочной полости
скопляется значительное количество
углекислоты, что и вызывает рефлекторное
открытие пневмостома. При нормальных
условиях у виноградной улитки (Helix
pomatia) пневмостом открывается и
закрывается приблизительно один раз в
минуту. При обилии углекислоты в
окружающей среде дыхательные движения
учащаются; по наблюдениям над нашими
слизнями (Agriolimax agrestis и Arion empiri-
corum) они доходят до 27 раз в минуту.
Кроме пары глаз, которые способны
различать не только степень интенсивности
освещения, но также, по-видимому, и
предметы на расстоянии до 1 еле, наземные
легочные улитки обладают хорошо
развитым химическим чувством,
или чувством обоняния.
Интересно, что органом этого чувства служат
не только концевые вздутия щупалец и
особенно задней их пары, как думали
раньше, но также и вся кожа передней части
тела, что было обнаружено рядом опытов
с ампутацией щупалец. Концевые вздутия
задней пары последних, а также участок
кожи, выстилающий вход в дыхательную
полость, обладают химическим чувством
в наибольшей мере, но оно свойственно
также коже передней части головы,
переднего края ноги и других участков тела.
Некоторые исследователи проделывали опыты с
целью выяснить, на каком расстоянии
6 Жизнь животных, т. 2
81
наземные улитки воспринимают
различные запахи и реагируют на них.
Оказалось, что многие сильно пахнущие
вещества, но такие, с которыми улиткам не
приходится сталкиваться в природных
условиях, вызывают реакцию с их
стороны только на очень близком расстоянии,
около 4 см. Так воспринимаются ими
керосин, бензин, хлороформ, аммиак.
Гораздо тоньше их обоняние, когда дело
касается запахов пищи. Виноградная
улитка чувствует запах зрелой дыни уже на
расстоянии 50 см, а запах капусты на
расстоянии 40 см, правда, при легких
дуновениях ветерка; в совершенно
неподвижном воздухе тот же запах действует на нее
всего лишь на расстоянии 6 см. Примерно
такой же остротой обоняния обладают и
некоторые слизни (Limax maximus), тогда
как другие ощущают запах пищи на
расстоянии около 2 м и даже более того (Arion
empiricorum). Хорошо развито у наземных
легочных улиток и чувство осязания,
тогда как чувство слуха у них,
по-видимому, вовсе отсутствует, и они не
воспринимают даже сильных шумов,
производимых на близком расстоянии. Пара ста-
тоцистов там, где она есть, имеет значение
лишь органа равновесия, дающего
возможность животному ориентироваться в
отношении силы тяжести, что проявляется,
например, в отрицательном геотропизме
некоторых форм (Agriolimax agrestis,
Helix), а также, по-видимому, дающего
животному правильную ориентацию в
передвижении: по крайней мере, перерезка
церебропедального нерва вместе с нервом
статоциста у виноградной улитки вызывает
у нее вращательное движение в ту
сторону, с которой была произведена операция.
Однако необходимо отметить, что общие
наши знания по физиологии наземных
легочных улиток, так же как и сведения
о различных сторонах образа жизни и
поведения многих из них, далеко
недостаточны.
Лучше изучены в этом отношении
некоторые представители семейства ге-
лицид (Helicidae) и некоторые более часто
встречающиеся в Европе голые слизни.
Те и другие имеют также и хозяйственное
значение, почему мы и займемся ими более
подробно. Семейство гелицид охватывает
свыше 50 родов и делится на несколько
подсемейств. Отдельные роды и виды
также крайне изменчивы, образуя множество
подродов, подвидов и рас, а в пределах
этих мелких систематических единиц
обычно наблюдается сильно выраженная
индивидуальная изменчивость, почему эти
улитки и стали, особенно в последнее
время, излюбленным объектом для изучения
явлений изменчивости. Ареал
распространения этого семейства в целом
ограничивается западной частью палеоарктической
области, охватывая страны Средней и
Южной Европы, Кавказ, Малую Азию,
Северную Африку и острова
Средиземного моря и Атлантического океана,
примыкающие к Европе и Северной Африке.
Есть указание на нахождение одного
из видов этого семейства на Алтае.
Виноградная улитка (Helix pomatia),
самая крупная из этого семейства, широко
распространена в странах Южной и
отчасти Средней Европы, в Передней Азии и
Северной Африке до Алжира. Раковина
этой улитки, кубаревидная по форме,
достигает высоты 50 мм при ширине около
45 мм и образует 42/2 быстро
расширяющихся оборота, которые оканчиваются
широким устьем. По светлому желто-
коричневатому фону обычно вдоль
оборотов завитка идут широкие коричневые
полосы, степень выраженности которых,
однако, очень изменчива, так что можно
встретить экземпляры и вовсе лишенные
этих полос, с однотонной окраской.
Виноградная улитка находит подходящие для
себя местообитания не только на
виноградниках, но также и в слабозатененных
садах, а иногда и на открытых местах. День
она проводит чаще всего спрятавшись в
свою раковину и выходит на кормежку
лишь ночью. Главную пищу ее
составляют зеленые части растений, и в связи с
этим она приносит иногда значительный
вред, объедая виноградные лозы.
Относительно размножения этих улиток
производились многочисленные наблюдения,
рисующие их поведение в период спаривания,
когда они приступают к характерной для
многих наземных легочных улиток так
называемой «любовной игре». Стремление
улитки к спариванию сразу
обнаруживается в ее поведении. Она медленно ползет,
82
как бы в поисках чего-то, часто
останавливается на полдороге и долго выжидает на
одном месте, слегка приподняв переднюю
часть своего тела. Если случайно
встретятся две такие улитки, они сейчас же
приступают к «любовной игре», которая
предшествует акту оплодотворения. Обе они
вытягиваются вверх одна против другой
и принимают характерное положение,
соприкасаясь участками подошвы и
ощупывая друг друга щупальцами и ротовыми
лопастями. Однако эти движения
прекращаются уже через короткое время,
животные падают и, плотно прижавшись
друг к другу подошвами, остаются
неподвижными примерно от четверти до получа-
Рис. 66. Виноградная улитка (Helix pomatia):
вверху — спаривание, внизу — кладка яиц.
са. После этого периода покоя снова
возобновляется прежняя игра, и весь этот
процесс длится около двух часов, когдаг
наконец, то из животных, которое
достигло большего возбуждения, не втыкает в
тело своего партнера любовную стрелу,
которая усиливает и его возбуждение.
После короткой паузы происходит самый
акт совокупления, причем каждая
улитка играет роль и самца и самки. Само
совокупление у виноградной улитки
продолжается всего несколько минут, тогда как
у некоторых других видов оно длится
часами. После этого, однако, проходит еще
продолжительное время, пока полностью
не произойдет обмен сперматофорами у
совершенно обессилевших животных.
Только после этого они расползаются в разные
стороны, но при этом вдоль ноги еще
долго проходят особенно сильные волны
сокращения, благодаря которым, вероятно,
сперматофоры, содержащие сперматозоиды,
легче проникают в семеприемник.
Оплодотворенные яйца, содержащие запас
питательного материала для будущего
зародыша и одетые каждое порознь защитной
оболочкой, откладываются в землю в
специально выкопанную улиткой ямку,
отверстие которой закрывается, когда
кладка окончена.
С наступлением осенних холодов
виноградная улитка подготовляет себе зимнее
убежище, выкапывая в земле ямку, куда и
залегает на зимнюю спячку. Ямку она
выкапывает ногой, подошва которой
плотно прижимается к земле, и производит с
силой движения, аналогичные ползатель-
ным. Если почва оказывается слишком
твердой, улитка опрокидывается на спину
и делает такие же движения ногой,
нагребая на себя сверху опавшую листву,
которой и пользуется как прикрытием.
Закопавшись, улитка втягивается в раковину
и выделяет мантийным краем содержащую
известь зимнюю к р ы ш е ч к у—э п и ф -
р а г м у. Изнутри выделяется затем еще
вторая крышечка из отвердевающей слизи
с пористым включением — «окошечком»,
приходящимся против дыхательного
отверстия. Улитка впадает в оцепенение,
но обмен веществ в ее теле не
прекращается, хотя и совершается очень медленно.
Число сердечных пульсаций падает до
6*
83
одного сокращения в минуту. Весной,
перед возвращением к активности, в первую
очередь набирается воздух в легкое, а
затем сбрасывается зимняя крышечка.
Общая продолжительность жизни
виноградной улитки 6—7 лет.
Хотя виноградная улитка и приносит
иногда вред виноградникам, она имеет
ценность как продукт питания и в ряде
стран употребляется в пищу в вареном
виде. Особенный cnpdc она находит в
Испании, во Франции, в Италии; здесь
уже во времена древнего Рима ее весьма
ценили как пищевой продукт. В связи
с этим в более позднее время она была
завезена монахами в ряд других стран,
как например в Северную Германию, где
ее разводили в монастырских садах.
Аналогичное значение имеет и другой
близкий, но более южный европейский вид
Helix aspersa; эта улитка была
акклиматизирована в США, Аргентине, Кейптауне,
Австралии, Тасмании и Новой Зеландии,
а также на Канарских островах, острове
Св. Елены и многих других. Однако в
некоторых местах массовое размножение
этой улитки приносит вред
плодоводческим хозяйствам, так как улитки
объедают цветы и листья на абрикосовых,
персиковых и других плодовых деревьях.
Почти тех же размеров, как и
настоящая виноградная улитка, достигает вид
Helix lucorum, встречающийся у нас в
области горного Крыма. Значительно
меньше ее по размерам вид Helix vulgaris,
широко распространенный на всем юге
Европейской части СССР. Из других
представителей семейства гелицид в состав
фауны СССР входит, с одной стороны, ряд
родов, имеющих широкий ареал
распространения; таковы, например, сухоустой-
чивые улитки Theba и Helicella, несколько
видов которых населяют южные районы
Европейской части СССР, тогда как
другие виды этого рода распространены на
запад вплоть до Атлантического
побережья Европы и до Марокко; широко
распространены также роды Trichia, Perforatella
и некоторые другие, входящие в состав
нашей фауны. С другой стороны, некоторые
роды гелицид эндемичны для фауны
Кавказа и не встречаются в других местах;
к таким относится улитка Caucasotachea
atrolabiata и многие другие. Также и
среди западноевропейских форм
семейства гелицид наряду с широко
распространенными родами встречаются роды,
захватывающие очень узкий ареал
распространения* например область
Пиренеев или другие отдельные горные
области Европы. Это находится в
прямой связи с характерной не только для
гелицид, но и вообще для наземных
пульмонат склонностью к образованию
большого числа локальных форм,
благодаря относительно весьма слабой
способности их к расселению по суше, что легко
создает условия изоляции в развитии
отдельных подвидов, видов и родов.
Несмотря на то что потребность в
достаточном количестве влаги имеет особенно
большое значение в жизни наземных
улиток, мы видим, что целый ряд
представителей гелицид, как равным образом и
некоторых других семейств легочных
улиток, приспособился к существованию в
засушливых областях, завоевав не
только сухие степи и полупустыни, но также
и настоящие пустыни. Так, например,
некоторые гелициды встречаются в Сахаре на
расстоянии до 4 км от оазисов, где уже нет
никаких следов растительности и где
температура в полдень достигает 43°. При
особенно сильном недостатке влаги эти
улитки впадают в спячку, закрывая устье
раковины эпифрагмой, и сохраняют
способность оживать через несколько лет
пребывания в покое. Интересно, что
представители тех же видов, но живущие в
условиях более влажного климата, например
на острове Мадейра, сохраняют
способность оживать после такой спячки всего
лишь в течение нескольких месяцев и
гибнут, если за это время не возвращаются
благоприятные для них условия. Во время
такой спячки происходит более или менее
значительная потеря влаги, и для
возврата к нормальной жизнедеятельности
необходимо ее пополнение. Поэтому, если
потерявшей часть своей влаги улитке,
которая также и голодала, предложить
пищу, она не сможет ее есть, пока не
пополнит запас воды в организме.
Будучи растительноядными формами,
гелициды в большинстве своем
малоразборчивы при выборе в качестве пищи того или
84
иного растения. В ряде
экспериментальных наблюдений было установлено, что
виноградная улитка, а также некоторые
другие европейские виды, как Сераеа
nemoralis и Helicigona arbustorum,
поедают самые различные растения, в том
числе и такие, как крапива; из этого
можно видеть, что жгучие волоски этого
растения не составляют надежной защиты
от улиток. Другие виды, например
садовая улитка (Сераеа hortensis), более
разборчивы и брезгают многими
растениями. В связи с многоядностью большинства
видов гелицид некоторые из них, обычно
питаясь дикорастущей растительностью,
нападают попутно также и на
культурные растения, принося существенный вред.
Так, например, улитка Helicella intersecta,
живущая в Англии, Бельгии, Франции и
Северной Испании, часто сильно вредит
зерновым хлебам; Helicella obvia,
распространенная на юго-востоке Европы,
вредит посевам клевера, люцерны и
эспарцета, а улитка Trichia rufescens вредит в
Западной Европе садовым культурам. В
качестве вредителей сельского хозяйства из
наземных раковинных улиток известен
также один представитель другого
семейства (Enidae) — улитка зебрина (Zebri-
na detrita), которая в годы своего
массового размножения приносит
значительный вред в Западной Европе как зерновым
культурам, так и виноградникам.
Семейство этих улиток характеризуется более
или менее высокой веретеновидной
раковинкой и охватывает значительное число
разнообразных видов, распространенных в
Европе, Азии и Африке. Ряд видов
семейства живет и в пределах СССР, причем у
многих из них наблюдается резко
выраженная внутривидовая изменчивость.
Однако наиболее значительный вред
сельскому хозяйству в культурных
странах Старого и Нового Света приносят
наземные легочные улитки, лишенные
раковины,— так называемые голые слизни;
наиболее известные из них относятся к
двум широко распространенным
семействам арионид (Arionidae) и лимацид
(Limacidae). Первое имеет более широкий
ареал распространения, охватывающий
Азию, Европу, Африку и Северную
Америку, тогда как естественный ареал
распространения лимацид ограничивается
палеоарктическои областью, если не считать,
что некоторые виды были случайно
завезены человеком и в другие части света. Что
касается южноамериканских голых
слизней, то они принадлежат уже к третьему,
хотя и близкому, семейству —
Philomycidae. Особую группу составляют
голые слизни, живущие в Новой Зеландии
и на некоторых островах Океании. Они
имеют сложно устроенное легкое,
состоящее из системы трубочек, открывающихся
в общую полость, чего нет ни у каких
других легочных улиток. Эти своеобразные
улитки, составляющие особое семейство
(Athoracophoridae), сильно вредят
культурным растениям в Новой Зеландии.
Само название голые с/шзшг, данное всем
этим улиткам, показывает на характерный
их признак — отсутствие у них наружной
раковины. Действительно, только у двух
калифорнийских улиток (Binneya и Нет-
philia), относящихся к семейству
арионид, имеется не закрытая мантией
раковина, прикрывающая внутренностный
мешок, и у одной палеоарктическои улитки
(Parmacella) из семейства
Limacidae только в молодом возрасте
сохраняется открытая раковинка в виде
маленькой пластинки с крошечным
спиральным завитком. У всех остальных
представителей этой группы раковина нацело
обрастает мантией и становится
рудиментарной, а у некоторых форм даже
полностью исчезает (африканский слизень
Oopelta). Что касается образа жизни
голых слизней, то они имеют много общего.
Будучи лишены раковины, эти улитки
гораздо в большей степени, чем, например,
гелициды, ограничены в своем
распространении лишь теми местообитаниями, где
они могут найти для себя подходящие
условия влажности и защиту как от
сильных холодов, так и от особенно
губительного для них сухого зноя. Защиту
от последнего они находят под корой
деревьев, особенно гниющих пней, во мху
и под опавшей листвой, под задерненными
камнями на лугах и тому подобными
прикрытиями, а на рыхлых почвах
забираются довольно глубоко под поверхность
земли, где сохраняется некоторое
количество влаги. Соответственно степи и
85
пустыни служат природными
препятствиями для расселения этих улиток. Однако
завоеванные ими области суши достаточно
обширны. Они проникли в область тундры,
заселяют хвойные и лиственные леса,
причем в последних находят более
благоприятную для себя обстановку, поскольку
опавшая хвоя представляет менее
удобную для них среду передвижения. В
своем вертикальном распространении
некоторые виды идут вплоть до зоны вечных
снегов, как показали, например,
исследования в Тирольских Альпах, где в
зоне до 2600 м было обнаружено 9 видов
наземных легочных улиток, ив том числе
3 вида голых слизней (Limax maximus).
Из значительного числа форм,
составляющих эту группу, мы остановимся лишь
на некоторых широко распространенных
европейских видах, которые имеют
большое значение как вредители сельского
хозяйства. На первом месте среди них
должен быть поставлен обычный в СССР
и в Западной Европе полевой слизень
(Agriolimax agrestis). Эта небольшая
улитка обычно держится днем в сырых и
темных уголках под гниющими листьями,
под досками, камнями и тому подобными
прикрытиями, прячась от яркого
солнечного света, а поздно вечером выходит
на кормежку. В этом отношении ее образ
жизни сходен с поведением и других
наших слизней. Наблюдения показали, что
максимум слизней на поверхности почвы
и на растениях бывает обычно между
9 часами вечера и 2 часами ночи. Потом
число их падает: если поблизости нет
подходящих для них прикрытий, то они
забираются под комки почвы, уходя на
глубину до 10—25 см. В засушливые
лета, когда почва сильно просыхает и
влажность ее падает до 30—35%, слизни
теряют в весе, становятся вялыми и плохо
принимают пищу, а при дальнейшем
падении влажности до 10—15% уже гибнут.
Иногда во время засух слизни пытаются
спастись, окружая себя коконом,
состоящим из частиц почвы, скрепленных
высохшей слизью. Зато влажное, хотя и
прохладное, лето особенно благоприятно для
жизни и размножения полевого слизня.
После спаривания полевой слизень
откладывает до 500 яиц кучками по 20—30
штук в землю или мох, и из отложенных
летом яиц через 2—3 недели выходит молодь,
которая уже через полтора месяца
становится половозрелой. При благоприятных
условиях число слизней возрастает поэтому
уже к осени, а из яиц, отложенных осенью,
весной выходит новое поколение, которое
вместе с теми взрослыми особями,
которые пережили зиму (большинство
взрослых гибнет), нападает на всходы
зерновых культур или на огородные растения.
Наоборот, при жарком засушливом лете
число слизней сильно сокращается в
результате большого процента гибели и
взрослых особей, и отложенных ими яиц.
Эта тесная зависимость размножения
слизня от климатических колебаний ведет
к периодическому массовому появлению
этой улитки, и в годы ее массового
появления вред, приносимый ею, становится
особенно ощутительным. Массовое
появление слизней сопровождается большими
повреждениями посевов, иногда на
десятках тысяч гектаров, причем часть посевов
гибнет полностью. Кроме того, слизни
уничтожают огромное количество
огородных культур.
Обычный полевой слизень — самый
главный, но не единственный среди
улиток вредитель наших полей и огородов.
Ближайший его родич, Agriolimax lae-
vis, также поедает озимые хлеба, но вред,
причиняемый им, относительно невелик.
Гораздо больший вред в этом
отношении приносит крупный слизень из рода
арион (Arion bourguignati), который тоже
в массе поедает всходы зерновых культур
или выползает на кормежку на
клеверные поля. Этот слизень иногда тоже
встречается в огромных количествах, и тогда
в подходящих для него убежищах,
например в сыром березовом лесу, под
каждым опавшим листом можно находить
и взрослых и молодых слизней этого вида.
Указанными видами не исчерпывается
перечень голых слизней — вредителей
культурных растений: к числу вредных
форм относятся также европейский
большой слизень (Limax maximus) и другой,
тоже европейский вид, Arion empirico-
rum, так как тот и другой наносят
существенный вред всходам древесных пород;
вредит и ряд других видов.
86
Внешний вид повреждений, наносимых
улитками растениям, весьма характерен:
на листьях они выгрызают большие
неправильно-округлые дыры, чаще всего
в середине листа и гораздо реже по краю;
на плодах, которыми они лакомились,
остаются широкие выеденные ямки, по краям
которых под лупой можно видеть следы
радулы. Обычно также кучки
экскрементов и оставленная на растениях слизь
выдают с наступлением дня ночных обжор.
Вред, причиняемый растениям слизнями,
усугубляется еще тем, что они
оказываются разносчиками многих заболеваний
растений, вызываемых различными
грибками, споры которых в неповрежденном
виде проходят через кишечник улиток.
В качестве мер борьбы против слизней-
вредителей употребляются или
различные заграждения, которые делают вокруг
растений и через которые слизни не могут
переползти (эти средства малодейственны),
или же проводятся мероприятия по массо-
Рпс. 67. Хищные улитки: тестацелла (Testacella
maugei), поедающая дождевого червя (вверху),
и Euglandina rosea.
вому уничтожению слизней, для чего
употребляются различные едкие вещества,
например негашеная известь и некоторые
другие, которыми опыляют или
опрыскивают зараженные моллюсками участки.
Кроме описанных вредных форм, среди
слизней есть целый ряд видов, которые
совершенно безвредны и питаются
преимущественно водорослями и лишайниками,
например живущий на деревьях Limax
arborum или обычный в наших лесах Arion
subfuscus, который питается грибами.
Наиболее крупные слизни, живущие в
пределах СССР, встречаются на Кавказе;
например, гигантский черный слизень
(Eumilax niger) достигает в вытянутом
состоянии длины 15 см.
Наряду с растительноядными формами
среди наземных легочных улиток
насчитывают значительное число хищников.
К хищникам относится, например,
живущая в средиземноморской области улитка
тестацелла (Testacella). Главную пищу
этой улитки составляют дождевые черви,
за которыми она охотится в их норах,
а также мелкие формы гелицид. Другим
примером хищной наземной улитки может
служить олеацина, или гландина (Glan-
dina), живущая в средиземноморской
области и в Вест-Индии. Она питается
другими наземными улитками, поедая цикло-
стом, гелицид и других улиток. К
хищникам относятся и другие роды
семейства Oleacinidae, распространенные
в Вест-Индии, улитки рода ритида
(Rhytida), живущие в Южной Африке,
Австралии и Новой Зеландии, и ряд
других. Этот хищный способ питания
характерен не для одной какой-нибудь
систематической группы, а развился у
представителей разных далеко стоящих друг от
друга семейств.
Не лишены хищности и типичные
слизни, для которых характерна
рудиментарная раковина, скрытая в коже. Так,
хищниками являются все виды слизней
из семейства тригонохламид (Tri-
gonochlamididae), распространенного у нас
на Кавказе. Они в «погоне» за своими
жертвами — почвенными червями—уходят
довольно глубоко в землю.
Совсем недавно был найден новый вид
этих хищных тригонохламид, получив-
87
ший название «пещерный разбойник»
(Troglolestes). Такое название как нельзя
более подходит к нему, так как этот
слизень — обитатель центральных отделов
одной пещеры на Черноморском побережье
Кавказа и к тому же хищник, питающийся
мелкими почвенными малощетинковыми
червями энхитреидами. Жизнь в вечном
мраке наложила свой отпечаток на
внешний облик и внутреннее строение
животного. Его тело лишено пигмента —
снежно-белое, с просвечивающими сквозь кожу
розоватыми внутренними органами. Глаза
рудиментарны и впячены под покровы
кожи; они не воспринимают дневной свет,
несмотря на то что сохранили остаток слоя
пигментных клеток и стекловидное тело
с частью хрусталика. Интересно то, что
в других пещерах не только иных
районов страны, но и Кавказа подобные слизни
не найдены, так же как не обнаружены
они и в других залах этой пещеры.
Продолжительность жизни весьма
различна у наземных легочных улиток; тогда
как виноградная улитка живет до 7 лет,
мелкие виды гелицид достигают своего
предельного возраста уже в 4—5 лет;
лимаксы живут от 1 до 3 лет. Характерно
при этом, что продолжительность жизни
некоторых лимацид может быть
увеличена, как показали опыты Кюнкеля, если
держать животных в изоляции и не давать
им спариваться, так как лимацидам,
подобно виноградной улитке, свойственна
так называемая «любовная игра»,
предшествующая половому акту и,
по-видимому, сильно истощающая силы
животного. При этом у разных видов слизней
манипуляции, в которых выражается эта
игра, имеют свои характерные черты.
«Любовные стрелы», которые служат для
возбуждения партнера, имеются не у всех
пульмонат: нет их, например, у
представителей семейства патулид
(Patulidae), нет их и у хищных улиток,
хотя они относятся к разным семействам.
При половом акте обычно, как мы видели,
происходит взаимное оплодотворение
партнеров, но из этого правила имеются
исключения, как например у улиток янта-
рок, которые характеризуются отчетливым
протерандрическим гермафродитизмом,
т. е. более ранним созреванием мужских
половых продуктов. Поэтому при
копуляции оплодотворяется лишь та улитка,
у которой уже созрели женские половые
продукты, тогда как мужские к этому
времени обычно бывают уже
израсходованы при предшествовавшей копуляции.
Долгое время составляло загадку,
почему у наземных пульмонат, у которых
мужские и женские половые протоки
соединяются у своего выхода, не происходит
самооплодотворения. Оказалось, что
сперматозоиды достигают полной зрелости и
способны оплодотворять яйцевые клетки
только через некоторое время после того,
как они будут находиться в особой части
полового аппарата, так называемом опло-
дотворительном мешочке, в который они
попадают после копуляции.
В то время как многие легочные улитки
откладывают яйца под естественные
прикрытия, не принимая особых мер к их
охране, у других, как мы видели это
на примере виноградной улитки, кладка
яиц обеспечивается искусственно
подготовленным прикрытием. Еще более
сложное прикрытие для своего потомства делает
улитка Cochlostyla leucophthalma,
живущая в Ост-Индии на острове Сангир; она
откладывает икру между склееннымиполо-
винками согнутого листа; все
манипуляции, необходимые для приготовления
такого домика, улитка проделывает своей
ногой, а выделенная слизь играет здесь
роль цемента. Так же поступают и
некоторые другие виды древесных улиток,
живущие на Филиппинских островах.
У одной маленькой улитки из семейства
патулид икра приклеивается к
собственной раковине и вынашивается матерью.
Наконец, известно много (около 80
видов) живородящих легочных улиток —
африканские ахатиниды, хищные улитки
из семейства Paryphantidae,
живущие в Новой Зеландии и на островах
Океании, Pupil]a muscorum и другие.
Наземные улитки имеют много врагов
среди различных групп животного мира.
Из млекопитающих за ними охотятся
йасекомоядные, их поедают свиньи и мышп.
Среди пернатых очень много видов,
которые уничтожают улиток наряду с другой
пищей: таковы вороны, галки, сороки,
скворцы, чибисы, некоторые дрозды, утки,
88
Рис. 69. Улитка Cochlostyla leucophthalma и
кладка ее яиц между согнутыми и склеенными
половинками листа.
фазаны, голуби. Последние иногда поедают
улиток в довольно большом числе: в зобу
одного почтового голубя было обнаружено
67 штук гелицид, причем все относились
к виду Helicella ericetorum.
Охотятся за улитками многие
пресмыкающиеся и земноводные: в содержимом
желудка лягушки Rana temporaria
наземные легочные улитки составляют
иногда около 9% заглоченной пищи. Из
насекомых к врагам улиток, особенно
голых слизней, относятся крупные
жужелицы и их личинки, жуки-стафилиниды
и другие. Некоторые личинки жуков
вонзаются челюстями в голову гелицид и
дают себя увлечь в раковину, где начисто
съедают моллюска; после этого они
окукливаются в пустой раковине. Наконец,
личинки некоторых мух (Onesia cognata,
виды Sarcophaga и др.) паразитируют
на улитках. В перечне врагов улиток
надо иметь в виду также и хищных
улиток, поедающих растительноядные формы.
Среди некоторых видов развит также
и каннибализм, т. е. поедание более
слабых особей своего вида.
Выше уже говорилось об
использовании виноградной улитки как продукта
питания для человека. Такое же значение
имеют и некоторые лимациды, особенно
в Италии. Молодых виноградных улиток
употребляют во Франции также и в
качестве корма для домашней птицы.
Использование наземных легочных улиток для
приготовления из них лекарственных
средств теперь уже имеет лишь
историческое значение. В древнегреческой
медицине ценилась рудиментарная раковина
Limax. Уже в новое время, вплоть до
середины XIX в., в ходу была мазь из
слизней, а также косметические средства,
приготовленные из гелицид.
Многие стебельчатоглазые моллюски
являются промежуточными хозяевами ряда
опасных гельминтов птиц и
млекопитающих. Иногда можно заметить, как ярко
окрашены и вздуты щупальца у янтарок:
это происходит потому, что щупальца
набиты личинками лейкохлорид —
трематод птиц. Брадибены и макрохламисы —
промежуточные хозяева дикроцелиума и
эуритремы, пу пи л лиды и гелициды —
многих нематод, паразитирующих в легких
у копытных.
В фауне нашей страны наземные
легочные моллюски представлены примерно
семьюстами видами. Особенно много видов
стебельчатоглазых обитает на Кавказе
и в Средней Азии. При всем
разнообразии видов стебельчатоглазых и несходстве
их в различных районах выделяется все
же группа раковинных улиток, виды
которых распространены почти повсеместно.
Они обитают во влажных местах — на
сырых лугах, в лесной подстилке.
К ним относятся некоторые виды
семейства пупилл (Pupillidae)—
очень маленькие башневидные улиточки,
встречающиеся на дерновинах. Вместе
с ними обитают маленькие же дисковидные
беловатые валлонии (Vallonia) и
стекловидно-прозрачные, маленькие
пузыревидные вертигины (Vertiginidae). В зарослях
крапивы по берегам рек, на верхней
стороне водных растений и в более сухих
участках леса многочисленны виды с е-
м е й с т в а янтарок (Succineidae),
похожие по своей раковине на
пресноводных прудовиков. По краям болот и в очень
сырых лесах часто встречаются стройные
прозрачные кохликопы (Cochlicopa),
плоские просвечивающие зонитиды (Zonitidae),
дисковидные, ребристые дискусы
(Discus) и веретеновидно-удлиненные клау-
зилии (Clausiliidae) с раковиной, большей
частью завитой влево. В сыром валежнике
лиственных лесов часто можно найти едва
ли не самого мелкого из брюхоногих
моллюсков — «маленькую точку» (Punc-
tum pygmaeum) из семейства
Entodontidae.
В пойменных лесах Европейской
части страны и в лесах Сибири на зарослях
крапивы обильна кубаревидная крупная
кустарниковая улитка (Bradybaena fru-
89
ticum) — единственный представитель в
лесной зоне южноазиатского
семейства брадибен г (Bradybaenidae). Этот
вид рассматривается как реликт древней
теплолюбивой фауны, в третичное время
распространенной на территории
современной Сибири и Северной Европы. В более
сухих лесах обитают среднего размера
улитки с крепкой раковиной, устье
которой несет мощные складки; это перфорател-
лы (Perforatella), относящиеся к с е-
мепству гелицид (Helicidae). Вместе
с башневидными крупными видами с е-
мейства энид, также характерными
для сухих лесов, гелициды являются
представителями групп, особенно широко
распространенных в странах
Средиземноморья.
Кроме отмеченных только что
чрезвычайно широко распространенных по всей
нашей стране раковинных легочных
улиток отряда стебельчатоглазых, есть и
несколько широко распространенных
безраковинных слизней. Тут следует назвать
сетчатого слизня (Deroceras reticulatus)
и полевого слизня (D. agrestis), в больших
количествах встречающихся в лесах, на
огородах и полях, особенно осенью2.
На Кавказе стебельчатоглазые весьма
разнообразны, но особенно обильны виды
следующих семейств: гелицид, энид, клау-
зилиид и нескольких семейств слизней,
преимущественно лимацид. Большинство
видов двух первых семейств ведет
открытый образ жизни, благодаря чему их легко
заметить на деревьях, на скалах и в траве.
Клаузилии и слизни выбирают более
скрытые убежища — в пнях, в мхе,
покрывающем стволы деревьев, среди валежника.
Как уже упоминалось, в Закавказье
обитают хищные слизни тригонохламиды и
даудебардии, питающиеся дождевыми
червями. Указывалось также, что в одной из
пещер Кавказа найден весьма интересный
представитель хищных тригонохламид,
сильно измененный в связи с жизнью
в вечном мраке.
1 Ранее это семейство называлось фрутици-
колами (Fruticicolidae).
2 Оба эти вида и ряд близких к ним ранее
называли родовым названием Agriolimax.
Правильнее же называть их Deroceras.
Общий облик фауны стебельчатоглазых
Кавказа свидетельствует, что природные
условия этой части нашей страны близки
к таковым в странах, окружающих
Средиземное море.
В этом отношении к Кавказу близок
Крым. Однако во многих районах этого
полуострова обитают моллюски, не
требующие для своего существования
высокой влажности воздуха. Среди многих
других видов в Крыму весьма обильны
сухоустойчивые гелициды — гелицеллы,
с плоской раковиной, по поверхности
которой идут многочисленные темные полоски.
Характерны также зебрины —
представители семейства энид с
цилиндрической ярко-белой раковиной. В некоторых
районах Крыма можно видеть, как эти
цилиндрические зебрины и полосатые
гелицеллы гроздьями висят под
палящими лучами солнца на ветках кустарника.
Карпаты тоже населены
разнообразными стебел ьчатоглазыми, однако без
значительного преобладания видов какого-
нибудь семейства. Некоторые обитающие
там виды— общие с Балканами, Альпами
и Пиренеями.
Чрезвычайно интересна и богата сте-
бельчатоглазыми фауна гор Средней Азии,
в то время как среднеазиатские пустыни
лишены их. В западной части
среднеазиатских гор преобладают сухоустойчивые виды
из семейства гелицид, что сближает эту
часть нашей страны со Средиземноморьем.
В горах восточной части преобладают виды
тропического семейства ариофантид
(Ariophantidae) — макрохламисы (Масго-
chlamys), с зеленоватой, просвечивающей
раковиной. Крайне разнообразны виды
восточноазиатского семейства
брадибен, для которых характерна раковина
с яркими коричневыми полосами. Весьма
любопытна изменчивость одного из
немногочисленных в Средней Азии видов энид —
яминии потанина (Jaminia potaniniana).
Этот вид состоит из большого количества
так называемых «морф», т. е.
отличающихся по облику раковины групп особей.
Каждая морфа связана переходными
особями с другой морфой, и каждая из
них обитает в своеобразных условиях.
Последнее обстоятельство и вызывает
различие в облике раковины.
90
Для лесов Дальнего Востока наиболее
характерны виды семейства брадибен,
достигающие там особенно больших
размеров. Вместе с ними в лесах Приморья
обитает несколько видов из тех семейств
и родов, которые представлены на
островах Тихого океана и в восточных районах
Северной и Южной Америки
(семейства стробилопсид и филомицид—Stro-
bilopsidae, Philomycidae).
КЛАСС ЛОПАТОНОГИЕ (SCAPHOPODA)
Класс лопатоногих охватывает
небольшую группу морских моллюсков,
в строении которых причудливо
сочетаются, с одной стороны, некоторые черты,
свойственные брюхоногим, и, с другой
стороны, ряд признаков, характерных
для представителей класса
двустворчатых {ракушек). Но тогда как ракушки,
а особенно брюхоногие, обнаруживают
большое разнообразие форм, лопатоногие
приспособились исключительно к
роющему образу жизни, населяют только
морское дно и встречаются лишь в весьма
ограниченном числе видов.
Наиболее характерен для всего этого
класса и лучше других изучен так
называемый морской зуб, или денталиум
(Dentalium), раковинка которого по своей
внешней форме напоминает слоновый
бивень или клык крупного хищника.
Заостренный узкий конец ее соответствует
верхней, или задней, части тела животного,
вогнутая поверхность — спинной, а
выпуклая — брюшной стороне. Сама
раковина представляет собой цельную трубку,
открытую с обоих концов, и нацело
охватывает со всех сторон тело животного.
Только небольшой придаток края мантии
может высовываться наружу из заднего
отверстия, а через переднее отверстие
животное высовывает свою голову и при
передвижении сильно вытягивающуюся
ногу. Этот орган прекрасно приспособлен
к копанию в грунте: концевая часть ноги
имеет форму тупого конуса, от основания
которого отходит пара боковых лопастей.
Их наличие и послужило основанием для
наименования всего класса
—«лопатоногие», хотя у некоторых других
представителей этой группы нога, сохраняя
значение копательного органа, приобрела
удлиненно червеобразную форму и
оканчивается круглым зубчатым диском.
Предполагают, что отверстие на
суженном, заднем конце раковины служит для
засасывания воды в мантийную полость
и для выбрасывания ее из этой полости.
Мантийная полость расположена вдоль,
от заднего до переднего конца животного.
Жабры отсутствуют, и дыхание
осуществляется с помощью тонких складок
мантии на стенке мантийной полости.
Отсутствует и сердце с кровеносными
сосудами. Циркуляция же крови,
наполняющей полости тела, происходит благодаря
ритмическим сокращениям ноги.
Совершая передвижения по грунту,
денталиум подбирает пищу — чаще всего
фораминифер — с помощью нитевидных
выростов, расположенных около ноги.
«Денталиум,— пишет Лаказ-Дютье, —
обитает во множестве на северном
побережье Бретани. Нет надобности это
принимать на веру, в этом можно с
легкостью убедиться, попав на берег.
Необходимо только знать, где и как он живет,
иначе можно проискать его тщетно и
найти в лучшем случае лишь выброшенные
морем пустые раковины. Так как у меня
было живейшее желание изучить это
животное, я терпеливо продолжал поиски
там, где находил наибольшее количество
выброшенных раковин, руководствуясь
этим признаком и считая это верным
знаком того, что на данном участке берега
должны жить денталиумы. Однако, как
ни были долги и прилежны мои поиски,
мне ничего не удавалось обнаружить.
Однажды не совсем спокойное море
выбросило мне живого денталиума, и это
дало мне возможность наблюдать его
поведение и привычки. Как только я по-
91
местил его в банку, я увидел, что он
старается закопаться на ее дне. Тогда я
высадил его обратно в лужицу, оставленную
отливом между водорослями и морской
травой, и наблюдал, как он снова и снова
закапывался в песок. Теперь я узнал, что
для животного непривычно находиться
вне грунта, как я его случайно нашел, и
что впредь мне следует искать его в
прибрежном песке.
Животное, закапываясь, принимает
положение приблизительно под углом
в 45°, но направление и глубина
закапывания зависят от свойства грунта. Ден-
талиум не может жить в черноватом и часто
зловонном илистом грунте, который
обыкновенно находится под верхним слоем
прибрежного песка. Если слой песка
тонок, животное принимает более
горизонтальное положение, и тогда его обычно
очень трудно найти, потому что моллюск
целиком зарылся и ничем не
обнаруживает своего присутствия. Обычно же он
высовывал на 1—2 мм край раковины
над поверхностью крупного песка в
сосуде, куда я его помещал. Отсюда легко
понять, что денталиум часто
выбрасывается прибоем, так как даже
небольшое волнение скоро вымывает его из
песка. Но не следует думать, что вымытое
из грунта животное и оставленное
отливом на суше не закапывается быстро
опять. Напротив, оно тотчас же
проделывает это; оно вытягивает ногу, погружает
ее в песок и через несколько минут
выпрямляется и кажется уже вросшим в
песок. При содержании в неволе бывает
трудно отличить среди находящихся на
поверхности грунта денталиумов живых
от мертвых, но я использовал их
стремление к закапыванию, чтобы отбирать
живые экземпляры. Выложив кучу
денталиумов на влажный песок, я
убеждался, что те, которые не закапывались,
оказывались мертвыми или полуживыми.
Когда при отливе песок обнажается,
денталиум зарывается в него целиком
и исчезает с глаз наблюдателя. Я
хочу к этому добавить одно наблюдение,
которое относится ко всем
закапывающимся животным и имеет как
естественнонаучный, так и практический интерес.
Наиболее благоприятный момент для
сбора во время отлива обитающих в
прибрежном песке животных — это момент,
непосредственно предшествующий началу
следующего прилива. Почему это так?
Когда вода отступает, вначале в песке
остается еще достаточное количество ее
и обитатели песка еще не ощущают в ней
недостатка. Но по мере того как море
отступает с отливом все дальше,
оставшаяся на песке вода стекает с него и во
время наиболее низкого стояния воды,
перед началом прилива, песок начинает
просыхать, а животные, ощущая недо-
Рис. 70. Лопатоногие моллюски:
1 — сифоноденталиум (Siphonodentalium lafotense); 2 — морской зуб (Dentalium vulgaris); 3 — денталиум;
извлеченный из раковины; 4 — Siphonodentalium lafotense, извлеченный из раковины; 5 — личинка ден-
талиума.
92
статок воды, начинают менять места в
поисках более влажных участков. Этот
момент наиболее удобен для сбора
зарывшихся в грунте животных; к какому бы
классу они ни принадлежали, они все
выдают свое местопребывание бороздами
и движением грунта. Тогда очень легко
обнаруживается большое число
обитающих в песке ракушек; я находил также
самых больших и красивых сипункулусов,
вылезающих из грунта, как раз в тот
момент, когда волна прилива уже гнала
меня и принуждала прекратить поиски;
так же и денталиумы обнаруживают себя
в это время взрыванием песка. Сначала
они делают маленькую отчетливую
борозду, по которой их можно легко принять
за маленькую ракушку пандору, но
борозда, оставляемая денталиумом,
представляет собой искривленный след, потому
что одна сторона раковины более выгнута,
чем другая. Зная этот признак, уже
невозможно ошибиться. Итак, денталиум
сначала обнаруживает свое
местонахождение бороздой в песке; затем
показывается его легко опознаваемая раковина,
как будто вросшая в песок; наконец, он
появляется на поверхности весь и падает
на песок. Ознакомившись с этими
обстоятельствами, я уже мог без труда во время
одного большого отлива собирать штук
по 200 интересовавшего меня моллюска.
Таким образом, денталиум — обитатель
более глубоких мест,и рассчитывать
встретить его можно только при сильных
отливах. Он предпочитает зарываться в
довольно крупный песок; в очень мелком
же я его никогда не находил. Экземпляры
денталиума, долго сохранявшиеся в
неволе живыми, по-видимому, чувствовали
себя довольно хорошо в песке из мелких
обломков раковин; в мелком же песке,
который бывает снизу илист и затхл,
животные очень скоро погибают.
Вышеприведенные факты ясно показывают, что
денталиум не малоподвижен, как
многие ракушки, и что, наоборот, он часто
меняет свое местопребывание. Для
закапывания он пользуется обеими боковыми
лопастями ноги, которые при этом играют
роль якорных крючьев, так, что, когда
животное, выбросив вперед ногу,
подтягивается ею, его тело подвигается вперед».
Большинство лопатоногих
относительно мелкие животные, размером от
одного до нескольких сантиметров, но
среди обитателей тропических морей
встречаются и более крупные формы,
достигающие длины 7—8 см, как,
например, слоновый морской зуб (Dentalium
elephantium), живущий у Молуккских
островов и у берегов Филиппин. С другой
стороны, известны и очень мелкие
представители класса, не превышающие 3 мм.
Как уже отмечалось, лопатоногие бедны
формами: насчитывается всего около 150
ныне живущих видов этого класса. В
большинстве своем они населяют умеренные
глубины моря, но некоторые виды
встречаются и на глубинах 3—5 тыс. м.
Распространение этой группы весьма широко,
и ее представители встречаются почти
во всех морях, не исключая арктических,
но наибольшее разнообразие их форм
приурочено к тропическим морям.
В морях СССР известен другой вид,
относящийся к тому же р о д у Dentalium
entalis; он живет у нас в Баренцевом море,
образуя иногда значительные скопления
на песчаных грунтах с примесью ракушки.
Одна драга приносит иногда до 100
экземпляров этого животного. Денталиум
встречается преимущественно на глубинах 50—
110 ле, где он наиболее многочислен, но
вообще вертикальное распространение
его значительно шире, и его можно
находить в Кольском заливе на глубинах 10—
350 м. Длина раковины у наших форм
достигает 48 мм. Окраска ее напоминает
цвет слоновой кости, иногда раковина
бывает молочно-белого цвета.
Другой вид лопатоногих, живущий у нас
в Баренцевом море, принадлежит к более
мелким формам и относится к роду
сифоноденталиум (Siphonodentalium
vitreum). Этот моллюск, длина тела
которого не превышает 18 мм, встречается
на глубинах 40—350 м, но наиболее
многочислен на глубинах 100—200 м. Хотя он
встречается и на песчаных грунтах, но
главными местообитаниями его являются
участки морского дна с илистым грунтом.
Раковина его, тонкая и
стекловидно-прозрачная, с вырезами на заднем конце,
гораздо менее прочна, чем крепкая
толстостенная раковина денталиума.
93
КЛАСС МОНОПЛАКОФОРЫ (MONOPLACOPHORA)
До недавнего прошлого моноплако-
форы были известны лишь в ископаемом
состоянии. Их раковины встречались
в морских отложениях кембрия, силура
и девона. Только в 1952 г. датская
морская глубоководная экспедиция,
работавшая на исследовательском судне «Гала-
тея», неожиданно подняла на борт
корабля с глубины 3950 м современных
представителей этого класса — несколько
экземпляров первого современного вида—
неопилины галатеи (Neopilina galatheae).
Это интереснейшее животное названо
в честь корабля экспедиции.
Шапочковидная раковина неопилины
со склоненной вперед вершиной и
многочисленными парными отпечатками мест
прикрепления мускулов оказалась такой
же, как раковины ископаемых видов из
древнейших отложений.
Раковина неопилины покрывает
довольно высокий внутренностный
мешок с небольшой и не резко
обособленной головой и округлую, присоско-
видную ногу. Края внутренностного
мешка оторочены складкой— м а н т и е й.
Широкая мантийная борозда
со всех сторон окружает ногу. По бокам,
на дне мантийной борозды, лежат
своеобразные жабры, по пяти с каждой стороны,
рядом с ними — отверстия почек. Аналь-
Рис. 71. Глубоководный моллюск Neopilina
galatheae.
ное отверстие расположено сзади. Между
передним краем ноги и ртом лежат два
пучка щупальцевидных выростов, видимо
органов обоняния. Рот
окружен лопастями. Анатомические
исследования показали, что неопилина обладает
обширными отделами вторичной полости
тела, сердцем с двумя парами предсердий.
Глотка снабжена радулой, а в
кишечник впадают две симметричные доли
печени. Органы выделения
представлены несколькими парами почек.
Таким образом, в наружном и
внутреннем строении неопилины видна метамер-
ность многих органов. Эта метамерность
прослеживается также в расположении
многих пар мышц, идущих от ноги
к раковине и прикрепляющихся к ее
внутренней поверхности. Нервная система
построена по типу нервной системы
панцирных моллюсков.
Организация неопилины до сего времени
вызывает оживленную дискуссию. Одни
исследователи в метамерии неопилины
видят метамерию кольчатых червей,
другие считают эту метамерию вторичной.
Нет также единого мнения и в
отношении образа жизни этого вида. Лемхе,
впервые исследовавший неопилину,
считает, что этот моллюск лежит на дне,
а его раковина обращена к субстрату.
В кишечнике неопилины обнаружены
радиолярии. Наличие мощной радулы,
видимо, опровергает представления Лемхе
и заставляет предполагать, что это
животное питается путем соскребывания или
собирания пищи.
Вслед за открытием неопилины, которое
как бы вернуло к жизни группу животных,
казавшихся вымершими еще в древнейшие
геологические периоды, на больших
глубинах океана были обнаружены и
другие представители этого класса. Но все
они близки к неопилине, а общее
количество видов едва ли более пяти.
94
КЛАСС ДВУСТВОРЧАТЫЕ МОЛЛЮСКИ (BIVALVIA)
Класс двустворчатых моллюсков1 имеет,
как известно, четыре различных
названия, каждое из которых в какой-то мере
отражает главнейшие черты их строения.
Название «двустворчатые» (Bivalvia)
было впервые предложено Линнеем A758)
и является наиболее правильным,
поскольку оно применимо ко всем
представителям этого класса. Безголовыми (Асе-
phala) они были названы Линком A807),
что отразило факт редукции у них
головного отдела тела по мере развития у них
в процессе эволюции двух створок
раковины и смыкания этих створок вокруг
тела моллюска. Третье название —
«пластинчатожаберные» (Lamellibranchia),
предложенное Блэнвилем в 1814 г.,
может быть полностью применимо лишь
к одному отряду данного класса,
поскольку остальные отряды имеют жабры иного
строения; таким образом, это название
неприменимо, как и четвертое — «топоро-
ногие» (Pelecypoda, Гольдфус, 1880),
так как строение ноги у двустворчатых
моллюсков весьма разнообразно. Таким
образом, наиболее правильным и
всеобъемлющим является первое, линнеевское
наименование, которое и следует сохранить
применительно к этому классу.
Двустворчатые широко
распространены в Мировом океане и его краевых
морях, в реках и озерах и даже в прудах.
Общее количество видов двустворчатых
составляет около пятнадцати тысяч, и
большая их часть связана в своем
обитании с солеными морскими водами, и лишь
около одной пятой общего числа их
видов населяет пресные воды. На суше
двустворчатые моллюски не встречаются.
В морских водах они распространены
чрезвычайно широко, встречаясь во всех
климатических зонах, от теплых вод тро-
1 В разделе «Двустворчатые моллюски»
читатель найдет после наименования рода некоторых
моллюсков латинские названия, заключенные в
скобки. Это названия под родов, которые
приобрели широкое распространение в систематике
моллюсков.
пических морей до Арктики и
Антарктики и до холодных глубин океанической
абиссали. Они населяют практически все
глубины Мирового океана — от приливо-
отливной зоны (литорали) и прибрежных
мелководий вплоть до огромных глубин
впадин Мирового океана, где они были:
найдены на глубине почти 10,8 км.
Особенно изобилуют двустворчатые
моллюски в прибрежных мелководных
районах морей (до глубины 100—200 м), где
они по своей биомассе (весу на 1 м*
площади дна) и по плотности поселений
составляют часто большую часть всей
обитающей здесь донной фауны.
В настоящее время количество видов
глубоководных двустворчатых
моллюсков, обитающих в абиссали Мирового
океана (т. е. на глубине более 2000 м)>
судя по неполным еще данным, составляет
около 400 видов, но и это число следует
считать сильно преуменьшенным.
Велико разнообразие размеров,
строения и окраски раковин двустворчатых
моллюсков. Так, гигант среди моллюсков
вообще, обитатель тропических морей
тридакна может достигать 200 кг веса,
а длина ее мощной раковины 1,4 де.
Наряду с этим размеры ряда обычных
глубоководных моллюсков не превышают
2—3 мм.
Пестрыми узорами и яркими красками
блещут раковины и края мантии многих
тропических мелководных моллюсков,
незаметных среди зарослей водорослей,
белых, розоватых, лиловых и
желтоватых кораллов, ярких звезд и других
беспозвоночных. Разнообразные выросты,
шипы, чешуйки и ребра украшают створки
этих моллюсков, что помогает им
укрепляться в этих зарослях и противостоять
действию морских волн и течений.
Более скромную окраску имеют
раковины моллюсков, живущих на песчаных
или илистых грунтах умеренной или
арктической области.
Глубоководные формы имеют, как
правило, бледно окрашенную раковину, часто
очень тонкую и полупрозрачную.
95
Скромно окрашена в зеленоватые и
коричневатые тона большая часть
пресноводных форм.
Большое разнообразие строения тела
и раковины двустворчатых моллюсков
тесно связано с образом их жизни, местом
их обитания, с глубиной и с качеством
грунта, на котором они живут, или
прикрепляются, или в который закапываются.
Это сказывается прежде всего на
строении их раковины; на наличии на ней ребер
о тем или иным вооружением ее так
называемого замка, при помощи которого
скрепляются створки; на наличии или
отсутствии сифонов — особых выростов
мантии (двух мягких лопастей,
окружающих тело моллюска и выделяющих
-его раковину); на форме и размерах ноги
и наличии в ней особой железы,
выделяющей нити так называемого б и с с у с а,
при помощи которого они могут
прикрепляться к грунту, а также и на многом
другом. У форм, закапывающихся
более или менее глубоко в мягкий грунт,
сзади развиваются особые выросты
мантии — сифоны, через которые
засасывается и выводится вода, необходимая для
дыхания и питания моллюска,
погруженного в грунт. Таковы различные ма-
комы, теллины, йольдии и др.
Моллюски, живущие на поверхности грунта,
ползающие или слегка закапывающиеся
в него, имеют лишь зачаточные сифоны
или вовсе лишены их (например, сердце-
видки, венусы, астарты и др.).
Моллюски, обитающие в прибрежных
мелководных районах на более жестких песчаных
грунтах с примесью камней, имеют более
крепкие, толстые раковины (например,
арки, скафарки, морские гребешки —
пектены и хламисы), а различные
обитатели мягких илистых грунтов имеют более
тонкие раковины (батиарки, морские
гребешки пропеамуссиумы и др.).
Многие формы, живущие на прибрежных
мелководьях, прикрепляются нитями бис-
суса к камням, скалам, друг к другу,
нередко образуя целые гроздья, сростки
(многие мидии), или даже прирастают
своими створками к камням, или
срастаются друг с другом (устрицы).
Очень крепкой раковиной с острыми
зубчиками на ребрах обладают многие мол-
люски-камнеточцы; некоторые из них
выделяют особый кислый секрет,
растворяющий известь прибрежных скал и
камней, в которых они протачивают себе
норки. Мягкое червеобразное тело
древоточца тередо (Teredo) прикрыто лишь
спереди маленькой сложной раковиной,
Таблица 6. Двустворчатые моллюски морей СССР.
Отряд Гребенчатозубые (Taxodonta):
1 — ореховидна Ацила (Acila divaricata);
2 — леда обыкновенная (Nuculana pernula costigera);
3 — йольдия северная (Yoldia hyperborea);
За — то же, вид с брюшной стороны;
4 — йольдия широкая Yoldia (Megayoldia) thraciaeformis;
5 — арка вздутая Anadara (Scapharca) broughtonii;
в — батиарка холодноводная (Bathyarca glacialis);
7 — лимопсис (Limopsis vaginatus);
8 — пектункулюс полосатый (Glycymeris albolineatus).
Отряд Связочнозубые (Dysodonta):
9 — модиола обыкновенная (Modiolus modiolus);
10 — модиола адриатическая (M. adriaticus);
11 — модиола фазеолиновая М. (Modiolula) phaseolinus;
12 — митилястер (Mytilaster lineatus);
13 — мускулюс черный (Musculus nigra);
14 — гребешок черноморский Chlamys (Flexopecten) glabra pontica;
15 — гребешок гренландский (Propeamussium groenlandicum);
16 — гребешок глубоководный Рг. (Hyalopecten) frigidus;
17 — лима северная Lima (Limatula) ftyperborea;
18 — аномия гигантская Pododesmus macrochisma;
19 — аномия шиповатая Anomia (Heteranomia) squamula aculeata.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
20 — астарта крената Astarte (Astarte) crenata;
21 — астарта атлантическая рыжая А. (А.) sulcata;
22 — астарта северная A. (Tridonta) boreaiis;
23 — астарта овальная A. (Elliptica) elliptica.
служащей ему для сверления, а не для
защиты тела; проводя всю жизнь в
прогрызаемых в древесине ходах, эти
моллюски не нуждаются в защите своего слабого
длинного тела раковиной. Наконец,
огромное разнообразие различных тропических
моллюсков, постоянных обитателей
коралловых рифов, тесно связано с их жизнью
в мелководной, крайне разнообразной по
характеру субстрата зоне.
Крепкие известковые раковины
двустворчатых, как и других моллюсков,
хорошо сохраняются в отложениях (глинах
и песках) в течение целых геологических
эпох. Остатки их поселений чрезвычайно
ценны для геологов и палеонтологов. Эти
остатки могут прекрасно характеризовать
не только гидрологические и
климатические условия, при которых эти отложения
образовались (т. е. при которых жили
найденные здесь виды моллюсков), но и
возраст данной толщи отложений. Так,
скопления ископаемых раковин ныне
живущего в арктических морях холодно-
водного моллюска портландии
арктической (Portlandia arctica) в отложениях
севера Европы прекрасно указывают на то,
что эти районы были ранее заняты
холодными, слегка распресненными водами
мелководного так называемого Йольдиева
моря. Это море с холодноводной фауной,
где руководящую роль играла
арктическая портландия, было связано с
периодом похолодания в послеледниковое время
(примерно около 8—10 тыс. лет назад).
И наоборот, отложения теплого Литто-
ринового моря, образовавшегося позднее
C—5 тыс. лет назад), характеризуются
присутствием остатков совсем других,
тепловодных моллюсков, как например
исландской циприни (Cyprina islandica),
съедобной сердцевидки (Cerastoderma edu-
1е), цирфеи гребенчатой (Zirfaea crispata)
и др. Эти виды живут теперь лишь
в Северной Атлантике, в наиболее теплых
районах Баренцева и отчасти Белого
морей, тогда как в эпоху Литторинового
моря они продвигались дальше на север.
Представители класса двустворчатых
моллюсков впервые появляются в
отложениях в палеозое, т. е. в древнейших
отложениях нашей планеты, а именно
в верхнекембрийских слоях,
образование которых насчитывает около 450—500
млн. лет. Найденные здесь первые
двустворчатые моллюски относились к четырем
родам, из которых такие, как Ctenodonta
и Paleoneilo, имели гребенчатый замок
и внешне напоминали современных орехо-
видок (Nuculidae) и маллеций (Malleti-
idae) из отряда гребенчатозубых (Тахо-
donta). Наибольшего видового
разнообразия двустворчатые моллюски
достигали в меловое время, т. е. за 100—
130 млн. лет до нашего времени.
Таким образом, класс двустворчатых
моллюсков является одной из самых
древних групп донных беспозвоночных.
Таблица 7. Морские двустворчатые моллюски:
1— Yoldia (Cnesterium) scissurata;
2 — Astarte (Nicania) montagui striata;
3 — Clinocardium californiense;
4 — Lithophaga lithophaga;
5 —Astarte (Tridonta) rollandi;
6 —Chlamys farreri nipponensis;
7 — Trachycardium flavum;
8 — Cyclocardia paucicostata;
9 — Corculum cardissa;
10 — Hemicardia hemicardlum;
11 — Venus (Circomphalus) casina;
12 — Chamelea (Clausinella) fasciata;
13 — Ensis ensis;
14 — Propeamussium (Cyclopecten)
imbriferum major;
15 — Papyri dea spinosa;
16 — Glycymeris glycymeris;
17 — Mya truncata;
18 —Chlamys (Pallidum) tigerina.
7 Жизнь животных, т. 2
С давних времен многие двустворчатые
моллюски используются человеком, они
служили и служат предметом добычи. Их
раковины постоянно встречаются в так
называемых «кухонных кучах»
доисторического человека, жившего у берегов
морей, рек, озер. В раскопках стоянок
человека палеолита в Крыму неизменно
находят большое количество раковин
устриц, мидий, гребешков и других
моллюсков, которых промышляют и в
настоящее время. Двустворчатых моллюсков
добывают из-за их вкусного, очень
полезного и легко усвояемого организмом человека
мяса (как, например, устриц, мидий,
морских гребешков, петушков-тапесов и
венеруписов, мактр, песчаную ракушку,
сердцевидок, арок, морских черенков я си-
новакул, пресноводных перловиц, лампси-
лин, беззубок, корбикул и др.).
По калорийности они могут даже
превосходить мясо многих рыб, как морских,
так и пресноводных. Пищевая ценность
мяса моллюсков обусловливается также
большим содержанием в них витаминов
А, В, С, D и др., высоким содержанием
таких редких в обычной пище человека
минеральных веществ, как йод, железо,
цинк, медь и др. Последние, как известно,
входя в состав ряда ферментов, гормонов,
играют исключительно большую роль
в окислительном, углеводном и белковом
обмене веществ, в регуляции
гормональной деятельности. Мясо и раковины
моллюсков широко используют для
изготовления кормовой муки, идущей на откорм
домашней птицы, а также для
изготовления удобрительных туков.
В последние десятилетия, ввиду того
что естественные природные запасы
наиболее ценных съедобных моллюсков (даже
в морях) истощены, а спрос на них
продолжает увеличиваться, во многих
странах их стали переселять в новые районы,
акклиматизировать, а также и разводить
искусственно как в морских, так и в
пресных водах, на «фермах»—специально
подготовленных отмелях и в охраняемых от
хищников небольших бухтах,
искусственных водоемах. Успешно разводят и
культивируют не только морских моллюсков
(устриц, мидий, морских петушков, та-
песов), но и пресноводных (лампсилин).
В настоящее время значительно более
половины добываемых двустворчатых
моллюсков получают в результате их
искусственного разведения. Вылов моллюсков
в местах их естественного обитания в
водоемах и их искусственное разведение
стали теперь в ряде стран доходной и
важной частью пищевой индустрии.
Двустворчатых моллюсков теперь
нередко добывают на больших судах
специально сконструированными орудиями лова;
широко используется отлов моллюсков
водолазами. Моллюски поступают на
рынок не только в свежем и сушеном виде,
но особенно в мороженом; сильно
развилось также приготовление различных
консервов из моллюсков.
Добыча двустворчатых моллюсков в
последние десятилетия резко возросла. Если
перед началом мировой войны ежегодная
добыча их составляла около 5 млн. ц,
то уже в 1962 г. она возросла примерно
до 17 млн. ц и стала составлять около 50%
мировой добычи всех морских
беспозвоночных, или 4 % от общей мировой добычи
D26 млн. ц) всех продуктов моря (рыбы,
китов, беспозвоночных, водорослей).
Наибольшее количество (около 90%)
двустворчатых моллюсков добывается
в северном полушарии — в Тихом и в
Атлантическом океанах. Промысел
пресноводных двустворчатых моллюсков дает
лишь несколько процентов от общей их
мировой добычи. Особенно большое
значение имеет промысел двустворчатых
моллюсков в таких странах, как Япония,
США, Корея, Китай, Индонезия,
Филиппинские острова и другие острова
Тихого океана. Так, в Японии добывается
около 90 видов двустворчатых моллюсков,
из которых около двух десятков видов
имеют большое промысловое значение, а
10 видов разводятся искусственно. В
европейских странах промысел и
разведение двустворчатых моллюсков
наиболее развиты во Франции, Италии и др.
В СССР промысловое значение имеют
главным образом крупный приморский
гребешок (Pecten (Patinopecten) yessoensis),
а также различные мидии, белая
ракушка (Spisula sachalinensis), песчаная
ракушка (Mya (Arenomya) arenaria), петушки
(Tapes, Venerupis) и некоторые другие.
98
Двустворчатые моллюски издавна
добывались и ради их раковин, дающих
не только прекрасное сырье для
перламутровых изделий (многие пресноводные
перловицы и жемчужницы, морские
жемчужницы — пинктады, птерии и др.)»
но и ценнейший жемчуг. В начале
текущего столетия были найдены
промышленные способы более быстрым
искусственным путем получать жемчуг (находки
которого являются в природных условиях
довольно редкой случайностью), не
отличимый от жемчуга, образовавшегося
естественным путем. Фермы для содержания
морских жемчужниц и выращивания в
них жемчуга особенно успешно работают
в Японии. Так, уже в 1936 г. здесь было
выращено 140 тыс. раковин морских
жемчужниц и получено 26,5 тыс. жемчужин.
В некоторых странах, особенно в
тропической части Тихого океана, добываемые
раковины двустворчатых моллюсков
широко используются для получения извести.
Почти все двустворчатые моллюски, за
исключением крупных форм с крепкой,
толстой раковиной, служат излюбленной
пищей придонных рыб — бентофагов (т. е.
питающихся донными животными), в том
числе и многих промысловых рыб, как
морских, так и пресноводных: камбал,
некоторых тресковых (пикши),
осетровых, многих карповых (лещ, сазан),
зубаток, бычков и др. Некоторые рыбы
благодаря преобладанию в их пище
мелких моллюсков получили название «мол-
люскоедов», как, например, каспийская
вобла. Районы, где наряду с другими
донными животными (многощетинковыми
червями, офиурами и др.) наблюдается
массовое развитие мелких двустворчатых
моллюсков, служат местами откорма
различных придонных промысловых рыб.
Моллюсков охотно поедают многие
крупные десятиногие раки (омары, раки-
отшельники, крабы), морские звезды—
исконные враги двустворчатых
моллюсков. Промысловые устричные банки
периодически очищают от морских звезд
при помощи особых швабр, которые
волокут по дну небольшие суда.
Немалую роль играют двустворчатые
в питании промысловых камчатских
«крабов» (Paralithodes kamtschatica).
Каковы же основные черты строения
двустворчатых моллюсков? Чтобы легче
понять их устройство, представим себе
книгу в переплете, поставленную
корешком вверх. Обе половинки переплета
будут соответствовать правой и левой
створкам раковины, охватывающим тело
моллюска с боков. Корешок книги будет
аналогичен эластичной наружной связке
(лигаменту), соединяющей обе створки
на спинной стороне раковины и
одновременно растягивающей их. Первый и
последний листы книги соответствуют двум
лопастям мантии, охватывающим тело
с правой и левой сторон, а следующие два
листа книги спереди и сзади будут
аналогичны двум парам жабр с каждой стороны
тела. И наконец, между обеими парами
жабр внутри расположено само тело и нога
— обычно довольно крупный мускулистый
топоровидный или клиновидный орган,
Рис. 72. Беззубка (Anodonta).
А — внешний вид раковины: 1 — передний край; 2 —
брюшной край; з — задний край; 4 — спинной, или
замочный, край; 5 — макушка; 6 — наружная
связка. Б — вид тела моллюска с левой стороны при
удаленной раковине: 7 — передний
мускул-замыкатель; 8 — задний мускул-замыкатель; 9 — передний
ретрактор ноги; 10 — протрактор; 11 — задний рет-
рактор ноги; 12 — элеваторы ноги; 13 — нога; 14 —
правая складка мантии; 15 — вводной (дыхательный)
сифон; 16 — выводной сифон; 17 — клоакальная
камера; 18 — спинной мантийный канал; 19 — спинное
мантийное отверстие; 20 — ротовое отверстие; 21 —
ротовые лопасти; 22 — левая наружная полужабра;
23 — левая внутренняя полужабра; 24 — область
Кеберова органа; 25 — область перикардия; 26 —
линия, по которой обрезана левая мантийная складка.
7*
99
направленный вперед; у прикрепленных
или малоподвижных форм нога может
превращаться в небольшой вырост, и, наоборот,
у активно двигающихся видов (например,
у сердцевидок) нога становится сильной,
слегка ко ленчатоизогнутой,
приспособленной для продвижения в мягком
песчанистом грунте.
Расположение частей тела
двустворчатого моллюска станет яснее, если
рассмотреть вскрытого моллюска, например
беззубку, обычную в наших пресноводных
водоемах с илистым дном и медленно текущей
или стоячей водой. Наиболее
распространенной является обыкновенная беззубка
(Anodonta cygnea) — довольно крупный
моллюск из отряда настоящих
пластинчатожаберных (Eulamellibranchia).
При осмотре моллюска важно определить
передний и задний концы раковины.
Передний конец у беззубки легко узнать по
более округлой форме раковины и по
направленной вперед ноге; на заднем,
несколько более суженном конце между
створок видны короткие выросты мантии—
сифоны. Вдоль верхнего спинного
края, позади макушек, расположена
довольно крупная наружная связ-
к а , или л и г а м е н т,— упругий
эластичный тяж, при «сокращении» которого
створки раскрываются. Он состоит из
волокнистого рогового вещества, близкого
к хитину,— конхиолина:
образуется он из наружного покрова самой
раковины (периострака). «Работа» лига-
мента определяется взаимодействием
различно расположенных волокон
конхиолина, из которых он состоит. Когда
мускулы-замыкатели, сокращаясь,
стягивают створки раковины, то волокна в
нижней части лигамента сжимаются, а в
верхней — растягиваются, а при расслаблении
мускулов — наоборот; поэтому у мертвых
моллюсков створки раковины всегда
полуоткрыты. У двустворчатых моллюсков
связка может быть наружной или
внутренней или той и другой вместе.
У анодонты нет замковых зубов и
спинной край гладкий, отсюда и ее название—
беззубка (Anodonta). У большинства же
двустворчатых моллюсков для более
прочного соединения створок между собой под
Макушкой, изнутри, на спинном или
замочном краю раковины, имеются
разнообразные (по форме, числу и расположению)
выросты, так называемые зубы, каждый
из которых входит в соответствующее
углубление на противоположной створке.
Все это, вместе взятое, образует замок
раковины. Устройство замка, характер,
количество и расположение его зубов
служит важным систематическим
признаком у двустворчатых моллюсков и
характерно для семейства, рода и вида. Лига-
мент также является частью замкового
аппарата двустворчатых моллюсков,
поскольку служит для соединения створок
между собой.
Поверхность раковины у большинства
двустворчатых моллюсков, в том числе
и у беззубки, покрыта различно
окрашенным наружным слоем, или п е-
риостраком. Он легко
соскабливается ножом, и тогда под ним
обнажается белый фарфоровидный, или
призматический, известковый слой (ostra-
cum). На нем хорошо видны
концентрические линии — следы нарастания
раковины, идущие параллельно ее краям.
Внутренняя поверхность раковины у
многих моллюсков, в том числе и у беззубки,
выстлана перламутровым слоем
(hypostracum).
Периострак, состоящий из
конхиолина, устойчив к внешним воздействиям
(как механическим, так и химическим)
и служит, таким образом, хорошей
защитой для внутреннего известкового слоя
раковины. Особенно хорошо заметна
стойкость периострака к действию
растворенной в морской воде угольной кислоты.
Она может накапливаться у дна, в самых
придонных слоях, и в грунте, где живут
моллюски (благодаря разложению
органического вещества, отчасти и дыханию
водных организмов), и возрастать по мере
увеличения глубины и давления. Так,
в Карском море часто встречаются совсем
мягкие отмершие раковины астарт, йоль-
дий или портландий с растворенной
известковой частью раковины, от которой
остался один неповрежденный мягкий роговой
слой — периостракум.
Оба других слоя раковины состоят из
известковых призмочек или пластинок,
соединенных небольшим количеством
100
конхиолина. В среднем (фарфоровом)
слое они располагаются перпендикулярно
к поверхности раковины, а во
внутреннем (перламутровом) — параллельно ей;
благодаря такому расположению
получается интерференция света, которая и дает
блеск и радужные переливы перламутра.
Чем тоньше пластинки этого слоя, тем
красивее и ярче этот блеск. Наиболее
красивый перламутр бывает у тех
моллюсков, у которых толщина пластинок
перламутра в слое составляет 0,4—0,6 мк.
Раковина моллюска образуется в
результате секреторной работы его мантии:
вдоль ее края расположено большое
количество железистых клеток,
вырабатывающих различные слои раковины. Так,клетки
особой мантийной бороздки, идущей
вдоль всего края мантии, образуют
наружный конхиолиновый слой, эпителиальные
клетки так называемой краевой
складки дают призматический слой раковины,
а наружная поверхность мантии выделяет
перламутровый слой.
Раковина двустворчатых моллюсков,
состоящая более чем на 90% из СаС03,
содержит его в виде кальцита или
арагонита, находящихся в различных
соотношениях. У тропических моллюсков
раковина содержит больше арагонита, а также
довольно много стронция.
Кристаллографическое изучение состава раковин
ископаемых моллюсков позволяет теперь
судить о температуре тех морей, в которых
жили эти моллюски.
Кальций, откладывающийся в раковину
мантией, поступает в нее не только через
кровь, куда она попадает из пищи через
кишечник, но, как показали последние
эксперименты с радиоактивным кальцием,
клетки мантии могут и сами извлекать
кальций из воды.
Рост раковины происходит как путем
общего утолщения створок в результате
наслаивания все новых и новых
известковых пластинок на внутренней
поверхности створок, так и путем нарастания
всей раковины по ее свободному краю.
При наступлении неблагоприятных
условий (в зимний период, при ухудшении
питания и т. д.) рост раковины
замедляется или даже приостанавливается, что
хорошо заметно на поверхности раковины у
многих моллюсков, где в это время
образуются характерные сгущения линий,
имеющих вид концентрических полос, идущих
параллельно брюшному кр-аю раковины.
По зимним кольцам — сезонным
остановкам роста — иногда можно определить
приблизительно возраст моллюска.
Однако у некоторых видов такие кольца
неразличимы; у тропических форм, где нет
сезонных явлений, такие кольца обычно
вообще не образуются. У наших
пресноводных перловиц и беззубок такие
сезонные зимние перерывы роста имеются,
поэтому годовые кольца выражены обычно
хорошо.
Чтобы открыть створки раковины
беззубки, надо прежде всего перерезать два
имеющихся у нее внутри спереди и сзади
довольно сильных мускул а-з а м ы-
к а т е л я, стягивающих обе створки
в поперечном направлении и закрывающих
раковину. У живой беззубки легче
сломать ее тонкую раковину, чем открыть ее,
не перерезав этих мускулов.
Когда мускулы перерезаны, то створки
сами свободно раскрываются,
растягиваемые лигаментом, и можно видеть две
мягкие полупрозрачные розоватые или
желтоватые лопасти — мантию,
охватывающую тело с боков. Края мантии слегка
утолщены. Этим местом она прикрепляется
к раковине, на внутренней поверхности
створок которой образуется так
называемая мантийная линия. Мантия
беззубок, срастаясь сзади, образует два
коротких сифона, опушенных короткими
чувствительными выростами.
У моллюсков, закапывающихся в грунт,
образуются длинные сократимые сифоны;
места прикрепления втягивающих их
мышц образуют на внутренней поверхности
раковины отпечаток, так называемый
синус. Чем глубже синус, тем длиннее
сифоны у моллюсков, тем глубже они
могут закапываться в грунт.
На брюшной стороне из-под края
мантии вперед выдается довольно крупная
клиновидная нога, направленная острым
концом вперед. Нога у беззубки очень
подвижна (как и у многих других
моллюсков), и ее действие легко наблюдать
в аквариуме. Как только анодонта
успокоится, ее раковина приоткрывается, пока-
101
зываются розовато-желтые края мантии
и наружу высовывается кончик ноги.
Если все кругом спокойно, нога
высовывается еще дальше (у крупных анодонт
на 4—5 см), погружается в песок, и
моллюск начинает двигаться вперед или
закапываться в грунт передним концом, слегка
приподнимаясь на своей ноге. На
пройденном ею пути остается след в виде
неглубокой бороздки.
Большая подвижность ноги беззубки
обусловливается главным образом
сокращением имеющихся в ней различных
групп гладких мышц. Имеются парные
передний и задний мускулы: ретра-
к т о р ы, втягивающие ногу косо вверх,
протракторы, выдвигающие ногу
вперед, и группа более мелких м у с к у-
л о в-п однимателей (элеваторов)
ноги кверху. Все эти мышцы
прикрепляются к внутренней поверхности
створок раковины, где отпечатки мест их
прикрепления довольно хорошо заметны
(у ретракторов вдоль замочного края
раковины). Кроме того, в ноге имеется
еще целый ряд более мелких мышц, не
прикрепляющихся к створкам и
расположенных в ноге послойно и поперечно.
Если отвернуть мантийную лопасть
кверху, то откроется мантийная полость
беззубки, где расположены ее основные
органы: ротовые лопасти, коричневатые
жаберные листки (по два с каждой
стороны тела), нога, основание которой
находится между правой и левой
жабрами. Спереди, в
углублении между ногой и передним
мускулом-замыкателем,
помещается ротовое отверстие,
окруженное двумя парами
небольших треугольных сократимых
околоротовых лопастей.
Каждая жабра у анодонты состоит
из двух листков полужабр,
которые в свою очередь сложены
двумя пластинками —
восходящей и нисходящей.
Рис. 73. Схема внутреннего
строения беззубки.
А — продольный разрез через тело, вид
слева; Б — поперечный разрез и схема
кровеносной системы. 1 — ротовое
отверстие; 2 — пищевод; 3 — желудок; 4 —
средняя кишка; 5 — прямая кишка; 6—
анальное отверстие; 7 — печень; 8 —
протоки печени; 9 — мешочек
кристаллического стебелька; 10 — перикардий; и —
желудочек сердца; 12 — предсердие;
13 — передняя аорта; 14 — задняя аорта;
15 — почка; 16 — половая железа
(гонада); 17 — полужабра; J я — нога; 19 —
передний мускул-замыкатель; 20 —
задний мускул-замыкатель; 21— задний рет-
рактор ноги; 22 — мантийная складка;
23 — вводной сифон; 24 — выводной
сифон; 25 — клоакальная камера; 26 —
спинной мантийный канал; 27 —
спинное мантийное отверстие; 29 —
внутренностная и ножная вена; 30 — венозная
система почек; 31 — жаберные артерии;
32 — приносящие жаберные сосуды; 33 —
выносящие жаберные сосуды; 34 —
жаберные вены; 35 — мантийные вены; 36 —
сосуды от Кеберова органа; 37 —
плевральные нервные стволы; 38 — нижняя
выдыхательная камера мантийной
полости; 39 — створки раковины; 40 —
связка; 41 — верхняя выдыхательная часть
(камера) мантийной полости.
102
Каждая жаберная пластинка состоит
из рядов отдельных нитей (ф и л а м е н-
т о в), а каждая нить соответственно
образует восходящее и нисходящее колена.
У анодонты между соседними нитями
и между образуемыми ими коленами
имеются сосудистые соединения (мостики),
что характерно для всего отряда часто-
ящих пластинчатожаберных. Каждая
полужабра представляет собой, таким
образом, решетчатую, сложно
продырявленную двуслойную пластинку.
У представителей других отрядов
двустворчатых моллюсков жабры имеют иное
устройство (о чем будет сказано ниже).
Мантийная полость и расположенные
в ней жабры все время омываются током
воды, который создается главным образом
мерцанием мельчайших ресничек
эпителия, покрывающего поверхность мантии,
жабр, ротовых лопастей и стенок тела.
Вода поступает в мантийную полость
беззубки через нижний (дыхательный)
сифон, попадая сначала в ее большую,
нижнюю часть — дыхательную
камеру, затем процеживается через
щели в жабрах и уходит в верхнюю
часть мантийной полости —в ыдыха-
тельную камеру, откуда наконец
и выходит наружу через верхний
(выводной, или анальный) сифон. Засасывание
воды через входной сифон происходит
в результате разницы в гидростатическом
давлении между поджаберным и
наджаберным пространствами мантийной
полости и между нею и окружающей моллюска
водой; эта разница вызывается не только
работой ресничного эпителия, но также
и сокращением жаберных нитей и
мускулатуры мантии и сифонов. Когда ток воды
замедляется, входя в большую
«вдыхательную камеру» мантии, грубые и
крупные частицы выпадают из него и оседают
на поверхности мантии, а затем удаляются
из моллюска. Легко убедиться в наличии
интенсивных токов воды, поступающей
в мантийную полость, если положить
беззубку в неглубокий сосуд с водой так,
чтобы вода лишь слегка покрывала
раковину. Дав ей успокоиться, надо
подсыпать в воду вблизи заднего ее конца какой-
нибудь порошок, остающийся
взвешенным в воде (например, тушь, кармин,
Рис. 74. Основные типы строения жабр у
двустворчатых моллюсков: А — первичножаберные
(Protobranchia); Б — нитчатожаберные (Filibran-
chia); В — настоящие пластинчатожаберные
(Eulamellibranchia); Г — перегородчатожабер-
ные (Septibranchia).
I — ось ктенидия; 2 — наружный лепесток ктенидия;
3 — внутренний лепесток ктенидия; 4 — наружная
жаберная нить, состоящая из нисходящего и
восходящего колен; 5 — внутренняя жаберная нить и
ее нисходящее и восходящее колена; 6 — наружная
полужабра; 7 — внутренняя полужабра; 8 —
мускулистая перегородка (септа), пронизанная
кровеносными сосудами; 9 — отверстия в септе; Ю — нога;
II — туловище; 12 — мантийная полость; 13 —
мантия; 14 — раковина; 15 — связка (лигамент).
сухие тертые водоросли). Тогда можно
увидеть, как крупинки порошка уходят
через нижний (вводной) сифон внутрь
раковины и через некоторое время с
сильной струей воды выбрасываются наружу
через верхний (выводной) сифон. Время от
времени беззубка, часто без всякого
внешнего раздражения, с силой захлопывает
створки раковины и выбрасывает наружу
струи воды, обновляя сразу всю воду,
содержащуюся в мантийной полости.
Вскоре створки раковины опять
приоткрываются и нормальная медленная
циркуляция воды возобновляется.
Чтобы убедиться в интенсивности
работы ресничного эпителия, можно вырезать
кусочек мантии беззубки и положить
103
Рис. 75. Фильтрационные токи воды на жабрах
двустворчатых моллюсков (схема): А — ктени-
дии нукулы; Б — нитчатожаберные (мидия);
В — настоящие пластинчатожаберные
(перловица); Г — строение и ресничное вооружение
конца жаберной нити мидии.
1 — боковые (латеральные) ряды ресничек; 2 — ла-
теральнофронтальные (передние) ряды ресничек;
3 — фронтальные реснички; 4 — пищевой желобок
(стрелками показано направление токов воды и
пищевых частиц; обозначения стрелок те же, что на
рис. 76).
его внутренней поверхностью книзу на дно
сосуда. Благодаря продолжающейся еще
некоторое время работе ресничек этот
кусочек будет слегка перемещаться и даже
слегка ползти по наклонной плоскости.
Засасывание воды и ее циркуляция
внутри мантийной полости обеспечивают
беззубку не только необходимым для ее
дыхания кислородом, но и пищей. Как
и все двустворчатые моллюски, беззубка
лишена головы и целого ряда связанных
с ней органов—обособленной глотки,
слюнных желез, жестких образований для
пережевывания пищи (как, например,
хитиновых пластиночек — терки,
имеющейся у брюхоногих моллюсков).
Поэтому беззубка не может питаться
крупными организмами. Она и большинство
двустворчатых моллюсков (Eulamellibran-
chia и Filibranchia) являются активными
фильтраторами. Такие моллюски
питаются взвешенными в толще воды детритом
(мельчайшими остатками отмерших
растений и животных) и микропланктоном
(одноклеточными водорослями,
бактериями и очень мелкими животными). При
помощи сложного ресничного механизма
жабр и околоротовых лопастей моллюски
отфильтровывают их из воды, отделяя
несъедобную минеральную взвесь и
крупные для них пищевые частицы.
Жаберные нити моллюсков имеют
расположенные в определенных местах ряды
различных по своим размерам ресничек,
которые и могут отфильтровывать и
отсортировывать пищевые частицы,
обволакивать их слизью, а затем направлять
в пищевые бороздки, расположенные
вдоль брюшного края полужабр (в местах
перехода нисходящих жаберных колен
в восходящие) или у их основания.
Ряды довольно крупных боковых,
наиболее интенсивно работающих ресничек на
жаберных нитях производят фильтрацию
воды через узкие щели между жаберными
нитями и обеспечивают прохождение ее
из «вдыхательной» в «выдыхательную»
камеру мантийной полости. Особенно
крупные латеральнофронтальные реснички,
сидящие по бокам жаберных нитей,
отцеживают пищевые частички из воды или
улавливают их обильно выделяемой
слизью и проталкивают на наружную сторону
жаберных нитей. Здесь расположены
фронтальные реснички, которые собирают
пищевые частички и направляют их вниз,
к пищевому желобку. Пищевые частицы,
собирающиеся в пищевых желобках,
также обволакиваются слизью, образуют
здесь комочки, уплотняются и благодаря
работе ресничек желобка направляются
к ротовым лопастям. Ротовые лопасти
моллюсков — очень эффективный
сортирующий аппарат, освобождающий пищу от
несъедобных частиц. Они вооружены
многими чувствительными элементами— хемо-
и механорецепторами. Они имеют ряды
поперечных бороздок, вооруженных особо
длинными ресничками; самые мелкие
частицы, пригодные для питания, по ряду
таких бороздок направляются к ротовому
желобку (расположенному у основания
обеих лопастей), по которому они далее
направляются к ротовому отверстию, где
и заглатываются. По другим бороздкам (с
ресничками, работающими в
противоположном, чему предшествующих, направлении)
более крупные частички и слизистые
комочки, непригодные для питания,
скатываются вниз и падают на мантию. Сильные
104
реснички мантийных краев гонят эти
частицы обратно к основанию вводного
сифона; по мере продвижения туда эти
частицы склеиваются, уплотняются и в
виде так называемых
псевдофекалий выбрасываются наружу.
У двустворчатых моллюсков из группы
Protobranchia (ореховидок, лед, йоль-
дий, портландий и т. п.), имеющих
наиболее просто устроенные лепестковидные
жабры-к т е н и д и и, ротовые щупальца
очень большие, сократимые и снабжены
длинным желобчатым выростом. При
помощи его они собирают с поверхности
грунта мелкие пищевые частички —
детрит, которые затем переносятся
ресничками по желобку на пластинки ротовых
щупалец, где и подвергаются сортировке;
жабры-ктенидии служат главным образом
для создания токов воды. Работа
отфильтровывающего и сортирующего аппарата
двустворчатых моллюсков довольно
совершенна. Так, мидии могут
отфильтровывать частицы размером от 40 до 1,5—2 мк
(лучше всего — в 7—8 мк), полностью
извлекая их из воды. Они задерживают
одноклеточные водоросли и жгутиковых;
более тяжелые частицы минеральных
взвесей, даже размером в 4—5 мк, не
задерживаются мидиями. Из смеси водорослей
и пурпурных бактерий устрицы извлекают
только водоросли; они обычно
задерживают жгутиковых, водоросли и
органические частицы размером более 2—3 мк
и пропускают все частицы размером
в 1 мк и меньше.
Двустворчатые моллюски
профильтровывают очень большие объемы воды. Так,
устрица может профильтровать за час
около 10 л воды; мидия — до 2—5 л
(при более высокой температуре воды
больше, при более низкой — меньше);
съедобная сердцевидка при температуре воды
в 17—19,5°С — от 0,2 до 2,5 л, в среднем
0,5 л воды за час; мелкие морские
гребешки фильтруют со скоростью 1 л в час
на 1 г их веса, а старые — только 0,7 л.
Пищеварительная система беззубки, как
и у всех двустворчатых моллюсков,
состоит из короткого пищевода, более или
менее округлого желудка, средней и
задней кишки; в желудок открываются
протоки парной пищеварительной железы —
Рис. 76. Фильтрация воды двустворчатыми
моллюсками для их питания и дыхания (схема).
А — настоящие пластинчатожаберные; Б — пер-
вичножаберные из гребенчатозубых.
1 — раковина; 2 — передний мускул-эамыкатель; 3 —
задний мускул-замыкатель; 4 — мантия; 5 — нога;
6 — вводной сифон; 7 — выводной сифон; 8 — жабры;
9 — околоротовые щупальца; ю — придатки
околоротовых щупалец; 11 — псевдофекалии. Стрелки:
толстые — направление основных токов воды, входящих в
мантийную полость и выходящих из нее; тонкие —
фильтрационные и отсортировывающие токи воды на
жабрах и ротовых щупальцах; пунктирные — путь
несъедобных частиц и выносящих их токов воды вдоль
мантии.
печени, а на брюшной стороне выступает
конец так называемого
кристаллического стебелька. Отходящий
от желудка кишечник (средняя кишка)
в основании ноги, в массе половых желез
образует 1—2 оборота, затем переходит
на спинную сторону и, пронизывая
нижнюю стенку перикардиальной сумки,
проходит через желудочек сердца, выходит
за пределы перикардия через его
спинную часть, проходит над задним замыка-
тельным мускулом и оканчивается
анальным отверстием, открывающимся в
клоакальную камеру мантийной полости с ее
выводным сифоном. Часть кишечника,
идущая от перикардия до анального
отверстия, обычно называется прямой или
задней кишкой. Кишечный тракт
двустворчатых моллюсков не имеет
мышечных волокон, и передвижение пищи
в нем происходит благодаря работе
выстилающего его ресничного эпителия.
Удалению непереваренных остатков
способствует мышечная связка, окружающая
анальное отверстие.
105
Попав через короткий пищевод в
желудок, пищевые частицы благодаря
деятельности ресничного тока и желудочного
желобка отсортировываются на мелкие и
крупные. Крупные пищевые частицы
попадают в кишечник, а более мелкие
разносятся по складкам желудка и собираются
у выступающего конца кристаллического
стебелька. Его выступающий конец все
время вращается, что способствует
перемешиванию пищевых частиц и их
сортировке. Кристаллический стебелек
образуется в особом мешковидном органе и
представляет собой стекловидную палочку
студенистого вещества, состоящего из
белка типа глобулина с адсорбированными
в нем ферментами (амилазой и др.),
способными переваривать углеводы
(крахмал, гликоген). Попадая в слабокислую
среду кишечника, он начинает
растворяться и выделять адсорбированные в нем
ферменты — единственные выделяемые
двустворчатыми моллюсками в кишечный
тракт для внеклеточного переваривания
пищи. Мелкие пищевые частицы,
обработанные энзимами кристаллического
стебелька, поступают из желудка в выросты
печени. Она состоит из очень большого
числа удлиненных слепых трубочек —
дивертикул и не является
пищеварительной железой в обычном смысле;
в ней не вырабатываются и не выделяются
в кишечный тракт какие-либо
пищеварительные ферменты, и она является органом
внутриклеточного (а не внеклеточного)
переваривания пищи и всасывания.
Внутриклеточное переваривание у
двустворчатых моллюсков производится главным
образом фагоцитирующими блуждающими
клетками — амебоцитами. Они во
множестве встречаются не только в
дивертикулах печени, но и в желудке и в
средней кишке. Амебоциты обладают
различными ферментами и способны переваривать
не только углеводы, но и белки и жиры
и т. д. Блуждающие клетки могут
проходить через эпителий кишечного тракта
в его просвет и обратно возвращаться
в ткани. Заглатывают и переваривают
пищевые частицы также и клетки печени;
они также могут блуждать по просвету
дивертикул и обратно возвращаться в
стенки печени. Блуждающим клеткам и
принадлежит основная роль в переваривании
пищи у двустворчатых моллюсков.
При отмирании амебоцитов и клеток
печени их пищеварительные ферменты
могут поступать в просвет кишечного
тракта. Поэтому следы различных ферментов
(протеаз, липаз) находят в вытяжках из
печени и желудка двустворчатых
моллюсков.
Далеко не все попавшие в кишечный
тракт организмы перевариваются
двустворчатыми моллюсками. Нередко,
особенно при большом количестве пищи,
в фекальных массах моллюсков находят
живых диатомей (одноклеточные водоросли
с кремниевым скелетом), мелких
веслоногих рачков и т. п. Поэтому при
искусственном разведении двустворчатых моллюсков
(устриц) в выростных бассейнах стараются
поддерживать определенную (не
избыточную) концентрацию планктонных
водорослей, которыми они питаются.
Из сказанного видно, что пищеварение
у двустворчатых моллюсков очень
своеобразно. Они могут переваривать внекле-
точно лишь углеводы, а белковые и
жировые компоненты их пищи перевариваются
фагоцитирующими блуждающими
амебоцитами и клетками их «печени». Таким
образом, двустворчатые моллюски
являются очень специализированной группой
животных, приспособленной к питанию
детритом, одноклеточными водорослями и
бактериями.
Кровеносная система у беззубок, как
у всех двустворчатых моллюсков,
незамкнутая, и кровь — гемолимфа —
циркулирует не только по кровеносным
сосудам — артериям и венам, но
и в пространствах между органами, и в
соединительной ткани по целой системе
лакун и синусов, не имеющих собственных
стенок. Артериальная кровь течет
главным образом по сосудам, а венозная
система имеет преимущественно лакунарный
характер. Кровь прогоняется через всю
систему благодаря сокращению сердца,
а также мускулатуры тела. Сердце у
двустворчатых моллюсков (у анодонты)
состоит из желудочка и двух предсердий
и лежит в перикардиальной полости, или
околосердечной сумке, расположенной на
спинной стороне тела. Перикардий пред-
106
ставляет собой вытянутый тонкостенный
мешочек, наполненный гемолимфой, и
является у двустворчатых моллюсков
частью сильно уменьшенной в объеме
их вторичной полости тела. Желудочек
обладает мощными мышечными стенками
и имеет вид мешка грушевидной формы,
своим широким концом обращенного
назад. Предсердия очень тонкостенны,
полупрозрачны и чаще всего имеют вид
вытянутых треугольников, вершины которых
открываются в желудочек; при входе
в последний они снабжены небольшими
лолулунными складками — клапанами,
пропускающими кровь только из
предсердия в желудочек.
У беззубки, как у большинства
двустворчатых, желудочек пронизан проходящей
через него задней кишкой, но его полость
лри этом вполне замкнута и отделена от
кишки своей стенкой. Из желудочка кровь
расходится по телу: к заднему концу —
через заднюю аорту, которая
разделяется на две артерии, питающие сосуды
задней части мантии и заднего замыкатель-
ного мускула; к переднему концу —
через переднюю аортуи
отходящие от нее артерии к ноге, к
внутренностям и к передней части мантии. Из
артериальных сосудов кровь изливается
в не заполненные тканями промежутки,
и через систему лакун кровь, ставшая
венозной, собирается по синусам и венам в
большой продольный венозный синус,
лежащий между выделительными органами.
Пройдя отсюда через венозную систему
почек, она вливается в приносящие парные
жаберные артерии, проходящие у
основания каждой жабры. Из них венозная кровь
по приносящим жаберным сосудам
нисходящих жаберных пластинок растекается
по жаберным нитям и их сосудистым
перемычкам. Окисленная в жабрах
артериальная кровь, насыщенная кислородом,
по выносящим сосудам восходящих
жаберных пластинок вливается в парные (с
каждой стороны моллюска) жаберные вены,
откуда поступает в предсердия. В
предсердия поступает и та часть крови,
которая, минуя жабры и почки, окислилась
в сосудах мантийных складок и по
мантийным венам поступила в наружные жабер-
лые вены. Мантия с ее сильно
разветвленными кровеносными сосудами играет
у двустворчатых моллюсков очень
большую роль в дыхании и обогащении крови
кислородом.
То, что у большинства двустворчатых
моллюсков желудочек сердца оказывается
пронизанным прямой кишкой, находит
свое объяснение в особенностях их
эмбрионального развития и в эволюции всей
этой группы. У ряда низших
представителей двустворчатых имеется не только два
предсердия, но и два раздельных
желудочка, лежащих по бокам кишки (у арок);
у других — непарный желудочек лежит
над кишкой (ореховидки, аномии, лимы),
у третьих — он лежит книзу от кишки
(устрицы, жемчужницы и др.). Все это
указывает на то, что расположение кишки
и сердца по отношению друг к другу
претерпевало большие изменения в
процессе эволюции двустворчатых моллюсков,
и на то, что первоначально у них было
два раздельных желудочка, которые затем
слились вместе. Частота сокращения
сердца у двустворчатых моллюсков,
являющихся в общем малоподвижными
организмами, невелика, обычно не более 15—30
раз в минуту, в то время как у таких
подвижных и активных моллюсков, как
головоногие, сердце сокращается 40—80
раз в минуту. Все отделы сердца
двустворчатых моллюсков могут сокращаться
автономно.
У двустворчатых моллюсков, как и
вообще у беспозвоночных с незамкнутой
кровеносной системой, кровяное
давление очень низкое и сильно изменчиво.
Кровь—гемолимфа—двустворчатых
моллюсков играет огромную роль в их жизни,
обмене веществ. Она выполняет целый
ряд функций: обеспечивает внутренние
органы и ткани кислородом и
питательными веществами, уносит продукты их
жизнедеятельности (углекислоту,
продукты азотистого обмена и т. п.), создает
и поддерживает постоянство их
внутренней среды (ионного состава,
осмотического давления). Наконец, играет очень
большую роль в создании
гидравлического механизма давления, необходимого
тургора (напряжения), а также при
движении моллюсков. Изучение циркуляции
крови в теле двустворчатых моллюсков
107
объяснило явление разбухания ноги, что
наблюдается при движении и закапывании
животного. Оно происходит благодаря
наполнению ее кровью, которая и
придает ноге необходимую упругость, создает
необходимый тургор. Когда нога
вытягивается и ножные мышцы расслабляются,
кровь поступает по артерии в ногу, а при
ее сокращении уходит обратно в тело. Так,
у морского черенка, который может очень
быстро закапываться, нога сначала
погружается в грунт и кровь быстро втекает
в нее, расширяя конец ноги в виде диска;
последний служит якорем, когда ножные
мышцы, сокращаясь, тянут моллюска
вниз. При подъеме моллюска из грунта
на поверхность ножные мышцы
расслабляются, а конец ноги снова расширяется,
заполняясь кровью; держась на таком
«якоре», нога вытягивается, так как
часть крови поступает в верхнюю часть
ноги и подталкивает моллюска вверх.
Приток, нагнетание необходимого для
набухания ноги количества крови и ее
отток регулируются так называемым
кеберовым органом,
играющим при этом как бы роль клапана.
В противоположность животным с
замкнутой кровеносной системой
двустворчатые моллюски, как и все беспозвоночные,
обладающие незамкнутой кровеносной
системой, имеют довольно значительный
объем крови — гемолимфы. У моллюсков
(кроме головоногих) он составляет 40—60%
(объемных процентов) к весу их тела без
раковины. У пресноводной жемчужницы
(Margaritifera) и у мидии (Mytilus califor-
nianus) на 100 г веса тела объем крови
составляет около 50 мл.
В крови двустворчатых моллюсков
имеется много форменных элементов,
главным образом различных форм амебоцитов
(лейкоцитов). Количество их колеблется
у разных видов от 6000 до 40 000 в 1 мм3.
У двустворчатых моллюсков имеются и
эритроциты; иногда их даже может быть
больше, чем некоторых форм лейкоцитов.
Гемоглобин обнаружен у довольно
многих видов (арки, морские черенки, тел-
лины, пектункулюсы, астарты и др.).
Важная для газообмена (для
снабжения органов и тканей кислородом)
способность крови насыщаться кислородом
у двустворчатых моллюсков очень мала
и составляет 1—5% к объему их крови.
Так, 100 мл крови беззубки способны
поглотить всего лишь 0,7 мл кислорода,
а у мидий — 0,3 мл. Беззубка потребляет
0,002 мл 02 на 1 г своего веса за один час
(при 10°С); устрицы соответственно —
0,006 мл 02 (при 20°С); мидии—0,055 мл 02.
Несколько больше обычно потребляют
виды более подвижные, как, например,
морские гребешки Pecten grandis,
потребляющие 0,07 мл 02 на 1 г своего веса за 1 час
(или 70 см3 02 на 1 кг веса). Мелкие формы
также нередко потребляют больше
кислорода, чем крупные. Например, шаровка
роговая при оптимальной для нее
температуре воды A8°С) потребляет 0,05 мг 02
на 1 г веса за час, но при снижении
температуры воды до 0,5°С потребление
кислорода у нее почти прекращается.
В потреблении кислорода, т. е. в
интенсивности обмена, у многих двустворчатых
моллюсков наблюдаются сезонные
колебания; так, мидии в летнее, наиболее
активное время их жизни потребляют
примерно в два раза больше кислорода,
чем в зимнее, холодное время года.
Многие двустворчатые моллюски могут
довольно долгое время жить при очень
малом содержании кислорода в воде или
даже при его отсутствии. Так, песчаная
ракушка (Муа arenaria) может жить в
бескислородных условиях при14сС до 8 дней, а
при 0° даже несколько недель;
виргинская устрица также выносит такие
условия в течение недели и более. Обмен
веществ в такие периоды анаэробиоза резко
снижается, но моллюски могут при этом
получать необходимый для их жизни
кислород путем интрамолекулярного
дыхания — гликолитического расщепления их
запасных веществ (углеводов и жиров)
по типу брожения. Такая способность ко
временному (факультативному)
анаэробиозу (аноксибиозу) особенно характерна
и необходима для видов, живущих на
литорали (как, например, для песчаной
ракушки, мидий, балтийской макомы,
съедобной сердцевидки). Во время отлива они
закрывают свои раковины, небольшое
количество кислорода в их мантийной
полости довольно быстро исчезает, и они
начинают жить за счет процессов аноксибио-
108
за. Во время прилива они раскрывают свои
раковины, непрерывно фильтруют воду
и дышат кислородом, растворенным в
воде; вначале у них резко (в несколько раз)
повышается интенсивность фильтрации
и потребления кислорода, а затем через
некоторое время приходит в норму,
характерную для их жизни в воде.
Органами выделения у
двустворчатых моллюсков являются почки,
а также, но в меньшей степени так
называемый кеберов орган;
последний представляет собой железистое
утолщение передней части и передне-боковых
стенок перикардиальной сумки.
Почки, или боянусовы
органы, открываются своими
внутренними концами в перикардий, а
наружными — в мантийную полость. Почки у
беззубки имеют вид двух темно-зеленых
изогнутых трубчатых мешочков; один конец
имеет железистые стенки и представляет
собственно выделительную часть почки,
другой имеет вид пузырька, где
накапливаются продукты обмена веществ,
подлежащие удалению из тела.
У двустворчатых моллюсков нет такой
концентрации частей центральной нервной
системы (нервных узелков, или ганглиев),
как у брюхоногих моллюсков. У беззубки,
например, имеется одна пара ганглиев над
ртом, немного позади него, другая пара —
глубоко в ноге и третья — позади заднего
замыкательного мускула. Между первой
и второй парой ганглиев, а также между
первой и третьей проходит по паре
нервных стволов, а каждая пара узелков
соединена между собой поперечными
перемычками (комиссурами).
Органы чувств у двустворчатых
по сравнению с другими классами
моллюсков развиты довольно слабо. Однако эти
органы у них довольно разнообразны по
своему строению и рассеяны в различных
частях тела: вдоль наружных краев
мантии, на концах сифонов, на первых
жаберных нитях, около ротового отверстия
на околоротовых щупальцах, на краях
заднего замыкательного мускула, в
выдыхательной камере, около задней кишки
и т. д. Эти органы чувств представляют
собой как довольно сложные
образования — глаза, или фоторецепторы, органы
Рис. 77. Свободноживущие личиночные стадии
двустворчатых моллюсков: Л — личинка-трохо-
фора, вид с брюшной стороны; Б —
личинка-парусник (велигер), вид спереди; В — осевшая
ползающая личинка дрейссены, с ямкой на
поверхности ноги; Г — осевшая ползающая личинка
устрицы; Д — молодые устрицы, приросшие
раковинкой к субстрату.
1 — теменная пластинка с теменным пучком ресничек;
2 — прототрох (венчик ресничек); 3 — рот; 4 — задний
пучок ресничек; 5 — парус (велюм); 6 — раковина;
7 — вход в мантийную полость; 8 —
мускулы-ретракторы; 9 — кишечник; ю — печень; 11 — мешочек
кристаллического стебелька; 12 — анальное отверстие;
13 — ножной ганглий; 14 — нога.
равновесия — статоцисты, или статорецеп-
торы, так и более простые — осфрадии,
различные чувствительные выросты,
а иногда и просто скопления
пигментированных чувствительных клеток.
Фоторецепторы у двустворчатых
моллюсков могут быть устроены очень
различно: от простых эпителиальных
пигментных (оптических органелл) до довольно
сложных глаз с хрусталиком и
сетчаткой. Такие глаза могут быть очень
многочисленными, особенно у свободноживущих
109
форм, как например мантийные глаза
у гребешков, у которых их
насчитывается иногда до 100 на обоих краях мантии.
Различно устроенные глаза и глазки
могут располагаться у двустворчатых
также на первых жаберных нитях (жаберные
глаза у арок, аномии), на коротких
выростах вокруг отверстий сифонов (у
некоторых сердцевидок и др.).
У многих двустворчатых имеются так
называемые оптические органеллы,
сферические или удлиненные, концентрирующие
свет на особом внутриклеточном иннерви-
рованном образовании (ретинелле). Такие
фоторецепторы разбросаны на концах
сифонов и в других частях тела
моллюсков.
Органы равновесия у
двустворчатых представляют собой
пузыревидное впячивание эпителия, хорошо иннер-
вируемое, выстланное изнутри ресничным
эпителием, замкнутое (статоцист)
или открытое (статокрипт). Они
содержат внутри твердое минеральное
зерно (с т а т о л и т) или мелкие песчинки
(с т а т о к о н и и). Обычно, например
у беззубки, статоцисты расположены не-
далеко от ножного ганглия или на
спинной стороне моллюска. Органы равновесия
хорошо развиты у свободноживущих
форм, например у морских гребешков.
Осфрадии представляют собой обычно
очень мелкие парные пигментированные
хорошо иннервируемые группы
чувствительных клеток. Они могут располагаться
в различных местах —- на ноге, в области
жабр, задней кишки и т. д. Их роль еще
недостаточно ясна: являются ли они хе-
морецепторами или органами осязания.
Анодонты, как и большинство
двустворчатых, раздельнополы, но в
условиях водоемов со стоячей водой могут
встречаться отдельные гермафродитные
особи или даже целые их колонии. При
этом во избежание самооплодотворения
сначала продуцируются мужские
половые продукты — сперматозоиды, а затем
женские — яйца. Парные, сильно
расчлененные половые железы (гонады) у
двустворчатых (в том числе и у анодонты)
залегают в спинной части ноги, где их
окружает петля кишечника и выросты
печени; выводные протоки гонад
открываются в мантийную полость рядом с
отверстиями выделительной системы.
Только у некоторых самых примитивных
двустворчатых моллюсков протоки половых
желез открываются общим отверстием
с выделительной порой. У некоторых
пресноводных двустворчатых половой
диморфизм выражен настолько резко, что
самцов и самок одного и того же вида
иногда относят к разным видам.
Развитие молоди у двустворчатых
моллюсков происходит разнообразно. Почти
все морские формы, обитающие на
мелководьях, откладывают яйца прямо в воду,
где и происходит оплодотворение, или
же оно происходит в мантийной полости
материнской особи. Яйца свободно
плавают в воде, реже склеиваются или
прикрепляются к раковинам, водорослям.
Исключение представляют живородящие
(точнее, личинкородящие) формы, как
некоторые устрицы, арки и др.
Оплодотворенные яйца двустворчатых
моллюсков, пройдя стадию дробления
спирального типа, образуют трохофорообраз-
ную личинку, сходную с личинкой много-
щетинковых червей (полихет). Однако
Рис. 78. Развитие
анодонты: А — разрез через
жабру беззубки,
наполненную зародышами;
Б — личинка глохидий с
полураскрытыми
створками, вид спереди; В —
глохидий, паразитирующие в тканях
плавника рыбы.
1 — раковина; 2 — краевой зуб с зубчиками по
краю; з — замыкательная клетка краевого зуба;
4 — мускулы-замыкатели; 5 — пучки
чувствительных щетинок; 6 — личиночная биссусная железа.
110
в процессе эмбрионального развития
двустворчатых моллюсков почти отсутствует
процесс сегментации, столь характерный
для развития личиночных стадий
кольчатых червей. У личинки двустворчатых
моллюсков имеется зачаток ноги и
первичной раковины (продиссоконха), которая
первоначально закладывается в виде
одной пластинки, расположенной на
спинной стороне личинки. После ряда
изменений трохофора, у которой возникает
парус (velum) — ресничный
теменной диск, двустворчатая раковинка и
зачатки других органов, превращаются в
велигер. Наличие свободноплавающей
личинки (велигер) представляет собой очень
важную жизненную стадию
моллюсков, так как обеспечивает им
возможности широкого расселения, поскольку
взрослые двустворчатые моллюски ведут
обычно малоподвижный или даже
прикрепленный образ жизни. Вместе с тем
эта личиночная стадия жизни, как и у
других беспозвоночных, является самой
чувствительной к неблагоприятным внешним
условиям, и лишь высокая плодовитость
двустворчатых моллюсков обеспечивает
сохранение вида и его расселение.
У некоторых видов двустворчатых эта
пелагическая личиночная стадия
утрачена и яйца развиваются внутри
мантийной полости материнского организма (как
у многих пресноводных форм), так что
из материнской раковины выходят уже
почти готовые молодые особи. У
некоторых пресноводных же форм цикл развития
осложняется своеобразным паразитизмом
личинок на теле рыб. Этот период следует
за периодом первоначального развития
на жабрах материнской особи, куда и
откладываются их яйца, при этом у
некоторых видов на жабрах образуются
специальные выводковые камеры (марзупиум).
Примером может служить развитие
личинки анодонты —глохидия,
протекающее, как и у всех Unionidae, в
наружных жабрах материнской особи. В
дальнейшем он становится паразитом рыб,
прикрепляясь к их коже или к жабрам
биссусом или зубцами раковинки.
Погружаясь в ткани рыбы, глохидий
окружается цистой, питается за счет соков тела
хозяина. Этот паразитический период
продолжается 1—2 месяца, после чего
моллюски выходят в воду и, опускаясь
на дно, переходят к свободному образу
жизни.
У ряда морских холодноводных и,
видимо, у многих глубоководных
океанических видов двустворчатых моллюсков
развитие может происходить с очень
укороченной стадией плавающей личинки
или совсем без нее. В последнем
случае образуются немногочисленные
крупные яйца, снабженные большим
количеством питательного вещества. Это
позволяет им развиваться вне
зависимости от наличия пищи в окружающей воде,
что особенно важно для глубоководных
форм, где количество пищи для молоди
у дна очень ограниченно.
Основываясь на строении раковины,
замка, жабр, на расположении и
количестве мускулов-замыкателей, связки и пр.,
различают отряды: Гребенчатозубые,
Связочнозубые, Настоящие
пластинчатожаберные и Перегородчатожаберные.
ОТРЯД ГРЕБЕНЧАТОЗУБЫЕ
(TAXODONTA)
К этому отряду относятся
двустворчатые моллюски, имеющие ряд
примитивных черт в своем строении, например
в первую очередь замок «таксодонтного
типа», т. е. состоящий из ряда мелких
зубчиков, расположенных вдоль спинного
(верхнего) края раковины. Особенно ясно
выражены такие признаки у видов из
надсемейства нукулид(Nuculacea),
являющихся наиболее древними среди
прочих гребенчатозубых. У нукулид
имеются наиболее просто устроенные жабры
типа ктенидиев, состоящие из ряда
мелких округлых листочков, по одному ряду
с каждой стороны тела, сидящие на
общей оси (Protobranchia); нога у них со
складывающейся подошвой, а полость
гонад соединяется с почками.
Виды второго большого
надсемейства арок (Агсасеа), хотя и имеют
существенные отличия от нукулид, но также
сохраняют ряд примитивных черт в
строении гребенчатого замка и жабр. Их жаб-
111
ры состоят из рядов тонких, свободных,
не сросшихся между собой нитей (Filib-
ranchia). Нога без подошвы, маленькая
и нередко образует биссусные нити.
Моллюски, относящиеся к отряду гре-
бенчатозубых, очень широко
распространены в Мировом океане и его
окраинных морях; они встречаются во всех
наших морях, кроме Азовского и
Каспийского озера-моря, обитают на самых
разнообразных глубинах, почти от уреза воды
вплоть до ультраабиссальных глубин
океанических впадин (более 6000 м).
Рассмотрим наиболее важные из них.
К надсемейству Nuculacea относятся
семейства: ореховидки (Nuculidae),
ледиды (Nuculanidae, или Ledidae) и малле-
циа (Malletiidae).
Представители первых двух широко
распространены в Мировом океане,
особенно в мелководных шельфовых морях,
где многие массовые виды входят в состав
биоценозов донной фауны. В семействе
же Malletiidae преобладают более
глубоководные виды, которые встречаются
обычно на больших глубинах, особенно в
абиссали — на ложе океана, до 4—6 км.
Из первого семейства наиболее известна
тонкая ореховидка-нукула (Nucula tenuis)—
одна из обычнейших форм морей северного
полушария, где обитает на илах до
глубин не более 100—150 м, а в северных
частях Атлантического и Тихого океанов
изредка встречается и на глубинах около
1000 м. Ее небольшая A—2 см) округло-
треугольная «орешковидная» блестящая
зеленоватая раковина изнутри имеет
перламутровый слой. Она массами попадается
в ряде мест прибрежной зоны, где
обитает в самом поверхностном слое мягкого
илистого грунта, поскольку не обладает
сифонами и не может глубоко
закапываться. Местами (например, в
Чукотском море) ее количество иногда
достигает 4000 экземпляров на 1 м2, при весе
около 700 г на 1 м2.
На мелководьях Баренцева моря и у
берегов Мурмана эта ореховидка, как и
другие двустворчатые, является
излюбленной пищей ряда донных промысловых рыб
(пикши, камбал, молоди тресковых), в
желудках которых встречается десятками.
Из семейства Nuculanidae
наиболее известны различные йолъдии, порт-
ландии, леды и др.
Виды рода Йолъдия (Yoldia) очень
обычны и многочисленны в морях
северного полушария. У настоящих йоль-
дий поверхность раковины гладкая, у
видов под род a Cnesterium она украшена
тонкими косыми линиями; у видов под-
рода Megayoldia раковина самая
крупная среди всех йольдий(до6—7 см), с
широким, как бы обрубленным сзади кон-
Таблица 8. Двустворчатые моллюски морей СССР.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
1 — циклокардия (Cyclocardia crebricostata);
2 — исландская циприна (Cyprina islandica);
3 — сердцевидка волосатая (Clinoeardium ciliatuiro,
4 — сердцевидка гренландская (Serripes groenlandicus);
5 — сердцевидка малоребристая (Cardium (Acanthocardia) pauci-
costatum);
6 — черноморский венус-петушок (Chione (Chamelea) gallina cor-
rugatula;
6a — то же, вид сверху;
7 — дозиния японская (Dosinia (Phacosoma) japonica);
8 — японский петушок (Venerupis (Amygdalum) japonica);
9 — тихоокеанская питария (Callista brevisiphonata);
10 — черноморский тапес (Venerupis (Polititapes) rugata proclivis);
11 — гульдия маленькая (Gouldia minima);
12 — гомфина (Liocyma fluctuosa);
13 — мерценария стимпсона (Mercenaria stimpsoni);
14 — прототака (Protothaca euglypta);
15 — камнеточец-петрикола (Petricola lithophaga);
16 — теллина желтая (Angulus (Peronidia) lutea);
17 — теллина ангулюс (Angulus (Moerella) donacinus);
18 — гастрана хрупкая (Gastrana fragilis);
19 — известковая макома (Macoma calcarea);
20 — балтийская макома (Macoma balthica);
21 — синдесмия овальная (Abra ovata);
22 — оливковая кровянка (Sanguinolaria (Nuttallia) olivacea).
цом. Для йольдий характерно наличие
пары длинных сросшихся сифонов,
широкие, сократимые ротовые лопасти и
сильная мускулистая нога со
складывающейся подошвой. При таком «вооружении»
они легко могут закапываться в мягкий
илистый грунт прибрежных мелководий,
выставляя над дном лишь задний конец
раковины, а также сифоны и ротовые
щупальца. При этом сифоны и околоротовые
лопасти могут вытягиваться далеко за
пределы раковины. Лопасти «ощупывают»
поверхность дна вокруг моллюска,
собирая детрит с его поверхности, а через
сифоны идет фильтрация воды из
придонного слоя; йольдий наиболее обычны на
глубине до 100 м, редко глубже.
В южных морях СССР йольдий не
встречаются. Наиболее обычны и широко
распространены в водах северного
полушария, в том числе в северных и
дальневосточных морях, йолъдия северная (Yoldia
hyperborea), а также Y. limatula, Y. mya-
lis, Y. (Cnesterium) scissurata, Y. (Mega-
yoldia) thraciaeformis и многие другие.
Особый интерес представляют широко
распространенные на мелководьях
арктических морей небольшие холодновод-
ные моллюски портландии, раньше также
называвшиеся йольдиями.
В этом типично арктическом роде Port-
landia имеется всего два вида: весьма
изменчивая по форме раковины и
экологии арктическая портландия (P. arctica),
образующая в северных морях несколько
подвидов, и более постоянная по
очертаниям раковины эсту арная портландия
(P. aestuariorum).
Арктическая портландия играла
большую роль в северных морях прошлых
геологических эпох. Она была
руководящей формой (характернейшей и массовой)
холодного «Йольдиевого моря»
(названного так по ее имени), которое около
8—10 тыс. лет назад покрывало в
межледниковый период весь север Европы вплоть
до Балтийского моря. Остатки массовых
поселений арктической портландии
указывают на пределы распространения
этого моря, образовавшегося вслед за
отступлением края ледника, и на условия,
при которых существовала населявшая
его в то время донная фауна.
Близки к портландиям виды из чисто
морского рода йолъдиелл (Yoldiella),
куда относится много глубоководных
океанических форм.
В составе настоящей морской фауны
моллюсков большую роль играют леды —
представители рода Nuculana, или Leda,
куда входит много видов. Для них
характерна удлиненная раковина с сильно
оттянутым задним ее концом, вследствие
чего ее длинные несократимые сифоны
Таблица 9. Морские промысловые двустворчатые моллюски:
1 — Crenomytilus grayanus;
2 — Mytilus edulis;
3 — Saxidomus purpuratus;
4 — Ostrea edulis;
5 — Crassostrea gigas;
6 — Crassostrea virginica;
7 — Ostrea denselamellosa;
8 — Pecten maximus;
9 — Pecten (Patinopecten) yessoensis.
8 Жизнь животных, т. 2
Рис. 79. Строение замка и тела у гребенчатозубых и разнозубых моллюсков. Гребенча-
тозубые — ореховидка нукула (Nucula tenius): A — правая створка со спинной стороны;
Б — вид тела при удаленной правой створке. Разнозубые: В—Г — гренландская сердцевидка
(Serripes groenlandicus) — вид тела при удаленной левой створке.
1 — гребенчатые замковые зубы; 2 — выросты краев мантии между зубами; з — ножные мускулы; 4 —
передний вамыкательный мускул; 5 — задний замыкательный мускул; 6 — обрезанная часть лопасти
мантии; 7 — нога; 8 — подошва ноги; 9 — внутренностный мешок; 10 — рот; 11 — ротовые лопасти;
12 — придатки ротовых лопастей; 13 — правый ктенидий; 14 — лопасть мантии; 15 — левая наружная
полужабра; 16 — левая внутренняя полужабра; 17 — вдыхательный сифон; 18 — выдыхательный
сифон; 19 — чувствительные выросты; 20 — наружная связка; 21 — кардинальные зубы; 22 — передний
боковой зуб; 23 — задний боковой зуб.
лежат в раковине, как в футляре. У
типичного и наиболее обычного для этого
рода вида обыкновенная леда (Nuculana
pernula) поверхность зеленовато-желтой
раковины покрыта тонкой
концентрической иечерченностью. Обыкновенная леда
обитает почти во всех морях северного
полушария, где образует несколько подвидов.
В третье семейство отряда
гребенчатозубых — Malletiidae—входит
несколько типично глубоководных
океанических родов, среди которых имеются
очень древние примитивные формы: Mal-
letia, Tindaria, Neilonella, Spinula и др.
Для всех маллеций характерна
небольшая светло окрашенная раковина
с наружным лигаментом и с одним
(выводящим) сифоном. Нога сильная, со
складывающейся подошвой, поверхность
раковины имеет обычно тонкую
концентрическую исчерченность.
Виды этих родов обычны в абиссали
ложа Мирового океана D—6 км
глубины), в том числе и в абиссали Северного
Ледовитого океана. Реже они встречаются
на склоне материковой ступени A—2 км).
Единичные виды маллеций встречаются
в глубоководных впадинах, как,
например, спинула витязя (Spinula vityazi),
найденная в Курило-Камчатской впадине
Тихого океана на глубине около 8 км.
(Она названа так в честь
исследовательского корабля Института океанологии
Академии наук СССР «Витязь».) В морях
маллеций редки (даже в абиссали
Охотского и Берингова морей), на
мелководьях почти неизвестны.
К Nuculacea (Protobranchia) также
относят очень интересное семейство
солемий (Solemyidae), представители
которого известны еще из отложений девона.
Как и нукулы, они обладают жабрами-
ктенидиями и ногой с подошвой; их
удлиненно-овальная раковина совсем лишена
зубов и покрыта тонким периостракумом,
выступающим в виде радиальных выростов
114
за брюшной край их раковины. Хотя они
и не имеют сифонов, но могут
закапываться в грунт. Многие солемии
глубоководные, имеются и литоральные виды.
Ко второму надсемейству Аг-
сасеа из отряда гребенчатозубых
относятся семейства арок (Arcidae), ли-
мопсисов (Limopsidae), пектункулюсов
(Glycymeridae), большая часть видов
которых обитает в теплых морях и на
тропических мелководьях.
У Агсасеа удлиненно-трапециевидная
или округлая раковина, с прямым или
слегка изогнутым замочным краем, с
многочисленными вертикальными или
скошенными зубами. Раковина с
радиальными ребрами. Нередко она покрыта
снаружи тонкими волосовидными выростами
периострака или гладкая. Между
спинным краем раковин и макушками имеется
ромбовидная площадка; нога слабая,
имеется биссус. Жабры нитевидные, несрос-
шиеся, сифонов нет.
В наиболее богатое семейство
арок (Arcidae) входит около 200 видов,
из которых в наших северных морях
широкое распространение имеют только три
североатлантических вида из рода
батиарок (Bathyarca) с сероватой
бархатистой опушкой поверхности раковины. В
Белом море батиарок нет. В Черном море
Рис. 80. Изменчивость видов рода Портландия
северных морей.
Подвиды арктической портландии (Portlandia arctica):
1 — a. arctica (основной подвид); 2 — a. siliqua;
3 — a. portlandica (из Белого моря); 4 — эстуарная
портландия (Portlandia aestuariorum). Левые створки;
а — ростр ум — задний суженный конец раковины.
8*
встречается лишь одна мелкая (около 1 см)
арка — Area (Arcopsis) lactea —
средиземноморско-атлантический вид.
Настоящего расцвета арки достигают
на мелководьях субтропической и
тропической зон, где обитают различные виды
из родов Area, Litharca, Barbatia, Ana-
dara, Trisidos, Cucullaea и др. Здесь часто
встречаются крупные ребристые формы
с крепкой, иногда пятнисто окрашенной
раковиной, делающей их незаметными
среди подводных зарослей (как,
например, арка полосатая — Area zebra).
Многие арки, особенно крупные,
съедобны, и их промышляют. Так, в Японии
добывают крупную (размером до 12 см)
вздутую арку (Anadara (Scapharca) brough-
tonii); она обычна и у наших берегов
Японского моря, в заливе Петра Великого,
на заиленных грунтах, на глубине до 20 м*
ОТРЯД СВЯЗОЧНОЗУБЫЕ
(DYSODONTA)
В этот отряд входит 13 семейств
моллюсков, весьма различных по устройству
своей раковины и тела, а также по
распространению и образу жизни. Вместе с тем
их объединяет большая или меньшая
редукция переднего
мускула-замыкателя, который либо может быть сильно
уменьшенным в размере (Неравномускуль-
ные — Anisomyaria), или имеется лишь
один крупный мускул-замыкатель, обычна
расположенный посредине раковины (Одно-
мускульные — Monomyaria).
Нередко весь этот отряд связочнозу-
бых называют неравномускулъными.
Общим для всего отряда является
отсутствие настоящего замка, однако у
некоторых (Mytilidae) под макушкой имеются
по краю раковины зубовидные выросты.
Соединение створок между собой
происходит главным образом при помощи ли-
гамента (связки). Жабры нитчатые (Fili-
branchia), состоят из двух складчатых
(наружных и внутренних) листков
(полужабр) с каждой стороны тела. Они
образованы длинными опушенными
ресничками, тонкими согнутыми жаберными
нитями, образующими сперва нисходящее,
а затем восходящее колена. Отдельные
жаберные нити не срастаются, а лишь
сцепляются между собой при помощи
жестких ресничек, сидящих на особых
дисках. Восходящее и нисходящее
колена жаберных нитей могут срастаться
соединительнотканными перемычками лишь
в местах их сгиба или у концов их
восходящих колен.
Почти все связочнозубые являются
формами эпифауны, т. е. живут на
поверхности грунта, прикрепляясь к нему своим
биссусом или даже прирастая створкой
к камням и скалам, или свободно лежат
на поверхности грунта и даже могут
плавать; некоторые из них живут частично
«воткнувшись» в грунт. Молодые формы
обычно свободно передвигаются при
помощи своей ноги, которая у взрослых
обычно редуцируется. Все они активные
фильтраторы.
Из этого отряда рассмотрим лишь
некоторых его представителей, интересных
с точки зрения их строения, образа
жизни или хозяйственного значения.
Ксемейству Mytilidae относится
много массовых видов: мидий, модиол,
мускулюсов, митилястеров и др., широко
распространенных в морях земного шара,
преимущественно в мелководных их
районах. Все они имеют клиновидную «мити-
лидного» типа раковину; передний
мускул-замыкатель маленький и узкий, а
задний — большой, округлый.
Наиболее широко известны мидии,
многие виды которых являются съедобными
и промысловыми: обыкновенная
съедобная мидия (Mytilus edulis),
дальневосточная гигантская мидия, или черная ракушка
(Crenomytilus grayanus), средиземноморско-
черноморская мидия (М. galloprovincialis),
калифорнийская (М. californianus), мидия
Магеллана (М. magellanicus) и др.
Мидии—обычные обитатели прибрежной
зоны, где они часто образовывают
массовые поселения. Срастаясь биссусами друг
с другом, они образуют на берегах так
называемые щетки; их большие
скопления на литорали и на подводных
мелководьях в более открытых частях моря
носят название мддиевых банок.
У мидий удлиненная клиновидная
раковина, суженная спереди, расширенная
сзади. В связи с их прикрепленным
образом жизни макушка сдвинута на передний
(приостренный) конец раковины. Окраска
раковины темная, часто иссиня-черная,
внутренняя поверхность имеет тонкий
перламутровый слой. Имеется несколько
мелких замковых зубчиков; биссус хорошо
развит.
Железа, выделяющая биссусные нити,
лежит у мидий в маленькой пальцевидной
ноге, которая у взрослых мидий
благодаря их неподвижному образу жизни
почти полностью утратила свою
двигательную функцию. Молодые мидии (менее
1—2 см длиной) могут хорошо
передвигаться, что можно видеть, наблюдая их
в аквариуме. Взрослые же мидии меняют
свое место обитания лишь при очень
неблагоприятных условиях, обрывая биссус
и переходя на новое место. Крепкие нити
биссуса дают им возможность
выдерживать действие даже сильного прибоя,
и только удары волн самого сильного
шторма могут обрывать связки мидий,
опустошая их поселения. Лишь сеть из
остатков биссусных нитей на скалах, остатки
раковин да битая ракушка указывают
на места их недавних густых поселений.
Биссусная железа состоит из двух
частей: одна лежит в глубине ноги, у ее
основания, а другая — на ее переднем
конце. Выделенное первой частью железы
вещество нити попадает в бороздку,
идущую вдоль нижней поверхности ноги до
ее вершины. Здесь она и приклеивается
к субстрату особым прикрепительным
диском, который выделяется другой частью
железы. Нити биссуса прикрепляются
к субстрату в определенном порядке и
своим натяжением, как якорями, удерживают
животное. Кроме того, у мидий для
втягивания ноги имеется несколько пар
мускулов, прикрепленных изнутри к
створкам. При их сокращения моллюск
подтягивается и прижимается к субстрату, а не
просто болтается на нитях биссуса,
поэтому даже при очень сильном прибое
мидии не разбиваются о скалы.
Мидии раздельнополы, но пол их можно
определить лишь у молодых форм по
цвету их созревающих половых желез,
частично залегающих в складках мантии:
у самцов обыкновенной съедобной мидии
116
Рис. 81. Свободнолежащие, плавающие и прикрепленные двустворчатые моллюски (схема):
1 — морские гребешки, свободнолежащие на дне и плавающие; 2 — лима, ползающая по дну и
плавающая; з — лима в «гнезде»; 4 — пинна; 5 — модиола; 6 — устрицы; 7 — мидии. Сплошные стрелки
показывают направление движения моллюска, пунктирные — токи воды, выходящие из мантийной полости.
мантия имеет кремовый, а у самок —
оранжевато-красный цвет, а у
дальневосточной гигантской мидии —
соответственно белый и розовый цвет. Размножаются
мидии в наиболее теплое время года.
Оплодотворение яиц происходит в воде, куда
выбрасываются половые продукты.
Плодовитость мидий очень велика и
повышается с возрастом. Обычно во время
каждой кладки самка мидии
выбрасывает от 5 до 12 млн. яиц, а крупные
экземпляры — до 25 млн. Самка гигантской
мидии начинает размножаться на шестом
году жизни (продолжительность жизни
этой мидии около 20 лет), откладывая
каждый раз около 20 млн. яиц.
Примерно через 20 часов после оплодотворения
из яйца появляется личинка т р о х о-
ф о р а, которая дня через два плавания
в толще воды превращается в личинку
парусник (велигер). При
неблагоприятных для оседания условиях
продолжительность планктонной (плавающей)
стадии жизни личинки парусника может
сильно затягиваться от нескольких дней
до нескольких месяцев. При размере 0,2—
0,3 мм велигер оседает на дно, имея уже
по 2 пары жаберных нитей. Некоторое
время молодая мидия еще ползает по дну,
но потом прикрепляется биссусом.
Молодь мидий в большом количестве обычно
встречается в зарослях литоральных
водорослей, которые предохраняют их от
высыхания во время отлива.
К концу первого года жизни молодь
мидии у берегов Европы достигает 3—4 см
длины, а в Белом море — всего 0,5 см;
живет мидия до 13—14 лет.
Пищей мидий является детрит (остатки
водных растений и животных), а также
живущие в толще воды одноклеточные
водоросли и мелкие планктонные
животные и бактерии. При температуре около
20°С одна мидия (в 5—6 см длины) может
профильтровать около 3 л воды в час;
плотное же поселение мидий на банке
может профильтровывать за сутки от 50 до
280 м3 воды. Таким образом, большие
поселения мидий представляют собой
мощный биофильтр, высасывающий из
окружающей воды большое количество взвеси,
как минеральной, так и органической,
и мелкого планктона. Пищевые частицы
при этом отсортировываются и
направляются в ротовое отверстие, а более тяжелые и
минеральные удаляются в виде
псевдофекалий. Таким образом, мидии, как и
многие другие двустворчатые моллюски (свя-
зочнозубые, подавляющее большинство
разнозубых и гребенчатозубых),
—активные фильтраторы. Они не только очищают
окружающую воду от взвеси, но своими
псевдофекалиями и фекалиями принимают
участие в образовании илистых грунтов.
Мидии обычно питаются круглый год,
но зимой менее интенсивно.
У мидий множество врагов среди морских
рыб, птиц и млекопитающих. Птицы
охотятся на них во время отлива на литорали.
Мидиевые банки в мелководных районах
сильно страдают от скатов, камбал и
трески, а в Черном море — от осетровых рыб.
Но постоянным их врагом, обитающим
обычно в районах поселений мидий,
являются крупные морские звезды, которые
ими питаются, как, например, в
Атлантике —Asterias rubens, а в
дальневосточных морях — A. amurensis, Patiria pecti-
nifera и др. Одна звезда съедает
ежедневно в среднем одну-две мидии размером до
2 см. Охотятся на них также крабы,
крупные брюхоногие моллюски и др.
Обыкновенная съедобная мидия (Myti-
lus edulis) — один из наиболее
широкораспространенных видов двустворчатых
моллюсков; это изменчивый вид, образующий
экологические и физиологические расы.
Она живет у Атлантического побережья
Европы, у берегов Исландии, Южной
Гренландии, по Атлантическому и
Тихоокеанскому побережью Канады и США,
обитает в Баренцевом, Белом и
Балтийском морях, в юго-западных участках
Карского и в дальневосточных морях.
Большие поселения обыкновенной
мидии наблюдаются главным образом в
литоральной (приливо-отливной) зоне, где
дважды в сутки эти поселения мидий
обсыхают во время отлива. Однако, крепко
смыкая в это время свои створки, они
обходятся в течение 5—6 часов тем
количеством воды, которое остается в их
мантийной полости. Плотность поселений
обыкновенной мидии может достигать
нескольких тысяч экземпляров, а биомасса—
нескольких килограммов на 1 м2.
118
Обыкновенная мидия — эвригалинная
форма, т. е. выносит значительные
колебания солености и опреснение до 3°/00.
Однако если она постоянно живет при
пониженной солености, как например
в Балтийском море, то она растет
медленнее и мельчает. Так, размеры взрослых
мидий при солености 2 и 3°/0о в
Ботническом заливе в 4—5 раз меньше, чем у
живущих при солености 15°/00 в датских
проливах и в Кильской бухте.
Обыкновенная мидия также очень эври-
термна, т. е. может выносить
значительные колебания температуры: летом на
литорали во время отлива она может сильно
прогреваться солнцем, а зимой в суровых
условиях Белого и Баренцева морей
мидии могут даже промерзать, оставаясь
живыми. Однако, по-видимому, из-за
истирающего действия льдов, покрывающих
зимой литораль в восточных участках
Баренцева и в прилегающих частях
Карского морей, мидии живут здесь не в осуш-
ной зоне, а на глубине нескольких метров.
Дальневосточная гигантская мидия (Сге-
nomytilus grayanus), достигающая 20—25 см
длины, обитает в Японском море, у берегов
Сахалина и Северного Китая. Она живет
на самых различных, преимущественно на
каменисто-песчаных грунтах, на глубине
до 50—60 м, нередко образует большие
поселения — банки, срастаясь биссусом
между собой в друзы; ее биомасса может
достигать 20 кг на 1 м2 грунта и более.
В 20—30-е годы гигантская мидия вместе
с устрицами была завезена из Японии
в Калифорнию, где и прижилась.
Средиземноморско-черноморская мидия
(Mytilus galloprovincialis) очень
изменчива по форме своей раковины,
образует ряд подвидов. В Черном море она
встречается при солености более 10°/00
на различных грунтах и глубинах—от
каменистых и от уреза воды вплоть до
мягких илистых грунтов и глубин около
80 м. На глубине 50—80 м в Черном море
широко распространен биоценоз «мидиево-
го ила», где одна из разновидностей
черноморской мидии (v. frequens) является
руководящей формой. Биомасса донной
фауны достигает здесь 0,5 кг на 1 м2.
С древнейших времен мидии имеют
для человека промысловое значение, на
Рис. 82. Изменчивость формы раковины у мидий
(правые створки): А — обыкновенная
съедобная мидия (Mytilus edulis), прикрепившаяся к
субстрату; разновидности (морфы) черноморской
мидии Mytilus galloprovincialis; Б — nominata;
В — dilatata; Г — frequens.
1 — биссусные нити; 2 — нога; 3 — макушка,
сдвинутая у мидий на передний конец раковины; 4 —
выросты заднего края мантии; 5 — выводной сифон.
119
что указывают остатки их раковин
(вместе с устрицами), образующие так
называемые «кухонные кучи» древних
поселений людей начиная с каменного века.
Ежегодная мировая добыча мидий
(главным образом съедобных) в последние годы
возросла до 2—2,5 млн. ц.
Благодаря большим запасам мидий в
наших морях развитие их промысла в СССР
имеет большие перспективы. Общие
запасы мидий у Черноморского побережья
СССР составляют по грубым подсчетам
около 65 млн. т, а доступных для
промысла около 7 млн. т, или в пересчете
па мясо около 800 тыс. т. Очень велики
запасы мидий в Японском и Охотском
морях и у берегов Камчатки, однако
промысел их здесь развит еще недостаточно.
В Белом море запасы мидий только в
районе Карельского и Поморского берегов
составляют около 3 тыс. т.
Мидий ловят особыми зубчатыми
драгами с небольших моторных судов или
собирают водолазы.
Ввиду того что во многих странах
естественные запасы мидий значительно
подорваны и грозят исчезновением, как
например во Франции, их начали
разводить искусственно. Такие «мидиевые
парки» имеются в Нормандии и других
районах. Там они часто растут лучше, чем на
естественных, «диких» банках.
Существует несколько способов искусственного
разведения мидий. Обычно их
воспитывают на специальных плетнях («бушо»),
стоящих в воде, куда переносят
собранную молодь. Здесь мидии растут 1,5—3
года до наилучших их вкусовых качеств.
На побережье Франции разводимые
таким образом мидии дают ежегодно около
8 т мяса с 1 га; в настоящее время
считают, что при интенсивной культуре можно
получать до 150 кг мидий с одного
погонного метра такого плетня. Так, в Италии,
в заливе Таранто, при продуктивной
культуре мидий получают в среднем свыше
1 т со 100 м2.
Мидий употребляют в пищу в жареном,
вареном и в консервированном виде
(сырые мидии быстро портятся и
становятся ядовитыми). Пищевая ценность мидий
высока: мясо свежих мидий содержит
около 10% белков, 1% жиров, 0,5% уг-
леводов(гликогена), а также витамины В и С
и много разных полезных для организма
человека солей.
Из мелких мидий делают также очень
питательную кормовую муку, которую
используют для корма домашних
животных.
Близок к мидиям род Modiolus;
интересны по своему образу жизни также виды
из рода мускулюс (Musculus, или Modi-
olaria) и камнеточцы (Lithophaga), часто
называемые также «морскими финиками».
Наиболее известна в северных частях
Атлантического и Тихого океанов крупная
тепловодная обыкновенная модиола
(Modiolus modiolus), до И еле длины (обычно
5—8 см). Она имеет светло-коричневую
выпуклую раковину, опушенную сзади
кожистыми выростами периострака.
Обитает главным образом на жестких
каменисто-песчаных грунтах; на глубине до
100 м в Баренцевом море образует очень
характерный богатый биоценоз эпифауны.
Фазеолиновая модиола—Modiolus (Modio-
lula) phaseolinus имеет небольшую
бобовидную коричневую раковину и хорошо
известна, например, в Черном море, где
образует массовые поселения на глубине 60—
120 м, благодаря чему эта зона получила
название «фазеолиновой», а грунты с
остатками раковин — «фазеолиновых илов».
Некоторые виды модиол могут сплетать
из своего биссуса подобие гнезда,
облепленного снаружи камешками и обломками
раковин. Подобные же «гнезда» могут
строить из нитей своего биссуса и
кусочков водорослей виды из близкого к модио-
лам рода мускулюс. В такое «гнездо»
Musculus discors прячет слизистые шнуры
своей яйцекладки. Подобное же «гнездо»
делает и черный мускулюс (М. nigra) из
кусочков морской травы зостеры (по
наблюдениям у берегов Дании). У обоих
видов зародыши развиваются в этих гнездах,
не проходя стадию свободноплавающей
личинки, как у других митилид. Таким
образом, постройка этих «гнезд»
представляет собой один из видов заботы о своем
потомстве.
Интересным примером активного
расселения морских организмов в новом
водоеме служит история появления в
Каспийском море моллюска митилястера
120
(Mytilaster lineatus, или Brachyodontes li-
neatus). Это небольшой моллюск около 2 см
длиной, с темной клиновидной («мити-
лидной») раковинкой, с сильно развитым
биссусом. Замок состоит из 2—3 зубов,
расположенных под макушкой; спинной
край изнутри мелко зазубренный.
Этот вид широко распространен в
Средиземном, Черном и Азовском морях.
В Азовском море он входит в состав ряда
биоценозов донной фауны; осенью его
количество здесь может достигать И тыс.
экземпляров на 1 м2 при биомассе 600 г на
1 м2.
В 20-е годы нашего столетия мити-
лястер случайно, возможно на
днищах деревянных судов, перевозимых
по суше из Азовского в Каспийское море,
был завезен сюда, где нашел весьма
благоприятные условия существования (как
в отношении солености — 10—12°/00, так
и питания), быстро размножился и
широко расселился. Теперь на глубинах до
50 м он стал самым многочисленным
компонентом донной фауны в большей части
Каспийского моря. Местами в Среднем
и Южном Каспии он составляет больше
двух третей общей биомассы донной
фауны, а его собственная биомасса может
достигать нескольких сот граммов на 1 м2,
а иногда даже более 1 кг на 1 м2. В ряде
районов Каспийского моря иногда
наблюдается даже вытеснение митилястером
некоторых местных видов моллюсков.
Любопытны по образу жизни морские,
или каменные, финики из родов Ли-
тофага (Lithophaga) и Ботула (Botula).
Они имеют низкую удлиненную
раковину с гладкими или исчерченными
створками, покрытыми толстым периостраком.
Они ведут «скрытый» образ жизни,
протачивая в известковых камнях и скалах
ходы и норки, где и живут, прикрепляясь
к их стенкам биссусом и выставляя
свои длинные сифоны. Поэтому находить
и собирать их очень трудно. Часто они
живут вместе с другими моллюсками,
например камнеточцами фоладами. Хорошо
известны такие виды, как
средиземноморская литофага (Lithophaga lithophaga),
8 см длиной, калифорнийская ботула
Botula (Adula) californiensis — до 4 см
длиной, и др. Морские финики
протачивают свои ходы, выделяя особый кислый
секрет (сок), растворяющий известковую
породу прибрежных скал. Железа,
выделяющая этот секрет, находится в
передней части края мантии; поэтому они имеют
тонкую раковину, которая не участвует
в сверлении. На берегах Средиземного
моря морские финики ценятся как
лакомое блюдо.
Очень интересны моллюски из
тропического надсемейстса птерий (Pte-
riacea), раковины у которых обладают
хорошо развитым перламутровым слоем,
передний мускул-замыкатель очень мал
или редуцирован, а нога обычно обладает
сильно развитым биссусом. Сюда
относятся различные пинны и атрины, или
раковины-перья (сем. Pinnidae), морские
молоточки (Malleus, сем. Isognomonidae)
и настоящие морские жемчужницы, или
крыловидки (сем. Pteriidae). Пинны и
атрины имеют очень характерную
клиновидную, нередко очень крупную
раковину без замковых зубов и с очень сильно
развитым биссусом. Поверхность
раковины обычно украшена радиальными
ребрами или множеством чешуек.
К родам Pinna и Atrina относится
около 20 видов, живущих в Средиземном
море, в Атлантическом, Тихом и
Индийском океанах. Благородная пинна (Pinna
nobilis), живущая в Средиземном море и
в Атлантике, достигает 30 см длины,
являясь самой крупной среди
двустворчатых моллюсков европейских морей.
Внутри раковины пинн красивого красноватого
цвета нередко встречаются красноватые
же или темные жемчужинки.
Биссус пинн, ради которого они
главным образом добываются, имеет вид
густого пучка тонких шелковистых нитей
желтоватого или коричневатого цвета.
Нити его эластичны, очень прочны и
имеют красивый блеск; в их составе имеется
белковое вещество, близкое к фиброину,
входящему в состав шелка. Пинны и
атрины живут на мелководьях теплых
морей, прикрепляясь к твердому грунту
или полупогружаясь узким концом в
песок; во всех случаях широкий конец
раковины торчит вертикально вверх.
Изделия из биссуса пинн высоко
ценились в прежние века. Из него делались
121
дорогие ткани и кружева, отличавшиеся
необычным блеском и красотой. Так,
в конце XVIII в. в Италии пара
перчаток из биссуса пинн стоила 20 золотых
дукатов.
Еще более интересны виды из
тропического индо-тихоокеанского рода
Malleus. За причудливую форму раковины
они получили название «морской
молоточек». Замковый край у раковин мал-
леуса сильно удлинен, створки же
толстой известковой раковины сужены и
вытянуты наподобие ручки молотка. Внутри
раковин М. malleus, M. albus и других
видов этого рода иногда также встречается
жемчуг, но реже, чем у обычных
жемчужниц.
Морские жемчужницы издавна славятся
высоким качеством своего перламутра
и способностью образовывать самый
лучший и ценный жемчуг. Для них
характерна крупная различной формы
раковина, с прямым замочным краем, обычно
вытянутым сзади в уховидный или
клювовидный выступ; замковый край с од-
ним-двумя зубовидными утолщениями
и передний мускул-замыкатель
редуцированы, биссус сильно развит. Раковина
птерий состоит из двух слоев: наружного
(призматического) и мощного
внутреннего (перламутрового) слоя. Наиболее
крупная из жемчужниц Pinctada margaritifera
может достигать 30 см в поперечнике и
веса 10 кг (обычно они мельче).
К морским жемчужницам-крыловидкам
относится много тропических видов,
распадающихся на два рода: птерий (Pte-
ria, или Avicula) и пинктады (Pinctada).
Наибольшее промысловое значение
имеют: Pinctada margaritifera,
мелководный индо-тихоокеанский вид,
живущий также у берегов Центральной
Америки и Австралии, P. martensii — у
берегов Японии; цейлонская жемчужница (Р.
vulgaris) и др. После постройки Суэцкого
канала цейлонская жемчужница
проникла и в Средиземное море.
Пинктады и птерий ведут сидячий
образ жизни, прикрепляясь своим густым
и прочным биссусом к подводным скалам
и камням. Часто они образуют банки —
колонии и грозди на глубине от 5—6 до
60 м (обычно на глубине 10—15 ле),
обитая здесь вместе с другими формами эпи-
фауны, т. е. животными, сидящими на
поверхности жесткого грунта, —
кораллами, губками, гидроидами и др.
Жемчужницы не выносят опреснения, являясь
типичными обитателями чисто морских
районов. Они растут очень медленно:
в возрасте трех лет они достигают
размеров всего 5—6 см.
Обитающие в различных районах
океана жемчужницы даже одного и того же
вида имеют большие различия в размерах,
толщине раковины и в качестве
перламутрового слоя, чем определяется их
промысловая ценность. Так, раковина
цейлонских жемчужниц имеет обычно 5—6,5 см
в длину и до 8 см в высоту и тонкие
створки; те же виды из Персидского залива
больше и толще, а из Красного моря еще
крупнее. Здесь вес их раковины может
достигать 1 кг, а перламутр — самого
высокого качества.
Жемчужины обычно образуются в
раковинах жемчужниц более или менее
случайно, когда по каким-либо причинам
мантийный эпителий, который обычно
выделяет раковину, попадает в соединительно-
тканый слой мантии. Этот попавший
внутрь участок эпителия, разрастаясь,
образует мешочек, который выделяет
внутрь вещество раковины, т. е.
жемчужину. Она растет, как и облегающий ее
эпителий. Попадание эпителия в глубь
мантии вызывается часто совершенно
случайными причинами, как, например,
песчинкой, или кусочком раковины, или даже
каким-нибудь животным-паразитом,
попадающим между створкой раковины и
мантией моллюска. Жемчужины могут
образовываться не только в мантии, но
и в мускулах моллюска; это явление в
целом представляет собой некоторую
защитную реакцию организма на попавшее
в него инородное тело, поэтому
жемчужины вообще встречаются в ничтожно малом
проценте раковин. Очень редко образуется
несколько жемчужин в одном моллюске;
известны случаи нахождения 67 и 150
жемчужин в одной особи птерий.
Жемчужины состоят обычно из чередующихся
слоев перламутра и конхиолина, т. е.
всех слоев раковины, выделяемых
мантийным эпителием внутрь мешочка. Поэ-
122
тому жемчужины имеют внутри
концентрически-слоистое строение и
разнообразный цвет. Наличие конхиолина под
тонким перламутровым слоем дает
голубовато-серый цвет. Японский и индийский
жемчуг имеет сильный блеск и розоватый
цвет, австралийский — белый, панам-
ский — золотистый. Крупные жемчужины
бывают в больших и старых раковинах.
У испанского короля Филиппа II была
белая жемчужина грушевидной формы,
3 см длиной. Самая крупная из
известных жемчужин была найдена на
Филиппинских островах в раковине Tridacna,
самого крупного из двустворчатых
моллюсков. Эта жемчужина имела 24 см
в длину и 14 см в поперечнике. Обычно
жемчужины, вынутые из моллюсков,
«живут» не больше 100—150 лет; они
постепенно высыхают и через 50—60 лет начинают
разрушаться. При ношении жемчуга на
теле он постепенно тускнеет, темнеет, так как
его конхиолиновое вещество разлагается
и высыхает.
Важнейшими районами добычи
жемчужниц в Тихом океане служат море Зулу,
австралийский Большой Барьерный риф,
Торресов пролив, побережье Панамы
и Японии; в Индийском океане —
Персидский залив, Красное море, побережье
о. Цейлон и Мадагаскар; в Атлантике—
Мексиканский залив. Знаменитые банки
жемчужниц облавливаются уже в
течение нескольких сот лет. Промысел
ведется довольно примитивно — местными
ныряльщиками; водолазные костюмы и
акваланги применяются редко. Многие
банки жемчужниц у о. Цейлон в настоящее
время уже опустошены, поскольку
добыча их здесь производится уже несколько
столетий. Добыча жемчужниц пловцами,
ныряющими без всякого снаряжения,—
дело очень тяжелое и даже опасное,
поскольку всегда возможна встреча с
акулами. Опытный ныряльщик может
пробыть под водой 53—57 секунд; обычно
через несколько лет такой работы
человек глохнет и становится инвалидом,
так как ему ежедневно приходится нырять
до 30—40 раз, собирая за день до 2000
раковин. В Персидском заливе на
богатых жемчужных промыслах ежегодно
работает до 30 тыс. ловцов. Однако банки
Рис. 83. Схема образования жемчужины у
двустворчатых моллюсков: А> Б, В —
последовательные стадии, Г — шлиф через жемчужину
нормальной структуры.
1 — перламутровый слой раковины; 2 — мантийный
эпителий; з — соединительная ткань мантии; 4 —
инородное тело; 5 — мантийный мешочек; 6 —жемчужина;
7 — перламутровые слои (заштрихован только первый
слой); * — ядро; 9 — конхиолиновые слои; ю—
призматические слои.
эти здесь находятся на глубине до 50 м,
поэтому работа ныряльщиков здесь
особенно трудна. Водолазные работы
требуют специально оснащенных судов—лю-
геров и дорогого оборудования, что
невыгодно для предпринимателей.
Поскольку хищнический вылов
жемчужниц подорвал их запасы во многих
промысловых районах, то были введены
охранные меры, а также начали разводить
жемчужниц искусственно. Японский
зоолог Митсукури в заливе Аго впервые
создал хозяйство для выращивания
жемчуга. Однако более успешный способ
выращивания жемчуга у пинктад был пред-
Рис. 84. Промысловый приморский гребешок
Pecten (Patinopecten) yessoensis: A — наружный
вид правой створки; Б—он же, вскрытый с
правой стороны.
1 — макушка; 2 — переднее ушко; 3 — передний край
раковины; 4 — заднее ушко; 5 — задний край
раковины; 6 — брюшной край раковины; 7 —
радиальные ребра; 8 — межреберные промежутки; 9 —
внутренний лигамент; 10 — наружный лигамент; 11 —
передний отдел мускула-замыкателя; 12 — задний
отдел мускула-замыкателя; 13 — нога; 14 — биссусная
бороздка ноги; 15 — ротовые лопасти; 16 — рот;
17 — внутренностный мешок; 18 — нисходящая
пластинка наружной полужабры; 19 — восходящая
пластинка наружной полужабры; 20 — верхний край
восходящей пластинки наружной полужабры; 21 —
жаберный сосуд; 22 — жаберная ось (основание
жабр); 23 — печень; 24 — околосердечная сумка с
сердцем; 25 — прямая кишка; 26 — свободный край
паруса; 27 — парус левой лопасти мантии; 28 —
щупальца мантии; 29 — глаза.
ложен позже и заключался во введении
кусочков мантийного эпителия в толщу
мантии для образования эпителиальных
мешочков. Освоив этот способ, в Японии
научились получать около 50 тыс.
жемчужин в год, для чего нужно ежегодно
оперировать и содержать в особых садках
около 1 млн. жемчужниц.
К группе одномускульных (Monomya-
ria) из отряда Dysodonta относятся
три богатых видами семейства
моллюсков. Таково в первую очередь
семейство Морские гребешки (Pectinidae),
насчитывающее много родов и видов,
широко распространенных почти во всех
морях и океанах, на самых разнообразных
глубинах вплоть до ультраабиссальных;
так, в Курило-Камчатской впадине
Тихого океана на глубине 8100 м «Витязем»
был найден крупный полупрозрачный De-
lectopecten, и это пока наибольшая
глубина встречаемости морских гребешков.
Особенно богат и разнообразен мир
морских гребешков в водах прибрежных
мелководий субтропической и умеренной
зон Мирового океана.
В морях Советского Союза встречается
сравнительно немного видов морских
гребешков, наибольшее их количество
обитает в морях Дальнего Востока. На
прибрежных мелководьях Японского моря,
до глубины около 50 м, от Кореи до
Сахалина и Южно-Курильских островов
обитают хорошо известные дальневосточные
гребешки: крупный (до 20 см в
поперечнике, реже — больше) приморский
промысловый гребешок Pecten (Patinopecten)
yessoensis, с белой радиальноребристой
раковиной, и очень красивый гребешок
Свифта — Chlamys (Swiftopecten) swifti.
В южной части Японского моря
встречается также и японский гребешок Фаррера
(Chlamys farreri nipponensis).
В Беринговом, Охотском морях и в
южной части Чукотского моря обитает бе-
ринговоморский гребешок (Chlamys berin-
gianus), а также ряд других видов из
рода Chlamys. Беринговоморский
гребешок наиболее обычен на глубинах от 50
до 100 ле, встречается также у
Тихоокеанского побережья Америки до Калифорнии.
В дальневосточных морях и особенно
в Баренцевом и Белом морях и в юго-
124
западной части Карского моря (куда
проникают с запада более теплые воды)
обитает довольно крупный (до 8 см в
поперечнике) красивый исландский гребешок
(Chlamys islandicus). Он обычен также
и у берегов Исландии, Норвегии, Южной
Гренландии, у Атлантического побережья
Северной Америки. Встречается на
глубинах до 100 м и входит в состав
некоторых биоценозов донной фауны. Мясо
исландского гребешка очень вкусно, но
промысла на него в наших морях нет.
На мягких илистых грунтах Баренцева,
Карского, Норвежского и Гренландского
морей на глубинах более 100 м живут
мелкие виды морских гребешков пропе-
амуссиумов, с тонкой хрупкой раковиной.
Это холодноводный гренландский гребешок
(Propeamussium groenlandicum) и
чешуйчатый гребешок (Pr. (Cyclopecten) im-
briferum), обитающий в Северной
Атлантике и на юго-западе Баренцева моря.
В Черном море живет лишь один черно-
морский гребешок Chlamys (Flexopecten)
glaber ponticus — подвид
средиземноморского гребешка. Его небольшая (до
5 см) раковина, ярко окрашенная в
желтый, розовый и другие цвета, с
небольшим количеством ребер, известна всем,
кто бывал на берегах Черного моря.
Черноморский гребешок обитает на глубине
50—60 м, главным образом в биоценозе
ракушечника, вместе с венусами,
талесами, модиолами и сердцевидками.
Морские гребешки имеют округлую
раковину с прямым замочным (спинным)
краем, выдающимся по бокам в виде
угловатых выступов — «ушек». Верхняя
створка обычно более уплощенная, а
нижняя — более выпуклая. Раковина
украшена радиальными или концентрическими
ребрами, часто несущими шипы или
чешуйки. У мелководных гребешков (Pec-
ten, Chlamys) раковина крупная,
крепкая, разнообразно окрашенная в
розовый, белый, лиловый, красноватый цвет,
часто с красивым пятнистым узором.
У более глубоководных форм (Amussium,
Propeamussium, Delectopecten) створки
раковин хрупкие, тонкие, часто
полупрозрачные, с тонкими наружными, а иногда
и внутренними ребрами. Замковых
зубов нет, но лигамент развит хорошо.
Замыкательный мускул крупный,
мясистый, расположен посредине раковины;
как и у устриц, он разделен на два
неравных отдела: большой, передний состоит
из поперечноисчерченных мускульных
волокон и способен к быстрым
энергичным сокращениям; меньший, задний отдел
мускула-замыкателя состоит из гладких
мускульных волокон. Нога маленькая,
пальцевидная, с бороздой, в которую
открывается биссусная железа. У
взрослых морских гребешков нога не служит
для передвижения. Жабры состоят из
двойных несросшихся жаберных нитей,
коленчатоизогнутых пополам. Края
обеих лопастей несросшейся мантии
утолщены и немного загнуты внутрь,
образуя так называемый «парус», играющий
большую роль при плавании гребешков.
Разноцветная мантия несет по краю
много тонких чувствительных выростов, у
основания которых расположены
многочисленные мелкие «мантийные глаза»,
которые у живых гребешков светятся
красивым зеленоватым светом. Количество и
расположение глаз у гребешков очень
различно, но может достигать ста; они
более многочисленны, и количество их
больше на верхней половине мантии.
Глаза гребешка могут «видеть» лишь на
небольшом расстоянии. Так, лишь когда
величайший враг морских гребешков
морская звезда приблизится к нему почти
вплотную, моллюск скачком «обращается
в бегство».
Мантийные глаза морских гребешков
довольно сложны: они представляют
собой замкнутые пузыревидные
образования, сидящие на небольших стебельках.
Глаза имеют роговицу,
светопреломляющий хрусталик и блюдцевидную
(инвертированную) сетчатку (ретину),
подостланную двумя слоями — так
называемым «зеркальцем» (tapetum), отражающим
свет (от чего и зависит зеленоватый блеск
их глаз), и пигментированным слоем.
Глазки иннервируются нервом, идущим
вокруг мантии. По своему происхождению
мантийные глаза гребешков отличаются
от головных глаз других моллюсков, так
как их сетчатка произошла путем впячи-
вания наружного (а не внутреннего) слоя
глазного пузырька.
125
Почти все гребешки могут плавать,
передвигаясь в воде короткими скачками,
при этом створки раковины сначала
открываются, а потом снова быстро
захлопываются, края паруса развертываются
и плотно сжимаются, так что вода
выталкивается из мантийной полости наружу
двумя сильными струями, выходящими
в области «ушек», куда края паруса не
доходят. Получающийся таким образом
толчок продвигает раковину над
грунтом брюшным краем вперед. Направление
скачка при этом противоположно
направлению струй воды, выталкиваемой через
«ушки». Скачок крупного гребешка
обычно достигает в длину полуметра или даже
больше. Так он может проплывать
значительное расстояние.
В движении гребешков большую роль
играют его парные органы равновесия—
статоцисты, расположенные вблизи
ножного нервного ганглия. Они представляют
собой маленькие замкнутые пузырьки,
выстланные внутри чувствительными
ресничными клетками; внутри этих
пузырьков находятся известковые образования
(статолиты). Левый статоцист развит
сильнее и содержит более крупный статолит,
а в правом, меньшем, статоцисте имеются
и более мелкие известковые зерна (стато-
конии). При обычном положении
гребешка — выпуклой створкой вниз — левый
статоцист располагается вверху. Если во
время плавания моллюск случайно упадет
на дно своей верхней (плоской) створкой
вниз, то он сейчас же толчком
переворачивается на 180°. Способность двигаться
таким образом позволяет мелководным
гребешкам в жаркое время года
переплывать в более глубокие прохладные
места, а зимой перемещаться ближе к
берегам.
Гребешки питаются детритом и
различными мелкими планктонными
организмами, извлекая их из воды, засасываемой
в мантийную полость. Один гребешок
размером 4 см может профильтровать за
час около 3 л воды, а гребешок размером
7 см — до 25 л воды в час.
У морских гребешков, как и у
других моллюсков, много врагов, из которых
самыми беспощадными являются морские
звезды и донные осьминоги: от них
гребешков не всегда спасает даже их
способность плавать. Кроме того, раковины
гребешков могут протачиваться
сверлящими губками, на них могут поселяться
различные водоросли, мшанки, балянусы
(морские желуди) и другие
беспозвоночные, затрудняющие передвижение
гребешков.
Из большого количества видов морских
гребешков особенно красивы обитатели
тропических морей, окраска раковины
и мантии которых бывает очень
разнообразных оттенков. Даже в Японском море
живые гребешки очень ярки и красивы.
Мясо морских гребешков (точнее, их
крупный мускул-замыкатель, а иногда
и мантия) издавна считалось вкусным
и лакомым блюдом, и даже древние греки
и римляне всегда высоко ценили его. В
настоящее время почти во всех странах мира,
особенно в приморских и островных
странах, гребешки употребляются в пищу как
в свежем, так и в мороженом,
консервированном и сушеном виде. Промышляют
почти все виды крупных прибрежных
гребешков (пектенов и хламисов). Так, в
Атлантическом океане используются:
большой гребешок (Pecten maximus), гребешок
св. Якова (P. jacobeus), гребешок Магеллана
P. (Placopecten) magellanicus и другие.
В 1962 г. было выловлено 1160 тыс. ц
морских гребешков и они заняли в
мировой добыче двустворчатых моллюсков
третье место (после устриц и мидий).
Морского гребешка ловят драгами,
сачками, или его собирают водолазы. Один
водолаз за 6 часов работы может собрать
несколько тысяч штук.
Промысел морских гребешков успешно
развивается в Советском Союзе, и их
свежезамороженное или консервированное
мясо находит широкий сбыт. Основным
объектом добычи у нас в Японском море
служит крупный приморский гребешок.
Приморский гребешок живет долго — до
15—16 лет, когда он достигает размера
18—20 см, однако такие старые особи
попадаются редко, обычно же встречаются
гребешки в возрасте 7—9 лет. Половая
зрелость наступает у них на третьем году
жизни (при размере 9—10 см),
размножение происходит летом (июнь — июль).
Пяти-, шестилетние самки A2—13 см) мо-
126
гут выметывать до 30 млн. яиц. Планнтон-
ная личинка велигер довольно скоро
оседает на дно, превращаясь в молодого
гребешка (малька). В большом количестве
мальки обитают в прибрежных зарослях
бурых и багряных водорослей, то
прикрепляясь к ним биссусом, то плавая, то
ползая при помощи ноги (которая затем
у взрослых гребешков редуцируется).
К концу осени мальки достигают размера
7-10 Плотность поселений
приморского гребешка у берегов Южного
Приморья достигает 7—10 экземпляров на 1 м2.
В Японии так интенсивно используют
запасы гребешков, что здесь начали
разводить приморского гребешка
искусственно, хотя это очень трудоемкое и
дорогое мероприятие. Для этого молодь
гребешков собирают в море после их
оседания и пересаживают на бамбуковые
палки-коллекторы, с которыми их потом
переносят для выращивания в
подходящие для этого участки морского дна.
Близки к рассмотренному выше
семейству морских гребешков моллюски из
семейств спондилюсов (Spondylidae)
и лим (Limidae).
Лимы имеют изящную
овально-ребристую раковину; замковые зубы не развиты,
мускул-замыкатель один. Мантийных глаз
нет, но имеются чувствительные выросты
по краю мантии, часто очень длинные
и ярко окрашенные. Имеется также
мантийный парус, поэтому лимы могут
плавать скачками, как и гребешки. Иногда
лимы образуют биссус, с помощью
которого некоторые виды, например
зияющая лима (Lima hians), могут строить
«гнезда», скрепляя биссусом куски
ракушки, мелкие камешки, обломки
кораллов и пр. После того как этот материал
скреплен сетями из биссусных нитей,
лима выстилает свой «дом» изнутри нежной
пряжей из этих нитей, и он становится
похожим на уютное птичье гнездо.
По-видимому, лима, живущая в таком гнезде,
редко его покидает, выпуская наружу
лишь густую подвижную бахрому краев
мантии, которые способствуют созданию
токов воды для ее питания и дыхания.
Однако многие лимы ведут и свободный
образ жизни, как, например, небольшие
белые северные лимы — Lima (Limatula)
hyperborea, обитающие в северных морях,
или многие более глубоководные виды,
встречающиеся в открытых частях морей
и реже на ложе океанов.
Моллюски из семейства Аномии
(Anomiidae) ведут прикрепленный образ
жизни. Только что осевшие на скалы и
камни молодые аномии прочно
прикрепляются к ним биссусом, который позже
пропитывается известью, превращаясь в
твердый стебелек, выходящий в отверстие
в нижней створке раковины. Зубов нет;
форма раковины неправильно округлая,
так как повторяет обычно неровности
камней, к которым плотно прилегает.
Обычна для морей теплой и умеренной
зон Атлантики Anomia ephippium, а в
дальневосточных морях гигантская
аномия — Pododesmus (Monia) macrochisma,
или Monia macrochisma. Размеры первой —
около 1—2 см, а второй — до 9—10 см.
Обе обитают на камнях и скалах
прибрежной зоны и небольших глубин. A.
ephippium встречается также в Средиземном
море, но в Черное не проникает.
К аномиям близка крупная
тропическая плакуна (Placuna placenta),
имеющая почти плоскую полупрозрачную
раковину 13—14 см в поперечнике. Она
живет на скалистых грунтах в Индийском
и в западной части Тихого океана. В
Индии, Китае, на о. Целебес и на
Филиппинах ее створки издавна употребляют
вместо стекол в окнах домов. Даже теперь
в одной только Маниле для этих и
других целей ежегодно добывается около
5 млн. штук плакун; в настоящее время
ее здесь даже разводят. Раковины плакун
размалывают также в серебристый
порошок, из которого приготовляют краску.
Мясо плакун нередко употребляют в пищу.
Устрицы (сем. Ostreidae) — одна из
самых популярных промысловых групп
двустворчатых моллюсков, которые
употребляются человеком в пищу с
незапамятных времен. Об этом свидетельствует
нахождение их раковин в так называемых
«кухонных кучах» — отбросах,
оставшихся от древних поселений людей, живших
еще в каменном веке на берегах морей
(Черного, западной части Балтийского
моря и др.). Упоминание о промысле
устриц находят еще у Плиния (т. е. за 150
127
лет до начала нашего летосчисления).
Все это говорит о том, что высокие
вкусовые и питательные качества устриц
были известны людям уже очень давно.
В результате долгого хищнического
истребления устричных банок уже в середине
XIX в. встал вопрос о необходимости
строгого регулирования промысла
устриц и об искусственном их разведении.
В Японии культура устриц возникла еще
в XVII в.
Устрицы имеют несимметричные грубо-
чешуйчатые створки раковин
изменчивой формы; при этом левая имеет блюдце-
образную форму, а правая (более плоская)
прикрывает ее, как крышечкой. Левой
(нижней) створкой устрица прирастает
к камням или скалам, а часто и к
створкам других устриц или иных сидячих
моллюсков. Нога и биссус у взрослых
устриц полностью редуцированы.
Известно около 50 видов устриц. Все они
тепловодны и не проникают на север
далее 66° с. ш. Основным промысловым
видом в разных морях служит обыкновенная,
или съедобная, устрица (Ostrea edulis),
обычная у берегов Европы — от
Норвегии до Алжира и в Средиземном море,
а в Балтийском море она отсутствует из-за
пониженной его солености. Издавна
промышляется и разводится: так, во
Франции устриц начали разводить в
семидесятых годах прошлого столетия.
Настоящая съедобная устрица — очень
изменчивый вид (как и вообще больший,
ство видов устриц) и образует различные
местные расы и морфы, нередко
считающиеся даже отдельными видами, как
например адриатическая устрица (О. е.
adriatica), богатые устричники которой
были известны издавна, скаловая устрица
(О. е. sublamellosa) и наша черноморская
грядовая устрица (О. е. taurica). У
атлантических берегов Франции промышляют
также португальскую устрицу (Crassostrea
angulata).
В Черном море устрицы встречаются как
в защищенных бухтах и на прибрежных
скалах, так и на мелководьях более
открытых частей моря, образуя в зоне
ракушечника биоценозы устричных гряд и
банок. Большие устричные банки у
Крымского и Кавказского берегов (например,
Гудаутская банка) почти не используются
промыслом. Кроме того, они сильно
пострадали от выедания устриц и мидий
хищным брюхоногим моллюском рапаной
(Rapana).
В США промышляют и разводят: по
Атлантическому побережью виргинскую
Таблица 10. Двустворчатые моллюски морей СССР.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
1 — морской черенок (Siliqua alta);
2 — мактра полосатая (Mactra sulcataria);
3 — мактра сахалинская, или белая ракушка (Spisula sachali-
nensis);
4 — мактра овальная S. (Mactromeris) voyi;
5 — мактра шизотерус (Schizothaerus keenae);
6 — саксикава арктическая, или обыкновенная (Hiatella arctica);
6а — она же, без шипов;
7 — саксикавида панопеа японская (Panope japonica);
8 — песчаная ракушка Муа (Arenomya) arenaria;
8а — то же, общий вид с телом;
9 — корбула амурская (Potamocorbula amurensis);
10 — камнеточец фолас (Pholas dactylus) — вид со спинной
стороны; Юа — правая створка; юб— левая створка;
11 — цирфея гребенчатая (Zirfaea crispata);
12 — цирфея Пилсбри (Z. pilsbryi);
13 — морское сверло фоладидея Pholadidea (Penitella) penita;
14 — песчаная лионзия (Lyonsia arenosa);
15 — пандора холодноводная (Pandora glacialis);
16 — трация (Thracia myopsis);
17 — морская ложечка периплома (Periploma fragilis).
Отряд Перегородчатожаберные (Septibranchia):
18 — глубоководная лионзиелла (Lyonsiella abyssicola);
19 — поромия (Poromya granulata);
20 — куспидария арктическая (Cuspidaria arctica).
устрицу (Crassostrea virginica), a no
Тихоокеанскому — калифорнийскую устрицу
(Ostrea lurida).
У берегов Японии встречается
несколько видов устриц, из которых промышляют
и разводят гигантскую устрицу
(Crassostrea gigas, или О. laperousi), японскую
устрицу (О. nippona), листоватую устрицу
(О. denselamellosa) и др. Обычная в
Японском море гигантская устрица встречается
у берегов Приморья и Сахалина, образуя
богатые скопления — устричники,
которые у нас еще не используются. В начале
текущего столетия эту устрицу перевезли
на Тихоокеанское побережье США (штат
Вашингтон), где ее широко промышляют.
Гигантская устрица очень изменчива
по форме — от длинной продолговатой
до овально-укороченной. Много других
видов устриц промышляется в Австралии,
Новой Зеландии, в Индии и многих
других странах.
Благодаря своему большому
промысловому значению и необходимости
искусственного разведения строение и биология
устрицы хорошо изучены в Японии, США
и других странах, особенно ранние стадии
ее развития. Искусственно
оплодотворенные яйца устриц хорошо развиваются
и выращиваются в специальных бассейнах
(«танках»); для их питания служит мелкая
водоросль зоохлорелла, также
искусственно культивируемая.
Устрицы обычно живут на жестких
грунтах — камнях, скалах или на
смешанных песчанисто-каменистых грунтах,
на небольшой глубине, от 1 до 50—70 м.
Различают устричные банки и береговые
поселения (устричники), которые в
Японском море, например, могут простираться
на 300—400 м от берега. Устричные
банки находятся на отдельных
мелководьях, в некотором удалении от берега»
Устрицы очень чувствительны к
температуре воды, особенно во время их
размножения, которое происходит при
температуре около 18—20° С.
Устрицы могут выносить некоторое
опреснение. Минимальная соленость, при
которой они могут существовать, не ниже
12°/00 A,2%).
Соленость воды отражается на росте
устриц и их вкусовых качествах. Наибо-
Рис. 85. Промысловые устрицы: А —
черноморская устрица (Ostrea edulis taurica); Б, В, Г—
верхние створки различной формы у
дальневосточной устрицы (Crassostrea gigas).
I — свободная (правая) верхняя створка, обросшая
морскими желудями; 2 — нижняя створка,
прирастающая к субстрату.
лее жирных и вкусных устриц собирают
при солености между 20 и 30°/00 B—3%),
там, где наблюдается небольшое
опреснение речными водами. При высокой
морской солености (до 3,5%) они растут
хорошо, но мясо их становится жестким и
неприятным на вкус; при еще большей
солености (около 3,7%) рост устриц замедля-
9 Жизнь животных, т. 2
129
ется. Эти их свойства были хорошо
известны даже древним римлянам, которые
выдерживали собранных в море устриц
в небольших опресненных водоемах.
Отмечалось, что самые вкусные устрицы
добывались лишь там, где имелся приток
пресных вод.
Имея открытую мантию и жабры,
устрицы очень чувствительны к чистоте воды
и к достаточному количеству в ней
кислорода для дыхания. Поэтому часто
наблюдается гибель целых устричных банок
при заносе их илом и песком после
штормов; ил засоряет жабры устриц и делает
невозможной фильтрацию ими воды,
необходимой для питания и дыхания.
На банках устрицы иногда живут
очень плотно, тогда их раковины стоят
вертикально, брюшным краем вверх;
часто устрицы живут друг на друге,
в несколько ярусов, образуя громадные
сростки—«щетки». Местами количество их
в среднем составляет около 20—50 штук
на 1 м2 площади дна. Кроме того, они
могут жить и одиночными экземплярами.
Почти все устрицы (Ostrea)
гермафродиты, мужские и женские доли гонад у
них перемешаны, а иногда яйца и
спермин производит одна и та же долька.
Возможность самооплодотворения
исключается т. к. разные половые продукты
созревают в разное время. У одной и той же
особи пол может периодически меняться.
Оплодотворенные яйца некоторое время
находятся в мантийной полости
материнской особи, и в воду попадают уже
сформированные личинки. Число яиц,
откладываемых обыкновенной устрицей,
изменяется от 300 тыс. до 6 млн.
Личинка парусник (veliger), размером около
0,2 мм, имеет уже маленькую
двустворчатую раковину. Оседая, личинка
выбирает для себя подходящий грунт, к
которому прикрепляется вначале при помощи
биссуса, а затем уже всей раковиной.
Промысловые виды Crassostrea
раздельнополы; гермафродитные особи крайне редки.
Оплодотворение наружное. Как
указывалось выше, культура устриц развита в
настоящее время очень широко.
Разработка и применение основных
методов выращивания устриц принадлежит
Франции, а именно Котэ, которому
Франция обязана существованием и развитием
современной устричной промышленности.
Первая задача, которую приходилось
разрешать для разведения устриц, это
сбор молоди устриц, так называемого
с п а т а, и пересадка ее на коллекторы,
которые ставятся на устричных банках
в период размножения устриц. Для этого
вначале применяли фашины (пучки
прутьев), черепицу или другие материалы.
В Японии применяют бамбуковые стебли.
Коллекторы со спатом стоят в воде
в течение года, пока молодь не подрастет.
Весной спат снимается с коллекторов
и переносится в особые ящики —
приемники для вскармливания. Приемники
держат в «устричных парках», т. е. на
специальных огороженных участках
морского дна. Здесь они содержатся около
двух лет, затем их продают на устричные
заводы для откорма. Здесь они живут
в мелких искусственных бассейнах, «клэ-
рах», до 4—5 лет, после чего их можно
продавать. Воду в этих бассейнах обычно
удобряют минеральными солями, что
приводит к быстрому развитию
одноклеточных водорослей, которыми и питаются
устрицы. Одна особь виргинской устрицы
нормально может профильтровать от
5 до 16 л воды за час. Перед продажей
устриц переносят в специальные
«очистительные» бассейны, где выдерживают без
пищи несколько дней в чистой свежей
воде, после чего моют щетками, а воду
спускают. Теперь они готовы к упаковке.
Промысел и разведение устриц
получили широкое развитие в очень многих
странах; искусственное разведение устриц
дало большое увеличение их уловов и
представляет целые особые отрасли устричной
промышленности. По величине вылова
устрицы занимают первое место среди
двустворчатых моллюсков.
ОТРЯД НАСТОЯЩИЕ
ПЛАСТИНЧАТОЖАБЕРНЫЕ
(EULAMELLIBRANCHIA)
К отряду настоящих
пластинчатожаберных моллюсков относится
наибольшее число видов, которые обитают как
в морских, так и в пресных и солоноватых
130
Рис. 86. Двустворчатые моллюски, обитающие на поверхности дна и неглубоко закапывающиеся
(схема):
А — нукула; Б — куспидария; В — йольдия; Г — дидакна; Д — пектункулюс; Е — астарта;
Ж — циприна; 3 — тапес и венерупис, различные стадии движения.
1 — нога; 2 — вводной сифон; з — выводной сифон; 4 — ротовые щупальца и придатки
(стрелки показывают направление движения воды).
Рис. 87. Двустворчатые моллюски, глубоко закапывающиеся в грунт (схема):
А — черенок солен, различные стадии закапывания; Б — мия; В — скробикулярия;
Г — теллина; Д — донакс; Е — макома; Ж — лорипес и тиазира.
1 — нога; 2 — вводной сифон; 3 — выводной сифон; 4 — трубчатый ход, выстланный сливью.
9*
водах. Встречаются во всех морях и в
океанах на самых различных глубинах — от
приливо-отливной зоны и почти до самых
больших глубин Мирового океана A0,8
км), в одной из глубоководных впадин
в южной части Тихого океана.
Основным признаком отряда является
строение их жабр, которые представляют
собой две двухслойные решетчатые, слож-
нопродырявленные полужабры с каждой
стороны тела. Каждая полужабра состоит
из рядов коленчатоизогнутых жаберных
нитей, соединенных между собой соедини-
тельноткаными или сосудистыми
спайками и образующих внутреннюю и
наружную жаберные пластинки с полостью
между ними. Спайки имеются и между
восходящими и нисходящими коленами
самих жаберных нитей.
Замок устроен различно, но у
большинства видов он так называемого гетеродонт-
ного типа и состоит из центральных
(кардинальных) и боковых зубов;
характер, количество и расположение зубов
могут варьировать, отдельные части
замка или весь замок в целом могут быть
редуцированы. Передний и задний замы-
кательные мускулы одинаково развиты.
Многие настоящие
пластинчатожаберные моллюски являются формами инфау-
ны, т. е. обитают в толще различных
грунтов, в большей или меньшей степени
закапываясь в них. Для глубоко
закапывающихся форм характерны уплощенные,
гладкие или очень слабо скульптирован-
ные раковины, сросшаяся мантия с
отверстием для выхода ноги, длинные
разделенные или сросшиеся сифоны.
Благодаря таким сифонам, находясь «в
укрытии» в толще грунта (спасаясь от
врагов всех двустворчатых моллюсков—
морских звезд или брюхоногих
моллюсков — букцинид, натицид и др.), они
могут засасывать необходимую им для
дыхания и питания воду из самых
придонных слоев, собирать детрит с
поверхности грунта.
Формы, ползающие по поверхности
грунта или лишь частично и неглубоко
закапывающиеся в него, относятся к так
называемой о н ф а у н е. Они обычно
обладают крепкой, вздутой ребристой
раковиной с хорошо развитым замком; у них
несросшаяся мантия, сильная нога,
небольшие или даже зачаточные сифоны.
Ребра укрепляют створки раковины, и
различные выросты на них препятствуют
«проваливанию» животных в грунт и
помогают им удерживаться в самых
поверхностных слоях грунта.
У форм, закапывающихся, вбуравливаю-
щихся в грунт, наблюдается ослабление
и даже полная редукция замковых зубов
и усиление роли внутренней связки,
а сверлящие формы лишены и замка
и связки.
Отряд Настоящие
пластинчатожаберные включает более ста семейств и
разделяется на четыре подотряда: Рас-
щепленпозубые (Schizodonta), Разнозубые
(Heterodonta) — самый большой по
количеству семейств, родов и видов, Адапе-
донты (Adapedonta) и Аномальносвязоч-
ные (Anomalodesmata).
Обширную группу настоящих
пластинчатожаберных моллюсков, целиком
приспособившихся к жизни в пресной воде,
составляют прежде всего моллюски из
большого подотряда расщепленнозубых',
из подотряда разнозубых к пресным
водам приурочены моллюски из надсемей-
ства Sphaeriacea, а также почти все
виды из семейства Dreissenidae.
Все эти указанные группы настоящих
пластинчатожаберных широко
распространены в пресных водах всех континентов,
особенно в водах Америки, где их видовое
разнообразие особенно велико.
Большинство видов расщепленнозубых
относится к надсемейству Unio-
nacea, или Najadacea), в которое входит
2семейства: пресноводные жемчужницы
(Margaritiferidae) и униониды (Unionidae).
У всех у них раковина имеет хорошо
развитый перламутровый слой, нога
большая, топоровидная, связка наружная,
позади макушек; мантия несросшаяся,
сифоны зачаточные. Замок, если он
имеется, очень изменчивого строения и
относится к особому унионидному («псевдогете-
родонтному») типу; зубы могут быть
более или менее расщепленными,
расходящимися и несут функции центральных
или боковых зубов.
Очень характерно развитие у Unionacea:
молодь развивается у материнских особей
132
Рис. 88. Обыкновенная пресноводная
жемчужница: А — вид снаружи (правая створка); Б —
вид изнутри (левая створка).
1 — передний конец раковины; 2 — задний конец;
3 — наружная связка; 4 — отпечаток места
прикрепления переднего мускула-замыкателя; 5 —
отпечаток заднего мускула-эамыкателя; 6 — ложнокарди-
нальные зубы; 7 — жемчужина; 8 — отпечаток края
мантии.
внутри жаберных полостей, играющих
роль выводковой сумки — марзупи-
у м а, где и формируется особая
личинка — глохидий, которая затем
выходит в воду и закрепляется на жабрах
или на коже различных рыб (окунь,
чехонь, язь, ерш и др.). Затем глохидий
инкапсулируются, паразитируя в
течение одного-двух месяцев на рыбах, где
и происходит их превращение в молодых
моллюсков. На одной рыбе может
«выкормиться» огромное количество глохидиев:
так, на жабрах одного небольшого окуня
длиной около 8 см было найдено около 400
глохидиев. Выйдя из мешочков (капсул),
молодь моллюсков оседает на дно.
Унионид употребляют в пищу, но
используют главным образом на откорм
домашней птицы и скота (свиней); раковины
многих их видов, а также и жемчужниц
используют для различных изделий из
перламутра. В 1963 г. мировая добыча
пресноводных двустворчатых моллюсков,
главным образом Unionidae (и отчасти
Corbiculidae), составляла 350 тыс. ц.
В семейство пресноводных ЖеМ/~
чужниц входит всего один род — Marga-
ritifera, или Margaritana, виды которого
живут в Северной Америке, Европе и на
севере Восточной Азии, в том числе на
Сахалине и в Японии.
Это наиболее примитивные формы из
Unionacea, имеющие редуцированные
боковые зубы; жабры их не срастаются
сзади с мантией. У нас известна главным
образом обыкновенная жемчужница (Marga-
ritifera margaritifera) длиной до 12 см.
Обитает в небольших реках севера нашей
страны, а также на севере Европы,
Америки и в Японии. Добывается ради
жемчуга давно. В реках Дальнего Востока живет
несколько видов жемчужниц, например:
даурская жемчужница (М. dahurica) длиной
до 18 см, обитающая в бассейне р. Амура
и в Приморье (ее издавна промышляли
ради перламутра и жемчуга); камчатская
жемчужница (М. middendorffi) длиной
9 см; овальная раковина этой жемчужницы
отличается от беззубок значительно
большей толщиной створок, мощным
перламутровым слоем, наличием замка с одним-дву-
мя центральными зубами.
Жемчужницы живут только в чистых
проточных водах (реках, ручьях), не
встречаясь в стоячих водоемах, так как
они особенно требовательны к чистоте
и аэрации воды. Современный ареал их
распространения сильно сократился по
сравнению с прошлым: так, они псчезлп
в реках, загрязненных сточными водами
городов и фабрик, лесосплавом, из-за
изменения химизма воды, а также при
заболачивании. Кроме того, хищническое
истребление жемчужниц в погоне за
жемчугом и перламутром, имевшее место
в прошлом, также подорвало запасы этих
ценных раковин. Их запасы трудно
восстанавливаются из-за медленного роста
этих моллюсков. На первом году жизни
жемчужницы достигают размера 0,5 смг
на пятом году — 2 см, к 7—8 годам —
3—4 см, а на десятом — 6 см, давая затем
ежегодно прирост лишь около 1 мм;
самые крупные экземпляры в 12—13 см
имеют возраст около 70 лет.
133
Соответственно медленно растут и
жемчужины: за 12 лет они вырастают с
горошину, а размера в 8 мм достигают за 30—
40 лет; жемчуг высокого качества.
Раковины жемчужниц используют для
изготовления главным образом
перламутровых пуговиц.
Обнаружить жемчужниц в реках, в
которых они сохранились, нелегко. Обычно
это реки с быстрым течением, с порогами,
валунами, где река бурлит и клокочет,
Рис. 89. Перловицы: Л — перловица
обыкновенная (Unio pictorum), правая и левая створки;
Б — перловица вздутая, или клиновидная (U.
tumidus); В — перловица толстая (U. cras-
sus); Г — ланцеолярия ножичковидная (Lan-
ceolaria grayana, левая створка).
1 — ложнокардинальные зубы; 2 — ложнобоковые
зубы.
а потом течет спокойно по спокойному
плесу до следующего порога. На плесах на
глубине нескольких метров жемчужниц
можно ловить только драгой, но зато
на мелких порожистых местах их можно
собирать руками. Наиболее густые их
поселения встречаются в области несколько
замедленного течения. Если место
выбрано удачно, то за час можно набрать
несколько десятков жемчужниц.
Плотность поселения их местами может
доходить до 60 экземпляров на 1 м2.
Оплодотворение, выход яиц и заражение
ими жабр у жемчужниц наблюдается
в июле — августе. В отличие от перловиц
и беззубок их яйца развиваются до глохи-
диев во всех четырех (наружных и
внутренних) полужабрах материнского
организма. Глохидии очень мелкие и лишены
клювовидного острого зубца; уже в
сентябре они встречаются на коже и жабрах
рыб (хариуса, горчака и др.).
В семейство Unionidae входит
около 80 родов, объединяемых в 6
подсемейств, из которых наиболее важны
перловицы (Unioninae), беззубки (Anodontinae),
а также самое обширное из них
подсемейство лампсилин (Lampsilinae),
куда входит более 20 родов и около 600
видов, обитающих исключительно в
Северной и Центральной Америке,
преимущественно в реках, впадающих в Тихий океан.
Перловицы широко распространены
в Европе, Азии, Африке, Северной и
Центральной Америке, Японии, на Малайском
архипелаге.
Наибольшее разнообразие перловиц
наблюдается в Восточной Азии, где известно
около тридцати их родов и большое
число видов; в Северной Америке известно
около 12 родов, в Африке 3 рода, а в
Европе 1 род — настоящие перловицы (Unio,
или Nodularia).
Настоящие перловицы (Unio) обладают
крепкой раковиной, хорошо развитым
замком с одним-двумя центральными и
хорошо развитыми пластинчатыми боковыми
зубами. Наружные полужабры сзади до
конца срастаются с мантией, а края
мантии срастаются только над верхним
(выводным) сифоном; верхний и нижний
(вводной) сифоны разделены
перегородкой.
134
Глохидии довольно крупные, с
шиповатым зубом, развитие их происходит
обычно только во внешних полужабрах.
Оплодотворение яиц и заражение ими
жабр у перловиц наблюдается с конца
апреля до июня; созревание глохидиев
в жабрах и выбрасывание их происходит
с конца мая по август. Вынашивание
яиц и развитие глохидиев продолжается
20—40 дней.
В Европейской части СССР хорошо
известны и широко распространены три
очень сильно изменчивых вида.
Перловица обыкновенная (Unio pictorum), длиной
до 14,5 см. Длинная узкая раковина ее
с почти параллельными спинным и
брюшным краями. Обитает в реках и озерах
всей Европейской части нашей страны,
в Средней и Северной Европе.
Перловица клиновидная, или вздутая
(U. tumidus), с выпуклой, суженной на
заднем конце раковиной, длиной до И см.
Ее распространение сходно с предыдущим
видом.
Перловица толстая (Unio crassus), с
овальной, довольно вздутой раковиной, нередко
с розовым перламутровым слоем внутри.
Обитает в реках Европы и Европейской
части СССР, в озерах обычно не встречается.
В реках Дальнего Востока, в озере Ханка,
в Приморье наиболее хорошо известна:
дальневосточная перловица (U. douglasiae),
а также ножичковидная ланцеолярия (Lan-
ceolaria grayana), с сильно вытянутой
почти цилиндрической раковиной,
закругленной спереди и суженной сзади. Оба
эти вида обитают также и в Китае,
а дальневосточная перловица — также и
в Японии.
В Западной и Центральной Сибири в
настоящее время перловиц нет; богатая их
фауна, бывшая здесь в плиоцене, вымерла
во время ледникового периода, а
физические преграды (горные цепи и большие
сибирские реки), видимо, препятствовали
проникновению в Сибирь амурских видов
перловиц. Отсутствие их в
среднеазиатских реках (Аму-Дарья и Сыр-Дарья)
объясняется, видимо, большой мутностью
воды этих рек.
Обнаружить перловиц в реке или в
озере нетрудно по их следам — бороздкам,
которые они оставляют на мягком грунте
Рис. 90. А — гребенчатка Cristaria plicata;
Б — схема изменчивости обыкновенной
европейской беззубки (Anodonta cygnea), контуры
раковины.
1 — Anodonta cygnea cygnea; 2 — беззубка
вытянутая (A. cygnea cellensis); 3 — беззубка рыбья (A.
cygnea piscinalis); 4 — беззубка утиная (A. cygnea ana-
tina); 5 — беззубка озерная (A. cygnea lacustris).
дна. Наиболее благоприятен для них
илисто-песчаный грунт; вязкого ила они
избегают.
Движения перловиц очень медленны и
вялы A—1,5 м в час). К осени перловицы
зарываются в ил, погружаясь в него почти
целиком; там они и зимуют, плотно
закрыв створки раковины и пребывая в
состоянии оцепенения. Перловицы весьма
чувствительны к недостатку кислорода в
воде: они отсутствуют там, где кислорода
мало. Питаются, как и все Unionacea,
детритом, взвешенным в воде, и мелкими
планктонными организмами; для питания
и дыхания профильтровывают большое
количество воды, способствуя тем самым
очищению ее от взвеси. Очень много
различных родов перловиц обитает в Южной
и Юго-Восточной Азии (Индии,
Индокитае, Китае и т.д.), островах Индонезии
(Суматре, Яве, Борнео и др.)-
Подсемейство беззубок (Anodon-
tinae) охватывает значительное
разнообразие форм пресноводных двустворчатых,
большинство родов и видов которых живет
в Северной и Центральной Америке, но
где они все же значительно уступают по
количеству видов лампсилинам (особенно)
и перловицам.
Беззубки лишены замковых зубов, их
раковины большей частью тонкостенные,
с довольно слабым перламутровым слоем;
края мантии образуют лишь зачаточные
сифоны; их молодь — эмбрионы —
вынашивается с осени в течение всей зимы (в
период так называемой жаберной
беременности) в наружных полужабрах. Перед
этим периодом у них развиваются особые
вентиляционные трубочки, проводящие
воду и значительно улучшающие снабжение
кислородом развивающихся в жабрах
эмбрионов. Выбрасывание обычно довольно
крупных глохидиев, вооруженных двумя
(по одному на каждой створке) крючками,
происходит весной.
У нас хорошо известна беззубка
обыкновенная европейская (Anodonta cygnea),
достигающая 20 см длины. Живет она в
Средней и Северной Европе, в
Европейской части СССР и в Западной Сибири,
обитая в прудах, озерах и речных заводях
и затонах. Это очень изменчивый вид,
образующий (чуть ли не в каждом большом
водоеме, районе и т. п.) различные морфы,
экологические расы, разновидности.
Некоторые из них считаются нередко даже
отдельными видами, как, например:
живущая в озерах и речных старицах
вытянутая беззубка (Anodonta cygnea cellensis),
обитающие в озерах и реках рыбья
беззубка (A. cygnea piscinalis) и утиная
беззубка (A. cygnea anatina).
В реках Европейской части СССР
обитает также и узкая беззубка (Pseudanodonta
complanata) с очень сжатой, удлиненно-
овальной раковиной; проникает в сильно
опресненные участки наших внутренних
морей. На Кавказе в реках, старицах,
озерах с прозрачной водой обитает
кавказская беззубка (Anodonta cyrea).
На Дальнем Востоке, в Приморском
крае, а также в Китае в реках и озерах
обитают наиболее крупные виды из рода
анодонт с сильно вздутой раковиной —
беззубка Вуда—Anodonta (Pteranodon) wo-
odiana, до 15 см длиной, и сводчатая
беззубка— Anodonta (Haasiella) arcaeformis,
до 10 еле. В бассейнах рек Амура и Уссури,
в озере Ханка, озерах Монголии и реках
Китая, в Японии обитает наиболее
крупный вид пресноводных двустворчатых
моллюсков гребенчатка (Cristaria plicata),
одна из разновидностей которой (v. hercu-
lea) достигает 34 см длины и веса 1 кг.
Встречается она на глубине 1,5—2 ле, на
мягких илистых грунтах, в хорошо
прогреваемых и защищенных участках озер
или в реках со слабым течением. В Китае
гребенчатку используют для добычи
перламутра. У гребенчатки широкая, довольно
толстостенная раковина, у которой
спинные края образуют высокий плоский
гребень. Замковые зубы рудиментарны. В
озере Ханка гребенчатка может встречаться в
количестве до 10 экземпляров на 1 м2 дна.
Лампсилин (Lampsilinae) в США не
только вылавливают, но довольно давно
искусственно разводят, особенно
некоторые быстро растущие виды из родов Lamp-
silis, Obovaria и др. При этом рыб
искусственно заражают их глохидиями. У
многих лампсилин наблюдается половой
диморфизм, выраженный даже в форме их
раковины. Выводковая камера образуется
в задней части наружных полужабр.
Кроме перловиц и беззубок,
постоянными обитателями наших пресноводных
водоемов являются мелкие моллюски из
подотряда разнозубых, из надсе-
м е й с т в a Sphaeriacea. Это прежде
всего такие широко известные и обычные
обитатели прудов, рек, озер и луж, как
шаровки (Sphaerium) и горошинки (Pisidium)
из семейства Sphaeriidae.
Шаровки — это мелкие двустворчатые
с округлой раковинкой, которая у самых
крупных видов— у речной шаровки
(Sphaerium rivicola) — достигает лишь 2,5 см
длины. Шаровки «живородящи»: яйца
развиваются у них в особых выводковых
камерах, которые образуются на внутренних
жабрах, так что из материнской раковины
выходят уже готовые молодые ракушки.
Интересно отметить, что количество этой
молоди обычно не превышает десятка, в то
время как глохидии у перловиц
выбрасываются сотнями. У шаровок забота о по-
136
томстве оказывается весьма эффективной,
так как обеспечивает им выживание и
расселение вида. Гермафродиты.
Шаровки встречаются в Европе, Азии,
Америке, Африке. Из обитающих в речных
водах в пределах СССР наиболее обычны
речная шаровка (Sphaerium rivicola),
прочная шаровка (S. solidum), некоторые
разновидности роговой шаровки — Sph. corneum;
остальные разновидности этой шаровки
обитают в стоячих водоемах.
По наблюдениям в реке Эльбе, шаровка
роговая может иметь две генерации в год:
моллюски, родившиеся под осень, после
зимнего покоя, в апреле — мае растут, в
июне размножаются, а в августе отмирают.
Летнее потомство этой генерации,
родившееся от нее в июне, в августе — сентябре
размножается, а в октябре отмирает, т. е.
живет всего 5 месяцев, в то время как
моллюски, родившиеся осенью, живут
почти год.
Самые мелкие представители
семейства горошинки (Pisidium) населяют
наши пресные водоемы, как стоячие, так и
проточные, почти повсеместно, от рек,
озер до прудов, болот и пещер.
Горошинки чрезвычайно разнообразны
и изменчивы и образуют большое
количество видов и подвидов, чему, видимо,
способствует изолированное их местообитание
в отдельных водоемах. Наиболее обычны
речная горошинка (Pisidium amnicum),
блестящая горошинка (P. nitidum),
болотная горошинка (P. casertanum) и др.
Кроме шаровок и горошинок, к Sphae-
riacea также относится семейство
Corbiculidae, виды которого обитают в
пресных и солоноватых водах субтропиков
и тропиков. Крупные виды корбикул
промышляют. У нас в Закавказье и в Средней
Азии в озерах, реках, оросительных
каналах, а также в заливах Южного Каспия
обитает западная корбикула (Corbicula
fluminalis), а в Приморье — восточная
корбикула (С. fluminea).
Своеобразную группу представляют
собой небольшие пресноводные и солонова-
товодные моллюски дрейссены (Dreissena,
или Dreissensia) и конгерии (Congeria) из
семейства Dreissenidae. Хотя
дрейссены по форме раковины напоминают
мелких митилид, однако они им не
родственны, а относятся к отряду настоящих
пластинчатожаберных (Eulamellibranchia).
Дрейссены имеют заостренный передний
конец раковины и расширенный задний.
Замковых зубов нет. Внутри, в самой
передней части раковины образуется
перемычка (септа), к которой прикрепляется
передний небольшой мускул-замыкатель.
Мантия имеет отверстие для выхода ноги
и образует два коротких сифона; нога с
биссусом. Развитие со свободно
плавающей личинкой.
Дрейссены раздельнополы. Яйца
выметываются прямо в воду в небольших
слизистых комочках, в которых содержится
много мелких яиц. После их
оплодотворения развивается личинка велигер,
которая .плавает короткое время, а потом
оседает на дно, после чего прикрепляется
биссусом к субстрату. Наличие
плавающей личинки — черта, свойственная чисто
морским моллюскам, и это указывает на
морское происхождение дрейссен.
Рис. 91. Корбикулы, шаровки и горошинки:
А, а — корбикула восточная (Corbicula
fluminea); Б, б — шаровка роговая (Sphaerium
corneum); В —горошинка речная (Pisidium
amnicum); а — замок с кардинальными и
боковыми зубами; б — раковина со спинной стороны.
137
Рис. 92. Дрейссены:
А — дреиссена речная (Dreissena polymor-
pha) — гроздь раковин, сросшихся биссусными
нитями; Б — дреиссена каспийская (Dreissena
caspia); В — дреиссена клювовидная (Dreissena
rostriformis).
Речная дреиссена (Dreissena polymorpha)
имеет зеленовато-желтую очень
изменчивой формы раковину до 3—5 см длиной,
нередко с рисунком из поперечных или
зигзагообразных коричневых полосок;
образует много подвидов.
Распространена речная дреиссена очень
широко, обитая во многих реках, некоторых
озерах Европы и Европейской части СССР
вплоть до Западного Казахстана,в лиманах
рек Черного моря, в опресненных
участках Азовского, Каспийского морей, а
также в Аральском море.
Речная дреиссена живет начиная от
уреза воды до глубины около 10 м;
встречается местами в громадном количестве,
образуя грозди, сростки, щетки, чаще
всего обрастая камни, сваи, различную
подводную растительность и т. п.
Очень сильно развились поселения
дрейссены в крупных водохранилищах Волги,
Днепра, Дона, где она обрастает остатки
затопленных деревьев, кустарников, сваи
и различные гидротехнические
сооружения — водотоки, трубы, проводящие воду
к турбинам и для технического и
питьевого водоснабжения, защитные решетки и
т. д. Все это затрудняет проход воды и
требует постоянной очистки от обрастаний
этих сооружений. Местами дреиссена
может сплошь и в несколько слоев покрывать
всякий подходящий субстрат; ее
количество здесь может достигать 10 тыс.
экземпляров на 1 м2, а вес — 7 кг на 1 м2 и более.
Большая жизнестойкость и высокая
плодовитость дрейссен сильно затрудняют
борьбу с ее обрастаниями. Приходится
применять не только механические, но и
различные химические, электрические и
биологические способы.
Дреиссена живет до 7—8 лет. Половые
продукты у нее созревают в апреле, после
чего начинается порционное икрометание,
которое продолжается все лето, благодаря
чему ее личинки (велигер) наблюдаются
в планктоне водохранилищ с конца мая
(когда температура воды повышается до
15—20°) до глубокой осени.
В период интенсивного размножения
дрейссен количество личинок велигер
может доходить до 50 экземпляров в 1 м3.
Молодь дрейссен, осевшая летом, хорошо
растет и достигает больших размеров,
чем молодь, осевшая осенью. Прирост
молоди за первый год жизни может
достигать 15 мм. Наилучший рост дрейссен
наблюдается на глубине 1,5—2 М/ В более
или менее защищенных местах, особенно
в южных районах.
Как и многие пресноводные моллюски,
речная дреиссена сохраняет свою
нормальную жизнедеятельность не только в
пресных водах, но и при повышении солености
до 2°/00, а ее разновидности из Северного
Каспия живут и при более высокой
солености. Дрейссены из Волги в опытах
начинали отмирать при солености 6—7°/00.
Кроме речной дрейссены, имеется еще
ряд видов дрейссен, свойственных только
Каспийскому или Аральскому морю (так
называемые эндемичные формы, т. е. нигде
больше не встреченные).
138
В Каспийском и Аральском морях все
дрейссены поедаются донными рыбами.
В Каспийском море обитают: каспийская
дрейссена (Dr. caspia), с более мелкой,
узкой, удлиненной раковиной, живущая при
солености 5—110/00> и морская клювовидная
дрейссена (Dr.rostriformis, или Dr. distin-
cta), более крупная, овально-каплевидной
формы, с маленькими острыми
отвернутыми в разные стороны носиками, живущая
при солености до 12,7°/00, и др. Считают,
что в настоящее время каспийская
дрейссена вымерла. Другие представители Dreis-
senidae живут в пресных или солоноватых
водах Америки, Африки, Индии.
Возвращаясь к рассмотрению чисто
морских моллюсков из отряда настоящих
пластинчатожаберных, начнем с наиболее
массовых форм подотряда разно-
зубых (Heterodonta).
Одной из обычных и массовых групп
двустворчатых моллюсков морей
северного полушария являются различные астар-
ты (сем. Astartidae). Для них характерна
большая внутривидовая изменчивость, как
и для других моллюсков, которые
образовывают массовые поселения. Астарты в
целом связаны с холодными или
умеренными водами северного полушария, где они
живут главным образом на шельфе (до
глубин 200 м). Несколько видов известно
также в Антарктике.
Замок у всех астарт имеет довольно
сходное устройство: 2 центральных
(кардинальных) зуба и по одному слабому
боковому в каждой створке.
Астарты не имеют сифонов, поэтому не
могут глубоко закапываться в мягкий
грунт, на поверхности которого они
обычно живут. Молодь всех астарт служит
пищей многим донным рыбам (камбалам,
треске, пикше). Астарты играют большую
роль в сообществах донной фауны
Северной Атлантики, наших северных и
дальневосточных морей.
Наиболее характерной для краевых
морей Северной Атлантики и в Баренцевом
море является астарта крената — Astarte
(Astarte) crenata.
Она имеет плотную желтоватую
концентрически-ребристую раковину, длиной
около 3 см, обычно мелкозубчатую по
внутреннему краю. Живет на мягких илисто-
песчаных грунтах средних глубин.
Отсутствует в Белом море и сибирских морях.
Северная астарта Astarte (Tridonta)
borealis имеет более крупную (до 5 см)
и более тяжелую раковину, покрытую
грубым коричневатым периостраком, почти
без ребер. Обитает на мелководьях всех
морей северного полушария, на мягких
илистых грунтах, в том числе в Белом,
Балтийском и в дальневосточных морях.
Близки к астартам кардиты, венерикар-
дии и другие кардитиды(СаМ1Шае), но они
обладают более мощным замком, обычно с
удлиненными задними кардинальными
зубами. Живут главным образом в теплых
и тропических водах. Очень красивые и
ярко окрашенные раковины у
представителей родов Cardita и Glans. На
мелководьях Берингова и Охотского морей
обитает несколько видов циклокардий:
Cyclocardia borealis, С. crebricostata, С.
paucicostata и др.
В Средиземном море и в Атлантическом
океане живет интересный моллюск из
семейства Glossidae — морское
сердце (Isocardia cor, или Glossus rubicundus),
по форме раковины действительно
напоминающий человеческое сердце. Раковина
тонкая, вздутая, размером до 5 см, имеет
выпуклые сильно закрученные внутрь
макушки.
Исландская циприна (Cyprina islan-
dica, или Arctica islandica) из семейства Аг-
cticidae представляет интерес как
единственный современный представитель
рода; тепловодный североатлантический
европейский вид, обитающий в наших
северных морях — в западной
прибрежной части Баренцева моря
Рис. 93. Морское сердце (Isocardia cor):
1 — правая створка; 2 — вид спереди.
139
Рис. 94. Моллюск комменсал лептонида Pseu-
dophytina rugifera, живущий на морском
десятиногом раке.
режья, а также и в наиболее теплых
районах Белого моря. Циприна имеет
крупную (до 12 см) выпуклую
округлую раковину, покрытую коричневым
лоснящимся периостраком. Замок
сложный, с хорошо развитыми кардинальными
и боковыми зубами.
Благодаря своей тепловодности
циприна служит хорошим показателем
распространения теплых атлантических вод в
Баренцевом море. В эпоху теплого Лит-
торинового моря она была распространена
шире, чем теперь, и заходила гораздо
дальше на восток (до полуострова Таймыр).
В настоящее время циприна является
массовой, руководящей формой сообществ
донной фауны с преобладанием моллюсков;
они распространены на песчаных грунтах
не только в наших морях, но и на
мелководьях Северной Атлантики, у берегов
Исландии, в датских проливах и т. д.
Мелкие циприны служат пищей ряду
промысловых донных рыб; крупными
питаются многие морские птицы, например
серебристая чайка. Птица, нырнув, достает
со дна крупную циприну, а затем, взлетев
в воздух, роняет ее на прибрежные
камни, о которые разбивается раковина даже
крупных циприн. После этого мясо
моллюсков поедается птицами.
Богатое видами (около 200)
семейство Люциниды (Lucinidae)
распространено главным образом в тропических морях.
Они играют здесь большую роль в
питании рыб, а некоторые виды
используются в пищу и человеком. Много крупных
и красивых видов с нежно окрашенными
раковинами, как например Codakia tigeri-
па, достигающая размера 12 см, обитаете
Атлантике, в Тихом и Индийском океанах.
В Черном море встречается лишь два
мелких вида — Divaricella (Lucinella) di-
varicata и Loripes lacteus.
В питании многих донных рыб заметное
место занимают мелкие моллюски из
близкородственного люцинам семейства
Thyasiridae—тиазиры (Thyasira, или Axi-
nus) и аксинопсис (Axinopsis). Они
обладают хрупкой округлой раковиной, нередко
с радиальной складкой, лишенной зубов,
и не имеют сифонов; нога червеобразная
с расширением на конце, способная очень
сильно вытягиваться на расстояние в
4—5 раз большее, чем сам моллюск. При
помощи такой ноги они закапываются в
мягкий грунт приблизительно на глубину,
равную длине своей вытянутой ноги. При
этом они строят два трубчатых хода до
поверхности грунта и выстилают их
слизью, выделяемой ногой. Эти ходы
выполняют для них роль вдыхательного и
выдыхательного сифонов и прочищаются ногой.
Примером моллюсков-комменсалистов
могут служить многие мелкие моллюски
из Leptonidae и Montacutidae, живущие
на жабрах и ножках некоторых
десятиногих раков, на иглокожих, в трубках
червей и т. п.
Очень красивы хамы (Chama) из
семейства Chamidae, которые живут в
тропических морях, прирастая одной из
створок раковины к скалам и камням.
Сердцевидки, или кардиумы (сем. Cardii-
dae),— очень большая группа
мелководных, преимущественно тропических и
субтропических, форм, распадающихся на 5
подсемейств, куда относится более 20
родов и много видов. Для них
характерна сердцевидная раковина с радиальными
ребрами, часто украшенными шипами или
чешуйками, или имеющая острое ребро
(«киль») на задней стороне раковины.
Замок хорошо развит, обычно с двумя
кардинальными и с передним и задними
боковыми зубами в каждой створке, или зубы
могут быть частично редуцированы, как
у группы солоноватоводных Limnocardii-
пае и др.
Сердцевидки обладают сильной
клиновидной ногой и с ее помощью не только
передвигаются и быстро закапываются в
грунт, но и подпрыгивают на несколько
сантиметров над дном, когда им грозит
140
опасность со стороны их врагов, в первую
очередь со стороны морских звезд. Из-за
своих коротких сифонов они не могут
закапываться глубоко, нередко выставляя
над грунтом заднюю половину раковины
с опущенными сифонами.
Сердцевидки, особенно мелкие или
молодые формы, охотно целиком поедаются
рыбами, а у крупных форм рыбы нередко
откусывают лишь их ноги.
Сердцевидки съедобны, и ряд их видов
промышляют. Так, в 1962 г. в Испании их
было добыто 99 тыс. ц, а в Голландии —
24 тыс. ц.
Особенно красивы и разнообразны по
форме, окраске и «вооружению» ребер
раковины сердцевидок, живущих в
мелководных тропических районах, на коралловых
рифах и у островов Тихого, Индийского
и Атлантического океанов.
Красивой, нередко нежно окрашенной,
неравносторонней, высокой треугольной
раковиной, с килеватой, усеченной задней
стороной, с ребрами, часто украшенными
шиповатыми выростами, отличаются
тропические сердцевидки из подсемейства
Fraginae. Сюда относятся различные виды
из родов Фрагум (Fragum), Коркулюм
(Corculum), Hemicardia и др. Не
менее красивы овально-треугольные
раковины Papyridea и Trachycardium с
многочисленными узкими ребрами,
усаженными шипиками или чешуйками
(подсемейство Trachycardiinae). Настоящие кардиумы
(Cardium) из подсемейства Cardii-
пае отличаются вздутыми раковинами с
резкими, широко раздвинутыми ребрами,
по гребню которых нередко проходит
дополнительное нитевидное ребрышко. Их
ребра вооружены шиповатыми, загнутыми
вверх выростами или чешуйками, а
широкие межреберные промежутки имеют
поперечную штриховатость. Обитают настоящие
кардиумы в морях южной части бореаль-
ной зоны и на тропических мелководьях,
известны в Средиземном море, у берегов
Японии и т. д.
В холодных северных морях и в
северной части бореальной зоны количество
видов сердцевидок резко уменьшается, а
их раковина имеет обычно почти гладкие
ребра и бледную окраску. Для северных
морей характерны виды из
подсемейства Laevicardiinae, куда относятся
роды Cerastoderma, Serripes, Clinocar-
dium.
Наиболее обычным видом у
европейских берегов Атлантики является
съедобная сердцевидка (Cerastoderma edule) с
небольшой, до 5—6 см, вздутой крепкой
сердцевидной раковиной, с уплощенными,
почти гладкими, покрытыми мелкими
чешуйками радиальными ребрами.
Изменчивый вид, образует ряд подвидов.
Основной подвид Cerastoderma edule edule —
более округлых очертаний, с более
уплощенными ребрами, особенно в средней
части раковины, с хорошо развитыми
чешуйками на них; обитает в обширном
районе, от берегов Мурмана до Средиземного
моря. Живет главным образом на
песчаных отмелях приливо-отливной зоны
вместе с другими типично литоральными
видами — с балтийской макомой, с песчаной
ракушкой, червем пескожилом и др.
Встречается также на глубине до 10 м. У берегов
Рис. 95. Движение сердцевидок Gardium (Acanthocardia) echinatum, спасающихся
от морских звезд.
141
Рис. 96. Сердцевидки:
А, Б и В — сердцевидка Ламарка — Cerastoderma edule lamarcki
(А — из Черного моря; Б — из Каспийского моря; В — из
Азовского моря); Г — съедобная сердцевидка (Cerastoderma edule edule).
Мурмана, на восточной и северной
границе своего распространения, С. edule edule
может еще не только жить, но и
размножаться, благодаря тому что летом в
мелких лужах, остающихся во время отлива,
вода достаточно прогревается.
Сердцевидка Ламарка (С. edule lamarcki)
обладает более косо-овальной или даже
округло-ромбической раковиной с
килевым перегибом в задней части раковины,
с менее уплощенными, а в задней части
даже угловатыми ребрами; обитает как у
берегов Европы, так и в Средиземном,
Адриатическом, Черном, Азовском,
Каспийском морях, часто в эстуариях,
лиманах, лагунах, при солености от 2—4 до
40°/00, на песчанисто-илистых и даже
смешанных грунтах, до глубины 10—15 м,
реже — глубже. По форме раковины и по
размерам сердцевидка Ламарка очень
изменчива: в Азовском, Каспийском и
Аральском морях она сильно мельчает. В
Балтийском море С. edule заметно мельчает при
понижении солености.
На илистых грунтах холодных
мелководий северных морей и в дальневосточных
морях хорошо известен массовый вид —
сердцевидка волосатая (Clinocardium cili-
atum). Она имеет довольно крупную (до
5—6 см) сероватую раковину, а ее при-
остренные ребра несут ряды волосков. В
Баренцевом море эта сердцевидка в
большом количестве встречается на
илисто-песчаных грунтах его юго-восточных
мелководий; охотно поедается донными рыбами.
Особого упоминания заслуживает
крупная (до 8—10 см длиной) холодноводная
гренландская сердцевидка (Serripes groen-
landicus), с гладкой округлой
раковиной и с малозаметными
ребрами на переднем и заднем ее
концах. Окраска взрослых темная,
сероватая, а молодых более
светлая, с зигзагообразным
рыжеватым рисунком; такие формы
ранее считали отдельным видом и
называли гренландской афроди-
той.
Сильная, довольно длинная,
коленчатоизогнутая снизу
зубчатая нога гренландской
сердцевидки может вытягиваться далеко за
пределы раковины. Ногу
крупных сердцевидок любят откусывать многие
донные рыбы Баренцева моря, поскольку
сам моллюск слишком велик для них;
нога у поврежденных моллюсков
впоследствии регенерирует. На севере
гренландских сердцевидок охотно поедают также
моржи.
Гренландская сердцевидка обитает на
глубине до 75—100 м, начиная от
юго-восточных мелководий Баренцева моря, во
всех наших сибирских морях, водах
арктического побережья Канады и в холодных
районах Охотского и Берингова морей.
Большой интерес представляет группа
сильно изменчивых видов сердцевидок,
обитающих в Каспийском, Аральском и
опресненных участках и лиманах
Азовского и Черного морей из р о д о в Дидакна
(Didacna), Монодакна (Monodacna) и Адак-
на (Adacna). Их нередко выделяют даже в
подсемейство Limnocardiinae,
поскольку они обитают в солоноватых водах;
у них нет боковых зубов и наблюдается
редукция кардинальных зубов. Это реликты
более богатой сарматской морской фауны,
обитавшей в районе наших южных морей в
прежние геологические эпохи. Уже в
плиоцене Понтическое озеро-море имело
богатую дидакнами и монодакнами фауну.
Все виды лимнокардиид, кроме крупных
дидакн, служат пищей многим каспийским
донным рыбам, в том числе осетровым.
Виды рода Didacna — ныне
эндемики Каспийского моря (т. е. обитают
только в нем). У них довольно крупная (до
5—6 см) треугольно-овальная или
овальная раковина, с ясным килеватым
перегибом перед задней частью раковины, с пло-
142
скими радиальными ребрами, с двумя,
реже одним, кардинальными зубами;
сифоны зачаточные, синуса нет. Из
нескольких ныне живущих в Каспийском море
(кроме сильно опресненных районов
Северного Каспия) дидакн наиболее обычны и
многочисленны Didacna trigonoides, D.
barbot de marnyi, D. crassa и D. protracta.
Первая встречается преимущественно в
мелководной прибрежной зоне, на
песчаных и смешанных твердых грунтах, а
последняя из указанных видов — на
мягких заиленных грунтах, на глубине
около 50 м и более.
Монодакны обладают округло-овальной,
зияющей сзади раковиной, обычно с
плоскими радиальными ребрами, с
небольшими сифонами и неглубоким синусом; у мо-
нодакн обычно один конусовидный
кардинальный зуб, а иногда и рудимент
второго (на правой створке). В Каспийском
море широко распространена М. caspia,
особенно изобилующая в северной его
части. Обитает при солености от 0,1 до
12—14°/оо- Встречается также в лиманах
Черного моря. В Таганрогском заливе
Азовского моря, при солености не более
4°/00, живет более крупная (до 4 см) М.
colorata, которую недавно нашли и в
Северном Каспии. Она
акклиматизировалась в Веселовском водохранилище, и ее
переселили в Цимлянское водохранилище
для улучшения его кормности для рыб.
Настоящие адакни (Adacna) имеют
также не килеватую, зияющую спереди и
сзади, почти гладкую, с очень низкими
плоскими ребрами раковину, обычно
лишенную зубов или с одним рудиментарным
зубом; сифоны длинные, синус глубокий.
В Каспийском морена илистых грунтах,
на глубине до 40 м и более, обитают адак-
ны с овально-треугольной, хрупкой,
нежной раковиной — Adacna vitrea, а также
A. minima, живущая и в Аральском море.
Наиболее крупная из адакн — A. laevi-
uscula (до 4 см), с хрупкой, косо-овальной
раковиной, живет главным образом в
Северном Каспии при солености до 10°/00.
В Каспийском море и в лиманах Черного
моря при солености до 8—14°/00 живет
небольшая, до 3—4 см, адакна с
многочисленными раздвинутыми резкими острыми
ребрами — A. (Hypanis) plicata.
Особый интерес представляют
близкородственные сердцевидкам виды тридакн
(сем. Tridacnidae). Это типичнейшие
обитатели тропических морей, особенно
характерные для зоны распространения
коралловых рифов в Индо-Тихоокеанской
и Австралийской областях. К этому
семейству относится два рода: собственно
тридакна (Tridacna) и гиппопус (Hippo-
pus), охватывающие около двух десятков
видов. Поистине «царь-ракушкой» среди
всех современных двустворчатых
моллюсков по своим размерам и мощности
раковины является гигантская тридакна
(Tridacna gigas), достигающая иногда
1,4 м длины и общего веса 200 кг, причем
вес только ее тела достигает 30 кг.
Однако обычные размеры тридакн меньше —
около 1 м, чаще 30—50 см.
Неподвижно лежащие среди скал и
кораллов, обычно обросшие различными
сидячими животными, кораллами и
водорослями, даже крупные тридакны не всегда
заметны в воде. Человек, неосторожно бро-
Рис. 97. Солоноватоводные сердцевидки из южны:?
морей СССР:
А — Adacna vitrea; Б — A. (Hypanis) plicata;
В — Monodacna caspia; Г — Monodacna colorata;
Д — Didacna trigonoides; E — Didacna protracta.
143
дящий по рифу, может не только тяжело
пораниться об ее острые края, но и
накрепко защемить ногу, попавшую между
створками. Однако морские
охотники-полинезийцы успевают просунуть внутрь
крупной раковины руку с ножом и
перерезать ее могучий мускул, прежде чем
тридакна захлопнет створки.
Обычно раковины тридакн с их
крупными чешуйчатыми ребрами хорошо видны на
рифе во время отлива, когда их крепко
сомкнутые раковины наполовину покрыты
водой. Под водой прилива раковина снова
медленно приоткрывает свои створки и
расправляет толстые края своей красивой
разноцветной мантии, охватывающей
острые, глубоко волнистые края раковины, и
фильтрация воды возобновляется.
Большой Барьерный риф в тропической
части Австралии представляет собой
настоящее царство тридакн. Огромные
мелководные пространства океана, занятые
здесь зарослями всевозможных кораллов,
прогреваемые тропическим солнцем и
кишащие жизнью, представляют для тридакн
прекрасное место обитания.
Однако тридакны могут жить и в
более глубоких местах, на внешних краях
рифов, постоянно омываемых океанским
прибоем: здесь они также живут среди
богатых зарослей кораллов и других
животных.
Тридакны интересны по своему
внутреннему строению и по симбиозу с
одноклеточными водорослями зооксантеллами,
что встречается у моллюсков крайне редко.
Тридакны обычно лежат на дне
макушкой и спинной, наиболее толстой и
тяжелой частью вниз, а брюшными краями
раковины и своей мантии вверх, т. е.
прямо противоположно тому, как живут
в грунте сердцевидки и остальные
двустворчатые моллюски. Поэтому у тридакн
по сравнению с другими моллюсками
наблюдается смещение ряда органов тела.
Такое положение тридакн на грунте
обеспечивает им наилучшие условия питания.
Ротовое отверстие у тридакн очень
маленькое; передний замыкательный мускул
редуцирован. Мантия сросшаяся и имеет
изнутри сверху лишь два отверстия—
вдыхательную щель спереди и
выдыхательное отверстие в ее средней части.
Нога и биссус выходят внизу через третье
отверстие мантии в щель под макушкой
между створками раковины. Лопасть
наружного края мантии очень сильно
утолщается, иногда даже не помещаясь из-за
этого в раковине, когда ее створки
смыкаются. Эта внутренняя лопасть края
мантии всегда сильно освещается солнцем
и ярко пигментирована. Она окрашена
в зеленый, синий, пурпурный и даже
черный цвет, часто с пятнами иной окраски.
Таблица И. Двустворчатые моллюски коралловых
рифов:
i — тридакна гигантская (Tridacna gigas);
2 — раковина-перо пинна (Pinna muricata);
3 — устрица петуший гребень (Ostrea crista-
galli);
4 — лима шероховатая (Lima scabra);
5 — морские жемчужницы пинктады (Pinctada
inartensii).
Интереснейшее явление представляет
собой симбиоз тридакны с
одноклеточной мелкой (около 7 |х) водорослью зоо-
ксантеллой. Ткань наружного края
мантии тридакн буквально набита этими
водорослями, они имеются также в других
частях мантии, в тканях мускула и даже
в крови, но в значительно меньшем
количестве. Хотя тридакна и является
фильтратором, она, несомненно, питается
и своими собственными зооксантеллами,
которые «разводит» главным образом в
хорошо освещенных тканях наружных
краев своей мантии. Зооксантеллы на
разных стадиях переваривания всегда
имеются в кишечнике и в желудке
тридакн.
С незапамятных времен раковины и
мясо тридакн использовались народами
Океании, полинезийцами и другими
жителями тропических островов. Крепкая
раковина тридакн заменяла им в какой-то мере
камень, которого эти народы, как жители
коралловых островов, не знали. Из нее
делались тесла, топоры, рыболовные
крючки и пр., сама раковина использовалась
как сосуд для воды, пальмового вина. Из
ее створок выпиливались круги, имевшие
значение монеты: на один такой круг
можно было купить несколько сот кокосовых
орехов. Теперь же тридакны служат лишь
любимым украшением садов и дворов
местных жителей на островах Фиджи,
Таити и др.
По своему образу жизни среди тридакн
различают свободно лежащих на дне,
куда относятся более крупные формы
(Т. derasa, T. gigas, Hippopus hippopus
и др.); все они могут сильно обрастать
другими животными и водорослями.
Другие, более мелкие, как, например, Trida-
спа сгосеа, имеют сильный биссус и
широкое отверстие для его выхода между
створками раковины на спинной стороне.
Погружаясь в расселины между
кораллами, механически «вбуравливаясь» в
щели, эти тридакны образуют углубления,
где и сидят макушкой вниз, время от
времени механически расширяя их и
прикрепляясь внутри этих углублений своим
биссусом.
Наш обзор будет неполным, если не
рассмотреть хотя бы вкратце такие
богатые видами, преимущественно
тропические и южнобореальные, группы разно-
зубых моллюсков, как семейства
Венериды (Veneridae), включающее более
400 видов, и Теллиниды (Tellinidae)—
более 200 видов.
Венериды (различные венусы, тапесы,
венеруписы, дозинии, прототаки, мерцена-
рии и т. д.) широко распространены
Таблица 12. Морские двустворчатые моллюски:
1 — арка полосатая (Area zebra);
2 — солемия бореальная (Solemya borealis);
3 — морской молоток (Malleus malleus);
4 — морская жемчужница (Pinctada margariti-
fera);
5 — спондилюс красивый (Spondylus pictorum);
6 — амусиум японский (Amussium japonicum);
7 — гребешок Свифта (Chlamys (Swiftopecten)
swifti);
8 — исландский гребешок (Chlamys islandi-
cus);
9 — кардита флоридская (Cardita floridana);
10 — хама чешуйчатая (Chama macerophylla);
11 — сердцевидка Фрагум (Fragum unedo);
12 — люцинида тигровая (Codakia tigerina);
13 — венерида—королевский гребень(РЦаг dione);
14 — венерида — сунетта (Sunetta meroe);
15 — теллина-фараонелла (Pharaonella perieri);
16 — донакс черноморский (Donax (Paradonax)
venustus);
17 — мидия зеленая (Mytilus viridis);
18 — куспидария кардиомия (Cardiomya goul-
diana);
19 — спинула клювовидная (Spinula calcar);
20 — черенок большой австралийский (Solen gran-
dis).
10 Жизнь животных, т. 2
в умеренно тепловодных и тропических
морях, обитая на мелководьях
прибрежной зоны, преимущественно на песчаных
грунтах. Не обладая длинными сифонами
или даже совсем лишенные их, венериды
не могут глубоко закапываться и живут
лишь в самом поверхностном слое грунта,
активно ползая при помощи своей
довольно сжатой клиновидно-топоровидной ноги.
Раковины венерид очень красивы,
различной величины и формы. Ребра раковин
иногда могут быть пластинчатыми с
бугорками и даже с причудливыми выростами,
как, например, у королевского гребня—
Pitar dione. Замковая пластинка крепкая,
с тремя кардинальными зубами,
удлиненными или в той или иной мере
расщепленными, могут быть и передние боковые
зубы.
Большинство венерид съедобно, и
многие их виды являются промысловыми, а
в последнее время венерид начали также
разводить искусственно и
акклиматизировать в новых для них районах.
В 1963 г. в Японии было выловлено
1370 тыс. ц различных венерид, главным
образом так называемого японского пе-
тушка-венеруписа — Venerupis (Amyg-
Рис. 98. Тридакна (Tridacna crocea). Вид
моллюска при удаленной левой створке раковины
и левой лопасти мантии:
I — правая створка раковины; 2 — макушка; з —
наружная связка; 4 — края правой лопасти мантии;
5 — вдыхательное отверстие; 6 — выдыхательное
отверстие; 7 — нога; 8 — биссусные нити, 9 — внутренняя
левая полужабра; ю — наружная левая полужабра;
II — рот; 12 — ротовые лопасти; 13 — анус; 14 — за-
мыкательный мускул; 15 — края срезанной
мантии. Стрелки указывают направление входящих
и выходящих токов воды.
dalum) japonica. Этот моллюск, с
продолговатой, удлиненно-овальной
раковиной (длиной до 6 см), распространен по
прибрежью нашего Приморья. Ряд
других венеруписов и тапесов добывается в
Японии, Китае, Америке, Франции и
других странах; так, в Испании в 1963 г.
было добыто 58 тыс. ц. В большом
количестве саксидомусов (Saxidomus nut-
talli и S. purpuratus) вылавливают в США
и Японии; в 1963 г. их было
добыто 445 тыс. ц. По Атлантическому
прибрежью США главным промысловым
объектом из венерид является мерцена-
рия (Mercenaria mercenaria); этого
довольно крупного моллюска, до 10—
12 см длиной, переселили из Атлантики
на Тихоокеанское побережье, где он
успешно акклиматизировался, и теперь
его здесь промышляют. В Японском море,
у берегов Японии и у нас в Приморье
на песчаных грунтах, на глубинах от
2 до 45 м, обитает очень близкий вид
мерценария Стимпсона (М. stimpsoni).
Крупные особи этих мерценарий
достигают 9 см, тяжелая раковина довольно
высокая, треугольно-округлая, грязно-
белого цвета, с многочисленными
концентрическими пластинчатыми ребрами.
Эту мерценарию также добывают.
Промысловым моллюском в Японии является
и обитающая в Приморье на песчаных
грунтах на глубинах 15—50 м
тихоокеанская питария (Callista brevisiphonata).
В Черном море живет несколько атлан-
тическо-средиземноморских видов венерид.
Они обитают у берегов, главным образом
в зонах биоценоза песка, илистых
прибрежий ракушечника (особенно) и мидие-
вого ила, на глубинах до 50—70 м (реже
несколько глубже).
Одним из самых обычнейших, известных
и массовых видов моллюсков является
черноморский венус-петушок — Chione
(Chamelea) gallina corrugatula.
He менее известны и обычны
черноморские венеруписы •— Venerupis (Polititapes)
rugata proclivis и V. (Polititapes) discrepans.
Очень близко к венеридам
семейство петрикол (Petricolidae), к
которому относится самый обычный камнето-
чец Черного моря, средиземноморско-
атлантический вид Petricola lithophaga,
146
сверлящий здесь известковые породы,
прибрежные скалы, а также плотные
глины. Благодаря такому образу жизни
раковина петриколы может быть сильно
деформирована. Мантия у петрикол
сросшаяся, с небольшим отверстием для
выхода небольшой ноги; сифоны длинные,
также частично сросшиеся. Местами
петриколы встречаются на берегах в
огромном количестве, и поэтому берега
здесь разрушаются волнами гораздо
быстрее. Для сверления моллюску служит
очень крепкая овально-треугольная
раковина, с тонкими многочисленными
мелкими радиальными гранулированными
ребрами; такая структура раковины
напоминает поверхность рашпиля;
закрепившись ногой в какой-нибудь точке
и делая вращательные движения, петри-
кола медленно соскабливает слой за
слоем породу этими зубчиками, постепенно
погружается в протачиваемый ею ход и
остается в нем.
Обширная группа умеренно
теплолюбивых и тропических форм,
объединяемых в надсемейство Tellinacea,
распадается на четыре семейства:
собственно Теллины (Tellinidae), Скро-
бикулярии (Scrobiculariidae), Донаксы
(Donacidae) и Псаммобии (Sanguinola-
riidae). Все они имеют относительно
тонкую и более легкую, чем у венерид,
овальную или удлиненную раковину.
Замок состоит в основном из двух
центральных кардинальных зубов в каждой
створке, из которых один нередко раздвоен
сверху, часто есть и боковые зубы. Нога
языковидная, подвижная, без биссуса.
Синус глубокий, сифоны длинные,
разделенные и очень подвижные. Последние
очень помогают им в собирании пищи—
детрита — с поверхности мягкого илисто-
песчаного или песчаного грунта, когда
моллюск глубоко закапывается в него.
Раковина теллинид часто бывает красиво
окрашена и снаружи и изнутри, как,
например, у ярко-розовой тропической
Pharaonella perieri или как у атлантиче-
ско-средиземноморских и живущих также
и в Черном море видов: желтая — у
хрупкой гастпраны (Gastrana fragilis), розовые—
у мелких ангулюсов — Angulus (Fabulina)
fabula и Angulus (Moerella) donacinus.
Благодаря небольшим размерам и
тонкой раковине почти все Tellinacea —
излюбленная пища придонных рыб
(камбал и др.); многие Tellinacea съедобны.
Как указывалось выше, в наших
северных морях из семейства Tellinidae
обитают только виды рода Macoma; у видов
этого рода отсутствуют боковые зубы,
и раковины их обычно белые, покрытые
очень тонким серым периостраком. В
морях северного полушария, в том числе
и в наших северных и дальневосточных
морях, наиболее широко
распространена умеренно холодноводная известковая
макома (Macoma calcarea). Она нередко
образует здесь массовые скопления на
мягких заиленных песках, на глубине
до 100—120 м; ее количество может
достигать нескольких сот экземпляров на
1 ж2, а биомасса — нескольких сот
граммов на 1 м2. М. calcarea входит в состав
многих сообществ донной фауны, являясь
их руководящей, массовой или одной из
самых характерных форм.
Едва ли не самой известной формой
среди маком является балтийская макома
(Macoma balthica).
Эта маленькая (до 2 см),
округло-треугольная, окрашенная внутри в розовый
цвет макома является массовой
руководящей формой биоценоза
илисто-песчаных пляжей приливо-отливной зоны.
Несмотря на сильное выедание ее рыбами,
плотность ее поселений на литорали может
доходить до нескольких тысяч экземпляров
на 1 м2, а биомасса до нескольких сот
граммов на 1 м2.
Балтийская макома имеет очень
широкое амфибореальное географическое
распространение. Это обычнейшая
литоральная форма в Белом море, у берегов Мур-
мана и далее на запад, вдоль
атлантических берегов Европы — до Франции; по
Атлантическому побережью Северной
Америки балтийская макома распространена
от Лабрадора до штата Джорджия, в
северной части Тихого океана — от
Берингова моря до Японского, а по
американскому побережью — до Калифорнии.
В юго-восточной части Баренцева моря
и в прилегающей части Карского моря
она не живет в литорали, а обитает на
глубине нескольких метров. В Балтий-
10*
147
ском море она заселяет всю основную
часть, далеко заходя во все его
заливы, и живет здесь до глубины более ДООле,
являясь руководящей формой ряда
сообществ его донной фауны.
Одной из самых крупных теллинид,
обитающих во всех наших
дальневосточных морях, а также в Чукотском море
и вдоль западных берегов Северной
Америки, является Angulus (Peronidia) lutea
(до 10 см длиной).
Очень красивы маленькие (не более
2—3 см) округло-треугольные, с
макушкой, сильно сдвинутой к заднему краю,
блестящие раковинки донаксов (семейство
Donacidae). Они бывают окрашены в
самые различные цвета, нередко с
лучистыми радиальными полосками, благодаря
чему их называют морскими бабочками.
Около 50 видов этого семейства —
обитатели песчаных мелководий умеренной
и тропической зон. У нас в Черном море
живут два вида донаксов, из которых
Donax (Paradonax) venustus наиболее
обычен.
Значительно более крупны (до 8—10 см
длиной) различные псаммобии (семейство
Sanguinolariidae, или Psammobiidae, или
Gariidae, или Asaphidae).
Около 100 видов этого семейства
обитает главным образом на небольших
глубинах и илистых грунтах,
преимущественно в тропиках. Их
удлиненно-округлые раковины обычно зияют спереди
и сзади и окрашены в коричневые и
фиолетовые тона. У берегов в южной части
Охотского моря и в Японском довольно
обычна оливковая кровянка (Sanguinolaria
(Nuttallia) olivacea).
Из семейства Scrobiculariidae
(или Semelidae) наиболее известны
различные синдесмий (Abra, или Syndesmya). Из
двух атлантическо-средиземноморских
видов синдесмий, обитающих в Черном
море, овальная синдесмия (Abra ovata)
обитает также и в Азовском море. Это
мелкий моллюск (до 2,5 см), с треугольно-
овальной тонкой, слегка приоткрытой
спереди и сзади раковиной; живет на
илистых грунтах и небольших глубинах
(не более 20—40 м); может выносить
значительное понижение солености (до 2°/00),
предпочитает жить даже в более
опресненных участках. Может также выносить
и недостаток кислорода в воде, перенося
даже временные заморы, наблюдающиеся
иногда в центральной части Азовского
моря (в районе, занятом биоценозом этой
синдесмий).
Для улучшения кормовой базы
осетровых рыб синдесмия была пересажена из
Азовского моря в Каспийское. Она здесь
хорошо прижилась, расселилась и теперь
вошла в пищевой рацион многих
каспийских донных рыб. Наибольшая плотность
поселений синдесмий в Каспии достигает
7000 экземпляров на 1 д2, а биомасса—
300 г на 1 м2.
К подотряду Adapedonta относится пять
надсемейств: Черенки (Solenacea)—
с одним семейством, Мактры (Mactra-
сеа) — с четырьмя семействами, Мии
(Муасеа) — с тремя семействами, Сакси-
кавы (Saxicavacea, или Hiatellacea) — с
одним семейством и Бессвязочные (Adesma-
сеа) — с двумя семействами. Их
объединяет большая или меньшая редукция
замка вплоть до потери всех зубов и
усиление роли внутренней связки. Многие
виды из основных семейств,
относящихся к первым трем группам, являются
объектами промысла, а также играют
большую роль в питании рыб.
Большинство видов из этих групп —
закапывающиеся формы, а бессвязочные
представляют собой специализированные
группы моллюсков — камнеточцев и
древоточцев.
Черенки (сем. Solenidae) — типичные
обитатели песчаных грунтов, в которые
они закапываются при помощи своей
сильной расширенной на конце ноги. Их
раковины очень характерной формы:
тонкостенные, вытянутые, узкие, с
параллельными спинным и брюшным краями,
почти прямоугольные (реже удлиненно-
овальные), зияющие с обоих концов,
с сильно сдвинутой на передний край
макушкой.
В Черном море на глубине до 10 м
живет атлантическо-средиземноморский вид
обыкновенный черенок (Solen vagina, или
S. marginatus), с очень удлиненной (до
12 см) узкой прямоугольной
раковиной; макушка на самом переднем конце
спинного края.
148
Настоящие мактры (Mactridae)
широко распространены на песчаных
мелководьях тепловодных морей бореальной
зоны, а также и в субтропической зоне;
в арктических морях их нет. Замковая
площадка довольно широкая, крепкая;
особенно характерен замок: с одним
раздвоенным лямбдовидным кардинальным
зубом в левой створке, а в правой — с
двумя сходящимися к макушке тонкими
кардинальными зубами; позади заднего
центрального зуба правой створки
располагается довольно большая треугольная
площадка для прикрепления внутренней
связки. Обычно имеются пластинчатые
(приближенные к центральным) боковые
зубы.
Мактры съедобны и многие служат
предметом промысла в США, Японии, Китае
и т. д. Их продают в свежем, мороженом,
сушеном и особенно в консервированном
виде.
Так, у Атлантического побережья США
в 1963 г. было добыто 860 тыс. ц мактр,
главным образом Spisula solidissima.
Эта крупная (до 12—15 см) мактра
обильно заселяет прибрежные песчаные
пляжи, начиная почти от уреза воды.
После сильных зимних штормов ее
миллионами находят в выбросах на берегу,
за что и называют прибойной мактрой.
В Японии очень интенсивно
добывают несколько видов крупных мактр,
больше всего овальную мактру — Spisula
(Mactromeris) voyi, затем сахалинскую
мактру, или белую ракушку — Spisula
sachalinensis), полосатую мактру (Мас-
tra sulcataria) и др.
В Приморском крае промысловые
запасы сахалинской мактры весьма
значительны, и при ловле драгой один
промысловый бот может добыть в среднем
2—3 тыс. штук в день. Эта крупная
(до 12 см) белая ракушка здесь типичный
обитатель мелкозернистых песков в
прибойных бухтах, где живет на глубине до
5 м или несколько глубже.
Размножается в июле — августе и быстро растет,
достигая к пяти годам среднего размера
около 8 см; затем ее рост замедляется;
живет 10—13 лет. Ее молодь развивается
на больших глубинах, а затем
откочевывает к берегам.
Овальная мактра обитает у берегов
всех наших дальневосточных морей до
глубины около 75 м, а также у
Тихоокеанского побережья Канады; полосатая
мактра, как и сахалинская, обитает в южной
части Охотского и Японского морей, а
последняя также и у берегов Китая.
Почти всесветное распространение
имеет обыкновенная, или арктическая, сакси-
кава (Hiatella arctica). Это небольшой
моллюск с грубой, толстой,
неправильной, более или менее
удлиненно-прямоугольной, зияющей с обоих концов
раковиной, лишенной зубов (может быть лишь
рудимент одного зуба). Мантия
сросшаяся, сифоны довольно длинные, сросшиеся.
Живет свободно на дне или в норках,
которые мажет высверливать в
различных грунтах. При сверлении саксикава
закрывает отверстия своих сифонов и
опирается сократившимися, сжавшимися
сифонами в стенки норки.
К надсемейству Муасеа
относится довольно большое число видов из
трех семейств: Мии (Myidae)—8
родов, Корбулы (Corbulidae, или Aloididae)—
4 рода и Gastrochaenidae — 3 рода.
У всех Myidae замок слабо развит,
раковина без зубов или с одним
зубовидным выростом в каждой створке; помимо
наружной, сильно развита внутренняя
хрящеватая связка, которая прикрепляется
одним концом к ложкообразному выросту
замочного края под макушкой в левой
створке, а другим концом—соответственно
к полукруглой выемке в правой створке.
Мантия сросшаяся, с отверстием для
выхода ноги и с длинными покрытыми
кутикулой сифонами.
К глубоко зарывающимся в мягкий
илисто-песчаный грунт прибрежных
мелководий формам относятся хорошо
известные в северных и дальневосточных морях
крупные моллюски мии: песчаная
ракушка — Муа (Arenomya) arenaria с овально-
яйцевидной раковиной (длиной до 10 см) и
Муа truncata с округло-угловатой, сзади
вертикально обрубленной раковиной (до
5—6 см длиной).
Песчаная ракушка — более тепловодный
амфибореальный вид, обитатель приливо-
отливной зоны и самых мелководных
районов прибрежной зоны (не более 10 м
149
Рис. 99. Моллюски, сверлящие плотные грунты
и известковые породы: А — тридакна (Tridacna
сгосеа), сверлящая коралловые рифы,
сидит макушкой вниз; Б — морской финик (Li-
thophaga), точит и растворяет известняки; В —
саксикава (Hiatella); Г — она же во время
сверления, сифоны сокращены; Д — гастрохена (Ga-
strochaena); E — морское сверло (Pholas).
1 — нога; 2 — биссус; 3 — вводной сифон; 4 —
выводной сифон; 5 — защитные отложения извести.
глубиной); в арктических морях не
встречается. Другой вид — Муа truncata —
живет несколько глубже первого и
распространен очень широко почти во всех
морях северного полушария.
Муа (Arenomya) arenaria легко
переносит опреснение воды; Муа truncata
придерживается более постоянной и более
высокой морской солености.
Все мии — типичные фильтраторы,
питающиеся взвешенными в воде
частицами (детритом) и мелкими планктонными
организмами. На обнажающихся во время
отлива песчаных отмелях, обильно
заселенных песчаной ракушкой, обычно видны
многочисленные отверстия,
представляющие собой входы в норки этих глубоко
закапывающихся (на 20—30 см)
моллюсков, по которым легко находить их
поселения. Норки и их края укреплены
слизью, выделяемой моллюсками. У
закопавшихся песчаных ракушек концы
сифонов могут слегка высовываться из грунта,
так что видны их отверстия, окруженные
чувствительными выростами. При
приближении опасности или даже при
раздражении сифоны могут сильно сокращаться;
в вытянутом состоянии они раз в пять
длиннее раковины.
Взрослые моллюски обычно не меняют
места своего обитания.
Песчаная ракушка размножается
летом. Ее личинка парусник уже через
несколько дней оседает на дно. Первое
время молодой моллюск не зарывается,
а лежит на дне, прикрепляясь биссусом
к камням, и может переползать с одного
места на другое. Через некоторое время
молодые моллюски начинают пробовать
закапываться, пока не выберут
окончательно место своего обитания.
Рост молодых мий в первые годы
жизни происходит очень быстро, но сильно
зависит от условий обитания, в
первую очередь от температуры и
солености воды. При сильном опреснении рост
может задерживаться; при хороших
условиях молодая раковина достигает
величины 2—3 см.
Быстрота размножения, высокий темп
роста, эвригалинность, легкая
приспособляемость к новым условиям делают
песчаную ракушку удобным объектом для
переселения и акклиматизации.
Больше всего промысел песчаной
ракушки развит в США, где в 1963 г. ее
было добыто 215 тыс. ц. В США ее не
только добывают, но и искусственно разводят.
На Тихоокеанское побережье США
песчаная ракушка была завезена случайно.
Здесь она расселилась и теперь служит
объектом промысла.
Песчаная ракушка поступает на рынок
в свежем или мороженом виде, реже —
сушеная и копченая, но большей частью
в консервированном виде; особенно много
из нее приготовляют бульона-экстракта.
Песчаную ракушку используют также
для наживки рыболовной снасти.
150
Большой интерес представляют камне-
точцы и древоточцы из группы лишенных
связок моллюсков (Adesmacea) из с е-
м е й с т в морских сверлильщиков —
фоладид (Pholadidae) и древоточцев (Теге-
dinidae). Это формы, приспособившиеся
к обитанию в ходах, проточенных ими в
скалах, известняках, плотных породах
или в дереве. Изменения в строении тела
и раковины, которые претерпела эта
группа моллюсков, оказались настолько
глубокими, что многие из них утратили свой
исходный внешний вид двустворчатых
моллюсков, став неузнаваемыми, как,
например, корабельный червь (Teredo).
Его длинное голое тело напоминает
червя, а не моллюска; маленькая
своеобразная раковина, играющая роль
сверла, располагается на самом переднем
конце тела; поверхность высверливаемого в
дереве хода покрыта известковыми
выделениями мантии моллюска и образует
как бы трубку, защищающую нежное
тело.
В связи с приспособлением к
механическому сверлению твердых или плотных
пород грунта раковины фоладид
(Pholadidae) очень крепкие, сложной формы,
со своеобразной скульптурой
поверхности, неравносторонние (макушка сдвинута
к переднему, реже — к заднему концу),
со значительным переднебрюшным и
задним зиянием. Части створок, граничащие
с верхним (замочным) краем, в большей
или меньшей степени отогнуты наружу,
образуя так называемую
макушечную складку. Передняя большая
часть раковины вооружена резкими
шиповатыми ребрами.
Изнутри под макушкой створки
раковин имеют довольно узкий, ножевидный,
слегка изогнутый, направленный вниз
особый вырост — апофиз, служащий для
прикрепления ножных мышц. Раковина
фоладид лишена связки (иногда может
быть очень небольшая внутренняя
связка), и ее створки скреплены только
мускулами-замыкателями, что значительно
увеличивает подвижность створок.
Передний мускул перемещен на макушечную
складку раковины и не несет функций
замыкателя, а является антагонистом
заднего аддуктора. При сокращении
переднего мускула раковина фоладид
раскрывается сзади, а при сокращении заднего —
спереди; они сокращаются не
одновременно (как у всех других двустворчатых
моллюсков), а поочередно, что делает
движения раковины фоладид при
сверлении более разнообразными. При такой
работе мускулов происходит как бы
качание створок вверх, вниз и по дуге назад.
Благодаря наличию на створках
острых, снабженных шипами и бугорками
ребер и вращению раковины и
происходит сверление фоладидами грунта.
Сифоны у фоладид мощные, сросшиеся,
нередко покрытые хитинизированным пе-
риостраком. Нога довольно хорошо
развитая, короткая, на конце усеченная, с
подошвой, служит для прикрепления к
стенкам норки во время сверления.
Некоторые фоладиды светятся, как, например,
встречающийся в Черном море камнеточец
фолас — Pholas dactylus. У него светятся
пять хорошо ограниченных участков,
включая и оба сифона; светящаяся слизь
выделяется особыми железами с
длинными протоками и при раздражении
выпускается через сифон.
Фоладиды довольно крупны, некоторые
достигают 10—12 см длины. Живут
главным образом в прибрежной зоне, на
малых глубинах. Фоладиды могут
повреждать не только известковые породы,
песчаник, мергель, но и дерево и даже бетон,
что способствует более быстрому
разрушению последнего в морской воде.
Рис. 100. Глубоководный моллюск ксилофага,
сверлящий дерево (Xylophaga grevei, вид со
спинной стороны):
1 — правая створка; 2 — левая створка; з —
переднее зияние; 4 — нога; 5 — сифон.
151
Некоторые виды фоладид проделывают
свои ходы также и в плотном иле или в
глине: атлантические и
средиземноморские Pholas dactylus и Barnea Candida,
или В. spinosa; обитающая по европейскому
и американскому побережьям цирфея гре-
бенчатая (Zirfaea crispata); по
Тихоокеанскому побережью Северной Америки—Z.
pilsbryi и в Японском море—морское
сверло— Pholadidea (Penitella) penita.
Не менее известны и обычны сверлящие
дерево фоладиды из рода Xylophaga—
ксилофаги, как, например, X. dorsalis,
живущая в Атлантическом океане и
Средиземном море. Они часто встречаются в
кусках плавающей и затонувшей
древесины, в верхней оболочке кокосовых
орехов и пр. и могут сверлить различные
предметы вплоть до изоляции подводных
кабелей. Много новых видов ксилофаг
было найдено датской экспедицией на
«Галатее» в глубоководных впадинах
Тихого океана, например X. grevei,
найденная на глубине более 7 тыс. м, что
является пока наибольшей глубиной
нахождения морских сверлильщиков.
Хотя ксилофаги точат свои ходы в
древесине и пропускают большое количество
древесных опилок через свой
приспособленный к этому кишечник, они не
могут переваривать древесину. Таким
образом, проточенные ходы в древесине служат
ксилофагам лишь для укрытия, как и всем
другим сверлящим норки фоладидам.
Наиболее совершенные приспособления
к протачиванию ходов в древесине
наблюдаются у морских древоточцев — весьма
специализированных моллюсков из
семейства Teredinidae — опасных вредителей
деревянных свай, пристаней, подводных
частей деревянных судов. Эти моллюски
наносят огромный материальный ущерб
в теплых и особенно в тропических водах.
Морские древоточцы очень широко
распространены и встречаются почти во всех
морях обоих полушарий, отсутствуя лишь
в холодных арктических водах.
Большинство известных в настоящее
время видов древоточцев (около 70)
относится к двум основным родам-— Тередо
(Teredo) и Банкия (Bankia).
Обычным местом их обитания служат
куски дерева, сваи пристаней,
деревянные суда, корневища
морских трав, а в тропиках
также заросли мангровых
деревьев, затонувшие в море
кокосовые орехи и др.
Лишь единичные виды
древоточцев безвредны, как,
например, живущий на песчаных
мелководьях и прибрежных
пляжах Kuphus polythalamia;
он обитает в Тихом океане у
Филиппин, на Малайском
архипелаге, у Северной Австралии;
строит в песке довольно тол-
Рис. 101. Схема строения
древоточца банкия (Bankia): A — общий вид
моллюска сбоку; Б — палетка; В —
левая створка раковины снаружи;
Г — левая створка раковины
изнутри; Д — положение сифонов и
палеток при фильтрации воды; Е — при
сверлении палетки закрывают
входное отверстие — сифоны втянуты.
1 — раковина; 2 — нога; з — палетки;
4 — сифоны; 5—переднее ушко; 6 —
средняя часть створки; 7 — заднее ушко;
8 — апофиз.
152
стые известковые трубки более метра
длиной. Безвредны и древоточцы Zachsia,
обитающие среди корневищ морской травы
зостеры и филлоспадикс.
Общее строение древоточцев служит
ярким примером приспособительных
изменений морских животных. Как
указывалось выше, они обладают червеобразным
удлиненным телом, на заднем конце
которого имеется два длинных тонких сифона —
вдыхательный и выдыхательный. Они
могут сильно вытягиваться из норки и
служат для связи моллюска с внешней
средой, в первую очередь для дыхания и
питания. Подсчитывая число пар таких
сифонов, выставленных наружу из
мелких отверстий их ходов в древесине,
можно определить, сколько штук «червей»
ведет в данном месте свою
разрушительную работу. На заднем же конце тела у
основания сифонов имеется пара
известковых хитиновых пластинок —
палеток. Они служат для защиты заднего
конца тела древоточцев и закрывают при
сверлении ими дерева очень маленькое
входное отверстие, сифоны при этом
втягиваются внутрь норки. Палетки у рода
Teredo очень небольшие, простые,
ложкообразные, а у Bankia более сложные,
колосовидные и состоят из многих мелких
входящих друг в друга воронкообразных
образований.
Небольшая неправильно округлая,
крепкая, сильно зияющая спереди и сзади
раковина покрывает только самый
передний конец тела моллюска; у взрослого
древоточца раковина занимает лишь
1/зо"_1/4о часть длины его тела. Между
широко раздвинутыми створками
раковины спереди выходит небольшая
округлая нога, а сзади удлиненное мягкое тело
моллюска.
Створки раковины служат прекрасным
инструментом для сверления древесины.
Ее передняя часть небольшая,
треугольная, выступающая в виде ушка, несет
горизонтальные тонко зазубренные
гребни; средняя, основная часть вооружена
мощными грубозубчатыми гребнями,
идущими под прямым углом к гребням
передней части. Задняя часть раковины
гладкая. На створках изнутри имеется
длинный вырост — апофиз. У терединид,
Рис. 102. Древоточец тередо (Teredo) в куске
источенного им дерева:
1 — ходы, проточенные моллюском в дереве; 2 —
известковая выстилка ходов (ложная раковина); з —
тело моллюска; 4 — раковина; 5 — нога; 6 —
сифоны; 7 — палетка.
как и у фоладид, сокращение заднего и
переднего мускулов-замыкателей
происходит поочередно. При этом передние
концы створок раздвигаются с такой силой,
что острые зубчики раковины сдирают
слой древесины, как рашпилем. Нога при
этом играет роль присоски, которая
удерживает моллюска в протачиваемом им
ходе; сверлящие движения происходят
ритмически — 8—12 раз в минуту. Ходы
древоточцев выстланы внутри тонким
слоем известковых отложений; поперечный
разрез куска дерева, источенного ими,
имеет вид пчелиных сот.
Интересно отметить, что у древоточцев
их соседние ходы в древесине никогда
153
не пересекаются, как бы густо они ни
заселяли зараженную ими древесину.
Повороты и изгибы ходов, которые приходится
делать моллюскам, чтобы избежать друг
друга, несомненно, приводят к еще
большему «изрешечиванию» древесины.
Морские древоточцы питаются, как и
другие моллюски-фильтраторы,
взвешенными в воде мелкими планктонными
организмами, засасывая воду через входной
сифон, и древесными опилками, которые
они соскабливают при сверлении.
В связи с питанием древесиной их
вытянутый желудок имеет особый большой
слепой мешковидный вырост, всегда
набитый у них древесными опилками. В этом
выросте скапливаются частицы древесины
двух сортов: очень мелкие, образованные
в результате соскабливания древесины
тонкими зубчиками передней части
раковины, и удлиненные, волокнистые,
соскобленные более крупными зубчиками
среднего отдела раковины.
Мельчайшие частички дерева
заглатываются амебоцитами в многочисленных
открывающихся в желудок
пищеварительных дивертикулах (выростах) печени и
перевариваются ими внутриклеточно. У дре-
Рис. 103. Стадии развития древоточца тередо:
А — свободноплавающая личинка велигер;
Б — только что осевшая личинка; В — молодой
тередо.
J — раковина; 2 — ресничный пояс и пучок
теменных ресничек; з — нога; 4 — биссус; 5 — сифоны;
6 — зачатки палеток; 7 — тело моллюска.
воточцев обнаружен фермент целлюлаза,
переваривающий клетчатку, гидролизую-
щий ее до глюкозы. Наблюдения
физиологов над питанием древоточцев показали,
что в переваривании древесины
принимает участие несколько ферментов и что
клетчатка (целлюлоза) используется ими
в большей степени, чем гемицеллюлоза
(углеводы клеточных стенок водорослей).
Все кусочки древесины, соскобленные
раковиной, заглатываются через рот,
проходят через пищеварительный тракт, а
непереваренные остатки древесины
выводятся через выдыхательный сифон
наружу.
В средних широтах древоточцы
размножаются только летом, а в тропиках
круглый год.
Терединиды — гермафродиты, но при
этом каждая особь сначала производит
сперматозоиды, а затем яйца; такое
чередование полов исключает
самооплодотворение. Иногда яйца развиваются и
без оплодотворения (партеногенетически).
Оплодотворенные яйца вначале довольно
быстро развиваются внутри жаберной
полости материнской особи: через 20—30
часов образуется личинка трохофора, а еще
через 36—48 часов — личинка велигер;
только через 2—3 недели велигер выходит
наружу, в воду, где 2—2,5 недели
плавает свободно, после чего отыскивает для
себя кусок дерева или сваю, в которую
внедряется.
Молодь осевших древоточцев быстро
растет. Так, тропическая Bankia gouldi
через 12 дней после оседания имеет
длину 3 мм, через 30 дней — 6 см, через
36 дней — 10 см. Протачивая ход,
молодые тередо сначала идут вдоль волокон
древесины, а позднее они могут точить ее
и поперек.
Размеры тела и трубок древоточцев
очень различны, часто очень значительны,
особенно у тропических видов. Так,
обычный обитатель мангровых зарослей Dicya-
thifer manni имеет раковину более 2 см
диаметром, а его трубки достигают длины
более 2 м.
В морях СССР обитает несколько видов
древоточцев, из которых наиболее
известны: в Черном и Балтийском морях —
Teredo navalis (до 25 см длиной), распро-
154
Рис. 104. Морская леечка (Вге-
chites giganteus):
j — раковина; 2 — ложная
раковина — защитная известковая трубка;
3 — нижний конец; 4— верхний
открытый конец.
страненныи также в водах
Северной Атлантики и
северной части Тихого океана;
в юго-западной части
Баренцева моря — Nototeredo
norvagica (до 50 см длиной)
и еще более крупный Т.
megotara. Последний также
обитает в Северной
Атлантике, Средиземном море и в
Тихом океане. В Японском
море у наших берегов
обитает крупная Bankia setacea
(длиной до 60 см) и
несколько более мелких видов
Teredo, в том числе
обитающая в зарослях морских трав
Zachsia zenkewitchi.
Меры борьбы с морскими
древоточцами очень
разнообразны, но сводятся в
основном к покрытию подводных
сооружений слоем
специальных красок и к пропитыванию
древесины токсическими для моллюсков
веществами, в первую очередь
креозотом, каменноугольным лаком,
карболовой кислотой и др. По отношению к
деревянным судам давно практикуется
регулярный заход на 3—4 дня в пресную
воду в период размножения тередо и
оседания его личинок, которые очень
чувствительны к сильному опреснению,
осушка судов сроком от 5 до 30 дней и
некоторые другие методы борьбы.
К подотряду аномальносвязочных
(Anomalodesmata) относится небольшое
число видов, обитающих в различных
морях; раковины со слабым замком, часто
даже лишенные зубов, с более или менее
обызвествленной внутренней связкой;
наружная полужабра нередко загнута вверх
или редуцирована.
В морях СССР обитают виды только
из четырех семейств этого
подотряда: Лионзии (Lyonsiidae), Пандоры (Pan-
doridae), Tрации (Thraciidae) и Морские
ложечки (Periplomatidae). К отряду
аномальносвязочных относят также очень
интересных и необычных по своему
внешнему виду тропических моллюсков,
известных под названием «морских леечек»,
из семейства Clavagellidae. Их
маленькие тонкостенные раковинки
срастаются одной створкой (Clavagella) или
обеими (Brechites, или Penicillus) с длинной
известковой трубкой — ложной
раковиной, выделяемой сифонами. Края мантии
сросшиеся, с отверстием для ноги;
сифоны длинные, сросшиеся. Нижний конец
такой трубки расширен в виде диска,
вооруженного по краям мелкими
трубочками, при помощи которых они
закрепляются в грунте, а сама трубка торчит
вверх. Верхний конец трубки открытый
и несет по краям листовидные выросты.
ОТРЯД ПЕРЕГОРОДЧАТОЖАБЕРНЫЕ
(SEPTIBRANCHIA)
К этой интересной, но еще мало
изученной группе моллюсков относятся
небольшие, преимущественно
глубоководные формы. У многих из них жабры
маленькие, редуцированные или
преобразованные в мускулистую поперечную
перегородку — жаберную септу с
несколькими отверстиями в ней. Нога без биссуса;
внутренний лигамент имеет известковое
ядро (литодесму). Наиболее
распространенными из этого отряда являются роды
и виды семейства куспидарий (Cus-
pidariidae). Их раковина оттянута сзади
в виде более или менее длинного рост-
рума (выроста), где лежат сифоны.
К отряду перегородчатожаберных
относятся также еще два семейства:
Поромии (Poromyidae) и Вертикордии
(Verticordiidae), среди которых имеется
много глубоководных видов, обитающих
в открытых частях многих морей при
высокой (морской) солености и в океанах.
Виды из отряда
перегородчатожаберных не встречаются во внутренних морях
СССР и при пониженной солености.
Перегородчатожаберные моллюски по
способу питания, вероятно, мелкие
хищники.
155
КЛАСС ГОЛОВОНОГИЕ МОЛЛЮСКИ
(CEPHALOPODA)
Головоногие — самые необычные,
крупные, хищные и самые совершенные из
моллюсков. Головоногие достигли
высокой степени развития. Это своего рода
приматы среди беспозвоночных
обитателей моря.
Внешне, на первый взгляд, головоногие
моллюски ничем не напоминают своих
родичей — улиток и ракушек. Посудите
сами: ведь речь идет об осьминогах,
кальмарах и каракатицах — именно их
зоологи называют головоногими моллюсками.
Что общего у осьминога с устрицей или
виноградной улиткой? У него ведь даже
и раковины нет, столь типичной для
других моллюсков. Однако наукой доказано,
что общего у них много. Общее
происхождение, много общего и в анатомии.
Головоногими эти животные были
названы потому, что на голове у них
расположены щупальца, или «руки»,
которые называют также и «ногами», потому
что осьминоги (да и кальмары тоже)
часто ходят на них по дну, как на ходулях.
Кроме того, эмбриологами было
установлено, что щупальца головоногих
моллюсков развились из «ноги» их древнего
предка — первобытного моллюска.
(«Нога», напомним,— тот языковидный или
похожий на подошву вырост тела, на
котором ползают улитки и ракушки.)
Размеры головоногих моллюсков очень
разнообразны. Некоторые из них
принадлежат к числу самых мелких
представителей нектона, другие же — самые
крупные беспозвоночные животные
вообще. Размеры Decapoda колеблются от 1 см
(Idiosepius) до 18 м (гигантские кальмары
рода Architeuthis).
Анатомические признаки. По числу
щупалец и другим признакам класс
головоногих моллюсков делят на два
подкласса. Осьминоги (подкласс Octo-
poda) имеют 8 щупалец, а раковину,
вернее ее недоразвитый остаток (две хряще-
видные палочки или хрящевые
образования другой формы), они носят не
снаружи, а под кожей спины.
У кальмаров и каракатиц (п о д-
класс десятиногих — Decapoda) не
восемь, как у осьминогов, а десять
щупалец и тело снабжено плавниками (у
обычных осьминогов плавников нет).
Туловище каракатицы плоское, как
лепешка; у кальмара оно конусовидное,
похожее на ракету. На узком конце
«ракеты», где полагалось бы быть хвосту,
расходятся в стороны ромбовидные
плавники.
Раковина головоногих моллюсков
рудиментарная: у каракатицы —
известковая пластиночка, у кальмара —
хитиновое перышко, похожее на римский меч
гладиус. Гладиусом и называют
недоразвитую раковину кальмара.
Щупальца головоногих моллюсков
венчиком окружают рот. На них в один или
в два ряда, реже в три или четыре, сидят
присоски *. В основании щупальца
присоски мельче, в середине расположены
самые большие, а на концах — совсем
крошечные.
Рот у головоногих небольшой,
глотка мускулистая, а в глотке —
черный роговой клюв (у
кальмара коричневый), кривой, как у попугая.
От глотки к желудку тянется тонкий
пищевод. По пути он насквозь пронзает
мозг. У осьминогов довольно большой
мозг: в нем четырнадцать долей. Мозг
осьминога покрыт зачаточной корой из
мельчайших серых клеток —
диспетчерский пункт памяти, а сверху он защищен
настоящим хрящевым черепом.
Хрящевой скелет головоногих моллюсков по
своему составу близок к хрящу
позвоночных животных. Хрящ чрезвычайно
редко встречается у беспозвоночных
животных, а у головоногих моллюсков
хрящевой внутренний скелет хорошо развит.
Он окружает не только головной мозг, но
и поддерживает чашевидными выростами
глаза. Опорные хрящи развиты также
1 У осьминога триоксиопуса три ряда
присосок.
156
в основании щупалец, плавников и в
кнопках запирающего мантийного аппарата.
Нервная система головоногих
моллюсков сложнее, чем у всех других
беспозвоночных животных. Ганглии ее
очень велики и так плотно сближены друг
с другом, что по существу образуют
единую нервную массу. Лишь на разрезах
можно различить составляющие ее узлы.
Нервная система головоногих
моллюсков высокоспециализированна. По
тонкости своих чувств, точности восприятия
и сложности ответных реакций и
поведения головоногие превосходят многих
морских животных. Некоторые
исследователи, которые произвели наиболее
совершенные эксперименты над поведением
головоногих моллюсков, считают, что
имеется много общего в образовании
условных рефлексов и процессов торможения
в головном мозгу человека и осьминога,
хотя центры, заведующие этими
функциями, и не гомологичны по своему
происхождению. Клетки мозга со всех сторон
плотно облегают пищевод. Поэтому
осьминоги (кальмары и каракатицы тоже),
несмотря на очень хищные аппетиты, не
могут проглотить добычу крупнее
лесного муравья. Но природа наделила их
теркой, которой они приготавливают
«пюре» из крабов и рыб. Мясистый
язык головоногих покрыт
полусферическим роговым чехлом. Чехол усажен
мельчайшими зубчиками, которые
перетирают пищу, превращая ее в кашицу.
Пища смачивается во рту слюной и
попадает в желудок, затем в слепую кишку;
а это по сути дела второй желудок.
Есть и печень и
поджелудочная железа.
Пищеварительные соки, которые они выделяют, очень
активны — быстро, за четыре часа,
переваривают пищу.
У других холоднокровных животных
переваривание затягивается на многие
часы, у камбалы, например, на 40—60
часов.
У головоногих не одно, а три сер д-
ц а: главное, состоящее из одного
желудочка и двух предсердий, гонит кровь
по телу, а два других (жаберные
сердца) проталкивают ее через жабры.
Главное сердце бьется 30—36 раз в минуту.
Рис. 105.
Анатомический разрез тела
каракатицы (А) и
осьминога (Б):
1 — глотка с клювом; 2 — мозг;
3 — хрящевая капсула; 4 —
остаток раковины; 5 —
чернильный мешок; б — жабры; 7 —
воронка; 8 — вороночные хрящи;
9 — внутренностный мешок;
10 — хрящевые кнопки
мантии; J J — мантия.
В отличие от других моллюсков у
головоногих кровеносная система
почти замкнутая: во многих местах (в
коже, мускулатуре) артерии через
капилляры переходят непосредственно в вены.
У них кровь необычная — голубая!
Темно-голубая, когда насыщена
кислородом, и бледная — в венах.
Цвет крови животных зависит от
металлов, которые входят в состав кровяных
телец (эритроцитов) или веществ,
растворенных в плазме. Вместо гемоглобина
кровь головоногих моллюсков содержит
гемоцианин. Медь, входящая в
состав гемоцианина, и придает их крови
синеватый цвет.
Мы переходим теперь к описанию
самого интересного органа головоногих
моллюсков — «реактивного двигателя».
Обратите внимание, как просто, с какой
минимальной затратой материала решила
природа сложную задачу.
Внизу, у «шеи» кальмара (рассмотрим
в качестве примера этого моллюска),
заметна узкая щель — мантийное
отверстие. Из нее, словно пушка из
амбразуры, торчит наружу какая-то
трубка. Это воронка, или сифон,— «сопло»
реактивного двигателя.
И щель и воронка ведут в
обширную полость на «животе»: это мантийная
полость — «камера сгорания» живой
ракеты. Всасывая в нее воду через широкую
мантийную щель, моллюск с силой
выталкивает ее затем через воронку. Чтобы
вода не вытекала обратно через щель,
кальмар ее плотно замыкает при помощи
особых «застежек-кнопок», когда «камера
сгорания» наполнится забортной водой.
По краю мантийного отверстия
расположены хрящевые грибовидные бугорки.
На противоположной стороне щели им
соответствуют углубления. Бугорки
входят в углубления и прочно запирают, как
кнопки, все выходы из камеры, кроме
одного — через воронку.
Когда моллюск сокращает «брюшную»
мускулатуру (мускулатуру брюшной
стенки мантии), сильная струя воды бьет из
сифона. Реактивная сила, возникающая
при этом, толкает кальмара в
противоположную сторону. Воронка направлена
к концу щупалец, поэтому головоногий
моллюск обычно плывет задним концом
тела вперед.
Реактивные толчки и всасывания воды
в мантийную полость с неуловимой
быстротой следуют одно за другим, и кальмар
ракетой проносится в синеве океана.
Движение. Перекачивая через
себя воду, головоногий моллюск скользит
в лазурных волнах, точно ракета.
Высшего совершенства в реактивной
навигации достигли кальмары. У них даже тело
своими внешними формами копирует
ракету (или, лучше сказать, ракета
копирует кальмара, поскольку ему
принадлежит в этом деле бесспорный
приоритет).
Тело у кальмара длинное,
цилиндрическое, предельно обтекаемое, спереди и
сзади заостренное. На хвосте живая
ракета несет стабилизаторы —
ромбовидные плавники. Щупальца на
наружной, противоположной присоскам
стороне вооружены мощными
продольными килями. Когда щупальца сложены
вместе, они напоминают хвостовое
оперение стрелы или авиационной бомбы.
Изгибая сложенные пучком щупальца
вправо, влево, вверх или вниз, кальмар
поворачивается в ту или другую сторону.
Поскольку такой руль по сравнению с
самим животным имеет очень большие
размеры, то достаточно его
незначительного движения, чтобы кальмар даже на
полном ходу легко мог увернуться от
столкновения с препятствием. Резкий
поворот руля — и пловец мчится уже в
обратную сторону. Вот изогнул он конец
воронки назад и скользит теперь головой
вперед. Выгнул ее вправо — и
реактивный толчок отбросил его влево. При
быстром передвижении воронка всегда торчит
прямо между щупальцами и кальмар
мчится хвостом вперед, как бежал бы
рак-скороход, наделенный резвостью
скакуна.
Если кальмар или каракатица плавают
не спеша, ундулируя плавниками,
миниатюрные волны пробегают по ним спереди
назад, и животные грациозно скользят,
изредка подталкивая себя также и струей
воды, выброшенной из-под мантии.
Октопусы и некоторые другие
осьминоги (Eledone, Agronauta) могут
передвигаться по дну на «руках». Для этого
обычно служат боковые пары рук, которые,
как правило, длиннее прочих. Поэтому
осьминоги передвигаются по дну боком,
как крабы. Они приподнимаются на
руках, опускают туловище вниз к
вентральной паре рук и поворачивают отверстие
воронки вбок, обычно вправо. В такой
позе осьминоги удивительно напоминают
уродливого головастого человечка.
Некоторым исследователям приходилось
наблюдать передвижение осьминогов по дну
«на цыпочках» — на самых кончиках
вытянутых вертикально вниз щупалец.
В критические минуты осьминоги, как и
кальмары, могут развивать большую
скорость. Обычно же осьминоги плавают
сравнительно медленно. Джозеф С а й н л,
изучавший миграции спрутов, подсчитал:
осьминог размером в полметра плывет
по морю со средней скоростью около
15 км в час. Каждая струя воды, выбро-
158
шенная из воронки, толкает
его вперед (вернее, назад,
так как осьминог плывет
задом наперед) на 2—2,5 м.
Органы чувств.
«Если, — пишет один
ученый,— попросить зоолога
указать наиболее
поразительную черту в развитии
животного мира, он назвал бы
не глаз человека (конечно,
это удивительный орган) и
не глаз осьминога, а
обратил бы внимание на то, что
оба эти глаза, глаз человека
и глаз осьминога, очень
похожи». Похожи они не
только своим строением, но
часто даже и выражением —
странный факт, который всегда поражал
натуралистов.
Глаз осьминога мало отличается от
глаза млекопитающего животного или даже
человека. Но есть и некоторые отличия:
например, роговица у большинства
головоногих моллюсков не сплошная, а
пронзена спереди небольшим (каракатицы) или
довольно широким (кальмары)
отверстием. Хрусталик не эллиптический, а
круглый, разделенный пополам тонкой
эпителиальной пластинкой.
«Кроме того,— пишет известный
советский исследователь В. А. Д о г е л ь,— в
глазу Cephalopoda имеются
приспособления к видению как при более сильном, так
и более слабом освещении. В клетках
сетчатки заключается бурый зернистый
пигмент. Последний при ярком дневном
свете распространен по всей клетке и,
таким образом, отчасти защищает клетку
от чересчур сильного освещения. Ночью
же весь пигмент сосредоточивается лишь
в основании клетки, чувствительность
которой вследствие этого повышается».
Аккомодация (установка зрения
на разные дистанции, фокусировка) у
человека достигается изменением кривизны
хрусталика, а у головоногих моллюсков —
удалением или приближением его к
сетчатке, подобно тому как в фотоаппарате
движется объектив.
Ни у кого из обитателей моря нет таких
зорких глаз, как у осьминога и его роди-
Рис. 106. Схематический рисунок глаза кальмара, каракатицы
и осьминога:
1 — головной хрящ; 2 — передняя полость глаза; 3 — сетчатка; 4 —
хрусталик; 5 — радужина; 6 — зрительный ганглий; 7 — отверстие в
роговице; 8 — роговица; 9 — веко.
чей. Только глаза совы, кошки да
человека могут составить им конкуренцию.
На одном квадратном миллиметре
сетчатки глаза осьминога насчитывается
около 64 тыс. воспринимающих свет
зрительных элементов, у каракатицы еще
больше — 150 тыс., у кальмара — 162 тыс.,
у карпа — 50 тыс., у кошки — 397 тыс.,
у человека — 400 тыс., а у совы даже
680 тыс.
И размер глаз у головоногих моллюсков
рекордный. Глаз каракатицы лишь в
десять раз меньше ее самой, а у гигантского
спрута глаза величиной с небольшое
колесо: 40 см в диаметре! У
тридцатиметрового голубого кита глаз не
превышает в длину 10—12 см (в 200—300 раз
меньше самого кита).
Даже ослепленные осьминоги видят свет.
Вернее, ощущают его всей поверхностью
тела. Оно у них очень чувствительное:
в коже рассеяны осязательные,
светочувствительные, обонятельные и вкусовые
клетки.
Вкус пищи, предлагаемой
экспериментаторами, осьминоги распознавали не
только языком. И даже главным образом
не языком, а «руками». Вся внутренняя
поверхность щупалец (но не наружная)
и каждая присоска участвуют в
дегустировании пищи. Чтобы узнать,
соответствует ли его вкусу предлагаемое блюдо,
осьминог пробует его кончиком щупалец.
Чувство вкуса у осьминога настолько
159
тонко, что он, видимо, и врагов
распознает на вкус.
Мак-Гинити, американский
океанолог, выпустил из пипетки около спрута
капельку воды, которую набрал в другом
аквариуме поблизости от мурены —
злейшего врага осьминогов. Спрут поступил
соответственно имитированной ситуации:
испугался, побагровел и пустился наутек.
Впрочем, еще неизвестно, каким
чувством он распознал врага — вкусом или
обонянием. Разница между этими
чувствами невелика, а у осьминога и вовсе,
возможно, ее нет. Мы уже знаем, что
органы вкуса, способные отличить
сладкое от кислого, горькое от соленого,
расположены у осьминога, помимо языка
и губ, еще на внутренней стороне
щупалец. Но щупальцами осьминог отлично
распознает и запахи: запах мускуса и
других пахучих веществ. Какое чувство
оповещает, например, лишенного зрения
спрута о том, где лежит мертвая рыба?
Он безошибочно находит ее даже на
расстоянии полутора метров. Вкус?
Обоняние?
Сытый осьминог не проявляет обычно
интереса к пище — он не обжора, но
отрезанное у того же осьминога щупальце,
лишенное контроля головного мозга,
упорно ползет за лакомым кусочком.
По-видимому, у осьминогов (и у
кальмаров и каракатиц) вкус и обоняние
неразделимы.
Осталось упомянуть еще об одном
чувстве — о слухе. Слышат осьминоги
или они ко всему глухи?
Наверное, немного слышат, если
крикнуть им в самое ухо. Впрочем, сделать
это непросто: снаружи осьминожье «ухо»
найти нелегко. Никаких внешних
признаков, которые указывали бы на его
существование, нет. Но если разрежем
хрящевой череп осьминога, внутри найдем
два пузырька с заключенными в них
кристалликами извести. Это статоцисты —
органы слуха и равновесия. Удары
звуковых волн (но только, пожалуй, лишь
сильные удары) колеблют известковые
камешки, они касаются чувствительных
стенок пузырька, и животное
воспринимает звук, очевидно, как неясный гул.
Кристаллики извести сообщают
осьминогу также о положении его тела в
пространстве. Осьминоги с вырезанными ста-
тоцистами теряют ориентировку: плавают
спиной вниз, чего нормальные животные
никогда не делают, а то начнут вдруг
вертеться волчком или путают верх и низ
бассейна.
Образ жизни. Все головоногие
моллюски — исключительно морские животные.
Таблица 13. Головоногие моллюски:
1 — стайка кальмаров Ommastrephes sloanei-
pacificus;
2 — осьминог Octopus vulgaris ;
3 — рбссия (Rossia glaucopis);
4 — каракатица Sepia officinalis.
Обитают они только в океанах и
полносоленых морях. Особенно многочисленны
они в тропической и субтропической
области и в умеренных широтах.
Содержание солей в морской воде должно быть
не менее 33°/0о- (Поэтому головоногие
моллюски не встречаются у нас в Черном
море, соленость которого почти вдвое
меньше океанской.)
Прибрежные виды головоногих
моллюсков лучше переносят опреснение и
могут жить, по-видимому, в воде с
соленостью, пониженной до 33°/оо» Но
обитатели открытого моря, например кальмар
Brachioteuthis nisei, плохо себя
чувствуют уже при солености ниже 35°/оо-
В океане головоногие моллюски
встречаются от абиссальных глубин и до
поверхности, от тропиков и до полярных
морей. Одни из них, как и многие рыбы,
свободно плавают в толще воды, другие
предпочитают прятаться на дне.
Большинство кальмаров и пелагических
осьминогов — типичные представители
нектона, но есть среди них и
планктонные организмы. Таков,
например, палочковидный Doratopsis ver-
micularis, обитающий в открытом море
в сообществе медуз и сальп. Этот кальмар
совершенно прозрачен и напоминает
плавающий в воде кусочек льда.
Планктонный образ жизни ведут личинки почти
всех головоногих моллюсков, некоторые
из них на этой стадии напоминают своей
формой D. vermicularis. Таковы молодые
особи Chiroteuthidae, некоторых Сгап-
chidae.
Мелких аргонавтов (Argonauta hias)
тоже следует отнести к представителям
макропланктона Их самки,
обремененные раковиной, в своих
перемещениях в значительной степени зависят
от воли ветра и волн.
Наконец, головоногие моллюски (Se-
piolidae, Octopodidae, Cirroteuthidae) ведут
придонный, прибрежный и
малоподвижный образ жизни. Каракатицы
предпочитают песчаные или илистые грунты.
Здесь они подолгу лежат на дне,
наполовину закопавшись в песок, и
подстерегают добычу. Опустившись на грунт, сепия
волнообразными движениями своих
плавников взмучивает его; осаждаясь, ил
покрывает ее тонким слоем и отлично
маскирует. Способность изменять окраску в
тон окружающим предметам помогает ей
оставаться незаметной на любом грунте.
У самого дна держатся и многие
другие представители подотряда
каракатиц (Sepiella, Sepiola, Rossia).
Подавляющее большинство осьминогов —
тоже донные и прибрежные животные.
Таблица 14. Головоногие моллюски:
1 — осьминог адский вампир (Vampyroteuthis
infernalis);
2 — глубоководный кальмар кранхия (Cranchia
scabra);
3 — глубоководный кальмар батотаума (Batho-
thauma lyromma);
4 — безглазый глубоководный осьминог цирро-
таума (Cirrothauma murrayi);
5 — кальмар чудесная лампа (Thaumatolampas
diodema);
6 — глубоководный блинообразный осьминог
Opistoteuthis depressa.
11 Жизнь животных, т. ?
Рис. 107. Присоски Octopoda G, 2) и присоски
с роговыми зубцами Decapoda C, 4).
Октопусы предпочитают каменистые
грунты. Они затаиваются среди камней или
в расщелинах скал. Некоторые из них
(Paroctopus conispadiceus, Eledone mos-
chata) поселяются и на песчаных грунтах.
Как показали наблюдения за
осьминогами, содержавшимися в аквариумах,
даже на песчаном грунте они устраивают
себе убежище из камней, которые
приносят издалека.
Кальмары — обитатели главным
образом открытых морей.
Питание. Основная пища
головоногих моллюсков — рыбы, крабы и
ракушки. Но многие виды (особенно
глубоководные) едят и падаль. Едят они
и друг друга. Мелкие кальмары и
осьминоги живут в постоянном страхе за свою
жизнь, которой угрожает алчность их
более крупных собратьев. Это одно из
обстоятельств, затрудняющих содержание
осьминогов в аквариумах: более крупные
спруты съедают мелких. И не всегда
голод служит причиной каннибализма.
Поэтому еще Аристотель, раздумывая над
дурными обычаями полипу сов, решил, что
они едят друг друга, чтобы поддерживать
в себе жизненную силу: осьминог, не
отведавший осьминожьего мяса, будто бы
хиреет и умирает.
Еще более странная привычка
спрутов — автофагия,
самопожирание.
Натуралисты наблюдали иногда, как
содержавшиеся в неволе осьминоги вдруг
без всякой видимой причины начинали
себя есть! Обкусывали начисто щупальца
и... умирали.
Хотя головоногие моллюски и очень
прожорливы, при необходимости они
могут подолгу голодать. В аквариумах
осьминоги иногда жили без пищи несколько
недель, а насиживающие самки ничего
не едят около двух месяцев, иногда и
больше, пока не выведут детенышей.
Защитные приспособления.
Все без исключения головоногие
моллюски—хищники, и хищники весьма
прожорливые. Габариты жертвы их не
смущают: головоногие нападают даже на
животных, в несколько раз превышающих их
своими размерами. Это, без сомнения,
самые агрессивные и воинственные
обитатели морей. Невидимыми, но прочными
нитями биологических взаимоотношений
связаны цефалоподы со всеми
обитателями океана. Они поедают множество рыб и
крабов и сами дают пищу миллионам
пожирающих их хищников: тут и рыбы —
акулы, мурены, тунцы, макрели, треска;
тут и птицы — альбатросы, поморники,
пингвины и морские звери — киты,
дельфины, тюлени.
Что и говорить — врагов много. Но
головоногие не сдаются без борьбы: они
отлично вооружены. Их щупальца
усажены сотнями присосок, а у многих
кальмаров — также и когтями, острыми и
кривыми, как у кошек. Зубов нет, но есть
клюв. Роговой, крючковатый, он без труда
прокусывает рыбью кожу и панцири
крабов, протыкает насквозь даже прочные
раковины двустворчатых моллюсков х.
Каракатица может раздробить клювом
панцирь большого рака или череп рыбы,
вдвое более крупной, чем сама. Четырех,-
шестикилограммовые кальмары легко
перекусывают проволочную леску
спиннинга, и поэтому опытные спиннингисты,
желая «поудить» спрутов, применяют
прочную стальную жилку.
1 У осьминогов и кальмаров «бульдожий
прикус»: надклювье короче подклювья, нижняя
челюсть заходит за верхнюю и крючком
загибается вверх.
162
Пронзенный острогой, кальмар грызет
ее клювом с такой яростью, что только
щепки летят. Дозидикусы охотятся на
четырехпудовых тунцов и объедают
гигантскую рыбу дочиста, не трогая лишь
голову.
О силе щупалец головоногих говорит
следующий эпизод.
В Брайтонском аквариуме в Англии
экспериментировали с небольшим,
размером в фут, осьминогом. Служитель
опускал в бассейн краба, привязанного к
бечевке, а натуралист наблюдал внизу, сидя
у стекла. Как только краб коснулся воды,
осьминог пулей выскочил из своего угла
и схватил его, вырвав бечевку из рук
служителя.
— Давайте другого краба,— сказал
натуралист.— И держите крепче.
Опустили второго краба. Осьминог с
вожделением посматривал на него, но не
хотел расстаться с первой добычей. Краб,
покачиваясь, приближался. В осьминоге,
казалось, боролись два чувства —
жадность и благоразумие. Жадность
восторжествовала. Удерживая пойманного
краба семью руками, он протянул восьмую
вверх и схватил новое угощение. Рывок,
еще рывок. Осьминог дернул третий раз,
и бечевка лопнула!
Автотомия1 — древнейшее
средство страхования жизни — есть в
арсенале защитных приспособлений и у
осьминога. Восемь длинных рук, которые
исследуют каждую пядь незнакомого
пространства, когда осьминог выходит на
добычу, чаще других частей тела
подвергаются опасности.
Щупальца прочные — ухватившись за
одно, можно всего осьминога вытащить
из норы. Вот тут спрут «автотомирует»
себя: мышцы попавшего в плен щупальца
спазматически сокращаются.
Сокращаются с такой силой, что сами себя
разрывают. Щупальце отваливается, словно
отрезанное ножом. Хищник получает его
в виде выкупа за жизнь.
Осьминог Octopus defilippi в
совершенстве постиг искусство автотомирования.
Схваченный за руку, он тотчас расста-
1 Автотомия — самокалечение.
ется с ней. Щупальце отчаянно
извивается — это ложный маневр: враг
бросается на него и упускает главную цель.
Отверженное щупальце долго еще
дергается и, если отпустить его на свободу,
пытается даже ползти и может
присасываться.
Головоногие моллюски в процессе
эволюции приобрели еще более удивительное
чудо-оружие — чернильную бомбу.
Вместо куска живой плоти кальмар
выбрасывает перед раскрытой, чтобы сожрать
его, пастью грубую подделку собственной
персоны. Кальмар как бы раздваивается
на глазах и недругу оставляет своего
бесплотного двойника, а сам быстро
исчезает.
В минуту опасности головоногие
выбрасывают из воронки струю черной
жидкости. Чернила расплываются в
воде густым облаком, и под прикрытием
«дымовой завесы» моллюск более или
менее благополучно удирает, оставляя
врага блуждать в потемках.
В чернилах содержится органическая
краска из группы меланинов, близкая по
составу к пигменту, которым окрашены
наши волосы. Оттенок чернил не у всех
головоногих одинаков: у каракатиц он
сине-черного тона (в сильном разведении
цвета «сепии»), у осьминогов — черный,
у кальмаров — коричневый.
Чернила вырабатывает особый орган —
грушевидный вырост прямой кишки. Его
называют чернильным мешком.
Это плотный пузырек, разделенный
перегородкой на две части. Верхняя половина
отведена под запасной резервуар, в нем
хранятся чернила, нижняя — заполнена
тканями самой железы. Ее клетки набиты
зернами черной краски. Старые клетки
постепенно разрушаются, их краска
растворяется в соках железы — получаются
чернила. Они поступают на «склад» —
перекачиваются в верхний пузырек, гд&
Хранятся до первой тревоги.
Не все содержимое чернильного мешка
выбрызгивается за один раз.
Обыкновенный осьминог может ставить «дымовую
завесу» шесть раз подряд, а через
полчаса уже полностью восстанавливает весь
израсходованный запас чернил. Красящая
способность чернильной жидкости необы-
И*
163
чайно велика. Каракатица за пять секунд
окрашивает извергнутыми чернилами всю
воду в баке вместимостью в 5,5 тыс. л.
А гигантские кальмары извергают из
воронки столько чернильной жидкости, что
морская вода мутнеет на протяжении
сотни метров!
Головоногие моллюски рождаются с
мешком, наполненным чернилами. Одна
крошка каракатица, едва выбравшись из
оболочки яйца, тут же окрасила воду
пятью чернильными залпами.
И вот какое неожиданное открытие было
сделано биологами в последнее
десятилетие. Оказалось, что традиционное
представление о «дымовой завесе» головоногих
моллюсков следует основательно
пересмотреть. Наблюдения показали, что
выброшенные головоногими чернила
растворяются не сразу, не раньше чем на что-
нибудь наткнутся. Они долго, до десяти
минут и больше, висят в воде темной
и компактной каплей. Но самое
поразительное, что форма капли напоминает
очертания выбросившего ее животного.
Хищник вместо убегающей жертвы
хватает эту каплю. Вот тогда она
«взрывается» и окутывает врага темным облаком.
Акула приходит в полное замешательство,
когда стайка кальмаров одновременно,
как из многоствольного миномета,
выбрасывает целую серию «чернильных бомб».
Она мечется из стороны в сторону,
хватает одного мнимого кальмара за другим,
и вскоре вся скрывается в густом облаке
рассеянных ею чернил.
В 1956 г. доктор Д. X э л опубликовал
в английском журнале «Нейчур»
интересные наблюдения над маневрами, к
которым прибегает кальмар, подменяя себя
чернильным макетом *.
1 Правда, еще в 1878 г. Фредерик писал,
что каракатица сепиола выбрасывает чернильные
капли, похожие на нее по форме, и благодаря
такой имитации спасается от хищников. Но
этому наблюдению не придали значения. Так
нередко случалось и с другими открытиями,
идущими вразрез с общепринятым мнением.
Прошло почти сто лет, прежде чем наблюдение
Фредерика было вновь «открыто». Исследованием
этого вопроса наука обязана многим
натуралистам, и в первую очередь должны быть
упомянуты немецкий зоолог Вильгельм Ш е ф е р
и английский океанолог Д. Хэл.
Зоолог посадил кальмара в кадку и
попытался поймать его рукой. Когда его
пальцы были уже в нескольких дюймах
от цели, кальмар внезапно потемнел и,
как показалось Хэлу, замер на месте.
В следующее мгновение Хэл схватил...
чернильный макет, который развалился
у него в руках. Обманщик плавал в
другом конце кадки.
Хэл повторил свою попытку, но теперь
внимательно следил за кальмаром. Когда
его рука вновь приблизилась, кальмар
снова потемнел, выбросил «бомбу» и тут
же стал мертвенно бледным, затем
невидимкой метнулся в дальний конец кадки.
До чего тонкий маневр! Кальмар ведь
не просто оставил вместо себя свое
изображение. Нет, это сцена с переодеванием.
Сначала он резкой сменой окраски
привлекает внимание противника. Затем тут
же подменяет себя другим темным
пятном — хищник автоматически фиксирует
на нем свой взгляд — и исчезает со сцены,
переменив «наряд». Обратите внимание:
теперь у него окраска не черная, а белая.
Чернила головоногих моллюсков
обладают еще одним удивительным защитным
свойством. Мак-Гинити,
американский биолог, провел серию
экспериментов над калифорнийским осьминогом и
муреной. И вот что установил: чернила
осьминога, оказывается, парализуют
обонятельные нервы хищных рыб!
После того как мурена побывает в
чернильном облаке, она утрачивает
способность распознавать запах притаившегося
моллюска, даже когда натыкается на него.
Больше часа длится парализующее
действие осьминожьего наркотика!
Чернила головоногих моллюсков в
большой концентрации опасны и для них
самих. Джильпатрик сделал
такой опыт: посадил небольшого спрута
в ведро с морской водой и добавил в нее
чернила, извлеченные из пяти таких же
моллюсков. Минуты через три осьминог
был мертв.
Подобный же эксперимент проделал и
Ли-Сюер: пустил в пятилитровый
сосуд двух маленьких осьминогов. Они
быстро окрасили воду в черный цвет,
опорожнив свои чернильные мешки, и...
умерли через десять минут.
164
В море, на воле, осьминог избегает
вредоносного действия своего оружия, быстро
покидая отравленное место. В
ограниченном пространстве ему нелегко это сделать.
В бассейнах с плохой сменой воды
концентрация чернил быстро превышает
допустимую норму, отравляет пленников,
и они гибнут.
Опасны ли чернила головоногих для
человека?
Ответить на этот вопрос попросим
такого знатока подводной охоты, как Джеймс
Олдридж1. Он говорит: «Я
настолько свободно вел себя с осьминогом, что
получил струю чернил прямо в лицо.
А так как я был без маски, то жидкость
попала мне в глаза и ослепила.
Окружающий мир от этого, правда, не потемнел,
а окрасился в чудный янтарный цвет. Все
вокруг казалось мне янтарного цвета до
тех пор, пока пленка этих чернил
держалась у меня на глазах. Это длилось минут
десять или около того. Этот случай не
повлиял на мое зрение».
В той же книге Олдридж пишет:
«Осьминоги удивительно быстро и гармонично
окрашиваются под цвет окружающей их
местности, и, когда вы, подстрелив одного
из них, убьете или оглушите его, он не
сразу потеряет способность менять
окраску. Это я наблюдал однажды сам,
положив добытого осьминога на газетный лист
для разделки. Убитый осьминог
моментально изменил окраску, сделавшись
полосатым, в белую и черную полоску!»
Ведь он лежал на печатной странице
и скопировал ее текст, запечатлев на
своей коже чередование черных строк и
светлых промежутков. По-видимому,
осьминог этот не был совсем мертв, глаза его
еще воспринимали оттенки меркнущих
красок солнечного мира, который он
навсегда покидал.
Даже среди высших позвоночных
животных немногие обладают бесценным
даром изменять по прихоти или
необходимости окраску кожи, перекрашиваться,
копируя оттенки внешней декорации.
1 Джеймс Олдридж — английский писатель и
спортсмен, написал превосходную книгу
«Подводная охота», изданную в 1960 г. издательством
«Физкультура и спорт» на русском языке.
У всех головоногих моллюсков под
кожей расположены эластичные, как
резина, клетки. Они набиты краской, словно
акварельные тюбики. Научное название
этих чудесных клеток — хроматофоры.
Каждый хроматофор —
микроскопический шарик (когда пребывает в покое)
или точечный диск (когда растянут),
окруженный по краям, будто солнце лучами,
множеством тончайших мускулов—
дилататоров, т. е.
расширителей. Лишь у немногих хроматофоров
только четыре дилататора, обычно их
больше — около двадцати четырех. Дила-
таторы, сокращаясь, растягивают
хроматофор, и тогда содержащаяся в нем
краска занимает в десятки раз большую, чем
прежде, площадь. Диаметр хроматофора
увеличивается в шестьдесят раз: от
размеров иголочного острия до величины
булавочной головки. Иными словами,
разница между сократившейся и растянутой
цветной клеткой столь же велика, как
между двухкопеечной монетой и
автомобильным колесом.
Когда мускулы-расширители
расслабляются, эластичная оболочка
хроматофора принимает прежнюю форму.
Хроматофор растягивается и сокращается
с исключительной быстротой. Он
изменяет свой размер за 2—3 секунды, а по
другим данным еще быстрее — за 112—111
секунды.
Каждый дилататор соединен нервами
с клетками головного мозга. У
осьминогов «диспетчерский пункт»,
заведующий сменой декораций, занимает в
мозгу две пары лопастевидных долей.
Передняя пара контролирует окраску головы
и щупалец, задняя — туловища. Каждая
лопасть управляет своей, т. е. правой или
левой, стороной. Если перерезать нервы,
ведущие к хроматофорам правой стороны,
то на правом боку моллюска застынет
одна неизменная окраска, в то время
как его левая половина будет «играть»
колерами разных тонов,
Какие органы корректируют работу
мозга, заставляя его изменять окраску
тела точно в соответствии с фоном
окрестностей? Глаза. Зрительные впечатления,
полученные животным, по сложным
физиологическим каналам поступают к нерв-
165
ным центрам, а те подают
соответствующие сигналы хроматофорам: растягивают
одни, сокращают другие, добиваясь
сочетания красок, наиболее пригодного для
маскировки. Слепой на один глаз
осьминог теряет способность легко менять
оттенки на безглазой стороне тела.
Удаление второго глаза приводит к почти
полной потере способности менять окраску.
Исчезновение цветовых реакций у
ослепленного осьминога неполное, потому
что изменение окраски зависит также и
от впечатлений, полученных не только
глазами, но и... присосками. Если лишить
осьминога щупалец или срезать с них
все присоски, он бледнеет и, как ни
пыжится, не может ни покраснеть, ни
позеленеть, ни стать черным.
Хроматофоры головоногих содержат
черные, коричневые, красно-бурые,
оранжевые и желтые пигменты. Самые
крупные — темные хроматофоры, в коже они
лежат ближе к поверхности. Самые
мелкие — желтые. Каждый моллюск наделен
хроматофорами только трех каких-нибудь
цветов: коричневыми, красными и
желтыми либо черными, оранжевыми и
желтыми. Их сочетание, конечно, не может
дать всего разнообразия оттенков,
которыми знамениты головоногие моллюски.
Металлический блеск, фиолетовые,
серебристо-голубые, зеленые и голубовато-
опаловые тона сообщают их коже клетки
особого рода — иридиоцисты. Они лежат
под слоем хроматофоров и за прозрачной
оболочкой прячут множество блестящих
пластиночек. Иридиоцисты заполнены,
словно комнаты смеха в парках, рядами
«зеркал», целой системой «призм» и
«рефлекторов», которые отражают и
преломляют свет, разлагая его на великолепные
краски спектра.
Раздраженный осьминог из пепельно-
серого через секунду может стать черным
и снова превратиться в серого,
продемонстрировав на своей коже все тончайшие
переходы и нюансы в этом интервале
красок. Бесчисленное разнообразие оттенков,
в которые окрашивается тело осьминога,
можно сравнить лишь с изменчивым
цветом вечернего неба и моря.
Если бы кому-нибудь пришло в голову
устроить всемирное состязание
хамелеонов, первый приз наверняка получила бы
каракатица. В искусстве маскироваться
никто не может с ней соперничать, даже
осьминог. К любому грунту каракатица
приспосабливается без труда. Только что
была она полосатой, как зебра,
опустилась на песок и тут же перекрасилась —
стала песочно-желтой. Проплыла над
белой мраморной плитой — побелела. Вот
лежит она на гальке, освещенной солнцем,
ее спину украшает узор из светлых (в тон
солнечным бликам) и серо-бурых пятен.
На черном базальте каракатица черная,
как ворон, а на пестром камне пегая.
Исследователь Холмс описал
девять цветовых образцов, которыми
пользуется каракатица для выражения чувств
(три образца) и маскировки (шесть
образцов).
Полосатая или пятнистая окраска,
составленная из резко контрастирующих
элементов (черные полосы на белой шкуре,
либо белые на черной, или черные пятна
на желтом фоне), встречается у многих
животных: у тигра, леопарда, ягуара,
оцелота, жирафа, антилопы куду и бонго,
у окапи, рыб, змей, бабочек.
Обратили ли вы внимание, что у всех
перечисленных животных полосы и пятна
идут рядами поперек тела? Ведь это не
случайно. Дело в том, что поперечные
полосы, достигая границ силуэта,
внезапно обрываются. Сплошная линия
контура при этом расчленяется
чередующимися то белыми, то черными полями
расцветки, и животное, теряя свои
привычные глазу очертания, сливается с фоном
местности. К такому же способу
маскировки прибегают и люди, когда
раскрашивают военные объекты светлыми и
темными пятнами, расчленяющими контуры
маскируемого сооружения.
Если же черные и белые полосы идут
не поперек, а вдоль контуров тела, то они
не расчленяют, а, наоборот, подчеркивают
их. Хорошо заметная окраска выгодна
ядовитым или обладающим дурным
запахом существам, чтобы хищники не
хватали их по ошибке. Таковы, например,
саламандра и скунс: у них действительно
полосы идут вдоль тела.
Контрастирующие полосы, расчленяя
силуэт каракатицы, помогают ей слиться
166
с окраской любого грунта. Ведь зеброид-
ный рисунок — универсальный
камуфляж.
Даже новорожденные осьминоги не
остаются безоружными. Пока не развились
еще их собственные боевые средства,
малютки вооружаются «ядовитыми
стрелами» медуз.
Медузы жалят, как крапива. Их
щупальца усажены микроскопическими
батареями стрекающих пузырьков — н е-
м а т о ц и с т а м и.
Немецкий ученый Адольф Н э ф ловил
в Средиземном море личинок г тремок-
шопусов — миниатюрных пелагических
осьминогов — и с удивлением обнаружил,
что каждая личинка держит перед собой
в слабеньких «ручонках» заграждение из
обрывков щупалец медуз.
Нэф решил, что стрекающие немато-
цисты, которыми усажены щупальца
медузы, служат осьминожьим младенцам
оружием защиты.
Владеет ли еще какое-нибудь живое
существо таким разнообразием защитных
инстинктов и столь совершенной «боевой
техникой», как головоногие моллюски?
Органы свечения. Жан
Батист В е р а н и любил приходить на
берег моря, когда рыбаки возвращались
с уловом. Диковинных животных
привозили их лодки. Однажды недалеко от
Ниццы он увидел на берегу толпу людей.
В сети попалось существо совершенно
необычное. Тело толстое — мешком, как
у осьминога, но щупалец десять, и
связаны они тонкой перепонкой, словно
зонтиком.
Верани опустил причудливого
пленника в ведро с морской водой и «в тот же
момент,— пишет он,— я был захвачен
удивительным зрелищем сверкающих
пятен, которые появились на коже
животного. То это был голубой луч сапфира,
который слепил меня, то опаловый —
топаза, то оба богатых оттенками цвета
смешивались в великолепном сиянии, ок-
1 Молодь головоногих моллюсков условно
называют личинками, хотя никакого
превращения эти животные не совершают и их личинки
(за немногими исключениями) отличаются от
взрослых особей лишь меньшими размерами.
Рис. 108. Гистиотевтис бонеллиана (Histioteuthis
bonelliana).
ружавшем ночью моллюска, и он казался
одним из самых чудеснейших творений
природы».
Так Жан Батист Верани, молодой
французский натуралист, открыл в 1834 г.
биолюминесценцию
головоногих моллюсков. Он не ошибся, когда
решил, что многочисленные голубоватые
точки на теле животного — светящиеся
органы (фотофоры). У глубоководного
кальмара гистиотевтиса, которого
исследовал Верани, около двухсот таких
«фонариков»; некоторые из них достигают в
диаметре 7,5 мм — настоящие
прожекторы! Фотофор по конструкции
напоминает прожектор или автомобильную фару.
И форма у него приблизительно такая
же — полусферическая. Орган покрыт со
всех сторон, кроме обращенной наружу
светящейся поверхности, черным,
светонепроницаемым слоем. Дно его выстлано
блестящей тканью. Это зеркальный
рефлектор. Непосредственно перед ним
расположен источник света — фотогенное
тело, масса фосфоресцирующих клеток.
Сверху «фара» прикрыта прозрачной лин-
167
зой, а поверх нее — диафрагмой (слоем
черных клеток — хроматофоров).
Наползая на линзу, хроматофоры закрывают
ее — свет гаснет.
Светящиеся органы кальмаров наделены
еще целым рядом других оптических
устройств.
У каллитевтиса, например, исходящий
от фотогенной массы свет пересекает косо
поставленное «зеркало». Особые мускулы
поворачивают «зеркало» в разные стороны,
и луч света меняет свое направление.
Есть в фотофорах и светофильтры —
экраны из разноцветных клеток. Иногда
роль светофильтра выполняет цветной
рефлектор. Нередко один моллюск
обладает осветительными средствами десяти
различных конструкций.
Некоторые кальмары буквально
усеяны крупными и мелкими фотофорами, и
не только снаружи, но и изнутри. Многие
носят под мантией «пояс огненных
драгоценных камней». Свет от сияющих
«камней» проникает наружу через прозрачные
«окна» в коже и мускулатуре этих
животных.
Часто фотофоры сидят на глазах —
на веках или даже на самом глазном
яблоке, а иногда они сливаются в
сплошные полосы, окружающие глазную
орбиту светящимся полукольцом.
У таксеумы и батотаумы,
причудливых обитателей глубин, глаза сидят на
длинных стебельках и каждый глаз
наделен мощными фотофорами. У этих
кальмаров, замечает Фрэнк Л е й н, сразу
два оптических прибора — дальномеры и
прожекторы.
Фотофоры на глазах обнаружены не
только у кальмаров, но и у некоторых
глубоководных раков и рыб. Очевидно,
приближенный к самым глазам источник
света помогает рассматривать близко
находящиеся предметы. О дальнем зрении
на глубинах не может быть и речи.
Светящиеся органы каракатиц иного
строения, чем у кальмаров: в них нет
твердой массы фотогенных клеток.
Светящиеся фонарики каракатицы —
самые экономные в мире лампочки. Без
перезарядки горят они годами. Дающее
свет горючее размножается быстрее, чем
успеваег сгорать. Каракатицы носят в
особой капсуле внутри тела целый мирок
светящихся бактерий.
«Пузырек» с бактериями погружен в
углубление чернильного мешка. Дно
углубления выложено, словно перламутром,
слоем блестящих клеток. Это зеркальный
рефлектор. Есть и линза-коллектор у
«карманного фонарика» каракатицы.
Студневидная и прозрачная, лежит она
сверху — на мешочке с бактериями.
Есть и выключатель у «фонарика».
Когда нужно «потушить» свет, каракатица
выделяет в мантийную полость несколько
капелек чернил. Чернила покрывают
тонкой пленкой мешочек с бактериями, как
бы набрасывают на него черное
покрывало, и свет гаснет.
Двурогой сепиолой назвали зоологи чо-
чин-ику — миниатюрное создание,
размером с ноготь большого пальца, которое
охотится за рачками в водах океана
вблизи берегов Японии и Курильских
островов. Ночью сепиола светится. Лучезарный
нимб окружает ее крошечное тельце, и
сияющая малютка парит над черной
бездной моря, как живая звездочка.
Поймать сепиолу нетрудно. Годится для
этого простой сачок на длинной палке.
Перевернув ее на спину и осторожно
отогнув край мантии, мы увидим большой,
двурогой формы (отсюда и название
малютки) «пузырек». Он лежит на
чернильном мешке, покрывая его целиком, и
наполнен слизью. Это мицетом1 —
«садок» для светящихся бактерий.
Наблюдения показали, что чочин-ика,
спасая свою жизнь, мечет во врага
«жидкий огонь» — мгновенно вокруг
животного вспыхивает светящееся облако.
Хищник, пытавшийся схватить каракатицу,
слепнет. Тем временем моллюск спешит
укрыться в безопасном месте.
Однако лучших результатов в
«огнеметном» искусстве добился гетеротевтис —
«пиротехник», о котором писал еще
Аристотель. Гетеротевтис живет в
Атлантическом океане и Средиземном море на
небольших глубинах — до 500—1000 м.
1 Мицетомы — камеры со светящимися сим-
биотическими бактериями; найдены только у
каракатиц и некоторых кальмаров (Loliginidae),
которые по анатомическим признакам близки
к каракатицам, хотя внешне и не похожи на них.
168
М и ц е т о м гетеротевтиса
снабжен большим резервуаром.
Стенки резервуара эластичны,
н, когда окружающие их
мускулы сокращаются, миллионы
бактерий извергаются наружу,
вспыхивая в глубинах моря
ярким фейерверком.
Светящиеся органы
головоногих моллюсков работают очень
экономно: 80 и даже 93%
излучаемого ими света
составляют лучи с короткой волной и
только несколько процентов —
тепловые лучи. В
электрической лампочке лишь 4%
подведенной энергии
преобразуется в свет, а 96% — в тепло.
В неоновой лампе коэффициент
полезного действия несколько
выше — до 10%.
Размножение. Во
время размножения самцы
головоногих моллюсков одним из
щупалец — его называют г е к -
токотилем — достают из-
за «пазухи» (из мантийной
полости) упакованную в «пакеты»
сперму и переносят ее в
мантийную полость самки. «Пакеты» со
спермой носят название сперматофоров.
Форма их разнообразна, но обычно
напоминает бутыль, трубочку, «казацкую
шашку». Размеры их — от 3 мм до 115 см
(у осьминога Octopus dofleini). Сперма-
тофоры хранятся в особом вместилище —
нидхэмовом органе. Они лежат
компактной пачкой параллельно друг
другу. Во время размножения струи
воды выносят их наружу через воронку.
Здесь моллюск подхватывает их одной из
своих рук и «преподносит» самке.
Конструкция сперматофора довольно
сложна и несколько напоминает
устройство мины. Основной частью ее
«взрывного» аппарата служит свернутая в
большое число витков упругая пружина и
особая пробочка — «запал» биологической
«мины». После того как сперматофор
попадет в мантийную полость самки, пробочка
сперматофора набухает и лопается,
словно взрывается, пружина с силой
разворачивается и выбрасывает сперму.
Рис. 109. Две стадии
развития гектокотиля
аргонавта.
Румынский биолог Эмиль Р а-
к о в и ц а был первым
исследователем (не считая, может
быть, Аристотеля), которому
удалось в конце прошлого века
наблюдать спаривание
осьминогов. Животные сидели на
некотором расстоянии друг от
друга. Самец удерживал самку
одной из восьми рук, а щу-
пальцем-гектокотилем доставал
из своей мантийной полости
сперматофоры и переносил их
в мантийную полость самки.
У многих видов осьминогов
гектокотиль имеет форму
гибкой руки, снабженной двумя
пальцами, из которых один
очень длинный (лигула),
второй очень короткий (калямус).
Этими пальцами и
захватываются сперматофоры. Однако
механизм действия гектокотиля
еще недостаточно ясен. У
самцов глубоководного осьминога
Vampyroteuthis infernalis
вообще нет гектокотилей.
В высшей степени
замечательное приспособление к
оплодотворению наблюдается у мелких
пелагических осьминогов из группы Argonau-
tidae. Очень крупный гектокотиль у
самцов аргонавтов и тремокотопусов
развивается в особом мешке между
четвертой и второй руками левой стороны.
Зрелый гектокотиль отрывается от тела
самца и, извиваясь, уплывает на
розыски самки своего вида. Найдя ее,
гектокотиль заползает к ней в мантийную
полость, там содержащиеся внутри его
один или два сперматофора «взрываются»
и оплодотворяют яйца. Подробнее об этом
будет рассказано в главе об аргонавтах.
У более крупных осьминогов рода
Ocythoe наполненные спермой гектоко-
тили тоже отрываются от тела самца,
плывут самостоятельно и, найдя самку,
заползают в полость ее мантии и
присасываются там.
Сперматофоры других головоногих
моллюсков обычно переносятся самцом
непосредственно в мантийную полость самки
и помещаются там вблизи от входа в яйце-
169
Рис. 110. Яйца головоногих
моллюсков:
7 — Eledone; 2 — Girroctopus;
3 — Loligo; 4 — Sepia.
вод. Однако это наблюдается не у всех
Cephalopoda, а главным образом у
осьминогов. У более примитивных форм
(Nautilus, Sepiidae, большинства Loliginidae,
некоторых Sepiolidae и кальмаров, в
частности Ommastrephidae) сперматофоры
прикрепляются самцом к складкам или семе-
лриемникам ротового конуса самки.
Оплодотворение яиц головоногих
моллюсков большей частью происходит при
их откладке, когда они выходят из
яйцевода и попадают в мантийную полость
или же когда, вынесенные током воды
через воронку, проходят мимо рта. При
этом сперма захватывается студенистой
оболочкой яиц или слизистой массой,
покрывающей их. Только у Argonautidae
яйца оплодотворяются еще в яйцеводе,
так что в нем находят далеко зашедшие
стадии развития яиц, которые к моменту
откладки прошли уже по крайней мере
дробление. У Ocythoe процесс развития
яиц во время их пребывания в яйцеводах
заходит настолько далеко, что этот
осьминог, по некоторым сведениям, рождает
уже живых детенышей.
Достоверно установлено, что
живородящие формы были уже среди аммонитов.
В жилой камере раковины верхнеюрской
Oppelia sterospis было найдено 60
маленьких а п т и х и й, т. е. остатки раковин
60 детенышей. Известны отпечатки
других аммонитов (Dactylioceras commune,
Harpoceras lythense) вместе с молодью
внутри раковины.
Принято считать, что у головоногих
моллюсков оплодотворение яиц
внутреннее. Но это неверно. У многих
головоногих моллюсков, у которых сперматофоры
прикрепляются к ротовому конусу самки,
оплодотворение происходит заведомо вне
тела животного. Но даже у тех видов,
у которых прикрепление сперматофоров
и оплодотворение яиц происходит внутри
мантийной полости, мы не можем
последнее считать внутренним, так как
мантийная полость не является внутренней
полостью тела. Она постоянно наполнена
водой, и находящиеся в ней яйца
пребывают, по существу, в условиях внешней
среды. В этом отношении ее можно
сравнить с сумкой сумчатых животных, хотя
она и имеет совершенно иное значение *.
Лишь у тех видов головоногих моллюсков,
у которых оплодотворение яиц происходит
в яйцеводах (Agronauta ocythoe), оно
является действительно внутренним.
Таким образом, в пределах класса
Cephalopoda мы встречаемся с тремя
различными типами размножения,
представляющими собой последовательное
развитие одного и того же процесса: внешним
оплодотворением, оплодотворением
внутренним и живорождением.
Яйца кальмаров еще в яйцеводах самки
«упаковываются» в длинные студенистые
нити, которые выталкиваются наружу
через воронку. Затем самка
переворачивается вниз головой, встает почти
вертикально и, быстро дергая хвостовыми
плавниками, рывками передвигается по дну
на руках, не выпуская, однако, из них
яиц. Так, балансируя на кончиках
щупалец, идет вниз головой до тех пор, пока
не наткнется на какой-нибудь
выступающий предмет — на раковину, например,
1 Впрочем, у некоторых видов Cephalopoda
в мантийной полости вынашиваются яйца,
следовательно, в этом отношении она является уже
полным аналогом сумки сумчатых
млекопитающих (см. об этом ниже).
170
или камень. Тогда самка в течение двух-
трех секунд ощупывает этот предмет,
словно исследуя его пригодность в качестве
якоря для яиц, после чего прикрепляет
к нему яйцевую нить.
Некоторые каракатицы в пору
размножения выделяют, по-видимому,
светящуюся слизь. Самки плавают у поверхности,
самцы устремляются к ним из глубины,
точно светящиеся стрелы.
Способы, которыми каракатицы
прикрепляют свои яйца к подводным
предметам, повергали в недоумение многих
натуралистов, находивших их
яйцекладки. Каждое яичко висит на длинной
ножке — стебельке. Стебельки всех яиц
настолько тщательно переплетены друг с
другом и прочно обернуты вокруг
водоросли, что, кажется, и человек с его
ловкими пальцами не смог бы проделать это
более аккуратно. Прикрепление яиц
требует очень сложных движений щупалец
моллюска.
Число видов и геологическое прошлое.
Зоологи описали уже около шестисот
видов головоногих моллюсков (каждая
крупная группа — кальмары, каракатицы,
осьминоги — имеет приблизительно по
двести видов). Каждое новое исследование
моря приносит, как правило, неизвестные
науке новые виды этих животных. Так
что в действительности на Земле
обитает, наверное, значительно больше
головоногих моллюсков, чем открыто в
настоящее время.
А было время, когда моря и океаны
нашей планеты буквально кишели
головоногими моллюсками. Палеонтологам
известно уже более 11 тыс. ископаемых
видов.
Древнейшими из головоногих
моллюсков были наутилоидеи. От них
произошли аммониты, названные так по имени
древнеегипетского бога Аммона, которого
жрецы изображали с головой барана.
Свернутый спиралью бараний рог,
похожий на раковину аммонита, был эмблемой
бога-барана.
И наутилусы и аммониты жили в
массивных спиральных или прямых
раковинах. Медленно ползая по дну,
животные с трудом таскали их на себе.
Постепенно в процессе эволюции в раковине
Рис. 111. Аммонит.
Рис. 112. Плывущий белемнит.
развились обширные камеры,
наполненные газом,— дом сразу стал легким, как
воздух, из грузила превратился в
поплавок. Животные, словно надувные лодки,
теперь свободно дрейфовали по волнам
и расселились по всем морям и океанам.
Были среди древних наутилусов и
аммонитов и малютки не больше горошины.
Другие таскали раковины-блиндажи
величиной с небольшой танк. Моллюск эндо-
церас жил, например, в раковине,
похожей на пятиметровую шишку. В ней
свободно могли разместиться три взрослых
человека.
Раковина аммонита пахидискуса —
чудовищное колесо диаметром в 3 м\ Если
раскрутить все витки, то из нее можно
было бы соорудить лестницу до
четвертого этажа. Никогда и ни у кого ни
прежде, ни теперь не было таких огромных
раковин.
Четыреста миллионов лет безмятежно
плавали по волнам аммониты и
наутилусы. Затем вдруг вымерли. Случилось это
восемьдесят миллионов лет назад, в конце
171
мезозойской эры. Наукой с точностью
не установлено, когда и как произошли
от наутилусов белемниты — ближайшие
родичи кальмаров и каракатиц. Двести
миллионов лет назад они уже бороздили
моря.
Белемниты почти не отличались от
кальмаров. Разве только удельным весом
своей раковины: она была тяжелая,
пропитанная известью г. Как это случилось,
неизвестно, но раковина постепенно
переместилась с поверхности моллюска внутрь
его.
Белемниты ее словно бы проглотили,
или, лучше сказать, поглотили.
Раковина со всех сторон обросла складками
тела и оказалась под кожей. Теперь это
был уже не дом, а своего рода
позвоночник. Но раковина-позвоночник долго еще
сохраняла древнюю форму — полый,
разделенный на камеры конус с массивным
наконечником. Внешне она напоминала
копье или дротик.
Вот откуда белемниты получили свое
странное название: belemnon (греч.) —
дротик2.
Белемниты вымерли чуть позже
аммонитов. От белемнитов произошли
кальмары. Царство динозавров еще не
достигло своего величия, а кальмары уже жили
в море. Осьминоги появились позднее —
сто миллионов лет назад, в конце
мелового периода 3.
Ну а каракатицы совсем молодые (в
эволюционном смысле) создания. Они начали
свое развитие в одно время с лошадьми
и слонами — всего каких-нибудь
пятьдесят миллионов лет назад.
Как выглядели предки осьминогов, мы
можем судить не только по их
окаменевшим раковинам, но и по живым образцам:
шесть видов из старейшего рода морских
патриархов дожили до наших дней.
Пережившие свою эпоху наутилусы обитают
на юго-западе Тихого океана: у
Филиппин, Индонезийских островов и у
Северной Австралии.
В строении тела современных
наутилусов сохранились многие примитивные
черты, свойственные предкам всех
головоногих.
Они, например, живут в раковинах,
словно улитки, у них очень
несовершенные глаза, воронка состоит из двух
свернутых в трубку лопастей, края которых
не срослись. У наутилусов 4 почки
(нефридия), 4 жабры и 4 жаберных сердца.
Поэтому зоологи выделили их в
подкласс четырехжаберных головоногих
моллюсков, а осьминогов, кальмаров и
каракатиц, у которых лишь по две жабры,—
в подкласс двужаберных.
ПОДКЛАСС ЧЕТЫРЕХЖАБЕРНЫЕ (TETRABRANCHIA)
НАУТИЛУСЫ, ИЛИ ЖЕМЧУЖНЫЕ
КОРАБЛИКИ
Это очень примитивные головоногие
моллюски с внешней многокамерной
раковиной. Раковина спиральная, закручена
экзогастрически, т. е. не на брюшную
(как у улиток), а на спинную сторону
животного, разделена поперечными
перегородками на несколько камер. Животное
помещается в последней, самой большой
камере, все прочие камеры наполнены
газом. (Подрастая, наутилус передвигается
в новую, более обширную камеру, которая
нарастает со стороны устья раковины.)
Перегородки между камерами пронзены
отверстием и пронизаны тонким сифоном—
задним выростом тела животного. Вокруг
отверстий края септ (перегородок) заги-
1 Правда, советский палеонтолог Г. К.
Кабанов предполагает, что раковина
белемнитов пропитывалась известью уже после их
смерти, в процессе окаменения.
2 В народе ископаемые раковины белемнитов
называют «чертовыми пальцами».
3 Пока найден (в меловых отложениях Сирии)
всего лишь один ископаемый осьминог Palaeocto-
pus newbaldi. У него большие плавники и
одинарный (не двойной) ряд присосок на каждом
щупальце.
172
баются назад или вперед и частично
облекают сифон, образуя сифонные дудки.
Внутренняя поверхность раковины
выложена перламутром, внешняя по виду
напоминает фаянс. Верхняя часть ноги нау-
тилид образует массивную лопасть —
головной капюшон, прикрывающий сверху
плывущее животное и закрывающий
входное отверстие раковины, когда животное
прячется в ней. (У аммонитов такую роль
выполняла, по-видимому, крышечка
раковины.) Воронка состоит из двух несрос-
шихся лопастей, которые свернуты в
узкую спереди и расширяющуюся сзади
трубку. В ее середине имеется языкооб-
разный, направленный вперед клапан.
Большая голова несет бокаловидные
глаза, лишенные хрусталика и
стекловидного тела, и многочисленные лишенные
присосок ногощупальца, которые
собраны вокруг рта в два венчика — внешний
и внутренний. Количество ногощупалец
у разных полов различно, в среднем около
90 штук. Последние снабжены большими
влагалищами, в которые могут
втягиваться. Влагалища дорзальных рук
значительно увеличены и срослись друг с другом,
образуя упомянутый выше головной
капюшон.
Рот снабжен парой мощных роговых
челюстей и широкой радулой. Чернильная
железа отсутствует. Большие ребристые
яйца прикрепляются на дне.
Многочисленные виды этого подкласса
(более 7,5 тыс.) населяли древние моря
нашей планеты на протяжении около 500 млн.
лет от среднего кембрия и до мелового
периода. С мелового периода и до наших
дней сохранился лишь один род этой
обширной группы моллюсков — Nautilus,
несколько видов которого обитает, как
упоминалось уже, на юго-западе Тихого
океана.
Рис. ИЗ. Наутилус (Nautilus pompilius).
В биологии разных видов наутилусов
много общего. Они живут в раковинах,
разделенных перегородками. Когда
наутилус хочет опуститься на дно, он наполняет
раковину водой, она становится тяжелой
и легко погружается. Чтобы всплыть на
поверхность, наутилус нагнетает в свои
гидростатические «баллоны» газ, который
вытесняет воду, и раковина всплывает.
Жидкость и газ находятся в раковине
под давлением, поэтому перламутровый
домик не лопается даже на глубине
в 700 м, куда наутилусы иногда
заплывают.
Наутилусы почти всегда держатся ниже
сублиторали, в тихих водах, где нет
волнения. Здесь они медленно плавают или
ползают по дну, цепляясь своими гибкими
щупальцами за неровности грунта.
Охотятся на небольших раков, не
отказываются и от падали.
Жители Индонезии ловят наутилусов
в верши с дохлыми крысами или раковым
отваром вместо приманки. Из раковин
жемчужных корабликов делают
украшения, посуду, пуговицы. Мясо их едят на
многих островах Тихого океана.
ПОДКЛАСС ДВУЖАБЕРНЫЕ (DIBRANCHIA)
Это высшие головоногие моллюски
с внутренней (а не внешней, как у
наутилуса) недоразвитой раковиной. Она имеет
обычно форму узкой пластинки,
хитиновой (у кальмаров) или известковой (у боль-
шинства каракатиц) либо двух
заостренных хрящевых палочек (у обычных
осьминогов). Лишь у одной спирулы (из
группы каракатиц) раковина спиральная, но
тоже внутренняя, спрятана под покро-
173
вами мантии, которая покрывает снаружи
все тело головоногого моллюска. В
мантийной полости лежат два нефридия, две
жабры, пульсируют два жаберных сердца.
Края образующих воронку лопастей
полностью срослись в цельную трубку,
которую и называют воронкой или с и-
фоном.
Глаза двужаберных почти такие же
совершенные, как у высших позвоночных
животных. У некоторых глубоководных
кальмаров глаза стебельчатые или
телескопические. Лишь у одного вида, у осьминога
цирротаумы (Cirrothauma murrayi), глаза
полностью редуцированы.
Позади глаз лежат особые
обонятельные органы. Щупалец 8 или 10. Они
вооружены присосками или крючьями,
напоминающими своей формой когти кошек.
В мускулистой глотке—роговые челюсти.
Радула в подавляющем большинстве
хорошо развита.
У некоторых глубоководных осьминогов
радула редуцирована. Яйца обычно
гладкие, иногда покрыты небольшими ребрами
или иной скульптурой, прикрепляются
гроздьями к подводным предметам. Дву-
жаберные — пелагические и придонные
хищники. Число вымерших и доживших
до нас видов не превышает 1000; они
появились на Земле около 200 млн. лет назад,
начиная с триаса.
Подкласс двужаберных
систематики делят на несколько отрядов и
подотрядов, наименования и классификация
которых не у всех авторов одинакова. Мы
возьмем за основу старую классическую
систему. Она по многим причинам удобнее
новейших подразделений, которые требуют
еще многих уточнений.
По новейшей классификации, например,
предложенной некоторыми современными
авторами, лжекальмары, или лолиго,
отнесены к настоящим кальмарам, с
которыми они внешне действительно очень
сходны (явно конвергентное сходство,
поскольку образ жизни у лолиго и кальмаров
одинаков), но в строении и развитии
внутренних органов (глазного яблока,
яйцеводов и пр.) обнаруживают отчетливые черты
родства с каракатицами.
ОТРЯД ОСЬМИНОГИ (OCTOPODA)
К этому отряду принадлежат двужабер-
ные головоногие моллюски с восемью
щупальцами, которые называют также р у-
к а м и или ногами. Правда, у
некоторых глубоководных осьминогов есть еще
одна пара тонких бичевидных выростов
умбреллы — перепонки, соединяющей
основания щупалец.
У осьминогов есть целый ряд и других
анатомических особенностей, которые
дали право систематикам выделить их в
отдельный отряд.
Присоски осьминогов, например,
лишенные стебельков, прямо плоским дном
прирастают к щупальцам (у каракатиц и
кальмаров присоски расположены на
стебельках, которыми они прикрепляются к
внутренней поверхности рук). Присоски без
опорных роговых колец (у каракатиц и
кальмаров зазубренные роговые кольца, словно
обручи, растягивают изнутри устье
каждой присоски). Поэтому у осьминогов
никогда не бывает когтей, или крючьев,
которые развиваются из роговых колец
присосок.
Присоски сидят на руках в один или два
продольных ряда.
Тело короткое, мешковидное, у
настоящих осьминогов без плавников (у
глубоководных осьминогов— одна или две пары
плавников).
Можно упомянуть еще и о таких
признаках, характерных для осьминогов:
передний край мантии на затылке сращен
у них с головой; «замыкающий
аппарат» — хрящевые кнопки на брюшном
краю мантии и в основании воронки,
запирающий мантийную щель, после того
как в нее будет набрана забортная вода,
не развит; хрящевой остаток раковины
имеет стилетообразную, подкововидную,
седловидную форму или полностью
отсутствует.
Все осьминоги нектобентические и
пелагические хищники. С мелового
периода и до наших дней специалистам
известно около двухсот вымерших и ныне
живущих видов.
174
ПОДОТРЯД БЕСПЛАВНИКОВЫЕ,
ИЛИ НАСТОЯЩИЕ, ОСЬМИНОГИ
(APTEROTI, или INCIRRATA)
Этот подотряд составляют обычные
прибрежные осьминоги, которых объединяют,
кроме сходного образа жизни, еще и
некоторые общие им всем анатомические черты:
у них нет плавников и присоски всегда
без усиков (у осьминогов следующего
отряда справа и слева от крупных
присосок растут осязательные, по-видимому,
усики), а остаток раковины имеет вид
двух хрящевых палочек, спрятанных под
кожей спины, либо отсутствует полностью.
Присоски обычно располагаются на
щупальцах в два продольных ряда.
Живут бесплавниковые, или настоящие,
осьминоги обычно недалеко от берега и
неглубоко, кроме представителей
подсемейства Bathypolypodinae, т. е.
батиальных осьминогов, которые
встречаются в океанах вплоть до глубин в 8 тыс. м.
Предпочитают каменистое дно, но
поселяются иногда и на илистых и песчаных
грунтах, например обычный у нас на
Дальнем Востоке песчаный пароктопус
(Paroctopus conispadiceus).
Типичный и наиболее крупный
представитель всего отряда и рода настоящих
осьминогов — обыкновенный осьминог,
которого называют также и обычным
спрутом (Octopus vulgaris). Он обитает в
тропических, субтропических и в умеренных
морях и океанах. К югу его
распространение простирается до острова св. Павла
и Австралии, к северу — до берегов
Шотландии.
В наших морях, на Дальнем Востоке,
он встречается очень редко. Здесь самые
обычные осьминоги: уже упомянутый
песчаный пароктопус и шершавый пароктопус
(Paroctopus asper).
Все осьминоги подсемейства
Octopodinae (к которому принадлежат
и обычный спрут, и многие наши
дальневосточные виды) по повадкам и образу
жизни похожи друг на друга. Многие
из этих животных еще очень плохо
изучены.
Но даже те немногие факты, которые
уже известны исследователям,
превзошли всякие ожидания.
Однажды в морском аквариуме в
Калифорнии осьминожиха по кличке Мефиста
отложила яйца — маленькие студенистые
комочки. Свои восемь рук Мефиста
сплела наподобие корзины. Это было гнездо.
Два месяца, пока осьминожиха
вынашивала в нем яйца, она ничего не ела.
Если кто-нибудь из служителей
осмеливался подбросить кусочек мяса к самой
ее голове, Мефиста вспыхивала в гневе
кирпично-красным цветом, освобождала
руку из импровизированной корзины и
отбрасывала прочь любимую прежде пищу:
ведь этот «мусор» мог попасть на ее
драгоценные яйца!
Когда Мефисту не тревожили, она
нежно перебирала яйца, покачивала их,
словно баюкая, и поливала водой из воронки.
Другая осьминожиха из Брайтонского
аквариума отложила яйца в углублении
искусственной скалы (близко от стекла,
так что за животным легко было
наблюдать). Свое гнездо она окружила
«крепостным валом», притащив несколько
десятков живых устриц и нагромоздив их
друг на друга. За этой баррикадой
устроилась сама, только выпуклые глаза
выглядывали из «крепости», зорко осматривая
окрестности. Два самых длинных щупальца
осьминожиха вытянула за укрепление, их
концы постоянно извивались в разных
направлениях, словно выискивая
возможных врагов.
Бдительная мать не позволяла ни
одному живому существу приблизиться к ее
гнезду х. Свободное от сторожевой вахты
время она отдавала «домашним делам»:
нежно поглаживала яйца, легонько
встряхивала их, полоскала водой из воронки.
Ее присоски, как крохотные пылесосы,
очищали яйца от мелкого мусора и
паразитов.
Лишь редкие осьминожихи решаются
принять немного пищи вблизи от
охраняемых яиц. Обычно же они ничего не
едят месяц и два, и даже четыре месяца,
пока длится насиживание. Аскетизм этот
1 Одна осьминожиха, рассказывает
американский натуралист Джильберт Клинджел,
столь ревностно оберегала свои яйца, что убила
самца, неосторожно приблизившегося к ее
логову.
175
вызван стремлением предохранить яйца
от загрязнения. Даже взрослые осьминоги
не переносят несвежую воду. Поэтому
насиживающие самки постоянно
поливают яйца струей из воронки —
промывают их. Все, что может гнить,
изгоняется осьминогами из гнезда. Вода должна
быть чистой. Ради этого самки голодают:
боятся уронить даже крошки со своего
стола на драгоценные яйца, в которых
заключено будущее их вида.
Фанатичная преданность своим
материнским обязанностям, продиктованная
суровым инстинктом, часто наносит
непоправимый вред здоровью осьминогов.
Большинство из них, по-видимому,
погибает, дав жизнь новому поколению.
Страсть осьминогов к посуде, их
стремление забираться в различные полые
предметы, в том числе в пустые раковины,
известна давно. Сто двадцать лет назад
об этом писал французский зоолог О р-
б и н ь и. Но еще раньше и с большой
выгодой для себя использовали эту осьми-
ножью страсть рыбаки с берегов
Средиземного моря. Как использовали —
расскажу несколько позже.
Когда нет готовых квартир, осьминоги
строят их сами. Стаскивают в кучу
камни, раковины и панцири съеденных
крабов, сверху в куче делают
глубокий кратер, в котором и устраиваются.
Часто осьминог не довольствуется лишь
«крепостным валом» из камней, а
прикрывает себя сверху большим камнем, словно
щитом.
Предпринимая небольшие вылазки,
осьминог не оставляет щит дома, а держит
его перед собой. При тревоге выставляет
его в сторону, откуда грозит опасность,
одновременно обстреливая врага струями
воды из воронки, словно из брандспойта.
Отступая, осьминог пятится назад, за
«крепостной вал», прикрывая свой тыл
каменным щитом.
Градостроительством осьминоги
занимаются по ночам. До полуночи они обычно
не предпринимают никаких вылазок, а
потом, словно по команде, отправляются
на поиски камней. Восьмирукие
труженики тащат камни непомерной
величины — в пять, десять и даже двадцать раз
превышающие их собственный вес.
В некоторых местах, особенно
приглянувшихся осьминогам, водолазы находили
на дне моря целые осьминожьи города —
один каменный дом недалеко от другого.
«На плоском дне отмели к
северо-востоку от Поркерольских островов,—
пишут Кусто и Дюм а,— мы напали
на город осьминогов. Мы едва верили
своим глазам. Научные данные,
подтвержденные нашими собственными
наблюдениями, говорили о том, что спруты обитают
в расщелинах скал и рифов. Между тем
мы обнаружили причудливые постройки,
явно сооруженные самими спрутами.
Типичная конструкция имела крышу в виде
плоского камня полуметровой длины,
весом около восьми килограммов. С одной
стороны камень возвышался над грунтом
сантиметров на двадцать, подпертый
меньшим камнем и обломками строительного
кирпича. Внутри была сделана выемка
в двенадцать сантиметров глубиной.
Перед навесом вытянулся небольшой вал
из всевозможного строительного мусора:
крабьих панцирей, устричных створок,
глиняных черепков, камней, а также из
морских анемонов и ежей. Из жилища
высовывалась длинная рука, а над валом
прямо на меня смотрели совиные глаза
осьминога. Едва я приблизился, как рука
зашевелилась и пододвинула весь барьер
к входному отверстию. Дверь закрылась.
Этот дом мы засняли на цветную
пленку... Тот факт, что осьминог собирает
стройматериал для своего дома, а потом,
приподняв каменную плиту, ставит под
нее подпорки, позволяет сделать вывод
о высоком развитии его мозга».
Развитие у животных органов,
способных использовать простейшие орудия,
приводит к образованию более сложного
мозга, к расширению сферы его
деятельности, к формированию разнообразных
Таблица 15. Головоногие моллюски:
Сверху — новорожденные осьминоги; снизу — самка осьминога; и днем и ночью
бессменно дежурит ока у своих яиц, которые висят гроздьями в глубине ниши.
приспособительных рефлексов и
врожденных инстинктов.
«Интеллигентность» животного в первую
очередь определяется способностью его
мозга к запоминанию опыта, т. е., говоря
языком науки, к образованию стойких
условных рефлексов.
Один ученый выработал у осьминогов
условный рефлекс на свет. Осьминога
легонько кололи куском проволоки и
одновременно включали лампочку.
Осьминог «мрачнел»: расширял черные хрома-
тофоры, кожа приобретала темную
окраску. Обучение продолжалось шестнадцать
дней. На семнадцатый день включили свет,
но проволокой осьминога не коснулись.
Однако он потемнел, как прежде.
Восемьдесят один день мозг осьминога сохранял
память об уколе, который должен был
следовать за вспышкой света.
Постепенно, лишь в конце третьего
месяца, исчез условный рефлекс на свет,
ни разу не подкрепленный за это время
раздражителем — уколом.
В результате своих опытов английские
ученые Бойкот и Юнг пришли к
выводу, что осьминоги наиболее
«одаренные» из всех беспозвоночных и даже
некоторых позвоночных животных,
например рыб.
Бойкот и Юнг установили также, что
осьминоги поддаются дрессировке. Не
хуже слонов и собак различают они
геометрические фигуры — маленький квадрат
отличают от более крупного,
прямоугольник, поставленный вертикально, от
поставленного горизонтально, белый круг
от черного круга такого же размера, крест
от квадрата, ромб от треугольника.
За правильно сделанный выбор
животных награждали пищей, за ошибку они
получали слабый удар электрическим
током.
Бойкот и Юнг кормили своих
пленников крабами, привязанными за нитку.
Когда подопытный осьминог привыкал
к такому угощению, в бассейн с крабом
опускали металлическую пластинку.
Осьминог, ничего не подозревая,
появляется из своего убежища. Как всегда,
смело атакует краба и вдруг
отскакивает. Бледнеет, выбрасывает струю воды
в предательского краба и спешит назад
в свое логово. Через краба с пластинкой
экспериментаторы пропустили слабый
электрический ток. Его удар напугал
осьминога.
Через два часа краб с пластинкой снова
в аквариуме. Осьминог не бросается на
него, как обычно, из своего убежища,
словно ракета. Он ведет себя теперь
совсем иначе. Осторожно появляется из
расщелины. Идет по дну с вытянутыми
вперед щупальцами. Каждую секунду
готов повернуть обратно. Борются два
чувства — желание получить лакомый
кусочек и страх. Разрешая мучительное
противоречие, осьминог то приближается
к крабу с вытянутыми вперед
щупальцами, то боязливо удаляется в свой угол,
чтобы через минуту снова вернуться.
Наконец чувство голода побеждает.
Осьминог хватает краба. Удар! Он бледнеет
и удирает в нору.
Условный рефлекс теперь прочно
закреплен в его мозгу. Осьминог берет
обычных крабов. Но краб с пластинкой не
вызывает у него никакого интереса. Он
лишь высовывает голову из своего
убежища, чтобы понаблюдать за трюками
странных существ» Когда опасный краб
приближается к его логову, осьминог
багровеет и пускает в него серию залпов из
своего водяного пистолета.
Осьминоги с удаленными кусочками
мозга, в которых замыкаются условные
рефлексы, действовали так, словно
никогда в жизни не получали электрических
ударов. Заметив краба с пластинкой,
оперированный осьминог торопливо
выбирался из своего убежища, хватал краба,
получал удар, убегал и тут же возвращался
вновь. Память о боли, полученной
только что, совершенно отсутствовала —
условный рефлекс на краба с пластинкой не
образовывался. Один осьминог в течение
тридцати пяти дней, пока длился
эксперимент, с неистощимой яростью бросался
на краба. Даже в конце дня, в течение
которого он получил пятнадцать ударов
током, осьминог с большой готовностью
появлялся из убежища, чтобы напасть
на краба, который причинял ему только
боль.
Юлиус К о л ь м а н наблюдал за
жизнью осьминогов на биологической стан-
12 Жизнь животных, т. 2
177
ции близ Неаполя. Его работы вошли
в литературу о поведении этих животных
как классический образец, но, к
сожалению, давно стали библиографической
редкостью. С наблюдениями Кольмана
познакомил широкий круг читателей Фрэнк
Л е й н в книге «Царство осьминога».
Осьминоги, которые жили на
неаполитанской станции, стали совсем ручными.
Они знали в лицо сторожа аквариума
и любили его. Если он протягивал к ним
руку, животные обвивали ее щупальцами
и нежно гладили. Они явно принимали
участие в игре, когда сторож
поддразнивал их. Он прятал кусок мяса в руке,
и осьминоги терпеливо пытались
разжать пальцы, боролись, но ни разу не
причинили боль человеку.
В 1875 г. британский натуралист Генри
Л и издал книгу «Осьминог—дьявольская
рыба, правда и вымысел». Это было
первое научно-популярное сочинение об
осьминогах. В ней рассказал он о забавном
происшествии в Брайтонском аквариуме.
Однажды по непонятной причине из
аквариума стали пропадать редкие рыбы
циклоптерусы (пинагоры) г. Ежедневно
исчезало по одной рыбе, но никаких следов,
которые указывали бы на их похитителя,
не оставалось.
И вот, наконец, вор был пойман с
поличным. Им оказался осьминог, который
жил в той же комнате, но в другом
аквариуме. Обнаружив, что в соседнем
аквариуме находятся рыбы, осьминог
регулярно совершал на них налеты по
ночам. К утру он всякий раз возвращался
в свой бассейн. В то утро, когда его
поймали, разбойник, видно, решил для
разнообразия переночевать неподалеку от
места пиршества.
После разоблачения за
осьминогом-похитителем было установлено тщательное
наблюдение. Повторит ли он свои ночные
странствия? Генри Ли пишет: «...наш
подозреваемый пират не отправлялся в свою
грабительскую экспедицию до тех пор,
пока кто-либо находился в помещении.
1 У пинагоров 1 или круглоперов (Cyclopteri-
dae), брюшные плавники образуют крупную
воронкообразную присоску, которой эти рыбы
прочно присасываются к камням.
Как будто он знал, что за ним
наблюдают, и около недели спокойно оставался
дома».
Затем, когда засада была снята, сразу
два осьминога выбрались ночью из
аквариума. Один из них был как раз тем
самым спрутом, которого уличили в
грабеже.
А вот что пишет Ч. Дарвин о
тактической хитрости спрутов.
Однажды он застал на мелководье
небольшого осьминога. Заметив человека,
моллюск осторожно и, как показалось
Дарвину, осмысленно пытался уйти в
безопасное место. Сначала он замер на месте,
затем украдкой, очень скрытно, словно
кошка за мышью, «короткими
перебежками» переползал вперед на несколько
сантиметров и вновь замирал в
неподвижности. Всякий раз, заняв новую позицию,
осьминог в соответствии с окружающим
пейзажем изменял свою окраску.
Так продолжалось, пока осьминог не
добрался до более глубокого места. Тогда
он внезапно рванулся вперед, оставляя за
собой густое облако чернил, чтобы скрыть
от преследователя расщелину, в которой
спрятался.
Как спят осьминоги?
Первым описал спящего осьминога Жан
В е р а н и. Он наблюдал в аквариуме за
мускусным спрутом. Осьминог спал
«сидя», присосавшись ко дну основаниями
щупалец и приподняв вверх тело. При
малейшем всплеске воды по телу животного
пробегали, словно вспышки, темные
пятна.
При более глубоком сне все щупальца,
кроме двух нижних, прижаты к телу. Две
вытянутые в стороны руки несут
сторожевую вахту. Осьминог погружен в
глубокий сон. Он ничего не видит и не
слышит. Можно подойти к аквариуму
вплотную, кричать над ним, шуметь как
угодно — осьминог не проснется. Но стоит
лишь слегка сотрясти воду или чуть
прикоснуться к сторожевым щупальцам —
животное сейчас же вскакивает. Тело его
раздувается и бледнеет. Бородавки и
черные пятна покрывают кожу. Если тревога
не была ложной и угроза реальна,
осьминог окутывает себя облаком чернил и,
не раздумывая, ищет спасения в бегстве.
178
Когда спрут спит, глаза он не
закрывает, лишь сильно сокращает зрачки,
дыхание замедляется, а окраска становится
желто- или буро-серой. Иногда
сторожевые щупальца вытягиваются вверх и
медленно кружатся над спящим осьминогом,
словно антенна радара.
«Спруты обладают необыкновенным
любопытством,— говорит Теодор Руссо.—
Если вы часто посещаете какой-нибудь
отдаленный пляж и осьминоги к вам
привыкнут, то, влекомые любознательностью,
они иногда приближаются к вам. Сначала
неуверенно, потом смелеют, подплывают
близко и, случается, усаживаются на
вашей ноге или руке».
Руссо несколько раз вытаскивал
одного осьминога из щели в скале. Сначала
тот отчаянно сопротивлялся и
выбрасывал в гневе чернила. Но постепенно
привык к странному человеку и завязал с ним
дружбу. Он сам покидал свое убежище,
спокойно плавал вокруг. Не шарахался
в испуге, когда человек протягивал к
нему руку, позволял почесать спину и,
казалось, получал от этого удовольствие.
«У меня сложилось впечатление,—
заключает Руссо,— что, если бы я мог
проводить с ним больше времени, он стал бы
совсем ручным».
Страшные рассказы об осьминогах,
опубликованные газетами, усугубили
недоверие, которое по незнанию испытывали
люди к необычным на вид обитателям
чуждой стихии. Рожденное невежеством
предубеждение питалось баснями
недобросовестных сочинителей, и вот осьминог,
создание не более опасное, чем крупная
треска, но гораздо более
доброжелательное и понятливое, превратился в
чудовище непомерной силы и свирепости.
Это, конечно, не так.
Описано более ста видов осьминогов, но
все они — животные мелкие, длиной не
более полуметра. Лишь три-четыре вида
заслуживают внимания как возможные
противники человека: это обыкновенный
осьминог, осьминог Дофлейна, осьминог
аполлион и близкий к нему гонконгский
осьминог. Первый обитает во всех
тропических, субтропических и тепловодных
морях и океанах. Второй обычен у
берегов Японии и изредка встречается у
южных Курильских островов и в заливе
Посьета. Осьминог аполлион живет в
скалах у побережья Аляски, Западной Канады
и Калифорнии (описанный мною близкий
к нему вид Parocpus asper обитает у
берегов Камчатки и северных Курил).
Обыкновенный осьминог и осьминог
Дофлейна — массивные, коренастые
создания с недлинными и толстыми
щупальцами. В длину они достигают 3 м и весят
при таких размерах около 25 кг.
Гигантом среди осьминогов можно
было бы назвать аполлиона, но гигант этот
весьма субтилен.
В конце прошлого века у берегов
острова Ситха рыбаки поймали осьминога,
который пропорциями своими напоминал
паука-сенокосца: маленькое туловище на
длинных и тонких ногах-щупальцах.
Общая длина его была около 5 м (в размахе
щупалец около 8,5 м), но тело вместе с
головой не превышало в ширину 15, а в
длину — 30 см.
Позднее еще несколько осьминогов этого
вида, но меньшего размера попались в сети
у берегов Калифорнии, Канады и Аляски.
Североамериканские «субтильные»
осьминоги аполлионы уступают своим
собратьям двух упомянутых выше видов
и в силе и в весе. И уступают при
одинаковых размерах примерно вдвое.
Опасны ли двух-,трехметровые
осьминоги?
Попросим ответить на этот вопрос
людей, которые с ними встречались.
Один водолаз недалеко от Мельбурна
расчищал устье реки. Он заложил
динамит между двумя камнями и взорвал его.
Затем спустился вниз, чтобы проверить,
какие разрушения причинил взрыв.
Большой камень не был сдвинут с места.
Водолаз лег на него и подсунул правую
руку под камень — хотел заложить еще
один заряд.
«Вдруг я почувствовал,— рассказывает
водолаз,— что кто-то держит руку...
Когда муть рассеялась, я увидел, к своему
ужасу, щупальце большого осьминога,
обвившееся вокруг моей руки подобно
удаву. Бол$> была нестерпимой: словно тело
мое разрывали на куски, и, чем отчаяннее
я пытался освободиться, тем сильнее
становилась боль.
12*
179
Мне нелегко было удержать свои ноги
внизу, так как воздух скапливался под
одеждой и раздувал ее. Если бы ноги
поплыли вверх, я бы скоро потерял
сознание, вися вниз головой.
Нельзя дать и сигнал тревоги,
попросить, чтобы меня подняли наверх. Эта
гнусная тварь меня бы не отпустила,
и скорее всего я оказался бы со
сломанной рукой».
Позади лежал железный лом, и водолаз
стал осторожно подтягивать его ногой.
Вот схватил лом рукой. И началась
борьба. «Чем я больше бил по спруту,
тем сильнее он сжимал мою руку. Она
совсем онемела, но скоро я почувствовал,
что хватка стала ослабевать. Однако
животное еще сопротивлялось, пока я не
изрубил его на куски, тогда присоски
ослабли.
Могу вас уверить, что за двадцать
минут этой борьбы я был совершенно
измучен. Мы подняли осьминога, вернее то,
что от него осталось, наверх.
Распластали его: в поперечнике восемь футов1.
Я совершенно уверен, что это животное
может удержать на дне пять или шесть
здоровых мужчин».
Второй рассказ принадлежит человеку,
несравненно более компетентному в
биологии,— известному знатоку моллюсков
Жану В е р а н и. В книге «Головоногие
Средиземного моря» он говорит, что
самые крупные осьминоги, которые живут
в Средиземном море, бывают длиной до
3 ж и весят 25 кг.
«Старый рыбак, очень ловкий и
опытный, встретил такого спрута напротив
портового мола Ниццы». Рыбак решил
пощекотать осьминога, поиграть с ним,
как с котенком. Но спрут был старый
и недружелюбный — из породы
закоренелых пиратов, которые могут
«удержать на дне пять или шесть здоровых
мужчин».
Он, конечно, утащил рыбака на дно?
И не подумал. Может быть, и хотел
бы это сделать, но, увы, сила была не на
его стороне. Спрут трагически
заламывал руки, отчаянно барахтался, он очень
1 Около 2,5 м. (Прим. автора.)
хотел вырваться из цепких лап двурукого
страшилища, но не мог. Рыбак со смехом
обнимал насмерть перепуганного
осьминога, перевернул его вверх брюхом и
кончил тем, что «поцеловал в нос» и отпустил
восвояси. Спрут удрал, багровый от
волнения, а человек тяжело перевел дух:
после возни с полуторапудовой
«зверюгой» он изрядно устал.
В. К. А р с е н ь е в, наш славный
исследователь дальневосточного края,
встретился однажды в Приморье с большим
осьминогом.
«Китайцы далеко разбрелись по
берегу,— пишет он.— Я сел на камень и стал
смотреть в море. Вдруг слева от меня
раздались какие-то крики. Я повернулся в ту
сторону и увидел, что в воде происходила
борьба. Китайцы палками старались
выбросить какое-то животное на берег,
наступали на него и в то же время боялись
его и не хотели упустить. Я побежал туда.
Животное, с которым боролись китайцы,
оказалось большим осьминогом».
Наконец осьминога вытащили на берег.
Арсеньев измерил его. Тело у спрута
было длиной 0,8 ж, голова — 28 еж, а
щупальца — 1,4 ж. Весь моллюск,
следовательно, был длиной около 3 ж, но «пять-
шесть здоровых мужчин» без особого
труда выкинули его палками на берег.
Два противоречивых свидетельства, два
разных мнения о силе спрутов: водолаз
из Мельбурна утверждает, что двух-,трех-
метровый осьминог может справиться
с пятью-шестью мужчинами, а рассказы
Верани и Арсеньева убеждают нас, что
осьминоги такой величины уступают
человеку и в силе и в агрессивности.
Раз мы должны сделать выбор, то
предпочтем, конечно, свидетельство
натуралистов — Верани и Арсеньева, чем басни
мельбурнского водолаза и других
подобного же сорта сочинителей, без меры
расписавших атлетические свойства
осьминогов.
Рассуждая о силе осьминогов, мы
совершенно упустили из виду другое
оружие этих хищников — их укус. Родичи
беззубых улиток и ракушек, они
приобрели в процессе эволюции очень острые
челюсти, роговые и крючковатые, по
форме похожие на клюв попугая.
180
«Ядовитыми канаваями» называют
индийские рыбаки некоторых маленьких
осьминогов и очень боятся этих тварей. Если,
говорят, осьминога, попавшего в лодку
вместе с рыбой, не выбросить немедленно
за борт, то он сам может напасть на
человека и укусить его в ногу или руку. Боль
такая, словно ужалил скорпион. Нога
распухает, человек несколько недель
чувствует слабость и головокружение.
Яд выделяет задняя пара слюнных
желез, но это не пищеварительный
фермент, а особая вирулентная жидкость,
близкая по химическому составу к
алкалоидам. Яд осьминогов вводили в тело
крабов, рыб и лягушек. Он действовал
парализующе на центральную нервную
систему. У крабов немедленно наступали
судороги, и через несколько минут они
умирали.
Яд осьминогов опасен и для человека.
Однажды сотрудник Калифорнийского
аквариума был укушен в ладонь небольшим
аполлионом. В ту же ночь рука так
распухла, что не видно стало суставов,
прошло четыре недели, прежде чем опухоль
спала. Признаки болезни напоминали
симптомы змеиного укуса.
Яд осьминогов, даже мелких, опасен.
Правда, эти животные редко и неохотно
пускают в ход свое ядовитое оружие.
М а к-Г и н и т и, известный океанолог,
говорит, что в его руках побывало
несколько тысяч осьминогов и ни один из
них его не укусил. И все же такие случаи,
как мы видели, бывают.
Достоверных сообщений о встрече под
водой с очень крупными осьминогами
известно очень мало. Но мелкими
осьминогами морское дно местами буквально
кишит. Ныряльщик, попавший в их
общество, не должен забывать, что имеет
дело с существами ядовитыми и отнюдь
не кроткими.
Особую группу настоящих осьминогов
составляют представители подсемейст-
в a Bathypolypodinae. (Все описанные
ранее разновидности принадлежат к
подсемейству Octopodinae.) Это
миниатюрные осьминожики, длиной не больше
детской ладони. У них очень маленькие
присоски и совсем нет чернильного мешка
(у головоногих это бывает редко).
Рис. 114. Аргонавт (Argonauta argo).
Почему у батиполипусов нет
чернильного мешка, вполне понятно: это
глубоководные животные, обитатели лишенных
света океанских глубин. Раз живут они
во мраке, то, понятно, дымовое облако
как средство защиты никакой пользы
им не принесет: ведь его в темноте не
видно. За ненадобностью чернильная железа
атрофировалась.
Батиполипусы — одни из немногих
головоногих моллюсков, приспособившихся
к жизни в холодных водах. Это
единственные осьминоги, обитающие в наших
северных морях: Карском, Восточно-Сибирском
и Чукотском.
Совершенно необычные повадки и образ
жизни у осьминогов семейства Argo-
nautidae — у аргонавтов. Было описано
довольно много видов этого своеобразного
семейства.
Большинство из них едва ли
существует в действительности, а
представляет скорее всего разновидности
шести бесспорных видов. Из них три имеют
циркумтропическое распространение, т. е.
обитают в тропических зонах трех океанов
(Argonauta argo, A. hians и A. nodosa),
два встречаются в Индийск о-Западно-
Тихоокеанской тропической области (Ин-
до-Вест-Пацифической): на западе Тихого
океана и на востоке Индийского (А. сог-
nuta и A. buttgeri), и лишь один вид живет
только в Тихом океане (A. nourgi).
181
В наших водах, у берегов
Южнокурильских островов, изредка встречается,
насколько пока известно, один вид этих
очень интересных животных — A. hians.
Их своеобразие особенно проявляется
в пору размножения.
Во время размножения самцы
головоногих моллюсков, как мы уже знаем,
одним из щупалец достают из-за «пазухи»
(из мантийной полости) сперматофоры
и переносят их в мантийную полость
самки. Лишь осьминоги аргонавты
поступают иначе. У них щупальце
самостоятельно, без помощи самца, исполняет
эти обязанности. Захватив пару спермато-
форов, оно отрывается и уплывает на
поиски самки, словно торпеда с
дистанционным управлением.
«Когда аргонавт предлагает «руку»
даме своего племени, она принимает ее и
сохраняет, унося с собой», и рука кавалера
«становится подвижным существом,
живущим своей жизнью еще и некоторое время
после того, как перешла во владение
дамы» — так несколько напыщенно, но
вполне благопристойно выражался Генри
Ли, первый натуралист, который
сумел найти популярные слова для
описания самых интимных сторон жизни
спрутов.
Еще Аристотель изучал аргонавта. Но
античные натуралисты ошибались,
полагая, что этот крошка может плавать,
словно под парусами.
Дело в том, что аргонавты вернулись
к традициям предков — вновь стали жить
в раковинах. Раковинки тончайшие,
будто пергаментные. Животные скользят
в них по волнам, как в миниатюрных
лодках. На верхней паре рук у
маленьких мореплавателей есть расширенные
лопасти. В древности думали, что при
благоприятной погоде аргонавты поднимают
вверх эти лопасти, ветер ударяет в них,
и раковинки плывут, словно под
парусами.
Но оказалось, что лопасти на
щупальцах не навигационные, а строительные
органы: они выделяют жидкое вещество,
которое, застывая, образует раковину.
А в ней аргонавты вынашивают свое
потомство. Значит, раковина у них не
только лодка, но и колыбель.
Когда говорят об аргонавтах, обычно
имеют в виду их самок: это существа более
импозантные, чем самцы, и только они
живут в раковинах. Самцов и заметить-то
трудно — такие они карлики. Ведь самка
аргонавта раз в двадцать больше самца.
Самый крупный известный науке
аргонавт-самец уместился бы на ногте
большого пальца человека, в то время как
длина одной самки, хранящейся в
Британском музее, целых 310 мм.
Когда наступает пора размножения,
одно щупальце самца аргонавта начинает
вдруг быстро расти. Достигнув
допустимого предела, оно отрывается от головы
животного и уплывает, предварительно
захватив с собой один-два сперматофора.
Щупальце, извиваясь, рыщет в морских
джунглях—ищет самку своего вида. Найдя ее,
заползает к ней в мантийную полость, там
сперматофоры «взрываются» и
оплодотворяют яйца.
Своим видом
щупальце-путешественник напоминает червя с двумя рядами
«ножек»—недоразвитых присосок. Не
удивительно, что первые натуралисты
считали это странное щупальце паразитом,
живущим в мантийной полости самки
аргонавта.
Известный французский анатом и
палеонтолог Жорж Кювье принял мнимого
паразита за самостоятельное существо
и дал ему научное название Hectocotylus—
«обладатель ста присосок».
Аргонавты представляют единственную
среди головоногих моллюсков группу с
ясно выраженным половым
диморфизм ом. Самки у них значительно
крупнее самцов, наделены наружной
раковиной и большими лопастевидными
выростами на паре дорзальных (т. е.
самых верхних) щупалец.
У других видов головоногих моллюсков
самцов от самок очень трудно отличить
по внешности (если не считать гектокоти-
ля, которого самки лишены). Лишь у
самцов некоторых осьминогов заметны более
крупные присоски, чем у самок того же
вида.
182
ПОДОТРЯД ГЛУБОКОВОДНЫЕ ОСЬМИНОГИ
(PTEROTI, ИЛИ CIRRATA)
Эти некрупные и малоизвестные
неспециалистам животные обитают только на
больших глубинах. От обычных
осьминогов отличаются они главным образом
плавниками. Их одна или две пары;
плавники не ромбовидные, как у
кальмаров, а напоминают своей формой тупые
крылья. Кроме того, щшсоски на
щупальцах у глубоководных осьминогов
располагаются всегда в один ряд, а справа и
слева от присосок на щупальцах же растут
чувствительные усики — ц и р р ы. Сами
щупальца этих осьминогов почти до самых
концов соединены перепонкой —
умбре л л о й. Чернильного мешка нет.
Часто и радула бывает недоразвитой,, У
некоторых недоразвита и мантийная полость
и воронка. Передвигаются такие
осьминоги выталкивая воду не из воронки, а из-
под колокола умбреллы.
Некоторые виды наделены светящимися
органами, например упоминавшийся уже
черный глубоководный осьминог адский
вампир (Vampyroteuthis infernalis).
Несмотря на свой страшный вид, существо
это довольно безобидное. У другого очень
странного представителя этого отряда —
цирротаумы (Cirrothauma murrayi) —
фотофоры скрыты, по-видимому, в сильно
утолщенных и вздутых основаниях
присосок. Этот осьминог знаменит также
своей слепотой: глаза у него атрофированы—
единственный известный науке пример
редукции глаз у головоногого моллюска.
Обычно у этих животных глаза всегда
хорошо развиты.
Некоторые глубоководные осьминоги
приспособились к жизни на вязких илах
океанских впадин. Формой тела они
похожи на раскрытые зонты или даже на
Рис. 115. Глубоководный осьминог
Amphitretus pelagicus.
блины. Большой поверхностью
соединенных умбреллой и раскинутых широко
в стороны рук ложатся они на зыбкий
грунт и не проваливаются. Подобный же
тип адаптации к особым условиям среды
развит у некоторых глубоководных звезд,
офиур и других животных.
ОТРЯД ДЕСЯТИНОГИЕ ГОЛОВОНОГИЕ
МОЛЛЮСКИ (DECXPODA)
В отличие от осьминогов у десятиногих
головоногих моллюсков (у каракатиц и
кальмаров) не восемь, а десять щупалец.
От осьминогов десятиногие отличаются
еще и другими признаками: присоски со
стебельками и снабжены опорными
роговыми кольцами, которые у многих
кальмаров превращаются в когти или крючья.
На щупальцах присоски располагаются
в два или в четыре продольных ряда
(поперечных рядов может быть больше сотни).
По бокам тело десятиногих головоногих
моллюсков всегда несет пару ромбовидных,
сердцевидных, веслобидных или
каемчатых плавников. Так называемый
замыкающий аппарат, т. е. хрящевые «кнопки»,
запирающие мантийное отверстие перед
реактивным толчком, почти у всех декапод
хорошо развит.
Все десятиногие — пелагические или
нектобентические хищники. С триасового
периода мезозойской эры и до наших дней
науке известно около 700 вымерших и
ныне живущих видов этого отряда.
ПОДОТРЯД КАРАКАТИЦЫ (MYOPSIDA)
Внешне типичную каракатицу, или
сепию, легко отличить от кальмара. У нее
уплощенное сверху вниз тело (у кальмара
оно цилиндрическое, кеглевидное),
отороченное по бокам узкой каймой плавников.
Щупальца короткие, а тентакули,
или ловчие руки, в покое и в
движении втянуты внутрь головы, в особые
карманы. Когда каракатица хватает
добычу, она с силой, словно из катапульты,
выбрасывает их из карманов и
присасывается ими к жертве.
183
В анатомии каракатиц систематики
выделили следующие черты, несвойственные
кальмарам (назовем из них только
главные): роговица у каракатиц не пронзена
в центре широким круглым отверстием,
как у кальмаров, т. е. глазные камеры
у каракатиц, как говорят зоологи,
замкнутые; раковина у них обычно
известковая (у кальмаров без примесей извести,
чисто хитиновая); роговые (вернее, тоже
хитиновые) кольца присосок никогда не
изменяются в крючья; светящиеся
органы состоят только из мицетомов с симбио-
тическими бактериями (фотофоров не
бывает), а из двух яйцеводов функционирует
только один, левый.
Интересно, что в процессе эволюции
некоторые каракатицы, которые ведут
сходный с кальмарами образ жизни,
приобрели и типичную внешность этих
животных, хотя и сохранили характерные
для своего подотряда анатомические
признаки. Этот однообразный отпечаток,
который одинаковый образ жизни
накладывает на разных по происхождению
животных, называют конвергентным
сходством.
Калъмароподобные каракатицы, или
лолиго, обитают в тропических и теплых
морях (тоже характерная черта типичных
каракатиц). Это очень подвижные и
хищные животные.
Настоящие каракатицы, или сепии
(Sepia), не так подвижны, как лолиго, но не
менее их хищны. Сепии не живут в
открытых просторах океанов. Они держатся
обычно у берегов и неглубоко (до 200 м,
глубже не опускаются).
Рис. 116. Россия (Rossia glaucopis).
Эти животные, говорит британский
натуралист Фрэнк Л е й н, «буквально
оставили след в человеческой культуре», ведь
в течение веков люди писали их
чернилами.
Знаменитая краска «сепия» получила
свое название от научного названия
каракатицы — Sepia, из чернильной
жидкости которой она изготовляется. Краска
очень ценная, необыкновенно чистого
коричневого тона. Конечно, химики в
производстве современной «сепии» научились
обходиться без каракатиц. Но
по-прежнему еще натуральная «сепия» в большом
количестве потребляется
промышленностью для приготовления краски, которая
носит ее название.
Не менее древнее и разнообразное
употребление находит у людей и «кость»
каракатицы. Так называют остаток
недоразвитой внутренней раковины этого
моллюска. Она имеет перовидную форму и
состоит, как известно, из извести.
«Кость» каракатицы
(«морская пена» — samudra phena
по-индийски) очень популярна среди парфюмеров
и ювелиров, любителей птиц и
животноводов, аптекарей и медиков.
Ее собирают на берегу моря, главным
образом на побережьях Португалии,
Северной Африки и Индии. В 1954 г. из одного
лишь Туниса вывезли 175 т «костей»
каракатиц.
Животноводы ценят «кость»
каракатицы за высокое содержание в ней кальция,
необходимого для скелета растущих
животных; чертежники — за превосходные
качества промокательной бумаги и
ластика, которыми она обладает. Ювелиры
приготавливают из нее формы для
отливки. Годится «кость» каракатицы и для
шлифования металла, полировки
дерева, добавляют ее в политуру и даже
в зубной порошок, чтобы придать блеск
зубам.
«Костью» каракатицы лечили и лечат
кожные и ушные заболевания,
воспалительные процессы и многие другие
болезни.
А недавно три японских исследователя
открыли еще одно неоценимое качество
каракатицы: ее жир, оказывается,
обладает свойствами антибиотика.
184
ПОДОТРЯД КАЛЬМАРЫ (OEGOPSIDA)
Переходим теперь к описанию одной
из самых интересных групп морских
животных — кальмаров. Все они обитатели
открытых морских просторов, отличные
пловцы, едва ли не самые быстрые в море
животные, а некоторые и отличные
летуны!
Кальмары, даже сравнительно
некрупные, могут развивать скорость до 55 км
в час. Об этом можно судить по скорости
и дальности полета так называемых
летающих кальмаров.
Лучший пилот среди моллюсков —
кальмар стенотевтис. Английские моряки
называют его флайнг-сквид (летающий
кальмар)г.
Это небольшое животное размером с
сельдь. Кальмар преследует рыб с такой
стремительностью, что нередко
выскакивает из воды, стрелой проносясь над ее
поверхностью. К этой уловке он прибегает
и спасая свою жизнь от хищников —
тунцов и макрелей.
Развив в воде максимальную
реактивную тягу, кальмар-пилот стартует в
воздух и пролетает над волнами более 50 м.
Апогей полета живой ракеты лежит так
высоко над водой, что летающие кальмары
нередко попадают на палубы океанских
судов. Четыре-пять метров не
рекордная высота, на которую поднимаются
в небо кальмары. Иногда они взлетают
еще выше.
Английский исследователь моллюсков
доктор У. Р и с описал в научной статье
кальмара (длиной всего в 16 еле), который,
пролетев по воздуху изрядное
расстояние, упал на мостик яхты,
возвышавшийся над водой почти на 7 м.
Случается, что на корабль сверкающим
каскадом обрушивается множество
летающих кальмаров.
Осьминоги тоже умеют летать.
Французский натуралист Жан В е р а н и
видел, как обычный осьминог разогнался
в аквариуме и вдруг задом вперед нео-
1 Кальмары других видов — Onychoteuthis
banksii, Ommastrephes sagittatus, Symplectoteu-
this oulaniensis — тоже могут совершать полеты
над морем.
Рис. 117. Обыкновенный лолиго — Loligo
vulgaris (слева) и кальмар Ommastrephes sp.
(справа).
жиданно выскочил из воды. Описав в
воздухе дугу длиной метров в пять, он
плюхнулся обратно в аквариум. Набирая
скорость для прыжка, осьминог двигался не
только за счет реактивной тяги, но и греб
щупальцами.
Глубоководные кальмары отличаются
часто очень причудливой внешностью.
Особенно поражают их глаза. У одних
они торчат вверх телескопами, у других
на тонких стебельках вынесены далеко
в стороны, а есть и такие кальмары,
у которых (небывалое дело!) глаза
асимметричные: левый в четыре раза больше
правого.
Как плавают эти животные — ведь
голова у них не уравновешена?
Немалые, наверное, приходится им
прилагать усилия, чтобы плыть вперед и не
переворачиваться.
Профессор Джильберт В о с с из
Океанографического института в Майами
(США) думает, что большой глаз
приспособлен к глубинам, он собирает своей
мощной оптической системой рассеянные там
крохи света. Маленьким же глазом каль-
Рис. 118.
Чудесная лампа (Lyco-
teuthis diadema).
мар обозревает окрестности,
всплывая на поверхность.
Это вполне возможно
(асимметричные глаза типичны
для представителей
семейства Histioteuthidae).
У кальмаров есть и совсем особенные
глаза, ни у кого в природе больше не
встреченные,— термоскопические г. Они
«видят»... тепло. На плавниках кальмара
мастиготевтиса около тридцати
миниатюрных термолокаторов, способных,
очевидно, воспринимать тепловые лучи. Темными
точками они рассеяны в коже. Под
микроскопом видно, что орган состоит из
шаровидной капсулы, наполненной прозрачным
веществом. Сверху капсула прикрыта
толстым слоем красных клеток — это
светофильтр, он задерживает все лучи, кроме
инфракрасных.
По-видимому, в термоскопиче-
ских глазах кальмаров происходят
фотохимические процессы такого же типа,
как и на сетчатке обычного глаза или на
фотопластинке. Поглощенная органом
энергия приводит к перекомбинации
светочувствительных (у кальмаров теплочувст-
вительных) молекул, которые
воздействуют на нерв, вызывая в мозгу
представление о наблюдаемом объекте.
У гремучих змей Америки и
щитомордников, которые водятся у нас в Сибири,
тоже есть на голове своеобразные
термолокаторы, но устроены они иначе — по
принципу термоэлемента. Змеи при
помощи термолокаторов разыскивают в
темноте теплокровных грызунов и птиц,
которые, как и всякое нагретое тело,
испускают инфракрасные лучи.
1 Термоскопические глаза открыты у
глубоководных кальмаров Chiroteuthis bomplandi, Masti-
goteuthis grimaldii, Abralia sp. Enoploteuthis sp.
А зачем термоскопические глаза
кальмарам? Ведь на глубинах, где они
обитают, нет теплокровных животных.
Нет ли? А кашалот! Этот прожорливый
кит ныряет очень глубоко и охотится
в морской бездне на кальмаров. Съедает
их в день несколько тонн. 95% его меню
составляют глубоководные кальмары.
Сотни тысяч кашалотов пожирают
ежедневно сотни миллионов кальмаров,
преимущественно глубоководных. Поэтому,
наверное, развились у жителей холодной
пучины глаза, которые «видят» тепло.
Местных теплокровных животных там
нет — это правда, зато сверху, с
сияющей лазури моря, вторгаются в царство
вечного мрака огромные прожорливые
звери. Сигналы о их приближении подают
кальмарам термолокаторы.
Многие из кальмаров невелики по
размерам. Таков, например, очень обычный
в наших дальневосточных морях
тихоокеанский кальмар Ommastrephes sloanei
pacificus, которого японцы добывают по
700 тыс. т в год. Но есть среди кальмаров
и настоящие гиганты, встреча с которыми
не сулит человеку ничего хорошего. О них
сложено много легенд, но долгое время
гигантские кальмары, или архитевтисы
(Architeuthis), не попадались в руки
зоологов, и ученые уже начали было
сомневаться, существуют ли они в
действительности.
С древнейших времен люди, жизнь и
труд которых тесно связаны с морем,
верили, что в морской пучине живут
странные огромные существа, непохожие
ни на рыб, ни на медуз, ни на раков, ни
на других обитателей океана. Почти у всех
приморских народов, живущих по
берегам океанов и открытых морей, есть свои
мифы об этих загадочных животных. Одни
называют их полипусами, другие — кра-
186
кенами, третьи — пульпами. У чудовищ
много и других названий.
О крупных морских животных,
вооруженных многочисленными щупальцами
с присосками, писал еще Аристотель. Он
называет их большими «тевтисами» и
добавляет, что «в Средиземном море они
достигают размеров в пять локтей (около
2,5 м) и отличаются красноватым цветом
и округлыми плавниками на хвосте».
Шли столетия, и странные морские
чудовища выросли в устах молвы до поистине
сказочных размеров: уже не локтями
измеряли их рассказчики, а милями...
Рассказы о них, сообразно со вкусами
и уровнем познаний эпохи, приобретают
еще более фантастичный и курьезный
характер.
Норвежцы, для которых море стало
родной стихией, больше других европейцев
страдали от этих чудовищ. Упоминания
о кракенах особенно часто встречаются
Рис. 119. Хиротевтис верании (Chiroteuthis
veranyi).
в сочинениях средневековых
скандинавских писателей и летописцев.
Олаус Магнус, архиепископ Упса-
лы и известный хроникер, в своей
«Истории северных народов» (издана в 1555 г.)
рассказывает о «чудовищной рыбе»,
появляющейся у берегов Норвегии. «Рыба»
эта в длину не меньше мили и похожа
скорее на остров, чем на животное.
Вполне естественно, что такое
страшилище могло «потопить много больших
кораблей со множеством сильных
матросов».
Вид «рыбы» ужасен. У нее непомерно
большая квадратная голова, усаженная
уродливыми бородавками и буграми, и
огромные глаза — отныне традиционные
признаки кракена.
В конце XVIII и начале XIX в. в
Парижском музее естественной истории
работал известный французский зоолог Д е-
ни де Монфор. Он написал много
научных трудов о животных, но
знаменитым его сделала книга «Естественная
история моллюсков», изданная в 1801—1802 гг.
Де Монфор собрал все известные ему
рассказы о кракене, которого называл
«колоссальным пульпом». Немало добавил
и от себя. Получилось удивительное
сочинение — суррогат науки и фантастики.
Его «Естественная история» смело могла
претендовать на звание
научно-фантастического романа, и даже в наше время этот
«роман», наверное, считался бы одним из
самых увлекательных.
«Колоссальный пульп» де Монфора —
чрезвычайно опасное и агрессивное
чудовище, настоящий пират океанских
просторов. Однажды он, неожиданно
вынырнув из глубины, схватил своими
щупальцами трехмачтовое судно и потащил его
ко дну. В другом месте де Монфор
рассказывает о том, как стая этих чудовищ
потопила за одну ночь десять военных
кораблей.
30 ноября 1861 г. произошло следующее
знаменательное событие: французский
пароход «Алектон» встретил в море...
кракена.
В два часа пополудни недалеко от
Канарских островов было замечено с
корабля огромное и странное существо, которое
медленно плыло у самой поверхности.
187
На голове чудовища, словно клубок змей,
копошился десяток длинных щупалец.
Корабль был быстро приведен в
боевую готовность. На чудовище навели
жерла пушек, но сильная бортовая качка
помешала бомбардирам точно прицелиться.
Ни один из десяти снарядов не попал
в цель. Тогда «Алектон» подошел
вплотную к диковинному «зверю», и несколько
удачно брошенных гарпунов вонзилось
в его тело.
Чудовище словно очнулось от
оцепенения и рванулось к кораблю с широко
раскрытым клювом, затем отплыло в
сторону. «Алектон» вновь настиг «зверя»,
и новые гарпуны вонзились в него.
Сражение продолжалось больше трех часов.
Матросам не удалось поймать кракена.
Достался им лишь небольшой кусок его
хвоста весом около 20 кг. Но судовой
художник успел сделать с животного
цветной рисунок; он хранится сейчас во
Французской академии наук.
В 1873 г. наконец-то одно из чудовищ
попалось живым в руки людей. Кракен
попал в сеть!
Ньюфаундлендские рыбаки вытягивали
на берег закинутый в море невод. Он
казался необычно тяжелым и вел себя очень
странно: яростно рвался из стороны в
сторону. С трудом сеть подтянули к берегу.
Но когда она показалась на поверхности,
люди едва не выпустили ее из рук — в
сети оказался кракен.
Рыбаки увидели массу извивающихся
щупалец, а в центре — два сверкающих
глаза. Несколько длинных щупалец
протянулось сквозь дыру в сети, стараясь
достать людей. С минуту они извивались
в воздухе, разыскивая свою жертву, но
расстояние было слишком велико, и змеи-
щупальца вернулись обратно.
Испуганные рыбаки готовы были обрубить невод,
но не вступать в бой с таким врагом.
Однако нашелся смельчак. Выбрав
удобный момент, он подскочил сзади к чудо-
Рис. 120. Кальмары:
слева — Enoploteuthis leptura (молодой самец); посредине — Onychoteuthis banksii;
справа — Bathothauma lyromma (самка).
188
вищу и вонзил длинный нож между
огромными глазами кракена, а затем
быстро отсек голову от туловища.
Этот кракен целиком достался Р. Г а р-
в е ю. Он измерил кракена — морское
чудовище достигало в длину 10 м — и
положил в огромный бак, наполненный
соленой водой. Позднее
законсервированное животное доставили в Лондон в
Музей естественной истории.
По-видимому, какая-то тяжелая болезнь
свирепствовала в семидесятых годах среди
кракенов. Много гигантских животных
погибло тогда. Их часто находили на
поверхности океана, полуживых и мертвых,
и даже на берегу, выброшенных морем.
Рыболовные суда, промышлявшие у
берегов Ньюфаундленда, подобрали в море
около сотни полумертвых чудовищ.
Рыбаки разрубали кракенов на куски и
наживляли их мясом снасти для ловли
трески. Много животных попало и в руки
ученых. Вот тогда их изучили как следует.
Одним из первых исследовал кракенов
американский зоолог Эдиссон В е р рил.
Он дал животным этого рода название,
присвоенное им датчанином С т е е н с т-
р у п о м — Architeuthis (архитевтис). Вер-
рил составил описание животного по всем
правилам зоологической науки.
Гигантские кальмары обитают в
открытом море, по-видимому, на глубине 200—
1000 ле. Известно около десяти видов
этого рода, распространенных по всем
океанам.
Питаются эти животные крупными
рыбами, возможно, даже дельфинами: на коже
некоторых дельфинов находили следы
присосок крупных спрутов г.
Исполнилось уже сто лет, как с
достоверностью было установлено
существование гигантских кальмаров, однако
немного узнали мы о их жизни и повадках 2.
Известно, что цвет их кожи обычно темно-
1 Слово «спрут» скандинавского
происхождения. Норвежцы и датчане обозначают им (sprute
или blekksprute) гигантских головоногих. В
русском языке спрутом иногда называют также
обыкновенного осьминога, но лучше сохранить это
название только для очень крупных головоногих.
2 Даже в наиболее объемистом описании жизни
животных — в четырнадцатитомном сочинении
А. Б рема — отведена для гигантских кальмаров
лишь одна страница.
зеленый (в покое) или кирпично-красный,
когда спруты раздражены, что у них
самые большие в мире глаза (почти
полметра в диаметре), что длина архитевтисов
нередко достигает 10—15 м. Самый
большой кальмар, точно измеренный
зоологами, был длиной в 18 м и весил, по-
видимому, около 8 т. Однако время от
времени поступают сообщения о еще
более огромных спрутах, и это вполне
возможно.
Удивительно ли, что, обладая такими
размерами, спруты оставляют на теле
своих врагов — китов кашалотов —
глубокие шрамы и царапины? Сражение
продолжается и внутри кита: на стенках
желудков убитых кашалотов исследователи
находят иногда отпечатки присосок
проглоченных спрутов.
Титанические битвы, которые
разыгрываются между кашалотом и спрутом,
принадлежат к числу самых грандиозных
битв в природе.
Советский специалист по китам
Б. А. Зенкович в книге «Киты
и китобойный промысел» пишет, что
однажды в море его внимание привлекло
необычное поведение кашалота. Кит,
точно в предсмертной агонии, то
выскакивал из воды, то вертелся у поверхности.
Моряки заметили, что тело его опутано
щупальцами огромного кальмара.
Кашалот схватил моллюска в пасть и пытался
проглотить, но мешали щупальца,
присосавшиеся к голове кита. Чтобы сбросить
их, кашалот дико вертелся и выпрыгивал
из воды. Ему удалось освободиться от
цепких «арканов», и он разорвал и
проглотил кальмара. Когда корабль
подошел к месту битвы, кашалот нырнул под
воду. Несколько потерянных им щупалец
растерзанного кальмара медленно тонуло
в пенящихся волнах.
Трудно, конечно, поверить, что спруты
питаются стотонными китами. Никто не
находил в их желудках остатков китов.
Правда, кальмары настолько тщательно
«пережевывают» свою пищу, что в пищевод
и желудок она попадает в виде мелко
перетертой кашицы. Кроме хищного клюва,
во рту у кальмаров есть ведь и «терка» из
сотен мельчайших роговых зубчиков,
которой они и приготавливают себе «пюре»
189
из рыбы или крабов. Поэтому
очень трудно судить о составе
пищи кальмаров по содержимому
их желудков.
Напротив, меню кашалота
наглядно представлено в его
желудке. Кашалот не пережевывает
пищу, он лишь разрывает на части
крупную добычу, а мелкую
глотает целиком. В желудке убитого
кашалота почти всегда можно
найти несколько десятков
глубоководных рыб и полтысячи
мелких кальмарчиков, обычно целых
и неповрежденных (вполне
пригодных для музейных коллекций).
Однако очень редко даже в
желудках тех кашалотов, которые
носят на теле свежие рубцы
ранений, полученных в битве со
спрутами, попадаются остатки
гигантских кальмаров.
Невольно возникает вопрос: не
говорит ли это о том, что в
непримиримой вражде спрута с
кашалотом нападающей стороной
чаще бывает не кит, а чудовищный
властелин мрачного подводного
царства, во владения которого
вторгается кашалот в поисках
пищи?
В декабре 1946 г. норвежский
журнал «Природа» опубликовал
интересное сообщение: танкер
«Брунсвик», океанское судно
водоизмещением в 15 тыс. т и
длиной в 150 м, подвергся в Тихом
океане нападению... гигантского
кальмара. Это произошло среди
бела дня между Гавайскими
островами и Самоа.
Автор сообщения, капитан
норвежского флота Арне Греннингзе-
тер, с интересными подробностями
описывает это необычное
происшествие.
Огромный кальмар длиной более 20 м
неожиданно вынырнул из глубины,
быстро нагнал корабль, который шел со
скоростью 12 узлов B1,6 км в час). Скорость
самого кальмара была около 32—40 км
в час. Некоторое время кальмар плыл
параллельным с кораблем курсом на рас-
Рис. 121. Galiteu-
this armata.
стоянии приблизительно 30 м от
его левого борта. Затем описал
полукруг, обогнав корабль, зашел
справа и, стремительно
бросившись в атаку, вцепился в борт,
нанося сильные удары по
обшивке.
Таким образом, мы вновь
возвращаемся к вопросу: кальмар —
жертва или охотник? Возможно,
что в природе эта альтернатива
решается либо тем, либо другим
образом в зависимости от
размеров спрута. Хорошо известно,
что кашалот охотится не только
на мелких, но и на очень
крупных кальмаров длиной до 10—
15 м. А разве исключено, что
вдвое более крупные кальмары
могут быть опасны и для самого
кашалота?
Сообщение капитана Греннинг-
зетера заставляет с меньшим
недоверием отнестись к легенде о
кальмарах—«пожирателях китов».
Бесспорно, рассказы о краке-
нах, огромных, как остров,
неправдоподобны. И все-таки
действительные размеры этих
животных превзошли всякие
ожидания. Гигантские кальмары
длиной в 10—15 м совсем нередки.
С семидесятых годов прошлого
века и до настоящего времени
море выбросило на побережья
Ньюфаундленда, Англии, Исландии,
Норвегии, Дании, Японии и
Новой Зеландии более 80
гигантских кальмаров. Почти все они
имели в длину 10—15 м. Самый
крупный из измеренных
экземпляров — кальмар, изученный Вер-
рилом. Исполинское животное
достигало в длину 55 футов, т. е.
около 18 м. Несколько длиннее
A9 м) был кальмар, найденный
на побережье Новой Зеландии. Этот
кальмар вошел в науку под
названием «длиннорукого спрута» Architeuthis
longimana. Его щупальца отличались
невероятной длиной; лежа на земле,
кальмар мог дотянуться ими почти до
шестого этажа!
190
Рассказывают, что на побережье
Ньюфаундленда несколько десятилетий назад
был выброшен
двадцатичетырехметровый кальмар. Однако у ученых нет
полной уверенности в том, что этот спрут был
правильно измерен.
Исследовав рубцы на теле кашалота
от присосок спрутов, установили, что
некоторые из чудовищ, оставивших на коже
китов отпечатки своих «пальцев», были
длиной около 30 м.
О весе наиболее крупных спрутов нам
известно немного. Вес «дьявольской
рыбы», с которой вступил в сражение
французский корабль «Алектон», был
определен командой этого судна в 2—3 т.
Другой кальмар был найден мертвым на
поверхности моря у берегов
Ньюфаундленда. Усилиями всей команды с трудом
удалось поднять его на палубу шхуны.
Кальмара разрубили и взвесили по
частям. Оказалось, что при длине тела
всего в 5 м (без щупалец) он весил 907 кг.
ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ
ГОЛОВОНОГИХ МОЛЛЮСКОВ
Головоногих моллюсков издавна
употребляют в пищу. В Китае, Корее и
Японии использование этих животных в
качестве пищевых продуктов уходит в глубь
веков; в средиземноморских странах оно
имеет также очень давнюю историю.
Античные писатели, в частности Аристотель,
Плутарх и Плиний, сообщают, что в
древней Греции и Риме искусно
приготовленный осьминог был обычной пищей.
В Португалии каракатиц и осьминогов
варят в их собственном соку, затем
консервируют в жестяных банках и
отправляют на экспорт. Еще более
значительную статью португальского и испанского
экспорта составляют сушеные осьминоги
и кальмары. Перед сушкой их разрезают
вдоль, извлекают гладиус и
внутренности, затем растягивают на палках и
высушивают на солнце.
В Китае, Корее, Японии и на островах
Полинезии головоногих моллюсков
употребляют в сыром, сушеном, маринованном,
печеном, вареном и жареном виде. Короче
говоря, этих животных там едят во
всяком виде, и никакие части их тела не
пропадают. Даже глаза и присоски сушат на
сковороде и затем едят, как орехи,
внутренности идут на вытопку жира, а
выжимки — на корм домашней птице.
По основным показателям
питательности — калорийности и белковому
составу — кальмары, особенно
консервированные, превосходят всех других
употребляемых в пищу моллюсков и даже
некоторых рыб. Они незначительно
уступают говяжьему мясу и телятине.
За последние двадцать лет мировая
добыча головоногих моллюсков возросла
в несколько раз. До второй мировой
войны основными поставщиками этих
животных были Япония и Тунис. В настоящее
время головоногих моллюсков добывают
и экспортируют многие страны. За
двадцать пять лет японцы намного увеличили
добычу этих моллюсков. В 1925 г. в
Японии было поймано 250 тыс. т
головоногих моллюсков, а в 1952 г.—701100 т.
Из них 600 900 т — кальмара «surume-
ika» (Ommastrephes sloanei pacificus) и
и 37 500 m осьминога «tako» (Octopus
vulgaris).
Валовая добыча головоногих
моллюсков составляет около V6 всей морской
продукции Японии, включая водоросли,
устриц и других раковинных моллюсков,
и около V4 годовой добычи рыбы.
На втором месте в мире по промыслу
головоногих стоит Китай: в 1956 г. здесь было
добыто 80 тыс. т одних только каракатиц.
В Соединенных Штатах в 1945 г. на
Тихоокеанском побережье было поймано и
заморожено для отправки на
Филиппины и в средиземноморские государства
(эти страны — основные потребители
американского экспорта) около 8 тыс. т
головоногих моллюсков, преимущественно
кальмара Loligo opalescens.
Во всех странах мира добывают
ежегодно около миллиона тонн головоногих
моллюсков. Эта цифра позволяет нам
сделать, весьма приблизительный правда,
подсчет общего количества этих животных,
обитающих во всех морях и океанах
земного шара. Промыслом берется едва ли
сотая часть всей биомассы головоногих
моллюсков. Если же учесть, что им охвачено
191
всего лишь несколько наиболее
многочисленных видов кальмаров, осьминогов
и каракатиц, а виды мелкие, редкие и
глубоководные совсем не промышляются, то
скорее всего мы должны предположить,
что на Земле обитает в тысячу раз больше
головоногих моллюсков, чем добывают
их в настоящее время люди, т. е. около
миллиарда тонн. Количество пищи,
необходимое этому неисчислимому
множеству хищников, должно быть в 15—20 раз
больше — около 15—20 млрд. т.
Из шестисот видов головоногих
моллюсков объектами интенсивного промысла
служат всего лишь один-два десятка
видов, обычно формы, обитающие в
поверхностной зоне и образующие массовые
скопления. Среди них на первом месте стоят
представители семейства Ommast-
rephidae, главным образом Ommastrephes
sloanei pacificus, Shenoteuthis bartrami,
Illex illecebrosus. На долю
первого—кальмара, интенсивный промысел которого
ведется в Японии, приходится почти 75%,
больше 600 тыс. т мировой добычи всех
головоногих моллюсков. На втором месте
за ним стоит обыкновенный осьминог
Octopus vulgaris, которого в одной
Японии ежегодно добывают около 40 тыс. т.
В значительном количестве добывают его
и во всех средиземноморских странах.
Основными орудиями лова головоногих
моллюсков служат обыкновенные н е в о-
д а. Впрочем, в обычное время эти
быстрые животные лишь случайно попадают
в сети, и их промысел бывает
добычливым только в весенние и раннелетние
месяцы, когда у них наступает период
размножения (в тепловодных бассейнах)
и кальмары, собираясь большими стаями,
приближаются к берегам. Случается, что
в эту пору после сильного шторма сотни
тонн кальмаров оказываются
выброшенными на берег.
Известны и другие способы лова
головоногих моллюсков. В Японии используют
для этой цели простые глиняные кринки,
которые на длинных бечевках опускают
на дно. Осьминоги охотно в них
забираются и весьма неохотно покидают, даже
когда их вместе с кринкой поднимают на
поверхность. Этот способ лова бывает
особенно добычлив опять-таки в период
размножения осьминогов, когда они
собираются у берегов в больших
количествах и ищут подходящих убежищ для
откладки своих яиц. Так же ловят этих
животных и на северном побережье
Африки, особенно в Тунисе. Случается, что
ловец из каждого кувшина извлекает не
одного, а 8—10 осьминогов.
В Италии «кувшинный перемет»
называют муммареллой. Применяют
его и в Индии, но вместо кувшинов
индийцы подвешивают на лине крупные
раковины морской улитки птероцеры —
семьсот — девятьсот раковин. И
ежедневно добывают такой снастью двести-три-
ста осьминогов.
Кубинским рыбакам муммарелла тоже
известна. Здесь осьминогов «соблазняют»
раковинами улитки стромбуса. Проверяют
ловушки каждый день, приблизительно
четверть раковин бывает заселена пуль-
по, т. е. осьминогами.
В Тунисе для ловли осьминогов
применяют еще более добычливое орудие —
глиняные дренажные тру-
б ы, которые опускают на дно, и осьминоги
забираются в эти трубы не менее охотно,
чем в горшки.
Кальмаров ловят иначе. Самая
распространенная для них снасть — это
джиг— многоякорный крючок с
фарфоровой бляхой вместо приманки. Его
дергают в воде вверх—вниз, вверх—вниз.
Кальмары бросаются на блеск фарфора
и попадаются на один из десяти или
тридцати крючков. Французы называют
джиг турбутом, а итальянцы —
л о н т р о. Джиг применяют и японцы, и,
пожалуй, успешнее, чем в других
странах. Один японский рыбак, используя
джиг, добывает тысячу кальмаров в час
(когда погода хорошая и кальмаров
много). Каждые три секунды — кальмар!
Мелких кальмаров в Японии не
выбрасывают, а используют для удобрения,
Таблица 16. Осьминог, присосавшийся к скале.
Хорошо видны темные пятна растянутых до д редел а хроматофоров,
корма скота и птицы или в качестве
наживки для рыболовных снастей. В
Канаде и Ньюфаундленде почти всех
добытых кальмаров употребляют для
наживки. В районе Ньюфаундлендской банки
половину всей трески ловят на наживку
из кальмара Illex illecebrosus. Для этой
же цели употребляют и гигантских
кальмаров рода Architeuthis, когда их
находят выброшенными на берег. В годы,
когда к берегам Канады подходит мало
кальмаров, улов трески сильно падает,
и, наоборот, треска бывает особенно
упитанной, когда в море много кальмаров.
Драгоценная амбра, которую
находят в кишечниках и фекалиях кашалотов,
представляет собой, по существу, тоже
продукт, получающийся из головоногих
моллюсков. Прежде считали, что амбра
образуется в кишечнике кашалота из кожных
желез головоногих моллюсков в
результате болезненного нарушения пищеварения.
Теперь же полагают, что амбра образуется
из уплотненных и прошедших
определенную стадию разложения клювов
кальмаров.
А велик ли вред, который головоногие
моллюски причиняют людям?
В некоторые годы по причинам еще не
вполне ясным осьминоги сильно
размножаются и появляются у берегов в
большом количестве. О «страшном нашествии»
обыкновенных осьминогов на северное
побережье Франции и южное побережье
Англии сообщал в 1899 г. Гарстэнг.
В окрестностях Шербурга с любого камня
у берега можно было увидеть одного или
нескольких осьминогов, беспокойно
переползающих по дну с места на место.
Местами с одного камня можно было
насчитать в море до 68 осьминогов. После
шторма сотни тонн осьминогов были
выброшены на берег. В некоторых местах
их разлагающиеся тела,
нагроможденные большими грудами, угрожали
здоровью населения. Сотни возов этих
выбросов были вывезены на поля для удобрения.
За два года A899—1900) своего
исключительного изобилия осьминоги нанесли
большой ущерб краболовству. Сильно
пострадали от них и устричные хозяйства.
В качестве примера Гарстэнг ссылается
на одного рыбака из Плимута, который,
поставив в море 180 верш на омаров,
поймал в них только три живых краба и 15
живых омаров. 44 краба и 41 омар были
убиты и изувечены осьминогами; 64
осьминога не успели убежать, пока поднимали
верши, и были вытащены наверх.
Омары и крабы, преследуемые
осьминогами, покидали родную стихию и
вылезали на берег. В сентябре 1900 г. в течение
нескольких дней на восточной стороне
плимутской гавани находили сотни
больших крабов и омаров, спасающихся от
осьминогов на берегу.
Подсчитали, что в течение шести
летних месяцев (апрель — сентябрь) 1900 г.
на южном побережье Англии было добыто
омаров на 69 тыс., а крабов на 281 тыс.
меньше, чем за тот же период в
предыдущем году, т. е. улов омаров упал на 18,
а крабов на 32%. Понесенные убытки
были бы еще большими, если бы они
сравнивались не с 1899 г., когда уже
началось нашествие осьминогов, а с 1898 г.
В 1922 и 1950 гг. нашествие
осьминогов на Ла-Манш повторилось. Опять в
окрестностях Шербурга осьминоги
буквально кишели у берега. Они были так
голодны, что жадно хватали всякие крохи
пищи, которые падали в воду. Тут же
разрывали на куски и своих собратьев,
попавшихся на крючок.
Кальмары тоже иногда появляются у
берегов огромными стаями, но вред,
который они наносят рыболовству и крабо-
водству, менее велик. Они объедают рыбу,
попавшую в ставные сети. Даже в годы
своего чрезмерного изобилия кальмары
пожалуй, приносят больше пользы, чем
вреда: их стаи привлекают крупные
косяки рыбы и дельфинов.
В странах, занимающихся промыслом
осьминогов, исследуются сейчас вопросы
не более рационального использования
продуктов, которые дает осьминожий
промысел (это уже пройденный этап), а
способы охраны и увеличения естественной
численности этих животных.
Первые опыты сделаны в Японии.
В префектуре Хиого несколько лет назад
был организован осьминожий заповедник.
В период размножения осьминогов
охраняли и даже подкармливали. В результате
уловы «тако» значительно повысились.
13 Жизнь животных, т. 2
193
Иглокожие
Echinodermata
Щетинкочелюстные
Chaetognatha
Вторичноротые (Deuterostomia) — двустороннесимметрич-
ные животные с вторичной полостью тела и кровеносной
системой, в отдельных случаях редуцированной.
В противоположность первичноротым у вторичноротых
ротовое отверстие формируется не на месте расположения
первичного личиночного рта (бластопора), а на противоположном
конце тела, т. е. на том месте, где у первичноротых образуется
анальное отверстие взрослого животного. На месте же
ротового отверстия личинки образуется дифинитивное анальное
отверстие, почему вся группа и получила название
вторичноротых.
Полость тела и мезодерма образуются
энтероцельным путем, т. е. из энтодермы, в виде слепых
карманов первичного кишечника, развивающихся в три пары
следующих друг за другом полостей.
Спиральный тип дробления отсутствует. Гаструла
образуется путем впячивания внутрь бластулы части
эктодермы, образующей энтодерму и формирующей кишечник (фаго-
цитобласт).
Кишечник почти полностью энтодермический.
Скелет у вторичноротых, в противоположность
первичноротым, внутренний, т. е. образуется из мезодермы,
наружный скелет (эктодермический) выражен слабо.
Нервная система неганглиозного типа и
закладывается в виде тяжа или трубки, закладка ее
происходит путем погружения внутрь зародыша эктодермальной
пластинки с образованием продольного желобка, а затем и
канала (невроцеля).
Покровный эпителий, хотя бы на отдельных
участках тела, многослойный.
В подраздел вторичноротых входят следующие типы:
Иглокожие (Echinodermata), Щетинкочелюстные (Chaetogna-
tha, Погонофоры (Pogonophora), Полу хордовые (Hemichordata),
Оболочниковые (Tunicata), Головохордовые (Cephalochordata) и
Позвоночные (Chordata), а всего 7 типов. Пять последних могут
быть объединены в н а д т и п Хордовые (Chordaria).
Первые 6 типов обитают только в морях. Круглоротые и рыбы
обитают в морских и пресных водах, амфибии населяют
пресные воды и воздушную среду, остальные три класса
позвоночных стали обитателями воздушной среды, и некоторые их
представители вторично переселились в пресные и морские
воды.
Среди вторичноротых очень редко проявляется склонность
к паразитизму, очень широко развитому у первичноротых.
тип hiviok©jkhe(echinodermata)
Иглокожие представляют собой
самостоятельный и весьма своеобразный
тип животного мира. По плану своего
строения они совершенно не сравнимы
ни с какими иными животными и
благодаря особенностям своей внешней
организации и оригинальной форме тела,
напоминающей звезду, цветок, шар, огурец
и пр., очень давно привлекают к себе
внимание. Название «иглокожие» дано еще
древними греками. В античное время уже
различали среди животного мира морских
звезд, голотурий и морских ежей, и
история изучения этих животных связана
с именами Аристотеля и
Плиния. В XVIII и в начале XIX в. ими
занимались такие видные ученые, как
Клейн (Kiein, J.), Линней
(Linnaeus, С), Л а м а р к (Lamarck, J. В.) и
Кювье (Cuvier, G.). Однако
большинство старых зоологов соединяло
иглокожих либо с червями, либо с
кишечнополостными.
Название лучистые, или зоофиты,
сохраняется за иглокожими вплоть до
1847 г., когда X. Ф р е й и Р. Лей-
карт (Н. Frey und R. Leuckart)
выделили их в отдельный самостоятельный тип,
указав на значительные отличия
иглокожих от лучистых, в частности от
кишечнополостных животных.
Последующие работы таких ученых,
как Людвиг (Ludwig, Н.), А г а с с и ц
(Agassiz, А), А. Ковалевский,
И. Мечников, Тиль (Theel, H.),
Мэк Брайд (Mac Bride, E. W.), Л и-
б е р к и н д (Lieberkind, I.), Фишер,
(Ficher, W. К.), Мортенсен, (Мог-
tensenT.H.) и др., значительно расширили
представление о систематике, анатомии
и эмбриологии иглокожих, о
родственных отношениях между отдельными
представителями этого типа. Русский ученый
Мечников своими работами по
развитию иглокожих установил их родство с
кишечножаберными, показав, что ветвь
иглокожих родственно близка ветви
хордовых животных. Датским ученым Мортен-
сеном была не только разработана
система для современных и ископаемых классов
иглокожих, но и дана классификация
личинок параллельно системе взрослых. Это
особенно важно, так как личинки
иглокожих очень своеобразны, весьма отличны
от взрослых форм, ведут пелагический
образ жизни и долгое время считались
самостоятельными организмами.
Среди советских исследователей,
особенно много сделавших для развития
знаний об иглокожих животных, прежде
всего необходимо отметить А. М. Д ь я-
конова и Д. М. Федотова,
которые своими работами по анатомии,
морфологии, развитию, регенерации, филогении
и палеонтологии данной группы
способствовали выяснению положения
иглокожих среди вторичноротых.
В настоящее время нет сомнений в том,
что тип иглокожих относится к группе
наиболее высокоорганизованных
беспозвоночных животных — к группе
вторичноротых.
197
Рис. 122. Ископаемые иглокожие.
Иэ класса гетеростелей: 1 — Trochocystites bohemicus (кембрий), 2 — Rhipidiocystis baltica
(нижний силур); из класса цистидей: з — Aristocystites bohemicus (нижний силур), 4 — Pleuro-
cystis filitextus (нижний силур); из класса бластоидей: 5 — Anthoblastus brouweri (пермь); из
класса эдриоастероидей: в — Agelacrinites ephraemovianus (девон), 7 — Cyathocystis plautinae (нижний
силур); из класса морских лилий: 8 — Platycrinus symmetricus (карбон), 9 — Gilbertsocrinus
tuberosus (карбон), 10 — Acrocrinus amphora (карбон); из класса офиоцистий: 11 — Volchovia vol-
borthi (нижний силур); из класса морских звезд: 12 — Macroporaster matutinus (нижний силур);
из класса офиур: 13 — Bundenbachia beneckei (нижний девон); из класса морских ежей: 14 —
Bothriocidaris pahleni (нижний силур).
Иглокожие появились на Земле давно,
свыше 520 млн. лет назад. Остатки этих
животных находят уже в отложениях
раннего кембрия. Обладание
известковым скелетом способствовало довольно
хорошему сохранению иглокожих в
ископаемом состоянии, поэтому их история
нам известна лучше, чем история других
беспозвоночных, тем более что особенности
скелета иглокожих в значительной
степени позволяют судить о всем строении
животного.
Древние иглокожие достигали в
палеозойскую эру огромной численности и
большого разнообразия форм. Это были
преимущественно двустороннесимметричные,
главным образом прикрепленные
животные с мешковидным, шаровидным илича-
шечкообразным телом, покрытым
пластинками в виде панциря.
Большинство животных относилось к
подтипу прикрепленных
иглокожих — пелъматозоа (Pelmatozoa), к пяти
его классам1: гетеростелей (Hetero-
stelea), цистидей, или цистоидей (Cystidea),
бластоидей (Blastoidea), эдриоастероидей
(Edrioasteroidea) и морских лилий (Crinoi-
dea). А из подтипа свободноподвижных
иглокожих—Eleuterozoa—только один
небольшой ископаемый класс Ophiocistia,
насчитывающий всего 5 родов и менее чем
1G видов, был ограничен палеозоем,
остальные же классы этого подтипа
достигают расцвета и в настоящее время.
Представители класса Ophiocistia (рис. 122)
обладали дисковидным или шаровидным
телом, лишенным выступающих лучей и
сплошь покрытым пластинками. Рот
помещался в центре стороны, обращенной
к субстрату, и был снабжен крепкими
подвижными челюстями. Эти животные
появились в среднем силуре и уже в
среднем девоне исчезли.
Наиболее древние и примитивные
формы, ближе всего стоящие к предкам
иглокожих животных, относились
к классу гетеростелей (рис. 122).
Они имели мешковидное сдавленное тело,
лишенное радиальной симметрии, и
обладали рядом признаков, сближающих
их с низшими хордовыми животными
1 Согласно сводке Хайман (Hyman, 1955).
и с ланцетником. Характерные черты
типа иглокожих получают значительное
развитие у цистидей, которые
господствовали в морях 500—400 млн. лет назад.
В связи с прикреплением к субстрату
у них развилась и закрепилась
радиальная симметрия, причем уже можно было
наблюдать появление пятилучевой
симметрии. Цистидей были чрезвычайно
разнообразной формы (рис. 122), процветали
в силуре и вымерли в девоне. Вполне
вероятно, что от них произошли
бластоидей и морские лилии. Бластоидей (рис.
122) являлись очень обособленными
формами, которые вымерли в пермский
период, не оставив потомства. Напротив ^
некоторые представители класса морских
лилий дожили до наших дней.
Ископаемые морские лилии (рис. 122),
представленные в силуре, девоне и в
нижнем карбоне, согласно Фишеру, 5 тыс.
видами, относившимися к 5 отрядам,
почти полностью вымерли к концу палеозоя.
Лишь немногие их представители перешли
в мезозойскую эру и постепенно исчезли.
Однако в начале мезозоя, приблизительно
180 млн. лет назад, появилась новая группа
морских лилий — отряд артикулята
(Articulata), представители которого не
только сохранились до наших дней, но
и достигают наибольшего расцвета в
настоящее время.
Эдриоастероидей (рис. 122), известные
с раннего кембрия до карбона, вероятно,
тоже происходили от ранних цистидей,
так как наиболее примитивные
представители их по уровню своей организации
были близки к цистидеям. Однако они
уже обладали целым рядом признаков,
сближающих их с морскими звездами,
поэтому их часто называют сидячими
морскими звездами. Эдриоастероидей
располагались на субстрате ртом кверху.
Большинство из них прирастало к
субстрату широкой поверхностью тела и
только очень немногие имели стебелек, а
некоторые формы, очевидно, вообще не
прирастали к субстрату, хотя лежали
неподвижно на дне. Таким образом,
эволюция древних классов иглокожих п о д т и-
п а палъматозоа почти полностью прошла
в палеозое, и до нашего времени дошел
только один отряд класса морских лилий.
199
Ныне живущие иглокожие, которых
насчитывается около 6000 видов, за
исключением класса морских лилий, относятся
к подтипу свободноподвижных
иглокожих — элеутерозоа (Eleutherozoa).
Среди современных иглокожих (табл. 17)
различают класс Морские звезды (Astero-
idea) со звездообразным или многолучевым
телом и ртом, расположенным в центре
нижней стороны, обращенной к
субстрату; класс Офиуры, или Змеехвостки
(Ophiuriidea), очень похожие на морских
звезд, но с лучами, сильно отшнурован-
ными от диска; класс Морские ежи
(Echinoidea)—почти шарообразные
животные, без вытянутых лучей; класс
Морские кубышки, или Голотурии (Holothu-
rioidea),— мешковидные или
червеобразные животные, рот и анальное
отверстие у которых почти всегда находятся
на разных концах тела; класс
Морские лилии (Crinoidea) — напоминающие
цветы, с сильно ветвящимися лучами, у
которых рот и анальное отверстие
сближены и лежат на одной стороне,
обращенной вверх.
Размеры тела иглокожих обычно
колеблются в пределах от 5 до 50 см, но имеются
виды, длина которых не превышает
нескольких миллиметров, а у других,
наоборот, может достигать даже 2 м.
Известные нам классы иглокожих очень
своеобразны и весьма отличаются друг
от друга, но, несмотря на большое много-
Рис. 123. Схема строения амбулакральной
системы морской звезды:
1 — кольцевой канал; 2 — радиальный канал; з —
боковые ветви радиального канала; 4 —
амбулакральные ножки; 5 — ампулы амбулакральных
ножек; 6 — мадрепоровая пластинка; 7 — каменистый
канал; 8 — полиевы пузыри.
образие форм, они имеют целый ряд
признаков, свойственных только данным
животным, которые указывают как на
общность происхождения всей этой группы,
так и на своеобразный путь ее эволюции.
Действительно, лишь иглокожие
обладают особой водоносной
(амбулакральной) системой, состоящей из серии
тонкостенных каналов, наполненных
жидкостью, имеют лучистый план строения,
обычно кратный 5, и сильно развитый
скелет, который по своему происхождению,
в отличие от других беспозвоночных
животных, является внутренним.
Амбулакральная система
у большинства форм служит для
движения, осязания, а у морских лилий и
некоторых ежей выполняет дыхательную
функцию. Она в наиболее типичном
случае состоит из кольцевого кана-
л а, окружающего ротовое отверстие
с внутренней стороны, и 5радиаль-
ных каналов (рис. 123), отходящих
от кольцевого и заканчивающихся слепо
или небольшим чувствительным
отростком. Радиальные каналы, проходя по
внутренней стороне луча, дают попарно
боковые веточки к ножкам, которые
называются амбулакральными
ножками. Ножки представляют
собой тонкие цилиндрические очень
растяжимые трубочки, снабженные обычно на
конце присоской, реже лишенные ее.
У основания каждой ножки находится
тонкостенный пузырек, или ампула.
Между радиальными каналами (и н т е р-
радиально) от кольцевого канала
амбулакральной системы отходят
тонкостенные, сильно растяжимые, часто очень
больших размеров мешки — полиевы
пузыри, служащие резервуарами
жидкости амбулакральной системы. Число
полиевых пузырей сильно варьирует,
иногда их отходит несколько даже в
одном интеррадиальном промежутке, у
других форм они вообще могут отсутствовать.
Межрадиально с кольцевым каналом
соединяется непарный каменистый
канал, названный так из-за твердости
своих стенок, в которых содержатся
отложения углекислого кальция. На
противоположном своем конце каменистый канал
соединяется с лежащей у большинства
200
форм на поверхности животного
пластинкой, которая называется мадрепоро-
вой пластинкой или м а д р е-
п о р и т о м. Он пронизан мельчайшими
отверстиями — порами, через которые
в амбулакральную систему поступает
морская вода. Детали строения и функция
амбулакральной системы могут
претерпевать ряд изменений как у различных
классов, так и в пределах одного класса,
отряда и даже семейства иглокожих. Так,
у части голотурий радиальные каналы
сильно укорачиваются до степени
коротких обрубков или вообще исчезают. У
некоторых звезд, офиур и голотурий бывает
несколько каменистых каналов и
несколько мадрепоровых пластинок, причем
у большинства голотурий каменистый
канал теряет сообщение с наружной средой
и вместе с мадрепоритом свисает в полость
тела. У морских звезд, ежей и
представителей одного отряда офиур кольцевой
канал имеет особые гроздевидные придатки,
состоящие из простых или разветвленных
трубочек. Это тидемановы тельца,
назначение которых точно не установлено.
Особенно большие изменения в строении
и функции претерпевают
амбулакральные ножки. Например, у морских лилий
они лишены ампул, присасывательных
дисков, остроконечны, никогда не служат
органами движения, а выполняют
чувствительную и дыхательную функцию. Две
пары ножек, расположенные около рта,
превращаются в ротовые щупальца,
помогающие при захвате пищи. У многих
голотурий у одного и того же экземпляра
встречаются как двигательные ножки,
снабженные присосками, так и ножки,
лишенные присосок, превращенные в
сосочки и выполняющие уже другие
функции, а у представителей двух отрядов
голотурий ножки на теле вообще
редуцируются и исчезают. Ротовые щупальца
голотурий представляют собой частично
видоизмененные ножки.
Среди морских ежей тоже встречаются
виды с очень разнообразными по форме
и функции амбулакральными ножками.
У них бывают наряду с двигательными
ножками конические, розеткообразные,
кистевидные ножки, которые несут уже
другие функции. Несмотря на такое
изменение ножек, основное их значение у
морских звезд, большинства голотурий и
ежей — передвижение животного по дну
морей. В этом случае жидкость
амбулакральной системы гонится из кольца в
радиальные каналы, откуда по боковым
ветвям поступает в ампулы ножек.
Благодаря сокращению мышц жидкость из
ампул перегоняется в ножки, которые
от этого сильно вытягиваются, и подошвы
ножек соприкасаются с субстратом. От
этого соприкосновения сокращаются
мышцы подошвы (присоски), последняя
вдавливается, и между подошвой и
субстратом образуется разреженное
пространство, обусловливающее прикрепление
ножки к субстрату. Прикрепление ножек к
субстрату благодаря давлению воздуха
и воды и действию клейкой слизи,
выделяемой железистыми клетками присоски,
столь прочно, что при попытке достать
прикрепившееся животное ножки скорее
обрываются, чем отстают от субстрата.
Последующее сокращение
прикрепившихся ножек подтягивает все тело животного,
а жидкость из сократившейся ножки
вновь поступает в расслабленную к этому
времени ампулу. Клапаны, находящиеся
в ампулах, регулируют наполнение их
жидкостью.
Другой характерной чертой иглокожих
является лучистое строение
тела, обычно кратное 5, которое
сказывается в том, что тело их поделено на
10 частей (секторов). Секторы, в
которых расположены амбулакральные
ножки и радиальные каналы
амбулакральной системы, называются а м б у л а-
к р а м и или радиусами. Они
чередуются с межрадиальными
секторами, которые называются
интеррадиусами или и н т е р-
амбулакрами. У разных
классов иглокожих радиусы и
интеррадиусы выражены различно. У одних они
слиты с общим телом животного
(голотурии и морские ежи), у других радиусы
могут сильно вытягиваться, образуя
подвижные или неподвижные лучи, иногда
называемые руками. В таких случаях
животные приобретают звездообразную
форму (звезды, офиуры). Иногда лучи
могут древовидно разветвляться, что на-
201
блюдается у ветвящихся офиур, или
делиться перистообразно, как у морских
лилий. У звездообразных форм число
лучей тоже может меняться от 5 до 16
и даже до 50. Изредка среди обычных пяти-
лучевых форм могут попадаться четырех-
и шестилучевые экземпляры.
Не менее характерной чертой типа
иглокожих наряду с амбулакральной
системой и лучистым планом строения
является наличие у них известкового
скелета, заложенного в соединитель-
нотканом слое кожи. Кожа иглокожих
состоит из однослойного покровного
эпителия, толстого слоя кутиса, или собственно
кожи, и снова слоя ресничного эпителия,
ограничивающего собой вторичную
полость тела, или целом.
Наружный покровный эпителий у
большинства форм (исключение составляют
голотурии) снабжен ресничками,
расположенными неравномерно. Действие этих
ресничек вызывает возникновение на
поверхности тела животного токов воды, идущих
в определенных направлениях и
служащих для подачи пищи, дыхания и
очищения тела от грязи. В наружном эпителии
кожи имеется большое количество
железистых клеток, выделяющих слизистый,
клейкий или ядовитый секрет, а у
некоторых видов встречаются железистые
клетки, секрет которых вызывает свечение
организмов. Здесь также находятся
скопления пигментных клеток, придающих
удивительно красивую и яркую окраску
иглокожим животным. Кутис,
подстилающий наружный эпителий, состоит из
соединительной ткани, в которой можно
различить три слоя. В его нижнем,
внутреннем, слое располагаются мышцы, сильно
развитые у голотурий, у которых они
образуют кожно-мускульный мешок, и
значительно редуцированные у морских
лилий, ежей и большинства офиур в
связи с развитием у них прочного скелета.
В наружном слое кутиса, под эпителием,
образуются скелетные элементы, поэтому
по своему происхождению из мезодермаль-
ного слоя они скорее соответствуют
покровным костям позвоночных животных,
а не раковине улиток.
Для построения своего скелета
иглокожие используют углекислый кальций,
растворенный в морской воде. По своему
происхождению скелет всех иглокожих
является внутренним и возникает в виде
мелкого, едва заметного под микроскопом
зернышка углекислого кальция внутри
клеток соединительной ткани. Это
зернышко растет, приобретает трехлучевую
форму, выпадает из давшей ему начало
клетки и становится межклеточным
известковым тельцем. При дальнейшем
развитии лучи известкового тельца начинают
ветвиться, соседние ветви соединяются
друг с другом, замыкая отверстия и
образуя мелкие продырявленные пластиночки.
Более крупные пластинки скелета
получаются путем слияния многих мелких
пластиночек своими краями, поэтому они
всегда имеют губчатую структуру. Форма
образующихся скелетных элементов
специфична и передается по наследству. Она
зависит от той роли, которую выполняют
скелетные элементы в организме. Хотя
форма скелетных элементов иглокожих
весьма разнообразна, но всем им присущи
оптические и кристаллографические
свойства одного кристалла известкового
шпата. Поэтому подобно последнему каждый
скелетный элемент иглокожего обладает
двояким лучепреломлением. Однако
скелетные элементы иглокожих не являются
просто кристаллами известкового шпата,
так как их удельный вес и показатель
преломления меньше, чем у кристаллов
известкового шпата. Это б и о к р и-
с т а л л ы, которые возникают в плазме
клеток главным образом из кальцита с
незначительным добавлением
органической массы. Поэтому они не ограничены
плоскостями кристалла, а форма их
определяется деятельностью внутриклеточной
образовательной плазмы. Известковый
скелет иглокожих может быть развит
слабо и состоять из отдельных очень мелких
пластинок чрезвычайно разнообразной
формы, что мы наблюдаем почти у всех
голотурий и в амбулакральных ножках
морских ежей, либо достигать мощного
развития (все остальные классы
иглокожих). В последнем случае он состоит из
хорошо заметных пластинок, которые
могут скрепляться неподвижно друг с
другом в виде плотной скорлупы, или
образовывать более или менее рыхлую или плот-
202
ную сеть, или соединяться друг с другом
подобно позвонкам. Пластинки скелета
довольно часто несут на своей
поверхности разнообразные иглы, бугры, мелкие
гранулы или особые хватательные
органы — педицеллярии.
Обладание таким скелетом со
всевозможными выростами послужило
основанием для того, чтобы назвать данных
животных «иглокожими», хотя не все
представители этого типа имеют иголки.
Среди скелетных элементов, пожалуй,
самыми примечательными являются
хватательные органы — педицеллярии.
Они встречаются у морских звезд и ежей
иногда в очень большом количестве и
представляют собой видоизмененные
иглы. Часто педицеллярии достигают
такой сложности, что трудно бывает себе
представить их происхождение от самой
обычной иглы скелетной пластинки. Так,
у морских звезд наряду с самыми
простыми педицелляриями, сидящими
непосредственно на теле животного и
представляющими собой 2 иглы, наклоненные друг
к другу, либо две (двустворчатые
педицеллярии) или несколько
створок, часто расположенных в ряд
(гребенчатые
педицеллярии), встречаются педицеллярии с
более удлиненными очень подвижными
створками (рис. 142), снабженными
зубцами, выростами и сидящими на особом
основании. Такие педицеллярии часто
образуют пучки или валики вокруг игл
и могут прикрепляться к телу животного
при помощи мягкой шейки или
стерженька. Они очень подвижны, снабжены
мышцами, захлопывающими и
раскрывающими створки. Основная функция таких
педицеллярии — очистка кожных покровов
от посторонних частиц, защита мягких
частей звезд от повреждений различными
мелкими животными. Часто
педицеллярии служат и для захвата добычи, но
иногда они используются как органы
нападения или защиты и тогда бывают
снабжены ядовитой железой. Ядовитые
педицеллярии встречаются у морских ежей
вместе с другими типами педицеллярии,
получившими дальнейшее развитие и
усложнение своей организации по
сравнению с педицелляриями морских звезд.
Скелетные пластинки
иглокожих очень часто имеют лучистое
расположение. Лучистое строение у
иглокожих проявляется не только во внешних
формах или в размещении пластинок
скелета, но и в расположении внутренних
органов. Так, нервная и кровеносная
системы иглокожих, подобно
рассмотренной выше амбулакральной системе,
построены по лучистому типу и имеют
сходное с ней расположение.
Нервная система иглокожих
довольно примитивна, состоит из трех
отделов, два из которых, называемые ги-
поневральной и апикальной системами,
являются преимущественно двигатель,
ными. Они развиты весьма различно и
подчас очень слабо в разных классах
данного типа животных, тогда как третий,
чувствительный отдел (эктоневраль-
ная система) очень постоянен и
хорошо развит у всех иглокожих. Все три
отдела нервной системы тесно связаны
между собой, так как при таком
распределении функций без этой связи не было
бы возможным осуществление передачи
воспринятого извне раздражения к
мышцам, железам и другим органам. Каждый
из названных отделов нервной системы
состоит из нервного кольца и отходящих
от него радиальных нервных стволов,
число которых соответствует числу амбу-
лакров (рис. 124). Кольцо и нервные
стволы состоят из нервных клеток и
нервных волокон и представляют собой нерв-
Рис. 124. Схема тройной нервной системы
морской звезды:
7 — эктоневральный, 2 — гипоневральный и
3 — апикальный отдел.
203
Рис. 125. Схемы строения органов чувств
иглокожих:
А — сферидий морского ежа (в разрезе): 1 —
ресничный эпителий; 2 — известковое тело сферидия; з —
система каналов; 4 — чувствительные клетки; 5 —
стебелек сферидия, сочленованный с бугорком
панциря. Б — отоцисты голотурии (в поперечном
разрезе): б — радиальный нерв; 7 — нерв отоциста; 8 —
отоцист; 9 — отолиты; ю — радиальный канал
водоносной системы. В — глазок морской звезды
(продольный разрез): 11 — линза; 12 — глазная ямка;
13 — чувствительные клетки; 14 — нервные волокна;
15 — опорные клетки.
ные центры, воспринимающие и
проводящие раздражения. Наиболее
поверхностно на ротовой стороне животного
расположен чувствительный (эктоневраль-
ный) отдел нервной системы, залегающий
у морских лилий и звезд непосредственно
в наружном эпителии амбулакральных
желобков, а у змеехвосток, морских ежей
и голотурий — в эпиневральных
каналах, образованных погружением стенок
амбулакральных желобков внутрь и
замыканием их в трубки. Нервное кольцо
эктоневральной системы окружает глотку
или пищевод, а радиальные стволы у
большинства форм доходят до конца амбу-
лакров, повторяя сложные разветвления
лучей и посылая веточки к различным
органам. Глубже этой системы и отделяясь
от нее только тонким слоем
соединительной ткани находится второй,
двигательный отдел нервной системы — г и п о-
невральная система. У
голотурий эктоневральная и гипоневральная
системы так тесно примыкают друг к
другу, что долгое время их даже не
различали. Наиболее значительно этот отдел
нервной системы развит у морских лилий
и офиур в связи с усиленной
подвижностью лучей, причем у офиур радиальные
нервы образуют ганглиозные утолщения,
посылающие нервы к мышцам позвонков
лучей. Третий отдел — апикальная
энтоневральная система -
расположен еще глубже предыдущего, на
стороне, противоположной ротовой, и
связан с эпителием полости тела. Эта
двигательная система отсутствует у голотурий,
слабо развита у морских звезд, ежей,
офиур и только у морских лилий
получила достаточное развитие. У морских
лилий она ведает движением лучей и их
частей и имеет сложное строение.
Благодаря такому строению нервной
системы, когда преобладающее развитие
получает эктоневральный отдел, многие
иглокожие становятся весьма
чувствительными к внешним раздражениям, хотя
органы чувств у них в целом развиты
слабо. Многочисленные
чувствительные клетки, одиночные или
собранные в группы, имеющиеся на
амбулакральных ножках морских звезд, офиур,
ежей и голотурий, коже рук морских
лилий, ротовых щупальцах и т. д.,
служат органами осязания.
Специальные органы зрения
встречаются сравнительно у немногих
иглокожих, хотя все эти животные обладают
значительной светочувствительностью.
Наиболее развитые глаза, построенные
по типу глазных ямок (рис. 125), имеются
у большинства морских звезд. Они
расположены на конце каждого луча у
основания концевого щупальца радиального
канала водоносной системы. Более слабо
развитые глаза встречаются в передней
части тела некоторых голотурий. Такие
глаза определяют только степень
яркости, но не различают отдельных предме-
204
тов. Пигментные пятна
морских ежей, расположенные, подобно
глазкам морских звезд, над концами
радиальных каналов водоносной системы, тоже
весьма чувствительны к свету и могут
рассматриваться как редуцированные
глаза. Несмотря на то что световое
раздражение проводится главным образом через
глаза, удаление их не лишает животное
светочувствительности. Так, весьма
обычная у нас в Белом море морская звезда
Asterias rubens с отрезанными вместе с
глазками кончиками лучей продолжает
уползать от очень яркого света и очень
сильной темноты. Да и остальные
иглокожие, например офиуры, у которых нет
органов зрения, сильно реагируют на
изменение освещенности, так как вся кожа
иглокожих обладает диффузной
восприимчивостью к свету.
Чувства обоняния и вкуса
у иглокожих развиты хорошо. Они могут
воспринимать вкусовые раздражения даже
на значительных расстояниях, хотя
органы обоняния и вкуса у них развиты слабо
и представлены многочисленными
чувствительными клетками, расположенными
на амбулакральных ножках, щупальцах
офиур, ротовых ножках некоторых ежей,
на щупальцах отдельных голотурий.
Некоторые зоологи считают органами вкуса
также небольшие, едва заметные глазу
образования — сферидии,
встречающиеся почти у всех морских ежей. Они
обычно расположены близко ко рту на
середине амбулакров и состоят из
плотного, сильно преломляющего свет
вещества, покрытого ресничным эпителием.
Сферидии (рис. 125) соединены с
панцирем коротким стебельком, в основании
которого находятся чувствительные
клетки и сплетения нервов. Окончательно
функция этих органов не выяснена.
Сходство их со статическими органами медуз
заставляет предположить, что у морских
ежей они скорее могут играть роль органов
чувства равновесия, чем органов вкуса.
Статическими органами,
позволяющими животному определять,
где верх и где низ, являются о т о ц и-
с т ы (рис. 125), встречающиеся лишь
у двух семейств голотурий, представители
которых либо живут на больших
глубинах, либо зарываются в песок. Отоцисты
имеют форму пузырька, содержащего
несколько или много мелких отолитов, и
связаны непосредственно с радиальными
нервами.
Кровеносная система
иглокожих также построена по лучистому
типу. Она состоит из околоротового
кольца и пяти радиальных кровеносных
сосудов, идущих в амбулакры между
радиальными нервными стволами и каналами
водоносной системы (рис. 126). У морских
звезд, ежей и офиур на стороне,
противоположной ротовой *, имеется второе
кольцо кровеносной системы, которое дает
боковые ветки к половым железам и
соединяется с первым кольцом особым
осевым органом, характерным только длг
иглокожих животных. Строение его буде^
рассмотрено дальше. Однако кровеносна}
система у иглокожих представляет собой
систему скорее лакун, чем кровеносных
сосудов. Она лишена собственных стенок
и образована просветами в рыхлой
соединительной ткани. Жидкость кровеносной
системы по своему составу близка к
жидкости полости тела или амбулакральной
системы. Кровеносная система иглокожих
скорее сравнима с лимфатической
системой позвоночных животных, ибо основное
ее назначение — разносить по телу
питательные вещества. Поэтому в толще
стенки кишки имеются кровеносные лакуны
или, как у голотурий и морских ежей,
хорошо развитые спинной и брюшной
кровеносные сосуды, способные
периодически сокращаться, что вызывает слабое
движение «крови». Пульсации, обычной
для кровеносной системы, у иглокожих
нет, но имеется пульсация перикардия,
стенки которого, как и у хордовых
животных, образуют впячивание — сердце.
Пульсация перикардия вызывает слабое
движение кровеносной жидкости, однако
правильного движения «крови» у
иглокожих нет, да и газового обмена, столь
характерного для кровеносных систем,
у них не установлено. Функцию газового
обмена выполняет жидкость полости тела.
1 Ротовая сторона иглокожих называется
оральной, а противоположная ей сторона —
аморальной.
205
Очень тесное отношение к кровеносной
и нервной системе имеет система
каналов, выстланных эпителием и
развивающихся из обособленных участков общей
вторичной полости тела. Эта система
каналов окружает кровеносные сосуды,
поэтому называется псевдогемаль-
ной (перигемальной)
системой. Она состоит из кольца вокруг
глотки или пищевода, расположенного
между амбулакральным кольцом и
околоротовым нервным кольцом, и пяти
радиальных каналов, заходящих в амбулак-
ры и проходящих между радиальными
нервами и амбулакральными каналами.
Жидкость псевдогемальной системы
соответствует жидкости полости тела,
содержит большое число амебоцитов, а
близкое расположение этой системы к нервной
дает возможность предположить, что она
служит для питания нервных тяжей и
предохранения их от сдавливания. Псев-
догемальная система развита неодинаково
у разных иглокожих. Так, у морских ежей
и голотурий имеются только радиальные'
каналы этой системы, а у морских лилий
она вообще, как правило, отсутствует.
Лучистый план строения иглокожих
проявляется отчасти и в расположении
Рис. 126. Кровеносная система морской звезды: А — общая
схема; Б — кольцевые каналы и осевой орган.
1 — ротовой (оральный) кольцевой канал; 2 — аборальный
кольцевой канал; 3 — сосуды осевого органа; 4 — радиальные
каналы; 5 — сосуды кишечника; 6 — сосуды половых желез.
половых органов. Половая система
состоит из полового тяжа и половых желез,
образующихся в качестве выростов
первого. Наиболее своеобразное и
примитивное строение имеет половая система
морских лилий. У них половые тяжи в виде
пяти плотных клеточных лент проходят
в лучи и ветвятся, повторяя
разветвления последних. Концевые веточки их
заходят в особые выросты на лучах,
называемые пиннулами, и принимают вид
полых слепых мешков. Половые
продукты развиваются из клеток половых
мешков внутри пиннул и выводятся наружу
через разрыв их стенок. У морских звезд,
офиур и ежей половой тяж имеет вид
пятиугольного кольца, стенки которого,
разрастаясь, дают начало половым
железам, расположенным интеррадиально.
Половые железы у морских звезд имеют
вид пяти пар ветвящихся гроздевидных
мешочков, помещающихся в основании
лучей или заходящих в них далеко. Они
открываются наружу короткими
каналами между лучами. У офиур
многочисленные мелкие половые железы расположены
на внутренней стороне пяти пар особых
выводковых сумок (бурс). Половые
железы открываются непосредственно в
половые сумки, которые сообщаются
с внешней средой через узкие
щели, находящиеся на ротовой
стороне диска в основании лучей. У
морских ежей пять гроздевидных
половых желез помещаются в
верхней части панциря, причем у
многих неправильных ежей
пятимерное строение нарушается
исчезновением задней железы. У
голотурий же правильное лучистое
расположение элементов половой
системы совершенно утрачивается.
У них имеется единственная
половая железа, состоящая из пучка
простых или ветвящихся трубочек.
Все трубки соединяются в один
половой проток, который у
большинства форм открывается наружу в
передней части тела, в спинном
интеррадиусе.
Нарушение пятилучевой
симметрии у современных иглокожих
встречается довольно часто. Так,
206
все голотурии и неправильные морские
ежи являются билатеральносимметричны-
ми животными и ясно обладают
обнаруживающимися передним и задним концами
тела. Да и у всех остальных иглокожих,
кроме некоторых многолучевых звезд,
ветвистых офиур и лилий, у которых
наблюдается увеличение числа
каменистых каналов и мадрепоровых пластинок,
фактически можно провести только
одну плоскость симметрии, несмотря на
лучистое строение, из-за положения
осевого комплекса органов,
расположенных лишь в одном из интеррадиусов. Эта
черта современных иглокожих
унаследована ими от билатеральносимметричных
предков, тогда как двусторонняя
симметрия голотурий и неправильных ежей
скорее должна рассматриваться как
вторичное явление, связанное с утратой
радиальной симметрии предков в связи с
переходом их к ползающему и роющему
образу жизни.
Осевой комплекс органов
очень характерен для строения
иглокожих и представляет собой
пространственное объединение весьма разнородных
органов. Он образован амбулакральной,
кровеносной и половой системами, а также
участками вторичной полости тела.
Основными составными частями этого
комплекса являются каменистый канал, мадре-
поровая пластинка с поровыми каналами,
осевой орган, идущий вдоль каменистого
канала в виде продолговатого мешка,
два обособленных участка вторичной
полости тела (левый и правый
осевые синус ы), заключающие
в себе осевой орган, и, наконец, половой
тяж, помещенный в особом участке
полости тела — половом синусе.
Осевой орган образован сплетением
кровеносных сосудов, отходящих от ротового
кровеносного кольца, перикардием
(правый осевой синус) и значительным
количеством соединительной ткани, а также
подвижными клетками —
амебоцитами. Функция осевого комплекса
довольно многообразна, но еще
недостаточно выяснена. С помощью пульсирующего
перикардия вызывается движение
жидкости в кровеносной системе; путем притока
и оттока морской воды через мадрепоро-
вую пластинку происходит регулирование
постоянного гидростатического давления
в амбулакральной системе и удаление
ненужных веществ; осевой орган служит,
с одной стороны, местом накапливания
и выведения из организма продуктов
распада, с другой стороны,
лимфатической железой, образующей
амебоциты — клетки, свободно
передвигающиеся в жидкости полости тела.
Возможно, что осевой орган, связанный с
половым тяжем, является также органом
внутренней секреции. Осевой комплекс
органов имеется у морских звезд, ежей
и офиур, хотя его положение и
соотношение органов в нем неодинаковы у
разных классов в связи со смещением мадре-
порита. У морских лилий и голотурий
осевого комплекса органов нет, а имеются
лишь каменистые и поровые каналы,
которые очень часто открываются
непосредственно в общую полость тела.
Целом иглокожих у большинства
форм достигает значительного объема,
выстлан ресничным эпителием, заключает
в себе кишечник и большую часть
внутренностей, покрытых этим эпителием. Лишь
у морских лилий в связи с развитием
значительного количества соединительно-
тканых мезодермальных тяжей,
заполняющих полость тела, да у ряда офиур,
у которых имеются особые эктодермаль-
ные сумки (бурсы), глубоко
вдающиеся в полость тела, объем его резко
уменьшается. Вторичная полость тела
возникает из двух задних целомов личинки,
причем мезентерий, отделяющий правый
и левый целомы личинки, у многих
взрослых иглокожих сильно редуцируется и
сохраняется только у голотурий, следуя
за изгибами кишечника. Полость тела
заполнена полостной, или пери-
висцеральной жидкостью,
очень прозрачной и близкой по своему
составу к морской воде, но содержащей
примесь белков и большое количество
клеточных элементов. Эта полостная
жидкость находится в постоянном движении,
вызванном ресничным эпителием,
выстилающим полость тела, и играет очень
важную роль в процессах обмена веществ.
Клеточные элементы (амебоциты),
содержащиеся в перивисцеральной жидкости,
207
участвуют в распределении питательных
веществ и в выделении продуктов распада.
Полостная жидкость имеет
также существенное значение и в
процессах дыхания: у некоторых иглокожих в
перивисцеральной жидкости имеются
красные кровяные клетки с
гемоглобином; кроме того, многие органы дыхания
образованы стенками полости тела. Так,
наружные околоротовые, сильно
разветвленные (перистомальные) жабры
большинства морских ежей представляют собой
выпячивания обособленного участка
вторичной полости тела — ротового, или
перифарингсального, целома,
расположенного вокруг пищевода. Через стенки
жабр, покрытых чрезвычайно тонким
слоем кожи, растворенный в воде кислород
проникает в полостную жидкость.
Кожные жабры, или папулы, в виде
тонкостенных сократимых очень мелких
пузырьков имеются у морских звезд.
Наряду с жабрами существенную роль в
дыхании, очевидно, играют амбулакральные
ножки и другие тонкостенные придатки
амбулакральной системы, так как у ряда
форм в клетках жидкости
амбулакральной системы найден пигмент, близкий к
гемоглобину. Большинство иглокожих
весьма нуждается в кислороде и быстро
погибает в плохо вентилируемых
аквариумах. Однако органы дыхания у них
развиты слабо и химизм дыхания изучен
еще мало. Настоящие органы дыхания —
водные легкие — имеются только
у голотурий. У тех голотурий, у которых
водные легкие отсутствуют, дыхательную
функцию, очевидно, могут выполнять
ротовые щупальца, кишечник, сравнительно
тонкая кожа тела и, наконец, спинные
придатки амбулакральной системы, через
которые проникает кислород. У офиур
дыхание происходит через тонкостенные
мешковидные камеры—бурсы,
расположенные в основании лучей и периодически
заполняющиеся морской водой благодаря
сокращениям спинной стенки диска и
наполнениям и опорожнениям желудка.
Физиология дыхания иглокожих изучена
недостаточно, но вполне вероятно, что
различные системы органов могут принимать
участие в дыхательных процессах наряду
с выполнением ими других функций.
Органы пищеварения
развиты не одинаково в разных классах
иглокожих, что связано с различием в
способах питания и с различным планом
строения представителей отдельных классов.
Пищеварительная система начинается
ртом, который в зависимости от способов
питания может быть снабжен скелетом
или лишен его. У голотурий рот,
окруженный щупальцами, помогающими при
ловле добычи, переходит, как у морских
ежей и лилий, сначала в короткую экто-
дермальную глотку, а затем в длинный
узкий энтодермальный кишечник, длина
которого значительно превышает длину
тела животного. Кишечник поэтому
образует петли (рис. 127), лежащие во
вторичной полости тела и подвешенные к
стенке тела при помощи мезентериальных
перепонок. У представителей этих трех
классов можно различить передний,
средний и задний отделы кишечника. Задний
конец кишки открывается наружу
выводным (анальным) отверстием, которое у
морских лилий расположено вблизи рта,
у ежей — на стороне, противоположной
ротовой, или на краю скорлупы. У
голотурий задняя кишка расширяется в
довольно объемистую мускулистую клоаку,
которая у большинства видов открывается
наружу на заднем конце тела. У морских
звезд и офиур глотка отсутствует и рот
ведет непосредственно в объемистый
мешковидный желудок, прикрепленный к
стенке тела мезентериальными тяжами.
Желудок офиур складчатой формы, с
радиальными и интеррадиальными
выростами, заканчивается слепо, так как
офиуры не имеют ни печеночных
придатков, ни задней кишки, ни анального
отверстия. У морских звезд желудок
имеет 5 пар печеночных выпячиваний,
заходящих в лучи и обильно выделяющих
пищеварительные соки. Таким образом,
кишечный канал иглокожих сохраняет
особенности, унаследованные
современными животными от двустороннесиммет-
ричных предков. Он почти целиком
развивается из первичного кишечника и не
имеет лучистого строения. Влияние пяти-
лучевой симметрии сказывается только
на кишечнике морских звезд, обладающем
пятью парами печеночных отростков.
208
Процессы пищеварения иглокожих
изучены недостаточно. Известно, что стенки
кишечника содержат большое количество
цилиндрических клеток, выделяющих
пищеварительные соки, а также много
свободных амебовидных клеток. У них
найдены ферменты: липаза, амилаза, маль-
таза, инвертаза, трипаза и некоторые
другие, но не установлено, где различные
ферменты образуются.
Весьма мало известны и процессы
выделения иглокожими продуктов распада,
особенно процессы выделения жидких
веществ. У иглокожих нет специальных
органов выделения, а функцию
выделения выполняют многочисленные
амебоциты, находящиеся в большом числе
всюду в целомической жидкости, поэтому
выделение носит диффузный характер.
В одном случае амебоциты выбрасываются
наружу путем разрыва стенок тела,
причем избираются места с наиболее тонкими
покровами. В другом случае амебоциты
вместе с продуктами распада
откладываются в разных местах тела, образуя
крупные окрашенные массы.
Из только что изложенной
характеристики иглокожих вытекает, что их
лучистое строение — до некоторой степени
кажущееся явление, так как положение
осевого комплекса органов, строение
кишечного канала, половой системы
голотурий свидетельствует о первичной дву-
боковой симметрии этих животных.
Изучение развития иглокожих также дает
основание считать, что они произошли
от двустороннесимметричного свободножи-
вущего предка.
Развитие иглокожих всегда
сопровождается сложным превращением с
прохождением стадии пелагической двубокой
личинки. У всех классов развитие на
ранних стадиях протекает более или менее
сходно, но затем наблюдаются
значительные различия. У большинства форм раз-
Рис. 127. Типы кишечника иглокожих: А —
морской лилии; Б — голотурии; В — морской
звезды; Г — морского ежа; Д — офиуры.
I — рот; 2 — кишечник; з — клоака; 4 — анус; 5 —
целом; 6 — печеночные придатки; 7 — водные
легкие голотурий; 8 — амбулакральные ножки; 9 —
амбулакральное глоточное кольцо; 1 о — мадрепорит;
II — кровеносные сосуды; 12 — полиев пузырь; 13 —
половая железа; 14 — бурсы.
множение только половое и яйца
откладываются прямо в воду, где они
оплодотворяются и развиваются. После
дробления, которое, как правило, полное,
радиального типа, образуется типичная
бластула, покрытая ресничками. На одном
полюсе этой бластулы происходит
глубокое впячивание (инвагинация), дающее
начало энтодермальной средней кишке,
и бластула превращается в гаструлу.
Первое важное изменение в строении гаст-
рулы происходит с образованием по бокам
первичной кишки выпячиваний целоми-
ческих карманов. Порядок их
образования весьма изменчив у различных
личинок, но наиболее обычным и первичным
14 Живнь животных, т. 2
209
для них является независимое
образование трех пар боковых выпячиваний,
последующее отшнуровывание которых от
первичной кишки дает 3 пары целомиче-
ских мешочков. На этой стадии развития
личинки иглокожих более всего сходны
с гребневиками, сравнение с которыми
было сделано впервые Мечниковым
A874). В дальнейшем, в отличие от
большинства червей, членистоногих и
моллюсков, первичный рот личинки, или бла-
стопор, превращается в анальное
отверстие, а ротовое отверстие образуется
вторично путем впячивания эктодермы
брюшной стороны навстречу
изгибающемуся концу первичной кишки. Подобное
образование рта свойственно также
погонофорам, полухордовым и хордовым
животным, что сближает их с иглокожими
и позволяет отнести последних к группе
вторичноротых животных.
После смещения анального отверстия на
брюшную сторону личинка иглокожих
становится вполне двустороннесимметрич-
ной, с выпуклой спинной и вдавленной
брюшной стороной, обладает
околоротовым венчиком ресничек и тремя парами
целомических мешков. Такая стадия
развития личинок называется д и п л е у-
р о л о й и является общей для
большинства иглокожих. При дальнейшем
развитии в разных классах иглокожих
диплеурола испытывает значительные
изменения, проявляющиеся особенно в
видоизменении внешней формы и формы
околоротового венчика ресничек. Меньше
Рис. 128. Личинки иглокожих:
1 — долиолярия морских лилий; 2 — аурикулярия голотурий; з — бипинария морских
ввезд; 4 — эхиноплутеус морских ежей; 5 — офиоплутеус офиур.
всего изменяется форма диплеуролы у
голотурий. Их личиночная стадия,
называемая аурикулярией (рис. 128),
имеет овальную форму, рот, лежащий
в глубине брюшного вдавления и
окруженный одним венчиком мерцательных
ресничек, а также скелет в виде мелких
микроскопических телец. Личинки
морских звезд называются бипинариями,
офиур— офиоплутеусами, а морских
ежей—эх иноп л у теу сами. Все они
характеризуются присутствием на теле
особых шнуров, снабженных ресничками, при
помощи которых могут свободно плавать.
Личинка морских лилий — долиоля-
р и я — отличается от остальных тем, что
она лишена рта и имеет бочонкообразное
тело, опоясанное не сплошным ресничным
шнуром, а расположенным в виде пяти
ресничных колец. В дальнейшем все
лилии проходят еще стадию личинки типа
пентакринус, прикрепленной стеблем к
субстрату.
Размеры личинок иглокожих очень
незначительны, длина их менее 1 мм. Период
их жизни в планктоне весьма различен
и определяется не только видом
животного, но и окружающими условиями. Так,
некоторые личинки одного и того же вида
при размножении летом находятся в
планктоне 2—3 недели, но зимой эта
стадия может затянуться до 2—3 месяцев.
Затем, чтобы превратиться в животное,
сходное внешне со своими родителями,
личинки иглокожих претерпевают еще
большие изменения. У морских ежей,
звезд и офиур большая часть тела
личинки и ее органов разрушается и погибает.
Но у голотурий при развитии
атрофируется лишь небольшая передняя часть
личинки, а остальная часть переходит в
тело взрослой особи. Растут иглокожие
очень медленно, в течение всей жизни,
половой зрелости достигают через
несколько лет. Рассмотренные выше типы личинок
являются основными для различных
классов иглокожих, однако следует заметить,
что внешний вид личинок и способ их
превращения во взрослый организм
меняются не только от семейства к семейству,
но и от вида к виду.
Находясь в планктоне, личинки могут
относиться на громадные расстояния от
места обитания взрослых форм. Это, с
одной стороны, положительный факт,
способствующий расселению вида. С другой
стороны, как планктонные организмы,
личинки иглокожих поедаются другими
животными, но компенсируется это
производством иглокожими чрезвычайно
большого количества яиц, чем
обеспечивается выживание вида.
Очень часто иглокожие проявляют
заботу о своем потомстве
(табл. 21). В таких случаях мать
прикрывает телом отложенные яйца или
вынашивает их. Обычно молодь вынашивается
на определенных местах тела матери, реже
ползает по всему организму. У
иглокожих известны случаи живорождения,
когда зародыш развивается внутри
материнского организма, помещаясь в полости
тела, либо в половых трубках, либо в
бурсальных впячиваниях (у офиур) и
получает от него питание, необходимое для
развития. Живорождение особенно
распространено у арктических и
антарктических видов. В этом случае развитие
прямое, без стадии планктонной личинки.
Наряду с половым размножением
у иглокожих существует в отдельных
случаях и бесполое размножение, когда
тело взрослого организма делится
пополам или на несколько кусков. Каждая
половина или кусок восстанавливает
утраченные части. Такой способ
размножения наблюдается у некоторых морских
звезд, офиур и отдельных голотурий.
В известной связи с бесполым
размножением стоит очень характерное для
иглокожих явление регенерации, т. е.
способность восстановления утраченных
отдельных органов или частей тела. Очень часто
при неблагоприятных условиях или при
нападении врага иглокожие отбрасывают
(автотомируют) лучи или части тела,
выбрасывают внутренности, а иногда даже
распадаются на куски. Скорость
восстановления утраченных частей различна и
очень зависит от температуры. Так, у
тропических форм и летом регенерация
протекает быстрее, чем у арктических
и бореальных форм или в зимнее время.
Восстановление утраченных частей у
молодых особей идет быстрее, чем у
старых, однако личинки иглокожих обла-
14*
211
дают очень незначительной регенераци-
онной способностью. Это защитное
полезное приспособление в очень слабой
степени развито у морских ежей. Тело их
покрыто прочным панцирем,
предохраняющим от внешних повреждений,
поэтому морские ежи могут при
неблагоприятных условиях отбрасывать и затем
восстанавливать только педицеллярии, иглы
и амбулакральные ножки.
Относительно продолжительности
жизни иглокожих имеется мало сведений,
однако вполне вероятно, что живут они
долго. На пластинках скелета некоторых
иглокожих установлены годичные кольца
роста. По подсчетам этих колец возраст
крупных морских ежей исчисляется
35 годами. Некоторые морские звезды
только к 14 годам достигают своих
обычных размеров. Офиуры, возможно, живут
от 3 до 4 лет или немного больше.
Все иглокожие обитают исключительно
в морях и весьма чувствительны к
малейшим опреснениям воды. Они совершенно
отсутствуют в Каспийском море, в
Балтийском представлены всего 1 видом, а
в Черном — только 8 видами, зато в
Баренцевом, Карском, Чукотском и
Охотском морях составляют основную часть
донных животных. Сравнительно
немногие виды способны переносить довольно
значительные понижения солености — до
20—24 г солей на 1 кг воды B0—24°/00)
и даже иногда встречаются в эстуарных
участках; напротив, другие виды
прекрасно чувствуют себя в воде с
чрезвычайно высокой соленостью. Примером может
служить Красное море, обладающее
соленостью 43—47 г солей на 1 кг воды и
обильно населенное самыми
разнообразными иглокожими. Иглокожие живут в
морях и океанах всех широт земного
шара, на самых разнообразных грунтах и
глубинах — от приливо-отливной зоны до
максимальных глубин океанских впадин,
которые бывают глубже 10 000 м. Чаще
они приурочены к определенным
глубинам и встречаются либо исключительно
на литорали, либо в сублиторальной или
батиальной зоне, либо обитают только
в абиссальной зоне, причем в пределах
отдельных групп иглокожих видовое
разнообразие иногда увеличивается с
глубиной. Однако среди иглокожих
значительное количество видов оказывается
эврибатными1 формами, диапазон
вертикального распространения которых
иногда превышает 5—7 тыс. м. Таких видов,
которые могут встречаться на самых
различных глубинах, особенно много среди
офиур. Для ряда иглокожих были
отмечены сезонные миграции из глубин к
берегам и, наоборот, связанные с периодами
размножения, с изменениями температур.
Степень приуроченности иглокожих к
определенным грунтам различна. Многие
голотурии, морские ежи и офиуры живут
исключительно на илистом грунте или
песке, но некоторые представители этих
классов предпочитают каменистый грунт.
Однако вполне вероятно, что характер
грунта не играет существенной роли в
распространении иглокожих. Более
важную роль, очевидно, в их расселении
играет соленость и солевой состав воды,
а также глубина, течения, температура,
история развития морей и океанов и целый
ряд биологических факторов. Среди
иглокожих встречаются виды,
предпочитающие определенную температуру. Так,
арктические формы встречаются только
при температурах близких к 0° С, а
высокоарктические — исключительно при
отрицательных температурах, тогда как
арктическо-бореальные виды являются
более эвритермными, т. е. могут
переносить значительные колебания
температуры. Среди бореальных видов тоже
имеются эвритермные формы, которые даже
зимой остаются на литорали, в условиях
снежного покрова, но основная масса
видов с наступлением хюлодов уходит в
более глубокие воды. Особенно пышно
представлены иглокожие в фауне
тропических морей и прибрежных зон, что
связано с более постоянными высокими
температурами и с наиболее разнообразными
условиями обитания в этих районах по
сравнению с холодными или умеренными
морями или с большими глубинами. Тем
не менее даже и в последних двух
районах иглокожие довольно многочисленны
и нередко принимают участие в образова-
1 Эврибатность — широкое
распространение в водоеме по вертикали.
212
нии определенных сообществ, или
биоценозов, являясь руководящими формами
большинства из них. Богата иглокожими
фауна Тихого океана, тогда как фауна
Атлантического—много беднее. Некоторые
виды иглокожих имеют довольно широкое
распространение по всем океанам, другие—
обладают разорванными ареалами
(биполярные, амфибореальные и амфипацифи-
ческие виды), третьи — приурочены только
к тропическим, бореальным арктическим
или антарктическим областям. Многие
виды, роды и семейства иглокожих эндемичны
для определенных районов, они
прекрасно характеризуют как литоральные, так
и глубоководные фауны различных
областей. Поэтому зоогеографическое
значение иглокожих весьма велико.
Иглокожие играют важную роль и в
общей экономике моря, являясь
многочисленными и типичными обитателями
его. Подобно моллюскам и другим
животным, они участвуют в поддержании
определенного солевого состава морской воды,
потребляя в процессе питания большое
количество солей, особенно углекислого
кальция, из которого строится скелет.
Иглокожие обладают способностью
извлекать из воды радиоактивные вещества,
которые прежде всего накапливаются у
них в жидкости полости тела. Кроме того,
иглокожие служат пищей различным
животным. Например, треска, пикша,
камбала, зубатка и другие рыбы поедают
значительное количество офиур и морских
ежей. Морских ежей употребляет в пищу
и морская выдра—калан. Охотятся за
иглокожими и некоторые крупные моллюски
и ракообразные. С другой стороны,
многие иглокожие являются хищниками и
сами поедают различных животных.
Особенно хищными слывут морские звезды,
которые могут съедать рыбу,
превышающую размерами их тело, и часто нападают
на устричные банки и рыболовные сети.
Некоторые иглокожие используются в
пищу человеком. Среди них наиболее
важное значение имеют съедобные
голотурии — трепанги, которых насчитывается
свыше 40 видов и разновидностей.
Наиболее старыми потребителями трепангов
являются жители Китая, Японии,
островов Индо-Малайского архипелага.
Однако и в этих странах трепанги скорее
лакомое блюдо, так как высокая цена
и ограниченная производительность мест
лова препятствуют более широкому
потреблению этих животных. В Китае по
своим целебным свойствам трепанги
сравниваются с корнем женьшеня и
называются «корнем моря». Кроме голотурий,
по берегам Средиземного моря добывают
отдельные виды морских ежей, икра
которых используется в пищу. Иглокожие
являются прекрасным объектом для
экспериментальных эмбриологических
исследований, и в последние годы яйца их,
особенно морских ежей, часто использовались
в космических полетах для выявления
влияния космических и других лучей на
живые организмы. Полезными для
человека являются и ископаемые иглокожие,
так как некоторые строительные
материалы, такие, как дербоширский и бельгийский
мрамор, трахитовый известняк, состоят
главным образом из остатков данных
животных. Кроме того, ряд ископаемых
иглокожих, особенно морских ежей,
используется в качестве руководящих форм,
важных для стратиграфии.
Почти все иглокожие ведут свободный
образ жизни, паразитов среди них,
можно сказать, нет, хотя питание молоди
некоторых видов офиур на колониях
кораллов и последующее их переселение
на диск и в полость тела взрослых особей
собственного вида, а также обитание
карликовых самцов нескольких видов офиур
постоянно около ротового отверстия самки
и высасывание ими с помощью специально
заложенных в глотке мышц пищевых
соков изо рта самки, пожалуй, можно
подвести под понятие паразитизма. Зато
комменсалы среди иглокожих известны
давно. Часто офиуры поселяются между
иглами морских ежей, где, очевидно,
находят себе пищу и защиту, а
представители одного из родов тропических
офиур (рис. 129) постоянно живут на ротовой
поверхности морских лилий около,
амбулакральных борозд и пользуются пищей,
собираемой этими животными, не
причиняя хозяевам заметного вреда. Целый
ряд других животных — простейших,
гидроидов, ракообразных, моллюсков и
даже рыб — являются комменсалами
213
иглокожих и обитают прикрепившись к
иглам, поверхности тела морских ежей
или звезд либо поселяются в полости тела,
кишечнике и других органах различных
иглокожих, делясь с ними пищей и
получая от них некоторую защиту. Однако
существуют и настоящие паразиты
иглокожих. Правда, они немногочисленны, но
относятся к разным группам животных.
Очень распространенными и наиболее
древними паразитами иглокожих,
известными еще с каменноугольного периода,
являются своеобразные кольчатые черви
семейства Myzostomidae. Объектом
интенсивного заражения этими паразитами
становятся морские лилии, как более или
менее прикрепленные животные (рис. 129).
Часто в голотуриях паразитируют
некоторые моллюски, а в офиурах — различные
паразитические ракообразные и
неполовозрелая форма трематоды, являющейся
паразитом зубаток.
Среди современных иглокожих только
стебельчатые морские лилии неподвижно
прикреплены к субстрату, а все остальные
животные свободно передвигаются.
Обычно иглокожие ползают по грунту как по
горизонтальным, так и по вертикальным
поверхностям, реже зарываются в грунт,
и лишь единичные виды голотурий ведут
пелагический образ жизни и могут
встречаться в толще воды. Закапывание в грунт
распространено довольно широко во всех
группах иглокожих, кроме лилий, у
которых в связи со своеобразным способом
питания амбулакральные борозды
должны быть всегда чистыми. Большинство
форм зарывается в грунт на небольшую
глубину, оставляя часть тела или часть
лучей над грунтом. Закапывание в грунт
у многих иглокожих связано с питанием.
Такие животные заглатывают грунт без-
выборочно, причем захват грунта с
находящимися в нем питательными
веществами происходит механически вместе с
поступательным движением животного
вперед в грунте. Грунтоеды встречаются
среди многих голотурий, морских ежей
и некоторых звезд. Большинство
иглокожих питается животной пищей, но
некоторые из них питаются исключительно
прибрежными водорослями или даже
остатками наземной растительности.
По поводу происхождения
иглокожих и филогенетических
взаимоотношений внутри этого
типа до сих пор существуют только
многочисленные догадки и гипотезы. Это
вызвано до некоторой степени тем, что
появление иглокожих относится к глубокой
древности, так как в конце кембрия и
в начале силура, свыше 500 млн. лет
назад, уже имелись представители почти
всех известных нам классов иглокожих,
за исключением голотурий, достоверные
остатки которых обнаружены только с
карбона. Предполагают, что иглокожие
возникли в докембрии и наиболее древ-
Рис. 129. А — офиура Ophiomaza, живущая на морской лилии Comanthus; Б —
паразитические черви семейства Myzostomidae на морской лилии; В — мизостомида, снятая
с морской лилии, вид с брюшной стороны;
1 — лучи морской лилии; 2 — мизостомиды.
214
ние из них, вероятно, имели
слаборазвитый скелет, что не способствовало их
сохранению в ископаемом состоянии.
На основании данных эмбриологии,
сравнительной анатомии и палеонтологии
считают, что предки иглокожих, очевидно,
имели двустороннесимметричное,
вытянутое тело, снабженное пятью парами
щупалец, трубковидным кишечником и тремя
парами целомов. Рот у них находился
на передней части тела, анус — на
задней.
Эти животные вели придонный образ
жизни, который привел их к некоторой
асимметрии в строении. Наиболее
близкими к такому предку были представители
примитивных классов подтипа
пельматозоа — гетеростелей и цистидей, в
пределах которых уже был намечен переход
от двусторонней симметрии к
радиальной. Прикрепление к субстрату правой
стороной переднего конца тела
животного, которое имело место у
представителей этого подтипа, вызвало смещение
рта влево и вверх, ануса — вправо и
приобретение лучистого плана строения.
Таким образом, эволюция пельматозоа
шла в направлении развития радиальной
симметрии, и у высших представителей
этого подтипа, ряда морских лилий, у
которых наблюдается увеличение числа
каменистых каналов, она охватывает
почти всю их организацию, за исключением
кишечника и способа отхождения
приводящих бороздок от рта.
Относительно происхождения
подтипа элеутерозоа единого мнения нет.
Однако большинство зоологов считает,
что они произошли от пельматозоа, так
как в организации последних имеется
много черт, из которых можно вывести
строение первых. Эволюция подтипа
элеутерозоа шла в направлении
восстановления двусторонней симметрии в связи
с атрофией стебля, с переходом к
подвижному образу жизни и активному
захвату пищи.
Вызывает разногласия и вопрос о
происхождении и взаимоотношении классов
этого подтипа. Вполне вероятно, что
различные классы элеутерозоа не имеют
общего предка, а произошли из разных
групп пельматозоа. Среди элеутерозоа
наиболее близки между собой морские
звезды и офиуры. Первые известны с
кембрия, вторые — с силура. Из всех классов
пельматозоа ближе всего к звездам и офиу-
рам стоят эдриоастероидеи по строению
своих амбулакров. Поэтому, исходя из
данных сравнительной анатомии и
эмбриологии, предполагают, что звезды и
офиуры в качестве двух близких и
частично параллельных стволов отделились от
раннекембрийских эдриоастероидеи.
Среди современных морских звезд наиболее
примитивными считают явно
пластинчатых звезд (Phanerozonia). Какой из
отрядов современных офиур более
примитивен, пока точно не выяснено, но многие
считают, что наиболее примитивен о т-
р я д ветвистых офиур (Еuryale).
Что касается класса морских ежей,
первые представители которых известны
с раннего силура, то такие признаки, как
наличие осевого набора органов и
полового тяжа, очень сближают их со
звездами и офиурами. О сходстве их с офиу-
рами свидетельствуют морфология
личинок и некоторые биохимические свойства.
Поэтому вполне возможно, что ежи
произошли от форм, близких к ранним эдрио-
астероидеям. Однако некоторые зоологи,
в частности Мортенсен, считают,
что они имеют дифинитическое
(двойное) происхождение.
Наиболее древних ежей семейства
Bothriocidaridae связывают с цистидеями,
а остальных ежей — с эдриоастероидея-
ми. Среди современных ежей наиболее
примитивными считают отряд
эластичных ежей (Lepidocentroida),
большинство форм которых вымерло.
Голотурии стоят несколько
обособленно от других иглокожих, но наличие у
них ряда примитивных черт, например
строение половой системы,
свидетельствует о их древнем происхождении и
сходстве с цистидеями. Благодаря слабому
развитию скелета палеонтологическая
история голотурий почти неизвестна.
Многие авторы сходятся лишь в том, что
голотурии относятся к подтипу элеутерозоа,
но произошли они независимо от других
классов этого подтипа. Более
примитивными считают отряд голотурий с
древовидными щупальцами (Dendrochirota).
215
ПОДТИП СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
(PELMATOZOA)
КЛАСС МОРСКИЕ ЛИЛИИ (CRINOIDEA)
Название класса греческого
происхождения и в переводе на русский язык
означает «подобный лилиям». Действительно,
представители этого класса обладают
причудливой формой тела, напоминающей
цветок. Великолепная же пестрая или яркая
окраска большинства из них еще больше
увеличивает это сходство. Они являются
настоящим украшением подводных садов.
Живут морские лилии исключительно в
морях и океанах, прикрепившись к
подводным предметам.Одни из них—стебельчатые
лилии — всю жизнь проводят в
прикрепленном состоянии, раскачиваясь на своем
стебле. Другие — бесстебельчатые
лилии — перешли к свободному образу
жизни, лишились стебелька и приобрели
способность отрываться от субстрата и
проплывать небольшие расстояния, двигая
своими лучами, как плавниками. Однако
каждая бесстебельчатая лилия в своем
развитии проходит прикрепленную
стебельчатую стадию, что говорит о близости
обеих групп современных морских лилий.
Строение морских лилий весьма
своеобразно. Тело их имеет вид чашечки,
обращенной расширенной стороной вверх,
от которой отходят перисто
разветвленные лучи, или руки. Лучи — чрезвычайно
характерное для данного класса
образование, и все многообразие морских лилий
в значительной степени связано с
особенностями строения лучей.
И стебельчатые и бесстебельчатые
морские лилии в противоположность
остальным иглокожим направлены ротовой
(оральной) стороной вверх, а к
субстрату — противоположной, аборальной
стороной. Все они имеют прекрасно
развитый известковый скелет, состоящий из
крупных пластинок разнообразного
размера и формы, часто пронизанных
отверстиями для прохождения нервов или
каналов амбулакральной системы. Хотя
скелетные пластинки закладываются в коже
животного, они хорошо различимы
снаружи, так как у взрослых лилий их
поверхность полностью оголяется. Абораль-
ная сторона чашечки покрыта панцирем,
состоящим из двух
(моноциклическая чашечка) или трех (д и ц и-
клическая чашечка)
венчиков, чередующихся пластинок,
расположенных по радиусам и интеррадиусам
вокруг центральной (основной) пластинки
чашечки, по 5 пластинок в каждом
венчике. У стебельчатых морских лилий с
основанием чашечки, точнее с ее
центральной пластинкой, соединен гибкий
стебель, также служащий для прикрепления
животного к субстрату. Следует заметить,
что способы прикрепления морских лилий
к субстрату различны. У некоторых форм
конечная пластинка стебля расширяется
в виде диска или зацепки, у других
от основания стебля отходят небольшие
корешки, у третьих подвижные
отростки (цирры) расположены кольцами вдоль
всего стебля на некотором расстоянии
друг от друга. У бесстебельчатых лилий,
у которых от стебля остается одна
конечная пластинка, сливающаяся с
центральной пластинкой чашечки, временное
прикрепление к субстрату осуществляется
членистыми корешками (циррами),
снабженными на конце коготками. Цирры
соединены со скелетными элементами
чашечки, притом часто, как это можно
наблюдать у нашей северной лилии Heliometra
glacialis (рис. 130), центральная
пластинка чашечки разрастается и образует так
называемый центральный конус, несущий
особые ямки для прикрепления цирр.
На дне каждой такой ямки имеется
отверстие, через которое в цирру проходит
нервный ствол. Цирр может быть более ста.
216
Рис. 130. Строение морской лилии Heliometra glacialis: А — внешний вид лилии; Б —
детали чашечки и части лучей; В — ротовой диск молодой лилии (увеличено).
1 — центральный конус чашечки; 2 — цирры; 8 — места прикрепления цирр; 4 — лучи; 5 — первый
членик (позвонок) лучей; 6 — второй членик лучей; 7 — радиальная пластинка; 8 — пиннулы; 9 —
ротовое отверстие, усаженное папиллами; 10 — амбулакральные бороздки; и — анальное
возвышение; 12 — анус; 18 — саккулы; 14 — поры, ведущие в ресничные воронки; 15 — рудиментарные
пластинки ротового скелета.
Руки морских лилий тоже обладают
прекрасно развитым поддерживающим
скелетом, состоящим из отдельных
члеников, или позвонков, называемых б р а-
хиальными пластинками.
Первые из брахиальных пластинок при-
членяются к радиальным пластинкам
последнего венчика чашечки, расположенным
близ границы ротовой стороны (рис. 130).
Скелетные пластинки соединены между
собой при помощи мышц, обеспечивающих
их чрезвычайную гибкость и
подвижность. Такое сочленение позвонков лучей
заметно с наружной стороны в виде
довольно широкой косой щели между ними.
Однако в некоторых местах соединение
брахиальных пластинок происходит без
мышц, тогда границы между ними менее
заметны и представляются в виде тонкой
поперечной бороздки. Эти сочленения
называются сизигиальными, и
способность лилий обламывать свои лучи
при неблагоприятных условиях,
например при высокой температуре, недостатке
кислорода, нападении врагов, связана в
значительной степени с таким менее
прочным способом соединения позвонков.
Исследования показали, что от 75 до 90%
лилий обламывают свои лучи на сизи-
гиальных швах и сравнительно редко —
на мускульных сочленениях.
Естественная автотомия (отламывание) рук—
весьма распространенное явление среди
морских лилий, причем утраченные части
очень быстро восстанавливаются
(регенерируют). Обычно регенерированный луч
в течение некоторого времени легко
можно отличить от остальных лучей по более
светлой окраске и меньшим размерам.
Как правило, сизигиальные швы
чередуются с мускульными и встречаются
через 3—4 позвонка. Почти к каждому
позвонку луча попеременно то справа,
то слева причленяются боковые веточки—
п и н н у л ы, также состоящие из
отдельных члеников, или позвонков,
расположенных на аборальной стороне. Эти пин-
нулы и придают лучам перистый вид.
Лучи морских лилий сравнительно редко
не ветвятся и сохраняются в числе пяти.
Обычно же начиная со второй брахиаль-
ной пластинки они раздваиваются, тогда
их становится уже 10, или многократно
делятся, и тогда их число может
доходить до 200. По оральной стороне луча,
включая все его разветвления вплоть до
пиннул, проходит соответствующим
образом ветвящаяся амбулакральная
бороздка, усаженная двойным рядом
амбулакральных ножек. У основания лучей эти
борозды соединяются вместе и переходят
на оральный диск чашечки, где по
радиусам направляются к ротовому отверстию,
расположенному у большинства форм в
центре орального диска (рис. 130).
Оральный диск чашечки покрыт только мягкой
кожей и почти совершенно лишен
скелетных элементов. Кожа его бывает
пронизана многочисленными порами, которые
ведут в ресничные воронки и далее в
полость тела и служат для наполнения
амбулакральной системы водой. Ближайшие
ко рту амбулакральные ножки
превращаются в околоротовые щупальца,
снабженные чувствительными папиллами.
Первая пара пиннул, лишенная борозд,
часто заворачивается на оральную
сторону и подобно ротовым щупальцам
помогает при приеме пищи. Анальное отверстие
размещается на небольшом возвышении,
расположенном у большинства видов в
одном из интеррадиусов орального диска,
ближе к его краю. Рот морских лилий
ведет в пищевод, переходящий в
желудок, затем в кишечник, образующий одну
или несколько петель.
Пищей лилиям служат мелкие
планктонные организмы, небольшие частицы
детрита. Способ их питания весьма
примитивен по сравнению со способами питания
других иглокожих. Они питаются
пассивно. Пища доставляется в рот при помощи
амбулакральных ножек и благодаря
действию многочисленных ресничек
покровного эпителия амбулакральных борозд.
Немалую роль при этом играет слизь,
выделяемая железистыми клетками
борозд. Она обволакивает пищевые частицы,
образует пищевые комки, которые с
токами воды, вызываемыми действием
ресничек, по амбулакрам направляются в рот.
Эффективность такого метода питания в
значительной степени зависит от длины
борозд. Чем больше разветвлены лучи,
тем больше длина борозд, тем больше,
следовательно, пищи может быть достав-
218
лено в рот. Было подсчитано, что у
стебельчатой морской лилии Metacrinus го-
tundus, имеющей 56 лучей, общая длина
борозд 72 м, а у 68-лучевой тропической
Comantheria grandicalix длина борозд
может достигать 100 м.
Такая большая поверхность лилий по
сравнению с их относительно небольшими
общими размерами исключает
необходимость развития у них специальной
дыхательной системы. Дыхание лилий
осуществляется, вероятно, через кожу,
амбулакральные ножки и анальное отверстие.
Морские лилии — весьма
малоподвижные животные. Стебельчатые лилии могут
Рис. 131. Стадии развития морской лилии:
1 — долиолярия морской лилии; 2 — морская лилия
на стадии цистоидеи; 3 — стадия пентакринуса; 4 —
пиннула стебельчатой живородящей морской лилии
Phrixometrus nutrix с пентакриноидами,
развившимися в выводковой сумке.
двигать только руками, некоторые
многолучевые бесстебельчатые тропические
формы очень медленно переползают с места на
место, представители же семейства
Antedonidae могут проплывать небольшие
расстояния (до нескольких метров в один
прием). Плавают антедоны не очень
часто. Если условия позволяют, то они
несколько месяцев могут находиться на
одном месте, прикрепившись своими цир-
рами. Малая подвижность лилий делает
их доступными для заражения другими
организмами, вплоть до настоящих
паразитов. Так, червей семейства Myzosto-
midae (рис. 129) можно найти более сотни
на одной лилии, причем ведут они себя
по-разному: одни свободно ползают по
поверхности лилии, другие образуют на
пиннулах, лучах, особые вздутия, где и
помещаются, третьи становятся
настоящими внутренними паразитами.
Среди самых страшных врагов морских
лилий надо назвать мелких хищных
моллюсков семейства Melanellidae.
Ползая по лилиям, они своими хоботками
буравят твердые скелетные части, влезают
в мягкую ткань и пожирают ее. Лилии
нередко поражаются различными мелкими
ракообразными, поселяющимися либо в
пищеварительном тракте, либо в
анальном конусе, либо на диске среди цирр.
Все морские лилии раздельнополы.
Половые продукты развиваются в
ближайших к чашечке пиннулах. Очень
часто самцы выпускают сперму первыми
через особые отверстия, образующиеся
в пиннулах к моменту созревания
половых продуктов. Это стимулирует
выбрасывание яиц самками. Последние не имеют
никаких специальных половых протоков,
и яйца выводятся наружу путем разрыва
стенок пиннул. Яйца большинства видов
оплодотворяются прямо в воде. Из
оплодотворенного яйца сначала образуется
бочонкообразная личинка
долиолярия, которая имеет довольно короткую
жизнь в планктоне по сравнению с
личинками других иглокожих. Через 2 или
3 дня она опускается на дно и
прикрепляется к субстрату или к каким-нибудь
твердым предметам, включая своих
родителей. Прикрепление долиолярии
осуществляется передним концом, после чего
219
она теряет реснички и становится
неподвижной. Тело личинки начинает
удлиняться и дифференцироваться на стебелек
и чашечку, на вершине которой затем
образуется рот. Это цистоидейная
стадия личинки (рис. 131). Вскоре
чашечка обнаруживает пятилучевое строение,
по краю ротовой стороны развиваются
руки, стебель продолжает удлиняться,
разрастается прикрепительный диск, и
личинка становится похожей на
маленькую морскую лилию, раскачивающуюся
на своем стебельке. Это уже стадия пен-
такринуса. Такое ее название
связано с тем, что ранее, когда не было еще
исследовано развитие атлантической
бесстебельчатой лилии Antedon bifida,
подобные личинки принимались за
самостоятельный вид стебельчатых лилий,
названный Pentacrinus europeus. Размеры
пентакринусов сравнительно небольшие—
от 4 мм до 1 см, но в холодных
антарктических водах могут встречаться более
крупные формы, до 5 см длиной.
Дальнейшее развитие обеих групп
современных морских лилий протекает
различно. У стебельчатых морских лилий,
остающихся всю жизнь прикрепленными,
со стороны чашечки образуются все новые
и новые членики стебелька. Стебель все
более увеличивается в размерах. Он
состоит из отдельных члеников (позвонков),
расположенных один над другим,
напоминая стопку монет. Членики стебля,
соединенные между собой подвижно с
помощью мышц и пронизанные в центре
каналом, по которому проходят нервы
и другие органы, развивают у некоторых
видов боковые цирры, расположенные
вдоль всего стебля, у других — только
в его основании. Морская лилия
становится совершенно похожей на цветок.
Длина стебля современных лилий
достигает 75—90 см, а ископаемые формы были
настоящими гигантами, длиной до 21 м.
Иначе протекает развитие пентакринуса
бесстебельчатых морских лилий.
Приблизительно через месяц-полтора их чашечка
самопроизвольно отламывается от стебля
и переходит к свободному образу жизни,
а стебель постепенно отмирает.
Стебельчатые морские лилии — самые
древние животные среди современных
иглокожих, но обнаружены они в морях
сравнительно недавно. Первый их
экземпляр был найден в 1765 г. у острова
Мартиника (Атлантический океан) и описан
под названием «морская пальма». В
настоящее время известно 75 видов
живущих стебельчатых лилий,
распространенных главным образом на больших
глубинах, вплоть до 9700 м. Напротив,
бесстебельчатые морские лилии
предпочитают более мелкие воды, могут встречаться
даже на литорали, поэтому известны
зоологам значительно раньше стебельчатых.
Упоминание о средиземноморских видах
Antedon можно встретить уже в конце
XVI в. Свободноживущие морские лилии
развиты более пышно. В современных
морях их известно 540 видов,
встречающихся как в тропической области, так
и в водах Антарктики и Арктики. Однако
главная область распространения этих
животных — тропические районы
Индийского и Тихого океанов. Все современные
лилии относятся к одному отряду
членистых лилий (Articulata) и четырем
подотрядам, три из которых
объединяют стебельчатых лилий и только
один — бесстебельчатых (Comatulida).
Из числа стебельчатых лилий наиболее
известны представители подотряда
изокринида (Isocrinida). Они имеют
длинный почти пятигранный стебель, несущий
на всем своем протяжении кольца
крупных цирр, по пять цирр в каждом,
расположенных на некотором расстоянии
друг от друга. Лучи лилий сильно
разветвлены, и крона их чрезвычайно
похожа на цветок. Эти лилии почти всегда
при драгировках добывались
обломанными, поэтому способ их прикрепления к
субстрату долгое время оставался
неизвестным. Сравнительно недавно целые
экземпляры были обнаружены на
телеграфных кабелях. Оказалось, что морские
лилии этого подотряда имеют на
основании стебля незначительное расширение,
которым прикрепляются к субстрату.
Прикрепление к субстрату довольно
непрочное, лилии часто обламываются и
ведут более или менее подвижный образ
жизни, временно прикрепляясь циррами
стебля к подходящему предмету.
Удавалось наблюдать лилий, поднятых со дна
220
обломанными, у которых кольцо цирр,
ближайшее к излому, было завернуто
внутрь, т. е. находилось в положении
хватания. Большая часть видов этого
подотряда относится к роду Metacrinus,
представленному главным образом в Индо-
Малайской области. Здесь можно
встретить Metacrinus nobilis (табл. 17),
обитающего на глубинах около 250 м. Эта
лилия обладает почти белым стеблем со
светло-желтой или
красновато-оранжевой кроной.
На глубинах 145—400 м у берегов
Японии можно обнаружить другой вид —
Metacrinus interruptus. Он легко цепляет-
Рис. 132. Стебельчатые морские лилии:
I — Rbizocrinus lofotensis; 2 —- Но lop us rangii.
ся за любой предмет, так как имеет
членистые цирры, снабженные коготочками.
В наших водах можно встретить
представителей другого подотряда
стебельчатых лилий — подотряда миллери-
кринида (Millericrinida), отличающихся
более мелкими размерами, менее
разветвленными лучами, а также округлым
стеблем, несущим цирры у своего основания.
Из них прежде всего следует упомянуть
некоторые формы глубоководного рода
Bathycrinus, насчитывающего 9 видов,
распространенных на больших глубинах
тропических и умеренных вод.
В Тихом океане в районе Командорских
островов на глубине 2840 м был найден
Bathycrinus complanatus. Эта
сравнительно маленькая, длиной в несколько
сантиметров, хрупкая лилия прикрепляется
к субстрату короткими корешками,
расположенными только у основания стебля.
Остальная часть стебля лишена цирр.
Очень близок к предыдущему виду
Bathycrinus pacificus, найденный к югу от
Японии на глубинах 1650 м. Размеры его
небольшие, чашечка и лучи
бледно-желтого цвета (табл. 22).
Более крупного размера
североатлантический вид Bathycrinus carpenteri.
Длина его стебля—27 см, а рук — 3 см.
Стебель оканчивается немногими довольно
грубыми корешками, прикрепляющими
животное к субстрату. Найден батикри-
нус карпентера близ Исландии,
Гренландии, Норвегии и Шпицбергена на
глубинах 1350—2800 м.
Весьма широкое распространение в
Атлантическом океане имеет Rhizocrinus
lofotensis. Его ареал простирается от
Норвегии до Бискайского залива в восточной
части Атлантического океана и от
пролива Девиса до Флориды — в западной
части. Маленький, изящный лофотенский
ризокринус, несущий на 7-сантиметровом
тоненьком стебельке пятилучевую
(иногда 4- и 7-лучевую) головку, обладает и
большим диапазоном распространения по
глубинам от 140 до 3 тыс. м. Он
прикрепляется к субстрату, подобно
предыдущим видам, тонкими, сильно
разветвленными корешками (рис. 132).
Несколько иной способ прикрепления
у представителей других семейств милле-
221
рикринид. Например, Proisocrinusruber-
rimus, относящийся к семейству
Apiocrinidae, укрепляется на грунте при
помощи простого расширенного
основания стебля. Встречена эта лилия на
глубине 1700 м близ Филиппинских островов.
Характерной ее особенностью является
удивительно яркая алая раскраска.
Существует предположение, что эти лилии
могут обламываться и некоторое время
плавать над субстратом.
Еще более своеобразен способ
прикрепления представителя третьего
подотряда стебельчатых лилий — Cyrtocri-
nida. Единственный ныне живущий вид
этого некогда обширного подотряда —
Holopus rangi — был обнаружен в 1837 г.
в районе Карибского моря на глубине
180 м. С тех пор добыто всего около
дюжины экземпляров холопу с, найденных
в том же районе на глубинах от 10 до
180 ж. Это живое ископаемое внешне
напоминает кулак в рыцарской перчатке
(рис. 132,2). Стебель у него укорочен,
и прикрепление к субстрату
осуществляется основанием чашечки. При этом все
пластинки чашечки, возможно, и
некоторые пластинки стебля, а также первый
и второй позвонки луча сливаются вместе
и образуют трубку, нижний конец
которой расширяется, обхватывая часть скалы
и прочно прикрепляясь к ней. Таким
образом, внутренние органы и ротовой диск
лилии помещаются внутри трубковидной
чашечки. Рот открывается в центре диска
и окружен пятью крупными
треугольными пластинками. Все десять рук лилии
разного размера, с одной стороны они
крупнее, чем с другой, поэтому при
сворачивании их в виде улитки животное
приобретает кривобокий вид. Пиннулы
на руках, в отличие от других лилий,
подворачиваются внутрь, заходят друг
за друга, образуя почти непрерывную
трубку вдоль каждого луча. Питается
холопус, подобно другим лилиям,
планктонными организмами, которые
доставляются ко рту через образованные пин-
нулами трубки токами воды, вызванными
действием ресничек амбулакров.
Холопус — одна из наиболее мелких
современных лилий. Длина его самого
крупного экземпляра едва достигает 6 см.
Все 540 видов бесстебельчатых лилий
относятся к одному подотряду ко-
матулида (Comatulida). Коматулиды
ведут свободный образ жизни, они плавают
или ползают, держа ротовую поверхность
всегда вверх. Если какую-нибудь кома-
тулиду перевернуть ртом к субстрату, то
она снова быстро принимает правильное
положение. Большинство коматулид
(исключение составляют представители
семейства Comasteridae) постоянно
отрываются от опоры и некоторое время
плавают, грациозно поднимая и опуская
то одни, то другие лучи. Многолучевые
особи при плавании используют
попеременно разные секции своих лучей, пока
все руки не примут участия в движении.
Двигаются коматулиды примерно со
скоростью 5 м в минуту, делая при этом около
100 ударов, но никогда они не проплывают
сразу больших расстояний. Плавание у
них носит пульсирующий характер, т. е.
плавают они с остановками, так как
быстро утомляются и некоторое время
отдыхают. Считают, что за один прием
коматулиды проплывают не более 3 м, но
после отдыха они снова плывут, пока
не найдут подходящего места для
прикрепления . Прикрепляются коматулиды
к субстрату с помощью цирр, число,
внешний вид, длина и характер которых
весьма зависят от мест обитания
различных видов лилий. Например, живущие
на мягких илах коматулиды имеют
длинные тонкие почти прямые цирры,
способные покрыть большие пространства
грунта и обеспечивающие хорошую якорную
стоянку. Напротив, лилии, живущие на
камнях, снабжены короткими, сильно
изогнутыми циррами, крепко
обхватывающими какие-либо твердые предметы.
В движении большинства лилий цирры
участия не принимают.
Лишь немногие коматулиды
безразличны к свету, как, например, Tropiometra
carinata. Значительная же их часть
предпочитает жить в затененных местах и
избегает действия прямого солнечного
света.
Если глыбу, к которой прикреплены
лилии, перевернуть к свету, то они
довольно быстро переселяются снова на ее
нижнюю, затененную часть.
222
Самое обширное семейство данного
подотряда — семейство антедонид(Ап-
tedonidae) — насчитывает 130 видов,
относящихся к 46 родам. Встречаются анте-
дониды всюду, от литорали до 6000 м,
и весьма обычны вне тропиков. Среди них
преобладают 10-лучевые особи, тогда как
многолучевые весьма редки. Очень
известный и ранее весьма обширный род
Antedon сейчас включает только 7
европейских видов. Все эти виды очень близки
между собой и отличаются главным
образом характером лучей, длиной и
толщиной цирр и пиннул.
В Атлантическом океане у берегов
Англии, Ирландии, Франции, Португалии,
вплоть до Азорских островов на глубинах
от 5 до 450 м можно встретить Antedon
bifida (рис. 133). Эта лилия часто
прикрепляется своими короткими, сильно
изогнутыми циррами к прутьям корзин,
спущенным для ловли крабов, а у берегов
Франции в больших количествах
поселяется на корневищах и стеблях морских
водорослей. Окраска ее чрезвычайно
варьирует: наряду с интенсивно
пурпурными особями встречаются розовые,
желтые или оранжевые, а иногда и пятнистые.
Тонкие, гибкие лучи ее бывают длиной
до 12,5 см. Они очень хрупкие и при
малейшем прикосновении легко
обламываются. Подобно многим другим видам
Antedon bifida легко обламывает свои лучи
при малейшем раздражении или
опасности. Очень редко можно встретить
экземпляр, у которого имелись бы все 10 рук
в полной сохранности, почти всегда один
или несколько лучей находятся в
состоянии регенерации. Регенерационная
способность у антедон столь велика, что если
разрезать его на 2 части, то каждая
половина развивается до целого экземпляра,
а содранный с чашечки оральный диск
вскоре замещается новым, с ротовым,
анальным отверстиями и приводящими
бороздками. Регенерации не происходит
только тогда, когда от лилии отрезаны
все руки. В этом случае животное
лишается возможности питаться и погибает.
При питании антедон прочно
прикрепляется циррами к субстрату и вытягивает
свои руки с расправленными под прямым
углом пиннулами в стороны, образуя
своеобразную сеть. Способ приема пищи
этими лилиями был исследован Г и с л е-
н о м (Gislen Т.).
Гислен наблюдал в аквариуме за
североатлантическим видом Antedon petasus.
Голодные антедоны сидели с
раздвинутыми лучами, расправленными пиннулами
и чрезмерно выпрямленными
амбулакральными ножками. Как только пища
попадала в аквариум, вся лилия
приходила в активное состояние: обычно
закрытые амбулакральные борозды
раскрывались, замкнутый до этого рот становился
округлым, амбулакральные ножки
наклонялись к борозде и сбрасывали в нее
попавшую на них пищу. Стоило лишь
частицам пищи и мелким организмам
попасть в борозду, они сразу начинали
обволакиваться клейкой слизью, выделяемой
железистыми клетками борозды, и вместе
с ней благодаря движению ресничек
направлялись по бороздам в рот. Гисленом
было замечено, что на ротовом диске
антедон имелось и обратное движение
ресничек в интерамбулакрах, которое было
направлено к краю диска. Этот ресничный
поток гнал остатки пищи к краю дискау
откуда их сбрасывал и тем самым очищал
диск от загрязнений. Исследование пищи
показало, что она состояла из смеси
детрита, планктона и мелких бентосных
организмов. Встречается Antedon petasus у
берегов Норвегии, Исландии, Англии на
глубинах 20—325 м. В отличие от других
Рис. 133. Бесстебельчатая морская лилия
Antedon bifida.
223
близких видов он откладывает яйца прямо
в воду, не прикрепляя их к пиннулам рук,
как делают, например, средиземноморский
(Antedon mediterranea) и адриатический
антедон (Antedon adriatica). У обоих
видов, размножение которых начинается
весной или летом в зависимости от места
обитания, оплодотворенные яйца при
помощи слизи подвешиваются к пиннулам
самки, где находятся около 5 дней. Из яиц
вылупляется вполне развитая личинка,
обладающая пятью ресничными шнурами.
В Атлантическом океане часто
встречаются представители и другого рода ко-
матулид. Так, на илистом грунте на
глубине около 50 м у берегов Англии живет
Leptometra celtica, легко отличающаяся
по зеленой или синеватой окраске и очень
длинным, тонким «корешкам» — циррам.
Такие длинные цирры, вытянутые по
субстрату, дают лептометре возможность
жить на мягких, вязких грунтах, не
проваливаясь в них.
В наших морях весьма обычна холодно-
водная гелиометра (Heliometra glacialis).
Эта крупная десятилучевая желтоватого
цвета лилия (рис. 130) распространена на
глубинах от 10 до 1300 м во всех
арктических морях, в северной части
Атлантического океана, а также в Японском и
Охотском морях. Дальневосточные
экземпляры очень крупные, длина их лучей
может достигать 35 см; местами они
образуют настоящие заросли на глубинах от
150 до 600 м.
Такие же крупные лилии, очень близкие
к холодноводной гелиометре, живут в
Антарктике, например Florometra antarctica.
Среди антарктических лилий имеются
виды, проявляющие заботу о своем
потомстве. У лилий рода Phrixometra
эмбрионы развиваются в выводковых
камерах, причем у разных видов степень
развития эмбрионов неодинакова. Так, у
самок Phrixometra longipinna выводковые
камеры располагаются вдоль пиннул и
в них размещаются многочисленные
эмбрионы, находящиеся все на одной стадии
развития. Как только у них образуются
ресничные шнуры, они покидают
материнский организм и плавают в воде до
прохождения пентакринусной
стадии. Напротив, у другого
антарктического вида семейства Bathymet-
ridae — живородящей фриксометры
(Phrixometra nutrix) — эмбрионы в выводковых
сумках матери проходят все стадии
развития, включая пентакринусную стадию.
На самках этого вида можно видеть
маленьких пентакринусов, прикрепленных
стеблем к выводковым сумкам матери
(рис. 131). Молодь покидает материнский
организм вполне сформированной
маленькой коматулидкой.
Таблица 17. Современные иглокожие.
Морские лилии:
1 — Metacrinus nobilis.
Голотурии:
3 — Cucumaria Japonica;
4 — Trochostoma arcticum.
Морские звезды:
2 — Ceramaster patagonicus;
7 — Asterias forbesi.
Морские бжи:
5 — Rotula orbiculus;
9 — Stylocidaris affinis.
О ф и у р ы:
в — Gorgonocephalus caryi;
8 — Ophiura sarsi.
Вынашивание молоди приводит к
развитию полового диморфизма. У
представителей семейства Isometridae,
живущих в антарктических водах,
половые пиннулы самок, вынашивающих
молодь, расширяются в виде свода, а у
самцов остаются нормальными. По этим
признакам сразу можно различить пол,
например, такого вида, как Isometra vivi-
рага. В крупных сводчатых пиннулах
живородящей изометры развиваются
богатые желтком яйца вплоть до
образования личинкой ресничных шнуров. Затем
личинка выходит из выводковой камеры,
но период ее плавания чрезвычайно крат-
ковремен: она тут же оседает на цирры
взрослой особи, где и проходит
следующую, пентакринусную стадию развития.
В связи с заботой о потомстве
количество производимых яиц резко
сокращается, поэтому у антарктического вида
Notocrinus virilis в выводковых сумках
удается обнаружить всего двух-трех
эмбрионов, находящихся на одной стадии
развития. Выводковые сумки этого вида
имеют форму кармана, помещающегося в
основании пиннул. Яйца попадают в них
уже оплодотворенными, путем разрыва
стенки между яичником и выводковой
сумкой, однако способ оплодотворения
яиц остается до сих пор не выясненным.
Подобную заботу о потомстве
проявляют представители и других семейств
коматулид, но мы здесь хотим обратить
внимание только на наиболее интересные
с точки зрения их биологии или
распространения виды.
Чрезвычайно привлекательны своим
внешним видом лилии семейства
Comasteridae. Это обширнейшее семейство
насчитывает около 100 видов,
относящихся к 19 родам. Среди них преобладают
многолучевые формы с длиной рук до 20—
25 см, живущие в прибрежных водах
тропиков. Их пестрая или яркая окраска
усиливает сходство этих животных с
цветами (табл. 18—19). Представители этого
семейства отличаются от других свобод-
ноживущих лилий тем, что рот у них
сдвинут к краю диска, а анус занимает
центральное положение. Другой их
отличительной чертой являются своеобразные
ротовые пиннулы. Они длинные, состоят
из многочисленных коротких, сжатых с
боков сегментов, на верхней стороне
которых расположены зубцы, придающие
концам пиннул пилообразный внешний вид.
Это, очевидно, приспособление для
захвата или даже разрезания мелких
объектов, но наблюдений за использованием
пиннул очень мало. Гислен высказал
предположение, что комастериды благодаря
Таблица 18. Иглокожие тропических мелководий.
Морские лилии:
1 — Comatella stelligera;
2 — Pterometra pulcherrima.
Голотурии:
4 — Brandtothuria arenicola;
7 — Stichopus chloronotus;
10 — Ludwigothuria atra.
Морские звезды:
5 — Linckia laevigata;
11 — Oreaster nodosus.
Морские ежи:
6 — Heterocentrotus mammillatus;
8 — Colobocentrotus atratus.
О ф и у р ы:
3 — Ophiotrix coerulea;
9 — Ophiomastix annulosa.
15 Жизнь животных, т. 2
таким пиннулам имеют дополнительный
способ питания. Они пользуются не
только пищей, пассивно поступающей в рот
по их желобкам, но, в отличие от других
коматулид, могут активно захватывать
пильчатыми пиннулами мелких животных
и передавать их на приводящие желобки.
Это предположение согласуется еще с тем,
что амбулакральная система у комасте-
рид несколько редуцирована, а кишечник
в несколько раз длиннее, чем у прочих
бесстебельчатых лилий.
Довольно часто среди комастерид
попадаются лилии с разной длиной рук. Такие
руки разделены на передние (ловчие) и
задние (короткие), несущие половые
продукты. Подобные лилии, как Comatula
pectinata (рис. 134), прочно
прикрепляются ко дну и расправляют веером
перпендикулярно течению длинные ловчие
руки с хорошо развитыми
амбулакральными бороздками.
Комастерид очень редко замечали
плавающими, это медлительные животные.
Их жизнь наблюдал Кларк (Clark, H.)
й Торресовом проливе. Он заметил, что,
когда комастериды отрываются от
субстрата, они медленно и трудно ползут
путем вытягивания некоторых рук и
схватывания подходящего объекта вершинами
пиннул, выделяя при этом клейкий сек-
Рис. 134. Морская лилия Comatula pectinata
(вид с аборальной стороны).
рет. Затем зацепившиеся руки
сокращаются и лилия подтягивается, отталкиваясь
в то же самое время от субстрата
противоположными руками. Это ползание может
продолжаться часами со скоростью 40 м
в час, пока лилия не найдет
благоприятное место для прикрепления. Если лилия
имеет лучи разной длины, что
наблюдается и у тропической Comatula purpurea,
то более длинные руки всегда
используются для вытягивания и прикрепления
к объекту, а короткие — для
отталкивания от субстрата при подтягивании тела.
Обычно большинство комастерид
прикрепляется к грунту с помощью цирр, но
у некоторых видов, живущих на
коралловом песке, цирры редуцируются,
центральный конус чашечки превращается в
плоский пятиугольник, лежащий почти
в одной плоскости с лучами. Такие лилии,
как, например, Comatula rotolaria,
распространенная на коралловых рифах в
районе Индо-Малайского архипелага,
просто лежат на песке.
Полную редукцию цирр можно
наблюдать и у 190-лучевой Comathina schlege-
Ш, обитающей близ Филиппинских
островов.
Количество лучей у многолучевых
комастерид может варьировать у разных
экземпляров одного и того же вида.
Весьма обычная на литорали Индо-Малайского
архипелага пестрая Comatella stelligera
(табл. 18) имеет от 12 до 43 лучей.
Примечательно, что у некоторых
тропических комастерид вымет половых
продуктов связан с фазами луны. Было
замечено, что живущий на литорали южной
части Японии японский комантус (Со-
manthus japonicus) откладывает икру один
раз в год в первой половине октября,
когда луна находится в первой или
последней четверти. Вымет половых
продуктов всегда происходит вечером, самцы
первые выпускают сперму, что
стимулирует самок к откладыванию яиц. Яйца
выводятся наружу путем разрыва
наиболее тонких возвышенных мест пиннул,
причем все лучи многолучевой лилии
выпускают половые продукты одновременно.
Оплодотворенные яйца заключаются в
оболочку, часто снабженную различными
шипами, иглами и т. д. В этой мембране
226
происходит развитие яиц до стадии
личинки, снабженной ресничными шнурами.
Прекрасно раскрашенных красивых
лилий, живущих в тропиках, можно встретить
и среди других семейств бесстебельчатых
лилий. Очень красива Amphimetra discoi-
dea, широко распространенная от Японии
до Австралии на глубинах 5—35 м. Этот
представитель большого семейства
Himerometridae, насчитывающего около
50 видов, имеет 10 чрезвычайно
правильно расположенных крупных лучей,
окрашенных в коричнево-желтые тона, a Ste-
phanometra spicata (табл. 19) из семейства
Marimetridae имеет 20 лучей,
раскрашенных в красно-желтые тона.
ПОДТИП СВОБОДНОЖИВУЩИЕ
ИГЛОКОЖИЕ (ELEUTEROZOA)
ЕЛАСС МОРСКИЕ КУБЫШКИ (HOLOTHURIOIDEA)
Морскими кубышками или морскими
огурцами называют животных, тело
которых при малейшем прикосновении
сильно сжимается, после чего оно у
многих форм становится похожим на
старинную кубышку или свежий огурец. Их
известно около 900 видов.
Название «морские огурцы» дано этим
животным еще Плинием, а описание
некоторых видов принадлежит
Аристотелю, так давно эти животные
привлекали к себе внимание.
Голотурии, или морские кубышки,
не только интересны своими внешними
особенностями, яркой окраской,
занятным образом жизни и некоторыми
повадками, но и имеют довольно существенное
хозяйственное значение. Свыше 40 видов
и разновидностей голотурий используется
в пищу человеком. Съедобные голотурии,
которых называют трепангами, давно
ценились как очень питательное и
целебное блюдо, поэтому промысел их
практикуется со времен глубокой древности.
Главные промыслы трепангов
сосредоточены преимущественно в тропических
районах: в водах Индо-Малайского архипелага,
островов Тихого океана, на
Филиппинских островах, у берегов Китая и Японии.
Менее значительные промыслы ведутся
в Индийском океане, в Красном море,
у берегов Америки, в районе Африки
и Италии. У нас в дальневосточных
морях добываются два вида съедобных
голотурий, которые идут на приготовление
консервов и сушеных продуктов. В пищу
чаще употребляются голотурии в виде
бульонов и рагу и вареная кожа их,
предварительно подвергнутая длительной
обработке и сушке. Некоторые современные
европейские фирмы изготавливают из
голотурий различные консервы, пользующиеся
большим спросом. В Италии рыбаки едят
жареных голотурий, не подвергая их
предварительной сложной обработке, а жители
островов Тихого океана съедают в сыром
виде икру и водные легкие этих животных»
Добыча трепангов в Тихом океане
составляет около 10 тыс. ц в год.
Голотурии представляют собой довольно
крупных животных, средние размеры
которых бывают от 10 до 40 см. Однако среди
них встречаются и карликовые виды, едва
достигающие нескольких миллиметров, и
настоящие гиганты, длина тела которых
при сравнительно небольшом диаметре,
около 5 см, может достигать 2 м и даже
больше. По форме тела голотурии весьма
15*
227
отличаются от других классов
иглокожих. Большинство из них скорее
напоминает крупных червей, но некоторые имеют
почти цилиндрическое или
веретенообразное, а иногда шарообразное или
несколько уплощенное тело, несущее на спине
различные выросты (табл. 23).
Несмотря на такую форму тела, у
голотурий почти всегда довольно хорошо можно
определить спинную и брюшную стороны,
правда, их брюшная сторона
морфологически не соответствует таковой других
двустороннесимметричных животных.
Они фактически ползают на боку,
ротовым концом вперед, поэтому название
«брюшная» и «спинная» стороны условно,
но вполне оправдано. У многих форм
брюшная сторона бывает более или менее
сильно уплощена и приспособлена для
ползания. К брюшной стороне относится
3 радиуса и 2 интеррадиуса, поэтому ее
часто называют тривиум, а спинная
сторона, или б и в и у м, состоит из 2
радиусов и 3 интеррадиусов. Расположение
ножек на теле морских кубышек еще более
усиливает разницу между спинной и
брюшной сторонами, так как сильно
сократимые ножки тривиума, сосредоточенные на
радиусах или иногда встречающиеся и на
интеррадиусах, снабжены присосками
и служат для движения животного, тогда
как ножки бивиума часто теряют
двигательную функцию, лишаются присосок,
становятся более тонкими и несут уже
чувствительные функции.
Никакого обособления головы у
голотурий нет, хотя у некоторых форм,
особенно у глубоководных представителей
отряда боконогих голотурий
(Elasipoda), можно заметить некоторое
отделение переднего конца от остального тела,
поэтому иногда его называют головным.
Рот, лишенный каких-либо
приспособлений для размельчения пищи и замкнутый
околоротовым сфинктером, расположен
на переднем конце тела или слегка сдвинут
на брюшную сторону; анус помещается
на заднем конце. У сравнительно
немногих форм, которые зарываются в ил или
прикрепляются к скалам, рот и анальное
отверстие передвигаются на спинную
сторону, придавая животному шарообразную,
колбообразную или сводчатую форму.
Очень характерны для всех голотурий
щупальца, окружающие рот. Число их
колеблется от 10 до 30, а строение и
внешний вид определенны для каждого отряда
данного класса (рис. 135). Они бывают дре-
Рис. 135. Схема строения
ротовых щупалец различных
отрядов голотурий:
1 — древовидные щупальца (отряд
Dendrochirota); 2 — щитовидные
щупальца (отряд Asp i doc hi rota);
3 — руковидные щупальца
некоторых представителей отряда
Elasipoda; 4 — пальцевидные
щупальца (отряд Molpadonia); 5 —
перистые щупальца (отряд Apoda).
вовидно разветвленными и сравнительно
большими, охватывающими большое
водное пространство при ловле добычи, либо
более короткими, щитовидными,
напоминающими небольшие цветочки и
предназначенными главным образом для сбора
питательного материала с поверхности грунта,
либо простыми пальцевидными с разным
количеством пальцевидных отростков,
либо перистыми, помогающими при
зарывании голотурии в грунт. Все они подобно
амбулакральным ножкам связаны с
каналами водоносной системы и имеют
существенное значение не только для питания,
движения, но и для осязания, а в
отдельных случаях и дыхания.
Другой отличительной чертой морских
кубышек являются мягкие кожные
покровы большинства форм. Только
немногие представители отряда
древовиднощупальцевых голотурий (Dendro-
chirota) имеют заметный невооруженным
глазом наружный скелет в виде
пластинок, плотно прилегающих друг к другу
и образующих подобие панциря. Скелет
кожи остальных голотурий состоит из
микроскопических пластинок весьма
причудливой и удивительно красивой формы
(рис. 136). Мы можем встретить наряду
с гладкими пластинками, содержащими
небольшое количество отверстий, ажурные
корзиночки и бокалы, палочки, пряжки,
теннисные ракетки, башенки, крестики,
колесики и даже настоящие якоря. Кроме
кожи тела, известковые пластиночки
могут быть найдены в щупальцах, около-
Рис. 136. Скелетные элементы голотурий:
I—2 — части глоточного кольца древовиднощупальцевых голотурий; 3—25 — пластинки кожи: 3—8 —
древовиднощупальцевых голотурий; 9—17 — щитовиднощупальцевых голотурий, 18—19 — боконогих
голотурий; 20—21 — бочонкообразных голотурий; 22—25 — безногих голотурий.
ротовой мембране, амбулакральных
ножках, половых органах. Только у редких
видов они отсутствуют совершенно, а для
большинства весьма характерны и играют
важную роль при их определении.
Самое крупное скелетное образование
находится внутри тела животного и
окружает глотку (рис. 136). Глоточное
известковое кольцо голотурий бывает
разнообразной формы: с отростками или без
них, цельным или мозаичным и т. д.,
но, как правило, состоит из 10
кусочков, 5 из которых соответствуют
радиусам животного, а другие 5 —
интеррадиусам. У ряда форм оно служит местом
прикрепления пяти лентовидных мышц
(мускул ов-р етракторов),
втягивающих передний конец тела внутрь вместе
со щупальцами. Расправление переднего
конца тела и вытягивание щупалец
обусловливается действием других пяти
лентовидных мышц (мускулов-протракто-
р о в), прикрепленных к глоточному
кольцу рядом с ретракторами.
Мускулатура у морских кубышек
достаточно развита и усиливает прочность
их покровов. У них имеется кожно-мус-
кульный мешок, состоящий из слоя
поперечных мышц и пяти пар продольных
мышечных лент, располагающихся по
радиусам. При помощи такой сильной
мускулатуры некоторые голотурии двигаются,
зарываются в грунт и сильно сокращают
тело при малейшем раздражении.
Внутреннее строение морских
кубышек уже было рассмотрено при
характеристике типа (рис. 127, Б). Следует,
пожалуй, только обратить внимание на особое
защитное приспособление — к ю в ь е-
ровы органы, имеющиеся у
отдельных групп этих животных, затем на
специальные органы дыхания — водные
легкие.
Кювьеровы органы имеются
у различных представителей отряда
щитовиднощупальцевых голотурий (Aspido-
chirota). Они представляют собой
железистые клейкие трубковидные образования,
впадающие в расширенный задний отдел
кишечника — клоаку, и при раздражении
животного способны выбрасываться через
клоаку наружу, прилипая к
раздражающему предмету.
С клоакой общим протоком соединены
иводные легкие, отсутствующие
только у боконогих голотурий (Elasipoda)
и у некоторых форм безногих голотурий
(Apoda). Они представляют собой два
сильно разветвленных ствола,
расположенных слева и справа от клоаки и
связанных со стенкой тела и с петлями
кишечника очень тонкими мускулистыми
и соединительноткаными тяжами.
Водные легкие могут быть ярко окрашены
в оранжеватые тона и занимают
значительную часть полости тела животного.
Конечные боковые веточки легочных стволов
образуют тонкостенные ампулообразные
расширения, и довольно часто левое водное
легкое бывает опутано сетью кровеносных
сосудов. Стенки водных легких снабжены
значительно развитой мускулатурой,
расслабление которой ведет к расширению
полости легкого и втягиванию морской
воды через клоаку внутрь, а сокращение—
к выталкиванию воды из легкого. Таким
образом, благодаря ритмичным
сокращениям и расслаблениям клоаки и водных
легких морская вода заполняет
мельчайшие разветвления последних, а
растворенный в воде кислород через их тонкие
стенки проникает в жидкость полости
тела и разносится ею по всему организму.
Очень часто водные легкие становятся
местом выделения ненужных организму
веществ. Их тонкие стенки легко
разрываются, и амебоциты, нагруженные
продуктами распада, выводятся наружу.
Почти все голотурии раздельнополы,
гермафродиты очень редки, причем
встречаются они чаще среди отряда безногих
голотурий. Обычно наблюдается так
называемый протерандрический
гермафродитизм, когда
половые железы вырабатывают сначала
мужские половые элементы — сперматозоиды,
а затем женские — яйца; но имеются виды,
у которых в одной половой железе
развиваются одновременно и мужские и женские
половые продукты. Например, безногая
голотурия Labidoplax buskii, живущая в
северных районах Атлантического океана,
размножается у берегов Швеции осенью,
с октября по декабрь. В ее гермафродит-
ной половой железе в это время года
находятся одинаково зрелые и женские и муж-
230
ские половые элементы, но каждая
голотурия сначала выпускает в воду яйца,
а через день-два — сперму, или наоборот.
Выпуск половых продуктов в воду может
происходить с интервалами небольшими
порциями. По многим наблюдениям,
морские кубышки выметывают икру вечером
или ночью. Видимо, темнота является
стимулом для икрометания. Чаще
размножение происходит весной или летом и
связано с температурой, но известны виды,
у которых спелые половые продукты
могут быть найдены в течение круглого года,
однако максимальное их развитие,
например у Holothuria tubulosa,
наблюдается в августе или сентябре. Сроки
икрометания различны не только для разных
видов, но и для одного и того же вида,
если он широко распространен, по
различным морям. Так, очень часто
встречающийся в Баренцевом, Карском морях
морской огурец (Cucumaria frondosa) мечет
в этих морях икру в июне—июле, а у
берегов Англии и Норвегии в феврале—
марте.
Обычно половые продукты выводятся
в воду, где яйца оплодотворяются и
развиваются. После их дробления образуется
очень характерная свободноплавающая
личинка— аурикулярия (рис. 128).
Многие аурикулярии сравнительно
крупного размера. Так, Auricularia antarctica
имеет длину более 4 Auricularia
nudibranchiata — 6 мм. У ряда морских
кубышек личинки, прежде чем стать
похожими на взрослый организм, проходят
еще одну личиночную бочонкообразную
стадию — долиолярию, а затем уже
последнюю личиночную пятищупальцевую
стадию, называемую пентактулой.
Однако не все голотурии развиваются
таким путем. Сейчас известно свыше 30
видов морских кубышек, которые
проявляют заботу о своем потомстве и
вынашивают молодь на материнском организме
или внутри его. У таких видов,
распространенных главным образом в
антарктических водах, стадия свободноплавающей
личинки утрачивается и яйца развиваются
либо за счет большего количества желтка,
либо получая питание непосредственно от
материнского организма. В самом простом
случае яйца и молодь развиваются на
поверхности тела матери, например под
защитой разросшихся скелетных
пластинок, или в набухших кожных валиках
спины, или просто прикрепившись к пол-
зательной подошве (табл. 21). Дальнейшая
эволюция в этом направлении приводит к
образованию кожных углублений,
внутренних выводковых камер, вдающихся во
вторичную полость тела, а у ряда ветви-
стощупальцевых и безногих голотурий
наблюдается развитие молоди до поздних
стадий даже непосредственно в полости
тела. Во всех этих случаях легко можно
различить пол голотурий, тогда как
обычно это сделать почти невозможно,
так как у большинства морских кубышек
нет полового диморфизма.
Известны случаи бесполого
размножения голотурий, когда животные
делились пополам и восстанавливали
утраченные части, но такие случаи единичны.
Живут голотурии, как и все иглокожие,
исключительно в морях, но среди
остальных классов этой группы животных они
менее чувствительны к опреснениям воды.
Так, из иглокожих в малосоленом
Черном море встречаются главным образом
голотурии, а некоторые представители
безногих голотурий могут жить даже в
слабосоленой воде мангровых болот.
Морские кубышки — донные
животные, они обычно медленно ползают по
грунту с помощью амбулакральных ножек,
щупалец или мускульных сокращений
тела, реже закапываются в грунт.
Известны и случаи плавания над поверхностью
грунта, но только для очень немногих
форм, а несколько видов боконогих
голотурий семейства Pelagothuriidae
всю свою жизнь проводят плавая в воде,
правда, на довольно значительных
глубинах, являясь тем самым настоящими
пелагическими формами.
Питаются морские кубышки мелкими
животными и растениями или донным
детритом. Это сравнительно
малоподвижные животные, поэтому они почти
беззащитны против различных паразитов
и комменсалов. Разнообразные
инфузории, грегарины поселяются на
поверхности тела, в водных легких, в кишечнике,
в полости тела или даже в кровеносных
лакунах морских кубышек, получая от
231
них кров, пищу, кислород без «оплаты»
этих услуг в свою очередь. Но не только
простейшие организмы используют
голотурий в качестве объектов своего
поселения. Различные черви, моллюски,
ракообразные и даже рыбы могут быть
встречены на поверхности морских кубышек
или в полости их тела, в кишечнике, или
в полиевых пузырях, или в различных
других органах, где они иногда причиняют
существенный вред.
Голотурии разделяются на пять
отрядов: древовиднощупальцевых (Dendro-
chirota), щитовиднощупалъцевых (Aspido-
chirota), боконогих (Elasipoda), бочон-
кообразных (Molpadonia) и безногих
(Apoda).
ОТРЯД ДРЕВОВИДНОЩУПАЛЬЦЕВЫЕ
ГОЛОТУРИИ (DENDROCHIROTA)
Очень обширный и широко
распространенный отряд
древовиднощупальцевых голотурий объединяет наиболее
примитивных представителей данного
класса, обладающих лучше всего
выраженной пятилучевой симметрией,
древовидными щупальцами и амбулакральными
ножками, имеющими присоски и
служащими для движения. Основная масса
видов этого отряда обитает главным
образом в прибрежных водах и на глубинах
до 500—1000 м, и только единичные
виды найдены на настоящих
абиссальных глубинах до 4—5 тыс. м. В пределах
этого отряда можно наблюдать голотурий
с весьма разнообразной формой тела,
чрезвычайно приспособленной для обитания
в различных условиях. Например,
представители семейства морских
огурцов, живущие на поверхности песчаного
грунта, имеют червеобразное либо
удлиненно-цилиндрическое тело с ротовым
отверстием на переднем конце и анальным
на заднем. А формы, закапывающиеся
в грунт, приобретают изогнутую, U-образ-
ную форму тела, выставляя над
поверхностью ротовое и анальное отверстия.
К отряду древовиднощупальцевых
голотурий принадлежит промысловый
японский морской огурец (Cucumaria
japonica). Это крупная голотурия,
длиной до 30—40 см (табл. 17), темно-бурой
или темно-фиолетовой окраски разных
оттенков, ведет малоподвижный образ
жизни. Ее можно встретить на глубинах
от 5 до 50 м и даже до 200 м у берегов
Японии, Сахалина, Курильских
островов и в других районах Японского,
Охотского и Берингова морей. Молодые особи
японского морского огурца предпочитают
заросли водорослей и прогреваемые летом
мелководья, а более старые — большие
глубины, от 40 до 90 м, и сравнительно
открытые участки илистого или скалистого
дна. Название этой голотурии очень
соответствует ее внешнему виду. Тело ее по
форме напоминает огурец. Кожа очень
толстая. Торчащие ножки придают ей
мелкобугристый вид. Брюшная сторона
слегка приплюснута и несколько светлее
спинной. Многочисленные
амбулакральные ножки на брюшной стороне
расположены по радиусам в 2—4 ряда, а на
спинной стороне могут находиться как на
радиусах, так и на интеррадиусах. Ножки
брюшной стороны крупнее спинных,
снабжены присосками и служат для движения.
При движении животного, которое
происходит ротовым концом тела вперед, они
вытягиваются, прикрепляются к субстрату
и, сокращаясь, несколько продвигают
голотурию. Вообще амбулакральные
ножки японского морского огурца сильно
сократимы и могут полностью втягиваться
внутрь. Рот у него окружен кольцом из
10 длинных, сильно ветвящихся щупалец.
Если животное находится в покое, то
можно наблюдать, как оно медленно
расправляет свои щупальца и расставляет их
подобно сетям для ловли добычи. Но стоит
его немного потревожить, как моментально
весь передний конец тела вместе со
щупальцами втягивается внутрь благодаря
сокращению втяжных мускулов (ретрак-
торов). Спустя некоторое время, когда
опасность миновала, животное с помощью
мускулов (протракторов) расправляет
щупальца и вытягивает передний конец
тела. При участии мускулатуры
осуществляется и дыхание. Когда животное
спокойно, можно заметить, как
периодически через анальное отверстие
затягивается и выталкивается вода благодаря
232
сокращению и расслаблению мускулатуры
клоаки и водных легких. Так вместе
с морской водой поступает в организм
кислород, а затем через тонкие стенки
водных легких проникает в жидкость полости
тела и разносится ею по всему организму.
Мускулатура у японского морского
огурца столь сильно развита и сила сжатия
при сокращении настолько велика, что
покровы животного не выдерживают
давления полостной жидкости и лопаются.
Через разрывы выбрасываются наружу
ярко-оранжевые водные легкие,
зеленоватые в виде густого пучка длинных
трубок половые органы, длинный трубковпд-
ный кишечник, заполненный мелкими
организмами, детритом и песчинкамп грунта.
Иногда вместе с внутренностями выкидывается
большой, до 1 м длиной, червеобразный
брюхоногий моллюск Parenteroxenos dogeli,
паразитирующий в полости тела этой
голотурии и прикрепленный одним концом к ее
кишечнику.
Плотные мускулистые стенки тела
японского морского огурца, толщина которых
более 5 мм, и являются сырьем для
приготовления пресно-сушеной или
варено-соленой кукумарии, а также
кормовой муки и консервов. Но несмотря на
огромные запасы японского морского
огурца в наших дальневосточных морях,
специального его промысла до сих пор
не существует. Он добывается в качестве
дополнительного продукта при ловле
донных рыб, так как попадается в больших
количествах в тралах и снюрневодах.
В водах нашего Приморья на отдельных
участках за одно траление поднимали
12—15 ц этой голотурии, а близ Южного
Сахалина — 2—4 ц. Средний вес каждого
животного 350—500 г, но попадаются
экземпляры весом в 1 «г и больше.
Пойманных голотурий разрезают, очищают
от внутренностей, промывают и варят
30—40 минут в пресной или соленой воде.
Когда голотурии остынут, их сушат в
течение 9—14 дней. Хорошо высушенный
продукт содержит большое количество белков
(по данным Иванова A955), до 81,8%),
минеральные вещества, немного жира и
может длительное время храниться. В
сырых тканях японского морского огурца
обнаружены витамины В, С,
рибофлавин, тиамин. Недавно было
предложено готовить из него кормовую
муку для кормления домашних животных.
Опыты, проведенные в данном
направлении, показали, что эта кормовая мука
отличается от рыбной большим количеством
белков и минеральных веществ, поэтому
ее приготовление, если учесть большие
запасы кукумарии в наших морях, имеет
большой практический смысл.
Очень похож на японский морской
огурец североатлантический морской огурец
(Cucumaria frondosa), встречающийся на
глубине от литорали до 200 м у берегов
Норвегии, Англии, Северной Америки, в
районе Шпицбергена, в Баренцевом, Карском
морях. Эти два вида отличаются только
по форме известковых телец кожи,
остальные внешние признаки, вплоть до окраски
и образа жизни, у них очень совпадают.
Удивительно красив и чрезвычайно
приметен по своей окраске трехцветный
морской огурец (Cucumaria tricolor) х,
обнаруженный в прибрежных водах островов
Ару (Индо-Малайский архипелаг). Бочон-
кообразное тело его, длиной 15 см и
шириной 9,5 см, сине-фиолетового цвета
с пятью интенсивно алыми радиусами,
окаймленными сбоку белыми полосами
(табл. 19). Рот окружен десятью сильно
разветвленными беловато-красными
щупальцами, из которых два брюшных
немного меньше остальных.
Не менее красив Colochirus luteus
(табл. 19), встречающийся в районе Индо-
Малайского архипелага на глубине 55—
90 м. Тело его лимонно-желтое, папиллы
розовые, щупальца оранжевые, а самые
маленькие зеленоватые. Такая яркая
окраска отличает его от очень близкого
вида Colochirus quadrangularis (табл. 46),
известного из района Австралии, Индо-
Малайского архипелага, Филиппинских
островов и Южно-Китайского моря.
Способ питания, свойственный древо-
виднощупальцевым голотуриям, был
исследован благодаря наблюдению за гладким
1 Мы относим многие виды семейства Cucu-
mariidae к старому роду Cucumaria, чтобы
избежать дальнейшей путаницы, так как
предложенная Паннингом (Panning, 1949) новая система
для этого семейства содержит целый ряд ошибок,
требует проверки и доработки.
233
морским огурцом (Cucumaria lubrica),
живущим на рифах западного побережья
Калифорнии среди скоплений моллюсков.
Эта небольшая голотурия, длиной с
обыкновенную спичку или чуть меньше,
серовато-коричневого цвета, прячется среди
створок ракушек, выставив только свои
разветвленные щупальца, которыми она
улавливает мелкие планктонные организмы
и взвешенные в воде органические частицы.
Когда к клейким веточкам щупалец
прилипнет достаточное количество пищи, то
они одно за другим начинают сокращаться
и заворачиваться в глотку. Как только
щупальца целиком будут засунуты в рот,
он замкнется, а щупальца начинают
медленно вытягиваться обратно через
суженное ротовое отверстие и таким образом
освобождаться от улова. Два брюшных
щупальца, которые у многих видов
бывают короче остальных, помогают
животному сберечь пойманную добычу. Они
не только способны прикрыть рот, но и
«вытирают» вынутое изо рта щупальце,
если на нем остались пищевые частицы.
Движение щупалец происходит почти
непрерывно.
Подобный способ питания свойствен
многим представителям семейства
морских огурцов. Однако вытянутый
морской огурец (Cucumaria elongata) ведет
несколько иной образ жизни. Его можно
встретить на глубинах 4—65 м в
Средиземном море, у берегов Англии, Дании,
Швеции. Эта темно-коричневая
голотурия, длиной до 15 см, имеет изогнутое
U-образное тело (табл. 23), сильно
суживающееся к заднему концу. Она обычно
живет на илистом грунте, зарывшись
в него своим передним концом и выставив
анальный конец над поверхностью грунта.
Наблюдения за ней в аквариуме показали,
что голотурия длительное время может
жить зарытой в грунт и питаться не только
планктонными организмами и детритом,
но и заполняя кишечник непосредственно
грунтом.
Обычно зарываются в грунт близкие
«родственники» морских огурцов,
относящиеся к р о д у тионе (Thyone). Наиболее
известен среди них Thyone briareus,
обитающий в прибрежных водах
Атлантического побережья Северной Америки.
Эта зеленоватая или коричневатая
голотурия длиной в 10—12 см имеет бочон-
кообразное гибкое тело, снабженное
многочисленными заостренными ножками,
расположенными как на радиусах, так и на
интеррадиусах и создающими впечатление
волосистых покровов. Живет она обычно
на илах или песке, зарываясь в грунт
так, что над поверхностью торчат только
передний и задний концы тела.
Закапывание в грунт происходит очень медленно
и удивительно своеобразно. Путь в
субстрате прокладывается средней частью
тела, которое благодаря сокращению
мускулатуры суживается в виде клина
и втискивается в грунт. Последующее
расслабление мускулов ведет к
расширению животного и оттеснению песка
либо ила в обе стороны. Круговые
и продольные сокращения мускулатуры
чередуются до тех пор, пока вся
средняя часть тела не будет зарыта.
Закапывание продолжается 2—4 часа и
заканчивается прикреплением ножек к
твердым объектам. Для обеспечения
гибкости и подвижности не только
внешней формы тела, но и внутренних органов
при таком образе жизни большое
значение имеет известковое глоточное кольцо
этого животного, которое становится
узким, мозаичным, снабженным длинными
членистыми отростками, состоящими из
отдельных кусочков скелета.
Наблюдения над представителями рода
тионе показали, что они уклоняются от
яркого света и ведут более активный
образ жизни вечером или ночью (особенно
в зимнее время года), а в утренние часы
держат щупальца в сжатом состоянии.
Размножаются они, как большинство
древовиднощупальцевых голотурий, путем
вывода половых продуктов прямо в воду.
Период икрометания у Thyone briareus
начинается в июне. В момент вывода
половых продуктов животное вытягивается
и шевелит своими распущенными
щупальцами, помогая яйцам лучше рассеиваться.
Сначала самцы выпускают в воду сперму,
затем самки откладывают яйца. Процесс
икрометания длится около 30 минут, но
может быть затянут в зависимости от
окружающих условий до 4 часов. Опыты,
проводимые Г о л в и н (Golwin) над жи-
234
вотными этого вида, показали, что вымет
половых продуктов начинается при
повышении температуры на 4°С по сравнению
с обычной, причем икрометание чаще
происходит ночью или вечером даже в
условиях аквариума. После оплодотворения
яйца развиваются с прохождением
стадии планктонной личинки.
Но не все голотурии семейства морских
огурцов развиваются подобным образом.
Некоторые имеют прямое развитие и
проявляют заботу о своем потомстве.
Примером наиболее простого случая,
когда молодь развивается просто на теле
животного, может служить небольшая
антарктическая голотурия Cladodactyla
crocea (табл. 21). У данного гермафродит-
ного вида яйца погружаются в
утолщенную губчатую ткань на радиусах спины
и, по-видимому, придерживаются там
присасывательными дисками невытягиваю-
щихся ножек.
Дальнейшая эволюция в этом
направлении ведет к образованию специальных
выводковых камер. Такие камеры имеются
у ледяного морского огурца (Cucumaria
glacialis), очень обычного в морях
Арктики, а также в Беринговом, Охотском
и Японском морях. Эта голотурия, длиной
до 8 см, имеет в передней части тела
брюшной стороны две глубокие
мешковидные выводковые камеры, снабженные
специальными выводковыми отверстиями,
открывающимися наружу. В этих
камерах развиваются яйца, но способ их
попадания туда еще до сих пор не выяснен.
Подобные выводковые камеры имеются
у рода антарктических видов, например
Cucumaria lateralis, С. vaneyi, а также
у северотихоокеанских видов С. ijimai
Stereoderma lamperti, Y. Thyone imbri-
cata.
Еще более выраженную заботу о
потомстве можно наблюдать у маленькой
красной голотурии Pachithyone rubra,
живущей на литорали и вблизи берегов в заливе
Монтерей (Калифорния). У нее яйца
развиваются прямо в полости тела, причем
можно встретить эмбрионов на разных
стадиях развития, размером от 2 до 7 мм.
У самых крупных эмбрионов уже имеется
10 щупалец, известковое глоточное
кольцо, хорошо развитые ножки, известковые
пластинки в коже, которая, в отличие
от взрослых животных, совершенно
лишена пигмента и бесцветна. Выход
развившихся эмбрионов происходит через
разрывы в покровах матери, но взрослый
организм от этого не погибает, так как
раны очень быстро заживают.
Среди представителей семейства
морских огурцов известны случаи бесполого
размножения. Небольшая, до 4 см
длиной, голотурия Cucumaria lactea, живущая
на ракушечном грунте среди известковых
водорослей у берегов Англии и Норвегии,
может размножаться делением тела
надвое с последующим восстановлением
утраченных половинок.
Если обитатели континентального плато
семейства морских огурцов имеют
сравнительно небольшие ареалы, то
глубоководные представители этого семейства—
Abyssocucumis abyssorum и Ipsilothuria
bitentaculata — широко распространены
на больших глубинах (до 5 тыс. м)
Тихого, Индийского, а первый вид — п
Атлантического океанов. Глубоководный
морской огурец (Abyssocucumis
abyssorum) имеет обычную для морских
огурцов форму тела, но очень тонкие
прозрачные покровы и весьма редкие, мелкие
ножки, расположенные только на
радиусах. Зато жесткая голотурия
(Ipsilothuria bitentaculata) мало похожа на своих
ближайших родственников (табл. 22).
Тело ее почти шарообразно и покрыто
наподобие панциря крупными
прозрачными, снабженными шипами пластинками.
Пластинки имеют отверстия для
прохождения амбулакральных ножек,
сосредоточенных преимущественно у ротового и
анального отверстий. Внешний вид у этой
маленькой голотурии, размеры которой
не превышают 2 см, напоминает изделие
из тонкого фарфора: сквозь прозрачные
покровы просвечивают внутренности.
Спинная сторона укорочена, а ротовое и
анальное отверстия сближены и помещаются
на вершине двух небольших конусов.
Такая форма тела помогает голотурии
жить на мягких вязких грунтах больших
глубин.
Семейство листоносных
голотурий (Phyllophoridae) очень близко к
предыдущему. Листоносные голотурии отли-
235
чаются от морских огурцов главным
образом числом щупалец, которых может быть
от 12 до 30 в виде двойного или тройного
кольца вокруг рта,а также большим
количеством крупных ножек, расположенных
как на радиусах, так и на интеррадиусах.
Эти мягкие ножки, рассеянные по всему
телу, создают впечатление листьев, что
и послужило основанием для такого
названия семейства. Известно свыше 30
родов и около 100 видов листоносных
голотурий, встречающихся на умеренных
глубинах в самых различных районах
Мирового океана. Однако подавляющее
большинство видов найдено в тропических
районах Тихого и Индийского океанов.
Среди родов этого семейства наибольшим
количеством видов представлен Phyllo-
phorus, насчитывающий 15 видов,
обитающих исключительно в тропических или
субтропических районах. Наиболее
характерным видом является урнообразная голо-
турия (Phyllophorus urna), обычная на
литорали и в прибрежных водах западной
части Средиземного моря. Эта среднего
размера голотурия, длиной около 10 см,
окрашена в фиолетово-пурпурные тона
разного оттенка. Особи этого вида
живородящие, вынашивают молодь в полости
тела до очень поздних стадий развития.
В советских водах листоносные
голотурии представлены несколькими видами,
в том числе прозрачным тионидиумом
(Thyonidium pellucidum). Эта голотурия
имеет амфибореальное распространение
и встречается на глубинах от 10 до 500 м
в северных районах Атлантического и
Тихого океанов. Размеры ее 10—12 см,
окраска розовато-коричневатая или лило-
ватая. Особи этого вида предпочитают
селиться на песчаном либо илистом грунте
и обычно прикрывают себя ракушками
или другими обломками. Они очень часто
служат пищей различным рыбам,
особенно тресковым.
Среди листоносных голотурий имеется
небольшая, но очень интересная группа
видов, у которых наблюдается сильное
укорочение спины и усиленное
разрастание брюшной стороны. Форма тела
становится совершенно булавовидной или кол-
бовидной (табл. 23). На узком вытянутом
конце почти рядом лежат рот и анальное
отверстие, а между ними помещается
половое отверстие. У животного
получается 10 радиусов и 10 интеррадиусов,
так как 5 радиусов спускаются по горлу
колбообразного тела вниз и переходят
в 5 радиусов, которые поднимаются по
колбе вверх. Известно 6 видов таких
животных, обитающих у берегов Африки.
Относятся они к двум родам: Rhopa-
lodina и Rhopalodinopsis.
Западноафриканский вид Rhopalodinopsis capensis
(табл. 23) и родственные ему формы живут
зарывшись в ил, откуда выставляют в воду
только длинные «горлышки» своих тел.
Такой неподвижный образ жизни вызвал
у этих животных возврат к радиальной
симметрии, так как взаимная близость
и правильное радиальное расположение
всех десяти концов радиусов как на
выступе тела, так и на значительной части
туловища придают им радиальносиммет-
ричный облик. Однако эта радиальная
симметрия достигается иным путем, чем
у остальных иглокожих.
Немного более 12 видов, относящихся
к довольно обширному семейству
чешуйчатых голотурий (Psolidae), можно
встретить в наших водах. Эволюция в
этом семействе шла в направлении
выработки приспособлении к жизни на
твердых грунтах и скалах. Тело этих
животных приобретает отчетливый двусторон-
несимметричный облик. Оно сплющивается,
и часть брюшной стороны превращается
в плоскую ползательную подошву,
покрытую мягкой кожей, а выпуклая часть
тела покрывается крупными, сильно
разросшимися пластинками скелета,
образующими прочный свод. Спинная сторона
укорачивается, передний и задний концы тела
несколько приподнимаются над
субстратом, ротовое и анальное отверстия
смещаются на спинную сторону. У наиболее
богатого видами рода Psolus
амбулакральные ножки имеются только на полза-
тельной подошве, а у двух небольших
родов — Thyonepsolus и Psolidium—
они встречаются и на спинной стороне,
просовываясь в отверстия между
чешуйками или в чешуйках.
Наиболее известна таинственная
чешуйчатая голотурия (Psolus phantapus),
обитающая на глубинах 20—540 м в север-
236
ных районах Антарктики, в Баренцевом,
Белом, Карском морях (табл. 23).
Молодые особи плотно прикрепляются своей
подошвой к различным предметам,
а более взрослые зарываются в ил,
выставив наружу анальный и ротовой конусы.
Цвет голотурии от желто-бурого до
почти черного, длина может достигать
15 см. Тело высокое, с сильно
выступающим анальным конусом. Рот, окруженный
десятью ветвящимися щупальцами,
размещается на менее высоком конусе. Полза-
тельная подошва имеет вид
прямоугольника, по краям которого вдоль середины
расположены ножки. Чешуйки спины
покрыты густыми гранулами, поэтому
трудно различимы. Период размножения
продолжается с марта до середины июля.
Развитие сопровождается прохождением
стадии бочонкообразной личинки красного
цвета. Чешуйчатая голотурия (Psolus
chitonoides), известная из района
Командорских и Алеутских островов, побережья
Калифорнии с глубины 50—70 м (табл. 25),
отличается от предыдущего вида сильно
уплощенным телом, покрытым крупными
грубыми чешуями с утолщенным краем.
Размеры чешуи заметно уменьшаются
к ползательной подошве, имеющей почти
круглую или овальную форму. Рот и
анальное отверстие сдвинуты на спинную
сторону, но не возвышаются сильно над
остальной поверхностью. Благодаря такой
форме тела ярко-желтая с розовым
оттенком голотурия может так сильно
присасываться к подводным скалам
и другим предметам, что ее трудно
оторвать от них.
Антарктические виды чешуйчатых
голотурий очень часто проявляют заботу о
своем потомстве. Так, у антарктической
чешуйчатой голотурии (Psolus antarcticus)
на ползательной подошве можно найти до
20 штук молоди, а у самок Psolus ephippi-
fer (табл. 21) чешуйки спины сильно
разрастаются и под их защитой находят
себе убежище развивающиеся эмбрионы.
Очень маленькая створчатая
чешуйчатая голотурия (Psolus valvatus),
размеры которой едва достигают 2 см в
длину, интересна тем, что у нее известно
только бесполое размножение. Тело
делится на две части поперечным делением,
и каждая половина восстанавливает
утраченные части. Эта голотурия белого или
желтоватого цвета обнаружена только
у побережья Норвегии и западного берега
Гренландии на глубине от 20 до 500 м.
Назвали ее «створчатой» потому, что
ротовой конус при сокращенных щупальцах
прикрыт пятью крупными треугольными
пластинками — створками.
Представители двух других родов
данного семейства — Thyonepsolus и Psoli-
dium — очень небольшого размера,
длиной в 2—3 см. Большая часть их видов
обнаружена в Тихом океане и в
антарктических водах. Среди видов этих родов
наиболее примечательным является
живородящий тионепсолюс (Thyonepsolus nut-
riens). Молодь у этого вида развивается
в коже спины, причем попадает она туда
очень занятно. При икрометании самки
изгибают свои распущенные щупальца так,
что они образуют сеть, улавливающую
икринки. С помощью щупалец и двух
сильно вытянутых ножек икринки
переносятся на спину и затем действием ряда
других ножек проталкиваются к
углублениям в коже, где застревают и
развиваются. Обитают эти голотурии в
прибрежных водах Калифорнии и в районе
Галапагосских островов.
ОТРЯД ЩИТОВИДНОЩУПАЛЬЦЕВЫЕ
ГОЛОТУРИИ (ASPTDOCHIROTA)
Характерный признак представителей
данного отряда указан в самом его
названии: голотурии обладают короткими
щитовидными щупальцами (рис. 135), которые
не втягиваются внутрь, так как у них
отсутствуют мускул ы-р етракто-
р ы, вворачивающие передний конец
тела. Своим внешним видом эти
животные напоминают очень крупных червей,
а иногда толстую подошву. У многих
форм брюшная сторона заметно
отличается от спинной и приспособлена для
ползания. Щитовиднощупальцевых
голотурий можно встретить на литорали и в
прибрежных водах, а также на больших
глубинах, причем среди них имеются
настоящие глубоководные виды, относя-
237
щиеся к таким родам, как Paelopatides
и Paroriza. Эти виды живут
преимущественно на глубинах более 2 тыс. м и даже
глубже 6 тыс. м. Так, Paroriza grevei
была найдена датской экспедицией 1950—
1952 гг. на судне «Галатея» в Тихом
океане во впадине Банда на глубине 7280 м.
Среди морских кубышек отряд
щитовиднощупалъцевых наиболее богат
видами. К нему принадлежит и основная
масса съедобных животных—трепангов.
Из четырех его семейств самое обширное
и известное — семейство настоящих
голотурий (Holothuriidae). Настоящие
голотурии — обитатели теплых морей.
Они весьма обычны в тропиках и
соседних с ними районах, но отсутствуют в
наших водах. Их часто можно обнаружить
на коралловых отмелях, где они местами
образуют значительные скопления и
играют весьма существенную роль в
уплотнении кораллового песка, так как
питаются пропуская через кишечник грунт
и усваивая находящиеся в нем
органические частицы. Наблюдения, проводимые
на Бермудских островах, показали, что
животные этого семейства, обитающие на
площади около 6 км2 A,7 кв. миль),
пропускают через кишечник от 500 тыс.
до 1 млн. кг грунта в год. При этом они
заглатывают грунт без особого выбора,
сгребая его своими 20 щупальцами, и
очень медленно продвигаются вперед при
помощи мускулатуры тела и
амбулакральных ножек. Многие настоящие голотурии
имеют пеструю окраску и довольно
большие размеры. Длина крупных
экземпляров ряда видов может быть 35—45 см.
Живут эти голотурии около 10 лет. Они
малоподвижны, иногда прикрыты сверху
кусками ракушки или растительными
остатками и служат объектом для
поселения различных паразитов или
комменсалов. Очень своеобразным и весьма
обычным сожителем различных видов
щитовиднощупалъцевых голотурий является
жемчужная рыбка — Carapus. Эта маленькая
лентообразная рыбка длиной 15—-19 см
использует в качестве убежища водные
легкие различных представителей
данного отряда, в том числе и видов
семейства настоящих голотурий. Она
долго шныряет между голотуриями, пока
не найдет заднего конца животного. При
очередном дыхательном движении
голотурии карапус быстро вставляет хвост или
голову сначала в ее клоаку, откуда при
последующих дыхательных движениях
голотурии проникает все глубже и
глубже в водные легкие, пока не останется
торчать наружу одна голова. Голотурия
первоначально сопротивляется входу
рыбки, сжимает тело, замыкает анальный
сфинктер, но необходимость дыхания
заставляет ее раскрыть анальное отверстие,
чем и пользуется карапус. Если его
насильно вытащить из голотурии, то он
начинает безостановочно искать себе
новое убежище и не успокоится до тех пор,
пока не найдет его снова. Весьма
любопытно, что эта рыбка может залезать
в мертвые особи голотурий либо в
разрывы стенок тела данных животных, но
никогда не использует в качестве убежища
другие объекты, весьма сходные внешне
с ее хозяином. Покидает карапус свою
голотурию только ночью для ловли
мелких рачков, служащих ему пищей, и вскоре
снова возвращается в свое убежище.
Голотурия же, очевидно, не имеет никакой
пользы от такого сожительства, так как
очень часто крупные рыбки разрывают
водные легкие и оказываются в полости
тела голотурии.
Самым характерным родом семейства
настоящих голотурий, широко
распространенным в тропической области,
является род Holothuria. Старый род Holo-
thuria насчитывал около 120 видов,
многие из которых в настоящее время
выделены в самостоятельные близкие роды.
Приблизительно V4 часть этих видов,
главным образом родов Holothuria,
Bohadschia, Actinopyga, Microthele и др.,
употребляется в пищу человеком.
Наиболее обычной промысловой голотурией на
островах Индо-Малайского архипелага,
в Японии, на Коралловых рифах в
Австралии, на островах Тихого и Индийского
океанов, у восточных берегов Африки
является шероховатая голотурия
(Holothuria scabra). Это животное, длиной до
30 см с коричневой спиной и
зеленоватой брюшной стороной (табл. 19),
имеет многочисленные ножки,
превращенные на спине в особые чувствитель-
238
ные папиллы, придающие всей
поверхности шершавый вид, что послужило причиной
такого названия животного. Хорошо
развитые кювьеровы органы, выбрасываемые
через анальное отверстие при опасности,
защищают голотурию от врагов.
Встречаются шероховатые голотурии на
литорали или в прибрежных водах теплых
морей. Добывают их чрезвычайно
примитивным способом, собирая во время
отлива на обнажающихся рифах или
отмелях либо ловя с лодок при помощи
острог (бамбуковых палок длиной до 5 де,
снабженных зазубренными остриями).
Добыча острогой возможна только в
тихую погоду. При этом ловец, лежа на
борту лодки, высматривает через ящик
со стеклянным дном голотурий.
Замеченные животные накалываются на зубья
остроги и втаскиваются в лодку. При
таком способе лова за светлое время
суток один ловец может достать 200—
300 голотурий. Ловля с помощью
ныряльщиков распространена меньше. В пищу
употребляется кожно-мускульный мешок
голотурии, предварительно подвергнутый
варке (для удаления соли) и длительной
сушке или копчению. Из сушеного
продукта готовят различные супы и рагу.
Иногда добывают песчаную голотурию
(Brandtothuria arenicola), которая еще
более широко распространена в тропиках—
от Красного моря и Занзибара на западе
до Японии и Галапагосских островов
на востоке. Эта крупная голотурия
(табл. 18), длиной до 30 см, часто
зарывается в прибрежный песок. Ее легко
можно отличить от других видов по бело-
желтой окраске с красно-бурыми пятнами
на спине. Довольно тонкая кожа
животного делает его не очень ценным при
трепанговом промысле.
Зато весьма высоко ценятся два других
вида: Microthele nobilis и Actinopyga
echinites. Первый вид можно встретить
в тропической части Индийского и Тихого
океанов на глубинах от 1 до 18 м.
Коричневая Actinopyga echinites,
распространенная также широко в Индийском и
западной части Тихого океана, особенно
ценится на Филиппинских островах, так
как ей приписывают
кровоостанавливающие свойства.
Среди представителей семейства
настоящих голотурий имеются ядовитые виды.
В 1947 г. Смит (Smith) сообщал, что
жители Маршалльских островов
употребляют раздавленных черных голотурий
для глушения рыбы. При этом они
разрезают голотурию пополам и выдавливают
в мелкие водоемы, после чего рыбы
всплывают на поверхность. Яд черной
голотурии (Ludwigothuria atra, табл. 18)
применяется при рыбной ловле жителями
и других островов Тихого океана.
Человек невосприимчив к яду этой голотурии,
и ее промышляют на островах Тихого
океана для переработки на трепангов.
Кроме того, туземцы едят совершенно без
всякого для себя вреда рыбу,
оглушенную ядом черной голотурии.
Недавно были исследованы токсичные
свойства другого вида, Actinopyga agas-
sizi, обитающего в западной части
Атлантического океана в районе Карибского
моря. Эти красноватые или
темно-коричневые голотурии хорошо видны сквозь
толщу воды. Они медленно передвигаются
с помощью многочисленных
амбулакральных ножек по коралловому песку
прибрежных вод. Было замечено, что
помещение данных животных в аквариум с
рыбами приводило к быстрой гибели
последних. Гибель рыбы можно было вызвать
и водой, в которой ранее находились
голотурии. Выяснением токсических
свойств Actinopyga agassizi много
занимался Нигрелли (Nigrelli, 1955). Было
выяснено, что кожа и внутренности этой
голотурии выделяют слабый яд, а
кювьеровы органы — весьма сильный. Одна
унция B7 г) экстракта из кювьеровых
органов, разведенная в 750 галлонах
A галлон равен 3,8 л) морской воды,
убивала рыбу за 23 минуты. Этот яд был
назван голотурии н. К голотурпину
оказались чувствительными не только рыбы,
но и другие животные,в том числе
различные простейшие, моллюски и некоторые
растения. Сейчас ведутся работы по
выяснению возможности применения голоту-
риина в медицине.
Промысловых голотурий можно
встретить и среди видов семейства стихоподид
(Stichopodidae). Так, Thelenota ananas,
одна из наиболее крупных голотурий
239
отряда щитовиднощупальцевых, считается
самым ценным видом и перерабатывается
на трепанг на островах Индо-Малайского
архипелага, в Австралии, Японии, на
Филиппинских островах.
В этих же районах, а также в
Индийском океане добывают Stichopus chlorono-
tus (табл. 18), S. variegatus и другие
виды семейства стихоподид.
Наша промысловая голотурия —
дальневосточный трепанг (Stichopus japonicus) —
тоже относится к данному семейству
(табл. 25). Тело дальневосточного
трепанга червеобразной формы с хорошо
различимыми брюшной и спинной сторонами,
но при раздражении оно сильно
сжимается и становится почти шарообразным.
Окраска трепанга отчасти зависит от
мест обитания и варьирует от
темно-зеленой до темно-коричневой. Наиболее
ценные голубые трепанги попадаются очень
редко. Считают, что трепанг, обитающий
преимущественно на скалистых грунтах
и в более глубоких местах,
темно-коричневый, а особи, живущие на песчаных или
илисто-песчаных мелководьях, более
светлые с зеленым оттенком. Как правило,
у всех трепангов брюшная сторона
значительно светлее спинной. Она снабжена
амбулакральными ножками,
расположенными на радиусах, спинная же сторона
несет различной величины мягкие
конусовидные выросты — спинные папиллы,
цвет которых на концах совершенно белый.
Рот животного несколько сдвинут на
брюшную сторону и окружен кольцом
из 18—20 щупалец. Живут
дальневосточные трепанги на небольших глубинах, от
1 до 60 ле, в защищенных от штормов
бухтах и заливах. Очень хорошее
исследование биологии дальневосточного трепанга
было проведено А. И. Савиловым.
По его наблюдениям, выполненным в
водолазном костюме, трепанги в тихую погоду
массами выползают на илисто-песчаные
площадки, расположенные рядом с
каменистыми россыпями или с зарослями
морской травы или водорослей и питаются,
собирая с помощью щупалец самый
поверхностный слой грунта, богатый различными
мелкими организмами. На твердых
грунтах среди камней и в зарослях морской
травы или водорослей они находят себе
убежище во время штормов и летних
спячек. Часто трепанги образуют большие
скопления, называемые «трепанговыми
полями». Ареал дальневосточного
трепанга простирается от вод Южного
Сахалина и Курильских островов на юг до
Сянгана. Особенно обилен он в
прибрежных районах Сахалина, Приморья, залива
Петра Великого, Японии и в Желтом море.
Таблица 19. Иглокожие тропических мелководий.
Морские лилии:
1 — Stephanometra spicata;
2 — Lamprometra protectus
Голотурии:
4 — Holothuria scabra;
7 — Thyone mirabilis;
10 — Cucumaria tricolor;
11 — Colochirus luteus.
Морские звезды:
5 — Acanthaster planci;
6 — Luidia quinaria;
12 — Culcita novae-guineae.
Морские ежи:
8 — Toxopneustes pileolus;
9 — Phyllacanthus imperialis;
13 — Diadema setosum.
О ф и у р а:
3 — Opliiothrichoides pulcherrima.
Запасы его в наших водах изучались
3 а к с о м, который подсчитал, что
общий сырьевой запас трепанга в заливе
Петра Великого примерно составляет
150 млн. экземпляров.
Следует добавить, что трепанг обладает
огромной плодовитостью: количество яиц
в половых железах одной особи достигает
2 млн. Трепанги раздельнополы, но
самцы и самки внешне совершенно
неразличимы. С повышением температуры в
прибрежных участках моря до 12—18°С
трепанги приступают к размножению,
причем старые и более крупные особи
несколько раньше молодых. По данным С а-
в и л о в а, вымет половых продуктов
в заливе Петра Великого начинается
в начале июля и происходит небольшими
порциями в течение одного — трех дней.
Сразу после этого голотурии прячутся
в убежище среди скал, водорослей, в
пустые раковины моллюсков, перестают есть
и впадают в «спячку». Это состояние
оцепенения у них продолжается около месяца,
после чего сильно ослабевшие голотурии
выползают из своих убежищ и начинают
усиленно питаться. Личинки же к этому
времени успевают развиться в маленьких
голотурий длиной до 2—4 см. Рост и
питание голотурий продолжается всю зиму,
и к концу первого года жизни они уже
бывают до 15—20 см в длину. К концу
второго года жизни молодь невозможно
отличить от взрослых особей. Живут
дальневосточные трепанги около 5 лет,
размножаться начинают на третьем году
жизни, зиму проводят в тех же самых
местах, что и лето.
Основным способом лова трепангов
в наших водах является водолазный
промысел. Водолаз двигается по дну и при
помощи острого крючка собирает
трепангов в подвешенную к поясу сетку.
Обычно водолазы добывают
одновременно и трепанга и гребешка. Каждый
водолаз работает не более двух часов.
Водолазы ловят трепанга на самых
разнообразных грунтах, которые бывают
недоступны для лова драгой. В среднем в
водах Приморья добывалось по 5400 ц
дальневосточного трепанга в год.
Промысел трепангов связан в значительной
степени с особенностями их жизни. У нас
в заливе Петра Великого он начинается
в марте и продолжается до июля, затем
снова возобновляется с конца августа
и длится до начала ноября. Пойманных
трепангов подвергают очень длительной
обработке (чистке, варке, сушке и т. д.),
на которую уходит почти 2—3 недели.
Только после этого получают продукт,
годный к длительному хранению и тран-
Таблица 20. Иглокожие северных морей СССР.
Морская лилия:
1 — Heliometra glacialis.
Морские звезды:
2 — Asterias rubens;
3 — Hippasteria phrygiana;
4 — Urasterias lincki;
7 — Ctenodiscus crispatus;
9 — Henricia sanguinolenta.
Морской еж:
8 — Strongylocentrotus
droebachiensis.
О ф и у р ы:
5 — Ophiopleura borealis;
6 — Ohiopholis aculeata.
16 Жизнь животных, т. 2
спортировке. Всякие разнообразные блюда
готовят уже из варено-сушеного трепанга.
Если представители двух первых семейств
отряда щитовиднощупальцевых
голотурий были распространены главным
образом на литорали или в прибрежных водах,
то виды третьего семейства Synal-
lactidae проникают уже в абиссальные
глубины. В 1950 г. экспедиция Института
океанологии АН СССР на
экспедиционном судне «Витязь» обнаружила в
Беринговом море на глубинах 2220—2400 м
интересных голотурий, относящихся
к роду Paelopatides. Крупные экземпляры
этого вида, длиной до 20 см, были
окрашены в темно-фиолетовый цвет. Довольно
плоское их тело с хорошо различимой
спиной и брюшной стороной напоминало
толстую подошву, поэтому вид назван
подошвообразным пелопатидесом
(Paelopatides solea, табл. 22). Paelopatides
в свою очередь в переводе на русский
язык означает «бродящий по илу».
Действительно, животных этого рода,
известных как в Тихом, так и в Атлантическом
океане, всегда находили только на
илистых грунтах. Однако «бродить» по вязкому
илу весьма трудно, поэтому у этих
голотурий в процессе эволюции выработались
интересные приспособления,
позволяющие им жить в таких условиях. У всех
у них внутренний слой кожи сильно
утолщается и принимает вид студенистой
массы, ножки редуцируются, а на границе
между спинной и брюшной сторонами
развивается своеобразная складка кожи
(кайма). Кайма имеет волнистый вид
и образуется видоизмененными
амбулакральными ножками боковых радиусов
брюшной стороны. Она немного лучше
развита в задней и передней части тела,
чем с боков, и нависает над ротовым
отверстием, сдвинутым на брюшную сторону.
Вполне вероятно, что при помощи этой
складки животные могут некоторое время
плавать над грунтом и таким образов
передвигаться на мягких вязких илах. Такое
предположение еще больше
подтверждается тем, что у плавающего представителя
данного отряда, Galatheathuria aspera,
обнаруженного в Южно-Китайском море
и Бенгальском заливе, имеется подобная
складка кожи, позволяющая ему плавать.
Внешне Galatheathuria aspera немного
напоминает подошвообразного пелопати-
деса. Это крупная фиолетового цвета
голотурия, длиной до 20 см, с хорошо
развитой краевой складкой и заметно
обособленной передней частью тела. Голотурия
прекрасно приспособлена к плаванию и
может плавать длительное время,
удаляясь на значительные расстояния от
грунта. Как сообщает Хансен (Hansen В.),
пойманные голотурии продолжали плавать
в сосуде с водой, двигая своей боковой
кожной складкой. Однако они не перешли
к обитанию исключительно в толще воды,
как это сделали некоторые формы боконо-
гих голотурий. Голотурии могут
опускаться на дно и отдыхать на грунте.
Изучение содержимого их кишечника
показало, что они питаются как донным
материалом, так и пелагическими видами.
Добыты они в толще воды на больших
глубинах, около 3—4 тыс. м.
Среди представителей четвертого с е-
мейства — Gephyrothuriidae —
наибольший интерес представляют виды
рода Pseudostichopus. Их
цилиндрическое тело почти всегда покрыто иглами
губок и других посторонних частиц,
придающих животному совершенно
несвойственный им «мохнатый» вид.У нас в
Японском море на глубине свыше 1000 м
встречается Pseudostichopus trachus,
15—18-сантиметровое тело которого густо покрыто
песчинками и иглами губок, торчащими
перпендикулярно к поверхности тела
животного и глубоко внедряющимися в его
кожу. Вначале даже бывает трудно
решить, что это за животное перед вами.
Такая удивительная способность
обволакивать свое тело посторонними частицами,
по мнению Мортенсена, является
защитным приспособлением против рыб
и других животных, поедающих голотурий.
ОТРЯД БОКОНОГИЕ ГОЛОТУРИИ
(ELASIPODA)
Боконогие голотурии во многих
отношениях весьма интересные животные,
среди которых имеются и подлинно
глубоководные виды, не встречающиеся на глу-
242
бинах менее 2 тыс. м, и настоящие
пелагические формы, всю жизнь плавающие
в воде.
У представителей этого отряда
особенно ясно выражена двусторонняя симметрия
тела. У них брюшная сторона сильно
уплощена и заметно отличается от более
выпуклой спинной стороны. Рот, окруженный
короткими щитовидными, не
втягивающимися внутрь щупальцами, сдвинут на
брюшную сторону, анальное отверстие — также.
Крупные редко расположенные
амбулакральные ножки имеются на боках тела,
а на спине они либо редуцированы, либо
превращены в сложные выросты,
направленные вперед или назад. Эти «хвосты»,
«антенны», «парусы», «плавники» еще
сильнее подчеркивают двустороннюю
симметрию животных и придают им нередко
причудливую внешность (табл. 22, 23).
Отсюда и удивительные названия этих
голотурий, например: Deima в переводе
означает «страшилище», Oneirophanta—
«фантастическая», Scotoplanes —
«блуждающий во тьме», Psychropotes —
«пьющий холодную воду» и т. д.
К данному отряду относятся 5
семейств, распространенных главным
образом на больших глубинах, поэтому
об образе жизни их представителей
известно очень мало.
К настоящим обитателям больших
глубин (свыше 2—4 тыс. м) относятся виды
семейства Psychropotidae. Один из
них, Psychropotes raripes (табл. 22),
предпочитает глубины свыше 3—4 тыс. м.
Он найден близ Курильских островов
и в восточной части Тихого океана в районе
Галапагосских островов. Подобно многим
глубоководным животным этот психропо-
тес имеет пурпурную окраску тела. Его
довольно легко отличить от других видов
по длинному хвостовому придатку,
образовавшемуся за счет разрастания папилл
спины, окруженных складкой кожи, а
также по воротнику, прикрывающему
щупальца. Воротник представляет собой тоже
складку кожи, окружающую
видоизмененные спинные папиллы.
На больших глубинах океанов и морей
распространены виды другого
глубоководного семейства — Elpidiidae, самый
обширный род которого — Peniagone—
насчитывает около 30 видов,
встречающихся на глубинах свыше 3 тыс. м
в Атлантическом, Тихом и Индийском
океанах. Всех их перечислить невозможно.
В качестве примера можно привести
голотурию с характерным парусом в
передней части тела — Peniagone lugubris,
обнаруженную в Атлантическом океане
на глубинах более 5 тыс. м (табл. 23).
Своеобразным местообитанием
отличается полярная эльпидия (Elpidia glacialis).
В отличие от других представителей
глубоководного рода эльпидия она
населяет арктические моря, где по склонам
желобов может подниматься на
чрезвычайно малые для глубоководных видов
глубины, равные 60—70 м. Тело
полярной элъпидии овальной формы, с довольно
прозрачной, но плотной кожей, густо
набитой мелкими скелетными
пластинками в виде разветвленных
палочек и колесиков. По бокам тела
имеется 4 пары крупных невтяжных ножек,
а на спине 3—4 пары более мелких
папилл. Рот загнут на брюшную сторону
и окружен кольцом из 10 щупалец.
Удивительно своеобразно устроены ротовые
щупальца этих голотурий. По форме они
напоминают ладонь с пальцеобразными
отростками (рис. 135,5), число которых
сильно варьирует. Размеры особей этого
вида небольшие, крупные экземпляры
едва достигают 2 см в длину.
В Тихом океане на больших глубинах
живут очень близкие к полярной эльпидии
формы, которые долгое время считались
тем же самым видом. Однако последние
исследования показали, что это близкие
подвиды или вполне самостоятельные виды,
отличающиеся размерами, расположением
и числом спинных папилл, формой тела
и очертанием скелетных пластинок кожи.
Они приурочены к определенным
желобам и впадинам Тихого океана. Так,
во впадине Кермадек на глубине 6620—
8300 м обнаружена Elpidia glacialis ker-
madecensis, в Ново-Британской впадине на
глубине 8940 м — Elpidia glacialis solo-
monensis, в Курильской впадине — свой
самостоятельный вид Elpidia sp. (табл. 22)
и т. д. Эльпидия из Курильской впадины
с глубины 8100 м отличается крупными
размерами, иногда превышающими 5 см.
16*
243
Внешне весьма сходны с эльпидиями
представители рода Scotoplanes. Они
тоже имеют овальное с плоской брюшной
стороной тело, несколько пар крупных
боковых ножек и три пары спинных папилл.
Длинные спинные папиллы очень
характерны для данных животных и отличают
их от близких форм. На больших
глубинах в Беринговом море была обнаружена
Scotoplanes murrayi (табл. 22), известная
ранее в южной части Индийского океана
и у берегов Перу.
Среди видов третьего семейства —
Deimatidae — имеются эврибатные формы,
встречающиеся как на глубинах менее
1000 м и даже менее 500 ж, так и на
больших глубинах, свыше Зтыс. м. Примером
широкого распространения по глубинам
может быть Pannychia moseleyi,
найденная на глубинах от 210 до 2600 м. Эта
довольно крупная голотурия (табл. 23)
известна как в западной, так и в
восточной части Тихого океана и в различных
районах своего обширного ареала
представлена самостоятельными
географическими расами. Так, у берегов Америки,
от Калифорнии до Галапагосских
островов, обитают типичные представители
вида; в Беринговом море, в районе
Алеутских островов, у северных берегов Японии
и о. Сахалин встречается подвид
virgulifera, а в Японском море —
подвид mollis. Внешне подвиды мало
отличаются друг от друга, основное их
различие заключается в строении скелетных
элементов кожи. Средняя величина
Pannychia moseleyi около 15—20 см,
внутренний слой кожи желатинообразной
консистенции, спина несет длинные отростки.
Еще более длинные и многочисленные
спинные папиллы, придающие животному
совершенно фантастический вид,
встречаются у видов рода Oneirophanta. Они
так и называются «фантастическими»
голотуриями. Несколько видов этого рода
обнаружено во всех океанах, включая
антарктические воды, за исключением Арктики.
В Атлантическом океане, между
Азорскими островами и Бискайским заливом,
на глубинах свыше 4 тыс. м
распространена Oneirophanta alternata, наиболее
крупные экземпляры которой могут
достигать в длину 12 см. Тело этой
голотурии удлиненно-овальное, с выпуклой
спинной стороной, несущей два ряда длинных
отростков (табл. 23).Более крупные
спинные отростки чередуются с несколько
более мелкими, что является основным
отличительным признаком данного вида.
Длина наиболее крупных спинных папилл
достигает 5—7 см. Цвет живых
экземпляров почти белый.
Представители четвертого семейства
боконогих голотурий — семейства
Laetmogonidae — не встречаются глубже
4 тыс. м. Тело их скорее
удлиненно-цилиндрическое, с хорошо развитыми
амбулакральными ножками, расположенными на
боковых радиусах брюшной стороны. Они
распространены в Тихом, Индийском и
Атлантическом океанах и особого интереса с
точки зрения своего строения или образа
жизни не представляют.
Наиболее интересны с этой точки
зрения виды семейства пелагических
голотурий (Pelagothuriidae). Форма тела
их мало напоминает морских кубышек
и чрезвычайно приспособлена для
плавания. По своему внешнему виду они скорее
похожи на маленьких медузок, так как
прежде всего у них заметен хорошо
развитый студенистый плавательный диск.
Это удивительно изящные и нежные
животные, часто окрашенные в розоватые
или фиолетовые тона. Всю жизнь они
проводят в толще воды, плавая всегда
ротовой стороной вверх. Было описано
восемь видов этих интересных голотурий,
обитающих главным образом в глубоких
слоях толщи воды, но исследования X е-
д и н г a (Hebing) показали, что число их
следует сократить до трех.
Общий план строения этих животных
очень сходен, все они похожи на Pelago-
thuria natatrix (табл. 23), известную
в тропических районах Тихого океана.
Мягкое студенистое тело голотурии
совершенно лишено всяких скелетных
элементов, что уменьшает вес тела. Вокруг рта,
окруженного 12—16 короткими
щупальцами, развит мощный плавательный диск,
образованный разрастанием стенки тела.
К наружному краю плавательный диск
становится все тоньше и несет довольно
длинные отростки, число которых равно
количеству щупалец. Каждый такой отрос-
244
ток имеет канал, соединенный с
амбулакральным каналом щупальца. Питаются
пелаготурии мелкими планктонными
организмами, например радиоляриями,
диатомовыми водорослями.
ОТРЯД БОЧОНКООБРАЗНЫЕ
ГОЛОТУРИИ (MOLPADONIA)
Как интересную особенность этих
голотурий следует отметить их
приспособление к жизни в грунте. Тело их бочонко-
образной либо веретенообразной формы,
с вытянутым в виде короткого или
длинного хвоста задним концом, лишено
амбулакральных ножек и других неровностей
поверхности, мешающих животному
закапываться в грунт. В качестве единственных
остатков различных придатков водоносной
системы у них сохраняются щупальца и
небольшое число папилл около анального
отверстия. Щупальца сильно укорочены,
снабжены часто пальцеобразными
отростками или парными перистыми выростами
(рис. 135,4) и приспособлены для
разрывания грунта. Число их чаще равно 15.
Они окружают рот, расположенный на
дисковидном переднем конце тела. В коже
этих голотурий наряду со скелетными
пластинками в виде различных якорей,
башнеобразных тел, пластинок с прямыми
или крючкообразными отростками
встречаются слоистые, округлой формы,
красного либо бурого цвета тельца,
содержащие фосфор.
Живут бочонкообразные голотурии на
илистых грунтах и питаются, заглатывая
грунт с находящимися в нем питательными
веществами. При этом они разгребают
грунт щупальцами и благодаря круговым
сокращениям тела продвигаются вперед
под некоторым углом к поверхности.
Скорость такого продвижения достигает
1 см в минуту, и через 10—20 минут
голотурия целиком закапывается в ил.
Но для дыхания животным нужна чистая
вода, поэтому они выставляют над
поверхностью грунта хвост, на конце
которого находится анальное отверстие. Через
это отверстие в их развитые водные
легкие поступает вода, богатая кислородом.
До сих пор во взглядах на
систематическое положение отдельных групп этого
отряда нет полной согласованности.
Многие виды и роды трудно различимы между
собой, их часто смешивают друг с другом,
поэтому в настоящее время почти
невозможно себе представить более или менее
точную картину распространения целого
ряда форм. По лучше обоснованной, на
наш взгляд, теории к этому отряду
принадлежат три семейства, наиболее
характерным из которых является
семейство Molpadiidae, куда относится и
широко известный вид Molpadia musculus
(табл. 23). Его красновато-коричневое
с фиолетовым оттенком тело заканчивается
длинным хвостом. Ареал его охватывает
все тропики, а также значительную часть
бореальных областей северного и южного
полушарий.
В Норвежском, Баренцевом и Карском
морях встречается крупная розоватого или
беловатого цвета арктическая трохостома
(Trochostoma arcticum), у которой тело
сигарообразной формы с коротким слабо
обособленным хвостом (табл. 17).
В арктической области можно встретить
и представителей семейства Eupyr-
gidae. Это все мелкие формы с бочонкообраз-
ным телом и коротким хвостом. Их
каменистый канал сообщается с внешней средой
через тонкий поровый канал,
открывающийся наружу близ полового отверстия.
Широко распространенный в Арктике
Eupyrgus scaber имеет в длину всего 1 см.
Представители семейства Caudi-
nidae отличаются очень длинным, но не
слишком резко обособленным от тела
хвостом. Длина хвоста может составлять
почти половину длины тела. Часто в
штормовых выбросах в Японском и Желтом
морях попадается розоватого цвета
голотурия Paracaudina ransonetii, длина
которой может достигать 14 см.
ОТРЯД БЕЗНОГИЕ ГОЛОТУРИИ
(APODA)
Характерной особенностью
представителей этого отряда является полное
отсутствие у них амбулакральных ножек.
В связи с этим у них исчезают и радиаль-
245
ные каналы водоносной системы.
Единственным придатком амбулакральной
системы остаются щупальца, которые
сообщаются непосредственно с кольцевым
каналом. Число и форма их различны. Они
бывают простыми, пальцевидными,
перистыми (рис. 135). Тело безногих
голотурий почти червеобразной формы. Кожа
тонкая, довольно прозрачная, но
шершавая или бородавчатая из-за скоплений
известковых телец. Последние удивительно
своеобразны и изящны. Это либо
колесики с разным числом спиц, либо сигмоиды,
либо настоящие якоря,соединенные с
якорными пластинками.
Крупные якоря помогают при
движении животного: располагаясь косо в коже
и тесно примыкая к якорным пластинкам,
они создают выступы, на которые
опираются голотурии, как змеи на концы
своих ребер.
Дыхание у безногих голотурий
осуществляется через тонкие кожные покровы,
водные легкие у них отсутствуют. Зато
имеются специальные органы выделения —
ресничные воронки,
образующиеся за счет выпячиваний ресничного
перитониального эпителия и
соединительной ткани. Они имеют вид урн либо
воронок и располагаются на стенке тела или
на мезентерии. В них накапливаются
твердые продукты распада, которые затем
и выводятся из организма.
Безногие голотурии лучше других
иглокожих переносят опреснение воды.
Некоторые тропические виды живут даже
в слабосоленой воде мангровых болот.
Многие виды зарываются в грунт или
прячутся под камнями, некоторые
тропические формы живут среди кораллов.
Коралловые виды имеют яркую
раскраску, чем сильно отличаются от своих
закапывающихся в грунт родственников,
которые окрашены в бледные тона или
почти бесцветны.
Этот отряд включает три семейства,
различающихся строением скелетных
элементов. Самое большое из них—
семейство синаптид (Synaptidae). Виды его
распространены по всем морям, но больше
в тропических водах.
К этому семейству относятся самые
крупные и самые мелкие формы иглокожих.
Чрезвычайно крупная пятнистая си-
напта (Synapta maculata) может быть
длиной до 200 см при максимальном
диаметре в 5 см. Она живет на коралловых
рифах и отличается от прочих
голотурий весьма быстрыми движениями.
Название голотурии —«пятнистая»— связано
с тем, что на зеленом или
голубовато-сером фоне ее покровов выделяются
светлые или темные пятна, часто
расположенные рядами.
В противоположность этому гиганту
малая лептосинапта (Leptosynapta minuta),
найденная в Северном море, в 400 раз
меньше. Ее длина едва достигает половины
сантиметра. Она интересна еще тем, что
вынашивает молодь в полости своего тела.
Большинство синаптид живет в грунте;
если их вытащить из нор, то они быстро
забираются туда снова. К бурильщикам
грунта относится и цепляющаяся голотурия
(Leptosynapta inhaerens),
распространенная от берегов Норвегии до Средиземного
моря. Она большую часть жизни проводит
зарывшись в грунт. Закапывание, с одной
стороны, представляет собой защитное
приспособление, с другой — является
способом добывания пищи.
Осуществляется оно при помощи щупалец и
мускульных сокращений тела. Если голотурию
положить на дно, то она сначала
погружает в грунт щупальца и делает
углубление, затем при помощи мускульных
сокращений тела расширяет его и таким
способом втискивается в грунт. При этом
якорьки кожи цепляются за неровности
субстрата и предохраняют животное от
скольжения назад при мускульных
сокращениях тела. Закапывание происходит
довольно быстро: через 5—10 минут
животное может скрыться в песке.
Проделанные в песке ходы предохраняются от
осыпания слизью, выделяемой
железистыми клетками голотурии. Цепляющиеся
голотурии достигают в длину 30 см.
Сквозь их розовую прозрачную кожу
просвечивает совершенно прямой
кишечник и 5 мышечных лент. Размножается
этот гермафродитный вид у берегов
Англии весной либо летом. Яйца очень
мелкие, развитие у них прямое без стадии
пелагической личинки. Особи вида
чрезвычайно чувствительны к малейшим раз-
246
дражениям и легко распадаются на
отдельные куски при загрязнении воды,
прикосновении и т. д. Такой способ защиты
от врагов широко распространен среди
представителей данного отряда.
Распадается на куски и встречающаяся в
Черном море Labidoplax thomsoni, при этом
конец, несущий щупальца, зарывается
в грунт и восстанавливает утраченные
части, остальные куски погибают. Эта
голотурия известна также в
Средиземном море и на Атлантическом побережье
Франции.
Очень интересна своим образом жизни
Synaptula hidriformis (табл. 23),
встречающаяся в Атлантическом океане в районе
Бермудских островов. Она не закапывается
в грунт, а живет среди водорослей или
разветвлений кораллов, цепляясь за них
своими щупальцами или якорными
пластинками кожи. Часто ее окраска зависит
от водорослей, среди которых она обитает:
среди красных водорослей цвет ее особей
коричнево-красный, а среди зеленых —
зеленый. Питается эта голотурия только
водорослями. Оригинален способ ее
передвижения. Она цепляется
щупальцами за выступающие предметы и с
помощью сокращений мускулатуры
подтягивает тело. Представители этого вида живут
целыми сообществами. Они вынашивают
молодь в полости своего тела, куда яйца
попадают через разрывы стенок яичника.
Оплодотворение происходит, очевидно,
прямо в полости тела попавшей вместе
с водой спермой. Молодь в полости тела
матери часто находится на разной стадии
развития, наиболее крупные личинки
достигают в длину 15—20 мм и уже похожи
на взрослых. Они выходят наружу через
разрывы тела матери, которые быстро
заживают.
Виды семейства Chiridotidae
отличаются присутствием в коже
колесиков и сигмоидных тел (рис. 136). Подобно
представителям предыдущего семейства
они имеют почти прозрачные покровы и
живут на мягких илистых грунтах, часто
зарываясь в них.
В прибрежных водах Баренцева,
Карского, Белого морей до глубины в 100 м
встречается Chiridota laevis. Эта
бесцветная голотурия длиной до 20 см часто
используется в пищу различными донными
рыбами.
Среди видов этого семейства имеются
интересные комменсалы морских ежей.
Например, Taenogirus cidaridis в районе
Индо-Малайского архипелага обнаружен
на ежах, где он живет, зацепившись
своим задним концом за их длинные
иглы.
Ксемейству Myriotrochidae
относится самый глубоководный из
современных иглокожих вид. Это Myriotrochus
bruuni, найденный датской экспедицией
на судне «Галатея» в Филиппинской
впадине и в желобе Новая Британия на
глубинах от 8920 до 10 240 м.
Род Myriotrochus весьма эврибатен.
Он включает виды, встречающиеся
исключительно в прибрежной зоне, а также
несколько настоящих глубоководных форм.
Имеются и виды, обладающие большим
диапазоном распространения по глубинам.
Так, Myriotrochus mitsukurii в Японском,
Охотском морях и у берегов Камчатки
обнаружен на глубинах от 70 до 1500 м.
КЛАСС МОРСКИЕ ЗВЕЗДЫ (ASTEROIDEA)
Название класса Asteroidea
(морские звезды) происходит от формы тела
объединяемых в этом классе
представителей иглокожих, имеющих вид звезды,
чаще всего пятиконечной, или плоского
правильного пятиугольника.
Среди животных, доживших до наших
дней, морские звезды — одна из наиболее
древних групп. В ископаемом состоянии
они известны с нижнего палеозоя — с
ордовикского периода, отстоящего от нашего
времени примерно на 400 млн. лет.
247
Рис. 137. Современный представитель древней
группы Somasteroidea — Platasterias latiradiata.
В морях с нормальной соленостью
различных морских звезд всегда можно
увидеть у самого берега — на литорали, т. е.
в зоне, заливаемой водой во время прилива
и обнажающейся при отливе. Поэтому
морские звезды были хорошо известны
человеку уже в глубокой древности.
Изображения их были обнаружены на
фресках 4000-летней давности, найденных
при раскопках на острове Крит. Само
название Aster, т. е. звезда, было дано этим
удивительным животным еще древними
греками. Аристотель более 2 тыс. лет
назад включил морских звезд в
созданную им классификацию животных, и ему
было известно, что «они, нападая,
высасывают многие раковины».
Класс морских звезд включает два
отряда — Platyasterida и Hemizonida,
полностью вымершие еще в палеозое и
изученные только по находкам их скелетных
остатков, и три отряда, многочисленные
представители которых существуют и в
настоящее время: 1) Phanerosonia (от
греческих слов phaneros — явный и zone—
пояс), или явнопластинчатые звезды,
получившие свое название от хорошо
заметных снаружи рядов известковых
пластинок, окаймляющих края звезды;
2) Spinulosa (от латинского spinula —
небольшая иголочка, шип),
или игольчатые звезды, и 3) Forcipulata
(от измененного латинского forceps —
щипцы), или педицелляриевые звезды,
т. е. звезды, у которых на поверхности
тела всегда имеются многочисленные
скелетные придатки в виде сидящих на
ножках крохотных щипчиков-педицеллярий.
При общем весьма однотипном плане
строения всех морских звезд они очень
разнообразны как по внешней форме,
размерам, окраске и характеру известковых
скелетных образований (пластинок, игл,
шипов, педицеллярий), от которых очень
зависит внешний облик животного, так
и по своему образу жизни и условиям,
в которых они обитают. Известно более
1500 современных видов морских звезд,
относящихся примерно к 300 родам и
29 семействам.
Морские звезды — типичные обитатели
океанов и морей, воде которых присуща
нормальная океаническая соленость
(около 35°/00). Ни один из представителей
морских звезд не сумел приспособиться
к жизни в пресных водах. В сильно
опресненных морях звезды или отсутствуют,
как, например, в Азовском и Каспийском
морях, или в них проникают лишь
единичные виды, представленные в этих условиях
явно угнетенными формами. Так,
например, взрослые особи звезды Asterias
rubens проникают в западную часть
Балтийского моря до районов с соленостью
8°/оо (У острова Рюген), но, по-видимому,
не способны размножаться при таком
сильном опреснении, и популяция этой звезды
поддерживается здесь за счет приноса
готовых к оседанию личинок из более
осолоненных районов. А единственная
звезда, проникшая из Средиземного моря
в Черное (Marthasterias glacialis), живет
лишь в его наиболее осолоненном юго-
западном углу вблизи пролива Босфор
и не смогла расселиться по остальному
морю. Такая соленолюбивость морских
звезд связана с тем, что все они относятся
к пойкилосмотическим
животным, т. е. к таким, которые не способны
регулировать общую концентрацию солей
в жидкостях, заполняющих полость тела.
Поэтому концентрация солей в полостных
жидкостях, омывающих все ткани и
внутренние органы звезд, всецело зависит
от солености окружающей воды, и сильное
248
опреснение оказывается губительным для
этих типично морских животных.
При благоприятных условиях некоторые
виды морских звезд могут размножаться
в очень больших количествах и
образовывать популяции большой плотности.
В океанах и полносоленых морях морские
звезды распространены повсеместно—от
Северного Ледовитого океана и вод,
омывающих побережья Антарктиды, до
тропических и экваториальной зон океана.
Они обильны и на литорали наших
северных морей, где выдерживают резкие
сезонные колебания температуры, а в
зимнее время могут даже промерзать, и на
постоянно прогретых тропических
мелководьях, где многочисленные виды
обитают среди пышных зарослей рифообразу-
ющих кораллов. По мере увеличения
глубины разнообразие морских звезд сильно
убывает, но и на дне ложа океана с его
постоянно однородными условиями —
полным отсутствием света, низкой
температурой и громадным давлением — они
встречаются повсюду, а представители трех
семейств (Porcellanasteridae, Pteras-
teridae и Brisingidae), по одному из каж-
дого отряда, проникают до глубин более
6 км. При работах советского
экспедиционного судна «Витязь» в Тихом океане
два вида звезд из р о д о в Porcellanaster
и Hymenaster были найдены в северной
части Марианского глубоководного
желоба на глубине 7600 м, что, видимо,
составляет предел проникновения морских звезд
в глубины океана.
Все морские звезды во взрослом
состоянии ведут донный образ жизни, ползая
по поверхности дна или закапываясь в
песок или ил. Многие звезды, особенно из
числа обитающих на мелководьях,—
активные хищники, питающиеся различными
моллюсками, ракообразными,
кишечнополостными и другими беспозвоночными,
в том числе и иглокожими. Наряду с этим
среди глубоководных звезд очень
распространены и илоядные формы,
заглатывающие грунт, в котором они живут,
и использующие в пищу содержащиеся
в нем органические остатки.
По-видимому, морские звезды были
очень многочисленны в морях прошлых
геологических эпох, но целые, хорошо
сохранившиеся экземпляры встречаются
Рис. 138. Морская звезда
Astropecten, вскрытая со
спинной стороны:
I — кожа; 2 — верхние
краевые пластинки; 3 — нижние
краевые пластинки; 4 — мадре-
поровая пластинка; 5 —
полнев пузырь; 6 — ампулы
амбулакральных ножек; 7 —
амбулакральные ножки; * —
желудок; 9 — печеночные
выросты; 10 — половая железа.
.?№
в ископаемом состоянии довольно редко.
Известковые пластинки морских звезд не
образуют единого прочного скелета, а
скреплены друг с другом соединительной
тканью и мускулами. Поэтому чаще в
ископаемом состоянии находят лишь
разрозненные пластинки или отпечатки звезд.
Наиболее древние и примитивные звезды
из группы Somasteroidea были в 1951 г.
описаны В. Спенсером (W. Spencer) из
нижнеордовикских отложений Южной
Франции. А несколько лет назад
новозеландским зоологом Б. Феллом (В. Fell)
к этой же группе была отнесена
современная звезда Platasterias latiradiata,
обитающая на мелководьях Тихоокеанского
побережья Америки у Южной Мексики и
Никарагуа. Спенсер и Фелл рассматривают
группу Somasteroidea как подкласс к л а с-
с а звездчатых иглокожих (Stelleroidea),
к которому они в качестве двух других
подклассов относят настоящих морских
звезд и офиур. Однако здесь мы
рассматриваем морских звезд и офиур как
отдельные классы в соответствии с
классификацией, общепринятой в
современных руководствах.
Рис. 139. Шаровидная звезда Podosphaeraster
polyplax, вид со спинной стороны:
1 — анальное отверстие; 2 — амбулакральная борозда.
Пунктирные линии проведены по радиусам,
соответствующим спинной стороне лучей.
Морские звезды обычно имеют более
или менее уплощенное тело с центральным
диском, постепенно переходящим в ради-
ально расходящиеся от него лучи, или
руки (рис. 138). Обращенная книзу
сторона звезды с ротовым отверстием в
центре называется оральной, т. е. ротовой,
а верхняя сторона — аборальной. Иногда
оральную сторону условно называют
брюшной, а аборальную — спинной.
У звезд, которые имеют анальное
отверстие, оно располагается вблизи центра
аборальной стороны диска. Посередине
нижней стороны каждого луча проходит
борозда, в которой находятся
многочисленные мягкие подвижные выросты —
амбулакральные ножки, служащие звезде
для передвижения.
Типичным для морских звезд является
пятилучевое строение, но довольно
многочисленные виды имеют шесть или больше
лучей. У обитающих в антарктических
водах звезд из рода Labidiaster число
лучей может достигать 45, а у солнечной
звезды Heliaster даже 50. Иногда число
лучей бывает различно у разных особей
одного вида. Так, у обычной в наших
северных и дальневосточных морях звезды
Crossaster papposus число лучей
колеблется от 8 до 16. В некоторых случаях
число лучей у звезд одного и того же
вида может меняться в зависимости от
района обитания, в других оно может
увеличиваться по мере роста звезды.
Соотношение размеров лучей и диска также
бывает очень различно. Для
характеристики формы тела морских звезд обычно
используют показатель соотношения
длины радиуса (R), т. е. расстояния от
центра диска до конца одного из лучей,
и интеррадиуса (г) — расстояния от
центра диска до его края в промежутке между
лучами. Наиболее часто длина лучей
в 3—5 раз превышает радиус диска. У
наиболее длинно лучевых звезд, например
у некоторых глубоководных Brisingidae
с очень маленьким диском и
многочисленными очень тонкими и хрупкими лучами,
R может в 20—30 раз превышать г, а
у многих звезд из отряда Phanerozonia
и у некоторых Spinulosa лучи могут лишь
немного выдаваться за края широкого
диска и тело имеет форму правильного
250
пятиугольника, в одних случаях
настолько уплощенного, что звезда похожа на
плоское печенье, в других — более или
менее выпуклого. Известны даже звезды,
внешний вид которых так изменен, что
с первого взгляда их трудно признать
за морских звезд. У обычной обитательницы
коралловых рифов Culcita (табл. 19) сильно
вздутое тело напоминает по форме почти
круглую подушку или булку. Однако
такая форма тела развилась вторично и
характерна лишь для взрослых особей,
а совсем молодые кульциты выглядят как
обычные пятиугольные звездочки. Еще
сильнее изменен недавно описанный вид
Podosphaeraster polyplax (рис. 139).
Единственный экземпляр этого необычного
представителя морских звезд был найден
на глубине около 80 м в
Южно-Китайском море. Подосферастер имеет вид
шарика диаметром немного более сантиметра
и внешне больше похож на морского ежа,
чем на звезду. Однако пять
амбулакральных борозд проходят у него лишь по
нижнему полушарию, соответствующему
оральной стороне прочих звезд, тогда как
у морских ежей полосы пластинок с
амбулакральными ножками переходят и на
верхнюю половину тела. Ближайшие
родственники этой звезды, составляющие
вместе с ней семейство Sphaerasteridae,
известны только в ископаемом состоянии
из отложений юрского и мелового
периодов. Размеры морских звезд очень
различны. У самых мелких видов диаметр, или,
точнее, наибольшее расстояние между
концами противоположно направленных
лучей, не превышает 1—1,5 см, тогда
как у наиболее крупных особей некоторых
видов — Acanthaster и Pycnopodia (табл.
19 и 25) —- оно может достигать 50 и даже
80 см, а у бризингиды (Freyella remex)
длина каждой из ее десяти рук равна
45 см.
У многих звезд, главным образом у
видов, обитающих на литорали и на
небольших глубинах, верхняя сторона тела часто
окрашена в очень яркие цвета, тогда как
нижняя обычно имеет бледную
желтоватую окраску. В окраске аборальной
стороны преобладают разнообразные оттенки
оранжевого, розового и красного цветов,
но встречаются также звезды, окрашен-
Рис. 140. Схема расположения скелетных
пластинок морской звезды на поперечном разрезе
луча:
1 — амбулакральная; 2 — адамбулакральная; з —
нижняя краевая; 4 — верхняя краевая пластинка;
5 — пластинки аборальной стороны; 6 — паксиллы.
ные в фиолетовый, синий, зеленый,
коричневый и даже черный цвет. Иногда
расцветка бывает пятнистой и различные
яркие цвета могут образовывать
причудливый рисунок. Звезды, населяющие
глубины океана, иногда окрашены в
красноватые тона, но чаще лишены яркой
пигментации и выглядят желтоватыми или
грязно-серыми. Окраска звезд зависит
от пигментных включений, находящихся
в клетках кожного эпителия.
При первом же взгляде на морскую
звезду прежде всего замечаешь
многочисленные, расположенные на поверхности
тела элементы известкового скелета —
пластинки иглы, мелкие шипы, бугорки
и т. п. Однако в действительности скелет
морских звезд по своему происхождению
не наружный, как, например, у
моллюсков или членистоногих, а внутренний,
образующийся в соединительнотканом
слое стенки тела. Все заметные снаружи
части скелета одеты покровным
эпителием и тонким слоем соединительной
ткани, но у взрослых животных эти
покровы на наиболее выступающих частях
могут стираться, обнажая скелетную
основу.
Основной опорный скелет звезд (рис. 140)
состоит из многочисленных отдельных
известковых пластинок, связанных между
собой соединительной тканью и
мускулами. Скелет оральной стороны развит
сильнее. Вдоль всей нижней стороны
каждого луча проходят два ряда амбула-
251
кральных пластинок, имеющих вид
лежащих поперек луча продолговатых
брусочков. Наклонно расположенные
противолежащие пластинки каждого ряда
соединены своими верхними концами, образуя
как бы крышу идущей вдоль луча
амбулакральной борозды. К этим пластинкам
с каждой стороны борозды примыкает
продольный ряд адамбулакральных
пластинок, за которыми следует по два ряда
краевых (магистральных) пластинок,
образующих боковые края лучей и диска.
У одних звезд краевые пластинки
крупные, хорошо заметные снаружи (Phanero-
zonia), у других они развиты слабо и мало
отличаются от скелетных пластинок
верхней стороны звезды. У большинства звезд
в той или иной мере развиты
дополнительные ряды пластинок, особенно в
интеррадиальных пространствах нижней стороны
диска. Скелет верхней стороны тела у
разных звезд развит в очень различной
степени. Чаще всего он образует
неправильную сеть из многочисленных
соприкасающихся своими концами пластинок
(рис. 141, Б).
В одном из интеррадиусов верхней
стороны диска, обычно вблизи его края,
находится особая мадрепоровая пластинка,
бороздчатая поверхность которой
пронизана мелкими порами. Через мадрепоро-
вую пластинку осуществляется связь
амбулакральной системы с внешней средой.
Кроме опорного скелета, у морских
звезд есть разнообразные и многочислен-
ные скелетные придатки, чаще всего в
виде игл или шипов, имеющих защитное
значение. Обычно они подвижно
сочленены с пластинками основного скелета.Иглы
могут быть простыми и в виде паксилл,
представляющих собой известковые
столбики, увенчанные на вершине группой
мелких шипиков или иголочек (рис. 142,
1—4), У некоторых звезд паксиллы сидят
на спинной поверхности так тесно, что
их шипики образуют сплошной покров.
Часть игл видоизменена в различной
формы мелкие щипчики —
педицеллярии (рис. 141,Л и 142,5—4),
сидящие непосредственно на пластинках
основного скелета или на иглах или
поднимающиеся над ними на гибких стебельках.
Подвижные и очень многочисленные
педицеллярии очищают поверхность тела от
загрязнения, схватывая и отбрасывая
попадающие на нее посторонние частицы.
Наряду с этим они могут выполнять и
защитную функцию.
В промежутках между скелетными
пластинками спинной стороны, а у некоторых
звезд и на оральной стороне наружу
выступают многочисленные нежные
тонкостенные выросты, в которые заходит
полость тела,— папулы, или кожные
жабры, служащие главным образом
для дыхания (рис. 141,-4). В значительной
степени дыхание происходит и через
стенки амбулакральных ножек, а иногда и
через другие участки поверхности тела.
Передвигаются звезды при помощи
многочисленных
амбулакральных ножек, попарно
отходящих от тянущихся до конца
каждого луча радиальных
каналов амбулакральной
системы (рис. 123). Через
промежутки между
амбулакральными пластинками
скелета ножки выходят наружу
в амбулакральную борозду.
Располагаются ножки
большей частью двумя, а у
некоторых звезд четырьмя
рядами вдоль всей длины луча.
Общее их число в каждом из
лучей может доходить до
нескольких сотен. Каждая
ножка соединена с находя-
Рис. 141. Поверхность кожи аборальной стороны (А) и абораль
ный скелет (Б) морской звезды Asterias rubens:
l — пластинки; 2 — иглы; 3 — педицеллярии; 4 — кожные жабры.
252
Рис. 142. Паксил-
лы и педицелля-
рии морских звезд:
1 — з — паксиллы
явнопластинчатых
звезд; 4 — паксиллы
игольчатых звезд из
семейства Pterasteri-
dae,
поддерживающие
супрадореальную мембрану; 5—
10 — педицеллярии
явнопластинчатых
звезд: 5 — Luidia;
6 — Hippasteria; ?—
8 — открытая и
закрытая педицелляр ии
Hacelia (семейство
Linckiidae); 9 —
пучковидная педицелля-
рия Astropectinidae;
10 — гребенчатая
педицелляр ия Bentho-
pectinidae; 11—12 —
прямые и 13—14 —
скрещенные
педицеллярии педицел-
ляриевых звезд.
щейся внутри луча ампулой —
мускулистым мешочком в виде небольшого
пузырька, способного сокращаться и
растягиваться. Сами ножки также могут
сильно вытягиваться и сжиматься, а также
изгибаться в любом направлении. У
большинства звезд ножки снабжены на конце
присосками. Одна присосавшаяся к
какой-либо поверхности ножка, сокращаясь,
может развивать усилие до 30 г, а при
совместном координированном действии
многочисленных ножек нескольких лучей
сила тяги, развиваемая звездой, может
доходить до нескольких килограммов.
Этого, оказывается, более чем достаточно
даже для передвижения звезды вверх по
вертикальной плоскости, а также для
удержания довольно крупной добычи и
для открывания плотно сомкнутых
раковин двустворчатых моллюсков. Любопытно,
что сила сцепления присоски ножки с
субстратом превышает атмосферное давление
и более 40% этой силы обусловлено не
присасывательным действием
образуемого присоской вакуума, а клейкой
слизью, постоянно выделяемой железистыми
клетками эпителия присоски.
Амбулакральные ножки снабжены только
продольной мускулатурой, обеспечивающей
их сокращение, а растягивание их
вызывается гидростатическим давлением
жидкости, нагнетаемой в ножки из
сжавшихся ампул.
Мускулатура у морских звезд
развита относительно слабо.
Мускульный тяж, проходящий вдоль спинной
стороны каждого луча, служит для
изгибания луча кверху. Система мышц,
связывающих скелетные пластинки оральной
стороны, обеспечивает сужение и
расширение амбулакральной борозды и боковое
изгибание лучей. Мускульными волокнами
снабжены наружные скелетные придатки,
амбулакральные ножки, кожные жабры.
Звезды глубоководного семейства Bentho-
pectinidae (Phanerozonia), для которых
характерны длинные и гибкие лучи,
отличаются от всех других звезд парными
мускульными тяжами, проходящими по
сторонам каждого луча вдоль его
спинной поверхности. По-видимому, звезды
этого семейства способны с помощью
таких мускулов совершать движения,
позволяющие им всплывать и проплывать
небольшие расстояния вблизи дна.
Хотя большинство звезд активные
хищники, каких-либо приспособлений для
разрывания добычи и для разжевывания
пищи у них нет. Специализированные
ротовые пластинки скелета приспособлены
лишь для проталкивания пищи в рот.
Окруженный мягким кожистым
пространством — перистомом, рот способен
широко растягиваться. Коротким пищеводом
он соединен с объемистым желудком,
заполняющим большую часть внутреннего
пространства диска. Отходящая от
верхней части желудка короткая и узкая
задняя кишка открывается на спинной
стороне вблизи центра диска анальным
отверстием. Однако последнее настолько мало,
что практически не может служить для
удаления непереваренных остатков пищи,
и они выбрасываются наружу через рот.
В верхней части желудка от него в
каждый луч отходят парные ветвистые слепые
выросты, так называемые пилорические,
или печеночные, придатки, во
внутренние каналы которых обильно выделяются
пищеварительные соки. В стенках
печеночных придатков происходит
всасывание переваренной пищи и накапливаются
запасы питательных веществ, которые
могут расходоваться в тех случаях, когда
звезда длительное время не питается,
например в период перед размножением
или при вынашивании молоди. Голодание
в течение 6—8 недель приводит к полному
расходованию таких запасов.
У некоторых звезд задняя кишка,
анальное отверстие и радиальные
выросты желудка отсутствуют и весь
пищеварительный тракт состоит лишь из
мешковидного желудка. Такие звезды обычно
относятся к илоядным формам.
Многие звезды, обладающие длинными
и гибкими руками, главным образом из
семейства Asteriidae, способны
выворачивать наружу желудок (его
нижнюю часть), обволакивать им свою жертву
и переваривать ее, не заглатывая. В
желудках таких звезд никогда не находят
остатков пищи. Другие звезды с
короткими или с недостаточно гибкими руками
не способны длительно удерживать
добычу. Они заглатывают целиком иногда
довольно крупных животных и после
переваривания их мягких частей выбрасывают
через рот наружу непереваренные
остатки, как, например, раковины моллюсков
или скорлупу морских ежей.
Пищевой спектр многих крупных
хищных звезд очень широк. Хотя они обычно
предпочитают питаться теми или иными
определенными видами животных, но
зачастую поедают и многих других или их
трупы. Но у некоторых звезд питание
строго избирательное. Одни из них
поедают только губок, другие питаются
коралловыми полипами, а пищу Coscinaste-
rias calamaria составляют главным
образом брахиоподы, раковины которых эта
звезда открывает так же, как другие
звезды открывают двустворчатых
моллюсков.
Нервная система морских
звезд очень примитивна. Никаких четко
выраженных мозговых ганглиев у них нет.
Несмотря на примитивность нервной
системы морских звезд, проведенными
рядом исследователей опытами было
показано, что у некоторых звезд могут
вырабатываться условные рефлексы. У обычной
на литорали Мурманска звезды Aste-
rias rubens многократным сочетанием
пищи с определенной поверхностью
субстрата, по которому ползает звезда,
удавалось выработать избирательную реакцию
на этот тип поверхности. Звезды этого же
вида, запутанные в сетку, постепенно
«научались» высвобождаться из нее
значительно быстрее, чем при первых
опытах. Выработанные у звезд условные
рефлексы могут сохраняться без
подкрепления до пяти суток.
Из морфологически обособленных
органов чувств у звезд развиты только глаза.
Последняя непарная амбулакральная
ножка каждого луча лишена присоски и
представляет собой короткое щупальце, у
основания которого находится красный
глазок, состоящий из многочисленных
отдельных глазных бокальчиков. С помощью
своих глаз звезда не может «видеть»
в собственном смысле этого слова, а
способна лишь различать интенсивность
освещения и направление света.
Кроме непарного концевого щупальца,
несколько соседних с ним
амбулакральных ножек также не имеют присосок и
выполняют функцию осязания. Ползущая
звезда всегда вытягивает их вперед и
ощупывает ими поверхность субстрата. В
эпидермисе стенки тела звезд повсюду встре-
254
чаются чувствительные нервные клетки.
Они особенно многочисленны в присосках
амбулакральных ножек, у оснований игл
и педицеллярий и по сторонам
амбулакральных борозд. В таких участках их
число может достигать 70 тыс. на площади
в 1 мм2. По-видимому, действовать как
органы осязания могут не только
концевые щупальца, но и присоски остальных
ножек. Кроме того, скопления нервных
клеток способны воспринимать вкусовые
и другие химические раздражения. Звезды
некоторых видов реагируют на пищу,
находящуюся от них на расстоянии до
полуметра. Описан даже случай, когда звезда
Asterias vulgaris в течение двух суток
двигалась прямо по направлению к пище,
находившейся от нее на расстоянии 12 м.
Рыбакам, ставящим ловушки на крупных
морских раков лангустов, хорошо
известно, что звезды часто сползаются к
ловушкам и скапливаются в них, поедая
предназначенную для лангуста приманку.
Однако иногда хеморецепция практически
отсутствует. В ряде случаев голодные
звезды проползали в нескольких сантиметрах
Рис. 143. Развитие морской звезды Asterias: А — Д — бипиннария; Е — Ж — брахиолярия;
3 — только что сформировавшаяся звездочка (вид с брюшной стороны).
1 — рот; 2 — анальное отверстие; 3 — желудок; 4 — руки брахиолярии; 5 — присоска; 6 —
формирующаяся звездочка; 7 — амбулакральные ножки.
от своей излюбленной пищи, не замечая
ее, и поедали пищу лишь тогда, когда
случайно на нее наталкивались.
Специальных выделительных органов
у звезд нет. Удаление из тела конечных
продуктов обмена веществ производится
главным образом подвижными
амебоидными клетками, многочисленными в
жидкости, заполняющей полость тела.
Нагруженные продуктами обмена амебоидные
клетки движутся к поверхности тела и
выбираются наружу в основном через
тонкостенные кожные жабры. Кроме того,
конечные продукты обмена выделяются
специальными секреторными клетками в
каналы печеночных придатков, откуда
попадают в желудок и удаляются наружу.
Большинство морских звезд
раздельнополы. Но нередки случаи и
гермафродитизма в различных его проявлениях. Так,
например, у Marthasterias glacialis наряду
с особями того или другого пола
встречаются и звезды с мозаичными гермафро-
дитными гонадами. А у обычной у
Атлантических побережий Европы Asterina
gibbosa в популяциях некоторых районов
преобладает так называемый протерандри-
ческий гермафродитизм, когда молодые
звезды функционируют только как самцы,
а выросшие до определенных размеров —
только как самки. Но в других районах
звезды этого же вида могут быть самцами,
самками или гермафродитами независимо
от размера.
В типичных случаях в каждом луче
вблизи его основания находятся парные
половые железы, протоки которых
открываются наружу между лучами или иногда
на оральной стороне тела. Но у ряда
видов многочисленные гонады располагаются
внутри лучей серийно и открываются
наружу каждая своим отверстием.
По внешнему виду мужские и женские
особи обычно неразличимы, за
исключением тех случаев, когда самки проявляют
заботу о потомстве и вынашивают
развивающуюся молодь. Но иногда звезды
разного пола могут несколько отличаться по
размерам или по окраске в период
размножения. У уже упоминавшейся Asterina
gibbosa самцы в период размножения
скапливаются вокруг самки и прижимают
к ее лучам свои таким образом, чтобы
выпускаемая сперма попадала прямо на
выметываемые в воду яйца. А
мелководные тропические Archaster при
размножении даже объединяются попарно.
Эмбриональное развитие звезд может
протекать со стадией свободноплавающей
личинки, из которой в результате
метаморфоза формируется молодая звезда, или
быть прямым. У большинства звезд
половые клетки выпускаются прямо в воду,
где и происходит оплодотворение яиц и их
Таблица 21. Забота о потомстве у различных видов
иглокожих:
1 — морская звезда Pteraster obsurus с молодью,
вылезающей через покровы;
2 — морская звезда Trophodiscus uber с
молодью на спине;
3 — самка Abatus philippii с маленькими ежами
на спине;
4 — морской еж Ctenocidaris nutrix с
маленькими ежами вокруг рта;
5 — голотурия Cladodactyla сгосеа с молодью
на спине;
6 — голотурия Psolus ephippifer с крупными
пластинками на спине, прикрывающими
выводковые камеры.
дальнейшее развитие в двустороннесим-
метричную личинку бипиннарию
(рис. 143,А—Д). Такая симметрия
личинки—след происхождения морских звезд
от далеких предков всех современных
иглокожих, также имевших двустороннюю,
а не радиальную симметрию. После
нескольких недель жизни в толще воды, во
время которой личинка активно питается
микроскопическими водорослями, на
переднем конце бипиннарии образуется три
выроста и она превращается в личинку
следующей стадии — брахиолярию
(рис.143, Е — Ж). Брахиолярия вскоре
оседает на дно, прикрепляется
находящейся между выростами переднего конца
присоской и претерпевает метаморфоз,
при котором выполняющая роль
стебелька передняя часть брахиолярии
дегенерирует, а из задней части, содержащей
желудок, формируется молодая звездочка.
При прямом развитии сравнительно
немногочисленные крупные и богатые
желтком яйца развиваются без стадии
свободноплавающей личинки. Многие звезды
с таким типом развития в той или иной
мере проявляют заботу о потомстве. У As-
terina gibbosa и некоторых других звезд
эта забота ограничивается тем, что они
прикрепляют яйца к различным
подводным предметам, чаще всего к нависающей
над дном нижней поверхности камней,
а затем уползают. Однако у большинства
обитателей холодных вод полярных
областей или больших глубин развивающиеся
зародыши и даже уже сформировавшиеся
молодые звездочки вынашиваются на теле
матери до тех пор, пока они не становятся
способными вести самостоятельное
существование. В частности, вынашивают свою
молодь все звезды, населяющие
Антарктическую область, независимо от того,
к какому отряду они принадлежат.
У звезд из отрядов Spinulosa и
Forcipulata с более или менее длинными
и гибкими лучами вынашивание молоди
чаще всего происходит следующим
образом. Материнская особь опирается на дно
концами лучей, а их основания и диск
изгибает так, что они образуют подобие
колокола, внутри которого находится рот.
В полости этого колокола и помещаются
развивающиеся звездочки, соединенные
общим тяжом, к которому каждая из них
прикреплена своей ротовой частью. В
течение всего времени вынашивания своего
потомства мать не питается. У живущей
в холодных водах Арктики небольшой
звезды Leptasterias groenlandica молодь
развивается внутри желудка матери —
в специальных сумчатых выростах его
передней части.
Таблица 22. Иглокожие больших глубин Тихого океана.
Морская лилия:
1 — Bathycrinus pacificus.
Голотурии:
2 — Paelopatides solea;
3 — Psychropotes raripes;
4 — Lpsilothuria bitentaculata;
5 — Scotoplanes murrayi;
6 — Elpidia sp.
Морские звезды:
7 — Astrocles actinodetus;
8 — Styracaster horridus.
Морские ежи:
9 — Aerop^is fulva;
10 — Pourtalesia tanneri.
17 Жизнь животных, т. 2
Звезды с короткими широкими лучами
и менее гибким телом не способны
образовывать выводковую камеру под ротовой
частью диска, и развитие молоди может
происходить у них на спинной стороне
тела, или непосредственно на ее
поверхности между паксиллами (некоторые
представители Phanerozonia), или в
специальной выводковой камере (семейство
Pterasteridae из Spinulosa). Более
подробное описание таких звезд будет дано
ниже.
Плодовитость звезд, у которых
развитие протекает в толще воды, может быть
исключительно велика. По подсчетам
английского исследователя Геммилла
(J. Gemmill), Asterias rubens только в
течение двух часов выпустила в воду около
2,5 млн. яиц, а в течение сезона
размножения выметывание яиц у этого вида
может происходить несколько раз.
Известный специалист по иглокожим датчанин
Т. Мортенсен (Т. Mortensen)
установил, что еще более плодовита другая звез-
да — Luidia ciliaris, в яичниках
которой может находиться до 200 млн. яиц.
Плодовитость звезд, вынашивающих свое
потомство, несравненно меньше. У звезд,
вынашивающих молодь на нижней стороне
тела около рта, развивающихся зародышей
обычно бывает не более 200, хотя иногда
их число может доходить до 1000. А число
молодых звездочек, развивающихся на
спине или в специальных выводковых
камерах, не превышает нескольких десятков.
У некоторых звезд наряду с половым
может происходить и бесполое
размножение путем деления диска на две части.
Линия деления обычно проходит в
определенном направлении через интеррадиусы,
так что лучи каждой образовавшейся
в результате деления части остаются
неповрежденными. Обе части звезды
начинают существовать самостоятельно, и у
них восстанавливаются недостающие лучи
и участки диска. Такой тип деления
встречается у ряда видов семейства
Asteriidae (Forcipulata), обычно у тех,
которые имеют более пяти
лучей, а также у некоторых
представителей семейства
Asterinidae из отряда
Spinulosa, например у
распространенных в тропических водах
Австралии звезд из рода
Nepanthia (рис. 144, Б). У
одних видов размножаться г.еле-
нием могут только молодые
звезды, у других способность
Рис. 144. Бесполое размножение морских звезд: А — образование звезды из
отделившегося луча у Linckia; Б — образование двух звезд после поперечного»
деления у Nepanthia (отряд Spinulosa).
258
к делению сохраняется на протяжении
всей жизни. Звезд, размножающихся
таким способом, редко удается найти с
лучами одинаковой длины: отрастающие
заново лучи одной из половин тела
значительно мельче лучей другой половины,
доставшейся звезде в наследство от
материнской особи. У всех видов рода
Linckia (Phanerozonia) бесполое
размножение происходит другим способом (рис.
144,Л), о котором мы расскажем при
описании этих звезд.
Со способностью к размножению путем
деления связана и очень развитая у звезд
способность к регенерации, т. е. к
восстановлению частей тела, утраченных
вследствие ранения. Некоторые звезды
могут восстанавливать все недостающие
части даже после разрезания их на
несколько кусков, однако обычно лишь
в тех случаях, когда такой кусок состоит
хотя бы из одного луча и прилежащего
к нему участка диска. А у Linckia вся
звезда может вырасти заново даже из
части луча. Известны случаи, когда
звезды могут при опасности самопроизвольно
отбрасывать лучи, а затем отращивать их
заново. Если конец луча оказывается в
результате ранения расщепленным в
продольном направлении, то каждая часть
может продолжать расти самостоятельно
и образуется своеобразный луч,
раздвоенный на конце в виде вилки. Интересно,
что у звезды, лишившейся всей руки,
регенерация протекает с большей скоростью,
чем при потере только кончика луча. У
молодых звезд утраченные части обычно
восстанавливаются быстрее, чем у
старых, а у тепловодных тропических видов
быстрее, чем у обитателей холодноводных
районов.
На поверхности тела морских звезд
и в их внутренних органах нередко
встречаются паразиты, главным образом
представители многощетинковых червей из
семейства Myzostomidae,
различные переднежаберные брюхоногие
моллюски и ракообразные, преимущественно
из целиком паразитического отряда Asco-
thoracida.
Среди самих морских звезд формы,
паразитирующие на других животных,
неизвестны.
ОТРЯД ЯВНОПЛАСТИНЧАТЫЕ ЗВЕЗДЫ
(PHANEROZONIA)
Для звезд этого отряда очень
характерны хорошо заметные снаружи верхний
и нижний ряды краевых пластинок,
окаймляющих диск и лучи (рис. 145, 7, 3).
Граница между этими рядами
соответствует границе между верхней и нижней
сторонами звезды. Строение педицелля-
рий у Phanerozonia простое: они никогда
не имеют подвижной ножки, и их створки
сидят непосредственно на скелетных
пластинках. Phanerozonia наиболее древний
и примитивный отряд из доживших до
наших дней отрядов морских звезд.
Ископаемые представители этого отряда
известны начиная с нижних слоев
ордовикского периода. Из трех современных
отрядов звезд отряд Phanerozonia наиболее
богат видами, объединяющимися в 14
семейств.
Особенно разнообразны Phanerozonia на
тропических мелководьях Тихого и
Индийского океанов. Более двух третей всех
встречающихся здесь видов звезд
принадлежит к этому отряду.
Наиболее характерные среди морских
звезд обитатели больших океанических
глубин относятся к семейству
Porcellanasteridae. Ни один из видов
этого семейства не встречен на глубинах
менее 900 ле, а 7 видов найдены глубже
6 км.
Порцелланастериды — пятилучевые,
обычно небольшие звезды
желтовато-серого цвета, чаще всего с относительно
крупным диском и короткими лучами.
Живут они на мягком илистом дне,
зарываясь в ил при помощи своих лишенных
присосок ножек. Для порцелланастерид
очень характерны своеобразные так
называемые гребневидные органы (рис. 145,5).
Каждый такой орган состоит из
многочисленных параллельных вертикальных
рядов тонких, покрытых ресничным
эпителием пластиночек, сидящих на краевых
пластинках диска в промежутках между
лучами (интеррадиусах). В каждом иа
пяти интеррадиусов может быть один
или несколько гребневидных органов.
Кожные жабры у порцелланастерид раз-
17*
259
виты слабо или их нет совсем и
гребневидные органы служат главным образом
для дыхания. Возможно также, что они
играют вспомогательную роль в питании
звезды, направляя создаваемыми ими
токами воды сверху вниз комочки
оседающего из толщи воды детрита, которые
далее транспортируются ножками ко рту.
Однако в основном эти звезды питаются
илом.
Печеночных выростов, задней кишки
и ануса у порцелланастерид нет и вся
полость диска заполнена объемистым
слепым желудком. Часто он бывает до такой
степени набит илом, что спинная сторона
звезды вздувается, как подушка, а
покрывающая ее кожа растягивается до тонкой
пленки. Вес проглоченного грунта часто
равен весу самой звезды, а иногда может
даже вдвое превышать его. Судя по тому,
что в желудке вместе с илом встречаются
и довольно крупные камни, звезды
заглатывают все, что попадается им по пути,
без какого-либо выбора. После
переваривания находящихся в грунте
органических остатков и мелких животных все
содержимое желудка выбрасывается
наружу и звезда начинает заглатывать новую
порцию ила, подгребая его ко рту
ножками и сидящими по краям
амбулакральных борозд иглами, нередко
видоизмененными в маленькие лопаточки или
совки.
Один из наиболее обычных
представителей порцелланастерид Eremicaster te-
nebrarius с тремя гребневидными
органами в каждом из пяти межлучевых
участков диска широко распространен в
абиссали Тихого и Индийского океанов. Очень
своеобразный облик имеют звезды другого
рода этого семейства — Styracaster
(табл. 22, рис. 145). Их довольно сильно
вытянутые, жесткие и хрупкие лучи одеты
сплошным панцирем известковых
пластинок, а по спинной стороне каждого луча
тянется ряд длинных острых
неподвижных шипов, по которым стиракастера
сразу можно отличить от всех других
морских звезд.
Размножение порцелланастерид
не изучено, но, судя по относительно
крупным и немногочисленным яйцам,
которые находили у них в яичниках,
развитие у этих звезд прямое, без
свободноплавающей личинки. По-видимому, они в той
или иной форме проявляют заботу о
потомстве подобно большинству других
глубоководных животных.
Близкородственная и похожая на порцелланастерид
звезда Ctenodiscus australis (семейство
Рис. 145. Элементы скелета у звезд семейства Porcellanasteridae: А и Б — Styracaster; A — вид
луча сбоку; Б — спинная сторона интеррадиальной части диска; В и Г — паксиллы Vitjazas-
ter; В — вид сверху; Г — вид сбоку; Д — педицеллярия Porcellanaster.
1 — верхние краевые пластинки; 2 — их шипы; 3 — нижние краевые пластинки; 4 — мадрепоровая
пластинка; 5 — паксиллы; 6 — столбик паксиллы; 7 — концевые шипики паксиллы; 8 — гребневидные органы.
260
Goniopectinidae) вынашивает молодь на
спинной стороне тела между паксиллами.
Одно из обширных семейств отряда
Phanerozonia составляют гребенчатые
звезды (Astropectinidae). Это плоские пятилу-
чевые звезды, у которых крупные
краевые пластинки обычно вооружены
игловидными шипами, окаймляющими края
диска и лучей. В окраске многих звезд
этого семейства преобладают оранжево-
красные тона. Звезды рода Astropecten
(рис. 138) предпочитают селиться на песке,
в который они зарываются, быстро
отбрасывая его в стороны от лучей ножками,
не имеющими присосок. Таким способом
астропектен может полностью скрыться
в песке в течение одной минуты. Астро-
пектены обычно чувствительны к
изменениям освещенности, причем реагируют на
такие изменения даже в случае удаления
у них глаз на всех пяти лучах.
Распространенный в тропических водах Тихого
и Индийского океанов A. polyacanthus
проводит большую часть времени
зарывшись в песок и остается неподвижным
и в темноте, и при ярком дневном свете.
Звезды выползают на поиски добычи лишь
на непродолжительное время дважды
в сутки — утром и в конце дня при
определенной и довольно слабой
освещенности дна. Рот астропектенов способен очень
широко растягиваться, и они питаются
преимущественно разными моллюсками,
заглатывая их целиком и буквально
набивая ими свой желудок. Так, например,
в желудке одного средиземноморского
золотистого астропектена (A. aurantiacus)
было обнаружено 10 гребешков и 6
других двустворчатых моллюсков, 5 лопа-
тоногих и несколько брюхоногих
моллюсков.
В амбулакральных бороздах
астропектенов часто можно найти поселяющихся
тут многощетинковых червей из родов
АсЫое и Harmothoe (семейство Aph-
roditidae). Иногда до 5—6 червей находят
приют на одной звезде, располагаясь не
только в амбулакральной борозде, но и
в желобках между краевыми
пластинками, и на спинной стороне между
паксиллами. Польза такого сожительства для
червя очевидна — звезда обеспечивает его
пищей. Голова находящегося в
амбулакральной борозде червя всегда направлена
в сторону рта звезды. Удавалось даже
наблюдать, как червь, не довольствуясь
остатками пищи, которые он подбирает
вокруг рта звезды, засовывает свою
голову через ее рот в желудок без какого-либо
вреда для себя. Какую пользу получает
звезда от такого сожительства, точно не
выяснено, но, видимо, черви, очищая
околоротовое пространство звезды,
выполняют роль мусорщиков.
По-видимому, астропектен выделяет в воду какие-то
вещества, привлекающие червей. Снятый
со звезды и выпущенный неподалеку от
нее, червь возвращается к своему
хозяину. А специально проведенными опытами
установлено, что червя привлекают
даже вырезанные из тела звезды кусочки
тканей.
К этому же семейству относятся
обитающие только в Охотском море небольшие
коротколучевые ярко-красные звездочки
Trophodiscus (табл. 21), вынашивающие
свою молодь, как и упоминавшийся выше
Ctenodiscus australis, на спинной стороне
между паксиллами. Иногда на одной
материнской особи можно видеть до 30
молодых звездочек от 3 до 8 мм в поперечнике.
Подрастая, звездочки все более
раздвигают окружающие их паксиллы, а
достигнув определенного размера,
выползают наружу, а на их месте остаются
покрытые голой кожей углубления. Такие
углубления часто видны на спинной
стороне матери рядом с еще вынашиваемой
молодью.
На тропических мелководьях Индо-Ма-
лайского архипелага и у побережья
некоторых островов Индийского океана живут
внешне похожие на астропектена звезды
из р о д a Archaster (семейство Arcfr-
asteridae). Архастеры интересны тем, что
среди всех звезд только у них при
размножении происходит спаривание. На
песчаных пляжах бесчисленных островков
Яванского моря у самого берега
многочисленны небольшие пятилучевые звезды
A. typicus. Обычно они лежат
полузарывшись в коралловый песок и едва
заметны из-за своей почти сливающейся
с песком окраски. В период размножения
самцы и самки этого вида объединяются
попарно таким образом, что лучи находя-
261
щегося сверху самца чередуются с
лучами несколько более крупной самки.
Обе особи одновременно выпускают в
воду половые продукты, и благодаря этому
яйца оплодотворяются тотчас же после
вымета.
Обычные на небольших глубинах
теплых и тропических морей Luidia
(семейство Luidiidae) с длинными и гибкими
лучами, окаймленными многочисленными
иголочками, часто окрашены в тусклые
серовато- или зеленовато-бурые тона и
достигают значительных размеров (табл. 19).
Эти звезды селятся на песчаном дне
и могут ползать не только по его
поверхности, но и зарывшись в толщу песка.
Питаются они главным образом офиурами
и морскими ежами, заглатывая их
целиком. В желудке луидий нередко находили
по нескольку десятков офиур. Известен
случай, когда луидия сумела проглотить
такого крупного плоского ежа Mellita,
что, переварив его мягкие части, не
смогла выбросить наружу его панцирь и в
результате погибла. Для одного из видов
луидий описано любопытное явление,
происходящее при метаморфозе личинки би-
пиннарии. Молодая звездочка развивается
лишь из небольшой части личинки,
отрывающейся от нее и оседающей на дно.
Сама же бипиннария, точнее ее большая
часть, остается в толще воды и еще
некоторое время продолжает жить
самостоятельно.
Среди коралловых рифов тропических
морей часто можно видеть довольно
крупных звезд из семейства Oreasteridae.
Одетых жестким скелетом,
неповоротливых оранжево-красных Oreaster nodosus
(табл. 18) легко узнать по большим
неподвижным коническим
темно-коричневым шипам, увенчивающим их высокий
диск. К этому же семейству принадлежит
и описанная выше, похожая на
небольшую подушку Culcita (табл. 19). Кульцита
замечательна не только своей необычной
для звезд формой, но также тем, что в ее
полости тела иногда находят небольшую
так называемую жемчужную рыбку Сага-
pus (известную также под более старым
названием Fierasfer). К арапу с обычно
держится вблизи некоторых голотурий и в
случае опасности использует в качестве
временного убежища их водные легкие.
По-видимому, в кульциту карапус
проникает, когда при опасности поблизости
не оказывается его обычного хозяина.
Но в полость тела звезды карапус,
вероятно, может проникнуть, только
пробравшись через ее рот в желудок и
затем пробуравив его стенку. Удается ли
рыбке снова выбраться на свободу из
такого необычного убежища, пока
неизвестно.
Весьма обычная на тропических
мелководьях Тихого и Индийского океанов
ярко-синяя звезда с пятью длинными,
почти цилиндрическими лучами — Linc-
kia laevigata (семейство Linckiidae,
табл. 18). Как для этой звезды, так и для
всех других видов рода Linckia очень
характерен особый тип бесполого
размножения, не встречающийся у других звезд.
Линкии обладают способностью
периодически аутотомировать, т. е.
самопроизвольно обламывать свои лучи. Процесс
этот начинается с отделения друг от
друга скелетных пластинок, чаще всего на
определенном расстоянии от диска.
Затем отделяющаяся часть руки начинает
отползать от материнской особи, еще
будучи соединенной с ней мягкими тканями
и кожей. В течение трех-четырех часов
ткани эти все более растягиваются (иногда
до 5 см) и наконец разрываются, после
чего оторвавшаяся рука начинает
самостоятельную жизнь. Через некоторое время на
месте облома у такой руки начинает
развиваться новая звезда, в результате чего
сначала образуется так называемая ко-
метная форма звезды с группой крохотных
лучей на конце единственной крупной
руки (рис. 144, А). В дальнейшем новые
лучи все более вырастают и звезда
приобретает нормальный облик. У материнской
звезды на месте оторвавшейся руки
вырастает новая. В местах, где линкии
многочисленны, нередко встречаются как
звезды-кометы, так и звезды, регенерирующие
одну или несколько рук. Если у аутото-
мированной руки линкии дополнительно
отрезать и ее кончик, то иногда
регенерация может начаться с обоих концов, и
таким образом могут образоваться две
молодые звездочки, соединенные толстым
участком материнской руки.
262
ОТРЯД ИГОЛЬЧАТЫЕ ЗВЕЗДЫ
(SPINULOSA)
У большинства звезд, входящих в
отряд Spinulosa, краевые пластинки
развиты слабо и не образуют резкой
границы между верхней и нижней сторонами
тела.
Педицеллярий у звезд этого отряда
обычно не бывает. Некоторые
представители имеют признаки, переходные между
Phanerozonia и Spinulosa, и отряд в целом
не очень резко отграничен от
предыдущего. В состав отряда входит 11 семейств.
Широко распространены в умеренных
и холодных водах обоих полушарий
многочисленные виды рода Henricia (с е-
м е й с т в о Echinasteridae). Это обычно
небольшие пятилучевые звезды с узкими,
почти цилиндрическими лучами и
маленьким диском. Скелетные пластинки
образуют на спинной стороне хенриции —
узкопетлистое сплетение — и покрыты
мелкими иголочками, сплошной покров
которых создает впечатление бархатистой
поверхности. Хенриции обычно однородно
окрашены в один из оттенков красной
гаммы цветов — от светлых
желто-оранжевых до темных вишнево-красных и
малиновых тонов.
В Арктике и северной Атлантике
обычна так называемая кровяная звезда
(Henricia sanguinolenta), получившая
название за свою сочную красную окраску.
Питается эта звезда исключительно
различными видами морских губок. При
этом она может распознавать посредством
хеморецепции предпочитаемые ею виды
губок, даже находясь на значительном
расстоянии от них.
Все виды хенриции живородящи.
Клубок соединенных общим тяжом
развивающихся зародышей звезда вынашивает на
нижней стороне тела подо ртом. У двух
видов хенриции обнаружены
приспособившиеся к существованию на этих
звездах комменсалы из группы морских nay-
ков, или многоколенчатых (Pantopoda).
А обитающий в Индо-Малайском
архипелаге представитель того же
семейства Echinaster luzonicus интересен
тем, что только на поверхности его тела
встречается своеобразный ползающий
гребневик Coeloplana astericola, ярко
выделяющийся на коричневом фоне
звезды своей окраской из чередующихся
белых и красных пятен.
Очень эффектны многолучевые звезды
из семейства Solasteridae:
фиолетово-розовые Solaster endeca и малиново-
красные Crossaster papposus (табл. 25).
Они широко распространены в
сублиторали северных районов Атлантического
и Тихого океанов. Обе эти звезды могут
достигать крупных размеров — до 40—50
см в диаметре. Это активные и очень
прожорливые хищники, питающиеся
преимущественно другими иглокожими,
особенно звездами, но поедающие также
различных моллюсков и других беспозвоночных.
Solaster endeca может нападать на морских
ежей и выедать их внутренности, a
Crossaster papposus поедает офиур, голотурий
и других звезд, иногда даже
превосходящих размерами его самого. Жертвами
кроссастера постоянно становятся такие
звезды, как Asterias, сами являющиеся
активными хищниками. Ханкоком
(Hancock) было установлено, что снижение
численности Asterias rubens на одном из
участков побережья Англии может в
значительной мере обусловливаться
уничтожением этих звезд кроссастером. Самим
кроссастерам не грозит опасность быть
съеденными другими животными, так как
их ткани обладают ядовитыми
свойствами. Паркером (С. Parker) описан
случай, когда два домашних кота,
полакомившиеся выловленными Crossaster
papposus, погибли в результате сильного
отравления. Crossaster — рекордсмены среди
звезд по скорости движения. Молодые
звезды этого вида могут проползать до 2 м
в минуту, у большинства же других
звезд скорость обычно не превышает 5—15
см в минуту. Кроссастер откладывает
крупные, богатые желтком яйца, и их
развитие проходит без стадии
свободноплавающей личинки. Молодые звезды довольно
быстро растут, увеличиваясь в диаметре
в среднем на 1 мм в неделю.
У кроссастера, хенриции и некоторых
других представителей отрядов
Spinulosa и Forcipulata в полости тела
встречаются паразитические ракообразные
263
Dendrogaster astericola из близкого к усо-
ногим ракам отряда Ascothoracida.
Строение этих рачков чрезвычайно
упростилось и видоизменилось в результате
паразитического образа жизни. Взрослый
дендрогастер выглядит как
прикрепленный к внутренним тканям звезды
оранжевый мешок с дольчатыми выростами, не
имеющий никакого сходства со свободно-
живущими ракообразными. Лишь в
процессе развития дендрогастера сохранилась
типичная для ракообразных личиночная
стадия.
Представители еще одного
семейства игольчатых звезд — Pterasteridae—
в основном обитатели холодных вод
полярных и умеренных зон обоих
полушарий и больших глубин океана. Звезды
этого семейства обычно имеют форму
вздутого (Pteraster) или сильно уплощенного
(Hymenaster) пятиугольника с крупным
диском и широкими короткими лучами,
лишь немного выступающими за края
диска. Hymenaster окрашены в нежные
розоватые, голубоватые тона или имеют
почти бесцветное полупрозрачное тело.
А цвет арктической сублиторальной
звезды Pteraster obscurus с 6—12 лучами
(табл. 21) темно-бурый, нередко почти
черный. Звезды этого семейства
живородящи. Развитие зародышей
происходит у них в весьма своеобразной
выводковой камере, ничего похожего на
которую нет у других звезд. В качестве такой
камеры используется пространство между
собственно спинной стороной тела и
расположенной над ней кожистой
оболочкой — супрадорзальной
мембраной, поддерживаемой вершинами
сидящих на спинной поверхности паксилл
(рис. 142,^). В центре мембраны находится
окруженное подвижными иглами широкое
отверстие — оскулум, а сама мембрана
пронизана многочисленными более
мелкими отверстиями — спиракулами.
Кроме того, находящаяся под
мембраной полость сообщается с внешней средой
отверстиями по бокам лучей. Ритмичными
движениями несколько раз в минуту
пространство между спинной поверхностью и
мембраной сжимается и расширяется, и в
нем происходит постоянная смена свежей
воды, входящей через боковые отверстия
и спиракулы и выходящей главным
образом через оскулум. Циркулирующая в
полости вода постоянно омывает и снабжает
кислородом находящиеся на ее дне кожные
жабры и развивающихся зародышей. Таким
образом, эта полость не только служит для
вынашивания молоди, но функционирует
и как орган дыхания звезды. Поскольку
супрадорзальная мембрана есть не только
у самок, но и у самцов, можно
предполагать, что первоначально она
сформировалась как приспособление для дыхания,
а в дальнейшем стала использоваться
и как выводковая камера. Одновременно
в выводковой камере развивается до 20
зародышей. Питание глубоководных
видов птерастерид не изучено.
Сублиторальный же вид Pteraster tesselatus
питается главным образом губками и
актиниями.
К паразитированию в звездах из р о д а
Pteraster приспособилась описанная
В. Н. Беклемишевым турбелля-
рия Pteraster icola fedotovi. Она была
найдена только в кишечнике трех видов
звезд этого рода, обитающих у побережий
Мурманска, и не обнаружена в каких-либо
других звездах.
На литорали Японского моря очень
обычна исключительно эффектно
окрашенная небольшая звезда Patiria pectinifera
(семейство Asterinidae), имеющая вид
правильного пятиугольника (табл. 25). На
верхней стороне этой звезды
ярко-оранжевые пятна разбросаны на фоне сочного
чисто синего цвета, а оральная сторона
имеет однородную палевую окраску. К
этому же семейству принадлежит
распространенная у Атлантического побережья
Западной Европы и в Средиземном море Anse-
ropoda placenta. Анзеропода —
зарывающаяся в песок звездочка, около 10 см
диаметром, замечательна своим
чрезвычайно уплощенным телом,
бледно-розовая или голубоватая поверхность
которого сплошь покрыта пучками очень
мелких иголочек. По фактуре
поверхности и ничтожной толщине тела
анзеропода напоминает вафлю. Тело ее
настолько тонко, что верхняя и нижняя
стороны кажутся вплотную
прижатыми друг к другу, не оставляя места
для каких-либо внутренних полостей*
264
Тем не менее анзеропода ухитряется
заглатывать целиком небольших крабов и
раков-отшельников, а также моллюсков и
иглокожих.
На коралловых рифах Тихого и
Индийского океанов нередко встречается
крупная звезда Acanthaster planci (табл. 19),
достигающая 40—50 см в диаметре (с е-
м е й с т в о Acanthasteridae). Обычно
принято считать, что все морские звезды
совершенно безвредны для человека,
однако неосторожное обращение с акантасте-
ром может причинить серьезные
неприятности. От широкого уплощенного диска
акантастера отходят многочисленные
короткие лучи. Однако молодые звездочки
имеют типичное для большинства звезд
пятилучевое строение, и число лучей
увеличивается лишь по мере роста звезды.
Акантастер — одна из немногих звезд,
имеющих не только большое число лучей,
но и многочисленные мадрепоровые
пластинки, количество которых также
увеличивается с возрастом. У наиболее
крупных звезд этого вида число лучей может
достигать 18—21, а мадрепоровых
пластинок — 16. Вся спинная поверхность
диска и лучей вооружена сотнями
сидящих на подвижных ножках крупных
и очень острых игл длиной 2—3 см,
концы которых по своей форме
напоминают наконечник копья. За свою форму,
обилие и остроту шипов эта звезда
получила название «терновый венец».
Окраска тернового венца может
меняться от голубовато- или зеленовато-серых
тонов до фиолетово-пурпурного и
малинового цвета. Изменение цвета в
значительной мере зависит от того, насколько
сокращены или расправлены
многочисленные на верхней стороне диска кожные
жабры — папулы. Питается акантастер
главным образом коралловыми полипами.
Звезды ползают среди рифов, оставляя
позади себя белую полосу известковых
скелетов кораллов с начисто
объеденными мягкими частями. Изменчивая
окраска тернового венца хорошо маскирует
его среди ярких и разнообразных красок
кораллового рифа, и звезду нелегко
заметить с первого взгляда.
Терновый венец пользуется дурной
славой среди жителей многих тропических
островов. Его невозможно взять в руки,
не получив причиняющих жгучую боль
уколов острых игл. Сборщикам жемчуга
на атолле Тонгарева в центральной части
Тихого океана нередко приходится иметь
дело с этими звездами. Минер пишет,
что, если ныряльщик случайно наступит
на одно из этих ужасных созданий, иглы
пронзают ступню и обламываются,
заражая кровь ядовитыми выделениями.
Местные жители считают, что получивший
такое ранение должен тотчас же палкой
перевернуть звезду ротовой стороной
кверху и прижать ногу к ее рту. Они
утверждают, что звезда с силой присасывается
к ноге и высасывает обломки игл и яд,,
после чего раны быстро заживают. Б ар-
нес и Эндин (J. Barnes a. R. Enbean)
недавно описали клиническую картину
отравления мальчика с острова Йорк в
Торресовом проливе, наступившего на
терновый венец. Пораженные участки ног
были сильно воспалены, и в течение
четырех суток через каждые 3—4 часа
начиналась продолжительная рвота.
Мальчик поправился только на шестые сутки.
Такая картина отравления хорошо
известна жителям острова Йорк, считающим
эту звезду очень опасной. Отравляющее
действие, видимо, вызывается
оранжевым и красным пигментом кожи,
покрывающей иглы звезды, но возможно, что»
ядовиты также и ее педицеллярии.
ОТРЯД ПЕДИЦЕЛЛЯРИЕВЫЕ ЗВЕЗДЫ
(FORCIPULATA)
Звезды этого отряда отличаются от всех
остальных характерными педицел-
л я р и я м и, состоящими из трех
подвижно сочлененных скелетных
элементов — основания и двух створок. Обычно
педицеллярии сидят на мягких
подвижных стебельках. Створки педицеллярии
или соединены у своих нижних концов
наподобие пинцета, или перекрещиваются,
как ножницы (рис. 142, 11—14). Обычно
педицелляриевые звезды имеют небольшой
диск и длинные гибкие лучи. Из-за
отсутствия морфологически обособленных
краевых пластинок верхняя сторона тела,
265
постепенно закругляясь, переходит в
нижнюю и четкой границы между обеими
сторонами нет. Снабженные присосками
амбулакральные ножки у многих Forci-
pulata расположены в
амбулакральных бороздах в 4 ряда. В состав отряда
входит 4 семейства, но большинство его
представителей относится к одному очень
обширному семейству — Asteriidae
(табл. 17, 20 и 25).
Типичные представители этого
семейства — виды рода Asterias — обычны
на морских и океанических мелководьях
умеренной зоны северного полушария.
На побережьях Баренцева и Белого морей
и северо-восточной Атлантики среди
камней и зарослей водорослей, обнажающихся
во время отлива, почти повсеместно
можно увидеть многочисленную в этих
районах пятилучевую звезду Asterias rubens
(табл. 20), длина рук которой может
достигать 30 см. Окраска этой звезды очень
изменчива. Наряду со светлыми особями
желто-оранжевого цвета встречаются
звезды, окрашенные в красноватые и
коричнево-бурые тона, и еще более темные
звезды буро-черного цвета. На литорали
наших дальневосточных морей обычны
звезды близкого к предыдущему вида — А.
amurensis, большей частью окрашенные
в фиолетовый цвет. У Атлантического
побережья Северной Америки на севере
распространена A. vulgaris, а южнее, у
берегов США, ее сменяет A. forbesi (табл. 17).
Все эти виды очень сходны по своему
внешнему облику и образу жизни.
Биология Asterias forbesi изучена наиболее
детально и всесторонне, и поэтому на
описании этой звезды мы остановимся
подробнее.
Asterias forbesi — небольшая пятилу-
чевая звезда, расстояние между концами
противолежащих лучей обычно не
превышает у нее 20 см, но чаще всего
встречаются звезды диаметром около 10 см.
Окраска A. forbesi варьирует от оранжево-
красных до зеленовато-черных тонов.
Питается A. forbesi главным образом
устрицами и мидиями, но поедает также
других моллюсков, мелких ракообразных,
червей и мертвых рыб, а при случае
нападает и на живых, особенно на больных или
запутавшихся в сети. При недостатке пищи
у A. forbesi отмечены и случаи
каннибализма — более крупные звезды поедают более
мелких особей своего вида. A. forbesi
причиняет огромный вред устричным
хозяйствам. Поэтому американские ученые Галь-
цов и Лузанов (P. Galtsoff, V. Loossa-
noff) специально посвятили ряд лет
изучению биологии этой звезды и разработке
мероприятий по борьбе с ней. По данным
этих авторов, прожорливость A. forbesi
настолько велика, что одна звезда средних
размеров может ежедневно уничтожать
нескольких годовалых устриц. При этом
A. forbesi очень плодовиты и при
благоприятных условиях размножаются в
громадных количествах, буквально
опустошая и разоряя устричники. В 20-х годах
нашего столетия морские звезды ежегодно
уничтожали у Атлантического побережья
США в среднем около 500 тыс. бушелей
устриц (бушель — мера объема,
составляющая около 35 л), что причиняло
убытки на сумму около полумиллиона
долларов в год.
Размножение A. forbesi обычно
происходит несколько раз в течение лета. При
этом стимулом для начала размножения
может служить даже небольшое
повышение температуры воды. Звезды обоих
полов приподнимают свое тело над дном
на концах лучей и выметывают половые
продукты в воду через парные отверстия
у основания каждого луча. Остатки
половых желез после вымета дегенерируют,
а осенью начинается формирование
новых гонад, которые быстро растут и к
началу следующего лета снова оказываются
наполненными зрелыми яйцами и
сперматозоидами. Личинки после трех-четырех
недель свободного существования в воде
оседают и превращаются в крохотных
звездочек диаметром около 1 мм, которые
вскоре начинают питаться недавно осевшей
на дно молодью моллюсков и других
животных. Поедают молодые звездочки и
друг друга, в результате чего
численность их за первый месяц после оседания
сильно уменьшается. В течение жизни
в планктоне личинки не относятся далеко
от места вымета яиц, и наиболее массовое
оседание молоди происходит обычно
именно там, где особенно многочисленны
и взрослые звезды.
266
Рост звезд очень зависит от условий
питания. При хорошем питании A. for-
besi за полгода вырастает до 8—10 см
в диаметре, а половозрелость наступает
уже к концу первого года жизни. При
плохом питании звезды растут медленнее
и созревают только к двухлетнему
возрасту. Общая продолжительность жизни
звезд из рода Asterias не превышает
5—6 лет.
В половых железах A. forbesi и других
видов этого рода иногда встречаются
паразитические инфузории Orchitophrya
stellarum. Массовое развитие этих
инфузорий может приводить к полному
разрушению гонад.
A. forbesi обычно медленно ползают
по дну посредством координированного
движения ножек, расположенных в четыре
ряда в каждом луче. На горизонтальной
поверхности звезда использует свои
ножки как рычаги, не подтягивая тело
сокращением присосавшихся ножек, а скорее
толкая его вперед, т. е. в прямом смысле
ходит на своих многочисленных ножках.
Подтягивание тела ножками происходит
лишь тогда, когда звезде приходится
подниматься по наклонной или
вертикальной поверхности.
Средняя скорость движения составляет
около 10 см в минуту, но иногда в течение
короткого времени звезда может двигаться
значительно быстрее — до 25—30 см в
минуту.
По-видимому, звезды в течение своей
жизни не могут совершать больших
миграций. Это было установлено опытом по
своеобразному мечению звезд безвредной
для них синей краской. Погруженные
на короткий срок в слабый раствор
краски, звезды окрашиваются в голубой цвет,
который сохраняется у них более 9
месяцев. Около 12 тыс. таких окрашенных
звезд было выпущено на одной устричной
банке у побережья штата Коннектикут.
В течение 10 месяцев все сборщики
устриц в радиусе 10 миль от места выпуска
должны были отмечать все находки
необыкновенных голубых звезд. К концу этого
срока удалось обнаружить около 300
меченых звезд, но ни одна из них не была
найдена дальше чем за полтора километра от
места выпуска.
Питается A. forbesi, как мы уже
указывали, главным образом различными
двустворчатыми моллюсками. Найдя
моллюска, звезда наползает на него и принимает
очень характерную позу, приподнимая
срединную часть тела и присасываясь
многочисленными ножками лучей к
створкам моллюска (табл. 26). Затем звезда
начинает растягивать створки, развивая
при этом силу до 4,5 кг. Только очень
крупным устрицам иногда удается
противостоять в течение длительного
времени такой силе, и звезда бывает
вынуждена оставить свою жертву и отправиться
на поиски более легкой добычи. В
подавляющем же большинстве случаев мускул,
запирающий створки моллюска, в конце
концов несколько расслабляется и между
створками образуется щель, в которую
звезда всовывает свой вывернутый наружу
желудок и постепенно переваривает все
тело моллюска внутри его раковины.
После этого желудок втягивается обратно
и звезда уползает в поисках новой
жертвы, оставляя после себя пустую
раковину.
Несмотря на медлительность своих
движений, звезды способны поедать не только
неподвижных моллюсков. Жертвами
морских звезд становятся даже такие
моллюски, как гребешки (Pecten). Обычно
спокойно лежащие на дне гребешки при
приближении к ним их главного врага —
хищной морской звезды — сразу же
начинают хлопать створками раковины, с
силой выталкивая из нее воду. Используя,
таким образом, ракетный принцип
движения, они всплывают и, продолжая
открывать и захлопывать раковину,
могут отплывать на значительное
расстояние. Если же гребешок почему-либо не
успевает спастись бегством, звезда
обхватывает его своими руками, открывает
и поедает (табл. 26).
В результате массового размножения
звезд промысел гребешка в заливе Баз-
зардс в штате Массачусетс с 1929 по 1931 г.
снизился с 750 тыс. долларов до 142 тыс.,
т. е. более чем в пять раз.
Ущерб, причиняемый звездами,
настолько велик, что оказалось целесообразным
содержать специально оборудованные
небольшие суда для вылова и уничтожения
267
звезд, на что только в штате
Коннектикут затрачивается ежегодно более 100
тыс. долларов. Отлов звезд производится
драгой или протаскиваемыми по дну
швабрами, в которых звезды запутываются.
Изобретена даже специальная, так
называемая сосущая, драга—соединенная с
трубой воронка, опускаемая на небольших
глубинах на дно и по мере движения
судна всасывающая при помощи помпы
морские звезды. Приведем некоторые
цифры, позволяющие судить об огромном
количестве звезд, скапливающихся на
устричных банках. В местах обильных
скоплений одна сосущая драга за 10 минут
работы может поднять на борт до
нескольких центнеров звезд. В 1931 г. только
с площади около 600 га было выловлено
и уничтожено 7 млн. звезд, а после
ущерба, причиненного промыслу гребешка в
заливе Баззардс, за три года в этом заливе
было выловлено более 60 млн. звезд.
У представителей рода Asterias,
подобно большинству других звезд, хорошо
развита способность к регенерации
утраченных частей тела. Известен
любопытный случай, когда при массовом
развитии одной из хищных звезд,
уничтожавшей ценных моллюсков, звезд
вылавливали, рубили на куски и выбрасывали
обратно в море. Результат получился
противоположный ожидаемому. Подобно
тому как у легендарных драконов на месте
отрубленной головы появлялось
несколько новых, так и из разрубленной звезды
вырастало несколько звезд, и численность
их еще более увеличилась. Поэтому при
отлове звезд их обычно убивают
погружением в горячую воду и в дальнейшем
перерабатывают на удобрение.
Способ, которым морским звездам
удается открывать двустворчатых моллюсков
и поедать их, был хорошо изучен К р и-
стинсеном на другой звезде из того
же семейства — Evasterias troschelii,
обитающей на мелководьях у Тихоокеанского
побережья Северной Америки. У
двустворчатых моллюсков Protothaca
перерезали запирающий мускул и после этого
стягивали их створки резиновым пояском,
представляющим собой своеобразный
динамометр. Наблюдая, как звезды поедают
таких моллюсков, удалось установить,
что звезда с лучами длиной 20 см может
растягивать створки с силой более 5 кг.
При этом звезде достаточно лишь чуть-
чуть приоткрыть створки. Даже в щель
шириной в несколько десятых долей
миллиметра она способна просунуть свой
растягивающийся, как резина, желудок.
У мидий в месте выхода из раковины
тонких нитей биссуса, которыми моллюск
прикрепляется к субстрату, есть незакры-
вающаяся щель шириной около 0,1 мм.
Для просовывания желудка внутрь
раковины звезде оказывается достаточно и
такого ничтожно малого отверстия, и,
чтобы полакомиться мидией, ей даже не
приходится тратить усилий на открывание
раковины. Чтобы выяснить, на какую же
длину звезда может вытягивать свой
вывернутый наружу желудок, звездам
предлагали мидий, помещенных внутрь
пластмассовых трубок на разном
расстоянии от их конца. Оказалось, что звезда
способна уничтожить мидию,
находящуюся в 10 см от отверстия, вытягивая свой
желудок на расстояние, равное половине
длины луча, а в некоторых случаях и на
всю его длину.
До сих пор окончательно не выяснено,
выделяют ли звезды какие-либо ядовитые
для моллюсков вещества, вызывающие
расслабление запирающего мускула. Для ряда
видов доказано, что звезда открывает
раковину только посредством механической
силы. Но не исключено, что у некоторых звезд
используются одновременно оба способа.
Очень интересные наблюдения были
проведены Смитом (L. S. Smith) над
крупной хищной пятилучевой звездой Pisaster
breyispinus. Ползая по дну, эта звезда
безошибочно останавливается над местом,
где находится один из закапывающихся в
грунт моллюсков из родов Saxidomus и
Protothaca. После этого звезда начинает
разрывать грунт, отбрасывая своими ножками
в стороны песок и мелкие камешки размером
до 2 см. Эта работа продолжается двое или
трое суток, причем рытье происходит только
ночью, а днем звезда неподвижно лежит
на месте производимых ею раскопок.
В конце концов звезда выкапывает нору,
равную по диаметру размерам ее тела (до
70 см) и глубиной около 10 см.
Добравшись до моллюска, который всегда оказы-
268
вается в самом центре норы, как раз
против рта звезды, звезда присасывается
находящимися вблизи рта ножками к
вершине раковины. Затем она приподнимает,
опершись на концы лучей, центральную
часть своего тела и вытаскивает
моллюска наружу, после чего расправляется
с ним обычным для астериид способом,
открывая раковину и всовывая внутрь
ее желудок.
Иногда звезды одного и того же вида из
различных местообитаний значительно
отличаются друг от друга по своей биологии,
в частности по характеру питания и
связанному с ним поведению. Так, Pisaster brevi-
spinus, обитающие у побережий
Калифорнии, поедают главным образом плоских
ежей, Dendraster, а севернее — в заливе
Пьюджет-Саунд проползают среди
поселений этих ежей, не обращая на них
внимания, и питаются моллюсками,
выкапывая их, как было описано выше.
Соответственно этому различна и реакция
Dendraster в том и другом районе на близость
этой звезды. Калифорнийские ежи сразу
же начинают закапываться в песок,
когда вблизи них проползает опасная для
них звезда, а ежи из Пьюджет-Саунд не
реагируют на звезд даже на расстоянии
нескольких сантиметров и начинают
закапываться, лишь потревоженные случайно
наползшей на них звездой.
Защитные реакции на прикосновение или
близость хищных звезд вырабатываются и у
многих других животных. Большей частью
это реакция бегства от звезды. Ф е д е р
(Н. М. Feder) очень красочно описывает
такую реакцию у крупного брюхоногого
моллюска морское ушко (Haliotis). При
соприкосновении со звездой Pisaster ochra-
ceus моллюск приподнимает раковину
на своей толстой ноте и начинает
стремительно поворачивать ее на 180°
попеременно в одну и другую сторону.
Освободившись такими стряхивательными
движениями от присосавшихся к раковине
ножек звезды, халиотис поворачивается
и уползает от хищника «аллюром,
напоминающим галоп». Нога его при этом резко
сокращается и вытягивается, производя
движения, более свойственные пиявке или
гусенице пяденицы, чем крупной улитке.
Подобным же образом реагируют на
хищных звезд брюхоногие моллюски —
блюдечки (Acmaea).
Другая морская улитка — Natica
catena — при прикосновении к ней звезды
Asterias rubens обволакивает всю свою
раковину способной сильно
растягиваться ногой, к скользкой поверхности
которой не могут присосаться ножки звезды.
Это спасает натику, так как звезды
начинают выворачивать желудок, чтобы
переварить свою добычу, лишь после того,
как захватывают и прочно удерживают
ее своими ножками. А морской еж Рага-
centrotus lividus реагирует на
нападающих на него хищных звезд, ощетинивая
и быстро раскачивая на стебельках свои
многочисленные ядовитые педицеллярии,
которые схватывают и отрывают
ощупывающие ежа ножки звезды.
Одновременно еж стремится уползти от звезды.
Подобной реакции не происходит, если
к ежу прикасаются звезды безвредных
для него видов.
На мелководьях Тихоокеанского
побережья Северной Америки, от Калифорнии
до острова Ванкувер, живет небольшая
пятилучевая звезда Astrometis sertulifera,
(табл. 25), предпочитающая селиться в
защищенных от яркого света местах.
Длина лучей астрометис обычно не
превышает 8 см. Ее спинная поверхность окрашена
в сочный темно-зеленый цвет и усажена
многочисленными шипами с
ярко-красными кончиками и темно-синими или
пурпурными у оснований. Нижняя
поверхность звезды соломенно-желтая, а
амбулакральные ножки яркого канареечного
цвета. Основания спинных шипов
окружены у нее розетками из многочисленных
мелких педицеллярии, а более крупные
одиночные педицеллярии рассеяны по
поверхности тела. По наблюдениям
Дженни н г с а (Н. Jennings), основное
назначение педицеллярии—защита расположенных
между шипами нежных кожных жабр. При
раздражении поверхности кожи
заползшими на звезду мелкими рачками или другими
животными папулы сокращаются и
втягиваются, а педицеллярии начинают
открывать и закрывать свои щипчики, пока
им не удастся схватить вызвавшее
раздражение животное или попавшую на кожу
чужеродную частицу. Захваченных мел-
269
ких рачков педицеллярии могут держать,
не выпуская, более двух суток. Все
схваченное педицеллярии удерживают
настолько прочно, что можно, например,
поднять звезду из воды за педицеллярии,
ухватившие волоски руки.
К семейству Asteriidae относится
и настоящий гигант среди морских
звезд — Pycnopodia helianthoides (табл.
25), живущая на скалистых участках дна,
покрытых зарослями бурых водорослей,
у северо-восточного побережья Тихого
океана от Калифорнии до Алеутских
островов. Спинной скелет у этой звезды
практически отсутствует, и ее многочисленные
лучи отличаются чрезвычайной гибкостью
и подвижностью. Наиболее крупные
звезды достигают 80 см в диаметре и 4,5 кг
веса. Когда такая звезда ползет,
распластав по дну два десятка своих лучей,
ее тело покрывает площадь около 0,5 м2.
Красно-коричневая поверхность тела
покрыта многочисленными группами серо-
фиолетовых разветвленных папул, между
которыми разбросаны гроздья
педицеллярии.
Известный специалист по морским
звездам У. Фишер (W. Fisher) так
описывает поведение пикноподии: «Питается она
главным образом морскими ежами,
раками-отшельниками и другими
животными, которых ей удается поймать,
нападает на крупных голотурий и поедает
мертвых или ослабленных рыб.
Последних она ловит своими лучами, почти столь
же подвижными, как руки осьминога.
Возбужденная близостью пищи, она
движется очень быстро и более активна,
чем все другие звезды, которых мне
приходилось наблюдать. Когда эта звезда
быстро ползет с тысячами своих
извивающихся ножек, она производит
внушительное впечатление, а ее многочисленные
помпоны из цепких педицеллярии и
широкое, гибкое тело делают ее грозным
орудием уничтожения. В борьбе с
сопротивляющейся рыбой или крабом она может
привести в действие более 15 тыс. ножек
с присосками. Пикноподия целиком
заглатывает крупных морских ежей Stron-
gylocentrotus, а спустя некоторое время
выбрасывает наружу лишенную игл
чистую скорлупу ежа. После сражения
с морским ежом ножки пикноподии
бывают обильно усажены педицелляриями
ежа, ярко выделяющимися своей
пурпурной окраской на светло-желтом фоне
ножек. Иногда пикноподии попадаются даже
на удочки рыбаков, схватывая наживку
из мяса рыбы или моллюска».
Интересна пикноподия не только своими
крупными размерами и хищностью. У этой
звезды вторично развились некоторые
черты двусторонней симметрии дополнительно
к таковым, унаследованным звездами от
их предков. Свою жизнь на дне
пикноподия начинает в виде небольшой пятилуче-
вой звездочки, у которой вскоре вырастает
шестой луч, занимающий, как правило,
строго определенное положение по
отношению к интеррадиусу с мадрепоровой
пластинкой. Дальнейшее увеличение
числа лучей происходит путем образования
по обеим сторонам шестого луча все
новых и новых пар симметричных лучей,
число которых может в конце концов
достигнуть 24. Проявляется двусторонняя
симметрия и в физиологии звезды.
Пикноподия обычно двигается направляя
вперед одни и те же определенные лучи и эти
же лучи использует в первую очередь для
переворачивания в нормальное
положение, если ее положить ротовой стороной
кверху.
Из звезд отряда Forcipulata следует
упомянуть еще о типичных обитателях
больших глубин — бризингидах (Brisin-
gidae), составляющих наиболее
примитивное семейство этого отряда. Бризин-
гиды — изящные звезды с маленьким
круглым диском и четко отграниченными
от него многочисленными длинными
тонкими и гибкими лучами (табл. 22,7). Число
лучей — от 7 до 15, а их длина может
достигать 45 см. Лучи окаймлены по бокам
иглами, часто снабженными на конце
пучком скрещенных педицеллярии.
Скелет спинной стороны развит у бризингид
слабо и скелетные пластинки разделены
широкими промежутками мягкой кожи.
Лучи обычно настолько хрупки, что
почти всегда ломаются у звезд,
попадающихся в трал или драгу при глубоководных
ловах. Поэтому поднять на палубу
корабля целых звезд почти никогда не удается.
Для многих бризингид характерны се-
270
риальные половые железы,
расположенные двумя рядами внутри каждого луча
вдоль его боковых стенок. По-видимому,
у всех бризингид развитие происходит
без свободной личиночной стадии.
Обитающая в антарктических водах Odinella
nutrix вынашивает свою молодь в свое-
образных выводковых камерах между
сильно вздутыми основаниями лучей.
Иглы соседних лучей в этих местах
переплетаются друг с другом, образуя как бы
защищенные со всех сторон корзиночки,
в каждой из которых находится 5—9
развивающихся зародышей.
КЛАСС МОРСКИЕ ЕЖИ (ECHINOIDEA)
Весьма красочное описание
представителей этого класса дано К л и н д ж е-
л о м в его популярной книге «Остров
в океане». Вот что он пишет: «Морского
ежа можно назвать дикобразом
подводного царства. Его невозможно взять в
руки, разве что найдется человек с
пальцами, вылитыми из металла; все тело у
морских ежей защищено длинными, острыми
иглами, крепящимися к телу при помощи
хитроумно устроенных шарниров.
Наступить на такого ежа и больно и опасно:
его иглы покрыты слизью, насыщенной
бактериями, которые вызывают тяжелое
нагноение». Однако не все ежи так
опасны и страшны, как описывает их Клинд-
жел. Значительная часть видов, в том
числе и тех, которые встречаются в наших
морях, совершенно безобидна для
человека. Их свободно можно брать в руки,
безо всякой для себя опасности.
Некоторые же плоские ежи, такие, например,как
Echinarachnius parma, покрыты столь
мелкими иголками, что их поверхность
скорее кажется бархатистой, чем колючей.
Тело морских ежей лишено выдающихся
лучей и имеет почти шарообразную, либо
плоскую дисковидную, либо
сердцевидную, либо яйцеобразную форму. Такая
форма у большинства видов остается
постоянной, так как тело их заключено
в панцирь, образованный крупными
пластинками, скрепленными между собой
неподвижно. Только небольшие площадки
вокруг рта (перистом, ротовое
поле) и вокруг анального отверстия
(перипрокт) остаются мягкими.
Подвижность пластинок сохраняется лишь
у представителей древнего семейства
Echinothuriidae.
Панцирь всех современных ежей
устроен по одной и той же схеме. Он состоит
из 20 рядов пластинок, располагающихся
по меридианам. Если, например,
очистить от игл скорлупу атлантического ежа
Echinus acutus (рис. 146, А), то сразу
станет заметным пятилучевой план его
строения. В центре несколько приплюснутой
стороны, всегда обращенной к субстрату
и часто называемой брюшной, ибо
животное ползает на ней, помещается рот.
Противоположный, аборальный
(апикальный) полюс несет в своем центре
анальное отверстие. От ротового поля к
анальному отверстию по меридианам шара идут
пять парных рядов амбулакральных
пластинок. Таких пластинок может
насчитываться несколько десятков в каждом ряду,
и каждая пластиночка несет одну или
несколько парных дырочек — пор, через
которые проходят амбулакральные
ножки. Каждой паре пор соответствует одна
амбулакральная ножка, так как каналы,
соединяющие ножки и ампулы
водоносной системы, у морских ежей двойные.
Общее количество ножек у морских ежей
огромно. У некоторых видов на одном
амбулакре их может быть несколько
сотен, а на всех пяти амбулакрах даже
больше тысячи. Это самое многоногое
животное на свете. С помощью таких
эластичных, способных вытягиваться или
сокращаться цилиндрической формы
образований, снабженных на конце присоской,
животное может не только передвигаться
с места на место и ползти даже по отвес-
271
ным скалам, но и прочно прикрепляться
к субстрату в самых прибойных местах
или добывать себе пищу.
Каждый амбулакральный ряд
заканчивается на своем аборальном конце одной
небольшой глазной пластинкой, на
которой помещается маленький глазок. С
амбулакральными рядами чередуются пять
парных интерамбулакральных рядов,
состоящих из более крупных пластинок,
лишенных пор для прохождения
амбулакральных ножек. Каждый
интерамбулакральный ряд заканчивается на своем
аборальном конце одной половой
пластинкой, имеющей половое отверстие. Одна
из пяти половых пластинок пронизана
еще множеством пор, через которые
поступает в водоносную систему вода, т. е.
является также и мадрепоровой
пластинкой. Таким образом, чередующиеся друг
с другом глазные и половые пластинки
образуют вокруг перипрокта венчик
пластинок, составляющих апикальное поле.
Сравнивая форму тела морских ежей с
формой тела звезд, можно видеть, что
первые обладают как бы укороченными
лучами, которые загнуты концами к
аморальной стороне и сходятся вокруг
анального отверстия. При этом, подобно
голотуриям, у них происходит сильное
увеличение ротовой (оральной)
поверхности за счет уменьшения
противоположной — аборальной.
Ротовое поле правильных морских ежей,
затянутое мягкой кожей, у большинства
форм имеет только 5 пар округлых
ротовых пластинок, снабженных ротовыми
ножками. По краю ротового поля, в
особых десяти выемках интерамбулакров
располагаются древовидно разветвленные
наружные кожные жабры. В центре поля
в глубине ротового отверстия помещается
особый жевательный аппарат, так
называемый аристотелев фонарь
(рис. 147, 7). Он состоит из 25
известковых перекладин и пластинок, подвижно
соединенных между собой при помощи
мышц. Главной составной частью его
являются 5 парных пирамидок, внутри
которых помещается по одному длинному
зубу. Кончики этих зубов и торчат из
ротового отверстия. Сам фонарь подвижно
соединен с помощью мышц с особыми
дуговидными выростами
интерамбулакральных пластинок, называемыми ушками.
С помощью острых зубов аристотелева
фонаря морские ежи могут соскребать с
поверхности камней водоросли, захватывать
другую пищу, а иногда и более быстро
передвигаться, отталкиваясь от
субстрата при смыкании зубов.
Вся скорлупа морских ежей несет на
своей поверхности многочисленные
различно устроенные бугорки, служащие для
прикрепления игл. На одной и той же
скорлупе бугорки и иглы бывают самой
Таблица 23. Различные представители класса голотурий:
i — Cucumaria elongata;
2 — Colochirus quadrangularis;
3 — Psolus phantapus;
4 — Rhopalodinopsis capensis;
5 — Pelagothuria natatrix;
6 — Psychropotes kerhervei;
7 — Pannychia moseleyi;
8 — Peniagone lugubris;
9 — Oneirophanta alternata;
10 — Molpadia musculus;
11 — Synaptula hydriformis.
разнообразной величины, наиболее
крупные из них называются первичными.
У правильных ежей первичные бугорки
и иглы располагаются более или менее
правильными меридиональными рядами
от апикального поля до ротового.
Основание каждой иглы имеет суставную ямку,
в которую входит полушаровидной формы
бугорок. Такое сочленение иглы с
бугорком охвачено суставной сумкой с сильными
мышцами, дающими возможность иглам
вращаться в разных направлениях.
Форма игл различна (рис. 147). Они могут
быть гладкие, ребристые, шиповатые и
даже с отростками. Иногда их длина в
несколько раз превышает диаметр тела ежа.
Часто они используются не только как
защитное приспособление, но и для
движения. В последнем случае животное
опирается на них, как на ходули.
Среди обыкновенных игл разбросаны
особые хватательные органы — педи-
целлярии. Они устроены в виде
мельчайших подвижных щипчиков, сидящих
на бугорках скелетных пластинок, к
которым они прикреплены при помощи
известкового стебелька и мышц. Таким
образом, каждая педицеллярия морского ежа
состоит из стебелька и головки,
заключающей обычно 3 или 4 известковые створки,
форма которых характерна для
определенного вида морского ежа (рис. 147). Очень
часто головка соединяется со стебельком
посредством мягкой шейки. У самых
крупных педицеллярий, называемых
трезубыми (тридентными), длина
головки едва превышает 2—3 мм, тогда
как вместе со стебельком и шейкой они
бывают до 10 мм длиной и даже больше.
Другие педицеллярий ежей, например
змееголовые (офицефаль-
н ы е), получившие название за сходство
с головой змеи, и трилистные
(трифильные), имеющие створки
в виде листочка, гораздо мельче трезубых
педицеллярий. Лишь шароносные
(г л о б и ф е р н ы е) педицеллярий
вместе с мягкой оболочкой могут
конкурировать своими размерами с трезубыми
педицелляриями. Головка шароносных
педицеллярий бывает более 3 мм длиной,
она сильно расширена и заключает в себе
ядовитые железы, а створки этих
педицеллярий заканчиваются в верхней части
острыми зубцами. Железы могут
помещаться и на стебле педицеллярий. Вся
педицеллярия покрыта ресничным
эпителием и слоем соединительной ткани и
поэтому имеет мясистый вид. По внутренней
стороне створок проходят чувствительные
нервные волокна, а в стебельке находится
нервный центр, регулирующий
деятельность педицеллярий. Мышечные волокна,
проходящие в различных направлениях,
обеспечивают движение головки и работу
створок. Железы шароносных педицелля-
Таблица 24. Современные морские ежи и офиуры.
Морские ежи:
1 — Acanthocidaris maculicollis;
2 — Salenocidaris miliaris;
3 — Arbacia spatuligera;
4 — Echinosigra paradoxa.
О ф и у р ы:
5 — Astrochlamys bruneus;
6 — Ophiocoma delicata;
7 — Ophiura multispina.
18 Жизнь животных, т. 2
Рис. 146. Схема строения скорлупы морских
ежей: А — правильного (сводчатозубого); Б —
неправильного (сердцевидного) морского ежа
(со спинной стороны); В — того же
сердцевидного ежа (с брюшной стороны).
1 — апикальное поле; 2 — амбулакральные пластинки
с отверстиями для прохождения ножек; 3 —
амбулакральные ряды; 4 — интерамбулакральные ряды
пластинок; 5 — бугорки на пластинках для
прикрепления игл; 6 — иглы; 7 — мадрепоровая
пластинка; 8 — петалоиды; 9 — ротовое поле; 10 —
губа, или скребок; 11 — непарные перистомальные
интерамбулакральные пластинки; 12 — грудные
пластинки (стернум); 13 — эпистернум; 14 — анус; 15 —
фасциола.
рий вырабатывают ядовитую жидкость,
которая у отдельных видов столь
токсична, что доожет вызвать гибель различных
животных даже при сильном разведении.
Действие яда морских ежей на человека
было прослежено японским
исследователем Фудживара (Fujiwara),
уколовшим палец при исследовании морского ежа
Toxopneustes pileolus. Он сразу
почувствовал боль, которая стала
распространяться до сердца, затем наступил паралич
губ, языка и всего лица, потом
последовало онемение конечностей. Все это
длилось 6 часов, затем паралич стал
постепенно проходить. Такое действие яда
было вызвано уколом всего одного пальца,
а более значительные дозы яда могут
вызвать более сильные параличи, поэтому
японские рыбаки называют подобных ежей
«убийцами». С помощью ядовитых педицел-
лярий морские ежи борются с врагами,
например с морскими звездами, с педи-
целляриями которых они часто вступают
в борьбу при нападении последних.
Педицеллярии предохраняют также
поверхность ежа от загрязнения. В этом
случае, если частицы грунта или грязи
застрянут между иглами ежа, то они
моментально извлекаются оттуда рядом
расположенными щипчиками (педицеллярия-
ми), которые передают их соседним
щипчикам, а те все дальше и дальше, пока
удаляемые частицы не достигнут
амбулакральных ножек, которые и выталкивают
их в воду. Поэтому, несмотря на то что
многие ежи живут в прибойных местах,
где в воде много песка, обломков и
другого мусора, они всегда имеют
удивительно опрятный внешний вид.
Педицеллярии используются и при захвате добычи.
Стоит только какому-нибудь животному
коснуться ежа, как педицеллярии сразу
приходят в движение и пытаются его
схватить. Как только одному из щипчиков
удалось поймать жертву, он крепко
сжимает ее и держит до тех пор, пока она
не погибнет. После этого добыча
передается от педицеллярии к педицеллярии,
от амбулакральной ножки к
амбулакральной ножке, пока не достигнет рта.
Биологическое значение педицеллярии столь
велико, что они развиваются у ежей очень
рано, еще на личиночной стадии.
274
У всех правильных ежей хорошо
выражен лучистый план строения, поэтому
у них невозможно различить передний
и задний концы тела. Напротив, у
неправильных ежей в связи со смещением
анального отверстия из центра
апикального поля на край скорлупы или даже на
брюшную сторону животное приобретает
более или менее билатеральное строение
с хорошо различимым передним и задним
концами тела. При этом интеррадиус,
куда происходит смещение анального
отверстия, считается задним, а
противоположный ему радиус — передним.
У большинства неправильных ежей
амбулакральные ряды на спинной стороне
расширяются в особые образования —
пета л о и д ы, имеющие вид листочков
(рис. 146, Б). У некоторых групп
неправильных ежей они помещаются в
углублениях скорлупы и отличаются тем,
что несут парные поры, тогда как
остальные пластинки амбулакров имеют всего
одну непарную пору. Рот, обычно
помещающийся в центре брюшной стороны,
у неправильных сердцевидных ежей
смещается по переднему радиусу вперед.
К нему примыкают непарные пластинки
интерамбулакров, называемые п е р и-
стомальными. Одна из этих
пластинок, расположенных в заднем
интеррадиусе, имеет вид скребка или губы, при
помощи которой животное копается в
мягком грунте. Позади губы помещаются
крупные пластинки, образующие щит,
или пластрон (стернум).
Все пластинки панциря неправильных
ежей, подобно пластинкам правильных
ежей, несут бугорки, на которых
размещаются многочисленные иглы и педи-
целлярии. Местами на скорлупе
сердцевидных ежей можно обнаружить
скопление мелких бугорков в виде извилистых
полос, называемых фасциолями. На
мелких бугорках фасциолей сидят
маленькие, очень густо расположенные иголочки.
Несмотря на такую мощную защиту, как
панцирь, иглы, ядовитые педицеллярии,
морские ежи могут стать добычей крабов,
некоторых моллюсков, морских звезд,
крупных рыб, млекопитающих и птиц.
Весьма интересно обходится с ежами
морская выдра (калан). Она собирает в при-
Рис. 147. Скелетные элементы морского ежа:
1 — аристотелев фонарь; 2 — трезубая педицеллярия;
3 — трилистная педицеллярия; 4 — шароносная
педицеллярия; 5 — змееголовая педицеллярия; 6,7 —
створки трезубых педицеллярии; 8 — створка
змееголовой педицеллярии; 9 — створка шароносной
педицеллярии; ю — различные типы игл.
18*
275
брежных водах несколько морских ежей,
берет их в передние лапы и плывет на
спине, держа добычу на груди перед собой,
затем разбивает о скалы или другие
твердые предметы скорлупу ежей и съедает
половые железы.
Песцы и птицы тоже добывают
морских ежей во время отлива. Было
замечено, что птицы бросают собранных ежей
с высоты на камни, разбивают их и
выклевывают мягкие части, главным
образом икру.
Зрелые половые продукты некоторых
видов морских ежей очень питательны
и употребляются в пищу человеком в
сыром, жареном либо подсоленном виде.
Главный промысел съедобных ежей
производится по берегам Средиземного моря,
Америки, Новой Зеландии и в тропиках.
В Средиземном море их добывают около
10 млн. штук в год.
Ежи обладают высокой плодовитостью.
Так, самка съедобного морского ежа
Echinus esculentus в течение одного периода
размножения дает около 20 млн. яиц.
Развитие большинства морских ежей
сопровождается прохождением
плавающей личиночной стадии. В отличие от
личинок других классов иглокожих личинка
ежа (эхиноплутеус) имеет 12 очень
длинных отростков (рук) и сильно
развитый известковый скелетв виде
удлиненных палочек. Тело его билате-
ральносимметрично, но при дальнейшем
развитии претерпевает сложные
изменения, превращаясь в радиальносиммет-
ричное тело взрослого животного. При
этом только часть личинки,
преимущественно ее левая сторона, развивается в
зародыш, тогда как большая часть правой
стороны отмирает и отбрасывается.
Переход от билатеральной личинки к радиаль-
носимметричному взрослому организму
происходит через асимметричную стадию
и сопровождается полной
переориентировкой внутренних органов личинки,
значительная часть которых разрушается
и отбрасывается. Зародыш ежа как бы вы-
почковывается на теле личинки. Левая
сторона личинки становится
оральной стороной зародыша, а правая —
аборальной. Зародыш ежа на этой
стадии имеет равномерно развитую
оральную и аборальную стороны и напоминает
маленькую звезду. Но в дальнейшем
происходит отставание в развитии
аборальной стороны зародыша и разрастание
оральной стороны, что приводит к
образованию ежа, вполне сходного со
взрослым животным.
Растут ежи довольно медленно, полных
размеров достигают в течение
нескольких лет. В лабораторных условиях
наблюдения проводились над Psammechi-
nus miliaris, широко распространенным
в восточной части Атлантического океана.
Было замечено, что сразу после
метаморфоза маленький ежик имел скорлупу
диаметром всего в 1 мм. В месячном
возрасте его скорлупа уже была 10 мм
в диаметре, к году — 20 мм, в 2 года —
26 мм, в 3 года — 29 мм, в 4 года — 30 мм
и к 6 годам — 38,7 мм в диаметре.
Другие ежи могут расти быстрее. Так, Stron-
gylocentrotus droebachiensis, наиболее
широко распространенный в северных и
дальневосточных морях вид, к 6 годам
имел диаметр скорлупы около 60 мм,
a Echinus esculentus уже в 4 года
превышал 110 мм в диаметре.
В естественных условиях ежи растут
быстрее, чем в аквариумах, и размножаться
начинают уже к концу первого или в
начале второго года жизни. Самки и самцы
внешне совершенно сходны между собой.
Только те особи, которые проявляют
заботу о потомстве, обладают временным
половым диморфизмом, так как у самок
этих видов в период размножения на теле
помещается молодь, которую они
вынашивают. Такая забота о потомстве чаще
проявляется у видов, живущих в
Антарктике. У сердцевидного ежа Abatus philip-
pii (табл. 21) два передних петалоида
самки сильно углублены и превращены в
выводковые камеры, где развивается молодь.
У некоторых копьеносных морских ежей,
распространенных в Антарктике, местом
вынашивания молоди становится ротовое
поле, например у Ctenocidaris nutrix
(табл. 21). У близких к нему форм, тоже
обитающих в Антарктике (Stereocidaris
nutrix), молодь сначала вынашивается
около рта матери, а на более поздних
стадиях — на поверхности тела, где она
ползает во всех направлениях.
276
Живут морские ежи на различных
грунтах, но правильные ежи чаще встречаются
на скалах, камнях, коралловых рифах
и других более твердых грунтах, тогда как
неправильные ежи предпочитают мягкие
грунты, в которые они часто закапываются.
В удобных местах побережья морские
ежи собираются в таких количествах, что
иглы их соприкасаются друг с другом.
Некоторые виды занимают различные
углубления в скалах, другие же способны
сами сверлить себе укрытия, которые
служат им защитой от волн. Некоторые ежи
способны сверлить даже стальные сваи.
Нередко ежи прикрывают себя обломками
раковин, кусками водорослей или
мелкими камнями, чтобы, очевидно,
предохранить себя от действия прямого света или
замаскироваться от врагов. Есть виды,
которые целый день прячутся под камнями
и выходят питаться только ночью.
Правильные морские ежи едят главным
образом водоросли, но не брезгают и
животной пищей.
Экспериментальное изучение питания
Strongylocentrotus intermedins,
проведенное японскими исследователями у
северных берегов Японии, показало, что эти
ежи могут поедать разнообразные
водоросли и морскую траву. В период развития
половых продуктов, с июля по октябрь,
наблюдалось уменьшение потребления
пищи.
Отдельные группы ежей отдают
предпочтение определенному сорту пищи. Так,
копъеносные ежи (Cidaroidea) едят
животных с твердым внешним скелетом: мшанок,
корненожек, моллюсков и т. д., которых
разгрызают своими мощными зубами.
Представители семейства Diademati-
dae, живущие большей частью на
коралловых рифах, питаются кораллами с
сидящими на них животными. При питании
все правильные ежи придерживают пищу
иглами, подталкивают ее в рот и
откусывают небольшие куски зубами аристотелева
фонаря. Зарывающиеся в грунт
сердцевидные ежи собирают клейкими ножками
пищевые частицы с поверхности грунта,
поэтому их кишечник почти всегда
бывает набит диатомовыми водорослями, фо-
раминиферами, кусочками червей,
кишечнополостных, моллюсков.
Передвигаются морские ежи при
помощи амбулакральных ножек и игл. На
вертикальные стены они забираются только
благодаря своим ножкам, а по
горизонтальным поверхностям двигаются на
иглах, как на ходулях. Движение с
помощью игл совершается быстрее, чем при
помощи ножек. Так, Centrostephanus lon-
gispinus, «шагая» на своих длинных иглах,
продвигается на 150—200 см в минуту,
a Arbacia punctulata, двигающаяся при
помощи ножек, всего на 2 еле в минуту
и даже если очень спешит, то не более
чем на 4 еж в минуту. Скорость видов
рода Echinus при таком способе
передвижения несколько больше и может
достигать 15 см в минуту.
Двигаются морские ежи сравнительна
мало. Это довольно медлительные
животные, поэтому они часто становятся
объектом для поселения различных других
организмов, вплоть до паразитов.
Нередко на длинных иглах копьенос-
ных ежей можно встретить различных
моллюсков или усоногих раков. На рисунке
468 изображен один из представителей
этих ежей — Notocidaris gaussensis,
несущий на своих иглах целое скопище
ракушек. Многие животные находят себе
убежище на поверхности панциря и между
иглами ежей. Как было указано выше,
некоторые безногие голотурии встре-
Рис. 148. Морской еж Notocidaris gaussensis
с многочисленными моллюсками на иглах.
277
чаются на скорлупе копьеносных ежей,
а офиуры иногда поселяются близ рта
отдельных видов рода Strongylocent-
rotus. Часто, видимо, «воруют» пищу
у ежей различные многощетинковые
черви, постоянно встречающиеся на морских
ежах. В обильном количестве в кишечнике
правильных ежей встречаются
инфузории. На ежей нападают и мелкие
паразитические брюхоногие моллюски из с е-
м е й с т в Melanellidae и Stiliferidae. Они
могут внедряться в основание крупных
первичных игл копьеносных ежей,
образуя галлы, часто очень причудливой
формы, мешающие росту иглы. Некоторые
виды этих моллюсков прикрепляют свои
кладки к иглам Strongylocentrotus droe-
bachiensis и видам рода Echinus. На одном
еже может быть до 40 таких кладок.
Развивающиеся моллюски просверливают
скорлупу и просовывают свои хоботки
внутрь, питаясь тканями ежа.
Благодаря своему твердому панцирю
морские ежи прекрасно сохранились в
ископаемом состоянии. По этим остаткам
можно судить о том, что морские ежи
процветали уже около 500 млн. лет
назад, когда в морях еще не было рыб,
а на суше — наземной растительности.
Целые большие группы ежей известны
только в ископаемом состоянии.
Современные морские ежи, которых
насчитывается 800 видов, объединяются в
два подкласса: Правильные ежи
(Regularia) и Неправильные ежи (Irregu-
laria) — и относятся к 9 отрядам.
ПОДКЛАСС ПРАВИЛЬНЫЕ ЕЖИ (REGULARIA)
ОТРЯД LEPIDOCENTROIDA
К этому отряду относятся наиболее
примитивные формы современных ежей.
Основная масса его видов вымерла еще
в отдаленные геологические эпохи, и до
наших дней дошли лишь представители
семейства кожистых ежей (Echino-
thuriidae).
В отличие от других современных ежей
они обладают эластичной
скорлупой, способной спадаться, так как
пластинки их скелета не соприкасаются
друг с другом, а разобщены участками
кожи. Поэтому пластинки подвижны,
могут надвигаться друг на друга и менять
форму тела животного. Обычно
шаровидная в естественных условиях форма этих
животных становится совершенно диско-
видной при их вылове. Многие виды
кожистых ежей являются глубоководными
формами, но в районе Индо-Малайского
архипелага отдельные виды были найдены
в прибрежных водах. Это все крупные
формы с удивительно красивой
окраской. Как протекает развитие этих ежей,
неизвестно, но большие яйца ряда
глубоководных форм дают возможность пред-
положить, что развитие у них прямое, без
стадии пелагической личинки.
В Индийском океане на глубине свыше
1000 м была найдена Sperosoma biseria-
tum. Цвет ее скорлупы голубовато-серый,
а кожистые промежутки между
пластинками беловатые, амбулакральные
ножки у основания черно-фиолетовые, а на
вершине желтоватые.
Близкий вид, Araeosoma thetidis, живет
в прибрежных водах Новой Зеландии
и не заходит глубже 70 м. Взрослый еж
имеет сферическую скорлупу диаметром
около 20 см. Цвет ее красно-пурпурный.
Стоит только вытащить ежа на
поверхность, как он сразу спадается и
превращается в блин из-за потери телом
жидкости, выходящей через естественные
отверстия. На кончиках его игл имеются особые
железы. Укол такой иглой может вызвать
паралич и смерть. Примечательная
особенность этих ежей заключается в том,
что они питаются листьями наземных
растений, далеко уносимых ветром в море.
Любопытен своими защитными
приспособлениями жгучий кожистый еж (Asthe-
nosoma urens), обитающий в гавани Три-
комали (Цейлон). Прикосновение к ежу
вызывает жгучую боль, которая быстро
278
проходит. Дело в том, что
острые иглы этих ежей,
расположенные на интерамбулак-
рах, заключены в особые
утолщения, клетки которых
вырабатывают вещества,
вызывающие при
соприкосновении с иглой ощущение
укуса пчелы. Окраска особей
данного вида
красновато-коричневая с ярко-синими или
зеленоватыми возвышениями
и фиолетовыми иглами.
Весьма примечательно
строение игл третьего
представителя этого удивительного
семейства. У Hygrosoma luculentum (рис. 149),
найденного в районе Филиппинских
островов, близ Новой Гвинеи и в Индийском
океане у Андаманских островов на
глубинах от 200 до 400 м, ротовая сторона
плоская. На ней на крупных первичных
бугорках имеются иглы длиной в 35 мм
с характерным копытообразным
расширением на конце. Сам еж темно-пурпурный
с белым рисунком и белыми «копытами».
ОТРЯД КОПЬЕНОСНЫЕ ЕЖИ
(CIDAROIDEA)
Чрезвычайно эффектны виды этого
отряда, обладающие толстыми, очень
длинными, редко расположенными иглами
интерамбулакров, заметно
отличающимися от мелких, невзрачных игл
амбулакров. Форма этих игл сильно изменчива.
Они могут иметь грани, выросты,
бугорки и т. д. и могут быть похожими на
маленькие копья. Отсюда и название отряда
«копьеносные». Характерным признаком
его представителей является то, что
ротовое поле у них не затянуто мягкой
кожей, а сплошь покрыто
амбулакральными и интерамбулакральными
пластинками. Амбулакральные пластинки
простые и обычно с одной парой пор для
выхода амбулакральных ножек. Наружные
жабры, сферидии, змееголовые и три-
листные педицеллярии у них отсутствуют.
Отряд содержит одно семейство —
Cidaridae, включающее около 30 родов
Рис. 149. Мягкий еж Hygrosoma luculentum.
и свыше 120 видов, распространенных по
всем океанам и морям, за исключением
Арктики. Они встречаются как на
литорали, так и на больших глубинах около
4 тыс. м, но основная масса видов
предпочитает глубины менее 1000 м и даже менее
500 м. Наиболее известен из этого
семейства обыкновенный копьеносный морской еж
(Cidaris cidaris). Его шарообразная
скорлупа, диаметр которой может достигать
65 мм, вооружена длинными
копьевидными иглами, превышающими почти в 2 раза
размер скорлупы. Они находятся
исключительно на интерамбулакральных
пластинках по 5—9 игл в каждом ряду. Вдоль
всей длины этих копий имеются ребра,
образованные рядами мельчайших
бугорков. Окраска таких игл ярко-розовая,
а мелких иголочек амбулакров
желтоватая либо серовато-белая. Распространен
этот вид вдоль европейского берега
Атлантического океана от Норвегии до
Азорских островов, а также в Средиземном
море. Его часто добывают рыбаки на
глубинах от 50 до 1800 м. Живет он на
самых разнообразных грунтах, питается
донными животными, имеющими хорошо
развитый скелет. В его кишечнике часто
находили фораминифер, губок, мшанок,
горгонид, обломки ракушек. При питании
еж придерживает добычу иглами ротовой
стороны и откусывает кусочки зубами.
Очень интересно передвигается этот еж по
субстрату. Он опирается на подвижные
околоротовые иглы и поддерживается
более длинными боковыми иглами скорлупы.
Амбулакральные ножки при движении
используются только молодыми особями.
279
Очень сходен с предыдущим видом Sty-
locidaris affinis (табл. 17), известный в
Средиземном море, тропических районах
Атлантики, как по побережью Африки, так
и в Мексиканском заливе и близ Флориды.
Этого изящного ежа находили на
глубинах от 30 до 1000 м. Примечательна
окраска его крупных игл с хорошо заметными
коричневатыми кольцами на светлом фоне
и красная расцветка вторичных игл и игл
амбулакров.
Еще более красив Acanthocidaris macu-
licollis (табл. 24), встречающийся в Тихом
океане от Японии до Индо-Малайского
архипелага, а также в Индийском океане
на глубинах от 40 до 225 м. Его крупные
толстые иглы, длина которых может быть
более 10 см и равна двум-трем диаметрам
скорлупы, имеют в своем основании очень
примечательный орнамент. Это украшение
состоит из нескольких гребней,
снабженных зазубринками, окрашенными в
светлые тона, и серии красных пятен,
разделяющих такие зазубренные возвышения.
Подобной раскраски нет ни у одного вида
данного семейства.
Очень крупными размерами отличается
еще один представитель копьеносных
ежей. Громадные экземпляры Phyllacant-
hus imperialis (табл. 19) часто
встречаются на коралловых рифах в
тропической части Индийского и Тихого океанов.
Его толстые, грубые, но малочисленные
иглы часто бывают усажены
паразитическими брюхоногими моллюсками. Очень
часто эти ежи днем прячутся под глыбами
кораллов, а ночью выходят на своих
ходулях для промысла пищи.
ОТРЯД АУЛОДОНТА (AULODONTA)
Основная особенность представителей
данного отряда заключается в том, что
зубы у них простые, без килеобразного
выступа, не имеют на внутренней стороне
характерного желобка. К этому отряду
относится широко известное с е м е й с т-
в о диадемовых (Diadematidae), виды
которого пользуются дурной славой из-за
своих весьма опасных игл. Скорлупа ежа
диадемы средних размеров снабжена
чрезвычайно длинными тонкими
многочисленными иглами, длина которых может
достигать 30 см. Иглы являются прекрасным
защитным приспособлением. Они очень
подвижны, моментально реагируют на
раздражение, включая простое затемнение,
и сразу направляются в сторону
раздражителя. При этом животное даже
складывает несколько игл вместе, образуя опас1-
ную пику. Темный, почти черный цвет
игл делает их малозаметными, ибо концы
игл даже тоньше кончика булавки. Но
стоит только случайно коснуться этого
конца, как игла моментально впивается
в тело, и вытащить ее нет никакой
возможности.
Все иглы полые внутри, легко
обламываются, а по всей длине усажены
спирально расположенными шипиками,
острие которых направлено назад,
поэтому игла легко и глубоко входит в тело»
К этому необходимо добавить, что кожа
вокруг игл имеет железистые клетки,
выделяющие ядовитый секрет красноватого
цвета, который при ранении иглой еще
больше усиливает боль. Часто такая
заноза осложняется тяжелым нагноением,
поэтому освободиться от нее лучше сразу,
вырезав иглу из тела.
Виды семейства диадемовых широко
распространены в тропиках и
прилежащих к ним районах Индийского, Тихого
и Атлантического океанов. На небольших
глубинах Индийского и Тихого океанов
встречаются Diadema setosum (табл. 19),
Diadema savignyi. Они предпочитают
селиться на коралловых отмелях,
образованных обломками кораллов, и поражают
великолепием своей окраски. Их темная
бархатистая скорлупа темно-коричневого
и темно-пурпурного оттенка украшена
изумительными по красоте голубовато-синими
пятнами. Эти пятна снабжены
чувствительными клетками и помогают животному
воспринимать малейшую тень, падающую от
приближающегося врага. Обычно
животное спокойно лежит в коралловом песке
и медленно вращает свои длинные иглы,
охраняя себя от врагов.
Очень интересное явление наблюдается
у Diadema antillarum, встречающейся в
западной части Атлантического океана в
районе Флориды. Она способна менять
280
свою окраску в зависимости от освещения.
Так, совершенно темная при дневном
свете, за каких-нибудь полтора часа в
темноте она становится бледно-серой с белой
звездой вокруг апикального поля,
окаймленной радужным синим краем,
продолжающимся на амбулакры.
Подобное явление было замечено и
у другого представителя данного
семейства, у Centrostephanus longispinus,
распространенного в Средиземном море и
прилегающих к нему частях Атлантического
океана. Этот удивительно красивый еж
с темно-пурпурной скорлупой и длинными
тонкими фиолетовыми иглами,
опоясанными белыми узкими колечками, в
темноте за полчаса становится бледно-серым.
В отличие от своих близких
родственников, передвигающихся с помощью
амбулакральных ножек, он ходит на длинных
иглах, причем довольно быстро,
передвигаясь на 100—150 см в минуту.
ОТРЯД КОПЬЕВИДНОЗУБЫЕ ЕЖИ
(STIRODONTA)
К этому отряду относится 8 семейств,
включающих современные и ископаемые
формы.
Сюда принадлежат виды с компактным,
неподвижным панцирем, покрытым
длинными, прочными иглами без полости
внутри. Зубы у них с внутренним килем,
копьевидные. Из педицеллярий развиты
трезубые и змееголовые.
Древнее семейство Salenidae,
расцвет которого относится к юрскому
периоду, удаленному от наших дней
приблизительно на 155 млн. лет, имеет своих
представителей и в современных морях.
Они относятся к двум родам: Salenia
и Salenocidaris. SaJenia pattersoni
известен в западной, тропической части
Атлантического океана, отличается характерной
окраской своих длинных игл, состоящей
из чередующихся красных и белых колец.
Salenocidaris miliaris (табл. 24)
распространен на больших глубинах от 1200 до
3 тыс. м в центральной части Тихого
океана, от Японии до Галапагосских островов.
Как все представители этого семейства,
он имеет шиповатые первичные иглы
только на интерамбулакральных
пластинках. Длина этих игл может быть в 4 раза
больше горизонтального диаметра.
Вторичные же иглы короткие и плоские.
Более известны виды семейства
Arbaciidae (табл. 24), которые снабжены
сравнительно короткими, гладкими,
крепкими первичными иглами. Их может быть
больше одной на каждой
интерамбулакральной пластинке.
Почти у всех видов наружный слой
(чехол) на первичных иглах исчезает и
сохраняется лишь в виде небольшой «чашечки»
на иглах ротовой стороны. Интересным
распространением обладают виды рода
Arbacia. Они встречаются только у
западного и восточного берегов Америки и по
Атлантическому побережью Африки, от
Анголы до Средиземного моря, включая
последнее. Все 6 видов этого рода обитают на
мелководьях и преимущественно на
каменистых грунтах.
Весьма обычен в Средиземном море,
у берегов Африки и у Азорских островов
черный морской еж (Arbacia lixula). Живет
он в прибойных местах на скалах и
питается известковыми водорослями. Он часто
забирается в узкие расщелины между
камнями, где бывает недосягаем для
своих врагов — морских звезд. Ножки
его снабжены чрезвычайно сильными
присосками, с помощью которых он может
взбираться по совершенно отвесным
скалам в поисках пищи. Ежи эти, диаметром
до 5 см, густо покрыты черными иглами
длиной в 2 см или немного больше, но
в темноте они изменяют свой цвет и
становятся коричневыми.
ОТРЯД СВОДЧАТОЗУБЫЕ ЕЖИ
(CAMARODONTA)
Один из обширнейших отрядов морских
ежей, включающий несколько семейств,
широко распространенных в современных
морях, в том числе и семейство
Strongylocentrotidae, виды которого
встречаются и в нашей фауне.
Представители этого отряда найдены
в отложениях еще мелового периода, тем
281
не менее и в настоящее время они весьма
обычны среди обитателей морского дна.
Название отряда связано с наиболее
характерным отличительным признаком
его представителей. Все они имеют зубы,
снабженные внутренним килем, и
замкнутые, в виде свода, отверстия зубных
пирамидок аристотелева фонаря.
Значительная часть его видов весьма интересна
с точки зрения биологии,
распространения или хозяйственного использования.
К нему относится и основная масса
съедобных форм.
Самое известное семейство Stron-
gylocentrotidae, распространенное в
основном в арктических и умеренных районах
северного полушария, представлено в
наших морях восемью видами. Среди них
прежде всего следует назвать
обыкновенного ежа (Strongylocentrotus droebachien-
sis), очень обычного в Арктике и в
северных частях Тихого и Атлантического
океанов от литорали до глубин свыше 1000 ле
(табл. 20). Его часто используют в пищу
морская выдра, песцы и птицы.
На литорали южной части Японского
моря, вдоль северо-западного берега
Японии и в Желтом море можно видеть
крупных темных ежей, медленно шагающих
на своих длинных черных иглах. Это
Strongylocentrotus nudus (табл. 25).
Его ближайший родственник — пурпур-
ный стронгилоцентротус
(Strongylocentrotus purpuratus), по сообщению И р в и-
н a (Irvin), проделывает большое
количество нор в стальных сваях портовых
сооружений Тихоокеанского побережья
Калифорнии. Этот средних размеров еж
(табл. 25) покрыт значительным
количеством крепких, длинных, пурпурного цвета
игл, вращая которыми он высверливает
себе углубление. Очевидно, и зубы помогают
ему в этой работе.
В районе Калифорнии и к югу от нее
распространен самый крупный вид из
современных ежей — Strongylocentrotus
franciscanus. Размеры его скорлупы
примерно равны детской голове (превышают
16 см в диаметре). Икра его
употребляется в пищу человеком, поэтому промысел
его с каждым годом увеличивается.
В больших количествах у берегов
Японии добывают другого представителя
данного семейства — Hemicentrotus pul-
cherrimus. Размеры его много меньше
предыдущего вида: диаметр его скорлупы
едва достигает 4 см, однако промысел
составляет около 16 тыс. т в год.
Значительное количество добываемых
ежей относится к следующему
семейству данного отряда — Echinidae,
отличающемуся главным образом строением
створок шароносных педицеллярий.
Наиболее характерный род семейства —
Echinus — содержит 17 видов, половина
из которых распространена в
Атлантическом океане. К этому роду относится
и съедобный морской еж (Echinus escu-
lentus), добываемый у берегов Португалии
и в некоторых районах Англии, в
Северном море. Распространен он от Баренцева
моря до берегов Испании, предпочитает
селиться в прибрежных водах на
глубинах от литорали до 40 м, реже до 100 ле,
но известны и случаи его нахождения
до 1200 ле. Внешний вид его очень красив.
Он обладает крупной, до 16 см в диаметре,
шарообразной скорлупой красноватого
цвета, покрытой короткими тонкими
красноватыми иглами с фиолетовыми
кончиками и большим количеством
педицеллярий, при помощи которых животное
содержит в чистоте скорлупу, а также добывает
себе пищу. Съедобный морской еж почти
всеяден. Кишечник его всегда плотно
набит различными водорослями, особенно
морской капустой (Laminaria), а также
остатками различных мелких животных:
усоногих раков, гидроидных полипов,
мшанок и даже морских ежей. Благодаря
этому его легко держать в аквариумах.
В спокойном состоянии он долго может
сидеть на дне аквариума, вытянув вверх
целый лес своих амбулакральных ножек.
При помощи ножек, игл и педицеллярий
он доставляет пищу ко рту. Любопытно,
что двигается этот еж довольно часто
с помощью своих зубов. При этом зубы
погружаются в субстрат, смыкаются и
приподнимают ежа, затем он
подталкивается иглами и продвигается вперед.
Двигаясь с помощью амбулакральных
ножек, он может проходить 15 см за одну
минуту.
Вдоль европейского берега
Атлантического океана от Норвегии до Марокко
282
можно встретить берегового морского ежа
(Psammechinus miliaris). Он довольно
обычен на устричных банках и в
прибойных местах. Сильные волны ему не
страшны, так как с помощью своих грубых
коротких игл он проделывает углубление
в грунте, где и скрывается от них.
Диаметр его скорлупы не более 50 мм, цвет
зеленоватый, иглы зеленые с фиолетовыми
кончиками. Питаясь всевозможной
животной пищей (гидроидами, сидячими по-
лихетами, молодыми устрицами и т. д.),
он, подобно морским звездам, наносит
вред устричным хозяйствам. Этот еж так
всеяден, что в аквариумах ел асцидий,
дохлую рыбу, икру, сырое мясо, раков,
дохлых крабов, мягкие части моллюсков,
мшанок, червей, гидроидов, губок,
различные водоросли, в том числе и
известковые, а также пустые раковины. Были
случаи, когда он жил в аквариуме 3 года.
При кормлениях в неволе пищу кладут
прямо на спину животному, тогда он
быстро начинает передвигать ее в рот с
помощью ножек и игл.
Наиболее известным сверлильщиком
скал является скальный морской еж (Рага-
centrotus lividus), распространенный от
берегов Англии до Африки, включая
Средиземное море. Он образует часто
огромные скопления на наклонных скалистых
поверхностях и в зарослях морской
травы. Его можно встретить от литорали
до глубины в 30 м. Любопытно, что
средиземноморская раса этих ежей несколько
отличается своим поведением от
атлантической расы. Так, особи, живущие в
Атлантике, поселяются в углублениях скал,
проделанных ими с помощью игл и зубов.
Напротив, в Средиземном море они
никогда не сверлят скалы, а селятся на
слабонаклонных поверхностях, прикрыв себя
кусками ракушки, морской травы и
другими предметами. Это связано, очевидно,
с тем, что в Средиземном море прибой
слабый, а в Атлантике очень сильный,
поэтому ежи и делают себе защиту от
волн. Иногда они оказываются
замурованными в своих убежищах, так как вход
в норку становится уже тела ежа.
Спасаясь от волн, маленький еж делает себе
в скале убежище и остается там на
длительное время. Тело его растет, он
расширяет вокруг себя углубление, но вход
оставляет прежним и через некоторое
время становится пленником своего дома,
питаясь только тем, что принесут ему
волны в норку. Ежи эти растительноядны,
они питаются различными водорослями
и морской травой. Скорлупа их достигает
7 см в диаметре. Окраска ее варьирует
от темно-фиолетового до
зеленовато-коричневого цвета. По некоторым наблюдениям,
самцы и самки отличаются по цвету:
самцы более темные, а самки более яркие, их
скорлупа красновато-фиолетовая. Половой
диморфизм проявляется и в очертании
панциря, более плоского у самок.
Икрометание у них происходит небольшими
порциями в течение всего лета.
Для многих животных этот еж опасен.
Его педицеллярии ядовиты. Вытяжка из
30 педицеллярии быстро убивала краба
длиной в 4—5 см. Однако иглокожие
любого класса оказались невосприимчивыми
к этому яду. Для человека этот яд тоже
безопасен. Икру скального морского ежа
употребляют в пищу. Основной его
промысел ведется на Средиземном море.
По берегам Чили и Эквадора добывают
белого локсехинуса (Loxechinus albus),
икру которого едят в жареном виде.
В Атлантическом океане рыбаки
острова Мартиника ловят на корраловых
рифах, ограничивающих большую лагуну,
морского ежа — Tripneustes ventricosus,
относящегося к семейству Тохор-
neustidae. Добывают его либо
ныряльщики, либо с плотов при помощи
бамбуковой палки, расщепленной на конце.
Собранных ежей на берегу вскрывают,
вынимают из скорлупы икру и варят ее в
котле на слабом огне, пока она не будет
иметь вид густой массы цвета пчелиного
воска, после чего ее снова закладывают
в очищенные скорлупки. Эта вареная икра
продается разносчиками поштучно.
Ежегодно креольское население потребляет
такое большое количество ежей, что
местами на острове их панцири образуют
целые горы.
Наряду со съедобными ежами в этом
семействе имеются виды, яд которых
опасен не только для различных животных,
но и для человека. Все они
распространены в прибрежных водах тропических
283
частей различных океанов. Наиболее
обычен на литорали в прибрежных водах
Индийского и Тихого океанов, от
Красного моря и восточного берега Африки до
Японии, Австралии и островов Фиджи,
Toxopneustes pileolus. Действие его яда
на человека было описано выше. Его
крупная скорлупа покрыта зеленоватыми
иглами с белыми или красными
кончиками, хотя окраска может сильно
варьировать и бывает иногда даже красной или
пурпурной. Среди этих игл разбросано
большое количество мелких глобиферных
педицеллярий, створки которых всегда
раскрыты (табл. 19). Крупные глобифер-
ные педицеллярий встречаются реже
мелких, створки их всегда сомкнуты. Длина
их головки достигает 2—3 мм. Под
головкой на стебельке имеются железы,
которые выделяют беловатый секрет,
чрезвычайно ядовитый как для животных, так
и для человека. Ядовитая жидкость этого
секрета была названа токсопнеус-
тес-токсин.
В Средиземном море и в прилегающих
к нему районах Атлантического океана
живет темно-фиолетовый морской еж
(Sphaerechinus granulans). Этот вид,
распространенный преимущественно на
литорали, очень красив. Его крупная,
диаметром в 13 см, скорлупа имеет пурпурную
окраску с более светлыми зонами на амбу-
лакрах и зеленоватым апикальным полем.
На скорлупе расположены фиолетовые
или пурпурные иглы, совершенно белые
на своих концах. Он часто забирается в
расщелины между скалами, но сам
никогда не делает норок. Подобно многим
мелководным животным, часто
прикрывает себя кусками водорослей, ракушек
или другими предметами. Обычно он
медленно ползает среди зарослей водорослей,
питаясь ими. Иногда собирает детрит с
находящимися в нем мелкими
организмами. Его ядовитые глобиферные
педицеллярий являются защитным
приспособлением против нападения главных врагов —
морских звезд. Ему удается спастись, если
нападает лишь одна звезда, но при
нападении нескольких хищников даже
ядовитые педицеллярий не спасают.
Среди представителей семейства
Echinometridae особенно интересны ежи
Heterocentrotus mammillatus (табл. 18)
и Colobocentrotus atratus (табл. 18). Оба
вида широко распространены на литорали
тропического района Индийского и Тихого
океанов и удивительно приспособлены для
жизни в этих своеобразных условиях
сильного прибоя.
Heterocentrotus mammillatus обладает
очень толстыми грубыми иглами,
помогающими ему рыть пещерки на литорали
коралловых рифов. Проделывает он это
главным образом иглами ротовой стороны,
концы которых снабжены тонкими
зубцами. Норка эта столь мала, что
животное в ней еле-еле поворачивается. Часто
растущий еж остается на всю жизнь
замурованным в своей пещере и питается
только тем, что заносит ему в убежище
морской прибой, поэтому норки этого ежа
бывают буквально вылизаны.
Colobocentrotus atratus (табл. 18) более
своеобразно приспособился к жизни в
полосе сильного прибоя. Скорлупа его
низкая, овальная, вооружена короткими
многоугольными иглами. По краю ротовой
стороны расположены лопатовидные
иголки. Такая плоская ротовая поверхность
панциря вместе с направленными косо
вниз лопатовидными краевыми иглами и
многочисленными амбулакральными
ножками создает столь мощный
присасывательный диск, что отцепить ежа от скалы
можно только с помощью ножа. Низкая
же аборальная поверхность скорлупы,
вооруженная короткими многоугольными
иглами, прекрасно противостоит действию
волн. Питается этот еж различными
организмами, живущими в тех же условиях,
например известковыми водорослями.
Под ним часто прячется планария Сега-
topleana colobocentroti, которая без него
не удержалась бы в прибое.
Комменсалами его являются также маленький
крабик Proechinoecus dimorphicus и
один вид моллюсков.
Интересен сожитель морского ежа
Caenocentrotus gibbosus,
распространенного на литорали Перу и Чили. В прямой
кишке ежа поселяется краб Pinaxodes
chilensis, который, разрастаясь, может
деформировать кишку и причинить ежу
вред, хотя сам получает прекрасную
защиту.
284
ПОДКЛАСС НЕПРАВИЛЬНЫЕ ЕЖИ (IRREGULARIA)
Два отряда этого подкласса —
Holectypoida и Cassiduloida,
представители которых процветали в юрский и
меловой периоды мезозойской эры, почти
нацело вымерли. Из многочисленных
представителей неправильных ежей до наших
дней дошли только немногие, довольно
редко встречающиеся виды.
ОТРЯД ПЛОСКИЕ ЕЖИ
(CLYPEASTROIDA)
Это обширный отряд, представленный
многочисленными ископаемыми и
современными формами.
Отличительным признаком видов этого
отряда является плоская, почти дисковид-
ная скорлупа, иногда имеющая по краям
вырезы, как у Rotula orbiculus (табл. 17),
или даже сквозные отверстия, как,
например, у Mellita sexiesperforata.
На спинной стороне у представителей
этого отряда имеется характерная фигура
в виде пяти лепестков цветка петалои-
ды, или петалодии, образованная
амбулакральными рядами. Анальное
отверстие перемещается на край скорлупы
или на брюшную сторону, в центре
которой расположен рот, снабженный
крупными зубами.
Весь панцирь плоских ежей густо покрыт
мелкими иголочками, придающими ему
щетинистый либо бархатистый вид.
Этот отряд, насчитывающий большое
число видов, распространен почти
исключительно в тропических морях. В наших
водах встречаются только виды рода
Echinarachnius, в том числе и амфибо-
рально распространенный Echinarachnius
parma.
Большинство плоских ежей живет в
мягком грунте, где может двигаться во
всех направлениях, как, например,
Echinarachnius parma. Он при помощи своих
иголок накапывает на себя грунт и за. 10—
15 минут может скрыться из виду.
Сведения о питании плоских ежей
весьма скудны. Видимо, питательные частицы
и микроорганизмы направляются
действием ресничек по бороздам брюшной
стороны в рот. В кишечнике плоских ежей
можно обнаружить мелкие песчинки,
кусочки растений, диатомовых водорослей,
корненожек, мелких копепод и т. д.
ОТРЯД СЕРДЦЕВИДНЫЕ ЕЖИ
(SPATONGOIDA)
Ежи этого отряда отличаются своей
причудливой и удивительно разнообразной
формой, напоминающей то яйцо, то
бутылку, то вазу, то сердце. У них нет
аристотелева фонаря, а рот снабжен губами
или одной губой, приспособленной для
копания грунта.
Этот обширнейший отряд,
представленный в современной фауне 12 семействами,
имеет широкое распространение по всем
морям. Виды его встречаются как в
прибрежной полосе, так и на больших
глубинах. Рассмотрим наиболее интересных
его представителей.
Самый обычный на глубинах от
литорали до 230 м в умеренных широтах
Атлантического и Тихого океанов вид
настоящий сердцевидный еж (Echinocardium cor-
datum), относящийся к семейству
Loveniidae.
Этот еж живет зарывшись в песчаный
грунт (рис. 150, 2), где делает ходы,
укрепляя их стенки слизистыми выделениями.
Он зарывается вертикально в грунт с
помощью своих боковых игл
приблизительно на 20 см в глубину. Когда еж сидит
в грунте, то с поверхностью его связывает
вертикальный ход, сцементированный
слизью. Через этот ход в яму поступает
свежая вода, необходимая для дыхания.
Кистеобразные передние ноги животного
сильно вытягиваются и высовываются
через трубку наружу. Ими еж собирает с
поверхности грунта мельчайшие пищевые
частицы (рис. 150). Липкие пальцеобраз-
285
Рис. 150. Закапывание морских ежей в грунт:
1 — Echinocardium cordatum; 2 — Mellita longifissa.
ные выросты этих ножек довольно быстро
собирают нужное количество пищи и,
втягиваясь обратно в норку, передают
эти частицы иглам на верхней губе,
которые направляют пищевые частицы в рот.
В то же самое время задние ножки
растягиваются на несколько сантиметров назад
в заднюю трубку и способствуют лучшему
удалению экскрементов.
Сердцевидный еж в поисках пищи
медленно ползет в грунте через песок,
отталкиваясь своими веслообразными
брюшными иглами. При этом задняя трубка
осыпается, а верхняя (дыхательная)
трубка делается заново. Движение игл
вызывает круговорот воды в ямке,
помогающий поступлению все новых порций,
чистой содержащей кислород воды,
необходимой для дыхания.
Этот еж редко появляется на
поверхности, так как рискует быть унесенным
волнами ближайшего прилива.
Пурпурный сердцевидный еж (Spatangus
purpureus), относящийся к семейст-
в у Spatangidae, делает менее глубокие
ходы. Он живет чаще на битой ракушке
и углубляется всего на 5 см от
поверхности, не образуя никаких
дыхательных ходов. Этот крупный
еж, достигающий в длину 12 см,
имеет пурпурный панцирь и
более светлые, даже белые,
изогнутые иглы верхней стороны.
Распространен он в северной
Атлантике вдоль европейского берега до
Азорских островов и
Средиземного моря и может опускаться до
глубины в 900 м.
Размножается этот еж, как
большинство его собратьев,
откладывая яйца в воду, где они
проходят личиночную стадию
эхиноплутеуса,
характеризующуюся длинным задним
отростком. Такие личинки в
северной Атлантике встречаются в
летние месяцы.
В Тихом океане, в Беринговом
море, вдоль американского берега
от полуострова Аляска до Калифорнии
распространен Brisaster townsendi (табл.
25). Этот вид, относящийся к семейст-
в у Schizasteridae, характеризуется сильно
углубленными петалодиями и тем, что
самая широкая часть скорлупы находится
впереди. Живет он на глубинах от 120 до
1043 м.
Виды отряда, встречающиеся часто на
больших глубинах океанов и морей,
принадлежат к семейству Pourtale-
siidae, характеризующемуся вытянутой
формой тела, имеющей на конце хобото-
образный выступ.
В Атлантическом океане на глубинах
свыше 1500 м обитает интересный, почти
бутылкообразной формы еж — Echinosig-
ra paradoxa. Его хрупкая скорлупа едва
достигает в длину 3,5 см (табл. 27).
На глубинах Тихого океана от 2150
до 5200 м распространен яйцевидный
морской еж (Aeropsis fulva). Его хрупкое
овальное тело, заметно суживающееся к
заднему концу, имеет сильно
выступающий передний амбулакр. Вся скорлупа
покрыта нежными иглами, расширенными
на конце (табл. 22).
286
КЛАСС ОФИУРЫ,
ИЛИ ЗМЕЕХВОСТКИ (OPHIUROIDEA)
Офиуры внешне сходны с морскими
звездами. Тело их звездообразной формы,
но, в отличие от последних, с сильно
обособленными от диска и очень подвижными
членистыми лучами, напоминающими
хвосты змей. Отсюда и название этого класса
«змеехвостки». Не только внешнее
сходство, но и ряд черт внутренней
организации сближает офиур со звездами. На
основании этого некоторые зоологи
объединяют их в один класс. Однако в
наиболее известных руководствах офиуры
рассматриваются как самостоятельный класс,
представленный в современной фауне
двумя отрядами — Euryalae и Ophiurae,
насчитывающими свыше 250 родов и около
2 тыс. видов.
Ныне живущие офиуры отличаются от
морских звезд прежде всего тем, что у них
отсутствуют открытые амбулакральные
борозды, так как в процессе
филогенетического развития у них появились
пластинки, закрывшие их. Кроме того, лучи
у офиур столь резко обособлены
от диска и разрастание
пластинок лучей такое сильное, что
полость тела в лучах весьма
сокращена и, в отличие от звезд,
отростки желудка не заходят туда.
Змеехвостки — обычные
обитатели морского дна. Их можно
встретить как в прибрежной зоне,
так и на больших глубинах* Самые
глубоководные из них найдены на
глубине в 6720 м. Некоторые из них
являются настоящими абиссальными
видами и не встречаются выше 2000—2500 м,
но большинство форм обладает большой
амплитудой распространения по
глубинам, у отдельных видов даже
превышающей несколько тысяч метров. Живут они
на самых разнообразных грунтах. Многие
литоральные и более мелководные виды
часто прячутся под камнями, в губках
и кораллах или среди водорослей.
Некоторые из них, особенно представители
семейства Amphiuridae,
зарываются в песок либо в ил, выставляя над
поверхностью грунта лишь кончики своих
лучей. Местами офиуры встречаются в
огромных массах и являются руководящими
формами некоторых донных сообществ
(биоценозов). Так, например, в Японском
море, вдоль берегов острова Сахалин,
на глубинах от 100 до 200 м огромные
пространства заняты биоценозом, в
котором первостепенную роль играет
змеехвостка Ophiura sarsi. Встречаясь в боль-
Рис. 151. Схема строения наружного
скелета офиур.
А — вид с ротовой стороны: 1 — рот; 2 —
челюсть; з — ротовые папиллы; 4 — зубы;
5 — адоральный щиток; 6 — ротовой щиток;
7 — чешуйки диска; 8 — бурсальная щель;
9 — луч; ю — брюшные щитки лучей;
11 — боковые щитки лучей; 12 — иглы луча;
13 — амбулакральные поры для выхода
ножек; 14 — амбулакральные чешуйки.
Б — часть диска и луча со спинной стороны:
15 — радиальные щитки; 16 — спинные
щитки лучей. В — различные типы строения
ротового угла офиур; 17 — непарная и 18 —
парная инфрадентальные папиллы; 19 —
зубные папиллы.
287
ших количествах, офиуры имеют
существенное значение в общей экономике моря,
поедая множество мелких морских
организмов и являясь в свою очередь пищей
различных донных рыб.
Тело большинства змеехвосток
уплощено и состоит из округлого или
пятиугольного диска и пяти, реже шести или
семи—девяти лучей (табл. 24). Лучи часто
очень длинные и в несколько раз (а у
наиболее длиннолучевых форм даже в 20 раз)
могут превышать диаметр диска, средние
размеры которого около 1—2 см, но у
отдельных видов ветвистолучевых офиур
достигают 10 см. Обычно лучи простые,
суженные к концу, однако у части офиур,
таких, например, как голова горгоны
(табл. 17), они ветвятся.
Характерной особенностью змеехвосток
является сильное развитие у них не только
внешнего (в виде тонких пластинок), но
и внутреннего скелета лучей, состоящего
из отдельных позвонков, скрепленных
между собой специальными мышцами и
связками. У одних офиур сочленовная
поверхность позвонков, гладкая, в виде
часового стекла, поэтому лучи их
чрезвычайно подвижны, могут сворачиваться
на брюшную сторону и обвиваться вокруг
различных предметов. У других офиур
позвонки имеют более сложные
соединения в виде ямок и бугорков, в таких
случаях лучи менее подвижны и могут
изгибаться главным образом в горизонтальной
плоскости. Снаружи позвонки прикрыты
тонкими скелетными пластинками,
образующими 4 ряда вдоль каждого луча.
Среди них различают брюшные щитки,
прикрывающие амбулакральные борозды,
спинные щитки, противоположные
брюшным, и два ряда боковых щитков, несущих
различно устроенные иглы лучей
(рис. 151). Иногда пластинки лучей могут
быть недоразвиты или вообще
отсутствовать.
На брюшной стороне луча в
каждом членике имеются два отверстия для
выхода амбулакральных ножек. Ножки
офиур лишены присосок и ампул и
выполняют главным образом функцию дыхания
или осязания.
Диск змеехвосток, подобно лучам,
снаружи покрыт пластинками, имеющими
вид чешуек. Среди них на спинной
поверхности у основания лучей выделяются
более крупные парные пластинки —
радиальные щитки. Иногда чешуйки
диска бывают снабжены различными
иголочками, гранулами, бугорками, а
радиальные щитки имеют вырез, несущий
мелкие иголочки, так называемые
папиллы дискового выреза.
Таблица 25. Иглокожие северной части Тихого океана.
Морские звезды:
1 — Pycnopodia heliantoides;
2 — Strongylocentrotus nudus;
3 — Crossaster papposus;
4 — Distolasterias nipon;
5 — Evasterias retifera;
6 — Patiria pectinifera.
Офиуры:
7 — Strongylocentrotus purpuratus;
8 — Amphiodia rossica;
9 — Psolus chitonoides.
Морские ежи:
10 — Ophiacantha bidentata;
11 — Brisaster townsendi;
12 — Stichopus japonicus .
Голотурии:
13 — Astrometis sertulifera;
14 — Amphiophiura ponderosa.
В центре диска на стороне, обращенной
к субстрату, помещается рот. Форма его
звездообразна, так как интеррадиально
в него вдаются пять треугольных
выступов — челюстей (рис. 151). На
вершине челюсти располагаются челюстные
(ротовые) пластинки, снабженные по краю
чешуйчатыми иголочками —
ротовыми папиллами, одна или две самые
верхние из которых называются и н ф-
радентальными. За ними в
глубине рта помещаются зубы, а у
некоторых видов — зубные папиллы
(рис. 151, 19). Рядом с челюстными
пластинками находятся узкие а д о р а л ь-
ные щитки, обхватывающие с боков
крупную пластинку — оральный
щиток. Один из оральных щитков
пронизан порами, через которые в
водоносную систему поступает вода. Челюсти
у офиур подвижны и помогают не только
удерживать и размельчать пищу, но
также и захватывать ее.
Большинство змеехвосток
раздельнополы. Случаи гермафродитизма отмечены
сравнительно для немногих форм.
Наиболее полный список гермафродитных
форм, составленный Т. Мортенсеном
в 1936 г., включает 36 видов. Как правило,
все гермафродитные виды проявляют
заботу о потомстве и вынашивают молодь,
однако не все вынашивающие молодь виды
являются гермафродитами.
Обычно офиуры откладывают половые
продукты прямо в воду. Размножение
чаще происходит весной либо летом, при
этом многие офиуры приподнимаются на
концах своих лучей и выметывают
половые продукты за короткий промежуток
времени с небольшими паузами. Чаще
первыми приступают к размножению
самцы. Оплодотворенные в воде яйца
развиваются в пелагическую личинку — офиоп-
лутеус (рис. 128), отличающуюся
длинными отростками и своеобразным
скелетом, состоящим из двух симметричных
половинок. На образование офиоплутеуса
требуется около трех недель с
момента оплодотворения яйца. Все дальнейшие
превращения личинки во взрослый
организм происходят в толще воды, пока офио-
плутеус плавает. В отличие от звезд
сидячая стадия в развитии офиур отсутствует.
Претерпев сложные изменения, маленькая
офиура опускается на дно и начинает вести
донный образ жизни.
Не все виды выметывают половые
продукты прямо в воду. В районе Новой
Зеландии, поданным Фелла(Н.Fell),имеется
вид, который прикрепляет от 20 до 200 яиц,
богатых желтком, к нижней поверхности
камней или в расщелинах. Эту
любопытную особенность можно считать уже
некоторой заботой о потомстве, получающей
дальнейшее развитие у отдельных видов
змеехвосток, распространенных главным
образом в антарктических и
субантарктических районах. Сейчас уже известно
около 60 видов, вынашивающих молодь
либо на своем теле, либо в бурсах, либо
прямо в яичниках. В самом простом
случае яйца откладываются в воду, но затем
молодь оседает на тело матери и долгое
время ползает по его поверхности. При
внутреннем вынашивании молоди яйца
задерживаются в бурсах, в которые через
щели все время поступает свежая вода,
а вместе с ней легко может проникать и
сперма, оплодотворяющая яйца. Обычно
в бурсе имеется всего несколько
экземпляров молоди, но у вида Astrochlamys
bruneus (табл. 24) Мортенсеном было
обнаружено в каждой бурсе до 200 эмбрионов.
Виды, вынашивающие молодь в бурсах,
имеют более растянутый период
размножения, чем виды, откладывающие икру
в воду. Так, очень широко
распространенная почти по всем морям Amphipholis
squamata имеет в своих бурсах
развивающуюся молодь в течение круглого года.
У этого вида, возможно, имеется случай
подлинного живорождения, когда
эмбрион развивается за счет питательного
материала, получаемого от матери. Фелл
сообщает, что в стенках бурс, которые
содержат эмбрионы, развиваются особые
лакуны кровеносной системы. Кроме того,
яйца этой офиуры очень мелкие и,
вероятно, не содержат достаточного на весь
период развития желтка, так как,
вынутые из бурс, они быстро погибают, если
не получают дополнительной пищи. У
целого же ряда других видов,
вынашивающих молодь, яйца крупные, богаты
желтком и, проходя в бурсах весь путь
развития, очевидно, не получают дополни-
19 Жизнь животных, т. 2
289
тельного питания за счет материнского
организма. Некоторые авторы
высказывают предположение, что во многих
случаях более развитые эмбрионы могут как-
либо использовать для своего питания
яйца, находящиеся на более ранних
стадиях развития. С подлинным случаем
живорождения мы встречаемся у
некоторых антарктических видов, у которых
местом развития молоди становятся
яичники, например у Ophionotus hexactis.
У большинства видов офиур пол
различить невозможно, если не считать
самок, вынашивающих молодь. Только 4
вида имеют вполне выраженный
половой диморфизм. У Amphilycus andro-
phorus (рис. 152), Ophiosphaera insignis,
Ophiodaphne materna и Astrochlamys bru-
neus известны карликовые самцы, всегда
сидящие на более крупных самках.
Некоторые виды змеехвосток могут еще
размножаться поперечным делением.
Случаи подобного бесполого размножения
чаще всего имеют место у мелких шести-
лучевых видов, диаметр диска которых
всего около 3 мм. При этом диск может
делиться в любом направлении, но всегда
каждая часть остается с тремя лучами
и восстанавливает недостающие. Поэтому
у таких форм часто попадаются
экземпляре. 152. Самец Amphylicus androphorus, napa-
ззпирующнй на ротовой стороне самки того же
вида.
ры с тремя длинными и тремя более
короткими лучами. Наиболее распространено
бесполое размножение среди видов рода
Ophiactis из отряда настоящих офиур
(Ophiurae).
Со способностью восстанавливать
недостающие части, столь широко
распространенной среди офиур, связана и другая их
особенность: они легко не только
обламывают свои лучи, но некоторые змеехвостки
могут временно лишаться спинной части
диска или внутренностей. Все это
довольно быстро восстанавливается
(регенерирует), если не повреждено околоротовое
нервное кольцо и диск сохранил хотя бы
один луч.
Растут офиуры в течение нескольких
лет и достигают максимальных размеров
на 4—5—6-м году жизни, хотя многие
из них начинают размножаться уже на
2—3-й год.
Офиуры — наиболее подвижные
животные среди иглокожих. Они передвигаются
с помощью лучей и редко используют для
этого амбулакральные ножки. Несмотря
на ясно выраженное лучистое строениег
при движении офиуры ведут себя как
билатерально симметричные животные.
При этом 2 или 4 луча волнообразно
изгибаются, тогда как непарный луч
направлен вперед либо назад, а диск приподнят
над субстратом. Вообще офиуры могут
использовать и другие способы
передвижения. Некоторые виды подтягиваются,
захватывая предметы одним или двумя
лучами и подталкиваясь остальными.
Несмотря на отсутствие ампул и присосок,
виды используют при движении
амбулакральные ножки, упираясь ими в
неровности грунта. Имеются также
наблюдения, что змеехвостки могут ползать по
стеклянным стенкам аквариума при
помощи амбулакральных ножек, но в этих
случаях железистыми клетками офиур
выделяется особая слизь, как бы
приклеивающая ножки к поверхности.
Амбулакральные ножки играют весьма важную
роль при закапывании офиур в грунт.
Новейшие анатомические и
гистологические исследования показали, что у офиур
имеются особые приспособления,
заменяющие ампулы ножек и дающие им
возможность вытягиваться.
290
Очень часто лучи и ножки офиур
принимают активное участие в захвате пищи
и подведении ее ко рту. Питаются
змеехвостки различными мелкими животными,
донным детритом, а отдельные виды
исключительно водорослями. При этом
мелкие частицы подносятся ко рту ножками,
а более крупные захватываются лучами,
которые, изгибаясь, доставляют их в рот.
Исследование желудков европейских
офиур показало, что они на 75—90%
заполнены донным детритом, перемешанным с
остатками мелких рачков, полихет,
моллюсков, молоди иглокожих и других
мелких морских организмов. В аквариумах
офиур можно кормить рыбой. Многие
офиуры чувствуют добычу на расстоянии
и ползут за ней.
Опыты, проведенные над офиурами,
показали, что чувствительными к
пищевым раздражениям являются в
значительной степени амбулакральные ножки,
которые, даже отделенные вместе с частью
луча от диска, продолжали направлять
пищевые частицы в сторону бывшего рта,
а посторонние частицы быстро
отбрасывать, если они, например, не были смочены
рыбьим соком.
Офиуры весьма чувствительны к
световым раздражениям и реагируют на
различные механические воздействия, однако
о их органах чувств почти ничего
неизвестно.
Некоторые офиуры способны светиться.
Следует заметить, что светятся только
лучи и их иглы, а иногда еще и оральные
щитки. Довольно сильное свечение
желтовато-зеленоватым светом часто является
ответом на малейшее прикосновение к
офиуре.
Свечение офиур часто привлекает к себе
различных веслоногих рачков, которые
поселяются на их теле или забираются
во внутренние органы. Некоторые рачки
рода Cancirilla были найдены на
ротовой стороне Amphipholis squamata,
другие виды копепод паразитируют в бурсах
офиур, откладывая в них массу яиц и тем
самым мешая размножению хозяина. Иногда
копеподы вызывают значительные галлопо-
добные вздутия в основании лучей
змеехвосток. Следует, однако, отметить, что
паразиты и комменсалы офиур
немногочисленны, хотя и известны из разных
групп животных. Подвижный образ жизни
и прочный панцирь, покрывающий тело,
до некоторой степени предохраняют их от
нападения мелких врагов. Тем не менее
в офиурах можно встретить различных
инфузорий, паразитических моллюсков,
ракообразных, червей. Сами же офиуры
чрезвычайно склонны жить на других
организмах. Некоторые змеехвостки
обитают постоянно среди игл морских ежей.
Например, очень мелкая офиура Nanno-
phiura lagani, диаметр диска которой едва
достигает 0,5мм, приспособилась к жизни
на брюшной стороне плоского морского
ежа Laganum depressum, где перелезает
с одной иглы на другую, как маленькая
обезьянка с дерева на дерево. Весьма
часто офиуры встречаются на губках и
кораллах.
Почти все 5-лучевые представители
отряда ветвистых офиур (Euryalae)
обычно сидят на различных кораллаху
а некоторые даже питаются их
щупальцами.
В литературе отмечено несколько
случаев сожительства офиур с морскими
лилиями. Так, офиуры рода Ophio-
maza живут в сообществе с морскими
лилиями. Ophiomaza cacaotica (рис. 129)
неоднократно была найдена на ротовой
стороне тропической морской лилии р о-
д a Comanthus, чашечку которой она
крепко обвивает своими лучами.
Раскраска этой офиуры довольно пестрая и очень
соответствует раскраске хозяина.
Окрашены офиуры более скромно, чем
остальные иглокожие. Отдельные виды,
правда, имеют ярко-красный или
малиновый цвет, но они малозаметны из-за
сравнительно небольших размеров. В
окраске змеехвосток преобладают кремовые,
желтые, зеленоватые, оливковые, бурые-
тона, часто оживляемые различными
пятнами контрастного цвета.
Ископаемые остатки офиур
немногочисленны и изучены далеко не полно. Офиуры,
сходные с современными, известны с
конца палеозоя, но офиуроподобные
организмы, отличающиеся от современных
открытой амбулакральной бороздой и
сближающие змеехвосток со звездами, найдены уже
в силуре, т. е. в раннем палеозое.
19*
291
ОТРЯД ВЕТВИСТЫЕ ОФИУРЫ
<EURYALAE)
Представители этого отряда не имеют
сплошного чешуйчатого или
пластинчатого покрова на диске и лучах; диск и
лучи у них затянуты толстой кожей, часто
покрытой гранулами. К этому отряду
относятся все офиуры с разветвленными
лучами, а также некоторые формы,
имеющие простые, но загибающиеся на
брюшную сторону лучи. Последние
чрезвычайно подвижны, могут сворачиваться в
спираль и обвиваться вокруг различных
предметов. Всего в отряде 4 семейства
и около 50 родов, известных как в
прибрежных водах, так и на больших
глубинах. Многие из них приурочены только
к тропикам, другие довольно широко
распространены по всем океанам и морям.
Один из наиболее известных
представителей отряда—Asteronyx loveni (рис. 153)—
широко распространен в теплых и
умеренных водах Мирового океана на глубинах
от 100 до 3 тыс. м. Его обычно находят
обвившимся вокруг морских перьев (Реппа-
tulidae). Перес (Peres), спускаясь в 1959 г.
в батискафе по западному склону
Яванской впадины, на глубине 1650 м видел
астерониксов, которые двумя лучами,
скрученными, как виноградная лоза,
держались за субстрат, а тремя вытянутыми
вперед лучами делали волнообразные
движения и таким образом ловили пищевые
частицы, взвешенные в воде. Цвет
астерониксов кораллово-красный. Размеры их
довольно значительны: диаметр диска
около 4 см, а длина лучей около 40 см.
Молодь астерониксов ползает по дну и
питается детритом, но, достигнув размеров
около 5—-6 мм в диаметре диска,
взбирается на морское перо и переходит,
подобно взрослым животным, на питание
планктоном или взвешенными в воде
частицами, однако вполне возможно, что
не брезгает и полипами, на которых
поселяется.
Не менее известны, чем предыдущий
вид, представители семейства гор-
гон (Gorgonocephalidae), включающего
свыше 30 почти исключительно
тропических родов. В наших водах встречаются
Рис. 153. Змеехвостка Asteronyx loveni
на морском пере.
представители всего трех родов. Лучи
этих офиур очень сильно ветвятся и
чрезвычайно подвижны. Поэтому когда они
двигаются, то действительно напоминают
голову мифического чудовища со змеями
вместо волос на голове, тем более что диск
292
этих офиур довольно крупный, часто
достигающий 10 см в диаметре.
По всем нашим дальневосточным морям
на глубинах от 25 до 785 м распространен
восточный горгоноцефал (Gorgonocephalus
caryi). Эта, обычно оранжевого или
желтовато-буроватого цвета, змеехвостка
достигает 5 см в диаметре диска (табл. 17).
Весьма близок к нему
североатлантический горгоноцефал (Gorgonocephalus euc-
nemis), распространенный в северной
части Атлантики и в западном секторе
Арктики вплоть до Восточно-Сибирского
моря. Взрослые особи горгоноцефала
питаются мелкими животными, которых они
захватывают широко разветвленными
лучами либо собирают вместе с грунтом;
молодь же на ранних стадиях развития
живет паразитически на коралловом
полипе, наружными покровами которого она
питается. Затем, когда на руках молодой
офиуры появится уже несколько
ветвлений, молодые особи переходят на спину
взрослых горгоноцефалов, на которых они
проводят некоторое время, питаясь
мелкой добычей, застревающей в
разветвлениях их рук. Случается, что они наносят
повреждения и взрослому организму, на
котором поселяются, соскабливая с
поверхности его кожу.
Среди отряда ветвистых офиур известен
только один вид, проявляющий заботу
о потомстве. Это Astrochlamys bruneus
(табл. 24), также относящийся к
семейству горгон. Яйца у этого вида
развиваются в бурсах. Распространен астрохла-
мис в Антарктике на глубинах до 3 тыс. м.
Диаметр его диска едва превышает 1 см,
но лучи в 7—8 раз длиннее диаметра
диска. Обычно астрохламисов находят с
маленьким самцом, сидящим на аборальной
поверхности диска самки.
ОТРЯД НАСТОЯЩИЕ ОФИУРЫ
(OPHIURAE)
Подавляющее большинство известных
змеехвосток относится именно к этому
отряду. Диск у этих офиур обычно
покрыт чешуйками, а лучи—пластинками.
Лучи никогда не ветвятся и менее
подвижны, чем у представителей предыдущего
отряда. Они изгибаются только в
горизонтальной плоскости, так как
сочленение позвонков у них более сложное —
при помощи бугорков и ямок.
Одно из наиболее обширных семейств
этого отряда — Ophiacanthidae —
содержит большое число видов, широко
распространенных по океанам, причем
многие из них часто обитают на больших
глубинах. У офиакантид диск на
спинной стороне сплошь затянут покровом
из тесно расположенных низких бугорков,
шипиков или иголочек, маскирующих
чешуйки диска. Лучи снабжены большим
числом часто очень длинных и шиповатых
иголочек. Наиболее типичным
представителем семейства является двузубая офиа-
канта (Ophiacantha bidentata, табл. 25),
очень часто встречающаяся в Арктике,
Атлантике и в Тихом океане на глубинах
от 10 до 4500 м. Эта довольно мелкая
буроватая офиурка, с диаметром диска
приблизительно до 12 мм, обладает
способностью светиться.
Однако эта способность в основном
распространена среди представителей другого
семейств а—амфиурид(Amphiuridae),
отличающихся главным образом строением
своего ротового угла, на вершине
которого сидят две инфрадентальные ротовые
папиллы (рис. 151). Наиболее интенсивно
светятся Ophiopsila annulosa и Ophiop-
sila arenea, распространенные в
Средиземном море на глубинах до 100 м. Эти
офиуры начинают светиться при
малейшем их раздражении. Достаточно
дотронуться палочкой либо пинцетом до луча
данной офиуры, как она моментально
вспыхивает ярким светом сначала в месте
прикосновения, затем и на других лучах.
При более длительном раздражении эти
змеехвостки светятся ярким зеленовато-
желтым светом, причем кажется, что свет
исходит от всей поверхности офиур.
Однако гистологические исследования
данных видов показали, что железистые
клетки, секрет которых вызывает свечение,
расположены лишь на определенных
местах тела офиур. Светиться могут иглы,
брюшные и боковые пластинки лучей.
Такая любопытная особенность этих
офиур была использована при изучении
293
образа жизни данных животных. Оба
вида активны ночью, а при дневном
освещении они прячутся в песок. Способность
светиться дала возможность наблюдать
этих животных ночью без
дополнительного освещения. Оказалось, что офиуры
питаются ночью, выставив из убежища
три луча и поставив их перпендикулярно
течению и таким образом улавливая и
отфильтровывая взвешенные в воде
питательные частицы.
Способны светиться и другие виды
семейства амфиурид. Интересно напомнить,
что свечение офиур впервые было
обнаружено более 150 лет назад у Amphipholis
squamata. Оказалось, что светиться
способны только живые особи, причем
основной свет исходит от основания игл, а
ножки, как правило, не светятся. Какое
значение имеет свечение для змеехвосток, до
сих пор не выяснено. Вполне возможно,
что яркие вспышки, возникающие при
прикосновении к ним, пугают рыб,
питающихся многими офиурами, и дают
возможность данным животным лучше
спрятаться. Любопытно, что эта весьма
маленькая офиурка с диском всего около
4—5 мм в диаметре сумела
приспособиться к чрезвычайно разнообразным
условиям обитания и распространиться почти
по всем морям тропических и умеренных
районов. Ее можно встретить и в западной
части Баренцева моря. Цвет офиуры
голубоватые или серовато-белый. Живет она
от приливной зоны до глубины 250 м.
Она относится к одному из немногих гер-
мафродитных видов змеехвосток. Яйца
у нее развиваются в бурсах, причем
период размножения очень растянут и
эмбрионы могут быть обнаружены в бурсах
в течение круглого года. Очень редко
удается найти экземпляры этого вида с
неповрежденным диском, так как
чрезвычайно часто при неблагоприятных
условиях они отбрасывают спинную часть
диска, иногда даже вместе с желудком
и половыми железами, и очень скоро их
вновь регенерируют (восстанавливают).
Проявляет заботу о потомстве и
маленькая черноморская амфиура (Amphiura
stepanovi). Молодь в бурсах ее можно
обнаружить в начале лета и осенью.
Исследуя внутреннюю организацию
черноморской амфиуры, Д. М. Федотов
пришел к заключению, что она является
не только живородящей, но и гермафро-
дитной. Живет она зарывшись в песок
или скрывшись в раковинах устриц. Эту
офиуру можно встретить по всему
Черному морю до глубины в 250 м, а также в
Мраморном море.
В Средиземном море, вдоль
европейского берега Атлантического океана и у
западного берега Африки обитает очень
близкий к предыдущему вид — Amphiura
chiajei. Положенная на песок, эта
змеехвостка быстро закапывается в него с
помощью своих ножек, оставляя торчать
над поверхностью грунта лишь кончики
своих лучей. Стенки проделанных в песке
углублений офиура укрепляет слизью,
чтобы они не осыпались, а волнообразные
движения лучей и различные сокращения
диска способствуют циркуляции воды в
углублениях и тем самым улучшают
дыхательные процессы офиуры. Наблюдения
над экземплярами, содержащимися в
аквариумах, показали, что они около 18
месяцев могли жить зарытыми в песок и
по своей воле оттуда никогда не
вылезали. Они только немного перемещались в
грунте, притягивая к себе лучи и затем
выставляя их в новом месте и слегка
подтягивая вслед за лучами тело. Но и эти
передвижения были незначительны.
Питались амфиуры находящимися в воде
питательными частицами, которые
улавливались выставленными над поверхностью
грунта кончиками лучей и продвигались
ко рту с помощью амбулакральных
ножек. Более крупные частицы
улавливаются и подносятся ко рту самими лучами.
Оказалось, что живой добычи этот вид
никогда не ловит. Но не все частицы
пищи, подведенные ко рту, поступают
в него. Околоротовые ножки как бы
сортируют пищу и часть ее отбрасывают.
Они же отбрасывают в сторону и не
переваренные офиурой частицы пищи.
Представителей рода амфиура
можно встретить в самых разнообразных
районах всех океанов. Например, в
антарктических водах встречается Amphiura
antarctica (рис. 154), а в тропической
части Атлантического океана Amphiura
stimpsoni, вынашивающая свою молодь.
294
В наших дальневосточных морях, в
Японском море, в Татарском проливе, в
Охотском море, на мелководьях южных
Курильских островов, очень часто попадается
довольно крупная темно-красная офиу-
ра — Amphiodia rossica (табл. 25),
предпочитающая небольшие глубины. Ее часто
используют в пищу различные донные
рыбы.
Не менее обычна в наших
дальневосточных морях и в северной части
Атлантического океана очень эффектная колючая
змеехвостка (Ophiopholis aculeata,
табл. 20), относящаяся к семейству
Ophiactidae. Она часто встречается среди
скоплений губок, камней, на известковых
водорослях на глубинах от 5 до 500 м.
Представители этого вида имеют пеструю
окраску различных красно-фиолетовых
оттенков, иногда с зеленоватым узором.
Диск усажен мелкими иголочками, часто
пестро окрашенными.
Среди семейства офиактид встречаются
виды, способные размножаться бесполым
путем. Маленькая шестилучевая офиурка
Ophiactis virens, известная в Средиземном
море и Атлантическом океане,
размножается делением тела на две части. После
деления каждая половина быстро
восстанавливает утраченные части и снова
становится 6-лучевой, однако очень часто
их лучи отличаются своими размерами.
Рост лучей происходит за счет
прибавления новых члеников на их концах,
поэтому более старые членики и оказываются в
основании лучей.
В тропических водах, на коралловых
рифах, очень часто встречаются
представители семейства Ophiothrichidae,
отличающиеся главным образом тем, что
у них отсутствуют ротовые папиллы, а
на вершине челюстей имеется группа
зубных папилл (рис. 151). Диск у них покрыт
чешуйками, которые могут быть
замаскированы густым покровом из очень низких
бугорков и иголочек. Они очень часто
пестро окрашены и имеют причудливый
рисунок.
К этому семейству относится Ophiomaza
cacaotica, живущая на морских лилиях
и часто окрашенная под цвет хозяина.
Великолепный темный фиолетово-синий
цвет имеет Ophiothrix coerulea (табл. 21).
Она была найдена в Тихом океане вблизи
островов Лиу-Киу. Спинная сторона этой
офиуры имеет характерный рисунок, так
как радиальные щитки обрамлены белым,
а лучи — с темными кольцами, между
которыми выделяются белые точки,
расположенные на спинных щитках лучей.
Иглы лучей Ophiothrix coerulea с
фиолетовым оттенком.
Не менее красив и вид Ophiothrichoides
pulcherrima, найденный там же.
Название ее в переводе на русский язык
означает «прекраснейшая». Она темно-синего
цвета с желтыми полосами. Диск с
чередующимися синими и желтыми кольцами.
Лучи с чередующимися синими и
желтыми спинными щитками. Ротовые щитки у
нее тоже желтые с широким синим
бордюром. Иглы лучей стекловидные с
голубоватыми концами (табл. 19).
Единственный представитель семе й-
с т в а офиотрихид в наших водах
встречается иногда в южной части Черного
моря. Это ломкий офиотрикс (Ophiothrix
fragilis), очень широко распространенный
в Атлантическом океане вдоль берегов
Европы и Африки, а также в Средиземном
Рис. 154. Amphiura antarctica со спинной
стороны.
295
море. Вид встречается от литорали до
глубины в 1200 м; отличается тем, что
лучи его несут длинные зазубренные иглы.
Питается эта офиура главным образом
донными животными, червями,
моллюсками и даже мелкими морскими ежами.
Живет офиотрикс обычно в разных
укрытиях, среди камней, в пустых раковинах
моллюсков.
В тропиках в основном распространены
и представители семейства Ophio-
comidae и Ophiodermatidae. Среди них
также преобладают пестроокрашенные
формы. Весьма привлекательна красивая
офиокома (Ophiocoma delicata, табл. 24),
найденная на глубине около 35 м у юго-
восточного побережья Австралии. Ее пя-
Рис. 155. Astrophiura marione:
l — вид с ротовой стороны; 2 — вид со спинной
стороны.
тиугольный маленький очень плоский
диск бледно-коричневого цвета с
несколькими круглыми или овальными пятнами,
каждое с широкой белой каймой.
Пластинки лучей тоже двухцветные: одна их часть
темно-пурпурная, а другая почти белая;
поэтому лучи выглядят
поперечнополосатыми.
На коралловых отмелях и других
местах тропической зоны Тихого океана
можно часто встретить и другого
представителя офиокомид — Ophiomastix annu-
losa (табл. 18).
На литорали тропической части
Атлантического океана в районе Мексиканского
залива и Карибского моря обитает корот-
коиглая офиодерма (Ophioderma brevi-
spina). Эта офиура характерна тем, что,
в отличие от многих офиур,
передвигающихся с помощью лучей, она при
движении использует амбулакральные ножки,
цепляясь ими за неровности субстрата.
В наших морях чаще попадаются виды
одного из самых обширных семейств
змеехвосток — семейства настоящих
офиур (Ophiuridae). Наиболее
характерным представителем этого семейства
является змеехвостка Сарса (Ophiura sarsi,
табл. 17), очень часто встречающаяся в
арктических морях, в северной части
Атлантического и Тихого океанов от
прибрежной зоны до глубины в 3000 м.
Подобно остальным представителям данного
семейства, она имеет довольно толстый
диск, имеющий в основании лучей на
спинной стороне вырез, усаженный папил-
лами, и сравнительно короткие лучи.
Род офиур очень обширен, его
представителей можно встретить во всех
морях. Так, у берегов Австралии обитает
Ophiura multispina, близкая к описанной
выше офиуре Сарса. Диск у нее также
покрыт крупными пластинками и лучи
относительно короткие, что придает
офиуре коренастый вид.
Еще Солее грубого сложения
представители рода Amphiophiura. Диск у них
высокий, толстый, покрытый толстыми,
часто вздутыми пластинками, лучи
крепкие, почти треугольные в поперечном
разрезе. Виды этого рода чаще встречаются
на больших глубинах. В Охотском море,
и у южных Курильских островов, и у бе-
296
регов Японии, а также в районе Аляски
и Калифорнии на глубинах от 130 м и
глубже обитает Amphiophiura ponderosa
(табл. 25). Эта кораллово-красная амфио-
фиура одна из самых крупных среди
видов отряда настоящих офиур. Диск ее,
покрытый грубыми вздутыми
пластинками, иногда бывает около 5 см в
диаметре, а лучи в 4—5 раз длиннее диаметра
диска.
В противоположность предыдущему
виду представители рода Ophiopleura
имеют диск, затянутый мягкой кожей,
скрывающей мелкие чешуйки. Самым
обычным видом в наших северных морях
является Ophiopleura borealis (табл. 20).
Поверхность ее диска кажется совершенно
атласной. Это тоже одна из наиболее
крупных офиур, с диаметром диска порой
свыше 4 см. Цвет офиуры красный или
оранжевый.
Представителей семейства настоящих
офиур можно встретить и в
антарктических водах. Некоторые из них
проявляют заботу о своем потомстве. Особенно
интересен среди них Ophionotus hexactis,
молодь которого развивается в яичниках
матери. Эта шестилучевая красновато-
фиолетовая офиура с диаметром диска
около 3 см встречается на небольших
глубинах у острова Кергелен, Диск у
самок часто бывает растянут несколькими
большими эмбрионами, диаметр диска
которых может быть около 1 см, а длина
лучей — 2—2,5 см. В каждом яичнике
офиуры содержится по одному эмбриону.
Яичники этого вида имеют вид полых
пузырьков, причем в каждом из них
созревает только одно яйцо, которое
выпадает в просвет яичника и там развивается
после оплодотворения в маленькую офи-
урку. Остальные яйца дегенерируют,
возможно, что они служат питанием для
развивающегося эмбриона. Как происходит
оплодотворение яиц, до сих пор неясно.
Очевидно, сперма попадает в бурсы вместе
с водой, постоянно туда поступающей.
После развития маленькие офиуры
выходят наружу, очевидно, через бурсальную
щель, освобожденный яичник сильно
сокращается, а затем регенерирует заново.
Не менее любопытны и офиуры, своей
оригинальной формой напоминающие
нечто среднее между звездой и офиурой.
Отсюда происходит и их название —
Astrophiura, т. е. звездоофиура. Однако
внимательное изучение этих животных
показало, что они ничего общего со
звездами не имеют, а являются настоящими
офиурами. У них ряд пластинок диска
и луча сильно разрастается и образует
нечто вроде щита (рис. 155), а свободные
части лучей становятся очень хрупкими.
Они лишены брюшных и спинных щитков,
а также амбулакральных пор для
прохождения ножек. Образ жизни этих
интересных офиур неизвестен. Этот маленький
род, насчитывающий несколько видов,
встречается в прибрежной зоне у острова
Мадагаскар, у берегов Японии. Вдоль
побережья Калифорнии на глубинах
менее 1000 м обнаружена Astrophiura mario-
пе (рис. 155).
Самый глубоководный вид этого рода,
Astrophiura chariplax, был обнаружен
экспедиционным судном «Витязь» к
северу от Командорских островов на
глубине 2440 м.
ТИП ЩЕТИНКОЧЕЛЮСТНЫЕ,
ИЛИ „МОРСКИЕ СТРЕЛКИ"
(CHAETOGNATHA)
Это небольшая группа (около 40 видов)
типично морских планктонных
организмов. В систематическом отношении
хетогнаты резко обособленная группа вторич-
норотых животных, отличающаяся
большой древностью.
Представители этой группы, весьма
сходные с современными, известны со
среднего кембрия.
Размеры хетогнат — 1—9 см, тело их
прозрачно, имеет удлиненную форму и
двустороннесимметричное строение,
покрыто кутикулой с небольшим
количеством ресничек. По бокам тела
расположены одна или две пары плавников, а
на заднем конце — непарный хвостовой.
Однако плавники не способны к
самостоятельному движению и только усиливают
изгибательный эффект. Хетогнаты быстро
изгибают тело и производят молниеносные
скачкообразные движения.
Кишечник у морских стрелок прямой,
на переднем конце тела с брюшной
стороны расположено ротовое отверстие, а
анальное — на расстоянии четверти
длины тела от его конца.
Голова хетогнат вооружена сильным
ловчим аппаратом из правого и левого
пучков сильных щетинок — крючьев,
способных при помощи сильных мышц быстро
раздвигаться и захлопываться,
захватывая добычу, нередко более крупную, чем
сам хищник.
Внутри тело хетогнат разделено двумя
поперечными перегородками на три отдела
и одной продольной — на правую и левую
стороны.
Органы дыхания, выделения и
кровеносная система отсутствуют. Нервная
система развита хорошо и состоит из трех
ганглиев: головного, вестибулярного и
брюшного. На голове имеются два глаза.
Мускулатура хетогнат
поперечнополосатая, что дает им возможность совершать
быстрые движения. Они гермафродиты,
и им свойственно прямое развитие, без
личиночной стадии.
Щетинкочелюстные распространены во
всех морях и океанах. В отдельные годы
численность их резко возрастает, и тогда
хетогнаты истребляют большое количество
планктона, в основном из семейства кала-
нид, и даже личинок рыб.
Хетогнаты способны совершать
суточные и сезонные вертикальные миграции
в десятки и сотни метров.
298
Рис. 156. Строение щетинкочелюстных (Chaetognatha).
А — сагита, общий вид: 1 — голова; 2 — передние боковые плавники; з — задние
боковые плавники; 4 — хвостовой плавник; 5 — кишечник. Б — нервная система, вид
с левой стороны: 1 — ротовое отверстие; 2 — кишка; з — головной мозг; 4 —
висцеральный ганглий; 5 — вентральный ганглий; 6 — глаз. В — голова морской стрелки
с раздвинутыми B) и сомкнутыми B) ловчими щетинками.
Погонофоры
Pogonophora
Полухордовые
Hemichorda
Оболочники
Tunicata
ТИП ПОГОНОФОРЫ (POGONOPHORA)
Погонофоры интересны во многих
отношениях. Открытие их явилось одним из
самых неожиданных событий в
современной зоологии. Еще недавно казалось, что
время описания новых классов и типов
давно миновало, что все группы
животных высокого систематического ранга
давно хорошо известны. История изучения
погонофор опровергла это представление.
Этот тип, объединяющий несколько
десятков своеобразных морских животных, был
установлен лишь в середине нашего
столетия, а детально изучен лишь в
последние 10 лет.
Погонофоры относятся ко вторичноро-
тым (Deuterostomia) — большой
эволюционной ветви животного мира, к которой
принадлежат также иглокожие,
полухордовые и хордовые (в том числе
позвоночные). О родстве погонофор со всеми
этими типами особенно убедительно
говорит их эмбриональное развитие. Многие
примитивные особенности погонофор
позволяют по-новому оценивать первые
этапы эволюции вторичноротых.
Погонофоры интересны также
отсутствием кишечника, который в процессе
эволюции исчез у них полностью. Этим они
резко отличаются от всех остальных сво-
бодноживущих многоклеточных животных
(не считая очень примитивных губок).
Известные случаи полной утраты
пищеварительного канала касаются исключительно
паразитических животных (скребни,
ленточные черви, некоторые эндопаразитиче-
ские брюхоногие моллюски и ракообразные).
Как только отсутствие кишечника у
погонофор было доказано, встал сложный
вопрос о том, как они питаются.
Погонофоры интересны своим образом
жизни. Это животные, ведущие почти
неподвижную жизнь на дне моря внутри
длинной защитной трубки. Подобным же
образом в трубках обитают многие много-
щетинковые черви (Polychaeta, Sedenta-
ria), относящиеся к типу кольчатых
червей. Одинаковый образ жизни этих
двух неродственных друг другу групп
животных привел к выработке у них
сходных, но возникших независимо
приспособлений. Благодаря этому погонофоры и
живущие в трубках полихеты приобрели
большое внешнее сходство, настолько
смущавшее некоторых зоологов, что они
ошибочно считали погонофор своеобразно
измененными кольчатыми червями (Л и-
ванов и Порфирьева, 1962).
Таким образом, погонофоры дают
возможность изучить важные вопросы о
закономерностях развития у неродственных
животных сходных приспособлений (к о н-
вергенций) под влиянием
одинаковых условий жизни.
Наконец, любопытна и история
изучения погонофор. Эти своеобразные
обитатели океанических глубин, еще недавно
казавшиеся большой зоологической
редкостью, в действительности принадлежат
303
к самым обычным элементам морской
фауны. Не удивительно поэтому, что
погонофоры уже давно попадали в руки
ученых. Однако они не привлекали к себе
внимания или причислялись к давно
известным группам животных. В одном из
музеев естественной истории в США
многие десятки лет хранились неопознанные
погонофоры, собранные еще в прошлом
столетии в Тихом океане американскими
океанографическими экспедициями. Лишь
теперь, когда мы уже знаем, что такое
погонофоры, эти уникальные экземпляры
«дождались» своих исследователей.
Недавно английский зоолог Д.
Карлайл (Carlisle) писал: «Когда в 20-х
годах английское исследовательское судно
«Дискавери II» работало в Антарктике
и Субантарктике, одна из обычнейших
неприятностей при добывании донных
животных с больших глубин заключалась
в том, что после 10 или 12 часов работы
драга поднималась, забитая массой
каких-то волокон. Очень часто мешок драги
был настолько заполнен этой противной
массой «gubbins», что исключалась
возможность получения какого бы то ни было
материала, интересного в биологическом
отношении, и весь улов после быстрого
и грубого осмотра выбрасывался за борт.
Это случалось так часто, что биологи на
борту судна употребляли шуточное
наименование «Gubbinidae» в качестве
названия «семейства» для этого досадного
материала».
Вспоминая этот период на борту
«Дискавери II», профессор сэр Алистер
Харди сказал мне, что «Gubbinidae»,
несомненно, были погонофоры. Таким
образом, некоторые из ведущих морских
биологов того времени выбросили за борт
много тонн погонофор. Пикантность этой
истории состоит также в том, что на
«Дискавери II» было строгое правило —
не выбрасывать никакого собранного
биологического материала. Он должен быть
сохранен целиком и привезен в Англию.
Это правило существовало для того, чтобы
предотвратить как раз подобные случаи
уничтожения материала первостепенного
научного интереса только потому, что он
не был сразу правильно определен.
Совершенно очевидно, что биологи того времени
не подозревали о биологической природе
«Gubbinidae».
Честь открытия первого представителя
погонофор принадлежит французскому
зоологу М. Коллери (Coullery). В
1914 г., изучая кольчатых червей, добытых
с большой глубины в индонезийских морях
голландской морской зоологической
экспедицией на судне «Зибога», он обнаружил
нитевидное животное, обитающее в
тонкой кольчатой трубочке. Коллери
исследовал его и назвал Siboglinum weberi
(Siboga — название судна, linum — в
переводе с латинского нить; weberi —
видовое название в честь зоолога Вебера,
начальника экспедиции). Хотя Коллери
в 1944 г. переисследовал Siboglinum,
выяснить полностью план строения этого
животного и его систематическое
положение не удалось. Вторая находка была
сделана почти 20 лет спустя советским
зоологом П.В.Ушаковым в
Охотском море. В трале, поднятом с глубины
3500 м, оказалось несколько длинных
бурых трубок, содержащих животных с
пучком щупалец на переднем конце.
Ушаков описал их в 1933 г. под названием
Lamellisabella zachsi (от латинского
lamella— пластинка, Sabella — родовое
название многощетинкового червя;
zachsi — видовое название в честь русского
зоолога И. Г. 3 а к с а) и отнес к много-
щетинковым червям (Polychaeta) — классу
уже давно и хорошо известному,
объединяющему множество видов, широко
распространенных во всех морях.
Однако шведский зоолог К. Э. И о-
г а н с с о н (Johansson), исследовав
Lamellisabella на разрезах, установил, что
это животное ничего общего с многощетин-
ковыми червями не имеет, и в 1937 г.
выделил его в новый класс, названный
Pogonophora (от греческого pogon — б о р о-
да и phoro — несу). Так в системе
животного мира среди примерно семидесяти
классов появился еще один, в то время
установленный для одного вида.
Природа этого класса оставалась темной,
а родственные связи и происхождение
неясными, поскольку ни строение, ни
эмбриональное развитие Lamellisabella не
были еще изучены. Но интерес был
возбужден, и в специальной зоологической
304
литературе началось оживленное
обсуждение систематического положения нового
класса, которое, однако, без нового
фактического материала не могло привести
к определенным результатам. Интересно,
что долгое время никому не приходило
в голову сравнить животных, описанных
Ушаковым и Коллери, и только в 1951 г.
была доказана принадлежность Sibogli-
num к классу Pogonophora.
С 1949 г. начался новый период
изучения погонофор — в морях Дальнего
Востока приступил к исследованиям
океанических глубин «Витязь», флагман
советского исследовательского флота,
экспедиционное судно Академии наук СССР.
Первые же зоологические сборы «Витязя»,
сделанные на больших глубинах в
Охотском и Беринговом морях и в Курило-
Камчатской глубоководной впадине,
содержали множество различных погонофор.
За 15 следующих лет почти непрерывной
экспедиционной работы «Витязь»
совершил 35 исследовательских рейсов,
участники экспедиций изучили фауну Тихого
и Индийского океанов и собрали обильный
уникальный материал по погонофорам.
Эти животные, против всяких ожиданий,
оказались весьма широко
распространенными во многих океанах и морях. Новый
материал дал возможность советским
зоологам открыть более 60 новых видов,
построить классификацию погонофор,
изучить строение и эмбриональное развитие
и сделать наблюдения над образом жизни
и географическим распространением.
Сравнение погонофор позволило
В. Н. Беклемишеву еще в 1944 г.
высказать убеждение, что эта группа животных
заслуживает ранга типа. Однако
обоснованная характеристика нового типа
погонофор и доказательства
принадлежности его к эволюционной ветви вторич-
норотых были даны лишь в 1955 и 1957 гг.
А. В. Ивановым, изучившим
главные особенности строения и
эмбрионального развития этих животных. К числу
других существенных открытий относится
объяснение способа питания погонофор,
сделанное на основе изучения их щупалец
и кровеносной системы (Иванов, 1955),
установление особенностей раннего
развития зародыша (Иванов, 1957),
выяснение химической природы трубки и
пигмента крови (Brunet a. Carlisle, 1958;
Е. Southward a. A. Southward, 1963) и,
наконец, обнаружение особого заднего
отдела тела (Webb, 1964). Последнее
открытие явилось совсем неожиданным.
Оказалось, что на заднем конце длинного
тела животного имеется особый,
снабженный щетинками участок, который до сих
пор оставался неизвестным, несмотря на
то что через руки исследователей прошли
тысячи погонофор. Это объясняется тем,
что этот отдел тела помещается в заднем,
очень нежном, тонкостенном участке
трубки, который очень легко обрывается и или
теряется при промывке улова трала, или
вообще не попадает в трал, оставаясь на
дне моря в глубоких слоях ила.
В настоящее время погонофоры
найдены почти во всех морях, даже там, где
донная фауна хорошо изучена. Так,
недавно они обнаружены в Баренцевом море
близ Мурманского побережья, в проливе
Скагеррак у берегов Швеции, в западной
части Атлантического океана, в фиордах
Норвегии. Около 15 лет назад мы знали
лишь 3 вида погонофор, ныне описано
около 100 видов. Судя по тому, что все
время в новых местах открываются все
новые виды, и учитывая слабую
изученность фауны Мирового океана, естественно
предположить, что действительное
количество видов погонофор выражается
четырехзначным числом.
Географическое распространение
погонофор имеет некоторые особенности. Они
очень обычны в глубоководных районах
Тихого, Индийского и Атлантического
океанов, более или менее прилегающих
к материкам, а также в окраинных морях,
широко сообщающихся с океанами.
Напротив, в большом удалении от островов
и материков на обширных пространствах
океанического дна они отсутствуют. Нет
их также во внутренних морях, т. е.
в морях, изолированных от Мирового
океана или связанных с ним лишь
сравнительно мелководными проливами. Так,
например, очень обильной и
разнообразной фауной погонофор отличаются
Охотское и Берингово моря, тогда как
соседнее с ними Японское море лишено
погонофор. Отсутствуют они и в сильно опрес-
20 Жизнь животных, т. 2
305
ненном Балтийском море, а также в
Каспии, в Азовском и Черном морях. Недавно
погонофоры обнаружены в Средиземном
море.
Погонофоры в большинстве обитатели
больших глубин. Почти половина
известных видов живет глубже 3000 м.
Исключительно глубоководными являются лишь
20 видов B5%), большей частью это
эндемичные обитатели глубоководных
впадин — Курило-Камчатской, Японской,
Бугенвильской в Тихом океане и
Яванской впадины в Индийском океане.
Двадцать пять видов (около 30%) ни разу
не встретились глубже 1000 м, а 7 видов
(около 10%) живут на мелководье не
глубже 300 м. Исключительно широким
вертикальным распространением
отличается Siboglinum caulleryi — форма,
обычная в Охотском море, где она встречается
даже на глубине около 20 му но найденная
также в пучине Курило-Камчатской
впадины на глубине 8100 м.
Необходимыми условиями
существования погонофор являются низкая
температура (от —1,3 до +13°) и нормальная
соленость воды. По-видимому, «глубоко-
водность» погонофор обусловливается
прежде всего тем, что в море наиболее
холодная вода занимает более глубокие
горизонты. С другой стороны,
мелководные виды погонофор распространены в
северных областях, где прибрежные воды
подвержены сильному охлаждению.
Таковы Siboglinum caulleryi в Охотском море,
Nereilinum murmanicum в Баренцевом
море, Galathealinum arcticum и
Siboglinum hyperboreum в северном полярном
бассейне.
Погонофоры всегда встречаются на
илистом грунте. Нередко на твердых и
длинных трубках Polybrachia, Lamellisabella
и некоторых других поселяются
различные сидячие беспозвоночные: форамини-
феры, губки, гидроидные полипы,
актинии, многощетинковые черви, мшанки,
асцидии и даже мелкие стебельчатые
морские лилии. Расположение этих «обраста-
телей» показывает, что верхняя часть
трубки свободно торчит над поверхностью
грунта, располагаясь более или менее
вертикально, тогда как остальная,
нижняя, часть ее глубоко погружена в ил.
Впрочем, у многих мелких форм трубка
настолько нежна и эластична, что,
несомненно, стелется своей верхней частью на
поверхности грунта. Вероятно, в
поселениях погонофор особи расположены
довольно тесно. Необычайное обилие
особей было отмечено в некоторых хорошо
изученных точках северо-западной части
Тихого океана, в Северном море, в
Скагерраке, в антарктических водах и в
других местах. Тралы приносят здесь массу
населенных и пустых трубок погонофор,
забивающих иногда мешок трала и даже
повисающих на раме и на тросе. Если
судить по обилию пустых и
разлагающихся трубок погонофор, часто
примешивающихся к илу, они играют важную роль
в формировании грунта во многих местах
океана.
Недавно было обнаружено, что
некоторые погонофоры ведут сверлящий образ
жизни.
Таковы описанный недавно Веббом
(Webb) из одного фиорда в Норвегии Scle-
rolinum brattstromi и найденный в 1959 г.
экспедицией на судне «Витязь» на дне
Яванской впадины (около 6 тыс. м
глубины) Sc. javanicum. Тонкие буроватые
трубочки этих видов, содержащие
животных, пронизывают кусочки затонувшего
дерева, например обломки древесных
веток. У Sclerolinum javanicum трубочки
проникают сквозь очень плотную
древесину, так что нет никаких сомнений в
способности животного каким-то образом
сверлить дерево, но остается совершенно
загадочным, как это происходит.
Погонофоры, отфильтровывающие из
воды мелкие частицы детрита и
планктонные организмы, зависят от количества
этого взвешенного в морской воде
пищевого материала и от развивающейся в нем
бактериальной флоры. Поэтому они
наиболее многочисленны именно в тех
местах, где имеются более или менее
постоянные скопления придонной органической
взвеси, зависящей от направления и силы
глубинных течений и от рельефа дна.
По-видимому, этим и объясняется тот
факт, что в морях и глубоководных
впадинах фауна погонофор гораздо богаче,
чем в очень удаленных от континента
открытых пространствах океана.
306
Погонофоры не способны покидать свою
трубку, так как не приспособлены к
передвижению на открытом месте. Внутри
трубки они, однако, могут быстро
перемещаться, то высовывая наружу передний
конец тела с щупальцами, то уходя
глубоко в трубку. В этом отношении они
напоминают живущих в трубках много-
щетинковых червей. По длине трубка
всегда в несколько раз превосходит
животное и не препятствует таким
движениям. Для опоры на внутреннюю
поверхность трубки служат хитиноидная
уздечка, многочисленные прикрепительные
сосочки, или папиллы, зубчатые щетинки
поясков и щетинки заднего отдела
туловища (см. ниже). Тело погонофор может
сильно вытягиваться и сокращаться.
Высовывание переднего конца тела из трубки
наружу осуществляется путем сильного
вытягивания туловища, причем задняя
часть последнего в это время прочно
прицепляется к стенкам трубки посредством
щетинок поясков и заднего отдела. При
раздражении или при опасности
высунувшееся наружу животное мгновенно
исчезает в глубине трубки вследствие простого
сокращения мощной продольной
мускулатуры, а прочное прикрепление заднего
конца облегчает, конечно, это «движение
бегства».
Погонофоры, поднятые на палубу с
большой глубины, не обнаруживают
никаких движений, хотя остаются некоторое
время живыми, если их содержать в
холодной воде. Английские зоологи супруги
Саусворд (Southward) и датчанин К и р-
к е г а а р д (Kirkegaard) успешно
содержали живых погонофор (представителей
рода Siboglinum) в аквариуме при
температуре в + 5° С. Однако животные не
проявляли никакой активности в
течение многих дней. Затем Карлайлу
(Carlisle) удалось установить, что
Siboglinum, подобно многим морским животным,
активен лишь в темноте и что красный
свет не препятствует его активности.
Прибегнув к красному свету, Карлайл
наблюдал движения животного и собирание
пищи щупальцем.
Очень древнее происхождение
погонофор не оставляет сомнений. Однако об
ископаемых погонофорах до самого
последнего времени ничего не было известно,
хотя, казалось бы, их остатки в виде
трубок должны встречаться начиная с
нижнего палеозоя.
Погонофоры относятся к числу самых
длинных животных. Их сильно вытянутое
тело нитевидно или шнуровидно. Длина
его обычно в 100—500 раз превышает
ширину. Размеры тела варьируют очень
широко. К числу миниатюрных форм
относится, например, Siboglinum minutum из
Берингова моря, у которого тело имеет
не более 5,5 еле в длину при ширине менее
0,1 мм. Таких мелких видов очень много,
особенно в роде Siboglinum. К крупным
погонофорам принадлежат Spirobrachia
grandis, длиной не менее 25 см при
диаметре туловища до 2,5 мм, и более тонкая
Zenkevitchiana longissima, достигающая
36 см в длину. Однако самой большой
формой является обитающий в Полярном
бассейне у берегов Канады Galathealinum
arcticum. Одна передняя часть самки
этого вида имеет в длину 50 см и 1,33 мм
в диаметре, все животное, вероятно,
достигает 1 м. Сравнительно крупными
являются и другие виды этого рода.
Мелкие и среднего размера формы
полупрозрачны, бесцветны или беловаты;
сквозь стенку тела просвечивают
наполненные красной кровью кровеносные
сосуды, придающие щупальцам розовый или
красноватый оттенок. У самцов в
туловище обычно просвечивают беловатые
половые органы, а у самок в средней части
тела — желтоватые или красноватые
яйца. Крупные Spirobrachia grandis и
Galathealinum arcticum окрашены довольно
ярко, первая в кирпично-красный,
вторая форма в розовато-коричневый цвет.
Внешний вид животных весьма
своеобразен. Червеобразное цилиндрическое
тело их состоит из трех главных отделов,
или сегментов: двух передних,
сравнительно коротких, и заднего, необычайно
длинного. На самом коротком, первом
сегменте располагаются очень длинные
щупальца и спинная головная лопасть.
Несколько более длинный второй сегмент
несет пару косых хитиноидных ребрышек,
или килей, называемых уздечкой; они
служат для опоры передней части тела
на стенку трубки. Третий сегмент, или
20*
307
туловище, значительно меняется от
переднего конца к заднему. В нем ясно
различимы два главных участка: передний —
преаннулярный,
простирающийся до особых поясков, и задний, лежащий
позади поясков, —п останнуляр-
н ы й. Последний в свою очередь
подразделяется на длинный (передний) участок,
несущий прикрепительные папиллы, и на
задний отдел, снабженный
палочковидными щетинками.
По-видимому, все наиболее примитивные
формы имеют только 4—6 очень длинных
щупалец, но у большинства число
щупалец в процессе эволюции увеличилось до
10, 20 и даже до 250 и более. С другой
стороны, у некоторых форм (Siboglinum)
произошла редукция многих щупалец и
осталось лишь одно правое. На щупальцах
обычно располагаются ряды очень
нежных и тонких п и н н у л, или
ворсинок.
У большинства погонофор все
щупальца свободно прилегают друг к
другу, но у Lamellisabella и Spirobrachia
спаяны между собой. У Lamellisabella,
которая имеет круговое расположение
щупалец, при этом образуется полый
щупальцевый цилиндр, а у Spirobrachia
формируется щупальцевая пластинка,
свернутая спирально наподобие рулона бумаги.
Чрезвычайно характерными
образованиями туловища являются
прикрепительные сосочки, или папиллы. Это небольшие
округлые или грушевидные подушечки,
обычно снабженные маленькими хитино-
идными пластинками. Замечательно, что
у наиболее примитивных погонофор они
слабо развиты, представлены
неопределенно большим числом и неправильно
рассеяны на поверхности туловища. У более
развитых родов и видов передние
папиллы принимают правильное расположение,
а именно лежат на брюшной стороне
попарно и метамерно. Общее число папилл
на туловище при этом сильно
сокращается* В ряду высших форм количество
передних метамерных папилл постепенно
возрастает, и, наконец, почти все
папиллы туловища становятся метамерными.
Преаннулярная область
туловища отличается еще наличием
широкой ресничной полоски на спинной
стороне. Судя по связи со спинным
нервным стволом, это орган чувств (может
быть, орган химического чувства, или
водного обоняния).
В средней части туловища находятся
2 или 3 пояска очень мелких зубчатых
щетинок, которые, несомненно, служат
для прикрепления тела к внутренней
поверхности трубки. Позади поясков
находится длинный участок туловища,
отличающийся развитием метамерных папилл
на брюшной стороне. За ним, наконец,
следует небольшой задний отдел,
несущий метамерные щетинки, которыми
животное закрепляется в глубине трубки.
Характерные нарушения метамерии
щетинок показывают, что первоначально они
располагались на этом участке тела
беспорядочно.
Покровы погонофор состоят из
однослойного эпителия, богатого
одноклеточными железами и почти сплошь
покрытого кутикулой. Лишь
незначительные участки на щупальцах и на спинной
стороне туловища одеты мерцательным
эпителием. Под кожным эпителием лежит
тонкий слой кольцевых мускульных
волокон, за которым располагается мощный
продольный мускульный слой. Имеются
многочисленные многоклеточные железы,
открывающиеся протоками на поверхности
тела; они выделяют вещество трубки.
Хорошо развитая полость тела
имеет собственные эпителиальные стенки
и, следовательно, представляет собой
вторичную полость тела, или целом.
Впереди, у основания щупалец, лежит
непарный мешковидный или подковообразный
целом первого сегмента. От него отходят
целомические каналы щупалец. Кроме
того, пара выделительных каналов
открывается своими внутренними
концами в этот целом, что указывает на
принадлежность их к первому сегменту. Во
втором и третьем сегментах имеются
свои парные целомические мешки.
Нервная система крайне
примитивна и целиком залегает в толще
кожного эпителия. Она представлена почти
сплошным нервным сплетением, клетки
и волокна которого образуют сгущения
на спинной стороне тела — мозг — в
области первого сегмента и спинной нерв-
308
Рис. 157. Погонофора Spirobrachia beklemischevi, извлеченная из трубки. Вид с
брюшной стороны. Туловище сзади оборвано.
J — щупальцевая крона, состоящая из 39—72 щупалец, спаянных вдоль в виде спирально свернутой
щупальцевой пластинки; 2 — передний отдел тела, состоящий из первого и второго сегментов;
3 — уздечка; 4 — брюшной желобок; 5 — прикрепительные папиллы с хитиноидными
пластинками; 6 — метамерная часть туловища; 7 — неметамерная часть туловища; 8 — пояски зубчатых
щетинок; 9 — участок туловища позади поясков с метамерными рядами папилл. (По Иванову.)
ный тяж, проходящий вдоль всего тела.
От мозга отходят нервы щупалец.
Кровеносная система
состоит из продольного брюшного сосуда, по
которому кровь течет сзади наперед, и
спинного сосуда, несущего кровь назад.
Брюшной сосуд у основания щупалец
снабжен мускулистым сердцем,
сокращения которого проталкивают кровь в
приносящие сосуды щупалец. У
представителей отряда Athecanephria к сердцу
Рис. 158. Передний
конец тела самца Oligo-
brachia dogieli: A — со
спинной стороны, В —
с брюшной стороны.
1 — щупальца; 2 — первый
сегмент; з — уздечка; 4 —
второй сегмент; 5 —
спинная ресничная полоска;
6 — туловище; 7 —
железы, просвечивающие сквозь
стенку тела; 8 — мужское
половое отверстие; 9 —
головная лопасть. (По
Иванову.)
Рис. 159. Задний щетинконосный
отдел тела Siboglinum caulleryi:
2 — щетинки; 2 — просвечивающие
мускульные пучки. (По Иванову.)
со спинной стороны прилегает
мешковидный участок целома —
околосердечная сумка, или перикардий. В спинной
сосуд кровь поступает из выносящих
сосудов щупалец. В задней половине
туловища оба продольных сосуда соединяются
многочисленными поперечными ветвями.
Кровь погонофор имеет красный цвет
и содержит гемоглобин.
Органами выделения
служат целомодукты первого
сегмента. Это длинные ресничные каналы, по
степени развития и по локализации
которых все погонофоры делятся на две
группы. У представителей отряда
Athecanephria они широко расставлены и
прилегают к задним участкам боковых
ветвей спинного продольного сосуда. У форм,
принадлежащих к отряду Thecanep-
hria, целомодукты очень длинны, извиты
и сближены друг с другом медиально,
причем прилегают к спинному сосуду,
глубоко впячивая его брюшную стенку.
Погонофоры раздельнополы, при этом
самцы и самки не отличаются по внешнему
виду. У самцов сперматозоиды развиваются
в паре очень длинных семенных мешков,
лежащих в задней половине туловища. От
семенных мешков отходит пара
семяпроводов, тянущихся вперед до передней
границы туловища, где они открываются
наружу на брюшной стороне тела. В них
формируются многочисленные уплощенные
или веретеновидные перепончатые спер-
матофоры с очень длинной тонкой нитью.
У самок яичники занимают переднюю
половину туловища. Зрелые яйца
сначала попадают в полость тела через
разрыв стенки яичников, затем выводятся
наружу по яйцеводам.
Самка откладывает яйца в переднюю
часть своей собственной трубки, в
которой и протекает все эмбриональное
развитие. Дробление яйца происходит, как
у низших вторичноротых, по радиальному
типу. Образование стенок полости тела
(мезодермы) осуществляется так
называемым энтероцельным путем, при котором
мешковидные зачатки вторичной полости
тела сперва возникают как боковые
выпячивания первичного кишечника, а потом
отделяются от него. Этот способ
развития целома чрезвычайно характерен для
310
вторичноротых. У зародыша сперва
развивается энтодермальная кишка, ее
крупные клетки переполнены запасными
питательными материалами — желтком и
жиром. При превращении зародыша в
молодое животное первичная кишка,
выполнив свою роль временного резервуара
запасных питательных веществ,
полностью рассасывается. Настоящей
планктонной личинки погонофоры не имеют.
Молодое животное выбирается из
материнской трубки и после очень короткого
периода свободной жизни оседает на дно
и выделяет свою собственную трубку*
Для решения вопроса о том, как
питаются погонофоры, эти неподвижные
обитатели морского дна, полностью
лишенные обычной пищеварительной системы
и неспособные активно разыскивать
пищу, рассмотрим строение их
щупальцевого аппарата.
Несмотря на все разнообразие в числе
и особенностях прикрепления щупалец,
в щупальцевом аппарате погонофор
всегда проявляются одни и те же
характерные особенности. Прилегая друг к другу,
щупальца ограничивают
межщупальцевое пространство, у некоторых форм,
как уже было сказано выше, щупальца
даже соединяются друг с другом. При
этом у одних (Lamellisabella) образуется
полый цилиндр, а у других (Spirobrachia)
спирально свернутая щупальцевая
пластинка.
Таким образом, наличие временной
(при свободных щупальцах) или
постоянной (при спаянных щупальцах)
межщупальцевой полости той или иной
конфигурации очень характерно. Она
образуется даже у Siboglinum вследствие што-
порообразного закручивания единственного,
но зато необычайно длинного щупальца.
Каждое щупальце является длинным
выростом тела и содержит целомический
канал. На внутренней поверхности
щупальца у большинства погонофор
расположены ряды пиннул—длинных и очень
тонких выростов. Каждая пиннула
представляет собой одну очень вытянувшуюся
клетку эпителия щупальца, содержит одно
ядро и пронизана двумя параллельными,
соединяющимися у вершины пиннулы
тончайшими кровеносными капиллярами —
приносящим и выносящим. Пронизывая
толщу эпителия щупальца, эти
капилляры сообщаются с соответственными
сосудами самого щупальца. У основания пин-
нул располагаются ряды мерцательных
клеток. При соприкосновении соседних
щупалец они образуют временные (у боль-
Рис. 160. Схема строения переднего конца тела
погонофор: А — вид с брюшной стороны; Б — вид
с правой стороны.
1 — целомические каналы щупалец; 2 — щупальца;
3 — приносящий сосуд щупальца; 4 — целом первого
сегмента; 5 — первый сегмент; 6 — наружное
выделительное отверстие; 7 —целомодукт первого сегмента;
8 — второй сегмент; 9 — целом второго сегмента;
10 — целом туловищного сегмента; 11 — туловищный
сегмент; 12 — спинной кровеносный сосуд; 13 —
брюшной кровеносный сосуд; 14 — семяпровод; 15 —
мужское половое отверстие; 16— сердце; 17—перикардий;
18 — основание щупальцевых нервов; 19 — выносящий
сосуд щупальца; 20 — головной сосуд; 21 — мозг;
22 — боковой головной сосуд; 23 — продольный
нервный ствол. (По Иванову.)
311
Рис. 161. Поперечный разрез кроны щупалец
Lamellisabella zachsi:
I — щупальца; 2 — целомический канал щупальца;
3 — кутикула; 4 — нерв щупальца; 5 — ресничные
бороздки между щупальцами; 6 — кровеносные
сосуды; 7 — пиннула; 8 — межщупальцевое пространство.
(По Иванову.)
шинства форм), а при срастании
щупалец — постоянные продольные ресничные
бороздки. Между основанием пиннул и
мерцательными клетками иногда имеются
ряды железистых клеток. Характерно,
что пиннулы обращены в
межщупальцевое пространство, где образуют густое
сплетение.
Одной из функций щупалец,
несомненно, является дыхание.
Однако перечисленные выше детали их
строения показывают, что они служат также
для собирания пищи, ее переваривания
и всасывания продуктов пищеварения.
Вероятно, мерцание ресничных бороздок
на щупальцах гонит воду в
межщупальцевое пространство. Вода входит между
концами щупалец и выходит между их
основаниями. Взвешенные в воде
микроскопические организмы и детрит застревают
в густой сети пиннул, которая, таким
образом, играет роль фильтра. В
межщупальцевом пространстве выделяются
пищеварительные ферменты. Здесь же
происходит переваривание пищи.
Питательные вещества всасываются пиннулами,
играющими роль своеобразных
кишечных ворсинок, и поступают в кровь их
сосудов, откуда с током крови попадают
через выносящие сосуды в спинной сосуд
и разносятся по всему телу. Возможно,
фильтрация воды щупальцами и
переваривание пищи происходят не
одновременно, т. е. сперва накапливается
некоторый запас пищи, потом животное
переваривает ее, втянув крону щупалец
в трубку.
Относительно способа питания Sibogli-
num, обладающего всего одним
щупальцем, сначала на основании изучения его
строения было сделано предположение
(Иванов, 1960), что животное
подбирает пищевые частицы с поверхности ила,
«исследуя» окружающее пространство дна
своим щупальцем. Затем щупальце обви-
Рис. 162. Поперечный разрез трех соседних
щупалец Lamellisabella zachsi:
l — нерв щупальца; 2 — выносящий сосуд щупальца;
3 —выносящий кровеносный капилляр пиннулы; 4 —
приносящий капилляр; 5 — пиннула; 6 — ее ядро;
7 — ресничная клетка; 8 — приносящий сосуд
щупальца; 9 — целомический канал щупальца; ю —
продольные мускульные волокна; 11 — эпидермис;
12 — кутикула. (По Иванову.)
312
Рис. 163. Трубки различных погонофор:
Л — Siboglinum fedotovi; Б —
Lamellisabella johanssoni; В — Polybrachia
annulata; Г — устье трубки Spirobrachia
beklemischevi. (По Иванову.)
Рис. 164. Трубочка сверлящей
погонофоры Sclerolinum,
торчащая из кусочка дерева,
поднятого тралом со дна
Яванской впадины. (По Иванову.)
вается вокруг пищевого комочка и
вытягивается в трубку, где пища и
переваривается. В самое последнее время
Карлайлу удалось полностью
подтвердить это предположение прямыми
наблюдениями над животными, содержащимися
в аквариуме.
Трубки погонофор имеют почти
цилиндрическую форму, так как очень
постепенно расширяются от заднего конца к
переднему. Длина их колеблется от нескольких
сантиметров до 1,5 м у Zenkevitchiana
longissima, а у Galathealinum arcticum,
вероятно, достигает нескольких метров.
Строение трубки в общем у всех
погонофор сходно; однако у одних форм она
гибка и нежна и обладает характерной
кольчатостью, у других имеет беловатые
пергаментообразные стенки, у третьих
отличается прочностью и твердостью и
темным бурым цветом. Детали строения
трубок часто используются как хорошие
видовые признаки. У большинства
погонофор передний конец трубки имеет
особое строение. Здесь стенки ее особенно
тонки, обычно совсем прозрачны и легко
спадаются. Напротив, у Lamellisabella
и Spirobrachia передний конец трубки
обладает толстыми непрозрачными
стенками, а устье трубки имеет форму
воронки. Трубки погонофор состоят из
хитина.
Тип погонофор содержит один класс,
который делится на два отряда. К
первому отряду — Athecanephria —
относятся погонофоры со свободными
щупальцами и с перикардиальным мешочком.
Сперматофоры у представителей этого
отряда веретеновидные, а передняя часть
трубки всегда пленчатая, со
спадающимися стенками. Второй отряд — Checa-
nephria — характеризуется свободными
или спаянными щупальцами, отсутствием
перикардия и листовидными спермато-
форами. У некоторых относящихся сюда
форм трубка с воронковидным устьем.
ОТРЯД ATHECANEPHRIA
К этому отряду, содержащему три
семейства, относятся примитивные формы,
иногда лишенные метамерии туловищных
папилл.
Семейство Склеролиниды (Sclero-
linidae) — особое семейство, установлен-
313
ное недавно Веббом (Webb) для рода
Sclerolinum, виды которого ведут
сверлящий образ жизни в затонувших кусках
дерева.
К сожалению, эти, вероятно очень
примитивные, погонофоры еще слабо
изучены. Первый представитель рода (Scle-
rolinum sibogae) и сам род были описаны
И. Саусвордом (Е. Southward) в 1961 г. по
материалам, собранным еще голландской
экспедицией на судне «Зибога» в морях
Малайского архипелага. Лучше
известен, однако, изученный Веббом Scleroli-
num brattstromi из Хардангер-фиорда
в Норвегии. Интересный материал по
новому виду Sclerolinum собран «Витязем»
в Яванской впадине. Еще один вид, по-
видимому, обитает в Тихом океане.
Sclerolinum отличается от других погонофор
не только своей экологией, но также
целым рядом особенностей строения.
Имеются лишь два щупальца, лишенные пин-
нул, туловищные папиллы отсутствуют,
так же как пояски зубчатых пластинок;
вероятно, все эти признаки следует
считать первичными. На этом основании Вебб
считает невозможным включать Sclero-
linidae в какой-либо из двух известных
отрядов. Так ли это, покажут будущие
исследования.
Семейство Олигобрахииды (Oli-
gobrachiidae) объединяет три рода, два
из них отличаются отсутствием
метамерии туловищных напил л, которые в
передней части туловища беспорядочно
сближены по обе стороны брюшного желобка.
Nereilinum murmanicum с двумя
щупальцами, лишенными пиннул, обычен в
Баренцевом море и встречается в северной части
Атлантического океана. Oligobrachia
имеет 4—9 щупалец, снабженных пиннулами,
и интересна своим амфибореальным
распространением: один вид — О. dogi-
eli — обитает в Охотском море и в
Тихом океане у берегов Камчатки, другой—
О. ivanovi — в восточной части
Атлантического океана. Birstenia vitjasi с 12
щупальцами и с хорошо развитой
метамерией туловищных папилл известна из
Берингова моря.
Семейство Зибоглиниды (Sibogli-
nidae) объединяет многочисленные мелкие
формы с двумя или с одним щупальцем
и с гибкой кольчатой трубкой. Более
примитивным родом является двущупальце-
вый Siboglinoides с двумя видами — S.
dibrachia (Индийский океан) и S. colum-
biensis (Карибское море). Однощупальце-
вый Siboglinum объединяет более 50 видов,
это самый обширный и наиболее широко
распространенный род. В мелководной
западной части Охотского моря обширные
пространства илистого дна на глубине
100—150 ж заняты густыми поселениями
Siboglinum caulleryi.
ОТРЯД THECANEPHRIA
Из трех семейств, принадлежащих к
этому отряду, самыми примитивными являются
полибрахииды. Они характеризуются
хорошо отграниченным первым сегментом
тела, не связанными друг с другом
щупальцами и пленчатым передним
участком трубки. Другие два семейства
отличаются слиянием первых двух сегментов
и срастанием щупалец, образующих более
или менее сложный щупальцевый
аппарат, а также имеют воронковидное устье
трубки.
Семейство Полибрахииды (Polybra-
chiidae) объединяет семь родов. Из них
Diplobrachia — наиболее примитивный
род — имеет от двух до четырех щупалец.
Четыре вида Diplobrachia обитают в
Японской глубоководной впадине, в
центральной части Индийского океана, в Аравийском
море и в Атлантическом океане. Zenkevi-
tchiana (названная так в честь
крупнейшего советского зоолога и океанолога
проф. Л. А. Зенкевича)
представлена одним видом Z. longissima — очень
характерным обитателем больших
глубин (до 10 тыс. м) Курило-Камчатской
впадины. Беловатые трубки Z. longissima,
достигающие 1,5 м длины, при удачном
тралении массами заполняют мешок
трала. Polybrachia объединяет три вида. Из
них P. annulata является очень обычной
формой в юго-западной части Берингова
моря (на глубинах от 1400 до 5 тыс. м)у
Galathealinum — очень крупные формы
с большим (до 268) количеством щупалец.
Первый вид этого рода ~ G. bruuni —
314
был добыт датской зоологической
глубоководной экспедицией на судне «Галатея»
в Целебесском море на глубине более
5 тыс. м. Второй вид — G. brachiosum,
обнаруженный экспедицией «Витязя» у
берегов Канады и США в Тихом океане, по-
видимому, является одной из наиболее
обычных глубоководных форм в этом
районе. Наконец, G. arcticum найден в
Полярном бассейне у берегов Канады, где
обитает на глубине 36 м. Это самая
крупная из известных погонофор,
достигающая, по-видимому, не менее 1 м в длину.
Семейство Ламеллисабеллиды (La-
mellisabellidae) включает два рода Lamel-
lisabella и Siphonobrachia. Его представители
отличаются полой цилиндрической
щупальцевой кроной, состоящей из спаянных
щупалец. Характерный представитель — L.
zachsi, имеющий 30—32 щупальца, был
описан П. В. Ушаковым в 1933 г. В 1937 г.
именно для этого вида К. Э. Иоганссон
установил класс Pogonophora. Lamelli-
sabella zachsi обитает в двух, по-видимому
изолированных, областях — в
глубоководной части Охотского моря и в
юго-западной части Берингова моря. Другим
видом этого рода является L. johanssoni
с 18 щупальцами — эндемичная форма
Японской глубоководной впадины, L. iva-
novi с 22 щупальцами из Панамского
залива в восточной части Тихого океана
и маленькая L. minuta с 10 щупальцами,
живущая у берегов Южной Индии.
Семейство Спиробрахииды (Spiro-
brachiidae) включает один род Spirobrachia
с двумя видами. Характерной
особенностью Spirobrachia является спирально
закрученная щупальцевая крона,
состоящая из многочисленных (до 223)
спаянных друг с другом щупалец. Известная
из Берингова моря S. grand is относится
к числу крупных форм, диаметр ее
трубки достигает 3 мм. Другой вид — S. bek-
lemischevi — свойствен Курило-Камчат-
ской глубоководной впадине.
ТИП ПОЛУХОРДОВЫЕ
(HEMICHORDATA)
Полу хордовые животные, к которым
относятся кишечнодышащие и крыложабер-
ные,— небольшая группа исключительно
морских организмов. Они живут на дне
моря. Одни из них по форме тела очень
похожи на червей и ведут свободнопод-
вижный образ жизни, роясь в иле,
Другие же являются прикрепленными
сидячими организмами, образующими весьма
своеобразные колонии. Больше всего по
внешнему виду они напоминают мшанок.
Группа кишечнодышащих представляет
собой исключительный интерес с точки
зрения истории возникновения и
развития всего раздела так называемых вто-
ричноротых животных, куда вместе с
иглокожими входят все хордовые животные,
в том числе и позвоночные.
Первый представитель кишечнодышащих
был открыт в 1821 г. И. Ф. Эшшоль-
ц е м (I. F. Eschscholtz), врачом
экспедиции О. Е. К о ц е б у, плававшим на
русском корабле «Рюрик». В районе Map-
шалловых островов в Тихом океане он
нашел крупное червеобразное животное,
названное им Ptychodera flava и принятое
за своеобразную и очень необычную
голотурию. Немного позже, в 1829 г., в
Средиземном море итальянскими
учеными был найден Balanoglossus clavige-
rus. Находки этих необычных животных
привлекли к себе большое внимание
зоологов. В прошлом веке большинство зоологов,
даже таких крупных ученых, как В. К ю-
кенталь (W. Kiikenthal), относило их
к червям. А. Агасс иц (A. Agassiz),
например, предполагал, что баланоглосс
является переходной формой между немерти-
нами и червями. Другие же просто
принимали их за червей, имеющих малое
количество сегментов тела. Эта точка зрения
продержалась очень долго. Даже после
замечательных работ крупнейшего русского
зоолога и эмбриолога А. О. Ков а л ев с к о-
г о A866), детально изучившего анатомию
баланоглосса и нашедшего у него
совершенно необычный жаберный аппарат,
устроенный по такому типу, который не имеет
никаких аналогов среди других
беспозвоночных животных и является одним из
признаков хордовых, многие, даже
известные, зоологи продолжали ставить их
в системе животных вместе с червями.
Личинка кишечнодышащих — торна-
рия, свободно плавающая в воде, также
уже давно была известна науке. Ее открыл
в 1850 г. И. Мюллер (I. Muller). Но по
своему общему виду она настолько похожа
на личинок иглокожих, что ее долгое
время считали личинкой какой-то морской
звезды. И только в 1870 г. другой
замечательный русский ученый и друг А. О.
Ковалевского И. И. Мечников доказал,
что торнария является личинкой
баланоглосса. Он высказал предположение о
родстве последнего с иглокожими. В 1912 г.
профессор Н. А. Холодковски й—
также один из известных русских зооло-
316
гов — предложил рассматривать кишеч-
нодышащих как дополнение к типу
иглокожих, с которыми их нередко и
объединяли в дальнейшем некоторые ученые.
Кроме сходства личинок, во внутреннем
строении самого баланоглосса имеются
некоторые черты, общие с иглокожими. Но
расхождения в их строении все же
чрезвычайно велики. Более внимательное
изучение кишечнодышащих позволило
обнаружить такие особенности их строения,
которые сближают их с хордовыми
животными. Еще в 1884 г. В. Бэте он указал на
возможность родства кишечнодышащих
с позвоночными животными. Некоторые
зоологи — В. М. Ш и м к е в и ч, В. А.
Догель и др.— еще совсем недавно
помещали их в системе животных в
начале типа хордовых, Баланоглосс
представляется тем самым как бы прообразом
хордовых животных—«хордовых на заре
своей юности».
В настоящее время точка зрения на
положение кишечнодышащих в системе
животных, видимо, установилась
окончательно. Академик И. И. Шмальгау-
зен и В. Н. Беклемишев
помещают класс Enteropneusta среди
раздела вторичноротых животных в
составе особого типа полухордовых —
Hemichorda, в который вместе с кишечно-
дышащими входят и почти нацело
вымершие в настоящее время граптолиты.
Кишечнодышащие стоят посредине между
двумя основными ветвями
вторичноротых животных — типом иглокожих,
в древние времена перешедших к
сидячему образу жизни и в связи с этим
подвергшихся кардинальной перестройке
всего плана строения тела своих предков,
и типом хордовых, активно
двигающихся животных, с активным питанием
и прогрессивным развитием различных
систем органов, в том числе нервной
системы. Кишечнодышащие не могут быть
отнесены ни к тому ни к другому типу.
Однако современных кишечнодышащих
нельзя рассматривать и как прямых предков
позвоночных животных. Таким образом,
нам предстоит познакомиться еще с
одним типом животных — Hemichorda, в
который включаются два класса
животных: 1 — палеозойские вымершие Gra-
ptolithoidea с современными их
представителями— Pterobranchia и 2—Enteropneusta.
Кишечнодышащие (Enteropneusta)
представлены в настоящее время 64 видами,
относящимися к 12 родам и 4 семействам.
Группа крыложаберных (Pterobranchia)
включает 21 вид, 3 рода и 2 семейства.
Представители этих двух классов
животных сильно отличаются друг от друга
и по внешнему виду, и по образу жизни.
Но и те и другие исключительно морские
животные, не выносящие даже
небольшого опреснения. Живут на дне моря.
Кишечнодышащие — свободноподвиж-
ные червеобразные животные, с очень
сильно вытянутым в длину мягким,
обильно покрытым слизью телом. Большинство
из них живет в мягком иле либо илистом
песке в прибрежной приливо-отливной
зоне океана, заселяя тихие куты морских
губ и заливов. Грунт в таких заливах в
некоторых случаях бывает даже сильно
заражен сероводородом и издает
неприятный сильный запах. И все-таки
животные обитают в нем. Другие виды
предпочитают песок с галькой, которые не
обнажаются во время отлива, а некоторые,
в частности уже известная нам Ptychode-
га, встречаются на коралловых рифах.
Роясь в грунте, кишечнодышащие
заглатывают огромное его количество и
извлекают из него мелких животных,
растительные остатки и другие органические
вещества, которыми они питаются. Они
являются, таким образом, в основной
своей массе грунтоядными животными, без-
выборочно заглатывающими грунт.
Однако некоторые крупные виды (Ptycho-
dera) могут быть и хищниками. Есть
среди них, по-видимому, и такие,
которые питаются мелкими планктонными
водорослями, взвешенными в воде (рис. 165).
Поселения энтеропнейст на
мелководьях бывают иногда очень значительны,
но, видимо, непостоянны. Бывают годы,
когда эти животные появляются на
некоторых участках литорали в огромном
количестве, а в другое время в том же
районе моря можно встретить лишь единичные
экземпляры. Например, живущий у нас
на литорали Белого и Баренцева морей
Saccoglossus mereschkowskii иногда
образует такие массовые скопления, при кото-
317
Рис. 165. Разные виды кишечнодышащих:
1 — Balanoglossus (Ptychodera) clavigerus; 2 — Schizocardium brasiliense;
3 — Saccoglossus pusillus; 4 — Protobalanus koeleri; 5 — Glandiceps malayanus.
рых в 1 дмъ грунта встречается 15—20
экземпляров животных размером до 20 см
и более. Видимо, их массовое развитие
зависит от каких-то особо благоприятных
комбинаций условий—температуры,
солености, состава грунта и других факторов.
Кишечнодышащие обитают в основном в
поверхностной—литоральной зоне, и к ней
приурочено в своем распространении
большинство их видов. Но они тем не менее
встречаются в океане на всех глубинах,
хотя количество глубоководных форм
очень невелико. До недавнего времени
наиболее глубоководным представителем
Enteropneusta считался Glandiceps abyssi-
cola, пойманный на глубине 4500 м.
Однако среди коллекций глубоководных
животных, собранных на советском
экспедиционном судне «Витязь» в Тихом
океане, есть кишечнодышащие, которые были
318
обнаружены в Курило-Камчатской и
Алеутской впадинах на глубине 700—800 м.
Большинство видов кишечнодышащих
F0%) обитает в теплых морях, 30% —
в морях умеренной зоны. И только
некоторые представители этого класса
встречаются в арктических областях.
Saccoglossus mereschkowskii, например, имеет
циркумполярное распространение. Он
найден у нас в Белом, Баренцевом,
Беринговом, Охотском и Японском морях.
Наиболее богата видами кишечнодышащих
тропическая зона, причем виды,
обитающие в тропиках, распространены, как
правило, очень широко. Но среди них есть
и такие, которые имеют ограниченный
ареал. Некоторые обитают только в
Индийском океане и в морях Малайского
архипелага.
Известны также свои виды для Ла-
Манша, Канады, Южной Африки и
других мест. Ни один вид Enteropneusta
неизвестен в Антарктике. Только Saccoglossus
otagoensis был найден на
северо-восточном побережье Новой Зеландии.
Представители класса
Enteropneusta имеют четкий план строения тела,
не измененный под влиянием сидячего
образа жизни, как у Pterobranchia.
Размеры тела Enteropneusta очень
варьируют и в среднем составляют 10—15 см
в длину. Самые мелкие из них имеют 3—5
см. Но известны и очень длинные кишечно-
дышащие. Balanoglossus cornosus, В. cla-
vigerus и др. могут достигать в длину
1,5 м. А самый крупный экземпляр В. gi-
gas имел в длину 2,5 м. Диаметр его тела
составлял при этом 2 см. Этот гигантский
представитель Enteropneusta был найден
впервые на Атлантическом побережье
Южной Америки, а затем в значительном
количестве был отмечен у побережья Бразилии.
В 1962 г. в тех же районах была найдена
другая гигантская энтеропнейста. Размеры
наиболее крупных экземпляров имели
в длину более 2 м. Животные относились
к новому виду редкого рода Willeya.
Окраска кишечнодышащих также
варьирует, чаще всего от серовато-белых
до светло-желтых тонов. Часто
встречаются и желтовато-коричневые и
коричневые особи. Но среди них есть и ярко
окрашенные виды — ярко-красные
Saccoglossus pusillus, оранжевые Ptychodera
flava, темно-коричневые Glandiceps и
черно-фиолетовые Pt. erythraea. Обычны
разные вариации окраски, и разные части
тела животных могут быть окрашены
различно. Наш северный Saccoglossus
mereschkowskii имеет хобот ярко-кирпичного
цвета. Воротник и передняя часть тела
темнее хобота и другого оттенка —
кроваво-алого цвета. К заднему концу
окраска постепенно слабеет и тело становится
более прозрачным с желтоватыми или
зеленоватыми оттенками. Животные,
добытые из глубоководных океанических
впадин, имели грязную розовато-бежевую
окраску и достигали в длину 10—15 см.
Длинное червеобразное тело
кишечнодышащих четко дифференцировано на три
отдела — хоботок, воротничок и
собственно туловище, где располагается
большинство органов животного (рис. 166,Л).
Хоботок представляет собой
мускулистое вытянутое или желудевидное
образование. Основная функция хоботка —
роющая. Основание его сужено в тонкую
шейку, которая охватывается
следующим отделом тела — коротким, также
мускулистым воротничком,
несколько приподнимающимся над
поверхностью тела в виде валика. Внутри
воротничка, как и внутри хобота, имеется
полость, соединяющаяся двумя
канальцами через особые воротничковые поры
с внешней средой. Следом за воротничком
идет длинное туловище животного,
составляющее 9/10 всей длины тела. В передней
части туловище с боков прорезано
двумя рядами многочисленных узких
жаберных щелей, расположенных поперечно. Все
тело животного одето нежным
однослойным эпителием, несущим движущиеся
реснички и называемым поэтому
мерцательным. Кроме того, сверху животное
обильно покрыто слизью, вырабатываемой
клетками воротничка. Внутри следуют два
слоя мышечных волокон: наружных —
кольцевых и внутренних — продольных.
На брюшной стороне тела у основания
хобота, ближе к границе последнего с
воротничком, лежит рот (рис. 166,i>). Рот
ведет в короткую глотку, лежащую на
протяжении воротничка. От спинной
стороны глотки отходит внутрь хоботка сле-
319
Рис. 166. Строение баланоглосса.
А — общий план строения: 1 — хоботок; 2 —
воротничок; з — туловище; 4 — жаберные отверстия;
5 — половые поры; 6 — печеночные железы. Б —
строение баланоглосса: 1 —продольные мышцы
хоботка; 2 — полость хоботка; з — клубочек; 4 — сердце;
5 — канал нервной трубки; 6 — спинной
кровеносный сосуд; 7 — жаберные щели глотки; 8 —
кишечный отдел глотки; 9 — брюшной кровеносный сосуд;
10 —полость воротничка; 11 — рот; 12 — скелетная
пластинка; 13 — нотохорд.
пое выпячивание. Этот вырост
называется нотохордом и считается
зачатком спинной струны, или хорды,
позвоночных животных. Нотохорд
доходит до середины длины хоботка, прилегая
к его брюшной стороне. Между ним
и брюшной стенкой хоботка развивается
плотная, хрящеобразная пластинка,
которая называется скелетом хобот-
к а. Вместе с нотохордом эта пластинка
придает хоботку известную плотность.
Далее глотка переходит в пищевод,
или дыхательный отдел кишечника.
В то время как спинная и брюшная
стенки пищевода сплошные, боковые стенки
пронизаны двумя рядами подковообразных
жаберных щелей. Через эти щели полость
пищевода сообщается с внешней средой.
Вода, проходящая через рот, поступает в
жаберный отдел и через жаберные щели
выходит наружу. В перегородках между
щелями проходят многочисленные
кровеносные сосуды, и кислород, растворенный
в воде, передается прямо в кровь
животного. Такое строение жаберного аппарата —
связь его с кишечником — неизвестно для
представителей беспозвоночных животных.
Только среди хордовых этот тип строения
дыхательного аппарата получает
широкое распространение. Благодаря
строению дыхательного аппарата эти животные
и стали называться кишечнодышащими.
По брюшной стенке пищевода проходит
продольный желобок, образованный
частично клетками, выделяющими секрет,
а частично клетками, несущими реснички.
Это эндостиль — образование,
которое считается гомологом щитовидной
железы позвоночных животных. Его
более подробное описание приводится при
разборе анатомии оболочников.
Мелкие частицы пищи, попадающие
с водой в пищевод, склеиваются слизью
эндостиля и гонятся его ресничками вдоль
желобка, в пищеварительный отдел
кишечника. За жаберным отделом
кишечника следует печеночный отдел, несущий
на спинной стороне многочисленные
печеночные придатки. Затем в виде трубки
идет средняя кишка, которая переходит
в заднюю. На заднем конце тела она
открывается наружу анальным отверстием.
Кровеносная система
состоит из продольных кровеносных
сосудов — брюшного и спинного. Брюшной
сосуд залегает лишь в туловище, тогда
как спинной продолжается в хоботке.
В хоботке образуется большая
центральная кровеносная лакуна — полость, куда
поступает кровь. По спинному сосуду
кровь течет вперед. Она частично
поступает в отходящие от спинного сосуда
парные приносящие жаберные сосуды,
проходящие через жаберные стенки, и
окисляется. Затем окисленная кровь
через уносящие жаберные сосуды
возвращается назад в спинной кровеносный
сосуд. Из центральной кровеносной ла-
Таблица 27. Иглокожие мелководий Японского моря.
Сверху — морской еж; снизу — морская звезда Patiria pectinifera.
320
куны кровь но двум окологлоточным
сосудам, окружающим глотку в области
воротничка, возвращается в брюшной
сосуд и направляется к заднему концу тела.
Через сеть лакун в области кишечника
она вновь поступает в спинной сосуд.
Циркуляцию крови обеспечивает сердечный
пузырь. Но кровь не проходит через
сердце. Этот полый замкнутый мускулистый
мешок, лежащий у спинной стенки
хобота над нотохордом, своим сокращением
проталкивает кровь из центральной
кровеносной лакуны по сосудам. Чтобы кровь
двигалась в нужном направлении,
имеется система специальных клапанов.
Выделительная система
состоит из двух пар особых трубочек,
посредством которых полости хобота и
воротничка, где накапливаются продукты
выделения, сообщаются с внешней средой.
Эти трубочки называются нефридия-
м и. Как правило, в хоботе сохраняется
только один левый нефридий, а в
воротничке их имеется два. Кроме того, в
хоботке, в центральной кровеносной лакуне,
в перегородке между нею и полостью
хоботка, имеется особый складчатый
орган, называемый клубочком. Этот
клубочек является как бы своеобразным
фильтром, через который продукты
обмена поступают из крови в полость хобота
и оттуда выводятся наружу.
Нервная система состоит из
двух нервных стволов — брюшного и
спинного. Брюшной нервный ствол
развит значительно слабее спинного. Он
залегает по средней брюшной линии туловища.
Спинной нервный ствол продолжается
в хоботок. В области воротничка оба
ствола соединяются нервным воротничковым
кольцом. В области воротничка нервный
ствол залегает в виде толстостенной
нервной трубки, или цилиндра. Эта часть
нервной системы наиболее развита и
называется иногда спинным мозгом.
Наличие хорошо развитой нервной системы
на спинной стороне тела является очень
важным признаком хордовых животных.
Кишечнодышащие и по этому признаку
также принципиально отличаются от
остальных беспозвоночных. Но, с другой
стороны, они отличаются от хордовых
наличием брюшного нервного ствола.
Специальных органов чувств
у кишечнодышащих нет, но под
наружным покровом их тела расположено
множество светочувствительных клеток.
Кишечнодышащие — раздельнополые
животные. Самцы и самки у них по
внешнему виду неразличимы, хотя иногда их
окраска не вполне одинакова. Половая
система устроена очень просто. Половые
железы образуют от 16 до 100 пар
овальных мешочков, лежащих по бокам
кишечника в средней части туловища вслед за
жаберным его отделом. Каждая гонада
открывается наружу коротким протоком,
через который половые продукты
выводятся в окружающую воду.
Кишечнодышащие, за единственным исключением,
размножаются половым путем. Почти все
они яйцекладущие животные.
Представители обоих полов у большинства их видов
встречаются в норках, где и происходит
спаривание. Оплодотворение наружное.
Яйца через половые отверстия или
выбрасываются в воду, или прикрепляются
к стенкам подземных ходов. Только у
тропического рода Glandiceps самцы и самки
во время периода размножения покидают
дно и плавают некоторое время у
поверхности воды, образуя плотные скопления
размером от 1 :до 5 м в поперечнике и
толщиной до 2 м. Животные беспорядочно
двигаются в таких скоплениях, видимо
отыскивая партнеров для размножения.
Однако среди них присутствуют и
неполовозрелые особи. Явление это довольно
загадочно, но оно безусловно способствует
широкому расселению вида в океане.
В зависимости от большего или
меньшего количества желтка, содержащегося в
яйце, развитие у разных видов
кишечнодышащих может происходить двумя
путями — без превращения и с
превращением, или метаморфозом. И у тех и у других
видов кишечнодышащих имеется в
принципе сходная свободноживущая личинка, но
у первых она мало отличается от взрослого
организма (Saccoglossus kowalevskii, Do-
lichoglossus). Развитие у этих видов
сильно сокращено. Из яйцевой оболочки
выходит уже готовая личинка, состоящая
из трех сегментов, каждый из которых
соответствует хоботку, воротничку или
туловищу взрослых форм (рис. 167). Та-
21 Жизнь животных, т. 2
321
кая личинка очень тяжела для того, чтобы
свободно плавать в воде. Она держится
на дне, где ползает или роется в иле. При
помощи своих ресничек она может только
немного плавать в придонных слоях воды.
И хотя эта личинка очень похожа на
молодое животное, она все-таки остается
настоящей личинкой. У нее имеются
временные личиночные органы —
чувствительная теменная пластинка с султаном
ресничек, движущим органом ей служит
прианальный венчик ресничек —
мерцательный поясок. После утери этих
органов превращение ее во взрослое животное
происходит путем вытягивания в длину
главным образом третьего сегмента,
формирования воротничка и внутренних
органов. Появляется нотохорд,
закладываются жабры, оформляется нервная
система.
У тех же Enteropneusta, яйца которых
бедны желтком,— у Ptychodera, Spenge-
lia и др. —развитие происходит через
стадию свободноплавающей пелагической
личинки. Эта личинка очень характерна и,
как уже указывалось выше, чрезвычайно
похожа на пелагических личинок морских
Рис. 167. Превращение личинки балано-
глосса во взрослое животное:
1 — теменной султан ресничек; 2 —
мерцательный поясок; 3 — жаберные отверстия;
4 — прикрепительный сосочек. 1 — хоботок,
II — воротничок, III — туловище.
Рис. 168. Торна-
рия:
1 — теменная
пластинка с султаном
ресничек; 2 —
мерцательный шнур;
з — сердце; 4 —
рот; 5 — желудок;
6 — задний проход;
7 — задний
мерцательный венчик
ресничек.
звезд (рис. 168). Развитие здесь
сопровождается довольно сложным метаморфозом.
Вследствие своего постоянного вращения
вокруг оси личинка называется торнарией.
Зрелая торнария по существу устроена по
тому же плану, что и рассмотренная выше
личинка. Она имеет кишечник с ротовым
отверстием на брюшной стороне и
анальным на заднем полюсе тела. На
переднем полюсе имеется теменная пластинка
с венчиком длинных чувствующих
ресничек. Внешний вид этой прозрачной
личинки усложнен сильно извитым
мерцательным шнуром, проходящим по
наружной ее поверхности. Имеется и
прианальный венчик ресничек. На первых стадиях
своего развития личинка активно плавает
в воде султаном вверх и вращается вокруг
своей оси. Постепенно объем ее тела
несколько уменьшается, утолщаются стенки
и личинка теряет свою прозрачность. Она
начинает расти в длину, и у нее возникает
5—6 пар жаберных отверстий. А затем
наступает резкое превращение — тело ее
делится на верхнюю и нижнюю половину.
Верхняя часть в дальнейшем
растягивается в хобот и обособляется перетяжкой
от остального тела, которое формируется
из нижней части. Позже около хобота
образуется валик — зачаток воротничка,
происходит упорядочивание органов, за
счет роста заднего конца тело удлиняется,
животное приобретает облик взрослой
формы и садится на дно.
Среди кишечнодышащих есть один
живородящий вид — Xenopleura vivipara,
а один вид — Balanoglossus proliferans—
способен размножаться как половым, так
и бесполым путем. В результате простого
поперечного деления тела взрослого жи-
322
вотного, у которого совершенно
отсутствуют половые железы, получаются новые
особи, способные к половому
размножению. Таким образом, здесь происходит
настоящее чередование поколений —
полового и бесполого. Причем оба эти
поколения внешне настолько резко отличаются
друг от друга, что первое давно было
известно науке под названием Balanoglos-
sus capensis, а второе — В. proliferans.
Как уже говорилось, большинство
видов кишечнодышащих роет в грунте
длинные ходы или постоянные норы. Другие же
живут на поверхности дна. Все эти виды
образуют за счет своей обильной слизи
трубки, в которых они и живут. На
поверхности грунта обитает большинство
Ptychodera, Spengelia, Glandiceps и Glos-
sobalanus. Все они прячутся под камнями,
обломками кораллов, иногда глубоко
внедряясь в блоки мадрепоров, в зарослях
водорослей или под корнями высших
водных растений в прибрежной зоне.
Некоторых из них находили в мертвых
раковинах My а, наполненных песком. Все
эти виды образуют за счет своей обильно
выделяемой слизи трубки, в которых они
живут. Смешиваясь с песком, слизь
быстро застывает в плотную коркообразную
массу. Животные могут свободно
двигаться в своих трубках. Постройка
трубки происходит следующим образом. Хобот
животного внедряется в песок, и песчинки,
окружающие его, прилипают и
вкрапливаются в слизь, обильно покрывающую
все тело животного. На конце хобота
получается маленький поясок толщиной в
в 1—2 песчинки. С помощью бьющих
в одном направлении назад ресничек
эпидермиса (наружного покрова тела) целым
потоком один за другим такие поясочки
песчинок, смешанных со слизью,
передвигаются к концу тела животного.
Образуется плотный кожух, охватывающий, как
рубашка, все тело животного. Очень
быстро он засыхает в прочную трубку.
Большинство видов Enteropneusta роет
в грунте длинные U-образные норы (рис.
169). Как правило, такая норка вначале
уходит вертикальной шахтой вниз на
довольно большую глубину. У Balano-
glossus и нового гигантского вида Willeya
эти ходы достигают 60—80 см. Затем
следует изгиб норы, и она снова выходит
на поверхность. Постоянно сам ход
выстилается быстро засыхающей слизью. Эта
слизь склеивает материал, из которого
состоят стенки хода, и образует гладкую
внутреннюю поверхность. Животные
почти никогда не покидают норок. Во время
прилива они поднимаются до поверхности
грунта и выставляют наружу или
передний конец тела, или задний, для того
чтобы выбросить фекалии, которые лежат
у выхода из норки горкой спирально
закрученных колбасок высотой в 3—6 см
так, как это наблюдается у червя
пескожила (Arenicola marina). По этим кучкам
закрученных экскрементов и по
небольшим дырочкам входов и можно
обнаружить домики Enteropneusta на литорали
морей.
При малейшем сотрясении грунта
животные молниеносно прячутся в свои
норки (или в трубки) и уходят на
спасительную глубину. Таким образом они
спасаются от врагов, и прежде всего от
хищных рыб, питающихся донной фауной.
В качестве защиты у многих видов,
возможно, служат содержащиеся в слизи
йодистые выделения кожных желез. Неко-
Рис. 169. Норка Balanoglossus clavigerus с
животным:
1 — U-образная норка с выходом и дополнительными
боковыми ходами; 2 — баланоглосс; 3 — кучка
экскрементов; 4 — грунт.
21*
323
торые Enteropneusta, например Balano-
glossus clavigerus, издают столь резкий
запах йодоформа, что можно, идя на этот
запах, найти их в песке.
Слизь кишечнодышащих может
светиться. Свечение прежде всего появляется
при сотрясении грунта. Оно менее
интенсивно в дневное время. Glossobalanus
minutus светится в течение всего дня,
а у Ptychodera имеется четкий суточный
ритм свечения, который сохраняется
неизменным до 8 суток, если содержать
животное в темноте. Химизм и биологический
смысл их свечения неизвестны.
Видимо, в связи с роющим образом
жизни, при котором нежное тело
животных может быть легко поранено, а также
в качестве защиты от врагов у
кишечнодышащих развилась необычайная
способность к регенерации, т. е. к
восстановлению тела (при его повреждении или
разрыве) из отдельных, даже небольших
частей. Например, маленький хвостовой
кусочек тела Ptychodera способен
восстановить целый организм. Интересно, что
передняя часть тела не может достроить
заднюю, а задние части легко
восстанавливают передние.
Мы уже знаем, что специальных
органов чувств у кишечнодышащих не
имеется. Но все части тела этих животных
отчетливо реагируют на механическое
раздражение и на свет. Хобот всегда
избегает света, и его поведение позволяет
установить, что животные могут
воспринимать не только сам свет, но и его
направление. Свет и даже небольшое
механическое раздражение всегда вызывают
бегство, уход, закапывание и сжатие
тела. Сильнее всего на эти раздражения
реагирует хобот.
Как же происходит закапывание
животных? За счет прилива и отлива крови
к хоботу, а также за счет напряжения его
мышц хоботок может становиться то более
упругим и сжиматься в жесткий кулак, то
расслабляться. Ptychodera, например,
косо внедряет в грунт его раздутый передний
конец. Потом она сжимает продольные
мускулы и перистальтическая волна
вздутия движется к заднему концу хобота.
Она доходит до его конца за 5 секунд.
Когда вздутие доходит до конца хобота,
животное прочно зацепляется в грунте,
а утонченный передний конец хобота за
счет сокращения кольцевых и продольных
мышц внедряется глубже в песок.
Ставшая в это время тонкой задняя часть
хобота подтягивается вперед, к передней
его части. При рытье хобот опирается на
воротничок, служащий для него в данном
случае твердой опорой.
Внутреннее напряжение воротничка при
рытье также сильно увеличивается за счет
сокращения его толстых мышечных
стенок. Кроме того, в полость воротничка
через воротничковые поры нагнетается
вода. Видимо, при рытье эти поры могут
герметически закрываться и тургор
воротничка за счет несжимаемой воды
увеличивается еще сильнее. Остальное тело
совершенно не принимает участия в
передвижении. Оно просто подтягивается вслед
за передними двумя отделами, при
необходимости сильно утончаясь (до 1 мм
у некоторых видов) и вытягиваясь.
Зарывание происходит довольно быстро. За
одну минуту происходит до 12
перистальтических сжатий хоботка. В это же время
реснички покровов тела синхронно и
постоянно уводят песчинки от кончика
хоботка ко рту и к концу тела. Эти
песчинки расширяют проход и облегчают
внедрение в него туловища. Одновременно
стенки хода обрабатываются
выделившейся слизью.
Двигаясь вперед, кишечнодышащие
заглатывают грунт. Причем заглатывание
грунта происходит не просто механически,
как у других грунтоедов. Реснички хобота
бьют по направлению назад и переносят
прилипшие к слизи песчинки и другие
частицы длинными параллельными
потоками назад. У основания хобота удар
ресничек направлен ко рту, так что нити
слизи и песка у рта сливаются и
создается один мощный поток. Край воротничка
при этом отодвигается далеко в сторону
от основания хобота и толстый поток
пищи проникает в рот. В пищеварительном
канале частицы грунта гонятся сначала
реснитчатыми клетками эндостиля по
жаберному отделу и далее по кишечнику.
Пищевой поток все время
переворачивается и вращается. Переваривание
происходит в печеночном отделе. Вместе с по-
324
ступившим грунтом в кишечник
попадают микроскопические
животные и водоросли,
которыми, собственно, и питаются
Enteropneusta. У некоторых
видов кишечник столь сильно
набит грунтом, что их нежное
тело рвется от тяжести при
поднимании животного. Фекалии
выбрасываются через задний
конец тела, который, высовываясь
из норки, совершает коловра-
щательное движение у
поверхности грунта. При этом и
получаются те спирально
закрученные колбаски экскрементов,
о которых мы говорили выше.
Близкие к Enteropneusta
представители другого класса
полухордовых животных—-Ярм-
ложаберные (Pterobranchia) —
ведут сидячий образ жизни.
Как мы уже знаем, эта очень
небольшая группа животных
включает в себя всего лишь
три рода: Cephalodiscus,
Rhabdopleura и
Athubaria.Представители рода Cephalodiscus
ограничены в своем
распространении южным полушарием. Они
встречаются главным образом
в Антарктике и на субантарктических
островах. Виды Rhabdopleura распространены
широко и встречаются во всех климатических
зонах от Арктики до Антарктики, a
Athubaria известна только у побережий Японии.
Pterobranchia живут в основном на
глубине от 100 до 650 м, и число их видов,
найденных на самом мелководье, очень
невелико. Размеры их тела колеблются от
0,2 до нескольких миллиметров.
Наиболее крупный представитель этой группы —
Cephalodiscus densus — имеет 1,4 еж в
длину. Все они, за исключением Athubaria,
строят колонии, очень напоминающие
колонии мшанок, с которыми их нередко
путали. Размер колоний обычно
составляет несколько сантиметров. Наиболее
крупные из них имеют 19 еж в
поперечнике и 25 см в высоту. Окраска зооидов
разнообразна. Часто на хоботе имеются
пигментированные пояса. У некоторых
видов тело окрашено в черный цвет, тог-
Рис. 170. Строение крыло-
жаберных: А — Athubaria;
Б — Rhabdopleura; В —
Cephalodiscus.
1 — руки; 2 — щупальца; з —.
лишенные щупалец концы
одной пары рук; 4—головной щит
(хобот); 5 — рот; 6 — задний
проход; 7 — жаберные щели;
8 — боковые складки
воротничка; 9—туловище; ю —
стебель; 11 —трубка; 12 — почки.
да как щупальца и ножка остаются
белыми. Колонии обычно светло-бежевые.
Строение отдельных особей во всех
главных чертах напоминает
Enteropneusta (рис. 170,А—В). Тело взрослых кры-
ложаберных, как и тело балаяоглосса,
расчленено на хобот, воротник и
туловище со стебельком, или ножкой. Хобот
имеет форму щитка и содержит полость,
открывающуюся наружу двумя порами.
От глотки в хобот отходит нотохорд,
над которым лежит сердце. На границе
хобота и воротника с брюшной стороны
лежит рот. Воротник, в отличие откишеч-
ножаберных, несет на спинной стороне
руки с перисто расположенными на них
щупальцами. Бывает от 1 до 8 пар рук.
Щупальца покрыты мерцательным
несущим реснички эпителием. Позади
воротника у Cephalodiscus и Athubaria имеется
по одной паре жаберных щелей. У
Rhabdopleura жаберных щелей нет. В туловище
325
Рис. 171. Колония Rhabdopleura.
А — часть колонии: 1 — трубочки-теки с
сократившимися зооидами; 2 — трубочки-теки с выступающим зо-
оидом; з — ножка зооида; 4 — черный столон. Б —
общий вид колонии на раковине моллюска.
помещается кишечник, изогнутый таким
образом, что заднепроходное отверстие
лежит недалеко от рта. Половые железы
представлены парой мешочков,
открывающихся протоками на спинной стороне
тела впереди ануса. Личинка их похожа
на личинку кишечнодышащих. Она
сплошь покрыта ресничками, имеет
теменной чувствительный орган и кишечник.
Черты упрощенного строения крыложа-
берных и различия в строении тел их
и Enteropneusta вызваны прежде всего
сидячим образом жизни этих животных
и малыми размерами их тела. Роющий
образ жизни кишечножаберных вызвал
сильное развитие хобота, исчезновение
щупальцевого аппарата, разрастание
туловища в длину. У крыложаберных в связи
с сидячим образом жизни хобот развит
умеренно, прекрасно развит щупальцевый
аппарат, а туловище искривлено. Но все
эти отличия лишь внешнего порядка.
Образ жизни этих животных
малоизвестен, поскольку они очень редки и
плохо переносят условия аквариума. Athubaria
живет главным образом на колониях
гидроидов и ведет свободноподвижный образ
жизни, не строя никаких домиков (рис.
170,Л). Она цепляется хвостиком за ветви
колоний и передвигается ползая по ним
при помощи грудного выроста и кончиков
своих рук. Особи ее имеют тенденцию
собираться группами. Rhabdopleura (рис.
171,Л, Б) являются настоящими
колониальными животными, зооиды которых
соединены между собой длинным
столоном черного цвета, в то время как Cepha-
lodiscus псевдоколониален. Отдельные
члены его колоний независимы друг от
друга и организуются в так называемый це-
ноций, внутри которого они могут
свободно передвигаться. Колонии или ценоции
Pterobranchia прикрепляются к подводным
камням, раковинам и другим твердым
предметам.
Форма и строение отдельных домиков
у разных видов весьма разнообразны.
Они являются продуктом выделения
железистых клеток головных щитов
животных, т. е. хобота. У Cephalodiscus домики
устроены наиболее простым образом и
называются ценоцием. У разных видов он
состоит из разветвленных, как дерево, или
неразветвленных, стелющихся или
приподнимающихся толстых ветвей с
помещениями для отдельных животных (рис. 172, А,
Б). Эти помещения могут быть или
полностью изолированными, или сообщаться
друг с другом неправильными каналами
и иметь одно выходное отверстие для
нескольких полостей. Иногда может быть
и единая полость для многих членов
сожительства, открывающаяся наружу
многими отверстиями. Особи сидят там
присосавшись кончиками стебельков, но могут
свободно ползать внутри и выползать
наружу. При ползании они пользуются
головным щитком. У некоторых видов
стебелек остается в домике, но он очень
сильно растягивается, так что животное
может выползать наружу и передвигаться
по поверхности домика на значительное
расстояние. При малейшем раздражении
все животные прячутся в домике.
Некоторые виды способны полностью вылезать
из домиков, и поверхность ценоция
покрывается как будто многочисленными
черненькими булавочными головками (рис.
172,/7). Иногда ценоций представляет собой
дерновину плотных нитей, по
поверхности которой ползают отдельные особи.
Колонии Rhabdopleura устроены более
сложно. Прежде всего их колонии
являются истинными и все особи внутри их
связаны воедино длинным стелющимся
столоном характерного черного цвета.
Столоны могут ветвиться, и от них
поднимаются вертикально вверх трубчатые
домики, или теки, отдельных зооидов (рис.
171 ,Л). Столоны покрыты тонкой и очень
нежной хитиноидной оболочкой и
заключены в цилиндрические трубки. Они могут
достигать 10 см длины и стлаться по
поверхности камней и раковин.
Развивающиеся на основном стелющемся столоне
новые молодые зооиды выделяют попереч-
Рис. 172. Колония ценоций Cephalodiscus: Л — Cephalodiscus nigrescens; Б — С. densis;
В — С. dodecalophus; Г — цефалодискусы, ползающие по разветвлениям ценоция.
J — выходы из камер; 2 — стебель животного; 8 — туловище; 4 — руки; 5 — головной щит.
ные перегородки, так что каждая особь
оказывается в отдельном отсеке столона.
Затем она продырявливает наружную
стенку трубки и строит вверх свою
собственную трубочку — теку, состоящую
как бы из вложенных друг в друга
колечек.
Rhabdopleura выползают из своих
домиков наружу с помощью головного
щитка только до устья трубки. Они
высовывают из нее только щиток, так что рот
лежит как раз на уровне края трубки.
Обе руки их изгибаются снаружи трубки
к спине. В случае испуга животные очень
глубоко втягиваются внутрь трубки за
счет сокращения стебелька. В желудках
крыложаберных находили кусочки
диатомовых водорослей, радиолярий и личинок
планктонных ракообразных. Все эти
пелагические организмы, видимо, были
пойманы с помощью ветвистых щупалец в
окружающей воде* И у Cephalodiscus и у
Rhabdopleura самцы и самки редко
объединяются в одном и том же ценоции. Чаще
они строят себе отдельные колонии.
Различия в строении самцов и самок
выражены слабо, но часто самцы бывают мельче
самок. Количество рук у них меньше,
и они не имеют отростков. О
размножении крыложаберных известно очень мало.
Кроме полового способа размножения,
они обладают и бесполым, путем
почкования. У Cephalodiscus на брюшной
стороне ножки может образовываться сразу
от 2 до 14 почек (рис. 170,5), почки рано
отделяются от материнского организма
и развиваются в новую взрослую особь.
У Rhabdopleura почки образуются на
черном столоне, и развившиеся из них
особи сохраняют связь с материнским
организмом. Они могут образовывать и
покоящиеся почки, заключенные в особые
мешочки, способные переживать
неблагоприятные условия.
Характер колоний крыложаберных
указывает на близкое родство этой группы
с граптолитами — обширной группой
ископаемых колониальных животных,
известных с кембрия до карбона, т. е.
примерно 250—550 млн. лет назад. Иными
словами, Pterobranchia является очень
древней группой животных. Но до
настоящего времени их окаменелые остатки
были крайне редки. Палеонтологам
известны формы, близкие к Rhabdopleura и
Cephalodiscus, только из верхнемеловых
отложений мезозоя — примерно 125 млн.
лет назад. Сходство их с граптолитами
так сильно, что, если бы трубки
крыложаберных были впервые найдены в
палеозое, их несомненно отнесли бы к грапто-
литам. Несомненно, что граптолиты, так
же как и Rhabdopleura, принадлежат к
типу полухордовых животных.
Если вспомнить то, что известно в
отношении самых общих черт строения всех
беспозвоночных животных, с одной
стороны, и хордовых — с другой, можно
сказать, что представители типа Hemichorda
отличаются от беспозвоночных животных
прежде всего присутствием жаберных
щелей, связанных с кишечным каналом,
а также наличием спинной нервной
системы и нотохорда. С другой стороны, они
отличаются от настоящих хордовых
присутствием брюшного нервного ствола,
мерцательным покровом тела и
некоторыми другими чертами. Иными словами,
в типе Hemichorda объединены признаки
обеих главных групп животного царства—
беспозвоночных и хордовых животных.
Они стоят в родословном древе где-то
в самом основании ствола группы
хордовых. В этом и заключается исключитель-
нейший интерес очень маленькой группы
полухордовых животных для изучения
истории развития жизни на Земле.
ТИП ОБОЛОЧНИКИ (TUNIGATA)
Оболочники, или туникаты, к
которым относятся асцидии, пиросомы,
сальны и аппендикулярии,— одна из наиболее
удивительных групп морских животных.
Свое название они получили за то,
что тело их одето снаружи особой
студенистой оболочкой, или туникой.
Туника состоит из вещества, чрезвычайно
близкого по своему составу к целлюлозе,
которая встречается только в царстве растений
и неизвестна ни для одной другой группы
животных. Оболочники —
исключительно морские животные, ведущие частично
прикрепленный,частично
свободноплавающий пелагический образ жизни. Они
могут быть или одиночными, или образуют
удивительные колонии, возникающие при
чередовании поколений в результате
почкования бесполых одиночных особей.
О способах размножения этих
животных — наиболее необычайных среди всех
живых существ на Земле — мы
специально будем говорить ниже.
Очень интересно положение
оболочников в системе животного царства.Природа
этих животных долго оставалась
загадочной и непонятной, хотя они были
известны еще Аристотелю более двух с
половиной тысяч лет назад под именем Tethya.
Только в начале XIX столетия было
установлено, что одиночные и колониальные
формы некоторых оболочников — сальп —
представляют собой лишь различные
поколения одного и того же вида. До тех
пор их относили к разным видам
животных. Эти формы отличаются друг от друга
не только по внешнему виду. Оказалось,
что только колониальные формы
обладают половыми органами, а одиночные —
бесполы. Явление чередования
поколений у сальп было открыто
поэтом и естествоиспытателем
Альбертом Шамиссо во время его плавания
в 1819 г. на русском военном корабле
«Рюрик» под командованием Коцебу. Старые-
авторы, в том числе и Карл Линней,
относили одиночных туникат к типу
моллюсков. Колониальные же формы были
отнесены им к совершенно другой группе—
зоофитам, а некоторые считали их
особым классом червей. Но на самом деле эти
внешне очень просто устроенные
животные не так примитивны, как они кажутся.
Благодаря работам замечательного
русского эмбриолога А. О.
Ковалевского в середине прошлого века было
установлено, что оболочники близки к хордовым
животным. А. О. Ковалевский установил,
что развитие асцидий идет по тому же
самому типу, что и развитие ланцетника,
представляющего, по меткому выражению
академика И. И. Шмальгаузена, «как бы
живую упрощенную схему типичного
хордового животного». Группа хордовых
животных характеризуется целым рядом
определенных важных черт строения.
Прежде всего это наличие спинной струны,,
или хорды, представляющей собой внут-
329
ренний осевой скелет животного. Личинки
туникат, свободно плавающие в воде,
также имеют спинную струну, или хорду,
которая совершенно исчезает при
превращении их во взрослую особь. Личинки
и по другим важнейшим признакам
строения стоят гораздо выше
родительских форм. По филогенетическим
причинам, т. е. по причинам, связанным с
происхождением группы, большее значение
у оболочников придается организации их
личинок, чем организации взрослых форм.
Такая аномалия неизвестна более ни для
одного другого типа животных. Кроме
наличия хорды, хотя бы в личиночной
стадии, с настоящими хордовыми
животными оболочников сближает целый ряд
других признаков. Очень важно, что
нервная система оболочников расположена
на спинной стороне тела й представляет
собой трубку, имеющую внутри канал.
Нервная трубка оболочников
образуется как желобообразное продольное впя-
чивание поверхностных покровов тела
зародыша, эктодермы, как это имеет
место у всех других позвоночных животных
и у человека. У беспозвоночных
животных нервная система всегда лежит на
брюшной стороне тела и образуется иным
способом. Основные сосуды кровеносной
системы оболочников, наоборот,
располагаются на брюшной стороне, в
противоположность тому, что характерно для
беспозвоночных животных. И наконец,
передний отдел кишки, или глотка, пронизан
у оболочников многочисленными
отверстиями и превратился в орган дыхания.
Как мы видели в других главах, у
беспозвоночных животных органы дыхания
очень разнообразны, но никогда
кишечник не образует жаберных щелей. Это
признак хордовых животных.
Эмбриональное развитие туникат также имеет много
общих черт с развитием Chordata.
В настоящее время считают, что
оболочники путем вторичного упрощения, или
деградации, произошли от каких-то
форм, очень близких с позвоночными
животными.
Вместе с другими хордовыми и
иглокожими они образуют ствол вторичноро-
тых животных — один из двух основных
стволов эволюционного древа.
Оболочники рассматриваются или как
отдельный подтип типа хордовых
животных — Chordata, куда вместе с ними
относятся еще три подтипа животных,
в том числе и позвоночные (Vertebrata),
или же как самостоятельный тип —Tuni-
cata, или Urochordata. К этому типу
относятся три класса:
Аппендикулярий (Appendiculariae, или Copelata),
Асцидий (Ascidiae) и Салыгы (Salpae).
Прежде асцидий делили на три отряда:
простые, или одиночные, асцидий (Monas-
cidiae); сложные, или колониальные, асцидий
(Synascidiae) и пиросомы, или огнетелки
(Ascidiae Salpaeformes, или Pyrosomata).
Однако в настоящее время деление на
простых и сложных асцидий потеряло
систематическое значение. Асцидий
делятся на подклассы по иным признакам.
Сальпы делятся на два отряда —
бочоночников (Cyclomyaria) и собственно
салъп (Desmomyaria). Иногда этим
отрядам придают значение подклассов. К саль-
пам же, видимо, относится и очень
своеобразное семейство глубоководных
донных оболочников — Octacnemidae, хотя
до сих пор большинство авторов считало
его сильно уклонившимся подклассом
асцидий.
Очень часто сальпы и пиросомы,
ведущие свободноплавающий образ жизни,
объединяются в группу пелагических
оболочников Thaliacea, которой придается
значение класса. Класс Thaliacea делят
тогда на три подкласса: Pyrosomida,
или Luciae, Desmomyaria, или Salpae,
и Cyclomyaria, или Doliolida. Как видно,
взгляды на систематику высших групп
Tunicata весьма различны.
В настоящее время известно более
тысячи видов оболочников. Громадное
большинство их падает на долю асцидий,
аппендикулярий насчитывается около 60
видов, около 25 видов сальп и примерно
10 видов пиросом (табл. 28—29).
Как уже говорилось, оболочники
обитают только в море. Аппендикулярий,
сальпы и пиросомы плавают в толще вод
океана, в то время как асцидий ведут
прикрепленный образ жизни на дне.
Аппендикулярий никогда не образуют колоний,
в то время как сальпы и асцидий могут
встречаться и в виде одиночных оргдниз-
330
мов7 и в виде колоний. Пиросомы же всегда
колониальны. Все оболочники —
активные фильтраторы, питающиеся или
микроскопическими пелагическими водорослями
и животными, или взвешенными в воде
частицами органического вещества —
детритом. Прогоняя воду через глотку и
жабры наружу, они отфильтровывают
мельчайший планктон, иногда пользуясь
при этом очень сложными
приспособлениями.
Пелагические оболочники обитают в
основном в верхних 200 м воды, но иногда
могут опускаться и глубже. Пиросомы
и сальпы редко встречаются глубже
1000 м, аппендикулярии известны до
3000 м. При этом специальные
глубоководные виды среди них, видимо,
отсутствуют. Асцидии в основной своей массе
также распределены в приливо-отливной
литоральной и в сублиторальной зонах
океанов и морей—до 200—500 м, однако
значительное количество их видов
встречается и глубже. Максимальная глубина
их нахождения — 7230 м.
Оболочники встречаются в океане то
единичными экземплярами, то в виде
колоссальных скоплений. Последнее
особенно характерно для пелагических форм.
В целом оболочники довольно обычны
в морской фауне и, как правило,
попадаются в планктонные сети и донные тралы
зоологов повсеместно. Аппендикулярии
и асцидии распространены в Мировом
океане на всех широтах. Они столь же
характерны для морей Северного
Ледовитого океана и Антарктики, как и для
тропиков. Сальпы и пиросомы, наоборот,
в основном приурочены в своем
распределении к теплым водам и лишь изредка
встречаются в водах высоких широт,
главным образом будучи занесенными
туда теплыми течениями.
Строение тела почти всех туни-
кат до неузнаваемости сильно отличается
от общего плана строения тела в типе
хордовых. Ближе всего к исходным формам
стоят аппендикулярии, и в системе туни-
кат они занимают первое место. Однако,
несмотря на это, строение их тела является
наименее характерным для оболочников.
Знакомство с оболочниками, видимо,
лучше всего начать с асцидии.
Строение асцидии. Асцидии — это
донные животные, ведущие прикрепленный
образ жизни. Многие из них одиночные
формы. Размеры их тела в среднем
составляют несколько сантиметров в диаметре
и столько же в высоту. Однако среди них
известны некоторые виды, достигающие
40—50 см, например широко
распространенная Cione intestinalis или
глубоководная Ascopera gigantea. С другой
стороны, есть очень мелкие асцидии,
размером менее 1 мм. Кроме одиночных
асцидии, существует большое количество
колониальных форм, у которых отдельные
мелкие особи, размером в несколько
миллиметров, погружены в общую тунику.
Такие колонии, очень разнообразные по
форме, обрастают поверхности камней и
подводных предметов.
Больше всего одиночные асцидии
похожи на продолговатый раздутый мешок
неправильной формы, прирастающий своей
нижней частью, которая называется
подошвой, к различным твердым предметам
(рис. 173, А). На верхней части
животного хорошо заметны два отверстия,
расположенные или на небольших бугорках,
или же на довольно длинных выростах
тела, напоминающих горлышко бутылки.
Это сифоны. Один из них —
ротовой, через который асцидия всасывает
воду, второй — клоакальный.
Последний обычно бывает несколько сдвинут
на спинную сторону. Сифоны могут
открываться и закрываться при помощи
мускулов — сфинктеров. Тело асцидии
одето однослойным клеточным
покровом — эпителием, который выделяет на
своей поверхности особую толстую
оболочку — тунику. Внешняя окраска
туники бывает различна. Обычно асцидии
окрашены в оранжевые, красноватые,
буро-коричневые или фиолетовые тона.
Однако глубоководные асцидии, как и
многие другие глубоководные животные,
теряют свою окраску и становятся грязно-
белыми. Иногда туника бывает
полупрозрачной и через нее просвечивают
внутренности животного. Часто туника
образует по поверхности морщины и складки,
зарастает водорослями, гидроидами,
мшанками и другими сидячими животными.
У многих видов ее поверхность покрыта
331
песчинками и мелкими камешками, так
что животное бывает трудно отличить от
окружающих предметов.
Туника бывает студенистой, хрящевид-
ной или желеобразной консистенции.
Замечательной ее особенностью является
то, что она более чем на 60% состоит
из целлюлозы. Толщина стенок туники
может достигать 2—3 см, обычно же она
значительно тоньше.
Часть клеток эпидермиса может
проникать в толщу туники и заселять ее.
Это оказывается возможным только
благодаря ее студенистой консистенции. Ни
в одной другой группе животных клетки
не заселяют образований аналогичного
типа (например, кутикула у нематод).
Кроме того, в толщу туники могут
врастать и кровеносные сосуды.
Под туникой залегает собственно стенка
тела, или мантия, в состав которой
входит однослойный эктодермический
эпителий, покрывающий тело, и соединитель-
нотканый слой с мышечными волокнами.
Наружные мышцы состоят из продольных,
а внутренние из кольцевых волокон.
Такая мускулатура позволяет асцидиям
производить сократительные движения
и при необходимости выбрасывать воду
из тела. Мантия покрывает тело под
туникой, так что оно свободно лежит внутри
туники и срастается с нею только в
области сифонов. В этих местах располагаются
сфинктеры — мышцы, замыкающие
отверстия сифонов.
Твердого скелета в теле асцидий нет.
Только у некоторых из них в различных
частях тела разбросаны мелкие
известковые спикулы разной формы.
Пищеварительный канал
асцидий начинается ртом, расположенным
на свободном конце тела на вводном, или
ротовом, сифоне (рис. 173, Б). Вокруг рта
располагается венчик щупалец, иногда
простых, иногда довольно сильно
разветвленных. Количество и форма щупалец
различны у разных видов, однако их не
бывает менее 6. Ото рта свешивается
внутрь огромная глотка, занимающая
почти все пространство внутри мантии.
Глотка асцидий образует сложный
дыхательный аппарат. По стенкам ее в строгом
Рис. 173. Строение
асцидий.
А — внешний вид асцидий:
1 — вводной, или ротовой,
сифон; 2 — выводной, или
клоакальный, сифон; 3 —
туника; 4 — подошва. Б —
схема строения одиночной
асцидий: 1 — ротовой
сифон; 2 — ротовые
щупальца; з — глотка с
жабрами; 4 — эндостиль; 5 —
спинная пластинка; 6 —
начало пищевода; 7 —
желудок; 8 — пищеварительная
железа; 9 — кишка; ю —
задний проход; 11 —
сердце; 12 — яичник и
семенник; 13 — клоака; 14 —
клоакальный сифон; 15 —
нервный ганглий;
16—спинной нерв; 17 —
околонервная железа; J я — туника.
В — часть стенки глотки
с жаберными щелями:
1 — жаберные щели; 2 —
продольные и поперечные
кровеносные сосуды.
332
порядке в несколько вертикальных и
горизонтальных рядов располагаются
жаберные щели, иногда прямые, иногда
изогнутые (рис. 173, В). Часто стенки глотки
образуют по 8—12 довольно больших
свисающих внутрь складок,
расположенных симметрично на двух ее сторонах
и сильно увеличивающих ее внутреннюю
поверхность. Складки также пронизаны
жаберными щелями, причем сами щели
могут принимать очень сложные
очертания, закручиваясь спиралями на
конусовидных выростах на стенках глотки и
складок. Жаберные щели покрыты
клетками, несущими длинные реснички. В
промежутках между рядами жаберных щелей
проходят, также правильно располагаясь,
кровеносные сосуды. Количество их
может достигать 50 на каждой стороне глотки.
Здесь кровь обогащается кислородом.
Иногда тонкие стенки глотки содержат
небольшие спикулы, поддерживающие их.
Жаберные щели, или стигмы, асцидий
незаметны, если рассматривать
животное снаружи, сняв только тунику. Из
глбтки они ведут в особую полость,
выстланную энтодермой и состоящую из
двух половин, сросшихся на брюшной
стороне с мантией. Эта полость называется
околожаберной, атриаль-
н о й или перибранхиальной
(рис. 173, Б). Она залегает с каждой
стороны между глоткой и наружной
стенкой тела. Часть ее образует клоаку. Эта
полость не является полостью тела
животного. Она развивается из особых впячи-
ваний наружной поверхности внутрь тела.
Околожаберная полость сообщается с
внешней средой при помощи клоакального
сифона.
Со спинной стороны глотки свисает
тонкая спинная пластинка, иногда
рассеченная на тонкие язычки, а по брюшной
стороне проходит особый поджаберный
желобок, или эндостиль. Биением
ресничек на стигмах асцидия гонит воду
так, что устанавливается постоянный
ток через ротовое отверстие. Далее вода
прогоняется через жаберные щели в
околожаберную полость и оттуда через клоаку
наружу. Проходя через щели, вода отдает
кислород в кровь, а различные мелкие
органические остатки, одноклеточные
водоросли и пр. захватываются эндостилем
и гонятся по дну глотки к ее заднему
концу. Здесь расположено отверстие,
ведущее в короткий и узкий пищевод.
Загибаясь на брюшную сторону, пищевод
переходит во вздутый желудок, из
которого выходит кишка. Кишка, изгибаясь,
образует двойную петлю и открывается
анальным отверстием в клоаку.
Экскременты выталкиваются из тела через
клоакальный сифон. Таким образом,
пищеварительная система асцидий очень
проста, но обращает на себя внимание
наличие эндостиля, являющегося частью их
ловчего аппарата. Клетки эндостиля двух
родов — железистые и реснитчатые.
Реснитчатые клетки эндостиля улавливают
пищевые частицы и гонят их к глотке,
склеивая выделениями железистых
клеток. Оказывается, эндостиль является
гомологом щитовидной железы
позвоночных и выделяет органическое
вещество, содержащее йод. Видимо, это
вещество близко по своему составу к
гормону щитовидной железы. У некоторых
асцидий у основания стенок желудка
имеются особые складчатые выросты и
дольчатые массы. Это так называемая
печень. Она соединяется с желудком
особым протоком.
Кровеносная система
асцидий незамкнутая. Сердце расположено на
брюшной стороне тела животного. Оно
имеет вид небольшой вытянутой трубки,
окруженной тонкой околосердечной
сумкой, или перикардием. От двух
противоположных концов сердца отходит по
крупному кровеносному сосуду. От переднего
конца начинается жаберная артерия,
которая тянется посередине брюшной стороны
и посылает от себя многочисленные ветви
к жаберным щелям, дающие между ними
побочные маленькие веточки и окружающие
жаберный мешок целой сетью продольных
и поперечных кровеносных сосудов. От
задней спинной стороны сердца отходит
кишечная артерия, дающая ветви к
внутренним органам. Здесь кровеносные сосуды
образуют широкие лакуны—пространства
между органами, не имеющие
собственных стенок, очень похожие по своему
строению на лакуны двустворчатых
моллюсков. Кровеносные сосуды заходят также
333
в стенку тела и даже в тунику. Вся система
кровеносных сосудов и лакун открывается
в жаберно-кишечный синус, иногда
называемый спинным сосудом, с которым
соединяются и спинные концы поперечных
жаберных сосудов. Этот синус значителен
по размерам и тянется посередине
спинной части глотки. Всем туникатам, в том
числе и асцидиям, свойственна
периодическая смена направления тока крови,
так как их сердце поочередно сокращается
в течение некоторого времени то сзади
наперед, то спереди назад. Когда сердце
сокращается от спинного отдела к
брюшному, кровь движется по жаберной
артерии к глотке, или жаберному мешку,
где она окисляется и откуда поступает
в кишечно-жаберный синус. Затем кровь
проталкивается в кишечные сосуды и
снова к сердцу, так же, как это имеет
место у всех позвоночных животных.
При последующем же сокращении сердца
направление тока крови меняется на
обратное, и она течет, как у большинства
беспозвоночных. Таким образом, тип
кровообращения у туникат является переходным между
кровообращением беспозвоночных и
позвоночных животных. Кровь у асцидий
бесцветная, кислая. Замечательной ее
особенностью является наличие ванадия,
который принимает участие в переносе
кислорода кровью и заменяет железо.
Нервная система у взрослых
асцидий чрезвычайно проста и развита
значительно слабее, чем у личинки.
Упрощение нервной системы происходит
благодаря сидячему образу жизни взрослых
форм. Нервная система состоит из
надглоточного, или мозгового, ганглия,
расположенного на спинной стороне тела
между сифонами. От ганглия берут начало
2—5 пар нервов, идущих к краям ротового
отверстия, глотке и к внутренностям —
кишечнику, половым органам и к сердцу,
где имеется нервное сплетение. Между
ганглием и спинной стенкой глотки
имеется небольшая околонервная железка,
проток которой впадает в глотку на дне
ямки в особом мерцательном органе. Эту
железку считают иногда гомологом
нижнего придатка головного мозга
позвоночных животных — гипофиза. Органы
чувств отсутствуют, но, вероятно, ротовые
щупальца имеют осязательную функцию.
Но тем не менее нервная система
оболочников в сущности не примитивна. Личинки
асцидий имеют спинномозговую трубку,
лежащую под хордой и образующую
вздутие на своем переднем конце. Это
вздутие, по-видимому, соответствует
головному мозгу позвоночных животных и
содержит личиночные органы чувств —
пигментированные глазки и орган
равновесия, или статоцист. Когда личинка
превращается во взрослое животное, вся
задняя часть нервной трубки исчезает, а
мозговой пузырь вместе с
личиночными органами чувств распадается; за счет
его спинной стенки и образуется
спинной ганглий взрослой асцидий, а
брюшная стенка пузыря образует
околонервную железу. Как отмечает В. Н.
Беклемишев, строение нервной
системы оболочников является одним из
лучших доказательств происхождения их
от высокоорганизованных подвижных
животных. Нервная система личинок
асцидий по своему развитию выше, чем
нервная система ланцетника, у которого
отсутствует мозговой пузырь.
Специальных выделительных
органов у асцидий нет. Вероятно,
в какой-то степени в выделении принимают
участие стенки пищеварительного канала.
Однако у многих асцидий имеются особые
так называемые рассеянные почки
накопления, состоящие из особых клеток —
нефроцитов, в которых накапливаются
продукты выделения. Эти клетки
располагаются характерным рисунком, часто
группируются вокруг кишечной петли
или гонад. Красновато-бурая окраска
многих асцидий зависит именно от
накопившихся в клетках экскретов. Только
после смерти животного и распада тела
продукты выделения освобождаются и
выходят в воду. Иногда во втором колене
кишки располагается скопление
прозрачных пузырьков, не имеющих выводных
протоков, в которых накапливаются
конкреции, содержащие мочевую кислоту.
У представителей семейства Mol-
gulidae почка накопления усложняется
еще больше и скопление пузырьков
превращается в один крупный
изолированный мешочек, полость которого содержит
334
конкреции. Большое своеобразие этого
органа заключается в том, что почечный
мешок мольгулид у некоторых других
асцидий всегда содержит симбиотические
грибки, которые не имеют даже
отдаленных родственников среди других групп
низших грибов. Грибки образуют
тончайшие нити мицеллы, оплетающие конкреции.
Среди них присутствуют более толстые
образования неправильной формы, иногда
образуются спорангии со спорами. Эти
низшие грибы питаются уратами —
продуктами выделения асцидий, и их
развитие освобождает последних от
накопленных экскретов. Видимо, эти грибки
необходимы асцидиям, так как даже ритм
размножения у некоторых форм асцидий
связан с накоплением экскретов в почках
и с развитием симбиотических грибков.
Как происходит перенос грибков от одной
особи к другой, неизвестно. Яйца
асцидий стерильны в этом отношении, а
молодые личинки не содержат в почке грибков,
даже когда экскреты уже
накапливаются в них. Видимо, молодые животные
вновь «заражаются» грибками из морской
воды.
Асцидий — гермафродиты, т. е. у одной
и той же особи имеются одновременно
и мужские и женские половые железы.
Яичники и семенники лежат по одному
или по нескольку пар с каждой стороны
тела, обычно в петле кишечника. Протоки
их открываются в клоаку, так что
клоакальное отверстие служит не только для
выхода воды и экскрементов, но и для
выведения половых продуктов.
Самооплодотворения у асцидий не происходит,
так как яйца и сперма созревают в
разное время. Оплодотворение чаще всего
происходит в околожаберной полости, куда
с током воды проникают сперматозоиды
другой особи. Реже оно бывает снаружи.
Оплодотворенные яйца выходят через
клоакальный сифон, но иногда яйца
развиваются в околожаберной полости и
наружу выходят уже сформированные
плавающие личинки. Такое живорождение
характерно особенно для колониальных
асцидий.
Кроме полового размножения, асцидиям
свойствен и бесполый способ
размножения путем почкования. При этом обра-
Рис. 174. Строение сложных асцидий.
А — внешний вид колонии: 1 — ротовые сифоны
отдельных особей; 2 — выход общей клоакальной
полости. Б — схема строения сложной асцидий: 1 —
ротовой сифон; 2 — глотка; з — пищевод; 4 —
желудок; 5 — кишка; 6 — задний проход; 7 — семенник;
8 — яичник; 9 — нервный ганглий; 10 —
околонервная железка; 11 — эндостиль; 12 — сердце с
выростами перикардия; 13 — общая клоака с отверстиями
клоакальных сифонов отдельных зооидов.
зуются разнообразные колонии асцидий.
Строение асцидиозооида-— члена
колонии сложных асцидий — в принципе
не отличается от строения одиночной
формы. Но размеры их много меньше
335
и обычно не превышают нескольких
миллиметров. Тело асцидиозооида вытянуто
в длину и разделено на два или три отдела
(рис. 174, А): в первом, грудном, отделе
располагается глотка, во втором —
кишечник, а в третьем — половые железы и
сердце. Иногда разные органы располагаются
несколько иначе.
Степень связи между отдельными
особями в колонии—асцидиозооидами—может
быть различной. Иногда они вполне
самостоятельны и соединяются только
тонким столоном, который стелется по грунту.
В других случаях асцидиозооиды
заключены в общую тунику. Они могут быть
или рассеяны в ней, и тогда наружу
выходят и ротовые и клоакальные отверстия
асцидиозооидов, или располагаются
правильными фигурами в виде колец или
эллипсов (рис. 174, Б). В последнем
случае колония состоит из групп особей,
обладающих самостоятельными ртами, но
имеющих общую клоакальную полость
с одним общим клоакальным отверстием,
в которую открываются клоаки отдельных
особей. Как уже указывалось, размеры
таких асцидиозооидов составляют всего
лишь несколько миллиметров. В случае
же, когда связь между ними
осуществляется только при помощи столона,
асцидиозооиды достигают более крупных размеров,
но обычно меньших, чем одиночные асци-
дии.
Развитие асцидий, их бесполое и
половое размножение будут описаны ниже.
Строение пиросом. Пиросомы, или
огнетелки,— это свободноплавающие
колониальные пелагические оболочники.
Свое название они получили из-за
способности светиться ярким фосфорическим
светом.
Из всех планктонных форм туникат они
ближе всего стоят к асцидиям. По
существу, это плавающие в воде колониальные
асцидий. Каждая колония состоит из
многих сотен отдельных особей —
асцидиозооидов, заключенных в общую, часто
очень плотную тунику (рис. 175, А).
У пиросом все з о о и д ы равноценны
и независимы в отношении питания и
размножения. Колония образуется путем
почкования отдельных особей, причем почки
попадают на свое место, передвигаясь
в толще туники с помощью особых
странствующих клеток — фороцитов. Колония
имеет форму длинного вытянутого
цилиндра с заостренным концом, имеющего
внутри полость и открытого на своем
широком заднем конце (рис. 175, Б). Снаружи
пиросома покрыта небольшими мягкими
шиловидными выростами. Важнейшее
отличие их от колоний сидячих асцидий
Таблица 28. Пелагические оболочники:
1 — колония сальп Thalia sp.;
2 — Salpa maxima (агрегатная форма);
3 — бочоночник Doliolum rarum;
4,5 — Pyrosoma sp.;
6 — Thalia democratica;
7 — колония сальп Cyclosalpa bakeri;
8 — Salpa maxima (одиночная форма).
заключается еще и в строгой
геометрической правильности формы колонии.
Отдельные зооиды стоят перпендикулярно
к стенке конуса. Их ротовые отверстия
обращены наружу, а клоакальные
находятся на противоположной стороне тела
и открываются внутрь полости конуса.
Отдельные маленькие асцидиозооиды
захватывают своими ртами воду, которая,
пройдя через их тело, попадает в полость
конуса. Движения отдельных особей
согласованы между собой, причем эта
согласованность движений возникает
механическим путем при отсутствии мышечных,
сосудистых или нервных связей. В тунике
пиросом от одной особи к другой
протянуты механические волокна, соединяющие
их двигательные мышцы. Сокращение
мышцы одной особи дергает при помощи
волокон туники другую особь и передает
ей раздражение. Сокращаясь
одновременно, маленькие зооиды проталкивают воду
через полость колонии. При этом вся
колония, похожая по форме на ракету,
получив обратный толчок, движется
вперед. Таким образом, пиросомы избрали
для себя принцип реактивного движения.
Такой с-дособ движения используется не
только пиросомами, но и другими
пелагическими туникатами.
Туника пиросом содержит такое
большое количество воды (у некоторых
оболочников вода составляет 99% от
веса тела), что вся колония становится
прозрачной, как будто стеклянной, и
почти невидима в воде. Однако встречаются
и окрашенные в розовый цвет колонии.
Такие пиросомы гигантских размеров —
длина их достигает 2,5 и даже 4 м, а
диаметр колонии 20—30 см —
неоднократно ловились в Индийском
океане. Их название — Pyrosoma spinosum.
Туника этих пиросом обладает настолько
нежной консистенцией, что, попадая
в планктонные сети, колонии обычно
распадаются на отдельные куски. Обычно же
размеры пиросом значительно меньше —
от 3 до 10 см длиной с диаметром от
одного до нескольких сантиметров.
Недавно описан новый вид пиросом — P. vi-
tjasi. Колония этого вида также имеет
цилиндрическую форму и размеры до
47 см. По описанию автора, сквозь
розоватую мантию как темно-коричневые
(вернее, темно-розовые у живых экземпляров)
включения просвечивают внутренности
отдельных асцидиозооидов. Мантия имеет
полужидкую консистенцию, и при
повреждении поверхностного слоя вещество
ее растекается в воде в виде тягучей
слизи, а отдельные зооиды свободно
распадаются.
Строение асцидиозооида
пиросом мало чем отличается от строения
Таблица 29. Оболочники.
Одиночные асцидии:
1 — Phallusia mammillata;
2 — Halocynthia roretzi;
4 — Stye la clavata;
5 — Ascidia virginea;
6 — Boltenia ovifera;
8 — Distomus variolosus;
9 — Boltenia echinata;
11 — Clavelina lepadiformis;
14 — Halocynthia aurantium;
16 — Ciona intestinalis;
17 — Ghelyosoma macleayanum;
18 — Pyura setosa;
19 — Pyura discoveryi;
20 — Styela plicata.
Колониальные асцидии:
3 — Gynandrocarpa placenta;
7 — Sarcobotrylloides racemosum;
10 — Ecteinascidia tortugensis;
12 — Sycozoa umbellata;
13, 15 — Botryllus schlossery.
22 Жизнь животных, т. 2
Рис. 175. Строение пиросом.
А — внешний вид колонии: J — отверстие полости колонии; 2 — околоротовые выросты
туники. Б — продольный разрев колонии: 1 — околоротовые выросты туники; 2 —
полость колонии; з — эооиды; 4 — отверстие полости колонии. В — строение зооида
пиросом: 1 — околоротовой вырост туники; 2 — ротовой сифон; 3 — глотка с
жаберными щелями; 4 — спинная пластинка; 5 — эндостиль; 6 — светящиеся органы; 7 —
кишечник; 8 — клоака; 9 — клоакальные отверстия; 1 о — столон; 11 — волокнистые тяжи
туники; 12 — общеколониальная туника.
одиночной асцидии, за исключением того,
что сифоны его располагаются на
противоположных сторонах тела, а не
сближены на спинной стороне (рис. 175, В).
Размеры асцидиозооидов обычно
составляют 3—4 мм, а у гигантских пиросом—
до 18 мм длины. Тело их может быть
сплющенным с боков или овальным.
Ротовое отверстие окружено венчиком
щупалец, или же может присутствовать только
одно щупальце с брюшной стороны тела.
Часто мантия перед ротовым отверстием,
также с брюшной стороны, образует по
небольшому бугорку или же довольно
значительному выросту. За ртом следует
крупная глотка, прорезанная жаберными
щелями, количество которых может
достигать 50. Эти щели располагаются
или вдоль, или поперек глотки.
Приблизительно перпендикулярно к жаберным
щелям проходят кровеносные сосуды,
число которых также варьирует от одного
до трех-четырех десятков. В глотке имеются
эндостиль и спинные язычки,
свешивающиеся в ее полость. Кроме того, в
передней части глотки по бокам расположены
светящиеся органы, представляющие собой
скопления клеточных масс. У некоторых
видов светящиеся органы есть еще и у
клоакального сифона. Органы свечения
пиросом заселены симбиотическими
светящимися бактериями. Под глоткой
лежит нервный ганглий, имеется и
околонервная железка, канал которой
открывается в глотку. Мышечная система
асцидиозооидов пиросом развита слабо.
Имеются довольно хорошо выраженные
кольцевые мышцы, расположенные вокруг
338
ротового сифона, и незамкнутое кольцо
мышц у клоакального сифона.
Небольшие пучки мышц — спинной и брюшной —
располагаются в соответствующих местах
глотки и лучеобразно расходятся по бокам
тела. Кроме того, есть еще пара
клоакальных мышц. Между спинной частью глотки
и стенкой тела имеются два кроветворных
органа, представляющих собой
продолговатые скопления клеток. Размножаясь
путем деления, эти клетки превращаются
в различные элементы крови —
лимфоциты, амебоциты и пр.
Пищеварительный отдел кишечника
состоит из пищевода, отходящего от
задней части глотки, желудка и кишки.
Кишка образует петлю и открывается
анальным отверстием в клоаку. На
брюшной стороне тела лежит сердце,
представляющее собой тонкостенный
мешочек. Имеются семенники и яичники,
протоки которых также открываются в
клоаку, которая может быть более или менее
вытянута и открывается клоакальным
сифоном в общую полость колонии. В области
сердца у асцидиозооидов пиросом имеется
маленький пальцевидный придаток —
столон. Он играет важную роль при
формировании колонии. В результате деления
столона в процессе бесполого размножения
от него отпочковываются новые особи.
Строение сальп. Так же как и пиросомы,
сальпы являются свободноплавающими
животными и ведут пелагический образ
жизни. Они делятся на два отряда:
бочоночников, или долиолид (Cyclomyaria),
и собственно сальп (Desmomyaria). Это
совершенно прозрачные животные в
форме бочонка или огурца, на
противоположных концах которого расположены
ротовое и анальное отверстия — сифоны.
Только у некоторых видов сальп
отдельные части тела, например столон и
кишечник, бывают окрашены у живых
экземпляров в голубовато-синий цвет.
Тело их одето нежной прозрачной
туникой, иногда снабженной выростами
разной длины. Сквозь стенки тела
хорошо просвечивает маленький, обычно
зеленовато-бурый кишечник. Размеры сальп
колеблются от нескольких миллиметров
до нескольких сантиметров в длину.
Самая крупная сальпа — Thetys vagina —
была поймана в Тихом океане. Длина
ее тела (вместе с придатками)
составляла 33,3 см.
Одни и те же виды сальп встречаются
или одиночными формами, или в виде
длинных цепочкообразных колоний.
Такие цепочки сальп представляют собой
соединенные друг с другом в ряд
отдельные особи. Связь между зооидами
в колонии сальп, как анатомическая, так
и физиологическая, чрезвычайно слаба.
Члены цепочки как бы слипаются друг
с другом прикрепительными сосочками,
и по существу колониальность их и
зависимость друг от друга едва выражены.
Такие цепочки могут достигать длины
более одного метра, но они легко
разрываются на части, иногда просто при ударе
волны. Отдельные особи и особи,
являющиеся членами цепочки, настолько
сильно отличаются друг от друга и по величине
и по внешнему виду, что даже
описывались старыми авторами под разными
видовыми названиями.
Представители же другого отряда —
бочоночники, или долиолиды, — наоборот,
строят чрезвычайно сложные колонии.
Один из крупнейших современных
зоологов В. Н. Беклемишев назвал бочоночни-
ков одними из самых фантастических
существ в море. В отличие от асцидий, у
которых образование колоний происходит
благодаря почкованию, возникновение
колоний у всех сальп строго связано с
чередованием поколений. Одиночные сальпы — это
не что иное, как появившиеся из яиц
бесполые особи, которые, почкуясь, дают
начало колониальному поколению.
Как уже говорилось, тело отдельной
особи, будь она или одиночной, или
членом колонии, одето тонкой прозрачной
туникой. Под туникой подобно обручам
бочонка просвечивают белесоватые ленты
кольцевых мышц. Таких колец у них
имеется 8. Они опоясывают тело
животного на определенном расстоянии друг
от друга. У бочоночников мышечные ленты
образуют замкнутые обручи, а у
собственно сальп они не смыкаются на брюшной
стороне. Последовательно сокращаясь,
мышцы проталкивают поступающую через
рот воду по организму животного и
выталкивают ее через выводной сифон. Как и:
22*
339
Рис. 176. Строение бочоночника:
1 — туника; 2 — мышечные кольца; 3 — ротовой
сифон; 4 — щупальца; 5 — глотка; 6 — жаберные1
щели; 7 — эндостиль; 8 — пищевод; 9 — желудок; ю —
кишка; 11 — задний проход; 12 — клоака; 13 —
клоакальный сифон со щупальцами; 14 — яичник; 15 —
семенник; 16 — сердце; 17 — нервный ганглий; 18 —
нервы; 19 — околонервная железка.
Рис. 177. Строение сальны.
А — внешний вид: 1 — туника; 2 — мышечные тяжи;
3 — ротовой сифон; 4 — глотка; 5 — эндостиль; 6 —
жаберное отверстие; 7 — кишка; 8 — клоака; 9 —
клоакальный сифон. Б — строение сальпы: 1 —
туника; 2 — мантия; 3 — ротовой сифон; 4 — глотка;
5 — эндостиль; 6 — жабра; 7 — пищевод; 8 —
желудок; 9 — пищеварительная железа; 10 — кишка;
11 — клоака; 12 — клоакальный сифон; 13 —
сердце; 14 — столон; 15 — нервный ганглий; 16 — глаз.
пиросомы, все сальпы движутся,
используя реактивный способ движения.
В отряде долиолид бочонки широко
открыты на обоих концах (рис. 176).
На одном конце располагается ротовое
отверстие, на противоположном —
анальное. Оба отверстия окружены
чувствительными бугорками. Внутренность бочонка
поделена косой перегородкой или
спинным выростом на две полости. Передняя
полость — глотка, задняя — клоака. Рот
ведет прямо в огромную глотку,
занимающую почти весь объем тела. В
отличие от асцидий боковые стенки
глотки бочоночников сплошные и только
задняя стенка, которая отгораживает
полость глотки от клоаки, пронизана
двумя сходящимися рядами жаберных
щелей. Щели соединяют глотку прямо
с клоакой, а особые околожаберные
полости, которые имеются у асцидий, здесь
отсутствуют. От них остается только одна
клоакальная полость. На дне глотки
имеется эндостиль, а по спинной стороне,
как и у других рассмотренных нами
туникат, проходит продольный вырост —
спинная пластинка. Эндостиль ведет
из глотки в кишечник, очень
укороченный, расположенный на брюшной части
перегородки между двумя полостями.
Кишечник состоит из короткого пищевода,
переходящего в колбообразный желудок,
к спинкам которого примыкает
пищеварительная железа, и кишки. Кишка
открывается анальным отверстием в клоаку.
Нервная система состоит из
расположенного над глоткой мозгового
ганглия, от которого отходят нервы.
Рядом с желудком лежит сердечный мешок.
От сердца отходят кровеносные сосуды,
которые, как и у всех оболочников,
образуют незамкнутые лакуны,
расположенные неправильной сетью.
Как и все оболочники, бочоночники—
гермафродиты. У них имеется один
яичник и один семенник. Половые железы
лежат по одну сторону от желудка и также
открываются протоками в клоакальную
полость. В яичнике развивается
одновременно только по одному крупному
яйцу.
Органы выделения
отсутствуют. Вероятно, их функцию выпол-
340
няют некоторые клетки крови, в которых
найдены желтовато-коричневые конкреции.
Током крови эти конкреции переносятся
в область желудка, где они
концентрируются, проникают затем в кишку и
выбрасываются вон из организма. У
некоторых сальп, например у Cyclosalpa,
найдены скопления ампуловидных клеток,
очень похожих на таковые у асцидий.
Они располагаются также в области
кишечника и, видимо, играют роль почек
накопления. Однако это еще не
установлено окончательно.
Только что описанное строение тела
относится к половому поколению бочо-
ночников. Бесполые же особи не имеют
половых гонад. Для них характерно
наличие двух столонов. Один из них, почкород-
ный, как и у пиросом, располагается на
брюшной стороне тела и называется
брюшным столоном; второй столон—спинной.
Собственно сальпи по своему строению
очень похожи на бочоночников и
отличаются от них только в деталях (рис. 177,
А, Б). По внешнему виду это также
прозрачные цилиндрообразные животные,
сквозь стенки тела которых хорошо
просвечивает компактный, обычно
оливкового цвета, желудок. Туника сальп может
давать разнообразные выросты, иногда
довольно длинные у колониальных форм.
Как уже указывалось, мышечные обручи
у них не замкнуты, а количество их
может быть больше, чем у бочоночников.
Кроме того, клоакальное отверстие
несколько сдвинуто на спинную сторону,
а не лежит прямо на заднем конце тела,
как у бочоночников. Перегородка между
глоткой и клоакой пронизана всего лишь
двумя жаберными щелями, но зато эти
щели огромны по размерам. И наконец,
мозговой ганглий у сальп развит несколько
сильнее, чем у бочоночников. У сальп
он имеет сферическую форму с
подковообразным вырезом на спинной стороне.
Здесь помещается довольно сложно
устроенный пигментированный глазок.
Сальпы и бочоночники обладают
способностью светиться. Органы свечения
у них очень похожи на органы свечения
пиросом и представляют собой скопления
клеток, расположенных на брюшной
стороне в области кишечника и содержащих
симбиотические светящиеся бактерии.
Особенно сильно органы свечения развиты
у видов рода Cyclosalpa, светящихся
более интенсивно, чем другие виды. У них
образуются так называемые «боковые
органы», расположенные по бокам с
каждой стороны тела.
Как уже неоднократно указывалось,
сальпы являются типичными
планктонными организмами. Однако существует
одна очень небольшая группа
своеобразных донных оболочников — Octacnemidae,
насчитывающая всего четыре вида. Это
бесцветные животные до 7 см в диаметре,
обитающие на морском дне. Тело их
покрыто тонкой полупрозрачной туникой,
образующей вокруг ротового сифона
восемь довольно длинных щупалец. Оно
уплощено и по внешнему виду
напоминает асцидий. Но по внутреннему
строению октакнемиды близки к сальпам.
В зоне прикрепления к субстрату туника
дает тонкие волосовидные выросты, но,
видимо, эти животные слабо укреплены
в грунте и могут плавать над дном на
небольшие расстояния. Некоторые ученые
считают их особым, сильно уклонившимся
подклассом асцидий, другие же склонны
рассматривать их как вторично осевших
на дно сальп. Octacnemidae —
глубоководные животные, найденные в
тропических районах Тихого океана и у берегов
Патагонии, а также в Атлантическом
океане южнее Гренландии, главным
образом на глубине 2000—4000 тыс. м.
Строение аппендикулярий. Аппенди-
кулярии — очень мелкие прозрачные
свободноплавающие животные. В отличие
от остальных оболочников они никогда
не образуют колоний. Размеры их тела
колеблются от 0,3 до 2,5 см. Личинки
аппендикулярий не подвергаются в своем
развитии регрессивному метаморфозу,
т. е. упрощению строения тела и утрате
целого ряда важных органов, например
хорды и органов чувств, вызванному
превращением свободноплавающей
личинки в неподвижную взрослую форму,
как это имеет место у асцидий.
Взрослая аппендикулярия по своему
строению очень похожа на личинку асцидий.
Как уже говорилось, такая важная черта
строения их тела, как наличие хорды,
341
которая всех оболочников ставит в одну
группу с хордовыми животными,
сохраняется у аппендикулярии в течение всей
жизни, и именно этим они отличаются
от всех остальных оболочников, по
внешнему виду абсолютно непохожих на своих
ближайших родственников.
Тело аппендикулярии распадается
на туловище и хвост (рис. 178, А).
Общий вид животного напоминает
головастика лягушек. Хвост, длина которого
в несколько раз превышает длину
округлого тельца животного, прикрепляется
к брюшной стороне в виде длинной тонкой
пластинки. Аппендикулярия держит его
повернутым на 90° вокруг его длинной
оси и подогнутым на брюшную сторону.
Вдоль середины хвоста по всей его длине
проходит хорда — эластичный тяж,
состоящий из ряда крупных клеток.
По бокам от хорды имеются 2 мышечные
ленты, каждая из которых образована
всего лишь десятком гигантских клеток.
На переднем конце туловища лежит
рот, ведущий в объемистую глотку
(рис. 178, Б). Глотка сообщается
непосредственно с внешней средой двумя продол-
говатоовальными жаберными
отверстиями, или стигмами. Околожаберной
полости с клоакой, как у асцидий, здесь нет.
По брюшной стороне глотки проходит
эндостиль, на противоположной, спинной,
стороне заметен продольный спинной
вырост. Эндостиль гонит комочки пищи
к пищеварительному отделу кишечника,
который напоминает подковообразную
изогнутую трубку и состоит из пищевода,
короткого желудка и короткой задней
кишки, открывающейся наружу анальным
отверстием на брюшной стороне тела.
На брюшной стороне тела под желудком
лежит сердце. Оно имеет форму про-
долговатоовального баллона, плотно
прилегающего своей спинной стороной к же-
Рис. 178. Строение аппендикулярии.
А — домик аппендикулярии: 1 — домик; 2 —
аппендикулярия; з — фильтр у входного отверстия домика;
4 — воронкообразное приспособление для лова
добычи; 5 — выходное отверстие домика; 6 — стенка
домика; 7 — отверстие для бегства. Б — строение
аппендикулярии (вид сбоку и со спинной стороны):
J — рот; 2 — глотка; з — эндостиль; 4 — жаберное
отверстие; 5 — пищевод; 6 — желудок; 7 — кишка;
8 — задний проход; 9 — сердце; 10 — нервный
ганглий и спинной нерв; 11 — статоцист; 12 —
мерцательная ямка; 13 — семенник; 14 — яичник с
яйцами; 15 — мышцы хвоста; 16 — хорда; 17 — нервные
ганглии хвоста. В — внешний вид животного без
домика (вид сбоку и со спинной стороны): 1 —
тельце; 2 — хвост.
Стрелки показывают направление тока воды и
движения домика.
342
лудку. К переднему отделу туловища от
сердца идут кровеносные сосуды —
брюшной и спинной. В переднем отделе глотки
они соединяются при помощи кольцевого
сосуда. Имеется система лакун, по
которым, как и по кровеносным сосудам,
идет циркуляция крови. Кроме того, по
спинной и брюшной сторонам хвоста
также проходит по кровеносному сосуду.
Сердце аппендикулярий, как и у
остальных оболочников, периодически меняет
направление тока крови, сокращаясь
в течение нескольких минут то в одном,
то в другом направлении. При этом оно
работает очень быстро, делая до 250
сокращений в минуту.
Нервная система состоит из
крупного надглоточного мозгового ганглия,
от него отходит назад спинной нервный
ствол, достигающий конца хвоста и
проходящий над хордой. У самого основания
хвоста нервный ствол образует вздутие—
небольшой нервный узелок. Несколько
таких же нервных узелков, или ганглиев,
имеется на протяжении всего хвоста.
К спинной стороне мозгового ганглия
тесно примыкает маленький орган
равновесия — статоцист, а на спинной стороне
глотки имеется небольшая ямка. Ее
обычно принимают за орган обоняния.
Других органов чувств у
аппендикулярий нет. Специальные органы
выделения отсутствуют.
Аппендикулярий — гермафродиты, у
них имеются и женские и мужские
половые органы. В задней части туловища
располагается яичник, тесно сжатый с
двух сторон семенниками.
Сперматозоиды выводятся из семенников наружу
через отверстия на спинной стороне тела,
а яйца попадают в воду только после
разрыва стенок тела. Таким образом, после
откладки яиц аппендикулярий погибают.
Все аппендикулярий строят
чрезвычайно характерные домики, являющиеся
результатом выделения их кожного
эпителия (рис. 178, В). Этот несколько
заостренный спереди домик — толстостенный,
студенистый и совершенно прозрачный —
сначала вплотную прилегает к телу, а
затем отстает от него так, что животное
может свободно двигаться внутри домика.
Домик и есть туника, но у
аппендикулярий она не содержит целлюлозы,
а состоит из хитина, вещества,
близкого по строению к роговому. На
переднем и заднем конце домик снабжен
несколькими отверстиями. Находясь
внутри, аппендикулярия производит
хвостом волнообразные движения, благодаря
чему внутри домика образуется ток воды,
и вода, выходя из домика, заставляет его
двигаться в противоположную сторону.
На той же стороне домика, в которую он
движется, наверху имеются два отверстия,
затянутые очень частой решеткой с
длинными узкими щелями. Ширина этих щелей
9—46 мк, а длина 65—127 мк. Решетка
представляет собой фильтр для
поступающих с водой внутрь домика пищевых
частиц. Аппендикулярий питаются только
тем мельчайшим планктоном, который
проходит через отверстия решетки. Обычно
это организмы размером 3—20 мк. Более
же крупные частички, рачки, радиолярии
и диатомовые водоросли, не могут
проникнуть внутрь домика.
Ток воды, поступив внутрь домика, по
падает в новую решетку, по форме
напоминающую вершу и заканчивающуюся на
конце мешковидным каналом, за который
держится своим ртом аппендикулярия.
Бактерии, мельчайшие жгутиковые,
корненожки и другие организмы, прошедшие
через первый фильтр, собираются на дне
канала, и аппендикулярия питается ими,
делая время от времени глотательные
движения. Но тонкий передний фильтр
быстро засоряется. У некоторых видов,
например у Oikopleura rufescens, он
перестает работать уже через 4 часа. Тогда
аппендикулярия покидает испорченный
домик и выделяет вместо него новый.
На постройку нового домика уходит всего
лишь около 1 часа, и снова она начинает
отфильтровывать мельчайший наннопланк-
тон. За время своей работы домик
успевает пропустить примерно 100 см3 воды.
Для того чтобы покинуть домик,
аппендикулярия пользуется так называемой
«калиткой для бегства». Стенка домика
в одном месте очень сильно утончена и
превращена в тонкую пленку. Пробив ее
ударом хвоста, животное быстро покидает
домик, для того чтобы тут же выстроить
новый. Домик аппендикулярий очень
343
легко разрушается при фиксации или
при механическом воздействии, и его
можно видеть только у живых организмов.
Характерной чертой аппендикулярий
является постоянство
клеточного состава, т. е. постоянство
количества клеток, из которых строится
все тело животного. Причем разные органы
строятся также из определенного числа
клеток. Такое же явление известно для
коловраток и нематод. У коловраток,
например, число клеточных ядер и
особенно их расположение всегда постоянны
для определенного вида. Один вид состоит
из 900 клеток, другой — из 959. Это
происходит в результате того, что каждый орган
формируется из небольшого числа клеток,
после чего размножение клеток в нем на всю
жизнь останавливается. У нематод не все
органы обладают постоянством
клеточного состава, а только мышцы, нервная
система, задняя кишка и некоторые
другие. Количество клеток в них невелико,
но зато размеры клеток могут быть
огромными.
Размножение и развитие оболочников.
Размножение оболочников представляет
собой удивительный пример того, какие
необычайно сложные и фантастические
жизненные циклы могут существовать в
природе. Всем оболочникам, кроме
аппендикулярий, свойствен как
половой, так и бесполый способ
размножения. В первом случае новый
организм образуется из оплодотворенного
яйца. Но у оболочников при этом развитие
до взрослой особи происходит с
глубокими преобразованиями в строении
личинки в сторону ее значительного
упрощения. При бесполом же размножении
новые организмы как бы отпочковываются
от материнской особи, получая от нее
зачатки всех основных органов.
Все половые особи оболочников —
гермафродиты, т. е. они обладают как
мужскими, так и женскими половыми
железами. Созревание мужских и женских
половых продуктов всегда происходит
в разное время, и поэтому невозможно
самооплодотворение. Мы уже знаем, что
у асцидий, сальпипиросом протоки
половых желез открываются в клоакальную
полость, а у аппендикулярий
сперматозоиды выходят в воду через протоки,
открывающиеся на спинной стороне тела, тогда
как яйца могут выйти наружу только
после разрыва стенок, что ведет к гибели
животного. Оплодотворение у
оболочников, кроме сальп и пиросом, наружное.
Это значит, что сперматозоид встречается
с яйцеклеткой в воде и там
оплодотворяет ее. У сальп и пиросом образуется
только по одному яйцу, которое
оплодотворяется и развивается в теле матери.
У некоторых асцидий оплодотворение яиц
тоже происходит в клоакальной полости
матери, куда с током воды через сифоны
проникают сперматозоиды других особей,
а оплодотворенные яйца выводятся через
анальный сифон. Иногда же зародыши
и развиваются в клоаке и лишь потом
выходят наружу, т. е. имеет место своего
рода живорождение.
Размножение и развитие
аппендикулярий. У аппендикулярий живорождение
неизвестно. Отложенное яйцо (около 0,1 мм
в диаметре) начинает дробиться целиком,
и вначале дробление идет равномерно.
Все стадии своего эмбрионального
развития — бластулу, гаструлу
и др.— аппендикулярий проходят очень
быстро, ив результате развивается
массивный зародыш. У него уже имеется тельце
с глоточной полостью и мозговым
пузырьком и хвостовой придаток, в котором
располагаются в ряд друг за другом 20 клеток
хорды. К ним примыкают мышечные
клетки. Затем из четырех клеток формируется
и нервная трубка, залегающая
вдоль всего хвоста над хордой.
На такой стадии личинка покидает
яйцевую оболочку. Она еще развита
очень мало, но в то же время у нее имеются
зачатки всех органов. Пищеварительная
полость зачаточна. Нет ни рта, ни
анального отверстия, но мозговой пузырек со
статоцистом — органом равновесия — уже
развит. Хвост личинки располагается
в продолжение передне-задней оси ее
тела, и его правая и левая стороны
обращены соответственно вправо и влево.
Далее следует превращение личинки
во взрослую аппендикулярию.
Формируется кишечная петля, которая растет
к брюшной стенке тела, где она
открывается наружу анусом. В это же время
344
глотка растет вперед, достигает внешней
поверхности и прорывается ротовым
отверстием. Образуются бронхиальные трубки,
которые открываются с двух сторон тела
жаберными отверстиями наружу и также
соединяют полость глотки с внешней
средой. Развитие пищеварительной петли
сопровождается оттеснением хвоста с
самого конца тела на его брюшную сторону.
В это же время хвост поворачивается
вокруг своей оси на 90° влево, так что
его спинной гребень оказывается с левой
стороны, а правая и левая стороны хвоста
обращены теперь вверх и вниз. Нервная
трубка вытягивается в нервный тяж,
формируются нервные узелки, и личинка
превращается во взрослую аппендикулярию.
Все развитие и метаморфоз личинок
аппендикулярий характеризуется
большой скоростью всех процессов,
происходящих во время этого развития. Личинка
вылупляется из яйца еще до окончания
своего формирования. Такая скорость
развития не вызывается воздействием
каждый раз каких-то внешних причин.
Она определяется внутренней природой
этих животных и является
наследственной.
Как мы увидим дальше, взрослые
аппендикулярий по своему строению очень
похожи на личинок асцидий. Только
некоторые детали строения отличают их друг
от друга. Существует точка зрения, что
аппендикулярий всю свою жизнь остаются
на личиночной стадии развития, но
личинка их приобрела способность полового
размножения. Такое явление известно
в науке под названием неотении. Широко
известен пример амфибии амбистомы,
личинки которой, называемые
аксолотлями, способны к половому размножению.
Живя в неволе, аксолотли никогда не
превращаются в амбистом. Они имеют
жабры и хвостовой плавник и живут
в воде, прекрасно размножаясь и давая
потомство, подобное себе. Но если их
кормить препаратом щитовидной железы,
аксолотли заканчивают превращение,
теряют жабры и, выходя на сушу,
превращаются во взрослых амбистом.
Неотения отмечена и у других земноводных—
тритонов, лягушек, жаб. Из
беспозвоночных животных она встречается у
некоторых червей, ракообразных, пауков и
насекомых.
Половое размножение на личиночных
стадиях иногда бывает выгодным для
животных. Неотения может быть не у всех
особей данного вида, й лишь у тех,
которые живут в особых, возможно
малоблагоприятных для них, условиях, например
при пониженной температуре. В
результате получается возможность
размножения в необычной обстановке. При этом
животное не затрачивает много энергии на
завершение полного превращения личинки
во взрослую особь и скорость созревания
увеличивается.
Неотения, вероятно, сыграла большую
роль в эволюции животных. Одна из
наиболее серьезных теорий о
происхождении всего ствола вторичноротых
животных — Deuterostomia, к которым
относятся и все хордовые животные, в том
числе и позвоночные, выводит их из
свободноплавающих кишечнополостных кте-
нофор или гребневиков. Некоторые
ученые считают, что предки
кишечнополостных были сидячими формами, а
гребневики произошли от плавающих в воде
личинок древнейших кишечнополостных,
приобретших способность полового
размножения в результате прогрессивной
неотении.
Размножение и развитие асцидий.
Развитие асцидий происходит более сложным
путем. Когда из яйцевой оболочки выходит
личинка, она вполне похожа на взрослую
аппендикулярию (рис. 179, Л). Она так же,
как и аппендикулярий, напоминает по
внешнему виду головастика, удлиненно-
овальное тельце которого несколько
сжато с боков. Хвостик вытянут и окружен
тонким плавником. По оси хвоста
проходит хорда. Нервная система личинки
образована нервной трубкой, залегающей над
хордой в хвосте и образующей на
переднем конце тела мозговой пузырек со ста-
тоцистом. В отличие от аппендикулярий
у асцидий есть еще пигментированный
глазок, способный реагировать на свет.
На передней части спинной стороны
располагается рот, ведущий в глотку, стенки
которой пронизаны несколькими рядами
жаберных щелей. Но, в отличие от
аппендикулярий, жаберные щели даже
345
у личинок асцидий открываются не прямо
наружу, а в особую околожаберную полость,
зачатки которой в виде двух впячивающихся
с поверхности тела мешочков хорошо видны
с каждой стороны тела. Они называются
нерибранхиальными впячиваниями. На
переднем конце тела личинки видны три липких
прикрепительных сосочка.
Вначале личинки свободно плавают
в воде, двигаясь при помощи своего хвоста.
Размеры их тела достигают одного или
нескольких миллиметров. Специальные
наблюдения показали, что личинки
плавают в воде недолго — 6—8 часов. За
это время они могут преодолевать
расстояния до 1 км, хотя большинство их оседает
на дно сравнительно недалеко от родителей.
Однако и в этом случае наличие
свободноплавающей личинки способствует
расселению неподвижных асцидий на значи-
Рис. 179. Строение и превращение личинки асцидий.
А — строение личинки асцидий: 1 — прикрепительные сосочки; 2 — эндостиль; з — глаза;
4 — перибранхиальная полость; 5 — хорда; в — нервная трубка; 7 — отверстие пери-
бранхиальной полости; 8 — мозговой пузырь; 9 — рот; 10 — глотка. Б—Ж —
превращение личинки асцидий во взрослую особь: J — прикрепительные сосочки; 2 — рот; з —
перибранхиальная полость; 4 — отверстие перибранхиальной полости; 5 — жаберные щели
в глотке; 6 — эндостиль; 7 — пищевод; 8 — желудок; 9 — задний проход; 10 — рудимент
хвоста; и — сердце; 12 — нервная трубка с остатками мозгового пузыря.
346
тельные расстояния и помогает им
распространяться по всем морям и океанам.
Оседая на дно, личинка прикрепляется
при помощи своих липких сосочков к
различным твердым предметам. Таким
образом, личинка садится передним концом
тела, и с этого момента она начинает
вести неподвижный, прикрепленный образ
жизни. В связи с этим происходит
коренная перестройка и значительное
упрощение строения тела (рис. 179, Б—Ж).
Хвост вместе с хордой постепенно
исчезает. Тело принимает мешковидную
форму. Статоцист и глаз исчезают, а вместо
мозгового пузырька остается только
нервный ганглий и околонервная железка.
Оба перибранхиальных впячивания
начинают сильно разрастаться по бокам
глотки и окружают ее. Два отверстия
этих полостей постепенно сближаются и
наконец сливаются на спинной стороне
в одно клоакальное отверстие. В эту
полость и открываются образующиеся вновь
жаберные щели. Кишка также открывается
в клоаку.
Садясь на дно своей передней частью,
на которой находится рот, личинка
асцидии попадает в очень невыгодное
положение в смысле захвата пищи.
Поэтому у осевшей личинки наступает еще
одно важное изменение в общем плане
строения тела: рот ее начинает медленно
перемещаться снизу вверх и в конце
концов располагается на самом верхнем
конце тела (рис. 179, Г—Ж).
Перемещение происходит по спинной стороне
животного и влечет за собой смещение всех
внутренних органов. Перемещающаяся
глотка толкает перед собой мозговой
ганглий, который в конце концов ложится на
спинной стороне тела между ртом и
клоакой. На этом заканчивается превращение,
в результате которого животное
оказывается совершенно непохожим по
внешнему виду на собственную личинку.
Образовавшаяся таким образом асцидия
может размножаться и иным, бесполым
путем, посредством почкования. В
наиболее простом случае от брюшной стороны
тела у его основания вырастает колбасо-
видное выпячивание, или почкород-
ный столон (рис. 180). Этот столон
окружен наружным покровом тела асци-
Рис. 180. Размножение сложных асцидий:
1 — асцидиозооид; 2 — столон; з — почки; 4 —
ротовой сифон; 5 — клоака; 6 — клоакальный сифон;
7 — глотка с жаберными щелями; 8 — эндостиль; 9 —
кишка; 10 — желудок; 11 — сердце; 12 — туника;
13 — корневые отростки столона с зачатками тканей.
дий (эктодермой), в него продолжается
полость тела животного и, кроме того,
слепое выпячивание задней части глотки.
Длинный отросток в столон дает и сердце.
Таким образом, в почкородный столон
заходят зачатки важнейших систем
органов. На поверхности столона образуются
небольшие бугорки, или почки, в которые
все перечисленные выше зачатки органов
также дают свои отростки. Путем сложной
перестройки эти зачатки образуют новые
органы почки. Из выроста глотки
развивается новый кишечник, из сердечного
выроста — новый сердечный мешок. В
покровах тела почки прорывается ротовое
отверстие. Путем впячивания эктодермы
снаружи внутрь образуется клоака и пери-
бранхиальные полости. У одиночных форм
такая почка, вырастая, отрывается от
столона и дает начало новой одиночной асцидии,
а у колониальных — почка остается сидеть
на столоне, вырастает, вновь начинает
почковаться, и в конце концов образуется
новая колония асцидий. Интересно, чю
347
почки у колониальных форм с общей
студенистой туникой всегда отделяются
внутри нее, но не остаются на месте, где
они образовались, а передвигаются сквозь
толщу туники на свое окончательное
место. Их почка всегда прокладывает себе
дорогу к поверхности туники, куда и
открываются ее ротовое и анальное
отверстия. У одних видов эти отверстия
открываются независимо от отверстий других
почек, у других — наружу открывается
только один рот, тогда как клоакальное
отверстие открывается в общую для
нескольких зооидов клоаку (рис. 174, Б).
Иногда при этом могут образовываться
длинные каналы. У многих видов зооиды
образуют тесный кружок вокруг общей
клоаки, а те, которые не поместились
в нем, выталкиваются прочь и дают начало
новому кружку зооидов и новой клоаке.
Такое скопление зооидов образует так
называемый кормидий.
Иногда такие кормидий устроены очень
сложно и имеют даже общую
колониальную сосудистую систему. Кормидий
окружается кольцевым кровеносным сосудом,
в который впадает от каждого зооида по
два сосудика. Кроме того, такие
сосудистые системы отдельных кормидиев
сообщаются и между собой, и возникает
сложная общеколониальная кровеносная
сосудистая система, так что все асцидиозооиды
оказываются связанными между собой.
Как мы видим, связь между отдельными
членами колоний у разных сложных ас-
цидий может быть или очень простой,
когда отдельные особи вполне
самостоятельны и только погружены в общую
тунику, а почки обладают, кроме того,
способностью передвигаться в ней, или
же сложной, с единой кровеносной
системой.
Кроме почкования посредством
столона, возможны также и другие виды
почкования — так называемые
мантийное, пилорическое,
постабдоминально е,— в
зависимости от тех частей тела, которые дали
начало почке. При мантийном почковании
почка возникает в виде бокового выступа
стенки тела в области глотки. Она состоит
всего лишь из двух слоев: наружного —
эктодермы и внутреннего — выроста
околожаберной полости, из которого
формируются в дальнейшем все органы нового
организма. Как и на столоне, почка
постепенно округляется и отделяется от
материнской особи тонкой перетяжкой,
которая затем превращается в стебелек.
Такое почкование начинается еще на
стадии личинки и особенно ускоряется после
того, как личинка садится на дно.
Личинка, дающая начало почке (она в данном
случае называется оозооид), погибает, а
развивающаяся почка (или бластозоид) дает
начало новой колонии. У других асцидий
почка образуется на брюшной
поверхности кишечного отдела тела, также очень
рано, когда личинка еще не вылупилась.
В этом случае в состав почки, покрытой
эпидермисом, входят ответвления
нижнего конца эпикарда, т. е. наружной
стенки сердца. Первичная почка удлиняется,
подразделяется на 4—5 частей, каждая
из которых превращается в
самостоятельный организм, а личинка — оозооид,—
давшая начало этим почкам, распадается
и служит для них питательной массой.
Иногда в почку могут входить части
пищеварительной системы желудка и задней кишки.
Такой способ почкования называется пило-
рическим. Интересно, что в этом очень
сложном случае почкования целый
организм возникает в результате слияния
двух почек в одну. Например, у Trididem-
num в состав первой почки входят
выросты пищевода, а во вторую — выросты
эпикарда. После того как обе почки
сольются, из первой образуется пищевод,
желудок и кишечник дочернего организма,
а также сердце, а из второй — глотка,
пронизанная жабрами, и нервная
система. После этого дочерний организм,
обладающий уже полным набором органов,
отшнуровывается от материнского. Однако
и другие части тела могут дать начало
почке. В некоторых случаях даже
выросты хорды личинки могут входить в
почку и из них формируется нервная
система и половые железы дочерней особи.
Иногда же процессы почкования бывают
настолько похожи на простое деление
организма на части, что трудно сказать,
какой способ размножения имеется в
данном случае. Кишечный отдел тела при
этом сильно удлиняется, в нем накапли-
348
ваются питательные вещества, которые
получаются в результате распада
грудного отдела. Затем происходит разделение
брюшного отдела на несколько
фрагментов, обычно называемых почками, из
которых возникают новые особи. У Атаго-
ucium вскоре после прикрепления
личинки на заднем конце ее тела образуется
длинный вырост. Он увеличивается в
размере, и у асцидии в результате этого
сильно развивается задняя часть тела —
постабдомен, в который смещается
сердце. Когда длина постабдомена намного
начинает превышать длину тела личинки,
происходит его отрыв от материнской
особи и разделение на 3—4 части, из
которых образуются молодые почки — бла-
стозоиды. Они перемещаются из
постабдомена вперед и располагаются рядом
с материнским организмом, сердце в
котором образуется заново. Развитие бласто-
зооидов происходит неравномерно, и,
когда одни из них уже закончили его,
другие только начинают развиваться.
Процессы почкования у асцидий
чрезвычайно разнообразны. Иногда даже
близкие виды одного рода обладают
разными способами почкования. Некоторые
асцидии способны образовывать
покоящиеся, остановившиеся в своем развитии
почки, которые позволяют им переживать
неблагоприятные условия.
При почковании у асцидий наблюдается
следующее интересное явление. Как
известно, в процессе эмбрионального
развития различные органы организма
животного возникают из разных, но совершенно
определенных частей эмбриона
(зародышевых листков) или слоев тела зародыша,
составляющих его стенку на самых
первых стадиях развития.
У большинства организмов имеется три
зародышевых листка: наружный, или
эктодерма, внутренний, или энтодерма,
и средний, или мезодерма. У зародыша
эктодерма покрывает тело, а энтодерма
выстилает внутреннюю кишечную полость
и обеспечивает его питание. Мезодерма
осуществляет связь между ними. В процессе
развития из эктодермы, как общее
правило, образуется нервная система,
кожные покровы, а у асцидий и
околожаберные мешки, из энтодермы —
пищеварительная система и органы дыхания, из
мезодермы — мышцы, скелет и половые
органы. При различных же способах
почкования у асцидий это правило
нарушается. Например, при мантийном
почковании всем внутренним органам (в том числе
и желудку и кишечнику, возникающим
из энтодермы эмбриона) дает начало
вырост околожаберной полости,
являющийся по своему происхождению образованием
эктодермическим. И наоборот, в случае,
когда в состав почки входит вырост
эпикарда (а сердце у асцидии в процессе
эмбрионального развития образуется как
вырост энтодермической глотки),
большая часть внутренних органов, в том
числе и нервная система и
околожаберные мешки, образуется как производное
энтодермы.
Размножение и развитие пиросом. Пиро-
сомы также обладают бесполым способом
размножения путем почкования. Но у них
почкование происходит с участием
специального постоянного выроста тела — почко-
родного столона. Оно характеризуется
также и тем, что происходит на очень ранних
стадиях развития. Яйца пиросом очень
крупны, до 0,7 мм и даже до 2,5 мм,
и богаты желтком. В процессе их
развития образуется первый индивид — так
называемый циатозооид. Циатозооид
соответствует оозооиду асцидий, т. е. он
является бесполой материнской особью,
развившейся из яйца. Он очень рано
перестает развиваться и разрушается. Всю
основную часть яйцеклетки занимает
питательный желток, па котором и
развивается циатозооид.
У описанного недавно вида Pyrosoma
vitjazi на желточной массе располагается
циатозооид, который представляет собой
вполне развитую асцидию размером в
среднем около 1 мм (рис. 181, А). Есть
даже маленькое ротовое отверстие,
которое открывается наружу под яйцевой
оболочкой. В глотке расположено 10—13 пар
жаберных щелей и 4—5 пар
кровеносных сосудов. Кишечник сформирован
полностью и открывается в клоаку,
сифон, который имеет форму широкой
воронки. Имеется также нервный ганглий
с околонервной железой и сердце, которое
энергично пульсирует. Кстати, все это
349
говорит о происхождении пиросом от ас-
цидий. У других же видов в период
максимального развития циатозооида можно
различить лишь зачатки глотки с двумя
жаберными щелями, зачатки двух
околожаберных полостей, клоакальный сифон,
нервный ганглий с околонервной
железкой и сердце. Рот и пищеварительный
отдел кишечника отсутствуют, хотя эндо-
стиль и намечается. Развита также клоака
с широким отверстием, открывающаяся в
пространство под яйцевыми оболочками.
На этой стадии еще в яйцевой
оболочке у.пиросом уже начинаются процессы
бесполого развития. На заднем конце
циатозооида образуется столон —
эктодерма дает вырост, в который заходят
продолжения эндостиля, околосердечной
сумки и околожаберных полостей. Из
эктодермы столона в будущей почке
возникает нервный тяж, независимый от
нервной системы самого циатозооида. В это
время столон разделяется поперечными
перетяжками на четыре участка, из которых
и развиваются первые почки—бластозоои-
ды, которые являются уже членами новой
колонии, т. е. асцидиозооид а-
м и. Столон постепенно становится
поперек оси тела циатозооида и желтка и
закручивается вокруг них (рис. 181, Б—Е).
Причем каждая почка становится
перпендикулярно к оси тела циатозооида.
По мере развития почек материнская
особь — циатозооид — разрушается, а
желточная масса постепенно
используется в качестве питания для первых
четырех почек—асцидиозооидов —
родоначальников новой колонии. Четыре
первичных асцидиозооида принимают
геометрически правильное крестообразное
положение и образуют общую клоакальную
полость. Это настоящая маленькая колония
(рис. 181, Е—Ж). В таком виде колония
выходит из тела матери и освобождается от
яйцевой оболочки. Первичные асци-
диозооиды в свою очередь
образуют столоны на своих задних
концах, которые, перешнуровываясь,
дают начало уже вторичным ас-
цидиозооидам, и т. д. Как
только асцидиозооид изолируется, на
его конце образуется новый
столон и каждый столон образует
цепочку из четырех новых почек.
Колония прогрессивно растет.
Каждый асцидиозооид становится
половозрелым и имеет мужские
и женские половые железы.
У одной группы пиросом асциди-
озооиды сохраняют связь с материн-
Рис. 181. Размножение и развитие
пиросом.
А — строение циатозооида: 1 — яйцевая
оболочка; 2 — циатозооид; з — туника; 4 —
клоакальный сифон; 5 — ротовой сифон;
6 — глотка с жаберными щелями и эндо-
стилем; 7 — пищеварительный отдел
кишечника; 8 — клоака; 9 — нервный
ганглий; 10 — сердце; 11 — желток; 12 —
первичный столон с почками. Б — Ж —
схема образования первичной колонии:
1 — циатозооид; 2 — столон с четырьмя
первичными почками; 3 — желток; 4 —
первичная колония с четырьмя асцидиозоои-
дами; 5 — полость колонии. 3 —
последовательность развития асцидиозооидов в
колонии пиросом (в центре — четыре первичных
асцидиозооида): 1—5 — вторичные асциди-
08ооиды в порядке их образования. / —
IV — кольцевые ряды асцидиозооидов.
350
ской особью и остаются на том месте, где
они возникли. В процессе образования
почек столон удлиняется и почки
соединяются между собой тяжами. Асцидиозооиды
располагаются друг за другом по
направлению к закрытому, переднему, концу
колонии, в то время как первичные
асцидиозооиды отодвигаются к ее задней,
открытой, части.
У другой же группы пиросом, куда
входит большинство их видов, почки не
остаются на месте. Как только они
достигают определенной стадии развития, они
отделяются от столона, который никогда
не удлиняется. В это же время они
подхватываются особыми клетками — форо-
цитами. Фороциты — это крупные, похожие
на амеб клетки. Они обладают способностью
передвигаться в толще туники при помощи
своих ложноножек, или псевдоподий, так
же, как это делают амебы. Подхватывая
почку, фороциты переносят ее через
тунику, покрывающую колонию, на строго
определенное место под первичными ас-
цидиозооидами, и, как только конечный
асцидиозооид отрывается от столона,
фороциты переносят его вдоль левой стороны
к спинной части его производителя, где
он окончательно устанавливается таким
образом, что старые асцидиозооиды все
дальше отодвигаются к вершине колонии,
а молодые оказываются на ее заднем
конце.
Каждое новое поколение асцидиозоои-
дов переносится с геометрической
правильностью в строго определенное место
по отношению к предыдущему поколению
и располагается этажами (рис. 181,3).
После образования трех первых этажей
между ними начинают возникать
вторичные, затем третичные и т. д. этажи.
Первичные этажи имеют по 8 асцидиозооидов,
вторичные — по 16, третичные — по 32
и т. д. в геометрической прогрессии.
Диаметр колонии увеличивается. Однако с
ростом колонии четкость этих процессов
нарушается, некоторые асцидиозооиды
путаются и попадают в чужие этажи.
У тех же самых индивидуумов в колонии
пиросом, которые размножались
почкованием, в дальнейшем развиваются
половые железы, и они приступают к половому
размножению. Как мы уже знаем, у
каждого из многих асцидиозооидов пиросом,
образовавшихся при помощи почкования,
развивается только по одному крупному
яйцу.
По способу образования колоний, а
именно по тому, сохраняется ли у
асцидиозооидов связь с материнским организмом
в течение долгого времени или нет, пиро-
сомы делятся на две группы — Pyrosoma
fixata и Pyrosoma ambulata. Первые
считаются более примитивными, так как
перенос почек при помощи фороцитов
является более сложным и более поздним
приобретением пиросом.
Образование первичной колонии из
четырех членов считалось настолько
постоянным для пиросом, что этот признак
вошел даже в характеристику всего
отряда Pyrosomida. Однако в
последнее время были получены новые данные
по развитию пиросом. Оказалось,
например, что у Pyrosoma vitjazi почкородный
столон может достигать очень большой
длины, а количество почек, одновременно
образующихся на нем,— около 100.
Такой столон образует под яйцевой
оболочкой неправильные петли (рис. 181, А).
К сожалению, пока еще остается
неизвестным, каким образом происходит у них
формирование колонии.
Размножение и развитие бочоночников
и сальп. У бочоночников процессы
размножения еще более сложны и интересны.
Из яйца у них развивается хвостатая,
как у асцидий, личинка, имеющая в
хвостовом отделе хорду (рис. 182, ^4).
Однако хвост в скором времени исчезает, а
туловище личинки сильно разрастается и
превращается во взрослого бочоночника,
который по своему строению заметно
отличается от половой особи, которую мы
описали выше. Вместо восьми
мускульных обручей у него имеется девять, есть
маленький мешковидный орган
равновесия — статоцист, жаберных щелей вдвое
меньше, чем у половой особи. У него
совершенно отсутствуют половые железы
и, наконец, посередине брюшной стороны
тела и на спинной стороне его заднего
конца развиваются два особых выроста—
столоны (рис. 182, Б). Эта бесполая
особь носит особое название —
корми л к а. В нитевидный брюшной сто-
351
Рис. 182. Размножение и развитие бочоночников.
А — личинка бочоночника: 1 — яйцевая оболочка; 2 — туловище личинки; 3 — хвост; 4 — хорда.
Б — внешний вид почкующегося бесполого бочоночника (кормилки) со спинным столоном: 1 —
кормилка; 2 — брюшной столон; 3 — спинной столон с почками; 4 — передвигающиеся почки.
В — перенос почек фороцитами с брюшного столона на спинной: 1 — брюшной столон; 2 —
спинной столон; з — путь почек по телу кормилки. Г — фороциты, несущие почки: 1 — фороциты;
2 — ядро клетки; 3 — ложноножки; 4 — почка с зачатками разных органов. Д — фрагмент спинного
столона: 1 — боковые почки, или гастрозоиды; 2 — рот; 3 — срединные почки, или форозоиды;
4 — кровеносные лакуны. Е — стебелек форозоида с половыми предпочками, или гонозоидами:
I — гонозоиды. Ж — свободноплавающий форозоид с гонозоидами: I — форозоид; 2 — гонозоиды.
л он кормилки, который является почко-
родным столоном, заходят выросты
многих органов животного — продолжение
полости тела, глотки, сердца и др.,— всего
восемь разных зачатков. Этот столон
очень рано начинает отпочковывать
крошечные первичные почки, или так
называемые предпочки. В это время у его
основания толпится множество уже
знакомых нам крупных фороцитов. Фороциты
по двое, по трое подхватывают почки
и несут их сначала по правому боку
кормилки, а потом по ее спинной стороне
к спинному столону (рис. 182, В, Г).
Бели при этом почки собьются с дороги,
они погибают. Пока почки передвигаются
и переходят на спинной столон, они все
время продолжают делиться. Оказывается,
что почки, образовавшиеся на брюшном
столоне, не могут развиваться и жить на
нем.
Первые порции почек усаживаются форо-
цитами на спинном столоне в два боковых
ряда на его спинной стороне. Эти боковые
почки очень быстро развиваются здесь
в маленьких бочоночников, имеющих
форму ложки, с огромным ртом, хорошо
развитыми жабрами и кишечником
(рис. 182, Д). Другие же органы у них
атрофируются. Они прикрепляются к
спинному столону кормилки собственным
спинным столоном, имеющим форму
отростка. Спинной столон кормилки в это
время сильно растет — удлиняется и
расширяется. В конце концов он может
достигать 20—40 см длины. Он представляет
собой длинный вырост тела, в который
заходят две большие кровеносные лакуны
кормилки.
Между тем подползают все новые и новые
фороциты с почками, но теперь эти почки
усаживаются уже не по бокам, а
посередине столона, между двумя рядами
описанных выше особей. Эти почки называются
срединными или форозоидами.
Они мельче боковых, и из них
развиваются бочоночники, похожие на половозрелых
особей, но не имеющие половых гонад.
Эти бочоночники прирастают к столону
кормилки особым тоненьким стебельком.
Все это время кормилка снабжает
питательными веществами всю колонию. Они
поступают сюда через кровеносные лакуны
спинного столона и через стебельки почек.
Но постепенно кормилка истощается. Она
превращается в пустой мускулистый
бочонок, который служит лишь для
передвижения уже значительной колонии,
образовавшейся на спинном столоне.
По поверхности этого бочонка
продолжают двигаться все новые и новые
почки, которые продолжает образовывать
брюшной столон. С того момента, как
кормилка превращается в пустой мешок,
роль ее в питании колонии берут на себя
большеротые боковые особи, которые
называются гастрозоидами (питающими
зооидами). Они захватывают и
переваривают пищу. Усвоенные ими
питательные вещества не только используются
ими самими, но и передаются срединным
почкам. А фороциты все еще подносят на
спинной столон новые поколения почек.
Теперь эти почки усаживаются уже не на
самом столоне, а на тех стебельках,
которыми прикрепляются срединные почки
(рис. 182, Е). Вот эти-то почки и превра--
щаются в настоящих половых
бочоночников. После того как на стебельке
срединной почки, или форозоида,
укрепилась половая предпочка, он отрывается
вместе со своим стебельком от общего
столона и становится свободноплавающей
маленькой самостоятельной колонией
(рис. 182, Ж). Задача форозоида состоит
в том, чтобы обеспечить развитие половой
предпочки. Иногда его называют корми-
лкой второго порядка. Во время
свободного периода жизни форозоидов половая
предпочка, осевшая на его стебельке,
делится на несколько половых почек—
гонозоидов. Каждая такая почка
вырастает в типичного полового бочоночника,
который уже был описан в предыдущей
части. Достигая зрелости, гонозоиды в
свою очередь отделяются от своего
форозоида и начинают вести жизнь
самостоятельных одиночных бочоночников,
способных к половому размножению. Надо
сказать, что и у гастрозоидов и у
форозоидов гонады также закладываются в
процессе их развития, но затем они исчезают.
Эти особи только помогают развитию
третьего настоящего полового поколения.
После того как от спинного столона
кормилки оторвутся все срединные почки,
23 Жиэнь животных, т. 2
353
кормилка вместе с боковыми почками
отмирает. Количество индивидуумов,
образующихся на одной кормилке,
чрезвычайно велико. Оно равняется нескольким
десяткам тысяч.
Как мы видим, цикл развития бочо-
ночников чрезвычайно сложен и
характеризуется сменой полового и бесполого
поколений. Краткая схема его такова: 1.
Половая особь развивается на брюшном
стебельке форозоида. 2. Половая особь
откладывает яйца, и в результате их
развития получается бесполая хвостатая
личинка. 3. Из личинки непосредственно
развивается бесполая кормилка. 4. На
спинном столоне кормилки развивается
поколение бесполых боковых
гастрозоидов. 5. Новое поколение бесполых
срединных форозоидов. 6. Появление и развитие
на брюшном столоне оторвавшегося от
кормилки форозоида половых гонозоидов.
7. Образование половой особи из гонозои-
да. 8. Откладывание яиц.
В развитии сальп также происходит
смена поколений, однако она не имеет
столь удивительной сложности, как у бочо-
ночников. Личинки сальп не обладают
хвостом, содержащим хорду. Развиваясь
из единственного яйца в теле матери, в ее
клоачной полости, зародыш сальп
вступает в тесную связь со стенками
материнского яичника, через которые к нему
поступают питательные вещества. Это
место соединения тела зародыша с тканями
матери называется детским местом или
плацентой. Свободноживущая
личиночная стадия у сальп отсутствует, и у их
зародыша имеется только рудимент
(остаток, не получивший полного развития)
хвоста и хорды. Это так называемый элео-
бласт, состоящий из крупных богатых
жиром клеток (рис. 183, А). Только что
развившийся зародыш, по существу еще
эмбрион, вышедший через клоакальный
сифон в воду, имеет на брюшной стороне
вблизи сердца и между остатком плаценты
и элеобластом маленький почкородный
столон. У взрослых форм столон
достигает значительной длины и обычно
закручен спирально. Эта одиночная сальпа
также представляет из себя такую же
кормилку, как и образовавшийся из
личинки бочоночпик (рис. 183, Б). На
столоне из боковых утолщений образуются
многочисленные почки, расположенные
двумя параллельными рядами. Обычно
сначала захватывается почкованием какой-то
определенный участок столона, дающий
начало определенному числу одновозраст-
ных почек. Количество их бывает
различно — у разных видов от нескольких
единиц до нескольких сотен. Затем
начинает почковаться второй участок, третий
и т. д. Все почки — бластозоиды —
каждого отдельного участка или звена
развиваются одновременно и равны по
величине. В то время как в первом
участке они достигают уже значительного
развития, бластозоиды второго участка
развиты гораздо менее и т. д., а на
последнем участке столона почки только
намечаются (рис. 183, В).
В процессе своего развития
бластозоиды испытывают перегруппировку,
оставаясь при этом связанными друг с другом
столоном. Каждая пара зооидов
приобретает определенное положение по
отношению к другой паре. Она поворачивается
своими свободными концами в
противоположные стороны. Кроме того, в каждой
особи происходит, как у асцидий,
смещение органов, приводящее к изменению их
первоначального взаиморасположения. На
образование почек идет все вещество
столона. У сальп все развитие почек
протекает на брюшном столоне, и они не
нуждаются в специальном спинном
столоне. Почки отрываются от него не
поодиночке, а целыми цепочками, соответственно-
тому как они возникли, и образуют
временные колонии (рис. 183, Г). Все особи
в них совершенно равноценны, и каждая
развивается в половозрелое животное.
Интересно, что, в то время как нервная
трубка, половой тяж, околожаберные
полости и пр. у разных индивидуумов
уже разделились, глотка остается общей
в пределах одной цепочки. Таким
образом, члены цепочки сначала органически
соединены между собой столоном. Но-
отрывающиеся зрелые отрезки цепочки
состоят из особей, соединенных между
собой только слипшимися
прикрепительными сосочками. Каждая особь имеет по
восемь таких присосок, что
обусловливает связь всей колонии. Эта связь и ана-
354
Рис. 183. Размножение и развитие сальп.
А — личинка сальп: 1 — тельце личинки; 2 — ротовой сифон; 3 — глотка;
4 — клоака; 5 — клоакальный сифон; 6 — ганглий; 7 — кишка; 8 — столон;
9 — рудимент хорды, или элеобласт; 10 — плацента. Б — внешний вид
почкующейся бесполой сальпы: 1 — сальпа; 2 — брюшной столон с почками. В —
схема почкования столона: I — III — участки столона, на которых одновременно
развиваются почки; 1 — рот; 2 — глотка; з — эндостиль; 4 — клоака; 5 —
нервный ганглий; 6 — кишечник; 7 — половые железы; 8 — сердце; 9 — элеобласт.
Г — схема расположения бластозооидов сальп в колонии. Вид сверху и
сбоку: 1 — ротовой сифон; 2 — нервный ганглий; 3 — зародыш; 4 —
прикрепительные папиллы.
23*
томически и физиологически крайне слаба.
Колониальность таких цепочек в сущности
едва выражена. Линейно вытяпутые
цепочки—колонии сальп — могут состоять
из сотен отдельных индивидуумов. Однако
у некоторых видов колонии могут иметь
форму кольца. В этом случае особи
соединены между собой выростами туники,
направленными, как спицы в колесе,
к центру кольца, по которому
расположены члены колонии. Такие колонии
состоят лишь из нескольких членов:
у Cyclosalpa pinnata, например, из восьми-
девяти особей (табл. 29).
Если теперь сравнить способы бесполого
размножения разных туникат, то,
несмотря на большую сложность и
разнородность этого процесса в разных группах,
нельзя не заметить и общих черт. А именно:
у всех них наиболее распространенным
способом размножения является деление
почкородного столона на большее или
меньшее число участков, дающих начало
отдельным особям. Такие столоны есть
и у асцидий, и у пиросом, и у сальп.
Колонии всех туникат возникают как
следствие бесполого способа размножения.
Но если у асцидий они появляются просто
в результате почкования и каждый зооид
в колонии может развиваться как
бесполым, так и половым путем, то у пиросом
и особенно у сальп их появление связано
со сложным чередованием полового и
бесполого поколений.
Образ жизни туникат. Посмотрим
теперь, как живут разные оболочники и
какое практическое значение они имеют.
Мы уже говорили выше, что часть их
живет на дне моря, а часть в толще воды.
Асцидий являются донными животными.
Всю свою жизнь взрослые формы
проводят неподвижно, прикрепившись к какому-
нибудь твердому предмету на дне и
прогоняя через свою пронизанную жабрами
глотку воду, чтобы отфильтровать из нее
мельчайшие клетки фитопланктона или
мелких животных и частицы
органического вещества, которыми асцидий
питаются. Они не могут передвигаться, и,
только испугавшись чего-либо или
проглотив что-нибудь слишком крупное, ас-
цидия может сжаться в комочек. При этом
вода с силой выбрасывается из сифона.
Как правило, асцидий просто
прирастают к камням или другим твердым
предметам нижней частью своей туники. Но
иногда тело их может приподниматься
над поверхностью грунта на тонком
стебельке. Такое приспособление
позволяет животным «облавливать» больший
объем воды и не тонуть в мягком грунте.
Оно особенно характерно для
глубоководных асцидий, которые живут на тонких
илах, покрывающих дно океана на
большой глубине. Для того чтобы не тонуть
в грунте, у них может быть еще и другое
приспособление. Отростки туники,
которыми в обычном случае асцидий
прикрепляются к камням, разрастаются и образуют
своего рода «парашют», удерживающий
животное на поверхности дна. Такие
«парашюты» могут появляться и у типичных
обитателей твердых грунтов, обычно
селящихся на камнях, при переходе их к жизни
на мягких илистых грунтах.
Корневидные выросты тела позволяют особям одного
и того же вида выйти в новую и не
обычную для них среду обитания и расширить
границы своего ареала, если другие
условия благоприятны для их развития.
В последнее время асцидий были
обнаружены среди очень специфической
фауны, населяющей тончайшие ходы между
песчинками грунта. Такая фауна
называется интерстициальной. Теперь уже
известно семь видов асцидий, избравших
местом своего обитания столь необычный
биотоп. Это чрезвычайно мелкие
животные — размер их тела всего лишь 0,8—
2 мм в диаметре. Некоторые из них
подвижны.
Одиночные асцидий образуют иногда
большие сростки, которые разрастаются
в целые друзы и селятся большими
скоплениями. Как уже говорилось, очень многие
виды асцидий колониальны. Чаще
других встречаются массивные
студенистые колонии, отдельные члены которых
погружены в общую довольно толстую
тунику. Такие колонии образуют
корковые наросты на камнях или же
встречаются в виде своеобразных шаров,
лепешек и выростов на ножках, иногда
напоминающих по форме грибы. В других
случаях отдельные особи колоний могут
быть почти самостоятельными.
356
Некоторые асцидии, например Clave-
iina, обладают способностью легко
восстанавливать, или регенерировать, свое
тело из разных его частей. Каждый из
трех отделов тела колониальной клаве-
лины — грудной отдел с жаберной
корзиной, отдел тела, содержащий
внутренности, и столон,— будучи вырезанным,
способен воссоздать целую асцидию.
Удивительно, что даже из столона вырастает
цельный организм с сифонами, всеми
внутренностями и нервным ганглием. Если
выделить у клавелины кусок жаберной
корзины одновременно проводимыми
двумя поперечными разрезами, то на
переднем конце превратившегося в округлый
комочек фрагмента животного образуется
новая глотка с жаберными щелями и
сифоны, а на заднем — столон. Если же
сделать сначала разрез сзади, а потом
спереди, то удивительным образом глотка
с сифонами образуется на заднем конце,
а сифон на переднем и передне-задняя
ось тела животного поворачивается на
180°. Некоторые асцидии способны в
каких-то определенных случаях сами
отбрасывать части своего тела, т. е. они
способны к аутотомии. И подобно тому
как заново вырастает у ящерицы ее
оторвавшийся хвост, из оставшегося куска
тела вырастает новая асцидия.
Способность асцидии к восстановлению
утраченных частей тела особенно ярко выражена
во взрослом состоянии у тех видов,
которые могут размножаться путем почкования.
Виды же, размножающиеся лишь
половым путем, например одиночная Giona
intestinal is, обладают регенеративной
способностью в гораздо меньшей степени.
Процессы регенерации и бесполого
размножения имеют много сходных черт,
и, например, Ч. Дарвин утверждал, что
эти процессы имеют единую основу.
Способность восстанавливать утраченные
части тела особенно сильно развита у
простейших, кишечнополостных, червей и
оболочников, т. е. у тех групп
животных, которым особенно свойственно
бесполое размножение. И в каком-то
смысле само бесполое размножение можно
рассматривать как проявляющуюся в
естественных условиях существования и
локализованную в определенных частях тела
животного способность к регенерации его
из фрагмента тела.
Асцидии широко распространены как
в холодных морях, так и в теплых. Они
есть и в Северном Ледовитом океане,
и в Антарктике. Они были найдены даже
непосредственно на побережье
Антарктиды при обследовании советскими
учеными одного из фиордов «оазиса» Бангера.
Фиорд был отгорожен от моря
нагромождениями многолетнего льда, и
поверхностная вода в нем была сильно опреснена.
На каменистом и безжизненном дне этого
фиорда удалось обнаружить лишь комочки
диатомовых и нити зеленых водорослей.
Однако в самом куту залива были
найдены остатки морской звезды и большое
количество крупных, до 14 см длиной,
розовато-прозрачных студенистых
асцидии. Животные были оторваны от дна,
вероятно, штормом и прибиты сюда
течением, но желудки и кишечники их были
целиком заполнены зеленой массой
несколько переваренного фитопланктона.
Вероятно, они питались незадолго до того,
как были выловлены из воды у самого
берега.
Особенно разнообразны асцидии в
тропической зоне. Имеются данные о том, что
число видов оболочников в тропиках
примерно в 10 раз больше, чем в умеренных
и полярных областях. Интересно, что
в холодных морях асцидии значительно
крупнее, чем в теплых, и поселения их
более многочисленны. Они, так же как
и другие морские животные, подчиняются
общему правилу, по которому в умеренных
и холодных морях обитает меньшее
количество видов, но они образуют
значительно большие поселения и биомасса их на
1 м2 поверхности дна во много раз
больше, чем в тропиках.
Большинство асцидии живет в самой
поверхностной литоральной или приливо-
отливной зоне океана и в верхних
горизонтах материковой отмели или
сублиторали до глубины 200 м. С увеличением
глубины общее количество видов их
уменьшается. В настоящее время глубже
2000 м известно 56 видов асцидии.
Максимальная глубина их обитания, на которой
были найдены эти животные,— 7230 м.
На этой глубине асцидии были обнару-
357
Рис. 184. Глубоководные оболочники:
1 — Culeolus murrayi; 2 — Ascopera gigantea;
3 — Octacnemus bythius.
жены во время работ советской
океанологической экспедиции на корабле
«Витязь» в Тихом океане. Это были
представители характерного глубоководного рода
Culeolus. Округлое тело этой асцидии
с очень широко открытыми сифонами,
совершенно не выступающими над
поверхностью туники, сидит на конце длинного
и тонкого стебелька, которым кулеолюс
может прикрепляться к мелким камушкам,
спикулам стеклянных губок и другим
предметам на дне. Стебелек не может
выдержать веса довольно крупного тела,
и, вероятно, оно плавает, колеблясь над
дном, уносимое слабым течением.
Окраска его беловато-серая, бесцветная, как
у большинства глубоководных животных
(рис. 184).
Асцидии избегают опресненных
участков морей и океанов. Громадное
большинство их живет при нормальной
океанической солености около 35°/00.
Как уже говорилось, наибольшее
количество видов асцидии обитает в океане
на небольшой глубине. Здесь же они
образуют и наиболее массовые поселения,
особенно там, где в толще воды достаточно
взвешенных частиц — планктона и
детрита,— служащих им пищей. Асцидии
селятся не только на камнях и других
жестких естественных предметах.
Излюбленным местом их поселения являются
также днища кораблей, поверхность
разных подводных сооружений и т. д.
Поселяясь иногда в огромных количествах
вместе с другими организмами-обраста-
телями, асцидии могут причинять
большой вред хозяйству. Известно, например,
что, поселяясь на внутренних стенках
водоводов, они развиваются в таком
количестве, что сильно сужают диаметр труб
и засоряют их. При массовом отмирании
в определенные сезоны года они настолько
забивают фильтрационные устройства, что
подача воды может прекращаться совсем
и промышленные предприятия терпят
значительный ущерб.
Одна из наиболее широко
распространенных асцидии — Ciona intestinalis,—
обрастая днища кораблей, может
поселяться в таких огромных количествах,
что скорость корабля значительно
снижается. Потери транспортного
судоходства в результате обрастаний очень велики
и могут составлять миллионы рублей
в год.
Однако способность асцидии
образовывать массовые скопления благодаря одной
удивительной их особенности может
представлять известный интерес для людей.
Дело в том, что в крови асцидии вместо
358
железа содержится ванадий, который
выполняет ту же роль, что и
железо,—служит для переноса кислорода. Ванадий-
редкий элемент, имеющий большое
практическое значение, растворен в морской
воде в чрезвычайно малых количествах.
Асцидии обладают способностью
концентрировать его в своем теле. Количество
ванадия составляет 0,04—0,7% веса золы
животного. Следует вспомнить еще, что
и туника асцидий содержит другое
ценное вещество— целлюлозу. Количество ее,
например, в одном экземпляре наиболее
широко распространенного вида Giona
intestinalis равно 2—3 мг. Эти асцидии
поселяются иногда в огромных
количествах. Число особей на 1 м2
поверхности достигает 2500—10 000
экземпляров, а их сырой вес — 140 кг на 1 м 2.
Появляется возможность обсуждения,
как можно практически использовать
асцидий в качестве источника этих веществ.
Не везде есть древесина, из которой
добывается целлюлоза, а залежи ванадия
немногочисленны и рассеянны. Если же
устроить подводные «морские огороды»,
то асцидий в огромных количествах
можно выращивать на специальных
пластинах. Подсчитано, что с 1 га площади моря
можно получать от 5 до 30 кг ванадия и от
50 до 300 кг целлюлозы.
В толще воды океана обитают
пелагические оболочники — аппендикулярии, пи-
росомы и сальпы. Это мир прозрачных
фантастических существ, обитающих в
основном в теплых морях и в тропической
зоне океана. Большинство их видов
настолько сильно приурочено в своем
распространении к теплым водам, что они
могут служить индикаторами изменений
гидрологических условий в различных
районах океана. Например, появление
или исчезновение пелагических
оболочников, в частности сальп, в Северном море
в определенные периоды связано с
большим или меньшим поступлением в эти
районы теплых атлантических вод. То же
самое явление неоднократно отмечалось
в районе Исландии, Ла-Манша, около
полуострова Ньюфаундленд и было
связано как с ежемесячными, так и с сезонными
изменениями в распространении теплых
атлантических и холодных арктических
вод. В эти районы заходят только три
вида сальп — Salpa fusiformis, Jhlea asym-
metrica и наиболее широко
распространенная в океане Thalia democratica.
Появление всех этих видов в большом
количестве у побережий Британских
островов, Исландии, Фарерских островов
и в Северном море редко и связано с
потеплением вод. У побережья Японии
пелагические туникаты являются
показателем пульсаций течения Куросио.
Ппросомы и сальпы особенно
чувствительны к холодным водам и предпочитают
не покидать тропическую зону океана,
в которой они распространены очень
широко. Области географического
распространения большинства видов сальп,
например, охватывают теплые воды всего
Мирового океана, где их встречается
более 20 видов. Правда, описаны два вида
сальп, обитающих в Антарктике. Это Sal-
pa thompsoni, распространенная во всех
антарктических водах и не выходящая
за пределы 40° ю. ш., т. е. зоны
субтропического опускания холодных
антарктических вод, и Salpa gerlachei, обитающая
только в море Росса. Аппендикулярии
распространены более широко, имеется около
десяти видов, обитающих, например, в
морях Северного Ледовитого океана, но и они
более разнообразны и многочисленны
в тропических районах.
Пелагические оболочники встречаются
при нормальной океанической солености
в 34—36°/00. Известно, например, что
в районе впадения реки Конго, где
температурные условия являются весьма
благоприятными для сальп, они отсутствуют
вследствие того, что соленость в этом
месте африканского побережья равна
всего лишь 30,4°/00. С другой стороны,
сальпы отсутствуют и в восточной части
Средиземного моря у Сирии, где соленость,
наоборот, слишком высока — 40°/00.
Все планктонные формы туникат —
жители поверхностных слоев воды, главным
образом от 0 до 200 м. Ппросомы, видимо,
не идут глубже 1000 м. Сальпы и
аппендикулярии в основной массе тоже не
опускаются глубже нескольких сотен метров.
Однако в литературе имеются указания
о нахождении пиросом на глубине 3000 ж,
бочоночников — 3300 м и сальп даже
359
до 5000 м. Но трудно сказать, обитают
ли на такой большой глубине живые
сальны, или же это были просто их мертвые,
но хорошо сохранившиеся оболочки.
На «Витязе» в ловах, сделанных
закрывающейся сеткой, пиросомы не были
обнаружены глубже 1000 м, а бочоночники —
2000—4000 м.
Все пелагические оболочники в общем
широко распространены в океане. Часто
они попадаются в сети зоолога
единичными экземплярами, но столь же
характерны для них и большие скопления. Аппен-
дикулярии попадаются в значительном
количестве — 600—800 экземпляров в
лове с глубины до 100 м. У берегов
Ньюфаундленда количество их значительно
больше, иногда свыше 2500 экземпляров
в таком лове. Это составляет
приблизительно 50 экземпляров в 1 л*3 воды. Но
благодаря тому что аппендикулярии очень
мелки, биомасса их при этом бывает
незначительна. Обычно она составляет 20—
30 мг на 1 м2 в холодноводных районах
и до 50 мг на 1 м2 в тропических районах.
Что касается сальп, то они способны
собираться иногда в огромнейших
количествах. Известны случаи, когда скопления
сальп останавливали даже большие
корабли. Вот как описывает один такой
случай участник советской антарктической
экспедиции зоолог К. В.
Беклемишев: «Зимой 1956—1957 гг. теплоход
«Кооперация» ( водоизмещением более
5000 т) доставлял вторую смену
зимовщиков в Антарктику, в поселок Мирный.
Ясным ветреным утром 21 декабря 1956 г.
в южной части Атлантического океана
с палубы судна на поверхности воды было
замечено 7—8 рыжеватых полос,
вытянувшихся по ветру почти параллельно
курсу корабля. Когда судно
приблизилось, полосы уже не казались рыжими,
но вода в них все же была не синей (как
вокруг), а беловато-мутной от
присутствия массы каких-то существ. Ширина
каждой полосы была больше метра.
Расстояние между ними — от нескольких
метров до нескольких десятков метров.
Длина полос — около 3 км. Едва
«Кооперация» начала пересекать под острым углом
эти полосы, как вдруг машина
остановилась и судно легло в дрейф. Оказалось,
что планктон забил машинные фильтры
и прекратилась подача воды к двигателю.
Во избежание аварии машину пришлось
застопорить, чтобы прочистить фильтры.
Взяв пробу воды, мы обнаружили в ней
массу продолговатых прозрачных существ
величиной около 1—2 см, называемых
Thalia longicaudata и относящихся к
отряду сальп. В 1 м3 воды их было не менее
2500 экземпляров. Ясно, что решетки
фильтров были ими совершенно
заполнены. Водоразборные кингстоны
«Кооперации» расположены на глубине 5 м и 5,6 м.
Следовательно, сальпы находились в
большом количестве не только на поверхности,
но и на глубине по крайней мере 6 м».
Массовое развитие оболочников и их
доминирование в планктоне, по-видимому,
характерное явление для краёв
тропической области. Скопления сальп отмечаются
в северной части Тихого океана, массовое
их развитие известно в зоне смешения вод
течений Куросио. и Оясио, у западного
Алжира, к западу от Британских
островов, у Исландии, в северо-западной
Атлантике в прибрежных районах, вблизи
южной границы тропической области в Тихом
океане, у юго-восточной Австралии.
Иногда сальпы могут преобладать в планктоне,
в котором уже нет других типичных
тропических представителей.
Что касается пиросом, то в столь
огромных количествах, как описывалось выше
для сальп, они, видимо, не встречаются*
Однако в некоторых краевых районах
тропической области обнаружены и их
скопления. В Индийском океане на 40—
45° ю. ш. во время работы советской
антарктической экспедиции было встречено
огромное количество крупных пиросом.
Пиросомы располагались на самой
поверхности воды пятнами. В каждом пятне
насчитывалось от 10 до 40 колоний,
которые ярко светились голубым светом.
Между пятнами расстояние было 100 м и
больше. В среднем на 1 м2 поверхности воды
приходилось 1—2 колонии. Аналогичные
скопления пиросом отмечались и у
берегов Новой Зеландии.
Пиросомы известны как исключительно
пелагические животные. Однако
сравнительно недавно в проливе Кука у Новой
Зеландии удалось получить несколько
360
фотографий с глубины 160—170 м, на
которых ясно были видны большие скопления
Pyrosoma atlanticum, колонии которых
просто лежали на поверхности дна.
Другие же особи плавали в
непосредственной близости от дна. Было дневное
время, и, возможно, животные ушли на
большую глубину, чтобы спрятаться от
прямых солнечных лучей, как это делают
многие планктонные организмы.
Судя по всему, они чувствовали себя
хорошо, так как условия среды были
благоприятны для них. В мае эта пиро-
сома обычна в поверхностных водах
пролива Кука. Интересно, что в том же
районе в октябре дно на глубине 100 м
бывает покрыто мертвыми, разлагающимися
пиросомами. Вероятно, это массовое
отмирание пиросом связано с сезонными
явлениями. Оно в какой-то степени дает
представление о том, в каких
количествах могут встречаться эти животные
в море.
Пиросомы, что в переводе на русский
язык означает «огнетелки», получили свое
название от свойственной им способности
к свечению. Было установлено, что свет,
возникающий в клетках органов свечения
пйросом, вызывается особыми симбиоти-
ческими бактериями. Они поселяются
внутри клеток светящихся органов и,
видимо, там и размножаются,
поскольку неоднократно наблюдались бактерии
со спорами внутри их. Светящиеся
бактерии передаются из поколения в
поколение. Током крови они переносятся
к яйцам пиросом, находящимся на
последней стадии развития, и заражают их.
Затем они поселяются между бласто-
мерами дробящегося яйца и проникают
в зародыш. Светящиеся бактерии
проникают вместе с током крови и в почки
пиросом. Таким образом, молодые пиросомы
получают светящихся бактерий в
наследство от материнских особей. Однако не
все ученые согласны с тем, что пиросомы
светятся благодаря
бактериям-симбионтам. Дело в том, что свечение бактерий
характеризуется своей непрерывностью,
а пиросомы испускают свет только после
какого-нибудь раздражения. Свет асци-
диозооидов в колонии может быть
удивительно интенсивным и очень красивым.
Кроме пиросом, светятся сальпы и аппен-
дикулярии.
Ночью в тропическом океане вслед за
идущим кораблем остается светящийся
след. Волны, бьющиеся о борта судна,
тоже вспыхивают холодным пламенем —
серебристым, голубоватым или
зеленовато-белым. В море светятся не только
пиросомы. Известны многие сотни видов
светящихся организмов — различные
медузы, ракообразные, моллюски, рыбы.
Нередко вода в океане горит ровным
немерцающим пламенем от мириад
светящихся бактерий. Светятся даже донныз
организмы. Мягкие кораллы горгонии
в темноте горят и переливаются, то
ослабляя, то усиливая свечение, разными
огнями — фиолетовым, пурпурным, красным
и оранжевым, голубым и всеми оттенками
зеленого цвета. Иногда их свет похож
на раскаленное добела железо. Среди всех
этих животных огнетелки по яркости
своего свечения, безусловно, занимают
первое место. Иногда в общей светящейся
массе воды как отдельные яркие шары
вспыхивают более крупные организмы.
Как правило, это бывают пиросомы,
медузы или сальпы. Арабы называют их
«морскими фонарями» и говорят, что их
свет похож на свет месяца, слегка
закрытого облаками. Овальные пятна света на
небольшой глубине часто упоминаются
при описании морского свечения.
Например, в выписке из журнала теплохода
«Алинбэк», приводимой Н. И.Тарасо-
в ы м в его книге «Свечение моря»,
в июле 1938 г. в южной части Тихого
океана были отмечены пятна света по
преимуществу правильной прямоугольной
формы, величина которых была
приблизительно 45 х Ю см. Свет пятен был
очень ярким, зеленовато-голубым.
Особенно заметным стало это явление при
наступившем шторме. Этот свет испускали
пиросомы. Большой знаток в области
свечения моря Н. И. Тарасов пишет,
что колония пиросом может светиться до
трех минут, после чего свечение
прекращается сразу и полностью. Свет
пиросом обычно голубой, но у утомленных,
перевозбужденных и гибнущих животных
он становится оранжевым и даже
красным. Однако не все пиросомы могут све-
361
титься. Описанные выше гигантские пиро-
сомы из Индийского океана так же, как
и новый вид Pyrosoma vitjazi, не имеют
органов свечения. Но не исключено, что
способность к свечению у пиросом
непостоянна и связана с определенными
стадиями развития их колоний.
Как уже говорилось, сальпы и аппенди-
кулярии тоже могут светиться. Свечение
некоторых сальп заметно даже днем.
Знаменитый русский мореплаватель и
ученый Ф. Ф. Беллинсгаузен, проходя
в июне 1821 г. мимо Азорских островов
и наблюдая свечение моря, писал, что
«море было усеяно светящимися морскими
животными, они прозрачны, цилиндрооб-
разны, длиною в два с половиною и в два
дюйма, плавают соединенные одно с
другим в параллельном положении,
составляя, таким образом, род ленты, длина
коей нередко аршин». В этом описании
легко узнать сальп, которые встречаются
в море как поодиночке, так и колониями.
Чаще светятся только одиночные формы.
Если у сальп и пиросом имеются
специальные органы свечения, то у аппенди-
кулярий светится все туловище и
некоторые места студенистого домика, в котором
они живут. При разрыве домика внезапно
возникает вспышка зеленого свечения по
всему туловищу. Светятся, вероятно,
желтые капельки особых секреторных
выделений, имеющиеся на поверхности тела
и внутри домика. Аппендикулярии, как
уже говорилось, распространены шире,
чем другие оболочники, и чаще
встречаются в холодных водах. Нередко именно
они обусловливают свечение воды в
северной части Берингова моря, а также
и в Черном море.
Свечение моря — необыкновенно
красивое зрелище. Можно часами любоваться
искрящимся буруном воды за кормой
идущего корабля. Нам неоднократно
приходилось работать в ночные часы во время
экспедиции «Витязя» в Индийском
океане. Большие планктонные сети,
приходившие из глубины моря, часто бывали
похожи на мерцающие голубоватым
пламенем большие конусы, а куты их, в
которых скапливается морской планктон,
напоминали какие-то волшебные фонари,
дающие такой яркий свет, что при нем
вполне можно было читать. Вода стекала
с сетей и с рук, падала на палубу
огненными каплями.
Но свечение моря имеет и очень
большое практическое значение, не всегда
благоприятное для человека. Иногда оно
сильно мешает кораблевождению, слепит
и ухудшает видимость в море. Яркие
вспышки его могут быть приняты даже
за свет несуществующих маяков, не
говоря о том, что светящийся след
демаскирует в ночное время военные корабли и
подводные лодки и наводит на цель
флот и самолеты противника. Свечение
моря часто мешает морскому промыслу,
отпугивая рыб и морских зверей от
облитых серебристым сиянием сетей. Но,
правда, и большие скопления рыб могут
быть легко обнаружены в темноте по
вызванному ими свечению моря.
Оболочники иногда могут вступать в
интересные взаимоотношения с другими
пелагическими животными. Например,
пустые оболочки сальп часто используются
планктонными рачками гипериидами-фро-
нимами в качестве надежного убежища
для выведения потомства. Так же как и
сальпы, фронимы абсолютно прозрачны
и незаметны в воде. Забравшись внутрь
сальпы, самка фронимы выгрызает все
внутри туники и остается в ней. В океане
часто можно найти пустые оболочки сальп,
в каждой из которых сидит по одному
рачку. После того как в своеобразном
родильном доме выведутся маленькие
рачки, они прицепляются к внутренней
поверхности туники и сидят на ней довольно
продолжительное время. Мать же,
усиленно работая своими плавательными
ножками, прогоняет сквозь пустой бочонок
воду, чтобы ее детям было достаточно
кислорода. Самцы, видимо, никогда не
поселяются внутри сальп. Все оболочники
питаются взвешенными в воде
мельчайшими одноклеточными водорослями,
мелкими животными или же просто частицами
органического вещества. Они являются
активными фильтраторами. У
аппендикулярии, например, выработалась
специальная, очень сложно устроенная система
фильтров и ловчих сетей для отлова
планктона. Об устройстве их уже писалось
выше. Некоторые сальпы обладают спо-
362
собностью скапливаться в огромные стаи.
При этом они могут настолько сильно
выедать фитопланктон в тех районах
моря, где они скапливаются, что серьезно
конкурируют из-за пищи с другим
зоопланктоном и являются причиной резкого
уменьшения его количества. Известно,
например, что у Британских островов
могут образовываться большие скопления
Salpa fusiformis, покрывающие площади
до 20 тыс. квадратных миль. В районе
своего скопления сальпы
отфильтровывают фитопланктон в таком количестве,
что почти нацело выедают его. При этом
зоопланктон, в основном состоящий из
мелких рачков Copepoda, также сильно
уменьшается в количестве, поскольку
Copepoda, как и сальпы, питаются
плавающими микроскопическими
водорослями.
Если такие скопления сальп в течение
долгого времени держатся в одной и той
же массе воды и такие воды сильно
обедненные фито- и зоопланктоном,
вторгаются в прибрежные области, они могут
оказать серьезное воздействие на местное
животное население. Выметанные
личинки донных животных гибнут из-за
отсутствия корма. Даже сельдь становится
очень редкой в таких местах, возможно,
из-за недостатка пищи или из-за
большого количества растворенных в воде
продуктов обмена оболочников. Однако
столь большие скопления сальп — явление
недолговечное, особенно в более холод-
новодных районах океана. При
похолодании они исчезают.
Сами сальпы, так же как и ппросомы,
могут иногда использоваться в пищу
рыбами, но только очень немногими видами.
К тому же туника их содержит очень
незначительное количество усвояемых
органических веществ. Известно, что в годы
наиболее массового развития сальп в
районе Оркнейских островов ими питалась
треска. Летучие рыбы и желтоперые
тунцы поедают сальп, а в желудках меч-
рыбы были найдены пиросомы. Из
кишечника другой рыбы — мунуса — размером
в 53 см однажды было извлечено 28 пиро-
сом. Аппендпкулярпп также встречаются
иногда в желудках рыб, причем даже
в значительных количествах. Сальпами
и пиросомами могут, очевидно, питаться
и те рыбы, которые поедают медуз и
гребневиков. Интересно, что крупные
пелагические черепахи каретты и некоторые
антарктические птицы поедают одиночных
сальп. Но большого значения в качестве
пищевого объекта оболочники не имеют.
Членистоногие
Arthropoda
ТИП ЧЛЕНИСТОНОГИЕ (ARTHROPODA)
ВВЕДЕНИЕ
Среди всех групп беспозвоночных
животных тип членистоногих выделяется
наибольшим разнообразием приспособлений
к самым различным условиям
существования, изумительным богатством форм и
огромным числом видов. Число видов
членистоногих приближается к 3 миллионам
и намного превышает число видов всех
остальных типов животных и растений,
вместе взятых, включая микроорганизмы.
Особенно обильны видами насекомые, на
долю которых падает более 90%
известных видов членистоногих.
Именно тонкими приспособлениями к
использованию всевозможных
местообитаний, всяких источников ппщи, часто
недоступных для других животных,
объясняется многообразие и обилие видов
членистоногих.
Трудно найти такие места, где не было
бы членистоногих. Они живут во всех
морях и океанах — и в толще воды, и на
дне, и в грунте на разных глубинах; они
обитают и во всевозможных пресных
водоемах — не только в реках и озерах, но
и в мелких пересыхающих лужах, в
подземных водах и пещерах, в скоплениях
воды в дуплах деревьев. Некоторые виды
приспособились к жизни в очень соленых
водоемах, другие — в горячих
источниках. Кажется, все разнообразие водной
среды на поверхности нашей планеты
освоено членистоногими.
Большая армия представителей этого
типа приспособилась и к жизни вне воды —
на суше, где членистоногие особенно
многообразны. Весь слой почвы,
пронизанный корнями растений, обильно
заселен и членистоногими — во многих
лесных и луговых почвах миллион особей
клещей и ногохвосток на 1 м2 не редкость.
Огромное количество членистоногих
обитает на поверхности различных частей
растений и внутри растительных тканей.
Многие членистоногие — паразиты,
постоянные или временные, наружные или
внутренние, как других видов
членистоногих, так и животных, относящихся к
другим типам, причем, например, среди
насекомых число видов паразитов не
многим, вероятно, уступает числу остальных
видов. Иногда членистоногие столь
изменяются под влиянием паразитического
образа жизни, что даже перестают быть
похожими на членистоногих.
Какими только способами, какими
только веществами членистоногие не
питаются. Одни фильтруют воду, потребляя
мелкие планктонные организмы и мелкие
взвешенные частицы разлагающихся
остатков организмов; другие роются в грунте
или почве, пропуская через кишечник
большое количество минеральных частиц,
чтобы усвоить рассеянные между ними
органические вещества; есть
членистоногие, которые питаются такими труднопе-
367
ревариваемыми веществами, как рог и воск;
есть существующие за счет клетчатки.
Некоторые способны усваивать с
помощью симбиотических микроорганизмов
атмосферный азот. Огромное число
представителей разных классов
членистоногих — хищники и паразиты; для наземных
членистоногих, особенно насекомых, один
из основных источников пищи — ткани
живых высших растений.
Во всех климатических зонах можно
встретить членистоногих. Огромно их
количество и велико многообразие в
тропиках, но есть они и в высоких широтах
Арктики и на побережье Антарктиды, где
недавно были обнаружены новые виды
клещей и ногохвосток. Членистоногих
можно встретить и в самых сухих пустынях.
Образно можно сказать, что
членистоногие — это тип животных, который по
полноте овладения поверхностью земного
шара не имеет равных.
По многим признакам строения
членистоногие близки к кольчатым червям,
особенно к полихетам. Замечательный
зоолог начала прошлого века Жорж Кювье,
предложивший деление животных на
типы, объединил кольчатых червей и
членистоногих в единый тип членистых
животных (Articulata). Знаменательно, что
и в наши дни некоторые авторитетные
зоологи, например В. Н. Беклемишев,
ратуют за восстановление типа членистых.
Это показывает, что членистоногие во
всяком случае очень близки к многощетин-
ковым кольчатым червям. От близких
к ним форм когда-то давно, еще в те
геологические эпохи, в отложениях которых не
сохранилось ископаемых остатков,
отделились предки современных членистоногих.
Членистоногие, как и кольчатые
черви, — это двустороннесимметричные
сегментированные животные. Сходство
членистоногих с кольчецами проявляется не
только в наружной сегментации.
Центральная нервная система у более
примитивных членистоногих очень сходна с
лестничной нервной системой полихет.
Специфические признаки членистоногих
связаны с тем, что у них тело покрыто
очень плотной и прочной кутикулой,
которая нередко образует очень твердый
панцирь. Кутикула у членистоногих
выделяется наружным слоем клеток, так
называемой гиподермой, имеет очень
сложное строение и сложный химический
состав. Раньше считали, что у
членистоногих кутикула состоит только из
хитина. Теперь выяснилось, что, кроме
хитина, в состав ее входят многие другие
соединения, в основном задубленные
белки, и что у большинства членистоногих
на долю хитина падает не больше трети
веществ, из которых состоит кутикула.
Хитин — это сложный полисахарид, в
состав молекулы которого входят атомы
азота. Вероятно, это не определенное
соединение, а группа близких соединений,
формулы которых точно не установлены. Хитин
эластичен, прочен и устойчив к
химическим воздействиям. Нередко в кутикуле
содержится много углекислого кальция.
Прочность и твердость кутикулы
членистоногих настолько велики, что в тех
местах, где она достигает большой
толщины, покровы становятся негибкими.
Подвижность сегментов тела друг
относительно друга, обеспечивающая движение,
у членистоногих достигается тем, что
между твердыми участками (склеритами)
сохраняются перепончатые участки с очень
тонкой кутикулой; такие перепонки на
сгибах образуют складки,
расправляющиеся при растягивании тела. Далеко не
все сегменты у членистоногих подвижны
друг относительно друга, очень часто
наблюдается полное слияние соседних
сегментов, особенно в передних отделах тела.
В теле членистоногих обычно можно
различить 3 отдела — г о л о в у, на
которой находятся основные органы чувств,
служащие для ориентировки в
пространстве (глаза, усики и др.), и органы
захватывания пищи. Сегменты головы у
членистоногих слитные. Далее следует
грудь — тот отдел тела, на
котором располагаются основные
двигательные придатки — ноги, а у насекомых и
крылья. Задний отдел тела называется
брюшком. У некоторых
членистоногих (например, у многоножек, а из
ископаемых — у трилобитов) все туловищные
сегменты более или менее однородны, и
тогда в теле животного различают только
2 отдела — голову и туловище. У многих
членистоногих (у паукообразных, десяти-
368
ногих раков) сливаются головной и
грудной отделы, образуя головогрудь. А у
клещей вообще утрачиваются границы
между отделами тела, тело слитное.
Передвигаются членистоногие с помощью
конечностей.
Поскольку конечности у них, как и все
тело, покрыты кутикулой, их движение
возможно тоже только при разделении ног
на членики. По строению конечностей,
на которые опираются членистоногие при
движении по твердой поверхности или
с помощью которых они плавают, весь
тип и получил свое название.
Некоторое количество пар конечностей
на голове у всех членистоногих
превращается в ротовые органы — челюсти,
использующиеся хотя бы частично для
захватывания или размельчения пищи. Это
тоже признак, характерный для типа
членистоногих. Ротовые части бывают
настолько видоизменены, что их трудно даже
признать конечностями, например,
хоботок комара или клопа. Число пар ног,
которые служат для передвижения,
неодинаково у разных членистоногих. Для
насекомых характерно развитие 3 пар
двигательных конечностей, для
паукообразных — 4 пар, а у некоторых
многоножек число ног превышает сотню.
Нередко туловищные конечности служат
не только как ноги, но и как жабры,
присоски и другие органы.
У членистоногих двигательная
мускулатура состоит из отдельных мышц,
концы которых присоединяются к разным
утолщенным участкам покровов,
отделенным друг от друга мягкой перепонкой.
Движение членистоногих обеспечивается
не изгибами тела при сокращении
всей мускулатуры туловища, как у
червей, а сокращениями обычно немногих
мышц, связанных с конечностями или
другими двигательными придатками
(крылья насекомых, вилочка ногохвосток
и др.). Мышцы членистоногих обладают
исключительно высокой способностью к
сокращению. Все они
поперечнополосатые, причем обнаруживаемая под
микроскопом поперечная исчерченность
мышечных волоконец у членистоногих выражена
больше, чем у позвоночных,— мышцы
членистоногих сильнее наших, если,
конечно, учитывать размеры. Так как панцирь
членистоногих служит и местом
прикрепления двигательных мышц, он является
и скелетом, называемым «наружным» в
отличие от привычного нам внутреннего
скелета позвоночных.
Панцирь членистоногих лишь в
результате долгой эволюции стал скелетом. А
сначала утолщение и упрочнение покровов
у обитавших в морях далеких
червеобразных предков членистоногих
служило, надо полагать, приспособлением к
защите внутренних органов от
повреждений. Утолщение и уплотнение кутикулы
делает ее менее проницаемой. Поэтому
понятно, что только очень мелкие
членистоногие, имеющие тонкие покровы,
живущие в воде или в очень влажных
местах, могут дышатьвсейповерхностью тела.
У всех более крупных членистоногих,
имеющих и более плотные покровы,
обязательно развиваются специальные
дыхательные органы, поверхность которых
покрыта такой тонкой кутикулой, что через
нее легко может поступать кислород.
У живущих в воде членистоногих это
жабры. Для дыхания в воде важно,
чтобы поверхность жабр была возможно
большей. Поэтому жабры всегда
представляют собой тонкие листовидные выросты
или пучки пальцевидных отростков.
Обычно жабрами у членистоногих становятся
конечности или части конечностей. В
воде благодаря большой ее плотности даже
тоненькие жаберные листочки не
слипаются друг с другом. Поэтому на
небольшом участке тела (в остальных местах
защищенного плотными покровами) может
разместиться много листовидных жабр.
Для доставки кислорода от жабр ко
всем удаленным от них органам служит
кровеносная система и полостная
жидкость. У членистоногих кровеносная
система представлена расположенным в
спинной части тела пульсирующим, обычно
продолговатым сосудом — сердцем. Кровь
поступает в сердце через закрывающиеся
клапанами боковые отверстия, а
выталкивается из него в полость тела или
непосредственно, или через немногие крупные
сосуды — система кровообращения
незамкнутая. У крупных морских членистоно-
гдх, имеющих толстый панцирь, — у ома-
24 Жизнь животных, т. 2
369
ров, у мечехвостов — в крови есть
дыхательный пигмент гемоцианин, сходный по
химическому составу с нашим
гемоглобином, но придающий крови не красный,
а синеватый цвет: в состав гемоцианина
входит не железо, как в гемоглобине,а
медь. Гемоцианин легко присоединяет
кислород при прохождении крови через жабры
и отдает его внутренним органам. Так
как у членистоногих кровеносная система
незамкнутая, правильнее называть их
«кровь» «гемолимфой», но и термином
«кровь» зоологи широко пользуются.
На открытой поверхности суши ни
кожное дыхание, ни дыхание с помощью жабр
невозможно -т- легкопроницаемые участки
покровов пропускают воду и быстро
подсыхают, теряя проницаемость. Поэтому водные
организмы, извлеченные на сушу, погибают
от потери воды или от недостатка
поступающего кислорода. На суше только скрыто-
живущие мелкие членистоногие,
обитающие в таких местах, где воздух всегда
насыщен водяным паром, например в
почве, могут дышать всей поверхностью тела
(мелкие клещи, ногохвостки, симфилы).
Жизнь на суше вне влажных
укрытий требует развития непроницаемых для
испарения покровов, а такие покровы не
пропускают и кислород.
У наземных членистоногих развиваются
особые дыхательные органы — либо
«легкие», представляющие глубокие
мешковидные впячивания со складчатыми
стенками, открывающиеся наружу лишь
небольшими отверстиями (у пауков, у
скорпионов), либо (у большинства наземных
членистоногих) трахеи. Трахеи
представляют собой тонкие, обычно обильно
ветвящиеся трубочки, открывающиеся
наружу маленьким входным отверстием
(«дыхальцем») на поверхности тела. Ветви
этих трубочек омываются полостной
жидкостью, а самые мелкие ответвления
доходят до отдельных клеток. Из тонких
трубочек трахей вода через дыхальца
испаряется очень медленно, и потери ее
незначительны, а кислород по ним
проникает быстро и легко.
Уже далекие предки разных групп
членистоногих приспособились к жизни
в разных средах. Можно выделить группу
водных членистоногих, к которым
относятся ракообразные, выделяемую в подтип
жабродышащих. Правда, среди
современных ракообразных есть и такие, которые
живут на суше (мокрпцы), но по
всем признакам они очень близки к
своим водным родичам. Другой подтип —
трахейнодышащие — представлен типично
наземными членистоногими — различными
группами многоножек и насекомых. Они
прекрасно приспособлены к жизни на
суше. В их покровах содержатся воско-
и жироподобные вещества, защищающие
организм от высыхания, а все системы
органов, связанных с расходованием
воды,— выделительная и дыхательная —
действуют так, что влага расходуется
минимально. Среди насекомых есть
отдельные виды, проводящие всю жизнь или часть
ее в воде, но строение таких водных
насекомых явно показывает их
происхождение от обитавших на суше предков.
За длившуюся сотни миллионов лет
историю эти группы обособились в связи
с приспособлениями к жизни в двух
разных средах — в воде и на суше.
Третий подтип членистоногих — хе-
лицеровые — очень существенно
отличается и от ракообразных, и от трахейноды-
шащих. Среди хелицеровых есть и водные
представители — мечехвосты, обитающие в
морях, и наземные — паукообразные.
В ископаемом состоянии сохранилось
много остатков хелицеровых. Их изучение,
дополняющее изучение современных
представителей, показывает, как шло
разделение классов в результате приспособления
к жизни в разных средах—в воде и на суше.
Изучение всех групп членистоногих
убеждает в том, что переход их к жизни на
суше был связан со способностью к
выработке предохраняющих от испарения
влаги покровов, к сохранению в теле
воды, без которой невозможна жизнь.
У членистоногих, хорошо
приспособившихся к жизни на суше, панцирь бывает
нередко тоньше, чем у обитающих в воде,
но зато на его поверхности отлагается
очень тонкий слой воскоподобных и жиро-
подобных веществ, не пропускающих пары
воды. Этот слой защищает организм от
высыхания и препятствует испарению.
Поэтому у всех членистоногих, живущих
открыто на поверхности суши, есть специ-
370
альные органы дыхания. Те
членистоногие, у которых специальных органов
дыхания нет, которые дышат всей
поверхностью кожи, вынуждены обитать во
влажных укрытиях — в почве, в лесной
подстилке и т. д., только иногда по
ночам и после сильных дождей показываясь
на поверхности.
У наилучшим образом
приспособившихся к жизни на суше членистоногих чаще
всего развивается трахейная система. Не
только дыхательная, но и другие системы
органов меняются у членистоногих с
переходом к жизни на суше.
Выведение растворенных вредных
продуктов разрушения белков у водных
членистоногих происходит непосредственно
во внешнюю среду через отверстия
выделительных органов, открывающиеся у
основания конечностей головного или
грудного отдела. А у всех членистоногих,
наилучшим образом приспособившихся к
жизни на суше, — у пауков, многоножек,
насекомых — выделительные трубочки
(«мальпигиевы сосуды») открываются в
начале задней кишки. По мере
прохождения по ней непереваренных остатков пищи
и продуктов выделения и из тех и из
других стенки задней кишки всасывают
воду, столь труднодоступную для
жителей суши, а экскременты и продукты
обмена выводятся из кишечника в
обезвоженном состоянии.
Приспособившиеся к жизни на суше
членистоногие выработали способность
запасать много жира в особых образованиях—
в жировом теле. Главная роль жирового
тела состоит в том, чтобы в критические
периоды, когда организм не может
получить воду извне, снабжать его... водой!
При окислении жиров образуется много
воды (так называемая «метаболическая
вода»), которая и идет на покрытие расхода
влаги животным. В этом отношении
насекомые сходны с верблюдами, у которых при
длительных переходах через пустыню
используется вода, образующаяся при
окислении запасов жира в горбу. У водных групп
членистоногих (ракообразных,
мечехвостов) жирового тела не бывает.
Исключительно многообразны у
членистоногих типы питания и характер пищи,
от которых зависит строение ротовых
органов и кишечника. При переходе к
жизни в воздушной среде большую роль в
завоевании членистоногими суши
сыграло приспособление к питанию высшими
растениями. Цветковые растения черпают
влагу из почвы и сами становятся
источником воды для наземных животных.
Представители многих групп насекомых,
некоторые клещи и совсем немногие
многоножки и мокрицы способны к
питанию живыми тканями высших растений.
Многие наземные членистоногие питаются
разлагающимися тканями растений, а
многие - хищники или паразиты. Но все
они, завоевывая сушу, следовали за
высшими растениями — не только
создателями поедаемых питательных веществ, но
и живыми резервуарами влаги.
Наружный панцирь членистоногих
настолько плотен, что в затвердевшем
состоянии не растягивается. Поэтому рост
членистоногих сопровождается
линьками. У растущего животного старый
панцирь становится тесным, лопается,
обычно сперва на спине, и сбрасывается. И
только в короткий период, пока новый
панцирь, образовавшийся под ним, не
затвердеет, животное может увеличиваться
в размерах. Во время сбрасывания старых
покровов, которые сходят не только со
всей наружной поверхности тела, но и
со всех конечностей, в том числе и с
ротовых органов, а также с передней и
задней кишки и со всех дыхательных
органов, членистоногие бывают вялыми,
малоподвижными, не питаются и стараются
спрятаться в каких-либо укрытиях.
Рост у членистоногих — не непрерывный
процесс, а ступенчатый, размеры тела
увеличиваются скачкообразно — сразу
после линьки. У многих членистоногих
во время линьки происходит и большее
или меньшее изменение строения тела.
Часто взрослое животное приспособлено
к совсем иному образу жизни, чем
молодое, и потому особи разных возрастов
(возрастом у членистоногих называется
период между линьками) бывают даже
совсем непохожи друг на друга. В таком
случае развитие сопровождается
метаморфозом. Превращение гусеницы в куколку,
а куколки в бабочку может служить
общеизвестным примером.
24*
371
Образ жизни разных членистоногих
неодинаков. Многие из них отличаются очень
сложным поведением, что всегда связано
с высоким развитием органов чувств и
центральной нервной системы.
У большинства членистоногих хорошо
развиты глаза. Глаза у членистоногих
бывают простые, имеющие одну линзу,
и сложные, или фасеточные. У высших
представителей всех подтипов глаза
сложные, фасеточные, состоящие из множества
расположенных плотно рядом друг с дру-
ном маленьких глазков. Иногда число
глазков (так называемых омматидиев) в
фасеточном глазу измеряется
несколькими тысячами (рис. 185). Фасеточные глаза
характерны для высших ракообразных,
мечехвостов и насекомых. Такие глаза
были и у живших в палеозойскую эру
трилобитов — членистоногих, полностью
вымерших в конце этой эры. Каждый омма-
тидий воспринимает только одну точку
находящегося передним предмета (рис. 186).
В результате сложный глаз дает
изображение, состоящее из большого числа
отдельных точек, т. е. мозаичное (как
тоновое клише в книге) и прямое (рис. 187,
см. т. 3, табл. 18). Вспомним, что
изображение, получаемое в нашем глазу,
перевернутое и цельное!
Глаза членистоногих, оказывается,
могут воспринимать не только те лучи,
которые видимы для нашего глаза, но и
ультрафиолетовую часть спектра.
Например, многие цветы, которые нам кажутся
равномерно белыми, для насекомых
оказываются пестрыми; у таких цветов есть
не видимый нами ультрафиолетовый
рисунок (см. т. 3, табл. 19).
И наземные и водные членистоногие
способны различать обычный свет и так
называемый поляризованный (те световые лучи,
электромагнитные колебания которых
совершаются в одной плоскости).
Поскольку для рассеянного света неба
характерна определенная степень
поляризации лучей в меридиональной плоскости,
членистоногие по небу, как по компасу,
могут ориентироваться и в ясные и в
пасмурные дни. Таким образом,
членистоногие различают не только частоту, но
и плоскость колебаний световых лучей.
Мы не ощущаем магнитные волны. А
например, самки термитов в гнезде лежат
так, что ось их тела направлена по
магнитному меридиану, а если поместить
рядом сильный магнит, меняют свое
положение. Как воспринимают членистоногие
магнитные волны, пока не выяснено.
Отмечено, что членистоногие могут
реагировать и на короткие радиоволны.
Хорошо развит и слух у членистоногих.
Звуковые волны определенной длины (чаще
короткие) они воспринимают с помощью
тонких слуховых волосков на
поверхности тела, особенно на усиках, с помощью
Рис. 185. Сложный глаз насекомого с вырезанным
сектором (схема):
1 — линзы роговицы (фасетки); 2 — кристаллические
конусы; 3 — главные пигментные клетки; 4 — ядра
зрительных клеток; 5 — рабдомы.
специальных слуховых органов,
напоминающих по строению струну, натянутую
над резонаторной ямкой (как струна на
балалайке), и других приспособлений.
Часто наземные членистоногие издают
ультразвуки, которых мы не слышим, с
помощью различных «стридуляционных»
органов, по принципу действия
напоминающих струну и смычок.
Особенно замечательна способность
членистоногих воспринимать запахи —
ничтожные концентрации многих веществ,
растворенных в воде или взвешенных в
воздухе. Никакие наши химические
приборы не дают возможности уловить такие
слабые концентрации вещества, а
насекомые, например, могут по ним
ориентироваться. Бабочки совки могут прилетать
за километр на тарелку с бродящей
патокой, а самцы шелкопрядов летят за
километры на не ощущаемый нами и вблизи
запах коробки, в которой сидела самка.
Не менее тонко восприятие и запахов,
и содержания солей водными
членистоногими. Многие наземные членистоногие
(мокрицы, личинки насекомых) способны
улавливать разницу в относительной влажности
воздуха порядка 1—2%.
У водных членистоногих прекрасно
развиты органы равновесия.
Наиболее универсальное чувство,
присущее всем членистоногим,—осязание.
Осязательную функцию выполняют
многочисленные волоски на поверхности тела
членистоногих. Для многих членистоногих,
особенно для донных, почвенных и
живущих в тканях растений и животных,
характерно стремление к наибольшей
поверхности контакта с твердыми
предметами — «тигмотаксис».
Многообразная «информация»,
получаемая из внешнего мира органами чувств
членистоногих, требует «обработки
информации» центральной нервной системой.
Головной мозг — парные скопления
нервных клеток над пищеводом — у высших
активных членистоногих очень хорошо
развит, в нем различается несколько
отделов, связанных с основными органами
ориентировки в пространстве,
расположенными в головном отделе. Чем
сложнее поведение членистоногого, чем
многообразнее те условия, с которыми животное
Рис. 186. Схематический разрез через фасеточный
глаз насекомого:
I — линзы роговицы; 2 — кристаллические конусы;
3 — главные пигментные клетки; 4 — ядра
зрительных клеток; 5 — рабдомы; 6 — нервные клетки
сетчатки; 7 — рассматриваемый предмет; 8 — мозаичное
изображение предмета в глазу.
Рис. 187. Изображение пламени свечи в глазу
жука-светляка (Lampyris^:
1 — в наружном слое фасеточного глаза; 2 — на
сетчатке.
373
сталкивается, и чем сложнее ответы
организма на воздействия среды, тем более
сложно строение головного мозга. Головной
мозг у членистоногих связан
окологлоточным нервным кольцом с брюшной
нервной цепочкой. Исходным у
членистоногих, по-видимому, является
соответствие каждому сегменту тела пары
ганглиев брюшной нервной цепочки. Но при
слиянии отдельных сегментов и даже
отделов тела друг с другом происходит
уменьшение числа нервных узлов, частично
сливающихся друг с другом, частично
исчезающих. Степень расчленения или
слитности брюшной цепочки связана со
степенью расчленения или слитности тела,
и концентрация узлов брюшной нервной
цепочки совсем не может служить
показателем сложности поведения.
Поведение членистоногих, особенно в
период размножения (спаривание, забота о
потомстве), бывает очень сложным, в чем
можно убедиться из знакомства с
приводимыми ниже описаниями образа жизни
разных представителей этого типа. Очень
часто поведение определяется прямой
реакцией на раздражение, например
приближение к источнику света
(«положительный фототаксис») или уход от света
(«отрицательный фототаксис»). Еще более
распространены реакции на раздражение
путем усиления или ослабления
подвижности («кинезисы»). Например, многие
насекомые активнее двигаются при более
сильном или слабом освещении и
малоподвижны при среднем. Таким образом,
после ненаправленного быстрого
движения они, наконец, попадают в
благоприятные для себя условия освещения и
перестают двигаться.
Кроме непосредственных стандартных
ответов на раздражения (безусловных
рефлексов), для членистоногих очень
характерны и сложные цепи наследственно
закрепленных рефлекторных действий
(инстинкты). Инстинкты представляют такое
сложное поведение, какое в нормальных
для данного вида условиях является
целесообразным, но оно остается
неизменным и при изменении условий и поэтому
может становиться нецелесообразным.
Например, роющие осы вырывают
норку, приносят в нее парализованное
насекомое, которым должна питаться
личинка, откладывают на добычу яйцо и
заделывают норку. Все действия осы
«целесообразны». Но если в процессе
деятельности осы, когда она приступает
к заделыванию норки, вынуть и добычу
и яйцо, оса завершает свою уже ненужную
работу по замуровыванию норки.
Наряду со сложными формами
инстинктивного поведения у членистоногих
(высшие ракообразные, пауки, высшие
насекомые) прослеживаются и индивидуальные
навыки; у них можно выработать и
условные рефлексы. Мало того, например,
пчелы могут передавать друг другу
приобретенные навыки, обмениваться информацией.
Определенными телодвижениями рабочая
пчела сообщает другим пчелам в улье, в
каком направлении надо лететь за взятком.
Членистоногим, несомненно, свойственны
многие проявления высшей нервной
деятельности, которую еще недавно считали
характерной только для позвоночных.
Все членистоногие размножаются
половым путем — обычно с оплодотворением,
но иногда и без оплодотворения (парте-
ногенетически). Большинство
членистоногих раздельнополые, но некоторые,
ведущие неподвижный образ жизни
(например, усоногие ракообразные) или
паразитические формы (например,
некоторые равноногие
ракообразные),—гермафродиты.
Типы оплодотворения у членистоногих
разнообразны.
У водных членистоногих, например
мечехвостов, бывает настоящее наружное
оплодотворение, когда самка откладывает
яйца в воду, а самец туда же выводит
семенную жидкость. Проникновение
сперматозоида в яйцо происходит при
наружном оплодотворении во внешней среде —
в воде.
Часто при наружном оплодотворении
у водных членистоногих самец
прикрепляет «пакеты» с семенной жидкостью (спер-
матофоры) к выводным половым
отверстиям самок — откладываемые яйца
оплодотворяются во внешней среде, но
вероятность оплодотворения каждого яйца
в таком случае увеличивается.
С переходом к жизни на суше низшие
членистоногие, обитающие во влажной
374
среде,— в почве, в гнилой древесине и
т. п. (многие клещи, скорпионы, ложноскор-
пионы, многоножки, ногохвостки и др.)»
выработали новый способ оплодотворения.
Самцы откладывают сперматофоры во
внешнюю среду, как при наружном
оплодотворении (на землю, на остатки растений), а
самки подбирают их своими половыми
отверстиями, и, таким образом, проникновение
сперматозоидов в яйцо осуществляется
внутри организма самки, как при
внутреннем оплодотворении. Такое
оплодотворение называется наружно-внутренним.
У обитающих на поверхности суши в
открытой атмосфере членистоногих
оплодотворение внутреннее: самцы вводят
семенную жидкость непосредственно в
половые отверстия самок. Проникновение
сперматозоида в яйцо происходит в
защищенных от высыхания условиях
внутри материнского организма.
В отдельных группах водных
членистоногих, например у усоногих раков,
также выработалась способность к
внутреннему оплодотворению, повышающая
плодовитость.
У большинства членистоногих
наблюдается откладка яиц, живорождение
наблюдается редко, например у некоторых
двукрылых насекомых. Чаще
наблюдается яйцеживорождение: самка рождает
живых детенышей, однако развитие
детенышей происходит за счет питательных
запасов яйца, но не во внешней среде,
а в яйцеводах самок (например, у тлей).
Как упоминалось, развитие многих
членистоногих происходит со сложным
превращением. В тех случаях, когда
взрослая особь и форма, вышедшая из яйца,
резко различаются по строению, молодые
особи называются личинками.
У малоподвижных или неподвижных
морских ракообразных расселяются
подвижные личинки. У очень подвижных во
взрослом состоянии крылатых насекомых
личинки—в основном питающаяся стадия.
В тех случаях, когда яйца богаты
желтком, из них выходят молодые
членистоногие, уже похожие на взрослых. Такое
развитие характерно для многих
пресноводных ракообразных, паукообразных и
некоторых насекомых.
Членистоногие — древние животные.
Самые достоверные данные, на
основании которых можно судить о
происхождении тех или иных групп животных, дают
исследования ископаемых окаменевших
остатков организмов или их отпечатков в
слежавшихся и окаменевших наслоениях
ила, песка или глины, под наносами кото-
Рис. 188. Смена сред обитания в эволюции основных групп членистоногих:
сплошные линии — филогенетические связи; пунктирные линии, заканчивающиеся стрелкой без
кружка,— выход отдельных представителей группы в новую среду; +—ископаемые формы;
в скобках — редкие случаи для данной группы; в кавычках — сборное название группы, не
имеющей четкой систематической характеристики.
рых трупы животных были погребены.
Но не от всех прошедших эпох, когда
уже существовала на Земле жизнь,
сохранились ископаемые остатки организмов.
Отложения самых древних периодов
развития жизни на Земле подвергались в
течение последующих эпох таким сильным
изменениям, что точно установить,
какие организмы обитали на Земле около
миллиарда лет тому назад, невозможно,
сколько-нибудь уцелевших остатков
животных из отложений более древних, чем
палеозойские, не сохранилось. А уже в
самых глубоких («нижних») осадочных
породах кембрийского периода
палеозойской эры (отлагавшихся более
полумиллиарда лет назад) встречаются трилобиты
(см. т. 3, рис. 3)— животные с явно
выраженными признаками членистоногих,
имевшие развитой наружный скелет,
расчлененные конечности, сложные глаза и другие
свойственные представителям типа Arthro-
poda признаки организации. Трилобиты—
класс членистоногих, полностью вымерший
в конце палеозойской эры (длившейся
300—350 миллионов лет).
Из кембрийских отложений известны
и ракообразные, некоторые из которых
относятся к современным отрядам, с
силурийского периода известны родичи
мечехвостов (см. т. 3, табл. 1) и даже
скорпионов (см. т. 3,табл. 3, i, 2), в девонских
отложениях были найдены остатки ногохвосток,
а недавно московский палеонтолог Б. Б.
Родендорф установил, что в девоне
жили и крылатые насекомые. Все эти данные
свидетельствуют о том, что тип
членистоногих отделился от каких-то животных,
похожих на кольчецов, очень давно.
Поэтому точно установить родственные
связи различных членистоногих очень
трудно. Несомненно, что одной из первых
оформившихся групп членистоногих были
трилобиты. В палеозое трилобиты
достигли расцвета, они буквально кишели в
прибрежной зоне морей и океанов 500
миллионов лет назад. Трилобиты
сохранились в ископаемом состоянии
настолько хорошо, что по некоторым
остаткам удалось не только исследовать их
внутреннее строение, но на шлифах
изучить даже микроскопическую структуру
тканей этих животных, а также
проследить их развитие. А применяя некоторые
методы современной химии, удалось даже
выяснить, что в их крови был гемо-
цианин, как у современных высших
ракообразных.
Трилобиты, обычно выделяемые в
отдельный класс членистоногих,
заслуживают выделения в самостоятельный подтип
Trilobita.
Из современных групп членистоногих
наиболее близки к трилобитам,
по-видимому, мечехвосты (см. т. 3,табл. 1), личинки
которых несколько сходны с трилобитами,
а зародыши—с личинками трилобитов («про-
таспис»—см. т. 3, рис. 2).
Мечехвосты и паукообразные имеют ряд
общих черт строения, позволяющих их
объединить в единый подтип хелицеровых
(Chelicerata).
По характеру органов дыхания и
некоторым другим признакам некоторые
общие с трилобитами черты имеют и
ракообразные (класс Crustacea). Однако
непосредственных родственных связей между
трилобитами и ракообразными установить
нельзя. Ракообразные представляют
особый подтип жабродышащих (Branchiata)
в типе членистоногих. А так как среди
современных ракообразных многие имеют
более примитивное строение, чем хели-
церовые, принято рассмотрение типа
членистоногих начинать с них.
Последний подтип — трахейнодышащие
(Tracheata)—объединяет различных
многоножек и насекомых, т. е. типично
наземных членистоногих.
В настоящее время не вызывает
сомнений происхождение насекомых от каких-
то древних членистоногих, близких
к многоножкам, а многоножек — от
примитивных червеобразных членистоногих.
Вероятно, водные хелицеровые
произошли от общих предков с трилобитами —
это была другая ветвь эволюционного
развития членистоногих. А
приспособившиеся к жизни в водной среде ракообразные,
сохранившие много черт древних
членистоногих, представляют третью основную
ветвь развития членистоногих (рис. 188).
Знакомство с типом и с его
многообразными представителями удобнее всего
начинать с подтипа жабродышащих, с
класса ракообразных.
376
ПОДТИП ЖАБРОДЫШАЩИЕ
(BRANCHIATA)
КЛАСС РАКООБРАЗНЫЕ (CRUSTACEA)
Бесконечно разнообразны водоемы,
существующие на нашей Земле,—от
необозримого океана, глубина которого достигает
почти 11 км, до маленьких
пересыхающих луж и капиллярных вод,
пропитывающих морской и речной песок. Различны
их температура, соленость, кислотность,
газовый режим и все остальные условия
обитания в них живых организмов.
Немногие группы многоклеточных
животных приспособились к жизни во всех
этих столь разнообразных условиях.
Большинство обитает или в морской, или в
пресной воде, или на дне, или в толще
воды, ведет или свободный, или
паразитический образ жизни. Этого никак нельзя
сказать о ракообразных.
Ракообразные населяют любые
существующие на Земле водоемы,
пресноводные и морские, маленькие и большие,
живут на дне и в толще воды, проникают
в подземные воды, опускаются на
предельные глубины Мирового океана.
Большинство ракообразных свободно
передвигается по дну или в толще воды, но среди
них есть и сидяче-прикрепленные виды,
например морские желуди и морские
уточки, есть и паразиты, прикрепляющиеся
к рыбам и другим водным животным.
Водная среда — царство ракообразных,
однако некоторые из них приспособились
и к жизни на суше. Таковы
общеизвестные мокрицы, некоторые тропические
крабы и раки-отшельники.
Многие ракообразные чрезвычайно
многочисленны. Морской планктон под всеми
широтами и на всех глубинах в
основном (не менее чем на 90% по весу)
состоит из ракообразных. Очень велико
значение ракообразных и в планктоне
пресных вод. Донные ракообразные также
нередко встречаются в массовом
количестве.
Ракообразные играют очень важную
роль в экономике природы. Органическое
вещество в водоемах создается главным
образом за счет жизнедеятельности
микроскопических водорослей. Ракообразные
поедают эти водоросли и сами, в свою
очередь, поедаются рыбами. Они выступают
в качестве посредников, делающих
создаваемое в водоемах органическое
вещество доступным для рыб. С другой
стороны, они используют в пищу огромные
массы погибших водных животных,
обеспечивая таким образом очищение
водоемов.
Существование почти всех рыб, как
морских, так и пресноводных, в
значительной степени зависит от ракообразных.
Некоторые рыбы, как например сельдь, всю
свою жизнь питаются планктонными
ракообразными, другие используют их
только вскоре после выхода из икринки, а
затем переходят на какую-нибудь другую
пищу. Для гигантов моря — беззубых
китов — ракообразные служат основной
пищей.
Многие ракообразные
непосредственно используются человеком в качестве
высокоценного пищевого продукта. Во многих
странах развит промысел креветок,
крабов, омаров, лангустов и некоторых
других съедобных видов. В последнее время
проводятся успешные опыты
использования морских планктонных
ракообразных для добывания витаминов, жиров
и других важных веществ. На
рыбоводных заводах разводятся некоторые виды
ракообразных, необходимые для
кормления молодых рыб.
Размеры тела ракообразных
колеблются в очень широких пределах — от
долей миллиметра до 80 см. Столь же
разнообразны форма их тела, строение его
частей, окраска и другие признаки.
377
В настоящее время известно более
20 000 видов ракообразных.
Тело ракообразных, так же как и
других членистоногих, обычно покрыто
хитиновым панцирем. Он отсутствует или
слабо выражен только у паразитических
и сидяче-прикрепленных видов. Панцирь
прекрасно защищает животных от
всевозможных вредных для них физических и
химических воздействий, но, с другой
стороны, препятствует их беспрерывному
росту. Поэтому время от времени
ракообразные линяют — сбрасывают старый
панцирь и, пока не затвердел новый, быстро
растут. Многие виды после линьки
оказываются совершенно беспомощными и
вынуждены укрываться от врагов в каких-
нибудь убежищах.
Тело ракообразных состоит из
отдельных сегментов, частично срастающихся
друг с другом. Каждый сегмент снабжен
парой членистых конечностей, но нередко
в процессе эволюции некоторые
конечности исчезают. Сегменты тела
группируются в три отдела, имеющиеся и у
других членистоногих,— голову, грудь
и брюшко.
Голова представляет собой результат
слияния головной лопасти и 5 следующих
за ней сегментов и снабжена
соответственно 5 парами придатков — передними
п задними антеннами, находящимися
у взрослых особей перед ротовым
отверстием (только у Cephalocarida задние
антенны расположены позади рта),
жвалами, или мандибулами, и двумя
парами челюстей, или м а к с и л л.
Обе пары антенн служат в первую
очередь органами чувств. Они богаты
различными чувствительными
щетинками, осуществляющими функции
обоняния, осязания и чувства химического
состава воды. Жвалы размельчают пищу,
челюсти фильтруют пищевые частицы и
участвуют в процессе подачи пищи ко
рту.
На голове помещаются также глаза.
Обычно у ракообразных есть пара
сложных фасеточных глаз, устроенных так же,
как у других членистоногих. Иногда эти
глаза сидят на стебельках, иногда
непосредственно на поверхности головы. У
некоторых ракообразных, например у вет-
вистоусых, сложные глаза сливаются в
один непарный глаз. У личинки
ракообразных — науплиуса — сложные глаза
еще не развиты, но имеется один
непарный простой, так называемый науплиаль-
ный глаз. У многих ракообразных этот
глаз сохраняется и у взрослых, а у
веслоногих он служит единственным
светочувствительным органом в течение всей
их жизни. У глубоководных и подземных
видов глаза исчезают.
Задний край головы многих
ракообразных снабжен покрытой хитином складкой,
которая охватывает сверху и с боков весь
грудной отдел тела или его переднюю часть.
Это — карапакс, имеющий форму
щита, двустворчатой раковины или
полуцилиндра. В тех многочисленных случаях,
когда карапакс отсутствует (жаброногие,
веслоногие, бокоплавы, равноногие и др.),
он исчез вторично. Карапакс не только
служит защитным образованием, но и
определяет направление токов воды,
необходимых для питания и дыхания.
Грудной и брюшной отделы тела у
разных ракообразных состоят из различного
количества сегментов. Нередко один или
несколько передних грудных сегментов
срастаются с головой и их конечности
преобразуются в так называемые ного-
челюсти, принимающие участие в
захватывании пищи.
Брюшко заканчивается тельсоном,
соответствующим пигидию многощетинко-
вых червей.
У наиболее примитивных ракообразных
грудные ножки выполняют три основные
функции — двигательную, дыхательную
и функцию подачи пищи ко рту. Каждая
ножка снабжена несколькими лопастями,
расположенными на ее внутреннем и
наружном краях. Некоторые из наружных
лопастей превращены в жабры, другие
лопасти вызывают ток воды ко рту, третьи
служат веслами. У более
высокоорганизованных ракообразных наблюдается
разделение функций между разными
конечностями: одни используются только для
передвижения, другие — только для
дыхания, третьи — только для
захватывания пищи.
Чрезвычайно характерным для
ракообразных следует считать способ их дыха-
378
ния. Как все настоящие водные
животные, они дышат жабрами, через тонкие
стенки которых растворенный в воде
кислород проникает в кровь. Даже
сухопутные ракообразные—мокрицы имеют
жабры и используют кислород смачивающего
их тонкого слоя влаги. Обыкновенно
жабрами служат выросты грудных ног, но
у равноногих и ротоногих ракообразных
в жабры превращены брюшные
конечности. У части ракообразных (веслоногие,
мистакокариды, усоногие, ракушковые)
жабр нет и дыхание совершается всей
поверхностью их тела.
Кровь приводится в движение биением
сердца, которое помещается недалеко от
жабр. У ракообразных, жабры которых
находятся на брюшном отделе, сердце
помещается сзади, у остальных оно
расположено в грудном отделе.
Кровеносные сосуды есть только у представителей
подкласса высших ракообразных, у
остальных кровь течет по полостям между
внутренними органами (рис. 189).
В крови часто растворены
дыхательные пигменты — красный гемоглобин и
синий гемоцианин, в котором железо
заменено медью. Дыхательные пигменты
могут связывать большое количество
кислорода.
Питаются ракообразные различной
пищей. Одни фильтруют имеющуюся в воде
взвесь, включающую и мелкие
организмы — бактерии, одноклеточные водоросли
и т. д., другие своими жвалами
откусывают куски мертвых и живых животных
или растений. Пища перетирается
жвалами и попадает в рот, а оттуда в
пищевод. Задняя часть пищевода у
большинства ракообразных преобразована в
желудок, в котором иногда имеются
хитиновые зубы, завершающие измельчение
пищи. Экскременты выбрасываются через
анальное отверстие, расположенное на
заднем конце тела.
Органами выделения служат железы,
открывающиеся наружу или при основании
задних максилл (максиллярные
железы), или при основании задних
антенн (антеннальные железы).
Эти железы соответствуют метанефридиям
кольчатых червей. Выделяемая ими моча
у многих пресноводных ракообразных
Рис. 189. Кровеносная система разных
ракообразных:
1 — веслоногих; 2 — ветвистоусых; 3 — ракушковых;
4 — равноногих; 5 — бокоплавов; 6 — десятиногих.
очень бедна солями. Животные удаляют
из полости тела воду, и концентрация солей
в их полостной жидкости оказывается
выше, чем в окружающей среде.
Почти все ракообразные
раздельнополы, только усоногие и некоторые
десятиногие — гермафродиты.
Обыкновенно самцы резко отличаются от
самок своим внешним видом или размерами.
У одних ракообразных самцы крупнее,
у других мельче самок. Часто у самцов
есть какие-нибудь приспособления,
помогающие им удерживать самку во время
спаривания. Спаривание заключается в
том, что самец прикрепляет близ половых
отверстий самки сперматофор, т.е.
кучку сперматозоидов, одетую оболочкой.
Когда самка начинает откладывать яйца,
вместе с ними выделяется вещество,
растворяющее оболочку сперматофора;
сперматозоиды оказываются на свободе и
оплодотворяют яйца. В других случаях
сперма вводится при помощи специальных ко-
пулятивных органов непосредственно в
половые пути самки. Яйца
представителей подкласса жаброногих ракообразных
способны развиваться партеногене-
379
тически, т. е. без оплодотворения.
Имеются виды, состоящие из одних
самок, дающих начало новому поколению
самок.
Сравнительно редко яйца
откладываются прямо в воду. Обычно самка носит
их на себе в специальной выводковой
камере или прикрепляет их к
поверхности тела или к конечностям.
Из яйца развивается характерная для
всех ракообразных личинка науплиус.
Тело науплиуса внешне не
сегментировано. Он имеет три пары конечностей —
передние и задние антенны и мандибулы.
У некоторых ракообразных науплиус
выходит из яйца и приступает к
самостоятельному существованию, у других эта
стадия формируется внутри яйцевой
оболочки и личинка покидает яйцо,
достигнув более поздней стадии развития.
Наконец, есть ракообразные с прямым
развитием, у которых все личиночные стадии
проходят в яйце, а из него рождается уже
сформировавшийся маленький рачок.
Личинки ракообразных часто
непохожи на взрослых и нередко ведут совсем
иной образ жизни. Так, например,
личинки многих донных ракообразных плавают
в толще воды.
Сравнение между собой по многим
признакам различных ракообразных
позволяет разделить этот огромный класс на
5 подклассов, к рассмотрению которых мы
и переходим.
ПОДКЛАСС ЖАБРОНОГИЕ РАКООБРАЗНЫЕ
(BRANCHIOPODA)
Жаброногие ракообразные считаются
самыми примитивными представителями
всего класса ракообразных. Их голова не
срастается с грудными сегментами.
Грудные ножки листовидные, слабо хитини-
зированные и служат одновременно для
передвижения, дыхания и направления
пищи ко рту. При их взмахах создается
только один ток воды, который
используется и для движения, и для питания,
и для дыхания.
Число грудных сегментов непостоянно,
строение половой, нервной и
кровеносной систем примитивнее, чем у других
ракообразных.
Жаброногие обитают преимущественно
в пресных водах, морские виды
немногочисленны и имеют пресноводное
происхождение.
Все жаброногие ведут свободный
образ жизни, среди них нет ни паразитов,
ни сидяче-прикрепленных видов. Они не
выработали приспособлений к
существованию в подземных водах и на больших
океанических глубинах.
В этот подкласс объединяются два
отряда ныне живущих ракообразных —жаб-
роноги (Anostraca) и листоногие (Phyllo-
poda).
ОТРЯД ЖАБРОНОГИ (ANOSTRACA)
Бурное таяние зимних снегов
закончилось. На полях и в лесах еще остались
лужи, заполненные талыми водами. Эти
лужи недолговечны: через несколько дней
они высохнут. Они существуют
настолько непродолжительное время, что не
успевают заселиться обычными
пресноводными животными. Тем удивительнее
наблюдать в некоторых из них сравнительно-
крупных (около 1 см длиной), изящных,
полупрозрачных рачков, плавно
двигающихся в воде брюшной стороной вверх.
Это — жаброноги, принадлежащие к
одной из наиболее примитивных групп
современных ракообразных.
Жаброноги обитают не только во
временных весенних водоемах. Условия их
существования весьма разнообразны. Среди
них есть виды, живущие круглый год
в мелких холодных озерах Арктики,
виды, населяющие слабосолоноватые
водоемы степей и полупустынь, есть виды,
приуроченные только к высокогорным
водоемам, и, наконец, один вид
замечателен тем, что в огромном количестве
развивается в пересоленных, даже в
самосадочных озерах; он выдерживает более вы-
380
сокую концентрацию солей, чем все
остальные многоклеточные животные.
Всего известно около 180 видов жабро-
ногов.
Казалось бы, что при такой
способности приспосабливаться к самым
различным условиям среды, жить при
высокой и низкой температуре и солености,
жаброноги должны быть очень обычными
и широко распространенными животными.
В действительности это далеко не так. Они
встречаются сравнительно редко и в очень
немногих водое::ах.
В чем же дело? По-видимому,
основным условием возможности их
существования надо считать отсутствие их врагов
и конкурентов. Как правило, в водоемах,
населенных жаброногами, не бывает ни
рыб, ни других сильных хищников. В
процессе эволюции жаброноги выработали
тонкие приспособления к жизни в крайних,
неблагоприятных для других животных
условиях и только таким образом смогли
выжить в жестокой борьбе за
существование, избегая столкновений с более
высокоорганизованными животными.
Тело жаброногов четко разделяется на
голову, грудь и брюшной отдел. На голове
можно заметить шов, отделяющий
переднюю ее часть с антеннулами,
антеннами и глазами от задней, несущей жвалы
и челюсти. Передние антенны, или антен-
нулы, обычно довольно длинные, но не
разделены на членики, задние гораздо
более мощные, особенно у самцов.
Самцы удерживают своими задними
антеннами самок во время спаривания. У
самцов некоторых жаброногов антенны
снабжены многочисленными выростами и
имеют весьма причудливую форму. По
бокам головы располагается пара сидящих
на стебельках сложных фасеточных глаз,
а на лбу находится один непарный, так
называемый науплиальный глаз.
Жвалы мощные, но лишены щупиков, а
челюсти развиты сравнительно слабо.
Перед жвалами находится имеющая форму
козырька складка кожи — верхняя губа,
задерживающая направляемый вперед
грудными конечностями ток воды.
У большинства жаброногов грудь
состоит из 11 и только у немногих из 17—
19 сегментов. Каждый сегмент несет пару
листовидных ножек. Все они устроены
одинаково, и каждая имеет с наружной
стороны 2—3 лопасти, служащие для
дыхания, и одну лопасть, играющую роль
весла, а с внутренней стороны 6 лопастей,
служащих для направления тока воды
вперед к ротовому отверстию (рис. 190,2).
Между левыми и
правыми ногами вдоль брюшной
стороны тела проходит
глубокая бороздка, по
которой ток воды и
устремляется ко рту.
Грудные ножки
действуют последовательно.
Каждая из них начинает
изгибаться назад только
после того, как это
проделала предыдущая.
Двигаясь назад, ножки
отталкивают тело рачка от
воды, а загибаясь вниз,
они возбуждают ток воды,
направляющийся между
ногами к голове (рис.
Рис. 190. Жаброноги:
1 — общий вид Branchipus stag-
nalis сбоку; 2 — грудная ножка;
3 — движения грудных ножек
(стрелками обозначены токи воды).
Рис. 191. Размножение
жаброногов:
1 — спаривание Branchipus;
2 — яйцо Chirocephalus; 3—
науплиус Branchinecta.
190, 5). Таким образом, каждая
ножка выполняет одновременно три
функции: служит для передвижения рачка,
направляет своими внутренними
лопастями ток воды с пищевыми частицами ко
рту и своими жаберными лопастями
усваивает растворенный в воде кислород.
За грудью следует длинное
цилиндрическое брюшко, в состав которого входят
обычно 8 сегментов и концевая пластинка—
тельсон. Первые два брюшных
сегмента частично сливаются между собой. Здесь
у самцов находятся два направленных
вперед трубковидных выроста, играющих
роль мужского копулятивного аппарата,
а у самок к передним брюшным
сегментам прикрепляется яйцевой мешок. С тель-
соном сочленены ветви фурки, или
вилочки, снабженные многочисленными
щетинками.
Пищей жаброногам служат
микроскопические водоросли и мелкие частицы
разлагающейся растительности и другие
органические частицы. Обитая всегда
только в стоячих водоемах, жаброноги
вынуждены весьма интенсивно фильтровать
воду, пропуская ее через щетинки
внутренних лопастей своих грудных ног и
челюстей, с тем чтобы добыть себе
достаточное количество пищи. Поэтому каждая
грудная ножка совершает от 140 до 400
взмахов в минуту в зависимости от
температуры. Собирающаяся перед верхней
губой пища перетирается жвалами и
только после этого поступает в рот.
Железы верхней губы выделяют вещество,
слепляющее мелкие пищевые частицы в
пищевой комок.
Когда наступает время спаривания,
самец подплывает под самку и хватает ее
задними антеннами. Удерживая ее этими
придатками, он изгибает свое тело так,
чтобы его трубковидные отростки
соприкасались с половым отверстием самки
(рис.191 ,2). Сперма попадает в половые
пути самки, где происходит
оплодотворение яиц. Весь этот процесс продолжается
несколько минут, после чего рачки
расходятся. Яйца еще в половых путях самки
окружаются желтком и покрываются очень
плотной оболочкой, снабженной у
некоторых видов шипами или бугорками (рис.
191,2). Яйца выметываются самкой через
2—3 суток после спаривания и попадают
в яйцевой мешок, но затем выбрасываются
в воду, опускаются на дно и там
развиваются.
Эти яйца отличаются необыкновенной
стойкостью. Они переносят полное
высыхание, резкие колебания температуры (в
условиях опыта нагревание до 80°С). Они
сохраняют жизнеспособность в течение 3—
4 лет. Если водоем, в котором жили
жаброноги, высох, яйца рачков могут
остаться на месте в высохшем иле до
следующей весны, когда это место снова
заполнится водой. Вместе с пылью яйца могут
переноситься ветром из водоема в водоем,
что служит основным способом
распространения жаброногов. Некоторые
исследователи считали, что высыхание и
промерзание необходимы для нормального
развития жаброногов. Однако
специально поставленные опыты не подтвердили
этого предположения. В аквариумах,
постоянно содержащих воду, отложенные жаб-
роногами яйца развивались на
следующий год после откладки вполне нормально.
Яйца испытывают полное дробление, и
из них выходит первая личинка —
науплиус (рис. 191,5). В течение 2—3 недель
происходит 7—8 линек личинки. С каж-
382
дой линькой увеличивается количество
сегментов тела рачка и относящихся к
ним конечностей. Продолжительность
жизни взрослых рачков зависит от
температуры. Так, например, Chirocephalus gru-
bei при 5,2° С живет 90 дней, при 7,9°С—
78 дней, а при 13,5°С — всего 5 дней.
По характеру размножения
резко отличается от остальных жаброногов
обитатель соленых озер артемия (Arte-
mia salina). Во-первых, нередко мы
находим в водоемах только самок артемий,
которые откладывают яйца, способные
развиваться без оплодотворения, т. е.
партеногенетически. Наряду с этим столь
же обычно присутствие самцов,
спаривание их с самками и развитие
оплодотворенных яиц, так же как у других
видов. Во-вторых, в одних случаях самки
артемий откладывают яйца подобно
прочим жаброногам, а в других — рождают
вышедших из яиц в яйцевых мешках
науплиусов. Способность выносить очень
высокую концентрацию солей,
изменчивость, связанная с жизнью в различных
по солености водоемах, полиплоидность
и важное практическое значение
заставляют несколько подробнее остановиться
на биологии этого рачка.
Артемия распространена очень широко,
но приурочена к водоемам определенного
типа и к определенным климатическим
условиям. Она обитает в пределах
степей и полупустынь Европы, Азии и
Америки, в морских осолоненных лиманах
и в соленых материковых озерах. Обычно
она живет при солености от 40 до 230°/00,
но может встречаться и при более низкой
(вплоть до полного опреснения), и при
более высокой солености. Наиболее
благоприятно для артемий преобладание в
растворе ионов натрия, но необходимо
присутствие и других солей.
Проф. Н. С. Гаевская содержала
артемий в растворах химически чистых
хлористого натрия, хлористого магния,
сернокислого магния и некоторых
других солей, причем оказалось, что в таких
условиях рачки погибают через 16—44
часа, а в смеси этих солей живут гораздо
дольше. В данном случае ядовитое
действие какого-нибудь одного иона
нейтрализуется действием его иона-антагониста.
В пресной воде большинство особей
артемий погибает через 2—3 суток. Перед
этим они прекращают размножаться.
Артемии, живущие при разной
солености, заметно отличаются друг от друга
по строению заднего конца тела. У рачковf
существующих при очень высокой
солености, ветви фурки совсем не выражены
или если они есть, то очень маленькие,
не отчлененные от тельсона и
вооруженные всего 1—3 щетинками. Рачки из
водоемов несколько меньшей солености имеют
более сильно развитую и богатую
щетинками фурку, и только у рачков из
сравнительно слабо осолоненных водоемов фур-
ка отчленена от тельсона и вооружена
многочисленными щетинками (рис. 192).
Кроме того, параллельно уменьшению
солености изменяется соотношение длины
грудного и брюшного отделов тела и
степень развития задних антенн.
Раньше все эти формы считались
особыми видами. Однако еще в 1875 и 1877 годах
одесский биолог В. Шманкевич,
воспитывая артемий при различной
солености, показал, что именно соленость
определяет эти морфологические
особенности рачков. Он считал, что при
уменьшении солености артемия приближается по
своему строению к пресноводному роду
Branchipus и даже переходит в него. В
действительности такого перехода,
конечно, не происходит. Просто артемий при
Рис. 192. Строение Artemia salina,
живущей при разной солености воды.
383
различной солевой концентрации
достигают половозрелости на разных стадиях
развития — при более высокой солености
на более ранних стадиях, чем при
пониженной. Формы артемии, развивающиеся
при пониженной солености, приобретают
лишь поверхностное сходство с Branchi-
pus.
Существуют ли в природе водоемы
настолько высокой солености, что артемии
не могут в них жить?
Некоторые наблюдения показывают, что,
возможно, верхний солевой предел
жизни артемии лежит около 300°/00.
Прежние исследователи Каспия, посещавшие
огромный осолоненный залив этого моря
Кара-Богаз-Гол, где в результате очень
высокой концентрации солей идет
выпадение мирабилита, были поражены
обилием здесь артемии. Из-за массы рачков,
кишевших в воде, она была красноватого
цвета. После падения уровня Каспия
обмелел пролив, соединяющий море с Кара-
Богаз-Голом, и соленость залива еще
повысилась. Живые рачки исчезли. Теперь
на поверхности залива плавает масса яиц
артемии, которые при такой высокой
солености развиваться не могут.
Для выяснения особенностей
физиологии артемии, позволяющих рачку
переносить изменения солености от очень
низких ее показателей до величины около
300°/00, была изучена соленость крови
артемии при помещении рачков в воду
различной солевой концентрации.
Обнаружилась удивительная картина: оказалось,
что артемия сохраняет постоянную
соленость крови независимо от колебаний
солености во внешней среде. Эта
неизменная концентрация солей в крови артемии
такая же, как у других жаброногов,
живущих в обычной пресной воде. Такое
совпадение не случайно. Оно свидетельствует
о пресноводном происхождении артемии
и о вторичном характере ее
приспособления к существованию в осолоненных
водоемах.
Независимость солености крови
артемии от солености внешней среды связана
со способностью рачка выводить из
организма поступающие в него соли, причем
в разном количестве при жизни в разных
концентрациях солей. Благодаря этому
концентрация солей в крови артемии
почти во всех населенных ею водоемах
оказывается ниже, чем в окружающей среде.
Таких совершенных способностей к
регуляции солености крови мы не знаем
у других беспозвоночных.
Основной пищей артемии служит
несколько видов микроскопических
водорослей, так же, как она,
приспособленных к жизни в соленых водоемах, и,
кроме того, бактерии грунта. В Сиваше,
где артемии очень много, они поедают
диатомовые (Amphora, Navicula) и зеленые
(Dunaliella) водоросли. Если водорослей
в воде недостаточно, рачки опускаются на
дно, взмучивают придонную воду и
отфильтровывают бактерий.
Есть указания на существование в
некоторых соленых озерах в большом
количестве только артемии и зеленой
водоросли дуналиеллы. Рачки питаются ду-
налиеллой, а она, в свою очередь,
усваивает некоторые продукты
жизнедеятельности рачка, в частности соединения азота.
Таким образом, в водоеме
устанавливаются необыкновенно простые
взаимоотношения между двумя организмами,
составляющими единую пищевую цепь.
Дыхание артемии нередко бывает
затруднено тем, что при высокой
концентрации солей и малочисленности зеленых
водорослей содержание кислорода в воде
падает. Но в таких случаях артемия
вырабатывает гемоглобин, растворенный
в ее крови и связывающий значительное
количество кислорода.
Период размножения артемии весьма
продолжителен и определяется
подходящей для этого температурой. Так,
например, в Сиваше артемия размножается с
мая до сентября — октября. При этом
иногда она откладывает «зимние яйца»,
способные к развитию только на
следующий год, а иногда рождает живых науп-
лиусов, развивающихся без перерыва во
взрослого рачка. Обычно рождение науп-
лиусов происходит весной и летом, а
откладка яиц осенью, поскольку такие яйца
хорошо переносят неблагоприятные
зимние условия, однако оба процесса могут
происходить и одновременно. По
наблюдениям А. Ф. Гунько, при резких
изменениях температуры, солености и пита-
384
ния у артемии преобладает
живорождение, а при постоянстве этих условий —
откладка яиц. Замечательно, что
оплодотворение не влияет на судьбу яйца: как
оплодотворенные, так и неоплодотворен-
ные, развивающиеся партеногенетически,
яйца могут либо превращаться в зимние,
либо сразу давать науплиусов. За один
вымет самка производит до 70 яиц или
науплиусов, а размножаться она может
три раза в сезон, с промежутками от 6
до 28 суток. Через 10—16.суток,
перелиняв за это время 7 раз, личинка
превращается в молодого рачка, а через 20—29
суток после рождения, перелиняв еще 4—5
раз, рачок достигает половозрел ости.
Замечательной особенностью артемии
следует считать существование недавно
открытых у нее полиплоидных рас.
Полиплоидия, довольно обычное явление
среди растений, в мире животных
встречается очень редко. В Италии обнаружены
ди-, три-, тетра-, пента- и октоплоидные
расы артемии, т. е. расы, состоящие из
рачков с двойным, тройным, четверным,
пятерным и восьмерным набором
хромосом. Среди диплоидных и тетраплоидных
рас есть как способные к обоеполому
размножению, так и состоящие только из
размножающихся партеногенетически самок.
Остальные расы включают
исключительно партеногенетически размножающихся
самок. В отличие от того что
наблюдается у растений, расы артемии не
различаются ни по размерам, ни по внешнему
виду. К сожалению, это явление у
артемии еще недостаточно изучено; в
частности, остается неизвестным, отличаются
ли эти расы артемии по жизнеспособности,
отношению к солености и т. д.
Как уже указывалось, при
благоприятных условиях артемия развивается в
водоемах в массовом количестве. По
подсчетам В. П. Воробьева, в Сиваше
насчитывается в среднем 13,6 г артемии на
1 ле3, что для всего залива составляет
огромную величину — 14,8 тысячи тонн.
Известно, что в отдельных случаях эти
рачки служили пищей человеку. Так,
например, американские индейцы, жившие
на берегах Соленого озера в штате Ута
(США), охотно ели артемию. Арабы,
кочевавшие к западу от дельты Нила,
вылавливали в соленых озерах артемии,
приготавливали из них пасту, напоминавшую,
по словам известного географа Петермана,
соленую селедку, и охотно использовали
эту пасту вместо мяса.
Широкое распространение получила
артемия среди аквариумистов, которые
выводят из яиц рачков, а затем
скармливают их аквариумным рыбкам.
Высушенные яйца артемии пересылаются по почте
из страны в страну и служат
предметом торговли. В последнее время и у
нас начали их заготовку для экспорта
за границу. Эти яйца массами
выбрасываются волнами на берега Сиваша и
других богатых артемией водоемов, и их сбор
не представляет особых трудностей.
Однако наиболее существенно значение
артемии в рыбоводстве. Она представляет
собой первоклассный, высокопитательный
корм для молоди ценных промысловых
рыб, разводимых на рыбоводных
заводах, в первую очередь для осетровых.
Строительство электростанций на
наших реках отрицательно влияет на
запасы проходных рыб, т. е. таких, которые
для нереста мигрируют в реки. Плотины
преграждают рыбам путь на нерестилища.
Для того чтобы сохранить запасы
ценных промысловых проходных рыб, и
прежде всего осетровых, необходимо в
широких масштабах проводить их
искусственное разведение на рыбоводных заводах.
Самым трудным делом при этом оказалось
снабжение вышедших из икры мальков
живым кормом. Молодые рыбки должны
некоторое время выкармливаться в
специальных бассейнах, прежде чем они
достаточно окрепнут, чтобы их можно было
выпустить на волю. Они едят только
живых животных, и артемия по своей
калорийности и химическому составу
оказалась одним из лучших видов корма для
молоди осетровых.
В настоящее время разработаны и
внедрены в производство методы разведения
артемии в заводских масштабах. На Рогож-
кинском и Ачуевском рыбоводных
заводах, в низовьях Дона, сооружены
прямоугольные цементные бассейны размером
11,5x20x1 м каждый. В них налит
рассол соленостью около 40—60°/0о»
изготовленный из добытой на Сиваше соли. По
25 Жизнь животных, т. 2
385
мере испарения воды в бассейны
подливается пресная вода. В качестве корма
для рачков в бассейны периодически
вносятся гидролизные дрожжи. В таких
условиях артемии все летнее время
производят живых науплиусов и развиваются в
таком количестве, что их бывает
достаточно для откорма молоди разводимых
на этих заводах осетровых рыб. Осенью
рачки дают покоящиеся яйца, а весной
из них выходит новое поколение.
По мере необходимости артемий
вылавливают из бассейнов с соленой водой и
переносят в бассейны с пресной водой, в
которых содержится молодь осетровых. В
пресной воде рачки остаются живыми в
течение нескольких часов и за это время
усиленно поедаются осетрятами.
Выкормленная на артемии молодь осетровых
отличается быстрым темпом роста и высокими
биохимическими показателями. Нет
никакого сомнения в том, что разведение артемий
будет организовано и на других
многочисленных рыбоводных заводах нашей страны
и что эти замечательные рачки сыграют
важную роль в трудном деле сохранения
запасов наших ценнейших промысловых
рыб — осетровых.
ОТРЯД ЛИСТОНОГИЕ РАКООБРАЗНЫЕ
(PHYLLOPODA)
Этот отряд отличается от отряда жабро-
ногов присутствием головогрудного щита,
или карапакса, который представляет собой
разросшуюся назад двойную складку
покровов заднего сегмента головы, покрытую
хитином. У части листоногих карапакс
имеет вид настоящего щита,
прикрывающего тело сверху, у других этот щит как бы
перегнулся так, что обе половины его
превратились в створки двустворчатой
раковины. Сложные глаза лишены стебельков
и расположены непосредственно на
поверхности головы. Грудные ножки
листовидные и похожи на ножки жаброногов.
Листоногие обитают главным образом
в пресных водах, морских видов очень
мало. Отряд включает в себя три
подотряда, различающихся строением, образом
жизни и характером местообитания.
ПОДОТРЯД ЩИТНИ (NOTOSTRACA)
Вернемся к весенним лужам, в которых
мы находили изящных полупрозрачных
жаброногов. Если нам повезет, то в
некоторых лужах мы обнаружим гораздо более
крупных (до 5 см длиной)
темно-коричневых рачков, то плавающих в воде,
повернувшись брюшной стороной вверх, то
опускающихся на дно и энергично
взмучивающих поверхностный слой грунта.
Нередко щитни попадаются в водоемах,
казалось бы совсем не подходящих для
жизни водных животных, например в
заполненных грязной водой колеях
проселочных дорог. Местные жители иногда
наивно объясняют внезапное появление этих
крупных рачков тем, что они падают из
туч вместе с дождем.
По своему строению щитни резко
отличаются от жаброногов. Их плоский
овальный щит покрывает голову, грудь и
переднюю часть брюшка рачка. Близ переднего
края щита на его поверхности находится
глазной бугорок, на котором слева и
справа расположено по темному, состоящему
из многих фасеток сидячему (т. е.
лишенному стебелька) глазу. Непосредственно
перед сложными глазами между ними виден
непарный науплиальный глаз, а за ними
так называемый четырехклеточный орган.
Функции этого органа еще не выяснены.
Предполагается, что он служит органом
внутренней секреции. По бокам
головогрудного щита просвечивают крупные
каналы выделительных желез,
открывающихся при основании второй пары
челюстей. Задний край щита имеет вид
полукруглой выемки, оставляющей непокрытой
заднюю часть брюшка. Оно заканчивается
тельсоном, снабженным парой длинных
членистых нитей — фуркой.
Рассматривая щитня с брюшной
стороны, можно разобраться в строении его
сегментов и конечностей. Головогрудной
щит спереди загибается на брюшную
сторону, и здесь с ним сочленяется большая,
почти квадратная верхняя губа. Передние
и задние антенны очень маленькие, зато
жвалы крупные, с многочисленными
зубцами. Позади верхней губы, между
жвалами помещается рот, а позади жвал две
пары маленьких пластинчатых челюстей
386
(максилл). Далее следуют 10
грудных сегментов, каждый из
которых несет по паре ножек. Эти
ножки устроены приблизительно
одинаково и во многом похожи на
рассмотренные ранее ножки
жаброногов. Они, так же как у жаброногов,
имеют с внутренней стороны 6
лопастей, служащих для подачи
пищи ко рту, а с наружной
стороны — одну большую плавательную
лопасть и одну (а не две, как у
жаброногов) дыхательную лопасть,
превращенную в жабру.
Детальные исследования мускулатуры
показали, что полного
соответствия между листовидными
ножками жаброногов и щитней нет, и
возможно, что у тех и других
листовидные ножки развились
независимо, но функционируют они
сходным образом, осуществляя
одновременно дыхание,
передвижение и подачу пищи ко рту.
Первая и в меньшей степени
вторая пара грудных ножек щитня
отличаются тем, что четыре
внутренние их лопасти удлинены,
расчленены и превращены в жгутики,
выступающие наружу за края го-
ловогрудного щита. Эти жгутики
принимают на себя функцию
чувствительных органов и даже
внешне напоминают жгуты антенн
других ракообразных. Такое
видоизменение внутренних лопастей
передних грудных ножек щитней,
несомненно, связано с редукцией
их антенн.
На одиннадцатом сегменте
туловища щитней открывается
парное половое отверстие, и этот сегмент
считается последним грудным. У самок
его конечности имеют весьма
своеобразное строение: большая наружная
лопасть, служащая на других ножках для
плавания, на одиннадцатой паре ног
превращена в круглую яйцевую капсулу,
в которой откладываются и
вынашиваются яйца. У самцов ножки
одиннадцатой пары не отличаются от остальных.
В строении щитня наблюдается еще одно
удивительное явление. На каждом сег-
Рис. 193. Щитень (Tri-
ops cancriformis):
l, 2 — вид со спинной и
с брюшной стороны; з —
грудная ножка; 4 —
спаривание щитней.
менте, начиная с тринадцатого, имеется
не одна, а от 4 до 6 пар ног. Поэтому общее
число пар ног рачка нередко достигает 70.
Таким огромным количеством ног не
обладает ни одно другое ракообразное.
Размеры ножек постепенно уменьшаются
по направлению спереди назад, а задние
сегменты тела совсем лишены ножек.
Существенным отличием ножек щитней
от ножек жаброногов следует считать
присутствие у первых на основной части
передних конечностей направленных внутрь
25*
387
шипов. Это связано со способностью щит-
ней захватывать шипами сравнительно
крупные пищевые частицы и затем
передавать их от ноги к ноге вперед, ко рту.
Фильтровать мелкие взвешенные в воде
частицы, как это делают жаброноги,
щитни, по-видимому, не могут. Их задние
ножки совершают преимущественно
дыхательные движения. Удается наблюдать,
как во время недолгих остановок рачка
задние его ножки продолжают действовать,
а передние остаются неподвижными. При
плавании щитня все его многочисленные
ножки последовательно сгибаются и
разгибаются так, как будто по ним пробегает
волна. Шведский зоолог Лундблад
пипеткой вносил около задних ног щитня
немного воды, подкрашенной кармином. Эта
вода медленно двигалась вперед по
желобку между задними ножками рачка, но как
только она достигала уровня десятой пары
ног, она начинала струиться вперед, что
говорит о важном значении передних
ножек в процессе подачи пищи ко рту.
При плавании щитней важную роль
играет зрение. Освещение дна аквариума
в темной комнате заставляет рачков
немедленно повертываться брюшной
стороной вверх: при таком положении
расположенные на спинной стороне тела глаза
воспринимают свет. Тот же эффект
получается даже после того, как сложные глаза
покрывают непрозрачным лаком и рачок
лежит брюшной стороной на дне
аквариума. По-видимому, он реагирует на свет
благодаря тому, что глазной бокал науп-
лиального глаза обращен вниз и
продолжается в прозрачный соединительнотканный
канал, который заканчивается на брюшной
стороне тела, перед верхней губой,
лишенным пигмента «окошком». Благодаря этому
щитень может одновременно воспринимать
свет, направленный как сверху, так и снизу.
Поиски пищи осуществляются не с
помощью зрения, а с помощью
химического чувства, органами которого
служат жгутовидные внутренние отростки
первой пары грудных ног. Щитень легко
находит в аквариуме и поедает куски
тела земляного червя, но, если к
такому куску добавлен хинин, рачок
чувствует добавку жгутами и отказывается от
невкусной пищи.
В естественных условиях щитни
питаются частицами грунта, растениями и
мелкими животными. Опускаясь на дно, они
взмучивают грунт и при помощи
внутренних лопастей передних ног гонят воду
с частицами грунта по срединному
желобку вперед ко рту. В пищу используются
сравнительно крупные частицы, а мелкая
взвесь обратным током воды направляется
назад. Наряду с этим щитни нападают на
жаброногов, которых истребляют
беспощадно, на хирономид и других мелких
насекомых, не имеющих твердых
хитиновых покровов, и даже на головастиков
лягушек и на мальков рыб. Наконец, они
могут обгрызать нежные части растений.
С этим связаны сравнительно
немногочисленные случаи нанесения щитнями
вреда хозяйству человека. На рыбоводных
заводах на Дону после заливания
предназначенных для выращивания мальков
осетровых бассейнов в них появлялись
массы щитней, которые набрасывались на
рыбок и поедали их. Есть также указания
на то, что щитни размножались на
залитых водой рисовых полях и объедали
побеги риса.
В наших широтах, а также на севере
щитни размножаются преимущественно
при помощи неоплодотворенных яиц.
Население наших прудов и луж состоит
почти исключительно из самок щитней. На
1000 самок приходится не более 10
самцов, а иногда их не бывает совсем. По
направлению с севера на юг относительное
количество самцов увеличивается, и в
тропиках самцы по числу особей
превосходят самок.
Спаривание щитней из-за редкости
самцов удалось наблюдать только дважды
(рис. 193, 4). Самец сверху прикреплялся
к самке и резко сгибал свое тело так, что
значительная его часть оказывалась под
телом самки. Своей одиннадцатой парой
ног он удерживал одиннадцатую пару
ног самки, и, вероятно, в этот момент его
сперма попадала в половые пути самки.
Как оплодотворенные, так и неоплодот-
воренные яйца откладываются самкой в
уже упоминавшуюся яйцевую камеру,
образованную наружными лопастями
одиннадцатой пары ног. Там яйца
находятся короткий срок, а затем выпадают
388
на дно водоема. Интересно, что никаких
отличий в строении, жизнеспособности
и развитии оплодотворенных и неопло-
дотворенных яиц заметить не удается.
Эти мелкие яйца снабжены очень
прочной оболочкой. Они переносят высыхание,
промерзание и сохраняют
жизнеспособность в течение 7—9 лет, в сухом
состоянии выдерживают повышение
температуры до 80° С. Без всякого вреда для себя
они проходят через кишечник лягушки.
Они легко переносятся ветром, причем,
по-видимому, даже на очень далекие
расстояния. Благодаря таким качествам
своих яиц щитни широко распространены
по всему свету. Внезапное их появление
в каком-нибудь мелком водоеме
объясняется попаданием в него их покоящихся
яиц, а вовсе не выпадением взрослых
рачков из дождевых туч, как думают жители
некоторых районов.
В случае попадания яйца в подходящие
условия на следующий год из него
выходит личинка. У широко
распространенного щитня Triops cancriformis эта личинка—
типичный науплиус, с тремя парами
конечностей и без всяких следов
сегментации. У других видов щитней личинка
выходит из яйца на более поздней
стадии — метанауплиуса,
характеризующегося сегментацией заднего конца тела.
Для достижения половозрелости Triops
cancriformis должен слинять около 40 раз,
причем эти линьки быстро следуют одна
за другой и при летней температуре все
развитие от науплиуса до взрослого рачка
заканчивается в течение двух недель.
Весенний щитпень (Lepidurus apus) после
17 линек достигает в длину 12 мм,
причем после 14-й линьки у него появляются
половые различия.
Несмотря на очень широкое
распространение в стоячих пресных водоемах всего
земного шара от приполярных озер до
жарких тропических луж, щитни
представлены чрезвычайно малым количеством
видов. Современные зоологи, учитывая
изменчивость этих ракообразных,
различают всего 9 видов щитней — 4 вида рода
Triops и 5 видов рода Lepidurus.
Некоторые из них, как, например, Triops
cancriformis и Lepidurus apus, встречаются почти
повсеместно, другие, как, например,
Lepidurus arcticus, приурочены только к
водоемам Крайнего Севера. Есть виды,
обитающие только в Африке, и виды,
живущие только в Австралии.
Чрезвычайно интересной особенностью
щитней следует считать их
необыкновенную геологическую долговечность.
Современный вид Triops cancriformis был
обнаружен в ископаемом состоянии в
триасовых отложениях Германии. Детальный
анализ многочисленных отпечатков
хорошо сохранившихся рачков и их
конечностей показал, что триасовый щитень
вполне сходен с современными. При этом
он жил в водоемах такого же типа, какие
и сейчас населены щитнями. Современные
африканские виды щитней также были
найдены в ископаемом состоянии, но в
юрских и в более молодых отложениях.
Мы не знаем других подобных случаев
существования современных видов в столь
отдаленные от нас геологические периоды.
Щитней можно с полным правом называть
«живыми ископаемыми».
Как можно представить себе
неизменность вида на протяжении 200 миллионов
лет, отделяющих нас от триасового
периода? Английский ученый Лонгхерст
предполагает, что это объясняется главным
образом постоянным разносом
покоящихся яиц щитней на огромные пространства,
благодаря чему не создается изоляции
отдельных поселений рачков. Однако
следует учесть и другие не менее важные
биологические особенности щитней, ставящие
их в особое положение в борьбе за
существование. В период их активной жизни в
населяемых ими водоемах нет ни их врагов,
ни их конкурентов. К тому же
естественный отбор действует на активных рачков
всего в течение каких-нибудь двух недель
на протяжении целого года. Все остальное
время рачки пребывают в стадии
покоящихся яиц. Совершенно очевидно, что
существование видов щитней зависит в
первую очередь от стойкости таких яиц.
Если допустить, что их яйца приобрели
свои замечательные свойства еще за
несколько миллионов лет до наших дней,
а рачки в активном состоянии не
подвергаются естественному отбору, то становится
понятной поразительная консервативность
видов этих удивительных ракообразных.
389
ПОДОТРЯД КОНХОСТРАКИ (CONCHOSTRACA)
В мелких, иногда пересыхающих
водоемах как средней, так и северной части
нашей страны, а нередко и высоко в горах
обитают рачки, менее заметные и не
достигающие столь крупных размеров, какщит-
ни. Они обычно окрашены в буроватые,
розоватые или зеленоватые тона. Все тело
их покрыто двустворчатой хитиновой
раковинкой, которая у большинства видов
имеет концентрические полосы прироста,
несколько напоминающие кольца на
раковинах двустворчатых моллюсков,
например беззубки и перловицы. Эти полосы
у рачков возникают после каждой линьки,
поскольку при линьке наружная часть
старой раковины сохраняется, а затем
раковина начинает расти по периферии.
Число полос соответствует числу линек.
Обе створки скреплены одним
поперечным сильным мускулом-замыкателем.
Рачки реагируют на опасность, сокращая
этот мускул, причем створки раковины
плотно сжимаются и рачок, как мертвый,
остается неподвижно лежать на дне.
Раскроем створки раковины, чтобы
рассмотреть строение тела рачка. Голова
многих видов уплощена сверху вниз. Сидячие
фасеточные глаза сближены и
расположены на лобной ее части, а у одного рода
(Cyclestheria) даже соединяются между
собой. Перед ними находится простой
науплиальный глазок. Передние антенны
развиты слабо и не расчленены. Зато
задние антенны длинные, двуветвистые,
причем обе ветви снабжены многочисленными
щетинками. Рачки плавают
преимущественно за счет взмахов задних антенн,
действующих как весла. Лишь у очень обычного
рода Lynceus, способного широко
раскрывать створки своей шаровидной раковины,
не только задние антенны, но и грудные
ножки совершают веслообразные движения.
Число сегментов, составляющих
грудной отдел, колеблется от 10 до 32, и
соответственно этому изменяется число пар
грудных ножек. Они имеют
приблизительно такое же строение, как у щитней, и их
размеры также постепенно уменьшаются
по направлению спереди назад. У самцов
часть внутренних лопастей первой, а
иногда и второй пары грудных ног
преобразована в крючковидные придатки, которыми
самец удерживает самку при спаривании
(рис. 194, Б). У самок наружные лопасти
нескольких средних ног превращены в
длинные, загнутые вверх нитевидные
отростки, приспособленные для удерживания
яиц на спинной стороне тела под
раковиной. Однако основные функции грудных
ног — это подача пищи вперед ко рту и
дыхание. Жабрами служат
ближайшие к основанию наружные
лопасти всех грудных ног. Для
обеспечения этих функций
грудные ножки все время находятся
в движении, благодаря чему вода
между створками раковины
беспрерывно обновляется. Тело
заканчивается крупным хитинизи-
рованным тельсоном, который
или сжат с боков и в таком
случае снабжен парой крупных ког-
Рис. 194. Строение и биология Con-
chostraca:
А — строение самки Limnadia; Б —
копуляция Cyzicus; В — яйца разных
видов; Г — метанауплиус Lynceus; 1 —
передняя антенна; 2 — задняя антенна;
3— фасеточный глаз; 4 —
мускул-замыкатель раковины; 5 — яичник; в —
нитевидные отростки грудных ножек.
390
тей, либо сплющен в спинно-брюшном
направлении и несет короткие коготки.
Большинство Conchostraca — донные
животные. Наиболее крупные из них —
Limnadia, достигающие в длину 17 лш,
лежат боком на дне, Cyzicus роется в
грунте, взмучивая ил движением брюшка и
отчасти ножек. Представители этих и
других родов плавают лишь в период
спаривания. Только сравнительно мелкий D—
5 мм) Lynceus brachyurus, отличающийся
шаровидной раковиной без полос
прироста, обычно плавает спиной кверху, раскрыв
створки раковины, и лишь изредка
опускается на дно. Он питается главным
образом планктонными организмами, в то
время как Cyzicus ест грунт с находящимися
в нем организмами, a Limnadia объедает
микроскопические водоросли,
находящиеся на дне и на поверхности водных
растений. Крупные несъедобные частицы
Limnadia склеивает специальным секретом,
выделяемым железами ножек, и
выбрасывает через заднюю щель своей раковины.
Размножение Conchostraca происходит
по-разному. Все представители семейства
Limnadiidae размножаются только парте-
ногенетически, самцы среди них
неизвестны. У других семейств самцы встречаются
регулярно, причем, чем южнее обитают
рачки, тем больше относительная
численность самцов. При спаривании самец
удерживает самку крючковидными отростками
своих задних антенн и просовывает задний
конец тела в заднюю щель между
створками раковины самки. Яйца
откладываются в пространство между спинной
стороной тела самки и' ее раковиной. У Cycles-
theria часть задних грудных сегментов
расширена таким образом, что перед ними
образуется настоящая выводковая сумка.
Проносив некоторое время яйца на спине
под раковиной, самка линяет, и яйца
выпадают на дно водоема.
Эти яйца окружены плотной и прочной
оболочкой и способны переносить
высыхание, замерзание и другие
неблагоприятные условия. Нередко они имеют
причудливую форму, у многих видов они
снабжены выростами, способствующими их
переносу ветром и различными животными
(рис. 194, В). Яйца некоторых видов
сохраняют жизнеспособность в течение 7 лет.
После того как яйцо попадет в
благоприятные для его развития условия, из него
выходит личинка, находящаяся на стадии
науплиуса. Науплиус Conchostraca
отличается очень маленькими передними
антеннами. На следующей стадии — метанауп-
лиуса — у личинки развивается зачаток
раковины. У большинства Conchostraca
эта зачаточная раковина уже
двустворчатая и очень напоминает раковину ветвисто-
усых ракообразных (см. ниже). У Lynceus
она имеет вид плоского щита, похожего
на карапакс щитней (рис. 194, Г), и только
на более поздних стадиях развития она
перегибается пополам и становится
двустворчатой. Все развитие происходит очень
быстро. Так, например, Cyzicus достигает
половозрелости через 19 дней после
выхода личинки из яйца.
Современные Conchostraca но имеют
никакого практического значения. Однако
вымершие виды этих ракообразных
привлекают к себе внимание геологов. В
пресноводных отложениях, вообще очень
бедных ископаемыми, хорошо
сохраняющиеся раковинки Conchostraca встречаются
довольно часто. По ним удаемся
определять геологический возраст
соответствующих слоев.
В предыдущие геологические периоды
Conchostraca были, по-видимому, более
многочисленны и шире распространены,
чем ныне. Вымерших видов известно около
300, а современных вдвое меньше. Наиболее
древние Conchostraca найдены в девонских
отложениях. Замечательно, что среди
древних вымерших видов, особенно триасовых,
многие жили в море, где сейчас
Conchostraca полностью отсутствуют.
ПОДОТРЯД ВЕТВИСТОУСЫЕ РАКООБРАЗНЫЕ
(CLADOCERA)
Почти в каждом пресноводном водоеме,
начиная с огромных глубоких озер и
кончая мелкими прудками, любой лов сачком
или специальной сеткой из мельничного
шелкового газа приносит десятки и сотни
мелких рачков, часто называемых
водяными блохами (табл. 30). Они оживленно
снуют в толще воды, делая характерные
взмахи своими задними антеннами, а некоторые
391
из них передвигаются по дну или по
подводным растениям. Их хорошо знают не
только ученые-зоологи, но и все любители
аквариумов, которые кормят ими,
живыми или сушеными, своих рыбок. Их
знают и рыбоводы, занимающиеся их
разведением для кормления молоди рыб,
выращиваемой на рыбоводных заводах. Многие
из этих рачков всю жизнь проводят в
толще воды и составляют значительную часть
пресноводного планктона. Иногда они
развиваются в таком количестве, что вода
приобретает буроватый оттенок. Наряду
с планктонными есть и донные виды. Есть
и такие, которые то прикрепляются к
растениям или опускаются на дно, то
всплывают в толщу воды. Подавляющее
большинство видов ветвистоусых обитает в
пресных водах, но есть и несколько мор-
Рис. 195. Строение дафнии:
1 — передняя антенна; 2 — задняя антенна; 3 —
науплиальный глазок; 4 — фасеточный глав; 5 —
кишечник; 6 — сердце; 7 — яичник; 8 —
эмбрионы в выводковой сумке; 9 — брюшко; 10 —
грудная ножка.
ских видов. Размеры этих рачков
незначительны, обычно меньше 1 мм, редко до
5 мм и в исключительных случаях до
10 мм. Поэтому для изучения их
строения приходится пользоваться
микроскопом.
Тело большинства ветвистоусых
ракообразных, например дафнии (рис. 195),
заключено в хитиновую полупрозрачную
двустворчатую раковинку, а голова
вытянута в направленный вниз клюв, или рост-
рум. На лобной части головы расположен
непарный фасеточный глаз, возникший в
результате слияния двух боковых глаз,
а перед ним — простой науплиальный
глазок, отсутствующий у некоторых видов.
Число фасеток, образующих сложный
глаз, различно у разных видов и зависит
от способа их питания. В глазу
отфильтровывающей мелкие частицы дафнии
насчитывается всего 22 фасетки, а у хищных
Evadne и Leptodora соответственно 80 и
300. При нападении на мелких животных
глаза хищных ветвистоусых имеют важное
значение. Под рострумом прикрепляются
палочковидные обыкновенно
относительно очень маленькие передние антенны.
Зато задние антенны непропорционально
велики по сравнению с телом. Они дву-
ветвисты, причем каждая ветвь снабжена
длинными перистыми щетинками. Задние
антенны служат основным органом
передвижения ветвистоусых. Одновременно
взмахивая обеими задними антеннами,
рачки отталкиваются ими от воды и таким
образом плывут короткими скачками. На
нижней поверхности головы находится
рот, окруженный спереди верхней губой,
с боков жвалами и сзади челюстями.
Грудной отдел ветвистоусых укорочен
и, судя по количеству пар ножек, состоит
из 4—6 сегментов. Он, как уже
указывалось, помещается внутри двустворчатой
раковины, но у самок между спинной
поверхностью тела и спинным краем
раковины имеется обширная полость,
выполняющая роль выводковой сумки. В эту
сумку откладываются яйца, и при
благоприятных условиях там же они
развиваются. У хищных ветвистоусых раковина
уменьшена в размерах, оставляет
непокрытым почти все туловище и прикрепляется
к его спинной стороне (табл. 30, 2, 5, б, 8У
392
14, 18); в таких случаях она служит
только для вынашивания яиц.
Строение грудных ножек различно у
разных видов и зависит от характера их
питания. Эти ножки в общем напоминают
листовидные ножки жаброногов, щитней
и Conchostraca. У большинства ветвисто-
усых ракообразных грудные ножки
служат для отфильтровывания мелких
взвешенных в воде частиц, которыми рачки
питаются. В соответствии с этим лопасти
грудных ножек снабжены
многочисленными перистыми щетинками,
образующими вместе фильтрационный аппарат.
Ножки совершают до 300—500 взмахов в 1
минуту.
В простейшем случае, например у Sida
(рис. 196,2), вода входит между створками
в передней части раковины. При
одновременном движении всех 6 пар ножек вперед
между ними возникает как бы ряд
всасывательных камер, куда просачивается
вода. При этом щетинки отфильтровывают
всю мелкую взвесь. Движение ножек назад
сжимает камеры и выгоняет воду через
заднюю часть раковины.
Отфильтрованные частицы направляются при помощи
специальных щетинок в продольный
желоб, расположенный на нижней стороне
туловища между основаниями грудных
ножек. По этому желобу находящимися
на его стенках щетинками пищевой комок
проталкивается вперед, по направлению
ко рту.
У Daphnia этот аппарат усложнен и
усовершенствован (рис. 196, 2). Из пяти пар
ее грудных ножек фильтруют только
третья и четвертая. При движении первой
пары ножек вперед вода поступает в
фильтрующую камеру. В это время третья и
четвертая пары изгибаются вперед,
обеспечивая дальнейший ток воды.
Отфильтрованные пищевые частицы поступают
в брюшной желобок. Пятая пара ног
ограничивает сзади фильтрующую камеру.
Делая взмах назад, она вместе с четвертой
парой вызывает выводящий ток воды.
Таким образом, между грудными ножками
дафнии функции разделены, что говорит
о большей их специализации.
Ближайшие к основанию наружные
лопасти грудных ног фильтрующих ветвисто-
усых ракообразных превращены в жабры.
Рис. 196. Движения ветвистоусых:
1, 2 — токи воды (обозначены стрелками), вызываемые
движениями конечностей Sida A) и Daphnia B); 3,4 —
движения в воде Daphnia C) и Bosmina D).
По системе полостей кровь поступает в
жабры и обогащается в них кислородом.
В плазме крови ветвистоусых растворен
гемоглобин, от которого в значительной
степени зависит окраска рачков. Его
концентрация меняется. Если содержание
кислорода в воде низкое, кровь
приобретает темно-красный цвет, если кислорода
в воде много, кровь окрашена значительно
светлее. При изменении содержания
кислорода в воде меняется окраска рачков,
но для этого требуется около 10 дней.
393
Дафнии с темпо-красной кровью в воде с
низким содержанием кислорода живут
значительно дольше, чем дафнии, имеющие
светлую, бедную гемоглобином кровь.
Очевидно, гемоглобин, как и у
позвоночных животных, связывает кислород.
Кровь приводится в движение
мешковидным сердцем, взбалтывающим ее в
полости тела (рис. 195, 6). Настоящих
кровеносных сосудов нет. Сердце дафнии бьется
очень часто. При температуре 10° С оно
делает 150, а при температуре 20° С даже
500 ударов в минуту.
Совсем иначе устроены и действуют
грудные ножки хищных ветвистоусых
ракообразных (табл. 30). Их боковые
лопасти недоразвиты или отсутствуют, жабр
нет совсем, а хорошо развитые внутренние
ветви снабжены сильными шипами и
шиловидными щетинками. При помощи
грудных ног рачки схватывают других мелких
животных, служащих им пищей. Хищные
ветвистоусые воспринимают кислород
скоплением железистых клеток,
находящихся на голове.
Задний брюшной отдел тела
ветвистоусых укорочен, часто изогнут под прямым
углом и может прятаться между створками
Рис. 197. Прикрепляющиеся ветвистоусые:
1 — Simocephalus vetulus на нити водоросли; 2 —
Chydorus sphaericus на нити водоросли; з — Scaphole-
beris mucronata у поверхности воды.
раковины. Он обычно заканчивается
двумя когтями. Однако у многих хищных
видов задний отдел имеет форму трубки,
сохраняет сегментацию и иногда
продолжается в длинный шип, назначение
которого остается непонятным.
Наблюдая за поведением ветвистоусых
рачков в водоеме или в аквариуме,
нетрудно убедиться в том, насколько различны
повадки разных видов. Особенно
многочисленны рачки, проводящие всю свою
жизнь в толще воды. Планктонные дафнии
и некоторые другие ветвистоусые после
взмаха задними антеннами и вызванного
этим движением скачка некоторое время
неподвижно парят в воде. Благодаря тому
что центр тяжести их тела находится ниже
места прикрепления антенн, в момент
удара антенн тело рачка наклоняется, а
в период покоя снова выпрямляется.
Распростертые антенны препятствуют
погружению рачка (рис. 196, 5).
Планктонные ветвистоусые из рода Bos-
mina плавают иначе. Они беспрерывно
машут своими задними антеннами, причем
передняя часть их тела при каждом ударе
несколько приподнимается. Однако
длинные хоботообразные передние антенны
служат балансиром, возвращающим тело
в горизонтальное положение (рис. 196, 4).
Если босмине отрезать передние антенны,
она начинает беспомощно кувыркаться
и теряет способность к плаванию.
Среди зарослей подводных растений в
прибрежной части озер и прудов часто
попадается крупный (до 4 мм) ветвисто-
усый рачок Sida crystallina, который, как
бы устав беспрерывно плавать, время от
времени прикрепляется к растениям при
помощи специальной присоски,
находящейся на задней части его головы. Другой
обитатель зарослей — Simocephalus —
цепляется зг растения щетинками задних
антенн. Маленький шарообразный хидо-
рус (Chydorus sphaericus), поплавав
некоторое время, садится на водоросли и
начинает ползать по ним, удерживаясь
щетинками края раковины и отталкиваясь
второй парой грудных ног и брюшком.
Наиболее своеобразен способ
передвижения Scapholeberis mucronata, который
благодаря присутствию несмачиваемых
щетинок на нижнем крае раковины может при-
394
крепляться снизу к поверхностной пленке
натяжения воды и быстро скользить по ней
спинной стороной вниз (рис. 197).
Наряду с такими постоянно или
временно плавающими формами существует
большое количество видов, плавающих очень
плохо и постоянно ползающих по дну.
Некоторые из них способны даже
зарываться в поверхностный слой ила.
С жизнью многих ветвистоусых
ракообразных в толще воды связано одно
любопытное явление, получившее название
цикломорфоза. Сущность его
заключается в том, что форма и величина
раковины определенных видов
закономерно изменяются по сезонам года. В
летнее время у некоторых видов дафнии на
голове появляется так называемый «шлем»,
а у босмин на спинной стороне — «горб».
Кроме того, у дафний удлиняется задний
шип на раковине, а у босмин — передние
антенны (рис. 198).
Эти изменения происходят параллельно
изменению температуры воды. В данном
случае важна, однако, не сама
температура, а ее воздействие на плотность воды.
Прогретая вода обладает меньшей
плотностью, чем охлажденная. Для движения
и парения в менее вязкой прогретой воде
необходимы более длинные выросты, а
иногда и иная форма тела. Поэтому летние
формы дафний и босмин развивают
специальные приспособления для
передвижения и парения в таких условиях.
Значение плотности воды для изменений формы
тела рачков подтверждается
наблюдениями Е. Ф. Мануйловой над дафниями
озера Балхаш.
В этом озере соленость по направлению
с востока на запад постепенно
увеличивается и вместе с тем вода становится все
более плотной. Местная планктонная
дафния — Daphnia balchashensis, живущая
при сравнительно высокой солености,
имеет совершенно круглую голову, а
живущая в опресненной части озера — голову,
снабженную загнутым назад шлемом.
Большинство ветвистоусых
ракообразных, как уже указывалось, питается,
отфильтровывая мелкую, находящуюся в
воде взвесь. Основной их пищей служат
бактерии, одноклеточные водоросли и мелкие
органические остатки — детрит. Наиболь-
Рис. 198. Цикломорфоз планктонных ветвисто-
усых:
1 — Daphnia cucullata; 2 — D. cristata; 3 — Bosmina
coregoni (римскими цифрами обозначены месяцы, в
течение которых существует каждая форма).
шее значение имеют бактерии. В опытах
многих исследователей удавалось
воспитывать ветвистоусых при кормлении их
одними бактериями, а американский зоолог
Банта получил в таких условиях около
1600 поколений дафний. В природных
водоемах ветвистоусые прекрасно живут и
размножаются при концентрации
бактерий не менее 1 миллиона в 1 см3 воды.
Если численность бактерий меньше 500
тысяч на 1 см3, наступление половозрело-
сти дафний задерживается; если же
численность бактерий падает ниже 200 тысяч
на 1 см3, жизнь рачков становится
невозможной. С другой стороны, избыточное
количество бактерий (более 3—5
миллионов на 1 см3) также вредно влияет на рач-
395
ков и вызывает прежде всего замедление
или приостановку их размножения, а
иногда даже их гибель. Это объясняется
выделением бактериями продуктов обмена
веществ, ядовитых для ветвистоусых.
От количества бактерий в водоеме
зависит численность ветвистоусых.
Неоднократно наблюдалось уменьшение числа
ветвистоусых в период массового
развития сине-зеленых водорослей. Некоторые
исследователи предполагали вредное
воздействие сине-зеленых на рачков. Однако,
по данным Е. Ф. Мануйловой, бурное
развитие сине-зеленых само по себе
никакого влияния на рачков не оказывает. Оно
подавляет развитие бактерий и именно
таким образом косвенно воздействует на
ветвистоусых. Часто наблюдающиеся периоды
резкого увеличения численности
ветвистоусых обычно следуют за периодами
размножения бактерий.
Помимо бактерий, фильтрующие ветви-
стоусые используют в пищу
одноклеточные водоросли. Они могут заглатывать
только самые мелкие виды протококковых
и зеленых водорослей, которые обычно
развиваются в большом количестве в
прудах, но не в крупных озерах. Обычные,
особенно колониальные, планктонные
водоросли слишком велики для того, чтобы
быть проглоченными рачками. Во многих
случаях водоросли приобретают большое
значение для ветвистоусых не потому, что
поедаются ими, а потому, что после
отмирания вызывают массовое развитие
бактерий — основного корма рачков.
При фильтрации пищи рачки не могут
отсортировывать съедобные частицы от
несъедобных. Они способны только
отбирать частицы по их размеру, отбрасывая
слишком крупные. С этим связаны, по
наблюдениям В. М. Рылова, те
изменения в составе планктона, которые
наблюдаются при проникновении планктона
стоячих водоемов в текучие. Эти
изменения сводятся главным образом к резкому
уменьшению численности ветвистоусых
ракообразных, в то время как численность
других планктонных животных, например
коловраток, существенно не меняется.
Оказывается, что речной поток несет
мелкие минеральные частицы и попавшие в
него планктонные ветвистоусые
отфильтровывают их и заполняют ими свой
кишечник. Рачки идут ко дну и погибают, а
коловратки, захватывающие пищу иным
способом, и притом еще более мелкую,
остаются невредимыми.
Фильтрация пищи рачками происходит
беспрерывно. Время, нужное для
заполнения кишечника, колеблется от 10 до
240 минут и зависит от величины рачков,
размеров пищевых частиц, их
концентрации и температуры. Мелкая босмина в
культуре водоросли Ankistodesmus
наполняет кишечник в течение 10 минут, а
в культуре бактерии Bacillus coli тратит
на это 2—2,5 часа. В течение суток одна
дафния поглощает от 4,8 до 40,8 миллиона
бактерий (в зависимости от их
концентрации). Даже после наполнения кишечника
рачки продолжают фильтровать, и
отфильтрованные частицы двигаются вперед по
их брюшному желобку. Но в рот они не
попадают: ротовые придатки и задний
отдел тела выталкивают их снова в воду.
Рачки начинают заглатывать пищу только
после того, как кишечник хотя бы
частично освободится от ее остатков.
Совсем иначе добывают себе пищу
хищные виды. Самый крупный совершенно
прозрачный ветвистоусый рачок Leptodo-
ra kindtii (табл. 30, 14) неподвижно парит
в воде и своим громадным глазом
высматривает добычу. Достаточно нескольких
сильных взмахов его задних антенн для
того, чтобы он настиг какого-нибудь более
мелкого планктонного рачка или
коловратку, схватил жертву своими далеко
вынесенными вперед грудными ногами и
вонзил в нее острые зазубренные жвалы.
Приблизительно так же охотится и другой
крупный хищник — Bythotrephes (табл.
30,2). Оба рачка сначала размельчают
жертву жвалами, а затем всасывают жидкую
пищу. Они нападают почти на всех
планктонных животных, включая собственную
молодь, но не трогают сидячих и даже
временно прикрепляющихся рачков,
например сиду и симоцефалюса.
Лептодора и битотрефес весьма
прожорливы. При температуре 20—25° С каждая
лептодора съедает за сутки более 50, а
битотрефес около 25 рачков.
Соответствующие расчеты показывают, что в некоторых
водоемах, например в Рыбинском водохра-
396
нилище, хищные ветвистоусые выедают
заметную часть зоопланктона (более 40%
всех планктонных животных). Наносимый
ими таким образом вред не так велик,
поскольку они в свою очередь интенсивно
потребляются многими рыбами.
В некоторых водоемах планктонные
ветвистоусые ракообразные подобно другим
планктонным животным совершают
регулярные суточные вертикальные миграции.
В темное время суток они поднимаются
в поверхностные слои воды, а в светлое
время уходят вниз. Однако во многих
озерах и водохранилищах большая часть
рачков круглые сутки остается близ
поверхности.
До тех пор пока условия жизни ветви-
стоусых рачков благоприятны, в водоемах
встречаются только самки,
откладывающие и вынашивающие в своих выводковых
камерах неоплодотворенные яйца. Число
этих яиц различно не только у разных
видов, но даже у различных особей одного
вида. Разные виды дафний образуют до
50 или даже до 100 яиц на самку, босми-
ны — до 16 яиц, а мелкие донные виды —
не больше 2 яиц. Яйца развиваются в
выводковой камере матери, где из них
образуются сначала подвижные эмбрионы, а
затем вполне сформировавшиеся
маленькие рачки, покидающие выводковую
камеру и приступающие к самостоятельному
существованию. Летом, при достаточном
количестве пищи, рождение молоди и
появление в камере новых яиц происходят
каждые 1—2 дня, благодаря чему
численность ветвистоусых может очень быстро
увеличиваться. Обыкновенно рождение
молоди сопровождается линькой самки.
У хищных ветвистоусых, например у леп-
тодоры и битотрефеса, выводковая камера
изолирована от окружающей среды и при
выходе молоди каждый раз разрушается,
а затем восстанавливается вновь.
Вышедшая на свободу молодь часто
линяет и быстро растет. Первые три линьки
происходят через 1—1,5 суток, а
следующие через 2—3 суток. Большинство
видов достигает половозрелости после
третьей-четвертой линьки, т. е. через 2—
6 дней после рождения. Так получаются
новые поколения самок, продолжающие
размножаться партеногенетически.
Но вот условия существования
ветвистоусых рачков ухудшились вследствие
понижения температуры, уменьшения
количества пищи или загрязнения водоема.
Это ухудшение сейчас же сказывается на
судьбе яиц, находящихся в половых путях
самки. Из неоплодотворенных яиц после
их откладки в выводковую камеру
выходят не самки, а самцы. Кроме того, часть
яиц в половых путях самки подвергается
второму делению созревания, в
результате которого количество хромосом в яйце
уменьшается вдвое. Такие яйца могут
развиваться только после
оплодотворения. При слиянии яйца и сперматозоида
восстанавливается полное количество
хромосом.
Замечательно, что даже
кратковременные отклонения от нормальных условий
жизни могут прервать процесс партено-
генетического размножения. Судьба яйца
определяется за 15 минут до его выхода
из половой системы самки. Если в этот
момент рачки подвергнутся каким-нибудь
неблагоприятным воздействиям, из их
яиц выведутся самцы или эти яйца будут
непременно нуждаться в оплодотворении.
Самцы ветвистоусых ракообразных —
карлики по сравнению с самками. Часто
они в несколько раз меньше самок
(рис. 199). Их передние антенны удлинены,
а первая пара грудных ножек
снабжена коготками, помогающими удерживать
самку.
Обыкновенно самцы появляются в
водоемах осенью, после значительного
понижения температуры. Они прикрепляются
к самкам при помощи коготков передних
ног и при помощи передних антенн,
причем у дафний нередко к одной самке
прикрепляются два самца. Сперма проникает
в выводковую камеру близ отверстий
яйцеводов, а потом и в них.
Оплодотворенные яйца во многих
отношениях отличаются от
неоплодотворенных. Их гораздо меньше (редко больше
двух), но зато они значительно богаче
желтком и поэтому крупнее, чем партено-
генетические. Вокруг них развиваются
защитные оболочки. У Sida, Diaphanoso-
ma, Holopedium и хищных видов
оплодотворенные яйца снабжены двумя
оболочками — плотной внутренней и клейкой
397
наружной. Такие яйца откладываются
в воду и опускаются на дно.
У большинства ветвистоусых
оплодотворенные яйца откладываются в
выводковую камеру, а в образовании их защитных
оболочек принимает участие материнская
раковина. Ее спинной край становится
толще, под раковиной собираются клетки,
плотным слоем окружающие яйцо или
яйца. У дафний поверх этих клеток
выделяется хитиновая оболочка и в выводковой
камере возникает так называемое
седлышко, или эфиппиум, содержащее обычно два
яйца. Формирование эфиппиума
продолжается 2—3 дня, и самка носит его в
течение 1—2 дней. Затем она линяет, и
эфиппиум оказывается на свободе. Благодаря
присутствию воздухоносного слоя эфип-
пиумы дафний плавают на поверхности
воды. У некоторых других ветвистоусых
наружной оболочкой эфиппиума служит
вся раковина или значительная ее часть,
сброшенная при линьке (рис. 199).
Главная биологическая особенность
оплодотворенных яиц заключается в том,
что они могут развиваться только после
более или менее продолжительного
периода покоя и поэтому называются
покоящимися. Такие яйца помогают рачкам
переносить всевозможные неблагоприятные
условия; они вмерзают в лед, высыхают,
сохраняя в течение длительного времени
жизнеспособность. В то же время они
служат средством расселения ветвистоусых,
легко переносясь из водоема в водоем
ветром, вместе с илом, присохшим к лапкам
водоплавающих птиц, и т. д. Благодаря
своим покоящимся яйцам ветвистоусые
широко распространены во всех стоячих
водоемах нашей планеты.
Многие виды зиму проводят в стадии
покоящегося яйца, которое начинает свое
развитие весной и дает начало самке,
приступающей к партеногенетическому
размножению. У всех ветвистоусых, за
исключением лептодоры, из яйца выходит
вполне сформировавшийся рачок. Лепто-
дора — единственный представитель
ветвистоусых, имеющий личинку; она
вылупляется из покоящегося яйца на стадии
метанауплиуса (табл. 30, 17).
Число известных видов ветвистоусых
достигает 380, в СССР найдено 164 вида.
Рис. 199. Размножение ветвистоусых:
1 — спаривание дафний; 2 — поперечный разрез через
яйцо Daphnia magna; 3 — 13 — эфиппиумы разных
ветвистоусых: 3 — Daphnia magna, 4 — D. balchashen-
sis, 5 — Leydigia acanthocercoides, 6 — Oxyurella te-
nuicaudis, 7 — Bosmina longirostris, 8 — Eurycercus
lamellatus, 9 — Drepanothrix dentata, 10 — Ilyocrip-
tus sordidus, 11 — Macrothrix hirsuticornis, 12 —
Alonella excisa, 13— Moina macrocopa.
Некоторые из них распространены очень
широко и встречаются в самых
разнообразных водоемах, другие приурочены к
определенным районам или населяют
водоемы какого-нибудь особого типа. Во
всех странах света обитают обыкновенная
дафния (Daphnia pulex), планктонная
дафния (D. longispina), босмина (Bosmina
398
longirostris), xudopyc (Chydorus sphaeri-
cus) и многие другие. В то же время
существуют виды, живущие только на
Крайнем Севере (Daphnia arctica, Eurycercus
glacialis). Преимущественно в северной
части СССР и Америки встречается
своеобразный Holopedium gibberum, имеющий
поверх раковины студенистую оболочку,
придающую рачку вид прозрачного
шарика (табл. 30, 16). Этот рачок изредка
встречается и в средней части СССР, в водоемах,
связанных с моховыми болотами. Наконец,
есть и тропические виды, в нашей стране
обитающие на залитых водой рисовых
полях Узбекистана, в Закавказье, в водоемах
дельты Волги. Высказывается
предположение о том, что их покоящиеся яйца
переносятся на колоссальные расстояния
воздушными потоками.
Некоторые ветвистоусые рачки
населяют только те водоемы, вода которых имеет
кислую реакцию и бедна минеральными
солями, но большинство видов способно
существовать в самых разнообразных,
в том числе и неблагоприятных условиях,
например в условиях недостаточного
содержания кислорода в воде.
Немногочисленные морские
представители ветвистоусых ракообразных также
распространены очень широко, но
приурочены преимущественно к прибрежным
водам. Только в Каспийском море обитает
несколько чрезвычайно своеобразных
видов (табл. 30, 6, 18), частично присущих
только Каспию, частично встречающихся
также в Азовском море и в опресненных
районах Черного моря. Среди них
особенно причудливы виды рода Cercopagis,
снабженные необыкновенно длинной
хвостовой иглой, в 5—6 раз превосходящей
по длине их тело. Каспийские
ветвистоусые близки к некоторым пресноводным
северным видам (например, Bythotre-
phes); предполагается, что при таянии
великого ледника эти виды были внесены
в Каспийское море и, приспособившись
к новым условиям, дали начало
каспийским эндемикам.
Значение ветвистоусых
ракообразных в жизни стоячих пресноводных
водоемов очень велико. Они служат
излюбленным и весьма питательным кормом для
многих рыб, как взрослых, так и молоди,
и истребляются рыбами в огромном
количестве. В кишечнике рипуса из уральского
озера Увильда оказалось 47 тысяч босмин
и 8,5 тысячи дафний. Чудской сиг, снеток,
ряпушка, молодь судака, амурского
сазана и многих других рыб питаются главным
образом ветвистоусыми ракообразными.
Питательность этого корма очень высока:
содержание белка в теле дафнии достигает
50%, а жира 11 % по весу. Многие морские
рыбы также в большом количестве
поедают ветвистоусых.
Высокие пищевые качества
ветвистоусых ракообразных в полной мере оценены
рыбоводами. На рыбоводных заводах
нашей страны осуществляется Массовое
разведение дафний и моин, которыми кормят
молодь осетровых и лососевых рыб.
Перед рыбоводами, занимающимися
этим трудным делом, стоит задача
обеспечить беспрерывное партеногенетическое
размножение разводимых рачков. Для
этого необходимо создать в бассейнах, в
которых разводятся рачки, постоянные
благоприятные для них условия. В случае
хотя бы временного ухудшения условий
существования рачков их яйца, как было
описано раньше, оплодотворяются и
превращаются в покоящиеся. Подойдя к
бассейну, рыбовод обнаруживает вместо
массы живых копошащихся рачков
плавающие на поверхности воды эфиппиумы,
которых рыбки есть, конечно, не будут.
Дело может кончиться гибелью от голода
сотен и тысяч маленьких осетрят и сев-
рюжат.
Дафний и моин разводят в
цементированных или просто выкопанных в земле
бассейнах, воду которых удобряют
навозом, отваром с жиротопки и
азотобактерином. Эти вещества благоприятствуют
массовому развитию бактерий, используемых
в пищу рачками. Иногда бассейны делают
проточными, что позволяет регулировать
их температурный и газовый режим.
Регулярно следя за состоянием рачков,
удается получать достаточное их количество
для откорма молоди разводимых на
заводах рыб.
399
ПОДКЛАСС ЦЕФАЛОКАРИДЫ (CEPHALOCARIDA)
Несколько лет тому назад американский
зоолог Сандерс занимался изучением
морской фауны близ Океанографического
института в Вудс-Холе, на берегу
Атлантического океана. Его интересовали не
только крупные, но и мелкие обитатели
дна. На небольшой глубине (9—20 м) он
обнаружил небольших (около 3 мм)
рачков, которые оживленно двигались среди
частиц грунта, изгибая свое длинное
тонкое тело. Внимательно рассмотрев
добычу, Сандерс установил, что эти рачки
непохожи на всех известных до сих пор, но
в то же время имеют те или иные общие
признаки почти с каждым из известных
подклассов ракообразных. Это заставило
изучить их более подробно, так как при
исследовании их строения можно было
надеяться выяснить родственные
отношения главных групп класса ракообразных.
Эти рачки (рис. 200) имеют крупную
голову, задний край которой нависает над
первым грудным сегментом. Глаз у них
нет. Задние антенны расположены позади
рта. У всех остальных ракообразных во
взрослом состоянии обе пары антенн
находятся перед ртом, и только у личинок
они занимают такое же положение, как
у взрослых цефалокарид. За большой
верхней губой открывается рот, по краям
которого помещаются жвалы. Передние
челюсти маленькие, а задние имеют такое
же строение, как следующие за ними
грудные ножки: большая основная часть,
снабженная внутренними выростами,
служащая для передвижения, расчлененная
внутренняя ветвь и две листовидные
наружные лопасти — псевдоэшшодит и эк-
зоподит. Сходство в строении и функциях
челюстей и грудных ножек может
считаться признаком примитивной
организации; челюсти еще не специализировались
так, как это произошло у других
ракообразных.
Грудной отдел состоит из 10 сегментов.
Конечности двух задних грудных
сегментов недоразвиты, и последняя их пара,
вероятно, выполняет половую функцию.
Грудные ножки совершают такие же
движения, как у жаброногов. Они
сгибаются последовательно, вызывая ток воды,
направленный вперед, ко рту. При
изгибании ножек назад вода с боков
устремляется между ними (рис. 200,1—6), а при
их изгибании вперед вода проталкивается
также вперед (рис. 200, 7—0). Оседающие
при этом на щетинках взвешенные
частицы попадают в рот и поедаются.
Таблица 30. Ветвистоусые ракообразные:
1 — Daphnia pulex;
2 — Bythotrephes longimana;
3 — Diaphanosoma brachyurum;
4 — Bosmina longirostris;
5 — Evadne nordmanni;
6 — Cercopagis socialis;
7 — Chydorus sphaericus;
8 — Polyphemus pediculus;
9 — Camptoccrcus rectirostris;
10 — Sida crystallina;
11 — Macrothrix hirsuticornis;
12 — Ceriodaphnia reticulata;
13 — Moina rectirostris;
14 — Leptodora kindtii;
15 — Simocephalus vetulus;
16 — Holopedium gibberum;
17 — метанауплиус Leptodora kindtii}
18 — Caspievadne maximovitchi.
Брюшко лишь незначительно уже, но
несколько длиннее грудного отдела. В
него входит 9 сегментов приблизительно
равных размеров, и заканчивается оно
тельсоном с длинными фуркальными
ветвями, вооруженными необычайно
длинными щетинками.
Цефалокариды раздельнополы, но
половые различия внешне не проявляются.
О процессе их спаривания ничего не
известно.
Самка носит яйцевой мешок снизу на
последнем грудном сегменте. Из яиц
вылупляются личинки, напоминающие
взрослых, но имеющие всего 3 грудных
сегмента; их антенны и жвалы развиты полностью,
а обе пары челюстей зачаточны (рис. 200,Г).
Дальнейшее развитие происходит очень
плавно: с каждой последующей линькой
добавляется 1—2 сегмента тела и так же
постепенно увеличивается количество
конечностей. Через 18 линек рачки
достигают половозрелости.
Личинки, так же как и взрослые, ведут
донный образ жизни. Они непохожи на
науплиусов жаброногих ракообразных,
так как имеют сравнительно длинные
расчлененные передние антенны, короткоз
основание задних антенн и двуветвистыз
жвалы. Эти признаки свойственны науп-
лиусам других ракообразных.
Таблица 31. Веслоногие ракообразные:
1 — Sapphirina ovatolanceolata:
2 — Oithona plumifera;
3 — Calanus finmarchicus;
4 — Oncaea mediterranea;
5 — Phoenna spinifera;
6 — Calocalanus pavo;
7 — Sapphirina vorax;
8 — Gorycaeus obtusus;
9 — Euchirella maxima;
10 — Pontella mediterranea;
11 — Gopilia vitrea;
12 — Calanus plumchrus.
Первый открытый Сандерсом вид цефа-
локарид — Hutchinsoniella macracantha—
оказался весьма многочисленным. На
глубине 9—30 м насчитывается в среднем 176
рачков на 1 м2 поверхности дна. Этот
вид найден и на глубине 300 ле, но на
глубине живет всего около 7 рачков
на 1 м2.
За последнее время в прибрежных водах
удалось найти еще 3 вида цефалокарид —
Lightiella serendipita в гавани
Сан-Франциско, L. incisa у берегов Барбадоса и
Пуэрто-Рико и Sandersella acuminata у
берегов Японии. Следовательно, эти рачки
обитают не только в Атлантическом, но
и в Тихом океане. Можно думать, что они
будут впоследствии обнаружены и в
других местах.
Lightiella incisa развивается в общем так
же, как Hutchinsoniella, но число грудных
сегментов у ее ранних личинок больше, чем
у личинок соответствующих стадий
Hutchinsoniella, в процессе развития у нее
редуцируется щупик жвал и внутренний вырост
задней антенны. Этот вид размножается в
течение круглого года.
Сравнение признаков цефалокарид и
других ракообразных показывает, что
цефалокариды занимают центральное
положение в этом классе. Из конечностей
цефалокарид можно вывести все типы конеч-
26 Жизнь животных, т. 2
Рис. 200. Цефа-
локариды:
А — общий вид
Hutchinsoniella;
Б — ее грудная
ножка; В —
движения
конечностей и вызываемые
ими токи воды
(обозначены
стрелками; цифрами
сверху показана
последовательность взмахов
ног); Г — первая
личинка.
Рис. 201. Движения Hutchinsoniella на
поверхности грунта.
ностей, свойственные разным подклассам
и отрядам ракообразных. В особенности
сходны конечности цефалокарид и жабро-
ногов, как по форме, так и по функциям.
Строением головы и заднего конца тела
цефалокариды напоминают веслоногих,
много общего у них и с личинками высших
ракообразных. По всей вероятности,
цефалокариды сохранили многие признаки,
которыми обладали некогда
существовавшие общие предки всех
ракообразных.
402
ПОДКЛАСС МАКСИЛЛОПОДЫ (MAXILLOPODA)
Грудной отдел у Maxillopoda состоит
из определенного числа сегментов, обычно
из 6 или реже оно сокращено до 5 или 4.
Грудные ножки служат только для
передвижения или для возбуждения токов
воды, но не несут дыхательной функции
и лишены внутренних отростков,
подающих пищу ко рту. Ротовые придатки
хорошо развиты, часто служат для фильтрации.
Сложные глаза и жабры обычно
отсутствуют, кровеносная система сильно
упрощена или отсутствует. Развитие со
сложным метаморфозом.
В состав этого подкласса наряду со сво-
бодноживущими рачками, сохраняющими
типичное для Maxillopoda строение,
входят ракообразные, ведущие
сидяче-прикрепленный и паразитический образ
жизни, сильно изменивший их организацию.
Maxillopoda распространены
чрезвычайно широко. Они населяют как море,
так и пресные воды, живут как на дне, так
и в толще воды, обитают в подземных и в
капиллярных водах, пропитывающих
песок, опускаются на огромные океанические
глубины, паразитируют на других
животных, в том числе и на других
ракообразных,— словом, они эволюционировали в
различных направлениях и
приспособились к существованию в самых
разнообразных условиях.
В этот подкласс включают 5 отрядов —
веслоногих, карпоедов, мистакокарид,
аскоторацид и усоногих.
ОТРЯД ВЕСЛОНОГИЕ РАКООБРАЗНЫЕ
(COPEPODA)
При изучении жизни каждого водоема—
от маленькой лужи до океана —
исследователь встречается с этими рачками.
Планктонная сеть, протянутая на любой
глубине океана, в жарких тропиках или
в холодных приполярных водах, приносит
улов, состоящий преимущественно из
веслоногих рачков. В ручьях и озерах
глубоких темных пещер также обитают эти
ракообразные. Они массами плавают во всех
обычных пресноводных водоемах, часто не
уступая по численности ветвистоусым.
Некоторые из них паразитируют на рыбах
и других водных животных, причем
нередко под влиянием паразитизма меняются
настолько, что не сразу можно угадать их
истинную природу. Веслоногие массами
поедаются рыбами. Сельдь, иваси и
многие другие рыбы питаются почти
исключительно веслоногими. Большинство этих
ракообразных — животные
микроскопические, длина их тела от 1 мм и меньше
до 5 мм; лишь отдельные глубоководные
и паразитические виды достигают в длину
1—3 см.
Тело свободноживущих веслоногих
разделяется на головогрудь, грудь и брюшко
(рис. 202). Голова слитная» без всяких
следов сегментации, срастается с первым
грудным сегментом, образуя головогрудь.
Передний конец головы часто вытянут в
загнутый вниз клюв, или рострум. Очень
характерно отсутствие парных
фасеточных глаз; на лобной части головы
расположен только науплиальный глазок. Именно
это обстоятельство позволило датскому
натуралисту Мюллеру в свое время
назвать обычных пресноводных веслоногих
«циклопами» в честь одноглазого гиганта
греческой мифологии.
Голова снабжена 5 парами придатков.
Передние антенны часто очень длинные,
иногда длиннее тела, и участвуют в
плавании и парении рачков. Кроме того, они
выполняют и функции органов чувств: на
них сидят чувствительные щетинки и
цилиндрические чувствительные придатки.
Задние антенны короткие, обычно двувет-
вистые. Жвалы мощные и имеют двувет-
вистый щупик. Их жевательная сильно
хитинизированная часть обладает
острыми зубцами, помогающими раздроблять
пищу. При внимательном рассмотрении
зубцов жвал некоторых морских
веслоногих выяснилось, что эти зубцы покрыты
кремневыми коронками, увеличивающими
их прочность (рис. 203). Открытие
кремневых коронок интересно в двух
отношениях. Во-первых, оно указывает на
способность веслоногих ракообразных усваивать
26*
403
и концентрировать кремний; такой
способности лишены почти все высшие
беспозвоночные — черви, моллюски, а также
другие членистоногие. Во-вторых, можно
надеяться найти в геологических
отложениях кремневые коронки древних
веслоногих, почти совсем не сохранившихся
в ископаемом состоянии.
Передние челюсти веслоногих устроены
очень сложно, так как снабжены
внутренними и наружными лопастями и
многочисленными перистыми щетинками.
Задние челюсти имеют только внутренние
лопасти и также многочисленные щетинки.
К головным придаткам присоединяется
пара одноветвистых ногочелюстей,
принадлежащих переднему слившемуся с
головой грудному сегменту.
Задние антенны, щупики жвал и
передние челюсти фильтрующих веслоногих
совершают частые и беспрерывные взма-
Рис. 202. Общий вид самок трех подотрядов
веслоногих:
1 — Galanoida, 2 — Cyclopoida; 3 — Harpacticoida.
Рис. 203. Край жующей пластинки Eucalanus
bungii и один зубец при большом увеличении.
хи, создающие круговороты воды,
которые приносят взвешенные пищевые
частицы. Эти частицы отфильтровываются
главным образом щетинками задних
челюстей.
Грудной отдел состоит из 5 сегментов
с ясно видимыми границами между ними.
Все 5 пар грудных ножек у примитивных
веслоногих устроены одинаково. Каждая
ножка состоит из 2-члениковой основной
части и двух обычно 3-члениковых
ветвей, вооруженных шипами и щетинками.
Эти ножки совершают одновременные
взмахи, действуя как весла и отталкивая тело
рачка от воды. У многих более
специализированных видов пятая пара ножек
самца преобразована в аппарат,
приспособленный для удерживания самки во время
спаривания и прикрепления сперматофо-
ров к ее половым отверстиям. Нередко
пятая пара ног редуцирована.
Брюшной отдел состоит из 4 сегментов,
но у самок часто их число меньше, так как
некоторые из них сливаются между собой.
На переднем брюшном сегменте
открывается парное или непарное половое
отверстие, и у самки этот сегмент часто
крупнее остальных. Брюшко заканчивается
тельсоном, с которым сочленены фуркаль-
ные ветви. Каждая из них вооружена
несколькими очень длинными, иногда
перистыми щетинками. Эти щетинки
особенно сильно развиты у планктонных видов,
у которых они приспособлены для
парения в воде, так как препятствуют
погружению рачка.
Строение паразитических видов сильно
отклоняется от описанного. Их головные
придатки превращаются в органы
прикрепления к хозяину и в связи с этим часто
имеют вид хитиновых крючков,
погруженных в кожу рыб или других водных
животных. Плавательные ножки иногда
исчезают совсем, или число их сокращается.
В большей или меньшей степени исчезает
сегментация, покровы становятся
мягкими. Специализированные паразиты
приобретают червеобразную форму тела и
теряют всякое сходство с ракообразными.
Однако можно подобрать полную серию
переходов между такими предельно
упрощенными видами и нормальными свобод-
ноживущими. Кроме того, яйца прикреп-
404
ляются к телу паразитических видов
совершенно так же, как к телу свободно-
живущих, и из яиц выходят такие же
личинки.
Дыхание веслоногих осуществляется
всей поверхностью тела, жабры
отсутствуют. С этим, возможно, связано и слабое
развитие или даже отсутствие
кровеносной системы. Сердце есть только у
представителей подотряда Calanoida, да и у
них оно невелико, хотя и бьется очень
часто: например, у морского рачка Labi-
docera оно совершает более 150 ударов
в минуту. У остальных веслоногих
полостная жидкость приводится в движение
сокращениями кишечника.
Все веслоногие раздельнополы. Самцы
обычно меньше самок и у свободноживу-
щих видов отличаются от самок
искривленными (одной или обеими) передними
антеннами, а также иным строением пятой
пары грудных ножек. Особенно резко
различны самцы и самки многих
паразитических видов. У некоторых из них
прикрепленные к хозяину самки полностью
теряют сегментацию и конечности, а
маленькие самцы сохраняют облик поздней
личинки и свободно плавают. У других
(семейство Lernaeopodidae) маленькие
самцы, также сходные с поздней личинкой,
своими клешневидными челюстями и но-
гочелюстями прикрепляются к крупным
червеобразным паразитическим самкам.
При спаривании самец удерживает
самку пятой парой грудных ножек и первыми
антеннами и при помощи той же пятой
пары ног приклеивает колбасовидный спер-
матофор близ ее половых отверстий, т. е.
к нижней стороне первого брюшного
сегмента. У некоторых видов одна из ветвей
пятой пары ног самца снабжена на конце
щипчиками, захватывающими спермато-
фор и переносящими его на нужное
место (рис. 204). Из сперматофора сперма
попадает в семеприемник самки. При
выметывании яиц они оплодотворяются.
Большинство морских планктонных
видов откладывает яйца прямо в воду, но
у всех пресноводных и у всех
паразитических, а также у морских донных и
прибрежных видов яйца склеиваются между
собой специальным секретом и
прикрепляются близ половых отверстий самки.
Рис. 204. Спаривание Calanoida:
1 — прикрепление сперматофора к генитальному
сегменту самки у Diaptomus; 2 — пятая пара ног
Pareuchacta glacialis; последний членик левой ноги
с «щипчиками», удерживающими сперматофор.
У одних веслоногих таким образом
формируется один, у других — два яйцевых
мешка, которые самка носит на себе до выхода
из яиц личинок.
Из яйца выходит личинка науплиус.
Личинка многократно линяет и
постепенно приближается по своим признакам к
взрослому рачку. Различают 12
личиночных стадий веслоногих. Первые две
стадии — ортонауплиуса — характеризуются
присутствием только обеих пар антенн
и пары жвал, у следующих четырех
стадий — метанауплиуса — закладываются
и развиваются остальные ротовые
придатки, но тело остается несегментирован-
ным. Последние 6 стадий называются
копеподитными и отличаются
сегментацией заднего конца тела и постепенным
развитием грудных ножек. Для
завершения метаморфоза разным веслоногим
требуется различное время, да и биология
личинок далеко не у всех видов
одинакова.
Образ жизни, способ питания и среда
обитания веслоногих рачков настолько
разнообразны, что лучше рассмотреть
этот отряд не целиком, а каждый из
входящих в него подотрядов в отдельности.
В пределах отряда веслоногих принято
различать 5 подотрядов, из которых 2
объединяют исключительно паразитиче-
405
L^-
Рис. 205. Движения Calanoida:
j — движение Diaptomus в воде; 2 — токи воды,
образуемые конечностями калануса.
ские виды1. Свободноживущие веслоногие
относятся к подотрядам Calanoida, Cyclo-
poida и Harpacticoida (рис. 202).
Calanoida — исключительно
планктонные животные. Их голова и грудь
значительно длиннее узкого брюшка, передние
антенны очень длинные, превосходят
голову и грудь, а нередко и все тело рачка,
если есть яйцевой мешок, то один.
Cyclopoida обитают преимущественно на
дне или в придонных слоях воды. Среди
них есть и паразиты. Их голова и грудь
относительно всего тела гораздо короче,
чем у Calanoida, передние антенны
короткие, не достигают и середины головогруд-
ного отдела, брюшко уже груди. Самки
всегда носят два яйцевых мешка.
Harpacticoida, за единичными
исключениями, живут на дне и больше ползают,
чем плавают. Тело их червеобразно
благодаря тому, что брюшной отдел почти не
отличается по ширине от грудного. Передние
1 Некоторые исследователи разделяют
паразитических веслоногих на 5 или 6 подотрядов.
антенны очень короткие, самки
большинства видов образуют один яйцевой мешок.
Представители всех трех подотрядов
населяют как моря, так и пресные воды.
ПОДОТРЯД КАЛАНИДЫ (CALANOIDA)
Вся организация Calanoida превосходно
приспособлена к жизни в толще воды.
Длинные антенны и перистые щетинки
фуркальных ветвей позволяют морскому
Calanus или пресноводному Diaptomus
неподвижно парить в воде, лишь очень
медленно погружаясь. Этому способствуют
находящиеся в полости тела рачков капли
жира, уменьшающие их удельный вес.
Во время парения тело рачка
располагается вертикально или наклонно, причем
передний конец тела расположен выше
заднего. Опустившись на несколько
сантиметров вниз, рачок делает резкий взмах
всеми грудными ножками и брюшком и
возвращается на прежний уровень, после
чего все повторяется сначала. Таким
образом, путь рачка в воде рисуется
зигзагообразной линией (рис. 205, 1). Некоторые
морские Calanoida, как например
приповерхностный вид ярко-синего цвета Pontel-
lina mediterranea, совершают настолько
резкие скачки, что выпрыгивают из воды и
пролетают какое-то пространство по
воздуху наподобие летучих рыб.
Если грудные ножки действуют время
от времени, то задние антенны, щупики
жвал и передние челюсти вибрируют
беспрерывно с очень большой частотой,
совершая до 600—1000 ударов ежеминутно.
Их взмахи вызывают мощные
круговороты воды с каждой стороны тела рачка
(рис. 205, 2). Эти токи проходят через
образованный щетинками челюстей
фильтрационный аппарат, и отфильтрованные
взвешенные частицы проталкиваются вперед
к жвалам. Жвалы размельчают пищу,
после чего она поступает в кишечник.
Пищей фильтрующим Calanoida служат
все организмы и их остатки, взвешенные
в воде. Рачки не заглатывают только
сравнительно крупные частицы, отталкивая
их ногочелюстями. Основой питания
Calanoida следует считать планктонные
водоросли, потребляемые рачками в огромном
406
количестве. Eurytemora nirundoides в
период массового развития водоросли Nitz-
schia closterium поедала до 120000 особей
этих диатомовых в день, причем вес пищи
почти достигал половины веса рачка. В
случаях такого избыточного питания
рачки не успевают усвоить все органическое
вещество пищи, но все-таки продолжают
ее заглатывать.
Для определения интенсивности
фильтрации Calanus применялись водоросли,
меченные радиоактивными изотопами
углерода и фосфора. Оказалось, что один рачок
за сутки пропускает через свой
фильтрационный аппарат до 40 и даже до 70 см3
воды, причем питается он
преимущественно ночью.
Питание водорослями необходимо для
многих Calanoida. Так, например, половые
продукты Calanus finmarchicus созревают
только при достаточном потреблении
рачком диатомовых водорослей.
Помимо фильтраторов, среди Calanoida
есть и хищные виды, большинство
которых обитает на значительных или
больших океанических глубинах, где
планктонные водоросли не могут существовать
из-за отсутствия света. Задние челюсти
и ногочелюсти таких видов снабжены
крепкими острыми шипами и
приспособлены для схватывания жертв. Особенно
интересны приспособления для добывания
пищи у некоторых глубоководных видов.
Винкстед наблюдал, как
глубоководная Pareuchaeta неподвижно висит
в воде, расставив в стороны свои
удлиненные ногочелюсти, образующие нечто вроде
капкана (рис. 206). Как только между ними
оказывается жертва, ногочелюсти
смыкаются, капкан захлопывается. При
крайней разреженности организмов на
больших океанических глубинах такой способ
охоты оказывается наиболее
целесообразным, так как затрата энергии на активные
поиски жертв не окупается их поеданием.
С особенностями движения и питания
Calanoida связана сложная проблема их
суточных вертикальных миграций. Уже
давно было замечено, что во всех водоемах,
как пресных, так и морских, огромные
массы Calanoida (и многих других
планктонных животных) ночью поднимаются
ближе к поверхности воды, а днем
опускаются вглубь. Размах этих суточных
вертикальных миграций различен не только
у разных видов, но даже у одного вида
в разных местах его обитания, в разные
сезоны года и у разных возрастных стадий
одного вида. Нередко науплиусы и
младшие копеподитные стадии держатся всегда
в поверхностном слое, а старшие
копеподитные стадии и взрослые рачки
мигрируют. В северной части Атлантического
океана диапазон вертикальных миграций
Calanus finmarchicus составляет 300—
500 м. Такие же громадные расстояния
ежесуточно преодолевают
дальневосточные Metridia pacifica и М. ochotensis. В
то же время другие массовые
дальневосточные Calanoida — Calanus plumchrus, С.
cristatus, Eucalanus bungii — мигрируют
не более чем на 50—100 м.
Скорость передвижений рачков при их
вертикальных миграциях измеряется
величинами порядка 10—30 см в минуту.
Если принять во внимание длину их тела
(для Calanus finmarchicus, например,
около 2 мм), то такую скорость приходится
считать значительной. При этом не толькЬ
подъем кверху, но и опускание вниз
осуществляется за счет активных движений
рачков, а не за счет их пассивного
погружения.
Не следует думать, что, совершая
вертикальные миграции, все рачки
одновременно двигаются в каком-нибудь
определенном направлении. Английский ученый
Бэйнбридж опускался под воду и
проводил наблюдения над мигрирующими
Рис. 206. Раскрытый «ловчий капкан»
Pareuchaeta.
407
Рис. 207. Вертикальные миграции Calanus в
Баренцевом море в условиях
круглосуточного дня A) и смены дня и ночи (-?).
веслоногими. Он видел, как в одном и
том же слое воды часть рачков движется
вверх, а другая — вниз. В зависимости
от того, какое движение преобладает,
перемещается вверх или вниз вся масса
животных.
Вопрос о причинах вертикальных
миграций до сих пор окончательно не выяснен.
Совершенно очевидно, что стремление
рачков подняться в поверхностные слои
объясняется изобилием там планктонных
водорослей, которыми питаются веслоно-
гие-фильтраторы. Менее понятны причины,
заставляющие рачков покидать эти
богатые пищей слои. Многие исследователи
считают, что свет вредно влияет на рачков
и, избегая его, они утром начинают
уходить вниз. Важное значение света
подтверждается наблюдениями В. Г. Б о г о -
р о в а над вертикальным распределением
веслоногих в Баренцевом море летом, т. е.
в условиях круглосуточного освещения.
Оказалось, что в это время Calanus fin-
marchicus неизменно находится на одной
глубине, там, где условия освещения для
него наиболее благоприятны. В этом
районе морй в толще воды наблюдаются
внутренние волны, которые должны то
несколько приподнимать, то несколько опускать
рачков. Очевидно, рачки активно
передвигаются в противоположном
направлении, так как они в течение круглых суток
не выходят за пределы определенного
горизонта. Осенью, когда
восстанавливается смена дня и ночи, возобновляются
обычные вертикальные миграции (рис. 207).
Не только солнечный, но и лунный свет
заставляет рачков уходить из
поверхностных слоев воды в более глубокие.
Однако далеко не во всех случаях
вертикальные миграции удается связать
непосредственно с действием света.
Существуют наблюдения, показывающие, что
рачки начинают опускаться вниз задолго
до восхода солнца. Эстерли содержал
веслоногих рачков Acartia tonsa и A. clausi
в полной темноте, и, несмотря на это, они
продолжали совершать регулярные
вертикальные миграции.
По мнению некоторых ученых, уход
рачков утром из освещенного слоя воды
следует считать защитной реакцией,
помогающей избегать выедания рыбами.
Доказано, что рыбы видят каждого рачка,
на которого они нападают. Опустившись
в глубокие темные слои воды, рачки
оказываются в безопасности, а в богатых
водорослями поверхностных слоях ночью
рыбы их также не могут рассмотреть. Эти
представления не могут объяснить многие
хорошо известные факты. Так, например,
целый ряд веслоногих рачков совершает
регулярные миграции небольшой
протяженности, не выходя из освещенной зоны
и оставаясь, следовательно, доступными
для планктоноядных рыб.
Помимо суточных вертикальных
миграций, морские веслоногие совершают и
сезонные миграции. В Черном море летом
температура поверхностного слоя
повышается, и обитающий там Calanus helgolan-
dicus опускается приблизительно на 50 ле,
а зимой снова возвращается на меньшую
глубину. В Баренцевом море молодые
стадии С. finmarchicus весной и летом
держатся в поверхностных слоях. После того
как они подрастут, осенью и зимой рачки
опускаются вниз, и перед весной
достигающие половозрелости особи начинают
подниматься к поверхности, где и отрождается
новое поколение. Особенно
многочисленны в поверхностных слоях рачки,
находящиеся на IV—V копеподитных стадиях
и известные под названием «красного ка-
лануса», так как содержат большое
количество жира буровато-красного цвета.
408
«Красный каланус» — излюбленная пища
многих рыб, в частности сельди.
Подобный характер сезонных миграций, т. е.
подъем в поверхностные слои воды для
размножения, обнаружен у многих других
массовых видов, например у Calanus gla-
cialis, С. helgolandicus, Eucalanus bungii
и т. д. Самки этих видов нуждаются для
развития половых продуктов в обильном
питании водорослями, а возможно, и в
освещении. Другие виды (Calanus cristatus,
С. hyperboreus), наоборот, размножаются
в глубоких слоях, а к поверхности
поднимается только их молодь. Взрослые рачки
С. cristatus совсем не питаются; у
половозрелых особей даже редуцируются жвалы.
Протяженность сезонных миграций
обычно бывает больше, чем суточных.
Первые иногда захватывают 3—4 тысячи
метров, а вторые — самое большее несколько
сотен метров.
Представители подотряда Calanoida
преимущественно морские животные. В
настоящее время известно около 1200
морских видов этих рачков, принадлежащих
к 150 родам и 26 семействам. В пресных
водах обитает всего около 420 видов,
распределяющихся между 12 родами и 4
семействами.
Проведенные в последнее время
подробные исследования фауны морских каланид
показали, что прежние представления о
широком распространении многих видов
этих ракообразных неправильны. В
каждой части океана обитают в основном
присущие только ей виды. Каждый вид
морских каланид расселяется благодаря
несущим рачков течениям. Так, например,
поступающие в Полярный бассейн
ответвления Гольфстрима заносят туда каланид
из Атлантического океана. В
северо-западной части Тихого океана в водах
теплого течения Куросио обитают одни виды,
а в водах холодного течения Оясио —
другие. Нередко по фауне каланид
удается определить происхождение тех или
иных вод в определенных частях океана.
Особенно резко различаются по своему
составу фауны вод умеренных широт и вод
тропиков, причем тропическая фауна
богаче видами.
Каланиды обитают на всех океанических
глубинах. Среди них ясно различаются.
поверхностные виды и виды
глубоководные, никогда не поднимающиеся в
поверхностные слои воды. Как уже указывалось,
на больших глубинах преобладают
хищники, а на малых — фильтраторы.
Наконец, существуют виды, совершающие
вертикальные миграции огромного
диапазона, то поднимающиеся к поверхности, то
опускающиеся на глубину до 2—3 км.
Некоторые мелководные виды каланид
в умеренных водах развиваются в
огромном количестве и по весу составляют
преобладающую часть планктона. Так,
например, планктон Баренцева моря
приблизительно на 90% состоит из Calanus
finmarchicus (табл. 31, 3). Эти рачки
характеризуются высокой питательностью: в
их теле содержится 59% белков, 20%
углеводов и часто более 10—15% жиров.
Многие рыбы, а также усатые киты
питаются главным образом каланидами.
Таковы, например, сельдь, сардина,
скумбрия, анчоус, шпрот и многие другие.
В желудке одной сельди было обнаружено
60 000 проглоченных ею веслоногих
рачков. Из китов активно потребляют
огромные массы каланид финвал, сейвал, синий
кит и горбач.
Calanoida внутренних водоемов по своей
биологии напоминают морские виды. Они
также приурочены только к толще воды,
также совершают вертикальные миграции
и питаются так же, как морские
фильтраторы. Они обитают в самых
разнообразных водоемах. Некоторые виды, как
например Diaptomus graciloides и D.
gracilis, живут почти во всех озерах и прудах
северной и средней части СССР. Другие
приурочены только к Дальнему Востоку
или к южной части нашей страны. Весьма
интересно распространение Limnocalanus
grimaldii, который населяет многие озера
севера нашей страны (в том числе
Онежское и Ладожское) и Скандинавии. Этот
вид близок к прибрежному солоновато-
водному L. macrurus, обитающему в пред-
устьевых пространствах северных рек.
Озера, населенные L. grimaldii, некогда
покрывались холодным Иольдиевым морем.
В Байкале в огромном количестве обитает
свойственный только этому озеру рачок
Epischura baicalensis, служащий
основной пищей омулю. Очень своеобразны,
409
хотя и еще мало известны, условия
существования недавно открытого единственного
подземного представителя каланид — Spe-
odiaptomus birsteini. Этот слепой рачок
найден в глубоких и узких заполненных
водой расселинах нижнего этажа Скель-
ской пещеры, расположенной в Крыму,
недалеко от Севастополя. За ним удалось
наблюдать в аквариуме, причем оказалось,
что он плавает так же, как обычные
пресноводные каланиды. Остается загадкой, чем
он питается, фильтруя чистую, полностью
лишенную водорослей и, вероятно, очень
бедную бактериями воду подземного
бассейна. По-видимому, он может считаться
единственным настоящим подземным
планктонным животным.
Некоторые пресноводные каланиды
появляются в водоемах только в
определенное время года, например весной. В
весенних лужах часто попадается сравнительно
крупный (около 5 мм) Diaptomus amblyo-
don, окрашенный в яркий красный или
синий цвет. Этот вид и некоторые другие
широко распространенные пресноводные
каланиды способны к образованию
покоящихся яиц, стойко переносящих высыхание
и вымерзание и легко разносящихся
ветром на далекие расстояния.
ПОДОТРЯД ЦИКЛОПЫ (CYCLOPOIDA)
Другой подотряд веслоногих
ракообразных — Cyclopoida — наибольшим
количеством видов представлен в пресных
водах.
Пресноводные циклопы живут во
всевозможных водоемах, от мелких луж до
крупных озер, причем нередко встречаются
в очень большом количестве экземпляров.
Основной зоной их обитания служит
прибрежная полоса с зарослями водных
растений. При этом во многих озерах к зарослям
определенных растений приурочены
определенные виды циклопов. Так, например,
для Валдайского озера в Ивановской
области описано 6 группировок растений
с соответствующими им группировками
видов циклопов.
Сравнительно немногие виды могут
считаться настоящими планктонными
животными. Некоторые из них,
принадлежащие преимущественно к роду Mesocyclops,
постоянно обитают в поверхностных слоях
воды, другие (Cyclops strenuus и другие
виды того же рода) совершают регулярные
суточные миграции, опускаясь днем на
значительную глубину.
Плавают циклопы несколько иначе, чем
каланиды. Одновременно взмахивая
четырьмя парами грудных ножек (пятая
пара редуцирована), рачок делает резкий
прыжок вперед, вверх или вбок, а затем
при помощи передних антенн может
некоторое время парить в воде. Поскольку
центр тяжести его тела смещен вперед,
во время парения его передний конец
наклоняется и тело может принять
вертикальное положение, причем погружение
замедляется. Новый взмах ножками
позволяет циклопу подняться. Эти взмахи
молниеносны — они занимают 1/60
секунды.
Много занимавшийся биологией
циклопов Л. Исаев описывает их движения
следующим образом: «Передвигаясь
ритмическими скачками, циклоп может
хорошо держаться на одном уровне,
подниматься вверх и опускаться вниз под углами
различной крутизны. Циклоп может
плавать с одинаковой легкостью,
перевернувшись на спину. Циклоп хорошо
описывает дуги, делает мертвые петли,
одинарные и множественные, прямые и
обратные. Циклоп может делать поворот под
углом 90°, вращаться вокруг оси не только
со снижением, напоминающим витки
«штопора» самолета, но и с поднятием вверх.
Циклоп может скользить на антенну,
делать через нее переворот, пикировать вниз
головой под углом в 90° и скользить на
хвост. Характер «фигур», выполняемых
циклопом, очень сходен с фигурами
высшего пилотажа. Обладание фигурами
высшего пилотажа, необходимыми для
самолетов-истребителей, несомненно
облегчает циклопу — активному хищнику —
возможность обеспечить себе
существование охотой за водными обитателями,
служащими ему пищей».
Большинство циклопов хищники, но
среди них есть и растительноядные виды.
Такие обычные, широко распространенные
виды, как Macrocyclops albidus, M. fuscus,
410
Acanthocyclops viridis и многие другие,
быстро плавают над дном или среди
зарослей в поисках добычи. При помощи своих
антенн на очень коротком расстоянии они
чуют мелких олигохет и хирономид,
которых хватают вооруженными шипами
передними челюстями. В передаче пищи жвалам
участвуют задние челюсти и ногочелюсти.
Жвалы совершают быстрые режущие
движения в течение 3—4 секунд, за
которыми следует минутная пауза. Циклопы
могут поедать олигохет и хирономид более
крупных, чем они сами. Скорость поедания
жертв зависит от их размеров и от
твердости их покровов. Для размельчения и
заглатывания мотыля длиною 2 мм
требуется 9 минут, а личинка длиною 3 мм
уничтожается уже в течение получаса.
Более нежный, хотя и более длинный
D мм), малощетинковый червь Nais
поедается всего за 3,5 минуты.
Хищные циклопы нападают не только
на таких малоподвижных животных, как
сидящие на дне или на растениях олигохеты
и мотыли, но и на плавающих в воде
других рачков, в том числе и на себе
подобных, а также на коловраток и т. д. Они
охотно заглатывают и личинок
паразитических червей — корацидиев широкого
лентеца и личинок ришты (см. том 1) —
и становятся, таким образом,
промежуточными хозяевами этих паразитов человека.
Неоднократно наблюдалось нападение
циклопов на икринки и даже личинок рыб.
Растительноядные циклопы, в частности
обычные Eucyclops macrurus и Е. macru-
roides, питаются главным образом
зелеными нитчатыми водорослями (Scenedes-
mus, Micractinium), захватывая их
приблизительно так же, как хищные
захватывают червей и мотылей; кроме того,
используются различные диатомовые, пери-
диниевые и даже сине-зеленые водоросли.
Многие виды могут поедать только
относительно крупные водоросли. Mesocyclops
leuckarti быстро набивает себе кишечник
колониями Pandorina (диаметр колонии
50—75 мк) и почти совсем не заглатывает
мелких Chlamydomonas.
Пресноводные циклопы распространены
очень широко. Некоторые виды
встречаются почти повсеместно. Этому
способствуют прежде всего приспособления к
перенесению неблагоприятных условий, в
частности способность рачков переносить
высыхание водоемов и в виде цист пассивно
расселяться по воздуху. Кожные железы
многих циклопов выделяют секрет,
обволакивающий тело рачка, часто вместе с
яйцевыми мешками, и образующий нечто
вроде кокона. В таком виде рачки могут
подвергаться высыханию и вмерзанию в
лед, не теряя жизнеспособности. В
опытах Камерера циклопы быстро
выводились при размачивании сухого ила,
сохранявшегося около 3 лет. Поэтому нет
ничего удивительного в появлении
циклопов в весенних лужах, возникающих при
таянии снега, в только что залитых
рыбоводных прудах и т. д.
Второй причиной широкого
распространения многих видов циклопов следует
считать стойкость рачков, находящихся
в активном состоянии, по отношению к
недостатку кислорода в воде, кислой ее
реакции и многим другим факторам,
неблагоприятным для остальных пресноводных
животных. Cyclops strenuus в течение
нескольких дней может жить не только при
полном отсутствии кислорода, но даже в
присутствии сероводорода. Некоторые
другие виды также хорошо переносят
неблагоприятный газовый режим. Многие
циклопы превосходно существуют в воде
с кислой реакцией, при высоком
содержании гуминовых веществ и крайней
бедности солей, например в водоемах,
связанных с верховыми (сфагновыми) болотами.
Тем не менее известны виды и даже роды
циклопов, ограниченные в своем
распространении какими-нибудь определенными,
в частности температурными и солевыми,
условиями. Так, например, род Ochrido-
cyclops обитает только в озере Охрид в
Югославии, род Bryocyclops — в Юго-
Восточной Азии и в экваториальной
Африке. К последнему роду близок
исключительно подземный род Speocyclops, виды
которого найдены в пещерах и грунтовых
водах Южной Европы, Закавказья, Крыма
и Японии. Эти слепые мелкие рачки
считаются остатками некогда более широко
распространенной теплолюбивой фауны.
Некоторые циклопы приспособились
к жизни в солоноватых и даже в очень
соленых водоемах. Род Halicyclops, на-
411
пример, весьма обычен в Каспийском море
и не встречается в пресной воде. Micro-
cyclops dengizicus широко распространен
только в солоноватых и соленых водоемах
зоны пустынь (Ирак, Индия, о. Гаити,
Египет, Калифорния, в СССР — в
Карагандинской области, в Муганской степи) и
прекрасно живет даже при солености,
превосходящей морскую (до 41°/оо)- Многие
обычные пресноводные виды могут
существовать и в солоноватой воде, как например
Mesocyclops leuckarti в Финском и
Ботническом заливах.
Значение циклопов в жизни наших
внутренних водоемов очень велико. С одной
стороны, они служат пищей многим рыбам
и их молоди, но, с другой стороны,
выступают в качестве конкурентов тех же
самых рыб, поедая множество мелких
животных, как планктонных, так и донных.
Не следует забывать и о том, что циклопы
способствуют заражению человека
некоторыми паразитическими червями, в
первую очередь широким лентецом (Diphyl-
lobothrium latum) и риштой (Dracunculus
medinensis). Циклопы заглатывают
плавающих в воде личинок этих червей. Кора-
цидий широкого лентеца, попав в
циклопа, сбрасывает свой
мерцательный покров, проникает из
кишечника в полость тела рачка
и превращается там в стадию
процеркоида. Если такой
циклоп будет проглочен рыбой,
процеркоид перейдет в стадию
плевроцеркоида, а затем, при
поедании рыбы человеком,
кошкой и другими
млекопитающими, из плевроцеркоида
возникнет взрослый червь, часто
вызывающий тяжелое
малокровие. Личинки ришты,
проглоченные циклопами, также
проникают в полость их тела и
остаются там в течение 1—1,5
месяцев. При питье сырой воды
человек проглатывает
циклопов и заражается риштой.
Морские представители
подотряда Cyclopoida менее
разнообразны, чем пресноводные.
Среди них обычны и нередко
многочисленны в морском планк-
Рис. 208. Циклопы —
промежуточные хозяева
паразитов человека:
1 — Paracyclops с процер-
коидами лентеца широкого
в Полости тела; 2 — Meso-
cyclops с личинками
ришты в полости тела.
тоне виды рода Oithona. Очень
характерны также крупные (до 8 мм) уплощенные
виды рода Sapphirina, поверхность тела
которых отливает яркими синими,
золотистыми или темно-красными тонами
(табл. 31, 1). Другой близкий морской
род — Опсаеа (табл. 31, 4)—обладает
железами, выделяющими светящийся
секрет, и нередко, совместно с другими
организмами, вызывает свечение моря.
ПОДОТРЯД ГАРПАКТИЦИДЫ
(HARPACTICOIDA)
Об образе жизни представителей
третьего подотряда — Harpacticoida —
известно гораздо меньше. Эти
червеобразные, большей частью очень мелкие рачки,
чрезвычайно разнообразны как в морских,
так и в пресных водах, но никогда не
встречаются в массовом количестве. Описано
более 30 семейств и несколько сотен видов
Harpacticoida.
Большинство гарпактицид ползает по
дну и донным растениям. Лишь единичные
виды способны продолжительное время
плавать и входят в состав
морского планктона (Microsetella).
Гораздо более характерны
целые группы родов и видов
гарпактицид,
приспособившиеся к обитанию в особых,
необычных условиях, в частности
в капиллярных ходах
между песчинками морских пляжей
и в подземных пресных водах.
Всего несколько лет тому
назад зоологи применили очень
простой прием, позволяющий
изучать население
капиллярных ходов морского песка. На
пляже, выше уровня моря,
выкапывается яма. В ней
постепенно накапливается вода,
заполнявшая капилляры песка.
Эту воду фильтруют через
планктонную сеть и таким образом
добывают представителей
своеобразной фауны, получившей
название интерстици-
а л ь н о й.
412
Гарпактициды составляют заметную
часть интерстициальной фауны. Их
обнаружили всюду, где проводились
соответствующие исследования,— на пляжах
Англии, вдоль европейского и
американского берегов Атлантического океана, на
Средиземном и Черном морях, у берегов
Африки и Индии, на островах
Мадагаскар, Реюньон и Багамских. Большинство
интерстициальных гарпактицид
принадлежит к особым, обитающим только в
таких условиях родам, отличающимся
необыкновенно тонким и длинным телом
(рис. 209), позволяющим рачкам двигаться
в узких капиллярных ходах.
Замечательно, что некоторые из этих
специализированных видов были найдены в очень
отдаленных друг от друга местах. Так,
например, на Багамских островах оказались
Arenosetella palpilabra, ранее известная
только из Шотландии, и Horsiella trisae-
tosa, ранее известная только из Кильской
бухты. Трудно объяснить подобное
распространение, поскольку покоящихся яиц
у интерстициальных гарпактицид нет.
Гарпактициды пресных подземных вод
также представлены рядом
специализированных родов — Parastenocaris, Elapho-
idella, Ceuthonectes и других, частично
очень широко распространенных, частично
имеющих узкие и разорванные ареалы.
Так, например, два вида рода
Ceuthonectes обитают только в пещерах Закавказья,
Югославии, Румынии, Италии и Южной
Франции. Эти удаленные друг от друга
местонахождения считаются остатками
некогда гораздо более обширной области
распространения древнего рода. В
некоторых случаях можно допускать
тропическое происхождение подземных
гарпактицид Европы. Среди многочисленных
видов рода Elaphoidella есть как
тропические, так и европейские виды. Первые
обитают в наземных, вторые (за
единичными исключениями) в подземных водах.
По всей вероятности, под землей
сохранились остатки древней тропической фауны,
погибшей на поверхности земли под
влиянием климатических изменений. В
тропических наземных водоемах некоторые
гарпактициды приспособлены к условиям
существования, напоминающим условия
жизни в подземных водах. Известны тро-
Рис. 209. Характерные интерстициальные
гарпактициды:
1 — Nitocra chelifer; 2 — Arenosetella spinicauda; 3 —
Goffinella stylifer; 4 — Paraleptastacus brevicaudatus;
5 — Emertonia gracilis.
пические виды Elaphoidella, живущие
в своеобразных микроводоемах,
образующихся в пазухах листьев водных растений
из семейства Bromeliaceae. В таких же
условиях живет тропическая Viguierella
соеса, обнаруженная на этих растениях
в ботанических садах почти всех стран.
Чрезвычайно богата видами
гарпактицид своеобразная фауна Байкала. Она
состоит из 43 видов, из которых 38
эндемичны для этого озера. Особенно много
этих рачков в прибрежной части Байкала,
на камнях и водных растениях, а также
на растущих здесь губках. По-видимому,
они питаются за счет губок и в свою
очередь становятся жертвами также
ползающего по губкам бокоплава Brandtia
parasitica.
Некоторые виды гарпактицид
приурочены только к водоемам, очень бедным
солями, характеризующимся повышенной
кислотностью, т. е. связанным с
верховыми, сфагновыми, болотами. Таков,
например, Arcticocamptus arcticus, биология
которого подробно изучена Е. В.
Бору ц к и м.
413
Рис. 210. Стадии вылупления науплиуса Arcticocamptus
arcticus из яйца.
A. arcticus широко распространен на
севере Европы, от Болыпеземельской
тундры до Скандинавии, на западном
побережье Гренландии, на Новой Земле.
Кроме того, он найден в Альпах и в
нескольких пунктах средней полосы
Европейской части СССР, в том числе в
Косине под Москвой, в Звенигороде, под
Ярославлем и т. д. Повсюду он обитает
в водоемах, связанных со сфагновыми
болотами.
Из многочисленных водоемов,
расположенных близ Косина, рачок живет только
в двух болотах и в Святом озере, лежащем
среди сфагнового торфяника. Очевидно,
условия существования в других
разнообразных соседних водоемах
неблагоприятны для A. arcticus. При этом даже в
немногих населенных им водоемах рачок
в активном состоянии существует только
в течение 1г/2—2 месяцев весной,
остальную часть года, т. е. 10—10V2 месяцев,
он проводит в стадии покоящегося яйца.
Жизненный цикл A. arcticus тесно
связан с изменениями
растительного покрова болота. Е. В. Б о-
р у ц к и й пишет: «Как только с
первыми весенними теплыми
лучами рыхлый снег начинает таять и
на поверхности болота образуются
лужи — все животные, проведшие
суровую зиму в тех или других
стадиях в ледяном заключении,
начинают реагировать на первые
весенние лучи. A. arcticus одним из первых
выходит из состояния анабиоза и
появляется в водоеме. Уже в
небольших лужах, еще среди снега, там,
где поверхностные слои сфагнума
оттаяли, можно найти его личинок,
медленно и неуклюже
передвигающихся среди листочков мха в
поисках пищи. Личинки — на первой
науплиальной стадии и, очевидно,
только что вылупившиеся из яйца.
С каждым днем науплиус крепнет,
движения его становятся более
уверенными и быстрыми, наконец,
наступает момент первой линьки — он
меняет старую узкую оболочку на
новую, более просторную. За
первой линькой следует вторая, третья
и т. д., и вот спустя две-три недели
мы уже встречаем взрослые экземпляры
или личинок на последней копеподитной
стадии. Но они уже не пользуются тем
простором, который имели в своем раннем
личиночном состоянии: вместо обширных
луж, полных воды, в которых они свободно
переплывали с одного сфагнового
кустика на другой, теперь только влажный мох
и незначительное количество воды.
Вместо жалких голых ветвей теперь нежные
розовые цветы Кассандры и клюквы,
белые чашечки андромеды и шапки
цветущего богульника. Болото изменилось —
яркий зеленый сфагновый ковер пестрит
розовыми и белыми пятнами цветов. И это
изменение картины болота как нельзя
лучше совпадает с определенным
моментом в биологии A. arcticus, именно с
периодом копуляции. В продолжение
нескольких дней мы встречаем почти
исключительно копулирующие пары. Но эти
цветы — роковые для A. arcticus: с
постепенным увяданием их наблюдается по-
414
степенное уменьшение численности
рачков, копуляция встречается все реже и
реже, чаще попадаются самки с
яйцевыми мешками и, наконец, к середине
или к концу июня A. arcticus совсем
исчезает из водоема, и лишь только
запоздалые экземпляры встречаются в
незначительном количестве в июле или в начале
августа».
Рачок оставляет в водоеме свои
яйцевые мешки, имеющие форму двух
соединенных вместе шаров, покрытых общей
«мешковой» оболочкой, осуществляющей
механическую защиту яиц, а также
защищающей их от высыхания. Кроме того,
каждое яйцо имеет свою более тонкую
прозрачную оболочку. Она
непроницаема как для солей, так и для воды. К
осени в каждом яйце формируется науплиус,
причем науплиусы двух соединенных
яиц всегда направлены своими передними
концами в противоположные стороны.
Науплиусы одеты еще одной очень
тонкой и эластичной внутренней оболочкой,
снабженной различными тяжами и
нитями. Эта оболочка проницаема для воды,
но не для солей.
Когда наступает время вылупления
науплиусов, т. е. в период весеннего
таяния льда, в мешковой оболочке с одной
стороны образуется трещина, через
которую начинает выпячиваться эластичная
внутренняя оболочка. Сначала этот
процесс идет очень медленно, но после того,
как около половины окруженной
оболочкой личинки окажется снаружи,
происходит резкий толчок и личинка,
заключенная в полый шар, как бы «выстреливает»
из яйцевого мешка и отскакивает в
сторону или задерживается за края щели
придатками эластичной оболочки.
Замечательно, что сам науплиус при этом
остается почти все время совершенно
пассивным. Только в самом начале процесса
вылупления науплиус производит
несколько довольно слабых движений,
приводящих, по-видимому, к разрыву яйцевой
оболочки. Основную роль здесь играет
полупроницаемая эластичная оболочка,
через которую диффундирует вода,
вызывающая ее разбухание, что сначала
заставляет лопнуть мешковую оболочку, а
затем выпятиться из нее окруженному
эластичной оболочкой науплиусу. Тяжи
и нити этой оболочки действуют как
пружины, причем расположены они таким
образом, что науплиус внутри
образованного оболочкой полого шара
всегда выходит своим головным концом
вперед. За первым науплиусом через ту
же щель в мешковой оболочке
выпячивается таким же способом второй, или оба
«выстреливают» одновременно. Первым
толчком к разбуханию эластичной
оболочки служит, по-видимому, разрыв
науплиусом яйцевой оболочки (рис. 210).
Только через некоторое время
новорожденный науплиус, находящийся
внутри полностью раздутой, принявшей
шаровидную форму эластичной оболочки,
начинает двигаться и пытается ее разорвать-
Это удается ему не сразу, после чего
оболочка спадается и личинка оказывается на
свободе. Утомленная тяжелой работой,
она сначала почти не способна быстро
передвигаться, но это ей и не нужно, так
как она находит достаточное количество
пищи на поверхности только что
покинутой мешковой оболочки, которая обычно
за многомесячное пребывание в водоеме
обрастает водорослями и покрывается
частицами детрита.
При помещении яиц A. arcticus в воду
неблагоприятных для его существования
водоемов науплиусы внутри яиц
развивались нормально, но их вылупления не
происходило. Специально поставленными
опытами Е. В. Боруцкий доказал, что
при сравнительно высоком содержании
солей в воде не происходит диффузии воды
Рис. 211. Цисты гарпактицид:
1 — Canthocamptus staphylinus;
2 — Attheyella wulmeri.
415
через эластичную внутреннюю оболочку
и она не разбухает. Если вода не имеет
кислой реакции, яйцевая оболочка
частично не растворяется и не размягчается,
что также исключает возможность вы-
лупления науплиуса. Таким образом, обе
эти оболочки препятствуют вылуплению
науплиуса при попадании яйца в
неблагоприятные для рачка условия,
обрекающие его на гибель. Действительно,
личиночные стадии и взрослые рачки
погибали в воде низовых (несфагновых) болот,
а также других водоемов, содержащей
обычное количество солей и имеющей
нейтральную или щелочную реакцию.
Все это делает понятным строгую
приуроченность A. arcticus к верховым,
сфагновым, болотам с их специфическим
гидрохимическим режимом.
Если A. arcticus в активном состоянии
существует весной, то некоторые другие
виды пресноводных гарпактицид
встречаются только зимой или только летом.
При этом известны виды, проводящие
период покоя не в стадии покоящегося
яйца, как A. arcticus, а в стадии цист,
несколько напоминающих описанные
выше цисты циклопов. У Canthocamptus
staphylinus такие цисты округлые, у
Attheyella wulmeri и A. northumbrica
они овальные, причем фуркальные
щетинки рачка торчат из оболочки (рис* 211).
Среди пресноводных гарпактицид есть
виды, способные к партеногенетическому
размножению, не свойственному всем
остальным веслоногим ракообразным. У
широко распространенной в Европе Е1а-
phoidella bidens самцы вообще неизвестны,
а в условиях эксперимента у этого вида
удалось получить 5 поколений партено-
генетических самок. К
партеногенетическому размножению оказался способным
и Epactophanes richardi, хотя в
естественных условиях он представлен как
самками, так и самцами. По-видимому, и
некоторые другие виды гарпактицид могут
размножаться партеногенетически.
Практическое значение гарпактицид
несравненно меньше, чем каланид и
циклопов. В некоторых водоемах они
составляют заметную часть пищи рыб, в
особенности их молоди.
ПАРАЗИТИЧЕСКИЕ ВЕСЛОНОГИЕ
Паразитические веслоногие
ракообразные прикрепляются к самым различным
рыбам, как морским, так и
пресноводным, а также ко многим морским
беспозвоночным — сифонофорам, кораллам,
моллюскам, полихетам, эхиуридам,
высшим ракообразным, иглокожим и к
некоторым другим. Как правило, каждый
Т а б л и ц а 32. Усоногие ракообразные:
1 — Lepas anatifera;
2 — L. fascicularis;
3 — Verruca stromia;
4 — Mitella mitella;
5 — Conchoderma auritum на Coronula
dema;
6 — Scalpellum Japonicum;
7 — Balanus improvisus;
8 — B. eyermanni;
9 — B. balanus;
10 — Chthamalus stellatus;
^1 — Xenobalanus globicipitis;
12 — Cryptolepas rajchianecti.
вид паразита паразитирует на
определенном виде хозяина, однако есть и
менее разборчивые, способные использовать
разные виды рыб.
Большинство паразитических
веслоногих представляет собой эктопаразитов.
Они поселяются на наружных покровах,
на жабрах, на глазах, реже в ротовой
полости рыб. Среди паразитов морских
беспозвоночных много эндопаразитов. Так,
например, Monstrillidae живут в
кровеносных сосудах полихет, Mytilicola —в
кишечнике моллюсков, несколько родов—
в кишечнике иглокожих.
Паразитические веслоногие
принадлежат к трем подотрядам — Cyclopoida,
включающему наряду с паразитическими
и только что рассмотренные свободножи-
вущие виды, Caligoida и Lernaeoida, в
состав которых входят исключительно
паразитические виды.
Паразитические Cyclopoida отличаются
от свободноживущих прежде всего
строением своих головных придатков. Задние
антенны крючковидные, приспособленные
для прикрепления к хозяину. Жвалы,
если есть, не бывают жующего типа,
другие ротовые части также видоизменены
или отсутствуют. Тело некоторых видов
сегментировано так же, как у свободно-
живущих, но у других сегментация в той
или иной степени исчезает. У таких
специализированных видов редуцируются и
грудные ножки. К этому подотряду
относится большое количество паразитов
пресноводных и морских рыб, а также
морских беспозвоночных.
Одними из самых обычных паразитов
как пресноводных, так и морских рыб
можно считать представителей рода Ег-
gasilus, прикрепляющихся к жабрам
линей, щук и многих других рыб. Нередко
к одной рыбе прикрепляется по
нескольку десятков, а иногда даже до 3000
рачков, что приводит ее к гибели. Замечено,
что, чем старше рыба, тем больше рачков
на ней паразитирует. Рачки охватывают
жаберные стебельки своими крючковид-
ными задними антеннами, при смыкании
образующими кольцо, на котором
паразит, по выражению В. А. Догеля,
болтается, как замок на скобе (рис. 212, 5).
Постепенно эпителий жабр разрушается.
Вышедшие из яиц личинки Ergasilus ведут
свободный образ жизни. Через 2—21/2
месяца рачки достигают половозрелости
и спариваются. Оплодотворенные самки
активно двигаются против течения. Это
помогает им оседать на жабры рыб,
поскольку из-под жаберной крышки
направляется ток воды.
Таким же путем жабры рыб
поражаются глохидиями перловиц (см. выше).
Интересно, что между эргаз ил идами и
глохидиями существует антагонизм: одни
вытесняют других и совместно на жабрах
одной рыбы не встречаются.
Если эргазилиды паразитируют во
взрослом состоянии и ведут свободный
образ жизни на ранних стадиях своего
развития, то представители семейства
Monstrillidae, паразитирующие в морских мно-
гощетинковых червях, имеют совсем
другой жизненный цикл. Взрослые рачки
оживленно плавают подобно обычным сво-
бодноживущим веслоногим.
Замечательно, однако, что они полностью лишены
кишечника и совсем не питаются. Из яиц
этих самок выходят также
свободноплавающие науплиусы, как и взрослые,
лишенные кишечника. Науплиусы
отыскивают определенные виды полихет (из
Рис. 212. Ergasilus sieboldi:
1 — рачок на жабре линя; 2 — рачок на жаберном
лепестке; 3 — то же, вид сверху; 4 — жабры
линя, пораженные Ergasilus.
27 Жизнь животных, т. 2
417
Рис. 213. Развитие Наетосега danae (сем. Monstrillidae):
Л — свободноплавающий науплиус; Б — науплиус после проникновения в червя; Б, Г —
две стадии зародыша в кровеносном сосуде червя; Д — многощетинковый червь Salmacina
danae с почти взрослой самкой рачка в спинном кровеносном сосуде; Е — самка из
кровеносного сосуда Salmacina; Ж —свободноплавающая самка; 1 —отростки зародыша; 2—
шипы на ваднем конце его тела; 3 — самка; 4 — всасывающие пищу отростки самки; 5 —
глаз; 6 — яичник; 7—зачатки грудных ножек; 8 — яйцевой мешок; 9 — передняя антенна.
Рис. 214.
Самка
Mytilicola.
родов Polydora, Salmacina и др.) и
внедряются в них, проникая в спинной
кровеносный сосуд. Здесь науплиус
линяет, превращаясь в многоклеточное
овальное тельце. Впоследствии у этого
зародыша на переднем конце вырастают два
длинных придатка, служащих для
всасывания пищи. Зародыш линяет еще раз
и преобразуется в длинное колбасовидное
тело, внутри которого формируется
взрослый рачок с хорошо развитыми
половыми органами. Он прорывает стенку
кровеносного сосуда и покровы хозяина и
приступает к активному существованию
(рис. 213).
Из паразитов беспозвоночных
наибольшее хозяйственное значение имеет Му-
tilicola (рис. 214). Взрослые самки рачка
достигают 8 мм длины и живут в
кишечнике мидий и устриц. Тело паразита
вытянутое, червеобразное, но следы
сегментации сохраняются; грудные ножки
редуцированы. Науплиусы и метанауп-
лиусы живут свободно, первая копепо-
дитная личинка внедряется в кишечник
моллюсков и превращается там во
взрослого рачка* Заражение Mytilicola
вызывает массовую гибель мидий. Запасы этих
промысловых моллюсков у берегов
Голландии в результате заражения рачками
в 1949 и 1950 годах уменьшились в 10 раз.
Представители подотряда Caligoida
характеризуются расширенным,
сплющенным в спинно-брюшном направлении
телом, сегментация грудного отдела в той
или иной степени утеряна, самки имеют
очень крупный передний брюшной (ге-
нитальный) сегмент, к которому
прикрепляются два яйцевых мешка, ротовые
придатки образуют хоботок, позволяющий
высасывать кровь хозяина. Самки и
самцы мало различаются по величине и по
строению.
Эти рачки паразитируют главным
образом на рыбах, как морских, так и
пресноводных, но род Archicaligus живет на
замечательном головоногом моллюске
Nautilus. Взрослые особи могут на некоторое
время покидать хозяина, плавать, а
затем садиться на другого. Науплиусы ве-
418
дут свободное существование. Личинка
на копеподитной стадии снабжена
специальной лобной нитью, при помощи
которой она прикрепляется к хозяину.
После этого она переходит в стадию
куколки, помещающейся в кутикулярной
оболочке предыдущей стадии, неподвижной и
непитающейся. Куколка линяет и
превращается в также прикрепленную
последнюю личиночную стадию — хали-
мус, которая преобразуется, наконец, во
взрослого рачка (рис. 215, Б).
Некоторые калигиды, будучи сами
паразитами рыб, подвергаются нападению
других паразитов. Так, например, на
Lepeophtheirus, живущем на многих
морских рыбах, паразитируют плоские черви
Udonella и инфузории Conidophrys,
выступающие в качестве «сверхпаразитов»,
т. е. паразитов паразита (рис. 215, А).
Последний подотряд паразитических
веслоногих — Lernaeoida — объединяет
наиболее сильно видоизмененных под
влиянием паразитизма рачков. Взрослые
половозрелые самки обычно имеют
червеобразное тело без всяких следов
сегментации или даже разделения на отделы
тела, с выростами на переднем конце,
погруженными в кожу хозяина, и с
двумя яйцевыми мешками на заднем конце.
Самцы гораздо мельче самок и по своему
строению напоминают копеподитные
стадии. Рачки паразитируют на рыбах, реже
на амфибиях и китах. Некоторые из них
достигают очень крупных размеров:
паразит китов Penella balaenoptera длиною
32 см может считаться крупнейшим
представителем веслоногих. Он погружен в
наружные покровы китов на 5—7 см.
Развитие этих рачков обычно
укорочено. Стадии науплиуса и метанауплиуса
они проходят в яйце. Из него
вылупляется свободная копеподитная личинка,
достигающая половозрелости. Вскоре
происходит копуляция, после которой самцы
умирают. Молодые самки семейства Ler-
naeidae сначала паразитируют на одних
рыбах, например на камбаловых, а затем
переходят на тресковых, теряя следы
сегментации, конечности и превращаясь
в червеобразные существа (рис. 216).
Первым хозяином гигантской Penella
служат головоногие моллюски, а вто-
Рис. 215. Калигоидные рачки:
А — Lepeophtheirus с гиперпаразитами — Udonella (I)
и Conidophrys B)\ з — яйцевой мешок рачка; Б —
личиночные стадии Caligus: 4 — науплиус, 5 —
копеподитная личинка, 6 — куколка, 7 — халимус.
рым — киты. У представителей другого
обширного семейства этого подотряда —
Lernaeopodidae — смены хозяев не
происходит и копеподитная личинка
совершает свой метаморфоз на одном хозяине.
Лернеиды нередко вызывают гибель
пораженных ими рыб, как морских, так и
пресноводных. Они вредят лососевым
рыбам, разрушая их жабры, описаны случаи
смерти морских рыб, вызванной этими
рачками. Глубоко внедряясь в тело рыб,
некоторые из лернеид достигают сердца и
печени своих хозяев, другие вызывают
образование кровоточащих ран на
поверхности тела хозяина. Кроме того, питаясь
кровью рыб, эти паразиты сильно их
истощают.
ОТРЯД КАРПОЕДЫ, ИЛИ
КАРПОВЫЕ ВШИ (BRANCHIURA)
На пресноводных рыбах паразитируют
не только веслоногие ракообразные.
Нередко на поверхности их тела, особенно
27*
419
Рис. 216. Стадии развития Lernaeocera branchi-
alis:
1 — взрослая самка с еще недоразвитыми яйцевыми
мешками; 2 — самец с жабр камбалы; 3 — неоплодотво-
ренная самка; 4 — копеподитная свободноплавающая
личинка; 5 — взрослая самка с развитыми яйцевыми
мешками с жабр трески.
там, где кожа тоньше и не так сильны
токи омывающей рыбу воды, т. е. за
жаберными крышками и за грудными
плавниками, можно найти сравнительно
крупных C—8 мм) очень плоских рачков,
прикрепленных к коже рыбы присосками и
жадно сосущих их кровь.
В отличие от веслоногих их нельзя
считать постоянными паразитами:
насытившись, карпоеды оставляют рыбу и с
огромной скоростью уплывают прочь.
Карпоеды живут не только в пресных
водах, но и в море. Всего их известно
около 60 видов.
По своему строению эти рачки
довольно резко отличаются от веслоногих
(рис. 217). Их плоское тело явственно
подразделяется на два отдела — передний и
задний. Передний, головогрудной, отдел
сверху покрыт широким головогрудным
щитом — карапаксом, на спинной
поверхности которого расположена пара
крупных фасеточных глаз, а между ними три
простых науплиальных глазка. Частично
этот щит покрывает и передний из
четырех относящихся к заднему отделу тела
свободных грудных сегментов,
снабженных каждый парой двуветвистых
плавательных ножек.
Тело заканчивается укороченным
брюшным отделом, слившимся с широкими фур-
кальными ветвями.
Придатки переднего отдела
приспособлены к паразитическому существованию,
а придатки заднего отдела — к
плаванию. Обе пары антенн очень короткие.
Жвалы образуют направленный назад
хоботок, вводимый в покровы рыб и
служащий для сосания их крови. По
направлению вперед от него отходит острый шип,
также помогающий рачку удерживаться
на коже рыб. По бокам от хоботка
расположены мощные присоски,
представляющие собой очень сильно измененные
передние челюсти. Задние челюсти одно-
ветвистые и служат для прикрепления к
хозяину. Четыре пары грудных ножек
несут многочисленные перистые щетинки
и совершают сильные взмахи, благодаря
которым рачок может быстро плавать
спинной стороной вверх. Кишечник кар-
поедов так же приспособлен к
паразитическому существованию, как и их
ротовые придатки. Средний его отдел снабжен
разветвленными слепыми выростами,
служащими резервуарами для высосанной
крови. Благодаря способности запасать
пищу впрок карпоеды могут до 3 недель
не питаться, постепенно расходуя свои
запасы.
При нападении рачка на рыбу зрение
не играет никакой роли. Рачки
руководствуются главным образом движением
воды и отчасти химическим чувством. Они
420
воспринимают соответствующие
раздражения многочисленными чувствительными
щетинками, расположенными на разных
частях тела. Если в аквариум с карпое-
дами опустить фильтровальную бумажку
и подвигать ею в воде, рачки бросаются
на бумажку, но, прикоснувшись к ней,
отплывают прочь. Если бумажкой
предварительно обтерта рыба, карпоеды
задерживаются на ней несколько дольше.
Карпоеды преследуют рыбу, быстро
двигаясь в том же направлении параллельно
ей, а затем садятся на голову жертвы.
Впоследствии они переползают на те части
тела рыбы, которые наименее сильно
омываются водою и где покровы
сравнительно тонкие, т. е. прикрепляются
преимущественно позади жаберных крышек и
грудных плавников. Присосавшись к
рыбе, рачки продолжают действовать
своими плавательными ножками, возбуждая
ими ток воды, необходимый для дыхания.
Оно осуществляется через тонкостенные
участки карапакса. Сердца у карпоедов
нет; кровь в полости тела двигается
благодаря сокращениям кишечника и
мускулатуры брюшного отдела.
Никакого предпочтения определенным
видам рыб карпоеды не оказывают.
Больше того, они могут нападать и на других
водных позвоночных — на тритонов и на
головастиков лягушек. Некоторые
морские виды карпоедов паразитируют на
головоногих моллюсках, например Argulus
arcussonensis на каракатицах.
Самец оплодотворяет присосавшуюся к
рыбе самку, удерживая ее задние грудные
ножки своими и подсовывая под нее свой
брюшной отдел так, чтобы отверстия его
семепроводов совпали с отверстиями ее
яйцеводов. Сперма попадает
непосредственно в половые пути самки. Перед
откладкой яиц самка покидает рыбу и
плавает в поисках подводных растений,
камней или другого подходящего субстрата.
Она откладывает двойной ряд яиц
(обычный Argulus foliaceus от 20 до 250 яиц)
на этот субстрат, прикрепляя их
специальным секретом. Обычно яйца
выметываются не сразу, а с перерывами, во
время которых самка снова нападает на
рыб и сосет их кровь. В этом отношении
карпоеды отличаются от большинства
других ракообразных, самки которых
носят яйца на себе в яйцевых мешках или
в специальной выводковой сумке.
Через 3—5 недель из яиц Argulus
foliaceus выходят молодые, но еще не
вполне сформировавшиеся рачки (рис. 217, 3).
Они имеют длинные задние антенны, с
помощью которых плавают, их жвалы
снабжены щупиками, передние челюсти еще
не превращены в присоски,
плавательные ножки недоразвиты, а карапакс
укорочен. Личинки прикрепляются к рыбам,
используя для этого передние антенны и
концевые шипы передних челюстей. В
течение первых 8 дней пребывания на рыбе
рачок дважды линяет, причем его задние
антенны и мандибулы укорачиваются, ко
зато полностью развиваются
плавательные ножки. После третьей линьки за счет
передних челюстей начинают
образовываться присоски; этот процесс
заканчивается только после пятой линьки. Так,
постепенно, в результате линек,
происходящих каждые 3—4 дня, формируется
взрослый рачок. У некоторых других
видов карпоедов развитие сокращено еще
больше: из яйца выходит вполне
сформировавшаяся, но еще неполовозрелая
особь.
Карпоеды довольно широко
распространены как в пресных водах, так и в морях
Рис. 217. Карпоеды:
I, 2 — Argulus foliaceus, вид спереди (i) и снизу B);
3 — вышедшая из яйца молодая особь Argulus japoni-
cus.
421
и могут считаться весьма обычными
паразитами рыб, в том числе и хозяйственно
ценных. Однако вред, приносимый ими
рыбам, невелик. В очень редких случаях
они вызывают гибель рыб, да и то только
молодых.
ОТРЯД МИСТАКОКАРИДЫ
<MYSTACOCARIDA)
Песчаные морские пляжи южных морей
привлекают массу отдыхающих.
Искупавшись, любители загорать
растягиваются на песке и подставляют свое тело под
жаркие лучи солнца. Но вряд ли
кто-нибудь из них знает, что в толще песка, на
котором он расположился, идет
интенсивная жизнь мелких своеобразных существ.
Только поверхностные слои песка, да
и то на некотором расстоянии от уреза
воды, лишены влаги. На той или иной
глубине песок пропитан морской водой.
Она заполняет узкие капиллярные
пространства между песчинками, и в этих
извилистых ходах, диаметр которых обычно
меньше 1 мм, обитает особая фауна,
получившая название интерстициальной. В
нее входят представители разных групп
животных — простейшие,
кишечнополостные, различные черви, водяные клещи и
многие другие, но особенно
многочисленны приуроченные к таким своеобразным
условиям ракообразные. Выше уже
упоминались интерстициальные гарпактици-
ды (рис. 209), однако представители этого
подотряда веслоногих встречаются не
только в пропитывающей песок воде, но и
в других весьма различных условиях.
Совместно с гарпактицидами в капиллярных
ходах пляжей Средиземного моря и
некоторых участков Атлантического и
Индийского океанов живут мелкие (до 0,5 мм)
рачки, принадлежащие к особому
отряду мистакокарид, который приспособлен
к существованию исключительно в интер-
стициальных водах.
Мистакокариды были открыты только в
1943 году на американском побережье
Атлантического океана. Впоследствии они
были обнаружены на Атлантическом
побережье Европы и Африки и вдоль почти
всех берегов западной половины
Средиземного моря, далее, в Южной Африке и,
наконец, в Чили. При предварительных
исследованиях интерстициальной фауны
Черного и Каспийского морей найти этих
рачков пока не удалось, так что для СССР
они еще не указаны. Все они
принадлежат к одному роду Derocheilocaris,
включающему всего 3 вида — американский
D. typicus, европейско-африканский D.
remanei и чилийский D. galvarini.
Подобно другим представителям
интерстициальной фауны мистакокариды имеют
длинное и тонкое червеобразное тело,
позволяющее им ползать по узким ходам
между песчинками. Голова удлиненная и
устроена своеобразно. Ее передняя часть —
акрон, несущая передние антенны,
отделена швом от задней части, на которой
расположены задние антенны и хорошо
развитые ротовые придатки. С
задней частью головы частично
сливается первый грудной сегмент, конечности
которого преобразованы в ногочелюсти.
Далее следуют 4 свободных грудных
сегмента. Каждый из них снабжен парой
коротких нерасчлененных придатков с
щетинками на концах, представляющих
собой редуцированные грудные ножки.
Брюшной отдел состоит из 6 сегментов,
подобных грудным, но лишенных
конечностей. Он заканчивается длинными ког-
тевидными фуркальными ветвями. На всех
грудных и брюшных сегментах, за
исключением последнего, с боков расположены
щелевидные углубления. Назначение этих
органов неизвестно (рис. 218, 2).
Наиболее характерной особенностью
мистакокарид можно считать сильное
развитие головных и редукцию грудных
придатков, чего не наблюдается у других
ракообразных. В соответствии с этим
движения мистакокарид весьма своеобразны:
если другие ракообразные передвигаются
преимущественно при помощи грудных
ножек, то мистакокариды используют в
качестве органов движения головные
придатки и ногочелюсти. Рачки перебирают
попеременно задними антеннами,
щупиками жвал, челюстями и ногочелюстями,
отталкиваясь ими или сидящими на них
щетинками от стенок хода, по которому
ползут. При этом передние антенны на-
422
правлены вперед и нащупывают свободное
пространство между песчинками, а
грудные ножки остаются прижатыми к телу
и никакого участия в движении рачка не
принимают. Кроме ползания, мистакока-
риды способны передвигаться прыжками.
Они круто изгибают свое тело на
брюшную сторону так, что конец фурки
достигает уровня задних антенн и касается
верхней губы, а затем распрямляются.
Опираясь на заднюю часть тела, они
проталкивают его вперед. Необычайная
подвижность грудных и брюшных сегментов
друг относительно друга позволяет
рачкам не только изгибать тело в разных
направлениях, но и производить
«телескопические» движения, при которых каждый
последующий сегмент слегка погружается
в предыдущий, причем тело укорачивается,
а затем, при возвращении сегментов к
нормальному положению, снова
удлиняется.
Пищей мистакокаридам служат мелкие
органические частицы, которые
направляются сзади наперед щетинками
внутренних краев челюстей и ногочелюстей.
Перед ртом находится необыкновенно
длинная, загнутая назад верхняя губа,
задерживающая пищевые частицы, по бокам
рта расположены жевательные лопасти
жвал, размельчающие пищу, а челюсти
благодаря удлинению головы отодвинуты
далеко назад от ротового отверстия, и
только их щетинки достигают рта.
Таким образом, в отличие от других
ракообразных, мистакокариды пользуются
челюстями больше как органами
передвижения, чем как ротовыми придатками.
Мистакокариды раздельнополы, но
самцы и самки внешне совершенно сходны.
Процессы спаривания и откладки яиц этих
ракообразных наблюдать не удалось.
Изучение их личинок показало, что из яйца
выходит метанауплиус длиною 0,15 мм.
Личинка линяет 9 раз, причем при каждой
линьке добавляется по одному сегменту
и постепенно развиваются недостающие
конечности. Такое равномерное и
постепенное превращение личинки во
взрослое животное и значительное количество
личиночных стадий говорит о
примитивном характере постэмбрионального
развития мистакокарид.
Рис. 218. Derocheilocaris remanei:
1 — со спинной стороны; 2 — сбоку, перед
проталкиванием тела вперед; видна верхняя губа; 3 — первая
личинка.
Далеко не каждый песчаный пляж
населен мистакокаридами. Они могут жить
только в очень мелком песке, частицы
которого имеют диаметр около 0,2 мм.
Как в более, так и в менее
крупнозернистом грунте они не встречаются. Гораздо
менее чувствительны эти рачки к
солености и температуре. Наблюдения на пляжах
Средиземного моря показали, что
мистакокариды без всякого вреда для себя
переносят колебания солености от 10 до
32°/00 и температуры от 10 до 26°С. Все
известные их местонахождения как в
северном, так и в южном полушарии
приурочены к зонам, температура которых даже
в самое холодное время года не опускается
ниже 10° С (рис. 219).
Может быть, именно этим объясняются
неудачи поисков мистакокарид в наших
морях.
Все мистакокариды найдены на
пляжах выше уровня моря. Только
чилийский Derocheilocaris galvarini обнаружен
на глубине 25 м. На этом основании можно
предполагать, что интерстициальная
фауна населяет и толщу морских грунтов
423
Рис. 219. Географическое распространение мистакокарид и температура наиболее холодного месяца
года (для северного полушария — февраля, для южного — августа).
в тех случаях, когда диаметр частиц
грунта оказывается подходящим для
составляющих ее животных.
Распространение мистакокарид ставит
перед исследователями некоторые
трудные и еще не решенные вопросы. На
рисунке 219 видно, что D. remanei образует
отдельные колонии, удаленные друг от
друга на сотни и тысячи километров.
Никаких покоящихся стадий у
мистакокарид нет, возможности их расселения
крайне ограничены, и, следовательно, эти
колонии полностью изолированы одна от
другой. В таких случаях обычно
представители различных изолированных
колоний эволюционируют в разных
направлениях, что приводит в конце концов к
возникновению разных видов. Между тем
мистакокариды Средиземного моря и
атлантического побережья Южной Европы
совершенно идентичны рачкам с
атлантического побережья Африки.
Приходится признать, что эволюция обитателей
интерстициальных вод идет очень
медленно; поэтому, несмотря на давнюю
изоляцию отдельных колонии мистакокарид,
строение составляющих их рачков
осталось неизменным.
Если предположение о замедленном
темпе эволюции обитателей
интерстициальных вод правильно, то можно
рассчитывать встретить среди них древние, мало
изменившиеся виды. К таким видам
следует причислить и мистакокарид, которым
свойственны многие чрезвычайно
примитивные признаки (например, строение
нервной системы и ротовых придатков) и
примитивный характер развития.
ОТРЯД МЕШКОГРУДЫЕ
РАКООБРАЗНЫЕ (ASCOTHORACIDA)
В 1887 году на организованной
незадолго до этого на Соловецких островах
биологической станции проводились
исследования фауны Белого моря. Известный
русский зоолог Н. Вагнер вскрывал
морских звезд. В полости тела одной из
424
них он обнаружил разветвленное мягкое
тело какого-то паразита. Вскоре будущий
академик В. М. Шимкевич нашел
такое же существо в полости тела
многолучевой морской звезды Solaster. В 1889
году он писал про свою находку:
«Настолько загадочная форма, что почти
ничего невозможно сказать относительно ее
природы». Этой «загадочной формой»
заинтересовался другой крупный зоолог и
будущий океанограф — Н. М. К н и п о-
в и ч, детально ее изучивший. В
результате кропотливых многолетних
исследований он установил, что это существо
принадлежит к группе паразитических
ракообразных, которую в то время относили
к отряду усоногих, а теперь считают
самостоятельным отрядом мешкогрудых
ракообразных. Н. М. Книпович назвал
изученное им существо Dendrogaster, что
значит «разветвленный живот».
Организация Dendrogaster очень
сильно изменена под воздействием
паразитизма. Тело самки состоит главным
образом из двух боковых многократно
ветвящихся отростков. Эти два ответвления
отходят от срединной непарной части, на
самом переднем конце которой находятся
передние антенны, а также сложенные в
виде пирамиды жвалы и челюсти, очень
сильно видоизмененные. Никаких других
признаков ракообразных (сегментации,
туловищных конечностей и т. д.) животное
не сохранило (рис. 220, А). В ответвлениях
тела самки помещаются карликовые
паразитические самцы, более сходные с
обычными ракообразными. Тело их
сегментировано, голова снабжена такой же
ротовой пирамидой, как у самок, но грудной
отдел несет 5 пар двуветвистых ножек.
Необычный вид самцу придает пара очень
длинных задних выростов, в которые
заходят ветви кишечника и семенники
(рис. 220, В).
Не все мешкогрудые видоизменены в
такой степени, как самки Dendrogaster.
Среди них есть и виды, сохранившие облик
нормальных ракообразных, есть и менее
измененные, чем Dendrogaster.
Представителей этого отряда по степени
упрощения организации можно расположить в
определенный ряд, по которому можно
судить о том, как шла эволюция
мешкогрудых ракообразных в связи с
приспособлением к паразитическому
существованию и с переходом с одних хозяев на
других.
На поверхности колоний коралла Ап-
tipates larix в Средиземном море живет
мешкогрудый рачок Sinagoga mira. Его
самцы и самки имеют одинаковое строение,
могут свободно передвигаться с места на
место и даже плавать. Покровы головы
образуют два боковых листка,
покрывающих тело не целиком. Шестисегментный
грудной отдел снабжен шестью парами
двуветвистых плавательных ножек.
Сегментированное брюшко заканчивается
снабженной перистыми щетинками фур-
кой. Близкий вид — S. metacrinicola —
паразитирует на морской лилии Meta-
crinus rotundus у берегов Японии. У самок
этого вида мантия покрывает все тело, у
самца она несколько короче.
По-видимому, самки малоподвижны, но все же могут
иногда менять место своего
прикрепления. Другие роды того же семейства Si-
nagogidae — Parascothorax и Ascothorax —
обитают внутри мешковидных
опухолей (цист), которые под их
воздействием образуются на поверхности диска
зараженных ими офиур или в бурсальных
полостях этих иглокожих. Самки этих
родов гораздо крупнее самцов,
неподвижно прикреплены клешневидными
передними антеннами к хозяину, мантия их
расширена за счет увеличения объема и
количества выводковых камер,
сегментация и грудные конечности имеются, но не
все они двуветвисты, брюшко подогнуто -
под грудь. Грудными ножками самки
возбуждают ток воды, направленный в
бурсу и необходимый им для дыхания.
Самцы живут в тех же полостях, что и самки
(рис. 220, 5, Г).
Рассмотренное раньше семейство Den-
drogastridae, представители которого
паразитируют в полости тела морских звезд
и морских ежей, отличается от семейства
Sinagogidae тем, что мантия самок
чрезвычайно сильно разрастается с боков,
принимая разветвленную древовидную
форму или имея вид мешка, иногда
раздвоенного. Собственно тело рачка
помещается в самой передней части непарного
срединного ствола мантии; оно не сегмен-
425
Рис. 220. Мешкогрудые ракообразные и их личинка:
А, Б — самка и самец Dendrogaster dichotomus; В, Г — самка и самец
Ascothorax; Д — аскоторацидная личинка Ascothorax, внешний вид и характер
сегментации; 1 — передние антенны; 2 — ротовая пирамида; 3 — задняя кишка; 4 —
ответвления кишечника.
тировано, лишено грудных ножек или они
не членистые. Маленькие самцы
паразитируют в мантийной полости самок.
В несколько ином направлении
эволюционировало семейство Lauriidae.
Относящиеся к нему рачки паразитируют на
кораллах, причем самки — эндопаразиты,
сидящие в цистах под сарколеммой, а
самцы — эктопаразиты тех же хозяев,
внутри которых живут самки. У самок
мантия разрастается вверх и вниз и
превращается в толстую пластинку, внутри
которой разветвляются яичники и
печеночные выросты; грудные ножки в той
или иной степени недоразвиты,
сегментация сохранилась. Самцы
приблизительно такие же, как у других мешкогрудых,
значительно мельче самок.
Таким образом, при переходе
мешкогрудых от эктопаразитизма к эндопара-
зитизму отличия между самцами и
самками становились все более резкими,
самки теряли сегментацию, их грудные
ножки редуцировались, а мантия
разрасталась. Самцы изменялись гораздо меньше
самок.
Все мешкогрудые ракообразные
прикрепляются к хозяину при помощи
передних антенн, концевые членики которых
образуют подобие клешни. Задние антенны
отсутствуют. Жвалы и челюсти сложены
вместе и составляют колющий хоботок.
Этим хоботком эктопаразитические Si-
nagoga прокалывают ткани своих хозяев
и сосут их соки. Ascothorax питается
разрушенными им клетками стенок бурсы
офиур, а паразит неправильных морских
ежей — Ulophysema — поедает половые
железы хозяина. Разветвленные самки
Dendrogaster заглатывают клетки
полости тела морской звезды, одевающие
паразита сплошным слоем.
Обычно присутствие паразита вызывает
кастрацию хозяина. Если Ulophysema
непосредственно разрушает половые
железы морских ежей, то Ascothorax и
Dendrogaster подавляют действие половых
желез офиур и морских звезд
механически.
Специальных органов дыхания у
мешкогрудых ракообразных нет. Кислород
усваивается стенкой мантии. Весьма
вероятно, что разветвленная мантия
Dendrogaster представляет собой
приспособление, позволяющее использовать
наибольшее количество кислорода, растворенного
в полостной жидкости морской звезды,
благодаря значительному увеличению
усваивающей поверхности. Молодые самки,
двигая ветвями мантии, могут медленно
перемещаться в полости тела хозяина.
Все мешкогрудые ракообразные
раздельнополы, за исключением паразита
одиночных кораллов Petrarca bathyactidis,
которому свойствен гермафродитизм. Чем
более специализированы роды
мешкогрудых, тем сильнее выражены различия
между самцами и самками. У всех
внутренних паразитов самки гораздо крупнее
самцов и гораздо сильнее видоизменены
под воздействием паразитизма. Обычно
совместно с одной самкой живет по
нескольку самцов, а у Dendrogaster они
паразитируют в выростах мантии самки.
Оплодотворенные самки откладывают
яйца в выводковые камеры,
расположенные с внутренней стороны мантии. У
Dendrogaster в этих же камерах
паразитируют самцы. Число откладываемых
самкой яиц различно у разных родов,
причем, чем глубже приспособления к
паразитизму, тем плодовитость выше. Мало-
измененная Sinagoga откладывает всего
50-60 яиц, Ascothorax — 250—300, а
Dendrogaster — около 20000.
Из развивающихся в выводковых
камерах яиц выходят личинки, находящиеся
на стадии науплиуса, но лишенные нау-
плиального глаза. У
малоспециализированной Sinagoga, а также у Lauriidae на-
уплиусы покидают тело матери и
приступают к самостоятельному существованию.
У остальных мешкогрудых науплиусы
остаются в выводковых камерах и там
продолжают свое развитие. У Parascothorax
личинка выходит на волю на стадии мета-
науплиуса, а у Ascothorax и почти всех
Dendrogastridae личинки еще дальше
развиваются в выводковых камерах, достигая
в конце концов свойственной только меш-
когрудым стадии так называемой а с к о-
торацидной личинки. Именно на
этой стадии рачок выбирается наружу,
плавает и заражает другие особи своих
хозяев. У некоторых видов Dendrogaster
из яйца сразу появляется аскоторацидная
427
личинка, т. е. происходит сокращенное
развитие. Для этой личинки характерны
двустворчатая мантия, клешневидные
передние антенны, колюще-сосущие ротовые
придатки, шесть пар плавательных
грудных ножек, 5-сегментное брюшко и
плавательная фурка (рис. 220, Д). В общем
аскоторацидные личинки похожи на
взрослых Sinagoga и на самцов
большинства родов.
Аскоторацидные личинки проникают в
своих хозяев и там развиваются во
взрослых рачков. Если такая личинка окажется
в одиночестве, она становится самкой.
Если же она прикрепилась около вполне
сформировавшейся самки или на
поверхности ее тела, то развивается самец.
Процесс превращения аскоторацидной
личинки во взрослую самку Dendrogaster
весьма своеобразен. Личинка садится на
поверхность тела морской звезды, линяет,
теряет сегментацию и грудные ножки и
приобретает грушевидную форму. После
этого она проникает через стенку тела
хозяина в его половые железы.
Последующий рост происходит исключительно
за счет разрастания мантии; собственно
тело зародыша почти не увеличивается в
размерах (рис. 221).
Мешкогрудые ракообразные, в
особенности Sinagogidae, широко
распространены в Мировом океане. Dendrogastridae
обитают в умеренных и высоких широтах
северного полушария,
Lauriidae—преимущественно в тропиках. Они поражают не
Рис. 221. Формирование самки Dendrogaster.
Пять последовательных стадий (заштриховано тело
рачка).
только мелководных, но и глубоководных
животных. Так, например, в Беринговом
море в морских звездах Psilaster pectinatus
и Eremicaster tenebrarius с глубин 2300 и
3940 м найдены Dendrogaster astropectinis
и D. beringensis. Всего известно около
40 видов мешкогрудых, половина которых
принадлежит к описанному впервые
Н. М. Книповичем роду Dendrogaster.
ОТРЯД УСОНОГИЕ РАКООБРАЗНЫЕ
(CIRRIPEDIA)
Чарлз Дарвин в своей автобиографии
писал: «В октябре 1846 г. я начал
работать над «Усоногими ракообразными»... я
занялся изучением и анатомированием
ряда обычных форм, и это постепенно
привело меня к исследованию всей группы.
В течение восьми лет я непрерывно
работал над этим предметом и в конце концов
издал два толстых тома, содержащих
описание всех известных современных видов,
и два тонких in quarto о вымерших видах.
Не сомневаюсь, что сэр Э. Литтон-Бул-
вер, выведя в одном из своих романов
профессора Лонга, который написал два
увесистых тома о ракушках, имел в виду
меня».
Усоногие ракообразные, которым отдал
восемь лет своей жизни великий
натуралист, во многих отношениях замечательны,
и интерес к ним Ч. Дарвина вполне
понятен. Внешне они даже непохожи на
ракообразных. Во взрослом состоянии они
ведут сидячий образ жизни, прикрепляясь
к всевозможным подводным предметам —
скалам, камням, сваям, днищам судов, а
также к самым разнообразным морским
животным. Тело большинства этих
рачков заключено в твердую известковую
раковину, состоящую из различного
количества отдельных пластинок. Часть этих
пластинок подвижно сочленена между
собой. Рачок может их раздвигать, и в
образующуюся щель просовываются время
от времени грудные ножки, совершающие
характерные взмахи. При этом вода с
находящимися в ней организмами и их
остатками загоняется внутрь раковинки;
так осуществляется питание и дыхание.
428
Твердая известковая раковина,
постоянное прикрепление к субстрату и
отсутствие внешних признаков ракообразных
долго мешали выяснению истинной
природы усоногих. Даже такой зоолог, как
Ж. К ю в ь е, по примеру предыдущих
исследователей относил усоногих к
моллюскам. Ж. Б. Л а м а р к рассматривал
усоногих как переходную группу от
червей к моллюскам. Только в 1830 году,
после того как стали известны личинки
усоногих, сходные с личинками других
ракообразных, была доказана
принадлежность усоногих к этому классу.
В настоящее время в этом никто не
сомневается. Больше того, нетрудно
подметить многие признаки, сближающие
усоногих с внешне совершенно несходными
с ними веслоногими. Однако усоногие
эволюционировали в ином направлении,
чем подавляющее большинство остальных
ракообразных. Среди непаразитических
представителей этого класса только они,
вместо того чтобы совершенствовать свою
двигательную систему, перешли к
прикрепленному существованию и развили
мощное средство пассивной защиты —
толстую известковую раковину,
несколько напоминающую раковину моллюсков.
В связи с прикрепленным образом жизни
организация усоногих очень сильно
изменилась: исчезли наружная сегментация и
хитиновый панцирь, бесполезный при
наличии защитной известковой раковины,
перестроилась голова, которой рачки
прикрепляются к субстрату, укоротился и
почти исчез брюшной отдел. Изменилась
и функция конечностей. Они перестали
служить для передвижения и
приспособились к тому, чтобы загонять воду в
мантийную полость.
Часть усоногих ракообразных перешла
от сидяче-прикрепленного образа жизни
на поверхности тела различных морских
животных к паразитическому
существованию, сменив питание планктоном на
использование соков своих хозяев.
Можно наметить полную серию переходов от
свободного существования этих
ракообразных к паразитизму. Как и в других
подобных случаях, паразитизм вызвал ряд
изменений организации усоногих и эти
изменения отразились на
приспособлениях к прикрепленному образу жизни.
Среди паразитических усоногих известны
предельно упрощенные формы, утерявшие
все признаки не только отряда, но даже
класса ракообразных и типа
членистоногих. Только изучение строения личинок
позволяет понять их истинную природу.
Все усоногие —- морские обитатели,
причем некоторые из них могут выносить
значительное опреснение. Тело их окружено
двойной мантийной складкой,
соответствующей карапаксу других
ракообразных. Мантия непаразитических форм
выделяет различное количество известковых
пластинок, образующих защитную
раковину. Мягкое тело подразделено на
головной и грудной отделы. Передние
антенны служат для прикрепления к
субстрату, снабжены цементными железами
и очень сильно укорочены. Задние
антенны отсутствуют, ротовые придатки
упрощенного строения. На грудном отделе
расположено 6 пар двуветвистых ножек.
У сверлящих усоногих число грудных
ножек сокращено до 4 пар и они одноветви-
сты. У подотряда корнеголовых
ракообразных, все представители которого —
паразиты, ротовых придатков и грудных
ножек нет совсем.
Из яиц усоногих выходит типичный нау-
плиус, отличающийся от науплиусов
других ракообразных присутствием боковых
выростов на переднем конце тела, так
называемых «рожек». Перелиняв
несколько раз, науплиус переходит в стадию ме-
танауплиуса, для которой характерно
появление челюстей и зачатков других
конечностей и начало развития фасеточных
глаз по бокам от науплиального.
«Рожки» у метанауплиуса обычно сохраняются,
задняя часть тела часто снабжена
выростами. За метанауплиальной стадией
следует свойственная только усоногим стадия
циприсовидной личинки, иногда
называемая также «куколкой». Она обладает
двустворчатой хитиновой раковиной,
покрывающей все ее тело, 4-члениковыми
передними антеннами с
присасывательными дисками на предпоследнем
членике и с цементными железами при
основании, упрощенными жвалами и челюстями
и 6 парами грудных двуветвистых
плавательных ножек. На переднем конце ли-
429
Рис. 222. Личиночные стадии морской уточки
Lepas anatifera:
1,2 — первая и вторая стадии науплиуса; 3 — ципри-
совидная личинка.
чинки расположены науплиальный и
пара фасеточных глаз. Присутствие
двустворчатой раковины придает этой личинке
внешнее сходство с представителями
отряда ракушковых ракообразных (см.
ниже), в частности с относящимся к нему
родом Cypris, почему личинка и получила
название циприсовидной. Циприсовидная
личинка при помощи своих грудных ног
свободно плавает в воде, а затем
прикрепляется передними антеннами к
подходящему субстрату. Дальнейшее ее
превращение во взрослого рачка у разных
подотрядов происходит по-разному.
В отряд усоногих включают три
подотряда, резко различающихся как по
строению, так и по образу жизни.
ПОДОТРЯД ТОРАЦИКОВЫЕ РАКООБРАЗНЫЕ
(THORACICA)
Вновь построенный корабль покидает
гавань и начинает плавание. Скорость его
хода известна, и сначала он идет точно по
графику. Однако с каждым днем скорость
его уменьшается. На преодоление одного
и того же маршрута тратится все больше
времени и топлива, и рейсы корабля
обходятся все дороже и дороже. В чем тут
дело?
Днище корабля обросло различными
морскими животными, трение его о воду
увеличилось, и скорость хода
соответственно уменьшилась. Корабль
приходится ставить в док и удалять обрастания
его днища.
Подсчитано, что из-за обрастаний судов
и необходимости их периодического до-
кования владеющие значительным флотом
страны ежегодно терпят миллионные
убытки.
Основу обрастаний судов составляют
усоногие ракообразные — морские
желуди. Они нередко покрывают днище
корабля сплошным слоем. С китобойной
матки длиной 140 ле, плававшей в
течение 7 месяцев к югу от экватора, сняли
40 т морских желудей. На наших
дальневосточных судах за год плавания
нарастает 10—12 кг морских желудей на
на 1 м2 поверхности днища.
Морские желуди и относящиеся к тому
же подотряду морские уточки (табл. 32),
отличающиеся присутствием стебелька, на
котором расположено защищенное
известковыми пластинками тело рачка,
поселяются не только на кораблях.
Прибрежные скалы и камни усыпаны ими,
они прикрепляются к раковинам
моллюсков, к панцирям ракообразных, к
раковинам себе подобных, иногда к рыбам.
Некоторые виды приспособились к
существованию на коже китов и акул. Есть
морские желуди, сидящие на скалах
значительно выше уровня моря и
довольствующиеся брызгами, которые долетают
до них только при сильных штормах.
С другой стороны, известны усоногие,
обитающие на глубине до 6860 м. Всего
описано около 700 видов этих
ракообразных.
430
Тело морских желудей (семейство Ва-
lanidae) и веррук (семейство Verrucidae)
целиком находится внутри раковинки,
состоящей из нескольких известковых
пластинок. Часть этих пластинок
неподвижно соединена друг с другом и
составляет стенки домика. Другие образуют его
крышку. Они могут смыкаться и
размыкаться благодаря действию специальных
мышц. На дне домика спинной стороной
вниз лежит рачок. Передняя часть его
сильно видоизмененной головы подогнута
под тело так, что остатки передних антенн
оказываются приблизительно в центре
подошвы. У морских уточек (надсемейство
Lepadomorpha) передняя часть головы
вытянута в стебелек, голый или
снабженный известковыми чешуйками, и только
остальная часть тела покрыта
известковыми пластинками. Этим стебельком, на
конце которого помещаются остатки
передних антенн, рачки прикрепляются к
субстрату. Задняя часть головы всех
Thoracica вздутая. На ней расположен
обращенный кверху рот, окруженный
верхней губой, жвалами, передними и
слившимися воедино задними челюстями.
Строение всех ротовых придатков упрощенно,
так как они не служат для фильтрации.
Глаза очень слабо развиты и
представлены раздвоенным науплиальным глазом.
Грудной отдел несет 6 пар двуветвистых
ножек. Их размеры увеличиваются по
направлению спереди назад.
Многочисленные членики ветвей ножек снабжены
длинными щетинками на направленной
ко рту стороне. Эти ножки, высовываясь
из домика, расправляются, как веер, а
затем складываются. Они совершают,
таким образом, характерные взмахи,
загоняя воду и взвешенные в ней частицы в
полость домика.
Частота взмахов ножек зависит от
внешних условий, в первую очередь от
скорости течения воды и от температуры.
Обычный морской желудь Balanus balanoides
производит 6,2 биения в 10 секунд при
17° С и скорости течения более 0,1 см/сек.
При температуре ниже 1,8°С и выше
31,5° С, а также при скорости течения
больше 10 см /сек рачок смыкает створки
раковины и прекращает движения ножек.
Концентрация кислорода и углекислоты
в воде не влияет на ритм работы рачков.
Щетинки на передних ножках сидят
более, а на задних менее часто, благодаря
чему разные ножки отфильтровывают
частицы разного размера. Поэтому пища усо-
ногих рачков довольно разнообразна: это
водоросли, даже бактерии и целый ряд
планктонных животных, в первую
очередь веслоногих. Захватывают они и
собственных личинок, но, по имеющимся
наблюдениям, циприсовидные личинки в
кишечнике усоногих не перевариваются.
Морской желудь Chthamalus, живущий в
условиях сильного течения, держит свою
ловчую сеть расправленной до тех пор,
пока в ней не окажется достаточное
количество пищевых частиц.
Весьма своеобразны способ питания и
функции грудных ножек морской уточки
Lepas fascicularis (табл. 32, 2), широко
распространенной в тропической и
субтропической частях всех океанов. Эти рачки
прикрепляются к каким-нибудь плаваю-
Рис. 223. Строение раковины
и тела торациковых:
А — морского желудя (Bala-
nus); Б — морской уточки (Le-
pas); l — мускул крышки
раковины; 2 — ротовой конус;
3 — грудные ножки; 4 —
пенис; 5 — редуцированное
брюшко.
431
тцим предметам (кусочки дерева, пемза,
водоросли) или к выделяемому ими
пенистому поплавку и целыми группами
плавают на самой поверхности моря. Они
могут грести своими ножками и активно
подплывать к сифонофорам (Velella, Porpita),
которых обгрызают жвалами.
Есть морские уточки, добывающие себе
пищу не прибегая к помощи грудных
ножек, которые в таких случаях
укорачиваются и упрощаются. Malacolepas
conchicola живет в мантийной полости
двустворчатых моллюсков и использует
пищевые частицы, поступающие через
вводной сифон хозяина. Pagurolepas conchicola
селится в раковинах брюхоногих
моллюсков, занятых раками-отшельниками,
и питается экскрементами последних.
Некоторые морские желуди прикрепляются
к коже китов и в той или иной степени
питаются за счет своих хозяев. Н. И. Т а-
расов и Г. Б. Зевина пишут:
«Род Coronula не полностью перешел к
питанию за счет тканей и крови китов,
причем молодые особи, по-видимому, в
большей степени питаются биосестоном
[взвешенными в воде частицами], нежели
взрослые, сросшиеся с кожей хозяина,
проникшей в полости стенки их домика.
Tubicinella глубоко погружена в кожу
кита, а взрослых Cryptolepas невозможно
механически отделить от плотно и
полностью облекающей их кожи
единственного хозяина этого вида — серого кита
Eschrichtius gibbosus». У таких
специализированных паразитов грудные ножки
укорочены, а известковые пластинки
домика уменьшены и частично исчезли
совсем (табл. 32, 12).
Взмахи грудных ножек служат не
только для добывания пищи, но и для
обновления воды в полости домика. Это
необходимо для дыхания и для удаления из
полости домика продуктов выделения.
Дыхание совершается через тонкие покровы
тела; у некоторых видов имеются
нитевидные придатки, которым приписываются
функции жабр Сердца у усоногих нет и
кровь передвигается по полости тела
благодаря сокращению мышц.
Весьма своеобразны половая система и
особенности размножения Thoracica. В
отличие от подавляющего большинства
остальных членистоногих эти рачки, за
немногими исключениями, гермафродиты.
Каждая особь обладает и мужскими и
женскими половыми железами. Яйцеводы
открываются при основании передней пары
грудных ножек. Выходящие из них яйца
откладываются в мантийную полость. Се-
мепроводы впадают в длинный трубкооб-
разный мужской совокупительный
орган — пенис, представляющий собой
вырост редуцированного брюшного отдела и
Таблица 33. Равноногие ракообразные, или изоподы:
1, 2 — Gnathia dentata, самец и самка;
3 — Asellus aquaticus;
4 — Neophreatoicus assimilis;
5 — Mesidotea entomcm;
6 — Aega psora;
7 — Cyathura carinata;
8 — Haplomesus insignis orientalis;
9 — Serolis sp.;
10 — Bathynomus giganteus;
11 — Munnopsis typica;
12 — Storthyngura herculea:
13 — Microcharon sp.;
14 — Porcellio scaber;
15 — Sphaeroma serratum;
16 — Hemilepistus cristatus;
17 — Antarcturus ultraabyssalis.
432
помещающийся непосредственно позади
задней пары грудных ножек. При
спаривании пенис распрямляется, высовывается
из раковины наружу, проникает в
мантийную полость соседней особи и выделяет
туда сперму, оплодотворяющую яйца.
Поскольку обычно морские желуди и
морские уточки сидят в непосредственной
близости друг к другу, происходящий,
таким образом, процесс перекрестного
оплодотворения осуществляется без всяких
затруднений. В некоторых случаях может
происходить самооплодотворение, как это
следует из опытов по изоляции
отдельных особей, которые и в одиночестве
продолжали размножаться.
Только у морских уточек из семейства
Scalpellidae, обитающих на больших
океанических глубинах и часто сидящих
поодиночке, наблюдаются более сложные
явления, на которые впервые обратил
внимание Ч. Дарвин. Среди них известны
раздельнополые виды; их самки развиты
нормально, а самцы в той или иной
степени редуцированы (они маленькие,
иногда мешковидные, у некоторых
сохраняются остатки ножек и пластинок раковины,
у других — нет) и сидят по одному или
попарно, прикрепившись к стенке
мантийной полости самки. Есть и
виды-гермафродиты, в мантийной полости
которых находятся один или несколько де-
Таблица 34. Разноногие ракообразные, или бокоплавы:
1 — Brandtia parasitica;
2 — Niphargoides maeoticus;
3 — Axelboeckia spinosa;
4 — Eulimnogammarus virgatus;
5 — Pallasea bicornis;
6 — Panoploea eblanae;
7 — Acanthogammarus godlewskii;
8 — Leucothoe richiardii;
9 — Grubia crassicornis;
10 — Lysianassa punctata;
11 — Biancolina algicola;
12 — Ampelisca diadema;
13 — Cyphocaris richardi;
14 — Megaluropus agilis;
15 — Dexamine spinosa.
28 Жизнь животных, т. 2
градировавших «дополнительных» самцов.
Наконец, есть виды, нормальные особи
которых представлены как самками, так
и гермафродитами, причем и в тех и в
других живут карликовые самцы.
Некоторые исследователи предполагают,
что некогда все усоногие были
раздельнополыми. Те виды, которые имеют
мало редуцированных самцов, должны
считаться наиболее древними, а те,
которые совсем их лишены и
представлены только гермафродитными особями,—
наиболее молодыми.
Недавно дополнительные самцы были
неожиданно открыты и у одного из
представителей морских желудей — Balanus masig-
notus с западного побережья Центральной
и Южной Америки. По своему строению
они напоминают самцов некоторых видов
морских уточек, но располагаются снаружи
от мантии, в специальной камере,
образованной углублением внутренней стенки
передней непарной пластинки раковины, а не
близ мускула крышки раковины, как у
стебельчатых усоногих. Поскольку все
морские желуди гермафродиты, присутствие и
у них дополнительных самцов осложняет
понимание биологической сущности и
происхождения этого явления.
Весьма интересен, хотя и до настоящего
времени совершенно неясен, вопрос об
условиях, определяющих развитие личин-
ки в самца, в самку или в гермафродит-
ную особь. По некоторым наблюдениям,
в случае попадания личинки в
мантийную полость гермафродитной или
женской особи из нее развивается самец, а в
случае ее прикрепления к
неорганическому субстрату —• гермафродитная или
женская особь. Эта гипотеза нуждается в
дополнительной проверке.
Кучки яиц после оплодотворения в
мантийной полости родительской особи
объединяются тонкими хитиновыми
оболочками в так называемые яйценосные
пластинки и подвергаются дроблению.
У большинства видов из яиц выходят
науплиусы, у некоторых метанаупли-
усы.
У холодолюбивых морских желудей
(Balanus balanoides, В. hameri, В. balanus)
яйца образуются летом, оплодотворение
их происходит зимой, а личинки выходят
на волю весной; теплолюбивые виды
(В. improvisus, Elminius modestus, Chtha-
malus stellatus и др.) успевают отложить
яйца несколько раз в течение лета. Один
морской желудь (В. balanoides) в наших
северных морях за период размножения
выпускает 3—4 тысячи науплиусов.
Температура и освещение оказывают очень
сильное влияние на разные этапы
процесса размножения. Так, например,
круглосуточное освещение подавляет
созревание яиц.
Выход личинок из мантийной полости
родительской особи в воду зависит от
других причин. У Balanus balanoides он
определяется не календарными сроками
и не температурой воды, а массовым
развитием микроскопических водорослей,
которые служат пищей науплиусам морских
желудей. Когда эти водоросли
размножатся, науплиусы наших северных
морских желудей в огромном количестве
появляются в планктоне. Они интенсивно
питаются и трижды линяют, после чего
переходят в стадию метанауплиуса. Мета-
науплиусы ведут столь же активный образ
жизни и тоже трижды линяют, превращаясь
в конце концов в стадию циприсовидной
личинки. Она не питается, некоторое
время плавает, а затем, попав в
подходящие условия, прикрепляется к
субстрату при помощи своих передних антенн.
У тропических видов все личиночное
развитие протекает в течение 3—5 суток, у
холодноводных видов оно занимает 2
недели — месяц, а то и больше. У многих
преимущественно глубоководных Scalpel-
lidae из мантийной полости родителя
выходит сразу циприсовидная личинка.
Изучение хода оседания циприсовидных
личинок помогает понять особенности
распределения взрослых морских желудей и
морских уточек как на камнях, скалах
и других естественных субстратах, так
и на днищах судов, сваях, буях и т. д.
По-видимому, разные виды оседают при
разных скоростях течения. При скорости
течения более 50 см/сек оседание личинок
невозможно. Обычно личинки активно
движутся против течения, а оседание
происходит в момент относительного
покоя, т. е. тогда, когда скорость движения
течения сравняется со скоростью
движения личинки. Личинки Elminius modestus
садятся при большей, a Balanus
balanoides — при меньшей скорости. Личинки
В. crenatus лишены передних антенн и
прикрепляются при помощи специальных
липких тяжей; поэтому они оседают при
незначительной скорости течения.
Различные завихрения течений, образуемые
ими водовороты вызывают оседание
личинок плотными кучками. Кроме того,
доказано, что особенно охотно личинки
садятся на домики особей того же вида
или непосредственно рядом с ними.
Только соприкосновение личинки с домиком
морского желудя стимулирует ее
оседание. Выделяемые взрослым рачком в воду
вещества не имеют здесь значения.
Благодаря этому возникают плотные однови-
довые поселения морских желудей и
морских уточек, что для них очень выгодно,
так как обеспечивается возможность
перекрестного оплодотворения.
Однако существуют наблюдения,
показывающие, что личинки морских желудей
оседают таким образом, чтобы
распределиться равномерно по заселяемой
поверхности и благодаря этому избежать
ожесточенной внутривидовой конкуренции.
Н а й т-Д жонс и Мойз так описывают
оседание личинок Elminius на шиферные
плиты: «Самые первые особи, заселяющие
эти плиты, образуют скопления, руковод-
434
ствуясь, без сомнения, соответствующим
инстинктом. Затем промежутки между
скоплениями заполняются и в конечном
счете распределение по поверхности
становится равномерным. Все же плотность
заселения плиты составляет лишь 35—55
особей на 1 см2, тогда как циприсовид-
ные личинки эльминиусов, длина
которых не превышает 0,5 мм, а ширина еще
меньше, способны заселять ровные
поверхности с плотностью до 230 особей на
1 см2 даже в естественных условиях.
При тех плотностях, при которых
вступает в действие механизм, регулирующий
распределение по поверхности, еще
остается много места, так что запоздалым
пришельцам есть где прикрепиться и
передвигаться. Приходится сделать вывод,
что рачки продолжают передвигаться,
пока не найдут свободное место».
Выяснилось также, что личинки В. balanoides
распределяются равномерно по
отношению к молоди и взрослым особям только
своего вида и близких видов, но садятся в
непосредственной близости или даже
прямо на домик представителя другого
семейства— Chthamalus stellatus. Это
приводит к ожесточенной конкуренции между
Balanus и Chthamalus.
Личинки некоторых видов оседают
только на других животных. Есть виды,
живущие исключительно в губках (Acasta
spongicola, Balanus spongicola и др.) или
главным образом на раковинах
моллюсков и на панцирях ракообразных, в том
числе камчатского краба (В. hesperius в
Тихом океане).
В процессе дальнейшего роста молодых
морских желудей степень густоты их
поселения очень важна, так как определяет
форму их домика. В густых поселениях
часто он приобретает «лилиевидную»
форму, т. е. его диаметр в верхней части
больше, чем в нижней, в разреженных
поселениях он конический. Возможно, что
на форму домика действуют и другие
факторы.
Морские желуди растут довольно
быстро. В тропиках некоторые виды достигают
половозрелости через 8—16 дней после
оседания. Обычному в Балтийском,
Черном и Азовском морях В. improvisus,
а также дальневосточному В. cariosus для
этого требуется 3 месяца.
Продолжительность жизни разных видов колеблется от
1—2 до 5—7 и более лет.
Как уже указывалось, морские желуди
и морские уточки населяют самые
различные глубины. Некоторые виды хорошо
приспособлены к существованию в
приливо-отливной зоне. Они активны только
во время прилива, а когда вода
отступает, они плотно закрывают крышки
домика и погружаются в состояние, сходное с
анабиозом. При переносе в аквариум
рачки в течение нескольких дней сохраняли
тот же ритм, прекращая биение ножек в
часы отлива. Однако, даже находясь вне
воды, животные продолжают дышать,
хотя и менее интенсивно, чем под водой.
Только при наступлении резко
неблагоприятных условий они впадают в
состояние «скрытой жизни» и в течение
нескольких месяцев сохраняют
жизнеспособность в сухом состоянии, без вреда для
себя подвергаются длительному действию
пресной воды, а в условиях опыта
переносят заливание вазелиновым маслом,
содержание в азоте и водороде, т. е.
продолжают существовать при полном
отсутствии кислорода.
Этой способностью к «скрытой жизни»
объясняется, между прочим, появление
морских желудей (В. improvisus и В. ebur-
neus) в 1955 году в Каспийском море,
где их раньше не было. Они, несомненно,
переселились туда из Черного и Азовского
морей на днищах судов, прошедших по
Волго-Донскому каналу, и сумели
пережить длительное воздействие пресной
воды. Вероятно, несколько легче было
преодолеть еще более длительное расстояние
австралийскому морскому желудю Elmi-
nius modestus, появившемуся на
побережье Англии только в 1943—1944 годах.
Он также проделал этот путь на днищах
судов, но оставаясь все время в морской
воде, а затем стал быстро расселяться
вдоль берегов Европы, вытесняя местные
виды Balanus balanoides и В. improvisus.
Однако при пониженной солености
В. improvisus преобладает над новым все-
ленцем, а при низкой температуре
преимущество получает В. balanoides.
Выше остальных морских желудей
поселяются виды рода Chthamalus. В Чер-
28*
435
ном море Ch. stellatus (табл. 32, 10) часто
и в большом количестве (у Карадага в
среднем 28 экземпляров на 1 м2)
встречается на прибрежных скалах, выше
уреза воды, довольствуясь брызгами, которые
иногда до него долетают. Другие
морские желуди, в частности обычные виды
рода Balanus, образуют массовые
скопления в пределах приливо-отливной полосы
и во время прилива оказываются под
водой. Целый ряд видов морских желудей,
как например крупный (до 20 см высотой)
дальневосточный В. evermanni (табл. 32,
?), и морских уточек приурочен к
области материкового склона. На больших
океанических глубинах преобладают морские
уточки из рода Scalpellum, к которому
относится и наиболее глубоководный из
известных видов S. japonicum,
обнаруженный в Курило-Камчатской и Японской
океанических впадинах до глубины
6860 м.
Некоторые морские уточки
прикрепляются не к дну, а к плавающим в воде
предметам — кускам дерева, судам, а
также к различным животным. Таковы виды
родов Lepas и Conchoderma. Обычный
широко распространенный вид С. auritum
встречается и на кусках дерева, и на
днищах судов, и на китах, морских
черепахах и медленно плавающих рыбах. При
этом С. auritum всегда садится только на
твердую поверхность. На китах эта
морская уточка находится только на
прикрепившихся к их коже морских желудях
Coronula (табл. 32, 5) и на китовом усе,
а у кашалотов — на зубах, образуя целые
гроздья. Уховидные придатки рачка,
служащие, вероятно, для отвода
профильтрованной воды, сильнее развиты у особей,
сидящих на быстро движущихся
животных.
Практическое значение
Thoracica определяется прежде всего тем,
что морские желуди составляют основу
обрастаний судов. Как уже указывалось,
обросшее морскими желудями (и другими
животными) судно теряет исходную
скорость. Соответственно увеличиваются
время рейса, затраты на топливо и все
остальные эксплуатационные расходы. Корабли
периодически приходится очищать от
обрастаний, а это обходится очень дорого.
Подсчитано, что торговый флот США
ежегодно терпит убытки от обрастаний,
превосходящие 100 миллионов долларов.
Во многих странах усиленно работают
над проблемой защиты судов от
обрастаний, в частности над изобретением таких
красок для днищ кораблей, которые были
бы ядовиты для оседающих на суда
личинок морских желудей. Некоторые
успехи в этом направлении достигнуты, но
«необрастающие» краски очень дороги.
Морские желуди вредят не только
судам. Проникнув в Каспийское море, они
стали оседать на ставных сетях, неводах
и других рыболовных орудиях и принесли
немало неприятностей местным
рыбакам, которые раньше с ними не
сталкивались.
С другой стороны, несомненно и
положительное значение морских желудей и
морских уточек. Их личинки нередко
составляют существенную часть
прибрежного морского планктона и в большом
количестве поедаются некоторыми планкто-
ноядными рыбами. Так, например, у
берегов Норвегии весной сайда питается
преимущественно личинками у соногих,
поедает их и беломорская сельдь, а также
некоторые другие рыбы. Взрослые усоно-
гие в малой степени используются
рыбами, которые часто не могут оторвать их
от субстрата и раскусить. Тем не менее
морские желуди отмечены в качестве
объектов питания трески, пикши, зубатки,
осетра и др.
Человек также не брезгает усоногими.
Из крупного морского желудя Balanus
psittaceus в Чили делают консервы. В
Испании и Португалии добывают морскую
уточку Mitella pollicipes, которая
используется и в пищу, и для
приготовления тука. Многие другие усоногие также,
несомненно, съедобны. Тур Хейердал,
описывая свое замечательное путешествие
на «Кон-Тики», рассказывает, между
прочим, о том, как он и его спутники варили
суп из морских желудей, которыми
быстро обрастал их плот, и высоко оценивает
вкусовые качества этих рачков. Весьма
возможно, что в будущем человек станет
использовать усоногих в большей
степени, чем ныне.
436
ПОДОТРЯД АКРОТОРАЦИКОВЫЕ
РАКООБРАЗНЫЕ (ACROTHORACICA)
Занимаясь усоногими ракообразными,
Ч. Дарвин открыл один сильно
отличающийся по своему строению и образу
жизни от морских желудей и морских
уточек вид, сверлящий раковины моллюсков,
который он считал необходимым
выделить в особый подотряд. В настоящее
время таких видов известно более десятка.
Они точат ходы в раковинах брюхоногих
и панцирных моллюсков, а также в
стволах кораллов (рис. 224).
Будучи хорошо защищены известковой
толщей раковины моллюска или ствола
коралла, они, естественно, лишены
собственных известковых пластинок,
свойственных их только что рассмотренным
родичам. Только на передней части
головы самки есть хитиновая пластинка,
принимающая, вероятно, участие в процессе
сверления. Эта передняя часть головы
перетяжкой отделена от задней части и от
грудного отдела. Вокруг ротового
отверстия расположены пара жвал и две
пары челюстей, а позади рта пара
передних грудных ножек. На значительном
расстоянии от них находятся собранные в
одну группу остальные три пары одно-
ветвистых грудных ножек. Грудные
ножки совершают около 60 взмахов в минуту,
но не высовываются при этом за края
мантии, как это бывает у морских
желудей и морских уточек. Тем не менее
благодаря взмахам грудных ножек вода
внутри мантийной полости рачка все время
сменяется и приносит достаточное
количество мелких пищевых частиц. Они
поступают в кишечник, лишенный заднего
отдела и анального отверстия, но зато
снабженный многочисленными
ветвящимися слепыми выростами, заходящими в
голову, в грудные ножки и в другие
части тела.
Все Acrothoracica, за исключением
живущего в кораллах рода Lithoglyptes,
раздельнополы. Карликовые @,4—2 мм)
самцы живут поодиночке, прицепившись к
мантии самки, длина которой достигает
2 см. Они обычно сохраняют облик цип-
рисовидной личинки или имеют очень
упрощенное строение. Из оплодотворенного
яйца выходит науплиус, такой же, как
у Thoracica. Перелиняв три раза, он
превращается в метанауплиуса, который дает
начало типичной циприсовидной личинке.
Циприсовидная личинка садится на
подходящий субстрат, сбрасывает свою
раковину и превращается в
сформировавшегося рачка. Рачок приступает к
сверлению известняка, пользуясь хитиновой
пластинкой и выделяя вещество,
растворяющее соли кальция.
Обыкновенно Acrothoracica селятся в
пустых раковинах уже погибших
брюхоногих моллюсков (Buccinum, Fusus и
др.) или в раковинах, занятых раками-
отшельниками.
ПОДОТРЯД КОРНЕГОЛОВЫЕ РАКООБРАЗНЫЕ
(RHIZOCEPHALA)
Еще больше отличаются от обычных
усоногих ракообразных паразитические
корнеголовые, организация которых
предельно упрощена под воздействием па-
Рис. 224. Строение Tripetosa lampos:
А — взрослая самка с карликовым самцом;
Б — молодая самка, передняя часть тела
которой еще не превращена в диск; В —
самец при большом увеличении; 1 —
передний конец тела; 2 — грудной отдел с
ножками; з — самец; 4 — семенник; 5 —
передняя антенна; 6 — лопасти мантии; 7 —
пенис.
437
Рис. 225. Sacculina carcini:
A — взрослый рачок, паразитирующий в крабе Carci-
nus maenas (левая половина тела краба изображена
прозрачной); Б — развитие личинки: 1 — науплиус,
2 — циприсовидная личинка, з — ее прикрепление
и первая линька, 4 — строение личинки после линьки,
5 — вторая линька и формирование кентрогона, б —
миграция клеток кентрогона в тело краба, 7 —
молодой паразит на поверхности средней кишки краба.
разитизма. Взрослые представители этого
подотряда не сохраняют совершенно
никаких признаков членистоногих
животных. Они живут в десятиногих
ракообразных — в крабах, раках-отшельниках и
некоторых креветках.
Все тело краба, зараженного корнего-
ловым рачком Sacculina, пронизано
ветвящимися, как корни растений,
отростками паразита. Они оплетают все
внутренние органы хозяина — кишечник,
половые железы, нервную систему, внедряются
в мускулатуру. Этими отростками рачки
высасывают соки краба. Отростки
впадают в обширный мешок,
расположенный под брюхом хозяина на поверхности
его тела. В мешке находятся сильно
развитые яичники и маленькие семенники.
Все остальные органы —- кишечник,
органы чувств и органы выделения —
полностью отсутствуют.
Систематическое положение этого
существа, так же как и в других подобных
случаях (например, описанного выше ден-
дрогастера), выясняется при изучении его
развития (рис. 225, Б). Из яиц,
откладываемых в полость мешка, выходит типичный
для усоногих науплиус, выбирающийся
наружу и плавающий в воде. Через
8 дней он превращается в метанауплиуса,
который после 4—5 линек переходит в
столь же типичную стадию циприсовид-
ной личинки. Все эти личиночные
стадии корнеголовых отличаются от
соответствующих им стадий других усоногих
главным образом отсутствием кишечника.
Они питаются за счет содержащихся в их
теле запасов желтка. Проплавав 3—4 дня,
циприсовидная личинка прикрепляется
передними антеннами к основанию
щетинок конечностей или брюшной стороны
тела молодых крабов. После этого она
испытывает удивительные превращения.
При очередной линьке она теряет
большую часть своего тела со всеми
конечностями, но сохраняет мантию, раковину и
глаз, а затем линяет еще раз и
отбрасывает и их. От личинки остается
маленькое овальное мешковидное тельце, внутри
которого располагаются
недифференцированные клетки, имевшиеся еще у науплиу-
са. Эта стадия получила название стадии
кентрогона. Покровы и
передние антенны кентрогона образуют
пронизанный каналом вырост,
пробуравливающий покровы краба. Через этот канал
кучка недифференцированных клеток
проникает в тело краба и, вероятно, кровью
хозяина переносится на поверхность его
средней кишки.
Здесь начинается рост паразита.
Отростки его тела удлиняются, разветвля-
438
ются и начинают оплетать внутренние
органы краба. Они не разрушают ни
кишечника, ни половых желез хозяина,
всасывая питательные вещества из омывающей
их крови и выделяя в организм хозяина
отбросы своей жизнедеятельности.
Через 7—8 месяцев паразит образует
мешковидный вырост. Когда он достигнет
2 мм в поперечнике, он прорывает
покровы хозяина и выпячивается наружу,
а затем продолжает расти в течение 6
недель, увеличиваясь до 12 мм в диаметре.
К этому времени в мешковидном выросте
заканчивается созревание половых
желез, которые начинают функционировать.
Кроме того, здесь же формируется очень
слабо развитая нервная система. Всего для
развития взрослой саккулины требуется,
таким образом, 9 месяцев с момента
прикрепления к хозяину циприсовидной
личинки. Паразит живет около 3 лет, и
хозяин обычно переживает его и
продолжает существовать после смерти
саккулины.
Присутствие саккулины оказывает
мощное воздействие на организм краба.
Заражение саккулиной молодого самца
подавляет развитие его половых желез.
С каждой последующей линькой он
становится все более похожим на самку:
брюшной отдел его расширяется,
исчезают превращенные в совокупительный
орган передние брюшные конечности,
меняется характер щетинок брюшка. Японские
зоологи И ш и к а в а и Янагимачи
описали зараженного саккулиной
самца краба Eriocheir japonicus, семенники
которого под влиянием паразита
переродились в яичники, продуцировавшие
нормальные яйца. Если саккулина заразила
взрослого крупного самца или самку, это
не приводит к изменению вторичнополо-
вых признаков хозяина, но нарушает
деятельность его половых желез —
происходит паразитарная кастрация.
Нет сомнения в том, что причиной
таких изменений в организме краба
служит воздействие саккулины на его
органы внутренней секреции.
Непосредственно или выделяя специфические вещества,
паразит подавляет выделение гормонов,
действующих на формирование вторично-
половых признаков самца и
контролирующих деятельность половых органов самца
и самки.
Паразиты раков-отшельников Peltogas-
ter и Peltogastrella также вызывают
паразитарную кастрацию своих хозяев, но
заражение ими самцов не приводит к
развитию у них вторичнополовых
признаков самок. В отличие от саккулины у
этих паразитов есть дополнительные
самцы, сохраняющие облик циприсовидных
личинок. Семенники гермафродитных
особей очень малы и не функционируют.
Самцы прикрепляются передними
антеннами к мешковидной части
гермафродитных особей, и масса заполняющих их
недифференцированных клеток
переселяется в недоразвитый семенник
гермафродита. Часть этих клеток преобразуется
в сперматозоиды.
При линьке раков-отшельников
наружные мешковидные части их паразитов
отваливаются вместе с покровами хозяина.
Однако это не приводит к гибели
паразита: из корневидной, находящейся
внутри рака-отшельника части паразита
вновь отрастает его мешковидная часть.
У Chologaster наблюдалось
четырехкратное отпадение мешков и замена их
новыми.
Способность к почкованию особенно
ярко проявляется у Thompsonia —
паразита креветок из семейства
раков-щелкунов. На теле зараженной креветки
всегда находится большое количество
мешковидных паразитов, объединенных общей
корневой системой. Она возникает из
одной личинки, а затем образует много
мешковидных особей.
Корнеголовые ракообразные широко
распространены во всех морях и океанах,
за исключением холодных южных и
северных морей. Есть они и в Черном море,
и в наших дальневосточных морях. В
Северной Атлантике, Средиземном и Черном
морях около 6% представителей
массовых видов крабов заражено
саккулиной. В тропиках некоторые
корнеголовые вместе со своими хозяевами обитают и
в пресных водах. Всего известно около
120 видов этих удивительных
ракообразных.
439
ПОДКЛАСС РАКУШКОВЫЕ РАКООБРАЗНЫЕ (OSTRACODA)
К этому подклассу принадлежат
большей частью мелкие (обычно мельче 1 мм,
в редких случаях до 23 мм длиной)
рачки, обладающие двустворчатой, часто
пропитанной солями кальция раковиной,
целиком покрывающей все их тело. Между
створками раковины при движении
животных высовываются только антенны и
иногда концы ножек, а также фурка.
Тело очень сильно укорочено и потеряло
сегментацию. Число грудных ножек
сокращено до одной-двух пар, а иногда их нет
вовсе. Все головные придатки и грудные
ножки очень сильно специализированы:
каждая пара выполняет какую-нибудь
определенную функцию. Сердца и жабр
в подавляющем большинстве случаев нет.
Из яйца выходит науплиус, имеющий,
как и взрослые особи, двустворчатую
раковину.
Ракушковые обитают как в море, так
и в пресных водоемах. Большинство
видов этих рачков передвигается по дну и
водным растениям, но среди морских
видов есть и планктонные. Благодаря проч-
Рис. 226. Ракушковые ракообразные:
1 — Cypris pubera; 2 — Gigantocypris; 3 — Conchoecia
elegans.
ной раковине ракушковые хорошо
сохраняются в ископаемом состоянии и
используются геологами для определения
возраста различных отложений. Судя по
геологическим данным, это очень
древняя группа животных. Остатки ракушко-
вых известны начиная с кембрия.
Подкласс включает единственный
отряд, имеющий то же название.
ОТРЯД РАКУШКОВЫЕ
РАКООБРАЗНЫЕ (OSTRACODA)
Трудно найти водоем, где не жили бы
ракушковые рачки. По разнообразию
условий, к которым они приспособились, их
можно сравнить только с веслоногими.
В мелких лужах, в прудах, в крупных
озерах, в подземных водах, на всех
глубинах Мирового океана, как на дне, так
и в толще воды, можно обнаружить этих
невзрачных мелких животных. Больше
того, один вид ракушковых — Mesocypris
terrestris — перешел к сухопутному
существованию: он живет в лесной
подстилке влажных тропических лесов Южной
Африки. Известно около 2000 видов
ракушковых.
Несмотря на столь разные условия
существования и различный образ жизни
(среди ракушковых есть хищники, есть
и грунтовды, донные и планктонные
формы, зарывающиеся в грунт и ползающие
по растениям и т. д.), организация
ракушковых в общем довольно однообразна.
Все они обладают толстой двустворчатой
раковиной, соответствующей карапаксу
других ракообразных. Внутренняя ее
поверхность хитинизирована, а наружная
обычно известковая. Левая и правая
створки раковины соединены на
спинной стороне эластической связкой. У
многих видов есть и замок, состоящий из
выступов на одной створке и
соответствующих им углублений на другой.
Приблизительно в середине тела расположен
мощный мускул-замыкатель, прикреп-
440
ленный к правой и левой створкам. При
сокращении этого мускула раковина
плотно закрывается.
Форма и строение раковины зависят от
образа жизни. У рачков, живущих на
поверхности жесткого грунта, раковина
обычно толстостенная и снабжена
различными буграми и выростами; у видов,
роющихся в грунте, она толстостенная,
но гладкая, а у видов, ползающих по
растениям и живущих на мягком илистом
грунте, она тонкая и гладкая. В
особенности тонкой раковиной обладают морские
планктонные виды, удельный вес тела
которых приближается к удельному весу
воды.
Голова часто хитинизирована. На ней
обычно помещается непарный наупли-
альный глаз, а у представителей морского
семейства Cypridinidae имеется пара
сложных фасеточных глаз, достигающих
у глубоководного планктонного Gigan-
tocypris очень крупных размеров (рис. 226,
2). Участки раковины, находящиеся перед
глазами, прозрачны, так что рачки видят
через раковину.
Все пять пар придатков головы
хорошо развиты. Наиболее мощная из них —
задняя антенна, служащая основным
органом передвижения. Она богата
различными щетинками, приобретающими у
некоторых видов форму маленьких клешней.
У многих пресноводных ракушковых
передняя антенна также длинная, хотя и
тоньше задней. Они плавают, взмахивая
передними антеннами вперед и вверх, а
задними вперед и вниз, двигаясь при этом
по равнодействующей, т. е. прямо. У
представителей преимущественно морского
семейства Cytheridae на конце
редуцированной наружной ветви задних антенн
открываются протоки парной паутинной
железы. Вертикально опускаясь с
какого-нибудь подводного предмета, рачки
выделяют, как пауки, паутинную нить, а
затем могут ею пользоваться при
возвращении назад. Эта нить позволяет им также
удерживаться на грунте и так
противостоять течению.
Жвалы снабжены сильно развитым
щупиком, который нередко помогает
движению рачков. Некоторые Cypridinidae,
например Asterope, зарываются в грунт
Рис. 227. Ракушковые ракообразные:
А — строение Cypridopsis vidua; Б — движения Cypris;
В — науплиус; 1 — мускул-замыкатель раковины; 2 —
передняя антенна; 3 — верхняя губа; 4 — жвала;
5 — передняя челюсть; 6 — задняя челюсть; 7 — на~
ружная ветвь передней челюсти; 8 — задняя грудная
нога; 9 — передняя грудная нога; 10 — ползание по
дну; 11 — ползание по растениям; 12 — плавание.
при помощи щупиков жвал. Челюсти у
разных групп ракушковых устроены
различно. Обыкновенно наружная ветвь
одной из них превращена в широкую
пластинку, взмахи которой сменяют воду в
полости раковины. Дыхание
осуществляется через стенку тела, и только у Asterope
на заднем отделе тела есть специальные
выросты покровов, функционирующие как
жабры. У большинства ракушковых
челюсти снабжены
специализированными щетинками и участвуют в
захватывании пищи, но у Cytheridae задняя
челюсть служит для передвижения рачка
и представляет собой ходильную
конечность.
441
Первая, а у некоторых семейств (Су-
theridae, Darwinulidae) и вторая пара
грудных ног также ходильные, но у Сур-
ridae и у всех представителей морского
подотряда Myodocopa задние грудные
ножки загнуты на спинную сторону и
приспособлены для очистки мантийной
полости: они удаляют попавшие туда
мелкие посторонние частицы. Есть роды,
совсем лишенные обеих пар (Polycope) или
задней пары (Cytherella) грудных ножек.
Задняя часть тела в большинстве случаев
заканчивается хорошо развитой фуркой,
принимающей участие в движении рачка.
Эти движения очень разнообразны.
Многие пресноводные Cypridae могут и
плавать и ползать. При ползании они
цепляются щетинками задних антенн за
субстрат и отталкиваются от него передними
грудными ногами и фуркой; передние
антенны при этом протянуты вперед и
нащупывают путь (рис. 227, Б). Некоторые
виды (Notodromas monacha, Heterocypris
incongruens, Cyproides marginata) могут
передвигаться брюшной стороной вверх,
подвешиваясь снизу к поверхностной
пленке натяжения, как это делает
описанный выше ветвистоусый рачок Scapho-
leberis. Другие способны зарываться в
грунт на глубину до 7 см. Виды,
приспособленные к передвижению по
поверхности грунта и растений, часто отличаются
удлиненными задними антеннами и
передними грудными ножками. Морские
планктонные Conchoecia и Gigantocypris
плавают при помощи своих мощных
задних антенн. О способе передвижения
единственного сухопутного представителя
отряда — Mesocypris terrestris — ничего не
известно, так как наблюдать его живым
не удалось. Судя по тому, что задние
антенны очень крупные и вооружены
крепкими когтевидными щетинками, можно
предположить, что они служат основным
органом передвижения.
Также различны и способы питания.
Многие ракушковые питаются
преимущественно водорослями, остатками
растений и животных и органическим веществом
грунта (Cytheridae, Darwinulidae, Can-
donidae, Cypridopsis). Среди морских
видов есть хищники. В кишечнике Cypridina
castanea обнаружены остатки мизид и ки-
леногих моллюсков, в кишечнике С. пог-
vegica — остатки полихет. Планктонные
ракушковые поедают других
планктонных животных, в первую очередь
веслоногих, но иногда и щетинкочелюстных и
мелких рыб, а некоторые виды Conchoecia
наряду с зоопланктоном используют и
фитопланктон. Некоторые роды (морские
Asterope, Cyclasterope, Cytherella,
пресноводный Notodromas) — настоящие филь-
траторы с соответственно
устроенными ротовыми придатками. Asterope и
Cyclasterope зарываются в грунт,
выставляя над ним концы передних антенн,
образующие отверстие в грунте, через
которое проходит служащий для
фильтрации ток воды.
Все ракушковые раздельнополы. По
внешнему виду самцы почти не
отличаются от самок. Впрочем, у многих
пресноводных Cypridae самки размножаются
партеногенетически, а самцы вообще
неизвестны. В условиях аквариума Нег-
petocypris reptans содержали в течение
30 лет, причем все это время происходило
партеногенетическое размножение и
самцы ни разу не появились. Есть виды,
которые в северной части области
распространения размножаются
партеногенетически, а в южной встречаются и
самцы и самки и наблюдается процесс
оплодотворения.
Половые органы ракушковых устроены
очень сложно. Одной из самых
удивительных особенностей этих рачков можно
считать огромные размеры их сперматозоидов,
превосходящих по величине
сперматозоиды всех остальных животных. Так,
например, у ракушкового рачка Pontocypris
длиною 0,7 мм длина сперматозоида
достигает 6 мм, т. е. оказывается в 8 раз больше
самого животного. Длина сперматозоида
человека @,062 мм) почти в 100 раз
меньше, чем рачка.
Вмещающие такие гигантские
сперматозоиды половые пути самцов и самок
представляют собой сильно извитые каналы.
Самцы имеют сложный копулятивный
аппарат, при помощи которого через щель
раковины вводят сперматозоиды в половое
отверстие самки. Яйца оплодотворяются
при выходе из полового отверстия и
сначала оказываются в полости раковины,
442
в ее задней части. Через некоторое время
самка откладывает яйца на камни,
поверхность грунта или на растения.
Некоторые Cytheridae и Darwinulidae носят
яйца под раковиной до выхода из них
личинок, а у Cypridinidae под раковиной
полностью формируются молодые рачки,
после чего мать удаляет их наружу
своими грудными ногами.
У остальных ракушковых из яйца
выходит науплиус своеобразного строения.
Он снабжен двустворчатой раковинкой,
более короткой и сильнее суживающейся
к заднему концу, чем раковинка взрослых
рачков. Его задние антенны и жвалы одно-
ветвисты, причем последние заканчиваются
длинным когтем. Науплиус может не
только хорошо плавать, но и ползать при
помощи задних антенн и жвал. Личинка
линяет 5—8 раз, постепенно приобретает
недостающие конечности и превращается
во взрослого рачка. У пресноводных видов
продолжительность личиночного развития
колеблется от 5 недель до 4 месяцев. В
отличие от других ракообразных взрослые
ракушковые не линяют. Длительность их
жизни различна у разных видов.
Пресноводные виды, науилиусы которых
вылупляются из перезимовавших яиц, живут
обычно не более 4 месяцев, виды семейства
Candonidae — 9 и более месяцев, а морской
Philomedes globosus — не менее 2х/2 лет.
ПОДКЛАСС ВЫСШИЕ РАКООБРАЗНЫЕ (MALACOSTRACA)
Основные признаки видов подкласса —
постоянное количество грудных и
брюшных сегментов, а также присутствие
брюшных конечностей. Грудь состоит из 8, а
брюшко из 6 или 7 сегментов и
заканчивается тельсоном. Нередко передний или
несколько передних грудных сегментов
срастаются с головой и их конечности
превращаются в ногочелюсти. Иногда
срастаются между собой или с тельсоном
и брюшные сегменты. Мужские половые
отверстия всегда открываются на
последнем, а женские на шестом грудном
сегменте. Желудок подразделен на
жевательную и фильтрующую камеры. Всегда есть
сильно развитая пищеварительная железа,
сердце и кровеносные сосуды.
Название «высшие» не совсем точно.
Если постоянство числа грудных и
брюшных сегментов можно считать признаком
более высокой организации, чем их
непостоянство, присущее другим подклассам
ракообразных, то присутствие брюшных
конечностей, наоборот, говорит о меньшей
специализации, чем их исчезновение,
наблюдающееся у других подклассов. По
всей вероятности, высшие ракообразные
развивались независимо от других
подклассов, каждый из которых сохранил те
или иные примитивные признаки,
унаследованные от общих предков.
Представители высших ракообразных,
как правило, крупнее, чем рачки,
относящиеся к другим подклассам. Они
распространены чрезвычайно широко: населяют
самые разнообразные водоемы и частично
приспособились к обитанию на суше.
Систематика подкласса очень сложна: он
делится на надотряды, которые в свою
очередь подразделяются на отряды. Таких
надотрядов насчитывается пять.
НАДОТРЯД ФИЛЛОКАРИДЫ
(PHYLLOCARIDA)
Этот надотряд включает единственный
очень бедный видами отряд, сохраняющий
признаки примитивной организации.
ОТРЯД ТОНКОПАНЦИРНЫЕ
РАКООБРАЗНЫЕ (LEPTOSTRACA)
Тонкопанцирные ракообразные
обитают исключительно в море. Большая их
часть встречается в прибрежной области
и ведет придонный образ жизни и только
один вид — Nebaliopsis typica —
планктонное глубоководное животное. Известно
443
всего 8 видов этих своеобразных рачков,
но распространены они очень широко.
Карапакс тонкопанцирных имеет вид
тонкой двустворчатой раковинки,
покрывающей весь грудной и переднюю часть
брюшного отдела рачка. С ним подвижно
сочленен направленный вперед овальный
пластинчатый рострум, прикрывающий
сверху большие стебельчатые глаза и
основания антенн. Наклоняя или приподнимая
рострум, рачок регулирует силу тока воды,
возбуждаемого грудными ножками и
направляемого спереди назад между
створками раковины.
Передняя и задняя антенны довольно
длинные и богаты чувствительными
щетинками. Ротовые придатки обычного
строения и служат для фильтрации и
размельчения пищи. Кроме того, удлиненный
щупик передней челюсти очищает полость
раковины от попадающих туда
посторонних частиц. Грудной отдел состоит из
8 сегментов, каждый из которых снабжен
парой одинаково устроенных ножек,
приспособленных одновременно и для
передвижения, и для дыхания, и для подачи
пищи ко рту. Эти ножки пластинчатые,
листовидные. Их наружные лопасти —
эпи- и экзоподит — служат жабрами, а
снабженная длинными щетинками более
узкая и сильнее хитинизированная
внутренняя лопасть — эндоподит —
функционирует преимущественно как орган
движения. На внутреннем крае основной
части каждой ноги сидят щетинки двух типов:
одни более короткие и тонкие, но более
многочисленные отфильтровывают
пищевые частицы, которые передаются сзади
наперед другими более длинными и
малочисленными щетинками.
Грудные ножки сгибаются и
разгибаются последовательно. Их движения
напоминают движения ног жаброногов (см.
стр. 381). При встречном изгибании ножек
соседние пары образуют фильтрующие
камеры. Вода проходит спереди назад по
продольному желобку вдоль брюшной
стороны тела, оставляя между
фильтрующими щетинками мелкие взвешенные
частицы, а затем двумя потоками расходится
в стороны. Эпиподиты и частично экзопо-
диты ножек используют при этом
растворенный в воде кислород, но, кроме того,
он усваивается и тонкими покровами
внутренней стенки раковины. Даже когда
рачки неподвижно сидят на грунте, их ножки
продолжают совершать свои ритмичные
взмахи. Только при зарывании в грунт,
когда рострум загибается вниз и закрывает
спереди вход в раковину, работа грудных
ножек прекращается.
В состав длинного цилиндрического
брюшка входит 7 сегментов. Из них 4
передних несут длинные двуветвистые
ножки, богатые щетинками и служащие для
плавания. Ножки следующих 2 сегментов
короткие, редуцированные, а последний
брюшной сегмент совсем лишен
конечностей. За ним следует удлиненный тельсон
с двумя длинными ветвями фурки.
Самцы по внешнему виду и по своему
поведению несколько отличаются от
самок. Их раковинка уже, брюшной отдел
с тельсоном относительно головы и груди
длиннее, глаза крупнее, а число
чувствительных щетинок на передних антеннах
гораздо больше. У донных видов самцы
быстрее и более продолжительное время,
чем самки, плавают, сильно взмахивая
своими передними брюшными ножками,
и активно отыскивают менее подвижных
самок.
Самки носят откладываемые яйца на
себе, удерживая их щетинками внутренних
краев задних грудных ножек. Из яиц
выходят молодые, еще не вполне
сформировавшиеся рачки, первое время
остающиеся на ножках матери. После двух линек
они переходят к самостоятельному
существованию.
Из немногочисленных видов
тонкопанцирных наиболее широко распространена
Nebalia bipes, длиною 6—11 мм,
живущая у нас в Баренцевом море, в
дальневосточных морях, а также почти во всем
Мировом океане, избегая опресненных его
частей. Она встречается близ берегов, на
глубине от 10 до 400 м, среди водорослей,
между камнями и т. д. Рачки иногда
часами лежат на дне, фильтруя мелкую взвесь
или поедая более крупные частицы
(остатки растений и животных), которые они
находят при помощи антенн и щупиков
жвал. Иногда они зарываются в мягкий
ил, орудуя при этом своим
ложкообразным рострумом. Тело рачка может круто
444
Рис. 228. Nebalia bipes:
1 — самец; 2 — самка; 3 — грудная ножка; 4 — схематичный
продольный разрез через три грудные ножки (стрелками обозначен путь
пищевых частиц ко рту).
изгибаться на брюшную сторону так, что
тельсон касается конца рострума. Это, по-
видимому, защитная реакция.
Совсем иной образ жизни ведет самый
крупный (до 40 мм длиной) представитель
тонкоцанцирных — Nebaliopsis typica. Он
обитает в толще воды на больших
глубинах почти во всем Мировом океане, в
том числе в Беринговом море. Взрослые
половозрелые особи попадаются только
на глубине более 4—5 км и
лишь в Беринговом море
поднимаются на глубину менее
200 м, молодые могут жить и
на меньшей глубине. По мере
роста рачки опускаются все
глубже и глубже и могут
размножаться только на огромных
глубинах. Кишечник N. typica
устроен весьма своеобразно.
Его передний отдел снабжен
выростом, который может очень
сильно растягиваться, что
позволяет рачку заглатывать
крупную пищу.
В растянутом кишечном
мешке Nebaliopsis нашли крупный
комок, представлявший, как
предполагают, желток икринки
какой-то глубоководной рыбы.
При крайней бедности пищевых
ресурсов больших океанических
глубин способность заглатывать
сразу большое количество пищи
очень важна, поскольку
подобный счастливый случай
представляется не часто и его
следует использовать в полной
мере.
Однако Nebaliopsis может
питаться и фильтруя мелкую
взвесь.
Тонкопанцирные
ракообразные, сохраняющие многие
примитивные признаки, только
сравнительно недавно были
найдены в ископаемом состоянии.
Из пермских отложений
Германии описали Nebalia bentzi,
очень близкую к
обыкновенной N. bipes. Это
подтверждает глубокую древность
отряда.
В палеозое существовал близкий к
тонкопанцирным отряд Archaeostraca,
принадлежащий к тому же надотряду Phyllo-
carida. Среди Archaeostraca известны
виды, которые считаются самыми
примитивными из всех высших ракообразных, как
в частности Nahecaris из девонских
отложений Германии. Однако систематическое
положение многих Archaeostraca не вполне
ясно.
445
НАДОТРЯД СИНКАРИДЫ
(SYNCARIDA)
Этот надотряд также характеризуется
примитивностью организации и
глубокой древностью. Замечательно, что
сначала были открыты его ископаемые
представители (в каменноугольных
отложениях) и только через 40 лет были найдены
ныне живущие.
Все современные Syncarida обитают в
пресной воде. Они населяют высокогорные
озера и ручьи Тасмании и подземные воды
юго-восточной части Австралии, а также
подземные воды других материков, за
исключением Северной Америки.
Рис. 229. Анаспидовые:
1 — Anaspides tasmaniae; 2 — Paranaspides lacustris;
9 — Koonunga cursor; 4 — Micraspides calmani.
В отличие от тонкопанцирных они
полностью лишены карапакса. Все их
грудные и брюшные сегменты имеют сходное
строение и приблизительно одинаковые
размеры. Грудные ножки двуветвисты.
Конечности последнего брюшного
сегмента преобразованы в рулевые ноги — так
называемые у р о п о д ы. Левая и правая
жвалы симметричны. Самки откладывают
яйца в воду и не носят их на себе.
Надотряд включает 3 отряда — Anaspi-
dacea, Bathynellacea и Stygocaridacea.
ОТРЯД АНАСПИДОВЫЕ
РАКООБРАЗНЫЕ (ANASPIDACEA)
Ныне живущие представители анаспи-
довых принадлежат всего к 5 видам. Они
обитают только в высокогорных озерах
и ручьях Тасмании, и лишь один из них
живет в грунтовых водах близ г.
Мельбурна в Австралии. В палеозойское время
анаспидовые были распространены
гораздо шире. Их ископаемые остатки найдены
во многих местах Европы и Северной
Америки, причем древнейшие из этих
ракообразных были морскими животными.
Современные виды можно считать
последними остатками древней, некогда
процветавшей группы, дожившими до наших дней
в очень ограниченной области, в
водоемах, где нет или очень мало их
конкурентов и врагов.
На голове этих рачков находятся
стебельчатые (у Anaspides и Paranaspides)
или сидячие (у Koonunga) глаза, совсем
отсутствующие у подземного Micraspides.
Антенны довольно длинные. В основном
членике передней антенны расположен
орган равновесия — статоцист, весьма
своеобразно устроенный. В нем нет ста-
толита, как это обычно бывает, но на его
стенках сидят булавовидные щетинки,
утолщенные концы которых сгибаются
под действием земного притяжения.
Передний грудной сегмент срастается
с головой, однако его конечности
сохраняют характер ходильных. Только у
Koonunga они приспособлены к рытью грунта,
а у остальных родов имеют такое же
строение, как последующие, если не считать
446
присутствия внутренних
выростов, участвующих в
подаче пищи ко рту. Грудные
ножки при основании
снабжены тонкостенными
наружными выростами — эпиподи-
тами, выполняющими
функцию жабр. На передних 6—
7 парах грудных ножек есть
и наружные ветви — эк-
зоподиты. Они совершают
беспрерывные взмахи (у Апа-
spides 100, а у Paranaspi-
des 250—300 раз в минуту),
благодаря чему жабры все
время омываются свежей
водой. Две передние пары
брюшных ножек самцов
преобразованы в копулятивный
аппарат. Они имеют вид
трубок и направлены вперед.
Следующие три пары
снабжены сильно развитой мно-
гочлениковой, богатой
щетинками наружной и
короткой, иногда совсем
отсутствующей внутренней ветвью.
При помощи наружных
ветвей этих ножек Anaspides и
Paranaspides могут
некоторое время плавать, держась
спинной стороной вверх.
Последние брюшные ножки — уроподы —
пластинчатые, направлены назад и вместе
с тельсоном составляют хвостовой веер.
Резко изгибая тело и отталкиваясь
хвостовым веером от воды, рачки способны
передвигаться скачками задним концом
вперед.
Самка Anaspides откладывает крупные
(диаметром около 1 мм) яйца на
поверхность водных растений или на другие
подводные предметы. Из яиц выходят молодые
рачки длиной 2,7 мм, отличающиеся от
взрослых недоразвитыми стебельчатыми
глазами, вместо которых имеется
непарный глаз, отсутствием внутренних ветвей
передних брюшных конечностей и
расщепленным на конце тельсоном, задне-боко-
вые отростки которого напоминают ветви
фурки других ракообразных. После
нескольких линек молодые рачки становятся
вполне подобными взрослым.
Рис. 230. Bathynella natans
Anaspides tasmaniae —
самый крупный представитель
отряда, достигающий в
длину 5 еле, живет
преимущественно в ручьях и мелких
речках нагорного плато
Тасмании. Значительно более
мелкие A. spinulae и
Paranaspides lacustris обитают там же,
но в высокогорных озерах.
Все они питаются главным
образом донными
водорослями и бактериями и
органическими остатками грунта,
который они взмучивают
грудными ножками и
отфильтровывают задними
челюстями. Anaspides может
захватывать и более крупную
пищу, например червей и
других водных животных.
Сравнение
физико-химических условий водоемов,
населенных этими тремя
видами, показало, что они
выносят довольно значительные
колебания температуры,
различное содержание в воде
кислорода и различный
химический состав ее.
Очевидно, ограниченность области
их распространения
объясняется не какими-нибудь особыми,
присущими ей физико-химическими
условиями, а геологической историей
нагорной части Тасмании, позволившей
сохраниться только там «живым ископаемым».
Четвертый тасманийский вид — Mic-
raspides calmani длиной около 8 MfMf —
обитает в полостях торфяника. Он лишен
глаз, что указывает на его подземный
образ жизни. То же можно сказать о
зарывающемся в грунт единственном
австралийском представителе анаспидовых—
Koonunga cursor.
ОТРЯД БАТИНЕЛЛИЕВЫЕ
РАКООБРАЗНЫЕ (BATHYNELLACEA)
Не только капиллярные, пропитанные
соленой водой ходы морских пляжей
населены специально приспособленными к
447
жизни в таких условиях ракообразными —
гарпактицидами и мистакокаридами. На
значительном удалении от моря на
обширных пространствах суши, особенно там,
где имеются отложения известняка, под
сравнительно тонким слоем почвы
циркулируют подземные воды. Они
пропитывают горные породы, протекают через
узкие пространства между частицами
грунта, заполняют колодцы и образуют
водоемы в глубине известковых пещер. В этих
водах также обитает своеобразное
население, в том числе некоторые
ракообразные. Наиболее характерными из них
следует считать представителей отряда
Bathynellacea.
Микроскопические размеры (обычно до
2 мм, Baicalobathynella magna до 5,4 мм),
тонкое червеобразное тело и короткие
конечности позволяют этим рачкам
пробираться по узким подземным ходам и
проползать между частицами грунта. Вместе
с грунтовыми водами они проникают в
колодцы, пещерные водоемы, а иногда
и в придонные слои воды крупных озер
и живут и в таких условиях, в том
случае если температура воды достаточно
низка. Первый представитель
Bathynellacea — Bathynella natans — был
обнаружен чешским зоологом В. Вейдов-
с к и м в колодце в Праге в 1886 году, а
затем в течение 28 лет никому не
удавалось находить этих рачков. Впоследствии,
после того как методы исследования
подземных вод были усовершенствованы и на
их фауну стали обращать серьезное
внимание, Bathynellacea были открыты во
многих странах и оказались очень широко
распространенными. Теперь они известны
из ряда мест Европы, экваториальной
Африки, Мадагаскара, полуострова Малак-
ка, Японии и Южной Америки. В СССР
их нашли в пещерах Кавказа, в колодцах
близ Дона, в озере Байкал, в пещерах
Средней Азии и Дальнего Востока. Всего
известно 64 вида и подвида Bathynellacea.
Тело Bathynellacea удлиненное,
цилиндрическое. В связи с подземным образом
жизни глаза отсутствуют. Антенны
довольно короткие, с многочисленными
чувствительными щетинками. Все 8 грудных
сегментов свободные. Их конечности дву-
ветвистые и снабжены эпиподитами,
участвующими в процессе дыхания. Однако
газообмен происходит, по-видимому, и
через стенки тела. У самцов конечности
последнего грудного сегмента сильно
видоизменены и превращены в
совокупительный аппарат, а у самок они могут
отсутствовать. Брюшные сегменты по длине и
ширине не отличаются от грудных. По-
Таблица 35. Креветки:
1 — Pandalus latirostris;
2 — Palaemon elegans;
3 — Alpheus brevicristata;
4 — Sclerocrangon salebrosa;
5 — Lebbeus sp.;
6 — Hetairus groenlandicus;
7 — Penaeus japonicus;
8 — Notostomus sp.;
9 — Macrobrachium sp.;
10, 11 — Hippolyte varians, различно
окрашенные;
12 — Troglocaris anophthalmus;
13 — Stenopus hispidus.
следний брюшной сегмент срастается с
тельсоном, на конце которого
расположены фуркальные ветви. Короткие
редуцированные ножки есть только на одном или
двух передних брюшных сегментах, или
их нет совсем, но на последнем сросшемся
с тельсоном брюшном сегменте всегда
имеются сильно развитые двуветвистые уро-
поды.
При помощи грудных ножек Bathynel-
lacea медленно ползают по узким ходам
между частицами грунта, изгибая при
этом свое тело. Попав в более обширное
пространство, они могут некоторое время
плавать, последовательно взмахивая
своими грудными ножками и оставляя тело
выпрямленным. Однако их движения в
толще воды настолько слабы, что рачки
совсем не могут противостоять течению
любой силы, которое легко их сносит.
Обыкновенно они находятся на грунте
или в самом придонном слое воды, и их
удается поймать планктонной сеткой
только в том случае, если она зачерпнет
поверхностный слой грунта.
Размножение изучено только у Bathy-
nella natans. Самка откладывает на грунт
поодиночке довольно крупные (диаметром
0,11 мм) по отношению к ее телу яйца.
При температуре 11°С из них через
2 месяца выходят малоподвижные личинки
длиной около 0,45 мм. У личинок уже
имеются антенны, все ротовые придатки
и передняя пара грудных ножек, а также
10 туловищных сегментов. Они еще не
питаются, используя находящиеся в их
теле запасы желтка. После первой
линьки добавляется вторая пара грудных
ножек, еще один туловищный сегмент,
а также возникают зачатки недостающих
конечностей. Личинки становятся более
активными и питаются самостоятельно»
С каждой последующей линькой (а их до
окончания превращения личинки
насчитывается пять) личинки постепенно
приближаются по своему строению к взрослым.
Через 9 месяцев после рождения
заканчивается формирование взрослого
животного, но для достижения половозрелости
самцы должны перелинять еще 4 раза,
а самки 5 раз. В общем жизнь рачков
при температуре 12—14°С длится не
менее 2 лет.
Большинство Bathynellacea
обнаружено в различных выходах грунтовых вод
при температуре не выше 12—14°С.
Несколько видов обитает в воде,
пропитывающей песок близ берегов озер или
лагун, как например Parabathynella сараг-
ti, найденная в песке на расстоянии 1 м
Т а б л и ц а 36. Крабы:
1 — Callinectes sapidus;
2 — Pachygrapsus marmoratus;
3 — Pugettia quadrifrons;
4 — Carcinus maenas;
5 — Hyas coarctatus;
6 — Chionoecetes opilio;
7 — Cancer pagurus;
8 — Maja squinado;
9 — Calappa sp.;
10 — Randallia eburnea;
11 — Lyreidus tridentatus;
12 — Lambrus validus;
13 — Dromia vulgaris;
14 — Pinnoteres boninensis.
29 Жизнь животных, т. 2
от уреза воды озера Танганьика в
Африке, или виды того же рода с острова
Мадагаскар. Bathynella baicalensis и Baica-
lobathynella magna живут на
значительной глубине (до 1440 м) в озере Байкал.
К совершенно необычным для Bathynel-
lacea условиям приспособились Thermo-
bathynella adami и Th. amyxi. Первый
вид населяет теплые E5°С) подземные
воды близ озера Упемба в экваториальной
Африке, а второй — пропитывающие
песок солоноватые воды в дельте Амазонки.
Широкое и разбросанное
географическое распространение Bathynellacea при
полном отсутствии у них покоящихся
стадий и, следовательно, способности к
пассивному расселению до сих пор остается
нерешенной загадкой. Некоторые
исследователи допускают давнее морское
происхождение этих рачков, предполагая, что
они сохранились в участках суши,
некогда покрывавшихся морем, однако
прямых доказательств правильности этой
гипотезы пока не приведено. Во всяком
случае, несомненна глубокая древность
этих мельчайших высших ракообразных.
ОТРЯД СТИГОКАРИДЫ
(STYGOCARIDACEA)
Представители отряда Stygocaridacea по
своему внешнему виду, размерам и по
образу жизни напоминают Bathynellacea.
Они были открыты только в 1963 году
при обследовании фауны грунтовых вод
Южной Америки. Известно всего 2 рода
и 4 вида Stygocaridacea.
По характеру сегментации тела этот
отряд довольно резко отличается от
Bathynellacea. Передний грудной сегмент
Stygocaridacea полностью срастается с
головой, но задний брюшной сегмент
свободный, отчлененный от тельсона. Тельсон
снабжен рудиментарной фуркой. В
основании передних антенн имеется орган
равновесия — статоцист. Передние брюшные
ножки самца преобразованы в копулятив-
ный аппарат. У обоих видов рода Sty-
gocaris отсутствуют наружные ветви
грудных конечностей.
Рис. 231. Parastygocaris.
Все Stygocaridacea живут в интерсти-
циальных пресных водах Аргентины и
Чили, причем Parastygocaris andina
известен в Андах на высоте 2000 м над
уровнем моря. Биология этих рачков не
изучена. Сходство их с Bathynellacea
объясняется одинаковой средой обитания (ин-
терстициальные воды), к которой
представители обоих отрядов приспособились
независимо друг от друга.
НАДОТРЯД ГОПЛОКАРИДЫ
(HOPLOCARIDA)
Гоплокариды — крупные морские
ракообразные, пять передних грудных
сегментов которых срастаются с головой. Глаза
стебельчатые. Короткий карапакс не
покрывает трех задних грудных сегментов.
Брюшко очень длинное, передние его
конечности несут жаберные придатки. Самки
вынашивают яйца под грудью. К этому
надотряду относится всегЬ один отряд.
ОТРЯД РОТОНОГИЕ РАКООБРАЗНЫЕ,
ИЛИ РАКИ-БОГОМОЛЫ
(STOMATOPODA)
На мелководьях тропических и
субтропических морей, а также среди
коралловых рифов, в норах или в расселинах
коралловых массивов укрываются
крупные (от 1—2 до 34 см длиной) хищные
раки. Наружу они выставляют только
самый передний конец тела со
стебельчатыми глазами, чувствительными
антеннами и мощными ловчими ногами. Если
мимо них проплывет креветка или
мелкая рыба, они молниеносно выскакивают
из норы и хватают жертву своими ловчими
ногами, последний членик которых воору-
450
жен многочисленными острыми шипами и
прижимается к предпоследнему, как
лезвие перочинного ножа к его ручке.
Пронзенная шипами жертва оказывается
зажатой между последним и предпоследним
члениками. По способу охоты и строению
ловчих ног эти ракообразные несколько
напоминают широко известных хищных
насекомых — богомолов, почему они и
получили название раков-богомолов.
Нередко они сами оказываются добычей рыб
или человека. В Средиземном и
Адриатическом морях, а также в Индийском и
Тихом океанах существует регулярный
промысел ротоногих ракообразных,
считающихся очень вкусными.
Строение их весьма своеобразно. Голова
и передняя часть их грудного отдела
сверху покрыты карапаксом, имеющим вид
щита, лишь в малой степени защищающего
бока животного. Передний край кара-
пакса сочленен с подвижным рострумом,
по бокам которого располагаются
сидящие на длинных стебельках фасеточные
глаза. Между ними находится науплиаль-
ный глаз. Фасеточные глаза
раков-богомолов отличаются необыкновенной
подвижностью. Они могут поворачиваться
вслед за проплывающими мимо
животными и в процессе ловли добычи играют
очень важную роль. Каждая из
передних антенн состоит из длинной 3-члени-
ковой основной части и трех
прикрепленных к ней жгутов. На жгутах расположены
многочисленные органы химического
чувства и вкуса — цилиндры и щетинки.
Перед тем как схватить замеченную глазами
добычу, рак пускает в ход передние
антенны и только после того, как убедится в
съедобности объекта, приступает к его
ловле. В аквариуме рак-богомол
пристально следил за привязанным к нитке
шариком из пластилина, которым
экспериментатор двигал перед ним, но после
исследования шарика передними
антеннами не делал никаких попыток его
поймать. Совсем иначе рак вел себя, когда
к нитке вместо пластилинового шарика
привязывали кусок рыбы. Задние ан^-
тенны короче передних. Их наружная
ветвь преобразована в чешуйку, как
у многих десятиногих ракообразных
и у мизид.
Конечности пяти передних слившихся
с головой и прикрытых сверху
карапаксом грудных сегментов превращены в
ногочелюсти. Первая из них короткая,
тонкая, густо покрыта щетинками и
служит органом осязания. При помощи этих
конечностей рак также удаляет со
своего тела посторонние частицы.
Остальные четыре пары замечательны тем, что
их последний и предпоследний членики
образуют так называемую подклешню
за счет способности последнего
членика пригибаться к предпоследнему. Так
возникает приспособление к
захватыванию добычи или, в случае
необходимости, комочков грунта. Из этих
конечностей особенно сильно развита передняя
(т. е. вторая ногочелюсть), превосходящая
остальные по размерам не менее чем вдвое,
с вооруженным острыми шипами
последним члеником. Именно эта пара ног и
осуществляет схватывание добычи.
Остальные ногочелюсти передают
пойманную добычу вперед, ко рту, и, кроме того,
выполняют основную работу по рытью
грунта при постройке нор. Все пять пар
ногочелюстей лишены наружных ветвей,
но несут маленькие эпиподиты, в
незначительной степени участвующие в
процессе дыхания.
Три задних грудных сегмента
подвижно сочленены между собой и не покрыты
карапаксом. Их конечности
функционируют как ходильные ноги. Они имеют
длинные внутренние и короткие
наружные ветви и лишены эпиподитов.
Брюшной отдел длиннее головы и груди
вместе взятых. Все его 6 сегментов сильно
развиты, они толще и длиннее, чем
предшествующие им задние свободные
грудные сегменты. Пять передних пар
брюшных конечностей довольно крупные, дву-
ветвистые, листовидные, причем обе
ветви по краям снабжены многочисленными
перистыми щетинками. При помощи
зубцов, имеющихся на внутреннем крае
основания каждой ножки, левая и правая
конечности каждой пары могут
соединяться между собой и производить взмах как
единое целое.
Функции передних брюшных ножек
весьма разнообразны. Благодаря их
взмахам ротоногие плавают. Кроме того,
29*
451
на всех: передних брюшных ножках
располагаются жабры, имеющие вид
тонкостенных, многократно ветвящихся придатков.
Так, например, у обыкновенного рака-
богомола Средиземного моря — Squilla
mantis длиной 15 см — на брюшных
ножках насчитывается в общей сложности
около 10 000 жаберных придатков
(основной придаток каждой из 10 ножек
разветвляется на 30 вторичных ветвей, каждая
из которых дает 25—30 третичных
ответвлений) длиной около 1 см при диаметре
2 мм. Дыхательная поверхность всех жабр
составляет огромную величину порядка
600 см2. Брюшные ножки непрерывно
совершают последовательные взмахи со
скоростью 25—40 раз в минуту, возбуждая
таким образом ток воды, достаточный для
дыхания. Когда рак сидит в норе, эти
взмахи приводят к постоянной смене воды
в его жилище.
Наконец, у самцов внутренние ветви
первой и второй пар брюшных ножек
преобразованы в копулятивный
аппарат. Последняя пара брюшных ножек —
уроподы — плоские, сильно хитинизиро-
ванные и направлены назад, располагаясь
по бокам крупного лопатообразного тель-
сона. Его задний конец снабжен зубцами,
а спинная поверхность — различными
килями и буграми. Продольные кили есть и
на поверхности брюшных сегментов. Они
устроены так, что вода поступает в
нору при движении брюшных ножек
определенным образом направленными
струями.
Большинство ротоногих роет норы в
морском грунте. Мелкие виды родов Go-
nodactylus и Coronida прячутся в
расселинах и щелях между ветвями кораллов,
а мелкий B,7 см длины) Lysiosquilla vi-
cina использует для жилья норы Bala-
noglossus, причем оба животных обитают
в одной норе, по-видимому не мешая друг
другу.
Раки-богомолы зарываются в грунт
передним концом тела, орудуя рострумом
и ногочелюстями и выбрасывая грунт
из норы брюхом, тельсоном и уроподами.
Готовая нора обычно имеет два выхода,
и вода, направляемая взмахами передних
брюшных ножек, свободно протекает
через нее. Норы Lysiosquilla excavathrix
достигают глубины 1 м, Squilla
mantis обычно зарывается на глубину около
15 см.
Большую часть времени раки-богомолы
проводят в норах. Вылезая наружу, они
ползают по поверхности грунта при
помощи задниг грудных ног, а также
ловчих ногочелюстей, которые при этом
сгибаются и на которые рак опирается, как
Рис. 232. Ротоногие ракообразные:
1 — Squilla oratoria в ожидании добычи; 2, з — личиночные стадии: первая пелагическая стадия псевдозоэа
Squilla B) и первая пелагическая стадия антизоэа Lysiosquilla C).
452
на костыли. Раки могут и довольно
быстро плавать. При этом они гребут
передними брюшными ножками, ногочелюсти
сложены и прижаты к телу, а чешуйки
задних антенн, тельсон и уроподы
изгибаются под разными углами и служат рулями
глубины. Кроме того, животные могут
передвигаться скачками вперед и назад,
резко изгибая и выпрямляя тело.
Squilla mantis и, вероятно, другие
виды обладают очень хорошим зрением.
Лучше всего рак видит объекты, находящиеся
от него на расстоянии 10 см. Доказано,
что число зрительных элементов,
участвующих при этом в зрении, у него такое
же, как у человека, смотрящего на тот
же объект с дистанции 1,2 — 1,5 м. Рак
свободно различает на таком расстоянии
креветок длиною 3—5 см, но обращает
внимание преимущественно на
двигающиеся объекты. Лучше всего Squilla видит
при сравнительно слабом освещении A0—
20, до 100 люксов), соответствующем тому,
которое господствует на глубине его
обитания F0—100 м).
Как уже указывалось, зрение имеет
важное значение при добывании пищи
раками-богомолами.
Большинство этих животных охотится
на других ракообразных, прежде всего
на креветок, но Lysiosquilla охотно
нападает также на рыб и моллюсков, a Gono-
dactylus поедает актиний.
Икрометание продолжается около 4
часов. При этом самки сгибают брюшко и
вместе с яйцами выделяют липкий
секрет, который склеивает яйца в крупные
яйцевые мешки. Формирование яйцевого
мешка осуществляется главным образом
тремя задними ногочелюстями. Они
придают ему форму эллипса, длиной у S.
mantis до 14 и шириной до 10 см. В таком
мешке у этого вида содержится до 50 000
яиц, расположенных в три слоя. У мелких
Gonodactylus длина мешка всего 8—14 мм.
Яйцевой мешок помещается между
ногочелюстями самки, и она носит его в
течение 10—11 недель. В это время она не
питается.
Из яиц выходят личинки, разные у
разных родов. У большинства родов,
в том числе у Squilla, из яйца появляется
так называемая псевдозоэа. Она
обладает плоским карапаксом с длинным
рострумом и с ддумя парами длинных
шипов на переднем и заднем краях,
стебельчатыми глазами, сформированными
головными придатками и двумя передними
парами ногочелюстей, из которых вторая
уже имеет характерное строение ловчих
ног. Брюшные сегменты также хороша
развиты, за исключением последнего, еще
слитого с тельсоном, и несут
плавательные ножки (рис. 232,2). Псевдозоэа сначала
живет на дне, но после двух линек,
происходящих у Squilla в течение 2—
3 дней, при длине 2—2,5 мм переходит
к планктонному существованию. В толще
воды она линяет еще 4 раза и
превращается в стадию с и н з о э а длиною 8 деле,
у которой развиты все грудные конечности
и которая и по другим признакам более
сходна с взрослыми раками-богомолами.
Однако синзоэа также плавает в толще
воды и, только перелиняв еще 4 раза,
опускается на дно и дает начало
маленькому ротоногому рачку.
У родов Lysiosquilla и Coronida
личиночное развитие начинается с
планктонной стадии антизоэа. В отличие от
псевдозоэа ее глаза сидячие, брюшной
отдел короткий и нерасчлененный, зато уже
обособлены все грудные сегменты (рис.
232,5). Пять пар передних грудных ножек
двуветвисты и служат для плавания.
После пяти линек, в результате которых у
личинки развиваются брюшные сегменты
и недостающие конечности, антизоэа
переходит в только что описанную стадию
синзоэа. У Lysiosquilla в Неаполитанском
заливе личиночное развитие занимает 2—
3 месяца.
Личинки ротоногих вскоре после
рождения начинают вести хищный образ
жизни. Псевдозоэа ловит мелких животных
ловчими ножками, а антизоэа сначала
шипами передних челюстей; после третьей
линьки у нее появляются ловчие ноги
и функции добывания пищи переходят
к ним. Достаточно продолжительный
период планктонного существования
обеспечивает разнос личинок течениями и
способствует расселению раков-богомолов.
Из 170 известных видов этого отряда
подавляющее большинство обитает в
тропических и субтропических морях на не-
453
большой глубине* В отдельных случаях
раки-богомолы опускаются до глубины
760 ле. Как правило, они избегают
сколько-нибудь значительного опреснения, но
некоторые тропические виды могут
обитать и в солоноватых водах, а один из
них даже проникает в низовья рек на
острове Тайвань. За пределами
тропической и субтропической зон раки-богомолы
встречаются в Средиземном и
Адриатическом морях (Squilla mantis, Lysiosquilla
maculata и др.).
В морях СССР зарегистрирован всего
один вид — S. oratoria, попадающийся в
Японском море и на мелководьях залива
Петра Великого.
НАДОТРЯД ПЕРАКАРИДЫ
(PERACARIDA)
Этот обширный надотряд объединяет
очень различных как по внешнему виду,
так и по образу жизни ракообразных,
обитающих в море, в пресных водах и на
суше, ползающих по дну и плавающих
в толще воды, большей частью свободно-
живущих, но частично и паразитирующих
на других животных. Однако все они
обладают некоторыми общими
признаками, указывающими на их родство между
собой.
Голова всех перакарид срастается с
одним или изредка с двумя передними
грудными сегментами, конечности которых
превращаются в ногочелюсти. Правая и левая
жвалы устроены различно: на левой
обычно есть подвижная пластинка,
отсутствующая на правой. Самки
вынашивают яйца под грудью в выводковой
сумке, образованной направленными внутрь
листовидными отростками грудных ножек
(только у отряда Thermosbaenacea
выводковая сумка помещается на спинной
стороне тела, под карапаксом). Из яиц
выходят почти вполне сформировавшиеся
молодые рачки, так что настоящих
личинок у перакарид нет.
К надотряду Peracarida относятся
отряды мизид, кумовых, клешненосных
осликов, спелеогрифовых, термосбеновых,
равноногих и разноногих ракообразных.
ОТРЯД МИЗИДЫ (MYSIDACEA)
По внешнему виду мизиды несколько
напоминают маленьких креветок. Тело
их вытянуто в длину, глаза стебельчатые,
голова и грудь покрыты цилиндрическим
карапаксом, брюшко тонкое и
заканчивается хвостовым веером, состоящим из
пластинчатого тельсона, по краям
которого расположены плоские двуветвистые
уроподы. Подобно креветкам они плавают
у дна или в толще воды.
Однако при более внимательном
рассмотрении мизид нетрудно заметить их
важные отличия от креветок. Карапакс
мизид срастается не больше чем с тремя
передними, а не со всеми грудными
сегментами. Над задними грудными
сегментами он просто нависает. У мизид всего
одна пара ногочелюстей, да и то очень
слабо специализированных, а не три, как
у креветок, клешней, как правило, нет.
Самки мизид вынашивают яйца под грудью
в выводковой сумке, а не на брюшных
ножках, как креветки. По-разному устроены
и жвалы обеих групп ракообразных.
Трудно сомневаться в том, что мизиды и
креветки приобрели общий облик
независимо друг от друга. Это объясняется их
сходным способом передвижения.
Мизиды распространены чрезвычайно
широко. Всего их известно около 500
видов. Они обитают во всех морях и
океанах, как у дна, так и в толще воды.
Некоторые виды проникают из морей в
реки, причем поднимаются по ним очень
высоко. Каспийские и азовско-черномор-
ские мизиды заселили не только Волгу,
Днепр, Дон, Донец и их притоки, но и
водохранилища, построенные на этих
реках. Есть и чисто пресноводные виды,
как например обычная в крупных озерах
северной части Европы и Америки Mysis
relicta. Несколько своеобразных видов
открыто в пресных подземных водах.
Многие мизиды служат излюбленной
пищей рыб, в том числе промысловых. Для
улучшения условий питания рыб как во
вновь созданных водохранилищах, так и
в ранее существовавших водоемах у нас
в стране были в широких масштабах
произведены искусственные переселения ми-
454
зид. Их перевезли в Аральское море, в
озеро Балхаш и во многие водохранилища
Украины. В последнее время и в США
осуществляется заселение мизидами водоемов,
в которых раньше этих рачков не было.
Длина тела большинства мизид 10—
20 мм, но среди глубоководных видов
многие гораздо крупнее (до 18 см). Тело
мелководных видов полупрозрачное.
У многих мелководных видов через
тонкие покровы просвечивают находящиеся
в соединительной ткани крупные
звездчатые клетки — хроматофоры,
заполненные темным пигментом. Они расположены
по одной на верхней стороне брюшных
сегментов и тельсона, а более мелкие
разбросаны по другим частям тела. На
свету или на светлом грунте зерна пигмента
собираются в центре хроматофора и рачок
светлеет, в темноте или на темном грунте
пигмент рассеивается по всей клетке, что
приводит к потемнению рачка. Эти
передвижения пигмента в хроматофорах, так
же как у десятиногих ракообразных (см.
ниже), регулируются деятельностью
находящихся в глазных стебельках органов
внутренней секреции. Глубоководные ми-
зиды, как правило, лишены хроматофоров
и окрашены в красный цвет.
Карапакс мизид обычно гладкий, без
выростов, с коротким рострумом или без
него, и только у видов глубоководного
рода Gnathophausia снабжен длинным
рострумом, продольными килями и
оттянутыми в шипы задне-боковыми углами (рис.
233,3). Стебельчатые глаза с
многочисленными фасетками на конце, но у многих
глубоководных и пещерных видов фасетки
и пигмент отсутствуют, а у
глубоководного рода Pseudomma оба слепых глаза
срастаются между собой, образуя единую
пластинку. Передняя и задняя антенны
очень длинные. Передняя антенна с двумя
жгутами, богатыми чувствительными
щетинками и цилиндрами, а у самцов
между обоими жгутами находится
специальный чувствительный придаток,
помогающий отыскивать самок. Наружная ветвь
задней антенны превращена в чешуйку,
а внутренняя имеет вид длинного жгута.
Из ротовых придатков наибольший
интерес представляют передние челюсти: их
наружная ветвь преобразована в широкую
пластинку, движения которой
обеспечивают смену воды под краем карапакса,
что необходимо для процесса дыхания.
Внутренние лопасти обеих челюстей и
щупик задней челюсти снабжены
многочисленными щетинками и, пригибаясь друг к
ДРУГУ* образуют фильтрующую камеру,
ограниченную спереди большой верхней
губой. Большинство мизид питается
отфильтровывая мелкие взвешенные в воде
частицы, но, кроме того, они могут
захватывать и крупные куски отмерших
растений и животных и даже трупы себе
Рис. 233. Мизиды:
1,2 — самка и самец Mysis relicta; 3 — Gnathophausia
gigas; 4 — Eucopia unguiculata.
455
Рис. 234. Токи воды, возбуждаемые Hemi-
mysis lamornae:
1 — при плавании; 2 — при опускании на дно.
подобных, которые они разрывают и
размельчают жвалами.
Все 8 пар грудных ножек снабжены
наружными ветвями — экзоподитами.
Передняя из них — ногочелюсть —
отличается от остальных меньшими размерами и
присутствием внутренних лопастей при
основании. Она принимает участие в
процессе фильтрации и захватывания
пищевых частиц. Остальные служат для
передвижения. Их экзоподиты совершают
взмахи, благодаря которым по бокам тела
рачка возникают два круговорота воды,
проталкивающие животное вперед (рис. 234).
Перебирая внутренними ветвями
грудных ножек, мизиды ползают по дну. У
некоторых видов часть этих внутренних
ветвей расширена и приспособлена для
рытья грунта. Такие виды могут очень
быстро зарываться в поверхностный слой
грунта. У подотряда Lophogastrida,
который объединяет почти исключительно
глубоководные виды, на основных частях
грудных ножек находятся разветвленные
жабры (эпиподиты), остальные
многочисленные мизиды относящиеся к другому
подотряду — Mysida, лишены жабр;
газообмен у них совершается через тонкие
стенки карапакса, в полостях которых
циркулирует кровь. Некоторые хищные
глубоководные мизиды имеют длинные
ловчие ноги, приспособленные для
захватывания других животных. У половозрелых
самок всех мизид к основаниям грудных
ножек прикреплены направленные внутрь
пластинки (оостегиты), образующие
выводковую сумку.
Длинный и тонкий брюшной отдел
мизид состоит из 6 цилиндрических, очень
подвижных друг по отношению к другу
сегментов, из которых последний самый
крупный. У рода Gnathophausia на нем
ясно заметен поперечный шов,
свидетельствующий о том, что этот сегмент двойной,
представляющий результат слияния
шестого и седьмого сегментов. Действительно,
у эмбрионов мизид шестой и седьмой
брюшные сегменты самостоятельны и
срастаются только на поздних стадиях развития.
Очевидно, некогда мизиды имели 7
брюшных сегментов подобно тонкопанцирным
(см. выше).
У всех мизид хорошо развиты
конечности шестого брюшного сегмента —уро-
поды. Они двуветвистые, плоские и
расположены по бокам крупного
пластинчатого тельсона, образуя вместе с ним
хвостовой веер. В основании внутренней ветви
уроподов у всех мизид, за исключением
Lophogastrida, находится орган
равновесия — статоцист. На стенках полости ста-
тоциста расположены чувствительные
щетинки (у Praunus около 60), на которые
опирается статолит, состоящий из
органического вещества и известковых
наслоений. Он давит на те или иные
чувствительные щетинки, и рачок, ощущая это
давление, повертывает свое тело так,
чтобы спинная его сторона была обращена
вверх. Конечности передних пяти
брюшных сегментов хорошо развиты только
у Lophogastrida и рода Boreomysis, у
которых они служат основным органом
плавания. У остальных мизид они в той
или иной степени редуцированы, особенно
у самок. У самцов одна из них (часта
четвертая пара) имеет удлиненную ветвь,
помогающую при оплодотворении яиц.
456
Спаривание мизид происходит ночью.
Оно наблюдалось лишь у немногих видов.
Самцы при помощи химического чувства
отыскивают недавно слинявших самок и
плавают под ними брюшной стороной
вверх. Приблизившись к самке, самец
Mesopodopsis просто выпускает
сперматозоиды в воду, а затем направляет их
удлиненными внутренними ветвями
четвертых брюшных ножек в выводковую
камеру самки. Самец Hemimysis u Praunus
схватывает самку на 1—2 секунды
внутренними ветвями грудных ножек и
вводит сперму в ее выводковую сумку при
помощи трубкообразных отростков
последнего грудного сегмента, в которые
открываются его семепроводы. Такой способ
спаривания, не связанный с
необходимостью длительно удерживать самку,
объясняет одну своеобразную особенность
мизид: в отличие от большинства других
высших ракообразных их самцы мельче
и слабее самок.
Через 20—30 минут после
проникновения спермы в выводковую сумку самка
откладывает туда же яйца. Число яиц
различно у разных видов и, кроме того,
зависит от размера, т. е. от возраста
самки. Так, например, самка пресноводной
Mysis relicta длиной 13—15 мм
откладывает 10—20, а длиною
17—21 мм — 25—40 яиц;
крупные, пережившие зиму самки Neo-
mysis vulgaris в районе г. Киля
носят до 55 яиц в выводковой
сумке, а мелкие A0 мм) самки,
родившиеся летом, — всего 9 яиц.
В Северном Каспии
перезимовавшие самки Paramysis baeri
откладывают весной в среднем 89,
максимально 156 яиц.
Яйца развиваются в
выводковой сумке, причем составляющие
ее пластинки двигаются так, что
вода в сумке все время сменяется.
Вышедшие из яиц молодые
рачки также первое время остаются
в выводковой камере матери, а ок-
репнув, переходят к
самостоятельному существованию.
Вынашивание яиц и эмбрионов у разных
видов длится разное время, при
высокой температуре оно заканчи-
вается гораздо раньше, чем при низкой,
В прибрежных водах Индии, близ
Мадраса, при температуре воды 25—29°С
вынашивание яиц и эмбрионов Mesopodopsis in-
dica продолжается всего 4 дня, а обитатель
холодных (до 14°С) озер Mysis relicta
носит яйца и эмбрионов в течение 2—3
месяцев. В холодном Белом море у Mysis
oculata, M. mixta и М. litoralis
вынашивание яиц и эмбрионов растягивается на
6—7 месяцев.
Рост молоди, вышедшей из выводковой
камеры матери, также зависит от
температуры. В хорошо прогреваемом
мелководном Северном Каспии родившиеся в мае
рачки уже в июне достигают половозре-
лости и приступают к размножению. До
наступления зимы северокаспийские ми-
зиды успевают дать 4 поколения. За весь
период размножения одна пара Paramysis
baeri может произвести на свет 430 000
потомков.
Neomysis integer в Северном море дает
два поколения. Родившиеся весной особи
живут в течение 4—51/2 месяцев и через
1—1% месяца после рождения достигают
половозрелости. За свою жизнь самки 4—
6 раз откладывают яйца, правда,
немногочисленные (в среднем 20 яиц). Другое по-
Рис. 235. Спаривание Praunus flexuosus.
457
коление состоит из рачков, родившихся
в конце лета — осенью. Они
перезимовывают и становятся половозрелыми только
к июню следующего года, размножаются
всего 3, реже 4 раза в жизни, но зато
откладывают в среднем по 40 яиц.
Продолжительность их жизни достигает 10—11
месяцев. Мизиды Северного Каспия
живут 12—14 месяцев, а другие виды
несколько дольше: пресноводная Mysis ге-
licta и морские виды рода Praunus —до
1Уг лет, арктическая М. oculata — более
2 лет, беломорские М. litoralis, M.
oculata и М. mixta — около 3 лет.
Подавляющее большинство мизид
обитает в море, где они встречаются на всех
глубинах, от приливо-отливной зоны до
огромных глубин — 7260 м (Amblyops
magna в Курило-Камчатской впадине).
Литоральные виды — Gastrosaccus sanctus
европейских морей и Archaeomysis greb-
nitzkii Японского моря — на время
отлива зарываются в песок, а в прилив ведут
активное существование. Многие виды
живут главным образом среди зарослей
растений (Praunus flexuosus), другие среди
камней и на жестком грунте (Neomysis
integer u Siriella jaltensis в Черном и
Средиземном морях).
Многие виды совершают регулярные
горизонтальные и вертикальные миграции.
Neomysis integer, Mesopodopsis slabbed
и Praunus flexuosus осенью откочевывают
от берегов, зиму проводят на
сравнительно больших глубинах, а весною
возвращаются на мелководье. В Рижском
заливе зимой мизиды также концентрируются
на глубине, а весной переходят на
мелководье. При этом более теплолюбивая
Neomysis vulgaris занимает глубины до 15 м,
на глубине от 15 до 25 м преобладает
Mysis mixta, а на большей глубине
обитает холодолюбивая М. relicta.
Мигрирующие виды нередко образуют
стаи или косяки. Мигрирующие стаи
Neomysis integer достигают 1 км в длину и от
одного до многих метров в ширину.
Не менее характерны для мизид и
вертикальные миграции. Многие мелководные
виды днем живут на дне, а ночью
поднимаются в поверхностные слои воды.
Gastrosaccus sanctus зимой появляется в
поверхностных слоях воды в 4 часа, а
летом в 9 часов вечера, т. е. в обоих случаях
в сумерки. Суточные вертикальные
миграции широкого диапазона свойственны
каспийским мизидам Mysis microphthalma,
М. amblyops и Paramysis loxolepis. Ночью
они поднимаются в поверхностный 50-
метровый слой, а днем опускаются до 600—
750 м. Четвертая пелагическая мизида
Каспия —М. caspia—совершает суточные
вертикальные миграции между 150 и 400 м.
Значительный интерес представляют
вертикальные миграции многих планктонных
глубоководных мизид, не связанные со
сменой дня и ночи. Крупные
половозрелые особи родов Gnathophausia и Eucopia
всегда встречаются только на большой
глубине, а молодые, неполовозрелые особи
поднимаются довольно высоко. Так,
например, в северо-западной части Тихого
океана молодь G. gigas живет на глубине
до 200 — 500 м, а половозрелые особи
обитают только на глубине 4—7 км.
Очевидно, по мере роста рачки опускаются
все глубже и глубже и размножение их
происходит на предельной для каждого
вида глубине. Там же обитает
новорожденная молодь, которая потом начинает
подниматься вверх. Так происходят так
называемые «онтогенетические» миграции,
свойственные и некоторым другим
планктонным глубоководным животным, в
частности рассмотренному ранее
тонкопанцирному рачку Nebaliopsis typica
(см. стр. 445).
Мы не знаем, каким образом на
глубине нескольких километров в полном
мраке самцы и самки таких животных
находят друг друга. Может быть, этому
помогает удивительная способность
Gnathophausia выбрасывать струю
светящейся жидкости, вырабатываемой железами
задних челюстей. Другие мизиды не
способны к свечению.
Среди глубоководных мизид некоторые
также мигрируют на несколько
километров, но их миграции имеют характер
кормовых, а не онтогенетических. В
северозападной части Тихого океана кишечники
Boreomysis plebeja, В. curtirostris и Нурег-
amblyops nana, пойманных на глубине
200—500 м, были набиты планктонными
водорослями. В кишечнике тех же видов
с глубин более 500—1000 м водоросли
458
были в значительной степени переварены i
а у особей с еще большей глубины в
кишечниках удавалось обнаружить только
остатки водорослей. Из этих данных
следует, что рачки вынуждены подниматься
в верхние слои воды, богатые
фитопланктоном, заполнять им свой кишечник, а
затем опускаться вниз, постепенно
переваривая запасенную пищу. Как часто они
это делают и сколько времени им
требуется для того, чтобы преодолеть огромное
расстояние в 4—6 км, остается пока
неизвестным. Можно только предполагать, что
планктонные мизиды родов Boreomysis и
Hyperamblyops на больших океанических
глубинах не находят для себя
достаточного количества пищи.
Другие глубоководные, как
планктонные, так и донные, мизиды — хищники.
У многочисленных в толще глубинных
слоев Eucopia внутренние ветви всех
грудных ножек снабжены подклешнями,
причем три задние пары грудных ног
удлиненные и, вероятно, действуют как
ловчие ноги. Есть ловчие ноги и у придонного
глубоководного Pethalophthalmus.
Представители этих родов лишены
фильтрующих щетинок на челюстях и, очевидно,
могут питаться только за счет
добывания других животных. Таким образом,
для мизид, так же как и для веслоногих,
отмечается повышение относительного
количества хищников с увеличением
глубины.
Среди мизид, обитающих в наших
южных морях, многие способны существовать
при различной солености, причем
несколько видов выдерживает даже полное
опреснение. Такие виды, как Paramysis ullskyi,
P. lacustris, P. intermedia, P. baeri, Lim-
nomysis benedeni и некоторые другие,
распространены не только во всем
Каспийском море и в опресненных участках
Черного и Азовского морей, но заходят и во
впадающие в них реки — Волгу, Урал,
Терек, Дон, Днепр, Донец, Прут, Дунай
и др. Особенно широко расселилась по
речным системам P. ullskyi, поднявшаяся
по Волге выше Рыбинска. Некоторые из
речных мизид сильно размножились и в
построенных на наших реках
водохранилищах, благодаря чему улучшились
условия питания обитающих там рыб. Как
уже указывалось, мизид переселили в
такие водохранилища и другие водоемы,
куда они не могли проникнуть
самостоятельно. Так, например, понто-каспийские
мизиды Paramysis lacustris, P. intermedia,
Limnomysis benedeni успешно
размножились во многих водохранилищах Украины
и Литвы, а также в озере Балхаш и в
Аральском море и используются там в пищу
промысловыми рыбами.
Не только понто-каспийские мизиды
вселяются в пресные воды. В северной части
Тихого океана некоторые виды рода Neo-
mysis заходят в низовья рек и связанные
с ними озера, есть такие виды и в Юго-
Восточной Азии,, на Зондских островах,
в Австралии и т. д.
Особый интерес представляет
распространение северной пресноводной мизиды
Mysis relicta. Она живет в крупных
холодных чистых озерах Карелии, в
Ладожском и Онежском озерах, в нескольких
озерах Белоруссии и Смоленской области,
в устьях северных рек, в озерах северной
Германии и скандинавских стран, в
некоторых озерах Канады и США. Расселение
М. relicta по всем этим озерам связано
с четвертичным оледенением, когда перед
краем наступающего или отступающего
ледника возникали огромные водоемы,
фауна которых частично содержала и
виды морского происхождения. Вместе с М.
relicta в таких остаточных озерах
обычно обитают веслоногий рачок Limnocala-
nus grimaldii и некоторые виды бокопла-
вов. М. relicta, как правило, живет при
температуре не выше 14°С, при высоком
содержании кислорода в воде. Рачок
совершает регулярные суточные миграции,
поднимаясь ночью в поверхностные слои
воды.
Почти ничего не известно об образе
жизни немногих подземных мизид. Однг
из них —¦ Troglomysis vjeternicensis —
найдена в большой пещере Ветреница в
Югославии. Она близка к понто-каспий-
ским видам, от которых отличается,
между прочим, полной слепотой. Другие виды
принадлежат к родам Stygiomysis и Le-
pidops. Они настолько своеобразны, что
выделяются в особые семейства Stygio-
mysidae и Lepidopidae. Виды первого
семейства живут на огромных расстояниях
459
Рис. 236. Кумовые ракообразные:
А, Б — Diastylis rathkei, самка и самец; В —
строение передней ногочелюсти; 1 — сифон; 2 —
эпиподит; 3 — жаберный придаток; 4 — шипы
внутреннего края.
друг от друга — в Италии, на Антильских
островах и в Индии, второго — в
Занзибаре, Италии и Мексике.
Практическое значение ми-
зид довольно велико, поскольку они
служат излюбленной пищей многих рыб, в
том числе промысловых. Снеток и
некоторые сиги питаются преимущественно Му-
sis relicta, молодь судака в большом
количестве поедает мизид, прежде чем
перейти к потреблению рыб. Мизиды
усиленно поедаются салакой в Балтийском
море, кильками и некоторыми сельдями
в Каспийском море, многими рыбами в
наших северных и дальневосточных
морях.
В некоторых странах Юго-Восточной
Азии мизид охотно едят и люди. Из
рачков приготовляют пасту или соус,
которыми сдабривают рис. На острове Ява
это кушанье носит специальное название
«джембрет».
ОТРЯД КУМОВЫЕ РАКООБРАЗНЫЕ
(CUMACEA)
Кумовые ракообразные по некоторым
особенностям своей организации близки
к мизидам, но по образу жизни довольно
резко от них отличаются. Все кумовые
приспособлены к зарыванию в
поверхностный слой морского грунта и лишь на
короткие промежутки времени покидают
грунт и плавают или ползают
по его поверхности. Несмотря
на сравнительно большое
количество относящихся к этому
отряду видов (около 600), все они
имеют сходное строение,
соответствующее их однообразной
биологии. Они обитают почти
исключительно в море, и лишь
несколько понто-каспийских
видов и один дальневосточный
вид встречаются в пресных
водоемах. Длина тела этих
рачков обычно не достигает 1 смг
и лишь немногие виды крупнее
(до 35 мм).
Тело кумовых резко
разделяется на более широкую
переднюю часть, включающую голову и
грудь, и тонкую заднюю часть,
состоящую из брюшка и тельсона. Передняя
часть тела покрыта коротким карапак-
сом, оставляющим свободными 5, реже
меньше задних грудных сегментов.
Строение карапакса весьма своеобразно.
Передне-боковые его углы вытянуты
вперед и сближены между собой так, что
образуют часто довольно длинный так
называемый «ложный рострум». За
основанием ложного рострума на поверхности
карапакса находится непарный глаз,
состоящий из двух соприкасающихся друг
с другом фасеточных глаз. Число
фасеток в каждом из них очень невелико по
сравнению с тем, которое свойственно
другим высшим ракообразным, обычно 3—4Г
самое большее И, и зрение не играет
существенной роли в жизни кумовых
ракообразных. Боковые края карапакса сзади
и с боков плотно прилегают к стенкам
тела, и только по бокам от ложного
рострума остаются отверстия, ведущие слева
и справа в пространства между краем
карапакса и стенкой тела; эти
пространства представляют собой жаберные
полости. Такое их строение позволяет рачку
дышать, когда он сидит, зарывшись в грунт
и выставив над его поверхностью только
самый передний конец тела. Грунт не
попадает в защищенные краями карапакса
жаберные полости, а вода проникает в них
через отверстия по бокам рострума»
Передние антенны очень короткие, каж-
460
дая с двумя жгутами, как у мизид.
Задние антенны у самок также очень короткие,
но у половозрелых самцов они удлинены,
нередко превосходят по длине тело и
богаты чувствительными щетинками. С их
помощью самцы отыскивают самок. Жва-
лы лишены щупиков. На обеих парах
челюстей с внутренней стороны находятся
выросты с фильтрующими щетинками.
Три передних, покрытых карапаксом
сегмента грудного отдела срастаются с
головой, и их конечности превращены в ного-
челюсти. Первая из них короткая, но
устроена особенно сложно и выполняет
несколько ответственных функций. Левая
и правая ногочелюсти сцеплены воедино
шипами на внутреннем крае и направлены
вперед. Крупный эпиподит каждой ножки
разделен на две части. Передняя из них
также направлена вперед, и конец ее
свернут так, что образует нечто вроде
трубки; это сифон, по которому вода
проходит в жаберную полость. Задняя часть
направлена назад, она тонкостенная,
листовидная и снабжена жаберным
придатком, на котором сидят в ряд 4—12, иногда
до 40 пальцевидных жаберных листочков.
Задняя часть эпиподпта первой
ногочелюсти служит основным органом
дыхания, но в дыхании участвует и внутренняя
поверхность стенки карапакса. Ток воды
в жаберную полость вызывается
движениями пластинчатой задней части эпипо-
дита, совершающей около 40 взмахов в
минуту. Остальные две пары ногочелю-
стей гораздо длиннее и служат главным
образом для захватывания пищевых
частиц и передачи их вперед, к ротовому
отверстию. Задняя из них снабжена
многочисленными перистыми щетинками, при
помощи которых она очищает жаберные
полости от попадающих туда
посторонних частиц.
На пяти свободных задних грудных
сегментах расположено по паре ходильных
ножек. Все они, обычно за исключением
последней пары, помимо внутренних,имеют
многочлениковые, несущие
многочисленные щетинки наружные ветви, при помощи
которых рачки плавают. Внутренние
ветви служат для разгребания грунта, а
передние, кроме того, загребают пищевые
частицы. У половозрелых самок на
последней паре ногочелюстей и на трех
передних ходильных ножках возникают
пластинки, образующие выводковую сумку.
Брюшко тонкое, длинное,
цилиндрическое, и составляющие его 6 сегментов
подвижно сочленены между собой. Поэтому
брюшко может легко изгибаться как на
спинную, так и на брюшную сторону.
Передние брюшные конечности у самок
отсутствуют, а у половозрелых самцов
частично или полностью развиваются. Они
короткие, обычно двуветвистые, с
длинными щетинками и служат для плавания.
К последнему брюшному сегменту у обоих
Рис. 237. Зарывание в грунт кумовых
ракообразных:
1 — Iphinoe trispinosa; 2 — Leucon nasica; 3 —
Diastylis rathkei; 4 — передний конец тела
зарывшейся D. rathkei в момент захватывания
пищи; 5 — Pterocuma pectinata.
461
Рис. 238. Самец и самка Diastylis glabra в
брачном наряде.
Рис. 239. Спаривание кумовых:
1 — Lamprops fasciata; 2 — Pseudocuma longicornis;
3 — конец второй грудной ножки самца Lamprops,
закрепленный за край карапакса самки.
полов прикреплены направленные назад
длинные тонкие двуветвистые уроподы.
Загибая брюшко на спинную сторону,
рачок может очищать ими спинную
поверхность карапакса и свободных грудных
сегментов. Расположенный между
основаниями уроподов тельсон имеет различную
форму у разных видов, а у некоторых
срастается с последним брюшным сегментом.
Большую часть жизни, во всяком случае
светлое время суток, кумовые проводят
зарывшись в поверхностный слой грунта.
Опускаясь на дно, рачок загибает вверх
брюшко и передний конец тела и начинает
разгребать грунт внутренними ветвями
трех пар задних грудных ног. Им
помогает изгибающееся в разные стороны
брюшко. В конце концов все тело рачка
погружается в грунт и только его самый
передний конец оказывается на уровне
поверхности грунта. У некоторых видов над
грунтом торчат и концы уроподов.
Изогнув брюшко на спинную сторону, рачок
спокойно сидит в грунте. При помощи
задних ногочелюстей и передних
ходильных ног он вырывает перед своим ротовым
отверстием воронку и приступает к
питанию (рис. 237).
Виды, зарывающиеся в песчаный грунт
(Lamprops, Pseudocumidae, Cumopsis и
др.), удлиненными внутренними ветвями
передней пары ходильных ног подгребают
к ротовому отверстию, находящемуся на
дне воронки, отдельные песчинки. Ногоче-
люсти передают их челюстям и жвалам,
которые счищают с песчинок налет из
водорослей и бактерий, служащих пищей
рачкам. Некоторые кумовые могут
использовать таким образом частицы грунта, не
уступающие по размерам их карапаксу.
«Обработанные» частицы отбрасываются
назад через голову. Другие виды,
живущие на мягком илистом грунте
(Diastylis, Cumella и др.)> разводят в стороны
внутренние ветви своих последних
ногочелюстей и начинают пропускать ил
через щетинки передних ногочелюстей и
челюстей. Бактерии и находящиеся в
грунте разные органические частицы поедаются,
а минеральные частицы направляются
вперед между основаниями передних антенн.
Вылезая из грунта, кумовые плавают;
самки гребут наружными ветвями
грудных ножек, а самцы, кроме того,
брюшными ножками. Плаванию помогают и
изгибание и распрямление брюшка.
Благодаря более мощному двигательному
аппарату самцы передвигаются вдвое быстрее
самок. При температуре воды 12°С самец
Pseudocuma longirostris длиной 3 мм
всплывает вверх со скоростью 1 м в 45
секунд.
462
Большая быстрота движений самцов и
их длинные богатые чувствительными
органами задние антенны служат
приспособлениями, позволяющими им активно
отыскивать самок. Кроме того, глаза и
наружные ветви грудных ног у самцов часто
бывают развиты сильнее, чем у самок.
Все эти признаки появляются у самцов
в результате линьки, наступающей после
созревания половых продуктов. Тогда же
усложняется и скульптура их карапакса—
конфигурация его килей, бугорков и т. д.
Все это позволяет применять к таким
готовым к половому процессу рачкам
условный термин — самец в брачном наряде.
Брачный наряд самок гораздо
скромнее. Он выражается в появлении на
задних ногочелюстях и передних грудных
ногах пластинок, составляющих
выводковую сумку.
Спаривание происходит в ночное или
сумеречное время в толще воды, вне
грунта, после линьки самки. Самец Lamprops
прикрепляется к спинной стороне тела
самки, держась за края ее свободных
грудных сегментов своими грудными ножками,
снабженными специально
приспособленными для этого коготками и щетинками,
а также жгутами задних антенн. Остается
пока неизвестным, оплодотворяются ли
яйца в выводковой сумке или в половых
путях самки. Самка откладывает в
выводковую сумку яйца, число которых
различно у разных видов (от 3 до 200) и,
кроме того, зависит от размера самки.
Так, например, в выводковой сумке
самок Diastylis rathkei длиной 10 мм
насчитывается от 12 до 49 яиц, а у самок того
же вида длиной 15 мм — от 85 до 102 яиц.
Яйца и выходящая из них молодь
остаются продолжительное время в
выводковой сумке матери, причем новорожденные
рачки линяют там 3 раза. После этого
они приобретают облик взрослых, но
отличаются от них отсутствием задней пары
грудных ножек. Такая стадия развития,
свойственная и некоторым другим пера-
каридам (клешненосным осликам,
равноногим), получила название стадии «манка».
На стадии манка рачки покидают
выводковую сумку матери и переходят к
самостоятельному существованию. Они линяют
еще раз, но остаются пока на той же
стадии. У берегов Швеции Diastylis rathkei
после этого линяет еще 4 раза перед тем,
как после пятой линьки «надеть брачный
наряд», т. е. быть готовой к размножению.
Продолжительность жизни этого вида —
4 года, но многие другие кумовые живут
не больше года.
Длительность пребывания яиц и молоди
в выводковой сумке матери, а также меж-
линечных периодов зависит от
температуры. При содержании D. rathkei в
аквариуме при температуре 10—13°С молодь
выходила из выводковой сумки через 55—
60 дней после откладки яиц, при
температуре 3—8,5°С — через 76 дней, а при
температуре 3,5°С — через 90—101 день.
При этом формирование молодых рачков
происходит не одновременно и иногда
первые детеныши вылупляются на две недели
раньше последних.
Период размножения большинства
кумовых очень растянут. В Белом море у
арктических видов (Diastylis scorpioides,
D. sulcata и др.) он длится с мая до
октября, но бореальные виды (Lamprops fas-
ciata, Brachidiastylis resima)
размножаются только в наиболее теплое время —
в июле — августе. В дальневосточных
морях некоторые виды (Eudorella pacifica,
Eudorellopsis integra), наоборот,
размножаются в наиболее холодное время
(февраль — май и сентябрь — декабрь).
Кумовые ракообразные широко
распространены во всех морях и океанах.
Некоторые семейства, как например Diastyli-
dae, Leuconidae, Lampropidae,
приурочены к холодным водам и встречаются
только в высоких и умеренных широтах
северного и южного полушарий,
отсутствуя в тропиках, другие, как Bodotriidae
и Nannastacidae, наоборот, приурочены
к тропической и субтропической областям.
Многие виды способны жить как на
малых, так и на больших глубинах. Так,
например, Leucon fulvus и L. pallidus.
населяют глубины от 9—10 до 2560 м,
Eudorella emarginata — от 9 до 2000 м
и т. д. Однако большинство кумовых (в
наших дальневосточных морях 70%
известных видов) не опускается глубже
материковой отмели. С другой стороны,
известен один род (Makrokylindrus),
объединяющий только глубоководные виды.
463
Весьма своеобразна фауна кумовых
Каспийского моря. Она состоит из 17 видов,
принадлежащих к семейству Pseudocumi-
dae. Из них 6 ограничены в своем
распространении Каспием, а остальные
встречаются, кроме того, в опресненных частях
Черного и Азовского морей. Некоторые
из этих видов заходят в русла рек и
поднимаются по ним вверх на 100—150 и
иногда даже на 350 км от устья.
Единственным чисто пресноводным видом
кумовых можно считать Lamprops korroensis,
обитающего в озерах и низовьях рек
Охотского моря, Сахалинского залива,
залива Де-Кастри, Сахалина и Курильских
островов.
Кумовые ракообразные используются
в пищу многими рыбами, в частности
промысловыми. В Каспийском море их
поедают молодь осетровых, вобла, некоторые
бычки и особенно охотно лещ, в Азовском
и Черном морях — тарань, камбалы,
бычки. В Баренцевом море и в
дальневосточных морях они входят в состав пищи
пикши, трески, камбал, морского окуня
и других рыб.
Учитывая важное кормовое для рыб
значение кумовых, их акклиматизируют
в некоторых водохранилищах. Так,
например, Pseudocuma cercaroides была
перевезена в Днепровское водохранилище,
где прижилась и размножилась.
ОТРЯД КЛЕШНЕНОСНЫЕ ОСЛИКИ
(TANAIDACEA)
При сборах донных морских животных
наряду с кумовыми, бокоплавами,
равноногими изредка попадаются мелкие
высшие ракообразные, принадлежащие к
отряду клешненосных осликов, или танаи-
довых. Они ведут скрытый образ жизни,
укрываясь в построенных ими трубках,
зарываясь в грунт или прячась среди
водорослей, камней и т. д. Клешненосные
ослики встречаются как на мелководье,
так и на больших океанических глубинах,
но нигде не образуют массовых скоплений»
На Курильских островах и в Южной
Америке один или два вида обитают и в
пресных водах, но, как правило,
клешненосные ослики не переносят опреснения и
могут считаться настоящими морскими
животными. Длина их тела составляет
обычно от 2 до 5 мм, и только некоторые
глубоководные виды достигают значительно
более крупных размеров (Herpotanais kir-
kegaardi до 26 мм). Из-за сравнительной
редкости и немногочисленности этих
рачков их биология мало изучена. Описано
около 250 видов клешненосных осликов.
Тело этих ракообразных удлиненное,
цилиндрическое или уплощенное в спинно-
брюшном направлении., Короткий
щитовидный карапакс покрывает голову и
слившиеся с нею два передних грудных
сегмента. Между его боковыми краями и
стенкой тела, так же как у кумовых, с каждой
стороны находится дыхательная полость.
Передний край карапакса часто образует
плоский треугольный или полукруглый
рострум. У многих мелководных видов по
бокам от основания рострума на
специальных выростах головы располагаются
маленькие глаза с очень небольшим
количеством фасеток (у Heterotanais — 12, у
Apseudes —8). Глазные выросты Tanaida-
сеа существенно отличаются от глазных
стебельков тонкопанцирных, мизид и
десятиногих, поскольку они не сочленены с
головой и не могут поворачиваться.
Обе пары антенн обычно короткие. У
представителей подотряда Monokonophora
передние антенны с двумя жгутами, а
задние с чешуевидной наружной ветвью,
у представителей другого подотряда —
Dikonophora — на передних антеннах
всего один жгут, а задние лишены
наружной ветви. Ротовые придатки
хорошо развиты только у самок, а у
самцов многих видов они частично или даже
полностью редуцируются. Жвалы у
Monokonophora несут щупик, отсутствующий
на жвалах Dikonophora. Челюсти
снабжены внутренними лопастями,
многочисленные щетинки которых образуют филь-
Таблица 37. Камчатские крабы (Paralithodes camtscha-
tica) на дне Охотского моря.
464
трующий аппарат. На передней ногоче-
люсти, так же как у кумовых, имеется
крупный эпиподит, приспособленный для
возбуждения тока воды в дыхательной
полости, под боковым краем карапакса. Вода
заходит под край карапакса сзади,
омывает дыхательную полость и выходит
позади места прикрепления передней ного-
челюсти. Кислород усваивается
внутренней поверхностью боковых краев
карапакса, а также стенкой тела, прилегающей
к дыхательной полости. Здесь
располагается густая сеть капиллярных
кровеносных лакун, впадающих близ заднего края
карапакса в околосердечную сумку.
Конечности второго, слившегося с
головой грудного сегмента крупнее остальных
и снабжены на конце настоящей клешней
или у некоторых видов подклешней.
Благодаря характерному строению этих ножек
клешненосные ослики и получили свое
название. У самцов клешненосные ноги
обычно бывают крупнее, чем у самок и
нередко имеют несколько иное строение,
но биологическое значение этих различий
пока остается неизвестным. Остальные 6
пар грудных ножек устроены одинаково
и представляют собой ходильные
конечности. Все они одноветвисты и только у
некоторых Apseudidae u Kalliapseudidae
на передней из них, а также на клешне-
носных ногах есть короткие наружные
ветви, напоминающие соответствующие
придатки мизид и кумовых. Кроме того, у
рода Apseudes ноги переднего свободного
грудного сегмента отличаются от
остальных тем, что их последние членики
расширены и приспособлены для рытья
грунта. У самок перед размножением на
четырех, а у некоторых видов всего на одной
паре грудных ножек развиваются
пластинки, образующие выводковую сумку.
Так же как у кумовых, они исчезают
после линьки, следующей за окончанием
вынашивания яиц и молоди, а затем,
перед новым периодом размножения,
возникают вновь.
Брюшной отдел всегда значительно
короче и обычно несколько уже грудного.
Пять передних составляющих его
сегментов свободные, а последний, шестой
сегмент срастается с тельсоном, образуя так
называемый плеотельсон.
Конечности передних пяти брюшных сегментов
короткие, но двуветвистые и снабжены
по краям многочисленными щетинками.
Они служат плавательными ножками и
обычно у самцов развиты сильнее, чем
у самок. Самки некоторых видов совсем
лишены этих ножек. К плеотельсону
прикрепляется последняя пара брюшных
ножек — уроподы. Они обычно двуветвисты,
причем и ствол и обе ветви тонкие,
палочковидные. У некоторых клешненосных
осликов, например у Apseudes, ветви уропо-
дов удлиненные и расчлененные.
Большинство клешненосных осликов
живет в выкапываемых ими норах или в
трубках. Внутреннюю поверхность норы рачки
обкладывают песчинками, остатками
растительности и другими частицами,
которые они собирают и перетаскивают клеш-
неносными ногами. Весь этот материал
скрепляется специальным секретом. На
боках трех передних свободных грудных
сегментов располагаются железы,
вырабатывающие этот секрет. Протоки желез
очень длинные; они тянутся через
каждую из трех пар передних ходильных
ножек и открываются в их когтевых
члениках. Этими ходильными ножками рачки
и скрепляют обкладку своих нор и стенки
своих трубок. Сидя в норе или в трубке,
рачки энергично машут брюшными
ножками и таким образом обновляют воду
в своем убежище.
Время от времени они вылезают из нор
и трубок и передвигаются по дну,
частично ползая при помощи грудных,
частично плавая над дном при помощи
брюшных ножек. Кроме того, они могут
двигаться скачками, сгибая и разгибая тело.
Пищей клешненосным осликам служат
находящиеся в грунте остатки отмерших
растений и другие органические частицы.
Некоторые виды отфильтровывают их
щетинками челюстей, используя ток воды,
направляемый в дыхательные полости.
Таблица 38. Омар Homarus gammarus и лангуст
ч- Palinurus vulgaris.
30 Жизнь животных, т. 2
Рис. 240. Клешненосные ослики:
1 — Apseudes spinosus; 2 — Herpotanais kirkegaardi;
3 — Tana is stanfordi; 4 — две фазы брачного танца
Heterotanais oerstedi.
Самцы клешненосных осликов
отличаются от самок большим количеством
чувствительных щетинок на жгутах антенн
и сильнее развитыми плавательными
ножками, что позволяет приписывать им
активную роль в поисках самок. Наряду
с этим самцы многих видов частично или
полностью лишены ротовых придатков,
следовательно, не могут питаться и,
очевидно, недолговечны.
Механизм определения пола у
клешненосных осликов очень интересен. Еще
недавно предполагалось, что некоторым из
них свойственна протогиния
—явление, довольно редкое в животном мире.
При протогинии созревающий организм
превращается в самку, которая затем
переходит в самца.
Новые наблюдения Рамиреца над
Heterotanais oerstedi позволяют считать
по крайней мере этот вид потенциальным
гермафродитом. У него самцы могут
возникать либо непосредственно из молодых
особей, либо из половозрелых самок,
закончивших вынашивание молоди или даже
еще не отметавших яиц. Это зависит от
условий существования рачков. При
совместном содержании самца и самки в
одном сосуде перемены пола не происходит.
При содержании молодой особи вместе
с самцом из нее получается самка и,
наоборот, если ее подсадить к самке, то —
самец. Содержание молодой особи в
одиночестве приводит к превращению ее в
самку. Таким образом, пол у Н. oerstedi
не связан с присутствием определенного
гена.
Гермафродитные особи известны не
только для Н. oerstedi, но также для крупного
A5 мм) антарктического Apseudes specta-
bilis, Tanais stanfordi и глубоководного
Neotanais serratispinosus. При этом у
широко распространенного Т. stanfordi
(рис. 240,5) гермафродитизм наблюдается
только в отдельных частях его обширного
ареала, в частности на Курильских
островах, а в других местах этот вид
представлен нормальными, не переходящими
друг в друга самцами и самками.
Замечательно, что все известные до сих пор
гермафродитные танаидовые живут при
низкой температуре.
Размножение клешненосных осликов
удалось изучить лишь недавно,
сконструировав для наблюдений специальные
микроаквариумы. Самец Heterotanais oerstedi
забирается в трубку самки, и в течение
11 часов рачки исполняют там брачный
танец. Сначала самец преследует самку,
затем она переворачивается через голову,
и оба партнера оказываются «лицом к
лицу», соприкасаясь антеннами (рис. 240, 4).
После этого они поворачиваются друг к
другу задними концами тела и контакти-
руются уроподами, для чего самцу
приходится предварительно переворачиваться
через голову. Наконец, самец
оказывается лежащим на спине, а самка
располагается над ним. В таком положении (в
течение 3 секунд) происходит копуляция, при
которой сперматозоиды попадают в вы-
водковую сумку самки. Вслед за этим
самец покидает трубку самки, а она
закрывает вход в трубку. Через некоторое время
самка откладывает в выводковую сумку
яйца.
Число откладываемых самкой в
выводковую сумку яиц незначительно: у
мелких видов обычно от 3 до 8, у крупного
Apseudes antarcticus — 16. Яйца и молодь
все время омываются током воды,
вызываемым движениями составляющих
сумку пластинок. Молодые рачки, оставаясь
в сумке, линяют 2 раза и выходят из
нее на стадии манка. У Heterotanais oers-
tedi при температуре 17°С это происходит
через 18—19 дней, а при температуре
20°С — через 14 дней после откладки яиц.
Первая свободноживущая стадия манка
достигает в длину 0,6 мм, в то время как
длина ее матери около 2 мм. Через 10—
11 дней наступает первая на свободе
линька молодого рачка. Вскоре после
освобождения от потомства самка также линяет
и теряет выводковую сумку, с тем чтобы
Рис. 241. Головогрудь Apseudes talpa
(снизу); стрелками обозначены токи
воды:
1 — передняя антенна; 2 — задняя антенна;
3 — глаз; 4 — жвала; 5 — нижняя губа;
6 — передняя челюсть; 7 — ногочелюсть;
8 — основание клешненосной ноги.
приобрести ее вновь после 3 линек,
последняя из которых предшествует
следующему периоду размножения.
Дальнейшее развитие молоди
прослежено у антарктического вида Heterotanais
antarcticus. После выхода из выводковой
сумки и до возникновения у самок
выводковой сумки рачки линяют 7 раз, причем
первые 4 стадии развития должны
считаться стадией манка, поскольку у них
отсутствует или недоразвита последняя
пара грудных ножек. Как уже
указывалось, эта стадия, помимо клешненосных
осликов, свойственна только кумовым и
равноногим ракообразным.
Распространение клешненосных
осликов изучено далеко не достаточно.
Наибольшее количество их видов приурочено
к небольшим глубинам полносоленых
морей умеренной зоны и высоких широт.
Можно отметить четкие различия в
составе фауны этих ракообразных на разных
по характеру грунтах — песчаных,
илистых и каменистых. На больших и
предельных океанических глубинах обитают
преимущественно представители
семейства Neotanaidae, характеризующегося
многими примитивными признаками.
ОТРЯД СПЕЛЕОГРИФОВЫЕ
РАКООБРАЗНЫЕ
(SPELAEOGRIPHACEA)
Столовая гора близ Кейптауна в
Южной Африке богата различными
подземными ходами и пещерами. В некоторых
из них есть водоемы. При обследовании
этих пещер в одном пещерном ручье,
температура воды которого равнялась 10°С,
были обнаружены небольшие G,5 мм)
слепые и лишенные пигмента рачки. Они были
названы Spelaeogriphus lepidops. Их
строение оказалось настолько своеобразным,
что их пришлось выделить в особый
отряд, имеющий некоторые общие признаки
с клешненосными осликами и кумовыми.
Больше нигде Spelaeogriphacea найти
пока не удалось.
Об образе жизни спелеогрифовых ничего
не известно.
30*
467
Тело Spelaeogriphus длинное, немного
уплощенное сверху вниз, с параллельными
боковыми краями.
Голова срастается только с одним
передним грудным сегментом и вместе с ним
покрыта коротким карапаксом с
направленным вперед коротким треугольным
рострумом. Под боковыми краями кара-
пакса расположены, как у кумовых и
клешненосных осликов, дыхательные
полости. Большой ложковидный эпиподит
ногочелюсти функционирует, очевидно, так
же как у кумовых и клешненосных
осликов, возбуждая ток воды, проходящий
мимо сочлененных с головой подвижных
глазных стебельков, полностью лишенных
фасеток и пигмента.
Конечности семи свободных грудных
сегментов устроены приблизительно
одинаково и представляют собой ходильные
ножки.
Все они, за исключением ножек
последней пары, снабжены короткими
наружными ветвями, а у самок на передних
пяти парах есть направленные внутрь
пластинки, образующие выводковую
камеру.
Все шесть брюшных сегментов
свободные. На передних четырех брюшных
сегментах находится по паре двуветвистых
плавательных ножек. Их одночлениковые
ветви несут по краям многочисленные
щетинки, и, судя по их строению, рачок
может не только ползать, но и плавать.
Пятый брюшной сегмент *лишен конечностей,
а на шестом помещаются направленные
назад двуветвистые уроподы. Тельсон
короткий, овальный.
Судя по содержимому кишечника, рачки
заглатывают грунт.
Рис. 242. Spelaeogriphus lepidops.
ОТРЯД ТЕРМОСБЕНОВЫЕ
РАКООБРАЗНЫЕ
(THERMOSBAENACEA)
Среди перакарид есть еще один отряд,
представители которого обитают только
в подземных водах. Сначала они были
открыты в горячем солоноватом ключе
в Тунисе, но потом близкие виды удалось
обнаружить в пещерах южной Италии,
в солоноватых интерстициальных водах
морского побережья Югославии, а в
последнее время в пещерах США. По своему
строению термосбеновые во многих
отношениях сходны с кумовыми и клешненос-
яыми осликами, но по способу
размножения они резко отличаются от всех
перакарид: выводковая сумка, в которой самки
вынашивают яйца, образуется у них на
спинной стороне под краем карапакса, как
у ветвистоусых ракообразных, а не под
грудью за счет отростков грудных ножек,
как у всех остальных перакарид. На
этом основании некоторые исследователи
считают возможным выделять термос-
беновых в особый надотряд Pancarida,
но другие, учитывая строение их жвал
и многие детали анатомии, включают их
в состав надотряда Peracarida. Всего
известно 6 видов этих ракообразных, из
которых один принадлежит к роду
Thermosbaena, а остальные к роду Мопо-
della.
Длина их тела не превышает 3,5 мм.
Туловище цилиндрическое, грудной и
брюшной отделы одинаковой ширины.
Будучи исключительно подземными
животными, они совершенно слепы и
полностью лишены пигмента. Передний
грудной сегмент объединен с головой.
Цилиндрический карапакс срастается с головой
и передним грудным сегментом, но
нависает и над несколькими следующими
грудными сегментами. Особенно сильно
он разрастается назад у половозрелых
самок, у которых он может покрывать
все грудные сегменты. У них, как уже
указывалось, между карапаксом и
спинной стороной тела образуется выводковая
сумка, куда откладываются яйца.
Внутренняя поверхность карапакса служит
органом дыхания.
468
На всех свободных грудных сегментах
у Monodella или только на пяти передних
у Thermosbaena находится по паре дву-
ветвистых ходильных ножек. Брюшной
отдел, как обычно, состоит из 6 сегментов,
но у Thermosbaena последний брюшной
сегмент срастается с крупным широким
тельсоном, образуя плеотельсон. На двух
передних брюшных сегментах имеются
короткие нерасчлененные ножки.
Последний брюшной сегмент снабжен
направленными назад двуветвистыми уроподами.
Thermosbaena может только ползать
по грунту, перебирая своими грудными
ножками, Monodella argentaria из
итальянских пещерных озер, кроме того, плавает,
часто поднимаясь к поверхности воды,
М. halophila, как и другие обитатели
интерстициальных вод, пробирается,
изгибая свое узкое тело, по капиллярным
ходам между песчинками. Все эти рачки
заглатывают грунт и усваивают
находящееся в нем органическое вещество.
Перед откладкой яиц карапакс самки
удлиняется. У самок Monodella от
заднего конца каждого яичника вверх
отходит канал, по которому яйца, вероятно,
проходят в выводковую сумку. Кроме
него, есть и другой канал,
открывающийся в основании ноги пятого грудного
сегмента половым отверстием. Можно
предполагать, что именно по последнему
каналу проникает сперма после спаривания
и яйца откладываются в выводковую
сумку уже оплодотворенными, однако ни
спаривания, ни откладки яиц у термо-
сбеновых еще никто не наблюдал. У
Monodella в выводковой сумке насчитывается
от 4 до 16 яиц. Из них там же выходят
почти полностью сформированные рачки,
отличающиеся от взрослых отсутствием
ног на двух последних грудных сегментах.
Они покидают выводковую сумку матери
и начинают жить самостоятельно. Через
неделю они достигают в длину 2 мм, и
к этому времени у них появляется
сначала шестая, а потом и седьмая пара
ходильных ног.
Условия существования
немногочисленных известных видов термосбеновых
весьма разнообразны. По-видимому, исходной
средой их обитания был морской грунт.
В наиболее близких к исходным усло-
Рис. 243. Термосбеновые:
1 — Thermosbaena mirabilis, самка с яйцами;
2 — Monodella stygicola.
виях существует Monodella halophila,
населяющая заполненные солоноватыми
водами интерстициальные ходы адриати-
ческого побережья Югославии. В
солоноватых водах живет и М. relicta из
ключей близ Мертвого моря в Израиле; она
считается реликтом морей, некогда
покрывавших этот район. Два вида Monodella
(М. argentaria и М. stygicola) из пещерных
озер южной Италии, по всей вероятности,
проникли в эти сравнительно крупные
подземные водоемы из системы грунтовых
вод, поскольку их местонахождения
расположены недалеко от моря и им
приписывалось морское происхождение.
Однако недавнее открытие М. texana в
пещерах США, находящихся на расстоянии
200 км от ближайших морских берегов,
опровергает сложившиеся ранее
представления о связи распространения Monodella
со Средиземным морем и сравнительно
недавними этапами его геологического
прошлого. Вероятно, род древнее, а исто-
469
рия его расселения сложнее, чем это
ранее предполагалось.
В совершенно необычайных условиях
существует первый по времени открытия
представитель термосбеновых, по имени
которого назван весь отряд — Thermos-
baena mirabilis. Теплый ключ в Тунисе—
единственное место обитания этого вида—
круглый год имеет температуру 45°С.
Рачок узко приспособлен к жизни при
такой высокой температуре и при
понижении температуры до 30°С погибает.
М. texana и М. argentaria живут в
пресной, а остальные виды отряда в
солоноватой воде.
Обширное и разорванное
географическое распространение, а также условия
обитания термосбеновых позволяют видеть
в их современных немногочисленных
представителях остатки древней, в
значительной степени исчезнувшей фауны. В этом
отношении они подобны цефалокаридам,
анаспидовым и батинеллиевым.
ОТРЯД РАВНОНОГИЕ РАКООБРАЗНЫЕ
(ISOPODA)
Равноногие ракообразные (табл. 33)—
один из немногих отрядов этого класса,
многочисленные D500 видов)
представители которого приспособились к самым
различным условиям существования.
Большинство относящихся к нему видов
обитает в море, причем населяет все глубины,
начиная от приливо-отливной зоны и
кончая предельными океаническими
глубинами порядка 11 км. Среди морских
равноногих есть обитатели дна, но есть и
немногочисленные планктонные виды; есть
также и паразиты других ракообразных
и рыб. Чрезвычайно широко
распространены равноногие и в пресных водах, как
поверхностных, так и подземных;
пресноводные водяные ослики могут считаться
очень обычными и многочисленными
представителями фауны самых различных озер
и прудов северного полушария. Наконец,
богатый видами подотряд равноногих —
мокрицы — выработал весьма совершенные
приспособления к жизни на суше. Они
встречаются почти во всех климатических
зонах земного шара — от холодных
приполярных районов до влажных тропических
лесов и раскаленных пустынь.
Очевидно, в процессе эволюции равноногие
приобрели какие-то важные особенности,
позволившие им впоследствии
специализироваться в разных направлениях и освоить
различные среды обитания. Одной из таких
особенностей можно считать строение
брюшного отдела и его конечностей,
выполняющих различные функции —
половую, двигательную и дыхательную — и
хорошо защищенных от неблагоприятных
внешних воздействий, в частности у
сухопутных рачков от высыхания.
Тело равноногих обычно уплощено
сверху вниз, реже цилиндрическое или
даже сжатое с боков (у подотряда Phrea-
toicidea, табл. 33, 4). Его покровы очень
разнообразны, часто гладкие, но нередко
снабжены килями, шипами или
различными бугорками. У некоторых мокриц
бугорки поверхности тела образуют
сложную водопроводную систему. Если рачок
прикоснется к капельке росы, влага под
действием капиллярных сил начинает
распространяться в промежутках между
бугорками, достигая заднего конца тела
и орошая находящиеся там жабры. Также
разнообразна окраска равноногих.
Обычно она бурая или желтоватая, но у видов,
живущих на морских растениях, она
зеленая, под цвет субстрата, а подземные и
глубоководные виды лишены пигмента,
белые или иногда розоватые.
Карапакс полностью отсутствует.
Голова срастается с одним или изредка
с двумя передними грудными сегментами.
На ней с боков находятся сидячие глаза.
Число составляющих их фасеток
колеблется в очень широких пределах: у самого
крупного вида равноногих — Bathynomus
giganteus (табл. 33, 10), достигающего
в длину 27 см, в каждом глазе
насчитывается 3000 фасеток, у более мелких
хищных морских Aega (табл. 33, 6) —
около 400, у мокриц — обычно 20—30,
а у водяных осликов—4. У глубоководных
и подземных видов глаза не развиты
совсем. Обе пары антенн имеют разную длину
у различных видов. У многих
глубоководных равноногих задние антенны
значительно длиннее тела. Строение ротовых
470
придатков зависит от способа питания.
У большинства равноногих они
жующего типа, но у некоторых хищников и
паразитов жвалы и челюсти превращены в
шиловидные острия и сложены вместе в виде
пирамиды, так что ротовой аппарат
приобретает характер колюще-сосущего.У
взрослых особей из подотряда Gnathiidea
ротовые придатки, за исключением жвал
самцов, редуцированы, и рачки вовсе не
питаются (табл. 33, 2, 2).
Семь свободных грудных сегментов
несут по паре ходильных ног, всегда
лишенных наружных придатков. У многих
равноногих эти ножки имеют приблизительно
одинаковое строение и одинаковую длину,
почему отряд и получил свое название.
Однако у других они специализированы
в разных направлениях. Нередко на одной
передней или даже на трех передних
ножках развивается хватательное
приспособление — подклешня. У многих
представителей подотряда Asellota, в
особенности часто у глубоководных, конечные
членики трех задних пар грудных ножек
расширены и снабжены по краям
многочисленными щетинками. Такие ножки
служат для плавания (табл. 33,11,12). У
половозрелых самок на передних пяти
или четырех (у некоторых Arcturidae на
одной) парах грудных ножек с
внутренней стороны возникают пластинки,
образующие выводковую сумку.
Особенно характерно строение
брюшного отдела и его конечностей. Он всегда
гораздо короче грудного, и количество
составляющих его сегментов сокращено
за счет того, что один, несколько или даже
все сегменты срастаются с полукруглым
или треугольным тельсоном, образуя
крупный плеотельсон. Только у
подотряда Anthuridea этого не происходит и
последний брюшной сегмент и тельсон
сохраняют самостоятельность.
Конечности пяти передних брюшных сегментов
состоят из короткого основания и двух
широких листовидных, часто
тонкостенных ветвей. Эти ветви накладываются друг
на друга, как страницы книги, и
направлены назад таким образом, что они
находятся под плеотельсоном (рис. 244, 2).
Основная функция передних брюшных
ножек дыхательная. Они служат жабрами.
Рис. 244. Брюшные ножки равноногих:
1,2 — плеотельсон Mesidotea снизу: 1 — с уроподами,
2 — без уроподов (видны жабры); 3 — брюшная ножка
мокрицы Porcellio scaber с псевдотрахеями (вид сверху);
4 — продольный разрез через эту ножку.
Через их тонкие стенки растворенный
в воде кислород проникает в проходящие
в ножках кровеносные лакуны и окисляет
кровь. В связи с этим, как уже
указывалось, сердце у равноногих ракообразных
в значительной степени смещено в
брюшной отдел и расположено близко от жабр.
Наружные ветви одной из пар брюшных
ножек обычно сильно хитинизированы и
служат крышкой, покрывающей
остальные нижние жаберные листочки, которые,
таким образом, оказываются сверху
защищенными плеотельсоном, а снизу этой
крышкой (рис. 244,2).
Такое строение дыхательного аппарата
послужило, несомненно, предпосылкой,
позволившей части равноногих перейти
к сухопутному существованию. Их
жабры оказались хорошо защищенными от
высыхания и не потребовалось
дополнительных усовершенствований для того,
чтобы полностью их обезопасить в новых
условиях существования. Большинство
мокриц, несмотря на то что они живут на
471
суше, дышат при помощи своих брюшных
ножек, как настоящие водные животные:
они используют кислород, растворенный
в тонком слое покрывающей их жабры
влаги. Следовательно, их жабры должны
все время увлажняться. Это достигается
у разных мокриц разными путями. У
одних, как уже упоминалось, приток воды
к жабрам обеспечивается благодаря
своеобразной водопроводной системе,
образованной скульптурой покровов. У других
задние брюшные ножки — уроподы —
устроены таким образом, что при
соприкосновении с капелькой воды она, также
под влиянием капиллярных сил, поступает
в жабры по уроподам. Наконец, третьи
орошают свои жабры жидкостью,
выделяемой из анального отверстия и
добываемой из поедаемой рачками сочной
растительности. Однако есть мокрицы,
способные при дыхании использовать не
только растворенный в воде, но и
атмосферный кислород. У таких мокриц в толще
наружных ветвей передних брюшных
ножек находится полость, соединяющаяся
щелью с наружной средой, так что в эту
полость свободно проникает воздух. От
нее отходят ветвящиеся,
заканчивающиеся слепо каналы, напоминающие трахеи
насекомых и других наземных
членистоногих и поэтому называемые
псевдотрахеями (рис. 244, 5, 4). Через них
кислород воздуха проникает в кровь,
циркулирующую в лакунах в толще
наружных ветвей брюшных ножек, а
кислород, растворенный в смачивающей жабры
воде, используется внутренними ветвями.
Судя по наблюдениям над туркменскими
мокрицами рода Protracheoniscus,
размеры трахейного поля наружных ветвей
брюшных ножек зависят от условий
существования. Виды, обитающие в пустыне
и на других открытых сухих местах, имеют
относительно крупное трахейное поле,
а виды, существующие при
повышенной влажности, наоборот, относительно
небольшое. Совсем маленькое трахейное
поле свойственно виду P. birsteini,
который вторично перешел от жизни на суше
к жизни в воде и был обнаружен среди
зарослей водных растений в обширных
мелководных озерах Делили в бассейне
реки Атрек.
У некоторых морских равноногих
дыхательная поверхность брюшных ножек
увеличивается за счет образования складок,
как у некоторых Sphaeromidae,
пальцевидных выростов, как у Bathynomus,
или продольного расщепления ветвей, как
у некоторых представителей
паразитического подотряда Epicaridea.
Однако передние брюшные ножки
равноногих служат не только для дыхания.
У самцов некоторые из них частично
преобразованы в совокупительные органы.
Эту функцию у многих выполняют
внутренние ветви второй пары. У водяных
осликов, например, они представляют
собой нечто вроде шприцев весьма сложного
строения. При спаривании самец сначала
набирает в этот шприц свою сперму, а
затем впрыскивает ее в половые отверстия
самок. Первая пара брюшных ножек
также принимает участие в этом процессе.
Сходным образом, при помощи двух пар
передних брюшных ножек, производят
перенос спермы в половые пути самок
мокрицы. Понятно, что по строению
передних брюшных ножек самцы равноногих
всегда отличаются от самок, у которых
первая пара брюшных ножек нередко
совсем отсутствует. Некоторые группы
равноногих используют брюшные ножки не
только как органы дыхания и
совокупления, но и как органы движения. Anthu-
ridea при плавании над дном прижимают
грудные ножки к бокам тела и гребут
брюшными. Также при помощи трех пар
передних брюшных ножек плавают
плоские антарктические Serolidae (табл. 33, 9)>
некоторые Cymothoidae и др.
Последняя пара брюшных ножек
—уроподы — имеет разное строение у разных
подотрядов равноногих. Так, например,
у Asellota и мокриц они прикреплены
к заднему краю плеотельсона и
направлены назад, у Valvifera они с боков
подвернуты под брюшную сторону
плеотельсона и, как крышки, прикрывают
передние нежные дыхательные брюшные
ножки (рис. 244, 2, ?), у Anthuridea
и Flabellifera они сочленены с основанием
плеотельсона и вместе с ним образуют
хвостовой веер.
У паразитических Epicaridea, в
особенности у их самок, организация всего тела
472
обыкновенно очень упрощена. Они
нередко теряют сегментацию и конечности и
превращаются в бесформенный мешок,
набитый яйцами. Однако самцы и
личинки полностью сохраняют признаки
равноногих ракообразных.
Основным способом передвижения всех
равноногих ракообразных, за
исключением паразитических, следует считать
ползание по дну, почве или растениям при
помощи грудных конечностей.
Большинство видов двигается только таким
способом. Среди морских видов многие, кроме
того, могут плавать. Обыкновенно они
проплывают небольшие расстояния над
дном, с тем чтобы потом снова перейти
к ползанию. При этом представители
некоторых семейств Asellota гребут
специально приспособленными к плаванию задними
грудными ножками, a Anthuridea, Cymo-
thoidae и Serolidae— брюшными ножками.
Idoteidae орудуют при плавании и
задними грудными и брюшными ножками.
Для некоторых равноногих плавание
играет важную роль в их жизни.
Полупаразитические полухищные виды Aega
и некоторых других Cymothoidae вплавь
нападают на рыб, прикрепляются к ним
и начинают сосать их кровь. Личинки
паразитических Epicaridea плавают при
помощи задних антенн и брюшных ножек
до тех пор, пока не найдут своих хозяев—
различных ракообразных, после чего
начинается превращение личинок во
взрослые формы. Так же ведут себя личинки
равноногих подотряда Gnathiidea, так
называемые праницы (рис. 249, 1),
которые сначала плавают, а потом
паразитируют на рыбах, с тем отличием от
Epicaridea, что взрослые ведут свободный,
а не паразитический образ жизни.
Наконец, известно несколько планктонных
видов равноногих, которые плавают всю
жизнь. Это преимущественно
глубоководные животные, как например широко
распространенная Munneurycope murrayi и
Anuropus bathypelagicus, свойственный
только северной половине Тихого океана.
Idotea algirica живет на плавающих по
поверхности моря саргассовых и других
водорослях, кусках дерева и вместе с
этими предметами пассивно переносится с
места на место. Однако, как и другие виды
этого рода, она хорошо плавает и сама,
перебираясь с одного скопления
«плавника» на другое.
Среди донных морских видов многие
способны зарываться в грунт. Очень
быстро зарываются, например, крупная Mesid--
tea (табл. 33, 5) и мелкие представители
подотряда Asellota — Desmosoma, Mac-
rostylis и др., имеющие специальные копа-
тельные грудные ножки. Cyathura carinata
и, вероятно, другие виды подотряда
Anthuridea делают в илистом грунте
направленные вертикально глубокие ходы, которые
они покидают только ночью. Если
экспериментальный аквариум с начавшим
строительство хода рачком повернуть на 90°,
придав ходу горизонтальное направление,
рачок продолжает рыть ход, также
изгибая новый его отрезок под прямым углом
к прежнему. Способность
ориентироваться по отношению к силе земного
притяжения объясняется присутствием у
Cyathura в основании тельсона парного стато-
циста. Оперативное удаление статоциста
нарушает способность рачка рыть ходы
в определенном направлении. Самцы
Gnathiidea своими жвалами вырывают ходы
длиной по 15—20 см.в илистых крутых
склонах дна. В конце ход расширяется
в камеру, в которой живет около десятка
самок.
Некоторые пресноводные равноногие
также могут глубоко зарываться в грунт
и таким образом спасаться от
пересыхания водоемов. Так поступает, например,
обычный американский водяной ослик
Asellus communis, а также представители
подотряда Phreatoicidea, обитающие в
болотах Южной Африки. В засушливыэ
периоды Phreatoicidea вырывают во
влажном мхе ямки глубиной 2,5—5 см и
массами сидят в них, прикрывая задними
грудными ножками жабры и ожидая
появления воды.
Однако наиболее совершенные норы
строят приспособившиеся к жизни в
пустынях мокрицы из рода Hemilepistus
(табл. 33, 16). В пустыне Каракум норы
Н. cristatus достигают глубины 60—100 см.
Температура в них круглый год не падает
ниже 10° и не поднимается выше 26°С,
относительная влажность воздуха близка
к 100%. На поверхности почвы днем тем-
473
пература достигает 60°С, а влажность
уменьшается до 15%. Летом мокрицы
покидают норы только утром и вечером,
а зимой впадают в спячку и совсем не
выходят из нор. Там же в конце мая самки
рождают по 60—80 молодых рачков.
Родители не выпускают их из норы и защищают
их от врагов, закрывая вход в нору
своими передними грудными сегментами. Для
питания детей родители затаскивают в
нору растительные остатки. Через 21А
месяца молодь вырастает, но продолжает
жить в норе до следующей весны, и только
в марте рачки выходят на поверхность
и приступают к строительству
собственных жилищ. В старой норе остаются самец
и самка, обзаводящиеся снова детьми.
Таким образом, в каждой норе всегда
живет одна семья и эти семьи не
смешиваются между собой.
Питание растениями присуще очень
многим равноногим, как морским, так и
пресноводным и сухопутным. Виды рода Idotea
питаются водорослями, на которых они
живут, а также обрастающими их
диатомовыми. Преимущественно водорослевыми и
бактериальными обрастаниями питаются
литоральные Jaera, Sphaeroma, Eurydice,
Ligia и некоторые другие. Интересно, что
изменчивая окраска широко
распространенной Idotea viridis, обычной, в
частности, у нас в Балтийском и Черном
морях, зависит от ее пищи. Живущие на
зостере и зеленых водорослях рачки —
зеленые, на бурых — коричневые, на
красных — красные. Пигмент растений
окрашивает не только содержимое их
кишечника, но также кровь и полостную
жидкость, просвечивающие через покровы тела.
Если рачков пересадить с одних растений
на другие, их окраска через 10—15 дней
соответственно изменится. Поедая
растительную пищу, Idotea не брезгает и
попадающимися ей среди обрастаний
животными — гидроидами, мшанками,
личинками хирономид и т. д.
Обычные обитатели пресных вод —
водяные ослики — также типичные
растительноядные организмы. Помимо водных
растений, живых и отмирающих, они в
большом количестве поедают листья деревьев,
падающие в водоемы. За всю свою жизнь
каждый водяной ослик потребляет 168 мг
листьев (в сыром весе). В водоемах, в
которые попадает много древесных листьев,
водяные ослики обыкновенно живут в
большом количестве. В пруду на
территории парка Тимирязевской академии
насчитывается 5000 рачков на 1 л*2.
Африканские и австралийские Phrea-
toicidea едят растительные остатки и
водяные мхи.
Почти все мокрицы питаются
растениями, которые служат для них источником
не только пищи, но и влаги, необходимой
для увлажнения жабр. Поглощая
растительный материал, массовые виды мокриц,
например некоторые виды родов Oniscus
и Porcellio, выступают в качестве важных
факторов почвообразовательного
процесса. Их экскременты частично
непосредственно входят в состав почвенного слоя,
частично подвергаются дальнейшей
переработке ногохвостками, которые их
поедают. Особенно важное значение имеет
почвообразовательная деятельность
мокриц рода Hemilepistus в некоторых
пустынях, в частности в пустыне Каракум
в Туркмении. При рытье нор они выносят
на поверхность частички глубже
лежащего грунта. Кроме того, они
откладывают вокруг нор свои экскременты,
которые вместе с частичками грунта образуют
вокруг отверстия норы своеобразные
кольцевые валики. На 1 га площади обычно
приходится более 1 миллиона мокриц.
В течение 3 месяцев своей ежегодной
активной деятельности они дают около г/2 т
богатых водорастворимыми солями
частичек почвы глубинных слоев и до 1 т
экскрементов. На образованных ими
плодородных выбросах хорошо растут
растения. Кроме того, благодаря
многочисленным B0 000 на 1 га) глубоким норам
мокриц повышается скважность почвы
и улучшаются условия существования
почвенных бактерий. Все это приводит
к значительному повышению плодородия
почвы.
К питанию зелеными частями растений и
их остатками близко использование в
пищу древесины. Среди равноногих
известны древоточцы, вредящие деревянным
сооружениям в море и питающиеся
древесиной. Это, во-первых, многочисленные
виды рода Limnoria и, во-вторых, некото-
474
рые виды рода Sphaeroma. Виды Limno-
ria встречаются почти во всех морях.
Только в наших дальневосточных и
северных морях известно 4 вида, в Черном
море — 2 вида и т. д. Некоторые из них
живут на водорослях и питаются ими, но
большинство поселяется на сваях
пристаней, на стоящих у причалов
деревянных судах и т. д. Острыми жвалами они
жуют древесину, причем разрушают
преимущественно более мягкие летние слои,
не трогая зимних. Под поверхностью сваи
на глубине 1—2 см они выгрызают ходы,
превосходящие по диаметру ширину их
тела, следуя по летним слоям
нарастания дерева. Каждый такой ход одним
или двумя отверстиями соединяется с
наружной средой. В нем обычно обитает
самка (длиной до 5 мм) и более мелкий самец,
но точит дерево главным образом самка.
При сильном повреждении на 1 смг
поверхности древесины насчитывается 12—15
отверстий, ведущих в ходы рачков.
Источенная рачками поверхность сваи
становится рыхлой, губчатой, иногда сваи
истончаются и теряют первоначальную
прочность. Их приходится заменять
новыми.
Рачки заглатывают опилки,
образующиеся при бурении древесины, и
пропускают их через свой кишечник. При этом
клетчатка переваривается, а лигнин
выделяется с испражнениями. При
прохождении через кишечник рачка древесные
опилки теряют 50—95% своего исходного
сухого веса.
В проточенных в дереве ходах
происходит и размножение Limnoria. Летом
каждая самка приносит 8—10, изредка до
30, осенью 1—4 молодых рачка. Вылезая
из ходов, рачки при помощи брюшных
ножек довольно быстро плавают спинной
стороной вниз, но не удаляются дальше
чем на 1 м от места своего рождения.
Расселение лимнорий на далекие расстояния
происходит пассивно, главным образом
путем переноса течениями зараженных
рачками бревен, стволов, кусков дерева
и т. д.
Заглатывание грунта и использование
содержащегося в нем органического
вещества распространено среди морских
равноногих не менее широко, чем питание расте-
Рис. 245. Лимнория и вызываемые ею
повреждения:
1 — Limnoria lignorum; 2 — разрез ее хода в дереве;
3 — свая, источенная лимнорией.
ниями. Многие представители Asellota,
как мелкие, так и крупные, а также
многие Valvifera, в частности из семейства
Arcturidae, питаются именно таким
образом. В особенности характерно питание
грунтом для глубоководных равноногих.
По мере увеличения глубины количество
растительноядных и хищных видов
равноногих постепенно уменьшается, и,
начиная с 3—4 км и кончая предельными
океаническими глубинами порядка 11 км, все
равноногие оказываются грунтоедами. При
этом некоторые из них способны
сортировать грунт, заглатывая преимущественно
наиболее мелкие его частицы с несколько
повышенным содержанием органического
вещества. Приблизительно таким же
образом питаются обитатели подземных
пресных вод, в частности подземные водяные
ослики и Cirolanidae.
Немало среди морских равноногих и
хищников. Mesidotea при помощи
чувствительных щетинок и цилиндров своих
задних антенн разыскивает зарывшихся в
грунт бокоплавов, полихет, личинок хиро-
475
Рис. 246. Astacilla pusilla:
1 — в охотничьей позе сбоку; 2 — передняя
часть тела рачка снизу.
номид и других мелких животных, хватает
их подклешнями передних грудных
ножек и поедает. Нередко она нападает
на рыб, в особенности попавших в
рыболовные сети, и местами, например в
низовьях наших северных рек, приносит
заметный ущерб промыслу, так как
портит улов. По-видимому, она может иногда
распространяться вверх по рекам,
прицепившись к мигрирующим рыбам.
Astacilla охотится на различных мелких
животных следующим образом: она держится
задними грудными ножками за водоросли,
ветви гидроидов или мшанок и
приподнимает вытянутую переднюю часть тела
так, чтобы длинные, богатые
крепкими щетинками передние грудные ножки
и чувствительные задние антенны были
направлены вперед; в таком положении
она легко хватает передними ножками
свои жертвы, прежде всего разных мелких
рачков (рис. 246).
Многочисленные представители
семейства Cirolanidae образуют ряд постепенных
переходов от хищничества к паразитизму.
Виды родов Cirolana и Conilera нападают
на рыб, правда преимущественно больных
или попавших в сети, и на недавно
слинявших десятиногих раков. Особенно
охотно они поедают трупы рыб, оставляя
от них буквально только кожу да кости.
У этих родов ротовые придатки жующего
типа. Виды рода Aega — временные
паразиты рыб. Они быстро плавают, прочно
прикрепляются тремя парами крючковид-
ных передних грудных ног к коже рыб,
там, где она тонкая (жабры, плавники,
жаберные крышки), прокалывают ее
колюще-сосущим ротовым аппаратом и сосут
кровь рыб. Однако, наевшись, они
оставляют своего хозяина. Наконец, виды
подсемейства Cymothoinae (Cymothoa, Mei-
nertia, Livoneca, Anilocra и др.) постоянно
живут на рыбах, причем не только на
жабрах, но и во рту, а также на
поверхности тела. Они плотно прикрепляются
к рыбе, благодаря тому что все (иногда за
исключением последней пары) их грудные
ножки крючковидны и своими концами
внедряются в кожу хозяина. Иногда
прикрепившиеся рачки обрастают кожей рыбы,
образуя на ее поверхности опухоли.
Самки Cymothoinae теряют двустороннюю
симметрию, и часто тело их искривляется.
Они паразитируют как на морских, так
и на пресноводных рыбах. В бассейне
Амура на теле многих рыб можно найти
вздутия, в которых помещаются попарно
крупная асимметричная самка и более
мелкий симметричный самец Livoneca amu-
rensis.
Помимо Cirolanidae, на рыбах
паразитируют личинки (праницы) Gnathiidea
(рис. 249, 1). Они подплывают к рыбам,
прикрепляются к их плавникам крючко-
видными передними ногами и начинают
разрывать ими ткани хозяина, погружая
все глубже свой колюще-сосущий
ротовой аппарат, пока не доберутся до
кровеносных сосудов. При этом они
удерживаются в ране жвалами, как гарпунами,
а сосущий хоботок составляют челюсти.
Они сосут кровь чрезвычайно жадно.
Средний отдел их кишечника
переполняется кровью, и задняя часть груди
раздувается настолько, что сегментация ее
исчезает. Впоследствии кровь переходит
в пищеварительные железы. Ее запасов
хватает на всю жизнь рачка. После
линьки личинок возникшие из них взрослые
рачки больше не питаются, их ротовые
придатки, за исключением жвал самца,
редуцируются, а кишечник у некоторых
родов частично даже зарастает.
476
Наиболее глубокие приспособления к
паразитическому существованию
свойственны подотряду Epicaridea, все
представители которого паразитируют на
различных ракообразных. Их организация,
в особенности самок, в различной
степени упрощена вплоть до полной потери
сегментации и конечностей и превращения
тела в бесформенный мешок. Самки
представителей семейств Bopyridae, Dajidae
и Entoniscidae, поселяющиеся в
жаберной полости креветок и крабов, а иногда
(Dajidae) и мизид, в той или иной степени
сегментированы, но обычно
асимметричны, ротовые придатки их превращены
в сосущий хоботок, которым они сосут
кровь своих хозяев. Выводковая сумка
очень крупная и содержит большое
количество яиц. Самцы двустороннесиммет-
ричны, гораздо мельче самок и живут на
них (рис. 247, А), Виды семейства
Cryptoniscidae паразитируют на самых
различных ракообразных — на ракушковых, усо-
ногих, мизидах, кумовых, других
равноногих, бокоплавах, клешненосных
осликах. Половозрелые самки Cryptoniscidae
имеют мешковидное тело, без всяких
следов сегментации и конечностей, с
хоботком на переднем конце, которым они
протыкают покровы хозяина и сосут его кровь.
Такие самки образуются из маленьких
самцов, сохраняющих основные признаки
равноногих ракообразных. Таким
образом, Cryptoniscidae, в отличие от других
раздельнополых Epicaridea, представляют
собой протерандрических гермафродитов:
при созревании они становятся самцами,
а затем преобразуются в самок.
Среди Cryptoniscidae особенно
интересны роды Dunalia и Liriopsis,
паразитирующие на корнеголовых ракообразных Saccu-
lina и Peltogaster, которые в свою
очередь паразитируют на крабах и раках-
отшельниках (стр. 438). Самка Dunalia
помещается в жаберной полости
пораженного саккулиной краба, но своим
длинным хоботком через покровы краба
достигает ответвлений «корней» саккулины и
питается за ее счет. Срок
продолжительности жизни Dunalia B—3 месяца)
значительно меньше, чем саккулины, так что
наступает момент, когда паразит
освобождается от своего сверхпаразита. Молодые
самки Liriopsis овальные, прикрепляются
к мешковидному телу Peltogaster и сосут
его соки. Затем животное проникает в
мантийную полость Peltogaster, округляется,
тело его перешнуровывается на две
части, оно прекращает питаться и достигает
половозрелости (рис. 247, Б—Г).
Все Epicaridea имеют
свободноплавающих личинок, при помощи которых они
расселяются и заражают своих хозяев.
Размножение подавляющего
большинства равноногих происходит так же, как
у других перакарид. Однако, помимо
раздельнополых видов, среди
паразитических видов, как уже упоминалось, есть
протерандрические гермафродиты (Суто-
thoinae, Cryptoniscidae), а для Cyathura
carinata описана протогиния. Наконец,
среди мокриц известно несколько видов,
которым свойственно партеногенетическое
размножение — явление исключительно
редкое у высших ракообразных.
Половой диморфизм у раздельнополых
видов, если исключить Epicaridea,
проявляется главным образом в строении
передних брюшных ножек, приспособленных у
самцов для копуляцпп, и нередко в разли-
Рис. 247. Паразитические равноногие:
А — Cancricepon elegans, самка с выводковой сумкой
и карликовым самцом (I); Б, В, Г — Liriopsis, молодая
питающаяся самка (Б, В) и половозрелая нспитаю-
щаяся самка (Г); 2 — выводковая сумка; з — плеоподы;
4 — уроподы.
477
чиях размеров самцов и самок:
у водяных осликов, например,
самцы крупнее самок, но у
некоторых других видов
наблюдаются обратные соотношения. У
Gnathiidea самцы имеют жвалы,
отсутствующие у самок (табл. 33,
2, 2). У ряда Sphaeromidae у
самцов две передние пары
грудных ножек снабжены подклеш-
нями, а у самок только одна.
Половому процессу
предшествует длительное спаривание.
Самец удерживает самку
грудными ножками, располагаясь
на ее спинной стороне и ожидая
ее линьки. У водяных осликов и,
вероятно, у других видов
перенос спермы возможен только
в течение 10 часов после
линьки самки, когда ее половые
отверстия расширены и не
образовались пластинки выводковой
сумки. Линька всегда
происходит в два приема: сначала
старый панцирь сбрасывается с
задней, а потом с передней
части тела. В промежуток между
этими двумя линьками самец
своими копулятивными
брюшными ножками переносит
сперму в половые пути самки.
Через 2—10 часов у самки
образуется выводковая сумка и
туда откладываются
оплодотворенные в половых путях яйца.
Число их различно у разных
видов, причем по мере развития
оно уменьшается, так как часть
яиц вываливается из
выводковой сумки и погибает. Так,
например, у водяного ослика из 150
откладываемых сначала яиц к концу
развития остается 80, у Idotea из 30—40—
только 4—6, у Jaera albifrons из 15—16
не больше 12, а обычно 3—4 яйца.
Наибольшее количество яиц, как и у других
животных, образуется у паразитических
видов. В выводковой сумке Cymothoa
oestrum насчитывается 2450 яиц.
Наоборот, очень низкой плодовитостью
отличаются подземные и глубоководные виды.
Мелкие интерстициальные виды Micro-
Рис. 248. Спаривание
водяного ослика и строение
внутренней ветви второй
брюшной конечности самца
(самец зачернен).
charon носят в сумке всего 1—2
яйца. Из всех равноногих
только некоторым Gnathiidea, в
частности роду Paragnathia,
свойственно живорождение. Из
половых отверстий самок сразу
выходят подвижные личинки на
стадии манка, и выводковой
сумки не образуется совсем.
У всех остальных яйца
развиваются в выводковой сумке.
Движения ее пластинок
приводят к тому, что вода все
время омывает яйца и молодь.
У мокриц полость выводковой
сумки увлажняется за счет
водопроводной системы покровов,
но у некоторых видов на
брюшной стороне тела яйценосных
самок образуются
пальцевидные выросты, выделяющие
влагу в выводковую сумку. Из яиц
в выводковой сумке выходят
молодые рачки, находящиеся,
так же как у кумовых и
некоторых других отрядов, на
стадии манка. Обычно эти
молодые рачки отличаются от
взрослых главным образом
недоразвитием последней пары
грудных ножек, но у Gnathiidea
и Epicaridea стадия манка
имеет ряд специальных
приспособлений к активному
существованию и поискам хозяина, на
котором ей или последующим
стадиям предстоит паразитировать.
О стадии манка Gnathiidea,
носящей специальное название
праница, уже говорилось выше
(стр. 473). У Epicaridea эта
стадия называется эпикаридиум. Эпи-
каридиум обычно достигает в длину
0,2—0,35 мм и активно плавает при
помощи крупных задних антенн и брюшных
ножек. Грудные ножки его крючковидные
(рис. 249,5). Он питается только за счет
желтка, унаследованного от яйца. Судя по
наблюдениям над Portunion (для других
видов жизненный цикл не установлен),
эпикаридиум прикрепляется к планктонным
веслоногим и после линьки превращается
в паразитическую стадию м и к р о н и с-
478
ц и у м, которая теряет членистые
конечности и плавательные щетинки, но,
слиняв несколько раз, дает начало новой
свободноживущей стадии —к р и п т о-
нисциуму. Криптонисциум имеет
нормальную сегментацию, 7 пар крючко-
видных грудных и 6 пар плавательных
брюшных ножек, хорошо развитые
антенны (рис. 249, 4). Он отыскивает раков,
которые служат окончательными
хозяевами данному виду паразита, прикрепляется
к ним, линяет и начинает
преобразовываться во взрослую стадию. У
раздельнополых Epicaridea криптонисциум,
прикрепившийся к еще не зараженному
хозяину, превращается в самку. Если же
личинка садится поблизости от уже имеющейся
самки, она становится самцом,
причем ее облик меняется незначительно:
редуцируются только глаза, антенны,
плавательные ножки, но характер
сегментации и строение грудных ножек остаются
прежними. У протерандрических
гермафродитов Cryptoniscidae личинка
криптонисциум, садясь на хозяина, без всякой
линьки превращается в самца, так как
в ней развиваются семенники. Затем в
половых железах, помимо сперматозоидов,
начинают созревать яйца, паразит
несколько раз линяет, постепенно теряя
при этом сегментацию, конечности и
другие признаки ракообразных, и в конце
концов приобретает мешковидную форму,
характерную для половозрелой
самки.
Партеногенетическое
размножение известно у некоторых видов
мокриц из рода Trichoniscus (Т.
elisabethae, T. provisorius) и
также у обычных широко
распространенных мокриц Armadillidium
vulgare и Cylisticus convexus. Все
эти виды распадаются на
отдельные расы, различающиеся по
числу хромосом. Расы с обычным
диплоидным числом хромосом
включают и самцов и самок, и у них
размножению предшествует
оплодотворение яиц. Наряду с ними
существуют расы с триплоидным
числом хромосом (у Trichoniscus
диплоидное число хромосом 16,
триплоидное 24), представленные
исключительно самками, к которым
иногда добавляются в ничтожном количестве
(менее 1,6%) самцы. Яйца триплоидных
самок при созревании не испытывают
второго редукционного деления и
оказываются способными к развитию без
оплодотворения. Такие размножающиеся
партеногенетически расы приурочены к
сравнительно неблагоприятным условиям
существования, в частности к засушливым
местам, и обычно обитают близ границ
области распространения каждого
данного вида.
У свободноживущих равноногих, как
водных, так и сухопутных, вышедшие из
яиц и находящиеся на стадии манка
молодые рачки некоторое время остаются в
выводковой сумке матери и линяют там два
или три раза. После этого, будучи еще на
той же стадии, рачки покидают
выводковую сумку матери. У морских видов из
семейства Arcturidae молодь перебирается
из выводковой сумки матерп на ее
длинные задние антенны и некоторое время
живет на них (рис. 250). Позднее молодые
рачки время от времени линяют и в
конце концов достигают половозрелости.
Промежутки между линьками и число
их до достижения половозрелости
различны у разных видов. Так, например,
у мокрицы Porcellio эмбриональное
развитие длится 35 дней, первая стадия
манка продолжается 21 день, из которых 16
Рис. 249. Личинки равноногих:
1 — праница Gnathia; 2 — ее голова; з — эпикаридиум
Clypeoniscus; 4 — криптонисциум Portunion.
479
приходится на пребывание в выводковой
сумке, вторая и третья — по 23 дня,
а всего до возникновения вполне
сформированного, но еще неполовозрелого рачка
требуется 102 дня. У морского рачка
Sphaeroma rugicauda при температуре
15°С яйца в выводковой сумке развиваются
45 дней, у обычного водяного ослика под
Москвой — от 14 дней в мае до 22 дней
в августе. Самки водяного ослика
достигают половозрелости через 50—55 дней
после рождения, перелиняв за это время
13—15 раз.
Общая продолжительность жизни
установлена для немногих видов. Мокрица
Oniscus asellus живет 3—4 года, Aega —
не менее 2L/4 лет> водяной ослик — около
2 лет, Cryptoniscidae — менее полугода.
Как уже указывалось, большинство
известных видов равноногих ракообразных
обитает в море, где они встречаются от
приливо-отливной зоны до предельных
океанических глубин. Литораль
населена небольшим количеством видов,
из которых наиболее характерны
представители родов Jaera и Sphaeroma.
J. albifrons и J. marina широко
распространены в северной части Атлантического
океана, а первая из них обычна также
в Баренцевом и Белом морях. Окраска
этих рачков очень изменчива: некоторые
особи целиком темные, у других вдоль
тела проходит темная полоса, у третьих
темная голова или плеотельсон и т. д.
(рис. 251). Соотношение особей с разным
типом окраски в различных поселениях
рачков различно. Специальные
исследования по скрещиванию между собой
различно окрашенных особей показали, что
каждый тип окраски зависит от
определенного гена. Одни из них доминантны,
другие рецессивны. Есть гены некоторых
типов окраски, сцепленные с генами пола.
Это значит, что определенный тип окраски
встречается только у самцов или,
наоборот, только у самок. Некоторые поселения
рачков обеднены генами по сравнению
с другими, и соответственно разнообразие
окраски составляющих их особей меньше.
Таким образом, окраска Jaera
определяется совершенно иначе, чем у Idotea
(см. стр. 474).
На морских мелководьях, глубже
приливо-отливной зоны, богатство фауны
равноногих возрастает. Среди водорослей,
в зарослях гидроидов и мшанок
преобладают виды с узким телом, позволяющим
рачкам пролезать между ветвями,
например различные виды Idotea, Arcturidae
и др. На мягких илистых грунтах
обитают уплощенные сверху вниз равноногие
вроде Mesidotea, Synidotea, Serolidae и
Таблица 39. Личинки десятиногих ракообразных:
1 — зоэа краба Rhithropanopeus;
2 — глаукотоэ Pagurus bernhardus;
3 — личинка Porcellana longicornis;
4 — эрионеикус Eryooidae,
5 — зоэа Palaemon longirostris;
в — поздняя личинка Sergestes cornutus;
7 — филлозома лангуста Panulirus inflatus;
8 — мегалопа краба Hyas araneus;
9 — мизисная стадия Nephrops norvegicus;
10, 11 — глаукотоэ и зоэа камчатского краба
(Paralithodes camtschatica);
12 — науплиус Trachypenaeus constrictus.
др. Зарывающиеся в мягкий грунт виды
часто имеют удлиненное цилиндрическое
тело (Anthuridea, Gnathiidea, Ischnome-
sidae и др.)- На больших океанических
глубинах обитают почти исключительно
представители подотряда Asellota. Этот
подотряд явно тяготеет к холодным водам.
Его виды многочисленны на мелководьях
в высоких и умеренных широтах
северного и южного полушарий, а также на
больших глубинах всего Мирового
океана, но почти не встречаются на
тропических мелководьях.
Для многих глубоководных Asellota
характерны удлиненные грудные ножки,
препятствующие погружению их тела в
мягкий грунт (табл. 33,8,11,12), а также
антенны, богатые чувствительными
щетинками и цилиндрами. Однако особенно
интересна четко выраженная у них
тенденция к увеличению размеров тела по
мере увеличения глубины. Чем глубже
обитает какой-нибудь вид определенного
рода, тем крупнее его размеры по
сравнению с близким видом с меньшей глубины.
Так, например, длина тела видов рода
Haplomesus с глубины менее 3 км не
достигает 5 мм, а Н. gigas с глубины 8330—
8430 м имеет в длину 15 мм.
Специальные исследования показали,
что увеличение тела глубоководных
равноногих происходит главным образом за
счет увеличения размеров составляющих
их организм клеток, а не за счет
возрастания их количества. Это явление не
связано с понижением температуры среды
обитания. Предполагают, что в качестве
основного фактора, вызывающего
«глубоководный гигантизм», выступает
повышенное на больших глубинах
гидростатическое давление, действующее на обмен
веществ.
Среди морских мелководных
равноногих многие способны переносить
значительные колебания солености. Jaera albif-
rons и Cyathura carinata (табл. 33, 7)
часто встречаются в опресненных
участках моря и даже заходят в пресные воды.
Mesidotea entomon (табл. 33, 5) обитает
преимущественно в солоноватых или даже
в пресных водах, причем при большом
опреснении она достигает более крупных
размеров. В западной части Балтийского
моря наибольшая длина самцов 50 мм,
а в опресненной восточной — более 80 мм.
Этот вид населяет Средний и Южный
Каспий и некоторые озера,
покрывавшиеся во время великого оледенения морем.
Фауна пресноводных равноногих
значительно беднее морской. Наиболее широко
распространено и обильно видами чисто
пресноводное семейство водяных осликов
(Asellidae). Обыкновенный водяной ослик
(Asellus aquaticus, табл. 33, 3) обитает
Таблица 40. Пальмовый вор (Birgus latro)
и его жизненный цикл:
1 — 5 — последовательные стадии
развития;
6 — взрослая особь»
31 Жизнь животных, т. 2
во множестве стоячих водоемов всей
Европы, исключая Пиренейский полуостров.
На Кавказе, кроме него, известен близкий
вид A. monticola. На значительном
протяжении Азии водяные ослики отсутствуют.
Только в Байкале живет пять присущих
лишь этому озеру видов. Кроме того, по
одному эндемичному виду есть в Ципо-
ципиканских озерах (бассейн Витима), в
бассейне Иртыша и в бассейне Уссури.
Местонахождения этих видов могут
считаться остатками некогда более широкого
распространения рода Asellus. Далее,
вдоль восточного берега Азии, включая
северную Японию и северный Китай,
широко распространен близкий к A.
aquaticus вид водяного ослика — A. hilgendorfi.
В Западной Европе наряду с A. aquaticus
встречаются и некоторые другие виды
водяных осликов, в частности A. meridia-
nus, A. banyulensis и т. д. Как правило,
A. aquaticus и A. meridianus, обладающие
чрезвычайно сходной биологией, тем не
менее не живут вместе в одном водоеме.
Между ними существуют
антагонистические отношения. A. aquaticus постепенно
вытесняет A. meridianus, но механизм
этого вытеснения, так же как и в других
подобных случаях (стр. 494, 525),
разгадать пока не удалось. Есть особые виды
водяных осликов в наземных водоемах
Северной Америки и Северной Африки.
Много видов водяных осликов
приспособилось к жизни в подземных пресных
водах. Такие виды известны из
Закавказья, Европы (особенно много их на
Балканском полуострове), бассейна Амура
и Японии. Только в подземных водах
обитают особые роды этого семейства—
Stenasellus, Stygasellus и Johanella.
Несколько видов Stygasellus населяет
только Португалию, Johanella найдена лишь
в одном ключе в Северной Африке, a
Stenasellus имеет очень широкое, но
разорванное распространение. Наибольшее
количество видов этого рода приурочено к
пещерам стран, лежащих к северу от
Средиземного моря (южная Франция, Италия,
Пиренейский полуостров, Балканы), 4
вида открыто в подземных водах
экваториальной Африки, а один вид обитает
в теплом источнике Ходжа-Койнар,
температура воды которого круглый год
равняется 20—22°С и который находится
в горной системе Кугитангтау, в
восточной Туркмении. По всей вероятности,
Stenasellus — очень древний род
тропического происхождения, сохранившийся
только в подземных водах. Ему
свойственны многие чрезвычайно примитивные
морфологические признаки (чешуевидная
наружная ветвь на задних антеннах,
сравнительно крупные передние брюшные
сегменты и т. д.).
Помимо водяных осликов, в пещерах
Европы обитают своеобразные роды Typh-
locirolana, Caecosphaeroma, Cirolanides,
Typhiosphaeroma, принадлежащие к
морским семействам Sphaeromidae и Cirola-
nidae. Вместе с некоторыми другими
животными они образуют группу видов
морского происхождения в составе
пресноводной подземной фауны.
Морское происхождение свойственно
также чрезвычайно мелким равноногим,
обитающим в интерстициальных пресных
и морских водах. Как и другие
представители интерстициальной фауны, они имеют
длинное червеобразное тело и
укороченные конечности, что позволяет им
активно передвигаться по узким пространствам
между песчинками и другими частицами
грунта. Род Microcharon (табл. 33, 13)
объединяет 16 видов и 5 подвидов, боль-
Рис. 250. Arcturus ulbani, самка с молодью.
шинство которых обитает в пресных
грунтовых водах бассейна Средиземного моря,
но один вид известен из Закавказья, а
два из Средней Азии, причем М. halo-
philus, известный из пещеры Каптархана
в восточной Туркмении, живет в соленом
подземном озере, соленость воды которого
равняется 12°/00. Кроме этих обитателей
внутренних водоемов, к роду Microcharon
принадлежат еще два вида из
интерсоциальных морских вод Ламанша. Нет
особых сомнений в том, что виды этого
рода переселились из системы морских
в систему пресных интерстициальных вод.
Сходная история расселения
приписывается и интерстициальному семейству
Microcerberidae, принадлежащему к
подотряду Anthuridea. Только к пресным
водам приурочен ныне подотряд Phreatoi-
cidea. Относящиеся к нему 47 видов
распространены в Австралии, Тасмании,
Новой Зеландии, Южной Африке и Индии
как в наземных, так и в подземных водах.
Однако в ископаемом состоянии они
известны из морских отложений пермского
периода в Архангельской области, Англии
и Германии. Очевидно, в конце палеозоя
они жили в море и населяли северное
полушарие, а до наших дней сохранились
только в пресных водах некоторых стран
южного полушария (рис. 252).
К жизни на суше приспособился только
один подотряд равноногих — Oniscoidea,
или мокрицы. Они обитают во всех
ландшафтных зонах земли, но особенно
многочисленны и разнообразны в степях.
Наиболее примитивные из них —
представители рода Ligia — еще связаны с морем.
Ligia живет на морских берегах и может
длительное время существовать под водой.
В условиях эксперимента эти мокрицы
оставались под водой более 80 дней,
причем нормально питались в таких условиях.
Так же тесно связаны, но с пресными
водами виды рода Ligidium, часто
находимые в ручьях и ключах. Для всего
семейства Ligiidae, к которому принадлежат
оба названных рода, характерно
отсутствие приспособлений к дыханию
атмосферным воздухом. Все остальные
мокрицы существуют совершенно независимо от
водоемов и представляют собой
настоящих сухопутных животных. Однако они
Рис. 251. Изменчивость окраски Jaera albifrons*
могут жить только в условиях
повышенной влажности, обеспечивающей их
нормальное дыхание и размножение. Поэтому
они выработали ряд приспособлений,
благодаря которым они заселили даже такиеt
казалось бы, не подходящие для них ме~
стообитания, как пустыни.
Эти приспособления направлены
главным образом на то, чтобы свести к
минимуму потерю влаги организмом, что
достигается разными способами. Все
мокрицы активны только ночью, что защищает
их от действия солнечных лучей. Светлое
время суток они проводят зарывшись в
поверхностный слой почвы, спрятавшись
под камнями, в различных других
укрытиях или в норах, которые выкапывают
и в которых поддерживается повышенная
влажность. Есть мокрицы, живущие в
норах грызунов и муравейниках. Ночью
они поедают растения, получая не только
31*
483
Рис. 252. Географическое распространение ныне живущих
и ископаемых Phreatoicidea.
пищу, но и влагу. В условиях
эксперимента, да и в природе они отрицательно
фототропичны: активно стремятся уйти
от света, чтобы избежать высыхания. Для
большинства мокриц смертельно
пребывание в условиях влажности воздуха менее
86% насыщения, причем Oniscus с менее
совершенными ложными трахеями
погибает раньше, чем РогсеШо, ложные трахеи
которого более сложны.
При помещении мокриц в условия
пониженной влажности они сначала очень
быстро теряют воду, но затем этот процесс
замедляется, благодаря тому что из крови
выделяются некоторые вещества,
увеличивающие непроницаемость покровов. С
другой стороны, испарение влаги
позволяет мокрицам в известных пределах
регулировать температуру своего тела,
избегать перегрева. Разница между
температурой нагретого солнцем до 36°С пляжа
и температурой тела Ligia равняется 8°,
для Oniscus и Armadillidium эта разница
составляет 4—5°, для РогсеШо — 2—3°.
Способность регулировать
температуру своего тела
свойственна только живым мокрицам.
Температура тела мертвых
животных совпадает с
температурой субстрата. Все эти
морфологические и физиологические
особенности позволяют
утверждать, что из всех
ракообразных мокрицы наиболее полно
приспособились к жизни на
суше.
Практическое
значение равноногих, несмотря
на их широкое
распространение и видовое разнообразие,
довольно ограниченно. С одной
стороны, они приносят
некоторый вред человеку, поскольку
Limnoria u Sphaeroma
разрушают деревянные сооружения в
море, а мокрица Hemilepistus
zachvatkini объедает луговую
растительность в Узбекистане.
Но, с другой стороны,
мокрицы, перерабатывая
значительное количество растений,
способствуют увеличению
плодородия почвы. В особенности
важную роль в этом деле играют виды рода
Hemilepistus в пустынях, где они
выполняют такую же огромную работу, как
земляные черви в других климатических зонах.
ОТРЯД РАЗНОНОГИЕ РАКООБРАЗНЫЕ,
ИЛИ БОКОПЛАВЫ (AMPHIPODA)
Бокоплавов хорошо знают не только
зоологи, но и все рыбаки. В разных
частях СССР местные жители называют их
по-разному: «стонога» на Каспии, «мор-
мыш» или «мормышка» на Урале и в
Западной Сибири, «бармаш» на Байкале и
в Восточной Сибири. На Байкале
существует подледный зимний промысел омуля—
«бармашенье»: бокоплавов из окрестных
озер живьем привозят в бочках на Байкал,
пробивают во льду проруби и горстями
швыряют туда рачков, привлекая таким
образом омуля, которого ловят на
поддев. Широко известно кормовое значение
484
бокоплавов для рыб. Они служат
удильщикам прекрасной наживкой. Рыбоводы
перевозят их вместе с мизидами во вновь
созданные водохранилища для улучшения
условий откорма рыб. Делались попытки
искусственного разведения этих рачков
на рыбоводных заводах. В естественных
условиях многие рыбы используют в
пищу бокоплавов, а некоторые, как
например форели, питаются исключительно ими.
Наряду с этим также хорошо известно,
что бокоплавы в отдельных случаях
портят рыболовные сети и объедают
попавших в них рыб.
Бокоплавы подобно равноногим
распространены очень широко. Наибольшее
количество их видов обитает в море, где
они населяют все глубины и живут как
на дне, так и в толще воды. Только в море
встречается и небогатая видами группа
бокоплавов, приспособившаяся к пара-
зитированию на китах — так называемые
китовые вши (семейство Cyamidae). В
пресных водах видовое разнообразие
бокоплавов гораздо меньше, чем в море;
по необычайному богатству видами из всех
пресных водоемов выделяется
замечательное озеро Байкал, где живет около 240
видов бокоплавов, причем почти все они
за пределы Байкала не выходят.
Довольно богата и разнообразна фауна
бокоплавов подземных пресных вод. Однако, в
отличие от равноногих, бокоплавы не
сумели приспособиться к сухопутному
существованию. Правда, есть виды
бокоплавов, большую часть жизни проводящие
на суше, но они живут только на морских
пляжах или в других местах в
непосредственной близости от моря и остаются
с ним очень тесно связанными. Всего
в настоящее время известно около 4500
видов бокоплавов.
По своему строению они во многом
сходны с равноногими ракообразными,
однако тело их часто бывает сжато с боков,
а не сверху вниз, как у равноногих.
Впрочем, и среди бокоплавов есть виды с
уплощенным в спинно-брюшном направлении,
а также с цилиндрическим телом. Голова,
так же как у равноногих, срастается с
первым, изредка с двумя первыми
грудными сегментами, а карапакс
отсутствует. Глаза сидячие и расположены по
бокам головы. У пелагических Phronima
каждый глаз разделен на две, а у
семейства Ampeliscidae даже на 3 части (табл.
34, 12). С другой стороны, у Oedicerotidae
оба глаза на спинной стороне
соединяются так, что образуется один громадный
непарный глаз. Глубоководные и
подземные виды, как это бывает обычно, слепы,
но у некоторых из них на месте глаз
находятся темные, лишенные фасеток
«глазные пятна», назначение которых остается
неизвестным. Под покровами головы,
близ ее спинной стороны, у многих
бокоплавов имеется пара статоцистов, с 1—3
статолитами в каждом. Обе пары антенн
обычно длинные и снабжены
чувствительными цилиндрами и щетинками.
Ротовые придатки жующего типа.
Конечности всех семи свободных
грудных сегментов устроены по-разному, что
и отражено в названии отряда «разноно-
гие». Первые две пари ножек обычно
снабжены подклешнями, в редких случаях
имеются настоящие клешни или эти ножки:
не специализированы как хватательные.
У следующих двух пар ножек когти
обращены назад, а у последних трех пар —
вперед. У некоторых планктонных
глубоководных видов подклешни есть не
только на передних, но и на следующих
за ними или даже на всех парах грудных
ножек. При помощи этого приспособления
рачки временно прикрепляются к
медузам и гребневикам, которыми питаются.
У видов, ползающих по водорослям,
гидроидам и т. д., когти задних грудных
ножек заметно удлинены; особенно сильно
они развиты на задних ногах
паразитических Cyamidae, которые вонзают их
в кожу китов и таким образом прочно
прикрепляются к хозяину. У этого
семейства, а также у семейства «морских
козочек» (Caprellidae) третья и четвертая пары
грудных ног редуцированы.
У всех бокоплавов эпиподиты грудных
ножек, за исключением ног первой пары,
а иногда и некоторых других,
превращены в листовидные тонкостенные жабры.
Во многих случаях жабры складчатые,
благодаря чему увеличивается их
дыхательная поверхность, а иногда они
снабжены пальцевидными выростами.
Поскольку дыхание осуществляется придат-
485
Рис. 253. Строение бокоплавов:
1— Gammarus; 2 — Caprella anatifera; 3 — Рагасуа-
mus boopis.
ками грудных ножек, сердце целиком
помещается в грудном отделе. У
половозрелых самок к некоторым из грудных
ножек с внутренней стороны от жабр
прикреплены пластинки выводковой
сумки. В отличие от равноногих, кумовых
и других выводковая сумка бокоплавов
не исчезает после конца каждого периода
размножения.
Брюшной отдел состоит из 6 сегментов.
Обычно он несколько короче грудного,
но имеет такую же ширину. Однако у
многих планктонных бокоплавов он сужен,
благодаря чему все тело приобретает
каплевидную форму (рис. 255,1). Конечности
трех передних брюшных сегментов
приспособлены для плавания. Их ветви многочле-
никовые и снабжены многочисленными
плавательными щетинками. Конечности
трех задних брюшных сегментов
направлены назад, и ветви их не расчленены
(за исключением наружных ветвей
последней пары, которая часто состоит из 2
члеников). Это прыгательные ножки, или
уроподы. У представителей подотряда
Laemodipodea, в который объединяются
морские козочки и китовые вши, брюшной
отдел очень сильно укорочен и лишен
сегментации, а брюшные ножки
редуцированы и нередко отсутствуют совсем.
У бедного видами подотряда Ingolfielli-
dea плавательные ножки превращены в
маленькие нерасчлененные пластинки. За
брюшным отделом следует короткий тель-
сон, имеющий форму треугольника, овала
или расщепленный выемкой на две лопасти.
Покровы тела бокоплавов часто
гладкие, но во многих случаях вооружены
различными килями, зубцами и шипами.
Такая скульптура покровов иногда имеет
защитное значение. Среди
многочисленных бокоплавов Байкала часть гладкие,
а часть «вооруженные». В кишечниках
байкальских бычков, питающихся
преимущественно бокоплавами, резко
преобладают гладкие виды. Очевидно,
«вооруженные» в какой-то мере защищены от
нападения бычков.
Бокоплавы, как правило, окрашены
довольно однообразно в буроватый,
зеленоватый и желтоватый тона. Исключение
составляют байкальские виды, среди
которых есть пестрые, синие, красные,
зеленые (табл. 34, 1, 4,5,7). Глубоководные
и подземные виды бесцветны, но среди
планктонных глубоководных видов есть
и красные, как например Cyphocaris
(табл. 34,13), Paracyphocaris и ряд близких
родов.
Зеленоватая окраска пресноводного
Gammarus вызывается каротиноидами,
вырабатываемыми из содержащихся в поеда-
мых рачком растениях каротинов. В
аквариуме представителей лишенной
пигмента подземной расы Gammarus
содержали в течение длительного времени в
полной темноте и в условиях постоянного
освещения. Независимо от этого они
приобретали нормальную зеленоватую
окраску, если получали в пищу растения.
Однако есть и наследственные факторы,
определяющие окраску. Изредка наряду
с зеленоватыми встречаются красные особи
бокоплавов. Эксперименты по
скрещиванию их между собой и с нормальными
особями показали, что окраска зависит
от трех пар генов, причем ген
зеленоватой окраски резко доминирует.
В соответствии с различным строением
ног движения разноногих весьма
разнообразны. Большинство этих рачков может
ползать по дну и растениям, перебирая
грудными ногами, плавать при помощи
передних брюшных ножек и прыгать,
отталкиваясь от субстрата задними
брюшными ножками. Следует иметь в виду, что
название всего отряда «бокоплавы»
неточно. Только в очень мелких ручьях
или у самого берега водоема рачки
действительно плавают на боку, а там, где
глубина это позволяет, они плавают
спиной вверх, но нередко лежат на грунте
на боку. Однако поскольку их легче всего
заметить на самой ничтожной глубине,
за ними и утвердилось заведомо неверное
наименование.
Большинство бокоплавов
передвигается всеми тремя упомянутыми способами,
переходя с одного на другой в
зависимости от обстоятельств. Даже морские
козочки, несмотря на отсутствие плавательных
брюшных и средних грудных ножек, могут
не только ползать по водорослям и
гидроидам, но и плавать, изгибая свое тело.
Полусухопутные Talitridae прекрасно
прыгают на расстояние до 30 см и даже больше.
Но наряду с такими донными,
придонными и полусухопутными обитателями
существуют и настоящие планктонные
бокоплавы, плавающие всю жизнь. Это,
во-первых, все многочисленные виды
подотряда Hyperiidea и, во-вторых,
отдельные представители самого обширного
подотряда бокоплавов — Gammaridea.
Планктонные бокоплавы
характеризуются очень тонкими, часто прозрачными
покровами и присутствием в теле
жировых включений, что уменьшает их
удельный вес и облегчает парение в воде. У
Hyperiidea тело обычно каплеобразное,
благодаря тому что передняя его часть
широкая, вздутая, а задняя сужена.
Интересно, что одно также планктонное, но
совершенно неродственное им семейство
Hyperiopsidae, принадлежащее к
подотряду Gammaridea, имеет очень сходное
строение тела. Вероятно, при такой его
форме сопротивление воды при движении
Рис. 254. Движения Caprella.
487
Рис. 255. Планктонные бокоплавы:
1 — Hyperia galba (сбоку и сверху); 2 — Rhabdonectes
armatus; S— Macrohectopus branickii.
Рис. 256. Niphargoides similis, зарывшийся в грунт
и поедающий комочки детрита.
рачка вперед минимально. У некоторых
гипериид тело, наоборот, тонкое,
стрелообразное (рис. 255, 2). Обычно у планктон-
ных бокоплавов сильно развиты
плавательные ножки, а прыгательные
выполняют функцию рулей глубины. У
мелководных и полуглубоководных Hyperiidea
почти всегда увеличены размеры глаз,
которые могут покрывать всю или почти
всю голову. Прямых наблюдений над
функциями этих громадных глаз не
производилось, но можно предполагать, что с их
помощью рачки находят свою добычу.
Кроме того, глаза имеют немаловажное
значение при суточных вертикальных
миграциях, весьма характерных для
мелководных гипериид. Все планктонные
бокоплавы, за одним исключением, обитают
в море и совершенно не переносят
опреснения. Единственный пресноводный
планктонный представитель этого отряда —
Macrohectopus branickii — живет в
Байкале.
Донные бокоплавы нередко также
плавают довольно продолжительное время.
Так, например, половозрелые самцы
широко распространенной в озерах северной
части Европы и Америки, в опресненных
участках Балтийского моря и в Каспии
Pontoporeia affinis отличаются от самок
удлиненными задними антеннами и
большую часть жизни проводят в толще воды*
отыскивая ползающих по дну самок.
Некоторые донные виды (Bathyporeia, Coro-
phium) ночью покидают дно и поднимаются
к поверхности воды.
Многие бокоплавы охотно и быстро
зарываются в грунт. Обычно они втыкают в
грунт задние, а иногда и передние
антенны и начинают разгребать его грудными
ножками, отбрасывая частицы грунта
хватательными передними ножками.
Иногда это происходит с очень большой
быстротой. На песчаных побережьях Азовского
и средней и южной части Каспийского
моря можно наблюдать, как каждая
набегающая волна выносит на берег массы
бокоплава Niphargoides (Pontogammarus)
maeoticus. Когда она начинает отступать,
рачки зарываются в грунт до появления
следующей волны, заставляющей их
вылезать из грунта, а потом повторяется все
сначала. Способность зарываться в грунт
488
облегчает некоторым понто-каспийским
видам распространение вверх по рекам,
так как рачки, таким образом, могут
противостоять течению и не сноситься вниз.
Так, например, Niphargoides (Pontogam-
marus) sarsi населяют всю Волгу вплоть
до ее верхнего течения, закапываясь в
песчаный грунт реки.
Другие виды вырывают в грунте
настоящие норы, а некоторые строят из грунта
трубки или убежища иной формы.
Некоторые виды подземного бокоплава Niphar-
gus роют в мягком грунте подземных озер
довольно сложные туннели с несколькими
входами и с расширениями — «жилыми
камерами». Представители многих семейств
подотряда Gammaridea (Ampeliscidae,
Corophiidae, Aoridae, Amphithoidae, Pho-
tidae и др.) обладают одноклеточными
железами, расположенными в средних
члениках задних грудных ножек или в
боковых пластинках грудного отдела и
открывающимися в когтях грудных ног. Эти
железы вырабатывают секрет, при помощи
которого рачки при строительстве трубок
и домиков скрепляют частицы грунта,
обрывки водорослей и т. д.
Все Corophiidae живут в построенных
ими трубках. Corophium volutator
цементирует секретом стенки вырываемого им
туннеля длиною 4—8 см, а перед
наступлением зимы углубляет туннель до 20 см.
С. curvispinum прикрепляет
свои трубки к поверхности
грунта, камней, раковин
моллюсков, а также к днищам судов.
Благодаря своим
прикрепленным к судам домикам этот
каспийский вид распространился
очень широко: суда развезли
его по всей Волге и другим
русским рекам, он проник в
бассейн Балтийского моря и
даже в Англию. При
строительстве убежищ корофииды
пользуются удлиненными задними
антеннами, которыми они
захватывают подходящий
строительный материал (рис. 257).
Массовые морские
Ampeliscidae строят из песка или ила
небольшие тонкостенные
мешковидные домики,вмещающие толь-
ко тело рака, а его голова с антеннами
торчит наружу. Leptocheirus сооружает из
частиц грунта или обрывков растений нечто
вроде купола над веткой гидроида или
водоросли, служащей как бы полом его
жилища. Microdeutopus, Microprotopus и
другие из того же материала, иногда с
добавлением собственных экскрементов,
делают трубки с входным и выходным
отверстиями. При этом внутри трубки им
приходится многократно поворачиваться
вокруг собственной оси, поскольку
цементные железы, как уже упоминалось,
открываются в когтях грудных ног, и рачок
может цементировать какой-нибудь
участок хода, будучи обращен к нему только
брюшной стороной тела.
Донные бокоплавы, не зарывающиеся
в грунт и не делающие нор и домиков,
обыкновенно прячутся среди водорослей,
зарослей гидроидов и губок или под
камнями, в расселинах скал и т. д. Многие
представители семейства Leucothoidae
живут в каналах губок и в мантийной
полости асцидий, пользуясь этими животными
только как местом поселения. Морские
козочки ползают по водорослям и ветвям
гидроидов и, держась за них тремя
парами задних грудных ножек, приподнимают
остальное тело так, чтобы иметь
возможность схватывать проплывающих мимо
животных хватательными передними нож-
Рис. 257. Трубки Corophium.
Две особи вадними антеннами собирают детрит для строительства
трубки.
489
ками. Их охотничья поза напоминает
соответствующее положение тела равноногих
рачков Astacilla (рис. 246, стр. 476).
Большинство бокоплавов можно
считать всеядными животными, это значит,
что они способны использовать
органическое вещество в разных формах.
Пресноводные и многие морские бокоплавы
едят растения, как живые, так и
отмершие, грунт, трупы и остатки животных,
а при случае и мелких живых животных.
Они откусывают жвалами куски пищи
и измельчают их, причем челюсти
задерживают мелкие частицы, не давая им
вывалиться из сферы действия ротовых
придатков. Некоторые виды могут наряду
с этим добывать пищу путем фильтрации.
Массовый бокоплав побережий
Каспийского и Азовского морей — Niphargoides
maeoticus — пассивно профильтровывает
взвесь, приносимую волнами. Когда
волна начинает отходить от берега, рачки
сидят в грунте, высунув из него передний
конец тела, при обнажении грунта они
зарываются в него целиком.
Фильтрация служит основным способом
добывания пищи Leptocheirus, Corophiidae
и Ampeliscidae. Эти животные, сидя в
своих домиках, возбуждают сильный ток
воды взмахами передних брюшных ножек,
пропуская воду через густую сеть
щетинок, расположенных на передних
грудных ножках. При этом Corophiidae
взмучивают поверхностный слой грунта удли-
Рис. 258. Hyperia galba, прикрепившаяся
к краю колокола медузы Суапеа.
ненными задними антеннами. Диатомовые
водоросли, бактерии и мелкие
растительные остатки усваиваются рачками. Другие
виды, как например многие представители
семейства Haustoriidae, соскребают с
частиц грунта водорослевые и бактериальные
обрастания. Chelura terebrans, подобно
лимнории и сфероме (стр. 475), точит
дерево и питается, вероятно, древесными
опилками.
Обитатели подземных вод заглатывают
грунт, который всегда обнаруживается в их
пищеварительном тракте. Однако
длительные наблюдения над Niphargus orcinus
virei показали, что содержащееся в
грунте органическое вещество не может
полностью обеспечить всех жизненных
отправлений рачка, в частности его роста
и размножения, а только поддерживает
его существование. Время от времени
в подземные водоемы заносятся остатки
растений и животных, и только такая
более питательная пища дает возможность
нифаргусам расти и размножаться. С этим
связано строение ротовых придатков ни-
фаргуса, сохраняющих характер жующих.
Донный бокоплав наших северных морей
Anonyx nugax питается преимущественно
в ночные часы. Интенсивность его
питания различна и в разные сезоны: она
повышается осенью и зимой и падает весной
и летом.
Хищников среди донных бокоплавов
сравнительно мало. К ним относятся
морские козочки. Молодые рачки едят
гидроидных полипов, а подросшие ловят
передними хватательными ножками
веслоногих рачков, мелких бокоплавов,
червей и т. д. Среди планктонных бокоплавов
хищничество преобладает. Судя по
постоянному присутствию в кишечниках
Hyperiidea стрекательных клеток
кишечнополостных, гиперииды нападают на медуз
и питаются ими. Под колоколом ушастой
медузы часто живет по нескольку
экземпляров Hyperia galba, которая, вероятно,
находит там и стол и дом. Гиперииды
прицепляются не только к медузам, но
и к гребневикам. Phronima выедает тело
сальп и пирозом, а затем забирается в
пустую мантию отдельных особей этих
животных и в ней плавает. Планктонные
гаммариды также, по-видимому, питаются
490
преимущественно кишечнополостными. В
их кишечниках, так же как у гипериид,
были обнаружены стрекательные клетки
медуз, а строение ротовых придатков и
грудных ножек с подклешнями многих
планктонных Lysianassidae позволяет
предполагать их полупаразитическое
существование на медузах и, возможно, на
гребневиках.
Настоящими паразитами среди бокопла-
вов следует считать только китовых вшей
(Cyamidae). Огромными массами они сидят
на китах, обычно близ полового и
анального отверстий, и грызут их кожу,
нередко вызывая глубокие язвы, доходящие
до жирового слоя. В отличие от других
паразитических ракообразных у китовых
вшей нет никаких свободноплавающих
стадий. Вероятно, они переходят с одной
особи хозяина на другую во время
спаривания китов, а китята заражаются при
рождении или при кормлении молоком.
Все бокоплавы раздельнополы.
Половой диморфизм часто бывает хорошо
выражен, но по-разному у различных
семейств и родов. У представителей
семейства Gammaridae самцы, как правило,
крупнее самок, но у представителей
семейства Lysianassidae наблюдаются обратные
соотношения размеров. У некоторых боко-
плавов Байкала, принадлежащих к
семейству Gammaridae, самцы настолько
мельче самок, что их называют
карликовыми. Они значительно раньше самок
достигают половозрелости, после чего их
рост прекращается. Так, например, длина
половозрелых самцов планктонного Мас-
rohectopus branickii не превосходит 5,5 мм,
в то время как длина
половозрелых самок колеблется между 14 и 30 мм.
У самцов многих Gammaridae и всех Talit-
ridae подклешни передних грудных ножек
сильнее развиты, чем у самок. Нередко
самцы обладают более длинными
антеннами с более многочисленными
чувствительными органами на них. У многих
видов подземного рода Niphargus самцы
резко отличаются от самок удлиненным
конечным члеником наружной ветви
задних уроподов, а иногда, кроме того,
удлиненными ветвями одной или двух пар
передних уроподов. Половозрелые самки
всегда имеют выводковую сумку.
Рис. 259. Спаривание и копуляция Gammarus
pulex.
Исследования последних лет показали,
что развитие мужских вторично-половых
признаков у бокоплавов определяется
гормоном специальных желез внутренней
секреции, так называемых андрогенных
желез, лежащих вдоль семепроводов, но
с ними не связанных. Этот гормон
выделяется в кровь. Пересадка андрогенных желез
молодым самкам Orchestia gammarella
приводила к возникновению у них
характерных для самцов хватательных ножек и
даже к перерождению их яичников в
семенники. В некоторых случаях
определение пола зависит от внешних условий,
в частности от температуры. У солонова-
товодного бокоплава Gammarus duebeni
при созревании яиц при температуре
ниже 5°С из них выходят самцы, а при
температуре выше 6°С—самки. Благодаря
этому все рожденные зимой рачки
оказываются самцами, а самки появляются
на свет только весной.
Спаривание продолжается обычно
несколько дней. Самец располагается на
спинной стороне самки, удерживаясь
своими подклешнями за передний край
491
ее первого и за задний край ее
пятого свободного грудного сегмента и
ожидая ее линьки. После линьки самки
самец передвигается под ее брюшную
сторону, складывает вместе свои передние
брюшные ножки, несколько раз
просовывает их между задними пластинками ее
выводковой сумки и в это же время
выделяет из своих половых отверстий сперму.
При помощи передних брюшных ножек
сперма переносится в выводковую сумку,
куда через 134—4 часа (у Gammarus)
откладываются яйца, которые здесь же
оплодотворяются.
Нормальная откладка яиц может
происходить только тогда, когда в
выводковой сумке есть сперма. В опытах с
Gammarus duebeni удавалось закупоривать
половые отверстия самцов. После
спаривания с такими самцами,
происходившего нормально, если не считать того, что
они не выделяли сперму, половина самок
вообще не отложила яиц, а остальные
отложили их не полностью, в малом
количестве.
Число откладываемых самками боко-
плавов яиц различно у разных видов
и, кроме того, в пределах каждого вида
определяется величиной самки. Обычно
оно колеблется от 4 до 100, изредка,
например у Gammarus oceanicus,
достигает 177. При этом плодовитость видов,
размножающихся несколько раз в
течение года, к концу лета и к осени
уменьшается. У некоторых бокоплавов в
разных частях области их распространения
плодовитость оказывается различной: на
севере она больше, чем на юге. Весьма
плодовиты некоторые каспийские виды
(Amathillina spinosa — до 251, Niphar-
goides robustoides — до 239, Gammara-
canthus loricatus caspius — до 336 яиц).
Большая плодовитость
зарегистрирована у антарктической Chevreuxiella
obensis. В выводковой сумке единственной
пойманной до сих пор самки этого вида
оказалось 344 эмбриона. Однако еще выше
плодовитость крупных D6 мм) самок
беломорского Anonyx nugax, вынашивающих
до 950 зародышей.
Находящиеся еще в яйцевых
оболочках зародыши бокоплавов изогнуты на
брюшную сторону, чем они
отличаются от зародышей других перакарид,
которые изогнуты, наоборот, на спинную
сторону. Другим важным отличием
бокоплавов от большинства близких к ним
отрядов следует считать присутствие у
вышедших из яиц молодых рачков всех
грудных конечностей. Таким образом, стадии
манка у бокоплавов нет.
Молодые рачки обычно через 20—30 дней
покидают выводковую сумку матери.
Длительность инкубационного периода
зависит от температуры. Так, например,
у берегов Англии молодь Gammarus obtu-
satus остается в сумке матери в течение
12—14 дней, а в Белом море не меньше
21 дня. У пещерного Niphargus orcinus
virei, живущего при постоянной
температуре около 11° С, инкубационный период
продолжается 2%—3 месяца.
Вышедшие из выводковой сумки
молодые рачки растут довольно быстро и
равномерно, периодически линяя. До
достижения половозрелости молодь Gammarus
и Niphargus должна слинять 13 раз, но
у разных видов и при различной
температуре на это требуется различное время.
В прибайкальских озерах G. lacustris
достигает половозрелости через 3 месяца
после выхода из выводковой сумки, в
озерах Западной Сибири и в озере Севан
тот же вид становится половозрелым на
следующий год после рождения, a
Niphargus orcinus virei — только через 2гЛ
года.
Период размножения бокоплавов обычно
весьма продолжителен и приходится на
наиболее теплое время года. Так,
например, в Южном Каспии у большинства
видов он начинается в феврале—марте и
заканчивается в сентябре—октябре, в
Белом море литоральные виды Gammarus
(кроме G. setosus) размножаются в июне—
августе. Обычный пресноводный G.
lacustris начинает размножаться в апреле—мае
и заканчивает размножение к концу лета
или осенью (в зависимости от
температуры).
Бокоплавы рода Anisogammarus с
литорали Курильских островов вынашивают
яйца и молодь в течение всей зимы, но
молодые рачки покидают выводковую
сумку только весной или летом, когда
температура достигнет определенной величины,
492
различной для разных видов. У двух
видов молодь выходит на волю при 2—4°С,
у четырех видов—при 4—8°С и у одного—
при 7—10°С. Если температура среды
обитания остается более или менее
постоянной, размножение бокоплавов
может продолжаться круглый год. В
ручьях и ключах Германии G. pulex
размножается с января по октябрь. Однако
для пещерного Niphargus orcinus virei,
несмотря на постоянство температуры его
обитания, отмечена периодичность в его
размножении, которую не удается
связать с факторами внешней среды. С
другой стороны, литоральные бокоплавы
северной Атлантики — G. zaddachi и,
вероятно, G. finmarchicus — испытывают
значительные колебания температуры, но
тем не менее размножаются круглый год.
В течение периода размножения каждая
самка дает от двух до 5—6 пометов.
Поскольку некоторые из молодых рачков в
тот же сезон успевают достигнуть полово-
зрелости и в свою очередь дать потомство,
численность бокоплавов может очень
быстро возрастать. Продолжительность их
жизни обычно от 1 до 2 лет, но Niphargus
orcinus virei живет в среднем 6 лет,
достигая иногда 30 лет.
Огромное большинство бокоплавов
населяет морские водоемы, в которых эти
рачки повсеместно широко
распространены и очень многочисленны. В пределах
приливо-отливной зоны, а во многих
случаях и на некотором отдалении от
покрываемой максимальным отливом полосы
обитают полусухопутные «морские блохи»—
бокоплавы из семейства Talitridae. Свое
название они получили благодаря тому,
что на суше они часто прыгают,
отталкиваясь брюшком и уроподами от
поверхности грунта. Днем морские блохи
закапываются в песок, прячутся под камнями
или под выбросами водорослей и т. д.,
а ночью они активно двигаются по
пляжам и другим побережьям, отыскивая
отмершие водоросли, которыми питаются.
Они дышат жабрами и могут существовать
только в достаточно влажной атмосфере.
В условиях эксперимента морские блохи
некоторое время выдерживают
пребывание под водой, но всегда стремятся
выбраться на сушу.
На Командорских островах они зимуют
высоко над уровнем моря, под толстым
слоем снега, впадая в анабиоз. На Шан-
тарских островах морские блохи с
наступлением заморозков переселяются с
побережья в леса и иногда забираются на
чердаки домов, а весной возвращаются
к морю.
Замечательна их способность
ориентироваться по солнцу. Итальянские
исследователи Папи и Парди проделали такой
опыт: они взяли круглый кристаллизатор и
разделили его радиальными планками на
16 секторов. Этот нехитрый прибор был
снабжен магнитной стрелкой. В центр круга
помещали по сотне бокоплавов. Через
некоторое время подавляющее
большинство рачков собиралось в том секторе,
который был обращен к морю. Оказалось,
что в каждый час суток рачки двигаются
под определенным углом к солнцу
(а ночью—к луне). В темноте они не
способны ориентироваться. При этом рачки,
живущие в различных участках берега,
приспособлены к ориентации по
отношению к солнцу под разными углами в
зависимости от направления берега. Эта
поразительная способность не зависит от таких
внешних условий, как температура.
Закономерные изменения угла между источником
света и направлением движения животного
в течение суток могут считаться одним
из лучших примеров существования так
называемых «биологических часов», т. е.
закономерных суточных изменений
особенностей организма, управляемых
внутренними факторами.
Обычными обитателями
приливо-отливной полосы наших северных и
дальневосточных морей можно считать несколько
видов Gammarus и Anisogammarus. Во
время отлива они прячутся среди
водорослей или под камнями, а в прилив
оживленно передвигаются в поисках пищи.
Некоторые из них хорошо выдерживают
значительное или даже полное опреснение. На
литорали наших северных морей нередко
насчитывается несколько тысяч особей
этих рачков на 1 м2.
Наиболее богата и разнообразна фауна
бокоплавов материкового склона. В
Баренцевом море здесь обитает около 260
видов, в Японском — 250 видов. Некото-
493
рые из видов бокоплавов материкового
склона встречаются в огромном количестве.
В Чукотском море на 1 ле2 дна приходится
до 24 тысяч экземпляров Pontoporeia и до
14 тысяч экземпляров Lembos. Трал в этом
море приносил такую массу бокоплавов,
что, высыпанные на палубу, они
образовывали копошащиеся кучи высотой до
полуметра.
С глубиной видовое разнообразие и
количество бокоплавов уменьшаются,
однако даже с огромных океанических глубин,
более 6000 м, в настоящее время известно
около 300 видов. Большая часть их
относится к широко распространенным родам,
встречающимся и на меньших глубинах,
но среди них есть и очень своеобразные
представители. Так, например, в Курило-
Камчатской впадине живет пелагический
бокоплав Vitjaziana gurjanovae,
выделенный в особое семейство и не
поднимающийся на глубины менее 6000 м.
Пресные воды населяет сравнительно
небольшое количество видов бокоплавов.
В северном полушарии чрезвычайно
широко распространен озерный бокоплав Gam-
marus lacustris, обитающий в самых
различных озерах нередко в громадном
количестве. Он может существовать как в
пресных, так и в сильно минерализованных
водоемах и переносить различные
неблагоприятные условия, в том числе зимнее
уменьшение содержания кислорода в воде.
При наступлении зимнего замора массы
рачков скапливаются под нижней
поверхностью льда. В Сибири бокоплавов
добывают, пробивая во льду проруби и разными
способами облавливая нижнюю его
поверхность. В текучих водах обитают другие
виды того же рода — G. pulex, G. balcani-
cus и т. д.
Необыкновенно богата и своеобразна
фауна бокоплавов озера Байкал,
состоящая из 240 видов. Они живут на дне или
у дна, от уреза воды до предельных
глубин, т. е. до 1620 м, и лишь один вид —
Macrohectopus branickii — ведет
планктонный образ жизни. Разные виды
приурочены к различным глубинам и грунтам.
Многие из них снабжены килями, шипами
или буграми, придающими им весьма
причудливый облик. Считается, что все эти
виды произошли в Байкале от немногих
исходных предков в сравнительно
короткое по геологическим масштабам время.
Лишь 52 вида проникают из Байкала
в вытекающую из него Ангару, и около
20 из них распространилось дальше по
Енисею до Енисейской губы. После
создания на Ангаре Иркутского водохранилища
численность байкальских бокоплавов в
новом водоеме уменьшилась, а некоторые
виды исчезли совсем.
В реках, впадающих в Каспийское,
Черное и Азовское моря, живут бокоплавы
морского происхождения, обитающие
также в самом Каспийском море и в
опресненных частях Азовско-Черноморского
бассейна. Некоторые из них поднимаются
высоко вверх по течению, например по
Волге до Ярославля, где встречаются
Dikerogammarus haemobaphes, Corophium
curvispinum и Niphargoides sarsi. Еще выше
они заходят по Оке и Каме, удаляясь на
3200 км от моря. Иногда в реках они
развиваются в очень большом количестве.
В нижнем течении Оки насчитывается
до 168 тысяч экземпляров Corophium на
1 м2 дна.
Один из бокоплавов каспийского
происхождения — Gammarus ischnus —
проник из Понто-Каспийского бассейна в
Вислу, принадлежащую к бассейну
Балтийского моря, a Corophium curvispinum
расселился еще шире благодаря способности
прикреплять свои домики к днищам судов.
Замечательно, что бокоплавы
каспийского происхождения, вселяясь в реки,
вытесняют древне-пресноводные виды,
почти никогда не встречаясь совместно
с ними. Такие же антагонистические
отношения отмечены и для некоторых других
видов и родов разноногих. В Англии
Gammarus pulex вытесняет G. duebeni,
в Молдавии G. balcanicus и G. kischinef-
fensis также исключают один другого.В
Крыму, в Румынии и в Германии виды
рода Gammarus никогда не встречаются
совместно с подземными бокоплавами из
рода Niphargus, даже если проникают в
подземные воды.
Пока еще непонятно, каким образом
осуществляется это вытеснение. В
аквариуме некоторые из видов-антагонистов
мирно живут совместно. Только в одном
случае удалось выяснить механизм вытес-
494
нения одного вида, а именно G. duebeni
другим — G. salinus. Оказалось, что
самцы G. salinus охотно спариваются с
самками G. duebeni, в то время как самцы
G. duebeni спариваются только с самками
своего вида. После спаривания с самцом
другого вида самки G. duebeni
откладывают неоплодотворенные, неспособные к
развитию яйца. Благодаря этому в местах
соприкосновения обоих видов
численность G. duebeni все время уменьшается.
Бокоплавы обычны не только в
водоемах поверхности земли, но и в подземных
водах. В пещерах, колодцах и ключах
Западной Европы, Кавказа и Западной
Украины живет очень богатый видами род
Niphargus. В подземных реках и ручьях
Закавказья встречается распространенный
только там особый poflZenkevitchia.
Представители родов Crangonyx и Synurella
найдены в отдельных выходах грунтовых
вод на огромном протяжении всего
северного полушария. Среди остальных
многочисленных подземных бокоплавов особый
интерес с точки зрения их
распространения имеют представители подотряда Ingol-
fiellidea. В настоящее время известно
всего 11 видов этого подотряда. Из них 7
обитают в подземных пресных водах
Южной Европы, экваториальной Африки и
Южной Америки, 3—в капиллярных ходах
морского песка в Ламанше, Сиамском
заливе и у берегов Перу, а один — на
глубине 3521 м в Дэвисовом проливе. Такое
разбросанное распространение этих
примитивных рачков, способных
существовать в столь разнообразных условиях,
остается пока не разрешенной загадкой.
Практическое значение
бокоплавов, как уже указывалось, очень
велико и определяется использованием их
в пищу многими рыбами, в том числе и
промысловыми. Так, например, в Каспии и
Азовском море они составляют
значительную часть пищи леща, молоди осетровых,
на Дальнем Востоке — многих камбал,
в устьях северных рек — муксуна,
омуля, ряпушки, в пресных озерах —
различных сигов, форелей и т. д. Для
улучшения условий откорма ценных рыб
бокоплавов перевезли во многие вновь
созданные водохранилища и в озерал
где их раньше не было.
ИАДОТРЯД ЭВКАРИДЫ
(EUCARIDA)
У представителей надотряда эвкарид
всегда имеется карапакс, он покрывает
все головные и грудные сегменты и
срастается со спинной стороной последних*
Глаза стебельчатые, жвалы симметричные,
без подвижной пластинки. Самки носят
яйца прикрепленными к брюшным
ножкам или реже откладывают их прямо в
воду. Из яиц обычно выходят личинки,
непохожие на взрослых, у немногих видов
развитие прямое. Надотряд объединяет
два отряда — эвфаузиевых и десятиногих
ракообразных.
ОТРЯД ЭВФАУЗИЕВЫЕ
РАКООБРАЗНЫЕ (EUPHAUSIACEA)
Каждый год наши китобойные флотилии
промышляют китов в Антарктике. Туда же
направляются китобои из многих других
стран. Хорошо известно, что наибольшее
количество усатых китов сосредоточено
в антарктических морях. Киты находят
там огромные запасы своей основной пи-
щи — скопления «криля», т. е.
эвфаузиевых рачков Euphausia superba. Эти
скопления настолько велики, что в настоящее
время делаются попытки непосредственно
использовать рачков для изготовления
из них пищевых продуктов для людей,
кормовой муки, жира, витаминов и т. д.
Эвфаузиевые рачки живут не только
в Антарктике. Они широко
распространены во всем Мировом океане, избегая
лишь его опресненных районов. Повсюду,
в частности в наших северных и
дальневосточных морях, они служат важным
объектом питания рыб и китообразных.
Всего известно 85 видов эвфаузиид.
Все эвфаузиевые — планктонные
животные. Тело их прозрачное, у некоторых
глубоководных видов красное. Его длина
колеблется от 7 до 96 мм. По внешнему
виду эвфаузииды напоминают небольших
креветок, однако отличаются от них
многими признаками. Боковые края карапак-
са эвфаузиид короткие и оставляют
непокрытыми кустистые жабры,
прикрепленные к основаниям грудных ног. Ни одна
495
из 8 пар грудных ног не превращена в но-
гочелюсти. Задние грудные ножки
укорочены. На заднем крае тельсона, по
бокам от его заостренного конца,
располагается пара пластинок, отсутствующая у
десятиногих ракообразных. Чрезвычайно
характерны для эвфаузиид органы
свечения — фотофоры. Обычно их бывает 10:
одна пара на глазных стебельках, по
одному на основных члениках второй
и седьмой пар грудных ножек и по
одному на нижней поверхности
каждого из 4 передних брюшных
сегментов. Они испускают желтый или
желто-зеленый свет, причем светятся как
при раздражении, так и сами по себе,
однако не беспрерывно, то вспыхивая, то
затухая. Каждая вспышка продолжается
3—6 секунд, после чего фотофоры
медленно гаснут. Иногда вспыхивают только
наиболее яркие фотофоры глазных
стебельков, иногда все вместе. Когда
фотофоры не действуют, рачки в темноте
становятся невидимыми благодаря
прозрачности своего тела. Фотофоры помогают
рачкам собираться в стаи, а самцам
находить самок. Только у одного вида —
глубоководной Bentheuphausia amblyops—
фотофоры отсутствуют, но у нее, в
отличие от других эвфаузиид, редуцированы
и глаза. У остальных эвфаузиид,
наоборот, глаза хорошо развиты. Они
стебельчатые и очень крупные: нередко их длина
равняется 1/6—V8 длины тела рачка.
У многих хищных видов каждый глаз
подразделен на две части: одна, более
крупная, направлена вперед, а другая —
вверх. Такое строение глаз, по всей
вероятности, увеличивает кругозор рачка и
помогает ему охотиться на движущихся
животных. Особенно эффективными
оказываются двураздельные глаза при
слабом освещении. Способность видеть при
слабом освещении достигается еще
высокой концентрацией в теле эвфаузиид
витамина А, который на 90—95%
сосредоточен в их глазах, а также астаксантина.
Эти вещества участвуют в образовании
светочувствительных соединений, в
частности определяющих возможность
сумеречного зрения. По содержанию в теле
витамина А и астаксантина эвфаузииды
превосходят остальных ракообразных.
Рис. 260. Эвфаузиевые:
1 —Meganyctiphanes norvegica; 2 — Bentheuphausia
amblyops; 3 — Thysanoessa raschii; 4 — Nematoscelis
atlantica.
У видов с двойными глазами есть еще
одно приспособление, свидетельствующее
об их хищных повадках: грудные ноги
второй или третьей пары, а иногда и те
и другие у них очень сильно удлинены
и нередко на одной из этих пар развиты
клешни. Это так называемые ловчие
или разбойничьи ноги, которыми рачки
хватают свои жертвы (рис. 260, 4).
Эвфаузииды — прекрасные пловцы. На
пяти передних брюшных сегментах у них
расположено по паре сильных двуветви-
стых плавательных ножек — п л е о п о-
д о в. Эти ножки уплощены по направ-
лению спереди назад, а их ветви снабжены
по краям перистыми щетинками. На
основаниях ножек имеются направленные
внутрь крючковидные шипы, при помощи
которых левая и правая ножки каждой
пары могут сцепляться и действовать как
единое целое. Своими плавательными
ножками эвфаузииды гребут. При этом Mega-
nyctiphanes norvegica передвигается со
средней скоростью 271 м в час. Но, кроме
того, резко сгибая и распрямляя брюшко
и отталкиваясь тельсоном и
пластинчатыми уроподами от воды, они совершают
молниеносные прыжки. Судя по
наблюдениям в аквариуме, разные виды в толще
воды ведут себя различно.
Растительноядная Euphausia беспрерывно двигается
во всех направлениях, а хищные Stylochei-
ron и Nematoscelis в течение длительного
времени неподвижно парят в воде, держа
тело в вертикальном положении и
расправив свои длинные ловчие ноги в
ожидании добычи.
Подвижность эвфаузиид позволяет
многим из них совершать регулярные
вертикальные миграции. Мелководные виды
ночью поднимаются к поверхности и там
питаются, а днем опускаются в более
глубокие слои. Однако, по-видимому, не все
особи каждые сутки проделывают такое
путешествие; значительная часть остается
на обычной глубине их обитания, так как
отдельные особи поднимаются в
поверхностные слои через сутки или даже через
двое суток, а самые крупные мигрируют
только для размножения. В Баренцевом
море суточные миграции наблюдаются
только весной и осенью.
Кроме того, в некоторых морях
происходят сезонные миграции эвфаузиид. Так,
например, в Баренцевом море рачки зимой
опускаются и образуют на мелководьях и
банках мощные придонные скопления.
Весной половозрелые рачки для размножения
поднимаются в поверхностные слои воды.
Это происходит начиная с середины апреля
и совпадает с весенним развитием
водорослей, которые служат пищей личинкам
эвфаузиид. Подъем начинается под
воздействием усиливающегося освещения.
Поднимающиеся для размножения
эвфаузииды образуют стаи, хорошо видимые
с борта корабля. Э. А. Зеликман
следующим образом описывает «роение»
Thysanoessa raschii в Баренцевом море:
«Рачки концентрировались на площади,
приблизительно равной 9 кв. милям.
Эвфаузииды плавали на этом участке как
рассеянно, поодиночке, так и стайками.
Стайки были двух типов — шаровидные,
от 0,5 до 2 м диаметром, и лентовидные,
длиною около 5 м и шириной около 1 м.
Основная масса рачков придерживалась
верхнего двухметрового слоя. С борта
судна стайки ярко-розовых рачков в воде
выглядели красновато-рыжими пятнами. Кое-
где под стайками эвфаузиид шла мойва,
которая питалась рачками и преследовала
их; рачки уходили от рыбы в
поверхностный слой, и многие выпрыгивали из воды.
Эвфаузиид пожирали и многочисленные
чайки, главным образом моевки. За два
часа наблюдений с корабля каждая из
стаек рачков сохранялась как целое».
Массовый подъем антарктической
Euphausia superba также связан с
размножением. Огромные скопления половозрелых
рачков приурочены к зоне так называемой
антарктической дивергенции, где
происходит поднятие к поверхности глубинных
вод, богатых солями фосфора и азота. Эти
соли дают возможность развиться
огромному количеству водорослей,
используемых в пищу рачками. Самки Е. superba
выметывают яйца на прибрежных
мелководьях. Затем яйца погружаются на
глубины 1800—3000 ле, где из них выходят
личинки. По мере развития личинки
постепенно поднимаются в поверхностные
слои. Все эти вертикальные перемещения
приводят к тому, что разные личиночные
стадии переносятся то поверхностными,
то глубинными течениями в различных
направлениях, однако в результате все
рачки остаются в пределах обычной и
наиболее благоприятной для них области
распространения, окружающей довольно
узким кольцом берега Антарктиды.
Поднимающиеся к поверхности воды
самки всех эвфаузиид уже оплодотворены.
Процесс оплодотворения у них происходит
еще во время пребывания у дна.
Непосредственно наблюдать его не удавалось, но
несомненно, что при этом важную роль
играет сложно устроенный придаток на
передних брюшных ножках самца, так
32 Жизнь животных, т. 2
497
Рис. 261. Личинки Thysanoessa raschii:
I — науплиус; 2 — метанауплиус; 3 — первый калиптопис; 4 — второй
калиптопис.
называемая петазма. У самок на
брюшной поверхности шестого грудного
сегмента имеется карманообразный
орган — телику м, в который
открываются половые отверстия. При помощи пе-
тазмы самец прикрепляет два
образующихся в его половых путях сперматофора
к теликуму самки. Иногда наблюдались
самки не с двумя, а с четырьмя и даже
с шестью сперматофорами,
оплодотворенные, очевидно, дважды и трижды.
Одни эвфаузииды (Euphausia,
Thysanoessa, Meganyctiphanes и др.)
выметывают оплодотворенные яйца прямо в воду,
а другие носят их на себе в виде яйцевых
мешков, прикрепленных к задним
грудным ногам. У Nyctiphanes образуется два
удлиненных грушевидных мешка с 60
яйцами в каждом, у Nematoscelis и Sty-
locheiron — один мешок, содержащий
у первого рода 36—60, а у второго — всего
18 яиц.
Плодовитость эвфаузиид,
откладывающих яйца прямо в воду, значительно
больше. Каждая самка антарктической
Euphausia superba длиною 33—45 мм
выметывает в зависимости от размера от 200
до 3600, а по некоторым данным до 11 000
яиц, Thysanoessa inermis, Т. raschii, T.
longicaudata и Euphausia pacifica в наших
северных и дальневосточных морях — от
1000 до 2000 яиц.
Из яиц эвфаузиид выходит
типичный науплиус. Он линяет,
оставаясь на той же стадии, а
затем, после второй линьки,
переходит в стадию метанауп-
лиуса. Через 2—3 дня (у
Thysanoessa Баренцева моря)
метанауплиус превращается в
присущую только эвфаузиидам
стадию — калиптопис, у
которой брюшко удлинено и
четко отграничено от грудного
отдела, а глаза находятся под ка-
рапаксом. Калиптопис имеет
сквозной кишечник и начинает
питаться водорослями.
Калиптопис трижды линяет; после
третьей линьки возникает
последняя стадия — ф у р ц и -
лия, характеризующаяся
постепенным развитием всех
грудных и брюшных ножек и освобождением
из-под карапакса стебельчатых глаз. Фур-
цилия линяет несколько раз и постепенно
переходит во вполне сформированного
рачка. Число ее линек у разных видов, по-
видимому, различно. Фурцилии старших
возрастов, задние антенны которых уже не
служат для плавания, иногда выделяются
в особую стадию — циртопию. У
некоторых эвфаузиид, в особенности у
носящих яйца на себе и населяющих
прибрежные воды, число личиночных стадий
уменьшается и метаморфоз оказывается
сокращенным.
Яйца, науплиусы и метанауплиусы
мелководных эвфаузиид наших северных и
дальневосточных морей находятся в
поверхностных слоях воды. Калиптопис
и фурцилии опускаются несколько
глубже, а взрослые особи начинают совершать
вертикальные миграции.
Через год после рождения эвфаузииды
наших морей, а также некоторые другие,
в частности Euphausia superba, достигают
половозрелости и приступают к
размножению.
На следующий год эвфаузииды
размножаются вторично, после чего погибают,
в значительной своей части становясь
добычей рыб. Небольшая часть рачков
достигает трехлетнего возраста и
размножается еще раз. Крупные глубоководные
498
виды, несомненно, долговечнее, но их
жизненный цикл еще не изучен.
Пища эвфаузиид разнообразна.
Некоторые виды питаются главным образом
планктонными водорослями,
отфильтровывая их из воды. Е. superba поедает
преимущественно диатомовые водоросли,
диаметр которых не превосходит 0,04 мм.
Грудные ножки рачка с внутренней
стороны снабжены длинными, густыми
перистыми щетинками, образующими
фильтрующий аппарат. Если в эту ловчую сеть
попадают более крупные частицы, рачок
удаляет их щетинками последних
члеников грудных ног. Большинство других
эвфаузиид питается сходным образом, но
наряду с водорослями потребляет и
планктонных животных — веслоногих и ра-
кушковых ракообразных, глобигерин,
тинтинноидей и т. д. Молодые Meganycti-
phanes norvegica длиной до 19 мм едят
детрит и диатомовые водоросли,
рачки длиной 21—29 мм —
преимущественно детрит, а крупные особи, длиной
31—39 мм,— планктонных ракообразных.
Глубоководные виды рода Thysanopoda —
хищники, нападающие исключительно на
животных, в том числе и на довольно
крупных (щетинкочелюстные, медузы).
Среди мелководных видов хищниками
могут считаться виды Nematoscelis, Tessa-
robrachion, Thysanoessa longipes и др.
Своими ловчими ногами они хватают
жертву. Эти ноги обычно вооружены
крупными шипами, которые прокалывают
пойманное животное. Глубоководная Веп-
theuphausia amblyops, помимо животных,
в большом количестве потребляет
водоросли, для чего время от времени
поднимается в поверхностные слои воды.
Эвфаузииды служат основной пищей
многим морским млекопитающим и рыбам.
Усатые киты питаются главным образом
этими рачками. Как уже указывалось,
скопления китов в Антарктике связаны
с зонами массового развития Euphausia
superba. Этот вид распространен на
глубинах до 40—50 м вокруг берегов
Антарктиды; зимой и весной он держится среди
льдов, летом и осенью — в открытой воде,
образуя стаи самых разных размеров —
площадью от нескольких до 2000 м2.
Каждая стая состоит из одноразмерных
рачков. Е. superba составляет основную,
а часто и единственную пищу финвала,
синего кита, горбача и сейвала. В течение
года они выедают 38 миллионов тонн
рачков. Однако количество Е. superba остается
значительным. На одну квадратную
морскую милю приходится около 14
миллионов рачков, что достаточно для
наполнения желудков более чем 50 китов.
В северной части Тихого океана
желудки китов также бывают обычно набиты
эвфаузиидами. Вес содержимого желудка
финвала, наполненного одними
эвфаузиидами, достигает 360 кг. Для Северной
Атлантики доказана зависимость
присутствия китов от распространения
скоплений эвфаузиид. Столь же охотно поедают
этих рачков некоторые ластоногие,
например акиба, миграции которой
связаны с районами концентрации эвфаузиид,
а также многие птицы — пингвины, чайки
и др.
В огромном количестве поедают
эвфаузиид в Баренцевом море сельдь (до 60%
по весу от всей пищи), морской окунь
E0%), треска, молодь многих
других рыб; в дальневосточных морях —
скумбрия, сардина, сельдь, лососевые г
минтай, молодь трески и т. д. Места
обитания многих из этих рыб в период их
откорма приурочены к районам
концентрации эвфаузиид. Интенсивное выедание
рыбами рачков может уменьшить их
численность в море. После резкого
сокращения в 1957 и 1958 гг. запасов одного из
основных потребителей эвфаузиид —
трески — численность этих рачков в
южной части Баренцева моря увеличилась
в 4 раза.
ОТРЯД ДЕСЯТИНОГИЕ
РАКООБРАЗНЫЕ (DECAPODA)
Из всех ракообразных десятиногие
издавна пользуются наиболее широкой
известностью. Популярный представитель
этого отряда — речной рак —
неизменный герой бесчисленных сказок и басен.
Он фигурирует в числе знаков Зодиака,
а с крабами связаны многие мифы инков,
ацтеков и древних египтян. К десятино-
32*
499
гим принадлежат крупнейшие из
ракообразных: длина омаров иногда превосходит
80 см, расстояние между когтями
вытянутых в стороны средних ног японского
краба Macrocheira составляет 3 м.
Благодаря своим крупным размерам
и отличным вкусовым качествам
некоторые десятиногие ракообразные служат
важным объектом промысла. В 1962 году
во всем мире было выловлено около
1 миллиона тонн ракообразных —
креветок, крабов, омаров, лангустов и т. д.,
т. е. приблизительно в 2 раза больше, чем
всех лососевых. В СССР промысел
камчатского краба производится огромными
судами — плавучими консервными заводами,
на которых работает несколько тысяч
человек, и существует специальная крабо-
консервная промышленность.
Десятиногие ракообразные
распространены чрезвычайно широко. Они населяют
все океаны и моря, от уреза воды до
глубины около 5 км. Особенно богата видами
десятиногих тропическая мелководная
фауна. В пресных водах обитают
общеизвестные речные раки, а в тропических
Рис. 262. Разные десятиногие ракообразные:
1 — креветка Pandalus borealis; 2 — краб Eriphia verrucosa; 3 — рак-отшельник Pagurus
bernhardus; 4 — норвежский омар Nephrops norvegicus.
и субтропических странах — крабы и
креветки. Наконец, в тропиках обычны
сухопутные десятиногие — крабы и сильно
видоизмененные раки-отшельники.
По образу жизни и по внешнему виду
десятиногие весьма разнообразны. К
этому отряду принадлежат плавающие
креветки с длинным мускулистым брюшком,
ползающие по дну крабы, брюшко
которых подвернуто под грудь,
раки-отшельники, прячущие свое спирально извитое
брюшко в пустые раковины брюхоногих
моллюсков, также ползающие по дну
речные раки, омары и лангусты,
обладающие подобно креветкам длинным
симметричным брюшком, и некоторые другие.
Общее число видов десятиногих достигает
8500; их приблизительно столько же,
сколько видов птиц.
Что же заставляет объединять столь
различных животных в один отряд? Всем
им свойственны некоторые общие
признаки, свидетельствующие об их тесном
родстве и едином происхождении.
У всех десятиногих ракообразных три
передних грудных сегмента срастаются
с головой, а их конечности превращены
в ногочелюсти. Пять пар задних грудных
ног, принадлежащих к свободным
грудным сегментам, служат для передвижения,
почему весь отряд и получил название
десятиногих. Часто на некоторых из этих
ног есть клешни или иногда подклешни.
Жабры целиком покрыты боковыми
краями карапакса и снаружи не видны.
Между боковыми краями карапакса и стенкой
тела расположено с каждой стороны по
жаберной полости. Самки вынашивают
яйца, прикрепляя их к брюшным
конечностям, и только самые примитивные
креветки просто откладывают яйца в воду.
Из яиц выходят планктонные личинки,
совсем непохожие на взрослых, у многих
пресноводных и глубоководных видов
развитие прямое.
Голова и грудь десятиногих покрыты
карапаксом, плоским и коротким у крабов
и цилиндрическим, удлиненным у
остальных представителей отряда. Передний
конец карапакса часто вытянут в
шиловидный, острый на конце рострум.
Поверхность карапакса иногда снабжена килями,
буграми или шипами, имеющими защитное
Рис. 263. Схема расположения A — 5) и:
направления воздействий F — 8) органов
внутренней секреции краба:
J — орган X; 2 — мозг; з — синусная
железа; 4 — орган Y; 5 — подглоточный
ганглий; 6 — линька; 7 — содержание сахара в
крови; 8 — хроматофоры.
значение. По бокам от основания ростру-
ма прикрепляются стебельчатые глаза.
Они могут поворачиваться в разные
стороны и обеспечивают широкий кругозор.
Каждый глаз состоит обычно из
большого (у креветки Palaemon serratus —
3020) количества фасеток, число которых
увеличивается с возрастом. Фасетки
отделены друг от друга пигментными
клетками, и каждая фасетка воспринимает лишь
лучи, падающие перпендикулярно к ее
роговице. Она видит только небольшую
часть объекта, на который смотрит рак,
а другие фасетки видят иные части этого
объекта. Таким образом осуществляется
«мозаичное зрение». Ночью пигмент
расходится к концу и к основанию глаза,
и косые лучи могут достигать сетчатки —
рак видит объект целиком, но довольно
туманно. В воде десятиногие видят только
501
близко расположенные предметы. Рак
Galathea на расстоянии 1 гА м не замечает
даже крупные движущиеся объекты, но
на расстоянии 40 см хорошо различает
белые квадраты площадью 2 см2. Поэтому
водные десятиногие в поисках пищи,
самок, убежища пользуются не столько
зрением, сколько обонянием, осязанием
и химическим чувством. Зато в жизни
сухопутных видов, обладающих особенно
длинными глазными стебельками, зрение
играет очень важную роль.
В глазном стебельке находится
несколько органов внутренней секреции. Их
выделяющиеся в кровь гормоны регулируют
расположение пигмента в пигментных
клетках, процесс линьки, обмен веществ,
содержание в крови сахара и кальция
и взаимодействуют с другими органами
внутренней секреции(рис. 263).
Передние и задние антенны у крабов
очень короткие, а у остальных
десятиногих длинные, нередко, особенно у
глубоководных креветок, превосходящие по
длине тело. У многих «длиннохвостых»,
т. е. обладающих длинным брюшком,
десятиногих на каждой из передних антенн
находится по два или даже по три жгута.
Эти жгуты богаты чувствительными
щетинками и цилиндрами, выполняющими
функции органов обоняния, осязания
и химического чувства. В основном
членике каждой из передних антенн
расположен орган равновесия — статоцист.
У большинства десятиногих полость
статоциста соединяется с наружной средой
узкой щелью. На стенках полости
находятся чувствительные щетинки двух
типов — крючковидные и нитевидные. На
крючковидные щетинки опираются один
или несколько статолитов. Большинство
десятиногих использует в качестве
статолитов песчинки, которые после линьки
рак запихивает в полость статоциста, у
некоторых статоцист образуется за счет
затвердевания выделяемого специальными
железами вещества; у многих крабов
статолитов нет. Ощущая давление статолита
на те или иные чувствительные щетинки,
животное ориентируется по отношению
к силе земного притяжения.
Для выяснения механизма действия ста-
тоцистов был проделан такой
остроумный опыт. Креветок Palaemon содержали
в аквариуме с грунтом из железных
опилок. После линьки рачки вынуждены были
вложить в свои статоцисты вместо
песчинок частицы металла. При приближении
сверху к аквариуму магнита креветки
дружно поворачивались брюшной
стороной вверх, поскольку этого требовали
их притягиваемые магнитом железные ста-
толиты. Поднося магнит к стенке
аквариума, можно было легко заставить
животных повернуться боком и т. д. Действия
нитевидных щетинок не связаны со ста-
толитами. Они регулируют повороты тела
десятиногих по отношению к
вертикальной оси.
Жвалы десятиногих всегда мощные
и служат для перетирания и разрывания
пищи. Передние челюсти маленькие, а на
задних имеется крупная наружная
лопасть — скафогнатид, благодаря
движениям которой осуществляется смена
воды в жаберной полости. Она совершает
от 120 до 200 взмахов в минуту, причем
в жаберной полости в течение 10 минут
сменяется от 2 до 25 объемов воды, что
зависит от температуры воды, содержания
в ней кислорода и видовой
принадлежности рака.
Из 8 пар грудных конечностей 3
передние превращены в ногочелюсти. Они
принимают участие в захватывании пищевых
частиц и в передаче их к ротовому
отверстию. Остальные 5 пар грудных ног
служат главным образом для ползания и,
кроме того, передние из них — для
хватания пищи. На таких хватательных ногах
развиваются клешни. У крабов, раков-
отшельников и других близких к
последним форм клешни есть только на передней
паре ходильных ног, у большинства
креветок — на двух передних парах, а у
речных раков, омаров и креветок из семейств
Penaeidae (табл. 35, 7) и Sergestidae — на
трех передних парах, причем клешни
первой пары значительно крупнее
остальных. У креветок из семейства Crangonidae
на передних ходильных ногах настоящие
клешни заменены подклешнями (табл. 35,
4). У лангустов, раков-медведей и
некоторых других клешней нет совсем.
Нередко левая и правая клешни первой
пары имеют разное строение: одна из них
502
крупнее или они различны по форме.
Так, например, у омаров одна (то левая,
то правая) более массивная клешня
приспособлена для раздавливания панцирей
морских ежей, раковин моллюсков,
домиков морских желудей и носит название
дробящей, а другая, более тонкая, режущая
клешня помогает делить на куски тело
рыб, креветок и т. д. Пресноводные
родственники омаров — речные раки
унаследовали от своих предков асимметрию
клешней. Особенно резко выражена
асимметрия клешней у самцов тропических
полусухопутных так называемых
манящих крабов, обладающих маленькой левой
и колоссальной правой клешней, сгибая
которую они производят своеобразные
манящие движения (рис. 284). У раков-
щелкунов (сем. Alpheidae) одна из двух
передних клешней также значительно
крупнее другой и устроена таким
образом, что подвижный палец может с силой
прижиматься к неподвижному, причем
раздается резкий звук (рис. 278).
Задние грудные ноги в некоторых
случаях также имеют какие-нибудь
специальные приспособления. У
крабов-плавунцов (сем. Portunidae) последние их
членики уплощены, ими крабы
отталкиваются от воды, когда плавают (табл.
36, 1). У раков-отшельников задняя пара
грудных ног укорочена и служит для
удерживания раковин брюхоногих, в которых
они живут. Родственные им Lithodidae
в процессе эволюции утеряли способность
жить в раковине, но сохранили
укороченные задние грудные ноги. Эти конечности
у Lithodidae подвернуты под край кара-
пакса и используются для очистки жабр.
С грудными конечностями связаны
жабры. Они прикрепляются или к основному
членику ногочелюстей и ходильных ног,
или к сочленению между этим члеником
и телом, или, наконец, к стенке тела над
местом прикрепления той или иной
грудной конечности. Часто на каждом грудном
сегменте расположены жабры всех трех
или двух типов. Как уже указывалось,
жабры прикрыты боковым краем карапак-
са, ограничивающим снаружи жаберную
полость.
У плавающих креветок вода свободно
проникает в жаберные полости и,
направляемая движениями скафогнатида задней
челюсти, омывает жабры. У ползающих
десятиногих нижние края карапакса
плотнее прижимаются к стенке тела,
основные членики ног развиты сильнее и
благодаря этому жаберные полости почти
полностью изолированы от внешней среды,
что препятствует загрязнению жабр
грунтом. Для проникновения воды в жаберные
полости имеются специальные отверстия,
расположенные у крабов по одному перед
основанием клешней, а у
раков-отшельников, речных раков, омаров и других
между основаниями ходильных ног. Из
жаберной полости вода выходит из-под
нижнего края карапакса в его передней
части.
Зарывающийся в грунт краб Corystes
складывает свои задние антенны так, что
они образуют трубку, которую краб
выставляет над поверхностью грунта. По
этой трубке вода достигает жаберных
полостей краба. Близкая к
ракам-отшельникам Albunea имеет сходное
приспособление, но ее дыхательная трубка
образуется не задними, а передними антеннами.
Краб Calappa , зарываясь, разгребает
клешнями грунт так, что перед входами
в жаберные полости остается свободное
пространство, заполненное водой.
Многие закапывающиеся крабы имеют на всем
крае головогрудного щита от глаз до
основания клешней ряд щетинок,
действующих как фильтр, мешающий песчинкам
попасть в жаберную полость при дыхании.
У многих сухопутных десятиногих
дыхательная система сильно изменяется. На
внутренней поверхности стенок
жаберных полостей у сухопутных потомков
раков-отшельников — пальмового вора
(Birgus latro) и Coenobita — развиваются
гроздевидные складки кожи, в которых
разветвляются многочисленные
кровеносные сосуды. Это — настоящие легкие,
позволяющие использовать кислород
заполняющего жаберные полости воздуха.
Легкие вентилируются благодаря
движениям скафогнатида, а также вследствие
способности животных время от времени
приподнимать и опускать карапакс, для
чего служат специальные мышцы.
Замечательно, что при этом сохраняются и
жабры, правда, относительно малых размеров
503
(рис. 264, Б). Удаление жабр нисколько не
вредило дыханию BirgusnCoenobita; с
другой стороны, оба рака полностью
потеряли способность дышать в воде.
Погруженный в воду пальмовый вор погибал через
4 часа. Остаточные жабры, по-видимому,
не функционируют.
За грудным отделом тела следует
брюшной, строение которого различно у разных
десятиногих и тесно связано с их образом
жизни. У плавающих десятиногих —
креветок — брюшко сильно развито, длиннее
головы и груди, часто сжато с боков
и снабжено относительно крупными
плавательными ножками— плеоподами.
По паре плеоподов прикреплено к
каждому из пяти передних брюшных сегментов,
а конечности последнего брюшного
сегмента — уроподы — широкие,
пластинчатые и образуют вместе с тельсоном
хвостовой веер. Ползающие по дну
длиннохвостые ракообразные (омары, речные
раки, лангусты и др.) также имеют длинное
брюшко, но оно более или менее сжато
в спинно-брюшном направлении, а пле-
оподы относительно короткие и слабые
и для плавания не служат. У
большинства раков-отшельников покровы брюшка
мягкие, кожистые и оно спирально
завернуто, так как помещается внутри пустой
раковины брюхоногих моллюсков и
следует за ее оборотами. Асимметрия брюшка
раков-отшельников проявляется и в
строении его конечностей: плеоподы
развиваются только на стороне брюшка,
прилегающей к наружной стенке раковины, а уро-
под с этой стороны гораздо крупнее, чем
с противоположной. У некоторых
родственных ракам-отшельникам десятиногих,
например у представителей семейства Li-
thodidae, к которому, между прочим,
принадлежит камчатский краб, а также у
сухопутных пальмового вора и Coenobita
брюшко укорочено и подвернуто под
грудь, но и у них в расположении
хитиновых щитков, покрывающих брюшко, и
плеоподов заметны признаки асимметрии,
свидетельствующие об их происхождении
от форм со спирально закрученным
брюшком. Наконец, у настоящих крабов
брюшко сильно уменьшено в размерах и всегда
подвернуто под грудь, число
составляющих его сегментов, особенно у самцов,
сокращено, плеоподы недоразвиты, а
уроподы отсутствуют.
Первичная функция плеоподов
плавательная, как это свойственно креветкам.
Однако даже у креветок плеоподы, кроме
того, участвуют в процессе размножения.
У самцов креветок первая или вторая
пара плеоподов частично преобразована
в совокупительный орган, а самки, за
исключением представителей семейств Ре-
naeidae и Sergestidae, прикрепляют к пле-
оподам яйца. У ползающих десятиногих
половая функция плеоподов стала
основной, самки используют их только для
вынашивания яиц, самцам передние плео-
Рис. 264. Органы дыхания десятиногих:
А — схематичный поперечный разрез через правую половину грудного отдела краба, видна
жаберная полость и жабра в ней; Б — поперечный разрез через грудной отдел пальмового вора на
уровне четвертой ходильной ноги; J — дыхательные выросты стенок жаберной полости (легкие); 2 —
остаточные жабры; 3 — задние грудные ноги, служащие для очистки жаберной полости.
504
поды служат копулятивным органом.
У самцов крабов сохраняются только
первая или две первые пары плеоподов,
превращенные в копулятивный аппарат.
Десятиногие — весьма подвижные
животные. По способу передвижения их
можно разделить на плавающих и
ползающих, причем к первым принадлежат
почти все креветки, а ко вторым
остальные. Резкой границы между этими
группами нет. Большинство креветок может не
только плавать, но и ползать по дну или
растениям, а среди таких типичных
ползающих десятиногих, как крабы,
известно обширное семейство крабов-плавунцов
(Portunidae), представители которого
приспособились к плаванию.
Креветки плавают при помощи плео-
подов, отталкиваясь ими от воды, причем
прижимают антенны и грудные ноги к
телу и выпрямляют брюшко. Планктонные
виды держатся в воде вертикально,
работая не только плеоподами, но и
наружными ветвями грудных ног. При этом они
вытягивают в стороны свои длинные
антенны и внутренние ветви грудных ног
и таким образом препятствуют
погружению. При приближении врага креветки
резкими скачками уплывают задом
наперед, порывисто сгибая брюшко и
отталкиваясь тельсоном и уроподами от воды.
Ползание цо дну осуществляется при
помощи четырех пар задних грудных ног.
Десятиногие сгибают и разгибают их
последовательно, причем никогда левая и
правая ноги одной пары не действуют
одновременно. Омары, лангусты, раки-
отшельники ползают передним концом
вперед, но скорость их передвижения
невелика. Гораздо быстрее бегают
десятиногие с укороченным грудным отделом и
подогнутым под него брюшком, т. е. крабы
и крабовидные родичи
раков-отшельников— камчатский краб (табл. 37),
пальмовый вор (табл. 40), Coenobitan др. Все они
передвигаются боком, достигая иногда
очень большой скорости. Обычный
«травяной краб» Carcinus maenas бегает
со скоростью 1 м в секунду, а сухопутный
краб-привидение Ocypode мчится на своих
вытянутых ногах так быстро, что
умудряется ловить даже мелких птиц. Крабы-
плавунцы также плавают боком. При этом
Рис. 265. Краб-привидение Ocypode rotundata.
вторая — четвертая пары грудных ног
делают 630—780 взмахов в минуту, а
последняя пара работает еще более
интенсивно.
Вошедшее в басни и поговорки мнение
о том, что «рак пятится назад» не совсем
справедливо. В нормальной для него
обстановке, на дне водоема, он этого не
делает. Когда же пойманного рака
выпускают обратно в воду, он начинает, так же
как креветки, сгибать брюшко и
действительно, плывя толчками задом наперед,
опускается на дно.
Окраска десятиногих разнообразна.
Большинство донных ползающих видов
бурого или зеленоватого цвета,
напоминающего цвет грунта, на котором они
живут. Обитающие среди водорослей или
ползающие по ним креветки и крабы
зеленого цвета. Прикрепляющийся к саргас-
совым водорослям и к другим плавающим
по поверхности моря предметам краб
Planes minutus окрашен по-разному:
особи, сидящие на водорослях, бурые или
зеленоватые, а сидящие на голубых сифоно-
форах — синеватые. Очень пестра
окраска многочисленных крабов — обитателей
коралловых рифов, но и она делает
рачков незаметными на фоне многокрасочного
кораллового известняка. Тропический
сухопутный краб-привидение Ocypode имеет
цвет песка, по которому он быстро бегает
на своих длинных, почти выпрямленных
ногах. При этом на ярком солнце его
тело отбрасывает на светлый песок тем-
505
ную тень. При приближении врага краб
подгибает ноги и прижимает плоский ка-
рапакс к песку, делаясь совершенно
незаметным.
Наряду с этими и многими другими
примерами покровительственной окраски
десятиногих можно указать и случаи
существования у них предупреждающей
окраски. Карапакс африканского сухопутного
краба Sesarma meinerti пурпуровый
и окаймлен желтым кантом, а клешни
ярко-красные. При приближении врага
Sesarma не прижимается к песку, как
краб-привидение, а, наоборот, приподнимается на
своих ходильных ногах и потрясает в
воздухе клешнями. У самцов манящих крабов
цвет их крупной клешни обычно резко
контрастирует с цветом карапакса, а также
грунта, что делает манящие движения
клешней еще более заметными. Так,
например, карапакс Uca batuenta золотисто-
коричневый, а клешня совершенно белая,
у U. beebi карапакс зеленый, большая
часть клешни желтая или розовая, а ее
пальцы белые.
Все эти типы окраски определяются
присутствием в покровах пигментов,
большая часть которых представлена кароти-
ноидами. Особенно обычен из них астак-
сантин, в чистом виде имеющий красный
двет. Соединяясь с белками, он образует
пигменты синеватых и буроватых тонов.
При повышении температуры эти
соединения разрушаются, астаксантин
освобождается. Поэтому раки при варке краснеют.
Помимо десятиногих, цвет которых
остается неизменным, существуют виды,
меняющие свою окраску в зависимости
от цвета грунта, на котором они
находятся, или в зависимости от времени суток.
Креветки Hippolyte, Palaemon и Palae-
monetes на светлом грунте приблизительно
через 1 ?% часа светлеют, а на темном
темнеют (табл.35, 10,11). Наряду с этим
ночью они светлеют, становясь прозрачными,
а днем приобретают ту или иную окраску.
Креветка Crangon, наоборот, ночью
окрашена темнее, чем днем. Манящие крабы
темнеют во время отлива, когда они
активны, и светлеют во время прилива,
когда они сидят в своих норах.
В толще покровов таких десятиногих
расположены многочисленные снабженные
Рис. 266. Хроматофор в стадии
наибольшего распространения зерен
темного (внутри) и светлого
(снаружи) пигмента по всем отросткам.
длинными разветвленными отростками
органы, называемые хроматофорами
и содержащие огромное количество зерен
пигмента. У некоторых креветок в
каждом хроматофоре находится несколько
пигментов, причем каждый из них
занимает свои ответвления хроматофора. Хро-
матофоры Crangon имеют ответвления с
черным, белым, желтым и красным пигмента-
ми,хроматофоры Penaeus устроены так же, но
черный пигмент в них заменен синим.
У крабов и других ползающих
десятиногих каждый хроматофор содержит один
или самое большее два пигмента. Когда
все пигментные зерна концентрируются
в середине хроматофора, животное
светлеет, когда же пигментные зерна
распространяются по всем отросткам, оно
темнеет. Если в отростках находятся
пигментные зерна одного цвета, а остальные
собраны в центре хроматофора, креветка
оказывается окрашенной в цвет
распыленного пигмента.
К хроматофорам не подходят нервы.
Как показано многочисленными опытами,
их деятельностью управляют органы
внутренней секреции, находящиеся главным
образом в глазном стебельке, а также в
головном ганглии и в отходящих от него
окологлоточных нервных тяжах. Световое
раздражение, воспринимаемое глазами,
передается этим органам, а они выделяют
в кровь различные гормоны. Одни гор-
506
моны заставляют зерна того или иного
пигмента распространяться по отросткам
хроматофора, другие, наоборот, вызывают
их концентрацию. Отрезание глаз или
тугая перевязка глазных стебельков
полностью нарушает движение зерен пигмента
в хроматофорах, так как преграждает
путь гормонам в кровь.
Замечателен автоматизм движений
пигмента в хроматофорах некоторых
ракообразных. У манящих крабов эти
движения, как уже указывалось, точно
согласованы с приливо-отливным ритмом. У
крабов Uca pugnax из двух соседних
участков, заливающихся приливом из-за
разной конфигурации берега в разное время,
периодичность изменения окраски
различна. Эти крабы вместе помещались в
темную комнату, где каждый из них
сохранял присущий ему ритм передвижений
пигмента в хроматофорах. Круглосуточное
освещение или, наоборот, помещение
крабов в условия постоянной темноты, а
также содержание их при различной
температуре (от 6 до 26° С) не нарушает этого
четкого ритма. Учитывая разницу в часах
прилива и отлива в разных частях земного
шара, крабов Uca pugilator на самолете
перевезли с Атлантического побережья
Америки на Тихоокеанское. Невольные
путешественники сохранили в новом
местообитании выработавшийся у них на родине
ритм распыления и концентрации
пигментных зерен в хроматофорах. Все это
позволяет считать миграции пигмента в
хроматофорах манящих крабов одним из
лучших примеров существования
«биологических часов», идущих точно и правильно
независимо от условий внешней среды.
Все же некоторые внешние факторы
влияют на хроматофоры манящих крабов.
Замечено, например, что при усилении
космической радиации степень
распыленности темного пигмента в хроматофорах
увеличивается; крабы таким образом
защищаются от вредного действия
коротковолновых излучений. Повышение
атмосферного давления уменьшает
интенсивность космической радиации, и можно
подумать, что крабы реагируют на
колебания атмосферного давления, а не
непосредственно на излучение. Для проверки этой
гипотезы крабов помещали в камеру с
постоянным атмосферным давлением,
причем оказалось, что расположение пигмента
в их хроматофорах меняется в
соответствии с интенсивностью космической
радиации независимо от атмосферного
давления.
Способы и объекты питания
десятиногих довольно разнообразны. Большинство
этих ракообразных можно считать
хищниками. Морские ползающие виды находят
различных животных — полихет,
моллюсков, иглокожих, других ракообразных,
разрывают или раздробляют добычу
клешнями, перетирают ее жвалами и поедают.
Многие способны расправляться с
крупными и сильными жертвами. Краб Loxorhyn-
chus grandis (с карапаксом длиной 26 см
и шириной 21 см) разрывал клешнями
осьминогов и больших морских звезд.
Некоторые десятиногие наряду с животными
поедают и богатый органическим
веществом грунт, а также растения.
Раки-отшельники охотно едят разных беспозвоночных,
но наряду с ними и грунт, который
они захватывают своей меньшей
(обычно левой) клешней. Креветки Pandalus,
Palaemon, Crangon и др. также едят
главным образом мелких животных, но
заглатывают и грунт и водоросли. Наши речные
раки питаются преимущественно
водными растениями — урутью,
роголистником, рдестом, харовыми водорослями, но
при случае охотно поедают моллюсков,
личинок насекомых, а также трупы любых
животных. Чрезвычайно широк
ассортимент объектов питания некоторых
сухопутных десятиногих. Пальмовый вор ест
падающие на землю плоды различных
пальм, особенно охотно маслянистые
плоды панданового дерева, грунт, а также
нападает на других сухопутных
ракообразных, в том числе на больных особей
своего вида. Мнение о питании его
кокосовыми орехами неверно. Несмотря на
огромную силу своих клешней, он не может
разгрызть скорлупу зрелых кокосовых
орехов и поедает только разбитые.
Помещенный в садок с кокосовыми орехами,
пальмовый вор через несколько недель
погибал от голода. Неверно также мнение,
что пальмовый вор забирается на
кокосовые пальмы, чтобы сбросить орехи вниз.
Он не может влезать высоко на деревья и
507
Рис. 267. Путь
камчатского краба к пище в
цементном бассейне.
Схематичный рисунок
краба — его исходное
положение, в конце пути —
моллюск литторина,
пунктирная линия — путь краба,
цифры на ней — время
прохождения пути.
совсем не способен слезать с них. Вообще
никакой связи между ним и кокосовыми
пальмами нет, поскольку он обитает на
многих островах, на которых отсутствуют
кокосовые пальмы.
Некоторые десятиногие используют в
пищу грунт. Тропические и
субтропические пресноводные креветки из семейства
Atyidae на концах пальцев клешней имеют
кисточки щетинок, при помощи которых
захватывают ил и передают его в рот. Так
же питаются и пещерные представители
этого семейства, в том числе живущая в
пещерах Закавказья, Балканского
полуострова и южной Франции Troglocaris
anophthalmias. Полусухопутные манящие
крабы также питаются грунтом, поглощая
его в огромном количестве. Вокруг
отверстий их нор всегда располагаются комочки
их экскрементов, состоящие из
пропущенного через кишечник грунта. Морские
Galathea метут дно щетинками задних
ногочелюстей, причем частицы грунта,
разлагающихся водорослей и т. д.
застревают между щетинками, а затем счищаются
вторыми ногочелюстями и отправляются
в рот. Более крупные пищевые частицы
раки хватают клешнями.
Фильтраторы среди десятиногих
немногочисленны. Живущие в норах Callianas-
sidae энергично машут плеоподами,
вызывая таким образом сильный ток воды,
направленный в нору. Этот ток проходит
через сложенные в виде корзинки передние
грудные ноги, густо обсаженные
щетинками, которые задерживают взвешенные
в воде частицы. Время от времени раки
резко сгибают брюшко, и получающийся
при этом обратный тек воды промывает
фильтрующую камеру, освобождая ее от
попавших в нее крупных частиц. Крабо-
видный рак Porcellana машет задними
ногочелюстями, снабженными
многочисленными щетинками, отфильтровывая ими
мелкие взвешенные частицы.
При добывании пищи основное значение
у большинства водных десятиногих имеют
чувства обоняния и осязания. После
внесения в аквариум с крабами или раками-
отшельниками пищи животное реагирует
на нее характерными движениями
передних антенн, богатых чувствительными
цилиндрами, и, почуяв добычу, приступает
к ее поискам. Дальнейшее поведение
камчатского краба Д. Н. Логвинов и ч
описывает так: «В выборе направления (к
пище) краб руководствовался тщательным
ощупыванием бетонного дна бассейна
концами клешней. Краб опускал клешни
вертикально вниз и, касаясь грунта концами
пальцев клешней, быстро закрывал и
открывал их, постепенно обследуя площадку
перед собой. Эти щупающие движения
чрезвычайно характерны, энергичны,
«нервны». Краб искал как бы вслепую,
описывая чрезвычайно запутанную
маршрутную линию. По мере приближения к
корму краб приходил в большое возбуждение,
что выражалось в усилении щупающих
движений клешней по грунту. Однако
даже в непосредственной близости от корма
(например, на расстоянии 1 см от конца
клешней до корма) краб неоднократно
промахивался и удалялся от него. Это
указывает на то,что осязание концов
клешней не выправлялось обонянием или
зрением. Наступает момент, когда положение
кормового объекта точно определяется
крабом, что выражается в стремительном
схватывании добычи, которое
производится правой или левой клешней или
обеими сразу». В опытах Д. Н. Логвинович
из-за неспособности направиться прямо
к предлагаемой пище краб терял на путь
к ней несообразно много времени. В одном
случае краб достиг корма, положенного
от него на расстоянии 75 см, через 23
минуты, причем исколесил 930 см, в другом
случае для достижения корма, лежавшего
в 260 см, потребовалось всего 17 минут,
а в третьем случае, когда корм находился
всего в 5 еж от краба, последний искал
его в течение 65 минут.
Важная роль органов обоняния и
осязания при поисках пищи подтверждается
и опытами над креветкой Palaemon. После
удаления глаз креветки отыскивали
пищу в течение 4—5 минут, но когда у
них были удалены и передние антенны,
время разыскивания увеличилось до 20
минут, однако даже лишенные обеих пар
антенн креветки в конце концов все-таки
находили пищу, чему помогали
чувствительные щетинки ротовых придатков и
пальцы ходильных ног.
Несколько иначе происходит охота за
движущимися объектами, при которой
основное значение приобретает зрение.
Хищные крабы неподвижно лежат на дне
и высматривают добычу. При виде
движущейся жертвы они бегут, а
крабы-плавунцы быстро плывут прямо к ней и обычно
ее настигают. Многие крабы, в особенности
крабы-плавунцы, таким образом успешно
ловят рыб.
Все сухопутные десятиногие при
поисках пищи руководствуются в первую
очередь зрением. Пальмовый вор видит пищу
с расстояния в несколько метров и прямо
направляется к ней. Он успешно
преследует других сухопутных десятиногих,
например краба Cardysoma.
Крабы-привидения (Ocypode) благодаря своим
необыкновенно длинным глазным
стебелькам и сложно устроенным глазам (рис. 265)
издали видят добычу и умудряются, как
уже указывалось, ловить даже птиц.
Будучи в большинстве хищниками,
десятиногие сами подвергаются нападению
многочисленцых врагов — рыб,
головоногих моллюсков и других десятиногих, а
сухопутные виды — птиц и грызунов. Для
защиты от врагов десятиногие выработали
разнообразные и совершенные
приспособления. К таким приспособлениям
принадлежат разобранные ранее примеры
покровительственной окраски, свойственной
некоторым креветкам и крабам, и
угрожающие позы многих крабов. Той же цели
служат и другие особенности строения и
поведения десятиногих.
Многие ползающие десятиногие, как
например омары, лангусты (табл. 38),
крабы, в том числе обычные в Черном море
Pachygrapsus и Carcinus (табл. 36, 2, 4),
укрываются между камнями и раковинами
моллюсков, в расщелинах скал и т. д.
Существуют крабы и креветки, которые,
стремясь к безопасности, закапываются
в грунт. Креветки Crangon, Nectocrangon
и, вероятно, другие представители
семейства Crangonidae, а также донные
креветки из семейства Penaeidae днем зарываются
Рис. 268. Рак-крот Callianassa и его норы:
1 — Callianassa tyrrhena; 2 — норы Callianassa.
509
Рис. 269. Манящий краб Uca,
закрывающий вход в нору A) и сидящий в ней B).
в грунт и сидят в нем горизонтально,
выставив наружу глаза и антенны, а ночью
выходят на охоту. Так же поступают
крабы-плавунцы, Galatheidae, некоторые
раки-отшельники. Многие крабы, зарывшись
в грунт, располагают свое тело
вертикально.
Различные десятиногие строят более
или менее совершенные норы, в которых
проводят значительную часть жизни.
Представителей семейства Callianassidae
называют «раками-кротами» за то, что они
вырывают глубокие ходы, часто имеющие
форму латинской буквы U, а иногда и
более сложную. Рачок роет грунт
клешнями, выбрасывая его из норы задними
грудными ногами. Стенки хода
обмазываются илом или выделениями кожных
желез рачка (рис. 263).
Менее совершенны норы речных раков,
которые они вырывают клешнями в
откосах берегов рек и ручьев. В этих норах
раки проводят дневное время, а кроме
того, зимуют. Некоторые виды
американских речных раков из рода Cambarus
строят целые туннели длиной до 75 см.
Расселившийся в недавнее время по берегам
морей и рекам Западной Европы
китайский мохнаторукий краб Eriocheir sinensis
делает норы в откосах берегов низовьев
рек, куда еще доходит прилив. Эти норы
наклонные и построены так, что в их низко
расположенной концевой части остается
морская вода после отлива. Е. sinensis
своими норами повреждает плотины,
оросительные каналы и другие
гидротехнические сооружения.
Почти все десятиногие,
приспособившиеся к жизни на суше, сооружают норы,
имеющие в их жизни большое значение.
Обычно норы сухопутных крабов
настолько глубоки, что достигают уровня
грунтовых вод и их обитатели могут пить
находящуюся в их домах воду. В других случаях
норы расположены так, что они
заливаются во время прилива. Часто норы имеют
два выхода. Некоторые манящие крабы,
влезая в нору, закрывают вход в нее
крышкой из грунта. В каждой норе обитает
один краб. Рытье нор осуществляется
ходильными ножками одной стороны тела
и меньшей клешненосной ногой, причем
краб располагается в строящейся норе
боком (рис. 269, 2).
Манящие крабы образуют на илистых
прибрежных отмелях тропиков густые
поселения. На 1 м2 нередко насчитывается
50 и более их нор. При приближении
опасности все население такого поселка
дружно и молниеносно скрывается в своих
убежищах. Крабы замечают летящую птицу
на расстоянии 7 ле, а человека — на
расстоянии 9 м. По новым данным, они
оповещают своих соседей об опасности
ударами клешней о грунт. Благодаря
колебаниям грунта сигнал принимается даже в
том случае, когда крабы не видят друг
друга.
Пальмовый вор выкапывает в почве
неглубокие норы, которые выстилает
волокнами кокосовых орехов. Ч. Дарвин
рассказывает, что туземцы на некоторых
островах выбирают из нор пальмового
вора эти волокна, нужные им в их
немудреном хозяйстве. Иногда пальмовый вор
довольствуется естественными убежищами—
расщелинами в скалах, полостями в
осушенных коралловых рифах, но и в таких
случаях использует для их обкладки
растительный материал, сохраняющий в
жилье повышенную влажность.
Некоторые крабы сами нор не роют, а
используют в качестве жилища норы или
трубки других животных, например
трубки сидячих полихет. В трубках червя
Chaetopterus поселяются разные
представители семейств крабов Pinnoteridae
(табл. 36,14) и крабовидных Porcellanidae.
510
Последние живут также в ходах,
выточенных в известняке моллюском-камнеточ-
цем Pholas.
Раки-отшельники используют пустые
раковины брюхоногих моллюсков, пряча
в них свое длинное, лишенное твердых
покровов брюшко. Они подбирают себе
раковины по размерам и плотно в них
удерживаются уроподами и укороченными
задними грудными ногами. На врагов
рак-отшельник реагирует, забираясь
целиком в раковину и закрывая ее устье
своими клешнями. Раки-отшельники
одного вида могут заселять раковины
разных видов моллюсков. Так, например,
Pagurus prideauxi использует раковины
не менее 25 видов брюхоногих. Однако
раки-отшельники, по-видимому, отдают
предпочтение раковинам определенных
видов. В аквариуме Pagurus acadianus
выбирал в первую очередь раковины Вис-
cinum, во вторую — Thais и в третью —
Littorina, что соответствует частоте
использования раковин этих видов раком-
отшельником в естественных условиях.
Предпочтение, оказываемое одним
раковинам перед другими, связано с
различным отношением внутреннего объема
раковины к ее весу. При выборе раковины
раки-отшельники руководствуются не
зрением, а осязанием. Насильно
извлеченные из своего убежища, они стремятся
возвратиться в старую раковину, несмотря
на возможность занять предлагаемую им
другую того же вида и размера. Однако
после линьки старая раковина
оказывается тесной для ее владельца и он
подыскивает новую, более вместительную.
Искусственно лишенный раковины
рак-отшельник чувствует себя крайне неспокойно,
напоминая, по выражению Тэйлора,
купальщика, у которого украли одежду.
Некоторые виды раков-отшельников
поселяются не в спирально извитых
раковинах брюхоногих моллюсков, а в почти
прямых конусовидных раковинах лопато-
ногих моллюсков или даже в кусках
бамбука. У таких видов брюшко вполне
симметрично. Близкий к ракам-отшельникам
сухопутный рак Coenobita прячет свое
мягкое брюшко в пустые раковины
наземных моллюсков. Обитающий на
Бермудских островах С. diogenes использует в
качестве жилья только раковины Livona
pica. Этот вид здесь вымер, и рак
разыскивает его полуископаемые остатки.
Других подходящих моллюсков на
Бермудских островах нет, и чрезвычайно
интересно, какая судьба ждет С. diogenes в
дальнейшем, когда запас пустых раковин
Livona истощится.
Рис. 270. Рак-отшельник Pagurus pubescens,
меняющий раковину.
511
Рис. 271. Краб Pinnixa chaetopterana, живущий
в трубках червя Chaetopterus.
Рис. 272. Маскировка крабов:
1 — Dorippe granulata, прикрытый раковиной Масота
calcarea и без нее; 2 — Oregonia gracilis, обросший
мшанками, гидроидами, губками, водорослями (вид
спереди) и очищенный от обрастаний (вид сверху).
От поселения в чужих ходах и пустых
раковинах один шаг к жизни на
поверхности тела и в полости тела живых
животных, которые могут быть также
использованы как убежища. Такой шаг многие
десятиногие ракообразные сделали.
Существуют креветки, проводящие всю
жизнь внутри губок. В двух губках Spe-
ciospongia vespara общим объемом 50000сде3
было обнаружено 18 000 экземпляров
двух видов раков-щелкунов Synalpheus.
Креветки из рода Spongicola сидят
попарно (самец и самка) в полостях стеклянных
губок, куда они попадают еще будучи
личинками и откуда затем, увеличившись
в размерах, выбраться уже не могут.
Другие креветки и некоторые крабы обитают
под колоколом медуз, есть десятиногие,
живущие среди щупалец актиний и
других коралловых полипов. В этих случаях
рачки оказываются защищенными от
врагов стрекательными клетками щупалец
кишечнополостных. Маленькие (самки до
5—6 деде, самцы обычно до 1,2 деде) краби-
ки из семейства Hapalocarcinidae
образуют на стволах рифообразующих
кораллов известковые галлы, в полости
которых обитают парами рачки. Молодой
крабик выедает тело полипа из его
известковой чашечки и занимает его место.
Растущая колония постепенно замуровывает
крабика, но остается отверстие, через
которое в полость галла попадает вода, а
вместе с ней кислород для дыхания и
органическая взвесь для питания. Среди игл
морских ежей укрываются другие мелкие
крабики, в некоторых случаях
объедающие мускулы, приводящие в движение иглы
и педицеллярии ежа. Ряд креветок
из подсемейства Pontoniinae, остальные
представители которого также тесно
связаны с другими животными (губками,
оболочниками, кишечнополостными),
приспособлен к жизни в чашечках морских
лилий, под цвет которых креветки
окрашены. Среди крабов такими же обычными
сожителями других животных можно
считать так называемых «крабов-горошинок»
(сем. Pinnoteridae), характеризующихся
малыми размерами и относительно очень
широким карапаксом. Различные виды
этого семейства обитают внутри самых
разных животных —- в задней кишке мор-
512
ских ежей и голотурий, в мантийной
полости двустворчатых моллюсков, в асци-
диях, а иногда в трубках полихет и в
норах различных червей и раков (рис. 271).
Десятиногие ракообразные используют
других животных не только в качестве
убежища, но и для маскировки
собственного тела, а также в качестве оружия,
направляемого против своих врагов.
Мелководные крабы из семейства Dorippidae
при помощи загнутых на спинную
сторону грудных ножек последней пары
таскают над собой створку или обломок
раковины какого-нибудь двустворчатого
моллюска, полностью закрывая свое тело,
за что п получили меткое название
«стыдливых крабов». Крабы из семейства Majidae
срезают своими клешнями различных
обрастающих камни животных — губок,
гидроидов, мшанок, колониальных асцидий —
и помещают их на спинную сторону кара-
пакса, где все эти животные продолжают
расти. Этому способствуют
специальные крючковидные щетинки
поверхности карапакса, к которым хорошо
прикрепляется живой наряд краба, а также
секрет желез задних ногочелюстей,
склеивающий в кучки оторванных клешнями
животных.
Крабы из семейства Dromiidae
покрывают свое тело губкой, выкраивая из нее
кусок, совершенно подходящий для них
по размеру. В аквариуме удалось
проследить во всех подробностях, как краб это
делает. Он выбирает губку, сидящую на
круглом камне или на раковине брюхо-
ногого моллюска, т. е. выпуклую сверху
и вогнутую снизу. Если губка сидит в
раковине, занятой раком-отшельником, краб
вступает с ним в борьбу и обычно
побеждает. Затем краб залезает на губку сверху,
как бы отмеривая, какой кусок ему нужен,
и начинает клешнями резать губку,
сначала с одной, потом с другой стороны.
После этого он переворачивает губку
вогнутой стороной вверх, переворачивается
сам на спину и начинает надевать на себя
готовое платье, передавая кусок губки
задним грудным ногам, специально
приспособленным для ее удерживания.
Наконец, закончив одевание, краб становится
на голову, опрокидывается через нее и
принимает нормальное положение — спи-
Рис. 273. Краб Drdmia caput-mortuum^
надевающий на себя губку.
Рис. 274. Краб Lybia tesselata с актиниями
в каждой клешне.
ной вверх. Такой маскарад имеет
глубокий смысл: осьминоги не трогают крабов,
покрытых губками, но нападают на
крабов, с которых губки удалены. Некоторые
представители семейства Dromiidae
используют в качестве защитного покрова
не губок, а сложных асцидий, поступая
с ними сходным образом, другие, подобно
Dorippe, прикрываются створками
раковин двустворчатых моллюсков. Многие
раки-отшельники и некоторые крабы
пользуются в качестве средства защиты
от врагов жгучими щупальцами актиний.
33 Жизнь животных, т. 2
513
Обычные обитатели коралловых рифов —
крабики Lybia и Polydectus всегда
держат своими специально для этого
приспособленными клешнями мелких актиний
Bunedeopsis, которых выставляют вперед
лри нападении врага. Их клешни служат
только для удерживания актиний, а пищу
они захватывают задними ногочелюстями.
На поверхности левой клешни
рака-отшельника Diogenes edwardsi обычно
прикреплена актиния; когда он закрывает
клешней отверстие своей раковины,
актиния остается снаружи и как бы
охраняет вход в жилище. Другой
рак-отшельник — Paguropsis typica — держит над
собой защищающую его актинию Mammi-
lifera.
Сожительство раков-отшельников с
актиниями — обычное явление. Для раков-
отшельников актинии служат средством
защиты от врагов, актинии питаются
остатками пищи своих хозяев. Поскольку
как раки-отшельники, так и актинии
получают от совместной жизни обоюдную
пользу, это сожительство считается одним из
наиболее ярких примеров симбиоза.
Некоторые виды раков-отшельников и
актиний могут жить как вместе, так и
порознь. В таких случаях раки-отшельни-
ки при смене раковины оставляют свою
сожительницу на старой брошенной
раковине. Другие виды раков-отшельников
(Pagurus arrosor) иногда существуют без
актиний, но если рак находит актинию,
то немедленно сажает ее на свою раковину,
а при смене жилища пересаживает на
новую. Наконец, рак-отшельник Pagurus
prideauxi и актиния Adamsia palliata
всегда живут только совместно. Лишь
молодые актинии сидят на камнях, а взрослые —
только на раковинах раков-отшельников,
иногда по нескольку экземпляров на одной
раковине. После поселения на
раке-отшельнике актинии ее подошва
разрастается, покрывает почти всю раковину и
часто даже продолжается за устье. Если
на раковине находятся две или несколько
актиний, они располагаются так, чтобы
уравновесить друг друга и не свалить на
бок раковину и ее обитателя. Меняя
раковину, рак-отшельник осторожно и
заботливо клешнями переносит на новую
раковину свою сожительницу.
Искусственно разделенные рак-отшельник и актиния
порознь потребляют меньше кислорода,
чем когда они живут совместно.
Рак-отшельник постоянно поедает пищу,
обстрелянную стрекательными клетками
Рис. 275. Рак-отшельник Pagurus bernhardus
с актиниями Sagartia parasitica на раковине.
Рис. 276. Рак-отшельник Pagurus arrosor,
сажающий на свою раковину актинию.
514
актинии. В связи с этим он приобретает
иммунитет по отношению к ее яду.
Впрыскивание этого яда переносится
раком-отшельником безболезненно, в то время как
такая же доза вызывает гибель краба Саг-
cinus.
Третьим сожителем рака-отшельника и
актинии часто бывают многощетинковые
черви. Nereilepas fucata обитает в
раковине, занятой Pagurus prideauxi, a Nereis
cyclurus — в раковинах, занятых P. ocho-
tensis и P. armatus. Эти черви иногда
встречаются и в других условиях — в норах
различных животных и в пустых
раковинах. Замечательно, что рак-отшельник не
трогает «своего» червя, хотя охотно поедает
других. Больше того, при переселении
в новую раковину он переносит с собой
и червя. Черви принимают участие в
трапезах рака-отшельника, высовываясь в
это время из раковины и подбирая куски
разрываемой хозяином пищи. Они
приносят пользу своему сожителю, очищая
полость его раковины и объедая паразитов
с его мягкого брюшка.
Раки-отшельники сожительствуют не
только с актиниями, но и с
представителями другой близкой группы
кишечнополостных — зоантариями, играющими в
этом симбиозе такую же роль, как
актинии. Некоторые виды актиний и зоанта-
рий живут только на определенных видах
раков-отшельнпков, другие, как например
Sagartia parasitica, сожительствуют с
многими видами.
Помимо кишечнополостных, раковины
раков-отшельников часто обрастают
губки. При этом очень часто губка почти
полностью растворяет раковину, и рак-
отшельник оказывается окруженным
губкой.
Защитное значение имеет, по-видимому,
и способность некоторых десятиногих
издавать звуки. В этом отношении особенно
интересны раки-щелкуны (сем. Alpheidae),
широко распространенные на тропических
и субтропических мелководьях и
встречающиеся также у нас в Черном и Японском
морях. Они обычно прячутся между
стволами кораллов, среди колоний мшанок,
водорослей и камней, а также в губках.
Одна из клешней этих рачков очень
крупная и имеет своеобразное строение. На
Рис. 277. Симбиоз рака-отшельника Pagurus
ochotensis и червя Nereis cyclurus.
внутренней поверхности неподвижного
пальца расположено углубление, а на
подвижном пальце — соответствующий ему
выступ. Перед щелчком подвижный палец
отгибается, а затем благодаря сокращению
сильного мускула прижимается к
неподвижному. Выступ подвижного пальца
входит в углубление неподвижного, и
через канал, возникающий при смыкании
пальцев, выбрызгивается струя
выдавливаемой из углубления воды. При этом
раздается громкий щелчок, усиливающийся
благодаря пустым пространствам внутри
клешни, которые служат резонаторами
(рис. 278). Поскольку альфеусы обычно
образуют густые скопления, одновременное
щелканье сотен рачков создает сильный
шум. По описанию Н. И. Тарасова, «при
извлечении губки, густо населенной альфеу-
сами, слышна долго не утихающая громкая
трескотня, напоминающая горение
хвороста», а в других случаях при щелканье
альфеусов «возникают шумы,
напоминающие звуки, которые раздаются при
заклепывании котла».
При щелчке возникают не только
звуковые, но и ультразвуковые колебания. По
всей вероятности, биологический смысл
щелканья заключается в том, что мелкие
ракообразные и рыбы получают сильный,
а иногда и смертельный ультразвуковой
удар. Таким образом, щелканье можно
считать защитной реакцией: недаром
альфеусы отвечают им на всякого рода
раздражения.
33*
515
Рис. 278. Щелканье альфеуса:
А — щелкающий рачок, видна возникающая
при щелканье струя воды; Б, В — щелкающая
клешня альфеуса перед щелчком (Б) и после
него (В); 1 — выступ подвижного пальца; 2 —
углубление на неподвижном пальце.
Лангусты также издают громкие звуки,
отпугивая ими своих врагов. На первом
членике задних антенн у лангустов есть
лопасть, которая скребет о килевидный
выступ передней части карапакса, причем
возникает трескучий звук.
Калифорнийский лангуст перед стрекотаньем
угрожающе машет задними антеннами,
усаженными острыми шипами. Рыбы отскакивают
от лангуста и, по наблюдениям М о у л т о-
на, иногда теряют при этом свою добычу,
которую подбирает лангуст. Известны
и некоторые другие стрекочущие и
скрипящие десятиногие, но биологическое
значение издаваемых ими звуков остается
пока неясным.
К защитным приспособлениям
десятиногих принадлежит также присущая
многим из них способность к автотомии —
произвольному отрыванию своих
конечностей. Когда враг хватает за ногу какого-
нибудь рака, тот оставляет свою
конечность врагу, а сам на оставшихся ногах
спасается бегством. Привязанный за ногу
краб отрывает ее при виде осьминога.
Таким образом можно лишить краба шести
ног, но, если краба ослепить, автотомии
не происходит. Отрывание конечностей
осуществляется всегда в строго
определенном месте, недалеко от их основания, при
помощи специальных мышц — автотоми-
заторов, которые загибают ногу вверх до
тех пор, пока она не обломится. В месте
отрыва внутри конечности находятся два
расположенных поперечно
соединительнотканных листка, помогающих быстро
приостановить кровотечение. Некоторые
десятиногие могут в случае необходимости
отрывать себе конечности своими клешнями
или задними ногочелюстями.
Способность к автотомии всегда
сопровождается способностью крегенерации.
Утраченная конечность постепенно
отрастает вновь, тем скорее, чем чаще
животное линяет, поскольку рост
регенерирующей конечности происходит только после
линьки. У крупных омаров полная
регенерация клешней происходит только через
2 года после их потери, у самок речных
раков — через 3—4 года, а у чаще
линяющих самцов — через 1%—2 года. Если
при утере конечности или глазного
стебелька были повреждены связанные с
ними нервные центры, то на их месте может
вырасти придаток ненормального
строения. Так, у многих креветок взамен
глазного стебелька появляется усик,
напоминающий переднюю антенну. Такое
явление называют гетероморфозом.
Почти все десятиногие ракообразные
раздельнополы, причем самцы внешне
хорошо отличаются от самок. У креветок,
как правило, самцы мельче самок, а у
ползающих десятиногих наоборот, и только
у крабов, живущих в других животных
(Hapalocarcinidae, Pinnoteridae), самцы,
так же как у креветок, уступают по своим
размерам самкам. Часто самцы имеют
более сильно развитые клешни,
помогающие удерживать самку при спаривании.
Иногда, как например у краба-плавунца
Neptunus, самец и самка различаются по
цвету, а у травяного краба Carcinus по
форме карапакса.
516
Особенно четко проявляется половой
диморфизм в строении брюшка и его
придатков. Как уже указывалось, передние
брюшные ножки самцов целиком или
частично преобразованы в органы, при
помощи которых самец переносит
сперматофоры, прикрепляя их к половым отверстиям
самки, а у крабов вводя их в ее половые
пути. Нередко брюшко самок бывает шире,
чем у самцов (самки носят под ним икру),
а у крабов оно, кроме того, часто состоит
из большего числа сегментов, так как у
самцов брюшные сегменты частично
сливаются между собой.
У некоторых крабов наблюдается
полиморфизм самцов. Так, например, среди
самцов Inachus различаются три
категории: мелкие крабики с толстыми клешнями,
несколько более крупные, но с плоскими,
как у самок, клешнями и самые крупные,
опять с толстыми клешнями, причем
семенники хорошо развиты только у крабов
первой и третьей категории.
Гермафродитизм свойствен лишь
немногим креветкам. Таковы все представители
семейства Pandalidae, в том числе и
промысловые виды Pandalus borealis и P. lati-
rostris. Это типичные протерандрические
гермафродиты. При наступлении полово-
зрелости они становятся самцами, а на
третий год жизни превращаются в самок.
Сходным образом происходит смена пола у
Lysmata seticauda и Calocaris macandrae.
Самцы водных десятиногих находят
самок, руководствуясь чувством обоняния
и осязания. На жгутах передних антенн
самцов креветок особенно много
чувствительных щетинок, помогающих, вероятно,
в поисках самок. Сухопутные крабы
пользуются для этого зрением. Самцы издали
видят самок и быстро бегут к ним.
Непосредственно перед спариванием
самка линяет. У многих крабовидных
отшельников, например у камчатского краба
и у некоторых настоящих крабов, в том
чпсле у мохнаторукого краба, линьке
самки предшествует так называемое
«положение рукопожатия», при котором
самец и самка стоят друг перед другом и
самец крепко держит в своих клешнях
клешни самки. При этом оба партнера
приподнимаются на своих грудных ногах.
«Положение рукопожатия» может длиться
довольно долго, у камчатского краба от
3 до 7 дней, и заканчивается линькой
самки, причем, по некоторым наблюдениям,
самец помогает самке слинять, стаскивая
с нее старый панцирь. Другие крабы ждут
линьки самки, взобравшись на нее сверху.
После линьки самки наступает
спаривание. Самец при помощи передних
брюшных ног прикрепляет сперматофоры близ
половых отверстий самки и только у
крабов вводит их в семеприемники самки.
Самцы некоторых раков-отшельников
приклеивают сперматофоры к раковине, в
которой живет самка.
У разных видов спаривание
осуществляется по-разному. Креветки Sicyonia cari-
nata и Athanas nitescens располагаются
перпендикулярно друг к другу, причем
самец оказывается лежащим на спине
снизу, а самка спиной кверху над ним. У
речных раков, омаров, лангустов, Galatheidea
и некоторых других самка лежит на спине,
а самец находится над ней спиной вверх.
У многих крабов и у крабовидных
отшельников самец сидит сверху, на спинной
стороне самки. Молодые самцы (длиной 10 мм)
Athanas nitescens в течение нескольких
минут спариваются с разными самками
4—5 раз, более крупные A7—20 мм) —
один раз. Самцы других креветок,
китайского и камчатского крабов, речного рака
Cambarus также часто спариваются с
несколькими самками. Самкам некоторых
крабов для оплодотворения яиц
достаточно одного спаривания в несколько лет,
так как сперматозоиды в семеприемниках
остаются живыми. Самка американского
Рис. 279. «Положение рукопожатия» у
мохнаторукого краба Eriocheir sinensis.
517
Рис. 280. Спаривание десятиногих:
1 — американского речного рака Cambarus
affinis; 2 — креветки Sicyonia carinata (самец
светлый).
голубого краба (Callinectes) спаривается
один раз в жизни, в возрасте 3 лет, а
живет 7 лет и откладывает яйца ежегодно.
Промежуток между спариванием и
откладкой яиц у разных видов различен.
Самки Athanas и других креветок
начинают откладку яиц через несколько минут
после спаривания, речные раки—через 10—
45 дней, краб Carcinus—через несколько
месяцев. Для откладки яиц омары,
речные раки и раки-отшельники
опрокидываются на спину, а креветки ложатся на
бок и подгибают брюшко так, чтобы между
ним и нижней поверхностью груди
образовалось пространство, куда и попадают
яйца. Вместе с яйцами выделяется секрет,
растворяющий оболочки сперматофо-
ров, сперматозоиды освобождаются и
оплодотворяют яйца. Самки лангустов
разрывают оболочку сперматофора когтем
задней ходильной ноги. Секрет цементных
желез, находящихся у крабов в семепри-
емниках, а у остальных десятиногих на
нижней поверхности брюшка и на
брюшных ножках, образует наружную
оболочку каждого яйца, а также тяж, который
прикрепляет его к брюшным ножкам
матери. Крабы Carcinus при откладывании яиц
зарываются в грунт, без чего не
происходит прикрепления яиц. В дальнейшем яички
остаются на плеоподах вплоть до выхода
личинок или молодых рачков. Только у
самых примитивных креветок,
объединяемых в семейство Penaeidae, процесс
откладывания яиц происходит иначе: они
откладывают яйца прямо в воду.
Плодовитость десятиногих колеблется
в широких пределах. При этом чем
больше яиц, тем меньше их размеры. Очень
плодовиты некоторые крабы: число яиц,
выметываемых Cancer pagurus и Carcinus
maenas, достигает 3 миллионов (но
в Черном море у последнего не более
180000 яиц), у Callinectes sapidus —2
миллионов, у Eriocheir sinensis оно колеблется
от 270 000 до 920 000 и т. д. Гораздо ниже
плодовитость наших северных крабов
(Hyas coarctatus —1 000—16 000 яиц, Н.
araneus — до 110 000 яиц). Очень высока
плодовитость лангустов — от 500 000 до
1,5 миллионов яиц. У раков-отшельников
число яиц обычно не доходит до 10 000,
но у близкого к ним камчатского краба
оно значительно больше — от 70 000 до
270 000. Среди креветок наиболее
плодовиты Penaeidae, откладывающие яйца
в воду: Penaeus trisulcatus мечет около
800 000, a Parapenaeopsis longirostris около
400 000 яиц. Креветки, вынашивающие
яйца на себе, откладывают их в меньшем
количестве: Pandalus latirostris — 150—
450, P. borealis—400—3 000, Palaemon ad-
spersus — 160—3600, P. elegans — 280—
1600, Crangon crangon — 1 500—15 000.
Сравнительно невелика и плодовитость
европейского и норвежского омаров (8 000—
32 000 и 1300-4 000 яиц).
У всех пресноводных и глубоководных
десятиногих плодовитость значительно
меньше, чем у морских мелководных. Наши
речные раки откладывают от 60 до 500 —
600 яиц, тропические пресноводные
крабы — 20—150, глубоководные
планктонные креветки — 15—60, глубоководный
B000—2600 м) Munidopsis rostrata — 230.
Из их крупных яиц выходит не личинка,
а уже почти сформировавшийся рачок.
Яйца остаются прикрепленными к
брюшным ножкам матери в течение продолжи-
518
тельного времени. Камчатский краб и
европейский омар вынашивают их почти
целый год, норвежский омар — 9 месяцев,
краб Cancer pagurus — 7—8 месяцев,
речные раки—около полугода, Pandalus bore-
alis—5 месяцев, Carcinus maenas — летом 2
месяца, зимой 5—6 месяцев, китайский
мохнаторукий краб и креветки рода Palae-
mon—1—1У2 месяца, пальмовый вор —
3—4 недели, краб-плавунец Portunus tri-
tuberculatus — 2—3 недели. Эти
различия зависят в первую очередь от
температуры, которая влияет на скорость
эмбрионального развития.
У большинства десятиногих из яйца
выходит личинка, резко отличающаяся
от взрослых как по своему строению, так
и по образу жизни. Только у многих
пресноводных, глубоководных и обитающих
при низкой температуре видов развитие
прямое, т. е. все личиночные стадии
проходят под покровом яйцевой оболочки и
из яйца появляется маленький почти
сформировавшийся рачок.
Различают несколько стадий
плавающих личинок десятиногих ракообразных
(табл. 39). Первая из них — науплиус —
свойственна только примитивному
семейству креветок Penaeidae (табл. 39, 12).
Стадия метанауплиуса есть также только
у них и у некоторых представителей
близкого семейства Sergestidae. У большинства
морских мелководных десятиногих из яйца
выходит личинка, находящаяся на стадии
зоэа (табл. 39, 1,5). Для нее характерно
разделение тела на покрытую карапаксом
головогрудь и сегментированное брюшко,
причем последний его сегмент еще не
отделен от тельсона. Глаза зоэа стебельчатые,
очень сильно развиты ногочелюсти,
наружные ветви которых служат основными
органами плавания, а задние грудные и все
брюшные ножки еще не образовались или
зачаточны. У зоэа крабов передняя часть
тела вздутая, на карапаксе расположены
длинные шипы — один на спине и два по
бокам, а кроме того, имеется длинный
рострум (табл. 39, 1). У зоэа
раков-отшельников и близких к ним групп задний
край карапакса выемчатый, задне-боко-
вые его углы оттянуты в шипы и также
есть длинный рострум (табл. 39, 11). Все
эти выросты помогают личинкам
существовать в толще воды, так как затрудняют
их погружение.
За стадией зоэа у длиннохвостых
десятиногих следует стадия м и з и с. На этой
стадии все грудные конечности хорошо
развиты, снабжены наружными ветвями
и служат для плавания, задний брюшной
сегмент отделен от тельсона, начинается
развитие брюшных конечностей. У
большинства креветок стадия мизис сходна
с мизидами, почему она и получила свое
название, но у некоторых других она
имеет причудливую форму. Таковы,
например, листовидные, совершенно
прозрачные мизисные личинки лангустов, так
называемые филлозомы (табл. 39, 7),
или шаровидные личинки глубоководных
Eryonidae, так называемые эрионеи-
кусы (табл. 39, 4). У омаров личинка
выходит из яйца на этой стадии (табл. 39, 9).
Мизисной стадии длиннохвостых
соответствует стадия метазоэа крабов, раков-
отшельников и близких к ним групп.
Метазоэа отличается от мизис отсутствием
наружных ветвей грудных ног.
Последняя личиночная стадия носит
название декаподитной. На этой
стадии карапакс уже почти такой же, как
у взрослого ракообразного, но брюшко
часто имеет иное строение. Декаподитная
стадия крабов—м егалопа и декаподитная
стадия раков-отшельников и близких к
ним форм — глаукотоэ
характеризуются еще не подогнутым под грудь и
вполне симметричным брюшком,
снабженным сильными плеоподами, при помощи
которых личинка плавает (табл. 39, S, 10).
Затем она садится на дно, линяет и
превращается в молодую особь с нормально
устроенным брюшком, подогнутым под
грудь у крабов или спирально
закрученным у раков-отшельников.
Продолжительность существования
личинок в планктоне различна у разных
видов: у креветок Palaemon adspersus и
Crangon crangon и краба Carcinus
maenas — 4—5 недель, у пресноводной
креветки Atyaephyra desmaresti и у
европейского омара — 2—3 недели, у креветки
Pandalina brevirostris — 2 месяца, у
обыкновенного лангуста Palinurus — 3 месяца,
у креветки Sergestes arcticus — 4—5
месяцев, а у лангуста Panulirus — до 6 ме-
519
сяцев. За время своего пребывания в
планктоне филлозомы лангустов
разносятся течениями на далекие расстояния.
С этим связывают циркумтропическое
распространение некоторых лангустов.
Многие планктонные личинки
десятиногих — хорошие пловцы. Когда они
плавают при помощи ногочелюстей, т. е. на
стадии зоэа, они двигаются задним концом
вперед или, как многие зоэа крабов,
спинной стороной вперед. При этом зоэа Ga-
Jathea проплывает 1 Ль за 45—56 секунд,
Porcellana — за 65—92 секунды. После
превращения в последующие стадии,
когда органами движения становятся плео-
поды, личинки двигаются передним
концом вперед. Пищей личинкам служат
разные планктонные водоросли и животные.
Огромное количество личинок погибает,
становясь жертвами различных хищников
или попадая в неподходящие условия. По
расчетам японского зоолога Марукава,
из 200 000 зоэа первой стадии, которых
дает одна самка камчатского краба, до
второй стадии зоэа доживает 13, а до
стадии глаукотоэ 7 личинок.
У большинства пресноводных,
глубоководных и холодноводных десятиногих
яйца крупные, богатые желтком, и всё
превращение личинки проходят под
покровом яйцевой оболочки. Из яйца
выходит маленький рачок, который
отличается от взрослых главным образом
отсутствием последней пары брюшных
конечностей. Новорожденные рачки
удерживаются на плеоподах матери при помощи
своих конечностей. У речных раков се-
Рис. 281. Движения зоэа Muni da A) и крабов B).
верного полушария (сем. Astacidae) для
этого служат клешни, а у речных раков
южного полушария (сем. Parastacidae) —
задние грудные ножки. Сходным образом
ведут себя вылупившиеся из яиц
пресноводные крабы (сем. Potamonidae),
полуглубоководная креветка Sclerocrangon ferox
и глубоководная S. zenkevitchi.
С размножением связаны миграции
многих десятиногих. Они стремятся к
моменту выхода из яиц личинок попасть в
условия, наиболее благоприятные для
существования своих новорожденных потомков.
Мохнаторукий краб, поднимающийся
довольно высоко вверх по рекам Западной
Европы, для размножения переселяется
в солоноватую воду предустьевых
пространств, что необходимо для его личинок.
Точно так же мигрируют в море для
размножения некоторые Penaeidae, остальное
время проводящие в солоноватой или
пресной воде, как например Penaeus carinatus
и Penaeopsis dobsoni из индийского соло-
новатоводного озера Чилька или Penaeus
plebejus из рек Австралии. Многие
морские креветки и некоторые крабы, когда
приходит время выхода личинок из яиц,
перекочевывают на прибрежные
мелководья, где вода сильнее прогрета.
Наиболее интересны нерестовые миграции
сухопутных десятиногих. Самки сухопутных
раков-отшельников — пальмового вора
и Coenobita, а также сухопутных крабов
из семейства Gecarcinidae, несущие на
брюшных ножках яйца, в которых
сформировались зоэа, массами стремятся к морю.
Они идут иногда издалека, преодолевая
по нескольку километров. Переселения
обычно совершаются ночью или днем во
время дождя, поскольку период
размножения сухопутных десятиногих
приходится на сезон тропических ливней.
Достигнув морского берега, самки заходят в
воду или забираются на камни и, как
пальмовый вор, стряхивают яйца в воду
либо, частично погрузившись в воду, ждут,
когда из яиц вылупятся личинки. В
дальнейшем личинки развиваются в море, там
оседают на дно и только через некоторое
время выбираются на сушу.
Развитие личинок пальмового вора дает
довольно полное представление об истории
формирования этого вида (табл. 40, 1—5).
520
Рис. 282. Вылупившиеся из яиц
мальки широкопалого рака:
1 — рачки, прикрепившиеся к пле-
оподам матери (кроме мальков,
видны пустые яйцевые оболочки);
2 — малек со спинной стороны.
Планктонные зоэа и глаукотоэ пальмового
вора такие же, как у раков-отшельников.
После оседания на дно, т. е. через 4—6
месяцев после вылупления из яйца, личинка
пальмового вора имеет спирально
изогнутое мягкое брюшко, которое она прячет
в пустую раковину какого-нибудь
морского брюхоногого моллюска. В это время
она совершенно сходна с обычным раком-
отшельником. Затем она перебирается на
сушу и использует раковины наземных
моллюсков, напоминая взрослые
экземпляры близкого сухопутного рода Coenobita.
Наконец, после очередной линьки, она
оставляет раковину, в результате
нескольких следующих линек ее брюшко
постепенно укорачивается и подгибается под
грудь, и таким образом она превращается
из рака-отшельника в пальмового вора.
Вероятно, так же, но в течение очень
длительного времени шла эволюция этого
вида и Coenobita, происшедших от морских
раков-отшельников.
Закончившие превращение молодые
десятиногие ведут такое же существование,
как взрослые, с каждой линькой
увеличиваясь в размерах. Сначала они линяют
часто, потом промежутки между
линьками удлиняются, и крупные особи таких
видов, как омар и камчатский краб,
линяют один раз в два года. Линька
контролируется органами внутренней секреции,
находящимися в глазных стебельках.
Удаление глазных стебельков вызывает
более частые линьки, так как в кровь
перестает поступать тормозящий их гормон.
Перед линькой под старым панцирем
формируется новый, еще мягкий, не
содержащий солей кальция. Старый
панцирь в процессе линьки лопается на
границе между грудью и брюшком, а у
крабов, кроме того, по бокам грудного отдела.
Длиннохвостые десятиногие вытаскивают
через образовавшуюся щель сначала
переднюю, а затем заднюю части тела,
крабы сначала брюшко, потом голову
и грудь, ходильные ноги и, наконец,
клешни. Речные раки при этом, как впервые
описал еще Реомюр в 1712 году,
опрокидываются на спину и начинают резко
сгибать брюшко и конечности, вследствие
чего и образуется щель в старом панцире.
Возможно, что при вытаскивании из
старого панциря тела и конечностей они
механически вытягиваются и рост
осуществляется непосредственно в процессе
линьки. Она занимает обычно 10—30 минут,
но затвердевание нового панциря требует
гораздо более продолжительного времени:
у камчатского краба 10 дней, у омара
20—50 дней, у мохнаторукого краба
больше месяца, у краба Cancer pagurus
2—3 месяца. При этом соли кальция
поступают из крови в новый панцирь и
пропитывают его. Перед линькой они
накапливаются либо в печени (у большинства
десятиногих), либо в так называемых гас-
тролитах — двух овальных твердых
образованиях, находящихся в желудке
речных раков.
До тех пор пока не затвердел новый
панцирь, десятиногие остаются
совершенно беззащитными. Кроме того, они
не могут питаться, так как все их
ротовые придатки и перетирающий пищу
аппарат желудка тоже мягкие. Многие
десятиногие на это время забиваются под
камни или в другие убежища, где они
оказываются в безопасности, и там
мужественно переживают тяжелый для них
период жизни.
Некоторые десятиногие живут очень
долго: американский омар до 50 лет,
широкопалый речной рак до 20 лет,
камчатский краб до 23 лет, лангуст более 15
лет, мохнаторукий краб до 6, иногда
до 10 лет. Пальмовый вор длиной
более 10 см имеет от роду не менее 5 лет.
Продолжительность жизни креветок обыч-
521
Рис. 283. Глубоководные светящиеся креветки:
А — Systellaspis, спасающийся от рыбы; Б — Sergestes
corniculum; В — фотофор Sergestes; Г — схема
расположения фотофоров на брюшной стороне; 1 — линза;
2 — светящиеся клетки; 3 — рефлектор.
но не превышает 4 лет (Crangon crangon,
Pandalus), а мелкие виды не доживают
и до года.
Подавляющее большинство десятиногих
обитает в области морского мелководья.
Особенно богата ими фауна тропиков.
Коралловые рифы населены
бесчисленным множеством самых
разнообразных крабов, креветок и
раков-отшельников, часто очень пестро окрашенных
и имеющих причудливую форму. По
направлению на север и на юг от
тропической зоны фауна десятиногих беднеет.
В пределах умеренных широт по
богатству десятиногими выделяется северная
часть Тихого океана, где обитает
наибольшее количество родов и видов таких
семейств креветок, как Crangonidae, Hippo-
lytidae, Pandalidae, а также крабовидных
отшельников Lithodidae. В
Беринговом море известно 117, в Охотском 113,
в Японском 82 вида десятиногих. Из
северной части Тихого океана шло
расселение некоторых групп десятиногих в
Полярный бассейн и даже в северную часть
Атлантического океана. Очень бедна
десятиногими ракообразными Антарктика,
откуда указано всего 8 видов.
Морские десятиногие обитают от
приливо-отливной полосы до глубины 5300 м.
В этом отношении они уступают многим
другим отрядам ракообразных (бокопла-
вам, равноногим и др.)> которые
приспособились к жизни и на больших
глубинах, вплоть до максимальных (И км).
Среди глубоководных донных
десятиногих много древних родов, отсутствующих
на мелководьях, но известных в
ископаемом состоянии (Eryonidae, крабы Prosopo-
nidae и Homolidae и др.). Очень
характерны глубоководные планктонные креветки,
принадлежащие к семействам Oplopho-
ridae (табл. 35, S), Penaeidae, Serges-
tidae и др. Обычно они темно-красного
цвета, их конечности удлинены, и многие
из них обладают способностью к свечению.
У многих глубоководных Sergestidae
на конечностях, вдоль боковых краев
карапакса и на брюшной стороне тела
располагается до 150 фотофоров. У
некоторых видов часть фотофоров снабжена
линзами. Среди глубоководных креветок
известны виды, лишенные фотофоров, но
тем не менее светящиеся. Их покровы
очень тонкие, и сквозь них
просвечивают участки печени, превратившиеся в
органы свечения — так называемые
органы Песта. Наконец, некоторые кревет-
522
ки, преимущественно из рода Systella-
spis, выбрасывают струю светящейся
жидкости и при помощи «огневой завесы»
скрываются от врагов. Присутствие фо-
тофоров, расположение и число которых
для каждого вида строго постоянно,
помогает в полном мраке океанических
глубин самцам находить самок и всем вместе
собираться в стаи.
Приливо-отливная полоса тропических
морей населена очень своеобразными
десятиногими, из которых наиболее
характерны манящие крабы. Это мелкие
(ширина карапакса 0,8—3,5 см) крабики,
живущие на илистом грунте большими
колониями и замечательные тем, что их
самцы своей огромной клешней или
обеими клешнями совершают сложные
манящие движения, ритмически разводя
клешни в стороны, поднимая и опуская
их и располагая их перед собой. Манящие
движения в деталях различны у разных
видов. У видов рода Uca с узким лобным
краем преобладают движения клешней в
вертикальной, у видов с широким лобным
краем — в горизонтальной плоскости.
Приближение самки или другого самца
вызывает более энергичные манящие
движения. Изучение их у разных видов
(с применением киносъемки и иных
современных методов) указывает на
многообразие биологического значения этих
движений. Во-первых, таким образом самцы
отпугивают других самцов, сообщая им
о том, что данный участок уже занят. Во-
вторых, в период спаривания манящие
движения самца служат для привлечения
самки. В промежутки между этими
движениями самец постепенно
приближается к самке, которая сначала принимает
позу защиты и отступает, а затем сама
начинает двигаться навстречу самцу. При
этом оба партнера размыкают и смыкают
пальцы своих клешней. Если
какой-нибудь самец не обратит внимания на
предупреждение и вторгнется на чужую
территорию, между ее хозяином и
пришельцем возникает ожесточенное сражение
(рис. 284). Если перед самцом поставить
зеркало, он бросается в бой со своим
отражением, что указывает на ведущую роль
зрения во всех этих явлениях. У манящих
крабов рода Dotilla победитель после
Рис. 284. Поведение
манящих крабов:
слева — манящие
движения самца Uca annulipes;
справа — драка между
двумя самцами Uca marionis
из-за самки
(схематизированные кадры кинофильма,
снятого со скоростью 16
кадров в секунду).
523
окончания драки исполняет
триумфальный танец. Нередко драки между двумя
самцами происходят из-за самки, которая
в таких случаях сама удаляется и ждет
исхода боя. Замечательно, что совершенно
сходные повадки свойственны некоторым
представителям другого семейства
крабов—Grapsidae, живущим также в приливо-
отливной полосе тропических морей.
Сравнительное исследование манящих
движений приводит к выводу о том, что они
развились независимо у представителей двух
разных семейств из поз защиты и
нападения, причем впоследствии к
первоначально исключительно агрессивному их
назначению добавилось сексуальное.
Некоторые морские десятиногие могут
переносить значительное опреснение; сре-
Рис. 285. Крабы-переселенцы:
1 — китайский мохнаторукий краб Eriocheir sinensis,
2 — Rhithropanopeus harrisi.
ди них есть виды, обитающие
преимущественно в опресненных участках моря и
способные заходить в совершенно пресную
воду. Таковы, например, мохнаторукие
крабы рода Eriocheir. Один из них —
Е. japonicus — живет перед устьями рек
от Амурского лимана до Гонконга и
Тайваня, а также Северо-Восточного
Сахалина, другой — Е. sinensis — в таких же
условиях в Китае. Китайский
мохнаторукий краб в балластных цистернах
судов был случайно перевезен около 1912
года в бассейн Эльбы. Оттуда он начал
быстро расселяться вдоль западных
берегов Европы и сейчас многочислен во
многих реках и их устьевых частях в
Германии, Бельгии, Голландии,Франции, Дании,
Швеции и Финляндии; он разрушает
своими норами плотины, рвет рыболовные
сети и портит пойманную в них рыбу.
Другой солоноватоводный крабик—
Rhithropanopeus harrisi — таким же образом попал
из предустьевых пространств американских
рек в Голландию, а затем проник в
Балтийское, Азовское и Каспийское моря,
где сильно размножился и используется
в пищу рыбами.
В пресных водах обитает
сравнительно немного видов десятиногих. Среди них
широко известны речные раки,
принадлежащие к трем семействам — Astacidae,
Parastacidae и Austrastacidae. Astacidae
живут в умеренной и отчасти южной
полосе северного полушария,
Parastacidae — в умеренной полосе южного
полушария — в Южной Америке,
Австралии, а также на острове Мадагаскар,
Austrastacidae — только в южной части
Австралии. Почти на всем протяжении
тропиков речных раков нет. В пределах
СССР речные раки представлены двумя
родами — Astacus и Cambaroides. Род
Astacus занимает почти всю Европу и
западную часть Азии, род Cambaroides —
бассейн Амура, северную Японию, Корею
и Сахалин.
Наиболее широко распространены и
имеют довольно важное промысловое
значение два наших вида речных раков —
широкопалый (A. astacus) и узкопалый
(A. leptodactylus). Широкопалый рак
живет главным образом в водоемах,
относящихся к бассейну Балтийского моря. Он
Рис. 286. Распространение речных раков в северном полушарии.
обычен в Латвии, Литве, Эстонии, в
Ленинградской области, в некоторых
районах Украины и Белоруссии. Область
распространения узкопалого рака гораздо
шире и охватывает водоемы бассейнов
Каспийского, Черного и Азовского морей,
Каспийское и Азовское моря, а также
реки и озера Западной Сибири.
Широкопалый и узкопалый раки
совместно не встречаются, и их ареалы не
перекрывают друг друга. В прошлом веке
узкопалый рак начал расселяться вверх
по бассейну Волги, вытесняя
широкопалого. Известны многочисленные случаи
посадок раколовами узколапого рака в
озера, населенные широкопалым.
Проходит 10—20 лет, и широкопалый рак
исчезает, полностью замещаясь узкопалым.
Оба вида сходны по своей биологии.
Они живут в воде со сравнительно
высоким содержанием кислорода и
минеральных солей, питаются преимущественно
растительной пищей, имеющейся обычно
в избытке. Узкопалый рак несколько
более плодовит и более вынослив по
отношению к химизму воды и ее кислородному
режиму. Однако это еще не объясняет,
каким образом он неизменно вытесняет
широкопалого. Не удалось получить
ответ на этот вопрос и при совместном
содержании обоих видов в бассейнах. Время
от времени раки дрались, но драки
между особями разных видов происходили
не чаще, чем между особями одного вида.
В последнее время в Литве, где
встречаются и широкопалый и узкопалый
раки, удалось заметить, что на стыке их
ареалов обитают особи, по своим признакам
занимающие промежуточное положение
между обоими видами. Я.М. Цукерзис
считает их гибридами и допускает
возможность постепенного исчезновения
признаков того или иного вида у потомков этих
гибридов. В искусственных условиях
удалось скрестить между собой оба вида
и получить гибридную молодь, но она не
доросла до взрослого состояния. Следует
при этом иметь в виду, что узкопалый и
широкопалый раки различаются не
только по форме клешней, рострума,
вооружению карапакса и т. д., но и по числу
хромосом: у узкопалого рака их 184, а
у широкопалого около 100. Для растений
давно известно, что полиплоиды
характеризуются повышенной
жизнеспособностью, плодовитостью и другими
преимуществами; по-видимому, это
подтверждается и на примере речных раков.
На численность и распространение
речных раков большое влияние оказали их
болезни, в первую очередь рачья чума.
В 70-х годах она появилась во
Франции,затем перекинулась в Германию, а в 1890
году — в западную часть России. К 1898
году эпизоотия рачьей чумы захватила
всю Россию, включая Западную Сибирь.
Во многих местах раки вымерли
поголовно и исчезли, в других их запасы посте-
525
пенно начали восстанавливаться. Но в
1919—1920 годах возникла новая
вспышка рачьей чумы, опять уничтожившая
огромное количество раков.
Заболевшие животные ходят на
выпрямленных ногах, выползают из нор днем, чего
здоровые раки никогда не делают, через
некоторое время у них начинаются судороги,
п в течение недели раки погибают.
Возбудителем болезни служит грибок Aphanomy-
ces astaci, развивающийся в покровах и в
центральной нервной системе больных
раков. Болезнь распространяется посредством
ооспор этого грибка.
В пресных водах тропиков взамен
отсутствующих там речных раков водится
много различных крабов и креветок. К
пресным водам целиком приурочено
богатое видами семейство крабов Potamo-
nidae, включающее около 20 родов и более
200 видов. Некоторые виды этого
семейства встречаются даже в высокогорных
водоемах (до 2100 м над уровнем моря в
Индии). Один из видов Potamonidae — Ро-
tamon potamios — широко распространен
в бассейне Средиземного моря и обычен
у нас на Кавказе, в Крыму и в Средней
Азии. В сырых лесах Закавказья он
встречается и во влажной земле, в которой
роет свои норы. Среди креветок есть
также одно целиком пресноводное семейство—
Atyidae, характеризующееся
присутствием пучков щетинок на пальцах клешней,
при помощи которых рачки захватывают
грунт. Разбросанное географическое
распространение этого семейства, а также его
морфологические особенности указывают
на его глубокую геологическую древность.
В бассейне Амура водится два вида атиид,
представляющих вместе со многими
рыбами и моллюсками тропический элемент в
амурской фауне. Еще одна креветка из
этого семейства — Troglocaris
anophthalmia (табл. 35, 12)—живет в пещерных
водоемах Закавказья. Кроме этих двух
семейств, в тропических пресных водах
обитают виды крабов и креветок,
относящиеся к преимущественно морским родам.
Особый интерес представляют
пещерные десятиногие. Известно около 20 сле-
Рис. 287. Речные раки:
1 — широкопалый рак (Astacus
astacus); 2— узкопалый рак (А.
leptodactylus).
526
пых пещерных видов Atyidae и 10 видов
Palaemonidae, разбросанных по
различным континентам и островам. В пещерах
США живет 18 видов слепых речных
раков, а в Мексике — слепой пещерный
краб Typhlopseudothelphusa mocinoi,
относящийся к семейству Potamonidae.
Наиболее удивительным следует считать
существование представителя рода Muni-
dopsis (сем. Galatheidae) в солоноватом
подземном озере на одном из Канарских
островов; все остальные многочисленные
виды этого рода обитают на больших
океанических глубинах. Очевидно, и они и
подземный М. polymorpha некогда
произошли от вымерших мелководных морских
предков и дожили до наших дней в
подземных и глубинных водах, как в
убежищах, сохранивших много различных
остатков древних фаун.
Десятиногие используются в пищу
людьми и интенсивно добываются почти во всех
странах. В СССР особенно большое
экономическое значение имеет промысел
камчатского краба (Paralithodes camtschatica),
из мускулов ног которого делают
консервы. Он принадлежит к семейству Litho-
didae, т. е. крабовидных
раков-отшельников, и населяет Японское, Охотское и
Берингово моря.
Ширина карапакса самцов этого вида
в среднем 16 см, а у некоторых
экземпляров достигает 25 см. Расстояние между
концами средних ходильных ног таких
крупных особей — 1V2 м, а общий вес
их тела — 7 кг. Самки значительно
мельче (ширина карапакса до 16 см), и их
использование для изготовления консервов
нецелесообразно.
Всю свою долгую жизнь камчатские
крабы проводят в странствованиях,
причем каждый год повторяют один и тот
же маршрут. Их миграции в значительной
степени зависят от колебаний
температуры. Для самцов наиболее благоприятна
температура между 2 и 7° С, хотя они
выдерживают некоторое время и колебания
от —2 до +18° С. Оптимальная
температура для самок несколько выше. Если
температура повышается или понижается
за пределы оптимальной, крабы
перемещаются в районы, более подходящие для
них по тепловому режиму. Другой
причиной миграций служит размещение
кормовых животных: крабы кочуют в
поисках пищи, пригодной для той или иной
фазы их жизненного цикла.
Наибольшие скопления крабов
сосредоточены у западной Камчатки, где в
основном и ведется их промысел. Здесь
крабы проводят зиму вдали от берегов,
на значительной глубине — 110—200 м,
куда зимнее охлаждение еще не успевает
распространиться и где господствуют
низкие положительные температуры. Весной
начинается прогрев воды на мелководьях,
а охлаждение достигает мест зимовки
крабов. Это и заставляет крабов начинать
переселение с больших глубин к берегам.
Скорость передвижения крабов
невелика. Наблюдая с высокой скалы за
движением одного из них по прямой,
Н. П. На возов-Лавр о в определил ее
приблизительно в 1 милю A,8 км) в час.
Но крабы всегда передвигаются зигзагами,
и путем мечения удалось показать, что
за сутки они проходят по прямой не
более 5—7 миль. При этом разные особи
бродят в разных направлениях, и все
стадо в целом обычно перемещается за сутки
всего на 1—2 мили. Каждое стадо кочует в
довольно ограниченном районе, площадь
которого составляет у Камчатки около
100, а в Приморье около 40 кв. миль. У
охотоморских берегов Камчатки
различаются 4 таких района и соответственно
4 стада, из которых наиболее
многочисленно северное, приуроченное к Хайрю-
зовскому району.
С мест зимовки к берегам крабы
двигаются отдельными косяками — крупные
самцы отдельно от самок и молоди. Самки
несут на своих брюшных ножках яйца,
содержащие уже вполне
сформировавшихся личинок. По дороге на мелководья
личинки выходят из яиц, а самки
продолжают свой путь.
Приблизительно через месяц после
начала миграции косяки самцов и самок
встречаются на мелководьях E—30 м),
где температура воды в это время
достигает 2—4 °С, и перемешиваются. Самцы
располагаются перед самками и
держатся своими клешнями за их клешни. Часто
удается багром с берега вылавливать
такие парочки в «положении рукопожатия».
527
Рис. 288. Расселение молоди камчатского краба вдоль западного берега Камчатки в различные годы.
Оно длится от 3 до 7 суток. Затем самцы
помогают самкам слинять, стаскивая с
них загрязненный старый панцирь,
после чего происходит спаривание. Самец
отбрасывает старый панцирь и
прикрепляет к основаниям третьей пары ходильных
ног только что слинявшей самки сперма-
тофор — лентовидную массу спермы,
покрытую нерастворимой в воде оболочкой,
после чего оба партнера расходятся.
Через несколько часов, а иногда и дней
после спаривания самка откладывает на
свои брюшные ножки икру, которая при
этом оплодотворяется и которую самка
носит до следующей весны, т. е.
приблизительно 11 V2 месяцев,
После спаривания косяки самцов и
самок опять мигрируют раздельно. Если
весеннюю миграцию можно назвать
нерестовой, то летние должны считаться
кормовыми. Перед ними самцы линяют,
но, в отличие от самок, в одиночестве,
среди каменистых россыпей, обычно против
далеко выдающихся в море мысов.
Со второй половины июля и до конца
сентября у Камчатки и с конца мая —
начала июня до ноября в заливе Петра
Великого крабы совершают миграции,
переходя с одного кормового поля на другое.
Самцы держатся глубины 20—60 м и
температуры 2—7°С. Самки перемещаются
несколько ближе к берегам. Осенью
прибрежные воды начинают охлаждаться, и
крабы отступают от берегов, пока не
достигнут мест своей зимовки. Следующей
весной все повторяется сначала.
Вышедшая из яйца первая личинка —
протозоэа — линяет через несколько
минут после вылупления и превращается в
зоэа, которая живет в толще воды около
2 месяцев, линяет 4 раза и переходит в
глаукотоэ. Глаукотоэ оседает на дно и в
течение 20 дней ползает в зарослях
водоросли анфельции, среди гидроидов и
других зарослей.
После линьки она превращается в
молодого краба — малька, вполне сходного
со взрослыми, но имеющего относительно
528
более длинные шипы на панцире. Ширина
его тела 2 мм. Малек живет так же и в
таких же условиях, как глаукотоэ, и
несколько раз линяет. Только через 3 года
он покидает заросли водорослей и
гидроидов и переходит на участки песчаного
грунта. К 6—7 годам, достигнув ширины
панциря около 7 см, молодые крабы
начинают собираться в косяки и мигрировать.
Самки растут несколько медленнее, чем
самцы, но зато раньше достигают долово-
зрелости. Уже на восьмом году жизни они
начинают откладывать икру, а самцы
становятся половозрелыми только в
возрасте 10 лет. В это время они достигают
ширины панциря 12,5 см и годятся для
изготовления консервов.
Вследствие позднего наступления по-
ловозрелости и медленного роста
камчатского краба его запасы, подорванные
интенсивным промыслом,
восстанавливаются с большим трудом. Из огромного
количества ежегодно рождающихся
личинок до взрослого состояния доживают
очень немногие. Остальные становятся
жертвами различных хищников или
уносятся течениями. Заросли, подходящие
для оседания личинок, есть только в Хай-
рюзовском районе охотоморского
побережья Камчатки. Отсюда молодь краба
заселяет более южные участки. Однако
в годы с пониженной температурой воды
на мелководьях такая миграция
невозможна, и южные стада камчатского краба
совсем не получают пополнения (рис. 288).
Уже более 30 лет тому назад было
сделано предложение об акклиматизации
камчатского краба в Баренцевом море. В
настоящее время есть сторонники и
противники этой идеи. Методика перевозки
крабов и их личинок с Дальнего Востока
разработана, перевозка осуществлена, и
сейчас ведутся наблюдения в аквариаль-
ных и естественных условиях над
поведением камчатского краба в новом для него
водоеме.
Краба ловят ставными сетями,
которые стремятся поставить так, чтобы
перегородить основные пути миграции.
Небольшие суда привозят сети с пойманными
крабами на краболов, где производят вы-
путку крабов, отрывают у них ноги и
клешни, варят их, рубят, вытряхивают
мясо и изготовляют из него консервы.
Помимо камчатского краба, в наших
дальневосточных морях в меньшем
количестве добывают креветок из рода
Pandalus. Этот промысел может быть
расширен, так как запасы различных
креветок в наших морях очень велики.
В пресных водах, преимущественно на
Украине, ведется промысловый лов
речных раков.
За рубежом добывают много различных
десятиногих, из которых на первом месте
стоят креветки. В 1962 году их было
поймано 700 тысяч тонн, причем особенно
развит этот промысел в Китае, США,
Индии, Японии. Наибольшее значение в
уловах имеют семейства Penaeidae, Рап-
dalidae и Crangonidae. В некоторых
странах Южной и Юго-Восточной Азии
ведется искусственное разведение креветок,
преимущественно пенеид. Прибрежные
низменные пространства при помощи дамб
превращают в цепь прудов,
соединяющихся каналом с морем. В прилив вода
заливает пруды и остается там благодаря
тому что шлюзы во время отлива
закрывают. В такие пруды креветки заходят
сами, или туда выпускают пойманных в
море личинок. Креветок подкармливают
рисовыми отрубями. При таком ведении
хозяйства удается получать до 500 кг
креветок с гектара.
Меньшее значение имеет промысел
настоящих крабов, особенно развитый в США.
Их продают свежезамороженными,
вареными и в виде консервов, причем высоко
ценятся недавно слинявшие особи. Из
других десятиногих считаются
деликатесом лангусты и омары. Только на Кубе
ловом лангуста занято более 1000 судов,
вылавливающих ежегодно свыше 7500 тонн,
что составляет около 1/4 всего улова
республики. Запасы европейского омара
подорваны промыслом, и Канада
экспортирует в Европу довольно значительное
количество американских омаров. Для
восстановления запасов омара
производится его разведение в специальных
садках.
Наша страна использует десятиногих
еще совершенно недостаточно. Нет
сомнения в том, что в будущем их промысел
у нас значительно возрастет.
34 Жизнь животных, т. 2
УКАЗАТЕЛЬ ТЕРМИНОВ
(Звездочкой отмечены страницы, на которых помещены рисунки; жирным шрифтом — цветные или
тоновые таблицы с изображением данного животного; в скобках—синонимы.)
А
Абиссаль 114
Абиссальные двустворчатые 95,
112, 114
— иглокожие 242, 260, 287
Аборальная сторона 205, 216,
218, 250, 272, 276
Аборальный канал кольцевой
206*
— полюс 271
Автотомизатор 516
Автотомия (самокалечение) 163,
211, 218, 262, 357, 516
Автофагия (самопожирание) 162
Аддуктор (мускул раковинный) 27
Аккомодация 159
Акрон 422
Амбра 193
Амбулакральная система 200,
200*, 246, 252
Амбулакральное глоточное
кольцо 209*
Амбулакральные бороздки 218,
223, 250*, 251, 252, 255, 261,
266, 287
— желобки 204
— каналы 206, 245
— ножки 200, 200*, 204, 208,
209*, 212, 218, 223, 230, 231,
232, 235, 236, 249*, 251, 252,
253,255,255*,266,269, 271, 274,
277, 283, 288, 290, 294, 296
— пластинки 271, 274*, 279
— поры 287*
— ряды 274*, 285
Амбулакры (радиусы) 201, 204,
205, 206, 218, 271
Амебоидные клетки 209, 256
Амебоциты 106, 108, 154, 207,
209 230 338
Ампулы 200, 200*, 201, 249*, 253
Амфибиотический образ жизни 73
Анабиоз 414, 435, 493
Анальное возвышение 217*
— отверстие (анус)
дефинитивное 196, 218
иглокожих 196, 210, 235,
245, 250*, 254, 255*, 271,275,
285
Анальное отверстие моллюсков
23*, 102*, 105
— — оболочников 278, 339
— — хетогнат 299*
Анальный конус 237
— сфинктер 238
Анаэробиоз 108, 109
— факультативный (аноксибиоз)
108, 109
Аноксибиоз 109
Антеннулы (передние антенны) 381
Антенны 378, 381, 440, 446, 448,
485
— задние 380, 381, 383, 386,
390, 390*, 392, 392*, 400, 403,
404, 417, 420, 422, 427, 429,
441, 444, 451, 455, 461, 464,
467*, 470, 489*, 502
— передние (антеннулы) 380, 381,
386, 390, 390*, 392, 392*, 403,
418*, 420, 422, 425, 426*, 427,
428, 429, 431, 437*, 441, 441*,
444, 451, 455, 460, 464, 467*,
470, 502
Антизоэа 452*, 453
Анус (анальное отверстие) 208,
209, 217*, 225, 228
Аорта 21
— задняя 102*, 107
— передняя 102*, 107
Апикальное (теменное) поле 272,
273, 274*, 275
Апикальный полюс 271
Апофиз 151, 152*, 153
Аппарат дыхательный 471
— копулятивный
(совокупительный) 442, 447, 448, 450, 452,505
Ареал разорванный 213
Аристотелев фонарь 272, 275*, 277
Артерии 107, 157
Артерия кишечная 333
Астаксантин 496, 506
Асцидиозооид 335, 336, 337, 338,
347*, 350, 350*
— вторичный 350, 350*
— первичный 350, 350*
Атриальная полость 333
Аурикулярия 210*, 211, 231
Б
Бесструктурная мембрана 43*
Бивиум 228
Билатеральносимметричные
животные 7, 207, 276, 286
Биокристаллы 202
Биологические часы 493, 507
Биолюминесценция (свечение) 167
Бипиннария 210*, 211, 255*, 257,
262
Биполярность 213
Биссус 96, 97, 116, 121, 128, 130,
144, 150, 150*, 154*
Биссусная железа 116, 125, 127
— — личиночная 111*
Биссусяые нити 116, 119*, 121,
127, 146*
Бластозоид 348, 349, 350, 354
Бластопор (рот первичный) 196,
210
Бластула 209, 344
Блуждающие клетки (амебоциты)
106
Боянусовы органы (почки) 109
Брахиальные пластинки 218
Брахиолярия 255*, 257
Брачный наряд 462*, 463
— танец 466
Бронхиальные трубки 345
Брюшко 368, 378, 382, 386, 392*,
401, 403, 428, 431*, 443, 444,
450, 460, 461
Брюшной желобок погонофор
309*, 314
Бугорки ежей 272, 273, 274*,
275
— офиур 288, 293
— первичные 279, 287
Бурса (выводковая сумка) 206,
207, 208, 209*, 289, 293, 294,
297
Бурсальная щель 287*
В
Веко 159*
Велигер (парусник) 10, 22, 45,
46*, 52, 56, 110*, 111, 118, 137,
138, 154
530
Велум (парус) 10
Вена внутренностная 102*
Вены 107, 157
Ветви фуркальные 401, 404, 420,
422, 449
Вилочка (фурка) 369, 382, 425
Внутренностный мешок 20, 21,
22—41, 43, 114*, 157*
Вода метаболическая 371
Водные легкие 230, 233, 254
Водоносная (амбулакральная)
система 200, 204, 229, 245, 246,
272
Воронка головоногих 157*, 158,
173, 174, 176, 183
Вороночные хрящи 157*
Воротник голотурий 243
— кишечнодышащих 319, 320*,
321, 322, 325
Воротничковые поры 319, 324
Вторичноротые 196, 197, 303, 305,
317, 345
Выводковая камера 18, 111, 224,
225, 231, 235, 236, 258, 264,
271, 276, 380, 425, 427, 468
— сумка 23*, 206*, 219*, 224, 225,
391, 392, 392*, 421, 454, 456,
461, 467, 468, 469, 471, 477*,
486, 492
Выделительные каналы 310
Выдыхательная камера 102*, 103
Вымершие животные 171, 172, 248
Г
Галлоподобные вздутия 291
Галлы 278
Ганглии моллюсков 8, 21, 22, 23,
23*, 72, 109, НО, НО*, 157, 159*
— оболочников 332*, 334, 335*,
338, 340, 340*, 342*, 343, 347,
349
— хетогнат 298, 299*
Ганглий головной 506
— подглоточный 501*
Ганглиозные утолщения 204
Гастрозоиды 352*, 353
Гастролиты 521
Гаструла 196, 209, 344
Гектокотиль 169, 169*, 182
Гемоглобин 108, 157, 208, 370,
379, 384, 394
Гемолимфа 106, 107, 370
Гемоцианин 157, 370, 376, 379
Генитальная складка 39
Генитальный желобок 56
Гермафродитизм брюхоногих 13,
23, 28,38,40, 44,58, 61, 67, 72
— двустворчатых НО, 134, 154
— иглокожих 230, 246, 256, 289,
294
— моллюсков 10
— оболочников 335, 340, 343, 344
— протандрический (протеранд-
рический) 13, 29, 40, 44, 67,
88, 230, 256
— протерандрический
(протандрический) 477, 479, 517
— хетогнат 298
Гермафродитизм членистоногих
374, 379, 427, 432, 433, 466,
517
Гермафродитная железа 73*
Гетероморфоз 516
Гидростатический аппарат 65
Гиперпаразиты (сверхпаразиты)
419*
Гиподерма 368
Гипостракум (известковый слой)
8, 25
Гладиус 156, 191
Глаз (ок) науплиальный 378, 381,
386, 390, 392, 392*, 403, 420,
427, 430, 441, 451
Глаза брюхоногих 8, 23, 43, 48,
56, 72, 76*, 81
— головоногих 159*, 172, 174, 183,
185, 189
— двустворчатых 109
— иглокожих 204, 254
— мантийные 109, 125
— моллюсков 8
— оболочников 340*, 346*
— панцирных 16
— простые 372
— фасеточные (сложные) 372,
372*, 373*, 378, 381, 386, 390,
390*, 392, 392*, 403, 420, 441,
451, 460, 464, 470
— термоскопические 186
— членистоногих 372, 400, 41.8*,
444, 448, 450, 454, 455, 467*, 485
Глазки (омматидии) иглокожих
205, 272
— оболочников 334, 341, 345
— членистоногих 372
Глазная пластинка 272
Глазные пятна 485
— стебельки 496, 502, 507
— ямки 204, 204*
Глаукотоэ 480, 519, 520, 521, 528,
529
Глотка головоногих 156
— двустворчатых 68, 72
— иглокожих 204, 208
— моллюсков 9
— моноплакофор 94
— оболочников 330, 332, 332*,
333, 335*, 338*, 339, 340*,346*,
347
— полухордовых 325
Глоточное кольцо известковое
229*, 230, 234, 235
Глохидий 111, 111*, 133, 136,417
Глубоководные брюхоногие 43
— головоногие 169, 174, 183, 185
— двустворчатые 95, 111, 114,
124, 132, 155
— иглокожие 212, 220, 228, 242,
250, 253, 259, 264, 278, 287,
292 296
— оболочники 330, 341, 356, 358
— панцирные 16, 17
— погонофоры 306, 314, 315
— полухордовые 318, 319
Гнезда 117*, 120, 127
Голова моллюсков 8, 21, 21*,
23*, 29*, 67. 68, 70, 77, 94
Голова ракообразных 378, 381, 390,
392, 400, 403, 422, 437, 441,
451, 454, 460, 468, 470, 485,
501
Головной капюшон 173
— мозг головоногих 156, 157,
157*, 165, 177
— хрящ 159*
— щит 325, 325*, 326, 327*, 328
Головные лопасти 307, 308, 310*
Головогрудь 369, 403, 467*
Голотурии 239
Гонады (половые железы) 10, 110,
266, 321
Гонозоиды 352*, 353
Гребневидные органы 259, 260,
260*
Грудь 368, 378, 381, 392, 403,
443, 460, 501
Грунтоеды 214, 234, 317
Губа (скребок) 274*, 275, 285, 286
— верхняя 381, 386, 392, 400,
423, 431, 441*, 455
д
Двустороннесимметричные
животные 196, 199, 208, 215, 228, 236,
298
Дексиотропная (правозакручен-
ная) раковина 20
Деление иглокожих 211, 235, 237,
258, 259, 290
— полухордовых 322
— поперечное 258*, 290, 322
Детритоеды 12, 13, 24, 234
Дивертикулы печени 106
Диморфизм половой НО, 136,
182, 225, 276, 283, 477, 491, 517
Диплеурола 210
Диски иглокожих 257, 259, 262,
265, 270, 287*, 288, 292, 296
Дифинитическое происхождение
215
Долиолярия 210*, 211, 219,
219*, 231
Домики полухордовых 323, 326,
326*, 328, 341, 342*, 343
Донные брюхоногие 39, 63
— двустворчатые 112, 121, 125,
161
— иглокожие 231, 249
— оболочники 330, 331, 341, 356
— панцирные 14
— полухордовые 316, 317
Древние животные 97, 220, 250,
259, 281, 307, 328
Древоточцы 97, 148, 151, 151*,
152, 152*, 153*, 154, 154*, 306
Дробление яйца 10
— радиального типа 209, 310
— спирального типа 110
Дыхальце 370
Дыхание интрамолекулярное 108
Дыхательная камера мантийной
полости 103
— полость 465, 468
Дыхательный аппарат
полухордовых 320
34*
531
•ль?
Жаберная полость 23, 27, 35, 59,
460, 461, 501, 503, 504*
— септа 155
Жаберно-кишечный синус
оболочников 334
Жаберные артерии 102*, 333, 334
— вены 102*, 107
— глаза 109
— крышки 420, 421
— листочки 369, 461, 471
— листки 102
— нити 103, 103*, 104, 107, 1Q9,
132
— отверстия 320*, 322, 322*, 341,
342*, 345
— придатки 461
— сердца 157
— сосуды выносящие 102*, 107,
320
приносящие 102*, 107, 320
— щели 319, 320*, 325, 325*,
328, 332*, 333, 338, 340, 340*,
345, 347*, 349
Жаберный аппарат 316, 320
— мешок 333, 334
— отдел кишечника 320, 321
Жабры брюхоногих 21, 23, 27,
38,40, 43, 62, 70, 72, 76*
— вторичные 9, 23, 28, 29*, 33,
38, 58, 76*, 78
— головоногих 157, 157*, 172, 174
— двустворчатых 99, 103, 104*,
105, 105*, 111, 130, 155
— моллюсков 7, 9
— моноплакофор 94
— наружные 111, 279
— нитчатые 115
— оболочников 340*
— остаточные 504, 504*
— панцирных 14, 16
— первичные 28, 31
— перистомальные 208
— полухордовых 322
— ракообразных 378, 387, 390, 403,
405, 432, 440, 441, 447, 452,
456, 471, 471*, 485, 495, 501,
503, 504*
Жвалы (мандибулы) 378, 381,
386, 392, 400, 403, 417, 420,
422, 425, 431, 437, 441*, 454,
464, 471, 495, 502
Жгутики (жгуты) 387, 388, 502
Железа белковая 73. 73*
— лимфатическая 207
— ножная 40, 45, 56, 81
— синусная 501*
— скорлуповая 73
— ядовитая 21, 57, 203, 273,
274
Железистые клетки двустворчатых
101
иглокожих 202, 246, 253,
280, 290, 293
— — оболочников 333
— — погонофор 312
— — полухордовых 326
Железка околонервная 334, 335*,
338, 340, 349
Железы андрогенные 491
— антеннальные 379
— выделительные 386
— выделяющие кислоту 21
— клейкие 70
— максиллярные 379
— паутинные 441
— пищеварительные 14, 58,332*,
443
— сверлильные 52
— слизистые 44
— слюнные 9, 21, 49, 52, 58, 72,
181
— цементные 429, 489, 518
Желудок брюхоногих 21, 58, 72
— головоногих 156
— двустворчатых 102*
— иглокожих 208, 218, 249*,
254, 255*, 261, 267, 287
— моллюсков 9
— оболочников 332*, 333, 335*,
340, 340*, 341, 342*
— полухордовых 322*, 328
— ракообразных 379, 443
Желудочек сердца 21, 72, 102*,
105, 107, 157
Жемчуг И, 99, 121, 122, 123,
126
—, выращивание 123
Жемчужины образование 122, 123,
123*
Жемчужный промысел 123
Живорождение 211, 289, 335,
344, 375, 385, 478
Живородящие 18, 35, 39, 44, 88,
НО, 136, 170, 237, 263, 294,
322
Зародыш 258, 264,271, 276, 311,
330, 344, 354, 355*
Земноводный образ жизни 39
«Зеркальце» 125
Зияние заднее 151, 151*
— переднебрюшное 151*
Зияющая раковина 143, 148, 149,
151, 151*, 153
Зоб 21
Зооиды 325, 326, 326*, 327, 335*,
336, 337*, 339, 348
Зоэа 480, 519, 520, 520*, 521, 528
Зубные папиллы 287*, 289, 295
Зубовидные выросты 115, 149
Зубцы радулы (терки) 9, 14, 24*, 72
Зубы боковые 114*, 130, 132,
137*, 139, 140, 146, 147, 149
— двустворчатых 100, 114*, 132
— кардинальные 114*, 130, 137*,
139, 140, 143, 146, 149
— ложнобоковые 134*
— ложнокардинальные 133*, 134
— морских ежей 272, 281, 282,
285, 287*, 289
— устья легочных 71
И
3
Забота о потомстве у иглокожих
211, 224, 231, 256, 256*, 260,
276, 289, 293, 297
моллюсков 12, 40, 88, 120
Завиток 20, 21*, 41, 43, 46, 47,
75, 82
Закапывающиеся (зарывающиеся)
беспанцирные 13
— брюхоногие 24, 49, 52, 56
— двустворчатые 96, 97, 102, ИЗ,
130, 140, 148
— иглокожие 214, 230, 234, 236,
245, 261, 277, 294
— лопатоногие 92, 93
— полухордовые 324
Замковые зубы 100, 114*, 135
— — рудиментарные 136
Замок двустворчатых 96, 100,
114*, 132*, 137, 139, 140, 147,
149, 155
— гетеродонтного типа 132
— таксодонтного типа 111
— унионидного типа 132
Замочный край 100
Замыкающий аппарат
головоногих 183
Иглы вторичные 280, 281
— иглокожих 212
— морских ежей 273, 274*, 275*,
277, 278, 280, 284, 286
звезд 252*, 260, 265, 270,
271
— офиур 287*, 295
— первичные 281
Известковое тело сферидия 204*
Известковые пластинки
моллюсков 13, 14, 20
— тельца голотурий 246
— — межклеточные 202
— шипики беспанцирных 12, 12*
Известковый слой раковины 100
— стебелек иедицеллярии 273
Икра 17, 35, 39, 39*, 227, 231,
276, 282, 283, 289
— неритическая 35
Икрометание 231, 234, 235, 237,
283
Инвагинация 209
Интерамбулакральные пластинки
271, 272, 279, 281
Интерамбулакры 223
Интеррадиусы 201, 206, 216, 228,
232, 236, 250*, 252, 258, 270,
275
Инстинкт 374
Ископаемые иглокожие 198*,
251, 278, 285, 291
— моллюски 10, И, 97
— оболочники 328
— погонофоры 307
К
Кайма голотурий 242
Калиптопис 498, 498*
Калямус 169
532
Каменистый канал 200, 200*, 201,
207, 245
Камеры раковины головоногих
171, 172
Камнеточцы 96, 121, 146, 147, 151
Каннибализм 44, 89, 162, 266
Капилляры 157, 312*
Карапакс (головогрудной щит)
378, 386, 420, 421, 429, 440, 444,
446, 450, 451, 454, 455, 460, 462*,
465, 468, 470, 485, 495, 501, 506
Каротиноиды 486, 506
Каротины 486
Кастрация паразитарная 439
Каури 25
Кеберов орган 99*, 108
Кентрогон 438, 438*
Кинезис 374
Кишечник иглокожих 208, 209*,
218, 234, 246
— моллюсков 9, 94, НО, 110*
— оболочников 330, 335*, 336, 339,
349
— первичный 196, 311
— полухордовых 320, 322, 324, 326
— ракообразных 392*, 417, 420,
426*, 437
Кишка 9, 14, 21, 72
— задняя 23*, 105, 254, 311
— первичная 209, 333, 335*
— средняя 102*, 105
Кладка 36*, 55, 70, 75, 83*, 88, 89*
Клапан воронки 173
Клаузилий 71, 81
Клешни 454, 465, 485, 496, 501,
502, 503, 516*
Клоака иглокожих 209*, 230, 238
— оболочников 332*, 333, 335*,
338*, 339, 339*, 340*, 344, 347
Клоакальная камера 102*, 105
— полость 336, 340, 342
Клоакальное отверстие 335, 336,
338*, 340
Клоакальный сифон 333, 335,
335*, 338, 339*, 340*, 347*
Клюв головоногих 156, 162, 193
— (рострум) ракообразных 392, 403
Кнопки замыкающего аппарата
головоногих 157, 158, 183
Когти головоногих 183
Кожа брюхоногих 43
— голотурий 227, 236, 237, 239,
242, 243, 246
— иглокожих 202
— морских ежей 280
звезд 249*, 252*, 270
— офиур 297
Кожная складка голотурий 242
Кожное дыхание 9, 28, 81, 219
Кожно-мускульный мешок
иглокожих 202, 230, 239
Кожные включения 15
— жабры (папулы) 208, 252, 252*,
253, 256, 259, 264, 265, 269
— железы 29, 193, 323
— скелетные пластинки 229*, 235
Кожный скелет 63
— эпителий 308
Коконы брюхоногих 44, 45, 75, 86
Колонии оболочников 329, 331,
335*, 336, 337, 337*, 339
— полухордовых 316, 325, 326,
326*, 327*, 328
Колонка (столбик) брюхоногих 20
Кольца роста годичные 18, 101,
212
Кольцевой канал иглокожих 200,
200*
Комменсалы иглокожие 213, 231,
238, 247, 263, 284, 291
— моллюски 140
Конвергенция (конвергентное
сходство) 184
Конхин 15, 16
Конхиолин 8, 100
Копулятивный орган 37, 73*
Копуляция 390*, 415, 466, 491*
Коралловых рифов, обитатели 17,
64, 143, 226, 238, 246, 251, 261,
265, 277, 283, 295, 317
Корацидий 411, 412
Корешки морских лилий 221
Кормидии 348
Кормилка 351, 352*, 353
Кость каракатицы 184
Криптонисциум 479, 479*
Кристаллический стебелек
двустворчатых 105, 106
Кровеносная лакуна централь-
лая 320, 321
— система головоногих 157
— — замкнутая 157
— — иглокожих 205, 206*
— — моллюсков 9, 15, 106
— — незамкнутая 21
— — оболочников 330
— — погонофор 310
— — полухордовых 320
членистоногих 369,379*, 380,
403, 405
Кровеносные лакуны 205, 289,
333, 340, 352*, 471
Крона морских лилий 220
Крышечка (оперкулюм)
брюхоногих 33, 37, 41, 43, 45, 48, 52,
73, 83, 184
— головоногих 173
Крючья головоногих 183
Ктенидии (жабры) брюхоногих 21,
23, 23*, 58, 73, 105
— двустворчатых 103*, 104*, 105,
111
— моллюсков 9
Куколка 371, 419, 429
Кутикула моллюсков 12
— оболочников 332
— погонофор 308, 312*
— членистоногих 368, 369
Кутис 202
Кухонные кучи 24, 97, 120, 127
Кювьеровы органы 230, 239
Л
Лакун система 9, 106, 107, 205,
321 342
Легкие 39,72,73, 85, 370, 503, 504*
— водные 208, 209*, 227, 230
Легочная полость 72, 72*, 76, 81
Лейкоциты 108
Леотропная (лев озакр ученная)
раковина 20
Лигамент брюхоногих 70
— двустворчатых 99, 100, 101,
103*, 115, 155
Лигула 169
Лимфоциты 339
Линза 16, 44
Линька 371, 378, 382, 397, 401,
449, 453, 463, 467, 478, 492,
498, 517, 521, 529
Литодесма 155
Литоральные брюхоногие 29,35, 64
— двустворчатые 70, 109, 115,
118, 141, 147
— иглокожие 212, 235, 249, 264,
266, 279, 282, 287, 296
— оболочники 331, 357
— панцирные 14
— полухордовые 317, 318, 323
Личинка брюхоногих 22, 40, 42,
47, 52, 68, 70
— голотурий 231, 237, 241
— двустворчатых НО, НО*, 130,
132 154
— иглокожих 197, 210, 210*, 211
— моллюсков 13
— морских ежей 276, 286
звезд 248, 257, 266
лилий 219, 224, 225, 227
— оболочников 330, 334, 341,
344, 345, 346*, 348, 352*, 354,
355*
— полухордовых 321, 322, 322*,
326
— ракообразных 401, 402*, 405,
414, 421, 423, 423*, 426*, 427,
429, 433, 434, 438, 443, 449,
453, 454, 473, 479, 479*, 480,
495, 497, 498*, 501, 519, 520
аскоторацидная 426*,427, 428
копеподитная 418,419, 420*
циприсовидная 429, 430*,
431, 434, 437, 438, 438*, 439
Личинкородящие двустворчатые
Личиночное закручивание
брюхоногих 22
Лобная нить 419
Ловчий аппарат крылоногих 69
— — оболочников 333
хетогнат 298
Лучи ветвистые 288, 292
— морских звезд 250, 258, 259,
264, 265, 270
лилий 216, 217*, 218, 220, 223
— офиур 287, 287*, 290, 295
— простые 288
М
Мадрепорит (мадрепоровая
пластинка) 207, 209*
Мадрепоровая пластинка
(мадрепорит) 200*, 201, 207, 249*,
252, 260*, 265, 272, 274
Максиллы (челюсти) 378, 387
35 Жизнь животных, т. 2
533
Макушечная складка 151
Макушка 99*, 110, 116, 119*,
146*, 148, 151
Мандибулы (жвалы) 378, 380
Мантийная борозда 101
— жидкость 109
— линия 101
— лопасть 102, 144
— полость брюхоногих 22, 26,
33, 39, 40, 72, 91
— — головоногих 169, 170, 183
двустворчатых 102, 103,
103*, 104
— — моллюсков 7, 9, 10
ракообразных 427, 429, 432,
437, 442
— складка 102*, 429
— щель 158
Мантийное отверстие 158
спинное 99*, 102*
Мантийные вены 102*
— глаза 109, 125
Мантийный аппарат головоногих
157
— парус 125, 126, 127
— спинной канал 99*, 102*
Мантия брюхоногих 21, 26, 29*,
39, 47, 58, 69, 71, 91
— головоногих 157*
— двустворчатых 96, 99, 99*, 101,
103, 105*, 107,132,144,146,147
«— моллюсков 7
— моноплакофор 94
— оболочников 338
— панцирных 14, 15
— ракообразных 425, 427, 428,
429, 437, 437*
Марзупиум (выводковые камеры)
111, 132, 133
Маскировка 166
Мегалопа 480, 519
Мезентериальные перепонки 208
— тяжи 208
Мезентерий 246
Мезодерма 196, 310, 359
Мелководные двустворчатые 96,
114, 116, 121, 125, 126, 139, 148
— иглокожие 220, 249, 261, 266,
281, 287
— погонофоры 306
— полухордовые 325
Меридианы 271
Мерцательное поле 68
Мерцательные клетки 312
Мерцательный венчик 322
— поясок 322, 322 *
— шнур 322, 322 *
Метазоэа 519
Метамерия (сегментация) 7,10, 94,
308, 314
— внутренняя 10
— наружная 10
Метамерное строение 7
Метаморфоз иглокожих 256, 257,
262, 276
— моллюсков 110
— оболочников 341, 345
— полухордовых 321, 322
— членистоногих 371, 403,498
Метанауплиус 389, 390*, 391,
398, 405, 418, 423, 427, 429,
434, 437, 438, 498, 498*. 519
Метанефридии 379
Метаподиум 24, 45
Механорецепторы 104
Миграции вертикальные 298, 407,
408 *, 409, 458, 488, 497
— горизонтальные 458
— кормовые 458, 528
— нерестовые 520, 527
— онтогенетические 458
— сезонные 408, 409, 497
Мидиевые банки 116, 118, 119
— парки 120
Мидиевый ил 119
Мизис 519
Микронисциум 478
Митилоидная раковина 116, 121
Мицетом 168, 169, 184
Мозг головной 373, 374
Морские беспанцирные 13
— брюхоногие 24, 32
— вторичноротые 196
— головоногие 156, 160, 185
— двустворчатые 95
— иглокожие 232, 248
— моллюски И
— оболочники 329
— панцирные 14, 19, 20
— погонофоры 310
— полухордовые 316, 317
— щетинкочелюстные 298
Мускул-втягиватель 74*
—замыкатель 390, 390*, 440,
441*
задний 99*, 102*, 105, 105*,
114*, 116, 132
передний 99*, 102*, 105*,
114*, 116, 122
ножной 114*
раковинный (аддуктор) 27
—ретрактор ноги задний 99*,
102*
передний 99*
Мускулатура (мышцы, мускулы)
иглокожих 158, 230, 246, 253
— моллюсков 17, 43, 72
— оболочников 338, 339
— погонофор 307, 308, 312*
— полухордовых 319, 320*
— хетогнат 298
Мускулы (мышцы) - дилататоры
165
—замыкатели 70, 100, 101,
111, 151
—подниматели 102
—протракторы 99*, 102, 230, 232
—ретракторы 102, 230, 232, 237
И
Наземные животные 20, 24, 33, 71,
78, 81
Наннопланктон 343
Науплиус 378, 380, 382, 383, 384,
385, 389, 391, 401, 405, 407, 414,
414*, 415, 418, 418*, 427, 429,
430*, 434, 437, 438, 438*, 440,
441*, 443, 480, 498, 498*, 519
Невроцель 196
Нектон 161
Нематоцисты 167
Неотения 345
Нервная система иглокожих 196,
203, 204*, 205, 254
— — моллюсков 8, 15, 21, 72,
109
оболочников 330, 334, 340,
343
— — погонофор 308
— — полухордовых 321, 322
ракообразных 380, 438
Нервная трубка 346*
— цепочка брюшная 374
Нервное кольцо (кольцевая ко-
миссура) 14, 203
— — окологлоточное 8, 14, 72
Нервные клетки 8
— комиссуры 109
— стволы моллюсков 8, 14, 23,
102*, 109
погонофор 308, 311*
— — полухордовых 321, 328
— узлы 8, 21, 109
Нервных клеток скопление 21, 308
— стволов перекрест 23, 58
Нефридии 172, 174, 321
Нефроциты 334
Нидхэмов орган 169
Нога брюхоногих 2.1, 24, 26, 42,
44, 56, 68, 70, 73
— головоногих 156, 173
— двустворчатых 94, 99, 99*,
101, 102*, 105, 110*, 123, 125,
140, 144, 151, 151*, 153
— лопатоногих 91
— моллюсков 7, 11, 14
Ногочелюсти 378, 404, 422, 443,
451, 452, 454, 456, 460*, 461,
465,467*, 468, 496, 501, 502
Ножки (конечности) брюшные 444,
447, 449, 450, 452, 456, 461,
468, 471, 471*, 478*, 486
— грудные 380, 381, 381*, 386,
387, 387*, 388, 390, 393, 394,
400, 402*, 403, 404, 417, 418*,
420, 422, 425, 427, 428, 429,
431, 431*, 432, 437, 440, 441*,
444, 445*, 446, 447, 449, 450,
451, 456, 461, 462*, 465, 468,
469, 471, 485, 495, 496, 501,
502, 503, 504*
Ножная вена 102*
Норы двустворчатых 150, 269
— морских ежей 282, 283, 284, 286
— полухордовых 321, 323, 323*
Нотохорд 320, 320*, 321, 322,
325, 328
Нотум 62
О
Обонятельные клетки 159
Обрастатели 138, 306, 358
Околожаберная (перибранхиаль-
ная) полость оболочников 333,
335, 346, 346*
534
Околоротовой сфинктер 228
Околосердечная сумка
(перикардий) 9, 15, 310, 333
Омматидии (глазки) 372
Онфауна 132
Оозоид 348
Оостегиты 456
Оперкулюм (крышечка) 43, 45
Оплодотворение внешнее
(наружное) 170, 344, 375
— внутреннее 22, 170, 335, 375
— головоногих 169, 170
— иглокожих 297
— моллюсков 18, 83, НО, 118,
133
— наружно-внутреннее 375
— оболочников 335
— перекрестное 10, 29
— полухордовых 321
Опорные клетки 204*
Оптические органеллы 110
Оральная (ротовая) сторона 205,
216, 218, 250, 276
Оральный диск 218, 223
— кольцевой канал 206*
Орган копулятивный 37,73,379,505
— совокупительный 472, 504
— четырехклеточный 386
Органическое вещество
раковины 8, 15
Органы вкуса 205
— внутренней секреции 455, 502,
506, 521
— выделения 109, 256, 310, 340,
371, 379, 438
— дыхания 427, 468, 472
— обоняния 22, 58, 81, 94, 205,
343, 509
— осязания (химического
чувства) 22, 44, 58, 204, 509
— Песта 522
— половые 418, 442, 472
— равновесия (статоцисты) 43,
43*, 44, 109, НО, 205, 334, 343
— статические 205
— стридуляционные 373
— химического чувства (осфра-
дии) 22, 81, 308
— чувств головоногих 159
иглокожих 204
— — моллюсков 15, 16, 22, 109
— — полухордовых 321
Ортонауплиус 405
Осевой орган кровеносной
системы 205, 206*, 207
Оскулюм (устье) 264
Остракум 8, 25
Осфрадий (орган химического
чувства) 9, 23, 53, 58, 72, 109, НО
Осязательные клетки 159
— усики 31
Отверстие анальное 379, 437
— половое 379, 382, 387, 404,
442, 443, 469, 498
— ротовое 437
Отдел брюшной 394, 404, 420, 422,
429, 451, 456, 465, 468, 469,
471, 486, 504
— головной 429
Отдел головогрудной 420
— грудной 392, 400, 403, 404,
418, 425, 429, 431, 437,
437*, 444, 451, 461, 468, 486,
504, 504*
Отолит 43*, 204*
Отоцисты 204*, 205
Офиоплутеус 210*, 211, 289
II
Паксиллы 251*, 252, 253*, 258,
260*, 261
Палетки 152*, 153, 153*, 154*
Панцирь иглокожих 211, 216,
271, 275, 281, 291
Папиллы иглокожих 218, 239,
243, 287*, 288, 293, 295, 296
Папулы (кожные жабры) 208,
252, 265, 269, 270
Паразитизм внутренний 24
— иглокожих 196, 213
— моллюсков 24, 51, 111
— наружный 24, 40
Паразитические иглокожие 264,
290*, 293
— моллюски 24, 40, 50, 278, 280,
291
— черви 214*
Паразитический образ жизни 51,
264
Параподии 58, 61
Партеногенез (партеногенетиче-
ское размножение) 154, 374, 379,
383, 385, 397, 416, 442, 477,
479
Парус 10, 45, 46*, 47, 70
Парусник (велигер) 10, 110*, 111,
118, 130, 150
Педицеллярии 203, 212, 252*,
253*, 259, 265, 269, 273,
275
— гребенчатые 203, 253*
— двустворчатые 203
— змееголовые (офицефальные)
273, 275*, 281
— пучковидные 253*
— скрещенные 270
— трезубые (тридентные) 273,
275*, 281
— трилистные (трифильные) 273,
275*
— шароносные (глобиферные) 273,
275*, 282, 284
— ядовитые 203, 248, 269, 274,
275, 283, 284
Пелагические брюхоногие 41, 42,
44
— головоногие 174, 183
— иглокожие 231, 242, 244
— оболочники 329, 336, 336*, 337,
359
— полухордовые 328
Пелагический образ жизни 44,
66, 214, 338
Пенис 431*, 432, 433, 437*
Пентакринусная стадия 219*, 220,
224, 225
Пентактула 231
Перибранхиальная
(околожаберная) полость 333, 346*, 347
Перивисцеральная (полостная)
жидкость 207, 208
Перигемальная (псевдогемальная)
система 206
Перикардиальная сумка 105
Перикардий (околосердечная
сумка) иглокожих 205, 207
— моллюсков 9, 15, 21, 58, 72,
99*, 102*, 105, 107
— оболочников 333, 335*
— погонофор 333, 335*
Периостракум 8, 15, 20, 80, 100,
115, 139, 151
Перипрокт 271, 272
Перистом 254, 271
Перистомальные пластинки 274*,
275
Перламутр 11, 27, 28, 31, 98,
101, 122, 133, 136, 173
Перламутровый слой 21, 26, 32,
100, 101, 112, 122, 123*, 132,
133, 135
Петазма 498
Петалодии (петалоиды) 285, 286
Петалоиды (петалодии) 274*, 275,
276, 285
Печеночные выросты 208, 209*,
249*, 260, 427
— железы 220*
— (пилорические) придатки 254,
256, 320
Печень 9, 21, 23*, 58, 63, 72, 102*,
106, НО*, 157, 333
Пещерные брюхоногие 36, 38,
73, 88
Пигидий 378
Пигмент 16, 44, 262
Пигментные клетки 202
Пиннулы 206, 217*, 218, 219*,
223,225, 308, 312*
Пищеварение внутриклеточное
106, 154
Пищеварительная система
иглокожих 208
— — моллюсков 105
— — оболочников 333
— — погонофор 311
Пищевод головоногих 156
— двустворчатых 102*, 105
— иглокожих 204, 218
— оболочников 332*, 333, 335*,
339*, 340*, 342*, 346*
— полухордовых 320
Пищевой желобок 104, 104*
Плавательные лопасти 68
Плавательный диск 244
Плавники 42, 43, 67, 69, 156, 158,
174, 183, 299*
Планктонные животные 34, 41,
161, 211, 298, 359
Планктонный образ жизни 24, 34,
58, 67
Пластрон (щит) 275
Плацента 354, 355*
Плевроцеркоид 412
Плеоподы 477*, 496, 504, 521*
35*
535
Плеотельсон 465, 469, 471, 471*,
480
Пневмостом 81
Поджаберный желобок 333
Поджелудочная железа 157
Подклешнд 451, 465, 471, 485,
501, 502
Подошва брюхоногих 29, 40, 42
— двустворчатых 112, 113, 114,
151
Позвонки (членики) 217*, 218,288,
293
Пойкилосмотические животные
248
Полиевы пузыри 200, 200*, 209*,
232, 249*
Полиморфизм 517
Полиплоидность 383
Половая пластинка 272
— система брюхоногих 22
— — иглокожих 206, 215
кишечнодышащих 321
ракообразных 380, 432
Половое отверстие 39, 236, 272
— — женское 42
мужское 310*, 312
Половой синус 207
Половые железы (гонады)
иглокожих 206, 209*, 230, 249*,
266, 271, 276, 279
моллюсков 10, 15, 58, 72,
102*, НО, 116
— — оболочников 335, 336, 344
полухордовых 321, 323,
326
— поры 320*
— тяжи 206
Положение рукопожатия 517,
517*, 527
Полостная жидкость 207, 208, 233,
248, 369 ч
Полость тела вторичная (целом)
94, 196, 310
— — сегментированная 10
Полужабры 99*, 102, 102*, 103,
103*, 115, 132, 133, 134, 146*,
155
Поплавок 44
Поровые каналы 207, 245
Постабдомен 349
Почки брюхоногих 15, 21, 23,
58, 72, 126
— головоногих 172
— двустворчатых 102*, 109
— моллюсков 10
— моноплакофор 94
— накопления 334, 340, 341
— полухордовых 325*
Почкование мантийное 348
— оболочников 329, 335, 339, 349,
351, 354, 356
— пилорическое 348
— постабдоминальное 348
Практическое значение
ракообразных 377, 399, 416, 436, 460,
464, 484, 485, 495, 499, 524,
527 529
Праница 473, 476, 478, 479*
Превращение сложное 375
Предпочки 352*, 353
Предсердие 15, 21, 23, 72, 94,
102*, 107, 157
Пресноводные брюхоногие 20, 24,
32, 36, 38, 71, 78
— двустворчатые 95, 101, 108,
132, 137, 138
— панцирные И
Прибойные (обитающие в зоне
прибоя) 14, 16, 30,93, 272, 281,
283
Прибрежные брюхоногие 20, 29,
34, 49
— головоногие 161, 175, 184
— двустворчатые 95, 97, ИЗ,
116, 124, 130, 139, 146,149,
152
— иглокожие 225, 232, 236, 242,
278, 283, 287, 292
— погонофоры 306
— полухордовые 317
Призматический слой 100, 101,
122
Прикрепительный диск 220
— сосочек 322*, 346, 346*, 354
Прикрепленные формы 199, 220,
316, 329
Прикрепленный образ жизни 39,
НО, 117*, 137, 216, 220, 222,
347
Прирастающая створка 128,129*,
140
Присасывательный диск 201, 284
Присоска ротовая 40
Присоски головоногих 156, 159,
162*, 166, 174, 181
— иглокожих 201, 228, 232, 253,
255*, 281
Продиссоконх (первичная
раковина) 110
Промысел головоногих 191, 192,
193
— гребневиков 267
— двустворчатых 120, 126, 128,
130, 150, 267
— иглокожих 227, 241, 276, 282,
283
Промысловые брюхоногие 25, 27,
30, 52, 59
— головоногие 191
— двустворчатые 98, 115, 119,
126, 129*, 146, 148
— иглокожие 232, 238, 239
Проподиум 24, 44, 45, 56
Протаспис 376
Протогиния 466, 477
Протозоэа 528
Процеркоид 412, 412*
Псевдогемальная (перигемаль-
ная) система 206
Псевдозоэа 452*, 453
Псевдоколониальность 326
Псевдоконха 69
Псевдотрахеи 471*, 472
Псевдофекалии 105, 105*, 118
Псевдоэпиподит 400
Пупок 20, 44
Пурпурная железа 54
Пурпурное вещество 55
р
Радиальная пластинка 217*
Радиальные выросты желудка 254
кишечника 208
— каналы водоносной системы
200, 200*, 204, 205, 206, 206*
Радиусы (амбулакры) 201, 216,
228, 230, 232, 234, 236, 250, 275
Радужина 159*
Радула (терка) брюхоногих 21,
24*, 33, 51, 58, 67, 87
— головоногих 157, 173, 183
— моллюсков 9, 17
— моноплакофор 94
— панцирных 17
Разведение моллюсков 98, 106,
120, 123, 127, 128, 130
Развитие иглокожих 209, 219*,
220, 224, 235, 246, 257, 276, 289
— моллюсков 10, 13*, 110
— оболочников 336, 345
— полухордовых 321, 322
— прямое 380, 495, 501, 519
Размножение иглокожих 231,241,
256, 266, 286, 289, 294
— моллюсков 18, 38, 82, 86, 199
— оболочников 343, 347*, 350*,
352*, 355*
— ракообразных 391, 397, 398*,
432, 449, 466, 467, 477, 497
— бесполое иглокожих 211, 231,
235, 258*, 262, 290, 295
кишечнодышащих 322, 328
оболочников 335, 339, 344,
349, 356
— половое иглокожих 211, 258
кишечнодышащих 321, 322,
328
оболочников 335, 336, 343,
345, 351, 356
Раковина брюхоногих 20, 26, 28,
29, 30, 37, 44, 68*, 70, 78, 80
— внешняя (наружная) 172
— внутренняя 46, 69, 172, 184
— вторичная 69
— головоногих 156, 171, 172,
182
— двустворчатых 96, 97, 99, 101,
103*, 105*, 111, 137*, 139, 145,
147, 153*, 154, 155*
— левозакрученная 20, 39, 71,
74, 74*, 76*, 77
— личиночная 10, 42, 68, 71
— ложная 153*, 155, 155*
— лопатоногих 91
— минеральная 7
— митилидная 116
— моллюсков 7, 8, 11
— моноплакофор 94
— наружная (внешняя) 70, 85,
172, 182
— панцирных 14, 15
— правозакрученная 20, 39, 71
— ракообразных 390, 392, 428,
431, 431*, 440, 441
— роговая 184
— рудиментарная 85, 156
— хитиновая 184
536
Раковинные пластинки
панцирных 15, 16, 18
Растительноядные 58, 60, 72, 85,
87, 283
Расы полиплоидные 385
Рачья чума 525
Реактивное движение 158, 185,
337, 339
«Реактивный двигатель»
головоногих 157
Ребра 132, 146, 151, 174
Регенерация (самовосстановление)
брюхоногих 64
— иглокожих 211
— морских звезд 259, 262, 268
лилий 218, 223
— оболочников 357
— офиур 290, 294, 297
— полухордовых 324
— ракообразных 516
Редукция амбулакральной
системы 226
— амбулакральных ножек 201,
242 243
— глаз 88, 174, 183, 205
— головы 95
— жабр 28, 33, 67, 155
— заднепроходного отверстия
114
— замка 132, 148
— замковых зубов 132
— зубов 140, 148
— ктенидия 9, 22, 23
— мускула-замыкателя 115, 121,
144
— мускулатуры 202
— ноги 40, 58, 67, 116, 127
— полужабры 155
— почки 23, 51
— предсердия 23
— радулы 174
— раковины 7,15, 21, 50,57, 62,
67
— сердца 24, 51, 58
— слюнных желез 9
— цирр 226
— щупалец 308
Реснички иглокожих 202, 285
— моллюсков 11, 69, 104, 104*
— полухордовых 324, 326
Ресничная полоска спинная 308,
310*
Ресничные бороздки 312, 312*
— воронки 217*, 218, 246
— клетки 22, 43*, 333
— кольца 69
— шнуры 224, 225
Ресничный аппарат 69
— пояс 154*
Ретина 125
Рибофлавин 233
Ринофоры 58, 62, 67
Рифы коралловые 17
Роговица 44, 125, 159, 159*,
184
Рост раковины 101
Рострум (клюв) 392, 403, 444,
451, 455, 464, 468, 501
— ложный 460
Рот брюхоногих 58, 69, 70
— головоногих 156, 173
— двустворчатых НО*, 114*, 146*
— иглокожих 199, 208, 209*, 222,
228, 232, 255*, 271, 287*, 289
— моноплакофор 94
— оболочников 342*, 346*
— панцирных 14
— первичный (бластопор) 196, 210
— полухордовых 320, 320*, 322*,
325, 325*
— ракообразных 392, 400, 403,
423, 431
Ротовая полость (стома) 9, 21
— пирамида (конус) 425, 426*,
431*
Ротовое отверстие иглокожих 210,
217*, 235, 272
моллюсков 9, 48, 99*, 102,
102*, 144
оболочников 333, 337, 338,
339
полухордовых 322
— — хетогнат 298
— поле (перистом) иглокожих
254, 271, 272, 274*, 276, 279
Ротовой диск 217*, 222, 223
— конус 170
Ротовые лопасти 76*, 99*
— ножки 272
— пластинки 272
— придатки 420, 422, 428, 429,
444, 464, 471, 485
Роющий образ жизни иглокожих
207
кишечнодышащих 324,
326
Руки головоногих 156, 158, 174,
183
— крыложаберных 325*, 327*, 328
— морских звезд 250, 255*
лилий 218, 222
с
Саккулы 217*
Самоампутация 56
Самокалечение (автотомия) 163
Самооплодотворение 154, 335
Самопожирание (автофагия) 162
Сверлильная железа 45
Сверлящие губки 126
— моллюски 19, 54, 97, 121, 132,
147, 150*, 151, 151*, 152
— морские ежи 277, 282, 283
— погонофоры 306, 313*, 314
Сверхпаразит 419, 477
Светочувствительность
диффузная 205
Светочувствительные клетки 16,
159, 321
Светящиеся бактерии 168, 341,
361
— брюхоногие 44, 67
— головоногие 167
— двустворчатые 51
— органы (фотофоры) 167, 183,
338, 338*, 341, 361
Светящиеся офиуры 291, 293, 294
— кишечнодышащие 321
— оболочники 360, 361
Свечение (люминесценция) 202,
321, 361, 362
— моря 362
Свободноживушде иглокожие 200,
220 222 225
— оболочники 329, 336, 338, 341
— полухордовые 316, 326
Связка (лигамент) 101, 102*, 103*,
132, 146*, 151
— внутренняя 148, 149, 151
Сегментация (метамерия) 10
Сегменты погонофор 308, 309*,
310*, 311*, 314
— наружные 100, 131*, 133*
— полухордовых 316, 321
Седлышко (эфиппиум) 398
Секрет слизистый 71
Секторы 201
Семенники 332*, 335, 335*, 338,
339, 340*, 343, 437*, 438
Семенные мешки 310
Семеприемник 73, 73*, 170
Семепровод (ы) 73*, 311*, 432
Септы 103*, 172
Сердце брюхоногих 21, 23*
-— головоногих 157
— двустворчатых 106
— иглокожих 205, 310
— моллюсков 9
— моноплакофор 94
— оболочников 332*, 335*, 336,
339, 339*, 340*, 346*, 347*, 350*
— погонофор 311*
— полухордовых 321, 322*, 325
— ракообразных 379,392*, 394, 405,
421, 432, 440, 443, 471, 486
— членистоногих 369
Сетчатка 44, 109, 125, 159, 159*
Сидячий образ жизни 7, 122, 316,
317, 325, 326, 334
Сизигиальные швы 218
Симбиоз 144, 145, 514, 515*
Симбиотические грибки 335
Симметрия двусторонняя 8, 63,
215, 243, 257, 270
— пятилучевая 199, 206, 208, 232
— радиальная 199, 215, 236, 276
Синзоэа 453
Синус венозный 107
— осевой 207
Синусы 9, 107, 143, 147
Сифон (ы) брюхоногих 21, 45, 45*,
48, 51
— вводной (дыхательный) 99*,
102*, 103, 105*
— выводной (выдыхательный) 99*,
102*, 114*, 119*, 140, 150, 158,
331, 338
— головоногих 172, 174
— двустворчатых 96, 100, 114,
131*, 132, 151, 151*, 154*
— клоакальный 331, 335*, 340*,
347*, 350*
— ротовой 331, 332*, 335*, 338,
340*, 347*, 350*
— ракообразных 460*, 461
537
Сифонные дудки 73
Скафогнатид 502, 503
Скелет аборальный 252*
— внутренний 200, 251, 288
— иглокожих 199, 202, 203, 216,
251, 270, 276, 285
— кожи голотурий 229
— наружный 196, 229, 287, 369
— ротовой морских лилий 217*
— хоботка кишечнодышащих 320
Скелетные пластинки 212, 218,
236, 245, 251*, 259, 270, 278,
288 320*
— элементы 216, 218, 229*, 243,
245 275*
Скорлупа 271, 272, 274*, 275, 276,
278, 279, 282, 283, 284, 286
— эластичная 278
Скребок (губа) 274*, 275
Сожительство 261, 291, 326
Солоноватоводные животные 36,
38, 137, 140
Сосуды кровеносные 443
— мальпигиевы 371
Спаривание 379, 382, 382*, 387*,
388, 391, 398*, 401, 405*, 423,
433, 457, 457*, 462*, 463, 478*,
491, 491*, 517, 518*, 528
Спат 130
Сперма 154, 169, 226, 231, 247,
289, 310, 335
Сперматозоид(ы) 15, 72, 73, 83,
169, 182, 230, 266, 310, 313,
335, 342, 344, 374, 375, 379,
442
Сперматофор (ы) 73, 83, 169, 182,
310, 313, 374, 375, 379, 404,
405, 405*, 498, 517, 528
Спикулы 332
Спинная пластинка 333, 337, 340
— струна 329, 330
Спиракулы 264
Спячка 39, 84
Стадия декаподитная 519
— кентрогона 438
— копеподитная 405, 407, 408,
414, 418, 419
— манка 463, 467, 478, 479, 492
— мизисная 480, 519
Статоконии 109, 126
Статокрипт 109
Статолпт 446, 456, 485, 502
Статоцисты головоногих 160
— моллюсков 8, 22, 58, 67, 72,
82, 109, 126
— оболочников 334, 342, 342*
— ракообразных 446, 450, 456,
473, 485, 502
Стебель крыложаберных 325,
325*, 326, 328
— морских лилий 216, 220, 222
Стернум 274, 275
Стигмы (жаберные щели) 333, 341
Столбик (колонка) 20, 21*
Столон 326, 326*, 339, 341, 350*
— почкородный 347, 349, 351,
353, 356
Стрекательная капсула 65
Стрекательные клетки 65
Супрадорзальная мембрана 264
Суставная сумка 273
— ямка 273
Сферидий 204*, 205, 278
Сфинктеры 331, 332
Съедобные двустворчатые 98, 116,
120, 127, 141, 146, 149
— иглокожие 227, 238, 276, 282
— панцирные 19, 27
Т
Тека 327
Теликум 498
Тело жировое 371
Тельсон 378, 382, 383, 386, 390,
401, 404, 443, 444, 447, 449,
450, 452, 454, 456, 460, 462,
468, 469, 471, 486, 496, 497,
504
Теменная пластинка 322
Теменной султан 322, 322*
— чувствительный орган 326
Тентакули 183
Тепловодные формы 44, 97, 139,
147, 238, 319, 331, 357
Терка (радула) 157, 189
Тиамин 233
Тигмотаксис 373
Тидемановы тельца 201
Токсопнеустес — токсин 284
Торнария 316, 322, 322*
Трахеи 370
— ложные 484
Трепанговые поля 240
Тривиум 228
Тропические брюхоногие 33, 34
— головоногие 179, 181, 184
— двустворчатые 95, 126, 139,
143, 147, 152, 154
— иглокожие 236, 246, 256, 280,
292 294
— оболочники 341, 357, 359, 360
— полухордовые 319, 320*
Трохофора 10, 110, НО*, 118
Трубки (трубочки)
кишечнодышащих 323
— крыложаберных 325*, 327
— погонофор 303, 305, 306, 311,
3i3 313*
Трубочки-теки 326*, 328
Туника 329, 331, 332, 333, 336,
338*, 339, 340*, 347*, 350*
У
Уздечка 307, 309*, 310*
Ультрабиссальные 112
Умбрелла 174, 183
Уроподы 446, 447, 449, 452, 454,
456, 462, 466, 468, 469, 471*,
472, 477*, 486, 497, 504
Условные рефлексы 157, 254
Устричники 128, 266
Устричные банки 99, 128, 130,
213, 268, 283
— парки (хозяйства) 130, 193,
266
Устье 21, 21*, 35, 41, 54, 56, 71,
75, 81
Ушки двустворчатых 125, 126
— морских ежей 272
Ф
Фагоцитобласт 196
Фазеолиновая зона 120
Фазеолиновые илы 120
Фарфоровидный слой 20
Фасетки 372*, 392, 501
Фасциоли морских ежей 274*, 275
Фасциолез 76
Фауна интерстициальная 412, 413,
413*, 422
Филлозома 480, 519, 520
Филаменты (жаберные нити) 103
Фильтраторы брюхоногие 24
— двустворчатые 104,116,145,150
— моллюски 11
— оболочники 331, 356, 362
Форозоид 352*
Фороциты 336, 351, 352*, 353
Фотогенное тело головоногих 167
Фотогенные клетки головоногих
168
Фоторецепторы 109
Фототаксис отрицательный 374
— положительный 374
Фотофоры (светящиеся органы)
167, 168, 183, 496, 522*, 523
Фурка (вилочка) 382, 383, 386,
425, 428, 440, 442, 444, 450
Фурцилия 498
X
Халимус 419, 419*
Хвост 243, 245, 342, 342*, 344,
346*
Хвостовой веер 447, 454, 456,
472, 504
Хеморецепторы 104
Хитин 313, 368
Хищные брюхоногие 24, 51, 53*,
58, 66, 69, 87*
— головоногие 156, 162, 174, 180,
184
— двустворчатые 128, 155
— иглокожие 213, 249, 263, 267,
269
— моллюски 9
— полухордовые 317
Хобот (хоботок) моллюсков 40,
45, 48, 51, 69
— полухордовых 319, 320*, 323,
326
— ракообразных 418, 420
Хозяин промежуточный 76, 89
Хозяйственное значение
головоногих 191
— — иглокожих 227
— — моллюсков 24
Холодноводные иглокожие 224,
264
— моллюски 12, 95, 97, 111,
114, 139, 141
— оболочники 357, 359, 360
538
Хорда 320, 341, 342, 342*, 344,
346*, 352*
Хроматофоры 165, 166, 168, 177,
190*, 455, 501*, 506, 506*
Хрусталик 88, 109, 125, 159, 159*
Хрящевая капсула 157*
Хрящевой скелет 156
— череп 156
Хрящи опорные 156
ц
Целлюлоза 359
Целом (вторичная полость тела)
196
— иглокожих 202, 207, 209*
— моллюсков 9, 10
Целомическая жидкость 207
Цел омические каналы 311*, 312*
— карманы 209
— мешки 210, 310
Целомодукты 310, 311*
Ценоций 326, 327, 327*, 328
Центральная пластинка 216
Центральный диск 250
— конус 216, 217*,, 226
Цефалоконии 70
Циатозооид 349, 350, 350*
Цикломорфоз 395, 395*
Цирры головоногих 183
— иглокожих 216, 217*, 220, 221,
224
Циртопия 498
Цисты 411, 415*, 416
Цистоидейная стадия 219*, 220
Ч
Чашечка 216, 217*, 220, 222
— дициклическая 216
— моноциклическая 216
Челюсти 68, 77, 287*, 289, 369, 378,
381, 386 392, 422, 425, 437, 455,
464
— задние 400, 404, 420, 431, 441*,
455, 502
— передние 400, 404, 420, 431,
441*, 455, 467*, 502
— роговые 173, 180
Чередование поколений
оболочников 329, 339, 354, 356
— — полухордовых 323
Чернила головоногих 163, 165,
168, 178, 179, 184
Чернильная железа 173, 181
— жидкость 184
Чернильный мешок 157*, 163,
164, 168, 181, 183
«Чертовы пальцы» 172
Членики (позвонки) 216, 217*,
218, 220, 295
Чувствительные клетки 22, 43*,
58, 109, НО, 204, 204*, 205,255
— папиллы 239
Ш
Шипы 260, 260*, 262, 269
щ
Щетинки 307
Щит (пластрон) 275
— головогрудной (карапакс) 386,
420, 503
Щитки адоральные 287*, 289
— боковые 287*, 288
— брюшные 287*, 288
— радиальные 287*, 288, 295
— ротовые (оральные) 287*, 289,
295
Щупальца брюхоногих 22, 44,
56, 70, 72, 81
— головоногих 156, 158, 175,
178, 183
— двустворчатых 105
— иглокожих 232, 233, 237, 245
— моллюсков 8
— оболочников 332, 338, 340*
— погонофор 307,308, 310*, 311*,
312*, 315
— полухордовых 325, 325*
Щупальцевая крона 309*, 315
— пластинка 308, 309*, 311
Щупальцевидные придатки 23, 31,
31*, 38, 40, 56, 58, 72
Щупальцевый аппарат 311, 314,
326
Щупик 403, 422, 441, 444, 455,
461, 464
Э
Эврибатные животные 212, 244,
247
Эвригалинные 150
Эвритермные 212
Экзогастрическое закручивание
раковины 172
Экзоподит 400, 444, 447, 456
Эктодерма 196
— оболочников 330, 332, 347,
349
Эктопаразиты (паразиты
наружные) 50, 51*, 417, 427
Элеваторы (мускулы-поднимате-
ли) 99*, 102
Эмбрионы 224, 235, 237, 289,
294, 297
Эндемичные брюхоногие 84
— двустворчатые 138, 142
— иглокожие 213
— погонофоры 306, 315
— формы 84, 138, 142, 213, 306,
315
Эндопаразиты 417, 427
Эндоподит 444
Эндостиль 320, 332*, 335*, 338,
338*, 340*, 346*, 347*
Энтодерма 196, 349
Эпиневральные каналы 204
Эпикаридиум 478, 479*
Эпиподиальные осязательные
усики 31
Эпиподит 444, 447, 448, 451, 456,
460*, 461, 465, 468, 485
Эпиподиум 24, 26, 31
Эпистернум 274*
Эпителий 103, 311
Эпителий мерцательный И, 308,
319, 325
— многослойный 196
— однослойный 202, 308, 319,
331, 332
— покровный 196, 202, 204, 218,
251
— полости тела 204
— ресничный 41, 103, 204*, 207,
246, 259, 273
Эпифрагма 71, 83
Эрионеикус 480, 519
Эритроциты 108
Эстеты 16, 19
Эфиппиум (седлышко) 398, 398*
Эхиноплутеус 210*, 211, 276, 286
Я
Яд брюхоногих 57
— головоногих 181
— голотурий 239
— морских ежей 274, 283, 284
Ядовитые голотурии 239
— морские звезды 263, 265
Ядовитый секрет 9, 202, 226, 280,
284
Язык головоногих 157
— панцирных 14
Яичники иглокожих 247, 258,
289* 290 297
— оболочников 332*, 335, 338,
339, 340*, 342*, 343
— погонофор 310
— ракообразных 390*, 392*, 418*
Яйца беспанцирных 13
— брюхоногих 38, 46, 51, 56,
66, 68, 75, 83, 86
— головоногих 170, 170*, 173
— двустворчатых НО, 118, 129,
134, 154
— зимние 384, 385
— иглокожих 211, 219, 225, 226,
241, 257, 266, 286, 299
— оболочников 335, 336, 340, 344
— панцирных 15, 18
— партеногенетические 379, 383,
385, 397
— погонофор 310
— покоящиеся 386, 389, 398, 410,
413, 414, 416
— полухордовых 321, 322
— ракообразных 382, 382*, 384,
385, 389, 390*, 391, 397, 398*, 421
Яйцевая капсула 36*, 45, 51, 56,
387
Яйцеводы брюхоногих 73*
— головоногих 170
— погонофор 310
Яйцевой (ые) мешок (ки) 382, 401,
405, 415, 418*, 419, 420*, 421,
453, 498
Яйцевые шнуры 68, 69, 75
Яйцеживорождение 375
Яйцеклетка оболочников 344
Яйценосные пластинки 434
Якорные пластинки кожи 246,
247
Якорьки 246
539
УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ
А
Ддакна 142
Адакны настоящие 143
Адапедонты 132
Акантастер 265
Акантодорис 64*
Акмеа 30
Аксинопсис 140
Актеон 59, 60*
Актинии (Морские анемоны) 306,
453, 512, 513, 513*, 514, 514*,
515
Альфеус (ы) 515, 516*
Аммониты 11, 170, 171
Ампулярии 39, 39*
Амуссиум японский 145
Амфибол иды 73
Амфиофиура 297
Амфиура 294
Амфиуриды 293
Анадара выпуклая 96
Ангария 31
Ангулюсы (теллины) 147
Анзеропода 264
Анодонта (Беззубка) 99*, 102,
102*, 109, НО, 110*
Анома 80
Аномал ьносвязочные 132, 155
Аномии 107, 127
Аномия гигантская 96, 127
— шиповатая 96
Антедон 223
— адриатический 224
— средиземноморский 224
Антедониды 223
Анцилиды 78
Аплекса 74, 74*
Аплизия (Морской заяц) 61,
61*
Аппендикулярии 329, 330
Аргонавты 161, 169, 169*, 181,
181*, 182
Арион 86
Ариониды 85
Ариофантиды 90
Арка вздутая 115
— полосатая 145
Арки 96, 98,107,108, 111,115
Артемия 383, 384, 385, 386
Артикулята 199
Арфа 56
Архастеры 261
Архитевтис (Гигантский кальмар)
186, 189
Аскоторациды 403
Астарта настоящая 96, 139
— овальная 96
— рыжая атлантическая 96
— северная 96, 139
Астарты 96, 108,131*, 139
Астерииды 269
Астероникс 292
Астрометис 269
Астропектен золотистый 261
Астропектены 261
Астрохламис 293
Асцидии 306, 329, 330, 331, 332*,
345, 346*, 349, 356, 357
— простые, или одиночные 330,
331, 344
— сложные, или колониальные
330, 335*, 344, 347*
Атииды 526
Атланта 41, 42, 42*
Атрины (Раковины-перья) 121
Аулодонта 280
Ахатиниды 88
Ахатины 79, 80, 81*
Ацера (Улитка-шарик) 59, 60,
60*
Ацила 96
Б
Байкалия 38
Баланоглосс 316, 317, 320*, 322*
Банкия 152, 152*
Батиарка 96, 115
— холодноводная 96
Батикринус карпентера 221
Батиполипусы 181
Батотаумы 161, 168
Безголовые (Двустворчатые,
Пластинчатожаберные, Топо-
роногие) 7, И, 91
Беззубка (Анодонта) 98, 99*, 100,
102*, 109, 111*, 390
— вуда 136
— вытянутая 135*, 136
— кавказская 136
— обыкновенная европейская 100,
135*, 136
— озерная 135*
— рыбья 135*, 136
— сводчатая 136
Беззубка узкая 136
— утиная 135*, 136
Беззубки 133, 136
Белемниты 171*, 172
Бертелиния 70, 70*
Беспанцирные И
Бессвязочные 148
Битинии 36, 36*, 37, 78
Битотрефес 396, 397
Бластоидеи 198*, 199
Богомолы 451
Б окоп лавы (Разноногие
ракообразные) 375*, 378, 379*, 413,
433, 459, 464, 475, 477, 484,
485, 486, 486*, 487, 488, 488*,
489, 490, 491, 492, 493, 494,
495, 522
Бороздчатобрюхие И, 16
Босмина (ы) 394, 395, 396, 397,
398, 399
Ботула 121
— калифорнийская 121
Бочеяочники (Долиолиды) 330,
336, 338, 339, 340*, 340,351,
352*
Брадибены (Фрутициколы) 90
Бризингиды 251, 270
Брюхоногие 7, 11
— древние 25
Букциниды 52
Букцинум 52, 53, 53*, 56
Буссикон 25
В
Валлония 89
Вальваты 38, 38*, 78
Велутина 46
Венериды 145
Венерикардия 112
Венерупис(ы) 98, 131*, 145
Венус петушок 146
— черноморский 112
— королевский гребень 145
— сунетта 145
Венусы 97, 145
Верметиды 40
Верметус 40, 41*
Верруки 431
Вертигины 89
Вертикордии 155
Витрея 80
Водяные блохи 391
540
г
Галиотис (Морское ушко) 28, 269
Гаммариды 490
Гарпактициды 412, 413, 413*,
415*, 416, 422, 448
Гастрана хрупкая 112, 147
Гастрохена 150*
Гелиометра 224
Гелициды 82, 84, «7, 89, 90
Гелициниды 33
Гетеростелии 199, 215
Гетеротевтис 168, 169
Гиббула 31
Гидробии 36, 36*
Гидроидные полипы 490
Гидроиды 474, 476, 480, 487, 489,
512* 513
Гиперииды 362, 488, 490, 491
Гиппопус 143
Гистиотевтис 167
— бонеллиана 167*
Гландина (Олеацина) 87
Глаукус 65
Глеба 69, 69*
Глобигерины 499
Глоссодорис 64
Голова Горгоны 2?8
Головоногие 7, 11, 156
Головохордовые 196
Голожаберные 62, 63
Голотурии (Морские кубышки)
200, 201, 205, 207, 208, 209*,
210*, 215, 224, 225, 227, 228*,
229*, 256, 257, 272, 288, 513
— безногие 228, 229*, 230, 232, 245
— боконогие 228, 229*, 230, 232,
242
— боченкообразные 229, 232, 245
— древовиднощупальцевые 229,
229*, 232
— листоносные 235, 236
— настоящие 238
— пелагические 244
— фантастические 244
— чешуйчатые 236
— щитовиднощупальцевые 229*,
230, 232, 237, 238
Голотурия антарктическая 237
— жесткая 235
— красная 235
— песчаная 239
— створчатая 237
— урнообразная 236
— цепляющаяся 246
— черная 239
— чешуйчатая 237
— шероховатая 238
Гомфина 112
Гоплокариды 450
Горация 38
Горгона 292
Горгоноцефал восточный 293
— североатлантический 293
Горошинка блестящая 137
— болотная 137
— речная 137, 137*
Горошинки 136,137
Граптолиты 317
Гребенчатка 136
Гребенчатозубые 96, 97, 105*, 112,
114*, 114
Гребешки 267
Гребешок беринговоморский 124
— большой 126
— глубоководный 96
— гренландский 96, 125
— исландский 125, 145
— Магеллана 126
— приморский 98, 124, 124*, 126
— Свифта 124, 145
— св. Якова 126
— Фаррера японский 124
— черноморский 96, 125
— чешуйчатый 125
Гребневики (Ктенофоры) 345, 485,
490, 491
Губки 413, 435, 489, 512, 512*, 513,
513*, 515
Гульдия маленькая 112
д
Даудебардия 80
Дафния (и) 392, 392*, 393, 394,
395, 396, 397, 398, 398*, 399
— обыкновенная 398
— планктонная 398
Двужаберные 172, 173, 174
Двустворчатые (Безголовые,
Пластинчатожаберные, Топоро-
ногие) 7, И, 91
Декапода 183
Дендрогастер 264, 438
Дендронотус 64, 65*
Денталиум (Морской зуб) 25, 91,
92. 92*, 93
Десятиногие 156, 183, 499, 500,
501, 505, 519, 521, 527
Диадема 280
Дидакна 131*, 142
Дикроцелий 89
Дозидикусы 163
Дозиния 145
— японская 112
Долииды (Улитки-бочонки) 49
Долиолиды (Бочоночники) 339,
340
Долиум 49
Донакс черноморский 145
Донаксы 131*, 147, 148
Дориды 63
Дорис 63, 64*
Древоточцы 151
Дрейссена каспийская 138*, 139
— клювовидная 138*, 139
— речная 138, 138*
Дрейссены 137, 138*
Жабы 345
Жабродышащие 370, 376, 377
Жаброноги 380, 381*, 382*, 386,
388, 393, 400, 402, 444
Жемчужная рыбка (Карапус) 238
Жемчужница даурская 132
— камчатская 133
— морская 145
— — цейлонская 122
— обыкновенная 132
— пресноводная 98, 104, 134,
133*
Жемчужницы морские настоящие
(Крыловидки) 98, 99, 122, 123,
144
— пресноводные 98, 132
Жемчужные кораблики
(Наутилусы) 173
Живородка лужанка 37, 37*
— уссурийская 37
Живородки 37
— настоящие 37
г
Заднежаберные 22, 58
Звездоофиура 297
Зебрины 79,85, 90
Зибоглиниды 314
Змеехвостка колючая 295
— Сарса 296
Змеехвостки (Офиуры) 200, 287
Зоантарии 515
Зонитиды 80, 89, 90
Зонитоидес 80
Зооксантеллы 145
Зостера 155
Зубатка 214
И
Игл обок 19
Иглокожие 195, 196, 197, 198*,
199, 202, 204*, 210*, 212, 257,
303, 317, 417, 425, 507
— свободноживущие (Элеутеро-
зоа) 199, 227
— стебельчатые 216
Идулия 64, 65, 65*
Изокринида 220
Изометра 225
Изоподы (Равноногие
ракообразные) 432
Инфузории 231, 419
Ишнохитон малый 17
й
Йолъдиелла ИЗ, 114
Йольдия 96, 105, 112, ИЗ, 131*
— северная 96, ИЗ
— широкая 96
К
Каволиния 68
Калан (Выдра морская) 275
Каланиды 406, 409, 410, 416
Каланус 406*
— красный 408, 409
Калигиды 419
Калиптреи 40
541
Каллиоетома 31, 31*
Каллитевтис 168
Кальмар гигантский (Архитевтис)
156, 186
— летающий (Стенотевтис) 185
— чудесная лампа 161, 186*
Кальмары 156, 157, 159*, 160,
162, 171, 173, 184, 185, 188*
— настоящие 174
Камбала 213
Камнеточец-петрикола 112
— фолас 128, 151
Камнеточцы (Морские финики,
Фоладиды) 120, 121
Кантокамптус 375*
Капулиды 40
Каракатицы 156, 157, 157*, 159,
159*, 160, 161, 172, 183, 184,
191, 421
— кальмарообразные
(Лжекальмары, Лолиго) 184
— настоящие (Сепии) 184
Карапус (Жемчужная рыбка) 238,
262
Кардита флоридская 145
Кардиомия 145
Кардиты 108
Кардиумы (Сердцевидки) 140
— настоящие 141
Каринарии 41, 43
Карихиумы 73
Карповые вши (Карпоеды) 419
Карпоеды (Карповые вши) 403,
419, 420, 421, 421*
Кассидария 48
Кассиды (Улитки шлемовидные)
48
Кассис 47*
Катушка роговидная 77, 77*
Катушки 77
Кашалот 190
Киленогие 41, 43
Китовые вши 485, 486, 491
Кишечнодышащие 316, 317, 318*,
321, 324, 326
Кишечнополостные 345, 375*, 422,
491, 512, 515
Клаватор 80*
Клаузилииды 71
Клаузилия 80, 89, 90
Клещи 367, 368, 369, 370, 371,
375
— водяные 422
Клионе 69, 69*
Клоп 369
Коловратки 396, 411
Кольчецы многощетинковые (По-
лихеты, Черви многощетинковые)
19
Комар 369
Комастериды 225, 226
Коматулида 222, 224, 226
Коматус японский 226
Конгерии 137
Конус 33, 57, 57*
Конхостраки 390
Копеподы 291
Кораллы 416, 425, 427, 437, 512,
515
Коралловые полипы 512
Корбикула восточная 137, 137*
— западная 137
Корбикулы 11, 98, 137
Корбула амурская 128
Корбуломии 149
Корбулы 149
Корифелла 65
Коркулюм 141
Корофииды 489
Кохликопы 89, 90
Краб(ы) 375*, 377, 438, 438*,
439, 449, 477, 480, 499, 500,
500*, 501, 501*, 502, 503, 504,
507, 508, 509, 510, 512, 512*,
513, 513*, 515, 516, 517, 518,
519, 520, 522, 523, 526, 527
— голубой 518
— -горошинки 512
— камчатский 435, 464, 480, 500,
504, 505, 508*, 517, 518, 519,
520, 521, 527, 528*, 529
— манящие 503, 506, 507, 508,
510, 510*, 523, 523*
— мохнаторукий 510, 517, 517*,
519, 520, 521, 524, 524*
плавунцы 503, 505, 509, 510,
516, 519
— -привидение 505, 505*, 509
— стыдливые 513
— травяной 505, 516
Кранхия 161
Креветка (и) 377, 438, 439, 448,
450, 453, 454, 477, 495, 500,
500*, 501, 502, 503, 504, 505,
506, 507, 508, 509, 512, 516, 517,
518, 518*, 519, 520, 522, 522*,
526, 529
Крезеис 68
Крепидула 39*, 40
Криптоплакс 15
Криптохитон 15
— Стеллера 17, 19
Кровянка оливковая 112, 148
Кроссастер 263
Круглоперы (Пинагоры-циклоп-
терусы) 178
Крыловидки (Жемчужницы
морские настоящие) 121
Крыложаберные 316, 317, 325,
325*, 326, 328
Крылоногие 59, 67, 68, 69
— голые 69
— раковинные 69
Ксенофоры 41
Ксилофага 151*, 152
Ксилофаги 152
Кульцита 251, 262
Куспидария 131*, 155
— арктическая 136
Л
Ламеллисабеллиды 315
Ламеллярииды 46
Лампсилины 98, 133, 134, 136
Лангуст (ы) 255, 377, 465, 480,
500, 501, 502, 504, 505, 509,
516, 517, 518, 519, 520, 529
Ланцелярия ножичковидная 134,
134*, 135
Ланцетник 329
Легочные (Пульмонаты) 21, 22, 71
Леда 105, 113, 115
— обыкновенная 96, 114
Ледиды 112
Лентец широкий 411, 412, 412*
Лептодора 396, 398
Лептометра 224
Лептосинапта малая 246
Лернеиды 419
Лжекальмары (Каракатицы
кальмарообразные, Лолиго) 174
Лима 107, 117*, 127, 144
— зияющая 127
— северная 96, 127
Лимапонция 70
Лимациды 85, 89
Лимацина (Спирателла) 68, 68*,
69
Лимнокардииды 142, 143*
Лимнория (и) 475, 475*, 490
Лимопсис 96, 115
Линкии 262
Лионзиелла глубоководная 128
Лионзия 155
— песчаная 128
Литотамний 19, 28
Литторины (Улитки береговые)
35, 508*
Литофага (Камнеточец) 120, 121
— средиземноморская 121
Ложноскорпионы 375
Локсехинус белый 283
Лолиго (Каракатицы кальмаро-
подобные, Лжекальмары) 174
— обыкновенный 185*
Лопатоногие 7, 91, 92*, 94
Лорипес 131*
Лоттия гигантская 30
Лофотенский ризокринус 221
Лужанка 23
Лучистые 197
Люцина тигровая 145
Люциниды 140
Лягушки 345
М
Макома 96, 131*
— балтийская 109, 112, 141, 147
— известковая 112, 147
— обыкновенная 112, 147
Макрохламисы 90, 91
Максиллоподы 403
Мактра 98, 148
— овальная 128, 149
— полосатая 128, 149
— сахалинская (Белая ракушка)
98, 128, 149
— шизотерус 128
Мактры настоящие 149
Маллеция 97, 112, 114
Маргарита 31
Мастиготевтис 186
Медузы 485, 490, 490*, 491, 499, 512
Меланелла 50
— пещерная 51*
Меланеллиды 50
542
Меланииды 38
Мелания 38
Меланопсис 38
Мелибе 66
Мерценарии 145, 146
Мерценария Стимпсона 112, 146
Мечехвосты 370, 371, 372, 374, 375*,
376
Мешкоязычные 70
Мидии 11, 268, 418
Мидия 11, 98, 104*, 117*
— гигантская (Черная
ракушка) 116, 119
— зеленая 145
— калифорнийская 116
— Магеллана 116
— обыкновенная съедобная 118,
119*
—
средиземноморско-черноморская 116, 119, 119*
Мизиды 442, 451, 454, 455, 455*,
456, 458, 459, 460, 477, 485, 519
Мизомения 12*, 13*
Мизостомиды 214*
Мии 148, 149, 150
Миллерикринида 221
Миратеста 76*
Мистакокариды 379, 403, 422,
423, 424, 424*, 448
Митилястер 96, 120
Митриды 56
Мия 131*
Многоножки 368, 369, 370, 371,
375, 375*, 376
Моднола 117*
— адриатическая 96
— обыкновенная 96, 120
— фазеолиновая 96, 120
Моины 399
Мокрицы 370, 371, 373, 375*, 377,
379, 470, 471, 471*, 472, 473,
474, 477, 478, 479, 480, 483, 484
Моллюски (Мягкотелые) 7, 232,
404, 417, 418, 429, 430, 435,
453, 503, 507, 509, 511, 526
— беспанцирные 8, 10, И, 12*, 13
— брюхоногие (Улитки) 7, И,
20, 23, 32, 33, 259, 303, 432,
437, 501, 503, 504, 511, 521
— головоногие И, 156, 160, 161,
170*, 176, 418, 419, 421, 509
— двустворчатые
(Пластинчатожаберные) 7, И, 95, 96, 103*,
104*, 106*, НО, 131*, 136, 144,
390, 432, 513
— киленогие 442
— легочные (Пульмонаты) 71, 79,
82, 84
— лопатоногие 91, 92*, 511
— панцирные 8, И, 14, 19
— переднежаберные 23*, 25, 50,
55, 259
— пластинчатожаберные
(Двустворчатые) 91, 95, 96
Монодакны 142, 143
Моноплакофоры 7, 9, И, 94
Морская бритва 98
— выдра (Калан) 213, 275
— звезда кровяная 263
Морская леечка 155*
— ложечка периплома 128
— пальма 220
Морские блохи 493
— гребешки И, 98, 108,117*, 124
— ежи 198*, 200, 202, 204, 205,
206, 209*, 210*, 211, 212, 213,
215, 224, 225, 240, 256, 257,
271, 273, 288, 425, 427, 503,
512, 513
— — диадемовые 280
— — кожистые 278
— — копьевиднозубые 281
копьеносные 277, 279
неправильные 206, 207, 276,
277, 278, 285
— — плоские 285
правильные 277, 278
— — сводчатозубые 281
— — сердцевидные 277, 285
— — эластичные 215
— желуди 19, 377, 430, 431, 431*,
432, 434, 435, 436, 437, 503
— звезды 198*,, 200, 200*, 201,
203*, 204, 206*, 209*, 210*, 212,
215, 224, 225, 240, 241, 247,
249*, 250*, 251*, 252*, 255*, 256,
257, 258*, 259, 425, 427, 428, 507
— — гребенчатые 261
игольчатые 248, 263, 264
педицелляриевые 248, 265
явнопластинчатые 248, 259
— козочки 485,486,487,489,490
— кубышки (Голотурии) 200, 227,
231
— лилии 198*, 199, 207, 209*,
210*, 214,214*, 216, 217*, 219*,
224, 225, 240, 291, 425, 512
бесстебельчатые 216, 219,
220, 223
стебельчатые 216, 219, 220,
221, 221*, 222
— — членистые 220
— ложечки 155
— молоточки 121, 122
— огурцы (Голотурии) 227, 232
— пауки (Многоколенчатые) 263
— перья (Пеннатулярии) 292
— стрелки (Щетинкочелюстные,
хетогнаты) 298
— уточки 377, 430, 430*, 431,
431*, 432, 435, 436, 437
— финики (Камнеточцы) 121, 150*
Морское сверло (Фоладидея) 128,
150*, 152
— сердце 139, 139*
— ушко (Галиотис) 25, 26, 32, 269
Морской еж береговой 283
— — кожистый жгучий 278
копьеносный обыкновенный
279
— — обыкновенный 282
сердцевидный 286
— настоящий 285
— — скальный 283
съедобный 276, 282
— — темно-фиолетовый 284
черный 281
— — яйцевидный 286
Морской заяц (Аплизия) 61, 61*
— зуб (Денталиум) 91, 92*
— — слоновый 93
— молоточек 145
— огурец 231
— — вытянутый 234
— — гладкий 233
— — глубоководный 235
— — ледяной 235
— — североатлантический 233
— — трехцветный 233
— — японский 232
Мотыль 411
Мукроналия 51
Мурекс 54*
Мускулюс 120
— черный 96, 120
Мшанки 19, 306, 474, 476, 480,
512*, 513, 515
Мягкотелые (Моллюски) 7
II
Насекомые 345, 367, 368, 369,
370, 371, 372, 372*, 373, 373*,
374, 375, 376, 388, 507
— двукрылые 375
— крылатые 375, 375*
— первичнобескрылые 375*
Насса (Улитка верша) 25, 53
Натика (Улитка пупочная) 44,
45*, 46*
Наутилусы (Жемчужные
кораблики) 171, 173
Наутилоидеи 171
Нематоды И, 375*
Немертины 316, 375*
— древесные 375*
— наземные 375*
Неомения 12*
Неопилина 94*, 95
— галатеа 94
Нептуниды 53
Нептуния 53*
Неравномускульные (Связочнозу-
бые) 115
Неритиды 32
Нитчатожаберные 103*, 104*
Нифаргусы 490
Ногохвостки 367, 368, 369, 370,
375, 376, 474
Нукула 112, 131*
Нукулиды 112
О
Оболочники (Оболочниковые, Ту-
никаты) 196, 301, 329, 330, 337,
362, 363, 512
Оболочники донные
глубоководные 330, 341, 358*
— пелагические 330, 359, 360
Оболочниковые (Оболочники, Ту-
никаты) 196, 329
Огнетелки (Пиросомы) 329, 330,
331, 336, 337, 338, 349, 350*,
351, 356, 359
Одномускульные 124
Октопусы 158, 162, 162*
Олеацина (Гландина) 87
543
Оливы 55, 56, 56*
Олигобрахииды 314
Олигохеты 411
Омар (ы) 369, 377, 465, 500, 501,
502, 503, 504, 505, 509, 516,
517, 518, 521, 529
— американский 521, 529
— европейский 518, 519, 529
— норвежский 500*, 518, 519
Онихофоры 375*
Онцидииды 79
Ореховидка Ацила 96
— нукула 114*
— тонкая 112
Ореховидки 97, 105, 107, 112
Орхестия 375*
Ослики водяные 470, 472, 473,
474, 475, 478, 478*, 480, 481,
482
—клешненосные (Танаидовые) 454,
464, 465, 466, 466*, 467, 468, 477
Осы роющие 374
Осьминог адский вампир 161,183
— аполлион 179
— гонконгский 179
— Дофлейна 179
— обыкновенный (Спрут
обычный) 175, 179, 189, 192
Осьминоги 156, 157, 157*, 159,
159*, 160, 161, 171, 172, 174,
176, 191, 192, 507, 513, 516
— бесплавниковые 175
— глубоководные 175,183
— настоящие 175, 183
Офиаканта двузубая 293
Офиактиды 293, 295
Офиодерма короткоиглая 296
Офиокома 296
Офиокомиды 296
Офиотрикс 296
— ломкий 295
Офиотрихиды 295
Офиоцистии 198*
Офиура темно-красная 295
Офиуры (Змеехвостки) 200, 201,
204, 205, 206, 209*, 210*, 213,
214 ,214*, 215, 224, 225, 273,
287, 287*, 425
— ветвистые 215, 291, 292
— настоящие 290, 293, 296
П
Пальмовый вор 375*, 481, 503,
504, 504*, 505, 507, 510, 519,
520, 521
Пандора холодноводная 128
Пандоры 93, 155
Панопеа японская 128
Панцирные 7, 14, 16
Пароктопус песчаный 175
— шершавый 175
Парусник 66
Пателла (Улитка-блюдечко) 29,
29*, 32
— обыкновенная 29
— черноморская 30
Пате л л иды 30
Патулиды 88
Пауки 345, 370, 371, 374, 375*
Паукообразные 368, 370, 375,
376
Пахидискус 171
Пектены 96
Пектункулюс 115, 131*
— полосатый 96
Пелаготурии 245
Пелопатидес подошвообразный
242
Пельматозоа 199, 215
Пенеиды 529
Пеннатулярии (Морские перья)
292
Пентакринус 224
Перакариды 454, 463, 468, 477, 492
Первичножаберные 103*, 105*
Первичноротые 196
Перегородчатожаберные 103*, 111,
128, 155
Переднежаберные 22, 23, 23*, 24,
24*, 25, 48
Периплома (Морская ложечка)
128
Перловица 98, 104*
— вздутая 135, 135*
— дальневосточная 135
— клиновидная 134*, 135
— обыкновенная 134*, 135
— толстая 134*, 135
Перловицы 98, 134, 390
— настоящие 134
Перфорателла 90
Пескожил 141
Петрикола 112, 146
Петушки-тапесы 98
Петушок японский 112, 146
Пикноподия 270
Пикша 213
Пинагоры (Круглоперы-циклоп-
терусы) 178
Пинктада 99, 122, 124
Пинна 117*, 121, 124
— благородная 121
Пинны (Раковины-перья) 121
Пиросомы (Огнетелки) 329, 330,
331, 336, 337, 338, 338*, 349,
350*, 351, 356, 359
Пирула (Грушевидная улитка) 49,
50*
Питария тихоокеанская 112, 146
Пиявки 375*
Плакуна тропическая 127
Планарии наземные 375*
Пластинчатожаберные
(Безголовые, Двустворчатые Топоро-
ногие) 95
— настоящие 96, 103, 103*, 104*,
105*, 130, 132
Плевробранхея 62
Плевробранхус 62, 63*
Плеуротомария 26
Погонофоры 196, 301, 303, 305,
306, 307, 309*, 310, 311*, 313*
Подосферастер 251
Позвоночные 196, 317, 330
Покрытожаберные 58, 59, 62, 62*
Полибрахииды 314
Полипусы 162
Полихеты (Черви многощетинко-
вые) 259, 261, 368, 417, 442, 475,
507, 510, 513
Полухордовые 196, 301, 303, 316,
317, 325
Поромия 128, 155
Портландия 105, 113, 115
— арктическая 97, 113, 115*
— эстуарная 113, 115*
Порцелланастериды 259, 260
Пронеомения 12
Пропеамуссиум 96, 125
Простейшие 422
Прототака 112, 145
Прудовик болотный 75
— малый 75, 78
— обыкновенный 74, 74*, 75, 75*,
76*
— ушастый 75
— яйцевидный 75
Прудовики 74, 76, 78
Псаммобии 147, 148
Психропотес 243
Птерии 99, 121, 122
Птерозома 41, 43
Птеротрахея 42*, 43, 43*, 59
Птероцера 192
Пульмонаты (Моллюски
легочные) 80, 84
Пунктурелла, 23*, 28
Пупиллы 89
Пчелы 374
Р
Разнозубые 114*, 132, 136, 139
Раковины-перья (Атрины и
пинны) 121
Рак (и) 511, 513, 516
— -богомолы (Ротоногие
ракообразные) 450, 451, 452, 453,
454
кроты 509*, 510
медведи 502
отшельники 377, 432, 437,
439, 477, 500*, 501, 502, 503,
504, 505, 507, 510, 511, 511*,
513, 514, 514*, 517, 518, 519,
520, 521, 522
— речной (ые) 499, 500, 501, 502,
503, 504, 507, 510, 517, 518,
519, 521, 524, 525*, 526, 526*,
527
— — американский 510, 518*
узкопалый 524, 525, 526*
— — широкопалый 521, 521*,
524, 525, 526*
щелкуны 439, 503, 512, 515
Ракообразные 232, 303, 345, 371,
375, 375*, 376, 377, 507
— акроторациковые 437
— анаспидовые 446, 446*, 470
— батинеллиевые 447, 470
— веслоногие 375*, 378, 379,
379*, 401, 402, 403, 404, 404*,
405, 406, 416, 419, 420, 429,
431, 440, 442, 459, 478, 499
544
Ракообразные ветвистоусые 378,
379*, 391, 392, 393, 393*, 394,
394*, 395, 395*, 396, 397, 398,
398*, 399, 400, 403, 442, 468
— высшие 372, 374, 375*, 376,
402, 416, 443, 445, 450, 457,
464, 477
— десятиногие 369, 375*, 379*,
438, 451, 455, 476, 496, 499,
500, 500*, 501, 502, 503, 504*,
505, 506, 507, 508, 509, 510,
512, 513, 515, 516, 517, 518,
518*, 519, 520, 521, 522, 523,
524, 526
— жаброногие 378, 379, 380, 401
— корнеголовые 429, 437, 438,
439, 477
— кумовые 454, 460, 460*, 461*,
462, 464, 465, 467, 468, 477,
478, 486
— листоногие 380, Э86
— мешкогрудые 424, 425, 426*,
427, 428
— равноногие (Изоподы) 374,
375*, 378, 379, 379*, 432, 454,
463, 464, 467, 470, 471, 471*,
472, 473, 474, 475, 477, 477*,
478, 479, 479*, 480, 481, 482,
485, 486, 490, 522
— разноногие (Бокоплавы) 433,
454, 484, 487
— ракушковые 379, 379*, 440,
440*, 441, 441*, 442, 477, 499
— ротоногие (Раки-богомолы) 379,
450, 451, 452*, 453
— спелеогрифовые 454, 467
— термосбеновые 454, 468, 469,
469*, 470
— тонкопанцирные 443, 444, 445»
446, 456, 458
— торациковые 430, 431*
— усоногие 375, 379, 403, 416,
425, 428, 429, 430, 433, 436,
437, 438, 477
— эвфаузиевые (Эвфаузииды) 495,
496*
Ракушка белая (Мактра
сахалинская) 149
— песчаная (Миа) 108, 128, 141,
149
— черная (Мидия гигантская)
116, 119
Рапана 55, 128
Расщеп л еннозубые 132
Рачки калигоидные 419*
Реклюзия 44
Ретуза 62
Риссоиды 36
Ритида 87
Ришта 411, 412, 412*
Ропаломения 12
Россия 160, 184*
Рыбы 7
— летучие 363
С
Саккулина 439, 477
Саксидемус 146
Саксикава(ы) 148, 150*
— арктическая или
обыкновенная 128, 149
Сальпы 329, 330, 331, 336, 339,
340*, 351, 355*, 356, 360, 490
— собственно 330, 339
Связочнозубые (Неравномускуль-
ные) 96, 112, 115, 116
Сепиола двурогая 168
Сепии (Каракатицы настоящие)
161, 183, 184
Сердцевидка волосатая 112, 142
— гренландская 112, 114*, 142
— Ламарка 142, 142*
— малоребристая 112
— съедобная 97, 109, 136*, 141,
142*
— Фрагум 145
Сердцевидки (Кардиумы) 96, 98,
109, 140, 141*
Сида 396
Симоцефалюс 396
Симфилы 370
Синапта пятнистая 246
Синаптиды 246
Синдесмия 148
— овальная 112, 148
Синкариды 446
Синовакула 98
Сипункулус 93
Сифоноденталиум 92*, 93
Сифонарииды 73
Сифоноротые 46
Сифонофоры 416, 432
Скафандер 62
Скафарки 96
Склеролиниды 314
Скорпионы 370, 375, 376
Скребни 303
Скробикулярии 147
Скробикулярия 131*
Скутигеры 375*
Слизень большой 73*, 86
— пещерный 87*
— полевой 49, 86, 90
— сетчатый 90
— черный 87
Слизни голые 72, 85, 86
Совки 373
Солемии 114, 115
Солемия 145
Соляриелла 31
Спинула витязя 114, 115
— клювовидная 145
Спирателла (Лимацина) 68, 68*
Спиробрахииды 315
Спирула 173
Спондилюс 127, 145
Спрут гигантский 159
— длиннорукий 190
— обычный (Осьминог
обыкновенный) 175
Стенотевтис (Летающий кальмар)
185
Стигокариды 450
Стилифер 50, 51, 52*
— брихиус 51*
Стиракастер 260
Стихоподиды 239
Стробилопсиды 91
Стромбус 47*, 192
Стронгилоцентротус пурпурный
282, 286
Строфохилус 80, 80*
Сферома 490
Т
Таксеумы 168
Танаидовые (Ослики клешненос-
ные) 464, 466
Тапес черноморский 112
Тапесы 145
Теллина 96, 108, 131*
— ангулюс 112
— желтая 112
— фараонелла 145
Теллиниды 145
Те л лины 96, 147
— собственно 147
Тенагодус 41
Теребра 57
Терединиды 154
Тередо 96, 152, 153*, 154*
Термиты 372
Терновый венец 265
Тестацелла 87, 87*
Тетис 66
Тиазира 131*
Тиазиры 140
Тика 40
Тинтинноидеи 499
Тионе 234
Тионепсолюс живородящий 237
Тионидиум 236
Тифобия 39, 39*
Тли 375
Топороногие (Безголовые,
Двустворчатые,
Пластинчатожаберные) 95
Трахейнодышащие 370, 376
Трации 155
Трация 128
Трематоды (Сосальщики) 214
Тремоктопусы 167, 169
Трепанг дальневосточный 240
Трепанги 227, 238, 240, 241
Треска 213
Тригонохламиды 87
Тридакна 95, 143, 144, 145, 146*,
150*
— гигантская 143
Трилобиты 368, 372, 375*, 376
Триоксиопус 156
Тритониум 49, 50, 54*
Тритоны 345
Трохиды (Улитки-волчки) 31
Трохостома арктическая 245
Трохус 25, 31
Туникаты (Оболочники, Оболочни-
ковые) 329, 330, 356
Тунец желтоперый 363
Турбо 32
У
Улитка актеон 59, 60*
— арфа 56
545
Улитка блюдечко (Пателла) 29,
29*, 32
— верша (Насса) 53, 53*
— виноградная 11, 72*, 80, 81,
82, 83*, 84
— грушевидная (Пирула) 49, 50*
— древовидная 64
— епископская шапка 56
— зонтик 62, 63*
— кустарниковая 49, 90
— пеликанья нога 49, 50*
— пупочная (Натика) 44
— пурпурная 54, 55
— ручьевая 36
— садовая 85
— туфелька (Крепидула) 40
— устричное сверло 55
— фарфоровая (Ципрея) 47
— цекум 41
— шарик (Ацера) 60
— шишак 48
Улитки (Моллюски брюхоногие)
20
— безъязычные 50
— береговые (Литторины) 35
— бочонки (Долииды) 47*, 49
— винтовые 56
— волчки (Трохиды) 31
— голожаберные 62, 63
— игольные (Церитии) 46
— киленогие 41
— легочные (Пульмонаты) 79, 84,
88
— сидячеглазые 73
— стебельчатоглазые 78, 79, 90
— сухолюбки 84
— червеобразные (Верметиды) 40
— шлемовидные (Кассиды) 48
Униониды 132
Усоногие 19
Устрица адриатическая 128
— виргинская 128
— гигантская 128, 129*
— калифорнийская 128
— листоватая 128
— обыкновенная, или съедобная
128
— петуший гребень 144
— португальская 128
— скаловая 128
— черноморская 128
— японская 128
Устрицы 11, 98, 107, 110, 117*,
127, 418
Ф
Фасциола печеночная 76
Феретима индийская 375*
Физа 74, 74*
Физиды 74
Филины 62
Филлирое 66*, 67
Филлокариды 443
Филомициды 91
Фоладиды (Камнеточцы-морские
финики) 150*, 151
Фолады 121
Фрагум 141, 145
Фриксометра 224
Фронимы 362
Фрутициколы (Брадибены) 90
X
Хама чешуйчатая 145
Хамы 140
Хелицеллы 89, 90
Хелицеровые 370, 375*, 376
Хенриция 263
Хетогнаты (Морские стрелки, Ще-
тинкочелюстные) 298
Хидорус 394, 399
Хирономиды 388, 411, 474, 475
Хиротевтис верании 187*
Хитон 17
— белый 19
— изящный 17*
— красный 16
Хламисы 97
Холопус 222
Хордовые 196, 303, 316, 319, 330
Хэтодерма 12*, 13
Хэтоплевра 17
ц
Цекум 41
Церитии (Улитки игольные) 46
Цестоды (Черви ленточные) И
Цефалодискусы 327
Циклокардия 112
Цефалокариды 400, 401, 402, 402*,
470
Циклоптерусы (Круглоперы,
Пинагоры) 178
Циклопы 403, 410, 411, 412, 412*,
416
Циклостома 34, 34*
Цилихна 62
Цимбулия 69
Ципрея (Улитка фарфоровая) 25,
47
Циприна 131*
— исландская 97, 112, 139
Цирротаума 161, 174, 183
Цирфея гребенчатая 97, 128, 152
— Пилсбри 128
Цистидеи 198*, 199, 215
Ч
Черви 232, 316, 345, 404, 422, 490,
513, 515*
— дождевые 375*
— кольчатые (Аннелиды) 10, 95,
303, 375*, 379
¦— ленточные (Цестоды) 303
Черви малощетинковые 375*
— многощетинковые (Полихеты)
259, 261, 303, 304, 368, 375*,
378, 417, 418*, 515
— плоские 375*, 419
Червь дождевой обыкновенный
87, 87*
— корабельный 151
Черенки 148
Черенок большой австралийский
145
— морской 108
— обыкновенный 148
— солен 131*
Четырехжаберные 172
Членистоногие 367, 368, 369, 370,
371, 372, 373, 374, 375, 375*,
376, 404
Членистые 368
Ш
Шаровка 136
— прочная 137
— речная 136, 137
— роговая 137, 137*
Шелкопряды 373
щ
Щетинкочелюстные (Морские
стрелки, Хетогнаты) 195, 196,
298, 299*, 499
Щитень весенний 389
Щитни 386, 387* 387*, 388, 389,
390, 391, 393
Э
Эвкариды 495
Эвфаузииды (Эвфаузиевые
ракообразные) 496, 497, 498, 499
Эдриоастероидеи 199, 215
Элеутерозоа (Иглокожие свобод-
ноживущие) 200, 215
Эллобииды 73
Эльминиусы 435
Эльпидия 243
— полярная 243
Эндоцерас 171
Эниды 90
Энтеропнейсты 317» 319
Энтоконха 51, 52*
Эолиды 64, 67
Эолис 63, 64, 65, 65*
Эргазилиды 417
Эуритрема 89
Эхиуриды 416
Я
Яминия потанина 91
Янтарка 49, 79, 79*, 80, 88, 89
Янтина 44
546
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ
А
Abatus philippii 256, 276
Abra (Syndesmya) 148
— ovata 112, 148
Abralia sp. 186
Abyssocucumis abyssorum 235
Acanthaster 265
— planci 240, 265
Acanthasteridae 265
Acanthinula aculeata 80
Acanthocardia 112, 141*
Acanthochites fascularis 19
Acanthochiton rubrolineatus 16
Acanthocidaris maculicollis 273,
280
Acanthocyclops viridis 411
Acanthodoris pillosa 64, 64*
Acanthogammarus godlewskii 433
Acanthopleura 19
Acartia clausi 408
— tonsa 408
Acasta spongicola 435
Acepbala (Lamellibranchia,
Bivalvia, Pelecypoda) 95
Acera bullata 59, 60*
Aceridae 59
Achatina 80
— fulica 80
— mauritana 81*
Achatinidae 79
Achloe 261
Acila divaricata 96
Acmaea 269
— cassis 32
— lineata 34
— polita 34
— rubella 30
— testudinalis 30, 32
— virginea 30, 31
Acmaeidae 28, 30
Acoela 62
Acrocrinus amphora 198*
Acroloxus lacustris 77*, 78
Acrothoracica 437
Actaeon 59
— tornatilis 59, 60*
Actaeonidae 59
Actinopyga 238
— agassizi 239
— echinites 239
Adacna 142, 143
— laeviuscula 143
— minima 143
Adacna (Hypanis) plicata 143,143*
— vitrea 143, 143*
Adamsia palliata 514
Adapedonta 132, 148
Adesmacea 148, 151
Adula 121
Aega 470, 473, 476, 480
— psora 432
Aeolis 63, 64
— papillosa 64, 65, 65*
Aeropsis fulva 257, 286
Agelacrinites ephraemovianus
198*
Aglossa 50
Agriolimax 81, 90
— agrestis 81, 82, 86
— laevis 86
Albunea 503
Aloididae 149
Alonella excisa 398*
Alpheidae 503, 515
Alpheus brevicristata 448
Amathillina spinosa 492
Amblyops magna 458
Ampelisca diadema 433
Ampeliscidae 485, 489, 490
Amphibolidae 71, 73, 102
Amphilicus androphorus 290,
290*
Amphimetra discoidea 227
Amphiodia rossica 288, 295
Amphiophiura 296
— ponderosa 288, 297
Amphipeplea (Myxas) 76
Amphiperasinae 48
Amphipholis squamata 289, 291,
294
Amphipoda 484
Amphithoidae 489
Amphitretus pelagicus 183*
Amphiura antarctica 294, 295*
— chiajei 294
— stepanovi 294
— stimpsoni 294
Amphiuridae 287, 293
Ampullaria gigas 39*
Ampullariidae 39
Amygdalum 112, 146
Amussium 125
— japonicum 145
Anadara 115
— broughtonii 96, 115
Anaspidacea 446
Anaspides 446, 447
— spinulae 447
Anaspides tasmaniae 446*, 447
Ancylidae 77*, 78
Ancylus fluviatilis 77*, 78
Anilocra 476
Anisogammarus 492, 493
Anisomyaria 115
Angulus fabula 147
— donacinus 112, 147
— lutea 112, 148
Anodonta 13, 99*, 100
— arcaeformis 136
— cygnea 100, 135*, 136
anatina 135*, 136
cellensis 135*, 136
— — cygnea 135*
— — lacustris 135*
piscinalis 135*, 136
— cyrea 136
— woodiana 136
Anodontinae 134, 136
Anoma tricolor 80
Anomalodesmata 132, 155
Anomia ephippium 127
— squamula aculeata 96
Anomiidae 127
Anonyx nugax 490, 492
Anostraca 380
Anseropoda placenta 264
Antarcturus ultraabyssalis 432
Antedon 223
— adriatica 224
— bifida 220, 223, 223*
— mediterranea 224
— petasus 223
Antedonidae 219, 223
Anthoblastus brouweri 198*
Anthuridea 471, 472, 473, 481, 483
Antipates larix 425
Anuropus bathypelagicus 473
Aoridae 489
Aphroditidae 261
Apiocrinidae 222
Aplacophora 7, 11
Aplexa 74
— hypnorum 74, 74*
Aplysia 61
— depilans 61*
Apoda 228*, 230, 232, 245
Appendiculariae (Copelata) 330
Aporrhais pes pelicani 49, 50*
Apseudes 464, 465
— antarcticus 467
— spectabilis 466
— spinosus 466*
— talpa 467*
547
Apseudidae 465
Apteroti (lncirrata) 175
Araeosoma thetidis 278
Arbacia 281
— lixula 281
— punctulata 277
— spatuligera 273
Arbaciidae 281
Area 110, 1J5
— lactea 115
— zebra 115, 145
Arcacea 111, 115
Archaeogastropoda 25, 34
Archaeomysis grebnitzkii 458
Archaeostraca 445
Archaster 256, 261
— typicus 261
Archasteridae 261
Archicaligus 418
Architeuthis 156, 186, 189, 193
— longimana 190
Arcidae 115
Arcopsis 115
Arctica (Cyprina) islandica 139
Arcticidae 139
Arcticocamptus arcticus 413, 414,
414*, 415, 416
Arcturidae 471, 475, 479, 480
Arcturus ulbani 482*
Arenicola marina 323
Arenomya 98, 108, 128, 149, 150
Arenosetella palpilabra 413
— spinicauda 413*
Argonauta 158
— argo 181, 181*
— cornuta 181
— bottgeri 181
— hians 181, 182
— nodosa 181
— nourgi 181
— ocytnoe 170
Argonautidae 169, 170, 181
Argulus arcussonensis 421
— foliaceus 421, 421*
— japonicus 421*
Arion bourguignati 86
— maximus 49
— empiricorum 49, 81, 82, 86
— subfuscus 49, 87
Arionidae 85
Ariophantidae 90
Aristocystites bohemicus 198*
Armadillidium 484
— vulgare 479
Artemia salina 383, 383*
Arthropoda 367, 376
Articulata 199, 220, 368
Asapbidae (Sanguinolariidae,
Psammobiidae, Gariidae) 148
Ascidia virginea 337
Ascidiacea 330
Ascidiae salpaeformis (Pyrosomata)
330
Ascopera gigantea 331, 358*
Ascothoracida 259, 264, 424
Ascothorax 425, 426*, 427
Asellidae 481
Asellota 471, 472, 473, 475, 481
Asellus aquaticus 432, 481, 482
Asellus banyulensis 482
— communis 473
— hilgendorfi 482
— meridianus 482
— monticola 482
Aspidochirota 228*, 230, 232, 237
Astacidae 520, 524
Astacilla 476, 490
— pusilla 476*
Astacus 524
— astacus 524, 526*
— leptodactylus 524, 526*
Astarte 96, 139
— borealis 96, 139
— crenata 96, 139
— elliptica 96
— montagui striata 97
— rollandi 97
— sulcata 96
Astartidae 139
Asterias 255*, 263, 267, 268
— amurensis 118
— forbesi 224, 266, 267
— rubens 118, 205, 241, 248, 252*,
254, 258, 263, 266, 269
— vulgaris 255, 266
Asteriidae 254, 258, 266, 270
Asterina gibbosa 256, 257
Aterinidae 258, 264
Asteroidea 200, 247
Asteronyx loveni 292, 292*
Asterope 441, 442
Asteropecten 249*, 261
— aurantiacus 261
— polyacanthus 261
AsteropeGtinidae 253*, 261
Asthenosoma urens 278
Astrochlamys bruneus 273, 289,
290, 293
Astrocles actinodetus 257
Astrofiura 297
— chariplax 297
— marione 296*, 297
Astrometis sertulifera 269, 288
Astropecten 249*, 261
— aurantiacus 261
— polyacanthus 261
Astropectenidae 261
Athanas 518
— nitescens 517
Athecanephria 310, 313
Athoracophoridae 85
Athubaria 325, 325*, 326
Atlanta 42
— peronii 42*
Atlantidae 41, 42
Atrina 121
Attheyella nortbumbrica 416
— wulmeri 415*, 416
Atyaephyra desmaresti 519
Atyidae 508, 526, 527
Aulodonta 280
Auricularia antarctica 231
— nudibranchiata 231
Austrastacidae 524
Avicula (Pteria) 122
Axelboeckia spinosa 433
Axinopsis 140
Axinus (Thyasira) 140
B
Baicalia 38
— stiedea 38*
Baicalobathynella magna 448, 450
Balanidae 431
Balanoglossus 323, 452
— capensis 323
— clavigerus 316, 318*, 319, 323*,
324
— cornosus 319
— gigas 319
— proliferans 322, 323
Balanus 431*, 435
— balanoides 431, 434, 435
— balanus 416, 434
— cariosus 435
— crenatus 434
— eburneus 435
— evermanni 416, 436
— hameri 434
— hesperius 435
— improvisus 416, 434, 435
— masignotus 433
— psittaceus 436
— spongicola 435
Bankia 152, 152*, 153
— gouldi 154
— setacea 155
Barbatia 115
Barnea Candida 152
— spinosa 152
Basommatophora 73
Bathothauma lyroma 161, 188*
Bathyarca 115
— glacialis 96
Bathycrinus 221
— carpenteri 221
— complanatus 221
— pacificus 221, 257
Bathymetridae 224
Bathynella baicalensis 450
— natans 447*, 448, 449
Bathynellacea 446, 447, 448, 449,
450
Bathynomus 472
— giganteus 432, 470
Bathypolypodinae 175, 181
Bathyporeia 488
Benedictia 36
— fragilis 36*
Bentheuphausia amblyops 496,
496* 499
Benthopectinidae 253, 253*
Berghia coerulescens 48
Bertelinia 70, 70*
Biancolina algicola 433
Binneya 85
Birgus latro 481, 503
Birstenia vitjasi 314
Bithinia 24*, 36
— leachi 36*, 37
— striata 37
— tentaculata 36, 37
Bivalvia (Lamellibranchia, Pele-
cypoda, Acephala) 7, 95
Blastoidea 199
Bodotriidae 463
Bohadschia €38
548
Boltenia echinata 337
— ovifera 337
Bopyridae 477
Boreomysis 456, 459
— curtirostris 458
— plebeja 458
Bosmina 393*, 394
— coregoni 395*
— longirostris 398*, 399, 400
Bothriocidaridae 215
Bothriocidaris pahleni 198*
Botryllus schlossery 337
Botula 121
— calif orniensis 121
Brachidiastylis resima 463
Brachiodontes lineatus 121
Brachioteuthis rusei 161
Bradybaena fruticum 49, 90
— maacki 49
Branchiata 376, 377
Branchinecta 382*
Branchiopoda 380
Branchipus 382*, 383, 384
— stagnalis 381*
Branchiura 419
Brandtia parasitica 413, 433
Brandtothuria arenicola 225, 239
Brechites 155
— giganteus 155*
Brisaster townsendi 286, 288
Brisingidae 249, 250, 270
Bryocyclops 411
Buccinidae 52
Buccinum 52, 437, 511
— undatum 21*, 52, 53*
Bulla lignaria 33
Bundenbachia benecki 198*
Bunedeopsis 514
Bythinella adsharica 36
Bythotrephes 396, 399
— longimana 400
С
Caecosphaeroma 482
Gaenocentrotus gibbosus 284
Calanoida 404*, 405, 405*, 406,
406*, 407, 409
Calanus 406, 407, 408*
— cristatus 407, 409
— finmarchicus 401, 407, 408, 409
— glacialis 409
— helgolandicus 408, 409
— hyperboreus 409
— plumchrus 401, 407
Calappa 449, 503
Caligoida 417, 418
Caligus 419*
Callianassa 509*
— tyrrhena 509*
Callianassidae 508, 510
Callinectes 518
— sapidus 449, 518
Calliostoma 31
— zizyphinus 31*
Callista brevisiphonata 112, 146
Calocalanus pavo 401
Calocaris macandrae 517
Calyptraea sinensis 40
Calyptraeidae 40
Camarodonta 281
Cambaroides 524, 525*
Cambarus 510, 517, 525*
— affinis 518*
Camptoceras 77
Camptocercus rectirostris 400
Cancer pagurus 449, 518, 519, 521
Cancirilla 291
Cancricepon elegans 477*
Candonidae 442, 443
Canthocamptus staphylinus 415*,
416
Caprella 487*
— anatifera 486*
Caprellidae 485
Capulidae 40
Carapus (Frierasfer) 238, 262
Carcinus 509, 515, 516, 518
— maenas 438*, 449, 505, 518, 519
Cardiidae 140
Cardiinae 141
Cardiomya gouldiana 145
Cardita 139
— floridana 145
Carditidae 139
Cardium 141
— edule 97
— echinatum 141*
— paucicostatum 112
Cardysoma 509
Carinariidae 41, 43
Caspievadne maximovitchi 400
Cassidaria 48
Cassididae 48
Cassidula 73
Cassiduloida 285
Cassis cameo 25
— cornuta 33, 47*, 48
— rufa 25
Caucasotashea atrolabiata 49, 84
Caudinidae 245
Carichium minimum 73
Cavolinia gibbosa 69
— tridentata 68, 68*, 69
Centrostephanus longispinus 277,
281
Cepaea hortensis 85
— nemoralis 49, 85
Cephalocarida 378, 400
Cephalochordata 196
Cephalodiscus 325, 326, 327*, 338
— densis 325, 327*
— nigrescens 327*
— dodecalophus 327*
Cephalopoda 7, 156, 159, 170
Cephalopyge 67
Ceramaster patagonicus 224
Cerastoderma 141
— edule 141
edule 141, 142, 142*
lamarcki 142, 142*
Ceratoplana colobocentroti 284
Cercopagis 399
— socialis 400
Ceriodaphnia reticulata 400
Cerithiidae 46
Cerithiolum reticulatum 46
Cerithium 47
Cerithium ponticum 46
— vulgatum 46
Certonardoa semiregularis 52*
Ceuthonectes 413
Chaetoderma nitidulum 12*, 13
Chaetodermatidae 13
Chaetognatha 195, 196, 298, 299
Chaetopleura bullata 17
Chaetopterus 510, 512*
Chama 140
— macerophylla 145
Chamelea 112, 146
— fasciata 97
Chamidae 140
Chelicerata 376
Chelura terebrans 490
Chelyosoma macleayanum 337
Chevreuxiella obensis 492
Chione gallina 112. 146
Chionoecetes opilio 449
Chiridota laevis 247
Chiridotidae 247
Chirocephalus 382*
— grubei 383
Chiroteuthidae 161
Chiroteuthis bomplandi 186
— veranyi 187*
Chiton elegans 17*
— fulvus 17
— polii 19
— rubicundus 16
— tuberculatus 18, 19
Chlamys 124
— beringianus 124
— farreri nipponensis 97, 124
— glabra pontica 96, 125
— islandicus 125, 145
— swifti 124, 145
— tigerina 97
Choanomphalus 77
Chologaster 439
Chordaria 196
Chordata 196, 330
Chromodoris coerulea 48
Chthamalus 431
— stellatus 416, 434, 435
Chydorus sphaericus 394, 394*,
399, 400
Cidaridae 279
Cidaris cidaris 279
Cidaroidea 277, 279
Ciona intestinalis 331, 337, 357,
358, 359
Circomphalus 97
Cirolana 476
Cirolanidae 475, 476, 482
Cirolanides 482
Cirrata (Pteroti) 183
Cirripedia 428
Cirroctopus 170*
Cirroteuthidae 161
Cirrothauma murrayi 161, 174, 183
Cirsotrema perpexum 32
Cladocera 391
Cladodactila crocea 235, 256
Clausilia laminata 80
Clausiliidae 71, 90
Clausinella 97
Clavagella 155
36 Жизнь животных, т. 2
549
Clavagellidae 155
Glavator clavator 79, 80*
Clavelina 357
— lepadiformis 337
Glinocardium 141
— californensis 97
— ciliatum 112, 142
Glione limacina 69, 69*
Clypeastroida 285
Glypeoniscus 479*
Gnesterium 97, 112, 113
Gochlicopa 90
— lubrica 49
Gochlitoma 80
Gochlostyla leucophthalma 88, 89*
Godakia tigerina 140, 145
Coeloplana astericola 263
Goenobita 503, 504, 505, 511, 520,
521
— diogenes 511
Colobocentrotus atratus 225, 284
Colochirus luteus 233, 240
— quadrangularis 233, 272
Comantheria grandicalix 219
Comanthus 214*, 291
— japonicus 226
Comasteridae 222, 225
Gomatella stelligera 225, 226
Comathina schlegelii 226
Comatula pectinata 226, 226*
— purpurea 226
— rotolaria 226
Gomatulida 220, 222
Conchoderma auritum 416, 436
Gonchoecia 442
— elegans 440*
Conchostraca 390, 390*, 391, 393
Conger ia 137
Gonidae 57
Conidophrys 419, 419*
Gonilera 476
Conus 24, 57
— ebraeus 33
— marmoreus 33
— mediterraneus 57
— textilis 57*
Gopelata (Appendiculariae) 330
Gopepoda 363, 403
Gopilia vitrea 401
Gorbicula fluminalis 137
— fluminea 137, 137*
Gorbiculidae 137
Corbulidae 149
Gorculum 141
— cardissa 97
Goretus corneus 77, 77*
Goronida 452, 453
Goronula 432, 436
— diadema 416
Gorophiidae 489, 490
Corophium 488, 489*, 494
— curvispinum 489, 494
— volutator 489
Gorycaeus obtusus 401
Coryphella rufibranchialis 48, 65
— sarsi 65
Gorystes 503
Goscinasterias calamaria 254
Granchia scabra 161
Cranchidae 161
Crangon 506, 507, 509
— crangon 518, 519, 522
Crangonidae 502, 509, 522, 529
Crangonyx 495
Crassostrea 130
Crassostrea angulata 128
— gigas 113, 129, 129*
— nippona 129
— virginica 113, 129
Grenomytilus grayanus 113, 116,
119
Crepidula 40
— farnicata 39*
— grandis 32
Creseis 68
Grinoidea 199, 200, 216
Gristaria plicata 135*, 136
— plicata var. herculea 136
Crossaster 263
— papposus 250, 263, 288
Crustacea 376, 377
Gryptochiton stelleri 16, 17, 19
Cryptolepas 432
— rachianecti 416
Cryptoniscidae 477, 479, 480
Gryptoplacidae 17
Cryptoplax 17, 19
— japonica 16
Gtenocidaris nutrix 256, 276
Ctenodiscus australis 260, 261
— crispatus 241
Gtenodonta 97
Cucullaea 115
Gucumaria 233
— elongata 234, 272
— frondosa 231, 233
— glacialis 235
— ijimai 235
— japonica 224, 232
— lactea 235
— lateralis 235
— lubrica 234
— tricolor 233, 240
— vaneyi 235
Cucumariidae 233
Culcita 251, 262
— novae-guineae 240
Culeolus 358
— murrayi 358*
Gumacea 460
Cumella 462
Gumopsis 462
Cuspidaria arctica 128
Cuspidariidae 155
Gyamidae 485, 491
Gyathocystis plautinae 198*
Cyathura carinata 432, 473, 477,
481
Cyclasterope 442
Gyclestheria 390, 391
Cyclocardia borealis 139
—- crebricostata 112, 139
— paucicostata 97, 139
Gyclomyaria (Doliolida) 330, 339
Gyclopecten 97, 125
Cyclopoida 404*, 406, 410, 412, 417
Cyclops strenuus 410, 411
Cyclopteridae 178
Cyclosalpa 341
— bakeri 336
— pinnata 356
Gyclostoma (Pomatias) elegans 34*
Cylichna 62
— alba 62*
— reinchardti 62
— scalpta 62
Gylisticus convexus 479
Cymbium amporus 33
Cymbulia peronii 69
Cymothoa 476
— oestrum 478
Gymothoidae 472, 473
Cymothoinae 476, 477
Cyphocaris 486
— richardi 433
Cypraea 47
— annulus 25
— caurica 32
— moneta 25
— tigris 32
Gypraeidae 47
Cypridae 442
Cypridina castanea 442
— norvegica 442
Gypridinidae 441, 443
Cypridopsis 442
— vidua 441*
Cyprina islandica 97, 112, 139
Gypris 430, 441*
— pubera 440*
Cyproides marginata 442
Cyrtocrinida 222
Gystidea 199
Cytherella 442
Cytheridae 441, 442, 443
Cyzicus 390*, 391
D
Dactylioceras commune 170
Dajidae 477
Daphnia 393, 393*
— arctica 399
— balchashensis 395, 398*
— cristata 395*
— cucullata 395*
— longispina 398
— magna 398*
— pulex 398, 400
Darwinulidae 442, 443
Daudebardia 80
Decapoda 156, 162*, 183, 499
Deima 243
Deimatidae 244
Delectopecten 125
Delphinula rugosa 32
Dendraster 269
Dendrochirota 215, 228*, 229, 232
Dendrogaster 269, 425, 427, 428,
428*
— astropectinis 428
— astericola 264
— beringensis 428
— dichotomus 426*
Dendrogastridae 425, 427, 428
Dendronotus 64
— arborescens 48
550
Dendronotus frontosus 65*
Dentalium 91
Dentalium elepbantium 93
— entalis 93
— vulgaris 92*
Derocersa 90
— argestis 49, 90
— reticulatus 90
Derocheilocaris 422
— galvarini 422, 423, 424*
— remanei 422, 423*, 424, 424*
— typicus 422 , 424*
Desmomyaria (Salpae) 330, 339
Desmosoma 473
Deuterostomia 196, 303
Dexamine spinosa 433
Diadema antillarum 280
— savignyi 280
— setosum 240, 280
Diadematidae 277, 280
Diapbanosoma 397
— bracbyurum 400
Diaptomus 405*, 406, 406*
— amblyodon 410
— gracilis 409
— graciloides 409
Diastylidae 463
Diastylis 462
— glabra 462*
— rathkei 460*, 461*, 463
— scorpioides 463
— sulcata 463
Dibranchia 173
Dicyathifer manni 154
Didacna 142
— barbot de marnyi 143
— crassa 143
— protracta 143, 143*
— trigonoides 143, 143*
Dikerogammarus baemobaphes 494
Dikonopbora 464
Diogenes edwardsi 514
Dipnyllobothrium latum 412
Diplobrachia 314
Discus 90
— (Patula) solarius 80
Distarsio (Murex) anus 32
Distolasterias nipon 288
Distomus variolosus 337
Divaricella divaricata 140
Dolabella 61
Dolicboglossus 321
Doliidae 49
Doliolida (Cyclomyaria) 330
Doliolum rarum 336
Dolium 49
— perdix 47*
Donacidae 147, 148
Donax venustus 145, 148
Doratopsis vermicularis 161
Dorioconus aulicus 33
Dorippe 513
— granulata 512*
Dorippidae 513
Doris 63
— bobretzkii 64
— pilosa 48
— verrucosa 64*
Dosinia japonica 112
Dotilla 523
Doto (ldulia) 64
— corona ta 48
Dracunculus medinensis 412
Dreissena (Dreissensia) 137
— caspia 138*, 139
— distincta 139
— polymorpha 138, 138*
— rostriformis 138*, 139
Dreissenidae 132, 137, 139
Dreissensia (Dreissena) 137
Drepanothrix dentata 398*
Dromia caput-mortuum 513*
— vulgaris 449
Dromiidae 513
Dunalia 477
Dysodonta 96, 115, 124
E
Echinarachnius 285
— parma 271, 285
Ecbinaster luzonicus 263
Ecbinasteridae 263
Echinidae 282
Echinocardium cordatum 285, 286*
Ecbinodermata 195, 196, 197
Echinoidea 200, 271
Ecbinomenia corallophila 16
Echinometridae 284
Echinosigra paradoxa 273, 286
Ecbinothuriidae 271, 278
Ecbinus 278, 282
— acutus 271
— esculentus 276, 282
Ecteinascidia tortugensis 337
Edrioasteroidea 199
Elaphoidella 413
— bidens 416
Elasipoda 228, 228*, 230, 232, 242
Eledone 158, 170*
— moschata 162
Eleutherozoa 199, 200, 227
Elliptica 96
Ellobiidae 73
Elminius modestus 434, 435
Elpidia glacialis 243
— — kermadecensis 243
— — solomonensis 243
— sp. 243, 257
Elpidiidae 243
Emertonia gracilis 413*
Ena obscura 49
Enidae 85
Enoploteutbis leptura 188*
— sp. 186
Ensis ensis 97
Enteropneusta 317,319,322,323
Entocolax ludwigi 51
Entoconcha 51
— mirabilis 52*
Entoconchidae 51
Entodontidae 90
Entoniscidae 477
Epactophanes richardi 416
Epicaridea 472, 473, 477, 478, 479
Epischura baicalensis 409
Eremicaster tenebrarius 260, 428
Ergasilus 417
Ergasilus sieboldi 417*
Eriocheir 524
— japonicus 43", 524
— sinensis 510, 517*, 518, 524,524*
Eripbia verrucosa 500*
Eryonidae 480, 519, 522
Eschrichtius gibbosus 432
Eubranchus (Galvina) flava 48
Eucalanus bungii 404*, 407, 409
Eucarida 495
Eucbirella maxima 401
Eucopia 458, 459
— unguiculata 455*
Eucyclops macruroides 411
— macrurus 411
Eudorella emarginata 463
— pacifica 463
Eudorellopsis integra 463
Eulamellibranchia 96, 100, 103*,
104, 128, 130, 137
Eulimax niger 87
Eulimnogammarus virgatus 433
Euphausia 497, 498
— pacifica 498
— superba 495, 497, 498, 499
Euphausiacea 495
Eupyrgidae 245
Eupyrgus scaber 245
Euryalae 215 287, 291, 292
Eurycercus glacialis 399
— lamellatus 398*
Eurydice 474
Eurytemora hirundoides 407
Evadne 392
— nordmanni 400
Evasterias retifera 288
— troschelii 268
F
Fabulina 147
Fasciola hepatica 76
Filibrancbia 103*, 104, 112, 115
Flabellifera 472
Flexopecten 96, 125
Florometra antarctica 224
Forcipulata 248, 257, 258, 263,
265, 270
Fraginae 141
Fragum 141
— unedo 145
Freyella remex 251
Frierasfer (Carapus) 262
Friticicolidae 90
Fusus 437
G
Galathea 502, 508, 520
Galathealinum 314
— arcticum 306, 307, 315
— brachiosum 315
— bruuni 314
Galatbeatburia aspera 242
Galatheidae 510, 527
Galatheidea 517
Galba truncatula 75, 76*, 78
Galiteutbis armata 190*
36«
551
Galerita hungarica 32
Gammaracanthus loricatus cas-
pius 492
Gammaridae 491
Gammaridea 487, 489
Gammarus 486, 486*, 492, 493
— balcanicus 494
— duebeni 491, 492, 494, 495
— finmarchicus 493
— ischnus 494
— kischineffensis 494
— lacustris 492, 494
— obtusatus 492
— oceanicus 492
— pulex 491*, 493, 494
— salinus 495
— setosus 492
— zaddachi 493
Gariidae (Sanguinolariidae,
Psammobiidae, Asaphidae) 148
Gasterosiphon 50
Gastrana fragilis 112, 147
Gastrochaena 150*
Gastrochaenidae 149
Gastropoda 7, 20
Gastrosaccus sanctus 458
Gecarcinidae 520
Gephyrothuriidae 242
Gibbula 31
— albida 31
Gigantocypris 440*, 441, 442
Gigantomilax kollyi 49
Gilbertsocrinus tuberosus 198
Glandiceps 319, 321, 323
— abissicola 318
— malayanus 318*
Glandina 87
Glans 139
Glaucus 65
Gleba coronata (Tiedemania
neapolitana) 69, 69*
Glossidae 139
Glossobalanus 323
— minutus 324
Glossodoris 64
Glossus rubicundus (lsocaridia
cor) 139
Glycymeridae 115
Glycymeris albolineatus 96
— glycymeris 97
Glyptopbysa resvoi 77
Gnatbia 479*
— dentata 432
Gnathiidea 471, 473, 476, 478,481
Gnatbophausia 455, 456, 458
—gigas 455*, 458
Goffinella stylifer 413*
Goniopectinidae 261
Gonodactylus 452, 453
Gorgonocephalidae 292
Gorgonocephalus carryi 224, 293
— eucnemis 293
Gouldia minima 112
Grapsidae 524
Graptolithoidea 317
Grubia crassicornis 433
Gymnosomata 69
Gynandrocarpa placenta 337
Gyraulus gredleri var. borealis 77
H
Haasiella 136
Hacelia 253*
Haemocera danae 418*
Halicyclops 411
Haliotidae 26
Haliotis 26, 269
— fulgens 28
— gigantea 28
— lamellosa 28
— rufescens 28
— tuberculata 26, 28, 32
Halistemma 67
Halocynthia aurantium 337
— roretzi 337
Halopsychae 68
Hanleya adyssorum 17
Hapalocarcinidae 512, 516
Haplomesus 481
— gigas 481
— insignis orientalis 432
Harmothoe 261
Harpa 56
— ventricosa 33
Harpacticoida 404*, 406, 412
Harpoceras lythense 170
Haustoriidae 490
Heliaster 250
Helicella 84
— candacharica 49
— ericetorum 89
— intersecta 85
— obvia 85
Helicidae 82, 90
Helicigona arbustorum 85
Helicinidae 33
Heliometra glacialis 216, 217*,
224, 241
Helix 82
— aspersa 84
— lucorum 84
— pomatia 72*, 81, 82, 83
— vulgaris 49, 84
Hemicardium 141
— hemicardium 97
Hemicentrotus pulcherrimus 282
Hemichorda 301, 316, 317, 328
Hemicbordata 196
Hemilepistus 473, 474, 484
— cristatus 432, 473
— zachvatkini 484
Hemimysis 457
— lamornae 456*
Hemizonida 248
Hemphilia 85
Henricia 263
— sanguinolenta 241, 263
Hermaenia maculosa 48
Hermissenda crassicornis 48
Herpetocypris rep tans 442
Herpotanais kirkegaardi 464, 466*
Hetairus groenlandicus 448
Heteranomia 96
Heterocentrotus mammilatus 225,
284
Heterocypris incongruens 442
Heterodonta 132, 139
Heterostelea 199
Heterotanais 464
— antarcticus 467
— oerstedi 466, 466*, 467
Hiatella 150*
— arctica 128, 149
Hiatellacea 148
Himerometridae 227
Hippasteria 253*
— phrygyana 241
Hippolyte 506
— varians 448
Hippolytidae 522
Hippopus 143
— hippopus 145
Histioteuthidae 186
Histioteuthis bonelliana 167*
Holectypoida 285
Holopedium 397
— gibberum 399, 400
Holopus rangi 221*, 222
Holothuria 238
— scabra 238, 240
— tubulosa 231
Holothuriidae 238
Holothurioidea 200, 227
Homarus gammarus 465
Homolidae 522
Hoplocarida 450
Horatia 38
Horsiella trisaetosa 413
Hutchinsoniella 401, 402*
— macracantha 401
Hyalopecten 96
Hyas araneus 480, 518
— coarctatus 449, 518
Hydrobia 36
— steini 36, 36*
— ventrosa 36
Hydrobiidae 36, 36*
Hydrosoma luculentum 279, 279*
Hymenaster 249
Hypanis 143, 143*
Hyperamblyops 459
— nana 458
Hyperia galba 488*, 490, 490*
Hyperiidea 487, 488, 490
Hyperiopsidae 487
I
Idiosepius 156
Idotea 474, 478, 480
— algirica 473
— viridis 474
Idoteidae 473
Idulia (Doto) 64
— coronata 65, 65*
Illex illecebrosus 192, 193
Ilyocriptus sordidus 398*
Inachus 517
Incirrata (Apteroti) 175
Ingolfiellidea 486, 495
Iphigena ventricosa 49
Iphinoe trispinosa 461*
Ipsilothuria bitentaculata 235, 257
Irregularia 278, 285
Ischnochiton exiquus 17
— hasodadensis 16
— magdalensis 17
552
Ischnomesidae 481
Isidora 77
Isocardia cor (Glossus rubicundus)
139, 139*
Isocrinida 220
Isognomonidae 121
Isometra vivipara 225
Isometridae 225
Isopoda 470
J
Jaera 474, 480
— albifrons 478, 480, 481, 483*
— marina 480
Jaminia potaniniana 49, 91
Janthina 44
— janthina 44
Janthinidae 44
Jhlea asymmetrica 359
Johanella 482
K
Kalliapseudidae 465
Kobeltocochlea 36
— maxima 36*
Koonunga 446
— cursor 446*, 447
Kuphus polythalamia 152
L
Labidiaster 250
Labidocera 405
Labidoplax buskii 230
Laemodipodea 486
Laetmogonidae 244
Laevicardiinae 141
Lambrus validus 449
Lamellaria 46
Lamellariidae 46
Lamellibranchia (Acephala, Bival-
via, Pelecypoda) 95
Lamellisabella 304, 306, 308, 311,
313, 315
— ivanovi 315
— johanssoni 313*, 315
— minuta 315
— zachsi 304, 312*, 315
Lamellisabellidae 315
Laminaria 282
Lamprometra protectus 240
Lampropidae 463
Lamprops 462, 463
— fasciata 462*, 463
— korroensis 464
Lampsilinae 134, 136
Lampsilis 136
Lampyris 373*
Lanceolaria grayana 134*, 135
Lapidoplax digitata 52*
— thomsoni 247
Lartetia (Paladilhia) 36
Lauriidae 427, 428
Lebbeus 448
Leda (Nuculana) 113
Ledidae (Nuculanidae) 112
Lembos 494
Lepadomorpha 431
Lepas 431*, 436
— anatifera 416, 430*
— fascicularis 416, 431
Lepeophtheirus 419, 419*
Lepetidae 28
L'epidocentroida 215, 278
Lepidochiton aleuticus 16
Lepidopidae 459
Lepidopleurus 19
— arcticus 19
Lepidops 459
Lepidurus 389
— apus 389
— arcticus 389
Leptasterias groenlandica 257
Leptocheirus 489, 490
Leptodora 392
— kindtii 396, 400
Leptometra celtica 224
Leptonidae 140
Leptostraca 443
Leptosynapta inchaerens 246
— minuta 246
Lernaeidae 419
Lernaeocera branchialis 420*
Lernaeoida 417, 419
Lernaeopodidae 405, 419
Leucon fulvus 463
— nasica 461*
— pallidus 463
Leuconidae 463
Leucothoe richiardii 433
Leucothoidae 489
Leydigia acanthocercoides 398*
Lightiella incisa 401
— serendipita 401
Ligia 474, 483, 484
Ligidium 483
Ligiidae 483
Liliopa 36
Lima hians 127
— hyperborea 96, 127
— scabra 144
Limacidae 85
Limacina (Spiratella) 68, 68*
— helicina 68, 69
— retroversa 68
Limapontia 70
Limatula 96, 127
Limax 89
— arborum 87
— maximus 49, 73*, 82, 86, 87
— variegatus 73
Limidae 127
Limnadia 390*, 391
Limnadiidae 391
Limnaea abyssicola 76
— foreli 76
— peregra 76
— — var. geysericola 76
— profunda 76
— stagnalis 74, 74*, 75*
Limnocalanus grimaldii 409, 459
— macrurus 409
Limnocardiinae 140, 142
Limnomysis benedeni 459
Limnoria 474, 475, 484
— lignorum 475*
Limopsidae 115
Limopsis vaginatus 96
Linckia 258*, 259, 262
— laevigata 225, 262
Linckiidae 253*, 262
Liocyma fluctuosa 112
Liriopsis 477, 477*
Litharca 115
Lithodidae 503, 504, 522, 527
Lithoglyphus naticoides 36
Lithoglyptes 437
Lithophaga 120, 121, 150*
— lithophaga 97, 121
Littorina 511
— littorea 35, 36*
— obtusata 35, 36*
— palliata 35
— rudis 35
Littorinidae 35
Livona pica 511
Livoneca 476
— amurensis 476
Loliginidae 170
Loligo 170*
— opalescens 191
— vulgaris 185*
Lophogastrida 456
Lorica (Callistochiton) viviparus 18
Loricata 7, 14
Loripes lacteus 140
Losechinus albus 283
Lottia gigantea 30, 140
Loveniidae 285
Loxorhynchus grandis 507
Luciae (Pyrosomida) 330
Lucinella 140
Lucinidae 140
Ludwigothuria atra 225, 239
Luidia 253*, 262
— ciliaris 258
— quinaria 240
Luidiidae 262
Lybia 514
— tesselata 513*
Lycoteuthis diadema 186*
Lynceus 390, 390*, 391
— brachyurus 391
Lyonsia arenosa 128
Lyonsiella abissicola 128
Lyonsiidae 155
Lyreidus tridentatus 449
Lysianassa punctata 433
Lysianassidae 491
Lysiosquilla 452*, 453
— excavathrix 452
— maculata 454
— vicina 452
Lysmata seticauda 517
M
Macoma 147
— balthica 112, 147
— calcarea 112, 147, 512*
Macrobrachium 448
Macrocheira 500
Macrochlamis 91
Macrocyclops albidus 410
— fuscus 410
553
Macrohectopus branickii 488, 488*,
491, 494
Macroporaster matutinus 198*
Macrostylis 473
Macrothrix hirsuticornis 398*, 400
Mactra sulcataria 128, 149
Mactracea 148
Mactridae 149
Mactromeris 128, 149
Maja squinado 449
Maiidae 513
Makrokylindrus 463
Malacolepas conchicola 432
Malacostraca 443
Malletia 114
Malletiidae 97, 112, 114
Malleus 121, 122
— albus 122
— malleus 122, 145
Mammilifera 514
Margarita 31, 32
Margaritana 133
Margaritifera 108, 133
— dahurica 133
— margaritifera 133
— middendorffi 133
Margaritiferidae 132
Marimetridae 227
Marsenia 46
Marseniopsis 46
Marthasterias glacialis 248, 256
Mastigoteuthis grimaldii 186
Maxillopoda 403
Megaluropus agilis 433
Meganyctiphanes 498
— norvegica 496*, 497, 499
Megayoldia 96, 112, 113
Meinertia 476
Melanella 50
— troglodytes 50, 51*
Melanellidae 50, 219, 278
Melania 38
— amurensis 39
Melaniidae 38
Melanopsis 38
— acicularis 39
Melibe rosea 66
Mellita 262
— longifissa 286*
— sexiesperforata 285
Mercenaria mercenaria 146
— stimpsoni 112, 146
Mesidotea 471*, 473, 475, 480
— entomon 432, 481
Mesocyclops 410, 412*
— leuckarti 411, 412
Mesocypris terrestris 440, 442
Mesogastropoda 33, 34, 39, 41, 50
Mesopodopsis 457
— indica 457
— slabberi 458
Metacrinus 221
— interruptus 221
— nobilis 221, 224
— rotundus 219, 425
Metridia ochotensis 407
— pacifica 407
Micraspides 446
— calmani 446*, 447
Microcerberidae 483
Microcharon 432, 478, 482, 483
— halophilus 483
Microcyclops dengizicus 412
Microdeutopus 489
Micromelanidae 38
Microprotopus 489
Microsetella 412
Microthele 238
— nobilis 239
Millericrinida 221
Miratesta 77
— celebensis 76*
Mitella mitella 416
— pollicipes 436
Mitra 24*
— cruentata 33
— episcopalis 33, 56
— strictica 33
Mitridae 56
Modiolaria (Musculus) 120
Modiolula 96, 120
Modiolus 120
— adriaticus 96
— modiolus 96, 120
— pbaseolinus 96, 120
Moerella 112, 147
Moina macrocopa 398*
— rectirostris 400
Molgulidae 334
Mollusca 5, 7
Molpadia musculus 245, 272
Molpadiidae 245
Molpadonia 228*, 232, 245
Monascidiae 330
Monia macrochisma (Pododesmus
macrochisma) 96, 127
Monodacna 142
— caspia 143, 143*
— colorata 143, 143*
Monodella 468, 469
— argentaria 469, 470
— balophila 469
— relicta 469
— stygicola 469, 469*
— texana 469, 470
Monokonophora 464
Monomyaria 115, 124
Monoplacophora 7, 94
Monstrillidae 417, 418*
Montacutidae 140
Mopalia middendorffii 16
— schrenkii 16
— seta 16
Mucronalia 50
Munida 520*
Munidopsis 527
— polymorpha 527
— rostrata 518
Munneurycope murrayi 473
Munnopsis typica 432
Murex 32, 54*
— erinaceus 53*
— fortispina 54
— saxatilis 32
— scarpio 32
— ternispina 32
Muricidae 54
Musculus (Modiolaria) 120
Musculus discors 120
— nigra 96, 120
Mya arenaria 98, 108, 128, 149, 150
— truncata 97, 149, 150
Myacea 148, 149
Myidae 149
Myodocopa 442
Myopsida 183
Myriotrochidae 247
Myriotrochus 51, 247
— bruuni 247
— mitsukurii 247
Mysida 456
Mysidacea 454
Mysis amblyops 458
— caspia 458
— litoralis 457, 458
— microphthalma 458
— mixta 457, 458
— oculata 457, 458
— relicta 454,455*,457,458,459,460
Mystacocarida 422
Mytilaster lineatus 96, 121
Mytilicola 417, 418, 418*
Mytilidae 115, 116
Mytilus calif ornianus 108, 116
— edulis 113, 116, 118, 119*
Mytilus galloprovincialis 116, 119,
119*
dilatata 119*
— — frequens 119, 119*
— — nominata 119*
— magellanicus 116
— viridis 145
Myidae 149
Myxas (Amphipeplea) 76
— glutinosa 76
Myzomenia 12*
Myzostomidae 214, 214*, 259
N
Nahecans 445
Nais 411
Najadacea (Unionacea) 132
Nannastacidae 463
Nannophiura lagani 291
Nassa 53
— reticulata 46, 53, 53*
Natica 44
— catena 269
— josephina 45*
— vitellus 32
Naticidae 44
Nautilus 170, 173, 418
— pompilius 173*
Nebalia bentzi 445
— bipes 444, 445, 445*
Nebaliopsis typica 443, 445, 458
Nectocrangon 509
Neilonella 114
Nematomenia flavens 16
Nematoscelis 497, 498, 499
— atlantica 496*
Neomenia 12*
Neomysis 459
— integer 457, 458
— vulgaris 457, 458
Neophreatoicus assimilis 432
554
Neopilina galatheae 94, 94*
Neotanaidae 467
Neotanais serratispinosus 466
Nepanthia 258, 258*
Nephrops norvegicus 480, 500*
Neptunea despecta 53, 53*
Neptunidae 53
Neptunus 516
Nereilepas fucala 515
Nereilinum murmanicum 306
Nereis cyclurus 515, 515*
Neritidae 32
Neritina 24*
Nerita spinosa 32
Nicania 97
Niphargoides (Pontogammarus)
maeoticus 433, 488, 490
— robustoides 492
— sarsi 489, 494
— similis 488*
Niphargus 489, 491, 492, 494, 495
— orcinus virei 490, 492, 493
Nitocra chelifer 413*
Nodularia (Unio) 134
Notocidaris gaussensis 277, 277*
Notocrinus virilis 225
Notodromas monacha 442
Notostomus 448
Notostraca 386
Nototeredo norvagica 155
Nucula tenuis 112, 114*
Nuculana (Leda) 113
— pernula 114
— — costigera 96
Nuculacea 111, 112, 114
Nuculanidae (Ledidae) 112
Nuculidae 97, 112
Nudibranchia 63
Nuttallia 112, 148
Nyctiphanes 498
O
Obovaria 136
Ochridocyclops 411
Octacnemidae 341
Octacnemus bythius 358*
Octopoda 156, 162*, 174
Octopodidae 161
Octopodinae 175, 181
Octopus defilippi 163
— dofleini 169
— vulgaris 160, 175, 191, 192
Ocypode 505, 509
— rotundata 505*
Ocythoe 169, 170
Odinella nutrix 271
Oedicerotidae 485
Oegopsida 185
Oicopleura rufescens 343
Oithona 412
— plumifera 401
Oleacinidae 87
Oligobrachia 314
— degieli 310*, 314
— ivanovi 314
Oligobracbiidae 314
Oliva 56
— maura 56*
Oliva mustelina 33
— nana 25
Olividae 56
Ommastrephes sagittatus 185
— sloanei pacificus 160, 186, 191,
192
Ommastrephes sp. 185*
Ommastrephidae 170, 192
Oncaea 412
— mediterranea 401
Onchidoris 64
— bilamellata 64
— muricata 64
Oncidiella 79
Oneidiidae 79
Oneirophanta 243, 244
— alternata 244, 272
Onesia cognata 89
Oniscoidea 483
Oniscus 474, 484
— asellus 480
Onychoteuthis banksii 185, 188*
Oopelta 85
Ophiacantha bidentata 288, 293
Ophiacanthidae 293
Ophiactidae 295
Ophiactis 290
— virens 295
Ophiocistia 199
Ophiocoma delicata 273, 296
Ophiocomidae 296
Ophiodaphne materna 290
Ophioderma brevispina 296
Ophiodermatidae 296
Ophiomastix annulosa 225, 296
Ophiomaza 214*, 291
— cacaotica 291, 295
Ophionotus hexactis 290, 297
Ophiopholis aculeata 241, 295
Ophiopleura 297
— borealis 241, 297
Ophiopsila annulosa 293
— arenea 293
Ophiosphaera insignis 290
Ophiothrichoides pulcberrima 240,
295
Ophiothrichidae 295
Ophiothrix coerulea 225, 295
— fragilis 295
Ophiura multispina 273, 296
— sarsi 224, 287, 296
Ophiurae 287, 290, 293
Ophiuriidae 200, 296
Ophiuroidea 287
Opisthobranchia 22, 57
Opistoteuthis depressa 161
Oplophoridae 522
Oppelia sterospis 170
Orchestia gammarella 491
Orchitophrya stellarum 267
Orconectes 525*
Oreaster nodosus 225, 262
Oreasteridae 262
Oregonia gracilis 512*
Ostracoda 440
Ostrea 110, 130
— cristagalli 144
— denselamellosa 113, 129
— edulis 113, 128, 129
Ostrea edulis adriatica 128
sublamellosa 128
taurica 128, 129*
— laperousi (Crassostrea gigas) 129
— lurida 129
Ostreidae 127
Otina otis 73
Oxyurella tenuicaudis 398*
P
Pachygrapsus 509
— marmoratus 449
Pachythyone rubra 235
Paelopatides 238, 242
— solea 242, 257
Pagurolepas conchicola 432
Paguropsis typica 514
Pagurus acadianus 511
— armatus 515
— arrosor 514, 514*
— bernhardus 480, 500*, 514*
— ochotensis 515, 515*
— prideauxi 511, 514, 515
— pubescens 511*
Paladilhia (Lartetia) 36
Palaemon 502, 506, 507, 509, 519
— adspersus 518, 519
— elegans 448, 518
— longirostris 480
— serratus 501
— squilla 54
Palaemonetes 506
Palaemonidae 527
Palaeoctopus newboldi 172
Paleoneilo 97
Palinurus 519
— vulgaris 465
Pallasea bicornis 433
Palliolum 97
Pancarida 468
Pandalidae 517, 522, 529
Pandalina brevirostris 519
Pandalus 507, 522, 529
— borealis 500*, 517, 518, 519
— latirostris 448, 517, 518
Pandora glacialis 128
Pandoridae 155
Pannychia moseleyi 244, 272
— — mollis 244
— — vergulifera 244
Panope japonica 128
Panoploea eblanae 433
Pantopoda 263
Panulirus 519
— inflatus 480
Papyridea 141
— spinosa 97
Parabathynella caparti 449
Paracaudina ransonetii 245
Paracentrotus lividus 269, 283
Paracyamus boopis 486*
Paracyclops 412*
Paracypbocaris 486
Paradonax 145, 148
Paragnathia 478
Paraleptastacus brevicaudatus 413*
Paralithodes camtschatica 99, 464,
480, 527
555
Paramysis baeri 457, 459
— intermedia 459
— lacustris 459
— loxolepis 458
— ullskyi 459
Paranaspides 446, 447
— lacustris 446*, 447
Parapenaeopsis longirostris 518
Parascothorax 425, 427
Parastacidae 520, 524
Parastenocaris 413
Parastygocaris 450*
— andina 450
Parenteroxenos dogieli 233
Pareuchaeta 407, 407*
— glacialis 405*
Parmacella 85
Paroctopus asper 175, 179
— conispadiceus 162, 175
Paroriza 238
— grevei 238
Patuia 80
Paryphantidae 88
Patella 24*, 29
— algira 29*
— coerulea 30
— granulans 32
— pellucida 30
— pontica 30
— vulgata 29, 30
Patellidae 28, 29
Patinopecten 98, 113, 124, 124*
Patiria pectinifera 118 264, 288
Patulidae 88
Pecten 125, 267
— glaber ponticus 96
— grandis 108
— jacobeus 126
— magellanicus 126
— maximus 113, 126
— yessoensis 98, 113, 124, 124*
Pectinidae 124
Pelagothuria natatrix 244, 272
Pelagothuriidae 231, 244
Pelecypoda (Lamellibranchia,
Bivalvia, Acephala) 95
Pelmatozoa 199
Peltogaster 439, 477
Peltogastrella 439
Penaeidae 502, 504, 509, 518,
519, 520, 522, 529
Penaeopsis dobsoni 520
Penaeus 506
— carinatus 520
— japonicus 448
— plebejus 520
— trisulcatus 518
Penella balaenoptera 419
Peniagone 243
— lugubris 243, 272
Penicillus (Brechites) 155
Penitella 128, 152
Pennatulidae 292
Pentacrinus europaeus 220
Peracarida 454, 468
Perforatella bidens 49
Periploma fragilis 128
Periplomatidae 155
Peronidia 112, 148
Perotrochus birasei 32
Petbalophthalmus 459
Petrarca bathyactidis 427
Petricola lithopbaga 112, 146
Petricolidae 146
Phacosoma 112
Phalium strigatum 33
Phallusia mamrnillata 337
Phanerozonia 215, 248, 250, 258,
261, 263
Pbaraonella perieri 145
Philomedes globosus 443
Philomycidae 85, 91
Philomycus bilineatus 49
Phoenna spinifera 401
Pholadidae 151
Pholadidea penita 128, 152
Pbolas 150*, 511
— dactylus 128, 151, 152
Photidae 489
Phreatoicidea 470,473,474,483,484*
Phrixometra 224
— longipinna 224
— nutrix 224
Phronima 485, 490
Pbyline 62
— lima 62, 62*
Phyllacanthus imperialis 240, 280
Pbyllirrhoe 67, 68
— bucephalum 66*
Phyllirrhoidae 67
Phyllocarida 443, 445
Phyllophoridae 235
Phyllophorus 236
— urna 236
Pbylomicidae 91
Phyllopoda 386
Pbysa 74
— acuta 74
— fontinalis 74, 74*
Physidae 74
Pinaxodes chilensis 284
Pinctada 122
— margaritifera 122, 145
— martensi 122, 144
— vulgaris 122
Pinna 121
— nobilis 121
— muricata 144
Pinnidae 121
Pinnixa chaetopterana 512*
Pinnoteres boninensis 449
Pinnoteridae 510, 512, 516
Pisaster brevispinus 268, 269
— ochraceus 269
Pisidium 136, 137
— amnicum 137, 137*
— casertanum 137
— nitidum 137
Pitar dione 145, 146
Placiphorella stimpsoni 16
— uschakovi 16
Placopecten 126
Placuna placenta 127
Planes minutus 505
Planorbidae 77
Planorbis leucostoma 77
— pfeifferi 78
— planorbis 77
PlanorLfg rossmaessleri 77
— vortex 77
Platyasterida 248
Platycrinus symmetricus 198*
Pleurobranchus 62
— testudinalis 62, 63*
Pleurocystis filitextus 198*
Pleurotomaria 26
— hirasei 32
Pododesmus macrochisma (Monia
macrochisma) 96, 127
Podosphaeraster polyplax 250*, 251
Pogonophora 196, 301, 303, 304,
305, 315
Polititapes 112, 146
Polybrachia 306, 314
— annulata 313*, 314
Polybra biidae 314
Polychaeta 303, 304
Polycope 442
Polydectus 514
Polydora 418
Polyphemus pediculus 400
Pomatias elegans (Cyclostoma
elegans) 34*
Pontella mediterranea 401
Pontellina mediterranea 406
Pontocypris 442
Pontoniinae 512
Pontoporeia 494
— affinis 488
Porcellana 508, 520
— longicornis 480
Porcellanidae 510
Porcellanaster 249, 260*
Porcellanasteridae 249, 260*
Porcellio 474, 479, 484
— scaber 432, 471*
Poromya granulaa 128
Poromyidae 155
Porpita 44, 432
Portlandia 113
— aestuariorum 113, 115*
— arctica 97, 113, 115*
— — arctica 115*
— — portlandica 115*
— — siliqua 115*
Portunidae 503, 505
Portunion 478, 479*
Portunus trituberculatus 519
Potamocorbula amurenis 128
Potamon potamios 526
Potamonidae 520, 526, 527
Pourtalesia tanneri 257
Pourtalesiidae 286
Praunus 456, 457, 458
— flexuosus 457*, 458
Proechinoecus dimorphicus 284
Proisocrinus ruberrimus 222
Proneomenia sluiteri 12
— thiilensis 12
Propeamussium 125
— frigidus 96
— imbrifer 125
— imbriferum major 97
— groenlandicum 96, 125
Prosobranchia 22, 23
Prosoponidae 522
Protobalanus koeleri 318*
556
Protobranchia 103*, 111, 114
Protopecten 96
Protothaca 268
— euglipta 112
Protracheoniscus 472
— birsteini 472
Psammechinus miliaris 276, 283
Psammobiidae (Sanguinolariidae,
Gariidae, Asaphidae) 148
Pseudanadonta complanata 136
Pseudancylastrum 78
— dybowskii 77*
Pseudocuma cercaroides 464
— longicornis 462*
— longirostris 462
Pseudocumidae 462, 464
Pseudomma 455
Pseudophytina rugifera 140*
Pseudostichopus 242
— tranchus 242
Psilaster pectinatus 428
Psolidae 236
Psolidium 236, 237
Psolus 236
— antarcticus 237
— chitonoides 237, 288
— ephippifer 237, 256
— phantapus 236, 272
— valvatus 237
Psychropotes 243
Psychropotidae 243
Pteranodon 136
— kerhervei 272
— raripes 243, 257
Pteraster 264
— obscurus 256, 264
— tesselatus 264
Pterastericola fedotovi 264
Pterasteridae 249, 253*, 258, 264
Pteria (Avicula) 122
Pteriacea 121
Pteriidae 121
Pterobranchia 317, 319, 325, 326,
328
Pterocera 48
— rugosa 33
Pterocuma pectinata 461*
Pterocyclus 34
Pterometra pulcherrima 225
Pteropoda 67
Pterosoma 43
Pteroti (Cirrata) 183
Pterotracheidae 41, 43
Pterotrachea 43
— sp. 42*
Ptychodera 317, 322, 323, 324
— erythraea 319
— flava 319
Pugettia quadrifrons 449
Pulmonata 22, 71
Punctum pygmaem 90
Puncturella noachina 23*, 28
Pupilla muscorum 49, 88
Pupillidae 89
Purpura lapillus 32
Pycnopodia 251, 270
— helianthoides 270, 288
Pyrosoma ambulata 351
— atlanticum 361
Pyrosoma fixntn 351
— sp. 336
— spinosum 337
— vitjazi 337, 349, 351
Pyrosomata (Ascidiae salpaefor-
mis) 330
Pyrosomida (Luciae) 330, 351
Pyrula 49, 50*
— pernessa 33
Pyura discoveryi 337
— setosa 337
R
Radix auricularia 75, 76*
— ovata 75, 76*
— peregra 76*, 78
Rana temporaria 89
Randallia eburnea 449
Rapana 55
— thomasiana 33
Recluzia 44
Regularia 278
Retusa 62
— partenius var. turrita 62*
Rhabdonectes armatus 488*
RhajDdopleura 325, 325*, 326, 326*,
328
Rhipidiocystis baltica 198*
Rhisocrinus lofotensis 221, 221*
Rhithropanopeus 480
— harrisi 524*
Rhizocephala 437
Rhopalodina 236
Rhopalodinopsis 236
— capensis 236, 272
Rhopalomenia aglaopheniae 12*
Rhytida 87
Rissoidae 36
Rissoa costata 36*
Rodope 58
Rossia 161
— glaucopis 160, 184*
Rotula orbiculus 224, 285
s
Saccoglossa 70
Saccoglossus kowalevskii 321
— mereschkowskii 317, 319
— otagoensis 319
— pusillus 318*, 319
Sacculina 438, 477
— carcini 438*
Sagartia parasitica 514*, 515
Salenia 281
— pattersoni 281
Salenidae 281
Salenocidaris 281
— miliaris 273, 281
Salmacina 418
— danae 418*
Salpa fusiformis 359, 363
— gerlachei 359
— maxima 336
— tompsoni 359
Salpae (Desmomyaria) 330
Sandersella acuminata 401
Sanguinolaria (Nuttalia) olivacea
112, 148
Sanguinolariidae (Psammobiidae,
Gariidae, Asaphidae) 147, 148
Sapphirina 412
— ovatolanceolata 401
— vorax 401
Sarcobotrylloides racemosum 337
Sarcophaga 89
Saxicavacea 148
Saxidomus 268
— nuttali 146
— purpuratus 113, 146
Scalaria 24*
Scalpellidae 433, 434
Scalpellum 436
— japonicum 416, 436
Scaphander 62
Scapharca 96, 115
Scapholeberis 442
— mucronata 394, 394*
Scaphopoda 7, 91
Schistosomum bovis 78
Schizasteridae 286
Schizocardium brasiliense 318*
Schizochiton 19
Schizodonta 134
Schizothaerus keenae 128
Schizoglossa novoselandica 80
Sclerocrangon ferox 520
— salebrosa 448
— zenkevitchi 520
Sclerolinidae 313, 314
Sclerolinum 313*, 314
— brattsomi 306, 314
— javanicum 306
— sibogae 314
Scotoplanes 243, 244
— murrayi 244, 257
Scrobiculariidae (Semelidae) 147,
148
Sedentaria 303
Segmentina hilida 77
Semelidae (Scrobiculariidae) 148
Sepia 170*, 184
— officinalis 160
Sepiella 161
Sepiidae 170
Sepiola 161
Sepiolidae 161, 170
Septibranchia 103*, 128, 155
Sergestes 522*
— arcticus 519
— corniculum 522*
— cornutus 480
Sergestidae 502, 504, 519, 522
Serolidae 472, 473, 480
Serolis 432
Serripes 141
— groenlandicus 112, 114*, 142
Sesarma meinerti 506
Siboglinidae 314
Siboglinoides 314
— columbiensis 314
— dibrachia 314
Siboglinum 304, 305, 307, 308,
311, 312, 314
— caulleryi 306, 310* 314
— fedotovi 313*
557
Siboglinum hyperboreum 306
— minimum 307
— weberi 304
Sicyonia carinata 517, 518*
Sida 393, 393*, 397
— crystallina 394, 400
Sigaretus 46
Silicodermata 8
Siliqua alta 128
Simocephalus 394
— vetulus 394*, 400
Sinagoga 427, 428
— metacrinicola 425
— mira 425
Sinagogidae 425, 428
Sinusigera 55
Siphonariidae 73
Siphonodentalium lafotense 92*
— vitreum 94
Siriella jaltensis 458
Solariella 31
Solaster 425
— endeca 263
Solasteridae 263
Solemya borealis 145
Solemyidae 114
Solen grandis 145
— marginatum (vagina) 148
— vagina (marginatus) 148
Solenacea 148
Solenidae 148
Solenogastres 11
Somasteroidea 250
Spatangidae 286
Spatangoidae 285
Spatangus purpureus 286
Speciospongia vespara 512
Spelaeogriphacea 467
Spelaeogriphus lepidops 467, 468*
Spengelia 322, 323
Speocyclops 411
Speodiaptomus birsteini 410
Sperosoma biseriatum 278
Sphaerasteridae 251
Sphaerechinus granularis 284
Sphaeriidae 136
Sphaerioidea 132, 136, 137
Sphaerium 136
— corneum 137, 137*
— rivicola 136, 137
— solidum 137
Sphaeroma 474, 475, 480, 484
— serratum 432
— rugicauda 480
Sphaeromidae 472, 478, 482
Spinula 114
— calcar 145
— vityazi 114
Spinulosa 248, 250, 257, 258,
258*, 263
Spiratella (Limacina) 68, 68*
Spirobrachia 308, 311, 313, 315
— beklemischevi 309*, 313*, 315
— grandis 307, 315
Spirobrachiidae 315
Spisula (Mactromeris) voyi 128, 149
— sachalinensis 98, 128, 149
— solidissima 149
Spondylidae 127
Spondylus calcifer 17
— pictorum 145
Spongicola 512
Squilla mantis 452, 453, 454
— oratoria 452*, 454
Stagnicola palustris 75, 76*, 78
Stelleroidea 250
Stenasellus 482
Stenoglossa 51, 55
Stenopus hispidus 448
Stephanometra spicata 227, 240
Stereocidaris nutrix 276
Stereoderma lamperti 235
Sthenoteuthis bartrami 192
Stichopodidae 239
Stichopus chloronotus 225, 240
— japonicus 240, 288
— variegatus 240
Stilifer 50
— brychius 50, 51*
— celebensis 52*
Stiliferidae 278
Stirodonta 281
Stomatopoda 450
Storthyngura herculea 432
Strohilopsidae 91
Strombidae 48
Strombus 48
— auris-dianae 33
— gigas 25, 32
— lentiginosus 47*
Strongylocentrotus 270, 278
— droebachiensis241,276,278,
282
— franciscanus 282
— intermedins 277
— nudus 282, 288
— purpuratus 282, 288
Strongylocentrotidae 281, 282
Strophochilus popelairianus 80, 80*
Styela clavata 337
— plicata 337
Stygasellus 482
Stygiomysidae 459
Stygiomysis 459
Stygocaridacea 446, 450
Stygocaris 450
Stylocheiron 497, 498
Stylocidaris affinis 224, 280
Stylommatophora 78
Styracaster 260, 260*
— horridus 257
Succinea putris 49, 79, 79*
Succineidae 80, 89
Sunetta meroe 145
Sycozoa umbellata 337
Symplectoteuthis oulaniensis 185
Synallactidae 242
Synalpheus 512
Synapta maculata 246
Synaptidae 246
Synaptula hydriformis 247, 272
Synascididae 330
Syncarida 446
Syndesmya (Abra) 148
Synidotea 480
Synurella 495
Systellaspis 522*, 523
Swiftopecten 124, 145
T
Taenogiras cidaridis 247
Talitridae 487, 491, 493
Tanaidacea 464
Tanais stanfordi 466, 466*
Tapes 98
Taxodonta 96, 97, 111
Tectibranchia 59
Tectus maximus 32
Tellinacea 147
Tellinidae 145, 147
Tenagodus 41
Terebra 57
— subulata 32
Terebridae 56
Teredinidae 151, 152
Teredo 96, 151, 152, 153, 153*
— megotara 155
— navalis 154
Tessarobrachion 499
Testacella 87
— maugei 87*
Tethya 329
Tethys (Fimbria) 66
— fimbria 48
— leporina 66
Tetrabranchia 172
Thais 511
Thalia democratica 336, 359
— longicaudata 360
— sp. 336
Thatcheria mirabilis 33
Thaumatolampas diodema 161
Theba 84
— fruticum 49
Thecanephria 310, 313, 314
Thecosomata 68
Thelenota annanas 239
Theodoxus fluviatilis 32*, 33
— pallasi 32, 32*
— transversalis 32*
Thermobathynella adami 450
— amyxi 450
Thermosbaena 468, 469
— mirabilis 469*, 470
Thermosbaenacea 454, 468
Thompsonia 439
Thoracica 430, 431, 432, 436, 437
Thracia myopsis 128
Thraciidae 155
Thyasira (Axinus) 140
Thyasiridae 140
Thyca 40
Thyone 234
— briareus 234
— imbricata 235
— mirabilis 240
Thyonepsolus 236, 237
— nutriens 237
Thyonidium pellucidum 236
Thyrophorella 81
Thysanoessa 498
— inermis 498
— longicaudata 498
— longipes 499
— raschii 496*, 497, 498, 498*
Thysanopoda 499
Thyasiridae 140
558
Tiara praestantissima 33
Tiedemania neapolitana (Gleba
coronata) 69*
Tindaria 114
Tiphobia 39
— horei 39*
Tonicella granulata 16
— marmorea 16, 19
Toxopneustes pileolus 240, 274,
284
Toxopneustidae 283
Tracheata 376
Trachycardium 141
— flavum 97
Trachycardiinae 141
Trachydermon 19
— albus 19
— ruber 19
Trachypenaeus constrictus 480
Trematoda 78
Trichia 84
— rufescens 85
Trichoniscus 479
— elisabethae 479
— provisorius 479
Tridacna 143
— crocea 145, 146*, 150*
— derasa 145
— gigas 143, 144, 145
— mutica 143
Tridacnidae 143
Trididemnum 348
Tridonta 96, 97, 139
Trifora 47
Trigonochlamididae 87
Trigonochlamys imitatrix 49
Trilobita 376
Triops 389
— cancriformis 387*, 389
Tripetosa lampos 437*
Tripneustes ventricosus 283
Trisidos 115
Tritonia hombergi 48
Tritonium 46, 49, 54*
— undatum 53*
Trivia 47
Trochidae 31
Trochocystites bonemicus 198*
Trochostoma arcticum 224, 245
Trochus 31
— adriaticus 31
— niloticus 31
— occidentalis 31
Troglocaris anophthalmus 448, 508,
526
Troglolestes 87*, 88
Troglomysis vjeternicensis 459
Trophodiscus 261
— uber 256
Trophonopsis breviatus striata 55
Tropiometra carinata 222
Tubicinella 432
Tunicata (Urochordata) 196, 301,
329, 330
Turbinidae 32
Turbo 32
— olearius 32
Turitella terebra cerea 32
Typhlocirolana 482
Typhlopseudothelphusa mocinoi
527
Typhiosphaeroma 482
U
Uca 510*, 523
— annulipes 523*
— batuenta 506
— beebi 506
— marionis 523*
— pugilator 507
— pugnax 507
Udonella 419, 419*
Ulophysema 427
Umbrella botanicum 62
— mediterranea 63*
Unio (Nodularia) 134
— crassus 134*, 135
— douglasiae 135
— pictorum 134*, 135
— tumidus 134*, 135
Unionacea (Najadacea) 132, 133
Unionidae 111, 132, 133, 134
Unioninae 134
Urasterias lincki 241
Urochordata (Tunicata) 330
Urosalpinx cinerea 55
V
Vallonia 89
— costata 49
Valva valva 33
Valvata brandti 38*
— cristata 37*, 38*
— piscinalis 38, 38*
— fluviatilis 38*
Valvatidae 38
Valvifera 472, 475
Vampyroteuthis infernalis 161,
169, 183
Velella 44, 66, 432
Velutina 46
Veneridae 145
Venerupis 98
— japonica 112, 146
— discrepans 146
— rugata proclivis 112, 146
Venus casina 97
— fasciata 97
Vermetidae 40
Vermetus 40
— lumbricalis 41*
Verruca stromia 416
Verrucidae 431
Vertebrata 330
Verticordiidae 155
Vertiginidae 89
Vharonia tritonis 32
Viguierella coeca 413
Vitjazaster 260*
Vitjaziana gurjanovae 494
Vitrea 80
Viviparidae 37
Viviparus 37
— contectus 37, 37*
— ussuriensis 37
— viviparus 23*, 38
Volchovia volborthi 198*
Voluta 33*
W
Willeya 319, 323
X
Xenobalanus globicipitis 416
Xenophora 41
Xenophoridae 41
Xenopleura vivipara 322
Xerophila 84
Xylophaga 152
— dorsalis 152
— grevei 151*, 152
Y
Yoldia 112
— hyperborea 96, 113
— limatula 113
— myalis 113
— scissurata 97, 113
— thraciaeformis 96, 113
Yoldiella 113
Z
Zachsia 153
— zenkewitchi 155
Zebrina cylindrica 49
— detrita 85
Zenkevitchia 495
Zenkevitchiana 314
— longissima 307, 313, 314
Zonitidae 80, 90
Zonitoides 80
— nitidus 80
Zospeum spelaeum 73
Zyrfaea crispata 97, 128, 152
— pilsbryi 128, 152
ИМЕННОЙ УКАЗАТЕЛЬ
А
Агассиц А. 197, 316
Аристотель 162, 168, 169, 182,
187, 191, 197, 227, 248
Арсеньев В. К. 180
Б
Банта 395
Барнес 265
Бауер 46
Беклемишев В. Н. 368
Беклемишев К. В. 360
Беллинсгаузен Ф. Ф. 362
Берг Л. 46
Блэнвилль 19, 95
Богоров В. Г. 408
Бойкот 177
Боруцкий Е. В. 413, 414, 415
Брем А. 189
Бруне 305
Бэйнбридж 407
Бэссон 317
В
Вагнер Н. 424
Вебб 305, 306, 314
Вебер 304
Вейдовский В. 448
Верани Ж. Б. 167, 178, 180,
185
Веррил Э. 189
Винкстед 407
Воробьев В. П. 385
Восс Дж. 185
Г
Гаевская Н. С. 383
Гальцов 266
Гарвей Р. 189
Гарстэнг 193
Гегенбауер 69
Гемилл 258
Гислен 223, 225
Голвин 234
Гольдфусс Г. А. 95
Грей 35
Греннингзетер А. 190
Гунько А. Ф. 384
д
Дарвин Ч. 178, 357, 428, 433, 437,
510
Дерюгин К. М. 35, 64
Дженнингс 269
Джильпатрик 164
Догель В. А. 159, 317, 417
Дьяконов А. М. 197
3
Закс И. Г. 241, 304
Зевина Г. Б. 432
Зеликман Э. А. 497
Зенкевич Л. А. 314
Зенкович Б. А. 189
Зимрот 17, 26, 29, 47, 57, 64, 68
И
Иванов А. В. 233, 305, 313
Иоганссон К. Э. 304, 315
Ирвин 282
Исаев Л. 410
Ишикава 439
К
Кабанов Г. К. 172
Камерер 411
Карлайл Д. 304, 305, 307, 313
Киркегаард 307
Кларк Г. 226
Клейн 197
Клинджел Дж. 175, 271
Книпавич Н. М. 425, 428
Ковалевский А. О. 52, 197, 316,
329
Коллери М. 304, 305
Кольман Ю. 177
Котэ 130
Коцебу О. 317, 329
Крумбах 66
Кун 67
Кусто 176
Кювье Ж. 182, 197, 368, 429
Кюкенталь В. 316
Л
Лаказ-Дютье А. Ж. Ф. 55, 91
Ламарк Ж. Б. 197, 429
Лейкарт Р. 197
Лейн Ф. 168, 178, 184
Ли Г. 178, 182
Либеркинд 197
Ливанов Н. А. 303
Линк 95
Линней К. 95, 197, 329
Ли-Сюер 164
Литтон-Булвер Э. 428
Логвинович Д. Н. 508, 509
Лонгхерст 389
Лузанов 266
Лундблад 388
Людвиг 197
М
Мак Гинити 160, 164, 181
Мануйлова Е. Ф. 395, 396
Марукава 520
Мейер Э. А. 53, 59, 60
Мебиус К. 53, 59, 60
Мечников И. И. 210, 316
Миддендорф А. Ф. 17
Минер 265
Митсукури 123
Мозелей 16
Мойз 434
Монфор Д. 187
Мортенсен Т. 197, 215, 242, 258,
289
Моултон 516
Мэк Б. 197
Мюллер И. 316
Мюллер О. Ф. 403
II
Навозов-Лавров Н. П. 527
Найт-Джонс 434
Некрасов П. Д. 38
Нигрелли 239
Нэф А. 167
О
Олаус М. 187
Орбиньи А. Д. 170
II
Паннинг 233
Папи 493
Парди 493
560
Паркер X. 263
Пельзенеер 55
Перес 292
Петерман 385
Плиний 128, 191, 197, 227
Плутарх 191
Порфирьева Н. А. 303
Поулсен 307
Р
Раковица Э. 169
Рамирец 466
Реомюр 521
Рис У. 185
Родендорф Б. Б. 376
Румф Г. 47
Руссо Т. 179
Рылов В. М. 396
С
Савилов А. И. 240, 241
Сайнл Дж. 158
Сандерс 400, 401
Саусворд А. 305, 307
Саусворд И. 305, 307, 314
Смит 239, 268
Спенсер В. 250
Стеенструп 189
Т
Тарасов Н. И. 432, 515
Тиль 197
Трошель Ф. Г. 49
Тэйлор 511
Ф
Федер 269
Федотов Д. М. 197, 294
Фелл Б. 250, 289
Фишер 17, 197, 199, 270
Фредерик Л. 164
Фрей 197
Фудживара 274
X
Ханкок Д. 263
Хансен 242
Харди А. 304
Хединг 244
Хейердал Т. 436
Холмс 166
Холодковский Н. А. 316
Хэдлей 17
Хэл Д. 164
ц
Цукерзис Я. М. 525
Ш
Шамиссо А. 329
Шефер В. 164
Шимкевич В. М. 317, 425
Шмальгаузен И. И. 317, 329
Шманкевич В. 383
Э
Эндин 265
Эстерли 408
Эшшольц 316
Ю
Юнг 177
Я
Янагимачи 439
СОДЕРЖАНИЕ
ТИП МОЛЛЮСКИ (MOLLUSCA) 7
Общий очерк —
КЛАСС БЕСПАНЦИРНЫЕ (APLA-
COPHORA). Б. В. Властов, П. В.
Матекин И
КЛАСС ПАНЦИРНЫЕ (LOR1CA-
ТА). Б. В. Властов, П. В.
Матекин 14
КЛАСС БРЮХОНОГИЕ
(GASTROPODA). Б. В. Властов, П. В.
Матекин 20
Подкласс Переднежаберные (Рго-
sobranchia) 23
Отряд Древние брюхоногие (Archa-
eogastropoda) 25
Отряд Mesogastropoda 33
Отряд Stenoglossa 51
Подкласс Заднежаберные (Opisthob-
ranchia) 57
Отряд Покрытожаберные (Tectib-
ranchia) 59
Отряд Acoela 62
Отряд Крылоногие (Pteropoda) 67
Отряд Мешкоязычные (Sacoglos-
sa) 70
Подкласс Легочные (Pulmonata) 71
Отряд улиток с глазами у
основания щупалец (Basommatophora) 73
Отряд улиток с глазами на концах
щупалец (Stylommatophora) ... 78
КЛАСС ЛОПАТОНОГИЕ (SCA-
PHOPODA). Б. В. Властов,
П. В. Матекин 91
КЛАСС МОНОПЛАКОФОРЫ (МО-
NOPLACOPHORA). Б. В.
Властов, П. В. Матекин 94
КЛАСС ДВУСТВОРЧАТЫЕ (B1VAL-
V1A), В. И. Зацепин,3. А. Филатова 95
Отряд Гребенчатозубые (Taxodonta) 111
Отряд Связочнозубые (Dysodonta) 115
Отряд Настоящие
пластинчатожаберные (Eulamellibranchia) . . . 130
Отряд Перегородчатожаберные
(Septibranchia) 155
КЛАСС ГОЛОВОНОГИЕ
(CEPHALOPODA). И. И. Акимушкин ... 156
Подкласс Четырехжаберные (Tet-
rabranchia) 172
Наутилусы, или Жемчужные
кораблики —
Подкласс Двужаберные (Dibran-
chia) 173
Отряд Осьминоги (Octopoda) . . 174
Подотряд Бесплавниковые, или
Настоящие, осьминоги (Apteroti,
или lncirrata) 175
Подотряд Глубоководные
осьминоги (Pteroti, или Cirrata) 183
Отряд Десятиногие головоногие
моллюски (Decapoda) —
Подотряд Каракатицы (Myopsida) —
Подотряд Кальмары (Oegopsida) 185
ТИП ИГЛОКОЖИЕ (ЕСНШО-
DERMATA) .197
ПОДТИП СТЕБЕЛЬЧАТЫЕ ИГЛОКОЖИЕ
(Pelmatozoa) 216
КЛАСС МОРСКИЕ ЛИЛИИ (CR1-
N01DEA). 3. И. Баранова . . . —
ПОДТИП СВОБОДНОЖИВУЩИЕ
ИГЛОКОЖИЕ (Eleuterozoa) .... 227
КЛАСС МОРСКИЕ КУБЫШКИ
(HOLOTHUR101DEA). 3. И.
Баранова —
Отряд Древовиднощупальцевые
голотурии (Dendrochirota) .... 232
Отряд Щитовиднощупальцевые
голотурии (Aspidocbirota) .... 237
Отряд Боконогие голотурии
(Elasipoda) 242
Отряд Бочонкообразные голотурии
(Molpadonia) 245
Отряд Безногие голотурии
(Apoda) —
КЛАСС МОРСКИЕ ЗВЕЗДЫ (AS-
TER01DEA). Г. М. Беляев .... 247
Отряд Явнопластинчатые звезды
(Phanerozonia) 259
Отряд Игольчатые звезды (Spinulo-
sa) 263
Отряд Педицелляриевые звезды
(Forcipulata) 265
КЛАСС МОРСКИЕ ЕЖИ (ЕСН1-
N01DEA). З.И.Баранова . . . . 271
Подкласс Правильные ежи (Regula-
ria) 278
Отряд Lepidocentroida —
Отряд Копьеносные ежи (Cidaroi-
dea) 279
Отряд Аулодонта (Aulodonta) . . 280
Отряд Копьевиднозубые ежи
(Stirodonta) 281
Отряд Сводчатозубые ежи (Camaro-
donta) —
562
Подкласс Неправильные ежи (lrre-
gularia) 285
Отряд Плоские ежи (Clypeastroi-
da) —
Отряд Сердцевидные ежи (Spa-
tongoida) —
КЛАСС ОФИУРЫ, ИЛИ
ЗМЕЕХВОСТКИ (OPH1UR01DEA). 3. И.
Баранова 287
Отряд Ветвистые офиуры (Euryalae) 292
Отряд Настоящие офиуры(Ордшгае) 293
ТИП ЩЕТИНКОЧЕЛЮСТНЫБ,
ИЛИ «МОРСКИЕ СТРЕЛКИ»
(CHAETOGNATHA). Л. А
Зенкевич 298
ТИП ПОГОНОФОРЫ (POGONO-
PHORA). А. Я. Иванов 303
Отряд Athecanephria 313
Отряд Thecanephria 314
ТИП ПОЛУХОРДОВЫЕ (НЕМ1-
CHORDATA). Я. Г. Виноградова 316
ТИП ОБОЛОЧНИКИ (TUNICA-
ТА). Я. Г. Виноградова .... 329
ТИП ЧЛЕНИСТОНОГИЕ
(ARTHROPODA) 367
Введение. М. С. Гиляров —
ПОДТИП ЖАБРОДЫШАЩИЕ (Вгап-
chiata). Я. А. Бирштейн 377
КЛАСС РАКООБРАЗНЫЕ
(CRUSTACEA) —
Подкласс Жаброногие
ракообразные (Branchiopoda) 380
Отряд Жаброноги (Anostraca) . . —
Отряд Листоногие ракообразные
(Phyllopoda) 386
Подотряд Щитни (Notostraca) . . —
Подотряд Конхостраки (Conchos-
traca) 390
Подотряд Ветвистоусые
ракообразные (Cladocera) 391
Подкласс Цефалокариды (Cepha-
locarida) 400
Подкласс Максиллоподы (Maxil-
lopoda) 403
Отряд Веслоногие ракообразные
(Copepoda) —
Подотряд Каланиды (Calanoida) . 406
Подотряд Циклопы (Cyclopoida) . 410
Подотряд Гарпактициды (Награс-
ticoida) 412
Паразитические веслоногие ... 416
Отряд Карпоеды, или Карповые
вши (Branchiura) 419
Отряд Мистакокариды (Mystacoca-
rida)
Отряд Мешкогрудые
ракообразные (Ascothoracida)
Отряд Усоногие ракообразные
(Cirripedia)
Подотряд Торациковые
ракообразные (Thoracica)
Подотряд Акроторациковые
ракообразные (Aero thoracica)
Подотряд Корнеголовые
ракообразные (Rhizocephala)
Подкласс Ракушковые
ракообразные (Ostracoda) 440
Отряд Ракушковые ракообразные
(Ostracoda) —
Подкласс Высшие ракообразные
(MalacoStraca) 443
Надотряд Филлокариды (Phylloca-
rida) —
Отряд Тонкопанцирные
ракообразные (Leptostraca) —
Надотряд Синкариды (Syncarida) 446
Отряд Анаспидовые ракообразные
(Anaspidacea) —
Отряд Батинеллиевые
ракообразные (Bathynellacea) 447
Отряд Стигокариды (Stygocarida-
сеа) 450
Надотряд Гоплокариды (Hoploca-
rida) —
Отряд Ротоногие ракообразные,
или Раки-богомолы (Stomatopoda) —
Надотряд Перакариды (Peracarida) 454
Отряд Мизиды (Mysidacea) ... —
Отряд Кумовые ракообразные (Си-
macea) 46(»
Отряд Клешненосные ослики (Та-
naidacea) 464
Отряд Спелеогрифовые
ракообразные (Spelaeogriphacea) 467
Отряд Термосбеновые
ракообразные (Thermosbaenacea) ...... 468
Отряд Равноногие ракообразные
(Isopoda) 470
Отряд Разноногие ракообразные,
или Бокоплавы (Amphipoda) . . 484
Надотряд Эвкариды (Eucarida) . . 495
Отряд Эвфаузиевые ракообразные
(Euphausiacea) —
Отряд Десятиногие ракообразные
(Decapoda) 499
Указатель терминов 530
Указатель русских названий . . 540
Указатель латинских названий . 547
Именной указатель 560
жизнь животных
в шести томах
2
том
Редакторы
В. И. СУЧИНСКАЯ, Е. Н. ХУНЦКАРИЯ
Редактор карт О. В. ПАЩЕНКО
Переплет художника М. В. СЕРЕГИНА
Суперобложка художников
В. И. ПРЕОБРАЖЕНСКОЙ, М. В. СЕРЕГИНА
Цветные и тоновые иллюстрации художников
Н. Н. КОНДАКОВА, А. В. КОНДРАТЬЕВА,
Н. Г. КОРОБОВОЙ, Т. Н. НАРБЕКОВОЙ,
В. И. ПРЕОБРАЖЕНСКОЙ, 0. Ф. ХЛУДОВОЙ
Макет и художественное редактирование
М. Л. ФРАМА
Технический редактор Н. Н. МАХОВА
Корректоры
В. А. ГЛЕБОВА, Т. М. ГРАФОВСКАЯ
Составители указателя
Э. С. КАРМОВСКАЯ,
Т. Л. МУРОМЦЕВА,
3. В. ЧАДАЕВА
Сдано в набор 5/VII 1966 г. Подписано к печати
17/VI 1968 г. 84Xl087ie. Янонис № 1
Печ. листов 59,22 C5,25) +вкл. 5,04C). Учетн.-изд. л.
57,6+вкл. 7,11. А03927. Тираж 300 тыс. экз.
•
Издательство «Просвещение» Комитета по печати
при Совете Министров РСФСР. Москва, 3-й проезд
Марьиной рощи, 41. Заказ № 1689.
Московская типография № 2 Главполиграфпрома
Комитета по печати при Совете Министров СС@Р. Москва,
проспект Мира, 105.
Цветные вклейки отпечатаны Калининским полиграф-
комбинатом Главполиграфпрома Комитета по печати
при Совете Министров СССР с диапозитивов,
изготовленных полиграфкомбинатом и типографией «Красный
пролетарий».
•
Цена 3 р. 60 к.
Таблица 1. Панцирные A—12 и бороздчатобрюхие
A3—14) моллюски:
1 — Placiphorella stimpsoni:
2 — Ischnochiton hasodadensis;
3 — Lepidochiton aleuticus;
4 — Tonicella granulata;
5 — Mopalia middendorffii;
6 — Mopalia schrenkii;
7 — Acanthochiton rubrolineatus;
8 — Placiphorella uschacovi;
9 — Cryptochiton stelleri;
10 — Tonicella marmorea;
11 — Cryptoplax Japonica;
12 — Mopalia seta;
13 — Nematomenia flavens;
14 — Echinomenia corallophila.
Таблица 2. Брюхоногие моллюски:
1 — Fissurella graeca,
2 — Acmaea cassis;
3 — блюдечко (Patella granulans);
4 — Acmaea testudinalis;
5 — Margarita;
6 — Perotrochus (Pleurotomaria)
hirasei:
7 — морское ушко (Haliotis tube-
rculata);
8 — Cypraea caurica;
9 — Tectus maximus,
10 — Cypraea tigris;
11 — Nerita spinosa;
12 — Delphinula rugosa;
13 — Purpura lapillus;
14 — Vharonia tritonis;
15 — Natica vitellus;
16 — Strorabus gigas;
17 — Turitella terebra cerea;
18 — Cirsotrema perpexum;
19 — Murex ternispina;
20 — Murex scarpio,
21 — Murex saxatilis,
22 — Distarsio (Murex) anus;
23 — Galerita hungarica;
24 — Grepidula grandis;
25 — Terebra subulata.
Таблица 3. Брюхоногие моллюски:
1 — Rapana thomasiana;
2 — Phalium strigatum;
3 — Bulla lignaria;
4 — Thatcheria mirabilis;
5 — Pyrula pernessa;
6 — Valva valva;
7 — Cassis cornuta;
8 — Pterocera rugosa;
9 — Strombus auris-dianae;
10 — Harpa ventricosa;
11 — Siliquaria cumingi;
12 — Конус (Conus marmoreus);
13 — Dorioconus aulicus;
14 — Conus ebraeus;
15 — Mitra episcopalis;
16 — Mitra cruentata;
17 — Oliva mustelina:
18 —Cymbium (Voluta) amporus;
19 — Tiara praestantissima;
20 — Mitra strictica.
Таблица 4. Голожаберные моллюски:
1 — Coryphella rufibranchialis;
2 — Hermissenda crassicornis;
3 — Eubranchus (Galvina) flava;
4 — Tethys fimbria;
5 — Dendronotus arborescens;
6 — Hermaenia maculosa;
7 — Tritonia hombergi;
8 — Berghia coerulescens;
9 — Doris pilosa с югадкой;
10 — Doto coronata;
11 — Chromodoris coerulea;
12 — Triopha carpenteri.
Таблица 5. Наземные моллюски:
1 — Jaminia potaniniana;
2 — Helicella candacharica;
3 — кустарниковая улитка (Bradybaena fruticum);
4 — янтарка (Succinea putris);
5 — Pupilla muscorum;
6 — Perforatella bidens;
7 — полевой слизень (Deroceras agrestis);
8 — Arion subfuscus;
9 — Arion empiricorum;
10— Limax maximus;
11 — Arion maximus;
12 — Cochlicopa lubrica;
13 — Theba fruticum;
14 — Ena obscura;
15 — Iphigena ventricosa;
16 — Gepaea nemoralis;
17 — Zebrina cylindrica;
18 — Gigantomilax kollyi;
19 — Bradybaena maacki;
20 — Caucasotachea atrolabiata;
21 — Helix vulgaris;
22 — Vallonia costata;
23 — Philomycus bilineatus;
24 — Trigonochlamys imitatrix.
Таблица 6. Двустворчатые моллюски морей СССР.
Отряд Гребенчатозубые (Taxodonta):
1 — ореховидка Ацила (Acila divaricata);
2 — леда обыкновенная (Nuculana pernula costigera);
S — йольдия северная (Yoldia hyperborea);
За — то же, вид с брюшной стороны;
4 — йольдия широкая Yoldia (Megayoldia) thraciaeformis;
5 — арка вздутая Anadara (Scapharca) broughtonii;
в — батиарка холодноводная (Bathyarca glacialis);
7 — лимопсис (Limopsis vaginatus);
8 — пектункулюс полосатый (Glycymeris albolineatus).
Отряд Связочнозубые (Dysodonta):
9 — модиола обыкновенная (Modiolus modiolus);
10 — модиола адриатическая (М. adriaticus);
11 — модиола фазеолиновая М. (Modiolula) phaseolinus;
12 — митилястер (Mytilaster lineatus);
13 — мускулюс черный (Musculus nigra);
14 — гребешок черноморский Chlamys (Flexopecten) glabra pontica;
15 — гребешок гренландский (Propeamussium groenlandicum);
16 — гребешок глубоководный Рг. (Hyalopecten) frigidus;
17 — лима северная Lima (Limatula) hyperborea;
18 — аномия гигантская Pododesmus macrochisma;
19 — аномия шиповатая Anomia (Heteranomia) squamula aculeata.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
20 — астарта крената Astarte (Astarte) crenata;
21 — астарта атлантическая рыжая А. (А.) sulcata;
22 — астарта северная A. (Tridonta) boreaiis;
23 — астарта овальная A. (Elliptica) elliptica.
Таблица 7. Морские двустворчатые моллюски:
1—Yoldia (Cnesterium) scissurata;
2 — Astarte (Nicania) montagui striata;
3 — Clinocardium californiense;
4 — Lithophaga lithophaga;
5 —Astarte (Tridonta) rollandi;
6—Chlamys farreri nipponensis;
7 —Trachycardium flavum;
8 — Cyclocardia paucicostata;
9 — Corculura cardissa;
10 — Hemicardia hemicardlum;
11 — Venus (Gircomphalus) casina;
12 — Chamelea (Clausinella) fasciata;
13 — Ensis ensis;
14 — Propeamussium (Cyclopecten)
imbriferum major;
15 — Papyri dea spinosa;
16 — Glycymeris glycymeris;
17 — Mya truncata;
18 —Chlamys (Pallidum) tigerina.
Таблица 8, Двустворчатые моллюски морей СССР.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
1 — циклокардия (Cyclocardia crebricostata);
2 — исландская циприна (Cyprina islandica);
3 — сердцевидка волосатая (Clinocardium ciliatum},
4 — сердцевидка гренландская (Serripes groenlandicus);
5 — сердцевидка малоребристая (Cardium (Acanthocardia) pauci-
costatum);
6 — черноморский венус-петушок (Chione (Chamelea) gallina cor-
rugatula;
ва — то же, вид сверху;
7 — дозиния японская (Dosinia (Phacosoma) japonica);
8 — японский петушок (Venerupis (Amygdalum) japonica);
9 — тихоокеанская питария (Gallista brevisiphonata);
10 — черноморский тапес (Venerupis (Polititapes) rugata proclivis);
11 — гульдия маленькая (Gouldia minima);
12 — гомфина (Liocyma fluctuosa);
13 — мерценария стимпсона (Mercenaria stimpsoni);
14 — прототака (Protothaca euglypta);
15 — камнеточец-петрикола (Petricola lithophaga);
16 — теллина желтая (Angulus (Peronidia) lutea);
17 — теллина ангулюс (Angulus (Moerella) donacinus);
18 — гастрана хрупкая (Gastrana fragilis);
19 — известковая макома (Macoma calcarea);
20 — балтийская макома (Macoma balthica);
21 — синдесмия овальная (Abra ovata);
22 — оливковая кровянка (Sanguinolaria (Nuttallia) olivacea).
Таблица 9. Морские промысловые двустворчатые моллюски:
1 — Crenomytilus grayanus;
2 — Mytilus edulis;
3 — Saxidomus purpuratus;
4 — Ostrea edulis;
5 — Crassostrea gigas;
6 — Crassostrea virginica;
7 — Ostrea denselamellosa;
8 — Pecten maximus;
9 — Pecten (Patinopecten) yessoensis.
Таблица 10. Двустворчатые моллюски морей СССР.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia):
1 — морской черенок (Siliqua alta);
2 — мантра полосатая (Mactra sulcataria);
3 — мантра сахалинская, или белая ракушка (Spisula sachali-
nensis);
4 — мантра овальная S. (Mactromeris) voyi;
5 — мантра шизотерус (Schizothaerus keenae);
6 — саксикава арктическая, или обыкновенная (Hiatella arctica);
6а — она же, без шипов;
7 — саксикавида панопеа японская (Panope japonica);
8 — песчаная ракушка Муа (Arenomya) arenaria;
8а — то же, общий вид с телом;
9 — корбула амурская (Potamocorbula amurensis);
10 — камнеточец фолас (Pholas dactylus) — вид со спинной сторо-
ны; Юа — правая створка; 106— левая створка;
И — цирфея гребенчатая (Zirfaea crispata);
12 — цирфея Пилсбри (Z. pilsbryi);
13 — морское сверло фоладидея Pholadidea (Penitella) penita;
14 — песчаная лионзия (Lyonsia arenosa);
15 — пандора холодноводная (Pandora glacialis);
16 — трация (Thracia myopsis);
17 — морская ложечка периплома (Periploma fragilis).
Отряд Перегородчатожаберные (Septibranchia):
18 — глубоководная лионзиелла (Lyonsiella abyssicola);
19 — поромия (Poromya granulata);
20 — куспидария арктическая (Cuspidaria arctica).
Таблица 11. Двустворчатые моллюски коралловых
рифов:
1 — тридакна гигантская (Tridacna ffigas);
2 — раковина-перо пинна (Pinna muricata);
3 — устрица петуший гребень (Ostrea crista-
galli);
4 — лима шероховатая (Lima scabra);
5 — морские жемчужницы шшктады (Pinctada
martensii).
Таблица 12. Морские двустворчатые моллюски:
1 — арка полосатая (Area zebra);
2 — солемия бореальная (Solemya borealis);
3 — морской молоток (Malleus malleus);
4 — морская жемчужница (Pinctada margariti-
fera);
5 — спондилюс красивый (Spondylus pictorum);
6 — амусиум японский (Amussium japonicum);
7 — гребешок Свифта (Chlamys (Swiftopecten)
swifti);
8 — исландский гребешок (Chlamys islandi-
cus);
9 — кардита флоридская (Cardita floridana);
10 — хама чешуйчатая (Chama macerophylla);
11 — сердцевидка Фрагум (Fragum unedo);
12 — люцинида тигровая (Codakia tigerina);
13 — венерида—королевский гребень(РЦаг dione);
14 — венерида — сунетта (Sunetta meroe);
15 — теллина-фараонелла (Pharaonella perieri);
16 — донакс черноморский (Donax (Paradonax)
venustus);
17 — мидия зеленая (Mytilus viridis);
18 — куспидария кардиомия (Cardiomya goul-
diana);
19 — спинула клювовидная (Spinula calcar);
20 — черенок большой австралийский (Solen gran-
dis).
Таблица 13. Головоногие моллюски:
1 — стайка кальмаров Ommastrephes sloanei-
pacificus;
2 — осьминог Octopus vulgaris ;
3 — рбссия (Rossia glaucopis);
4 — каракатица Sepia officinalis.
Таблица 14. Головоногие моллюски:
1 — осьминог адский вампир (Vampyroteuthis
infernalis);
2 — глубоководный кальмар кранхия (Cranchia
scabra);
3 — глубоководный кальмар батотаума (Batho-
thauma lyromma);
4 — безглазый глубоководный осьминог цирро-
таума (Cirrothauma murrayi);
5 — кальмар чудесная лампа (Thaumatolampas
diodema);
6 — глубоководный блинообразный осьминог
Opistoteuthis depressa.
Таблица 15. Головоногие моллюски:
Сверху — новорожденные осьминоги;
снизу — самка осьминога; и днем и ночью
бессменно дежурит она у своих яиц,
которые висят гроздьями в глубине ниши.
Таблица 16. Осьминог, присосавшийся к скале.
Хорошо видны темные пятна растянутых
до предела хроматофоров.
Таблица 17. Современные иглокожие.
Морские лилии:
1 — Metacrinus nobilis.
Голотурии:
3 — Cucumaria japonica;
4 — Trochostoma arcticum.
Морские звезды:
2 — Ceramaster patagonicus;
7 — Asterias forbesi.
Морские €!жи:
5 — Rotula orbiculus;
9 — Stylocidaris affinis.
Офиуры:
6 — Gorgonocephalus caryi;
8 — Ophiura sarsi.
Таблица 18. Иглокожие тропических мелководий.
Морские лилии:
1 — Comatella stelligera;
2 — Pterometra pulcherrima.
Голотурии:
4 — Brandtothuria arenicola;
7 — Stichopus chloronotus;
10 — Ludwigothuria atra.
Морские звезды:
5 — Linckia laevigata;
11 — Oreaster nodosus.
Морские ежи:
6 — Heterocentrotus mammillatus;
8 — Colobocentrotus atratus.
О ф и у р ы:
3 — Ophiotrix coerulea;
9 — Ophiomastix annulosa.
Таблица 19. Иглокожие тропических мелководий.
Морские лилии:
1 — Stephanometra spicata;
2 — Lamprometra protectus
Голотурии:
4 — Holothuria scabra;
7 — Thyone mirabilis;
10 — Cucumaria tricolor;
11 — Colochirus luteus.
Морские звезды:
5 — Acanthaster planci;
6 — Luidia quinaria;
12 — Culcita novae-guineae.
Морские ежи:
8 — Toxopneustes pileolus;
9 — Phyllacanthus imperialis;
13 — Diadema setosum.
О ф и у р а:
3 — OpHiothrichoides pulcherrima.
Таблица 20. Иглокожие северных морей СССР.
Морская лилия:
1 — Heliometra glacialis.
Морские звезды:
2 — Asterias rubens;
3 — Hippasteria phrygiana;
4 — Urasterias lincki;
7 — Ctenodiscus crispatus;
9 — Henricia sanguinolenta.
Морской еж:
8 — Strongylocentrotus
droebachiensis.
О ф и у р ы:
5 — Ophiopleura borealis;
6 — Ohiopholis aculeata.
Таблица 21. Забота о потомстве у различных видов
иглокожих:
1 — морская звезда Pteraster obsurus с молодью,
вылезающей через покровы;
2 — морская звезда Trop ho discus uber с мо-
лодью на спине;
3 — самка Abatus philippii с маленькими ежами
на спине;
4 — морской еж Ctenocidaris nutrix с малень-
кими ежами вокруг рта;
5 — голотурия Cladodactyla сгосеа с молодью
на спине;
G — голотурия Psolus ephippifer с крупными
пластинками на спине, прикрывающими
выводковые камеры.
Таблица 22. Иглокожие больших глубин Тихого океана*
Морская лилия:
1 — Bathycrinus pacificus.
Голотурии:
2 — Paelopatides solea;
3 — Psychropotes raripes;
4 — Lpsilothuria bitentaculata;
5 — Scotoplanes murrayi;
6 — Elpidia sp.
Морские звезды:
7 — Astrocles actinodetus;
8 — Styracaster horridus.
Морские ежи:
9 — Aerop^is fulva;
10 — Pourtalesia tannerL
Таблица 23. Различные представители класса голотурий:
1 — Cucumaria elongata;
2 — Colochirus quadrangularis;
3 — Psolus phantapus;
4 — Rhopalodinopsis capensis;
5 — Pelagothuria natatrix;
6 — Psychropotes kerhervei;
7 — Pannychia moseleyi;
8 — Peniagone lugubris;
9 — Oneirophanta alternata;
10 — Molpadia musculus;
11 — Synaptula hydriformis.
Таблица 24. Современные морские ежи и офиуры.
Морские ежи:
1 — Acanthocidaris maculicollis;
2 — Salenocidaris miliaris;
3 — Arbacia spatuligera;
4 — Echinosigra paradoxa.
О ф и у р ы:
5 — Astrochlamys bruneus;
6 — Ophiocoma delicata;
7 — Ophiura multispina.
Таблица 25. Иглокожие северной части Тихого океана.
Морские звезды:
1 — Pycnopodia heliantoides;
2 — Strongylocentrotus nudus;
3 — Crossaster papposus;
4 — Distolasterias nipon;
5 — Evasterias retifera;
6 — Patiria pectinifera.
Офиуры:
7 — Strongylocentrotus purpuratus;
8 — Amphiodia rossica;
9 — Psolus chitonoides.
Морские ежи:
10 — Ophiacantha bidentata;
11 — Brisaster townsendi;
12 — Stichopus Japonicus .
Голотурии:
13 — Astrometis sertulifera;
14 — Amphiophiura ponderosa.
Таблица 26. Охота морской звезды (Asterias vulgaris) за моллюсками (Pecten jacobeus).
Таблица 27. Иглокожие мелководий Японского моря.
Сверху — морской еж;
снизу — морская звезда Patiria pectinifera.
Таблица 28. Пелагические оболочники:
1 — колония сальп Thalia sp.;
2 — Salpa maxima (агрегатная форма);
3 — бочоночник Doliolum rarum;
4,5 — Pyrosoma sp.;
6 — Thalia democratica;
7 — колония сальп Cyclosalpa bakeri;
8 — Salpa maxima (одиночная форма).
Таблица 29. Оболочники,
Одиночные а с ц и д и е;
1 — Phallusia mammillata;
2 — Halocynthia roretzi;
4 — Styela clavata;
5 — Ascidia virginea;
6 — Boltenia ovifera;
8 — Distomus variolosus;
9 — Boltenia echinata;
11 — Clavelina lepadiformis;
14 — Halocynthia aurantiura;
16 — Ciona intestinalis;
17 — Chelyosoma macleayanum;
18 — Pyura setosa;
19 — Pyura discoveryi;
20 — Styela plicata.
Колониальные асцидии:
3 — Gynandrocarpa placenta;
7 — Sarcobotrylloides racemosum;
10 — Ecteinascidia tortugensis;
12 — Sycozoa umbellata;
13, 15 — Botryllus schlossery.
Таблица 30. Ветвистоусые ракообразные:
1 — Daphnia pulex;
2 — Bythotrephes longimana;
3 — Diaphanosoma brachyurum;
4 — Bosmina longirostris;
5 — Evadne nordmanni;
6 — Cercopagis socialis;
7 — Chydorus sphaericus;
8 — Polyphemus pediculus;
9 — Camptoccrcus rectirostris;
10 — Sida crystallina;
11 — Macrothrix hirsuticornis;
12 — Ceriodaphnia reticulata;
13 — Moina rectirostris;
14 — Leptodora kindtii;
15 — Simocephalus vetulus;
16 — Holopedium gibberum;
17 — метанауплиус Leptodora kindtii}
18 — Caspievadne maximovitchi.
Таблица 31. Веслоногие ракообразные:
1 — Sapphirina ovatolanceolata;
2 — Oithona plumifera;
3 — Calanus finmarchicus;
4 — Oncaea mediterranea;
5 — Phoenna spinifera;
6 — Calocalanus pavo;
7 — Sapphirina vorax;
8 — Corycaeus obtusus;
9 — Euchirella maxima;
10 — Pontella mediterranea;
11 — Copilia vitrea;
12 — Calanus plumchrus.
Т а б л и ца 32. Усоногие ракообразные:
1 — Lepas anatifera;
2 — L. fascicularis;
3 — Verruca stromia;
4 — Mitella mitella;
5 — Conchoderma auritum на Coronula dia-
dema;
6 — Scalpellum Japonicum;
7 — Balanus improvisus;
8 — B. eyermanni;
9 — B, balanus;
10 — Chthamalus stellatus;
H — Xenobalanus globicipitis;
12 — Cryptolepas rachianecti.
Таблица 33. Равноногие ракообразные, или изоподы:
1, 2 — Gnathia dentata, самец и самка;
3 _ Asellus aquaticus;
4 — Neophreatoicus assimilis;
5 — Mesidotea entomon;
6 — Aega psora;
7 _ Cyathura carinata;
8 — Haplomesus insignis orientalis;
9 — Serolis sp.;
10 — Bathynomus giganteus;
11 — Munnopsis typica;
12 — Storthyngura herculea:
13 — Microcharon sp.;
14 —. Porcellio scaber;
15 _ Sphaeroma serratum;
16 — Hemilepistus cristatus;
17 _ Antarcturus ultraabyssalis.
Таблица 34. Разноногие ракообразные, или бокоплавы:
1 — Brandtia parasitica;
2 — Niphargoides maeoticus;
3 — Axelboeckia spinosa;
4 — Eulimnogammarus virgatus;
5 — Pallasea bicornis;
6 — Panoploea eblanae;
7 — Acanthogammarus godlewskii;
8 — Leucothoe richiardii;
9 — Grubia crassicornis;
10 — Lysianassa punctata;
11 — Biancolina algicola;
12 — Ampelisca diadema;
13 — Cyphocaris richardi;
14 — Megaluropus agilis;
15 — Dexamine spinosa-
Таблица 35. Креветки:
1 — Pandalus latirostris;
2 — Palaemon elegans;
3 — Alpheus brevicristata;
4 — Sclerocrangon salebrosa;
5 — Lebbeus sp.;
6 — Hetairus groenlandicus;
7 — Penaeus japonicus;
8 — Notostomus sp.;
9 — Macrobrachium sp.;
10, 11 — Hippolyte varians, различно
окрашенные;
12 — Troglocaris anophthalmias;
13 — Stenopus hispidus.
Таблица 36. Крабы:
1 — Callinectes sapidus;
2 — Pachygrapsus marmoratus;
3 — Pugettia quadrifrons;
4 — Carcinus maenas;
5 — Hyas coarctatus;
6 — Chionoecetes opilio;
7 — Cancer pagurus;
8 — Ma]a squinado;
9 — Calappa sp.;
10 — Randallia eburnea;
11 — Lyreidus tridentatus;
12 — Lambrus validus;
13 — Dromia vulgaris;
14 — Pinnoteres boninensis.
Таблица 37. Камчатские крабы (Paralithodes camtscha-
tica) на дне Охотского моря.
Таблица 38. Омар Homarus gammarus и лангуст
Palinurus vulgaris.
Таблица 39. Личинки десятиногих ракообразных:
1 — зоэа краба Rhithropanopeus;
2 — глаукотоэ Pagurus bernhardus;
3 — личинка Porcellana longicornis;
4 — эрионеикус Eryonidae,
5 — зоэа Palaemon longirostris;
6 — поздняя личинка Sergestes cornutus;
7 — филлозома лангуста Panulirus inflatus;
8 — мегалопа краба Hyas araneus;
9 — мизисная стадия Nephrops norvegicus;
10, 11 — глаукотоэ и зоэа камчатского краба
(Paralithodes camtschatica);
12 — науплиус Trachypenaeus constrictus.
Таблица 40. Пальмовый вор (Birgus latro)
и его жизненный цикл:
1 — 5 — последовательные стадии
развития;
6 — взрослая особь.